




































































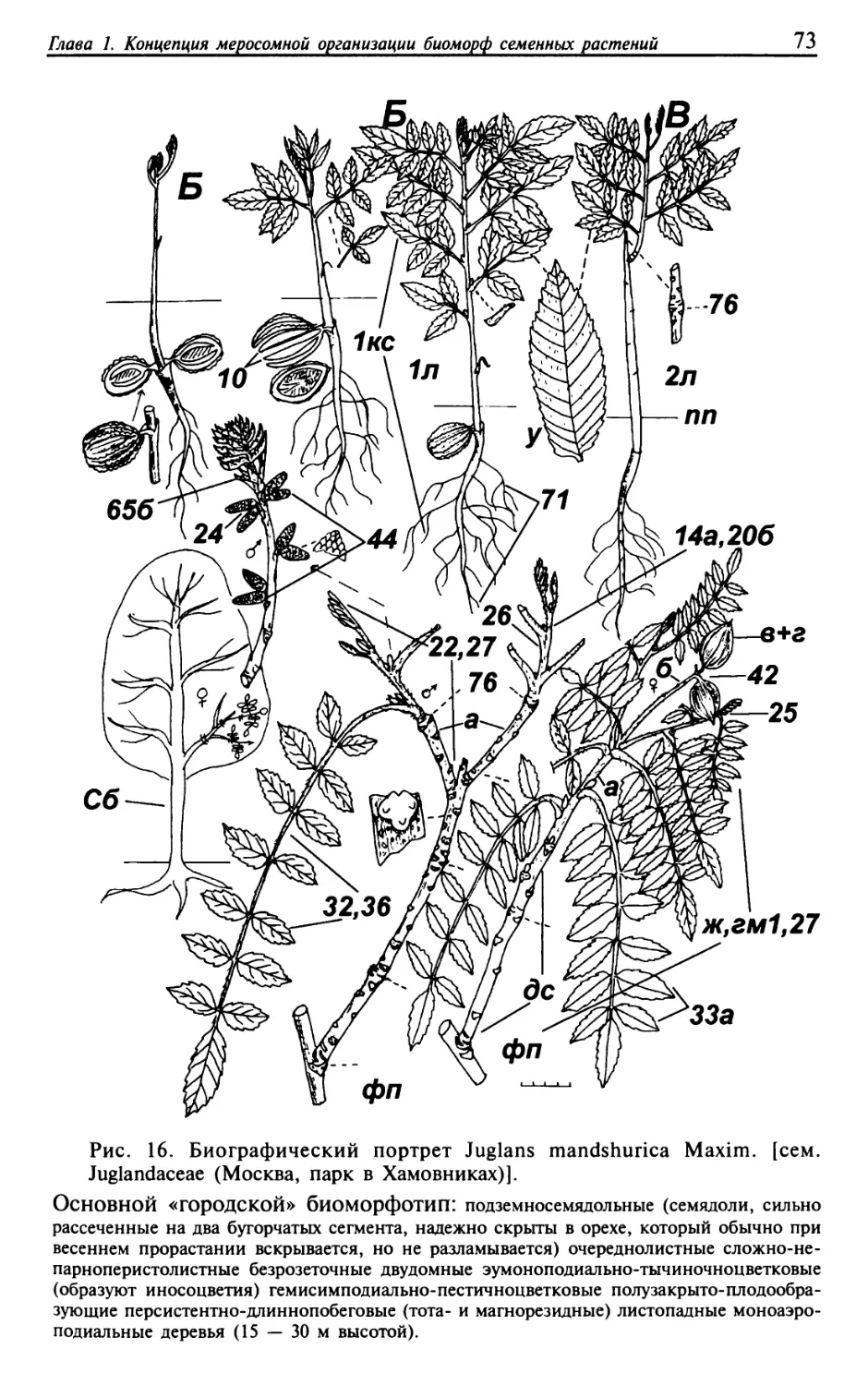
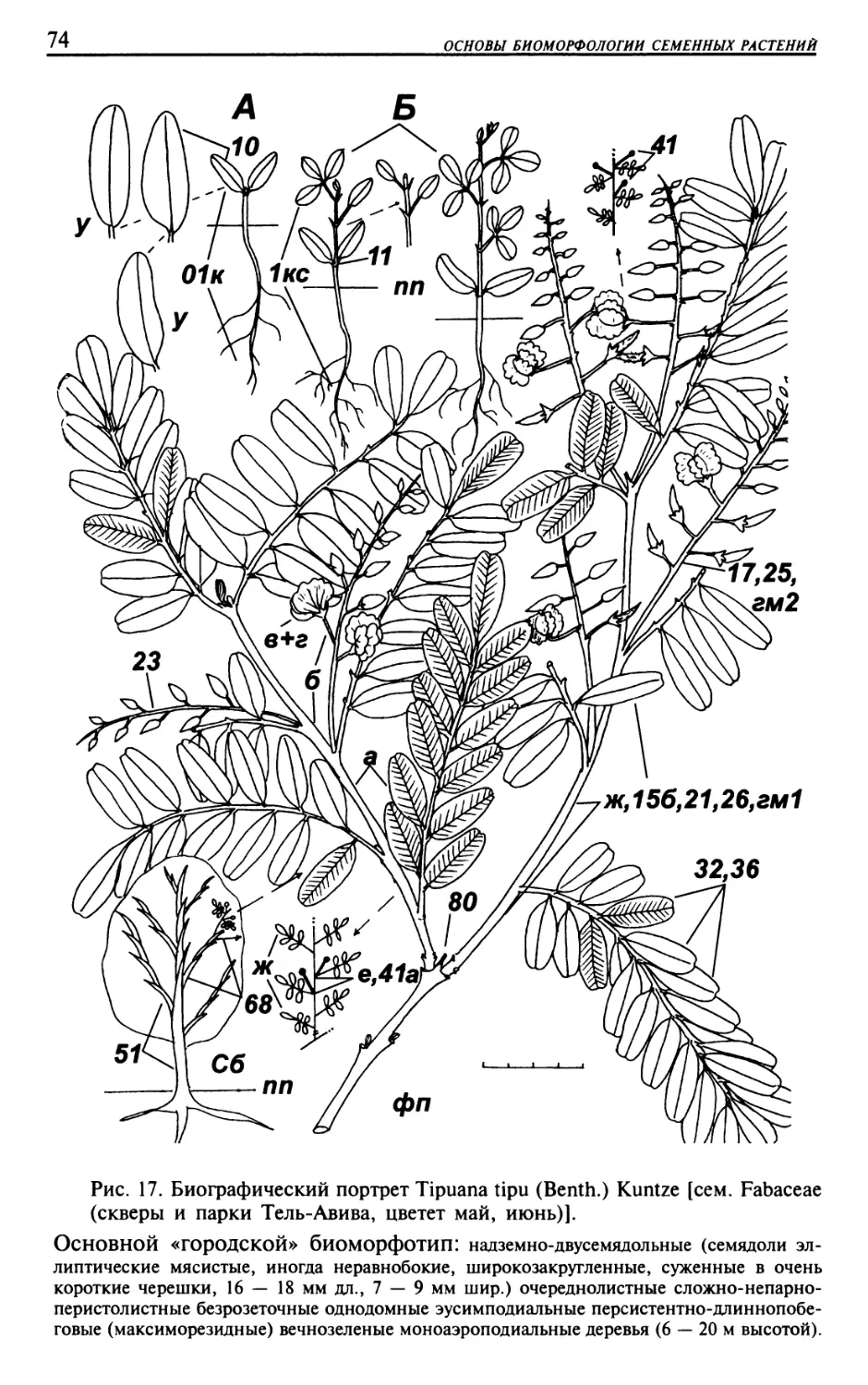
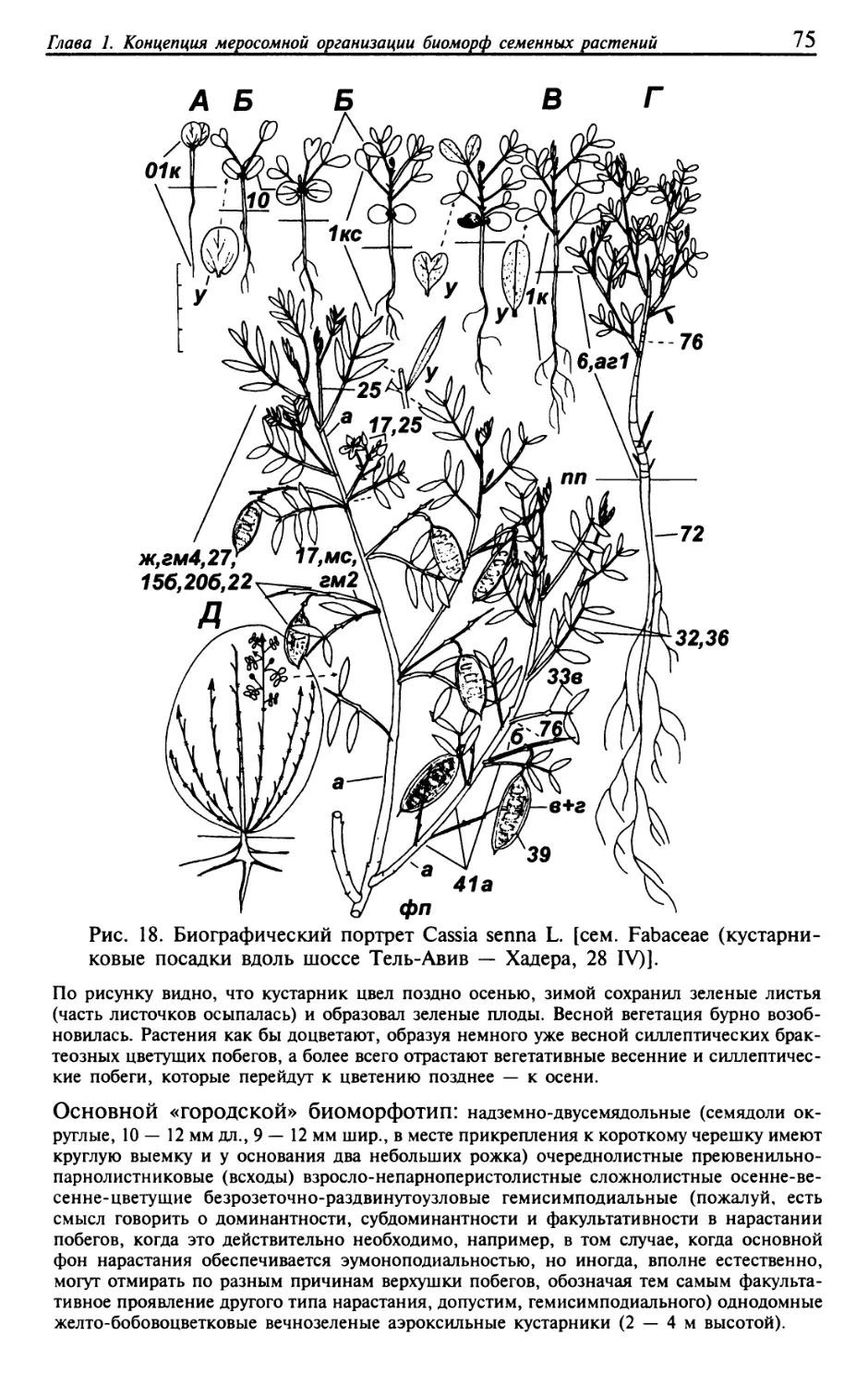

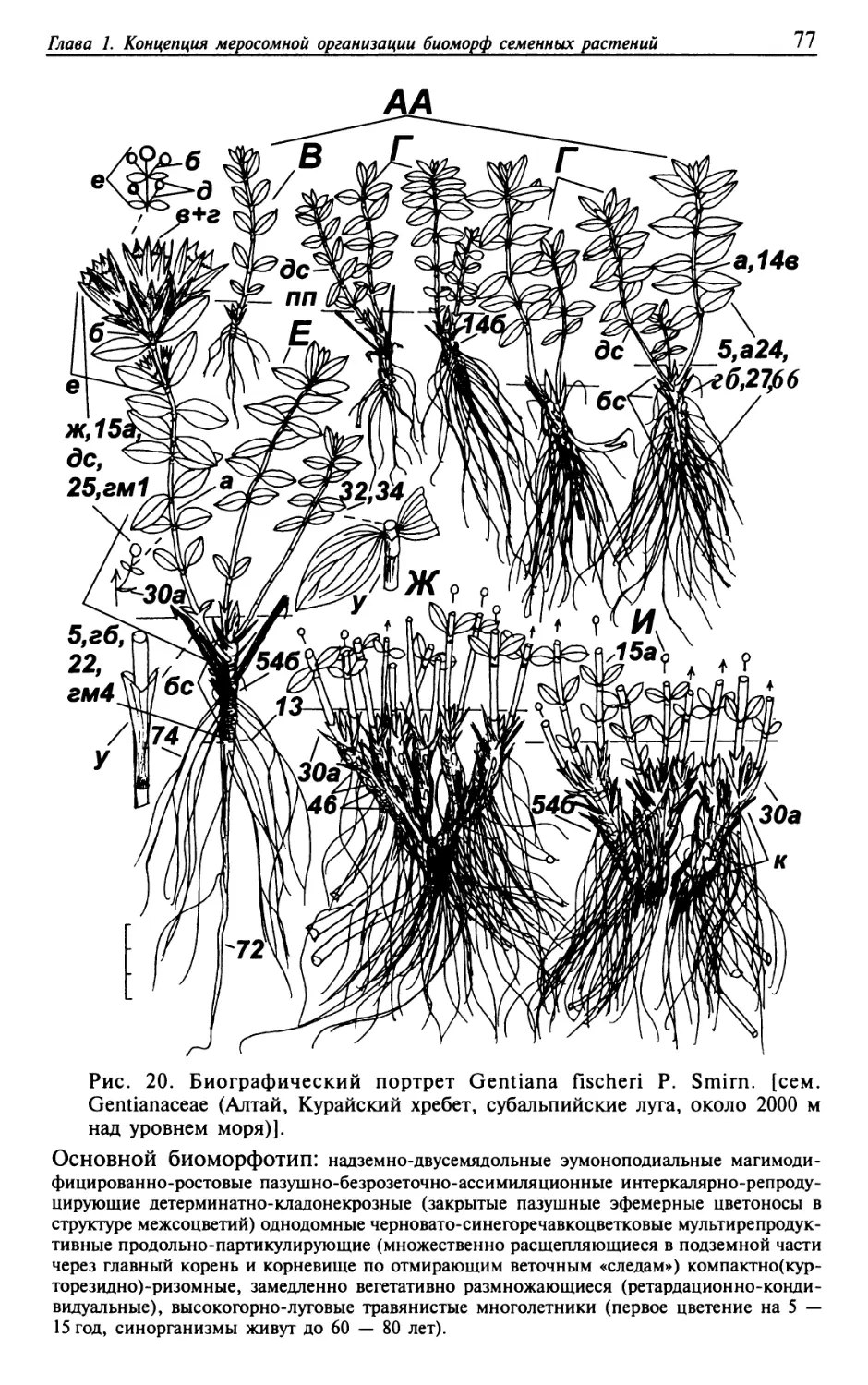
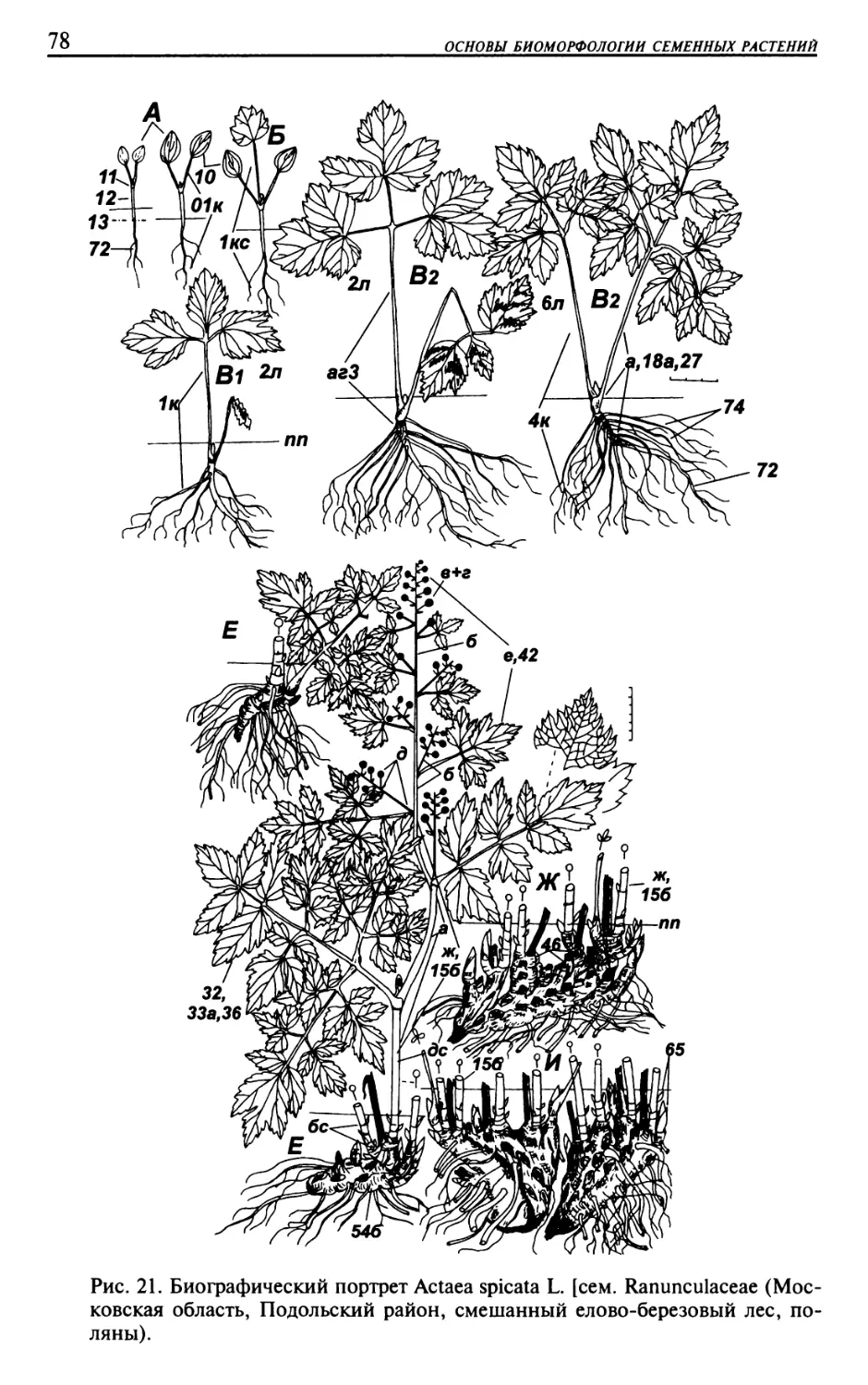

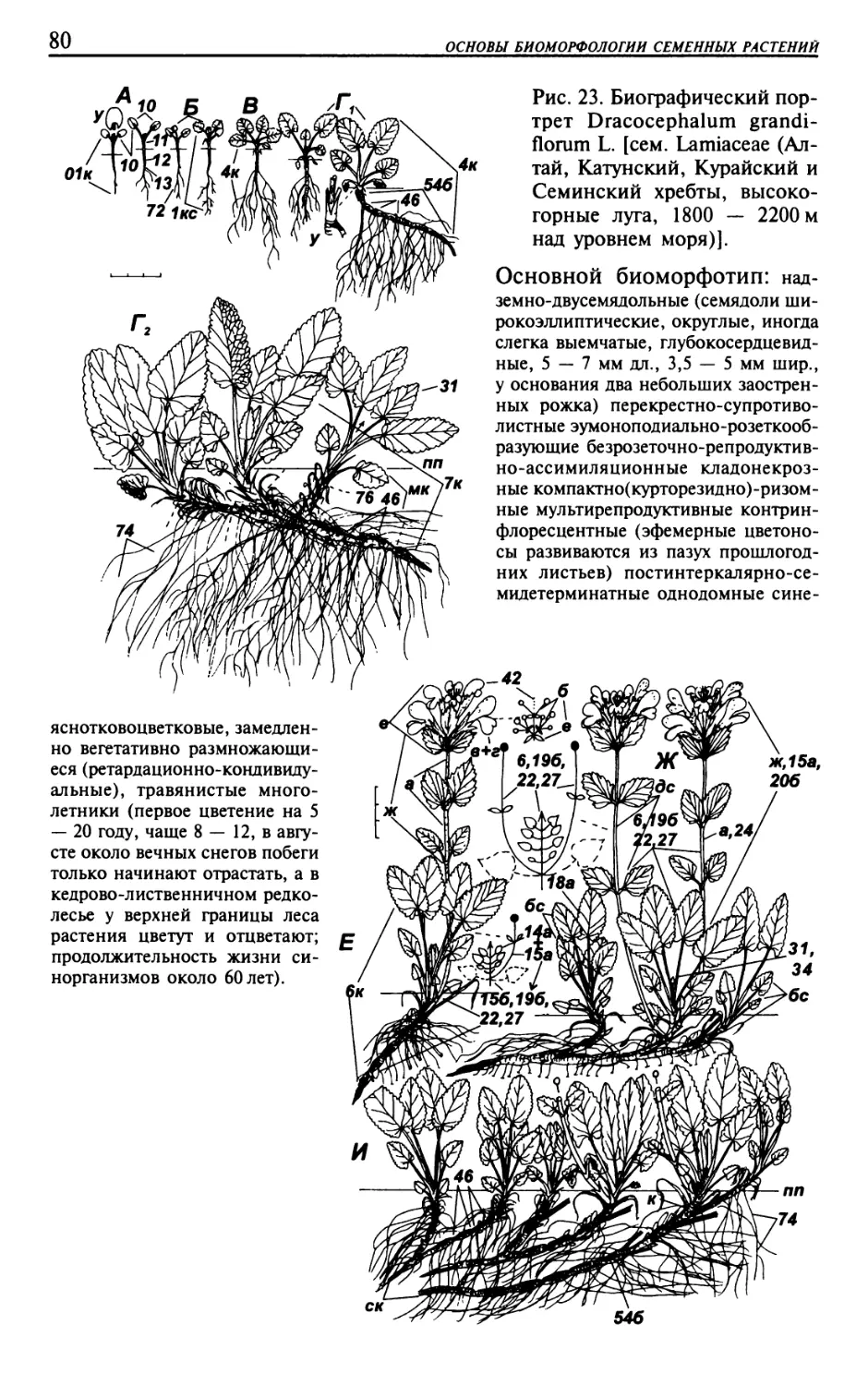
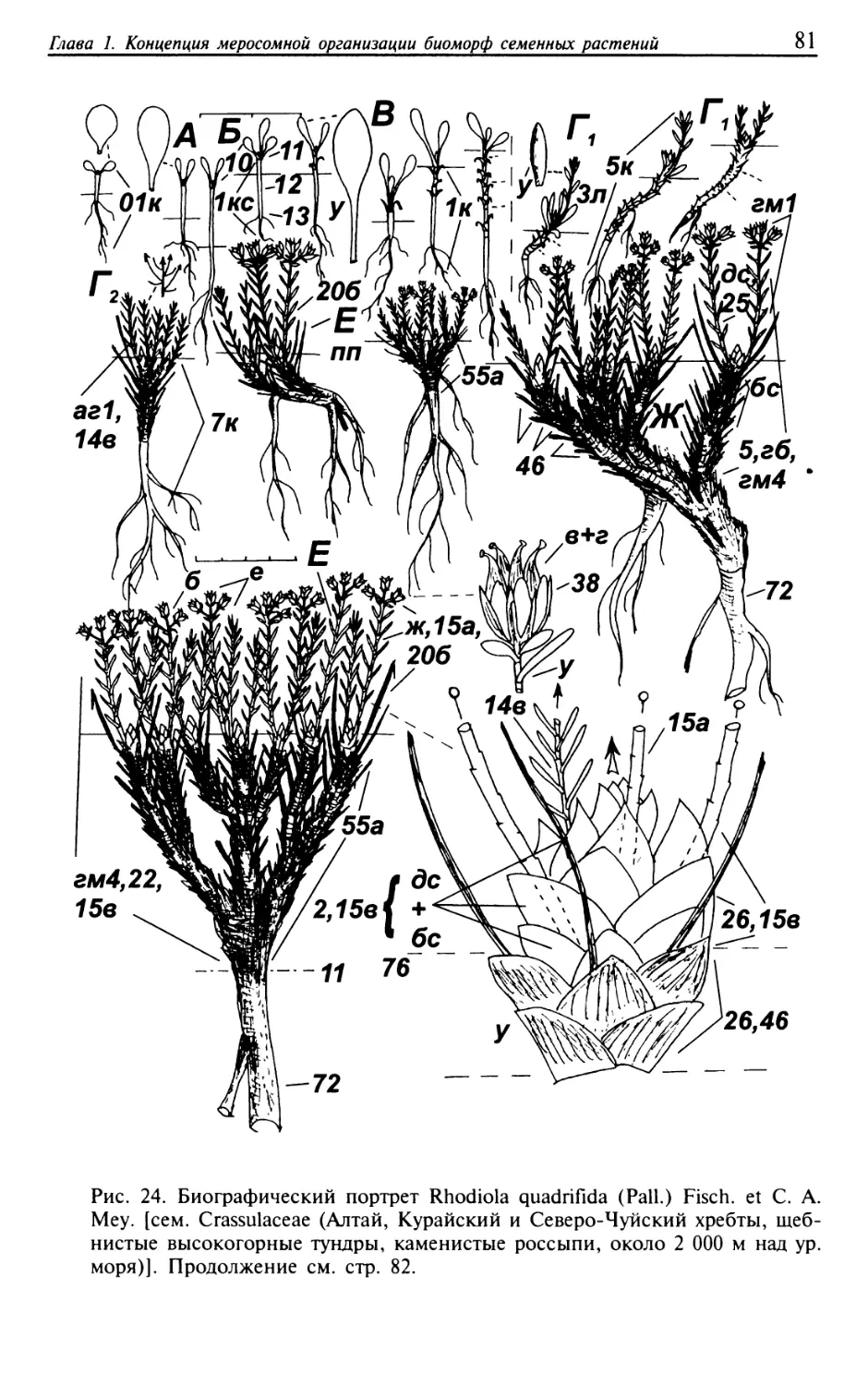
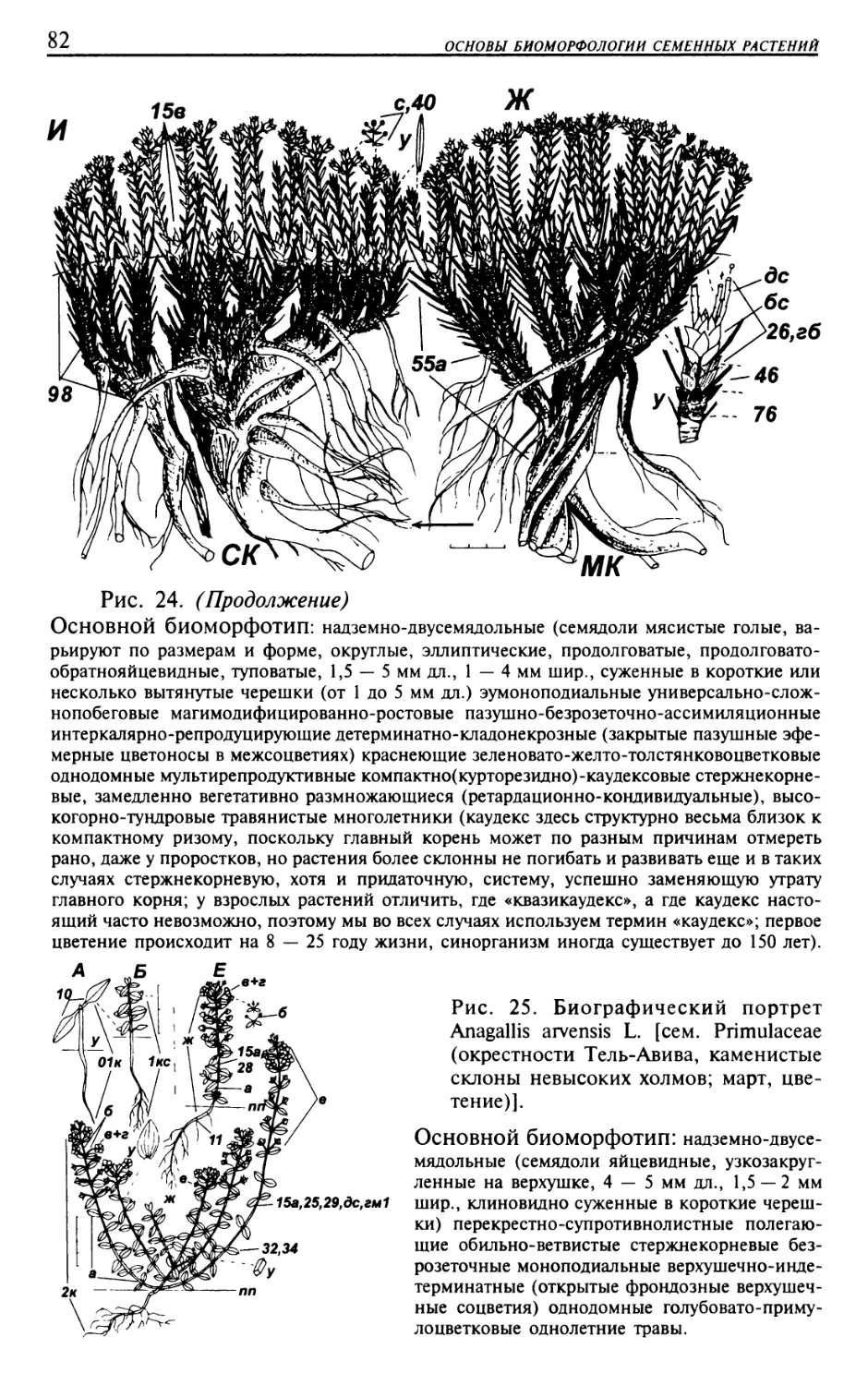
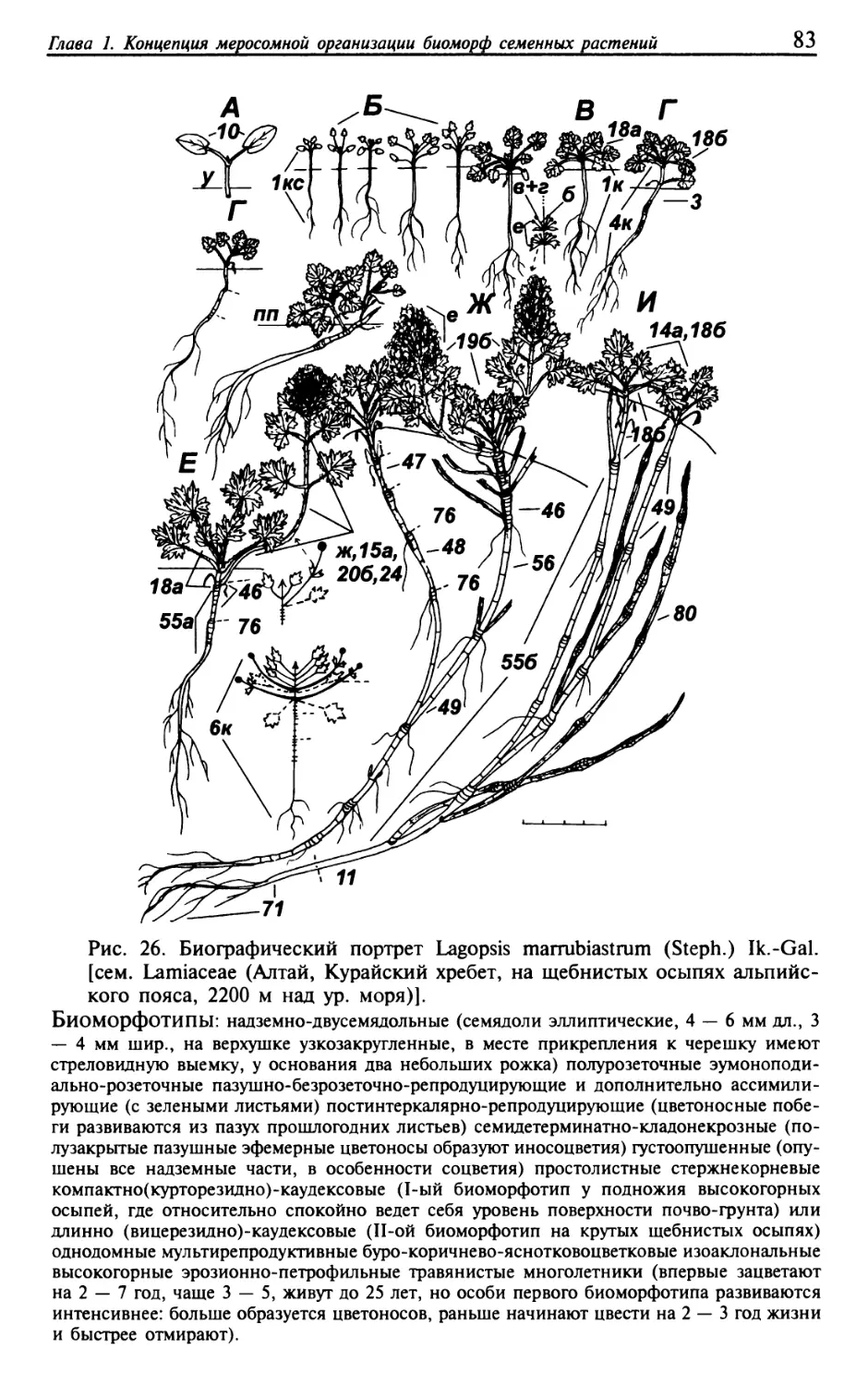
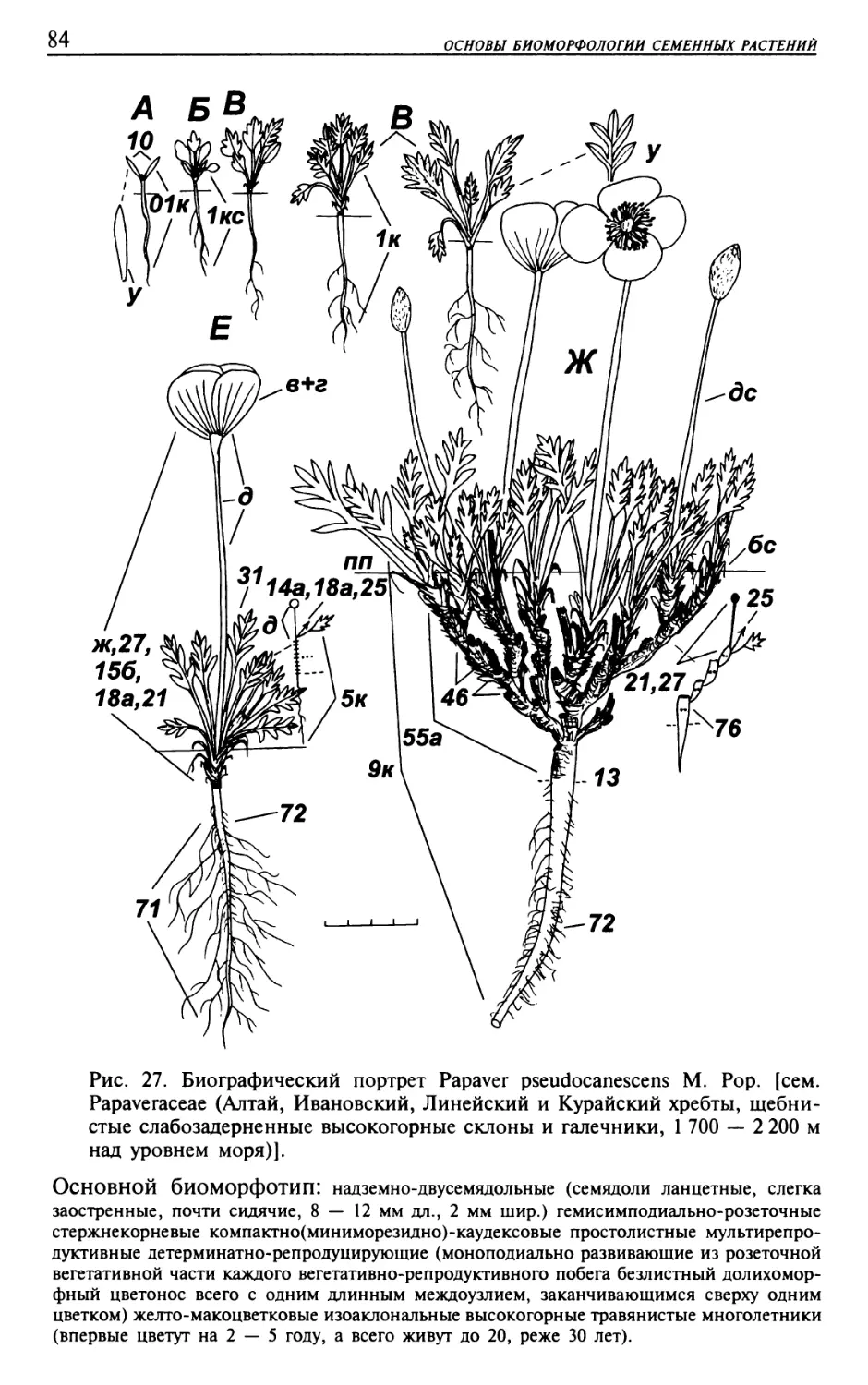
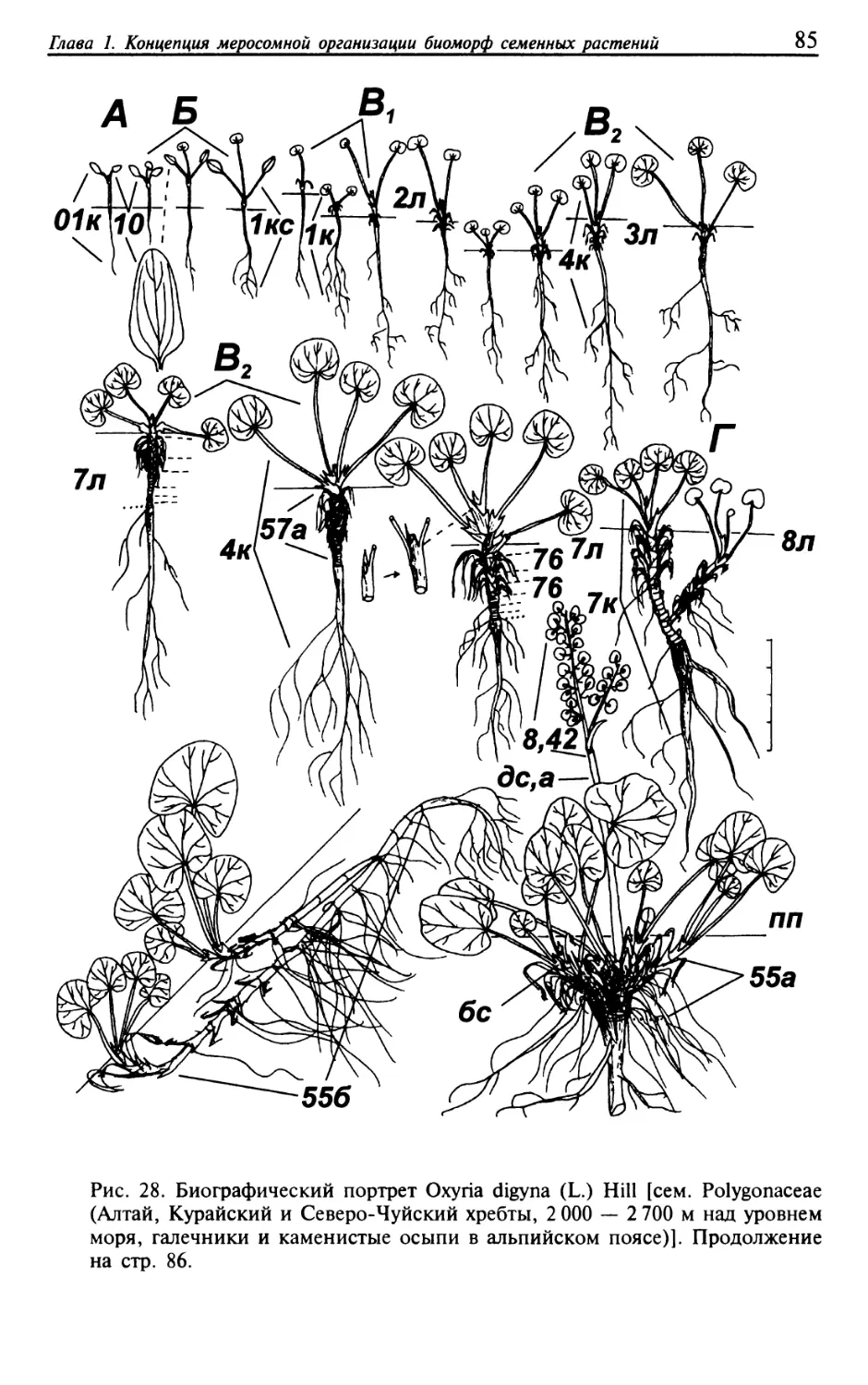
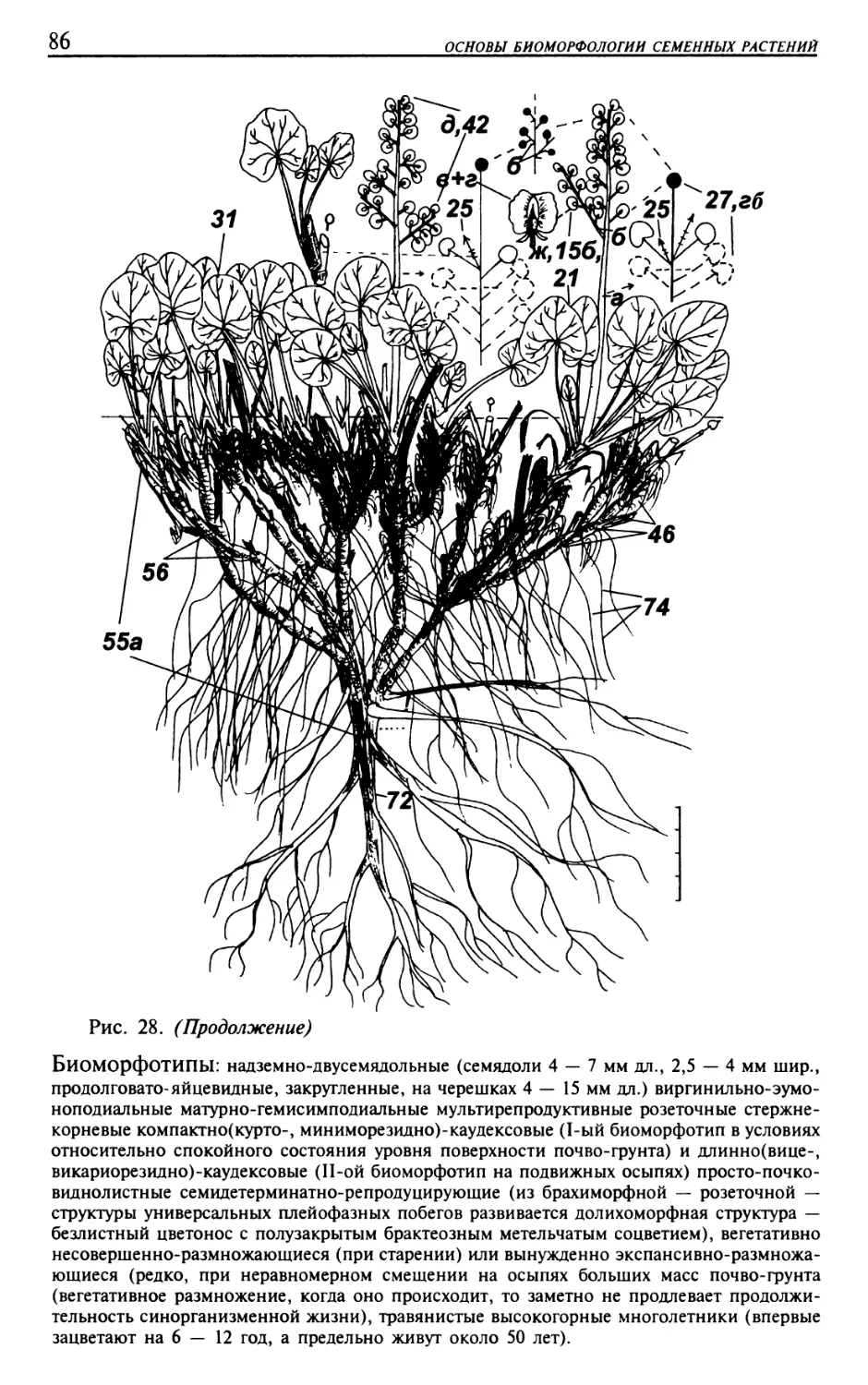





























