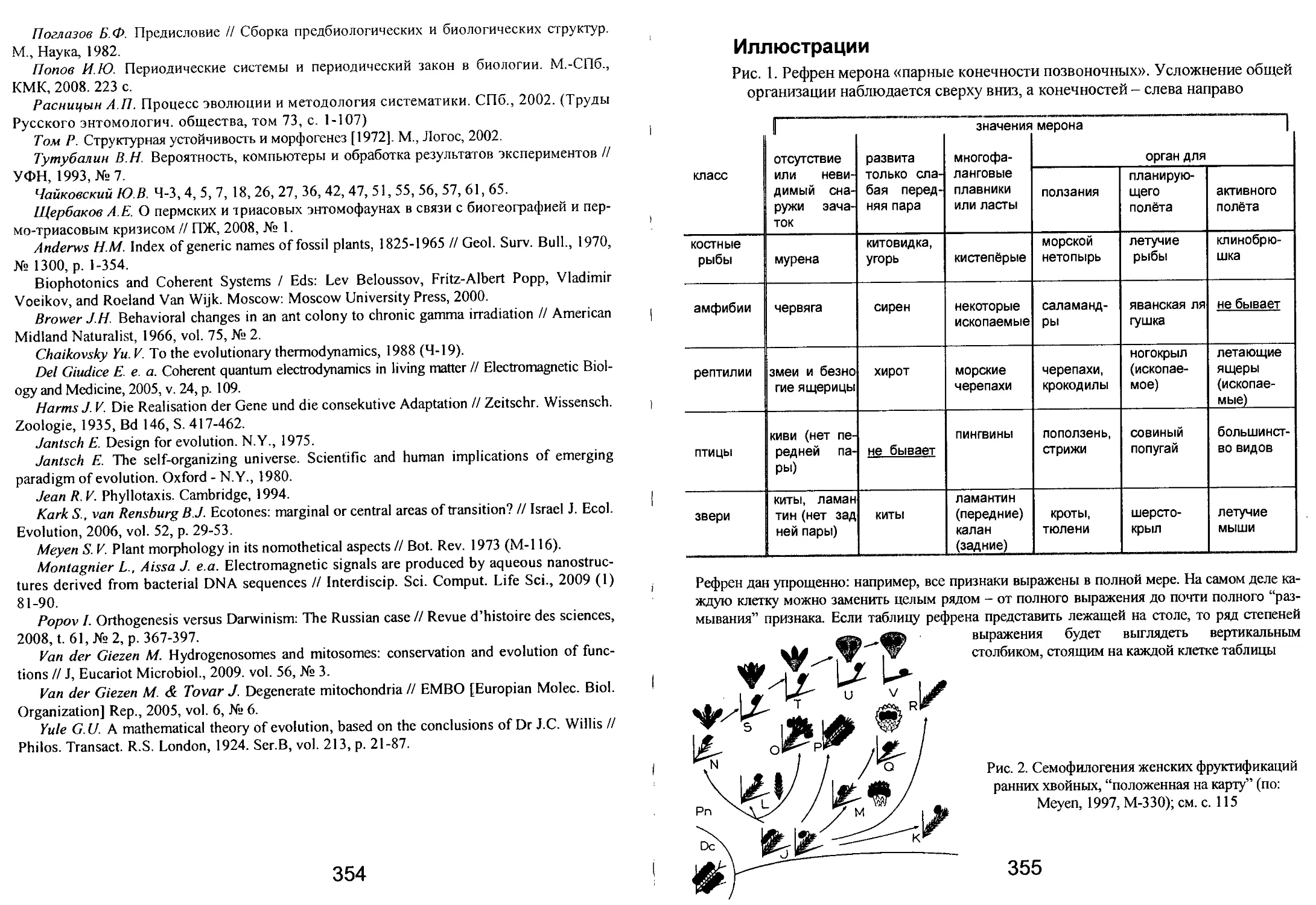
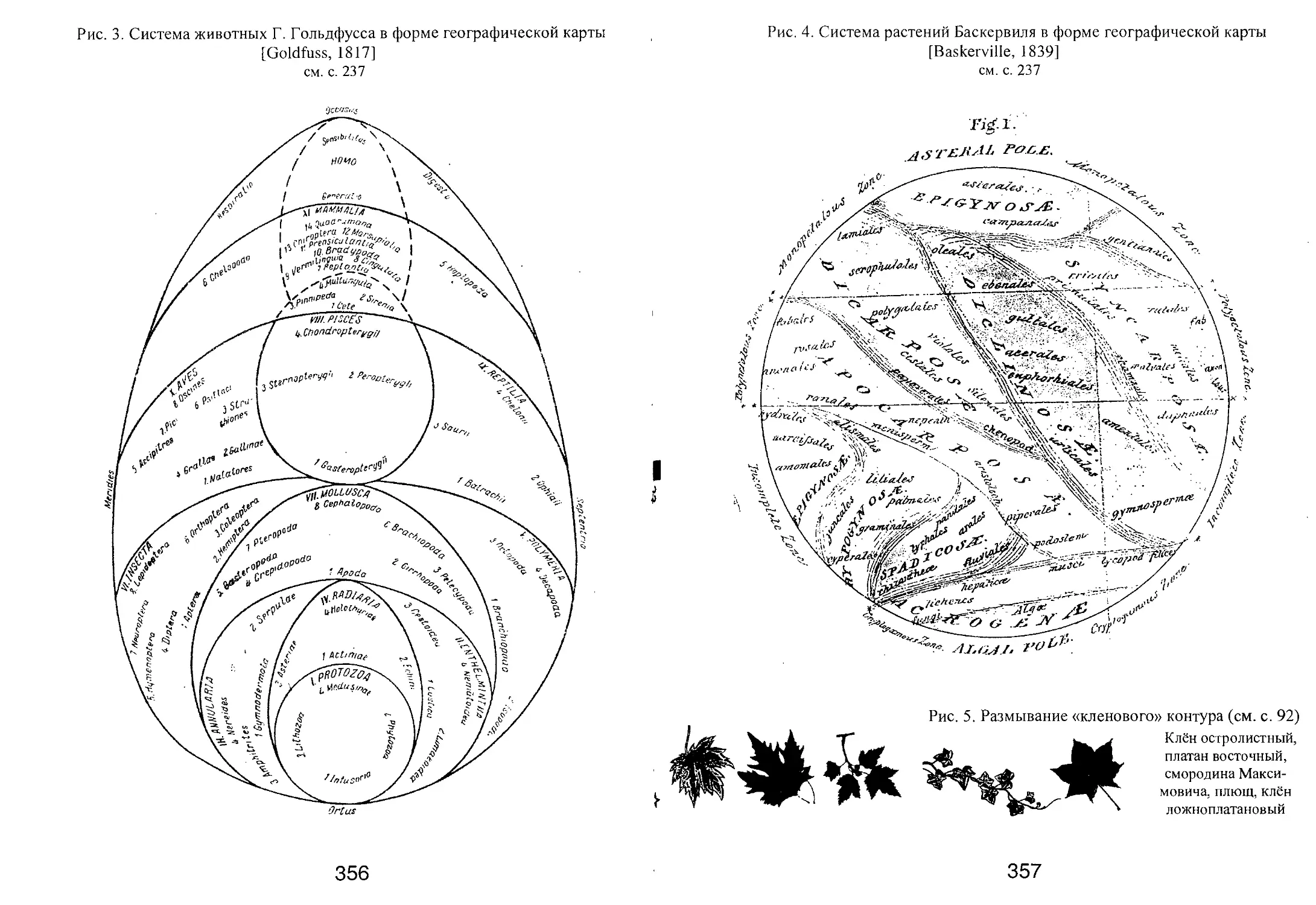
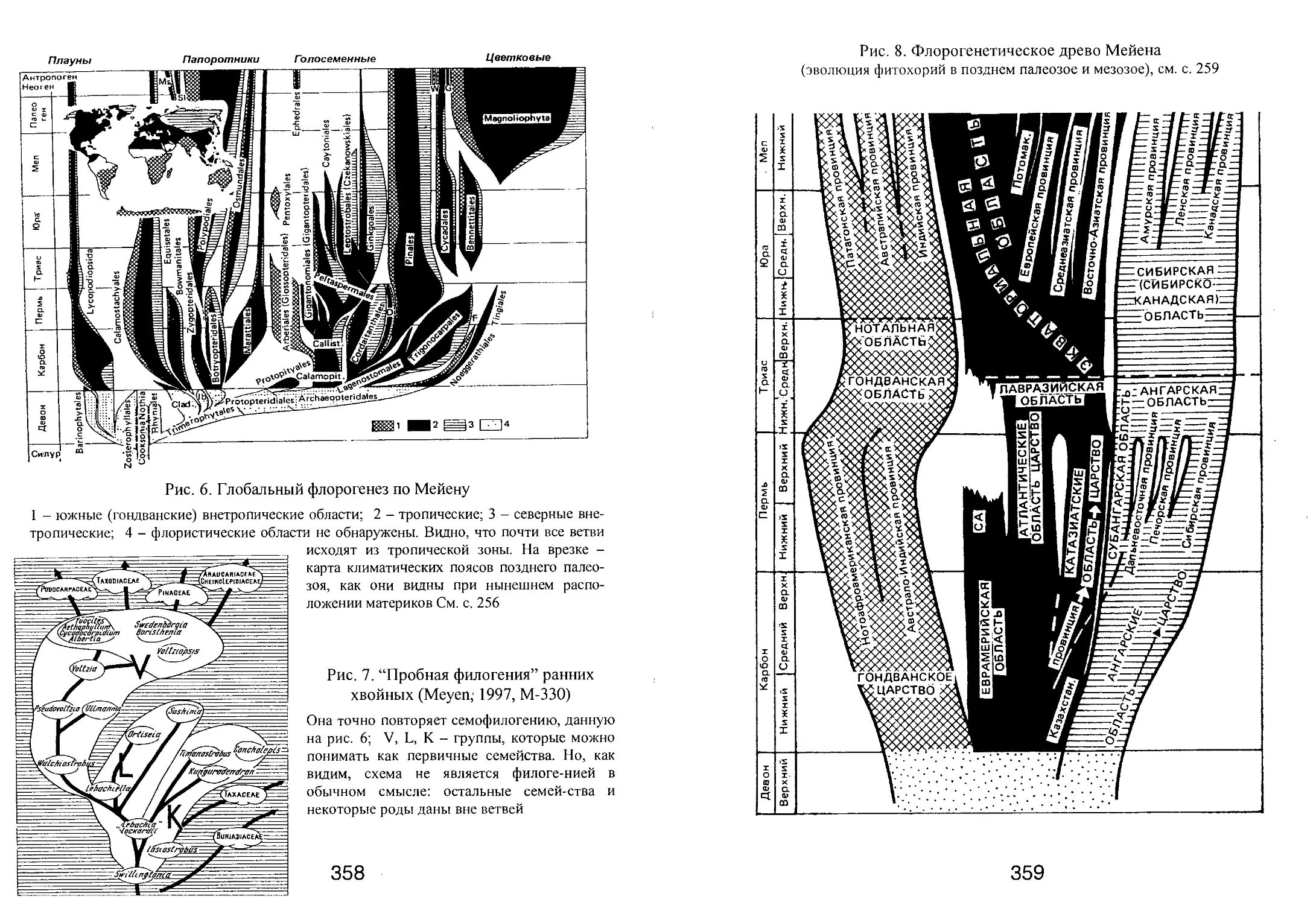
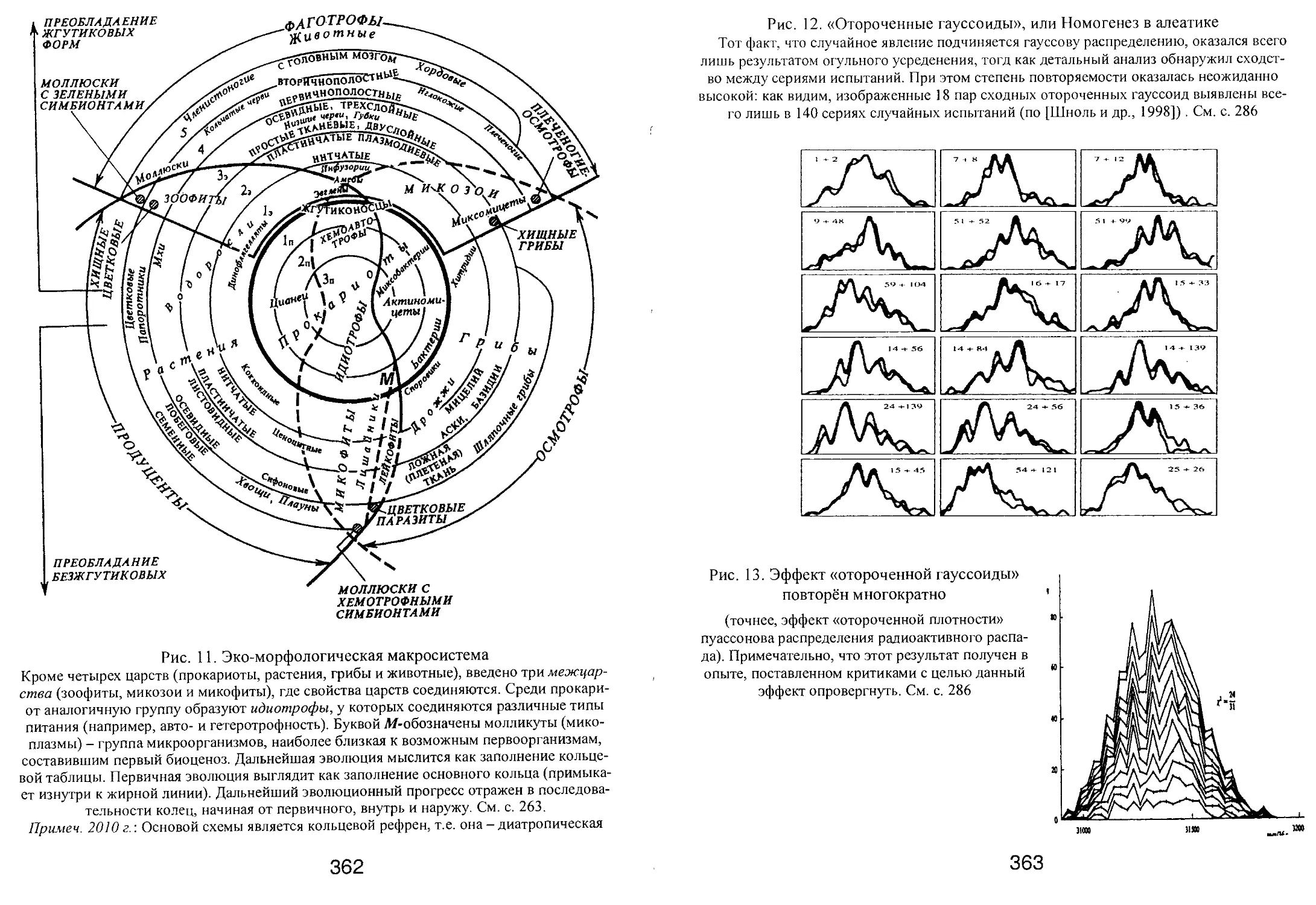
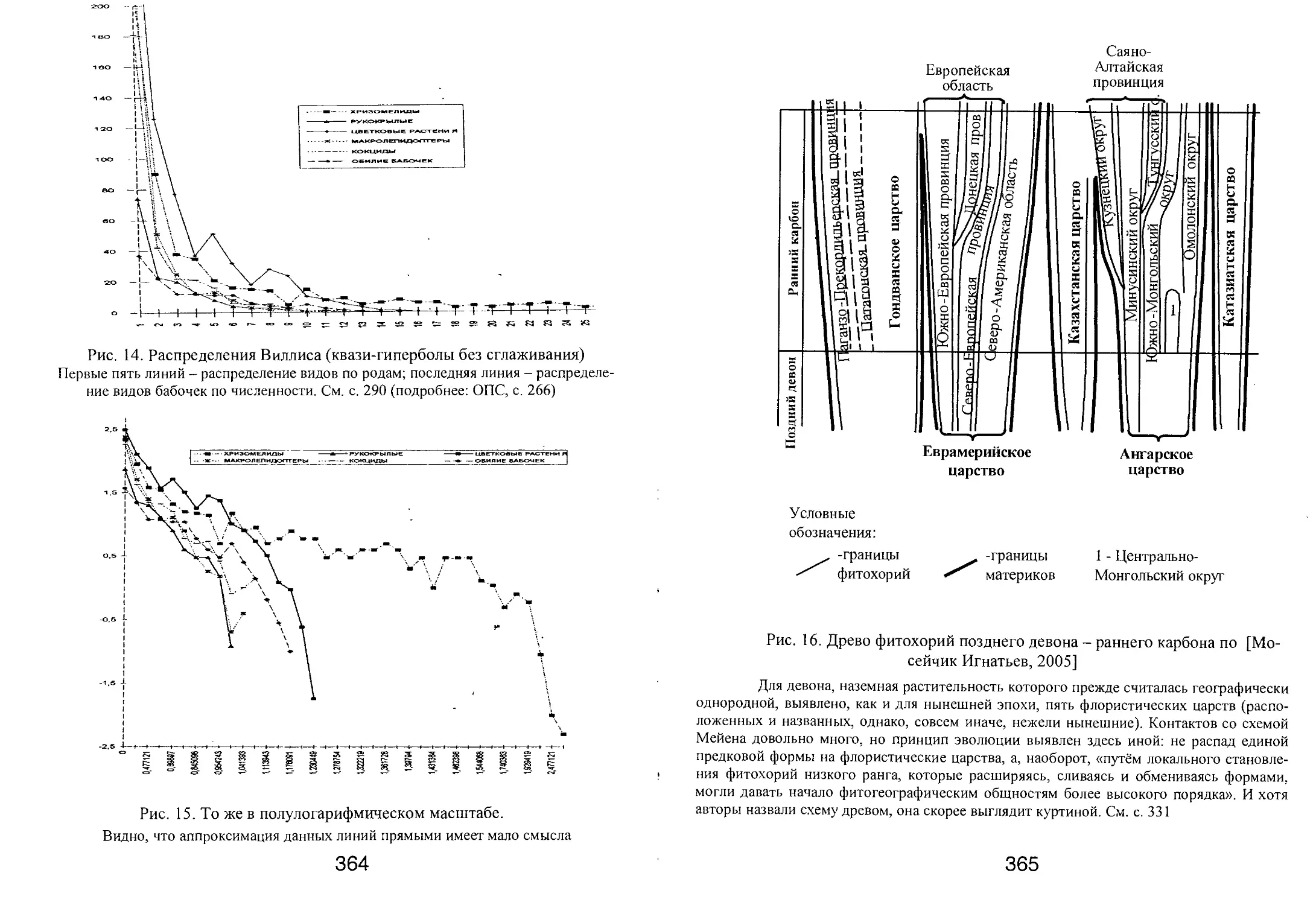
/









































































































































































































Автор: Чайковский Ю.В.
Теги: биологические науки в целом история естественных наук биология
ISBN: 978-5-87317-695-3
Год: 2010




Текст
Ю. В. Чайковский
Диатропика, эволюция
и систематика
К юбилею Мейена
Сергей Викторович Мейен (1935-1985)
Ю.В. Чайковский
Диатропика, эволюция
и систематика
К юбилею Мейена
Сборник статей
С приложением полного списка трудов
С.В. Мейена,
составленного И.А. Игнатьевым
Товарищество научных изданий КМК
Москва ❖ 2010
УДК 57(09)
ББК 20г
Ч 15
Чайковский Ю.В. Диатропика, эволюция и систематика. К юбилею Мей-
ена. Сборник статей. С приложением полного списка трудов С.В. Мейена
(составил И.А. Игнатьев). Москва: Товарищество научных изданий КМК.
2010. 407 с.
Сборник, приуроченный к 75-летию С.В. Мейена (1935-1987), палеобо-
таника, эволюциониста и философа, содержит работы Ю.В. Чайковско-
го, историка науки и эволюциониста, дружившего с Мейеном. Они по-
священы тем проблемам теоретической биологии, которые более всего
занимали Мейена. Заключительная статья написана специально для дан-
ного сборника и подводит итог работам этого круга. Остальные воспро-
изводимые здесь (целиком или частично) работы Чайковского публико-
вались прежде в различных изданиях, многие из которых в настоящее
время мало кому доступны. Для данного издания они снабжены новыми
комментариями.
На передней обложке:
Собственноручный рисунок С.В. Мейена, объясняющий гипотезу
преобразования фруктификаций беннетитовых в цветки
покрытосеменных. Из статьи: «Гипотеза происхождения
покрытосеменных от беннеттитов...» (М-286).
Перенос признаков с мужского пола на женский показан стрелкой;
синангии (сросшиеся спорангии) и пыльники показаны крапом;
семязачатки — черными кружочками, элементы обвёртки или
околоцветника заштрихованы. Подробнее см. заключительную статью.
На задней обложке:
Четырехцарственная диатропическая макросистема
(по 4-15, с уточнениями), в основе которой лежит кольцевой рефрен.
Подробнее см. с. 263
© Ю.В. Чайковский, текст, иллюст-
рации, 2010
© Товарищество научных изданий
ISBN 978-5-87317-695-3 КМК, издание, 2010
ОГЛАВЛЕНИЕ
Обозначения 7
Предисловие 8
ЧАСТЬ 1. О ЗАКОНАХ РАЗНООБРАЗИЯ 10
Многотрудный поиск многоликой истины 10
1. Разнообразие мнений и разнообразие явлений 10
2. Физика - образец науки? 12
3. Бывает ли вообще лженаука? 13
4. Наука не движется прямо 16
5. Прогресс науки - не в «охоте на ведьм», а в самокритике 18
Грамматика биологии 20
Параллелизмы и систематика 20
Как работает линнеевская система 21
«Свет без двойства являться не может» 23
Как должна работать типологическая система 26
Параллелизмы и «грамматика биологии» 28
Литература 32
Наука о разнообразии 33
Диатропика. Суждение о новизне 33
Тесный мир логики 35
Ядро и периферия 38
Разнообразие тоже наследуется 39
Берегите несоответствия! 40
ЧАСТЬ 2. ЭВОЛЮЦИЯ, ДИАТРОПИКА И НОВЫЙ НОМОГЕНЕЗ 42
Комбинативность. От Парацельса до школы Вавилова 42
Литература 46
Разнообразие, ряды и тенденции 47
2.1. О биологической грамматике 47
2.2. Мероно-таксономический анализ (типология по Мейену) 52
2.3. Ядро и периферия 57
2.4. Параллельные ряды 63
2.5. Другие тенденции 69
2.6. Гомология и аналогия взаимодополнительны 76
Литература 79
Новые тенденции в теории эволюции 81
Любищев, Мейен и плюрализм в эволюции 83
Литература 84
С.В. Мейен и теория биологической эволюции 85
Мейен как эволюционист и натурфилософ 101
3
Эволюция: с чем входим в XXI век? 102
1. Отношение к эволюции меняется 102
2. Начало жизни 105
3. Усложнение жизни 107
4. Механизмы эволюции 108
Феномен эволюции организмов в понимании С.В. Мейена ПО
1. Наука и идеология 110
2. Дарвинизм и марксизм 111
3. Место идей Мейена в эволюционизме 112
4. Мейен и эволюционная систематика 114
5. Флорогенез как свидетельство реальности эволюции 115
Литература 116
ЧАСТЬ 3. СОЦИАЛЬНАЯ ДИАТРОПИКА 117
Молодежь в разнообразном мире 117
Что такое разнообразие? 118
Ядро и периферия 121
Феникс разнообразия 122
Варианты ограничены. Рефрен Мейена 124
Место молодежи в обществе 125
Литература 129
Горизонт познания и познавательные модели 130
Г оризонт познания 131
Познавательные модели 134
Контуры будущей познавательной модели 137
Модели и усвоение нового 138
Ряды и модели 139
Историческое событие как пересечение рядов 143
Исключения уточняют правило 145
Поле предпочтений 147
Литература 149
ЧАСТЬ 4. ЮБИЛЕЙ ЛАМАРКА И ДАРВИНА 151
Дарвинизм, “Эволюция не по Дарвину” и близкий юбилей 151
1. В ожидании юбилея 151
2. Вера и наука могут меняться местами 153
3. Становление дарвинизма в свете юбилея 156
4. А что тогда было кроме дарвинизма? • 160
Литература 162
Пятьсот лет споров об эволюции 163
Вечно актуальная тема 163
1509. Европа открывает идею эволюции 165
1559. Идея эволюции начинает работать 166
1609. Активность и изменчивость 166
1659. Человек как модель эволюции 166
1709. Экологический взгляд на мир 167
1759. Эмбриология и эволюция 168
4
1809. Рождение ламаркизма 168
1859. Великое событие и его философские оценки 168
1909. Юбилей дарвинизма и рождение новой религии 171
1934. Малый юбилей. Гуманитарии об эволюции 172
1959. Столетие дарвинизма и рождение новых дисциплин 173
1979 Малый юбилей и большие ожидания 176
2009. Напрасные споры с креационизмом. Новое в теории эволюции 179
Литература 181
Ламарк, Дарвин и устройство науки 183
1. Ламарк и Дарвин 183
2. Учения Эразмовы и бэровы 185
3. Место отбора 189
4. О структуре науки и парадигмах 190
5. Нормальные и маргинальные компоненты эволюционизма 192
6. Что сказали методологи 194
7. К построению теории 197
Литература 199
ЧАСТЬ 5. ПРОИСХОЖДЕНИЕ ЧЕЛОВЕКА 201
Двускоростной механизм размножения и его роль в эволюции человека 202
Черты антропогенеза 202
Пример глубокой старины 204
Чисто биологические вопросы людской эволюции 206
Литература 209
Происхоадение человека в людском муравейнике 210
Если бы не штампы 213
Литература 214
ЧАСТЬ 6. ЭВОЛЮЦИЯ И СИСТЕМАТИКА 215
Зачем систематикам теория эволюции? 215
1. Является ли систематика эволюционной? 215
2. Дарвинизм и номогенез в систематике 218
3. В чем состоит использование систематики? 220
4. Как же строить систематику? 221
Литература 223
Теория систематики по Мейену 224
Табличные системы 225
Литература 226
Естественная система и таксономические названия 227
1. Названия неустойчивы 228
2. Названия не должны быть частью системы 231
3. Бытовые названия и народная таксономия 232
4. Искусственная и естественная системы организмов 235
5. Систематика и эволюционная идея. Тезис Короны 238
6. Почему Линней не построил естественной системы? 240
7. Различные понимания естественной системы 242
5
8. Место естественной системы в систематике 246
9. Является ли нынешняя систематика эволюционной? 251
10. Номогенез в систематике 253
11. Опыт грубой филогении (Мейен) 255
12. Опыт двух систем (бактериологи) 260
13. Опыт естественной макросистемы (Чайковский) 262
14. Опыт постоянных названий (Клюге) 263
15. Итог: третья система 264
Литература 266
ЧАСТЬ 7. ДИАТРОПИКА СЛУЧАЙНОГО 269
Разнообразие и случайность 269
3.1. Случайность и закономерность 269
3.2. Номотетико-идиографическое чередование 274
Случайность и самоорганизация 278
4-3. Рулетка Пуанкаре и вероятность как мера 279
4-4. От рулетки Пуанкаре к странным аттракторам 280
Организующая роль случайности 281
О математических моделях случайного 281
Случайность в игре 283
Случайность и тенденции 285
Эффект Шноля и диатропика случайности 286
Дарвинизм, ламаркизм и номогенез о случайности 289
Общие замечания 291
Литература к части 7 293
ЧАСТЬ 8. ЛИЧНОЕ 294
Серёжа Мейен, мой сосед 294
Некролог 304
ЧАСТЬ 9. СУДЬБА КОНЦЕПЦИИ 308
Диатропика С.В. Мейена: сегодняшний взгляд 308
1. Связь диатропики с типологией. Рефрен 309
2. Состав диатропики и ее практический смысл 311
3. Диатропика и эволюция. Филогения по Мейену 313
4. Диатропика и эволюция. Рефрены и блоки 314
5. Причина рефренов 316
Литература 317
Практичная теория (заключительная статья) 318
1. Эдванты 320
2. Практические достижения диатропики... 322
3. ...почти никому не известны 326
4. О флорогенезе 330
5. Три кита 334
6. О параллелизме бытия и мышления и об эффективности математики 337
7. О структуре пространства и природе рефрена 341
6
8. Место в общей теории 346
9. О диатропической картине мира 351
Литература 353
ИЛЛЮСТРАЦИИ 355
ПРИЛОЖЕНИЯ И ЛИТЕРАТУРА 366
1. С.В. Мейен. Легче вывести человека из современной свиньи 366
Пояснения к публикации 368
2. В.А. Брынцев. Рецензия 371
Пояснения к рецензии 374
3. Р.Г. Баранцев. Мини-реферат 376
Общий список литературы к сборнику 378
Список упомянутых в сборнике работ автора 388
Полный список опубликованных трудов С.В. Мейена 391
Обозначения
автогенез - эволюция, независимая от изменений среды (одна из форм номогенеза)
актиреф - активное движение вдоль рефрена (см. с. 96 и п. 7 заключит, статьи)
алеатика - общая наука о случайности (см.: «Организующая роль случайности»)
АСМ- Активный связный мир, 2008 (см. список работ автора)
ВИЕТ - Вопросы истории естествознания и техники (журнал, Москва)
ВФ - Вопросы философии (журнал, Москва)
ГИН - Геологический институт РАН (прежде АН СССР), Москва (место работы
С.В. Мейена)
диасеть - диатропическая сеть (рефренная структура разнообразия), с. 8, 187
ЖОБ - Журнал общей биологии (Москва)
ЗЭ - Зигзаги эволюции, 2010 (см. список работ автора)
ИНЕТ - Институт истории естествознания и техники РАН (прежде АН СССР), Мо-
сква (место работы автора)
ИМ - Игнатьев И. А., Мосейчик Ю.В. Творческая случайность... (см. Общий список)
ЛЧ - Любищевские чтения (ежегодник, Ульяновск)
МОИП - Московское общество испытателей природы
МСП - Материалы симпозиума, посвященного... (см. Общий список лит-ры)
ОПС - О природе случайности, 2004 (см. список работ автора)
ортогенез - эволюция в одном направлении (одна из форм номогенеза)
ПЖ - Палеонтологический журнал (Москва)
ПМ - познавательная модель, с. 134
РАН - Российская Академия наук
ТВ - теория вероятностей (наиболее разработанный раздел алеатики)
транпол - транзитивный полиморфизм - с. 39, 69, 81, 95, 100, 121, 200, 304, 310
ТСО - теория стабилизирующего отбора, с. 185
УФН - Успехи физических наук (журнал, Москва)
ЭКЭ - экосистемная концепция эволюции, с. 189
ЭЭД - Элементы эволюционной диатропики, 1990 (см. список работ автора)
7
Предисловие
Сборник приурочен к 75-летию со дня рождения Сергея Викторовича Мей-
ена и составлен из моих статей и отрывков, прямо касающихся его идей и их
продолжения. В основном, это темы, которые после его безвременной кончины
были собраны мной под именем диатропики.
Есть много наук, изучающих разнообразие: химия изучает разнообразие ве-
ществ, биология - организмов, лингвистика - языков и т. д. Для всех них разно-
образие - один из аспектов исследования. Наоборот, для диатропики разнообразие
представляет единственный объект исследования, зато интерес ее - не в описании
самих различий между предметами, а в выявлении и формулировке закономерно-
стей каждого разнообразия и в поисках того, что нескольким или даже всем раз-
нообразиям обще. Это, прежде всего, рефренная структура разнообразий, на ко-
торую первым указал, насколько знаю, Мейен. Позже вместо этого длинного тер-
мина мной был предложен короткий - диасеть (диатропическая сеть).
Первичный материал и понятийный аппарат Мейену дал номогенез - учение о
биологической эволюции, созданное в 1913-1928 гг. Д.Н. Соболевым, Л.С. Бергом
и, отчасти, Н.П. Вавиловым и А.А. Любищевым. После работ Мейена можно го-
ворить о новом номогенезе, каковой и стал основной темой данного Сборника. От
прежнего номогенеза он отличен тем, что включил в себя диатропику.
Диатропика отнюдь не является теорией ни эволюции, ни систематики, она,
как мне видится, призвана быть одной из основ для обеих этих дисциплин. К
ней непосредственно примыкают этические взгляды Мейена, без которых, по-
моему, диатропика не могла бы родиться. Поэтому мне представилось естест-
венным начать сборник со статьи о поиске истины.
Сама по себе диатропика составляла лишь небольшую часть обширной об-
ласти интересов Мейена, поэтому в сборнике довольно мало ссылок на работы
его коллег, однако данные работы представляются мне важными для понимания
тематики сборника. Укажу прежде всего схему флорогенеза, которую наметил
Всеволод Андреевич Вахрамеев (руководитель лаборатории), частично реализо-
вал, пользуясь данными лаборатории, Мейен (возглавивший после него лабора-
торию), а завершили уже после его смерти. Это были Михаил Алексеевич Ах-
метьев, возглавивший лабораторию после Мейена, а также (в несколько ином
смысле) Игорь Анатольевич Игнатьев, близкий ученик Мейена, и его ученица
Юлия Владимировна Мосейчик. О них пойдет речь в заключительной статье.
Отмечу также работу по обоснованию гипотезы Мейена о происхождении
покрытосеменных от беннетитов, которую провел Алексей Борисович Герман,
тоже близкий ученик Мейена, руководящий той же лабораторией ныне. Об этом
тоже скажу в заключительной статье. Он любезно предоставил мне в 2005 году
презентацию своей работы, из которой, между прочим, взята редко публикуемая
замечательная фотография Мейена, помешенная перед титулом сборника.
Исключений (выходов за рамки диатропики) в сборнике всего три. Первые
два - статьи «Многотрудный поиск многоликой истины» и «Дарвинизм, “Эволю-
8
ция не по Дарвину”...», где речи о диатропике нет. Однако их темы не только бы-
ли чрезвычайно важны для самого Мейена, но и представляются мне прямо-таки
необходимыми для понимания основного смысла других статей Сборника. Это -
параллель дарвинизма с марксизмом, уверенность, что важнее знать эволюцио-
низм до Дарвина, нежели споры вокруг дарвинизма, и, главное, этика учёного.
Третье исключение - список трудов Мейена (который составил Игнатьев) -
он дан целиком. Ссылки на него снабжены в Сборнике буквой «М», предваряю-
щей номер работы Мейена. В этом списке на 69 названий больше, чем в преды-
дущем (помещенном в книге М-318), а также указаны переиздания.
Аналогично, ссылки на список моих работ предваряются буквой «Ч».
Заключительная статья написана специально для Сборника и призвана подвес-
ти итог 40-летнего1 развития диатропики, дабы показать, что теория давно и успеш-
но работает. Остальные статьи уже были опубликованы прежде и здесь воспро-
изведены с восстановлением редакционных купюр и снятием той редакторской
правки, которая была внесена без моего ведома. Исправлены явные ошибки (в
основном, в датах и списках литературы) а также опущены абзацы и отдельные
фразы, оказавшиеся повторами или малоосмысленые. Эти купюры отмечены от-
точиями в квадратных скобках. Что касается новых фактов и суждений, то они,
в случае необходимости, приводятся в форме сносок, с обозначением года.
Все рисунки помещены в конце книги, чтобы избежать их повторения.
В качестве приложения публикуются отрывки из статьи Мейена «Врачу, исце-
лися сам», выпавшие из публикации М-162. Название подборке дано (по фразе из
отрывка) мной. Она, на мой взгляд, приоткрывает ту обстановку, в которой Сергей
Викторович почти всю жизнь работал. А именно, из нее видно, каковы были его
«непричёсанные мысли» и как их «причёсывали» даже благожелательные редакто-
ры. Статья, в печатном виде явно страдающая конформизмом, вышла из-под пера
автора, как оказывается, вовсе не ориентированной на какого-либо цензора.
В виде приложений даны также рецензия на мою книгу «Активный связный
мир» (ее написал теоретик ботаники Владимир Альбертович Брынцев) и краткий
реферат этой книги (его составил и передал мне Рэм Георгиевич Баранцев, близкий
друг Мейена и давний глава питерского клана любищевцев). В них затронута, в ос-
новном, лишь натурфилософская сторона книги (а не эволюция живого как тако-
вая), зато понять эту сторону тут легче, чем по самой книге. При этом роль диатро-
пики в становлении нового понимания эволюции в них вполне выявлена.
Обоим, рецензенту, и референту, приношу глубокую признательность. Игнать-
ев и Мосейчик очень помогли советами, и им я также рад выразить благодарность.
1 Если начинать отсчет со статьи «Корова и сено. К общему предку» (М-73), где , по сути,
Мейеном уже были введены рефрен (на примере параллельной эволюции клетки в разных
царствах) и блочность (на примере феномена многоклеточное™) - основные понятия диа-
тропики, а с тем и нового номогенеза.
9
Часть 1. О законах разнообразия
Многотрудный поиск многоликой истины'
Еще недавно отношение к методологии естествознания вполне резюмирова-
лось двумя расхожими афоризмами - немецким: «Кто не способен ни на что
путное, берется за методологию» и русским: «Кто умеет, тот делает, кто не уме-
ет, тот учит, как делать». Со студенческих пор страдал подобным скептицизмом
и автор этих строк, так что однажды, обнаружив, что сам давно занимается ме-
тодологией, был крайне удивлен.
1. Разнообразие мнений и разнообразие явлений
«Живущие в настоящее время бактерии являются, возможно, реликтовыми
формами, уцелевшими до наших дней от раннего периода биологической эволю-
ции. Они либо сохраняют старый образ жизни в каких-либо крайних условиях,
либо нашли для себя новые экологические ниши». Это - заключительный абзац
прекрасного учебника «Общая микробиология» Ганса Шлегеля (Москва, 1972).
Автор - знаток микробов и в своем труде ясно показывает, что бактерии в
целом - никак не изгои жизни, что они занимают в балансе природы ряд ключе-
вых пунктов, причем эти пункты не «крайние» и не «новые», а, наоборот, цен-
тральные и вечные (например, только они усваивают молекулярный азот из ат-
мосферы). Откуда же этот пессимизм по отношению к своему объекту? Отве-
тить просто: эти фразы некритически переписаны из какой-нибудь книги про
«дарвинизм XX века», где такие фразы обычны - их пишут авторы, не исследо-
вавшие экологию микробов. Непонятно другое. Разве микробиология не знает
уже более полувека, что биохимическая мощь царства бактерий гораздо выше,
чем в царстве животных?
Вот тут и приходится вспомнить о методологии. Микробиолог доверился
«мнению специалистов» (чужой дисциплины), не смутившись радикальным
противоречием с его собственной наукой. Он не подозревал, что у представите-
лей разных наук разное понимание истины: для него истина - всегда результат
хорошо согласующихся экспериментов; вряд ли он думал, что у эволюциони-
стов может быть не так. Однако, как известно, эволюционисты опытов не ставят
(а если иногда и ставят, то не на своем объекте, а на объекте генетиков), поэтому
для них истина - не столько доказанный факт, сколько правдоподобное выска-
зывание; правдоподобность же сама по себе зависит от объекта исследования и
допустимых приемов рассуждения, и правдоподобное для одних нередко лише-
но всяких оснований для других.
Ошибка Шлегеля в том, что выводы теории, основанной на идее взаимного
вытеснения, он приложил к проблеме равновесия биосферы, то есть к вопросам
взаимной подстройки. Идею конкуренции, которую сам Дарвин считал сущест-
* Химия и жизнь, 1980, № 10, с. 15-20. Подзаголовки даны заново в 2010 г.
10
венной для близких видов, он, вслед за дарвинизмом XX века, распространил на
взаимодействие разных царств. Инфузория, с ее сложнейшей структурой и изо-
бретательным поведением, способна синтезировать всего 10 аминокислот из
двадцати; как же ей конкурировать с «примитивной» бактерией, делающей все,
что ей нужно, из глюкозы и азота? Разумеется, они не конкурируют, а сотрудни-
чают (когда бактерия поселяется внутри инфузории), и экологи давно это знают.
Всю экологию можно назвать наукой о взаимоподстройке, которую нарушает
лишь конкуренция видов, наделенных близкими потребностями. Тем не менее
многие эволюционисты до сих пор видят в конкуренции главный движущий
фактор биологического прогресса. Кто же прав?
В сущности, вопрос не нов, он регулярно возникает в различных науках.
Считать ли протон твердым шариком или чем-то расплывчатым? Считать ли,
что возникновение жизни было предрешено в момент рождения Вселенной или
что жизнь отобрала сама себя из множества случайных вариантов развития ма-
терии? Считать ли исторические события уникальными или всего-навсего пово-
ротом одних и тех же стекляшек в калейдоскопе? Все подобные вопросы иллю-
стрируют, если хотите, принцип дополнительности Нильса Бора: свойства объ-
екта в значительной мере определяются тем, с какой точки зрения на него смот-
реть. Однако если физика достаточно точно указывает, в каких феноменах про-
тон удобно считать шариком, а в каких - волновым пакетом, то другие вопросы
- предмет бесконечных споров. Заметив это, физики уже встали в гордую позу:
мы-де свои утверждения доказываем, а вы ограничиваетесь описанием.
Однако дело не только в этом. Известный науковед Эдуард де Боно подчер-
кивает («Рождение новой идеи». М., 1976): никакая новая идея недостижима ло-
гически, она всегда рождается интуитивно, но затем может быть логически дока-
зана или опровергнута. При этом роль логического доказательства де Боно оцени-
вает невысоко, считая, что оно само в конечном счете подчинено эмоциональным
мотивам. Он приводит такой пример: гипнотизер внушает пациенту, что он пре-
вратился в собаку, и тот становится на четвереньки и лает; на вопрос, зачем это,
гипнотизируемый начинает доказывать логичность своего поведения: он - соба-
ка, следовательно... Такова сила рационализации! - заключает де Боно.
Физика знает множество таких доказательств-рационализаций. Например,
курс оптики издавна строился на «твердо доказанном» факте невозможности
синхронного излучения хотя бы у двух атомов. Надо было построить лазер (ге-
нератор синхронного излучения), чтобы заметить, что все «доказательства не-
возможности» просто-напросто относились к объектам, не допускающим син-
хронизации. Более того, оказалось, что еще лет за двадцать до появления лазера
была теоретически обоснована его осуществимость. Выходит, и здесь долгое
время сосуществовали исключающие друг друга «доказательства».
В строгом смысле слова доказательства возможны только в математике, и
не потому, что математики умнее других, а потому, что они сами создают все-
ленную для своих опытов, все же остальные вынуждены экспериментировать во
11
Вселенной, созданной не ими. Доказательство означает неопровержимую де-
монстрацию невозможности какого-то события (любая теорема допускает фор-
мулировку: «такое-то множество пусто») но утверждать невозможность бес-
смысленно, если в реализации события могут сыграть роль неизвестные обстоя-
тельства. Это и губит рано или поздно любое физическое «доказательство».
2. Физика - образец науки?
Однако, слов нет, физика гораздо точнее других наук, например, она боль-
ше, чем какая-либо другая наука, математизирована. Физики опять-таки видят в
этом плод своих трехсотлетних усилий и предлагают другим идти по их стопам.
Методологи возражают: напрасная трата сил. Математизация физики оказалась
возможной потому, что разнообразие физических объектов не препятствует
почти полному их охвату (то есть от опыта к опыту новые обстоятельства прак-
тически почти никогда не появляются), оттого и рассуждения могут быть «поч-
ти доказательствами». А в других сферах знания этого нет: геолог не встречает
двух месторождений (а биолог - двух организмов), настолько сходных, чтобы
говорить об их взаимозаменяемости. Организмы, как говорят вдумчивые биоло-
ги, неперечислимо разнообразны*. Та математика, что создана для нужд физики,
здесь вряд ли полезна.
Именно это обстоятельство - взаимозаменяемость объектов в рамках иссле-
дования - позволяет представлять явление природы в виде модели, которая в
свою очередь поддается математизации. Если геолог или биолог могут соста-
вить модель не иначе, как опустив что-то важное, то физик часто создает очень
полные модели, в которых отражено все явление, кроме «малых возмущений».
Причем нередко он даже не отдает себя отчета в том, что его уравнения описы-
вают, собственно говоря, не объект, а только модель объекта. Когда в середине
XIX века было установлено, что планета Меркурий ведет себя не вполне по
Ньютону (орбита поворачивается на 43 угловых секунды за столетие), разгадку
стали искать в чем угодно - в сопротивлении межпланетной среды, в том, что на
Меркурий влияет неизвестная планета, даже в неправильности закона тяготения.
Суть же дела в том, что ньютонов закон есть закон поведения ньютоновой мо-
дели Вселенной, но не самой Вселенной.
Тогдашняя методология утверждала, что малые искажения следует объяс-
нять малыми возмущениями, каковые следует вводить в уравнения в виде малых
поправок. Почти всех удовлетворило такое объяснение парадокса Меркурия: за-
кон тяготения следует записывать не в виде формулы F ~ */г2, где F - сила тяго-
1, 2,00000016
тения, г - расстояние между телами, а чуточку иначе: F ~ /г
Среди тогдашних парадоксов физики был и другой: в 1881 году А. Май-
кельсон нашел, что скорость света не зависит от того, куда направлен луч - по
ходу орбитального движения Земли или навстречу ему. Если бы кто-нибудь ска-
зал, что здесь есть что-то общее с парадоксом Меркурия, его подняли бы на
* Имелся в виду биолог К.Х. Уоддингтон (Англия). - Примеч. 1991 г.
12
смех: данные явления несопоставимы, они определяются разными законами
природы . Понадобился гений Эйнштейна, чтобы всего через 35 лет убедить
вдумчивых коллег (а через 50 лет - почти всех), что оба факта имеют одну и ту
же причину, но что уяснение ее требует новой модели мира.
На вопрос, почему переворот в умах произошел так быстро и относительно
гладко, ответить непросто - ведь большинство подобных идей входят в науку
столетиями. Дело не столько в удачном толковании наблюдений (разве в про-
шлом наука не игнорировала многие эмпирически наблюдаемые факты, такие
как падение метеоритов, гипнотизм, биологический эффект космических про-
цессов - вместе с их толкованиями?), сколько в благоприятных методологиче-
ских обстоятельствах. Одно из них легко заметить: первый результат Эйнштей-
на (искажение законов механики при возрастании скоростей) легко восприни-
мался на языке «малых возмущений». Казалось, что новые представления каса-
ются лишь узкого круга вопросов и практически не меняют установленных прежде
законов механики и электродинамики сред, движущихся с обычными скоростями.
Теперь-то мы знаем иное - что тогда произошла научная революция, пере-
вернувшая все наши представления о мире: оказывается, пространство и время
сплетены в нечто единое, и этот факт приводит к самым неожиданным послед-
ствиям в обычных земных условиях. Например, мучившее ученых со времен
Ньютона совпадение массы как меры веса с массой как мерой ускорения стало
просто новой аксиомой естествознания.
3. Бывает ли вообще лженаука?
Дискуссия натуралистов на методологические темы ширится у нас на гла-
зах; сошлемся на выступление в научно-популярной печати двух ведущих уче-
ных - палеоботаника Мейена** и физика-теоретика Мигдала***. Несходство про-
фессий в данном случае вряд ли имеет значение, так как речь идет о самых об-
щих вопросах. Существеннее, что оба специалиста, известные своими крупными
научными результатами, придерживаются едва ли не противоположных взгля-
дов на логику и методологию естествознания.
Оба автора - внимательные и благожелательные люди, у них не видно об-
щего с примитивными «охотниками на ведьм», ежеминутно готовыми бить лже-
ученых - спасать науку. Оба признают, что противоборство мнений в науке не-
избежно, что взаимное непонимание может быть вызвано объективными причи-
нами, преодоление которых требует специальной работы, сравнимой с собст-
Набранные рубленым шрифтом слова здесь и далее заимствованы из критикуемой ниже
статьи А.Б. Мигдала. -Примеч. 1991 г.
Мейен С.В. Врачу, исцелися сам... // Знание - сила. 1978. № 6; Может ли быть победи-
тель в дискуссии о номогенезе? // Природа. 1979. № 9.
Мигдал А.Б. От догадки до истины... //Химия и жизнь. 1979. № 12. - Примеч. 1991 г.:
академик Мигдал был крайне возмущен тем, что мой ответ на его статью не был ему
предварительно показан, и отказался впредь иметь дело с журналом.
13
венно научными исследованиями. Тем удивительнее, насколько различны их ре-
комендации.
А.Б. Мигдал берет, так сказать, быка за рога: «Единственный убедительный
способ установить истину - поставить... эксперимент, проведенный специали-
стами, дающий повторяющиеся результаты и подтвержденный независимыми
опытами других исследователей». Показаниям очевидцев он не только не дове-
ряет, но прямо заявляет: настоящий ученый, увидев что-либо сверхъестествен-
ное, обязан прежде всего исключить преднамеренный обман, обман зрения или
гипноз. Подобные феномены должен исследовать консилиум специалистов, в
числе которых должны быть и цирковые фокусники.
Вот и методологический тезис: «Задача науки - отбирать наиболее правдо-
подобные объяснения и придерживаться их до тех пор, пока опыт не заставит от
этого отказаться»*. Правдоподобное предположение служит для Мигдала пово-
дом к отказу от дальнейших исследований проблемы, и выходит, что все его от-
личие от упомянутых охотников состоит лишь в несколько большей терпимо-
сти: он не хочет ничего запрещать другим.
По-видимому, формулировки Мигдала не следует все же понимать букваль-
но: ведь математик требует не повторности результатов, а одного-единственного
доказательства; в биологии повторность обычно неточна, а палеонтолог вообще
не ждет второй находки археоптерикса, чтобы описать новый таксон, да и экспе-
риментов не ставит; не ставит их и астрофизик, хоть он и физик. Наконец, разве
Эйнштейн искал смещения орбит разных планет, а не только Меркурия? Разве
кто-нибудь не поверил первому же сообщения о запуске первого спутника? Одна-
ко не будем придираться: мысль автора ясна, и эта мысль достаточно здравая -
науке необходим некоторый консерватизм. Другое дело, нуждается ли этот кон-
серватизм в особой защите (и так ведь большинство научных работников заняты
только перепроверкой и правдоподобными объяснениями), но в принципе консер-
ватизм нужен в науке так же, как наследственность - в эволюции.
Гораздо хуже дело с очевидцами, обманом и гипнозом. Слов нет, обман бы-
вает повсюду, но разве приходило в голову самому Мигдалу, едучи в институт,
где совершено научное открытие, приглашать с собой эксперта-фокусника? Де-
ло, разумеется, не в том, что физический эксперимент подделать трудно; просто
немыслимо демонстрировать коллегам столь явную недоброжелательность. Та-
ким «экспертам» коллеги вряд ли что покажут, и будут правы.
Далее: разве, сославшись на возможный гипноз, автор что-нибудь объяснил
себе или нам? Навряд ли гипноз понятен ему больше, чем телепатия; такие
ссылки - просто выражение веры в правоту всякой академической науки. По-
Это - так называемый метод презумпций. Мейен относился к нему насмешливо-
критически. Беда метода в том, что, несмотря на уверения его приверженцев (что они-де
всегда готовы отказаться от презумпции под давлением фактов), он ведет к фактическому
прекращению поисков истины (подробно об этом см. в АСМ). - Примеч. 2010 г.
14
этому стоит напомнить, что гипноз прошел весь тернистый путь от лжеучения
до стандартного терапевтического метода и что в 1774 году врач Антон Месмер,
пытавшийся пользовать пациентов методом, который сейчас можно квалифици-
ровать как гипнотерапию, был ошельмован «консилиумом специалистов», то
есть группой авторитетных медиков, назначенной французским правительством
для рассмотрения вопроса, в котором они, увы, ничего не смыслили.
Характерно, что аргументы, некогда выдвинутые против гипноза, были те
же, что всегда выдвигаются против всевозможных «лжеучений», старых и но-
вых: отсутствие хорошей повторяемости (гипнозу поддаются не все), противо-
положные эффекты (средство, помогающее одним, ухудшает состояние других),
необъяснимость с точки зрения физики (Месмер отождествлял гипнотизм с маг-
нетизмом, а такой связи не найдено и поныне), наконец, неудачи контрольных
сеансов в присутствии комиссии. Что же касается множества больных, которым
гипноз все-таки помог, то эти случаи, разумеется, допускали «более правдопо-
добные объяснения».
Предрешать исход конкретного научного спора - не дело методологии, но
она обязана указывать спорщикам, когда они выходят за рамки компетенции
своих наук. Нельзя требовать от непонятного феномена, чтобы он подчинялся
тем же критериям истинности, что и задачи студенческого практикума.
Физика почти не умеет работать с потоком несходных уникальных наблю-
дений, поэтому не от нее следует ждать раскрытия тайн психики - она не смогла
подступиться даже к относительно простой, во всяком случае привычной, про-
блеме гипноза.
Прошло сто лет после Месмера, и гипнотизм как-то сам собой, без помощи
комиссий, стал достоянием лечебной медицины. Дело, как видим, не в новых
фактах или физических теориях, а в освобождении от власти придирчивой не-
доброжелательности. Истина не рождается в комиссиях, зато там легко наклеи-
вается ярлык «лженауки».
Именно об этом и размышляет С.В. Мейен. По его убеждению, наука отли-
чается от лженауки не арсеналом фактов и теорий, а этической атмосферой ра-
боты. Много раз уже предпринимались попытки дать формальное определение
лжеученого, чтобы облегчить работу научных инстанций, но до сих пор пользы
не вышло. Научный шедевр поначалу способны оценить немногие, а они-то и не
любят заседать в комиссиях. Однако у всех ученых должно быть то общее, что
необходимо им в работе и может быть без труда распознано при общении: уче-
ный готов слушать, чтобы понять, а не чтобы возражать; он хочет, чтобы его
творение совершенствовалось, ищет у себя просчеты и жадно слушает критику.
И наоборот: тот, кто изобретательно защищает свое творение и рад найти про-
счет у оппонента, тот выступает не как ученый.
«Степень самосознания, рефлективности - главный критерий зрелости нау-
ки», - пишет Мейен и тоже делает попытку определить границу науки и лже-
науки: «Рефлексия в науке подразумевает прежде всего такт и самоконтроль
15
ученого. Тогда на противоположном, “нерефлектирующем” и “лженаучном” по-
люсе будет отсутствие и такта, и самоконтроля. Каждый, кто хоть немного стра-
дает недостатком того или другого, приближается к тому, что стало принято на-
зывать лженаукой».
И здесь, пожалуй, хвачено через край: разве не знает наука крупных лично-
стей, которые слушать не умели абсолютно, но сами умели придумывать полез-
ное? Не относить же их всех к лжеученым! Однако Мейен безусловно прав в
том смысле, что эти добровольно оглохшие гении сами себя обкрадывали, огра-
ждаясь от неугодных мыслей, то есть поступали антинаучно. Вряд ли можно
найти хотя бы одного из таких добровольно оглохших, в чьих трудах жемчужи-
ны мысли не перемешаны с банальностями и ерундой. Эти-то места эпигоны ус-
воят легче всего - вот и подлинная лженаука.
Как же быть? Рекомендация Мейена и проста, и трудна: ничего не отвергая
сразу, размышлять над разными вариантами, а «природа рано или поздно ука-
жет, какие теоретические варианты бессмысленны. Важно только не принять за
эти указания собственные субъективные симпатии и антипатии, традиции науч-
ных школ».
4. Наука не движется прямо
Итак, согласно одному взгляду, наука - это то, что хорошо проверено, со-
гласуется с прежним опытом («фундаментом научных завоеваний прошлого») и
принято специалистами; согласно другому, наука - то, что «отрефлектировано»,
то есть прилажено само к себе. Разумеется, теория относительности научна в
обоих смыслах: она покорила мир цельностью (именно цельностью не обладали
«более правдоподобные» поправки); однако Эйнштейны рождаются редко и в
наших советах не нуждаются. Важнее понять, как быть с едва намечающимися
идеями, для проверки которых еще не рожден синклит знатоков.
Бросается в глаза: «наука по Мигдалу» направлена шипами въедливой крити-
ки наружу, не доверяет сенсациям, зато легко согласна пройти мимо клада; напро-
тив, «наука по Мейену», оснащенная шипами самокритики внутрь и, уязвимая для
чужой критики, согласна искать жемчужины в навозной куче и не дает своим
адептам почивать на достигнутом. Следовательно, вторая пригодна поставлять
новые идеи (ведь легче прилаживать их друг к другу, чем к опыту прошлого), а
первая хороша для отбраковки, развития и внедрения.
Между прочим, так обстоит дело с биологическим эволюционизмом, где бо-
рются два типа учений: первые исходят из того, что эволюция - это отбраковка
неудачных и распространение удачных организмов, возникающих случайно (то
есть как бы за рамками учения); вторые озабочены проблемой: откуда появляются
ростки нового?
Конечно, первый взгляд на науку удобнее для учебников. «Мы твердо зна-
ем, - продолжает А.Б. Мигдал, - что дальнейшее развитие науки не отменит ус-
тановленных соотношений, а только выяснит область их применимости». Пожа-
луй, учебник можно выдержать в таком духе (ведь мало кому из будущих уче-
16
ных доведется совершить научный переворот, а бороться с вечными двигателя-
ми всем придется), но давайте все же признаем, что сама наука до сих пор раз-
вивалась не так. Разве геоцентрическая система или теория флогистона не от-
вергнуты? А ведь когда-то они были респектабельной академической наукой.
Разве ограничение области применимости не есть отмена тех соотношений,
которые были предусмотрены до выяснения ограничений? Людвиг Больцман
прекрасно знал, что закон возрастания энтропии верен только для изолирован-
ных систем, и все же был уверен, что Вселенная деградирует; Илья Пригожин
(лауреат Нобелевской премии 1977 года), наоборот, показал, что в открытых
системах возможен самопроизвольный синтез структур*, - формально это не
противоречит Больцману, но как меняет все наше мировоззрение! Только после
Пригожина затихли речи о «термодинамической невероятности живого».
До этого физик не умел объяснить, почему эта формулировка бессмыслен-
на: оказывается, практическое понимание границы применимости достигается
лишь тогда, когда хоть немного приоткрывается то, что лежит по ту сторону
этой границы. Короче говоря, мы понимаем закон ровно в той мере, в какой зна-
ем границы его применимости, рассуждать же о будущей несокрушимости како-
го-то закона - значит, поддаться искушению той самой неоправданной рациона-
лизации, о которой мы толковали выше.
В целом можно сказать, что утверждение, будто наука движется путем са-
модостройки, не выдерживает критики. Далеко не новым и не самым смелым
является следующий вывод науковедов: естествознание движется вперед не
столько путем добавления новых фактов и законов к прежним, сколько регуляр-
ным пересмотром парадигм (о парадигмах в науке говорилось в статье В.В. На-
лимова в № 1 «Химии и жизни» за 1978 г.). Ньютонова парадигма неограничен-
ного пространства и абсолютного времени не подправлена, а отменена парадиг-
мой Эйнштейна, которая тоже, как уже сейчас видно, не вечна.
Аккуратные факты сами по себе, действительно, не отменяются дальней-
шим прогрессом науки, но науку образуют не факты, а модели, которые меня-
ются - приятно нам это или нет. То, что вчера было академической наукой, зав-
тра может быть сдано в архив (и потом снова извлечено лет через сто), а вче-
рашняя лженаука может стать наукой - примеров тому не счесть.
Именно так сложилась судьба одной из самых известных идей мировой нау-
ки - идеи биологической эволюции. Она претендовала на академизм с середины
XVIII века, но свыше ста лет прозябала в качестве лженауки, вплоть до появле-
ния книги Дарвина (подробно об этом в наших статьях: «Химия и жизнь», 1978,
№ 12; 1979, № 12). Можно подумать, что Дарвин представил именно тот ком-
плект фактов, который, наконец, убедил «специалистов», но это не так: недоста-
точность фактического обоснования нового учения отмечали не только его кри-
тики, но и многие из числа сочувствующих.
* Вернее сказать, что результат принадлежит всей школе Пригожина. - Примеч. 2010 г.
17
Вряд ли «Происхождение видов» вышло бы в свет без поддержки знамени-
того геолога Чарлза Лайеля, авторитет которого помог обойти «мнение специа-
листов»; а ведь сам ЛаЙель вовсе не был уверен в обоснованности дарвинизма:
он ценил идеи своего друга за их яркость. Признание дарвинизма в конечном
счете оказалось переосмыслением дано известного, а недостающие аргументы
стали поступать позже .
Все сказанное приводит нас к выводу: историю науки, пожалуй, и можно
излагать как «поступательное развитие», но это так же скучно, как казенная
биография ученого: получал результаты, дипломы, чины и... старел. Смена дня
и ночи, моменты взлета и полосы неудач, любовь и ненависть, успехи и катаст-
рофы, а главное - вечный поиск ускользающей истины, - все это остается по ту
сторону «поступательного варианта» и вне послужного списка, так что биогра-
фии ярких людей пылятся на полках нечитанные.
Разве в том дело, что Эйнштейн ошибся в формуле и писал об этом Фридма-
ну? Такие курсы истории науки студенты неизменно и справедливо прогулива-
ют. Насколько интереснее было бы для них узнать, что, например, идея устой-
чивости движения планет родилась у Джорджа Дарвина (сына Ч. Дарвина) по
аналогии с идеей естественного отбора, что общая теория относительности в
чем-то соприкасается с древнеиндийскими представлениями, что до Максвелла
электричество хотели объяснить механикой, а после него механику стали объяс-
нять электричеством. Тогда и нынешняя претензия видеть в биологии одно
лишь взаимодействие валентных электронов (плюс водородных связей) нашла
бы свое историческое место: ведь триста лет назад любили физику выводить из
биологии (считали, например, что кристалл растет из семени). Сейчас этот на-
строй мысли возрождается: кое-кто среди физиков говорит о прапсихике атома.
5. Прогресс науки - не в «охоте на ведьм», а в самокритике* **
Изложенного, по-видимому, достаточно, чтобы сформулировать тезис: ис-
тина не может быть безапелляционно отличаема от лжи с помощью «специали-
стов», поскольку любые каноны формулируются задним числом, в порядке ра-
ционализации пройденного. Либо нужно отказаться от термина «лженаука» и
ему подобных, либо придется признать, что лженаука - такой же феномен куль-
туры, как и привычная нам школьная наука (недавно, кстати, вышла книга В.Л.
Рабиновича, которая так и называется: «Алхимия как феномен средневековой
культуры»).
Среди исторических корней любой науки всегда найдется корешок «лже-
науки» (алхимия и химия - хрестоматийный пример). Но этого мало: наука и
лженаука то и дело меняются местами. Исследовать поверья и приметы - удел
лженауки (о чем прямо пишет и Мигдал), но вот Мейен считает, что респекта-
В то время мне казалось, что таковые аргументы существуют - Примеч. 2010 г.
** В статье М-162 тема развита подробно. Особенно, в снятом редакцией фрагменте 12
(см. Приложение Примеч. 2010 г.
18
бельный эволюционизм основан на утверждениях, которые иначе как научными
приметами не назовешь.
В тех дисциплинах, где разнообразие объектов много шире возможностей
описания, из разных «примет» можно складывать разные учения, равно убеди-
тельные для специалистов с разными вкусами, и они будут враждовать долгие
годы, разумеется, аттестуя друг друга «лженаучными». Те ученые, которые не
знают об этой конфронтации, могут строить чудовищные химеры по советам
«специалистов» (см. первую главу этой статьи), однако достаточно осознать
многоликость проблемы, чтобы понять: в этой дискуссии никогда не будет по-
бедителя. Прогресс - удел того, кто более самокритичен.
Заметим, что полуподпольный статус лженауки не мешает идеям получать
выход в практику: гипноз, иглоукалывание, гомеопатия и многое другое вошло
в практическое врачевание задолго до официального одобрения и вопреки от-
сутствию удовлетворительного научного объяснения. Влияние магнитных полей
на жидкости и организмы не объяснено до сих пор, но это не отталкивает тех,
кто с успехом использует это влияние. Что же касается экономичности самих
научных исследований, то здесь лженауке можно только позавидовать: считан-
ными людьми, самодельными приборами и почти без финансирования она под-
час делает очень многое*.
Впрочем, не будем пытаться дискредитировать альтернативный образ мыс-
лей, пусть по нашему убеждению и консервативный. У каждого подхода свои
достоинства, и если стоит над чем задуматься, так это над той же проблемой
контакта: не только разные специальности, но и разные методологии заставляют
ученых попусту враждовать и наделять друг друга нелестными эпитетами. Чуть
перефразируя Мигдала: можно понять даже своих коллег, если приложить к
этом усилия того же порядка, какие тратятся на занятия самой наукой**.
В наши дни тема, увы, опять стала актуальной: чиновники вновь призывают, ссылаясь
на недобросовестность отдельных лиц, громить «лженауку», под которой, как и при со-
ветской власти, понимают всё то, что противоречит решению чиновной Комиссии. См.
далее, п. 4 статьи «Ламарк, Дарвин и устройство науки» . - Примеч. 2010 г.
Надо отдать должное ныне покойному академику Мигдалу - несмотря на все мои вы-
пады и своё возмущение ими, он повел себя вполне достойно: не поддержал (а возможно,
и прямо пресёк) попытку моего тогдашнего директора С.Р. Микулинского (тоже ныне
покойного) учинить мне за эту статью «персональное дело», чтобы, возможно, потом
уволить. Тот факт, что журнал 8 лет меня не печатал, связан лишь с тем, что сам я ничего
им не предлагал (чтобы не ставить редакцию в неловкое положение) вплоть до разгара
«перестройки». - Примеч. 2010 г.
19
Грамматика биологии*
В языковом словаре каждое слово дано один раз, с указанием, если нужно,
типа его изменения. Например, существительное дается в именительном падеже
единственного числа с указанием типа склонения, а остальные варианты (слово-
формы) всякий, кто знает грамматику, восстановит сам. Пусть у правил граммати-
ки много исключений, но все же их запоминание проще, чем запоминание каждой
словоформы в качестве отдельного слова. Г. Гельмгольц ставил словарь и грамма-
тику в пример «естественноисторической системе», видя в них принципиальное
сходство, но следует признать, что и сейчас биологические «словари» (системати-
ческие сводки) сильно отстают от языковых: не только школьник, но и специалист
вынужден описывать виды, роды и т.д. только путем полного перечисления. Ни-
каких «таблиц склонения», по которым можно было бы, например, установить
возможный перечень подвидов, зная определение вида, до сих пор нет.
Потребность в такой рационализации особенно очевидна сейчас, когда чис-
ло описанных видов превысило 2 млн. и когда к материалу описательной биоло-
гии обращается множество неспециалистов. Однако в какой мере такая рацио-
нализация возможна вообще?
Параллелизмы и систематика
В 1834 г. немецкий ботаник И. Бернгарди заметил, что многие растения, в
других отношениях несходные, дают сходные ряды типов листа [1, S. 14]. В
1868 г. Ч. Дарвин добавил к этому наблюдению много других, и в частности та-
кое: гладкокожий персик выведен из немногих форм бархатистого, но «между
сортами бархатистых и гладких персиков наблюдается замечательный паралле-
лизм» - общие варианты цвета плода и его мякоти, характера косточки, цветков,
листьев и т.д., хотя «признаки каждого сорта гладкого персика получены отнюдь
не из соответствующего сорта бархатистого» [2, с. 716]. Через 50 лет Н.П. Вави-
лов, изучая злаки Памира, установил, что характерные признаки разновидностей
обыкновенной пшеницы с удивительной точностью повторяются у разных форм
ржи. Позже такой же четкий параллелизм (гомологические ряды) был обнаружен
у других злаков, а потом и у других растений и некоторых животных. Вавилов
справедливо решил, что видовое разнообразие в целом упорядочено.
Исследуя разновидности одного вида, мы как бы двигаемся по коридорам
одного этажа, а при исследовании другого, похожего вида - по коридорам дру-
гого этажа. Естественно, что в планировке разных этажей одного здания мы на-
ходим много общего, хотя положение отдельных комнат и их убранство доста-
точно различны. Можно сказать и по-другому: ряд, составленный из форм пше-
ницы, можно уподобить склонению какой-то одной видовой формы, и те же са-
мые «падежи» мы видим у форм ржи.
Вестник АН СССР, 1986, № 3, с. 47-58. Статья написана к 50-летию С.В. Мейена. Попу-
лярный ее вариант см.: Знание - сила, 1985, № 12, с. 30-32. Подробнее о различном пони-
мении искусственных и естественных систем см. далее, статьи части 6. - Примеч. 2010 г.
20
Однако, как писал в 1957 г. А.А. Любищев, вавиловские ряды - «только ма-
ленький отрезок грандиозной проблемы биологической системы» [3, с. 252]. Эту
проблему он видел в создании системы организмов, в которой «адрес» вида ука-
зывает все его существенные свойства. Такую систему Любищев называл естест-
венной. Подробнее см. [4].
Обычная (линнеевская) система, при всех ее достоинствах, этим свойством
не обладает: в ней близкие, почти идентичные «адреса» могут принадлежать
очень несходным организмам, а далекие - сходным. Например, неботанику уди-
вительно, почему столь несхожие для него виды, как картофель и баклажан,
«проживают» в одном роде, а ближайший к ним род - непохожий на них поми-
дор; что по соседнему с ними «адресу» находится овощной перец, тогда как ос-
тальные перцы «живут» в другом подклассе. Бывают даже случаи, когда морфо-
логически сходные виды относятся к трем разным царствам, например плазмо-
диевые грибы, плазмодиевые амебы и плазмодиевые водоросли. (Плазмодий -
это гигантская бесформенная многоядерная клетка.)
Конечно, ботаник объяснит, что в основе объединения видов в группы ле-
жат фундаментальные сходства, более важные, чем те, что видны постороннему.
Однако настораживает тот факт, что сами ботаники не всегда единодушны в
своих выводах о близости конкретных групп, да и в тех случаях, когда они еди-
нодушны, новые данные то и дело заставляют их перегруппировывать виды. В
частности, помидор прежде входил в один род с картофелем, а про овощной пе-
рец неизвестно даже, на сколько видов его следует подразделять: в начале века
их считалось около 30, а теперь разные систематики оценивают их количество в
интервале от 20 до 50. Даже число царств колеблется ныне от 4 до 19 и больше.
Поэтому давно, со времен Линнея, идет разговор о том, что нужны по край-
ней мере две системы - искусственная, удобная для нахождения вида, и естест-
венная, отражающая, как говорил Линней, суть вещей (имея в виду, что и сама ес-
тественность - понятие относительное). Любищев, как уже говорилось, видел ес-
тественность системы в информативности адресов, английский логик В. Уэвелл
(1840) - в устойчивости (в том, чтобы такую систему не надо было пересматри-
вать при обнаружении новых видов и признаков), а последние 100 лет принято
считать вслед за Э. Геккелем естественной систему, построенную на идее истори-
ческого родства. Все эти толкования не исключают, а дополняют друг друга, но
построить пока что удалось только одну - родословное древо. Любищев указывал,
что древо не может учесть отмеченные выше параллелизмы, он хотел соединить в
одной системе информативность («суть вещей») и удобство поиска, а это вряд ли
возможно: для поиска удобнее всего именно древо, то есть линнеевский принцип
иерархической классификации, проведенной по единому основанию.
Как работает линнеевская система
Вы поймали птицу и хотите определить ее вид. Ход и успех работы решаю-
щим образом зависят от того, что вы знаете заранее. Прежде всего, чтобы взять
определитель птиц, вы должны быть уверены, что поймали именно птицу, а это
21
предполагает какое-то знакомство с морфологией: не все ведь птицы летают, ле-
тать могут не только они, да и перья иногда на перья непохожи. Далее, к какому
отряду она принадлежит? Если вы взяли определитель впервые, то с удивлением
обнаружите, что различение даже отрядов (не говоря уже о меньших таксонах)
идет по признакам, ничего вам не говорящим. Например, принадлежность к от-
ряду воробьиных определяется по характеру щитков на задней стороне цевки
(часть ноги, от которой отходят пальцы).
Пусть вам уже удалось установить, что птица относится к роду синиц се-
мейства синицевых отряда воробьиных, и осталось только определить ее вид. В
определителе читаем: «Вдоль средней линии груди и брюха проходит черная
или черноватая полоска» - если это найдено, перед нами синица большая (Parus
major), если нет, - то надо читать дальше. Приходится читать: если на зашейке
имеется светлое пятно, то надо смотреть на концы кроющих перьев (если концы
белые, то это синица московка (Parus ater), если нет - читать дальше), если же
пятна нет, то надо смотреть на цвет спины и т.д.
Какой кропотливый 400-летний труд скрыт за этими диагнозами! Причем
эта удобная дихотомическая форма (если да, то читай одно, если нет - другое)
пришла в систематику только с «Французской флорой» Ламарка (1778), хотя
предложена была еще до Линнея. Самое интересное, что ее прекрасно знал Ари-
стотель - знал и отвергал, поскольку, определяя вид по дихотомической табли-
це, мы не познаем совокупности самых его существенных свойств, его сущно-
сти. Поясню: ведь большая синица - птичка яркая, а московка - грязноватая.
И все-таки, следуя дихотомии, мы почти всегда доходим до определенного
вида - потому, что всякая дихотомическая таблица делается теперь по готовой
системе. А сама система была придумана без дихотомии: систематик, желая
упорядочить известное ему разнообразие (например, воробьиных), следует лин-
неевскому афоризму - не признаки задают род, но род задает признаки - то есть
не пользуется формальным ключом, а основывается на непосредственном вос-
приятии, на критерии самоочевидности.
Только создав хорошую систему, можно начинать ее формализацию - хотя
бы составление дихотомической таблицы, и недаром Ламарк впервые достиг в
этом успеха лишь в год смерти Линнея. При Аристотеле же никакой писанной
системы не существовало, дихотомическое классифицирование рассматривали
как путь ее построения, и ничего, разумеется, не получалось.
Итак, предпосылкой дихотомии Ламарка была готовая линнеевская система
растений; но ведь, с другой стороны, и эта система была возможна лишь потому,
что еще давным-давно были выявлены и закреплены словами разговорного язы-
ка такие дихотомии, как живой - неживой, животное - растение, мужской -
женский, наземный - водный и т.п. Не менее четки и важны были также деления
организма на части: корень, стебель, лист, цветок, плод и т.п. Долиннеевские
систематики уже исходили из этих делений как из данных. Грубо говоря, общие
для данной группы свойства выявляет морфология, а различия между группами
22
- систематика, и друг без друга они развиваться не могут. Это звучит банально,
но фактически организовать взаимодействие внутри этой пары непросто.
«Свет без двойства являться не может»
Наличие двух взаимодополнительных точек зрения - самое удобное, что
можно пожелать, если хочешь углубляться в проблему путем последовательных
шагов (итераций). Недаром еще в 1827 г. молодой московский ботаник М.А. Мак-
симович, размышляя над теми же, что и мы сейчас, вопросами, написал: «В при-
роде везде двойство: свет без двойства являться не может» [5, оборот тит. листа].
Это «двойство» - взаимодополнительность двух альтернативных подходов, точек
зрения, путей, сходную с принципом дополнительности Н. Бора - он видел во
всяком познании. Выискивая «двойства», он даже подошел к гомологическим ря-
дам, то есть заметил, что если группировать виды по «сродству» (по гомологии) в
столбцы, то образующиеся при этом строки (соседние виды разных столбцов) то-
же будут демонстрировать сходства (аналогии). Что, однако, делать со всем этим
множеством пар? Ответ удалось дать только через 150 лет, когда С.В. Мейен
предложил первый эскиз того, что уже можно назвать грамматикой биологии.
Речь идет о выявлении структурных инвариантов разнообразия организмов.
В других науках такие инварианты успешно выявляются. Такова в этноло-
гии структура мифа, которая сохраняется неизменной при всех вариациях, про-
исходящих во времени и пространстве, в филологии - понятие порождающей
грамматики, то есть некоторого устройства, остающегося структурным инвари-
антом бесконечных вариаций речевой деятельности, и т.д.
В биологии для выявления инвариантов естественно обратиться к вавилов-
ским рядам. Почему открытия с их помощью так редки? Почему определители
не делаются в форме таблиц, где все таксоны одного ранга представлены по
единым правилам, пусть и с исключениями, подобно таблицам склонения (пол-
ным наборам падежных форм в языке)? Некоторые причины видны из аналогии
с грамматикой: одно слово может входить в разные таблицы, и ни одна грамма-
тика не сводится к одним лишь таблицам, зато каждая грамматика создавалась
столетиями, и была такая научная профессия - грамматик.
Слова обычно образуют большие группы грамматически сходных форм, та-
ких как пол и стол, где почти все падежные формы идентичны. Однако встре-
чаются слова, к исходной форме которых не подобрать пары, сходной и грамма-
тически, и по звучанию конца: существительному хрящ созвучны только крат-
кие причастия типа говорящ. Зато, просклоняв хрящ, найдем, что его производ-
ным формам созвучна группа слов того же склонения типа хвощ’, немного хуже
соответствует праща - другого склонения. Кроме них, много разрозненных
сходств - совпадений той или иной падежной формы с отдельными словофор-
мами других частей речи без какого бы то ни было совпадения рядов в целом:
хрящ-говорящ, хряще-вообще и т.д.
В языках, не имеющих падежей, ту же закономерность легче всего увидеть
на глагольных формах. Так, словоформы французских глаголов devenir, revenir,
23
souvenir, tenir, venir и т.п. (образующих свою небольшую группу спряжения: ve-
nir - venant - viens - vient - venu - vins) сходны с существительными: инфинитив
этих глаголов сходен с существительными avenir и souvenir, настоящее время -
с soutien и revient, активное причастие - с revenant и tenant, пассивное причас-
тие - с avenu и revenu, простое прошедшее - с vin и devin. Есть сходства с прила-
гательными (avenant, tenu), местоимением tien, междометием tiens и т.п.
Подобным же образом изменяются признаки биологических видов, родов и
т.п. Так, если в ботанике часты довольно полные параллельные ряды, то для
зоологии более характерны разрозненные сходства. Возьмем для примера тех же
синиц. Собственно синицы (род Parus семейства Paridae) образуют 12 видов (в
фауне СССР), но термин «синица» применяется и к другим воробьиным: ремезу,
длиннохвостой синице, усатой синице и др. Уже это говорит о параллелизме:
птицы других родов и даже семейств имеют характерный «синичий» облик. Од-
нако параллелизм наблюдается гораздо шире. Почти все признаки, отраженные
в названиях синиц, представлены и у других птиц. Так, характерная полоска
вдоль щеки, давшая видовое название усатой синице (из семейства толстоклю-
вых синиц отряда воробьиных), наблюдается у некоторых овсянок и одного дят-
ла (сирийского пестрого). Естественно, что в пределах отряда воробьиных па-
раллелизм полнее: здесь мы видим аналогов и хохлатой синице (свиристель, жа-
воронок хохлатый) и рыжешейной (овсянка рыжешейная) и ремезу (овсянка-
ремез) и усатой (некоторые овсянки) и длиннохвостой (личинкоед серый, отчас-
ти сороки) и другим синицам.
Перейдем к описанию того эскиза «грамматики биологии», который С.В.
Мейен называет типологией (прежде так иногда называли то, что мы выше упо-
минали как морфологию). Главная его идея в следующем: чтобы познавать разно-
образие путем попеременного обращения - то к науке о различиях, то к науке о
сходствах, надо их понимать иначе, чем до сих пор было принято. Науку о разли-
чиях он называет таксономией, то есть учением о таксонах, о том, как их образо-
вывать. Однако таксономия у него не синоним систематики, поскольку само поня-
тие таксона он использует широко - как любое множество объектов, достаточно
определенно отграниченное от других множеств (так, если обсуждается приспо-
собительная окраска, то зеленые гусеницы - таксон). Науку о сходствах Мейен
тоже представляет шире, чем морфологию, и дает ей название «мерономия».
В основу кладется понятие мерона. Меро* н (от греческого meros - часть) -
это «класс частей». Когда мы говорим, например, о конечности, это в равной
мере может относиться к руке, ноге или лапке насекомого, а это значит, что мы
умеем мысленно расчленять тела животных, выделяя части естественным обра-
зом. При этом слово конечность означает результат мысленного объединения
таких частей разных организмов, которые обладают некоторыми очевидными
общими свойствами, - это подвижные придатки тела, имеющие собственную
мускулатуру и служащие для передвижения или хватания (да простит нам мор-
фолог некоторое упрощение!). Мерой «конечность» - не конкретная рука или
24
лапка, а множество всевозможных конечностей. Иными словами, мерой - это
обобщенный класс однородных в каком-то отношении признаков.
Отсюда следует, что конкретный организм может обладать органами, частями
тела, свойствами, признаками, но не меронами; меронами обладают только груп-
пы объектов. (Понятие мерона обобщает введенное Р. Оуэном в 1855 г. понятие
гомологии.) Часто, выявляя мерой, мы называем некоторый таксон систематики
(например, мерой «позвоночник» характеризует подтип позвоночных в типе хор-
довых, а мерой «цветок» - отдел цветковых растений), но еще чаще мерой объе-
диняет в одну группу организмы, которые ни один систематик не признает таксо-
ном, например: мерой «пищеварение» объединит животных с насекомоядными
растениями, а мерой «голова» объединит (в частности) людей с осьминогами.
Одно лишь понятие мерона еще не ведет к пользе для таксономии, нужно
2
некое учение о меронах, то есть мерономия .
Еще в 1858 г. зоолог, более известный сегодня как публицист славянофиль-
ского направления, Н.Н. Страхов отметил, то сравнительная анатомия класси-
фицирует органы так же, как зоология классифицирует организмы. Развивая эту
мысль, Мейен пришел к фундаментальному выводу: гомологизация (классифи-
кация частей) и классификация организмов являются двумя логически само-
стоятельными процедурами, и их связь осуществляется итеративно, то есть по-
следовательными обращениями от одной процедуры к другой. Мы уже отмеча-
ли, что для определения птицы надо заранее знать, что такое птица, а это значит
не только уметь выявлять характерные для птиц мероны (определенная форма
тела, клюв, крылья, перья и т.д.), но и уметь мысленно складывать из них обоб-
щенный образ птицы, то есть знать, какое сочетание комбинаций меронов дела-
ет животное птицей. Этот образ - то самое, что Оуэн назвал архетипом. Если
сам организм можно мысленно расчленять на части тела, органы и т.д. и гово-
рить, что организм из них состоит, то архетип состоит из меронов.
Однако организмы можно сравнивать не только по частям их тел, но и по
всем их качествам: по способности летать или образовывать брачные пары, по
цветным узорам на теле, по роли в экосистеме и даже по размерам. Архетип по-
лучается куда богаче, чем по Оуэну, и строить его нужно по-новому.
Если прежде его изображали простеньким рисунком, содержащим только
самые общие свойства таксона (например, архетип позвоночного - это очень
схематические голова, позвоночник и хвост, столь же схематические органы
пищеварения и т.д.), то, по Мейену, архетип таксона - это обширная таблица,
где каждая строка - мерой, а каждый столбец — возможная в принципе особь.
Так, архетип позвоночных включает строку «конечности», в которой друг за
другом записаны всевозможные значения мерона: отсутствие, плавники, ласты,
лапы и т.д., причем каждый вид конечности может иметь различные степени
2 Взгляды С.В. Мейена изложены им в обширных статьях в периодике: [6; 7]. (В послед-
ней им развито понятие мерономии).
25
развития, число и индивидуальные особенности (например: шесть пальцев на
левой руке). Во избежание путаницы следует четко различать архетип по Оуэну
(рисунок) от архетипа по Мейену (таблицы).
Чаще всего эволюция - это реализация нового столбца, но прогрессивная
эволюция всегда включает появление новой строки (и связанных с нею новых
столбцов). До появления птиц в таблице позвоночных не было, например, стро-
ки «перья», а в строке «способность летать» почти все клетки были пусты, по-
скольку прежние позвоночные хорошо не летали и в воздухе не охотились.
Новые типы особей почти всегда являют лишь новые комбинации прежних
признаков, так что большинство признаков фигурируют в таблице по многу раз, а
большинство клеток пусто. Поэтому фактически выписывать таблицу смысла нет
- она важна как принципиальная иллюстрация. Биология к таким мысленным таб-
лицам не привыкла, зато они обычны в математике (например, теория матриц),
где их свойства умеют изучать, не выписывая самих таблиц в явном виде.
Как должна работать типологическая система
В архетипе позвоночных (если понимать этот термин по Мейену) архетип
птиц займет некоторые строки и столбцы, их можно вырезать из прежней таб-
лицы и склеить в меньшую, но тоже огромную таблицу; из нее можно вычле-
нить таблицу «архетип воробьиных», «архетип страусов» и т.д., то есть строить
систематику. У различных архетипов окажется довольно много общих строк
(меронов), поэтому встает вопрос: не попадут ли при таком вычленении птицы в
один класс с летучими мышами, не окажутся ли воробей и страус в разных клас-
сах? Мейен уверен, что этого не произойдет, и вот почему.
Ж. Кювье недаром брался восстановить облик животного по нескольким зу-
бам - одни свойства организмов влекут за собой другие. И понятие архетипа
призвано выразить эту взаимосвязь. Дело в том, что строки таблицы резко не-
равноправны: сравнительно немногие почти не содержат пустых клеток (при-
знак присутствует почти у всех особей) и образуют ядро таксона по Бэру3, а в
большей части строк пустые клетки преобладают - это периферические призна-
ки. Чтобы разумно вырезать из большей таблицы меньшую, надо так выбрать
столбцы, чтобы у них было свое ядро (почти сплошь непустые клетки в некото-
рых строках). Так, ядро класса птиц образуют летающие формы, имеющие
крепкие маховые перья и киль (гребень грудины), но ядро (в данном случае без
периферии, если не считать уродливых особей) отряда страусов - нелетающие
птицы без киля, с мягкими декоративными перьями на месте маховых. И все же
страус останется среди птиц при любом способе классификации, так как перья и
3 К. Бэр в 1825 г. говорил о том, что крупный таксон должен иметь свое ядро из харак-
терных форм (например, кошки и собаки среди хищных, воробьи и синицы среди во-
робьиных), в котором тип организации группы выражен четко, и периферию - нетипич-
ные, пограничные с другими таксонами формы (например, калан среди хищных, ворон
среди воробьиных) [8, с. 401-402]. О том же писал М.А. Максимович [5, с. 41].
26
многие другие его признаки принадлежат ядру класса птиц. Наоборот, при по-
пытке объединить каких-либо птиц и летучих мышей никакого ядра не получит-
ся - сходства будут слишком разрозненны.
Главной задачей типологии Мейен считает построение «естественной» сис-
темы организмов, столь же устойчивой к новым данным, как система химиче-
ских элементов. (Пока что споры о родстве конкретных групп не собираются за-
тухать и все родословные древа недолговечны.) Более того, Мейен не считает
обязательным и линнеевский принцип иерархии, напоминая, что Любищев
справедливо указывал: форма системы должна выявляться, а не постулировать-
ся. Что естественная система вообще-то возможна, ясно ему из того, что и в
обыденной жизни и в науке мы постоянно предсказываем свойства организмов:
по отдельным представителям таксона мы судим о других его представителях,
что было бы невозможно, если бы самим природным объектам не была свойст-
венна некая не зависящая от исследователя типологическая упорядоченность.
Поясним. У всякой птицы мы уверенно ожидаем найти перья, у каждого раз-
личимого глазом организма - много клеток, у всякого организма вообще - ДНК. И
дело здесь не в общем предке, о котором обычно ничего не известно, а в том, что
без этих меронов мы не в состоянии сконструировать соответствующие архетипы.
«Все современные птицы имеют перья, а ископаемые, для которых перья неиз-
вестны... немыслимы без них», - пишет зоолог А.С. Раутиан [9, с. 109]. Анало-
гично, единый архетип организма немыслим для нас без мерона «ДНК». Однако
типологический прогноз, как и всякий прогноз, может не оправдаться. Например,
возможен крупный организм, не состоящий из клеток (таковы плазмодии), и даже
универсальность генетического кода у некоторых одноклеточных нарушена.
Эволюционная схема (древо), показывающая происхождение данной груп-
пы видов от предполагаемого общего предка, называется филогенией (букваль-
но - «происхождение семьи»). Фактически всякая филогения, даже при наличии
ископаемого материала, строится путем группировки таксонов по сходным ме-
ронам, причем, разумеется, таксоны могут оказаться близки по одним меронам
и далеки по другим. Приходится говорить о селюфилогениях, то есть отделённых
филогениях признаков, например об эволюции плавника в лапу. Филогения дос-
товерно строится только тогда, когда все семофилогении сходятся к единой
предковой форме, но этого-то и не бывает. К. Фогт, один из первых немецких
дарвинистов, в 1863 г. горячо возражал против идеи общего предка (ее, считал
он, приписывают Дарвину противники, чтобы скомпрометировать новое уче-
ние). По Фогту, если производить строение человека от обезьяньего, то надо
взять голову ревуна, руку шимпанзе, скелет туловища гиббона, ногу гориллы и
мозг оранга; в этом он видел свидетельство того, что эволюция идет не путем
расхождения от общего предка, а путем скрещиваний. Дальнейший прогресс на-
ук, в том числе находки ископаемых, не улучшил положения: в 1977 г. ведущие
английские антропологи отказались указать филогению человека, хотя для неко-
торых костей есть «непрерывная временна* я последовательность в одной и той
27
же географической области», то есть хорошая семофилогения, но она не согла-
суется с представлениями о ходе эволюции человека в целом [10].
Причина подобных трудностей, по Мейену, в том, что сходные мероны мо-
гут быть открыты у существенно разных таксонов, а потому эволюцию можно
представить скорее не как появление разнообразия на месте однообразия, а как
преобразование одного разнообразия (полиморфизма) в другое.
Итак, биологическому языку предложено нечто вроде грамматики: словом в
ней служит мерой, осмысленной фразой - архетип (то есть таксон), а связным
текстом может служить или таксон более высокого ранга, или экосистема, или, к
примеру, последовательность сменяющих друг друга видов. По Мейену, описать
состав таксона и его архетип (то есть указать мероны) - это и значит собрать
«необходимые сведения о разнообразии объектов». Это верно, но нуждается в
оговорке: такие сведения необходимы, но недостаточны. В частности, получае-
мая система вряд ли заменит линнеевскую в отношении адресации: все-таки для
диагноза древо наиболее удобно.
Зато есть задачи, которые система-древо вообще решать не может, напри-
мер, обрисовать общую картину живой природы с ее переплетением сходств и
связей. Это не должно удивлять: в других отраслях знания тоже одной системы
не хватает. Грамматическое руководство не может заменить словарь, схематиче-
ский план города - адресную книгу. Покажем, как «грамматика биологии» по-
зволяет усвоить и упорядочить тот же феномен параллелизма, не учитываемый
обычной систематикой.
Параллелизмы и «грамматика биологии»
Если положить головами в одну сторону рыбу, лягушку, ящерицу, воробья и
крысу, то легко увидеть ряды сходных частей: ряд черепов, ряд глаз, ряд желудков
и т.д. В наше время сходство объясняют только общностью происхождения всех
позвоночных, но вопрос не так прост: ведь к тем же животным можно подложить
и рака, и жука - параллелей будет меньше, но все-таки много. Если здесь и можно
говорить об общем предке, то уж у него-то не было ни мозга, ни сердца, ни раз-
дельнополости, ни многого другого, чем сходы все высшие животные. Эту общую
тенденцию к появлению сходных свойств и называют параллелизмом.
Если взять много разных видов рыб, птиц, насекомых и т.д., то легко уви-
деть параллели в окрасках - не только покровительственные или контрастные,
но и просто сходные рисунки. Картина еще усложнится, если углубиться внутрь
организма: скелет руки поразительно сходен со скелетом ноги, хотя рука и нога
всегда имели отдельных «предков» - переднюю и заднюю лапы.
Когда удается обследовать много тысяч членов одного вида, открывается
еще один тип морфопараллелизма - уродства. Уродства часто реализуют при-
знаки, нормальные для других видов, и эта параллель очень важна для осозна-
ния морфологического единства природы. Еще в 1935 г. ботаник Н.П. Кренке
обосновал на ботаническом материале правило: признак, являющийся уродст-
вом у данного вида, обычно нормален в каком-то ином виде.
28
Обычно у миндаля плод сухой, а у персика - окруженный мясистым около-
плодником, но изредка бывает наоборот, причем необычная форма может суще-
ствовать и как индивидуальная вариация, и как устойчивая раса [11, Табл. 41,
пример 227]. Эта пара видов проявляет целый ряд сходств изменчивости, и его
можно сопоставить с падежным рядом существительных одного склонения. Как
и в грамматике, ряды сходств довольно полны для близкородственных видов
(персик и миндаль обычно включают в один род Amigdalus L.) и довольно обры-
вочны для форм далеких; но как и в грамматике, определенных границ для прояв-
ления сходств нет. Н.П. Кренке назвал этот феномен «законом родственной из-
менчивости». Вне ботаники прекрасным примером того же закона является мута-
ционная изменчивость дрозофил, почти одинаковая у 150 видов этого рода.
Это оказалось общим правилом: чем значительнее отклонение от нормы,
тем реже оно наблюдается и тем дальше в таксономическом отношении вид, у
которого соответствующее свойство можно встретить в качестве нормального.
С древности и до наших дней параллель между индивидуальными и таксо-
номическими различиями привлекалась для эволюционных гипотез (на уровне
расовых и видовых различий она легла в основу учения Дарвина), но здесь она
важна для другого. Она показывает, что формообразование у всех организмов
подчиняется некоторым общим законам, которых мы еще почти не знаем. Уже
на приведенных примерах можно видеть, насколько сложно явление паралле-
лизма, так что классифицировать разнообразные типы параллелей было очень
трудно без «Грамматики биологии». В ее же свете видно, что все типы паралле-
лизма подчиняются простой схеме: у разных таксонов - общие мероны.
Самое замечательное то, что этой формуле можно действительно придать
вполне грамматический вид, и недаром раздел грамматики, изучающей строение
слов, тоже называется морфологией. Слово делится на основу и окончание, ко-
торое, изменяясь, указывает грамматическую форму (род, число, падеж, время,
лицо). Изменяя окончания, мы получаем разные формы данного слова, то есть
термин «слово» выступает как род, где отдельные формы этого слова являются
видами. Основа в свою очередь может состоять из корня и аффиксов (то есть
приставки и суффикса), и группа слов с общим корнем образует гнездо, которое
играет здесь роль семейства.
Слова можно группировать по-разному - просто в алфавитном или алфа-
витно-гнездовом порядке (обычные словари), в алфавите окончания (рифмовни-
ки), по частоте употребления (частотные словари), по смыслу (словари синони-
мов), по происхождения (этимологические словари) и т.п., и какой бы способ ни
выбрать, обнаружится параллелизм слов - просто потому, что другие, не ис-
пользованные для классификации признаки будут заявлять о себе на каждой
странице. Точно так же и в биологии: по каким бы правилам ни создавать сис-
тему, другие, неучтенные закономерности выступят как параллелизм.
С.В. Мейен предлагает видеть в каждом таксоне как архетип по Оуэну (то
есть нечто коренное), так и изменчивое множество, причем это последнее ока-
29
зывается упорядоченным, как упорядочены падежные или личные формы. Эту
упорядоченность можно наблюдать на таксонах всех уровней - от подвида (бар-
хатистые и гладкокожие персики) до надцарства (водоросли повторяют свойства
цианобактерий), но мы поясним ее на одном примере - характере изменений в
пределах мерона «конечность».
Этот мерой вообще может отсутствовать в архетипе таксона (губки, черви,
растения), а может принимать самые различные значения. Если говорить только о
парных конечностях позвоночных и избегать деталей, то обнаруживается сле-
дующая картина. Позвоночные делятся на пять классов: рыбы, амфибии (земно-
водные), рептилии (пресмыкающиеся), птицы и звери (млекопитающие). Мерой
же принимает девять основных значений: простой плавник - только у рыб; плав-
ник-ласт (с суставчатым скелетом) - встречается у всех пяти классов (кистеперые
рыбы, ископаемые амфибии и рептилии, пингвины и киты); ласт (для плаванья и
ползания по земле) - у вымерших амфибий и рептилий, у черепах и млекопитаю-
щих (ластоногие); рыбы, как ни странно, ползают с помощью простых плавников;
ползательная лапа - у многих амфибий, рептилий и некоторых млекопитающих,
часто снабжена плавательной перепонкой; «.планер» (лапа или плавник с лета-
тельной перепонкой) - у некоторых рыб, лягушек, ящериц и мелких зверей; крыло
для активного полета - у вымерших птицеящеров, у птиц и летучих мышей; как
редкое исключение - у рыбок (из семейства харциновых) наблюдаются элементы
активного полета; нога для хождения и стояния без касания брюхом земли - у не-
которых вымерших амфибий, у рептилий, птиц и зверей; хватательная лапа - из-
вестна у всех классов, кроме рыб; рука - только у приматов*.
В этих параллельных рядах наблюдаются одни и те же тенденции. Как у
всякого мерона, у конечности бывают разные степени выражения: полное отсут-
ствие, рудимент (зачаток, невидимый снаружи или видимый, но бесполезный),
слабое развитие, полное развитие. Во всех классах мы видим тенденцию к реа-
лизации самых разных степеней выражения конечностей: полное отсутствие на-
блюдается у рыб (семейство угрей - отсутствует передняя или обе пары), у ам-
фибий (червяги) и рептилий (змеи); рудименты бывают у пещерных рыб, мно-
гих хвостатых амфибий и ящериц, у китов (задняя пара) и у некоторых птиц
(передняя пара). Оформленные, но совсем слабые конечности тоже хорошо из-
вестны (у всех пяти классов), так что можно выстроить пять параллельных ря-
дов по степеням их развития.
Другую тенденцию отметил еще Г. Брони (1858): по мере перехода от ти-
пично морских к типично наземным организмам наблюдается усложнение
строения организмов. Он назвал это террипетным (то есть устремленным к зем-
ле) развитием - developpement terripete, но мы можем говорить о большем: о
террипетной тенденции в организации разнообразия, как ныне существующего,
Таблица этого рефрена приведена в конце Сборника (рис. 1).- Примеч. 2010 г.
30
4 т'»
так и развивающегося в ходе эволюции . В частности, она хорошо видна и на
конечностях: всякое возвращение в воду (превращение конечностей в ласты и
плавники) приводит к упрощению морфологии конечностей, что видно во всех
четырех наземных классах.
«То, что разные члены рядов изменчивости объединяются одной тенденци-
ей, одним правилом преобразования, дает возможность предсказывать и целена-
правленно искать недостающие члены. Эту повторяющуюся, подчиненную од-
ному правилу преобразования последовательность состояний мерона назовем
рефреном», - пишет Мейен [7, с. 501]. Рефрен можно, пусть и вольно, сравнить
с набором форм спряжения глаголов или склонения существительных, причем
неполнота рядов в грамматике тоже обычна - как для отдельных слов («сутки»
не бывают в единственном числе), так и для целых классов (в русском языке со-
слагательное наклонение не имеет временны» х форм: «Если бы завтра был мо-
роз...»). И точно так же, как правила грамматики позволяют нам лучше пони-
мать язык при меньшей затрате сил, так и рефрен дает нам лучшее и экономное
средство для ориентировки в разнообразии.
Те же конечности позвоночных: можно ли как-то упорядочить их разнооб-
разие, выявить, что здесь возможно, а что нет? Оказывается, это разнообразие
складывается за счет наложения небольшого числа рефренов разного уровня.
Легче всего заметен тот рефрен, который привел Бронна к идее террипетно-
сти: переход с суши в болото и воду, с поверхности под землю, из толщи вод на
дно - все это вызывает редукцию (упрощение, уменьшение, утрату) конечно-
стей, преимущественно задних; если остается одна пара конечностей, то перед-
няя. Среди рыб мы видим это у угрей (зарываются в дно), у водных форм, при-
надлежащих наземным классам - у ихтиозавров и китов. Очень характерен и ка-
лан: у этого прибрежного морского зверя спереди еще лапы, а сзади уже ласты.
Хотя у современных амфибий и рептилий ласт нет (кроме некоторых черепах),
однако рефрен ясно виден и у них: у подземных, болотных и водных форм
уменьшаются или утрачиваются лапы. В обоих классах есть небольшие группы
«двуруких» (хвостатые амфибии си* рены и ящерицы хиро* ты), сохранившие
крошечные, но вполне развитые передние лапки. Объяснить их как приспособ-
ление никто и не пытается (другие виды в тех же условиях несут по четыре
крошечные лапки или вообще безноги), так что рефрен можно уверенно тракто-
вать как чисто типологический, а не экофизиологический феномен.
Донный угорь, болотный сирен, подземный хирот, морской хищник ихтио-
завр и поедатель планктона кит - все они встали в один ряд, реализовав в реф-
рене мерона «конечность» состояние «развита только слабая передняя пара».
Это состояние расположено на периферии всех пяти классов, занимая в рефрене
«террипетное преобразование конечностей» одну из последних позиций (далее
уже следует полное вторичное исчезновение конечностей).
4 О взглядах Бронна см. [12] (а также в ЭЭД и АСМ. - Примеч. 2010 г.).
31
Есть несколько примеров, казалось бы, противоречащих найденному ряду: у
нескольких видов змей и безногих ящериц заметны только рудименты задних
конечностей (змеями называют рептилий, не имеющих даже следов плечевого
пояса), а у нелетающих птиц развита только задняя пара конечностей. Однако
эти примеры сами образуют ряд, и он связан не с террипетностью, а с совсем
другой тенденцией, которая образует свой рефрен.
А именно, у амфибий, рептилий и зверей наблюдается тенденция к росту
задних конечностей и переносу на них тяжести тела, вплоть до прямосидения и
прямохождения, а затем и летания (при этом, в частности, рептилии эволюцио-
нировали в птиц). Естественно, что передняя пара при этом упрощается и срав-
нительно легко редуцируется*, а в случае полной утраты конечностей задняя па-
ра отстает, отчего на периферии классов и наблюдается упомянутый ряд**. В
данном рефрене мерона «конечности» он реализует группу состояний «более
редуцирована передняя пара». Факт, что задние рудименты выражены чаще и
лучше, отметил еще в 1931 г. морфолого-эволюционист А.Н. Северцов.
Грамматика, когда начинаешь ее изучать, подчас кажется надуманной и из-
лишней, и многие призывали «писать как слышится», однако фактически без
грамматики никто не пишет. Всякая наука рано или поздно составляет себе об-
щепринятую «грамматику»; недалеко, вероятно, то время, когда систематик,
предлагая объединить какие-то виды в один род, будет предъявлять таблицы их
«склонения», то есть единообразно описанной изменчивости.
Литература
1. Bemhardi J.J. Uber den Begriff der Pflanzenart und seine Anwendung. Erfurt, 1834.
2. Дарвин Ч. Собр. соч. M.; Л., 1951. Т. 4.
3. Любищев А.А. Проблемы формы, систематики и эволюции организмов. М., 1982.
4. Мейен С.В., Соколов Б.С., Шрейдер Ю.А. Классическая и неклассическая биоло-
гия: Феномен Любищева// Вестник АН СССР. 1977. № 10 (М-150).
5. Максимович М.А. О системах растительного царства. М., 1987.
6. MeyenS.V. 1973. Plant morphology... (М-116).
7. Мейен С.В. 1978. Основные аспекты типологии... (М-165).
8. Бэр К.М. О сродстве животных// Анналы биологии. М., 1959. Т. 1.
9. Раутиан А.С. Уникальное перо птицы... // ПЖ, 1978. № 4, с. 109.
10. Харрисон Дж. и др. Биология человека. М.: Мир, 1979.
11. Феногенетическая изменчивость/ Под ред. Н.П. Кренке. М., 1933-1935. Т. 1.
12. Чайковский Ю.В. 1984. «Происхождение видов»: Загадки первого перевода (4-10).
Эта тенденция носит ясный физиологический характер: при потере функции орган под-
вергается снятию давления нормы - явлению, которое описала зоолог Е.Е. Коваленко
(см. АСМ). Наоборот, у террипетности в целом такого характера не видно - она носит
преимущественно (а может быть, целиком) диатропический характер. - Примеч. 2010 г.
" Эволюция как движение вдоль рефрена позже получила название актиреф - см. далее,
п.7 заключительной статьи, а также ЗЭ, с. 56. - Примеч. 2010 г.
32
Наука о разнообразии*
Диатропика. Суждение о новизне
Разбирая публикации Сергея Викторовича и размышляя над кругом идей, ко-
торые они породили, я пришел к выводу, что можно говорить о новой науке,
предмет которой - разнообразие. Подыскивая для нее название из подходящих
греческих терминов, я выбрал слово диатропика, от диатропос - разнообразный,
разнохарактерный, поскольку само понятие тропос (поворот, способ, образ мыс-
лей, нрав, обычай, слог, стиль, направление) наиболее многозначно и хорошо со-
ответствует широте спектра задач, решаемых новой наукой. Исследуя те сходства
различных объектов и те различия сходных, на которые в других науках редко об-
ращают внимание, диатропика имеет отношение ко всему на свете.
Это значит, что очерчивая круг ее задач, важно соблюсти определенный такт,
чтобы наука не превратилась в пустой разговор на научные темы, как то не раз слу-
чалось с другими направлениями мысли. Иными словами, надо ясно определить не-
который аспект рассмотрения, в котором можно рассчитывать на содержательные
результаты и в то же время не пытаться подменить уже существующие науки (не
выдавать их результаты за собственные). Далее попробую очертить этот аспект.
Рождение действительно новой науки - вещь крайне редкая. Возникающие то и
дело новые названия дисциплин обычно отражают либо детализацию предмета
(так из науковедения выделилась наукометрия), либо появление нового объекта
исследований (селенология - геология Луны), либо стык наук (биогеохимия), а
чаще всего - все три феномена сразу (промышленная экология, внеатмосферная
астрономия). Это, так сказать, новые поколения научных дисциплин, вторичные
дисциплины, рожденные новыми фактами - прямо или опосредованно.
Несравненно реже мы видим рождение нового знания, основанное на переос-
мыслении старых фактов и представлений, где новые факты играют роль пусть и
существенную, но подчиненную, где дается новое воззрение на прежний мир. Так
около 1900 г. родилась генетика, когда Г. Де Фриз нашел, что наследование - не-
что большее, чем «физиология размножения». И уж совсем редко такое рождение
затрагивает все науки сразу, как это случилось с появлением общей теории сис-
тем, когда А.А. Богданов (1913) указал на то, что система имеет свои законы, не-
сводимые к свойствам входящих в нее элементов. Даже молекулярная биология и
кибернетика с этой точки зрения вторичны. У нас же пойдет речь о рождении дей-
ствительно новой, первичной науки, для которой общая теория систем дает фор-
мальный аппарат - как математика дает аппарат для физики или статистики.
Известность разных идей Мейена далеко не одинакова. Если как палеобота-
ник он был признан научным сообществом в роли высшего авторитета, если как
геолог он считался ведущим стратиграфом, если как философ и методолог он
был желанным участником конференций и сборников, то среди теоретиков эво-
Химия и жизнь, 1989, № 1, с. 40-48. Здесь опущены введение, взятое из Некролога (см.
далее), и малоосмысленный абзац в конце, отмеченный знаком [...].
33
люционизма его признавали лишь в качестве главы нетрадиционного направле-
ния, едва упоминаемого в руководствах, а как теоретик общей биологии Мейен
остался одинок, подобно Архимеду.
Я далеко не все понимаю у Мейена и не во всем, что мне понятно, с ним со-
гласен. Что могу, стараюсь изложить популярно, тем же заняты некоторые его
ученики, но чувствуется, что этого мало. В принципе, так и должно быть: кого
современники сразу поняли, тот не первопроходец, а скорее выразитель мнений,
к которым общество готово. Однако науковедение для того и родилось, как мне
кажется, чтобы избавлять развитие науки от ложных шагов, одним из которых
всегда было непризнание первопроходцев. Привычная для историка науки си-
туация - когда приходится констатировать, что ныне господствующие взгляды
много раз выдвигались в прошлом и отвергались современниками, - должна, по-
моему, перестать быть обычной, поскольку слишком дорого обходится людям.
Опыт показывает, что если ученый был настоящим корифеем (его лидерство не
было следствием его административного положения или научной моды) в своей
области, то и его размышления на общие темы не бывают совсем пустыми.
Шесть десятилетий о Л.С. Берге, создавшем учение о номогенезе*, говорили,
что он прекрасный ихтиолог и географ, но дарвинизма не понимает, и только
теперь, когда дарвинизм, похоже, прекратил свое существование как направле-
ние биологического исследования, сохранившись лишь как объект историческо-
го анализа, подражания и преподавания, - только теперь выясняется, что Берг
видел дальше своих критиков. Вдруг стали замечать давно известное - что среди
его приверженцев были такие знаменитые наши эволюционисты, как Ю.А. Фи-
липпенко, Н.И. Вавилов и А.А. Заварзин; что среди его гонителей, наоборот, ко-
рифеев не числится. Хотелось бы, чтобы с идеями Мейена этого не произошло.
Мейен ушел в эволюционизме гораздо дальше Берга и Любищева. Если у них
основным пафосом была критика, то Мейен, с самого начала усвоив ее, сосредо-
точил усилия на конструктивной теории. Если оригинальные построения номо-
генетиков мыслились как альтернатива дарвинизму, как объяснение эволюции
«не по Дарвину», то Мейен стремится к синтезу, к работоспособной теории’*.
Помните, как сказано у Маркса? Философы до сих пор объясняли мир, хотя
задача в том, чтобы его переделать. Перефразируя, можно сказать: эволюциони-
сты до сих пор спорили о том, какие факторы привели мир в наблюдаемое нами
состояние, хотя задача - в том, чтобы спасти его от этого состояния. Сейчас, ко-
гда экологический кризис ставит вопросы об эволюции биосферы не в порядке
гипотетического объяснений, а с целью найти способы жить в ней, когда преж-
ние натурфилософские построения (ламаркизм, жоффруизм, дарвинизм, номо-
генез) непременно должны уступить место конструктивным схемам, допускаю-
щим хоть в какой-то мере прогноз и практические рекомендации, - сейчас идеи
Мейена нельзя оставить вылеживаться до лучших времен.
Характеристику номогенеза Берга см.: ИМ, с. 106-110; АСМ; ЗЭ. - Примеч. 2010 г.
Что такая теория не будет синтезом прежних, мне стало ясно позже. - Примеч. 2010 г.
34
До сих пор в истории науки преобладает выявление предшественников ны-
нешних теорий, отчего альтернативные пути волей-неволей затаптываются. Вот
уже 300 лет вокруг стелы эволюционизма водит хоровод пяток одних и тех же
фигур, по очереди высвечиваемых лучом научной моды - Ламарк, Жоффруа,
Дарвин, Берг и Вернадский* (похожие дискуссии велись не раз; первая, извест-
ная мне, происходила в середине XVII века). Чтобы прочесть, наконец, что на
стеле написано, надо прекратить хоровод. Необходим синтез, к которому впер-
вые призвал Мейен и к которому он, насколько успел, приблизился.
Тесный мир логики
Исходное для диатропики понятие - ряд, она оперирует им так же, как опыт-
ные и наблюдательные науки - понятием факта. И так же, как факт не имеет
смысла (а подчас и места) вне объясняющей схемы, так и ряд бессмыслен без
сопоставления с другими. Смысл ряда радикально зависит от того, с какими ря-
дами он сравнивается. Именно этой произвольностью объясняется, по-моему,
тот факт, что диатропика до сих пор служила в качестве Madchen fur alles (де-
вочка для всего - нем.), а не заводила собственный дом.
Ряд может быть задан общим свойством его членов — например, ряд зеленых
стульев: из множества стульев извлечены обладающие свойством зелености.
Можно задать ряд способом его построения - ряд простых чисел, алфавитный
порядок слов. Наконец, можно задать ряд путем сопоставления с другим рядом
- например, в англо-русском словаре русская часть сопоставлена по смысловому
принципу английскому алфавитному ряду. Существенно, что ряд не может быть
задан перечислением, что он является подмножеством, извлекаемым из заданно-
го множества посредством определенного правила. Слово «ряд» не очень удач-
но, поскольку ассоциируется с линейной упорядоченностью (которой в действи-
тельности может и не быть), но оно уже прижилось, и менять его. поздно.
Наиболее частым является третий способ построения - сопоставление рядов,
связанное с параллелизмом. Упорядочивая элементы по одному принципу, мы
то и дело видим проявление какого-то другого принципа. Так, в англо-русском
словаре на буквы A, S видим массу эллинизмов, а по всему словарю - массу ла-
тинизмов в окончаниях на -tion (-ция) типа evolution - эволюция. Здесь паралле-
лизм проявляется поэлементно, но это необязательно. Так, во всех частотных
словарях зафиксировано одно и то же гиперболическое распределение слов по
частотам их употребления в текстах - это параллелизм системный.
Наиболее четким (правда, редким) случаем параллелизма является периодич-
ность - например, периодическая система химических элементов, где элементы
упорядочены в строки по заряду ядра, а сходства реализуются в форме столбцов.
Как сформулировал философ Ю.А. Урманцев, системы «действительно обна-
руживают определенный шаблон - повторяющиеся от системы к системе строй
В.И. Вернадскому в то время приписывали концепцию эволюции биосферы (включая
возникновение жизни), каковой у него нет (см. ЭЭД; 4-28; АСМ). - Примеч. 2010 г.
35
и порядок». С точки зрения логики, это - следствие известного принципа Ди-
рихле: объектов в природе больше, чем логических возможностей, вот свойства
объектов и повторяются. Мир логики тесен для мира феноменов. Встает вопрос
о законах этой повторности, который и решал на своем материале Мейен.
Материал этот в сущности стар, как сама ботаника. В «Инвентаре растений»
Каспара Баугина (1623) зарегистрировано много повторностей - как в форме
синонимов, так и при группировке видов, но нет никаких свидетельств осоз-
нания параллелизма как феномена. У Линнея (1735) феномен ясно осознан, и
даже упомянута потребность в таблице, но таблицы нет. Первую попытку за-
дать повторность в форме таблицы, пересечением строк и столбцов, дал, на-
сколько мне известно, шведский ботаник Элиас Фрис (1825), и почти тогда же
параллелизм описан в гуманитарной сфере, но тоже ботаником: Александр
Гумбольдт (1814) обнаружил типологическое сходство мифов Центральной
Америки и Африки. Однако только после формулировки Д.И. Менделеевым
(1869) периодического закона стали появляться различные «периодические
системы организмов».
Беда этих систем состояла в том, что они были или слишком общи, или в таб-
лицах преобладали пустые клетки. Именно на этом основании их отвергало
большинство систематиков, но вот Мейен понял главное: пустота клеток озна-
чает лишь избыточность табличного принципа записи, а не порочность самого
описания посредством пересечения строк и столбцов.
Этот принцип описания прокламировал Н.И. Вавилов в своем знаменитом за-
коне гомологических рядов (1920), но у него ряды задавались только сопостав-
лением друг с другом. В 1935 г. московский ботаник Н.П. Кренке предложил
систематизировать формы внутри каждого ряда с помощью своего «закона род-
ственных уклонений»: таблицами можно упорядочивать не только сами свойст-
ва, но и пути их изменений. С этого пункта и начал свое построение Мейен.
Другим исходным моментом было для него давно забытое замечание Н.Н.
Страхова (1858) о том, что сравнительная анатомия классифицирует части тела
так же, как систематика - организмы. По аналогии с понятием таксона (вид, род,
семейство, отряд, класс...) Мейен ввел понятие мерона - «класса частей». Орга-
низму свойственны признаки, а таксону - мероны. Позвоночное имеет две пары
конечностей (плавники, ласты, лапы, крылья, ноги, руки), а таксон «позвоноч-
ные» имеет мерой «парные конечности».
В этих терминах Мейен сформулировал два простых и важных утверждения.
Первое', классификационная наука состоит из таксономии (исчисления таксонов) и
мерономии (исчисления меронов), причем процедура классификации всегда состоит
в попеременном обращении то к одной, то к другой. Иными словами, новая система
всегда исходит из прежней (а в самом начале была интуиция), тогда как исследова-
тель порой думает, что строит систему сам. Второе: при переходе от одного таксона
к другому всегда наблюдается сходный (а иногда и тождественный) ряд меронов;
эту повторяющуюся последовательность Мейен назвал рефреном.
36
Наиболее ясные примеры рефренов дает грамматика в виде правил склонения
или спряжения. Очевидно, что слова «пол» и «стол» преобразуются по одному и
тому же закону, но самое интересное не в этой поверхностной симметрии, а в
том, что все русские существительные преобразуются по одному и тому же за-
кону, именуемому русской падежной парадигмой. Вместо того, чтобы запоми-
нать все формы каждого существительного, можно запомнить его исходную
форму и номер склонения. Сказавши, что в каждом склонении есть шесть па-
дежных форм (в единственном и множественном числах), мы кратчайшим обра-
зом опишем разнообразие форм существительных, если не захотим углубляться
в маловажные детали.
Увы, в биологии такой классификации нет, каждую видовую форму прихо-
дится описывать и заучивать отдельно. А ведь материал явно проявляет регу-
лярность. Возьмем хотя бы конечности позвоночных: в пяти главных классах
(костистые рыбы, амфибии, рептилии, птицы, звери) наблюдается один и тот же
рефрен - от полного отсутствия одной или обеих пар, через зачаток или слабо-
развитую пару, полноценный плавник (ласт), лапу и планирующую поверхность
- к органу активного полета (крылу). Правда, некоторых вариантов не бывает (у
амфибий не бывает крыльев, не бывает зверей без передних конечностей), но в
остальном параллелизм удивительно полон. Достаточно отметить, что у четырех
классов наблюдается хиротностъ (наличие крохотных «ручек» при полном от-
сутствии «ножек»): среди рыб это китовидка и угорь, среди амфибий - си* рен,
среди рептилий - хирот, среди зверей - киты (см. рис. 1 в конце книги).
У всех пяти классов мы видим ласты, способность к планированию, утрату
одной пары (или обеих пар) конечностей и тому подобное, причем формы от-
нюдь не связаны родством, как не связаны родством существительные первого
склонения. Китовидка удивительно похожа на гренландского кита, только кро-
шечного, хотя принадлежит к другому классу и живет в других условиях.
Сходство часто не связано ни с происхождением, ни с приспособлением. Вы-
вод Мейена: следует разработать две теоретические процедуры - исторических
реконструкций и адаптивных интерпретаций. Наряду с ними следует признать
всеобщность рефренной структуры, которая реализуется в силу чисто систем-
ных (диатропических) свойств всякого архетипа (см. предыд. статью - примеч.
2010 г.); приспособительный характер рефрена выражается в одном: плохо при-
способленные редки, а совсем неспособных к жизни, понятно, нет.
Это - совершенно новый для биологии принцип. Рядом с приспособлением,
господствовавшим у Ламарка и Дарвина, встает не менее важный феномен -
разнообразие. Пока биология имела дело только с фактами, а не с их рядами, за-
метить это было невозможно. Естественно встает вопрос: а законно ли вообще
рассматривать приспособление как главный принцип организации живого? Если
от поддакивающей истории обратиться к истории напоминающей, то окажется,
что этот вопрос давно задавали умнейшие эволюционисты, в том числе Анри
Бергсон (1907). Он видел в приспособлении к определенной среде лишь вторич-
37
ный эффект, а первичным полагал внутренний импульс к развитию. По этой
идеологии жизнь подобна фонтану: вверх струя взлетает, руководствуясь «внут-
ренним импульсом», вниз же капли разлетаются, приспосабливаясь ко внешним
обстоятельствам. В эволюции, как она представляется мне, равноправны три
феномена: прогресс, приспособление и разнообразие - но до сих пор ни одно
учение не претендовало на их синтез. Дарвинизм категорически игнорировал
прогресс, номогенез едва упоминает приспособление, а в ламаркизме приниже-
но разнообразие (Мейен сумел наметить синтез схем дарвинизма и номогенеза).
Ядро и периферия
Не имея дела с отдельными фактами, диатропика не может давать утвержде-
ний, справедливых абсолютно для всех объектов. Изучая какое-либо множество,
она всегда выявляет в нем ядро типичных объектов, для которых формулируе-
мые закономерности выполняются очевидным образом, и периферию - сравни-
тельно немногочисленные объекты, на которых закономерности данного мно-
жества видны плохо, вплоть до, может быть, полного отсутствия. Так, позво-
ночным свойственны две пары конечностей, точнее, это свойство ядра позво-
ночных, а на периферии типа мы видим три пары у кистепёрых рыб, одну пару у
хиротных и даже отсутствие обеих пар: один крохотный класс (круглоротые),
один отряд (змеи) и несколько более мелких таксонов.
Ядро и периферию можно выделить почти всюду (заметьте: без «почти» не
обойтись - такова уж диатропика), и это позволяет по-новому взглянуть едва ли
не на все науки. К примеру, классификация: вместо вековых бесплодных споров
о том, сколько в живой природе царств - 2, 4, 7 или 23, - следует задать вопрос:
сколько можно выявить ядер примерно равного ранга? Исследовав этот вопрос
подробнее, легко понять: царств всего четыре (бактерии, растения, грибы и жи-
вотные). Всё же остальное - периферические группы, не образующие ядер, но
легко объединимые в три межцарства, где попарно комбинируются свойства
царств растений, грибов и животных (см. табл, на обложке - примеч. 2010 г.).
Так же упрощается дело на всех уровнях систематики. Нечего спорить о том,
к енотам или собакам отнести енотовидную собаку, ибо она - периферический
вид между семействами псовых, енотовых и виверровых. Ее отнесение к псовым
- дело вкуса, и лучше уж не ломать установившуюся традицию, оставить это
существо среди псовых. Вместо спора о границах, надо четко выявлять ядра.
Вообще же можно сказать, что ядро и периферия есть не только у каждого так-
сона, но и у каждого закона5. Во всякой большой системе есть объекты (ядро за-
кона), на которых данная закономерность действует как бы в чистом виде, тогда
как у других ее тоже можно заметить, но она как бы смазана действием других
законов. Если ядро можно выявить, то закон налицо, если же оно теряется в сво-
ей периферии, то о законе лучше не говорить - вот и всё.
5 Под законом подразумевается формулировка (правило) и объясняющий это правило ме-
ханизм, а под закономерностью - фактически наблюдаемая в природе тенденция (ряд).
38
И все-таки ученые спорят, столетиями повторяя одни и те же аргументы и за-
ставляя вхолостую работать новейшую и сложнейшую технику - лишь потому,
что им не приходит в головы сперва договориться, какого типа закономерности
вообще можно надеяться наблюдать на данном множестве фактов.
Например, планетные астрономы спорят так о реальности и роли резонансов
- совпадений (или кратностей) периодов, и исход спора важен для понимания
процессов в Солнечной системе. Резонансы бывают разной степени точности -
от справедливых абсолютно (Луна всегда повернута к Земле одной стороной в
силу совпадения периодов ее вращения и обращения вокруг Земли), до весьма
сомнительных (приблизительное совпадение периодов солнечной активности и
обращения Юпитера). Взгляд с точки зрения диатропики здесь, по-моему, силь-
но упрощает дело: как качественная закономерность резонансы достоверны, но
как точные равенства они, вероятно, - лишь редкие исключения.
В сущности, всякая наука имеет дело не с самими законами, а с закономерно-
стями, описывающими их ядра. Мейен не раз напоминал, что самый респекта-
бельный и точный закон на поверку имеет периферию. Например, двудольные
плоды не всегда имеют две семядоли - надо лишь просмотреть достаточное число
экземпляров исследуемого вида. Совсем другой пример. Всем вроде бы известно,
что падежей в русском языке шесть, но это - лишь простейший способ описать
ядро именных словоформ. При попытках детализировать или уточнить само поня-
тие падежа их оказывается в русском языке до девяти. То же и в других языках.
Сколько раз превозносилась универсальность генетического кода! А потом
периферию нашли и тут: «пятая буква», нетипичные значения триплетов и т.д.
Разнообразие тоже наследуется
Если говорить о вкладе диатропики в эволюционизм, то надо начать с феноме-
на, который Мейен назвал транзитивным полиморфизмом. В череде поколений
разнообразие восстанавливает себя почти независимо от того, какая его часть бе-
рется для размножения. Яркий пример приводил Дарвин в 1868 г. Известно ог-
ромное разнообразие бархатистых персиков - по вкусу, форме и цвету плодов, по
форме косточки и т.д. Среди них возникла вариация (мутант) с гладкой кожицей,
и вот она повторила своими (вторичными) вариациями чуть ли не все формы бар-
хатистых персиков. Глядя на это вторичное разнообразие, нельзя понять, из какой
формы оно получено - какая форма послужила для транзита. Рефренная структура
вида была воспроизведена заново, и вновь появились свойства, которыми исход-
ный мутант с виду не обладал. Разнообразие было передано по наследству.
Транзитивный полиморфизм, как и все законы разнообразия, воспринимается с
трудом, несмотря на свою эмпирическую очевидность. Поскольку человек сам
производит отдельные действия и следит за отдельными объектами, то ему трудно
усвоить ту мысль, что природа оперирует с разнообразиями, взятыми целиком. В
частности, и эволюцию принято рассматривать как появление и изменение от-
дельных свойств у отдельных видов. Особенно грешат этим ламаркизм и дарви-
низм. Селекция пород, полезных в каком-то отношении, давно стала моделью
39
эволюции, хотя Спенсер еще в 1864 г. заметил (в книге «Основания биологии»),
что на самом деле эволюция представляет собой целостный процесс, в котором
нельзя выделить отдельные эволюционирующие по своим законам объекты.
Это, в сущности, библейская традиция - следить за генеалогией отдельных лиц
или их качеств, идея преследовать «до седьмого колена», основанная на уверенно-
сти, что «вор рождает вора». В действительности же общество само порождает всё
свое разнообразие, в том числе и воров. Пусть вор и происходит чаще всего от во-
ра, но это вовсе не значит, что после ликвидации всех воров их не будет через од-
но-два поколения ровно столько же - сколько соответствует системе. Передача
через поколения (транзит) может происходить и поэлементно, но важно понять,
что суть транзита не в этом, а в порождении разнообразием разнообразия. Именно
поэтому, в частности, бессмыслен всякий индивидуальный террор, если речь идет
об эволюционном масштабе времени. Однако краткосрочные перемены он поро-
ждать может - ведь транзит требует смены поколений.
Кстати, Мейен часто пользовался термином «индивидуальный террор» для
шутливого обозначения объяснений каждого факта в отдельности. Он призывал
эволюционистов сменить «индивидуальный террор фактов» на выявление тен-
денций, а затем - и их механизмов (законов). В отношении транзитивного поли-
морфизма он указывал на то, что в рамках возможностей данного рефрена каж-
дый признак реализуется сам собой - пусть и редко, но достаточно регулярно.
Выражение «сам собой» означает, в частности, что любое состояние мерона
может, в принципе, реализоваться у любого таксона, то есть у разных таксонов
бывают общие мероны. И наоборот: разные мероны (точнее: состояния одного и
того же мерона - примеч. 1998 г.) могут быть в общем таксоне. Мейен кратко
называл это обстоятельство мероно-таксономическим несоответствием, уста-
новление же самого феномена связывал с работами Урманцева.
Важно отметить, что такое несоответствие скрепляет разнообразное множество
(сообщество) в систему, обеспечивая в ней связь всех элементов со всеми. Мас-
штабы несоответствия могут быть очень различны. Там, где несоответствие слиш-
ком велико, бывает трудно выявить ядро и периферию, а значит - ввести класси-
фикацию объектов (элементов). В ботанике таксоны с такими свойствами издавна
называют crux botanicorum (крест ботаников). Там же, где несоответствие слиш-
ком мало, наблюдается распад системы на слабо связанные подсистемы с малым
разнообразием в каждой. Эволюционисты давно описали это как специализацию и
установили, что она дает краткосрочные выгоды, а затем ведет к вымиранию.
Более всех преуспели в специализации насекомые, например, те тли, у кото-
рых каждый вид может питаться одним-единственным видом растений. Остав-
шись без него, все или почти все особи мрут от голода при изобилии пищи. Эво-
люция таких видов невозможна без катастроф.
Берегите несоответствия!
Ценность диатропики - в ее междисциплинарности, в применимости к любым
системам, в том числе и социальным. Так, сейчас много пишут о том, что в нашем
40
обществе нарушены многие соответствия, необходимые для его успешного разви-
тия: цен и спроса, дохода и производительности, прав и обязанностей и т.д.
Все это верно, но никто еще, кажется, не написал, что у нас опасно нарушено
одно несоответствие - разнообразия общественных институтов с разнообразием
их функционеров. Всякий общественный институт организован по функцио-
нальному принципу, но люди, его наполняющие, демонстрируют разнообразие,
а оно подчиняется другим законам, законам диатропики. Им же подчиняется и
разнообразие самих общественных институтов - таксонов. Каждый человек об-
ладает множеством признаков (каждый признак есть состояние мерона), причем
далеко не все активны, не все используются. Специализация института выраже-
на в том, что он заполняется людьми с определенными активными качествами, в
результате чего падает мерономическое разнообразие этого таксона.
Разные таксоны при этом теряют значительную долю общности меронов, что
понемногу приводит их к невозможности прямого функционального контакта, к
потере общего языка. Обычно об этом говорят как о ведомственности и бюро-
кратизме, но понимания сути тут еще мало, что видно в безуспешности методов
борьбы с этим злом путем «координации». В одной из недавних публикаций6
сказано (со ссылкой на «Детей Арбата» Рыбакова): «Топор репрессий был... на-
правлен не на людей - на связи между ними». Хотя Сталина и беспокоили, как
отмечено там же, связи между руководителями, но достигнуто было гораздо
большее: мероны разместились по таксонам, и параллельные (недолжностные)
связи между общественными институтами стали распадаться. Кастовая замкну-
тость во многих из них намного превысила дореволюционную. О придворных,
сенаторах, министрах, главных фабрикантах и фельдмаршалах обычно было из-
вестно, что они любят, чем их можно заинтересовать или отпугнуть, где они бы-
вают и кто бывает у них. Сходная картина была до середины тридцатых годов, и
как курьез напомню, что Бабель водил компанию «с главными милиционерами»
(и чуть ли не с Ежовым). Сейчас этого нет, пресечено большинство каналов не-
официальной информации. Во всех отраслях нашей жизни аппарат живет обо-
собленно от управляемых и, насколько мне известно, столь же обособлены друг
от друга работники аппарата разных этажей и разных отраслей.
Указывать выход - задача не диатропики, но она может и здесь дать рекомен-
дации, неучет которых будет (как свидетельствует исторический опыт) губителен.
В частности, проводимые сейчас попытки сменить одни регламентации на другие
заведомо бесполезны, поскольку не восстановят полноценного разнообразия. [...]
Симпатии российского общества, к сожалению, всегда клонились в сторону
регламентации. Эта сторона дела, увы, - вне сферы действия диатропики. Я
лишь хотел подчеркнуть, что возможности новой науки чрезвычайно обширны,
а ее исследовательская программа очевидна.
6 Ефимов А. Элитные группы, их возникновение и эволюция// Знание - сила. 1988. № 1.
41
Часть 2. Эволюция, диатропика и новый номогенез
Комбинативность. От Парацельса до школы Вавилова'
Парацельс - алхимик, врач, фантазер, мыслитель и практик, послуживший
связующим звеном между средневековой и новой наукой, в 1530-е годы раз-
мышлял о разнообразии. Он воскресил гиппократову идею образования заро-
дыша слиянием материнских и отцовских частиц, поступающих ото всех частей
их тел. Он полагал, что внешние условия, в которых развивается организм, не
формируют его сами, но лишь реализуют те или другие его качества, потенци-
ально имеющиеся у зародыша данного вида. (Вот где источник позднейших
мыслей о прототипе* **.) Тем самым, разнообразие организмов - это прежде всего
разнообразие их потенций (заданных прототипами), а потом уже - условий су-
ществования. Эта простая мысль, проходя через всю историю биологии, всегда
понималась с большим трудом.
Парацельс сравнивал природу с аптекой, где Бог - главный провизор, дав
этим афоризмом начало целой веренице сравнений. Лейбниц сравнивал мир с
часами, позже мир видели как фабрику, как систему уравнений, как организм. А
самый организм видели как механическую машину (Декарт), как химическую
фабрику, как ЭВМ. «Мысль - не более, как ток электричества по проводам нер-
вов», - писал Томас Гексли, один из первых дарвинистов; вслед за ним мышле-
ние уподобляли работе парламента, телефонной станции, ЭВМ-и даже проводи-
ли параллель между памятью и голографией. Каждое такое объяснение казалось
очень убедительным - пока не проходила мода на соответствующую новинку
техники. Это нам надо постоянно иметь в виду: будем стараться не путать моду
с выявлением чего-то постоянного и важного.
С другой стороны, всегда есть опасность упустить важную мысль только
потому, что она выражена в ненаучной, с нашей нынешней позиции, форме. Па-
рацельс, например, высоко ценил астрологию, а ее развитие наук отвергло. Сто-
ит, однако, вдуматься в его рассуждение о человеке как «микрокосме», чтобы
понять, что Парацельс в своих фантазиях (врач «постигает пульс небесного сво-
да, физиономию звезд, хиромантию в минералах, дыхание в ветрах, лихорадку в
землетрясениях») искал общие законы природы, те, которые много позже Лейб-
ниц характеризовал как «единство в многообразии».
Парацельс решил, вопреки Библии, что каждая раса «имела своего Адама».
(Вопрос стоял тогда очень остро: обязательно ли считать полноправными людь-
ми вновь открытое население Америки, не упомянутое в Библии? Несмотря на
папскую буллу 1512 г., объявлявшую всех людей потомками Адама и Евы, кон-
кистадоры обращались с туземцами, как правило, до абсурда жестоко.) Вопрос о
независимом происхождении рас обсуждается, с перерывами, до сих пор.
* Пункт 1.5 из главы 1 книги ЭЭД, с. 27-32. Сохранены ссылки на параграфы книги.
" Прототип по Бюффону и Гёте - не исходная форма, но носитель всех форм (ЭЭД с. 23).
42
Первым собрал сведения о заселении Америки Мэттью Хэйл, правовед, бо-
гослов и натурфилософ, и пришел к заключению, что здешние высшие живот-
ные, равно как и люди, произошли от мигрантов из Старого Света, причем их
своеобразие - результат накопления изменений (возникших случайно или под
влиянием местных условий жизни) и скрещиваний измененных форм друг с
другом [Hale, 1677, с. 196-199]. Что касается низших животных, то их он считал
попросту порождением сочетаний «семенных молекул», витающих в земле, воде
и воздухе [там же, с. 274]. Он же провел замечательную аналогию между скре-
щиванием организмов и взаимовлиянием западных языков [там же, с. 197].
Значительно дальше продвинулся физик и философ Пьер Луи Мопертюи
[Maupertuis, 1756]. Он писал, что комбинацией частиц отцовского и материнского
«семени» можно объяснить не только сходство ребенка с обоими родителями, но
и феномены уродства (как недостаток или избыток частиц определенного типа), и
происхождение новых видов, и, естественно, - обычное разнообразие особей
внутри вида. Эти «частицы» не были для него абстрактной натурфилософской
идеей - он пришел к ним на основе реальных, пусть и скромных, наблюдений.
В 1747 г. он напоминал, что великий Ньютон видел целесообразность (боже-
ственный замысел) в орбитах планет (они движутся по почти круговым орбитам
все в одну сторону) и в строении животных - в их «единообразии» и «чудесной,
полной пользы, организации». Но, возражал Мопертюи, «если единообразие, на-
блюдаемое нами во многом, является доказательством, то не опровергается ли это
доказательство бесконечным разнообразием, которое мы наблюдали во многом
другом?» Так, некоторые видят целесообразность в складках кожи носорога (ина-
че она не могла бы сгибаться), но забывают при этом о черепахе, панцирь которой
действительно не сгибается. Можно, правда, искать целесообразность в строении
организма как целого, но зачем сложнейшие формы и инстинкты даны насекомо-
му, «которое пожирает первая птица»? Для Мопертюи явно односторонни и фи-
лософы, которые ищут целесообразность только в природе как целом, и Лейбниц
(мир просто невозможно сделать лучше), и Мальбранш (мир создан простейшим
из возможных способов), и Поп («все, что существует, хорошо»), и Декарт (мир
попросту движется по законам механики).
Сам он предлагал искать «первичные законы» - те, которым подчиняется
природа, но которые обычно не удается найти в феноменах, «являющихся только
слишком сложными следствиями этих законов». Существует общий закон творе-
ния, такой, что «движение животных, произрастание растений, вращение звезд
являются только его следствиями». Это - принцип наименьшего действия, откры-
тый самим Мопертюи, одна из основ механики [Мопертюи, 1959].
Здесь ясно выражена важная мысль: для понимания явлений надо выявить в
чистом виде те элементы, комбинация которых («слишком сложные следствия»)
создает разнообразие; так, единый набор частиц наследственности (через 150 лет их
назвали генами), комбинируясь, может вызывать к жизни, согласно Мопертюи, все
разнообразие организмов. Пусть мы не считаем (как считали первые генетики), что
43
организм сводится прямо-таки к набору генов, но так не думал и Мопертюи: его
«частицы» наделены атрибутами целостности - «сродством» и «памятью».
Бюффон включил понимание «частиц» Мопертюи в свою концепцию, кото-
рую усвоил Эразм Дарвин, дед Ч. Дарвина. (В 1868 г. Ч. Дарвин назвал ее панге-
незом, а еще через 20 лет ботаник Гуго де Фриз в статье «Гипотеза внутрикле-
точного пангенеза» дал набросок основ генетики.) Бюффон ничего не добавил к
идее «частиц» Мопертюи, но его идею комбинативности он блестяще развил как
морфолог, отметив, что она приводит к параллелизмам, к повторению свойств
от группы к группе: у одного конца группа млекопитающих сходна с птицами
(летучие мыши), у другого - с рыбами (киты, тюлени), в ее середине обезьяны
сходны с человеком. С другой стороны, группа птиц как бы протягивает ветвь в
сторону млекопитающих - это нелетающие птицы. С удивительной проница-
тельностью Бюффон писал: «...Ветви, которые простираются к другим отрядам,
всегда довольно коротки и образуют лишь очень маленькие роды»; эти редкие и
малые роды важны, однако, тем, что должны «дать понять философам, что она
[природа] не может быть сжата оковами наших классификаций...» (Последние
слова - упрек Линнею, системы которого он не признавал.) От этих «ветвей»
всего шаг до рефренов и периферии (см. гл. 2).
Интересный поворот той же мысли мы находим у Мильн-Эдвардса, в его
«законе повторений»: природа повсюду скупа на первичные элементы, повсюду
применяя одни и те же конструкции по многу раз, - это и многочисленные рес-
нички инфузории, и сходства планов строения позвоночных, и идентичность
плана руки и ноги. Все это он выводил из более общего «закона экономии».
Экономия на новшества - это, по существу, переосмысление принципа наи-
меньшего действия Мопертюи. Такое же переосмысление, но уже со ссылкой на
Мопертюи, произвел и Дарвин: «...Взгляд, согласно которому первоначально
были созданы немногие формы или лишь одна... согласуется с философской ак-
сиомой Мопертюи о “наименьшем действии”» [Дарвин, 1951, с. 107].
Другой подход к комбинативности составило изучение параллельных рядов.
Еще Баугин в своей «Описи растительного мира» фактически регистрировал па-
раллели форм. Так, род Plantago (подорожник) он разделил на три группы видов:
широколистные (Р. latifolia), узколистные (Р. angustifolia) и водные (Р. aquatica),
причем среди водных (вероятно, обозначавших нынешний род Littorella) тоже
выявил широколистную и узколистную формы [Bauhin, 1623, с. 189-190]. К
большинству названий видов Баугин давал списки синонимов, из которых вид-
но, что уже тогда параллелизм свойств вызывал путаницу в названиях. Чтобы
ликвидировать ее, Линнею пришлось постулировать иерархию диагностических
признаков; например, он полагал, что число тычинок цветка определяет класс, а
пестиков - порядок в этом классе. Тем самым Линней искусственно изобразил
логическим древом то, что само по себе {естественно) было таблицей: порядок
точнее мог быть задан пересечением строки (число тычинок) и столбца (число
пестиков). В результате параллелизм выпал из понятийных рамок систематики.
44
В XIX в. внимание к параллельным рядам возникало не раз, но оставалось
уделом отдельных авторов. Так, Иоганн Якоб Бернгарди в 1834 г. отмечал, что
растения, в других отношениях несходные, дают сходные ряды типов листа. Из-
ложив его мысль, Дарвин в 1868 г. добавил: гладкокожий персик выведен из не-
многих форм бархатистого, но «между сортами бархатистых и гладких персиков
наблюдается замечательный параллелизм» - общие варианты цвета плода и его
мякоти, характер косточки, цветков, листьев и т.д., хотя «признаки каждого сор-
та гладкого персика получены отнюдь не из соответствующего сорта бархати-
стого» [Дарвин, 1951, с. 716].
В 1825 г. шведский ботаник Элиас Фрис предложил систему растений, в ко-
торой отмечались сходства между таксономически далекими растениями, де-
монстрирующими сходные «степени развития», например, между однодольны-
ми и двудольными. Ботаник М.А. Максимович (более известный ныне как энту-
зиаст украинской словесности) значительно развил мысли Фриса. В своих выво-
дах он писал: «В природе везде двойство: свет без двойства являться не может».
Это «двойство» он видел во всяком познании, призывая идти попеременно то от
высших форм к низшим, то наоборот; то от общий свойств организмов к част-
ным, то наоборот; то от органов к плану строения, то наоборот; то от анатомии к
физиологии, то наоборот. Выискивая «двойства» (взаимодополнительные аспек-
ты - сказали бы мы сейчас, зная принцип дополнительности Бора), он отметил
замечательную мысль Фриса: если группировать виды по «сродству» (т.е. по
гомологии) в столбцы, то образующиеся при этом строки (соседние виды, обра-
зующие разные столбцы) тоже будут образовывать сходства - «аналогии» [Мак-
симович, 1827]. Через 93 года Н.И. Вавилов назвал этот феномен гомологиче-
ским рядом наследственной изменчивости. Он показал, что один и тот же видо-
вой комплекс может встречаться в различных родах настолько закономерно, что
его можно предсказывать. Так, изучая злаки Памира, он заметил, что характер-
ные признаки разновидностей пшеницы удивительно повторяются у разных
форм ржи. Следовательно, решил Вавилов, видовое разнообразие упорядочено:
«Мутации, идущие как бы случайно в разных направлениях, при объединении
их обнаруживают общий закон» [Вавилов, 1987, с. 97].
Теперь вопрос о параллелизмах приобрел совсем новый аспект; откуда они
берутся - создаются требованиями среды независимо, реализуются за счет
свойств, унаследованных от общего предка, прямо передаются от вида к виду
подобно инфекции, или виды, эволюционируя, могут развиваться не как попало,
но только в рамках ограниченного числа логических возможностей (как говорил
Дарвин, в рамках «природы организма»)? Два таксона могут как бы двигаться на
разных этажах, но по одинаковым коридорам. Дарвин писал: «...Члены одного и
того же класса, независимо от их образа жизни, сходны между собой по общему
плану организации» [Дарвин, 1939, с. 623], причем «гомологичные части склон-
ны изменяться одинаковым образом» [Дарвин, 1951, с. 697]. Тогда же (1868) па-
леонтолог Эдуард Коп, пытаясь истолковать параллелизмы у ископаемых, пи-
45
сал, что родовые признаки могут меняться со временем без обязательного изме-
нения видовых [Филипченко, 1977]. В целом же, как констатировал Любищев,
вопросы параллелизма мало занимали Дарвина и его последователей, поскольку
не гармонировали ни с его теорией, ни со взглядами эпохи.
После работы Вавилова некоторые стали смотреть на систематику и генети-
ку (а частично и на морфологию) как на науки о комбинациях. Среди них оказа-
лись такие крупные наши биологи, как ихтиолог Л.С. Берг, гистолог А.А. Завар-
зин, энтомолог А.А. Любищев, палеонтолог Д.Н. Соболев и генетик Ю.А. Фи-
липченко. Последнему принадлежит, кстати, первая обзорно-теоретическая ста-
тья о «законе Вавилова», где сформулированы три вида параллелизмов - гено-
типический, обусловленный сходными генами и наблюдаемый обычно у близ-
ких видов, анатомический, «вытекающий из одинаковых возможностей разви-
тия», и гистологический, «который выражается в сродстве гистологической
структуры одинаково функционирующих частей у очень далеких в смысле сво-
его систематического положения организмов» [Филипченко, 1924, с. 255]. Вер-
ный своему времени, автор предлагал применять термин «комбинация» только к
генотипическому параллелизму (там же. С. 254), поскольку только для генов
мог представить механизм комбинирования. (Впоследствии стало ясно, прежде
всего - на основе работ Мейена, что комбинироваться могут любые факторы,
ответственные за параллелизм.) Зато он как бы предвосхитил наше нынешнее
системное понимание сродства (которое будет изложено далее, в п. 3 главы 9) -
это видно из того, сколь необычно для тогдашней биологии он употребил под-
черкнутое мной слово «сродство». На эту замечательную работу Филипченко
мое внимание обратил В.В. Бабков.
В наше время понемногу выясняется, что познание разнообразия прежде
всего состоит именно в выяснении законов параллелизма (пользуясь аналогией
- плана коридоров, общего для разных этажей), и важное место среди них зани-
мает комбинативность. Только узнав эти и прочие законы, по которым упорядо-
чено разнообразие живого, можно всерьез заняться проблемами эволюции, ко-
торая предстает при этом как процесс исторического преобразования разнообра-
зия - экосистем, таксонов, организмов и составляющих их структур.
Литература
Дарвин Ч. Соч, т. 3, 1939; т. 4, 1951.
Максимович М.А. О системах растительного царства. М., 1827.
Мопертюи П. -Л. Законы движения и покоя, выведенные из Метафизического прин-
ципа// Вариационные принципы механики. М., 1959 .
Филипченко Ю.А. О параллелизме в живой природе И Успехи эксперимент, биоло-
гии, 1924, вып. 6.
Филипченко Ю.А. Эволюционная идея в биологии. М., Наука, 1977. 227 с.
Bauhin С. Pinax theatri botanici. Basel, 1623. XXIV + 567 p.
Hale M. The primitive origination of mankind... London, 1677.
Maupertuis P.L. Oeuvres. Lyon, t. 2, 1756.
46
Разнообразие, ряды и тенденции'
Основная цель этой главы - познакомить читателя с диатропической концеп-
цией С.В. Мейена в той мере, в какой это необходимо для понимания данной кни-
ги. Поэтому, в частности, опущен геологический материал, бывший для него ос-
новным [Мейен, 1977], зато привлечены другие примеры, на мой взгляд, более
доступные большинству читателей, а также проведен ряд историко-методоло-
гических параллелей. Мейен не оставил единого изложения своей концепции, од-
нако прочел первый вариант предлагаемого ниже текста и сделал много письмен-
ных замечаний, которые я постарался учесть со всею возможною полнотой.
2.1. О биологической грамматике
Учения о различиях (систематика), о сходствах (морфология) и о комбина-
тивности форм (например, элементы генетики) вошли в общее употребление с
интервалами примерно в сто лет (первая треть XVII, XIX и XX вв. соответст-
венно), хотя и начали свое развитие в одно время (вторая четверть XVI в.) и не
испытывали недостатка в талантливых исследователях. Очевидно, уже эти три
круга проблем достаточно различны по степени трудности их усвоения научным
сообществом, тогда как сейчас у нас речь пойдет о более трудном, о четвертой
линии познания разнообразия - об упорядочении материала, даваемого первыми
тремя линиями, о биологической грамматике'* 7 Введение этого термина долж-
Глава 2 книги ЭЭД. Сохранены ссылки на параграфы книги.
7 Прочтя это место в первом варианте рукописи, Мейен записал следующее: «Мне не
очень нравится такое разбиение: систематика - различие, морфология - сходство. Со
времен Жюссье систематика строится по конгрегационному принципу, т.е. по сходству;
мы идем путем последовательного объединения сходного. Кроме того, я считаю морфо-
логию (отчасти) систематикой частей, причем здесь действует (как показал Е.С. Смир-
нов) тот же конгрегационный принцип. Комбинативность же не рядоположена этой паре,
она указывает на один аспект распределения признаков в системе. Твоя четвертая линия
- результат соединения систематики и морфологии (это типология в.мое.м понимании)».
Здесь все верно, но противоречия между нами я не вижу. Все разрешается, по-моему,
взятым у Мейена в скобки словом «отчасти»: у меня речь идет о другой «части» классифи-
кационной науки. Ведь систематика тоже лишь отчасти строится по сходствам - уже тот
факт, что ни один объект не может остаться вне системы, говорит о том, что первичен здесь
принцип разбиения заданного множества (см. гл. 6), а принцип объединения вторичен. Если
же рассматривать только разграничительный аспект систематики (вольно говоря, система-
тику по Линнею) и только объединительный аспект морфологии (вольно говоря, морфоло-
гию по Юнгу), то вне их останется именно комбинативность, точнее - учение о паралле-
лизмах, которое в этом смысле им рядоположено. На базе этой тройки и нужен синтез, да-
ваемый далее схемой, которую Мейен называл типологией, а я - биологич. грамматикой.
Примеч. 2010 г.: Уверенность Мейена в том, что «со времен Жюссье систематика стро-
ится по конгрегационному принципу, т.е. по сходству» представляется мне теперь чрез-
мерной. А.-Л. Жюссье известен единицам, как и сам принцип конгрегаций. Равнодушие
почти всех систематиков к принципам классификации поразительно - как у нас, так и на
Западе (см. далее, часть 6), а презрение к мыслям предтеч даже демонстративно.
47
но подчеркнуть сходство познавательных процедур: если общность словоформ
рыбе и яме мы легко устанавливаем как «дательный падеж единственного числа
1-го склонения существительных», то нельзя ли так же обойтись, например, со
сходными формами описанных Дарвином персиков или Вавиловым - злаков?
Трудность усвоения связана с тем, что первые три линии познания никак не
назовешь последовательными ступенями - углубляясь в познание разнообразия,
все время приходится опираться, как при спуске в горную расщелину, то на од-
ну стену, то на другую, то на удачный выступ спереди или сзади. Чтобы понять
это, напомним, как пользуются системой организмов.
[Далее опущен текст, повторяющий первые два обзаца параграфа «Как ра-
ботает линнеевская система», приведенные выше, на с. 21-22.]
Какой кропотливый 400-летний труд скрыт за этими диагнозами!^. Однако,
определяя вид по дихотомической таблице, мы не познаём самых еще сущест-
венных свойств («завершающих различий»), мы ведем себя, словно комиссия в
музее, сверяющая инвентарные бирки ярких полотен и равнодушная к самим
полотнам. Ведь большая синица - птичка яркая, а московка - грязноватая.
И все-таки, вопреки великому Аристотелю, мы почти всегда доходим, сле-
дуя дихотомии, до определенного вида. Почему? Да просто потому, что всякая
дихотомическая таблица делается теперь по готовой системе, а сама система
была придумана без дихотомии; систематик, желая упорядочить известное ему
разнообразие (например, воробьиных), следует линнееву афоризму (не признаки
задают род, но род задает признаки), т.е. пользуется не формальным ключом, а
интуицией. Только создав хорошую систему, можно начинать ее формализа-
цию - хотя бы составляя дихотомическую таблицу, - и недаром Ламарк достиг
впервые в этом успеха лишь в год смерти Линнея. В то же время при Аристоте-
.. . о
ле никакой писанной системы не существовало, дихотомию рассматривали как
путь ее построения, и ничего, разумеется, не получалось.
