/
























































Автор: Чайковский Ю.В.
Теги: развитие органического мира (филогенез) эволюционное учение эволюция
ISBN: 978-5-904129-10-1
Год: 2010




Текст
БИБЛИОТЕКА_Ж V Р Н АЛА
НАУКА И ЖИЗНЬ
Ю. В. Чайковский
ЗИГЗАГИ
ЭВОЛЮЦИИ
РАЗВИТИЕ ЖИЗНИ И ИММУНИТЕТ
НАУКА И ЖИЗНЬ» Москва 2010
Ю. В. Чайковский
РАЗВИТИЕ ЖИЗНИ И ИММУНИТЕТ
«НАУКА И ЖИЗНЬ» Москва 2010
ISBN 978-5-904129-10-1
ББК 28.02
Ответственный редактор Е. Л. Лозовская
Макет, верстка М. Н. Михайлова, Т. Н. Черникова
Оформление обложки 3. А. Флоринская
Редактор Н. В. Костецкая
Корректоры Ж. К. Борисова, В. П. Канаева
Чайковский Ю. В.
Зигзаги эволюции. Развитие жизни и иммунитет. - М.: Наука и жизнь,
2010. (Библиотека журнала «Наука и жизнь». Серия «Об основах наук»),
ISBN 97В-5-904129-10-1
В основе этой небольшой книги - статьи Ю.В. Чайковского, опублико-
ванные в журнале «Наука и жизнь» в 2007 - 2009 годах. Книга рассказыва-
ет о возникновении, развитии и нынешнем состоянии науки о биологиче-
ской эволюции. В отличие от других книг, здесь новейшая наука подается
через ее историю: история призвана показать, что наши новые теории
обычно коренятся в прежнем знании, в традициях и в мировоззрении.
Эволюционная идея вошла в науку задолго до Ч. Дарвина, но прежний
эволюционизм был после появления дарвинизма надолго забыт. Сейчас
выясняется, что он во многом сходен с новейшим. Поэтому, чтобы ис-
пользовать прежний опыт и не повторять ошибок прошлого, надо не про-
сто знать о «предшественниках», но уметь понимать ход их мысли и суть
их споров. Это показано в книге на самых различных примерах. Многие из
них загадочны и потому не попадают в учебную литературу.
Представлен взгляд на эволюцию не как на «происхождение видов» и
прочих групп организмов, а как на единый процесс становления - и самих
организмов, и тех частей, из которых они образованы, и тех экосистем, в
которые они входят. Рассмотрены клеточные и молекулярные механизмы
эволюции, причем особый акцент сделан на нерешенных проблемах. В
качестве основного материала, поясняющего теорию, рассмотрено ста-
новление иммунитета.
Книга рассчитана на учителей биологии, студентов биологических
специальностей и старшеклассников школ биологического профиля, но
будет интересна всем, кто близок биологии, а особенно, всем специа-
листам, кому по роду их занятий надо знать новые эволюционные идеи
- биологам, экологам, инженерам, экономистам и обществоведам.
ББК 28.02
ISBN 978-5-904129-10-1
кгануугп гйтгЫ
© Ю. В. Чайковский, 2010, текст
© АНО «Редакция журнала «Наукаи жизнь», издание
© 3. А. Флоринская, оформление обложки,2010
ОГЛАВЛЕНИЕ
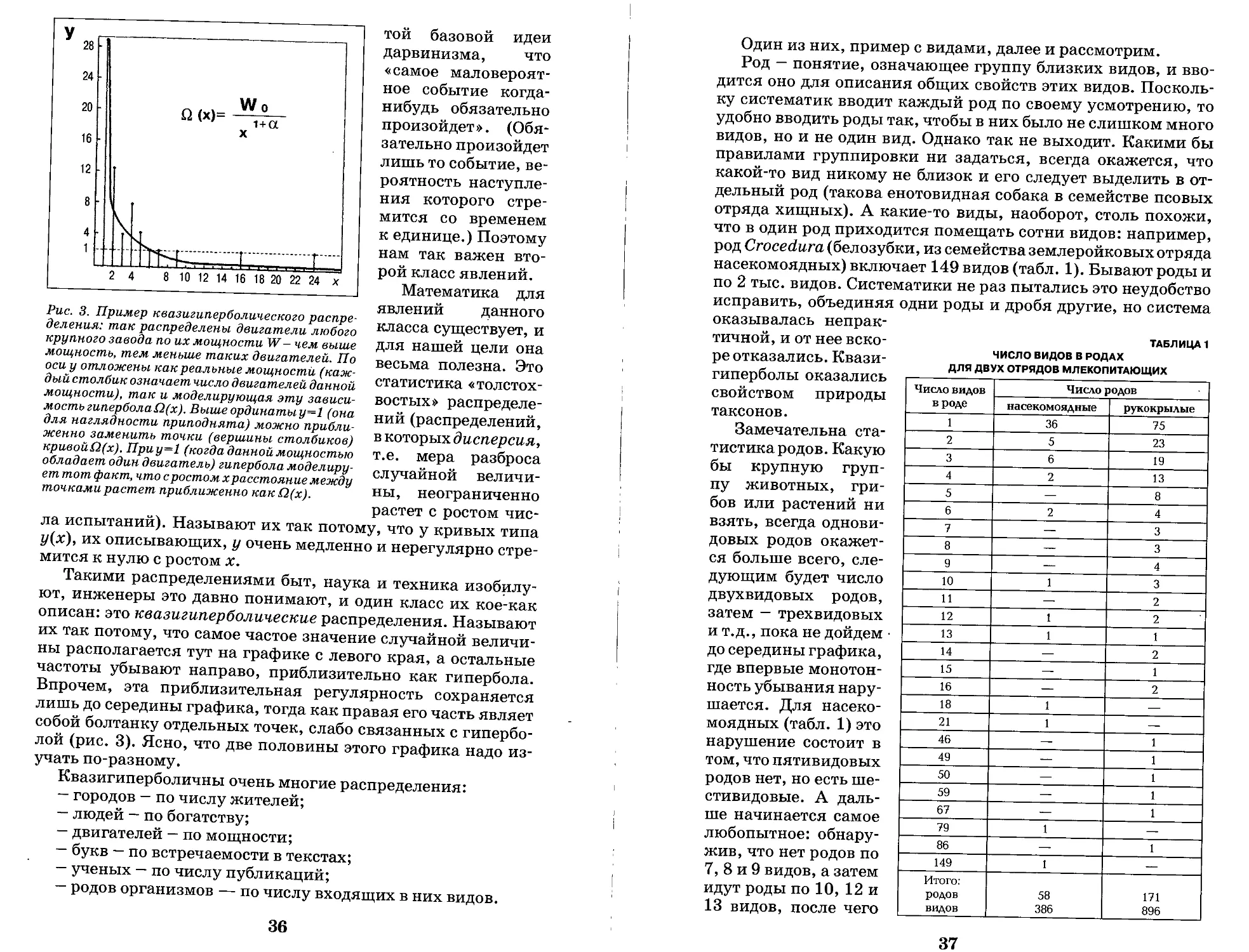
Введение. Была ли эволюция? Незаданный вопрос
и каверзный ответ...............................5
РАЗДЕЛ I. КАК ПОНИМАЮТ
ЭВОЛЮЦИЮ ЖИВОГО
Часть 1. Прежние теории эволюции
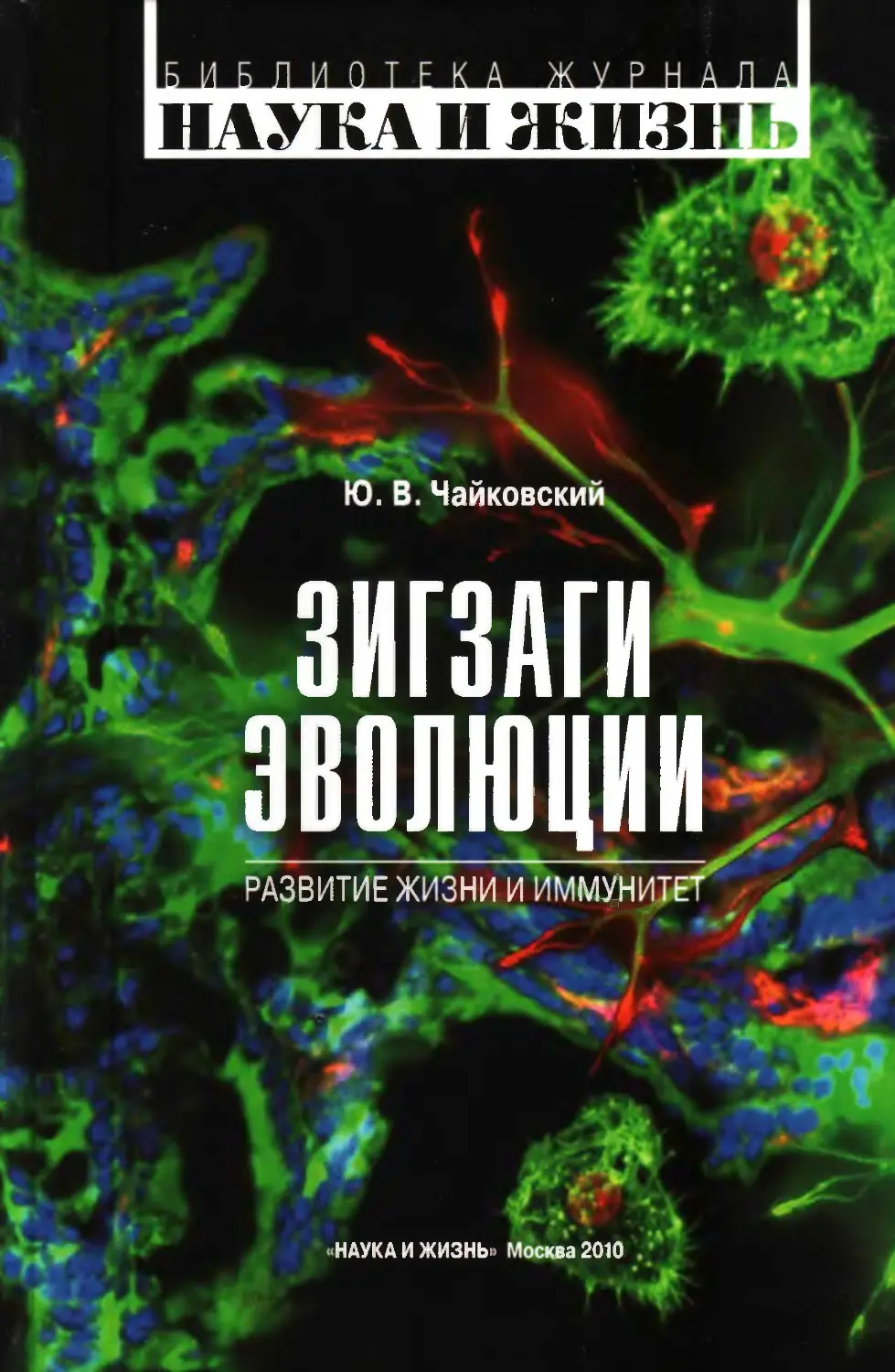
1. Первичный номогенез............................7
2. Ламаркизм.....................................13
3. Жоффруизм......................................9
4. Дарвинизм.....................................10
5. Метод толкований..............................13
6. Классический номогенез........................16
7. Новые достижения прежних учений. Адаптивный
компромисс ....................................19
8. Как быть, если отбора «по Дарвину» в природе нет?.22
Часть 2. Что же движет эволюцию?
9. Принцип активности и новый ламаркизм. Генетический
поиск и акт эволюции...........................26
10. Как появляются новые гены. Генетика XXI века.29
11. Активность, мышление и эволюция — они возможны
вместе.........................................32
12. Вопрос Ламарка—Дарвина. Экосистемная концепция
эволюции.......................................32
13. О случайности................................35
14. Новый номогенез и основа эволюции............39
15. Отбор (selection) и подбор (delectus).
Новый дарвинизм................................42
16. Делектус, математика и самоорганизация. Intelligent
Design.........................................44
17. Номогенез снежинок и организмов. Новый
жоффруизм......................................48
18. Активность и цель............................53
19. Нанотехнология эволюции......................54
Литература к разделу I.........................56
РАЗДЕЛ II. ЮБИЛЕЙ ЛАМАРКА - ДАРВИНА
И РЕВОЛЮЦИЯ В ИММУНОЛОГИИ
Часть 1. Иммунитет как борьба за существование
1. Эволюция сквозь призму иммунологии.............58
2. Мечников и борьба клеток......................59
3. Эрлих: активность и селекция антител..........60
4. Ландштейнер и искусственные антигены...........61
5. Попытка соединить теории.......................62
6. Долой арифметику! ............................64
7. Возвращение к арифметике......................67
8. Стил и ламаркизм в иммунологии................69
Часть 2. Иммунитет как активность организма
9. Поппер и «демон Ламарка»......................71
10. Каннингэм, Тонегаваи «Дарвинов микрокосм»....73
11. Вентребер и рождение молекулярного ламаркизма.76
12. Эдельман: морфогенез и иммунитет.............78
13. Джаневей и революция в иммунологии...........80
14. Революция в эволюции.........................82
15. Ламарк и прогресс............................83
16. Разум как активность иммунной системы........85
Часть 3. Иммунитет как упорядоченность
17. Вездесущий иммунитет и несовместимость тканей.86
18. Самонесовместимость и совместимость чуть подроб-
нее .............................................88
19. Иммунный механизм онтогенеза.................88
20. Иммунитет — это эволюционный ограничитель....92
21. 0 случайности в работе иммунной системы......93
22. Иммунитет и мозг.............................94
23. Презентация антигена. Отбор уступает «демону
Ламарка» ........................................95
Часть 4. Иммунитет как экосистема. Что возьмем
у классиков?
24. Иммунные рефрены...........................98
25. Три пары главных рефренов..................99
26. Экология и номогенез иммунного мира.......101
27. Не будем забывать классиков...............104
Заключение.................................. 105
Литература к разделу II.....................108
Словарик....................................109
Сокращения..................................110
ВВЕДЕНИЕ.
Была ли эволюция? Незаданный вопрос и каверзный ответ
Жан-Батист Ламарк впервые выступил с эволюционной
идеей на лекции в мае 1800 года в Париже. Через 200 лет, в
марте 2000 года, там же вышла книга [Nottale, 2000], авторы
которой (космолог, биолог и экономист) возродили тради-
цию сквозного описания эволюции - от космоса до общества.
Каждый написал свой раздел, а затем все трое описали еди-
ную закономерность эволюции: по их мнению, мир является
многомерным фракталом (что это такое, мы узнаем далее), а
эволюция мира - единым многофрактальным ростом.
Одни согласны с таким построением, другие протестуют
(«упрощение!»), третьи просто не понимают, зачем биологам
нужны фракталы, но нам важнее иное — то, что в науку воз-
вращается идея сквозного эволюционизма, то есть понима-
ние ее как процесса, единого для всей природы. До воцарения
дарвинизма такое понимание эволюции было основным, и,
на мой взгляд, именно его забвение побудило биологический
эволюционизм погрязнуть в деталях, упуская его суть. Исто-
рию забывать не следует.
Автор биологического раздела Жан Шалин (Chaline) почти
не упоминает ученых прошлого, нам же придется поступать
иначе - выводить каждую нынешнюю тему из работ прошло-
го и показывать сложное к ней отношение в разные времена.
А зачем?
У Владимира Высоцкого есть фраза:
А мы все ставим каверзный ответ
И не находим нужного вопроса.
В этом суть — чтобы задать нужный вопрос, надо уметь ви-
деть негодность каверзного ответа и прямо признать, что от-
вета нет. Такими каверзными ответами, в частности, полны
нынешние разговоры о биологической эволюции. Прежде
всего, это ходячие противоположные утверждения: «эволю-
ция — достоверный факт» и «эволюции нет и быть не может».
У каждого утверждения много защитников (тех, кто отрицает
эволюцию и уверен, что мир создан Богом в том виде, какой
мы наблюдаем ныне, именуют креационистами), и уже одно
это наводит на мысль, что тут не задан какой-то главный во-
прос, а ответы уже даны.
Для меня этот главный вопрос звучит так: « Была ли эволю-
ция?» Его никто, насколько знаю, не обсуждает. Пришел он
мне в голову отнюдь не сразу, а после внимательного чтения
доводов немногих умных креационистов. Оказалось, что «до-
казательства эволюции», которыми полны учебники, ничего
не доказывают (если уметь сомневаться), что они лишь пока-
зывают, как разнообразие организмов выглядит и выглядело
в различные эпохи. Доказательства надо искать.
В самом деле, наиболее очевидным доказательством эво-
люции считаются данные палеонтологии, однако сами по себе
они говорят лишь, кто за кем жил, поэтому для креациониста
5
они всего лишь — «следы Творения». А какие данные могли
бы убедить его (если, конечно, он способен вникать в чужой
строй мысли, то есть умный), что ископаемые свидетельству-
ют об эволюции, о естественном процессе?
История науки дает вполне определенный ответ: в про-
шлом ученые (в том числе верующие) переставали призывать
имя божие для объяснения своих данных тогда, когда могли
формулировать общее правило, действующее в данных усло-
виях всегда. Для появления новых форм организмов (как
ныне живущих, так и вымерших) таких правил не найдено,
оттого креационизм и продолжает существовать.
Ответ на наш главный вопрос будет дан положительный,
хотя на сегодня и в узких рамках (см. п. 16, раздел I и п. 1, раз-
дел II). Почему он для одних положителен вне всяких рамок,
а для других - отрицателен, притом тоже вне всяких рамок?
Нетрудно понять - именно из-за неумения ставить рамки от-
ветам, т. е. из-за того, что их ответы каверзны.
Один из самых известных каверзных ответов - ссылка на
естественный отбор. Ее используют для «объяснения» всего,
что не имеет объяснения, т.е. того, что биология не умеет опи-
сать конкретно. Это относится не только к эволюции (той, какую
никто не наблюдал), но и к постоянно наблюдаемым явлениям,
если механизм их неизвестен. Например, совершенно непонят-
но, каким образом аксон (ответвление нервной клетки) прорас-
тает через всю длину организма к месту своего прикрепления
(к нужному синапсу нужной нервной клетки). Недавно пред-
ложено (и принято на ура) такое «объяснение»: те, что растут
иначе, гибнут в силу «дарвиновского отбора» (подробнее см.
п. 15, раздел I).
Однако, как только механизм явления оказывается понят,
такие ссылки на отбор исчезают, и это явственно говорит о том,
что они ненаучны. Вместо них надо уметь честно сказать: «Не
знаем» - только тогда может открыться путь к знанию. И это
начинают понимать: уже появляются книги вообще без упоми-
наний отбора - например [Голубевы, 2009].
Таких загадок, как механизм роста аксона, в биологии мно-
го и одна, иммунная, будет рассмотрена в разделе II - это вы-
работка антитела, парализующего данный антиген (ее тоже
приписывают отбору именно в тех пунктах, где механизм не-
известен). Все вместе эти загадки ясно говорят, что мир дале-
ко не столь понятен, как пишут в учебниках.
Словом, само состояние науки требует всюду указывать на
каверзные ответы. Если так не делать, многие удивятся на-
писанному далее: зачем эти сложности, коли все можно объ-
яснить много проще? Да, можно: дарвинизм говорит «все от
отбора», а креационизм — «все от Бога». Такие объяснения ни
к какому знанию не ведут, но ум большинства людей устроен
так, что какое-то объяснение нужно, пусть и откровенно неле-
пое (подробнее см. статью [Чайковский, 2010]). И отказаться
от него почти никто не может, пока не узнает, что есть нечто
иное. Об этом «нечто» и пойдет речь в книге.
6
РАЗДЕЛ I. КАК ПОНИМАЮТ ЭВОЛЮЦИЮ ЖИВОГО
Часть 1. Прежние теории эволюции
Эволюционная идея принадлежит к основам европейского
видения мира, она пронизывает европейскую науку с первых
ее шагов, однако мы сможем далее коснуться лишь тех не-
скольких исторических фактов, которые необходимы для по-
нимания нынешних воззрений на эволюцию.
1. Первичный номогенез
Ранние намеки на идею исторического развития организ-
мов носили характер параллелей с онтогенезом (см. слова-
рик): лучшим примером в течение многих веков служило пре-
вращение личинки в куколку и затем во взрослое насекомое.
Ранние эволюционные фантазии полны рассказов о рыбах,
обратившихся в зверей и людей, и т.п. Все подобные схемы
исходили из той мысли, что существует некий заданный Бо-
гом механизм таких превращений, которые тем самым проис-
ходят не как попало, а согласно раз навсегда установленным
законам. Впоследствии, уже в XX веке, идея закономерно те-
кущей эволюции получила название номогенез. В ней, однако,
ссылки на Бога нет.
Первым автором, важным для темы номогенеза, был
французский натурфилософ и физик Пьер-Луи де Мопертюи.
В 1744 году он поставил вместе три вопроса: почему дитя по-
хоже на обоих родителей? как
происходит развитие зароды-
ша? откуда берутся уродства?
Исходной для него была та
античная мысль, что при
оплодотворении происходит
смешение мужской й жен-
ской «семенных жидкостей».
К ней Мопертюи добавил, что
плавающие в этих жидкостях
частицы соединяются в за-
родыш точно так же, как в
ходе химической реакции со-
бираются в единое вещество
элементы - по законам, ана-
логичным закону химическо-
го сродства. Свойства «ча-
стиц», аналогичные этому
сродству, он охарактеризовал
как «нечто аналогичное же Пьер-Луи Моро де Мопертюи
ланию, неприязни, памяти».
7
Эта параллель явила собой намек на принцип активности, о
котором речь будет далее.
Мопертюи предлагал искать «первичные законы», кото-
рые трудно уловить лишь потому, что они действуют совмест-
но (и тем создают наблюдаемое разнообразие явлений). Так
что задача ученого - выявлять их и исследовать поодиночке.
Одну из таких процедур он видел в выявлении «частиц» и по-
лагал, что «частицы» организма одного вида могут служить
для сборки организма другого вида, в качестве примера он
приводил кишечных паразитов, появление которых толковал
(говоря нашим языком) как самосборку частиц организма-хо-
зяина, т.е. самосборку прежних частиц в новый, более про-
стой организм.
Объяснение для нас наивно, но это - первый намек на то,
что ныне именуют блочной эволюцией. Оно навело физика на
один из самых глубоких вопросов биологии: откуда берется
сходство? Например, почему так похожи земляные черви,
личинки насекомых и глисты? Мопертюи склонен был видеть
здесь сходство законов формообразования - так по законам
химии и физики растет кристалл (добавлю: при одних усло-
виях углерод кристаллизуется в графит, при других - в ал-
маз, но не в кристалл вроде соли). Сходство итогов развития
он объяснял наличием общего закона. Самым ясным из таких
законов он полагал открытый им в механике принцип наи-
меньшего действия - простейший вариационный принцип.
Подробнее об этом см. книгу [Чайковский, 1990].
2. Ламаркизм
Жан-Батист Ламарк
(1744—1829).
Ламаркизмом в наше вре-
мя почти все именуют на-
следование приобретенных
свойств, но это - просто итог
утраты культуры. Ни сам
Ламарк, ни его последовате-
ли, читавшие его труды, не
видели в наследовании при-
обретенных свойств основы
учения. Наследование приоб-
ретенных свойств признава-
ли все, в том числе и Чарльз
Дарвин, вплоть до появления
в 1876 году работ германско-
го биолога Августа Вейсмана,
выдвинувшего принцип нена-
следования приобретенных
признаков.
Может быть, нужно сми-
риться с такой ошибкой по-
8
вседневных речей? (Говорим же мы «Солнце встало», хотя
знаем, что на самом деле это Земля повернулась.) Нет, такая
ошибка мешает думать. Дело в том, что третирующая ламар-
кистов доктрина дарвинизма всегда «предъявляла к ламар-
кизму гораздо более высокие требования, чем к себе самой»
[Аронова, 1997]. Так, принято отрицать всякий факт насле-
дования приобретенных свойств, если он нерегулярен, тогда
как в основу самого дарвинизма положена идея мутаций - не-
регулярных, да еще и редких.
Лучше вспомнить, что утверждал великий француз на
самом деле и что взяли у него последователи, а не пресле-
дователи. Главное у него — принцип активности особи как
фактор эволюции вида. Активность проявляется, по Ламар-
ку, тремя способами. Во-первых, активное использование
органа или функции вызывает их усиление, а неиспользова-
ние — ослабление (по-английски это именуют: use — disuse).
Во-вторых, результат данной процедуры наследуется (вот
единственное, что усвоено у Ламарка ныне). И, в-третьих,
организмы обладают стремлением к усложнению строения
и функций (к совершенствованию, к прогрессу — кто как вы-
ражается).
Американский биограф Ламарка писал: «Идея наследова-
ния приобретенных признаков стала известной как ламарков-
ский механизм благодаря чему-то вроде исторической шут-
ки». Он отмечал, что главный у Ламарка творческий фактор
эволюции — «внутренняя активность организма» — просто не
был понят учеными, что он никогда «не был опровергнут экс-
периментально» [Burkhardt, 1977].
3. Жоффруизм
Этьен Жоффруа Сент-Илер, ученик Ламарка, превратил в
теорию то, что до него было набором голых и несвязных идей.
Только его взгляд упомянут в книге, избегающей истории
[Nottale, 2000, с. 176]. Наш историк науки Б.Е. Райков писал:
«Природа, по Сент-Илеру, как бы штампует животных по одно-
му идеальному образцу, а разнообразие в их внешнем устрой-
стве зависит от разнообразных влияний со стороны внешних
условий» [Райков, 1969, с. 226]. Отличие от Ламарка видно у
Жоффруа1, во-первых, в признании «идеального образца» (его
называли еще и архетипом), а во-вторых — в том, что внешние
влияния действуют не на взрослый организм, а на растущий,
или даже на его зародыш. Тем самым эволюция выступает как
следствие преобразования зародышей под прямым действием
среды обитания.
1 Жоффруа — не второе имя, а первая фамилия ученого. Отсюда и термин «жоф-
фруизм» , предложенный зоологом и переводчиком Н.А. Холодковским в 1915
году.
9
Главную эволюционную
работу Жоффруа издал в
1833 году. В ней есть фраза:
«Если эти изменения приво-
дят к вредным последствиям,
то животные, у которых они
возникают, гибнут и заменя-
ются другими животными со
слегка измененными форма-
ми, притом измененными та-
ким образом, что они соответ-
ствуют новым условиям».
Здесь требуется пояснение:
о гибели неприспособленных
писали многие, начиная с
древности, но у Жоффруа мы
Этьен Жоффруа Сент-Илер впервые видим утверждение,
(1772—1844). что «слегка измененные» вы-
тесняют прежних. Никаких
примеров в подтверждение столь важного тезиса у него нет.
Однако, читая именно эту статью в феврале 1838 года, моло-
дой Дарвин сформулировал в записной книжке свой «закон
малых различий, производящих более плодовитое потомство»
(Notebook В, р. 236 - см. [Дарвин, 1959, с. 123]), - будущую
основу дарвинизма.
4. Дарвинизм
Вторым источником дарвинизма стал труд Томаса Мальту-
са «Очерк о населении», попавший в руки Дарвину в сентябре
того же 1838 года. Отсюда он почерпнул тот тезис, что боль-
шинство особей гибнет из-за нехватки дефицитного ресурса
(чаще всего — пищи).
Чтобы увязать эти два тезиса, надо увериться, что за счет
гибели одних выживают именно те самые другие, которые
слегка изменились. Из природы таких сведений извлечь не
удалось (ни до Дарвина, ни после), и молодой Дарвин взял их
из трудов селекционеров: лучшим вариантам, заявил он, да-
ется возможность оставить потомство.
Аналогия получилась слабая — ведь селекционер своей
волей выбирает нужных самца и самку, принудительно спа-
ривает их и полностью исключает возможность нежелаемого
спаривания как их самих, так и их потомства. Природа ниче-
го такого не умеет, поэтому Дарвину пришлось ввести в нее
воображаемого селекционера. Вот как он появился.
В рукописном очерке (1844 год, опубликован в XX веке)
Дарвин прямо писал: «Предположим теперь, что некое Суще-
ство, одаренное проницательностью, достаточной, чтобы пости-
гать совершенно недоступные для человека различия в наруж-
10
ной и внутренней организа-
ции, и предвидением, прости-
рающимся на будущие века,
сохраняло бы с безошибочной
заботливостью и отбирало бы
для какой-нибудь цели потом-
ство организма... я не вижу ни-
какой причины, почему бы оно
не могло создать новую расу»
[Дарвин, 1939, с. 133].
Рассуждение похоже по
форме на дарвинизм, но на
самом деле это — естествен-
ное богословие (эту дисци-
плину юный Дарвин изучал
на богословском факультете
- см. далее, п. 5). Божество,
как и селекционер, может от-
бирать желаемое свойство не-
зависимо от величины плодо-
витости его носителей.
В другом месте того же
очерка (с. 86) Дарвин заме-
тил: «Природа не позволяет
Чарльз Роберт Дарвин (1809—1882).
Зарисовка 1853 года - в то время он
писал книгу «Усоногиераки», до сих
пор настольную у специалистов.
На портрете мы видим серьезного
ученого, не отягченного той знамени-
тостью, что видна на более поздних
портретах.
своей расе портиться от скрещивания с другой расой». Но это
явное заблуждение: природа (если она не Бог) позволяет спа-
риваться всем, кто может. Для пресечения вредных скрещи-
ваний как раз и нужно то самое «Существо». Оно ко дню напи-
сания «Происхождения видов» из записей Дарвина исчезло,
его сменила избирательная размножаемость, хотя никто ни
до Дарвина, ни после не показал на реальных примерах, что
новое размножается лучше старого. Тем меньше оснований
верить, что лучше размножаются «слегка измененные».
Однако Дарвин верил. В «Происхождении видов» есть па-
раграф «Примеры действия естественного отбора», и в нем
читаем: «Растения, производящие цветы с самыми большими
нектарниками, выделяющими наибольшее количество некта-
ра, будут чаще посещаться насекомыми и чаще подвергаться
скрещиванию и, в конце концов, осилят своих соперников и
образуют местную разновидность».
Это оказалось не так: опыт показал, что если ничто на цвет-
ке не мешает, то «колибри и бабочки застревают надолго, вы-
пивая весь нектар, и не очень стараются сменить кормушку»
и переносить пыльцу [Никотин и колибри // Наука и жизнь,
2009, № 5, с. 32].
Еще один пример говорил о воображаемых волках, бегу-
щих за воображаемыми оленями. Остальная часть параграфа
убеждала читателя, что эти примеры (чья слабость видна само-
11
му автору) всё же можно принять во внимание. Конечно, мож-
но, но не более того: надо еще выяснить, в самом ли деле гибель
одних влечет перемену свойств у других.
Содержание параграфа огорчило даже друзей Дарвина. Ге-
олог Чарльз Лайель, прочтя книгу в корректуре, писал ему,
что в новом издании следует «кое-где опереться на реальные
случаи вместо абстрактных предположений»; Дарвин обещал
описать требуемые случаи, но так и не сделал этого, хотя писал
еще 22 года. Хуже, что этого так и не сделал никто за 150 лет.
Так же, как принцип отбора, в дарвинизме важен прин-
цип филогении, согласно которому новые виды происходят
исключительно путем разделения прежнего вида. На самом
деле новый вид может происходить и путем слияния видов,
например горизонтального переноса генов (см. словарик),
чего Дарвин знать не мог.
Нам интересен тот особый вариант дарвинизма, который
у западных историков науки именуется «Дарвин без Маль-
туса» и связан с российской наукой и общественной мыслью
[Tories, 1989]. Он отрицал те формы борьбы за существование,
которые возникают при перенаселенности (она делает какие-
то ресурсы дефицитными). И в самом деле, все биологические
факты, приведенные позже, в XX веке, в качестве примеров
естественного отбора, касались ситуаций, когда успех вида не
зависел от плотности его населения.
Тему «Дарвин без Мальтуса» породил ботаник и педагог
С.А. Рачинский, первый русский переводчик трудов Дарви-
на. Он дал такое понимание дарвинизма: «Из многочисленно-
го потомства всякого поколения
Сергей Александрович Рачинский
(1833—1902), первый русский пере-
водчик Дарвина, понимал подбор
как самоорганизацию живого.
органических существ выжи-
вает лишь малейшая доля, вы-
живают лишь те представители,
которые вполне и до последней
подробности приспособлены ко
всему строю внешней природы»;
она поступает «как искусный
заводчик, дорожащий чистотою
крови в своем стаде. Она строго
подбирает в каждом поколении
те особи, которые она предна-
значает для продолжения пле-
мени» [Рачинский, 1863].
Мы уже знаем, что это - не
дарвинизм в его обычном по-
нимании, а скорее естественное
богословие, но такое толкова-
ние Дарвина вскоре получило
широкое распространение, за-
тем было забыто, а в наши дни
12
становится главным довольно близкое понимание отбора.
Суть его в том, что под естественным отбором теперь всё чаще
понимают не преимущественную размножаемость, а соответ-
ствие изучаемого объекта той системе, частью которой он яв-
ляется (см. далее, пп. 15 и 16).
У Дарвина был младший друг, Томас Гексли. Работая на-
учным обозревателем газеты «Таймс», он более всех помог
продвижению нового учения в массы, и именно он предложил
термин «дарвинизм» - через два месяца после выхода «Про-
исхождения видов». У него есть изречение: «Дарвин или про-
рок Моисей — третьего не дано», и пафос его молчаливо на-
правлен против Ламарка.
Предлагаемая книга отвергает как раз это «третьего не
дано». Кстати, иногда меня упрекают в неразличении старого
дарвинизма и нового (который якобы преодолел недочеты ста-
рого). Но, к сожалению, пафос «третьего не дано» по-прежне-
му царит (см. п. 5). В частности, креационистам кажется, что
падение дарвинизма будет означать падение идеи эволюции
вообще. Мы увидим, что это не так.
5. Метод толкований
Толкование — неотъемлемый атрибут богословия: посколь-
ку в исходных догматах сомневаться нельзя, то всякому про-
тиворечию с ними или между ними богословы вынуждены
придумывать приемлемую для их религии трактовку. За это
им дают ученые степени.
Данный прием проник и в науку — в те времена, когда она
была тесно связана с религией. Таковым было, например, при-
думывание полезности наблюдаемым свойствам организмов,
разработанное в XVI—XVIII веках в рамках естественного
богословия. (Именно оно дало Дарвину, как в XX веке отмети-
ли западные историки науки, основной рабочий прием — при-
писывать каждому свойству организма полезность.) Ограни-
чусь одним примером из учебника «Natural Theology» (1802)
Уильяма Пэйли (Paley). Это вероятностный аргумент: если
пользы свойства не видно, можно предложить непроверяемую
гипотезу и назвать ее вероятной. Например: «А в рыбьем глазу
радужка неспособна к сужению. Это большое отличие, вероят-
ный смысл которого в том, что ослабленный водой свет никогда
не бывает слишком резким на сетчатке», - писал Пэйли.