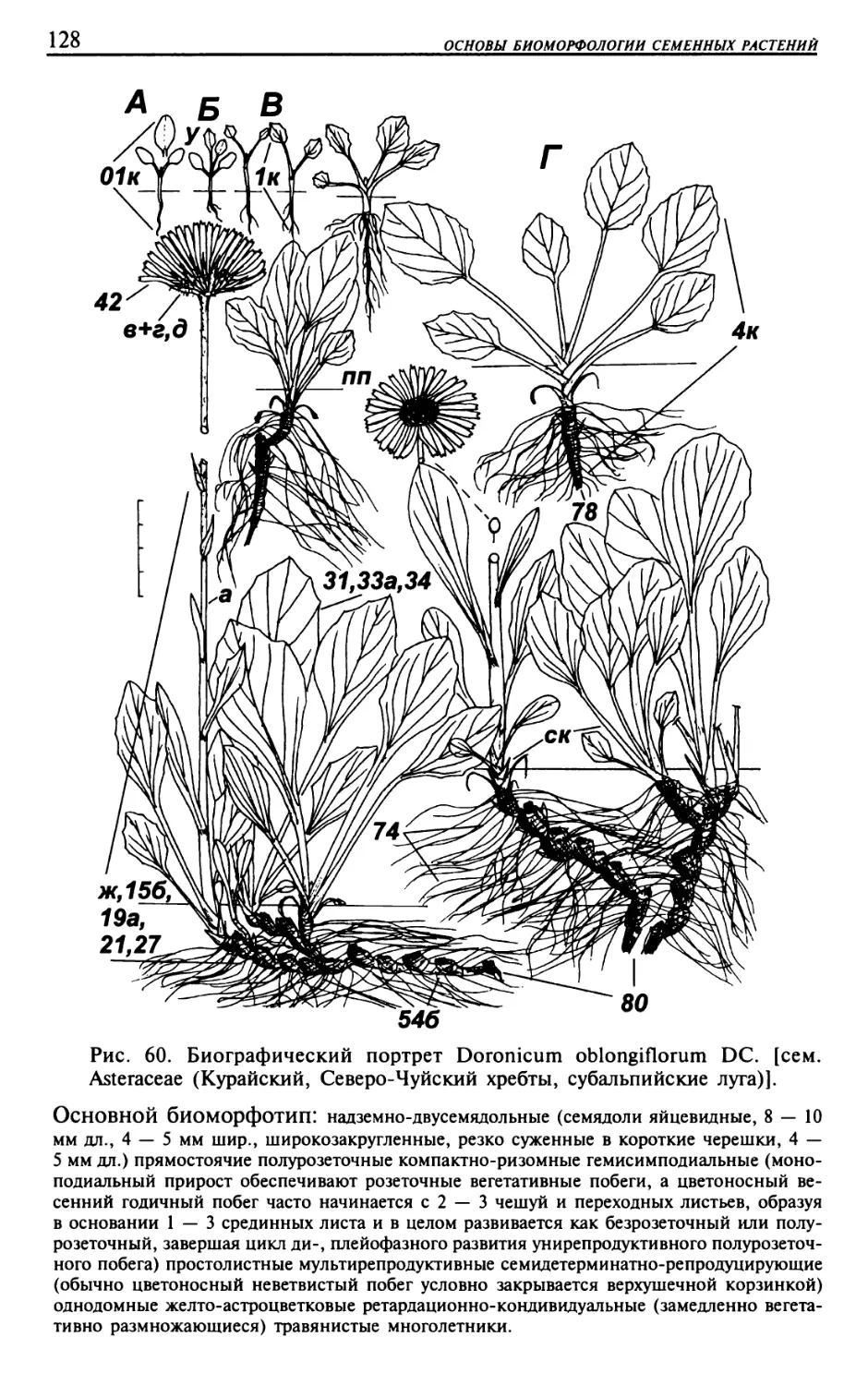

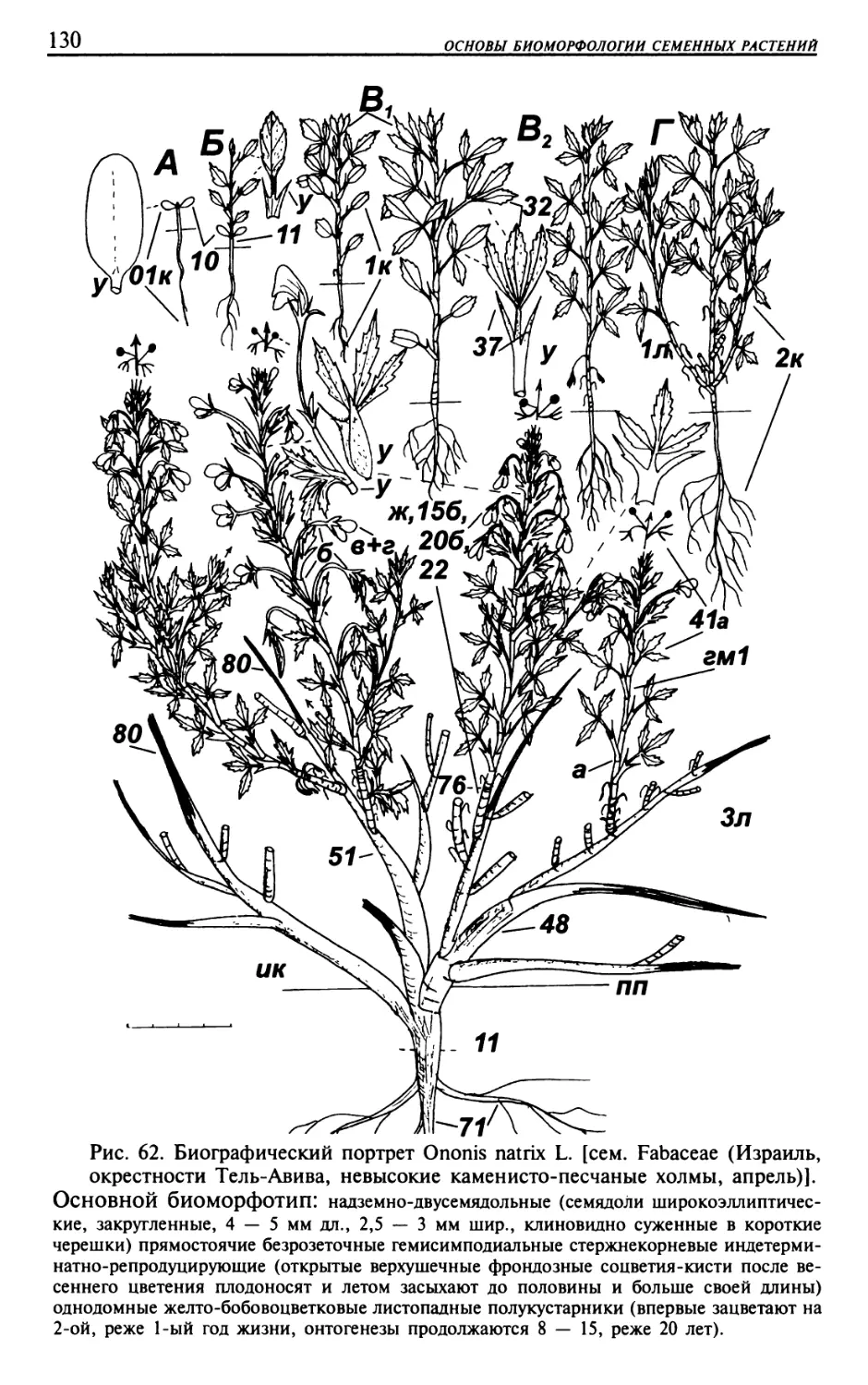
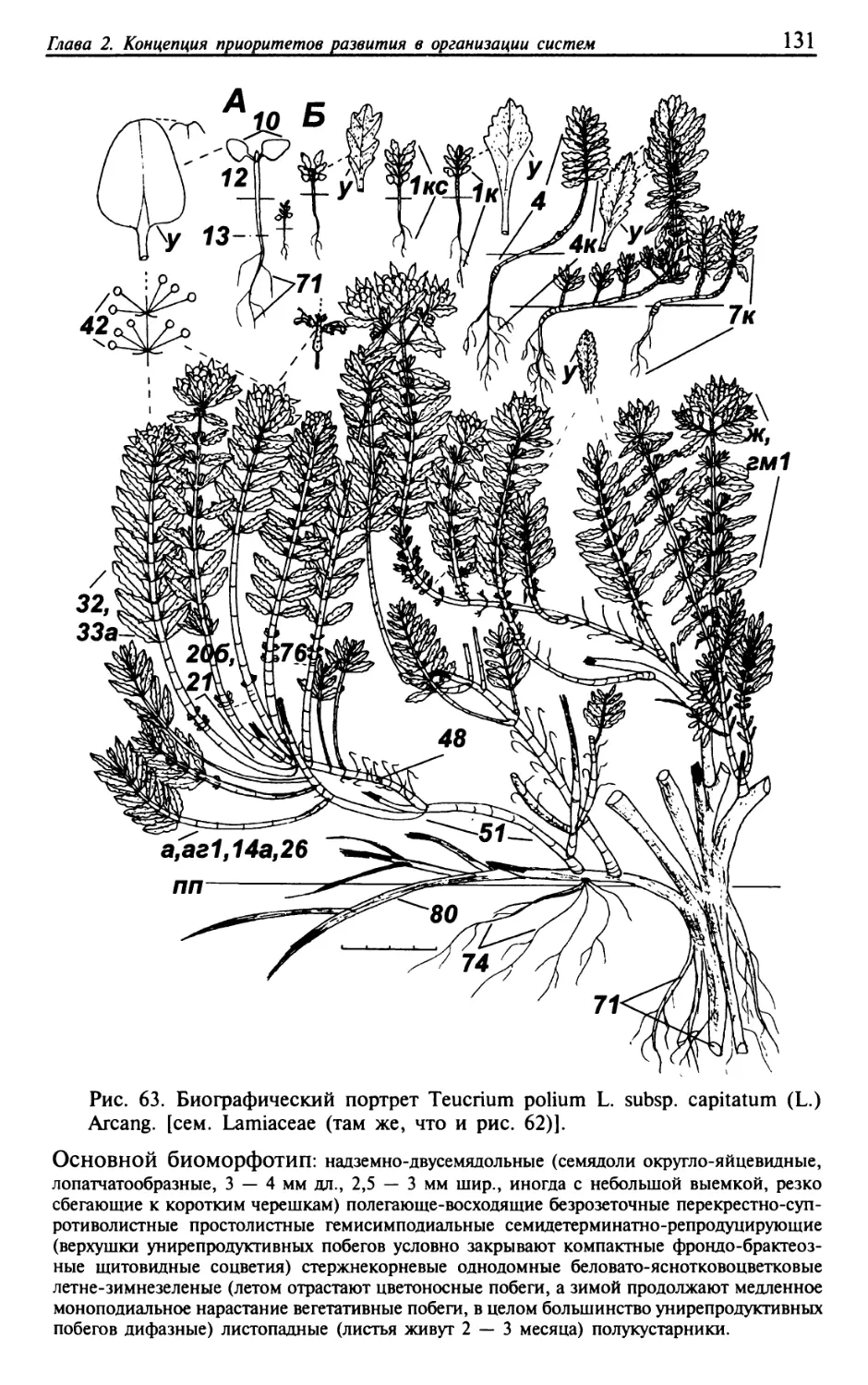
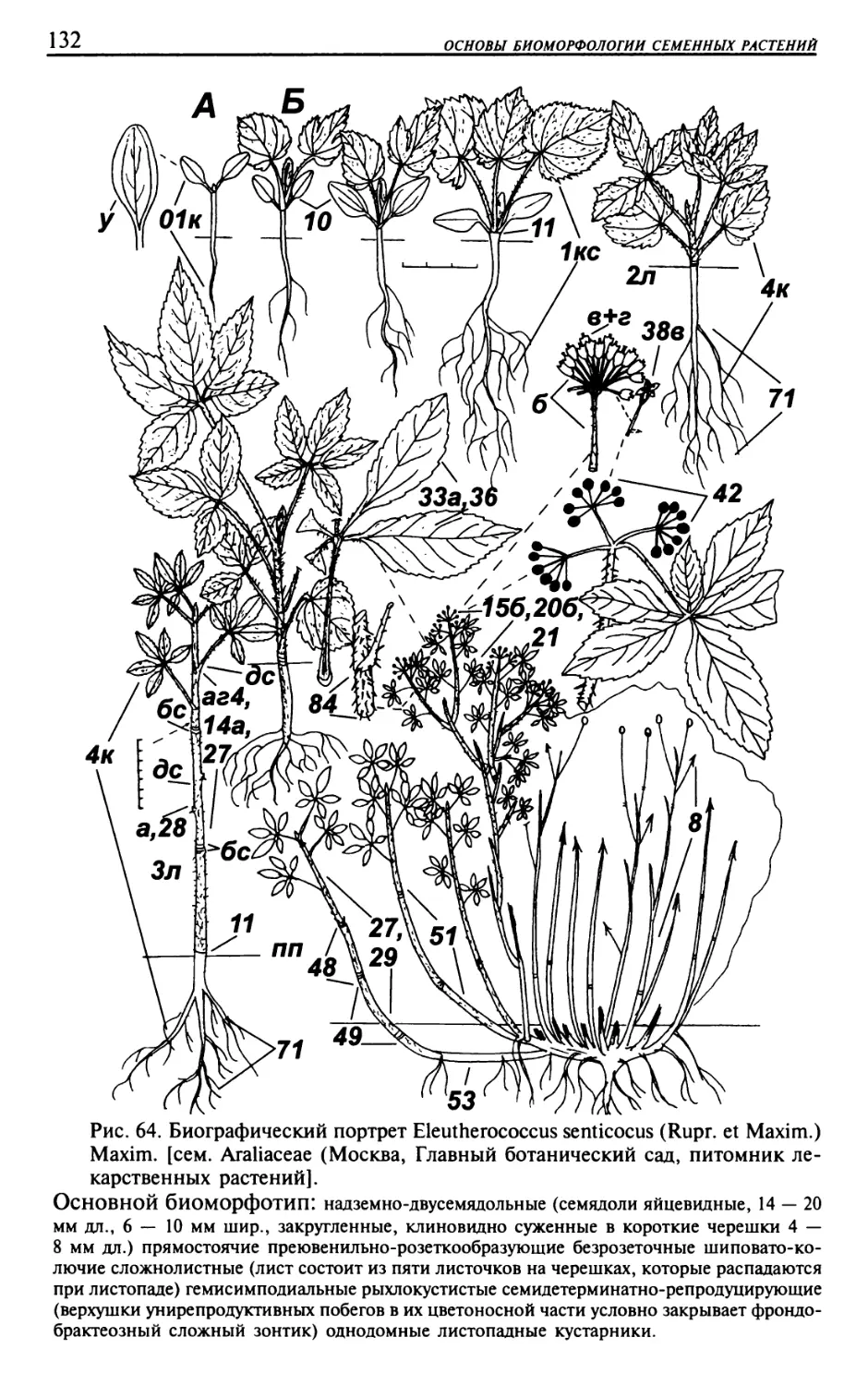

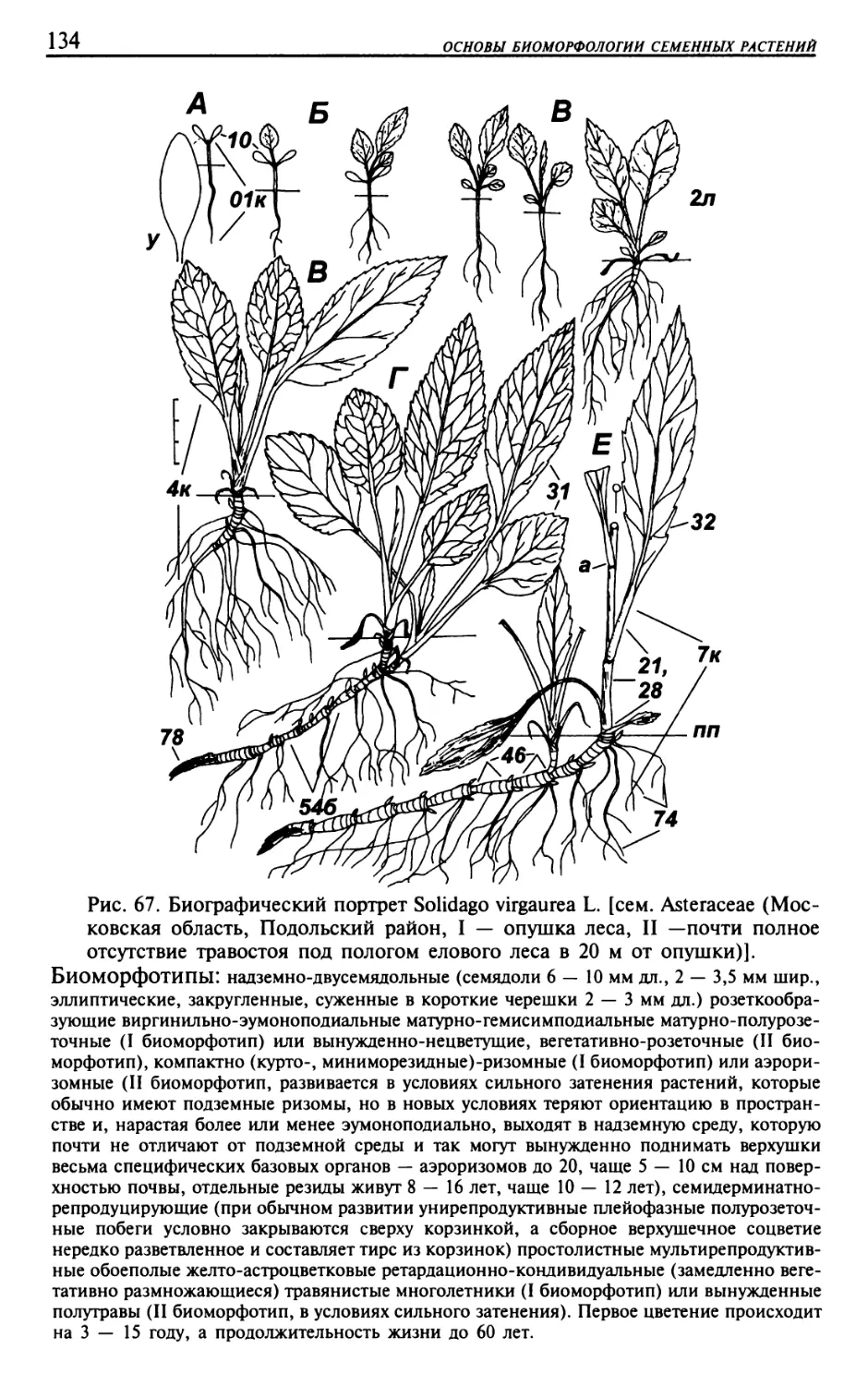
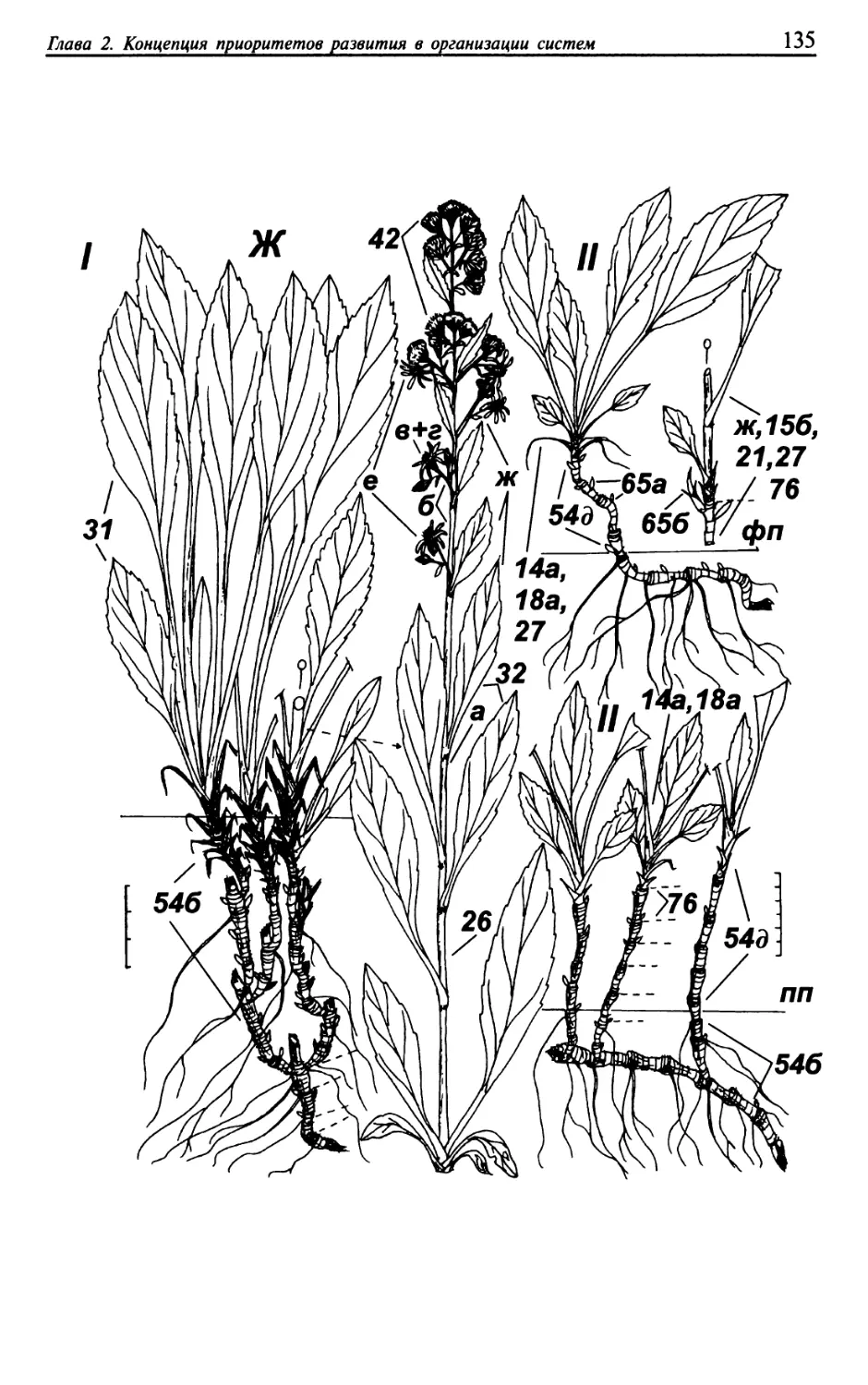
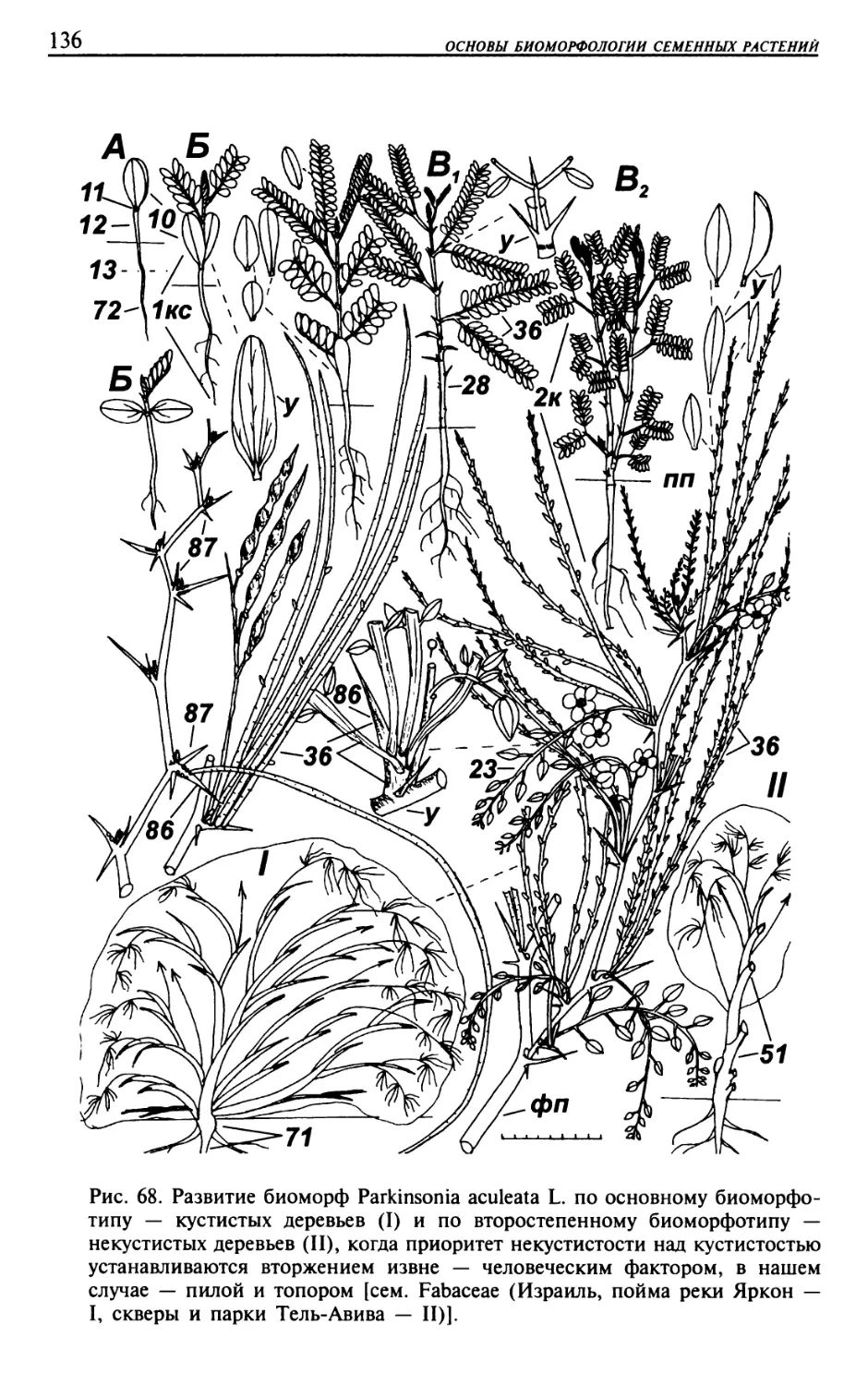




