На этом простом примере можно убедиться в очень важном факте: познание
разнообразия состоит как в его интуитивном уяснении, так и в его упорядоче-
нии, причем интуитивное понимание и формальное упорядочение невозможны
друг без друга. Да, дихотомия Ламарка построена на системе растений Линнея,
8 Удобная дихотомическая форма (если да, то читай одно, если нет - другое) пришла в
систематику только в 1778 г. с «Французской флорой» Жан-Батиста Ламарка, хотя пред-
лагали ее еще до Линнея (например, Мартин Иорений в 1710 г. [Максимович, 1827, с.
17]); ее прекрасно знал и Аристотель, который писал, что путем дихотомии «приходишь
к последнему признаку, но не к завершающему различию и виду» и что поэтому «тем,
кто применяет дихотомию, невозможно дойти до какого-нибудь отдельного животного»
[Аристотель, 1937, с. 47].
9 Мейен не раз подчеркивал, что в основе классификационных построений очень долго
лежали бытовые («кухонные») таксономические и морфологические представления, что
без них невозможно начать работу [Мейен, 1977, с. 29].
48
но ведь и система стала возможна потому, что еще давным-давно были выявле-
ны и закреплены словами разговорного языка такие дихотомии, как живой - не-
живой, животное - растение, мужской - женский, наземный - водный, и многие
другие. Не менее четки и важны были также деления организмов на части (на-
пример, корень, стебель, лист, цветок, плод и т.п.), Долиннеевы систематики
уже исходили из этих и подобных делений (морфологических, физиологиче-
ских, экологических и т.п.) как данных. Наука о разнообразии должна строиться
последовательными шагами: каждый успех морфологии позволяет улучшить
систематику, и обратно. Такой прием называется итеративным.
Одна из первых задач диатропики - сделать итеративный характер познания
разнообразия явным, эксплицировать итеративные процедуры. Это давно проделано
в математике, где известно, что итеративная процедура либо приводит к определен-
ному пределу, либо оказывается бесконечным блужданием. Предел (данной проце-
дуры на данном множестве) является инвариантом множества в том смысле, что ко-
нечная точка процедуры не зависит от того, с какой точки множества итерации на-
чаты. Встает задача выявления структурных инвариантов разнообразия организмов.
В других науках подобные инварианты успешно выявляются^0, а в биологии для их
выявления естественно обратиться к вавиловским рядам. Почему открытия с их по-
мощью так редки? Почему определители не делаются в форме таких таблиц, в кото-
рых все таксоны одного ранга были бы представлены по единым правилам, пусть и
с исключениями, подобно таблицам склонения (наборам падежных форм) или
спряжения (наборам личных и временны* х форм) в языке?
Выяснение законов «биологической грамматики» едва начато. Здесь дело
сдвинулось только после переосмысления закона Вавилова: «в законе Вавилова
в его обычно формулировке обращается внимание на повторность отдельных
признаков от таксона к таксону, но опущена повторность в правилах преобразо-
вания варьирующих признаков», причем «запомнить порознь все разновидности
и относящиеся к ним самостоятельные термины невозможно (это все равно, что
запоминать порознь все склонения [правильнее: падежные формы. - Ю.Ч.\ каж-
дого существительного, не познакомившись с правилами склонения)» [Мейен,
1978,с. 500-501]. Добавим, что падежи, когда их много, не просто перечисляют-
ся (как в латыни и русском), а упорядочиваются. Такова система локативов (ме-
стных падежей) в дагестанских языках.
10 Такова в этнологии структура мифа, которая сохраняется неизменной при всех вариа-
циях, происходящих во времени и пространстве, - на это обратил внимание еще Алек-
сандр Гумбольдт в 1814 г. [Гумбольдт, 1963, с. 293], а в XX в. - этнолог Клод Леви-
Строс. В филологии примерами инвариантов являются грамматические категории (па-
деж, время и т.д.): любое русское существительное принадлежит шестипадежному ряду,
именуемому склонением (если пренебречь рудиментарными падежами), любой русский
глагол - трехвременному трехличному ряду, именуемому спряжением, и т.д.
Примеч. 2010 г. : Общим для самых различных наук инвариантом разнообразий являют-
ся квази-гиперболические распределения (см. часть 7 Сборника).
49
Слова обычно образуют большие группы грамматически сходных форм (та-
ких, как бал и пол, где все падежные формы идентичны), т.е. подчиняются еди-
ной парадигме. Однако встречаются слова, к исходной форме которых не по-
добрать пары, сходной и грамматически, и по звучанию конца. Так, существи-
тельному хрящ нет созвучного ему существительного в том же числе и падеже
(оканчивающегося на -ящ). Зато, просклоняв хрящ, найдем, что его производ-
ным формам созвучна группа слов (того же склонения) типа хвощ', немного хуже
соответствует праща - в ходе развития языка слово попало в другое склонение.
Кроме них, много разрозненных сходств - совпадений той или иной падежной
формы с отдельными словоформами других частей речи (см. табл. 1)
Таблица 1. Пример фонетического параллелизма (совпадения ударных кон-
цов) между падежными окончаниями и прочими словоформами
Падеж Данное су- ществитель- ное Сходные по склоне- нию существит-ные Несходные части речи
Словоформа и ее исходная форма
им.-вин. род. дат. твор. предл. хрящ хряща хрящу хрящом хряще [пращ*] хвоща праща** хвощу пращу** хвощом пращом* хвоще праще горящ*** таща (деепр. от тащить) трещу (1-е лицо от трещать) [общо] (наречие от общий) вотще* (наречие от тщетный)
им. род. дат. твор. предл. хрящи хрящей хрящам хрящами хрящах хвощи пращи хвощей пращей хвощам пращам хвощами пращами хвощах пращах ищи (повелит, форма от искать) кощей (имен, падеж) овощам (дат. падеж от о* вощ) щами (твор. падеж от щи) Натощах* (наречие от тощий)
* архаическая форма
** несовпадение падежей (ныне праща - женского рода).
*** Краткое причастие от гореть
Прямые скобки означают неполное созвучие ударного слога
В языках, не имеющих падежей, ту же закономерность легче всего увидеть
на глагольных формах. Так, французские глаголы devenir, revenir, souvenir, tenir,
venir и т.п., образующие свою небольшую группу спряжения: ‘venir-venant-viens,
vient-venu-vins, сходны с существительными: инфинитив сходен с avenir и sou-
venir, настоящее время - с soutien и revient, активное причастие - с revenant и
tenant, пассивное причастие - с avenu и revenu, простое прошедшее - с vin и
devin. Есть сходства с прилагательными {avenant, tenu), местоимением tien,
междометием tiens и т.п.
Подобным же образом изменяются признаки биологических видов, родов и
т.п., но если в ботанике нередки довольно полные параллельные ряды, то для
50
зоологии более характерны разрозненные сходства. Возьмем для примера тех же
синиц. Собственно синицы (род Parus семейства Paridae) образуют в фауне
СССР 12 видов, но термин «синица» применяется и к другим воробьиным:
ре» мезу, длиннохвостой синице, усатой синице и др. Уже это говорит о паралле-
лизме: птицы других родов и даже семейств имеют характерный «синичий» об-
лик. Однако параллелизм наблюдается гораздо шире. Почти все признаки, отра-
женные в названиях синиц, представлены и у других воробьиных (см. табл. 2),
но, как и в примерах со словами, здесь нет таксономических границ для прояв-
ления разрозненных сходств. Так, многие птицы различных отрядов хохлаты
(крохали, турухтан, чибис и др.), а характерная полоска вдоль щеки, давшая ви-
довое название усатой синице (из семейства толстоклювых синиц отряда во-
робьиных), наблюдается и в отряде дятловых, у сирийского пестрого дятла (на
этот факт мое внимание обратил Е.Н. Панов). Параллелизм не ограничивается
классом птиц; например, хохлатыми называются некоторые виды млекопитаю-
щих. Естественно, что в пределах отряда воробьиных параллелизм полнее.
Таблица 2. Пример параллелизма внешних обликов организмов в рамках отряда
Синицы (отряд воробьиных) Другие воробьиные
хохлатая (Parus cristatus) рыжешейная (Parus rufonuchalis ремез (Remiz pendulinus) толстоклювые синицы (семейство) в том числе: усатая (Panurus biarmicus) длиннохвостая (Aaegithalos caudatus) Свиристель. Жаворонок хохла- тый Овсянка рыжешейная Овсянка-ремез Камышовка толстоклювая некоторые овсянки Личинкоед серый; сороки (отчасти
Оставляя профессионалу-орнитологу доведение таблицы 2 до максимально-
го сходства с таблицей 1, заметим, что обе таблицы и так проявляют одно сущест-
венное сходство - пересечение рядов. С одной стороны, слово хрящ входит в
обычный ряд существительных 2-го склонения, а с другой - в ряд слов на -ящ,
причем на пересечении этих рядов слово хрящ - единственное. Аналогично, хох-
латая синица - единственный вид на пересечении ряда синиц с рядом хохлатых
птиц. При этом сама принадлежность объекта ряду может определяться как одним
признаком (-ящ; хохлатость), так и комплексом признаков (принадлежность 2-му
склонению; комплекс признаков рода синиц). Как мы увидим, наиболее интерес-
ны пересечения комплексных рядов. Не раз высказывалась идея (В.М. Шимкеви-
чем, Н.И. Вавиловым, А.А. Еленкиным и др.), что саму систематику следует стро-
ить так, чтобы таксон (раса, подвид, вид, род и т.д.) задавался пересечением рядов.
В некоторых случаях это действительно удобно, но в любом случае надо знать,
возможно ли это в принципе, - чтобы понимать структуру разнообразия.
51
2.2. Мероно-таксономический анализ (типология по Мейену)
Главная идея анализа состоит в следующем: чтобы познавать разнообра-
зие, обращаясь итеративно то к различиям, то к сходствам, надо уметь различия
и сходства классифицировать сопряженным образом. В 2.1 мы видели, что объ-
ектами итерации служат систематика и морфология, поэтому их следует пред-
ставить единым (удобным для итерации) образом. Н.Н. Страхов (1858), зоолог,
более известный сегодня как публицист славянофильского направления, отме-
тил, что сравнительная анатомия классифицирует органы так же, как зоология
классифицирует организмы. Развивая эту мысль, Мейен пришел к фундамен-
тальному выводу: гомологизация (классификация частей) и классификация орга-
низмов являются двумя логически самостоятельными процедурами, но неявная
связь между ними существует - она существует итеративно', морфология обес-
печивает систематику «признаковым пространством и данными о соотношении
признаков у разных объектов» [Мейен, 1978, с. 496], а систематика дает морфо-
логии пространство объектов, допускающих соответствующие гомологизации.
Эту связь Мейен решил сделать явной.
В основу он положил понятие мерона.
[Далее опущен текст, повторяющий последний абзац на с. 24 и
первый абзац на с. 25.]
Предложено учение о меронах, мерономия, где основным является понятие
об архетипе. Мы видели, что для определения птицы надо заранее знать, что та-
кое птица, а это значит не только уметь выявлять характерные для птиц мероны
(определенная форма тела, клюв, крылья, перья и т.д.), но и уметь мысленно
складывать из них обобщенный образ (архетип) птицы, т.е. знать, какое сочета-
ние комбинаций меронов делает животное птицей. Если сам организм можно
мысленно расчленять на части тела и говорить, что организм из них состоит, то
архетип состоит в этом смысле из меронов.
Однако организмы можно сравнивать не только по частям их тел, но и по
всем их качествам: по способности летать или образовывать брачные пары, по
цветным узорам на теле, по роли в экосистеме и даже по размерам. Архетип по-
лучается куда богаче, чем по Оуэну, и строить его нужно по-новому. Прежде его
изображали простеньким рисунком, содержащим только самые общие свойства
таксона. Например, архетип позвоночного - это очень схематичный скелет,
правда, с очень сложными позвонками (см. рис. 22 в кн.: [Бляхер, 1976]), столь
же схематичные органы пищеварения и т.д., т.е. «инвариантная для всех членов
таксона совокупность меронов» [Мейен, 1977, с. 29]. Будем называть архетип-
рисунок архетипом по Оуэну. Архетипом по Мейену будем называть «архетип в
более широком смысле», когда «гомологизированные, то есть расклассифици-
рованные и ставшие меронами части организмов данного таксона в сумме со-
ставляют архетип таксона» [Мейен, 1978, с. 497]. Это обширная таблица, где
каждая строка - мерой, а каждый столбец - возможная в принципе для данного
52
таксона особь*. Для позвоночных таблица, например, включает строку «конеч-
ности», в которой друг за другом записаны всевозможные значения мерона: от-
сутствие, плавник, ласт, лапа и т.д., причем каждый вид конечности может
иметь разные степени развития. Грубо говоря, архетип по Оуэну - пересечение
множеств признаков, а по Мейену - их объединение. В терминах модальной ло-
гики [Целищев, 1977, с. 5-9] архетип по Мейену близок к понятию модельной
структуры (системы), а по Оуэну - общезначимому утверждению в ней.
Чаще всего эволюция - это реализация нового столбца, но прогрессивная
эволюция всегда включает появление новой строки (и связанных с нею новых
столбцов). До появления птиц в таблице позвоночных не было, например, стро-
ки «перья» или, если таблица менее детальна, - в строке «роговые покровы» бы-
ли пусты клетки, соответствующие всевозможным перьям. Вообще, большинст-
во клеток таблицы пусто всегда (поскольку особь задается комбинацией призна-
ков, а мерой - самими признаками), зато число строк и столбцов в ней необо-
зримо, и нет смысла выписывать ее в явном виде. Она - средство теоретическо-
го анализа, подобная матрицам в математике, которые можно изучать, не выпи-
сывая самих таблиц в явном виде.
Если положить головами в одну сторону рыбу, лягушку, ящерицу, воробья
и крысу, то легко увидеть ряды сходных частей: ряд черепов, ряд глаз, ряд же-
лудков и т.д. Их сходство объясняют общностью происхождения всех позвоноч-
ных, но вопрос не так прост: ведь сюда можно подложить и рака, и жука - па-
раллелей будет меньше, но все-таки много. Если здесь и можно говорить об об-
щем предке, то уж у него-то не было ни мозга, ни сердца, ни раздельнополости -
всего, чем сходны высшие животные. Эта общая тенденция к появлению сход-
ных свойств и есть параллелизм.
Параллелизмы бывают самые разные. Если взять много разных видов рыб,
птиц, насекомых и т.д., то легко увидеть параллели в окрасках - не только покро-
вительственные или контрастные, но и просто сходные рисунки. Скелет руки по-
разительно сходен со скелетом ноги, хотя рука и нога всегда имели отдельных
«предков» - переднюю и заднюю лапы. Когда удается обследовать много тысяч
членов одного вида, открывается еще один морфопараллелизм - уродства. В них
часто реализуются признаки, нормальные для других видов, и это важно для осоз-
нания морфологического единства природы.
Палеонтолог Д.Н. Соболев считал, что «почти для каждого из известных ти-
пов аномалий можно подобрать соответствующие примеры видообразования»
[Соболев, 1924, с. 161]. В 1935 г. П.Н. Кренке обосновал на ботаническом мате-
Здесь повторены устные разъяснения самого Мейена. Однако реальность выглядит
сложнее: плоские таблицы (вроде рис. 1) могут описать рефрен лишь изредка и то упро-
щенно. Реальные рефренные таблицы многомерны, однако все их координаты можно
разделить на «строки» (те, что описывают мероны) и «столбцы», определяющие свойства
особей, мыслимых в данном таксоне. - Примеч. 2010 г.
53
риале правило: признак, являющийся уродством у данного вида, обычно нормален
у какого-то иного вида [Мейен, 1978].
Обычно у миндаля плод сухой, а у персика - окруженный мясистым око-
лоплодником, но изредка бывает наоборот, причем необычная форма может су-
ществовать и как индивидуальная вариация, и как устойчивая раса. Эта пара ви-
дов проявляет целый ряд сходств изменчивости, и его можно сопоставить с па-
дежным рядом существительных одного склонения. Как и в грамматике, ряды
сходств довольно полны для близкородственных видов (персик и миндаль
обычно включают в один род) и довольно разрозненны для форм далеких; но
как и в грамматике, определенных границ для проявления сходства нет. Кренке
назвал этот феномен законом родственной изменчивости [Корона, 1987, с. 50—
53]. Вне ботаники прекрасным примером того же закона является мутационная
изменчивость дрозофил, почти одинаковая у многих видов данного рода.
Это оказалось общим правилом: чем значительнее отклонение от нормы,
тем реже оно наблюдается и тем дальше таксономически вид, у которого соот-
ветствующее свойство можно встретить в качестве нормального. Такой без-
обидный дефект, как нарушение пигментации, дает признаки другой расы (бе-
лый негр, описанный у Мопертюи); а уже «заячья губа» не встречается в качест-
ве нормы у человека и обезьян, зато обычна у лемуров (низшие приматы). Очень
редкие дефекты людей (нечто вроде чешуи) нормальны для низших позвоноч-
ных, а самые редкие (и тяжелые) уродства приходится искать в качестве нормы
еще дальше; так, две головы на одном теле и одна голова на два тела возможны
как норма у некоторых червей.
Отметим, что дело здесь не в атавизме: черви предками людей не считают-
ся. Не было и рогов у предков лошадей, а рогатые лошади иногда рождаются.
(Примеры сообщены мне С.В. Мейеном и А.С. Раутианом.)
Без «грамматики биологии» было почти невозможно сказать, что общего
есть между всевозможными параллелизмами, в ее же свете видно, что все они
подчиняются простой схеме: у разных таксонов общие мероны. Этой формуле
можно придать вполне грамматический вид, и недаром раздел грамматики, изу-
чающий строение слов, тоже называется морфологией.
Слово делится на основу и окончание, которое, изменяясь, указывает
грамматическую форму (падеж, время и т.д.). Изменяя окончания, мы получаем
разные словоформы, т.е. термин «слово» выступает как род, в котором отдель-
ные словоформы являются видами (диалектные варианты их выступают в каче-
стве рас). Основа может состоять из корня и аффиксов (приставки и суффикса),
и группа слов с общим корнем играет роль семейства. Слова можно группиро-
вать по-разному - просто в алфавитном порядке (обычные словари), в алфавите
окончаний (рифмовники), по частоте употребления (частотные словари), по
смыслу (словари синонимов), по происхождению (этимологические словари) и
т.п., и какой бы способ ни выбрать, обнаружится параллелизм слов - потому что
другие, не использованные для классификации признаки будут заявлять о себе
54
на каждой странице. Так же и в биологии: по каким бы правилам ни создавать
систему, другие, неучтенные закономерности выступят как параллелизмы.
Если в опытных и описательных науках исходным является понятие факта,
то в диатропике - понятие ряда^Л И так же, как факт не имеет смысла (а подчас
и места) вне объясняющей схемы, так и ряд бессмыслен без сопоставления (па-
раллели) с другими рядами. Тем самым, параллелизм выступает исходным тео-
ретическим элементом диатропики. С его помощью строятся архетипы. Архетип
по Оуэну сворачивает разнообразие в единый образ, архетип по Мейену макси-
мально разворачивает его с целью показать пределы возможного.
Мейен предложил видеть в каждом таксоне как архетип по Оуэну (т.е. не-
что коренное), так и изменчивое множество, причем это последнее оказывается
упорядоченным, как упорядочены падежные или личные формы (подробнее см.
2.4). Эту упорядоченность можно наблюдать на таксонах всех уровней - от под-
вида (бархатистые и гладкокожие персики) до надцарства (водоросли повторяют
свойства цианобактерий), но мы поясним ее на одном примере - характере из-
менений в пределах мерона «конечность».
Этот мерой вообще может отсутствовать в архетипе таксона (губки, черви,
растения), а может принимать самые различные значения. Если говорить только
о парных конечностях позвоночных и избегать деталей, то обнаружится сле-
дующая картина. Позвоночные делятся на пять классов: рыбы, амфибии (земно-
водные), рептилии (пресмыкающиеся), птицы и звери (млекопитающие). Мерой
же принимает девять основных значений: простой плавник - только у рыб;
плавник-ласт (с суставчатым скелетом) - встречается у всех пяти классов (кис-
теперые рыбы, ископаемые амфибии и рептилии, пингвины и киты); ласт (для
плаванья и ползанья по земле) - у вымерших амфибий и рептилий, у черепах и
млекопитающих (ластоногие); рыбы, как ни странно, ползают с помощью про-
стых плавников; ползателъная лапа - у многих амфибий, рептилий и некоторых
млекопитающих, часто снабжена плавательной перепонкой; «планера (лапа или
плавник с летательной перепонкой) - у некоторых рыб, лягушек, ящериц и мел-
ких зверей; крыло для активного полета - у вымерших птицеящеров, у птиц и
летучих мышей; как редкое исключение - у клинобрюшек (рыбок из семейства
харциновых) - наблюдаются элементы активного полета; нога для хождения и
стояния без касания брюхом земли - у некоторых вымерших амфибий, у репти-
лий, птиц и зверей; хватательная лапа - известна у всех классов, кроме рыб;
рука - только у приматов (рис. 1).
11 Слово «ряд» надо понимать широко, как латинское ordo, означающее и ряд и порядок.
Так, бытует выражение «падежный ряд», хотя, например, в языках Восточного Дагестана
лучше видеть систему таблиц: одни лишь локативы (падежи, соответствующие вопросам
где, куда, откуда), образуют здесь до 8 серий (верх, низ, глубь и т.д.) по 3 падежа [Маго-
метов, 1965, с. 117], т.е. таблицу. Порядок может означать не только линейную, но и лю-
бую организованность.
55
В этих параллельных рядах наблюдаются одни и те же тенденции. Как у
всякого мерона, у конечности бывают разные степени выражения: полное отсут-
ствие, рудимент (зачаток, невидимый снаружи или видимый, но бесполезный),
слабое развитие, полное развитие. Во всех классах мы видим тенденцию к реа-
лизации самых разных степеней выражения конечностей: полное отсутствие на-
блюдается у рыб (семейство угрей - отсутствует передняя или обе пары), у ам-
фибий (семейство червяг) и рептилий (змеи); рудименты бывают у пещерных
рыб, многих хвостатых амфибий и ящериц, у китов (задняя пара) и у некоторых
птиц (передняя пара). Оформленные, но совсем слабые конечности тоже хорошо
известны у всех пяти классов, так что можно выстроить пять параллельных ря-
дов по степени их развития (о других тенденциях см. 2.5).
«То, что разные члены рядов изменчивости объединяются одной тенденци-
ей, одним правилом преобразования, дает возможность предсказывать и целена-
правленно искать недостающие члены. Эту повторяющуюся, подчиненную од-
ному правилу преобразования последовательность состояний мерона назовем
рефреном», - писал Мейен [1978, с. 501]. Как и сами мероны, рефрены могут
быть выявлены в любой теории классификации.
Лучше всего обследованы языковые рефрены - формы спряжения глаго-
лов, склонения существительных и т.д. Неполнота рядов характерна и здесь -
как для отдельных слов («сутки» не бывают в единственном числе, «рожь» - во
множественном, хоть это и вынуждает строить громоздкие обороты речи), так и
для целых классов (в современном русском языке сослагательное наклонение
утратило формы времени). И всё же именно благодаря рефренам грамматика яв-
ляется полезным для всех инструментом - они помогают лучше понимать язык
при меньшей затрате сил: вместо заучивания каждой словоформы, мы запоми-
наем исходное слово и то правило, по которому изменяется данная группа слов
(о биологических рефренах см. 2.4).
Еще в 1862 г. Герман Гельмгольц, блестящий физик и физиолог, обращал
внимание на необходимость организации знания по грамматическому образцу.
Он говорил: «С помощью хорошего словаря гимназист может сделать теперь
кое-что в объяснении классиков, что должно быть так трудно было Эразму, не-
смотря на начитанность долгой жизни», так как требуется хорошая системати-
зация, «чтобы не потеряться безнадежно в лабиринте учености». Системы
«представляют, так сказать, основной фонд научного достояния человечества,
проценты с которого пускаются в хозяйство» [Гельмгольц, 1866, с. 15-16]. Он
указывал, что хорошая система - не просто каталог, а инструмент, способный к
предсказанию. (Примеч. 2010 г.: Подробнее см. далее, часть 6.)
Об этом писал и Мейен: возможность восстанавливать значения неизвест-
ных меронов на основании знания архетипа он называл типологическим прогно-
зом, возможность которого выводил из того свойства природы, которое называл
типологической упорядоченностью. В наших терминах это - диатропический
прогноз. (Примеч. 2010 г. : Подробнее см. далее, с. 337-338.)
56
У всякой птицы мы ожидаем найти перья, у каждого организма - ДНК с
четырехбуквенным кодом. И дело здесь не в происхождении их от общего пред-
ка, о котором обычно ничего не известно, а в том, что без этих меронов мы не в
состоянии сконструировать соответствующие архетипы. «Все современные пти-
цы имеют перья, а ископаемые, для которых перья неизвестны... немыслимы
без них», - пишет зоолог А.С. Раутиан [1978, с. 109]. Аналогично, единый архе-
тип организма немыслим для нас без мерона «ДНК». Однако, как и всякий про-
гноз, типологический прогноз может не оправдаться: даже универсальность ге-
нетического кода у некоторых одноклеточных нарушена (см. гл. 6).
Итак, биологическому языку предложено нечто вроде грамматики: словом в
ней служит мерой, осмысленной фразой - архетип (и соответствующий ему так-
сон), грамматическим правилом - рефрен, а связным текстом может служить или
таксон более высокого ранга, или экосистема, или, к примеру, последовательность
сменяющих друг друга видов - смотря о чем идет речь. Точнее, пока что намечен
аналог грамматической морфологии (теории построения слова), а аналог синтак-
сиса, т.е. теорию связи меронов в архетип (по Оуэну) еще предстоит создать.
Здесь уместно вспомнить, что по В.Н. Беклемишеву [1964, с. 20], «разделение це-
лого на части и описание частей составляют предмет тектологии\ синтез целого,
обратное его построение из частей, составляет предмет архитектоники». Архи-
тектоника и должна в будущем явиться искомым синтаксисом.
2.3. Ядро и периферия
Зоолог Карл Бэр в 1825 г. говорил, что всякий крупный таксон имеет ядро
из характерных форм (таковы, например, воробьи и синицы среди воробьиных,
кошки и собаки среди хищных) и периферию из сравнительно малочисленных
форм, у которых облик данного таксона выражен нечетко, образуя ряд переход-
ных форм, сближающих этот таксон с другими [Бэр, 1959, с. 401-404). Таковы,
например, ворона среди воробьиных и морской зверь калан среди хищных. О
том же, что и Бэр, вскоре писал Максимович [1827, с. 41], и в этом легко уви-
деть развитие идей Бюффона. В 1840 г. логик и историк науки Вильям Уэвелл
сформулировал на этой основе принцип классификации: «Класс установлен
твердо, хотя и не ограничен точно» (см.: [Мейен, 1978]). Напомнив это, вернем-
ся к «биологической грамматике».
Кювье недаром брался восстановить облик животного по нескольким зу-
бам - одни свойства организмов влекут за собой другие, и понятие архетипа
призвано выразить эту взаимосвязь. Дело в том, что строки таблицы резко не-
равноправны: сравнительно немногие почти не содержат пустых клеток (при-
знак присутствует почти у всех особей) и характеризуют ядро таксона по Бэру,
а в большей части строк пустые клетки преобладают - эти строки означают пе-
риферические признаки, т.е. признаки, характерные для периферических форм.
Чтобы строить естественную систему", надо разумно вырезать из большей таб-
Об этом понятии подробно рассказано далее, в части 6. - Примеч. 2010 г.
57
лицы меньшую, т.е. так выбирать столбцы, чтобы у них было свое ядро (почти
сплошь непустые клетки в некоторых строках). Так, ядро класса птиц образуют
летающие формы, имеющие крепкие маховые перья и киль (гребень грудины),
но ядро отряда страусов - нелетающие птицы без киля, с мягкими декоратив-
ными перьями на месте маховых. И все же страус останется среди птиц при лю-
бом способе классификации, так как перья, клюв вместо зубов, цевка, способ
прикрепления черепа к позвоночнику и многие другие его признаки принадле-
жат классу птиц.
Возможность строить систему связана с тем, что разнообразию свойствен-
на типологическая упорядоченность по Мейену или, что то же, с тем, что ядро
обычно сильно превосходит размерами периферию. Бэр пояснял это обстоятель-
ство на примере класса млекопитающих, которые в большинстве ходят на четы-
рех ногах, мало или совсем не покидают землю и голова их направлена вперед.
Это - ядро класса, значительное большинство его видов, «собственно четверо-
ногие», тогда как периферийные виды могут быть похожи на рыб (киты), на
птиц (летучие мыши), двигаться вертикально (обезьяны) и т.д.
Добавим, впрочем, что изредка (у некоторых крупных таксонов) ядро вы-
является плохо, не превышает периферии. Тогда классификация оказывается за-
труднительной, независимо от подходов тех или иных систематиков. Среди
цветковых такие таксоны издавна заслужили эпитет «crux botanicorum» (тяжкий
крест ботаников), например - порядок Salicales (ивовые), число видов которого
колеблется от 400 до 700 в различных системах, причем почти все они прихо-
дятся на один род Salix (ива). Такой же «сгих» являли собою все бактерии, пока
бактериологи не отказались от иерархических принципов систематики.
Однако для высших многоклеточных ядро обычно таково, как говорил Бэр.
К ядру зверей Бэр отнес хищных, а к их ядру - кошек и собак.
Рассмотрим ныне живущие виды собак чуть подробнее, чтобы выявить ло-
гику ядра и периферии. В семействе Canidae (псовые, или собачьи) ядром («цен-
тральным родом») все признают род Canis (собака), а ядром рода - вид Canis lu-
pus (волк), но в остальном это семейство подвергается непрерывной перегруп-
пировке (хотя новых его представителей в природе не обнаруживают с 1914 г.).
Причина - «промежуточные формы, которые не позволяют проводить четких
границ между ними, постулировать их дискретность» [Волк, 1985, с. 21].
Еще в 1880 г. Томас Гексли выявил в семействе два параллельных ряда
форм - Thooida (волкообразные) и Alopecoida (лисицеообразные), отличающие-
ся строением черепа и глаза (у волкообразных зрачок круглый, у лисицеобраз-
ных - вертикальный, эллиптический). Параллель заключается, в частности, в
том, что у южноамериканских псовых есть и свои «волки» и свои «лисы», хотя
эта группа произошла, как все признают, от своего общего предка.
Вывод Гексли остался без внимания, и в начале XX в. установилась очень
дробная и вычурная классификация семейства: 54 его вида были разделены на 4
рода, из которых 3 имели по 1 виду, зато род Canis делился на 13 подродов, объ-
58
единенных в 3 группы - волков, шакалов и лис [Hunde, 1908]. Затем число видов
уменьшили до 35-37, но о структуре семейства до сих пор договориться не мо-
гут: число родов то сокращают до 8, то увеличивают до 15.
Признаки видов семейства были изучены с помощью 3BM-[Clutton-Brock
е.а., 1976] и оказалось, что волк (вместе с динго и домашней собакой) - перифе-
рическая группа. Компактное ядро семейства образовано двумя тесно сомкну-
тыми субъядрами - группами лисиц и шакалов (к последней на некотором отда-
лении примыкает волк). Ближе всего к середине пространства признаков оказа-
лись две внешне сходные лисицы* - тибетская песочная (Vulpes ferrilata) и перу-
анская пустынная (Dusicyon sechurae).
Переоткрытие параллелизма не побудило авторов к существенному пере-
смотру системы Canidae - они лишь уменьшили число родов с 14 до 9. Харак-
терно, что для перегруппировки пришлось привлечь английские бытовые на-
звания видов псовых.
Однако в 1978 г. появилась новая классификация. Ее автор использовал в
основном те же материалы, но повторил построения прошлого века почти без
вариаций. Он предложил вернуться к широкому пониманию рода Canis - 8 под-
родов и 29 видов, - включающему упраздняемый им род Vulpes. (Тем самым
обыкновенной лисе предложено бинарное название Canis vulpes, и мы возврати-
лись на 220 лет назад, к Линнею). Остальные 6 родов семейства вновь содержат
по 1 виду [Van Gelder, 1978]. Такое блуждание будет бесконечно, если не заду-
мываться о теоретической базе (итеративной процедуре).
Как видим, детализация исследований не привела ни к единству мнений,
ни к их устойчивости, ни даже к существенно новым вариантам системы. Ана-
логичные примеры можно приводить из всех областей систематики. Среди мле-
копитающих «нельзя, пожалуй, указать ни одной группы, которая не нуждалась
бы в полной ревизии» [Павлинов, Россолимо, 1987, с. 4].
И авторы этих слов смело ревизуют «свою» группу: без объяснений ликви-
дировали отряд ластоногих, включив его виды в качестве двух отдельных над-
семейств в состав «естественного» отряда хищных. («Естественность» авторы
понимают в весьма искусственном смысле, который носит название кладисти-
ческого и оценку которому дал в свое время Любищев (1968); он указал, в част-
ности, что согласно кладизму надо бы отнести крокодилов к птицам, чего никто,
разумеется, не делает). Единственным поводом послужила ссылка на работу,
отмечавшую параллелизм моржей с псовыми, а тюленей - с куньими. Тот факт,
что с любой архетипической (целостной) точки зрения морж ближе к тюленю,
чем к собаке, для авторов несуществен, и сходство между ластоногими вообще в
* Уже одного этого факта достаточно, чтобы признать, что «середина пространства при-
знаков» не является информативной, поскольку архетипически ядром признан волк. Дело
здесь в том, что «середина» - понятие статистической ПМ, тогда как систематика, есте-
ственно, должна строиться на понятиях системной ПМ. - Примеч. 2010 г.
59
их «системе» никак не отражено. Как говорил Любищев, отказ теоретизировать
- худший вид теоретизирования: привычка игнорировать параллелизмы приво-
дит на деле к полной зависимости исследователей от любого частного паралле-
лизма, к невозможности выявить ядра таксонов.
При перетасовках часто страдают бинарные названия видов: пользуясь
разными трудами, не всегда можно понять, об одинаковых или различных видах
идет речь - если только не быть специалистом именно по данному семейству. В
отношении известных животных и растений спасают, как ни парадоксально, их
бытовые названия, не меняющиеся столетиями, - вопреки расхожему тезису,
что они вносят путаницу, устраняемую только ученой латынью. (К сожалению,
огромное большинство мелких организмов бытовых названий не имеет). Так,
виды фауны СССР, в прошлом относимые к роду Cams, значатся в общей слож-
ности под 16 родовыми названиями, хотя самих родов ни в один момент не на-
считывалось более пяти. Песец носил 4 родовых и 3 видовых названия, причем
нынешнее (Alopex lagopus) является комбинацией из третьего по времени родо-
вого и первого видового. И здесь, как видим, - блуждание вокруг старого.
Эти подсчеты оказались возможными потому, что недавно впервые вышла
сводка синонимов [Павлинов, Россолимо, 1987]. Она не содержит никаких опи-
саний или объяснений (опять-таки рассчитана только на узких специалистов),
охватывает едва ли тысячную долю всей мировой фауны и все-таки заняла 18
авторских листов. Ясно, что подобная сводка для всех видов животных, расте-
ний, грибов и микробов просто немыслима, да и бессмысленна, если не обуздать
словотворчество натуралистов. Таксономическая чехарда особенно недопустима
сейчас, когда спасение экосистем требует экологических обобщений, охваты-
вающих самые разные группы организмов и значительные промежутки времени.
В принципе задача стабилизации названий не так уж сложна, и ответ дан в
сущности самим Линнеем: не надо путать естественную систему с адресаций
(см. 1.3). От адреса требуются простота и постоянство, тогда как сам таксон мо-
жет быть устроен сложно, а взгляды систематиков на «естественность связей»
видов в нем могут то и дело меняться. Даже если допустить, что когда-нибудь
споры систематиков утихнут, то и тогда одной системой не обойтись: в силу на-
личия ядра и периферии естественная система не может быть удобна для ад-
ресации. Ведь даже нечеткие виды должны адресоваться четко. Методолог С.С.
Розова пишет, что «удобное для нахождения расположение материала» несо-
вместимо в одной системе с «выявлением внутренних системных отношений»
[Теория и методология... , 1983, с. 14].
Сравнение таксонов с адресами обнаруживает факт, неизвестный система-
тике организмов, - адрес не может заменить имя. (Если человек, переехав на
другое место жительства, меняет и имя, то найти его крайне трудно.) Латинское
название вида сможет служить ему именем только тогда, когда перестанет ис-
пользоваться в качестве адреса, т.е. если оно не будет меняться при переносе
вида в другой род. В отношении других таксонов систематика так и делает: пе-
ренос рода в другое семейство, семейства - в другой отряд и т.д. не обязано от-
60
ражаться в их названиях. Бинарное латинское название удобно как имя вида, ес-
ли не придавать совпадению первых слов у разных видов таксономического зна-
чения. Так, обыкновенную лису удобно навсегда назвать Canis vulpes L.72, а ее
принадлежность определенному роду обозначать отдельно. Например, можно
писать: вид Canis vulpes из рода Vulpes L. из рода Vulpes (точно так же, как сей-
час пишут: род Vulpes из семейства Canidae). До Линнея это допускалось: Бау-
гин мог, например, в составе рода Plantago назвать один вид Plantaginella palus-
tris (Подорожничек болотный); он счел это более точным и выразительным, чем
прежнее Plantago aquatica minima [Bauhin, 1623, с. 190]. Можно сделать и по-
другому: закрепить за словом, означающим род, определенное окончание (точно
так же, как окончание -idea означает семейство животных, а -асеае - семейство
растений, грибов или микробов), но не включать такие слова в имя вида. Это
имя не должно меняться, даже если вид будет признан подвидом или, наоборот,
разбит на несколько видов, как мы не перестаем называть «динго» австралий-
скую дикую собаку, хотя идут споры о ее видовом статусе.
В сущности, специалисты не так уж далеки от принятия такого принципа.
Так, в монографии, написанной по просьбе Международной ассоциации таксоно-
мии, признано, что «система номенклатуры, в которой названия изменяются при
изменении классификации, как справочное пособие имеет очевидные недостатки»
[Джеффри, 1980, с. 34]. В ее защиту приведен единственный аргумент: общее ро-
довое название у близких видов удобно, так как указывает на их большое сходст-
во. Этот аргумент сохранит полную силу и при навсегда закрепляемых бинарных
названиях. Понятие неизменных названий уже существует в таксономии (nomen
conservandum), и осталось только договориться о применении его к имени опреде-
ленного организма (голотипа), а не только к названию таксона.
Отделив имя от адреса, легко обнаружить, что в центре споров системати-
ков лежит именно вопрос о соотношении ядра и периферии таксона. Так, род
Canis «в узком смысле» (волк + ближайшие 5-8 видов) это и есть ядро рода, по-
нятого широко. А сам род Canis «в широком смысле» - ядро семейства Canidae.
(Те, кто хотят выявить это ядро более четко, склонны вводить особый таксон -
подсемейство Caninae.) Замечательно, что границы ядра рода CanisBce понима-
ют одинаково, а спорят только о том, какие формы внутри него считать отдель-
ными видами. Поэтому мы можем уверенно записать строение рода в форме:
группа «lupus» (волк, домашняя собака, динго и подвиды волка, иногда
выделяемые в виды) - внутреннее ядро рода Canis;
группа «thos» (койот, шакалы, включая эфиопского72) - внешнее ядро
рода Canis; 12 13 *
12 Символ L. должен означать, что это бинарное название дано Линнеем [Джеффри, 1980,
с. 59]. (Примеч. 2010 г.-. Практика, однако, бывает иной - виду дается нынешнее назва-
ние, а буква L. после него означает, что сам вид был узаконен Линнеем.)
13 Canis simensis. Он имеет ряд лисьих черт [Clutton-Brock е.а., 1976, с. 149], и его иногда
выделяют в особый подрод [Волк. 1985. с. 24].
61
группа «dusicyon» (южноамериканские лисицы, что приблизительно
соответствует роду Dusicyon - около 10 видов) - внутренняя периферия рода;
остальные Caninae - внешняя периферия рода.
Аналогично можно очертить ядро и периферию семейства: ядро - подсе-
мейство Caninae, т.е. род Canis в его широком понимании. Характерно, что Ван
Гельдер, строя систему Canidae, как раз и пользуется расстояниями видов (в
признаковом пространстве) от вида Canis lupus, т.е. описывает как бы слои ядра
и периферии. Следует только оговориться, что волк в качестве центра неуда-
чен*: центром семейства естественнее считать общую периферию родов Canis
(узкого) и Vulpes, где расположены тибетская и перуанская лисицы.
Тот факт, что периферии пересекаются, очень важен: если ядро, как бы его
ни понимать, может совпадать с каким-то таксоном, то периферия таксоном
быть не может, поскольку ее виды отличаются друг от друга больше, чем от ви-
дов соответствующих ядер, и поскольку какого-либо своего ядра (а только оно и
оправдывает введение таксона) в ней найти не удается. К сожалению, иногда
систематики собирают периферические виды в недолговечные таксоны (см. гл.
6). Важно и то, что между ядром и периферией нет четкой границы, что, однако,
по Уэвеллу, не лишает этих понятий строгости.
Ядро и периферия существуют объективно, а вовсе не являются следствием
плохого знания объектов. Поэтому, например, единственный способ определить
прочно место енотовидной собаки состоит в признании, что она - нечеткий род
между семействами псовых, енотовых и виверровых. А для адресации ее удобнее
оставить там, где она уже признана, - среди псовых, на их внешней периферии.
И наконец, отметим, что ядро и периферия - не особенность биологиче-
ских таксонов, а общее явление природы, одно из проявлений естественной ти-
пологической упорядоченности (по Мейену), свойственной группам как матери-
альных объектов, так и абстрактных понятий**. Например, в Солнечной системе
известны два правила, упорядочивающие движения небесных тел. Первое - пра-
вило планетных расстояний (закон Тициуса - Боде), которое гласит, что по мере
удаления от Солнца планетные орбиты располагаются всё реже и реже и что так
же располагаются вокруг планет орбиты почти всех спутников^. Второе прави-
ло - однонаправленность: почти все тела вращаются вокруг своих осей и обра-
щаются по орбитам в ту же сторону, в какую вращается Солнце. Хотя эти пра-
вила лежат в основе любой теоретической модели Солнечной системы, ряд ав-
* С позиции статистической ПМ, на которой стоял Ван Гельдер. - Примеч. 2010 г.
” Подробнее см. далее, с. 337. - Примеч. 2010 г.
14 Каждой орбите как бы соответствует свой дискретный энергетический уровень. По од-
ному уровню могут двигаться несколько мелких тел (пояс астероидов, троянцы, некото-
рые спутники). После открытия у планет-гигантов систем колец и связанных с ними мел-
ких спутников правило можно уточнить: оно действует снаружи от этих систем, т.е. вне
пяти планетных радиусов.
62
торов отрицает их познавательную роль, поскольку не все тела им подчиняются.
В сущности, предмета для спора нет: каждое правило имеет обширное ядро (те-
ла, подчиняющиеся ему с достаточной точностью) и скромную периферию
[Чайковский, 1978].
Другой пример являет система русских падежей. Обычно их называют 6, с
помощью которых упорядочивается подавляющее большинство (ядро) именных
словоформ, но есть еще как минимум 3 падежа. Так, например, звательный падеж
соответствует форме именительного (кто, что), но означает не подлежащее, а об-
ращение; кроме архаических словоформ (Батюшки! Боже! Владыко!), есть и вновь
возникшие, используемые в устной речи специально при обращении: бабусь,
браток, Вась, земе* ля и т.п. Этот падеж (как и другие архаизмы) нужен не только
для понимания изречений вроде «Вра* чу, исцелися сам», но и для выявления пара-
дигмы системы склонения в широком смысле, т.е. ее архетипа по Мейену. (Таб-
лицу архетипа для четырех индоевропейских языков см.: [Богородицкий, 1910].)
Парадигмой склонения в узком смысле именуют конкретный ряд вроде пол -
стол, пола - стола..., т.е. рефрен.) Тем самым, если не ограничиваться вариация-
ми слов «кто, что», а упорядочивать сами ряды словоформ, то выявляется пери-
ферия падежной системы. (О других малозаметных русских падежах см.: [Иванов,
1983, с. 268; Трубецкой, 1987, с. 72], а также: [Пялльидр., 1962, С. 93].)
Наконец, сложный пример - гомология по Оуэну (1.4). Мало кто принимал
ее в полном объеме, да и Оуэн защищал ее скорее как идею, чем как анатомиче-
ский факт. Однако место ее вполне ясно, если признать у этого понятия ядро -
то, что вошло в учебники (соответствие руки и ноги, руки и крыла, двух позвон-
ков), и периферию - те примеры, о которых долго спорили и не договорились.
Они не могут быть поняты сами по себе, так как в каждом из них мало от гомо-
логии (плечевой пояс не похож на позвонок), но находят свое место в общем ря-
ду гомологичных объектов, где на одном конце - почти полная идентичность
(два соседних позвонка), а на другом - полное отсутствие сходства. Поскольку
переходы в этом ряду достаточно плавны, мы можем искать общий механизм,
их порождающий, понимая, что его действие в одних случаях доминирует, в
других - маскируется действием других механизмов, а на пограничных объектах
(в смысле границ таксона «гомология») вообще исчезает.
2.4. Параллельные ряды
Параллельных рядов мы уже не раз касались, и в 2.2 были сформулирова-
ны три основных свойства параллелизма: он выражается правилом «у разных
таксонов - общие мероны», вызван невозможностью использовать для класси-
фикации сразу все существенные сходства объектов и фиксируется он наличием
у разных рядов общего для них рефрена. Теперь нам надо показать общебиоло-
гическую роль параллелизмов и как ими пользоваться.
Совокупность объектов образует ряд, если она обладает каким-то общим
свойством. Это свойство может быть либо заключенным в каждом объекте (на-
пример: ряд стульев), либо задавать способ построения ряда (например: ряд
63
четных чисел 2, 4, 6...), либо сопоставлять одну совокупность с другой (напри-
мер: англо-русский словарь). Поясним это на примерах, даваемых филлотакси-
сом (порядком расположения листьев на стебле/5.
Листья могут быть собраны в одном месте стебля, образуя розетку, или рас-
полагаться вдоль него регулярно - либо мутовками (т.е. группами), либо пооче-
редно. Если в мутовке только 2 листа, то листья называются супротивными. Тип
листорасположения может быть общим свойством крупного таксона (например, в
семействе губоцветных листья всегда мутовчаты и почти всегда супротивны, а в
семействе гвоздичных - почти всегда супротивны, но иногда очередны), однако
может меняться от вида к виду в одном семействе и даже роде, а иногда - и в рам-
ках одной особи (так, у можжевельника в нижней части обычно очередное, а в
верхней - мутовчатое расположение). Изредка листорасположение бывает беспо-
рядочно (например, у женьшеня). Самое же интересное в том, что очередные ли-
стья располагаются вдоль стебля по спирали (вообще спиральная симметрия рас-
тениям свойственна во многом) через равные промежутки, так что между двумя
листьями, расположенными на одной вертикали, помещается одно и то же число
листьев, характерное для данного вида. Спиральное расположение свойственно,
например, всему семейству злаков, причем у них всех (кроме одного австралий-
ского рода) на виток спирали приходится два листа.
Здесь естественно выявляются разные типы параллельных рядов. Так, для
большинства губоцветных и валериановых типично супротивное расположение
листьев, хотя эти семейства принадлежат разным порядкам. Это - пример па-
раллельных рядов с общим свойством.
Бук, вяз, липа, рожь и некоторые другие растения имеют по 2 листа на ви-
ток, хотя далеки таксономически. Столь же далеки друг от друга виноград, оль-
ха, орешник, осока и другие растения, имеющие по 3 листа на виток. Многие
растения имеют по 5 листьев на 2 витка (вишня, дуб, слива и др.) и по 8 на 3
витка (барбарис, малина, тополь и др.) - это, по-видимому, самые частые типы
спиралей. Гораздо реже встречаются 13 листьев на 5 витков (миндаль, облепиха)
и, как исключение, известны примеры, когда 21 лист приходится на 8 витков
(Linum регеппе - лен многолетний). Последовательность дробей
а, = 2/1, а2 = 3/1, а3 = 5/2, а4 = 8/3, а5 = 13/5, а6 = 21/8, ... (*)
где числители и знаменатели образованы числами Фибоначчи (каждое число -
сумма двух предыдущих), сходится ко квадрату золотого сечения [Петухов,
1981, с. 12]. Дальнейшие члены (а7 = 34/13 и т.д.) на листьях просто не удается
измерить, так как соседние члены последовательности (*) слишком мало разли-
чаются, но, например, на шишках они известны.
Сказанное дает возможность указать параллельные ряды, построенные по
обгцему закону. Так, представители порядка буковых - бук (аД ольха (а2) и дуб 15
15 Литературу можно найти по книге С.В. Петухова [1981].
64
(а<) имеют с розоцветными - вишня (а3), малина (аД и миндаль (а3) - то общее,
что являются членами последовательности (*). Заметим, что сама эта последова-
тельность - хороший пример рефрена.
Чаще, однако, рефрен не удается выразить в виде математической формулы, и
приходится описывать его путем непосредственного сопоставления параллельных
рядов. Так, если рассмотреть два порядка цветковых - норичниковые и сложно-
цветные, то в них обоих обнаружатся разные формы как мутовчатых, так и оче-
редных расположений, и, кроме того, есть розеточные формы (подорожники, чер-
тополох бесстебельный). Тем самым, можно построить два параллельных ряда.
Именно так, непосредственно сопоставлением, построены и ряды Вавило-
ва. В каждом таксоне Вавилов различал радикал, т.е. совокупность свойств, его
определяющих, и переменную часть, т.е. свойства, различные у разных предста-
вителей таксона. Если группировать таксоны по близким радикалам, то полу-
чится обычная систематика, но их можно сближать и по сходным переменным
частям. Два вида одного рода Вавилов представлял формулой
Lj(a + b + с + ...), L2(a + b + с + ...),
где - видовые радикалы, а а, Ь, с... - общая для них переменная часть. Не-
достачу одного из слагаемых в одном из рядов позволял Вавилову успешно ис-
кать новые формы растений, но это было лишь начало пути.
Мейен предложил (развивая мысли Кренке и Урманцева) более общее по-
нимание параллелизма: дело здесь не столько в повторности самих признаков,
сколько в повторности «законов преобразования» этих признаков. Другими сло-
вами, Мейен предложил рассматривать не столько те простейшие случаи, когда
а,Ь,с- признаки конкретных особей, сколько более общую ситуацию, когда а =
{at, а2...} - последовательность признаков а,.
Примером может служить последовательность листорасположений (*):
обычно растение данного вида имеет один тип а„, но иногда (например, как ска-
зано, у можжевельника) в разных частях растения наблюдаются разные типы.
Другие примеры сходств (расчленения листа, жилкования листа и др.) приведе-
ны у Мейена [Меуеп, 1973]; здесь они могут быть записаны как b = {bh b2...} {с
= С/, с2...} и т.д. Естественно, что прогнозировать недостающий признак а, по на-
личному ряду а можно гораздо увереннее, чем у Вавилова, где по признакам а и b
прогнозировался с. Однако нам здесь важнее другое: радикал-рефренная структу-
ра таксона допускает эффективную свёртку (компактное описание) информации о
нем, дает возможность увидеть в нем систему, а не случайный набор.
Поясним сказанное тем же примером парных конечностей позвоночных, о
котором шла речь в 2.2. Сведем в таблицу то, что касается изменения конечно-
сти от полного отсутствия до пригодности к активному полету (см. рис. 1, уже
упомянутый в 2.2). Для пяти классов позвоночных получим 5 параллельных ря-
дов, которые мало соответствуют последовательности исторических событий,
но именно этим и интересны. Киты не произошли прямо от рыб, тем более не
65
произошли они от рыб, лишенных задних (брюшных) плавников, но кит до того
похож на одну мелкую глубоководную рыбку, что ее так и называют - китовид-
ка (Cetomimus). Еще более похожи друг на друга хвостатая амфибия си* рен и
ящерица хиро* т, равно как безногая амфибия червяга похожа на змею.
При этом наблюдается сходство целых комплексов свойств. Так, китовидка
- черная, с голой кожей, огромными головой и ртом, с малюсенькими глазками,
не имеет брюшных плавников. Назвать сходство кита и китовидки независимым
приспособлением - значит не сказать ничего, поскольку условия их обитания
различны предельно. Аналогично, нет смысла толковать как независимое при-
способление крохотные, но вполне развитые «ручонки» сирена и хирота: другие
виды в тех же условиях несут по 4 крохотных лапки или вообще безноги. Зато и
здесь налицо целый комплекс сходств, в том числе неожиданных: и сирены, и
хироты живут только на юге Северной Америки, несут по 3-4 (сирены) и по 3-5
(хироты) пальцев на лапке и составляют семейства по три вида в двух родах ка-
ждое. (Правда, сейчас хиротов отдельным семейством не считают, но это, есте-
ственно, не умаляет их параллелизма с сиренами, а является лишь одним из
примеров той таксономической нестабильности, о которой мы говорили в 2.3).
Словом, сравнение сиренов и хиротов заслуживает особого исследования; сам
феномен - мелкие, слабые, но вполне развитые передние конечности при отсут-
ствии задних - назовем хиротностью.
Вывод из этого следует тот, что кроме сходств, порождаемых общностью
происхождения и функций, есть еще сходства чисто диатропической природы.
Обычно их связывают с еще неизвестными законами формообразования, но та-
кую догадку следует принимать с одной оговоркой: речь идет здесь не о путях
клеточной дифференцировки, а об общих законах построения форм. Эту разни-
цу Мейен любил пояснять таким примером: листья высших растений демонст-
рируют те же ряды форм, что и талломы сифоновых водорослей, хотя последние
не разделены на отдельные клетки. Спектр возможных вариантов формообразо-
вания достаточно широк, чтобы обеспечить наблюдаемое разнообразие, но все
же он ограничен (допускает классификацию), и это выражается в параллелиз-
мах. Если этого не учитывать, то неизбежны курьезы вроде упомянутого в 2.3
исчезновения ластоногих из системы млекопитающих.
Всеобщность феномена параллелизма привела Мейена к уверенности, что
делить свойства таксона на радикал и рефрен можно лишь условно, так как при-
знак, входящий в радикал, столь же изменчив в смысле диапазона возможных
изменений*, но только их гораздо реже удается наблюдать. Это различие частот
носит качественный характер и дает возможность практически выявлять ради-
кал, т.е. обеспечивает «естественную типологическую упорядоченность», как ее
понимал Мейен. В качестве примера изменчивости радикала он напоминал дав-
ние указания Д.Н. Соболева, который отмечал, например:
Так и должно быть, если система имеет фрактальную природу. - Примеч. 1997 г.
66
«Аномальное увеличение числа частей также может стать нормальным
отличительным признаком какой-либо группы... У коров, овец и коз в
виде аномалии вместо одной пары рогов появляются иногда две пары... и
если бы этот признак сделался стойким, то возникла бы четырехрогая
порода вроде азиатской четырехрогой антилопы (Tetraceros quadricornis),
которая, можно думать, так и возникла» [Соболев, 1924, с. 161],
Количество рогов входит в радикал, поэтому четырехрогость явно наруша-
ет радикал копытных. Столь же явным нарушением радикала червей является
наличие двух лентовидных тел при одной головке у Triplotaenia mirabilis [там
же, с. 162]. Редкость таких фактов не снимает вопроса о путях и причинах их
появления, точно так же, как редкость алмаза не делает его менее важным и ин-
тересным для кристаллографии, чем графит [Меуеп, 1973, с. 206, 233]. Редкие
варианты указывают на рамки возможного, что впервые использовал для эво-
люционной схемы Мопертюи (1756) и что ныне выступает в виде четкого тера-
то-таксономического параллелизма: то, что в одном таксоне выступает как
уродство, в другом является нормой.
Выявление принципиальных рамок формообразования приводит к воз-
можности эволюционного прогноза - только не в том смысле, какой обычно
вкладывают в этот термин. Появляющиеся изредка в литературе фантастические
изображения организмов будущего (человек с огромной головой на недоразви-
том теле и т.п.) вряд ли представляют научный интерес, но то внимание, которое
им уделяет критика, свидетельствует о некоем прогностическом вакууме, вызы-
вающем беспокойство, пусть и неосознанное, эволюционистов. Ведь способ-
ность к прогнозу обычно приводится в качестве основного критерия работоспо-
собности теории. Поэтому важно, что анализ рефренов и образуемых ими па-
раллельных рядов позволяет отсекать некоторые мысленные варианты эволю-
ции как практически невозможные. А это - первое условие для возможности
прогнозировать будущее развитие природы.
Так, мы можем уверенно сказать, что невозможен «ангел» (наземное по-
звоночное с третьим поясом конечностей), поскольку эта тенденция в соответ-
ствующих параллельных рядах отсутствует нацело. Однако иногда, очень редко,
мы в подобных негативных прогнозах все же будем ошибаться. Например, спо-
собность к планирующему полету у позвоночных обеспечивается конечностями
(иногда и перепонками между ними - ногокрыл, шерстокрыл) и хвостом, а дру-
гой возможности, казалось бы, нет. Однако в действительности существует род
летучих ящериц (Draco), у которых «планер» натянут на выростах ребер. Запрет,
казалось бы наложенный рефреном мерона «конечность», преодолен в рамках
другого параллелизма, о котором мы скажем в 2.5. Подобные казусы не должны
обескураживать, так как сопровождают прогнозы во всех науках.
Наконец, коснемся кратко множественного параллелизма. Он имеет ме-
сто между параллельными рядами А, В, С, D, ... тогда, когда один и тот же реф-
рен связывает любые два ряда (А, В), (А, С), (A, D), (В, С) и т.д.; т.е. когда мож-
67
но говорить о параллели между параллелизмами. В 1857 г. геолог и зоолог Луи
Агассиц описал 5 типов таких рядов: 1) геологическая последовательность по-
явления организмов, 2) их таксономическое положение, 3) их эмбриональное
развитие, 4) их «структурная градация» (т.е. сложность строения, о которой мы
говорили в 1.4), 5) их географическое распространение [Agassiz, 1857]. И сами
ряды, и их сходства между собой были обоснованы здесь далеко не одинаково
(что отметил сам автор), и в литературу вошло лишь обыкновение сопоставлять
первый, третий и четвертый ряды, получившее (после «Общей морфологии»
Геккеля) название метода тройного параллелизма.
До Агассица «теорией параллелизма» называлось сопоставление реально
наблюдаемых сходств зародышей высших животных со взрослыми формами
низших и, на этой основе гипотетически вывод в пользу идеи эволюции орга-
низмов. Вот что писал об этом Агассиц: «Столь поразительно сходство молоди
высших животных со взрослыми индивидами низших типов, что многими авто-
рами было допущено, что все высшие животные проходят, в ходе своего роста,
через фазы, соответствующие постоянной конституции низших классов. Эти до-
пущения, будучи результатом несовершенных исследований, даже стали осно-
вой системы философии Природы, которая представляет всех животных как
различные степени развития нескольких примитивных типов [там же, с. 118].
В своем отрицании примитивного эволюционизма Агассиц, формально го-
воря, был прав, так как общность законов развития сама по себе не указывает на
общность происхождения, но он, противник самой идеи эволюции, был бы, на-
верное, сильно удивлен, если бы узнал, что именно множественный паралле-
лизм, им открытый, дал впоследствии первое серьезное доказательство эволю-
ции. Это произошло через сто лет, когда данные по нынешним и вымершим так-
сонам были спроецированы на палеогеографическую карту (см. 7.3).
Замечательно и то, что обратиться к единой схеме параллелизма Агассица
побудила междисциплинарная параллель - возможность представить правило
планетных расстояний (о котором шла речь выше) той же последовательностью
(*), что и филлотаксис. Для него эта параллель была свидетельством божествен-
ного замысла [там же, с. 130], для нас же - общности системных законов.
Наиболее наглядный пример множественного параллелизма дают те же па-
дежные парадигмы. Родственные слова могут склоняться совсем по-разному
(например, дочь и дочка) или даже относиться к различным архетипам склоне-
ния (стул и немецкое der Stuhl), зато параллелизмы (пол - стол и т.д.) являют
собой самое тесное сходство склонений.
Однако главный интерес представляют параллели между параллелизмами.
Во-первых, все русские слова склоняются по единому архетипу, ядро которого
шестипадежно (см. 2.3); но, во-вторых, для уяснения его структуры надо знать,
что в основе его лежит древняя (санскритская) восьмипадежная парадигма [Бо-
городицкий, 1910], общая для славянских (кроме болгарского, где падежи почти
утеряны) и балтийских языков индоевропейской семьи. Парадигма здесь пора-
68
зительно устойчива (хотя отдельные падежи в отдельных языках утрачены), что
не мешает ей проявлять параллели с языками других семей (алтайской, угро-
финской и др.). Например, в самом устойчивом языке семьи - литовском, наряду
с обычными падежами в некоторых диалектах встречаются множественные ло-
кативы - падежи, основанные не на вопросах что, чего, чему..., а на вопросах
где, куда, откуда', для их описания потребовались термины угро-финской лин-
гвистики [Грамматика..., 1985, с. 90].
В русском языке что-то похожее видно в рамках родительного падежа. В
нем слились функции двух древних падежей - собственно генитива, означавше-
го принадлежность (вкус чая), и аблатива, означавшего отъединение (отказался
от чая), но кроме того, он выполняет функцию партитива, или частичного па-
дежа (немного чаю), свойственного, например, финскому языку.
Как и биологи, лингвисты склонны объяснять подобные сходства ссылкой
на общее происхождение и заимствование (так сказать, скрещивание); казалось
бы, ссылка носит эволюционный характер, но она весьма туманна как раз в эво-
люционном смысле. Почему диалекты воспроизвели не иноязычные окончания,
а сам локативный принцип, чуждый данной семье? Почему это произошло в зо-
не весьма слабых межсемейных влияний? Почему периферийные падежи в ли-
тературном русском языке путаются (на столе, но на полу) и в то же время вы-
являются четко в диалектах [Иванов, 1983, § 170]? Последнее естественно, если
считать периферийные падежи вымирающими, но Н.С. Трубецкой [1987, с. 214]
характеризовал их как новации.
Эти и подобные вопросы аналогичны биологическим и носят отчетливо
диатропический характер: всюду речь идет о рефренах, частых в одних таксо-
нах, но редких в других76. Историческое движение вдоль данного рефрена воз-
можно в любую сторону. Параллели с далекими языками - это именно паралле-
ли, подчас возникающие как безо всякого контакта их носителей, так и вне род-
ственных связей. Допускать сохранение диалектов с праиндоевропейских вре-
мен столь же абсурдно, как допускать ген двухголовости, унаследованной всеми
животными от безголового предка. Вернее сказать, что и в лингвистике, и в био-
логии мы видим феномен транзитивного полиморфизма, эволюционный фено-
мен, о котором речь пойдет в 7.3*.
2.5. Другие тенденции
Обычно за рефреном (правилом преобразования мерона) стоит какая-то
тенденция (массовая, не всегда четкая, но в каком-либо смысле бесспорная за-
16 Точнее, рефрены меронов «индоевропейский падеж» и «угро-финский падеж» совпадают
отдельными значениями в отдельных таксонах (языках). Совпадения часты, если принад-
лежат ядрам таксонов (именно так эстонские падежи терминатив и абессив соответствуют
русскому родительному: до чая и без чая [Пялль и др., 1962, с. 95]) и редки, если принадле-
жат перифериям или ядру и периферии (таковы упомянутые примеры диалектов).
В книге АСМ предложено короткое его обозначение: транпол - Примеч. 2010 г.
69
кономерность), хотя в рамках того же рефрена часто есть нечто далеко не мас-
совое и даже совсем редкое. Так, рефрен, обрисованный посредством рис. 1,
демонстрирует тенденцию превращения лап в ласты и в крылья, но он же дает
примеры редкостей - элементы активного полета у рыб, хиротность (отсутствие
задних конечностей при наличии передних) у некоторых амфибий и ящериц.
Эти редкости в совокупности также образуют тенденцию - редкое в одном так-
соне нормально в другом (правило Кренке). А пустые клетки таблицы свиде-
тельствуют о возможностях, вообще не реализованных известными на сегодня
организмами. Для упорядочения разнообразия надо у каждого рефрена выявить
по возможности все тенденции. Сделаем это для рефрена рис. 1.
В 1857 г. Брони сформулировал «закон террипетного развития» (le loi du
developpement terripete), т.е. развития, устремленного к земле. Согласно Бронну,
морфологический тип «стоит тем выше, чем более преобладают в нем наземные
классы и отряды» (подробнее см. [Чайковский, 1984]). Более того, по мере пере-
хода от типично морских к типично наземным организмам внутри крупного так-
сона тоже наблюдается усложнение строения организмов. В этом состоит тер-
рипетная тенденция организации разнообразия. В частности, она видна на ко-
нечностях: всякое возвращение в воду (превращение конечностей в ласты и
плавники) приводит к упрощению внешней морфологии конечностей, а выход
из воды - к ее усложнению. Таким образом, террипетная тенденция обратима.
Продолжая мысль Бронна, можно заметить, что переход с суши в болото и воду,
с поверхности под землю, из толщи вод на дно - все это вызывает редукцию
(упрощение, уменьшение, утрату) конечностей, преимущественно задних; если
остается одна пара конечностей, то передняя. Среди рыб мы видим это у угрей
(зарываются в дно), у водных форм, принадлежащих наземных классам, - у их-
тиозавров, китов, ламантинов. Очень характерен и калан: у этого прибрежного
морского зверя спереди еще лапы, а сзади уже ласты.
Хотя у современных амфибий и рептилий ласт нет (кроме некоторых чере-
пах), однако тенденция ясно видна и у них: у подземных, болотных и водных
форм уменьшаются или утрачиваются лапы. В обоих классах есть небольшие
группы (хвостатые амфибии си* рены и ящерицы хироты), сохранившие крошеч-
ные, но вполне развитые передние лапки. Объяснить их как приспособление ни-
кто и не пытается, так что рефрен можно в этой его части уверенно трактовать
как чисто диатропический, а не экофизиологический феномен. Донный угорь,
глубоководная китовидка, болотный сирен, подземный хирот, стремительный
морской хищник дельфин, неповоротливый прибрежный ламантин и поедатель
планктона гренландский кит — все они встали в один ряд, реализовав в рефрене
мерона «конечность» состояние «хиротность». Это состояние расположено на
периферии всех пяти классов, но может характеризовать ядра меньших таксонов
(например, отряда китообразных). Будучи редким в двух классах (амфибий и
рептилий), это состояние характеризует в рамках всего подтипа позвоночных
ясную тенденцию (входящую как часть в террипетную тенденцию).
70
Оказывается, что разнообразие конечностей позвоночных складывается за
счет наложения небольшого числа таких тенденций разного уровня. Обычно они
проявляются совместно и создают впечатление невозможности упорядочить
разнообразие, однако их в принципе можно выявить в качестве отдельных. В ча-
стности, для конечностей характерна тенденция к упрощению строения их ске-
лета по мере усложнения их функций: «пятипалая конечность развивалась не
путем усложнения, а путем упрощения из более богато расчлененной конечно-
сти», так что нормальная пятипалая конечность - итог развития из 7-10-лучевой
структуры кистеперых рыб, причем и у нынешних наземных форм наблюдаются
«остатки лишних лучей - элементы, лежащие впереди первого пальца... и эле-
менты, лежащие позади пятого пальца» [Шмальгаузен, 1938, с. 169-170].
В этом можно видеть «концентрацию» в смысле Бронна (см. 1.4) и «олиго-
меризацию» в смысле В.А. Догеля (см.: [Канаев, 1966]). Существенно, что об-
ратное превращение лапы в ласт (упрощение функции и внешней морфологии)
вновь увеличивает число фаланг пальцев, а иногда и число самих пальцев (их-
тиозавры), т.е. эта тенденция тоже обратима.
Не включаясь в давний спор о том, насколько прогрессивна олигомериза-
ция сама по себе, отметим только, что это - самостоятельная тенденция, не
имеющая отношения к террипетной (при которой преимущественно сохраняется
передняя пара). Олигомеризация может заходить так далеко, что будет уже оз-
начать явный морфофизиологический регресс - например, полная редукция ног
(а заодно - органов чувств и т.д.) у многих рептилий; и при этом редукция зад-
ней пары отстает, так что единственная рудиментарная пара здесь всегда - зад-
няя. Здесь есть свои закономерности. В частности, порядок утраты в точности
противоположен тому, по какому шло эволюционное приобретение соответст-
вующих структур [Северцов, 1945, с. 474, 496]. Характерно, что А.Н. Северцов,
отказавшись объяснить этот класс фактов по Ламарку или по Дарвину, т.е. как
чем-то полезных организмам, ограничился выводом, что редукция здесь являет-
ся самостоятельным «процессом, который, однажды начавшись, ведет к полно-
му исчезновению редуцирующегося органа» [там же, с. 500]; тем самым он кон-
статировал тенденцию, и притом необратимую. Замечательно, что «функциони-
рующие органы, несмотря на то, что они исправно функционируют, все же за-
частую редуцируются» [там же, с. 498].
Заметим на будущее (см. 4.4), что Любищев называл такую редукцию ру-
тинизацией (утерей способности к новизне) и видел причину ее в однообразно-
сти (рутинности) использования органа*. В качестве примера он приводил со-
хранение на ноге лошади лишь одного пальца: это называют приспособлением к
бегу, но ведь другие животные бегают подчас не хуже на обычных лапах, зато
остальные функции лапы копыто выполнять не может.
Тезис явно ламаркистский, хотя сам Любищев был к проблемам ламаркизма достаточно
равнодушен. - Примеч. 2010 г.
71
Третья тенденция исследуемого рефрена проявляется в том, что в ходе
эволюции наземных позвоночных регулярно наблюдался подъем головы и свя-
занный с этим перенос тяжести тела на заднюю пару конечностей. Это приводи-
ло к появлению прямосидения, прямохождения, а затем и летания (при этом, в
частности, рептилии эволюционировали в птиц). Тенденция видна еще у лягуш-
ки, но лучше всего выражена у птиц и у человека. В данном процессе неотъем-
лемой частью являлась, как ни странно, необратимая редукция. Так, первоптица
(археоптерикс) имела зубы и свободные пальцы на крыльях (которые тем самым
могли выполнять некоторые функции лап), но птицы утратили эти свойства и
притом необратимо: утеря способности к полету не возвращает крылу свойств
лапы, а наоборот - завершает редукцию крыла.
Комбинацией этих трех тенденций - террипетной, олигополимеризацион-
ной и подъема головы - разнообразие конечностей позвоночных, по-видимому,
в основном и упорядочивается. Каждой тенденции, когда она доминирует, соот-
ветствует свой пример нулевого значения мерона (утраты органа): первой - си-
рен (нет даже следов тазового пояса, но есть передние лапки), второй - змеи
(нет даже следов плечевого пояса, но часты рудименты задних лап), третьей -
киви (нет передних конечностей, но есть неуклюжие ноги).
Кроме трех тенденций, общих для позвоночных, есть еще частные тенден-
ции, выявляемые в рамках рефренов меньших таксонов. Так, мерой «плавники
костистых рыб» проявляет югулярную (от лат. jugularum - горло) тенденцию -
по мере снижения функциональной значимости брюшных плавников они сдви-
гаются вперед, вплоть до горла (то есть вперед от грудных), вырождаются (в де-
коративные лучи и т.п.) и подчас исчезают. Линней выделили горлопёрых (Jugu-
lares) в особую группу, а позже югулярность иногда рассматривали как паралле-
лизм [Бобрецкий, 1891]. Тем самым исчезновение брюшных плавников может
означать как отсутствие террипетности, так и последнюю ступень югулярности,
смотря в какой морфологический ряд ставится данный вид.
Югулярная тенденция замещает у рыб тенденцию подъема головы и отчасти
противоположна ей: плавание головой вверх освоили многие рыбы (вогмер, мор-
ской конек, сельдяной король и др.), но брюшные плавники при этом не развива-
ются, а наоборот, югулируют. Югулярности места нет у всех активно ползающих
рыб, даже у тех видов, которые не пользуются при ползании брюшными плавни-
ками. Причина этого сохранения бесполезных органов видится мне в том, что ак-
тивное движение по поверхности являет (в сравнении с пассивностью и с зарыва-
нием в субстрат) террипетность, а она связана с развитием конечностей. Если так,
то переход рыб к ползанию означает не специальное приспособление к какой-то
экологической нише, а преобладание террипетной тенденции над югулярной. Са-
мые неуклюжие из ползающих рыб, разучившись плавать, ничего еще не получи-
ли взамен, например, морской нетопырь Ogcocephalus (бывший Malthe).
Разнообразие любых объектов только кажется бесконечным, а в действи-
тельности часто видно, как оно довольно просто упорядочено и образуется за
72
счет комбинаций небольшого числа тенденций. Приведу пример из психологии,
который понадобится нам в главе 12. Это шесть тенденций, или типов мотива-
ций (жизненных установок), которые ввел в науку немецкий психолог Эдуард
Шпрангер [Spranger, 1914]: теоретический - стремление к познанию, экономи-
ческий - стремление к выгоде, эстетический - стремление к впечатлению и са-
мовыражению, социальный - стремление к общению и любви, политический (я
бы назвал его чиновным) - желание подчинять и подчиняться, религиозный -
поиск высшего смысла жизни. У отдельного человека обычно преобладает один-
два типа мотивации, но никогда не бывает, чтобы присутствовал лишь один.
Можно спорить о том, насколько перечень полон (я бы ввел седьмой тип:
животный - стремление сохранить себя и потомство, но кто-то может сказать,
что самосохранение - частный случай выгоды, а для кого-то забота о потомстве
- высший смысл жизни), однако несомненно, что тенденции очень удобны для
ориентировки в разнообразии. Становится ясным, например, что политэкономия
описывает не само поведение людей, а лишь те его аспекты, в которых домини-
рует второй тип мотивации (подробнее см. [Чайковский, 1988]).
Желая дать определение царствам животных и растений, философ Анри
Бергсон пришел к выводу, что нет не только ни одного признака, но и ни одного
комплекса признаков, позволяющего четко различить царства, зато указал на
четко выраженные у них «тенденции к развитию» [Бергсон, 1909, с. 94]. Напри-
мер, для растений характерно усвоение углерода из атмосферы, тогда как хищ-
ничество (хотя и представлено насекомоядными растениями) не получило здесь
развития, будучи животной тенденцией. Определение таксона через его тенден-
цию основано на выявлении ядра таксона. Мы воспользуемся этим в главе 6*.
Качественная тенденция, допускающая исключения, играет в описатель-
ных науках ту же роль, какую играет математически выразимых закон в науках
точных. Более того, к ней приходится прибегать и в самых, казалось бы, точных
науках - в тех случаях, когда закономерность налицо, но носит нечеткий харак-
тер. Таковы, например, три качественные тенденции в организации разнообра-
зия планет и спутников Солнечной системы [Чайковский, 1985; 1987].
В этом отношении описательные науки дают урок точным. В частности,
закон больших чисел (см. 3.6) является тенденцией, что важно для понимания
Недавно Анна Дамбикур-Маласэ пришла к выводу, что в происхождении человека ве-
дущую роль сыграла пара тенденций к развитию - двуногость (высвобождение передних
конечностей от функции передвижения) и цефализация (увеличение головы), а вовсе не
обезлесение, как полагали до сих пор [Dambicourt-Malasse, 1996]. На цефализацию ука-
зывали и раньше (Джеймс Дана, 1852 и многие позже), нам же важно указание на авто-
номность тенденций: они действовали сами по себе, а не как реакция на изменение сре-
ды. Это соответствует взглядам Бергсона, Берга и Мейена, особо упиравшего на негод-
ность общепринятого приёма - искать адаптивную ценность для любого наблюдаемого
свойства (см. например: М-263). - Примеч. 1997 г.
73
философских оснований ТВ: вероятностные законы нечетки не только тем, что
выполняются лишь на массовом материале, но и тем, что даже на нем выполня-
ются почти всегда, а не всегда. Для традиционной математики это - изъян, кото-
рый она пытается, не всегда удачно, преодолеть (анализ см.: [Чайковский,
1989]), диатропика же ищет качественные методы работы с подобными объек-
тами как таковыми.
Тенденция может проявляться не только во времени. Так, для последних
100 млн. лет эволюции характерно наступление цветковых растений, но в север-
ной флоре до сих пор господствует хвойный лес. Это - одна из тенденций в про-
странстве. Другая состоит в том, что разные группы обладают различными пре-
имущественными способами освоения новых зон обитания. Например, птицы
освоили водную среду не так, как звери.
С водой связаны водные (170 видов) и 4 - водные и околоводные (200 ви-
дов), но только 1 отряд (состоящий из одного семейства пингвинов - 17 видов)
обладает подлинно водным обликом (сигарообразное тело и ласты). В прошлом
сходным обликом обладала еще бескрылая гагарка. В других же отрядах собст-
венно водных форм нет, и более того, бывают водные виды, совсем лишенные
характерных водных черт строения. Самый яркий пример (его любил повторять
Любищев): оляпка (водяной воробей) плавает, ныряет и бегает по дну, не имея
даже перепонок на пальцах. Почти так же удивителен зимородок.
Совсем другую картину мы видим у плацентарных млекопитающих [То-
милин, 1977, с. 141-143]: здесь имеются три водных отряда - китообразные,
ластоногие и сирёнообразные (к ним относится ламантин) - причем все их 108
видов обладают водным обликом. Всего же плацентарных меньше, чем птиц:
около 4 тысяч видов в 17 отрядах. Даже отдельные представители неводных от-
рядов (например, калан из хищных) имеют водные черты строения. Только у бе-
лого медведя перемена в строении сильно отстает от образа жизни. Очевидна
тенденция: птицы легче других подстраивают поведение (вспомним, что только
птиц можно обучить имитации человеческой речи), но довольно жестко ограни-
чены в изменении строения. Плацентарные, наоборот, легко изменяют как
строение, так и привычки. Этим они резко отличаются от сумчатых, у которых
только 1 вид из 250 - водяной опоссум - связан, и то частично, с водой.
Змее летать не на чем, да и в истории всего отряда змей ничто не говорит о
такой возможности. Однако и летающие змеи бывают - в Индонезии. Там кто
только не летает! Мало того, что там летает самое крупное из рукокрылых мле-
копитающих (калонг - размах крыльев до 1,5 м), но только там летают амфибии
и рептилии (последние, так сказать, «залетают» частично и на материк).
О «летающем драконе» (ящерице) мы уже говорили, но в Индонезии лета-
ет и другая ящерица (из гекконов) - на перепонках, тянущихся от носа до хвоста
и захватывающих обе пары лап. При этом, как бы специально для иллюстрации
параллелизма, точно так же летает (и только здесь) млекопитающее - шерсто-
крыл. Наконец, здесь летает лягушка (растопыривая огромные лапы с перепон-
ками) - единственная летающая амфибия вообще.
74
После такого перечня как-то не столь удивительно узнать, что несколько
видов древесных ужей совершают здесь 10-метровые прыжки-перелеты между
ветвями, а для одного вида известно даже, как он добивается устойчивости в по-
лете: растопыривая костные щитки и втягивая живот, он превращается в нечто
вроде уголковой балки. Почему все они летают (точнее - планируют) именно
здесь (в далеком прошлом рептилии планировали повсюду), никто не знает, но
поразительно, что в сходной по климату зоне Центральной Америки вместо
страсти к полету царит страсть висеть на хвосте - здесь цепкохвосты и амфибии,
и рептилии, и звери. Того же сорта явление (хоть и помельче) - «хиротность»
амфибий и рептилий на юге Северной Америки, о которой мы уже говорили.
Этот географический стиль (термин Любищева) пусть и непонятен в привычных
терминах приспособления, но самый факт наличия ряда свидетельствует о тен-
денции. Таких рядов можно указать много.
Феномен географического стиля характерен не только для биологии. Так,
для языков Европы характерна тенденция к редукции падежной системы с востока
на запад, причем она захватывает языки разных семей с очень разными структу-
рами падежной парадигмы, так что объяснить это прямым заимствованием (скре-
щиванием) можно далеко не всегда. Языки пограничья Восточной Европы и Азии
многопадежны: наибольшее (в Европе и в мире) число падежей (более 40) имеют
языки Восточного Дагестана (кавказская семья), к северу от них калмыцкий язык
демонстрирует наибольшее число (11) падежей монгольской семьи, далее к северу
господствует угро-финская семья (иногда более 20 падежей). От Каспия и Урала к
Карпатам и Балтике данные семьи уступают место индоевропейской (до 7 паде-
жей) семье. Затем, к Пиренеям и Британии, падежи редуцируются в рамках этой
семьи, так что в португальском и валлийском практически отсутствуют. То же де-
монстрирует другая широтно ориентированная семья - кавказская, так что самый
западный ее язык (абхазский) падежей не имеет.
Тенденция редукции падежной системы в целом несомненна, хотя в ир-
ландском языке есть 4 падежа, а в румынском 3. Она столь же несомненно со-
пряжена с тенденцией к усложнению глагольных конструкций, хотя в самом
многопадежном языке (52 в табасаранском) глагольные конструкции - отнюдь
не самые простые [Взаимодействие лингвистических ареалов, 1980, с. 208-209].
Таксономические тенденции могут быть не проще географических. Почему
треть видов отряда кукушкообразных - гнездовые паразиты, тогда как среди дру-
гих птиц это редкость? Почему две группы насекомых (палочники и клопы-
хищнецы) почти сплошь состоят из видов, имитирующих чью-то окраску, тогда
как у других насекомых это бывает редко? Почему в одном отряде динофлагеллат
(панцирных жгутиконосцев) и только в нем встречаются почти все типы клеточ-
ного деления? Почему сумчатые избегают воды? Ответов на эти вопросы пока
нет, но, как мы увидим в главе 3, резкая неравномерность имеет свои причины, а
здесь заметим, что неравномерность задает тенденцию сама по себе, без выявле-
ния рефрена {примеч. 2010 г.’, если не видеть рефрен в повторе неравномерности).
75
2.6. Гомология и аналогия взаимодополнительны
Вавилов назвал свои ряды гомологическими (ссылаясь на гомологические
ряды в химии и на Дж. Майварта [Вавилов, 1987, с. 21, 51], но какова их связь с
понятием гомологии? И с каким именно из многих значений этого термина? А
можно ли назвать гомологическими те более широкие ряды, которые описаны
после Вавилова?
Дарвин называл подобные ряды аналогическими, т.е. не связывал с гомоло-
гией и даже как бы противопоставлял ей. Прямо противопоставлять их стало
обычным после Геккеля, без сомнений объяснявшего параллелизм независимым
приспособлением. Таким способом он объяснял, например, параллелизм между
отрядами сумчатых и плацентарных зверей [Haeckel, 1866, с. CXLIII]. Произошло
смешение трех понятий - гомологии, наследования и приспособления. Как спра-
ведливо отмечал в 1956 г. немецкий морфолог А. Ремане [см. Канаев, 1966, с.
117], нельзя путать объяснение гомологии (как эволюционного феномена) с опре-
делением самого понятия «гомология» (носящего структурный характер и воз-
никшего до идеи его эволюционного или физиологического толкования). Обычно
гомология противопоставляется аналогии - так поступал еще Оуэн (см. 1.3), затем
Дарвин и Геккель, а Карл Гегенбаур, один из последних приверженцев Оуэна,
уточнил данное противопоставление афоризмом: «Аналогия для физиолога - то
же, что гомология для морфолога» (подробнее см.: [Чайковский, 1987]).
В начале XX в. эта мысль поляризовалась в двух противоположных тези-
сах, когда морфолог-ботаник К. Гёбель утверждал: «Морфологично то, что еще
не может быть понято физиологически», а его ученик В. Тролль - наоборот:
«Морфологично то, что вообще нельзя понять физиологически [Канаев, 1966,
133]. Зажатая между этими антитезами, проблема гомологии не могла быть ре-
шена - хотя бы потому, что феномен параллелизма лежал как бы в другой плос-
кости. Прежде всего - считать ли параллельные ряды аналогиями (как Дарвин)
или гомологиями (как Вавилов)?
Гёбель еще в 1880-х годах считал их гомологиями [Stevens, 1984, с. 188]. В
1928 г. гистолог А.А. Заварзин писал, что морфология, пользующаяся понятием
гомологии для выяснения родственных связей, «становится в тупик перед сход-
ствами, часто поразительными, которые существуют между органами только
лишь аналогичными». Их «приходится рассматривать как случайность, как не-
которую игру природы и обозначать самый факт ничего не объясняющим тер-
мином конвергенции». Если же рассмотреть параллелизмы («конвергенции») в
их совокупности, то выясняется, «что они являются выражением каких-то зако-
номерностей, не менее важных, чем даже те, которые лежат в основе учения о
гомологических частях» [Заварзин А.А., 1986, с. 80-81]. Заварзин развил целое
учение о параллелизме в строении тканей, где объяснял его общностью тех
функций, которые выполняют ткани в разных органах и организмах. Однако
многие параллелизмы, как мы видели, этим не объясняются.
В 1930-е годы морфолог М.М. Новиков четко указывал, что вопрос о соот-
ношении гомологии и аналогии шире, чем вопрос о связи формы с функцией, и,
76
в частности, связан со статусом, приписываемым феномену параллелизма
[Бляхер, 1976, с. 201].
Ссылаясь на «Номогенез» Берга, Заварзин утверждал, что формы, имею-
щие различное происхождение, «в своем развитии не могут беспорядочно варь-
ировать, а эволюционируют в одном более или менее определенном направле-
нии, почему в различных параллельных рядах и развиваются конвергирующие
признаки. Таким образом, с этой точки зрения даже многие гомологические час-
ти обязаны своим сходством конвергенции», на что в частности указывает закон
Вавилова [Заварзин А.А., 1986, с. 81].
Конвергентное понимание гомологии звучит странно, но именно так по-
нимал ее Оуэн: он, в отличие от Жоффруа, выводившего гомологию из сходства
развития зародышей, видел в гомологии лишь соответствие самих структур. И
совсем отказаться от понимания Оуэна никто не пытается, ибо некоторое разли-
чие путей развития часто наблюдается даже там, где гомология несомненна для
всех (включая строгих последователей Геккеля). Наиболее наглядно это видно
на дроблении (первых стадиях развития многоклеточного зародыша).
У высших животных господствуют два типа дробления - спиральное у
первичноротых (в частности, у моллюсков и членистоногих) и радиальное у
вторичноротых (в частности, иглокожих и позвоночных), но ярко выражен «па-
раллелизм эволюции радиального и спирального типов» [Преснов, Исаева, 1985,
с. 62], так что приходится то и дело говорить о «вторичных утратах», «вторич-
ных приобретениях», «сборных и неоднородных группах» и т.д. То есть трудно-
сти здесь в точности те же, что и в любой систематике.
Сходство типа дробления с таксономическим типом может оказываться чис-
то номинальным, так что например, по дроблению млекопитающие сходны с пло-
скими червями, а рыбы - с осьминогами. Наоборот, с другими позвоночными и
даже некоторыми млекопитающими дробление у человека несходно. Тем самым,
налицо различие ранних стадий развития всех органов, которые гомологичны у
человека со всеми позвоночными (для некоторых первичноротых такое различие
еще очевиднее). Различие типов дробления осьминога и улитки могло бы поста-
вить под сомнение все гомологии моллюсков, но никто в них не сомневается.
В свете сказанного проясняется «понимание гомологизации как классифи-
кации частей» [Мейен, 1978, с. 496], а не как путей их развития. Те части, кото-
рые принято гомологизировать в руководствах (рука и нога, перо и чешуя и
т.п.), это - элементы радикала таксона по Вавилову (2.4). Но ведь радикал отли-
чается от рефрена лишь малой частотой вариации, а не ее возможными преде-
лами. Поэтому гомологизировать элементы параллельных рядов можно с тем же
основанием, что и другие регулярно реализуемые признаки организмов, в част-
ности - функциональные признаки с нефункциональными (чисто структурны-
ми). Грань между гомологией и аналогией стирается. Гомологизировать прихо-
дится не только основные свойства организмов (принадлежащие радикалу так-
сона), но и все регулярно встречающиеся признаки: «Принадлежность меронов
77
одному классу означает их гомологию» [Мейен, 1977, с. 30]. Более того, понятие
архетипа по Мейену (2.2) подразумевает, что гомологизировать в принципе
приходится не только наличные, но и принципиально возможные свойства - на
этом-то и основаны были прогнозы Вавилова. Но если любые сходства могут, с
некоторой точки зрения, оказаться гомологией, то что же такое аналогия?
Очевидно - то, что рассматривается с другой точки зрения, что дополняет
основной набор сходств. Если какой-то набор сходств признан основным, т.е.
особи, им обладающие, отнесены к одному таксону, тот этот набор признается
первичным (гомологичным), а все остальные сходства - вторичными (аналогия-
ми или параллелями). Так, весь комплекс псовых свойств енотовидной собаки
признан достаточным для включения ее в семейство псовых; при этом все ено-
товые и виверровые ее черты признаются параллелизмом. Это обосновано серь-
езными морфологическими соображениями, имеется ископаемый материал для
сравнений, и все же в 1959 г. зоолог С. Фрешкоп предлагал отнести ее к еното-
вым. Предложение не было принято [Van Gelder, 1978], но налицо тот факт, что
она «не имеет тесного сродства ни с какими другими псовыми» и внешне очень
похожа на енота [Clutton-Brock е.а., 1976, с. 165). Так что в принципе гомология
и аналогия могут меняться местами, и мы вправе говорить о взаимодополни-
тельности этих понятий (в смысле Бора).
Этот вывод звучит странно для всех, кто привык связывать гомологию с
наследованием от общего предка и полагать поэтому, что в принципе гомология
- нечто более важное, чем аналогия, и что в будущем всякая гомология будет
установлена или отвергнута окончательно. Сказанное выше приводит к другой
точке зрения: общий предок - лишь один из источников гомологии, источник
частной гомологии. Поскольку феномен множественного параллелизма носит
всепроникающий характер, то механизм, его обеспечивающий, тоже всеобщ, и
Вавилов справедливо связал его с гомологией. Столь же справедливо Оуэн на-
звал однотипность органов (вроде позвонков) серийной гомологией', онтогенез
носит характер серийного производства. Два соседних позвонка сходны, как две
детали, сошедшие с одного станка, а далекие позвонки могут быть совсем не-
сходны, и все же они гомологичны, принадлежат одной серии, созданы в прин-
ципе одним и тем же онтогенетическим механизмом.
Таковы же принципиальные основания для гомологизации глаз позвоночных
- не только между собой, но и со всеми типами камерных глаз (моллюсков, коль-
чатых червей, медуз) [Заварзин А.А., 1986, с. 81]. Это - свойство гомологии как
понятийного таксона (см. 2.3). Стоит вспомнить Дарвина, говорившего о гомоло-
гии как о свидетельстве родства, а не как о следствии родства [Чайковский,
1987]. В главе 9 мы увидим, что современный термодинамический эволюционизм
рассматривает «подлинную гомологию» как общность процессов, а не предков.
Итак, ряды сходств - следствие общности законов развития, и более важные
(для данного исследования) сходства обычно именуются гомологиями, а осталь-
ные - аналогиями. Такой подход позволяет снять одно старое противоречие, от-
78
меченное Мейеном [Меуеп, 1973, с. 244]. Он напомнил, что часто гомологические
ряды объясняют как результат сходных мутаций в сходных генах. Действительно,
простейшие примеры так объяснить можно (например, альбинизм всегда вызван
нехваткой пигмента), но если связывать ряды с родством вообще, то придется до-
пустить, например, что «общий предок» высших растений обладал способностью
к реализации всех типов расположения листьев вдоль стебля, хотя не имел ни ли-
стьев, ни стебля, а значит - и генов, управляющих их развитием*.
Мы еще почти не знаем, в чем состоят эти законы, но видим, что действие их
можно описать, выявляя общие закономерности. Точно так же, как грамматика
сформулирована и успешно применяется, хотя нет еще определенных представле-
ний о законах развития языка и лингвисты спорят даже о самом факте развития.
Да и геология успешно использовалась для поиска руд задолго до появления
сколько-нибудь убедительных теорий рудообразования.
Литература
Беклемишев В. И. Основы сравнительной анатомии беспозвоночных, т. 1. М., 1964.
Бергсон А. Творческая эволюция. М., Канон-Пресс, 1998. 383 с.
Бляхер Л.Я. Проблемы морфологии животных. М., Наука, 1976.
Богородицкий В.А. Очерки по языковедению и русскому языку. Казань, 1910.
Бэр КМ. Об искусственной и естественной классификациях животных и растений. О
сродстве животных. // Анналы биологии. М., МОИП, 1959. Т. 1, с. 367-383; 395-405.
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. М., 1987.
Взаимодействие лингвистических ареалов. Л., 1980.
Волк/ред. Д.И. Бибиков. М., 1985.
Гельмгольц Г. Популярные научные статьи. СПб. Вып. 1, 1866.
Грамматика литовского языка / ред. В. Амбразас. Вильнюс, 1985.
* Здесь высказано общее (но не Мейена) тогдашнее убеждение, что новое свойство вызыва-
ется появлением нового гена. Теперь, спустя 20 лет, известно, что это не так. Генов оказа-
лось слишком мало для задания посредством них даже основных свойств высшего орга-
низма, не говоря уж о мелких различиях. Различия свойств организмов (как мелкие, так и
крупные) приходится объяснять различием режимов фрактального роста при морфогенезе -
см. далее, с. 198-199. Хороша эта гипотеза или плоха, но другой пока мне неизвестно. При
этом эволюцию чисто условно (пока, за отсутствием фактов) можно принять как следствие
каких-то непонятных изменений в ДНК (напомню, что гены составляют ее малую часть),
изменяющих параметры этого роста. Если так, то появление стеблей и листьев (со всем
спектром листорасположений) могло осуществиться без появления новых генов - доста-
точно было, например, малых изменений в некодирующих участках ДНК (если вообще свя-
зывать эволюцию с изменением ДНК). Параллелизмы листорасположений в различных так-
сонах означают тогда лишь тот номогенетический факт, что в этих таксонах независимо
реализуются одни и те же логические возможности морфогенеза, определяемые различием
режимов фрактального роста. Такое сходство связано не с родством, а с малым числом ло-
гических возможностей (мы видим, что они заданы первыми членами рядов Фибоначчи,
каковых тоже совсем немного). Это, в сущности, и утверждал Мейен. - Примеч. 2010 г.
79
Джеффри Ч. Биологическая номенклатура. М., Мир, 1980.
Заварзин А.А. Труды по теории параллелизма и эволюционной динамике тканей. Л.,
Наука, 1986.
Иванов Валер. Вас. Историческая грамматика русского языка. М., 1983.
Канаев И И. Очерки из истории проблемы морфологического типа от Дарвина до на-
ших дней. Л., Наука, 1966.
Корона В. В. Основы структурного анализа в морфологии растений. Свердловск, Изд.
СГУ, 1987. 272 с.
Мейен С.В., 1977 Таксономия и мерономия // Вопросы методологии в геологиче-
ских... (М-148).
Мейен, 1978, Основные аспекты типологии организмов // Ж (М-165)
Павлинов И.Я., Россолимо О.Л. Систематика млекопитающих СССР. М., МГУ, 1987.
Петухов С.В. Биомеханика, бионика и симметрия. М., 1981.
Преснов Е.В., Исаева В.В. Перестройки топологии при морфогенезе. М., 1985.
Пялль Э., Тотсель Э., Тукумцев Г. Сопоставительная грамматика эстонского и русско-
го языков. Таллин, 1962.
Раутиан А.С. Уникальное перо птицы // ПЖ, 1978. № 4.
Северцов А. И. Собр. соч., т. 3. М.-Л., 1945.
Соболев Д.Н. Начала исторической биогенетики. Харьков - Симферополь, 1924.
Теория и методология биологических классификаций (Труды семинара в МОИП). М.,
Наука, 1983. 169 с.
Томилин А.Г. Снова в воду. М., 1977.
Трубецкой Н.С. Избранные труды по филологии. М., 1987.
Целищев В. В. Философские проблемы семантики возможных миров. Новосибирск, 1977.
Чайковский Ю.В., 1984. «Происхождение видов». Загадки первого перевода И Природа,
1984, №7, с. 88-96 (4-10).
Чайковский Ю.В., 1985 Разнообразие и случайность (4-13).
Чайковский Ю.В., 1987; История науки и обучение науке (на примере дарвинизма) //
ВИЕТ, № 2 (4-16).
Чайковский Ю.В., 1987; Нечеткие закономерности в планетной астрономии (4-18).
Чайковский Ю.В., 1988; Молодежь в разнообразном мире (4-20).
Чайковский Ю.В., 1989; История науки и обучение науке (на примере понятий «слу-
чайность» и «вероятность» (4-23).
Шмальгаузен И.И. Основы сравнительной анатомии позвоночных. М., 1938.
Agassiz L. Contributions to the natural history of USA. Pt 1. Boston, 1857.
Bauhin C. Pinax theatri botanici. Basel, 1623. XXIV + 567 p.
Clutton-Brock e.a. A review of the family Canidae // Bull. Brit. Mus. (Nat. Hist.) Zool.,
1976, vol. 29, № 3.
Haeckel E. Generelle Morphology der Organismen. Berlin, 1866. Bd 2.
Hunde // Brockhaus’ Konversations-Lexicon. Bd 9. Leipzig, 1908. S. 412-418.
Meyen S., 1973. Plant morphology in its nomothetical aspects // Bot. Rev. (M-l 16).
Spranger E. Lebensformen. Halle, 1914.
Stevens P.F. Metaphors and typology in the development of botanical systematics 1690-
1960...//Taxon, 1984, vol. 33, N 2.
Van Gelder R.G. A review of canid classification // Amer. Mus. Novitates, 1978, № 2646.
80
Новые тенденции в теории эволюции
Прошло двадцать пять лет без А. А. Любищева и десять лет без С.В. Мейена,
и настало время, чтобы подвести некоторые итоги. Насколько развитие науки
пошло по тем путям, какие они указывали? Здесь речь пойдет о теории эволю-
ции’*, где в последние годы произошел, по-моему, качественный сдвиг: многое
из того, в чем они были одиноки, сейчас, наконец-то, становится научной нор-
мой. Появилось, разумеется, и такое, чего они не предвидели.
Оба мыслителя настойчиво убеждали нас, что всякое эволюционное иссле-
дование должно сопровождаться понятийным анализом, а иначе может стать
беспредметным. Еще в первой статье об эволюции (1925) Любищев показал, на-
сколько важно понимать, что именуется эволюцией. Его 70 лет не слушали и
продолжали под именем эволюции описывать нечто иное. Так, Любищев разли-
чал эволюцию и эманацию, и лишь сейчас понято, что общепринятый «кладоге-
нез (дробление таксона на подтаксоны) объясняется принципом эманации, а не
эволюции» (Поздняков // Журнал общей биологии, 1996, N 1).
Любищев (1946) указал на необходимость принятия преадаптации любой
теорией, не признающей телеологии, причем «вопрос стоит только о том, как
ограничить рамки преадаптации». И вот преадаптация предложена как основной
фактор - видообразования и только (Gayon // Bull. Soc. zool. France, 1995, N 4).
Далее, вместо эволюции традиционно описывают внутривидовую изменчи-
вость. Мейен своим анализом множественного (он неудачно говорил - пяти-
кратного) параллелизма начал, а своим мероно-таксономическим анализом за-
вершил объяснение живучести этой досадной подмены: разные формы изменчи-
вости (внутри- и межвидовая, одновременная и историческая) действительно
организованы однотипно и потому легко путаются.
Однако путать их не следует: сдвиг изменчивости популяции не может рас-
сматриваться как шажок эволюции - в силу того, что такие сдвиги абсолютно об-
ратимы, поскольку члены популяции наследуют не признаки, а потенции, отчего
популяция наследует полиморфизм как целое (феномен транзитивного полимор-
физма*** по Мейену). Организация полиморфизма, как всю жизнь подчеркивал
Любищев, основана на собственных законах, не сводимых к влияниям среды.
Поэтому стоит вспомнить обоих мыслителей, когда читаешь новейшие мо-
дели разнообразия, основанные на теории фракталов (математических само-
подобных структур дробной размерности)”*’. Пусть Любищев и Мейен не знали
’ Любищевские чтения, 1997. Ульяновск, 1997, с. 31. Поехать я не смог из-за отсутствия
денег, и реферат был зачитан Р.В. Наумовым, председателем оргкомитета Чтений. - Все
примечания к тексту реферата сделаны в 2010 г.
” Характеристику эволюционных взглядов Любищева и их отличие от номогенеза Берга
см.: ИМ, с. 112-115.
”* В книге АСМ предложено короткое его обозначение: транпол - Примеч. 2010 г.
Точнее о затронутых проблемах диатропики, в том числе про мероно-таксономи-
ческий анализ, транпол и фрактальность разнообразия, см. книгу АСМ.
81
о фракталах и не могли предугадать, что систему организмов можно трактовать
как многомерный фрактал с неопределенностью в порождающем правиле, но
фрактальная модель действительно описывает указанные ими свойства системы.
В частности, Ю.Г. и А.Ю. Пузаченко (Журнал общей биологии, 1996, N 1) свя-
зали множественный параллелизм с единством фрактальной структуры над- и
внутриорганизменного уровней.
Там, где понятийного анализа нет, до сих пор лучшие опыты ничего не
могут дать теории. Недавно S.Via & A.Shaw (Evolution (USA), 1996, N 1) в
сущности повторили известные у нас опыты Г.Х. Шапошникова (1957 г.) по
мгновенной эволюции тлей в опытах со сменой кормового растения, получили
те же результаты и так же, как гонители Шапошникова, приписали их отбору.
Факт мгновенности, естественно, опять не объяснен.
Огромны успехи в понимании конкретного хода эволюции. Построены
схемы глобальной эволюции фаун, аналогичные "глобальному флорогенезу"
Мейена*; как и у него, эволюция понята как преобразование разнообразия
(Барсков, Жерихин, Раутиан // Журнал общей биологии, 1996, N 2).
К сожалению, о роли идей Любищева и Мейена обычно не говорится, хотя
авторы постоянно ими пользуются**.
* В действительности дело обстояло сложнее. Во-первых, Мейен фактически опирался в
своих построениях на флорогенетическую схему ленинградских ботаников Е.В. Вульфа
(1944) и М.Г. Попова (1963), по существу им не упомянутую (подробнее см. [Игнатьев,
2003]). Меня ввела в заблуждение его фраза: «О флорогенезе говорили и до меня, но
практически - по новым и четвертичным данным, т.е. схемы в целом получались ad hoc, а
это неинтересно» [Из бесед, 2003]. В действительности это было более чем интересно,
поскольку задало ему вектор поиска, однако мне стало ясно многб позже, после беседы с
И.А. Игнатьевым. Во-вторых, глобальной аналогии с фауногенезом не вышло, и мой то-
гдашний оптимизм был излишним. В-третьих, основной приём Мейена («наложение фи-
логении на географическую карту») задолго до него широко использовал биогеограф Ле-
он Круаза из Венесуэлы (1958 и позже), однако вряд ли Мейену могло быть о нем из-
вестно, как не было известно и мне. Подробнее см. АСМ и [Моггопе, 2000], а также за-
ключительную статью Сборника.
** Этот феномен («Избегание предтеч») подробно описан в работе 4-40. Стоит добавить,
что и сам Мейен попал под его влияние - см. предыдущую сноску.
82
Любищев, Мейен и плюрализм в эволюции
А.А. Любищев в "Уроках истории науки" видел главные достижения разума
вовсе не в универсальных схемах, а в «преодолении претензий на универ-
сальность». Тезис мало кому был тогда близок: лишь в наше время понемногу
распространяется та мысль, что в большинстве ситуаций ни одно решение,
пусть и лучшее, не может быть единственно правильным. Мысль равно
плодотворна при анализе как природы, так и общества.
Самое главное, на мой взгляд, упущение марксизма XX века состоит в
нежелании видеть разнообразие социальных устройств. В частности, одной
фазой капитализма - империализмом - именуется (в том марксизме, которому
нас учили) строй и Англии начала века, и фашистской Германии, и нынешних
США, и даже нынешней Швеции, в некотором смысле социалистической.
Естественно, при столь грубом взгляде на нынешнее социальное разнообразие
нельзя ожидать верных указаний на будущее, что, по-моему, и привело к
конфузу всех нынешних реформаторов (а они сплошь марксисты по воспитанию
и, как бы они это ни отрицали, по кругозору).
Не собираюсь топтать поверженных классиков - в свое время марксизм, как и
дарвинизм, сказал людям много нового и был не только полезен, но и неизбежен.
Параллель между дарвинизмом и марксизмом становится понятной, если признать,
что перед нами две идеологии, а вовсе не две научные (фактически обоснованные)
теории, причем - идеологии, выражающие одно и то же мировоззрение.
Что же это за мировоззрение? Мне уже не раз приходилось писать о нем как
о СТАТИСТИЧЕСКОМ мировоззрении. Оно стало безраздельно господствовать
в науке как раз при Дарвине и Марксе, а в наши дни уступает СИСТЕМНОМУ.
Последнее повсюду старается видеть функциональное единство, так что идея
всепроникающей борьбы (хоть видов организмов, хоть социальных групп
людей, хоть научных идей) теряет популярность. Теперь более популярны
указания на сотрудничество и взаимозависимость. Уход от устаревшей
идеологии отраден, но таит в себе новую опасность.
Вернемся к тезису Любищева. В его время системное мировоззрение едва
начинало прорубать себе путь в джунглях статистического, и трудно было
понимать человека, думавшего в рамках еще более нового способа видеть мир -
плюралистического. Думаю, что вскоре он станет обычным.
Все сказанное требует для своего осмысления нового теоретического
аппарата, каковой видится мне в методе познавательных моделей (ПМ),
который предложен методологом А.П.Огурцовым. См. об этом [1]. Статис-
тическая ПМ - одна из наиболее заметных в миропонимании ученых, всюду
видит средние и балансы. Она в XX в. во многом уступила место системной,
которая всюду видит единство и целесообразность. Ныне в общество проникает
* ЛЧ, 2000, с. 35-37. О социальных темах см. также часть 3 Сборника. - Примеч. 2010 г.
83
следующая ПМ, диатропическая, которая видит всюду организованное
разнообразие. В частности, основной прием дарвинизма (указание на полезность
каждого признака каждого организма) терпит крах, когда анализируется
разнообразие соответствующих признаков: многие из них необязательны для
исполнения предполагаемых функций, как и сами функции. Непревзойденным
мастером такого анализа был Любищев.
Если Любищев аппелировал к разнообразию в основном затем, чтобы пока-
зать несостоятельность как претензий на универсальность, так и дарвинистиче-
ских объяснений, то С.В. Мейен, вполне усвоив его метод, начал строить тео-
рию разнообразия, диатропику. В письме Любищеву от 29.11.1970 он писал "о
высокой степени изоляции сначала трех, а затем четырех крупных флористичес-
ких областей (царств) в течение доброй сотни миллионов лет. Получается как
бы несколько независимых моделей эволюции... и закономерности формообра-
зования, параллелизмы, ортогенез, выпирают особенно четко". Мейен собирался
"сформулировать модусы преобразований, разобраться со становлением во вре-
мени типов симметрии у высших растений и т.д." Вскоре он многое реализовал.
Мейеном отмечено, что эволюционный подход к живому привел к
"предпочтению анализа динамических сторон проявления тех или иных форм и
структур" в ущерб "складывающимся закономерным сочетаниям форм и
структур, безотносительно к их истории и систематической принадлежности".
Желая исправить положение, Мейен предложил тогда два пути: "Один путь,
очевидно, это установление некоторых констант, аналогичных закону
постоянства углов в кристаллах... Другой - изучение повторности конкретных
морфологических типов и их сочетаний в разных таксонах" [2, р. 206, 207]*.
Статья была посвящена второму пути, причем было введено и основное понятие
- "повторяющееся полиморфное множество", понятие, которому Мейен вскоре
нашел новое короткое обозначение - рефрен.
Первым путем он проследовал немного позже и отметил этот факт
введением еще более общего понятия - им стал мерон (класс частей) [3]. Тогда
рефрен - это "упорядоченная изменчивость однотипных меронов в разных
таксонах". Эволюция организмов при этом выступает как заполнение объек-
тивно существующих рефренных таблиц, т.е. как реализация потенциального
разнообразия. Но разные организмы занимают разные клетки таблицы, и потому
познать их удается лишь с позиции плюрализма**
Литература
1. Чайковский Ю.В. Ступени случайности и эволюция. // ВФ, 1996, № 9.
2. Меуеп S.V. Plant morphology in its nomothetical aspects. // Bot. Rev., 1973, vol. 39, № 3.
3. Мейен C.B. Основные аспекты типологии организмов. // Ж. общей биол., 1978, № 4.
Здесь наиболее ясно выражен структурализм воззрений Мейена. - Примеч. 2010 г.
** То есть их сходство нельзя определить в рамках единой таксономии. - Примеч. 2010 г.
84
С.В. Мейен и теория биологической эволюции*
Как эволюционист Сергей Викторович заявил о себе всерьез в конце 1970
года, в статье «Некоторые теоретические вопросы современной палеоботаники».
До этого он касался эволюции как минимум в дюжине публикаций, но только
теперь выступил с цельной оригинальной программой, которую затем разраба-
тывал много лет. Чтобы понять, как и почему она оказалась столь глубоко от-
личной от общепринятого тогда понимания эволюционной проблематики, надо
хотя бы бегло коснуться предшествующей подспудной работы мысли молодого
палеоботаника.
Как и А.А. Любищев (его будущий идейный наставник), Мейен еще в
школьные годы был удивлен явлением параллелизма, и (точно так, как и Люби-
щеву) ему долго никто не мог указать на литературу по этому вопросу. Была на
то в последние годы сталинизма своя веская причина: гомологические ряды Ва-
вилова, самое крупное и известное тогда достижение в области изучения парал-
лелизма, были под запретом - с тех пор, как в 1940 г. Н.И. Вавилов исчез в ста-
линском застенке. Однако смею уверить, что главное было не в страхе, а в не-
знании - и позже, когда имя Вавилова было возвращено в список классиков, а
его работа «Гомологические ряды» многократно переиздана, положение мало
изменилось: большинство палеонтологов и эволюционистов не знает, что с эти-
ми рядами делать. Не знают и на Западе, где Вавилова чтили всегда.
Насколько помню из рассказа С.В., он был первокурсником, когда геолог-
эрудит В.В. Меннер объяснил ему, наконец, про вавиловские ряды. С тех пор
Мейен многократно (и уже целенаправленно) фиксировал параллелизмы на ис-
копаемом материале, понемногу подходя к мысли, то их надо как-то учитывать
в систематике. Легко сказать - надо, но как? Ведь иерархическая систематика
обращаться с ними не умеет, а ни о какой другой прочесть было решительно не-
где. Все споры систематиков тогда (как и почти все теперь, 40 лет спустя) ве-
лись в рамках иерархической идеи. Поэтому подспудный период и оказался у
юного Мейена долог.
На Мейена-студента большое впечатление произвела кристаллография:
оказывается, разнообразие кристаллов жестко ограничено возможностями про-
странственной симметрии. Согласно С.Г. Смирнову, позже Мейен сопоставил
эту ограниченность с ограниченностью вариантов онтогенеза. Отсюда недалеко
до признания той мысли, что формообразование имеет свои собственные зако-
ны, несводимые к потребностям функционирования. Ю.А. Шрейдер описал это
приобретение в теоретическом багаже Мейена как признание им идеи Любище-
ва о «примате формы над функцией».
Все это так, но следует добавить, что к моменту знакомства с творчеством
Любищева Мейен уже защитил кандидатскую диссертацию, где написал: «В на-
* Написано в 1991 г. Опубликовано в МСП, 2001, с. 51-70. Популярное изложение напи-
сано в 1993 г. и опубликовано ранее данного: Химия и жизнь, 1994 (4-31).
85
стоящее время большинство исследователей придерживается убеждения, что в
основе стратиграфических подразделений должны лежать этапы эволюции основ-
ных групп организмов. Установить эти этапы без разработки естественной систе-
мы невозможно» (Автореферат, 1964 [М-17], с. 3). А естественная система, в свою
очередь, требовала описать такие, например, факты: род Rufloria «развивался дву-
мя параллельными ветвями, одна из которых (подрод Rufloria) доминировала в
каменноугольное и позднепермское время, а другая (подрод Neuburgia - в ранне-
пермское время»^^. То есть надо было понять, где эволюция объясняет систему, а
где - наоборот, и Мейен подошел к этой проблеме изнутри своей работы.
Подошел он к проблеме эволюции и извне, от общей проблематики. Еще на
3-м курсе он прочел «Номогенез» Л.С. Берга и «понял, что дарвинизм небеспо-
рочен». Берг не удовлетворил его - как тем, что «заменил понимание терми-
ном», так и своей позицией «или - или», стремлением предложить альтернативу
вместо синтеза. Столкновение дарвинизма с антидарвинизмом породило у юно-
го Мейена «скепсис, может быть даже агностицизм»^. Далее, Мейен высоко
ценил учебник палеоботаники А.Н. Криштофовича, а там был поразительный
параграф «факторы эволюции» - в нем не упоминался отбор, зато была фраза:
«Первым, хотя и менее энергичным фактором является само время». Из кон-
кретных учений в этом параграфе назван только катастрофизм, и притом сочув-
ственно: «преломленное в свете современных представлений и освобожденное
от вмешательства творческой силы извне, это учение само по себе не является
столь ошибочным, отражая коренные переломы в составе органического мира».
Именно это - внимание к течению времени как таковому и к резкой неравно-
мерности перемен стало основой эволюционных исследований Мейена.
Он всегда подходил к эволюции как палеонтолог, подход же Любищева
был иным: он считал все аргументы эволюционизма косвенными, хотя и при-
знавал, что палеонтологические свидетельства эволюции ближе к прямым дока-
зательствам, чем прочие. Поэтому Любищев как эволюционист прославился в
области критики, тогда как Мейен с самого начала своей деятельности подчинил
критику задачам конструктивной работы - реконструкции хода эволюции по
данным палеоботаники.
Наконец, знакомство Мейена с творчеством Любищева началось довольно
поздно - в 1968 г. В первом письме Любищеву, представившись адресату, он
смог уже выразить свою теоретическую позицию если не четко, то достаточно
явно. Он сослался на совместную с А.Ю. Розановым статью^, где понимание
параллелизма продвинуто несколько дальше, чем у самого Любищева. Кроме
17Мейен С.В. 1964. Ангарские кордаиты... (М-15).
18 Письмо А.А. Любищеву от 13 июня 1970 г. Переписка Любищева с Мейеном хранится
как в Петерб. отделении Архива РАН (фонд Любищева), так и в семейном архиве Мейена.
19 Мейен С.В., Розанов А.Ю. 1968. Среда: драматург или режиссер? (М-44).
86
нее, Мейен послал тогда Любищеву свою статью20 о роли неотении в эволюции,
„ 27
и рукопись популярной книги 1.
78-летний Любищев, едва начавший тогда немного работать после перелома
шейки бедра, сразу же ответил: «Разбор Вашей рукописи до известной степени
входит в мой план работы». Отметив: «Вы поняли неудовлетворительность со-
временной эволюционной теории, но Вы еще крепко держитесь за старое», - Лю-
бищев сделал ряд замечаний, из которых 7 носили характер конкретных упреков.
Мейен, однако, ни одного из них не учел, да и на письмо не ответил - вероятно, по
болезни. Но, нет худа без добра, у него «появилось время подумать, так как читать
и писать было трудно из-за глаз», - писал он почти через 2 года. Тогда же он за-
вершил монументальную докторскую диссертацию - палеонтологическую работу,
надолго оторвавшую его от общих проблем эволюции. Так и получилось, что вы-
шедшая в свет первая эволюционная книга Мейена в общем-то чужда тех люби-
щевских идей, которые позже стали для него очень и очень важными.
В книге поставлены почти все интересовавшие Мейена общеэволюционные
вопросы, но нет и намека на большинство из данных им впоследствии ответов.
Пожалуй, единственное исключение являет фраза: «Экспансия на незанятые
участки суши при незначительной конкуренции и богатом воздействии факто-
ров, каждый из которых был в диковинку выходцам из воды, - вот, по-
видимому, тот источник, которому обязан девонский мир своей сверхскорост-
ной эволюцией (указ, соч., с. 186). Пояснений не было, зато был дарвинистиче-
ский контекст, так что Любищев откликнулся восклицанием: «Вот так раз: не-
значительная конкуренция, т.е. слабый отбор - и сверхскоростная эволюция»
(письмо от 7 августа 1968 г.). В действительности, здесь налицо первый намек
на одну из позиций теории глобального флорогенеза, которая гласит: «С более
успешной макроэволюцией высших растений в экваториальной зоне можно свя-
зать лишь ослабление, а не усиление отбора в большинстве случаев»22.
По признанию Мейена, критика Любищева тогда еще не затронула основ
его мыслей, причем «с этими основами еще никто не знакомился, я больше вы-
нашиваю, чем пишу, а пишу только отдельные звенья и подготавливаю почву,
причем, скорее не для читателя (я не та персона, чьи публикации ждут с нетер-
пением и потом тщательно штудируют), а для себя» (письмо от 13 июня 1970).
* * *
В 60-е годы в литературе безраздельно господствовало (за вычетом несколь-
ких статей Любищева, еще не известных Мейену) убеждение, что теория эволю-
ции существует - очередной вариант дарвинизма - и что она носит характер
окончательной принципиальной истины, которую следует лишь уточнять и дета-
20 Мейен С.В. 1968. Дальновидная амблистома (М-50).
21 Мейен С.В. 1971. Из истории растительных династий (М-84).
22 Мейен С.В. 1987. География макроэволюции у высших растений (М-295), с. 307.
87
лизировать. Поэтому прежде всего надо было выявить проблемы эволюционизма.
Этот аспект книги Мейена «Из истории растительных династий» хорошо выражен
цитатой из Г. Эндрьюса: «Возможно, наибольший вклад, который сделала палео-
ботаника, это не заполнение пробелов в наших знаниях об эволюции растительно-
го царства, а показ нам того, как много пробелов существует» (указ соч., с. 20).
Большинство пробелов, указанных Мейеном, были одновременно и факти-
ческими, и теоретическими. Таков, например, первый из них: неизвестно, как,
когда и от кого произошли первые наземные растения. При этом, что особенно
важно, проблема прогресса сразу была разъединена от проблемы разнообразия:
«Откуда же берется все разнообразие девонской флоры? Добро бы только раз-
нообразие! Девонские растения имеют уже сложную внутреннюю структуру,
которая за всю последующую историю растительного мира менялась лишь по
линии второстепенных реорганизаций» (с. 21).
Значительно меньшей была ясность в отношении проблемы приспособле-
ния: с одной стороны, ясно сказано, что «приспособительный смысл большин-
ства свойств растений пока неизвестен» (с. 82) и что даже при катастрофических
изменениях среды никаких “эволюционных вспышек” не наблюдалось» (с. 181-
182), но с другой стороны, «вопрос о происхождении параллелизмов не может
быть окончательно решен, пока не выяснено приспособительное значение па-
раллельно появляющихся признаков» (с. 186). Та мысль, что разнообразие и
приспособленность - разные явления, мысль, которую я позже почерпнул имен-
но из бесед с Мейеном, в этой книге еще не выражена.
Приверженность старой системе взглядов видна и при обсуждении пробле-
мы вида. «Если бы биология могла точно сказать, что такое вид, то половина ее
проблем была бы решена» (с. 199) - эту мысль Мейена Любищев назвал в пер-
вом письме курьезной, заметив: «Современные дарвинисты продолжают повто-
рять мысли Дарвина (простительно в его время), что, разрешив проблему видо-
образования, мы разрешим проблему эволюции в целом». В действительности,
по Любищеву, в эволюции возможно образование высших таксонов помимо ви-
дообразования, а есть в биологии масса вопросов (в том числе - вся физиоло-
гия), к проблеме вида не относящихся.
Разумеется, Мейен уже и тогда не был дарвинистом в обычном смысле этого
слова. Его подспудную тогдашнюю тягу к приспособительным объяснениям можно
в равной мере назвать симпатией и к дарвинизму, и к ламаркизму, и к жоффруизму .
Если в чем он и отдавал предпочтение дарвиновой традиции, так это - в ее акценте
на внутривидовую изменчивость. Как и в схемах дарвинистов, так и у Мейена вид
определяется не столько типовым экземпляром, сколько набором рядов изменчи-
вости. В кандидатской диссертации23 он подробно рассмотрел «монотонные ря-
ды» ископаемых кордаитовых листьев. Монотонный ряд состоит из экземпляров,
* Об основных различиях этих учений см. далее, статью «Ламарк, Дарвин и...».
23 Мейен С.В. 1966. Кордаитовые верхнего палеозоя Северной Евразии... (М-31).
88
собранных в одном слое и связанных друг с другом постепенными переходами
(по морфологии и микроструктуре). Мейен показал, что можно избежать многих
ошибок в определении таксонов, «если больше внимания обращать на внутриви-
довую изменчивость спор и пыльцы, изучая ее как путем построения монотонных
рядов, так и анализируя содержимое спорангиев (пыльников)»2^.
Это исходное внимание к полиморфизму внутри вида имело два важных ре-
зультата. Во-первых, Мейен гораздо легче, чем номогенитики, контактировал с
дарвинистами. Если для Любищева современные дарвинисты (Э. Майр и др.)
были «чистые Угрюм-Бурчеевы от науки»2^, то Мейен легко и весело общался с
Майром (даже переводил его во время выступления того в Москве), а позже пи-
сал: «...из того, что Майр, как и его единомышленник Симпсон ... демонстри-
рует полное непонимание сущности типологии, вовсе не следует, что нет смыс-
ла рассматривать все взгляды этих эволюционистов»24 25 26. Именно Мейену при-
надлежит единственная известная мне серьезная попытка синтеза дарвинизма и
номогенеза. Во-вторых, сопоставление полиморфизма в пределах одной особи
(спорангия) с монотонным рядом (каковой есть упорядоченный полиморфизм
признаков особей одного вида или близких видов) вскоре привело Мейена к
формулировке правила множественного параллелизма - базового феномена при
выявлении рефренов. О том и другом речь пойдет позже.
О своих беседах с Майром он вспоминал, что тот - веселый, остроумный и
контактный человек, но «буквально звереет», когда ему показывают параллель-
ные ряды и говорят, что здесь скрыт какой-то особый закон эволюции. В 1970 г.
Майр издал руководство, в котором (вот правда, по-угрюм-бурчеевски) сказано:
«Типологическая концепция - другая крупная ошибочная концепция, которую
необходимо было полностью опровергнуть, прежде чем предлагать стройную
теорию эволюции». Приняв желаемое за достигнутое, он не назвал среди вопро-
сов и задач ничего, что относилось бы к морфологии и типологии, а паралле-
лизм помянул однажды: «Истинный параллелизм обусловлен реакцией орга-
низмов с общей наследственностью на сходные требования среды (сходные дав-
ления отбора)»27. Вся проблематика тем самым загонялась в словечко «истин-
ный» - что не обусловлено отбором, то вроде бы не истинно.
24 Мейен С.В. 1970. Некоторые теоретич. вопросы современной палеоботаники (М-76), с. 5.
25 Любищев пояснил: такие люди склонны «полагать нежелательные для селектогенеза
явления несуществующими» (первое письмо Мейену). Напомню соответствующее ме-
сто из характеристики глуповского градоначальника: «Но так как не было той силы в
природе, которая могла бы убедить Угрюм-Бурчеева “в неведении чего бы то ни было'’,
то в этом случае невежество являлось не только равносильным знанию, но, даже в из-
вестном смысле было прочнее его» (Салтыков-Щедрин М.Е. История одного города.
М., Художеств, литература, 1970, с. 201).
26 Мейен С.В. 1974. О соотношении номогенетического... (М-121) с. 353.
27 Майр Э. Популяции, виды и эволюция. М.: Мир, 1974. С. 12.
89
Поэтому прежде всего надо было показать, что есть другие параллелизмы, и
тем обосновать, хотя бы для себя самого, необходимость реанимации ошельмо-
ванной типологии. Эту задачу Мейен к 1970 г. блестяще выполнил, построив
ряды, несомненно игнорирующие «требования среды» и «общую наследствен-
ность». Делалось это и до него, причем иногда с блеском (школа Вавилова), но к
новой теории эволюции не привело. Почему?
Ответов можно дать много - смотря на какой точке зрения стоять, что знать в
теории и практике, что уметь понимать и какие цели ставить. Многоаспектность
проблемы раскрывалась Мейену постепенно и, пожалуй, первым крупным шагом
в ее понимании было сомнение в том, что сходство есть мерило родства. В рецен-
зии на книгу А.Л. Тахтаджяна, высоко оценив «глубокую проработку огромного
фактического материала», Мейен усомнился в ее основной гипотезе - что цветко-
вые и беннеттитовые произошли от общего предка (птеридоспермовых). На ос-
новной аргумент автора (невероятно независимое происхождение всех общих
признаков) Мейен возразил: «Мы привыкли верить теории вероятностей и ... опи-
раться на ее законы. Но при этом редко вспоминаем, что любое повышение
организации живых существ и, более того, само возникновение жизни представ-
ляют собой явления, вероятность которых обозначается числом с трехзначным
отрицательным показателем степени. А ведь жизнь когда-то возникла»25.
Разумеется, теория вероятностей здесь ни при чем, речь шла о другом: нельзя
принимать аргумент, когда он работает в пользу твоего убеждения, и игнориро-
вать, когда он работает против. Поняв это, Мейен получил в руки мощный инст-
румент, но тем самым и взвалил себе на плечи огромный груз: взялся за расчистку
вековых завалов, созданных теоретиками, подбиравшими аргументы в свою поль-
зу и отбрасывавшими все остальные как хлам. Весь корпус данных палеоботаники
высших растений надо было просмотреть с единой беспристрастной позиции, ни-
чего не упуская и не отбрасывая. Эта беспрецедентная работа была через много
лет щедро вознаграждена: родилась теория глобального флорогенеза, впервые в
науке* показавшая, как фактически, без домыслов шла эволюция.
Выход в свет рецензии на книгу А.Л. Тахтаджяна как бы подвел черту под
первым периодом работы Мейена по эволюции. Следующая публикация28 29 объ-
являла о начале построения конкретной теории.
* * *
Нехватка места не позволяет мне дальше описывать эволюционные воззре-
ния Мейена в их становлении, и я попробую просто дать эскиз его эволюцион-
ной концепции.
28 Мейен С.В. 1970. Отвратительная тайна раскрыта?// Природа. № 9 (М-75).
’ Ситуация сложнее - см. выше, статью «Новые тенденции...», где в сноске на с. 82 ска-
зано о предшественниках Мейена по этой части. - Примеч. 2010 г.
29 Мейен С.В. 1970. Некоторые теоретические вопросы... палеоботаники (М-76), с. 3-5.
90
Хорошо усвоив ту мысль Любищева, что систематику, морфологию и эволю-
ционизм надо строить вместе, Мейен добавил сюда еще экологию и биогеогра-
фию, но почти не коснулся физиологии, ибо такова уж специфика палеоботаники.
Как и Любищев, он исходил из того факта, что форма системы различна на разных
таксономических уровнях, причем на низшем (роды, виды, расы) уровне естест-
венна сетчатая (комбинативная) форма системы высших организмов, «хотя имен-
но на этом уровне, по доктрине Дарвина, можно ожидать наиболее четкую иерар-
хичность». То же относится и к уровням сложности организмов: «Если у низших
групп система имеет вид решетки свободно комбинирующихся признаков, то с
усложнением организмов решетка начинает вырождаться <.. > в иерархию» .
Налицо был загадочный параллелизм между двумя разделами систематики -
низших таксонов и низших уровней организации. Обратившись к общей теории
систем (вариант Ю.А. Урманцева), Мейен понял, что параллелизм - имманентное
свойство всякой большой системы и что поэтому бессмысленно искать отдельное
объяснение каждому конкретному сходству, а следует положить сам принцип па-
раллелизма в основу теории. До него в биологии было признано только одно на
сей счет правило - «правило тройного параллелизма», которое ввел Э. Геккель30 31
для сопоставления данных палеонтологии, эмбриологии и сравнительной анато-
мии. Грубое соответствие трех типов рядов Геккель широко использовал для эво-
люционных построений, и его родословное древо, по Мейену, - «уникальное и
непревзойденное собрание филогенетических анекдотов».
Мейен действовал иначе: вместо того, чтобы строить филогению (а по ней
и систему) на основе расплывчатых параллелей, решил сперва построить тео-
рию самих параллелизмов. Тройной параллелизм он дополнил до пятерного, до-
бавив к историческим, онтогенетическим и морфологическим параллелям меж-
ду далекими видами еще и параллели между особями близких видов (или даже
одного вида) и частями одного организма. На примере листьев это сделано в
конце работы'32. Вскоре и Мейену, и другим стало ясно, что сюда же надо доба-
вить гено-фенотический, географический и многие другие параллелизмы, так
что мы будем говорить сразу о феномене множественного параллелизма, хоть у
Мейена этого термина и не найдем.
Множественный параллелизм состоит в следующем. Если составить ряд ор-
ганизмов (или их частей, признаков), то, по какому бы принципу ряд не был со-
ставлен (лишь бы принцип вообще был), этому ряду найдутся соответствия в
рядах, составленных по другим принципам. К сожалению, сам Мейен нигде не
дал наглядного развернутого примера, иллюстрирующего его теоретическую
схему, и мне приходится конструировать пример самому.
30 Мейен С.В. 1982. Работы по проблемам системы, эволюции... (М-219), с. 41, 43-44.
31 Он опирался на множественный параллелизм, установленный Л. Агассицом в 1857 г.
Анализ см.: ЭЭД, с. 55.
32 Меуеп S. V. 1973. Plant morphology in its nomothetical aspects (M-l 16).
91
Составим ряд растений по принципу наличия «кленового контура» листа
(пальчато-лопастного с острыми концами лопастей). Сюда войдут многие (но не
все) клёны, платаны, ликвидамбары, смородины, аралиевые и т.п. Упорядочим
ряд: от остролистного клена (Acer platanoides), имеющего шилообразные концы
у глубоко вырезанных лопастей, до черной смородины (Ribes nigrum), у которой
и лопасти, и их острие едва намечены*. Затем упорядочим таким же образом род
Клён (Acer) - в этот ряд войдут все клёны из первого ряда плюс остальные клё-
ны, вплоть до клёна лаврового (A. laurinum), имеющего простой (без лопастей)
лист. Для третьего ряда возьмем все виды деревьев, имеющих лопастные остро-
конечные листья - сюда попадут многие члены первого ряда плюс, например,
некоторые дубы: так, дуб болотный (Quercus palustris) и дуб красный (Q. rubra)
имеют лист почти кленового контура, но не пальчатый (округлый, как растопы-
ренная ладонь), а слегка городчатый (вытянутый в длину).
Четвертый ряд построим совсем иначе: возьмем любое дерево (один экзем-
пляр) из первого ряда и упорядочим все его листья - от самых «кленовых» до
самых «тупых». Оказывается, в нем можно найти все или почти все контуры из
первого ряда (и не только из него), хотя крайних членов может оказаться мало.
В частности у молодых листиков лопасти если и выражены, то слабо.
Почему так - очень таксономически далекие листья имеют сходный контур, а
очень близкие (в том числе с одного дерева) - различный? Даже великий Люби-
щев прошел мимо этого (назвав в первом письме листья «недоорганами»), а вот
Мейен понял, что это - прекрасная модель для уяснения общей теории. Ведь так
устроено чуть ли не всякое разнообразие, только не всегда это столь заметно.
До Мейена ряды выявляли многие, особенно Вавилов, но они попросту со-
поставляли их почленно, не упорядочивая члены внутри ряда. Упорядочение
начал в 1930-е годы московский ботаник Н.П. Кренке, но он не выходил за рам-
ки предполагаемого родства, чем свел к минимуму общебиологический смысл
своих результатов. Наконец, в 60-е годы палеонтолог А.Ю. Розанов составил
периодическую таблицу археоциат, в которой «совершенно одинаковые струк-
туры появляются у разных групп археоциат в разное время. А последователь-
ность появления одинаковая», притом - практически не зависевшая от конкрет-
ных условий среды35. Поначалу Мейен старался объяснить это «по Шмальгау-
зену»: появление каждого свойства создает принципиальную возможность для
создания на его базе каких-то новых свойств, что и объясняет сходство последо-
вательностей. Однако вскоре он обнаружил на своем материале, что последова-
тельность появления свойств тоже может быть различной. Оставалось упорядо-
чивать, исходя из собственной логики рядов, что и привело к понятию повтор-
ного полиморфного множества5^, позже названного рефреном. 33 34
* Для примера см. рис. 5, заимствованный из АСМ, со с. 163. - Примеч. 2010 г.
33 Мейен С.В., Розанов А.Ю. 1968. Среда: драматург или режиссер? (М-44).
34 Meyen S. V. 1973. Plant morphology in its nomothetical aspects (M-116).
92
Известно, что Вавилова называли «ботаническим Менделеевым», однако
следует отметить, что у Вавилова нет периодических систем - их впервые наме-
тил Мейен. (О периодичности системы в биологии писал еще зоолог В.М. Шим-
кевич в 1906 г., но в его системе пантопод периодичности было не больше, чем в
системе цветковых К. Линнея^.) Некоторый аналог периодичности возникает
лишь тогда, когда в нескольких параллельных рядах, выстроенных по горизон-
тали в порядке возрастания (или убывания) некоторого модуса, обнаруживаются
параллельные ряды по вертикали.
Так, можно выстроить в порядке размывания «кленового контура» клены
(от остроконечного A. platanoides через округлый A. tegmentos и яйцевидный А.
tataricum до ланцетовидного A. laurinuin), под ним - смородины (от остроконеч-
ной R. horridium через округлую R. procumbes до яйцевидной R. villosum), а под
ним - аралиевые (от остроконечного Kalopanax septemlobium через округлый
Echinopanax elatum и яйцевидную Aralia mandschurica до почти ланцетовидного
Acantopanax sessiliflorum). При этом остроконечный формы образуют свой вер-
тикальный ряд, округлые - свой, яйцевидные - свой и т.д. Разумеется, некото-
рые клетки останутся пусты. Порядок расположения горизонтальных рядов (а с
тем - и порядок внутри каждого вертикального ряда) может отражать перемену
либо того же самого модуса, либо любого другого (например - тип плода).
Аналогично, периодические ряды можно построить для типов жилкования
листа. Они поразительно напоминают ряды типов жилкования крыльев насеко-
мых, так что нельзя говорить ни об общем предке, ни о приспособлении к об-
щим условиям существования: в обеих группах попросту реализованы все мыс-
лимые модусы.
«Таким образом, мы видим, что органическая форма (в данном случае лист)
распадается на несколько независимых систем с присущими целостными свойст-
вами, причем законы преобразования в пределах одной системы связаны с зако-
нами других систем, но не сводятся к ним»?^. Эти преобразования Мейен хотел
видеть (следуя идеям У. д’Арси Томпсона и Д.В. Наливкина) в форме преобразо-
ваний криволинейной симметрии - как типологических (среди ныне живущих
форм), так и исторических. «Развитие элементов симметрии у наземных растений
происходит со временем, причем раньше в микроструктуре, потом в макропри-
знаках, - я имею в виду геологическое время» (письмо Любищеву от 13.06. 1970).
Сравнив сказанное с монотонными рядами и с пятерным параллелизмом,
можно догадаться, что нащупан какой-то общий закон эволюции как преобразо-
вания разнообразия. Но чтобы ясно выразить его суть, Мейену пришлось при-
думать новое направление классификационной науки, взаимодополнительное к
обычной систематике - мерономию. * *
35 Любищев А.А. 1982. Проблемы формы, систематики и эволюции организмов. М, с. 249.
36 Мейен С.В. 1974. О соотношении номогенетического... (М-121), с. 357.
93
* * *
Мерой - это класс частей. Организм имеет признаки, органы и свойства, а
таксон имеет мероны. Так, позвоночное обычно имеет пары конечностей (плав-
ники, ласты, лапы, крылья, ноги, руки), а таксон «позвоночные» имеет мерой
«парные конечности». Мероном «цветок» характеризуется таксон (отдел цвет-
ковых), мероном «кленовый контур» - квазипериодическая система рядов рас-
тений разных таксонов, а мерой «пищеварение» объединяет животных с насеко-
моядными растениями. Рука и нога - это состояния мерона «конечности», ха-
рактеризующие таксон «человек».
По Мейену, классификационная наука (типология) состоит из таксономии
(исчисления таксонов), занятой классификацией как таковой, и мерономии (ис-
числения меронов), чья основная процедура (расчленение) взаимодополнитель-
на к классификации. Всякая классификация строится итеративно - путем попе-
ременного обращения то к таксономии, то к мерономии, так что всякая система
строится из какой-то прежней, в самом же начале строилась интуитивно. А ис-
следователь обычно думает простодушно, что строит систему сам.
При переходе от одного таксона к другому всегда наблюдаются сходства, а
как правило - сходные ряды. Так бывает даже при переходе от царства к царст-
ву, что подробно описал Г.А. Заварзин. Если можно указать то преобразование,
которое является общим для этих рядов, то налицо рефрен. Таков, например,
рефрен «кленовый контур», причем преобразованием является описанное выше
размывание синдрома «кленовый лист». Наиболее ярким рефреном является пе-
риодичность - когда сам переход от ряда к ряду тоже описывается как законо-
мерное преобразование. Так, в системе химических элементов преобразование
вдоль строки - заполнение одного уровня, а от строки к строке - появление но-
вого уровня электронов.
В 1976 г. Мейен формулировал: «Рефрен - это такое множество объектов,
принадлежащих разным таксонам, которые (объекты. - Ю.Ч.) могут быть сдела-
ны неразличимыми посредством одного и того же преобразования». Позже он
уточнил определение, выразив рефрен через мерой: «Эту повторяющуюся, под-
чиненную одному правилу преобразования последовательность состояний ме-
рона назовем рефреном»^7. Как видим, определение к тому же и смягчено: нет
ни требования неотличимости, ни «объектов» (рефрены строятся для свойств
объектов), ни требования разных таксонов. Однако мысль Мейена легче понять
по первому определению: именно оно показывает, что рефрен призван играть ту
роль, какую в математике играет формула.
В этих терминах многое упрощается. Множественный параллелизм выра-
жается правилом: у разных таксонов общие мероны. Онтогенез формулируется
как движение вдоль рефрена «индивидуальное развитие». А «основной биогене-
тический закон» Мюллера-Геккеля превращается из исторического анекдота в 37
37 Мейен С.В. 1978. Основные аспекты типологии организмов (М-165), с. 501.
94
инструмент познания: онтогенез вовсе не повторяет филогенез (это был бы фи-
зиологический абсурд), но похож на него в силу сходства рефренов. Причина
этого сходства - в том факте, что разнообразие организмов имеет единую реф-
ренную структуру, которую обычно называют законами морфологии.
Понимание данных законов выступает в качестве едва ли не главной задачи
биологии, тогда как до сих пор оно было занятием горстки чудаков. Чтобы при-
ступить к нему, надо переосмыслить в новых понятиях основную идею морфо-
логии - идею архетипа.
Со времен Р. Оуэна (1846) архетипом таксона называют обобщенный план
строения организмов этого таксона. В терминах типологии это «инвариантная
для всех членов таксона совокупность меронов»^, то есть, грубо говоря - схе-
матический рисунок, изображающий то, что есть у всех членов таксона. По-
скольку у Мейена каждый мерой задается не упрощенной схемой, а рядом (или
рядами), то архетип должен быть многомерной таблицей, где каждая строка -
мерой, а каждый столбец - особь (точнее - перечень возможных в принципе для
данного таксона свойств особи). Столь же грубо говоря: архетип по Оуэну - пе-
ресечение множеств признаков, а архетип по Мейену - их объединение. При
анализе эволюции оба понятия архетипа взаимодополнительны.
И опять многое становится понятнее. Например, если таксономия это попро-
сту «теория установления таксонов», то «теория выведения архетипов» - это ме-
рономия . Аналогично можно сказать, что в эволюции таксонов выявляются два
основных феномена - появление новых строк в архетипической таблице (которое
можно назвать прогрессивной эволюцией) и преобразование разнообразия в пре-
делах существующих строк (все прочие процессы). Теории (пусть и ущербные)
фактически предложены только для второго феномена. Однако главная проблема
- «в том, на основании каких критериев можно вычленить из процессов, происхо-
дящих в “кипящем слое”» (в ныне живущих популяциях. - Ю.Ч.\ устойчивые и
истинно эволюционные тенденции, отличив их от ... “броуновского” движения,
создающего лишь шумовое оформление эволюции». То есть встает вопрос - как
перейти «от разнообразия в синхронии к становлению этого разнообразия во вре-
мени. Ведь при желании можно любой современный полиморфизм прочитать как
временную последовательность» (там же), что и делается повседневно.
Чтобы осуществить этот переход, Мейену понадобились еще два понятия -
семофилогении и транзитивного полиморфизма^® Семофилогения - это эволю-
ционное преобразование мерона. Она часто бывает хорошо документирована
ископаемым материалом, который можно достаточно уверенно выстроить во
временной ряд, притом составленный из географически близких или даже моно-
38 Мейен С.В. 1977. Таксономия и мербномия (М-148), с. 29.
39 Мейен С.В. 1984. Принципы исторических реконструкций в биологии (М-263), с. 29.
40 Мейен С.В. 1978. Основные аспекты типологии организмов (М-165), с. 505.
95
топных форм. Говорить о филогении таксона можно только в тех случаях, когда
семофилогении основных меронов обнаруживают один и тот же эволюционный
путь. На практике, однако, мы чаще видим иную ситуацию: семофилогении
преобразуют разнообразие предкового таксона в разнообразие таксона (таксо-
нов) потомков так, что о филогениях остается лишь спорить. Часто у предка и у
потомка наблюдается один и тот же полиморфизм, который, тем самым, и вы-
ступает как нечто наследуемое. (Наследуются не столько сами свойства, сколько
их разнообразие). Этот феномен Мейен и назвал транзитивным полиморфизмом.
Там, где тот господствует, филогенетика теряет смысл, то есть нужна другая
процедура реконструкции прошлого. Теорию эволюции, понимаемую как пре-
образование разнообразия, Мейен называл номотетической*.
Для ее построения надо прежде всего понять те познавательные принципы,
на которых можно хотя бы приближенно выяснить, что есть истина в историче-
ском прошлом. До сих пор основным источником эволюционных построений бы-
ло наличие сходств между рефренами ныне живущих и ископаемых организмов, и
Мейену удалось показать, что на таком базисе ничего устойчивого построить
нельзя. Анализ рефренной структуры разнообразий сам по себе - чисто структу-
ралистский (в смысле К. Леви-Строса), а не исторический инструмент. Так назы-
ваемый «исторический метод в биологии» не умеет всерьез отличать диахронию
от синхронии, а потому и сводится к чехарде «филогенетических анекдотов».
Следовательно, для построения эволюционной теории надо разработать
принципы исторических реконструкций (ИР). Мейен посвятил им работы47, и ос-
новной его вывод состоял в следующем. Принципы ИР (он выявил их девять) об-
разуют познавательный инструмент: «Изолированное приложение любого из
принципов невозможно: все принципы так или иначе сцеплены друг с другом. По-
этому ИР всегда будет не разовым построением, выполняемым с одного захода, а
повторяющейся (рекурсивной, итеративной) процедурой, последовательно при-
ближающей к согласованию доступные факты и эмпирические обобщения»* 41 42.
Другими словами, ИР, как и система, строится на основе какой-то предыдущей
ИР, так что предыдущие построения не отбрасываются, а переосмысливаются.
Результатом работы должна быть система, которую не надо то и дело пере-
сматривать: «Устойчивость ИР на введение ... новых признаков и принимается
* Данный термин плохо воспринимается ввиду его явного тяготения к структурализму. В
данном Сборнике использован более эволюционный термин «новый номогенез». Сочета-
ние нового номогенеза с новым ламаркизмом приводит к идее активного движения вдоль
рефренов (актирефа), изложенной в последней статье Сборника. - Примеч. 2010 г.
41 Мейен С.В. 1978. О наиболее общих принципах исторических реконструкций в геоло-
гии (М-164); 1982. Специфика историзма и логика познания прошлого в геологии (М-
229); 1984. Принципы исторических реконструкций в биологии (М-263).
42 Мейен С.В. 1978. О наиболее общих принципах исторических реконструкций в геоло-
гии (М-164), с. 89-90.
96
критерием истинности»43 44 45. В этом очень важном пункте Мейен следовал за Лю-
бищевым. Кроме принципов ИР, Мейен рассмотрел еще четыре принципа адап-
тивных интерпретаций44, без помощи которых нельзя сделать серьезных выво-
дов о том, являлось ли данное приобретение приспособительным. Насколько
ему удалась эта работа, попробую рассказать на одном примере.
* * *
Попросту говоря, Мейену впервые* удалось показать, что эволюция была на
самом деле. Когда я сказал ему, что именно так воспринимаю его «глобальный
флорогенез», он удивился, но ничего не возразил.
Разумеется, отдельных, хорошо обоснованных филогений было предложено
много и до него, но если подвергнуть их рефренному анализу, то окажется, что об
эволюции они говорят мало. Их авторы (по-видимому, все без исключения) попа-
лись в одну методологическую ловушку: выстроив какие-нибудь ископаемые ор-
ганизмы в ряд от самых древних до самых молодых так, что соседние образцы
сильно похожи друг на друга, исследователь уверенно заявлял, что ряд является
генетическим45, хотя для уверенности нет оснований - в такие же ряды можно
выстроить одновременно живущие виды, а они заведомо не связаны отношением
«предок - потомок». Более того, родство ныне живущих организмов, известных
нам во всей полноте их свойств, приходится то и дело пересматривать, так что
общепризнанное близкое родство вдруг оказывается одень дальним. Вся система-
тика (как палео-, так и нео-) держится на святой вере в том, что таковое родство
заведомо имеет место, то есть сама идея родства проверке не подвергается.
Рефренный анализ рушит эту веру: если все сходства выстраиваются в реф-
рены, а рефрены в принципе одинаково устроены у живых и неживых форм, ес-
ли даже у живых форм идентичные рефрены могут ничего не говорить о родстве
(жилкование листьев и крыльев), то тезис «сходство есть мерило родства» теря-
ет смысл. Следовательно, Любищев был прав, отказывая современной ему па-
леонтологии в статусе эволюционного арбитра, и мы не имеем прямых доказа-
тельств эволюции. Те, кто не имел святой веры в родство, всегда заявляли, что
эволюция ничем не доказана, но до 1980 года их можно было не замечать. А в
43 Мейен С.В. 1982. Специфика историзма и логика познания прошлого в геологии (М-
229), с. 380.
44 Мейен С.В. 1984. Принципы исторических реконструкций в биологии (М-263).
* Нет, как сказано выше (в сноске на с. 82), это делали и до него. - Примеч. 2010 г.
45 То есть соседние формы связаны отношением «предок - потомок». Это отношение яв-
ляется логическим основанием всех филогений и нисколько не меняется от того, что их
авторы делают оговорки (заменяют слово «предковые» на «близкие к предковым» и т.п.).
Мейен решительно возражал против подобного самообмана: «Во имя осторожности
можно добавить к суждениям слово «по-видимому», оставив все остальное на своем мес-
те. В действительности же надо выявить постулаты и принцип ИР, раскрыть их содержа-
ние и граничные условия» (М-164, с. 82).
97
80-е годы начал (сперва среди протестантов-фундаменталистов США) возрож-
даться креационизм, т.е. святая вера в независимое творение каждого вида, и чи-
тающей публике не оставалось ничего другого, кроме как выбрать одну из вер.
Не ведая о рефренах, креационисты стали быстро набирать сторонников, в том
числе (с началом «перестройки») и в СССР. Так что «глобальный флорогенез»
оказался более чем актуален.
«Многое в истории разрешает география» - этой фразой из Гоголя Мейен на-
чал статью «География макроэволюции у высших растений» (1987). Задача новой
концепции - «совместить филогению таксонов с конкретным биогеографическим
фоном, как бы спроецировать ее на историю...», т.е. показать, как в своем кон-
кретном месте и времени один конкретный таксон преобразовывался в другой.
Вся теория, начиная с монотонных рядов'76 и кончая философией времени^7, ока-
залась работающей на эту концепцию. Ее тезисы опубликованы на английском и
русском^9, публикацию развернутого изложения палеоботанических аргументов
автор успел увидеть только по-английски^9, а самой концепции - только по-
русски в популярном изложении59. Научная публикация57 вышла в свет через 3
месяца после смерти автора, а в английском переводе только через 5 лет, в 1992 г.
Суть глобального флорогенеза в том, что «в последевонское время подавляю-
щее большинство порядков и семейств впервые появилось в экваториальном поя-
се, а далее они или не выходили за его пределы, или проникали в бореальные и
нотальные фитохории (сев. и юж. ботанико-географические зоны. - Ю. Ч.). Это
проникновение обычно происходило с задержкой, иногда на целый период, и, ве-
роятно, приурочивалось к эпизодам потеплений, далее судьба переселенцев из
низких широт была различной. Они или исчезали при очередном похолодании,
или же удерживались на новом месте, но с этого времени незначительно меняли
свою общую организацию, давали начало новым видам, реже родам и лишь в ис-
ключительных случаях таксонам более высокого ранга, но не выше порядка».
Другими словами, можно говорить о действии «экваториальной помпы»,
которая при потеплениях «закачивает» виды, возникшие у экватора, на юг и на
север. Эту картину автор назвал «фитоспредингом» - «по аналогии со спредин-
гом (“растеканием”) морского дна в рифтовых зонах» (там же, с. 298), т.е. по
аналогии с теорией дрейфа континентов. 46 47 48 49 50 51
46 Мейен С.В. 1966. Кордаитовые верхнего палеозоя Северной Евразии (М-31); Мейен
С.В. 1970. Некоторые теоретические вопросы современной палеоботаники (М-76).
47 Мейен С.В. 1982. Специфика историзма (М-229); Мейен С.В. 1983. Понятие о времени
и типология объектов: (На примере геологии и биологии) (М-246).
48 Мейен С.В. 1984. Филогения высших растений и флорогенез (М-256; М-257).
49 Meyen S. V. 1984. Basic features of gymnoperms systematics and phylogeny... (M-268).
50 Мейен С.В. 1986. Флорогенез и эволюция растений// Природа. № 11, с. 21-22 (М-280).
51 Мейен С.В. 1987. География макроэволюции у высших растений (М-295).
98
Менялись положение и очертания материков, менялись климаты и флоры,
но неизменно (в течение последних 350 млн. лет) действовал глобальный эво-
люционный механизм: новые формы наземных растений возникали в жарком
климате и, немного изменяясь, расселялись в более холодные зоны. Для таксо-
нов этот механизм фундаментально обоснован, но, кроме того, в виде догадки
заявлен он и для сообществ: «...наиболее продвинутые типы организации сооб-
ществ сначала появлялись в низких широтах и отсюда... смещались во все более
высокие широты»52 53 *. Образно говоря, Мейен показал, где, когда и каким образом
изменялись растения суши, постепенно преобразуясь из риний (первых робких
стебельков на суше) в хвойные.
Поскольку обнаружен и убедительно описан единый механизм преобразо-
вания всех (в указанных рамках) растений, то таксоны и флоры впервые пред-
стают перед нами в становлении. Говорить о том, что они каждый раз создава-
лись Богом сразу в законченном виде (как Адам и Ева, согласно книге Бытия,
созданы взрослыми), теперь нельзя. Верующий человек, изучивший основы па-
леоботаники, увидит теперь Бога не в примитивном облике кустаря, поштучно
изготавливающего всё на свете, а в облике величественного творца общих зако-
нов, в том числе - эволюции. Именно в такого Бога верили и Ламарк, и Дарвин,
и Мейен, продолжавшие этим традицию великих богословов-ученых, из кото-
рых вспомним хотя бы Альберта Великого и Фому Аквинского.