Так, замечу, можно «доказать» всё что угодно, и Пэйли
невольно внес лепту в будущий дарвинизм. По его учебнику
учился на богословском факультете Дарвин, и только этот
учебник он вспомнил в автобиографии с благодарностью.
В науке если и используют толкования, то неявно, и лишь
в дарвинизме они стали основным методом. Первым, кто пря-
мо заявил, что оборачивает в пользу дарвинизма богословские
1 ч
Сэмюэл Хаббард Скеддер (1837—
1911), толкованиям противопоста-
вил арифметику.
аргументы, был упомянутый
Гексли. Другие делали и де-
лают это непрямо.
Так, в 1867 году Альфред
Уоллес, сподвижник Дарви-
на, писал, что видел белых
бабочек на зеленом фоне и
склонен предположить, что те
предупреждают контрастной
расцветкой о своей несъедоб-
ности. Дарвин был в восторге:
«Никогда не слыхал ничего
более остроумного... Факт бе-
лой моли великолепен; кровь
загорается, когда видишь, что
истинность теории почти до-
казана», - писал он Уоллесу.
Ободренный Уоллес положил
метод в основу учения — в его
книге «Дарвинизм» 15 глав,
из них 4 посвящены толкова-
нию окрасок.
На самом деле «почти до-
казана» была отнюдь не «ис-
тинность теории», а правдоподобность частного толкования
частного примера. То есть предложен был еще один довод в
пользу своей правоты. Такой довод бесполезен в науке: он убе-
дителен только для тех, кто и без него думает «как надо», а для
думающих иначе выглядит нелепостью. И вскоре довод Уолле-
са об упреждающей окраске был отвергнут профессиональным
наблюдателем.
В декабре 1870 года в журнале «Nature» энтомолог Сэмюэл
Скеддер (Scudder) недоумевал: гусеница гротескной формы и
угрожающей окраски буквально кишит пожирающими ее па-
разитами, тогда как гусеницы близкого вида для них несъе-
добны. Почему оказался эффективным отбор столь сложной
формы и расцветки, если судьбу решают не птицы, а парази-
ты, съедающие 99,9% потомства на стадии яиц, гусениц и ку-
колок?
Этот вопрос в свое время заставил задуматься многих; но
для тех, кто верит в отбор, он просто неинтересен (если так
произошло, значит, отбор действовал - вот и толкование).
При размышлении над ним и подобными было найдено реше-
ние, изложенное далее. О Скеддере же мне пришлось недавно
вспомнить, читая книгу «Введение в палеоэнтомологию». Там
помещен большой портрет Скеддера - одного из основателей
данной науки, но на его еретические взгляды нет и намека.
Зато есть глава «Теоретические основы эволюционной биоло-
14
гии», по которой удобно выяснить, где в настоящее время про-
легает граница между наукой и толкованием.
Автор главы, наш ведущий палеонтолог А.П. Расницын,
привел две классификации теорий эволюции — мою (номо-
генез, ламаркизм, жоффруизм и дарвинизм - остальных он
не назвал) и свою, согласно которой есть всего два взгляда
на эволюцию. Первый - тот, что естественный: отбор дви-
жет эволюцию, в том числе прогрессивную; в него верит он
сам. Второй — «внеестественный (альтернативный отбору)».
Расницын пишет: «Поскольку привлечение внеестественных
факторов эволюции, на мой взгляд, законно лишь в рамках
веры, а с наукой несовместимо, выбор селектогенеза (теории
отбора. — Ю.Ч.) для меня неизбежен». Тем самым делавшие
иной выбор оказываются для него вне науки.
Такое юношеское самоутверждение конечно же радует (нам
обоим по семьдесят), но для дела важнее другое — тут есть осо-
бый взгляд на саму эволюцию. А именно, все взгляды, кроме
одного, аттестуются как несовместимые с наукой. Это черес-
чур смело - как быть, например, с номогенезом? Он ведь дает
верные прогнозы, самую возможность которых селектогенез
отрицает. Затем Расницын пишет и о толкованиях: «Именем
основателя логично называть учения, суть которых состоит
в толковании канонических текстов (христианство, буддизм,
марксизм), но не системы научных взглядов». Верно, и за-
мечу, что именно поэтому критики именуют теорию отбора
дарвинизмом. Это, кстати, признал и сам автор главы, когда
оценил мои взгляды как «яростный антидарвинизм»2 [Расни-
цын, 2008, с. 6]. Без имени основателя он, как видим, обой-
тись не смог, так что и мы будем использовать имена осно-
вателей. Это прежде всего Жоффруа, которого теперь, после
столетнего забвения, всё чаще вспоминают.
Так, например, Расницын видит в эволюции (как и Жоффруа)
преобразование онтогенезов, но толкует его как итог отбора. Ни-
каких данных в пользу такого толкования ни у кого нет, поэтому
оно не хуже и не лучше, чем у Жоффруа, толковавшего измене-
ние зародышей как результат прямого воздействия среды.
Теперь можно сформулировать сам метод толкований как
особый взгляд на эволюцию. Вот моя формулировка, немного
заостренная. Сперва вводится постулат', механизм эволюции
известен и не подлежит пересмотру, так что предметом изуче-
ния могут быть только его подтверждение и детализация. За-
тем - рабочий прием', все опровержения данного механизма
следует игнорировать, пока хотя бы одному из них не будет
2
Эта оценка вряд ли верна, что станет читателю видно далее. Моя цель — отправля-
ясь от дарвинизма, рассказать о достижениях более новых. Для сравнения отмечу
лишь, что никому не придет в голову сказать, будто теория Эйнштейна - «анти-
ньютонизм» или нынешняя космология — «антиэйнштейнизм».
15
найдено толкование, делающее возражение сомнительным в
рамках этого самого механизма. После чего объявляется вывод:
данное опровержение считается снятым. Это служит основани-
ем полагать снятыми все как-то связанные с ним (например,
принадлежащие тому же автору) прочие опровержения того
же механизма. Их опять следует игнорировать, а для внешнего
пользования (начальству, прессе, учащимся) предлагается за-
явление: опровержений механизма вообще нет.
Метод используется многими учениями. Еще Ламарк тол-
ковал длинную шею жирафа как результат активности при
обрывании листьев с высоких ветвей, игнорируя все противо-
речия (например, тот факт, что у других видов, тем же питаю-
щихся, шеи имеют обычный размер).
6. Классический номогенез
Термин «номогенез» предложил зоолог и географ Л.С. Берг
в книге «Номогенез» (1922). Он имел в виду создание учения
о биологической эволюции как о процессе, протекающем по
определенным законам, которые полагал не сводимыми к
воздействиям среды обитания. Этим он сам противопоставил
свое учение дарвинизму (а также ламаркизму и жоффруиз-
му). В зарубежной литературе термин не прижился.
Книга Берга довольно сумбурна, так что основные ее по-
ложения требуют разъяснения.
Первый закон учения Берга — «Изначальная целесо-
образность живого». Берг натерпелся за него изрядно, по-
скольку критики-коммунисты увидали в ней идеализм. На-
прасно Берг приводил чисто материалистические доводы
(например: Дарвин постулировал избыточное размноже-
ние и неопределенную изменчивость как первично целесо-
образные свойства щивого),
его не слушали и не слуша-
ют до сих пор.
Второй закон сам Берг вы-
разил так: «Эволюция в зна-
чительной степени есть раз-
вертывание уже существую-
щих задатков».В частности,
она подобна онтогенезу.
Третий закон Берга гласит:
«Наследственных вариаций
ограниченное число, и они
идут по определенным направ-
лениям». Так в дальнейшем и
Лев Семенович Берг (1876—1950).
Фото1920 года — в это время он
писал книгу «Номогенез».
16
оказалось (см. далее), но признали
это не всё.
Четвертый закон Берга — «Эво-
люция идет скачкообразно».
У Берга нет ни слова о концеп-
ции Жоффруа, хотя его второй за-
кон прямо к ней относится. Зато
ее использовал Д.Н. Соболев, ав-
тор второго варианта номогенеза
(1924).
Эволюция для Соболева — еди-
ный процесс «органического ро-
ста», идущий как при развитии
особи (онтогенезе), так и при исто-
рическом развитии (филогенезе).
«Органический рост» протекает в
череде поколений, так что каждое
поколение, вырастая, повторяет
онтогенез своих родителей не в
точности, а с ничтожными изме-
нениями.
История флор и фаун является
для него чередованием периодов
скачков и периодов постоянства,
поэтому в ископаемой летописи
почти нет переходных форм. О том
же самом писал еще в 1859 году
палеонтолог Освальд Геер (Heer)
в своей книге «Третичная флора
Швейцарии». Он обосновал тот
факт, ныне всем известный, что
нынешняя флора Европы произо-
шла из флоры третичного периода
быстро, после долгого постоян-
ства, и с тех пор снова как бы за-
стыла, мало меняясь до сих пор.
Хотя Геера и Соболева поддержи-
вали многие палеонтологи, боль-
шинство их не слушало. В 1972
году сходную мысль высказали
палеонтологи Стивен Гулд (Gou-
ld) с соавторами (США) и имели
большой успех.
По Соболеву, «земное населе-
ние» (теперь говорят: биосфера)
- единый организм, онтогенез ко-
торого основан на актах эволюции
таксонов. Ныне этими темами за-
Дмитрий Николаевич Соболев
(1872—1949), автор концепции
комбинаторного номогенеза и
первого наброска экосистемной
концепции эволюции.
Освальд Геер (1809—1883),
швейцарский естествоиспы-
татель, первым показал
быструю эволюцию земной
флоры.
17
нимается экосистемная концепция эволюции (см. далее, п. 12,
а также [Vestbroek, 1997]).
Часто мы видим, что сам факт эволюции вне сомнения,
но все конкретные пути сомнительны (и это служит козырем
креационизма). Почему так? Соболев отвечал: потому, что
организмы не несут в себе достаточной информации о своих
предках. Мысль не нова (ее высказал еще Мопертюи), но по-
нималась с трудом и немногими. Соболев искал способ упо-
рядочить ископаемые без детальных родословных. Лишь че-
рез полвека его нашел зоолог В.Ф. Мужчинкин как принцип
блочной эволюции. Простой его иллюстрацией является пер-
вая амфибия ихтиостега, которую шутливо зовут «рыба на
ногах»: конечности ее уже выглядят как наземные, а тулови-
ще еще имеет рыбий облик (рис. 1).
Рис. 1. Ихтиостега - первая настоящая амфибия.
Если прежние учения исходили из какого-то абстрактного
принципа, то номогенез - из анализа свойств реального раз-
нообразия организмов различных эпох. Это позволило обна-
ружить ряды изменчивости и на их основе дать несколько
примеров эволюционного прогноза. Не в смысле предсказа-
ния будущей эволюции (это фантазия), а в смысле указания
на эволюционную параллель, которая позже была обнаруже-
на фактически.
Первый прогноз предложил в старости еще Дарвин. Хотя
дарвинизм входил в моду, сам основатель разуверился в нем
и в 1872 году, в последний раз выпустив «Происхождение
видов», вернулся к своим любимым ботаническим работам.
Здесь его ждало открытие, значение которого выяснилось
только в XX веке, после появления номогенеза.
В книге «Усоногие раки» (1854) Дарвин описал странное
явление - дополнительных самцов у гермафродитных видов.
Описав, он добавил: «Ничего в точности похожего в животном
царстве неизвестно, но среди растений, в линнеевском клас-
се Polygamia3, есть много весьма сходных случаев» [Дарвин,
1936, с. 86]. Случаи оказались сомнительными, и теперь, в
книге «Различные формы цветов» (1877), он высказался осто-
К. Линней (Философия ботаники, п. 149) назвал четыре рода, в том числе Dios-
pyros (хурма) - прим. Ю. Ч.
18
рожнее: «Если существуют растения, особи которых являют-
ся гермафродитными и мужскими, их можно бы выделить
как андродиэцичные». Позже они были найдены, в том числе
хорошо известная полярникам куропаточья трава (Dryas oct-
opetala) [Дарвин, 1948, с. 16, 49].
Замечательно, что Дарвин не просто дал прогноз, но и как
бы поместил его в систему комбинаций блоков (типов размно-
жения). А именно, он противопоставил открытой им андроди-
эции более известную ботаникам гинодиэцию — когда у вида
представлены гермафродитные и женские особи. Некоторые
ботаники видели в этом приспособление (см. статью: [Карасе-
ва, 2009]), но опять вышло лишь толкование. По-моему, здесь
налицо ясный номогенез в смысле Соболева. Подробнее см.
книгу [Чайковский, 1990, с. 179].
7. Новые достижения прежних учений.
Адаптивный компромисс
Первую после появления дарвинизма попытку выйти за
рамки толкований в область теории (как ее понимают другие
науки) предпринял в старости сам Дарвин, и это, как мы только
что видели, оказался не дарвинизм, а первичный номогенез.
В XX веке в рамках ламаркизма удалось показать еще и
роль активности, и отличие прогресса от приспособления. В
рамках жоффруизма успех был еще большим: возникла новая
дисциплина, именуемая «evo-devo». Она возродила к жизни
старинную параллель между эволюцией и онтогенезом, од-
нако теперь данная параллель проводится на молекулярном
уровне. Обо всем этом речь пойдет далее.
Теперь о дарвинизме. Весь XX век он господствовал, и ты-
сячи писавших о нем, чтобы не повторяться дословно и, не
имея фактов, видеть себя исследователями, меняли формули-
ровки. Одна из них была смелой— «новый синтез» (1942). Как
позже показал анализ (см. обзор [Гилберт и др., 1997] - это
был первый обзор «evo-devo»), как раз синтеза там и не было.
Наоборот, из «синтетической теории эволюции» начисто ис-
чезли не только свойственные Дарвину ссылки на морфоло-
гию (которую развивал и развивает номогенез), физиологию
(ламаркизм) и эмбриологию (жоффруизм), но даже на эколо-
гию (борьба за существование). Ее заменило утверждение, что
каждому варианту организма можно приписать некий «коэф-
фициент отбора». Единственной дисциплиной, допущенной
в этот «синтез», была популяционная генетика - набор рас-
суждений о разнообразии генов, основанный на некоторых
фактах генетики 1930-х годов.
Но время шло, биология развивалась, дарвинизм отставал
(он быстро признал ДНК как синоним слова «ген», но РНК
ждала 30 лет), и иногда ему приходилось реагировать на не-
приятные открытия. Один пример надо привести, ибо он ясно
19
Георгий Христофорович Шапошни-
ков (1915—1997), первым получил
видообразование в эксперименте.
указывает на связь эволюци-
онных учений: ленинград-
ский энтомолог Г.Х. Шапош-
ников поставил в 1957 году
на своей родине (на Север-
ном Кавказе, близ Майкопа),
опыт, который справедливо
называют классическим. Вот
его суть, опуская всё, без чего
смысл еще виден.
Шапошников работал с
тлями рода Dysaphis. Эти
тли - монофаги, т.е. питают-
ся листьями растений только
одного вида. В течение лета
они производят 15—18 бес-
полых (точнее, партеногене-
тических) поколений, давая
одно половое поколение лишь
осенью, перед зимовкой. Ша-
пошников сажал бесполых
тлей на растение, для данного
вида почти или вовсе негод-
ное, и тли быстро эволюционировали. Точнее, было взято три
вида тлей: а, в и с, живущих на растениях видовА, В и С соот-
ветственно. Вид а на растении вида С не мог жить вовсе, но мог
кое-как жить на растении вида В, куда его и сажали.
Сперва в опыте падали численность тлей и их размеры, зато
сильно росли изменчивость и смертность. Но через несколько
поколений произошел «скачкообразный необратимый переход
в новое адаптивное состояние с ... резким снижением изменчи-
вости, прекращением естественного отбора, ... повышением
плодовитости; в 8—10-м поколениях возникли существенные
морфологические различия между исходной формой и обра-
зовавшейся новой, уже неспособной жить на старом хозяине»
[Шапошников, 1965, с. 23].
Выделенные мною слова звучат странно, но означают
лишь то, что ученый понимал под естественным отбором рост
смертности, достигавшей 75%. Он не исследовал отбора (луч-
шего размножения носителей некоторых качеств), хотя воз-
можность у него была: одна из сотен личинок вида а выжи-
ла прямо на виде С [Шапошников, 1961, с. 745]. Идеология
дарвинизма требовала обмерить ее (насколько сходна с видом
с), постараться получить расплод от нее и бросить силы на от-
ыскание случаев повторного выживания особей а на растени-
ях С.
Но Шапошникова привлекло иное - то, что он назвал «вос-
питанием» новой формы. В самые трудные для тлей моменты
20
автор возвращал их на сутки на исходное растение «для восста-
новления иссякающих жизненных сил». Эта ремарка - в духе
ламаркизма, но ламаркистом он себя не считал.
С 11 -го поколения тлей сажали на растение вида С, и теперь
они уже не гибли сплошь, а доживали до осени, до появления
полового поколения, дав новую форму. Замечательно, что она
не могла скрещиваться с исходным видом а, зато давала пло-
довитое потомство с видом с. Иными словами, Шапошников
получил новый вид животных за одно лето!
Сам он говорил осторожно: это новая «видовая форма»,
несущая черты как вида с, так и вида Ь. Это свойство, как мы
знаем, - блочность, т.е. элемент номогенеза по Соболеву. Но-
могенез по Бергу виден здесь в том, что изменения шли сразу
у многих особей в параллель и реализовали черты уже имев-
шегося вида. А эволюция путем прямого воздействия среды
на развитие личинок - ясный пример жоффруизма. Словом
- великолепное открытие.
Оно впоследствии было подтверждено другими исследо-
вателями на других видах насекомых и на клещах. Сходные
результаты получены и на растениях (злаках) [Агаев, 1978].
Взрывное видообразование, начинающееся примерно с вось-
мого поколения после начала резкого вредного воздействия на
исследуемую популяцию, оказалось обычным явлением.
Шапошников прожил еще 40 лет, но не мог ни продолжить
опыт, ни даже завершить публикацию сделанного. Его опыт
долго не поминали, а затем, забыв его суть, стали толковать
как пример эффективности отбора, т.е. дарвинизма. Вот но-
вейший пример. Уже известный нам автор, Расницын, писал:
«В опытах Шапошникова отбор явно был односторонним и
несбалансированным (инадаптирующим). Действительно, в
опыте способность питаться на новом растении-хозяине на ка-
кое-то время оказалась единственным жизненно важным ком-
плексом адаптации, а все остальные отошли на задний план»
[Расницын, 2008, с. 20].
Необходимо заметить, что ввести инадаптирующий (т.е.
разрушающий приспособленность) отбор — значит, отказать-
ся от отбора в смысле Дарвина. Дело в том, что если принять
первый как инструмент рассуждения, то рухнет второй — един-
ственный у Дарвина пример отбора животных: олени, убегая
от волков, стало быть, инадаптируются, ибо «единственным
жизненно важным» для них является спасение от волков. Эво-
люции как приспособления не получится.
К чести Расницына следует отнести тот факт, что он видит
коренное противоречие этого своего толкования со старин-
ной идеей отбора «по Дарвину» и предлагает вместо нее идею
адаптивного компромисса, столь же старинную. Состоит она
в том, что единицей отбора служит не признак, а весь комплекс
свойств, нужный организму для продолжения эволюции вида,
21
т.е. сразу всё, что требуется теоретику. Это - возрождение дар-
винизма по Рачинскому.
Здесь необходимо задать следующие вопросы:
1. Может ли комплекс свойств, бесполезных по отдельно-
сти, служить объектом отбора? Ведь наука не знает поисковых
систем, в которых улучшается всё сразу.
2. Если даже допустить, что комплекс мог быть отобран, то
куда исчезал на это время отбор отдельных свойств, гораздо
более мощный? Ведь коротконогие волки и олени гибнут на
самом деле.
3. Если и этот вопрос решен, то сколько нужно времени и
особей на отбор одного адаптивного компромисса? Но ариф-
метика, близкая сторонникам адаптивного компромисса в их
сокрушающей критике обычного дарвинизма, почему-то со-
всем чужда им, едва они переходят к своему толкованию эво-
люции (как множества разных адаптивных компромиссов).
Выходит, они просто сменили одно толкование на другое.
Но, может быть, в идее адаптивного компромисса есть более
серьезный смысл?
8. Как быть, если отбора «по Дарвину» в природе нет?
Смысл данного пункта отнюдь не нов. Фактов отбора нет4,
и всякое указание на это обижает дарвинистов. Но, по-моему,
им следует, согласно принятым во всех (кроме дарвинизма и
богословия) науках нормам, не придумывать толкования, а
выяснить суть дела хотя бы на одном реальном объекте. Если
это не удалось, надо либо смириться с упреками в религиозно-
сти дарвинизма, либо от данного понимания отбора отказать-
ся (что, как увидим, в нынешнем дарвинизме и происходит).
Принимать прежний «отбор» в качестве постулата нельзя,
демонстрация чего и является целью всех приводимых здесь
четырех групп примеров.
Во-первых, вне мира бактерий нет ни одного примера
получения нового свойства за счет различия размножаемо-
стей вариантов. Что касается бактерий, то 30 лет назад была
поставлена серия изящных опытов, показавшая реальный
предел действия отбора: для получения колонии мутан-
тов, в которых путем отбора произведена замена всего лишь
двух аминокислот в белке, бесполезных по отдельности и
полезных вместе, понадобилось провести через хемостат
4
Тот постулат, что естественный отбор (понимаемый по Дарвину: успешны те,
кто лучше размножается) служит движущим фактором эволюции, ни разу не
обоснован фактами, т.е. есть с цифрами и повторностью, с опытом и контролем.
До сих пор «доказательства действия естественного отбора» описывают либо рост
частот тех свойств, какие уже имелись к началу опыта, либо искусственный от-
бор, а при нем численное значение размножаемости роли не играет. Часто звучит
оправдание: «Отбор трудно наблюдать», но это значит лишь, что доказательств
нет. Кстати, трудно — не значит невозможно, и опыты, как увидим, проведены.
22
1013 (10 триллионов) штук бактерий. Авторы справедливо
заключили, что даже у бактерий эволюция путем отбора
нереальна [Hartley, 1976; Wu, 1978], что о более крупных
(даже одноклеточных) организмах говорить нет смысла.
Хотя эти работы изредка упоминаются, но суть их (как и
работ Шапошникова и Агаева) забыта: их, не читая, упо-
минают как подтверждение идеи эффективности естествен-
ного отбора.
Во-вторых, во всех аккуратно исследованных случаях ока-
зывалось, что новые варианты организмов поначалу долгое
время размножаются не лучше, а гораздо хуже исходных.
То есть новое формируется не потому, что лучше размножа-
ется, а потому, что успевает сформироваться прежде, чем
вымрет. Затем новые формы организмов ждут своего часа.
Известный пример — млекопитающие: они сформировались
при господстве динозавров и завоевали мир после их выми-
рания. Еще ярче (хотя и мало известен) пример преобразо-
вания кистеперых рыб в первых амфибий [Клэк, 2006]. Оно
заняло 20 млн лет, причем в толщах осадков, отложившихся
за это время, обнаружено всего 6 (шесть!) экземпляров пере-
ходных форм, тогда как находки исходных и конечных форм
вполне обычны.
В-третьих, многие организмы процветают при весьма
низкой размножаемости. Так, орхидеи часто называют вер-
шиной эволюции растений, ибо их цветки поражают красо-
той и сложностью, а семейство орхидных (ятрышниковых)
— самое обширное (35 тыс. видов). Пока мир растений опи-
сывали флористы (изучающие, что где растет) и системати-
ки (их интересует классификация организмов), всё казалось
просто. Однако в XX веке за дело взялись физиологи и эколо-
ги, и красивая картинка потускнела, а затем распалась.
Критерием успеха дарвинизм полагает успешность раз-
множения (многочисленность в череде поколений). Но орхи-
деи размножаются хуже всех цветковых®, и надо бы признать
их маргиналами, кандидатами на вымирание. Этого никто не
делает, т.е. все признают фактически, что их прогресс шел
иначе, помимо преимущества в размножении.
В-четвертых, хуже всего обстоит дело с естественным отбо-
ром как раз там, где его больше всего следует ожидать теоре-
тически, — там, где избирательная размножаемость наиболее
высока. Оказывается, наиболее жестко отбираемые организ-
мы вовсе не передают новых качеств потомкам. Один пример
(у Скеддера) мы уже рассмотрели, а еще лучший пример —
тропические подземные термиты.
° Орхидные не имеют двойного оплодотворения — главного приобретения цветко-
вых. У орхидей крайне затруднено опыление, а их семена не имеют эндосперма,
созревают медленно (год или два, как у хвойных) и только при содействии грибов-
симбионтов. Поэтому виды орхидей малочисленны.
23
Рис. 2. У термитов лишь один раз в
год на свет появляется поколение,
способное летать и размножаться.
Однако после образования брачных
пар термиты обламывают крылья
и становятся совершенно беззащит-
ными перед хищниками. На рисунке
из книги Карла Эшериха «Термиты
или белые муравьи» (пер. с нем., С.
Петербург, 1910) показаны стадии
развития крылатых термитов
Leucotermes lucifuges: а - имаго; б
- имаго после сбрасывания крыльев.
Хорошо видна бороздка для обламы-
вания крыльев.
Этот пример тоже стар и
тоже не опровергнут. Правда,
мне не раз заявляли (устно и
письменно, но не в печати),
что не учтены какие-то свой-
ства термитов. Выяснилось
много нового, но никто из воз-
ражавших не коснулся самой
процедуры отбора, т.е. сути
дела. Она проста и состоит
в том, что процесс, постули-
руемый всеми вариантами
дарвинизма, у этих термитов
не идет, зато идут процессы,
прямо противоположные. В
этом со мной никто не спорит.
Если взять руководство по
насекомым (например, том
3 «Жизни животных» изда-
тельства «Просвещение»),
то можно прочитать следую-
щее: в отряде термитов 2600
видов, в основном тропиче-
ских; все общественные, т.е.
живут огромными колония-
ми. В каждой размножение
осуществляет единственная
плодущая самка. Большин-
ство видов — подземные, они
живут весь год в закупорен-
ных термитниках, бесполые, слепые и бескрылые. Лишь раз
в году рождается и вылетает наружу зрячее крылатое половое
поколение (или каста) — самцы и самки. Это похоже на пчели-
ный рой, но есть три принципиальных отличия.
Первое: самок тут много. Второе: в момент образования
пар все особи обламывают себе крылья (для чего имеют осо-
бые инстинкт и борозду облома - см. рис. 2). Яркие, вкус-
ные и беспомощные (у данной касты изъяты все средства за-
щиты), они тут же гибнут от множества хищников, ждущих
их вылета (спасается одна пара из многих тысяч). Зато есть
третье: выжившая пара сразу основывает новую колонию
(термитник).
Как видим, самой природой из века в век ставится жесткий
селекционный опыт: если имеются, как гласит дарвинизм,
вариации всех свойств, то среди них должны быть вариации,
снижающие съедобность (в широком смысле: крепкие кры-
лья, колючий покров, ядовитость, отпугивающий запах или
вкус и т.д.), и они должны отбираться. То есть лучше раз-
24
множающиеся должны вытеснить хуже размножающихся (в
данном случае - несъедобные съедобных) — такова логика
дарвинизма, на которой построено всё учение, в том числе
- дарвиновы олени и волки. Проверять опытом надо ее, логи-
ку. Однако несъедобности у этих термитов не возникает.
Мало того, что ни на одном из видов подземных термитов
никто ее не наблюдал, но, главное, ее с позиции экологии и
не должно быть: половое поколение термитов обеспечивает
работу всей экосистемы тропического леса, о чем будет речь
далее (см. п. 12). Приспособленность всей экосистемы нали-
цо, и будем из этого исходить, но где и в чем могла тут быть
роль отбора? Ведь экосистемы не размножаются путем смены
поколений, а значит, отбор (в обычном смысле слова) для эко-
систем ввести нельзя. А в каком можно?
Да в очень простом: отбор - это иносказание. Каждая эко-
система являет собой экологический адаптивный компро-
мисс, точнее — компромисс между выеданием каждого вида
и его размножением. И с обычным отбором свойств одного
организма ситуация, как мы видели в п. 7, та же. Всюду уче-
ные видят взаимную подгонку свойств, иногда именуют ее
адаптивным компромиссом, но механизм ее формирования
додумывают, вводя метафору отбора (на что указал, уступая
критикам, еще сам Дарвин®). Однако на метафорах теорию не
построишь - ее и нет.
А что есть? Есть примеры приспособленности, которые
принято толковать как итог прежнего отбора. Однако толкова-
ния возможны и в иных рамках, в частности ламаркизм тол-
кует приспособленность как итог прежней активности особей
(выше был упомянут пример Ламарка: длинная шея жирафа
как итог его усилий в стремлении объедать высокие ветви).
Пример с термитами не толкуется никак и заставляет задать
главный вопрос: есть ли у нас вообще теория эволюции? То
есть можно ли обойтись без метафор?
Факт гибели одних организмов ничего не говорит о том, бу-
дут ли эволюционировать другие, выжившие. Поняв это, мы
избавляем себя от нужды вести бесконечные списки толкова-
ний и бесконечные споры с толкователями. Однако при этом
лишаемся основного прежнего инструмента объяснения, гла-
сившего: «Полезное выживает потому, что лучше размножа-
ется». Что можно предложить взамен? Попробуем ответить.
В декабре 1859 года в начале главы «Естественный отбор» он вставил во фра-
зу: «Можно метафорически сказать, что естественный отбор ежедневно, ежечасно
расследует по всему свету мельчайшие изменения...» слово «метафорически». В
1861 году он вписал перед этим разъяснение: «Естественный отбор - неправиль-
ное название (misnomer)», а в 1869 году усилил эту мысль: «Естественный отбор
- ложный (false) термин». Русский перевод смягчен (Дарвин, 1939, с. 328, 330].
Видимо, Дарвин сам потерял ясность в понимании явления отбора, и заменой тер-
минов помочь делу было нельзя. Ясность была внесена много позже — см. п. 15.
25
Часть 2. Что же движет эволюцию?
9. Принцип активности и новый ламаркизм.
Генетический поиск и акт эволюции
Если в самом деле новое возникает и формируется вопреки
своей низкой размножаемости, то весь процесс становления
нового должна вести какая-то иная сила, помимо размножае-
мости. Самые разные эволюционисты, пытаясь нащупать эту
силу, искали аналогии в физике и химии (вспомним Мопер-
тюи). В глаза бросались аналогии с гравитацией (пылинки
собираются в небесное тело) и с валентностью (простые веще-
ства соединяются в сложное; например, водород и кислород —
в воду). В обоих случаях новое обладает принципиально ины-
ми свойствами, нежели его компоненты (подробнее см. книги
[Чайковский, 2006; 2008]). Нет ли тут пути к общей законо-
мерности? Оказывается, есть.
Общим является то, что на всех уровнях сложности мы ви-
дим в основе развития какую-нибудь активность. Таковы гра-
витация в физике и валентность в химии — формы активности
неживой материи, самые простые для понимания. Элемен-
тарные частицы соединяются в атомы, атомы — в молекулы,
молекулы — в вещества и т.д. На каждом уровне действуют
свои формы активности, и, чем выше уровень, тем сложнее
эти формы. Пусть идут бесконечные споры о том, можно ли
вывести химию из физики, жизнь — из физики и химии, за-
коны общества — из биологии, но независимо от того, каков у
кого будет ответ, несомненно одно: этот ряд усложняющихся
активностей существует.
Та активность особи, которую положил в основу своего уче-
ния Ламарк, находит в данном ряду очевидное место, но не
стоит думать, что другие эволюционисты могли обойтись без
активности. Дарвин, например, ввел, никак не объяснив, две
формы активности живого: наследственную изменчивость и
стремление к размножению (притом к избыточному) — и счел
именно их ответственными за эволюцию. Иногда говорят (а
иногда нет) о третьей форме активности живого - борьбе за
существование.
Откуда они взялись? В печатных трудах Дарвина об этом
нет ни слова, зато в ранних рукописях сказано прямо - от
Бога (см. п. 4). В XX веке дарвинисты стали толковать на-
следственную изменчивость как случайные мутации (что ока-
залось ошибкой — см. далее, п. 10), а стремление к размноже-
нию так ничем и не объяснили.
Есть и другие формы активности живого - например, са-
мосборка макромолекул или рост тел. Словом, если будет ска-
зано, что в основе эволюции (как и любого развития - живо-
го и неживого) лежит принцип активности, мы не придем в
26
противоречие ни с какой теорией и ни с какими фактами, а
просто укажем на общеизвестное явление.
Саму активность ни из чего вывести нельзя — это одно из
первичных понятий (основное свойство мира), такое же, как
пространство, время, движение. Ее можно лишь описать. Вот
самое короткое описание: всякое развитие объекта происхо-
дит под действием некоторой формы активности; оно приво-
дит к появлению более сложных объектов, что в свою очередь
порождает более сложную форму активности, и так далее.
Уже само признание принципа активности оказывается
полезным. Например, стало ясно, что если нет активности,
то нет и возможности использовать никакой ресурс. Так, дав-
но было понятно, что инертный газ нельзя использовать как
реагент из-за отсутствия у него валентности. Таковы же все
попытки утолить нужды общества путем финансирования:
если нет активных (заинтересованных и имеющих силу) ис-
полнителей, то деньги пропадут, точнее, их присвоят носите-
ли иных активностей, чуждых данным нуждам.
Таков и мутационный ресурс полового поколения терми-
тов: мутации заведомо возникают (в том числе мутации к не-
съедобности), но если никакая активность их не использует,
то новое свойство не формируется.
Главный же толк от введения понятия активности виден
там, где можно выявить ее конкретные свойства. Выявление
их — дело долгое. Отошлю желающих к упомянутым книгам
[Чайковский, 2006; 2008], а здесь ограничусь парой приме-
ров и выводов.
Самый простой пример — принцип компенсации. Его ввел
в науку еще Аристотель. В трактате «О частях животных» он
писал: «Природа везде, взяв с одного места, отдает другой ча-
сти» и «Общих и многих средств защиты природа, однако, не
дала одному и тому же животному». В наших терминах эту
мысль можно выразить так: ни один вид организмов не об-
ладает активностью в таком количестве, чтобы развивать все
принципиально доступные ему качества.
С позиции этого принципа легко понять упомянутые
выше особенности орхидей: их активность настолько ушла
на усложнение цветка, что система размножения осталась на
уровне голосеменных. То же самое можно сказать об очень
многих биологических свойствах. Например, самые сложные
ткани (см. словарик) наблюдаются у низших червей, а самые
сложные типы деления клеток — у одноклеточных. Да и у
человека принцип хорошо виден: преимущество в мышлении
далось ценой явных недостатков — тяжкие роды, голое тело,
неспособность синтезировать витамин С и многое другое.
Пример посложнее — стресс. Попав в тяжелые (стрессо-
вые) условия, организм уменьшает активность по всем на-
правлениям, кроме одного-двух, которые должны вести к
27
снятию стресса и тем самым к выживанию. При этом проис-
ходит смена норм: если надо менять поведение, значит, преж-
нее поведение, нормальное прежде, перестало быть таковым,
а новое, которое приведет к снятию стресса, должно стать но-
вой нормой.