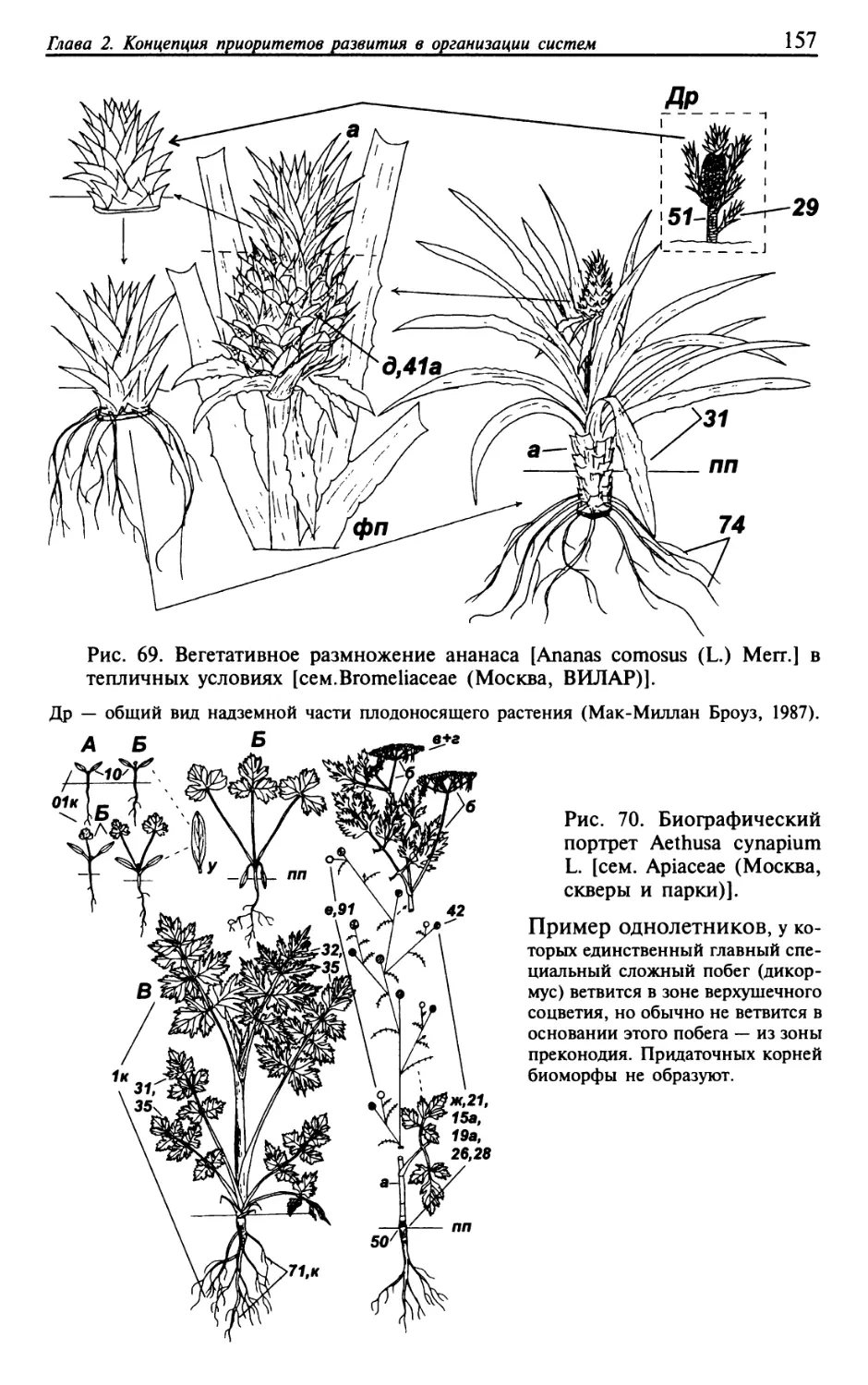










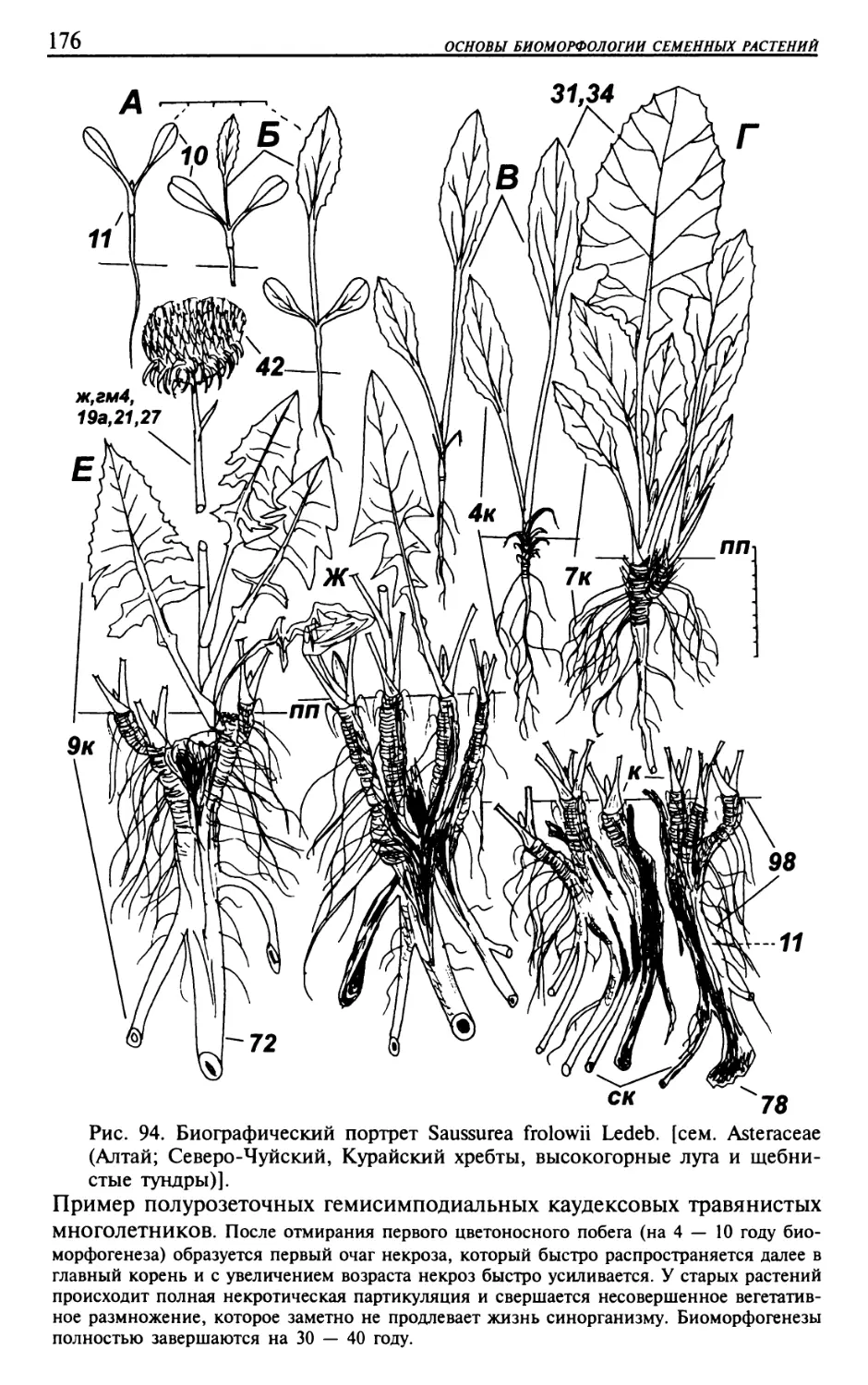




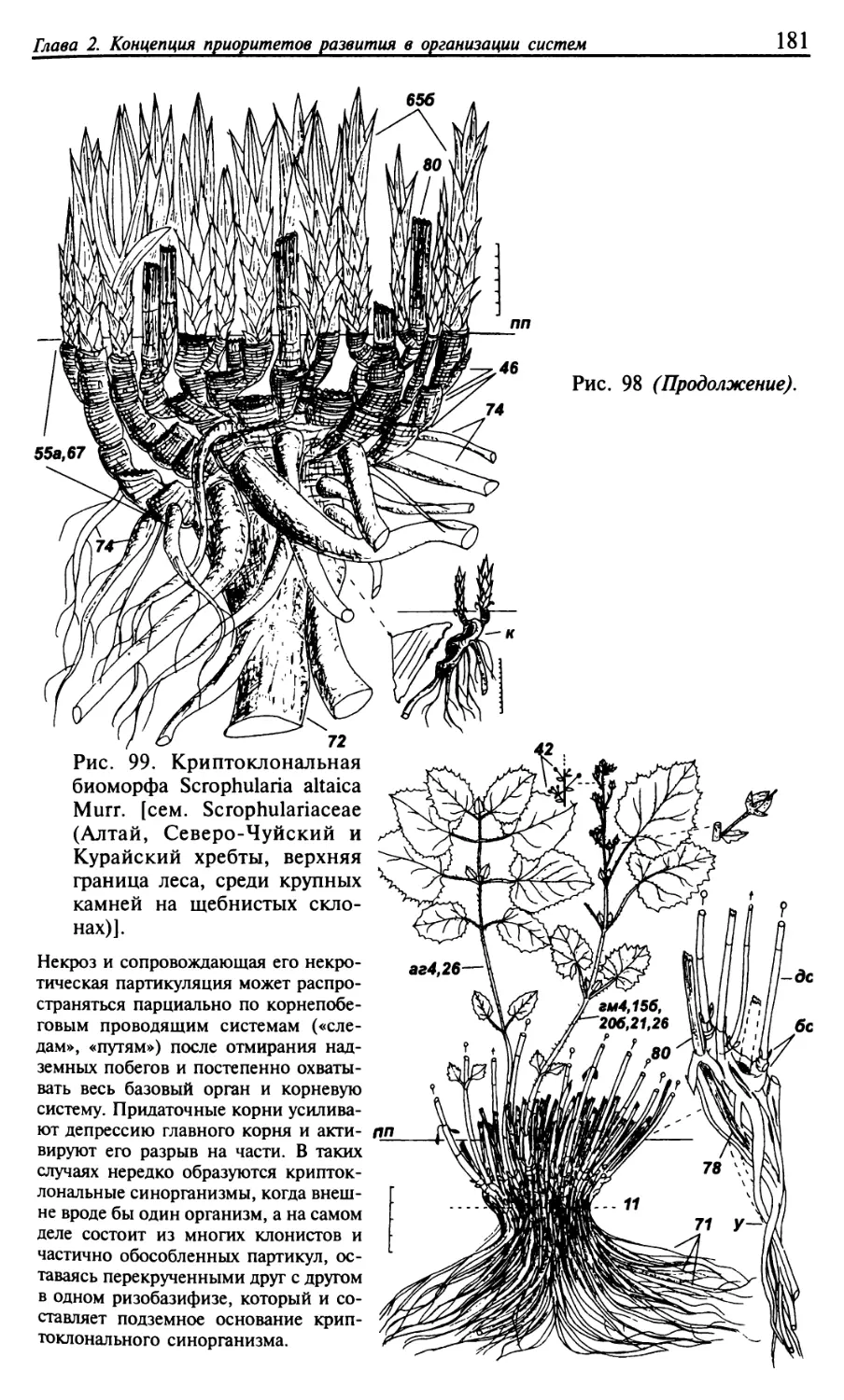



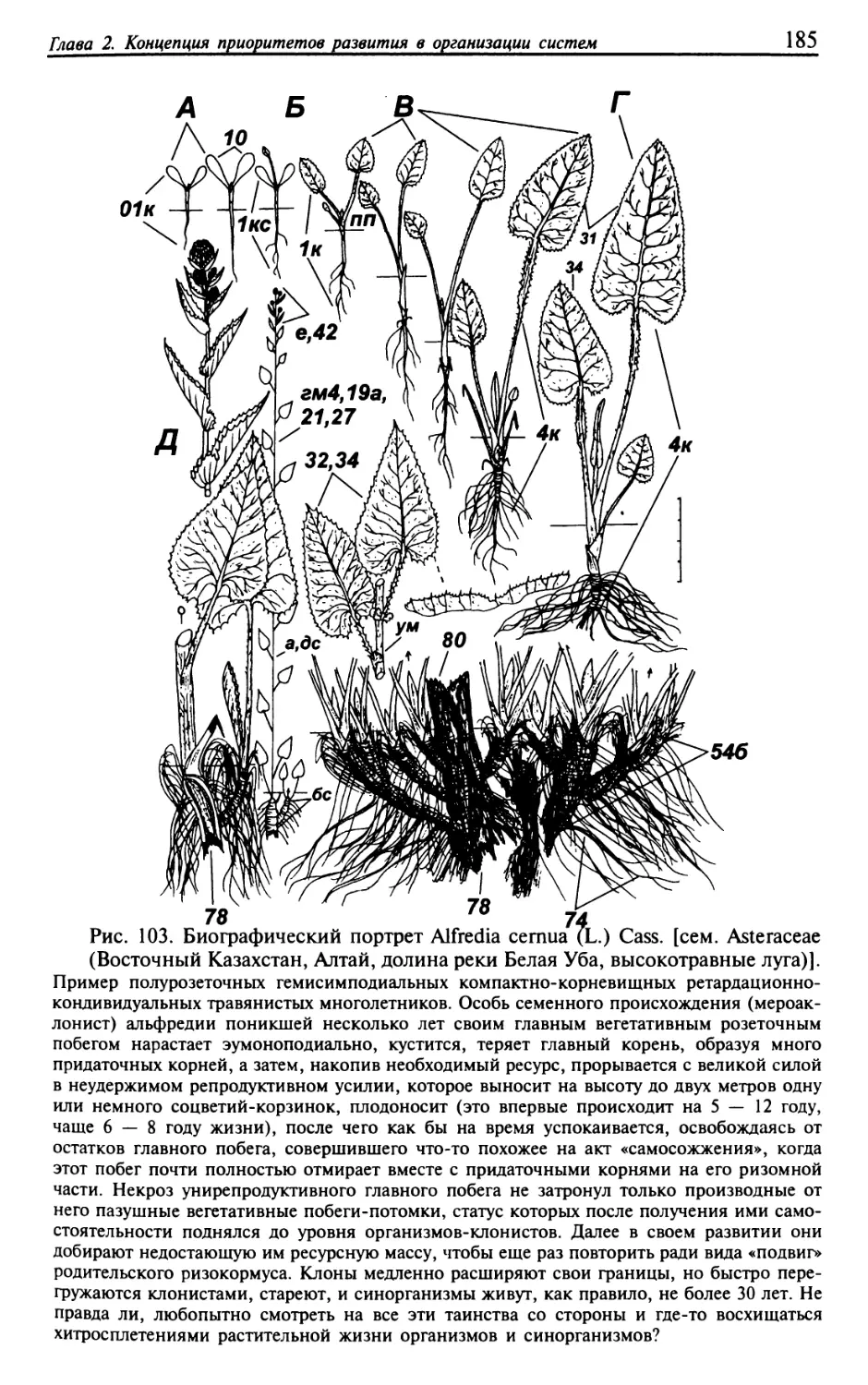
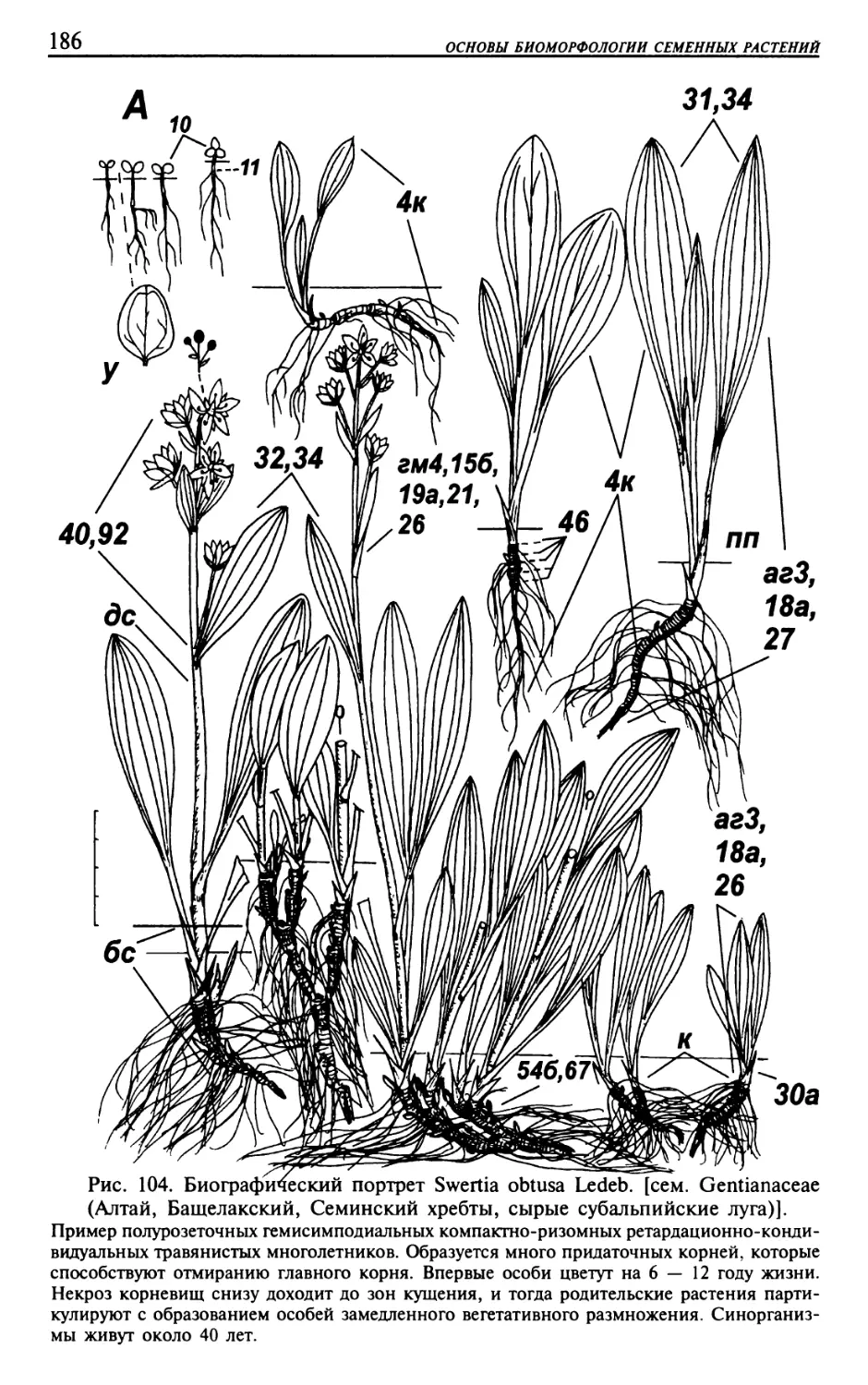
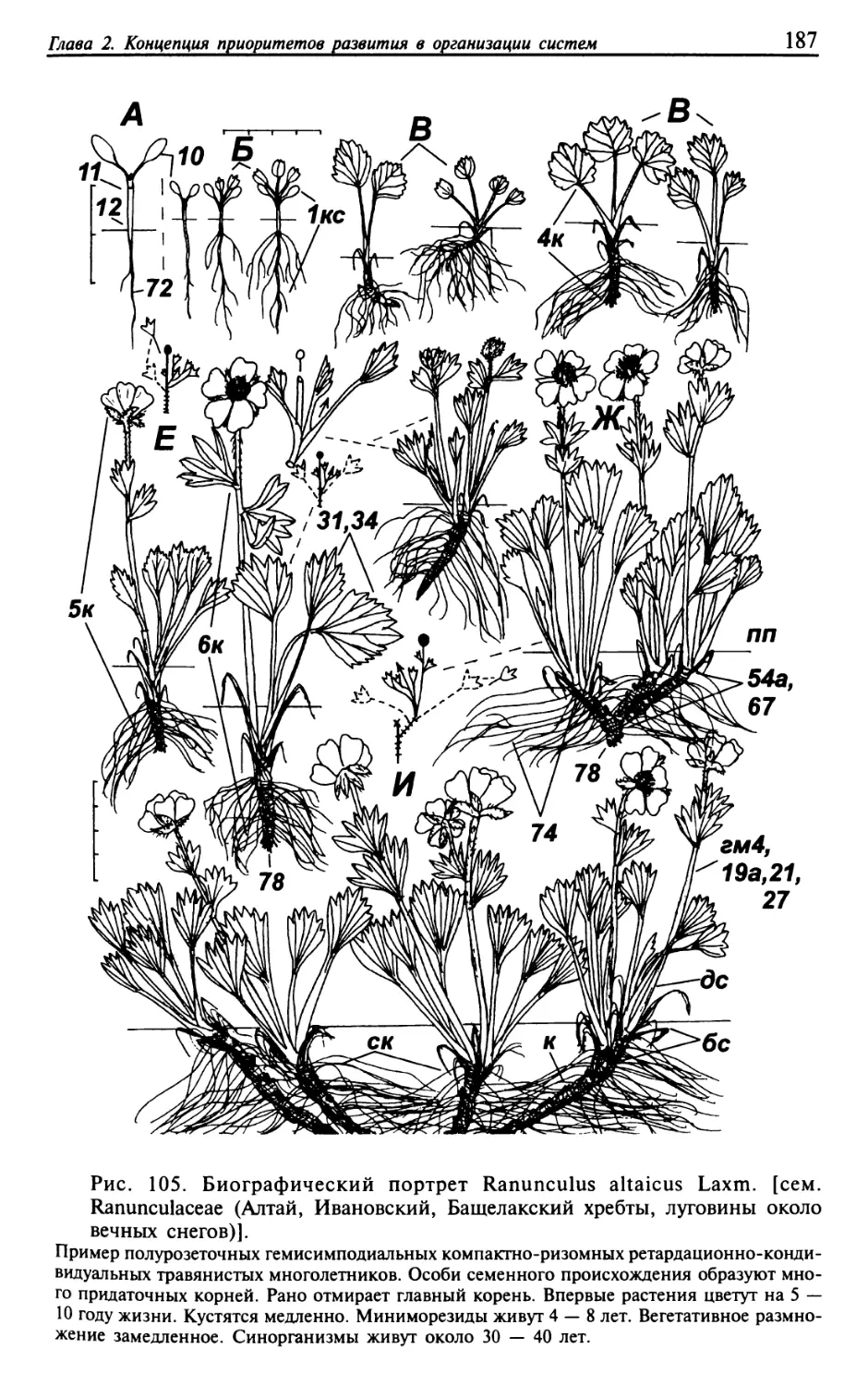
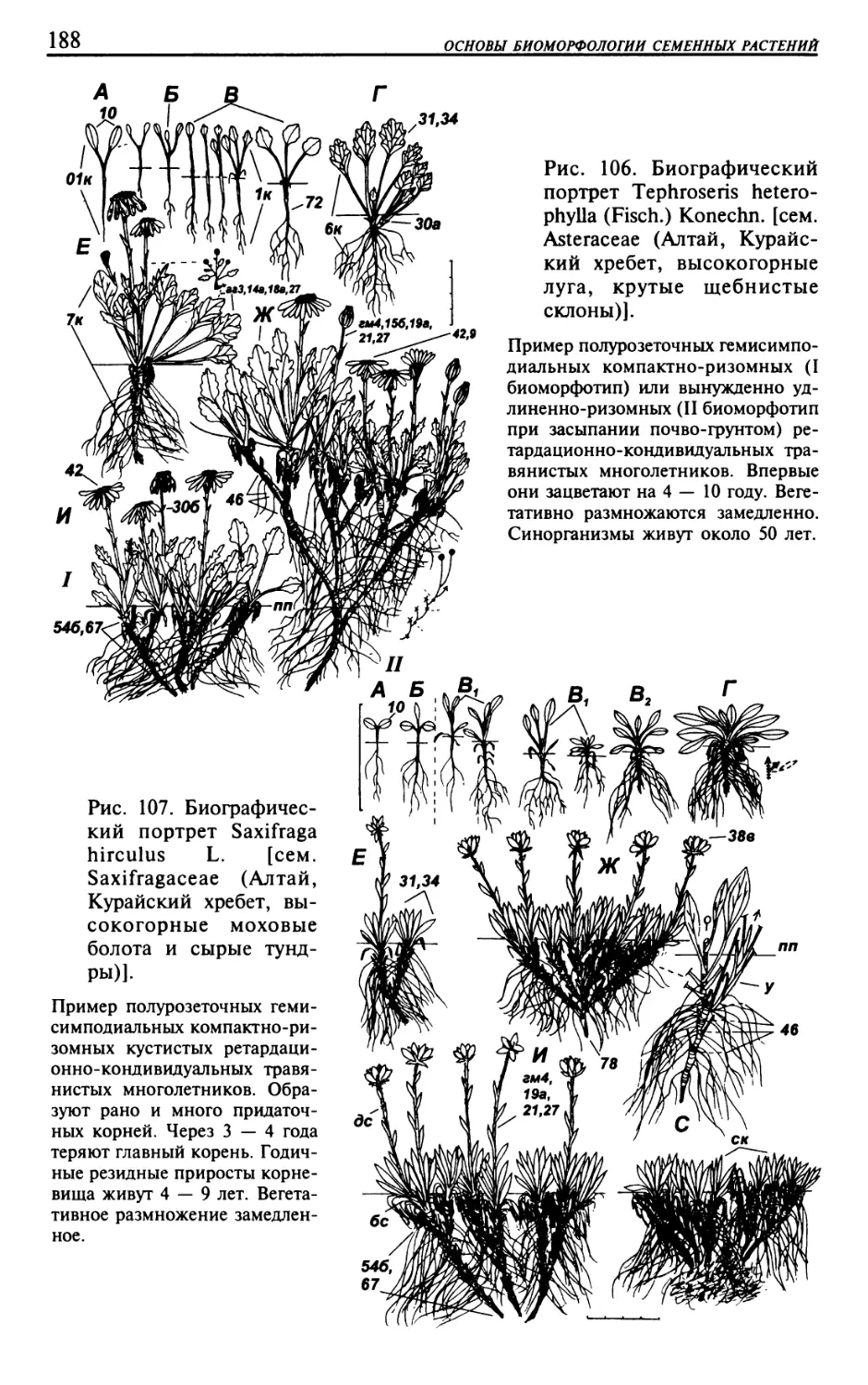
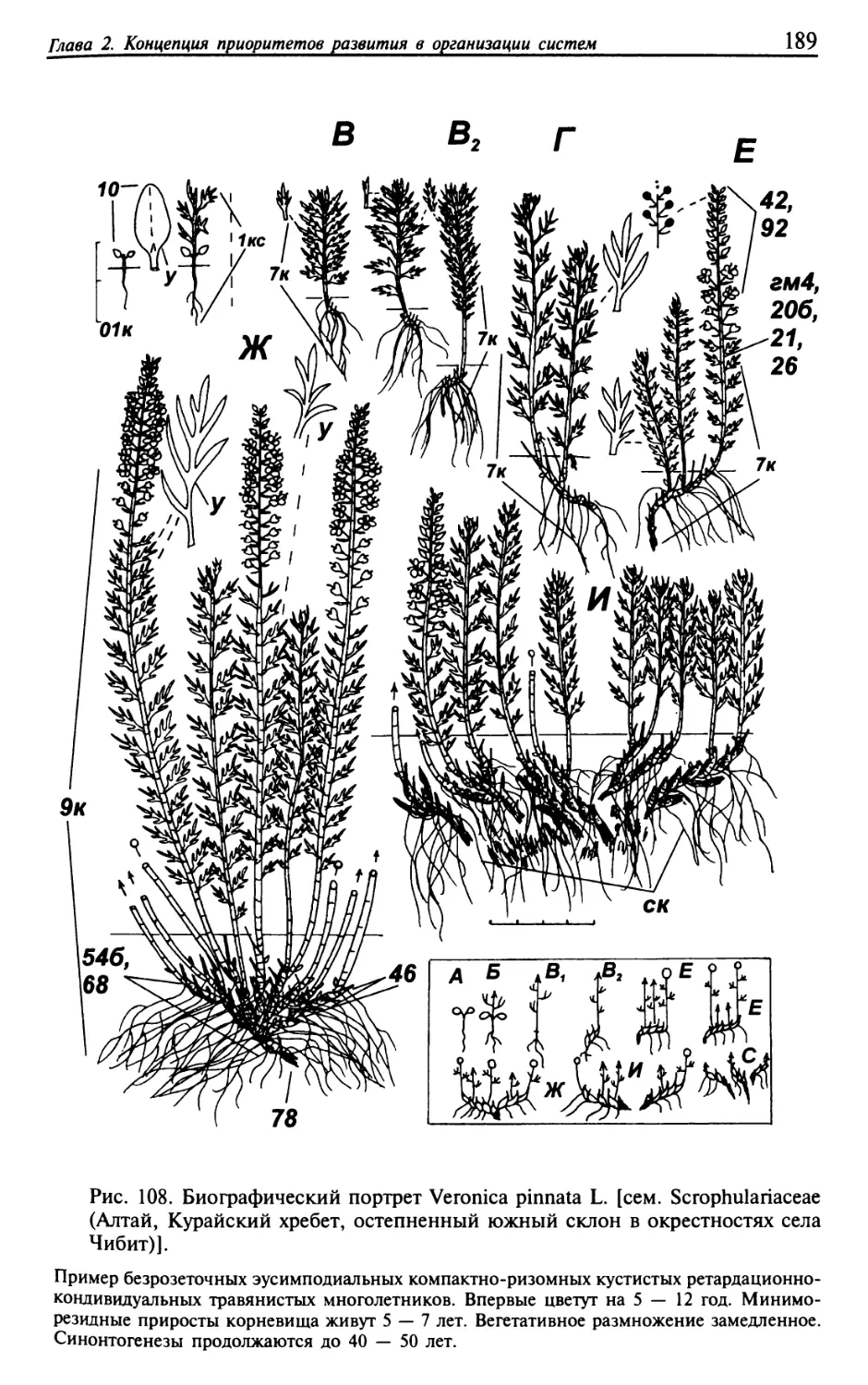
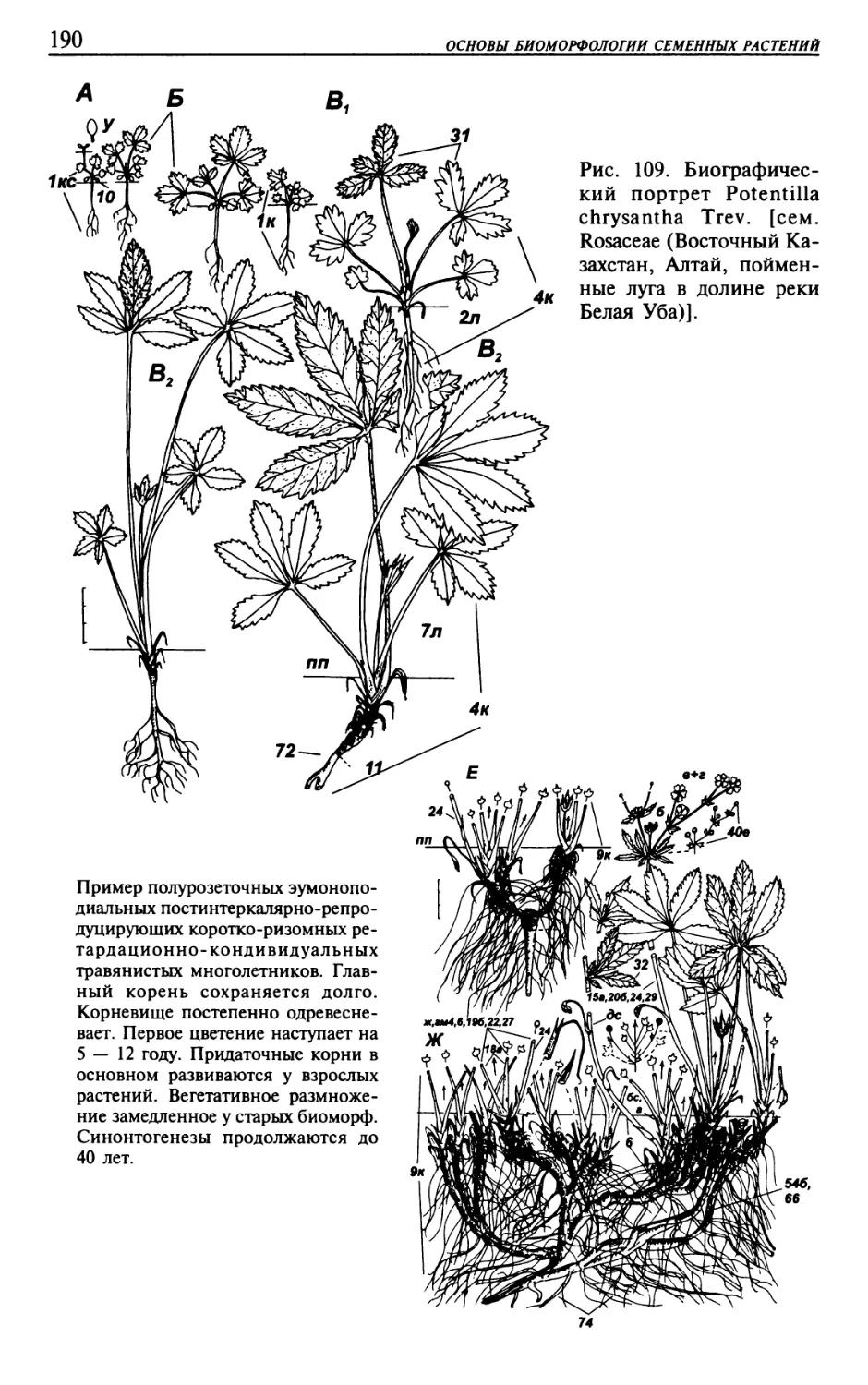