Основной результат Мейена, позволяющий мне уверенно говорить, что его
теория соответствует реальному ходу эволюции, состоит в следующем. Мейен
всю жизнь был противником априорного представления естественной системы в
форме древа, оставляя эту форму в качестве априорной только для систем ис-
кусственных (см., например, его «Основы палеоботаники»55), тогда как форму
естественной системы считал нужным выводить в ходе работы. Тем не менее,
его система высших растений, построенная как естественная, на основе данных
флорогенеза, оказалась именно древом (см. цветную таблицу в популярной ста-
тье «Флорогенез и эволюция растений», 1986). Он сам говорил мне об этом с
некоторым удивлением, но таков оказался итог: все семофилогении сошлась у
него в единую филогению’.
Мейен довольно точно указал таксономическую границу филогении: се-
мейства и выше. То есть налицо вовсе не возврат к геккелизму, не распростра-
52 Мейен С.В. 1986. Флорогенез и эволюция растений// Природа. № 11, с. 56 (М-280).
53 Meyen S.V. 1987. Fundamentals of palaeobotany (М-299), р. 12.
* Нет, смысл флорогенеза “по Мейену” состоит в ином: некоторые семофилогении непре-
рывно соответствуют географической основе флорогенеза, они и положены Мейеном в ос-
нову его «филогенетического древа». Остальные семофилогении либо разрывны (сальта-
ции, гетерогенез), либо это - параллелизмы, не имеющие отношения к данной семофилоге-
нии. - Примеч. 2002 г. (Более новые оценки концепции флорогенеза см. в заключительной
статье и в работах И.А. Игнатьева. - Примеч. 2010 г.)
99
нение дарвиновской идеи дивергенции на высшие таксоны, а обоснование
принципиально иного процесса: сетчато-параллельная эволюция низших (виды,
роды) таксонов сама собой вырождается на высших таксонах в древо.
Цельного описания схемы родо-видовой эволюции он дать не успел, но вот
частный пример: «В целом мы видим в эволюции позднепалеозойских хвойных
комплекс родов, в котором прихотливо переплетаются признаки, различающие-
ся по уровню примитивности и продвинутости. Как бы мы ни группировали эти
роды, будут получаться параллелизмы и реверсии примитивных состояний».
Это не помешало хвойным встать в систему-древо.
Вопрос о факторах эволюции он едва успел наметить*, но и здесь ясно ви-
ден его подход с позиции общей теории систем. Ограничусь одной, наиболее
четкой формулировкой: «Модель прерывистого равновесия, предложенная ра-
нее для видообразования (длительные периоды равновесия сменяются быстры-
ми перестройками), отражает то, что происходит с флорами. Периоды медлен-
ного изменения флор перемежаются короткими интервалами их перестроек (на
что палеоботаники обратили внимание еще в прошлом веке**). Основным фак-
тором, вызывающим быстрые флористические смены, было изменение клима-
та»^. Из других факторов он отмечал (как я уже сказал мимоходом в начале
статьи), ослабление отбора.
Прежние креационисты, умевшие спорить с эволюционистами всерьез, все-
гда отмечали главную трудность эволюционизма: никогда не удается указать
конкретного предка ни для одного вида. Теперь ясно, почему так: не вид преоб-
разуется в вид за счет дивергенции (как учил Дарвин), а семейство преобразует-
ся в семейство за счет транзитивного полиморфизма видов и родов. Быть может,
вообще бессмыслен вопрос о том, какой вид был предком данного.
Нельзя сказать, что эта мысль совсем уж нова - за 230 лет до Мейена нечто
похожее писал великий натурфилософ Пьер-Луи Мопертюи: «...Память о пер-
воначальном состоянии элементов исчезает, и от нас остается скрытым наше
происхождение». Но в биологии победила противоположная идея - во что бы то
ни стало выяснять родословные. Чтобы дать синтез этих, казалось бы, несовмес-
тимых идей, вновь понадобился великий натурфилософ. Им, как мне представ-
ляется, и стал Сергей Викторович, сумевший быть натурфилософом в науке, не
ставившей ни в грош натурфилософию.
Правильнее было бы сказать, что Мейен никогда всерьез не занимался факторами эво-
люции. В частности, здесь он всего лишь уменьшил роль отбора в пользу прямого влия-
ния среды, т.е. остался в рамках традиционных дискуссий. Более новые представления о
факторах эволюции можно найти, например, в книге АСМ. - Примеч. 2010 г.
" Первым, насколько знаю, был Освальд Геер (1859) - см. АСМ; ЗЭ. - Примеч. 2010 г.
54 Мейен С.В. 1987. Основы палеоботаники. М.: Недра, с. 377 (М-294).
100
Мейен как эволюционист и натурфилософ*
Как бы высоко ни оценивать конкретный вклад Мейена в теорию эволюции,
основная его роль в эволюционизме видится как натурфилософская. До него
эволюционисты лишь подбирали примеры под эмоционально приемлемые им
доктрины**, Мейен же заявил своим принципом сплошной просмотр доступного
корпуса фактов вне зависимости от того, в чью они пользу свидетельствуют.
Итогом стал "Глобальный флорогенез", заставивший Мейена во многом
изменить свои эволюционные установки. Результат представляется мне как
фактическим и методологическим, так и этическим. Почти полная его
невостребованность нуждается в уяснении.
Любищев называл линией Демокрита тенденцию произносить смутные
объясняющие фразы обо всем, вместо того, чтобы прямо заявлять - вот это я
знаю и умею объяснять, а вот этого - нет. Название неудачно, ибо данная
тенденция свойственна почти всем, а сутью жизни была отнюдь не для
Демокрита, а для софистов. Тенденцию смог на время пресечь Сократ,
показавший необходимость четко определять предмет обсуждения и введший в
философию нравственный принцип (поиск блага). Этот принцип усвоил Платон;
он же разработал концепцию, согласно которой наблюдаемые объекты - лишь
бледные копии идей, относительно которых только и может быть истинное
знание. Однако подлинную систему объектов тогдашнего знания смог дать
лишь Аристотель, сузивший понимание идеи и сменивший этический принцип
(заменивший всеблагое божество на всемудрое).
Если вместо любищевской дихотомии "линия Демокрита - линия Платона"
ввести дихотомию "линия софистов - линия Сократа", то сам Любищев окажется
Сократом эволюционизма, а софистами - эволюционисты от Дарвина до Майра,
искавшие доводы в свою пользу (сюда по сути должны попасть также ламарки-
сты, номогенетики и все прочие, но Любищев их мало касался). Отсутствие
собственной эволюционной концепции у Любищева выглядит тогда столь же
естественным, сколь и отсутствие своей системы у Сократа.
Но где новые Платон и Аристотель? Мне очевидно, что мероно-таксоно-
мический универсум Мейена эксплицирует концепцию мира идей Платона на
материале биоморфологии, а конкретные мейеновы классификации в страти-
графии и палеоботанике параллельны системе знаний Аристотеля.
Однако логика развития знания, по-видимому, не допускает столь крупных
скачков - хотя бы потому, что воззрения Платона и Аристотеля были отчасти
взаимодополнительны, а отчасти даже противоположны. Платон и Аристотель в
одном лице вряд ли может быть понятен сообществу, которому для этого нужно
время. Нужно оно и нам, чтобы осознать хотя бы путь, Мейеном указанный.
* Тезисы доклада 25 дек. 2000 г. в ГИН // МСП, 2001, с. 70-71.
** Нет, бывали исключения - например, Л. Круаза. См. сноску на с. 82. - Примеч. 2010 г.
101
Эволюция: с чем входим в XXI век?*
XX век подарил биологии несколько важнейших для понимания эволюции
инструментов - генетику, учение о биосфере и молекулярную биологию. Новую
жизнь обрела почти забытая морфология: если сто лет после Дарвина считалось,
что строение организмов нацело определяется как сумма их приспособлений, то
теперь биологи снова, как было до Дарвина, признают самостоятельные
морфологические законы. Кроме того, в толщах пород самых древних эпох,
прежде считавшихся безжизненными, в середине XX века обнаружены следы
первичной жизни, а затем была открыта совсем новая форма жизни -
сообщества организмов горячих источников.
Все это радикально преобразило эволюционную теорию, однако на препода-
вании эволюции почти не отразилось: ее по-прежнему преподают в понятиях ге-
нетики 1930-х годов, добавляя лишь несколько новых терминов. Такая ситуация
не может продолжаться долго, и в конце 1990-х гг. мы видим появление, в том
числе в солидных академических журналах, нескольких программных работ, ко-
торыми школьный преподаватель может и, по-моему, должен воспользоваться.
1. Отношение к эволюции меняется
Знаковым представляется мне факт публикации статьи красноярского
биолога Д.Л. Гродницкого "Критика неодарвинизма", появившейся в "Журнале
общей биологии" (1999, № 5), поскольку этот журнал до сих пор занимал строго
ортодоксальную позицию и считается выразителем взгляда академической
науки на теорию эволюции. Неодарвинизмом автор называет нынешний
дарвинизм как целое и разумно акцентирует внимание не на различиях его
вариантов (чему, увы, посвящены почти все дискуссии), а на трудностях
дарвинизма как такового. Большинство их, как пишет Гродницкий, вообще в
рамках дарвинизма не обсуждается.
По Гродницкому, "собственный язык неодарвинизма слишком беден: на нем
нельзя хотя бы сформулировать даже наиболее очевидные вопросы"; например
- почему у насекомых три пары ног, а у позвоночных - две; почему крупные
животные эволюционируют быстрее мелких. Причину он видит в том, что
строение и работа организма сами по себе не интересуют дарвинистов - они
заняты изучением только полезных отличий. Гродницкому же важны
"механизмы построения и деятельности организмов", поэтому для него "теория
эволюции должна быть основана на этих механизмах, тогда как неодарвинизм
относится к ним как к чему-то постороннему", рассматривая строение и
функционирование организмов "всего лишь как ограничения" выживаемости.
Поясню: мысль о том, что учение Дарвина само по себе не дает теории
эволюции, не нова, и высказывали ее многие, в том числе - многие почитатели
Дарвина. Так, в 1917 г. петроградский ботаник Р.Э. Регель писал (Труды Бюро
по прикладной ботанике, т. 10, № 1), что естественный отбор описан Дарвином
* Биология в школе, 2001, № 1, с. 9-14.
102
прекрасно, но он является не первичным фактором эволюции, а вторичным.
Регеля не услышали, хотя мысль его в сущности проста и очевидна: организмы
преобразуются по своим законам, которые должны быть предметом самосто-
ятельного изучения эволюционистов, а вот результат действия этих законов
демонстрируется отбором, и плохие результаты отметаются.
Вместо законов преобразования организмов дарвинизм XX века занялся
законами изменения генов и выражаемых ими признаков, что очень узко.
Напомню, что в XX веке механизмам функционирования традиционно уделял
внимание ламаркизм, а законам строения (точнее, формам) - номогенез (это
учение основали ленинградский ихтиолог Л.С. Берг в 1922 г. и харьковский
палеонтолог Д.Н. Соболев в 1924 г., а современный вид придал ему московский
палеоботаник С.В. Мейен в 1970-е гг.)
Как известно, дарвинизм во всех его формах утверждает непредсказуемость
эволюции и этим отводит любые упреки в отсутствии у него предсказательной
силы. Но Гродницкий заметил, что ряд базисных утверждений дарвинизма
носит характер прогнозов - таковы тезисы о необратимости эволюции и о
невозможности быстрого (в несколько поколений) видообразования; он
напомнил, что эти прогнозы часто опровергаются практикой. Один приводимый
им пример (эволюцию тлей) мы разберем далее.
Гродницкий уверен, что проблемы эволюции надо формулировать не на
языке генетики, а на языке, описывающем свойства организмов, т.е. на языке
морфологии, физиологии, экологии и этологии. Он призывает к новому синтезу
- этот призыв мы слыхали не раз, но теперь синтез начинает вырисовываться.
Гродницкий описал недавний синтез эмбриологии и генетики, но это, как он
признает - лишь часть дела. Поэтому надо рассказать про недавнюю попытку
общего синтеза, которую предпринял ульяновский зоолог А.Г. Зусмановский.
Доклад, с которым выступил проф. Зусмановский, так и назван: "На пути к
новому синтезу" (XII Любищевские чтения. Сб. докладов. Ульяновск, 2000).
Чуть ранее Зусмановский в своей небольшой книге развил ту мысль, что
полезные изменения развития обязаны не случайным мутациям, а приспосо-
бительным реакциям, которые становятся наследственными в ходе генетиче-
ского поиска - процесса, известного вот уже 30 лет, но совершенно не
упоминаемого в учебных курсах {Зусмановский А. Г. Механизмы эволюционной
изменчивости. Ульяновск, 1999). От дарвинизма им взята "непрогнозируемость
эволюционно значимых потребностей", от ламаркизма - активность особи как
фактор эволюции и "переход в наследственную форму" прижизненных
изменений, от номогенеза - "эмерджентность эволюции" (т.е. возникновение
принципиальных новшеств) и концепция формообразования, известная как
теория биологического поля. И дарвинизм, и ламаркизм, и номогенез тут новые:
Генетический поиск (генпоиск) состоит в активной перестройке генов (а не в пассивной,
как учит дарвинизм), когда организмы попадают в состояние стресса. Он подробно рас-
смотрен в книгах [Назаров, 2005; Зусмановский, 2007], а также в АСМ. -Примеч. 2010 г.
103
они соответствуют современным данным науки и служат базой для перехода к
экосистемной эволюции - в этом и состоит желаемый синтез.
Еще недавно подобные выступления клеймили как "антидарвинизм", но
ситуация меняется. Кроме названных работ, можно указать много других, из
которых в отношении влияния на учебные программы важны статьи бактери-
олога Г.А. Заварзина, поскольку они тоже помещены в официозном и
консервативном академическом журнале, а сам Заварзин недавно стал академи-
ком. В одной из них дано ясное разграничение эволюционной тематики:
«Дарвин решал вполне определенную задачу: происхождение видов. Она
распространяется на все множество живых объектов... Примененный им
принцип естественного отбора действительно имеет универсальное приложение
к очень широкому набору явлений, обозначая, что все неприспособленное
должно исчезнуть. Другое дело конкуренция как основной механизм
естественного отбора... Роль конкуренции как самоорганизующего систему
механизма преувеличена. Она действует лишь на уровне тонкого
приспособления компонента к системе. На других уровнях действуют
кооперация и взаимозависимость. Поэтому речь идет не об антидарвинизме, а о
недарвиновской области явлений» (Заварзин Г.А. Индивидуализм и системный
анализ - два подхода к эволюции. И Природа, 1999, № 1, с. 26).
Снова поясню: индивидуализмом Заварзин называет всякое понимание
эволюции, отправляющееся от свойств особи, популяции или вида (т.е. всякий
взгляд снизу), и противопоставляет его системному взгляду (т.е. взгляду сверху),
при котором свойства биосферы задают возможности экосистемам и входящим в
них видам. Первые шаги эволюции можно описать только системно, поскольку,
как ныне принято считать, жизнь возникла в форме экосистемы, позже
распавшейся на отдельные первоорганизмы. При дальнейшем описании жизни
приходится добавить взгляд снизу, который неминуемо должен играть все
большую роль по мере нашего движения вверх по геохронологической шкале, т.е.
по мере усложнения организмов. Традиционные учения (дарвинизм, ламаркизм,
номогенез и т.п.), равно как и упомянутый выше синтез эмбриологии и генетики,
все относятся к "индивидуализму". Наоборот, учение об эволюции биосферы, как
и всякий экологическиий эволюционизм, является взглядом сверху*.
Что касается идеи конкуренции, то ее Заварзин [1995] рассмотрел еще
раньше, и вывод его был тот, что она неправомерно заимствована в биологию из
политэкономии, где роль конкуренции тоже была в то время завышена.
Академик Заварзин принадлежит к тем эволюционистам, которые видят
развитие природы в виде ряда глобальных новшеств (кризисов, катастроф),
между которыми долго (иногда сотни миллионов лет) не происходит важных
изменений. Первым таким новшеством было само рождение жизни, затем
следовали появление фотосинтеза, кислородной атмосферы, ядерной
* См. ЭКЭ (экосистем, концепцию эволюции) далее, с. 189. - Примеч. 2010 г.
104
организации клетки, твердых скелетов и т.д., а последним - нынешний (отнюдь
не первый в истории Земли) экологический кризис. Прежние эволюционные
учения, либо не видевшие в кризисах ничего существенного, либо просто их
отрицавшие, оказались совершенно бесполезными перед лицом нынешнего
антропогенного кризиса - а ведь он не только преобразует живую природу (и,
следовательно, является эволюционным феноменом), но и срочно требует
работоспособной теории, дающей рекомендации.
Представление о резкой неравномерности хода эволюции тоже должно быть
включено в новый синтез. Далее я коротко расскажу о нем, причем рассказ
будет отчасти резюмировать серию статей, в которой читатель может найти и
ссылки на литературу (Чайковский Ю.В. Эволюция [Ч- 36]).
2. Начало жизни
Биохимические следы жизни обнаружены в самых древних породах
(возрастом более 3,8 млрд лет), так что неясно, была ли Земля безжизненной
хоть какое-то время после формирования суши, океана и атмосферы. Во всяком
случае, процесс возникновения жизни занял столь мало времени, что на
ожидание совпадения каких-либо редких случайностей его явно не было, а это
заставляет искать закономерные процессы превращения неживого в живое.
Дарвинизм тут ничем помочь не может, поскольку исходит из принципа
избыточности размножения, а размножения поначалу не было - оно само
должно было возникнуть по каким-то законам. Ламаркизм, номогенез и прочие
учения говорят не больше. Все ими выявляемые закономерности могут начать
работать, когда уже есть организмы.
Зато помогает новая термодинамика. Если прежняя термодинамика
утверждала, что сам собой может возникать только беспорядок, то теперь мы
знаем - в потоках вещества и энергии могут сами собой возникать и
усложняться структуры (геохимические циклы, залежи минералов, атмосфера,
гидросфера, затем биосфера). Это воспринимается наблюдателем как
собственная активность термодинамической системы. Такой системой явля-
ются и клетка, и организм, и любой биоценоз.
Ныне входит в обиход биоценотическая концепция абиогенеза', начавшаяся с
работы английского физика Джона Бернала (1957) и утверждающая, что жизнь
возникла в форме биоценоза, уже включенного в геохимические круговороты, но
еще не распавшегося на отдельные организмы. Источником первичной энергии
служил не солнечный свет, а восстановленные химические вещества из недр
Земли, т.е. первичная жизнь была не фотосинтетическая, а хемосинтетическая.
' Активность как ведущий фактор эволюции подробно рассмотрена в АСМ, а также да-
лее, в статье «Ламарк, Дарвин и устройство науки». Одной из главных форм активности
является самоорганизация, о которой см. заключит, статью. - Примеч. 2010 г.
* В книге АСМиспользован другой термин: ГЦК - гель-ценотическая концепция (проис-
хождения жизни) . - Примеч. 2010 г.
105
Представление о первых очагах жизни дают недавно открытые подводные
сообщества, живущие у гидротерм - выходов горячих вод и газов на дне океанов.
Новая концепция абиогенеза (происхождения жизни) гораздо оптимистич-
нее, чем теория "первичного бульона" А.И. Опарина (исходившая из идеи сов-
падения редчайших молекулярных событий) и идея "вечности биосферы" В.И.
Вернадского (фактически закрывавшая путь к анализу происхождения жизни).
Первый биоценоз не был биосферой, ибо не был замкнутым (как замкнута
биосфера): вещества из недр необратимо потреблялись, а некоторые продукты
накапливались в океане и в атмосфере. Замыкание произошло, и то не по всем
веществам (круговорота фосфора нет до сих пор), уже после того, как первичный
биоценоз распался на первичные организмы (примитивные бактерии).
Первичная жизнь была, вернее всего, построена на РНК: сперва
одноцепочечные РНК служили как ферменты (это были рибозимы) и как
структурные элементы. Затем появились двухцепочечные РНК, которые легли в
основу первичной наследственности - еще в рамках единой метаболической
системы, не поделенной на организмы. Впоследствии большинство функций
РНК отошло к белкам, липидам и ДНК, а уж затем стали появляться отдельные
носители живого - примитивные бактерии.
С какого момента следует вести историю жизни? С излагаемой точки
зрения, исходным свойством живого является не размножение, а возникшая
раньше собственная активность. О ней давно писали многие эволюционисты -
например, Анри Бергсон (1907) называл ее жизненным порывом. Новая
термодинамика трактует ее как суммарное превышение анаболизма (реакций с
удлинением молекул и усложнения структур) над катаболизмом (реакциями с
укорочением химических цепочек и упрощением структур), что впервые сфор-
мулировал физиолог И.А. Аршавский (1982). Кроме того, нынешняя биофизика
связывает собственную активность живого с биологическим полем - этим
термином называют фактор, организующий процедуру формообразования.
Биополе открыл в 1922 г. петроградский биолог А.Г. Гурвич. Самым простым
и наглядным его проявлением являются "митогенетические лучи" (термин Гур-
вича), стимулирующие митоз. Их природу (это оказался сверхслабый ультрафио-
летовый лазер) раскрыл недавно немецкий биофизик Фриц-Альберт Попп. Поле +
жизненный порыв представляются ныне Поппу ответственными за формирование
живых объектов на всех уровнях организации живого - от клетки до биосферы.
Биофизик В.Л. Воейков (биофак МГУ), развивая Поппа, обращает внимание на
тот мало известный факт, что без излучения (оно может быть не только ультра-
фиолетовым, но и, например, радиационным) никакой организм развиваться не-
способен. Поэтому Воейков полагает, что биополе появилось вместе с жизнью.
Почему-то считается, что дарвинизм обходится без идеи жизненного
порыва. Однако еще Дарвину указывали, что постулированное им стремление
организмов к избыточному размножению есть то первичное свойство жизни,
которое прежде всего нуждается в объяснении. Точно так же нуждается в
106
объяснении постулированное Ламарком стремление к прогрессу и
постулированное в ламаркизме и номогенезе стремление к приспособлению. Все
они могут быть объединены в одной концепции как различные формы
жизненного порыва. Любая теория эволюции обязана указать источник этой
первичной активности, и, на мой взгляд, лучше всего с этим справляется
эволюционная термодинамика, указывающая на неизбежность саморазвития
достаточно сложных и далеких от равновесия систем.
Обретшая свои первичные качества жизнь понемногу растекалась от
гидротерм по первичному океану, насыщая его органикой. С выходом жизни на
поверхность вод смог начать развиваться фотосинтез, который тоже не возник в
форме совпадения редких случайностей, а постепенно сформировался путем
замены хемосинтетических звеньев геохимических круговоротах на
фотосинтетические. Затем возник совершенный фотосинтез, протекающий с
выделением свободного кислорода, а появление кислорода в атмосфере
повлекло формирование привычной нам биосферы - это произошло около
миллиарда лет назад. Суша еще оставалась безжизненной.
3. Усложнение жизни
Первая биосфера была сформирована из бактерий, выполняла все свои функ-
ции, и зачем ей "понадобились" более крупные организмы, неясно. Еще Дарвину
задавали каверзный вопрос: если в борьбе за жизнь побеждают более совершен-
ные, то почему продолжают существовать низшие организмы, включая бактерий?
Ответ Дарвина (у низших-де просты условия жизни) вряд ли сейчас кого-то уст-
роит - жизнь "низших" отнюдь не проста; достаточно вспомнить фантастически
запутанные жизненные циклы некоторых низших червей или резкие перепады
температур и концентраций, в которых ухитряются выживать микробы. Но
важнее, что без бактерий мы просто не сможем существовать - это простейшее
экологическое обстоятельство в дни Дарвина еще не было сформулировано.
Гораздо лучшее объяснение факта существования организмов различной
сло-жности дает концепция жизненного порыва: то самое термодинамическое
свойст-во организмов, которое побуждает их активно жить и избыточно раз-
множаться (при этом изменяясь), ведет и к их усложнению. Однако разные ор-
ганизмы реализуют жизненный порыв по-разному - одни усложняют строение
клетки, другие - взаимодействие своих клеток, третьи усложняют химические
реакции внутри неизменных по форме клеток. Не будь такого различия, жизнь
была бы невозможна - как невозможна жизнь государства, в котором все люди
бросились бы осваивать одну профессию. Ни один организм не достигает успеха
сра-зу во всем - в этом состоит принцип компенсации Аристотеля. И ни один не
отстает сразу во всем, так что бактерии - отнюдь не низшие: биохимически они
успешнее нас. С данной позиции достаточно просто объяснить ученикам ход
эволюции за последний миллиард лет, когда образовались все, кроме бактерий,
формы жизни. Заодно естественно усваивается та мысль, что нынешние бакте-
рии вовсе не старше нас: если бацилла поражает только млекопитающих, есте-
ственно считать, что она младше класса млекопитающих.
107
4. Механизмы эволюции
Твердо установлено, что наследственность бактерий устроена проще, чем у
"высших", но и она столь сложна, что ясно говорит о долгой истории.
Нынешние бактерии заведомо гораздо сложнее первичных. "Случайные
ненаправленные мутации", о которых пишут учебники дарвинизма, хотя и
происходят в бактериях, но не играют в их эволюции существенной роли.
Гораздо важнее механизмы наследственного приспособления бактерий,
известные уже 60 лет, но лишь недавно начавшие получать признание.
Начало признанию положили уже ставшие знаменитыми опыты
американского генетика Джона Кэйрнса (Cairns), который в 1988 г. повторил
вошедшие во все учебники опыты, которые провели Сальвадор Лурия и Ганс
Дельбрюк; они в 1943 г. "доказали", что бактерии не могут приспосабливаться к
антибиотикам. По их (и многих других) мнению, могут выживать лишь те из
бактерий, в которых случайно возникла мутация, вызвавшая устойчивость) и
обнаружил удивительный факт: оказывается Лурия и Дельбрюк использовали
смертельные дозы антибиотиков, так что приспособление в принципе протекать
не могло. Достаточно было уменьшить дозы, и приспособление происходило, а
его результаты наследовались.
Механизм наследственного приспособления (генетический поиск) состоит в
том, что организм, попав в условия стресса, резко меняет режим работы своей
генетической системы. В частности, в популяции таких организмов у
большинства особей мутируют очень немногие гены и в очень немногих
направлениях каждый, зато очень часто. При этом основная масса особей
гибнет, но среди выживших часто находятся нашедшие приспособление,
позволяющее популяции выйти из стрессовых условий.
Как видно из сказанного ранее, на становление бактерий ушло в 2-3 раза
больше времени, чем на всю дальнейшую эволюцию. И вот что замечательно:
чем крупнее организмы, тем меньше их численность и дольше жизнь каждой
особи, однако никакого замедления эволюции у крупных форм не наблюдается.
Скорее наоборот: одни из самых быстро эволюционировавших зверей - слоны.
Это значит, что с усложнением организмов меняются и механизмы их
эволюции. Из них упомяну лишь один: с появлением мейоза и кроссинговера
главным поставщиком изменений при генетическом поиске становятся не
мутации, а рекомбинации блоков генетического материала.
Роль мутаций и рекомбинаций в генетическом поиске лучше всего изучена
на механизме иммуногенеза-, гены новых антител формируются за счет
перегруппировки блоков, а затем за счет мутаций происходит тонкая подгонка
антигенов. Иммуногенез оказался хорошей моделью эволюции.
У насекомых мы видим удачное сочетание высоко развитого генетического
поиска с высокой численностью особей и частой сменой поколений, что
приводит к удивительно быстрой эволюции. Самым ярким примером считаются
результаты классических опытов с тлями рода дизафис, которые ленинградский
108
энтомолог Георгий Христофорович Шапошников (1915-1997) публиковал в
1960-х гг. Вот как резюмирует их Гродницкий:
«Тли Dysaphis anthrisci majkopica Shap., в природе обычно встречающиеся на
купыре Anthriscus nemorosa, были перенесены на бутень клубненосный Chaero-
phyllum bulbosum - растение, на котором этот вид тлей редко встречается в при-
роде. После нескольких партеногенетических поколений тли адаптировались к
питанию этим мало пригодным растением и были перенесены на Chaerophyllum
maculatum - бутень пятнистый, кормиться на котором они были ранее не способ-
ны. В итоге экспериментальная популяция в течение 10-15 поколений не только
приспособилась к абсолютно новому растению-хозяину, но и изменилась морфо-
логически, обретя фенотипическое сходство с Dysaphis chaerophyllina Shap, - ви-
дом, который в естественных условиях питается на пятнистом бутне.
Эксперимент Шапошникова проходил в отсутствие полового размножения.
Однако когда в конце опыта были получены половые особи, то оказалось, что
экспериментальная популяция D. anthrisci обрела способность скрещиваться и
образовывать смешанные популяции с D. chaerophyllina; из яиц, отложенных
оплодотворенными самками, вышли личинки, способные к дальнейшему
размножению и предпочитающие питаться бутнем. В результате эксперимент
привел не просто к дивергенции... а ко вторичному возникновению вида, уже
существующего в природе».
К словам Гродницкого добавлю, что на эту тему существует обстоятельная
монография ленинградского ботаника М.Г. Агаева (1978), практически никем не
цитируемая, где данные Шапошникова сопоставлены с данными самого Агаева
по пшеницам. Замечательно: оба автора утверждали, без всякой аргументации,
что их опыты объясняются отбором случайных вариаций и что получившиеся
организмы являются вовсе не новыми видами, а лишь "видовыми формами", -
таковы были рамки советской цензуры. Недавно сходные результаты наконец-то
стали появляться на Западе и тоже почти не цитируются - научное сообщество,
как всегда, не может быстро менять убеждения.
Однако 40 лет - достаточный для сообщества срок, чтобы привыкнуть, ибо
за это время происходит смена научных поколений. Ныне начинают понимать,
что приспособление идет быстро и направленно, как утверждает ламаркизм (на
медленное просто нет времени, ибо среда тоже меняется быстро), и что
вариантов преобразования достаточно мало, как утверждает номогенез (в
сходных условиях может появиться тот же вид). И что все это нисколько не
противоречит идее естественного отбора.
Если данные по "экспериментальной эволюции" (их много) не замалчивать,
а ввести в преподавание, то нынешние ученики, став учеными, наверняка
завершат долгожданный "новый синтез" и дадут, наконец, обществу теорию
эволюции, которую можно будет не только заучивать, но и применять. То есть
теорию, полезную для выхода из нынешнего экологического кризиса.
109
Феномен эволюции организмов в понимании С.В. Мейена'
1. Наука и идеология
Малайская притча гласит, что трое слепорожденных захотели узнать, каков
слон. Попросили погонщика, он привел им слона и сказал: "Вот он, глядите".
Один из слепых оказался около хобота, другой под брюхом, третий у ноги сло-
на, и каждый ощупал, что смог. Когда слон был уведен, первый из слепых ска-
зал: "Слон подобен толстой веревке", второй - "огромной бочке", третий - "ко-
лонне". Притча кончается словами: "И так спорили они без конца".
Удивительно похоже на судьбу эволюционизма, особенно - биологическо-
го. Не знаю другой отрасли знаний, где так принято было бы выдавать свой ча-
стный опыт и личную склонность за общий закон природы. Едва ли не главным
приемом здесь является такой: изучить какую-то группу организмов с какой-то
точки зрения, подметить что-то общее, сопоставить его с одной из эволюцион-
ных доктрин и, если материал в нее уложился, объявить эту доктрину единст-
венно истинной теорией. Другие доктрины либо не упоминаются, либо аттесту-
ются как ошибочные. Ситуация скорее характерна для религии, чем для науки,
что отчасти отражается в названиях основных эволюционных доктрин - дарви-
низм и ламаркизм, обозначаемых именами тех, кто признан в качестве основа-
телей. Более, насколько знаю, нигде в естествознании так не принято.
Поэтому неизбежен вопрос: является ли эволюционизм отраслью естество-
знания или чего-то другого? Блестящий теоретик А.А.Любищев не раз прямо
говорил, что дарвинизм - не научная теория, а идеология**. Ему все всё прощали
(считая его чудаком), но из известных мне биологов долгое время один лишь
С.В. Мейен, столь же блестящий теоретик, развивал эту мысль далее. Ныне дар-
винизм аттестуют как идеологию многие. Среди ведущих теоретиков так пола-
гает биофизик В.Л. Воейков***.
Однако и другие учения тоже можно считать идеологиями - например,
ламаркизм и номогенез. Хотя главное признаваемое ими свойство историче-
ского преобразования организмов - тенденция к прогрессу, т.е. к усложнению
строения организмов - является наблюдаемым фактом, однако ламаркизм ис-
ходит из тезиса, что строение нацело определяется функционированием
("функция определяет форму"), номогенез же, наоборот, - из тезиса, что "фор-
ма определяет функцию", а это есть толкование фактов.
Мейен не только аттестовал в качестве идеологий дарвинизм и ряд других
эволюционных доктрин, но и предложил выход: надо не искать факты, рабо-
тающие на любимую доктрину, и не пытаться изложить чужую, эмоционально
неприемлемую доктрину как свою собственную (это предлагал Любищев, но это
’ В сборнике «Эволюция флор в палеозое». М., 2003, с. 99-105.
Что отражено в заглавии сборника [Теория эволюции..., 1998]. -Примеч. 2010 г.
Воейков В.Л. Правда Дарвина и ложь дарвинизма // Человек, 1997, № 3. См. о нем также
книги АСМ и ЗЭ. - Примеч. 2010 г.
110
не слишком получилось даже у него самого), а изучить весь корпус фактов, из-
вестных о какой-то крупной группе организмов, и основные доктрины, прила-
гавшиеся к этим фактам, а затем уже строить теорию. Такая программа доступна
лишь колоссальному уму, и Мейен сумел ее поставить и начал ее реализовывать
для ископаемых высших растений. Теория, к которой он пришел, оказалась
плюралистической - некоторые прежние доктрины выступили в качестве не
конкурентов, а аспектов общей теории.
2. Дарвинизм и марксизм
В 1977 году Мейен сказал мне в ходе домашней беседы, что "дарвинизм
конгениален марксизму" и что в этом-то кроется удивительная живучесть дар-
винизма. Что и говорить, живучесть удивительная: упустив эволюционную роль
едва ли не всех биологических открытий XX века, от дискретности гена (1901 г.)
до обсуждаемых ныне генов морфогенетического поля, оконфузившись со все-
ми своими практическими рекомендациями, от массовых стерилизаций конца
XIX в. до нынешних призывов проводить политические, хозяйственные и соци-
альные реформы по принципу отбора наиболее приспособленных, - дарвинизм
не склонен видеть в этом ничего зазорного. Всякое открытие дарвинизм сперва
яростно отрицает, затем молчит о нем, а после признания его научным сообще-
ством заявляет, что всегда признавал и само открытие, и следующие из него
идеи как свою неотъемлемую часть. Это действительно похоже на марксизм,
однако марксизм принято третировать, а дарвинизм, хоть уже и не господствует
в умах большинства, но преподается повсюду. В чем тут дело?
Прежде всего, не нужно видеть в этом ум одних и глупость других - просто у
разных людей различны исходные установки (вот чему научил меня Мейен). Од-
нако в разное время полезны (ведут к новому знанию) разные установки. Сказан-
ное требует для своего осмысления нового теоретического аппарата, каковой ви-
дится мне в методе познавательных моделей, который предложен в 1980 г. мето-
дологом А.П. Огурцовым и подробно описан в моей книге [Чайковский, 2001].
Дарвин, благопорядочный английский эсквайр (помещик), в качестве моде-
ли эволюции взял хорошо знакомую эсквайрам селекцию, т.е. выведение новых
пород. Даже не пытаясь исследовать природные механизмы, он назвал движу-
щий фактор эволюции естественной селекцией, т.е. положил в основу своего
учения аналогию, на что современники не раз обращали внимание. Успех дар-
винизма состоял, в этом смысле, в том, что аналогия оказалась злободневной.
Но, как мне уже случалось писать [Чайковский, 1988, с. 105], аналогия хо-
роша только для единомышленника, противника же она никогда не убеждает.
Наоборот, парируя аналогию, он уверен, что опроверг и сам критикуемый тезис.
Неспособность человека организовать нынешнее природопользование по анало-
гии с селекционной фермой и служит, на мой взгляд, одной из причин недове-
рия нынешних ученых к дарвинизму как модели эволюции, а это влечет повы-
шение внимания к суждениям, прежде игнорируемым. Другая причина видится
в неуспехах марксизма.
111
Самое главное, на мой взгляд, упущение марксизма XX века состоит в не-
желании видеть разнообразие социальных устройств. В частности, одной фазой
капитализма - империализмом - именуется (в том марксизме, которому нас учи-
ли), строй и Англии начала XX века, и фашистской Германии, и нынешних
США, и даже нынешней Швеции, в некотором смысле социалистической. Есте-
ственно, при столь грубом взгляде на нынешнее социальное разнообразие, нель-
зя ожидать верных указаний на будущее, что, по-моему, и привело к конфузу
всех нынешних реформаторов (а они сплошь марксисты по воспитанию и, как
бы они это ни отрицали, по кругозору).
Нет, я вовсе не собираюсь топтать поверженного классика - в свое время
марксизм, как и дарвинизм, сказал людям много нового и был не только полезен,
но и неизбежен. Это банально, как банально и то, что всякая идея рано или поздно
себя изживает. Мне только хочется понять самому и объяснить другим глубокий
смысл в отмеченной Мейеном параллели между дарвинизмом и марксизмом.
Параллель становится понятной, если признать, что перед нами две идеоло-
гии, а вовсе не две научные (фактически обоснованные) теории, причем - идеоло-
гии, выражающие одно и то же мировоззрение. Что же это за мировоззрение?
Мне уже не раз приходилось писать о нем как о статистическом мировоз-
зрении. Оно стало безраздельно господствовать в науке как раз при Дарвине и
Марксе, а в наши дни уступает системному. Последнее повсюду старается видеть
функциональное единство, так что идея всепроникающей борьбы (хоть видов ор-
ганизмов, хоть социальных групп людей, хоть научных идей) теряет популяр-
ность. Теперь более популярны указания на сотрудничество и взаимозависимость.
Уход от устаревшей идеологии отраден, но таит в себе новую опасность.
Тот же Любищев, в отличие от большинства своих современников, видел
главные достижения разума вовсе не в универсальных схемах, а в "преодолении
претензий на универсальность". Системный подход являет собой именно такую
претензию. В годы, когда системное мировоззрение едва начинало прорубать
себе путь в джунглях статистического, трудно было понимать Любищева, ду-
мавшего в рамках еще более нового видения мира - плюралистического. Лишь в
наше время становится хоть сколько-то популярной та мысль, что в большинст-
ве ситуаций ни одно решение, пусть и лучшее, не может быть единственно пра-
вильным. В этом мне видится суть нарождающегося диатропического мировоз-
зрения*, которое как раз и отвергает приемлемость единственных решений.
Замечу, что в эволюционизме зачинателем диатропичёского подхода был
Мейен. Подробно об этом рассказано в книге [Чайковский, 1990].
3. Место идей Мейена в эволюционизме
Любищев и Мейен замечательны не столько эрудицией, сколько необычны-
ми точками зрения на самые разные проблемы, ставшими отправными для мно-
О статистической, системной и диатропической познавательных моделях см. далее, ста-
тью «Горизонт познания и познавательные модели», 2007. - Примеч. 2010 г.
112
жества исследований. Обоим в этом помогало удивительное уменье общаться с
людьми чуждых взглядов. Оба по сути развивали номогенез (и во многом имен-
но они придали ему его нынешнюю форму), оба легко и охотно общались с дар-
винистами и ламаркистами, а Мейен, кроме того, стоял у истоков экологическо-
го эволюционизма, широко развившегося уже после его смерти.
Несмотря на отдельные фразы о целостности, дарвинизм исходит из идеи
зависимости судьбы организма от полезного признака, отсутствующего (или
слабее развитого) у конкурентов. Организм, тем самым, неизменно выступает в
дарвинизме как набор признаков. Аналогично, сообщество выступает как набор
организмов. Весь же остальной эволюционизм, наоборот, по сути является пред-
ставлением о преобразовании одних целостностей в другие, где элементы не за-
даны, а формируются вместе с целостностями. Сама идея происхождения вида в
форме приспособления его к некой среде выглядит теперь примитивной.
Вновь обретают популярность старые учения. В частности, приходится вновь
обращаться к основной идее номогенеза - к идее независимо ни от чего сущест-
вующих законов эволюции организмов. Но не стоит думать, что на этом пути
можно построить цельную теорию эволюции, поскольку на сегодня номогенез яв-
ляется чисто морфологической теорией, тогда как цельная теория должна вклю-
чать и функционирование организмов (физиологию, экологию и этологию), о чем
подробно говорится в моей книге [Чайковский, 2003]. И всё же, если речь идет о
вкладе Мейена, то надо говорить именно о номогенезе, добавляя к нему лишь не-
которые обстоятельства палеобиогеографии, несущие экологический смысл.
С диатропической точки зрения процесс эволюции состоит в заполнении объ-
ективно существующих рефренных таблиц. Они существуют, как и все законы
природы, самостоятельно, независимо от наличия или отсутствия конкретных ви-
дов (точно так же как теорема Пифагора была справедлива и до того, как кто-то
начертил первый прямоугольный треугольник); но таблица редко заполняется це-
ликом, и какие ее клетки заполнятся ныне, а какие останутся ждать пустыми, ни-
кто сказать не может. В этом случайность эволюции - согласно номогенезу.
Пусть рефренных таблиц мы почти никогда целиком выписать не можем, но
сама идея выражать разнообразие с помощью таблиц позволяет четко различать
в эволюции три совсем разных процесса:
1) заполнение прежде пустых клеток неизменной таблицы,
2) преобразование прежней таблицы и
3) появление совсем новой таблицы.
До сих пор именно смешение трех процессов в один и не давало, по-моему,
возможности построить работоспособную (дающую полезные рекомендации)
теорию эволюции. Стоит их разделить, как картина эволюции во многом прояс-
няется, и связано это с именем Мейена [Чайковский, 2003, гл. 6].
Живое предстает экологическим, физиологическим и морфологическим
единством. О последнем для растений писал Гёте, для животных - Бюффон и
Жоффруа, но только выявление Мейеном рефренной структуры любых разно-
113
образий позволило говорить о морфологическом единстве природы как целого.
Мы легко понимаем, что всякий вид занимает свое место в экосистеме (а именно
- экологическую нишу), но оказывается, что и его строение задано не менее же-
стко*. Именно этот факт позволяет строить систематику организмов.
4. Мейен и эволюционная систематика
Понимание эволюции как череды ветвлений родословного древа ведет свое
начало от иерархической систематики, возникшей задолго до К. Линнея. Оно
развито в XIX в. Э. Геккелем и господствует в литературе. Ей противостоит по-
нимание эволюции как преобразования разнообразия, идущее от взглядов П.-Л.
Мопертюи (ок. 1750 г.) и развитое в начале XX в. Л.С. Бергом и Д.Н. Соболе-
вым. Нынешнюю форму номогенезу придали Любищев, и, в основном Мейен.
Это направление в 1990 г. получило название эволюционной диатропики [Чай-
ковский, 1990].
Геккель фактически исходил из постулата: единственным модусом эволю-
ции является дивергенция (расхождение одного таксона на два) на всех уровнях,
от популяции до типа или даже царства. Целью систематики было признано по-
строение филогений (т.е. родословных древ - далее это слово употребляется
только в данном смысле, если не оговорено иное), а не упорядочение разнообра-
зия организмов, не выявление уровней их организации и способов взаимодейст-
вия организмов с миром. Филогенетик уверен, что система задается предшест-
вующей эволюцией, причем механизмом дивергенции служит конкуренция. Но
высшие таксоны не являются экологическими единствами, потому не конкури-
руют, и неясно, почему макроэволюция мыслится дивергентной.
Диатропика (у Мейена - типология) видит эволюцию как заполнение объ-
ективно существующей (подобно идее Платона) таблицы, в которой близкие
клетки (обладающие общим архетипом - это понятие по сути интуитивно) со-
ставляют общий таксон. Порядок заполнения клеток не задан жестко, хотя чаще
всего заполняются клетки, соседние с уже занятыми, что и создает видимость
дивергенции. После смерти Мейена появилось несколько изображений конкрет-
ных таблиц - например [Дьяков, 2000, с. 11].
Для диатропики система - это реализованная часть таблицы логических воз-
можностей данной группы организмов, мало зависящая от прежней эволюции
группы. Этим хорошо объясняются факты параллелизма, но плохо - иерархич-
ность общепринятой системы высших растений и животных. На сегодня синтез
этих двух подходов к систематике не виден, но можно идти по пути, намеченно-
му Мейеном. Сперва он отрицал филогенетику, а в 1984 г. сам построил схему
эволюции голосеменных, которую назвал филогенией [Меуеп, 1984; Мейен, 1992].
Но это - филогения в ином смысле. А именно, схема эволюции отдела голо-
семенных выглядит у Мейена древом, если рассматривать только порядки и
классы: они порождают новые порядки и классы; но при этом нельзя сказать,
* Подробнее о нишах в разном понимании см. АСМ, с. 372Примеч. 2010 г.
114
какое семейство (и тем более род) происходит из какого, т.е. разнообразие се-
мейств и родов переходит в разнообразие (а не элемент в элемент). Другие от-
делы детально им не рассмотрены.
Тем самым, “филогения по Мейену” - как бы грубый взгляд издалека на
процесс эволюции; если же вглядеться в нее детально, она предстает как сетча-
тая эволюция. Вместо прежнего понимания филогении как универсального де-
тального процесса эволюции путем дивергенции, Мейен дал совсем новое ее
понимание: эволюция выглядит как сеть, однако отдельные ее части связаны
слабо (между порядками и между классами сращений мало), схема их связей
похожа на расходящиеся ветви, и эволюцию в рамках этих таксонов можно гру-
бо представить древом. То есть у Мейена филогенетическое древо - грубая схе-
ма, не допускающая детализации-, виды, роды и многие семейства эволюциони-
руют “с широкой трансгрессией признаков” [Мейен, 1992, с. 97].
Поясню это на одном примере. На рис. 2 приведена семофилогения женских
фруктификаций ранних хвойных. Она распадается на три ветви: сверху экватори-
альные (N, S, Т, U, V), посреди северные (L, М, О, Р, Q, R) и внизу справа южный
(К) роды. Ее точно повторяет изображенная на рис. 2 “пробная филогения ранних
хвойных” [Меуеп, 1997, р. 422, 432]. Это значит, что “филогения” основана на
единственной семофилогении, на той семофилогении, какая “легла на карту”, т.е.
согласно которой сходные формы росли в одной зоне. Слово “филогения” я взял в
кавычки потому, что форма древа соблюдена тут весьма приблизительно (особен-
но в отношении семейств, названия которых помещены в “облаках”).
5. Флорогенез как свидетельство реальности эволюции
Конструируя свое понимание филогении голосеменных, Мейен открыл* яв-
ление глобального флорогенеза\ оказывается, все классы и почти все порядки
происходят в экваториальной зоне, а вне ее - только таксоны более низкого ран-
га. Это позволило ему поставить задачу - “положить филогению на карту”, т.е.
проследить, как таксон эволюировал, расселяясь. И тем самым отличать реаль-
ную эволюцию от вымышленной.
Для Мейена филогения реальна в той группе, где основные семофилогении
(филогении отдельных свойств или органов) легли на материал ископаемых
данных, причем этих семофилогений достаточно, чтобы увидеть архетип преоб-
разуемого таксона. Те семофилогении, которые выпали из этой процедуры, надо
счесть параллелизмами, а те свойства, которые появились вне семофилогений -
сальтациями.
Декларировав это как идею, на самом деле Мейен строил филогению по од-
ной семофилогении, если она ложилась на (грубый) палеогеографический фон.
Например, для ранних (верхнепермских) хвойных, рассмотренных выше, это
был абрис женской фруктификации, который оказался достаточно характерен
В действительности, он лишь развил эту хорошо изветную ему идею - см. сноску на с.
82 и заключительную статью- Примеч. 2010 г.
115
для каждой из трех палеогеографических зон - экваториальной, северной и юж-
ной. Этот факт истолкован Мейеном как свидетельство происхождения каждого
типа фруктификации в соответствующей зоне, а значит - друг от друга. Пусть
это и грубо, но все-таки гораздо тоньше, чем общепринятые “филогении”, во-
обще игнорирующие географию, а подчас и палеонтологию.
В целом открытие Мейеном флорогенеза и предварительное (эскизное) опи-
сание его механизма можно рассматривать как первое фактическое доказа-
тельство факта эволюции, тогда как прежние “доказательства” описывали
лишь разнообразие ныне существующих организмов или разрозненные разно-
образия ископаемых.
Напомню, что Любищев справедливо относил ископаемые к косвенным
свидетельствам эволюции (правда, считал их наиболее близким к прямым) и по-
лагал, что прямых доказательств эволюции вообще не бывает. Теперь можно
сказать точнее: разумеется, любую историческую последовательность ископае-
мых остатков можно представить и как свидетельство естественной эволюции, и
как итог последовательных актов творения, однако всю совокупность свиде-
тельств глобального флорогенеза можно признать свидетельством творения
только при допущении того, что Творец в процессе творения имитировал также
и процедуру естественного расселения таксонов. Творец в таком понимании
практически тождествен естественному процессу.
Литература
Дьяков ЮТ. Введение в альгологию и микологию. М., МГУ, 2000.
Мейен С.В. Основные черты систематики и филогении голосеменных по палеобота-
ническим данным (пер. с англ. изд. 1984 г., М-268), перевод 1992 года в книге М-328.
Чайковский Ю.В. О формировании концепции Ч. Дарвина, 1988. (4-21)
Чайковский /О.В.Элементы эволюционной диатропики. М., Наука, 1990. 4-24 (ЭЭД).
Чайковский Ю.В. О природе случайности, 2001. (4-43)
Чайковский Ю.В. Эволюция, 2003. (4-51)
Meyen S.V. Basic features of gymnosperm systematics and phylogeny as shown by the fos-
sil records // Botanical Review, 1984, vol. 50, № 1 (M-268).
Meyen S. V. Permian conifers of Western Angaraland // Review of Palaeobotany and Paly-
nology, 1997, № 96 (M-330).
Это неверно. Во-первых, до Мейена более детальную работу в данном направлении
провел Л. Круаза. В отличие от Мейена (который учёл географию лишь тем, что отнес
каждый рассмотренный таксон растений к одной из трех зон - южной, экваториальной и
северной) Круаза описал трэнды - т.е. пути расселения эволюирующих таксонов расте-
ний и животных. См. сноску на с. 82 и п. «1959» в статье «500 лет споров об эволюции»
(4-64). Во-вторых, существуют (правда, малочисленные) прямые доказательства эволю-
ции на уровне образования новых видовых форм, полученные экспериментально - на-
пример, в опыте Шапошникова (см. выше, с. 82, 109).- Примеч. 2010 г.
116
Часть 3. Социальная диатропика
Молодежь в разнообразном мире'
Документальный фильм Ю. Подниекса «Легко ли быть молодым»** пере-
сказать невозможно. Это надо увидеть: довольный взгляд, которым женщина-
прокурор провожает плачущего шалопая в колонию (откуда тот вернется злым
уголовником); назидательная брань, которой стая психиатров «успокаивает»
девушку, выбросившуюся из окна кабинета следователя (если от них она и вы-
рвется - то злобным инвалидом); кучка застенчивых мальчиков-калек среди
госпитальных коек; орден, стыдливо спрятанный в кулак («...я занимался чем-
то грязным, чуждым человеку») - это нельзя описать, как нельзя описать запах
или музыку.
«Ребенка родили, а теперь будем убивать, вместо того, чтобы воспиты-
вать» - такой упрек один из юных персонажей фильма бросает всем нам. Чем
мы можем на это ответить?
У некоторых картина вызывает протест: почему показаны лишь несчаст-
ные и заблудшие, почему нет среди ее героев обычной трудовой и учащейся
молодежи? При втором просмотре выясняется, что она есть, только не запоми-
нается - столь сильное впечатление производит молодежь неблагополучная.
На экране чудесный мальчик Геннадий. Он мечтает стать врачом и уверен,
что всякий врач глубоко сочувствует больному. Но тут же - цветы на могиле
Геннадия. Несчастный случай? И да и нет. Едва поступив в мединститут, он
поехал на картошку, простудил почки и умер в больнице, где его «как-то не-
удачно лечили» те, кого он так высоко оценивал.
Вопросы фильма адресованы в основном социологии. Но не только ей.
Скажем, в смерти Геннадия социологическая тема переплетается с экономиче-
ской, санитарной, организационной, правовой. Смерть или увечье студентов и
школьников на сельскохозяйственных работах - кто в этом повинен? [...]
Кто-то из персонажей фильма восклицает: «Нельзя ведь говорить о молоде-
жи как об однородной массе!» Действительно, подходить ко всем с одной мер-
кой - это значит медленно убивать многих. Если Геннадию был нужен только
заботливый организатор труда (а коли такого не нашлось, то, на худой конец, -
заботливый врач), то другому нужно с кого-то брать пример, третьему нужен за-
ботливый наставник, а четвертый от заботливого сбежит - ему нужен строгий. И
так до бесконечности, ибо до бесконечности разнообразны люди вообще и моло-
дые особенно. Но ведь я-то как раз и занимаюсь проблемой разнообразия!
Социологические исследования, 1988, № 1, с. 76-84.
** Рижская киностудия, 1986. Юрис Подниекс (Podnieks, 1950-1992) - латвийский кино-
режиссер-документалист. Фильм был первым «перестроечным» (наряду' с отснятым ра-
нее «Покаянием» Тенгиза Абуладзе), в Москве шел в декабре 1986 - январе 1987, с ог-
ромными очередями и, насколько помню, только в к/т «Москва». - Примеч. 2010 г.
117
Было бы самонадеянным в том, о чем пойдет речь далее, видеть решение
проблемы, но ее обсуждение представляется мне делом весьма важным.
Что такое разнообразие?
Если не гнаться за точным определением, то можно сказать так: наука о
разнообразии занимается теми различиями сходных объектов и теми сходства-
ми различных, какими обычно пренебрегают. Она ищет здесь закономерности.
Вряд ли найдется явление, которое было бы нам так же хорошо известно и
так же плохо понимаемо, как разнообразие. Все мы знаем, что нет двух одина-
ковых людей, что рассчитанная на «среднего ученика» программа одним скуч-
на, а другим непонятна, но все-таки живем так, словно нас окружают «сред-
ние». Мы привыкли усреднять, но в сложных системах средняя величина часто
оказывается просто статистической фикцией.
Так, в 1960 г. половина горожан СССР жила в городах с более чем 90 ты-
сячным населением, тогда как средняя численность советского города состави-
ла всего 50 тыс. человек. Сосредоточение большинства по одну сторону от сред-
ней непривычно и рушит саму «идеологию» средних величин. Рушат ее и дру-
гие обстоятельства. Именно там, где усреднение неинформативно, встает про-
блема разнообразия как таковая. Часто ли любой из нас может похвастаться,
что отнесся к другому не как к среднему, а как к единственной неповторимой
личности? Нет, даже к собственным детям мы прилагаем стадартные мерки.
И в то же время вся наша жизнь - эксплуатация разнообразия. Выбираем ли
мы себе спутника жизни или сотрудника, надеемся ли на восходящий талант или
болеем за спортсмена, консультируемся ли со специалистом или приглашаем в
дом грузчика - на первый план всегда выступают личные качества человека, ко-
торые в данный момент отличают его от других. То же можно сказать о разнооб-
разии животных, всяческих предметов. Сейчас, когда разнообразие организмов
стало угрожающе сокращаться, нельзя ограничиваться описанием сходства и
различий, самое детальное знание которых зачастую не помогает предотвратить
убыль в дикой природе, не говоря уж о восстановлении утраченных видов. Что
надо знать о разнообразии, чтобы контролировать его? Можно ли, хотя бы в
принципе, говорить об управлении разнообразием? Да и нужно ли это?
Может быть, мы способны разводить «полезные» виды, истребляя «вредные»
и игнорируя все прочие? Биологи дают отрицательный ответ, но в других отрас-
лях знания определенности пока нет. Зачем обществу убийцы, садисты или де-
билы? Появляются они (так же как, к примеру, чумные очаги) не потому, что их
кто-то культивирует, а потому, что огромное разнообразие организмов то и дело
реализует самые различные свои качества. Преследование чумных, прокажен-
ных, умалишенных дало для оздоровления общества ничуть не больше, чем пре-
следование еретиков и прочих инакомыслящих, - оздоровление достигнуто ис-
ключительно путем выяснения причин болезней и разработки методов лечения.
И победа над одними бедами сопровождается возникновением других, ранее ме-
заметных или даже вовсе неизвестных - таковы сюрпризы разнообразия.
118
Человечество оказалось не готово к тому, что общий рост благосостояния
породит много новых проблем, в частности, к тому, что молодежь, выровняв-
шись, если можно так сказать, «по сытости», вовсе не обнаружит подобной тен-
денции в других отношениях. Стремясь к равенству, никто не думал о том, надо
ли сохранять разнообразие и если надо, то какое и как. Оказалось же, что разно-
образие не сокращается, а лишь принимает новые формы, подчас зловещие.
Биосфера существует только целиком (хотя вроде бы каждый отдельный вид
можно из нее изъять), и только целиком может существовать общество. Попытки
ущемить отдельные национальные, религиозные или социальные группы приво-
дили к трагедии государства в целом. Одна из таких социальных групп - моло-
дежь, которую не раз пытались лишать ее собственных интересов и институтов.
Многие, наверное, думают, что в архаичных обществах молодежных институ-
тов не было. Это неверно. Достаточно обратиться к народным сказкам, чтобы убе-
диться: в незапамятные времена главным предметом повествования было социаль-
ное утверждение молодых до (или в ходе) создания семьи. Этот отрезок жизни на-
зывают «героическим» периодом [1], он завершался в сказке либо свадьбой, либо
тем, что молодые супруги доказывали свое право жить вместе. Мужские доблести
- силу, ловкость, отвагу - иногда демонстрируют и девушки, но чаще сказка отво-
дит им свою, чисто женскую роль: они проявляют завидную преданность, готов-
ность к самопожертвованию, мастерство и мудрость (обычно в форме волшебст-
ва). Именно «хитрость» невест спасает безрассудных женихов, а не наоборот, и
здесь есть над чем задуматься. Свадьба заканчивает сказку, переводя молодых в
разряд производителей («стали жить-поживать и добра наживать»). Кончался «ге-
роический период), кончались и законные молодежные формы досуга - совмест-
ные гулянки, девичьи хороводы, кулачные бои парней.
«Героический период» жизни остается главной темой любовного и при-
ключенческого жанров во всех обществах. Государства всегда пользовались им
(для войн, освоения новых территорий и т.п.), заодно эксплуатируя нетребова-
тельность молодежи к удобствам, комфорту и безопасности. И вот парадокс:
эксплуатируя специфику молодежи, общество старается не замечать ее специ-
фических требований, ее желания жить для себя, а не только для старших.
Обратите внимание: в древних текстах обычно отсутствует детство как та-
ковое. Адам и Ева возникли взрослыми, Ева родила Каина «и еще родила брата
его Авеля. И был Авель пастырь овец; а Каин был земледелец». Дети обычно
уподобляются взрослым: Геракл совершил первый подвиг новорожденным, а
герои Махабхараты изрекали мудрости еще из лона матери. До логического за-
вершения (фактически - до абсурда) отрицание роли становления личности до-
вел в середине XVI века Жан Кальвин, один из вождей Реформации: он учил,
что человек обречен на рай или ад уже в момент рождения. Тем самым роль
наставника сводилась к выявлению и разграничению. Только в Новое время
становление человека стали расценивать как нечто особое, отличное от роста.
Сперва детей перестали судить и казнить, после чего уже понемногу стали
119
признавать за ними особые права (на содержание, лечение, учение, развлече-
ние). Воспитание же еще и теперь нередко рассматривается просто как обуче-
ние взрослому делу.
У биологов тоже очень долго детский организм, и даже зародыш, рассматри-
вался как маленький сформировавшийся организм. Отсутствовало детство и в эво-
люционных учениях. В это трудно поверить (ведь параллель развития вида с раз-
витием организма - одна из самых давних в науке), но, например, дарвинизм об-
ращается к зародышу, личинке или детенышу лишь в двух планах - много ли их
рождается и борются ли они сами за существование. Только теперь в биологии на-
чинает утверждаться мысль (тоже очень давняя), что взрослый организм почти или
вовсе не участвует в эволюции вида, что эволюция состоит в изменении путей раз-
вития зародыша и отчасти - других незрелых форм.
Это важно знать и социологу: ведь социальные науки брали и берут эволюци-
онный элемент у биологии. Как в биологии, так и в социологии эволюция мало за-
висит от взрослых, поскольку мала их способность меняться. Поэтому-то всякое
социальное потрясение резко омолаживает слой активных деятелей, вплоть до
юных генералов революционных армий. Взросление человека и развитие общества
связаны так же неразрывно, как в биологии развитие зародыша и эволюция вида.
В застойном обществе и личность ущербна, если говорить не об отдельных
героях, а о «социальном характере». Найти место молодежи в обществе - значит
во многом направить его эволюцию, а потому нам следует думать не только о
влиянии на молодежь, но и о преобразовании взрослого общества для молодых.
Социология юношества сейчас только становится на ноги, пытаясь пре-
одолеть извечный «социологический вывод» - ворчание стариков на тему «в
наше время молодежь была лучше». Нет, молодежь не выродилась, она попро-
сту заведена в тупик и не знает, как ей быть со своим «героическим периодом».
И мальчики из фильма Подниекса на все лады повторяют гневный упрек
взрослым: это вы сделали нас такими. Отповедь младших извечному ворчанию
старших - знамение времени, но правы ли младшие?
По-моему, да. Младшие отстаивают свое отличие от старших, а те призы-
вают брать с них пример. Некоторые берут, но это - не решение проблемы:
молодежь разная, у одних «героический период» выражен слабо, у других на-
правлен в область, недоступную большинству. А что делать остальным, кого
природа не наделила особым талантом? В телевизионной передаче «Музы-
кальный ринг» (декабрь 1986 г.) девушка спросила бардов: «Что вы можете
сказать нам, восемнадцатилетним?» В ответ они спели свои песни про войну.
Тяжело и стыдно слушать, как сорокалетние воспевают от первого лица (будто
участники) подвиги 45-летней давности и разъясняют потом, что это - ответ
молодым на их тревогу.
По-моему, куда честнее тот бравурный голос, что поет из репродуктора:
«На всю оставшуюся жизнь нам хватит подвигов и славы». Молодым явно не
хватит; молодым нужно самим за что-то бороться. «Вы бороться не способны,
120
и это же хотите воспитать в нас», - резюмирует герой Подниекса, юный лидер
«металлистов».
Обрисовав положение, попробуем выяснить, чем тут может быть полезна
наука о разнообразии вообще и о биологическом в особенности.
Правомерен вопрос - допустимо ли закономерности, установленные в од-
ной науке, переносить на материал другой? Отвечу: вполне допустимо, если
рассматривать их не как аксиомы, а лишь как указание пути, на котором более
вероятен успех. В последние десятилетия возникла новая дисциплина - общая
теория систем, показавшая, что у сложных систем есть общие свойства, не за-
висящие от природы элементов, из которых сложена каждая система. Одним из
аспектов этой теории и является наука о разнообразии.
Ядро и периферия
Существует простой закон разнообразия, закон ядра и периферии. Суть
его состоит в следующем: всякая крупная группа в чем-то существенно сход-
ных объектов имеет не только типичные, легко отличимые объекты (ядро), но
и относительно небольшое число нетипичных, которые легко спутать с объек-
тами других групп. Это меньшинство составляет периферию. Так, звери (мле-
копитающие) обычно ходят на четырех ногах - это признак ядра класса зверей,
однако есть группы, представители которых имет вместо ног плавники (киты)
или крылья (летучие мыши) - они составляют периферию класса. Перифериче-
ские виды всегда вызывают споры, их то и дело объявляют отдельными груп-
пами или переносят из одной в другую, но это не должно мешать видеть об-
щую логику: система строится по типичным формам (по ядрам групп). Каждая
группа имеет право на существование, если удалось точно описать свойства ее
ядра: «Класс установлен точно, хотя и не ограничен четко» (В. Уэвелл, 1840).
Тем самым, классификация состоит не в проведении четких границ (как
многие думают), а в выявлении типичных форм. Что касается отнесения каж-
дой нечеткой формы к определенной группе, то это - вопрос воли и вкуса, он
не может быть решен раз и навсегда. Ядро можно выделить всюду. Э. Фромм
писал в 1941 г.: «Социальный характер содержит лишь выборку черт, сущест-
венное ядро структуры характера большинства членов группы. Хотя здесь все-
гда будут “отклоняющиеся” индивиды с совершенно иной структурой характе-
ра...» [2, с. 49]. Нам надо выявить ядро свойств той группы, которую обычно
именуют молодежью (хотя границ ее никто назвать не может).
С того времени, когда почти все младшие шли по стопам своих старших,
язык сохранил слово «стараться», первоначально означавшее «уподобляться
старшему». С тех пор слово «старший» имеет двойной смысл - «более старый» и
«распорядитель». Старание, старение и старшинство указывали человеку по сути
дела один и тот же жизненный курс, поскольку в течение жизни человека обще-
ство мало менялось, и с возрастом он обычно приобретал вес в своем кругу.
В раннем, доклассовом обществе юноша был воином, зрелый муж - про-
изводителем, старик - воспитателем, распорядителем, жрецом. Женский путь
121
был сложнее (сверстнице воина - 16 лет - полагалось уже включиться в изну-
рительную череду беременностей, родов, вскармливаний, т.е. стать «произво-
дительницей»; она, грубо говоря, проходила четыре стадии - невеста, произво-
дительница детей, воспитательница, распорядительница - и все время работала
по дому), но возрастное разнообразие в самом грубом виде соответствовало
общественному. Не имея возможности провести здесь эту схему рассуждений
через перипетии классовых обществ, отмечу только, что человек, успешно
прошедший «героический период», попадал на путь «старания-старения-
старшинства», куда его направлял отец или другой «старший». А если там он
не уживался, то оказывался на «периферии» общества. Упрек «у всех дети как
дети» имел тот смысл, что этими словами отец указывал на «ядро», к которому
принадлежал сам и которое видел единственно годным для сына. Ныне не так.
«Героический период» теперь предлагается просидеть за учебой, «произво-
дительницей» девушка становится поздно и ненадолго, различия полов во мно-
гом игнорируются обществом («равноправие»), братьев и сестер, прежде опре-
делявших микроструктуры общества, теперь мало, мать ведет хозяйство урыв-
ками. Старшие очень ограничены в возможности передавать опыт, поскольку
сами начинали жизнь не в той обстановке, что их дети и тем более внуки.
Поэтому старики теряют прежнюю роль наставников, и семья легко расста-
ется с ними, отселяется, а внуки отдаются на воспитание телевизору, школе,
улице. Только улица приучает их хоть к какой-то активности (увы, часто антисо-
циальной). Школа же готовит прежде всего служащих, способных о любом зада-
нии дать отчет, в котором все «гладко на бумаге», приучает «убивать время» на
уроках и вне их, отучает от инициативы и собственного мнения, культивирует
донос как форму общения со старшими. Все это молодежь усваивает и щедро
возвращает обществу в виде молодежных проблем и таких качеств взрослых, как
двуличие, бюрократизм, безответственность, склонность к анонимкам и т.п.
Естественно, что и прежде, когда роль главного воспитателя играла семья,
все эти проблемы существовали, но тогда они были периферийными, а сейчас
скорее характеризуют ядро городской молодежи. Пусть родители и принадлежат
«ядру» своего слоя, но если дети хотят последовать их примеру, им слишком
часто приходится оказываться на «периферии» своего, молодежного слоя, что
большинству неприятно. Вот лишь некоторые причины конфликтов.
Сказанное можно резюмировать так: разнообразие условий, в каких обще-
ство предлагает жить молодым, резко изменилось, поэтому прежнее разнооб-
разие самих молодых людей оказалось ему резко несоответствующим. В исто-
рии так бывало не раз.
Феникс разнообразия
Однако, осталось ли разнообразие молодых людей прежним? Или оно в
самом деле вырождается? Для ответа снова полезно обратиться к опыту теоре-
тической биологии. В работах недавно умершего С.В. Мейена, палеоботаника
и эволюциониста, сформулирован закон транзитивного полиформизма [3].
122
Точная его формулировка нам здесь не понадобится, поэтому отмечу лишь
следующее: существенные свойства крупных органических разнообразий вос-
производятся в череде поколений заново, независимо от того, какая часть
(лишь бы не слишком малая) исходного разнообразия была взята для размно-
жения. Наблюдается как бы самостоятельный переход (транзит) разнообразия
во времени, мало зависящий от признаков, которыми обладают конкретные
особи, послужившие основой для этого транзита.
Иногда удается наблюдать, как разнообразие рождается заново, загадоч-
ным образом повторяя свои прежние формы. Еще Дарвин удивлялся: гладко-
кожий персик выведен из бархатистого, и гладкие формы, изменяясь, повтори-
ли все существенные варианты бархатистых. За следующие сто лет подобных
примеров опубликовано множество, и многие рассуждали об их биологическом
смысле, но только с работами Мейена (особенно [4]) стал вырисовываться
фундаментальный закон природы: спектр возможных изменений организмов
ограничен и упорядочен, причем организм наследует свой спектр (свои тен-
денции) так же, как и свои наличные признаки.
Если сто лет биологи считали, что эволюция есть сплошной процесс при-
способления (и вымирания неприспособленных), то теперь выясняется, что
эволюция подчас идет туда, куда указывает определенный закон разнообразия
[3]. Приспособление состоит в таких случаях в том, что маложизнеспособные
варианты редки, а нежизнеспособных нет вовсе. Так, есть много видов хирот-
ных позвоночных (с передними конечностями, но без задних), хотя это никак
не приспособление. У рыб отсутствие задних плавников довольно обычно, у
китов - обязательно, у наземных животных хиротность - большая редкость, а
вот птиц таких просто нет. Хирот (буквально «рукач») - ящерица без задних
лап - произошел от обычной ящерицы, так что его хиротность - и не приспо-
собление, и не наследие от хиротного предка. Это - в чистом виде издержки
разнообразия, повтор одного и того же в разных условиях, транзит.
Разнообразие людей тоже весьма транзитивно. В частности, пожилые лю-
ди каждого поколения уверены, что они в молодости были лучше, чем нынеш-
няя молодежь. Человек с возрастом перемещается вдоль спектра разнообразия,
сам редко осознавая это во всей глубине. Размышляя над своим юношеским
дневником, Ромен Роллан изумлялся: «Я вижу перед собой странного мальчи-
ка, носящего мое имя и похожего - нет, не на меня (я не узнаю в нем себя!), а
на кого-то другого, мне знакомого».
Устойчивость транзитивного разнообразия ярко проявляется в том, что все
попытки изменить общество посредством преследования людей определенного
типа терпят провал. Древняя натурфилософская идея избавления от порока пу-
тем преследования порочных, идея, породившая социал-дарвинизм, террор и
многое другое, терпит крах при осознании феномена транзитивного полимор-
физма. В каждом поколении будут и грешники, и праведные, и безликие, хотя
не обязательно им наследовать такие качества от своих родителей.
123
Это не значит, что разнообразием вообще нельзя управлять. Можно, но
только надо действовать по законам его собственного движения, а не мешать
ему. Для поворота автомобиля надо вращать руль, а не толкать сбоку в корпус.
Чтобы изжить бюрократизм, надо ликвидировать бюрократический аппарат
управления (отсутствие эффективной обратной связи снизу вверх). Преследовать
конкретных чиновников, занятие, на мой взгляд, бесполезное. Да, бюрократизм
связан с определенными чертами характера, но было бы наивным полагать, буд-
то смена (даже полная) личного состава учреждения может надолго изменить
характер управления: произойдет транзит структуры аппарата, она сама отберет
себе тех, кто может в ней успешно существовать. Если же меняется принцип
управления, то люди прежнего склада понемногу перемещаются из ядра аппара-
та на периферию, а ядро заполняется за счет прежней периферии и посторонних.
Если мы хотим управлять структурой разнообразия молодежи, то надо восстано-
вить нарушенное соответствие последнего структуре общества.
Варианты ограничены. Рефрен Мейена
Казалось бы, всякое богатое разнообразие разнородно и хаотично, но это
не так. Мейен продемонстрировал на ботаническом материале, что в природе
господствуют рефрены - устойчивые ряды изменчивости, повторяющиеся в
разных группах, так что разнообразие создается, в основном, наложением неза-
висимых рядов. Рассмотренная выше хиротность позвоночных - пример зооло-
гического рефрена.
Сходное явление есть в грамматике: все русские существительные склоняют-
ся, т.е. реализуют один и тот же шестипадежный ряд, но независимо от этого они
изменяются в числе, следовательно налицо два независимых рефрена. У прила-
гательных - три независимо комбинирующихся рефрена (род, число, падеж) и
один особняком (степени сравнения). Столь же четки рефрены глаголов.
Рефрены сводятся в таблицы, что дает огромную экономию: каждое слово мы
должны запомнить только в одной форме (например, глаголы - в инфинитиве) с
указанием типа его изменчивости (номер спряжения и т.д.). Хотя есть много ис-
ключений, все же польза грамматики доказана опытом. Точно так же, отсутствие
безногих птиц не должно заслонять наглядности и экономичности табличного
описания вариантов изменчивости организмов.
Предложенная Мейеном параллель биологического и языкового разнообразий
достаточно перспективна: как не может быть существительного вне падежно-
числового ряда, так же нет надежды встретить «ангела» - позвоночное с тремя
парами конечностей; иными словами, «грамматика биологии» [5] способна к со-
держательному прогнозу. Такая установка в корне противоречит прежней, когда
считалось: путем приспособления можно получить все, что угодно, и прогнози-
ровать эволюцию в принципе невозможно.
Теперь уточним ранее сказанное: управлять наличным разнообразием
можно только - двигаясь вдоль рефренов, а не поперек, т.е. не следует пытать-
ся навязать таблице разнообразия клетку из чужого ряда (рефрена).
124
Не раз уже строились и рефрены людских качеств. Так, в 1914 г. Э. Шпран-
гер описал шесть «идеальных типов» личности, оговорив, что в чистом виде они
не встречаются, а являются лишь тенденциями. По-моему, лучше сказать, что он
выявил шесть рефренов, наложение которых в грубых чертах описывает разно-
образие людей в плане мотивации (жизненных установок). Вот они:
[Опущено перечисление, повторяющее первый абзац на с. 73]
Каждая из тенденций имеет, по Шпрангеру, много степеней и форм прояв-
ления. Так, касаясь религиозного типа, он подчеркивает, что это «может быть
человек, который не принадлежит ни к одной вере, никаких обрядов не выполня-
ет, а это именно человек, ищущий высший смысл, высшую правду, первопричи-
ну. И к этому типу он причислял Спинозу и Джордано Бруно» [6, с. 80].
Каждая тенденция у Шпрангера - обширный рефрен, и потому в его форму-
лировке свернуто очень много информации. Не берусь оценивать качество и
полноту типов Шпрангера; я взял их просто как четкий пример рефренов, к тому
же часто упоминаемый в литературе.
Личность определяют многие характеристики, например, темперамент, т.е.
«общее понятие, относящееся к тому аспекту поведения, который отвечает на
вопрос «как?». Он отличается от способностей, относящихся к тем аспектам
поведения, которые связаны с вопросами «что?» и «насколько хорошо?», а
также от мотивации, связанной с вопросами «зачем?» и «почему?» [7, с. 69].
Самую древнюю и простую классификацию темпераментов - холерик, мелан-
холик, сангвиник, флегматик - тоже сводят теперь к наложению двух характе-
ристик - скорости реакции и силы чувств. (Согласно другим авторам, число
рефренов темперамента бывает больше и может достигать семи.)
Далее нам понадобятся два свойства рефренов: во-первых, они налагаются
друг на друга почти независимо (например, темперамент человека ничего не
говорит о его мотивациях), а во-вторых, всякое развитие можно определить как
смену или состояний в рамках прежних рефренов, или самих рефренов. На-
пример, человек не находится в одной клетке таблицы разнообразия, а движет-
ся по ней вдоль некоторых (иногда и всех) рефренов (так, социальное начало в
нем может затухать, а теоретическое или экономическое - нарастать). Сущест-
венно, что эта смена состояний нерегулярна у индивида, но проявляет стати-
стическую регулярность в большом множестве индивидов.
Место молодежи в обществе
Является ли молодежь особым общественным слоем или она всего лишь
«переходный возраст» от детей ко взрослым? И.С. Кон допускает, что юность
имеет «определенный социальный статус, специфическое для данного возрас-
тного слоя общественное положение и деятельность» [8, с. 7], но не разъясняет,
что это конкретно значит, ограничиваясь замечаниями психологов, призван-
ными отделить юность от детства (в 15 лет «быстро нарастают индивидуаль-
ные различия»; молодость «характеризуется появлением потребности и спо-
собности к интимной психологической близости с другим человеком, включая
125
сексуальную») и от юности («важнейшее приобретение... взрослости - творче-
ская деятельность и сопутствующее ей чувство продуктивности»). Подобные
разрозненные замечания легко свести в единую систему, если воспользоваться
понятиями ядра и рефрена. По-видимому, особых молодежных социопсихоло-
гических рефренов нет, и это служит извечной причиной отрицания за моло-
дежью особого статуса. Однако за счет развития каждого человека в ходе жиз-
ни ядро свойств молодежи сильно отличается от ядер других социальных
групп; а этого, как мы знаем, достаточно для выявления группы.
Можно сказать, что для молодежи особо характерны социальные мотива-
ции, самоанализ (особенно для девушек) и быстрота реакций, зато ее гораздо
слабее, чем взрослых, заботят экономические мотивы, комфорт, безопасность и
ответственность за близких. Это и есть грубая характеристика молодежи как
особого слоя общества. Хотя такие свойства нередки и у взрослых, но сочета-
ние их у взрослых редко, а у юных обычно. Ядро социального типа молодежи
оказывается периферией типа взрослых и наоборот.
Жизнь, естественно, меняет соотношения ядра и периферии. Прежде, когда
молодежь в основном содержала себя сама, ее ядро сближалось с ядром взрос-
лых. Сам я, окончив школу в 1957 году, могу свидетельствовать: наши проблемы
до седьмого класса мало отличались от детских (акселерация стала заметна поз-
же, в 60-е годы), а в восьмом нас осталась половина. Работы и собственных денег
некоторые семиклассники ожидали с нетерпением. Слов нет, в 14 лет работать
рано, но вот результат: в старших классах почти не было лодырей - тон задавали
те, кто готовился в вуз. Как тут не вспомнить слова А.А. Любищева, сказанные
30 лет назад: «Обязательность среднего образования надо понимать в смысле
предоставления всем возможности учиться, но не в смысле обязательства для
учителей довести до аттестата зрелости каждого лентяя» [9].
Из кончивших десятый класс едва ли не половина моих сверстников опять
же пошла зарабатывать деньги, а большинство продолжавших учиться не могло
жить без стипендии. Тем самым проблемы наши сразу стали взрослеть.
Специфические молодежные проблемы начали нарастать в 60-е годы, ко-
гда родители в массе избавили молодежь от необходимости рано идти рабо-
тать, когда у молодых появились деньги на водку и на вещи. Поэтому многие
полагают, что молодежь бесится от сытости, но ранее сказанное приводит к
другому выводу: она бесится не от материальной сытости, а от умственного и
эмоционального голода.
Снятие экономического давления опасно для всякого, у кого не выражена
ярко какая-либо неэкономическая черта. А.Н. Леонтьев пишет: «...Личность не
может развиваться в рамках потребления, ее развитие необходимо предполага-
ет смещение потребностей на созидание, которое одно не знает границ» [10, с.
115]. И юные персонажи Подниекса, далекие от науки, фактически считают так
же, только преломляют это через призму своего мироощущения: «сегодня нет
ничего такого, ради чего стоило бы умирать», «хочется что-то перевернуть,
126
расколотить, чтобы чего-то нехватало, чтобы было к чему стремиться». Растут
наркомания и преступность.
Какие-либо запреты бессмысленны, бессмысленно и устрашать - ведь из-
вестно, что мальчишки во все времена любили рисковать жизнью и здоровьем,
не то что свободой. Надо, чтобы им самим хотелось не творить насилие над со-
бой и другими, а чего-то иного. Но чего именно? Что же предлагает общество
взрослых молодым для заполнения трудового вакуума?
Прежде всего - досуг, но ведь он хорош после работы, а как главное дело
досуг удовлетворяет далеко не всех. Разумеется, те из молодых, у кого уже яр-
ко выражены научные, технические, эстетические, спортивные, администра-
тивные или идейные устремления, распорядиться своим временем сумеют, но
не в них проблема, как и не в тех, кто ищет выгоду. Проблема - это молодые
люди, у которых нет устремлений или есть одна лишь тяга к общению.
Взрослые пытаются еще и ограничивать количество свободного времени
молодежи, увеличивая нагрузку на учащихся, расширяя объем «добровольно-
принудительных» работ, сокращая круг освобождаемых от военной службы и
т.п., причем без всякого учета разнообразия молодых. Результат тоже плачев-
ный: проблемы не решаются, а лишь переносятся в другое место и другую плос-
кость. Эти меры не делают молодых созидателями в их собственных глазах.
Конкретный выход я указать не могу, но в самом общем виде задача со-
стоит в восстановлении нарушенного соответствия двух разнообразий - соци-
ального и психовозрастного. В последнем молодежь все резче обособляется в
качестве отдельного слоя, тогда как общество не может предложить вступаю-
щим в жизнь поколениям особого места, если не считать принудительного
привлечения к тем работам, которые не удается обеспечить трудовыми ресур-
сами посредством свободного найма.
Это вопиющее нарушение законодательства громко именуется «трудовым
воспитанием», хотя, по-моему, воспитывается при этом не любовь к труду, а
безынициативность, апатия, неуважение к праву, закону. Углубляется разрыв
между молодежью и взрослыми. Ведь господствующая в XX веке тенденция
прямо противоположна: по мнению социологов, дети наконец-то перестают
быть источником дохода старших, который «повернул на 180 градусов, устре-
мился от родителей к детям», в связи с чем и распалась прежняя семья, которая
«не знает долга родителей по отношению к детям, напротив, дети - вечные
должники родителей» [11, с. 193].
Чтобы воспитание было действительно трудовым и успешным, молодых
надо не принудить, а увлечь. Но чем?
Ядро молодежи не видит связи производства с потреблением (так как по-
требляет на уровне, предлагаемом семьей, обычно - малодетной), работает
больше для «галочки» или для справки в вуз - так что обычный денежный сти-
мул тоже эффективным не будет, не увлечет. К тому же резкий рост производст-
ва, если и возможен, приведет к недопустимому ускорению разрушения окру-
127
жающей среды. Массовое молодежное дело надо искать на периферии нынеш-
ней системы «взрослого» народного хозяйства. Таково, например, восстановле-
ние природных и культурных ценностей, разрушаемых экономикой.
И опыт здесь уже есть: Подниекс показывает, как заблудшие «панки» и «ме-
таллисты» увлекаются экологией родного города. Возможны дела и покрупнее.
Например, более 30 тыс. молодых канадцев провели масштабную экологическую
операцию, в ходе которой были очищены от отходов берега рек и озер общей
длиной около тысячи километров. Это начинание молодых - прямой вызов
взрослым, их отношению к природе (Знание - сила. 1987, № 3, с. 41). Вызов этот
обществу принять придется, т.е. придется наконец обратить экологию из докуч-
ливого противника экономики в ее ведущую отрасль.
Связь молодежной и экологической проблем неслучайна - наряду с пробле-
мой разоружения все три проблемы ясно указывают, что прежняя ориентация
экономики неприемлема, но если в отношении разоружения это, в общем, всем
понятно, если экология начинает отвоевывать право диктовать граничные усло-
вия развитию экономики, то в отношении молодежи дело обстоит куда хуже.
Хотя традиционные затраты на нее (обучение, досуг, пропаганда) призна-
ются необходимыми, фактически понемногу возрождается тенденция быстрей-
шего получения от молодежи выгод - экономических и прочих. О принципиаль-
но новых затратах на нее и речи нет. Скорее наоборот, перестройка экономики
обострит проблему, поскольку еще более сократит контакты подростков с семь-
ей (за счет вечерних смен и прочих видов интенсификации труда - их и взрос-
лых). Однако новые затраты необходимы.
Народное хозяйство (не только экономика, но и охрана природы, и меди-
цина, и обучение) издавна основано на идее кредита: общество платит сейчас
за будущие предприятия, ресурсы, кадры. В наше время общество увеличило
аванс молодежи тем, что позволяет ей больше и дольше, чем прежде, пользо-
ваться доходами старших, но этот аванс не является кредитом, поскольку не
направлен на какое-либо развитие. Наоборот, такой аванс задерживает моло-
дых в детском статусе. Кредит молодым должен содействовать их развитию,
т.е. помогать им двигаться вдоль возрастных рефренов, а для этого надо знать
законы данного движения.
Во-первых, оно, как мы уже говорили, нерегулярно (конкретный путь
взросления человека почти непредсказуем), а во-вторых, при каждом его акте
юный субъект, как правило, ощущает желание проявить независимость от дик-
тата старших. Оба эти обстоятельства взрослые обычно игнорируют, полагая,
что сами лучше знают, что надо делать каждому из юных. Вот здесь-то и ну-
жен принципиально новый кредит (вспомним, что итальянское credito перво-
начально означало «доверие»): надо в максимально возможной мере доверить
юным самим решать, как использовать ту долю общественного внимания и
продукта, которую им кредитует общество. Это - не филантропия, а самый
дешевый способ сгладить накопившиеся противоречия.
128
В самом деле, сейчас, когда повсюду говорят о демократизации, странно
выглядит то, что выбор молодыми своего жизненного пути становится все ме-
нее и менее свободным, что его все более диктуют те, кому еще предстоит пе-
рестраиваться. Сперва одно начальство решает, кому идти в девятый класс, за-
тем другое раздает «комсомольские путевки», а третье объявляет, что все
должны увлеченно служить «в соответствии с потребностями общества».
Но давно пора признать, что увлеченность по указке невозможна, тем бо-
лее, что «потребности» начальство само и определяет. Если не хватает моло-
дых рабочих, то наивно думать: чем больше откроем новых ПТУ, тем больше
вчерашних школьников встанет к станку или конвейеру. Лучше попытаться
понять, почему их не хватает, то есть почему разнообразие молодых людей
стало плохо соответствовать разнообразию рабочих мест.
Вспомним, что в 60-е годы симпатии абитуриентов неожиданно поверну-
лись от точных и технических наук к биологическим и гуманитарным, - теперь
мы знаем, то был первый сигнал о неблагополучии технократического общест-
ва. Не о том же ли говорит теперь более широкое движение - поворот молоде-
жи от заводов и строек? Ведь всяческих сооружений становится все больше, а
природы - все меньше. Не хватает жилья в городах? Но на селе оно в избытке.
Не хватает машин на полях и заводов, делающих эти машины? Нет, и то и дру-
гое в избытке, но плохо работает.
Пусть мы еще не видим связи сумбурных претензий молодежи с будущим
устройством общества, но надо признать, что связь эта объективно существует,
что ее надо изучать и использовать. И потому сейчас, когда молодежь слабой
струйкой начала возвращаться в деревню, поддержать ее, по-моему, куда важ-
нее, чем возобновить на заводах двухсменку.
Поддерживать тех молодых, кто тянется перестраивать экономику и куль-
туру, защищать природу, - то же самое (в плане социально-экономическом),
что финансировать бесплатное обучение. Поколение в целом конечно же вер-
нет аванс, только не своим родителям, а своим детям. По-моему, такая страте-
гия куда перспективнее, чем нынешняя. Ведь молодежи принадлежит будущее,
а не наши представления о нем.
Литература
1. Берзин Э. Сивка-бурка, вещая каурка // Знание - сила. 1986. № 11.
2. Психология личности. Тексты. М.: Изд-во МГУ, 1982. С. 49.
3. Meyen S.V. Fundamentals of palaebotany. L., 1987 (M-299).
4. Мейен С.В. Основные аспекты типологии организмов, 1978 (М-165).
5. Чайковский Ю.В. Грамматика биологии, 1986 (Ч- 14).
6. Зейгарник Б.В. Теория личности в зарубежной психологии. М. 1982.
7. Стреляу Я. Роль темперамента в психологическом развитии. М., 1982.
8. Кон И.С. Психология юношеского возраста. М.: Просвещение, 1979.
9. Любищев А.А. О положении в средней школе// Эко. 1983. № 12.
10. Леонтьев А.И. Деятельность, сознание, личность. М.: 1975.
11. Вишневский А.Г. Воспроизводство населения и общество. М., 1982.
129
Горизонт познания и познавательные модели
Исторические параллели, и что с ними делать *
У историка Ю.В. Андреева есть замечательная мысль: "Историческая сти-
лизация всегда считалась одним из самых эффективных приемов политической
пропаганды" [Андреев, 1999, с. 3]. Мне тоже случалось писать, что с помощью
истории принято учить не столько тому, что было, сколько тому, что должно
(по мысли учителей) быть [Чайковский, 2001].
До каких пределов может доходить “стилизация”, показал нам блестящий
ученый и публицист биолог А.А. Любищев. Он известен широким кругозором,
слыл знатоком истории, ироническим скептиком и въедливым критиком, не
хотевшим ничего брать на веру. И, тем не менее, полвека назад он написал:
«Мы можем назвать великим государем лишь такого, деятельность которо-
го направлена в прогрессивную сторону; это - подлинные революционеры
на престоле, и к их числу мы бесспорно можем отнести нашего Петра Ве-
ликого». И далее: «Ближе всего к понятию демократического самодержца
подходил, конечно, Петр Великий» [Любищев, 1995, с. 130, 139].
Поразительно: ведь труд Любищева ставил целью доказать, что Ивана
Грозного нельзя счесть великим, поскольку он укрепил крепостное право; но
ведь именно это дело и завершил блестяще Петр через полтора века после
Ивана. Более того, Иван действовал в рамках шедшего тогда в центральной и
восточной Европе «вторичного закрепощения»* 55, Петр же, обратив крестьян в
рабов, явно тянул Россию назад, в “тёмные века” варварских королевств ранне-
го Средневековья. Заодно Петр свел к голой форме сам принцип коллективного
руководства, до него в России обычный.
Можно ничего не знать о жутко знаменитых “петровских казнях”; можно
не брать во внимание, что беспредельную власть чиновника, с которой Россия
не может совладать до сих пор, ввел именно Петр; можно не видеть в его мето-
дах террора (Тайный Приказ, узаконенные безнаказанные доносчики (фиска-
лы) и всевластие “гвардейской опричнины” [Гордин, 1994, с. 88]) связующего
звена от методов инквизиции56 к методам чрезвычайки. Можно даже признать
* Отрывки из статьи 4-60 и ее рукописи. Статья, слабая сама, испррчена еще и беспоря-
дочными сокращениями разных редакторов. Однако в ней содержится подход к истори-
ческой диатропике, что, надеюсь, видно из приводимых отрывков. В целом же выявление
исторических рефренов потребует сквозного знания одним учёным истории всех стран,
что весьма затруднительно. Замечу, что познавательные модели описаны еще в ЭЭД.
55 «С XV в. положение крестьянства от Эльбы до Волги не переставало ухудшаться... В
Польше, в Венгрии, в Чехии “вторичное закрепощение”... возникло к выгоде сеньеров...
В России главным действующим лицом было государство» [Бродель, 1992, с. 459].
56 Петр не скрывал корней своих методов: он прямо называл основанное на доносе и “ис-
креннем” признании следствие инквизицией [Ерошкин, 1983, с. 93-95].
130
(чего Любищев не делал), что высокая цель оправдывает мерзкие средства. Всё
это можно если не принять, то понять. Но как можно писать о вторичном за-
крепощении крестьян с негодованием и напрочь умолчать при этом, что имен-
но Петр сделал его всеобщим и уравнял крестьян с рабами юридически?
И ведь об этом Любищев мог прочесть в самых известных источниках, на-
пример, в словаре Брокгауза-Ефрона, где крупный историк права писал: «Ря-
дом указов 1719-1722 гг., вызванных введением подушной подати... для кре-
стьян водворилось формальное рабство» [Тарановский, 1915, стлб. 262-263].
Про то же упоминал даже учебник истории, по которому сам Любищев, веро-
ятно, учился в школе (Платонов С.Ф. Учебник русской истории для средней
школы. Несколько изданий в 1900-х гг.). Там же говорилось, что основные ре-
формы, приписываемые Петру, начались до него.
Далее, по Платонову: «при Петре началась продажа крестьян без земли не
только семьями, но и в розницу» [Платонов, 1907, с. 464]. Добавлю: Петр, на-
делив правом владеть людьми купечество, породил класс крепостных рабо-
чих57 58 (до этого и Грозный не додумался), сбор налогов отдал генералам, с это-
го кормившимся [Милюков, 1900, с. 165], чем обратил армию в оккупантов
(она при Петре стала жить “по квартирам”, вконец разоряя нищее население).
И ведь Любищев книги Платонова упоминал.
Перечень “демократических” деяний Петра можно продолжать почти бес-
конечно, но, думаю, сказанного достаточно, чтобы признать, что они остались
за любищевским горизонтом познания. Этот горизонт оказался тем самым, ка-
кой давала сталинская пропаганда. В те годы бытовал злой анекдот, который я
слыхал школьником: «Мама, мама, скажи - Иван Грозный, Петр 1, кто еще из
царей был за Советскую власть?»*
А ведь Любищев был одним из самых независимых умов ранней послеста-
линской поры. Он попытался расширить свой горизонт в направлении Ивана,
но в благости Петра был настолько уверен, что даже не пробовал ничего выяс-
нять. Вероятно, его заворожила формула “Петр прорубил окно в Европу”, хотя
и она весьма сомнительна5*. Его конфуз заставляет задуматься: откуда вообще
мы знаем то, в чем уверены?
Горизонт познания
Горизонт познания похож на географический горизонт: для видения вдаль
надо высоко подняться, а при этом не видишь, на чем стоишь. Большинство
57 Они составили костяк ткацких и горных заводов; участь их была еще тяжелее кресть-
янской и осталась таковой даже после отмены крепостного права, чем часто и объясня-
лась их ведущая роль в революции 1917 г.
В терминах диатропики, три эпохи являют параллелизм, и пропаганда это использует. В
частности, все процессы повторного закрепощения входят в один рефрен. - Примеч. 2010 г.
58 Характерны слова Петра: «Европа нужна нам еще на несколько десятков лет, а там
мы можем повернуться к ней спиной» [Ключевский. 1983, с. 16].
131
ученых решают эту проблему просто: всю жизнь “лежат пластом”, глядя на
ближайшее, и изредка “встают на четвереньки”, когда пишут обзоры; лишь в
конце жизни некоторые из них “встают во весь рост” (оглядывают свою науку)
или даже “подымаются на башню”, т.е. описывают свое запоздалое удивление
от мира, мало кому интересное. Не интереснее и те, кто всегда “живет на баш-
не” (методологи), ибо они обычно слабо знают суть того, о чем пишут.
[-.]
Чтобы очертить обычный горизонт научного познания, приведу примеры
массового неведения учеными того, что они, вроде бы, обязаны знать. Как и в
случае с Петром, не буду претендовать на знание истины, но хочу показать, что
истина лежит гораздо глубже, нежели принято считать.
Первый пример подсказан мне чтением Любищева: всю жизнь критикуя
марксистов за узость взглядов, он сам пользовался той же, что у них, схемой, по
которой история есть борьба классов - землевладельцев, буржуазии, крестьян и
рабочих. Как и марксисты, он едва упомянул “служилых людей”, притом в со-
ставе “населения, которое, как правило, не эксплуатировало других” [Любищев,
1995, с. 130]. А ведь старые книги по русской истории и энциклопедии прямо-
таки вопиют о господствующей роли чиновников.
Сто лет назад на слово “Бюрократия” все большие энциклопедии России59
давали антибюрократический памфлет, иногда с указанием литературы. В одних
бюрократия - класс, противостоящий остальным и “состоящий из агентов цен-
тральной правительственной власти”. В других она “внесословная и внеклассо-
вая”. Но и тут и там суть бюрократии заявлена в ее безнаказанности и в том, что
чиновники видят смысл работы не в пользе дела, а в исполнении воли свыше, в
отписках, что писание бумаг становится самоцелью. Бюрократия как метод
управления известна с глубокой древности, но особенно развилась в Европе к
концу Средних векоов с созданием громоздкой налоговой службы.
Там же мы узнаём, что бюрократизм слабо развит в Англии, но особо раз-
вит во Франции, где доведен до совершенства Бонапартом, и в России, где про-
цветает со времен Петра I и упорядочен (при Александре I) графом М.М. Спе-
ранским. Что он совершенно нечувствителен к смене законов, укладов и видов
правления, так что единственным средством против него считается рост местно-
го общественного самоуправления.
«Живучей бюрократию делает то, что она - общественная группа, наи-
более легко приспособляющаяся ко всякого рода новым порядкам. Она
на той стороне, за которой больше силы». Она первая изменит прежне-
му строю, так как она «внесословная и внеклассовая» [Дживелегов,
1911, стлб. 426].
59 А также знаменитая французская энциклопедия Ларусса [Bureaucratic, 1866]. Наобо-
рот, тогдашняя английская Britannica, столь же знаменитая, даже не упомянула этого
слова в указателе. Немецкий Брокгауз дал лишь подробное объяснение понятия.
132
«Характернейшая черта бюрократического режима - стремление умножать
число правительственных функций и, вместе с тем, увеличивать класс чи-
новников, иногда создавая почти фиктивные функции» [Водовозов, 1912].
Всё это мы вроде бы и так знаем, однако удивительно то, что в новых по-
литических трудах данная тематика почти отсутствует. Иногда петровскую сис-
тему именуют “военно-бюрократическим монстром” [Гордин, 1994, с. 10]; инрг-
да к ней подходят при обсуждении идеи “Нового Средневековья”, но останавли-
ваются как бы в испуге (см., например [Блехер, Любарский, 2003]). В последнее
время тема засилья чиновников стала популярна у российских журналистов, но
только - как атрибут нынешней российской ситуации. Почему же не исследуют
наше чиновничество как элемент вечного института?
Второй пример: связь капитализма с рынком. Рынок известен с глубокой
древности, Высокое средневековье сделало первые попытки его ограничить -
путем введения максимальных цен на товары первой необходимости и т.п. В
XVII в., с началом эпохи капитализма, европейские государства отказались от
этого, что теоретики запомнили как торжество рыночной экономики, однако
выпал из внимания более важный факт: именно с XVII века государство стало
фактически и юридически ограничивать рынок. Средством к успеху служит не
конкуренция, а уход от нее путем разграничения сфер деятельности; поэтому
капитализм надо отличать от рыночной экономики [Бродель, 1993; Чайковский,
1996а; 2004]. Поле действия конкуренции ныне радикально сузилось, она сохра-
нилась, в основном, в розничной торговле и обслуживании.
Главная сфера, где при капитализме видно расширение конкурентно-
рыночной практики, это - сфера наемного труда, но и это не всюду: в Японии,
цитадели капитализма, преобладает традиция пожизненного найма. Почему же
именно время капитализма трактуется как эпоха рыночной системы?
Третий пример связан со вторым - эволюция. Ученый и неученый люд
воспринимает ее одинаково: как то, что открыл Дарвин, как происхождение ви-
дов путем отбора случайных мутаций в процессе конкуренции. Всякому, кто чи-
тал критиков дарвинизма (на мой взгляд, самым блестящим из них был Люби-
щев), известно, что такой взгляд на эволюцию ничем не обоснован и, более того,
противоречит фактам. С критиками никто не спорит, однако все учебники (а по-
тому и почти все ученые труды) продолжают учить “по Дарвину”. Об этом мно-
го написано, в том числе по-английски (см. например [Meyer, 2005]), и с напи-
санным тоже никто не спорит. Его не замечают.
Ущерб для биологии от этого невнимания огромен [Чайковский, 2003], но
это полбеды. Хуже, что дарвинизм проникает в иные области, где ведет к самым
чудовищным действиям - вспомним хотя бы “евгенику” фашистов, снова ожи-
вающую [Глэд, 2005]. Удивительно, что полный неуспех прежней евгеники во-
все не принимается во внимание. Его тоже не замечают.
Недавно возникли “эволюционная экономическая теория” и “литератур-
ный дарвинизм”, авторы которых не цитируют ни одной биологической работы,
133
но уверены, что развивают идею Дарвина. Неведение снова грозит обернуться
бедами для миллионов людей. Почему “эволюционные экономы” и прочие про-
жектёры не хотят прочесть хоть что-нибудь новое об эволюции?
[...]
Примеры можно продолжать, но, по-моему, уже ясно, что ученым не дает
договориться различие их познавательных горизонтов: большинство просто не
видит того, что меньшинство (а оно-то и движет науку) видит как главную цель
познания. Эта массовая слепота - частный случай того, что Имре Лакатош (Ла-
катос) называл защитным поясом исследовательской программы [Лакатос,
1995]. Задолго до него великий Кант говорил об этой слепоте: “Там, где другие
проходят, не заметив препятствия, для меня встают Альпы проблем”.
Чем же объясняется различие горизонтов познания людей? Вряд ли можно
указать одну-единственную причину, но одной из ведущих причин представля-
ется мне различие их познавательных моделей. Понятие познавательной модели
ввел в 1980 г. методолог А.П. Огурцов, ввел устно, а опубликовал много позже,
в книге [Карпинская и др., 1995].
Познавательные модели
Будем понимать каждую познавательную модель (ПМ) как набор прие-
мов и утверждений, которые данному ученому (ученым) настолько наглядны и
самоочевидны, что через них принято объяснять (к ним сводить, ими моделиро-
вать) все остальные факты и понятия. Тем самым, ПМ— явление социальное по
определению. В любой данный исторический момент в обществе обычно гос-
подствует одна ПМ-(иногда две), формирующая научную парадигму (в смысле
Томаса Куна) в каждом разделе знания, а другие ПМ-оппозиционны ей. Хотя
каждая ПМ-удобна для описания лишь какого-то круга явлений, однако на прак-
тике ведущая модель привлекается для объяснения всего на свете, а то, чего она
описать не может, часто вообще не воспринимается. Методу ПМ-посвящена гла-
ва 5 книги [Чайковский 2004].
Понятие ПМ, сформулированное Огурцовым, было ориентировано на
классическую методологию науки, черпавшую поллвека назад материал из фи-
зики. Огурцов сделал шаг вперед: следует говорить о ПМ-данной эпохи в целом,
поскольку обычно ПМ-заимствуется в естественные науки из наук обществен-
ных и из техники. Насколько я понимаю мысль Огурцова, ПМ-отличается от па-
радигмы по Куну и исследовательской программы по Лакатошу своей всепрони-
кающей междисциплинарностью (выходящей даже за рамки 'науки как целого),
привязанностью к определенной эпохе и количеством самих ПМ-(их, в отличие
от программ, всегда немного, но, в отличие от парадигмы, всегда больше одной).
Парадигмой Кун в 1963 г. назвал господствующую в данной отрасли точку
зрения, претендующую на единственность, тогда как всякая ПМ-охватывает
науку в целом и конкурирует с другими ПМ. В отличие от парадигмы, ПМ-
может не осознаваться сообществом: так, статистическая ПМ-традиционно вос-
принимается в рамках механической картины мира, и мне известен всего один
134
автор, явным образом противоположивший "гиббсово мышление" (т.е. стати-
стическую физику) "ньютонову мышлению" - это французский социолог и фи-
нансист Жак Аттали [Attali, 1986, с. 17].
[...]
До рождения европейской науки в обществе царило не оформленное логи-
чески почитание природы как благого или как злобного начала. Это почитание
удобно описать как господство нулевой (донаучной) ПМ, которую можно на-
звать этико-эстетической (религиозной) моделью. В ее рамках мир (природа и
общество) понимался как храм. Считать ее нулевой следует еще и потому, что
она характеризует тот эмоциональный тип восприятия, который нельзя, строго
говоря, отнести к познанию.
Огурцов ввел две ПМ— семиотическую и механическую, основные при
рождении европейской науки. Семиотическая (знаковая') ПМ— тот тип описания
знания, при котором мир выступает как текст, а познание - как чтение, рас-
шифровка. Эта ПМ-была исходной для европейской науки, первой научной: ею
пользовались Высокое средневековье и Возрождение, когда познание понима-
лось как разгадывание замысла Творца. С ней в науку вошли понятие закона
природы и идея математизации науки. Хотя ныне наука, в общем, отошла от
знаковой трактовки знания, таковая всё еще присутствует в ней в форме семио-
тики - учения о знаках и знаковых системах. Первая ПМ-характерна для на-
чальных стадий формирования научных дисциплин.
Вторая (механическая) ПМ-сменила в XVI-XVII веках знаковую. Она
строит систему мира как механизм. Ее модельный объект - часы. В ее рамках
утвердились принцип причинности и идея эволюции (точнее, прогресса - как
социального, так и биологического). До сих пор мы говорим "понять механизм
явления", хотя бы явление было вовсе не механическим. Идея целостности за-
нимает мало места в данной модели, но всё же присутствует: каждый объект оп-
ределяется, как деталь, своим местом в целом механизме.
В XIX веке вторая ПМ-обогатилась идеей устойчивости движения - стали
считать реально интересными лишь те движения, которые при малых возмуще-
ниях не приводят к большим различиям в результатах. Ныне же основной инте-
рес представляют как раз неустойчивые движения, а их невозможно описать без
обращения к случайности.
Дальнейшее развитие рациональной европейской науки связано, на мой
взгляд, с тремя ПМ— статистической, системной и диатропической. Вся пя-
терка сменяющих друг друга научных моделей грубо, но в общем, по-моему,
верно описывает процесс европейского научного познания как социальное явле-
ние, каков он был в ходе ушедшего тысячелетия. Античную и восточную науку
таким образом описать не удается.
Третья (статистическая) ПМ-видит мир как совокупность балансов, сред-
них и инвариантов. Ее модельный объект - весы. С нею в науку вошли такие по-
нятия, как закон сохранения, торговый баланс, баланс природы, однородное и
135
изотропное пространство, процент. Возникла она впараллель со знаковой: впер-
вые понятие баланса родилось в бухгалтерии XV века, откуда идет традиция ви-
деть государство и природу как исконно сбалансированные Богом (прообраз
идей равновесия властей и экологии) и приводить доли к единой форме - про-
центной. Эта ПМ-завоевала науку лишь в XVIII-XIX веках; она до сих пор зани-
мает в науке центральное положение, и в ее терминах принято трактовать всё,
что связано со случайностью*.
Отличие ее от механической часто ускользает от внимания, поэтому пояс-
ню: если мы взвешиваем объект исследования на чашечных весах, то уравнива-
ем стрелку весов, кладя на другую чашу весов гири в нужном составе, то это -
механическая процедура; если же мы уравниваем сотню образцов килограммо-
вой гирей и ограничиваемся этим (зная лишь то, что средний вес образца равен
10 г), то это - статистическая процедура.
Четвертая (системная) ПМ-видит во всем целостность, уподобляет мир
организму. С нею в науку вошли идея оптимальности (экстремальные принци-
пы) и идея самоорганизации. В мировоззрение ученых эта ПМ-входит в настоя-
щее время, хотя отдельные ее положения утвердились давно (например, прин-
цип наименьшего действия). Несмотря на свои очевидные достоинства, систем-
ная ПМ-несет в себе радикальный изъян - уверенность в наличии единственно
правильного решения едва ли не каждой задачи .
Пятая (диатропическая, от греч. диатропос - разнообразный) ПМ-видит в
мире прежде всего разнообразие, видит природу как сад или как ярмарку (а не
как огород или рынок, где нет эстетического элемента), где ни один элемент не
обязателен, но в которой некая трудно уловимая целостность (часто - не функ-
циональная, а эстетическая) есть. Базой для этой ПМ-служит диатропика, наука
о закономерностях организации разнообразия, основу которой заложил палеобо-
таник и философ С.В. Мейен [Чайковский, 1991].
Первичным понятием диатропики является ряд объектов - он для нее то же,
чем для иных наук служит факт', первичной процедурой тут является сопоставле-
ние рядов, а первичным результатом - выявление их параллелей. Примеры рядов
приведены далее. К пятой ПМ-близка идея о “множественном порядке” (polyordre)
[Attali, 1986, с. 357], которую можно назвать социальной диатропикой. [...]
В наше время третья ПМ-повсюду уступает место четвёртой, а это значит,
что обе имеют достаточно много приверженцев, чем и объясняется нескончае-
мый спор вокруг дарвинизма и рынка. Факт смены ПМ-хорошо виден хотя бы в
том, что аргументы, прежде не привлекавшие внимания, вдруг попадают в центр
дискуссии. Системная ПМ-тоже, как уже сказано, имеет изъяны. Их пытается
* Ее явным образом держался и Мейен, поэтому он полагал, что для таксономии важны
частоты признаков, а итог своих пионерских наблюдений над рефренами формулировал
как «статистическую природу таксона». Идею ПМ-он узнать не успел, да и мне лишь
позже стало ясно, что фактически он работал в рамках четвертой ПМ. - Примеч. 2010 г.
136
исправить диатропический подход, но он еще не принят обществом, не завоевал
статуса ПМ. Диатропическая картина мира (одним из инструментов которой яв-
ляется теория фракталов), явственно заявившая о себе в конце XX века, претен-
дует на общественное внимание, но еще не завладела им. По всей видимости,
одна ПМ-может сменять другую только в определенном порядке. Подробнее см.
[Чайковский, 2004, гл. 5].
Основной интерес представляет пятая ПМ-как ныне входящая в науку.
Одиночное сходство не является предметом интереса диатропики, тогда как в
остальных науках оно занимает едва ли не главное место. В них обычны описа-
ние факта за фактом (анализ) и обобщение фактов (синтез), тогда как диатропи-
ка призвана исследовать ряды и выявлять параллели. Самым известным является
параллелизм сумчатых и плацентарных млекопитающих: известны сумчатые
крысы, кроты, барсуки, муравьеды, волки и т.д. [...]
Контуры будущей познавательной модели
В 1994 г. методолог Н.Ф. Овчинников высказал мне устно мысль о том,
что зародыш еще одной (шестой научной) ПМ-можно видеть в концепции пред-
расположенности Карла Поппера. Хотя тут еще нельзя всерьез говорить о соци-
альном явлении (каковым по определению является ПМ), однако мысль Овчин-
никова интересна, и параллели со взглядами других ученых видны.
В отличие от Овчинникова, композитор и методолог А.А. Кобляков [2000,
с. 309] полагает главным свойством будущей 'ПЪЛ-феномен творчества-.
«Какая же должна быть новая... познавательная модель, включающая субъ-
екта и связанную с ним категорию смысл? ... Какой же вид деятельности
наиболее полно репрезентирует интуитивное? Конечно, творчество!»
Тут уместно напомнить, что еще в 1907 г. французский философ-
эволюционист Анри Бергсон полагал феномен творчества главным в эволюции
[Бергсон, 1998], т.е. допускал его наличие вне сознания, в природе. Р.Г. Баран-
цев рассмотрев триаду “хаос - порядок - творчество”, приходит к выводу, что
“именно творчество, будучи достаточно глубоким понятием, объединяет поря-
док и хаос в целостную триаду” [Баранцев, 2002, с. 117].
Познавательную модель Кобляков понимает вне всякой связи с социаль-
ной проблематикой и для ее построения использует модальную логику (с кван-
тором уместности, отражающим контекст высказывания). То, что он предло-
жил, можно рассматривать не как ПМ, а лишь как заявку на будущую ПМ. В ней
нам интересны как отдельные соображения:
«У творчества и синергетики есть общая основа - гармония как взаимо-
дополнительность конкурирующих процессов»; «взаимодополнительность
процессов-антагонистов есть главный объединяющий принцип в поведе-
нии сложных систем»; «в неустойчивых нелинейных средах на уровне тон-
кой структуры фазовых переходов природа демонстрирует свою, если
угодно, психическую компоненту, способную самоорганизовываться»,
так и общие выводы:
137
1) «вся история последовательного расширения (обогащения) имеющихся
математических представлений... может рассматриваться не только гно-
сеологически (как акт познания), но и онтологически, как акт творения, как
акт творческого созидания нового» [Кобляков, 2000, с. 315].
2) «признание творчества в качестве новой парадигмы и модели устра-
нения противоречий в качестве новой познавательной модели неизбеж-
но влечет за собой следующую переформулировку закона-вектора твор-
ческого процесса, а именно: любая эволюция направлена в сторону
больших размерностей, больших степеней свободы. Структурное ус-
ложнение материи в процессе развития - результат действия этого зако-
на» [Кобляков, 2000, с. 319].
Эти смутные намеки не претендуют пока на то, чтобы формулировать бу-
дущую ПМ-(социальное явление), но осмелюсь допустить появление в будущем
шестой научной (креативно-пропенсивной60) ПМ. Свойства ее должны вытекать
из желания видеть мир как систему предпочтений, потенций и склонностей; в
данной системе всякое развитие должно радикально зависеть от актов свободного
выбора (а он, как мы увидим, иногда рассматривается как одна из форм случайно-
сти, самая трудная для понимания). В этих рамках однородное изотропное про-
странство (основное понятие физики) - не объективная реальность, а лишь одна
из моделей реальности, так что возможны другие ее модели, с предпочтительными
направлениями. Можно полагать, что эта ПМ-будет итогом синтеза системного и
диатропического подходов, т.е. будет брать за основу объект, образно говоря, ме-
нее организованный, чем организм, но более цельный, чем ярмарка.
Всякая ПМ-требует для ее уяснения указать модельный объект. Для шес-
той ПМ-назову железную игральную кость. Вне магнитного поля она падает,
как обычная кость - это поведение без предрасположенностей; если же вклю-
чить поле, то поведение изменится вплоть до возможности выпадения всегда
одной и той же грани или даже невозможности падения. Магнитное поле здесь
выступает как поле предрасположенностей (поле предпочтений, пропенсивное
поле).
Модели и усвоение нового
Как вообще возможно усвоение нового обществом, если люди мыслят
столь различно? Тезис Маркса “Практика - критерий истины” не дает ничего,
когда горизонты разных людей различны. Люди склонны не только не верить
практике, которая им неугодна, но и считать (что довольно странно) практикой
голое умозрение, даже грубо противоречащее практике, если оно угодно.
Самые простые и яркие примеры дает упомянутая выше эволюция. Ее сто-
летиями общество не замечало, хотя приводились те самые факты, которые те-
перь служат “доказательствами эволюции” во всех учебниках; и обратно: ныне
“научный креационизм” снова отрицает (и вполне успешно) идею эволюции,
60 От латинских creatio ex nihilo (творение) и propensity (склонность, расположенность).
138
пользуясь старыми аргументами, которые еще 30 лет назад выглядели несерьёз-
но почти для всех.
Когда в дни Дарвина идея эволюции утвердилась (в форме дарвинизма), то
ее противников сразу же перестали слушать, даже когда они говорили вполне
очевидные истины. Так, оставлены без ответа все расчеты, демонстрирующие
арифметическую нелепость идеи отбора малых вариаций.
[•••]
Разумеется, можно сказать, что эта смена одной слепоты на другую -
крайний пример как научной революции по Куну, так и защитного пояса иссле-
довательской программы по Лакатошу, но почему это фактически происходит и
нельзя ли научиться этого избегать?
Видимо, усвоение идет легко лишь тогда, когда новое предложено в поня-
тиях господствующей ПМ. Так, ранее [Чайковский, 2004] мне удалось показать,
что прогресс в понимании случайности шел об руку со сменой ПМ. И прогресс
генетики в XX веке выглядит естественно, когда поймешь, что он не требовал
новых ПМ. Успех идеи квантов потребовал новой (системной) ПМ, но она как
раз тогда и пришла в науку. Наоборот, недвижность “проблемы осуществления”
[Любищев, 2004], т.е. понимания механизма реализации линейной генетической
информации в трехмерную структуру организма, связан, на мой взгляд, с полной
непригодностью наличных ПМ-для этой цели.
[...]
Словом, в целом видно, как господствующая в умах ПМ-задает ученым их
горизонт познания. Почему люди так упорно держатся за марксову “пятичлен-
ку” (пять исторических формаций), если сам Маркс был вынужден для описания
многих государств Востока ввести “азиатский способ производства” (о нем см.
сб. [Феномен..., 1993]), не входящий в им же предложенный ряд? Чувствуется
связь конфуза исторического материализма с петровским конфузом Любищева;
от нее и попробуем двигаться.
Ряды и модели
Видна модельная природа приведенных исторических рассуждений: если
Маркс хотел дать единую модель истории хозяйства и марксисты оказались в ее
плену, то Любищев был в плену гораздо более старой модели “хороший прави-
тель”, без которой вряд ли строится хоть одна историческая картина.
Общее у обеих моделей то, что в основе каждой лежит ряд: в модели Мар-
кса - ряд сменяющих друг друга формаций (единый для многих государств), а в
модели “хороший правитель” - ряд правителей, служащих объектом “стилиза-
ции”. Правитель с эпитетом “Великий” нужен каждой культуре, и он находится,
какого бы насилия над истиной это ни стоило.
Безмерно возвеличенный при последующих царях (видимо, с бессозна-
тельной целью оправдания растущей бюрократизации государства и гниющего
крепостного режима), Петр I был совсем не нужен коммунистам в первые годы
их правления, и о нем официозное издание писало:
139
“всеми силами укреплял самодержавие и использовал крепостнические
отношения для промышленности, заполняя новые мануфактуры крепост-
ными рабочими... После удачной Северной войны... принял титул “Вели-
кий”, удержавшийся за ним в дореволюционной историографии... Как
личность Петр 1 соединял огромную волю, талант организатора и бога-
тую инициативу с крайней психической неуравновешенностью, жестоко-
стью, запойным пьянством и безудержным развратом” [Нечкина, 1930].
Новое возвеличение Петра началось в параллель с закрепощением кресть-
ян в колхозах и с самодержавными амбициями Сталина. Оно отмечено фильмом
“Петр I” (1937-39; премия в Париже, 1937; Сталинская премия, 1941), где царь
произносит длинную пламенную речь на фоне реющего стяга с двуглавым ор-
лом. В наше время Петр I опять возвеличен, ибо знаменует утраченное величие
нации. О его зверствах, разврате и сверх-крепостничестве не вспоминают. По-
чему именно Петр? Да больше, по сути, возвеличивать некого, а без Великого
нет великой державы. Так требует стилизация.
Екатерина Великая? Нет, она возвеличила дворянство62, почти никому
нынче не нужное, тогда как Петр I возвеличил вечное чиновничество. И тем, по-
лагаю, мил властям всех времен и мастей.
Удивительно невнимание к чиновникам как классу. Насколько знаю, лишь
Милован Джилас, югославский политик и публицист, написал книгу “Новый
класс” (1957), но он называл новым классом компартию, а не чиновничество. В
действительности же оно старо, как само государство, а компартия - лишь его
сравнительно новый инструмент. Ни в одну ходячую модель чиновничество не
попало, оставшись за горизонтом познания. Его принято лишь походя поминать,
тогда как роль его весьма велика и в разных культурах оно весьма сходно. То
есть его черты образуют ряды. Тем самым, полезна пятая ПМ.
Вот пример ряда, имеющий отношение к власти чиновников. В 1725-1825
годах смену власти в России вела гвардия: 6 раз удачно, при чем 5 раз приводи-
ла к власти женщину (чего ни до, ни после в русской истории не было, бывали
только опекунши царей-мальчиков), а 7-й раз неудачно - декабристы пытались
заменить Николая на Константина. Он не имел влияния в Петербурге, т.е., по их
мнению, не мог помешать реформам (и этим, добавлю, был для чиновников схо-
ден с женщиной). Подавив бунт декабристов, Николай сменил гвардейскую ох-
рану на мусульманский конвой, и гвардейских переворотов больше не возника-
ло. Из пяти женщин четыре возведены на престол в обход мальчика (наследника
или даже монарха), т.е. вопреки праву и обычаю.
В чем причина сходств этих переворотов? Сравнением единичных фактов
можно обосновать любую точку зрения, но пятая ПМ-предлагает иной путь. 61
61 Знаменитый указ "О вольности дворянства" (1762) подписал ее муж Петр III, но в
жизнь проводила, после его убийства, она. И столь же знаменит ее указ (1767), каравший
каторгой за любые жалобы на помещиков, независимо от их справедливости.
140
Ряд российских переворотов XVIII века параллелен ряду “солдатских импера-
торов” Рима: гвардия ставила слабого правителя (кто знал, что Екатерина П будет
сильной?), чтобы никому не подчиняться. (Исключением в данном ряду переворо-
тов было убийство Павла, притеснявшего саму гвардию.) Но для понимания сути
явления одной параллели мало.
Другую параллель русскому ряду переворотов являют ряды правлений
женщин. Удивительно: за 30 лет после Петра I в России правили (включая ре-
гентство Анны Леопольдовны) 5 женщин - столько же, сколько за 3 тыс. лет ис-
тории египетских фараонов (включая Клеопатру). Ряд это или не ряд? По-моему
- ряд: в древнем Египте 4 женщины-фараона из пяти
“получают трон в период упадка... Странно, но женщин допускали к престо-
лу только тогда, когда положение государства становилось совершенно без-
надежным и фараоновская власть ослабевала” [Томашевич, 1992, с. 313].
Примерно ту же, что в России (но совсем отличную от Египта), картину
видим в османской Турции. Сулейман Великий (правил в 1520-1566 гг.) сходен с
Петром Великим не только тем, что могущества империи достиг за счет роково-
го перенапряжения сил страны, но и тем, что 90 лет после него длилась нелепая
“власть гарема”: султаны были, в основном, слабы и за них правили их матери,
боровшиеся за власть с главными наложницами. Здесь ряд очевиден.
Важно, что в то время в Турции религиозно-сословная власть уступала ме-
сто военно-бюрократической (см.: Иванов Н.А. Организация шариатской вла-
сти... [Феномен... 1993]) - как в России при Петре. Ослабление царской власти
представляется мне основной причиной женских правлений.
Итак, один ряд может проявлять параллели с разными рядами.
Богаче всех на параллели этого рода был переворот 1730 года, когда гвар-
дия привела к власти Анну Иоанновну. Обычно пишут, что он был актом побе-
ды дворянства над аристократией. В самом деле, горстка аристократов (Верхов-
ный тайный совет, правивший при юном Петре II) поднесла Анне “Кондиции”,
т.е. условия принятия короны, которые резко ограничивали власть Анны в поль-
зу “верховников”, и Анна была вынуждена их подписать. Однако группа офице-
ров гвардии предложила ей взять власть во всей полноте, и Анна публично по-
рвала “Кондиции”. Власть верховников пала, в чем и состоял переворот.
Это, однако, половина правды. Вторая состояла в том, что в Москву на
свадьбу Петра II прибыло много дворян, но он неожиданно умер. Поэтому
«Собравшись на свадьбу и попав на похороны, дворяне очутились в во-
довороте политической борьбы. Проект верховников встречен был в об-
ществе глухим ропотом... Съехавшееся дворянство разбилось на кружки,
собиралось по ночам и повело оживленную оппозицию верховникам...
Французский уполномоченный Маньян писал тогда в депеше из Москвы:
“Здесь на улицах и в домах только и слышны речи об английской консти-
туции и о правах парламента”. Против чего же восстало дворянство?., оно
восстало против замысла верховников как олигархической затеи... было
141
против верховников потому, что их проект не давал дворянству доста-
точного обеспечения» [Ключевский, 1983, с. 36].
Более тысячи дворян (среди их глашатаев выделялся государственный дея-
тель и историк В.Н. Татищев) подписалось под проектами, поданными в Вер-
ховный тайный совет с требованием власти дворянству и городам. Верховники
пошли на внешние уступки дворянству, но успеха не имели, и в дело вмешалась
гвардия. Едва офицеры, охранявшие Анну, поняли, что ни верховники, ни дво-
рянство не имеют решающей силы, они предложили ей взять полную власть.
Чью волю выражала гвардия? Какой общественной группе было выгодно
отнять власть у аристократов и не допустить к ней дворянство? Подсказкой к
ответу служит тоже ряд', в 1730 г. на престол предлагалось 6 женских вариантов
и всего 1 мужской (будущий Петр III), причем его тогда никто, кроме иностран-
ных послов, даже обсуждать не стал [Гордин, 1994, с. 146-147] - возможность
сильного в будущем царя пугала всех*. Мы в России знаем: беспомощная власть
означает безнаказанность класса чиновников (а не класса дворян), тогда как при
сильной власти чиновники иногда несут наказания.
Замечательно, что дворяне выставили пункты о правах городов (а самая
большая группа добавила даже пункты о крестьянах и солдатах [Милюков, 1903,
с. 34]), т.е. пытались стать голосом всей нации, но прискорбно, что силы за со-
бой они не имели: она вся была у чиновного аппарата.
Все три реальные силы переворота были чиновничьими: верховники были
министрами и фельдмаршалами, на свои собрания звали военных и штатских
генералов (иногда полковников)6-2; гвардия тоже теснилась вокруг нескольких
высших чиновников, в том числе церковных, ущемленных верховниками (там
же, с. 40-42), и, вероятно, помнила свою полицейскую роль при Петре I; а в про-
екте Татищева прямо “вся власть сосредоточивалась в руках высшего чиновни-
чества” (там же, с. 32). Сама победа Анны стала возможна, когда сторонники
самодержавия объединились (увы, при посредстве Татищева) со сторонниками
конституции6,2 против верховников (там же, с. 45).
В перевороте 1730 г. есть и иные параллели с событиями русской истории:
в Смутное время претендентам на престол трижды (1606-1613) предлагались
“ограничительные записи”, на что и опирался Татищев [Гордин, 1994, с. 156]; в
1682 г. царевна Софья стала правительницей при малолетнем Петре I (и его хи- * 62 63
* Здесь напрашивается сопоставление одновременного и исторического рядов, что в диа-
тропике обычно (например, «биогенетический закон» в биологии). Законно ли такое со-
поставление в истории, сказать не берусь - недостаёт материала. - Примеч. 2010 г.
62 Лишь однажды, видя успех дворян, верховники вспомнили свою аристократическую
суть и призвали “из знатных фамилий шляхетство в рангах и без рангов” (там же, с. 32).
Это была явная параллель с Западом, но верховники тут же в ней разочаровались.
63 Первые обещали вторым принять их планы, что Анна частично исполнила (там же, с.
50). Дело не в ее честности, а в том, что некоторые идеи были признаны разумными.
142
лом брате Иване, отце Анны Иоанновны) по требованию бунта стрельцов; в
1725 г. гвардия возвела на престол Екатерину I вопреки наличию Петра II, пря-
мого наследника по мужской линии; в 1762 г. Екатерина II подписала манифест
об ограничении ее прав, но в тот же день порвала [Сватиков, 1905, с. 23] и т.д.
Это высвечивает истинно русский характер происходившего (ни право, ни обы-
чай, ни голос масс могут не приниматься в расчет), но одиночные параллели не
дают понимания явлений (произвольно комбинируя, можно доказать что угод-
но), поэтому надёжнее брать ряды параллелей.
Историческое событие как пересечение рядов
С позиции пятой (диатропической) ПМ-событие выступает как пересече-
ние рядов. Ранее [Чайковский, 2003, с. 402] пересечением рядов описано проис-
хождение человека. Попробуем применить этот прием к историческому акту.
Первый ряд для понимания событий 1730 года: гвардейские перевороты,
приводившие к власти женщин. Сам этот ряд - пересечение мирового ряда
“дворцовые перевороты” с русским рядом “опекунша лучше, чем опекун”, легко
перешедшим в ряд “царица лучше, чем царь”. Впервые вдовой, восшедшей на
престол вместо того, чтобы опекать малолетнего наследника (Петра II), стала
Екатерина I, а при следующем перевороте никто даже не хотел слушать, что ма-
лолетний наследник (будущий Петр III) вообще имеется.
Вторым можно взять ряд “Новое Средневековье”, который описывают вот
уже 200 лет [Блехер, Любарский, 2003, с. 492]. Суть его в том, что каждая эпоха
Нового времени содержит в себе важные средневековые черты. Петровская эпо-
ха (ей как раз и подражало правление Анны) и сталинская становятся в этот ряд,
имитируя эпоху Грозного (“Каждый век имеет свое средневековье” - грустно
шутил Станислав Ежи Лец, польский писатель, о власти коммунистов).
Третьим можно взять ряд “Парламентарные замыслы”. Русские дворяне в
1730 г. равнялись на польский, английский и шведский опыты, но для этого ряда
здесь не было никаких основ, так что сам собой (бессознательно) был разыгран
чиновный вариант на французский лад: переворот являет ясную параллель с Ге-
неральными штатами во Франции - чем-то вроде сословного парламента, кото-
рый был время от времени собираем королями в XIV-XVII веках, когда короли в
нем нуждались, и который игнорировали, едва в нем отпадала нужда (похоже на
земские соборы в допетровской России). Чиновники Франции и России одина-
ково использовали дворян как временную опору в борьбе с аристократией и тут
же о них забывали.
Четвертый ряд: “Русское дворянство как бюрократия”. Триста лет, от
Ивана III до Елизаветы, русское дворянство было подневольным классом, слу-
жившим там и столько, где и сколько скажет правитель. Единственным ограни-
чением этого служило местничество, т.е. право просить у правителя того места,
которое соответствовало родовитости просящего, но и оно было отменено в
1682 г. Если выписать свойства западных чиновников-дворян и китайских чи-
новников, что сделал китаист С.В. Волков [Феномен..., 1993], то русское дво-
143
рянство, до Указа о вольности дворянства (1762) сплошь чиновное, больше по-
хоже на китайское, чем на западное.
Переворот 1730 года предстает как пересечение этих рядов (а также дру-
гих, не названных). Если вместо третьего ряда (парламенты) взять ряд “опекун-
ша лучше, чем опекун”, то получим переворот 1740 г., приведший к власти Анну
Леопольдовну вместо Бирона. Если же третьим взять мировой ряд “Победа пат-
риотической оппозиции”, то получим воцарение Елизаветы в 1741 году. Этот
ряд, насколько знаю, имеет лишь один пример в русской истории.
Аналогично с другими заговорами. Так, чтобы обрисовать бунт декабри-
стов как пересечение рядов, надо вспомнить, что они, тоже в основном гвардей-
цы, обсуждали, в частности, возведение на престол вдовы Александра I.
Засилье чиновников в России Милюков [1900] объяснял тем, что всю
жизнь народа тут с давних пор формировало правительство, занятое только вой-
ском и средствами для его содержания; оно расселяло жителей по окраинам и
строило там города. На Западе было наоборот: земли были издревле заселены и
формировали правительство.
Ряд гвардейских переворотов (римских, русских и т.п.) - яркий пример яв-
ления, которое историк-структуралист Фернан Бродель (Франция) называл се-
рийным фактом, первичным материалом в его концепции. Для него серийным
фактом в равной мере мог служить и глобальный параллелизм, каковой он обна-
ружил в феномене «миров-экономик», и частный параллелизм типов помещичь-
их владений, наблюдаемый в разных культурах [Бродель, 1992, с. 17, 456].
Усвоив мысль Милюкова, Бродель пошел дальше него:
“Иван Грозный... не имел иного выбора, кроме как устранить старую ари-
стократию... а чтобы иметь в своем распоряжении войско и администра-
цию, создать новое служилое дворянство” [Бродель, 1992, с. 459].
Тем самым, победить в 1730 г. могло только чиновничество, и нельзя со-
гласиться с историком-публицистом Я.А. Гординым, полагающим, что успех
верховников означал бы иной путь развития России (вплоть до невозможности
прихода к власти большевиков через 187 лет). В подтверждение он не привел
ничего, кроме своих пожеланий, чтобы было так.
Да, годы Анны, с пытками и сажанием на кол недовольных, были злым
фарсом, имитировавшим совсем злое правление Петра, но ведь и верховники,
чьей победы Гордин желал бы, ратовавшие за справедливость, ничего, кроме
усиления своей власти, по-существу не предлагали. Кто-то йз них вполне мог
лично взять власть, но изменить ее не мог бы; да никто и не предлагал.
Зато сама Анна еще до въезда в Москву грубо нарушила “Кондиции” -
объявила себя командиром гвардии, чем показала знание того, на чьей стороне
сила. На гвардию должен был опереться любой победитель, и с тем же итогом.
Чтобы утверждать хотя бы принципиальную возможность падения власти
бюрократии, надо указать в рядах смен власти в России какую-то пригодную для
этого мысленную комбинацию реальных возможностей, а таковой не видно по
144
сей день. Наоборот, все антибюрократические движения в России кратки и вы-
глядят нежизненно, в том числе нынешние.
Как увидим в и. 9, Гордин рассуждает в понятиях третьей ПМ, что неверно
сразу в двух смыслах: во-первых, она тогда еще не пришла в Россию и потому
не могла быть реальным способом рассуждать, а во-вторых, ныне мы обладаем
лучшими ПМ, чем третья, и можем понять происшедшее тогда лучше, нежели
делают наивные приверженцы парламентаризма.
Зато Гордин прав в другом - в оценке состава общественных групп:
“выразителей интересов дворянства в природе, на мой взгляд, не сущест-
вует... в политических группировках оказывались люди самых разных
статусов и происхождений... Существует другой принцип деления...
принцип политико-психологических групп, образований межсословных и
межсоциальных... сложная игра которых и есть политическая борьба”
[Гордин, 1994, с. 191].
Одним из подтверждений служат Гордину данные о гвардейцах, привед-
ших к власти Елизавету: их было 308, и дворян среди них всего 17%, а основная
масса - выходцы из крестьян [Гордин, 1994, с. 318]. Добавлю, что долю дворян в
1730-х годах резко сократил Бирон, фаворит Анны, желая пресечь политическую
активность гвардии, но, как видим, не достиг ничего.
В 1730 г. людей объединили ненависть к прежней власти и надежда на но-
вую, а достигнута была новая победа прежней власти. В точности то же повто-
рялось много раз. Возможен ли выход из этого дурного круга?
Ответа у меня нет, но видно, где и как его искать: сперва надо выстроить
параллельные ряды, затем указать то пересечение рядов, которое будет означать
искомый выход (если он существует), и только после этого - те группы, которые
для этого должны объединить усилия. Первое относится к диатропике, второе -
к познавательным моделям и горизонту познания.
Надо пояснить второе. Для этого скажу о двух исключениях из правила
смены власти в России: обычно она является переворотом сверху, но в 1917 и
1991 годах в смене власти явно сыграли основную роль низы общества.
Исключения уточняют правило
Первую смену долго называли Великой Октябрьской социалистической ре-
волюцией, хотя к социализму привела не она, а коллективизация и индустриали-
зация 1930-х гг., перечеркнувшие главный тезис революции: “Земля - крестьянам,
заводы рабочим”. Называли потому, что лидером революции видели большеви-
ков, считая их одной из социалистических партий. Но чем она была на деле?
Известно, что большевики всегда скрывали профессиональный состав сво-
ей партии. До революции они простодушно называли всех членов партии “орга-
низованными рабочими”, хотя Л.Д. Троцкий полагал, что и большевики, и
меньшевики были чисто интеллигентскими организациями. Так ли это?
Мне пришлось устроить вождям большевиков небольшую перепись: по
Словарю Гранат [Деятели...] в первой сотне биографий оказалось:
145
рабочих (т.е. работавших таковыми более года) 23,
в том числе ушедших в служащие и публицисты 7;
батраков 4, в том числе 1 крестьянин (декханин);
служащих 39 и “интеллигентов” 30й4.
Выходит, что служащих уже до революции было больше, чем рабочих (да
и те редко были кадровыми рабочими). Видимо, Гордин прав: большевиков,
позже ставших известными, до революции объединяло отнюдь не социальное
положение, а способ думать.
После революции народ повалил в правящую партию массой, и масса эта
шла не к станку и не к пашне, а за должностью. Об этом впервые сообщил жур-
нал “Вопросы истории КПСС” (1968, №№ 7, 8, 9; серия публикаций, прерванная
после вторжения советских войск в Чехословакию). Оказывается, осенью 1918 г.
на 14 центральных губерний, где было более 750 крупных заводов, пришлось 3
тыс. партийных ячеек, из которых всего 156 были промышленными (притом ма-
лая часть - заводскими). Остальные были, в основном, учрежденческие и вой-
сковые (притом в большинстве уездов была партъячейка карательного отряда, а
то и две, не считая партьячейки ЧК)*.
Затем оказалось, что в 1918 г. московские наркоматы на 3/4 состояли из
петербургских чиновников [Ирошников, 1974]”.
Я стал искать что-нибудь об их партсоставе. Мне удалось найти всего одну
общую советскую работу - о партпереписи 1922 г. [Струмилин, 1964] и не-
сколько - о губернских организациях, где я и отыскал в хорошо зашифрованном
виде самое удивительное: в “рабоче-крестьянской” партии было около 10% ра-
бочих и крестьян64 65. Важно, что даже в рабочих и крестьянских ячейках рабочие
и крестьяне ничего не решали, поскольку и там 2/3 составляли заводские и воло-
стные чиновники. Но еще важнее, что в партию вступали (а это - способ ду-
мать), чтобы стать служащими: процент рабочих и крестьян по происхождению
втрое и больше превышал их процент по нынешнему занятию.
Революция предстала мне в неожиданном свете: получалось, что в 1917 г.
чиновники при содействии пишущих отняли власть у собственников, что про-
64 Собственно крестьян нет. Женщин три (интеллигентки). Рабочим и сыном рабочего
указали себя пятеро. Характерно, что, несмотря на запрос редакции, многие не указали
своего происхождения и/или дореволюционных занятий.
Всё это - серийные факты в смысле Броделя. Самый же общий для той поры серийный
факт видится в том, что партия большевиков каждый раз почти исчезала там, где теряла
власть, а получив ее снова, возникала заново (не из подпольщиков). - Примеч. 2010 г.
” Т.е. царские чиновники стали зародышем нового правящего класса. - Примеч. 2010 г.
65 По их работе в настоящее время (а вовсе не служащие “из рабочих и крестьян”). Гораздо
ниже был этот процент среди кандидатов: «Не служат (рабочие и крестьяне) - 4%» (Девя-
тый съезд РКП. Протоколы. М., 1960, с. 572). Редактор газеты "Беднота” Я.А. Яковлев при-
водил ходячее мнение: в партию шли, чтобы поправить хозяйство. Он же сообщал, что в
крестьянские ячейки входили волостные чиновники, имевшие надел [Яковлев, 1923].
146
изошла Великая Октябрьская бюрократическая революция. В годы “пере-
стройки” мои попытки (виноват, вялые) донести эту формулировку до интелли-
гентов вызывали у них удивление, но использовать ее в своей деятельности ни-
кто не захотел - неактуально. А зря: вскоре у нас перед глазами развернулась
вторая в XX веке смена власти, и прошла она с чертами дворцового переворота
и народной революции, которые вместе привели к новому расширению власти
бюрократии. Знание истории было бы в августе 1991 г. более чем актуально.
Два исключения заставляют уточнить правило: смена власти в России все-
гда есть бюрократический переворот сверху, но может использовать волну под-
держки снизу. Низы от этого ничего не выигрывают, зато многое теряют.
История смен власти предстает во многом как калейдоскоп, в котором кру-
тятся одни и те же стекляшки, т.е. как комбинация слабо зависимых блоков (се-
рийных фактов). Но во многом так же предстает и история смен форм живого;
там этот феномен именуется блочной эволюцией [Чайковский, 2003, ч. 3]. Эта
параллель неслучайна: формирование разнообразия путем комбинаций сравни-
тельно небольшого числа блоков - одно из основных свойств природы.
Пятая ПМ-помогает увидеть данный факт в самых разных науках. Для нее
важно, что блоки часто появляются задолго до того, как могут эффективно быть
использованы (в биологии это называется: преадаптация). Один из самых ярких
примеров являют “безмозглые глаза” кубомедуз: хотя оптическая конструкция
дает изображение, кубомедузе его нечем анализировать ввиду отсутствия у ме-
дуз соответственной нервной системы [Wehner, 2005]. Параллель с науками о
человеке и обществе может быть здесь весьма поучительна.
Общим для разных наук является и то, что ряды можно выявлять на всех
уровнях - вспомним Броделя. Его много порицали за подмену многогранной ре-
альной истории упрощенными «серийными фактами», хотя он и реальными дан-
ными владел виртуозно. На мой взгляд, выявление серий оправдано, когда исто-
рики ищут возможность диатропического прогноза (см. ниже). Ведь любая нау-
ка дает классификацию и прогноз не фактам многогранной реальности, а лишь
некоторым свойствам некоторой модели, причём этим как раз науки и ценятся.
[-.]
Поле предпочтений
Осталось выстроить общую цепочку познавательных моделей, грубо опи-
сывающую ход научного познания. Это уже делалось для идей экологии [Чай-
ковский, 1991], эволюции в биологии [Чайковский, 1996 а, б] и случайности
[Чайковский, 2004]. Приведу пример в отношении истории.
Старше всего идея “власть от Бога”, и ее выражает нулевая ПМ. Она лежит
в основе идеи наследственной монархии. Очень стара та античная идея, что для
хорошей власти необходимо и достаточно записать хорошие законы, она тоже
выражает первую ПМ. Второй (механической) ПМ-соответствует идея разумно-
го просвещенного правителя. Ее всегда эксплуатировали сторонники бюрокра-
тии, сравнивая государство с машиной. На Западе в XVIII веке утверждалась
147
третья (статистическая) ПМ: разумной считали, вслед за Шарлем Монтескье,
власть, которая опирается на баланс разнородных влияний (разделение властей,
коллективное руководство, выборы). В России данный ход мысли отвергали все,
кроме нескольких людей типа Татищева. В XIX веке бюрократию начали клей-
мить в печати. Критика велась с позиций той же третьей ПМ.
В XX веке пришла четвертая (системная) ПМ, и критика бюрократов по-
шла с другой стороны: они-де неспособны к самоорганизации, на что, по мне-
нию критиков, способны иные виды власти. В конце века пятая ПМ-добавила
требование плюрализма (разнообразия методов и целей управления), на что бю-
рократия тоже, якобы, мало способна. Однако бюрократия благоденствует, грозя
пережить все “более прогрессивные” системы. В чем же дело?
Дело, на мой взгляд, в том, что бюрократия вовсе не является ни продук-
том механистического взгляда на мир, ни механистической природы мира. Ме-
ханистична по своей сути командная система (как в армии), бюрократия же уст-
роена тоньше. Даже в армии на высших уровнях мы видим военный совет - ко-
мандующий обязан иметь при себе совещательный орган. В гражданской же
службе этот принцип всеобщ, и именно его сознательно ввел в 1803 году в рос-
сийскую систему управления Сперанский: начальник принимает решение сам,
но обязан обеспечить проработку вопроса во вверенном ему учреждении (отде-
ле) и выслушать мнения исполнителей задания [Яновский, 1891].
Приняв это к сведению, мы поймем, почему ныне угасла содержательная
критика бюрократии: прежние ПМ-оказались бесполезны, а новой подходящей
ПМ-еще нет. При господстве англоязычной науки иначе и быть не может, по-
скольку ей вопрос неинтересен, так что и выразить нашу нужду ей затрудни-
тельно (англ, слово bureaucracy означает “госслужащие” и не несёт ни характе-
ристики деятельности, ни, тем более, оценочного смысла). Я сам видел, как тол-
ковый американец не мог понять нашего смысла слова “бюрократия”, а потому
не смог и включиться (хотя хотел этого и свободно владел русским языком) в
нашу злободневную “перестроечную” беседу.
Тут-то и подходит шестая (будущая) ПМ: в ее терминах, учреждение фор-
мирует своему начальнику поле предпочтений. Эффективность бюрократии
радикально зависит от того, каким образом набор возможных альтернатив ис-
пользуется в качестве данного поля. Когда бюрократия чувствует себя в безо-
пасности, ее предпочтения направлены к собственной выгоде каждого, но в мо-
менты опасности она совершает чудеса самоорганизации, «плоть до полной
смены идеологии (1918 и 1992 годы).
Пользу от теории принято оценивать ее способностью к предсказанию. Это
не вполне удачно, поскольку оно возможно лишь в нескольких узких областях
науки (даже в небесной механике, где данный взгляд родился, оно возможно не
всюду). Более реалистичной мне видится идея диатропического прогноза [Чай-
ковский, 1991; 2003], который ставит целью лишь выявить возможные варианты,
но не вероятности их реализации. Если говорить о политике, то ученый дает
148
диатропический прогноз, бюрократия делает из него поле предпочтений, а пра-
витель принимает решение. При этом удачный прогноз вовсе не должен сбы-
ваться: наоборот, получив перечень возможностей, правитель должен сделать
всё возможное, чтобы не допустить катастрофических вариантов.
Аналогично, теория эволюции нужна не для фантазий о будущем облике
людей, а для принятия мер к их (и их окружения) сохранению. Если это удастся,
то апокалиптические предсказания, сделанные в видах спасения, не сбудутся. Но
именно для этого и нужно высказать их как прогноз. [...]*
Словом, общего ответа на вопрос об эффективности познания у меня нет,
но метод ПМ-кое-что проясняет: единая ПМ-обеспечивает ее обладателям общ-
ность горизонта познания и тем облегчает понимание текущих проблем.
Сама возможность упорядочения явлений природы и общества посредст-
вом ПМ-вызывает у многих сомнение - вспомним упрёки в адрес Броделя (его
можно назвать основателем исторической диатропики). Такие же упрёки разда-
вались и в адрес Мейена. Он полагал возможность упорядочения явлений путем
выявления параллельных рядов следствием «типологической упорядоченности»
мира и был уверен, что его метод универсален. Метод ПМ-был разработан уже
после его безвременной кончины и представляется мне одним из аспектов диа-
тропики. По-моему, здесь можно говорить о том, что в философии именуется
параллелизмом бытия и мышления**.
Литература
Андреев Ю.В. Тираны и герои. Историческая стилизация в политической практи-
ке старшей тирании // Вестник древней истории, 1999, № 1, с. 3-7.
Баранцев Р.Г. Синергетика в современном естествознании. М., УРСС, 2003. 142 с.
Бергсон А. Творческая эволюция. М., Канон-Пресс, 1998. 383 с.
Блехер Л.И., Любарский Г.Ю. Главный русский спор: от западников и славянофи-
лов до глобализма и Нового Средневековья. М., “Академический проект”, 2003. 605 с.
Бродель Ф. Время мира. Пер. с франц. М., Прогресс, 1992. 679 с.
Бродель Ф. Динамика капитализма. Пер. с фр. Смоленск, Полиграмма, 1993. 124 с.
Водовозов В.В. Бюрократия И Новый энциклопедический словарь Брокгауз-
Ефрон, т. 9, 1912, стлб. 160.
ГлэдДж. Будущая эволюция человека. Евгеника XXI века. М., Захаров, 2005. 169 с.
Гордин Я.А. Меж рабством и свободой. СПб., Лениздат, 1994. 380 с.
Деятели СССР и революционного движения России // Энцикл. словарь Русского
Библиографич. ин-та Гранат, т. 41. М., 1925-1926 (переиздано: М., Советская энцикло-
педия, 1989. 832 с.).
Дживелегов А.К. Бюрократия И Энцикл. словарь Русского Библиографич. ин-та
Гранат, 7-е издание, т. 10. М., 1911, стлб. 423-431.
* Далее приведен пример успешного диатропич. прогноза в политике: в 1938 г. Сталин, по-
лучив прогноз о возможной ведущей роли Польши в будущей войне, поспешил ее уничто-
жить. Прогноз считают неудачным (как несбывшийся), но это нелепо. - Примеч. 2010 г.
*’ См. об этом п. 6 заключительной статьи. - Примеч. 2010 г.
149
Ерошкин Н.П. История государственных учреждений дореволюционной России.
Учебник. М., Высшая школа, 1983. 352 с.
Ирошников М.П. Председатель Совета Народных комиссаров... Л., Наука, 1974.
Карпинская Р.С., Лисеев И К., Огурцов А.Л. Философия природы: коэволюцион-
ная стратегия. М., Наука, 1995. 250 с.
Ключевский В О. Неопубликованные произведения. М., Наука, 1983. 416 с.
Кобляков А. А. Синергетика и творчество // Синергетическая парадигма. М., Про-
гресс-традиция, 2000, с. 308-324.
Лакатос И. Фальсификация и методология научно-исследовательских программ.
М., Медиум, 1995. 236 с.
Любищев А.А. Понятие великого государя и Иван Грозный // Звезда, 1995, № 8, с.
117-156.
Любищев А.А. О природе наследственных факторов / Примеч. и послесл. Ю.В.
Чайковского; послесл. М.Д. Голубовского. Ульяновск, УГПУ, 2004. 211 с.
Нечкина М.В. Петр I // Малая Советская энциклопедия, т. 6. М., 1930.
Милюков П.Н. Очерки по истории русской культуры. Часть 1. СПб., 1900. 239 с.
Милюков П.Н. Из истории русской интеллигенции. СПб., 1903. 308 с.
Платонов С.Ф. Лекции по русской истории. Изд. 5. СПб., 1907. 651 с.
Сватиков С.Г. Общественное движение в России. Ростов н/Д., 1905. 400 + VI с.
Струмилин С.Г. Состав РКП к 1922 г. // Струмилин С.Г. Избранные произведе-
ния. Часть 1. М., 1964.
Тарановский Ф.В. Крестьяне в России до XIX в. // Новый энц. словарь Брокгауз-
Ефрон. Т. 23, Петроград, 1915, стлб. 257-264.
Томашевич О.В. Женщины на престоле Египта. Послесловие к книге: Жак К. Еги-
пет великих фараонов. М., Наука, 1992. 334 с.
Феномен восточного деспотизма. Структура управления и власти / отв. ред. Н.А.
Иванов. М., Восточная литература, 1993. 392 с.
Чайковский Ю.В. Познавательные модели, плюрализм и выживание // Путь. Фи-
лософский журнал. № 1, 1991, с. 62-108.
Чайковский Ю.В. О познавательных моделях // Исследования по математической
биологии. Сб. памяти А.Д. Базыкина. Пущино, 1996 а, с. 170-184.
Чайковский Ю.В. Познавательные модели в эволюции // Педагог-исследователь,
№ 1. Петрозаводск, 1996 б, с. 26-36.
Чайковский Ю.В. Экологический аспект преподавания истории // Философия эко-
логического образования. М., Прогресс-традиция, 2001, с. 385-397.
Чайковский Ю.В. Эволюция. Книга для изучающих и преподающих биологию.
М„ Центр системных исследований, 2003. 475 с.
Чайковский Ю.В. О природе случайности. М., Центр системных иссл., 2004. 278 с.
Яковлев Я. А. Деревня, как она есть. М., 1923.
Яновский А.Е. Бюрократические установления // Энцикл. словарь Брокгауз-
Ефрон, полутом 9, 1891.
Attali J. Les trois mondes. Pour une theorie de I’apres-crise. Paris, 1986.
Bureaucratie // Grand Dictionnaire universel du XIX ciecle / P., Larousse. T. 2, 1866.
Meyer S.C. The Origin of Biological Information and the Higher Taxonomic Catego-
ries // Proceedings of the Biological Society of Washington. August 20, 2005.
Wehner R. Brainless eyes // Nature, 2005, vol. 435, № 7039, p. 157-159.
150
Часть 4. Юбилей Ламарка и Дарвина
Дарвинизм, “Эволюция не по Дарвину” и близкий юбилей *
Предстояший в 2009 году юбилей «Происхождения видов» Ч. Дарвина за-
ставляет вспомнить, чем наука обязана ему, что было известно об эволюции до не-
го и как влияет дарвинизм на эволюционную науку сегодня. Последняя требует
совсем иных идей, и нынешнее возрождение креационизма есть реакция общества
на засилие дарвинизма, остающегося в понятийных рамках столетней давности.