Например, если исчез прежний источник питания, надо
искать новый. Процесс может быть очень болезнен и даже
привести к смерти основную часть популяции, поэтому его
любят называть «давлением отбора». Однако он может идти и
без роста смертности, так что суть не в том, сколько погибло,
а в том, что произошло с выжившими. Оказывается, они вы-
живают путем смены типа активности. Она происходит сразу
у заметной части популяции и должна быстро дать результат,
иначе все вымрут. Такова выработка фермента, нужного для
использования нового типа пищи (бактерии у Хартли и By
(см. п. 8) и, по всей вероятности, тли в опыте Шапошнико-
ва).
Подобные опыты ставили разные ученые на разных видах,
и всюду они давали сходные результаты. Новую литературу о
смене кормового растения у насекомых и клещей можно най-
ти в статье [Magalhaes, 2009]. В частности, у паутинных кле-
щей рода Tetranychus наследуемая адаптация к новому расте-
нию тоже шла около 15 поколений (8 месяцев). Но поскольку
эти клещи - полифаги (могут кормиться на различных рас-
тениях), то снижения приспособленности (adaptation costs) к
исходному растению вообще не наблюдалось.
Столь же быстро (по числу поколений) реагировали и дру-
гие объекты — например лисицы, которых отбирали на «до-
машнее» (т.е. неагрессивное) поведение, или пшеница, выра-
щенная М.Г. Агаевым в непригодном для нее климате [Агаев,
1978]. Всюду причиной резкой наследственной смены .типа
активности (поведения, физиологии) можно считать стресс7.
Ни о каком отборе случайных вариаций тут говорить нет
смысла, поскольку выжившие реагировали быстро и, глав-
ное, одновременно.
Наконец, самый сложный пример смены типа активности
— когда никакого стресса не наблюдается, а итог оказывается
примерно таким же. Уже Дарвин отмечал, что изменчивость
растений возрастает при улучшении ухода за ними. Еще уди-
вительнее, что самую высокую изменчивость показали те
лисята, которые лучше и быстрее всех приспосабливались
к жизни в неволе: у них проявились (кроме мирного поведе-
ния, на которое их только и отбирали) многие черты собак —
7 Можно привести большое количество работ о влиянии стресса на изменение ге-
нов. Особо интересны опыты генетика Джона Кэйрнса в США (Cairns, 1988), в ко-
торых снижение доз яда от смертельных до стрессовых привело к наследованию
приобретенных свойств - точнее, к наследованию устойчивости бактерий к анти-
биотикам (см. [Голубовский, 2001; Чайковский, 2006; 2008]).
28
висячие уши, хвост кольцом, короткая морда, размножение
вне сезонов, пегая окраска (последнее было досадно).
Получается такая схема акта эволюции. В состоянии нор-
мы изменчивость популяции низка. При утрате нормы актив-
ность организмов направляется на поиск. Он может быть по-
веденческим, физиологическим и генетическим. Активность
сначала тратится на первый тип поиска, при его неудаче — на
второй, затем на третий — тогда и растет изменчивость.
Например, если зверю холодно, он пробует уйти туда, где
теплее, затем повышает теплопродукцию (дрожь, бег и т.д.)
и, если прежние меры не дают эффекта, впадает в стресс, что
активирует генетическую систему. Начинается генетический
поиск (см. словарик). Простейший его случай — повышение
мутабильности в длительно неблагоприятных условиях, т.е.
при стрессе.
Однако известны и гораздо более сложные типы генетиче-
ского поиска. Например, упомянутые в следующем пункте
поиск антитела и весьма близкий к нему по сути сплайсинг.
Подробнее о поиске см. в книге: [Назаров, 2005] или в моих, а
также в статье [Голубовский, 2001].
Поиск нормы (он может выглядеть и как случайные му-
тации, и как обучение, и как наследование приобретенных
свойств8) поставляет основной материал для эволюции. Когда
новая норма найдена, изменчивость снова падает и перестает
быть фактором эволюции — вплоть до следующего акта утра-
ты нормы. Схема описана десятки раз, но те, для кого «выбор
селектогенеза неизбежен», ее просто не знают. Рассмотрим
пример — появление нового гена.
10. Как появляются новые гены. Генетика XXI века
В дарвинизме появление новых генов не рассматривается:
все рассуждения ведутся вокруг уже существующих генов —
либо их включения и выключения, либо замены в них отдель-
ных нуклеотидов. Эту несуразность можно было не замечать,
пока процесс формирования нового гена не был описан фак-
тически. Однако в 1965—1982 годах несколько выдающихся
генетиков из разных стран сумели расшифровать процедуру
формирования целой плеяды генов - тех генов, каждый из ко-
торых кодирует антитело (см. словарик).
У зародыша млекопитающих совсем немного генов, коди-
рующих иммуноглобулины, — около сотни, тогда как множе-
ство различных возможных антигенов (см. словарик) необо-
зримо велико. Поэтому в ходе развития и жизни организма
разнообразию генов иммуноглобулинов каждый раз прихо-
дится создаваться заново (точно так же, как заново создается
8 См. раздел «Приобретенные признаки наследуются», статья [Голубовский,
2001], где дана литература.
29
любой орган). Происходит это путем комбинирования фраг-
ментов существующих генов (это тоже блочность, о которой
мы говорили в п.7). Нужное антитело обычно не выбирается
из наличных иммуноглобулинов, а продуцируется в ответ на
конкретную заразу (на вторгшийся антиген).
В стрессовой ситуации, которую вызывает массовое вторже-
ние антигена, включается механизм перестройки иммуногло-
булиновых генов: по каким-то еще непонятным правилам гене-
тическая система режет и сшивает фрагменты генов и находит
приемлемый вариант — тот, что синтезирует антитело, которое
реагирует с вторгшимся антигеном, связывая его. Найденный
вариант гена интенсивно размножается (копируется).
Это стало известно в 1982 году, когда генетик Судзуми То-
негава (образование получил в Японии, работу начал в Швей-
царии и завершил в США) обнародовал итоговую статью по
данной теме (через пять лет он, и только он, получил Нобе-
левскую премию за расшифровку всего механизма — так уж в
Нобелевском комитете заведено).
Механизм комбинаций поставляет антитела, связывающие
антигены, но связывание идет пока довольно слабо. Поэтому
существует еще один механизм — соматический гипермута-
генез, который включается после создания нужной комбина-
ции фрагментов. Заключается он в том, что при копировании
гены найденного варианта мутируют с огромной частотой (тут
каждый тысячный нуклеотид заменяется, тогда как обычно
точковый мутагенез в миллионы раз менее интенсивен), так
что порождается масса чуть отличных антител, различающих-
ся одной аминокислотой или двумя, чем и достигается точная
подгонка антитела к антигену. Конечный вариант гена снова
копируется и запоминается иммуногенетической системой
организма, т.е. наследуется на время жизни особи. Мы рас-
смотрим данный круг вопросов подробнее в разделе II.
Открытия иммуногенетиков впервые показали, что вместо
случайных мутаций надо говорить о механизмах преобразова-
ния генетических текстов и искать данные механизмы. Ныне,
спустя четверть века после работ Тонегавы, многие начинают
понимать, что изменение гена - не источник изменений наслед-
ственной информации, а ее запись, производимая активными
процессами, текущими в клетке. Кратко см. об этом в книгах
[Чайковский, 2006; 2008], а более подробно - в обзоре [Griffiths,
Stotz, 2006]. Здесь остается заметить, что ламаркисты понимали
это давно, еще на заре молекулярной эры. О самом оригиналь-
ном из них, Вентребере, мы поговорим тоже в разделе II.
В указанном обзоре приведены новые данные о собствен-
ной активности генетической системы, некоторые из которых
весьма впечатляют. Таков транс-сплайсинг, т.е. сборка гена
из фрагментов, расположенных в различных местах генома.
Если при обычном сплайсинге (цис-сплайсинге) фрагменты
30
(экзоны) берутся из одного транскрипта, где расположены
линейно (друг за другом), то при транс-сплайсинге о линей-
ности говорить не приходится9, и, что называть словом «ген»,
становится совсем неясно.
Важно, что транс-сплайсинг тоже (как и цис-) может быть
альтернативным [Horiuchi, 2006], т.е. порождать целый
спектр генов на основе одного набора фрагментов. Теперь,
когда это открыто, стало видно, что данный набор практи-
чески бесконечен, а потому никакая случайная комбинация
не имеет шанса повториться. Тем самым любая повторность
транс-сплайсинга означает его направленность. Впрочем, это
относится и к иным генетическим механизмам (например, к
синтезу антител), только не всюду так заметно.
Тот факт, что новый ген может формироваться не путем
мутаций в прежнем гене, а путем конструирования (т.е. за-
ново), недавно показан на примере рисунков крыла бабочки:
в рамках одного тропического рода Heliconius одно и то же
изменение рисунка у вида Heliconius numata контролируется
одним геном, тогда как у других - четырьмя или пятью, при-
том на разных хромосомах [Baxter, 2008]. А эффект (рисунок
крыла) един.
Этот пример позволяет предполагать, что в опытах Ша-
пошникова и подобных тоже шло весьма интересное измене-
ние генов (к сожалению, генетический анализ таких опытов
не ведется до сих пор). Сам Шапошников допускал, что там
имели место длительные модификации (см. словарик). Их
механизм ныне отчасти расшифрован, причем оказалось, что
при долгом стрессе они могут включаться в постоянный ге-
ном [Голубовский, 2001].
Наоборот, для пшениц такой анализ недавно проведен, и
выяснилось, что быстрое видообразование при стрессе идет
вовсе не за счет мутаций в генах, а благодаря изменению со-
става массовых повторов некодирующей ДНК с помощью
транспозонов [Беляев, 2009]. (О транспозонах см. словарик.)
Активность этих массовых процессов очевидна уже сама по
себе, а их сверхбыстрое протекание в опытах Агаева (и им
подобных) убеждает в их активности еще более (точнее, речь
идет о генетическом поиске — см. п. 9).
Всё это подтверждает давнюю догадку ламаркистов, не раз
заявлявших, что изменение ДНК - не источник изменчиво-
сти, а лишь способ ее записи. Но если так, то вся эволюцион-
ная проблематика, связанная с генетикой, в корне меняется.
Далее обсудим генетику XXI века как базу нового понимания
эволюции.
9 Строго говоря, транс-сплайсинг - не новость. Он открыт давно (в 1985 году),
но его долго не признавали - видимо, неосознанно боясь расстаться с последним
классическим свойством гена (линейностью расположения текста). О сплайсинге
см. словарик.
31
11. Активность, мышление и эволюция —
они возможны вместе
Итак, механизм генетической изменчивости умеет ком-
бинировать не только нуклеотиды, но и составленные из них
блоки. Образно говоря — не буквы, а длинные слова и даже
фразы. Здесь нужно пояснение. Если перебрать все буквы во
всех коротких словах вполне реально (можно поочередно за-
менять в данном слове каждую букву и смотреть, осмысленно
ли получившееся слово), то перебор букв в длинных словах
просто невозможен. В разговорном языке всего лишь около
миллиона осмысленных слов (если считать всё словоформы
- различные падежи, времена, степени сравнения и пр.), тог-
да как комбинаций букв той же длины, что все эти слова, при-
мерно в квинтильон (1018) раз больше.
Далее, слова обретают смысл только в контексте, но даже
осмысленных словопар (не говоря уж о тройках слов и более)
в языке — миллиарды. Поэтому никто не мыслит путем пере-
бора - ни букв, ни слов, ни фраз - мышление устроено как-то
иначе.
И вот мы видим, что в механизме Тонегавы тоже идет не
перебор, а какая-то более сложная процедура, генетический
поиск. Пусть смысл ее нам пока неясен, но уже видно, почему
ее иногда называют внутриклеточным мышлением: рождает-
ся новизна — ген, какого никогда прежде не было (например,
антитела к искусственным антигенам).
Кстати, мышление непонятно тоже, но никто ведь не счи-
тает его набором случайных электрохимических импульсов.
Все признают, что мышление - процесс активный, причем
эту активность проявляет мыслящая особь. Давно пора ска-
зать то же о работе генов. О внутриклеточном мышлении не-
много сказано в книге [Чайковский, 2008, с. 386].
Наличие хотя бы одного-единственного примера изготов-
ления нового гена, кодирующего новое качество, дает уверен-
ность в возможности эволюции как таковой. И иммуногенез
такой пример нам дал. Кстати, все молекулярные генетиче-
ские процессы, текущие в иммуногенезе, порознь известны и
для других областей генетики.
12. Вопрос Ламарка—Дарвина.
Экосистемная концепция эволюции
И Ламарка, и Дарвина занимал вопрос: почему в ходе эво-
люции все низшие формы жизни не превратились в высшие?
Ламарк отвечал: потому, что низшие до сих пор заново воз-
никают в порядке самопроизвольного зарождения. Дарвин
же полагал иначе: низшие приспособлены к своим простым
условиям существования и потому не изменяются.
32
Нынешняя наука отрица-
ет оба объяснения — и воз-
можность самопроизвольно-
го зарождения жизни в наше
время, и то, что «низшие»
просто устроены и потому
в простоте живут. Бывает и
наоборот (см. п. 9). То есть
организмы, низшие в одном
смысле, могут быть высшими
в другом. Ныне принято объ-
яснять факт наличия низших
форм (как бы ни понимать
этот термин) тем, что каждая
форма жизни вписана в свою
экосистему.
Вспомним половую касту
термитов. Подземные терми-
ты — единственный в тропи-
ческом лесу существенный
канал, возвращающий отмершую подземную органику в на-
земный мир [Гиляров, 1969]. Половая каста обладает всеми
свойствами, чтобы быть тут же съеденной, самый суровый от-
бор ничего с этим поделать не может, и именно эту проблему
призвана решить экосистемная концепция эволюции (ЭКЭ).
Обратим внимание на всеобщность съедобности. Мало
того, что все что-то едят и всех кто-то ест, но многие средства
размножения, как и у термитов, почти нацело выедаются
(семена растений, икра рыб и т.п.), причем никто этому не
удивляется, хотя всюду одна мутация могла бы уничтожить
съедобность. Если бы отбор таких мутантов имел место, ника-
кая жизнь не была бы возможна. Вот его и нет. (Несъедобные
виды бывают, они служат кормовой базой лишь узкому кругу
видов, способных их поедать.)
Согласно ЭКЭ, вид вовсе не приспосабливается к некой за-
данной среде, как думали и Ламарк, и Жоффруа, и Дарвин,
но формируется вместе с нею. Приспосабливается не вид к
виду, а экосистема сама к себе — примерно так же, как любой
из нас приспосабливается к своим болезням. «В определен-
ном смысле поистине живой можно считать только экоси-
стему» [Лекявичюс, 2009, с. 237]. Представление о развитии
этой концепции дает книга [Назаров, 2005]. Там она названа
теорией, но, на мой взгляд, теории тут пока еще нет.
Согласно ЭКЭ, вид имеет ту численность, какую ему обе-
спечивает его место в экосистеме. Данную мысль впервые
высказал великий биолог Карл Бэр еще при жизни Дарвина.
Ту идею Дарвина, что вид имеет такую численность, какую
отбил у конкурентов в борьбе за жизнь, Бэр счел нелепой и
33
предложил другую: численность вида задается его местом в
экосистеме. Кого много едят, тот и многочислен.
С позиции Бэра нам следует считать, что, поскольку отсут-
ствие эндосперма у орхидей делает их семена невыгодными
для питания, они никому не служат пищевой базой, поэто-
му орхидеи могут выжить при малой численности и низкой
плодовитости. И термиты теперь понятны: круговорот ве-
ществ в экосистеме тропического деса замкнут посредством
почти сплошного выедания их полового поколения, и такая
экосистема жизнеспособна. Если что и удивительно, то вы-
едаемость лишь одного поколения (касты), но почти то же
мы видим у тех видов, где выедается почти нацело лишь их
огромная масса икры.
Разумеется, экосистема, как и организм, должна в каждый
момент быть жизнеспособной, а потому формируется быстро.
Такое понимание эволюции можно вести от того же 1859 года,
что и дарвинизм: тогда Геер (о нем шла речь в п. 6) впервые
привел свидетельства синхронного преобразования видов
швейцарской флоры.
Чтобы превратить ЭКЭ в теорию, прежде всего надо опре-
делить источник активности, формирующий экосистему.
Этого до сих пор не сделано, и остается заметить, что он вряд
ли может быть тем же самым, что в формировании вида, т.е.
собственной активностью особей. В самом деле, особь актив-
но стремится выжить сама и оставить потомство (если речь не
идет об общественных животных), что постоянно приводит
к уничтожению особей других видов. Активность оленя не
может быть направлена на обеспечение волка пищей, как и
активность волка - на спасение оленя. Активность же экоси-
стемы направлена и на то, и на другое, и на многое еще - всё
сразу. Вопрос об источнике ее активности пока что открыт, и
замечу, что примерно та же трудность встает при попытке по-
нять источник активности общественных животных: улей и
термитник выглядят то ли особями, то ли экосистемами. Вот
и предпочитают говорить про отбор: хотя его тоже не видно на
объектах, зато он «очевиден», пока на объекты не смотришь
слишком внимательно.
Но вернемся к «низшим». Бактерии — основа любой эко-
системы и выполняют функции, которые проще всего осу-
ществляются именно бактериями, а не организмами с более
сложным устройством тела. Царство бактерий выступает
как генетическое целое, поскольку между бактериями легко
происходит горизонтальный перенос генов. У него есть важ-
ное для ЭКЭ следствие, отмеченное Е.А. Ароновой [Аронова,
1997]: «Перенос функциональной информации от высших
форм жизни к примитивным практически не происходит
(хотя и здесь есть исключения) по причине невозможности
“понимания” генетической информации высших организмов
34
геномом низших. Так природа как бы законсервировала мир
примитивных форм, создав своеобразную генетическую базу
для экспериментов Эволюции». Иными словами, бактерии
являют собой базовый пласт эволюции, тот питомник генов,
из которых черпают материал.
Столь же важно, что горизонтальный перенос генов, как и
мутация, происходит случайно - в том смысле, что мобильно-
му гену удается встроиться в чужой организм редко. Но вжи-
вается он активно, это своего рода заражение. Далее рассмо-
трим, как можно (если можно) описать такую случайность.
13.0 случайности
Противники Дарвина всегда упрекали его в том, что он
преувеличил роль случайности в эволюции, заменив исследо-
вание ее реальных законов ссылкой на случайность наслед-
ственных изменений. Дарвинисты, наоборот, хвалят своего
основоположника за то, что тот ввел в рассмотрение «анти-
случайный фактор» (отбор) и тем самым указал случайности
ее место в эволюции. Однако ни те, ни другие не захотели ис-
следовать сам феномен случайности, так что долго оставалось
тайной, говорят ли спорщики об одном и том же явлении или
о разных. Оказалось, о разных. Более того, сплошь и рядом
один и тот же автор использует термин «случайность» в са-
мых различных, порой несовместимых, смыслах. Мы увидим
это в клонально-селекционной идее иммунологов (см. далее,
раздел II).
Мне пришлось в свое время потратить много времени
и сил на выявление разных типов и форм случайных явле-
ний, итогом чего явились книги [Чайковский, 1990; 2004].
Пересказать их смысл здесь не удастся, и остается лишь при-
вести несколько самых необходимых для нашей темы положе-
ний.
Случайным именуют то явление, которое имеет различные
исходы, т.е. может при данных условиях как произойти, так и
нет. Всё необозримое разнообразие случайных явлений мож-
но разделить на два класса:
1) имеющих устойчивую частоту появления каждого ис-
хода (ее интерпретируют как вероятность данного исхода);
2) не имеющих такой устойчивости.
Почти все случайные явления, с которыми наука умеет
обращаться, относятся к первому классу (их изучают теория
вероятностей и математическая статистика), а почти все био-
логические (в том числе эволюционные и иммунные) — ко вто-
рому. Однако на деле мало кто проводит это членение, поэто-
му эволюционизм и топчется на месте.
В частности, выражение «мутация происходит с вероятнос-
тью Р» смысла не имеет, поскольку у мутаций почти никогда
не наблюдается устойчивых частот. И тем меньше смысла у
35
Рис. 3. Пример квазигиперболического распре-
деления: так распределены двигатели любого
крупного завода по их мощности W- чем выше
мощность, тем меньше таких двигателей. По
оси у отложены как реальные мощности (каж-
дый столбик означает число двигателей данной
мощности), так и моделирующая эту зависи-
мость гипербола И (х). Выше ординатыу=1 (она
для наглядности приподнята) можно прибли-
женно заменить точки (вершины столбиков)
кривойП(х). Приу=1 (когда данной мощностью
обладает один двигатель) гипербола моделиру-
ет тот факт, что с ростом х расстояние между
точками растет приближенно какП(х).
той базовой идеи
дарвинизма, что
«самое маловероят-
ное событие когда-
нибудь обязательно
произойдет». (Обя-
зательно произойдет
лишь то событие, ве-
роятность наступле-
ния которого стре-
мится со временем
к единице.) Поэтому
нам так важен вто-
рой класс явлений.
Математика для
явлений данного
класса существует, и
для нашей цели она
весьма полезна. Это
статистика «толстох-
востых» распределе-
ний (распределений,
в которых дисперсия,
т.е. мера разброса
случайной величи-
ны, неограниченно
растет с ростом чис-
ла испытаний). Называют их так потому, что у кривых типа
у(х), их описывающих, у очень медленно и нерегулярно стре-
мится к нулю с ростом х.
Такими распределениями быт, наука и техника изобилу-
ют, инженеры это давно понимают, и один класс их кое-как
описан: это квазигиперболические распределения. Называют
их так потому, что самое частое значение случайной величи-
ны располагается тут на графике с левого края, а остальные
частоты убывают направо, приблизительно как гипербола.
Впрочем, эта приблизительная регулярность сохраняется
лишь до середины графика, тогда как правая его часть являет
собой болтанку отдельных точек, слабо связанных с гипербо-
лой (рис. 3). Ясно, что две половины этого графика надо из-
учать по-разному.
Квазигиперболичны очень многие распределения:
— городов — по числу жителей;
— людей — по богатству;
— двигателей — по мощности;
— букв — по встречаемости в текстах;
— ученых — по числу публикаций;
— родов организмов — по числу входящих в них видов.
36
Один из них, пример с видами, далее и рассмотрим.
Род — понятие, означающее группу близких видов, и вво-
дится оно для описания общих свойств этих видов. Посколь-
ку систематик вводит каждый род по своему усмотрению, то
удобно вводить роды так, чтобы в них было не слишком много
видов, но и не один вид. Однако так не выходит. Какими бы
правилами группировки ни задаться, всегда окажется, что
какой-то вид никому не близок и его следует выделить в от-
дельный род (такова енотовидная собака в семействе псовых
отряда хищных). А какие-то виды, наоборот, столь похожи,
что в один род приходится помещать сотни видов: например,
род Crocedura (белозубки, из семейства землеройковых отряда
насекомоядных) включает 149 видов (табл. 1). Бывают роды и
по 2 тыс. видов. Систематики не раз пытались это неудобство
исправить, объединяя одни роды и дробя другие, но система
оказывалась непрак-
тичной, и от нее вско-
ре отказались. Квази-
гиперболы оказались
свойством природы
таксонов.
Замечательна ста-
тистика родов. Какую
бы крупную груп-
пу животных, гри-
бов или растений ни
взять, всегда однови-
довых родов окажет-
ся больше всего, сле-
дующим будет число
двухвидовых родов,
затем — трехвидовых
и т.д., пока не дойдем
до середины графика,
где впервые монотон-
ность убывания нару-
шается. Для насеко-
моядных (табл. 1) это
нарушение состоит в
том, что пятивидовых
родов нет, но есть ше-
стивидовые. А даль-
ше начинается самое
любопытное: обнару-
жив, что нет родов по
7, 8 и 9 видов, а затем
идут роды по 10, 12 и
13 видов, после чего
ТАБЛИЦА 1
число видов в РОДАХ
ДЛЯ ДВУХ ОТРЯДОВ МЛЕКОПИТАЮЩИХ
Число видов вроде Число родов
насекомоядные рукокрылые
1 36 75
2 5 23
3 6 19
4 2 13
5 — 8
6 2 4
7 — 3
8 — 3
9 — 4
10 1 3
И — 2
12 1 2
13 1 1
14 — 2
15 — 1
16 — 2
18 1 —
21 1 —
46 — 1
49 — 1
50 — 1
59 — 1
67 — 1
79 1 —
86 — 1
149 1 —
Итого: родов видов 58 386 171 896
37
— опять провал, мы должны бы ожидать, что разнообразие
группы близко к исчерпанию, но оказывается наоборот. Впе-
реди еще 2/3 видов отряда, и свыше половины этого обилия со-
ставит один огромный род - упомянутые белозубки.
Похоже ведет себя отряд рукокрылых (в основном состоя-
щий из летучих мышей), но в нем вдвое больше видов, и пото-
му первый провал, достаточно крупный, расположен гораздо
дальше — нет родов по 17—45 видов. Отряд вдвое обширнее
предыдущего, и можно ожидать гигантского рода в конце та-
блицы, а его нет: тут самый большой род почти вдвое меньше,
чем в отряде землеройковых. Словом такие распределения
весьма хаотичны, и регулярно в них одно — наличие квази-
гипербол (максимум с краю и «толстый хвост»).
«Толстый хвост» выражен в данных распределениях тем,
что при у = 1 одиночные точки могут уходить по оси х нео-
пределенно далеко и с неопределенными интервалами между
ними. Именно поэтому дисперсия тут растет с ростом выбор-
ки и обычная («гауссова») статистика не работает. Это выра-
жается, прежде всего, в том, что нет смысла искать средние
величины, ибо они совсем неинформативны.
Средний рост людей или средний вес яблок полезно знать,
поскольку около этих величин группируется основная часть
объектов (людей, яблок и т.п.). А вот среднее число видов в
роде не говорит ничего: в обоих приведенных примерах оно
(43 и 75 видов в роде) приходится на провалы - родов с таким
или близким числом видов просто не имеется. Реальные роды
либо много крупнее, либо много мельче средней величины.
И никакой биологической специфики тут нет: таковы
почти все квазигиперболические распределения. Например,
около трети научных работников не оставляют после себя ни
одной публикации, затем следуют оставившие одну-две (.это
не только рано ушедшие из науки, но и те, кто рано понял, что
его стезя — обслуживание чужих исследований). Зато основ-
ную массу статей пишет та треть ученых, у которых больше,
чем по 30 статей на каждого. Авторов со средним числом пу-
бликаций (5—6 за жизнь) очень мало.
Оказывается, что почти все случайные явления мира укла-
дываются в одну из двух статистик — либо в обычную «гаус-
сову», либо в квазигиперболическую. У этого факта должна
быть общая причина, и она уже известна.
Если виды одинаково распределены по родам в самых раз-
ных семействах, это значит, что род — не просто набор видов,
а элемент единой системы организмов. Точно так же город
— элемент единой системы расселения народов. Например,
Большой Лондон, бывший сто лет назад крупнейшим горо-
дом мира (5,6 млн человек), уступил через полвека первен-
ство Нью-Йорку, а теперь занимает 20-е место (имея 8,7 млн
человек); хотя система народорасселения изменилась ради-
38
кально, однако форма квази-гиперболы населенности городов
осталась той же. Не вырос горб, не изменился тип хвоста и
т.п. (см. рис. 3).
Итак, если в опыте обнаружены квазигиперболы, это на-
водит на мысль о наличии какой-то нежесткой системы (не-
коего единства), которую надо выявить. В частности, их стоит
ожидать всюду, где отдельные части системы проявляют соб-
ственную активность.
Математическая статистика родилась в XIX веке, когда
была осознана важность того явления (известного тогда уже
200 лет), что многие случайные величины, в природе которых
не видно ничего общего, ведут себя сходно. Например, сред-
ний рост людей, средняя продолжительность жизни, средняя
смертность от инфаркта, среднегодовая температура и т.д.
- обладают устойчивостью средних значений (средние меня-
ются очень мало от опыта к опыту, от наблюдения к наблюде-
нию). Это - обычная «гауссова» статистика.
Затем, в XX веке, возникла нужда создать другую, квази-
гиперболическую, статистику, где вероятностей и устойчи-
вых средних величин нет. Только после этого разграничения
статистик удалось выяснить природу различий самих явле-
ний: оказалось, что первой статистикой описываются те, где
основную роль играет независимость между случайными ве-
личинами, а второй — те, где обязательна нежесткая зависи-
мость величин.
Для теории эволюции существенно, что «толстый хвост»
означает появление в системе всё более и более сложно орга-
низованных объектов (или, кому так понятнее, — подсистем:
ведь виды по родам распределены примерно так же, как роды
по семействам и семейства по отрядам). Так же распределены
мутации по частоте встречаемости и многое другое. Этого не
могли предвидеть ни Ламарк, ни Дарвин, ни Берг.
Забегая вперед, следует заметить, что две статистики - это
два общенаучных рефрена.
14. Новый номогенез и основа эволюции
В 1970-х годах на связь квазигипербол с системностью об-
ратил внимание палеоботаник С.В. Мейен. Главное его дости-
жение как теоретика — введение понятия рефрена.
Рефрен — это, упрощенно говоря, ряд направленных ря-
дов. У самого Мейена примеры рефренов относятся к анато-
мии растений и мало кому понятны.
Простой пример рефрена приведен в табл. 2: в классе кост-
ных рыб плавники могут отсутствовать, быть представлены
одной слабо развитой парой, одной сильной парой, двумя па-
рами лучевых лопастей, двумя-тремя парами «кистей» (ки-
степерые рыбы), планирующим крылом и, наконец, актив-
ным крылом (некоторые летающие рыбы). Этот ряд направ-
39
Сергей Викторович Мейен (1935—
1987), палеоботаник и философ,
более всех сделавший для нового по-
нимания номогенеза.
лен в том смысле, что вдоль
него наблюдается услож-
нение строения органа. Но
аналогичные направленные
ряды видны и на остальных
строках табл. 2, т.е. в других
классах позвоночных. Ряд,
составленный из этих рядов,
и есть рефрен.
Другой пример рефрена,
тоже простой, но более об-
щий, — тот самый мир ква-
зигипербол. Каждая квази-
гипербола - тоже направлен-
ный ряд(упорядоченный ряд
наблюдений), а всепроникаю-
щий феномен квазигипербол
— это рефрен, составленный
из рефренов, один из самых
общих рефренов в науке.
Через рефрены Мейен
пришел к новому варианту
номогенеза, которому после его смерти было дано название
«диатропика», т.е. наука о разнообразии (от греч. диатро-
пос - разнообразный). Мейен был уверен, что теория рефре-
ТАБЛИЦА2
РЕФРЕН «КОНЕЧНОСТИ ПОЗВОНОЧНЫХ»
(УСЛОЖНЕНИЕ ОБЩЕЙ ОРГАНИЗАЦИИ НАБЛЮДАЕТСЯ СВЕРХУ ВНИЗ,
А КОНЕЧНОСТЕЙ - СЛЕВА НАПРАВО)
Класс Описание свойств конечности
отсутствие или невиди- мый снаружи зачаток развита только слабая передняя пара много- фаланговые плавники или ласты орган для:
ползания планирую- щего полета активного полета
Костные рыбы Мурена Китовидка, угорь Кистеперые Морской нетопырь Летучие рыбы Клино- брюшка
Амфибии Червяга Сирен Некоторые ископаемые Саламан- дры Яванская лягушка Не бывает
Рептилии Змеи, безногие ящерицы Хирот Морские черепахи Черепахи, крокодилы Ногокрыл (ископаемое) Летающие ящеры (ископа- емые)
Птицы Киви (нет передней пары) Не бывает Пингвины Поползень, стрижи Совиный попугай Большин- ство видов
Звери Киты, ламантин (нет задней пары) Киты Ламантин (передние), калан (задние) Кроты, тюлени Шерстокрыл Летучие мыши
40
нов явится единым методом упорядочения разнообразия яв-
лений (как природы и общества, так и мышления), но, увы,
умер в начале этой работы. Более новое состояние диатропи-
ки освещено в статьях [Чайковский, 2006; 2010], а здесь будет
сказано только самое необходимое.
Совокупность рефренов образует диатропическую сеть (диа-
сеть), т.е. многомерную таблицу, где каждая клетка - воз-
можный вариант организма. Можно говорить как о диасети
каждого таксона (например, вида или рода), так и о диасети
экосистем, и даже о диасети всего живого мира. Чтобы пред-
ставить себе, например, диасеть позвоночных, надо к приве-
денному в таблице рефрену конечностей добавить рефрен го-
лов, рефрен позвоночников, рефрен пищеварительных трак-
тов, рефрен иммунитетов и т.д.
Понятие рефрена можно ввести для любого множества
свойств, но имеет смысл вводить только там, где у таблицы
оказываются интересные свойства. Например, табл. 2 инте-
ресна тем, что множества конечностей разных классов позво-
ночных не только высоко сходны сами, но и демонстрируют
сходную эволюцию. Иными словами, здесь налицо обширный
параллелизм — не только форм, но и процессов.
В таких понятиях эволюция таксона предстает как запол-
нение всё новых и новых клеток его диасети. Существенно,
что это - не толкование фактов, а их обобщение.
Встает вопрос: каков механизм заполнения новой клетки?
В рамках дарвинизма вопрос нельзя даже поставить, посколь-
ку дарвинизм толкует всякое сходство через допущение либо
общего предка, либо независимого приспособления к сходным
условиям среды. В нашем примере (см. табл. 2) любое из них
весьма ущербно. Например, о том, зачем клинобрюшке по-
лезен активный полет, есть только забавные фантазии, и уж
никто не станет выводить из ее «крыла» крыло птицы. Более
содержательный подход видится в том, чтобы признать как на-
личие самой диасети (в этом состоит новый номогенез), так и
активное ее заполнение (новый аспект ламаркизма). Но глав-
ное достижение видится мне в рамках жоффруизма, точнее, в
рамках «evo-devo»: в и. 17 мы узнаем, что есть гены, переклю-
чающие развитие зародыша с одного пути развития на другой.
Таким путем сразу несколько организмов могут попасть в дру-
гую клетку диасети и тем самым дать начало новому виду.
Процесс заполнения диасети назовем сменой норм. Суть его
в том, что в нормальном состоянии (когда объект соответствует
своей функции, своему назначению) изменчивость особей низ-
ка, а в ненормальном — высока. Мы уже касались данного явле-
ния в связи с генетическим поиском, а теперь добавим, что это
— основной акт эволюции (см. и. 9), запускающий эволюционное
изменение. После нахождения организмами новой нормы из-
менчивость снова падает, что первым показал Шапошников. Он
41
же показал, что новая норма берется из весьма ограниченного
набора (возможностей), каковой выше и назван диасетью.
Теперь можно описать основу эволюции так: «Процесс эво-
люции живого являет собой триединство: активность — диасеть
- смена норм» [Чайковский, 2008, с. 12, 609]. Слово «диа-
сеть» означает здесь, что эволюция понимается как заполне-
ние диасети.
Тут стоит задать вопрос: можно ли эту основу сообразовать
с идеей отбора?
15. Отбор (selection) и подбор (delectus).