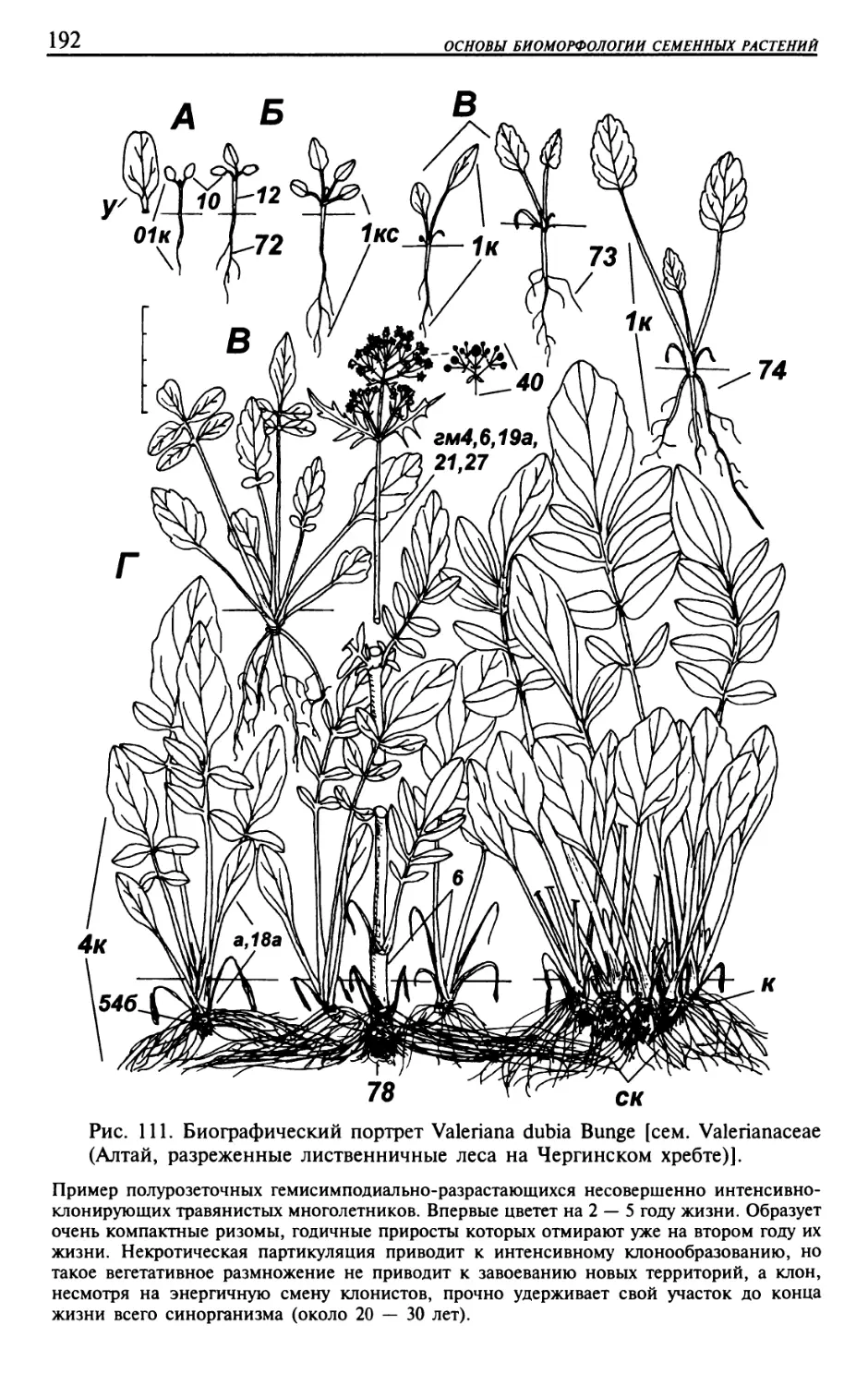

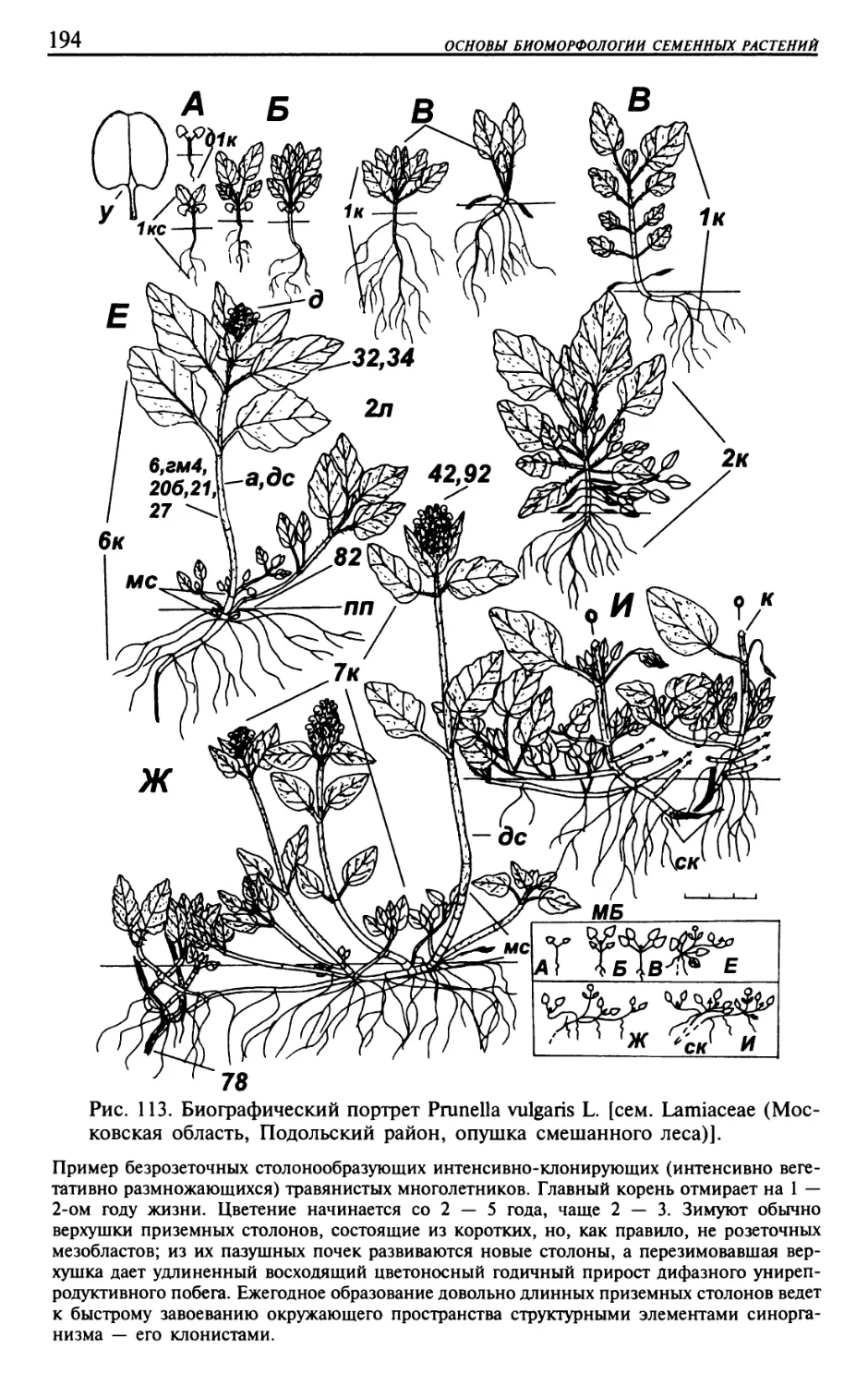
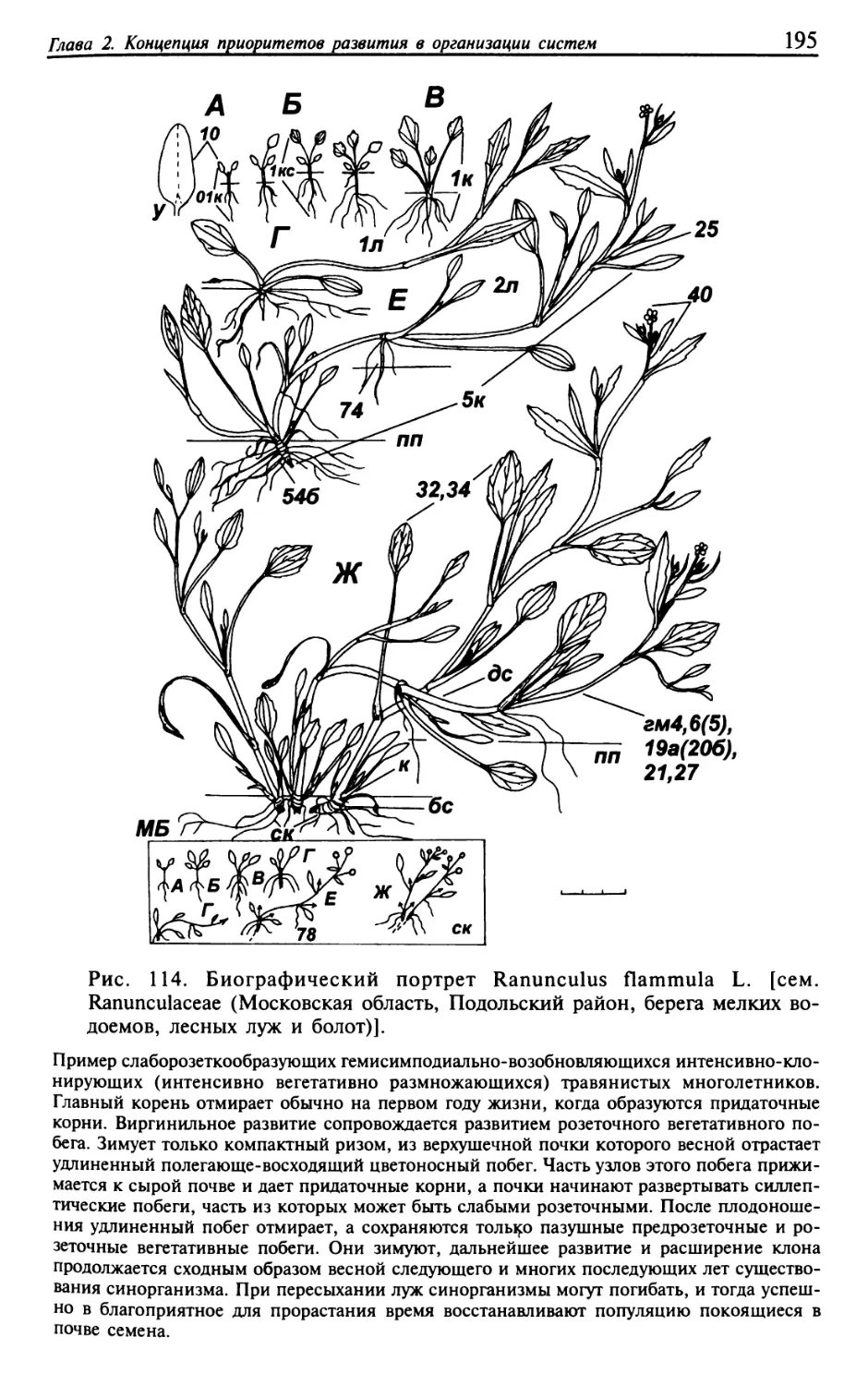
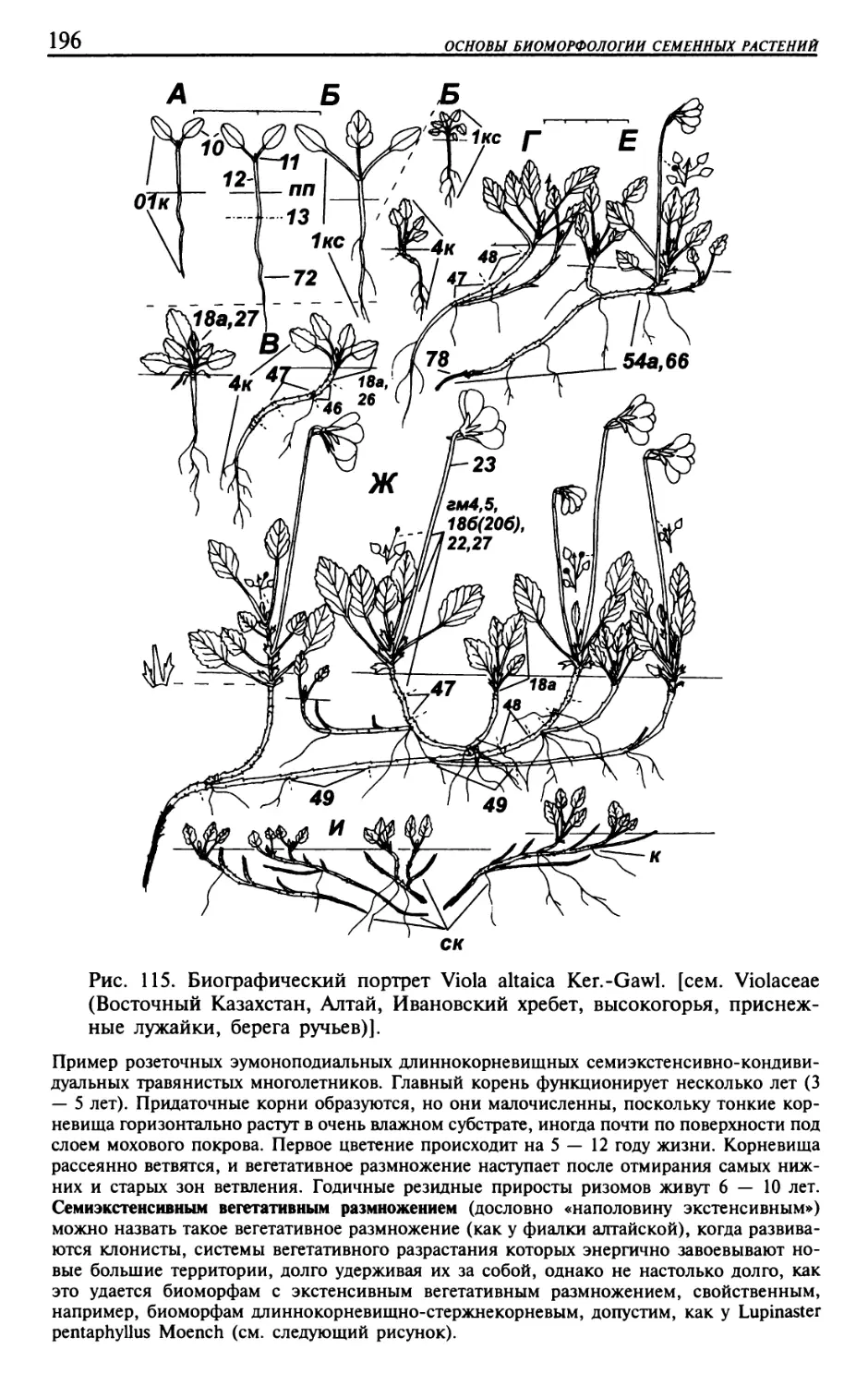









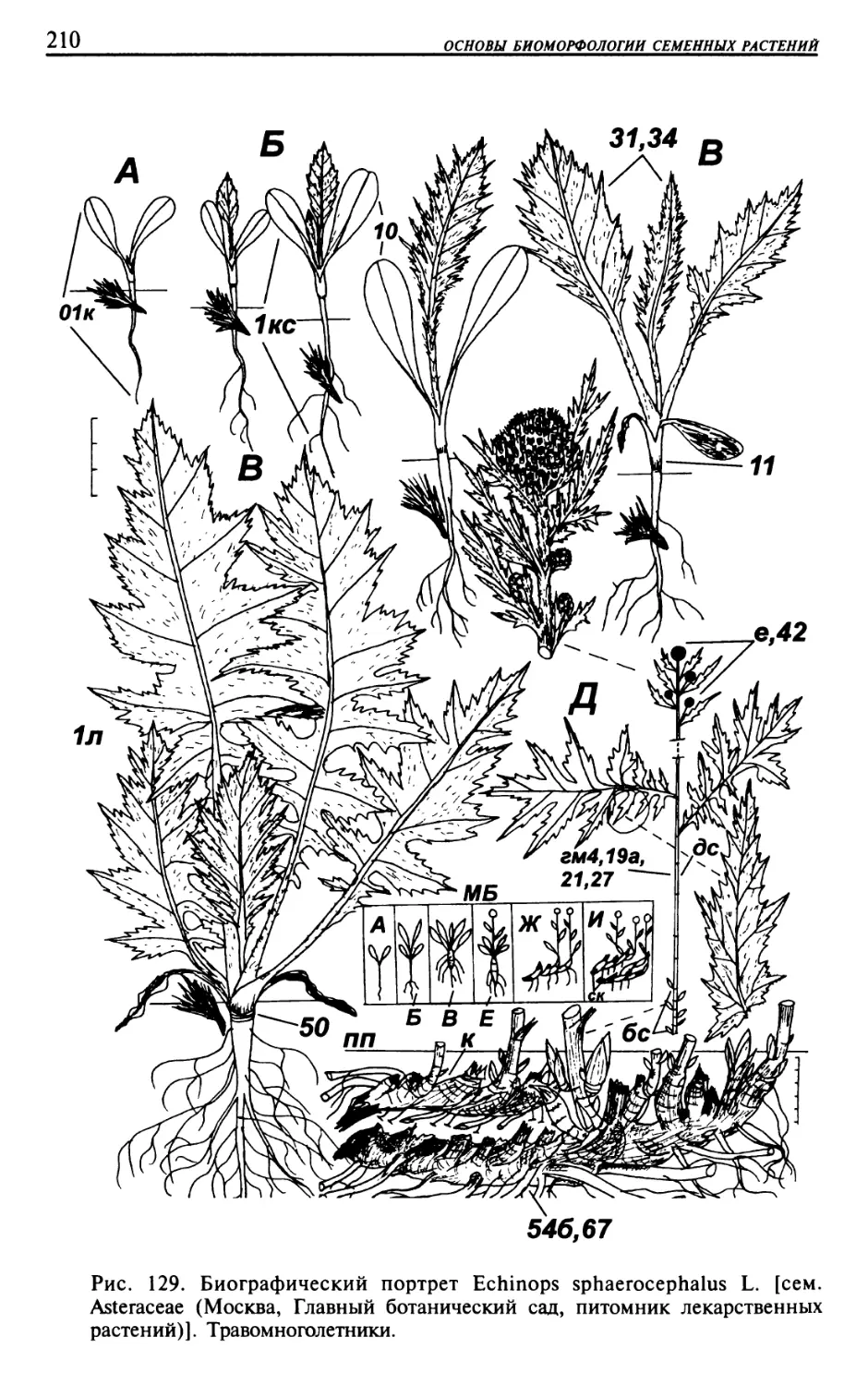
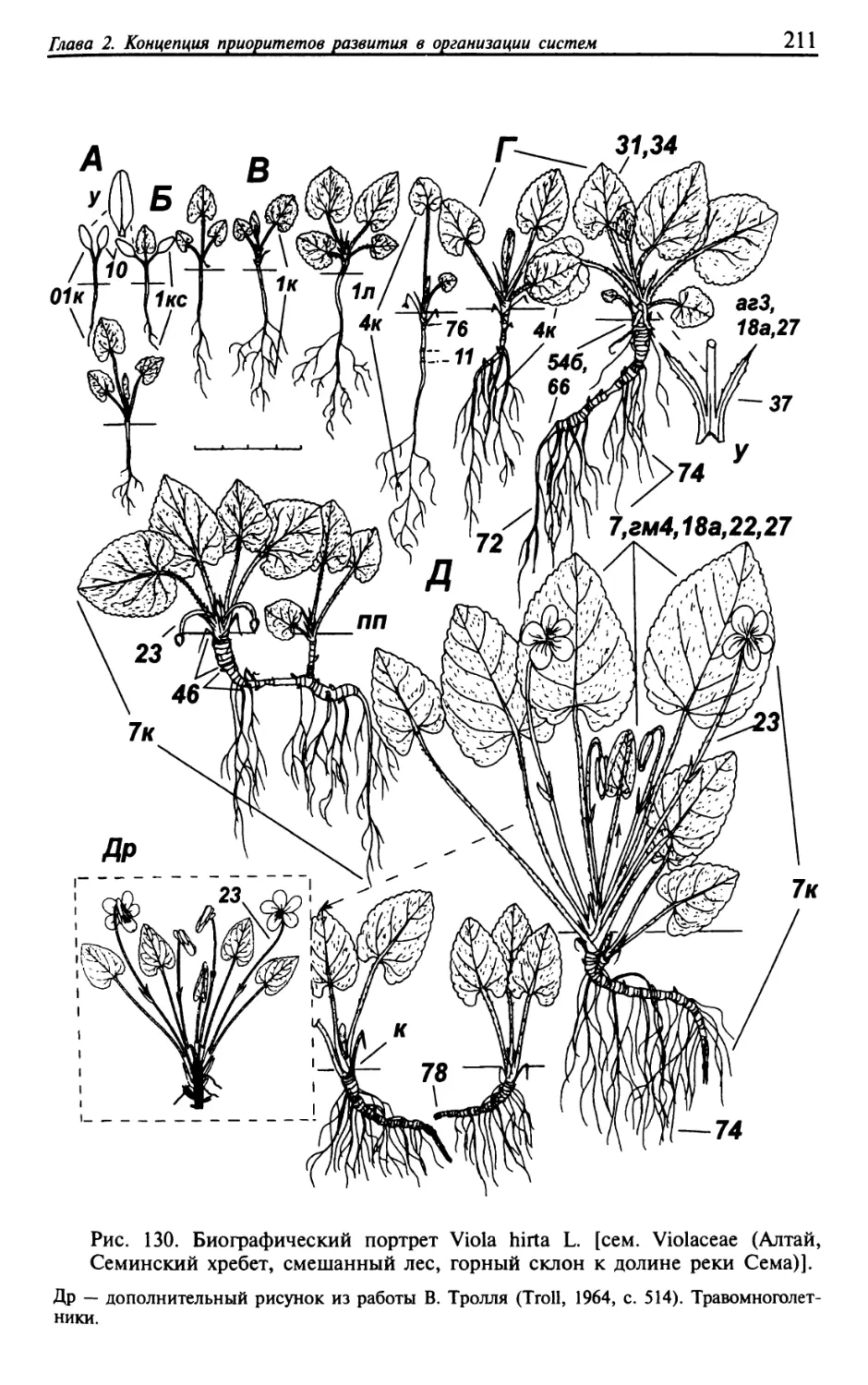


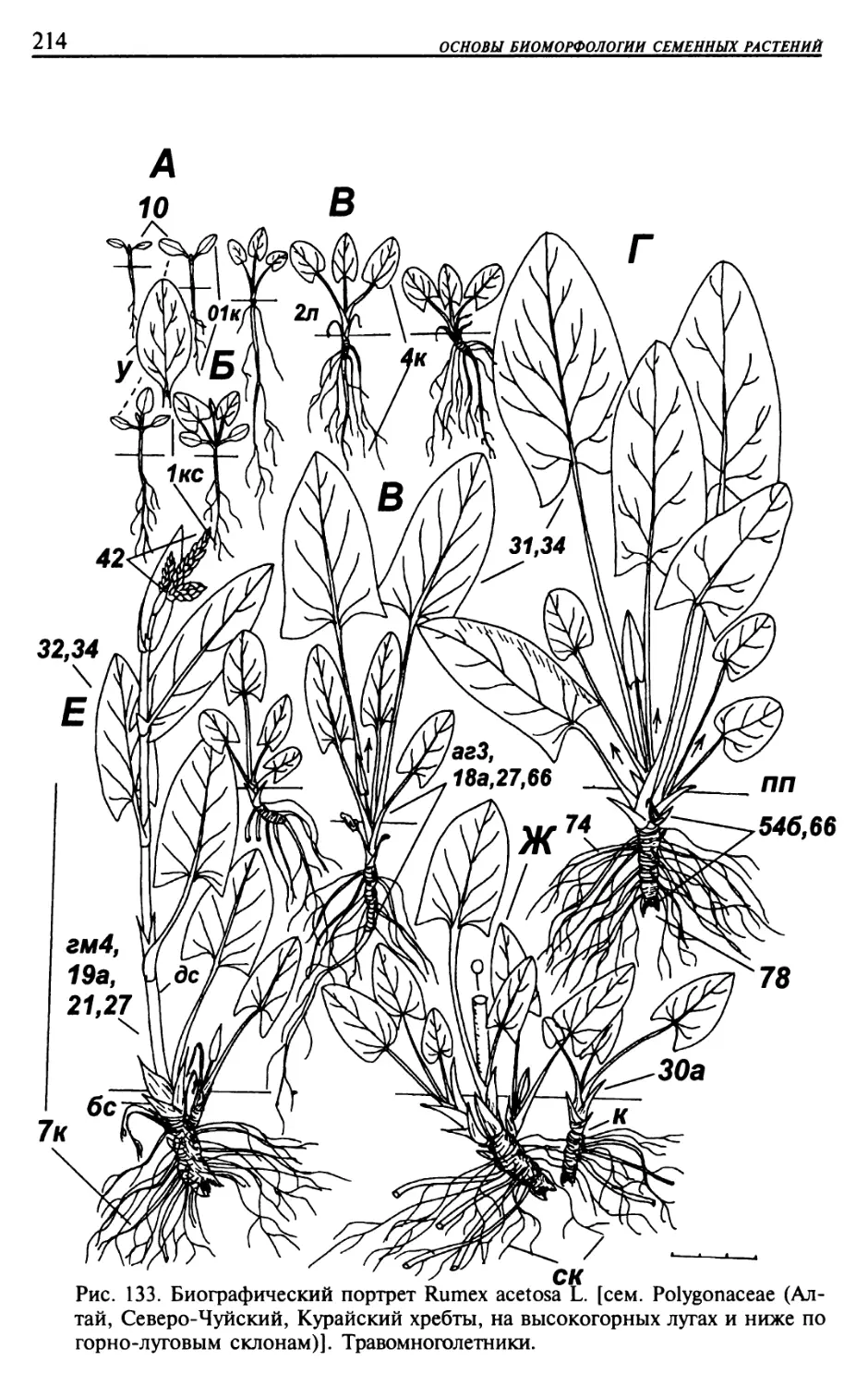



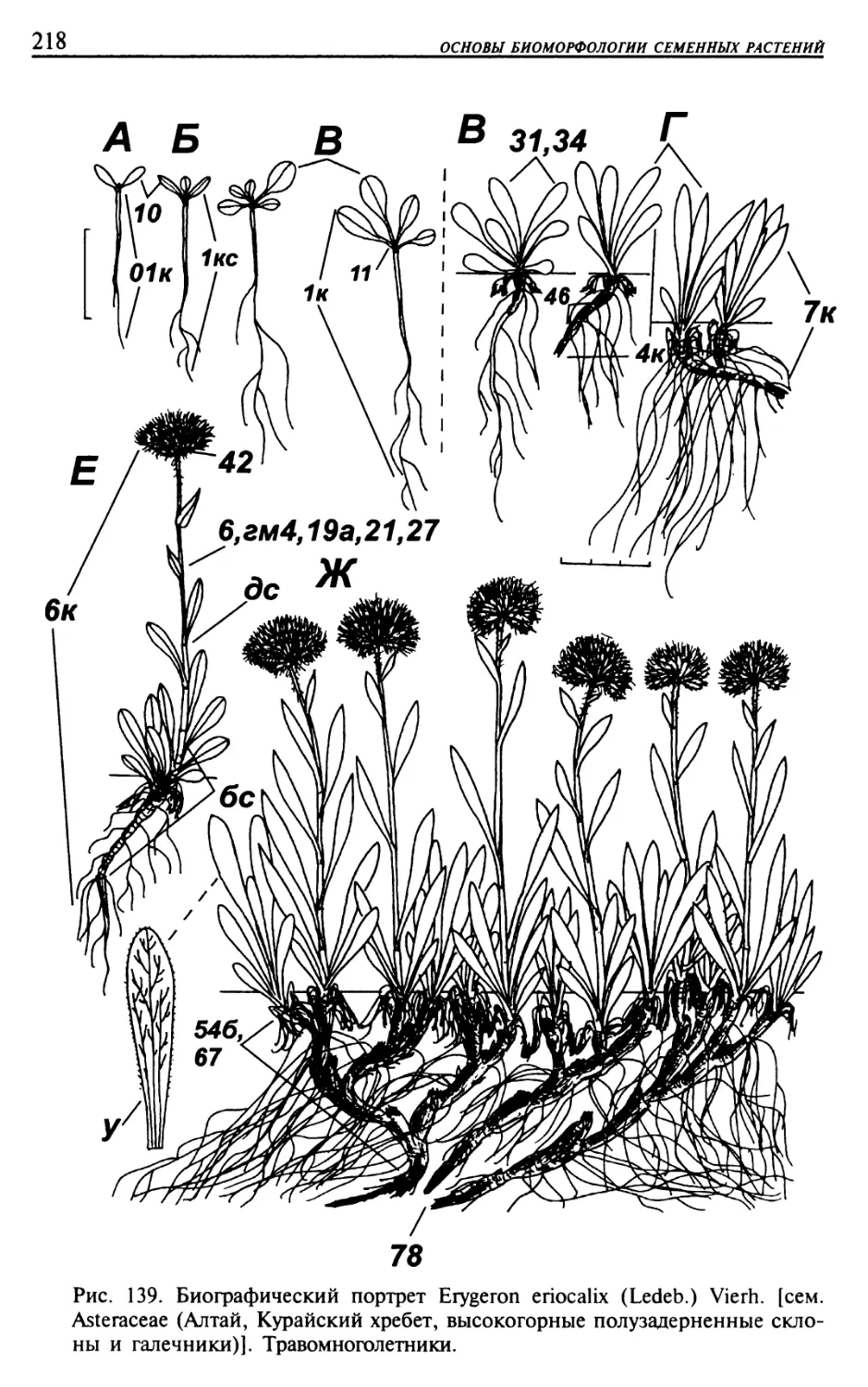
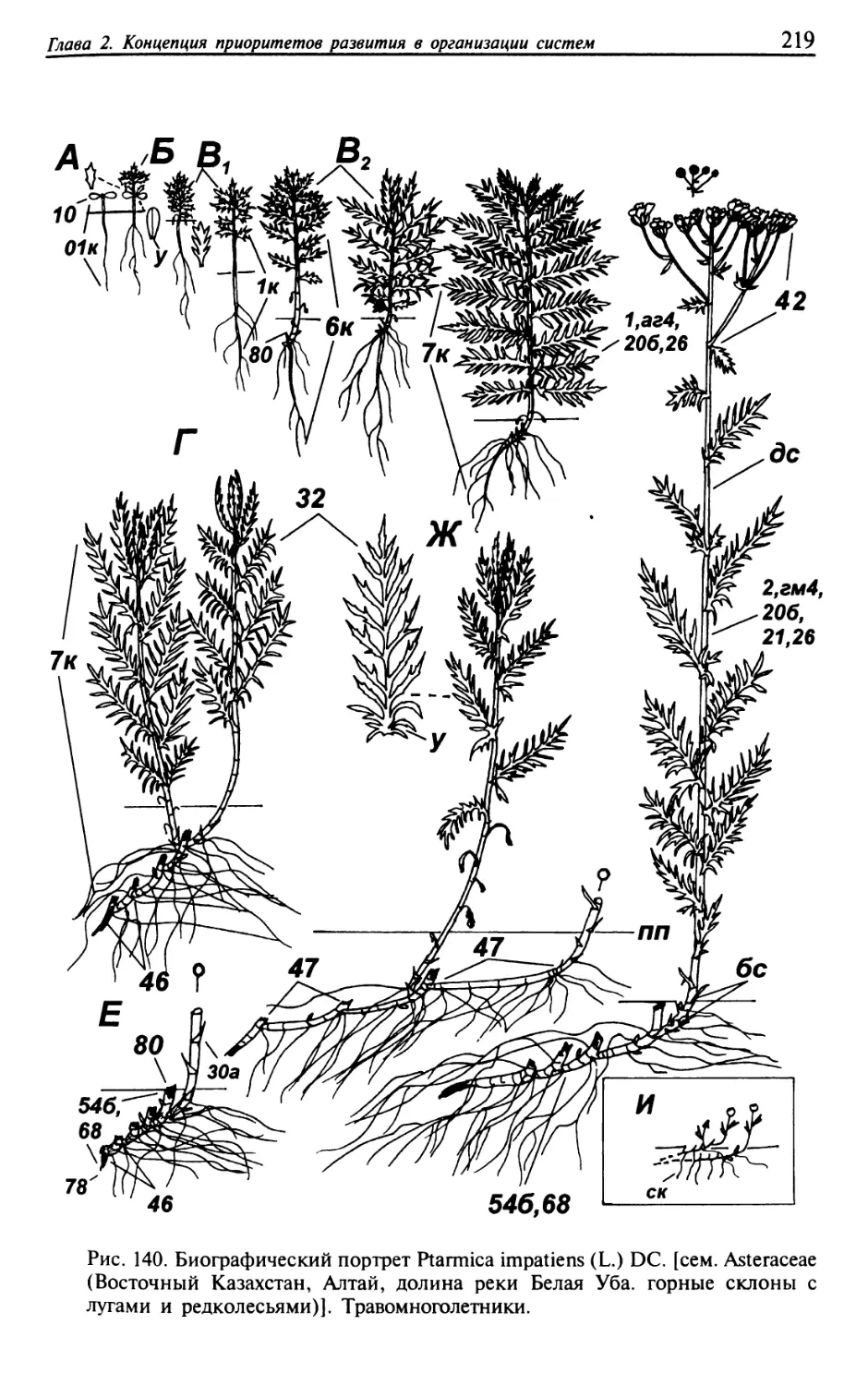