1. В ожидании юбилея
Взяться за эту статью меня побудило обсуждение книги Вадима Ивановича
Назарова [2005] в Институте философии РАН в феврале 2006 года (см. отчет о
нем [В помощь...]). Доклад самого Назарова назывался, как и книга, “Эволюция
не по Дарвину” и излагал новую (и не очень) эволюционную науку.
Кто-то крикнул с места: “Как вы можете спорить с Дарвином, если мы, гео-
логи, находим ископаемые остатки организмов?” А одна из слушательниц Наза-
рова, опытный исследователь англоязычной науки, выслушав доклад, даже вос-
кликнула в недоумении: “Так все-таки, есть эволюция или нет?” Видимо, она
так и не смогла понять, что эволюция - явление природы, тогда как дарвинизм
- одно из его толкований.
Странно - неужели вопрошавшие не слыхали прежде даже о ламаркизме? А
если и не слыхали, то как могли не заметить в докладе? Нет, конечно слыхали,
только в головах не удерживали. Потому и в докладе не заметили. Ведь это так
удобно: принадлежать господствующему учению и считать всех оппонентов
оного учения несуществующими.
Впрочем нет - одну группу оппонентов просто нельзя не заметить, даже
чисто внешне. Они слишком уж заметны, ибо претендуют на господство. Это
сторонники “шести дней творения” в его самой прямой форме. Они, кстати, то-
же делают вид, что эволюция и дарвинизм - одно и то же. Обе команды, так ска-
зать, играют в одни ворота, и нам надо понять - в чьи.
Первый из выступавших по докладу, философ В.Г. Борзенков, начал слова-
ми: “Возможно, от этой книги будут отсчитывать новую эру в учении об эволю-
ции”, а закончил словами: “Автор меня не переубедил, и я остаюсь дарвини-
стом”. Обезопасил себя на все предвидимые случаи.
О содержании книги Борзенков не сказал ничего (неясно даже, открывал ли
он ее), зато отметил, что первый юбилей дарвинизма (1909) прошел “и так, и
эдак”, что второй юбилей (1959) был полным апофеозом учения и что близкий
третий юбилей (2009) тоже, надо полагать, будет апофеозом.
В промежутке Борзенков зачитал длинную выдержку из статьи нынешнего
американского критика дарвинизма. Этот критик, профессор Рик Дейтон, обви-
* Общая и прикладная ценология, 2007, № 2, с. 24-30.
151
няет “эволюционную теорию” (так в США принято именовать дарвинизм) чуть
ли не во всех бедах XX века. Задав себе вопрос - что общего между всеми жес-
токими режимами XIX-XX веков, профессор отвечает:
“Их базовая идеология - философия эволюции! Катастрофические социаль-
ные последствия широко распространившегося дарвинизма оказались ужасны-
ми, почти невообразимыми” [В помощь, с. 139].
В таком негативном отношении к дарвинизму я согласен с критиком, но
нужны две оговорки: 1) виноват не сам дарвинизм (как биологическая идея), а
то мировоззрение, которое породило его и сделало популярным для пяти-шести
поколений читающей публики, в том числе ученых; 2) философия эволюции тут
ни при чем - все остальные, кроме дарвинизма, понимания эволюции отрицают
конструктивную роль вражды.
Как видим, путать дарвинизм и эволюцию ныне вполне принято. Начал это
еще Томас Гексли, младший друг Дарвина: “Либо Дарвин, либо пророк Моисей,
третьего не дано” - говорил он.
Столь же в российской науке принято (вот уже 85 лет, с появления книги
Л.С. Берга “Номогенез”) выражать уважение (а иногда и восхищение) к эволю-
ционным трудам “не по Дарвину” и, ничего в них не опровергнув и даже не ос-
порив, оставаться при господствующем учении.
Но гораздо важнее с апофеозом. Да, первый юбилей прошел “и так, и эдак”.
Значит, Борзенков что-то по истории дарвинизма читал, и тем удивительней его
оценка второго юбилея. Слов нет, в советской науке он был почти полным апо-
феозом^ - но лишь потому, что коммунистическая идеология обязывала сла-
вить Дарвина чуть ли не наравне с классиками марксизма. В около-философской
литературе эта традиция жива поныне. Так, доктор философских наук Э.И. Кол-
чинский (директор Петербургского филиала Института истории естествознания
и техники) пишет: “Юбилей 1959 г. стал годом триумфа дарвинизма”, в под-
тверждение чего ссылается на две американские конференции, и только [Кол-
чинский, 2002, с. 318].
Неловко напоминать, но историк (а в цитированной книге Колчинский вы-
ступал как историк) должен опираться не на торжественные речи, а на моногра-
фии, на весь корпус исследований. В них же всё гораздо сложнее.
Значительные деньги, отпущенные на тот юбилей в разных странах Запада,
позволили учёным впервые провести серьезные исследования старой литерату-
66 Почти, ибо были яркие исключения. Например, фитоценолог В.Н. Сукачев подверг
уничтожающей по сути (хотя и юбилейно корректной по форме) критике всю “теорию
борьбы за существование” Дарвина, противопоставив его “априорным умозаключениям”
эксперименты на высших растениях. Они показали, в частности, что борьба между близ-
кими формами (необходимая в схеме Дарвина) на деле почти отстутствует, и “дарвинов-
ское положение о более напряженной внутривидовой конкуренции не имеет места” [Су-
качев. 1959, с. 38]. Эту работу анализировать не полагается.
152
ры и архивов, и сообщество впервые узнало, что же, собственно, являл собой
дарвинизм в разные периоды. Конечно, во время самого юбилея хвалебных ста-
тей и речей было огромное большинство, но они не были исследованиями, даже
более того: по текстам видно, что никаких исследований дарвинизма авторы тех
выступлений не читали. Если же брать тогдашние документальные изыскания,
то ясно видно совсем иное: западные исследователи пришли в полное изумление
(см. далее).
Если в первые годы дарвинизма споры вокруг него носили, по-
преимуществу, научный характер (участники искали истину сами и пытались
объяснить свои установки оппонентам), то в последние сто лет этого не видно
вовсе. Стороны не только не испытывают желания кому-то что-то объяснить, но
чаще всего даже не желают знать, о чем, собственно, идет речь (что проявилось
и на обсуждении книги Назарова). А со стороны господствующей концепции
вообще налицо одно лишь стремление сохранить собственное господство. При-
меров не перечесть.
2. Вера и наука могут меняться местами
Вот самый из них поразительный. Американский физик-космолог Лоренс
Кросс (L. Krauss) по роду своей работы постоянно сталкивается с новыми идея-
ми об эволюции космоса и вынужден то и дело менять свои взгляды под давле-
нием новых фактов и новых сужений коллег. Он резюмирует:
“Кто-нибудь непременно должен выдвинуть совершенно новую идею.
Трудно предсказать, когда это произойдет. В 1904 г. нельзя было предвидеть,
что годом позже появится замечательная теория Альберта Эйнштейна”. И далее:
“Сегодня наука предлагает нам такие идеи, о которых не додумался бы ни один
фантаст. Я, например, сейчас занимаюсь космической антигравитацией: никто
не понимает, откуда в пустом пространстве может взяться энергия. В мире нет
ничего более сверхъестественного!” [Что беспокоит, с. 33].
Когда Кросса пригласили в Комитет стандартов обучения для обсуждения
программы школьного курса биологии, он решительно встал на защиту теории
эволюции и против идеи ввести курс креационизма. И
«мы выиграли, потому что не допустили теорию разумного замысла в учеб-
ные классы, и вместе с тем проиграли, поскольку “из чувства справедливости”
Комитет добавил... формулировку: “Школьники должны изучить, как ученые
критически исследуют эволюционную теорию”. Я настоятельно выступал про-
тив этого: в нормах должно быть сказано, что ученые вообще всё исследуют
критически!» (там же, с. 34-35).
Не странно ли? Зачем “настоятельно выступать против” - не проще ли было
настоять на действительном включении критического подхода в учебные про-
граммы? Нет, не странно:
«Как я и боялся, принятая формулировка открыла лазейку для креациони-
стов, утверждающих, что есть все основания сомневаться в справедливости эво-
153
люционной теории... Вы только подумайте, одно из занятий в учебном плане
было посвящено резкой критике дарвинизма!» (там же, с. 35).
Вот, оказывается, в чем дело: в своей области можно признавать всё, вплоть
до сверхъестественного, но дарвинизм - это святое, без всяких оговорок и кавы-
чек. Неумение Дейтона и Кросса (странное для профессоров) различить терми-
ны “дарвинизм” и “эволюционная теория” при этом выглядит просто мелочью.
Наши дарвинисты данную пару различают. Однако и у них есть свои высо-
ты, никем более не достигнутые. Об одной стоит рассказать, ибо достигнута она
профессором А.С. Северцовым, много лет возглавляющим главную в России
кафедру дарвинизма (Биофак Московского университета).
Собственный его учебник, много раз изданный, малоинтересен - в нем всё
застыло на уровне полувековой давности и ранее. В этом Северцов близок аме-
риканским дарвинистам. Зато интересен его отклик [Северцов, 2006] (приведен
также в обзоре [В помощь]) на книгу Назарова. То есть сама-то рецензия скучна,
поскольку является попросту пересказом оглавления книги, без попытки что-
либо объяснить читателю. Зато в ней есть 4 выпада против автора (два носят не-
уместно грубый характер), и вот они-то интересны. Не бранью, разумеется, а по-
зицией глухой обороны, занятой дарвинизмом.
1) Назаров привел давно известный экологам факт: в тропиках разнообразие
видов столь велико, что “чаще встретишь любой другой вид, чем вторую особь
того же вида” и сделал из этого вывод (тоже давно обсуждаемый), что внутри-
видовой конкуренции (на которой зиждется эволюционная схема Дарвина) в
тропиках практически нет. Но ведь
“именно в тропиках, как мы увидим в гл. 11, зародились все высшие систе-
матические группы. Вывод очевиден: естественный отбор является не мотором,
а тормозом эволюции” - заключил Назаров.
И рецензент парирует:
“По поводу этого высказывания надо отметить, что, прежде чем расписы-
ваться в собственной некомпетентности, В.И. Назарову стоило бы почитать
учебник экологии Э. Пианки” [Северцов, 2006, с. 312].
Помилуйте, дорогой профессор! Назаров - профессиональный историк нау-
ки, он пишет не по учебникам, он исследует научные труды. И изложил извест-
ную в литературе концепцию. Оспорьте именно ее, если это вам под силу.
2) Вторая грубость рецензента совсем коротка: “автор рецензируемой книги
грубо исказил представления И.И. Шмальгаузена” [Северцов, 2006, с. 316]. В
чем именно состоит искажение, не сказано, и мы остаемся в неведении.
Другие выпады корректны по форме, но не по сути.
3) В отличие от массы других наших дарвинистов, Северцов затронул опы-
ты американского генетика Джона Кэйрнса (1988 г.):
“Изложены выводы Кэйриса (sic!) и других авторов о наследовании благо-
приобретенных свойств у Е. coli, получившие некоторую известность..., но ока-
154
завшиеся слишком вольной интерпретацией экспериментальных данных, о чем
В.И. Назаров не упомянул” [Северцов, 2006, с. 315].
В чем состояла вольность, тоже не сказано, но тут как раз всё вполне ясно,
поскольку мне довелось спросить об этом самого Северцова.
В мае 2003 г. на эволюционном семинаре в МОИП случилось так, что А.С.
Северцов и двое других активных дарвинистов стали забавно убеждать друг
друга в том, что никаких данных о направленной наследственности не сущест-
вует, и при этом (еще более забавно) оперировали только данными 1930-х годов.
На мой вопрос - известно ли беседующим об опытах Кэйрнса, последовало об-
щее молчание. Тогда биохимик А.М. Словников объяснил несведущим суть
опытов и мягко добавил, что знать их необходимо, поскольку они рушат (он
сказал как-то мягче) базу дарвинизма. По наивности я ждал, что дарвинисты хо-
тя бы для виду спросят ссылки, но произошло совсем забавное: все трое взгля-
нули на часы, сказали, что спешат, и тут же исчезли, оставив председателя соб-
рания в полном замешательстве.
Очевидно, что тогда, через 15 лет после бурной дискуссии по опытам
Кэйрнса, Северцов ничего о “слишком вольной интерпретации данных” еще не
знал. Как не знал и самих данных. С тех пор он, видимо, что-то радикальное уз-
нал, и жаль, что он не поделился этой шокирующей новостью на страницах ре-
цензии.
Впрочем, самую шокирующую новость он выдал в конце рецензии.
4) Занятый “эволюцией не по Дарвину”, Назаров, естественно, не ставил це-
лью дать обзор работ дарвинистов, за что и получил упрек от рецензента. Осо-
бенно Северцова огорчило пренебрежение к работам сотрудников его кафедры.
В частности:
“Не упомянуты работы А.Г. Креславского, решившего трудную для селек-
ционизма проблему возникновения новых признаков, отсутствовавших у пред-
ков и потому недоступных отбору” [Северцов, 2006, с. 318].
Уточню: не трудную, а неразрешимую - недоступное отбору отобрано быть
не может, а других ведущих факторов дарвинизм не признаёт. Ни ссылки, ни
объяснения сути “решения” опять нет.
Дорогой профессор! Если ваш сотрудник действительно решил главную
проблему эволюционизма (проблему эмерджентности, то есть рождения новиз-
ны), проблему, которую дарвинизму вечно выставляет креационизм, то бросьте
на время писать скучные рецензии и архаичные учебники, обратите все силы на
пропаганду данного решения. Это же переворот в науке. Он даст главный ко-
зырь ученым в борьбе с мракобесием и воспрепятствует возвращению в школу
средневекового метода понимания мира.
Ну а если серьезно, то налицо лишь прискорбная параллель с Кроссом: кто
не верит в дарвинизм, тот не ученый, а кто верит, тому аргументы не нужны. И
не профессора это придумали. Нет, это извечная позиция всех религий: “Кто
знает истину, тот не доказывает, а кто доказывает, тот ее не знает”. Оба профес-
155
сора исповедуют то, чему их обучили в отрочестве, но Кросса отчасти извиняет
то, что он не биолог.
3. Становление дарвинизма в свете юбилея
Вернемся к прошлому юбилею дарвинизма.
Увы, как часто о Дарвине берутся судить, не прочтя его! И, что еще хуже -
его именем судить других. Сам Дарвин, пусть и не был в рассуждениях безупре-
чен, но всегда искал, как мог, истину и был против обращения его взглядов в дог-
му. Однако догма утвердилась, и до сих пор можно, например, прочесть такое:
«Согласно Дарвину... движущей силой эволюции является естественный от-
бор. Именно он и только он определил те пути, которыми шло развитие органи-
ческого мира от самых простых до самых сложных форм».
Если бы прекрасный генетик С.М. Гершензон, написавший эти слова в 1984
году*, поискал у Дарвина их подтверждения, он, разумеется, ничего не нашел
бы: Дарвин никогда не считал отбор единственным движущим фактором. Под-
робнее см.: [Чайковский, 2003, с. 120].
Почему, однако, столь широко принято апеллировать к Дарвину, не прочтя
его? Ответить просто, вспомнив, что точно так же обходятся и с другими идео-
логами, например, с Карлом Марксом. Если простых ученых, деятелей чисто
когнитивной истории науки, цитируют лишь те, кто ими интересуется, а потому
читал или хотя бы листал, то идеологов поневоле цитируют те, кому их труды
совершенно неинтересны.
Полвека назад юбилейные публикации, основанные на исследованиях, не-
смотря на их уважительный (кроме редких и маловажных исключений) тон и
стиль, начисто разрушили прежний образ Дарвина. Он предстал крупным, но
обычным ученым, продуктом своего века и класса, знатоком в нескольких от-
раслях науки, но редко читавшим литературу иных школ и почти не ходившим в
библиотеку. Главное же - по-новому предстало само учение.
Первым и, по-моему, самым значительным стал фундаментальный труд
шведского дарвиноведа Альвара Эллегора «Дарвин и простой читатель»
[Ellegard, 1958]. Выявив в английских журналах и газетах более трех тысяч ре-
цензий на Дарвина за 1859-1872 годы, Эллегор впервые показал, как пресса
* Цитология и генетика (журнал, Киев), 1984, № 1, с. 72. В этой заметке гл. редактора, за-
вершавшей разгром книги [Кордюм, 1982], разгром, о котором сам В.А. Кордюм писал:
«Принципиальностью здесь и не пахнет», Гершензон поносил его как раз за те результаты и
взгляды, которые сам получал и исповедовал, начиная с 1939 г. Эта тактика (демонстрация
верности официальной догме, выраженная в гротескной форме, призванной показать умно-
му читателю, что написанному верить не надо), оказалась вполне успешной как для Гер-
шензона, так и для Шмальгаузена (о нем см. 4-39). Разница в том, что Шмальгаузен, на-
сколько знаю, никогда никого не громил. Основная часть взглядов Кордюма вскоре стала
общим мнением, а сам Гершензон частично раскрыл свои взгляды (что основную часть из-
менчивости составляет направленная) много позже - в парадной статье [Гершензон, 1996]
по случаю собственного 90-летия. - Примеч. 2010 г.
156
формировала дарвинизм: ученые соглашались с фактом эволюции, а репортеры
и проповедники убеждали, что ей найден механизм - учение об отборе. Что
именно кем признано и что подтверждено фактами, простой читатель, естест-
венно, сам оценить не мог и не хотел. (Поясню: процесс шел приблизительно
так же, как в наши дни, когда идею благотворности "плановой экономики" сме-
няет идея благотворности "рыночной экономики"; на самом деле речь идет о
смене идеологий, имеющих равно малое отношение к реальной экономике, жи-
вущей по своим законам, плохо известным и довольно хорошо скрываемым.)
Вскоре американский дарвиновед Морз Пекхэм представил толстенную
книгу «Происхождение видов Чарлза Дарвина. Меняющийся текст» [The Origin,
1959], которую можно назвать анатомией «Происхождения видов». Фраза за
фразой воспроизведен весь путь книги от первого до последнего варианта. К
сожалению, изучены только лондонские издания, но счастливец, который имеет
возможность положить на стол книгу Пекхэма вместе с американским, немец-
ким и французским изданиями, где была дарвинская правка, а также всю пере-
писку Дарвина, может восстановить ход его мысли предельно полно. Тут-то и
выясняется, как Дарвин далек был от восторга своих почитателей и последова-
телей. В частности, выяснилась полная голословность приведенной цитаты из
Гершензона и ей подобных.
Позже мне удалось, включив в анализ нелондонские издания, показать, что
многие вставки, добавленные Дарвином в книгу, в том числе принципиальные
(например, всё понимание эволюционного прогресса), появились вовсе не в ходе
обстоятельного анализа им фактов и теорий, а в качестве разрозненных эмоцио-
нальных реплик на упреки критиков. Подробнее см. [Чайковский, 1988].
Книга Эллегора задала тон и самому юбилею, и последующим исследовани-
ям. Исследователи поняли, что прежнее голословное восхваление Дарвина и по-
ношение его оппонентов - не наука, что это мало почтенное занятие следует ос-
тавить огромной массе желающих и не обращать на них внимания. Равным об-
разом, ими были оставлены без внимания и голословные ниспровергатели Дар-
вина, включая “научных креационистов”. С тех пор серьезное дарвиноведение
тянется тонкой ниточкой (с заметным утолщением около года малого юбилея в
1984 гЛ7) среди мутного потока восхвалений и ниспровержений.
Выявить серьезные работы очень легко: их выдает научный аппарат (список
литературы, цитаты, численные данные). Столь же легко выявить голословные
публикации основного потока: общим местом в них является утверждение типа
“Дарвин в 1859 г. показал...”, за которым либо не следует ссылки, либо идет
ссылка на последнее издание “Происхождения видов” (1872), хотя у этих изда-
67 Из них нашей темы касаются, например, биография А.Р. Уоллеса [Brooks, 1984], где
показана его определяющая роль в становлении некоторых идей дарвинизма, очерк [11],
освещающий связь идей Дарвина с прежней натурфилософией, обычно отрицаемую, и
очерк о Г. Бронне, первом переводчике Дарвина [10].
157
ний совпадает лишь 1/3 фраз и общие выводы значительно различны (что как
раз и показал Пекхэм). Подробнее см. [Чайковский, 1988].
Юбилей 1959 года помог вспомнить "предшественников Дарвина", т.е. соз-
дателей эволюционизма. Оказалось, что ученые не знали не только тех, кто был
забыт задолго до Дарвина, но и тех, у кого черпал сам Дарвин. Вспомнили, что
лесовод Патрик Мэттью в 1831 г. сформулировал основные положения дарви-
низма. Был заново открыт знакомец молодого Дарвина зоолог Эдвард Блис, пер-
вый биолог, взявший (в 1835 г.) у Мальтуса идею "борьбы за существование" и
сам термин. Оба остались незамечены (см.: [Соболь, 1962]*55), и удивляться не-
чему - общество слышит лишь тех, кого готово слушать, а готово оно было к
1859 году. Вспомнить их важно не в видах защиты приоритета, а для уяснения
того простого факта, что Дарвин ничего не открыл. Он просто выступил тогда,
когда обществу это было нужно.
Парадоксально, но несомненно: без уяснения этого чисто исторического
факта не удается понять и саму нынешнюю концепцию эволюции. Ведь все спо-
ры о приоритете Дарвина были спорами об отборе малых ненаправленных из-
менений и дивергенции (видообразовании путем расхождения признаков). Ни
один эволюционист, видевший в эволюции нечто иное, не считался и не счита-
ется при этом предшественником Дарвина.
Но ведь главными, на кого равнялся (и от кого открещивался) сам Дарвин,
были его дед Эразм Дарвин, его знаменитый предшественник Ламарк и его со-
временник Роберт Чемберс. (Последний, кстати, был, в сущности, заново открыт
тоже в дни юбилея - книгой [Millhauser, 1959].) Они (как и почти все эволюцио-
нисты до рождения дарвинизма) видели эволюцию не как отбор малых вариаций,
а как закономерный процесс, никакого отношения к малым вариациям и отбору не
имеющий. И оказались ненужными обществу - ни при Дарвине, ни через сто лет.
Единственное, что признают за ними - что они отметили факт эволюции.
Это, конечно же, совсем не так. Именно они подробно рассказали обществу
про эволюцию в целом, чего Дарвин так никогда и не сделал. Более того, они
понимали эволюцию так же как мы, а Дарвину было тогда до этого еще далеко.
Его презрительная фраза из “Исторического очерка”: “мой дед Эразм Дарвин
предвосхитил взгляды и ошибочные мнения Ламарка” - хорошо известна, но
куда интереснее его замечание на полях книги Чемберса: “Идея, что рыба пере-
ходит в пресмыкающееся, чудовищна” [Дарвин, 1950, с. 298]. Напомню, что ам-
фибий тогда относили к рептилиям, так что этой репликой Дарвин отрицал эво-
люцию морских позвоночных в наземные*.
68 Где упомянуто 8 англоязычных работ 1958-1959 гг., касающихся идеи отбора до Дар-
вина. Работа С.Л. Соболя не закончена, поэтому Блис упомянут лишь на с. 17 и 22.
* При всей курьёзности данной ремарки (оказывается, «величайший эволюционист», раз-
рабатывая тему происхождения видов, отрицал эволюцию как происхождение классов),
Дарвин был по-своему прав: о превращении рыб в четвероногих серьёзных данных тогда
158
Заметьте: Дарвин сделал эту заметку уже после того, как завершил все свои
эволюционные записные книжки и все свои эволюционные очерки (1835-1844).
Он стоял тогда на позициях естественного богословия, а оно признавало лишь
небольшие приспособления, да и те - под божественным присмотром. Об этом
вспоминать не принято, и тем удивительнее, что советский дарвинист С.Л. Со-
боль осмелился, пусть и вскользь, отметить религиозный характер первой эво-
люционной концепции Дарвина. Сделал он это как раз в юбилейной статье [Со-
боль, 1959].
Соболь привел несколько цитат из молодого Дарвина. Например:
“Если один вид изменяется в другой, то это должно происходить путем
скачка”, чем и объясняется чёткое различие между видами. Виды “сотворены на
определенный срок существования” (с. 25), “Сотворение вовсе не связано с
только приспособленностью животных” (с. 26).
Соболь заключил: “Изложенная первая попытка Дарвина выяснить законо-
мерности эволюционного процесса носит, как мы видим, определенно идеали-
стический характер... а эволюционная концепция, созданная им ранней осенью
1836 г., сильно напоминает теории автогенетиков и мутационистов конца XIX и
первых десятилетий XX в.” (и, добавлю, нынешние). Соболь был уверен, что
молодой Дарвин уже через несколько месяцев смог “нацело отбросить создан-
ную им наспех ошибочную теорию”, что вскоре “пора натурфилософских иска-
ний закончилась и наступила пора серьезной исследовательской деятельности”,
каковую Соболь видел во всех текстах Дарвина, начиная с 1837 года, прямо
ведших к “Происхождению видов”.
Здесь маститый историк явно ошибся. Возможно, он не знал, что в начале
1839 г. Дарвин написал свой первый эволюционный очерк, вполне богословский
по сути (опубликован в [Gruber, 1974]), явившийся комментарием к свежему ес-
тественно-богословскому трактату Дж. Маккалока. Очерк служит естественным
звеном, связующим раннюю “идеалистическую” теорию с хорошо известными
Соболю очерками 1842 и 1844 годов, в которых принято видеть первые набро-
ски теории, легшей в основу “Происхождения видов”. Но и эти два очерка - то-
же богословские по сути. В обоих есть место:
"Предположим теперь, что некое Существо, одаренное проницательностью,
достаточной, чтобы постигать совершенно недоступные для человека различия в
наружной и внутренней организации и предвидением, простирающимся на бу-
дущие века, сохраняло бы с безошибочной заботливостью и отбирало бы для
какой-нибудь цели потомство организма... я не вижу никакой причины, почему
бы оно не могло создать новую расу" (V. Дарвин. Соч., т. 3. М.-Л., 1939, с. 133).
не было. Более того, нынешние данные, хотя и демонстрируют переход рыб в четвероно-
гих, но напрочь отвергают дарвинов механизм (приспособление путем ненаправленной
изменчивости): изменчивость при тетраподизации направленна и являет параллелизмы, а
приспособление обнаруживается лишь в конце процесса (см. 4-67) . — Примеч. 2010 г.
159
Это похоже с виду на дарвинизм, но это пока еще - естественное богосло-
вие: “Существо” явно наделено божественным сверхразумом. Однако дело даже
не столько в сверхразуме (для отбора он не нужен), сколько в малозаметном
словечке "безошибочно": в действительности выгодное качество дает выгоду не
“безошибочно”, а при сочетании (чаще всего редком) выгодных обстоятельств.
В другом месте (с. 86) Дарвин заметил, что «природа не позволяет своей расе
портиться от скрещивания с другой расой, и сельские хозяева знают, как трудно
это предупредить».
Но как раз природа (если она - не Бог) позволяет спариваться всем, кто мо-
жет, и новый вариант не рождается с запретом на встречу со старыми, а значит
не может находить себе пару "безошибочно". Для безошибочности нужно то са-
мое “Существо”. Ко времени написания «Происхождения видов» оно (вместе со
сверхразумом) из записей Дарвина исчезло. Чем “Существо” было заменено?
Исследование данного вопроса приводит к выводу, что обращение к Богу
было заменено у зрелого Дарвина порочным кругом: естественный отбор ос-
тался обоснованным аналогией с искусственным, а искусственный был заново
обоснован по аналогии с естественным [Чайковский, 1983, с. 98-99]. На этом
славном кругу дарвинизм и вертится плотораста лет.
Всех занимал один вопрос: как это можно себе представить? И на него Дар-
вин дал легкомысленный (а потому легко принятый публикой, несмотря на яро-
стное сопротивление всех крупных ученых той поры) ответ - путем отбора ма-
лых вариаций. Превращение рыбы в амфибию так не объяснялось (как не объяс-
нено и поныне [Чайковский, 2003, с. 393]) - ну и не надо.
Устойчивый интерес к естественному богословию сохранился у Дарвина до
старости, и в «Происхождении видов» мы видим сочувственные ссылки на бо-
гословские труды Батлера, Поуэлла, Пэйли, Уэвелла и Шпренгеля.
4. А что тогда было кроме дарвинизма?
Историк Мильтон Мильхаузер, вернувший полузабытого Чемберса в ряд
классиков эволюционизма [Millhauser, 1959], отмечал, что идея эволюции тогда
“носилась в воздухе”, к чему Дарвин не был причастен, поскольку ничего не
публиковал и с докладами не выступал. Вопрос, повторю, не в приоритете, а в
уяснении исторического процесса. Юбилей 1959 года ясно высветил тот про-
цесс, от которого нынешней публике остался известным только труд Дарвина:
если в дни его молодости царило религиозное понимание природы, то к 1859
году оно сменилось на механико-статистическое, и Дарвин йокорно сменил ар-
гументацию. Подробнее см. [Чайковский, 1983; 1984а, с. 177].
Самой же эволюции как процесса у Дарвина не было, и его успех стал воз-
можен лишь потому, что факт эволюции организмов был всем так или иначе из-
вестен, а известен он был от Ламарка, которого тогда все вспоминали, а также от
Чемберса, Бронна и других, об эволюционных трудах которых тогда вдруг все
забыли, хотя эти двое еще активно работали.
160
Немецкий ученый Генрих Георг Брони (1800-1862) был знаменит как зоолог
и палеонтолог, он первый взялся за перевод книги Дарвина, однако вместо эво-
люции он признавал непрерывное творение Богом новых видов и только в ходе
столетнего юбилея дарвинизма был назван эволюционистом. Тогда историк био-
логии Вальтер Барон писал: «Ясно, что слово “творение” означает у Бронна
ничто иное, как образование нового [вида]» и прямо относил его к ранним эво-
люционистам [Baron, 1961]. Исследовав этот вопрос, я пришел к выводу о пол-
ной правоте немецкого историка.
Исследуя нынешних и вымерших животных, Брони выявил конкретные ли-
нии их прогрессивного развития, чем в сущности и заложил основы того, что мы
именуем эволюционным учением. Однако душой он принадлежал к морфологи-
ческой школе Жоржа Кювье, которая порицала саму идею эволюции как нена-
учную. Личный выход он нашел в том, что взялся описать фактический ход эво-
люции, но самый факт порождения одним видом другого отрицал, полагая исто-
рию жизни непрерывной цепью творений. Позицию можно признать вполне на-
учной для того времени (первые прямые доказательства порождения одним ви-
дом другого получены лишь через сто лет [Чайковский, 2003, с. 195, 385]).
Англичанин Роберт Чемберс (1802-1871) был человеком совсем иным - не
ученым, а писателем и издателем. Идею эволюции он воспринял из творений
натурфилософов, в основном германских, и принял ее в самом широком смысле
- от космоса до людских обществ. В своей анонимной книге “Следы естествен-
ной истории творения” (1844) он и изложил ее как единый процесс, причем
впервые собрал вместе и основные данные об эволюции животных и растений, и
единую теорию тех творческих сил, которые эволюцию движут. Развивая Ла-
марка, он признавал две силы: одна ответственна за прогресс (усложнение
строения и жизнедеятельности организмов), а другая - за приспособление к ус-
ловиям существования.
Позже (1858) примерно те же силы признал Брони [Чайковский, 1984, с. 96].
Мы привыкли считать, что представление о постепенном развитии жизни
вошло в науку с Дарвином, но ведь у него нет ни описания истории флор и фаун
(зарождение в море, выход на сушу, появление деревьев и т.д.), ни анализа эво-
люционных изменений какой-либо группы организмов (например, изменения
скелета от рыб к человеку), ни экологического аспекта эволюции (связи разви-
тия жизни с составом атмосферы, животных с растенями и т.п.). Всё это стало
известно от Бронна, Чемберса и многих других до Дарвина.
Идея отбора малых вариаций тоже высказана до Дарвина - во Франции
(Этьен Жоффруа Сент-Илер, 1832), но ее, увы, ни одним фактом до сих пор под-
твердить не удалось. Все “доказательства отбора в природе” суть толкования -
те же факты можно истолковать и иначе, например, “по Ламарку”, полагавшему
основным фактором приспособления активную деятельность каждой особи.
Правда, часто можно услышать, что таким путем нельзя объяснить “пассивные
приспособления” (например, приспособительную окраску), но ее, как отмечал
161
еще Н.И. Вавилов, обычно нельзя объяснить и “по Дарвину”. Подробнее см.
[Чайковский, 2003].
Мы живем в совсем иную эпоху, нежели Дарвин, нам нужна настоящая
(дающая полезные рекомендации) теория эволюции, а не произвольные толко-
вания. Об этом см. подробно [Чайковский, 2003]. Дарвинизм же никаких реко-
мендаций не дает и даже гордится этим (“эволюция непредсказуема”). Поэтому
и из истории нам теперь важно извлечь не то, чем полны учебники дарвинизма,
а то, о чем они вот уже почти полтораста лет умалчивают - что эволюцию изу-
чали до Дарвина и помимо Дарвина, причем находили в ней закономерности.
Эти закономерности ныне прямо-таки бьют в глаза.
Разумеется, близкий юбилей дарвинизма протечет точно так же, как и пре-
дыдущий - почти все будут славить то, что принято славить, не заглядывая в
неприятные тексты, в том числе и в труды Дарвина. Однако в науке никогда не
остаются восхвалители (и редко ниспровергатели) - остаются исследователи.
Литература
1. В помощь аспиранту. Существует ли естественный отбор? (Материалы круглого
стола) // Высшее образование в России, 2006, №№ 7, 8.
2. Дарвин Ч. Избранные письма. М., 1950.
3. Колчинский Э.И. Неокатастрофизм и селекционизм... СПб., Наука, 2002.
4. Назаров В.И. Эволюция не по Дарвину. М., УРСС, 2005.
5. Северцов А.С. Рецензия на книгу: В.И. Назаров. Эволюция не по Дарвину. М., 2005
// Успехи современной биологии. 2006, № 3.
6. Соболь С.Л. Эволюционная концепция Чарлза Дарвина на первых этапах ее форми-
рования И Анналы биологии. Т.1. М., МОИП, 1959.
7. Соболь С.Л. Принцип естественного отбора в работах некоторых английских биоло-
гов... //История биологии, наук, вып. 9. М., 1962.
8. Сукачев В.Н. Новые данные по экспериментальному изучению взаимоотношения
растений// Бюллетень МОИП, Отдел биол., 1959, вып. 4.
9. Чайковский Ю.В. Рождение дарвинизма, 1983 (4-8).
10. Чайковский Ю.В. “Происхождение видов”. Загадки первого перевода, 1984 (4-10).
11. Чайковский Ю.В. Редкое и благородное спокойствие, 1984а (4-9).
12. Чайковский Ю.В. О формировании концепции Ч. Дарвина, 1988 (4-21).
13. Чайковский Ю.В. Эволюция, 2003 (4-51).
14. Что беспокоит физиков? // В мире науки, 2004, №11.
15. Baron W. Zur Stellung von H.G. Bronn in der Geschichte des Evolutionsgedankens //
Sudhoffs Archiv fur Geschichte der Medizin und Naturwissenschaften. Bd 45, Hefte 2. Wies-
baden, 1961.
16. Brooks J.L. Just before the Origin. Alfred Russel Wallace’s theory of evolution. N.Y., 1984.
17. A. Darwin and the general reader. Goteborg, 1958.
18. Gruber G.E., Barrett P.H. Darwin on man. N.Y., 1974.
19. Millhauser M. Just before Darwin. Robert Chambers and “Vestiges”. Middletown, Con-
neticut, 1959.
20. The Origin of species by Charles Darwin. A variorum text edited by Morse Peckham.
Philadelphia, 1959.
162
Пятьсот лет споров об эволюции
Двойной юбилей - 200-летие «Философии зоологии» Жана-Батиста Ламарка и 150-
летие «Происхождения видов» Чарлза Дарвина - хороший повод глянуть на исто-
ргло той идеи, которая в XX в. стала основой научного мировоззрения («Мысль об
эволюции мира - единственное абсолютное, за что еще можно держаться сознани-
ем» [6]), и оценить место, которое занимают в ней эти знаменитые книги.
Вечно актуальная тема
Если не углубляться в Античность и Средневековье, то можно сказать, что
всё началось с Христофора Колумба, который обнаружил в открытой им части
света новую расу людей. Сперва этот факт просто вызывал удивление, но когда
Америго Веспуччи убедил мир в том, что открыт именно новый материк (1503),
возник вопрос: могут ли его жители быть потомками Адама? Те, кто отвечал ут-
вердительно, позже стали именоваться моногенистами. Их позиция служила ба-
зой для защитников прав индейцев на человеческое существование. Их против-
ники - полигенисты, давали аргументацию тем, кто видел в индейцах всего
лишь «говорящий скот». Можно сказать, что моногенизм предварял идею эво-
люции человека. Как видим, эволюционное учение с самого своего появления
было радикально политизировано.
Католическая церковь сразу приступила к насильственному утверждению
«истинной веры» в Америке. Для основной массы миссионеров, далеких от бо-
гословия, но близких к отрядам конкистадоров, туземцы были «порождением
дьявола» [28, с. 29], но для иерархов Европы такая позиция была крайне не-
удобна, ибо лишала церковную идеологию главного стимулирующего тезиса —
обращать людей в истинную веру. И «верховная церковная власть Запада реши-
тельно отвергла возникшее было мнение, будто индейцы - низшие существа.
Было торжественно провозглашено, что они - потомки общечеловеческих пра-
родителей» [28, с. 118] Так гласила папская булла (1512). Словом, католическая
церковь заняла позицию будущего моногенизма.
Гуманно мыслившие миссионеры пользовались этой позицией церковных
властей в своих попытках противостоять чудовищным зверствам конкистадо-
ров. Увы, их усилия приносили лишь незначительный, краткий и локальный ус-
пех. В целом завоевание туземцев носило в Америке истребительный характер.
Он вполне оформился после того, как из Африки стали ввозить негров, которые
оказались более выносливыми, а потому и более удобными в качестве рабов.
Назову только такие знаменитые испанские имена, как историк Хуан Гинес
де Сепульведа (1490-1574) и его страстный противник мексиканский епископ
Бартоломе де Лас Касас (1474-1566) - оба числятся в справочниках как гумани-
сты. Однако первый восклицал: «Что может быть лучше для этих людишек, чем
покориться тем, кто их превосходит? На вполне законных основаниях можно
было бы истребить их, но вследствие свойственных испанцам гуманности и ре-
* Вопросы философии, 2009, № 2. с. 71-85.
163
лигиозности их всего лишь обратили в рабство, позволяющее вырвать их из лап
дьявола» [28, с. 30]. Второй негодовал: «Испанцы чуть что, начинают кричать...
об общении индейцев с дьяволом, дабы очернить их, воображая, что суеверие
этих людей дает больше права их грабить, убивать и угнетать. Это происходит
от великого невежества наших дюдей, ибо они не знают о слепоте, и о заблуж-
дениях, и суевериях, и идолопоклонстве древнего язычества, которого не мино-
вала и Испания» [28, с. 123].
Противоречие гуманности и жестокости породил еще Колумб, грубо подав-
лявший всякое сопротивление индейцев и водивший захваченных пленников по
городам Кастилии напоказ, словно диковинных зверей, но в письмах уверявший:
«Если у них попросить какую-нибудь вещь, они никогда не откажутся ее от-
дать» и даже писавший, что они похожи на «голых испанцев» [28, с. 149]. В ито-
ге их считали то людьми, то скотом, и обе позиции имели своих сильных защит-
ников в литературе.
В Латинской Америке, в полном согласии с моногенизмом, стали преобладать
метисы. Родилась новая этническая общность, которую Симон Боливар (1783-
1830), испанский дворянин и предводитель борцов за освобождение от Испании,
характеризовал так: «Мы не европейцы и не индейцы... Мы пребываем в кон-
фликте с самими собой. Ведь у коренных жителей мы оспариваем право на эту
землю и одновременно мы оспариваем право жить в стране, где мы родились, у
проитвостоящих нам захватчиков» [28, с. 149]. То есть - у испанских войск.
Совсем иным, хоть ничуть не лучшим, было положение индейцев в Север-
ной Америке, где завоевателями были, в основном, протестанты разных конфес-
сий. Они не так яро свирепствовали, истреблению чаще всего предпочитая вы-
теснение с удобных земель, и не стремились к тотальному насаждению христи-
анства. Зато они добились полного господства белой расы. Исключение соста-
вили земли, где колонизаторами были католики - например, Французская Кана-
да. Там «создалось население, до сих пор существующее - канадские метисы»
[12, с. 122].
К какой науке надо отнести анализ метисации людей? С одной стороны, ме-
тисы - новая раса, феномен явно биологический, причем эту расу некоторые ан-
тропологи давно признали [1, с. 284-285]. Параметры новой расы весьма внуши-
тельны: если европеоиды составляют 51% населения обеих Америк, негроиды -
7% и индейцы 5,5%, то метисы и мулаты (итог смешения европеоидов с негрои-
дами) - 36% [21, с. 24]. С другой стороны, фактором этой биологической эво-
люции выступает религия. К подобному «стыку наук» ни биологи, ни гуманита-
рии не готовы. Единственным известным мне исключением является концепция
историка и этнолога Л.Н. Гумилева (1912-1992), к которой мы еще обратимся.
Признание эволюции человеческих рас - торжество идеи моногенизма. Если
процесс един, то уместен вопрос: где он начался? Обычная схема, согласно ко-
торой мы все происходим от кроманьонцев Центральной Европы, казалась мне
очевидной и вполне доказанной фактами, пока я не узнал, что, например, китай-
164
ские антропологи считают ее наивным лепетом, ибо гораздо более древний си-
нантроп («пекинский человек») уже имел характерные монголоидные черты. Но
если каждая раса произошла отдельно, от разных праобезьян, то почему все ра-
сы свободно скрещиваются?
Бегло проследим судьбу эволюционной идеи в христианском мире. Чтобы
не потонуть в необъятном материале, коснемся только ключевых публикаций,
притом появившихся в юбилейные годы, и тех, что тесно с ними связаны.
1509. Европа открывает идею эволюции
В 1509 г., ровно 500 лет назад, в Венеции была впервые напечатана книга
позднеантичного автора, ныне известного как Псевдо-Плутарх. Она именова-
лась «Epitome» (Сокращение) и содержала краткое изложение научных мнений
античных ученых - в основном, о «физике», т.е. о естествознании. (Греческий
оригинал книги с латинским комментарием см. [40]. Там же описаны рукопис-
ная традиция и издания книги.) Эта книга до сих пор служит одним из основных
источников сведений о философии досократиков и ранних стоиков. Ею восполь-
зовался великий Коперник при обосновании идеи движения Земли, а Джордано
Бруно мог черпать из нее сведения о воззрениях античных авторов касательно
многочисленности обитаемых миров.
Есть там и места, которые позже историки науки отнесли к эволюционным.
Таковы фрагменты трудов Анаксимандра, Парменида и Эмпедокла - анализ см.
[34]. Например: «Анаксимандр: из влаги порождены первые животные, объем-
лемые щетинистой оболочкой; выходили же с возрастом превращения на сушу и
по облуплении оболочки жили недолго» [40, с. 430]. Его трактовка появления
жизни в форме брожения или гниения, а также догадка о превращении некоего
рыбообразного животного в человека стали основной построений для многих
мыслителей.
Тогда же (около 1509 г.) Леонардо да Винчи в своих записях обращался к
вопросам, которые мы именуем эволюционными. Первый - о природе ископае-
мых. В конце рукописной книги, состоящей из разрозненных записей (начата в
марте 1508 г.), Леонардо выразил изумление: «сколь много произошло событий
с тех пор, как эта удивительная порода рыб погибла» [22, с. 184]. Трудно ска-
зать, насколько эта реплика свидетельствует о понимании ее автором проблемы
вымирания видов, и одной ее мало для отнесения его к эволюционистам-
биологам. Зато к эволюционистам-геологам историки причисляют его уверенно,
так как в последующих записях Леонардо первый (после античных авторов) по-
нял значение ископаемых остатков животных и растений для воссоздания исто-
рии земных толщ. Это - второй вопрос.
Геология Леонардо изумительна: он не только дал набросок геологической
истории северной Италии и смежных областей, но и высказал прозрения. На-
пример, он был уверен в прежнем прорыве суши там, где ныне Гибралтарский
пролив: «И если мы находим устриц вместе огромными семьями, среди которых
видишь ты таких, у которых створки еще сомкнуты, это значит, что они остав-
165
лены здесь были морем еще живыми, когда прорезан был пролив Гибралтара»
[16, с. 238]. Более того, он счел живой и всю Землю, дав начало тому, что позже
было названо геофизиологией, а в XX веке - концепцией Геи (о ней см. [34]).
Третий вопрос - о единстве строения животных. Признав «человека, первого
среди животных» сходным с обезьянами, он дал себе задание изобразить «ноги
лягушек, которые имеют большое сходство с ногами человека как в костях, так и в
мышцах», а также «ноги медведя и обезьяны и других животных, с тем, чем отли-
чаются они от ноги человека» и «ноги какой-нибудь птицы» [16, с. 258-260].
После смерти Леонардо (1519) его записи ходили по рукам, были предметом
торга и даже краж. Они несомненно сыграли роль в развитии естествознания.
Вскоре Парацельс (1493-1541) писал о сходстве организма и мироздания, Р. Аг-
рикола (1544) - о началах исторической геологии, П. Белон (1555) - о началах
сравнительной анатомии. А затем появился собственно эволюционный труд.
1559. Идея эволюции начинает работать
Первую собственно эволюционную работу опубликовал Иоганнес Бутео, мо-
нах ордена св. Антония и геометр, в 1559 г., ровно за 300 лет до книги Дарвина. В
книге «Логистика», в главе «Ноев ковчег, его форма и вместимость», Бутео вы-
числил, что ковчег не мог вместить все виды известных животных. [37]. Отсюда
следовал тот вывод, что нынешнее разнообразие животных произошло от сравни-
тельно небольшого числа видов. Растения оставались еще вне поля зрения.
Книга Бутео дала спорщикам новый аргумент: изменение видов касается не
только людей, но и всех животных. Анализ проблемы см. [47].
1609. Активность и изменчивость
В этом году Галилей изобрел микроскоп, а Фрэнсис Бэкон издал книгу «De
sapientia veterum» (ее можно перевести и как «О мудрости древних», и как «О
мудрости стариков»). Оба достижения сходны тем, что дали позже эволюцио-
низму главные аргументы, но их авторам это в голову прийти не могло. С мик-
роскопом достижение понятно само по себе: узнав, что каждый организм начи-
нается с одной клетки, естественно допустить, что эволюция началась с одно-
клеточных. Отсюда знаменитая параллель развития зародыша с эволюцией мно-
гоклеточных. Такую же параллель видел и Бэкон, называя древность юностью
человечества, а Новое время - его умудренной опытом старостью. Но главной
для нас мыслью его книги было утверждение о движении и активности как ис-
конных свойствах материи. К собственно эволюции Бэкон,тоже обратился, но
позже: в «Новом органоне» (1620) заметил, что виды могут изменяться путем
накопления «ошибок природы», т.е. случайных изменений. В «Естественной ис-
тории» (1626, посмертно) он пояснял, что сам наблюдал - речные рыбы могут
как ухудшать форму и вес, так и улучшать их. Забегая вперед, замечу, что Ч.
Дарвин видел в такой изменчивости одну из двух форм эволюционной активно-
сти (второй формой было у него стремление к избыточному размножению).
1659. Человек как модель эволюции
В 1657 и 1659 гг. были опубликованы два тома писем покойного Декарта
166
(Париж), а в 1658 г. вышел трактат Т. Гоббса «О человеке» (Лондон). Этих книг
читатели давно ждали, мысли французского вольнодумца были хорошо извест-
ны, но ссылаться на него было опасно. В частности, Декарт объяснял все явле-
ния через движение частиц, и Гоббс, уже хорошо известный философ, писал те-
перь (1658) фактически по Декарту, его не называя: «благодаря движению при
акте совокупления возникает брожение, а затем и расширение сосудов. Оплодо-
творяющая жидкость прорывается внутрь и проникает в матку, где она посред-
ством своего специфического движения формирует из постепенно скопляющей-
ся крови человеческое тело». [9, с. 225].
Поскольку «Письма» Декарта появилсь в печати раньше его основного на-
турфилософского наследия, они поначалу были основным источником знания о
его картине мира. Нам важно, что в Англии нашелся его критик - юрист, финан-
сист и богослов Мэтью Хэйл. Споря с Декартом и Гоббсом и поневоле вторя
обоим, он создал первую концепцию биологической эволюции - происхождение
человеческих рас от единой предковой путем постепенного накопления мелких
изменений (см. [34]). Его «Происхождение человеческого рода» (1677, посмерт-
но) по сути ставило целью обосновать позицию моногенизма. В частности, Хэйл
высказал мысль об индейцах, господствующую ныне - что в Америку люди
пришли из Азии по сухопутному мосту, существовавшему в древности.
1709. Экологический взгляд на мир
У мыслителей Просвещения начала XVIII в. мы находим множество обра-
щений к теме баланса природы и места человека в нем [15, гл. 4]. Из массы
имен, рассмотренных в данной книге, упомяну двоих. Первым (1709) писал об
этом лорд Шефтсбери, философ-моралист. Он уверял (как делал до него и
Хэйл), что бедствия (эпидемии, голод и т.п.) предусмотрены Провидением в ви-
дах ограничения людской численности и что то же самое видно на всех других
организмах. Однако вольный философ шел много дальше царедворца Хэйла, ви-
дя мир неким единством - как рациональным, так и эстетическим. Резко протес-
туя против «войны всех против всех» (тезис Гоббса, позже легший в основу дар-
винизма), он уверял, что ее нет среди животных («волк волку - существо весьма
доброе и любящее»), а среди людей она неуклонно ослабляется культурой. Тут
практика, увы, опровергает моралиста ежечасно, но надо признать, что, в самом
деле, воюют отнюдь не все и не против всех.
Через 10 лет французский историк Жан-Батист Дюбо придал этой линии
мысли эволюционную окраску: люди разных стран различаются в одаренности,
причем разница меняется с поколениями и переселениями. «Во все времена за-
мечалось, что воздействие климата на человека сильнее, чем влияние крови и
происхождения». Он ярко описал (за 200 лет до Шпенглера!) экологическое бед-
ствие большого города. Еще через полвека в Германии, под пером натурфило-
софа И.Г. Гердера (хорошо знавшего данную линию мысли), всё это обратилось
в первую экологическую концепцию эволюции. Она рассмотрена как в книге
[15], так и в моей [34].
167
1759. Эмбриология и эволюция
В классическом труде Каспара Фридриха Вольфа «О зарождении» (1759)
[8], посвященном эмбриологии, нам важен тот тезис, что сходство между вида-
ми животных есть следствие сходства сил, действующих при формировании за-
родышей. Вольф развил тут идею Бэкона. Эволюционную трактовку придал те-
зису Карл Фридрих Кильмейер (1793): «Сила, благодаря которой происходит
развитие индивидуума, тождественна силе, которая производит различные орга-
низации на Земле». Отсюда пошла теория параллелизма зародышей и древних
форм. О ней см. [8].
1809. Рождение ламаркизма
«Философия зоологии» была уже третьим эволюционным трактатом Ламар-
ка. Первые два вышли в 1802 г., так что юбилей ламаркизма следовало отметить
раньше, что мной и было сделано [32]. Эволюционный аспект первых двух книг
состоял в утверждении самой идеи эволюции, а теперь третья обсуждала ее ме-
ханизмы. Здесь Ламарк изложил идеи Эразма Дарвина, положившего в основу
своего учения идею активности живого (Бэкон не назван). К ним Ламарк доба-
вил, что организмам свойственно усложняться (идея прогресса), чем и стал зна-
менит - подробнее см. [34].
Наоборот, мысль о наследовании приобретенных свойств была общеприня-
та и в его время, и позже (включая Ч. Дарвина), вплоть до пошатнувших ее ра-
бот А. Вейсмана, а потому счесть ее основой ламаркизма нельзя. Тем не менее, в
1930-х гг. прочно установилось именно такое его понимание (знаменит вопрос
Н.К. Кольцова: «С чем строить сельское хозяйство - с генетикой или ламаркиз-
мом?» [3, с. 553]).
Точно так же, нет оснований видеть в Ламарке творца первой эволюцион-
ной теории, что делают многие. Так, известный теоретик Грегори Бейтсон писал
40 лет назад (когда симпатия общества к Ламарку проходила через минимум):
«Ламарк (вероятно, величайший биолог в истории) перевернул лестницу объяс-
нения вверх ногами, заявив, что она начинается с инфузории, а изменения ведут
вверх к человеку» [4, с. 146]. Понимая его восхищение, замечу всё же, что «ле-
стницу существ» еще с 1760-х гг. рассматривали как в некотором смысле эво-
люционную Ш. Боннэ, Ж.-Ж.-Р. Робинэ и И.Ф. Гердер. Их эволюционизм, осно-
ванный на идее последовательных переворотов, менявших облик планеты [8],
сильно отличался от принятого ныне, но всё же это был эволюционизм, по-
скольку они признавали преемственность форм жизни. В, остальном Ламарк
следовал за Хэйлом и Э. Дарвином, тоже давшими связные учения. Историк
науки Ч. Гиллиспи (США) писал, что учение Ламарка так же относится к уче-
нию Ч. Дарвина, как диалектика Гегеля к диалектике Маркса [42, с. 268].
1859. Великое событие и его философские оценки
«Происхождение видов» вышло в свет в ноябре 1859 г. и ознаменовало по-
беду эволюционной идеи в биологии: до этого почти все полагали ее лженауч-
ной, теперь же ее стали повсеместно обсуждать, а вскоре и преподавать. Это, на
мой взгляд, необходимо помнить всем охотникам на ведьм.
168
Успех, как многие справедливо пишут, был вызван, прежде всего, тем, что
теперь эволюция толковалась как следствие всем понятной идеи - отбора. Лите-
ратура на данную тему необозрима, мы же сосредоточим внимание на том, что
гораздо менее известно.
1) Тема «Происхождения видов» созрела еще до выхода самой книги: боль-
шую дискуссию и в печати, и в собраниях вызвали журнальные заметки Ч. Дар-
вина и А. Уоллеса (1858) [31], чего со статьями обычно не бывает. Их первый
критик, зоолог Томас Бойд, удивлялся: разве можно строить столь широкую
теорию на столь узком фундаменте, как внутривидовая изменчивость? [36, с.
6357]. Добавим через 150 лет: в самом деле, куда девались у Дарвина все 300-
летние достижения эволюционизма (бегло затронутые выше) - морфологиче-
ские, эмбриологические, физиологические, поведенческие и экологические?
Учение построено без них и часто вопреки им - см. [34]. Пусть экологические
аспекты были чужды Дарвину6^, но и другим аспектам он уделил в книге лишь
по нескольку общих фраз, взятых из тогдашних руководств.
2) Одним из таких руководств была книга знаменитого палеонтолога Жана
Луи Агассица [35], где обстоятельно описан параллелизм зародышей и древних
форм - см. [34, с. 66, 90]. Дарвин в гл. 14 записал: «Так как зародыш часто ука-
зывает нам более или менее полно на строение... предка группы, то мы можем
понять, почему древние вымершие формы так часто похожи во взрослом со-
стоянии на зародыши ныне живущих видов того же самого класса. Агассиц ду-
мает, что это общий закон природы». Несмотря на прямую ссылку, принято ви-
деть здесь предвосхищение или даже открытие «биогенетического» закона. Это
наивно - Дарвин лишь честно повторял. Но важнее, что у него нет и намека на
вопрос - объяснимо ли это отбором?
3) Хотя палеонтологии отведено у Дарвина две главы, аргументов в пользу
учения и там нет, а есть лишь неуверенное допущение, что ее данные не рушат
идею эволюции. У других тогдашних авторов аргументация гораздо конкретней.
В том же 1859 г. вышла «Третичная флора Швейцарии» Освадьда Геера, где эво-
люция прямо описана как быстрая смена одной флоры на другую. В наше время
69 К экологии можно в дарвинизме отнести лишь идею конкуренции. Уже при Дарвине
было отмечено (П.А. Кропоткин и др.), что для эволюции важнее не она, а сотрудничест-
во. В XX в. выяснено, что формирование экосистем противоречит принципу конкуренции
[48]. Ныне можно указать и причину противоречия: дарвинизм исходит из того постула-
та, что новшество формируется путем лучшего размножения каждого следующего вари-
анта, но ни одного примера этому найти не удалось. На самом деле вид начинает запол-
нять экологическую нишу, уже сформировавшись. Поэтому реальная эволюция идет при
радикальном падении численности. Например, решающая стадия эволюции рыб в амфи-
бий описана по шести находкам из региона Зап. Гренландии (за 20 млн лет их эволю-
ции), тогда как исходные и конечные формы находят тысячами в каждом ярусе по всему
миру [34, с. 538]. По единичным костям из Вост. Африки изучают также ранний антропо-
генез. а ископаемых человекообразных обезьян не найдено вообще.
169
такой вывод почти общепризнан в рамках «теории прерывистого равновесия».
4) Многие критики отрицали схему Дарвина на том основании, что отбор не
может поддерживать начальные стадии формирования полезных свойств, ибо
зачатки не могут сразу быть полезными в борьбе за жизнь. Возражение, несмот-
ря на свою очевидность, долго оставалось без ответа, и Уоллес объяснял - по-
чему: оно относилось к весьма общим, а их Уоллес (как и Дарвин) не рассмат-
ривал: «Я напрасно доискивался у своих критиков чего-либо иного, кроме весь-
ма общих возражений» [46, с.415]. Он принимал как «настоящие» только те воз-
ражения, где давался конкретный пример - в нем всегда удавалось найти какую-
то неточность и успокоиться.
Однако в ноябре 1870 г. возражение о зачатках было высказано в самой
конкретной арифметической форме: ботаник Альфред Беннет подсчитал, что
для создания самого простого рисунка на крыльях бабочки, такого, какой можно
счесть приспособлением и тем самым - началом процедуры отбора, нужно, при
случайной изменчивости, как минимум 10 млн поколений. От такого фактора
никакой реальной пользы для эволюции не будет. Вызов был брошен, и Уоллес
его принял. Он вскоре же ответил: «Каждое успешное поколение под влиянием
наследственности получит всё большее преимущество, так что шанс воспроиз-
вести нужную вариацию будет всё выше». На это Беннет резонно возразил, что
Уоллес противоречит своей собственной теории: в нее вводится новый теорети-
ческий принцип - направленная наследственность, действующая до начала от-
бора. Как ни странно. Уоллес торжественно согласился с этой трактовкой: да,
новый фактор есть принцип наследственности (principle of heredity), дарвинизм
не может его игнорировать. Закончил он возгласом, что Беннет может-де не ут-
руждать себя «математической демонстрацией».
Эта дискуссия, шедшая на страницах журнала «Nature» в ноябре-декабре
1870 г., задала рамки всем последующим. Оказывается, арифметику (точный чи-
словой факт) можно отвергнуть презрительным возгласом, и никто из союзни-
ков не укажет на его нелепость. Поэтому противников, пишущих, что «прин-
цип» рушит теорию (в нынешних терминах, он является признанием номогене-
за), можно не слушать, а собственное признание можно забыть - в «Дарвиниз-
ме» Уоллеса (1889) нет ни слова о названной дискуссии. Хотя там есть ссылки
на тот самый номер «Nature», но «принцип наследственности» приписан (в пре-
дисловии) неким критикам дарвинизма, не стоящим внимания. Правда, в гл. 7
последнего издания «Происхождения видов» (1872) появились обширные встав-
ки, обсуждавшие возражения Беннета, но имя его отсутствует, и ответа на его
арифметический аргумент нет. Не появилось его и позже.
5) Естественно, такая иммунность к фактам поставила вопрос о теоретиче-
ском статусе учения Дарвина. Многие критики видели в дарвинизме новую
форму религии - первым, еще весной 1860 г., назвал его «новым Евангелием»
ирландский геолог Семюэл Хоутон [43, с. 226], но о религии поговорим чуть
позже. Опустив крайние мнения (Дарвин - величайший из ученых; учение Дар-
170
вина - вообще не наука [34, с. 93]), приведу один из множества философских
аспектов ранних откликов - тот, который одновременно (весной 1860 г.) выска-
зали Агассиц, отрицавший эволюцию вообще [43, с. 202], и Дж.Г. Луэс (Льюис,
Lewes), сторонник Дарвина [17, с. 286]. Оба отметили, что вид, как и всякий
таксон, - абстрактное понятие и потому в другой вид обратиться не может, что
он неизменен в том смысле, в каком формулируем. Луэс добавил, что вопрос
нуждается в «философском развитии», однако дарвинисты не увидали самого
вопроса, а противники эволюции (например, Хоутон [43, с. 210]) заявили, что
вид в принципе неизменен и что описанная Дарвином и Уоллесом изменчивость
- свойство, которым Бог наделил виды в целях их приспособления к условиям
обитания. (Вопрос решается в рамках номогенеза: вид есть компактная область
в пространстве свойств, и популяция занимает ее сразу, скачком.)
1909. Юбилей дарвинизма и рождение новой религии
В 1909 г. Уильям Бэтсон ввел понятие гена, и мир биологов раскололся на
два лагеря: один уверял, что дарвинизм обрел новый смысл (эволюция - процесс
изменений и новых комбинаций генов под действием отбора), а другой - что в
дарвинизме больше нет нужды, поскольку весь вопрос теперь не в отборе, а в
том, каков механизм появления новых генов (в нынешних терминах: каков ме-
ханизм молекулярного номогенеза). Ныне, через сто лет, почти все принадлежат
к первому лагерю, однако вопрос из второго лагеря остался без ответа: и поныне
дарвинизм умеет описывать только включение, выключение и мелкие модифи-
кации генов, уже существующих, но не умеет описывать появление новых ге-
нов. На это и указывают креационисты.
В юбилейном сборнике (1909) Вейсман заявил: («Мы должны признавать
естественный отбор... потому, что это единственно возможное объяснение» [39,
с. 61]): Интересен ответ Л.С. Берга: «Подобный ход мыслей пресекает дальней-
ший прогресс науки: селекционизм здесь выливается в догму, аналогичную ре-
лигиозным учениям» [5, с. 93]70. Позже дарвинизм аттестовали как религию
многие, но нам важнее то, что в рамках дарвинизма с тех пор «прогресса науки»
действительно не было. О новой религии шла речь и в положительном смысле -
например, в итоговом сборнике работ Фрэнсиса Гальтона, основателя евгеники
(«Очерки евгеники», 1909). Изложенная там программа развития науки включа-
ла, по словам историка генетики В.В. Бабкова, «превращение евгеники в орто-
70 Вскоре Вейсман заверил, что теорию естественного отбора «можно обосновать только
косвенным путем, прямое доказательство невозможно» [7, с. XIII]. Он был прав в том
смысле, что обнаружить отбор как фактор эволюции никому не удалось (попытки обна-
ружения с тех пор почти прекратились, уступив место «косвенным доказательствам», т.е.
произвольным толкованиям). Но еще до написания Вейсманом этих слов удадось найти
прямое опровержение идеи отбора - см. [34, с. 117, 150]. В таком странном состоянии
идея отбора и стала общепризнанной. Опять видим - равнодушие к фактам говорит о
догматической природе учения.
171
доксальный религиозный догмат будущего» [3, с. 19].
Библией этой религии евгеников стала «Генетическая теория естественного от-
бора» Р. Фишера (1930). Ее автор возглавил кафедру евгеники в Лондоне (1933) и
призывал к стерилизации «неполноценных» (которую вскоре развернули герман-
ские фашисты и, увы, не только они, но и ряд либеральных стран Запада [3, с. 483;
34, с. 639]). Как отметил Бабков, «евгеника заменяла им веру» [3, с. 18]. Мне остает-
ся уточнить: в религии обычно есть священная история (миф) и эсхатология (футу-
рология). В новой религии роль первой играет дарвинизм*, а второй - евгеника.
1934. Малый юбилей. Гуманитарии об эволюции
В 1934 г. вышли в свет знаменитые книги «Логика научного исследования»
Карла Поппера и «Исследование истории» Арнольда Тойнби (первый том). Да-
лекая от биологии, книга Поппера содержала, тем не менее, знаменательную
фразу: «Мы выбираем ту теорию, которая наилучшим образом выдерживает
конкуренцию с другими теориями, ту теорию, которая в ходе естественного от-
бора оказывается наиболее пригодной к выживанию» [23, с. 144]. С нее через 40
лет началась эволюционная эпистемология (термин ввел, обсуждая идею Поп-
пера, Д. Кэмпбелл в 1974 г.).
Замечу, что словами «естественный отбор» здесь названо совсем не то, что у
Дарвина - нет речи о преимущественом размножении. Недаром критики дружно
упрекнули Поппера в использовании вольных метафор вместо серьезного анали-
за и в «вопиющем антропоморфизме» [2, с. 708]. В том же, как известно, упре-
кали и Дарвина (например, Ф. Энгельс). Хотя сам Поппер и вспоминал, что пе-
ред написанием книги читал Дарвина и других эволюционистов, но историк
науки Е.А. Аронова, исследовавшая биологический аспект творчества Поппера,
нашла, что в этой его книге «обсуждение биологии... отсутствует полностью»
[2, с. 707]. Тем не менее, такое понимание отбора стало в наше время преобла-
дать, что говорит о фактическом отказе большинства биологов и всех гуманита-
риев от дарвинизма как такового.
В разделе «Мифологемы эволюционизма» статьи Ч-ЗО показан параллелизм структур
мифа и дарвинизма. Миф, как и дарвинизм, 1) равнодушен к противоречию, с ним нельзя
спорить логически (та идея, что вредные черты унаследованы, грубо противоречит идее
отбора); 2) не отличает объект от его знака (сколь угодно слабая выгодность признака
достаточна для его выявления отбором); 3) видит в природе культуру (естественный от-
бор списан с искусственного); 4) вместо объяснения свойств объекта говорит о его про-
исхождении (вся нынешняя систематика); 5) любое столкновение интересов выдает за
смертную вражду (борьба за существование); 6) выявление свойств объекта подает как
его расчленение (организм как состоящий из признаков, кодируемых генами; эволюция
как смена признаков); 7) всё важное помещает в прошлое, в сакральное время (для замет-
ной эволюции требует «миллионы веков», хотя все реально наблюдаемые процессы текут
быстро); 8) враг героя не может быть прав ни в чём (критику дарвинизма даже слушать
не надо). Этот параллелизм представляется мне важным для выявления рефренной струк-
туры общественного сознания,- Примеч. 2010 г.
172
Тойнби был еще дальше от биологии, но все же и он использовал некоторые
биологические понятия - например, «жизненный порыв» А. Бергсона [27, с.
138]. В своей концепции «вызов-и-ответ» Тойнби близко подошел к тому, что
теперь называют (см. [34, с. 272, 446]) ролью стресса в эволюции: «Человек дос-
тигает цивилизации... в ответ на вызов в ситуации особой трудности, вооду-
шевляющей его на беспрецедентное до сих пор усилие» (цит. по [11, с. 153]).
1959. Столетие дарвинизма и рождение новых дисциплин
К этому времени возникли научные дисциплины, осветившие всю пробле-
матику по-новому. Появилась биоценология, и ведущий фитоценолог В.Н. Су-
качев подверг уничтожающей по сути (хотя и юбилейно корректной по форме)
критике всю «теорию борьбы за существование», противопоставив «априорным
умозаключениям» дарвинизма свои эксперименты на высших растениях. Они
показали, в частности, что борьба между близкими формами (необходимая в
схеме Дарвина) на деле почти отстутствует, и «дарвиновское положение о более
напряженной внутривидовой конкуренции не имеет места» [26, с. 38]. Эту рабо-
ту не анализируют уже полвека.
В ходе юбилея само учение Ч. Дарвина оказалось во многом переосмысленн-
ным, поскольку критическое дарвиноведение, почти исчезнувшее со смертью со-
временников Дарвина, теперь как бы родилось вновь. Об этом уже говорилось в
работе [33]. (К сожалению, оно оказалось в советской науке под запретом.)
К юбилею Фишер переиздал «Генетическую теорию...» [41]. Этот странный
анахронизм (евгеника показала свою негодность и была забыта лет на сорок)
стоит упоминания, поскольку побудил биолога и философа А.А. Любищева дать
убийственный анализ позиции Фишера (показавший, что тот не хотел видеть
факты), вылившийся в небольшую книгу. Она ждала издания 25 лет [18], но до
этого ходила в Самиздате, где я и прочел ее. Она оказала большую услугу уче-
ным серьезной постановкой вопроса о связи биологии и истории, показав, что
уверение древних, будто вор рождает вора, а гений - гения (по сути близкое
Фишеру), противоречит фактам.
У Любищева, как и у Гумилева, основным двигателем истории цивилизаций
служит духовная элита общества. Если Тойнби полагал религию фактором, прямо
формирующим цивилизацию, то для Любищева прогресс требует значительной
(пусть и неполной) независимости власти и культуры от церкви. Оба, в отличие
от Гумилева, указывали на этносы, избегавшие упадка, зато обошли вниманием
биологическую сторону (пассионарность), ключевую для Гумилева.
В 1959 г. Гумилев издал 4 статьи по частным деталям востоковедения, а
вскоре - книгу о гуннах (Гумилев Л.Н. Хунну. М., 1960), где эти статьи служат
фрагментами его концепции. В ней он предложил новое видение истории.
Еще в сталинской тюрьме он задумался: как и почему маленький народ ста-
новится великим, а впоследствии перестает быть таковым? Из этих размышле-
173
ний родилось его учение по имени этногенез71. Гумилев первый у нас осмелил-
ся сказать, что эволюция человечества - процесс и социальный, и географиче-
ский, и биологический одновременно.
В 1959 г. Поппер опубликовал английскую версию своей книги 1934 года.
Теперь фраза об отборе теорий, приведенная выше, звучала по-новому, посколь-
ку еще в 1953 г. Поппер увязал воображаемый отбор с реальным понятием ки-
бернетики - методом проб и ошибок. Позже он добавил, что никакого иного
способа эволюции нет - ни в биологии, ни в теории познания [23, с. 260, 268,
406-407, 538]. Сейчас это звучит странно, но вполне отражает тогдашнюю ки-
бернетическую эйфорию.
По иронии судьбы в том же году родилась теория, позже начисто опроки-
нувшая доктрину отбора, понимаемого как метод проб и ошибок: тогда Л.А.
Растригин, один из творцов советской кибернетики, предложил метод случайно-
го поиска. В 1959 г. он получил патент [10, с. 10], а вскоре напечатал статью на
эту тему [24]. С них и началась теория, преобразившая в наше время эволюци-
олнную проблематику.
Метод содержит случайную компоненту, но отличается от проб и ошибок
наличием целевого (оптимизационного) критерия, памяти и многих уровней по-
иска [10, с. 28-29]. Показана невозможность эволюции «по Дарвину» не только
арифметически (что делалось еще при жизни Дарвина), но и функционально: ес-
ли одновременно эволюируют два признака и более, то их случайные вариации,
контролируемые лишь избирательной размножаемостью, вообще ведут не к
приспособлению, а к беспорядочным блужданиям в пространстве признаков.
Для приспособления нужен именно поиск, случайная компонента которого не-
обходима, но подчинена общей задаче.
Идею случайного поиска приложил к анализу эволюции С.Н. Гринченко,
ученик Растригина. Он пишет: «“Селектогенез” и “направленность” эволюции
71 Напомню основную идею Гумилева. Движущей силой этногенеза служит пассионар-
ность Р («способность к целенаправленным сверхнапряжениям») людей и, прежде всего,
вождей. Она противоположна инстинкту' самосохранения I. У пассионариев Р > I, у гармо-
ничных особей Р = /, у субпассионариев Р < I. Пассионарии есть в любом обществе всегда,
но в момент пассионарного толчка они становятся многочисленными и начинают опреде-
лять облик этноса. Он становится агрессивным и, мигрируя и смегйиваясь с иными этноса-
ми, порождает новый этнос или даже суперэтнос. Толчок всегда есть итог массовой мута-
ции. которая происходит под действием космической причины. Эта причина выглядит как
удар хлыста по дуге большого круга (круга на земной поверхности, имеющего центром
центр Земли). Новый этнос восприимчив к новациям. Затем, с падением пассионарности эт-
носа, в нем начинают преобладать гармоничные особи (таковы фазы надлома, инерции, го-
меостаза), а затем субпассионарии (фаза обскурации, т.е. распада этноса; часто она смазана,
а иногда даже сметена приходом и формированием нового этноса на месте старого). Фазу
инерции обычно видят как прогресс, что и произошло в XIX в. с Зап. Европой [11]. До Гу-
милева похоже писали Шпенглер и Тойнби, но без ссылок на биологию.
174
реализуются активными поисковыми процессами на всех уровнях интеграции
живого и “целевым” отбором на основных уровнях его интеграции по критери-
ям энергетического характера. То есть не Дарвиновскую селекцию... а селекцию
результатов адаптивного случайного поиска экстремальных значений всей ие-
рархической совокупности этих энергетических критериев. Другими словами,
отнюдь не по Спенсеровскому критерию “наибольшей приспособленности”, а
по вполне конкретному воплощению Берговского “эндогенного фактора” эво-
люции» [10, с. 35].
Если снять внутренние кавычки, по-моему, излишние, то смысл этой гро-
моздкой цитаты довольно ясен, что я попытаюсь показать далее. Пока же заме-
чу, что мне понять ее было легче, чем другим, потому что сам я пришел к похо-
жим выводам. Мой результат был не так общ, зато более нагляден: популяция
может одновременно улучшать два своих параметра, только если изменчивость
ее особей направленна72.
Осталось почти не замечено рождение панбиогеографии - учения, возво-
дившего увидеть эволюцию. Его автор, биолог Леон Круаза (1894-1982), италья-
нец, осевший в Венесуэле, впервые показал, что эволюция на самом деле проис-
ходит. Обычно в качестве «доказательств эволюции» принято составлять ряды
сходных ископаемых и утверждать, что ряды эволюционны (указывают на род-
ство); но разные ученые составляют их по-разному, а это позволяет креациони-
стам отрицать их эволюционность. Круаза, как позже говорили, «положил фило-
гению на карту», т.е. проследил, как исследуемая форма изменяется со временем
и одновременно распространяется по земной поверхности [38].
Позже такую же работу независимо провел палеоботаник и философ С.В.
Мейен. Оба нашли параллельные направленные ряды - то, что обычно именует-
ся номогенезом [34]. На мой взгляд, только после работ этих двух классиков
эволюционизм стал преображаться из набора догм в науку, имеющую факты.
Именно это надо бы усвоить креационистам и дарвинистам вместо споров друг
с другом.
Круаза сам издал и сам разослал свою увесистую книгу ведущим ученым,
но поначалу (1959) откликнулся только Дж. Симпсон, притом лишь письмом к
72
В дарвинизме рассуждают об отборе признаков в пассивной среде, но реальная среда
активна, ибо состоит из других видов. Поэтому даже простейшая задача адаптации вида к
среде должна быть формализована как игра. Если и вид, и среда могут изменять лишь по
одному параметру, то это - матричная игра 2X2, где каждый игрок - коллектив особей;
если же одновременно могут меняеться по два параметра, то это - матричная игра 3X3.
Если изменчивость ненаправленна, то каждый игрок знает лишь, выгодно ли выполняе-
мое им действие. Тогда процесс поиска оказывается неустойчивым - каждый игрок блу-
ждает далеко от оптимальной стратегии. Решение игры возможно, если игрок, выпол-
няющий невыгодное действие, знает, какое из двух других действий более выгодно [30].
Это и есть направленность изменчивости.
175
автору. В печати Симпсон, как и другие дарвинисты, никогда его не упоминал, а
в частных беседах аттестовал как автора ненаучного и даже экстремиста [44].
С 1959 г. этнолог Клод Леви-Строс читал в Париже лекции по структура-
лизму' (не опубликованы), где соединял подходы гуманитарный и биологический,
структурный и исторический [14]. Выявленные им параллели (например - между
мифами далеких друг от друга народов) оказались идейно близки к тем рядам, ка-
кие выявил в ботанике И.И. Вавилов. О структурализме в эволюции см. [34].
Если идею случайного поиска биологи не заметили, если книги Круаза ос-
тались уделом горстки специалистов, видевших больше, чем принято и удобно,
а структурализм был отвергнут, то концепция Гумилева вот уже полвека устой-
чиво владеет вниманием общества, и впору отметить ее юбилей тоже. Мы кос-
немся ее далее.
1979 Малый юбилей и большие ожидания
Целью моей статьи «Дарвин±120» (Химия и жизнь, 1979, № 12) было сооб-
щить о прежних (до Ч. Дарвина) эволюционистах, практически неизвестных то-
гда русскоязычному читателю, и показать важность их идей для понимания са-
мой эволюции. Ту же цель преследует и нынешняя моя статья, только погруже-
ние в прошлое идет теперь вдвое глубже (не на 240 лет, а на 500), и авторы, в
основном, названы иные.
Тогда же журнал «Природа» (1979, № 9) устроил дискуссию о номогенезе
Л.С. Берга. В ней ботаник А.К. Скворцов, пламенный дарвинист, утверждал, что
Берг не добавил ничего нового к ламаркизму, и был, конечно, прав (Дарвин
также ничего к ламаркизму не добавил, он предложил новое учение). Остальные
три участника говорили как раз о том новом, что нашли у Берга - концепцию,
альтернативную (или дополнительную) дарвинизму* **. Это обычно - каждый ви-
дит свое.
Один из участников, ботаник и философ Ю.А. Урманцев, привел два приме-
ра, когда разнообразие растительных структур (венчиков цветка барбариса и
жилок листа липы) не случайно и не приспособительно, а задается возможно-
стями соответствующих систем [29].
* Структурализм - метод гуманитарных наук, выявляющий ту устойчивую совокупность
отношений между элементами системы (структуру), которая оказывается важнее, чем
природа элементов, из которых система состоит. Пример языковой структуры: система
спряжений глаголов или склонений существительных данного языка. Обычно (в том чис-
ле у Леви-Строса) структурализм противостоит эволюционизму, однако существует по-
нимание эволюции как преобразования одной структуры в другую. Поскольку структуры
устойчивы, преобразование возможно только скачком. - Примеч. 2010 г.
" Ища место Берга в ряду эволюционистов, надо помнить, что он писал после Первой ми-
ровой войны и в брошюре [Берг, 1922] прямо связал своё учение ее опытом. Отрицая борь-
бу как фактор прогресса, он признал ее как фактор стабилизации и даже заявил: «Борьба с
природой и есть тот оселок, на котором должны заостряться способности человека» (с. 17).
Закончил он по Кропоткину: взаимопомощь полезна в борьбе. - Примеч. 2010 г.
176
Недавно канадский ботаник Р. Джан (R.V. Jean)[13] проследил конкрет-
ный номогенетический механизм эволюции растений, и стало понятно, что
Урманце верно объяснял тогда одно из свойств номогенеза - ограниченность
числа вариантов.
Всех участников этой дискуссии, как и ей подобных, объединяло одно -
уверенность, что отбор для эволюции необходим. Но способен ли этот постули-
руемый ими фактор выполнять возложенную на него теоретиками роль? В том
же году мы с Г.Г. Маленковым опубликовали статью [19], где ответ отрицателен
- селективная ценность сама по себе недостаточна для определения судеб обла-
дателей полезных мутаций. Вывод шел вразрез со всей традицией математиче-
ского дарвинизма, уверявшего, что отбор всесилен. Разница в методе - в статье
все допущения сделаны с учетом реальных черт эволюции (а не с целью обосно-
вать априорный тезис). В качестве помощника отбору в [19] упомянут генетиче-
ский поиск - понятие, введенное мною ранее (1976) и являющее собой частный
случай случайного поиска (анализ см. [20]).
Главным же для нашей темы событием того года представляется мне пуб-
ликация основного труда Л.Н. Гумилева [11]. Хотя начальство отказалось его
печатать и книга была лишь депонирована, но о концепции много говорили. Гу-
милева приглашали с лекциями, и на одной из них мне случилось самому его
слушать. Лектор говорил уверенно и отшлифованно, явно рассчитывая на об-
щий восторг, а не на критику. Описав пассионарность по дуге большого круга,
он бросил фразу: «Вы, биофизики, должны найти механизм этого явления». Бу-
дучи по образованию биофизиком, я, когда задавали вопросы, спросил его при-
близительно так: «Ваши примеры касаются довольно мелких событий, а для по-
иска механизма их мало. Можно ли уложить на большой круг, например, Вели-
кую французскую революцию?»
Ответ Гумилева меня поразил - она, мол, не заслуживает включения в число
этногенетических событий, поскольку тогда всего лишь «горстка санкюлотов
выгнала правительство». Сейчас мне понятно: выйдя из заключения, Гумилев
избегал актуальных тем (хотя именно ради актуальных аналогий его и пригла-
шали), и видимо привык осаживать неугодных оппонентов именно так - парали-
зуя их шокирующим тезисом. Мне бы смолчать и обдумать сказанное, но я па-
рировал: две главных революции (французская и русская) проявляют целый ряд
сходств и потому никакая теория не имеет права их игнорировать. То, что Гу-
милев ответил, было столь далеко от науки, что приводить это здесь не следует.
Лишь недавно, лет через 15 после его смерти, мне стало ясно, что тогда он вы-
ражал неприятие вовсе не мне (он не знал обо мне и даже не спросил, кто я), а
структурализму в духе Леви-Строса.
Словом, тогда «Этногенез» Гумилева прошел мимо меня. Прошел, насколь-
ко знаю, и мимо многих. Неприятие своей концепции Гумилев объяснял ее ори-
гинальностью, но вопрос куда сложнее. Давать ей оценку здесь не место, и я ог-
раничусь биологической стороной дела. Метисация - процесс биологический, и
столь же биологическим Гумилев полагал феномен роста пассионарности. Мно-
гократно сославшись на биогеохимическую энергию В.И. Вернадского, он не
177
только нигде не разъяснил ее роли в этногенезе вообще и в росте пассионарно-
сти в частности, но нигде толком не описал сам феномен этногенеза. Его описа-
ния - перечень множества не увязанных друг с другом примеров, никогда не из-
ложенных внятно и очень редко снабженных ссылками на более внятные изло-
жения. Хочется бросить чтение, что многие и делают; но, уверяю, так поступать
не надо. Разберем один пример.
Гумилев опирался на материал по Евразии, а этногенез, шедший в иных
местах, упоминал для сравнения. В своей главной книге он коснулся мимоходом
Африки и Америки в такой реплике: «Протестанты, попавшие в Южную Афри-
ку - голландцы, французские гугеноты и немцы - сложились в этнос, названный
“буры”. Они были наиболее нетерпимы к аборигенам. Рабство в Трансваале (од-
на из бурских республик - Ю. Ч.) было отменено только в 1901 г. А французы на
Гаити обучили негров-рабов французскому языку и католической религии... В
1792 г., когда английский флот блокировал революционную Францию, негры
подняли восстание против французских плантаторов... И убили всех находив-
шихся на острове французов» [11, с. 432].
Что хочет сказать автор и почему этногенез в Африке пояснен восстанием
негров в Америке, непонятно. Но мысль о влиянии религии на этногенез для Гу-
милева столь важна, что приходится вести поиски. В его итоговой книге читаем:
«...а как же Центральная Африка? А она была чужда и той и другой стихии. Доб-
ровольно она не принимала ни протестанство, ни католичество, но зато ислам там
распространялся с потрясающей легкостью, даже без каких-либо насилий... По-
пытка обратить негров в христианскую веру дала результаты райне мрачные. Ко-
гда французы стали заселять Гаити и построили там массу плантаций, великолеп-
ные гасиенды, они привезли туда негров-рабов и обратили их всех в свою католи-
ческую веру... Но случилась французская революция, и тогда негры сразу потре-
бовали, чтобы им дали свободу тоже. Об этом и речи, конечно, французы не до-
пускали. Свобода, равенство и братство были не для негров. Тогда негры восста-
ли, и... все белое французское население было вырезано. Подать помощь из
Франции было невозможно, так как... английский флот не пропускал француз-
ские корабли, и выехать с острова было нельзя. До сих пор там негритянская рес-
публика; официально религия там католическая» [12, с. 258-259].
От новых деталей суть не прояснилась, зато видно, что пояснение Африки
Америкой - не случайность, а теоретическая установка автора. Ищем дальше. В
1894 г. германский этнолог Ф. Ратцель, признав успешное распространение ис-
лама в черной Африке и роль мусульман-метисов, отмечал еще и совсем иное:
возник «новый народ либерийцев, состоящий из настоящих, вернувшихся из
Америки негров». Он напомнил их происхождение. В 1787 г. «английские дру-
зья человечества» купили у местых вождей полуостров Сьерра-Леоне (на западе
Африки) для расселения негров - то были беглые рабы и пленники, отбитые у
работорговцев. В 1820 г. опыт англичан повторили американцы, купившие мыс
Месурадо (к юго-востоку от Сьерра-Леоне) - эта земля и стала основой будущей
Либерии. Любопытно, что либерийцы расширили свою территорию «частью
178
мирным путем, частью с помощью той же грубой силы, какую по отношению к
ним употребляли белые» [25, с. 344, 388]. Следовательно, они в массе были (как
и положено беглецам) достаточно пассионарны. Либерия, республика негров,
существует с 1847 г. поныне, тогда как в Сьерра-Леоне община была вскоре раз-
громлена французами и в 1808 г. стала английской колонией.
Теперь ясно, что имел в виду Гумилев: за счет перемещения и смешения
племен родился «новый народ либерийцев», притом народ агрессивный, а это и
есть этногенез по Гумилеву. Но зачем Гумилев заменил Либерию на Гаити? Да
очень просто: таких действий протестантам, по его схеме, не полагалось, вот и
взят аналогичный пример с католиками^. К тому же в Гаити (в отличие от Ли-
берии) роль мулатов установлена документально: несколько лет (до 1820) там
агрессивной монархии негров противостояла республика мулатов, а восточная
часть острова (Доминиканская республика) состояла тогда (как и состоит ныне)
из мулатов на 3/4.
Итак, чтобы понять один абзац, вполне типичный для Гумилева, мне при-
шлось просмотреть более десятка книг и статей в энциклопедиях, на что цели-
ком ушло два рабочих дня. Читать так целую книгу (а у Гумилева книг много)
невозможно, и встает вопрос: как относиться к его концепции? Мое мнение -
надо пробовать понять каждую его идею и, если она выглядит здравой, то про-
верять каждый его аргумент и искать свои. Пусть связи с «дугой большого кру-
га» мне не видно, пусть в большинстве утверждений и понятий Гумилева мне не
удалось увидать реального смысла, пусть конкретные исторические реалии ока-
залось перепутаны, но рост пассионарности как фактор эволюции этносов пред-
ставляется мне мыслью верной и сходной с идеей роста активности как фактора
биологической эволюции (о нем см. [34]).
2009. Напрасные споры с креационизмом. Новое в теории эволю-
ции
Оценить нынешний юбилей можно будет, когда он закончится, однако глав-
ная его линия и сейчас очевидна: одни будут утверждать, что ничего, кроме
дарвинизма, нет и не нужно, а другие, указывая на ошибки дарвинизма - что
73 На самом деле специфика гаитянской революции была не столько в насаждаемой фран-
цузами религии, сколько в их манере колонизации. В испанской части острова Гаити
рабов было около 15% населения, 75% которого составляли мулаты, а на французской
рабов было 85%, и угнетались они более жестоко, причем мулатов было менее 10%. Вос-
стание началось, когда рабовладельцы решили игнорировать декрет Конвента Франции
об отмене рабства (1791) и пытались занять испанскую часть острова. Франция (вопреки
словам Гумилева) активно помогала им, но вскоре восстание негров повторилось. Уце-
левшие белые покинули остров вместе с католическим префектом (1804), после чего в
испанскую часть острова вторглась уже армия негров. Налицо пассионарный взрыв, хотя
и протекавший гораздо быстрее, чем требует схема Гумилева (видимо, поэтому он о ней
тут и нс сказал; напомню, что он не включал в свою концепцию даже великие револю-
ции). Литературу см. при статьях «Гаити» и «Либерия»: старую - в энциклопедии Брок-
гауза и Ефрона, новую - в Католической энциклопедии (М., 2002, 2005).
179
эволюции нет и быть не может. Это - спор двух религий (как стало мне ясно по
ознакомлении книгой В.В. Бабкова [3]). Участвовать в споре верующих нет
смысла, и вместо этого я закончу статью попыткой кратко ответить на вопрос:
что же появилось принципиально нового за последние полвека? И дарвинисты,
и креационисты об этом молчат.
Главными мне видятся реальное доказательство Круаза и Мейеном факта
эволюции и возврат идеи единства эволюционного процесса, обычной до появ-
ления дарвинизма и забытой с его воцарением. Здесь первой, насколько знаю,
явилась французская книга «Деревья эволюции» [45], где в основу положено
представление о единой фрактальной структуре мироздания. Оно же лежит в
основе моей книги [34], наряду с принципами активности (идеей Бэкона, заяв-
ленной ровно 400 лет назад) и сопряженности. Гринченко тоже кладет в основу
активность, но вместо фрактальности и сопряженности говорит об оптимально-
сти итогов случайного поиска. Активность как фактор эволюции видна под раз-
ными названиями и у других авторов.
Столь же важно понимание различия эволюции и приспособления, приспо-
собления и отбора, а также разъединение понятий отбора и размножения: из то-
го тривиального факта, что неспособное жить не живет, отнюдь не следует, что
избыточность размножения движет эволюцию, как был уверен Дарвин. Основ-
ные приобретения эволюции прошли при дефиците размножения (см., напри-
мер, сноску 1). Это - несомненное свидетельство в пользу номогенеза.
Общий процесс эволюции, единый для всего мироздания, обозначен у
Гринченко как «метаэволюция», а за термином «эволюция» он сохранил более
узкий смысл: «.Метаэволюция (процесс возникновения новых уровней... в
формирующейся иерархической системе) представлена в ней (книге [10] - Ю. Ч.)
как основополагающее свойство Мироздания во всех его проявлениях», так что
«понятие “метаэволюция” необходимо выделить из традиционно используемого
понятия “эволюция” - которое предлагается применять и далее, но относя его к
при этом лишь к приспособительным процессам, происходящим на уже сфор-
мировавшихся уровнях... системной иерархии (обычно “верхних”). Метаэволю-
ция же приспособительным процессом не является, а развивается по своим соб-
ственным законам» [10, с. 2, 8].
Проще говоря, Гринченко (вслед за Ламарком и многими после) различает
прогресс и приспособление, видя первый как создание новых уровней организа-
ции (которую понимает как управление), а второе - как пбиск на уже сущест-
вующем уровне. Лишь второе является для него эволюцией. Место дарвинизма
Гринченко определяет как анализ того единственного уровня природы, на кото-
ром «единицей эволюции является популяция, а переменными - гены» [10, с. 32]
(точнее - включение и выключение уже существующих генов). На этом уровне
ничего нового в принципе появиться не может - вот в дарвинизме ничего и нет,
кроме «кучи подробностей» (слова Френсиса Дарвина, сына Ч. Дарвина - см.
[34, с. 103]).
180
Зато у других направлений налицо явный успех. Ламаркизм и номогенез
(упорно поминаемые в учебниках дарвинизма лишь по работам 70-летней и бо-
лее давности) ушли за последние 30 лет далеко вперед [20; 32]. Так, многократ-
но показан факт направленности наследственных изменений, даны основы для
рациональной систематики [34, с. 217, 231, 603] и выяснены некоторые стороны
антропогенеза [33]. Даются практические рекомендации [34, гл. 12], отчего дар-
винизм уклонялся всегда.
На поставленный в начале статьи вопрос (почему все расы людей скрещи-
ваются) номогенез дает простой ответ: эволюция рас - не итог случайных мута-
ций; она образует параллельные ряды (в смысле Н.И. Вавилова), не выходящие
за рамки вида. Из иных достижений номогенеза укажу упомянутую выше книгу
[13]. Что касается ламаркизма, то теперь нет оснований отделять его от номоге-
неза: как следует из новейших работ биофизиков (В.Л. Воейков, Биофак МГУ;
Эмилио Дель Джудиче, Миланский университет), активность и упорядоченность
порождают друг друга [34].
Заново возник экологический эволюционизм - в частности, в форме этноге-
неза по Гумилеву. Последний требует теперь корректировки: в последние 150
лет с Дальнего Востока на запад движется, начиная с Японии, процесс рождения
активных этносов. Пассионарные взрывы здесь очевидны, и, хотя шли они без
массовой метисации и массовых нашествий чужеземцев, это не повод отрицать
саму идею пассионарного этногенеза. Даже креационизм, и тот иногда проводит
наблюдения и опыты, показывая, например, что наводнение («потоп») может
оставить следы, сходные со свидетельствами о долгой эволюции искпаемых уг-
лей [41а]. Даже будучи неправыми, такого рода креационисты повышают план-
ку эволюционных исследований.
Всё это давно нуждается в философском осмыслении.
Литература
1. Алексеев В.П. Человек. Эволюция и таксономия. М., 1985.
2. Аронова Е. А. Карл Поппер, «наука по Попперу» и дискуссии о ламаркизме в био-
логии // ВИЕТ, 2004, № 2.
3. Бабков В.В. Заря генетики человека. М., 2008.
4. Бейтсон Г. Шаги в направлении экологии разума. Избранные статьи по теории
эволюции и эпистемологии. М., 2005.
5. Берг Л. С. Труды по теории эволюции. Л., 1977.
6. Вавилов С. И. Из дневников 1939-1951 гг. // ВИЕТ, 2004, № 2.
7. Вейсман А. Лекции по эволюционной теории. Т. 1. Петроград, 1918.
8. Гайсинович А.Е. К.Ф. Вольф и учение о развитии организмов. М., 1961.
9. Гоббс Т. Сочинения в двух томах. Т. 1. М., 1989.
10. Гринченко С.Н. Метаэволюция (систем неживой, живой и социально-
технологической природы). М., 2007.
11. Гумилев Л.Н. Этногенез и биосфера Земли (1979). М., 2003.
12. Гумилев Л.Н. Конец и вновь начало. М., 2004.
13. Джан Р.В. Филлотаксис: системное исследование морфогенеза растений / пер. с
англ. М.-Ижевск, 2006.
181
14. Иванов Вяч. Вс. Послесловие // Леви-Строе К. Структурная антропология. М.,
2008.
15. Круть И.В., Забелин ИМ. Очерки истории представлений о взаимоотношении
природы и общества. М., 1988.
16. Леонардо да Винчи. Избранные произведения. Том. 1. М.-Л., 1935.
17. Льюис Г. Очерки животной жизни // Русский вестник, 1860, т. 29, сентябрь.
18. Любищев А.А. Расцвет и упадок цивилизаций (1969). Самара-Ульяновск, 1994.
19. Маленков Г.Г., Чайковский Ю.В. Катастрофический отбор // Генетика, 1979, № 10.
20. Назаров В.И. Эволюция не по Дарвину. М., 2005.
21. Народы мира. Историко-этнографический справочник. М., 1988.
22. Ольшки Л. История научной литературы на новых языках. Т. 1. Сретенск, 2000.
23. Поппер К. Логика и рост научного знания. Избр. работы. М., 1983.
24. Растригин Л.А. Экстремальное регулирование методом случайного поиска // Ав-
томатика и телемеханика, 1960, № 9.
25. Ратцель Ф. Народоведение / пер. со 2-го немец, изд. (1894). Т. 2. СПб., 1901.
26. Сукачев В.Н. Новые данные по экспериментальному изучению взаимоотношения
растений // Бюллетень МОИП, Отдел биол., 1959, вып. 4.
27. Тойнби А. Постижение истории. М., 1991.
28. Три каравеллы на горизонте. К 500-летию открытия Америки. М., 1991.
29. Урманцев Ю.А. Номогенез о сходстве в живой природе // Природа, 1979, № 9, с. 116.
30. Чайковский Ю.В. О непрерывном процессе решения матричной игры, 1971, 4-1.
31. Чайковский Ю.В. Перед выходом «Происхождения видов», 1981, ВИЕТ, Ч-7а.
32. Чайковский Ю.В. Ламаркизм умер. Да здравствует ламаркизм, 2002, ВИЕТ, Ч-46а.
33. Чайковский Ю.В. Философия дарвинизма против..., 2007, 4-62.
34. Чайковский Ю.В. Активный связный мир. 4-63 (АСМ).
35. Agassiz L. Contributions to the natural history of USA: First monograph: Essay on
classification. Boston, 1857. Pt 1.
36. Boyd Th. On the tendency of species to form varieties // Zoologist, 1859, № 1.
37. ButeoJ. Logistica. De quadrature circuli. Lyon, 1559.
38. Croizat L. Panbiogeography, or an introductory synthesis of zoogeography, phytogeogra-
phy and zoology; with notes on evolution, systematics, ecology, antropology etc. Caracas, 1958. 3
volumes.
39. Darwin and modem science. Cambridge, 1909.
40. Diels H. Doxographi graeci. Berlin, 1879 (репринт: 1929).
41. Fisher R.A. The Genetical Theory of Natural Selection. N.Y., 1958.
41a. Gastaldo R.A Debates on autochtonous and allochtonous origin of coal: Empirical
science versus the diluvialists // The Paleontological Society Papers, vol. 5, Octob. 1999.
42. Glass В. (ed.). Forerunners of Darwin. 1745-1859. Baltimore, 1959.
43. Hull D. Darwin and his critics. Cambridge (Mass.), 1973.
44. Nelson G. Review // Systematic Zoology, 1977, vol. 26, pp. 449-452.
45. Nottale L, Chaline J., Grou P. Les arbres de revolution. Univers, vie, sosietes. Paris, 2000.
46. Wallace .R. Contributions to the theory of natural selection. L., 1870.
47. Wichler G. Charles Darwin the founder of evolution and natural selection. Oxford, 1961.
48. Wynne-Edwards V.C. Ecology denies neo-Darwinism // Ecologist, 1991, vol. 21, № 3.
182
Ламарк, Дарвин и устройство науки
К недавнему юбилею «Философии зоологии» и «Происхождения видов»*
Прошедшее в прошлом году с большим размахом по многим странам праздно-
вание 200-летия со дня рождения Чарлза Дарвина огорчило меня отсутствием в пе-
чати всяких упоминаний о другой годовщине, о 200-летии «Философии зоологии»,
которую Жан-Батист Ламарк опубликовал в 1809 г.” (Была, правда, выставка о Ла-
марке в Дарвиновском музее в Москве, и за нее устроителям спасибо.) Как бы ни
относиться к учению Ламарка сейчас и к его месту в нынешней науке, невозможно
отрицать, что в дни Дарвина оно играло огромную роль [4, с. 93], что без него не
понять становление дарвинизма. И в прошлые юбилеи (1909, 1959) Ламарка поми-
нали много. Как говорили древние греки, «даже боги не властны сделать бывшее
небывшим». Впрочем, с той поры этому вместо богов научились люди.
1. Ламарк и Дарвин
Если от Дарвина осталось множество материалов, позволивших историкам
восстановить ход становления его учения, то учение Ламарка выглядит как Аф-
родита, вышедшая взрослой из пены морской. Однако историки давно (еще во
времена Дарвина) заметили, что оно содержанием и построением очень сходно с
«Зоономией» - объемистым трактатом, который написал и трижды (в 1794-1801
гг.) издал Эразм Дарвин, дед Ч. Дарвина. В наше время был найден и посредник,
у которого Ламарк, не читавший по-английски, узнал содержание «Зоономии».
Это был его хороший знакомый Жан Кабанис, врач и общественный деятель,
чью книгу7**, излагавшую, кроме прочего, мысли «Зоономии», Ламарк не раз
цитировал. Подробнее см. [23].
Не избежал влияния деда и внук - свою записную книжку по эволюции (1837)
он озаглавил: «Зоономия». Вскоре он в понимании эволюции с дедом разошелся,
но ряд положений Э. Дарвина присутствует в учениях как Ламарка, так и Ч. Дар-
вина. Поэтому общую схему эволюции, данную Э. Дарвином, стоит изложить.
* Вестник РАН, 2010, № 8.
О книге «Эволюция и систематика: Ламарк и Дарвин в современных исследованиях.
М., КМК, 2009 (Сб-к трудов Зоомузея МГУ, т. 51)» мне стало известно лишь недавно.
74 «Отношения между физической и нравственной природой человека». Первый вариант
вышел в журнале «Memoires de 1’Institut» (1798-99), книга в 1802 [28], а русский перевод
П.А. Бибикова - в 1865 г. Замечу, что Ламарк цитировал Кабаниса по любым темам, кроме
эволюции. Для нас это плагиат, но надо учесть, что другие вели себя сходно. В «Происхож-
дении видов» Ч. Дарвин ограничился небрежной фразой в сноске: «Любопытно, как широ-
ко мой дед Эразм Дарвин предвосхитил взгляды и ошибочные мнения Ламарка в своей
“Зоономии”». В других трудах он цитировал 1-е изд. «Зоономии», где эволюции отведено
всего 10 страниц, но нигде не коснулся их содержания, а иных изданий вообще не называл.
В работе «Жизнь Эразма Дарвина» (1879) он даже не упомянул, что у деда была идея эво-
люции. Примеч. после публикации-, «избегание предтеч» - феномен вполне обычный [4-40].
183
Глава 39 «Зоономии» названа «О порождении» («On generation»), занимает в
третьем издании 10 авт. листов (монография!) и излагает то самое, что мы ныне
называем учением об эволюции. Главное, чем должен бы стать знаменит Э.
Дарвин и чем стал знаменит Ламарк - та идея, что активность особи является
движущей силой эволюции. Активность Эразм понимал не только как поведе-
ние особи, но как всю совокупность текущих в ней процессов, поэтому его мож-
но назвать основателем физиологического эволюционизма, легшего в основу ла-
маркизма. «Три главных предмета желаний, которые изменяют формы большого
числа животных в поисках удовлетворения - половая страсть, голод и самосо-
хранение» (п. IV.8 главы 39).
Идею активности усвоили и Кабанис, и Ламарк, причем Кабанис ее углубил
- он допускал собственное, а не только связанное с внешней средой, развитие:
«Организмы могут к тому же без всякой видимой порчи своей природы подвергать-
ся глубоким изменениям в своих внутренних положениях; приобретать совсем но-
вую способность (aptitude) к новым впечатлениям и движениям» [18]. От слов Ка-
баниса «приобретать совсем новую способность» ведет начало всем известная
идея прогресса. Она едва затронута Э. Дарвином, но стала основной у Ламарка.
А слова: «без всякой видимой порчи» означают, надо полагать, что прогресс может
быть мысленно отделен от приспособления.
Напомню: Ламарк называл прогресс градацией, в противоположность де-
градации, т.е. упрощению организма, которое до него уже признал Жорж Бюф-
фон (полагавший, например, осла деградировавшей лошадью). Ламарк писал:
«Если бы... животные - все и всегда - жили в одинаковом климате, в однород-
ной по составу воде, на одной и той же глубине и т.д. - очевидно, что в органи-
зации этих существ наблюдалась бы правильная и равномерная градация», но на
деле «градация в развитии их организации оказалась во многих случаях иска-
женной до неузнаваемости» [9, с. 114, 115].
Другими словами, Ламарк видел приспособление как набор частных процедур,
нарушающих и маскирующих процедуру прогрессивного развития. Это в корне от-
лично от дарвинизма, видящего прогресс как совокупность приспособлений.
Основным механизмом, делающим активность фактором эволюции, у Эразма
названо упражнение органов (это нынешнее англ, use - disuse): то, что постоянно
упражняется, то развивается, усиливается и наследуется, а остальное пропадает.
Эта мысль проходит через все его труды и является также основной у Ламарка.
А вот в дарвинизме физиологии почти нет, ибо Ч. Дарвин взял из нее только
стремление к избыточному размножению. Зато он, вслед за Ламарком, отводил
(и с годами всё большее) место идее упражнения. В начале XX века она из дар-
винизма выпала, но затем И.И. Шмальгаузен ввел ее вновь в научный оборот:
«Мы видим, что непосредственное модифицирующее влияние измененной
среды, а также “упражнение” и “неупражнение” органов может привести и, ко-
нечно, приводит в новой обстановке к установлению новых, исторически нико-
гда не существовавших форм... Из семейства белок суслики живут на полях и
184
роют норы, белки перешли к лазанью по деревьям, а летяги лазают на деревьях
и перепархивают с ветки на ветку. Различная функция конечностей в этих слу-
чаях явно их преобразует... Функциональные изменения здесь должны играть
ведущую роль [...] Здесь активная работа органа должна была вести к некото-
рым модификационным изменениям, которые определяли затем направление
дальнейшей эволюции» [25, с. 156-157].
Эту ламаркову идею Шмальгаузен включил в дарвинизм следующим обра-
зом: назвал наследование итога упражнения органа его стабилизацией, а свою
концепцию - теорией стабилизирующего отбора (ТСО). Саму стабилизацию
легко наблюдать, но почему и как она отбирается, ни тогда, ни позже объяснено
конкретным материалом не было. Подробнее см. [23, с. 183]*. Позже он никогда
столь прямо об упражнении не писал, и через это умолчание ламаркова основа
ТСО сохранилась в науке до сих пор.
Ее роль в российском варианте дарвинизма радикальна: у ряда авторов она
остается основной связью с дарвинизмом. Вот как, например, выразил свое кре-
до зоолог А.С. Раутиан, известный теоретик дарвинизма:
«1) движущая форма отбора является разрушительным, а не созидающим
фактором; 2) созидательной функцией обладает стабилизирующая (канализи-
рующая) форма отбора... 3) эволюция - творческий процесс, но в основе твор-
чества лежит не пассивный отбор, а активный выбор; 4) наследственность неиз-
бежно порождает направленность и инерционность эволюции...» [14, с. 7-8].
Уже тут видно, что ламаркизм и дарвинизм тесно переплетены. Поскольку
таких связей много, есть нужда ввести для этих учений общее обозначение.
2. Учения эразмовы и бэровы
В религиоведении есть термин «Авраамические религии», которым обозна-
чают иудаизм, христианство и ислам вместе. Общее название подчеркивает поч-
тение всех трех религий к праотцу Аврааму, но важнее, что все они, несмотря на
вечную вражду, исповедуют монотеизм (единобожие) и основаны на общем ми-
фе о творении и о первых веках общественной жизни Палестины. Можно ска-
зать, что на сегодня авраамическими религиями монотеизм почти исчерпывает-
75
ся ; однако кроме них существует много политеистических религии - напри-
мер, индуизм.
Поступим аналогично: концепции, видящие эволюцию как цепь отдельных
удачных изменений в заданной среде обитания, будем именовать «эразмовы».
* Уточню: стабилизация - факт (такой же, как борьба между самцами, повлекшая идею
полового отбора), но связь обоих фактов с отбором ненаправленных наследственных ва-
риаций никем не отмечена. В "опытах по изучению стабилизирующего отбора" (напр.:
Зеликман А.Л. //ЖОБ, 1946, №4; Берг Р.Л. Ботанич. журнал, 1958, № 1) наблюдалась ста-
билизация признаков, а отбор и автономизация додуманы. - Примеч. после публикации.
75 Иногда к нынешнему монотеизму относят еще зороастризм, но в нем два начала - доб-
рое и злое. Полностью монотеизм не выдержан и в христианстве: вспомним хотя бы Бо-
городицу, исполняющую ряд функций женского божества (особенно в народной вере).
185
Хоть они и вечно враждуют, но во всех организм действует в некой среде, к ко-
торой ему надо приспособиться или, если это не удастся, умереть, не оставив
потомства. Рассмотрим их основные общие свойства.
У Э. Дарвина список свойств эволюции выглядит так.
1) Эволюция - очень медленный процесс, и «со дня сотворения» протекли
«миллионы веков». 2) Повышенная активность какого-либо органа вызывает его
усиление и усовершенствование. 3) Активность не означает сознательного по-
ступка; ею обладают и низшие животные и даже растения. 4) Результат измене-
ния, происшедшего в силу активности, наследуется. 5) Эволюционные новшест-
ва образуются путем изменения развития зародыша. 6) Причиной сходства ор-
ганизмов является происхождение от общего предка. 7) Причиной изменения
органов является изменение микроскопических «волокон», из которых они вы-
растают. (В одних группах «волокно» дало пальцы с ногтями, в других - когти,
в третьих - перепонки, в четвертых - копыта.) [30].
Эти положения вызывали и вызывают у читателей восхищение76, независимо
от того, считать ли их верными или нет. Ведь именно благодаря им эволюционизм
начал становиться наукой, тогда как до них он был лишь абстрактной идеей.
Ни ламаркизм, ни дарвинизм не используют по существу пятый тезис Эраз-
ма, поскольку оба оперируют со взрослыми организмами, полагая, что их свой-
ства определяют их эволюцию. Эразмовым учением, ставящим во главу угла
изменение развития зародыша, явилась концепция, которую в 1833 г. предложил
Этьен Жоффруа Сент-Илер. В ней главным процессом эволюции считается мас-
совое изменение хода онтогенеза, происходящее за счет изменений условий
среды обитания особей данного вида [1; 32]. В 1915 г. она получила в русской
литературе имя жоффруизм.
Эта тройка (ламаркизм, дарвинизм и жоффруизм) в целом, на мой взгляд, ис-
черпывает эразмову идею, но не проблематику эволюции. Что есть кроме тройки?
Тезис Ламарка: «потребности животных и употребление частей вызвали
развитие данных частей» [9, с. 187], более известный в наше время в краткой
форме «Функция определяет форму», до сих пор защищаем видными ламарки-
стами - см., например, книгу А.Г. Зусмановского [7, п. 6.1]. Тезис целиком усво-
ен дарвинизмом и, отчасти, жоффруизмом, но вызывает протест у многих уче-
ных. В самом деле, никто не смог указать тех различий функций, которые могли
быть ответственны за различие форм листьев или за различие форм колоний
бактерий. И вряд ли кто-то всерьез думает, что тысяча видов дрозофил имеет
столько же различий в их функциях.
76 Так, биограф Э. Дарвина видел тут гениальное предвосхищение роли ДНК и отметил,
что в 3-м изд. «Зоономии» (его Кабанис и Ламарк не использовали) автор наделил «во-
локна» не только наличными качествами, но и способностью к развитию (formative apti-
tudes) [31, с. 89], т.е. особой формой активности. Ныне можно уточнить эту параллель:
ДНК действительно активно формируется [6; 7; 23; 24] - например, в ходе иммуногенеза,
когда идет быстрое изготовление нового гена.
186
Поэтому в любом поколении биологов есть крупные ученые, полагающие,
что форма организма - это самодовлеющая сущность, к функциям организма
несводимая. Их главный аргумент состоит в том, что разнообразие форм значи-
тельно превышает (например, у семенных растений - в сотни раз) разнообразие
функций. Дисциплина, изучающая законы разнообразия - диатропика (от греч.
«диатропос» - разнообразный) показывает, что разнообразие форм организмов
имеет свои собственные законы, действие которых слабо связано (но связано) с
разнообразием их функций. Подробнее см. [19; 23; 24].
Если так, то должны существовать эволюционные учения, указывающие на
то, чего эразмовы не видят. Наиболее известен среди них номогенез - его созда-
ли в 1920-х гг. зоолог и географ Л.С. Берг и палеонтолог Д.Н. Соболев (у них,
конечно, были предшественники). Номогенез указывает на упорядоченность из-
менчивости: возникает только то, что может у данных организмов возникнуть.
Это исходное эмпирическое обобщение надо пояснить.
Эразмовы учения тоже, разумеется, признают некоторые границы изменчи-
вости. Например: «для того, чтобы изменилась кисточка хвоста, надо иметь
хвост» - любимая фраза дарвиниста Н.В. Тимофеева-Ресовского. Однако номо-
генез имеет в виду не столько ограниченность изменчивости, сколько ее упоря-
доченность. Как кисточки, так и сами хвосты, бывают не какие угодно - орга-
низмы таковы, что их можно классифицировать. Данный факт давно отмечен и
давно выставлен против дарвинизма. Отчего, спрашивали критики еще у самого
Ч. Дарвина, итогом эволюции является не каша форм, а дискретный набор ви-
дов, допускающий построение систематики?
Дарвин ничего ответить не смог (точнее, он ответил, что промежуточные
формы вымерли, но это - лишь пересказ вопроса; а именно, почему сохраняются
лишь дискретные формы?), не смогли и его последователи. В эразмовых учени-
ях зияет на сей счет провал - для Ламарка и Дарвина изменчивость организмов
ничем, кроме выживаемости вариантов, не ограничена. Что касается Жоффруа,
то он упорядоченность изменчивости видел, но в эволюционную схему не
включил, о чем скажем далее.
Палеоботаник С.В. Мейен, придавший в 1970-х гг. номогенезу его нынешний
вид [12], называл эту упорядоченность рефренной структурой разнообразия
(рефрен - это ряд направленных рядов изменчивости*), но оно, как слишком гро-
моздкое, не прижилось, и удобнее оказалось краткое ее название - диасеть (диа-
тропическая сеть) [23; 24]. Самым наглядным примером диасети служит периоди-
ческая таблица Менделеева, демонстрирующая структуру разнообразия химиче-
ских элементов: промежуточных форм элементов (кроме тех, что в таблице) не
бывает, зато между строками таблицы виден параллелизм.
Суть диасети была пояснена мною в «Вестнике» еще в 1986 г. на примере
сходства двух разнообразий - конечностей позвоночных и грамматических форм.
Точнее см. далее, часть 9, статью «Диатропика С.В. Мейена...». - Примеч. 2010 г.
187
Статья начиналась словами: «В языковом словаре каждое слово дано один раз, с
указанием, если нужно, типа его изменения. Например, существительное дается в
именительном падеже единственного числа с указанием типа склонения, а осталь-
ные варианты (словоформы) всякий, кто знает грамматику, восстановит сам.
Пусть у правил грамматики много исключений, но все же их запоминание проще,
чем запоминание каждой словоформы в качестве отдельного слова. Г. Гельмгольц
ставил словарь и грамматику в пример “естественноисторической системе”, видя
в них принципиальное сходство, но следует признать, что и сейчас биологические
“словари” (систематические сводки) сильно отстают от языковых» [18].
Фрагменты диасети приведены в книгах [19; 23]. Среди них - данный самим
Мейеном (структура разнообразия форм листьев). Сами они рисуют лишь упо-
рядоченность разнообразий и лишены эволюционного смысла, но дают понять
ту мысль Мейена, что эволюцию видов можно выразить как заполнение клеток
огромной диатропической таблицы. Она существует объективно и так же не за-
висит от наших знаний, как таблица Менделеева. Разница в том, что последнюю
мы видим целиком, а биологическую диасеть - лишь фрагментами. Кстати, эво-
люцию химических элементов тоже представляют как заполнение (в ходе эво-
люции Вселенной) менделеевской таблицы.
Первой попыткой выявить участок диасети организмов мне видится трактат
Жоффруа «Философия анатомии» (1818), где даны ряды сходств между скеле-
тами позвоночных - они приведены в табл. 1-Х книги И.Е. Амлинского [1]. Од-
нако в трактате нет эволюционной идеи (ее Жоффруа высказал позже, и как раз
при этом ничего похожего на тему диасети не высказал).
Кроме невнимания к порядку в изменчивости, в Эразмовых учениях зияет
провалом, как ни странно, пренебрежение к процедуре приспособления. Все они
повествуют о приспособлении данного вида к его заданной среде обитания, хотя
заданными можно полагать климат или соленость воды, но никак не свойства
окружающих видов, поскольку они могут меняться быстрее, чем он сам.
Простой пример: основной для данного вида источник питания (он тоже -
вид) может исчезнуть быстро, тогда как приспособление к новому источнику -
эволюционный процесс, требующий много времени. Это значит, что приспособ-
ление видов возможно только совместное, что оно - не поиск максимума некой
функции (как полагает, например, ТСО), а своего рода игра. Однако в теории
игр давно установлен простой математический факт: поиск решения даже про-
стейшей (матричной) игры может быть успешен только тогда, когда каждый иг-
рок в каждый момент знает, какое действие более выгодно [17], т.е. когда игро-
ки действуют целесообразно. Наделены ли виды таким свойством?
Хотя принято отвечать отрицательно, но на самом деле эразмовы учения не-
явно вводят целесообразность как первичное частное свойство: в ламаркизме
это - наследование полезного итога упражнения, в жоффруизме - изменение он-
тогенеза под воздействием среды («зародышевая изобретательность» [13, с.
195]), а в дарвинизме - «принцип наследственности» [23, с. 111]. На неявную
188
роль первичной целесообразности в дарвинизме впервые обратил внимание
Карл Бэр в 1873 г. [2, с. 70, 133], а позже Л.С. Берг согласился с ним и затем
ввел явным образом целесообразность живых систем как их общее первичное
свойство, ни из чего не выводимое [2, с. 101, 117].
Приведенный выше математический факт биологам неизвестен, однако ин-
туитивно многие из них чувствуют, что во взаимном приспособлении есть некая
трудность. Учением, обращенным к ней, является экосистемная концепция эво-
люции (ЭКЭ) [10; 13]. Она, как и номогенез, исходит из той идеи, что вид, воз-
никая, занимает некоторое место в пространстве свойств, именуемое здесь эко-
логической нишей. Основным путем эволюции ЭКЭ признает занятие пустую-
щей ниши, которая именуется лицензией. Напомню, что в дарвинизме, наоборот,
основным путем эволюции названо вытеснение новым видом прежнего из его
уже занятой ниши.
Сам факт признания лицензии как общего для экосистем феномена роднит
ЭКЭ с номогенезом. В частности, экосистема понимается как совокупность
ниш, некоторые из которых в данный момент пусты (то же мы видим в диасети).
Установлено, что ниши образуют (точно так же, как формы организмов) парал-
лельные ряды [10].
Бэр, один из главных защитников первичной целесообразности живого77 78,
всю жизнь размышлял об эволюции. Своей концепции не построил («ибо слиш-
ком хорошо знал биологию», как сказала мне в 1980 г. Майе Реммель, эстонская
исследовательница творчества Бэра). Зато он оказался одним из основателей как
диатропики [19], так и экологического взгляда на мир [5].
Поэтому естественно сказать, что номогенез и ЭКЭ (учения, противостоя-
щие эразмову пониманию эволюции ) - это бэровы учения. Два их главных от-
личия от эразмовых - признание первичной целесообразности живого и упоря-
доченности вариантов изменчивости - в качестве двух самостоятельных свойств
природы, в качестве ни из чего не выводимых сущностей.
3. Место отбора
Третье отличие бэровых учений от эразмовых видится в стремлении исхо-
дить, по возможности, из эмпирических обобщений. Сам Бэр отверг идею отбо-
ра за ее априорность, ничем тогда не подкрепленную. Еще до выступления Дар-
вина, в 1850 г., он выставил ее в смешном виде (животные имеют нужные свой-
ства потому, что не имеющих не существует7#) и отверг без серьезного анализа
77 Ее горячо отстаивал, ссылаясь в том числе и на Бэра, историк науки В.И. Назаров [13].
Общность номогенеза и ЭКЭ в том аспекте, что «поведение частей определяется зако-
нами целого», подчёркивал В.В. Жерихин [2003, с. 410]. - Примеч. после публикации.
78 Вольно говоря: Бэр отверг здесь биологический аспект «антропного принципа», сфор-
мулированного в XX веке. (Примеч. после публикации статьи: Антропный принцип гла-
сит: первичная согласованность свойств природы - например, мировых констант - за-
ставляет предположить, что мир уже при начале был задан таким, чтобы в нем мог суще-
189
[3, с. 434]. Это обычно для Бэра - так же легко и хлёстко он в 1828 г. разделался
с принципом упражнения-неупражнения [4, с. 30], тоже априорным. Вместо них
он указал факты быстрого, за 350 лет, одичания домашних животных, завезен-
ных в Америку [3, с. 452] (замечу, что столь быстрая эволюция противоречит
эразмовой идее «миллионов веков») и свой знаменитый принцип «идущей впе-
ред победы духа над материей» (1834) [4, с. 120]. Последний был для Бэра тоже
эмпирическим обобщением, но это понимание можно оспорить.
Путь эмпирических обобщений, трудный сам по себе, осложняется еще тем,
что сообщество легко принимает за них абстрактные аксиомы. Так, царит убеж-
дение, что Дарвин обосновал идею естественного отбора фактами. На самом де-
ле он ввел отбор как аксиому, и у него есть лишь два примера, ее пояснявших,
которые к тому же он сам назвал воображаемыми. Он признавал, что реальные
примеры следует искать, но сам их не нашел - наоборот, указал несколько
свойств (например, у цветков), которые не могут быть итогом отбора. Не появи-
лось доказательств отбора и позже.
Бэр же показал полную необоснованность идеи конкуренции как механизма
отбора79. Подробнее об отборе см. [13; 23; 24]. В основе учения Ч. Дарвина ле-
жит эразмова идея, из которой он пытался изъять принцип активности и заме-
нить его принципом отбора как ведущего фактора эволюции. Обнаружить этот
фактор в природе не удалось, тем не менее, учение утвердилось в науке. Как это
смогло произойти?
4. О структуре науки и парадигмах
Мейен любил говорить (не знаю, писал ли), что назначение методологов - не
столько научить ученого правильно работать, сколько «показать ему, что он, соб-
ственно говоря, делает». Ныне эта мысль стала привычной, входит в руководства
[16, с. 307], и мне представляется, что без нее теорию эволюции не построить.
Согласно теории развития науки Томаса Куна (1962), система принятых
обществом постулатов образует основу нормальной науки, тогда как на иных
постулатах базируются иные варианты науки - «экстраординарные» (марги-
нальные). Система постулатов данной отрасли нормальной науки образует ее
парадигму, которая, тем самым, в данной отрасли всегда одна. Когда нормаль-
ная наука перестает удовлетворять запросам общества, происходит, по Куну,
научная революция, причем новые постулаты берутся из маргинальных концеп-
ций. Поэтому, замечу, полное отсутствие изложения маргинальных положений в
преподавании является недальновидым.
ствовать наблюдатель, т.е. разумный человек. Не раз отмечено, что возможна и такая ин-
терпретация: мы существуем потому, что миров, где нас не может быть, не существует.
Не раз отмечена и идейная близость данной интерпретации дарвинизму - см. АСМ.)
79 Как показала Майе Реммель (Вальт) [5], высокая плодовитость, по Бэру, это не итог
победы в конкуренции, а требование экосистемы - плодовиты те, чья молодь служит пи-
щей многим. Гибнут же почти исключительно незрелые и престарелые особи, а вовсе не
худшие варианты.
190
Важно, что разница в статусе у бывшей и будущей парадигм радикальна:
первую обычно чтут как феномен истории науки, тогда как вторую чаще всего
преследуют как лженауку. Опыт обращения прежних «лженаук» в парадигмы,
увы, ничему не учит. Идея эволюции в любых ее формах (даже как простая до-
гадка о возможности появления новых видов) до победы дарвинизма восприни-
малась обществом как лженаука, и то же самое было с атомизмом, гипнозом и
многими другими концепциями. Это надо бы помнить всем «охотникам на
ведьм», однако не помнит этого почти никто*.
Из гражданской истории известно, что по свершении революции происхо-
дит переписывание курсов истории, и Кун отметил, что в истории науки
обычно то же самое. Он был далек от биологии, но и в ней видна та же зако-
номерность, а это говорит о важной роли теории Куна для науки вообще. Уход
Ламарка из поля зрения нынешних авторов есть самый заметный пример тако-
го переписывания^0.
Однако хуже то, что переписывают вклад Ч. Дарвина. В статье [21] говори-
лось о совсем невероятных воззрениях, какие приписывают ему нынешние уче-
ные. Теперь могу привести новое обстоятельство: утвердился термин «положи-
тельный (дарвинов) отбор», которым именуют фактор, по сути чисто номогене-
тический - сборку целостных систем, притом безо всякой смены поколений
(подробнее см. [24]). Это обращение Дарвина в номогенетика поначалу кажется
забавным, но смеяться вовсе не следует: опыт показывает, что расширительное
толкование классика (теми, кто не читал его трудов) препятствует развитию
науки, ибо новое объявляется давно известным, а потому не требующим иссле-
дования и уж совсем ненужным при обучении.
Подробно см. статью «Многотрудный поиск...» в наст. Сборнике. В наши дни ту же за-
бывчивость проявляет «Комиссия РАН по борьбе с лженаукой». Попытка понять, что же
имеют в виду под лженаукой ее гонители, привела меня к двум выводам. 1) Что касается
борьбы с идеями, то, что бы ни говорили гонители в свою защиту, на самом деле они пы-
таются изгнать тот пласт мысли, который инквизиторы именовали ересью. К счастью,
целиком это никогда не удавалось. 2) Что касается борьбы с мошенниками, то их-то как
раз нельзя считать лжеучёными с позиции Комиссии потому, что они действуют именно
в указанных ею рамках (имеют все нужные документы). - Примеч. после публикации.
ол
° Мне уже случилось писать, что из англоязычных энциклопедий давно ушло изложе-
ние сути ламаркизма (там его трактуют лишь как наследование приобретенных призна-
ков), но что русскоязычные энциклопедии сохраняют более высокий культурный уро-
вень [21]. Теперь, увы, не так - в переводном издании [27] вообще нет статьи «Ламар-
кизм», а в статье «Ламарк» с изумлением читаем лишь одно: «Представление Линнея о
наследовании приобретенных признаков (ламаркизм) было в 1930-х опровергнуто
большинством генетиков» (что неверно [6; 13; 23]). При этом в [27] даны ссылки на 3
статьи той же энциклопедии - «Ч. Дарвин», «Дарвинизм» и «Лысенко». В них о Ламар-
ке и ламаркизме нет ни слова, зато в статье «Лысенко» (с портретом, хотя портретов Ч.
Дарвина и Ламарка нет) ему приписано изобретение травопольного севооборота (что, к
счастью, редактор перевода тут же опроверг).
191
Здесь самым ярким является очень старый (но актуальный) пример - судьба
ТСО. Термин «стабилизирующий отбор» на Западе не прижился, а при обуче-
нии российских студентов он вводится как безобидная и очевидная частность -
как сохранение нормальных свойств организмов посредством преимуществен-
ного истребления носителей уклонений от нормы. Но затем он используется со-
всем иначе, что мы видели на примере доклада [14], - для обозначения некоего
универсального механизма, преобразующего цельные системы. Данного меха-
низма никто даже не пробовал наблюдать. Поскольку его тоже именуют отбо-
ром, создается впечатление, что ТСО - это дарвинизм, хотя в основе там видна
смесь ламаркизма с номогенезом.
Подобных примеров можно привести много, и такая подмена понятий тоже,
кстати, описана Куном. В итоге наблюдается полное нежелание разработать
идею отбора хотя бы приблизительно так же, как разработаны все другие науч-
ные идеи - детально, через конкретный материал. Вместо этого царят толкова-
ния (подробнее см. [24]), что ведет многих к ложному убеждению, будто эволю-
ционная наука - сплошное пустословие. Именно здесь черпает силу и сторон-
ников нынешний креационизм.
Общую причину застоя увидал еще Кун: он описал общее правило - пара-
дигма стареет и при этом обращается в «решение головоломок». Поначалу голо-
воломки еще научно содержательны, но затем начинают преобладать «приспо-
собления ad hoc». Поиск нового уступает спорам о частностях: «Хотя парадигма
все еще сохраняется, мало исследователей полностью согласны... о том, что она
собой представляет» [8, с. 133]. Это целиком относится к эволюции: не берусь
назвать двух авторов, принадлежащих к нынешней эволюционной парадигме,
которые бы одинаково понимали отбор или видообразование, не говоря уж о
понимании феномена эволюции вообще.
А как в иных науках? Ведь там есть выход в практику, а она, как нас учили
и учат, - критерий истины. Однако, занимаясь эволюцией почти полвека и по-
гружаясь в материал смежных дисциплин, я понемногу убедился, что в них си-
туация примерно та же: царит один подход (парадигма), заслоняющий всё ос-
тальное. Факты, противные парадигме, почти или вовсе не упоминаются, а ред-
кие упоминания уничижительны - приводим-де эту мелкую частность лишь для
объективности. Но объективным было бы отметить, что она - факт в пользу
иной концепции. Такие концепции часто дают вполне приемлемые методы,
пусть и не общие (что ставится им в упрек), но работающие там, где парадигма
оказалась бессильна. Увы, начинающему узнать это негде.
Тем же самым грешат и сами маргинальные концепции - они почти или во-
все не касаются парадигмы и причин ее господства. Научные публикации почти
сплошь являют прискорбное пристрастие авторов к «своей» теории. Ущерб для
науки огромный.
5. Нормальные и маргинальные компоненты эволюционизма
Приведу несколько таких противостояний, имеющих отношение к биоэволюции.
Не буду пытаться оценить истинность самих положений, но постараюсь показать,
192
что маргиналии всюду заполняют познавательный вакуум парадигм. Литературу
можно найти в книгах [13; 23; 24], и далее приведена лишь недостающая.
1) . В рассказах о появлении жизни царит идея А.И. Опарина (1924), под-
правленная данными 1950-х гг., по которой жизнь возникла в форме простой
клетки. Иногда добавляют, что первой молекулой жизни была случайно воз-
никшая короткая РНК, кодировавшая короткий белок. Идея противоречит как
принятым взглядам на биосферу (жизнь возникла в форме простого биоценоза)
и химической кинетике (требуемые молекулы не накапливаются), так и сама се-
бе: то, что она полагает исходным, было невообразимо сложным. Исходным ат-
рибутом клетки признана сложная мембрана, а значит, РНК первой клетки
должна была, кроме прочего, кодировать мембранные белки. Ту теорию, по ко-
торой внутреннюю среду клетки задает не мембрана, а структура ее собственно-
го геля [11], не принято даже упоминать. Если же ее учесть, то появляется внят-
ная картина рождения жизни и проясняется ранняя эволюция клетки.
2) . Господствует схема происхождения человека в Восточной Африке, по ко-
торой он расселился в Переднюю Азию около 100 тыс. лет назад, а в остальные
земли еще позже. Она хорошо соответствует большинству находок, но не всем.
Есть более древние (300-800 тыс. лет назад - новейшие данные см. [26]), и если их
учесть все вместе, то эволюция человека предстанет в совсем ином свете - как
процесс, в основных чертах не раз повторявшийся, т.е. по сути номогенетическйй.
3) . В любом учебнике и руководстве по эволюции можно прочесть, что ос-
новой ее понимания служит дарвинизм (или какая-то его форма). В англоязыч-
ной литературе обычно даже отождествление эволюции с дарвинизмом. Иные
теории едва упоминаются, да и то не всегда, хотя многие проблемы рассматри-
ваются только в них.
4) . Например, царит убеждение, что наследуются (и тем дают материал для
эволюции) лишь случайные ненаправленные мутации. Опытов в его пользу по-
ставлено мало, они стары (с тех пор само понятие мутации изменилось) и неко-
торые прямо опровергнуты [6]. Факты и концепции закономерного изменения
наследственных свойств (например, все виды активного генетического поиска -
о нем см. [6; 7; 13]) не упоминаются вовсе. Причина давно известна - боязнь об-
ращения к одиозному ламаркизму.
5) . Предыдущий вопрос упирается в анализ природы случайности. Таковой
в биологии отсутствует вообще, и мне пришлось потратить на него много лет
самому [22; 24]. Царит убеждение, что всякое случайное явление (в том числе,
всякая мутация) имеет устойчивую частоту, которую трактуют как вероят-
ность. Однако факты рисуют иную картину - отсутствие устойчивых частот
даже для мутаций, не говоря уж о более сложных изменениях. Теория неустой-
чивых частот существует, давно применяется на практике, но ни в одном руко-
водстве о ней не прочесть. Если хотя бы знать о ее наличии, фразы о мутациях
как основе эволюции становятся сомнительными.
6) . Выпадение из руководств речи о неустойчивых частотах вызвано, прежде
всего, тем, что преподаваемая ТВ является разделом теории меры, а та основана
193
на теории множеств, предложенной Георгом Кантором (1878). Эта теория уже 130
лет служит объектом споров. Ее главные теоремы доказаны лишь от противного,
и это вызвало протест логиков: противоречие бывает внешним и внутренним, чего
О 7
Кантор не знал и, смешав их, получил видимость доказательств [15] .
Если теорию множеств чуть изменить, то факт наличия устойчивых и неус-
тойчивых (в разной степени) частот становится естественным и наглядным - раз-
личные случайности опишутся множествами различной мощности, притом не та-
ких, как у Кантора: некоторые мощности промежуточны между мощностями ра-
циональных и вещественных чисел [22, с. 271]. Описывают они разные явления
природы, для чего в ТВ нет языка. Пример важен тем, что вековое противостояние
отмечено в математике, где «всё доказывается». Среди противников идеи Кантора
есть крупные ученые, и никто не ищет у них ошибок - просто «нам и так всё яс-
но». В точности как в эволюционизме.
Список можно продолжить - например, теорией дрейфа материков (царя-
щей в геотектонике): она замалчивает свои трудности, многие из которых есте-
ственно объяснимы в рамках таких концепций, как пульсация Земли или ее не-
давняя океанизация. Словом, для становления теории эволюции необходимо до-
полнить нормальную науку идеями из маргинальных концепций. Почему этого
так долго не происходит?
6. Что сказали методологи
Если парадигмы в одних дисциплинах довольно быстро падают под тяже-
стью опровергающих примеров, а в других не падают столетиями, то очевидно,
что схемы Куна недостаточно. Надо коснуться иных направлений. Литературу
можно отыскать по недавнему руководству Е.В. Ушакова [16], и далее приведе-
на лишь недостающая.
Еще до книги Куна, в 1960 г., Виллард Куайн отметил чисто эмоциональ-
ную черту деятельности ученых - их консерватизм, требующий ото всех но-
вовведений сохранения максимально возможной части старых учений: Причем
«консерватизм выражает нашу лень, но он же и стратегия открытий. И именно
он остается действующим, когда иссякает воображение» (цит. по [16, с. 321]).
Очевидно, что Кун имел основания для своего вывода, тоже по сути ирра-
ционального, о «психологической несоизмеримости» парадигмы и конкури-
рующих с ней концепций. Однако возникло целое течение мысли, полагавшее,
что смена взглядов ученых происходит на вполне рациональной основе.
В 1970 г. Имре Лакатос предложил модификацию схемы Куна: парадигму
81 Внешнее противоречие есть оценка теоремы как целого, а внутреннее - одной из фраз, ее
составивших. Допустив, что вещественные числа R можно перенумеровать, Кантор нашел
число, не попавшее в нумерацию, и тем получил противоречие (внешнее). Он счел ложной,
как принято, одну из фраз (исходное допущение), и заключил, что мощность множества {/?}
выше, чем у счетного множества. Но законно было лишь утверждать, что уравнять мощно-
сти этих множеств таким путём не удалось. Остальное надо доказывать отдельно.
194
рушат не опровергающие примеры, а новая теория; иррациональна лишь упор-
ная приверженность ученых старой парадигме (вопреки массе ее опроверже-
ний), тогда как смена парадигм идет рационально. Лакатос обрисовал и проце-
дуру охраны парадигмы от неприятных примеров, введя понятие исследова-
тельской программы, которое шире парадигмы.
Программа состоит из ядра - основных положений теории, признанных в
ней истинными, и защитного пояса - утверждений и приемов для защиты ядра
от опровержения, которые могут подлежать изменению, если защита ядра того
требует. Основную роль защиты играют эвристики - отрицательная, т.е. запрет
сомневаться в утверждениях ядра, и положительная, т.е. система приемов их
подтверждения. Ядро господствующей теории и есть парадигма. Для дарвиниз-
ма это - положения теории естественного отбора. Отрицательная эвристика -
запрет подвергать их сомнению (в либеральные времена разрешается сомне-
ваться в том, что отбор может всё), а положительная - уверение, что всякая при-
способленность есть итог отбора.
Схему Куна - Лакатоса уточнил в 1974 г. Куайн со своей позиции о консер-
ватизме: ученые легко соглашаются на те нововведения, которые не затрагивают
теории как целого (уточню: ядра по Лакатосу), и для ее сохранения широко
пользуются многозначностью терминов. При этом проверке на истинность под-
лежит вся теория, а не отдельные ее положения, отчего консерватизм и может
сохранять ее так долго.
Все названные авторы хотели объяснить феномен бесконечных споров уче-
ных. Наоборот, Ларри Лаудан (1977, 1984) решил обрисовать процедуру прихо-
да их к согласию. Среди споров ученых ему важны лишь те, где участники
стремятся понять друг друга и прийти к общему представлению о предмете их
исследования. Для начала следует выявить то, что у противников обще (таково,
замечу, объединение трех прежних учений в группу эразмовых), чтобы это об-
щее совместно использовать: «мы можем использовать те концепции, в отноше-
нии которых достигнуто согласие, для того чтобы уладить оставшиеся области
расхождения» (цит. по [16, с. 328]).
Лаудан назвал три уровня обсуждения (и несогласия): факты, методология и
ценности. Ему очевидно, что если ученые много лет не могут договориться,
подтверждена ли данная теория фактами или опровергнута, то следует обра-
титься к методологии - решить, что в данной теории является фактом для ее
сторонников, а что для противников, и найти единую базу надежных фактов.
Если это не удалось, то, значит, противники исходят из различных ценностных
установок. Их согласовать труднее всего, но Лаудан показал, что ученые спо-
собны менять и их, обсуждая факты и методологию.
По аналогии с исследовательской программой Лаудан ввел понятие иссле-
довательской традиции - это корпус подлежащих исследованию фактов и объ-
яснительных приемов теории. Различие традиций не дает сторонникам разных
теорий понять друг друга. Поясню: дарвинизм основан на фактах индивидуаль-
195
ной изменчивости и приспособленности, потому он не видит ценности в парал-
лелизмах свойств, на которых номогенез строит успешные прогнозы. Наоборот,
номогенез основан на анализе сходств, поэтому не видит ценности в указаниях
на беспорядочную изменчивость.
Если Лаудан обошел вопросы истинности, то Филип Китчер (1993) уверен,
что истина существует, что развитие любой дисциплины есть движение к ней и
что все ученые к ней стремятся. Он прав в том, что они сами так полагают и что
только с ищущими всерьез истину стоит иметь дело. Но его оптимизм видится
мне чрезмерным: по-моему, большинство людей, включая ученых, ищет ком-
форта, потому смена установок и идет лавиной (научные революции), когда
держаться за старое уже некомфортно.
В данной связи важно понять, что кем признаётся истиной. Еще в 1976 г.
Питер Кларк указал на то, что кинетическая теория теплоты была принята в
1860-е гг. как истинная, несмотря на противоречие опытным фактам. Причина в
том, что ее исходные постулаты (ненаправленность и равновозможность движе-
ний частиц газа) были наглядны и соответствовали мировоззрению эпохи [29].
Мне в 1993 г. удалось, идя по его стопам, показать, что аналогичные посту-
латы (ненаправленность и равновозможность наследственных вариаций) в силу
той же наглядности и в то же самое время явились достаточным основанием для
принятия дарвинизма тем же самым сообществом [20].
Главное у Кларка понятие - обратная эвристика (утверждение, которое
сперва предполагалось доказать, но которое вместо этого стало постулатом).
Легко видеть, что в дарвинизме обратной эвристикой оказался феномен естест-
венного отбора. В обеих теориях основным фактором, давшим апологетам ново-
го уверенность, был успех в смежной области: физики ссылались на успех у
биологов, а биологи - у физиков. (На данный взаимный процесс и указал, как
теперь вижу, Лаудан.) Замечу, что ни те, ни другие ни словом не обмолвились о
коренных противоречиях друг у друга - вернее всего, просто не знали о них, ибо
брали новшества из популярной прессы.
Идея Кларка о роли наглядности постулатов (наглядность же - частный
случай «принципа простоты») представляется мне одной из основ для понима-
ния успеха теорий: если исходные положения наглядны, большинство ученых
верит выводам - даже вопреки фактам.
Наконец, чтение гуманитарного раздела книги Ушакова [16] убедило меня в
том, что эволюционная наука во многом гуманитарна. Гуманитарным наукам
общ феномен интерпретации, для которого характерно, между прочим, объяс-
нение без понимания. Таковым является, например, ссылка на естественный от-
бор как на причину появления новых свойств организмов - ведь ни одного при-
мера того, как он реально действует, нет. Есть только объяснение приспособле-
ний как итога прежнего отбора, а нужно иное: показать, что они обязаны именно
отбору малых ненаправленных вариаций. Критики указывают на это вот уже 150
лет, но их (судя по редким ответам) просто не понимают («разве можно возра-
196
жать против наглядно очевидного?») и опытов для выявления роли отбора
(вполне, кстати, выполнимых) не ставят. Подробнее см. [13; 23; 24].
7. К построению теории
Теперь можно ответить на вопрос, поставленный в конце п. 2: как видим, дар-
винизм утвердился в ходе научной революции, а она есть социальное явление и
течет по законам социальной психологии, а не только рациональной науки. Хотя
последняя и дает материал, открывающий для революции поле действий, но куда
и как пойдет революция, заранее (как и в любой революции) не скажешь.
Поняв это, перейдем к ответу на вопрос, заданный в конце п. 4: смена эво-
люционных воззрений возможна лишь при изменении установок социальной
психологии, а таковое начинается лишь сейчас, с глобальным кризисом.
Хотя все идеологии XX века опирались на дарвинизм, однако их крушение
не повело к отказу от дарвинизма - именно в силу наглядной очевидности его
посылок для большинства людей. Наглядность не раз заводила человечество в
долгие тупики (идеи «Земля неподвижна», «вор рождает вора», «выжившие в це-
лом лучше погибших», «рынок рождает более разумную экономику, нежели
план» или наоборот и т.д.), но избавиться от ее магии, по-видимому, невозможно.
Остается понять, как с ней уживаться, и тут как раз помогает выявление общей ба-
зы давно враждующих учений.
Если мы признаем общую базу эразмовых учений, мы волей-неволей нач-
нем, по Лаудану, искать ее для всех концепций эволюции вообще. Это и есть
путь к искомой теории. Надо только не повторять ошибок прошлого, т.е. не
прельщаться наглядностью и не игнорировать противоречий. Зато надо внима-
тельно присмотреться к общим чертам развития науки, а они видны в исследо-
вательских программах и традициях.
В частности, маргинальные концепции всюду возникают по одной общей
причине - парадигма не может сама справиться с противоречиями. Сторонники
парадигмы, будучи в подавляющем большинстве, всегда блокируют продвиже-
ние за ее рамки, причем тоже по общей причине: то новое, что вне рамок, нару-
шает их комфорт (в широком смысле слова). По Куну, положение должно изме-
ниться само, и нам надо просто ждать научной революции, однако это дорогова-
то, да и нет сейчас на это времени. К счастью, мы знаем больше, чем во время
Куна, и можем действовать лучше.
Прежде всего, следует понять, что парадигма живет иначе, чем иные теории:
ее защитный пояс может намного превышать ядро, тогда как у иной теории, пока
она маргинальна, он мал - ее ведь третируют не за факты (их чаще замалчивают),
а за непривычность исходных постулатов. Пояс, разрастаясь, мешает увидеть вы-
рождение парадигмы из исследовательской программы в голую традицию. Поняв
это, можно обратиться к более сложному вопросу - какая часть парадигмы отно-
сится к естествознанию, а какая к мировоззрению (т.е. гуманитарна). К последне-
му обычно относятся как раз постулаты. Их следует сознательно привести в соот-
ветствие с нынешним пониманием строения природы и задач общества.
197
Проделав (в меру своих сил) эту работу для биоэволюции [18 - 24], я смог
убедиться, что ее парадигма осталась в рамках мировоззрения XIX века и даже
более ранних (в смежных науках она, как правило, много новее). Одна из при-
чин ее долговечности названа выше - это обманчивая наглядность ее постула-
тов, до сих пор позволяющая не видеть опровержений. Тем самым, наглядность
успешно служит отрицательной эвристикой по Лакатосу. Но почему физики по-
няли ее обманчивость еще сто лет назад (с появлением идеи квантов), а биологи
до сих пор не могут? Ответ на сегодня возможен только гипотетический, да и
тот потребовал бы отдельной статьи.
Зато новые эволюционные постулаты вполне просматриваются. Их не стоит
пробовать извлечь из старых учений, поскольку те никогда не работали (в том
смысле, в каком работали старые естественнонаучные теории) - иными словами,
очередной «новый синтез» наличных учений представляется мне утопией. Ис-
ходные положения теории следует черпать даже не просто из Эразмовой или бэ-
ровой идей, а из положений, совместимых с обеими (такова, например, роль ак-
тивности). В книге [23] это сделано в параграфах 11-1 («Исходные положения»
теории) и 11-2 («Стержень теории»), причем заняло с. 605-617. Изложить это
короче мне не под силу, и выскажу здесь лишь общую мысль.
Эразмова идея гласит, что организм создается постепенно (миллионы поко-
лений) таким, каким его требует среда; а сходство трактуется как итог родства.
Бэрова идея - что он создается сразу (несколько поколений) как комбинация
возможностей, заданных законами становления и устройства форм и функций.
Среда здесь - итог взаимодействия организмов, а сходство - итог общности за-
конов онтогенеза. Первая идея наглядна исходными допущениями (но нагляд-
ность эта сохраняется лишь в рамках видов одного семейства), вторая - наблю-
даемыми следствиями. Их предстоит согласовать.
Неожиданно то, что ни в одной смежной с биоэволюцией отрасли науки мне
не удалось ограничиться знанием ее парадигмы. Так, для описания хода эволю-
ции рептилий можно в целом применить теорию дрейфа материков (парадигму
геоэволюции), но несколько фактов («северные динозавры» мелового периода и
др.) парадигма объясняет лишь произвольными допущениями (вот ее защитный
пояс), тогда как маргинальные концепции успешно привлекают вполне реаль-
ную общую идею геологически кратких перегревов Земли.
Словом, «Маргиналы всех наук, соединяйтесь!» Но не для сокрушения сво-
их парадигм (хотя некоторые, слабые, и рухнут), а для построения единого на-
учного здания.
Однако зачем оно? Что мешает нам ограничиться отдельными теориями?
То, полагаю, что они сами по отдельности не дают мировоззрения, а без него не-
ясно, как нам жить дальше. Черный пессимизм экологов и розовый оптимизм
космоэволюционистов почти не увязаны, и вряд ли их можно увязать прежде,
чем будет увязана роль «антропного принципа» (см. сноску к п. 3) в теориях
космической и биологической эволюции.
198
Попыток увязать их было много, и одна из новейших [32] основана на тео-
рии фракталов. Если не гнаться за строгостью, то фрактал - это нелинейный (не
описываемый никаким линейным уравнением) объект, у которого часть устрое-
на в некотором смысле так же, как весь объект. Таково, например, любое фило-
генетическое древо систематики - по виду его рисунка (если он без надписей)
нельзя сказать, отображает ли оно деление отряда на семейства, семейства на
роды или рода на виды. Фрактал может быть получен как путем последователь-
ных ветвлений (такова филогенетическая система организмов), так и путем раз-
рывов, и путем изломов (fractus по-латыни - сломанный, разбитый). Пример ло-
манного фрактала - береговая линия на карте.
Каждый фрактал образуется посредством правила его построения, причем
правило может быть очень простым, а фрактал - очень сложным. При этом со-
всем малое изменение правила может приводить к качественному изменению
фрактала [23]. Фракталы открыл в 1975 г. Бенуа Мандельброт как математиче-
скую конструкцию, а недавно выяснено, что фрактальная структура характерна
для всех уровней мироздания [7а].
Антропный принцип выглядит теперь как совокупность фракталообразую-
щих правил .
Самое главное для нового понимания организма и его эволюции - тот фрак-
тальный процесс клеточных делений (формирование фрактала ветвлений), ка-
ким предстаёт ныне онтогенез52. С этой точки зрения эволюция предстаёт как
процедура изменения фракталообразующих правил, а эразмовы и бэровы кон-
цепции - как два взаимодополнительных аспекта изучения данной процедуры.
На этом пути мне и видится построение работоспособной (дающей практиче-
ские рекомендации - например, по сохранению природы) теории эволюции.
Литература
1. Амлинский И.Е. Жоффруа Сент-Илер и его борьба против Кювье. М., Изд. АН
СССР, 1955.
2. БергЛ.С. Труды по теории эволюции. Л., Наука, 1977.
Примеч. после публикации. Смысл фразы в том, что самосогласованность мира можно
теперь понимать не как предустановленную Творцом согласованность констант и на-
чальных условий, а как общее свойство единого многомерного фрактала. Факт, что мно-
гие фракталы проявляют самосогласованность, очевиден при разглядывании изумитель-
ных фрактальных узоров (см. АСМ, Дополн. 4). Он, однако, непонятен - в том смысле,
что не вытекает из каких-либо иных известных нам законов. Надо ли выводить эту само-
согласованность из чего-то более элементарного или разумнее счесть ее таким же пер-
вичным свойством мира, как активность (химическое сродство, самосборка макромоле-
кул и т.п.)? В книге [23] изложнена вторая позиция. См. также далее, пункт «Причина
рефренов» статьи 4-57 и п. 5 заключительной статьи сборника.
52 В частности, весьма вероятно, что мозг столь сложен (при столь малом числе генов) по-
тому, что в генах записаны не связи между его клетками, а параметры фракталообразующих
правил, реализуемых в ходе онтогенеза мозга.
199
3. Бэр К. Человек в естественноисторическом отношении // В книге: Симашко Ю.И.
Русская флора.... Часть I. СПб., 1850, с. 390-623.
4. Бэр К. Избранные работы / сост. и примеч. Ю.А. Филипченко. Л.. Госиздат, 1924.
5. Вальт М.Х. Экологические исследования К. Бэра и концепция борьбы за сущест-
вование // Петербургская Академия наук и Эстония. Таллин, Валгус, 1978, с. 102-119.
6. Голубовский М.Д. Неканонические наследственные изменения // Природа, 2001, №№
8, 9.
7. Зусмановский А.Г. Эволюция с точки зрения физиолога. Ульяновск, Ульяновская
Сельхозакадемия, 2007.
7а. Красный Л.И. Разномасштабная делимость // Вестник РАН, 2002, № 6.
8. Кун Т. Структура научных революций (1962). М., ACT Москва, 2009.
9. Ламарк Ж.-Б. Философия зоологии. Т. 1. М., Биомедгиз, 1935.
10. Лекявичюс Э. О некоторых аналогиях между эволюцией экосистем и развитием
экономики... // Эволюция: космическая, биологическая, социальная. М., Либроком, 2009.
И. Линг Г. Физическая теория живой клетки (незамеченная революция). СПб., Нау-
ка, 2008.
12. Мейен С.В. Основные аспекты типологии организмов, 1978, ЖОБ (М-165).
13. Назаров В.И. Эволюция не по Дарвину. Смена эволюционной модели. М., Либ-
роком, 2005.
14. Раутиан А. С. О вкладе С.В. Мейена в теоретическую биологию // Чтения памяти
С.В. Мейена (в ТИН). Сборник тезисов. М., ГЕОС, 1997.
15. Традиционная логика и канторовская диагональная процедура. М., Янус, 1997.
16. Ушаков Е.В. Введение в философию и методологию науки. СПб., КноРус, 2008.
17. Чайковский Ю.В. О непрерывном процессе решения матричной игры, 1971 (4-1).
18. Чайковский Ю.В. Грамматика биологии, 1986 (4-14).
19. Чайковский Ю.В. Элементы эволюционной диатропики, 1990. 4-24. (ЭЭД)
20. Чайковский Ю.В. Идея равновозможности в физике и биологии, 1993. 4-29.
21. Чайковский Ю.В. Иммунитет и эволюция, 2003. 4-49.
22. Чайковский Ю.В. О природе случайности, 2004. 4-53. (ОПС)
23. Чайковский Ю.В. Активный связный мир. М., КМК, 2008. 4-63. (АСМ)
24. Чайковский Ю.В. Зигзаги эволюции, 2010. 4-66. (ЗЭ)
25. Шмальгаузен И.И. Организм как целое в индивидуальном и историческом
развитии (1938). М., Наука, 1982.