Новый дарвинизм
Неизбежен вопрос: почему идея отбора, не имеющая опыт-
ного подтверждения, 150 лет господствует? По-моему, дело в
том, что она отражает два вечных свойства людского мышле-
ния, которые можно усмотреть на всех этапах развития евро-
пейской науки.
Первое — что всякий наблюдаемый порядок был получен из
исходного беспорядка или из первичной однородности. Ему
противостоит восточная идея — исходным является состояние
первичного порядка.
Второе — уверенность,
то лучше тех, кто вымер. Е
Александр Александрович Богданов
(Малиновский) (1873—1928) первым
понял подбор как самоорганизацию
неживого, живого и социального.
ныне живущие организмы чем-
тезис высказан в Библии («Мно-
го званых, мало избранных»),
увязан Дарвином с политэко-
номией Мальтуса и освобож-
ден от нее Рачинским. Мы
сейчас увидим, что ныне те-
зис воспринимается сам по
себе, без апелляции к выжи-
ваемости и размножаемости,
однако всё еще увязывается
с привычным термином из
дарвинизма — отбором.
Самым заметным в данной
традиции был А.А. Богданов,
революционер и основатель
общей теории систем (1913).
Он, как и Рачинский, приме-
нял для введенного им про-
цесса улучшения всего на
свете термин «подбор», при-
чем ссылался на Дарвина, но
- вообще, нигде не излагая
его по существу. У Богданова
подбор мыслится как универ-
сальный фактор, ведущий ту
подстройку частей к целому,
42
какая одинаково видна и в природе (неживой и живой), и в
обществе, а значит, не сводится к отбору «по Дарвину».
Чтобы не путать с естественным отбором (natural selec-
tion), подбор по Богданову пояснен мною в книгах [Чайков-
ский, 2006; 2008] через перевод следующим образом: на ла-
тынь - как delectus', на английский — как choice, self-assemb-
ling, tektological selection. Термином «делектус» мы и будем
пользоваться.
В недавние годы стало обычным понимать естественный
отбор (natural selection) как самоорганизацию, не связан-
ную с успехом размножения, т.е. как делектус. Первым это
сделал наш известный дарвинист С.Э. Шноль. Признав, что
общепринятая в новом (в те годы) дарвинизме идея отбора
точковых мутаций абсурдна, он ввел в виде движущего эволю-
цию фактора «блочно-иерархический принцип». Но блочный
принцип известен давно и является частью номогенеза (пп. 6,
7 и 10), ново же здесь было то, что в качестве механизма по-
строения блоков взято мышление: «Попробуем отыскать ме-
ханизмы возникновения наследственных текстов через линг-
вистические»; в языке «образуются и “шлифуются естествен-
ным отбором” относительно простые слова и правила грам-
матики», а «из них строятся всё более сложные конструкции.
Этот принцип блочно-иерархического совершенствования и
позволяет написать поэму». И далее: «Естественный отбор
тоже возможен лишь по блочно-иерархическому принципу»
[Шноль, 1990].
Слова во внутренних кавычках дают нам понять, что сам
автор не верит в серьезность объявления мышления формой
естественного отбора — действительно, преимущественного
размножения нужных мыслей в мозгу не допускал еще никто.
Однако далее, как видим, он назвал естественным отбором
(уже без кавычек) организмов тот же самый механизм, кото-
рый «позволяет написать поэму», т.е. мышление. Тем самым
к молекулярному номогенезу здесь добавлен молекулярный
ламаркизм, и всё это предложено понимать как молекуляр-
ный дарвинизм. Вскоре такой подход стал обычным.
В качестве яркого примера можно привести книгу «Прав-
доподобие жизни: решение дилеммы Дарвина» [Kirschner,
Gerhart, 2005]. Дилеммой там назван вопрос о том, каким об-
разом мутация порождает самоорганизацию, а предлагаемое в
книге ее «решение» таково: полезная случайная мутация (на-
пример, удлинение шеи жирафа) обязательно будет поддержа-
на естественным отбором, ибо все органы, клетки и даже их
микротрубочки подстраиваются к ней «методом проб и оши-
бок», причем все неудачные вариации тут же отмирают (shri-
nk) «в силу строгого отбора». Как всегда, вопрос о том, можно
ли проверить это толкование, даже не упоминается.
43
Наивно? И нет, и да. Нет, так как авторы вновь открыли
(в который раз!) делектус. Да, ибо не заметили этого и увере-
ны, что развивают дарвинизм, хотя всем известно, что каждая
микротрубочка растет направленно (а не методом проб и оши-
бок) и порождаются микротрубочки не размножением друг от
друга, а активностью клетки. Процесс их роста, по-моему, так
же загадочен, как и все прочие факты самоподстройки систем.
Нигде мы не видим случайных блужданий, и вот предложено
принять, что они есть, но пресекаются раньше, чем успевают
проявиться. То есть, что они столь же воображаемы, сколь во-
ображаем обычный отбор.
Другой способ переосмыслить отбор виден в ЭКЭ. Напри-
мер: «Естественный отбор принято определять как диффе-
ренциальное (избирательное. - Ю.Ч.) выживание и такую же
репродукцию особей. Не имею ничего против такого определе-
ния, однако суть отбора... сводится к неким ограничениям, на-
лагаемым на поле потенциальных возможностей, создаваемое
наследственной изменчивостью» [Лекявичюс, 2009, с. 254],
т.е., в наших терминах, диасетью. Здесь необходимо поясне-
ние — роль диасети в ЭКЭ играют экологические ниши.
А вот позиция американского генетика, пишущего в обзо-
ре «Ищут Дарвина совсем не там»: акты изменения генома,
которые ныне подают в печати как доказательства «поло-
жительного дарвиновского отбора», обычно демонстрируют
«отсутствие правдоподобного биологического механизма, ле-
жащего в основе воображаемого (авторами - Ю.Ч.) отбора»
[Hughes, 2007, с. 371]. Подробнее об этом обзоре см. в книге
[Чайковский, 2008].
Очевидно, что среди людей, именующих себя дарвиниста-
ми, происходит отказ от прежнего понимания отбора.
Есть мнение, что надо всё же сохранить этот термин, разъяс-
няя, что теперь отбор понимается как «начало, актуализиру-
ющее новообразования» [Пчеленко, 2009], но опыт показал,
что любые замены смысла привычных терминов порождают
путаницу. И лучше всё-таки ввести термин «делектус».
Новое понимание движущего фактора проявляется в виде
самых неожиданных концепций, именующих себя дарвини-
стическими. Приведу еще один пример.
16. Делектус, математика и самоорганизация.
Intelligent Design
Системы, внутри которых связи нежестки, подчиняются
той квазигиперболической статистике, о которой шла речь
в п. 13. Можно даже сказать: где есть самоорганизация, там
жди квази-гипербол, а где видны квазигиперболы, там ищи
самоорганизацию.
«Дарвину и в голову не могла прийти мысль о существова-
нии самоорганизации — недавно открытого природного свой-
44
ства, присущего некоторым природным системам», — писал
американский биокибернетик Стюарт Кауфман [Кауфман,
1991]. По его мысли, естественному отбору подвергаются от-
нюдь не случайные мутации, а цельные системы, т.е. итог са-
моорганизации. Термин «отбор» взят им из дарвинизма, но
давным-давно, еще при Дарвине, многие поняли, что если от-
бору может подвергаться только готовая система, то, значит,
саму систему сформировал не отбор, а какая-то иная сила.
Это сто лет не считалось дарвинизмом. В самом деле, ког-
да «иную силу» видят в самом организме, в его активности,
то это ламаркизм, а когда привлекают какие-либо законы
формообразования — это номогенез. Кауфман ничего тако-
го не знал — на Западе ламаркизмом именуют наследование
приобретенных свойств, а номогенез как явление там вообще
неизвестен (хотя книга Берга дважды издана на английском
языке). Там любят говорить лишь про ортогенез (см. п. 6), что
слишком узко. Мысль Кауфмана близка и к идее адаптивного
компромисса, изложенной в п. 7, однако решение проблемы у
него иное, без всяких толкований. Попросту говоря, Кауфман
предложил эскиз понимания делектуса.
Он совершил прорыв — нашел нужную для описания самоор-
ганизации математику и приложил ее к эволюции. Удивитель-
ное здесь то, что Кауфман нашел способ извлечь самооргани-
зацию из феномена хаоса: обнаружил ее на хвостах квази-ги-
пербол. Основная его идея состоит в том, что сложные системы
можно поделить на два класса — «газообразные» и «твердые»,
т.е. на хаотические и упорядоченные (на «облака» и «часы»,
как выразился еще до него философ Карл Поппер). При этом
между классами возможны переходы — система может как об-
рести жесткую структуру, так и утратить ее. Только при таком
переходе система совершает акт
эволюции, т.е. качественно изме-
няется. Его модель совершенно
абстрактна, она не имитирует ни-
какого биологического объекта, а
лишь демонстрирует роль необыч-
ной случайности.
Кауфман привел компьютер-
ные примеры, показавшие, что
система из многих тысяч связан-
ных функционально элементов
может быть довольно просто опи-
сана. А именно: она может обла-
дать совсем небольшим числом
устойчивых состояний. Для этого
нужно, чтобы элементы системы
были слабо связаны, т.е. чтобы
каждый имел мало (лучше всего
Стюард Кауфман (р. 1939 г.)
45
— два) «входов» и примерно столько же «выходов». Но ведь
слабая связность как раз и есть основное свойство объектов
квазигиперболической статистики.
Подвижку системы от порядка к хаосу или обратно (с точки
зрения физика, это фазовый переход) можно сравнить с воз-
гонкой и осаждением, т.е. с прямым переходом твердого тела
в газ и обратно. Яркий пример — осаждение кристаллов льда
из воздуха, образующее снежинки в пространстве и морозные
узоры на плоскости (стекле окна). И то и другое поражает зри-
теля разнообразием, а разнообразие — один из главных объ-
ектов исследования биологии.
Разнообразные снежинки — это различные устойчивые со-
стояния того порядка, который образуется из хаоса (здесь — из
охлаждаемого пара), т.е. различные формы фазового перехода
по Кауфману. Он писал: «Хаос, как бы он ни был интересен,
— это лишь часть поведения сложных систем. Существует так-
же не поддающееся интуитивному осознанию явление, кото-
рое можно было бы назвать антихаосом. Оно выражается в
том, что некоторые весьма беспорядочные системы спонтанно
“кристаллизуются”, приобретая высокую степень упорядо-
ченности. Я полагаю, что антихаос играет важную роль в био-
логическом развитии и эволюции».
Антихаосом Кауфман назвал феномен устойчивости не-
многих состояний, а эволюцией — смену таких состояний. Ее
он усмотрел на грани порядка и хаоса: «Высокохаотичные
сети будут настолько беспорядочными, что контролировать их
сложное поведение весьма трудно. С другой стороны, высоко-
упорядоченные сети слишком заморожены, чтобы координи-
ровать сложное поведение. Однако по мере того, как заморо-
женные компоненты расплавляются, становится возможной
более сложная динамика» - и онтогенеза, и эволюции. Вджно
отметить, что в схеме Кауфмана наблюдается сравнительно
небольшое число устойчивых вариантов развития и эволюция
предстает как смена режимов такого развития. Он не знал, что
в биоэволюции это называют номогенезом.
В недавно написанной книге Кауфман вместо антихаоса
пользуется термином «Дарвинова преадаптация», странным
хотя бы потому, что слова адаптация (т.е. приспособление)
в книге нет. У Дарвина преадаптаций тоже нет, это — понятие
XX века.
Преадаптацией именуют полезное свойство, возникшее
прежде, чем оно смогло быть использовано. Примером мо-
жет служить сложный скелет плавника-ласта кистеперой
рыбы (см. п. 6), ставший полезным только у наземных по-
звоночных, т.е. через 20 млн лет. Другой пример - человече-
ский мозг, способный решать задачи, каких не было в эпоху
становления человека как вида (такова способность к мате-
матике).
46
Кауфман, вернее всего, хочет сказать своим термином, что
решил ту же проблему, что и «дилемма Дарвина», - каким
образом возникает готовое приспособление. Кауфман предло-
жил совсем иное: структуры полезны сами по себе. Фактиче-
ски этим он вводит изначальную целесообразность (по Бергу).
Ее он тоже назвал странно: «порядок задаром» (order for free
[Kauffman, 2008, с. 106]). Желая понять место своих работ в
эволюционизме, Кауфман стал читать о корнях дарвинизма и
обнаружил, что Дарвин начал не с Мальтуса, а с естественно-
го богословия. В итоге свою книгу Кауфман назвал «Вновь от-
крывая священное». То есть, признавая дарвинизм, он указал
на то, что «отбор» истолкован теперь как всепроникающий
феномен самоорганизации (фактически это наш делектус),
что он является тем божеством, которое управляет миром,
включая общество. По-моему, он наконец-то разъяснил, что
значит «метафора» у Дарвина (см. п. 8). О связи идей эволю-
ции и религии см. в книге [Чайковский, 2006, с. 15—19].
Для Кауфмана «отбор» как нечто священное - важная
часть нынешнего научного видения мира.
Но главным у Кауфмана остается, на мой взгляд, указание
на эволюционную роль случайности без вероятности. К тому,
что сказано о ней в п. 13, надо добавить следующее.
С конца XX века на Западе широко известна концепция
разумного замысла (intelligent design), научная база креа-
ционизма. Ее главная мысль — наблюдаемый мир слишком
сложен, чтобы его можно было счесть возникшим самопро-
извольно, поэтому следует признать его продуктом высшего
ума. Вывод основан на том, что самопроизвольное приравне-
но к случайному, а «слепая случайность» противопоставлена
разумному замыслу. Но можно ли так делать? Единствен-
ный известный мне анализ этого вопроса провел Марк Перах
(США). Отметив, что серьезно обсуждать можно только ак-
куратные понятия, он предложил определения случайности
и сложности, тоже противопоставил «слепую случайность»
разумному замыслу и легко показал (в своих понятиях!) пол-
ную наивность всей концепции [Перах, 2001]. Его часто ци-
тируют, однако он даже не ставит вопроса: а что не наивно?
Здесь и надо вспомнить Кауфмана.
Перах показал одно: нельзя отрицать возможность одно-
кратного самопроизвольного возникновения сложного. Но
ведь загадка в другом - в ежесекундном появлении слож-
ного в каждой клетке. Ответ дал Кауфман: такое появление
сложного можно считать возможным в той же мере, что и его
первичное (при начале жизни) появление - если рассматри-
вать не вероятность (устойчивую частоту), а случайность на
грани порядка и хаоса. Эта случайность - не «слепая», она
активная.
47
Вольно говоря, если бы случайность была возможна лишь
в форме вероятности, жизнь была бы невозможна. Подроб-
нее см. книгу [Чайковский, 2004, с. 227, 230, 234]. Столь же
вольно: если возможна повседневная жизнь, то возможна и
эволюция. Но в какой мере? Появление человека из одной
клетки мы видим повседневно, а вот от четвероногого предка
- нет, этого никто не наблюдал, это умозаключение, это еще
надо доказать.
17. Номогенез снежинок и организмов.
Новый жоффруизм
Рассмотрим сперва снежинки (рис. 4). Чаще всего они
принимают форму плоского дендрита (ветвящейся звезды),
который мы в обиходе и называем снежинкой. Никогда не
было обнаружено двух одинаковых дендритов. Изредка (при
некоторых значениях температуры, влажности и давления)
образуются иглы, пластины, пирамиды и прочее. В основе
их форм — симметрия кристалла льда. Этот кристалл имеет
форму шестигранной призмы, т.е. его боковые грани прямо-
угольны (рис. 5). Поэтому на самом деле обычная снежинка
— это очень тонкая призма. При некоторых условиях замер-
зания получаются более толстые призмы, так что снежинка
может принять форму короткой колонны; возможны мелкие
изъяны лучей и как бы склеивание двух половинок звезд.
Но если в строении единого кристалла главное — запол-
нение пространства (замерзание сплошной массы воды дает
сплошную кристаллическую массу льда), то замерзание пара
и микрокапелек идет путем роста кристаллов в почти пустом
воздушном пространстве и потому дает массу отдельных сне-
жинок, каждая из которых устроена по-своему. Мы видим,
Рис. 4. Фотографии снежинок. Вид-
ны дендриты, пластинка и снежинка
с лучами в форме пластинок (комби-
нация блоков).
что они имеют различия, но
каждая весьма упорядочена.
Удивительно, что обычно
дендриты имеют примерно
одинаковый размер порядка
миллиметра. Что мешает сне-
жинке, пока она свободно па-
дает, расти и расти дальше?
Каждая предстает завершен-
ной симметричной формой
именно данного конкретного
размера. Еще удивительнее,
что эта симметрия одинако-
ва во всех частях снежинки.
Если дендриты столь раз-
личны и растут в свободном
пространстве, то что мешает
разным лучам одного ден-
48
дрита расти по различным законам? Например, иметь один
луч ветвящимся, а другой — в виде пластины? Очевидно, что
в снежинке, как целостной системе, идет какой-то процесс са-
моорганизации, вскоре завершающийся.
То же удивление возникает при изучении морозного узора.
Он возникает на внешней плоской опоре, а значит, имеет сво-
боду роста не в трех, а только в двух измерениях. Он замечате-
лен тем, что являет собой крупную картину, рисуемую неве-
домой рукой по единому правилу. В этом правиле не заметно
симметрии, зато давно отмечено другое его свойство: каждая
картина похожа на какое-нибудь растение.
Полвека назад биолог-теоретик А.А. Любищев провел у
себя в Ульяновске поучительный опыт. Он сфотографировал
несколько морозных узоров, показал снимки коллегам и за-
тем писал: «Один из рисунков квалифицированный ботаник
принял за фотографию чертополоха, в других — очевидное
сходство с ... листьями пальмы, корневищами растения с
отходящими от него листьями, подобием мхов и лишайни-
ков».
Кристаллизация идет не путем приспособления к среде и не
как следствие истории развития, а по законам осаждения твер-
дых тел из пара - это общее. А вот образование конкретного
рисунка есть следствие приспособления начального зародыша
кристалла к его микроусловиям — это частное. Аналогично и в
живом веществе должны, по Любищеву, играть роль собствен-
ные законы образования форм — общие и частные10. Их и следу-
ет искать.
Рис. 5. Развитие зародыша снежинки до стадии призмы.
Обычно снежинка имеет почти идеальную 6-гранную или
6-лучевую (редко 3-гранную или 12-лучевую) плоскую сим-
метрию. Тот факт, что каждый луч растет той же формы, что
и его братья, загадочен, но он ясно говорит о наличии общей
программы развития. В чем она состоит, где и каким кодом
записана, каким образом распределяется по шести лучам оди-
наково? Каким механизмом этот код реализуется в тело сне-
жинки? Ответов пока нет, однако того факта, что программа
есть, и притом без генов, вполне достаточно, чтобы обрисо-
вать задачи биологического номогенеза в его нынешнем по-
нимании.
10 Легко видеть параллель с прогрессом и приспособлением по Ламарку. У Люби-
щева о ней речи нет.
49
На первых стадиях роста снежинки из центра конденсации
(на пылинке или микрокапле в атмосфере) возникают струк-
туры всего нескольких типов — игла, столбик, пирамида,
пулька и др. Первые стадии развития для всех снежинок типа
пластины или дендрита одинаковы — от «точечного» центра
конденсации до призмы (см. рис. 5). Это похоже на ситуацию
в биологии: по первым стадиям развития зародыша опреде-
лить облик будущего организма невозможно — он выявляется
только тогда, когда появляются зачатки органов.
Еще больше сходства с развитием организма являет мороз-
ный узор. Картины его тоже неповторимы. Например, на рис.
6 видим нечто вроде снимка заснеженной заросли сорняков
на краю огорода. Каждая «ветвь» не просто растет на десятки
сантиметров от своего центра кристаллизации, но и образует
«листья» — все одного типа.
Считается, что у организма всё закодировано в его геноме.
Но генов у человека всего 30 тыс., для создания наличного
разнообразия их явно не хватит. Более того, даже некодиру-
ющих участков ДНК не хватит: их около миллиарда, а одних
лишь связей между нейронами — триллион. Нестыковка при-
водит в отчаяние, пока не узнаешь, что в снежинках и окон-
ных узорах великолепное огромное разнообразие создается
вообще без генов.
Следовательно, в каждом из этих случаев работает какой-
то механизм, порождающий бесконечно разнообразные боль-
шие структуры из однообразных крохотных. Этот механизм
известен уже более 30 лет как фрактальный рост. О его роли
в эволюции много написано (в частности, см. книги [Nottale,
2000; Чайковский, 2004; 2006; 2008]), и здесь необходимо до-
бавить: фрактальный механизм способен порождать крупные
закономерно устроенные формы из микроскопических заро-
дышей таким образом, что малое изменение фракталообразу-
Рис. 6. Морозный узор на стекле.
Каждый «стебель» тянется от
своего центра кристаллизации, со-
храняя форму (свою и «листьев»),
то есть свой тип развития. Фото:
www.nikvel.ru.
ющего правила может вызы-
вать радикальное изменение
крупной формы, не наруша-
ющее ее единства как цело-
го. Так, ничтожная разница
в зародыше, сложившемся
из первых молекул воды, на-
липших на пылинку, ведет
к появлению совсем разных
форм снежинки, причем из-
менение одинаково проявля-
ется на всех ее лучах. То же
— с морозным узором: из-за
ничтожного различия цен-
тров конденсации один узор
смотрится (по всей его дли-
50
не!) чертополохом, другой
— папоротником, третий —
пальмой.
В биологии акт переклю-
чения процесса развития (он
сходен с модификацией фрак-
тала) именуется гомеозисом.
Суть его в том, что из-за му-
тации одного гена онтогенез
может резко измениться —
например, у дрозофилы на
месте усика вырастет доба-
вочная ножка или на месте
жужжальца — добавочное
крылышко. Гомеозис был от-
крыт еще в 1894 году, но под-
робно его впервые исследова-
ла генетик Е.И. Балкашина,
сделавшая в 1928 году важ-
ное наблюдение: все четыре
известных тогда гомеозисных
гена дрозофил сидят рядом,
на одном коротком участке
третьей хромосомы. Она от-
метила, что гомеозисная му-
тация не только порождает
уродливый орган, но и видо-
Елизавета Ивановна Балкашина
(1899— 1981), замечательная ис-
следовательница, открывшая в
1928 году у дрозофил группу генов,
ответственных за ключевые стадии
онтогенеза (впоследствии у них был
найден почти одинаковый начальный
участок, названный гомеобоксом).
Вскоре была арестована и в науку
не вернулась.
изменяет другие органы (в наших нынешних терминах — как
бы меняет фракталообразующее правило), и сделала вывод,
что эти гены ответственны за переход к новым стадиям онто-
генеза.
Через полвека молекулярная генетика подтвердила вывод
Балкашиной и выявила поразительную общность гомеозиса:
все его гены имеют в начале участок (гомеобокс), почти или
совсем одинаковый для данного гена у самых разных организ-
мов — цветковых, червей, мух, позвоночных. Каждый гомео-
зисный ген играет во всех организмах одну и ту же роль: один
определяет закладку передне-задней оси зародыша (такая ось
есть и у листа), другой — закладку головы животных, третий
— закладку глаза и т.п.
И здесь следует поразмышлять: организмы устроены со-
всем различно (например, насекомые имеют внешний хитино-
вый скелет и фасеточный глаз, а позвоночные — внутренний
костный скелет и камерный глаз), но управляются одинако-
выми генами. Поэтому возврат интереса ученых к гомеозису
изменил всю эволюционную проблематику.
Во-первых, подтвердилась давняя догадка номогенетиков
- сходство само по себе еще не говорит о родстве. В самом
51
Александр Александрович Любищев
(1890—1972), сторонник номогенеза,
обеспечивший нам связь классической
научной культуры с нынешней.
деле, если даже допустить,
что общий предок насекомых
и позвоночных существовал,
то глаз у него не было, даже
примитивных.
Принято считать, что точ-
ное совпадение нуклеотид-
ного текста какого-либо гена
у разных организмов дока-
зывает их происхождение
от единого предка, так как
«случайные мутации не мо-
гут создать два идентичных
текста». Даже открытие гори-
зонтального переноса генов
эту уверенность не пресекло,
и на ней до сих пор строится
молекулярная систематика.
Однако предполагать всюду
случайные мутации не име-
ет смысла. Если ген — за-
пись итога решения какой-то
функциональной задачи, то
он и должен быть одинаков у всех, кто ее верно решил.
Приведем пояснение через аналогию: число л можно вы-
числять принципиально различными способами, но резуль-
тат окажется одинаковым до любого знака, кто бы и как его
ни вычислял. Таковы и гомеобоксы: гены (точнее, их началь-
ные участки), запускающие одну и ту же стадию онтогенеза,
оказались у самых различных организмов одинаковы просто
потому, что решают одну и ту же задачу, независимо от ис-
пользуемого материала.
Во-вторых, многие стали возвращаться к жоффруизму и
связанному с ним направлению «eoo-deoo» (см. п. 7), близко-
му к тому, что можно назвать молекулярным жоффруизмом.
Его авторы впервые (после полувекового перерыва) заявили,
что решение вопросов эволюции видится не на основе одной
лишь генетики: «Подобно тому как клетка (а не ее геном) слу-
жит единицей органической структуры и функции, морфоге-
нетическое поле (а не гены или клетки) представляется основ-
ной единицей онтогенеза, и его изменение влечет за собой из-
менения в эволюции».
Морфогенетическое поле — понятие, обозначающее фор-
мообразующую силу, полвека назад изгнанное из науки как
идеализм. Поле оказалось сходно устроенным у самых разных
организмов; вот почему гены, кодирующие морфогенетиче-
ские оси, одинаковы у разных животных и сходны даже с рас-
тительными. Очевидно, что мутация в таком гене может дать
52
организму совсем иную форму. Здесь номогенез (включающий
«evo-devo») естествен и материалистичен, так как объясняет
сходство объектов общностью законов формообразования - в
точности, как в химии. Но как при этом достигается приспо-
собленность к жизни? Тут с идеализмом так легко не раздела-
ешься, требуется тщательный подход.
18. Активность и цель
У фасеточного глаза насекомого и линзового глаза позво-
ночного общей является только идея зрения, у внешнего ске-
лета насекомого и внутреннего скелета позвоночного - идея
подвижного каркаса, а у их сердец - идея прокачки жидкости.
Материальные реализации совсем различны и от общего пред-
ка идти не могут. Общим предком оказывается не организм, а
его идея, что для дарвинистов неприемлемо, хотя именно они
привыкли вспоминать общего предка. Выход видится мне в
том, чтобы о нем больше не говорить.
Во-первых, стало ясно, что предка обычно указать нельзя
- как в силу блочности эволюции (номогенез по Соболеву), так
и в силу горизонтального переноса наследственности. Во-вто-
рых же, понять эволюцию всерьез — значит уметь с понима-
нием использовать, для чего надо знать механизм появления
важных нам свойств - например, чтобы уметь создать вид,
способный заполнить опустевшую экологическую нишу. Зоо-
лог Н.Н. Воронцов еще полвека назад ставил вопрос: как за-
полнить нишу стеллеровой коровы?
Для этого надо знать параллели свойств, они хорошо видны
из рефренов, а те поразительны и наводят на мысль о единстве
именно идеи, в них заложенной. В книгах [Чайковский, 2006;
2008] дано много примеров. Приведем один из них.
Черви, как известно, делятся зоологами на пять типов, т.е.
различаются не меньше, чем, например, рыбы, осьминоги И
мухи. И всё же личинки червей разных типов обладают удиви-
тельным общим свойством — умеют сменять поведение своих
промежуточных хозяев с адаптивного на самоубийственное,
активно мигрируя в их «мозг». Каждый такой случай фанта-
стичен и воспринимался скептиками как шутка, пока все вме-
сте они не выстроились в четкие ряды. Только эти ряды (ряды
идей) придают всему феномену реальность. В разделе II будет
приведен пример сходного ряда активных миграций, но - не
личинок, а молекул (презентация антигена).
Словом, феномен активности нам еще понятен мало, но
уже видно, что он подчинен некоторым правилам. Одно из
них - наличие цели в природе. В космологии, например,
ее именуют антропным принципом, и некоторые видят в
нем аналог дарвинизма. Пусть цель выглядит в неживом и
живом различно, но она и тут и там — итог активности ма-
терии.
53
Самый наглядный пример реализации дальней цели —
преадаптация (см. п. 16). Обычная адаптация - тоже реали-
зация цели, но ближней. Точнее, цель в мышлении, в наслед-
ственном приспособлении и в преадаптации достигается за
счет одного и того же явления — делектуса (или, согласно С.Э.
Шнолю, блочно-иерархического принципа - см. п. 15). Здесь
остается напомнить высказывания Берга, Любищева и дру-
гих классиков, видевших в целеполагании основное свойство
живого.
Хотя еще господствует точка зрения, что полагать себе
цель может лишь человеческое сознание (см., например, [Рас-
ницын, 2008, с. 8]), но этот наивный антропоцентризм яв-
ляется, на мой взгляд, лишь наследием царивших в XX веке
позитивизма и марксизма. Нам всем предстоит привыкнуть
к целеполаганию в природе как к одному из главных проявле-
ний активности материи — точно так же, как наши деды при-
выкли к относительности и квантам.
19. Нанотехнология эволюции
Кстати, квантовая теория имеет отношение и к пониманию
эволюции. Мне уже приходилось упоминать [Чайковский,
2008, с. 403] мнение итальянского физика Эмилио Дель Джу-
диче (Del Gludice), что квантовая электродинамика - «есте-
ственный концептуальный каркас для понимания материи и,
в частности, живой материи». Речь шла там об удивительных
свойствах комплекса (домена) молекул воды, ведущего себя как
единое целое, способное, в частности, испускать общий мощ-
ный квант, пригодный в качестве источника энергии для мно-
гих биохимических реакций. Они тем самым могут протекать
не только за счет разложения (понижения энергетического со-
стояния) органических молекул (прежде всего, глюкозы), но и
за счет понижения энергетического состояния доменов воды.
Иными словами, без изменения ковалентных связей.
Открытие такого вездесущего источника энергии, как до-
мен воды, позволяет понять многое в природе ферментатив-
ных реакций. Биофизик В.Л. Воейков (биофак МГУ) в своих
работах показывает, опираясь на идеи Дель Джу диче, что
вода служит как источником энергии, так и структурным кар-
касом самых разных биохимических объектов [Воейков, 2009;
Чайковский, 2008; Del Gludice, 2005].
Концепция Дель Джу диче и Воейкова задает «концепту-
альный каркас» будущей теории, но ничего не говорит о пу-
тях возникновения и развития конкретных биологических
структур (как термодинамика ничего не говорит о путях хи-
мических реакций). Поэтому очень важно и отрадно появле-
ние книги московских биофизиков Голубевых (2009), отца и
сына, прямо заявивших об определяющей роли квантового
вакуума (они называют его эфиром) в становлении разноо-
54
бразия биоструктур. Оценить справедливость их выводов не
берусь, ибо несведущ в новой квантовой теории, однако даже
если они все и окажутся неверными, за авторами останется та
огромная заслуга, что они нашли нужные вопросы, а не успо-
коились на общепринятых каверзных ответах.
Главным в концепции Голубевых является не вода, а угле-
род. Как известно, у него есть две основные кристаллические
формы — алмаз и графит. Голубевы в качестве блоков (единиц
комбинирования) структурного каркаса, формирующего ма-
кромолекулы, видят не домены воды, а кристаллы алмаза и
графита. Конечно, необходим синтез обеих названных кван-
тово-биологических концепций (уже видно, что первая тяго-
теет к ламаркизму, а вторая — к номогенезу), но его пока нет,
и нам остается использовать их порознь.
Для начала Голубевы рассмотрели микроскопические во-
доросли кокколшпофориды, составляющие основную часть
так называемого нанопланктона (планктона, ускользающего
из планктонной сети). Часть жизни клетка несет кальцито-
вый панцирь, фрагменты которого, кокколиты (в переводе с
греческого — камнезернышки), являют собой основную часть
залежей писчего мела. Кокколит состоит из наночастиц, ви-
димых лишь в электронный микроскоп, т. е. построение пан-
циря является естественной нанотехнологией.
Назначение панциря загадочно: хотя на его создание идет
половина энергии, добытой фотосинтезом, защитой он не слу-
жит (ведь животные глотают объем планктона целиком), зато
лишает клетку плавучести. И вот Голубевы пришли к выводу,
что он служит орудием антигравитации. Мысль выглядела
безумной, но вскоре антигравитацию открыли космологи, и
ныне ситуация меняется.
Разнообразие микронных кокколитов (оно частично пока-
зано на рисунках второй страницы обложки) сильно напоми-
нает разнообразие гораздо более крупных объектов - милли-
метровых снежинок. Нам важно, что в обоих случаях меха-
низмом формирования признан фрактальный рост кристал-
лических структур.
Не вдаваясь в предложенную Голубевыми теорию, скажу
только, что самосборку наноструктур они трактуют, по сути,
номогенетически, как комбинирование небольшого числа
блоков. Понимание устойчивости как самих блоков, так и про-
цессов сборки упирается у них в понимание структуры эфира.
Именно ею авторы объясняют всё то, что другие связывают с
полями, в том числе с гравитацией, антигравитацией и с упо-
мянутыми мною в п. 17 биополями. Структуру эфира они по-
лагают квазикристаллической, каковой является (что давно
известно) и структура воды.
В нынешних руководствах по биологии о механизмах эво-
люции не говорится ничего, кроме голых деклараций. Это,
55
по мнению Голубевых, вызвано пропастью «между широким
кругом биологических явлений и общепринятой физической
картиной мира». Без организующего эфира ни эволюционная
изменчивость (даже мелкая), ни и удачные варианты анти-
тел не могли бы появляться иначе, чем «на уровне единич-
ных курьезов» [Голубевы, 2009, с. 152, 154].
Вместо «организующего эфира» мы выше вели речь про
активность, но по сути дела речь шла о том же самом — мир
устроен так, что материя сама собой организуется, порож-
дая всё более и более сложные системы. На всех уровнях ор-
ганизации виден один и тот же эффект - актиреф (активное
движение вдоль рефрена; о рефренах см. п. 14). Если речь
идет о биологическом объекте, то актиреф имеет две формы:
развитие особи (онтогенез) и эволюцию. Поскольку рефрены
принадлежат общей диасети (о ней см. тоже п. 14), то эти две
формы сходны (известный из школы биогенетический закон).
Здесь мы видим соединение в одной схеме идей номогенеза
(наличие диасети), жоффруизма (активное изменение в раз-
витии особи) и ламаркизма (активная эволюция).
Давайте исследуем актиреф на примере. Обратимся, вслед
за Голубевыми, к «нанотехнологии» антител. Для этого при-
дется сперва рассказать кое-что про иммунологию вообще.
ЛИТЕРАТУРА К РАЗДЕЛУ I
Агаев М.Г Экспериментальная эволюция. Л., Изд-во ЛГУ, 1978.
Аронова Е. А. Неумирающий ламаркизм. Падения и взлеты // Биология
(прилож. к газете «Первое сентября»), 1997, № 41, 42.