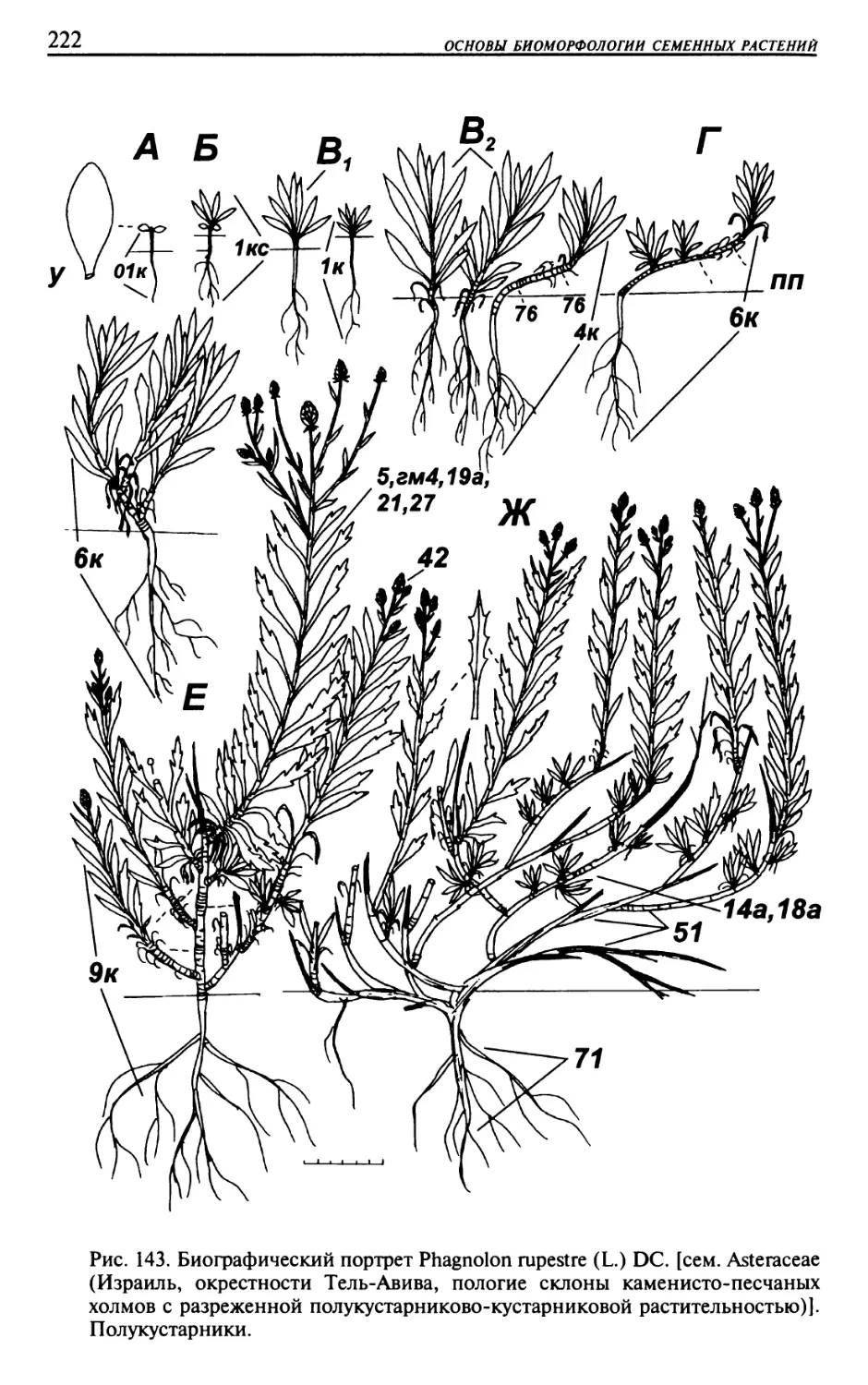
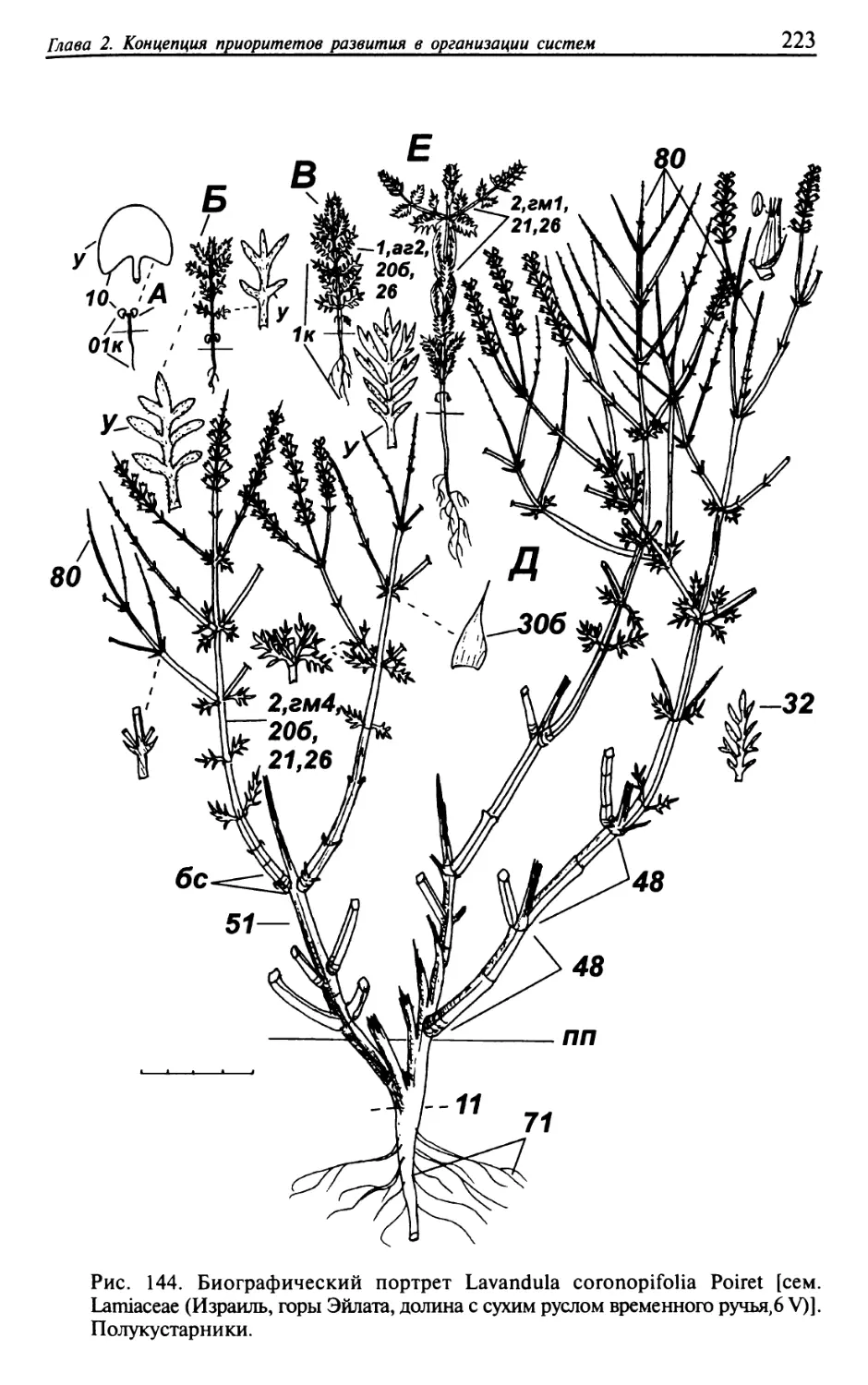


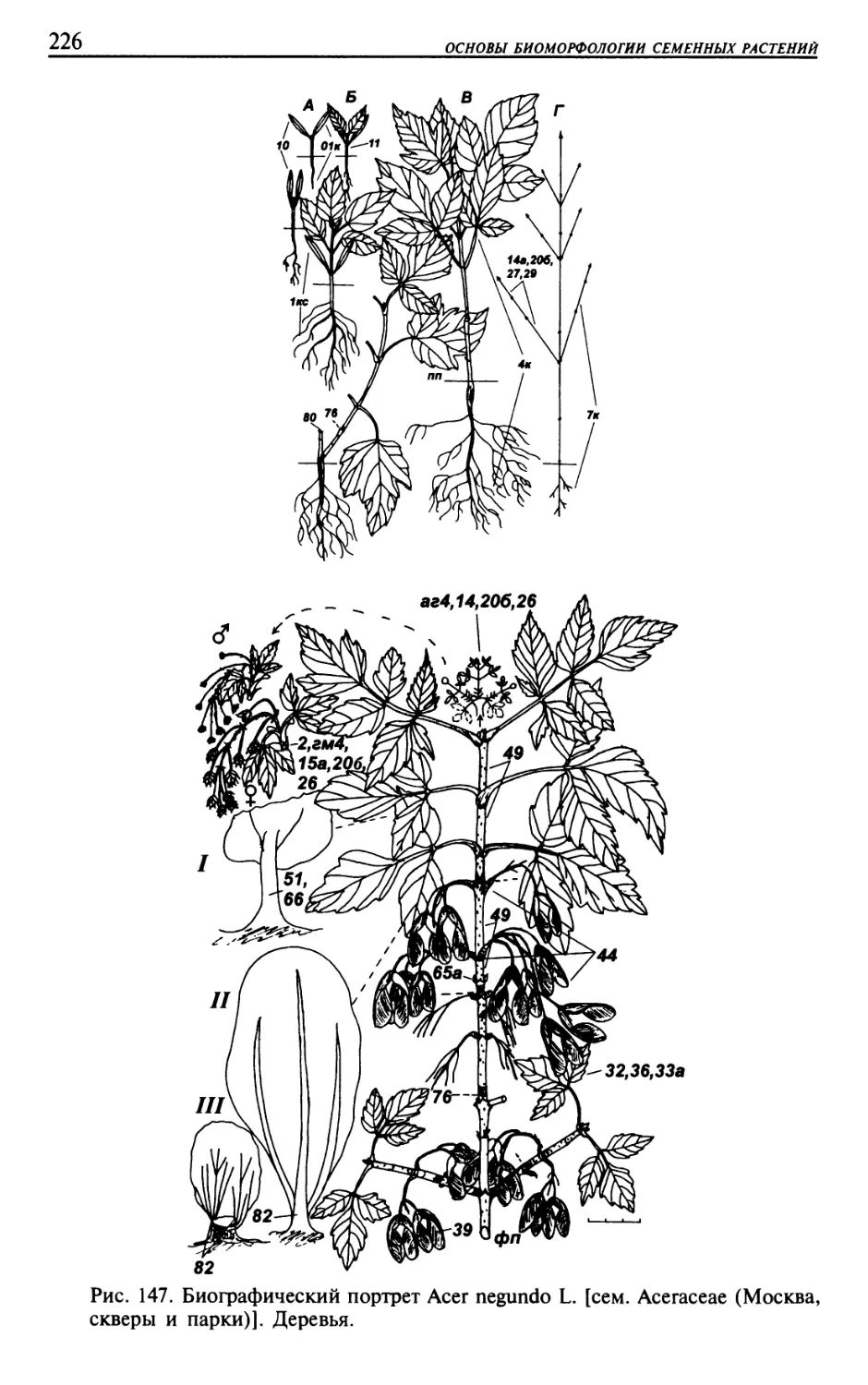








































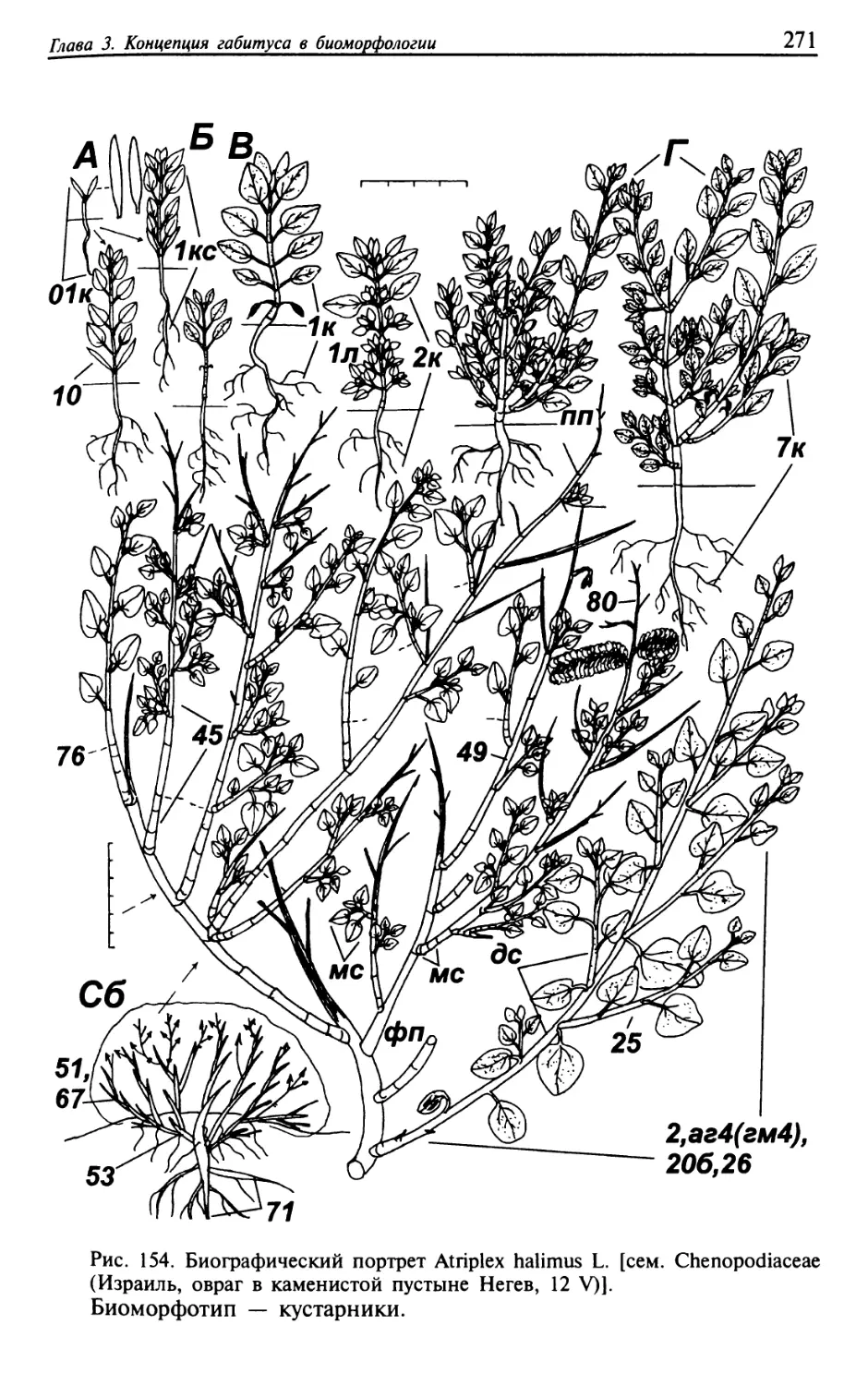
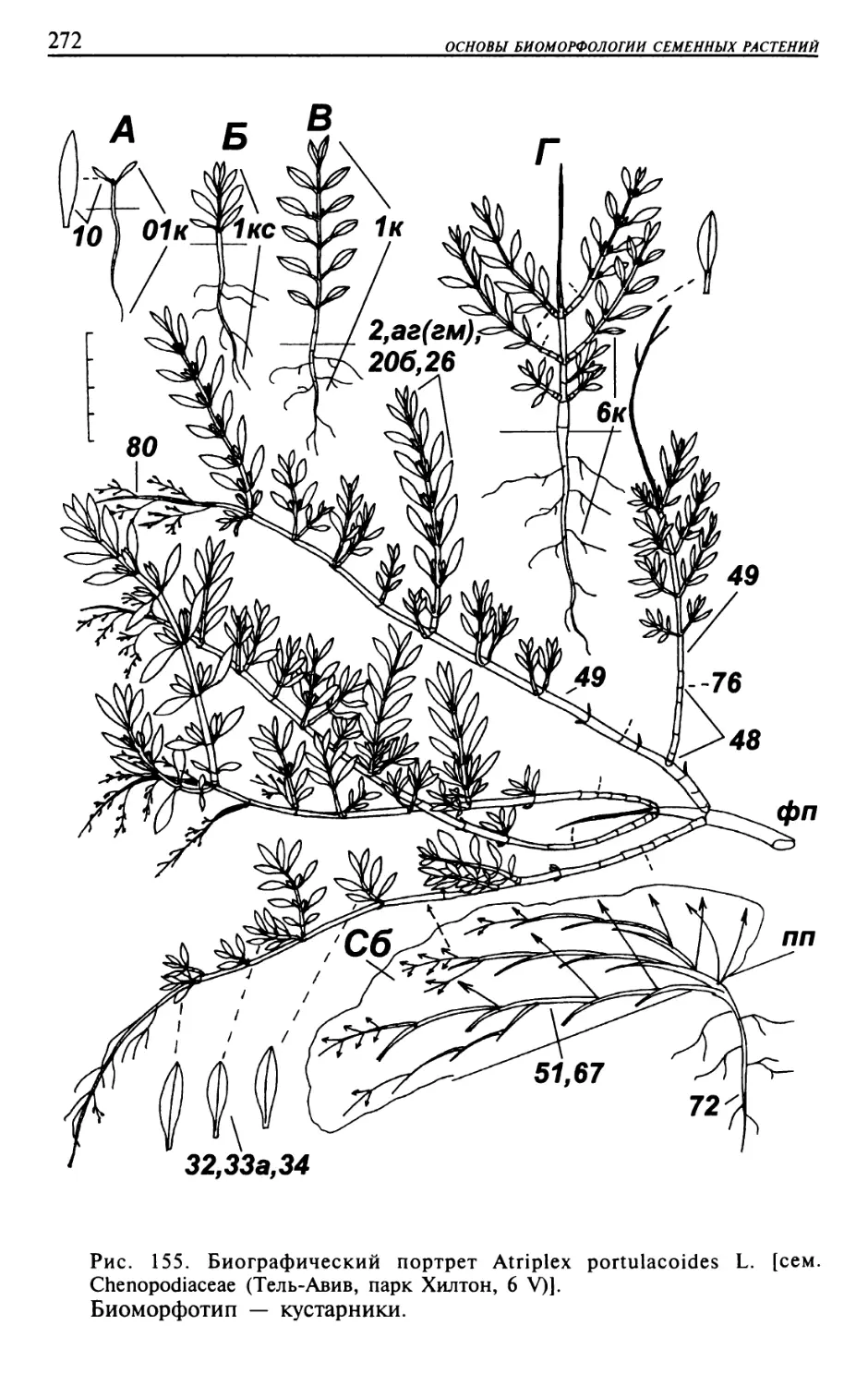


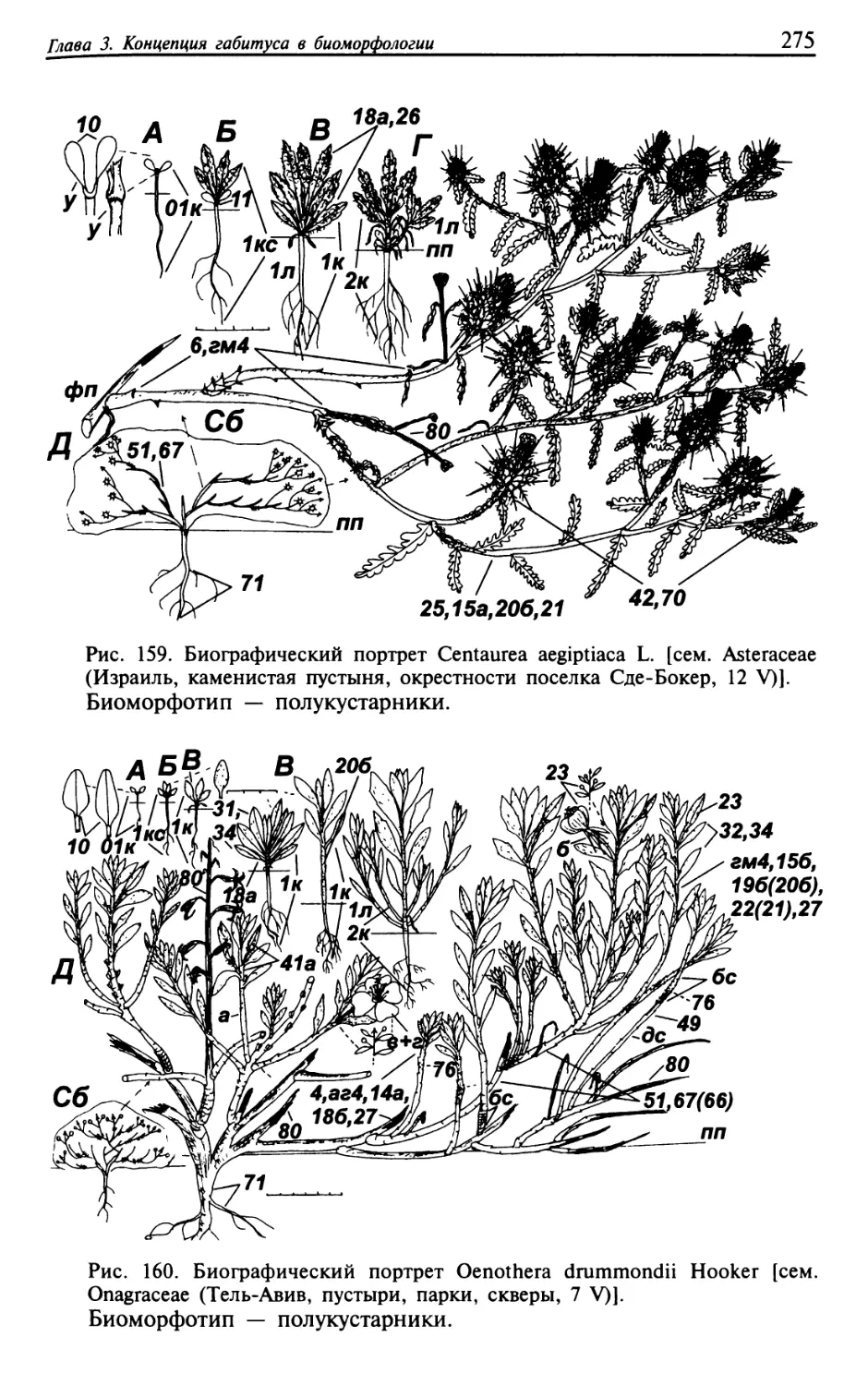




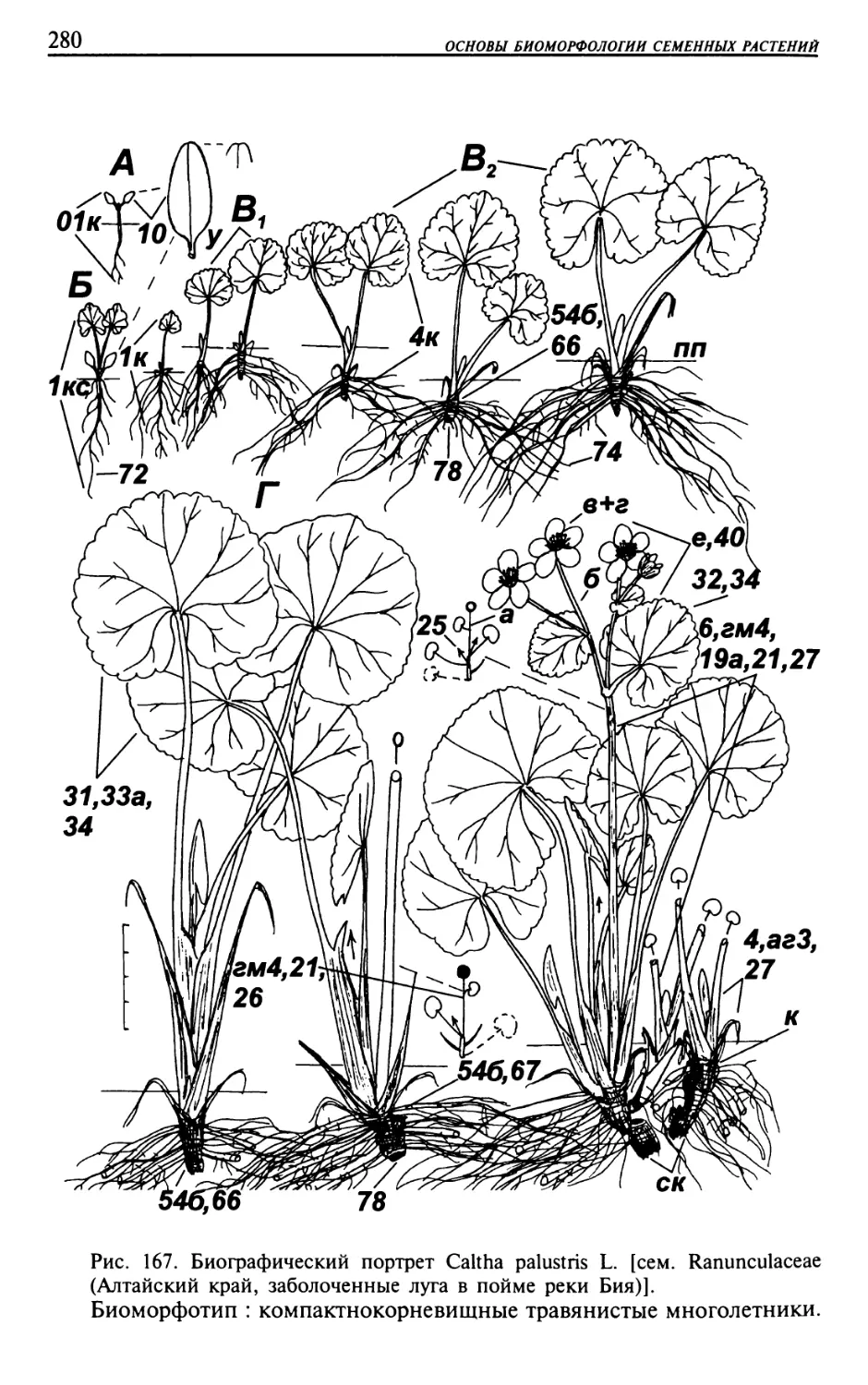


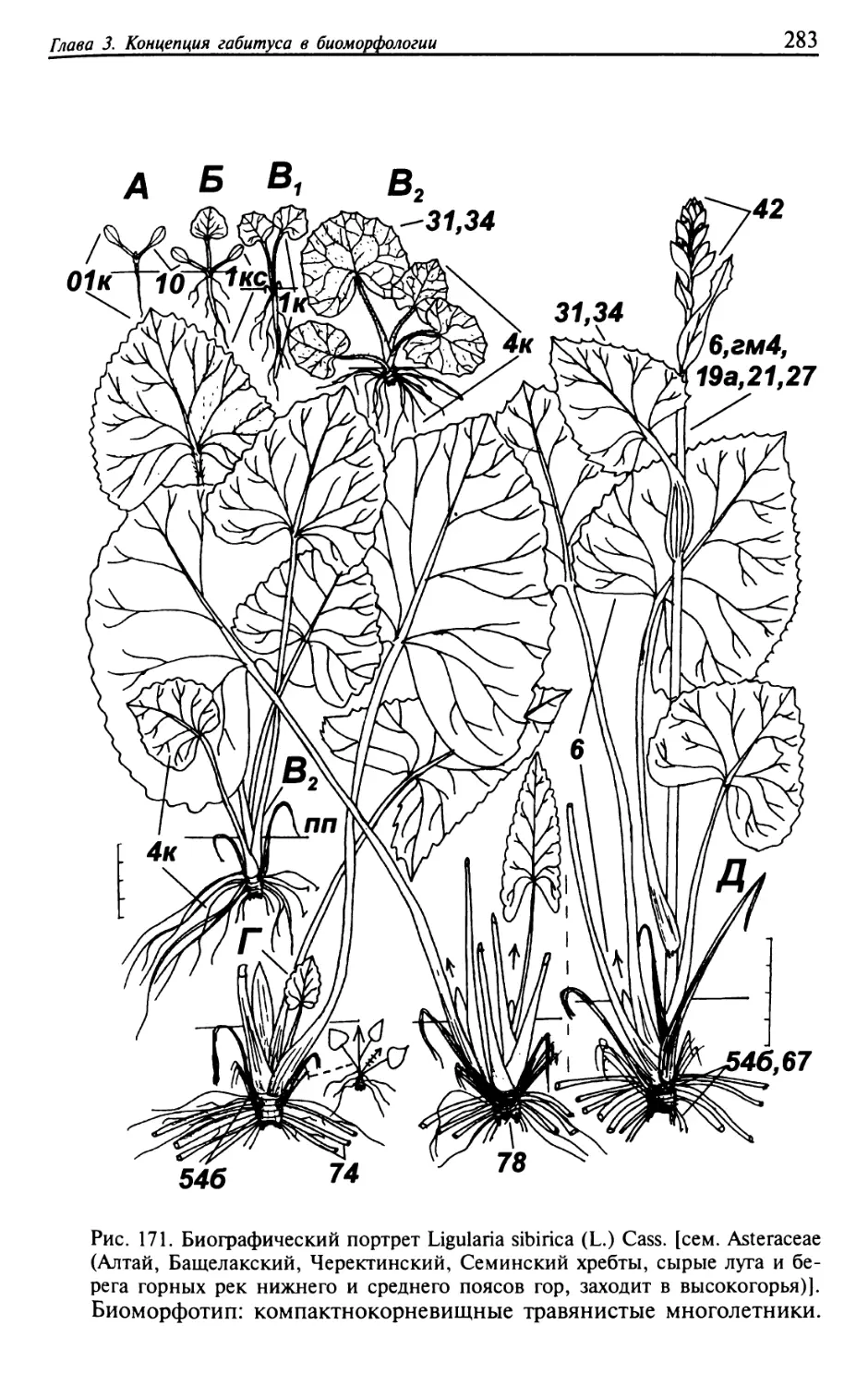
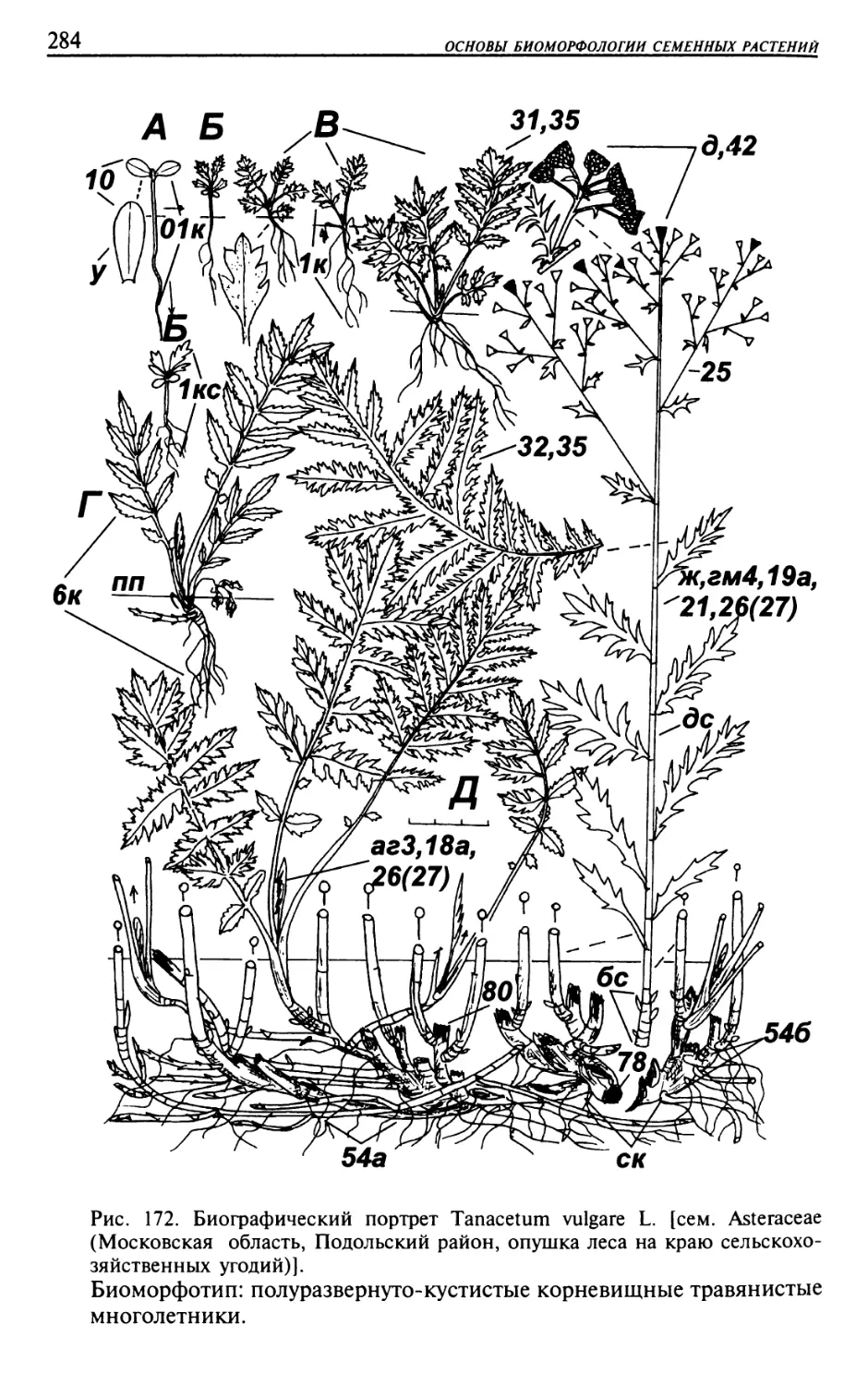
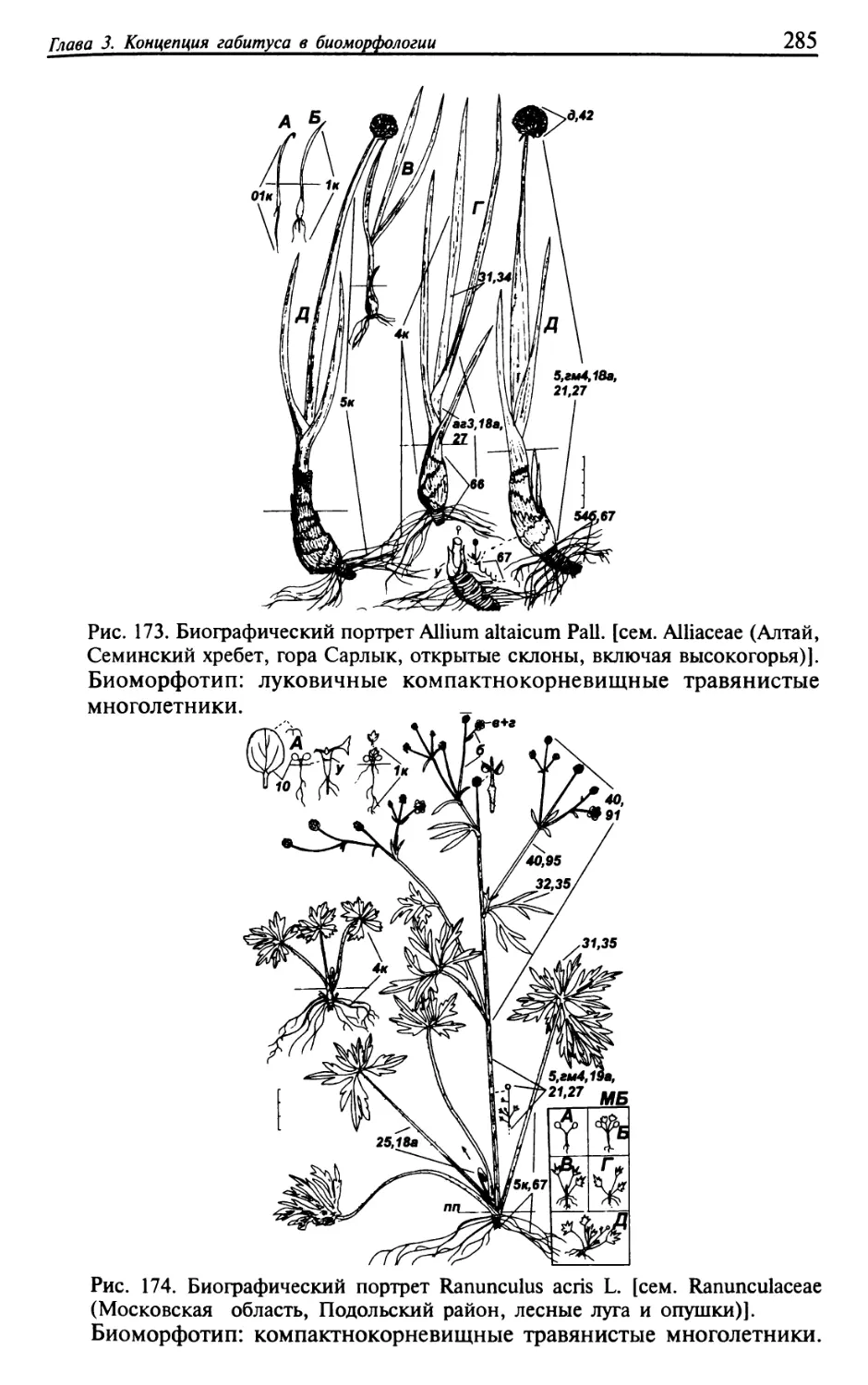
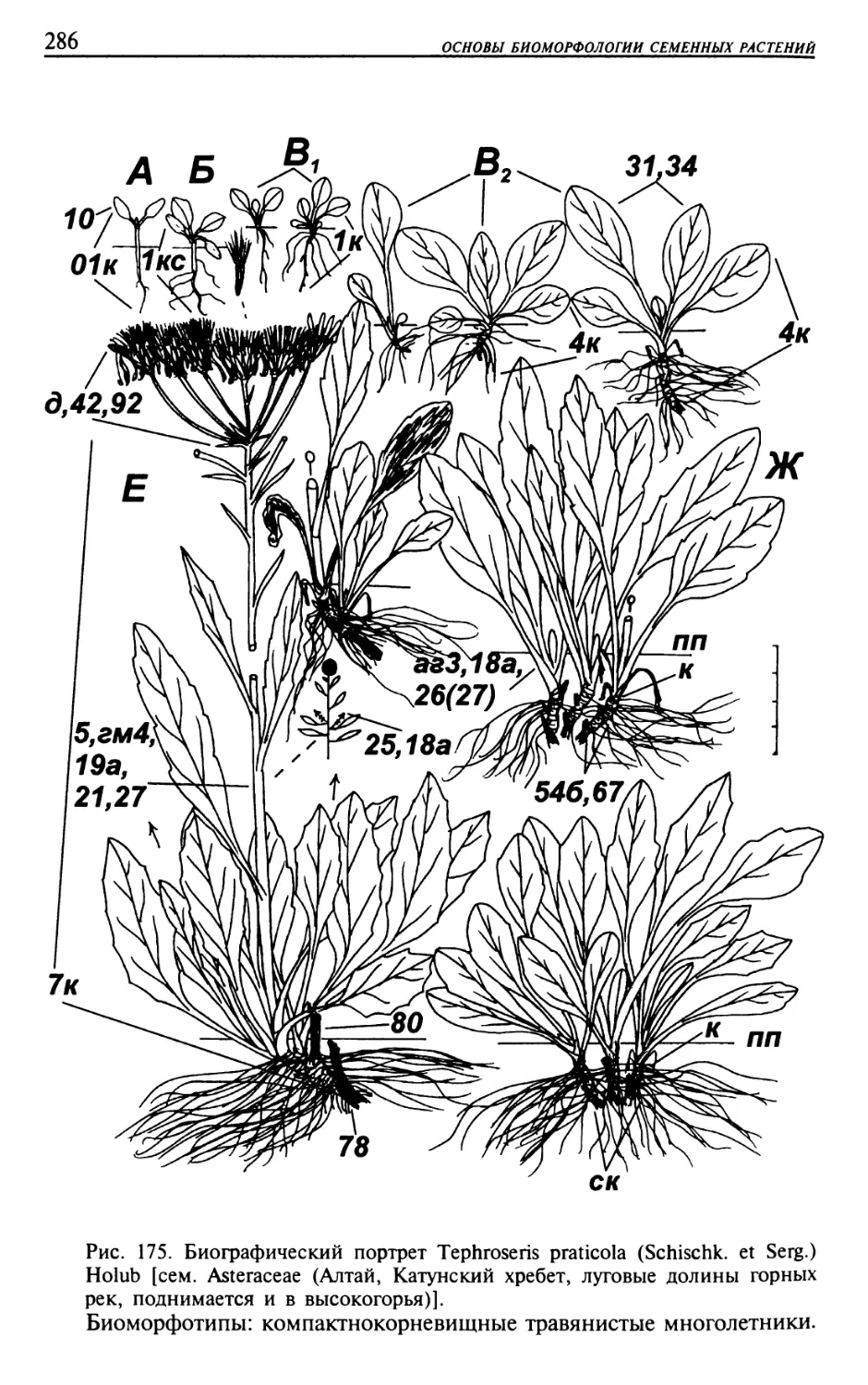