26. Alperson-Afil N. Continual fire-making by Hominins at Gesher Benot Ya’agov, Israel //
Quaternary Sci. Rev., 2008, vol. 27, p. 1733-1739.
27. Britannica. Настольная иллюстрированная энциклопедия. T. 1. М., ACT Астрель,
2009.
28. CabanisP.-J.-G. Rapports du physique et du moral del’homme. Vol. 2. P., 1802.
29. Clark P. Atomism versus thermodynamics // Method and appraisal in the physical sci-
ences. Cambridge, Cambr. Univ. Press, 1976.
30. Darwin E. Zoonomia or the laws of organic life. Third ed., t. 2, ch. XXXIX. L., 1801.
31. King-Hele D.J. Essential writings of E. Darwin. L., Worcester, 1968.
32. Nottale L., Chaline J., Grou P. Les arbres de revolution. Univers, vie, societe. P.,
Hachette, 2000.
200
Часть 5. Происхождение человека
{Проблема антропогенеза часто в наше время подается как проблема поиска
недостающих ископаемых «переходных форм», однако после уяснения блочного
характера эволюции она потеряла смысл. Неудача таких поисков служит важным
доводом креационистов против эволюционного объяснения возникновения людей
вообще. Приводимые далее статья и отрывок призваны показать, что к антропоге-
незу есть иные подходы, более содержательные, чем построениие филогений.
Главным мне видится понимание путей качественного усложнения психики.
Пока ее понимали примитивно (бихевиоризм, школа И.П. Павлова, биокиберне-
тика и т.п.), многим (но не всем) казалось, что психика могла возникнуть так же,
как любой признак. Ныне, когда утвердился системный взгляд (что система мо-
жет возникать только как целое), когда видна уже и его ограниченность, ясно,
что это не так. Вот новейшее на сей счет суждение: «Намечается новая картина
мира, в которой движение, изменение, активность являются основой и идут впе-
реди формы, функции, да и системы как таковой. Не эволюция систем, а эволю-
ция, рождающая системы, создающая их» [Брынцев, 2009, с. 1130].
Пример «эволюции, рождающей систему» и приведен далее. При опустыни-
вании Северной Австралии условия жизни ухудшились настолько, что люди
должны были там исчезнуть. Однако они сохранили зону обитания, освоив уди-
вительный общественный уклад. Вот его главные для нашей темы черты.
1. Все девочки вступали в брак предельно рано (8 лет) и почти все - со
старейшинам племени, которые все оказывались многожёнцами.
2. Все юноши были устранены от брака примерно до 20-и лет, после чего
могли взять женой старую вдову.
3. Фертильную жену мужчина обычно получал в 30 лет и позже.
4. Поскольку средняя длительность жизни не превышала там 35 лет, то
большинство мужчин не доживало до плодовитого брака. Это были основные
кормильцы племени. Зато остальные были основными производителями детей.
5. Кража жён была основной причиной конфликтов [Роуз, 1989].
Такая система обеспечивала повышенную выживаемость племени, поскольку
1) половое наслаждение было отдалено от размножения, в итоге чего
2) были низки как рождаемость, так и младенческая смертность (ок. 20%);
3) главные кормильцы работали не на свою семью, а на коллектив;
4) племя могло быстро повысить численность без изменения строя - про-
сто ослабив борьбу с кражей жён, если ее совершали крепкие кормильцы;
5) тем самым, основную массу детей производили мужчины высокого ста-
туса - старейшины и крепкие кормильцы.
Эта система позволяла выжить, но общество и хозяйство застыли на самом
низком уровне развития. Пример интересен как эволюционный, поскольку пока-
зывает две из принципиальных возможностей системогенеза, иначе не видных.
Первой посвящена статья, а второй - отрывок. - Вводное замечание 2010 г.}
201
Двускоростной механизм размножения и его роль в эволюции
человека'
Часто пишут, что с рождением цивилизации биологическая эволюция челове-
ка прекратилась, поскольку на него перестал действовать естественный отбор. В
действительности отбор на людей действует (поскольку они поддерживают свое
существование в историческом времени исключительно путем размножения),
только не по всем параметрам. [...] Другое дело, насколько отбор эффективен.
Оказывается, эффективность его очень низка, что подробно рассмотрено в кни-
ге[ 1], но это верно для любых организмов и не является особенностью людей.
Отказ видеть у человека естественный отбор связан, по-моему, просто с тем,
что статистика населения хорошо известна, поэтому многие высказывания об
отборе тут можно проверить, а проверки они обычно не выдерживают. Напри-
мер, отбор должен, по теории, уничтожать все свойства, ведущие к полному
бесплодию, однако он ничего не может поделать со многими из них. Таков аб-
солютный гомосексуализм: он поражает 4% населения, воспроизводясь в каж-
дом поколении заново. Тем меньше оснований считать итогом отбора те свойст-
ва, которые лишь изменяют долю выживающих.
Дело в том, что как у человека, так и других видов, наследуется вовсе не каж-
дый признак (как полагал Ч. Дарвин), а всё разнообразие свойств, членов популя-
ции как целое. С.В. Мейен называл такое наследование транзитивным полимор-
физмом. В каждом поколении ра'чообразие воспроизводится заново, независимо
от судьбы отдельных особей, и отбор не может влиять на отдельные признаки. Об
этом подробно сказано в книге [1].
Легко видеть, что эволюция человека как биологического вида идет, и притом
на разных уровнях. Например, жители разных широт и разных материков заметно
различаются как строением тела, так и функционально, причем в этих различиях
видны как разнообразие, так и итог эволюции (можно указать, какой вариант для
какого стал основой). Другое дело, что их эволюция не всегда носит характер при-
способления, но и эта черта эволюции характерна для организмов всех видов (о
чем тоже не раз писал Мейен). Однако есть и особые черты эволюции человека.
Черты антропогенеза
Первая черта -очень быстрая эволюция рода Ното, протекающая несмотря
на медленную смену поколений* ** и очень долгое взросление детей. У девочек
первые признаки взросления видны в 8-10 лет, а полностью взрослыми женщин
считают в 19-20 ле т [2]. Африканский слон демонстрирует примерно ту же ско-
рость эволюции, что и человек (1 род за 3 млн лет) при той же скорости смены
поколений (половая зрелость к 20 годам, долгота жизни 70 лет) и даже более
долгой беременности (22 месяца), однако слоны взрослеют так же быстро, как и
* Эволюция (журнал, ИИЕТ), № 2, 2005, с. 42-45. Подзаголовки даны заново.
" Если считать, как принято, по мужчинам (около 30 лет на поколение). Если считать по
женщинам, она шла примерно вдвое быстрее. Подробнее см. далее. - Примеч. 2010 г.
202
другие крупные млекопитающие, за несколько месяцев. Быстро взрослеют и са-
мые близкие к человеку обезьяны, например, шимпанзе. Это тем более странно,
что шимпанзе не уступают людям в общей длительности онтогенеза (периода от
зачатия до взрослости) - около 20% длительности жизни [3].
Взрослость людей принято оценивать не только как половую зрелость, тут
важна и душевная зрелость; без нее, например, нельзя быть сколько-то хорошей
матерью. Мне представляется, что уникально долгое душевное взросление лю-
дей необходимо для эволюции их интеллекта.
Вторая черта - внесезонное размножение. В п. 9-15 книги [1] было отмече-
но, что свойственный людям избыток половой активности можно рассматривать
и как итог внесезонного размножения, и как один из факторов антропогенеза.
Добавлю, что когда человек стал человеком, этот избыток не только не исчез, а
наоборот, стал одной из определяющих его поведение черт.
Третья черта - неугасающий в течение года инстинкт продления рода пере-
рос у людей в любовь, которая вышла за рамки процесса размножения. Она, види-
мо, столь же необходима для человеческого общества, сколь и избыток активно-
сти. Некоторые идейные течения учитывают это: есть религии, видящие половое
начало как энергетическое (особенно тантризм в Индии: он рассматривал половой
акт как священнодействие), а сто лет назад Зигмунд Фрейд положил феномен по-
ловой энергии (либидо) в основу понимания культурной деятельности .
Не будучи приверженцем этих учений, отмечу всё же, что нельзя вообще
игнорировать указанную ими черту культуры, как делает, например, христиан-
ство. В нем господствует идея греховности (порочности) полового акта, потре-
бовавшая даже ввести для божества “непорочное зачатие”. Как следствие, в
христианской литературе обычна мысль, что люди, совокупляющиеся не ради
размножения, а ради удовольствия, ведут себя “аки скоты”. Но на самом деле
соединяются лишь для размножения как раз “скоты”, а человек отличается от
них тем, что изобрел половую любовь, и данное отличие, как выше сказано, есть
культурообразующий фактор. Исподволь к половому фактору прибегает и хри-
стианство, лишь подвергая его сублимации (переводу во внешне неполовую
форму). Иногда сублимированная сексуальность христианских установлений
даже подчеркивается (“монахини - невесты Христовы”), но обычная сексуаль-
ность всегда горячо порицается.
Европейская наука выросла из идеологии христианства и сохраняет тот пе-
рекос мысли, что половой акт нужен лишь для размножения.
Народы резко различаются по степени активности, она носит, в основном,
мужской характер и ведет к огромным различиям в их численности (расселении) и
влиянии на ход истории. Народы, словно разные виды, различно расходуют свой
Его ученики Ф. Адлер и К. Юнг расширили рамки учения, фактически поняв либидо как
некую обобщенную активность. В основу АСМ положена именно активность в широком
смысле, а в данной статье сделан упор на ее половую составляющую. - Примеч. 2010 г.
203
жизненный порыв (понятие, которое ввел Анри Бергсон в 1907 г. для обозначения
главного фактора, движущего эволюцию). Некоторые народы попросту сбивают
его (например, путем уничтожения новорожденных девочек: этим достигается
снижение размножаемости, зато активность мужчин расходуется, в основном, на
истребление друг друга в борьбе за женщин83), но среди таких народов нет, на-
сколько знаю, исторически успешных. Есть более перспективные пути.
Четвертая черта, которая является главной для нашей темы - необычное
взаимодействие полов. Нам понадобятся четыре факта: 1) у женщин возраст раз-
множения наступает много раньше становления взрослой анатомии, физиологии и
психики, 2) их период размножения короче мужского, 3) самих женщин больше,
чем мужчин (если нет истребления девочек), 4) отец ребенка обычно старше мате-
ри. На этой основе попробую выделить одну линию в эволюции человека, линию,
носящую явно биологический характер.
С глубокой древности почти до наших дней было нормой выдавать замуж де-
вушек в 15 лет и ранее, тогда как жениться - в 30 лет и позже. Разница бывала
много выше двойной: возраст, с какого девочка считалась годной для мужчины,
часто бывал ниже 10 лет. Ислам провозгласил женским брачным возрастом 9 лет.
Античная литература не называет возрастов, но изобилует примерами, когда
мать взрослых детей становилась объектом любви мужчин и даже юношей (“царь
Эдип”), т.е. была молода. Пленницы и рабыни вступали в половую жизнь совсем
рано. С римских времен по всей христианской Европе был узаконен женский
брачный возраст с 12 лет, и эта норма сохранялась в праве очень долго, хотя на
практике уже в годы Возрождения ее стала теснить тенденция выдавать замуж
более взрослых. Однако Восток и Юг эксплуатируют малолетних жён поныне.
Пример глубокой старины
Наиболее разительна прежняя брачная практика у аборигенов Северной Ав-
стралии: старейшина брал в жёны 8-летних девочек, оставляя старух в качеств
жён для молодых мужчин. Этнограф Фредерик Роуз (F. Rose) еще в середине
XX века сам наблюдал остатки этого обычая. Он же приводил и более ранние
свидетельства, например: “Нередко... женой 50-летнего мужчины является 8-
летняя девочка. [...] Часто можно увидеть старых мужчин с женами-девочками и
молодых мужчин, состоящих в браке со старыми вдовами. Как правило, мужчи-
на не может получить жену по крайней мере до 30 лет” (Е.М. Сшт, 1887); или:
“18-25-летние мужчины либо совершенно не имеют жен, либо по возрасту жена
вполне годится им в бабушки” (Е. Eylmann, 1908) [5, с. 155-156].
Правда, естество брало верх и в тех юношах, чьи потенциальные жёны дос-
тались не им, а их дедам: в семьях аборигенов дед и внук считали себя как бы
OQ
«В ответ на просьбу рассказать о своем муже ни одна из взрослых женщин не назвала
единственного мужчину. Каждая повествовала, кто и как убил ее мужа, потом второго,
третьего...» - пишет о жителях Новой Гвинеи А.П. Назаретян [4], излагая данные амери-
канского этнографа Дж. Даймонда (J. Diamond).
204
братьями, вплоть до “шуточного” (joking) притязания внука на жён деда [6, с.
96-97]. Можно лишь гадать, во что оно обращалось вдали от посторонних глаз и
как существенно было его влияние на реальное отцовство, но бросается в глаза
сходство с токованием, где демонстрируют себя молодые самцы, а отцом боль-
шинства потомства является старый самец [7].
Будучи марксистом, Роуз дал этому укладу чисто экономическое толкова-
ние: муж, пользуясь властью в общине, брал девочку в свою семью ученицей к
старшим жёнам и работницей, вовсе не помышляя о любовных утехах.
“Доказательством” своей идеи Роуз счёл то обстоятельство, что не видал
беременных и матерей моложе 14 лет. Однако есть факты, разбивающие его
“доказательство”. Первый - так называемая подростковая стерильность: в го-
ды взросления девочек у них овуляции происходят гораздо реже, чем менструа-
ции, поэтому беременность наступает редко [2, с. 24], а Роуз наблюдал лишь од-
но небольшое племя. Второй - вряд ли этнограф мог много узнать об убитых и
умерших младенцах. Третий факт привел сам Роуз из своих наблюдений: “Са-
мому молодому женатому мужчине... было 22 года. Он являет собой чрезвычай-
но интересный и показательный случай. Этот абориген имел даже двух жен...
причем это были не старухи, а очень молодые девочки 10 и 8 лет соответствен-
но. Уже в этом возрасте этот абориген выделялся своей сообразительностью, ак-
тивностью и влиянием в общественных делах” [5, с. 156].
Разумеется, некий экономический элемент во всякой системе брака есть (так,
“бабушки” явно использовались молодыми мужчинами для ведения хозяйства),
но он никогда не бывает единственным. Очевидна потребность в более сложном
объяснении, с привлечением более сложных теорий общества, а также биологии.
Мне представляется, что в обществе, не знающем ни богатства, ни иерархии
власти, основным для старейшины средством повысить свой статус являются жё-
ны. А поскольку число возможных жён тоже ограничено (как размером племени,
так и трудностью их удержания и прокормления), то остается повышение их каче-
ства в глазах соплеменников, что и достигалось предельным снижением их воз-
раста. Причем делал так даже совсем молодой муж - поскольку его статус в пле-
мени дал ему такую возможность. Любопытно, что как раз взрослая женщина ему
оказалась ни к чему - видимо, девочки вполне сносно вели хозяйство. И уж со-
всем очевидно, что за отсутствием взрослой жены муж не мог ограничить оным
хозяйством свое общение с малолетними женами. Тут мы подходим к биологии.
Что бы ни думали о такой системе браков аборигены и этнографы, очевидна
ее биологическая функция: очень многие дети имели незрелых матерей и старых
отцов. Молодым же мужчинам оставалось утешать себя тем, что женщина “в
сорок пять ягодка опять”. Кстати, при всей дикости и странности для нас подоб-
ного уклада жизни, он недалек от уклада, обычного 200 лет назад в России.
Здесь практика часто была весьма далека от закона (по которому жениху
должно было быть как минимум 15 лет, а невесте 13). Известное признание няни
Татьяне (“Мой Ваня / Моложе был меня, мой свет, / А было мне тринадцать лет”
205
- Пушкин А.С. Евгений Онегин, 3, XVIII) удивляет само по себе, но главное ви-
дится в том, что оно оставляет без ответа вопрос - как мог мальчик справиться с
обязанностями мужа, да еще при столь юной жене.
Оказывается, вопрос обычно решал его отец (мог и дед): “Заинтересованные
в привлечении в семью лишней рабочей силы родители совершенно малолетне-
го жениха женили его на взрослой девушке, причем с ней вступал в сожительст-
во не муж, а кто-либо из старших членов семьи, - обычай, известный древнерус-
скому быту и крестьянскому обычному праву, содействовавший между прочим
происхождению и укреплению снохачества” [8, ста. 949].
Снохачество - открытое сожительство хозяина дома со снохой (женой сы-
на). Слово сноха (вероятно, связующая*) применяется только по отношению ее к
свёкру, тогда как свекровь называет жену сына невесткой [9]. Само слово про-
исходит, по всей видимости, от этого обычая. Налицо фактическое многоженст-
во в рамках христианства, причем младшие жёны всегда молоды. Вряд ли жена
(она же сноха) всегда достигала тринадцати лет: “бабушки середины XIX в.
помнили, как на другой день свадьбы бегали домой за куклами” [10, с. 48].
Позже тургеневский Базаров заявлял, что именно снохач есть пример без-
нравственности народной жизни. Судя по контексту (Тургенев И.С. Отцы и дети,
гл. X), речь шла не об извращенных индивидах, а об известном всем обычае. Да
и в конце XIX века снохач мог еще действовать открыто: “Ну и старик, чёрт!
Женил сына, отбил сноху и - прав!” (Горький А.М. На плотах).
Чисто биологические вопросы людской эволюции
1) Как молода могла быть мать? Известно, что подростковая стерильность не
абсолютна: как российское [2], так и американское [11] руководства упоминают
11-летних беременных. Ныне беременные младше 15 лет очень редки, однако при
прежней практике (12-летние невесты) доля юных матерей явно была значительно
больше нынешней, так что прорыв барьера подростковой стерильности был нере-
док. Возможно, что именно полудетский женский организм более восприимчив к
агрессии мужской наследственности, но пока данных на сей счет нет.
2) Важна и другая граница: 60-летний отец новорожденного - явление почти
обычное. Поэтому до тех пор, пока у людей были обычны 12-летние (на момент
зачатия) матери, размножение по женской линии шло у части популяции в 4-5
раз быстрее, чем по мужской**. Что это могло давать для эволюции?
Именно пятикратное различие скоростей размножения достигалось в одном
из опытов по переключению геронтогенов (генов старения), активностью которых
* Согласно этимологии, словарям, происходит от праиндоевропейского *sneu - вязать. В
частности, в немец, языке слово Schnur (бечёвка) еще сто лет означало также сноху. Эта
связь деда с поколением внуков, минуя сына, - вероятный след прежнего господства од-
ного из механизмов двускоростного размножения. Еще один пример такого следа дает
инцест, как он известен из мифов (см. АСМ, Дополн. 10). - Примеч. 2010 г.
** Речь идет о первенцах. Об их роли в древности см. АСМ, с. 557, 560. - Примеч. 2010 г.
206
определяется эволюционная стратегия видов: r-стратегия с быстрой сменой поко-
лений и Л'-стратегия с медленной их сменой [12]. Как видим, человек реализует
достоинства обеих стратегий: мужчины приобретают долгий опыт, а женщины
быстро передают* его потомству (быстрое наследование долгого обучения)**.
Тем самым, налицо двускоростной механизм размножения людей. Как
факт он вне сомнения, однако его роль в эволюции еще предстоит выяснить.
3) В Связи с этим вспомним известную идею В.А. Геодакяна (см. [1]) о разли-
чии ролей мужчин и женщин в эволюции, по которой женщина дает геному детей
лишь опыт поколений, а мужчина - еще и свой опыт. Если так, то есть эволюци-
онный смысл для позднего размножения мужчин, а учёным есть смысл искать на-
следование итогов обучения в механизмах мужской наследственности. К сожале-
нию, на сегодня данные о наследовании итогов обучения противоречивы.
4) С ростом возраста рожающих осложняются роды и учащаются патологии у
их детей. Напомню синдром Дауна: с одной стороны, он имеет явно генетическую
природу, а с другой - в десятки раз чаще поражает детей немолодых (40 лет и бо-
лее) матерей, т.е. явно зависит не только от наследственности. Вероятное объяс-
нение состоит в том, что женские половые клетки закладываются еще в эмбрионе
девочки и затем стареют в ожидании своего срока, тогда как мужские половые
клетки образуются непрерывно и потому всегда молоды [13, гл. 22]. [...]
5) Затем, есть мнение (не знаю, насколько обоснованное), что у выдающих-
ся людей обычны пожилые отцы и что возраст матери в этом плане безразли-
чен. Если и это верно, то естественно допустить, что для быстрой эволюции ин-
теллекта желательны молодые матери и старые отцы.
6) Наконец, встает вопрос о выживаемости юных рожениц и их детей. При-
нято писать, что 21 год - оптимальный возраст для первых родов, но это вряд ли
так: слишком уж многие специалисты уверены, что девочки рожают легче, чем
взрослые женщины [2; 11]. Есть, правда, противоположные мнения (там же)***.
К сожалению, статистики беременности и родов по возрастам внутри группы
“подростки”, по-видимому, не ведется, судя по тому, что каждый исследователь (и
у нас, и на Западе) исходит из своего опыта и отрывочных литературных данных.
При этом “подростками” именуют даже 19-летних девушек. Статистика патоло-
гий по возрастам беременных соединяет всех до 18 лет в одну группу [2; 11].
* Быстро реализуют само размножение, а не только передают опыт. - Примеч. 2010 г.
** Суть в различии физиологий взрослого отца и юной матери, находящейся в середине
взросления. Оно и вообще-то является радикальной перестройкой организма (его даже
сравнивают с метаморфозом, т.е.с превращением из личинки во взрослую форму - [Гил-
берт, 1995, с. 187]), а у беременной девочки оно проходит раз в 5-7 быстрее, чем в норме
[2, с. 130]. Тем самым, она находится в состоянии сильнейшего стресса (это отчасти вид-
но и в необычно высокой доле, 54%, мальчиков в потомстве юных матерей [2, с. 174]), а
стресс теперь уже прочно признан в качестве фактора эволюции [1] . - Примеч. 2010 г.
*** Противоречие тут, как обычно, вызвано невниманием к разнообразию (девочек).
207
Ныне, пока нет детальной статистики, можно высказать лишь общую догадку,
как бы впечатление от книг [2; 11]: в здоровых обществах роды у нормально раз-
витых девочек-подростков протекают легче и быстрее, чем у взрослых, а их дети
нормальны (“Существует даже мнение, что подростковый возраст является иде-
альным для вынашивания плода” [2, с. 136]); наоборот, в неустроенных слоях на-
селения на Западе и в целом по нынешней России девочки-подростки рожают по
всем показателям тяжелее, смертность и их самих, и новорожденных выше.
Можно понять, почему в прошлом царила тенденция совсем юных невест: в
здоровом обществе это спасало много жизней. Тех особых бед, которым подвер-
галась (и ныне подвергается) беременная девочка вне брака, вовсе не могло
быть, если она была женой или наложницей. А ее ребенок обычно поступал к ее
матери или в иные взрослые руки. До появления в последние 150 лет эффектив-
ных средств родовспоможения данная тенденция была, видимо, оптимальной
стратегией обществ. А старые отцы обычны в силу их высокого социального
статуса. Тем самым, двускоростной механизм размножения возник у людей без
умысла и отбора, просто как реакция на суровую реальность жизни.
Разумеется, нынешняя норма (взрослые невесты) имеет огромную ценность:
женщина выступает не как “родильная машина”, а как самоценное существо. Ра-
зумеется, девочка редко бывает хорошей мамой, а ее работающая мать редко
может целиком взять на себя ее заботы. Разумеется, возрождение прежних норм
(вполне реальное в условиях роста исламского экстремизма) было бы ужасно.
Но наши моральные оценки не должны мешать увидеть наш эволюционный ме-
ханизм. Его нельзя было увидеть, пока в умах царил неодарвинизм, считающий
единственным источником новаций случайные мутации и рекомбинации.
Однако теперь мы знаем [1, гл. 5], что организм изменяет свои гены активно
и что ведущим фактором эволюции служит изменение развития зародыша. Это
знание можно положить в основе предлагаемой эволюционной модели.
Суть ее видится так. В биологии размножение незрелого организма именуется
неотенией, а размножение со включением надставок старшего возраста - герон-
тогенезом. Его механизм (как свойства, возникшие в старшем возрасте, достают-
ся потомству) совсем непонятен, но сам факт давно обсуждается.
На сегодня известно, что в организме всегда имеются стволовые клетки (СК),
способные развиться в любую его часть или в целый организм; они способны миг-
рировать*. Как и все клетки, они могут обмениваться генами с иными клетками. По-
ловые клетки - частный случай СК Способны ли неполовые СК достигать половых
желёз и занимать место половых клеток, пока неизвестно, но если это замещение
допустить, то геронтогенез будет в принципе понятен.
У людей прошлого мы видим женскую неотению при мужском геронто-
генезе. Видимо, этой парой противоположных процессов и достигалась эффек-
тивность двускоростного механизма размножения людей.
* Есть они, что важно для нашей темы, и в мозгу [Корочкин, 2004]. - Примеч. 2010 г.
208
Хотя и эта сторона двускоростного механизма сама по себе налицо, роль ее в
эволюции тоже еще предстоит понять. Данный механизм соединяет достоинства
двух стратегий экологической эволюции (r-стратегия и /^-стратегия) и обходится,
по-видимому, без их недостатков (таковы краткость жизни при r-стратегии, вы-
званная ослаблением иммунитета, и вялость жизни при ^-стратегии, вызванная
низкой биохимической активностью [12]).
Если в резко двускоростном режиме постоянно живут хотя бы несколько
процентов какой-то локальной популяции, то в ней первенцы могут образовать
субпопуляцию, где мужчины экспериментируют долго, а женщины размножают
их достижения быстро, притом в полудетском состоянии.* [...]
Трудно сказать, когда в прошлом двускоростной механизм начал действо-
вать и насколько был всеобщ. Зато в будущем можно выяснить эксперименталь-
но, какие свойства могут приобретаться или теряться таким путем и прав ли
Геодакян, полагая, что гены переходят с Y-хромосомы на Х-хромосому. Если
да, то вполне возможно, что быстрый темп интеллектуальной эволюции челове-
ка был обеспечен двускоростным механизмом. Насколько механизм эффекти-
вен, можно выяснить популяционным генетическим исследованием, если уклад
вроде австралийского еще где-то сохранился.
Литература
1. Чайковский Ю.В. Эволюция, 2003 (4-51).
2. Гуркин Ю.А., Суслопаров Л.А., Островская Е.А. Основы ювенильного акушерства.
СПб., 2001.
3. Флиндт Р. Биология в цифрах / пер. с немецкого. М., 1992.
4. Назаретян А.П. Антропогенные кризисы: гипотеза техно-гуманитарного баланса
// Вестник РАН, 2004, N 4.
5. Роуз Р. Аборигены Австралии. Традиционное общество. М., 1989.
6. Radcliff-Brown A.R. On joking relations И Structure and function in primitive society.
L., 1958.
7. Панов E.H. Бегство от одиночества. M., 2001.
8. Нечаев В.М. Брачное право // Новый энц. словарь Брокгауза и Ефрона, т. 7, 1912.
9. Черных П.Я. Историко-этимологич. словарь современ. русского языка. М., 1993.
10. Бернштам Т.А. Молодежь в обрядовой жизни русской общины. Л., 1988.
11. Детская и подростковая гинекология / ред. Анна Гарден; пер. с англ. М., Мир, 2001.
12. Скулачев В.П. О биохимических механизмах эволюции и роли кислорода И Био-
химия, 1998, вып. 11.
* Дело не в быстроте как таковой, а в том, что эволюция психики, как всякий системоге-
нез, если и может происходить, то только скачком, за малое число поколений. Беремен-
ность в полудетском состоянии влияет на наследственность (см. сноску на с. 207, а также
АСМ, с. 558), т.е. на свойства первенцев. Особое к ним отношение во многих древних
культурах могло быть вызвано высокой долей необычных особей. Большинство их уми-
рало в детстве, зато лучшие из выживших могли часто выступать в роли племенных
(элитных для расплода) особейопрос требует изучения,- Примеч. 2010 г.
209
Происхождение человека в людском муравейнике*
Мы живем в совсем иную эпоху, нежели Дарвин, нам нужна настоящая
(дающая полезные рекомендации) теория эволюции, а не произвольные толко-
вания. Об этом см. подробно [3]. В учение об эволюции человека из дарвинизма
принято брать три идеи: 1) каждая стадия очеловеченья давала какое-то пре-
имущество и потому “подхватывалась отбором”5^; 2) с тех пор, как человек стал
человеком, его биологическая эволюция прекратилась, так как естественный от-
бор уступил место социальным факторам; 3) процесс становления нынешнего
вида людей есть линейная последовательность поколений, которую в принципе
можно приблизительно восстановить по ископаемым остаткам. Первые две - го-
лословные декларации (в огромном потоке литературы мне не попалось никаких
реальных аргументов в их пользу, и могу лишь напомнить, что сам Дарвин не
верил в их пригодность для объяснения происхождения человека), зато третья
вполне разработана, и итог оказался неожидан: линию эволюции человека найти
не удалось.
Об этом недавно вышла толстая книга [1]. Хотя ее автор не биолог, а исто-
рик, и не всегда ясно, что он имеет в виду, однако негативная сторона его выво-
дов вполне понятна и обоснована литературой. Автор убежден, что на Земле в
разные эпохи жили различные человечества и что они нам не предки; мы, евро-
пейцы, происходим от кроманьонцев, и ничего достоверного более о нас сказать
нельзя. Что касается монголоидов и негроидов, то о них нельзя достоверно ска-
зать и этого. Утверждение, что все люди произошли от общего предка (образно
говоря, от близкого предка кроманьонцев), всего лишь евроцентризм. По автору,
более древние антропоиды (образно говоря, питекантропы) тоже являют собой
различные линии эволюции.
Хотя в моей книге [3] тоже сказано, что дарвинизм не дает ключа для пони-
мания эволюции человека, однако только теперь мне стало ясно, что и я попался
в ловушку евроцентризма. А именно, вслед за большинством антропологов, я
исходил из той модели эволюции человека, где последние ее стадии идут при
чередовании холодных (ледниковых) и теплых периодов (этот процесс назван у
меня: “Дитя жары и мороза”). На самом деле, ледники были в Европе и Сибири,
* Конец статьи «Философия дарвинизма против философии эволюции» (4-62), с. 81-85.
ОД
Наиболее эффектный пример: прямохождение. Предки человека ходили вертикально 2
млн лет (в 200 раз дольше всей истории человечества), прежде чем начали изготовлять
орудия. Это рушит всю “трудовую теорию происхождения человека”, однако дарвинизм
придумал отговорку: поднятие головы над травой открывало обзор, позволявший рано
заметить опасность, а потому “было подхвачено отбором”. На это разумно возражают:
наоборот, прямохождение делало предка человека издали заметным для хищника, кото-
рый сам не ходил вертикально [1]. Ответ дарвинистов (предок выжил, значит польза была
выше вреда) являет собой логическую ошибку (именуемую в учебниках логики подменой
основания): идея отбора тут не подтверждается, а кладется в основание рассуждения.
210
и если они необходимы для очеловеченья, то надо признать, что кроманьонцы
заселили Африку, обратясь по дороге в негроидов, что нелепо (нет даже косвен-
ных данных). Вернее, что каждая раса ведет род от своих “питекантропов”, но
тогда единство нынешнего человечества (все со всеми скрещиваются и все всех
понимают ничуть не хуже, чем внутри рас) совершенно загадочно. Нужна со-
всем новая теория.
Та теория, абрис которой дан в книге [3], может несколько прояснить кар-
тину. В основе ее лежит диатропика (о ней см. [4]), о которой здесь достаточно
сказать, что эволюция в ней мыслится как заполнение клеток огромной много-
мерной таблицы свойств, и таблица эта содержит черты периодичности (как в
таблице химических элементов). Вопрос “откуда сама таблица?” в настоящее
время не имеет ответа, как не имел ответа вопрос “откуда таблица Менделеева?”
до рождения атомной физики. Это, однако, не мешает ею пользоваться, как
пользовался своей таблицей Д.И. Менделеев.
Приняв наличие таблицы как факт (он на сегодня обоснован самым разным
биологическим материалом, но пока еще не на людях), сразу можно понять, где
искать разгадку: различные расы могли в ходе эволюции заполнять клетки па-
раллельных столбцов диатропической таблицы, что и вызвало их фундамен-
тальное сходство. Польза от такого представления эволюции людей видна от-
нюдь не сразу, и я поясню ее примером, который без диатропики вовсе не виден.
Все знают, что Аристотель назвал человека “общественным животным”
(zoo/г politikori), но мы называем общественными совсем иных животных - насе-
комых, у которых разные касты выполняют разные функции; а функцию раз-
множения выполняет у них одна-единственная самка, оплодотворяемая малым
числом самцов (чаще одним). Это разделение функций заставляет считать орга-
низмом не особь, а рой, улей, муравейник, термитник и т.д. Но в каком смысле
надо общественным видом считать людей?
Да в том же самом: специфически человеческую деятельность может осу-
ществлять только общество. У нынешних людей этот факт замаскирован тем,
что они сами вступают в брак, сами выбирают себе работу и жильё, однако в ис-
тории было далеко не так, и до сих пор закрепленность человека видна в обяза-
тельности школы и военной службы. Французский социолог Эмиль Дюркгейм
сто лет назад утверждал, что человек стал человеком в нынешнем смысле тогда,
когда выделился из первобытного коллектива (стаи), т.е. стал мыслить и вести
себя самостоятельно.
Материалом ему послужили племена, не имеющие государственности (их
называют без должных оснований первобытными), где индивид почти лишен
самостоятельности. Обычно эта несвобода гораздо слабее, чем в улье, но в на-
стоящее время известны племена людей, где положение индивида близко к пче-
линому или муравьиному, и они могут пролить свет на эволюцию.
Наиболее разительна прежняя брачная практика у аборигенов северной Ав-
стралии: старейшина брал в жёны 8-летних девочек, оставляя старух в качеств
211
жён для молодых мужчин. Этнограф Фредерик Роуз (F. Rose) еще в середине
XX века сам наблюдал остатки этого обычая и приводил более ранние свиде-
тельства. В частности,
«мужчина получал жену, когда достигал 30 лет, а то и позже, в то время как
женщина переходила жить к своему первому мужу задолго до наступления
половой зрелости и была замужем на протяжении всей своей дальнейшей
жизни». Она «переходила к более молодому человеку, чем умерший муж...
Напротив, мужчина в конечном счете, если он жил достаточно долго, оказы-
вался состоящим в браке с несколькими женщинами». Смена супругов «под-
чинялась определенному социологическому закону, который совершенно не
зависел от воли и желаний отдельных лиц» [2, с. 146] (курсив мой - Ю. Ч.).
[..]
Мне представляется, что этот осколок доисторического быта интересен как
демонстрация такой возможности размножения, какая вряд ли может прийти
кому-то в голову, а если бы и пришла, никем не была бы принята как реальная
возможность. Но этнографы, к счастью, успели ее зафиксировать, и мы можем
проанализировать ее в свете данных нынешней биологии.
Это - двускоростной механизм эволюции, когда женские поколения сме-
няются много чаще мужских, причем наиболее подверженный изменениям не-
зрелый женский (девочка) геном соединяется с наиболее успешным мужским
(старейшина*). Идет быстрое наследование долгого обучения [3, п. 9-16].
Давно замечено (хотя не знаю, какой статистикой подтверждено), что со-
единение юных матерей и старых отцов производит выдающихся детей чаще
среднего. [...].
Могу добавить: этот положительный эффект сопутствует отрицательному -
старческим наследственным нарушениям (у мужчин они сказываются позже, чем
у женщин: отцом желательно быть до 60, а матерью до 40 лет). Но в древних об-
ществах, где детская смертность всегда превышала половину рождений, а мла-
денцев с явными уродствами убивали, носители нарушений гибли, так что дву-
скоростной механизм являл собой прекрасный инструмент эволюционного поис-
ка. Возможно, он** и привел к появлению Человека разумного. Если он и похож
внешне на отбор, то не на воображаемый естественный, а на реально известный
искусственный, так как в нем важна не плодовитость особей, а принудительное
спаривание одних и отстранение других (силой запретов) от размножения. По-
скольку средняя длительность жизни в таких обществах не превышает 35 лет,
большинство мужчин при данной организации общины не доживало до получе-
ния фертильной жены, т.е. исключалось из размножения, несмотря на полную их
пригодность - именно они поддерживали, в основном, жизнь общины.
* В оригинале (ВФ, 2007, № 9) здесь ошибочно значится: «вождь». Вождя в этих племе-
нах, очень слабо организованных, не было. - Примеч. 2010 г.
** В сочетании с инцестом, позже прекратившимся (см. АСМ). - Примеч. 2010 г.
212
Если бы не штампы
Налицо каста рабочих особей у людей, словно у общественных насекомых.
Параллель выглядела бы натянутой, если бы не наличие ясной переходной фор-
мы между данными формами общественного организма. Это - вид южноафри-
канских подземных грызунов по имени голый землекоп (Heterocephalus glaber).
Семья этих зверьков образует сверх-организм (иногда до 300 особей), похожий
на улей, муравейник или термитник. В частности, там имеется одна-един-
ственная плодущая самка, что, казалось бы, для млекопитающих невозможно
[5]. Все попытки объяснить данный феномен как приспособление являются про-
извольными отговорками, исходящими из привычных штампов мышления.
Подробнее см. [3, с. 329].
Диатропика указывает множество таких параллельных рядов, в которых
предлагаемые ламаркизмом и дарвинизмом объяснения (либо общее происхож-
дение, либо независимое приспособление) бессмысленны. Встает вопрос, откуда
подобные феномены берутся. Их приходится признать чисто диатропическими,
т.е. имманентным свойством разнообразия организмов, но это еще не само объ-
яснение, а лишь указание рамок поиска.
Философам данные рамки хорошо известны и привычны: это платонизм,
точнее, мир объективно существующих идей. Идея общественного организма реа-
лизована как у насекомых, так и у млекопитающих, только у последних она выра-
жена гораздо беднее, поскольку само разнообразие их гораздо меньше (4 тыс. ви-
дов против полутора млн видов насекомых). Точно так же, идея ядерной клетки
реализована у растений, грибов и животных. Примеров не счесть, но дело не в их
количестве и даже не в качестве, а в признании возможности реализации общей
идеи на разных материальных носителях. Если физики довольно легко (хотя и не
сразу) признали, что волна может быть как механической, так и электромагнит-
ной, то биологи до сих пор ищут всюду общий материальный носитель - либо в
форме общего предка, либо в форме сходного приспособления*. Таков штамп их
мышления.
Этот штамп именуется эволюционным, но как раз эволюции здесь нет. Факт
эволюции заимствован биологией из палеонтологии, где ни общее происхожде-
ние, ни приспособление почти никогда не обнаруживаются (а редкие исключения
таковы, что не дают основания для общих выводов). Наоборот, общее происхож-
дение и приспособление хорошо изучены биологией, но это изучение не показало
факта эволюции (за редкими примерами образования очень близких видов, о ко-
торых спорят, виды они или расы - таковы домашние собаки). В отождествлении
этих двух пониманий эволюции и состоит главный штамп обучения.
Даже происхождение человеческих рас таким путем понять не удалось: ни
общего предка, ни приспособления на ископаемых костях не видно, и рост числа
Это - следствие господства контактного детерминизма, неявно сохранившегося в умах
биологов со времен Декарта - см. заключительную статью. - Примеч. 2010 г.
213
находок лишь проясняет факт - налицо теоретический тупик [1]. Выход из него
видится мне в переосмыслении задач эволюционизма: вместо придумывания
общих предков и приспособлений, заняться, наконец, путями (как эволюция шла
[3, ч. 3]) и механизмами [3, ч. 2] эволюции.
Один пример выше рассмотрен: каждая раса людей прошла свой путь сама,
независимо от других, и все пришли к единому виду, поскольку реализовали од-
ну и ту же идею; на этом пути был, среди других, использован механизм, осно-
вой которому послужила идея, ранее реализованная насекомыми (а может быть,
и другими животными).
Каков механизм передачи идеи между разными (в том числе очень различ-
ными) видами? Вопрос отчасти рассмотрен в Дополнениях к книге [3]*. Даже
сейчас, в начале работы, видно, что для понимания эволюции нужны не преж-
ние штампы, а серьезный анализ**.
Литература
1. Маслов А. А. Другое человечество. Ростов на Дону, 2006.
2. Роуз Ф. Аборигены Австралии. Традиционное общество. М., 1989.
3. Чайковский Ю.В. Наука о развитии жизни, 2006. 4-58.
4. Чайковский Ю.В. Диатропика С.В. Мейена: сегодняшний взгляд, 2006. 4-57.
5. The Biology of naked mole-rat. Princeton, 1991.
*
Реализация одной и той же идеи на различных материальных субстратах в эволюции
обычна. Иногда она даже запускается с помощью одного и того же гена [3, п. 5-15*]. О
гипотезе, допускающей, что повторная реализация эволюционного достижения происхо-
дит быстрее и легче, нежели первая, см. Дополнение 7 к книге [3]. То же самое содержит-
ся (в одних и тех же местах) в книге [3] и в АСМ. - Примеч. 2010 г.
** Продолжение темы происхождения человеческих племён см. в Дополнении 10, как в
книге [3] так и в АСМ. - Примеч. 2010 г.
214
Часть 6. Эволюция и систематика
Зачем систематикам теория эволюции?
35 лет после Любищева*
Во всевозможных предисловиях часто пишут, что эволюционная идея - ос-
нова биологии, однако далее эволюцию обычно не поминают вообще или поми-
нают по мелочам. Хороший пример - систематика (таксономия): все признают,
что она основана на идее эволюции, но на деле строят систему организмов, мало
сообразуясь с данными об их эволюции**.
Например, про волка (Canis lupus) можно сказать, что у данного вида есть
точный адрес в системе: царство животных, тип хордовых, класс млекопитаю-
щих, семейство псовых (Canidae), род Canis (Собака). Это похоже на почтовый
адрес (страна, город, улица, дом, квартира), и можно сказать, что система орга-
низмов служит для их адресации. Эта же система указывает на сходство строе-
ния тела: все собаки более похожи друг на друга, чем другие псовые (например,
песец), псовые - нежели другие хищные и т.д. Такую общность строения по-
следние 140 лет трактуют эволюционно - общий предок рода Canis жил позже,
чем общий предок семейства псовых, этот предок жил позже, чем общий предок
хищных и т.д.
Однако так просто получается только на некоторых удобных примерах. Ес-
ли же мы посмотрим литературу, например, о семействе псовых, то увидим, что
структура его (в том числе состав рода Canis) то и дело радикально меняется,
хотя новых его видов не открывают уже почти сто лет. В частности, род Canis
понимают то очень узко (7 видов), то очень широко (до 30 видов, т.е. почти всё
семейство псовых). Добро бы при этом наблюдалось какое-то движение вперед,
так нет же: несмотря на новые методы исследований, систематика ходит по кру-
гу - ученые время от времени возвращаются к очень старым вариантам системы,
подчас к XVIII веку, к Карлу Линнею. Например, род Vulpes (Лисица) у Линнея
отсутствовал (его виды входили в состав рода Canis), и в наше время находятся
авторы, снова включающие (на основе компьютерного анализа!) лисиц в состав
рода Canis.
1. Является ли систематика эволюционной?
Нам важны не таксономические тонкости, а самые грубые эволюционные
обстоятельства. Так, в роде Vulpes есть вид корсак, водящийся в средней и юж-
ной Евразии, и вид американский корсак, водящийся в средней и южной части
Северной Америки; уже из их названий видно, что они очень сходны, а из отда-
ленности зон их распространения - что их общий предок, если он вообще суще-
ствовал, не мог жить недавно (жил до плейстоценового похолодания). Естест-
* Любищевские чтения. Ульяновск, 2007, с. 132-140.
" На что в течение полувека тщетно указывал А.А. Любищев. - Примеч. 2010 г.
215
венно допустить, что их тесное сходство не унаследовано, а возникло. Как бы
это проверить? Для этого надо прежде всего выстроить ряд аналогичных сход-
ных видов.
Оказывается, его являет собой всё семейство псовых: еще в 1880 г. Томас
Гексли разделил всех псовых на две группы - волкообразные и лисицеобразные,
причем обе представлены как в Евразии, так и в Америке, в основном, Южной.
Пары сходных видов (в каждой - один евразийский и один американский вид) и
образуют искомый ряд: естественнее допустить, что эти сходства возникли не-
зависимо, нежели то, что каждый евразийский вид каким-то образом перебрался
через океан, где породил сходный вид. И действительно, генетический анализ
показывает, что все южноамериканские виды ближе друг к другу, чем к евра-
зийским [1].
Выходит, что близкое сходство отнюдь не всегда означает близкое родство.
В книге [2] рассказано, что само понятие «ряд» было введено как раз для ото-
бражения этого факта - сходства без близкого родства.
Так в самом ли деле нынешняя систематика эволюционна в своей основе? В
новейшей программной статье «Таксономические новшества и проблемы поль-
зователей» [3] эволюция упомянута по сути всего один раз и вот в каком кон-
тексте: существует филогенетическая85 концепция вида, и дает она фактически
ту же систематику, что и прагматическая концепция, никакого отношения к
эволюции не имеющая. Согласно первой, вид есть множество организмов, кото-
рые можно уверенно отличить от членов других множеств и «внутри которого
существуют отношения предков и потомков» (что значит это странное словосо-
четание, не сказано). Согласно второй, вид есть просто множество организмов,
которые можно уверенно отличить от членов других множеств (с. 554).
Но ведь уверенно различаются и подвиды (например, аральская вобла, кас-
пийская вобла и тарань - подвиды плотвы обыкновенной) - почему бы не на-
звать их тоже видами? Оказывается, в новейшем каталоге рыб так и сделано, с
чем и спорит статья [3], резонно замечая, что на этом пути «невозможно оста-
новиться: практически каждую популяцию приходится признать самостоятель-
ным видом» (с. 555). Авторы поясняют это положение на роде Coregonus (Сиг),
в котором, например, название С. lavaretus новейшей сводкой закреплено ис-
ключительно «за сигом из оз. Буржэ во Франции» (там же). Разумеется, пользы
от такой системы, будь она целиком развита, окажется очень мало.
Пафос же статьи [3] состоит в следующем. Систематики заняты, в основ-
ном, изобретением новых видов, «выделение видов превращается в самоцель, а
85 Филогения - тот тип эволюции, который может быть представлен в форме родословно-
го древа, т.е. при котором у каждого таксона (вида, рода, семейства и т.д.) есть ровно
один таксон-предок. Например, все собаки считаются происшедшими от волка. Явным
нарушением филогении являются межвидовые гибриды (например, гибрид волка с соба-
кой или глухаря с тетеревом).
216
классификация становится бесполезной» (с. 557). Однако положение с видами
еще довольно сносно, поскольку у вида есть хоть какое-то определение, тогда
как у более высоких таксонов (родов, семейств и т.д.) нет и этого. Авторы при-
зывают «сначала исследовать и описать его (разнообразия организмов - Ю. Ч.)
структуру и лишь после этого дискретным группировкам (если таковые обна-
ружатся) придать таксономический статус и присвоить научные названия. Ины-
ми словами, предполагается продвижение от изучения объектов к их классифи-
кации... а не наоборот» (с. 556).
Прошу обратить внимание на подчеркнутое мной слово «дискретным» (то
есть четко отличным одна от другой). Откуда может взяться дискретность, если,
как утверждает господствующая эволюционная доктрина (дарвинизм), господ-
ствует плавная непрерывност всех наследственных преобразований? Для видов,
родов и более высоких таксонов дискретность свойств мыслится в дарвинизме
как итог вымирания промежуточных форм. И хотя никто этого вымирания на
практике не наблюдал, все-таки таковая возможность логически допустима. Од-
нако в цитированном рассуждении речь идет об исходной процедуре анализа,
когда объектом наблюдения являются не роды и не виды, а непосредственно на-
блюдаемые группы особей (популяции). В них-то и идет элементарный (а пото-
му, согласно дарвинизму, плавный) процесс обретения новых качеств - теорети-
ки дарвинизма в один голос утверждают, что здесь на уровне особей никаких
дискретностей быть не должно. Какой же теорией дискретность объясняется?
Авторы статьи [3] таких вопросов не ставят, однако признают, что никакой
теории у систематики нет, что каждый систематик творит, как хочет, а страдают
при этом пользователи, т.е. все остальные. Подобных заключений можно при-
вести много. Например: «Итак, естествоиспытатель вправе задать вопрос: за-
чем нужна такая классификация кому-либо, кроме филогенетиков?» [4, с. 526].
Поэтому неизбежны две группы вопросов. 1) Если никакой теории у сис-
тематики нет, то почему все-таки существует сама система организмов, кото-
рой, хорошо или плохо, но все пользуются? Почему, например, волком все
люди называют одну и ту же группу зверей, почему они при этом понимают
друг друга? Или, другими словами, почему виды и иные таксоны дискретны?
2) Если нынешняя биология в самом деле эволюционна, то почему не построе-
на эволюционная теория систематики? Строится ли она? Можно ли ее постро-
ить вообще, или удел эволюционного учения - ютиться в предисловиях?
На последний вопрос ответить проще всего: эволюционную систематику
вполне можно построить тогда, когда есть достаточный ископаемый материал.
Мы можем, например, подкрепить идею происхождения всех собак от волков
тем фактом, что собаки появились позже волков и появились там, где водились
волки. Другое дело, будет ли при этом система филогенетической, но об этом
скажу позже (п. 2), когда будет рассмотрена первая группа вопросов.
Там же, где ископаемого материала нет, всякие эволюционные рассуждения
голословны, а потому и недолговечны. Так, весьма скудны ископаемые данные
217
о происхождении южноамериканских псовых от остальных псовых, поэтому их
отнесение к одному семейству (если понимать каждый таксон как происшедший
от общего предка) то и дело подвергается сомнению, и молекулярные методы
помогают мало [1].
Точно так же со многими иными группами. Так, ныне в Африке водится 2
вида человекообразных обезьян - шимпанзе и горилла; они в ископаемом со-
стоянии вообще неизвестны (что с эволюционной точки зрения более чем
странно), поэтому приходится ограничиться сравнением нынешнего материала.
По строению тел считается, что они отделились от предков людей 15-20 млн лет
назад, по данным же молекулярной генетики выходит совсем иначе - они отде-
лились приблизительно 7 (горилла) и не более 5 (шимпанзе) млн лет назад. Рас-
хождение данных в 10 млн лет совершенно неприемлемо для целей эволюцион-
ной систематики, поскольку уже 7 млн лет назад в Африке существовали (со-
гласно ископаемым данным) предполагаемые предки людей, более близкие к
человеку строением скелета, чем шимпанзе. Это широко известно.
Но есть и более радикальная позиция: «Человек и шимпанзе по геному от-
личаются менее чем на 1%. Это значит, что время их расхождения не превышает
1,5 миллиона лет. Однако палеонтологические данные говорят о том, что уже
миллион лет назад на Земле существовал человек, близкий к человеку совре-
менного типа и отличающийся от него только на уровне подвида» [5, с. 15]. Тут
систематика неэволюционна в принципе.
В последние 30 лет появились молекулярные методы анализа сходств (пре-
жде всего - геносистематика [6]), которые претендуют на объективную исто-
рию. Однако объективный анализ должен, если он обоснован фактически, при-
водить разных авторов к одному и тому же выводу, на самом же деле разные ге-
носистематики приходят к весьма разным выводам, к тому же выводы эти почти
всегда противоречат выводам обычной систематики. Словом, на сегодня нет
эволюционной систематики, которая была бы настолько долговечна и общепри-
знанна, чтобы ею могли пользоваться остальные биологи.
2. Дарвинизм и номогенез в систематике
Читатель или слушатель, помнящий школьный курс биологии, удивится:
как же так? Неужели разнообразие организмов не есть итог эволюции? Оказы-
вается, на этот счет есть разные точки зрения. Мне ближе всего та, которую вы-
сказывал А.А. Любищев: не надо путать разнообразие организмов (явление при-
роды) и систему организмов (наше описание этого явления, т.е. плод человече-
ской мысли). Хотя первое и возникло в ходе эволюции, вторая может не иметь
(и, как оказывается, не имеет) к ней никакого отношения. Поясню этот тезис
аналогией с химией.
В книге [2] уже говорилось про гомологические ряды, сходные в биологии и
в химии. Продолжая тему, зададимся вопросом: является ли Периодическая сис-
тема элементов итогом эволюции? Разнообразие элементов таковым является:
сперва во Вселенной был один водород, затем появился гелий, а потом и более
218
тяжелые элементы. Но сами свойства элементов определяются отнюдь не их
происхождением, а характером энергетических уровней, на которых располага-
ются их электроны. Поэтому данный атом может появиться как путем соедине-
ния более легких частиц или за счет бомбардировки более легкого атома прото-
нами и нейтронами, так и путем распада более тяжелого атома.
Иными словами, ходом эволюции могло определяться лишь то, как и в ка-
ком порядке будут заполняться клетки менделеевской таблицы, но свойства са-
мой таблицы (в частности, ее периодичность) никакого отношения к эволюции
не имеют. Хотя весьма вероятно, что все элементы произошли из водорода, но
каждый из остальных элементов происходил заведомо различными путями, так
что принцип филогении тут бессмыслен.
Так вот, номогенез (эволюционное учение, о котором рассказано в книге [2])
утверждает, что эволюция организмов идет похожим образом: в ее ходе запол-
няются клетки некой огромной многомерной таблицы возможных форм орга-
низмов; каждая клетка может быть в ходе эволюции заполнена различными пу-
тями. Например, род Nyctereutes (Енотовидная собака) мог произойти как из
псовых, так и из енотовых. Он ныне отнесен к семейству псовых, причем это -
волевой акт, а не итог анализа эволюции.
Всю огромную таблицу окинуть взглядом невозможно, но некоторые ее
свойства легко видны: прежде всего, это - ее дискретность и гомологические
ряды. В химии дискретность выражается в том, что всякий атом занимает опре-
деленную клетку таблицы, а гомологические ряды в химии именуются перио-
дичностью, и номогенез трактует биологию так же. То есть утверждает как пер-
вичную реальность то самое, что дарвинизм пытается либо не замечать, либо
отрицать, либо считать итогом какого-то прошлого, никем не наблюдавшегося
процесса вымирания.
Согласно номогенезу, многообразие организмов (нынешних и вымерших) -
это множество заполненных клеток таблицы. Разнообразие организмов - это со-
вокупность всех различий и сходств между' организмами, занявшими разные
клетки таблицы. Если принять данный взгляд, то задача систематики окажется
не в исследовании путей заполнения таблицы, а в анализе свойств самой табли-
цы. Как в химии.
Так фактически систематика и строится - на выявлении сходств и различий
ныне живущих организмов, - а история додумывается. Видно это хотя бы из то-
го, что исторические связи нынешних групп то и дело пересматриваются, да и в
одновременно вышедших работах часто понимаются по-разному. Пользоваться
такой эволюционной систематикой невозможно, и, судя по приведенным репли-
кам специалистов (а также множеству сходных), ею никто и не пользуется. Вме-
сто нее пользуются тем фактом, что виды дискретны, не задумываясь о том, что
он по сути номогенетичен. По-моему, авторам статьи [3] стоило бы признать,
что они, как и остальные систематики, фактически исходят из идеи номогенеза,
когда признают дискретность в качестве исходного факта.
219
В отношении крупных организмов система достаточно устойчива (чтобы ею
можно было пользоваться) потому, что они имеют бытовые названия, к которым
и прибегают в своих спорах ученые, чтобы понять, о чем идет речь (см. [2, с.
566]; это же хорошо видно из статьи [3]).
Что же касается основной массы мелких организмов, не имеющих бытовых
названий, а также родов и семейств, то положение сложилось удручающее. Без
консультации с узким специалистом по данной группе тут не обойтись, а это
мало кому из практиков доступно. Об этом не раз писал Любищев, другие писа-
ли и позже.
3. В чем состоит использование систематики?
Практическое (то есть вне самой систематики) использование систематики
сводится, в основном, к двум процедурам: как определить вид (то есть как, пу-
тем исследования самого организма, узнать его положение в системе и научное
название) и как обрисовать разнообразие организмов.
Первая задача постоянно стоит перед самыми различными учеными и прак-
тиками. Ученый должен быть уверен, что работает с тем же объектом, что его
заморский коллега. Агроном, врач и ветеринар должны быть уверены, что обна-
ружили того самого возбудителя, который описан в руководстве. Эколог, гово-
рящий о возрастании числа видов в экосистеме в силу природоохранных мер,
должен исключить возможность того, что этот рост - просто итог разбиения
прежних видов новыми систематиками.
Задача определения видов никакого отношения к эволюции не имеет, даже
если определить надо ископаемый организм (биологи, отрицающие факт эволю-
ции, справляются с задачами определения не хуже прочих).
Поэтому, если систематики вдруг откажутся от эволюционного принципа
(для этого им потребуется немного: признать, что никаким эволюционным зако-
ном они всерьез не пользуются, а всего лишь именуют некоторые свойства раз-
нообразия организмов эволюционными), то процесс определения не пострадает
вовсе. Наоборот, он станет проще и яснее, поскольку системы перестанут ме-
няться вслед за сменой понимания того, какой вид к какому ближе эволюцион-
но. Такие смены понимания эволюции, увы, происходят в каждой крупной
группе чуть ли не ежегодно, и приближения к стабильности не видно.
А вот вторая задача радикально и по существу зависит от эволюционных
взглядов ученых. Так, дарвинист и ламаркист склонны описывать организм как
совокупность приспособлений - к иным организмам и к нежйвой природе; на-
оборот, номогенетик мало интересуется приспособлениями - ему интереснее
выявить общие закономерности, по которым разнообразие сложилось. Со сме-
ной эволюционных взглядов абрис живого мира должен сильно измениться.
Вместо нынешней уверенности, что всякий нынешний организм таков потому,
что приспособлен к среде лучше, чем вымершие, должно возобладать понима-
ние природы как целого, где каждый организм занимает свое место в системе
организмов.
220
Для тех, кто не видит большой разницы между обоими пониманиями систе-
мы организмов, поясню: первое исходит из идеи эволюции снизу (когда исход-
ным понятием является изменение признака), тогда как второе - из идеи эволю-
ции сверху (когда исходными понятиями являются изменение экосистемы и из-
менение таксона). Поскольку первое понимание явно зашло в тупик (эволюцио-
нисты потонули в деталях, так и не сумев внятно объяснить процесс эволюции
жизни), всё слышнее становятся голоса тех, кто предлагает понимать эволюцию
как цельный процесс. С этой позиции открывается надежда построения такой
системы организмов, которую можно будет не только заучивать, но и понимать.
4. Как же строить систематику?*
Едва в 1922 году появился номогенез, как Любищев выступил с парадок-
сальным заявлением: систематика вообще не должна быть эволюционной. Глав-
ный довод его, если использовать нынешние термины, был именно тот, что эво-
люция являет собой процесс заполнения объективно существующей таблицы
возможностей, причем систематика есть наука о свойствах данной таблицы и
способах ее построения учеными, а не о путях ее заполнения самою природой в
ходе истории жизни.
Любищев отметил, что изображение системы организмов в форме древа
(иерархия) взято отнюдь не из анализа процесса эволюции, а заимствовано из
логики систематики времен Линнея и раньше [2, гл. 10].
Систему организмов, говорил он, следует строить не в иерархической, а в
параметрической форме. (Примеры параметрических систем: система химиче-
ских элементов, где положение элемента задается одним параметром - зарядом
ядра; географическая карта, где пункт задается двумя параметрами - широтой и
долготой; почтовый адрес, где место жительства задается несколькими алфа-
витными параметрами.)
В конце жизни Любищев занял несколько более мягкую позицию, высказав
несколько общих положений о возможной роли номогенеза в развитии система-
тики; однако он не развил их [7].
Сейчас, через 85 лет после выхода «Номогенеза» Л.С. Берга и через 35 лет
после смерти Любищева, мы знаем о системе организмов гораздо больше. В
главе 10 книги [2] рассказано, что классификация в форме древ весьма удобна
для адресации и именно поэтому широко используется в разных отраслях зна-
ния, так что вряд ли стоит вообще от нее отказываться. Однако сама по себе она
ничего не говорит об истории, в том числе и об эволюции организмов. Более то-
го, в последние 30 лет выяснился феномен блочной эволюции', процесс эволюции
носит характер комбинации слабо зависимых блоков [2, с. 315]. Обычно этот
процесс удобнее изображать не древом, но сетью путей, а его итог - многомер-
ной таблицей.
* Подробнее см. следующую статью сборника, где, в частности, разобраны представления
С.В. Мейена, во многом вытекавшие из установок А.А. Любищева. - Примеч. 2010 г.
221
Поэтому, если система предназначена для целей адресации (в том числе для
определения видов), ее следует оставить в форме древа. Однако нельзя считать
это древо родословным и изменять его каждый раз при получении новых сведе-
ний или соображений об эволюции данной группы.
Если же мы хотим понять, как устроено разнообразие организмов, то древо
даст нам очень мало, независимо от того, будем ли мы считать ли его эволюци-
онным или нет. И свойства самих организмов, и их связи друг с другом образу-
ют не древо, а многомерную сеть. Итак, в биологии нужны две системы орга-
низмов, причем эта мысль идет от Линнея [2, с. 569].
Если говорить о системе царств (макросистеме), то, по мысли биолога Р.М.
Зелеева (Казань), она «видится в форме серии параметрических таблиц с указа-
ний траекторий развития...» [8, с. 86-87]. В частности, мне в 1986 г. удалось
изобразить ее в виде плоской кольцевой таблицы, которая многократно опубли-
кована (например: [2, с. 598], [9, с. 80]). Эволюция предстает в ней как движение
от кольца к кольцу и начинается с кольца «одноклеточные прокариоты». Такая
таблица удобна для единого взгляда на живую природу и ее эволюцию, но не
ставит целей адресации, каковая предполагается традиционной, в форме древа.
Фактически две систематики приняты на сегодня в бактериологии: одна,
филогенетическая, то и дело пересматривается и особой ценности не представ-
ляет [4], и вторая, параметрическая. В ней три параметра: группа (этот параметр
имеет всего 4 значения) и два параметра - названия рода и вида, задаваемые пе-
речнями в алфавитном порядке и находимые (в ходе определения вида) посред-
ством определительных ключей [10].
Обо всем этом не раз говорилось, в том числе - мною на Любищевских чте-
ниях в Москве [7] и Ульяновске [9]. По-моему, это имеет смысл рассказывать
студентам и даже старшеклассникам школ с биологическим уклоном, что неко-
торые преподаватели и делают. Что можно добавить нового?
Критические статьи типа [3; 4] достаточно ясно говорят, что осознание ту-
пика, в котором оказалась эволюционная систематика, становится всё более ши-
роким. Однако почти все систематики ищут решения в рамках тех же самых по-
нятий, которые оказались тупиковыми. Налицо вопиющее нежелание система-
тиков читать теоретическую литературу, в том числе - читать Любищева.
Моя кольцевая таблица теперь не одинока: в работах [2, 7] названы и другие
системы, в том числе в [2] приведена прямоугольная таблица из книги [11], изо-
бражающая вполне новую «Систему членистых» зоолога В.Я. Павлова (Моск-
ва), который прямо ссылается на понимание системы Любищевым как на свой
исходный пункт [11, с. 174-175]. Система Павлова является эволюционной (в
том же смысле, что и моя); это, полагаю, проясняет идею Любищева о роли но-
могенеза в построении систематики.
Наконец, стоит заметить, что главный пропагандист молекулярных эволю-
ционных древ Карл Вёзе (США) три года назад признал негодность своих преж-
них филогенетических построений (и самой идеи древа) для ранних стадий эво-
222
люции [12]. Его шаг вполне соответствует в теории тому, что на практике еще в
1994 г. сделали бактериологи [10]. Очередь за остальными биологами. Для это-
го им хорошо бы вспомнить о номогенезе .
Литература
1. Wayne R.K., Geffen Е. е.а. Molecular systematics of the Canidae // Syst. Biol., 1997,
vol. 46, № 4.
2. Чайковский Ю.В. Наука о развитии жизни. М., 2006.
3. Мина М.В., Решетников Ю.С., Дгебуадзе Ю.Ю. Таксономические новшества и
проблемы пользователей // Вопросы ихтиологии, 2006, № 4.
4. Заварзин ГА. Составляет ли эволюция смысл биологии? // Вестник РАН, 2006, № 6.
5. Мащенко Е.Н. Новые страницы каменной летописи // Наука и жизнь, 2006, № 5.
6. Медников Б.М. Геносистематика: проблемы и трудности И Биология в школе,
2000, № 1.
7. Чайковский Ю.В. Естественная система: 30 лет после Любищева // Любищев и
проблемы формы, систематики и эволюции. 30-е Любищевские чтения. М., 2003.
8. 3елеев Р.М. Проблемы построения системы организмов и некоторые критерии ме-
гатаксономии И XVIII ЛЧ. Ульяновск, 2004.
9. Чайковский Ю.В. Любищев и естественная система организмов // XVIII Любищев-
ские чтения. Ульяновск, 2004.
10. Определитель бактерий Берджи. М., Мир, 1997, тт. 1 -2.
11. Павлов В. Я. Периодическая система членистых. М., 2000.
12. Woese C.R. A new biology for a new century // Microbiology and Mol. Biol. Reviews,
2004, vol. 68, № 2.
i
I
* Речь идет о новом номогенезе - в том смысле, в каком это слово употребляется в на-
стоящем Сборнике. - Примеч. 2010 г.
223
Теория систематики по Мейену'
В комментариях Любищева к рукописи Кузина есть запись: “противоположе-
ние точек зрения систематики и сравнит, анатомии мне нравится, но центральным
пунктом систематики по-моему является не тип, а понятие формы и структуры
системы. Впрочем возможно, что это практически то же самое”. Из этого зароды-
ша мысли, равно как и из беглого признания Любищевым роли номогенетических
взглядов для систематики (см. п. 4), можно вывести основную линию рассужде-
ний Мейена - самого крупного, на мой взгляд, представителя школы Любищева.
Взгляды Мейена на систематику неоднократно излагались и комментирова-
лись, в том числе мною, так что здесь ограничусь выводом: Кузин и Любищев об-
ратили внимание на действительно центральный пункт теории системы, однако
структуры системы и типа оказались, по Мейену, не одним и тем же и не проти-
воположностями, а взаимодополнительной парой. Эта пара работает в теории, ко-
торую Мейен назвал типологией, а я (после его смерти) — основой диатропики.
Что же касается способа построения конкретной системы, то Мейен показал
(на примере голосеменных), как это фактически делается в рамках современного
ему номогенеза. Он рассматривал принятые в науке положения:
«(1) эволюционные преобразования постепенны, (2) преобразования орга-
нов идут уже известными нам способами (модусами), (3) разнообразие форм
должно быть сведено с помощью этих модусов к единственной исходной фор-
ме» [3, с. 303] - и пришел к выводу, что они верны не всегда. Наряду с ними на-
до рассматривать и противоположные им:
«1) эволюционное преобразование форм может быть дискретным; 2) коли-
чество и характер модусов преобразования не фиксировано; 3) разнообразие
форм, наблюдаемое в данном таксоне, выводится не из одной формы, а из раз-
нообразия же исходных форм» [3, с. 304].
По форме система Мейена является филогенией, но по сути она конгрегаци-
онна в смысле Е.С. Смирнова’*. В 1984 году Мейен писал (статья о систематике
голосеменных в журнале “Botanical Review”):
«Со времен A. de Jussieu система организмов строится по принципу, позже
(Smirnov, 1925) удачно названному конгрегационным. Конгрегационные таксо-
ны естественны в том понимании естественности, которое господствовало в ин-
дуктивной логике XIX века (W. Whewell, J. St. Mill) , а также в додарвиновской
биологии. Суть конгрегационной таксономии в том, что система строится “сни-
зу”, от таксонов низшего ранга к таксонам более высокого ранга. Таксоны по-
степенно группируются в конгрегации по признакам, выявляемым и оценивае-
мым в ходе работы. В конце концов для каждого таксона выводится некоторый
устойчивый комплекс признаков (синдром)... Допускается, что любой признак
может исчезнуть у каких-то представителей таксона..., тогда в диагностической * **
Из статьи «Естественная система. 30 лет после Любищева» (4-47), с. 44-47.
** См. п. 7 предыдущей статьи.
224
роли выступают другие признаки синдрома. Это значит, что вес признака не
считается фиксированным» [4, с. 42].
Суть (для нашей темы) системы голосеменных Мейена состоит в том, что
она построена как итог исследования эволюции (но не как объяснение заранее
заданной схемы). А итог состоял для Мейена в том, что принятая в науке схема
верна не для всех случаев, что в одних ситуациях прежнюю систему можно (с
модификациями) принять, а в других надо строить новую, причем различие си-
туаций выясняется при анализе эволюции конкретных групп голосеменных. Са-
ма же эволюция предстала Мейену как номогенез.
Мейен вовсе не отвергал идею филогении, но хотел убедиться, когда она вер-
на, а когда нет. И убедился в том, что она с приемлемой точностью верна для не
слишком давних времен и не слишком мелких таксонов. Конкретно говоря, мы не
должны требовать от филогении указаний родства точнее, чем до семейства в ме-
зозое и позже, точнее, чем до порядка в карбоне и перми, а для девона (когда го-
лосеменные появились) можно указать только родство классов', и дело вовсе не в
недостатке данных - когда их достаточно, они выявляют наличие “синтетических
типов”, т.е. сочетания свойств нескольких таксонов равного ранга [4, с. 94, 96].
Огорчает тот факт, что конкретная система построена Мейеном без явных
обращений к типологии и изложена упрощенно, без раскрытия методов; а статья
1984 года - еще и в расчете на англоязычных читателей: архетипом названа
“общая система признаков таксона”, типологией - перечень форм, взаимодо-
полнительность системы и формы опущена и т.д. Редкие упоминания положе-
ний подлинной типологии (диатропики) носят побочный характер, и неясно, на-
сколько они были нужны автору при разработке системы.
Однако среди англоязычных читателей нашлось трое проницательных, и
Мейену пришлось указывать, в ответ на их критику, теоретические корни своей
системы: “Все трое моих коллег хотели бы найти в моей статье какое-то недву-
смысленное объяснение тому, каким образом я провожу гомологии между орга-
нами ... и т.д. Нет сомнений, что понять выводы, сделанные автором, было бы
намного легче, если бы читатели познакомились с его позицией по этим теоре-
тическим проблемам”. Некоторые пояснения тут же даны - например, упомянут
“метод взаимного просветления”, в котором легко увидеть взаимодополнитель-
ность [4, с. 107, 110]. Словом, любищевская линия мысли налицо, но требуется
отдельный анализ, каковой может проделать только палеоботаник.
Табличные системы
В таблице Менделеева элементы упорядочены в строки, а сходства между строка-
ми (ряды) проявляются в форме столбцов. Кроме всем известной периодичности,
эта таблица содержит малоизвестную вторичную периодичность* [5]. Нельзя ли
Она имеет несколько форм. Наиболее заметны два параллельных ряда - лантаноиды и
актиноиды. Менее заметна периодичность свойств внутри побочных подгрупп (например, в
подгруппе редкоземельных элементов, куда входят и лантаноиды) [5]. - Примеч. 2010 г.
225
похожим образом упорядочивать организмы? После открытия периодического за-
кона (1869 г.) действительно появилось несколько «периодических систем» не-
скольких групп организмов, но успеха они не имели. Беда их состояла в том, что
они выражались громоздкими таблицами, в которых преобладали пустые клетки;
и только Мейен через сто лет дал нам понять, что запись в форме строк и столб-
цов полезна - не столько для построения конкретной системы, сколько для прояс-
нения логики систематики (и, добавлю, для уяснения хода эволюции).
Пустота большинства клеток означает всего лишь то, что для данной груп-
пы табличная форма записи очень избыточна, но она вовсе не означает порочно-
сти самого принципа описания объекта посредством пересечения строки и
столбца некоторой таблицы. Другими словами, таблица в принципе работает
всегда, однако конкретную таблицу имеет смысл строить лишь тогда, когда в
ней оказывается мало пустых клеток.
Яркий пример удобства таблиц являют определители, составленные по схеме
ботаника Б.Е. Балковского, например [1]”, но в них не видно ничего эволюцион-
ного. Пример эволюционной табличной систематики привел ботаник А.П. Хохря-
ков, чья система двудольных выражена им в форме таблицы из 128 клеток, среди
которых всего 16 пусто [6]. Автор нашел в ней любопытные тенденции, в том
числе эволюционные - например, подтверждение идеи Мейена о тропическом
происхождении таксонов высокого ранга. Есть тут и элементы периодичности; их,
если учесть частный характер системы (двудольные - подотдел отдела цветковых)
лучше всего сопоставить с упомянутой вторичной периодичностью элементов.
Ничего такого по обычной (филогенетической) системе увидеть невозможно.
Существует табличная система листовых пластинок [2]. К сожалению, дру-
гие известные мне примеры табличных систем гораздо менее удачны. Однако и
приведенных примеров достаточно, чтобы понять, что неразумно априори счи-
тать иерархию спасением.
Литература
1. Бородина И.А. и др. Деревья и кустарники СССР. Полевой определитель. М.,
1966.
2. Корона В.В. Основы структурного анализа в морфологии растений. Свердловск,
1987.
3. Мейен С.В. Органы размножения голосеменных..., 1982, (М-218).
4. Мейен С В. Эволюция и систематика высших растений , 1992, (М-328).
5. Мельников В.П., Дмитриев И.С. Дополнительные виды периодичности в периоди-
ческой системе Д.И. Менделеева. М., 1988.
6. Хохряков А.П. Табличное изображение системы двудольных // Общебиологич. ас-
пекты филогении растений. (Совещание в МОИП.) М., 1991.
** Подробнее см. АСМ, с. 323. - Примеч. 2010 г.
226
Естественная система и таксономические названия
Взяться за эту статью меня побудило появление программной работы [Мина
и др., 2006], авторы которой мягко, но ясно показывают, что профессионалы-
таксономисты совершенно равнодушны к проблемам пользователей, что ника-
кой теории в систематике по сути нет и что единственный серьезный совет, ка-
кой можно дать пользователям - надо избегать новых сводок, предпочитая им
старые просто потому, что они старые, устоявшиеся.
Да, они правы: несмотря на учёную оболочку, в систематике царит личное
мнение, и потому, в частности, безостановочно меняются названия. Факт доса-
ден, однако недавно он с гордостью декларирован в качестве нормы:
«Систематика является наукой и, следовательно, не может диктоваться
кем-либо; каждый исследователь вправе сам решать, какие принципы
систематики следует использовать и какой должна быть классификация:
он может основывать классификацию на филогении или на чем-либо ином
(на чем, не сказано - Ю.Ч.)... Согласно принципам, действующим в науке
начиная с эпохи Возрождения, никакой авторитет и никакая сколь угод-
но демократично избранная комиссия не может навязывать своего мне-
ния ученому, а может лишь пытаться переубедить его логическими до-
водами. В отличие от систематики, номенклатура не является наукой; т.к.
названия придумываются искусственно. Поэтому в номенклатуре могут
действовать правила, утвержденные комиссией» [Клюге, 2000, с. 43].
Здесь - цепь заблуждений, они легко заметны, и мне даже странно, что при-
ходится начинать статью с их разъяснения. Но приходится.
На самом деле номенклатура (учение о названиях) - тоже часть науки, толь-
ко не естественной, а гуманитарной, точнее - классификационной. В любой
науке есть своя классификация, и она нигде не терпит своеволия. В точных нау-
ках это очевидно (изменить систему элементарных частиц или химических эле-
ментов вряд ли возможно), но и в описательных ситуация по сути та же.
Так, лингвист отнюдь не вправе своевольно писать грамматику. Хотя она и
выглядит классификацией произвольно введенных правил, но на деле она отража-
ет закономерности данного языка. Правда, попытки видеть во всех языках латынь
делались (например, в XVII веке филологи находили в 14-падежном финском
языке всего 6 латинских падежей [Основы..., 1975, с. 160]), но то была заря фило-
логии. Она напоминает зарю ботаники, когда авторы травников XVI в. хотели ви-
деть в Германии лишь вариации средиземноморской флоры, описанной античны-
ми авторами, зато каждый группировал растения, как хотел.
Увы, в систематике происходит ныне откат назад: многие снова уверены,
что систематика для всех уровней и групп одинакова, зато каждый «вправе сам
решать». На самом деле ученый может допускать вольности лишь в рамках при-
нятых норм, и на их нарушение научное сообщество отвечает отнюдь не «логи-
* Линнеевский сборник/ред. И.Я. Павлинов. М., 2007, с. 381-436.
227
ческими доводами», а изгнанием его из сообщества (но, к счастью, не из жизни,
как бывало в эпоху Возрождения, да и позже).
Это, кстати, делает и сам Н.Ю. Клюге: из его книги, именуемой учебником,
изгнано всё, что выходит за рамки воззрений кладистов (той группы, к которой
принадлежит автор). Из иных взглядов помянуты лишь «традиционализм»,
единственным приверженцем которого назван Эрнст Майр (его книга 1971 г.),
хотя на самом деле он господствует у практиков, и «градизм» (ни имен, ни дат).
Притом замечу, что традиционализм, кладизм и градизм - филогенетические
идеи. Иные (нефилогенетические) взгляды Клюге даже не упомянул, хотя у него
сказано: «Подавляющее большинство современных исследователей сходится на
том, что классификация должна отражать филогению» (с. 24).
А остальные, подавляемое меньшинство? Почему оно есть, какие трудности
филогенетической систематики вынуждают его существовать? В известных мне
руководствах об этом нет ни слова (в отличие от эпохи Возрождения, когда спор
с оппонентами был нормой). Трудности же огромны.
Главная - поток новых названий не падает, а растет (см. далее). Это значит,
что ухватить суть системы до сих пор не удалось. Отсюда и странный лозунг:
«каждый вправе сам решать» - там, где все заняты общим делом (например,
строят одну и ту же систему), так не говорят.
Однако как раз в области номенклатуры биолог, вопреки Клюге, почти сво-
боден: правила задают допустимые рамки изменений названий, но, как показала
практика, почти не уменьшают числа самих изменений. Это выглядит странно,
поскольку назначение номенклатурных кодексов и комиссий - именно стабиль-
ность названий [Клюге, 2000, с. 44, 54].
Попробуем выявить (через анализ мнений меньшинства) причины несоот-
ветствий, чтобы понять, что же надо делать дальше. Как ни странно, основное
предложение по стабилизации названий мы найдем как раз у Клюге.
1. Названия неустойчивы
С проблемой названий мне пришлось столкнуться 35 лет назад, когда мы с
женой Риммой стали редактировать перевод «Иллюстрированной энциклопедии
животных» [Станек, 1972]. Оказалось, что текст содержал множество ошибок,
но отредактировать его во многих случаях было невозможно без консультации с
разными зоологами. Дело в том, что часто нам не удавалось соотнести описан-
ные там виды животных с видами из доступных для нас руководств - хотя бы с
видами из «Жизни животных», издававшейся тогда издательством «Просвеще-
ние». Не удавалось потому, что чешский автор ориентировался на немецкие
справочники разных лет издания, тогда как советское издательство поручало
каждую главу узкому специалисту, имевшему собственную «новейшую» точку
зрения и, соответственно, свои латинские названия.
Учёная латынь устаревает вообще очень быстро: любое название может в
новой сводке быть изменено. И будет затем охраняться номенклатурными пра-
вилами, если изменение совершено в соответствии с оными правилами.
228
Позже, занявшись историей науки, я смог убедиться, что чехарда латинских
названий - подлинный бич биологии, бьющий и по смежным наукам (экологии,
медицине, сельскому, лесному и рыбному хозяйству и т.д.). При необходимости
проследить судьбу видового ареала или способов борьбы с эпидемией, при вы-
яснении реального вклада в науку данного биолога или целой школы прошлого
- надо опять и опять искать специалиста по каждой узкой теме - подчас для того
лишь, чтобы сопоставить латинские названия.
Хорошо мне - в Москве, притом в кругу самых различных ученых, а каково
практикам на местах? Ну хотя бы: что делать эпидемиологу, если названия пара-
зита в определителе и в медицинском руководстве не совпали?
Сами систематики считают (насколько могу судить по нескольким беседам)
такое положение вполне удобным, поскольку чувствуют свою незаменимость,
но с этим, по-моему, пора кончать - как потому, что страдает практика, так и
потому, что в таксономической чехарде нет объективной нужды.
Выходит, неправду говорят учебники, в один голос утверждая, что учёная
латынь изобретена для удобства общения разноязычных биологов. Как раз язы-
ковый барьер все науки давно одолевают без латыни. Какую же роль на самом
деле играют латинские названия видов? Зачем изобретены?
Прежде всего, они вошли в биологию отнюдь не путем изобретения. Карл
Линней, как и почти все до него, писал свои труды по-латыни и называл виды их
латинскими именами (т.е. античными бытовыми), если таковые имелись; а в
иных случаях (вновь открытым или плохо описанным видам) давал названия,
тоже латинские по форме. Так, используя латинские бытовые имена canis, lupus,
vulpes (пёс, волк, лиса), Линней образовал род Canis с видами Canis familiaris,
Canis lupus, Canis vulpes и другими. А вот полярный зверёк песец латинского
имени иметь не мог, и Линней назвал его Canis lagopus, использовав греческое
слово lagopus (заячий), латинское по форме окончания.
Не Линней придумал обозначать вид двумя словами (род + вид) - то была
куда более давняя традиция. Бинарные латинские названия кое-каких видов на-
ходим еще у Отто Брунфельса (1530), и некоторые из них сохранились в науке
поныне: например, Plantago major (подорожник большой). Брунфельс был од-
ним из трех немецких «отцов ботаники» [Лункевич, 1960; Куприянов, 2005].
Через двести лет, ко времени Линнея, такая практика была почти всеобщей.
Но именно Линней сделал такое именование обязательным, составив единый
список всех известных ему видов растений (1753) и животных (1758). И именно
исчерпывающим списком (а вовсе не использованным для него языком, где вид
назван в два слова: род + вид) Линней добился краткого периода взаимопонима-
ния ученых. На данную работу никто его не уполномочивал, а признан ее ре-
зультат был (после жестоких споров) потому, что нужду в едином списке ощу-
щали все, а предложить иной общий список никому не было под силу.
Во времена Линнея обозначать вид при помощи указания рода было разум-
но, поскольку род был основной единицей классификации, выразителем сущно-
229
сти (т.е. именем для совокупности основных черт строения и функционирова-
ния) данного организма. Поэтому он и обозначался существительным в единст-
венном числе с заглавной буквы. Вид же рассматривался как частная реализация
идеи рода (одно из значений слова species - особенность; отсюда специя - при-
права). Семейств в системе Линнея не было, и запоминать предлагалось прежде
всего названия родов. Отрядов (порядков) у него было сравнительно мало, и
система держалась не на них, а на родах; это видно хотя бы из того, что теоре-
тические проблемы систематики он обсуждал, перечисляя роды, и в рамках рода
допускал то, что мы именуем эволюцией.
В наше время всё не так: латинские названия для ученых всех стран являют-
ся чужими, род стал рядовой единицей между семейством и видом, а вид, на-
оборот - центральным понятием систематики. Отрядов стало больше, чем у
Линнея было родов, так что общую сущность теперь скорее следует признать не
за родом, а за отрядом (порядком). Число родов близится к миллиону, запомнить
их невозможно, и в заглавии работы о каком-то роде и виде полагается указы-
вать семейство и даже более высокий таксон. Общность же (принадлежность
одному таксону) теперь принято выражать не через общую сущность, а через
предполагаемое родство. Этого достаточно, чтобы не считать род чем-то осо-
бенным (выразителем сущности, как бы ее ни понимать).
Если Линней полагал, что дает видам бинарные имена раз и навсегда, то те-
перь имена уверенно живут лишь до очередной ревизии группы. А ревизовать
«свою» группу хочет всякий таксономист - иначе он вроде как и не профессио-
нал. Обретая всё новые и новые методы исследований, систематика веками хо-
дит по кругу - ученые время от времени возвращаются к очень старым вариан-
там системы. В частности, и лисица, и песец побывали особыми родами (Vulpes
vulpes и Alopex lagopus', замечу: alopex по-гречески значит лисица}, вернулись в
род Canis и снова вышли из него. (Подробнее см. выше, с. 59 - примеч. 2010 г.)
Кстати, узнать об этом непросто. Род Canis разрастался и съёживался неод-
нократно. Сейчас его объём минимален (7 видов). Максимальный объём он имел
сто лет назад и включал тогда почти всё семейство псовых (51 вид из 54-х), в
том числе всех лисиц, песца и енотовидную собаку, но все-таки не включал гие-
новую собаку, до этого тоже относимую к роду Canis (Hunde, 1908). Даже в
справочнике (Павлинов, Россолимо, 1998), назначение которого - сопоставлять
названия, о перипетиях рода нет ни слова. В руководстве (Каталог..., 1981) тоже
не сказано про это, но здесь в рубрике «тип рода» можно хотя бы для каждого
вида псовых бывшего СССР проверить, входил ли он когда-нибудь в род Canis'.
’ К этому месту ред. И.Я. Павлинов дал примеч-е: «Здесь автор путает два рода справочни-
ков - систематические и номенклатурные. Первые призваны давать представление о теку-
щей классификации данной группы, вторые - об истории ее номенклатуры (они так и назы-
ваются - номенклаторы)». Примера номенклатора он не привел и сам согласился с моей
мыслью, что «хождение по кругу» не отражено нигде. - Примеч. 2010 г.
230
А тот факт, что систематика данного рода веками 8одит по кругу, выясняется
только в ходе длительных поисков по достаточно редким изданиям. В итоге сис-
тематик, поневоле не зная истории, подчас бывает уверен, что нашел новое
удачное решение, хотя на самом деле он лишь в очередной раз предложил ста-
рое, в своё время отвергнутое.
При таких поворотах разные латинские названия одного объекта подолгу су-
ществуют впараллель и впараллель же, вопреки друг другу, претендуют на обяза-
тельность. Это возникает чуть ли не при каждом появлении новых методов и мод
систематики. Словом, ученая латынь сама по себе вовсе не дает «единого языка
науки», как часто утверждают. Латынь тут не хуже и не лучше любого другого
языка - лишь бы названия были долговечными. Но этого как раз и нет. А почему?
2. Названия не должны быть частью системы
Во-первых, стабильности нет потому, что родовое название несет две плохо
совместимые функции - служит адресом (который надо найти в определителе
как итог поиска) и в то же время указывает место в нынешней системе (или,
точнее, «нынешнее мнение автора о систематическом положении вида» [Клюге,
2000, с. 68]). Когда-то так именовали (адресовали) и людей (Фалес Милетский,
Эразм Роттердамский - тут родовым служит второе слово), но давно от этого
отказались - неудобно. Система же организмов то и дело меняется, причем кон-
ца этому процессу ожидать не приходится, что признаёт и Клюге (с. 46). Но, как
бы ни сложилась судьба системы (будет ли найдено долговечное решение), про-
блему стабильности названий надо решать сегодня.
А во-вторых, как ни странно, стабильность названий совсем не интересует
тех, от кого она целиком зависит - систематиков. Теоретики ее всерьез даже не
обсуждают. Да и систематики-практики больше озабочены тем, как бы не от-
стать от модных названий, как бы выглядеть посовременнее.
Словом, систематика перестала выполнять полезную для остальных наук
функцию. Она работает сама на себя, на что ученые уже не раз обращали вни-
мание. Например: «Общий упрек (funding): таксономия неадекватна и во многом
уклоняется в изучение филогении, что не улучшает ни классификацию, ни но-
менклатуру» [Wheeler, 2004, р. 571]; «выделение видов превращается в само-
цель, а классификация становится бесполезной» [Мина и др., 2006, с. 557];
«Итак, естествоиспытатель вправе задать вопрос: зачем нужна такая класси-
фикация кому-либо, кроме филогенетиков?» [Заварзин, 2006, с. 526].
К сожалению, за этими упреками не следует ничего, кроме (иногда) призы-
вов улучшить инструменты той же самой филогенетики. Выход видится мне в
другом - в том, чтобы за дело взялись, наконец, сами «пользователи» таксоно-
мии. Так в истории наук бывало не раз. Например, натуралисты неоднократно
сами разрабатывали и математические, и философские основания своих дисцип-
лин и совершали прорыв, пока математики и философы погрязали в уточнении
устаревших схем. Лучшие примеры: механика Ньютона, квантовая теория и ста-
тистическая физика, а в биологии - статистика Виллиса, о которой будет речь в
231
п. 10. Сейчас такого же прорыва можно ждать от биосистематики [Wheeler,
2004]. Ее господствующее направление (филогенетика) отстало, на мой взгляд,
от классификационной науки и данных палеонтологии лет на сто.
Прежде чем взяться за дело, необходимо понять, каким образом система на-
званий (номенклатура - то единственное, что нужно пользователям) связана с
той системой организмов, какую предлагают теоретики таксономии.
3. Бытовые названия и народная таксономия
Легко соотносятся по всем изданиям лишь те виды, у которых есть бытовое
(обиходное, простое) название. Обычно оно переводится с языка на язык одно-
значно, не подвержено моде, не зависит от принадлежности автора к научной
школе и может измениться лишь с эволюцией языка (лет за 500 и больше), да и
это бывает редко. Слово волк является таковым даже в древнерусских летописях
(вълк), и примерно ту же картину мы видим в других языках. Поэтому не стоит
удивляться тому, что в таксономических работах мы то и дело видим апелляцию
к бытовым названиям как проясняющим суть дела.
Упомянутый в п. 1 термин «бытовое название» (nomen triviale) ввел в сис-
тематику Линней в 1749 году. Поначалу он имел в виду лишь слово, характери-
зующее вид в биномиальном (в два слова) латинском названии [Скворцов, 2007,
с. 8], но затем [Linne, 1753] применил данный термин ко всему биномиальному
названию. Позже он избегал биномиальных названий растений, поскольку науч-
ным считал такое бинарное (род + вид) название, в котором род именуется од-
ним словом, тогда как вид характеризуется полиномиально (в несколько слов)
[Бобров, 1958]. Только в XIX в., когда латынь перестала быть языком науки, ла-
тинские бытовые названия стали выглядеть как научные (а бинарная номенкла-
тура, прежде полиномиальная, стала биномиальной). Роль бытовых перешла к
названиям на новых языках, не всегда бинарным.
Латинские названия были бы скорее данью традиции, чем потребностью,
если бы все виды имели бытовые названия. Однако подавляющее большинство
видов их не имеет, и научные латинские являются единственными. Для этих ви-
дов и необходима единая международная номенклатура, так как переводить их
придуманные названия с разных языков было бы невозможно (их нет в языко-
вых словарях). А поскольку номенклатурные правила должны быть едины для
всех видов, то приходится применять латынь и для обиходных видов.
Другой вопрос - что для чего должно служить примером. По-моему, устой-
чивость бытовых названий разговорных языков должна и может быть примером
для организации латинских названий. При этом сам термин «бытовое название
вида» приходится понимать достаточно широко. Например, часто в качестве
бытового выступает прежнее научное название, которое нынешний систематик
отвергает, но без которого обойтись не может, ибо не может иначе объяснить, о
чем ведет речь.
Вот недавний пример: «Диагноз. Трофическая стадия представлена одно-
ядерными амебами, с... псевдоподиями или без них, или с псевдоплазмодиями
232
или одно- или многоядерными плазмодиями». Что это за группа? Какие у ее
членов еще есть стадии, кроме трофической? Этого не сказано.
Попробуем узнать группу по описанию ее размножения: «Половой процесс
имеется не всегда. Обычно бесполое размножение... редко оно отсутствует.
Жгутиковая стадия, если имеется, обычно в форме зооспоры; часто жгутики или
реснички отсутствуют на всех стадиях». Опять никакой ясности - любое свой-
ство необязательно (единственный четкий признак: «Хлорофиллы отсутству-
ют», - для амеб почти тривиален). Само по себе это затруднение довольно
обычно и легко преодолевается путем описания общего строения (т.е. архетипа
группы), но архетип (о нем см. п. 8), как и все вообще целостные характеристи-
ки, ныне у систематиков не в моде5*5, и диагноза не получилось.
О чем идет речь, осталось бы тайной, если бы не простодушное признание
авторов, что это - упраздняемые ими миксомицеты плюс еще ряд небольших
групп (например, энтамёбы), которые по невысказанным причинам авторы объ-
единяют с миксомицетами в одно царство [Кусакин, Дроздов, 1998].
Очевидно, что слово «миксомицеты» использовано здесь как бытовое на-
звание и что оно (а вовсе не приведенный «диагноз») несет всю информацию о
вновь вводимой группе. В оправдание авторов можно сказать лишь, что таких
«диагнозов» в печати появляется много, что их филема (филогенетическая сис-
тема) оказалась не хуже и не лучше других. Ни одна из таких схем не принята
никем, кроме их творцов, да и те вскоре обычно меняют свои взгляды.
Еще такой пример: в роде Vulpes есть вид корсак (V corsac) обитающий в
средней и южной Евразии, и вид американский корсак (V. velox), водящийся в
средней и южной части Северной Америки. Из самих этих бытовых названий
видно, что данные виды очень сходны, а из отдаленности зон их распростране-
ния - что их общий предок не мог жить недавно. Хотя в последний раз Берингия
(сухопутный мост с Чукотки на Аляску) ушла под воду не так уж давно, около
20 тыс. лет назад, но наивно думать, что из Евразии в Америку могли перейти
теплолюбивые звери вроде корсаков. Мост возникал только в ледниковые пе-
риоды, и из псовых перейти мог разве что песец. Следовательно, тесное сходст-
во корсаков не унаследовано, а возникло, и, с общепринятой точки зрения, тут
86 Противники архетипической систематики уверяют, что «у типологии нет критериев, с
помощью которых можно было бы обосновать введение типов» [Шаталкин, 2006, с. VI],
т.е. архетипов. Сторонники указывают такие критерии. Например, К.Г. Михайлов, излагая
Г.Ю. Любарского, пишет: «В качестве внутреннего критерия автор вводит соответствие
ранга таксона и уровня наиболее сильно измененного мерона. Так, изменения плана строе-
ния задают, как правило, тип животных, аппарата - класс, системы органов - семейство и
т.д. ... Внешние критерии - это просто сравнение изучаемого таксона с соседними, а также
с вышестоящими... На примере группы ellipticus рода Cryptophagus удалось проиллюстри-
ровать переход биоморфы в таксон,, стиля в архетип. Группу ellipticus можно интерпрети-
ровать как ныне формирующийся, причем явно не-монофилетический таксон родового
ранга» [Михайлов, 1997, с. 93].
233
налицо не родство, а параллелизм. Тем не менее, корсаков включают в общий
род, т.е. бытовое название оказалось соответствующим научному.
Более изощренный пример дает тот же род Canis. Он не поддается стабиль-
ному описанию [Павлинов, Россолимо, 1998], причем новые методы (молеку-
лярная генетика и компьютерный анализ массива признаков) не прояснили по-
ложения нисколько - систематика ходит по кругу [Чайковский, 1990, с. 45-46].
Знаменитый этнолог Клод Леви-Строс, один из основателей структурализма,
обрисовав таксономические успехи южноамериканских индейцев (включавшие
бинарную номенклатуру вид + подвид), писал в 1962 г.:
«скольких ошибок и заблуждений можно было бы избежать... если бы
прежние путешественники относились с доверием к туземным таксономи-
ям вместо того, чтобы измышлять новые. В результате 11 авторов дали од-
но и то же научное наименование - Canis azarae - трем различным родам,
восьми видам и девяти подвидам и, напротив, придали несколько разных
наименований одной и той же разновидности» [Леви-Строс, 1994, с. 147].
Поясню: термин Canis azarae использовался в XIX в. для обозначения са-
мых разных лисиц Южной Америки; например, так называли бразильскую ли-
сицу (ныне Dusicyon vetulus), а Дарвин в «Путешествии на Бигле» назвал терми-
ном Canis azarae одну из лисиц Огненной Земли, т.е. совсем другого зверя.
Фактически Леви-Строс обратился здесь к идее бытовых названий дважды.
Во-первых, призвал биологов предпочесть туземные названия европейским как
более естественные. Во-вторых, применил название Canis azarae, в его время
уже ушедшее из таксономической практики, как бытовое название, для обозна-
чения южноамериканских лисиц, в его время объединеных в род Dusicyon (око-
ло 10 видов). Досадно только, что он не привел тут самих туземных названий -
они помогли бы ему радикально прояснить его мысль.
Тем самым, Леви-Строс коснулся народной таксономии, которую оценил,
как видим, достаточно высоко. К сожалению, те труды, на которые он ссылался,
ныне забыты, а новые авторы приходят к иным выводам -что народы опериру-
ют единственной сущностью, той, которой канадский этнолог и историк науки
Скотт Атран [Atran, 1990] дал название «родо-вид». Если я правильно понял,
родо-вид может быть, с нынешней точки зрения, родом и видом сразу (таков в
русском языке бекас), и приметной группой видов (кулик как охотничий тер-
мин), и подвидом (такова вобла), и домашней породой (таков карп). Именно
господство родо-вида в массовом сознании, а вовсе не языковые обстоятельства,
является препятствием для использования бытовых названий в качестве науч-
ных - таков, если я правильно понял, вердикт ученых - см. [Куприянов, 2005].
Это, в общем, верно, но всё же не стоит совсем отрицать народную таксо-
номию. Если взять тех же бекасов, то увидим, что русским словом бекас уве-
ренно обозначается как род птиц Scolopax, введенный еще Линнеем, так и самый
известный его представитель Scolopax gallinago, известный также под именем
барашек (Словарь В.И. Даля). Аналогично с немецким Schnepfe (бекас).
234
Такое совмещение обычно и в научном языке - вспомним хотя бы волка: в
южной Сибири есть красный волк, прежде входивший в род Canis как С. pri-
maevus (затем выделен в род Сион) [Каталог..., 1981]. То же и вне биологии:
словом «эфир» именуют как класс соединений, так и самый известный его член
(этиловый эфир). Словом, видеть в таком совмещении изъян народной номенк-
латуры (а тем более, изъян «первобытного» сознания) незаконно.
Вероятно, различие в выводах ученых связано с различием доставшегося им
материала, и я позволю себе тоже привести пару примеров.
Несколько старых словарей позволили мне составить табличку:
бекас обыкновенный, или барашек (Scolopax gallinago)
лесной бекас, или вальдшнеп (S. rusticola, Waldschnepfe);
бекас-дупель, или доппельшнеп (S. major, Doppelschnepfe);
бекас стучик, или гаршнеп (S. gallinula, Haarschnepfe).
Род Scolopax выявил Линней - как видим, в соответствии с немецкой народ-
ной таксономией. В XIX веке эти виды были разнесены по разным родам, что,
однако, не должно мешать нам увидеть народную бинарную номенклатуру род
+ вид. То же являет собой род кроншнеп (Numenius) из семейства улитов, кото-
рый, заодно, выявляет указанный данным названием параллелизм кроншнепов с
бекасами, чего научная номенклатура избегает.
Ту же картину являют многие растения, например, род горчица, виды кото-
рого отнесены к одному роду и в русской народной таксономии, и Линнеем:
горчица дикая, или горчец (Sinapis arvensis);
горчица белая, или английская (S. alba);
горчица черная (S. nigra);
горчица сарептская (S. juncea; Сарепта - бывшее немецкое поселе-
ние, ныне в черте г. Волгограда).
Последние два вида были в XIX веке отнесены к роду Brassica (капуста).
Одной из заслуг Атрана мне видится тот факт, что он выявил господство
родо-вида не только в народной таксономии, но и в зоологических трудах Ари-
стотеля, и у натуралистов XVI-XVII веков. Однако было бы интересно оценить,
какую долю в народной таксономии составляют приведенные выше названия по
схеме род + вид. Пока же замечу, что систему бытовых названий таксономисты
то и дело рассматривают (пусть и неосознанно) как естественную.
4. Искусственная и естественная системы организмов
Естественной системой группы объектов принято называть такую их клас-
сификацию, в основу которой положены существенные свойства классифици-
руемых объектов, тогда как искусственной - такую, где для классификации ис-
пользуются произвольно выбранные (несущественные, побочные) свойства объ-
ектов, оказавшиеся удобными в данном исследовании для их различения и упо-
рядочения. Очевидно, что центральным тут является понятие существенности
свойства: то, что существенно для одного исследователя, может быть побочным
235
для другого, а это значит, что деление систем на естественные и искусственные
всегда субъективно и потому неоднозначно [Розова, 1986, с. 45].
Деление систем организмов на искусственную и естественную впервые на-
ходим у Линнея, однако и этим он также лишь завершил давнюю традицию.
Еще Иероним Бок, приятель Брунфельса, в своем «Новом травнике» (1538) рас-
положил рядом друг с другом растения, сходные строением, не давая этому
приему никакого объяснения. Зато расположил их столь удачно, что впоследст-
вии трем его группам был приписан статус семейств, сохранившихся поныне -
это губоцветные, крестоцветные и сложноцветные [Лункевич, 1960, т. 1].
Традиция обозначать существенные сходства компоновкой материала, а не
словесным описанием, надолго стала почти всеобщей, что вполне понятно, по-
скольку сходства выявлялись интуитивно; для их словесного описания еще не
был выработан понятийный язык. Так, Каспар Баугин широко пользовался ро-
дом и видом, но этого уже явно нехватало, и у него мы видим (труды 1596-1623
гг.) четырехступенчатую классификацию. Кроме рода и вида (которые он
обычно, но не всегда, обозначал, как и мы, двумя словами, первое с заглавной
буквы), он ввел для родов, близких по общему строению, термин «секция», а
близкие секции группировал в одну «книгу». Всего в трактате [Bauhin, 1623]
было 12 «книг», но названий им Баугин не дал.
Да и не у всех его «секций» есть названия. Так, в Книге I из пяти секций
лишь две имеют названия: «О травах» и «О зерновых». Причем в них даны да-
леко не все травянистые растения; заглавия остальных трех секций - просто пе-
речни родов. Часто в одну секцию включены растения, сходные лишь внешним
обликом (например, камыш и рогоз - мы в быту путаем их до сих пор).
Если одни секции похожи у Баугина на наши семейства (например, секция
зонтичных), то другие вызывают в наше время улыбку: так, в одной секции ока-
зались мхи и ряска (цветковое), в другой - хвощи и злаки. Это не должно нас
удивлять, поскольку способы размножения были известны далеко не для всех
растений, и если цветка не находили, то полагали, что он всё же есть, только
очень мелкий или очень недолговечный. Можно сказать, что естественные
группы выступали тогда из массы материала, упорядоченного каким-либо ис-
кусственным образом (чаще всего - по алфавиту латинских названий).
Уже у Баугина открылось то явление, которое с тех пор крайне затрудняет
работу систематиков: параллелизм. Например, при описании рода Plantago (по-
дорожник) описаны три группы видов: Р. latifolia (широколйстные, 13 видов), Р.
angustifolia (узколистные, 10 видов) и Р. aquatica (водные, 4 вида) [Bauhin, 1623,
р. 189-190]. В наше время ширину листьев почти не используют для различения
видов (исключение: подорожник ланцетолистный), поскольку она может сильно
меняться даже в пределах одного вида, а водные подорожники выделены в осо-
бый род Littorella (побережник).
И вот мы видим у Баугина такой прием: широколистные виды подорожни-
ков приведены подряд друг за другом (Plantago latifolia sinuata, Р. latifolia
236
glabra, P. major и т.д.), а затем то же самое сделано с узколистными. После этого
выписаны все 4 вида водных подорожников и отмечено, что среди них тоже есть
как широколистные, так и узколистные (Л aquatica latifolia и Р. aquatica
angustifolid). К сожалению, никаких пояснений Баугин не давал.
Может быть, надо было группировать иначе: отнести водный широколист-
ный к первой группе, а водный узколистный ко второй? Последующие система-
тики вовсю этим занимались - то и дело меняли (и меняют поныне) признаки
местами (по степени их важности для данного систематика) и получают разные
системы. Дело в том, что одни и те же синдромы часто встречаются у разных, в
том числе очень различных, видов (таковы, например, сходство формы плодов
разных перцев или клюв у утконоса) - это и есть параллелизм.
Качественно новый уровень понимания естественности и искусственности
видим у Линнея. Он явно и ясно заявил, что (если пользоваться нынешними
терминами) в биологии адресация требует одной системы (ее он для цветковых
растений построил, и ее с тех пор называют искусственной), а понимание уст-
ройства разнообразия организмов - другой. Последнюю он назвал «естествен-
ным методом» и едва наметил в «Философии ботаники» (1751) [Линней, 1989],
где назвал естественное сходство организмов сродством (affinitas).
Он дал понять (§§ 78 и 160), как видит сродство: им обладают члены одного
семейства. Эти 7 «семейств» (familiae): Грибы, Водоросли, Мхи, Папоротники,
Злаки, Пальмы, (прочие) Растения. Их не надо путать с семействами нынешней
систематики; скорее они близки к нашим типам (отделам).
Основной тут была у Линнея мысль: «Все растения проявляют друг к дру-
гу сродство, как земли на географической карте». Никакой «карты» он не
предложил, но впоследствии такая карта не раз строилась.
Например, в 1817 г. немецкий зоолог Георг Август Гольдфус расположил
группы животных от простейших (инфузории) на востоке до человека на запа-
де, причем холодных рептилий поместил на севере, а теплокровных птиц на юге
(рис. 3). Аналогичную попытку в 1839 г. предпринял английский хирург и бо-
таник-любитель Томас Баскервиль. Для низших растений он избрал южное по-
лушарие: у полюса - водоросли, выше них мхи, лишайники и пр.; для высших
же он отвел северное', у полюса - астровые, т.е. высший порядок двудольных,
пониже их помещены колокольчики и т.д. (рис. 4).
Такие системы-карты успеха никогда не имели, поскольку увлечение гео-
графической аналогией явно шло здесь в ущерб сути дела, однако вся тенденция
систем-карт весьма любопытна - она, вновь и вновь проявляясь в разные эпохи,
явственно говорит о наличии некой непреходящей потребности ума в наглядных
изображениях биоразнообразия. Главный положительный урок, который следу-
ет извлечь из систем-карт прошлого, тот, что наглядность может даваться соче-
танием различных форм симметрии, чего совсем не дает система-древо. А глав-
ный отрицательный - тот, что симметрия должна задаваться не симпатиями ав-
тора, но природой изображаемого объекта.
237
Во-первых, природа объекта включает его экологию, поэтому систему сле-
дует строить для всей природы, а не для отдельных царств.
Во-вторых, природа объекта должна включать упомянутый выше паралле-
лизм: сходные формы двух групп должны лечь рядом, с обеих сторон общей
границы групп. Узколистные впараллель с узколистными, перцы с перцами,
двухпестичные с двухпестичными и т.д. В отношении параллелизмов привер-
женцы всех форм филогенетической систематики ведут себя одинаково - упо-
минают их как главную трудность [Клюге, 2000, с. 19]:
«Наибольшие затруднения при реконструировании филогении вызывает
существование независимо возникающих гомологичных признаков (это
явление известно как закон гомологических рядов Н.П. Вавилова»)
- и затем игнорируют, ибо сам феномен параллелизма противоречит идее
древа (подробнее см. [Чайковский, 2006]). Поскольку параллелизм - один из са-
мых общих феноменов природы, постольку его обязана описывать (а не только
упоминать) любая система, претендующая отобразить биоразнообразие.
Как это сделать, будет сказано далее, а пока замечу, что факты параллелиз-
ма побудили многих правоверных дарвинистов (таким был в юности и А.А. Лю-
бищев) отказаться от уверенности, что филогенетическая систематика имеет
смысл, и задуматься над тем, что такое естественная система.
Существенно, что ни сам Линней, ни его последователи не пытались дове-
сти естественную систему до уровня видов, и это вполне разумно, коль скоро
она не служит для определения видов, а дает общий абрис живого мира.
Столь же важно, что Линней составил свои семь «семейств» отнюдь не из
классов своей искусственной системы, а из новых групп, которые назвал есте-
ственными классами. Для 7-го «семейства» он дал их примеры: Зонтичные, Му-
товчатые, Стручковые, Бобовые (мотыльковые), Сложноцветные и т.д. Естест-
венный класс Линнея - это примерно то, что именуют семейством в наше время.
Оба эти обстоятельства показывают, что естественную систему следует строить
не так, как систему, служащую для адресации.
Однако совет Линнея большинством не был услышан, и биологи вот уже
третий век движутся в тупик, пытаясь построить естественную систему, годную
для адресации. Хотят, так сказать, от одной коровы получить и мясо и молоко
сразу. Так не бывает: как мы увидим, понятие естественности радикально зави-
сит от моды (от взглядов, популярных в данную эпоху в данном кругу теорети-
ков), тогда как от адресации нужно прежде всего постоянство.
5. Систематика и эволюционная идея. Тезис Короны
Систематика сформировалась в XVI-XVIII веках без всякой связи с эволю-
ционной идеей, так что древовидная форма системы отражала не историю жиз-
ни, а царивший тогда способ классификации. (Другой способ, табличный, когда
объект задается пересечением строки и столбца, вошел в практику много позже,
с химической системой Д.И. Менделеева.) Именно эту систему-древо дарвинизм
и поставил целью рационально истолковать.
238
Недаром Чарлз Дарвин назвал свою главную эволюционную книгу «Проис-
хождение видов», а знаменитый палеонтолог Эдвард Коп назвал свою первую
эволюционную статью (1868) «Происхождение родов» - эволюцию тогда все
понимали как появление и вымирание таксонов. Сами таксоны брались для тео-
рии эволюции из традиционной систематики как нечто данное. Ныне ситуация
иная: система видов и прочих таксонов строится как итог эволюции.
От Эрнста Геккеля (1866) идет уверенность, что таксоны возникают в ходе
эволюции по единой схеме: все таксоны данной группы происходят путем ди-
вергенции (расхождения) от единого таксона-предка (т.е. монофилетически).
Только в том случае, когда группа таксонов считается монофилетической, ее
саму предлагают именовать таксоном (более высокого ранга).
Здесь сознательному принятию факта эволюции в качестве постулата сопут-
ствовало неосознанное принятие еще одного постулата: что эволюция имеет
вполне определенную форму - форму филогении, т.е. порождает родословное
древо (далее слово «филогения» используется лишь в этом смысле).
Постулат попал в биологию из логики иерархической классификации еще
до Линнея. В наши дни от нее отказалось большинство наук, но не биология, и
именно биологи вынуждены то и дело пересматривать свои системы.
Неизбежен вопрос: соответствует ли наличная систематика принятому по-
ниманию эволюции? Первым, насколько знаю, его задал А.А. Любищев в 1923 г.
и ответил отрицательно - на самом деле система строится на наличных свойст-
вах организмов, а происхождение додумывается. Такое додумывание общепри-
нято, хотя, по Любищеву, противоречит фактам:
«1) Можно было бы ожидать, что так как... дивергенция низших таксоно-
мических единиц произошла очень недавно, то всегда легче построить фи-
логению мелких таксономических групп; на самом деле именно в этой об-
ласти филогения... совершенно не удавалась... напротив, небольшие груп-
пы всегда легче классифицируются в виде решеток... 2) Постоянные пере-
стройки системы путем обнаружения “неестественности” групп, прини-
маемых ранее за естественные с филогенетической точки зрения; примеры
этого можно привести из любого класса и типа животного и растительного
мира... 3) Преобладание параллельного развития в хорошо изученных па-
леонтологических рядах» [Любищев, 1982, с. 25-26].
Такой вердикт (звучащий, кстати, вполне современно - см. далее) вовсе не
значит, что систематику надо тут же менять. Надо сперва выяснить, насколько
система таксонов - итог эволюции. Напомню, что есть иная точка зрения: номо-
генез видит эволюцию как процесс заполнения объективно заданной таблицы
свойств, которая сама от эволюции не зависит (от эволюции зависит лишь сте-
пень заполненности таблицы).
Ботаник В.В. Корона уловил самую суть дела, отметив, что древо, как фор-
ма системы, связано не с идеей эволюции, а с удобством, с тем, что
«наиболее рациональной формой упорядочения информации является
форма дихотомического ключа» и «Монофилия и дивергенция это логиче-
239
ское следствие принятых постулатов, а не собственная форма системы»
[Корона, 1987, с. 32].
Сформулируем данную мысль в такой форме: древо как форма классифи-
кации отражает свойства не объекта, а мышления.
Этот тезис Короны доказать или опровергнуть невозможно, так что прини-
мать его или нет - вопрос реальной помощи от него в работе. По-моему, он мно-
гое делает понятным. В нем, по словам Короны, кроется «причина устойчивости
филогенетической систематики» как явления науки, хотя она не решает ни од-
ной из стоящих перед нею задач. Их Корона видел три: «построить естествен-
ную систему органического мира, выяснить историю его развития и найти наи-
более упорядоченную форму представления существующего многообразия»
(там же). Могу добавить задачу создания устойчивой адресации.
Принять тезис Короны - значит признать, что сам способ изображать сис-
тему древом искусствен по существу. Другими словами, тот, кто полагает фило-
гению основой естественной системы, должен прежде всего доказать, что ре-
альный эволюционный процесс порождает именно филогению (древо).
К сожалению, филогения всегда постулируется, а не выводится из анализа
материала. В итоге «филогенетика - наука без собственного предмета исследо-
вания и занимается интерпретацией данных, которые представляет ей система-
тика» [Любарский, 1996, с. 31]. Материал же в последние годы явственно пока-
зывает, что итогом эволюции является не древо, а сеть. Обоснованию данного
факта посвящена значительная часть книги [Чайковский, 2006].
Недавно отрицание филогении стало появляться сразу во многих работах, и
я отмечу только сборник [Эволюционные...]. В нем даже энтузиаст фило-гении
зоолог И.Я. Павлинов пишет, что хотя новая филогенетика доминирует, близок
ее закат, поскольку в ней «слишком много от моды и конъюнктуры»; и указыва-
ет причину: она оперирует с отдельными признаками, что с точки зрения «орга-
низма как целого» бессмысленно (с. 21). Другие авторы сборника, палеонтологи,
прямо называют альтернативу филогенетике - древо должно быть заменено се-
тью. А именно, Э.И. Воробьева отмечает «параллельное приобретение сходных
признаков» в ходе блочной эволюции (с. 45-46), А.Г. Пономаренко видит «фи-
логенетический луг» (с. 149) вместо филогенетического древа, а С.В. Рожнов
спрашивает: «древо или луг?» (с. 162).
6. Почему Линней не построил естественной системы?
Линней описал свой «естественный метод» в духе Баугина - без всяких по-
яснений перечислил роды, входящие в «семейства». То есть завершил традицию
травников, где сходство отображалось соседством.
Но впараллель этой (по Линнею, географической) в систематике существо-
вала и другая первичная традиция - морфо-функциональная (у А.В. Куприянова
- методическая). Основателем ее стал итальянский ботаник Андреа Цезальпин
(Чезальпино) в дни юности Баугина, а завершил французский ботаник Жозеф
240
Питтон де Турнефор в дни младенчества Линнея. Она хорошо описана в брошю-
ре [Куприянов, 2005], и мне остается только сказать следующее.
По Куприянову, Цезальпин видел у растений две главные функции - пита-
ние (напомню, что фотосинтез был открыт через 200 лет) и размножение:
«Он полагал, что отличия в способе питания отражаются в более или менее
плотной консистенции корня и стебля, из которых первый отвечает за по-
глощение пищи из почвы, а второй - за распределение и усвоение этой пи-
щи». По словам Цезапьпина, «должна быть “вегетативная” субстанция, по-
зволяющая растениям воспроизводиться... На сей конец созданы были пло-
ды и части, обеспечивающие фруктификацию» [Куприянов, 2005, с. 33, 34].
Реже Цезальпин учитывал способность растения к движению и закреплению
(усики). Различия этих свойств и легли у него в основу классификации.
Турнефор тоже был достаточно физиологичен. Например: он «использовал
синюю бумагу, которая изменялась при воздействии на нее сока растения, указывая
на его кислое или щелочное качество» [Линней, 1989, с. 250].
Но чаще физиология Турнефора уже во времена Линнея могла вызвать
только улыбку: пестик у него служит для питания завязи, а тычинки - орган вы-
деления. Зато Турнефор не только развил и узаконил четырехступенчатую ие-
рархию таксонов, но и ввел два понимания рода: виды родов первого типа не
требуют для своего описания ничего, кроме цветка и плода, тогда как второй
тип требует привлечения других органов растения.
Упомянув данный факт, Куприянов не объяснил сути дела (что в его бро-
шюре видим часто), и надо обратиться к тексту самого Турнефора.
К родам второго типа Турнефор отнес, например, шафран (ныне род Crocus).
Его цветок весьма характерен - околоцветник похож на лилию, но вынесен вверх
на длинной ножке, тогда как завязь остается в земле; а плод - ничем не примеча-
тельная коробочка. Тем не менее, описав цветок и плод, Турнефор добавил
(«Элементы ботаники», 1694): «Корень у видов этого рода - мясистый клубень,
бородатый (barbu) в своей нижней части» [Tournefort, 1957, р. 173-174].
Как видим, дело не столько в том, что цветок и плод недостаточны для уве-
ренного определения вида, сколько в том, что «бородатый» клубень - самая ха-
рактерная часть данного растения (ныне ее именуют «клубнелуковица»).
Именно этот прием (выбирать для определения самую характерную часть
организма) и воспринял Линней. Но в искусственной системе, где выбор прово-
дился единый для всей системы, успех был неполон. Так, в окончательном ее
варианте пальмы пришлось описать вне системы, причем использовать для пер-
вого (грубого) их различения свойства листьев [Linne, 1753, t. 2]. До этого (в
«Философии ботаники») Линней избегал основывать классификацию на листьях
и там же назвал пальмы одним из семейств «естественного метода».
Очевидно, что теперь он прибег к фрагменту «естественного метода», по-
скольку осознал недостаточность искусственного. Что же помешало Линнею
продолжить строить конкретную естественную систему? Ведь он сохранял дее-
241
способность еще 20 лет после «Философии ботаники» и все эти годы много пе-
чатался. Самый простой ответ - задача оказалась ему не по силам. Ведь Линней,
блестящий систематик-практик^7, был довольно слаб как теоретик:
«приходится согласиться с большинством историков биологии, упрекаю-
щих Линнея в том, что он не обогатил науку о жизни ни одной подымаю-
щейся над средним уровнем идеей, ни одним важным в теоретическом
отношении открытием» [Лункевич, 1960, т. 2, с. 80],
а философом не был вообще. Его «Философия ботаники» - лучшее тому
подтверждение: ее афоризмы не увязаны общей мыслью и показывают лишь,
как он тонул в затронутых им проблемах. Есть даже мнение, что он вообще не
знал аристотелевой логики, основы тогдашнего образования [Winsor, 2006]. Од-
нако ответ «не по силам», пусть и верный формально, был бы поверхностным.
Достаточно заметить, что именно Линней первый увидел саму проблему
двух систематик. Более того, он вроде бы даже нащупал подход к ее решению
(см. ниже, п. 7). На мой взгляд, во времена Линнея просто еще не было аппарата
для ее решения. О нем и поговорим.
7. Различные понимания естественной системы
Что же такое естественная система? Должна ли она отражать историю (эво-
люцию)? Если да, то будет ли она древом? Если нет, то что она должна отражать
и какую форму иметь? Должна ли эволюционная система строиться на идее по-
лезности свойств? Если да, то почему на деле систематик предпочитает исполь-
зовать свойства, польза которых отрицается или сомнительна? Эти и подобные
вопросы почти не обсуждаются, но каждый систематик имеет на них свой осо-
бый ответ, поэтому систематика вязнет в противоречиях.
Далее изложена точка зрения на естественность системы, описанная в статье
[Любищев, 1972], в § 1.3 книги [Корона, 1987] и в §§ 6.1 и 6.2 книги [Чайковский,
1990]. Там есть ссылки на литературу, а недавние сведения можно найти в книгах
[Павлов, 2000; Эпштейн, 2002; Захаров, 2005]. В книге В.М. Эпштейна изложена
также идея Линнея о необходимости двух таксономий.
Линней понимал, как мы видели, что естественность системы - нечто более
глубокое, нежели удобство адресации, и предложил строить две системы расте-
ний: искусственную (для целей адресации - ее он построил) и естественную (для
уяснения «сродства» видов, которое он понимал как некую общую близость, но не
как родство - ее он едва наметил). [Линней, 1989],
Данный ход мысли напомнила юбилейная статья [Скворцов, 2007], к которой
позволю себе одно уточнение. Читая ее, можно подумать, что Линней хотел заме-
нить искусственную систему на естественную, хотя у Линнея на это нет и намека.
Линней ставил перед двумя системами совсем разные задачи и, в частности, нигде
87 «В отличие от современных систематиков, Линней был чужд тавтологии, и его диагно-
зы были настоящими, т.е. давали четкие различия от соседних таксонов» [Любищев,
1972, с. 53].
242
не доводил рассуждения о естественном методе до уровня вида. Идею замены ис-
кусственной системы на естественную впервые видим у Антуана Жюссье (1774 г.
и позже), а завладела умами она только в XIX веке.
Мишель Адансон, младший современник Линнея, видел естественной ту
систему, где рядом стоят таксоны, близкие по максимально возможному числу
признаков. В XX веке последователи Адансона дополнили его идею допущени-
ем, что признаки имеют различный вес (делятся на более и менее важные). Та-
кой подход именуют нео-адансоновским. Его новый вариант [Эпштейн, 2002, с.
55] тоже видит накопление знаний о признаках как естественность.
В 1819 г. Карл Бэр уточнил естественность как сходство общего строения, а
не отдельных, пусть и многих, частей или свойств. Он впервые прямо сопоста-
вил задачи двух систематик, воздал должное искусственным системам вообще, а
линнеевой в особенности, и заметил:
«Легко видеть, что в основу каждой системы положены различные за-
дачи, но... это редко понимается правильно, так как то стараются по
возможности заменить обе системы одной, то упрекают искусственную
за то, что она не дает того, что можно было бы ожидать только от есте-
ственной, или, наоборот, борются против этой последней, если она не
обеспечивает преимущества, которое дает первая» [Бэр, 1959, с. 367],
Прошло почти два века, а темы и упреки бытуют те же. В науке так практи-
чески не бывает, зато обычно в философии, и это надо помнить.
Саму естественную систему Бэр определил в 1825 г. так:
«В льве, в обезьяне легче узнать человека, чем в клеще. Результат сопос-
тавления организмов в соответствии с большим или меньшим сходством
между ними во всех отношениях называется естественной системой.
Степень сходства обозначается также выражением “сродство”. Спраши-
вается, можно ли установить в этом сродстве между животными всеоб-
щее правило?» [Бэр, 1959, с. 396-397].
Бэр полагал, что естественную систему следует то и дело пересматривать в
связи с новыми данными, и это мнение обычно до сих пор. Наоборот, англий-
ский логик и историк науки Вильям Уэвелл (Whewell) в 1840 г. предложил по-
нимать естественность системы как устойчивость: чтобы ее не требовалось пе-
ресматривать при появлении новых для классификации признаков (например,
при более детальном изучении прежних объектов).
Для Дарвина и Геккеля естественность системы состояла в общности предка
каждой группы, и это понимание стало господствующим в XX веке. Но тезис
«сходство свидетельствует о родстве» игнорирует ту истину, что само понятие
родства почти всегда основано на сходстве и ни на чем другом.
Сто лет назад математик Анри Пуанкаре в качестве естественной предло-
жил понимать параметрическую систему, в которой положение объекта задает-
ся одним (как в системе Менделеева), двумя (как на географической карте) или
несколькими параметрами. Любищев в 1923 г. соединил Пуанкаре и Уэвелла,
243
признав естественной ту систему, «где все признаки объекта определяются по-
ложением его в системе». Образцом он счел систему Менделеева, в которой но-
мер элемента задает его свойства.
В 1983 г. математик и философ Ю.А. Шрейдер уточнил это понимание: ес-
тественна та система, в которой существует универсальное правило, по которо-
му можно дать полное описание архетипа (здесь - плана строения) таксона по
его «короткому описанию» [Теория..., 1983, с. 97].
В 1926 г. зоолог Е.С. Смирнов, известный ламаркист, выступил с понимани-
ем систематики как анализа конгрегаций. Для него каждый таксон выглядел как
компактная область (скопление близких точек, конгрегация) в признаковом про-
странстве. Любищев высоко ценил этот подход прежде всего за то, что он по-
зволяет описывать даже те таксоны, в которых «крайние члены такой конгрега-
ции не имели ни одного общего признака». При этом, в отличие от Смирнова, он
видел такой принцип шире, чем иерархический: соседние конгрегации можно
сгруппировать в конгрегацию более высокого ранга и тем самым получить ие-
рархию, но можно выявить и кольцо конгрегаций [Любищев, 1975, с. 167].
Для Любищева широкая применимость принципа конгрегаций была свиде-
тельством ограниченности иерархического принципа. Но сам Смирнов, видя в
конгрегациях естественные группы, уверял: «В настоящее время следует счи-
тать доказанным, что иерархия таксонов отражает родственные отношения ор-
ганизмов» [Смирнов, 1969, с. 8]. Никаких ссылок на доказательство нет, и оста-
ется с горечью признать, что и тут налицо смешение знания с верой. Вообще же
ламаркизм, при всех его заслугах [Назаров, 2005; Зусмановский, 2007], проявля-
ет к своим основам так же мало внимания, как и дарвинизм.
Применимость конгрегационного принципа широка, но не абсолютна - по-
сле объединения сходных объектов в конгрегации вне их всегда остается неко-
торое количество объектов. Если их мало, то можно поступить двояко: либо на-
звать все их общей конгрегаций по имени «разное», либо счесть каждый такой
объект отдельной конгрегацией. Если же их много, то конгрегационная система
теряет наглядность. Это и произошло уже у Линнея.
В его «естественном методе» оказалось «неопределенными» 10% всех родов
[Линней, 1989, § 68]. Примерно столько же в его «Видах растений», хотя систе-
ма там - искусственная. Очевидно, что был нужен какой-то иной метод класси-
фикации, чтобы не отправлять каждый десятый род в общую кучу. И мы видим
у Линнея намек на него: в § 252 «Философии ботаники», говоря на другую тему
(о названиях), он заметил, что их в будущем надо давать «исходя из принципа
деления будущей системы». О делении (рассечении множества таксонов) как
методе классификации у нас речь пойдет далее, в п. 13.
В 1928 г. друг Любищева зоолог В.Н. Беклемишев, признав негодность
принципа родства для систематики, сохранил, однако, идею иерархии как един-
ственно годную для построения естественной системы. Эта вроде бы странная
позиция вызвана тем, что саму естественность он понимал отнюдь не как выяс-
244
нение какой бы то ни было «природы вещей», а лишь как краткую адресацию, и
считал идеалом понимание ее по Алансону [Беклемишев, 1994, с. 28]. С позиции
как Линнея, так и Любищева, и Короны, Беклемишев описал идеал вовсе не ес-
тественной, а искусственной системы.
Он показал невозможность построить работоспособную чисто комбинатив-
ную систему (сеть) - она слишком избыточна, потому громоздка и возможна
лишь как часть иной системы (что мы увидим). То же считал Смирнов. Т.е. для
них естественна компактная система. Этим они показали, что представляют
ее себе как нечто конструируемое, тогда как сама идея естественной системы
противоположна - почти все признают, что ее следует искать в природе.
Любопытно, что Любищев как раз общепринятой систематике предъявлял
упрек в огромной избыточности: из-за параллелизмов дивергентные определи-
тели наполнены огромным числом повторов тез и антитез. Добавлю: есть удоб-
ные определители, составленные по схеме ботаника Б.Е. Балковского, где в ос-
нове лежит не древо, а таблица (подробнее см. [Чайковский, 2006])*.
В 1930-е гг. Н.И. Вавилов и Н.П. Кренке видели естественность системы в
возможности прогнозировать ее элементы (находить новые виды).
В 1943-1950 гг. Любищев и его друг зоолог Б.С. Кузин обсуждали проблемы
систематики в письмах [Теория..., 1983, с. 143-165]. Любищев отдал предпочте-
ние «параметрической» форме (как на карте, где координаты точки суть 2 пара-
метра); а Кузин признал в качестве естественной иерархическую систему. Лю-
бищев стоял на логико-математической позиции (всякий таксон можно логиче-
ски определить, а различие таксонов измерить), а Кузин отстаивал (продолжая
Линнея) интуитивное и даже эстетическое понимание таксона.
В 1966 г. французский культуролог Мишель Фуко писал, что естественность
системы определяется ее соответствием тем представлениям о структуре мира и
порядке вещей в нем, которые в данное время господствуют в обществе. Ника-
кое накопление противоречий не заставит массу ученых отказать системе в ес-
тественности, пока не сменится картина мира.
Смирнов в те же годы видел естественность системы в ее общепризнанно-
сти, окончательности и единственности, «не допускающей вариантов». Эта по-
пытка развить Уэвелла неудачна: до сих пор всякое «окончательное» решение
оказывалось временным. Но позиция Смирнова стоит внимания, поскольку вся-
кий, предлагающий новую систему, строит ее как окончательную.
А недавно предложен совсем оригинальный критерий естественности: «Когда
я начинал заниматься функциональной морфологией... я и не помышлял о по-
строении какой-либо системы». Она «сложилась сама собой. И это, на мой взгляд,
является главным доказательством ее естественности» [Павлов, 2000, с. 6].
Систематику принято строить на устойчивых (долговечных) признаках, а они
часто бесполезны, что отметил еще Дарвин в гл. 14 «Происхождения видов».
См. также п. 2 заключительной статьи наст, сборника. - Примеч. 2010 г.
245
Палеоботаник и философ С.В. Мейен делал из этого вывод: «функциональ-
ность в принципе непригодна к объяснению всего разнообразия живого», и со-
глашался прилагать ее «лишь к отдельным особенностям организмов» [Чайков-
ский, 2003, с. 34]. Замечу: не будет противоречия, если перевернуть установку
Мейена, т.е. признать, что как раз отдельные особенности чаще всего бесполез-
ны, а как раз общие свойства обычно нужны его обладателям. Однако лучше
признать, что систему нельзя строить априорно (как на функциональных свой-
ствах, так и на отрицании их значения), что ее следует строить для каждого так-
сона на тех свойствах организмов, которые ему соответствуют.
Сказанное почти неизвестно практическим систематикам. Они чаще всего
попросту не знают, что такое естественная система, не говоря уж о различиях в
ее понимании. Соединяя Любищева с Кузиным, можно сказать, что искусст-
венную систему надо строить, а естественную - искать в природе.
Есть и другое понимание естественной системы: поскольку она «является
метафизическим конструктом, она не может быть определена единственным об-
разом без четкого указания того аспекта разнообразия, которому соответствова-
ло бы данное понятие» Поэтому «полное описание таксономического разнооб-
разия организмов может быть дано только совокупностью нескольких естест-
венных классификаций, разрабатываемых для разных аспектов биологического
разнообразия» (И.Я. Павлинов, см. наст. сб.).
Далее пойдет речь именно о поисках естественной системы, однако не ка-
кой-то единственной, а той, которая соответствует конкретной задаче - по-
строению как картины биоразнообразия, соответствующей нынешней картине
мира, так и достаточно постоянного перечня названий организмов.
8. Место естественной системы в систематике
Самым важным в рассуждениях Любищева о естественной системе мне
представляется та мысль, что каждый век имеет о ней свое представление: сна-
чала естественной была лестница существ, затем иерархическая система Линнея,
позже принявшая форму филогении, а в его время - комбинативные системы
(см. [Чайковский, 2003, с. 38]). Сам по себе комбинативный принцип Любищев
не считал пригодным для систематики ввиду избыточности получаемых таблиц,
полагая его лишь начальным этапом всякой систематизации.
Будущими этапами развития систематики он видел построение как парамет-
рической системы (см. выше, п. 7), так и коррелятивной, о которой высказался,
насколько знаю, лишь однажды, в рукописи 1947 года. Суть ее в следующем:
пустоты комбинативной таблицы предлагалось понимать не только как запреты
некоторых вариантов, но и в том смысле, что одними свойствами задаются дру-
гие. Например, членистость и хитин задают многие другие свойства насекомых
[Любищев, 1975]. Блестящим примером коррелятивной системы мне видится
грубая филогения по Мейену (см. п. 11).
Впоследствии было показано, что для каждой эпохи в развитии науки ха-
рактерна своя познавательная модель (ПМ) [Чайковский, 1990; 2006а]. Тем са-
246
мым, смена взглядов на естественность системы связана с общей сменой взгля-
дов на природу и общество, выражаемой сменой ПМ, обычных в данную эпоху*.
Лестница существ характерна для религиозной (этико-эстетической) ПМ; ие-
рархия - для двух ПМ, знаковой и механической; комбинативность - тоже для
двух ПМ, статистической (господствующей ныне) и диатропической (едва на-
мечаемой в общественном сознании); а коррелятивная - для системной ПМ,
входящей ныне в общее употребление [Чайковский, 2006а, п. 10-2*].
Любищев оказался прав: несмотря на появление компьютеров и геносисте-
матики, ни одна из общих проблем систематики не решена. Непрестанный пере-
смотр конкретных систем (а с тем и названий) продолжается на всех уровнях.
Только за счет дробления таксонов получают новые имена около 1% видов мле-
копитающих в год, а число видов приматов за 30 лет удвоилось [Павлинов,
2006]. Приближения к чему-то стабильному не видно, что говорит о познава-
тельном тупике. Этот конфуз является, на мой взгляд, следствием невнимания к
теории и, в частности, к разделению естественной и искусственной систем.
Хотя вопрос о стабильности названий иногда у филогенетиков ставится, но
не видно ни ответа, ни даже подхода к нему. Так, ими предлагалось
1) остаться при традиционной («линнеевской») систематике, поскольку та
«максимально проста, информативна и стабильна» [Wiley, 1979, р. 309];
2) отделить номенклатуру от систематики [Ereshefsky, 2001, р. 236];
3) улучшить номенклатурные кодексы [Knapp е.а., 2004].
Итог оказался нулевым, и вряд ли возможно иначе, пока без изменений ос-
тается сам принцип единственной системы. Действительно, если законсервиро-
вать единственную систему, то каким образом биологи смогут отражать измене-
ние своих представлений? Предложение разделить номенклатуру и систематику
означает, что первая должна обслуживать нечто, отличное от второй. Что же ка-
сается кодексов, то их улучшение сможет привести к стабильности названий
только тогда, когда именовать придется нечто слабо изменчивое. Однако все три
предложения имеют, как увидим, смысл, если их применить к схеме параллель-
но вводимых систематик - естественной и искусственной.
Разумеется, небрежение к теории ничего путного дать не может. В виде
примера небрежения Б.П. Захаров ссылается на Э. Майра, хотя можно бы со-
слаться и на нынешних авторов, пренебрегающих естественной системой без
попытки понять, что это такое. Например, в новом руководстве читаем;
«Систематика живых организмов традиционно строится как отражение их
филогении (или “естественной системы”, как ее называли до создания эво-
люционной теории), так что именно систематика позволяет объяснить суще-
ствование у тех или иных живых организмов тех или иных свойств»', «цен-
ность представляет только естественная классификация, основанная на фи-
логении» [Клюге, 2000, с. 7,43].
См. также в наст, сборнике статью «Любищев, Мейен и плюрализм в эволюции».
247
Неловко снова разъяснять, но приходится. Вам ценно одно, а мне другое. Но
важнее, что подчеркнутые мною слова Клюге выражают критерий естественно-
сти Любищева (положение организма в системе указывает его свойства), чуждый
самому Клюге. Более того, именно та идея филогенетиков, что родство и есть ес-
тественность, нацело игнорирует физиологию и экологию, а потому и ту идею,
что систематика должна объяснять основные свойства организмов.
Достаточно вспомнить, что тесное сходство свойств дельфина с ихтиозавром
не отражено (как и все прочие параллелизмы) в филеме никак55, зато в ней рядом
всегда помещаются китообразные и копытные, хотя они сходны (если вынести за
скобки общие свойства млекопитающих) только по тем признакам, какие взяты
филогенетиками для классификации. То же можно сказать о недавнем включении
ластоногих в отряд хищных [Павлинов, Россолимо, 1998, с. 86]. Более того, не
отражены целые ряды сходств, например, рода рожь и рода пшеница, хотя
именно этот пример, открытый Н.И. Вавиловым, все поминают.
Филогенетической систематике противостоит типологическая [Любарский,
1996; Михайлов, 1997; Захаров, 2005]. Обе именуют себя естественными, однако
адепты первой гордо демонстрируют, что не знают никаких пониманий естест-
венности, кроме собственного. Поэтому стоит напомнить, что первая рассматри-
вает лишь признаки, а вторая - сам организм, его целостность:
«Типология занимается упорядочением многообразия объектов - таксо-
номией, а также частей этих объектов - мерономией... Таксономический
признак (вне типологии - Ю.Ч.) в известном смысле произволен, тогда
как мерой - закономерно, естественно выделенная часть архетипа. Архе-
тип - это иерархическая система меронов и одновременно закон их изме-
нения. Не признаки определяют таксон, а, наоборот, таксон определяет
признаки... Гомология свидетельствует не о родстве таксонов, но о сход-
стве их архетипов» [Михайлов, 1997, с. 93].
Поясню: понятие гомологии введено в XIX в. именно для обозначения соот-
ветствия органа архетипу, а ее филогенетическое понимание (унаследование от
общего предка) относится только к частному случаю специальной гомологии и
всё равно неявно апеллирует к архетипу (подробнее см. [Чайковский, 2006]).
Вообще, типологии, как и всей нынешней науке, присущ интерес к целост-
ности, и только филогенетическая систематика всё еще строится на старинной
уверенности, что организм определяется признаками. Общую картину можно
иллюстрировать двумя новыми руководствами.
В руководстве [Протисты, 2000]59, в разделе «Систематика и ее задачи» ес- 88 89
88 Точнее, в ней есть единственный способ его учесть - признание полифилетической
природы того или иного таксона. Однако нынешняя филогенетика, особенно в ее клади-
стическом варианте, стремится начисто изгнать такие таксоны из системы.
89 Предвижу упрек: нельзя ссылаться на эту книгу как на руководство. Она, вопреки под-
заголовку, является сборником очерков, вопиюще неполным как по составу, так и по со-
248
тественная система упомянута трижды. На с. 88 читаем: «Классификация долж-
на быть естественной, т.е. должна строиться на генеалогически важных призна-
ках и, следовательно, отражать родственные отношения». Но на с. 91 тот же ав-
тор неявно использовал совсем иное свойство естественной системы -
возможность распространять выводы, полученные на отдельных экземплярах,
на все организмы данного таксона (С.В. Мейен назвал его типологической экс-
траполяцией). Редактор резонно заметил в сноске: «В этом смысле естественная
система - это система с максимально возможной прогностической силой». При-
ходится напоминать, что именно отсутствие прогностических свойств у филоге-
нии явилось одной из главных причин споров о сути систематики.
Третье упоминание (с. 91-92):
«Дарвин убедительно доказал, что естественные группы организмов разви-
вались путем расхождения признаков (дивергенции) из одного общего
ствола вследствие вымирания менее приспособленных промежуточных
форм. Отдельные группы возникали монофилетически... Все это очень хо-
рошо объясняло, почему естественная система организмов, отражающая
реально существующие в природе отношения, обладает иерархическим
строем... Однако это важнейшее положение дарвинизма решительно отвер-
гается его противниками, для взглядов которых характерен полный отрыв
филогении от системы, что, впрочем, не препятствует им упорно имено-
вать систему, предлагаемую ими самими, естественной».
Вероятно, имелась в виду типология. Это и есть мнение «подавляющего
большинства». Чтобы разобраться в приведенной мешанине несогласованных
мыслей, напомню: ничего такого у Дарвина нет. Он «убедительно доказал»
только наличие изменчивости и сам был смущен отсутствием фактических дока-
зательств остального, в том числе механизма эволюции. Что касается принципа
дивергенции, то он изложен Дарвином вообще без аргументов, даже мыслен-
ных, и это смущало самих дарвинистов. Например [Филипченко, 1977, с. 53].:
«Возвращаясь к принципу расхождения признаков, мы не можем не при-
знать в нем слабейшую часть всей теории, так мастерски развитой Дар-
держанию каждого. В книге отсутствуют важнейшие таксоны (инфузории, динофлагел-
латы и др.), и во второй, завершающей, части их тоже не предусмотрено [Протисты, с.
135]. Новых данных по цитологии, генетике и т.д., обещанных в предисловии, почти нет;
при описании разных групп уровень очень различен; лишь для немногих описан, и то
скудно, митоз и еще хуже - мейоз. Главы по эволюции и систематике никакого отноше-
ния к протистам не имеют и являют собой скорее упреки неназванным конкурентам, чем
главы руководства. Они, судя по содержанию, написаны в 1950-х гг. с добавлением ссы-
лок на ту литературу 1960-1990 гг., какая не может поколебать архаичную конструкцию.
Да, всё так, но книга принята “на ура” (даже теми, кто не мог не увидеть ее ущербность,
напр. [Малахов, 2002]) и вероятно будет служить руководством, а иных мне неизвестно.
Добавлю, что некоторые разделы (например, о молекулярных методах систематики про-
тает) довольно удачны. - Примеч. из [Чайковский, 2003, с. 44], добавлено в 2010 г.
249
вином... Он должен был прибегнуть для обоснования этого начала к со-
вершенно отвлеченным, чисто спекулятивным соображениям»
Всё это писано сотни раз, но оставлено дарвинизмом без ответа.
Вторую иллюстрацию дает статья из палеонтологического словаря, выра-
жающая линию любищевского направления:
«ЕСТЕСТВЕННАЯ СИСТЕМА, систематика... Привилегированная
(выделенная среди прочих) классификация (С.В. Мейен), отражающая ес-
тественный порядок природы (космос), врожденную (истинную) сущность
вещей (Аристотель), охватывающую в идеале все их сходства и различия...
Объекты общего рода в Е.С. находятся в большем соседстве между собой,
чем с объектами др. родов... Е.С. должна: а) быть устойчива, т.е. сущест-
венно не изменять своей структуры, к введению в нее новых объектов и
(или) новых признаков объектов... (В. Уэвелл); б) указывать по положению
(“адресу”) объекта в ее структуре все его существенные свойства; естест-
венность системы заключается в информативности ее “адресов” (А.А. Лю-
бищев). Если объекты возникают в процессе развития, т.е. обладают исто-
рией, как, напр., организмы, то их Е.С. неизбежно будет нести внушитель-
ную информацию о структуре их родственных отношений (Ч. Дарвин, Э.
Геккель). Преобладание центробежных тенденций (специализации) в раз-
витии науки над интегративными породило в XX в. нигилизм в отношении
сущностного (целостного) подхода к классификации объектов природы».
Далее - 4 примера отношения биологов к Е.С. [Палеонтология, 1995].
Об этой линии «подавляющее большинство» никогда по сути не говорит,
однако в русскоязычной литературе90 сложилось небольшое направление мыс-
ли, анализирующее общую проблематику. Центральной фигурой остается Лю-
бищев. Он не всегда упоминается, но сути дела это не меняет.
Так, морфолог растений Р.Е. Левина, хорошо знавшая Любищева, предло-
жила, не ссылаясь на него (на что имела в Ульяновске основания), «естествен-
ную морфологическую классификацию плодов». Вот ее замечания: «В ботани-
ческой литературе широко принято любую неэволюционную классификацию
снабжать эпитетом искусственной, независимо от объектов, задач и методов ис-
следования. Эта ошибочная точка зрения берет начало в систематике»; естест-
венной системой у Левиной названа «сущностная» система [Левина, 1987, с. 3,
18, 19]. Явно следуют Любищеву и названные мною в данной статье книги [За-
варзин, 1974; Корона, 1987; Павлов, 2000; Эпштейн, 2002; Захаров, 2005]. Все они
противостоят «подавляющему большинству», о чем любое руководство обязано,
на мой взгляд, сообщать пользователю.
90 К сожалению, насколько мне известно, только в ней. Русскоязычный характер обсуж-
даемой темы отмечают К.Г. Михайлов [1997, с. 95] и другие авторы. Связано это с тем,
что основой темы служит структурализм, чуждый господствующей ныне англо-
американской традиции. Подробнее см. [Чайковский, 2006, п. 6-11].
250
Есть, однако, еще более важный вопрос, почти никем не затрагиваемый: какая
классификация дает нам представление о картине биоразнообразия? Обычный от-
вет (что такова «система таксонов») - поверхностный. Ведь
«Помимо системы собственно таксонов, существуют, по меньшей мере,
еще две - система морфологических уровней и система жизненных форм»
[Михайлов, 1995, с. 42]. «Именно жизненные формы, а не таксоны, явля-
ются составляющими элементами биоценоза» [Михайлов, 1997, с. 93].
И в самом деле: общепринятая систематика говорит исключительно про род-
ственные связи видов, а не про свойства самих видов. Но «так ли это важно знать
- кто от кого и как произошел?» - спрашивал Мейен [Гоманьков, 1990, с. 76]. От-
вет может быть различным (кому важно, кому нет), но несомненно, что никакой
ответ о происхождении сам по себе не дает картины живой природы - ее дают
описания свойств организмов и их взаимодействий.
Поэтому не раз делались попытки построить эко-морфологическую систему.
Они не имели успеха, поскольку не доходят до уровня вида и неудобны для опре-
деления, но если этих задач перед ними не ставить, то надо признать, что они
весьма полезны - именно для уяснения картины мира (см. далее, п. 13).
9. Является ли нынешняя систематика эволюционной?
В п. 3 мы видели, что 2 вида корсаков (евразийский и американский) попали
в общий род, хотя произошли независимо. Оказывается, ряд таких же пар являет
собой всё семейство псовых: еще в 1880 г. Гексли разделил всех псовых на две
группы - волкообразные и лисицеобразные, и деление сохранилось в науке по-
ныне. Обе группы представлены как в Евразии, так и в Америке, в основном,
Южной. Пары сходных видов (в каждой паре один вид евразийский и один аме-
риканский) и образуют искомый ряд: естественнее допустить, что сходства воз-
никли независимо, нежели то, что каждый евразийский вид каким-то образом
перебрался через океан, где породил сходный вид. И действительно, в наше
время генетический анализ показал, что все южноамериканские виды псовых
ближе (в смысле родства) друг к другу, чем к евразийским [Wayne е.а., 1997]. То
есть иерархическая структура таксона здесь не отражает дивергентной эволю-
ции его видов. Вероятно, то же самое характерно и для корсаков. Данная ситуа-
ция типична и отражает тот тип эволюции, который известен как номогенез и
игнорируется «подавляющим большинством».
Неудивительно, что программная статья [Мина и др., 2006] упоминает эво-
люцию по сути всего один раз, и вот в каком контексте: существует филогене-
тическая концепция вида, она дает фактически ту же систематику, что и праг-
матическая концепция, никакого отношения к эволюции не имеющая. Согласно
первой, вид есть множество организмов, которые можно уверенно отличить от
членов других множеств и «внутри которого существуют отношения предков и
потомков» (что значит это странное словосочетание, не сказано). Согласно вто-
рой, вид есть просто множество организмов, которые можно уверенно отличить
от членов других множеств (с. 554).
251
Но ведь уверенно различаются и подвиды (например, аральская вобла, кас-
пийская вобла и тарань - подвиды плотвы обыкновенной) - почему бы не на-
звать их тоже видами? Оказывается, в новейшем каталоге рыб так и сделано, с
чем и спорит статья, резонно замечая, что на этом пути «невозможно остано-
виться: практически каждую популяцию приходится признать самостоятельным
видом» (с. 555). Систематики, согласно статье, заняты, в основном, изобретени-
ем новых видов, «выделение видов превращается в самоцель, а классификация
становится бесполезной» (с. 557).
Однако положение с видами, полагают авторы, еще довольно сносно, по-
скольку у вида есть хоть какое-то определение, тогда как у более высоких так-
сонов (родов, семейств и т.д.) нет и этого. Авторы признают, что никакой тео-
рии у систематики нет, что каждый систематик творит, как хочет, а страдают
пользователи, т.е. все остальные.
Поэтому, добавлю, неизбежны две группы вопросов. 1) Если никакой тео-
рии у систематики нет, то почему все-таки существует сама система организ-
мов, которой, хорошо или плохо, но все пользуются? Почему, например, вол-
ком все люди называют одну и ту же группу зверей, почему они при этом по-
нимают друг друга? Другими словами, почему почти все виды и иные таксоны
дискретны (четко различимы)? 2) Если нынешняя биология в самом деле эво-
люционна, то почему не построена эволюционная систематика? (Ее отличие от
филогенетической разъяснено в работе [Шаталкин, 2006].) Строит ли ее кто-
нибудь? Можно ли ее построить вообще?
На последний вопрос ответить проще всего: эволюционную систематику
можно построить там, где есть достаточный ископаемый материал. Мы можем,
например, подкрепить идею происхождения всех собак от волков тем, что соба-
ки появились позже волков и появились там, где водились волки.
Там же, где ископаемого материала нет, всякие эволюционные рассуждения
голословны, а потому и недолговечны. Так, весьма скудны ископаемые данные
о происхождении южноамериканских псовых от остальных псовых, поэтому их
отнесение к одному семейству (если понимать каждый таксон как происшедший
от общего предка) то и дело подвергается сомнению, и даже новейшие молеку-
лярные методы, как оказалось, помогают мало.
В последние 30 лет появились молекулярные методы анализа наследствен-
ных сходств (геносистематика), которые, будучи применены совместно с тра-
диционными, помогают уточнить систематику. Такова, например, упомянутая
статья [Wayne е.а., 1997]. Геносистематика претендует на объективное описание
истории, но анализ можно счесть объективным, только если он приводит разных
авторов к одному и тому же выводу. На самом же деле разные сторонники гено-
систематики приходят к весьма разным выводам, которые к тому же почти все-
гда противоречат выводам обычной систематики.
. Хорошей иллюстрацией служит упомянутое руководство [Протесты, 2000].
В главе «Система протестов» С.А. Карпов пишет: «Не было построено двух
252
одинаковых деревьев даже в одной лаборатории», а кладограммы на форзаце эту
мысль поясняют: на них человек изображен сразу между двумя видами дрозо-
фил и между протистом Plasmodium и многоножкой Schizophyllum.
Вообще же, систематика почти нацело игнорирует генетику - точно так же,
как и эволюцию. Например, статья [Мина и др., 2006] сетует, что новейшая
сводка по систематике рыб построена, как и двести лет назад, на морфологии
(изучении строения тел), а к данным генетики прибегает лишь изредка, когда
это почему-то удобно составителям. Разумеется, там, где в расчет принято раз-
личие генов, число видов сильно возрастает. Вопрос о том, осмысленно ли это,
не ставится. Словом, на сегодня нет эволюционной систематики, которая была
бы настолько долговечна и признанна, чтобы ее можно было преподавать и что-
бы ею, тем самым, могли пользоваться остальные биологи. А что же есть? На
чем в действительности основана систематика организмов?
10. Номогенез в систематике
Авторы упомянутой выше статьи [Мина и др., 2006] призывают «сначала
исследовать и описать его (разнообразия организмов - Ю. Ч.) структуру и лишь
после этого дискретным группировкам (если таковые обнаружатся) придать
таксономический статус и присвоить научные названия. Иными словами, пред-
полагается продвижение от изучения объектов к их классификации... а не на-
оборот» (с. 556). Они видят в этом приеме (движение от частного к общему)
суть новой систематики, однако мне придется напомнить, что именно так строи-
лась систематика до Линнея. Он же строил ее с обеих сторон, и его «естествен-
ный метод» как раз включает путь от общего (целого) к частному.
Главным новшеством у Линнея был знаменитый тезис - что не признаки за-
дают род, но род задает признаки, (родом тогда называли нечто более крупное
и важное, чем ныне.), усвоенный нынешней типологией (см. п. 8), но не филоге-
нетикой. Этот тезис означает, что сперва надо выявить план строения таксона
(то главное, что позволяет узнать его представителей), а затем уже искать при-
знаки, уверенно отличающие его от других таксонов. То есть в основе класси-
фикации у Линнея лежала интуитивная процедура выявления некоторой целост-
ности, а формальная процедура выявления признаков лишь ее обслуживала и
была вторичной. Подробнее см. [Любищев, 1972].