Беляев А.А., Раскина О.М. Высокоповторяющиеся фракции ДНК как
индикатор процесса видообразования у видов семейства Роасеае // Хромо-
сома 2009. Мат-лы междунар. конф. Новосибирск, 2009 (см. также: Беляев,
автореф. докт. дисс., C-Пб. унив-т, 2009).
Воейков В.Л. Ключевая роль устойчиво неравновесного состояния во-
дных систем в биоэнергетике // Российский химич. журнал, 2009, № 6.
Гилберт С.Ф., Опиц Д.М., Рэф Р.А. Новый синтез эволюционной биоло-
гии и биологии развития // Онтогенез, 1997, № 5.
Гиляров М.С. Отряд Термиты (Isoptera) // Жизнь животных, т. 3. М.,
1969.
Голубев С.Н., Голубев С.С. Взгляд на физический микромир с позиции
биолога. Владивосток, Дальнаука, 2009.
Голубовский М.Д. Неканонические наследственные изменения // Приро-
да, 2001, №8, 9.
Дарвин Ч. Сочинения. М.-Л., Изд. АН, 1936 (т. 2), 1939 (т. 3), 1948 (т. 7),
1959 (т. 9).
Карасева ТА. Самонесовместимость и гииодиэция... //XXIII Любищев-
ские чтения. Ульяновск, 2009, с. 47—58.
Кауфман С.А. Антихаос и приспособление // В мире науки, 1991, № 10.
КлэкДж. Трудный путь на сушу //В мире науки, 2006, № 3.
Лекявичюс Э. О некоторых аналогиях между эволюцией экосистем и раз-
витием экономики... // Эволюция: космическая, биологическая, социальная.
М., 2009.
Назаров В.И. Эволюция не по Дарвину. М., УРСС, 2005.
56
Перах М. Разумный замысел или слепая случайность? // Континент,
2001, № 107.
Пчеленко С.П. Естественный отбор: новые основания // XXIII Люби-
щевские чтения. Ульяновск, 2009, с. 144—157.
Райков Б.Е. Германские биологи-эволюционисты до Дарвина. Л., На-
ука, 1969.
Расницын А.П. Теоретические основы эволюционной биологии // Жери-
хин В.В. и др. Введение в палеоэнтомологию. М., КМК, 2008.
Рачинский С. А. Цветы и насекомые // Русский вестник, 1863, № 1.
Стил Э., Линдли Р., Бландэн Р. Что, если Ламарк прав? Иммуногенети-
ка и эволюция. М.: Мир, 2002.
Чайковский Ю.В. Элементы эволюционной диатропики. М., Наука, 1990.
Чайковский Ю.В. О природе случайности. М., Центр сист. исслед., 2004.
Чайковский Ю.В. Наука о развитии жизни. Опыт теории эволюции. М.:
КМК, 2006. 712 с.
Чайковский Ю.В. Диатропика С.В. Мейена: сегодняшний взгляд // Во-
просы философии, 2006а, № 5.
Чайковский Ю.В. Активный связный мир. Опыт теории эволюции жиз-
ни. М., КМК, 2008.726 с.
Чайковский Ю.В. Ламарк, Дарвин и устройство науки // Вестник Рос-
сийской Академии наук, 2010, № 4.
Шапошников Г.Х. Специфичность и возникновение адаптации... // Энто-
мологическое обозрение, 1961, №4.
Шапошников Г.Х. Морфологическая дивергенция и конвергенция в экспе-
рименте с тлями... // Энтомологическое обозрение, 1965, № 1.
Шноль С.Э. Хватает ли времени для дарвиновской эволюции? // При-
рода, 1990, № 11.
Baxter S. W. et al. Convergent evolution in the genetic basis of Mullerian
mimicry in heliconius butterflies // Genetics, 2008, vol. 108 (3), Nov.
Burkhardt R. W. The spirit of system. Lamarck and evolutionary bioligy. Ca-
mbridge (Mass.), 1977.
Del Gludice E e.a. Coherent quantum electrodynamics in living matter //
Electromagnetic Biology and Medicine, 2005, vol. 24, pp. 199-210.
Griffiths P.E., Stotz K. Genes in the postgenomic era // Theoretical Medici-
ne and Bioethics. 2006, vol. 6, p. 499—521.
Hartley B.S. et al. Experimental evolution of a xylitol dehydrogenase // Struc-
ture-function relationships proteins. Amsterdam, 1976.
Horiuchi T„ Aigaki T. Alternative trans-splicing: a novel mode of pre-
mRNA processing // Biol. Cell, 2006, vol. 98. p. 135—140.
Hughes A.L. Looking for Darwin in all wrong places: the misguided quest
for positive selection at the nucleotide sequence level //Heredity. 2007, vol. 99,
p. 364-373.
Kauffman S.A. Reinventing the sacred. A new view of science, reason and
religion. N.Y., 2008. 320 pp.
Kirschner M., Gerhart J. The plausibility of life: resolving Darwin’s dilem-
ma. Yale University, 2005.
Magalhaes S. et al. Are adaptation costs necessary to built up a local adapta-
tion pattern? // BMC Evolutionary Biology, 2009, vol. 9, August 3.
Nottale L., Chaline J., Grou P. Les arbres de revolution. Paris, Hachette,
2000.
Todes D. P. Darwin without Malthus. The struggle for existence in Russian
evolutionary thought. N.Y., 1989.
Westbroek P. La Terre est-t-elle un superorganisme? // La Recherche, 1997,
№ 295.
Wu T. T. Experimental evolution in Bacteria // CRCCrit. Rev. Microb. 1978,
vol. 6, № 1.
57
РАЗДЕЛ II. ЮБИЛЕЙ ЛАМАРКА - ДАРВИНА
И РЕВОЛЮЦИЯ В ИММУНОЛОГИИ
Рассмотрим подробнее одну иллюстрацию основы эволюции
— роль новых представлений об эволюции в понимании иммуни-
тета и, обратно, роль последнего в понимании самой эволюции.
Не зная места иммунитета в общей биологии (особенно - его
роли в онтогенезе и в нескрещиваемости), в наши дни можно
кое-как обсуждать пути эволюции, но никак не ее механизмы.
Точно так же, как сто лет назад нельзя было без генетики.
Конечно, лучше это сделал бы иммунолог, знающий эволю-
цию. Но где его найти? Поэтому для освещения данной темы
пришлось прибегнуть к помощи консультантов, котором ав-
тор книги выражает благодарность.
Часть 1. Иммунитет как борьба за существование
1. Эволюция сквозь призму иммунологии
Хотя в 1840—1858 годах Ламарк как эволюционист не упо-
минался в кругу Дарвина вовсе, однако его хорошо знали.
Почти все ранние комментаторы учения Дарвина — востор-
женные, критические и ниспровергавшие — сравнивали его
с учением Ламарка, и отнюдь не все видели в Ламарке про-
сто предшественника. Например, немецкий переводчик книг
Дарвина, знаменитый зоолог и палеонтолог Генрих Брони,
сравнивая концепции Ламарка и Дарвина, подчас отдавал
предпочтение первому — например, в вопросе: почему, несмо-
тря на эволюцию, до сих пор существуют низшие животные?
(поставлен в разделе I)
Словом, история науки отнюдь не так проста, как многим
кажется, и как раз в дни двойного юбилея — великих книг Ла-
марка и Дарвина — ее полезно вспомнить, посмотрев, как они
повлияли на развитие наук. Выберем иммунологию — она
формировалась в параллель с дарвинизмом11, черпая идеи из
него и из ламаркизма; а успехи и неудачи эволюционизма на
ней легче всего видны.
11 Роэдение иммунологии относят к 1876 году, когда Л. Пастер получил во Фран-
ции вакцину против куриной холеры, показав тем самым, что давно известный
пример лечения оспы посредством вакцины (Э. Дженнер в Англии, 1796 год) мож-
но распространить на другие болезни и что можно обосновать лечение теоретиче-
ски. То была иммунология растворов (гуморальная). В 1883 году И.И. Мечников
показал, работая в Италии, что реакцию иммунной защиты ведут особые клетки
- фагоциты, поедающие чужеродные клетки, и тем самым основал клеточную
иммунологию. А в 1901 году К. Ландштейнер в Австрии открыл группы крови и
их иммунную природу. Тогда же в обиход вошло ставшее основным в иммуноло-
гии понятие антитела - частицы, нейтрализующей чуждую частицу (антиген)
\Ульянкина, 1994]. В тот же год появилось понятие мутации, основное в генетике
и дарвинизме. Так начался для биологии XX век. Для физики он начался появле-
нием понятий кванта и относительности.
58
Генетика прекрасно объяснила, как из линейного текста
гена получается линейный текст белка, но ничего не смогла за
сто лет сказать (кроме совсем абстрактных предположений)
про то, каким образом из линейного текста генов получается
трехмерный орган — ну хотя бы капилляр. Поэтому вопрос:
«Можно ли наблюдать эволюцию?» на сегодня генетически
осмыслен только в одном плане: «Можно ли наблюдать по-
явление нового белка, такого, какого прежде в природе не
было?»
Да, можно, гласит иммунология. И это — огромный успех
всей биологии, успех, ключевой для эволюционной науки.
Однако на вопрос: «Как именно появляется нужный новый
белок?» ясного ответа нет, и это тоже видно из иммунологии.
2. Мечников и борьба клеток
Ознакомившись в 1863 году с «Происхождением видов» в
немецком переводе Бронна, юный Мечников (ему было всего
17 лет) тут же написал критическую статью о новом учении. В
ней много мальчишеского задора и лишней самоуверенности,
но черты будущего классика уже видны: это наблюдательность,
завидная эрудиция, умение сопоставлять факты из самых раз-
ных областей знаний и полная независимость в суждениях.
Приняв идею эволюции как возможность, юный автор напрочь
отверг предложенный Дарвином ее механизм — борьбу за дефи-
цитный ресурс как основу естественного отбора.
Впоследствии, в годы зрелости, Мечников признал учение
Дарвина, но опять-таки — без мальтузианской его компонен-
ты. Тем самым он, сам того не зная, внес солидный вклад в
особый вариант дарвинизма - «Дарвин без Мальтуса» (см. и.
4, раздел I).
Мечникова привлекала борьба за существование отнюдь
не между особями одного вида за ресурс (как у Дарвина), а
между особями разных- видов. Именно такую борьбу он нашел
в иммунной системе: фагоциты (рис. 7) поедают опасные для
организма клетки, не трогая клеток, ему полезных, каковые
и выступают победителями в борьбе за жизнь. Аналогичную
борьбу он увидел и в эволюции. Так в иммунологию вошел
дарвинизм.
В данной схеме мальтузианству действительно нет ме-
ста, но беда в том, что для объяснения эволюции она мало-
пригодна. В самом деле, дарвинизм объяснял и объясняет
процедуру постепенного формирования полезного свойства
исключительно как вытеснение каждым более удачным ва-
риантом каждого менее удачного, то есть борьбой между осо-
бями одного вида в условиях перенаселения. Отрицая ее роль,
Мечников должен был предложить что-то ей взамен, что он и
сделал в работе «Борьба за существование частей животного
организма» (1892). Там причиной гибели клеток при фагоци-
59
Рис. 7. Наше нынешнее представление о фагоците. Его функцию не удается
описать в терминах механики или химии, и ее всегда описывают на языке
поведения (то есть активности), словно фагоцит — это организм.
А — хемотаксис (движение в сторону повышения концентрации важного
для клетки вещества) фагоцита.
Б — фагоцитоз (поедание фагоцитом другой клетки); опсонины — общее
название для белков, которыми иммунная система покрывает клетку,
предназначенную к уничтожению.
тозе названа их неспособность работать на благо организма,
т.е. неупотребление органа по Ламарку. Так в иммунологию
вошел ламаркизм.
Эту причину Мечников видел и в эволюции. Например,
отмирание глаз у подземных животных он объяснял тем, что
глаза в почве легко воспаляются, тогда как пользы от них в
темноте нет. Неупотребление и отбор действуют здесь, по Меч-
никову, совместно. Но глаза столь же быстро отмирают и у пе-
щерных животных, где особых оснований к их воспалению нет,
т.е. неупотребление органа действует здесь само по себе.
3. Эрлих: активность и селекция антител
Новая наука притягивает выдающиеся умы. Двое великих
биологов заложили две ветви иммунологии: Луи Пастер — хи-
мическую, а Илья Мечников — клеточную. Сперва эти подхо-
ды конфликтовали, но на грани веков появился третий клас-
сик — немецкий врач и биохимик Пауль Эрлих, сумевший
взять от каждой ветви главное. У химической он заимствовал
понятие антитела (см. словарик), как раз тогда получившее
хождение среди ученых, а у клеточной — идею клетки как ак-
тивного иммунного деятеля.
Активность особи — основной фактор эволюции у Ламарка.
Употребление и неупотребление органа можно описать как
две крайние степени активности. Она имеет прямое отноше-
ние к иммунологии, поскольку позволяет говорить о поведе-
нии фагоцитов и других клеток, а также их частей, словно они
являются особями.
60
Главный вывод, к которому пришел Эрлих, состоял в том,
что антитело не может образовываться химически в ответ на
появление антигена (и в этом был, в рамках химии, прав). А
значит, решил он, «физиологические аналоги антител долж-
ны существовать заблаговременно в организме и в его клет-
ке». Если так, то попавший в клетку антиген вызывает всего
лишь усиленную выработку нужного антитела из предше-
ственника, т.е. (как стали говорить позже) производит свое-
образную селекцию. Здесь уже Эрлих был прав лишь отчасти:
конечно, какие-то предшественники антител необходимы, но
они никак не могут быть их «физиологическими аналогами»,
поскольку физиология будущих антител может быть весьма
различна, и заранее не предскажешь, какой аналог нужен.
4. Ландштейнер и искусственные антигены
В начале XX века в иммунологии появился еще и четвер-
тый классик — австрийский врач и биолог Карл Ландштей-
нер. У него много заслуг, но нам он интересен тем, как развил
ламаркизм в иммунологии. Ландштейнер усомнился в том,
что предшественники на все случаи жизни могут заранее су-
ществовать, и поставил, начиная с 1912 года, серию решаю-
щих опытов. Он вводил в кровь подопытных животных искус-
ственные антигены, т.е. такие химические вещества, каких в
прежней истории животных быть не могло. Антитела выраба-
тывались и на них!
«Специфичность антител оказалась настолько велика, что
оказалось возможным получать сыворотки, различающие
орто- и параизомеры одной и той же молекулы» [Аронова,
2006, с. 21]. Если число возможных вариантов антител столь
велико, то одновременное их наличие у каждой особи неверо-
ятно, и приходится признать, что иммунная система каким-то
образом узнает строение внедрившегося антигена и делает к
нему антитело. Другими словами, антиген как бы дает иммун-
ной системе инструкцию, по которой формируется антитело.
Так возник вековой спор двух школ (вернее, лагерей) в по-
нимании иммунного ответа — инструктивной и селективной.
Участники спора полагают, что основывают свои взгляды на
фактах иммунологии, не подозревая, что на самом деле про-
сто объявляют ученому миру свою принадлежность к опреде-
ленной натурфилософской позиции, что сами позиции гораз-
до раньше обозначены эволюционистами, а идут из антично-
сти [Lherminier, 1997].
Первая позиция исходит из убеждения, что все сущности
вечны и наблюдаемые тела - лишь реализация их идей (по-
зиция Платона), а вторая — что мы наблюдаем реальное по-
явление новых сущностей (позиция Аристотеля). Короче, ты-
сячелетний спор идет о том, что в природе старше — само тело
или его идея (его сущность, информация о нем)?
61
В наше время принято считать, что философским основа-
нием всех наук является принцип причинности, гласящий,
что всякое следствие следует во времени за своей причиной.
Поскольку антитело есть ответ организма на антиген, т.е.
следствие появления антигена в организме, то оно должно по-
являться позже, чем антиген. Инструктивная теория как раз
это и утверждает, тогда как селективная — наоборот. Выходит,
что с позиции принципа причинности первая теория научна,
а вторая — нет.
Так и утверждали инструктивисты, тогда как селекцио-
нисты отвечали им, что антитела возникают случайно и что
некоторые из них оказываются (случайно же) годными для
борьбы с данным антигеном. То есть принцип случайности
был выдвинут взамен принципа причинности, чтобы обойти
(сознательно или нет, неважно) вопрос о платонизме.
Законно ли это? Сделав допущение, что нужное антите-
ло появляется с определенной вероятностью (это далеко не
всегда так — см. раздел I), можно сказать: да, если случайные
события могут за приемлемое время (в примере с антителом
— около трех суток) породить в организме всё то, чего от них
ждут. Нет, если расчет покажет, что не могут. Словом, тогда
вопрос упирается в арифметику. Легко догадаться (ниже мы
это увидим), что арифметика высказалась решительно про-
тив селекционистов и в пользу инструктивистов.
Но Ландштейнера мало кто слушал. Получив в 1930 году
Нобелевскую премию, он с изумлением прочел в дипломе, что
его наградили лишь за весьма давнее (1901) открытие групп
крови, тогда как сам он считал своим главным делом жизни
получение антител к искусственным антигенам и анализ их
свойств [Ульянкина, 1994, с. 191].
5. Попытка соединить теории
Австралийский вирусолог Фрэнк Макферлейн Бернет
в 1941 году обратился к иммунологии и горячо поддержал
Ландштейнера, предложив свою модель выработки антител,
чисто инструктивную. В ее основе лежала модная в те годы
параллель между высокой специфичностью антител и высо-
кой специфичностью ферментов: если антитело столь высоко
специфично к антигену, то оно и устройством должно быть
похоже на фермент. Такие модели имели большой успех, но
он был недолог [Аронова, 2006; Ульянкина, 1994].
В 1955 году датский иммунолог Нильс Ерне высказал ги-
бридную (селективно-инструктивную) идею. Слабую сторону
селективной идеи (огромное число различных антител, зара-
нее обитающих в организме, что невероятно) Ерне укрепил
допущением, что каждого антитела в организме — всего одна
или несколько штук. Тогда различных антител в нем могут на-
ходиться миллиарды, так что на каждый антиген, попавший
62
внутрь организма, вроде бы может найтись нужное антитело.
Чтобы схема могла работать, надо, чтобы в ответ на попада-
ние антигена нужное антитело начинало взрывообразно раз-
множаться. Так бывает при автокаталитическом (самоуско-
ряющемся) процессе, если (вот второе допущение) антитело
служит само для себя матрицей, т.е. инструкцией.
Прочтя статью Ерне, Бернет понял: если инструкция
необходима лишь для размножения нужного варианта, то
от инструктивного принципа вполне можно избавиться —
достаточно допустить (третье допущение) вместо копирова-
ния белковой молекулы мутирование гена, кодирующего эту
молекулу. Вместо случайного нахождения антитела в орга-
низме Бернет ввел (в статье 1957 года) принцип его случай-
ного появления.
Даже единственной мутантной клетки, производящей
нужное антитело, будет достаточно для борьбы с заразой,
если (вот четвертое допущение) эта клетка создаст клон, т.е.
начнет, в отличие от остальных клеток, безудержно де-
литься и тем самым произойдет селекция нужного антитела.
Избирательное клональное деление иммунных клеток было
уже известно, но причина его оставалась загадкой.
Итак, инструктивную гипотезу Ерне (редкий белок -» ча-
стый белок) Бернет заменил гипотезой соматической (проис-
ходящей вне половых клеток) мутации: новый ген -» новый
белок. Напомним, что «центральная догма молекулярной
биологии» (ДНК -» РНК -> белок) была высказана позже, в
1958 году (Ф. Крик). Инструктивный компонент пропал, и ро-
дилась чисто селективная «клонально-селекционная теория
образования антител». Ее мгновенный успех побудил Берне-
та написать книгу «Клонально-селекционная теория приоб-
ретенного иммунитета». Она вышла в свет в 1959 году, в год
столетия «Происхождения видов», а через год Бернет стал но-
белевским лауреатом.
Разумеется, устав премии был соблюден: формально ее
дали не за «теорию», а за предсказание Бернетом вполне кон-
кретного опытного результата — приобретенной иммунной
толерантности. Результат был установлен в 1956 году ан-
глийской группой Питера Медавара (он получил премию вме-
сте с Бернетом) и состоял в следующем.
У большинства видов животных ткань, пересаженная от
одного организма другому того же вида, не приживается, а
отторгается. Причина в том, что у них идет непрерывная про-
цедура распознавания белков (своих и чужих) на предмет
выявления чужеродных. Последние уничтожаются, а свои —
нет; к ним организм проявляет толерантность. Оказалось,
что толерантность у теплокровных не наследуется, а фор-
мируется в каждой растущей особи заново. Поэтому Бернет
предположил, что можно сделать животное толерантным к
63
чужим белкам, если ввести их в кровь новорожденного. Так и
оказалось.
Предсказание следовало из уже отвергнутой самим Бер-
нетом инструктивной модели (ее еще держался в то время
Медавар), но это никого не занимало, и оба вошли в историю
иммунологии как гении, избавившие иммунологию от «ереси
ламаркизма» или даже от «ламаркистского психоза». Вскоре
появилась «центральная догма молекулярной биологии», гла-
сившая, что источником новой информации являются только
случайные мутации в половых клетках. Концепцию Бернета
впоследствии стали, по аналогии, именовать «центральной
догмой молекулярной иммунологии». В самом деле, обе дог-
мы утверждали одно и то же — отрицали активность поиска
новых вариантов, как ее отрицает дарвинизм.
Но догмы, как известно, относятся к религии, а не к науке,
и это вскоре дало о себе знать. В частности, концепция Берне-
та оставалась недвижной, хотя всё более и более расходилась
с арифметикой. В разделе I мы видели, что то же самое было
и с дарвинизмом.
6. Долой арифметику!
Арифметику тогда никто из иммунологов, кроме выми-
равшей горстки инструктивистов, не вспоминал, но вопрос:
«Почему у организма хватает времени и молекул для ответа
на инфекцию?» — оставался, вызывал упреки, и вот в 1969
году Бернет с высоты нобелевского Олимпа решился его про-
яснить, притом в самом слабом месте своей концепции — в
проблеме ответа организма на искусственные антигены.
Приведя возражение инструктивистов о невероятности
наличия в одном организме антител ко всем принципиально
возможным антигенам, включая искусственные, Бернет отве-
тил: «В основе этого возражения лежит совершенно ошибоч-
ная интерпретация концепции рандомизации», т.е. концеп-
ции случайности.
На самом деле вопрос не так прост, к одной случайности
его никак не свести, но послушаем доводы Бернета, они лю-
бопытны. Он пояснил: допустим, что в алфавите из 20 букв
(таково число различных кодируемых аминокислот) закоди-
рован активный центр антитела, состоящий из двух пепти-
дов по 10 аминокислотных остатков каждый. Всего возможно
2010 или примерно 1013 вариантов каждого пептида.
Это количество, по Бернету, «всёравно еще слишком вели-
ко», поэтому он допустил, что не все замены имеют функци-
ональное значение. И объяснил, что значит «не все». Но его
объяснение придется процитировать почти целиком (опустив
только уходы в сторону, отмеченные многоточиями), так как
при пересказе вряд ли кто поверит, что нобелевский лауреат
мог подобное написать.
64
«Селективная теория постулирует, что в индивидуальном
организме, скажем, человека или кролика, продуцирует-
ся около 100 тыс. подобных структур (коротких пептидов —
Ю. Ч.), однако в каждой клетке может функционировать толь-
ко одна такая структура... Возникнув в организме в резуль-
тате дифференцировки, любая такая клетка должна пройти
“сквозь строй” собственных самых разнообразных потенци-
альных антигенных детерминант, которых насчитывается
около 100 тыс., и, “случайно” прореагировав со структурой
соответствующей специфичности, может образовать с ней до-
статочно прочную связь. Если такое событие действительно
произойдет, то, согласно гипотезе, эта клетка будет элимини-
рована. Таким образом, может быть элиминировано до 90%
случайно возникших структур, несущих иммунологические
рецепторы. В результате в организме останется 10% первона-
чально возникших клеток, несущих, скажем, 10 тыс. иммуно-
логически специфичных структур, обладающих свойствами
антител... Эти 10 тыс. иммунологически специфичных струк-
тур способны реагировать с любым синтетическим веществом,
так же как и с любым природным...» И наконец: «Такой сво-
бодный подход к проблеме является большим достоинством
селективных теорий. Антитела или рецепторы не продуциру-
ются организмом по заказу, в соответствии с инструкцией
антигена, синтетического или природного. Организм ответит
на введение антигена иммунологической реакцией лишь в
том случае, если данный потенциальный антиген отыщет в
организме иммуноциты..., с которыми этот антиген способен
прореагировать» [Бернет, 1971, с. 256].
На этом рассуждение заканчивается, не давая отсылок
к другим местам книги или к другим трудам. Сравнив его с
остальными положениями книги, приходится заключить:
Бернет заявил, что природа устроена так, как ему нужно для
теории, а не иначе, причем он выдумал для этого недостающие
факты и числа. Не верите? Мне самому долго не верилось, и в
своих работах я даже ссылался на «клонально-селекционную
теорию». Прочтем еще раз внимательно приведенную цитату.
Давайте пройдемся по местам, которые выделены выше мною
курсивом.
1. Упрекнув инструктивистов (каким недавно был сам) в
непонимании случайности, Бернет интересно разъяснил нам
суть дела: сперва дал слово случайность в кавычках (дескать,
не придирайтесь, случайность — просто метафора), а затем —
без кавычек (будто термин уже введен). Именно тут и так он
отказался от принципа причинности, основы наук.
2. Как уже сказано случайностью можно заменять при-
чинность, если нужные события происходят достаточно ча-
сто. Чтобы убедить нас в достаточности, Бернет привел чис-
ла. Для их оценки он выбрал воображаемый активный центр
65
антитела, поскольку тогда уже было известно, что реальный
активный центр фермента (группа атомов, служащая катали-
затором ферментативной реакции) всегда невелик и оценка в
10 «букв» реальна. Настораживает только ремарка: «всёрав-
но еще слишком велико» — Бернет явно понимает, что провел
негласное занижение, но даже его недостаточно.
Занижение колоссально: ведь активный центр фермента
отвечает лишь за сам ход реакции, тогда как за ее специфич-
ность (за то, что будет допущено к реакции, а что нет) отвечает
остальная молекула, всегда огромная. Так что 20 надо бы воз-
вести не в десятую, а хотя бы в сотую степень. Но тогда полу-
чится нелепо огромное число вариантов, подлежащих апро-
бации, — даже число элементарных частиц во Вселенной не
превышает 1О80.
3. Трудность необъятного числа вариантов Бернет решил
обойти признанием, что не все варианты надо рассматривать.
Святая правда! Только вот какую долю всё-таки рассмотреть
надо? Тут он поразил читателей простодушием — вместо 1013
оставил 106, т.е. одну стомиллионную от им же принятого
(безбожно заниженного) числа вариантов.
4. И тут-то получилось всё, что надо? Если бы так! Да, Бер-
нет оставил к рассмотрению 10 тыс. вариантов антител на ор-
ганизм (как мы узнаем дальше, у мыши их примерно столько
и есть), но трудность не исчезла, а лишь сместилась в иную
плоскость. Различных бактерий известно около 100 тыс., та-
кого же порядка число известных видов вирусов, и каждый
может выделять по нескольку антигенных веществ. Приба-
вим к этому искусственные антигены, которых можно насин-
тезировать невесть сколько.
Значит, различных антител нужно одновременно больше
миллиона. Но каждая В-клетка (см. словарик) синтезирует
один вид антител, и клеток каждого вида нужно для борьбы с
заразой сразу очень много — миллионы. В мышке такое чис-
ло В-клеток просто не уместится: в ней меньше 108 иммунных
клеток, из которых В-клетки составляют едва ли пару процен-
тов. Чтобы В-клетки нужного вида смогли в ней поместиться,
нужно как минимум все остальные В-клетки изъять, но тако-
го явления не наблюдается. А иммунитет мыши работает, и
ничуть не хуже, чем у человека.
5. Здесь Бернет смог одно - провести манипуляцию со сло-
вом «любой». В первом употреблении оно значит у него «каж-
дый» (каждая возникшая клетка, т.е. каждая возможная), а
во втором — «тот, который будет выбран» (антиген, выбран-
ный в данный момент иммунной системой). Невнимательный
читатель, не увидев разницы, подумает, что задача решена:
для каждого возможного антигена найдется свое антитело.
Очевидно, что на самом деле иммунитет устроен как-то
иначе, что мы и увидим.
66
Как же могло так получиться, что этот бессвязный набор
произвольных утверждений почти всех устроил? Это трудно
осознать, но Бернет своим способом рассуждать в точности
повторял Дарвина. У того было всего два «примера действия
естественного отбора» (см. п. 4, раздел I), и оба - без всякой
арифметики; однако ученый мир его принял. Можно предпо-
ложить: знай Бернет, что отбора в эволюции не найдено, он
бы со своей «теорией» не выступил.
7. Возвращение к арифметике
В 1970-х годах в блестящей австралийской школе имму-
нологов нашелся исследователь совсем иного типа, Эдвард
Стил, тогда еще юный стажер. Его собственную концепцию
(не во всем оправдавшуюся) мы рассмотрим позже, а пока
только позаимствуем у него некоторые числа, которые никто
никогда не оспаривал и не оспаривает.
Что быстрее размножается — бактерия или В-клетка нуж-
ного типа? Учебники и руководства по иммунологии дружно
обходят данный вопрос, а вот у Стила написано: бактерия
обычно делится ежечасно, тогда как В-клетка — за 5—6 ч.
Наработка нужного количества данного варианта лимфо-
цита производится (как известно врачам) двое суток, т.е. за
это время произойдет всего 10 делений каждого лимфоцита.
Это значит, что если нужный вариант найден лишь однажды,
то появится всего лишь тысяча нужных клеток - ничтожно
даже для мыши. Наоборот, болезнетворные бактерии обычно
попадают в организм целыми колониями, а затем делятся в
пять раз быстрее В-клеток. Клонирование их никак не смо-
жет поспеть за размножением заразы.
Конечно, иммунологи знали это и раньше, но успокаивали
себя побочными соображениями. Например: каждая В-клет-
ка испускает по 3 тыс. антител — на всех хватит. Но за какое
время? Антитела составляют долю процента от массы поро-
дивших их клеток [Полетаев, 2008, с. 44], значит, антител,
вышедших из одной В-клетки, одновременно бывает всего
около сотни. Поскольку они облепляют жертву десятками, то
одна В-клетка может поразить за час всего три - четыре клет-
ки бактерий. Даже если допустить нереальное - что в какой-
то момент достаточно одного В-клона, то через час-два бакте-
рий всё равно станет слишком много, так как геометрическая
прогрессия обгонит арифметическую.
Можно услышать и такое: у каждой бактерии сотни точек,
атакуемых антителами, - значит, данную популяцию бакте-
рий может атаковать сразу много антител различных клонов.
Уловка столь же слаба численно, но главное другое — имму-
нитет так не работает. Если бы он работал так, он поражал бы
все бактерии, попавшие в организм, а на деле он весьма раз-
борчив, о чем мы узнаем в п. 26.
67
Словом, от арифметики никуда не деться, потому о ней
предпочитали молчать, пока верили в Бернета. Единичный
клон В-клеток сам по себе беспомощен, нужно, чтобы клонов
росло сразу много параллельно. И это у Стила отмечено: «Со-
ревнование может быть выиграно только при условии, что на-
чальное число В-клеток, связывающих эти бактерии, велико»
[Стил, Линдли, Бландэн, 2002, с. 111]. Но если размножение
начинается не с одной особи, а со многих (как оказывается на
практике — с миллионов), то его нельзя называть клональ-
ным. Тогда главное - понять, откуда сразу берутся миллионы
клонобразующих клеток.
Этот удар по клональной идее — не единственный. Каждый
В-лимфоцит синтезирует лишь один тип антител. Если бы
множество В-лимфоцитов, производящих данное антитело,
действительно было клоном единственной клетки, случай-
но нашедшей нужный ген антитела, следовало бы ожидать
огромного разброса сроков иммунного ответа больных — кому
как повезло с поиском. Но этого нет. Первичная иммунная
реакция организма наступает сразу, а затем несколько суток
(острый период инфекционной болезни) тратится на создание
«зародышевых центров», т.е. так называемых «фабрик анти-
тел» [Игнатов, 2002].
Столь же важно, каким числом различных типов антител,
присутствующих одновременно, может оперировать орга-
низм. Оказывается, их количество у мыши иммунологи оце-
нивают в 104, тогда как нужны миллиарды. И если иммунитет
справляется со своей задачей (для каждого или почти каждо-
го антигена находится антитело), то, значит, нужные антите-
ла продуцируются намного чаще, нежели «по Бернету». На-
сколько же чаще?
В наше время механизм формирования антител расшиф-
рован, и число возможных их вариантов, как у мыши, так и у
человека, оценивается в 1014. А присутствует их у мыши 104.
Разница, как видим, в 10 порядков (в 10 млрд раз). Вольно
говоря, лишь одна десятимиллиардная часть поиска прихо-
дится на случайность, в остальном же он целенаправлен. Вот
почему (наряду с другими причинами) Стил разочаровался в
схеме иммуногенеза «по Дарвину» (читай: «по Бернету»), В
поисках лучшей теории он обратился к ламаркизму, как он
его понимал тогда и как понимает ныне. Прав он или нет12, но
это интересно.
12 На самом деле вопрос сложнее. К ламаркизму относится феномен активного по-
иска структуры антитела, но если миллион лимфоцитов быстро нашел одну и ту же
структуру, то, по определению, налицо молекулярный номогенез. Иногда говорят:
«Номогенез нам не нужен, мы можем всё объяснить без него». Да, но «всё объяс-
нить» можно и без иммунологии вообще (вот обычное: «Бог оделяет нас терновым
венцом болезни и сам снимает его»), здесь же мы видим - опыт зафиксировал то
самое, что называется номогенезом.
68
8. Стил и ламаркизм
в иммунологии
Стил вовсе не отрицал
клонально-се лекционной
идеи, он лишь добавил к ней
допущение, что новый при-
знак, возникнув(например, в
результате той же соматиче-
ской мутации), может стать
наследственным. Способ пе-
редачи гена из соматической
клетки в половую был тогда
известен и даже уже обсуж-
дался как возможный фактор
эволюции иммунитета — это
был один из вариантов гори-
зонтального переноса наслед-
ственности (см. п. 12, раздел Эдвард Стил' Около 1980 года'
I) . Очередь была лишь за опытом, демонстрирующим факт на-
следования приобретенного иммунного свойства. Стил и его
сотрудник Per Горчинский, тоже тогда молодой, работая в То-
ронто (Канада), модифицировали тот опыт, который принес
успех Бернету и Медавару.
Схема получения приобретенной толерантности была
ими усложнена (рис. 8): «Мы показали, что если новорож-
денных самцов линии А повторно подвергать воздействию
большого числа лимфоцитов линии В, то толерантные сам-
цы могут передавать неко-
торые черты специфической
толерантности к антигенам
В-ткани своему потомству,
полученному от скрещива-
ния с нормальными самками
линии А... Положительная
Рис. 8. Опыт Стила и Горчинского по
обнаружению наследования иммуно-
толерантности, приобретенной от-
цом. Новорожденным самцам линии
А, приобретшим толерантность к
ткани линии В (опыт Медавара),
вводили ударную дозу лимфоцитов
той же линии В (инъекция изобра-
жена стрелкой от темной мыши во
второй строке). Родившееся от та-
ких самцов и обычных самок потом-
ство исследовано на толерантность
к клеткам линии В. У некоторых из
них толерантность на самом деле
проявилась.