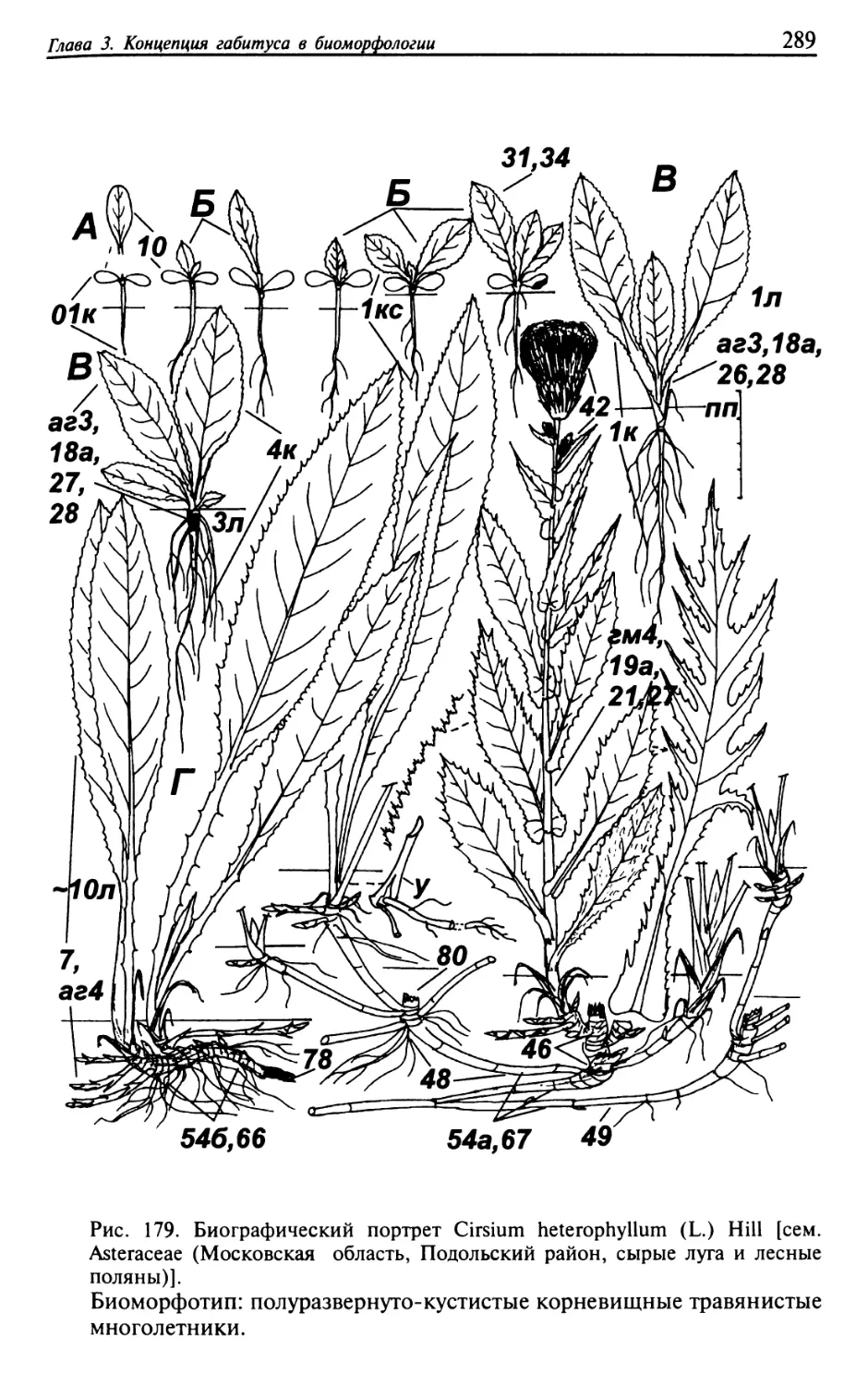

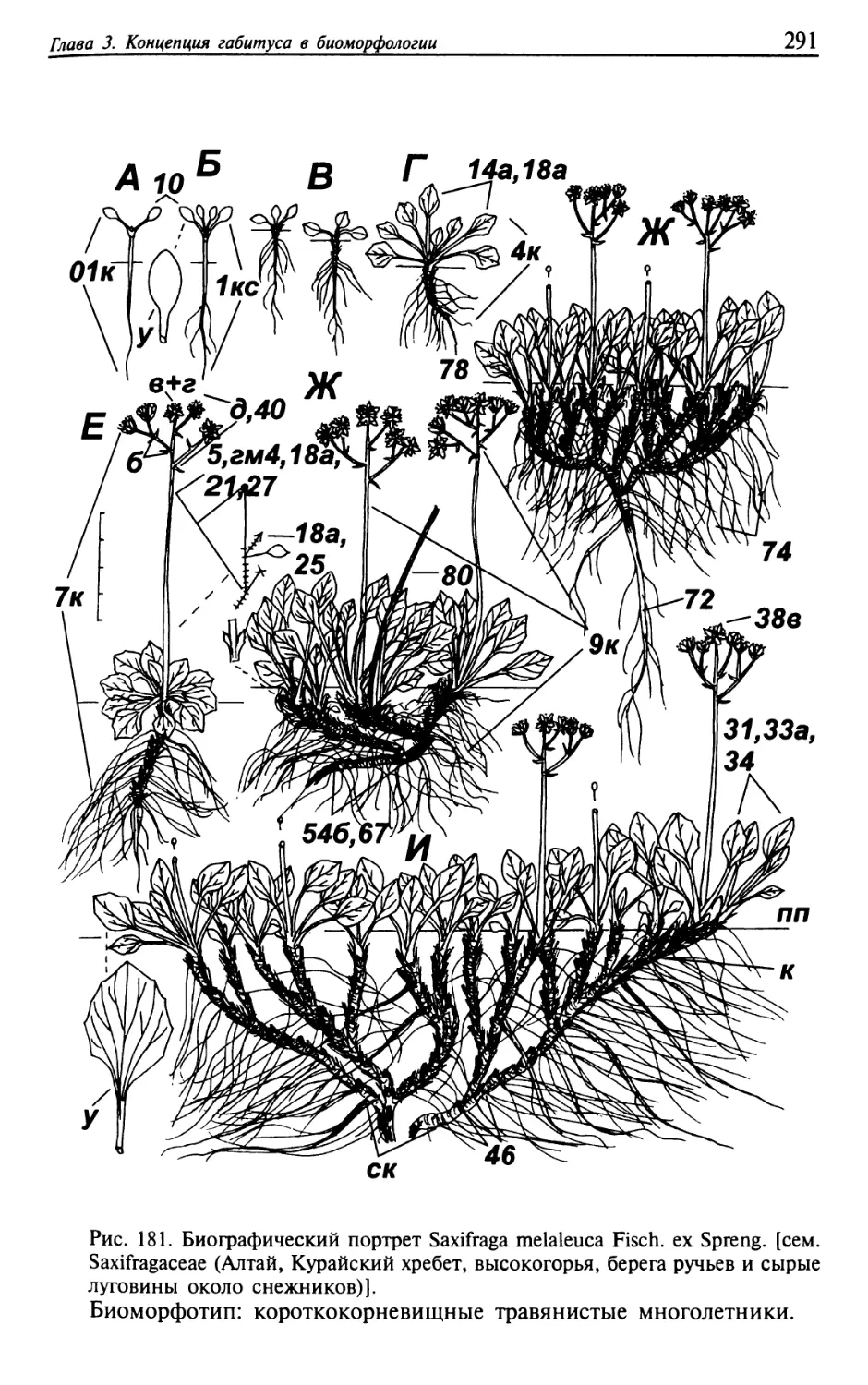



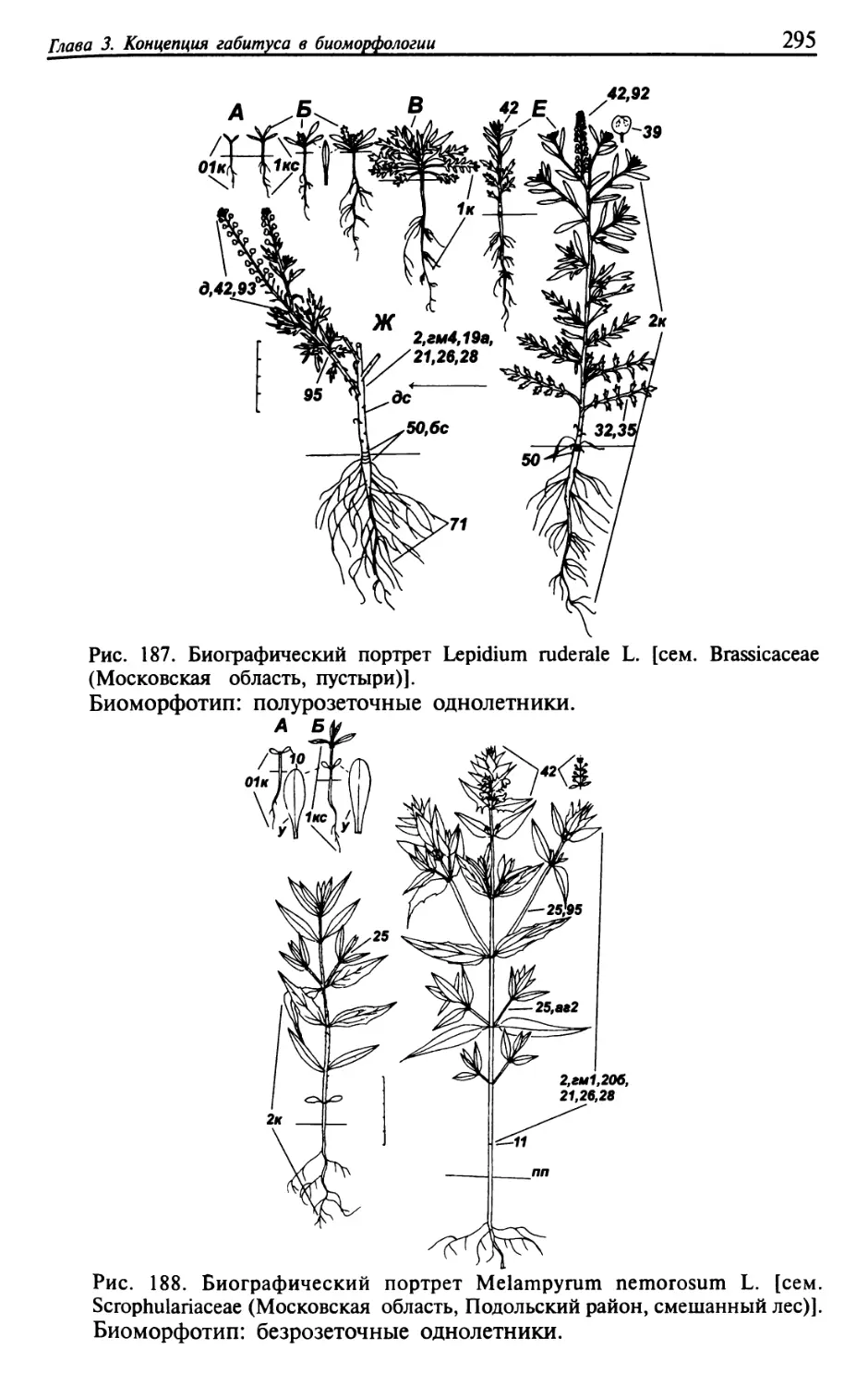
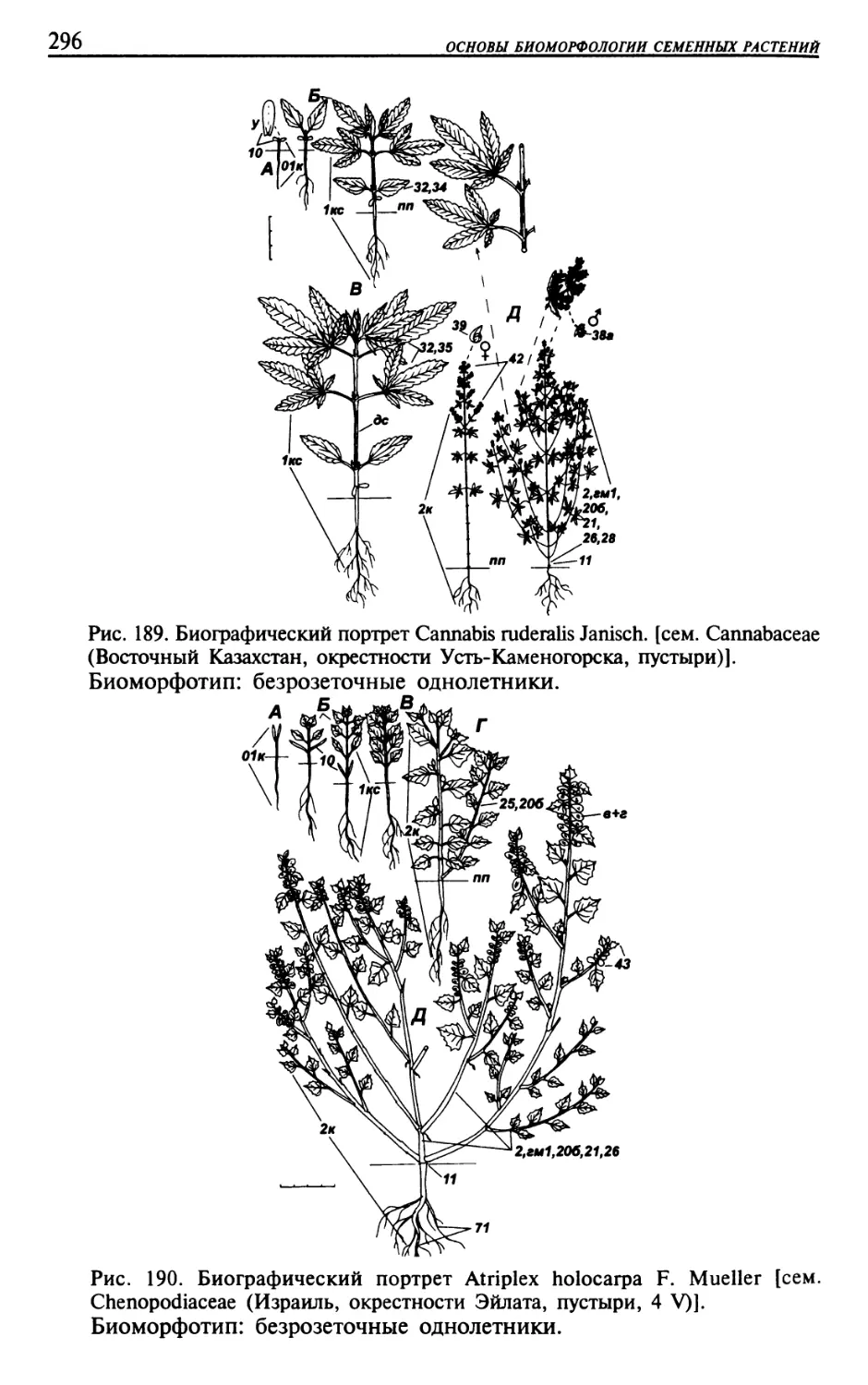













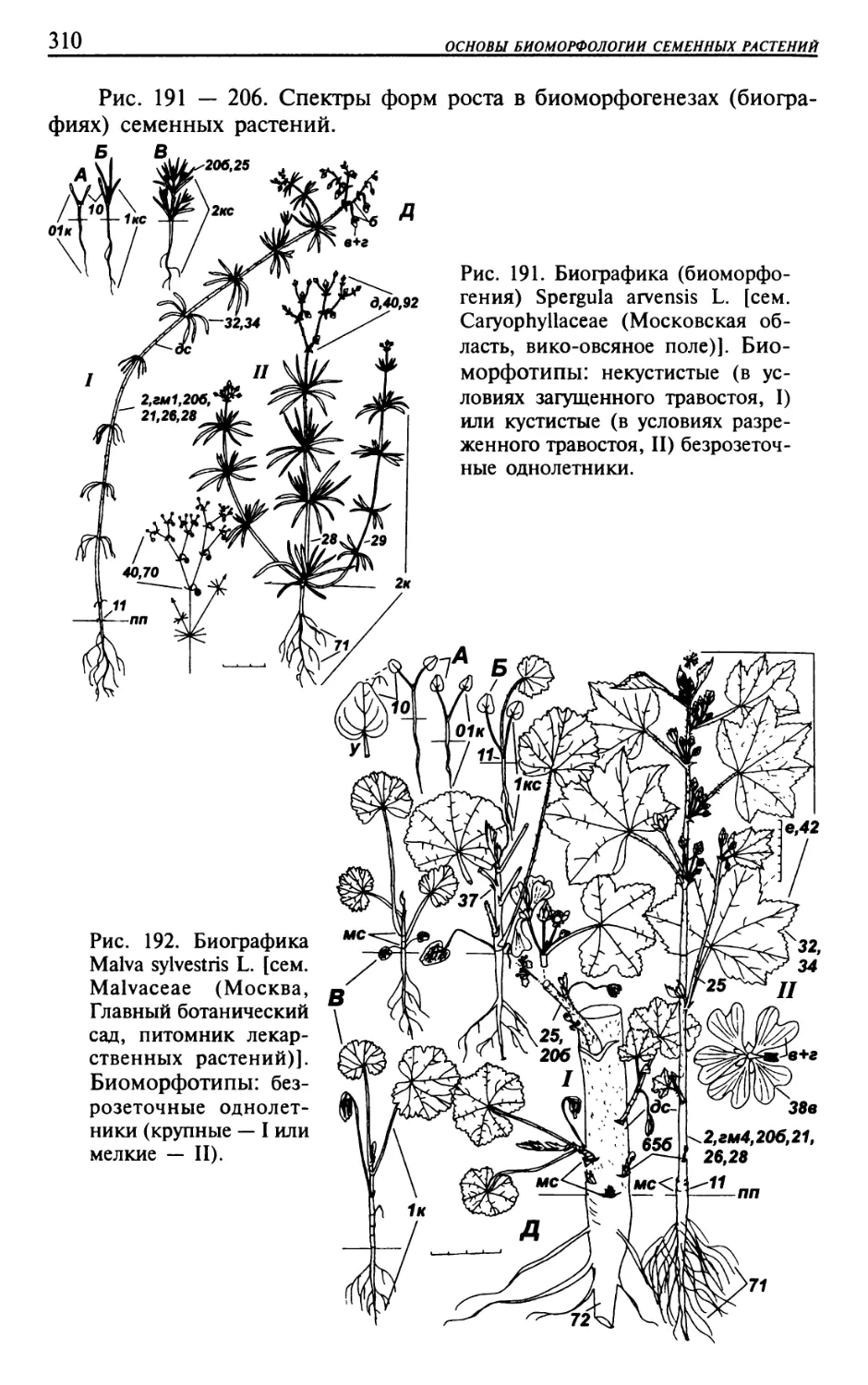
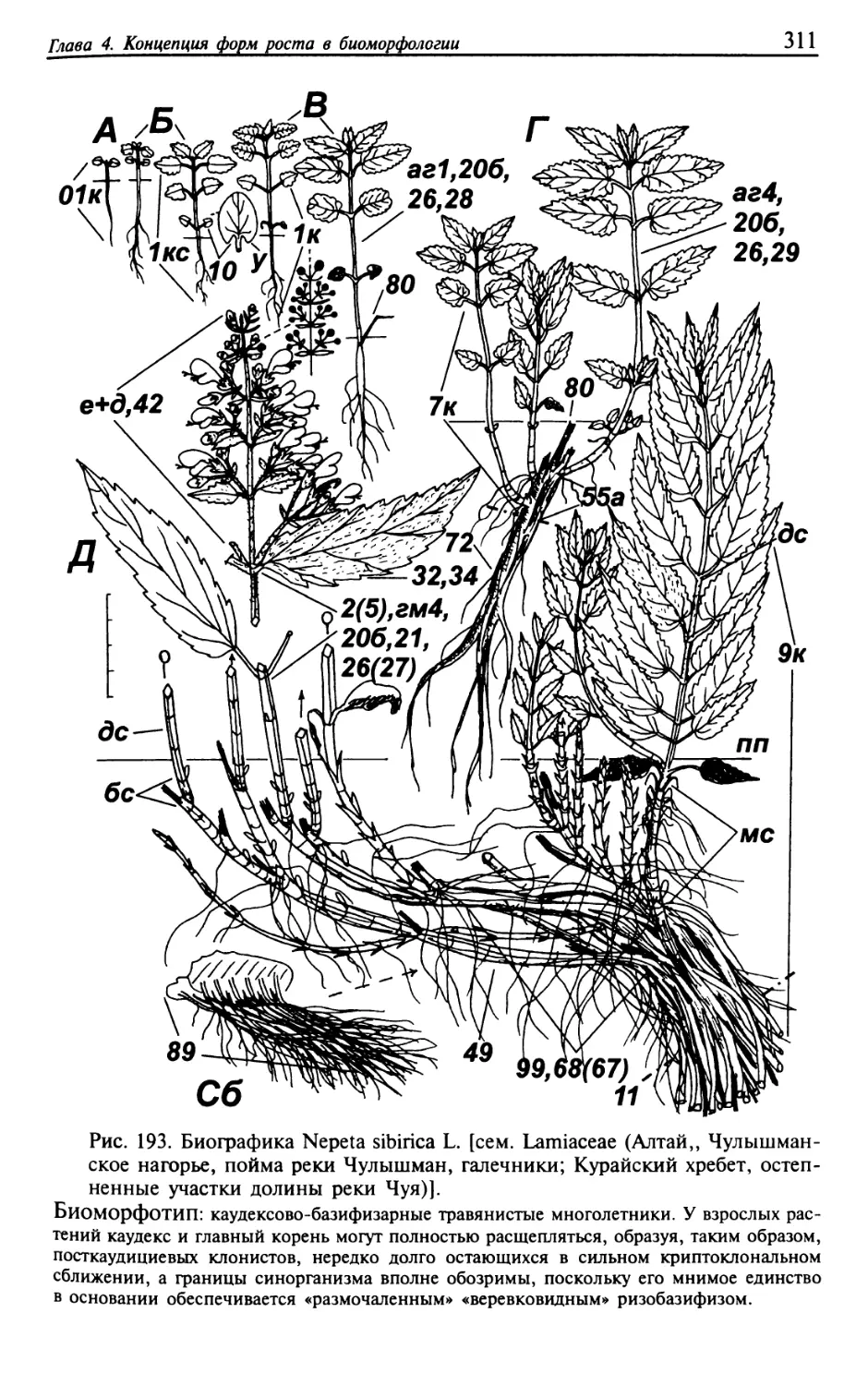
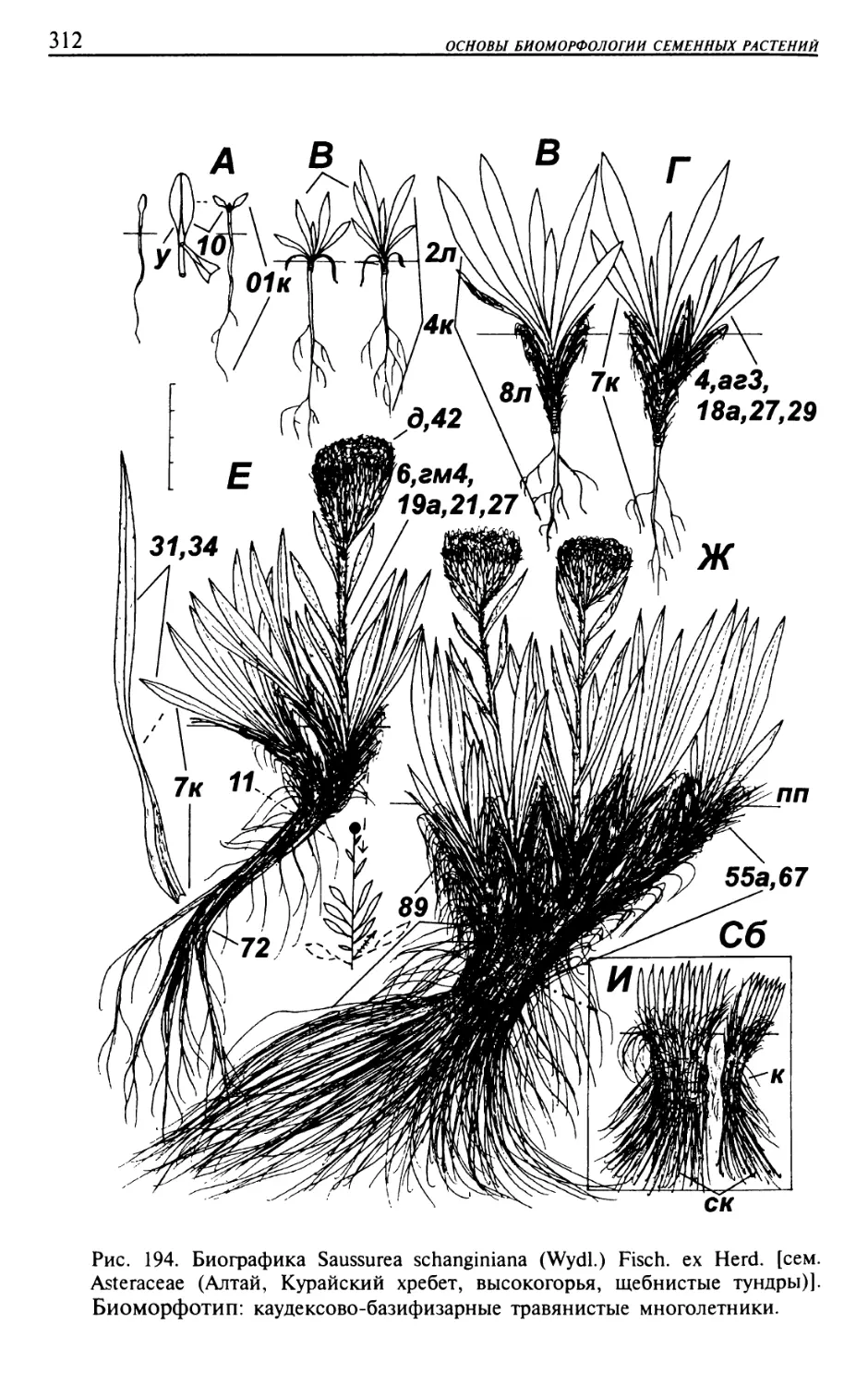
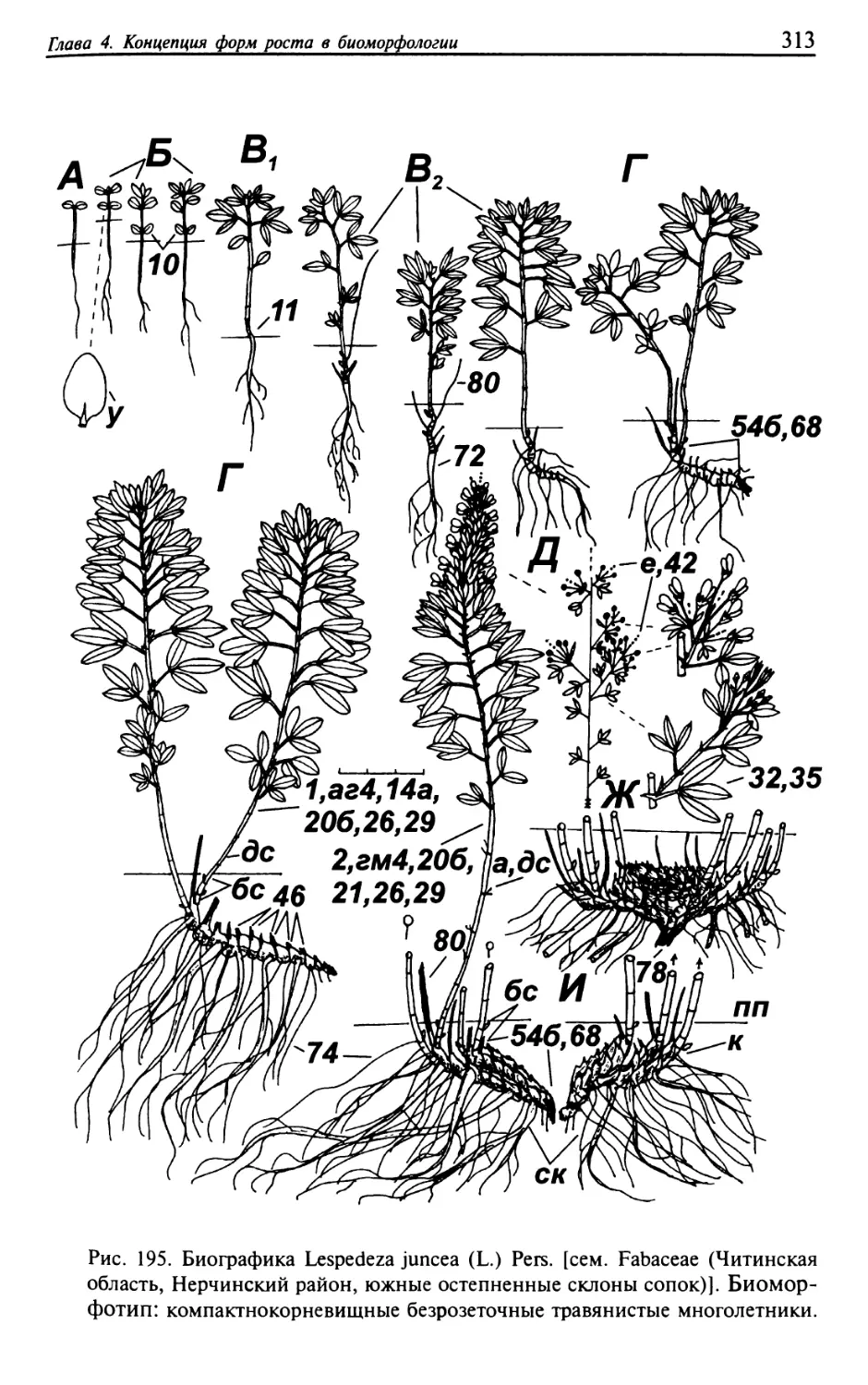
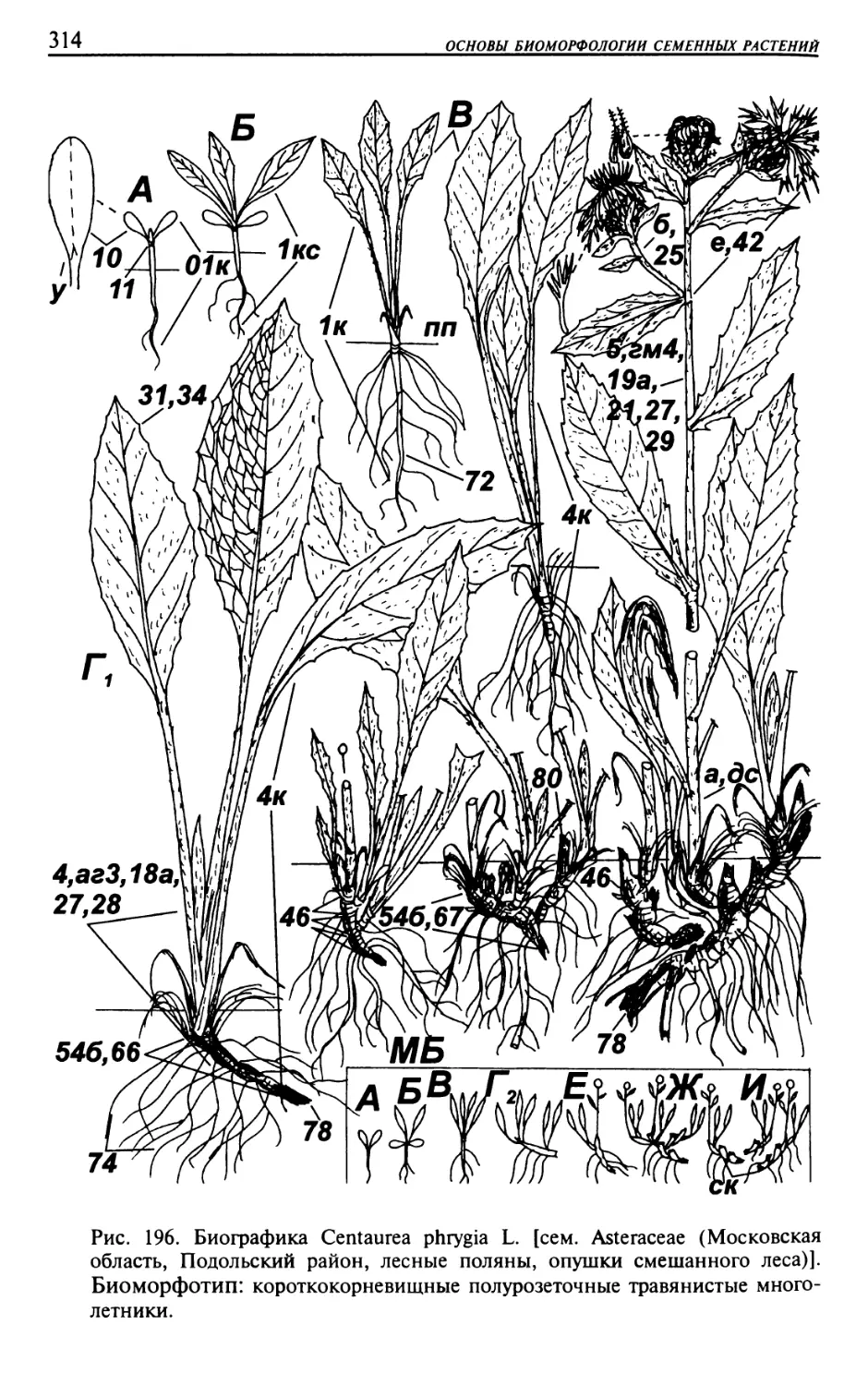