Позиция авторов статьи [Мина и др., 2006] (как и почти всех нынешних сис-
тематиков) противоположна: никаких интуитивных актов, двигаться надо от
учета признаков. Это - обычная для систематиков (и только для них) установка:
считать первичной реальностью не строение организма, а те признаки, которы-
ми различаются организмы. Из различий надо каким-то способом извлекать
классификацию. Каков способ, такова будет и теория. О способе авторы ничего
не говорят, оттого у них, как они сами признали, нет и теории.
Но при чем тут эволюция? А вот при чем. Единственное, что авторы отме-
тили - что для классификации нужна (подчеркнутая мною в цитате) дискрет-
ность, то есть четкое отличие исследуемой группы особей ото всех иных. Но от-
253
куда она берется? Откуда может взяться дискретность, если, как утверждает
господствующая эволюционная доктрина (дарвинизм), в природе царит плавная
непрерывность всех наследственных преобразований?
Дискретность свойств мыслится в дарвинизме как итог воображаемого вы-
мирания промежуточных форм. Никто этого процесса на практике не наблюдал
- повсюду наблюдают противоположную картину: новая организация (в отли-
чие от новых признаков) всегда возникает цельными блоками; в этом состоит
блочность эволюции, ныне всеми признанная. Но даже если вымирание проме-
жуточных форм где-то и удастся обнаружить, ясности не возникнет - надо будет
объяснить, почему эволюция сперва благоприятствовала промежуточным фор-
мам, а затем их истребила. Для этого дарвинизм тоже имеет лишь одно объясне-
ние - воображаемый стабилизирующий отбор, которого, однако, никто не толь-
ко не наблюдал, но и не искал. Он попросту постулирован*.
Другого объяснения в дарвинизме нет, и авторы статьи (дарвинисты) спра-
ведливо допускают, что таковой дискретности может и не обнаружиться. Одна-
ко все организмы удается классифицировать, то есть дискретность так или иначе
всегда место имеет* 91. Более того, она имеет место на всех уровнях, от биосферы
до подвида, что и отметили авторы, когда признали, что на пути выявления все
более мелких таксонов невозможно остановиться. Это, замечу, так называемая
фрактальность разнообразия [Чайковский, 2006].
Почему так? Ответа в дарвинизме нет, и приходится обращаться к другим
учениям. Ламаркизм, главный оппонент дарвинизма, ничего тут добавить не мо-
жет, поскольку обе доктрины объясняют разнообразие через приспособление к
каким-то своим особенным условиям существования, а почему оно дискретно, не
говорят. Тем более не говорят они о свойствах этой дискретности. Почему, к при-
меру, в семействе псовых роды «волк» и «лисица» имеют по многу видов, а боль-
шинство родов данного семейства имеют (как песец и енотовидная собака) лишь
по одному виду? Почему та же ситуация типична для всех крупных таксонов?
Но есть еще номогенез - эволюционное учение, которое принято относить к
одной старинной книге Л.С. Берга (1922), но которое на самом деле никогда не
исчезало и накопило огромную фактическую базу (новейший пример: [Мосей-
чик, 2005]). Ныне оно переживает взлет, о чем говорят новые книги: [Назаров,
2005; Попов, 2005; Чайковский, 2006; Зусмановский, 2007], но нигде не препо-
дается и «подавляющим большинством» не цитируется.
Номогенез дает предварительный ответ, исходя из дискретности форм орга-
низмов как основного факта, данного нам наблюдением, и на нем (а не на допу-
щениях) строит теорию. В этом огромное преимущество номогенеза, и для на-
шей темы им можно пользоваться, невзирая на его изъяны, о которых, кстати,
См. «Стабилизирующий отбор, или Святость веры» (4-39). -Примеч. 2010 г.
91 Исключение являют немногие роды-гиганты (такие, как лилия или дрозофила), внутри
которых классификация трудна; однако сами такие роды четко отличимы от иных.
254
тоже можно прочесть в указанных книгах.
Принять факт дискретности как основной приходится потому, что она уди-
вительно одинаково устроена у всех организмов: во всяком крупном таксоне
около 40% родов имеют по одному виду, около 15-17% родов - по два вида и
т.д., а основная часть видов содержится в одном-двух крупных родах. Это - так
называемое распределение Виллиса, открытое давным-давно, в 1918 году. Его
принято игнорировать, у нынешних филогенетиков нет о нем ни слова, а преж-
ние отвергали его на том «основании», что оно распространено и в неживой
природе, и в обществе. С позиции тех, кто изучает естественные системы, си-
туация противоположна: система, приблизительно подчиняющаяся распределе-
нию Виллиса, во многих науках признана как естественная (литературу см.
[Чайковский, 2006]). Биологам давно пора этим воспользоваться.
Объяснение природы данного распределения - важнейшая задача науки, но
она вовсе не является задачей номогенеза (и биологии вообще), поскольку точно
так же распределены самые различные разнообразия - города по их численно-
сти, буквы по их встречаемости в текстах, химические элементы по их встре-
чаемости в природе и многое другое. Тем самым, номогенез выступает как часть
общей теории эволюции природы, чего ни о ламаркизме, ни о дарвинизме ска-
зать нельзя - их не вписать в общую картину мира.
Если в основе ламаркизма лежит активность особи, а в основе дарвинизма -
ненаправленная изменчивость, то в номогенезе главное - план строения орга-
низма (подробнее см. [Чайковский, 2006]). Для систематики эта сторона номо-
генеза очень важна, и поэтому о нем приходится здесь много говорить, хотя ос-
тальные учения сами по себе не менее интересны. Так, для понимания становле-
ния функций необходим опыт ламаркизма [Зусмановский, 2007]. Однако в
большинстве отраслей биологии систематика физиологию игнорирует.
И. Опыт грубой филогении (Мейен)
Самым замечательным в систематике Мейена мне видится то, что она не
строилась в рамках заранее заданной формы - форма системы искалась в ходе ее
построения. Уйдя в юности от веры в дарвинизм, Мейен стал противником вся-
ческих филогений. В конце своей слишком короткой жизни он построил свою
систему голосеменных и обнаружил у нее, к собственному удивлению, черты
филогении, притом в монофилетическом варианте. Факт этот был отмечен его
учеником А.В. Гоманьковым, который выразил уверенность, что
«систему голосеменных он (Мейен - Ю. Ч.) перестраивает (причем весьма
кардинально), руководствуясь отнюдь не абстрактными “номотетически-
ми” идеалами Любищева, а на основе все той же реконструированной им
филогении, т.е. как правоверный дарвинист» [Гоманьков, 1990, с. 79].
Позже другой ученик Мейена, И.А. Игнатьев, выразился грубее:
«Эволюционные работы Мейена несомненно способствовали распростра-
нению антиселекционистских и номогенетических идей», однако он вовсе
не был номогенетиком. «Мейен развивал, причем только в молодости, не
255
номогенез, а дополненный историческим компонентом структуралист-
ский подход Любищева, пытаясь выстроить вслед за ним треугольник
“форма - система - эволюция”. Впоследствии он полностью перешел на
позиции эклектического плюрализма, понимая эволюцию существенно
тихогенетически» [Игнатьев, 2007, с. 48j.
Оба, как видим, декларировали возврат Мейена к дарвинизму. Оба, впро-
чем, не нашли никакого дарвинизма в последнем и главном труде Мейена
[Meyen, 1987], а увидали там его прежнюю позицию. Поскольку его работы по
эволюции и систематике голосеменных представляются мне важным достиже-
нием эволюционной систематики, ситуацию необходимо прояснить.
Дальнейшее изложено, если не указано иное, согласно п. 10-6 книги [Чайков-
ский, 2006], где можно найти подробности и ссылки на литературу.
Свои принципы классификации Мейен изложил в 1974 году так: хотя «в при-
знаковом пространстве растений есть свои регулярности», однако они не выра-
жаются общепринятой систематикой; нужна система, основанная на трех положе-
ниях: (1) признаки не связаны жестко, (2) любой признак может у какого-то члена
таксона отсутствовать и (3) параллелизмы должны быть учтены системой.
Сближение таксонов голосеменных Мейен проводил по органам размноже-
ния. Идя этим путем, он в 1986 г. пришел к такому выводу:
«В целом можно сказать, что в филогении высших растений, если говорить
о происхождении семейств и таксонов более высокого ранга до отдела
включительно, безраздельно господствует монофилия».
Монофилия является итогом филогении, а прежде Мейен полагал, что фило-
гению, как правило, построить невозможно и что эволюция представима в форме
не древа, а сети. В том же 1986 г. он писал: «Лично я считаю, что голосеменные
представляют собой монофилетический таксон», - но не привел фактических
обоснований филогении вообще и монофилии в частности. Есть у него схема
флорогенеза (она воспроизведена во многих учебных и научных работах - см.
рис. 6), которая названа в подписи при первой публикации (1984) филогенией.
Позже он называл ее осторожнее: «Географическое распределение главных фило-
генетических ветвей» [Мейен, 1988, с. 42]. Что в эти годы изменилось?
Рассмотрев схему, убеждаемся, что это никак не филогения: отдел цветковых
«повис», т.е. ни к какой ветви не отнесен; да и в отделе голосеменных также «по-
висли» 6 порядков. Идее монофилии тут противоречит и тот факт, что толстые
ветви часто переходят одна в другую без сужения. Мейен объяснил: это значит,
что порядок переходит в порядок, «с широкой трансгрессией признаков»; т.е.
нельзя сказать, какое семейство переходит в какое. Это значит, что о монофилии
на этом уровне говорить нет смысла.
Тем более, нет ее на уровне семейств и ниже: единственная у Мейена на сей
счет «филогения» (рис. 7) показывает, что большинство семейств и некоторые
роды повисли вне связей (подробнее см. [Чайковский, 2006а, с.99-100]).
В своей последней статье (М-324) Мейен пояснил:
256
«если учесть всю систему ранних хвойных и длительные тенденции в семо-
филогенетических преобразованиях органов92 93, то обнаруживаются более или
менее устойчивые состояния некоторых признаков и соответственно их раз-
ный вес в филогенетических и, следовательно, таксономических построени-
ях». Поэтому «в эволюции палеозойских хвойных намечается несколько ос-
новных тенденций... Это именно тенденции, а не отчетливые семофилогене-
тические ряды... Для растений характерна блочная (“модулярная”) структура,
причем в эволюции отдельные блоки легко меняются местами». И «мы видим
в эволюции позднепалеозойских хвойных комплекс родов, в котором прихот-
ливо переплетаются признаки... Как бы мы ни группировали эти роды, будут
получаться и параллелизмы, и реверсии примитивных состояний»
Как видим, он не нашел не только филогений, но и хороших семофилогений.
Два его отрицания филогении (раннее и позднее) смыкаются. В каком же смысле
Мейен «реконструировал филогению» голосеменных? А вот в каком.
«Филогенетическое древо» он строил на основе одной семофилогении (на-
пример, по женской фруктификации [Meyen, 1997, р 422]), причем выбор ее в ка-
честве базовой обосновывал «географически» - соединял родством те роды, кото-
рые, сменяя друг Друга, жили в общей географической зоне и притом сохраняли
значительное сходство по базовому синдрому (в данном случае, по женской фрук-
тификации). Конечно, тот факт, что эволюция женских фруктификаций ранних
хвойных в бореальной зоне средней и верхней перми приняла у Мейена форму
древа, информативен - могла бы получиться сеть. Однако для отождествления
этого древа с эволюцией самих растений как целостных единиц материала слиш-
ком мало. На вопросы скептика (достаточно ли число находок? Нельзя ли с дру-
гим синдромом получить другое древо? Можно ли для иных крупных таксонов
подобрать синдром и находки, дающие древо, или описана особенность данной
группы в данной местности? Законно ли принадлежность общей зоне, когда их
всего три92, называть географической близостью? И т.д.) ответить было бы, по-
моему, нечего. Но скептиков не нашлось, а сам автор себе их не задал (не успел?).
И тут мы подходим к ответу на вопрос: что изменилось?
Он и прежде призывал к синтезу учений и сочувствию в отношениях между
людьми (в этом - пафос воспоминаний Гоманькова и Игнатьева), теперь же Сер-
гею Викторовичу (он знал, что умирает) явно хотелось не споров с коллегами, а
сближения с ними. Он охотно сам вставал теперь на точки зрения коллег, порою
92 Семофилогении - историческое преобразование одного-единственного синдрома (цветка,
конечности и т.п.), если его можно изобразить в форме древа.
93 В текстах о происхождении высших таксонов растений у Мейена использовано всего три
зоны - северная, тропическая и южная. Иногда он добавлял еще две узких полосы - грани-
цы тропической зоны [Мейен, 1988, с. 41]. Глобальный флорогенез состоял, по Мейен}', в
том, что основная масса высших таксонов возникала в тропической зоне и затем распро-
странялась в северную и южную зоны. При этом сами таксоны претерпевали эволюцию.
257
плохо совместимые. Отсюда, если я верно понял, и упрёк ему в «эклектике», и
объяснение того, почему он назвал новое явление старым словом «филогения», и
того, почему при обсуждении теории он, тем не менее, остался на прежней анти-
филогенетической позиции. Такова душевная сторона дела.
Теперь - о биологической стороне. Мейен открыл (вернее, обозначил) новое
биологическое явление, которое можно назвать грубой филогенией. Суть дела в
том, что процесс эволюции в пределах целого отдела (голосеменные) предстал
ему как имеющий некоторые черты филогении, так что итог эволюции можно
счесть филогенией лишь при поверхностном взгляде. Точнее,
1) надо следить не за всеми (даже существенными) чертами организмов сразу,
а только за избранным комплексом (синдромом), полагая остальные свойства па-
раллелизмами; 2) избирается в качестве базового синдрома тот, который «лёг на
карту», т.е. для которого исторически близкие формы оказались близкими геогра-
фически; 3) надо ограничиться рассмотрением таксонов довольно высокого ранга
(классы, порядки и некоторые семейства)*.
Вместо привычного понимания филогении как единого детального процесса
эволюции путем дивергенции (расхождения одного таксона на два), Мейен дал но-
вое ее понимание: эволюция видов порождает некую сеть, отдельные части кото-
рой связаны слабо: как между классами, так и между порядками внутри каждого
из классов, сращений столь мало, что части сети похожи на ветви. Эволюцию в
рамках каждой части можно грубо представить древом. То есть тут древо - грубая
схема, не допускающая детализации: роды и многие семейства эволюируют, как
писал Мейен, «с широкой трансгрессией признаков».
Но если грубая филогения отсутствует на низших уровнях, значит, она явно
не является итогом дивергенции признаков - того, на чем строится обычная фило-
гения. Грубую филогению можно рассматривать как итог корреляции свойств, об-
разующих базовый синдром (благодаря чему сеть вырождается в почти древо), т.е.
как реализацию коррелятивной системы по Любищеву.
Грубая филогения может показаться чем-то вычурным и редкостным, но ведь
именно ее строили почти все, кто, вслед за Геккелем, описывал филогении: идея
обычной филогении иллюстрировалась только на семофилогениях высокого так-
сономического ранга. Когда же вставал вопрос о механизме дивергенции, то речь,
наоборот, переводилась на самый низший таксономический уровень - конкурен-
цию рас по Дарвину. Критики сотни раз обращали внимание на это явное несоот-
ветствие, но им не отвечали или упрекали их в неверии в науку.
Прим. И.Я. Павликова. Как представляется, идея грубой филогении близка к концепции
«широкой монофилии» Дж. Симпсона, которая при анализе таксонов высокого ранга также
не предполагает выяснения их происхождения от конкретных видов. Согласно названной
концепции, макротаксон (семейство, отряд) монофилетичен, если он произошел от другого
таксона того же или более низкого ранга.
258
Мейен вскрыл, в отличие от дарвинистов, свой метод, а это умеют очень не-
многие - из классиков, которыми я занимался, таким был только астроном Иоганн
Кеплер. Остальные попросту декларируют свое мнение в качестве установленной
истины (а на деле в основе у них чаще всего лежат удобные примеры) .
Словом, считать позднего Мейена филогенетиком можно только в смысле
указания им на грубую филогению. Как этим опытом воспользоваться?
Прежде всего, надо выяснить, насколько данный феномен реален. Мейен ста-
вил цель «положить филогению на карту», но фактически успел только положить
несколько семофилогений на картоид из трех зон. Для этого масштаба и материа-
ла грубая филогения - факт, но попытка его детализации не столь однозначна.
Дело в том, что Мейен извлек данные об эволюции и систематике таксонов
высших растений из построенной им же схемы флорогенеза (эволюции целых
флор, которую он представил в форме флорогенетического древа [Мейен, 1988,
с. 43]). Ее он успел довести до конца мезозоя (рис. 8).
Детально он смог рассмотреть только появление первых хвойных в Запад-
ной Ангариде (ныне - восток Восточной Европы) в кунгурском - татарском яру-
сах пермского периода. На его картосхеме [Меуеп, 1997, с. 353] можно видеть,
как хвойные, эволюируя, распространялись от самого раннего местонахождения
(к востоку от Камы) на северо-запад, запад и юго-запад (рис. 9).
В целом картина достаточно ясна, но на картосхеме есть два более северных
местонахождения (бассейн Печоры), и они выпадают из нее. (Возможно, они ука-
зывают на другой очаг эволюции ранних хвойных, но даже для гипотетического
суждения здесь недостает материала.) Тем не менее, они частично (!) включены в
общую схему, которую Мейен назвал «пробной филогенией» (рис. 6) [Меуеп,
1997, с. 433]. Если добавить, что в основании «пробной филогении» лежит род
Swillmgtonia, обнаруженный в вестфальском ярусе карбона (т.е. на 50 млн лет ра-
нее) и притом в Англии, то для иллюстрации единого флорогенеза камский при-
мер выглядит неудачным. Скорее его следует считать локальной вспышкой видо-
образования у группы (хвойные), известной с карбона.
После смерти Мейена (1987 г.) исследования были продолжены в его лабо-
ратории. В частности, М.А. Ахметьев продвинул флорогенетическое древо в
кайнозой (рис. 10) и получил подтверждение схемы флорогенеза со значитель-
ной детализацией [Ахметьев, 1999]. Наоборот, Ю.В. Мосейчик (ученица И.А.
Игнатьева), сузив рамки исследования во времени (ранний карбон) и простран-
стве (Подмосковный бассейн), а затем сравнив свои наблюдения с выводами
других ученых, пришла к иному выводу, чем Мейен. Вероятно, полагает она,
* Мысль этого абзаца (изъятого И.Я. Павлиновым из его издания) вызвала возражения у
палеоботаников школы Мейена. Они сомневаются в том. что Мейен действительно рас-
крыл свой метод: «На наш взгляд, монофилетизм построенной Мейеном филогении явля-
ется в значительной мере артефактом, обусловленным, с одной стороны, искусственной
монотетичностью семейств ископаемых растений, а с другой - построением системы
растений по конгрегационному принципу» (ИМ, с. 141). -Примеч. 2010 г.
259
«в разных местах Земли сходные эволюционные тенденции реализуются
при подходящих условиях. Л.С. Берг (1922) называл эти тенденции, ино-
гда проявляющиеся даже вопреки внешним условиям, автономическим
ортогенезом. Вопреки мнению С.В. Мейена, эти процессы, очевидно, не
были локализованы в пределах тропической зоны», и «можно предполо-
жить существование в каждом полушарии, по крайней мере, трех “поя-
сов” с... повышенной интенсивностью эволюционных процессов» [Мо-
сейчик, 2005, с. 102].
С этой позиции камский пример Мейена становится понятным - как один из
сходных случаев автономического ортогенеза. Однако глобальный флорогенез
следует подтверждать (или опровергать) более длинными рядами. Такую работу,
насколько мне известно, первым проделал биогеограф Леон Круаза (Croizat), ра-
ботавший в Венесуэле. В 1958-1962 годах он издал несколько огромных книг, в
которых скрупулёзно описал трэк (фактический ход эволюции, сопровождавшей-
ся расселением по суше или по морю) для многих таксонов животных [Попов,
2005]. К сожалению, его работы мне подробнее неизвестны*.
Полагаю, что понятие грубой филогении, если можно будет принять его как
общее для эволюции, в корне изменит всю проблематику таксономии - ведь
грубая филогения в принципе не может опуститься до уровня рода и вида. На
этом уровне царит, как отмечал Мейен, комбинативная эволюция, а с тем и сис-
тематика не может здесь строиться филогенетически.
Тут тоже встает вопрос о названиях: род Lebachiella, исходный на рис. 7 для
ветвей L и V, известен под четырьмя названиями: Lebachia, Walchia, Lebachiella
и Emporia, причем второе недавно предложено вновь - см. [Gomankov, 1997].
Еще важнее, что первое название всеми признается как «нелегитимное», однако
все обращаются к нему, чтобы объяснить, о чем идет речь. (В своем учебнике
Мейен даже уравнивал Lebachia с древнейшим хвойным Swillingtonia.') Самое же
важное то, что Мейен положил его в основу трех ветвей, лишь взяв в кавычки.
Можно сказать, что оно использовано как бытовое.
12. Опыт двух систем (бактериологи)
Единственная область биологии, где узаконены сразу две системы, это бак-
териология. Произошло это в 1994 г.: «В девятом издании “Определителя бакте-
рий Берджи” мы отступили от принципа прежних изданий - объединения,
обычно не очень успешного, сведений по систематике и идентификации» [Оп-
ределитель..., 1997, с. 6]. Оставив филогенетическую систематику в прежнем
«Bergey’s Manual of Systematic Bacteriology», авторы издали отдельный опреде-
литель («Bergey’s Manual of Determinative Bacteriology»), который «построен
только на фенотипических признаках, без попытки создать естественную клас-
сификацию... Такое подразделение представляется нам наиболее удобным для
диагностики».
* Подробнее см. статьи «500 лет...» (4-64) и [Моггопе, 2000] . -Примеч. 2010 г.
260
«Для практических целей» (т.е. для адресации) бактерии разделены на 4
«основные категории», категория делится на группы (всего их 35), группа - на
роды, а род, как обычно, на виды. Авторами подчеркивается, что такое членение
вызвано лишь удобством. Добавлю: во-первых, оно, как в систематике Линнея,
имеет всего четыре ступени (причем «основные категории» не используются);
во вторых, в нем главную роль, как в народной таксономии, играют род и вид,
иногда в форме родо-вида; а в-третьих, именно поэтому оно может претендовать
на долговечность.
Что касается естественной системы, то авторы сочли таковой филогению и
высказали уверенность, что ее таксоны существуют, но почему-то неудобны для
отыскания, отчего и пришлось отделить диагностику от систематики. Однако
материал говорил против этой уверенности. Например, в одном из семейств не
оказалось общих диагностических признаков [Определитель..., 1997, с. 295], т.е.
налицо не филогения по Геккелю, а конгрегация по Любищеву.
Это понимал редактор перевода Г.А. Заварзин, знаток любищевских тем, ав-
тор книги о естественной (в смысле Любищева) системе бактерий [Заварзин,
1974]. В предисловии к переводу «Determinative Bacteriology» он писал:
«Сейчас идентификация филогенетического положения прокариот... развива-
ется на основе нуклеотидных последовательностей 16S-pPHK. Усовершенст-
вованная методика... сделала этот подход практически безальтернативным при
определении родовой принадлежности новых организмов. Созданная филоге-
нетическая система имеет все достоинства и недостатки классификации, осно-
ванной на одном признаке. К достоинствам следует отнести почти полную со-
гласованность результатов, полученных в разных лабораториях... Недостаток
этой системы состоит в том, что она не дает представления о функциях бакте-
рий. Поэтому фенотипическая, или функциональная, система сохранит свое
значение для практических работников» [Определитель..., 1997, с. 5].
Это звучало странно: ведь род бактерий, как неоднократно отмечал сам За-
варзин, определяется своими функциями. Оставалось непонятным, на каком ос-
новании принято, что различие функций обязательно скажется в различии 16S-
рРНК (т.е. откуда уверенность, что такая филема не погрязнет в гомонимии). За-
то было ясно, что филеме бактерий тут отказано в статусе естественной по Лю-
бищеву (ибо она не указывает функций), а фенотипическая система по сути при-
равнена к диагностической.
И вот через 9 лет последовала легко ожидаемая реакция - уже известная нам
фраза: «зачем нужна такая классификация кому-либо, кроме филогенетиков?»
[Заварзин, 2006, с. 526]. Как обычно, о долговечности филогений говорить не
приходится, ибо она целиком зависит от модных сегодня методов.
Для меня этот опыт* состоит в том, что необходимость двух систем осоз-
нана, что естественная система бактерий ищется вне филогенетики и что это
* Признание Г.А. Заварзиным того, что прежде лишь подразумевалось. - Примеч. 2010 г.
261
тоже всё более осознаётся, пусть и немногими. К сожалению, конкретная фор-
ма естественной системы бактерий еще не видна, однако, как мы увидим да-
лее, она может быть найдена в рамках более широкой задачи - построения ес-
тественно макросистемы (и, на мой взгляд, только в этих рамках).
13. Опыт естественной макросистемы (Чайковский)
Основной упрек филогенетиков в адрес сторонников иных пониманий есте-
ственной системы состоит в том, что такую систему не удалось построить [Эп-
штейн, 2002, с. 46]. На самом деле есть несколько примеров успешного по-
строения таких систем, но надо понимать, что под системой в них имеется в ви-
ду не то, что принято у филогенетиков. Прежде всего, такие системы не ставят
целью адресацию и потому не доходят до уровня видов. Первой стала феноти-
пическая система бактерий [Заварзин, 1974], затем появилось еще несколько, в
том числе коррелятивная система Мейена, а вскоре табличные системы цветко-
вых [Хохряков, 1991] и членистых [Павлов, 2000].
Вопросы естественной системы наиболее разработаны в отношении макро-
системы, т.е. системы царств. Ботаник А.Б. Шипунов [2007, с. 33] пишет:
«Нам представляется, что на смену филогенетическому пониманию
царств постепенно приходит новое, основанное на системных принци-
пах... С этой точки зрения, всё разнообразие живого легко разбивается на
три группы в соответствии с уровнями структурной сложности: Bacteria,
Protista и два царства “многотканевых” - Vegetabilia и Animalia (два, по-
скольку их ткани негомологичны)... В целом смена концепции царств от
линнеевской механистической к системной вполне укладывается в идею
Ю.В. Чайковского [1990] о смене познавательных моделей**».
Показано, что для макросистемы требуется иной, кроме древа и конгрега-
ций, способ классификации. В качестве него предложен секционный метод, фак-
тически тоже идущий от Линнея. Многообразие организмов делится на основ-
ные группы (царства) по принципам, взятым из экологии и, отчасти, морфоло-
гии. Царства, подцарства и равные им группы задаются секционно, а типы (и
равные им группы) и таксоны меньших рангов - конгрегационно. В основу по-
ложены схема царств Уиттекера - Заварзина (1979 г.), и представление о парал-
лелизмах. Последнее легло вскоре в основу диатропики, которая была развита
Мейеном и, после его безвременной кончины (1987 г.), мною. Об этом см. [Чай-
ковский, 2006; 2006а].
Живой мир не только делится на царства, но царства еще и пересекаются.
Без учета этого факта картина живого мира будет в корне неверна. Поэтому в
макросистеме есть понятие «межцарство» (Interregnum), частный случай поня-
тия «межтаксон», вводимого вместо термина insertae sedis.
Помня тезис Короны, нет смысла избегать древ в классификации. От них
надо отказываться только там, где они явно противоречат материалу. Именно
так построена указанная макросистема: она выглядит как «парк», в котором
О них см. выше, статью «Горизонт познания...». - Примеч. 2010 г.
262
«рассажены деревья» - таксоны привычной систематики. Эволюционный аспект
системы виден в «планировке парка»: концентрические кольцевые «аллеи» - это
уровни организации, возникавшие в ходе эволюции последовательно. См.
рис. 11, где эта макросистема изображена*. Ее публикацию (с латинскими на-
званиями групп) см. [Чайковский, 1986]**. За истекшие годы она не потребовала
пересмотра и в этом смысле тоже показала свою естественность***.
Такой метод представляется мне единственным на сегодня возможным спо-
собом прекратить чехарду ежегодно предлагаемых макросистем, филогенетиче-
ских по форме, которые игнорируются всеми (включая и их авторов, вскоре
предлагающих новые макросистемы-однодневки). Однако гораздо важнее пре-
кратить чехарду родовых и видовых названий.
14. Опыт постоянных названий (Клюге)
Система биологических названий (номенклатура) подробно рассмотрена в
книге Клюге. Сам он держится довольно странного мнения - что номенклатура
нужна только потому, что систематика непрестанно меняется:
«Если бы существовала стабильная классификация, не нужны были бы пра-
вила номенклатуры, достаточно было бы раз и навсегда утвердить названия
для всех таксонов этой стабильной классификации» [Клюге, 2000, с. 46].
Что это не так, ясно из сравнения с иными классификациями. Так, в грамма-
тике тоже есть номенклатура (правила написания слов). Грамматика весьма ста-
бильна, но написание слов во многом кодифицировано: во-первых, удобнее за-
поминать правила плюс исключения, нежели запоминать написание каждого
слова в отдельности, во-вторых, надо уметь записать впервые услышанное сло-
во. То есть нужда в кодификации вызвана не столько переменами в классифика-
ции, сколько разнообразием классифицируемого объекта.
Зато Клюге предложил то, что нам нужно - неизменное имя вида:
«видовое название может быть записано следующим образом: вначале
пишется видовой эпитет, затем автор и год... затем в квадратных скобках
первоначальное родовое название (независимо от того, соответствует ли
оно современному представлению о систематическом положении данного
Другой вариант данной макросистемы см. на задней обложке Сборника, где добавлен
уровень 1и (интрон-экзонные прокариоты), более сложный, чем 1п. Нужен ли он в каче-
стве классификационного, т.е. существуют ли вообще в природе прокариоты без интро-
нов (если нет, то уровни In и 1и совпадают), пока сказать нельзя. - Примеч. 2010 г.
Таблицу с лат. названиями см. также в ст.: [Попов, 2008, с. 45]. -Примеч. 2010 г.
Подробное объяснение системы с цветными таблицами см. в статье «Система, не похо-
жая на другие» (4-38). Одна таблица там поясняет взаимное расположение царств и меж-
царств, где в центре находится «озеро» (царство прокариот), окруженное тремя «массивами
деревьев» (эвкариотными царствами), причем каждое «дерево» - это тип (отдел). Каждый
«массив» отделен от двух других «поляной» (межцарством) из малых типов. Другая табли-
ца там поясняет уровни организации: кольцевая сеть - это рефрен, где сложность организ-
мов возрастает от уровня 1п в обе стороны: в прокариотную (к центру «озера») и в эвкари-
отную. Цветная симметрия таблиц отражает главные параллелизмы. - Примеч. 2010 г.
263
вида); если первоначальная комбинация включала название подрода, оно
может быть записано в круглых скобках позади родового названия внут-
ри квадратных скобок. Такая или похожая форма записи широко исполь-
зуется в каталогах. Здесь родовое название выполняет такую же функ-
цию, как фамилия человека: первоначально присваивается на основании
родственных связей, но в дальнейшем не изменяется и используется
только для установления личности... Чтобы указать современное систе-
матическое положение вида, впереди видового эпитета можно написать
иерархическое название... любого из надвидовых таксонов». Оно «может
меняться по мере развития систематики» [Клюге, 2000, с. 68].
Клюге очень сложно понимает иерархическое название, и я предложу более
простой прием9^. Проще всего лисицу обыкновенную обозначить так:
первично Canis vulpes Linnaeus, 1758; ныне Vulpes vulpes Frisch, 1775 [Пав-
линов, Россолимо, 1998].
Литературная ссылка нужна, чтобы указать, сколь ново утверждение второ-
го (быть может, очень старого) названия в качестве нынешнего. Для пользовате-
ля желательна более краткая запись, например:
первично Canis vulpes, ныне Vulpes vulpes.
Вот и простейшее решение. Если его принять (например, если ведущий в
описательной биологии журнал обяжет авторов указывать первичное название
данного вида), то это, ввиду простоты, вскоре станет обиходной нормой, и вся-
кий сможет легко понять, о каком виде идет речь.
Читатель может спросить: если всё так просто, то к чему были все разгово-
ры про особую систему для адресации, про народную таксономию и про быто-
вые названия? Отвечу: дело в том, что множество первичных названий, если их
указание станет обязательным или хотя бы массовым, само собой обратится в
искусственную систему, параллельную нынешней, и будет исполнять роль на-
родной таксономии. Ее элементы будут играть роль бытовых латинских назва-
ний, т.е. продолжится традиция, начатая в «Species plantarum» Линнея.
Это потребует номенклатуры, простой и неизменной - правил, какое имя
считать первым (например, имя носителя голотипа, если он имеется); как быть,
если, кроме первого и нынешнего названий, есть общепринятое (лучше всего
назвать все три) и т.д. Номенклатуру лучше всего ввести сразу, не дожидаясь,
пока возникнут трудности, поскольку они легко предвидимы.
15. Итог: третья система
Итак, предлагается прекратить путаницу в названиях видов путем разделе-
ния функций систем. Перечень первичных названий - это искусственная (фор-
мальная) система, которая может оставаться постоянной и потому удобна для 94
94 Прим. ред. Такая номенклатура рекомендуется в Ботаническом кодексе: указывается
авторство исходного названия и принятого в данной сводке.
264
цели адресации. Она должна лишь пополняться вновь открытыми в природе ви-
дами и исправлениями найденных ошибок. Наоборот, система названий нынеш-
ней систематики постоянной быть не может, зато дает представление о текущих
взглядах на эволюцию*. Ее приверженцы полагают ее естественной, однако есть
и всегда будут иные понимания естественности.
Полагать, что их попросту нет, оказалось неразумно: сейчас, когда сами
адепты филогенетики начали признавать, что в ней не всё в порядке, им, адеп-
там, явно недостаёт знания альтернатив. В таких случаях полезно оглянуться на-
зад, чтобы увидать те развилки, где, быть может, и был избран тупиковый путь.
Выше было указано три развилки: после Линнея две системы соединили в одну,
после Дарвина филогению сочли единственной формой эволюции и системы, а
после В. Хеннига кладизм сочли единственной формой анализа филогении. На
всех трех были, по-моему, избраны ошибочные пути.
Ошибка во всех случаях видится общая: один из вариантов был объявлен
единственно верным; остальные могут даже не упоминаться при обучении и
быть забытыми научным сообществом (таковы первые две развилки). Лучше
всего это видно на примере понятий эволюция и естественная система: оба
термина смешали с филогенией. Теперь, когда многие видят, что эволюция - не
древо, а сеть, остальные искренне не понимают, что это значит.
Объяснить просто (но, увы, только тем, кто хочет и умеет слушать): фило-
гения - один из способов объяснить путь, которым создано разнообразие, и
только; все другие понимания естественной системы объясняют итог - как оно
устроено, т.е. какие в нем есть закономерности (инварианты). Инварианты вид-
ны в систематике как параллелизмы - они и образуют сеть. Идея филогении от-
казывается включать их в систему. Вольному воля, но поскольку в эволюции
обнаружено существенное сходство вне родства (горизонтальный перенос ге-
нов, блочность эволюции и т.п.), то филогению нельзя более признавать отра-
жающей эволюцию, т.е. нельзя считать естественной системой.
Приведенные в п. 13 примеры естественных систем построены на идее па-
раллелизма (но отнюдь не только на ней). Они не ставят цели описать разнооб-
разие с точностью до вида, как не ставил ее и Линней. Вместе с системой пер-
вичных названий (п. 14) они, на мой взгляд, полностью осуществят линнееву
традицию, если будут построены для всех групп организмов.
Наоборот, филема из этой традиции выпадает - идея непрестанно меняю-
щейся системы, не идущей к постоянству, противоречит всему, что делал Лин-
ней. Она - третий тип системы. Ее назначение с позиций первых двух - давать
Эту разницу Мейен в 1981 г. выразил так: «раскладка по ящикам для инвентаризации с
заранее заданными формальными требованиями или... организация материала таким об-
разом, чтобы можно было изучать таксоны по их выборкам» (МСП, с. 112). Таковое изу-
чение он в беседах называл типологической экстраполяцией (см. выше, п. 8) и полагал
систему, допускающую эту возможность, естественной. - Примеч. 2010 г.
265
им названия для новых таксонов. Но давать их может не только она: возможно,
что высшие таксоны (от семейства до класса) лучше опишет грубая филогения
(п. 11), а низшие - родо-видовая систематика в смысле п. 12.
Вне рассмотрения осталась геносистематика (п. 9). Отсутствие у нее проду-
манных основ и полная неясность того, какую роль какие изменения генов иг-
рают в эволюции, вполне гармонируют с несообразностью получаемых ею ре-
зультатов. Так что лучше ею пока пользоваться лишь для коррекций.
А.И. Шаталкин [2006, с. XIII] справедливо заключает:
«Сейчас эволюционное учение стоит на пороге радикальных изменений,
связанных с впечатляющими достижениями молекулярной биологии... и
биологии развития. Безусловно, новые эволюционные концепции вновь
пробудят интерес у систематиков к эволюционной проблематике».
Он уверен, что будет раскрыт архетип как генетическая программа укладки
белков (с. VI). Осмелюсь, однако, возразить: как раз новые факты делают эту
надежду сомнительной. Известно несколько примеров, когда один и тот же ген
управляет планом строения у позвоночного, насекомого и зеленого листа, т.е.
при полном различии молекулярных основ (см. [Назаров, 2005; Чайковский,
2006; Зусмановский, 2007]). Это значит, что он управляет не укладкой белка, а
формой, как ее понимают со времен Платона; так же ее понимает диатропика,
выступающая в систематике как типология [Чайковский, 2006а].
Если так, то от естественной системы в типологическом смысле никуда не
деться. Это, повторю, никоим образом не умаляет значения иных форм система-
тики, поскольку естественная система в этом понимании не претендует на де-
тальную (с точностью до вида и ниже) картину биоразнообразия.
Литература
А.хметьев М.А. Географическая дифференциация позднемеловых и кайнозойских
флор // Стратиграфия. Геол, корреляция. 1999, № 3, с. 42-62.
Беклемишев В.Н. Методология систематики (1928). М., КМК, 1994. 250 с.
Бобров Е.Г. Описательный метод Линнея, «Species plantarum» и современная но-
менклатура растений // Карл Линней. Сб. статей. М., Изд. АН, 1958, с. 78-100.
Бэр К.М. Об искусственной и естественной классификациях животных и растений. О
сродстве животных // Анналы биологии. М., МОИП, 1959. Т. 1, с. 367-383; 395-405.
Гоманьков А.В. Синтез или противоречие? // Природа, 1990, № 4, с. 73-79.
Заварзин Г.А. Фенотипическая систематика бактерий. М., Наука, 1974. 190 с.
Заварзин ГА. Составляет ли эволюция смысл биологии? // Вестник РАН, 2006, № 6.
Захаров Б.П. Трансформационная типологическая систематика. М., КМК, 2005. 165 с.
Зусмановский А.Г. Эволюция с точки зрения физиолога. Ульяновск, Ульяновская сель-
скохозяйственная академия, 2007. 394 с.
Игнатьев И.А. Воспоминания об С.В. Мейене. Об эволюционизме Мейена // Общая и
прикладная ценология. 2007, № 4, с. 47-48.
Каталог млекопитающих СССР. Плиоцен - современность / ред. И.М. Громов, Г.И.
Баранова. Л., Наука, 1981. 456 с.
Клюге Н.Ю. Современная систематика насекомых. СПб., Лань, 2000. 333 с.
Корона В.В. Основы структурного анализа в морфологии растений. Свердловск, Изд.
СГУ, 1987. 272 с.
266
Куприянов А.В. Предыстория биологической систематики. «Народная таксономия» и
развитие представлений о методе в естественной истории конца XVI - начала XVIII вв.
СПб., Европейский ун-т, 2005. 60 с.
Кувакин О.Г., Дроздов А.Л. Филема органического мира. СПб., Наука, 1998 (т. 2).
Левина Р.Е. Морфология и экология плодов. Л., Наука, 1987. 160 с.
Леви-Строс К. Первобытное мышление. М., Республика, 1994. 384 с.
Линней К. Философия ботаники. М., Наука, 1989. 452 с.
Лункевич В.В. От Гераклита до Дарвина. М., Учпедгиз, 1960. 2 тома.
Любарский Г.Ю. Архетип, стиль и ранг в биологической систематике М., КМК,
1996. 434 с.
Любицев А. А. К логике систематики // Проблемы эволюции. Т. 2., 1972, с. 45-68.
Любищев А.А. О некоторых постулатах общей систематики И Записки научных семинаров
Ленингр. отд. Математич. института АН СССР. Том 1. Л., 1975, с. 159-175.
Любищев А.А. Проблемы формы, систематики и эволюции организмов. М„ Наука, 1982.
278 с.
Мейен С.В. Методы палеофлористических исследований и проблемы флорогенеза//
Современная палеонтология, т. 2. М., Недра, 1988, с.31-44.
Мина М.В., Решетников Ю.С., Дгебуадзе Ю.Ю. Таксономические новшества и про-
блемы пользователей // Вопросы ихтиологии, 2006, № 4, с. 553-557.
Михайлов К.Г. Типология и практическая систематика И Теоретические проблемы
экологии и эволюции (Вторые Любищевские чтения). Тольятти, 1995, с. 40-46.
Михайлов К.Г. Рец. на книгу: Любарский Г.Ю. Архетип, стиль и ранг в биологиче-
ской систематике //Ж. общей биологии, 1997, № 4, с. 92-96.
Мосейчик Ю.В. Номогенетический характер эволюции растений (на примере флор
раннего карбона) И XVIII ЛЧ, т. 2. Ульяновск, 2005, с. 98-103.
' Назаров В.И. Эволюция не по Дарвину. М., УРСС, 2005. 520 с.
Определитель бактерий Берджи. / Предисл.: Г.А. Заварзин. М., Мир, 1997. 800 с.
Основы финно-угорского языкознания. Прибалтийско-финские, саамский и мордов-
ские языки. М., Наука, 1975. 348 с.
Павлинов И.Я. Рец. на книгу: Mammalian species of the World, 2 v. Baltimore, 2005 //
Зоологич. ж., 2006, № 11, с. 1386-1389.
Павлинов И.Я., Россолимо О.Л. Систематика млекопитающих СССР (дополнения).
М„ Изд. МГУ, 1998. 190 с.
Павлов В.Я. Периодическая система членистых. М., ВНИРО, 2000. 185 с.
Палеонтология и палеоэкология. Словарь-справочник / Редакторы: В.П. Макридин и
И.С. Барсков. М., Недра, 1995. 404 с.
Попов И.Ю. Ортогенез против дарвинизма. СПб., Изд. СПб. ун-та, 2005. 180 с.
Протесты. Часть 1. Руководство по зоологии. СПб., Наука, 2000. 679 с.
Розова С.С. Классификационная проблема в современной науке. Новосибирск, Нау-
ка, 1986. 223 с.
Скворцов А.К. У истоков систематики. К 300-летию Карла Линнея И Природа, 2007,
№ 4, с. 3-10.
Смирнов Е.С. Таксономический анализ. М., Изд. МГУ, 1969. 187 с.
Станек В.Я. Иллюстрированная энциклопедия животных. Пер. с чешского. Прага,
Артия, 1972. 612 с.
Теория и методология биологических классификаций (Труды семинара в МОИП).
М., Наука, 1983. 169 с.
267
Филипченко Ю.А. Эволюционная идея в биологии. М., Наука, 1977. 227 с.
Хохряков А.П. Табличное изображение системы двудольных // Общебиологические
аспекты филогении растений. М., Наука, 1991, с. 113-117.
Чайковский Ю.В. Опыт эко-физиологической макросистемы // Методы исследования
в экологии и этологии. Пущино, 1986, с. 6-33.
Чайковский Ю.В. Элементы эволюционной диатропики. М., Наука, 1990. 271 с.
Чайковский Ю.В. Естественная система. 30 лет после Любищева// Любищев и про-
блемы формы, эволюции и систематики организмов. М.. Центр системных исследований,
2003 (Ценологические исследования, вып. 23), с. 30-50.
Чайковский Ю.В. Наука о развитии жизни. М., КМК, 2006. 712 с.
Чайковский Ю.В. Диатропика С.В. Мейена: сегодняшний взгляд// Вопросы филосо-
фии, 2006а, № 5, с. 95-102.
Шаталкин А.П. Предисловие к книге: Симпсон Дж.Г. Принципы таксономии жи-
вотных М., КМК, 2006, с. Ш-ХШ.
Шипунов А.Б., 2007. «Systema Naturae»: 250 лет спустя. Судьба линнеевской концеп-
ции царств И Материалы конференции по морфологии и систематике растений, посвя-
щенной 300-летию со дня рождения Карла Линнея. М., КМК, с. 31-33.
Эволюционные факторы формирования разнообразия животного мира. М., КМК,
2005. 308 с.
Эпштейн В.М. Философия систематики. Книга 3. М., КМК, 2002. 235 с.
Atran S. Cognitive foundations of natural history: towards an antropology of science.
Cambridge: Cambr. Univ. Press, 1990, XII + 360 p.
Baskerville T. Affinities of plants: with some observations upon progressive developments.
London, 1839. X+ 114 p.
Bauhin C. Pinax theatri botanici. Basel, 1623. XXIV + 567 p.
Ereshefsky M. The poverty of the Linnean Hierarchy. Cambridge Univ. Press, 2001, 316 p.
Goldfuss G.A. Ueber die Entwickelungsstuffen des Thieres. Nuernberg, 1817. 58 S.
Gomankov A. V. Post-Script// Rev. Palaeobot. Palinol., 1997, vol. 96, p. 449-451.
Hunde // Brockhaus’ Konversations-Lexicon. Bd 9. Leipzig, 1908. S. 412-418.
Knapp S., Lamas G., Lughandha E.N., Novarino G. Stability or stasis in the names of organ-
isms: the evolving codes of nomenclature // Phil. Trans. R. Soc. Lond., B, 2004, p. 611-622.
Linne C. Species plantarum. Stokholm, 1753. T. 1, p. 1-560; t. 2, p. 561-1200.
Meyen S. V. Foundamentals of palaebotany. London - N. Y., Chapman & Hall, 1987. 432 p.
Meyen S.V. Permian conifers of Angaraland// Rev. Palaeobot. Palinol., 1997, vol. 96, p. 351-
447.
Toumefort. Paris, Museum nat. hist., 1957 (serie: Les grands naturalistes fran^ais). 321 p.
Wayne R.K., Geffen E., Girman D., Kopfelli K.P., Marshall C. Molecular systematics of
the Canidae // Systematic Biology, 1997, vol. 46, № 4, p. 622-649.
Wheeler Q.D. Taxonomic triage and poverty of philogeny// Phil. Trans. R. Soc. Lond., B,
2004, p. 571-583.
Wiley E. The annotated Linnean Hierarchy//Syst. Zool., 1979. vol. 128, p. 308-337.
Winsor M.P. Linnaeus's biology was not essentialist // Ann. Missouri Bot. Garden, 2006,.
vol. 93, p. 2-7.
268
Часть 7. Диатропика случайного
{Приводимые далее отрывки призваны показать, что диатропический взгляд
на мир дает процедуру, которая может, в принципе, работать со случайностью.
Мейен об этом и не догадывался: в работе ИМ-на с. 131-133 приведена на сей
счет подборка цитат. Из нее видно, что Мейен ограничивал анализ случайно-
сти в эволюции всего лишь двумя тезами: 1) преобразование разнообразия
есть перераспределение частот (иногда говорится - вероятностей) возможных
вариантов; 2) там, где такой подход не имеет смысла (при появлении принципи-
альной новизны, эмерджентности), царит полная, никак далее не анализируемая
случайность - она-то и есть эволюционное творчество в смысле Анри Бергсона.
В устах сторонника «системности» это звучит странно - тем более, что имен-
но от него я узнал о системной случайности (квази-гиперболах) и о ее всепрони-
кающем характере. Мне не раз приходилось спорить с Мейеном, пытаясь убедить
его, что природа таксона - не вероятностная, а системная. Однако Мейен нахо-
дился под влиянием В.В. Налимова, чей взгляд на мир не выходил за рамки веро-
ятностного (анализ см. ОПС). А понятием случайности без вероятности я еще не
владел. Неудача заставила меня развить метод ПМ-(см. выше, ст. «Горизонт...»),
так что мне роль Мейена очевидна. Увы, обсудить с ним метод не пришлось.
Налимов и Мейен понимали, что перераспределение частот - не эволюция:
«Если так, то в биологии мы вновь приходим к к идее неизменности видов, но
это... - неизменность потенций, на которые накладываюся разные вероятности
проявления» (ИМ, с. 132). (Уточню: потенции - это заданная диасеть.) В данных
понятиях подлинная эволюция - это появление новой строки в диасети, которое
происходит, по их мнению, абсолютно случайно. Можно ли сказать больше?
Налимов и Мейен оба рассуждали в рамках статистической ПМ, хотя область
их интересов требовала иных ПМ. В частности, их уверенность, что «абсолютное
творчество - это появление новых принципов, не сводимых к старым. Стало быть,
эти новые принципы абсолютно случайны» (там же), выглядит наивно уже с по-
зиции системной ПМ, не говоря о более новых. Новое случайно лишь при взгляде
снизу, с уровня прежней системы, но осмысленно (а возможно, даже детермини-
ровано) при взгляде сверху, с уровня новой системы (такова обычная философ-
ская трактовка теоремы Гёделя). Эволюция, как становится теперь ясно, детерми-
нирована как раз сверху (см. выше, п. 1 статьи «С чем входим...»). Впрочем, это
давно понимали многие, чем и воспользуемся. - Вводное замен. 2010 г.)
Разнообразие и случайность
3.1. Случайность и закономерность
Что значит - объяснить явление? Для одних - вывести его из очевидных по-
ложений, для других - найти его механизм, для третьих (особенно для биологов)
* Параграфы 1 и 2 из гл. 3 книги ЭЭД.Сохранены ссылки на параграфы этой книги.
269
- понять его полезность, для четвертых - указать ряд, к которому принадлежит
явление, тогда как для пятых достаточно, чтобы явление не вступало в явные
противоречия с другими, привычными (и в этом смысле понятными). То, что не
нашло приемлемого объяснения, чаще всего трактуется как случайность (см. ци-
тату из Вигнера в 1.1) и выпадает из дальнейших рассуждений, как выпал, на-
пример, географический стиль (2.5). На это выпадение обращали внимание мно-
гие методологи, например, А.А. Любищев и Имре Ла* катош. Всякая наука начи-
нается с упорядочения некоторых данных, до этого казавшихся хаотическими, а
наука о разнообразии ставит целью упорядочить сами эти процедуры частных
упорядочений, в ходе которых из случайного выявляется закономерное. Поэто-
му необходим анализ категорий «случайность» и «закономерность».
Ранее мы говорили о шести типах случайности, имеющих отношение к на-
шей теме [Чайковский, 1985]:
А. Случайность как непонятая закономерность.
Б. Случайность как скрещение несогласованных процессов.
В. Случайность как уникальность.
Г. Случайность как неустойчивость движения.
Д. Случайность как относительность знания.
Е. Имманентная случайность.
(Седьмой тип, Ж, будет введен в 3.2). В той же статье была приведена литера-
тура, на которой основан (если не оговорено иное) следующий краткий анализ.
О типах А и Б следует заметить только, что к ним относятся псевдослучай-
ные явления, т.е. явления, механизм которых в принципе может быть описан без
понятия случайности. Сейчас, после работы Мейена [1984], правильнее характе-
ризовать тип Б как скрещение несогласованных тенденций (процесс - это тен-
денция во времени). В качестве одного из главных источников разнообразия
Мейен указал мероно-таксономическое несоответствие (опираясь на работы
Ю.А. Урманцева); для нас это значит, что распределение меронов по таксонам и
наполнение таксонов меронами в определенной степени случайны. В прошлом
ученым было свойственно видеть во всех явлениях только случайности типа А и
Б, но нынешняя наука понимает случайность шире. Сложнее характеризовать
другие типы. Так, тип В может иметь в основе тот же механизм, что и типы А и
Б, но наблюдая явление только однажды, убедиться в этом невозможно. Тип В
бывает важен для понимания разнообразия, поскольку оно, как правило, уни-
кально: тогда как сами объекты появляются и исчезают многократно, их разно-
образие при этом не повторяется, а только все полнее разворачивается (ни пла-
нета, ни вид организмов, ни язык, ни поэт не появляются дважды). Выявить, что
в каждом таком разнообразии случайно, а что закономерно, нельзя без сопос-
тавления с другими разнообразиями. Случайность типа Г известна всем на при-
мере стока воды из ванны: симметричный поток сквозь отверстие в дне неус-
тойчив (несмотря на симметрию уравнения и начальных условий), и реализуется
водоворот, который закручивается вправо или влево. Случайность типа Г реали-
270
зуется, например, в играх (тасовка карт, бросание кости или монеты - здесь слу-
чайность вызвана нерегулярностью отображения множества начальных точек
движения во множество конечных). Она же лежит в основе механизма естест-
венного отбора. В детерминированных системах обычно возникает случайность
типов Б нГ [Заславский, 1984].
Достаточно очевидно, какую роль типы случайности А - Г играют в струк-
туре любого достаточно богатого разнообразия: случайность возникает за счет
сложности, неповторимости и нестабильности процессов, его породивших. Ко-
роче, случайность типов А - Г является в основном отражением истории мно-
жества, разнообразие которого исследуется. Наоборот, случайность типа Д от-
носится, как правило, не к истории, а к наличной структуре множеств. Об отно-
сительности знания говорят в разных смыслах и едва ли не первым, кто связал
ее с проблемой разнообразия, был Иоганн Кеплер, который в 1595 г. писал, что
истинное «заключение, выведенное из ложных посылок, является случайным; а
его внутренняя ложность немедленно обнаруживается, как только его применя-
ют к иному объекту, чем тот, для которого оно было выведено» [Kepler, 1938, с.
15]. Именно для того, чтобы преодолеть эту случайность, полезно исследовать
не отдельные факты, а их ряды.
Другой смысл относительности знания связан с принципом дополнительно-
сти. Первоначальная формулировка принципа - в форме соотношения неопре-
деленностей - прямо связала случайность с относительностью знания, но еще не
касалась разнообразия. Однако Нильс Бор, истолковав неопределенность как ча-
стный случай дополнительности и придав последней общефилософскую интер-
претацию, отмечал возникновение взаимодополнительных пар (аспектов) в са-
мых разных областях знания. Такова взаимодополнительность точек зрения,
терминологий, наук, культур и т.д. Тогда же Любищев, развивая так называе-
мую концепцию пробабилизма, писал, что в процессе познания метрическая
точность достигается в ущерб общности (достоверности) знания об объекте и
наоборот. Либо надо характеризовать разнообразие как некое качественное це-
лое (при этом каждый элемент описывается не своими признаками, а местом в
системе), либо детально (в том числе метрически) описывать отдельные элемен-
ты и мириться с тем, что свойства разных элементов выступают друг относи-
тельно друга как случайности.
Приведем пример. Всем известен тезис Дарвина, согласно которому эволю-
ция движется за счет естественного отбора, в нем никто не сомневается (и нико-
гда не сомневался), если понимать его в самом общем смысле - как апробацию
на жизнеспособность, которой среда подвергает организмы. Правда, в таком ви-
де тезис многих не удовлетворяет своей расплывчатостью, но оказывается, что
уточнить его можно только ценой уменьшения достоверности (отсюда и бесчис-
ленные споры о роли отбора в эволюции); наконец, если описать феномен отбо-
ра математически точно, то такое описание нельзя будет приложить ни к одному
природному процессу, поскольку оно оперирует с немногими наугад взятыми
271
параметрами организма так, словно они - единственные существенные для его
судьбы свойства (см. 3.4).
Можно уточнить мысль Любищева специально в аспекте анализа разнооб-
разия: то, что достоверно с одной позиции исследования, может быть сомни-
тельным, т.е. в некотором смысле случайны, с другой (дополнительной по Бору)
позиции. Так, общеизвестен спор о том, случайно ли то разнообразие мутаций,
из которого естественный отбор должен черпать материал, и разрешается этот
спор достаточно просто: если наблюдать за отдельными организмами популя-
ции, то каждая мутация предстает возникшей случайно (вне связи с конкретны-
ми условиями существования именно данного экземпляра); однако при анализе
механизма мутации (т.е. в эксперименте с препаратами, выделенными из клеток
большого числа особей) она предстает как цепь детерминированных физико-
химических процессов, в которых случайность (статистическая неопределен-
ность) играет роль только подчиненную. Неточность процедуры познания неиз-
бежна уже по той причине, что даже исчерпывающее теоретическое описание,
проведенное с одной позиции, окажется набором более или менее случайных
фактов с какой-то другой (в примере с мутацией таковыми оказались популяци-
онная и биохимическая позиции исследования).
В биологии случайность часть противополагают целесообразности, в дейст-
вительности же эта пара понятий тоже взаимодополнительна. Еще Бэр в 1876 г.
обращал внимание, что различные целесообразные поведения являются друг для
друга случайностями, когда не объединены в общую систему, обладающую сво-
ей собственной целесообразностью [Берг, 1977, с. 68]. Действительно, всякая
попытка точно указать место каждого элемент и цель, которую он себе ставит
(или ему ставят), приводит к толкованию всего разнообразия элементов как хао-
са; наоборот, при трактовке всей системы как целесообразного целого, ее разно-
образие оказывается понятным, хотя далеко не каждый элемент можно тракто-
вать при этом как целесообразный. Так, трудно назвать целесообразным пато-
генный микроб, уничтожающий свою жертву (и себя вместе с нею); но целесо-
образность биосферы в целом этим не умаляется: она не может существовать
без избыточности - как в отношении числа видов и индивидов, так и в отноше-
нии их разнообразия. Без этого она не была бы самодостаточна.
Если во всех случайностях типов А - Д оставалась хотя бы абстрактная воз-
можность видеть псевдослучайность, то в некоторых явлениях современное науч-
ное мировоззрение такой надежды нам не оставляет, и приходится говорить об им-
манентной случайности. Как математическое понятие она существует безусловно:
приходится признать имманентно случайной последовательность знаков любого
неконструктивного числа (числа, для записи которого в принципе не существует
алгоритма). Оказывается, что почти все действительные числа неконструктивны,
т.е. «случайны по Колмогорову», и если рассматривать этот математический факт
как модель реальности, то возникает мысль о преобладании в природе имманент-
но случайных процессов над детерминированными и псевдослучайными.
272
«Случайность по Колмогорову» из всех многочисленных определений слу-
чайности заслуживает наибольшего внимания. Принято считать, что закономер-
ности массовых случайных явлений должны устанавливаться теорией вероятно-
стей, и часто пишут даже (например, в недавней «Математической энциклопе-
дии»), что случайно то событие которое происходит с некоторой вероятностью.
Однако о вероятности можно говорить лишь в тех случаях, когда многократно
происходят однотипные события и вся случайность сводится к отсутствию види-
мого порядка их появления; тогда случайность можно описывать через частоту.
Мы же в этой книге должны прежде всего говорить о связи случайности со слож-
ностью, о системах в которых выявление каких бы то ни было частот - не главное
в понимании их структуры и функции. Примером подобной системы является
текст: чтобы выразить его суть, его надо понять. Анализ частот появления в нем
букв и слов, если и может при этом играть роль, то только подчиненную.
После появления в 1963 г. статьи А.Н. Колмогорова «О таблицах случайных
чисел» математики стали рассматривать связь случайности со сложностью в ас-
пекте теории информации: сложность последовательности знаков, записанных
по определенному алгоритму, оценивают по минимальной возможной длине
этого алгоритма. При этом максимально сложно называется последователь-
ность, которую нельзя выразить короче, нежели записав ее целиком. Такую по-
следовательность знаков и называют случайной, что связывают с интуитивным
пониманием случайности как отсутствия алгоритма. Полученные здесь резуль-
таты (например: последовательность знаков почти любого действительного чис-
ла случайна) важны для выявления оснований теорий информации и вероятно-
стей, но в других отношениях дают мало и могут даже заводить в познаватель-
ный тупик. Да, обоснованием применения вероятностных, а не каких-либо иных
методов к массовым цифровым данным действительно может служить тот факт,
что у наугад взятой последовательности чисел нельзя ожидать наличия жесткой
закономерности - алгоритма.
Но откуда берется сама посылка «наугад»? Ведь окружающий мир предла-
гает нам ряды чисел, образованные не наугад, а в результате совокупного дейст-
вия определенных законов; мы, извлекая из этих рядов конечные выборки, пы-
таемся угадать порядок их построения, но не всегда можем найти его (ведь за-
кон обычно проявляется не жестко, а демонстрирует ядро и периферию - см.
22.3). Далее, об алгоритме можно говорить, когда задан алфавит, на котором тот
записан, но его выявление уже предполагает понимание структуры изучаемого
множества. Так, прежде чем упорядочить звезды с помощью диаграммы Герцш-
прунга - Рассела, мы должны выявить реальные классы звезд, так как сам по
себе спектр конечным алфавитом не задается; а выявив классы, мы обнаружива-
ем, что распределение спектр - светимость неслучайно: оно обладает нечеткой,
но очевидной закономерностью (тенденцией).
Еще нагляднее уже упомянутый пример с текстом. Возьмем важный для нас
содержательный текст и сократим его настолько, чтобы из него уже нельзя было
273
изъять ни одной буквы без ущерба для смысла. Текст окажется идеалом инфор-
мативности (и в этом смысле - неслучайности), но он же будет «случаен по
Колмогорову». Здесь тип Е при ближайшем рассмотрении оказался типом Д,
«По сути дела, высокое разнообразие может трактоваться и как порядок и как
беспорядок» [Левин, 1980, с. 51].
В ходе неформальной по своему существу процедуры выявления алфавита
исследователь выражает свое, в основном интуитивное, понимание сложности
исследуемой системы, а вместе с тем - и ее случайности. Для дальнейшего, уже
более или менее формализованного изучения случайности остается обычно
только самый поверхностный уровень ее проявления - тот, который связан с по-
вторяемостью и вероятностью. (Однако есть ли на деле достаточная повторяе-
мость? Типично в этом отношении понимание биологической эволюции как
процесса естественного отбора удачных мутаций: если буквой считать нуклео-
тид, то длина минимального осмысленного текста оказывается много больше
числа актов чтения (поколений), а если буквы - возможный ген, то даже пере-
чень букв будет не короче длины возможных текстов. Здесь можно говорить о
случайностях типа В, но не о частотах и вероятностях.) Тогда закономерность
случайных явлений получается в ходе усреднения. Нас же, как уже отмечено в
1.2, больше будет интересовать та информация, которую можно извлечь из
обобщения95. Простейшими обобщениями являются ряд и тенденция.
Всякий конкретный пример случайности может быть, в принципе, с развити-
ем знаний, отнесен к одному из типов псевдослучайности: даже радиоактивный
распад, обычно приводимый в качестве примера чисто самопроизвольного (а по-
тому имманентного объекту) процесса, демонстрирует некоторую управляемость
(нейтрон, например, быстро распадается вне ядра). Поэтому необходимо особо
проанализировать типД- возникновение случайности в системе знаний.
Согласно представлениям, которые сформулировал философ Вильгельм
Виндельбанд на грани XIX и XX вв., каждая наука имеет свою номотетику, т.е.
общие правила, упорядочивающие в немногих словах большое число фактов, и
свою идиографию, т.е. описание конкретного материала. Виндельбанд утвер-
ждал, что науки сильно различаются по соотношению в них номотетического и
идиографического компонента, и он же заметил, что само разграничение номо-
тетики и идиографии зависит от позиции исследователя (анализ и литературу
см.: [Беклемишев, 1925]). Рассмотрим вопрос подробнее.
3.2. Номотетико-идиографическое чередование
Рассмотрим любое частное утверждение, хотя бы, например: «этот стол - де-
ревянный». Фраза, чисто идиографическая, указывает на частный конкретный
факт, но несет в себе в свернутом виде мощную номотетическую информацию.
95 Существенной особенностью диатропики (в отличие от формальной теории систем)
является акцент на итеративность процедуры обобщения и возможное отсутствие фор-
мальных выражений.
274
Ведь слово «стол» - символ принадлежности к некоторому абстрактному множе-
ству столов, указание на номотетику. Еще мощнее свертка информации в слове
«деревянный», которое указывает на все предметы, сделанные из дерева, причем
дерево может быть любое; т.е. проведена еще и свертка по всем видам деревьев,
тогда как, например, слово «дуб» уже означает свертку по всем видам и экземпля-
рам, принадлежащим к данному роду. Но более всех абстрактно слово «этот»,
формализующее не предметы или их качества, а отношение их к говорящему.
Оказывается, идиографический факт формулируется как пересечение номотетик.
Отметив это, можно перейти к вопросу о том, что номотетично, а что идио-
графично в той или другой научной дисциплине. Это зависит от позиции иссле-
дователя, целей, которые он ставит, и средств, которыми он располагает. Так,
для математика «Курс геометрии» - номотетический текст, а «Курс практиче-
ской систематики высших растений» - сугубо идиографический, и мало кто за-
мечает, что для ботаника ситуация прямо противоположна. Действительно, гео-
метрия для него, как правило, - набор теорем, которые нельзя усвоить иначе,
чем заучив их, и наличие доказательств не упорядочивает для него текст, но
только загромождает (надо заучивать еще и их); наоборот, инструкции «Практи-
ческой систематики» упорядочивают для него море растений, стоящее перед
глазами, дают метод свернуть необозримое множество признаков в компактное
множество диагнозов. Словом, номотетично то множество, в котором дан-
ный исследователь видит номотетику.
В этом отношении интересно вспомнить следующее определение полезно-
сти информации: информация тем полезнее для воспринимающего, чем более
она увеличивает его информированность. Другими словами, информация беспо-
лезна как для того, кто ее уже знает, так и для того, кто не может ее понять
[Шрейдер, 1965].
Приняв, что слово - это имя множества и тем самым указывает номотетику,
получаем следующую циклическую схему построения понятий, общую как для
разговорной и научной речи, так и для самой процедуры познания. Как уже го-
ворилось, любой конкретный идиографический факт задается пересечением
номотетик. Из идиографических фактов как из элементов строится всякая идио-
графия, т.е. всякое конкретное описание, составляющее первичный материал
науки и практической деятельности. Другими словами, идиография выступает
как объединение идиографических фактов. Наконец, номотетика оказывается
пересечением идиографий. Из того факта, что номотетика и идиография зада-
ют друг друга, следует, в частности, что в разговорном языке нет каких-либо ос-
новных слов, через которые определяются остальные; наоборот, язык определя-
ет сам себя, зато через него определяются искусственные языки. Поэтому в ос-
нове всякой формальной теории лежит словесное описание, причем почти вся-
кое слово разговорного языка может выступать как категория^. Очевидно, что
96 Это частный случай «категориальной когерентности» по Н. Гартману.
275
понятия номотетики и идиографии взаимно дополнительны - точно так же, как
понятия гомологии и аналогии. Нетрудно также увидеть сходство между номо-
тетико-идиографическим и мероно-таксономическим чередованием в процессе
познания соответствующих разнообразий.
Теперь можно сформулировать понятия случайности и закономерности,
удобные для наших целей. Именно, закономерностью назовем всякую возмож-
ность выявить и именовать некоторое номотетическое множество (т.е. множест-
во, элементы которого обладают существенным для наблюдателя общим свой-
ством). Тогда случайностью естественно назвать выбор элемента (или ряда
элементов) из какого-то множества (неважно, номотетического или нет), если
наблюдатель не может характеризовать этот выбор в номотетических терминах.
Другими словами, случайность разнообразия можно характеризовать как такой
взгляд на него, при котором мы отказываемся исчерпывающе характеризовать
отдельные объекты, но можем, хотя бы в принципе, характеризовать их множе-
ство. Удобно ввести седьмой тип случайности:
Ж. Случайность как произвольный выбор.
Он тесно связан с типами Б, ВиД, но не сводится к ним.
Вопрос о том, имеет ли данное множество номотетику (т.е. закономерно)
или не имеет (т.е. случайно), бессмыслен, пока не задан аспект, в котором жела-
тельно видеть упорядочение: ведь любую идиографию можно расчленить так,
что получатся отдельные идиографические факты, а можно так, что получатся
элементы, входящие в некоторые номотетики. Если же достаточно разнообраз-
ное множество не с чем сопоставить, то оно обычно выступает как идиография -
ведь номотетика создается пересечением различных идиографий. Такое понима-
ние случайности разнообразия характеризует процесс познания, а не само по-
знаваемое явление, и обычно для многих наук. Даже в такой точной науке, как
небесная механика, где царят формулы, работа началась с упорядочения.
Для первобытного человека звезды расположены беспорядочно, но уже в
глубокой древности было установлено, что все они, за исключением пяти пла-
нет, неподвижны на небесном своде. Тем самым понятия «звезда» и «планета»
породили два ряда. В древности же разнообразие наблюдений планет упорядо-
чили в систему кругов. Коперник заново упорядочил эти круги, положив цен-
тром Солнце (точнее, положив центр рядом с Солнцем; подробнее см. [Чайков-
ский, 19876], Кеплер упростил эту систему, заменив большое число кругов не-
большим числом эллипсов (т.е. найдя более простые ряды для известных на-
блюдений), после чего Ньютон объяснил их, найдя формулу закона тяготения. В
биологии до формул дело доходит редко, поэтому трудно решить, какой из воз-
можных рядов лучше («естественнее»), но упорядочение наблюдений проводит-
ся столь же неукоснительно.
Не следует объявлять какое-либо множество имманентно случайным, так
как вас может опровергнуть любой, кто укажет в нем ряд, вами не замеченный.
Даже для простейших множеств - числовых последовательностей нет способа
276
практически установить, что они случайны. Например, последовательность зна-
ков числа л случайна в том смысле, что ни один статистический критерий не
поможет предсказать (хотя бы в вероятностном смысле (/г + 7)-й знак по п
предшествующим знакам. Статистик вправе назвать эту последовательность
случайной, но это - случайность типа А и только, ибо любой знак числа п вы-
числяется. Тем более нельзя говорить, что какой-то опыт доказывает случай-
ность данного биологического явления (например, мутации).
Единственная бесспорная случайность, о которой можно всерьез говорить
при понимании ее как неупорядоченности, возникает в тех ситуациях, где вооб-
ще нельзя выстроить ряд - когда различных объектов слишком мало. Если, на-
пример, считать жизнь возникшей от единственной первобактерии, то это воз-
никновение можно считать случайным, но если допустить, что первых организ-
мов было два, то сразу обнаруживается пара богатых параллельных рядов (на-
следственная память, ее считывание, энергетика, мембраны, размножение и
т.д.), так что неслучайность налицо. Хиротность сирена и хирота (2.5) тоже
можно было считать случайным совпадением, пока они не встали в общий реф-
рен. Как видим, номотетико-идиографический подход оставляет случайности
очень скромное место.
И все же без идеи случайности разнообразие анализировать невозможно. Де-
ло в том, что при исследовании разнообразий часто бывает необходим один част-
ный атрибут случайности - независимость исследуемых событий. Если вам надо,
например, выяснить, действительно ли долгожительство является существенной
тенденцией разнообразия горцев, то самый простой выход - переписать возраст
всех горцев и установить процент долгожителей. Если же всех переписать невоз-
можно, то необходимо организовать выборку, в каком-то смысле случайную, и
есть довольно общий прием: обработать какой-нибудь обозримый ряд, заведомо
составленный без связи с возрастом (например, брать список жителей каждого де-
сятого поселения из алфавитного списка поселений). Вообще, основной источник
фактически удобной случайности (тип Б) - в том, что разные способы упорядоче-
ния одного и того же разнообразия - например, по возрасту и по алфавиту адресов
- можно считать несопоставимыми (хотя доказать это и невозможно).
Другой частый источник фактической случайности - неполнота комбина-
торики. Если виды одного рода различаются только десятью признаками, при-
чем ни один признак не влечет обязательно другого, и они принимают всего по
два значения (есть или нет), то даже тогда возможно 210, т.е. более тысячи видов.
А поскольку чаще всего род включает менее десяти видов (как и семейство
обычно включает менее десяти родов и т.д.), то фактически реализованной мы
видим небольшую долю комбинаций и можем, в зависимости от вкуса, или счи-
тать их случайными, или строить для них «родословные древа», или искать «за-
преты на определенные комбинации».
У двух голубоглазых родителей может родиться черноглазый ребенок - в
результате мутации. Случайна ли она? Да, случайна в том смысле, что мы не
277
можем предсказать, произойдет ли она у членов данной голубоглазой пары (как
не можем предсказать, что встретим сегодня на улице знакомого), но из этого
никак не следует, что мутация возникает без определенной причина (как нельзя
считать, что наши знакомые без цели бродят по улицам). Мутация - результат
очень сложной комбинации реакций (каждая из которых течет по четким зако-
нам), подчас несопоставимых друг с другом в таком же смысле, как несопоста-
вимы наши намерения с намерениями знакомого, которого мы встречаем на
улице. Замечательно, что Дарвин, ничего не зная о механизмах наследственно-
сти, понимал случайность наследственных изменений именно так: «Хотя каждое
изменение должно иметь собственную возбуждающую причину и хотя каждое
из них подчиняется закону, мы все-таки так редко можем проследить в точности
соотношение между причиной и следствием, что нам хочется говорить о вариа-
циях, как о проявляющихся произвольно. Мы даже можем назвать их случай-
ными...» [Дарвин, 1951, с. 770].
Это очень существенно: дарвинская установка означает готовность искать
закономерность там, где с виду есть только случайности. Наоборот, у позд-
нейших авторов мы чаще видим беспредметный спор - случайна ли та изменчи-
вость, которая поставляет материал для эволюции? Разумеется, она случайна в
одних смыслах и неслучайна в других.
Эту мысль легко пояснить техническим примером. При решении задач на
компьютере часто требуются таблицы случайных чисел, и для этой цели приду-
маны специальные короткие программы, всего в 5-7 арифметических действий.
Естественно, никакой случайности в работе программы нет, но в результате по-
следовательной ее работы получается таблица, содержащая миллионы чисел, и
любой статистический критерий подтвердит, что таблица - случайная.
Как говорится, статистике часто принадлежит первое слово, но последнее -
никогда. Она часто наводит на размышления, но никогда сама не дает ответов,
поскольку занимается лишь явлениями, а не их внутренней сутью. Первое слово
и у нас будет принадлежать статистике*.
Случайность и самоорганизация"
Как уже замечено во Введении, если подбросить монету один раз, то нельзя
предсказать, какой стороной она упадет, но если сделать это 500 раз, то извест-
но, что она упадет гербом приблизительно 250 раз, так как вероятность падения
любой стороной равна 1/2. И хотя математик может тщательно разъяснить, что
тут значит слово "приблизительно", он мало поможет в понимании равноверо-
ятности и совсем ничего не скажет насчет самого важного вопроса: как из не-
предсказуемых результатов складывается предсказуемый? Другими словами:
почему из совокупности неустойчивых процессов (полетов монеты) складывает-
Далее в ЭЭД следует изложение квази-гиперболич. статистики (о ней см. ЗЭ, с. 36).
” Отрывки из главы 4 книги ОПС. Сохранены ссылки на главы этой книги.
278
ся устойчивый (частота)? И всегда ли частота оказывается устойчивой?
"Все же здесь есть что-то парадоксальное. Мы можем предвидеть, что про-
изойдет в конечном итоге, но не можем предвидеть деталей" - писал венгеро-
американский математик и педагог Дьердь Пойа [1957, с. 307]. Аналогичную
ситуацию в квантовой физике ("законы квантовой физики можно понять, ... если
применить законы ТВ к большому числу частиц, однако эти законы не объясня-
ют поведения электрона или протона") венгерский математик Габор Секей тоже
назвал парадоксом [Секей, 1990, с. 209]. Но что делать с парадоксами, не сказа-
но. А ведь ситуацию прояснил еще сто лет назад Анри Пуанкаре.
4-3. Рулетка Пуанкаре и вероятность как мера
Пусть на вертикальной оси крутится горизонтальная стрелка, под нею поло-
вина плоскости белая, а половина черная. Слабо толкнув стрелку, легко подга-
дать, чтобы она остановилась над белым, но если она сделает хотя бы 10 оборо-
тов, подгадать трудно. Если же и секторов сделать не два, а 37, как в настоящей
рулетке, то подгадать практически невозможно. Говоря более строго: для любой
точности измерений существует число оборотов, после которого угадать цвет
остановки невозможно. То есть факт невозможности справедлив на том самом
языке "эпсилон - дельта", который принят за эталон строгости в математическом
анализе.
Сопоставив белое с гербом, а черное - с цифрой, получим прекрасную мо-
дель бросания монеты. В модели нет сложного полета, а главный результат на-
лицо: чем больше начальная скорость, тем меньше она влияет на итог, на цвет
остановки. Хотя точка остановки остается функцией начальной скорости и
близким начальным скоростям по-прежнему соответствуют близкие точки оста-
новки, но они всё чаще оказываются принадлежащими разным цветам.
Следовательно, необозримая сложность полета монеты не рождает случай-
ность, а лишь маскирует ее подлинный источник - неустойчивость процедуры
отображения бесконечного множества начальных состояний в конечное (в слу-
чае монеты в нем всего два элемента) множество итоговых. Эта неустойчивость
должна быть исследована, а не принята как данная опытом поколений. [...]
Вращение стрелки можно, не ограничивая общности рассуждения, всегда
начинать с одного и того же нулевого угла. Тогда угол а, на котором она оста-
новится, будет функцией начальной скорости и: а = а(и). Если v достаточно ве-
лика, то доля времени, проведенная стрелкой в белом секторе, а следовательно
и вероятность остановиться в нем, близка к той доле, какую составляет белая
дуга от всей окружности. Наоборот, зависимость угла остановки а как от v, так
и от закона, по которому совершается замедление, сказывается всё меньше и
быстро сходит на нет.
Иначе говоря, какова бы ни была монотонная непрерывная функция а(я),
искомая вероятность остановки стрелки в белом секторе задается не ею, а соот-
ношением длин дуг разных цветов. В этом смысле функция а(ц) произвольна.
Факт ее произвольности позволяет заняться исключительно соотношением длин
279
дуг, что мы и сделаем. Число белых дуг, их взаимное расположение и длина
каждой могут быть любыми, но если их .мера (суммарная длина) не меняется, не
изменится и вероятность остановки стрелки на белом. Поэтому естественно на-
звать вероятностью остановки стрелки на белом именно меру (суммарную дли-
ну) белых дуг. В этих терминах феномен существования постоянной вероятно-
сти вполне очевиден. Существенно, что белая часть круга может быть сколь
угодно "рваной" - это не мешает вероятности оставаться постоянной. (Эта идея
любой "рваности" отрезка и положена в основу концепции меры по Лебегу.)
Удивительно, сколь долго оставался незамеченным факт изоморфизма
свойств рулетки и монеты. У самого Пуанкаре рулетка рассмотрена с одной це-
лью - показать, что факт равновероятности справедлив при любом законе за-
медления стрелки, выражаемом произвольной функцией. В книге Бореля рулетка
Пуанкаре подробно описана, однако акт случайного бросания описан без связи с
ней и подан вполне традиционно, по Гоббсу: "Разве можно детально проанали-
зировать движение руки, бросающей кость?" [Борель, 1923, с. 63-66, 5]. На-
сколько знаю, первым рулетку с монетой сопоставил в 1935 году философ Шузо
Куки [Kuki, 1966, с. 150].
Но и после этого рулетка Пуанкаре осталась вне поля зрения ученых, и в
новейшей книге [Шрёдер, 2001, с. 207] читаем: чтобы регулярно выигрывать в
рулетку, “неоходимо прежде определить коэффициенты трения для шарика и
колеса”. Автор этого уверения не стал проверять его практически, а зря: хорошо
бы узнать, насколько прав был Пуанкаре, утверждая произвольность функции,
т.е. независимость цвета (но не угла) остановки от характера движения, в том
числе и от коэфициента трения.
4-4. От рулетки Пуанкаре к странным аттракторам
Обиходный взгляд на случайность исхода бросания монеты ("необозримо слож-
ный полет") выступает как итог ущербной философской позиции (именуемой
абсолютным детерминизмом), а вовсе не слабости фактических знаний. Со вре-
мен Ньютона и до работы Лоренца почти 300 лет ученые закрывали глаза на са-
модовлеющий характер феномена неустойчивости движения, полагая, что прак-
тический интерес представляют только устойчивые процессы. Оказалось не так.
Высказано убеждение [Растригин, 1969, с. 18], что источник неопределен-
ности имеет в рулетке квантовую природу (трение определяется хаотическим
движением атомов). Возможно, но он должен каким-то механизмом усиливать-
ся, и этим механизмом служит принципиальная неустойчивость акта остановки
рулетки.
Рулетка Пуанкаре оказалась простейшим примером динамического хаоса.
Теория динамического хаоса (появилась в 1960-х гг.) показала, что случайное по-
ведение является вполне обычным для широкого класса детерминированных сис-
тем. В настоящее время становится ясно, что в мире динамических систем господ-
ствуют два противоположных принципа - сжатых отображений и растянутых
отображений. Первый лежит в основе обычной теории динамических систем, за-
280
даваемых дифференциальными уравнениями, имеющими (кроме отдельных осо-
бых точек) единственные решения. Если правые части уравнений непрерывны, то,
в силу принципа сжатых отображений, единственный тип неустойчивости траек-
торий - расхождение от особых точек к асимптотам (замкнутые асимптоты име-
нуются предельными циклами) или на бесконечность. Если не рассматривать осо-
бые точки, то случайности тут места нет ("лапласов детерминизм").
В.П. Гачок [1989] рассматривает всякую (в том числе и не описываемую
дифференциальными уравнениями) самоорганизацию как сжатие отображений.
Г.М. Заславский [1984] связывает появление динамического хаоса с противопо-
ложной ситуацией - когда в динамической системе выполняется принцип растяну-
тых отображений; он имеет место в пространствах отрицательной кривизны; та-
ковы, например, "рассеивающие биллиарды" - т.е. системы с упругими столкно-
вениями шаров, когда производные непрестанно терпят разрыв, а углы расхожде-
ния со временем увеличиваются. В области действия этого принципа царит пере-
мешивание, а оно порождает простейшую случайность - стохастичность. Другими
словами, здесь возникают случайные события, повторяющиеся с регулярной час-
тотой, т.е. частотой, сходящейся к определенному пределу, именуемому вероят-
ностью. Иногда удается сформулировать и критерий стохастичности - условие,
при котором в динамической системе возникает перемешивание.
Сравнивая идеологию этих двух книг, приходишь к мысли, что стохастич-
ность динамических систем возникает там, где с течением времени происходит
расширение фазового объема системы, а самоорганизация, наоборот - там, где
происходит его сжатие. (Для сравнения: обычная статистическая физика исходит
из идеи сохранения фазового объема.) В больших системах возможно расширение
по одним фазовым областям или подпространствам и сжатие по другим, поэтому
траектория реального процесса может переходить из области сжатия в область
растяжения и обратно, что воспринимается как фазовые переходы. Поэтому, на-
блюдая за маргинальным участком (свойством, срезом) такой системы, естествен-
но ожидать встретить наиболее сложно устроеннную случайность.
Организующая роль случайности
О математических моделях случайного
...У нас вовсе не будет речи об имитационных моделях, когда ставится цель
имитировать какие-то желаемые свойства объекта. К ним относится подавляю-
щее большинство математических моделей. Речь пойдет только об ориентаци-
онных и мажорирующих моделях. Первые призваны показать какие-нибудь
новые логические возможности, до моделирования остававшиеся незамеченны-
ми, а вторые должны ограничить реальный круг приложимости таковых воз-
можностей. Однако сам я начал именно с имитационных моделей эволюции,
где, как и большинство авторов, "искал кошелек под фонарем".
* Отрывки из глав 8, 9 и 10 книги ОПС. Сохранены ссылки на ее главы.
281
Поэтому и путь к алеатике* оказался через оценку роли разных типов моделей.
Первый раз, когда мне пришлось исходить из наличного математического
аппарата, а не из реальных свойств изучаемого объекта природы, был связан с
желанием ответить на вопрос о том, может ли коллектив простых стохастиче-
ских объектов решить достаточно сложную задачу оптимизации (матричную иг-
ру). Ответ удалось найти [Чайковский, 1971]^7, и состоял он в том, что возмож-
ности таких коллективов скромны.
В этой работе было показано, что для нахождения оптимального поведения
в матричной игре игрок, являющийся однородным коллективом автоматов**
должен обладать как минимум следующим свойством: каждый член коллектива
должен знать номер хотя бы одного действия, к которому в данный момент t
следует переходить. Если же он знает чуть меньше - что выполняемое им дейст-
вие следует менять (но неизвестно, на какое), то при наличии трех и более дей-
ствий игра сводится к бесконечному перебору действий без приближения к ре-
шению. В данной работе еще господствует идеология предельного подхода,
позже мною не использованная.
Хотя исходная установка модели, позволившая мне применить аппарат тео-
рии игр, была заимствована из третьей ПМ-(мне тогда представлялось, что ре-
альные объекты в случайной среде способны оценивать средние величины, что
дискретные величины можно заменять непрерывными) и оказалась далекой от
биологической реальности, однако результат имел для меня решающее значе-
ние: начиная работу, я был уверен во всесилии естественного отбора (так нас
всех учили), а по окончании стал искать в литературе данные об иных механиз-
мах эволюции. Тем самым, модель модель оказалась ориентационной.
[...]
Еще один тип ориентационных моделей - это автоматы, которые не моде-
лируют никакого конкретного объекта, зато демонстрируют неизвестные ранее
возможности случайного поведения. Впервые их рассмотрели в 1960-х годах
московский кибернетик М.Л. Цетлин (увы, вскоре безвременно умерший) и
сложившаяся вокруг него группа исследователей. Они обнаружили ту форму
оптимального поведения, которая может реализоваться только в среде, случайно
Алеатика - общая теория случайности. Включает как ТВ и математич. статистику (т.е.
те разделы, где выполняется закон больших чисел), так и анализ тех видов случайности,
которые не обладают ни устойчивыми частотами (вероятностями в обыденном смысле
слова), ни вероятностью как мерой. Алеатика показывает, что случайные явления имеют
самую различ. природу; в т.ч. существует (вопреки мнению многих) подлинная, ни к че-
му не сводимая случайность. См. 4-35, а также ОПС, с. 18. - Примеч. 2010 г.
97 Публикации не повезло: на с. 1027 спутаны номера формул с номерами из списка ли-
тературы. Еще менее повезло ее переводу [Chaikovsky, 1971]: на с. 1246 он содержит
смысловые ошибки. Оба текста правлены в журналах без моего ведома.
** Здесь автомат - вычислительная система, минимизирующая штраф. - Примеч. 2010 г.
282
реагирующей на действие объекта. Эти работы служат отличной иллюстрацией
организующей роли случайности...
Вскоре американский биокибернетик Стюарт Кауфман открыл другой класс
автоматов, ничего конкретно не моделирующих, но коллектив которых спосо-
бен к самоорганизации. Вывод Кауфмана: эволюция может эффективно проте-
кать не в детерминированных и не в хаотических системах, на границе между
порядком и хаосом, причем приближение коллектива автоматов к этой границе
сравнительно легко достигается путем уменьшения числа связей между автома-
тами [Кауфман, 1991]* **. [...]
Независимо от того, считать ли случайность реальным феноменом или спо-
собом говорить о непонятном, нельзя отрицать, что она играет организующую
роль. Достаточно вспомнить, что неподвижно растущее растение не может ина-
че распространять свои семена, как предоставив их на волю случая. У опыляе-
мых ветром и опыляемых насекомыми эти случайности достаточно разноплано-
вы, но они есть всегда. Да и сознательно действующий человек не мог бы эф-
фективно размножаться, вовсе исключив из этого процесса случайность.
Говоря шире, случайность необходима для действия в разнообразном мире.
В качестве основной случайности, организующей элементы в целое, чаще
всего выступает мероно-таксономическое несоответствие . Этим термином
Мейен обозначил «отсутствие соответствия между классами частей и классами
самих индивидов» [Мейен, 1984, с. 18; М-254]. Чем значительнее это несоответ-
ствие, тем выше организующая роль случайности.
Организующая роль случайности хорошо выявлена в ряде физических работ
- см., например, пункт "Конструктивная роль динамической неустойчивости
движения" статьи [Климонтович, 1996], где отмечено, что разбегание траекто-
рий при динамическом хаосе обеспечивает физическую и информационную це-
лостность системы; и пункт "Тепловой шум в биосистемах - необходимый ком-
понент их работы" статьи [Иваницкий и др., 1998], где сказано: "Тепловой шум
в биосистемах... выполняет две полезные функции - поиск партнеров при моле-
кулярных взаимодействиях и адаптационную изменчивость". (Последняя, заме-
чу, далеко выходит за рамки шума, что мы увидим ниже, в п. 5.2.) Недаром с
древности важной частью многих социальных действий является жеребьевка, а в
наше время в науке и технике распространены методы случайного поиска. [...]
Такая ситуация достаточно типична: существует множество объектов, кото-
рые без случайностного компонента распались бы на отдельные бессмысленные
состояния или движения. [...]
Случайность в игре
...Простейший тип игры - матричная игра, т.е. игра двух игроков, в которой
первый должен выбрать одно из т действий, а второй - одно из п действий, и за-
* О Кауфмане и идее «На грани порядка и хаоса» см. АСМи ЗЭ. - Примеч. 2010 г.
** См. выше, статью «Наука о разнообразии» . - Примеч. 2010 г.
283
дание этой пары определяет выигрыш первого (он же - проигрыш второго). Вы-
игрыши образуют матрицу размером шХ/i. Решение игры в общем случае являет
собой пару оптимальных стратегий (два набора вероятностей, с которыми игро-
ки применяют свои действия), и реализующийся при этом средний выигрыш - це-
ну игры. Для матричной игры решение всегда существует (теорема фон Неймана,
1928 г.), причем игрок, применяющий свою оптимальную стратегию, гарантирует
себе выигрыш не менее цены игры, независимо от того, как ведет себя противник
(и получает больше цены, если противник ошибся).
Решение может быть найдено с помощью эффективной процедуры - реше-
ния системы дифференциальных уравнений. Эту процедуру я и использовал для
ответа на поставленный выше вопрос [Чайковский, 1971; Chaikovsky, 1971].
А именно, поиск решения игры был рассмотрен как взаимодействие двух
больших коллективов достаточно простых автоматов, когда каждый акт поиска
состоит в том, что каждый автомат первого коллектива играет с автоматом второ-
го согласно игровой матрице. Разумность игрока (коллектива) как целого достига-
ется сравнением выигрышей групп, применяющих в данный момент одно и то же
действие (применяющих одну и ту же ''чистую стратегию"). При этом распреде-
ление автоматов коллектива по действиям играет роль коллективной памяти.
Сама по себе приближенная оценка средних внутри коллектива несложна:
достаточно каждому автомату между актами игры несколько раз случайным об-
разом обменяться половиной своего текущего выигрыша с несколькими соседя-
ми, чтобы в коллективе быстро выровнялась величина, близкая к среднему вы-
игрышу коллектива. Столь же легко оценивается выигрыш, средний по автома-
там, выполняющим данное действие, а значит можно оценить и их разность -
среднюю выгодность данного действия. Этот случайный обмен и несет органи-
зующую функцию.
Оказалось, что для решения игры знать игровую матрицу необязательно. Дос-
таточно, чтобы каждый автомат знал лишь один из номеров действий, к которым
в данный момент выгодно переходить (причем каждый выгодный номер какой-то
части автоматов известен); наоборот, знание выгодности собственного действия
недостаточно: если каждый автомат знает лишь то, что номер выполняемого им
действия надо сменить на другой (неизвестно какой), то коллективы бесконечно
блуждают. Тем самым, граница эффективности случайного поиска в игре оказа-
лась четкой: если известно только, где плохо, игра не решается, а если каждому
члену коллектива известно хоть одно из направлений к лучшему, то решается.
[...]
Игровые задачи интересовали меня в одном отношении - можно ли их решать
методом проб и ошибок... [т.е.] встал вопрос: существует ли решение системы
уравнений, в которых вместо выгодностей взяты невыгодности (штрафы)? Ока-
зывается - нет. Удалось найти простой пример игры 3X3, в котором решение
системы уравнений не приближается к решению игры (траектории блуждают по
всему фазовому пространству). Это и было интерпретировано мной как недоста-
284
точность штрафов - в игре коллективов надо поощрять тех, чьи действия лучше
среднего. И наконец, оказалось, что численное значение выгодностей лишь за-
медляет процесс поиска: быстрее всего ищут те, кто знает лишь знак выгодно-
сти, т.е. лишь номер действия, к которому следует перейти.
Конечно, факт решаемости игры не несет существенного поведенческого
смысла, поскольку и сами автоматы, и способ их обмена информацией слишком
сложны и надуманны. Однако указание информационной границы, ниже которой
решение заведомо невозможно*, весьма осмысленно: самая простая конфликтная
ситуация (матричная игра) оказывается неразрешима методом случайного поиска,
притом - даже в самом грубом приближении. В этом состояло для меня ориента-
ционное значение игровой модели. Грубо говоря, рассмотрение ее привело сперва
к отказу от попыток решать эволюционные задачи методом проб и ошибок, а за-
тем и к пониманию ограниченности статистической ПМ-вообще”.
Случайность и тенденции
Тенденцией называется закономерность, вполне очевидная, но нечеткая и не
поддающаяся точной формулировке; чаще всего это происходит вследствие на-
личия противоречащих примеров. В 1907 г. философ Анри Бергсон ввел в обо-
рот эволюционной литературы тенденцию как эволюционное понятие, и глав-
ным его примером были тенденции царств природы: хотя всем известны поня-
тия животного и растения, однако нет ни одного признака, позволяющего четко
различить их; главное свойство растений (фотосинтетический механизм) явля-
ется лишь тенденцией - есть растения паразиты, а есть животные с фотосинте-
зом (многие жгутиконосцы и др.).
[...]•
Для нашей темы важна та тенденция, которую С.В. Мейен, палеонтолог,
эволюционист и философ, назвал мероно-таксономическим несоответствием.
Этим термином Мейен обозначил «отсутствие соответствия между классами
частей и классами самих индивидов» [Мейен, 1984, с. 18]; для нас это значит,
что распределение меронов по таксонам и наполнение таксонов меронами в оп-
ределенной степени случайны. Данная тенденция характерна для разнообразия
любых объектов. Так, для таксона "заводы" характерен мерой "высокие вытяж-
ные трубы", труба - как бы символ завода, но возможны заводы без них, зато
данный мерой характерен и для таксона "теплоцентрали". После работы Мейена
[1984], правильнее характеризовать случайность типа Б*** как скрещение несо-
гласованных тенденций (процесс - это тенденция во времени).
Возможность указать данную границу делает модель мажорирующей. - Примеч. 2010 г.
” Игровой подход к научным задачам означает применение к ним системной ПМ, а при-
водимое ниже использование тенденций - диатропической ПМ. Игра позволяет (отчасти)
моделировать случайность, включающую свободу выбора,- Примеч. 2010 г.
Классификацию типов случайности см. выше, в отрывке из гл. 2 книги «Элементы
эволюционной диатропики». - Примеч. 2010 г.
285
Организующая роль мероно-таксономического несоответствия выражается
во многом и, в частности, во всех процессах приспособления: для обретения
старым таксоном нового свойства чаще всего прежний мерой используется не
по назначению, а как положено мерону иного таксона. В биологии этот феномен
известен как принцип смены функций и является основным при объяснении по-
явления новых органов в эволюции. Так, рыба, оказавшись на суше, использует
плавники для ползания, что в ходе эволюции может привести к появлению лап.
Аналогично, тракторный завод может быстро перейти на выпуск подвижной
бронетехники, даже если это не планировалось при проектировании завода, т.е.
было случайным в смысле Б для его проекта. Случайность такого типа возможна
в качестве массовой именно потому, что пространство таксонов связано с про-
странством меронов не прямо (мерой - таксон), а как бы наискось, косой сеткой
- через мероно-таксономическое несоответствие.
Эффект Шноля и диатропика случайности
Как говорилось в главе 5, всякая ПМ-пытается описать со своей позиции все
явления и, в частности, всевозможные случайности, однако не все явления в
данную ПМ-хорошо укладываются. Поэтому нельзя говорить о знаковой или
диатропической случайности, но можно говорить о знаковом или диатропиче-
ском описании той или иной случайности. Более того, некоторые категории слу-
чайных явлений вообще не имели удовлетворительного описания до появления
соответствующих ПМ, о чем тоже в главе 5 шла речь.
В качестве яркого примера случайности, для описания которой не было
языка вплоть до появления пятой (диатропической) ПМ, можно привести ту, ко-
торую вот уже почти полвека исследует биохимик С.Э. Шноль. Его исследова-
ния коснулись самых разных природных явлений, и всюду было обнаружено,
что за экспериментальной "гауссоидой" скрывается сложная и достаточно ус-
тойчивая плотность распределения исследованной случайной величины, причем
локальные пики не исчезают с накоплением данных, а четко воспроизводятся. В
п. 7-5.1 этот феномен уже был кратко описан, и обнаруженная Шнолем плот-
ность была названа отороченной гауссоидой, а сейчас надо добавить, что сами
отороченные гауссоиды от опыта к опыту меняются, причем не как попало, а
выстраиваются в достаточно стройные ряды. Точнее говоря, если кривые сгла-
дить неким однообразным методом, то их оказывается всего около 30 вариантов,
тогда как самих опытов проведены тысячи. Таблицы см. [Шноль и др., 1998], а
одна из них воспроизведена на рис. 12.
Главное для нашей темы состоит в том, что "формы компьютерных гисто-
грамм, имитирующих статистику Пуассона, ничем не отличаются от гисто-
грамм, построенных по результатам измерения радиоактивности" [Шноль и др.,
1998, с. 1137]. К сожалению, компьютерные кривые не опубликованы, но при-
няв на веру, что какие-то из них существуют и чем-то схожи с природными,
можно заключить, что налицо не просто какой-то особенный материальный
процесс, но подлинно алеатический феномен.
286
Итак, можно уверенно говорить про эффект Шноля. Точнее, он состоит из
трех феноменов:
1. Феномен отороченной гауссоиды (простой эффект Шноля),
2. Ряды изменчивости отороченных гауссоид (диатропический эффект Шноля),
3. Космофизический эффект Шноля. В работах Шноля и его группы содер-
жатся материалы о существовании еще одного интереснейшего явления - зави-
симости рядов изменчивости от космофизических обстоятельств. Его мы касать-
ся не будем.
Даже въедливые критики, отвергнувшие всё в работах Шноля (в том числе и
космофизический компонент), тем не менее почти в точности воспроизвели эф-
фект отороченной гауссоиды и поместили соответствующие графики; один из них
приведен на рис. 13. В тексте их статьи этот факт (совпадение их собственных
данных с данными отвергаемой концепции) не упомянут, а в подписи к рисунку
сказано лишь: «По нашему мнению, наблюдаемые локальные максимумы и ми-
нимумы есть результат "статистической инерции"» [Дербин и др., 2000, с. 211].
Что имелось в виду, не сказано, и приходится признать, что простой эффект
Шноля - доказанный факт. Он ясно говорит о недостаточности языка и идеоло-
гии ТВ для описания самых обычных, хрестоматийных вероятностных явлений.
При этом важно иметь в виду, что и на кривых гиперболических распределений
также «существуют неизбежные выбросы (зубцы), отражающие не ошибку и не
случайность, а свойства ценоза, и требующие модификации... методов, экстра-
полирующих экспериментальные данные» [Кудрин, 1991а, с. 16].
Потребность в алеатике налицо95, а всеобщность феномена говорит о том, что
и причина столь же обща и что для описания эффектов, подчиняющихся стати-
стике, как стандартной, так и гиперболической, нужен новый общий подход.
Зацепкой к нему может оказаться факт, едва отмеченный авторами походя:
"Эта типичная фрактальность требует для своего объяснения дальнейших экспе-
риментов" [Шноль и др., 1998, с. 1133]. (Как историку науки мне очевидно иное -
накопление данных без руководящей идеи бесперспективно. На сегодня эффект
Шноля нуждается не в новых опытах, а прежде всего в рабочей теории, которая
подскажет, какие нужны опыты, и затем, вернее всего, уступит место иной тео-
рии.) Возможно, что дело окажется не во фрактальности, а в самоподобном
строении действительой оси [...]. Сама же группа Шноля, увы, ставит "развитие
теории" на последнее место в ряду своих задач [Шноль и др., 1998, с. 1139].
Вольному - воля, но вряд ли тогда стоит сетовать на косность оппонентов: на- 98
98 Это видят и Шноль с соавторами; они именуют алеатические аргументы арифметиче-
скими. Они надеются вывести отороченную гауссоиду из механизма многостадийного
химического (но, замечу, не радиоактивного!) процесса и выявления комбинаторики
взаимодействий [Шноль и др., 1998, с. 1137]. На мой згляд, надежда напрасна - таким
путем можно получить нечто вроде биномиальной плотности (как сделал 300 лет назад
Якоб Бернулли), а она одновершинна. Суть надо искать глубже.
287
сколько знаю, в истории науки нет примера, когда бы факт преодолел неприятие
ученого мира без объясняющей теории. Повторяю, сама теория может быть за-
тем отброшена, но факт без объяснения сообществом к рассмотрению не при-
нимается - такова логика социальной истории науки.
Разумеется, наличие рабочей теории лишь необходимо, но недостаточно для
принятия новых фактов научным сообществом. Достаточным условием пред-
ставляется, в терминах главы 5, соответствие теории бытующим в обществе
представлениям, т.е. господствующим ПМ. С позиции третьей (статистической)
ПМ-феномен отороченной гауссоиды нелеп. [...]
Исследование тут естественно начать с упорядочения разнообразия, т.е. с
выявления рядов изменчивости. Как сказано ранее, такие ряды сперва были вы-
явлены в химии, где получили название гомологических. С их помощью открыто
множество новых веществ (например, ряды углеводородов).
В 1920 г. Н.И. Вавилов с помощью данного метода предсказал существова-
ние одного вида злаков (безлигульная рожь) [Вавилов, 1987]. Параллель химии
и биологии в этом пункте достаточно очевидна [Урманцев, 1979], но игнориру-
ется почти всеми. Принято писать, что гомологические ряды в биологии - след-
ствие сходства генов у близких видов (например: «Связано это с тем, что от об-
щего предка может быть унаследован не фенотипический признак, а только ко-
дирующие его гены» [Клюге, 2000, с. 19]), хотя всем, это пишущим, известно,
что это не так. Например, общеизвестна параллель рядов жилкования у крыльев
насекомых и у листьев растений.
Можно полагать, что аналогично будут описаны ряды изменчивости оторо-
ченных гауссовд. Для принятия их научным сообществом понадобится то же
самое, чего пока нехватает рядам Копа - Вавилова, т.е. смена господствующей
ПМ, поскольку анализ разнообразия в терминах третьей ПМ-никогда не удается.
[...] Однако уже сейчас надо пытаться понять, что означает сам феномен
рядов, т.е. повторности форм плотностей. Если он реален, то приходится сделать
обескураживающий вывод: подлинно независимых событий не существует. Эта
мысль не раз уже высказывалась. Например, сто лет назад немецкий статистик и
философ Карл Марбе выступил с серией работ, в которых утверждал, что при-
рода обладает памятью и что поэтому, в частности, после 17 подряд выпадений
герба вероятность его дальнейших выпадений должна уменьшиться". В наше
время с похожей идеей выступил английский биолог и философ Руперт Шелд-
рейк [Sheldrake, 1981]. О нем будет немного сказано в следующих главах.
«Теория Марбе должна быть отвергнута, так как она не подтверждается
опытами» - писал Феллер [1964, с. 153], но вернее было бы написать, что она не
подтверждена и не опровергнута, поскольку для этого потребовалось бы бро-
сить реальную монету более 2 млн раз. И только теперь, с обнаружением груп-
99 Со своей теорией Марбе выступал и позже [Marbe, 1934] и вызвал обширную полеми-
ку, ныне забытую.
288
пой Шноля того факта, что эффект "памяти” или "статистической инерции" в
основе своей одинаков для физического процесса реализации случайностей и
для его имитации на компьютере, можно приступить к проверке компьютерного
аналога схемы Марбе.
В свете эффекта Шноля можно полагать, что идею Марбе будет трудно оп-
ровергнуть. Поэтому стоит повторить слова методологов "Мы на самом деле не
знаем, что такое независимость" [Кац, Улам, 1971, с. 72]. Как мы видели (п. 3-
3.1), понятие независимости было введено в ТВ просто для удобства построения
теории, и даже радиоактивный распад не вполне может считаться имманентно
случайным явлением (см. выше, п. 1, тип Е), т.е. даже для него независимость
отдельных актов нельзя постулировать. В ТВ независимость попросту постули-
рована, а всякое обоснование ТВ выводит феномен независимости из каких-то
конкретных допущений, которые иногда справедливы, а иногда нет.
Дарвинизм, ламаркизм и номогенез о случайности
Иногда дарвинисты "сдают позиции", говоря, что их учение не ставит целью
объяснить мелкие различия, а призвано лишь утвердить выгодность существен-
ных приобретений. Но, во-первых, это не так (вся литература по математиче-
скому дарвинизму состоит в исчислении выгодности малых различий, и прочий
дарвинизм видит в ней свое основное подтверждение), а во-вторых и в-главных,
если всерьез сравнивать крупные эволюционные приобретения, привлекая поня-
тие отбора только для оценки их (а не мелких различий), то основной интерес
представит не динамика их численности, а те законы, по которым эти приобре-
тения возникли. Данные законы изучаются отнюдь не дарвинизмом (он попро-
сту отрицает их наличие), а другими эволюционными учениями.
Из этих учений здесь надо назвать ламаркизм, понимаемый как совокуп-
ность всех представлений, в которых собственная активность особи рассматри-
вается как фактор эволюции, и номогенез, понимаемый как совокупность всех
представлений, основанных на признании закономерности появления новых
эволюционных приобретений, не сводимой к динамике выживания особей. Дан-
ные закономерности интересны здесь лишь в той мере, в какой включают слу-
чайность их появления. Обо всем этом шла речь в частях II и III книги [Чайков-
ский, 1990], и здесь стоит добавить немногое.
Во-первых, ориентационные автоматные модели... по сути являются ламар-
кистскими - в той же мере, как и “стабилизирующий отбор” Шмальгаузена. (Ра-
нее [Чайковский, 1998] я уже старался показать, что данная теория И.И. Шмаль-
гаузена являет собой явственный ламаркизм, по цензурным соображениям вы-
сказанный в дарвинистических терминах).
Во-вторых, решающую роль в эволюции играет свободный выбор. В ламар-
кизме это - свободный выбор поведения особью, а в номогенезе - самопроиз-
вольное появление новой эволюционной альтернативы (новой логической воз-
можности), о чем говорилось прямо в книге [Чайковский, 1990, с. 173]... С этой
позиции можно понять тех авторов, которых прежде считали мистиками и по-
289
нимать даже не пытались, - например, Бергсона, писавшего в 1907 г., что эво-
люция есть творчество: с позиции алеатики он всего лишь указал другую, неже-
ли дарвинисты, ступень случайности; если они имели в виду вероятность, то
Бергсон - произвольный выбор.
Тем самым Бергсон фактически ввел в оборот особый фактор эволюции, ко-
торый с позиции алеатики (это в-третьих) взаимодополнителен случайности, по-
стулируемой дарвинизмом, - в том же примерно смысле, в каком, по Мейену
[1974], взаимодополнительны дарвинизм и номогенез*. В связи с этим уместно
повторить, вслед за В.А. Красиловым, что противники дарвинизма обычно от-
вергают его как "сведение явлений на игру случайностей", а приверженцы, на-
оборот, главным в дарвинизме считают "представление о естественном отборе
как антистохастическом факторе"; и пусть основатель концепции номогенеза
Л.С. Берг отвергал случайность наследственных изменений, но на деле, наобо-
рот, "случайности Берг отводил гораздо большую роль, чем Дарвин", ибо пола-
гал большинство признаков организмов нейтральными по отношению к нуждам
адаптации [Красилов, 1979, с. 8, 7]. Это - тоже взаимодополнительность.
Наконец, и это в-четвертых, понимание эволюции требует рассмотрения
случайности как предпочтения, т.е. - привлечения шестой (креативно-пропен-
сивной) ПМ. Начало этому положили, как ни странно, дарвинисты. Так, один из
классиков дарвинизма XX века Эрнст Майр утверждал:
«Каждая группа животных "предрасположена" к изменчивости одних сво-
их структур и к удивительной стабильности других... У некоторых групп
млекопитающих имеется предрасположенность к развитию рогов на лбу, у
других - к развитию рогов на вершине головы, а у третьих - к тому, чтобы
вообще не иметь рогов... Лишь часть этих различий можно объяснить раз-
личиями в давлениях отбора» [Майр, 1974, с. 403].
Что касается остальных эволюционных школ, то все они, особенно номоге-
нез, включают анализ предпочтений, хотя сами эволюционисты и не думают,
что исследуют случайность. Ярким примером может служить свойственное
жоффруизму и номогенезу толкование появления новых видов через тератоге-
нез, который выше (п. 4.1) был описан как реализация редких предрасположен-
ностей. В этих терминах вся эволюция предстает как процесс изменения пред-
расположенностей. Вопрос о том, что управляет "пропенсивными полями", рас-
смотрен гипотезой Шелдрейка [Sheldrake, 1981, с. 93, 123] и выходит за рамки
данной книги. (См. АСМ- примеч 2010 г.)...
Какими законами управляется эта случайность, пока неизвестно, но можно
не сомневаться, что они сложнее, чем гиперболические. Достаточно рассмотреть
квази-гиперболы рис. 14 в логарифмическом масштабе, чтобы увидеть, что это
вовсе не прямые линии (рис. 15). На самой длинной из них (хризомелиды, т.е.
* Об этой работе (М-121) см. выше, статью «С.В. Мейен и теория биологической эволю-
ции». - Примеч. 2010 г.
290
жуки-листоеды - данные охватывают примерно четверть мировой фауны) хо-
рошо видно, что даже если принять среднюю часть за прямую, то правая часть
должна быть описана как-то иначе.
Общие замечания
В конце недавней французской книги о случайности приведена "не без
юмора" пара противоположных тезисов ("Последний закон мира - случай, и
весь возможный детерминизм обязан закону больших чисел" и "Последний за-
кон мира - полный детерминизм, и вся наблюдаемая случайность обязана де-
терминистическому хаосу"), из чего сделан вывод: на уровне бытия у нас нет
средств выбрать между этими моделями мира, а на уровне знания мы можем (и с
пользой) принять любую [Chaos, 1992, с. 405].
Сделать так можно, однако надо понимать, что кроме двух этих взглядов
(т.е. второй и третьей ПМ) есть и другие. Они нужны, в частности, для того,
чтобы осознать феномен случайности без вероятности, но для этого необходима
смена статистического мировоззрения, господствующего пока в обществе (его
яркие черты - вероятностное толкование текстов, подсчет всевозможных шан-
сов в обыденной жизни, вера в самодостаточность рынка и дарвинизма), на бо-
лее новое, основанное на новых ПМ.
Будущее алеатики видится мне в выявлении исторических линий мысли.
Линия от Кардано к Колмогорову представляется мне пройденной, а будущее - в
движении по забытой линии, т.е. по пути пропенсивности, от Августина и Пачо-
ли к Попперу, Шелдрейку и далее. Клеточные автоматы Кауфмана вселяют на-
дежду, что пропенсивность может быть вскоре понята вполне конкретно и даже
инструментально.
Однако... еще предстоит найти некоторый принцип - образно говоря, прин-
цип однородности событийного пространства-времени, в рамках которого при-
менима ТВ, и четко выявить эти рамки. В понятиях каких ПМ-это удобно будет
сделать? Однородное пространство - атрибут третьей ПМ, что же касается на-
рушений однородности, то тут, на мой взгляд, более всего подходит шестая (бу-
дущая) ПМ, охарактеризованная, насколько это сейчас возможно, как креатив-
но-пропенсивная (см. гл. 5). Случайность оказывается здесь почти нацело не-
стохастической (невероятностной), как, впрочем, и при пользовании остальны-
ми новыми ПМ, т.е. четвертой и пятой.
Однако, если о пропенсивности мы не можем на сегодня сказать ничего,
кроме нескольких абстрактных фраз, то системная и диатропическая случайно-
сти изучены лучше и допускают уже сейчас описание в довольно аккуратных
терминах (пусть и не столь точных, какими пользуется ТВ). А именно, в неже-
стких системах обычна сложная случайность с неустойчивыми частотами, кото-
рую нельзя характеризовать посредством вероятности-частоты, но, как правило,
удается описать через иные, менее жесткие, инварианты. Такими инвариантами
служат, например, плотности распределения неустойчивых частот и ряды из-
менчивости плотностей (см. выше, п. «Эффект Шноля»).
291
Наиболее сложна случайность тогда, когда система находится на грани ме-
жду порядком и хаосом. В таких системах обычен феномен спонтанной самоор-
ганизации, и случайность ему способствует (организующая роль случайности).
Поскольку "увеличивая долю канализирующих функций в сети, мы можем под-
талкивать систему к фазовому переходу между хаосом и порядком" и "устойчи-
вое ядро элементов идентично почти для всех аттракторов" [Кауфман, 1991, с.
64, 65], постольку самоорганизация может быть не только понята, но, в принци-
пе, и управляема. Здесь, по Кауфману, открывается также и путь к пониманию
эволюции .
Роль случайности во всякой эволюции оказывается при этом ключевой, од-
нако совсем не той, какую рисовал (для биологии) дарвинизм, видевший лишь
две ее функции - ненаправленность вариаций и непредсказуемость путей . Роль
ненаправленных вариаций оказалась несущественной, а пути развития - вполне
предсказуемыми, если понимать предсказание в диатропическом смысле - как
указание спектра возможностей.
В своей недавней книге [Kauffman, 2008] Кауфман, признавая дарвинизм, указал на то,
что «отбор» истолкован теперь в науке как всепроникающий феномен самоор-
ганизации. Это обстоятельство снимает, по Кауфману, противоречия между дарвиниз-
мом и креационизмом. Главным же вкладом Кауфмана в эволюционную науку остается,
на мой взгляд, указание на эволюционную роль случайности без вероятности - органи-
зующего феномена, возникающего на грани порядка и хаоса. Самоорганизация, возни-
кающая спонтанно, является новым законом развития природы (живой и неживой), от-
крытие которого радикально расширяет понимание активности как ведущего фактора
эволюции. - Примеч. 2010 г.
** Так, известный голландский глобальный эволюционист Петер Вестбрук говорил: «Есть
и другие возможные принципы организации, кроме естественного отбора. Например, ра-
бота Стюарта Кауфмана, показавшая, что порядок, структура может самопроизвольно
возникать из хаотической ситуации... Так, на грани между порядком и хаосом, поддер-
живает себя жизнь» [Westbroek, 1997, с. 101].
292
Литература к части 7
Беклемишев В.Н. Морфологические проблемы животных структур. Пермь, 1925.
Берг Л.С. Труды по теории эволюции. Л., Наука, 1977.
Борель Э. Случай. М. - П., 1923.
Вавилов И.И. Закон гомологических рядов в наследственной изменчивочти. М., 1987.
Гачок В.П. Странные аттракторы в биосистемах. Киев, 1989.
Дарвин Ч. Соч., т. 4, М.-Л, 1951 (Изменение животных и растений...).
Дербин А.В., Бакланов С.В., Егоров А.И., Муратова В.Н. Замечания к статье “О реа-
лизации дискретных состояний...” (Шноль С.Э. и др., 1998) // УФН, 2000, № 2.
Заславский Г.М. Стохастичность динамических систем. М. Физматлит, 1984. 272 с.
Иваницкий Г.Р., Медвединский А.Б., Деев А.А., Цыганов М.А. От “демона Максвелла”
к самоорганизации массопереноса в живых системах// УФН„ 1998, т. 168, № 11.
Кауфман С.А. Антихаос и приспособление // В мире науки, 1991, № 10.
Кац М., Улам С. Математика и логика. Ретроспектива и перспективы. М., 1971.
Климонтович Ю.Л. Критерии относительной степени упорядоченности открытых
систем И УФН, 1996, т. 166, № 11.
Клюге Н.Ю. Современная систематика насекомых. СПб., Лань, 2000. 333 с.
Красилов В.А. Роль случайности в эволюции // Эволюционные исследования. Влади-
восток, 1979, с. 7-19.
Кудрин Б.И. Проблемы создания и управления ценозами искусственного происхож-
дения И Кибернетические системы ценозов: синтез и управление. М., МОИП, 1991.
Левич А.П. Структура экологических сообществ. М., 1980.
Майр Э. Популяции, виды и эволюция. М., Мир, 1974.
Мейен С.В. 1974. О соотношении номогенетического и тихогенетического... (М-121).
Мейен С.В. 1984. Принципы исторических реконструкций в биологии (М-263).
ПойаД. Математика и правдоподобные рассуждения. М., 1957.
Растригин Л. А. Этот случайный, случайный, случайный мир. М., 1969.
Секей Г. Парадоксы в теории вероятностей и математической статитстике. М., 1990.
Урманцев Ю.А. Номогенез о сходстве в живой природе И Природа, 1979, № 9, с. 116.
Феллер В. Введение в теорию вероятностей и ее приложения. М., 1964.
Чайковский Ю.В.
Чайковский Ю.В.
Чайковский Ю.В.
Чайковский Ю.В.
Шноль С.Э., Коломбет В.А., Пожарский Э.В., Зенченко Т.А., Зверева И.М., Конрадов
А.А. О реализации дискретных состояний в ходе флуктуаций в макроскопических процес-
сах//УФН, 1998, № 10.
Шредер М. Фракталы, хаос, степенные законы. М. - Ижевск, 2001.
Шрейдер Ю.А. Об одной модели семантической теории информации // Проблемы ки-
бернетики, вып. 13. М., 1965.
Chaikovsky, 1971. On a continuous process of matrix game solution (4-2)
Chaos et determinisme / Sous la direction de A. Dahan Dalmedico. Editions de Seuil, 1992.
Kepler J. Gesammte Werke. Munchen, 1938, Bd 1.
Kuki Sh. Le probleme de la contingence (1935). Tokyo, 1966.
Sheldrake R. A new science of life. London, 1981.
1971. О непрерывном процессе решения матричной игры. 4-1.
1985. Разнообразие и случайность. 4-13.
1987 Нечеткие закономерности... 4-18.
1990. ЭЭД. 4-24.
293
Часть 8. Личное
Серёжа Мейен, мой сосед*
Познакомились мы с ним поздно - всего за 12 лет до его кончины, так что я
застал мало из того, чем он был известен в молодости. О его музыкальном обра-
зовании (виолончель), увлечении танцами (чарльстон) и итальянскими фильма-
ми узнал я лишь из воспоминаний в сборнике его памяти; в поле он ездил при
мне всего раз, ненадолго и без всякой радости (просто надо было увидеть свои-
ми глазами еще один разрез); и восходящей звездой его давно не считали - к со-
рока годам он был уже классиком. Долгое время мне и в голову не приходило,
что он может интересоваться религией, искусством или прекрасным полом. Во-
обще, при всей готовности помочь и внешней открытости, он пускал в свой
внутренний мир скупо. И правильно делал.
Как-то Юлий Анатольевич Шрейдер, математик и философ, рассказывал за
чашкой чая о Любищеве и на мое замечание, что есть вот в русле тех же идей
интересный эволюционист Мейен, откликнулся репликой: “Да он же ваш сосед!
Живет в вашем квартале”. Вскоре он пригласил меня на его доклад. Так, в апре-
ле в 1975 года, я впервые попал в Институт философии на Волхонке, где увидел
и услышал Сергея Викторовича.
Говорил он о том, что в теории эволюции следует допустить к рассмотре-
нию все воззрения, а не только дарвинизм. По тем временам это было смело
(ведь дарвинизм был частью партийной идеологии, а она не терпела альтерна-
тив), Мейен говорил очень аккуратно, чтобы никого лично не задеть, и мне по
наивности показалось, что он предлагает попросту улучшить дарвинизм. Поэто-
му в моем сумбурном выступлении главной мыслью был тот тезис, что хороши
все теории мироздания, что-то говорящие о мире, но не мифы вроде “земли на
трех китах”. Они не говорят о мире ничего, их включать в число альтернатив
познания не стоит, и к ним как раз относится нынешний дарвинизм. В ответ
один философ физики ответил внешне мирно, а по сути зло, что всё это верно “с
точностью до наоборот”, т.е. именно выступающего (меня) и надо игнорировать
- как желающего возродить “землю на трех китах”.
Было обидно - из-за моей несдержанности, так глупо контрастировавшей с
изящным докладом, Мейен конечно сочтет меня “чайником” и не воспримет
всерьёз. Каково же было мое удивление, когда он сам пригласил меня зайти к
нему, поскольку-де мы соседи. А ведь кроме моей выходки да слов Шрейдера,
он не знал обо мне ничего. И вот 1 мая мы встретились.
Помню, как мы с женой Риммой обсуждали - поздравлять ли Мейена с Пер-
вомаем. Римма посоветовала мне на всякий случай вопрос обойти - как бы не
было конфуза. С.В. вышел ко мне на улицу (быть может, дома готовили засто-
* Из сборника к 70-летию С.В. Мейена, с. 63-68 (4-56).
294
лье), и мы часа два гуляли по кварталу у метро “Речной вокзал”. Он был тогда
генеральным секретарем международного конгресса по карбону и с увлечением
рассказывал, как готовит конгресс, обходя препоны, то и дело выставляемые ор-
ганами партии и госбезопасности. Мне оставалось лишь изумляться - как он всё
это выносит, да еще с иронией.
На мое изумление он ответил просто и спокойно (чем изумил меня еще боль-
ше): да, три года для науки почти потеряно, но за это лаборатория получила обо-
рудование, и удалось даже взять еврея в штат института. И еще: поскольку урыв-
ками работать с образцами невозможно, он использует просветы во времени для
чтения, и прочел наконец-то старых натурфилософов и просто философов.
После этого я решился рассказать о наших сомнениях насчет поздравления,
и С.В. рассмеялся весело: неужто он похож на любителя Советской власти? (В
последующих беседах он величал ее Софьей Власьевной и радовался шутке не-
замысловатой: “старухе пора на пенсию”. Позже с восторгом рассказал мне
анекдот: грузин просит у друзей барана для свадьбы, каждый советует обратить-
ся к другому, и грузин взрывается: “Пачему все дают совет, никто нэ даёт ба-
ран?”; “А ты гдэ живёшь - в странэ Советов или в странэ баранов?”)
Мне тогда было решительно не с кем обсудить то, чем я занимался - как уст-
роено разнообразие одноклеточных и чем можно заменить нелепую “теорию”
симбиогенеза. Меня занимала новая идея “горизонтального переноса генов”, т.е.
обмена генами вне процедуры размножения. Обсуждать это Мейен тогда, на пер-
вомайской прогулке, не стал, зато (опять к моему изумлению) предложил мне дать
ему на прочтение мою рукопись на сей счет. Прочел очень быстро, опять мы гу-
ляли по кварталу в выходной день, и он снова меня огорошил: «По большому сче-
ту вы держетесь как раз за то старое, с чем пытаетесь спорить».
По его мнению, те сходства митохондрий и хлоропластов с бактериями, на
которых построена идея симбиогенеза, основаны вовсе не на обмене генами, а
носят типологический характер (другими словами: сходства не унаследованы от
общего предка, а вызваны общими законами формы). Сам он не занимался бак-
териями, но ряды сходств видел повсюду и предлагал объяснять их общим зако-
ном формы, а не обилием случайных актов горизонтального переноса. Правоту
С.В. я уяснил себе много позже. Вообще же он стал моим учителем, хоть и был
старше меня всего на четыре года.
Вскоре Мейена пригласили на зимнюю школу по теоретической биологии
для студентов Биофака МГУ, а он рекомендовал пригласить и меня. Школа была
в поселке Борок Ярославской области, в январе.
Выйдя из вагона в пять утра на тридцатиградусный мороз в осенних пальто,
мы тут же стали замерзать. Когда подали автобус, все мы бросились к нему, но
Мейена все, конечно же, пропустили вперед. Только устроитель (милейший че-
ловек и блестящий организатор), сам изрядно замерзший, загородил нам путь со
словами: «Тут есть постарше вас. Проходите, Алексей Меркурьевич». А.М. Ги-
ляров, совсем молодой кандидат наук, был сыном академика-секретаря Мерку-
295
рия Сергеевича Гилярова.
Препираться на морозе было бы глупо, и Меркурьич, как бы ничего не за-
метив, быстро поднялся в автобус, затем мы, лекторы, за нами толпа студентов,
Мейен тоже как бы ничего не заметил. Войдя в гостиницу, мы стали в ожидании
регистрации читать программу школы на стенде, и тут С.В. учинил “страшную
месть”. Отвел устроителя в сторонку и сказал тихо: «Исправьте, пожалуйста. Я
не кандидат биологических, а доктор геолого-минералогических». Устроитель
чуть не умер.
Мы все быстро подружились, и когда устроитель сделал доклад, а я поднял
руку, чтобы задать вопрос, Мейен одёрнул меня, ошибочно полагая, что я скажу
гадость. (Слова С.В., из лексикона царской семинарии, привести нельзя.) Позже
он пробовал устроить устроителя на хорошую работу, а вскоре сам устроитель
учредил свой семинар и открыть его предоставил Мейену. Еще много позже он
организовал вечер, увы, памяти Мейена, где доклад предоставил мне.
Зимняя школа в Борке - одно из лучших воспоминаний моей жизни. О ней
писали, и я ограничусь эпизодом. Биохимик С.Э. Шноль, дремучий дарвинист, в
своей лекции уверял, что генетика - основа всей биологии, а С.В. в реплике с
места недоумевал: как в терминах генетики описать ловлю кошками мышей?
Шноль отвечал в запальчивости, что и это станет ясно, когда будет целиком
прочтён геном кошки. Мейен промолчал, но по окончании сказал в коридоре,
что ни одну из общих проблем биологии прочтение генома решить не сможет.
Сейчас, через 30 лет, это начали понимать даже генетики.
Живя в одной комнате, мы с Серёжей перешли на “ты”, а по возвращении в
Москву стали общаться семьями. Рита Мейен поразила меня своим молодым
обликом (ведь она была его однокурсницей) и тихой ненавязчивой помощью во
всех его делах. Моя Римма тоже мне помогала, но ведь Рита вообще бросила ра-
боту и взвалила на себя всю секретарскую деятельность, а Мейен работал, как
целый институт. Да, без шутки: он с явной гордостью говорил мне, что у них в
дирекции два ящика с почтой - для него и для остального института.
Однажды в 70-х в Коми с его аспиранта потребовали оплаты за какую-то
научную услугу, Мейен пришел в ярость, взял журнал лаборатории и попросил
свою бухгалтерию выставить в Сыктывкар счёт - только за датировки образцов,
проведенные для них только им самим и только за истекший год; оттуда вскоре
же пришло извинение и просьба отозвать счёт как разорительный. А ведь это
малая толика: были еще бесчисленные консультации, как в Москве, так и с вы-
ездами в Сыктывкар и Ухту, и знаменитые лекции Мейена для одного слушате-
ля. Не только из Коми, а со всего Союза.
Встречались мы почему-то чаще у него, хотя нас было в трехкомнатной
квартире четверо, а их в точно такой же - пятеро (когда же их дочка Катя вышла
замуж и родила Петю - семеро). Как они ухитрялись жить мирно, ума не при-
ложу, но никаких следов скандала никогда не замечал.
Серёжа не ходил в магазины, это делали домочадцы. Когда в Москву приле-
296
тел его французский коллега и друг Ив Лемуань, Серёжа спросил меня: что бы
такое показать ему самое интересное? Я посоветовал отвести его в наш универ-
сам у метро после работы - пусть посмотрит драку у мясного или рыбного отде-
ла и поймет, что не политические свободы нас волнуют. Серёжа только усмех-
нулся: явно счёл негожим предлагать гостю “высматривать срам земли сей”.
Летом 1977 года Мейены гостили у нас на даче в Отдыхе. У меня там был
тайник с рукописями, и, чтобы развлечь гостей, я стал читать им вслух. Вскоре у
меня началась резь в глазах (нелепая манера: писать карандашом, чтобы легче
править - от нее после этого пришлось отказаться), затем отсох язык, а Серёжа
увлекся и просил читать еще. Чтение продолжила Римма.
Основной мой опус он отверг (ни слова не сказал по окончании, и всем всё
было ясно), но два поменьше, прочитанные вначале, ему понравились, и он зая-
вил, что их надо переслать на Запад для публикации под псевдонимом. Один на-
зывался «Декрет, которого не было» и объяснял, что большевики вовсе не даро-
вали, как они хвастают, независимости Финляндии, что они ограничились ниче-
го не значащим жестом, организуя в те же дни вторжение туда Красной гвардии.
Другой назывался «Адмирал» и повествовал о Колчаке как героическом поляр-
нике, талантливом адмирале и наивно-беспомощном правителе.
В январе ожидалось шестидесятилетие независимости Финляндии, и Мейен
взялся, если будет машинопись, сделать микрофильм и переправить его. Рита
героически смолчала, хотя понимала конечно, чем он рискует. Сам же он убеж-
дал нас всех, что всё безопасно. К осени я перепечатал «Декрет», и Серёжа с
гордостью показал мне, как мал (обрезана перфорация) и хорошо читаем мик-
рофильм. Затем то же было сделано с «Адмиралом». Один был отослан в “Но-
вый журнал” (США), другой в “Континент” (Париж) - какой куда, не помню.
Судьба их мне осталась неизвестна, что и к лучшему: готовя их к печати в годы
гласности, я понял, насколько они были сырыми.
Серёжа хвастался, что, насколько помню, у него 42 племянника (считал, ра-
зумеется, не он сам, а кто-то из троюродных братьев, спец по этой части). И вот
один из них осенью приехал из эмиграции родню проведать. Серёжа дал ему
«Адмирала» в полной уверенности, что тем самым отправляет его на Запад, но
племянник отверг текст с негодованием - это же советский пасквиль на белого
героя. Особо возмутило его мое выражение «в угасавшем уме адмирала». Серё-
жа был смущен, но, как учёный, спросил меня, откуда мне известно психическое
состояние Колчака в последние месяцы правления. Об этом писали многие, но я
по наивности назвал ему мемуары французского генерала Жанена, начальника
союзных войск в Сибири. Узнав это, племянник сменил ярость на презрение -
всем известно, что Жанен - предатель, выдавший Колчака красным.
Мне это был хороший урок: мемуаров избегать вообще и уж во всяком слу-
чае не ссылаться на них как на источник. Пришлось перечитать всё, что можно,
мемуарные сведения проверить по иным текстам, слова «в угасавшем уме адми-
рала» убрать. Итог был замечателен: очерк приобрел некую ценность.
297
(«Адмирал» был выправлен и опубликован^00, а повесть, сделанная из него
и «Декрета», лежит до сих пор. Большой опус - простым карандашом по серой
бумаге - теперь, полагаю, и прочесть уже невозможно. Да и ненужно. Мемуаров
стараюсь не писать, и данный текст - единственное исключение.)
В начале 80-х Мейену как минимум дважды удалось пресечь разгромные
собрания. Вообще-то они после смерти Сталина в естествознании не проводи-
лись, но нападки на дарвинизм все-таки преследовались. Первое собрание было
отведено генетику Л.И. Корочкину, но само появление Мейена, которого никто
не ждал (он был далек от генетики) напрочь расстроило все планы устроителей.
Забавно было слушать, как они друг за другом выходят к трибуне и объясняют,
что ничего против Леонида Ивановича не имеют.
Второе собрание было опаснее: героем был киевский генетик В.А. Кор-
дюм^0^, а Киев - не Москва, там уже вышли 2 рецензии погромного стиля^02, а
по “сигналу” из Москвы его могли с работы погнать. Да и сам Виталий Арноль-
дович своей иронией в адрес тогдашних дарвинистов разозлил многих. (То, что
он тогда писал как ересь - о роли горизонтального переноса в эволюции - сей-
час общепризнано.) У меня был заготовлен текст выступления под названием
«Нечитавшие критики», по окончании собрания я сделал запись впечатлений,
так что здесь у меня не мемуар, а как бы изложение дневниковой записи.
Мейен не выступал, действуя своим присутствием. Председатель требовал
говорить только о книге Кордюма, беря этим рецензентов под защиту. При-
шлось ждать повода, и он нашелся: все дружно называли Флеминга Дженкина
(критик Дарвина) Дженкинсом, и я смог начать с замечания, что все знают о нем
лишь по одной безграмотной ссылке. Затем прочел из своей заготовки абзац:
«То, что делают с Дарвином наши рецензенты, поразительно. Единственное,
что приходится допустить, что ни один из троих не читал Дарвина со студенче-
ских лет, открывал его с тех пор только в поисках цитат, но каждый уверен, что
суть дарвинизма знает хорошо. Это - лучшее, что я могу допустить».
Председатель предложил перейти к книге Кордюма, но я продолжил:
«То, что позже назвали мутацией, можно сопоставить с “самопроизвольной”
изменчивосью у Дарвина. Неопределенной изменчивости он придавал мало зна-
чения: в девятитомнике на нее всего 9 ссылок мимоходом, тогда как рубрика
“Изменчивость” занимает в указателе три больших колонки. Ключевое значение
он придавал определенной изменчивости и ей посвящал целые главы».
Председатель пригрозил лишить меня слова. Я успел еще процитировать
Гершензона: «Согласно Дарвину... движущей силой эволюции является естест-
Чайковский Ю.В. Гранит во льдах // Вокруг света, 1991, №№ 1, 2. Белогвардейская
часть там сведена к минимуму.
101 Кордюм В.А. Эволюция и биосфера. Киев, Паукова думка, 1982. 262 с.
102 Б.М. Медников и А.В. Яблоков; С.М. Гершензон // Цитология и генетика, 1984, № 1.
298
венный отбор. Именно он и только он...» и сел на место в ярости. Кордюм был в
восторге и шепнул мне: «Никогда не приходило в голову, что у Дарвина нет от-
бора случайных вариаций». Рецензент А.В. Яблоков выступал весь красный, го-
ворил что-то бессвязное, и я попросил Мейена спросить, где он это взял. «Брось
ты, ведь он же извиняется» - ответил Серёжа.
О Мейене обычно пишут в стиле “Житие святого”, и мне не хочется сбиться
на этот путь. Серёжа был изумителен, но именно потому, что святым не был. Он
более чем любил Риту, этой любви я поражался, но однажды мне пришлось ми-
рить их, ибо дело шло к разрыву. Ему тоже пришлось мирить меня с Риммой, но
без успеха. Не потому, что он был хуже как дипломат (он был и в этом гораздо
лучше меня), а потому, что мы оказались хуже, чем он и Рита.
Моя семья распалась, и мы с Серёжей с конца 1979 года больше не были со-
седями. Сохранив контакт с Риммой, он неохотно, но согласился познакомиться
с Наташей, моей новой женой, и после этого сказал мне, что от своей заочной
неприязни к ней отказывается. Однажды он даже был с Ритой у нас дома, но всё
же отношения стали скудеть. Хоть он и сердился на меня, но это прошло, зато
слабел контакт по-существу: Мейен понемногу стал понимать, что жить ему не-
долго, и понемногу же стал отбрасывать всё лишнее, в том числе эволюцию.
Летом 1982 года он первый раз попал в больницу. Мы с ним редактировали
тогда книгу Любищева, и он поделил работу так: «Ты сиди над рукописью, а по
кабинетам буду ходить я». Конечно, и над рукописью он корпел больше, чем я,
но главное - умело провел ее через враждебных чиновников и рецензентов. И
вот корректура, а он на койке.
Указатели мы стали делать порознь, но примерно поровну, а снимать вопро-
сы с редактором пришлось мне. Помню, как трогательно Серёжа умолял меня не
говорить редакторше резкостей («она такая благожелательная»), чего я, разуме-
ется, вовсе и не собирался. И в самом деле, всё было дружелюбно.
Когда-то в 60-е годы Любищев познакомился с инструктором^ ЦК КПСС,
тот оказался вполне порядочным человеком и, чем мог, Любищеву помогал.
Как-то в поезде Мейен тоже познакомился с инструктором ЦК и решил извлечь
из этого пользу - написал ему про беды в науке. В частности, примерно так: «В
биологии теоретики есть, но они вынуждены вести полуподпольное существо-
вание. Например, Чайковский». Вскоре меня вызвал директор института, куда
недавно (в 1980 г.) меня взяли с огромным трудом, и мрачно спросил, какую я
веду подпольную деятельность и с какой целью. В тот же вечер я рассказал это
103 Молодым поясню: в каждом партийном органе (ЦК, обком, горком, райком) инструк-
торами назывались рядовые штатные сотрудники. Однако вовне инструктор был грозным
начальником. Если он занимался делами института, то именовался “куратором институ-
та” и фактически мог приказывать директору, что, однако, бывало редко.
299
Серёже, мы посмеялись, и на том всё кончилось - возможно, он позвонил инст-
руктору, и тот дал отбой. Но не всегда было столь просто и мирно.
В начале 1983 года издательство научно-популярной литературы “Знание”
заключило со мной договор на книгу «По законам разнообразия». Не имея тогда
возможности издать книгу в научном издательстве, многие, в том числе Мейен,
пользовались случаем изложить свои мысли в книгах популярных. Рецензию на
мою рукопись писал Мейен, она была очень положительна и содержала фразу о
том, что особую ценность представляет изложение автором спорных и даже за-
бытых теорий. Редактор был болезненно труслив (на что, увы, имел основания)
и заявил, что рецензия далеко не положительна, что она призывает к бдительно-
сти. Он направил рукопись другому рецензенту, профессорше из Тимирязевской
сельхозакадемии. Та ответила почти так же, как Мейен, но добавила - не видно
политического лица автора, а устно пояснила: «Можно подумать, что автор пи-
шет в Нью-Йорке».
Редактор потребовал унавозить текст цитатами из Маркса и Ленина, убрать
или закутать в пелёнки пояснений цитаты из “не наших” авторов (например, из
Герберта Спенсера), а “отвергнутые” теории изъять или разоблачить. Разумеет-
ся, я отказался, и книга в свет не вышла.
Серёжа был явно смущен своим промахом (редактор был ему известен, а
рецензия явно повторяла ошибку письма к инструктору ЦК) и обещал «что-
нибудь придумать», но не сделал ничего, и я обиделся. Теперь мы встречались
только на учёных собраниях, но всё же обменивались дружескими репликами.
Вскоре, однако, последовало новое ухудшение.
В апреле 1985 года пожарная инспекция закрыла Библиотеку МОИГТ^4 по
причине аварийной перегрузки здания. Директор Библиотеки Н.В. Демьяненко
просила меня помочь, чем я и занимаюсь вот уже 20 лет. Нужно было найти
пустующее помещение (каковое и нашлось в том же дворе), но без активного
содействия Президиума МОИП получить его было невозможно, и я очень рас-
считывал на помощь Мейена, там весьма влиятельного. Он хорошо знал и лю-
бил эту библиотеку, со вниманием выслушивал мои донесения, но помощи не
предлагал, чем обидел меня снова.
(Осенью Библиотека была вновь открыта, но никакой помощи так и не полу-
чила, даже в юбилейные мероприятия 2005 года. Перегрузка возрастает*, 4 биб-
Мейен работал в МОИП с юности, а с 1975 г. был членом Президиума МОИП. Биб-
лиотека расположена в здании Библиотеки МГУ на Моховой, против Манежа, и имеет
фонд в полмиллиона единиц хранения. Примеч. 2010 г.', как позже выяснилось, фонд со-
ставляет около 300 тыс.; остальное (груду оттисков, списанных другими библиотеками и
свезённых в Библиотеку МОИП) покойная Н.В. Демьяненко когда-то приписала к фонду
в надежде повысить категорию своей библиотеки.
* Летом 2006 г. вопрос с помещением для хранения был решен. Библиотека МГУ вы-
везла часть фонда в новое помпезное (но с ничтожным объемом хранилища) здание на-
300
лиотекаря, даже когда все ставки заняты, обслужить огромный фонд не могут, он
приходит в негодность, и никаких надежд на ее спасение у меня не осталось.)
К этому добавилось наше расхождение по вопросам службы. Серёжа долго
помогал А.В. Яблокову войти в Академию наук, и наконец в 1984 году тот стал
член-кором. Беседуя об этом с Серёжей, я вдруг понял, что сам он тоже туда хо-
чет. «Зачем тебе в эту компанию?» - спросил я удивленно, на что он спокойно
ответил: «Разве я не имею права на хороший кусок мяса по пятницам?»^"’
Дело было, конечно, не в съестных благах (хотя тогда с ними стало еще ху-
же), а в служебных возможностях. Став в 33 года доктором наук, Мейен сразу
ощутил преимущество статуса, который был ему (в отличие от меня) был очень
нужен в общениях с начальством. Но эта регалия оказалась последней^, а к 50
годам ее для давления на начальство было мало. Еще в начале 80-х он пытался
получить диплом профессора (каковым фактически давно был), но внештатное
чтение лекций такой возможности ему не дало (хотя другим давало), а в науч-
ном институте для этого надо защитить пятерых аспирантов и обязательно быть
начальником. Мейен даже согласился стать заместителем директора, но райком
потребовал вступления его в партию, на чем эпопея и кончилась.
Были и другие попытки его повысить свой статус, но главное - продви-
нуть его в Академию, как он продвигал Яблокова, должен был кто-то другой,
ибо сам он был для этого слишком принципиален. Другой не появился, и это -
позор для самой Академии-^ Мейен был уязвлен, а я при редких встречах
только иронизировал.
против Главного здания. Освободилось пол-яруса рядом с хранением МОИП, и прошел
слух, что эту площадь отдадут Библиотеке МОИП. Не имея никаких подтверждений,
мы в один день заполнили своими книгами столько стеллажей, сколько смогли. Даже
А.М. Оловников (позже так цинично обойденный Нобелевским комитетом), случайно
зашедший, тоже возил тележки с книгами к лифту. Ни запрета, ни разрешения от рек-
тората не последовало, и ни денег, ни штатов для обслуживания этой площади Библио-
тека МОИП так и не получила. Библиотека умирает, однако книги и журналы все-таки
размещены (в этом большую роль сыграли И.А. Игнатьев и Ю.М. Мосейчик, расста-
вившие около полусотни стеллажей), так что можно надеяться, что они достоят до пе-
ремен к лучшему.- Примеч. 2010 г.
все Годы Советской власти, с 1918 г., действовала сеть закрытых распределителей,
после Сталина скрывшаяся (на нижних своих ступенях) в тени “отделов заказов”. Член-
корам полагался еженедельный “заказ” с киллограммом мяса, а также целый ряд услуг,
в том числе жилищных и медицинских.
106 pi3 советских. Зато Мейен был вице-президентом Международной организации па-
леоботаников; но это, насколько знаю, на чиновников не действовало.
107
и Показательно: так и не вошел в Академию А.Ф. Лосев, опальный филолог, историк
и философ. Многие ждали, что на “перестроечном” собрании Академии (1986 г.) три
отделения будут спорить за право ввести его в свой состав, а его даже не упомянули.
301
К его пятидесятилетию мне удалось напечатать две статьи о нем^. В день
юбилея хотел сообщить ему эту новость в виде подарка, но она так и осталась
при мне - Рита отказалась позвать его к телефону (“ты же знаешь, Сергей не
терпит юбилеев”). Позже, весной, при встрече где-то в коридоре, на мой вопрос,
читал ли он про себя, он молча наклонил голову и приставил каблук к каблуку -
как бы благодаря.
С апреля 1986 года мы с Сережей не виделись, и лишь в начале ноября мне
рассказали, что у него давний рак, что сейчас пошли метастазы и он едва ходит
на костылях. В тот же вечер, когда он вернулся с работы (как оказалось, в по-
следний раз), я приехал к нему на новую квартиру. Держался он (как и Рита) ве-
ликолепно, и вот начался наш последний срок общения.
Статусные темы потеряли смысл, их и не было (хотя он весною возглавил
лабораторию, в которой работал всю жизнь), почти не было и медицинских. Он
знал о безнадежности своего состояния и порой прямо говорил об этом («Ах,
Юрка, если б я знал, что мне жить год, разве стал бы я учить испанский?»), а по-
рой не хотел верить («У меня три вида на будущее: не встать, встать на костыли
и выздороветь полностью»).
То мы говорили про науку (теперь он с жадностью обсуждал эволюцию,
прежде им почти забытую ради чистой палеоботаники), то про начавшуюся то-
гда “перестройку” (в которую мы оба простодушно верили), то его скрючивала
боль, и иногда он (при всей-то его воле) начинал кричать. Анальгетики больше
не действовали, надо было колоть морфий, но Рита каждый раз медлила: и ам-
пулы в больнице давали скупо, и боялась она сделать его морфинистом (значит,
тоже надеялась на выздоровление - до, примерно, середины марта). Иногда об-
ходилось, и мы снова начинали разговор. Про болезнь он говорил мало (напри-
мер, с полуулыбкой: «как страшно, оказывается, услыхать слово “метастазы”,
когда это относится к тебе самому»).
Понемногу стало ясно, что обещания врачей не оправдались, пришлось ис-
кать знахарей. Кто-то (кажется, Рита) дал мне телефон питерского целителя, не
избегающего безнадежных раковых больных; он назначил мне встречу на 25 ян-
варя и, в питерской квартире, продиктовал какой-то сложный рецепт. В тот же
день я вернулся в Москву и увидал, что Серёжа ждет с большой надеждой. Пока
его родные покупали компоненты снадобья, мы беседовали про науку, и он
оживал еще до начала курса. По-моему, это была психотерапия^^ плюс обыч-
ное отступание болей при начале конца, но Серёжа решил, что выздоравливает.
Так или иначе, но 6 марта он именно в этом духе надписал мне экземпляр толь-
ко что вышедших «Основ палеоботаники». Надписал, сидя в постели, но позже,
насколько помню, уже не подымался. В последние встречи он еле шептал (“ше-
“Знание - сила”, 1985, № 12; “Вестник Академии наук”, 1986, № 3 (4-13; 4-14).
109 Беседы с философом Региной Карпинской он так и называл: “регинотерапия”.
302
лестел” - говорила Рита), и собеседникам приходилось опускаться перед его из-
головьем на колени. Первой, помнится, это догадалась сделать Р.С. Карпинская,
вскоре тоже умершая от рака и тоже державшаяся изумительно.
Сейчас, через 18 лет, приходится вновь и вновь спрашивать себя: можно ли
было иначе? Вылечить Серёжу - вряд ли. А вот отношения можно было выстро-
ить и по-другому. Не мне судить, был ли он прав, забросив туманную натурфи-
лософию в пользу конкретной науки: с одной стороны, этим путем он достиг
максимума возможного, а с другой - этого на сегодня, вероятно, достигли бы и
без него, а вот его “типологию” вряд ли кто достроит. Зато мне стоило бы по-
нять, что он видит дальше нас и не тянуть его на свою сторону.
Не успокоил он слишком трусливого редактора? Но ведь Мейен мог лишь
обезвредить свою неудачную фразу, а это вряд ли изменило бы судьбу книги. Да и
ей отказ пошел только на пользу: через 5 лет на ее основе была написана более
серьезная книга “Элементы эволюционной диатропики”.
Не вступился за библиотеку? Тут сложнее: тогда, на первой волне “пере-
стройки”, помещение получить было можно. Вероятно, Мейен прикинул, что в
Президиуме МОИП, где все любят только разговоры, добиваться придется ему,
а на это уже места не было на шагреневой коже его бюджета времени. Да и ясно
нам, еще живым, что в нашу эпоху не только бы отняли добавочное помещение,
но и саму библиотеку могли выбросить на улицу, как выбросили много чудес-
ных библиотек, музеев, школ и прочего. Эпоху Мейен предвидеть не мог, зато
вполне мог, зная нравы лучше, чем я, предвидеть месть обиженных*.
Хотел стать член-кором? Но ведь, доживи он до сего дня, заведомо был бы
академиком. Куда ни кинь, он всюду оказался прав.
Да и не в помещении, как теперь видно, была и осталась главная беда, а в отсутствии
интереса к старым книгам со стороны высокого начальства. - Примеч. 2010 г.
303
Некролог*
Сергей Викторович Мейен (17. XII. 1935 - 30. III. 1987)
С ним я имел счастье дружить. С того счастливого летнего дня 1971 г., ко-
гда я увидел на книжной обложке** его фамилию, дотоле мне неизвестную, и до
того страшного своей безнадежностью мартовского вечера нынешнего года, ко-
гда он в знак прощания шевельнул мне левой рукой (правая давно не двигалась),
- все эти годы я, как теперь понимаю, жил с потаенной мыслью: есть Сережа,
можно жить и что-то делать. И вот его нет.
Его жизнь потребует книги (не скучной «научной биографии», а книги), его
труды по биологии и геологии будут отмечены в некрологах в других журналах,
здесь же хочется вспомнить об ином. Прав был Н.Н. Воронцов, сказавши у гро-
ба, что роль Мейена - не только в естествознании или даже науке в целом, а во
всей нашей культуре, и что ясно это станет, вероятно, лишь через годы. Другие,
каждый на свой лад, досказали ту же мысль точно: мы потеряли гения. Это гово-
рилось и в рядах собравшихся, потом - на кладбище, назавтра - на Любищевских
чтениях в Институте философии. Так оно, я уверен, и есть на самом деле.
Многие удивятся: не через край ли хвачено? Почему же мы его не знали:
Почему он даже не претендовал на академическое кресло?
Да именно потому, что на него теперь надо претендовать, надо пробиваться.
Блестящий администратор, Мейен хорошо знал, как это делается, и не раз опи-
сывал унизительные подробности прохождения в академию. Как-то раз я даже
не выдержал: «Зачем ты помогаешь ему, заурядному, стать выше тебя в иерар-
хии?» Мейен спокойно разъяснил: ставши член-корреспондентом, заурядный
его приятель может помочь одному очень нуждающемся в помощи институту.
Нет, он не был равнодушен к регалиям, и с годами зависимость от их обла-
дателей стала его тяготить. Если в 70-е годы он мог заставить себя весело по-
смеяться над тем, как его пригласили в Париж или Лондон, а поехал другой, с
регалиями, то в 80-е юмора на это в себе не находил. Сил становилось все
меньше, а потребность видеть образцы ископаемых растений своими глазами -
все больше. Правда, теперь уже ни один академик не решался ступить куда-то
вместо Мейена, зато комом налипали бытовые трудности - в издательстве,
больнице, жилищном управлении, да и у себя в институте то и дело приходи-
лось доказывать что-то очевидное, доказывать потому, что не было в руках
единственного аргумента, убедительного для чиновников. И все же он подумать
не мог о том, чтобы самому себя продвигать.
ВИЕТ, 1987, № 3. Перепечатано в М-353. Это и следующие примечания - 2010 года.
Из истории растительных династий (М-84).
304
Он был более чем академик - был лучшим в своей науке. Зачем ему было
украшать себя членством в ученых обществах, если серьезные общества давно
почитали за честь украсить себя членством Мейена? В 40 лет он стал вице-
президентом Международной организации палеоботаников и был самым реаль-
ным кандидатом в ее президенты на ближайших выборах. Но какое это имеет
значение в сравнении с тем, что другого подобного палеоботаника попросту
нет? Дело не в том, что он поразительно знал весь свой фактический материал, и
даже не в том, что как бы между делом совершал открытия (например, указал
происхождение цветковых*, над которым бились более 100 лет), а в том, что дал
палеоботанике новый статус. Его концепция глобального флорогенеза - первая
эволюционная концепция, основанная на палеоботаническом материале (а не
только иллюстрированная им, как было прежде), причем этот материал в прин-
ципе не мог бы быть заменен палеозоологическим.