Линия А
источник
чужих клеток
Новорожденные самцы линии А,
которым введены лимфоидные
Линия А, толерантная Нормальная,
к В-антигенам не обработанная линия А
Потомство, проверенное на толерантность
Ожидаемое на основании
Наблюдаемое —
дарвинизма —
мыши нетолерантны
много толерантных
мышей
69
передача была непостоянной, проявляясь с большой часто-
той у одного-двух из десяти самцов» [Стил, Линдли, Блан-
дэн, 2002, с. 153].
Итак, была установлена высокая и достоверная, хотя и не-
устойчивая, частота наследования приобретенного иммунно-
го свойства: весьма заметная часть потомства воспринимала
в качестве своих те белки, которые были инъецированы во
младенчестве их отцам.
Неустойчивость частот прекрасно известна всем биологам,
поскольку таковы частоты едва ли не всех мутаций. Почти сто
лет из книги в книгу кочуют оценки частот порядка 10 7—106
на ген на особь, т.е. с разницей в сотни раз. К ним стоит до-
бавить явление, известное как «мода на мутации», когда для
данного вида оценка частоты мутаций вдруг подскакивает в
десятки раз, чтобы через несколько поколений упасть снова.
Это значит, что мы почти никогда не можем сказать, с какой
вероятностью мутирует интересующий нас ген: ведь вероят-
ность во всех прикладных науках— это устойчивая от опыта к
опыту частота. И тем не менее никто из-за этого не сомневает-
ся в реальности мутаций.
С открытием Стила вышло иначе. Сперва его окружал
успех, он печатался в самых престижных журналах, но вско-
ре (1980) им заинтересовался Медавар, ставший за четверть
века, истекшие со дня его собственного открытия, яростным
дарвинистом. (То же произошло тогда почти со всеми биоло-
гами, что выразилось в легкой победе «центральной догмы»,
ныне всерьез никем не защищаемой.) Его лаборатория повто-
рила опыт Стила и получила, по сути, тот же результат, одна-
ко вывод сделан был противоположный: если устойчивой ча-
стоты наследования нет, то нет и самого наследования. А что
есть? Есть, по Медавару, только случайные мутации (видимо,
он отнес к ним и рекомбинации) генов, кодирующих антите-
ла. Тот факт, что их частоты еще менее устойчивы и гораздо
более низки, во внимание принят не был и даже никому, ка-
жется, в голову не пришел.
Из данной истории, по-моему, ясно, что Бернету надо бы
в свое время не ставить и снимать кавычки у слова «случай-
ность», а исследовать феномен случайности всерьез. Об этом
мы немного поговорим в п. 21.
Медавар потребовал, чтобы Стил прекратил опыты и
сменил область деятельности. В 1981 году Стил писал: «Сэр
Питер Медавар и его коллеги сообщили мне... что я должен
сменить область своих научных интересов и не публиковать
ничего на тему сома -» зародышевая линия». Спорить с лау-
реатом было невозможно, Стил вернулся в Австралию, но и
там его не хотели брать на работу. Он был глубоко подавлен
и, вероятно, пропал бы для науки, если бы в дело не вмешал-
ся Философ.
70
Часть 2. Иммунитет как активность организма
9. Поппер и «демон Ламарка»
Нобелевских лауреатов появляется около десяти в год, а
крупных философов — несколько в столетие. Философов, изу-
чавших суть и структуру науки, за всю историю — единицы.
К ним относится Карл Поппер (1902—1994). Он родился в
Австрии и начал там работу, уехал от фашистов в Новую Зе-
ландию, а с 1946 года обосновался в Англии. Биологи знают
его в основном как автора принципа фальсифицируемости
(далеко не главного в его зрелом творчестве), согласно кото-
рому нельзя считать научной теорией концепцию, которая
в принципе не допускает опровержения. В виде примеров
ненаучных концепций он в молодости приводил марксизм,
фрейдизм и дарвинизм, но к последнему позже испытывал
симпатию и даже использовал его при пояснении своей идеи
развития науки.
Не считая дарвинизм теорией (ввиду невозможности ни
доказать его исходные положения, ни опровергнуть), Поп-
пер не видел в нем и эмпирики, т.е. обработки опыта. Мы уже
говорили, что нет ни одного примера действия естественного
отбора как фактора эволюции, но Поппер имел в виду иное
— противопоставлял эмпирике ситуационную логику. Дан-
ное понятие (на мой взгляд, еще более важное, чем фальси-
фицируемость) означает «объяснение различных процессов
в терминах ситуации, в которой они происходят». При этом
к основным принципам, общим для всего исследования, по
мере надобности «добавляется множество методологических
принципов, которые может использовать исследователь в той
или иной ситуации» [Аронова, 2006, с. 78]. Именно это, а не
анализ фактов видел Поппер в основе дарвинизма. С помощью
ситуационной логики можно доказать всё что угодно.
О ламаркизме Поппер имел довольно смутное представ-
ление, будучи всю жизнь уверен, что тот сводится к идее
наследования приобретенных свойств. Она заинтересовала
Рис. 9. Типичное
антитело состоит
из двух легких (L) и
двух тяжелых (Н)
белковых цепей. Ами-
нокислотный состав
«вилки» весьма из-
менчив (V — вариа-
бельные части цепей,
С — константные),
чем и достигается
соответствие данно-
го антитела своему
антигену.
71
о
о
X
5 юо-
LO
ПОЗИЦИЯ
Рис. 10. График By - Кэбота. Как видим, мутирует
лишь небольшая часть нуклеотидных оснований.
Поппера, ког-
да в 1979 году
Стил прислал
ему свою ру-
копись. Поп-
пера в идее
наследования
приобретен-
ных свойств
«привлекала
“фальсифици-
руемость”, т.е.
возможность
опровержения
(чего нет в дар-
винизме)», — пишет иммунолог А.А. Ярилин в предисловии к
книге [Аронова, 2006]. К его мыслям мы еще обратимся.
Поппер не был целиком уверен в точности допущений Сти-
ла и его выводов, однако решительно встал на его защиту, ког-
да тому пытались запретить работать. Поппер напомнил его
гонителям ту постоянно забываемую истину, что наука без
свободы поиска теряет смысл. Стила он характеризовал как
первопроходца, и письмо Поппера руководству Австралий-
ского университета (1984) стало решающим для принятия
Стила на работу.
Поппер полагал дарвинизм ущербным, но не видел ему
иных альтернатив, кроме «ламаркизма». В 1983 году он бе-
седовал с этологом (специалистом по поведению животных)
Конрадом Лоренцом. «Участники беседы сошлись в том, что,
если считать эволюцию сочетанием мутаций и отбора, невоз-
можно объяснить возникновение всего многообразия живо-
го... по крайней мере, ввиду недостатка времени». Слова «по
крайней мере» весьма важны — на самом деле главное отнюдь
не в дефиците времени, а в том, что процесс отбора случайных
изменений сам по себе никуда не ведет, что для эволюции ну-
жен иной, реальный, направляющий фактор (мы говорили об
этом в разделе I). В этой беседе двух классиков данный фактор
был обозначен (пишет Е.А. Аронова) как некий «сверхчелове-
ческий разум... но не Бог» и назван там «Демоном Ламарка»
[Аронова, 2006, с. 87].
«Демон Ламарка» к тому времени уже давно свирепство-
вал в иммуногенетике. Еще в 1970 году американские гене-
тики Тай Тэ By (Wu - тот самый, что цитирован в п. 8, раз-
дел I) и Элвин Кэбот (Cabat) опубликовали статью [Wu, Cabat,
1970], цитируемую поныне. В ней исследован результат му-
тационного изменения гена, кодирующего иммуноглобулин.
Мутации до этого всеми считались случайными, но оказалось
совсем иное.
72
Иммуноглобулин — это составной белок, образующий ан-
титело (рис. 9). Его активная часть являет собой вилку для
захвата антигена. Ее внутренняя часть и «ручка» образованы
двумя тяжелыми (т.е. длинными) цепями Н, а две внешних
части - легкими (т.е. короткими) цепями L. Наружная захва-
тывающая часть вилки (двузубец) образована вариабельными
(изменчивыми) участками тяжелых (Ун) и легких (VL) цепей.
By и Кэбот исследовали статистику мутаций вариабельных
участков. На рис. 10 приведен знаменитый график By—Кэбота
для участка VH одного из иммуноглобулинов человека. Как
видим, 27 из 116 аминокислотных остатков заменяются мно-
го чаще других, причем они образуют три компактные обла-
сти. Мутации тут далеко не случайны.
Позже было выяснено, что эти области «образуют поверх-
ность, которая вступает в тесный контакт с поверхностью
антигена» [Стил, Линдли, Бландэн, 2002, с. 128]. Чтобы по-
нять значение этого для эволюции и иммунологии, вернемся
в Австралию, знаменитую своими иммунологами, их спорами
о природе разнообразия иммунных клеток и о ламаркизме.
10. Каннингэм, Тонегава
и «Дарвинов микрокосм»
До отъезда в Канаду Стил работал в Канберре, в лаборато-
рии, которой руководил иммунолог Эл истер Каннингэм (A.J.
Cunningham). В чисто научном плане он был не ниже Берне-
та13, но сильно уступал ему в умении продвигать свои идеи.
Бернет начал как вирусолог и собрал в Мельбурне сильный
коллектив вирусологов, но, получив Нобелевскую премию
(1960), уволил их едва ли не всех, и они (кому удалось) были
вынуждены уехать в Канберру. Среди уволенных оказался и
Каннингэм. Это странно - и потому, что он был чистый им-
мунолог, и потому, что он-то как раз старался подтвердить
«клонально-селекционную теорию» Бернета, которую тот сам
полагал (до получения премии) гипотезой, требующей под-
тверждения. То ли Бернет, став лауреатом, не нуждался боль-
ше в аргументах, то ли предвидел, сколь опасны будут для
него аргументы Каннингэма, то ли просто сработал эффект
«два медведя в одной берлоге» - не знаю.
Бернет никак не объяснил, откуда берется огромное разно-
образие антител (см. и. 6). Это не помешало успеху его моде-
ли и утверждению ее в учебниках14, однако кое-кого всё-таки
13 Его работы по специфичности антигенов описаны в книге [Аронова, 2006]. В
параллель с ними в 1975 г. Э. Каннингэм и К. Лафферти постулировали процедуру
антиген-презентации (о ней см. далее, п. 23), затем открытую в опыте. Об этом за-
мечательном факте П. Матцингер писала: «Данное озарение показало, что теоре-
тическая биология и физика могут иметь больше общего, нежели подчас думают»
[Matzinger, 2002, примеч. 20].
14 За сто лет до того дарвинизм тоже вошел в науку и в учебники, не имея при-
73
беспокоило. Одним из них и был Каннингэм. Он фактически
открыл соматический гипермутагенез — процесс очень бы-
строго изменения гена, кодирующего вариабельную часть им-
муноглобулина. Этот процесс начинался в организме в ответ
на попадание антигена - до этого нужные клетки «начисто от-
сутствовали» [Аронова, 2006, с. 124]. (Это прямо опровергало
схему Бернета, но никто, включая Каннингема, видеть этого
не хотел, и в ход шли различные толкования, вряд ли нынче
интересные.) Обычно каждая В-клетка производит один-един-
ственный тип антител. Однако Каннингэм стал следить за от-
дельным клоном В-клеток и с удивлением обнаружил, что при
добавлении нового антигена около 10% потомков клетки-осно-
вателя начинают производить также и иные антитела. Если это
мутации, то — неимоверно интенсивные. Процесс идет в мил-
лионы раз быстрее, чем обычный мутагенез, достигая подчас
частоты в одну мутацию на каждую тысячу пар мутирующих
оснований. Очевидно, что, если бы такая частота относилась
ко всем нуклеотидным основаниям гена, он быстро бы распал-
ся, но вновь открытый процесс затрагивает лишь малую долю
оснований (рис. 10), причем прекращается столь же быстро,
сколь и начинается. Это и есть гипермутагенез.
Каннингэм понимал, что тут налицо параллель с эволю-
цией («эволюционный микрокосм»). В своих публикациях
(1974—1977) он настаивал, что развивает взгляды Бернета и
не выходит за рамки дарвинизма, ибо мутации в его опытах
остаются ненаправленными, но это далеко не всех убежда-
ло. В словах самого Каннингэма («организм “учится” от сво-
ей окружающей среды», клетки делают «активный выбор»
[Аронова, 2006, с. 125—126]) звучал ламаркизм, и многие, в
том числе Стил, видели тут именно то, что выше мы назвали
генетическим поиском (см. п. 9, раздел I). Его ведет иммун-
ная система в ответ на заразу. В самом деле, как уже сказано,
дальнейшие исследования показали, что в результате гипер-
мутагенеза изменяются как раз те точки молекулы антитела,
которые тесно контактируют с антигеном.
Это уже в дарвинизм не укладывалось никак. Хотя желаю-
щих его видеть и тут нашлось много, но их рассуждения вряд
ли интересны. Суть их всегда одинакова: поскольку всё в при-
роде есть итог отбора случайных вариаций, то во всем, что мы
наблюдаем, отбор имел место - достаточно лишь его поискать.
Кто должен искать? Тот, кто в этом сомневается, а нам и так
всё ясно. Но это уже не наука, это ближе к религии.
меров действия отбора в качестве фактора эволюции (зато есть опровергающие
наблюдения). Ни одно из возражений, предъявленных Бернету, не было оспорено
— их просто перестали упоминать. Учебники иммунологии перестроены на основе
идеи Бернета в течение 1970-х годов [Аронова, 2006]. Эта прискорбная параллель
еще ждет анализа методологов, каковой особо актуален сейчас, когда сомнения в
клональной селекции публикуются ежемесячно.
74
Стил был одним из немногих, кто прямо заявил, что
ки на отбор случайных вариаций здесь бессодержател
нужно искать механизм передачи информации. Так чт
его столкновения с Медаваром (см. п. 8) была отнюдь не
ных опыта (у обоих они были по сути одинаковы), а в то
Стилу данный механизм был нужен, а Медавару — не1
прасно Стил сокрушался, что «заключения были сдела
того, как были собраны какие-либо экспериментальные
зательства»15. Ведь и до этого Медавар был известен как
гонитель того, что относил к ламаркизму [Аронова, 20
98, 120].
Среди многих, занявшихся (под впечатлением раб
и Кэбота, а затем Каннингэма) проблемой разнообрази
мунных клеток, был молодой иммуногенетик Тонегава,
тором уже шла речь в п. 10, раздел I. Он исследовал пр
создания изменчивости генов, кодирующих иммуноглс
ны. Оказалось, что гипермутагенез — поздняя стадия им
генеза, прежде которой каждый такой ген уже должен
мироваться. Он заново собирается из блоков в ходе раз:
каждого организма (когда у плода, а затем у ребенка ф
руется иммунная система). Уже после этого идет гипер
генез (часто — при вторичной встрече с той же заразой),:
шающий точную подгонку антитела к антигену.
Рис. 11. Механизм Тонегавы. Ген тяжелой цепи антитела формирщ
одного V, одного D и одного J-фрагмента ДНК; полученная тройка
крибируется.
Сама сборка гена происходит так. Комбинируются
три типа генетических блоков — V, D и J. В геноме мле
тающего (как мыши, так и человека) есть около сотни б,
типа V, около 30 блоков типа D и шесть — типа J. Ген и
чивой части тяжелой цепи каждого антитела (см. рис.
бирается из одного V, одного D и одного J-элемента (рис
15 Опыт Стила занял два года, тогда как Медавар лишь в октябре 1980 го^
ложил сотрудникам ставить проверочный опыт, а уже в феврале 1981 год?
бовал, чтобы Стил «сменил область научных интересов».
С этой тройки (цепочки) и считывается первый вариант гена
нужного белкового домена антитела (второй вариант полу-
чится в ходе гипермутагенеза). Аналогично формируется ген
легкой цепи, только в нем нет блоков типа D.
Обнаружив это явление, Тонегава в 1982 году решил (а в
1987 году заявил в Нобелевской лекции), что выбор блока для
включения в данный ген происходит случайно, что удачная
комбинация блоков поддерживается отбором и, стало быть,
налицо «Дарвинов микрокосм». Правда, он оговорился, что
здесь имеет место « сложная неравномерная случайность », но
это никому тогда не было интересно.
Ситуация вполне обычна: если механизм явления поня-
тен, об отборе не вспоминают, а если непонятен, то в ход идет
единое толкование — «под давлением отбора». В следующие
20 лет механизм созревания В-клеток был расшифрован, и
его теперь описывают в терминах активного взаимодействия
клеток, а об их отборе все замолчали (см. п. 23).
Если бы механизм Тонегавы перебирал одну за другой все
возможные их комбинации, то, как показывает расчет [Стил,
Линдли, Бландэн, 2002], он наработал бы в одном организме
мыши за ее жизнь 3 млн различных антител. Но возможных
антигенов — миллионы миллиардов, и нет никакой гарантии,
что среди созданных были бы те самые антитела, какие в дан-
ное время для данной особи нужны. Поэтому естественно, что
процесс идет иначе: при комбинировании выбираются одни
варианты много чаще других. В частности, «около половины
И-элементов никогда не участвуют в образовании антитела»,
а реальное одновременное разнообразие антител — отнюдь не
3 млн: их всегда меньше 10 тыс. [Стил, Линдли, Бландэн,
2002, с. 111-112].
Позже стало ясно, что Тонегава открыл аналог альтерна-
тивного сплайсинга (см. словарик), но на уровне не РНК, как
обычно, а ДНК. Иначе говоря, в основе механизма Тонегавы
оказались не случайные вариации, а итог генетического поис-
ка, т.е. собственная активность генома. Налицо, кроме моле-
кулярного номогенеза, молекулярный ламаркизм.
11. Вентребер и рождение молекулярного ламаркизма
Французский зоолог Поль Вентребер, ведущий ламаркист,
сумел за свои 99 лет прожить две научные жизни: до выхода
на пенсию (1937) был эмбриологом и стал академиком имен-
но как эмбриолог, а затем стал эволюционистом. Как эмбри-
олог он известен своим стремлением понять физиологию за-
родыша (тогда, как и нынче, развитие зародыша понимали, в
основном, как становление формы). Он настаивал на том, что
форма частей зародыша (как и форма частей организма) зада-
ется их функцией. Тем самым он стоял у истоков эмбриофизи-
ологии, которая еще так и не сложилась поныне.
76
Поль Вентребер (1867-1966). Дал
новую жизнь ламаркизму, сопо-
ставив эволюцию и иммуногенез.
Наметил задачи молекулярного
ламаркизма.
Вентребер мог бы прямо
выйти на эволюционную про-
блематику через описание
изменений в развитии заро-
дышей, но он предпочел путь,
которым не ходил до него, ка-
жется, никто — представил
в качестве основного способа
эволюции иммуногенез, точ-
нее, выработку антител. Им-
мунитет, наследственность и
эволюцию сопоставляли дав-
но, но то были иммунологи,
и они ставили целью понять
иммуногенез, взяв за данное
какую-то эволюционную кон-
цепцию. Вентребер же хотел
понять саму эволюцию.
В 1949 году он сделал в
Парижской академии наук
доклад «Химический ламар-
кизм», где смело допустил,
что у иммуногенеза и эволю-
ции есть что-то общее. Он дал
свое понимание мутации, то есть сути молекулярного ламар-
кизма. А именно, Вентребер предложил следующую схему
адаптации организма к новой среде: если какой-то орган пере-
стает справляться со своей работой, он вынужден работать в
ненормальном режиме, в нем начинается производство како-
го-то вещества, вредного для организма, и это вещество слу-
жит сигналом для поиска адаптации.
Теперь все знакомы с таким пониманием запуска адапта-
ции: это концепция, согласно которой ее запускает стресс (за-
пускающие вещества именуют стрессорами). Австрийский
физиолог Ганс Селье развивал концепцию стресса с 1936 года,
но вряд ли Вентребер тогда знал о ней. Фактически он отож-
дествил стрессоры с антигенами и решил, что иммунный про-
цесс, ими запущенный, ведет к перестройке того гена, который
ответствен за работу данного органа. Здесь легко заметить то
понимание работы генетической системы, которое позже по-
лучило название «оперонная регуляция» и было предложено
французскими генетиками (Франсуа Жакоб и др., 1960). Как
это происходит, Вентребер, разумеется, сказать не мог: тогда,
в 1949 году, генетики еще не достигли согласия даже в том, что
служит веществом наследственности — ДНК или белок.
Однако Вентребер не только уверенно встал в ряд привер-
женцев ДНК, но и предположил механизм ее генетического
действия: нуклеиновая кислота, находясь в цитоплазме, как-
77
то узнает о строении антигена, а делает это знание наслед-
ственным та ДНК, которая попадает в клеточное ядро. Об этом
генетики стали говорить лишь в 1960-е годы.
Эти мысли Вентребер уточнял еще три года, а затем на де-
сять лет снова замолчал — писал книгу. За это время была
расшифрована двуспиральная природа ДНК, поставлена и
решена проблема генетического аминокислотного кода и от-
крыт оперон — основной элемент генетической регуляции у
бактерий. Однако книга Вентребера «Живое, творец своей
эволюции» (1962) оказалась вовсе не архаизмом, но, наобо-
рот, вполне актуальной. Читая книгу, никак не скажешь,
что автору 95 лет.
В те годы шло неумеренное увлечение генетикой как осно-
вой и даже сутью всей биологии (мы касались этой темы в п.
1), Вентребер же видел дело иначе: «Ген — продукт протоплаз-
мы. Собранный из ДНК и нуклеопротеина, он ... продукт, со-
творенный живым (веществом). Тем самым он — его делегат в
хромосомах, гормональная субстанция, стоящая в резерве и
используемая, когда надо» [Wintrebert, 1962, с. 137—138].
Данную форму активности живого он трактовал как им-
муногенез и видел в этом новое понимание ламаркизма. (На-
оборот, понимание ламаркизма как наследования приобре-
тенных свойств он считал недоразумением.) Вентребер не был
иммунологом и угадывал проблематику чисто интуитивно,
идя скорее от своей эмбриофизиологии. Однако в те же годы
сложилась настоящая экспериментальная эмбриоиммуноло-
гия, о которой теперь тоже надо немного сказать.
12. Эдельман: морфогенез и иммунитет
К 1960 году, т.е.к началу эры молекулярной биологии,
было известно, что при развитии зародыша запуск форми-
рования каждого органа сопровождается запуском синтеза
определенного вещества (индуктора) и что выявить этот ин-
дуктор можно иммунологическим приемом. Например, если
изготовить препараты зародыша головы цыпленка до и после
начала формирования глаза, а затем ввести пробы из этих
препаратов в кровь мыши, то у нее образуется много различ-
ных антител (ведь тело цыпленка чуждо телу мыши), причем
при второй пробе образуется такое антитело, какого не было
при первой пробе. Выделив это вещество, можно индуциро-
вать им запуск формирования глаза в зародыше цыпленка.
Такие вещества назвали антигенами дифференцировки [Гил-
берт, 1995, с. 21].
Затем, в конце 1960-х годов, подобный иммунологический
прием помог определить те вещества на поверхности клеток,
которые позволяют клеткам соединяться с аналогичными
(прилипать к ним) и не соединяться с чуждыми клетками.
Этим весьма разнообразным веществам позже было дано на-
78
Рис. 12. Некоторые иммуноглобулины (по Эдельману). Антитело плавает
в лимфе, остальные иммуноглобулины укоренены в клеточной мембране.
звание МКА — молекулы клеточной адгезии (адгезия — это
ученое название для прилипания).
Вскоре на МКА обратил внимание Джералд Эдельман,
американский биолог, получивший в 1972 году Нобелевскую
премию за расшифровку строения антитела (см. рис. 9). Вско-
ре он стал известен исследованиями взаимодействия клеток
многоклеточных организмов и роли этого взаимодействия в
развитии зародыша. В 1976 году он и соавторы обнаружили
среди МКА группу белков, которые оказались иммуногло-
булинами, весьма сходными с иммуноглобулинами антител
(некоторые изображены на рис. 12), и сумели вскоре рас-
шифровать их строение. Им удалось показать, что развитие
зародыша сопровождается многократными сменами типов
МКА на поверхностях его клеток. В итоге родилась концеп-
ция, согласно которой весь онтогенез запрограммирован по-
следовательностью синтезов МКА. Вряд ли она верна, но она
красива.
Так же, как и антитела, МКА весьма изменчивы, ив 1987
году Эдельман высказал предположение, что система имму-
нитета высших животных развилась в эволюции из системы
избирательной адгезии. Тут стоит добавить, что избиратель-
ная адгезия клеток наблюдается повсюду. Даже колонии бак-
терий имеют формы, особые для разных видов, что объясня-
ется различием в способах их слипания. Это можно рассма-
тривать и как простейший онтогенез, и как простейший им-
79
мунитет (распознавание родственными клетками друг друга).
Тем самым иммунология, поначалу служившая эмбриологам
лишь как инструмент исследований, теперь сама стала их те-
оретической базой: всё развитие многоклеточных предстало
как система смен иммунных предпочтений [Гилберт, 1995,
гл. 15]. Такой подход и можно назвать эмбриоиммунологией.
Всё это было красиво и занимательно, однако оставляло без
ответа как минимум два вопроса. Во-первых, чем заполнить
эволюционную пропасть между избирательной адгезией, ко-
торая есть у всех организмов, начиная с бактерий, и иммуни-
тетом высших животных? Во-вторых, какую роль играют на
поверхности клетки иные МКА, кроме иммуноглобулинов?
Таковых веществ было открыто много. Оба ответа последова-
ли вместе.
13. Джаневей и революция в иммунологии
В 1989 году, через два года после появления гипотезы
Эдельмана, американский иммунолог Карл Джаневей высту-
пил со статьей, смело названной «Эволюция и революция в
иммунологии» [Janeway, 1989]. Он не оспаривал блестящих
достижений в духе Тонегавы, он просто отметил, что иммуни-
тет ведет главные бои с заразой на ином фронте.
В самом деле, тот иммунитет, о котором мы говорили до
сих пор, — приобретенный, или адаптивный16, формирует-
ся, как и сам организм, в каждом поколении заново. Он есть
только у высших животных, тогда как с заразой успешно бо-
рются все — и животные, и растения. Этот всеобщий иммуни-
тет является врожденным (одну его форму, общую для мно-
гоклеточных животных, — фагоцитоз — открыл Мечников).
Врожденный иммунитет весьма эффективен, однако через сто
лет после открытия он оказался почти всеми забыт. Учебники
и руководства по иммунологии его едва упоминали (подчас
даже именуя «доиммунными формами» сопротивляемости),
а то и не поминали вовсе.
Конечно, вдумчивые иммунологи не раз отмечали, что
врожденный иммунитет отнюдь не прост и весьма важен, что
он тесно связан с адаптивным и даже является основой по-
следнего. Например, Г.И. Абелев писал: «Врожденный им-
мунитет... позвоночных функционирует не только самостоя-
тельно, но также и как первая и заключительная стадия им-
мунитета приобретенного» [Абелев, 1997]. Но в целом царило
пренебрежение.
Общность явлений врожденного иммунитета у самых раз-
ных организмов указывала на возможную общность их ме-
16 Адаптивный иммунитет - атрибут позвоночных, причем его сложность растет от
рыб к теплокровным [Галактионов, 2005]. Недавно открыты элементы адаптив-
ного иммунитета у высших насекомых и высших растений.
80
ханизма. И вот Джаневей
высказал предположение,
что на поверхности клеток,
ответственных за иммунную
реакцию (у любого организ-
ма), расположены «образ-
распознающие рецепторы»
(ОРР; или, по английски, pa-
ttern-recognition receptors -
PRR), которые он назвал так
потому, что они умеют разли-
чать, так сказать, не отдель-
ные тонкости, а самую суть
дела — классы молекул. Для
этого данный ОРР должен
узнавать ту часть распозна-
ваемой молекулы, которая
является общей для всех мо-
лекул данного класса.
Так вскоре и оказалось.
Например, для стенок бакте-
рий характерны липопроте-
иды — комплексы из липи-
Чарльз (Карл) Джаневей (1943-
2003) заложил основу нынешней
иммунологии.
дов и белков. Липопротеид
бактерий обычно содержит пару «пальмитиновая кислота —
цистеин». Первая — из числа жирных кислот, второй — ами-
нокислота, содержащая серу и тем самым служащая для
скрепления белковых нитей дисульфидными мостиками.
Именно эту пару распознает один из первоначально откры-
тых ОРР.
Хотя в 1990-е годы такие данные быстро накапливались и
идея Джаневея блестяще подтверждалась, в целом в иммуно-
логии царило пренебрежительное отношение к врожденно-
му иммунитету, так что Джаневею не пришлось дождаться
признания — оно пришло лишь через 15 лет. Вообще, это не
так уж много — подлинно новые идеи нередко ждут призна-
ния гораздо дольше, но Джаневей, увы, в январе 2003 года
умер.
На грани веков данные о врожденном иммунитете пошли
косяком, и с 2004 года стали появляться номера журналов,
посвященные его новому пониманию. Одним из них был № 6
французского журнала «Les Comptes Rendus de I’Academie des
Sciences de Paris. Biologies», где с удивлением читаем, что
«первичная роль врожденной иммунной системы — саморе-
гуляция.., а защитная вторична». Прежде так писали только
про иммунитет адаптивный (ту мысль, что главное назначе-
ние адаптивного иммунитета — контроль целостности тепло-
кровного организма, высказал еще Бернет). Но целостностью
81
обладают все организмы, и вот теперь выясняется, что ее, ве-
роятно, им обеспечивает иммунитет врожденный.
Наиболее простой и ясный обзор новой концепции [Мед-
житов, Джаневей, 2004] принадлежал самому Джаневею (в
соавторстве с Русланом Меджитовым, тогда ведущим сотруд-
ником, а ныне заведующим его лабораторией в Йельском уни-
верситете). Там сказано, что адаптивный иммунитет тепло-
кровных начинает бороться с заразой через несколько суток
после заражения, тогда как начальную реакцию организма на
заразу обеспечивает врожденный иммунитет. Он же запускает
в работу иммунитет адаптивный, а у тех, кто им не обладает,
ведет всю борьбу сам, с помощью ОРР. Они оказались весьма
сходными с МКА Эдельмана (см. для сравнения рис. 12 и 13),
а это значит, что его идею о происхождении иммунитета мож-
но с равным правом применить ко всем типам организмов.
Самые опасные инфекции губительны именно тем, что
рушат врожденную систему иммунитета и тем самым блоки-
руют включение адаптивной системы. Например, недавно
выяснено, что чумная бацилла Yersinia pestis своими ядами
отравляет популяцию фагоцитов жертвы, так что они теряют
способность вырабатывать молекулы, сигнализирующие о за-
разе, а затем и сами гибнут [Ruckdeschel et al., 2008].
В обзоре [Меджитов, Джаневей, 2004] впервые четко про-
ведена та мысль, что два типа иммунитета решают противопо-
ложные задачи: задача ОРР — общая, а множество задач анти-
тел — частные. (Впоследствии распределение данных ролей
оказалось сложнее, но здесь мы этого касаться не станем.) Все
вместе ОРР узнают основную массу чужеродных антигенов,
обеспечивая тем самым основную иммунную защиту, причем
безразличны к антигенам собственного организма. Антитела,
наоборот, могут возникать порознь на любые антигены, в том
числе на собственные. Поэтому синтез антител необходимо
жестко контролировать, что и делает система врожденного
иммунитета. С пониманием этого иммунология наконец-то
получила основной принцип, так что учебники и руководства
пора переписывать.
14. Революция в эволюции
Для эволюционистов же встала во весь рост проблема: если
врожденный иммунитет столь эффективен, то зачем возник
иммунитет адаптивный? Зачем антитела и прочее, если для
выживания вида его особям достаточно иметь ОРР? Если без
сигнала от ОРР не может начаться и синтез антител?
На своей проблемной лекции (биофак МГУ, ноябрь 2008
года) А.А. Ярилин задал эти вопросы и ответил, что адап-
тивный иммунитет нужен для иммунной памяти. Вероятно,
это верно, но зачем сама память? Ведь ее роль как быстрого
ответа (на повторную встречу с той же инфекцией) ничтож-
82
на в сравнении с бедами, какие адаптивный иммунитет несет
в форме иммунных болезней17. Ясно, что если она нужна, то
для чего-то иного.
Первое, что приходит в голову и не раз высказано, — что
иммунная память есть промежуточное звено между чисто
адаптивным иммунитетом, т.е. специфическим иммунным
ответом на заразу, попавшую в организм впервые, и врожден-
ным иммунитетом. Однако это любопытное допущение (из
него, кстати, исходил Стил) было умозрительным, и откры-
тие Джаневея его опровергло: врожденный и адаптивный им-
мунитеты оказались устроены принципиально различно, на
разных молекулах.
Не лучше и другие гипотезы. Например, что усложнение
иммунной системы нужно для устранения мутантных белков:
— чем крупнее организм и чем он дольше живет, тем больше
их накапливается и тем сложнее должна быть система их
устранения. Эту гипотезу (об иммунном надзоре) высказал
еще Бернет (см. выше, п. 6), и тогда она имела свой резон, но
вскоре был открыт куда более эффективный механизм устра-
нения подобных мутаций — разные типы репарации ДНК. К
тому же самые крупные и долговечные организмы (гигант-
ские деревья) ничего подобного иммунитету теплокровных не
имеют (а мыши, совсем мелкие, его имеют). У растений нет
даже фагоцитоза, обычного для животных.
Так зачем адаптивный иммунитет? Тут пора вспомнить,
что в ламаркизме есть основополагающий принцип — тенден-
ция к усложнению (прогрессу).
15. Ламарк и прогресс
«Да сохранит меня небо от Ламаркова нелепого “стрем-
ления к прогрессу”», — писал Дарвин в 1844 году, и «небо»
в самом деле сохраняло его от этого до самой смерти. Дарви-
низм обходит проблему до сих пор, но от прогресса никуда не
деться, ибо он в природе виден: в ходе эволюции появляют-
ся организмы, более сложно устроенные, чем прежние (так,
иммунитет комаров и зверей сложнее, чем у губок). Теория
должна его объяснять, а не постулировать, как дарвинизм,
что существует «отбор на более высокую организацию».
Ламаркизм, наоборот, не уходит от проблемы прогресса, а
прямо говорит, что прогресс — явление природы, отличное от
приспособления, и для своего осмысления требует введения
особого теоретического принципа. Многие поэтому относи-
ли ламаркизм к «мистическим учениям», но мистики тут не
больше, чем в остальной науке. Ведь всякая наука при своем
17 Иммунная память есть и у врожденного иммунитета: скорость отторжения по-
вторно пересаженной чужой ткани нарастает (по сравнению с пересаженной пер-
вично) от губок к позвоночным, достигая максимума у теплокровных [Галактио-
нов, 2005].
83
появлении объявляет какие-то принципы базовыми, а затем,
ища им обоснование, видоизменяет их.