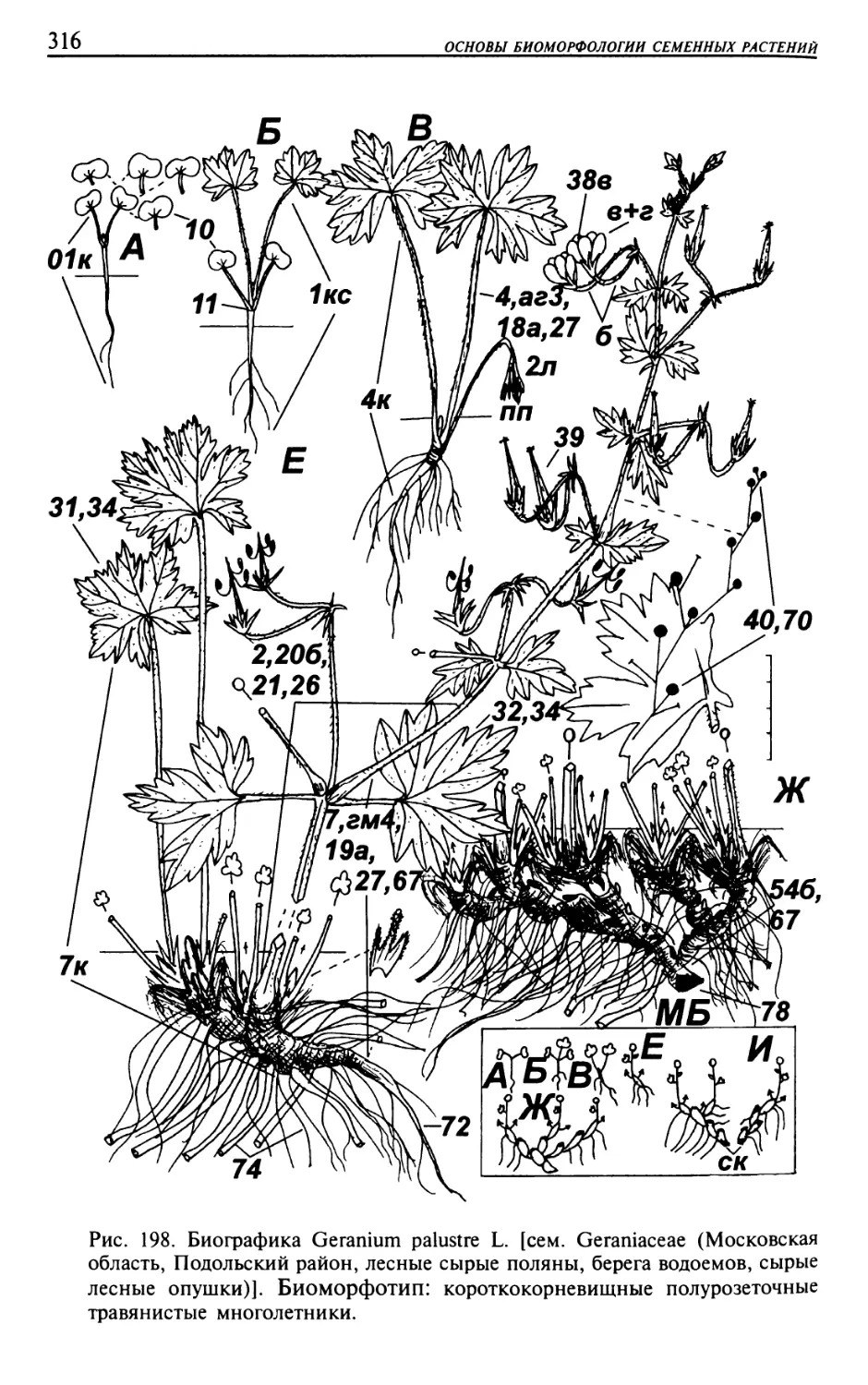
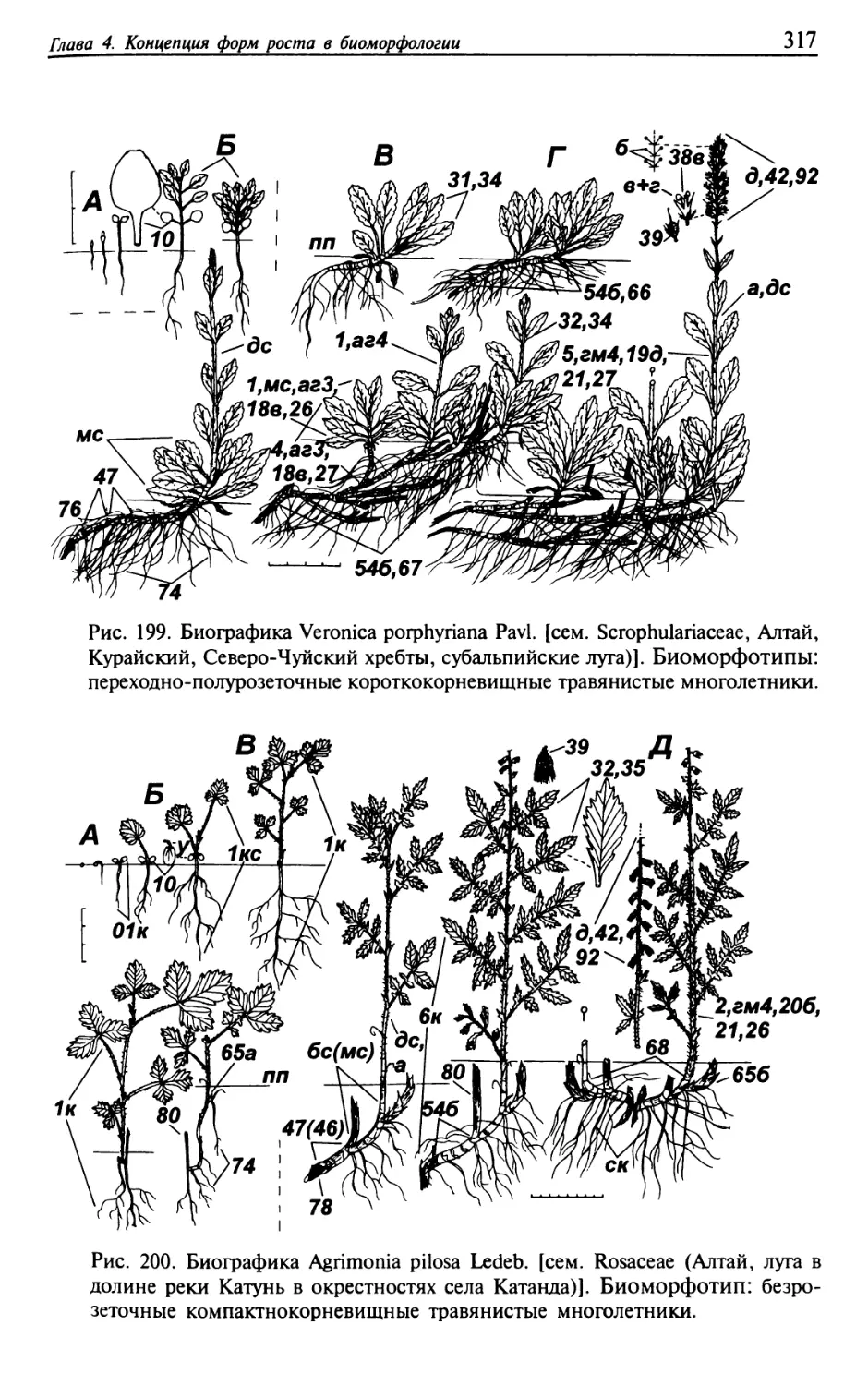




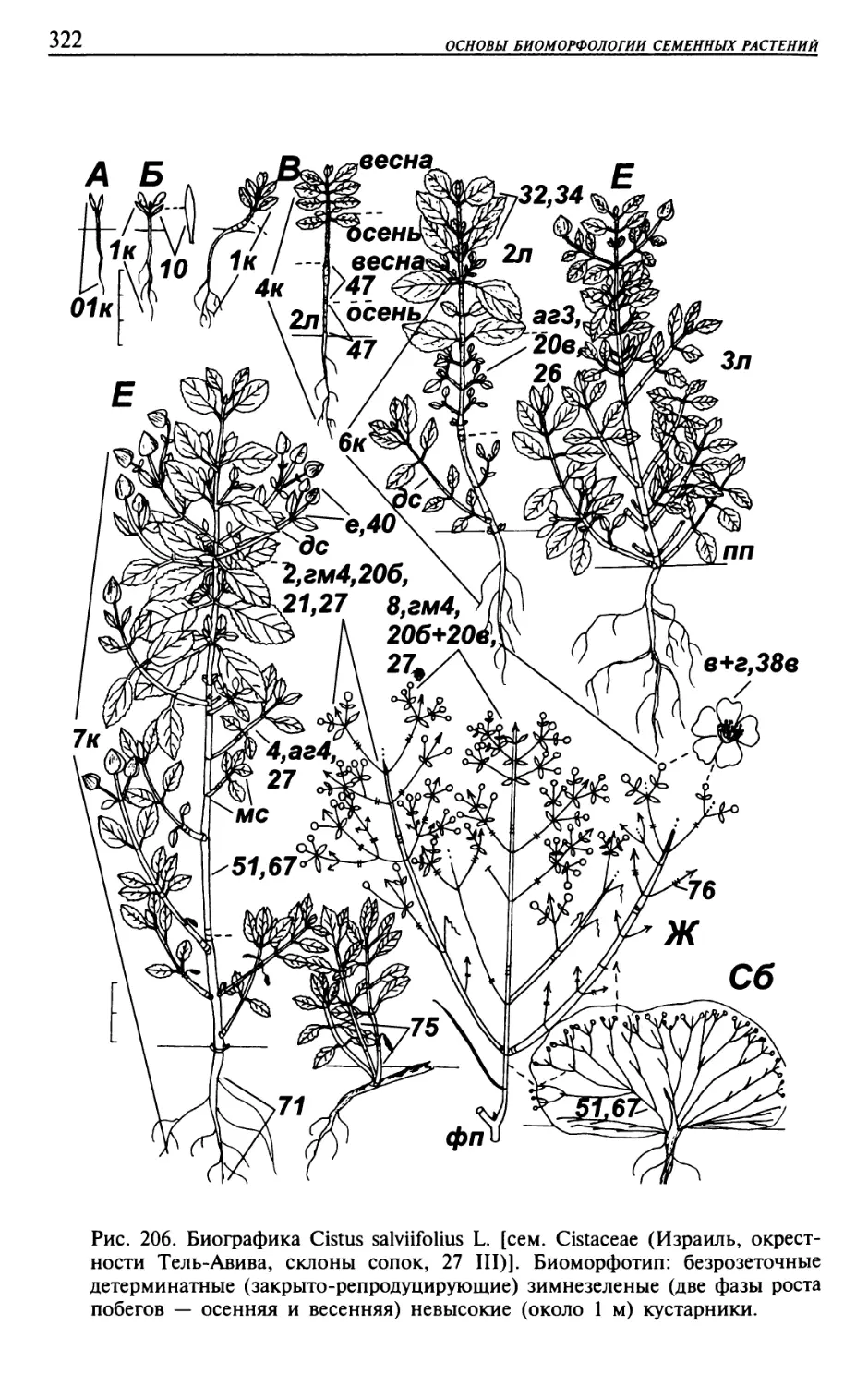










































































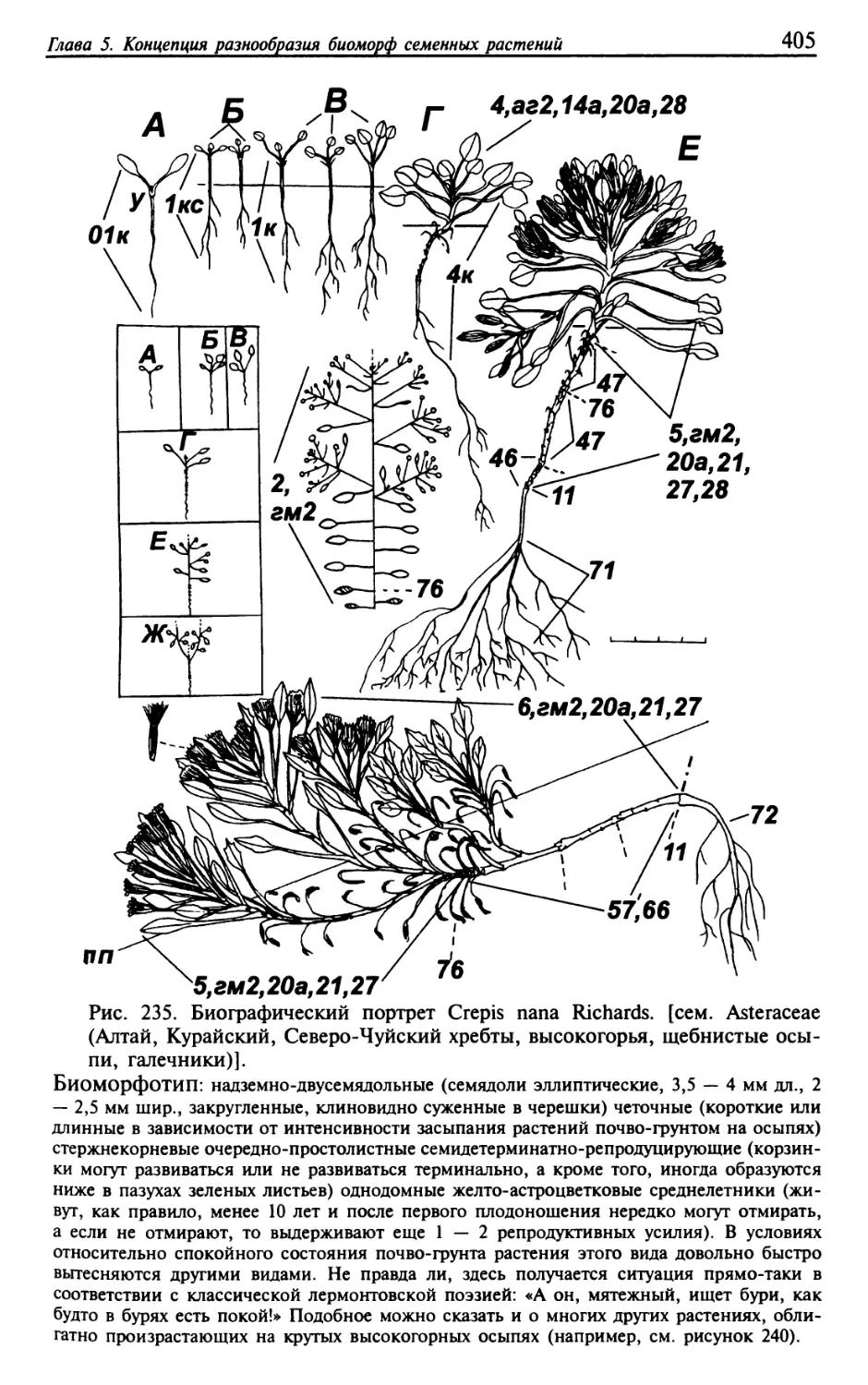
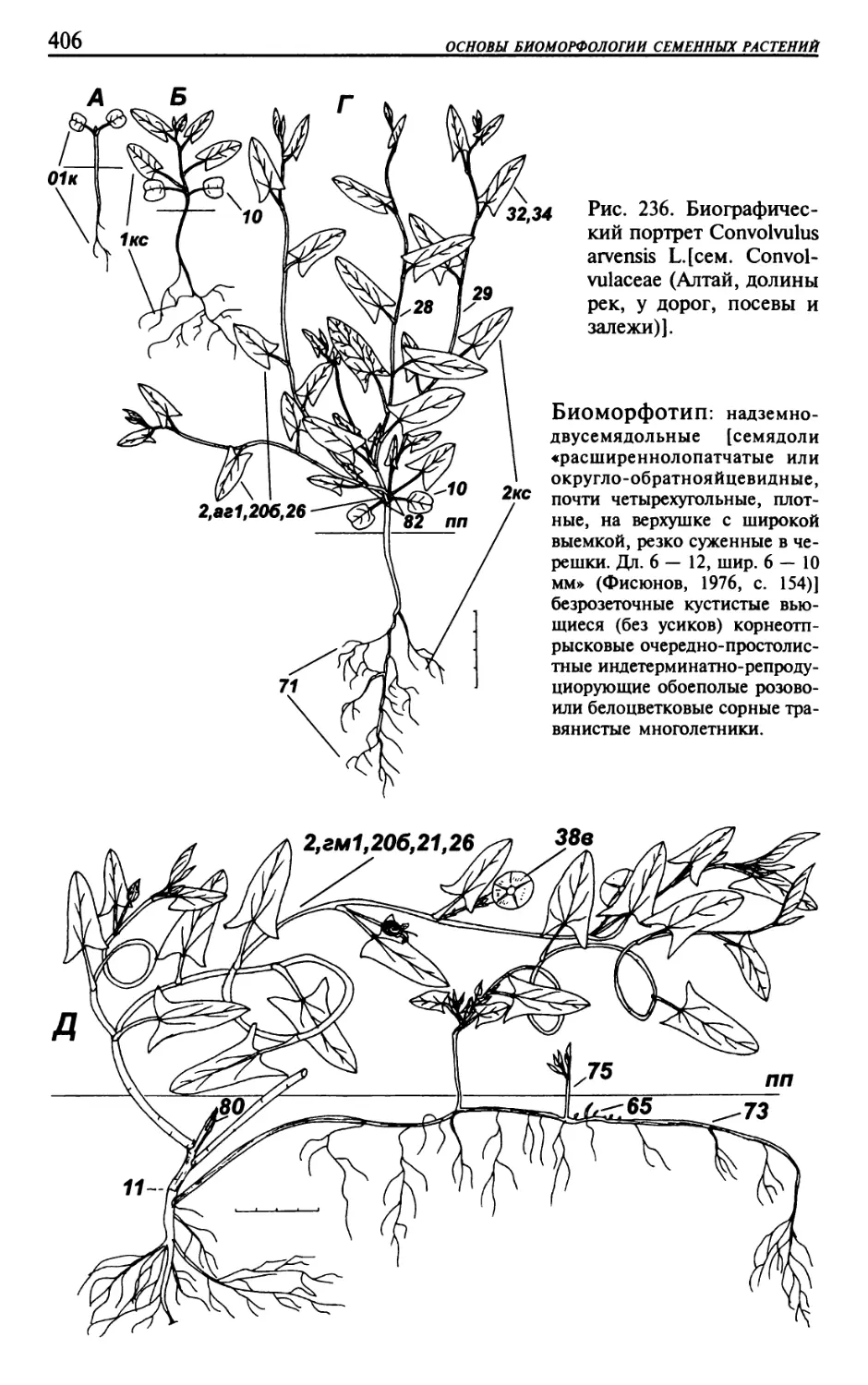
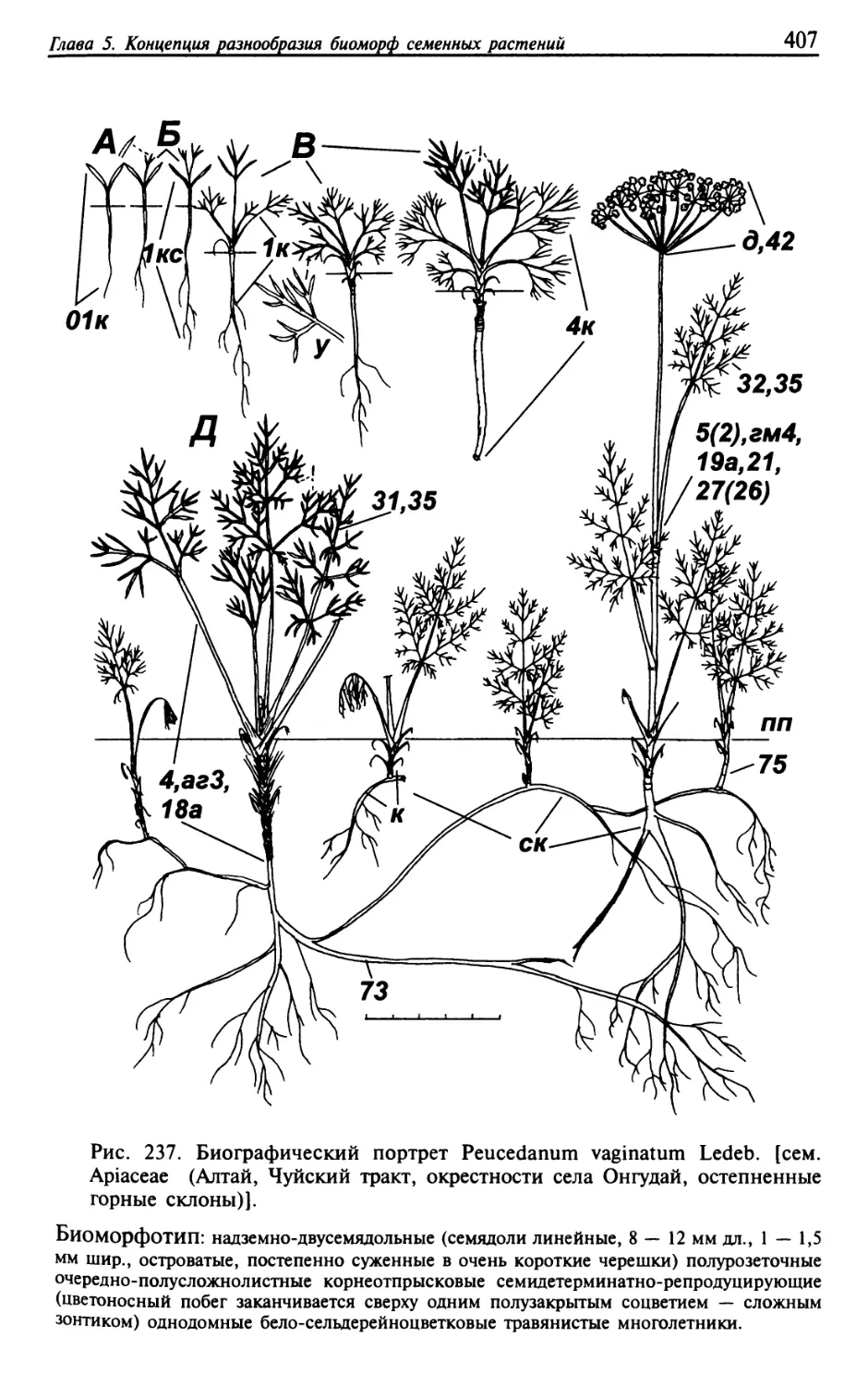
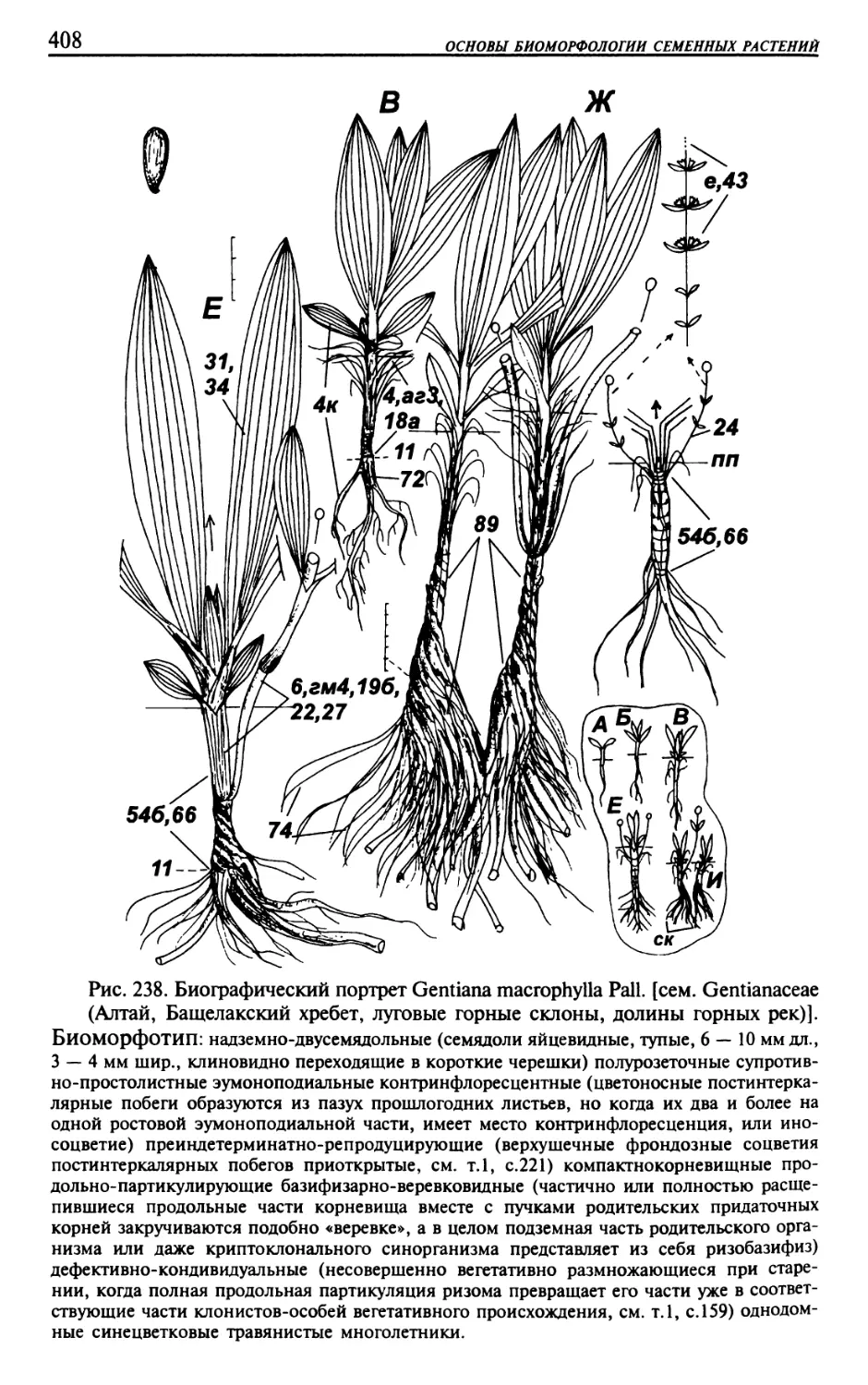