Такие результаты в каждой науке наперечет, и история склонна их авторов
объявлять основателями, до которых в этих науках якобы лишь копился факти-
ческий материал. Так Линней попал в основатели систематики, Кювье - палео-
нтологии, Бэр - эмбриологии, Дарвин - эволюционизма и т.д. Часто в этом ви-
новны сами «основатели», замалчивавшие или занижавшие роль своих предше-
ственников, поэтому спешу заявить: мало кто из ученых был так внимателен к
истории и так заботлив к ушедшим авторам, как Мейен, и если после «Основ
палеоботаники» потускнеет вся «домейенская» палеоботаника, то виноват будет
не он, а те, кто не захотят изучить другие труды Мейена, со всеми их историче-
скими экскурсами**.
Для нас, историков и методологов науки, Мейен был неким камертоном. В
отличие от других натуралистов он и не пренебрегал нами, и в то же время не
пытался повсюду искать выявляемые нами схемы, а указывал на их ограничен-
ность. Он любил говорить: «Подлинная задача методологов не в том, чтобы ука-
зать ученому верный путь исследования, а в том, чтобы объяснить ему, что же
он в действительности делает». Историко-философские поиски были ему столь
же необходимы, как анализ коллекций ископаемых. Без этих поисков не было
бы и схемы глобального флорогенеза. Натуралист Мейен немыслим без истори-
ка и методолога, точно так же, как был он немыслим без своей жены Маргариты
Алексеевны.
А.А. Любищев и Мейен в равной мере привлекли внимание натуралистов к
миру историко-методологических идей, открыли тогда, когда подавляющее
большинство их все еще наслаждалось самым примитивным презентизмом и
* Он сам сказал мне незадолго до смерти, что эту задачу решил. Однако в печати назвал
это не решением, а лишь гипотезой, что верно. Подробнее см. заключительную статью.
** Надеюсь, читатели простят мне преувеличения - ведь это начато до похорон и законче-
но сразу после них (поминок не было - измученные родственники не были на них спо-
собны, да и само это языческое действо было бы, по-моему, здесь неуместно.)
305
кумулятивизмом, то есть считало, что нынешняя наука - венец познания, во-
бравший в себя все нужное из прошлого. (Хотя большинство продолжает так
считать неявным образом и сейчас, однако я думаю, что всякий, кто вниматель-
но прочел хоть что-то из теоретических трудов этих двух классиков, понимает
роль старых, долгое время забытых или отвергаемых идей и неприятных фак-
тов.) Им обоим многие обязаны осознанием того, что новейшие концепции -
лишь звенья от науки прошлого к науке будущего. Именно беседы с Мейеном
впервые обратили меня к той мысли (о которой прочесть тогда было почти не-
чего), что путь к построению работоспособной теории биологической эволюции
лежит через историко-методологический анализ.
«Удел величия - одиночество», - так сказал о Канте В. Виндельбанд, один
из любимых философов Сергея Викторовича. Лишь время рассудит, кому суж-
дено попасть в великие, но скажу уверенно: у Сергея Викторовича Мейена были
все основания быть одиноким; но он сумел им не быть. В сущности ему не с кем
было обсудить свой взгляд на мир в целом, и все же он постоянно обсуждал раз-
ные его аспекты со всеми, кто мог понять хоть что-то. Его любимым делом было
проводить будни за образцами, а выходные и отпуск - за письменным столом, и
все. же он постоянно был окружен людьми со всего мира, активно включался во
все комиссии, коллегии и комитеты, в которых мог усмотреть хоть какую-то
пользу делу.
С удовольствием обсуждал он со мною механизмы и закономерности эволю-
ции, но я чувствовал себя не вполне полноценным собеседником, когда он начи-
нал обосновывать свои взгляды геологическим и ботаническим материалом, и уж
вовсе ощущал себя лишним, когда он пускался в палеоботанические рассуждения.
Не многим полезнее был я ему в философских вопросах, а ведь он и с философа-
ми беседовал едва ли не так же свободно, как с палеоботаниками. И в геологии он
был не только ведущим стратиграфом, но и собственно эволюционистом - один
из первых у нас развивал идеи мобилизма (глобальной тектоники плит), а в по-
следнее время один из первых в мире выражал уверенность, что классический мо-
билизм (который он считал лучшей из геологических теорий) должен уступить в
будущем место концепции расширяющейся Земли В.Н. Ларина.
Разумеется, главным делом для него в последние годы были его «Основы
палеоботаники» (первые экземпляры усеченного русского издания он успел
надписать, а экземпляр полного английского издания почтальон доставил ему
домой за час до смерти, доставил спешно, минуя сроки доставки, но автор уже
никак не дал знать, понял ли, что ему показывают), а в остальном его, кажется,
более всего волновали вопросы этики.
Они, эти вопросы, вышли теперь у него на первый план как в отношении к
семье и друзьям, так и в отношении к науке, природе и обществу. Начав со ста-
тьи «Научиться понимать» (Неделя. 1976. № 2)’, где впервые сформулировал
* М-133. Переиздано: М-353.
306
свой «принцип сочувствия»), теперь уже широко известный*, он понемногу раз-
вил целое учение о неразделимости познавательных и этических процессов; а
закончил статьей о проблемах перестройки в науке (ее он слабеющим голосом
произнес в диктофон), где показывал, к каким бедам приводят установившиеся в
Академии наук порядки**, и обсуждал приемлемые меры их преодоления. При-
веду пример его этической позиции.
Принесли недавнюю переписку писателей*** о состоянии нашей культуры,
где один «истинно русский» писатель мыслит «национальное возрождение рус-
ского народа» в полном отстранении «инородцев». Я спросил Мейена: как отде-
лить здесь обычный бесплодный шовинизм от естественного желания сохранить
национальное своеобразие, от неприятия безнациональных тенденций новейшей
культуры? И он ответил: индикатором должно служить отношение не к своей, а
к чуждой культуре - если «патриот» хоть чем-то принижает чужое, значит он
ратует не за своеобразие, не за разнообразие культур, а за свое господство, зна-
чит «возрождение» он видит в подавлении.
Уходил он из жизни героически (и едва ли не столько же героизма проявили
при этом его близкие). Как-то в январе, убедившись, что дикая боль не дает ему
повернуться на бок, он слабо улыбнулся: «Я стал подобен онтогенезу - каждая
новая стадия закрывает прежние степени свободы». Но работал еще 2 месяца. И
у нас, оставшихся, нет другой степени свободы, кроме как пытаться осознать,
что же он, собственно, для нас сделал.
* М-145. Переиздано: М-338; М-353.
** Академическая наука?.. (М-322). В сокращенном виде опубликовано в М-326 и МСП, с.
265-280.
Переписка Н.Я. Эйдельмана и В.П. Астафьева. Ходила, несмотря на уже набиравшую
силу «эпоху гласности», по рукам в машинописи (в «Самиздате»),
307
Часть 9. Судьба концепции
Диатропика С.В. Мейена: сегодняшний взгляд'
Сергей Викторович Мейен, высказал идею учения о разнообразии, исхо-
дившую из той давней посылки, что у формы есть собственные законы, не сво-
димые ни к функции, ни к материалу. Идея близка структурализму. Мейен не
дал своему детищу названия - лишь заметил, что «можно воспользоваться ста-
рым термином “типология”» [3, 497]. При этом он сослался на свою (с соавто-
рами) работу, где типология понята узко: «систематика, точнее, типология сла-
гается... из двух компонентов - таксономии (изучающей отношения между так-
сонами, причем не обязательно иерархическими) и мерономии (изучающей от-
ношения между архетипами и между меронами одного архетипа110)» [8, с. 120].
Получалось, что он предложил всего лишь новый способ строить систематику,
тогда как на самом деле речь у него шла о новом взгляде на всю биологию: «Я
думаю, что это - единственный рабочий способ выявления и исследования
уровней системности» [5, с. 6]. В этом несоответствии содержания концепции ее
названию мне видится первый просчёт ее автора.
На собрании в память Мейена (в Институте истории естествознания и тех-
ники АН СССР, дек. 1987) я сделал доклад “О теории разнообразия Мейена”,
после которого ко мне подошел методолог Николай Фёдорович Овчинников,
друживший с покойным, и сказал: вас никто не услышит, пока вы не дадите из-
лагаемому учению простое ясное название. Он пояснил: слово “типология” ни-
куда не годится, поскольку уже имеет много смыслов и тем запутывает слу-
шающих, а пара слов “теория разнообразия” ничего не говорит.
Овчинников полностью убедил меня, я сразу же взял в библиотеке грече-
ские и латинские словари, и к вечеру был готов термин диатропика - наука о
разнообразии (от греч. диатропос - разнообразный, разнохарактерный).
Термин ныне довольно широко употребляется гуманитариями (сотни ссылок
в Интернете), что же касается коллег Мейена, то почти все игнорируют как тер-
мин, так и саму концепцию. Кроме собственных работ, могу назвать всего одну
подборку материалов, где слово “диатропика” стоит в заголовке [2], и две работы,
где диатропика применена по сути: их авторы - зоологи Виктор Яковлевич Пав-
лов и Сергей Петрович Пчеленко. Да еще зоолог Кирилл Глебович Михайлов 10
лет назад обсуждал одну мысль из диатропики, именуя последнюю типологией.
Об этих трех зоологах скажу далее, пока же должен заметить, что круг идей
диатропики обсуждается, в том числе и в кругу коллег Мейена (хотя сам он при
этом не упоминается), однако с грустью добавить, что сдвига в понимании по-
ставленных Мейеном проблем не видно вовсе (кроме работ [10; 12; 15], прямо
* ВФ, 2006, № 5, с. 95-102. В основе доклад на Чтениях памяти Мейена в ГИН 21.12 2005.
1 *0 О структурализме см выше, с. 176, а об отношении мерона и архетипа см. [9].
308
опирающихся на работы Мейена). Последнее, на мой взгляд, есть прямое след-
ствие отказа видеть диатропику вообще и ее отличие от типологии в частности.
Первой темой будущей теории был у него феномен параллелизма, который
он хотел не просто описать, как делали до него, но и упорядочить. Даже можно
сказать - дать некое исчисление параллелизмов. Исчисления дано не было, а сама
идея осталась почти никем не понятой - прежде всего, из-за отсутствия содер-
жательных примеров. Это отсутствие - второй просчёт Мейена.
Единственный разобранный им самим пример (сходства в типах расчлене-
ния листовой пластинки у самых разных таксонов растений) интересен только
узкому кругу морфологов растений, но и у них, видимо, создает впечатление не-
кой частности, не имеющей общенаучного значения. На самом деле отмеченный
им феномен буквально пронизывает всю биологию и выходит далеко за ее пре-
делы, причем касается не только деталей, но и самой сущности явлений. Для его
описания Мейен ввел понятие рефрена.
1. Связь диатропики с типологией. Рефрен
Он сетовал, что никто идею рефрена не понимает [5], но на типах расчлене-
ния листа ее, по-моему, и нельзя понять. Перед мысленным взором Мейена
стояло несколько ярких рефренов (например: “совершенно одинаковые структу-
ры появляются у разных групп археоциат в разное время. А последовательность
появления одинаковая” [7, с. 282]), но все столь специальные примеры что-то
говорят только знатоку.
Мне пришлось начать анализ диатропики с построения всем ясного сущест-
венного примера рефрена. Им стал фрагмент рефрена “парные конечности по-
звоночных” [17, с. 52; 18, с. 240]. Он являет собой прямоугольную таблицу, в
ней 5 строк (для пяти основных классов - рыбы, амфибии, рептилии, птицы и
звери) и 6 столбцов. Каждая строка изображает изменение конечности от про-
стого плавника до сложной конструкции (крыло, рука и т.п.).
Легко видеть, что преобразование конечности шло во всех классах сходно.
Например, крыло, обеспечивающее активный полет, есть не только у птиц и ле-
тучих мышей, но и у тропической рыбки клинобрюшки. Было оно и у летающих
ящеров. Данный рефрен нельзя целиком объяснить ни унаследованием от обще-
го предка (он, если и был, не имел конечностей), ни приспособлением (в клетке
“имеется только слаборазвитая передняя пара” есть и совершенно негодные к
использованию крохотные “ручонки”).
Мейен дал, насколько знаю, три определения рефрена:
1) рефрен - это “повторное полиморфное множество”, причем определение
данного множества заняло журнальную страницу [21, 1973, с. 209]. Столь слож-
ную конструкцию довести до понимания читателей не удалось, и
2) чтобы дать сжатое определение, Мейену пришлось ввести понятие мерона
(о нем пойдет речь далее): “повторяющуюся, подчиненную одному правилу пре-
образования последовательность состояний мерона назовем рефреном” [3, с. 501];
309
3) “Рефрен - это такое множество объектов, принадлежащих разным таксо-
нам, которые могут быть сделаны неотличимыми посредством одного и того же
преобразования” [5, с. 5].
Последнее определение, самое красивое, Мейен не предал гласности, оста-
вив в рукописи; он понял, видимо, что таких рефренов вне физики нет. Нам ос-
тается использовать второе определение рефрена, через мероны.
Мерой - это класс частей. Если организм имеет части тела, свойтва и при-
знаки, то таксон имеет мероны. Например, организм позвоночного может иметь
плавник, ласт, лапу, крыло или руку, а таксон “позвоночные” имеет мерой “ко-
нечность”. Тем самым, Мейен предложил с помощью меронов классифициро-
вать части организмов точно так же, как систематика с помощью таксонов клас-
сифицирует организмы (данную идею высказал Н.Н. Страхов, зоолог и фило-
соф-почвенник, в 1858 г.).
Другими словами, классификацию самих организмов предложено строить
на базе классификации частей организмов, тогда как сами эти части невозмож-
но классифицировать без какой-то системы организмов; а она сама должна быть
перестроена после выявления меронов. Построение систематики оказывается
последовательной (итеративной) процедурой, и Мейен высказывал уверенность
(ничем не подтвердив ее), что такая процедура будет сходящейся, т.е. приведет к
устойчивой (долговременной) систематике.
Как видим, мерой здесь введен Мейеном не для анализа рефренов, а для целей
систематики, т.е. в этом пункте диатропика в самом деле есть часть типологии. Но
как раз в этом пункте никакого продвижения не последовало. Хотя сам Мейен
вскоре построил свою систему голосеменных растений, но предложенной им ите-
ративной процедуры не провел. Это наводит на мысль, что никакой надежды на
успех нового метода у него не было или же она иссякла по ходу работы. И в са-
мом деле: мы знаем массу примеров, когда с пересмотром системы меронов так-
сономия веками ходит по кругу (см. пример с семейством псовых: [17, с. 45]).
Есть ли тогда смысл в типологической части диатропики вообще? По всей
видимости, на сегодня он ограничен тезисом К.Г. Михайлова: «Главное, что да-
ет типология систематику - это снятие комплекса собственной неполноценно-
сти» [10, с. 95]. Точнее, смысл введения ее принципов (например, постулата об
устойчивости итеративно построенной систематики) видится мне примерно та-
ким же, каков он у основных принципов некоторых иных наук, т.е. как деклара-
ция мировоззрения - не более, но и не менее.
Таковы: принцип рыночной конкуренции, второй принцип термодинамики,
принцип естественного отбора и т.п. Они не имеют обоснования с позиции
обычных критериев науки* (графики, таблицы и формулы, демонстрирующие
Перечень примеров неудачен: отбор не имеет обоснования вообще, тогда как для ос-
тальных принципов лишь не указаны границы применимости. Но для всех трех верно то,
что их неадэкватное применение приводит к огромным потерям. - Примеч. 2010 г.
310
область применимости и точность действия принципа), но оказались достаточны
для формирования защитного пояса теории (термин И. Лакатоса)*, дающего
теории неуязвимость для критики со стороны иных теорий и практики.
А.С. Раутиан [16] рассмотрел типологию Мейена, опустив рефрены (т.е. всю
направленность преобразований), и пришел к тривиальному выводу, что она есть
сравнительный метод. В остальном диатропика от типологии резко отлична.
2. Состав диатропики и ее практический смысл
Ряды сходств, не связанные ни с родством, ни с приспособлением, были из-
вестны давно, еще до Дарвина, причем сам Дарвин признавал их, но пасовал пе-
ред этим феноменом и в своих построениях фактически игнорировал. Все по-
пытки построить какую бы то ни было теорию рядов (Эдвард Коп, Н.И. Вавилов
и др.) оказались тщетны - в том смысле, что не вошли в состав науки, несмотря
на громкие имена их авторов. Мейен решил исправить дело, введя вместо рядов
в рассмотрение рефрены.
Рефрен в рассматриваемом далее смысле - это ряд направленных рядов**.
Например, парные конечности позвоночных образуют в каждом из пяти основ-
ных классов позвоночных направленный ряд (от плавника к более сложным
формам), а вся пятёрка рядов образует рефрен. Он грубо изображается, как ска-
зано выше, таблицей, каждая строка которой - класс животных, а каждый стол-
бец - состояние мерона (плавник, ласт, лапа и т.д.). Для более точного выраже-
ния рефрена следует положить таблицу горизонтально и над каждой ее клеткой
построить столбец, отражающий степень выраженности свойства.
В целом каждый ряд таблицы, как горизонтальный, так и вертикальный, де-
монстрирует и прогресс (усложнение форм), и приспособление (каждая форма,
выражаемая столбцом таблицы, для чего-то полезна), однако для каждой от-
дельной позиции (клетки таблицы) этого не скажешь (зачем клинобрюшке ле-
тать? Ответы дать можно, и различные, но они будут произвольными, непрове-
ряемыми и не для всех биологов осмысленными). Сам же феномен рефрена на-
водит на мысль, что теорию разнообразия вовсе не надо искать в виде объясне-
ния свойств отдельных клеток или строк, что ее надо строить как единую.
О ней впервые немного сказал Карл Линней в “Философии ботаники”, но не
развил ее. Речь шла о двух способах упорядочивания организмов, которые он
назвал искусственной и естественной системами. Первая предназначена для
удобной адресации, а вторая - для понимания “сути вещей”. Последнюю он ни-
как не изобразил, но предлагал изображать в форме географической карты. По-
сле Линнея никто не вёл речи о необходимости двух систем - наоборот, все хо-
тели строить естественную систему, надеясь, что она сгодится и для адресации.
По-видимому, эта задача невыполнима, и мне это стало ясно как раз из диа-
тропики Мейена. Он писал: «Эмпирически установленный факт наличия и уни-
0 защитном поясе теории см. выше, с. 195. - Примеч. 2010 г.
Данное упрощение делает понятие рефрена практически применимым. - Примеч. 2010 г.
311
версальности рефренов делает обязательной большую или меньшую комбина-
тивность естественной системы» [3, с. 503]. Но ведь адресация не может стро-
иться на “большей или меньшей степени”, она должна быть чёткой и однознач-
ной. Таким свойством обладает принцип иерархии (последовательного подчи-
нения) таксонов, и не видно никаких оснований от него отказываться. А это зна-
чит, что удобная для адресации система не может быть дана естественной
системой организмов.
Чтобы прекратить вековую путаницу, здесь тоже надо ввести названия. Бу-
дем называть систему адресов видов адресацией, а естественную систему этих
видов будем, следуя Линнею, искать в форме карты. Адресация имеет форму
древа, а карта - форму плоского рисунка; его элементы обозначим словами, взя-
тыми из адресации. (Адресацию не следует путать с номенклатурой - так име-
нуется биологическая дисциплина, призванная давать названия видам и иным
таксонам [1]. К сожалению, обычно она следует за той систематикой, которая в
данный момент господствует.)
Если принять это, задача систематики радикально упростится: она сведётся
к построению удобной адресации, построенной на определительных ключах, ко-
торые весьма долговечны, а не на переменчивых суждениях о родстве. По сути
именно так поступают микробиологи [11], и мне представляется, что только не-
знание идей чужого круга мешает зоологам и ботаникам сделать то же самое.
Адресация может быть устойчивой, если будет полностью изолирована от спо-
ров о родстве и приоритетах, если изменения в ней будут допустимы только при
обнаружении г омонимии (если одним словом названы различные объекты) или
ошибки в ключе.
Наоборот, карта зависит от того, как понимать “суть вещей”, а потому не
может быть ни однозначной, ни вечной. Это отнюдь не значит, что она должна
меняться столь же часто, как системы меняются ныне - нет, если “суть вещей”
действительно найдена, то карта окажется устойчивой по Уэвеллу, т.е. не ме-
няющейся при появлении новых сведений. (Примеч. 2010 г.', см. [4-61, п. 7]).
Пример карты (точнее, система четырех царств живой природы) приведен
мною ранее [17; 18]. Это - кольцевая таблица основных рефренов живого, моя
попытка реализовать упомянутую выше идею Линнея. За 20 лет менять ее не
пришлось: новые сведения до сих пор лишь детализировали ее. Тем самым, она
оказалась устойчивой по Уэвеллу. (Наоборот, все известные мне системы
царств, построенные на идеях нынешней систематики, уже через 5 лет не упо-
минаются никем, даже их авторами.) Это - конкретный результат диатропики.
Другой результат диатропики являет собой карта, которую ее автор, В.Я.
Павлов, назвал: “Периодическая система членистых”. Это - прямоугольная таб-
лица основных рефренов членистых, в которой названия таксонов выступают
как данные, т.е. их можно рассматривать как заданные адресацией. Эволюция
предстает как заполнение этой таблицы [12, с. 173]. Этот замечательный резуль-
тат был замечен биологами, но остался без использования, и всё испортило, на
312
мой взгляд, как и у Мейена, неверное заглавие: на самом деле Павлов дал вовсе
не систему, а дополнительное к ней описание (карту в данном выше смысле),
дающее как абрис разнообразия, так и картину эволюции.
Третий результат диатропики - таблица рефрена форм маток коров, кото-
рую составил С.П. Пчеленко [15]. Она оказалась полезной для понимания про-
блем ветеринарного акушерства, от меня далеких.
3. Диатропика и эволюция. Филогения по Мейену
Из сказанного ясно, что диатропика нужна не столько для целей системати-
ки, сколько для работы с параллелизмами, для понимания хода эволюции. Диа-
тропика видит эволюцию как заполнение объективно существующей таблицы
(карты), которая подобна идее Платона и в которой близкие клетки (обладаю-
щие общим архетипом111) составляют общий таксон. Диатропика понимает
карту как реализованную часть множества возможностей данной группы орга-
низмов. Пути заполнения клеток не заданы жестко, хотя чаще заполняются
клетки, соседние с уже занятыми, что и создает видимость дивергенции видов
(расхождения вида на близкие виды).
Дивергенция была постулирована Ч. Дарвином в качестве единственного
механизма происхождения видов, а последующий дарвинизм сделал ее основ-
ным механизмом эволюции вообще: близкие роды мыслятся происшедшими из
общего рода, близкие семейства - из общего семейства и т.д.
Череду актов дивергенции именуют филогенией. При филогении каждому
виду можно указать единственный вид- предок, роду - род-предок и т.д. Обще-
принято выделять какие-то признаки вида А, именовать их ведущими, т.е. гово-
рящими о родстве с видами В и С, а все признаки, этого не говорящие, полагать
возникшими заново. Наоборот, Мейен предлагал прямо следить за эволюцией
меронов, т.е. за изменением каждого интересующего нас мерона при мысленном
движении вдоль рефрена. Такую эволюцию (семофилогению) можно наблюдать
всегда, а про филогению от таксона А к таксону В можно говорить только тогда,
когда все наблюдаемые семофилогении сходятся, т.е. ведут от Л к В.
Но этого никто еще не наблюдал. Естествен вопрос: как связана эволюция в
этом понимании с эволюцией, понимаемой как заполнение клеток таблицы?
В последней диатропической работе «Нетривиальные модусы морфологиче-
ской эволюции высших растений»222 Мейен писал: “понятия дивергенции, кон-
вергенции и параллелизма - мерономические (морфологические, физиологиче-
ские или экологические), а не таксономические” (с. 8). Иными словами, если мы
говорим, что вид А породил виды В и С, то всегда имеем в виду какие-то при-
знаки, за которыми следим, а не виды в целом.
Ш По Мейену, «Архетип - это структура частей и внешних связей (те и другие явля-
ются меронами), присущая всем объектам данного таксона» [9, с. 72]. На мой же взгляд,
это понятие по сути интуитивно.
1 12 Машинопись, 1986, 20 с. Третий экз. хранится у меня. (Примеч. 2010 г.: см. М-313).
313
Сам Мейен в 1984-1986 гг. писал и говорил, что построил филогению голо-
семенных, однако это была филогения в особом смысле: описаны только пере-
ходы одних порядков (порядок - таксон ботаники, равный отряду в зоологии) в
другие порядки и одних классов в другие классы. Относительно же низших так-
сонов этого сделать не удалось: о главном порядке Pinales (Сосновые) прямо
сказано: “порядок Pinales разделяется на 13 семейств. Их филогенетические от-
ношения практически неизвестны” [6, с. 94]. Не удалось указать ни одного пере-
хода рода в род (и, тем более, вида в вид), поскольку семофилогении вели от ро-
дов материнского семейства к родам дочернего без всякого порядка: одни семо-
филогении указывали на родство данных родов, другие же семофилогении это
родство опровергали.
Т.е. множество меронов от порядка А к порядку В наследовалось (а это - фи-
логения), но от одного семейства к иному семейству множество меронов, как пра-
вило, не наследовалось. Оказалось, что рефрен порядка (как и рефрен класса) на-
следуется целиком, но не по семействам. Мейен назвал этот феномен (сохранение
рефрена как целого в историческом времени) транзитивным полиморфизмом*.
Транзитивный полиморфизм всюду, где он имеет место, исключает возмож-
ность филогении в обычном смысле (от вида к виду, от рода к роду и т.д.). Вме-
сто нее следует говорить об эволюции как движении вдоль рефрена, которое
лишь на двух уровнях (порядок и класс) представимо в форме филогении77’.
Это тоже - диатропический результат.
Данное ограниченное сходство с филогенией следовало назвать новым тер-
мином, а не филогенией, но Мейен не сделал этого, в чем его третий просчёт.
Из-за этого сам результат остался неведом научному сообществу: сторонники
филогений сочли, что Мейен под конец жизни осознал их правоту (хотя его по-
смертные публикации прямо опровергают данную трактовку), а противники ли-
бо отрицают обоснованность системы Мейена, либо ссылаются на необъяснён-
ность самого явления рефрена (о которой скажу далее).
4. Диатропика и эволюция. Рефрены и блоки
Итак, эволюцию, вообще говоря, нельзя представить в форме филогении.
Зато она хорошо выражается через рефрены. То и дело публикуются новые реф-
рены, подчас весьма яркие, но авторам в головы не приходит, что это - рефрены.
В качестве примера приведу серию работ палеонтолога А.Г. Пономаренко, ко-
торую он никак не назвал. Он перечисляет: эвкариотизация, метазоизация, арт-
В книге АСМпредложено короткое его обозначение: транпол. - Примеч. 2010 г.
1 13
В том же смысле Мейен высказался устно [2, с. 133]. Поэтому его часто упоминаемую
фразу: “в филогении высших растений, если говорить о происхождении семейств и таксо-
нов более высокого ранга до отдела включительно, безраздельно господствует монофилия”
[4, с. 50] - следует считать опиской и читать: “о происхождении таксонов рангом выше се-
мейства”. Что касается группировки самих отделов (то же, что в зоологии типы) в рамках
царства растений, то это тоже удается только посредством рефренов [17, гл. 10].
314
роподизация, тетраподизация и т.д. протекали сходно. Я пока обозначу данный
феномен как «зация»*. У всех этих процессов Пономаренко видит общую суть:
а) процесс не только таксономический, а всебиосферный;
б) он начинается созданием подходящей для зации экосистемы и завершает-
ся осуществлением всех основных экологических потенций данной зации-,
в) может возникать независимо и неоднократно;
г) идёт самоускоряясь [14, с. 8].
В силу (в) всякая зация - рефрен, но важнее, что все вместе они образуют
рефрен высшего порядка (рефрен рефренов).
Тот феномен, на который указывает Пономаренко, является частным случа-
ем более общего, известного в настоящее время как блочность эволюции и ши-
роко обсуждаемого (литературу см. [18]). Например, первая амфибия ихтиосте-
га заслужила шутливое прозвище “рыба на ногах”, поскольку сочетала четыре
ноги с почти типичным рыбьим туловищем. Термин “блочность эволюции” во-
шел в употребление после смерти Мейена, однако у него можно найти весьма
глубокие мысли на сей счет. В упомянутой рукописи Мейен писал:
«считается, что лепестки имеют двойственное происхождение - от чашелисти-
ков (когда наблюдается постепенный переход от чашелистиков к лепесткам,
как у некоторых сложноцветных) или от тычинок (когда постепенный переход
связывает лепестки и тычинки, как у некоторых нимфейных). Если учесть вто-
114
ричность описываемого явления , то мы лишаемся возможности судить о
происхождении органов и об их гомологичности по наличию переходов между
ними... Важно то, что эти гомологизации не подкрепляются и ископаемым ма-
териалом. Допуская происхождение лепестков будь то от тычинок или чашели-
стиков, мы должны ожидать широкое развитие переходных органов у древ-
нейших покрытосеменных, особенно меловых. Между тем у всех наиболее
полно изученных меловых цветков никаких переходных форм между тычинка-
ми и членами околоцветника не наблюдается...
Возникновение вторичных переходов между органами, очевидно, связано
с перестройкой онтогенетических программ органов таким образом, что ра-
нее дискретные программы частично перекрываются... Этому явлению срод-
ни другое, при котором в каком-то органе потомка совмещаются онтогенети-
ческие программы разных органов предка, в результате чего получается су-
щественно новый орган (“гибридный” - Sachs, 1982)» (с. 11-12).
Через 2 года термин был легализован - в поздравлении Александру Георгиевичу читаем:
«Взяв за образчик маммализацию териодонтов, он проповедует артроподизацию, провоз-
глашает ангиоспермизацию и придает концепции “-зации” почти философское звучание»
(ПЖ, 2008, № 1, с. 106). Слово «почти» здесь, по-моему, излишне. - Примеч. 2010 г.
114 Мейен имел в виду, что переходы есть у нынешних видов, но отсутствуют у древних
представителей данных семейств цветковых, а потому не говорят о путях эволюции.
315
Мейен имел в виду статью [22], где была высказана важная для него мысль:
«возникшие в результате переноса органы занимают в системе морфологиче-
ских понятий то же место, что и гибриды [занимают] в таксономической систе-
ме». Речь фактически шла о блочной эволюции.
Далее Мейен привел палеоботанические примеры гибридных органов, по-
нятные лишь знатокам морфологии растений; вместо них могу назвать в качест-
ве типичного примера митохондрию - внутриклеточную энергетическую “фаб-
рику”. Она внешне похожа на бактерию и имеет свой небольшой геном. Для
объяснения этого сходства была предложена “теория”, по которой митохондрия
в самом деле является потомком бактерии, потерявшей самостоятельность.
Объясняя сходства, эта “теория” не может объяснить различий (например:
митохондрии есть у всех высших организмов и у многих совсем непохожи на
бактерий), но главное - остался совершенно непонятным тот факт, что все до
единой генетические программы построения и работы любой митохондрии за-
кодированы вперемешку в геномах самой митохондрии и клетки, ее содержа-
щей. Подробнее см. [18, с. 339]. Налицо именно гибрид.
Понятие гибридного органа и примеры таких органов из разных областей
биологии делают противоречивую “теорию” происхождения митохондрий из-
лишней и ликвидируют большой пробел в наших знаниях. Сходство бактерий и
митохондрий ложится в единый рефрен. Это тоже диатропический результат.
5. Причина рефренов
Поскольку феномен рефрена принято считать непонятным, в заключение
выскажу кратко свое мнение: рефрен вполне понятен как следствие фракталь-
ной структуры мира.
Фрактал - это математическая структура дробной размерности, для которой
характерно, что либо она сама, либо ее части обладают свойством самоподобия
(большое подобно малому, а то - еще меньшему и т.д.). На всех уровнях, от галак-
тик до атомов, наблюдается фрактальность, и причина этого тоже довольно оче-
видна: такова природа самих чисел, поэтому всё, что подчиняется арифметике,
должно быть фрактально. Любой отрезок вещественной прямой устроен точно так
же, как и любая часть его или любого иного отрезка. Это еще не фрактал, ибо вся-
кий отрезок имеет единичную размерность, но на отрезке легко получить (выбра-
сывая точки по очень простому закону) фрактал. Точнее, одномерный фрактал’.
Давно замечено, что обычная система таксонов (древо) тоже образует одно-
мерный фрактал [19], но возможны и многомерные фракталы (ведь самоподобие
касается и фигур на плоскости, и тел в пространстве). Ими определяются многие
свойства экосистем и их эволюции [20]. Многомерным фракталам свойственны
и параллелизмы, и мозаичность (достаточно взглянуть хотя бы на иллюстрации
в книге [13]), поэтому на них всюду выявляются рефрены.
Именуемый множеством Кантора. Это пример того, как манипуляция с числами помогает
обнаружить новые явления природы. См. п. 5 статьи 4-65, пример 6.
316
Может показаться странным, что объяснение биологического феномена да-
но не биологическое, а математическое. Но иначе и быть не может: если фено-
мен вездесущ, то и носители его вездесущи, а таковыми прежде всего являются
числа; арифметике подчиняется любое явление (кроме, насколько знаю, только
диалектического материализма и, как его варианта, дарвинизма).
Можно сказать, что теория фракталов и есть основа того будущего исчисле-
ния параллелизмов, о котором смутно мечтал Мейен. Он не успел ничего узнать
про фракталы, но его интуиция верно подсказала ему, что если феномен рефрена
вездесущ и внемасштабен, то он может лечь в основу теоретической биологии.
Литература
{.Джеффри Ч. Биологическая номенклатура. М., Мир, 1980.
2. Из научного наследия С.В. Мейена: к истории создания основ эволюци-
онной диатропики // Эволюция флор в палеозое. М., ГЕОС, 2003.
3. Мейен С.В. Основные аспекты типологии организмов, 1978 [М-165].
4. Мейен С.В. Флорогенез и эволюция растений // Природа, 1986 [М-280].
5. Мейен С.В. Нетривиальная биология (Заметки о...) [М-323].
6. Мейен С.В. Эволюция и систематика высших растений... [М-328].
7. Мейен С.В. Листья на камне. М., ГЕОС, 2001. [М-338].
8. Мейен С.В., Соколов Б.С., Шрейдер Ю.А. Классическая... [М-150].
9. Мейен С.В., Шрейдер Ю.А. Методологические аспекты... [М-140].
10. Михайлов К.Г. Рецензия на книгу: Любарский Г.Ю. Архетип, стиль и ранг
в биологической систематике. М., КМК, 1996 // Ж. общей биологии, 1997, № 4.
11. Определитель бактерий Берджи. М., Мир, 1997.
12. Павлов В.Я. Периодическая система членистых. М., ВНИРО, 2000.
13. Пайтген Х.-О., Рихтер П.Х. Красота фракталов. М., Мир, 1993.
14. Пономаренко А.Г. Артроподизация и ее экологические последствия // Эко-
системные перестройки и биосфера. Вып. 6, М., Палеонт. ин-т, 2004, с. 7-22.
15. Пчеленко С.П. К механизму явлений номогенеза // ЛЧ, 2005, с. 112-122.
16. Раутиан А.С. Апология сравнительного метода // Любищев и проблемы
формы, эволюции и систематики организмов. М., 2003.
17. Чайковский Ю.В. Элементы эволюционной диатропики. М., Наука, 1990.
18. Чайковский Ю.В. Эволюция. М., Центр системных исследований, 2003.
19. Burlando В. The fractal dimension of taxonomic systems // J. Theor. Biology,
1990, vol. 146, № 1.
20. Gisiger T. Scale invariance in biology: coincidence or footprint of a universal
mechanism? // Biological Reviews, 2001, № 2.
21. Meyen S.V. Plant morphology in its nomothetical aspects // Botanical Review,
1973, vol. 39, № 3.
22. Sachs T. A morphogenetic basis for plant morphology // Acta Biotheorethica,
1982, Ser A, vol 31, p. 118-131.
317
Практичная теория (заключительная статья)
Нет ничего практичнее, чем хорошая теория - говорил полтораста лет назад
Людвиг Больцман, знаменитый физик. Речь шла о том, что хороша та теория,
которая имеет выход в практику, оправдывается практикой. По-моему, это
вполне относится к номогенезу: будучи высказан Л.С. Бергом просто как логи-
ческая антитеза дарвинизму, он оказался, после модификаций Мейена (соста-
вивших основу диатропики), весьма полезен практически.
Правда, цельной теорией номогенез, в том числе новый, не стал, ибо разви-
вался почти безо всякой связи с физиологической (ламаркизм), эмбриологиче-
ской (жоффруизм) и экологической (ЭКЭ) эволюцией, однако составил сущест-
венную часть теории - ее морфологическую базу. Вырастающая сейчас теория
эволюции номогенезом не является и ранее (ЗЭ) получила название «акти-
реф»225 - активное движение вдоль рефрена (если понимать его как таблицу).
Но вернемся к новому номогенезу. Суть его видится мне в том, что многооб-
разие, из которого природа черпает конкретные варианты устройства организмов,
выявлено им как единое для всей природы и оно оказалось упорядоченным. Всё
собирается из одних и тех же блоков (т.е. существует единый резервуар разнооб-
разия), причем сходства выстраиваются в рефрены. На это, в сущности, и указы-
вал Мейен, скорее интуитивно, чем обоснованно, когда писал о рефренной струк-
туре всякого разнообразия и об эволюции любых объектов как преобразовании
разнообразий.
Об этом лучше всего говорит концепция ангиоспермизации (одновременно-
го становления цветковых в разных местах Земли и в разных таксонах), одна из
«заций» Пономаренко (см. выше, с. 315). В данном контексте гипотезу происхо-
ждения цветковых (М-286) надо понимать именно как мысленную сборку цветка
из нецветковых блоков. (Реальную форму, способную быть исходной, указал А.Б.
Герман - см. АСМ, с. 534). И в этом смысле она уже вполне обоснована, хотя в
более общем смысле ее еще предстоит «положить на карту».
Тот факт, что сочетаться могут почти любые свойства (некоторые, правда,
весьма редко - например, цветок и пищеварение) , знали и до Мейена, но боль- 115 * * * * * * * * * * *
115 Ее лучше бы назвать словом «хологенез» (греч.: порождение целого, целостное поро-
ждение), но данный термин давно занят под один аспект эволюции - как последователь-
ности меняющихся онтогенезов (см. АСМ, с. 181). Заняты оказались и подходящие ла-
тинские эквиваленты. Мне осталось извлечь название излагаемой концепции из главного
ее утверждения. Приведу формулировку: «...Мир устроен так, что материя сама собой
организуется, порождая всё более и более сложные системы. На всех уровнях организа-
ции виден один и тот же эффект - актиреф (активное движение вдоль рефрена...). Если
речь идет о биологическом объекте, то актиреф имеет две формы: развитие особи (онто-
генез) и эволюцию. Поскольку рефрены принадлежат общей диасети, то эти две формы
сходны (известный из школы биогенетический закон). Здесь мы видим соединение в од-
ной схеме идей номогенеза (наличие диасети), жоффруизма (активное изменение в разви-
тии особи) и ламаркизма (активная эволюция)» (ЗЭ, с. 56).
318
шинство полагало, что никакой теории тут построить невозможно. К идее Копа
- Вавилова о фундаментальном значении параллельных рядов изменчивости и к
указанию Урманцева на небходимость сопоставлять не факты, а ряды фактов,
Мейен добавил ту идею, что в рядах надо исследовать направленность парал-
лельных преобразований. Дело в том, что если само наличие сходных форм в
разобщенных флорах можно трактовать как унаследование от неизвестных
предков, то одинаковое их преобразование во времени (при явном отсутствии
контактов и в совсем различных средах) - невозможно. Здесь налицо номогенез,
что вполне понимал (судя по разрозненным репликам) Вавилов, прекративший
(можно полагать, именно поэтому) работы по анализу рядов. У него был факти-
ческий последователь - ботаник Николай Петрович Кренке, пытавшийся дви-
нутся дальше в рамках дарвинизма. Но материал, по сути номогенетический,
сопротивлялся, и сформулировать результат аккуратно не удалось, пока через 40
лет Мейен не ввел свои понятия (М-116).
Использовав идею Бэра о ядре и периферии, он дал обоснование как правилу
Кренке (признак, являющийся уродством у данного вида, как правило нормален у
какого-то иного вида - АСМ, п. 4-11), так и грубой филогении. В свою очередь,
принятие грубой филогении понуждало пересмотреть всю теорию эволюции.
Во-первых, пришлось задать вопрос: является ли нынешняя “эволюцион-
ная” систематика эволюционной? Оказалось - нет. См. об этом выше, часть 6.
Во-вторых, если в качестве механизма обычной филогении был деклариро-
ван (еще Дарвином) акт дивергенции (расхождение вида на два или несколько),
то для грубой филогении он не работает, и следовательно, нужен некий аналог
дивергенции. Мейен назвал его нетривиальной дивергенцией и в своей послед-
ней диатропической работе М-313 наметил соответствующий теоретический ап-
парат. Взяв три стандартных понятия - дивергенцию, параллелизм и конверген-
цию - и назвав их «тривиальными модусами эволюции», он предложил в каче-
стве новых единиц описания эволюционного процесса нетривиальные модусы,
т.е. механизмы эволюции. Их у Мейена тоже три.
1) «Нетривиальная дивергенция... это расщепление так называемых “синте-
тических типов”, которые широко распространены у высших растений. Под
“синтетическими типами” понимают также предковые таксоны, у которых по-
разному комбинируются признаки таксонов-потомков».
Данный модус, по-моему, напрочь рушит надежду понять происхождение
родов и выше как череду происхождений видов, однако Мейен на это, видимо,
внимания не обратил и сразу перешел к другим модусам. По Мейену, «для мор-
фологии гораздо интереснее нетривиальные конвергенции и параллелизмы».
2) Нетривиальный параллелизм - это, прежде всего, транзитивный полимор-
физм (у меня - транпол), т.е. «сохранение одной и той же упорядоченности поли-
морфизма (т.е. рефрена) в филогенетической последовательности таксонов». (За-
тем, к тому же модусу Мейен отнёс псевдоцикл - им является, например, эво-
люция цветка в соцветие и дальнейшая редукция соцветия в одиночный цветок.)
319
Данный модус напрочь рушит другую надежду - понять появление и утрату
каких-либо свойств путем истребления особей, их несущих.
3) Нетривиальная конвергенция - это, прежде всего, гамогетеротопия, т.е.
«перенос признаков с одного пола на другой, в результате чего половой димор-
физм... сменяется мономорфизмом»116. Мейен объяснял этим свою идею проис-
хождения цветковых из беннетитов. Как и при любой гетеротопии (вырастании
органа на новом месте), эволюция здесь возможна только скачком (сальтацией).
Подробнее об этом можно прочесть в книгах [Васильевы, 2009; АСМ].
Остальное сделано после безвременной кончины Сергея Викторовича.
1. Эд ванты
Прежде всего, следовало продолжить линию нетривиальных модусов. На-
пример, надо было исследовать все виды гетеротопии. Сам Мейен едва обозна-
чил тему, упомянув в статье М-286 допущение, что «игловидные листья хвой-
ных появились в результате распространения на вегетативные побеги онтогене-
тической программы чешуевидных листьев фертильных побегов». Следовало
указать иные типы гетеротопий.
Это и было сделано (без ввода самого термина) в книге 4-51, на с. 393. Как
пример перемещения самого органа указано изменение плоскости симметрии па-
ры плавников-ласт (вертикальная плоскость задней пары у кистеперых рыб пере-
шла в горизонтальную у четвероногих). А как пример переноса идеи органа на
иной материальный субстрат указано перемещение теменного отверстия (у рыб
оно расположено между лобными костямй, а у амфибий - между теменными).
Примеры гетеротопии можно множить почти бесконечно, если таковыми
считать все случаи, когда можно говорить о реализации прежней онтогенетиче-
ской программы на новом месте. Например, таковы серийная гомология (гомо-
логия в том смысле, в каком рука гомологична ноге) и югулярность (сдвиг
брюшных плавников под горло, вперед от грудных - см. выше, с. 72).
Важнее, однако, перенос не программы, а идеи (когда она реализуется на
новом материальном носителе). Наиболее поразительна презентация антигена'.
тремя разными способами достигается в разных иммунных клетках одного ор-
ганизма один и тот же результат - антиген, подлежащий узнаванию и (если
нужно) уничтожению, проносится сквозь клетку, препарируется и выставляется
на другой ее стороне, где узнающая клетка (Т-хэлпер) запускает некую (еще не
понятую учёными) операцию, приводящую к синтезу нового гена (ЗЭ, с. 95-96).
Если с одного способа тут списаны другие, то налицо гетеротопия, если же они
возникли независимо, то перед нами реализация параллельных строк одного
рефрена, т.е. другая форма нетривиальной конвергенции.
Однако громоздкий термин не может иметь хождения. Так, не смогло уко-
рениться «повторное полиморфное множество», так что Мейену пришлось за-
менить его словом «рефрен», и оно вскоре же стало использоваться - например,
Г.Ю. Любарским и С.В. Чебановым в связи с проблемой биологического стиля
116 Примеры гамогетеротопии приведены у Мейена (М-286) и у меня (АСМ, п. 9-12*).
320
(она в данном Сборнике не рассматривается). Поэтому предлагаю заменить тер-
мин «нетривиальный модус эволюции» термином эдвант (от англ, advance -
продвижение, улучшение; advanced - передовой, продвинутый).
Любящим математические аналогии замечу, что если счесть движения точек
по коническим сечениям (эллипсу, параболе и гиперболе) тривиальными моду-
сами теоретической механики, то ее эдвантами окажутся движения сплошных
сред согласно уравнениям эллиптического, параболического и гиперболическо-
го типов. Аналогия позволяет увидеть, что эдванты раскрывают глубинную
сущность тривиальных модусов. Так, парабола выглядит маловажным раздели-
телем между миром эллипсов и миром гипербол, тогда как уравнения параболи-
ческого типа описывают особый мир процессов (диффузию).
И, точно так же, параллелизм выглядит лишь разделителем между миром
дивергенций и миром конвергенций, тогда как эдвант параллелизма (транпол)
перевернул понимание задач эволюции и систематики. Это побуждает и в обыч-
ном параллелизме увидеть больше, чем видел Берг.
Статью М-313, а с тем и всю диатропическую деятельность, Мейен закон-
чил словами: «может быть, более продуктивным окажется отказ от тотального
адаптационизма в пользу той более гибкой точки зрения, на которой стоял Ч.
Дарвин». Это, конечно, верно, но нужен был ход и в противоположном направ-
лении - надо было отвести приспособлению определенное место в теории.
Если полагать, что адаптация - тривиальный модус эволюции (при «переносе
идеи» это, конечно, не так, но в простых случаях так считать можно), то следует
ожидать соответствующего нетривиального. Иными словами, должен существо-
вать адаптационный эдвант. Таковым мне представляется преадаптация, т.е. при-
способление, полезное не в момент своего появления, а для потомков, быть может
далёких. Понятие ввел Люсьен Кэно (1901) в чисто номогенетическом (в нашем
нынешнем понимании) смысле: он полагал, что при мутации реализуется одна из
потенций, уже существующих в организме.
Ламаркизм и дарвинизм не смогли сказать о преадаптации ничего определен-
ного [Георгиевский, 1974], зато еще Берг клал ее в основу учения, и в новом но-
могенезе она оказалась весьма к месту: указывает на тот факт, что эволюция в це-
лом течёт не в линейном, а в более сложном времени.
Адаптационными эдвантами можно считать также пост- и экстраадаптацию,
т.е. изменения, полезные в условиях уже исчезнувших или просто отсутствую-
щих. И если эти адаптации текут во времени, отличном от привычного (тривиаль-
ного), то естественно будет задуматься, в каком времени течет адаптация привыч-
ная. На тот факт, что реальное время устроено сложнее, чем время, указываемое
всевозможными часами, не раз указывал и Мейен. См.: М-229, М-246, М-281.
Полагаю, что каждому или почти каждому первичному эволюционному по-
нятию (тривиальному модусу по Мейену) можно найти свой эдвант. Так, реком-
бинация и гибридизация (модусы микроэволюции) имеют своим эдвантом блоч-
ность, наблюдаемую на всех уровнях эволюции.
321
2. Практические достижения диатропики...
Что касается систематики, то, как пишут супруги А.Г и И.А. Васильевы
[2009, с. 289], «мерономия С.В. Мейена уже прочно заняла нишу науки, изу-
чающей многообразие строения организмов и их частей».
Для Васильевых диатропика Мейена (они именуют ее популяционный ас-
пект популяционной мерономией) важна как теоретическое обоснование реаль-
ного способа построения систематики грызунов. Для классификации они берут
«неметрические», т.е. качественные (есть - нет, выпуклый - вогнутый, удвоен-
ный и т.п.) признаки черепа. Если для данной популяции все они вместе обра-
зуют в признаковом пространстве компактную область, и область эта почти или
вовсе не перекрывается с областями для других популяций, то данной популя-
ции можно придать таксономический статус. Обычно это - подвид или вид, но
метод годится и для выявления таксонов более высоких рангов.
Это старый «метод конгрегаций» (см. в наст. сб. часть 6). Его предложил
энтомолог-ламаркист Евгений Сергеевич Смирнов - тот самый, который еще до
Шапошникова ставил опыты по эволюции насекомых. Предложил еще в 1920-х
годах. Метод Смирнова не прижился - сперва из-за трудоемкости ручных расче-
тов, а затем, несмотря на появление ЭВМ-и «нумерической систематики», из-за
того, что был вместе с нею затоптан всепобеждающим учением (кладизмом).
Васильевы успешно прилагают свой метод также к диагностике уже выяв-
ленных видов: «Подставляя результаты классификации конкретной особи в
формулу данного дискриминантного ключа в виде значений 1, если признак
имеется, и 0, если он не обнаружен, можно диагностировать ее принадлежность
к соответствующему таксону» [Васильев, Васильева, 2009, с. 321].
Здесь тоже можно говорить о предшественнике - им был ботаник Б.Е. Бал-
ковский [1960]. У его последователей читаем (в популярном определителе):
«Обнаружив, что листья и веточки расположены супротивно, а на ветвях
отсутствуют колючки, запишите число 3; светло-бурой глубокопродольно-
морщинистой коре соответствует число 3, а молодые побеги, зелено-
чернеющие от железистых бородавочек, обозначены числом 8. Добавив
двойку, так как листья определяемого кустарника простые, яйцевидные,
голые или снизу по жилкам опушенные, получаем набор цифр 3382. Опре-
делить, какие были соцветия или цветки, осенью довольно трудно, поэто-
му переходим к рассмотрению плодов, поставив 2 прочерка вместо цифр,
соответствующих цветкам. Плоды определяемого кустарника - 4-
лопастные коробочки с семенами, окруженными оранжево-красными мя-
систыми присемянниками (кровельками); это соответствует индексу 16.
Окончательный шифр 3382-16 соответствует в таблице бересклету боро-
давчатому» (подробнее см.: АСМ, с. 323).
Нынешнее возрождение забытых методов объясняется, по-моему, тем, что
неметрические признаки легко трактуются теперь как состояния меронов. Так что
прав был К.Г. Михайлов, утверждая: «Главное, что дает типология систематику -
это снятие комплекса собственной неполноценности» - см. статью 4-57.
322
В той же моей статье, кстати, названы и другие практические достижения
диатропики: упорядочен и прояснен феномен параллелизма - через введение
понятия рефрена (п. 1); выявлена взаимодополнительность искусственной и ес-
тественной систем в таксономии, назначение которых оказалось одинаково важ-
но (п. 2); эволюция таксона представлена как движение вдоль рефрена - это по-
зволило понять, что такое грубая филогения (п. 3); дано понимание экосистем-
ной эволюции как рефрена рефренов, а также блочной эволюции; в частности -
как появления «гибридных органов» (п. 4). Наконец, в п. 5 был предложен под-
ход к пониманию природы самого феномена рефренной структуры разнообразия
(диасети) как фрактальной. К этому списку теперь следует добавить иные не-
давние успехи науки, тоже носящие явственно диатропический характер.
Прежде всего, надо сказать о работах школы Мейена. Кроме конкретных
работ, названных в предыдущих статьях Сборника, укажу на исторический об-
зор [Игнатьев, 2003], где «флорогенез по Мейену» погружен в исторический
контекст и стал выглядеть по-новому. Прежде многим (в том числе и мне) Мей-
ен казался в данной области первооткрывателем. См. сноску к докладу на с. 82.
Центральной фигурой во флорогенетике теперь мне представляется ботаник
Михаил Григорьевич Попов (1893-1955), который прежде других понял, что по-
строение теории эволюции невозможно на прежнем поле «происхождения ви-
дов», что вопрос о появлении новых видов, родов и т.д. может быть решен толь-
ко при совместном их рассмотрении. К сожалению, рассматривая механизмы
существования и размножения нынешних организмов, Попов почти не касался
ископаемого материала, т.е. процессов их реального появления в геологической
летописи и последующего расселения. О Попове см. ниже, п. 4.
Поэтому главная роль Мейена во флорогенетике видится в том, что он впер-
вые, имея сплошные данные по огромной группе вымерших растений, попытался
вплести их в ткань нового номогенеза (который именовал номотетической теори-
ей эволюции). Подробнее о флорогенезе будет сказано далее, тоже в п. 4.
Другой диатропический результат - поразительные открытия в иммуноло-
гии: эволюционное развитие иммунитета (от простейших актов ограниченной
совместимости тканей низших животных до адаптивного иммунитета высших)
образует у беспозвоночных и позвоночных два параллельных направленных ря-
да, т.е. рефрен. Данные ряды столь же разобщены, как северная и южная флоры
в построении Мейена. Значительный параллелизм наблюдается также с иммуни-
тетом животных и растений, так что скоро, по всей вероятности, можно будет
говорить о едином эволюционном рефрене иммунитета (подробнее см. ЗЭ). Од-
нако сейчас уже видно, что эти три параллельных ряда - ряды почти идентич-
ных идей, но не молекулярных механизмов, которые совсем различны.
И, так же, как ряды эволюции клетки (М-73, см. сноску к предисловию
Сборника) иммунные ряды могут быть исследованы почти исключительно на
нынешних организмах. И тут и там мы видим два основных направленных ряда,
тесно сходных, и третий ряд, сходный с ними меньше. В последнем случае речь
323
идет о клетках грибов. Мейен почти не коснулся грибов, но всё же отметил еще
один важнейший параллелизм, параллелизм межиагдцарственный - миксомице-
тов и миксобактерий.
Наконец, новый номогенез возродил интерес к систематике как науке, пока-
зав, что она может быть полезна самым разным биологам, да и не только им.
Еще до появления книги Берга российские зоологи П.И. Бахметьев [1903] и В.М.
Шимкевич (1906, см. [Попов, 2008]) пытались построить табличные формы сис-
тем отдельных групп животных по аналогии с периодической системой химиче-
ских элементов. С появлением номогенеза попытки приняли организованный
характер и, хотя построить такую систему тогда не удалось, но нынешний ана-
лиз проблемы позволил достичь понимания двух фундаментальных мыслей.
Во-первых, что если от табличной (в том числе периодической) системы не
ожидать того же, чем обладает иерархическая, а ставить ей собственные задачи,
то построить ее вполне можно, что для нескольких отраслей биологии уже сде-
лано - см. статьи Ч- 47, 61 в наст. Сборнике.
Во-вторых, что «поиски периодических систем и связанные с ними представ-
ления указывают на альтернативный механизм эволюции, а точнее, ее направлен-
ности, который можно охарактеризовать без всякой мистики» [Попов, 2008, с.
138]. Эту мысль историк науки Игорь Юрьевич Попов развил подробно, ссылаясь
на российских и зарубежных авторов. Работы по новому номогенезу (в том числе,
Мейена) ему известны (он привёл названия некоторых), и Попов признал превос-
ходство в этом смысле российской науки перед западной. Отметив (с. 42) порази-
тельное в этом пункте теоретическое невежество западных коллег, делающих
вновь и вновь самые убогие «открытия» по эволюции227, он пишет:
«Таким “открытиям” значительно труднее появиться в нашей стране, потому
что несмотря на все репрессии не забыты концепции направленной эволюции,
а в особенности номогенез Л.С. Берга. Некоторые работы сторонников этих
ересей связаны с поиском периодических систем» [Попов, 2008, с. 43].
Замечу: периодичность тут - примерно то же самое, что в данном Сборнике -
рефрен, и потому книга Попова может служить частичным перечнем названий
достижений старого и нового номогенеза (но, увы, не самих достижений, ибо ав-
тор почти нигде не старается понять суть описываемого, а иногда даже подменяет
изложение иронией над жертвами гонений, что вряд ли уместно).
117 Одним из важных примеров служит Попову книга Антонио Лима де Фариа [1992], где
заново «открыты» самые простые положения номогенеза. Пришлось писать об этом и мне
(4-26). Что же касается историков эволюционизма, то всё наоборот: у нас процветает, на-
пример, полное незнание творчества Дарвина и становления дарвинизма, в том числе и в
России, о чем мне тоже пришлось писать: «Потеряв культуру исторического поиска, позд-
несоветская... наука вынуждена опираться на западные исследования о России, а они сами
оказываются основанными на этой самой советской науке» (4-27, с. 145). С тех пор поло-
жение не изменилось: новое (2001) «академическое» переиздание «Происхождения видов»
демонстративно игнорирует всё западное (весьма, кстати, серьезное) дарвиноведение.
324
В частности, старая эволюционная концепция М.Г. Попова прослежена
здесь как номогенетическая (в смысле номогенеза по Соболеву, т.е. с упором на
ту комбинацию свойств, которая ныне именуется блочностью эволюции) и увя-
зана (не вполне явно) с более новыми взглядами на эволюцию. А именно, И.Ю.
Попов напомнил, что М.Г Попов полагал главной причиной появления новых
групп растений отдалённую гибридизацию - например, понимал первые цветко-
вые как гибриды гнетовых и цикадовых. Согласно М.Г. Попову, возникает такая
гибридизация крайне редко и только при радикальной смене условий жизни.
В качестве особого случая отдалённой гибридизации И.Ю. Попов рассмот-
рел симбиогенез 118. В этой связи он верно отметил [Попов, 2008, с. 30], что род-
ство митохондрий (главного нынешнего объекта учения о симбиогенезе) с бак-
териями весьма обманчиво:
«На срезах клеток эти органоиды выглядят похожими на бактерий. Надо за-
метить, правда, что срезы могут исказить картину. То, что кажется разрезан-
ными небольшими частицами, может быть срезом большой трубчатой еди-
ной структуры, что в некоторых случаях было установлено. Что касается
других черт сходства органелл и бактерий, то их можно рассмотреть и как
параллелизм или результат развития в одинаковом направлении».
В этом, замечу, суть: в большинстве своем (а отнюдь не «в некоторых слу-
чаях») митохондрии по форме резко отличны от бактерий, а похожи на бактерий
лишь те, что имеют размер бактерий. Они повторяют формы бактерий просто
потому, что число форм данного размера в природе ограничено, а потому по-
вторность здесь неизбежна, а всякая повторность приводит, как показала прак-
тика, к рефренной структуре. Родство же с бактериями видят те, кто не знает о
явлении «развития в одинаковом направлении», т.е. о рефренах.
Единственный серьезный аргумент симбиогенетиков - сходство мт-ДНК и
ДНК бактерий, имел силу полвека назад, когда о митохондриях мало знали. Уже в
1970-х годах стало известно, что и тут различий больше, чем сходств. Например, мт-
гены не кодируют сами ни одного белка, так что их гены были названы «якорями»,
к которым тянутся соответствующие хромосомные гены (аналогия с якорем, когда
118 Т.е происхождение принципиально нового организма путем объединения более про-
стых организмов. Реальных примеров симбиогенеза немного (лишайник как соединение
гриба и водоросли, несколько видов жгутиковых с фотосинтетическими бактериями, не-
сколько видов инфузорий, использующих жгутиковых эндосимбионтов для передвиже-
ния), однако концепция симбиогенеза претендует на объяснение главных актов эволюции
- прежде всего, происхождения эвкариотной клетки. Правда, о происхождении ее глав-
ных свойств (ядро, цитоскелет, митоз и тд.) концепция не говорит ничего существенного,
зато много говорит о происхождении митохондрий. А именно, несколько примеров сход-
ства некоторых митохондрий с бактериями (главное - сходство ряда свойств ДНК) заяв-
лено как основание утверждать, что все типы митохондрий произошли из бактерий-
эндосимбионтов и тем дали начало эвкариотной эволюции. Никакие факты, противоре-
чащие концепции, ее сторонниками даже не упоминаются. Подробно см. АСМ.
325
его завозят вдаль, чтобы лебёдкой снять судно с мели) [4-4]. И триплетный код ми-
тохондрии имеют не вполне тот, что у бактерий [Никоноров, 2010].
Тот довод , что митохондриальные гены устроены «по бактериальному типу»,
т.е. цепочками оперонов (а не экзон-интронно), тоже потерял силу, поскольку и
подобных генов много найдено также в ядре, а у некоторых митохондрий обна-
ружены интроны. И, наконец, само наличие митохондриальных генов оказалось
далеко не всеобщим:
«На Земле существует целый ряд организмов, лишенных типичных мито-
хондрий, продукция АТФ у которых происходит в гидрогеносомах и мито-
сомах, этих “остатках” митохондрий, главной отличительной чертой кото-
рых считается отсутствие собственного генома [Van der Giezen & Tovar,
2005]» [Никоноров, 2010, с. 156].
Слово «остатки» недаром взято автором в кавычки: никакие это не остатки
митохондрий, а совсем иные органеллы. Единственным доводом в пользу «ос-
татков» служит сама уверенность в идее симбиогенеза119, ставшая, тем самым,
типичной обратной эвристикой. Как сказано выше [4-65, п. 6], борьба с обрат-
ными эвристиками на уровне фактов и логики бессмысленна - нужна смена точ-
ки зрения. Ее и дает диатропика, видящая всё развитие разнообразия митохонд-
рий как рефрен, проявляющий параллелизм с рефреном бактериальным. Мейен
понимал это давно, хоть и был далёк от цитологии [4-56]. Позже стало понятно,
что митохондрия - обычный гибридный орган [4-57, п. 4].
3. ...почти никому не известны
Естествен вопрос: если всё сказанное верно, то почему диатропика совер-
шенно выпала из внимания биологов - за исключением узкого круга идейных
приверженцев Мейена? А если неверно, то где опровержения? Ведь работы
Мейена и его школы широко известны и пользуются общим уважением, так что
говорить о незнании их не приходится. Разумеется, играет роль элементарное
непонимание сути дела (надеюсь, авторы курьёзных высказываний - о диатро-
119 Ведущий митохондриолог Марк ван дер Гизен (Англия), больше всех, пожалуй, сде-
лавший для понимания природы митохондрий в их разнообразии, обнаружил недавно,
что некоторые организмы, не имеющие митохондрий, несут всё же в своих хромосомах
гены, характерные для митохондриальных белков [Van der Giezen, 2009]. Сам по себе
данный феномен скорее говорит против идеи симбиогенеза: ведь эти гены работают так
же, как и почти все хромосомные - кодируют белки, используемые вне ядра (митохонд-
рия, как известно, сама ни одного из своих белков целиком не кодирует). Однако Гизен
счел свое открытие доказательством всеобщности симбиогенеза, так как допустил, что
эти гены привнесены когда-то митохондрией, позже утерянной. Он как бы забыл, что сам
факт перенесения генов из митохондрии в ядро клетки - всего лишь допущение, выдви-
нутое для поддержки допущения о том, что митохондрия прежде имела автономный ге-
ном, который, в свою очередь, был додуман для поддержки того допущения, что мито-
хондрия когда-то произошла от бактерии. Хотя, повторю, многие митохондрии совсем
непохожи ни на какие бактерии, так что в нагромождении допущений нет нужды.
326
пике вообще и рефренах в особенности - не упрекнут меня за то, что здесь нет
ни имён, ни цитат), однако не может же оно быть причиной общей.
Чтобы подойти к возможному более общему ответу, обратимся к недавней
работе того же Попова (историка науки), но - адресованной западному читате-
лю, а не российскому. Тут автор перед нами предстаёт совсем иным. В обшир-
ной статье «Ортогенез против дарвинизма: русский пример» [Popov, 2008] он не
различает номогенез и ортогенез («номогенез-ортогенез», с. 387) и вообще ведет
читателя к мысли, что наука России вторична. Однако он выделяет эпоху работ
Любищева и Мейена как некое «возрождение», что здесь нам как раз и важно.
Подробно изложив некоторые взгляды Берга и Соболева и назвав имя Лю-
бищева, Попов перечислил еще несколько советских публикаций по «направ-
ленной эволюции». Об их содержании нет ни слова, кроме уверения, что все ци-
тировали Берга, многие - Любищева и лишь какие-то «палеонтологи-геологи» -
Соболева. Из последних назван только Мейен с одной его статьей М-127, но ее
Попов даже в руки не брал, судя по следующему поразительному заявлению:
«Зарубежные авторы ортогенеза не слишком известны (российским авто-
рам - Ю.Ч.) и не так влиятельны. Лишь одна из современных концепций
демонстрирует значительное европейское влияние, это концепция «конту-
ров эволюции» Эдуарда Ахназарова (1933-2001). Этот автор заявил о же-
лании переформулировать «феномен человека» Тейяра де Шардена в чис-
то научных терминах. Некоторые авторы решительно заявили, что пришли
к направленной эволюции без какого-либо теоретического влияния,
т.е.обосновали факты противоречия дарвинизму в процессе эмпирического
исследования» [Popov, 2008, с. 386-387].
И это всё? Неужели? Нет, есть еще перечень тех, кто после Вавилова писал
о гомологических рядах (с. 390). Трудно поверить, но ни Кренке, ни Мейена в
нем нет. Более о Мейене и новом номогенезе (к коему сам Попов приложил руку
основательно, предложив собственную периодическую систему [Попов, 2008, с.
88-118]) в статье - ни слова. Добавлю, что и Ахназаров тут не к месту: он был
системный эволюционист и факты биологии привлекал лишь как примеры.
Еще замечу, что если уж сказано о «европейском влиянии» на российскую
мысль, то надо назвать очень многих и, главное, хотя бы для приличия сказать
об обратном - о влиянии наших работ на западные. Например, о канадском фи-
томорфологе Роджере Джане, который прямо опирается на ранние диатропиче-
ские идеи Мейена [Jean, 1994] (русский перевод: [Джан, 2006]).
Сам Попов знает, что сказанное им о нашей науке - неправда (см. его преж-
ние работы), и встает вопрос, зачем эта ложь понадобилась. Простой ответ (ради
приглашений, публикаций и грантов) возможен, но неполон. Ведь всего того же
можно было добиться, не позоря страну и не позорясь самому - достаточно бы-
ло выразиться аккуратнее. Очевидна еще одна причина, чисто душевная.
Преподавая математику, я не раз пытался пояснять уравнения физическими
примерами и убедился, что многие ученики не могут видеть две науки сразу (бы-
327
вало и возмущение: «У нас же урок математики, а не физики»). А редактируя ста-
тьи для Советской энциклопедии, я видел, как легко их авторы лгут, даже если
правда безобидна. Спросив одного из них, зачем это, услыхал поразительный от-
вет: «А разве для энциклопедии надо иначе?» Видимо, и Попов, когда пишет для
западных коллег (точнее, для тех, кто уверен, будто русские хлебают щи лаптем),
то невольно хочет убедить в этом (в экзотике русской трапезы) и себя. На другом
«уроке» он такого не то что написать, но, полагаю, и подумать не сможет.
К чему разбирать это? К тому, что своей несуразной выходкой Попов лишь
выявил в уродливой форме то, что многие в приличной форме делают неявно. Но-
вый номогенез широко применяется, без него порой даже не обойтись, но упоми-
нать его и его классиков не принято, и предпочитают называть посторонних.
Для критики надо выбрать пример не слабый, как делают обычно, а (как при-
зывал Любищев) самый крепкий. Выберу работы, анализирующие феномен «за-
ции», открытый палеонтологом А.Г. Пономаренко (см. 4-57). Сам Александр Ге-
оргиевич встретил моё замечание, что зация - рефрен, снисходительным смехом
из зала (декабрь 2005 г., Чтения памяти Мейена в ГИНе), а в перерыве весело и
дружески дал мне понять, что ни в каких философиях не нуждается.
Но, как верно замечал Любищев, отказ от философии есть худшая из филосо-
фий (ведь какой-то философией пользуется каждый), и проблема продолжает о
себе заявлять. На зацию обратила внимание палеонтолог Эмилия Ивановна Во-
робьева, известная блестящими исследованиями палеозойских рыб и широким
кругозором. Она была хорошо знакома с Мейеном (см. М-302), они слушали док-
лады друг друга и обменивались оттисками публикаций. Позже она одна из пер-
вых заявила, что эволюция носит у ее объектов не столько филогенетический
(древо), сколько блочный (сеть) характер. Подробнее см. об этом в АСМ.
В ее недавней статье «Процессуальный подход к проблеме “тетраподиза-
ции”» [Воробьева, 2008] справедливо отвергается, как и у Мейена, «ретроспек-
тивный подход», т.е. подход, при котором система организмов строится на ма-
териале ныне живущих видов, а ископаемый материал призван, по сути дела,
лишь подтверждать общепринятую схему.
Вместо этого ущербного подхода в тексте сквозит новый номогенез. На-
пример, отодвигается на задний план построение родословных древ, так как
«акцент все больше перемещается с дискуссий о моно- либо полифилетическом
происхождении тетрапод на последовательность появления тетраподоподобных
признаков и... на процессы формирования тетраподной организации» (с. 13).
Предложено «отказаться от от термина “переходные формы”, взяв на воо-
ружение более соответствующий реальности термин “мозаичные формы”» (с.
20). «Идея поиска общего предка тетрапод, позволяющая построить единое фи-
логенетическое древо, все далее уходит вспять. Параллелизмы, конвергенции и
мозаичность в проявлениях тетраподных признаков уже на уровне рыбоподоб-
ных предшественников тетрапод придают процессам тетраподизации газонопо-
добный облик» (с. 21). Затем указана «общность морфогенетических основ» у од-
328
ного нынешнего и одного палеозойского родов (с. 23, подпись к рисунку), т.е.
обозначен общий резервуар разнообразия (о нем см. выше, начало статьи).
А в заключении статьи новый номогенез прямо-таки господствует:
«Исходной организацией для формирования первых... тетрапод служат
девонские кистеперые рыбы. Приобретение этими рыбами тетраподопо-
добных признаков происходило параллельно и локально мозаично, осуще-
ствляясь в разных филогенетических линиях и в геологически разные от-
резки времени, придавая их эволюции вид сальтаций. Тетраподоподобные
признаки носили скорее всего характер преадаптаций...120» (с. 25).
Такое течение событий характерно, как мы знаем, для всякой «зации».
Кто же из учёных всё это осознал? И почему «скорее всего»? Об этом не
сказано. В качестве теоретических работ по тетраподизации в статье указаны
только работы дарвинистов, вышедшие 35-70 лет назад, так что новая, противо-
речащая им, идеология подана как выведенная (кем-то не названным) прямо из
наблюдений (с. 13). Этого, как известно методологам, не бывает.
Мне могут возразить, что важно не то, кто был первым, а сам факт вхожде-
ния новых идей в научную практику. На это отвечу: нет, анализ источника не-
существен лишь тогда, когда спорят о приоритете, однако у нас речь идет о сути
концепции, а тут выявление исходных положений просто необходимо.
Если номогенезом называть одну лишь книгу Берга, а достижения нового
номогенеза приписывать авторам, ее отрицавшим, то понять концепцию тетра-
подизации (тем более - «зации» вообще) вряд ли можно, и уж во всяком случае
не удастся ввести новые идеи ни в практику исследований, ни в преподавание.
Ну как, к примеру, объяснить ученику тезис о преадаптации, если в том па-
раграфе, где она упомянута [Воробьева, 2008, с. 24-25], аргументов нет, а есть
только упоминания имен А.Н. Северцова, Догеля и Шмальгаузена? Ведь та пре-
адаптация, о которой у Воробьевой идет речь, при советской власти неизменно
отрицалась как идеализм, и ее оставалось либо поносить, либо не упоминать.
Вместо нее, как это обычно в дарвинизме, допущено было к рассмотрению
иное явление, благонадежное («смена и расширение функций»), которое и было
объявлено причиной преадаптации - в частности, причиной превращения плав-
ника в лапу. Все противоречия с фактами либо опускались (такова полная не-
функциональность многопальцевой структуры кистеперых рыб), либо прямо от-
рицались (таково участие психики в акте смены функций, третируемое как пси-
холамаркизм - см. Приложение 1, фр. 2). Как рассказать это нынешнему учени-
ку (читающему новую литературу), не вызвав у него горькой усмешки?
Итоги советской дискуссии о преадаптации в свое время подвел историк нау-
ки А.Б. Георгиевский. Подтвердив, что самим термином допустимо пользоваться
только в смысле смены или расширения функций, что преадаптацию в смысле
Кэно не следует даже обсуждать, ибо это - телеология, он, тем не менее, напом-
нил о поразительном опыте И. Гармса [Harms, 1935] над рыбами мангровых за-
120 О преадаптации как эдванте см. выше, в п. 1.
329
рослей. Считалось и считается, что ползанье рыб среди зарослей способствует об-
ращенью плавников в лапы, и вот мы читаем [Георгиевский, 1974, с. 126]:
«Опыты Гармса показали, что инъекция гормона щитовидной железы (ти-
роксина) молодому животному Periophthalmus вызвала утолщение эпидер-
миса, удлинение конечностей, уменьшение слуховых косточек и возник-
новение других признаков, необходимых для длительного пребывания на
суше и характерных для ближайших предков наземных позвоночных».
Георгиевский не стал этого комментировать, и не надо было ждать от него
ничего такого, но мы-то сейчас комментировать подобные факты обязаны.
Если написанное верно, то налицо шаг к кистеперости у рыбы другого отря-
да. Гормон лишь стимулировал этот шаг, тогда как сам шаг демонстрирует реф-
ренную структуру (диасеть) возможных вариантов. Не так уж важно даже, сколь
аккуратен был опыт - важнее то, что номогенетическая идея была высказана и
никого тогда не привлекла (Георгиевский привел лишь одну работу 1962 г.).
Если бы смысл опыта был тогда понят и использован научным сообщест-
вом, мы бы сейчас знали о диасети куда как больше, чем знааем. Но понимание
не извлекается прямо из наблюдений, а о номогенезе Гармс мог и не знать.
Сейчас обстановка совсем иная. Палеонтолог Дженнифер Клэк (Англия) по-
казала, что новейшие данные вновь заставляют видеть процесс превращения
плавника в лапу именно как преадаптацию в смысле Кэно. По ее свидетельству,
«было обнаружено много новых окаменелостей, которые... полностью пере-
вернули прежние представления о происхождении и эволюции ранних тет-
рапод... Акантостега имела конечности, оканчивающиеся пальцами, но эти
ноги не были приспособлены для наземного существования... акантостега
была проамфибией, т.е. лишь “зачатком” четвероногого животного, полно-
стью адаптированным] к жизни в воде... Всё меньше подтверждений нахо-
дила гипотеза, согласно которой выход на сушу сопровождался формирова-
нием конечностей со ступнями. Напротив... конечности тетрапод сформи-
ровались тогда, когда они обитали в воде» [Клэк, 2006, с. 53-54].
Именно об этом, т.е. о подлинной преадаптации в смысле Кэно, идет речь в
нынешней палеонтологии, именно такая преадаптация есть компонент нового
номогенеза, и именно о ней надо, на мой взгляд, говорить и писать прямо, если
мы хотим изучать природу, а не только царящие в учебниках мнения о ней.
4. О флорогенезе
Исаак Ньютон, основатель классической физики, полагал в старости глав-
ным своим достижением богословские труды и исправление библейской хроно-
логии. Мейен до старости не дожил, но и он успел сменить приоритеты: в по-
следние годы жизни ушел от общих проблем эволюции к конкретным пробле-
мам палеоботаники, а рефрены едва упоминал и вообще не печатал о них ничего
нового. В последний же месяц жизни даже назвал (беседуя со мной) главным
своим делом завершение каталога ископаемых растений.
И каков же итог? Что вошло в науку, что ожидает часа, а что определенно из
нее выпало? Каталог был в некотором смысле завершен и в таком же смысле из-
330
дан (М-325) - трудами его коллег, российских и английских. Правда, в таком
виде он оказался всего лишь приложением к известному каталогу [Anderws,
1970], да и в Россию почти не попал. Называть это главным итогом жизни вряд
ли можно. Если же говорить о достижениях Мейена в теоретической биологии,
то главным мне видится построение основ диатропики, как она изложена в дан-
ном Сборнике. Но она осталась бы мало кому интересным умствованием, если
бы не работала в практике и, прежде всего, в палеоботанике. А она работает.
Во-первых, детально описанные Мейеном две внетропические ветви флород-
рева - южное и северное (см. рис. 8), образуют параллель, которую не удается све-
сти к актам независимого приспособления (ни в форме отбора, ни в форме
прямого влияния среды обитания): «Роль естественного отбора и непосредствен-
ной адаптации в начале становления семейств и таксонов более высокого ранга у
высших растений сомнительна и, во всяком случае, не была убедительно показа-
на» (М-295, с. 307). Естественно видеть в этой параллели рефрен или совокуп-
ность рефренов, одну из самых общих в ботанике.
Во-вторых, Мейеном описаны на ископаемых растениях первые три эдван-
та, причем идея транзитивного полиморфизма (транпола) помогла Мейену вы-
явить самую суть феномена флорогенеза .
В-третьих, усмотрен в составе флорогенеза феномен, который был позже опи-
сан мною как «грубая филогения», тоже есть следствие транпола.
Мейен изобразил нижнюю часть древа флор121, оно было довершено М.А.
Ахметьевым и выглядит вполне солидно (рис. 10). Но изъян нового древа в том,
что оно достроено тем же самым ретроспективным методом, который справедли-
во осуждает Э.И. Воробьева (см. п. 3): от принятой автором нынешней системы
флористических царств оно уходит вниз (в прошлое), дабы соединиться в мезозое
с древом Мейена, построенным по совсем иному принципу - синхронистическо-
му. Естественно, вышли нестыковки. Например, «обломанная» в перми северо-
американская ветвь флородрева вырастает в олигоцене из совсем иного ствола.
Но в целом пользоваться древом можно.
Наоборот, эскизно намеченное Мейеном древо таксонов (рис. 6) никакого
развития не получило. Его воспроизводят, словно икону, хотя теперь, через чет-
верть века, это странно - ведь его предварительный, пробный характер очеви-
ден. Давно пора либо достроить его, либо признать, что оно из науки ушло. И в
этом не будет ничего для памяти Мейена обидного, ибо в науке такое обычно.
Ведь не перестаём же мы видеть в Копернике основателя нашей астрономии,
хотя от его системы (где Солнце - центр Вселенной) мало что осталось.
Достроить древо не удалось по многим причинам. Главное - почти ничего
не удалось с цветковыми, а они -сердцевина ботаники. Единственное, что уда-
121 На тот момент. Позже выяснилось [Мосейчик Игнатьев, 2005], что развитие флор может
быть выявлено еще ниже, в верхнем девоне, растительность которого прежде казалась гео-
графически однородной. Схема ранней эволюции флор не выглядит древом (рис. 16).
331
лось - А.Б. Герман подкрепил гипотезу Мейена об их происхождении от бенне-
титов (см. АСМ, с. 534), так что таксон покрытосеменных, висящий на схеме
рис. 6 без опоры, можно мысленно закрепить (при учете блочности). Независи-
мо от того, как сложится судьба гипотезы, на сегодня это явный успех.
Как отмечено в начале статьи, с диатропической точки зрения гипотеза уже
достаточно обоснована, так как указаны блоки, участвовавшие в процессе ангиос-
пермизации. Что же касается господствующей (филогенетической) точки зрения,
то данную гипотезу, по всей видимости, не удастся обосновать всерьез никогда,
как не удалось до сих пор обосновать никакую филогению вообще - все так назы-
ваемые филогении на поверку оказываются семофилогениями (см. с. 27, 95).
Надежда Мейена на то, что филогения, хотя бы грубая, окажется господ-
ствующей внутри таксона покрытосеменных, пока что не оправдалась (хотя от-
дельных семофилогений построено довольно много). Однако сам феномен гру-
бой филогении остается, на мой взгляд, очень существенным, и вот почему.
Дарвин хотел своей концепцией объяснить, как появилось наблюдаемое
многообразие организмов, которое представлял себе исключительно как иерар-
хическую систему таксонов. Имея до (работ Геккеля) самое смутное представ-
ление о возможных родственных связях животных, он составил свою уверен-
ность в эволюционном родстве групп животных именно на легко заметных фак-
тах грубой филогении. (Так, считалось, что кошачьи и псовые явно имеют об-
щего предка, а все кошачьи, в свою очередь - своего предка и все псовые - сво-
его.) Полагаю, что именно этот очевидный ему факт Дарвин и распространил
вниз, на свою воображаемую процедуру «происхождения видов». Вот почему
его единственный рисунок был древом, но не имел никаких названий.
С этой точки зрения понятно, почему дарвинизм так и не смог стать теори-
ей: единственная в нем наглядная картинка итога эволюции относилась к выс-
шим для той поры таксонам (семействам, отрядам и классам), тогда как единст-
венная наглядная картинка механизма эволюции - к внутривидовой изменчиво-
сти, т.е. к низшим таксонам (расам). Правда, Геккель в 1866 году на время спас
положение, наполнив родословное древо конкретными (местами вплоть до рода)
названиями. Это был титанический труд, однако его древо через сто лет стало
выглядеть, как любил говорить Мейен, собранием исторических анекдотов. По-
нятно теперь, почему: филогения реально описана толька как грубая. (Тот факт,
что грубая филогения - отнюдь не филогения в обычном смысле, мы с ним обсу-
дить успели (М-344, с. 133), а вот то, что она - единственный реальный источник
дарвинских представлений о дивергенции как механизме эволюции, не успели.)
Вернёмся к флорогенезу. Представления о фитоспрединге (возникновении
растительных таксонов у экватора и растекании их к полюсам) претерпели после
Мейена коренные изменения. Первоначальные увлечения этой схемой, выра-
зившиеся в попытках увидать тот же феномен у животных (зооспрединг - АСМ,
с 510), не дали сколько-то крупных результатов. В частности, в настоящее время
зоны таксонообразования обнаруживаются повсюду, а не только в низких широ-
332
тах, и не столь глобальны.
Палеонтолог Д.Е. Щербаков в недавней статье поместил параграф «Энто-
моспрединг», но в самом тексте этого слова нет, а приведенные примеры гово-
рят скорее против самой идеи зооспрединга. И он заключает:
«В перми и триасе низкие широты были заняты в основном аридными и
семиаридными зонами, поэтому важную роль в генерации новых таксонов
играли экотоны между семиаридными и умеренными зонами, располагав-
шиеся за пределами тропического пояса» [Щербаков. 2008, с. 20].
Экотон - это переходная полоса между флористическими областями. Те-
перь многие полагают, что он часто служил и как источник таксонообразования,
и как убежище (рефугиум) для сохранения древних таксонов:
«Экотоны во многих случаях характеризуются высоким разнообразием и
служат не только рефугиумами, но и центрами видообразования [Kark, van
Rensburg, 2006]» [Щербаков. 2008, с. 22].
На множественность зон интенсивного таксонообразования растений еще
ранее указывала Ю.В. Мосейчик [2005], выявившая в раннем карбоне северного
полушария три таких главных пояса - тропический, субтропический и умерен-
ный. Там же она отметила номогенетический аспект флорогенеза:
«В целом, развитие флор раннего карбона демонстрирует явную ортогене-
тическую направленность. В середине раннего карбона (приблизительно
335 млн лет назад) происходит глобальная смена растительности, связан-
ная с эволюционной радиацией древних голосеменных растений... По-
следняя идет практически одновременно в разных регионах тропической
области (современные Северная Америка, Европа, Казахстан, Китай)».
Замечу, что в ту эпоху эти 4 региона были разделены океаном.
Через три года эту «ортогенетическую направленность» Мосейчик предло-
жила использовать для целей практики стратиграфической корреляции:
«Основные трудности межрегиональных корреляций связаны, с одной сто-
роны, с высоким провинциализмом каменноугольных флор, который не
позволяет опираться на руководящие формы. С другой стороны, множест-
венные параллелизмы, характерные для растений, растущих в сходных
ландшафтно-климатических условиях, заставляют исследователей выде-
лять одни и те же роды и виды в географически разобщенных флорах, что
приводит к ложным стратиграфическим выводам. Выход из этой ситуации
видится в обращении к изучению эволюционных смен растительности, ко-
торые могут быть положены в основу выделяемых фитостратонов».
А именно,
«Причины многих из описанных флористических смен пока окончательно
не установлены. Для субглобальной флористической смены в середине ви-
зе222 показано, что она не связана с какими-либо заметными, едиными од-
нонаправленными изменениями условий среды... Вероятно, все три круп-
122 Визейский ярус раннего карбона (второй от его начала) - Ю. Ч.
333
ных рубежа в эволюции флор раннего карбона не зависели напрямую от
перемены окружающих условий и были связаны с автохтонной эволюцией
видов (автогенезом по Л.С. Бергу...). Это позволяет использовать их в ка-
честве основы для межрегиональных корреляций» [Мосейчик, 2008].
Признание хотя бы возможности того, что перестройки растительности бы-
вают независимы от изменений климата, выглядит странно и даже дико - ведь
связь растительности с климатом очевидна и самой Мосейчик в начале ее статьи
описана. Однако факт есть факт: увязать смены флор со сменами климатов ни-
кому еще всерьез не удалось (хоть попыток и было много), так что давно пора
признать эту связь неоднозначной и нечёткой - иными словами, признать собст-
венную направленность (автогенез) как одну из компонент развития флор.
Замечу, что на синхронность смен разобщенных флор Мейен тоже обращал
внимание, но в его дни совсем отрешиться от идеологии полной зависимости эво-
люции от влияния среды в пользу номогенеза было слишком трудно, если и воз-
можно (хотя до него такие попытки делались). Мейен настойчиво искал климати-
ческую подоплёку во всяком акте смены флор, но, по-видимому, без особого ус-
пеха. И вот Мосейчик сделала за него нужный шаг: автогенез представлен как од-
на из непременных компонент эволюции. Это - новый номогенез в практике.
5. Три кита
Сам по себе номогенез (в том смысле, в каком это слово используется в дан-
ном Сборнике) безразличен к проблеме движущих сил эволюции, поэтому спосо-
бен быть только базой теории, но не самой теорией. Любопытно выяснить, как
Мейен представлял себе то, чем сам не занимался - движущие силы эволюции. В
1970 году в письме Любищеву он выразился следующим образом:
«Итак, три основных кита: всеобщая кристаллография с двумя типами де-
терминации, психика, симбиоз+пол. Взаимодействие этих трех перемен-
ных определяет развитие живого» (М-352, с. 28123, письмо от 13.06.70).
Поясню: под всеобщей кристаллографией Мейен понимал законы морфогенеза,
где два типа детерминации (контактная и дистанционная) маркировали две идеи
эмбриологии - традиционную и ту, которая признаёт биологическое поле. Далее,
«психика» означала признание психоламаркизма, но Мейен не просто указал на
психику (одну из форм активности) как на фактор эволюции, а привлёк идею пан-
психизма (убеждения, что мир одушевлен как целое):
«Возможно, теперь самое время вспомнить идеи психического континуума,
в котором отдельные сознания - сгустки (об этом хорошо написано у Шре-
дингера в “Что такое жизнь”)».
А выражением «симбиоз+пол» Мейен обозначил ту идею, которая с XIX ве-
ка известна как социабилизм. Его он формулировал как ряд:
«Налипание органических частиц в первичном бульоне на минеральные
частицы..., образование плодового тела у миксомицетов и миксобактерий,
образование сверхорганизмов, симбиоз разных организмов - скорее всего,
123 Частично опубликовано ранее; данную цитату см.: М-348, с. 111; ИМ, с. 122.
334
звенья одного ряда. Мне кажется, что сюда же надо отнести конъюгацию,
а значит, и любовь...» (там же).
(Замечу, что этот ряд притяжений - от налипания до любви - в моих более
поздних работах именуется старым химическим термином «сродство».)
Независимо от того, насколько эти «три кита» описывают реальную эволю-
цию, схема - яркая и побуждала к поиску. В частности, видны активность и со-
пряженность всего со всем (лежащие теперь в основе АСМ), но видно и многое
другое, например, космизм сознания. К сожалению, через 5 лет, когда мы с Мейе-
ном познакомились, его занимала лишь одна сторона эволюции - преобразование
разнообразия (то, что выше он обозначил как всеобщую кристаллографию).
Почему, сказать не берусь. Есть такое мнение: «Представленная выше про-
грамма номогенетических исследований остановилась на первых исследователь-
ских шагах», поскольку «ее автор боялся и не хотел прослыть маргиналом»
(ИМ, с. 130). Некоторый резон тут есть, но вопрос глубже. Да, публично Мейен
избегал слишком смелых мыслей, и не раз упрекал меня в максимализме, когда
я поступал иначе. Естественно, если он в письме Любищеву сказал больше, чем
где-либо публично; однако о «трех китах» он ни разу не поведал и мне, хотя мы
часто говорили такое, за что «давали срок». Вернее, что тут причина была не
столько в осторожности, сколько в смене типа деятельности.
Тогда (1970) «три кита» явно занимали его всерьез, он очень ждал ответа
Любищева, но тот ответил на цитированное письмо кратко и не дал никакой
оценки «трем китам» (предпочтя, как обычно, увести разговор с явлений на по-
нятия), а иных собеседников в то время у Мейена, как он признавал, не было.
После смерти Любищева (1972) Мейен сам увлекся «метанауками» и стал
охотно переходить с обсуждения явлений на обсуждение понятий: «Я понял, что
во многих областях биологии меня интересует больше мета-компонента, чем
конкретные утверждения об об органическом мире. 1972 г. явился отрефлекси-
рованным началом моих мета-изысканий» - писал он в 1985 г. (МСП, с. 40).
Можно сказать даже больше: он взял на себя роль «методологизирующего скеп-
тика» (ИМ, с. 127), которую прежде столь блистательно играл Любищев.
Вполне вероятно, что при этом тема «трех китов» выпала из его планов124
как не стоящая требуемых затрат - Мейен решительно стремился сделать за
жизнь как можно больше и в этом преуспел. Нам же надо прояснить саму тему.
Еще до указанного письма Мейен писал в статье М-73:
124 Но не была им забыта. В 1978 г., в статье М-163, формально носившей обзорный ха-
рактер, Мейен писал: «Речь, разумеется, не идет о прямом наделении атомов или элемен-
тарных частиц “сознанием” или “свободой воли”, а лишь о “протопсихических свойст-
вах” атомов... Протопсихические свойства атомов преобразуются в психические свойст-
ва их агрегатов... При этом часто указывают на то обстоятельство, что по мере развития
исследований психические явления эмпирически наблюдаются на все более низких сту-
пенях биологической организации». Своего отношения к теме он не высказал, ограни-
чившись замечанием, что это - тоже своего рода редукционизм.
335
«Что толкнуло два организма навстречу друг другу - своеобразный канни-
бализм, прапраинстинкт или что-нибудь другое? Этого никто не знает. Но
уже то, что половой процесс широко распространен среди жгутиковых ор-
ганизмов, показывает, что половыми вопросами живые существа тоже “за-
интересовались” еще до разделения на два царства».
Сейчас мы знаем, что вопрос тут неверно поставлен: половой процесс стар,
как сама жизнь, так что спрашивать можно только о причине, путях и времени
возникновения раздельнополости, а ее-то у жгутиковых нет. Но важно не это, а
сам интерес автора к поведению на микроуровне. Об этом читаем там же:
«Если аккуратно перерезать проходящую в стебле жилку, то уже через не-
сколько дней можно видеть, как в обход надреза начинают образовываться
новые клетки сосудистого тяжа. Вскоре они соединят концы разорванной
магистрали. Во всех этих процессах клетки, игравшие совсем другую роль
и получившие свою специализацию, вдруг преображаются, начинают де-
литься и в конце концов становятся подобными утраченным клеткам».
Едва отправив письмо, Мейен отправился в Ленинскую библиотеку и про-
чел там знаменитую статью Владимира Яковлевича Александрова о клеточном
и внутриклеточном поведении [Александров, 1970], где говорилось о целесооб-
разности оного поведения и полной непонятности его механизмов, а потому
предлагалось учредить новую науку - цитоэтологию. (Как говорил баснописец
Иван Крылов, воз и ныне там.) Поскольку за данным журналом Мейен не сле-
дил, а статья лежала в «текущей периодике» второй месяц и о ней уже заговори-
ли, то можно смело утверждать, что он знал о ней, когда писал письмо.
Статья поразила Мейена, и назавтра он снова писал Любищеву (М-352):
«Она очень симптоматична, читая ее последние страницы, я все время ду-
мал, неужели лед скоро тронется. Жаль, что я прочел эту статью после, а
не до моего письма Вам, кое-что можно было бы добавить, кое-что иначе
сформулировать. Вообще, эта “цитоэтология”, выражаясь словами из Биб-
лии, “камень, который отвергли строители, сделался главою угла”».
Для уяснения тогдашних умонастроений ученых стоит еще добавить, что в
течение 1960-х годов появилось несколько работ, убедительно показавших, что
разумное поведение свойственно не только «низшим» организмам, но и клеткам
(АСМ, с. 679-681). Что же касается насекомых (вершины мира наземных беспо-
звоночных), то до сих пор часто цитируется тогдашняя статья [Brower, 1966],
описавшая, как муравьи дружно строят заслоны от источника гамма-излучения,
установленного людьми близ муравейника. (Это означает, что коллективная их
работа в данном случае сознательна и не определяется наследственными про-
граммами.) Всё это наводило на мысль, что психика распространена в природе
гораздо шире, чем принято думать, и, в частности, не обязательно связана с моз-
гом и даже с нервами. Так что внимание Мейена к панпсихике не в пустоте ро-
дилось. Способствовал ему и взгляд физиков на материю как на сгусток полей,
тогда уже обсуждавшийся среди любителей философии науки.
336
Осталось выяснить, насколько идея «трех китов», высказанная Мейеном до
его работ по диатропике, оказалась с ними в согласии. «Всеобщая кристалло-
графия» позже вся выстроилась в рефрены, это понятно. Она - то, на что дейст-
вует активность, то, что активностью формируется. Два других «кита» явно от-
носятся к самой активности, однако и тут налицо рефрены - например, «пове-
денческие рефрены» (АСМ, с. 679). Один из них демонстрируют паразиты, из-
меняющие поведение своих жертв с адаптивного на самоубийственное.
Панпсихизм, почти явно признанный Мейеном в указанном письме, сам
имеет рефренную природу, поскольку является аспектом параллелизма бытия и
мышления (о котором см. ниже, п. 6). Здесь же надо заметить, что чуть ранее, в
статье М-73, Мейен описал параллелизм строения клетки у животных и у расте-
ний - фактически тоже рефрен, притом столь же важный и общий125.
Сложней всего с тезисом «симбиоз+пол». Зачем Мейен объединил эти два
явления? Ориентировочный ответ отыскался в той же статье М-73: в половом
процессе, как и в симбиозе, происходит соединение того, что прежде существо-
вало порознь. Жизнь - не только непрерывное разделение при делении клеток и
размножении многоклеточных, но и регулярное соединение - как для размно-
жения (половой процесс), так и для совместной жизни.
Последнее общеизвестно в форме симбиоза и в форме жизни группами (се-
мейными, охотничьими и пр.), но Мейен обратил внимание еще и на третью
форму - соединение бесполых одноклеточных в единое плодовое тело у миксо-
мицетов и миксобактерий (есть и другие, которых он не упомянул).
Это весьма характерно для его способа мыслить - малозаметное (а потому
для большинства маловажное) явление взято как связующее звено между явле-
ниями, вроде бы никак не связанными. Речь, разумеется, не о родственной свя-
зи, а о едином законе преобразования, каковой позже Мейен надеялся увидеть в
каждом рефрене. (В данном случае он приблизительно нащупал ту общность в
проявлении активности, которая у меня фигурирует как сродство.)
Словом, фактически рефрены занимали Мейена уже тогда.
6. О параллелизме бытия и мышления и об эффективности ма-
тематики
Увы, сам Мейен мало рассказал о природе рефрена. В беседах он говорил о
«типологической упорядоченности мира» как о чем-то вполне естественном, но
125 Прочтя это место в рукописи, И.А. Игнатьев сделал замечание: «Выводя рефрен за об-
ласть морфологии, органической формы, Вы делаете это понятие настолько общим и абст-
рактным, что построение какой-либо содержательной теории, даже философской, ... дела-
ется фактически невозможным. У Вас рефрен абстрактнее фрактала». Сравнение странно
(фрактал так же абстрактен, как интеграл или вероятность - в чем упрёк?), а вот в отноше-
нии слишком широкого понимания рефрена резон есть, но тут я следовал Мейену, а он по-
лагал данное понимание годным всюду (М-165, с. 498), в чем прав был лишь отчасти. На-
пример, описание физиологии через рефрены хоть и возможно, но, по-моему, не более со-
держательно, чем описание В.В. Налимовым мышления через вероятности.
337
мне известна лишь одна на сей счет печатная его фраза: «Упорядоченность объ-
ектов в синхроническом срезе и во времени - лишь аспекты общей упорядочен-
ности, т.е. естественной типологии» (М-165, с. 505). Обращение Мейена к ана-
логии с грамматикой (см. ранее, с. 49) и его уверенность в том, что типология -
«единственный рабочий способ выявления и исследования уровней системно-
сти126» (М-323, с. 6), наводят на мысль, что он понимал рефренную структуру
природы как один из аспектов параллелизма бытия и мышления.
Эта почтенная идея пронизывает всю историю европейской философии. Она
вполне ясно выражена еще у Парменида и нашла изящное выражение у Декарта,
который был уверен, что понял устройство мира: мир разумен потому, что Бог
устроил его по законам математики. Вскоре Спиноза уточнил Декарта: если Де-
карт видел в математике прежде всего интуитивное озарение (таковым было,
например, введение им системы координат), то Спиноза - доказательство. В
этой (спинозовой) форме идея и стала господствующей.
Изъян данной концепции был в том, что замена интуиции доказательством
не оставляла в жизни человека места ни самосознанию, ни свободе воли. Вопрос
этот ясно высветил Вильгельм Виндельбанд [1913, с. 174, 184] (философ, кото-
рого, кстати, весьма ценил Мейен). Тем самым, принятие обществом позиции
Спинозы означало неминуемый разрыв естествознания с психологией. На эту
болевую точку науки как целого и указал, полагаю, Мейен.
Позже математика понемногу вытеснила идею Бога из умов почти всех учё-
ных, и проблема параллелизма бытует теперь в форме вопроса: почему матема-
тика, мысленная конструкция, столь непостижимо эффективна при описании ре-
ального мира? Формулу «непостижимая эффективность математики» высказал
полвека назад математик Юджин Вигнер, и с тех пор проблему по-разному ком-
ментируют популяризаторы. Так, математик Морис Клайн [1985], изложив раз-
личные западные точки зрения (восточные см. в конце п. 7), высказался в том
духе, что хотя общеприемлемого ответа нет и ждать его не приходится, однако
связь бытия и мышления налицо, чем и следует пользоваться.
Наоборот, другой популяризатор, физик В.В. Низовцев уверен, что «изо-
морфность математических моделей, описывающих процессы в физике, химии,
психологии, экономике и т.д., свидетельствуют о том, что эти модели не затра-
гивают существа изучаемых процессов», и делает «единственно возможный вы-
вод о том, что математические методы не могут служить носителями знания о
существе механизмов» [Низовцев, 2000, с. 64, 65]. Выражение «единственно
возможный» звучит здесь забавно (вопрос о системном сходстве объектов раз-
личной природы даже не упомянут), однако некоторый смысл тут есть.
126 Системность - частый у Мейена термин, появившийся под влиянием работ философа
Ю.А. Урманцева (см. ИМ, с. 122-123). Так обозначены те свойства целостности, которые
не зависят от природы составляющих ее элементов. Здесь эти слова означают, что, по
Мейену, рефренная структура неизбежна в любой системе.
338
Приведу пример из близкой мне области: подавляющее большинство слу-
чайных явлений подчиняется либо гауссовой (в широком смысле слова), либо
квази-гиперболической («виллисовой», «ципфовой» и т.д.) статистике, и это
часто служит примером «непостижимой эффективности». Однако принадлеж-
ность явления к первой статистике не говорит о его механизме ничего, а при-
надлежность ко второй - очень мало (точнее, она указывает на возможность на-
личия системной случайности, но не гарантирует ее - анализ см. в ОПС).
Да и сама «непостижимая эффективность» далеко не всегда имеет место
там, где ее декларируют. Так, третий популяризатор, В.Н. Тутубалин, профессор
ТВ, часто указывающий на трудности обоснования ТВ (и никогда не объясняю-
щий, в чем их суть), обратил внимание на то, что эффективность стандартной
ТВ сильно преувеличена. Он заявил, что устойчивость частот (без которой при-
менение ТВ теряет смысл) обычно в прикладных задачах места не имеет - экс-
перимент чаще всего показывает, что этой устойчивости нет. При этом
«за 200 лет, прошедших со времен Лапласа и Гаусса, наука не добилась
продвижения в фундаментальном вопросе - когда возникает статистиче-
ская устойчивость. Узнать, так обстоит дело или нет, мы можем только из
эксперимента» [Тутубалин, 1993, с. 98].
Однако еще важнее неявное признание Тутубалина в том, что ТВ неспособ-
на аккуратно описать даже свой исходный простейший опыт - бросание монеты:
оказывается, добиться хорошего приближения частоты выпадения герба (точно-
сти 0,1% при 24 тысячах бросаний) удалось только одним путем - путем пре-
кращения опыта в нужный момент, когда текущее значение частоты наиболее
приблизилось к ожидаемой, к 1/2 (анализ см.: ОПС, с. 98). Добавлю, что обычно
частота, даже там, где принято называть ее устойчивой (например, при бросании
монеты), сходится не к определенному числу, а к некоторому интервалу, како-
вой не сокращается с ростом количества испытаний выше некоторого.
В парадном примере сторонников математизации как основы теоретизации
естествознания - планетной механике - тоже часто приходится довольствовать-
ся малой точностью, около 10 процентов [4-18]. Всё это говорит о том, что «не-
постижимая эффективность математики» - отнюдь не факт, точно доказанный, а
всего лишь тенденция, одна из тех, о каких мы говорили во многих из выше-
приведенных статей. Без таких тенденций никакая наука не обходится.
Диасеть - одна из них, и рефренный анализ был задуман Мейеном как некое
обобщение математической физики (а может быть, и самой математики): мате-
матическая формула выступила у него как тот частный тип рефрена, когда зако-
номерность выражается строго - логически и количественно (точнее см. преды-
дущую статью). Еще важнее, что даже в области «точного» соответствия фактов
формулам вопрос об истинности теорий вызывает жестокие споры, которые ре-
шаются отнюдь не на языке формул и даже не фактов, а победой той или иной
философии природы - примеры см. в п. 2 первой статьи Сборника.
В биологии ситуация очень похожа: любое (даже предполагаемое) сходство
формул с опытными данными может быть объявлено доказательством биологи-
339
ческой теории (такова вся математическая генетика популяций, а также «адап-
- 727
тивныи компромисс» , приводимые в качестве доказательства справедливости
дарвинизма), тогда как полное (с точностью самих данных) совпадение форму-
лы с опытом просто никем не упоминается, если противоречит господствующей
философской установке. В качестве примера мне уже довелось напоминать (4-7;
ОПС) изящную математическую теорию ветвящихся процессов, которой мате-
матик Гаролд Юл объяснил в свое время гиперболическую статистику распре-
деления видов по родам, открытую незадолго до того Джоном Виллисом. Работа
Юла [Yule, 1924] исходила из неприемлемого тогда (но многим очевидного ны-
не) понятия видовой и родовой мутации, была встречена молчанием (хоть и
опубликована была в весьма престижном журнале), так что даже сам Юл позже
на нее не ссылался. Вновь в научный оборот она стала проникать в 1960-х годах,
в тех работах, где статистическая ПМ-уступала место системной127 128.
Зато те же ветвящиеся процессы, когда они подтверждали дарвинизм, быва-
ли признаны блестящим его доказательством, несмотря на произвольные исход-
ные допущения и вопиющее расхождение выводов с наблюдаемыми фактами.
Статьи на эту тему (Ч-З; 4-5) никем, насколько знаю, никогда упомянуты не бы-
ли, хотя появились в самом читаемом нашими генетиками журнале и некоторы-
ми (насколько знаю из бесед) были прочтены с огорчением.
Это значит, что и отмеченное выше пренебрежение большинства учёных к
концепции Мейена может быть связано не с ее низким качеством, а с его фило-
софской позицией, намного опережавшей таковую его коллег. Фактическая
обоснованность диатропики на сегодня очевидным образом превышает таковую
иных концепций - это фактически подтверждают и их приверженцы, когда ис-
пользуют ее, на нее не ссылаясь. Они полагают простодушно, что, отбрасывая
непонятную (а потому неприятную) им философию как хлам, «берут из теории
всё ценное». На самом деле так можно брать лишь крохи - те, что оказались
доступны пониманию. То есть, повторю, отказ от философии есть худшая из
философий, поэтому вопрос обоснования диатропики следует прояснить.
127 Так именуют то утверждение, что объектом отбора служит не признак, а сбалансиро-
ванная совокупность приспособлений. Основной апологет данного подхода палеонтолог
А.П. Расницын согласен с тем известным фактом, что «ни одна система не может быть
оптимизирована одновременно более чем по одному параметру» [Расницын, 2002, с. 14],
т.е. сам признал свой подход нелепым, однако положил его в основу своей модели эво-
люции, попросту назвав всю исследуемую совокупность искомым единственным пара-
метром. Пучение природы заменено словесным оборотом. Подробнее см. ЭЭД; ЗЭ.
128 Анализ см. в ОПС, а здесь замечу, что Мейен, видевший в статистике Виллиса общий
закон природы, тоже не хотел принимать теорию Юла как слишком частную: «Любопытно,
что сам Дж. Виллис не усмотрел в открытой им закономерности общего свойства таксоно-
мических систем и считал ее всего лишь следствием того, что новые таксоны появляются
не путем отбора мелких изменений, а “таксономическим прыжком”» (М-163, с. 156).
340
7. О структуре пространства и природе рефрена
Мир оказался устроен совсем не так, как нас учили и как учим мы. Принято
считать, что исходное состояние всего сущего - простота и однородность, что
сложность и разнообразие - всегда итог эволюции (позиция, идущая от филосо-
фа Герберта Спенсера, исповедовавшего ламарко-дарвинизм). Однако факты
давно в эту схему не укладываются. Да, в целом сложность и разнообразие рас-
тут с течением времени, но то тут, то там мы обнаруживаем в качестве исходно
наблюдаемых такие разнообразие и сложность, какие позже не достигаются.
Вот примеры: митозы одноклеточных, особенно низших, удивительно раз-
нообразны, а многоклеточных животных и растений - практически одинаковы; у
кистеперых кисть сложнее, чем у потомков; череп упрощается от древних че-
репных рыб к млекопитающим; у всех первобытных племён очень сложно и
различно устроена система браков; древнегреческий язык (самый древний евро-
пейский язык, известный нам из реальных текстов, а не реконструированный)
сложнее латыни, а она сложнее новых европейских языков.
Это обстоятельство многих приводило к вопросу: допустимо ли считать
всякое исходное состояние (которого мы почти никогда наблюдать не можем)
простым и неупорядоченным? В 1975 году Мейен писал (М-127):
«Это представление о вторичности и эктогенетичности (внешней обуслов-
ленности - Ю. Ч.) всякой упорядоченности унаследовано современной нау-
кой от донаучной метафизики и креационизма. Показательно, что пред-
ставления об эндогенетической (внутренне обусловленной - Ю. Ч.) приро-
де правильных форм природных кристаллов также не сразу были приняты.
В биологии же до сих пор господствует неявное убеждение, что первичная
упорядоченность форм живого привнесена извне... Это представление по
существу ни на чем не основано. Мы просто не знаем имманентных (внут-
ренне присущих объекту - Ю. Ч.) свойств биологического пространства,
его геометрию. Вполне правомерен как раз противоволожный постулат,
что специфика биологического пространства (Вернадский, 1965; Урман-
цев, 1971) имеет эндогенетический характер» (М-370, с. 256).
Итак, здесь предложено рассматривать биопроцессы (включая эволюцию)
как текущие в своем особом пространстве, заведомо неоднородном, и встал во-
прос о его структуре. Описать ее никто до сих пор не может, но некоторые свой-
ства этого пространства видны. Самым простым и наглядным выступает реф-
ренная структура разнообразия организмов (включая разнообразие их частей и
их сообществ). Мейен полагал это свойство основным, определяющим осталь-
ные. И главное, что он нам в этом плане (а не в плане конкретной палеоботани-
ки или стратиграфии) оставил, было понятие диасети, которую он (слишком
длинно) именовал рефренной структурой разнообразия.
Моё мнение не столь оптимистично (свойства объекта определяются не
только его положением в диасети), однако и мне ясно, что без анализа диасети
ни онтогенез, ни эволюцию не понять. Эволюция предстала не как порождение
разнообразия, а как преобразование одних разнообразий в другие.
341
В отличие от диасети химических элементов (плоской таблицы, открытой
Д.И. Менделеевым) всякая более сложная диасеть (в том числе химическая)
многомерна, так что обозревать мы можем лишь ее фрагменты, именуемые реф-
ренами (в качестве простого биологического примера на рис. 1 изображен уп-
рощенно рефрен конечностей позвоночных). Это обозрение приводит к выводу,
что диасеть многоуровнева, причем в процессе ее эволюции выявляются всё но-
вые и новые ее уровни. Например, в разных таксонах животных независимо по-
являются конечности, которые становятся из простых всё более и более слож-
ными, но не монотонно, а взлётами и падениями. Единой линии усложнения
указать нельзя, но сам тот факт, что усложнение имеет место, вне сомнения.
Словом, усложнение диасети - не точный закон, а тенденция (о тенденциях
см. с. 69), и понятно, почему надежда Мейена на формализацию его «типоло-
гии» оказалась напрасной: разнообразие и формализация совместимы плохо.
Даже в небесной механике, служащей примером формализации физики, точно
формализуема только Зато может оказаться вполне осмысленной противопо-
ложная задача - формализовать на манер биологии (то есть достоверно, но не-
точно, приблизительно) ту физику, где точные формулы дают мало. Именно та-
ков, если верно понимаю, пафос Низовцева (см. п. 6), который, кстати, процити-
ровал и рассмотренное выше письмо Мейена Любищеву. Взяв от Мейена «кон-
тактный детерминизм», но не «дистанционный», Низовцев вернулся, тем самым,
к Декарту, видевшему всякое взаимодействие как контакт - либо непосредст-
венный, либо через воображаемый эфир, заполняющий всякое пространство.
Отсюда Низовцев вывел (не будем обсуждать корректность вывода), что все
взаимодействия в природе должны быть непрерывными. В этом он по сути сле-
довал идее, которую высказал в 1972 году биоматематик Ренэ Том (Thom):
«Чтобы понять каждое частное микроскопическое явление внутри живого
существа, биолог должен для начала постулировать существование локаль-
ного детерминизма, а затем пытаться интегрировать все эти локальные де-
терминизмы в единую когерентную и устойчивую локальную структуру. С
этой точки зрения, основная проблема биологии является топологической,
поскольку топология это как раз та математическая дисциплина, которая по-
зволяет переходить от локального к глобальному» [Том, 2002, с. 116].
Интуиция у Тома была блестящая, но - классическая, дофрактальная. Через
три года (1975) Бенуа Мандельброт открыл (опираясь на идеи Гастона Жюлиа)
мир фракталов, и «переход от локального к глобальному» понемногу стали трак-
товать не в непрерывных понятиях топологии, а в дискретных, фрактальных. Это
была научная революция, равноценная той, что произошла, когда был открыт ма-
тематический анализ Ньютона - Лейбница, только теперь она прошла в обратном
направлении (тогда воцарилась идеология непрерывности и гладкости).
У нас о фракталах речь шла не раз, особенно - в предыдущей статье. О них у
342
Низовцева нет в указанной работе ни слова129, зато есть (и это важно) термин
«коллективное бессознательное», заимствованный у психолога Карла Юнга и
приложенный к мировоззрению современных физиков [Низовцев, 2000, с. 67].
Но у Юнга термин означает общую основу мышления людей, доставшуюся им
от глубокой древности, а у Низовцева - общую основу нынешнего научного
мышления, усвоенную при обучении. В чем она состоит, не сказано, но нужную
нам мысль давно уже сформулировал философ И.А. Акчурин [1974].
Речь идет об элементарном свойстве того пространства, в котором происхо-
дят интересующие нас события. Простейшим изученным классом событий были
механические движения, для которых основным свойством, изученным раньше
других, является предельный переход, позволяющий получить мгновенную ско-
рость (производную пути по времени) и мгновенное ускорение (производную
скорости по времени). Обладающие этим свойством движения именуются глад-
кими и замечательны тем, что на достаточно малом отрезке времени представи-
мы прямыми линиями. Пространство с таким свойством Акчурин назвал «ситэ
Декарта»130. Оно послужило бессознательной основой классической физики, а с
тем и всей науки того времени. Лишь сто лет назад, с рождением квантов и от-
носительности, понадобились новые ситэ - «ситэ Планка» для субмикромира и
«ситэ Эйнштейна» для сверхбольших скоростей, длин и масс. В них мир устроен
иначе: не допускает величин меньше (больше) некоторых. Эти ситэ сформули-
ровал тоже Акчурин.
Для описания фрактального мира мне тоже понадобилось ввести новое ситэ:
«Удобно ввести понятие ситэ Мандельброта, которое мы будем тракто-
вать как наблюдаемую в каждом масштабе структурированность. Но
структурировано - значит, не перемешано, т.е. такое ситэ вводится там, где
ни на каком уровне нет перемешивания, а значит, нельзя ввести вероятно-
сти - даже условные. Очевидно, что это понятие шире понятия фрактала,
поскольку не включает ни самоподобия, ни дробной размерности (хоть и
ориентировано на их описание)» (ОПС, с. 130).
Читателям, далеким от данной тематики, поясню: если трем предыдущим
ситэ свойственна идея масштаба (очень мелкие явления - ситэ Планка, средние
- Декарта, гигантские - Эйнштейна), то ситэ Мандельброта описывает внемас-
129 Хотя есть в предыдущей, написанной вместе с его аспиранткой Ольгой Панченко. Там
фракталы рассмотрены как аппарат для описания контактного детерминизма, а дистан-
ционный детерминизм назван «уступкой телеологии» со стороны Мейена [Низовцев,
Панченко, 1998, с. 149]. На мой же взгляд, фрактальное рассмотрение развития делает
оба детерминизма взаимодополнительными подходами к онтогенезу.
130 Само понятие «ситэ» ввел французский математик Александр Гротендик (1972), из-
вестный, в частности, умением вводить удачные понятия. Оно тоже выглядит весьма
удачным, поскольку французское la cite означает и город, и городок, и часть города, и
даже общежитие, а потому удобно для обозначения структуры, допускающей последова-
тельное изменение масштаба с сохранением исследуемого свойства. Наоборот, француз-
ское la ville означает город и ничего более.
343
штабные (масштабноинвариантные) структуры и явления. Одной из таких
структур является диасеть131, а одним из таких явлений - развитие (как индиви-
дуальное, так и эволюционное). Именно внемасштабность диасети позволяет
строить единую морфологию, единую систематику и выявлять параллелизмы.
В одном из параллелизмов Геккель простодушно увидал «повторение фило-
генеза в онтогенезе», хотя имел дело с внемасштабным параллелизмом развития
(онтогенеза и эволюции). Не будем упрекать его за это - то же самое многие
увидали и в других науках, едва эволюционная идея стала популярной. Напри-
мер, старинную параллель между взрослением человека и развитием общества
(см ранее, с. 166) тоже долго толковали по Геккелю [Иванов А.В., 1994].
Близок ко всему этому еще более общий параллелизм - бытия и мышления.
Разница в том, что его, кажется, никто еще не рассматривал как следствие эво-
люции. Наоборот, Гегель, например, видел саму эволюцию как следование при-
роды мыслям Высшего существа. Ни удивляться, ни возмущаться не стоит: с
более общей точки зрения, все акты параллелизма - итог актирефа. К актирефу
мы еще вернемся в п. 8, а здесь ограничусь аналогией: актирефу стоит удивлять-
ся не больше, чем одинаковому типу колебаний у маятника, электромагнитного
контура и химической системы (реакция Белоусова - Жаботинского). Если мир
устроен рефренно (а при его фрактальности это неизбежно - см. предыд. ста-
тью), то параллелизмы будут наблюдаться во всём.
Вообще, внемасштабность диасети позволяет выявлять «гомологические ряды»
на всех таксономических уровнях, что в ботанике впервые сделал Кренке, а выразил
в общенаучной форме Мейен. Именно внемасштабность - основа рефренов.
Меня нередко просят: «Нет, вы объясните, откуда на самом деле берутся
рефрены, объясните в понятных всем биологам терминах». Это - призыв объ-
яснить структуру мира, построенного на ситэ Мандельброта, тем «всем», кто
умеет мыслить только в терминах ситэ Декарта, в терминах контактного де-
терминизма. (На память приходит анекдот XIX века, где крестьянин просит
объяснить ему, где в паровозе «на самом деле» размещены лошади.) Для по-
нимания тут нужна новая картина мира (как для понимания работы паровоза
нужна идея механического, т.е. неживотного, двигателя). К ней призывает мой ре-
цензент (см. Приложение 2), и ей будет посвящен п. 9, здесь же надо сказать о ней
то, что требуется для завершения темы структуры событийного пространства.
А именно, сходство структур отмечено в пространствах самой разной приро-
ды. Приведу мнение четвертого популяризатора (трое названы в п. 6), психолога
А.В. Иванова, высказанное (что примечательно) в пособии для студентов-филосо-
фов [Иванов А.В., 1994]. Он убежден в необходимости соединения привычного
нам западного взгляда на мир с восточными и тем самым (что неожиданно) прямо
131 В частности, на всех уровнях таксономии говорят о включенности: и вид состоит из
подвидов, и надцарство - из царств; на всех уровнях морфологии говорят о рефренах и о
гомологии; на всех уровнях эволюции - о блочности и о прерывистом равновесии.
344
подходит к диатропическому пониманию параллелизма бытия и мышления:
«Идея о том, что нейродинамические структуры мозга соответствуют каким-
то важнейшим структурам материального мира, нам представляется заслу-
живающей внимания» (с. 29). «В отличие от фрейдовского психоламаркист-
ского объяснения совпадений между онто- и филогенезом мышления, кото-
рое сегодня не выдерживает критики, идеи К.Г. Юнга о существовании все-
общих психофизиологических структур мозга, ответственных за порожде-
ние сходных идеальных образований у разных индивидов в различные эпо-
хи и культуры, являются более перспективными для раскрытия параллелей
между онто- и филогенезом мышления» (с. 30).
Выстроив «примечательный философско-теоретический ряд аналогов» ан-
тичных, восточных, русских и западных взглядов на связь психики с космосом
(легко видеть, что это - довольно чёткий рефрен понятий), Иванов заключил:
«Таким образом, есть определенные... основания предположить, что дейст-
вительно существует какая-то единая протопсихическая энергийно-
субстанциальная основа бытия всех физических, биологических и низших
психических процессов» (с. 101). «Значит, есть какие-то... единые струк-
турные законы эволюции и инволюции, по которым развивается Космос,
природа, социальные и психические процессы. Но только в одном случае
они рационально-понятийно фиксируются в виде закономерностей диалек-
тического развития (как у Гегеля), а в другом - существуют в виде конкрет-
ных рецептурных психопрактических методик и символов (как, например,
буддийской иконописи и графической символике)» (с. 103). «Это предвидел
еще великий философ Индии Свама Вивекананда232, писавший: “тем, кто
ориентирован на познание внешнего мира, и тем, кто ориентирован на внут-
ренний опыт, суждено встретиться в единой точке...”» (с. 104).
Выглядит это непривычно, даже не очень научно, однако в терминах Сборни-
ка это можно понять так: отмеченный в разных культурах психо-физический реф-
рен (совокупность параллелей развития природы и сознания) имеет основой ре-
альную многоуровневую структуру мира. Ее внемасштабность позволяет модели-
ровать ее на языке фракталов233, причем в «единой протопсихической энергийно-
субстанциаль-ной основе» можно видеть вот что: в качестве энергиии - актив-
ность, а в качестве субстанции - диасеть. «Протопсихическую» суть структуры
мира отмечал, как мы видели, и Мейен. Теперь видно, что это - особое явление,
отличное от сродства (социабильности), и притом пока что гипотетическое.
8. Место в общей теории
Новый номогенез призван описать то, на что активные силы действуют -
рефренную структуру (диасеть), но не сами силы. Силы эти изучаются другими
132 Свами Вивекананда (в монашестве Нарендранатх Датта, 1863-1902), религиозный ре-
форматор, сторонник сближения религий (на базе индуизма) и освобождения Индии.
133 Возможно, обходясь без слишком частного языка генов - см. сноску на с. 79.
345
разделами эволюционной теории. Разные авторы называют данные силы по-
разному, и у меня они объединены общим и самым нейтральным термином «ак-
тивность». О нем см. подробно АСМ, а в данном сборнике - статьи 4-42; 4-55;
4-65. Поскольку Мейен этим кругом вопросов почти не занимался, активность
не могла занять много места в данном Сборнике, и здесь следует сказать о ней
еще тоже немногое.
Излагаемая в АСМ и ЗЭ концепция трактует эволюцию исследуемого объекта
как активное его движение вдоль своего рефрена, поэтому ей было дано в ЗЭ на-
звание актиреф. Можно говорить как о наблюдаемом актиреф-процессе (ЗЭ, с.
56), так и об актиреф-концепции, видящей в данном процессе самую суть эво-
люции. Но если процесс вполне очевиден (как способ описания фактического
хода дел) всякому, кому он удобен в работе, то концепция является объяснением
природы этого процесса и потому требует обоснования, чему и посвящен дан-
ный параграф. Удобно дать его в форме сопоставления с идеями Мейена - что
взято из них, что стало их развитием, что от них независимо, а что - против них.
Первое, что ясно - основными описанными на сегодня формами актирефа
являются эдванты, введенные в п. 1.
Столь же очевидно, но Мейеном не разработано - что прогресс (появление
нового уровня сложности организмов) можно описать как начало заполнения
рефренной строки, прежде пустовавшей. Когда эволюция протекает в рамках за-
данной диасети, ее издавна именуют плоской. Она носит чисто комбинативный
характер (комбинируются уже известные из более ранней эволюции блоки) и в по-
следние годы сама именуется блочной (см. п. 3 предыд. статьи). Наоборот, когда
движение происходит вдоль заново реализующихся рефренных строк, то такая
эволюция как раз и именуется прогрессом.
Как плоская эволюция, так и прогресс могут быть адаптивными или не быть.
Главным свойством активности является то, что она побуждает материю к ус-
ложнению форм, причем на каждом более высоком уровне сложности появляется
новая, более сложная форма активности, вновь вызывающая усложнение форм, и
так далее. Это и есть прогресс. Хотя именно «Мейен привнес в номогенез концеп-
цию уровней организации» (ИМ, с. 125), но сам он прогрессом не занимался, ог-
раничиваясь отдельными репликами. Одну из них (1984 г.) следует привести:
«Я довольно много размышлял над прогрессом, но потом интерес к этому
вопросу утратился. Недавно Чайковский выдвинул три ипостаси эволюции
- разнообразие, приспособление и прогресс. Это заставило меня снова за-
думаться над прогрессом... Я не претендую на решение вопроса и даже не
могу сформулировать достаточно ясно свою точку зрения» (ИМ, с. 135).
Далее он ушел от разбираемой здесь нами темы (что такое прогресс в биоэво-
люции), и мне остается объяснить упомянутые «ипостаси». Мейен имел в виду
«методический треугольник»
346
- мы как-то раз обсуждали его. Место его и роль в понимании эволюции рас-
смотрены в статье [Chaikovsky, 1988], откуда приведу цитату:
«Ламаркизм основан на идеях градации (усложнения, прогресса) и при-
способления, дарвинизм - на идеях приспособления и разнообразия, а но-
могенез - разнообразия и прогресса. Хотя и Ламарк упоминал о разнооб-
разии, и Дарвин - о прогрессе, и Л.С. Берг - об изначальной целесообраз-
ности..., но не в этом был пафос их учений. [...] Трем главным направле-
ниям эволюционизма соответствуют (нежестко) три главных эволюцион-
ных проблемы - разнообразия, прогресса и приспособления. Если поста-
вить их в вершины треугольника, то его стороны грубо представят собой
ламаркизм, дарвинизм и номогенез» [Chaikovsky, 1988, с. 69].
Впоследствии, при занятиях наукой о законах разнообразия (диатропикой),
мне пришлось отказаться от столь сильного утверждения - что дарвинизм осно-
ван на идее разнообразия в том же смысле, что и номогенез. На самом деле в ос-
нове дарвинизма лежит та установка, что у разнообразия нет собственных зако-
нов, а номогенез как раз эти законы и находит. Далее, на треугольнике среди
главных концепций нет жоффруизма (о нем см. выше, статью 4-65). В то время
мне достаточно было мысленно отнести преобразование онтогенеза под дейст-
вием среды к ламаркизму, а автономное - к номогенезу.
Однако «методический треугольник» оказался удобен для целей преподава-
ния, и после его публикации в газете для учителей (4-36, 2000, с. 5) некоторые
из них стали им пользоваться. Тем самым, он имеет право на существование.
Удивительно забвение Мейеном своего собственного прежнего «треуголь-
ника», изложенного выше, в п. 5, и, на мой взгляд, гораздо более интересного.
Там, в п. 5, уже было сказано, насколько он вошел в нынешнюю науку.
Активность порождает новые структуры, в тех возникают новые формы ак-
тивности и так далее. Для внешнего наблюдателя это чередование (спираль) вы-
ступает как многоуровневая самоорганизация. Факт этот описан неоднократно,
и я ограничусь одним ранним высказыванием об одном самом важном уровне:
«переход от химической эволюции абиогенных органических веществ к
эволюции биологической потребовал образования многомолекулярных
систем, сборка которых протекала спонтанно и могла быть многократно
воспроизведена. [...] Сейчас становится все более ясным, что самосборка
лежит в основе построения большинства простейших биологических
структур. И только дальнейшее усложнение этих структур, сопровождаю-
щееся образованием многокомпонентных систем, требует подключения
дополнительных регулирующих факторов» [Поглазов, 1982].
Подчеркнутые мною слова, а также отсутствие в цитируемой заметке слова «от-
347
бор» ясно говорят, что мыслящие люди уже тогда понимали суть проблемы, о
которой большинство предпочитает не думать до сих пор.
О спонтанной самоорганизации см. выше, часть 7, а здесь замечу, что вся
изложенная там проблематика выявлена уже после смерти Мейена. Не будем га-
дать, как бы он к ней отнесся, но сама идея системной случайности предстала
мне в ходе бесед с Мейеном и Б.И. Кудриным (о его роли см. в ОПС).
Теперь займемся упомянутым чередованием. При реализации нового уровня
организации рефренная структура обнаруживается регулярно, так что и этот
процесс явно описывается в терминах актирефа.
Одни спросят: каков механизм, творящий данную дивную спираль? Пока не
указано механизма, многим не хочется замечать и само явление. На этом пути
механизм, разумеется, не будет найден никогда.
Другим хватает такой аналогии: активность укладывает белок в нативную
структуру примерно так же, как она укладывает атомы в молекулу, а электроны
и нуклоны - в атом. Как химики приняли идею валентности без всякой оглядки
на физику (квантовая теория и поныне объясняет валентность весьма приблизи-
тельно), так следует принять и идею самосборки макромолекул - как особое
свойство природы. Это - вниз, в глубь микромира, но то же можно видеть и при
движении вверх, в просторы биосферы: как молекулы укладываются в клеточ-
ные структуры, клетки - в ткани и органы, а те - в макроорганизм, так же и ор-
ганизмы образуют экосистемы, а те - биосферу, структурированную не менее
очевидно (хотя и менее чётко), нежели периодическая таблица Менделеева. На
данном пути механизм каждого уровня фактически принято описывать, не пы-
таясь вывести его свойства из свойств нижележащих уровней.
Мейен призывал отказаться от таких попыток явным образом. Он писал, что
нужна «та концепция уровней, когда в качестве уровня выступает такой аспект
действительности, свойства которого не могут быть описаны в понятиях любого
иного аспекта» (М-163). В устных беседах он уточнял, что между различными
уровнями организации объективно существуют «барьеры несводимости».
Третьи (редукционисты), наоборот, пытаются это вывести. Например, пробу-
ют объяснить самосборку макромолекул в терминах физики и химии. Некоторые
детали при этом, конечно, выясняются (см. АСМ, с. 430), но вряд ли это позволит
понять суть дела, поскольку «барьеры несводимости» обнаруживаются всюду.
Поясню: в принципе соединение атомов в молекулу на языке квантовой
теории выразить можно, однако - только пока их мало. Когда элементов в сис-
теме становится достаточно много, вступают в силу собственные законы сис-
тем, а конкретные законы действия элементов играют всё меньшую роль, об-
ращаясь в ограничения (рамки) действия этих законов систем134. Данную
мысль, основную для науки, Мейен, насколько знаю, почерпнул у Урманцева.
Четвертым не нужно ничего, ибо им априори ясно, что все структуры мира
134 Пример: компьютер является системой, слабо зависящей от природы своих компонент.
Замена ламп на транзисторы, т.е. полная смена физики элементов, мало повлияла на тип
возможных вычислений. К революции привело уменьшение транзисторов (в интегральных
схемах), раздвинувшее рамки решаемых системой задач далеко за рамки математики.
348
сотворил Бог. Ныне почти все верующие ученые именуют Его термином «естест-
венный отбор» и лишь немногие (это креационисты) - божеством какой-либо из
старых религий. Вопрос, почему Он творит мир рефренами, дарвинистами не ста-
вится, зато сторонники понимания Бога как личности давно ответ дали - Ему эко-
номнее действовать по единому плану (архетипу), отчего мы и наблюдаем повсю-
ду ряды сходств. Забавно, что этот принцип экономии был признан (и как раз в
связи с вопросом об архетипе) самим Дарвином (см. выше, с. 44), но из аргумен-
тации дарвинизма выпал. Любопытно бы рассмотреть данную коллизию с пози-
ции Мейена, призывавшего к синтезу дарвинизма с номогенезом.
Теперь об активности. В книге АСМ и предыдущих публикациях актив-
ность подана, в основном, в описательном ключе. Тем читателям, кому интерес-
ны механизмы ее действия, могу предложить несколько новых работ. Биофизик
В.Л. Воейков [2009] рассматривает энергетический аспект активности, точнее -
источники свободной энергии, работающие в водных системах живой клетки и
обеспечивающие ее активность. Он работает в деятельном контакте с западны-
ми биофизиками (см. например: [Biophotonics..., 2000; Del Giudice, 2005]).
Информационному аспекту активности посвящено множество работ (на-
пример, упомянутые в АСМ книги П.П. Гаряева и А.Г. Зусмановского), однако
большинство научных работников считают их маргинальными и не читают. По-
этому укажу автора, которого даже дремучий ортодокс маргиналом назвать не
сможет - это нобелевский лауреат 2008 года Люк Монтанье (Франция), от-
крывший вирус спида. В недавней статье Монтанье с соавторами пишут, что от-
крыли совершенно новое явление: некоторые фрагменты ДНК активно излуча-
ют электромагнитные волны [Montagnier е.а., 2009]. Это подтверждает прежние
умозрительные построения Гаряева и Зусмановского, открывая тем самым со-
вершенно новые горизонты в понимании активности генетических процессов.
Активность, движущая эволюцию, расходуется между «ипостасями» (раз-
нообразием, приспособлением и прогрессом) очень различно в разных областях
живого - в одних группах идет преимущественно диверсификация (рост разно-
образия), в других - преимущественно усложнение организованности (про-
гресс), а третьи приспосабливаются к условиям, недоступным для остальных ор-
ганизмов. В этом - одна из форм реализации принципа компенсации Аристотеля
(см. п. 3 статьи 4-42). Тем самым, на передний план обычно выходит одна «ипо-
стась», и Мейена занимали, в основном, те процессы, где возрастало разнообра-
зие. Удерживать во внимании все три «ипостаси» сразу удается немногим. Од-
ним из них был системолог Эрих Янч (развивавший «философию процесса»,
предложенную за полвека до него Альфредом Уайтхедом).
Обычно эволюцию системы изучают после того, как сама система описана.
Янч же предлагал описывать ее через ее развитие. Западная наука, писал Янч,
привыкла исходить из неподвижных схем как чего-то исходного и строить вся-
кое движение как мысленную последовательность состояний (замечу: Бергсон
называл это кинематографическим методом), тогда как более естественно счи-
тать всякое состояние мысленно остановленным движением. Только тогда появ-
349
ляется надежда понять, почему движение может быть направлено к некоторой
цели. Это - системная динамика, с ее помощью легче понять целенаправленное
движение, как велосипед легче удержать в равновесии, когда он едет. Направ-
ленность движения, удивительная для стороннего наблюдателя, вполне естест-
венна для наблюдателя, движущегося вместе с системой [Jantsch, 1975].
В своей последней книге Янч заявил, что само понятие приспособления надо
переосмыслить. Как и Любищев, он видел в приспособлении не самоцель эволю-
ции, а инструмент системодинамики и, одновременно, аспект эволюции: «Эволю-
ция совершает эксперимент с целым филумом, эффективный сейчас» [Jantsch,
1980, с. 16]. (Тут нужна оговорка: вместо «эффективности сейчас» лучше гово-
рить об эффективности в течение всей истории «филума».) Эволюция идет по-
средством единого процесса коэволюции микро- и макроструктур, причем с тече-
нием времени неуклонно растет размер частиц микроуровня (от фотона до орга-
низма) и падает размер характерных объектов макроуровня (от протогалактик до
людских обществ). Подробнее см. [Chaikovsky, 1988; ЭЭД)].
Недавно у В.А. Брынцева традиция Янча получила продолжение. Он пишет:
«Я предлагаю новый системно-динамический подход. В основе его лежат
другие основания-сущности. В системно-динамической модели простран-
ство и время не являются сущностями - это эпифеномены. Сущностью яв-
ляется движение. Время и пространство порождаются системным (цикли-
ческим) движением. Каждая система (каждый цикл) создают свое про-
странство и свое время. В системно-динамическом подходе элементарная
система - это циклическое движение. Цикл, в свою очередь, — это само-
воспроизводящееся, порождающее себя движение. Данный взгляд посту-
лирует, что мир состоит из движения. [...]. Одно системное движение по-
рождает другое системное движение» [Брынцев, 2006, с. 77].
Легко видеть, что системным движением здесь названо как раз то, что у меня
называется активностью.
Для более нового (после выхода АСМ) понимания эволюции понадобилось
разделить традиционные эволюционные учения на эразмовы (организмоцентри-
ческие) и бэровы (объект интереса которых - пространство свойств) - см выше,
статью 4-65. Там сказано, что в эразмовых учениях организм мыслится как соз-
даваемый постепенно (миллионы поколений) таким, каким его требует среда; а
сходство трактуется как итог родства. И что в бэровых учениях новый организм
создается сразу (несколько поколений) как комбинация возможностей, заданных
законами становления и устройства форм и функций. Среда 6 бэровых учениях -
итог взаимодействия организмов, а сходство - итог общности законов онтогене-
за. Эразмова идея наглядна исходными допущениями (но наглядность эта со-
храняется лишь в рамках видов одного семейства), а бэрова - наблюдаемыми
следствиями. Эволюция предстаёт, как там сказано, процедурой изменения
фракталообразующих правил, а эразмовы и бэровы концепции выступают как
два взаимодополнительных аспекта изучения данной процедуры.
Мейен получил основные свои палео-результаты в рамках эразмовой тради-
350
ции, однако его главная эволюционная схема - бэрова: в самом деле, диасеть яв-
ляется, кроме прочего, описанием пространства свойств, а флорогенез (если
включать в это понятие и возникновение новых таксонов) - движений в нем.
Взаимодополнительное рассмотрение обеих традиций и представляется мне под-
ходом к построению работающей теории. Мейен любил называть взаимодополни-
тельный подход синтезом, однако, как теперь вижу, он противоречит тому пути к
синтезу, каким шёл он сам - попыткам соединить номогенез с дарвинизмом. От
последнего взять почти нечего.
9. О диатропической картине мира
О новой картине мира говорят часто, есть параграф о ней и в АСМ, напом-
нивший ту точку зрения, что прежняя картина мира, в основном физическая, ус-
тупает место нынешней, преимущественно биологической. В этой связи приве-
денные выше мысли Мейена о рефренах и более ранние (о «трех китах эволю-
ции») выглядят как одна из ранних попыток биологизировать (и даже далее -
психологизировать) научную картину мира. В его время это смотрелось дико,
поскольку царила противоположная установка - видеть в самой биологии лишь
очень сложную физику. Не стану занимать ничью точку зрения, но отмечу, что
чем шире круг физических (обычно - чисто механических) объяснений биоло-
гии, тем глубже пропасть между естествознанием и психологией. Эту пропасть,
полагаю, и пытался преодолеть (заполнить, снабдить мостом) Мейен.
Выше, в п. 7, сказано, что новая картина мира носит диатропический харак-
тер: эволюция, как и всякое развитие, мыслится текущей в пространстве, обла-
дающем собственной первичной структурой (диасетью) и носящей характер ак-
тирефа (активного заполнения диасети). Поскольку активность разработана в
ламаркизме, а рефрены - в номогенезе, актиреф выглядит, как их синтез, но на
самом деле он глубже, нежели синтез этих двух никогда не бывших работоспо-
собными концепций. Ламаркизм, как и дарвинизм, исходит из идеи «происхож-
дения видов» как самодовлеющего процесса, в чем противостоит бэровой идее.
Номогенез и ЭКЭ отличаются от них вниманием к бэровой идее, но не более то-
го. Подлинно бэров подход виден в системодинамике Янча.
Выше построение реально работающей теории эволюции намечено как син-
тез (точнее - осознание взаимодополнительности) эразмовой и бэровой тради-
ций.С этой позиции новая картина мира включает отказ от идеологии «проис-
хождения видов» как основы эволюции в пользу идеи эволюции природы как
целого посредством системодинамики по Янчу, т.е. сразу и сверху и снизу (см.
Вводное замечание к части 7).
О картине мира высказался и Брынцев:
«Намечается новая картина мира, в которой движение, изменение, актив-
ность являются основой и идут впереди формы, функции, да и системы как
таковой. Не эволюция систем, а эволюция, рождающая системы, создаю-
щая их» [Брынцев, 2009, с. 1130]. См. также Приложение 2.
Как видим, его привлекла в качестве мирообразующего начала активность, то-
351
гда как Мейена всегда больше занимало диатропическое начало. Роль этих на-
чал в будущей картине мира видится мне равно необходимой.
Разумеется, «впереди формы» эволюция не идет, коль скоро всякая новая
форма укладывается в какой-нибудь рефрен. Это видно как в большом (Мейен:
растительная и животная клетка усложнялись в эволюции параллельно - М-73) и в
среднем («зации» - см. выше, п. 3), так и в малом: ген антитела к данному впер-
вые возникшему антигену конструируется параллельно в тысячах штук в каждой
из множества зараженных особей (ЗЭ, с. 64-69; 95). Но главное даже не это.
Интереснее другое: чем «эволюция, рождающая системы» отлична от Твор-
ца? Вроде бы, эволюционист так ставить вопрос не должен, но он у меня возник и,
по сути, возразить мне Брынцеву нечего. Да, мир устроен так, что новые системы
рождаются (вот намечающаяся картина мира), но разговоры ученых о механизмах
превращения одной системы в другую не идут дальше пустых уверений (мол, это
понятно через диалектику перехода количества в качество), а о механизме проис-
хождения системы из чего-то несистемного и вовсе сказать нечего (кроме столь
же ценного уверения, что это обеспечивается обратной связью - см. АСМ, с. 617).
Зато о феномене эволюции можно сказать (и в п. 8 сказано) вполне опреде-
ленно: простое собирается в сложное, собирается регулярно (образуя рефрены)
и активно. А где видна регулярность, там наука может действовать. Тем самым,
эволюционная картина мира, притом диатропическая, вполне реалистична.
Реалистична в принципе, но еще не построена. Хотя примеры вроде эволю-
ции тлей Шапошникова (см. выше, с. 82, 109) достоверны и вселяют надежду на
объяснение широкого круга явлений, но всё это - в рамках уже реализованной
диасети. О появлении принципиально нового (такой строки диасети, которая
прежде пустовала, т.е. ни в одной ее клетке не значилось реально живущего или
жившего прежде организма) можно сегодня сказать очень немногое.
А именно, эмерджентность (см. с. 102) доказана только в процедуре форми-
рования гена антитела к искусственному антигену, прежде в природе не сущест-
вовавшему. К ней примыкает красивая, но пока что абстрактная аналогия мор-
фогенеза с ростом фрактала - см. с. 316, 199, а также сноску на с. 79.
Вот и всё, что пока понято. Остается верить (или не верить), что здесь-то и
будет вскоре совершен познавательный прорыв. Но верят все по-разному.
Одни верят, что всё создано разумным началом, другие - что рождение сис-
тем сможет быть описано, когда наука больше узнает о фрактальном строении
пространства-времени. Третьи верят, что обходятся вообще без веры, ибо знают
истину, но это вряд ли продуктивно, как всякий самообман.
Таков итог моего почти полувекового занятия проблемами эволюции. И по-
ловина его, увы, уже без Серёжи.
352
Литература
Акчурин И.А. Единство естественнонаучного знания. М., Наука, 1974.
Александров В.Я. Проблема поведения на клеточном уровне (цитоэтология) // Успехи
современной биологии, 1970, т. 69, № 2.
Волковский Б.Е. Цифровой политомический ключ для определения растений // Ботанич.
журнал, 1960, № 1; он же. Теза, антитеза и ряд признаков в диагностике растений // Бота-
нич. журнал, 1960, № 11.
Бахметьев П.М. Попытка установить периодическую систему бабочек // Труды Сара-
товского общества естествоиспыт., т. 4, 1903-1904.
Брынцев В.А. Филлотаксис. Опыт применения компьютерных моделей для решения
вопросов теоретической морфологии растений. Мос. гос. Университет леса. М., 2006.
112 с.
Брынцев В.А. Рец. на книгу «Активный связный мир» // Вестник РАН, 2009, № 12.
Васильев А.Г., Васильева И.А. Гомологическая изменчивость морфологических структур
и эпигенетическая дивергенция таксонов: основы популяционной мерономии. М., 2009.
Виндельбанд В. История новой философии. СПб., 1913. Том 1.
Воейков В.Л. Ключевая роль устойчиво неравновесного состояния водных систем в био-
энергетике // Российский химич. журнал, 2009, № 6.
Воробьева Э.И. Процессуальный подход к проблеме «тетраподизации» И ПЖ, 2008, № 2.
Георгиевский А.Г. Проблема преадаптации. Историко-критич. исследование. Л., 1974.
Дарвин Ч. Соч., т. 4. М., 1951 (Изменения животных и растений...)
Джан Р.В. Филлотаксис: системное исследование морфогенеза растений / пер. с англ.
М.-Ижевск, 2006.
Иванов А.В. Сознание и мышление (учебное пособие). М., Изд. МГУ, 1994.
Игнатьев И.А. Общие фитогеографические модели и эволюция позднепалеозойских
флор // Эволюция флор в палеозое. М., ГЕОС, 2003.
Клайн М. Математика. Поиск истины. М., Мир, 1985.
КлэкДж. Трудный путь на сушу // ВМН, 2006, № 3
Лима-де-Фариа А. Эволюция без отбора. Автоэволюция формы и функции. М., Мир,
1991.
Мейен С.В. М-73, 163, 165, 229, 246, 281, 286, 295, 313, 323, 325, 344, 352, 370.
Мосейчик Ю.В. Номогенетичесий характер эволюции растений (на примере раннего
карбона) // XIX ЛЧ. Ульяновск, 2005, с. 98-103.
Мосейчик Ю.В. О природе фитостратиграфических рубежей (на примере флористиче-
ских последовательностей раннего карбона Северной Евразии) // Био- и литостратигра-
фические рубежи в истории Земли. Труды международной конференции. Тюмень, 2008,
с. 208-214.
Мосейчик Ю.В., Игнатьев И.А. О начальных стадиях палеозойского этапа глобального
флорогенеза И Палеонтологическая летопись региональных и глобальных событий. Тезисы
докладов 51-ой сессии Палеонтологического общества при РАН (4-8 апреля 2004 г., СПб.)
СПб., 2005, с. 87-88.
Низовцев В.В. Время и место физики XX века. М., УРСС, 2000. 207 с.
Низовцев В.В., Панченко О.В. Контактный детерминизм при морфогенезе // Теория
эволюции: наука или идеология? Москва - Абакан, 1998, с. 147-164.
Ииконоров Ю.М. Барьерные функции митохондриального кода // XXIV ЛЧ. Улья-
новск, 2010, с. 156-161.
353
Поглазов Б.Ф. Предисловие // Сборка предбиологических и биологических структур.
М„ Наука, 1982.
Попов И.Ю. Периодические системы и периодический закон в биологии. М.-СПб.,
КМК, 2008. 223 с.
Расницын А.П. Процесс эволюции и методология систематики. СПб., 2002. (Труды
Русского энтомологии, общества, том 73, с. 1-107)
Том Р. Структурная устойчивость и морфогенез [1972]. М., Логос, 2002.
Тутубалин В.Н. Вероятность, компьютеры и обработка результатов экспериментов И
УФН, 1993, №7.
Чайковский Ю.В. Ч-З, 4, 5, 7, 18, 26, 27, 36, 42, 47, 51,55, 56, 57, 61,65.
Щербаков А.Е. О пермских и триасовых энтомофаунах в связи с биогеографией и пер-
мо-триасовым кризисом // ПЖ, 2008, № 1.
AnderwsН.М. Index of generic names of fossil plants, 1825-1965 // Geol. Surv. Bull., 1970,
№ 1300, p. 1-354.
Biophotonics and Coherent Systems / Eds: Lev Beloussov, Fritz-Albert Popp, Vladimir
Voeikov, and Roeland Van Wijk. Moscow: Moscow University Press, 2000.
Brower J.H. Behavioral changes in an ant colony to chronic gamma irradiation // American
Midland Naturalist, 1966, vol. 75, № 2.
Chaikovsky Yu. V. To the evolutionary thermodynamics, 1988 (4-19).
Del Giudice E. e. a. Coherent quantum electrodynamics in living matter // Electromagnetic Biol-
ogy and Medicine, 2005, v. 24, p. 109.
Harms J. V. Die Realisation der Gene und die consekutive Adaptation // Zeitschr. Wissensch.
Zoologie, 1935, Bd 146, S. 417-462.
Jantsch E. Design forevolution. N.Y., 1975.
Jantsch E. The self-organizing universe. Scientific and human implications of emerging
paradigm of evolution. Oxford - N.Y., 1980.
Jean R. V. Phyllotaxis. Cambridge, 1994.
Kark S., van Rensburg B.J. Ecotones: marginal or central areas of transition? // Israel J. Ecol.
Evolution, 2006, vol. 52, p. 29-53.
Меуеп S. V. Plant morphology in its nomothetical aspects // Bot. Rev. 1973 (M-l 16).
Montagnier L., Aissa J. e.a. Electromagnetic signals are produced by aqueous nanostruc-
tures derived from bacterial DNA sequences // Interdiscip. Sci. Comput. Life Sci., 2009 (1)
81-90.
Popov I. Orthogenesis versus Darwinism: The Russian case // Revue d’histoire des sciences,
2008, t. 61, №2, p. 367-397.
Van der Giezen M. Hydrogenosomes and mitosomes: conservation and evolution of func-
tions // J, Eucariot Microbiol., 2009. vol. 56, № 3.
Van der Giezen M. & Tovar J. Degenerate mitochondria // EMBO [Europian Molec. Biol.
Organization] Rep., 2005, vol. 6, № 6.
Yule G.U. A mathematical theory of evolution, based on the conclusions of Dr J.C. Willis //
Philos. Transact. R.S. London, 1924. Ser.B, vol. 213, p. 21-87.
354
Иллюстрации
Рис. 1. Рефрен мерона «парные конечности позвоночных». Усложнение общей
организации наблюдается сверху вниз, а конечностей - слева направо
значения мерона
класс отсутствие или неви- димый сна- ружи зача- ток развита только сла- бая перед- няя пара многофа- ланговые плавники или ласты орган для
ползания планирую- щего полёта активного полёта
костные рыбы мурена китовидка, угорь кистепёрые морской нетопырь летучие рыбы клинобрю- шка
амфибии червяга сирен некоторые ископаемые саламанд- ры яванская ля гушка не бывает
рептилии змеи и безно гие ящерицы хирот морские черепахи черепахи, крокодилы ногокрыл (ископае- мое) летающие ящеры (ископае- мые)
птицы киви (нет пе- редней па- ры) не бывает пингвины поползень, стрижи совиный попугай большинст- во видов
звери киты, ламан тин (нет зад ней пары) КИТЫ ламантин (передние) калан (задние) кроты, тюлени шерсто- крыл летучие мыши
Рефрен дан упрощенно: например, все признаки выражены в полной мере. На самом деле ка-
ждую клетку можно заменить целым рядом - от полного выражения до почти полного “раз-
мывания” признака. Если таблицу рефрена представить лежащей на столе, то ряд степеней
выражения будет выглядеть вертикальным
столбиком, стоящим на каждой клетке таблицы
Рис. 2. Семофилогения женских фруктификаций
ранних хвойных, “положенная на карту” (по:
Меуеп, 1997, М-ЗЗО); см. с. 115
355
Рис. 3. Система животных Г. Гольдфусса в форме географической карты
[Goldfuss, 1817]
см. с. 237
356
Рис. 4. Система растений Баскервиля в форме географической карты
[Baskerville, 1839]
см. с. 237
ASTEJiAL POES.
Рис. 5. Размывание «кленового» контура (см. с. 92)
Клён остролистный,
платан восточный,
смородина Макси-
мовича, плющ, клён
ложноплатановый
357
Плауны Папоротники Голосеменные Цветковые
Рис. 6. Глобальный флорогенез по Мейену
1 - южные (гондванские) внетропические области; 2 - тропические; 3 - северные вне-
тропические; 4 - флористические области не обнаружены. Видно, что почти все ветви
исходят из тропической зоны. На врезке -
карта климатических поясов позднего палео-
зоя, как они видны при нынешнем распо-
ложении материков См. с. 256
Рис. 7. “Пробная филогения” ранних
хвойных (Meyen,-1997, М-ЗЗО)
Она точно повторяет семофилогению, данную
на рис. 6; V, L, К - группы, которые можно
понимать как первичные семейства. Но, как
видим, схема не является филоге-нией в
обычном смысле: остальные семей-ства и
некоторые роды даны вне ветвей
358
Рис. 8. Флорогенетическое древо Мейена
(эволюция фитохорий в позднем палеозое и мезозое), см. с. 259
359
Рис. 9. Распространение и эволюция ранних хвойных в Печорской области
ранней перми (по М-ЗЗО).