Например, термодинамика поначалу была основана на
двух принципах: первым был закон сохранения энергии (об-
щий для всей физики), а вторым — закон стремления замкну-
той системы, не содержащей источников силовых полей, к
тепловому равновесию. Сто лет назад первый принцип при-
шлось изменить — по Эйнштейну, сохраняется не энергия, а
пара масса—энергия. Через полвека пришла очередь и второ-
го принципа: он оказался верен лишь вблизи теплового рав-
новесия, тогда как вдали от него могут самопроизвольно воз-
никать структуры. Простой пример: при слабом нагревании
идет лишь переток тепла от более теплого тела к менее тепло-
му, а при сильном может возникнуть пламя — структура, ко-
торая будет сама поддерживать себя, пока есть, чему гореть.
Не раз отмечено, что эта модификация второго принципа
открывает путь к объяснению прогресса, понимаемого как
усложнение. Выходит, что Ламарк, формулируя 200 лет назад
принцип прогресса, вовсе не был мистиком, а обогнал физику
своих дней лет на 160. Дарвинизм же ориентировался как раз
на физику своего времени — недаром Больцман ссылался на
Дарвина, а ранние дарвинисты — на Больцмана. Словом, при-
шла пора сменить старый призыв Дарвина к «небу» на еще
более старый Ламарков принцип прогресса. Именно его имел
в виду Лоренц, говоря Попперу про «Демона Ламарка».
Самый простой пример прогресса как усложнения — это
самосборка целого из частей. Первый пример самосборки
указал 90 лет назад К.Э. Циолковский: в замкнутой Вселен-
ной гравитация собрала пыль в звезды и в планеты, которые,
в свою очередь, обзавелись внутренними структурами. При-
мер противоречил старой термодинамике, но не новой.
Самосборку надо описывать, но нет нужды чем-то оправ-
дывать - разве что по Кауфману: тем, что так устроен мир — в
нем на всех уровнях обычна самоорганизация (см. пп. 15и 16,
раздел I) и самосборка - ее простейший вид. В мире нет места
перебору всех возможных вариантов - на это нет времени не
только в эволюции, но даже при самосборке белка: для пере-
бора конформаций одного короткого белка не хватит возраста
Вселенной [Савинов, 2006, с. 64, 68], а он собирается быстро,
т.е. почти или вовсе без «проб и ошибок».
В качестве наглядных примеров биологического прогресса
часто называют появление клеточного ядра, ткани и голов-
ного мозга. Столь же ясным примером служит и появление
адаптивного иммунитета. Ни одно из таких свойств не было
необходимо для выживания — ведь многие организмы в тех же
условиях обходятся без них. Например, бок о бок могут жить,
выполняя одну и ту же экологическую функцию, бактерия
(клетка без ядра) и высший организм (гриб или растение).
84
Важное приобретение сплошь и рядом сильно затрудняло
жизнь, так что его обладатели надолго становились малочис-
ленными. Зато в будущем открывались невиданные горизон-
ты для далеких потомков — обладателей новшеств. Именно так
произошло и с четвероногими, когда они освоили адаптивный
иммунитет. Подобную оптимизацию путей (а не конечных со-
стояний) эволюции физики именуют вариационным принци-
пом (первым тут был Мопертюи - см. п. 1, раздел I), а биологи
- преадаптацией.
16. Разум как активность иммунной системы
Разум принято видеть как форму активности нервной си-
стемы (точнее — головного мозга) людей. Многие добавляют:
«и высших животных». Иногда можно услышать, что мозг
лишь улавливает некую активность Космоса, однако при чем
тут иммунитет?
А вот при чем. Если врожденный иммунитет стар, как
сама жизнь, то адаптивный иммунитет явно произошел после
врожденного, и можно указать этапы его эволюции. Иммуно-
глобулины есть уже у низших вторичноротых (иглокожие),
а среди позвоночных они появляются впервые у некоторых
хрящевых рыб. Адаптивная система иммунитета позвоноч-
ных, постепенно усложняясь, целиком проявилась только у
млекопитающих [Галактионов, 2005], и естественно допу-
стить, что она связана с теми свойствами, которые более всего
отличают их от остальных животных. Это, прежде всего, их
интеллект.
Звери в целом умнее птиц, а птицы в целом умнее осталь-
ных животных, исключая головоногих моллюсков. Это озна-
чает (для всех, даже для тех оригиналов, кто считает, что мы
ловим мысли из Космоса), что мозг млекопитающих устроен
сложнее, чем у других. Как происходит мышление, мы не зна-
ем, но зато знаем, что усиление ума в эволюции означало уве-
личение числа нервных клеток и связей между ними.
Каждый нейрон (основная клетка нервной системы) связан
даже у пиявки с сотнями других нейронов, а у человека таких
связей нейрона — сотни тысяч. Связи эти возникают у заро-
дыша путем роста каждого аксона (ответвления нейрона) в
нужную сторону. Как он узнает, куда расти? Оказывается, он
ведет себя примерно так же, как подвижная клетка иммунной
системы, когда она ищет тот орган, где ей следует поселить-
ся. Причем, по Гилберту, у аксона пиявки видно сходство с
врожденным иммунитетом, а у аксона млекопитающего — с
адаптивным [Гилберт, 1995, с. 120—122].
Напрашивается мысль, что у каждого организма есть тот
тип иммунитета, который нужен для его онтогенеза. Когда
эта мысль впервые была высказана мною в научном журнале
(«наш с вами коварный иммунитет достался нам... потому, что
85
без него нас попросту не было бы — наш онтогенез невозможен
без сложной системы иммунологического надзора» [Чайков-
ский, 2003, с. 272]), она была почти лишена эволюционных
обоснований. С тех пор положение заметно изменилось, чем
и займемся далее.
Часть 3. Иммунитет как упорядоченность
17. Вездесущий иммунитет и несовместимость тканей
Иммунитет в какой-то форме есть у всех организмов, даже
у бактерий. Например, приспособление бактерий к антибио-
тикам тоже ведь иммунный акт. Это значит, что друг с дру-
гом борются различные иммунитеты и нужно говорить об
иммуноэкологии. Во многих отраслях биологии и медицины
иммунная сторона дела стала видна отнюдь не сразу. Так по-
лучилось, в частности, при объяснении неудачных попыток
пересадки тканей и органов от одного организма к другому.
Еще лет 60 назад все полагали, что эти неудачи (отторжение
чужих тканей и органов) носят чисто хирургический характер
— надо аккуратнее оперировать, и всё получится.
Английский иммунолог (родом из Бразилии) Питер Меда-
вар, о котором мы уже много говорили, вошел в историю на-
уки не столько своим частным успехом, быстро давшим ему
Нобелевскую премию (в 1960 году, вместе с Бернетом), сколь-
ко своим более давним достижением. А именно, ставя опыты
по пересадке органов животным, а также исследуя прижив-
ление тканей, пересаженных обожженным военным летчи-
кам, он в 1944 году доказал фундаментальный факт — оттор-
жение организмом чужой ткани является следствием работы
Рис. 13. В сканирующий микроскоп видно, что клетки покрыты множеством
выступов и ворсинок, которые, в частности, служат метками для их узна-
вания. Клетку узнают другие клетки — как соседние (с целью создания
тела) и иммунные (с целью уничтожения, если она опасна организму или
отслужила), так и чужеродные микробы (с целью ее повреждения и про-
никания внутрь).
а — клетки 8-клеточного зародыша; б — В-клетка иммунного ответа.
86
иммунной системы, и к пересадке пригодны только ткани
собственного организма или близкородственного. Через 10
лет американский иммуногенетик Джордж Снелл, исследуя
генетику отторжения, ввел понятие комплекса тканевой со-
вместимости (КТС) (по-английски: Major Histocompatibility
Complex — МНС). Данный комплекс как раз и делает иммуно-
логию такой же основой биологии, каковы, например, генети-
ка или экология.
КТС - это комплекс белков, кодируемых группой генбв,
совместно расположенных на одной хромосоме. У человека
ныне в данной группе известно 224 гена. Причем гены эти
удивительно разнообразны: некоторые имеют по 200 аллелей
- ничего подобного у других генов нет [Рабсон и др., 2006, с.
69]. Известно, что данные белки ответственны не только за
иммунное распознавание, но и за разнообразие рисунков кле-
точных поверхностей (рис. 13). Последнее необходимо для со-
единения клеток в ткань при онтогенезе.
Но зачем вообще нужно отторжение? Ведь в природе ни-
каких пересадок тканей не бывает. Очевидно, что отторжение
— побочное действие какого-то механизма, зачем-то организ-
му нужного. Его и надо искать.
Медавар и Снелл работали с теплокровными, поэтому уста-
новилась традиция видеть в КТС эволюционное достижение
высших животных. Но отторжение чужой ткани имеет место
у всех организмов (хотя в разных формах и в разной степени),
обладающих тканями. Губки, самые простые организмы, у
каких можно видеть некое подобие тканей, умеют отличать
свою «ткань» от чужой: если измельчить несколько губок
одного вида и перемешать фрагменты, то они вновь соберутся
в полноценных губок, тогда как фрагменты губок различных
видов к этому неспособны. У более сложных животных (на-
пример, медуз) оказываются несовместимыми уже ткани раз-
ных рас одного вида, а у высших организмов, имеющих раз-
витой адаптивный иммунитет, полностью совместимы только
ткани однояйцовых близнецов и особей лабораторных живот-
ных чистых генетических линий [Галактионов, 2005].
У растений вроде бы с совместимостью всё наоборот: ткани
близких видов вполне совместимы даже у самых высших форм
(у цветковых) — на этом построена прививка, основа плодо-
водства. Однако растения тоже кое-что распознают: яблоню
и грушу можно привить друг к другу или к рябине (одно се-
мейство), но не к липе. А главное, растения тоже способны к
индивидуальному распознаванию — всё те виды цветковых,
которые обладают обязательным перекрестным опылением,
обладают и самонесовместимостью — пыльца данной особи не
прорастает на рыльцах той же особи. То есть распознавание
«свой — чужой» идет у них столь же неукоснительно, как и
у нас с вами, только с противоположной целью. Самонесов-
87
местимость — это как бы толерантность навыворот. Словом,
явление несовместимости оказалось одним из самых общих
свойств жизни.
18. Самонесовместимость и совместимость чуть подробнее
Дело не в том, что иммунитет растений слишком прост и
чего-то не может, а в том, что растению нет нужды в аппарате
отторжения чужой ткани. Отторжение мы наблюдаем лишь
в лаборатории, в опыте, тогда как в природе чуждые ткани
встречаются друг с другом разве лишь в процессе пищеваре-
ния, а его у подавляющего большинства растений нет. (Оно
есть у хищных растений, и возможно предположить, что их
иммунитет окажется качественно сложнее, чем у других рас-
тений.) В ходе пищеварения чужие вещества разлагаются на
весьма мелкие части, теряя свою видоспецифичность, од-
нако этот процесс дает сбои, так что отдельные их не очень
мелкие фрагменты могут попадать из зоны пищеварения в
иные полости тела. Попадают туда также и целые микробы,
и вирусы, а это ведь тоже антигены. Может быть, КТС нужен
против них?
Нет, против чужих антигенов борется тот механизм врож-
денного иммунитета (ОРР), который умеет действовать вне
системы тканевой совместимости. Зато 224 гена, да еще по
200 аллелей у многих из них, наводят на иные мысли. Тут
можно ожидать (с учетом гетерозиготности) около 2 млрд
комбинаций, а это значит, что рисунки поверхностей клеток
могут быть практически неповторимы от человека к челове-
ку. Хотя у каждого человека этот рисунок довольно прост (за-
дается не более чем 12-ю вариантами молекул КТС [Полета-
ев, 2008, с. 38]), однако различие поверхностей клеток между
разными особями, создаваемое наличием КТС, огромно и
сильно затрудняет атаку организма микробами — к каждой
особи микробу нужен свой особый подход. Если бы поверх-
ности клеток у всех особей многоклеточных были одинаковы,
атаковать их микробам было бы намного легче. То есть КТС
действительно работает как защита от инфекций.
Это важно само по себе, но ничего не говорит про общие
причины усложнения иммунитета с ростом сложности орга-
низмов. Попробуем хотя бы подступиться к ним.
19. Иммунный механизм онтогенеза
В конце части 2 была высказана мысль, что у организма
есть тот тип иммунитета, какой нужен для его онтогенеза. В
обоснование можно привести несколько ярких, но всё же кос-
венных свидетельств. Прежде всего, это данные о сходстве
двух механизмов: разнообразие МКА, обеспечивающих со-
единение клеток друг с другом в единую ткань, устроено так
же, как разнообразие иммуноглобулинов (см. выше, п. 12).
88
Рис. 14. Галл в форме
шишки на ветке сосны.
Далее, простой иммунитет в са-
мом деле способен контролировать
сравнительно простую целостность.
Прекрасный пример приведен еще в
книге Л.С. Берга «Номогенез» (1922):
паразитирующие на растениях на-
секомые откладывают яйца в ткани
растений, а те защищаются, окру-
жая чужеродное тело оболочкой —
галлом, причем форму галла задает
растение. На хвойном это — недораз-
витая шишка (рис. 14). В такой реак-
ции растения мы видим сразу акт и
иммунной защиты, и онтогенеза.
Наконец, иммунологи приводят
такие примеры: если у низших жи-
вотных (губки и т.п.) масса живу-
щих в них бактерий может достигать
80% веса тела, то у человека - всего
2%. Зато при этом в нем живет око-
ло 500 видов микробов, образующих
устойчивую экосистему, в которую
невозможно внедрить новый вид.
«У беспозвоночных состав постоян-
ной микрофлоры обычно более узок
по составу, но положительные функ-
ции, которые они осуществляют для
организма хозяина, значительно более широки, вплоть до
управления морфогенезом и регуляции размножения орга-
низма-хозяина» [Лебедев, Понякина, 2008, с. 116]. Словом,
иммунитет и онтогенез явно переплетены, причем иммуни-
тет в ходе эволюции постепенно берет на себя новые функ-
ции, которые прежде исполнялись другими механизмами,
в том числе живущими внутри организма микробами. Но
для рождения теории нужно еще понять и молекулярную
сторону дела.
Понемногу становится ясно, что рассмотренная выше си-
стема КТС годна для контроля почти всего онтогенеза, одна-
ко — как раз кроме процесса формирования нервной системы.
Молекулы КТС присутствуют на поверхности почти всех кле-
ток организма, но это разные молекулы, и выполняют они
разные функции. У позвоночных, начиная с костистых рыб,
они бывают трех классов. Молекулы класса I имеются на всех
клетках, кроме нейронов и эритроцитов, и служат для кон-
троля состава собственных макромолекул организма. Моле-
кулы класса II представлены лишь на поверхностях некото-
рых иммунных клеток (например, В-клеток), и служат для
обнаружения чужеродных антигенов, предназначенных к
89
a
b
БЕСПОЗВОНОЧНЫЕ: DSCAM
ПОЗВОНОЧНЫЕ: ИММУНОГЛОБУЛИН
Рис. 15. Сопоставление элементов адаптивного иммунитета у малярийного
комара (а) и у человека (Ь). Хотя в первом случае налицо альтернативный
сплайсинг (преобразование РНК), а во втором — соматическая рекомбина-
ция ДНК, процессы и итоги весьма сходны. Рисунок из статьи [25].
уничтожению [Полетаев, 2008]. (Молекулы класса III нуж-
ны для запуска отдельных реакций иммунитета, и мы их ка-
саться не будем.)
Как видим, класс I осуществляет тот самый иммунный са-
моконтроль, на который указал когда-то Бернет, и этот класс
почему-то не касается нервной системы. А класс II ведет ту
самую борьбу с заразой, которой ограничивалось понимание
иммунитета до появления (полвека назад) идеи Бернета. Обе
части КТС заняты тем, что непрерывно прокачивают через
себя фрагменты макромолекул с поверхностей клеток, но
если цель работы класса II вполне понятна — борьба с заразой,
то назначение класса I довольно загадочно — зачем организму
ежечасно убеждаться в том, что он состоит сам из себя?
Первая мысль при ответе на этот вопрос — таким путем ведет-
ся контроль за перерождением здоровых клеток в раковые. Да,
иммунный надзор эффективен против одного типа опухолей,
именуемых высокоиммунными, но основная масса опухолей
избегает иммунного контроля. Причин этого много, и основная
- сбои в работе КТС [Рабсон и др., 2006, с. 256-258]. Создается
даже впечатление, что, не будь система КТС высших организмов
столь сложна, не было бы и основных типов рака. А значит, у
этой системы должна быть еще какая-то иная функция. Тут са-
мое время вспомнить, что нас интересует онтогенез.
На вопрос: «Управляет ли иммунитетом и онтогенезом
один и тот же механизм? » ответа и ныне дать нельзя, гипотеза
90
остается гипотезой. Но в последние годы появились примеры,
показывающие, что иногда это в самом деле так. Они, между
прочим, объясняют, почему КТС избегает касаться нейронов
(а с тем и построения мозга), — нейронами, оказывается, за-
нимается другая белковая система узнавания.
А именно, найдено несколько белков, каждый из которых
ответствен как за какой-то этап онтогенеза, так и за какую-
нибудь иммунную реакцию. В качестве недавнего примера
лучше всего указать на белок DSCAM, который ответствен не
только за рост аксонов, но и за иммунитет, причем обнаружен
и у человека, и у насекомых отряда двукрылых (дрозофила и
малярийный комар). Само название белка18 указывает, что он
контролирует нервную систему, а недавно выяснено к тому
же, что при повреждении его гена комар теряет часть своего
иммунитета — подвергается нашествию микробов, в том чис-
ле того самого плазмодия, что вызывает у людей малярию.
Выяснено, что в обоих случаях эффект действия DSCAM до-
стигается за счет чрезвычайной изменчивости данного белка.
Ее порождает альтернативный сплайсинг (см. п. 10). У насе-
комых DSCAM обеспечивает еще и некоторую адаптивность
иммунитета, похожую на нашу (рис. 15).
Особенно интересна статья «Разнообразие DSCAM суще-
ственно для нервной сети и для самораспознавания» [Hattory
et al., 2007]. Оказывается, что мутантные мухи, у которых
альтернативный сплайсинг выключен и потому синтезиру-
ется лишь одна форма DSCAM (испробовано три мутанта,
т.е. три формы белка), неспособны сформировать работаю-
щую нервную систему — концы аксонов крепятся как попало.
Единственное, что при этом аксоны делали правильно, — они
избегали отростков собственной клетки — проявляли самоне-
совместимость. Опять мы видим, что она нужна повсюду.
Но тут ученым может возразить внимательный читатель:
если иммунная система руководит онтогенезом, то почему
иногда рождаются вполне сформированные дети, лишенные
иммунной защиты и способные жить лишь в стерильных
условиях? Ответ известен врачам-иммунологам и достаточ-
но прост: у таких детей иммунитет сформирован почти нор-
мально, он содержит лишь один небольшой дефект: из-за от-
сутствия одного фермента врожденная система не посылает
адаптивной системе того сигнала, который должен запустить
процедуру размножения лимфоцитов. Онтогенезом же управ-
ляют отнюдь не лимфоциты, а, по всей видимости, рецепторы
МКА тканевых клеток.
18 DSCAM (Down syndrome cell adhesion molecule), т.е. молекула клеточной адге-
зии, ответственная за синдром Дауна. Дело в том, что у людей ухудшение психи-
ческого развития, известное как синдром Дауна, вызвано мутацией, портящей
именно данный белок.
91
20. Иммунитет - это эволюционный ограничитель
Вопрос Ламарка—Дарвина (см. раздел I, п. 12) имеет свою
иммунную сторону: если иммунитет полезен, то почему все
организмы не обзавелись в ходе эволюции вполне развитым
иммунитетом? Пусть рыбы остались рыбами в силу экоси-
стемных обстоятельств, но что помешало им за то же, что у
наземных позвоночных, время создать такой же развитой
иммунитет? Ведь обзавелись же все многоклеточные одним и
тем же (тонкости опускаем) типом клеточного деления, хотя
одноклеточные, от которых они, как принято считать, про-
изошли, делились весьма различно.
У всех организмов есть механизмы ограничения горизон-
тального переноса генов, оберегающие генетическое единство
каждого вида. Одним из таких ограничений и выступает им-
мунитет — ведь он препятствует, кроме всего прочего, проник-
новению чужих генов. Если бы иммунитет у всех организмов
был столь же сложным, как у высших, то эволюция была бы
сильно заторможена — за счет излишнего ограничения пото-
ков информации между организмами.
Иммунитет ограничивал эволюцию не только через горизон-
тальный перенос генов. Палеонтологи Роберт Кэрролл (Канада)
и Жан Шалин (Франция) независимо пришли к тому выводу,
что адаптивный иммунитет во многом определил эволюцию
теплокровных [Чайковский, 2008, с. 485]. Дело в том, что живо-
рождение возникало многократно, но только у теплокровных
вызвало иммунную проблему, и решили они ее по-разному.
Если у птиц и первых млекопитающих (яйцекладущих) жи-
ворождения нет; если сумчатые рожают ранее, чем можно ожи-
дать отторжения плода, то плацентарные окружают ранний
зародыш иммунным барьером. Это - трофобласт (слой клеток,
который затем становится плацентой), отделяющий плод от им-
мунных атак организма матери. Только отделенный от мира и
даже от матери иммунным барьером, зародыш человека может
достичь сегодняшнего итога всей эволюции — создать «себя не-
повторимого», способного мыслить и покорять мир.
Вот еще вопрос. В отличие от Ламарка и Дарвина мы зна-
ем, что царство бактерий — основа жизни для высших орга-
низмов и если бы оно исчезло, жизнь на Земле прекратилась.
Но одни бактерии породили, как принято считать, высших, а
остальные остались бактериями — иначе нас попросту не было
бы. Тоже — чем не объяснение? Вполне, кстати, в духе дарви-
низма. Но это не объяснение — на самом деле надо понять, как
и почему древние бактерии, не ведая о нуждах потомков (в
том числе и о наших с вами), удержались от того, чтобы всем
сплошь обратиться в высшие формы жизни и с тем погибнуть.
Ведь гибнут же, бездумно размножаясь, стая саранчи, рако-
92
вая опухоль или популяция чумной бациллы. Да и человече-
ство делает то же самое — так полагают многие.
О чумной бацилле стоит сказать особо. Зачем ей смертель-
ный яд, губящий «хозяина» (жертву), а с тем и ее саму? Как
недавно выяснилось, он вовсе не нужен ей самой (ее мутанты,
лишенные способности производить яд, выживают не хуже, а
лучше нормальных) и даже не служит отходом ее метаболиз-
ма. Он как бы специально сделан для убийства и затем - само-
убийства. Каким именно образом чумной яд убивает иммун-
ную систему жертвы, мы уже знаем (см. п. 13), но зачем?
Древние религии полагали, что чума — кара богов, но
сейчас такое объяснение существования чумы вряд ли кого
устроит, в том числе и верующих. Христианство, например,
утверждает, что Бог ничего плохого сам не творит. Тогда, мо-
жет быть, чумной яд творят черти? Нет, они, согласно христи-
анской догме, вообще ничего не творят, они могут только что-
то портить. Но ведь для синтеза яда особый оперон не только
создан (сотворен?), но и вписан в иммунитет бациллы — яд ее
саму не губит. Кем и как это сделано? Тут даже креационисты
ничего всерьез сказать не могут. Некоторые, правда, ссыла-
ются на божью кару, но эта идея прямо и грубо противоречит
догмату о благости божьей, так что им приходится погрязать
в толкованиях. Это - не путь к пониманию.
Не привлечь ли всемогущее божество другой правящей
религии - отбор? Но и отбор, даже если допустить его роль,
здесь бессилен, ибо обладатели яда гибнут много чаще других
бактерий, вместе с жертвами, для которых у них есть яд. По
этой же причине и Ламаркова активность работает здесь ни-
чуть не лучше. Может быть, яд возник случайно?
21. О случайности в работе
иммунной системы
Недаром Тонегава в нобелевской лекции оговорился, что
в исследованном им механизме формирования генов антител
имеет место «сложная неравномерная случайность»: действи-
тельно, теперь мы знаем, что основная часть иммуноглобули-
новых генов не мутирует вовсе, доля мутирующих убывает
примерно квазигиперболически, причем один из них мутиру-
ет втрое чаще основной массы мутирующих (см. рис. 10).
Вот почему у таких мутаций нет устойчивых частот. На-
сколько знаю, не найдено их и ни у каких иных мутаций. Более
того, сам термин мутация обрел столько смыслов, что лишь
запутывает дело, мешая строить теорию эволюции. Дело в
том, что утверждения типа «мутации объясняют эволюцию»
осмысленны только тогда, когда мутация понимается в самом
широком смысле - как любое наследуемое изменение. В то
же время случайными и ненаправленными являются только
мутации в весьма узком смысле - отдельные замены нуклео-
93
тидных пар в ДНК (точковые мутации) и другие изменения,
подобные им по простоте. Однако и у них случайность весьма
неравномерна, что выражается в их квазигиперболической
статистике.
Теперь видно, сколь наивен был окрик Медавара, чтобы
Стил либо показал регулярность частот (на гораздо более
сложном явлении, нежели точковый мутагенез), либо прекра-
тил исследование. Досадно, что квазигиперболическая стати-
стика разработана в 1920-е годы именно в Англии (основате-
лями явились ботаник Джон Виллис и математик Гаролд Юл
- G. U. Yule), где позже жил и работал Медавар, но она оста-
лась через полвека вне круга знаний остальных ученых, в том
числе и иммунологов.
Сейчас, спустя еще 30 лет, мы начинаем понимать, как с
такой случайностью можно работать — надо не говорить о ве-
роятностях (их тут нет), а выстраивать ряды — например, тех
же ядов и вызываемых ими инфекций, а с тем и эпидемий.
Эпидемии возникают вновь и вновь, пусть и без видимой
регулярности. А где есть повторность (ряды), там наука мо-
жет работать - в этом исходный тезис номогенеза. Если не
дарвинизм и не ламаркизм, то, может быть, номогенез? Ответ
положительный, см. часть 4.
22. Иммунитет и мозг
Как ни сложен узор стеклянной губки или цветка орхидеи,
эта сложность не идет ни в какое сравнение со сложностью со-
единений клеток в головном мозге, где каждый аксон каждо-
го нейрона растет туда, куда надо, чтобы мозг стал работать.
Разнообразие связей нервных клеток требует соответственно-
го разнообразия МКА. Еще в 1970 году генетик Сусуму Оно
обратил внимание на сходство мозга с иммунной системой:
«Каков был механизм, снабдивший геном Ното системой, в
которой не было непосредственной необходимости, но кото-
рая возникла как бы в предвидении будущих потребностей?..
У позвоночных известна такая система. Речь идет об иммун-
ной системе, дающей специфичные ответы на огромное ко-
личество антигенов, включая и искусственные, созданные в
пробирке» [Оно, 1973, с. 219]. Иммунолог Ерне (см. часть 1)
в своей Нобелевской лекции (1984) провел параллель даль-
ше: для Ерне иммунитет — аналог рассудочной деятельности.
В обеих системах (мозг и иммунитет) в каждом поколении на
основе небольшого числа генов разворачивается необозримое
разнообразие свойств. Это значит, что гены задают некое фор-
мирующее начало, а не конкретную схему устройства.
В наши дни параллель мышления с иммунитетом тоже
проводится, но уже на новых основаниях. Так, иммунологи
К.А. Лебедев и И.Д. Понякина отмечают: «После открытия
системы ОРР стало ясно, что функционирование иммунной
94
системы значительно сложнее и приближается к уровню
сложности работы ЦНС»; иммунная реакция в целом ныне
столь же мало понятна, сколь и появление в мозгу образа или
понятия [Лебедев, Понякина, 2008, с. 208].
Здесь требуется пояснение: если полвека считалось, что
одна сторона иммунитета (работа антител) понятна, то теперь
стало ясно, что антитело формируется не за счет случайной
мутации, а на основе анализа антигена в ходе его презента-
ции (см. п. 23). Информационная сущность этого анализа
совершенно непонятна (его-то и сравнивают с мышлением),
зато выяснилось, что синтез антител вплетен во всю работу
иммунной системы - как адаптивной, так и врожденной. И
вот парадокс - данное непонимание являет собой огромный
прогресс иммунологии, поскольку оно указывает на то, что
надо пытаться понять. Нужный вопрос дан отказом от каверз-
ного ответа.
А именно, надо понять, каким способом формируется спе-
цифический ответ организма на вызов среды - в данном слу-
чае на антиген. В точности та же задача стоит перед эволюци-
онистами, и они, в большинстве своем, так же плохо ее видят.
Разница в том, что в иммунологии можно сформулировать за-
дачу довольно четко, что и попробуем сделать.
23. Презентация антигена.
Отбор уступает «демону Ламарка»
Презентация - удивительная процедура, в ходе которой
антиген, обнаруженный иммунной клеткой, препарирует-
ся внутри нее и выставляется на ее поверхности (с другой ее
стороны) для опознания его другими иммунными клетками.
Удивительна она тем, что в разных имммунных клетках идет
тремя различными путями с использованием различных кле-
точных органелл и молекулярных механизмов переноса (т.е.
не могла произойти от одной предковой формы), однако всю-
ду приводит к одному и тому же результату.
На рис. 16 показана презентация антигена на трех типах
клеток (из числа изображенных на рис. 17):
1) макрофаг как любой фагоцит поглощает антиген неспе-
цифическим рецептором и доставляет в лизосому, где препа-
рирует;
2) В-клетка захватывает его антителом и переправляет на
свою другую сторону;
3) дендритная клетка попросту поедает его (этот способ
питания клетки называется пиноцитозом) и так же переправ-
ляет. В них во всех происходит активный перенос антигена
сквозь клетку.
Несмотря на эти различия, итог одинаков: антиген, под-
лежащий опознанию, соединенный с молекулой КТС, выстав-
ляется на другой стороне иммунной клетки. Здесь ее обна-
95
Рис. 16. Три типа презентации
антигена. В иммунных клетках
происходит активный перенос
антигена (показанного на рисун-
ке штриховкой) сквозь клетку.
Макрофаги поглощают антиген.
В-клетки захватывают антиген
с помощью иммуноглобулинов (Ig).
Дендритные клетки используют
для захвата пиноцитоз, то есть их
клеточная поверхность обволаки-
вает часть окружающей жидкости
вместе с содержащимися в ней веще-
ствами. Но, несмотря на различие
путей и молекулярных механизмов
переноса для клеток трех разных
типов, итог одинаков: антиген,
подлежащий опознанию в комплексе
с молекулой КТС, появляется на
другой стороне иммунной клетки
- для презентации.
руживает клетка-помощник (Т-хэлпер), назначение которой
- активировать те иммунные клетки, которым предстоит раз-
множаться и вести уничтожение заразы19.
По-видимому, именно в ходе активации В-клетки проис-
ходит процесс, который примиряет схему Бернета с арифме-
тикой (см. п. 6) - массовая подгонка гена антитела к антиге-
ну. Как именно она происходит, пока неизвестно, и, разуме-
ется, в ход идет ссылка на отбор. Например, Стил с соавтора-
ми пишут: «Вариабельные последовательности подвергаются
положительному дарвиновскому отбору путем связывания
с антигеном на уровне HL-гетеродимеров» [Стил, Линдли,
Бландэн, 2002, с. 129].
Тут нужно пояснить. В устах Стила эта апелляция к Дарви-
ну звучит странно, но здесь «положительный отбор» - это не
из Дарвина. Так иммунологи именуют действие химических
агентов, ускоряющих размножение иммунных клеток (а дей-
ствие тормозящих именуют отрицательным отбором). Далее,
HL-гетеродимеры - это обозначение тяжелых и легких цепей
(см. п. 9) вместе, т.е. отбор мыслится происходящим прямо
в вилке антитела. Или, иными словами, селективным терми-
ном названа инструктивная схема (см. п. 3), хотя в начале той
же книги она авторами отвергнута. В наших терминах вер-
нее будет сказать, что Стил и соавторы, говоря про «положи-
тельный отбор» фрагментов, имели в виду «делектус» (см. п.
15, раздел I). А если вспоминать имена классиков, то лучше,
по-моему, здесь подошел бы тот «демон Ламарка», о котором
шла речь в п. 9.
19 Кроме уже известных нам В-клеток, уничтожающих бактерии, это - тот основ-
ной массив Т-клеток, который призван уничтожать вирусы. Об этих клетках рас-
сказать здесь нет возможности (см. [Игнатов, 2002; Рабсон и др,, 2006]).
96
Как в действитель-
ности, никто не знает, и
любая догадка полезна,
пусть даже в будущем ее
опровергнут. Вредно (в
том числе для медици-
ны) только одно - делать
вид, что проблемы нет.
Проблема же, повторю,
состоит в том, что клон
иммунных клеток рас-
тет гораздо медленнее,
чем клоны их жертв,
т.е. существует процесс,
который быстро произ-
водит в параллель массу
клонобразующих кле-
ток. Быстро — значит,
без перебора огромного
числавариантов. «Демо-
на Ламарка» естествен-
но искать в процедуре
презентации антигена,
когда к нему приклады-
Рис. 17. Типы иммунных клеток и их про-
исхождение. Внизу - клетки системы
свертывания крови (мегакариоциты и
тромбоциты). Справа -В-и Т-клетки адап-
тивного иммунитета: Тх - Т-хэлперы (по-
мощники, они активируют другие клетки),
Тц - цитотоксические, то есть убивающие,
клетки (иначе - Т-киллеры). Остальные
изображенные на рисунке клетки относят-
ся к клеткам врожденного иммунитета и
клеткам-посредникам. Изображены не все
типы клеток.
ваются различные вари-
анты антител. Об этом, кроме группы Стила, говорят и другие
авторы. Их суждения ложатся в то общее русло мысли, которое
связано с делектусом. В качестве искомого «демона» некоторые
авторы указывают на обратную трансляцию — никем пока не
найденный механизм считывания информации полинуклеоти-
дом с белка - об этом см. [Чайковский, 2008, с. 422].
Сам Стил последние 20 лет занимается гипермутагенезом
и уверен, что тот включает в себя акты обратной транскрип-
ции. Она, в отличие от репликации ДНК, идет с высокой
(104 на пару оснований) частотой ошибок, что позволяет
понять огромную скорость гипермутагенеза [Steele, 2009].
Даже здесь (где идея случайных мутаций господствовала)
Стил видит, как и в объектах своих ранних работ, направ-
ленное изменение наследственности, т.е. ламаркизм в его
понимании (хотя теперь в его статьях ламаркизм не упоми-
нается).
Интересно проследить, как в течение 120 лет отступает идея
отбора случайных вариаций. До рождения иммунологии всё
было просто: полагали, что выживает тот, кто случайно оказал-
ся устойчив к данной заразе; он мыслился передающим данное
свойство потомкам. Затем, в годы Пастера и Мечникова, отбор
был перенесен с организма на его клетки: микроба стали пред-
ставлять как борющегося за существование с клетками орга-
97
низма (эритроцитами — у Пастера; фагоцитами — у Мечни-
кова). Открытие семейства лимфоцитов сузило идею отбора
до представления об отборе среди самих иммунных клеток
организма - успешно размножаются те из них, которые
производят нужные антитела. Это представление бытует
до сих пор, так что роль отбора сужается теперь далее в его
рамках.