Слева на таблице показаны позднепермские ярусы (татарский, казанский и уфимский) и
раннепермский (кунгурский) ярус, в середине - места находок, обозначенных на карте,
справа - виды ранних хвойных. Как видим, более поздние виды расходятся дугой на север,
запад и юг от двух самых древних кунгурских видов (обведенных кружком)
WESTERN ANGARALAND
Tatarian I 1 Upper 1 Vyatsky Aristovo, Titovo, Viled, Kalinovka, Luptyug, Dor Sashinia aristovensis, S. borealis, Quadrociadus dvinensis, Q. borealis, Dvinostrobus sagittalis, Geinitzia subangarica, Geinitzia sp. SVM—1
Severodvinsky Isady, Aleksandrovka Kargala Mines Kzyl—Bairak Kityak Sluda Vym —1, Vym—2 Dvinostrobus sagittalis, Quadrociadus schweitzeri, Geinitzia sp. SVM—2 "UHmannia" Steirophyllum biarmica, lanceolatum "Walchia" foiiosa Pseudovoltzia ?cornuta Quadrociadus (al. Steirophyllum) komiensis Timanostrobus muravievii Conchoiepis harrisii
Lower
| Kazanian I Upper Lower
Ufimian Sheshminsky Solikamsky Shapkina Kungurodendron ? sp, SVM—1 Cyparissidium entsovae
c ГО 3 Irensky Chekarda —1 Chekarda—2 Kungurodendron sharovii, Cyparissidium appressum, Taxodiella bardaeana
1 Kung Filippovsky Krutaya Katushka, Krasnaya Glinka, Matveevo—1 Kungurodendron sharovii, Cyparissidium appressum, Taxodiella bardaeana
360
Рис. 10. Эволюция фитохорий в мезозое и кайнозое, по М.А. Ахметьеву
За основу нижней части схемы (юра - нижний мел) взят вариант Мейена (изо-
браженный на рис. 8). Она соединена с верхней частью (верхний мел - современ-
ность), за основу которой взята система нынешних флористических царств.
См. с. 259
Главнейшие современные фитохории земного шара
361
ФАГОТРОФЫ
рвотные
ровным моэгбх;
МОЛЛЮСКИ
С ЗЕЛЕНЫМИ
СИМБИОНТАМИ
—ьг
нитчатые
ГнЛпор"
ПРЕОБЛАДАЕНИЕ
> ‘ ЖГУТИКОВЫХ
ФОРМ
икон
ХИЩНЫЕ
ГРИБЫ
МОЛЛЮСКИ С
ХЕМОТРОФНЫМИ
СИМБИОНТАМИ
Рис. 11. Эко-морфологическая макросистема
Кроме четырех царств (прокариоты, растения, грибы и животные), введено три межцар-
ства (зоофиты, микозои и микофиты), где свойства царств соединяются. Среди прокари-
от аналогичную группу образуют идиотрофы, у которых соединяются различные типы
питания (например, авто- и гетеротрофность). Буквой М-обозначены молликуты (мико-
плазмы) - группа микроорганизмов, наиболее близкая к возможным первоорганизмам,
составившим первый биоценоз. Дальнейшая эволюция мыслится как заполнение кольце-
вой таблицы. Первичная эволюция выглядит как заполнение основного кольца (примыка-
ет изнутри к жирной линии). Дальнейший эволюционный прогресс отражен в последова-
тельности колец, начиная от первичного, внутрь и наружу. См. с. 263.
Примеч. 2010 г.'. Основой схемы является кольцевой рефрен, т.е. она - диатропическая
М И'.К О з'о *
4 Я”’1
ПРЕОБЛАДАНИЕ
БЕЗЖГУТИКОВЫХ
рв«чнополостнь/л «
TrtJjbiE. ТРЕХОЮ
49ю, Гувки "Ые
3 Тканевые, двус„
а
о »
Е с
362
Рис. 12. «Отороченные гауссоиды», или Номогенез в алеатике
Тот факт, что случайное явление подчиняется гауссову распределению, оказался всего
лишь результатом огульного усреденения, тогд как детальный анализ обнаружил сходст-
во между сериями испытаний. При этом степень повторяемости оказалась неожиданно
высокой: как видим, изображенные 18 пар сходных отороченных гауссоид выявлены все-
го лишь в 140 сериях случайных испытаний (по [Шноль и др., 1998]) . См. с. 286
Рис. 13. Эффект «отороченной гауссоиды»
повторён многократно
(точнее, эффект «отороченной плотности»
пуассонова распределения радиоактивного распа-
да). Примечательно, что этот результат получен в
опыте, поставленном критиками с целью данный
эффект опровергнуть. См. с. 286
363
-1 во
Рис. 14. Распределения Виллиса (квази-гиперболы без сглаживания)
Первые пять линий - распределение видов по родам; последняя линия - распределе-
ние видов бабочек по численности. См. с. 290 (подробнее: ОПС, с. 266)
Рис. 15. То же в полулогарифмическом масштабе.
Видно, что аппроксимация данных линий прямыми имеет мало смысла
364
Саяно-
Европейская Алтайская
область провинция
царство царство
Условные
обозначения:
-границы
фитохорий
-границы
материков
1 - Центрально-
Монгольский округ
Рис. 16. Древо фитохорий позднего девона - раннего карбона по [Мо-
сейчик Игнатьев, 2005]
Для девона, наземная растительность которого прежде считалась географически
однородной, выявлено, как и для нынешней эпохи, пять флористических царств (распо-
ложенных и названных, однако, совсем иначе, нежели нынешние). Контактов со схемой
Мейена довольно много, но принцип эволюции выявлен здесь иной: не распад единой
предковой формы на флористические царства, а, наоборот, «путём локального становле-
ния фитохорий низкого ранга, которые расширяясь, сливаясь и обмениваясь формами,
могли давать начало фитогеографическим общностям более высокого порядка». И хотя
авторы назвали схему древом, она скорее выглядит куртиной. См. с. 331
365
Приложения и литература
1. С.В. Мейен. Легче вывести человека из современной свиньи*
1. Неожиданное разнообразие - главная проблема биологии. С ним ни физику,
ни химику почти не приходится иметь дела. Если требуется изучить свойства какого-то
химического элемента, то достаточно получить его в нужном количестве, позаботиться о
чистоте препаратов... Биолог находится в совершенно ином положении, хотя не всегда
это осознает.
2. Выступая с докладом в Москве, один из главных апостолов «синтетиче-
ской» теории эволюции Эрнст Майр проповедовал, что эволюционные сдвиги
начинаются с изменений в поведении. Это вызывает смену в давлении естест-
венного отбора и ведет к морфологическим переменам. Майр по профессии ор-
нитолог. Поэтому неудивительно, что в ответ на вопрос, как быть с растениями,
он лишь пренебрежительно махнул рукой. Его жест можно было понять только
так, что у этих дурацких растений все не как надо. Известный ботаник М.Г. Попов,
наоборот, заботился больше о растениях и предложил объяснять происхождение наибо-
лее крупных систематических групп гибридизацией. Ботаники не приняли этой гипотезы,
хотя и обсуждали ее, а зоологи не снизошли и до этого, так как скрещивание ужа с ежом
разрешается лишь в анекдотах. [См. пояснение].
3. Скорее их (эволюционные учения - Ю.Ч} можно сравнить с изложенны-
ми на научном языке собраниями примет, но уже принадлежащих не простому
люду, а ученым. Я готов всерьез отстаивать взгляд, что так называемые «эволюцион-
ные законы» вошедшие в сводки по теории эволюции, - не естественнонаучные законы,
а всего лишь приметы... Американский палеонтолог Дж.Г. Симпсон, один из соавторов,
«синтетической» теории эволюции, признался, что все эти «правила-законы» - стати-
стические обобщения со многими исключениями. [См. пояснение].
4. Ни один специалист по хвойным, зная признаки обыкновенной сосны и
сосны Станкевича, не смог бы предсказать, что первая войдет в число самых
многочисленных в мире деревьев, а вторая будет расти лишь в Крыму. Как по-
нять, что в травяном ковре некоторых мест Монголии и южного склона хребта Восточный
Танну-Ола прогрессивные цветковые растения потеснены плаунками (селягинеллами),
появившимися еще в каменноугольном периоде и, по всем эволюционным канонам,
весьма примитивными созданиями. [См. пояснение].
5. ...До сих пор нет теории, которая руководила бы исследователем адапта-
ций и указывала бы ему, как эти адаптации обнаружить. Я представляю отношение
физика к человеку, который пришел бы и сказал, что он открыл универсальный закон
природы, но вот только пока не знает, как его доказать или хотя бы показать его прояв-
ления... Чтобы не обижать биологов такой параллелью, скажу в их оправдание, что сами
* Места, выпавшие из публикации М-162. Фразы, публикуемые впервые, набраны рубленым
шрифтом, а сопровождавшие их журнальные - обычным (тайме). Подробнее см. Пояснения.
366
физики, увлекшись биологией, теряют требовательность и начинают рассуждать о роли
адаптаций в эволюции, отнюдь не заботясь о том, как эти адаптации вылавливать...».
6. Один из наиболее глубоких теоретиков филогении В. Генниг... признав
шаткость филогенетических гипотез во многих случаях, в конце концов делает
оптимистический вывод: все трудности преодолимы, и лучший тому пример -
успешное восстановление филогении человека, которая пользуется общим призна-
нием и подтверждается все новыми и новыми фактами. Это было написано в 1950 г., ко-
гда взгляды на происхождение человека действительно устоялись и опирались на из-
вестные находки питекантропов, синантропов, неандертальцев и кроманьонцев.
Прошло меньше десяти лет. В 1959 г. в Олдовайском ущелье (Танзания) Л.
Лики нашел череп человека вместе с галечными орудиями в слое, имеющем
возраст 1,75 млн лет. Это чуть ли не втрое больше возраста питекантропов и си-
нантропов. [См. пояснение].
7. А уж как из конкретных, известных палеоботанике, а не воображаемых
птеридоспермов вывести покрытосеменных, я совершенно не в состоянии себе
представить. Легче вывести человека из современной свиньи.
Существует и гипотеза, выдвинутая эмбриологами растений, что попытки филогене-
тически связать покрытосеменных с с известными группами высших растений безнадеж-
ны. Эмбриогенез покрытосеменных таков, что гораздо правдоподобнее считать их пред-
ками какие-то неизвестные пока растения, независимо происшедшие от водорослей.
[См. пояснение].
8. Тем не менее, многие эволюционисты с легкостью распространяют на все орга-
низмы то, что они наблюдают на собственном материале. Заключения, сделанные таким
способом, я совершенно серьезно отождествил с народными приметами (в чем вижу дос-
тоинство последних)... Другие же законы гораздо хуже примет, ибо выступают как непро-
веряемые догматы. Один из них - «эволюция это прогрессивная адаптация». Я поста-
рался показать, что с доказательством этого положения, как только речь заходит о кон-
кретном материале, дело обстоит из рук вон плохо... Эти трудности известны эволюцио-
нистам, и они попытались спрятаться за ссылку на предковые формы. У них, дескать, с
приспособительным значением признаков было все в порядке. Однако оказалось, что
найти предка еще трудней. [См. пояснение].
Я не буду раскапывать основания других эволюционных догматов, но заме-
чу, что с их обоснованием дело обстоит не лучше.
9. Теперь я сказал все, что было нужно, чтобы провести параллель между эволюци-
онной теорией и тем, что принято называть лженаукой.
[Приводятся обычные аргументы против существования телепатии и столь
же обычные ответы ее приверженцев - для них эти аргументы убоги - Ю.Ч.].
К теперь похожие упреки можно сделать какой-нибудь эволюционной теории. Возьмем
опять же самую популярную, которой учат студентов, - теорию естественного отбора.
Главный упрек: эта теория экспериментально не доказана. Ответ: эволюция не такая
вещь, которая может быть доказана экспериментально. Здесь нужны огромные промежутки
времени. [См. пояснение].
367
10. Последний упрек: если в эволюции нет ничего кроме случайных изменений и их
отбора, то совершенно непонятно становление сознания, психики. Придется допустить, что
вся материя подспудно сознательна, или считать сознание чем-то несущественным, пузы-
рями на поверхности потока. Ответ: вот и хорошо. Выбирайте. [См. пояснение].
Я опасаюсь, что меня поймут превратно. [Далее автор поясняет, что не сомне-
вается в факте эволюции, а лишь призывает эволюционистов не замыкаться в
рамках устаревших взглядов - Ю. Ч].
11. ...Один из сторонников «синтетической» теории эволюции, или СТЭ,
недавно провозгласил: «Вне и помимо СТЭ не существует никаких других на-
учных концепций эволюции». Можно взять переведенные на русский язык книги Э.
Майра. Они написаны в духе проповеди человека, не знающего никаких сомнений, знаю-
щего решительно все и задумывающегося лишь о мелочах. Насколько я знаю, отвергае-
мые редакциями рукописи сторонников телепатии порой написаны в том же стиле, и
именно он больше всего возмущает борцов с лженаукой. [См. пояснение]. Отсюда за-
манчиво сделать следующий шаг: те, кто против «синтетической» теории, вы-
ступают против науки... Эти обвинения выдвигались и против номогенеза.
12. Я не думаю, что наши представления о мире надо делить на научные и
лженаучные. В самых темных суевериях, не говоря уж о народных приметах, иногда со
временем обнаруживалась серьезная естественнонаучная подоплека. Наоборот, убеж-
дения виднейших ученых прошлого мы порой находим настолько наивными, что впору
считать их суевериями... Дарвин в «Происхождении видов» напоминает о нападках
Лейбница на теорию всемирного тяготения. Лейбниц был возмущен, что Ньютон выду-
мывает какие-то мистические силы тяготения. [См. пояснение].
Пояснения к публикации
У меня хранится ксерокопия (26 стр. машинописи) статьи, озаглавленной
С. Мейен, доктор геолого-минералогических наук
Врач, исцелися сам...
История ее такова. В течение 1976 - 1977 годов мы с Сергеем Викторовичем
много обсуждали вопрос о причинах коренного отличия эволюционной пробле-
матики ото всех иных научных дисциплин - о вековом повторении одних и тех
же споров и взаимных упреков. Непонимание учеными друг друга Мейен отно-
сил, согласно своим этическим принципам, за счет нежелания большинства лю-
дей встать на чужую точку зрения хотя бы на время ее анализа. Мне сейчас дело
представляется намного более сложным (большинство людей просто неспособ-
но выйти за рамки заученных штампов - см. конец части 4 наст, сб-ка), но тогда
позиция Мейена казалась мне безупречной. Когда он сообщил, что пишет на эту
тему статью и думает над названием, я предложил ему указанное выше.
Слово «врач» поставлено в именительном падеже, хотя в латинском ориги-
нале (medice, сига te ipsum) оно стоит в звательном (вокативе). Сделано это было
мной потому, что в русском языке звательный падеж известен плохо, а его
окончание -у вообще мало кому знакомо (кроме слова врачу, могу вспомнить
только плач петуха из сказки: «Котику-братику! Несёт меня лиса за синие ле-
368
са»), и сам я в юности, пока не узнал оригинал, читал его как врачу, считая по-
этому афоризм корявым. Однако редакция (в ней была тогда должность прове-
ряющего цитаты) предпочла хрестоматийное прочтение - см. М-162.
Статья была явно велика для журнала «Знание - сила», и сокращение ее бы-
ло неизбежно. Редактором была Татьяна Петровна Чеховская (тоже вскоре по-
сле Мейена умершая от рака), дама вдумчивая и благожелательная, так что сня-
то оказалось, в основном, то, что и следовало, - абзацы слабые и многословные,
а подчас и посторонние основному тексту, лишь затруднявшие его понимание.
Однако некоторые места изъяты, как мне кажется, за излишнюю прямоту, изъя-
ты из-за нежелания раздражать чиновников от науки. Они и помещены выше.
Кроме них, приведены также те, что удачно (на мой взгляд) поясняют мысль ав-
тора. Нумерация фрагментов дана мной.
Фраза, выбранная мной для заглавия всей подборки, в наши дни политиче-
ски безопасна, но тогда могла выглядеть крамольной, и, полагаю, за это и исчез-
ла из публикации. Достаточно заметить, что в том же журнале моя статья «Изу-
мительная асимметрия» (Ч- ) была так названа по требованию цензора, уви-
давшего в первоначальном заглавии («Изумительно кособокий мир») намёк на
кособокость советской власти.
Пояснения к фрагментам
фр. 2. Мейен переводил доклад Майра и дискуссию. Смена поведения как
фактор эволюции предлагалась не раз, но прежде вызвала решительный (и, в рам-
ках дарвинизма, обоснованный) протест самого Майра (АСМ, с. 182). Гибридиза-
ция как фактор эволюции животных на самом деле тогда обсуждалась, хотя и ма-
ло (так, в 1972 г. меня в ее роли безуспешно убеждал генетик Ю.П. Алтухов). Фак-
ты, заставлявшие говорить о ней тогда, ныне признаны, но в рамках другой идеи -
блочности эволюции (см. АСМ), а она - эдвант гибридизации (см. выше, с. 322).
Любопытно, что упомянут М.Г. Попов - основной предшественник Мейена в
теории флорогенеза (см. заключительную статью), обычно им не упоминаемый.
Как видим, Мейен не забывал о нем. Наконец, вот анекдот, упомянутый Мейеном:
«Что будет, если скрестить ужа с ежом? Полтора метра колючей проволоки».
фр. 3. Такие «законы» именуют тенденциями. Они лежат в основе наук, даже
точных (см. выше, с. 339), и в желании обойтись без них Мейен вряд ли был прав.
фр. 4. Упрек в адрес тезиса математика Роналда Фишера, писавшего: «Эво-
люция есть прогрессивная адаптация и больше ничего»).
фр. 6. Без изъятой части фрагмент довольно банален (нет ни одной филоге-
нии, которая не вызывала бы возражений, а после смерти Мейена сами филоге-
нетики всё чаще выражают сомнение в пользе своих занятий) и носит чисто
учебный характер. Восстанавливаемая же купюра призвана показать иное - что
даже многолетняя уверенность научного сообщества не гарантирует безопасно-
сти от крушения ни для какой научной концепции. См. АСМ, п. 9-15.
фр 7. Ситуация с животными примерно та же: так, тип ранних стадий разви-
тия у них часто совсем не согласуется с морфологическим типом (АСМ, с. 178).
фр. 8. Здесь Мейен совершил две ошибки, на которые сам не раз указывал.
369
Во-первых, использовал слово «эволюционисты», хотя имел в виду «дарвини-
сты», а во вторых, как бы согласился с тем, что польза в прошлом может объяс-
нить наличие признака в настоящем. Однако дарвинизм, позицию которого
Мейен вроде бы здесь излагает, утверждает, что отбор действует всегда и всюду
и что даже нейтральные признаки понемногу распадаются за счет мутаций.
фр. 9. Это - «главный упрек» отнюдь не с позиции Мейена (его возмущало
не отсутствие доказательства, а отсутствие желания искать его и придумывание
отговорок), а с позиции обывателя. Поэтому и ответ дан обывательский. Сам
Мейен вовсе не хотел унизить идею отбора (что видно из других его публика-
ций, например, М-121 или М-144; последняя издана в том же году, когда напи-
сана данная статья), ему важно было показать равенство познавательных стату-
сов идей отбора и телепатии с точки зрения проверки опытом. Здесь он, по-
моему, не совсем прав: телепатию отвергали и отвергают в силу отсутствия
«физического» ее объяснения, тогда как для отбора «физическое» объяснение
как раз есть, нет же другого - примеров конкретного его действия в природе.
фр. 10. Фрагмент выражает, в неудачной форме, мысль самого Мейена: если
есть только изменения наличных свойств, то психика выросла из каких-то своих
зачатков. Об этом (подспудной сознательности материи) в рамках дарвинизма
первым писал, насколько знаю, еще Геккель, признавший в конце жизни пра-
психику атома. В терминах данного Сборника лучше сказать, что материя, по
Мейену, на всех уровнях активна. О сознании как сгустке (примерно то же, что
здесь «пузыри») см. письмо Мейена Любищеву в п. 5 заключительной статьи.
фр. 11. Первая фраза принадлежит советскому автору, но она голословна,
тогда как у Майра есть ее развитие: «Немногочисленные противники этой тео-
рии (СТЭ - Ю.Ч.) - те, кто все еще оперирует ламаркистскими и финалистиче-
скими концепциями, - демонстрируют столь поразительное незнание основ ге-
нетики и всей современной литературы, что опровергать их было бы напрасной
тратой времени» {Майр Э. Зоологический вид и эволюция. М., Мир, 1968, с. 22).
Это, разумеется, верно - нелепо было бы Майру, вся эрудиция которого в облас-
ти современной ему генетики свелась к фразе: «ДНК ядра активно взаимодейст-
вует с клеточными белками через РНК» (там же, с. 148), тратить время на опро-
вержение профессионалов, державшихся молекулярного ламаркизма.
Последний имел в то время блестящих представителей. Таковым был, на-
пример, Поль Вентребер, один из тех, с кого началась нынешняя (а не мечни-
ковская) эволюционная иммунология (см. АСМи ЗЭ), о чем Майр, как и о номо-
генезе не имел, как видим, никаких знаний. Нет их и у его последователей.
фр. 12. Таковы, например, многие метео-астрономические приметы, над ко-
торыми обыватели, не понимая их значения, смеются до сих пор. Впрочем, глав-
ный смысл аргументации Мейена другой - этический: не осуждай ничего огуль-
но, можешь оскандалиться, пусть и после смерти.
370
2. В.А. Брынцев. Рецензия
на книгу: Чайковский Ю.В. Активный связный мир: Опыт теории эво-
люции жизни. М.: КМК, 2008. 726 с.*
Представляемая книга, объемный том по теории эволюции, предназначена для тех,
кто интересуется вопросами теоретической биологии и эволюционизма, не удовлетворя-
ясь простыми ответами на сложные вопросы.
Перед нами глобальный, от Анаксимандра до современных исследователей, научно-
исторический обзор эмпирических обобщений, теорий и гипотез по эволюционному направле-
нию. Особый интерес представляют внимательное отслеживание и анализ автором новых,
недавно появившихся в отечественной и зарубежной литературе эмпирических и теоретиче-
ских работ, связанных с эволюцией. Истории эволюционных идей полностью посвящена пер-
вая часть книги, но не только, ибо эта тема красной нитью проходит через всю работу.
Другая прослеживающаяся на протяжении всей книги тема - жесткая критика дарви-
низма, иногда несколько эмоциональная. В конце первой части приведен яркий пример с
размножением термитов, который, по мнению автора, является доказательством отсут-
ствия в природе естественного отбора. Взяв за основу опыт, поставленный самой приро-
дой, и используя попперовскую методику, Ю.В. Чайковский пытается фальсифицировать
отбор как научное теоретическое положение, видимо, забывая, что сам К. Поппер считал
дарвинизм феноменом, не вписывающимся в рамки научной теории.
Обширный материал книги помогает воспринять ее четкая структура: четыре части,
каждая по три главы, в них подглавы, представляющие собой логически законченный
текст. Работа снабжена подробным указателем терминов и имен, библиографией, иллю-
страциями, перекрестными ссылками. Издание относится к тем, с которыми приятно ра-
ботать. Однако создается впечатление, что перекрестными ссылками автор несколько
злоупотребляет. Часто в интересах смысла необходимо повторить мысль, а не отсылать
читателя за ней на другой конец книги.
Достаточное место в книге занимает науковедческая гносеологическая часть. Она
могла бы представлять отдельную главу, но здесь она вплетена в общий текст книги и
помещена в главу «Принцип активности, развитие особи и иммуногенез».
Ю.В. Чайковский объясняет методику своей работы. Она заключается в анализе вы-
двинутых за всю историю эволюционных принципов, отказе от тех, которые не работают,
и синтезе тех, которые признаются рабочими. При этом рассматриваются не только тео-
рии, а и отдельные принципы и идеи, охват которых невероятно обширен. Перед нами
предстает, наверное, один из самых полных обзоров эволюционных идей и их анализ.
В изложении материала автор использует исторический принцип, выводя каждую тему
из работ прошлого и показывая сложное к ней отношение в разное время. Многие продук-
тивные исторически сложившиеся эволюционные идеи, вытесненные господствующими
концепциями на периферию науки, вводятся им в круг научного анализа и обсуждения.
Формулируя требования к теории, автор отказывается от презумпций и «бритвы Ок-
кама», выражающейся в установке: «Не умножай сущностей сверх необходимого мини-
мума» (с. 213), что дает возможность широкого и всестороннего анализа материала, но
* Вестник Российской Академии наук, 2009, № 12, с. 1128-1130.
371
мешает построению своей строгой теории. Для анализа данный метод, бесспорно, под-
ходит. Не мешает он и синтезу из готовых теоретических блоков. Однако вырастить дей-
ствительно новую теорию невозможно без ограничений, которые достигаются посредст-
вом презумпций и «бритвы Оккама». Поэтому в качестве метода у автора остается под-
бор и крупноблочный синтез. При этом, как при любом синтезе, создается структура с но-
выми эмерджентными свойствами, но создать истинно новое таким путем не могут, ви-
димо, ни человек, ни природа. Единственная презумпция, принятая Ю.В. Чайковским, -
это «презумпция рационального объяснения явления», но в некоторых случаях автор от-
ходит от нее, находя опору в идеях платонизма. Платонизм помогает постановке задач,
но мешает их рациональному решению.
К методологической части относится глава о познавательных моделях. Этот подход,
развиваемый автором на протяжении двух десятков лет, позволяет оценить теории с точ-
ки зрения общедоступных для понимания моделей. При этом хорошо видны ограничения
каждой из них. Познавательные модели крайне продуктивны для исследования истории
теоретических знаний. Непонятно, почему автором не была выделена типологическая
познавательная модель; если она входит в диатропическую, то последняя должна быть
одной из старейших познавательных моделей.
Ведущим для эволюции в книге признан принцип активности. Проводя обзор и ана-
лиз новейших исследований по иммунологии, Ю.В. Чайковский приходит к выводу об ак-
тивности генома. Активность становится важным эволюционным фактором. На основе
анализа иммуногенетики, по аналогии с ней, строится эволюционная модель, в которой
ведущую роль играет активный выбор. В свою очередь, у автора активный выбор пред-
полагает наличие направленных механизмов. Согласно принципу активности, возрастает
значение онтогенеза для эволюции организмов. Онтогенез уже не является тенью фило-
генеза, а играет в эволюции свою активную самостоятельную роль. Эволюцию организ-
мов автор рассматривает как перестройку онтогенеза. Эволюция и онтогенез имеют сис-
темное сходство, поскольку в обоих случаях изменения определяются взаимодействием
активности сверху (от системы) с активностью снизу (от элементов). Следует отметить,
что при таком подходе главным становится не изучение изменения формы или функции,
а изучение законов самого процесса преобразования.
То, что эволюционисты начали понимать, что имеют дело с движением, хотя бы
только в форме активности, - большой шаг вперед. Однако в книге понятие активности
не формализовано. Пусть оно в целом неопределимо, но вполне может быть обсуждено
на формальном уровне. Автор же приводит многочисленные примеры, предоставляя де-
лать заключения читателю. При этом нередко возникают вопросы. Почему, скажем, спе-
циализация клеток относится к активности? При специализации клетка идет путем, жест-
ко прописанным ей организмом, - это, скорее, пример пассивного движения. То же может
относиться к заполнению диатропической сети. Поэтому хотелось бы понять, в чем про-
является активность: в расширении систем? в активном ответе на внешние вызовы? в
активном поиске путей своего развития?
Второй важнейший эволюционный принцип - принцип сопряженности. У сопряжен-
ности два основных вида: повторность форм и согласование явлений. Если второе по-
нятно, то с тем, что повторность форм - это не следствие, а именно вид сопряженности,
согласиться трудно. Тем не менее повторность форм составляет основу диатропической
372
познавательной модели, и ей в книге уделяется много места.
Ю.В. Чайковский, исследуя вопрос о феномене сходства, указывает, что прежде
было два ответа: грубо материалистический (родство, по Геккелю) и грубо идеалисти-
ческий (общая идея, по Платону). Автор говорит, что оба они маловразумительны и ос-
нованы только на вере. Вразумительным же ответом является следующий:
«...Сходные структуры и функции могут достигаться различными путями в силу упоря-
доченности диасети» (с. 327). По моему мнению, более понятны ответы Геккеля и Пла-
тона. Дело в том, что диасеть есть некая абстрактная (идеальная) конструкция, по-
строенная на эмпирических (материальных) обобщениях. Диатропика, как и любое на-
учное направление, не может находиться между материальным и идеальным, посколь-
ку устойчивого серединного положения здесь не просматривается. На материалисти-
ческих позициях диатропика имеет подчиненное положение, приводя в порядок полу-
ченные знания. Если диатропика - идеалистическая теория, то в упорядоченные ряды
встают «идеи» Платона. Как самостоятельное направление диатропика видится только
на основе платонизма. Только тогда «эволюцию можно рассматривать как достаточно
закономерное преобразование диасети» (с. 318). Такой подход возможен, интересен и
продуктивен, по нему шла, в частности, математика, но нужно всегда помнить, что при
этом находишься в идеальном мире теоретических знаний и связь с реальным процес-
сом эволюции будет весьма опосредованной.
Это же относится и к фрактальности, которой автор придает слишком большое зна-
чение. Масштабная инвариантность (одно из свойств фрактальности) в природе встреча-
ется достаточно часто и отражает создание подсистем по образу и подобию материнской
системы. Однако вложенность эта не очень глубокая и всегда прерывается. Фракталами
также описываются некоторые соотношения в процессах и явлениях, но это еще не зна-
чит, что природа использует фрактальные законы для построения своих форм. Фрак-
тальность есть некое идеальное отражение реальных процессов и, видимо, связана с
замкнутостью пространства движения.
Второму виду сопряженности - согласованности явлений, к сожалению, уделено
меньше внимания, хотя именно в ее изучении видится путь от системной к активностной
познавательной модели. Два вида сопряженности относятся к разным познавательным
моделям. В первом случае сопряженность в виде диасети направляет и определяет дви-
жение. Во втором случае «сопряженность - итог активности материи на всех уровнях».
Таким образом, два вида сопряженности - повторность форм и согласованность явле-
ний, смешанные автором в одной главе, относятся к разным познавательным моделям -
диатропической и активностной - и являются дополнительными одна к другой. Если пер-
вая не может без платонизма, то вторая вполне без него обходится. Перспективно разви-
вать обе модели, понимая, однако, что они не могут быть ни смешаны, ни объединены.
Общие выводы по прочтении книги следующие. Современные изменения в эволю-
ционных представлениях нуждаются не в замене неодарвинизма номогенезом, ламар-
кизмом или иным новым их синтезом, они требуют смены измерений, похожей на пере-
ход из плоского видения к объемному. Взгляды на эволюцию получили по крайней мере
три существенных изменения. 1. Значимым фактом для образования нового в эволюции
признается активность, которая направлена как снизу (от элементов к системе), так и
сверху (от системы к элементам). 2. Объектом эволюции видов становится не морфофи-
373
зиология, а онтогенез, то есть процесс. Изменяется размерность изучаемого. От позна-
ния изменений форм и функций наметился переход к изучению самого процесса измене-
ния. 3. Изучаемый объект не выделяется из системы, а рассматривается в ней и с ней.
Эволюция видов зависит от эволюции сообществ и биосферы в целом. Теория эволюции
превращается в теорию коэволюции систем одного и разного уровней организации. В
этом, на мой взгляд, суть принципа сопряженности.
Намечается новая картина мира, в которой движение, изменение, активность явля-
ются основой и идут впереди формы, функции, да и системы как таковой. Не эволюция
систем, а эволюция, рождающая системы, создающая их. Она не может быть описана ни
одной из существующих эволюционных теорий, ни их механическим синтезом. Но именно
развитие имеющихся эволюционных теорий приводит нас к новой картине мира.
У книги Ю.В. Чайковского есть большая и важная цель - дать рекомендации для че-
ловеческого общества, наметить общую стратегию его выживания, основанную на пони-
мании эволюционных процессов. Связывая прошлое великих идей и будущее человека,
книга возвращает нам настоящее - как время поиска и свершений.
Пояснения к рецензии
На мой взгляд, рецензент в целом прав, и позволю себе лишь несколько заме-
чаний. Они носят, по-моему, характер не столько возражений, сколько пояснений.
1) Отказ от использования презумпций действительно очень затрудняет рабо-
ту, и я сам пользовался ими, пока не убедился (в ходе бесед с Ю.А. Шрейдером
о главной презумпции европейской науки - «бритве Оккама»), что презумпции
не просто мешают исследованию, а прямо пресекают его в исходных пунктах.
На каком-то этапе прибегать к ним, видимо, можно, но на каком - не знаю.
2) Платонизм действительно мешает рациональному решению задач, если
полагать саму идею предсуществования мира идей иррациональной. Сам Пла-
тон так не считал, и математика служит подтверждением его правоты, прежде
всего - теория фракталов, независимость существования которых от нашего
сознания особенно очевидна.
Что касается «устойчивого серединного положения» между идеализмом и
материализмом, то таковым мне представляется позиция Аристотеля, полагав-
шего идеи не предсуществующими, а возникающими в ходе обобщения знаний
о мире. Этой позиции лучше всего и держаться, но не в отношении к фракталам,
где платонизм бьет в глаза. Тут более уместна позиция Платона. Напомню, что и
он полагал, что свою идею имеют отнюдь не все вещи.
3) Фракталы - тоже математика, так что применять их «на основе платониз-
ма» так же законно, как и любую математику. Связь их с биологией столь же
«опосредованна», как и классической математики с физикой. В частности, мате-
матический фрактал - структура бесконечная, воображаемая, тогда как реаль-
ные объекты конечны; поэтому введение фрактала - идеализация. Она, в прин-
ципе, столь же законна, как введение интегралов вместо сумм или вероятностей
вместо частот - осторожное введение полезно, а бездумное вредно и даже может
лишить работу всякого смысла (так произошло в популяционной генетике).
4) Гораздо большей, нежели фракталы, идеализацией реального мира являют-
ся ПМ. Достигаемая с их помощью схематизация знания (такая же, как с помо-
374
щью парадигмы, защитного пояса и т.п - см. выше, статью 4-65) оказалась для
многих ученых чрезмерна, но В.А. Брынцев к ним явно не относится и даже
предложил собственную ПМ [Брынцев, 2007]. Тем самым, рамки применимости
моделирования задаются, по-моему, не глубиной идеализации (абстрагирова-
ния), а тем, насколько модель ухватывает суть описываемого явления, т.е. выяв-
ляет инварианты исследуемого «движения» - в том широком смысле этого сло-
ва, каким пользуется рецензент.
5) Неизбежность построения теорий с помощью фракталов на сегодня очевид-
на, насколько знаю, всем, кто понимает, что такое фрактал. И не только потому,
что подсистемы в природе создаются «по образу и подобию материнской систе-
мы» и в связи с «замкнутостью пространства движения» (это рецензент отметил
верно), но и в силу более общей математической причины. Фрактальный подход
противоположен подходу Ньютона - Лейбница, породившему “гладкую” матема-
тику, которая служила 300 лет основой моделирования «движения» в широком
смысле. Первый удар ее господству был нанесен еще сто лет назад созданием тео-
рии функций действительного переменного (ТФДП) - первой “рваной” математи-
ки. (Недаром исторически первым фракталом оказалось множество Кантора, рас-
сматриваемое в ТФДП.) Однако ТФДП не посягала на анализ «движения» в ши-
роком смысле - это сделали фракталы Мандельброта. Самой простой и эффектной
демонстрацией силы нового метода явилось, на мой взгляд, распространение
принципа “рваности” на область комплексных переменных, которую прежде изу-
чали только средствами теории аналитических (предельно гладких) функций.
Именно тут открылись никем не рисованные узоры такой сложности и красоты,
что платонизм стал выглядеть не выдумкой гениального философа, а реальностью
мира, в котором мы живем (см. АСМ, дополнение 4).
6) На сегодня фракталы являются единственным аппаратом, обещающим опи-
сание онтогенеза. В рамках “гладкой” математики оно не удалось. Даже блестя-
щие попытки выразить усложнение зародыша в понятиях топологии [Том, 1972]
привели, на мой взгляд, лишь к уточнению проблематики, а попытки описать это
усложнение диф. уравнениями мат. физики обречены увязнуть в бесконечной
сложности тензоров высших порядков (см. 4-54). Упрощение, достигаемое язы-
ком фракталов, примерно так же радикально, каким было 300 лет назад упроще-
ние задач на экстремум, происшедшее после открытия производных.
Замечательно, что фракталы проявились и в попытках формализации система-
тики - еще один пример «непостижимой эффективности» (см. выше, с. 337 и д.).
7) На недоуменные вопросы рецензента («хотелось бы понять, в чем проявля-
ется активность: в расширении систем? в активном ответе на внешние вызовы? в
активном поиске путей своего развития?») можно ответить так: во всех трех ас-
пектах, особенно в последнем. Он почти не затронут в рецензии (достаточно от-
метить, что слово «самоорганизация», трижды упомянутое даже в приведенном
ниже мини-реферате, в ней отсутствует), откуда, по-моему, и недоумение.
8) Активность - не моё построение, а наблюдаемое свойство мира, не раз
отмеченное. Мне принадлежит лишь выбор самого нейтрального слова («актив-
ность») для обозначения этого удивительного свойства. Столь же удивительна
375
сопряженность, тоже наблюдаемая на всех уровнях мироздания. В рецензии
верно отмечено, что этим словом у меня обозначаются два разных понятия, и в
свое оправдание могу сказать лишь, что сам не умею (в отличие от В.А. Брын-
цева) сколько-то определенно их различить. Тот факт, что диасеть относится к
четвертой ПМ, а согласованность явлений - к шестой, проблему для меня не
снимает, поскольку любая ПМ-претендует на описание мира как целого. Состо-
ит же проблема в том, что сопряженность некоторых явлений выглядит такой же
первичной, как и диасеть. Взять хотя бы «проблему четырех Л» (АСМ, п. 7-12),
т.е. проблему первичной согласованности основных свойств наследственности.
За годы после ее формулировки она еще больше углубилась за счет открытия
параллели между редактированием РНК и конструированием ДНК в ходе имму-
ногенеза (ЗЭ, рис. 15). Параллель носит платоновский характер (идея повторена
на ином субстрате), так что без платонизма и здесь не обойтись.
3. Р.Г. Баранцев. Мини-реферат
книги «Чайковский Ю.В. Активный связный мир»
Биологическая эволюция неотделима от общей эволюции мира. Эволюция природы
- нечто единое, целостное. Направленность значимых изменений. Триединый принцип
эволюции живого: активность - диасеть - смена норм (с. 12).
Принцип сопряженности (когерентность? - Р.Б.) (с. 13).
Номогенез - эволюция на основе закономерностей (с. 161). Сходство - результат
общности законов развития (с. 164). Ряды Вавилова - номогенетические... Пять идей у
Берга: номогенетические ряды, аналогия с химией, ортогенез (длительная тенденция в
определенном направлении) преадаптация, пророческая фаза (с. 165).
Любищев: Основа природы - не польза, а гармония (с. 199). Форма - самодовлею-
щая сущность (с. 202).
Термин «активность» предпочтительнее (чем «жизненный порыв» А. Бергсона), ибо
подчеркивает общность феномена, его господство на всех уровнях организованности -
от элементарных частиц, через физику и химию, через жизнь и культуру, до космоса (с.
223). Эволюция - усложнение форм активности (с. 224). Познавательная модель (А.П.
Огурцов) - базисная метафора (с. 225).
На грани порядка и хаоса (с. 300). Любищев - «Морозные узоры»: законы образова-
ния форм (с. 301). Активность формирует варианты из возможных, реализуя сопряжен-
ность (лат. conjunction, самосогласованность. - Р.Б.) - атрибут целостности. Коэволюция
- сопряженная эволюция (с. 302).
Тенденция видна лишь на массиве (с. 306). Закономерности наблюдаются в форме
тенденций (с. 308). Через приспособление она не объяснима. Разнообразие служит од-
ной из форм сопряженности (с. 311). Разнообразие множества - это совокупность отно-
шений сходства и различия между элементами данного множества (с. 312).
Основной прием диатропики - сопоставление рядов. Таблицы живого многомерны
(с. 315). Мерой - класс частей (с. 316). Номогенетический механизм видообразования.
Рефрен - самый заметный вид сопряженности структур, а смена норм - сопряженности
Любезно передан мне самим референтом.
376
процессов (с. 325). Сходство нельзя сводить к родству. Сходные структуры и функции
могут достигаться различными путями в силу упорядоченности диасети (с. 327). В фено-
мене рефрена выявляется сопряженность разнородных явлений (с. 326).
Мозг живет в околокритическом режиме (с. 336). Наследование фракталообразующего
правила - механизм передачи разнообразия (с. 343)... Эти картины существовали всегда, а
мы только сейчас научились их видеть (с. 344). Фрактал - средство переноса больших ко-
личеств информации через маломощный канал (наследование). Многоуровневость коэво-
люции - нет собственного характерного масштаба в пространстве и времени (с. 345).
Ограниченность спектра возможных форм. Эволюция как движение по рядам, зада-
ваемым структурой разнообразия - это структурализм в эволюции, вариант номогенеза
(с. 358). Структурные инварианты (Гёте, А. Гумбольдт, Вернадский, Пропп, Леви-Строс,
Корона, Мейен) (с. 359). Инвариантом структуры у Мейена служит архетип (с. 360). Само-
организация - основное свойство природы на всех уровнях (с. 367). Феномен диасети го-
ворит о морфологическом единстве природы как целого (с. 371). Морфониша - совокуп-
ность сходных черт строения (с. 372).
Внутриклеточное мышление (? с. 386). Может быть, замысел записан вне особи?
Соответствие объекта системе, сопряженность (с. 387). Если принять платонизм, то эво-
люция предстает как активное движение вдоль заданной диасети (с. 390).
Мир не является иерархическим. Мир является сложным переплетением древовид-
ных и сетевидных фракталов (с. 574). Естественная система всегда субъективна. Приме-
ры в географии: естественная - по принадлежности к речной системе, политической или
хозяйственной структуре; искусственная - координаты (широта и долгота) (с. 582).
Свойства целостности, общие для всех уровней, - активность и сопряженность (ко-
герентность? фрактальность? - Р.Б.) (с. 605). В природе на каждом уровне видна самоор-
ганизация. Это процесс, ведомый активностью и приводящий к новым сопряженностям.
Другим аспектом активности является эмерджентность (создание совсем нового). Основ-
ной тезис общей теории эволюции: разнообразие - объект эволюции, активность - ее
движущая сила, богдановский подбор - ее регулятор, а сопряженность - ее итог. Этот те-
зис - философский постулат (с. 607).
Объект эволюции - многоуровневое разнообразие форм материи, взятое как целое.
Процесс эволюции - последовательность актов сопряжения частей в очередное целое (с.
608). Три формы активности живого: материальная, энергетическая, информационная. В
последней есть три аспекта сопряжения: узнавание, изобретение и наследование (с.
609). Биосфера - мультифрактал (с. 610). Три типа уровней: взаимодействия, классифи-
кации и сложности. И соответственно три аспекта эволюции: функциональный, структур-
ный и сложностный (с. 613).
Новое рождается в каждом онтогенезе, но без свободы выбора... Нет оснований
ожидать, что мышление и эмерджентность могут быть поняты по отдельности... Новое
рождается при сопряженном усложнении фрактальной структуры объектов, поэтому
эмерджентность так же неизбежна, как рост кристаллов (с. 617). Процессы самооргани-
зации должны служить первичными понятиями, а состояния - вторичными (с. 634). Взаи-
модополнительность - основа картины мира; мир - единая активная целостность, дости-
гаемая путем сопряжения процессов (в том числе сверху и снизу). Мир имеет вид много-
фрактальной диасети и непрестанно развивается... Взаимодействие активностей пред-
377
стает наблюдателю как силовое поле... Мысль о возможном наличии полей на всех
уровнях организации материи (с. 635).
Возможность давать практические рекомендации - главный критерий работоспо-
собности теории. Она должна дать спектр возможных вариантов события. Самая про-
стая и общая рекомендация - не путать эволюцию ни с отбором, ни с филогенией (с.
636). Дарвинизм во всех формах основан на идее обратной связи (именуя ее естест-
венным отбором), и начисто отрицает наличие прямых связей, а они-то и движут лю-
бой процесс. Прямой связью является формирующий принцип, а обратной - регули-
рующий (с. 637). Механизм становления сопряженности (прямую связь), увы, никто не
изучает. Теории его нет и до отказа от дарвинизма (шире: до смены ПМ и картины ми-
ра) явно не будет (с. 639).
Разнообразие организовано в диасеть (имеет рефренную структуру) всюду - от мак-
ромолекул и клеток до таксонов и (отчасти) сообществ. Выявление диасети как целого ви-
дится первой задачей для построения приемлемой систематики и работоспособной теории
эволюции. А также для диатропического прогноза, под которым понимается выявление (пу-
тем анализа рефренов) спектра возможных блоков - таких, что все огромное разнообразие
реальных особей есть набор комбинаций не очень большого числа блоков (с. 645).
Наука может жить только как часть культуры, а та культура, которая норовит стать
всемирной, с наукой сопрягается плохо... Я не вижу иных путей спасения нашей цивили-
зации, кроме тех, которые будут исходить из факта сопряженности всего со всем (the
consilience), из понимания того, что мы живет в связном мире (с. 655).
Типы активности организмов реализуются, как и формы их тел, в рамках достаточно
ограниченной диасети (с. 688). Источник эмерджентности таится в смене параметров
фракталов (с. 692). Проблема новаций требует иного взгляда на мир, а он, в свою оче-
редь, должен породить новую познавательную модель, вероятно - активностную (с. 693).
4 апреля 2010 г.
Общий список литературы к сборнику
Акчурин И.А. Единство естественнонаучного знания. М., Наука, 1974.
Александров В.Я. Проблема поведения на клеточном уровне (цитоэтология) // Успехи
современной биологии, 1970, т. 69, № 2.
Алексеев В.П. Человек. Эволюция и таксономия. М., 1985.
Амлинский И.Е. Жоффруа Сент-Илер и его борьба против Кювье. М., Изд. АН СССР,
1955.
Андреев Ю.В. Тираны и герои. Историческая стилизация в политической практике
старшей тирании // Вестник древней истории, 1999, № 1, с. 3-7.
Аронова Е. А. Карл Поппер, «наука по Попперу» и дискуссии о ламаркизме в биоло-
гии // ВИЕТ, 2004, № 2.
Ахметьев М.А. Географическая дифференциация позднемеловых и кайнозойских
флор // Стратиграфия. Геол, корреляция. 1999, № 3, с. 42-62.
Бабков В.В. Заря генетики человека. М., 2008.
Балковский Б.Е. Цифровой полигамический ключ для определения растений // Бота-
378
нич. журнал, 1960, № 1; он же. Теза, антитеза и ряд признаков в диагностике растений //
Ботанич. журнал, 1960, № 11.
Баранцев Р.Г. Синергетика в современном естествознании. М., УРСС, 2003. 142 с.
Барское И.С., Жерихин В.В., Раутиан АС. Проблемы эволюции биологического разно-
образия И ЖОБ, 1996, №2, с. 14-39.
Бахметьев П.М. Попытка установить периодическую систему бабочек // Труды Сара-
товского общества естествоиспытателей, т. 4, 1903-1904.
Бейтсон Г. Шаги в направлении экологии разума. Избранные статьи по теории эво-
люции и эпистемологии. М., 2005.
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных, т. 1. М., 1964.
Беклемишев В.Н. Морфологические проблемы животных структур. Пермь, 1925.
Беклемишев В.Н. Методология систематики (1928). М., КМК, 1994. 250 с.
Берг Л.С. Борьба за существование и взаимная помощь. П., Время, 1922. 35 с.
Берг Л. С. Труды по теории эволюции. Л., Наука, 1977. 387 с.
Бергсон А. Творческая эволюция. М., Канон-Пресс, 1998. 383 с.
Берзин Э. Сивка-бурка, вещая каурка // Знание - сила. 1986. № 11.
Бернштам ТА. Молодежь в обрядовой жизни русской общины. Л., 1988.
Блехер Л.И., Любарский Г.Ю. Главный русский спор: от западников и славянофилов
до глобализма и Нового Средневековья. М., “Академический проект”, 2003. 605 с.
Бляхер Л.Я. Проблемы морфологии животных. М., Наука, 1976.
Бобров Е.Г. Описательный метод Линнея, «Species plantarum» и современная номенк-
латура растений // Карл Линней. Сб. статей. М., Изд. АН, 1958, с. 78-100.
Богородицкий В.А. Очерки по языковедению и русскому языку. Казань, 1910.
Борель Э. Случай. М. - П., 1923.
Бородина НА. и др. Деревья и кустарники СССР. Полевой определитель. М., 1966.
Бродель Ф. Время мира. Пер. с франц. М., Прогресс, 1992. 679 с.
Бродель Ф. Динамика капитализма. Пер. с фр. Смоленск, Полиграмма, 1993. 124 с.
Брынцев В.А. Филлотаксис. Опыт применения компьютерных моделей для решения
вопросов теоретической морфологии растений. Мос. гос. Унивситет леса. М., 2006.
Брынцев В.А. Системно-динамическая познавательная модель // XXI ЛЧ, 2007, с.
72-77.
Брынцев В.А. Рец. на книгу «Активный связный мир» // Вестник РАН, 2009, № 12.
Бэр К. Человек в естественноисторическом отношении // Симашко Ю.И. Русская фло-
ра... . Часть 1. СПб., 1850, с. 390-623.
Бэр К. Избранные работы / сост. и примеч. Ю.А. Филипченко. Л., Госиздат, 1924.
Бэр К.М. Об искусственной и естественной классификациях животных и растений. О
сродстве животных. // Анналы биологии. М., МОИП, 1959. Т. 1, с. 367-383; 395-405.
В помощь аспиранту. Существует ли естественный отбор? (Материалы круглого сто-
ла) // Высшее образование в России, 2006, №№ 7, 8.
Вавилов Н.И. Закон гомологических рядов в наследственной изменчивости. М., 1987.
Вавилов С.И. Из дневников 1939-1951 гг. // ВИЕТ, 2004, № 2.
Вольт М.Х. Экологические исследования К. Бэра и концепция борьбы за существова-
ние // Петербургская Академия наук и Эстония. Таллин, Валгус, 1978, с. 102-119.
Васильев А.Г., Васильева НА. Гомологическая изменчивость морфологических структур
и эпигенетическая дивергенциятаксонов: основы популяционной мерономии. М., 2009.
Вейсман А. Лекции по эволюционной теории. Т. 1. Петроград, 1918.
Взаимодействие лингвистических ареалов, 1980
379
Виндельбанд В. История новой философии. СПб., 1913. Том 1.
Вишневский А.Г. Воспроизводство населения и общество. М., 1982.
Водовозов В. В. Бюрократия // Новый энциклопедия, словарь Брокгауз-Ефрон, т. 9,
1912, стлб. 160.
Воейков В.Л. Ключевая роль устойчиво неравновесного состояния водных систем в
биоэнергетике // Российский химич. журнал, 2009, № 6.
Волк / ред. Д.И. Бибиков. М., 1985.
Воробьева Э.И. Процессуальный подход к проблеме «тетраподизации» // ПЖ, 2008, № 2.
Гайсинович А.Е. К.Ф. Вольф и учение о развитии организмов. М., 1961.
Гачок В.П. Странные аттракторы в биосистемах. Киев, 1989.
Гельмгольц Г. Популярные научные статьи. СПб. Вып. 1, 1866.
Георгиевский А.Г. Проблема преадаптации. Историко-критическое исследование. Л.,
Наука, 1974.
Гершензон С.М. Избирательность мутагенного действия ДНК и других полинуклеоти-
дов//ЖОБ, 1996, №6.
Гилберт С. Биология развития. М., Мир, 1995, т. 3.
Глэд Дж. Будущая эволюция человека. Евгеника XXI века. М., Захаров, 2005. 169 с.
Гоббс Т. Сочинения в двух томах. Т. 1. М., 1989.
Голубовский МД Неканонические наследственные изменения // Природа, 2001, № 8,
№ 9.
Гоманьков А.В. Синтез или противоречие? // Природа, 1990, № 4, с. 73-79.
Гордин Я.А. Меж рабством и свободой. СПб., Лениздат, 1994. 380 с.
Грамматика литовского языка/В. Амбразас. Вильнюс, 1985.
Гринченко С.Н. Метаэволюция (систем неживой, живой и социально-технологической
природы). М., 2007.
Гумилев Л.Н. Конец и вновь начало. М., 2004.
Гумилев Л.Н. Этногенез и биосфера Земли (1979). М., 2003.
Гуркин Ю.А., Суслопаров Л.А., Островская Е.А. Основы ювенильного акушерства.
СПб'., 2001.
Дарвин Ч. Избранные письма. М., 1950.
Дарвин Ч. Соч.,тт. 1-9. М.-Л, 1935-1959 (т. 3, 1939; т. 4, 1951).
Де Боно Э. Рождение новой идеи. М., 1976
Дербин А.В., Бахланов С.В., Егоров А.И., Муратова В.И Замечания к статье “О реали-
зации дискретных состояний...” (Шноль С.Э. и др., 1998) // УФН, 2000, № 2.
Детская и подростковая гинекология / ред. Анна Гарден; пер. с англ. М., Мир, 2001.
Деятели СССР и революционного движения России // Энцикл. словарь Русского
Библиографич. ин-та Гранат, т. 41. М., 1925-1926 (переиздано: М., Совет, энциклопе-
дия, 1989. 832 с.).
Джан Р.В. Филлотаксис: системное исследование морфогенеза растений / пер. с англ.
М.-Ижевск, 2006.
Джеффри Ч. Биологическая номенклатура. М., Мир, 1980.
Дживелегов А.К. Бюрократия // Энцикл. словарь Русского Библиографич. ин-та Гра-
нат, 7-е издание, т. 10, 1911, стлб. 423-431.
Дьяков Ю.Т. Введение в альгологию и микологию. М., МГУ, 2000.
Ерошкин Н.П. История государственных учреждений дореволюционной России.
Учебник. М., Высшая школа, 1983. 352 с.
Ефимов А. Элитные группы, их возникновение и эволюция// Знание - сила. 1988. № 1.
380
Жерихин В.В. Избранные труды по палеоэкологии и филоценогенетике. М., КМК, 2003.
Заварзин А.А. Труды по теории параллелизма и эволюционной динамике тканей. Л.,
Наука, 1986.
Заварзин Г.А. Составляет ли эволюция смысл биологии? // Вестник РАН, 2006, № 6.
Заварзин Г.А. Фенотипическая систематика бактерий. М., Наука, 1974. 190 с.
Заславский Г.М. Стохастичность динамических систем. М. Физматлит, 1984. 272 с.
Захаров Б.П. Трансформационная типологическая систематика. М., КМК, 2005. 165 с.
Зейгарник Б.В. Теория личности в зарубежной психологии. М. 1982.
Зелеев Р.М. Проблемы построения системы организмов и некоторые критерии мега-
таксономии // XVIII Любищевские чтения. Ульяновск, 2004.
Зусмановский А.Г. Эволюция с точки зрения физиолога. Ульяновск, Ульяновская
Сельхозакадемия, 2007. 394 с.
Иваницкий Г.Р., Медвединский А.Б., Деев А.А., Цыганов М.А. От “демона Максвелла” к
самоорганизации массопереноса в живых системах И УФН„ 1998, т. 168, № 11.
Иванов А.В. Сознание и мышление (учебное пособие). М., Изд. МГУ, 1994.
Иванов Валер.Вас. Историческая грамматика русского языка. М., 1983.
Иванов Вяч. Вс. Послесловие // Леви-Строс К. Структурная антропология. М., 2008.
Игнатьев И.А. Общие фитогеографические модели и эволюция позднепалеозойских
флор // Эволюция флор в палеозое. М., ГЕОС, 2003.
Игнатьев И.А. Воспоминания об С.В. Мейене. Об эволюционизме Мейена // Общая и
прикладная ценология. 2007, № 4, с. 47-48.
Из научного наследия С.В. Мейена: к истории создания основ эволюционной диатро-
пики И Эволюция флор в палеозое. М., ГЕОС, 2003.
Ирошников М.П. Председатель Совета Народных комиссаров Вл. Ульянов (Ленин). Л.,
Наука, 1974.
Канаев И. И. Очерки из истории проблемы морфологического типа от Дарвина до на-
ших дней. Л., Наука, 1966.
Карпинская Р.С., Лисеев И.К., Огурцов А.П. Философия природы: коэволюционная
стратегия. М., Наука, 1995. 250 с.
Каталог млекопитающих СССР. Плиоцен - современность / ред. И.М. Громов, Г.И. Ба-
ранова. Л., Наука, 1981.456 с.
Кауфман С.А. Антихаос и приспособление // В мире науки, 1991, № 10.
Кац М., У лам С. Математика и логика. Ретроспектива и перспективы. М., 1971.
Клайн М. Математика. Поиск истины. М., Мир, 1985.
Климентович, 1996
КлэкДж. Трудный путь на сушу// ВМН, 2006, № 3
Клюге Н.Ю. Современная систематика насекомых. СПб., Лань, 2000. 333 с.
Ключевский В.О. Неопубликованные произведения. М., Наука, 1983. 416 с.
Кобляков А.А. Синергетика и творчество // Синергетическая парадигма. М., Прогресс-
традиция, 2000, с. 308-324.
Колчинский Э.И. Неокатастрофизм и селекционизм... СПб., 2002.
Кон И. С. Психология юношеского возраста. М.: Просвещение, 1979.
Кордюм В.А. Эволюция и биосфера. Киев, Наукова думка, 1982, 262 с.
Корона В.В. Основы структурного анализа в морфологии растений. Свердловск, Изд.
СГУ, 1987. 272 с.
Корочкин Л.И. Стволовые клетки в нейрогенетике // Генетика, 2004, № 6.
Красилов В.А. Роль случайности в эволюции // Эволюционные исследования. Влади-
381
восток, 1979, с. 7-19.
Красный Л.И. Разномасштабная делимость // Вестник РАН, 2002, № 6.
Кудрин Б.И. Проблемы создания и управления ценозами искусственного происхожде-
ния // Кибернетические системы ценозов: синтез и управление. М., МОИП, 1991.
Кун Т. Структура научных революций (1962). М., ACT Москва, 2009.
Куприянов А.В. Предыстория биологической систематики. «Народная таксономия» и
развитие представлений о методе в естественной истории конца XVI - начала XVIII вв.
СПб., Европейский ун-т, 2005. 60 с.
Кусакин О.Г., Дроздов А.Л. Филема органического мира. СПб., Наука, 1998 (т. 2).
Лакатос И. Фальсификация и методология научно-исследовательских программ. М.,
Медиум, 1995. 236 с.
Ламарк Ж.-Б. Философия зоологии. Т. 1. М., Биомедгиз, 1935.
Левина Р.Е. Морфология и экология плодов. Л., Наука, 1987. 160 с.
Леви-Строс К. Первобытное мышление. М., Республика, 1994. 384 с.
Левин А.П. Структура экологических сообществ. М., 1980.
Лекявичюс Э. О некоторых аналогиях между эволюцией экосистем и развитием эконо-
мики... И Эволюция: космическая, биологическая, социальная. М., Либроком, 2009.
Леонтьев А.Н. Деятельность, сознание, личность. М.: 1975.
Лима-де-Фариа А. Эволюция без отбора. Автоэволюция формы и функции. М., Мир,
1991.
Линг Г. Физическая теория живой клетки (незамеченная революция). СПб., Наука, 2008.
Линней К. Философия ботаники. М., Наука, 1989. 452 с.
Лункевич В.В. От Гераклита до Дарвина. М., Учпедгиз, 1960. 2 тома.
Любарский Г.Ю. Архетип, стиль и ранг в биологической систематике М., КМК, 1996.
Любищев А.А. К логике систематики//Проблемы эволюции. Т. 2., 1972, с. 45-68.
Любищев А.А. О некоторых постулатах общей систематики // Записки научных семинаров
Ленингр. отд. Математич. института АН СССР. Том 1. Л., 1975, с. 159-175.
Любищев А.А. Проблемы формы, систематики и эволюции организмов. М., Наука,
1982. 278 с.
Любищев А.А. О положении в средней школе//Эко. 1983. №12.
Любищев А.А. Понятие великого государя и Иван Грозный // Звезда, 1995, № 8.
Любищев А.А. О природе наследственных факторов / Примеч. и послесл. Ю.В. Чай-
ковского; послесл. М.Д. Голубовского. Ульяновск, УГПУ, 2004. 211 с.
Майр Э. Зоологический вид и эволюция. М., Мир, 1968.
Майр Э. Популяции, виды и эволюция. М., Мир, 1974.
Максимович М.А. О системах растительного царства. М., 1827.
Маслов А. А. Другое человечество. Ростов на Дону, 2006.
Материалы симпозиума, посвящённого памяти Сергея Викторовича Мейена (1935—
1987). Москва, 25-26 декабря 2000 г. М.: ГЕОС, 2001.
Мащенко Е.Н. Новые страницы каменной летописи // Наука и жизнь, 2006, № 5.
Медников Б.М. Геносистематика: проблемы и трудности // Биология в школе, 2000, № 1.
Мейен С.В. М-15, 17, 31,44, 50, 73, 75, 76, 84, 121, 127, 148, 163-165, 218, 219, 229; 246,
256, 257, 263, 268, 280, 281, 286, 294, 295, 299, 310, 313, 323, 325, 328, 338, 344, 352, 370.
Мейен С.В., Соколов Б.С., Шрейдер Ю.А. ...Феномен Любищева. (М-150).
Мейен С.В., Шрейдер Ю.А. Методологические аспекты... классификации// ВФ (М-140).
Мельников В.П., Дмитриев И.С. Дополнительные виды периодичности в периодиче-
ской системе Д.И. Менделеева. М., 1988.
382
Мигдал А.Б. От догадки до истины... // Химия и жизнь. 1979. № 12.
Милюков П.Н. Из истории русской интеллигенции. СПб., 1903. 308 с.
Милюков П.Н. Очерки по истории русской культуры. Часть 1. СПб., 1900. 239 с.
Мина М.В., Решетников Ю.С., Дгебуадзе Ю.Ю. Таксономические новшества и про-
блемы пользователей // Вопросы ихтиологии, 2006, № 4, с. 553-557.
Михайлов К.Г. Рец. на книгу: Любарский Г.Ю. Архетип, стиль и ранг в биологической
систематике // Ж. общей биологии, 1997, № 4, с. 92-96.
Михайлов К.Г. Типология и практическая систематика // Теоретические проблемы
экологии и эволюции (Вторые Любищевские чтения). Тольятти, 1995, с. 40-46.
Мопертюи П.-Л. Законы движения и покоя, выведенные из метафизического принци-
па// Вариационные принципы механики. М., 1959 .
Мосейчик Ю.В. Номогенетический характер эволюции растений (на примере флор
раннего карбона) // XIX Любищевские чтения, т. 2. Ульяновск, 2005, с. 98-103.
Мосейчик Ю.В. О природе фитостратиграфических рубежей (на примере флористиче-
ских последовательностей раннего карбона Северной Евразии) // Био- и литостратиграфи-
ческие рубежи в истории Земли. Труды междунар. конфер. Тюмень, 2008, с. 208-214.
Мосейчик Ю.В., Игнатьев И.А. О начальных стадиях палеозойского этапа глобального
флорогенеза // Палеонтологическая летопись региональных и глобальных событий. Тезисы
докладов 51-ой сессии Палеонтологического общества при РАН (4-8 апреля 2004 г., СПб.)
СПб., 2005, с. 87-88.
Назаретян А.П. Антропогенные кризисы: гипотеза техно-гуманитарного баланса //
Вестник РАН, 2004, № 4.
Назаров В.И. Эволюция не по Дарвину. Смена эволюционной модели. М., 2005.
Нечаев В.М. Брачное право // Новый энц. словарь Брокгауза и Ефрона, т. 7, 1912.
Нечкина М.В. Петр I // Малая Советская энциклопедия, т. 6. М., 1930.
Низовцев В.В. Время и место физики XX века. М., УРСС, 2000. 207 с.
Низовиев В.В., Панченко О.В. Контактный детерминизм при морфогенезе // Теория
эволюции: наука или идеология? Москва - Абакан, 1998, с. 147-164.
Никаноров Ю.М. Барьерные функции митохондриального кода // XXIV Любищев-
ские чтения. Ульяновск, 2010, с. 156-161.
Определитель бактерий Берджи. / Предисл.: Г. А. Заварзин. М., Мир, 1997. 800 с.
Основы финно-угорского языкознания. Прибалтийско-финские, саамский и мордов-
ские языки. М., Наука, 1975. 348 с.
Павлинов И.Я. Рец. на книгу: Mammalian species of the World, 2 v. Baltimore, 2005 //
Зоологии, ж., 2006, № 11, с. 1386-1389.
Павлинов И.Я., Россолимо О.Л. Систематика млекопитающих СССР. М., Изд. МГУ,
1987. Они же (дополнения). М., Изд. МГУ, 1998.
Павлов В.Я. Периодическая система членистых. М., ВНИРО, 2000. 185 с.
Пайтген X -О., Рихтер П.Х. Красота фракталов. М., Мир, 1993.
Палеонтология и палеоэкология. Словарь-справочник / Редакторы: В.П. Макридин и
И.С. Барское. М., Недра, 1995. 404 с.
Панов Е.Н. Бегство от одиночества. М., 2001.
Петухов С.В. Биомеханика, бионика и симметрия. М., 1981.
Платонов С.Ф. Лекции по русской истории. Изд. 5. СПб., 1907. 651 с.
Поглазов Б.Ф. Предисловие // Сборка предбиологич. и биологич. структур. М., 1982.
Пойа Д. Математика и правдоподобные рассуждения. М., 1957.
383
Пономаренко А.Г. Артроподизация и ее экологические последствия // Экосистемные пе-
рестройки и биосфера. Вып. 6, М., Палеонтол. ин-т РАН, 2004, с. 7-22.
Попов И.Ю. Ортогенез против дарвинизма. СПб., Изд. СПб. ун-та, 2005. 180 с.
Попов И.Ю. Периодические системы и периодический закон в биологии. М., 2008.
Преснов Е.В., Исаева В.В. Перестройки топологии при морфогенезе. М., 1985.
Протесты. Часть 1. Руководство по зоологии. СПб., Наука, 2000. 679 с.
Психология личности. Тексты. М.: Изд-во МГУ, 1982. С. 49.
Пчеленко С.П. К механизму явлений номогенеза И Любищевские чтения. XIX, т. 2.
Ульяновск, 2005, с. 112-122.
Пялль Э., Тотсель Э., Тукумиев Г. Сопоставительная грамматика эстонского и русско-
го языков. Таллин, 1962.
Расницын А.П. Процесс эволюции и методология систематики. СПб., 2002. (Труды
Русского энтомологич. общества, том 73, с. 1-107)
Растригин Л.А. Этот случайный, случайный, случайный мир. М., 1969.
Раутиан А.С. Уникальное перо птицы// ПЖ, 1978. № 4.
Раутиан А.С. О вкладе С.В. Мейена в теоретическую биологию // Чтения памяти С.В.
Мейена. Сб. тезисов. ГИИ. М., ГЕОС, 1997.
Раутиан А.С. Апология сравнительного метода // Любищев и проблемы формы, эво-
люции и систематики организмов. М., 2003.
Розова С. С. Классификационная проблема в современной науке. Новосибирск, Наука,
1986. 223 с.
Роуз Ф. Аборигены Австралии. Традиционное общество. М., 1989.
Сватиков С.Г. Общественное движение в России. Ростов н/Д., 1905. 400 + VI с.
Северцов А. Н. Собр. соч., т. 3. М.-Л., 1945.
Северцов А.С. Рецензия на книгу: В.Н. Назаров. Эволюция не по Дарвину. М., 2005 И
Успехи современной биологии. 2006, № 3.
Секей Г. Парадоксы в теории вероятностей и математической статитстике. М., 1990.
Скворцов А.К. У истоков систематики. К 300-летию Карла Линнея // Природа, 2007, №
4, с. 3-10.
Скулачев В.П. О биохимических механизмах эволюции и роли кислорода // Биохимия,
1998, вып. 11.
Смирнов Е.С. Таксономический анализ. М., Изд. МГУ, 1969. 187 с.
Соболев Д.Н. Начала исторической биогенетики. Харьков - Симферополь, 1924.
Соболь С.Л. Принцип естественного отбора в работах некоторых английских биоло-
гов... // История биологич. наук, вып. 9. М., 1962.
Соболь С.Л. Эволюционная концепция Чарлза Дарвина на первых этапах ее формиро-
вания // Анналы биологии. Т.Е М., МОИП, 1959.
Станек В.Я. Иллюстрированная энциклопедия животных. Пер. с чешского. Прага,
Артия, 1972, 612 с.
Стреляу Я. Роль темперамента в психологическом развитии. М., 1982.
Струмилин С.Г. Состав РКП к 1922 г. // Струмилин С.Г. Избранные произведения.
Часть 1. М., 1964.
Сукачев В.Н. Новые данные по экспериментальному изучению взаимоотношения рас-
тений И Бюллетень МОИП, Отдел биол., 1959, вып. 4.
Тарановский Ф.В. Крестьяне в России до XIX в. // Новый энц. словарь Брокгауз-
Ефрон. Т. 23, Петроград, 1915, стлб. 257-264.
Теория и методология биологических классификаций (Труды семинара в МОИП). М.,
Наука, 1983. 169 с.
384
Теория эволюции: наука или идеология? Труды Любищевских чтений. / отв. ред. Б.И.
Кудрин. Москва - Абакан, 1998. (Ценологич. исследования, вып. 7.)
Том Р. Структурная устойчивость и морфогенез [1972]. М., Логос, 2002.
Томашевич Ольга В. Женщины на престоле Египта. Послесловие к книге: Жак К. Еги-
пет великих фараонов. М., Наука, 1992. 334 с.
Томилин А.Г. Снова в воду. М., 1977.
Традиционная логика и канторовская диагональная процедура. М., Янус, 1997.
Трубецкой Н.С. Избранные труды по филологии. М., 1987.
Тутубалин В.Н. Вероятность, компьютеры и обработка результатов экспериментов
//УФН, 1993, №7.
Урманцев Ю.А. Номогенез о сходстве в живой природе // Природа, 1979, № 9, с. 116.
Ушаков Е.В. Введение в философию и методологию науки. СПб., КноРус, 2008.
Феллер В. Введение в теорию вероятностей и ее приложения. М., 1964.
Феногенетическая изменчивость / Под ред. Н.П. Кренке. М., 1933-1935. Т. 1.
Феномен восточного деспотизма. Структура управления и власти / отв. ред. Н.А. Ива-
нов. М., Восточная литература, 1993. 392 с.
Филипченко Ю.А. О параллелизме в живой природе // Успехи эксперимент, биологии,
1924, вып. 6.
Филипченко Ю.А. Эволюционная идея в биологии. М., Наука, 1977. 227 с.
Флиндт Р. Биология в цифрах / пер. с немецкого. М., 1992.
Харрисон Дж. и др. Биология человека. М.: Мир, 1979.
Хохряков А.И. Табличное изображение системы двудольных // Общебиологические
аспекты филогении растений. (Совещание в МОИП.) М., Наука, 1991, с. 113-117.
Целищев В.В. Философские проблемы семантики возможных миров. Новосибирск, 1977.
Черных П.Я. Историко-этимологич. словарь современного русского языка. М., 1993.
Что беспокоит физиков? И В мире науки, 2004, №11.
Шаталкин А. И. Предисловие к книге: Симпсон Дж.Г. Принципы таксономии живот-
ных М„ КМК, 2006, с. Ш-ХШ.
Шипунов А.Б. «Systema Naturae»: 250 лет спустя. Судьба линнеевской концепции
царств // Материалы конференции по морфологии и систематике растений, посвященной
300-летию со дня рождения Карла Линнея. М., КМК, 2007, с. 31-33.
Шлегель Г. Общая микробиология. М., 1972.
Шмальгаузен И.И. Основы сравнительной анатомии позвоночных. М., Учпедгиз, 1938.
Шмальгаузен И.И. Организм как целое в индивидуальном и историческом развитии.
М., Наука, 1982. (1-е изд.: 1938; оконч. вариант: 1942).
Шноль С.Э., Коломбет В.А., Пожарский Э.В., Зенченко ТА., Зверева И.М., Конрадов
А.А. О реализации дискретных состояний в ходе флуктуаций в макроскопических про-
цессах // УФН, 1998, № 10.
Шредер М. Фракталы, хаос, степенные законы. М. - Ижевск, 2001.
Шрейдер Ю.А. Об одной модели семантической теории информации // Проблемы ки-
бернетики, вып. 13. М., 1965.
Щербаков А.Е. О пермских и триасовых энтомофаунах в связи с биогеографией и
пермо-триасовым кризисом // ПЖ, 2008, № 1.
Эволюционные факторы формирования разнообразия животного мира. М., КМК,
2005. 308 с.
Эпштейн В.М. Философия систематики. Книга 3. М., КМК, 2002. 235 с.
Яковлев Я. Деревня, как она есть. М., 1923.
Яновский А.Е. Бюрократические установления // Энцикл. словарь Брокгауз-Ефрон, по-
лутом 9. СПб., 1891.
385
Agassiz. L. Contributions to the natural history of USA. Pt 1. Boston, 1857.
Alperson-Aftl N. Continual fire-making by Hominins at Gesher Benot Ya’agov, Israel //
Quaternary Sci. Rev., 2008, vol. 27, p. 1733-1739.
Anderws H.M. Index of generic names of fossil plants, 1825-1965 // GeoL Surv. Bull.,
1970, № 1300, p. 1-354.
Atran S. Cognitive foundations of natural history: towards an antropology of science.
Cambridge: Cambr. Univ. Press, 1990, XII + 360 p.
Attali J. Les trois mondes. Pour une theorie de 1’apres-crise. Paris, 1986. 416 c.
Baron W. Zur Stellung von H.G. Bronn in der Geschichte des Evolutionsgedankens // Sud-
hoffs Archiv fur Geschichte der Medizin und Naturwissenschaften. Bd 45, Hefte 2. Wies-
baden, 1961.
Baskerville T. Affinities of plants: with some observations upon progressive developments.
London, 1839. X + 114 p.
Bauhin C. Pinax theatri botanici. Basel, 1623. XXIV + 567 p.
Bernhardi J. J. Uber den Begriff der Pflanzenart und seine Anwendung. Erfurt, 1834.
Biophotonics and Coherent Systems I Eds: Lev Beloussov, Fritz-Albert Popp, Vladimir
Voeikov, and Roeland Van Wijk. Moscow: Moscow University Press, 2000.
Britannica. Настольная иллюстрированная энциклопедия. T. 1. М., Астрель, 2009.
Brooks J.L. Just before the Origin. Alfred Russel Wallace’s theory of evolution. N.Y., 1984.
Brower J.H. Behavioral changes in an ant colony to chronic gamma irradiation // American
Midland Naturalist, 1966, vol. 75, № 2.
Bureaucratic // Grand Dictionnaire universel du XIX ciecle / P., Larousse. T. 2, 1866.
Burlando B. The fractal dimension of taxonomic systems // J. Theor. Biology, 1990, vol.
146, № 1.
Cabanis P.-J.-G. Rapports du physique et du moral de 1’homme. Vol. 2. P., 1802.
Chaos et determinisme / Sous la direction de A. Dahan Dalmedico. Editions de Seuil, 1992.
Clark P. Atomism versus thermodynamics // Method and appraisal in the physical sciences.
Cambridge, Cambr. Univ. Press, 1976.
Clutton-Brock e.a. A review of the family Canidae // Bull. Brit. Mus. (Nat. Hist.) Zool.,
1976, vol. 29, N 3.
Dambicourt-Malasse A. Nouveau regard sur 1’origine de 1’homme // La Recherche, 1996,
№ 286.
Darwin E. Zoonomia or the laws of organic life. Third ed., t. 2, ch. XXXIX. L., 1801.
Del Giudice E. e. a. Coherent quantum electrodynamics in living matter // Electromagnetic Bi-
ology and Medicine, 2005, v. 24, p. 109.
Ellegard A. Darwin and the general reader. Goteborg, 1958.
Ereshefsky M. The poverty of the Linnean Hierarchy. Cambridge Univ. Press, 2001, 316 p.
Gisiger T. Scale invariance in biology: coincidence or footprint of a universal mechanism? //
Biological Reviews, 2001, № 2.
Goldfuss G.A. Ueberdie Entwickelungsstuffen des Thieres. Nuernberg, 1817. 58 S.
Gomankov A. V. Post-Script//Rev. Palaeobot. Palinol., 1997, vol. 96, p. 449-451.
Gruber G.E., Barrett P.H. Darwin on man. N.Y., 1974.
Haeckel E. Generelle Morphology der Organismen. Berlin, 1866. Bd 2.
Hale M. The primitive origination of mankind... London, 1677.
Harms J. V. Die Realisation der Gene und die consekutive Adaptation // Zeitschr. Wissensch.
Zoologie, 1935, Bd 146, S. 417-462.
Hunde// Brockhaus’ Konversations-Lexicon. Bd 9. Leipzig, 1908. S. 412-418.
386
Jantsch E. Design for evolution. N.Y., 1975.
Jantsch E. The self-organizing universe. Scientific and human implications of emerging
paradigm of evolution. Oxford - N.Y., 1980.
Jean R.V. Phyllotaxis. Cambridge, 1994.
Kark S., van Rensburg B.J. Ecotones: marginal or central areas of transition? // Israel J. Ecol.
Evolution, 2006, vol. 52, p. 29-53.
Kauffman S.A. Reinventing the sacred. A new view of science, reason and religion. N.Y.,
2008. 320 pp.
Kepler J. Gesammte Werke. Munchen, 1938, Bd 1.
King-Hele D.J. Essential writings ofE. Darwin. L., Worcester, 1968.
Knapp S., Lamas G., Lughandha E.N., Novarino G. Stability or stasis in the names of organisms:
the evolving codes of nomenclature // Phil. Trans. R. Soc. Lond., B, 2004, p. 611-622.
Kuki Sh. Le probleme de la contingence (1935). Tokyo, 1966.
Linne C. Species plantarum. Stokholm, 1753. T. 1, p. 1-560; t. 2, p. 561-1200.
Maupertuis P.L. Oeuvres. Lyon, t. 2, 1756 .
Meyen S. V. M-l 16, 268, 299, 330.
Meyer S.C. The Origin of Biological Information and the Higher Taxonomic Categories //
Proceedings of the Biological Society of Washington. August 20, 2005.
Millhauser M. Just before Darwin. Robert Chambers and “Vestiges”. Middletown, Conneti-
cut, 1959.
Montagnier L., Aissa J. e.a. Electromagnetic signals are produced by aqueous nanostructures
derived from bacterial DNA sequences // Interdiscip. Sci. Comput. Life Sci., 2009 (1) 81-90.
Morrone J. J. Between the taunt and the eulogy: Leon Croizat and the panbiogeography // In-
terciencia, 2000, N 1, p. 41-47. [Журнал изд. в Венесуэле]
Nottale L, ChalineJ., GrouP. Les arbres de 1’evolution. Univers, vie, societe. P., Hachette, 2000.
Popov I. Orthogenesis versus Darwinism: The Russian case // Revue d’histoire des sciences,
2008, t. 61, n 2, p. 367-397.
Radcliff-Brown A.R. On joking relations // Structure and function in primitive society. L., 1958.
Sachs T. A morphogenetic basis for plant morphology // Acta Biotheorethica, 1982, Ser A,
vol 31, p. 118-131.
Sheldrake R. A new science of life. London, 1981.
SprangerE. Lebensformen. Halle, 1914.
Stevens P.F. Metaphors and typology in the development of botanical systematics 1690-
1960... //Taxon, 1984, vol. 33, № 2.
The Biology of naked mole-rat. Princeton, 1991.
The Origin of species by Charles Darwin. A variorum text edited by Morse Peckham. Phila-
delphia, 1959.
Toumefort (serie: Les grands naturalistes fran^ais). Paris, Museum nat. hist., 1957. 321 p.
Van der Giezen M. & Tovar J. Degenerate mitochondria // EMBO [Europian Molec. Biol.
Organization] Rep., 2005, vol. 6, № 6.
Van der Giezen M. Hydrogenosomes and mitosomes: conservation and evolution of func-
tions // J, Eucariot Microbiol., 2009. vol. 56, N 3.
Van Gelder R.G. A review of canid classification // Amer. Mus. Novitates, 1978, № 2646.
Via S., Shaw A. J. Short-term evolution in the size and shape of pea aphids // Evolution. 1996, N 1.
Wayne R.K.. Geffen E., Girman D., Kopfelli K.P., Marshall C. Molecular systematics of the
Canidae // Systematic Biology, 1997, vol. 46, № 4, p. 622-649.
Wehner R. Brainless eyes //Nature, 2005, vol. 435, № 7039, p. 157-159.
387
Westbroek P. La Terre est-t-elle un superorganisme? // La Recherche, 1997, № 295.
Wheeler Q.D. Taxonomic triage and poverty of philogeny // Phil. Trans. R. Soc. Lond., B,
2004, p. 571-583.
Wiley E. The annotated Linnean Hierarchy // Syst. Zool., 1979. vol. 128, p. 308-337.
Winsor M.P. Linnaeus's biology was not essentialist // Ann. Missouri Bot. Garden, 2006,.
vol. 93, p. 2-7.
Worse C.R. A new biology for a new century // Microbiology and Mol. Biol. Reviews, 2004,
vol. 68, № 2.
Yule G.U. A mathematical theory of evolution, based on the conclusions of Dr J.C. Willis //
Philos. Transact. R.S. London, 1924. Ser.B, vol. 213, p. 21-87.
Список упомянутых в сборнике работ автора
Крестиком (+) обозначены публикации, вошедшие в данный Сборник частично,
двойным крестиком (++) - публикации, вошедшие целиком
4-1. 1971. О непрерывном процессе решения матричной игры // Доклады АН СССР, т.
199, №5, с. 1026-1028.
4-2. 1971. Chaikovsky Yu.V. On a continuous process of matrix game solution // Soviet
Math. Dokl., vol. 12, № 4, c. 1245-1247.
4-3. 1977. Выживание мутантного клона. Сообщ. 1, 2 // Генетика, т. 13, № 8, с. 1467-
1477, 1478-1488.
4-4. 1977. Генетическая интеграция клеточных структур как фактор эволюции I/ ЖОБ,
т. 38, №6, с. 823-835.
Ч-4а. 1978. Кошмар Дженкина, или Повесть об инженере, которого не могут забыть
биологи // Химия и жизнь, № 12, с. 109-117.
4-5. 1979 // Выживание мутантного клона. Сообщ. 3. Катастрофический отбор... //
Генетика, т. 15, № 10, с. 1809-1815 (совм. с Г.Г. Маленковым).
Ч-5а. 1979. Дарвин ± 120 // Химия и жизнь, № 12, с. 74-86
++Ч-6. 1980. Многотрудный поиск многоликой истины // Химия и жизнь, 1980, № 10, с.
15-20.
4-7. 1981. Изумительная асимметрия // Знание сила, № 2, с. 16-18, 48.
Ч-7а. Перед выходом «Происхождения видов», 1981, ВИЕТ, № 4, с. 79-86.
4-8. 1983. Рождение дарвинизма // Теоретич. проблемы современной биологии. Пущино,
с.94-103.
4-9. 1984. «Редкое и благородное спокойствие» // Книги, открывающие мир. М., Книга, с.
159-200. [О творчестве Ч. Дарвина]
4-10. 1984. «Происхождение видов». Загадки первого перевода//Природа, 1984, № 7, с.
88-96. [О творчестве Г. Бронна]
4-11. 1984. Анализ [понятий] эволюционной концепции [А.А. Любищева] // Систем-
ность и эволюция. М., Наука, с. 32-52.
4-12. 1985. Грамматика для биологии//Знание - сила, 1985, № 12, с. 30-32.
4-13. 1985. Разнообразие и случайность // Методы научного познания и физика. М.,
Наука, с. 149-168.
++Ч-14. 1986. Грамматика биологии // Вестник АН СССР, №> 3, с. 47-58.
388
Ч-15. 1986. Опыт эко-физиологической макросистемы // Методы исследования в эко-
логии и этологии. Пущино, с. 6-33.
4-16. 1987. История науки и обучение науке (на примере дарвинизма) // ВИЕТ, № 2, с.
92-101.
++Ч-17. 1987. Некролог//ВИЕТ, № 3, с. 169-171.
4-18. 1987. Нечеткие закономерности в планетной астрономии // Историко-астро-
номич. исследования, вып. 19. М., Наука, с. 69-86.
4-19. 1988. Chaikovsky Yu. V. То the evolutionary thermodynamics // Lectures in theoretical
biology. Tallinn, c. 65-72.
++Ч-20. 1988. Молодежь в разнообразном мире // Социологии, исследования, № 1, с. 76-84.
4-21. 1988. О формировании концепции Ч. Дарвина // Науки в их взаимосвязи. Исто-
рия. Теория. Практика. М.
++Ч-22. 1989. Наука о разнообразии//Химия и жизнь, 1989, № 1, с. 40-48.
4-23. 1989. История науки и обучение науке (на примере понятий "случайность" и "ве-
роятность") // ВИЕТ, № 4, с. 92-101
+4-24. 1990. Элементы эволюционной диатропики. М., Наука. (ЭЭД)
4-25. 1991 Познавательные модели, плюрализм и выживание// Путь. Филос. журнал.
№ 1, с. 62-108.
Ч-25а. 1991. Гранит во льдах// Вокруг света, № 1, № 2. [О Колчаке как полярнике]
4-26. 1992. Наука без истории - ремесло убогое // Химия и жизнь, № 10, с. 73-77. [Не-
лепые купюры редактора местами лишили текст смысла]
4-27. 1992. О классиках и комментаторах // ВИЕТ, № 4, с. 137-146. [О дарвиноведении]
4-28. 1992. Письмо в редакцию // ВИЕТ, № 2, с. 168-169. [Об эволюционных взглядах
В.И. Вернадского]
4-29. 1993. Идея равновозможности в физике и биологии // Физическое знание: его
генезис и развитие. М., Наука, с. 104-129.
Ч-ЗО. 1994. Междисциплинарность современного эволюционизма// Концепция самоорга-
низации в исторической ретроспективе. М., Наука, с. 198-237.
4-31. 1994. Преобразование разнообразия. Эволюционная теория Сергея Мейена //
Химия и жизнь, № 1, с. 20-29.
4-32. 1994. Становление статистического мировоззрения // Метафизика и идеология в
истории естествознания. М., Наука, с. 62-107
Ч-ЗЗ. 1996. О познавательных моделях // Исследования по математической биологии.
Сб. памяти А.Д. Базыкина. Пущино, с. 170-184.
4-34. 1996. Познавательные модели в эволюции // Педагог-исследователь, № 1. Пет-
розаводск, с. 26-36.
4-35. 1996. Ступени случайности и эволюция // ВФ, № 9, с. 69-81.
4-36. 1996-2000. Эволюция // Биология (приложение к газете "Первое сентября"). [Ран-
ний (журнальный) вариант книги «Эволюция», 2003; девять выпусков]
++Ч-37. 1997. Новые тенденции в теории эволюции // ЛЧ, 1997. Ульяновск, 1997, с. 31.
4-38. 1998. Макросистема, непохожая на другие // Биология в школе, № 6, с. 19-25.
4-39. 1998. Стабилизирующий отбор, или Святость веры // Теория эволюции - наука
или идеология? Москва - Абакан, с. 55-58.
4-40. 2000. Избегание предтеч // ВФ, № 10, с. 91-103.
++Ч-41. 2000. Любищев, Мейен и плюрализм в эволюции // ЛЧ, с. 35-57.
++Ч-42. 2001. С чем входим в XXI век? // Биология в школе, № 1, с. 9-14.
389
+4-43. 2001. О природе случайности. М., Центр системных исследований, 272 с. [Отли-
чий от ОПС, 2004 очень мало, номера страниц совпадают]
++Ч-44. 2001. С.В. Мейен и теория биологической эволюции // МСП, 51 -70.
++Ч-45. 2001. Мейен как эволюционист и натурфилософ // Тезисы доклада 25 дек. 2000
г. в ГИН // МСП, 2001, с. 70-71.
4-46. 2001. Экологический аспект преподавания истории // Философия экологии,
образования. М„ Прогресс-традиция, с. 385-397.
Ч-46а. 2002. Ламаркизм умер. Да здравствует ламаркизм, ВИЕТ, № 3, с. 430-447.
+4-47. 2003. Естественная система. 30 лет после Любищева // Любищев и проблемы
формы, эволюции и систематики организмов. М., Центр системных исследований,
2003 (Ценологические исследования, вып. 23), с. 30-50.
4-48. 2003. Из бесед с С.В. Мейеном // Эволюция флор в палеозое. М., ГЕОС, с. 132-134.
4-49. 2003. Иммунитет и эволюция: не впасть бы в другую крацность // Вестник РАН,
№ 3, с. 265-283.
++Ч-50. 2003. Феномен эволюции организмов в понимании С.В. Мейена И Эволюция
флор в палеозое. М., с. 99-105.
4-51. 2003. Эволюция. Книга для изучающих и преподающих биологию. М., Центр
системных исследований, 475 с. [Ранний вариант «Науки о развитии жизни», 2006]
4-52. 2004. Любищев и естественная система организмов // XVIII ЛЧ. Ульяновск.
+4-53. 2004. О природе случайности. М., Центр системных исследований, 2004. Изд-е
второе, исправл. и доп.; 278 с. (ОПС)
4-54. 2004. Неуслышанный совет. Послесловие к книге «О природе наследственных
факторов» (см.: Любищев, 2004), с. 161-176.
++Ч-55. 2005. Двускоростной механизм размножения и его роль в эволюции человека //
Эволюция, № 2, с. 42-45.
++Ч-56. 2005. Серёжа Мейен, мой сосед // Памяти С.В. Мейена (к 70-летию со дня рожде-
ния) Труды Междунар. палеоботанич. конференции 17-18 мая 2005 г., вып. 3, с. 63-68.
++Ч-57. 2006. Диатропика С.В. Мейена: сегодняшний взгляд // ВФ, № 5, с. 95-102.
4-58. 2006. Наука о развитии жизни. М., КМК. 712 с. [Ранний вариант АСМ]
++Ч-59. 2007. Дарвинизм, эволюция «не по Дарвину» и близкий юбилей // Общая и при-
кладная ценология, № 2, с. 24-30.
+4-60. 2007. Горизонт познания и познавательные модели. Исторические параллели, и
что с ними делать // Грани познания. Наука, философия и культура в XXI веке / ред.
Н.К. Удумян. Книга 2. М., Наука, с. 5-31.
++Ч-61. 2007. Естественная система и таксономические названия // Линнеевский сбор-
ник/ ред. И.Я. Павлинов. М., МГУ, с. 381-436
+4-62. 2007. Философия дарвинизма против философии эволюции // ВФ, № 9.
4-63. 2008. Активный связный мир. М., КМК. 726 с. (АСМ)
++Ч-64. 2009. 500 лет споров об эволюции // ВФ, № 2, с. 71-85.
++Ч-65. 2010. Ламарк, Дарвин и устройство науки // Вестник РАН, № 8, с. ... (в печати).
4-66. 2010. Зигзаги эволюции. Развитие жизни и иммунитет. М., «Наука и жизнь».
109 с. (ЗЭ)
++Ч-67. 2010. Практичная теория (заключительная статья) //данный Сборник, с. 318-353.
390
Полный список опубликованных трудов С.В. Мейена
(на август 2010). Составлен И.А. Игнатьевым
1955
1. Обрабатываем собранный материал // Моск, университет (газета). № 55 (1446).
С. 1. (Совместно с А. Соловьёвым).
1956
2. Надо быть осторожнее! // Моск, университет (газета). 24.02.56 г. С. 3.
3. Флора четвертичных известковых туфов Европейской части СССР // Бюлл.
МОИП. Отд. геол. Т. 61, № 3. С. 115.
1957
4. Новый способ фотографирования // Природа. № 10. С. 111-112.
1961
5. О палеоботанических вопросах, затрагиваемых в курсе «Палеогеография»
К.К.Маркова// ПЖ № 4. С. 172-173.
1962
6. Материалы к познанию морфологии вегетативного побега ангарских кордаитов //
ПЖ № 2. С. 133-144.
1963
7. Мария Фёдоровна Нейбург (1894-1962)//ПЖ № 1. С. 151-153.
8. Мхи в палеозое Ангариды И Природа. № 5. С. 73-76.
9. Об анатомии и номенклатуре листьев ангарских кордаитов // ПЖ № 3. С. 96-107.
10. О методах восстановления экологии ископаемых растений // Тезисы докладов на
IX сессии Всес. Палеонтол. о-ва. С. 42-43.
1964
11. Предисловие. Дополнительные данные к морфологии Tschernovia //
М.Ф.Нейбург. Пермская флора Печорского бассейна. Ч. II. Членистостебельные
(Sphenopsida). М.: Наука. С. 5-6, 77-78. (Труды ТИН. Вып. 111).
12. О морфологии, анатомии и номенклатуре ангаро-гондванского рода Noeggera-
thiopsis И Гондвана. М.: Наука. С. 87-100.
13. Генри Потонье (к 50-летию со дня смерти) // ПЖ № 1. С. 147-148. [Переиздано в
сб.: С.В.Мейен. Листья на камне: Размышления о палеоботанике, геологии, эволюции и
путях познания живого. М.: ГЕОС, 2001. С. 198-199; - см. №338.]
14. Альберт Чарльз Сьюорд (к 100-летию со дня рождения) И ПЖ № ЕС. 146-147.
[Переиздано в сб.: С.В.Мейен. Листья на камне: Размышления о палеоботанике, геоло-
гии, эволюции и путях познания живого. М.: ГЕОС, 2001. С. 194-196; - см. №328.]
15. Ангарские кордаиты (методы исследования, принципы систематики и основные
этапы эволюции) // Тезисы докладов на X сессии Всес. Палеонтол. о-ва. С. 26-28.
16. К вопросу об истории установления возраста угленосных отложений Кузбасса //
Изв. АН СССР. Сер. геол. № 5. С. 98-103. (Совместно с В.В.Меннером).
17. Кордаитовые верхнего палеозоя Северной Евразии (морфология, эпидермальное
строение, систематика и стратиграфическое значение): Автореф. дисс. ... канд. геол.-мин.
наук. М.: Наука. 19 с.
391
18. Изучение эпидермиса кордаитов из Кузнецкого бассейна (опыт подродовой клас-
сификации) И Ботан. журн. Т. 49, № И. С. 1533-1543.
1965
19. Предисловие. Дополнение об эпидермальном строении печорских кордаитов //
М.Ф.Нейбург. Пермская флора Печорского бассейна. Ч. III. Кордаитовые (Cordaitales), вой-
новскиевые (Vojnovskyales), семена голосеменных неясного систематического положения
(Semina gymnospermarum incertae sedis) // M.: Наука. С. 5-7, 39—41. (Труды ГИН. Вып. 116).
20. О классификации дисперсных кутикул (Автореф. докл., прочитанного 18.12.64 г.)
//Бюлл. МОИП. Отд. геол. Т. 40, № 3. С. 140- 141.
21. О некоторых проблемах палеофлористики палеозоя (Автореф. докл., прочитан-
ного 25.12.1964 г.)//Бюлл. МОИП. Отд. геол. Т. 40, № 3. С. 141-142.
22. О классификации дисперсных кутикул // ПЖ № 4. С. 75-87.
23. Дьюкинфилд Генри Скотт (к 110-летию со дня рождения и 30-летию со дня
смерти) И Палеонтол. журн. № 1. С. 162-163. [Переиздано в сб. №338.]
24. Вальтер Готан (к 10-летию со дня смерти) // ПЖ № 1. С. 163. [Переиздано в сб. №338.]
25. Йозеф Август Шенк (к 150-летаю со дня рождения) И ПЖ. № 3. С. 152. [Переиз-
дано в сб. №337.]8
26. Джон Линдли (к 100-летию со дня рождения) И ПЖ № 3. С. 153. [Переиздано в
сб. №337.]8
27. Рене Зейлер (к 50-летию со дня смерти) // ПЖ № 3. С. 153. [Переиздано в сб. №338.]
28. Дэвид Томас Гвин-Воган (к 50-летию со дня смерти) // ПЖ № 4. С. 109. [Переиз-
дано в сб. №338.]
29. Герман Зольмс-Лаубах (к 50-летию со дня смерти) // ПЖ № 4. С. 109-110. [Пере-
издано в сб. №338.]
30. Эдвард Уилбер Берри (к 90-летию со дня рождения и 20-летию со дня смерти) //
ПЖ № 4. С. 110. [Переиздано в сб. №338.]
1966
31. Кордаитовые верхнего палеозоя Северной Евразии (морфология, эпидермальное
строение, систематика и стратиграфическое значение) М.: Наука. 186 с. (Труды ГИН.
Вып. 150).
32. Рудольф Флорин (1894-1965)//ПЖ № 2. С. 141-142. [Переиздано в сб.: №338.]
33. О палеофлористическом районировании территории СССР в карбоне И ПЖ № 4.
С. 109-113.
34. О находке листостебельных мхов в пермских отложениях Южного Приуралья //
Докл. АН СССР. Т. 166, № 4. С. 924-927. (Совместно с В.П.Твердохлебовым)
35. Classification of dispersed cuticles // Intern. Geol. Rev. Vol. 8, № 8. P. 965-975. (Пе-
ревод на английский язык статьи № 20).
1967
36. О соотношении индийских и ангарских флор позднего палеозоя (Автореф. докл.,
прочитанного 9.12.66 г.) И Бюлл. МОИП. Отд. геол. Т. 42, № 2. С. 153.
37. Общие элементы в ангарской и гондванской флорах позднего палеозоя: паралле-
лизм или миграция? // Тезисы докл. к XIII сессии Всес. Палеонтол. о-ва. С. 32-34.
38. О некоторых методах восстановления экологии древних растений // Вопросы па-
леогеографического районирования в свете данных палеонтологии. Тр. IX сессии Всес.
Палеонтол. о-ва. М.: Недра. С. 102-106.
392
39. О сопоставлении разрезов верхнепалеозойских отложений Тунгусского и Куз-
нецкого бассейнов по кордаитам // Стратиграфия палеозоя Средней Сибири. Новоси-
бирск: Наука. С. 230-231.
40. Итоги изучения палеозойских флор СССР за 50 лет (1917-1967 гг.) // Ботан.
журн. Т. 52, № 12. С. 1701-1712.
41. Николай Александрович Шведов (1907-1966) И Палеонтол. журн. № 3. С. 139-
140. [Переиздано в сб. №338.]
42. О великой палеоботанической стене, Gondwanidium подлинном и поддельном и
ещё - о хорошем отношении к мелочам И Знание - сила. № 7. С. 23-26. (Совместно с
И.А. Добрускиной). [Переиздано в сб. №338.]
43. New data on relationship between Angara and Gondwana Late Paleozoic floras //1 Simp.
Intern, sobre Estratigr. V Paleontol. del Gondwana. Res. de trabaj. Mar Del Plata. P. 13-14.
1968
44. Среда: драматург или режиссёр? // Знание - сила. № 1. С. 16-19. (Совместно с
А.Ю. Розановым). [Переиздано в сб. №338.]
45. О возрасте острогской свиты Кузбасса и об аналогах намюра в континентальных
отложениях Северной Азии // Докл. АН СССР. Т. 180, № 4. С. 944-947.
46. О методике исследования и описания ископаемых растений И Палеонтол. журн.
№ 3. С. 103-112.
47. О некоторых общих вопросах систематики и эволюции хвойных в связи с откры-
тием семезачатков у Burladia // Палеонтол. журн. № 4. С. 28-31.
48. Об общих принципах палеофлористического районирования // Тезисы докл. на
XIV сессии Всес. Палеонтол. о-ва. Л. С. 28-30.
49. Основные этапы развития ангарской (тунгусской) флоры // Материалы к пленуму
постоянной стратиграфической комиссии МСК (29 января - 1 февраля 1968 г.). Л. С. 7-11.
50. Дальновидная амблистома, или шаг назад, два шага вперед // Знание - сила. № 2.
С. 38-40. [Переиздано в сб. №338.]
51. Где ты был, Адам? In statu nascendi И Знание - сила. № 11. С. 17-19. [Переиздано
в сб.: С.В.Мейен. Листья на камне: Размышления о палеоботанике, геологии, эволюции и
путях познания живого. М.: ГЕОС, 2001. С. 96-107; - см. №338.]
52. Paleofloristic zoning of the Carboniferous in USSR // Intern. Geol. Rev. Vol. 9, № 5.
P. 738-742. (Перевод на английский язык статьи № 32).
1969
53. О гипотезе перемещения континентов с точки зрения палеофлористики карбона и
перми // Геотектоника. № 5. С. 3-16.
54. Предисловие [и редактирование] сб. «Птеридоспермы позднего палеозоя и мезо-
зоя». М.: Наука. С. 5-6. (Тр. Геол, ин-та АН СССР. Вып. 190).
55. 0 некоторых проблемах в изучении птеридоспермов // Птеридоспермы позднего
палеозоя и мезозоя. М.: Наука. С. 7-13. (Труды ГИН. Вып. 190).
56. Эпидермальное изучение ангарских Callipteris и Compsopteris И Птеридоспермы
позднего палеозоя и мезозоя. М.: Наука. С. 59-84. (Труды ГИН. Вып. 190). (Совместно с
А.В.Мигдисовой).
57. О роде Zamiopteris Schmalhausen и его соотношении с некоторыми смежными
родами И Птеридоспермы позднего палеозоя и мезозоя. М.: Наука. С. 85-104. (Труды
ГИН. Вып. 190).
58. Новые роды Entsovia и Slivkovia из пермских отложений Русской платформы и
Приуралья // ПЖ № 4. С. 93-100.
393
59. Жажда в каменноугольной топи // Знание - сила. № 4. С. 21-23. [Переиздано в
сб.: №338.]
60. Карты палеофлористического районирования карбона СССР И Ред. В.М.Познер
(ред.). Атлас литолого-палео-географических карт СССР. 6 карт.
61. Владимир Петрович Маслов (1891-1968) И ПЖ № 2. С. 146-148. (Совместно с
И.Н. Крыловым).
62. Елена Степановна Рассказова (1911-1969) // ПЖ № 3. С. 163-164. (Совместно с
М.П. Долуденко).
63. Сравнительно-исторический анализ каменноугольных и пермских флор Евразии:
Автореф. дисс. ... докт. геол.-мин. наук. М.: Наука. 50 с.
64. Явления параллелизма и их значение для систематики ископаемых растений.
(Автореф. докл., прочитанного 23.5.69 г.) // Бюлл. МОИП. Отд. геол. Т. 44, № 6. С. 152.
65. The Angara members of Gondwana genus Barakaria and its systematical position //
Argumenta palaeobotanica. Heft 3. S. 1-14.
66. New data on relationship between Angara and Gondwana Late Paleozoic floras //
Gondwana stratigraphy. IUGS Symposium, Buenos-Aires, 1-15 October 1967. Paris:
UNESCO. P. 141-157.
1970
67. Понятие о флоре и растительности геологического прошлого. Принципы палго-
флористического районирования // В.А.Вахрамеев, И.А.Добрускина. Е.Д.Заклинская,
С.В.Мейен. Палеозойские и мезозойские флоры Евразии и фитогеография этого времени.
М.: Наука. С. 7-21. (Труды ТИН. Вып. 208).
68. Развитие взглядов на закономерности географического распределения растений в
геологическом прошлом // Там же. С. 22-36. (Совместно с В.А. Вахрамеевым).
69. Девонские флоры // Там же. С. 37-42.
70. Каменноугольные флоры // Там же. С. 43-110.
71. Пермские флоры // В.А.Вахрамеев, И.А.Добрускина, Е.Д.Заклинская, С.В.Мейен.
Палеозойские и мезозойские флоры Евразии и фитогеография этого времени. М.: Наука.
С. 111-157. (Труды ТИН. Вып. 208).
72. Когда перестает видеть глаз // Знание - сила. № 3. С. 48.
73. Корова и сено: к общему предку. // Знание - сила. № 2. С. 27-29; № 3. С. 25-28.
[Переиздано в сб. №338.]
74. Хвощ-прадедушка // Юный натуралист. № 3. С. 42-43.
75. Отвратительная тайна раскрыта? [рец.] // Природа. № 9. С. 112-114.
76. Некоторые теоретические вопросы современной палеоботаники // ПЖ № 4. С. 3-
15. [Переиздано в сб. № 320.]
77. Флоры и климаты Евразии в геологическом прошлом И Природа. № 11. С. 32-41.
(Совместно с В.А.Вахрамеевым).
78. Об основном делении каменноугольной системы (по палеоботаническим дан-
ным) // Проблемы стратиграфии карбона. Т. 4. С. 46-56.
79. Идея рождена! Что дальше? // Литературная газета. № 39. С. 2. (Совместно с В.В.
Меннером).
80. Epidermisuntersuchungen an permischen Landpflanzen des Angaragebietes //
Palaontol. Abhandl. Reihe B. Bd. 3. H. 3/4. S. 523-552.
1971
81. Об основных типах биостратиграфических ошибок // Бюл. МОИП. Отд. геол. Т.
46, № 2. С. 136-137. (Совместно с В.В.Меннером).
394
82. Современная палеоботаника и эволюционная теория И Природа. № 2. С. 48-57.
[Переиздано в сб. №338.]
83. Пермские флоры Русской платформы и Приуралья И Труды Палеонтол. ин-та АН
СССР. Т. 130. С. 294-308.
84. Из истории растительных династий. М.: Наука. 221 с. [В 1979 г. переведено на
эстонский язык Eesti loodus. №№ 1-7].
85. Новые данные о систематике филладодерм и их географическом распростране-
нии // Докл. АН СССР. Т. 198. № 3. С. 676-679. (Совместно с А.В. Гоманьковым).
86. Эволюция растений в документах // Природа. № 8. С. 111-114.
87. Концепция рода в палеоботанике // III Междунар. палинологическая конф. Пле-
нарный доклад. Новосибирск. 10 с.
88. Пермские отложения Карасорского прогиба (Центральный Казахстан) // Мате-
риалы по геологии Центрального Казахстана. Т. 10. М.: Изд-во Московск. гос. ун-та. С.
417—427. (Совместно с А.М.Курчавовым).
89. Genus concept in palaeobotany // III International Palynological Conference. Paper for
plenary session. Novosibirsk. 10 p. [Перевод на английский работы №87.]
90. Phyllotheca-like plants from the Upper Palaeozoic flora of Angaraland // Palaeonto-
graphica. Abt. B. Bd. 133. Lief. 1/3. S. 1-33. [Переиздана на русском языке в сб. № 320.]
91. Parallelism and its significance for the systematics of fossil plants // Geophytology.
Vol. 1, № 1. P. 34-47.
92. The Permian-Triassic boundary and its relation to the Paleophyte-Mesophyte floral
boundary // International Permian - Triassic Conference. August 23-26, 1971. Calgary; Al-
berta, Canada. Program with Abstracts. P. 339-340.
93. Some current theoretical problems of paleobotany // Paleontol. Joum. (Washington).
Vol. 4, № 4. P. 453-462. [Перевод на английский статьи №74.]
94. Morphology and systematics of Angara Carboniferous lepidophytes II B. Sahni Inst.
Palaeobot. Silver Jubilee. Palaeobot. Conf. December 1971. Abstracts. P. 35-36.
1972
95. Основные проблемы палеофлористики карбона и перми // Итоги науки и техни-
ки. Стратиграфия. Палеонтология. Т 3. М.: ВИНИТИ. С. 94-107.
96. Листья на камне. М.: Знание. 48 с. [Переиздано в сб. №338.]
97. Путь к новому синтезу, или куда ведут гомологические ряды // Знание - сила. №
8. С. 20-22. [Переиздано в сб. №338.]
98. Рецензия на книгу Л.Ш.Давиташвили «Эволюция накопления горючих ископае-
мых в связи с развитием органического мира» // Вести. АН СССР. № 2. С. 126-129.
99. Граница перми и триаса и ее отношение к границе палеофита и мезофита // Но-
вые данные по границе перми и триаса СССР. Л. С. 54-55.
100. К 60-летию В.А.Вахрамеева // Сов. геология. № 2. С. 152-153. (совместно с
М.А. Ахметьевым, М.П.Долуденко, Е.Л.Лебедевым).
101. Are there ligula and parichnos in Angara Carboniferous lepidophytes // Rev. Pa-
laeobot. Palynol. Vol. 14, № 1/2. P. 149-157.
102. On the origin and relationship of the main Carboniferous and Permian floras and their
bearing on general paleogeography of this period of time // 2nd Gondwana Symposium. South
Africa. 1970. Proc. a. Papers. Pretoria. P. 551-555.
1973
103. О систематическом положении некоторых верхнепермских мешковых миоспор
Ангариды // Основные вопросы палинологии перми и триаса СССР. Сыктывкар. С. 46-47.
395
104. 0 VIII Международном конгрессе по стратиграфии и геологии карбона // Изв.
АН СССР. Сер. геол. № 6. 1 с.
105. О соотношении мешковых миоспор верхнего палеозоя Ангариды и индийской
части Гондваны // ПЖ № 3. С. 108-118.
106. Биологические парадоксы А.А. Любищева// Природа. № 10. С. 38- 41. (Совме-
стно с Ю.А. Шрейдером). [Переиздано в сб. №338.]
107. Послесловие к книге: А.А.Гангнус «Через горы времени». М.: Мысль. С. 170-173.
108. Новейшие достижения в области изучения лепидофитов, членистостебельных,
птеридоспермов и кордаитов // Информационные материалы о деятельности Научного
совета по проблеме «Пути и закономерности исторического развития животных и расти-
тельных организмов» в 1972 г. М.: Наука. С. 45-50.
109. Жизнь в «безжизненную эру» [рец.] // Природа. № 4. С. 115-117.
110. Концепция рода в палеоботанике// Проблемы палинологии. М.: Наука. С. 65-70.
111. Об уровне нашего незнания // Знание - сила. № 5. С. 30-31. (Совместно с А. В.
Яблоковым). [Переиздано в сб. №338.]
112. Тип и таксон в палеоботанике (Автореферат доклада, прочитанного 23.05.72 г.)
// Бюлл. МОИП. Отд. геол. Т. 48, № 3. С. 147-148.
113. Кунсткамера для любознательных или инструмент современной науки? // При-
рода. № 12. С. 70-73. [Переиздано в сб. №338.]
114. Carboniferous and Permian floras of the Northern continents // A.Hallam (ed.) Atlas of
Paleobiogeography. Amsterdam: Elsevier Publ. Co. P. 169-186. (Совместно c W.G. Chaloner).
115. The Permian-Triassic boundary and its relation to the Paleophyte-Mesophyte floral
boundary // A.Logan, L.V.Hills (eds.). The Permian and Triassic systems and their mutual
boundary Canad. Soc. Petrol. Geologist. Mem. 2. Calgary. P. 662-667. [Переиздано на рус-
ском языке в сб. № 328.]
116. Plant morphology in its nomothetical aspects // Bot. Rev. Vol. 39, № 3. P. 205-260.
117. Ueber die Hypothese der Kontinentaldrift unter dem Aspekt der Palaeobotanik von
Karbon und Perm // Z. geol. Wiss. Berlin. Bd. 1, H. 4. S. 415-429.
1974
118. Закон? Есть закон? // Знание - сила. № 9. С. 8-9. [Переиздано в сб. №338.]
119. Морфология вегетативного побега ангарских каменноугольных лепидофитов /7
Палеонтол. журн. № 3. С. 97-110.
120. Об основных фитостратиграфических границах в карбоне и перми Тунгусского
бассейна//Тр. Томск, ун-та. Т. 232. С. 151-155.
121. О соотношении номогенетического и тихогенетического аспектов эволюции //
Журн. общ. биологии. Т. 35, № 3. С. 353-364.
122. Спорные вопросы теории стратиграфии // Природа. № 12. С. 16-22. [Переиздано
в сб. №338.]
123. Корреляция нижнепермских континентальных отложений СССР (по палеобота-
ническим данным) // Тезисы докладов к Пленуму Пермской комиссии МСК. Пермь. С.
31-32. (Совместно с М.В. Дуранте).
124. Введение в теорию стратиграфии // ВИНИТИ, деп. рук. № 1749-74. Деп. 186 с.
[Переиздано в сб. № 318.]
125. Понятие «естественность» и «одновременность» в стратиграфии // Изв. АН
СССР. Сер. геол. № 6. С. 79-90.
396
1975
126. Достаточен ли палеонтологический материал для суждений о движении конти-
нентов // Палеонтология, палеобиогеография и мобилизм. Тезисы докладов XXI сессии
Всес. Палеонтол. о-ва. 27-31 января 1975 г. С. 19-20.
127. Проблема направленности эволюции // Итоги науки и техники. Зоология позво-
ночных. Т. 7. М.: ВИНИТИ. С. 66-117.
128. Систематика и формализация // Биология и современное научное познание (ма-
териалы к конференции). Ч. 1. М.: Ин-т филос. АН СССР. С. 32-34.
129. Находка хвойных уникальной сохранности в нижнем триасе (?) Западной Яку-
тии //ПЖ № I.C. 131-133. (Совместно с В.В.Грицик).
130. Palaeobotanical taxonomy and nomenclature: the need for a new approach // Taxon.
Vol. 24, № 1. P. 251-254.
131. Comparison of the Late Palaeozoic saccate miospore assemblages of Angaraland and
Indian part of Gondwanaland // Geophytology. Vol. 5, № 2. P. 117-125.
132. Cladostrobus and the systematics of cordaitalean leaves // Lethaia. Vol. 8. P. 103-
123. (Совместно c H.K.Maheshwari).
1976
133. Научиться понимать // Неделя. № 2 (826). С. 4-5. [Переиздано в сб. № 353].
134. Эволюционная морфология и палеоботаника // Материалы Пятого Московского
совещания по филогении растений. М.: Наука. С. 116-117.
135. Принципы палеобиогеографического районирования // Палеонтология. Морская
геология. Междунар. геол, конгр. XXV сессия. Доклады советских геологов. М.: Наука.
С. 18-24.
136. Нижняя и верхняя граница карбона в континентальных отложениях // Границы
геологических систем. М.: Наука. С. 126—144.
137. Проблемы стратиграфии карбона и некоторые пути их решения // Сов. геология.
№8. С. 142- 147.
138. Ископаемые свидетели. Три «за» в пользу мобилизма // Техника - молодежи.
№ 11. С. 24-25. [Переиздано в сб. №338.]
139. «Головоломки» или проблемы эволюции [рец.] // Природа. № 8. С. 145-147.
(Совместно с Б.С. Соколовым). [Переиздано с сокращениями в сб. №338.]
140. Методологические аспекты теории классификации // Вопросы философии. №
12. С. 67-79. (Совместно с Ю.А. Шрейдером).
141. The concepts of «naturalness» and «synchroneity» in stratigraphy // Int. Geol. Rev.
Vol. 18, № 1. P. 80-88. [Перевод на английский язык статьи №125.]
142. Carboniferous and Permian lepidophytes of Angaraland// Palaeontographica. Abt. B.
Bd. 157, Lfgs. 5/6. S. 112-157. [Переиздано на русском языке в сб.: С.В.Мейен. Теорети-
ческие проблемы палеоботаники. М.: Наука, 1990. С. 76-124; - см. № 320.]
143. Permian conifers of the West Angaraland and new puzzles in the coniferalean phylog-
eny//Palaeobotanist. Vol. 25. P. 298-313.
1977
144. Классификация эволюционных теорий // Методологические и теоретические ас-
пекты биологии (Тезисы докладов Школы молодых учёных по теоретической биологии).
Петрозаводск. С. 44—45.
145. Принцип сочувствия // Пути в незнаемое. Сб. 13. М.: Сов. писатель. С. 401-430.
[Переиздано в сборниках: №338; № 353].
397
146. Олигомеризация и полимеризация в эволюции древнейших высших растений:
Сборник научных работ // Значение процессов полимеризации и олигомеризации. Л.: Зоо-
логический ин-т АН СССР. С. 75-77. [Переиздано в сб.: С.В.Мейен. Эволюция и система-
тика высших растений по данным палеоботаники. М.: Наука, 1990. С. 11-12; - см. № 328.]
147. Палеозойский этап в развитии наземной флоры И Жизнь на древних континен-
тах, ее становление и развитие. Тезисы докл. XXIII сессии Всес. палеонтол. о-ва (21-25
марта 1977 г.). Л. С. 51-52.
148. Таксономия и мерономия // Вопросы методологии в геологических науках. Ки-
ев: Наукова думка. С. 25-33.
149. Cardiolepidaceae - новое пермское семейство хвойных Северной Евразии // Па-
леонтол. жури. № 3. С. 128-138.
150. Классическая и неклассическая биология. Феномен Любищева // Вести. АН
СССР. № 10. С. 112-124. (Совместно с Б.С.Соколовым и Ю.А.Шрейдером).
151. Предложения по созданию Международной стратиграфической шкалы карбона
// Изв. АН СССР. Сер. геол. № 2. С. 5-24. (Совместно с А.Бурозом, Р.Г.Вагнером,
М.Гордоном, О.Л.Эйнором).
152. Relation of Angara and Gondwana floras: a century of controversies 11 Fourth Int.
Gondwana Symposium, Calcutta, India. January, 1977. Sec. 2. Gondwana flora. Key paper. 10
p. [Переиздана на русском языке в сб. № 320. С. 125-131.]
1978
153. Предисловие и редактирование т. 1-3 Трудов Международного конгресса по
стратиграфии и геологии карбона. М.: Наука. (1978, 1979).
154. Воображаемая или невообразимая биология // Знание - сила. № 3. С. 47-48.
[Переиздано в сб. №338.]
155. В поисках идеального геологического календаря [рец.] //Природа. № 4. С. 151-152.
156. Флоры прошлого Земли [рец.] // Палеонтол. журн. № 2. С. 148-150.
157. Первый каталог индийских ископаемых растений [рец.] // ПЖ № 3. С. 146.
158. /Геоботаника/. Рецензия // Новые книги за рубежом. Сер. В, биол., мед. и сельск.
хоз. № 4. С. 33-36
159. Неклассическая биология. Феномен Любищева И Химия и жизнь. № 6. С. 29-35.
(Совместно с Б.С.Соколовым и Ю.А.Шрейдером).
160. Послесловие (к кн. И.В.Круть. Введение в общую теорию Земли. Уровни орга-
низации геосистем). М.: Мысль. С. 360-365. (Совместно с Б.С.Соколовым).
161. Чтобы способности не пропали даром И Литературная газета. № 44. С. 13.
162. Врачу, исцелися сам ... // Знание-сила. № 7. С. 31-34. [Переиздано в сб. № 353].]
163. Проблема редукционизма в биологии // Диалектика развития в природе и науч-
ном познании (К 100-летию книги Ф.Энгельса «Анти-Дюринг»): Сборник научно-
аналитических обзоров. М.: ИНИОН АН СССР. С. 135-169.
164. О наиболее общих принципах исторических реконструкций в геологии И Изв.
АН СССР. Сер. геол. № 11. С. 79-91.
165. Основные аспекты типологии организмов И ЖОБ. Т. 39, № 4. С. 495-508.
166. Палеоботаническая характеристика стратиграфических подразделений верхне-
палеозойских отложений Средней Сибири И Новое в стратиграфии и палеонтологии
среднего и верхнего палеозоя Средней Сибири. Новосибирск. С. 111-125. (Совместно с
С.Г.Гореловой и С.В.Суховым).
167. Морфология проптеридофитов («псилофитов») И Бюлл. МОИП. Отд. биол. Т.
83, №. 2. С. 96-107. [Переиздано в сб. № 328.]
168. Систематика, филогения и экология проптеридофитов // Бюлл. МОИП. Отд.
биол. Т. 83, №. 4. С. 72-84. [Переиздано в сб. № 328.]
398
169. Рец-зия на «Биология моря» //ЖОБ. Т. 39, № 5. С. 793-795 (сонм, с В.В. Меннером).
170. An attempt of a radical improvement of suprageneric taxonomy of fossil plants //
Phyta. Vol. 1. P. 76-86.
171. Nomothetical plant morphology and the nomothetical theory of evolution: the need
for cross-pollination // Acta Biotheoretica. Vol. 27. Suppl.: Folia biotheoretica. № 7. P. 21-36.
172. Propositions pour une classification chronostratigraphique Internationale du Carbon-
ifere // Industrie Minerale (France). Vol. 60, № 10. P. 469-483. (Совместно c A. Bouroz, O.L.
Einor, M. Gordon, R.H. Wagner). (To же по-английски в т. I Трудов Междунар. конгресса
по стратиграфии и геологии карбона, а также по-русски - см. № 150).
173. Palaozoische und mesozoische Floren Eurasiens und die Phytogeographie dieser Zeit.
VEB Gustav Fischer Verlag, Jena. 300 S. (Совместно с V.A.Vakhrameev, I.A.Dobruskina,
E.D.Zaklinskaja).
174. A restatement of Taxon, 24, 251-254 // IOP Newsletter. № 5. P. 12.
175. Why “Neuberg” instead of “Neuburg”? // IOP Newsletter. № 6. P. 6.
1979
176. Вклад А.А.Любищева в теоретическую биологию // Доклады МОИП: Общ.
биол. I полугодие 1977 г. М. С. 109-110.
177. Предки высших растений // Природа. № 11. С. 40-47. [Переиздано в сб. №338.]
178. Может лн быть победитель в дискуссии о номогенезе // Природа. № 9. С. 114-
116. [Переиздано в сб. №338.]
179. О представителях семейства Peltaspermaceae из пермских отложений Русской
платформы // Палеонтол. журн. № 2. С. 124-138. (Совместно с А.В.Гоманьковым).
180. Вероятностный мир и вероятностный язык И Химия и жизнь. № 6. С. 22-27.
(Совместно с В.В.Налимовым). [Переиздано в сб. №338.]
181. USSR Colloquium on the systematics of U. Palaeozoic plants // IOP Newsletter. № 9.
P. 3-4.
182. Interpretation of Cardiolepis - an unfortunate error // IOP Newsletter. № 9. P. 11-12
(совместно c H.G. Smoller).
183. The North American Permian flora - an Angara palaeobotanisfs first impression //'
IOP Newsletter. № 10. P. 9-10.
184. Proposal on article 3: Remove «Form Genus» too! // Taxon. Vol. 28, № 5/6. P. 595-
598. (Совместно c A.Traverse).
185. Permian predecessors of the Mesozoic pteridosperms in Western Angaraland, USSR
// Rev. Palaeobot. Palynol. Vol. 28, № 2. P. 191-201.
186. Relation of Angara and Gondwana floras: a century of controversies // IV Int. Gond-
wana Symp., 1977. Calcutta, India. Hindustan Publ. Corporation. P. 45-50. [Ещё одно издание
статьи №148]
187. Probabilistic vision of the world // 6th Int. Congr. Logic. Methodol. and Phil. Sci. Ab-
stracts. Sect. 7. P. 253-257. (Совместно c V.V.Nalimov; русский вариант см. № 180).
1980
188. Экосистемы и принцип взаимозаменяемости признаков // Экосистемы в страти-
графии. Владивосток: ДВНЦ АН СССР. С. 16-21.
189. О стратиграфическом расчленении газоносных отложений перми центральной
части Вилюйской синеклизы И Стратиграфия нефтегазоносных отложений Якутии.
Якутск. С. 36-45. (Совместно с И.И.Голубевой, В.В.Граусман, Е.К.Петровой).
190. Мудрость науки и загадки жизни И Знание - сила. № 12. С. 26-28. [Переиздано
в сб. № 353].
399
191. Прогноз в биологии и уровни системности живого // Биология и современное
научное познание. М.: Наука. С. 103-120.
192. IX Международный конгресс по стратиграфии и геологии карбона // Изв. АН
СССР. Сер. геол. № 4. С. 151-154. (Совместно с П.П.Тимофеевым, Б.В.Полянским).
193. Новый палеоботанический справочник [рец.] // ПЖ № 2. С. 148-149.
194. Палеофлористика и стратиграфия // Стратиграфия в исследованиях Геологиче-
ского института АН СССР. М.: Наука. С. 106-107. (Совместно с В.А.Вахрамеевым).
195. Палеофлористика палеозоя // Стратиграфия в исследованиях Геологического
института АН СССР. М.: Наука. С. 107-117.
196. Проблемы систематики растительных остатков // Стратиграфия в исследованиях
Геологического института АН СССР. М.: Наука. С. 132-134.
197. Проблемы флористического районирования в геологическом прошлом // Стра-
тиграфия в исследованиях ГИН. М.: Наука. С. 134-137. (Совместно с В.А.Вахрамеевым).
198. Об организации хранения коллекций в Геологическом институте АН СССР // Ин-
формационные мат-лы о деятельности Науч, совета по проблеме «Пути и закономерности
исторического развития животных и растительных организмов» в 1979 г. М.: Наука. С. 79.
199. О соотношении комплексов растительных макро- и микрофоссилий в перми
Ангариды // ПЖ. № 4. С. 114-122. (Совместно с А.В.Гоманьковым).
200. Разведанные тропы, нерешённые загадки [Предисловие к книге А.А.Гангнуса
«Тропой времен»]. М.: Детская литература. С. 9-12.
201. Человек рождён для полёта И Литературная газета. № 7 (4761). С. 12. [Напеча-
тано полностью под названием «Есть ли рожденные ползать»» в сб. № 353].
202. Вымерший климат//Земля и люди. М.: Мысль. С. 18-25.
203. Пельтаспермовые птеридоспермы рода Tatarina // Палеонтол. журн. № 2. С.
116-132. (Совместно с А.В.Гоманьковым).
204. The «Correlation of coal-bearing formations» project // Newsletter on Carboniferous
Stratigraphy. № 1. P. 13-15.
205. Systematics & stratigraphical significance of Phylladoderma, USSR // IOP Newslet-
ter. № 12. P. 3-4.
1981
206. Рецензия на книгу: В.В.Плотников «Эволюция структуры растительных сооб-
ществ» // Экология. № 6. С. 102-104.
207. Палеогеография карбона и перми и проблема перемещения материков // Палео-
нтология, палеогеография и мобилизм. Труды 21-й сессии Всес. палеонтол. о-ва. Мага-
дан. С. 82-90 (совместно с В.И.Устрицким, Г.Е.Черняк).
208. От общей к теоретической стратиграфии И Советская геология. № 9. С. 58-69.
[Переиздано в сб. № 318.]
209. Исторические реконструкции в естествознании и типология // Эволюция мате-
рии и ее структурные уровни. М.: Ин-т философии АН СССР. С. 90-93.
210. Следы трав индейских. М.: Мысль. 159 с.
211. Extra-equatorial persistence // ЮР Newsletter. № 14. Р. 6-7.
212. Terminology of gymnosperm fructifications // IOP Newsletter. № 14. P. 7-8.
213. Correlation of coalbearing formations (C.C.F.) Palaeobotanical colloquium, Vorcuta,
March, 1981 // IOP Newsletter. № 15. P. 4.
214. Ginkgo - a living pteridosperm // IOP Newsletter. № 15. P. 9-11.
215. Some true and alledged Permotriassic conifers of Siberia and Russian platform and
their alliance // Palaeobotanist. Vol. 28/29. P. 161-176.
400
1982
216. Стратиграфия пермских отложений Вилюйской синеклизы // Изв. АН СССР.
Сер. геол. № 11. С. 57-67 (совместно с В.В.Граусман и В.Д.Матвеевым).
217. Фруктификации верхнепалеозойских кордаитантовых Ангариды // ПЖ № 2. С.
109-120.
218. Органы размножения голосеменных и их эволюция (по палеоботаническим
данным) // Журн. общ. биол. Т. 43, № 3. С. 303-323.
219. Работы по проблемам системы, эволюции и формы организмов // Александр
Александрович Любищев. М.: Наука. С. 38-52.
220. Типологические аспекты интеграции физического, биологического и социогу-
манитарного знания // Пути интеграции биологического и социогуманитарного знания
(сборник тезисов выступлений к методологическому семинару). М.: Ин- философии АН
СССР. С. 17-19.
222. Рецензия на книгу: С.В.Петухов «Биомеханика, бионика и симметрия» И Журн.
общ. биол. Т. 43, № 1. С. 239.
223. Ревизия индийских видов Glossoptei-is И ПЖ № 1. С. 137. (Совместно с
В. А.Вахрамеевым).
224. Теория и практика экосистемного подхода в стратиграфии // Палеонтология и
детальная стратиграфическая корреляция. Тезисы докладов XXVIII сессии Всес. палео-
нтол. о-ва. Ч. 1. Ташкент. С. 52-54.
225. Геоисторическая периодизация и геохронологическая шкала // Развитие учения
о времени в геологии. Киев: Наукова думка. С. 19-30. [Переиздано в сб. № 318.]
226. Концепция гомотаксиса и ее значение в геохронологии // Развитие учения о
времени в геологии. Киев: Наукова думка. С. 88-99. [Переиздано в сб. № 318.]
227. Проблема геохронологических границ / / Развитие учения о времени в геологии.
Киев: Наукова думка. С. 209-219. [Переиздано в сб. № 318.]
228. Палеоэкосистемный подход // Развитие учения о времени в геологии. Киев:
Наукова думка. С. 286-305. [Переиздано в сб. № 318.]
229. Специфика историзма и логика познания прошлого в геологии // Развитие уче-
ния о времени в геологии. Киев: Наукова думка. С. 361-381. [Переиздано в сб. № 318.]
230. Рецензия на книгу: T.N.Taylor. «Paleobotany. Introduction to fossil plant biology».
1981 // Ботан. журн. T. 67, № 12. С. 1694-1696.
231. Гинкго - современный птеридосперм (положение гинкговых в системе голосе-
менных) // Филогения высших растений. М.: Наука. С. 93-96.
232. Новые данные о спороношениях верхнепалеозойских папоротников Ангариды //
Палеонтол. журн. № 4. С. 108-112.
233. От редакции. Биографическая справка. Вводная статья: «О работах А.А.Любище-
ва по общим проблемам биологии». Комментарии И А.А Любищев. Проблемы формы, сис-
тематики и эволюции организмов. М.: Наука. С. 3-23, 257-262. (Совм. с Ю.В. Чайковским).
234. Divergent opinions on Orestovia and allied plants // IOP Newsletter. № 17. P. 16-17.
235. Trichopitys, Medullopteris and Autunia revisited // IOP Newsletter. №> 18. P. 4-6.
236. French-Soviet palaeobotanical cooperation // IOP Newsletter. № 19. P. 5. (Совмест-
но c Y.Lemoigne.)
237. Imperfection of the fossil record // IOP Newsletter. № 19. P. 6.
238. The Carboniferous and Permian floras of Angaraland (a synthesis) // Biol. Memoirs.
Vol. 7, № 1. P. 1-109. [Переиздана на русском языке в сб.: С.В.Мейен. Теоретические
проблемы палеоботаники. М.: Наука, 1990. С. 131-223; - см. № 320.]
239. Commentary on Dr.Cusset's paper// Acta biotheoretica. Vol. 31 A. P. 87-92. [To же
отдельным изданием: Axioms and Principles of Plant Construction. Proceedings of a sympo-
401
sium held at the International Botanical Congress, Sydney, Australia, August 1981. The Hague;
Boston; London: Martius Nijhoff/ Dr. W.Junk Publishers. P. 87-92.]
240. Ginkgo as a possible living pteridosperm // Studies on living and fossil plants. Special
issue of Phyta. Allahabad: The Society of plant taxonomists. P. 163-172.
1983
241. Thuringia - копролит. а не синангий // Докл. АН СССР. Т. 270, № 2. С. 431-432.
242. Систематика пельтаспермовых птеридоспермов и их место в филогении голосе-
менных // Бюлл. МОИП. Отд. биол. Т. 88, № 1. С. 3-14. [Переиздано в сб. № 328.]
243. Морфология, систематика и филогения растений геологического прошлого // Ос-
новные проблемы палеонтологических исследований в СССР. Материалы Всесоюзного со-
вещания. М.: Наука. С. 41-55. (Совместно с В.А.Вахрамеевым). [Переиздано в: сб. №339.]
244. Каузальная геология сейчас и в будущем // Будущее геологической науки. Тези-
сы докладов советских ученых. М. С. 21.
245. За кулисами доисторического ландшафта // Знание - сила. № 7. С. 23-26. [Пере-
издано в сб. №338.]
246. Понятие времени и типология объектов (на примере геологии и биологии) //
Эволюция материи и её структурные уровни. М.: Наука. С. 311- 316.
247. Не «кормите» меня, пожалуйста // Литературная газета. № 13. С. 14. [Напечата-
но полностью под названием «Самолечение духа» в сб. № 353].
248. Том Максвелл Харрис (1903-1983) // ПЖ № 4. С. 122- 123. (Совместно с В.А.
Вахрамеевым, М.П.Долуденко, В.А.Самылиной). [Переиздано в сб. №338.]
249. Сравнение принципов систематики современных и ископаемых организмов //
Математика и ЭВМ-в палеонтологии. Кишинёв: Штиинца. С. 10-27.
250. Систематика верхнепалеоэойских членистостебельных семейства Tchemo-
viaceae // Ботан. журн. Т. 68, № 6. С. 721-729. (Совместно с Л.В.Меньшиковой).
251. Thuringia: gymnosperm synangium or coprolite? // ЮР Newsletter. № 20. P. 6-7.
252. Two weeks in the Museum fuer Naturkunde (Berlin, DDR) // IOP Newsletter. №
20. P. 7.
1984
253. Корреляция пермских отложений Ангарской области // Основные черты страти-
графии пермской системы СССР. Л.: Недра. С. 226-229.
254. Наземная флора // Основные черты стратиграфии пермской системы СССР. Л.:
Недра. С. 270-272.
255. Среднесибирская провинция // Основные черты стратиграфии пермской систе-
мы СССР. Л.: Недра. С. 222-225. (Совместно с И.И. Голубевой).
256. Филогения высших растений и флорогенез // 27-й Междунар. Геол, конгресс.
Тезисы. Секция 01-03. Т 1. М.: Наука. С. 288-289. (То же на англ, языке).
257. Филогения высших растений и флорогенез // 27-й Междунар. Геол, конгр. Па-
леонтология. Секция. С.02. Доклады. Т. 2. Москва. 4-14 августа 1984. М.: Наука. С. 75-
80. (То же на англ, языке).
258. Происхождение главных групп высших растений // Актуальные проблемы био-
логической науки. М.: Просвещение. С. 128-165. [Переиздано в сб. №338.]
259. Высшие растения // Справочник по систематике ископаемых организмов (так-
соны отрядной и высших групп). М.: Наука. С. 153-163.
260. События на рубеже мезозоя и кайнозоя в развитии органического мира // Изв.
АН СССР. Сер. геол. № 1. С. 134-136.
402
261. Нужна ли теоретическая геология как некоторая консолидированная концепция
// Природа. № 1. С. 25-27. [Переиздано под названием «Зачем нужна теоретическая гео-
логия» в сб. №338.]
262. Верить ли геологической летописи? // Знание - сила. № 4. С. 7-10. [Переиздано
в сб. №338.]
263. Принципы исторических реконструкций в биологии // Системность и эволюция.
М.: Наука. С. 7-32 [Переиздано в сб.: In memoriam. С.В. Мейен: палеоботаник, эволю-
ционист, мыслитель. М.: ГЕОС. С. 319-345].
264. Типологические аспекты интеграции физического, биологического и социогу-
манитарного знания // Пути интеграции биологического и социогуманитарного знания.
М.: Наука. С. 88-99.
265. Будущее эволюционной теории - продолжение синтеза // Методологические
проблемы эволюционной теории. Тарту. С. 173-175. [Перепечатано в сб.: С.В. Мейен.
Принцип сочувствия: Размышления об этике и научном познании. М.: ГЕОС, 2006. С.
142-144;-см. № 353].
266. Сравнение темпов и форм эволюции высших растений в экваториальных и внеэк-
ваториальных биотах геологического прошлого // Макроэволюция. М.: Наука. С. 157-158.
267. Is Thuringia a gymnosperm synangium or a coprolite? // Z. geol. Wiss. Berlin. Bd.
12, H. 2. S. 269-270.
268. Basic features of gymnosperm systematics and phylogeny as evidenced by the fossil
record // Bot. Rev. Vol. 50, № 1. P. 1-111. [Переиздано в сб. № 328.]
269. The concept of assemblage-taxa// IOP Newsletter. № 23. P. 10-11.
270. Gamoheterotopy - a probable process in morphological evolution of higher plants //
IOP Newsletter. № 25. P. 4-5.
271. Nomenclature of early conifers // IOP Newsletter. № 25. P. 6.
272. A reappraisal of the Lower Carboniferous lepidophyte Eskdalia Kidston // Palaeon-
tology. Vol. 27, Pt. 4. P. 707-718. (Совместно c B.A.Thomas).
273. A system of form-genera for the Upper Palaeozoic lepidophyte stems represented by
compression-impression material // Rev. Palaeobot. Palynol. Vol. 41, № 3/4. P. 273-281. (Со-
вместно c B.A.Thomas).
274. Tunguskadendron borkii: gen. et sp. nov. a new Angaran lepidophyte // Palaeonto-
graphica. Abt. B. Bd. 193, Lfgs. 5/6. S. 121-126. (Совместно c B.A.Thomas).
1985
275. М.Ф.Нейбург - 40 лет служения «малой» науке // Страницы из истории Москов-
ской геологической школы. Очерки по истории геологических знаний. М.: Наука. С. 62-79.
276. Структура теоретической стратиграфии // Изв. АН СССР. Сер. геол. № 11. С. 8-
16. [Переиздано в сб. № 318.]
277. Sphenobaiera is under fire again // IOP Newsletter. № 26. P. 6.
278. TTiree weeks in palaeontological laboratories in France // IOP Newsletter. № 26. P. 8-9.
1986
279. Что мне скажет методолог? // Знание-сила. № 1. С. 21-22. [Переизд. в сб. № 338.]
280. Флорогенез и эволюция растений // Природа. № 11. С. 47-57. [Переизд. в сб. № 328.]
281. Время без часов, или похвальное слово создателям геохронологии И Знание -
сила. № 12. С. 33-34. [Переиздано в сб. №338.]
282. Голосеменные ангарской флоры. Деп. в ВИНИТИ, № 4080-В86. 54 с. [Переиз-
дано в сб. № 328.]
283. Пермские хвойные Западной Ангариды. Деп. в ВИНИТИ. № 3405- В86. 140 с.
403
284. Граница девона и карбона по макрофоссилиям растений // Граница девона и
карбона на территории СССР. Минск. С. 141-151.
285. Биологический энциклопедический словарь. М.: Советская энциклопедия. 23
статьи по палеоботанике.
286. Гипотеза происхождения покрытосеменных от беннеттитов путем гамогетеро-
топии (переноса признаков с одного пола на другой) // Журн. общ. биол. Т. 47, № 3. С.
291-309. [Переиздана в сборниках: № 296; № 320.]
287. Suchoviella - новый род руфлориевых из перми Западной Ангариды и основные
вопросы систематики и филогении кордаитантовых. Деп. в ВИНИТИ. № 5632-В86. 57 с.
(Совместно с И.А. Игнатьевым).
288. Gymnosperm systematics and phylogeny, a reply to commentaries by C.B.Beek,
C.N.Miller and G.W.Rothwell // Bot. Rev. Vol. 52, № 3. P. 300-320. [Переиздано в сб.:
С.В.Мейен. Эволюция и систематика высших растений по данным палеоботаники. М.:
Наука, 1990. С. 105-120; - см. № 328.]
289. The Anderson's «Prodromus»: continuing discussion // IOP Newsletter. № 30. P. 7-8.
290. Cladistics of marattialeans: another view // IOP Newsletter. № 31. P. 7-8.
291. The genus Mostochkla Chachlov (Upper Palaeozoic of Angaraland) and its bearing on
the characteristics of the order Dicranophyllales (Pinopsida) // Rev. Palaeobot. Palynol. Vol. 47,
№ 3/4. P. 205-223. (Совместно с Г.Г.Смоллер).
292. Dicksonites pluckenetii (Schlotheim) Sterzel and its affinity with Callistophytales //
Geobios. № 19, Fasc. 1. P. 87-97. (Совместно c Y. Lemoigne).
1987
293. Опять тройка // Знание - сила. № 1. С. 102-103.
294. Основы палеоботаники. М.: Недра. 403 с.
295. География макроэволюции у высших растений // ЖОБ. Т. 48, № 3. С. 291-310.
[Перепечатано в сб.: Общая палеоботаника - см. № 296. Деп. в ВИНИТИ. № 8673-В87. С.
149-177; - см. №288; перевод этой работы на английский язык см. № 315.]
296. Общая палеоботаника (Депонированное приложение к книге «Основы палеобо-
таники», М., Недра, 1987; - см. № 285). Деп. в ВИНИТИ. № 8673-В87. 202 с.
297. Стратиграфия верхнепалеозойских отложений Вилюйской синеклизы // Изв. АН
СССР. Сер. геол. № 10. С. 53-60. (Совместно с В.В.Граусман).
298. Кто первым бросит камень?.. // Знание - сила. № 12. С. 74-80. [Переиздано в сб.
№338; полностью в авторской редакции в сб. № 353].]
299. Fundamentals of Palaeobotany. London; New York: Chapman and Hall. 432 p.
300. Evolution of Ginkgoopsida: from Peltaspermales to Ginkgoales, Leptostrobales and
Caytoniales // Bull. Soc. bot. France. Vol. 134. Actualites bot. P. ...
Посмертные публикации
Звездочкой отмечены работы, сданные в печать самим С.В. Мейеном
1988
*301 -312. Главы // В.В.Меннер, В.П.Макридин (ред.). Современная палеонтология. Ме-
тоды. Направления. Проблемы. Практическое приложение. Справоч. рук-ство. М.: Недра.
404
Том 1
Введение. С. 8-25.
1.1. Морфологические исследования в палеонтологии. С. 80-123. (Совместно с
Э.И.Воробьёвой).
1.4. Реконструкции вымерших организмов. С. 159-197. (Совместно с Ю.М.
Смириным, В.М.Смириным, А Г.Пономаренко, И.С.Барсковым, Б.Т. Яниным).
2.5. Микроструктурные исследования в палеоботанике. С. 320-327.
3.3. Тафономические исследования. С. 416-434. (Совместно с В.А.Захаровым,
В. Г.Очевым, Б.Т.Яниным).
4.1. Принципы и методы палеонтологической систематики. С. 447-466.
4.2. Вопросы систематики в некоторых разделах палеоботаники (палеопалиноло-
гии и палеоальгологии). С. 446-479. (Совместно с С.И.Шумейко).
4.4 Проблемы филогенетической классификации организмов. С. 497-511.
Том 2
5.1. Палеобиогеографические исследования. С. 5-31 (совместно с В.П.Макридиным).
5.2. Методы палеофлористических исследований и проблемы флорогенеза. С. 31—44.
5.3. Методы палеогеоботанических реконструкций. С. 44-56.
8.1. Практическое значение палеонтологии и её связь с другими научными дис-
циплинами. С. 210-232. (Совместно с А.С. Алексеевым).
* 313. Нетривиальные модусы морфологической эволюции высших растений И Со-
временные проблемы эволюционной морфологии. М.: Наука. С. 91-103.
* 314. On the structure of theoretical biology// К. Kull, T. Tivel (ed.). Lectures in theoreti-
cal biology. Tallinn: Valgus. P. 15-21.
* 315. Gymnosperms of the Angara flora // Ch.B. Beck (ed.). Origin and evolution of
Gymnosperms //New York: Columbia Univ. Press. P. 338-381.
* 316. Origin of the angiosperm gynoeceum by gamoheterotopy // Bot. J. Lin. Soc. Vol. 97.
P. 171-178.
1989
* 317. Первые «сухопутные» растения // Природа. № 5. С. 17-20. [Переиздано в № 338.]
318. Введение в теорию стратиграфии. М.: Наука. 216 с.
*319. Suchoviella - gen. nov. from the Permian of Angaraland and a review of the sys-
tematics of Cordaitanthales // Rev. Palaeobot. Palynol. Vol. 57. P. 313-339. (Совместно c
И. А. Игнатьевым).
1990
320. Теоретические проблемы палеоботаники. М.: Наука. 287 с.
321. Палеоботаническая таксономия и номенклатура: необходимость нового подхода
// С.В.Мейен. Теоретические проблемы палеоботаники. М.: Наука, 1990. С. 49-76. [Ранее
опубликована со значительными сокращениями по-английски - см. № 128.]
322. Академическая наука?.. // Вопросы философии. № 9. С. 16-26. [Опубликовано
также с сокращениями: 1) № 327; 2) МСП. 2001. С. 265-280.]
323. Нетривиальная биология (Заметки о...) // Журн. общ. биол. Т. 51, № 1. С. 4-14.
(Фрагмент работы № 372).
324. Bardospermum - новый род хвойных из кунгура Приуралья и некоторые вопро-
сы эволюции ранних хвойных // Палеонтол. журн. № 2. С. 3-12.
325. Catalogue of fossil plant genera. London, Intemat. Org. Paleobot., XXII+57 pp.
(Текст на англ, и рус. языках, ред. А.В. Гоманьков).
405
1991
*326. Логико-методологические и теоретические стереотипы в биологии // Природа
биологического познания. М.: Наука. С. 21-29.
327. О перестройке в науке и в Академии наук // Знание - сила. № 1. С. 50-54. [Со-
кращенная публикация статьи №322].
1992
328. Эволюция и систематика высших растений по данным палеоботаники. М.: Нау-
ка. 174 с.
329. Geography of macroevolution in higher plants // Sov. Sci. Rev. G. Geol. Vol. 1. P.
39-70. [Перевод на английский язык статьи №295.]
1997
330. Permian conifers of Western Angaraland // Rev. Palaeobot. Palynol. Vol. 96. P. 351—
447. [Публикация на английском языке работы № 283.]
1999
331. Православие в духовной диаспоре // Записки Семинара по истории Церкви па-
мяти св. Стефана просветителя Пермского. С. 6-8. [Переиздано - см. № 353].
332. Креационизм и наука в книге Т.Хайнца «Творение или эволюция» // Той повеле,
и создашася: Современные учёные о сотворении мира. Клин: Фонд «Христианская
жизнь». С. 166-171. (Под псевдонимом С.Катюнин) [Переиздано - см. № 353].
2001
333. Заметки о редукционизме // Методология биологии: Новые идеи: Синергетика.
Семиотика. Коэволюция. М.: УРСС. С. 5-13.
334. М.Ф.Нейбург (заметки ученика) // Материалы симпозиума, посвящённого памя-
ти Сергея Викторовича Мейена. Москва, 25-26 декабря 2000 г. М.: ГЕОС. С. 241-258.
335. Ответы на анкету Ю.В.Линника «Александр Александрович Любищев» // Там
же. С. 78-84.]
336. Флорогенетика - интегративный подход в палеоботанических исследованиях //
Там же. С. 280-298.
337. Памяти А.А.Любищева // Александр Александрович Любищев: Творческий
портрет. Ульяновск: Симбирская книга. С. 4-71.
338. Листья на камне: Размышления о палеоботанике, геологии, эволюции и путях по-
знания живого. М.: ГЕОС. 493 с. (Труды ГИН. Научно-популяр. серия. Вып. 1).
2002
339. Памяти В.А.Вахрамеева И Сборник памяти члена-корреспондента АН СССР, про-
фессора Всеволода Андреевича Вахрамеева (к 90-летию со дня рождения). М.: ГЕОС. С. 62.
340. Современная палеоботаника: проблемы и перспективы // Там же. С. 106-110.
341. Концепция палеофлористических исследований // Там же. С. 119. (Совместно с
В. А.Вахрамеевым).
342. Основные проблемы палеофлористики // Там же. С. 119-121.
343. On the Subangara palaeofloristic area of the Permian // Там же. С. 232-246.
344. Флорогенез и стратиграфия //. С. 316-329. (Совместно с В.А.Вахрамеевым).
2003
345. Методологические установки А.А.Любищева // Эволюция флор в палеозое:
Сборник научных трудов. М.: ГЕОС. С. 106-107.
406
346. Типология // Там же. С. 107-108.
347. Тип // Там же. С. 108.
348. Из заметок, записей бесед и переписки по вопросам эволюционной диатропики
//Там же. С. 108-129, 131-134.
349. Главные недостатки и преимущества кладистики // Там же. С. 129-131.
350. На каком таксономическом уровне геологическая летопись полна? // Там же.
С. 134-135.
351. К вопросу о создании эволюционной биогеографии//Там же. С. 135.
2004
352. Из переписки [А.А. Любищева] с С.В.Мейеном // XVIII Любищевские чтения:
Современные проблемы эволюции. Ульяновск. С. 11-32.
2006
353. Принцип сочувствия: Размышления об этике и научном познании. М.:
ГЕОС. 212 с.
354. Докажи свою неправоту! // Там же. С. 58-64].
355. Наука и этика// Там же. С. 65-66].
356. Хотя бы синицу в руках // Там же. С. 67-75].
357. Из переписки с писателем Д.А. Граниным // Там же. С. 155-162.
358. Выступление о принципе сочувствия // Там же. С. 163-165.
359. Развитие науки и научная этика (I) И Там же. С. 166-168.
360. Развитие науки и научная этика (I) // Там же. С. 169-171.
361. Взаимодействие науки и этики // Там же. С. 172.
362. К вопросу о научном творчестве / Там же. С. 173-175.
363. Тривиальные слова // Там же. С. 176-180.
364. Наброски к статье «Кто первый бросит камень?» // Там же. С. 181-184.
2007
365. Форма - система - эволюция // In memoriam. С.В. Мейен: палеоботаник, эво-
люционист, мыслитель. М.: ГЕОС. С. 156-161.
366. Морфология растений в номотетическом аспекте // Там же. С. 162-222.
367. О путях снятия противоречий между селекционизмом и номогенезом // Там
же. С. 223-228.
368. О соотношении номогенетических и тихогенетических факторов в эволюции
//Там же. С. 229-233.
369. О статье А.А. Яценко-Хмелевского «Предначертана ли эволюция?» // Там же.
С. 234-243.
370. Роль направленных процессов в эволюции // Там же. С. 244-287.
371. Соотношение макро- и микроэволюции // Там же. С. 288-293.
372. Заметки о теоретической биологии // Там же. С. 294-318.
2009
373. Теоретические основы палеоботанических исследований (неизданные главы к
«Основам палеоботаники» [М., Недра, 1987]). М.: ГЕОС, 2009. 108 с.
2010
374. Легче вывести человека из современной свиньи // данный Сборник. С. 354-356.
407
Книга свёрстана автором
и печатается в авторской редакции
Обложка: С.М. Злочевский
Юрий Викторович ЧАЙКОВСКИЙ
Диатропика, эволюция и систематика.
К юбилею Мейена. Сборник статей.
С приложением полного списка трудов С.В. Мейена
(составил И.А. Игнатьев).
Москва: Товарищество научных изданий КМК. 2010. 407 с.
при участии ИП Михайлова КГ.
Для заявок:
123100 Москва, а/я 16, изд-во КМК
эл. почта: mikhailov2000@gmail.com
http://avtor-kmk. ги
Подписано в печать 28.09.2010. Формат 60x90/16. Объём 25.5 п.л.
Бумага офсетн. Тираж 500 экз. Заказ № 1806
Отпечатано в ГУП ППП «Типография «Наука» АИЦ РАН,
121099, Москва, Шубинский пер., 6.
Четырехцарственная
диатропическая
макросистема
ПРЕОБЛАДАНИЕ
ЖГУТИКОВЫХ
ФОРМ
фДГОГ₽Оф6/
Ж <в ° Utb/Z
£ головным Мозг5|
МОЛЛЮСКИ
С ЗЕЛЁНЫМИ
СИМБИОНТАМИ
ИчнополЗ
2э
1э
ы
кте
in
s
ПРЕОБЛАДАНИЕ
БЕЗЖГУТИКОВЫХ
3 э
СК^______
МОЛЛЮСКИ
С ХЕМОТРОФНЫМИ
СИМБИОНТАМИ
цереи
НИТЧАП,,^
оГ
С1
о
> в
5 St
й е
ЛЭ
е
<5
Ф
ЦВЕТКОВЫЕ
ПАРАЗИТЕ
ПЛЕЧЕНОГИЕ
ОСМОТРОФЫ
ХИЩНЫЕ
ГРИБЫ
8
О