А именно, если в годы Бернета отбор понимали как теку-
щий между В-клетками разной специфичности, то открытие
презентации и активации свело предполагаемый акт отбора к
отбраковке В-клеток, производимой Т-хэлперами и их окру-
жением. Смутная аналогия с естественным отбором уступила
место явной аналогии с отбором искусственным.
Что касается старого понимания отбора (лучше выживают
те, у кого совершеннее иммунитет), то оно почти выпало из
аргументации, когда выяснилось, что усложнение, связанное
с появлением адаптивного иммунитета, огромно, а массового
повышения выживаемости по сравнению с низшими организ-
мами вообще не видно. Наоборот, налицо целый спектр новых
болезней, и они часто — хуже для больного, чем смерть.
Вернемся внутрь клетки. Идея случайных мутаций, приво-
дящих к порождению нужных антител, уступила место пред-
ставлению о ступенчатом процессе, в котором случайность,
весьма ограниченная, сохранилась только за отдельными эта-
пами и притом в узком смысле — например, гипермутагенез
идет только в определенных точках ДНК. Эту случайность
вскоре тоже потеснили: одни замены происходят чаще, чем
другие [Steele, 2009].
Но главное, снова сузился отбирающий агент: теперь он
— не среда, а сам антиген. К нему, распростертому на презен-
тации, прикладываются варианты антител, тут происходит
их подгонка, и найденный вариант лимфоцита клонируется.
Это не отбор, а делектус.
Часть 4. Иммунитет как экосистема.
Что возьмем у классиков?
24. Иммунные рефрены
Итак, с Джаневеем иммунология обрела логический стер-
жень. Основой иммунитета служат у всех организмов врож-
денные механизмы, адаптивный же иммунитет высших жи-
вотных - надстройка, которая не только не обязательна, но и
не может работать без содействия иммунитета врожденного.
Если в основе врожденного иммунитета лежит распознавание
основных свойств опасных антигенов, то адаптивный постро-
ен на распознавании их индивидуальных особенностей.
В этой картине видны столь же ясные рефрены, как те, что
обсуждались в разделе I. Как мы знаем, молекулярные эмбри-
98
ологи были поражены сходством генов, запускающих главные
стадии онтогенеза у самых разных организмов (см. п. 17, раз-
дел I). Через 20 лет такой же сюрприз принесла иммунология,
когда обнаружилось фундаментальное сходство иммунитетов
у позвоночных, беспозвоночных и высших растений. В част-
ности, у растений и различных животных оказался одинако-
вым механизм разрушения микробных стенок посредством
антимикробных пептидов. «Особенно непонятно, а некото-
рым ученым кажется даже мистикой, что пептиды... никогда
не поражают клетки “хозяина”» [Белоконева, 2004].
Молекулярная основа иммунитета у трех групп (цветковые
растения, позвоночные, беспозвоночные) совсем различна, и
здесь самое время вспомнить о рефренах. Вернемся к табл. 2 —
Рефрен «Конечности позвоночных» (см. п. 14, раздел I).
Никто не станет выводить крыло птицы из плавника-крыла
рыбки клинобрюшки или ласт ихтиозавра из плавника-ласта
кистеперой рыбы. Так надо ли пытаться выводить иммунитет
высших из иммунитета низших? По-моему, полезнее обрисо-
вать ту диасеть, клетки которой заполнялись в ходе эволюции
иммунитета.
Для этого отметим, сколь удивительно сходство иммунных
процессов опознания, протекающих в самых разных клетках
одного организма. Несмотря на то, что различие клеточных
органелл и макромолекул, участвующих в опознании, ради-
кально, процессы протекают сходно (см. рис. 16). Сходство
процессов состоит в том, что в каждом из них распознаваемый
фрагмент молекулы активно проводится через распознаю-
щую клетку, выставляется на ее противоположной стороне,
где активно используется Т-хэлпером.
Такое молекулярное различие не дает вывести эти процес-
сы из общего предшественника — они тоже, как и иммуните-
ты трех групп организмов, явно произошли порознь. Тем са-
мым, процессы внутри одной особи тоже образуют рефрен. А
это значит, что перед нами некое глубинное сходство, которое
предстоит понять, чтобы изучать их (разные организмы, раз-
ные клетки) совместно.
Но есть общенаучные рефрены, более очевидные.
25. Три пары главных рефренов
Не раз отмечено — самые что ни на есть разные жизнен-
ные структуры могут обладать удивительным сходством,
которое нельзя понять как независимое приспособление
хотя бы потому, что не видно самого приспособления. Вот
примеры.
1. Очень похожи глаза (в форме камер с линзами) у позво-
ночных, головоногих, некоторых пауков и (трудно поверить!)
некоторых медуз. Причем у медуз нет мозга, способного при-
нять зрительную информацию, а в чем тогда польза от глаз?
99
2. Совсем различные механизмы обеспечивают у самых
разных организмов адаптивный иммунитет, и польза от него
видна не всем (вопрос обсужден в книге [Чайковский, 2008]).
3. Совсем различные объекты вдруг начинают безудержно
размножаться и вскоре от этого гибнут — таковы стая саран-
чи, чумная бацилла или раковая опухоль.
Однако если говорить не о пользе признаков, а о повторно-
сти явлений, то положение не выглядит столь уж безнадеж-
ным. Регистрация повторности — первый способ упорядоче-
ния разнообразия, вспомним рефрены Мейена (п. 14, раздел
I). Повторность — простейшая форма закономерности, а обна-
ружение закономерности — начало всякой науки. Объяснение
приходит позже.
Вспомним теперь содержание п. 13, раздел I. Там сказано,
что в статистической науке царит пара рефренов. Аналогично
в иммунологии тоже царит пара рефренов. Первый рефрен:
у позвоночных, беспозвоночных и высших растений незави-
симо развился сложнейший врожденный иммунитет. Второй:
у позвоночных есть еще более сложный иммунитет, адаптив-
ный, который независимо достиг своих высших форм у птиц
и зверей. Пары рефренов в науке обычны, и поищем третью,
нужную нам пару.
Единственный рисунок в «Происхождении видов» — ветвя-
щееся древо, коим Дарвин изобразил воображаемый процесс
расхождения признаков, различающихрасы. Этот процесс он
отождествил с происхождением видов и на этом схему завер-
шил. А его старший друг, геолог Чарльз Лайель, заявил, что
таким же образом можно объяснить и происхождение родов.
Заявление привело Дарвина в большой восторг («как глоток
спиртного» — писал он другу). Никакого примера образова-
ния нового вида или рода таким путем Дарвин и Лайель не
привели, на что критики конечно же указали.
Дальнейшее хорошо известно: в 1866 году германский зоо-
лог Эрнст Геккель изобразил в виде воображаемого древа всю
эволюцию — как животных, так и растений — от одноклеточного
предка. И доводов вскоре никто уже не спрашивал, поскольку
дарвинизм стал аксиомой. От древа Геккеля все давно отказа-
лись, но сам принцип древа господствует (под именем принци-
па филогении - см. п. 4, раздел I). Поскольку древом выглядит
и многое другое — схема роста клона клеток (как в онтогенезе и
в иммуногенезе, так и в раковой опухоли) и любой процедуры
роста численности особей, — то неизбежен вопрос: есть ли во
всем этом некий общий смысл? Оказывается, есть.
Всякое развитие есть направленный ряд, и, если направ-
ленные ряды проявляют существенное сходство, это рефрен.
Нам осталось понять, где рефрен-древо реален, а где вымыш-
лен (и где, следовательно, должны работать какие-то другие
рефрены).
100
При такой постановке вопроса ответ легко виден: рефрен-
древо реален там, где объекты исследования только расходят-
ся, но от него надо отказаться там, где объекты не только рас-
ходятся, но и сливаются; иначе говоря — там, где вместо древа
мы видим сеть. Слияние очевидно на уровне организмов — в
половом процессе и в горизонтальном переносе генов. Труд-
нее увидеть слияние объектов в экологии, но и там оно имеет
место: ведь экосистемы развиваются не делением, а проник-
новением друг в друга. Выходит, что кроме рефрена-древа
биологам необходимо рассматривать еще и рефрен-сеть. Вот
искомая третья пара рефренов — древо и сеть.
Все три пары сходны в одном: и обыденный опыт, и наука
заняты в основном одним членом каждой пары и почти не ви-
дят другого, а это сильно мешает как науке, так и практике.
Статистика царит «гауссова», иммунология — «адаптивная»,
биология - «древовидная». Рефрен-древо аккуратно описы-
вает лишь приведенную выше тройку — стаю саранчи, чумную
бациллу и раковую опухоль (а также подобные им объекты).
Подробно о такой однобокости науки см. статью (Чайковский
Ю.В. Ламарк, Дарвин и устройство науки // Вестник РАН,
2010, № 8). Поняв это, можно завершить наглядное описание
нынешней картины иммунитета.
26. Экология и номогенез иммунного мира
Если становление иммунной системы в онтогенезе можно
выразить в форме нескольких связанных между собой про-
цессов клонального роста, то иммунную систему в действии
лучше всего представить как экосистему (Ерне называл это
иммунной сетью).
Рассмотрим рис. 17. Как видим, все изображенные им-
мунные клетки клональным способом производятся из кле-
ток одного типа — стволовых кроветворных. Какую форму
примет данная иммунная клетка, зависит от условий: куда
она попадет и в какой момент жизни организма это случит-
ся. Наоборот, возникнув и развившись, иммунные клетки
действуют совместно, сообразно своему назначению (а не
своим наследственным свойствам — они у всех у них оди-
наковы). Это похоже чем-то на жизнь общества или на эко-
систему.
Проще всего понять с экологических позиций работу фа-
гоцитов: они пожирают те вещества и клетки, которые опо-
знаны как негодные или вредные. Это опознание ведут две
системы: сперва врожденный иммунитет, а затем включает-
ся и адаптивный — поэтому идет оно в две стадии. Их связь
обеспечивают некоторые фагоциты (моноциты), выделяя ве-
щества, оповещающие адаптивную систему о проникновении
заразы. Это — первый экологический аспект работы иммуни-
тета — внутренний.
101
Рис. 18. Нынешнее представление
о механизме порождения клона
В-клеток. Как видим, до начала
клональной стадии (внизу), приво-
дящей к массовому производству
антител, происходит сложный (и
во многом еще непонятный) обмен
с Т-хэлперами (Тх) информацией о
соответствующем антигене.
В-клетка (на свойствах В-
клеток строил свою концеп-
цию Бернет) активируется,
т.е. получает способность
производить нужный тип
антител вовсе не благодаря
случайной мутации, а в силу
помощи клеток, получивших
информацию об антигене.
Таковы Т-хэлперы и многие
другие клетки (рис. 18). Ин-
формационная роль Т-хэл-
перов в процессе активации
еще далеко не выяснена.
Ничто из этого в годы ра-
бот Бернета не было извест-
но, так что его увлечение
клонами можно понять, но в
наши дни ясно, что тайна воз-
никновения специфичности
антител существует и скрыта
именно во взаимодействии
клеток иммунной системы.
Второй экологический аспект — пограничный. В начале
части 3 мы говорили, что проникновение микроба в организм
сталкивает на границе (кожа, слизистая оболочка, рана) два
иммунитета, две активности: микроб пробует проникнуть,
жертва пробует не пустить, и оба используют иммунные при-
емы.
Третий экологический аспект — внешний. Такова работа
обоняния: животное узнает КТС-пептиды, исторгнутые из
других организмов, обнюхивая их самих и их отходы. Это по-
зволяет решать иммунные по существу задачи — избегать как
опасностей, так и инбридинга (родные братья и сестры пах-
нут сходно). Замечательно недавнее открытие: обонятельная
система использует для своей работы молекулы своего КТС
[Харченко, 2008].
Четвертый экологический аспект — общий (или, как теперь
любят говорить, системный). Животные питаются, поэтому
их иммунитет должен одновременно и бороться с заразой, и
обеспечивать сосуществование с нужными ему микробами
кишечника, и распознавать чужие молекулы, попавшие туда
с пищей. Всякий организм поедается, а значит, для кого-то
безвреден.
Недавно оренбургский патофизиолог А.П. Малышкин вы-
ступил со смелой гипотезой: как раз противоречие этих задач
и есть основной источник заразных болезней, ибо организм
обязан уметь поглощать нужные микробы и уметь с ними
102
сосуществовать (в частности, быть им пищей) [Малышкин,
2007]. Малышкин уверен, что патогенность не может быть
целью эволюции бактерии, поскольку снижает ее выживае-
мость — обычно популяция бактерий гибнет вместе со своей
жертвой. Эта уверенность, как мы знаем, справедлива лишь
с позиций дарвинизма, который полагает приспособление
единственным приемом и единственным итогом эволюции.
С этих позиций не видно решения загадки происхождения и
процветания смертельных инфекций, и Малышкин, сам того
не замечая, переходит от принятого им дарвинизма к идеям
ламаркизма и номогенеза.
А именно, он признал как собственную активность иммун-
ной системы - поглощение нужных микробов: его осущест-
вляют особые М-клетки (рис. 19), имеющиеся в слизистых
оболочках, так и ее упорядоченность (которую, по существу,
рассматривает как рефренную). Он справедливо полагает,
что смертельные инфекции не являют собой особого случая,
что поэтому теория иммунитета должна быть общей для всех
инфекций, а в чуме и т.п. надо видеть лишь патологию им-
мунного процесса. Сам же данный процесс у Малышкина —
вполне экологический, и эпидемия для него — такой же выход
экосистемы из-под контроля, как нашествие саранчи или (в
организме) раковая опухоль.
Начал Малышкин с того, что напомнил про таинственный
видовой иммунитет. Состоит он в том, что довольно близ-
кие виды могут не болеть болезнями друг друга. Человек и
грызуны подвержены чуме, однако человек, собака, свинья,
лошадь, корова и баран имеют каждый свою форму чумы. Хо-
лерой вообще болеет только человек. Среди бедствий крупных
эпидемий всегда находятся люди неболеющие, даже если они
тесно общаются с больными, а прежде с данной болезнью не
встречались. Всё это навело Малышкина на мысль, что пато-
генность — побочное свойство
микробов. А что главное?
Для него «функцией им-
мунной системы является
не “слепое” уничтожение
всего чужеродного..., а ста-
билизация инфекционного
процесса, предотвращение
его перехода в инфекцион-
ное заболевание». Как это
удается? Тут он обращает
внимание на поразительное
соответствие бактериальных
ядов и тех рецепторов орга-
низма, которые призваны их
распознавать. Ему ясно, что
Рис. 19. Активный перенос антигена
М-клеткой через слизистую оболоч-
ку. Антигеном (АГ) может быть
и молекула, и вирус, и бактерия. В
процессе участвуют Т- и В-лимфо-
циты, дендритные клетки (ДК) и
макрофаги (МФ). ЭК - эндоциты
(улавливающие клетки эпителия)
- см. [Рабсон и др., 2006, с. 104].
103
при этом можно представить себе заболевание как ошибку
в иммунном распознавании, но непонятно, откуда берется
сама патогенность, губящая популяцию микроба. Он лишь
приводит то соображение, высказанное до него, что яды и
метаболиты (нормальные участники обмена веществ) на-
столько сходны по строению, что яды «могут вмешиваться в
процессы метаболизма».
Остается напомнить, что понимание не всегда может до-
стигаться в терминах приспособления, иногда приходится
привлекать феномен повторности (рефрены), о чем, в сущно-
сти, и ведет речь Малышкин. В данном случае суть дела в том,
что опаснейшая бацилла Yersinia pestis имеет общий с осталь-
ными десятью видами рода Yersinia (из них девять незаразны
для людей) ген YopJ, ответственный за ее проникновение в
клетку млекопитающего [Zhou et al., 2004]. Поскольку почти
ко всем видам рода Yersinia у человека есть видовой имму-
нитет, его иммунная система легко может здесь ошибиться и
пропустить чуму.
Еще одна загадка видового иммунитета — в ВИЧ-инфекции
(СПИД). Пандемия СПИДа радикально отлична от всех пре-
дыдущих тем, что не склонна затухать, а лишь нарастает вот
уже четверть века. Эпидемиолог М.В. Супотницкий уверен,
что люди потеряли видовой иммунитет к СПИДу и что это —
акт экосистемной эволюции, способный убить человечество.
«СПИД-пандемия... “верхушка” более сложного природного
явления — эволюционного процесса, который представляется
нам в форме инфекционного» [Супотницкий, 2007]. Если в
самом деле так, то понимание эволюционной роли иммуни-
тета — главная и самая срочная задача, важней, по-моему,
всех нынешних кризисов.
27. Не будем забывать классиков
Ламарк смог сказать для своего времени главное: различил
основные формы активности - приспособление и прогресс.
Главное потому, что одним приспособлением эволюцию объ-
яснить не удалось — ни тогда, ни позже. К тому же геном в
очень малой мере изменяется за счет пассивной случайной
болтанки - наоборот, его активно (на грани порядка и хао-
са) изменяет сама клетка, а активность - это ламаркизм в его
классическом смысле.
Активность клетки видна во всем и хорошо исследована
в иммуногенезе. Пассивной болтанки не обнаружено нигде.
Таинство становления специфичности антитела в ходе пре-
зентации антигена (и связанных с ней процессов) будет рас-
крыто, едва отпадут ссылки на оную болтанку. Приложения,
прежде всего к медицине, ждать себя не заставят - если толь-
ко помнить, что ламаркизм всюду шествует об руку с номоге-
незом.
104
Но и Дарвина забывать не следует. Пусть основное поле
зрения его было слишком узко, так что о собственно эволюции
(появлении новшеств) он не смог сказать ничего существенно-
го, но его имя по-прежнему связано с натурфилософской иде-
ей - живущие лучше вымерших (см. п. 15, раздел I). И есть
пункт, в котором Дарвин ушел далеко вперед от Ламарка, —
вопрос о роли случайности в эволюции. Пусть Дарвин ее и
преувеличил, но Ламарк не увидал ее вовсе, а мы теперь зна-
ем, что эволюцию стоит ожидать именно на грани порядка и
хаоса (см. п. 16, раздел I).
Прогресс мы ныне в целом понимаем не по Ламарку (как
приближение к человеку), а шире. Как уже было сказано,
сложность «низших» организмов может быть намного выше
сложности «высших» — смотря как мерить сложность. На-
пример, у низших рачков, именуемых Полифемами, половая
структура (три пола) и общественное поведение куда сложнее,
чем у любых позвоночных (см. [Буторина, 2007]). Зато в пре-
делах подтипа позвоночных понимание прогресса довольно
близко к тому, что было у Ламарка: направление от «низших»
к «высшим» известно, никем не оспаривается (рыбы -> амфи-
бии -> рептилии —> птицы -> звери -> человек) и выражается
прежде всего в повышении интеллекта. В том же направле-
нии (от рыб к человеку) шло у позвоночных усложнение им-
мунитета. Естественно искать здесь связи, что выше и было
сделано.
До недавнего времени не было сведений о каком-то особом
характере иммунитета людей, но Супотницкий обращает вни-
мание на то, что особую роль в эволюции нашего иммунитета
играли транспозоны, доля которых особенно высока именно
в геноме людей [Stengel е. а., 2006]. Появление интеллекта
тоже следует связывать, кроме иммунитета, с транспозона-
ми. Для обоснования или отвержения этой гипотезы нужны
новые исследования.
Как мы видели, кроме ламаркизма в новом эволюциониз-
ме велика роль номогенеза — не столько книги самого Берга
(она играет примерно ту же роль, что «Философия зоологии»
и «Происхождение видов»: ее почитают, но не читают), сколь-
ко концепции рефренов. Особенно, по-моему, важны в эволю-
ции три типа рефренов, приведенные в п. 25.
Заключение
Итак, на грани веков снова, как и сто лет назад, произошел
прорыв в понимании устройства природы. Теперь это — пони-
мание места иммунитета в биологии (заставляющее строить
новую теорию эволюции с новой ее основой) и понимание ме-
ста темной энергии в космологии (разрушающее привычное
понимание роли гравитации [Ксанфомалити, 2005]). Темная
энергия равномерно разлита вокруг и внутри нас. Это тоже
105
форма активности (а именно, активности физического ваку-
ума), форма, о которой стало известно всего десять лет назад.
Последует ли за этим и прорыв в понимании эволюции мира?
Ведь понимание макро- и микромира обычно предполагает
совместное рассмотрение.
Свидетельств иного, нежели принято думать, устройства
сил природы накопилось в биологии тоже много (см. конец
раздела I). Самое время понять, что активность живого — это
целый ряд форм активности материи, что он продолжает тот
ряд (актиреф), который известен из наук о неживой приро-
де. Если живая материя активна во всем, на всех уровнях, то
естественно ожидать ее активности и на уровне наследствен-
ной изменчивости (что долгое время отрицалось).
Полтора века царило убеждение, будто на этом и только на
этом уровне материя пассивна (способна лишь на случайную
болтанку). Теперь такое убеждение — анахронизм: открытия
последних лет показали, что как гены, так и участки ДНК
между ними изменяются активно, по своим законам. Один
пример, иммуногенез в В-клетках, мы рассмотрели подроб-
нее, чтобы увидеть, что такое активность генетической систе-
мы на деле.
На этом примере легко понять, что являет собой «основа
эволюции» (п. 14, раздел I). Если врожденный иммунитет не
смог сам справиться с заразой и болезнь разрастается, то ор-
ганизм оказывается в тяжелом стрессе, ибо утратил норму.
Стрессом запускается поисковая активность иммунной си-
стемы20, одной из форм которой является идущий в В-клет-
ках генетический поиск (его обычно воспринимают как одну
из форм отбора по Дарвину, но на самом деле это делектус,
в частности он не требует смены поколений). Поиском рас-
ширяется изменчивость, которая за счет комбинации блоков
колоссальна. Она, однако, не выходит за рамки, задаваемые
диасетью. Когда нужный тип антитела найден, генетическая
система запоминает его в форме клеток иммунной памяти.
Тем самым фиксируется новая норма, генетический поиск
прекращается и изменчивость снова падает. Следует напом-
нить, что вне иммунологии основа эволюции видна лучше
всего на примере преобразования организмов в опыте Ша-
пошникова и подобных.
Вентребер и Стил предлагали, пусть и очень различно, ви-
деть в иммуногенезе модель эволюции. Хотя их модели ныне
выглядят наивно, но таков удел всех моделей, и мне предста-
вилось разумным развить эту линию мысли, ламарковскую
по сути. Тот факт, что иммунные механизмы управляют он-
20 В иммунологии эту мысль разработала Полли Матцингер [Matzinger, 2002], а в
зооморфологии близкую мысль о роли утраты нормы высказала Е.Е. Коваленко
(см. [Чайковский, 2008]).
106
тогенезом, вселяет надежду, что дисциплина «evo-devo» (по
сути - часть жоффруизма) всерьез поможет решить проблемы
эволюции.
Что касается ЭКЭ, то ей, на мой взгляд, еще предстоит стать
теорией и освоить для этого хотя бы азы иммуноэкологии. Но
самым важным вкладом иммунологии в теорию эволюции
видится демонстрация появления принципиальной новизны
- таково активное формирование антитела к искусственному
антигену. Именно этот факт впервые показал, что возникно-
вение новизны, а с тем и эволюция - реальность.
Такую эволюцию не могут отрицать даже креационисты.
Ведь вряд ли креационист, даже самый дремучий, станет
утверждать, что Бог сам, лично, руководит каждым процес-
сом, какие ежесекундно текут внутри всех клеток всех особей
всех видов организмов. Такая вечная рабская работа противо-
речила бы его величию, поэтому даже креационисты соглас-
ны, что эти процессы текут по единым законам природы, в ход
которых Бог, задав их, больше не вмешивается. Если в таком
процессе рождается новый ген, значит, новое действительно
может рождаться. А если так, то и эволюция, понимаемая
как преобразование генетических систем, происходить может
- даже с позиции креациониста.
По-моему, всем ученым, в том числе верующим в Творе-
ние, и надо ее исследовать, а не спорить о том, кто прав — Би-
блия или Дарвин. Общим для всех ученых исходным пунктом
может послужить осознание феномена активности. Ведь ак-
тивность действует в различных формах на всех уровнях ми-
роздания, и признать этот факт можно независимо от того,
считает ли данный автор ее источник божественным или нет.
Признав это, вспомним, что назначение теории — служить
людям. Не раз отмечено, что идея всеобщей борьбы служит
силам зла, и отрадно узнать, что она ничем, кроме веры в
зло, не обоснована. Можно надеяться, что иммунологам опи-
санная «основа эволюции» полезна уже сейчас (см. пп. 23 и
27), а о других приложениях прочитать в книге [Чайковский,
2008]. Там же можно узнать о тех немногих пока данных па-
леонтологии, которые не просто иллюстрируют разнообразие
древней жизни, как сказано во введении, а свидетельствуют
об эволюции всерьез.
Эти данные служат хоть и небольшим, но важным добавле-
нием ко всему, что сказано выше. А именно, в настоящее вре-
мя науке известно несколько существенных примеров того,
что эволюция живого протекает на самом деле. И пусть креа-
ционисты спорят с нынешней наукой, а не с учебниками про-
шлого века. Кстати, само возрождение креационизма, почти
исчезнувшего в середине XX века, вызвано, на мой взгляд,
тем, что эти учебники всё еще в ходу, хотя расхождение их с
достижениями науки разительно.
107
ЛИТЕРАТУРА К РАЗДЕЛУ II
Абелев Г. И. Загадка происхождения специфического иммунитета (По-
лемические заметки на книгу: Галактионов В.Г. Очерки эволюционной имму-
нологии. М., 1995) // Онтогенез, 1997, № 1.
Аронова Е.А. Иммунитет. Теория, философия и эксперимент. М., 2006.
Белоконева О. Иммунитет в стиле ретро // Наука и жизнь, 2004, № 1.
Бернет Ф. Клеточная иммунология. М., 1971.
Буторина Л. Сложная жизнь Полифема // Наука и жизнь, 2007, № 11.
Галактионов В.Г. Эволюционная иммунология. М., 2005.
Гилберт С. Биология развития. В 3 т. М., Мир, 1995. Т. 3.
Игнатов П.Е. Иммунитет и инфекция. Возможности управления. М., 2002.
Кокряков В.Н. Очерки о врожденном иммунитете. СПб., Наука, 2006.
Ксанфомалити Л. Темная энергия // Наука и жизнь, 2005, № 5.
Лебедев К.А., Понякина И.Д. Иммунология образраспознающих рецеп-
торов. Интегральная иммунология. М., 2008.
Малышкин А.П. Некоторые вопросы инфекции и видового (наслед-
ственного) иммунитета // Иммунология, 2007, № 3.
Меджитов Р„ Джаневей Ч. Врожденный иммунитет // Казанский
медиц. журнал, 2004, № 3.
Оно С. Генетические механизмы эволюции. М., 1973.
Полетаев А. Б. Иммунофизиология и иммунопатология. М., 2008.
Рабсон А. и др. Основы медицинской иммунологии. М., Мир, 2006.
Савинов А.Б. Биосистемология. Учеб, пособие. Ниж. Новгород, 2006.
Стил Э., Линдли Р., Бландэн Р. Что, если Ламарк прав? Иммуногенетика
и эволюция. М., 2002.
Супотницкий М.В. К вопросу о роли ВИЧ-инфекции...// Энвайронмен-
тальная эпидемиология (журнал, Киев), 2007, № 1.
Ульянкина Т.И. Зарождение иммунологии. М., Наука, 1994.
Харченко Е.П. Иммунное узнавание и иммунная привилегия // Имму-
нология, 2008, № 2.
Чайковский Ю.В. Иммунитет и эволюция: не впасть бы в другую край-
ность // Вестник Российской академии наук, 2003, № 3.
Чайковский Ю.В. Активный связный мир. М., КМК, 2008.
Hattory D. et al. Dscam diversity is essential for neural wiring and self-rec-
ognition // Nature, 2007, vol. 449, p. 223—227.24.
Janeway C.A., Jr. Approaching the asymptote? Evolution and revolution in
immunology // Cold Spring Harbor Symp. Quant. Biol., 1989, vol. 13.
Kurtz J. et al. Alternative adaptive immunity in invertebrates // Trends in
Immunology, 2006, № 11.
Lherminier Ph. Modeles instructifs et modeles selectifs de 1’origine des
especes // Jean-Baptiste Lamarck, 1744—1829. (119е Congres national des
societes historiques et scientifiques, 1994.) Paris, 1997, p. 409—424.
Matzinger P. The Danger model: a renewed sense of Self // Science, 2002,
vol. 296, p. 301-306 (April).
Ruckdeschel K. et al. Crosstalk of signalling process of innate immunity
with Yersinia Yop effector function // Immunobiology, 2008, № 3—4.
Steele E.J. Mechanism of somatic hypermutation: critical analysis of str-
and biased mutation signatures at A:T and G:C base pairs // Molec. Immunol.,
2009. vol. 46. p. 305-320.
Stengel A., Roos C. et al. Expression of endogenous retroviruses in Old Wo-
rld monkeys // Jour. Virology, 2006, vol. 80, № 9.
Wintrebert P. Le vivant createur de son evolution. Paris, 1962.
Wu T. T., Cabat E.A. An analysis of the sequences of the variable regions of ...
light chains and their implications for antibody complementarity // Journal
Exp. Med., 1970, vol 132, p. 211-250.
Zhou L. et al. Yersinia YopJ inhibits proinflammatory molecule expression in
human bronchial epithelial cells // Respiratory Physiology & Neurobiol. 2004,
vol. 140, p. 89-97.
108
СЛОВАРИК
Адаптивный (приобретенный) иммунитет - совокуп-
ность иммунных свойств и реакций, формирующихся
в ходе развития и дальнейшей жизни организма. Изве-
стен у всех групп позвоночных. Полностью представлен
у теплокровных (птиц и, особенно, млекопитающих) в
форме Т-клеток, управляющих распознаванием антиге-
нов и борющихся с вирусами, и В-клеток, борющихся с
бактериями. Приводится в действие сигналами системы
врожденного иммунитета (она есть у всех организмов и
вступает в действие сразу по проникновении заразы в
организм), причем он начинает действовать лишь через
несколько суток после заражения, если врожденный им-
мунитет не смог до этого справиться с заразой. Важней-
шая черта адаптивного иммунитета — иммунная память,
т.е. отсутствие или более легкая форма заболевания при
повторном заражении организма той же инфекцией. У
низших позвоночных представлен слабо. У насекомых и
высших растений имеет иную природу и известен мало.
У низших беспозвоночных не выражен вовсе (например,
у губок и одноклеточных).
Аллель — одно из возможных состояний гена. Напри-
мер, различие цвета глаз у людей вызвано тем, что ген
«цвет глаз» представлен у них различными аллелями.
Альтернативный сплайсинг — тип сплайсинга (см.
сплайсинг), при котором вырезаются не только все ин-
троны, но и некоторые экзоны (кодирующие участки
РНК). При таком сплайсинге на одной макромолекуле
РНК может быть синтезировано много различных бел-
ковых цепочек. Аналогичный процесс на уровне ДНК
именуется соматической рекомбинацией.
Антиген — молекула или более крупное тело (свое
или чужеродное), способное вызвать иммунный ответ.
Антитело — растворимый белок, нейтрализующий
антиген.
В-клетки — клетки адаптивной иммунной системы,
способные распознавать антигены и вырабатывать ан-
титела. Каждая В-клетка продуцирует только один вид
антител.
Генетический поиск — тип активности генетической
системы, при котором создаются новые генетические
тексты. При нем наследственная изменчивость возрас-
тает, давая материал для эволюции. Простейший при-
мер: рост мутабильности при стрессе.
Горизонтальный перенос генов — передача фрагмен-
тов ДНК от одного организма другому вне процесса по-
109
лового размножения (вертикального переноса) или в
пределах одного организма. Чаще всего происходит че-
рез посредство вирусов.
Гуморальный иммунитет — совокупность иммунных
реакций организма, связанных не с клетками, а с внекле-
точными жидкостями.
Длительная модификация — краткосрочное (на не-
сколько поколений) наследование приобретенных
свойств. Обычно идет за счет внехромосомного гена.
Если он встраивается в хромосому, наследование стано-
вится обычным (стойким).
Клон — потомство, полученное от одной особи (ею
может быть как клетка, так и больший организм) путем
последовательного многократного бесполого размноже-
ния.
Лимфоциты — клетки, плавающие в лимфе и ответ-
ственные за иммунный ответ.
Онтогенез — развитие особи от оплодотворения до по-
ловой зрелости.
Пептиды — короткие (короче белков) цепочки амино-
кислотных остатков. Содержат до нескольких десятков
остатков аминокислот.
Сплайсинг — процесс удаления интронов (некодирую-
щих участков) из мРНК с последующей сшивкой образо-
вавшегося в ней разрыва. Происходит после транскрип-
ции РНК, но до ее трансляции, т.е. до считывания с нее
аминокислотной цепочки.
Ткань — компактное множество клеток, предназна-
ченных для определенной функции.
Т-клетки — клетки адаптивной иммунной системы,
первично образующиеся в тимусе. Выполняют разные
функции: активируют В-клетки (Т-хэлперы), регулиру-
ют активность иммунной системы, уничтожают неугод-
ные клетки (Т-киллеры), уничтожают вирусы.
Транспозон — элемент генетической системы, способ-
ный перемещаться как целое в пределах генома организ-
ма или между геномами.
Фагоцит — иммунная клетка, предназначенная для
уничтожения других клеток (как чужеродных организ-
му, так и его собственных) путем их поглощения.
СОКРАЩЕНИЯ:
КТС — комплекс тканевой совместимости
МКА - молекулы клеточной адгезии
ОРР - образраспознающие рецепторы
ЭКЭ — экосистемная концепция эволюции
110
НАУКА И ЖИЗНЬ
Каталог «Гаэеты.Журналы»
агентства «Роспечать»,
индекс 70601
• Легендарный журнал -
более 100 лет с читателями!
• Уникальная энциклопедия для
всей семьи - интересная и
достоверная информация из
первых рук.
• О науке доступно, о жизни -
серьезно. Российским
достижениям - приоритет.
• Подписка на журнал - лучший
подарок к любому празднику.
• Умная реклама - для умных
людей. Нашим рекомендациям
доверяют миллионы!
www.nkj.ru
On-line версия журнала «Наука и жизнь»,
электронный архив на DVD и CD -
в интернет-магазине http.7/www.nkj.Rj/shop/
Тел.:(495)628-09-24
e-mail:re klama@nkj.ru
Юрий Викторович Чайковский
и I и
"в 11I И
Н 1г11 И
РАЗВИТИЕ ЖИЗНИ И ИММУНИТЕТ
Редакция журнала «Наука и жизнь»
Телефон: (495) 624-18-35; факс: (495) 625-05-90
101990, Москва, Мясницкая ул., д. 24, строение 1
E-mail: mail@nki.ru
www.nkj.ru
Подписано в печать 26.08.2010
Заказ 2441. Формат 60x90/16 Бумага офсетная
Печать офсетная
Печ.л. 7
Тираж 2 000 экз.
Отпечатано в соответствии с предоставленными материалами
в ЗАО “ИПК Парето-Принт”, г. Тверь, www.pareto-print.ru