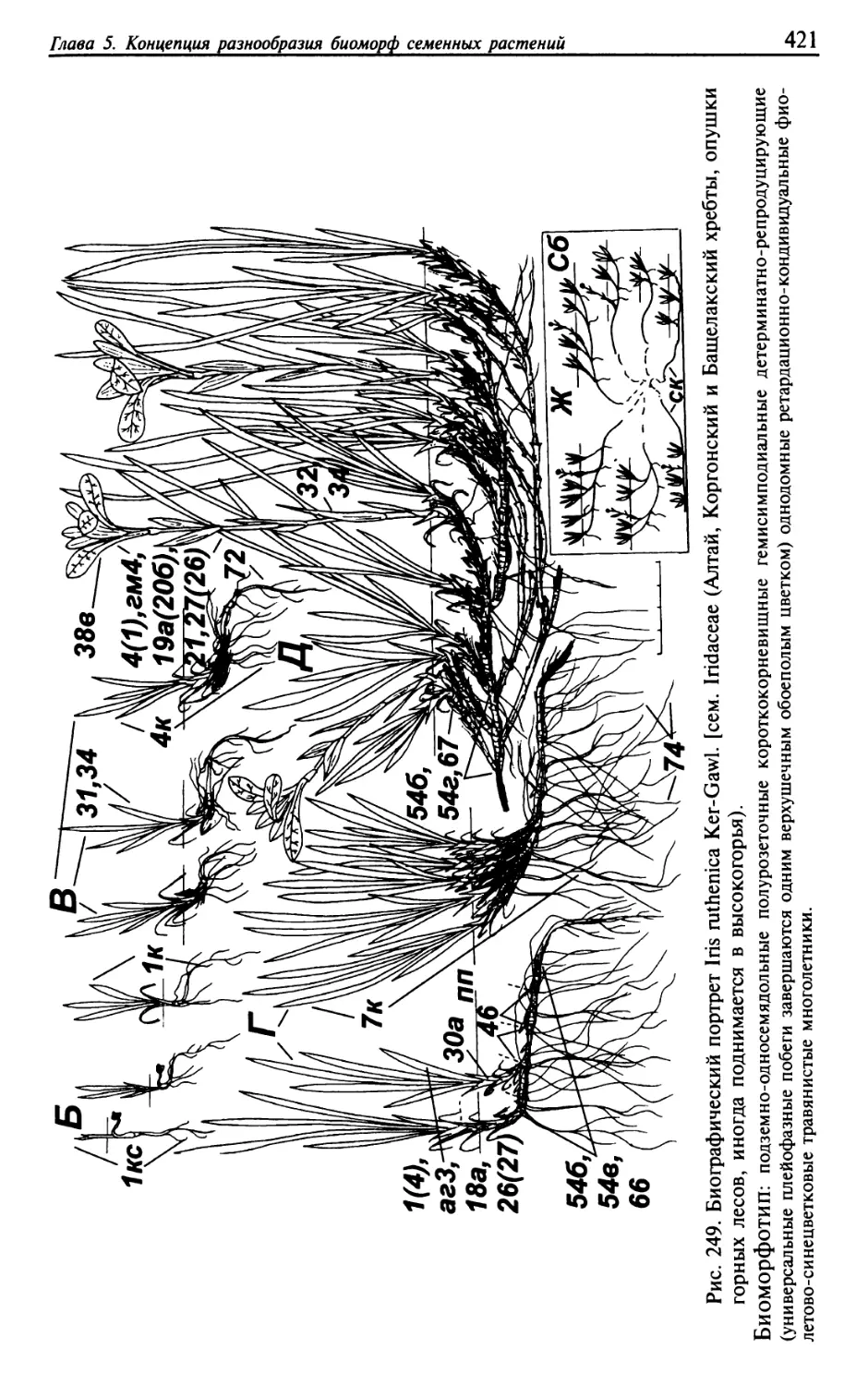
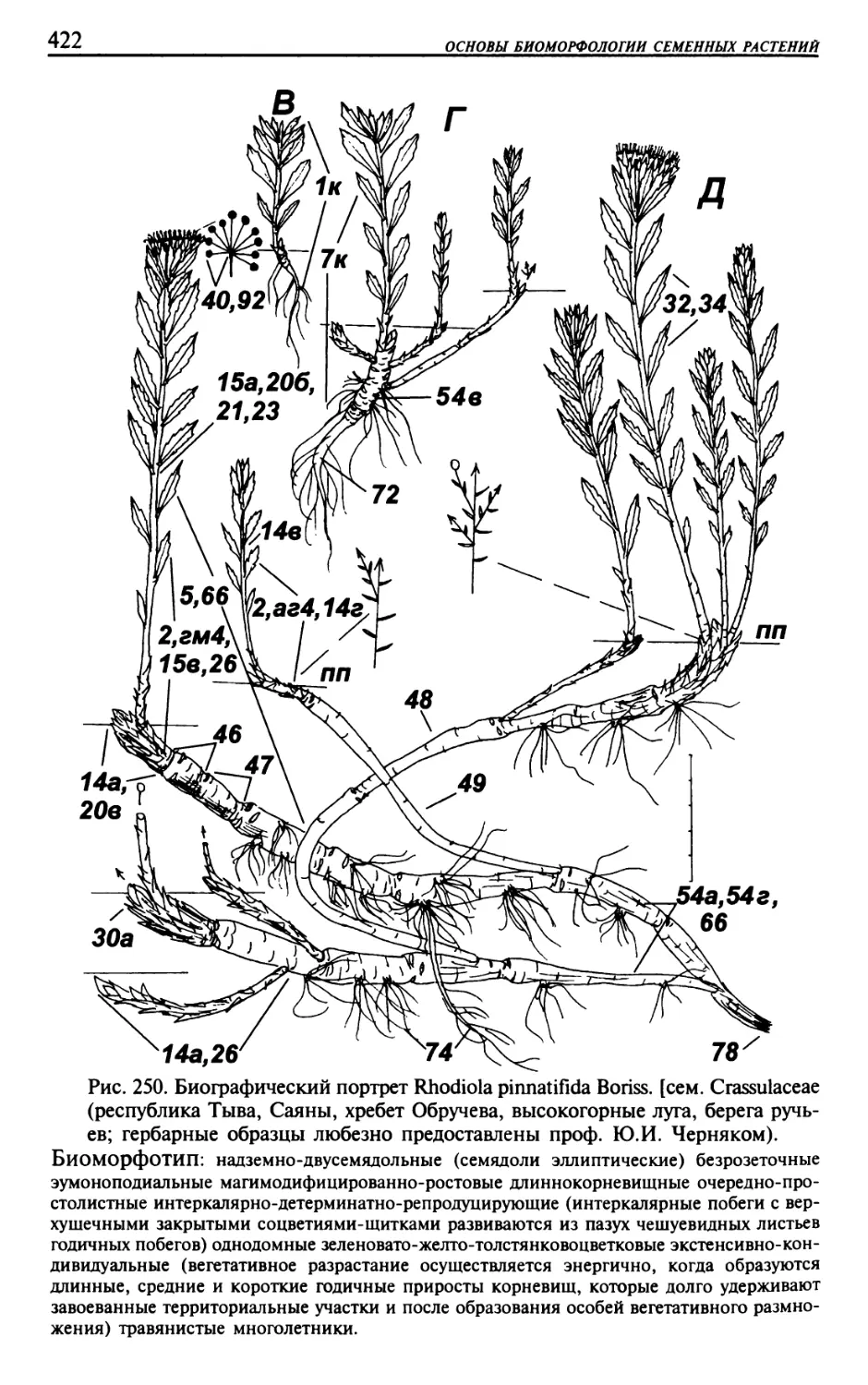
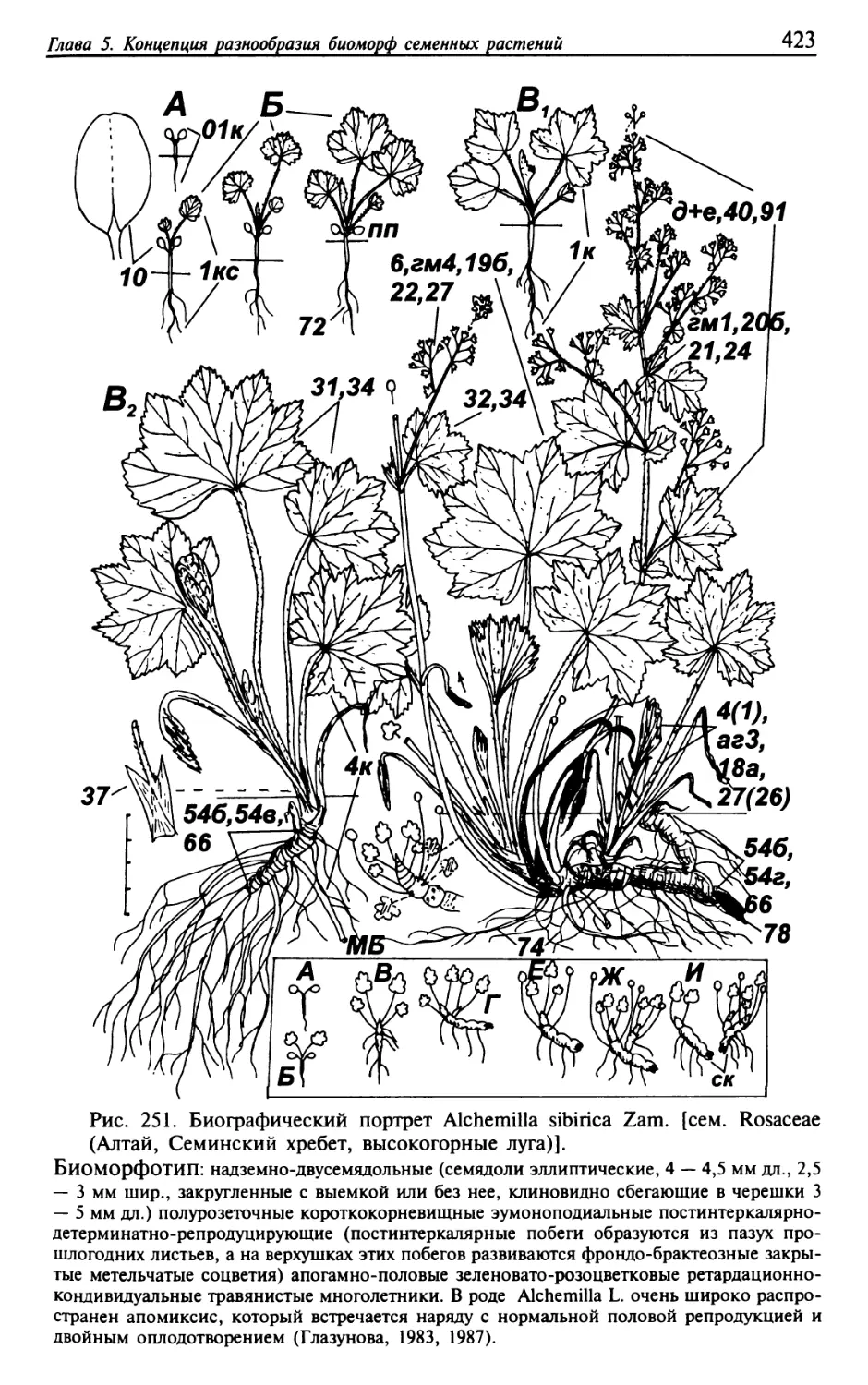
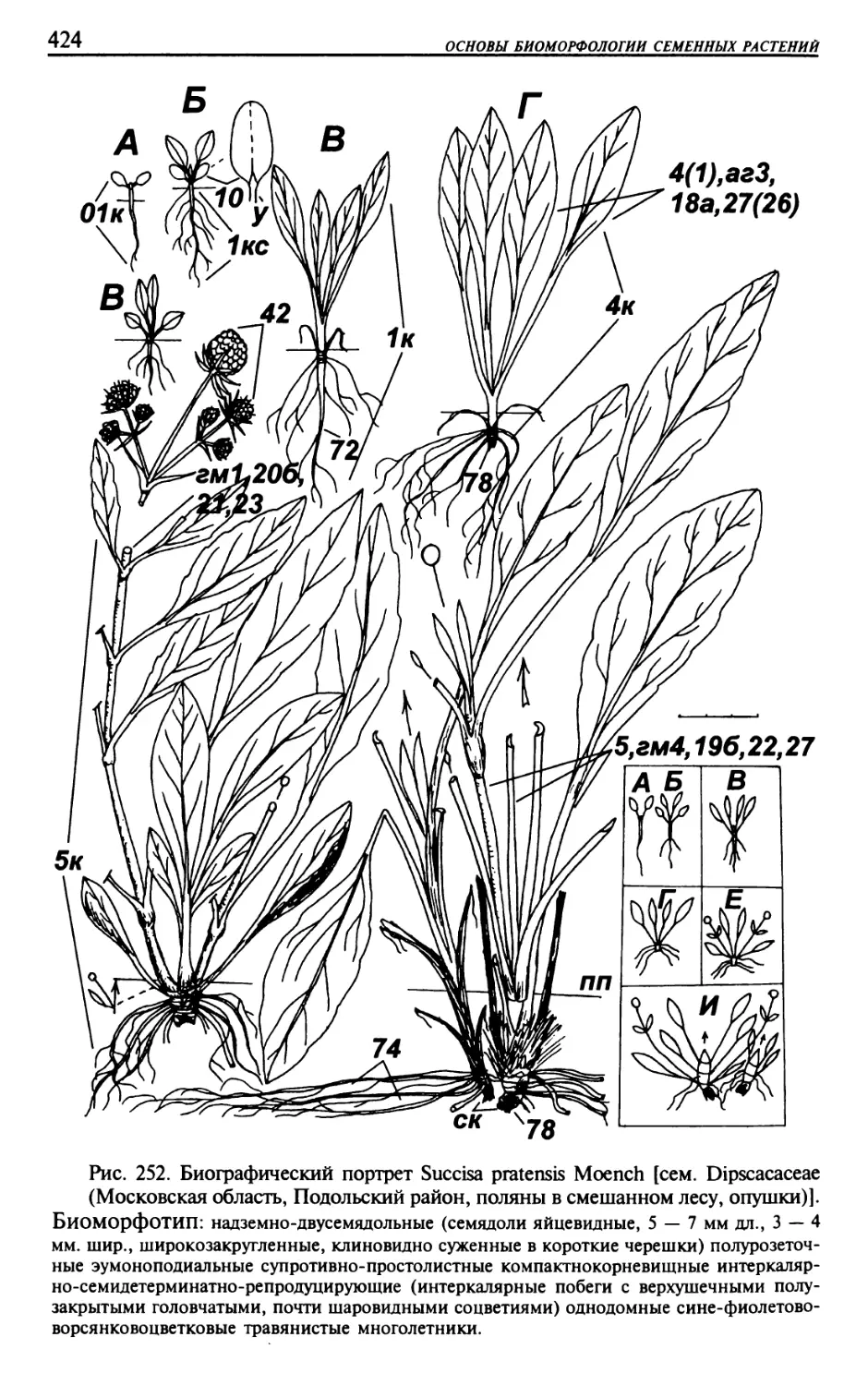
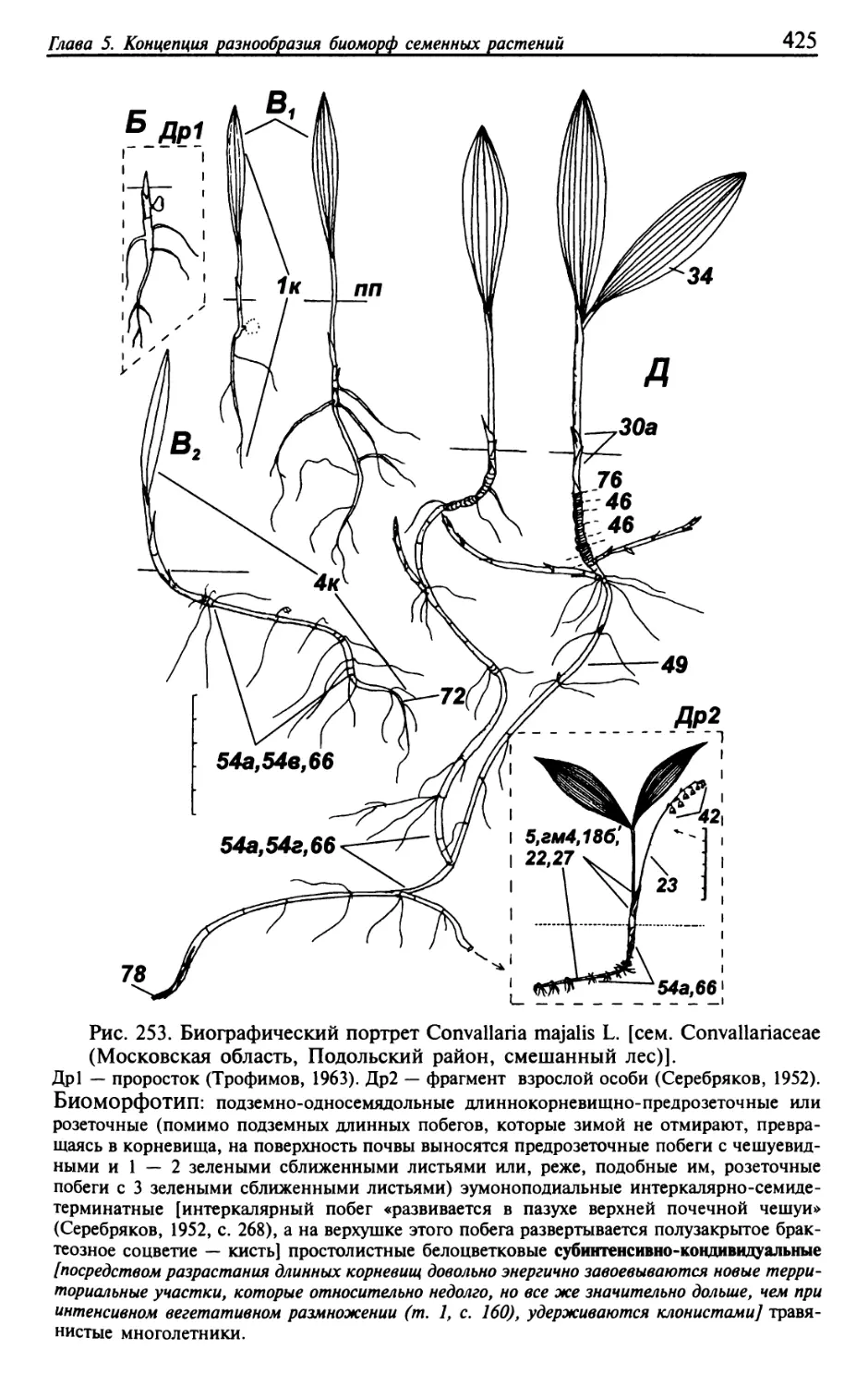
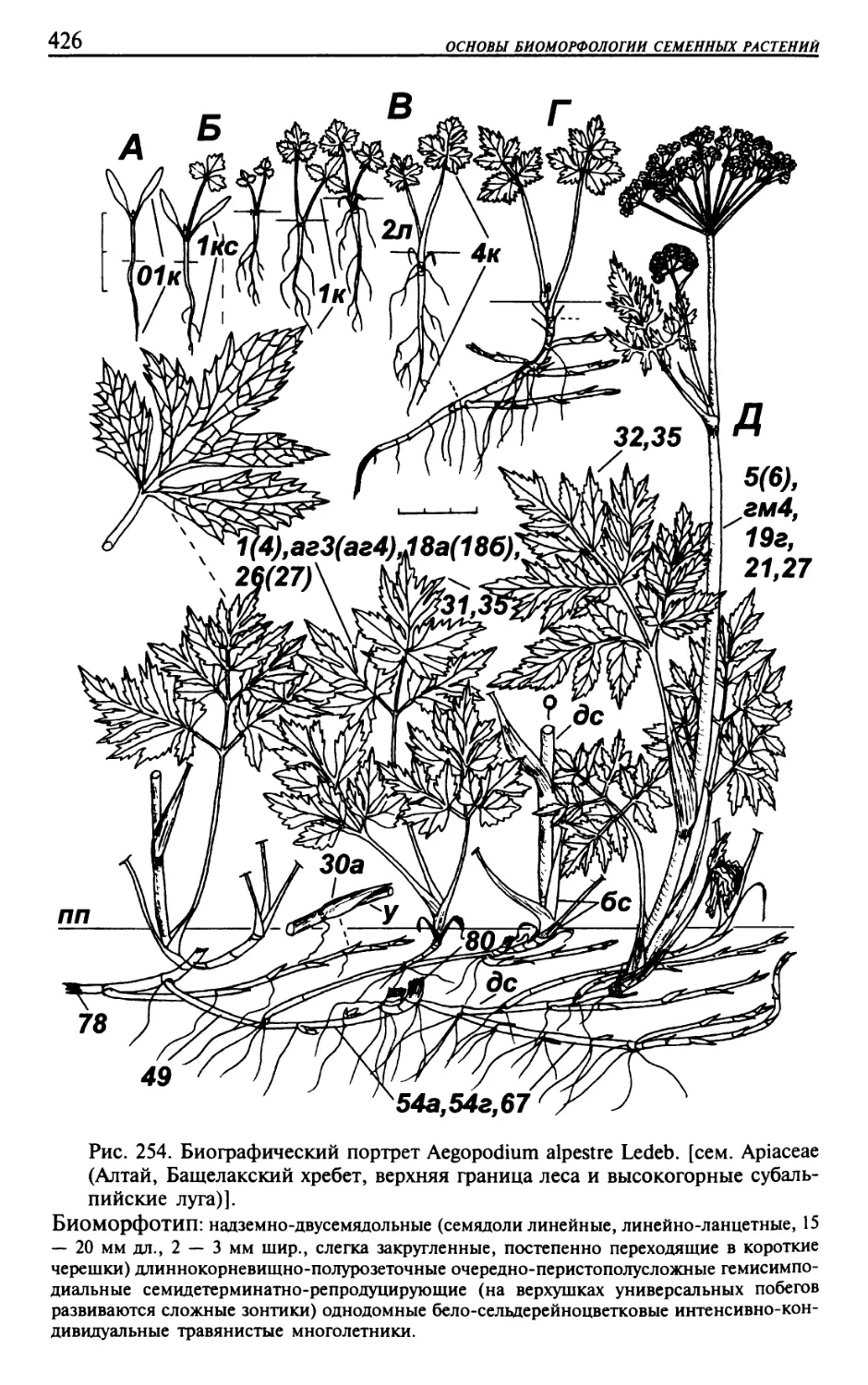

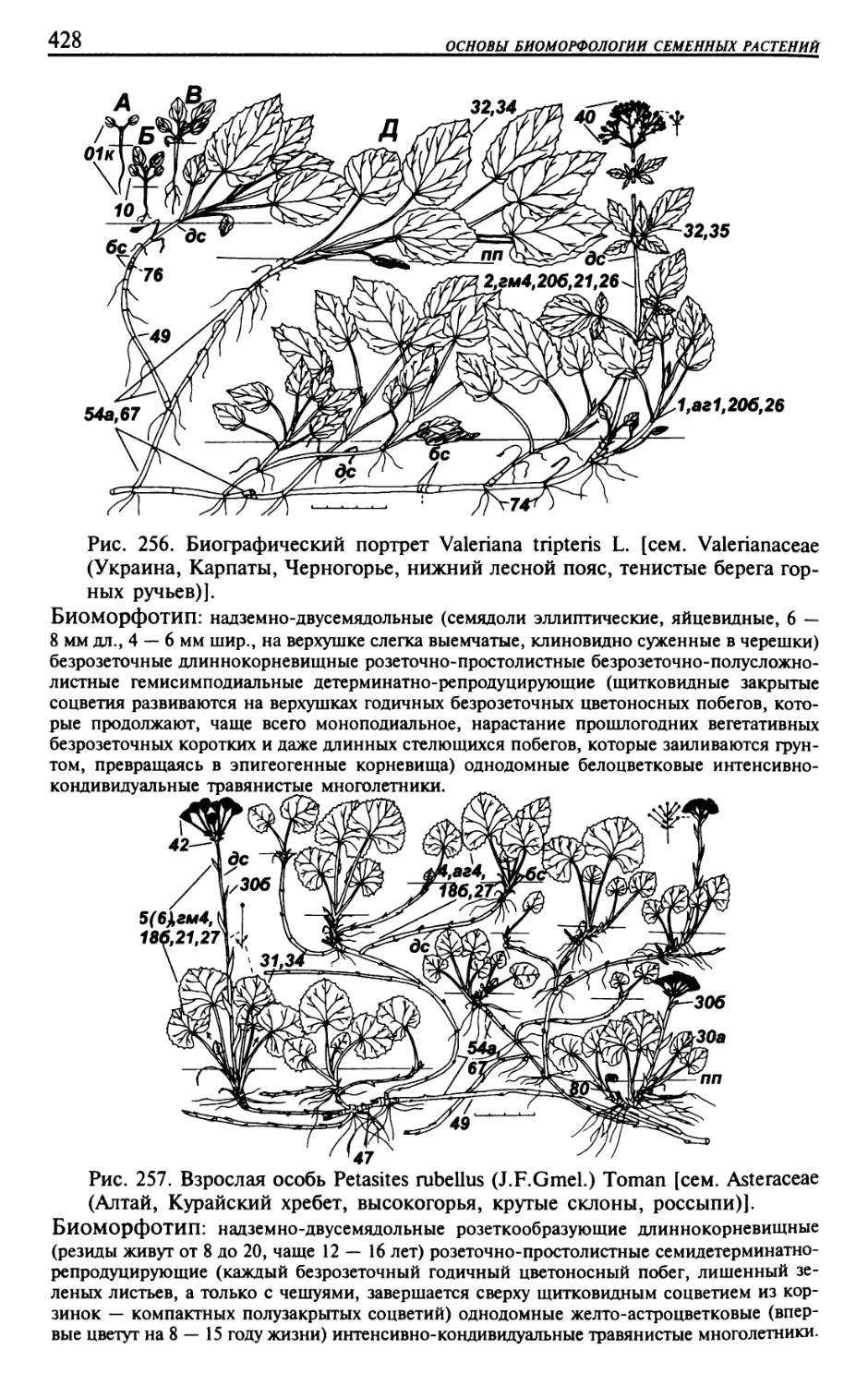
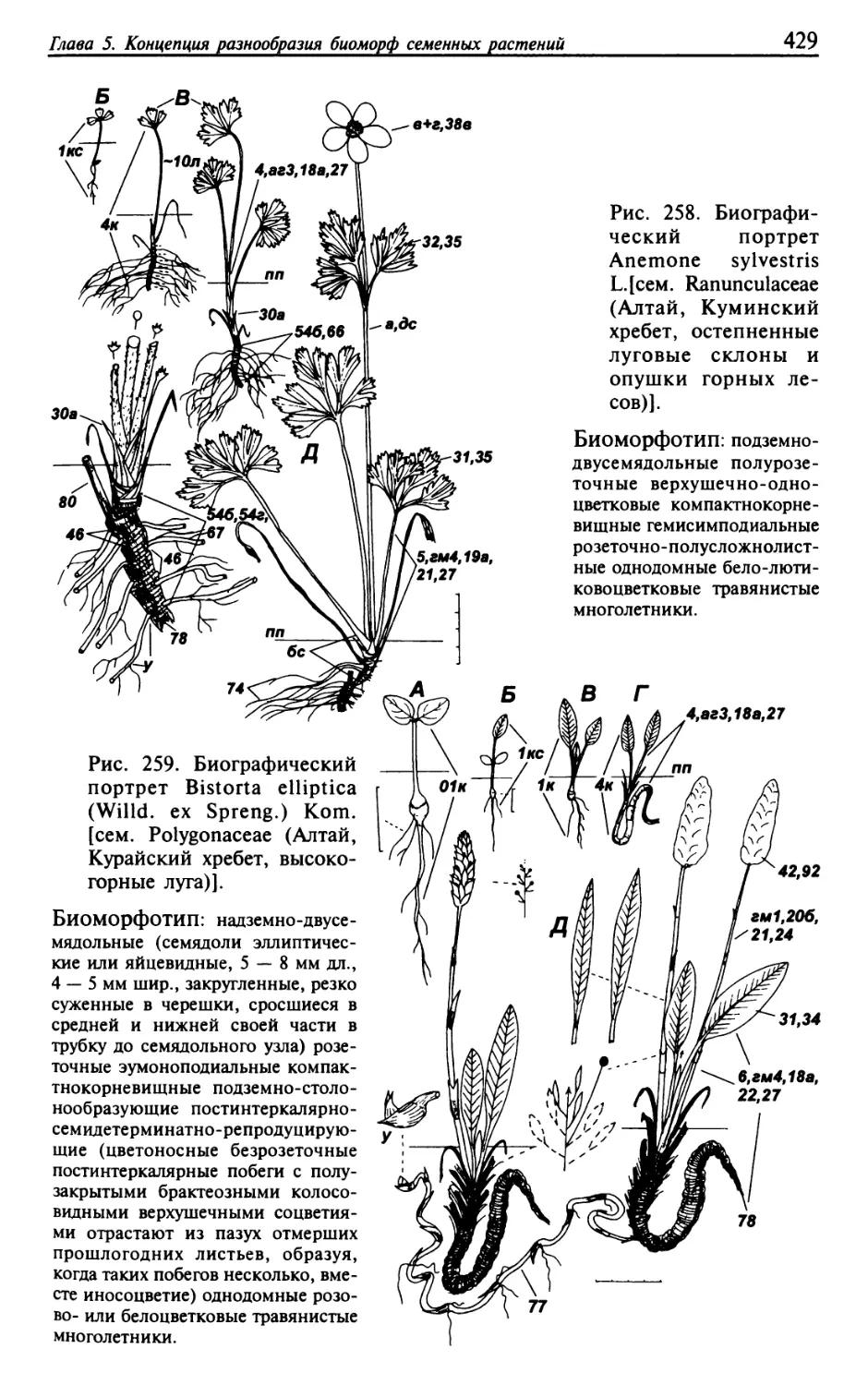
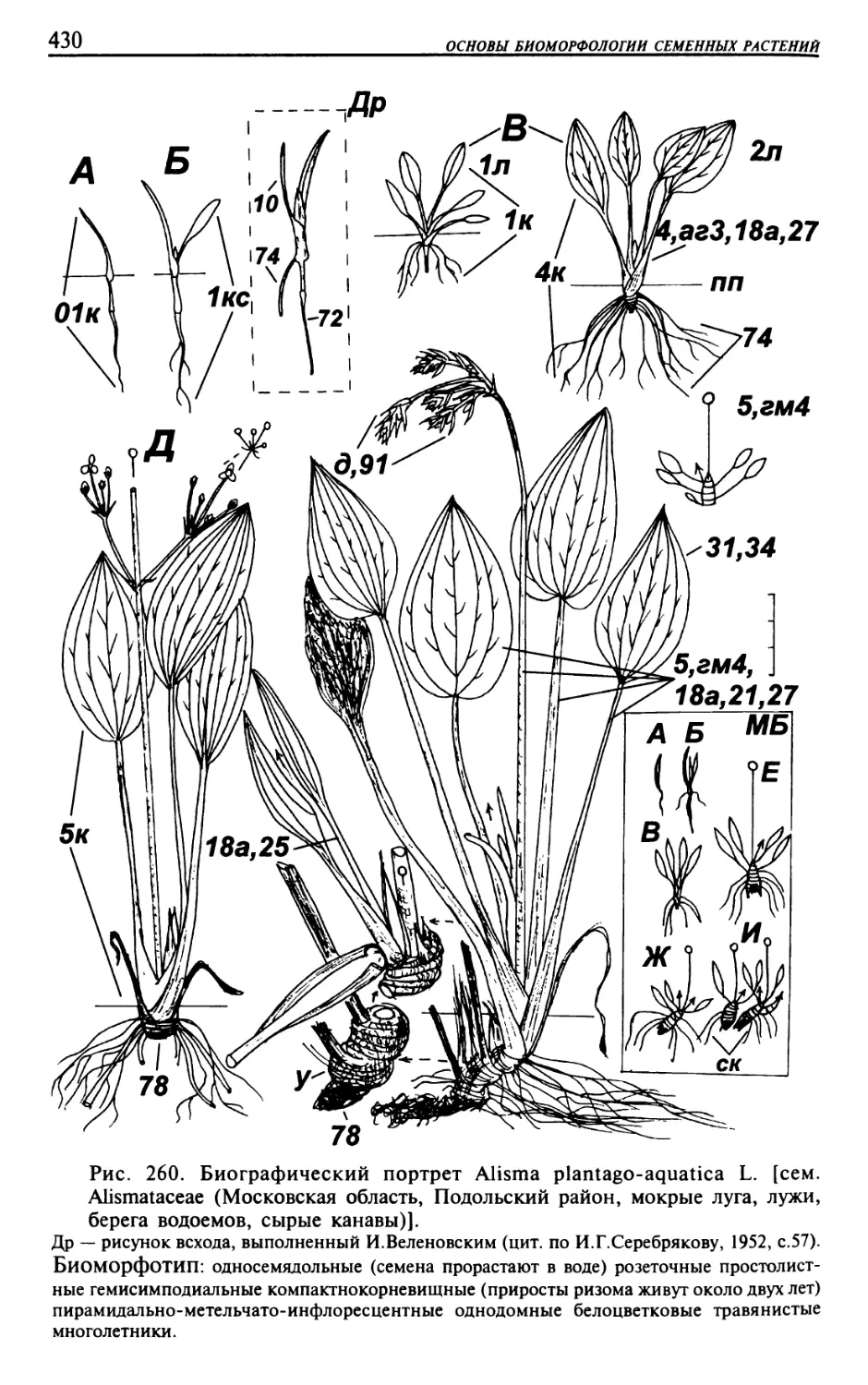
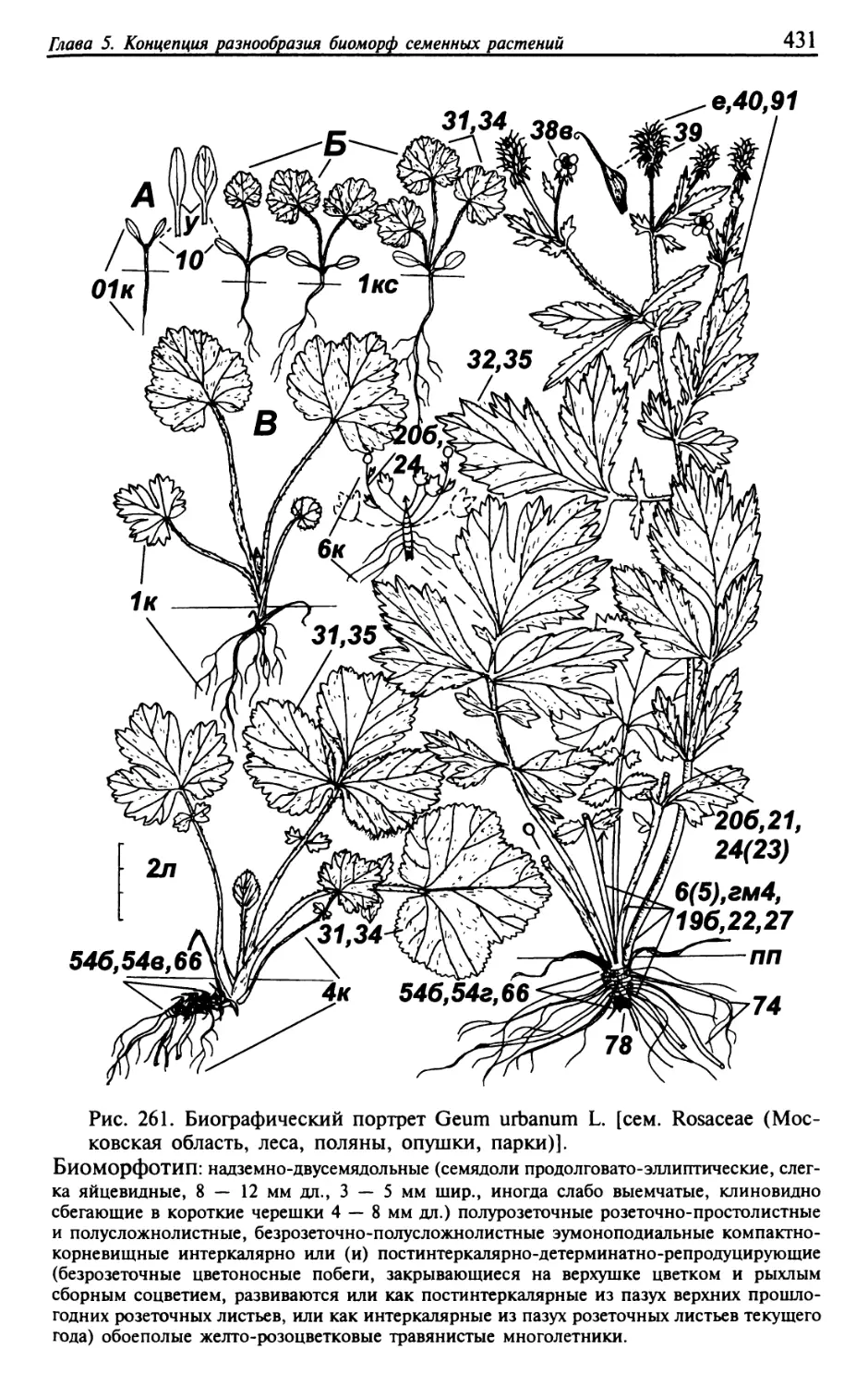

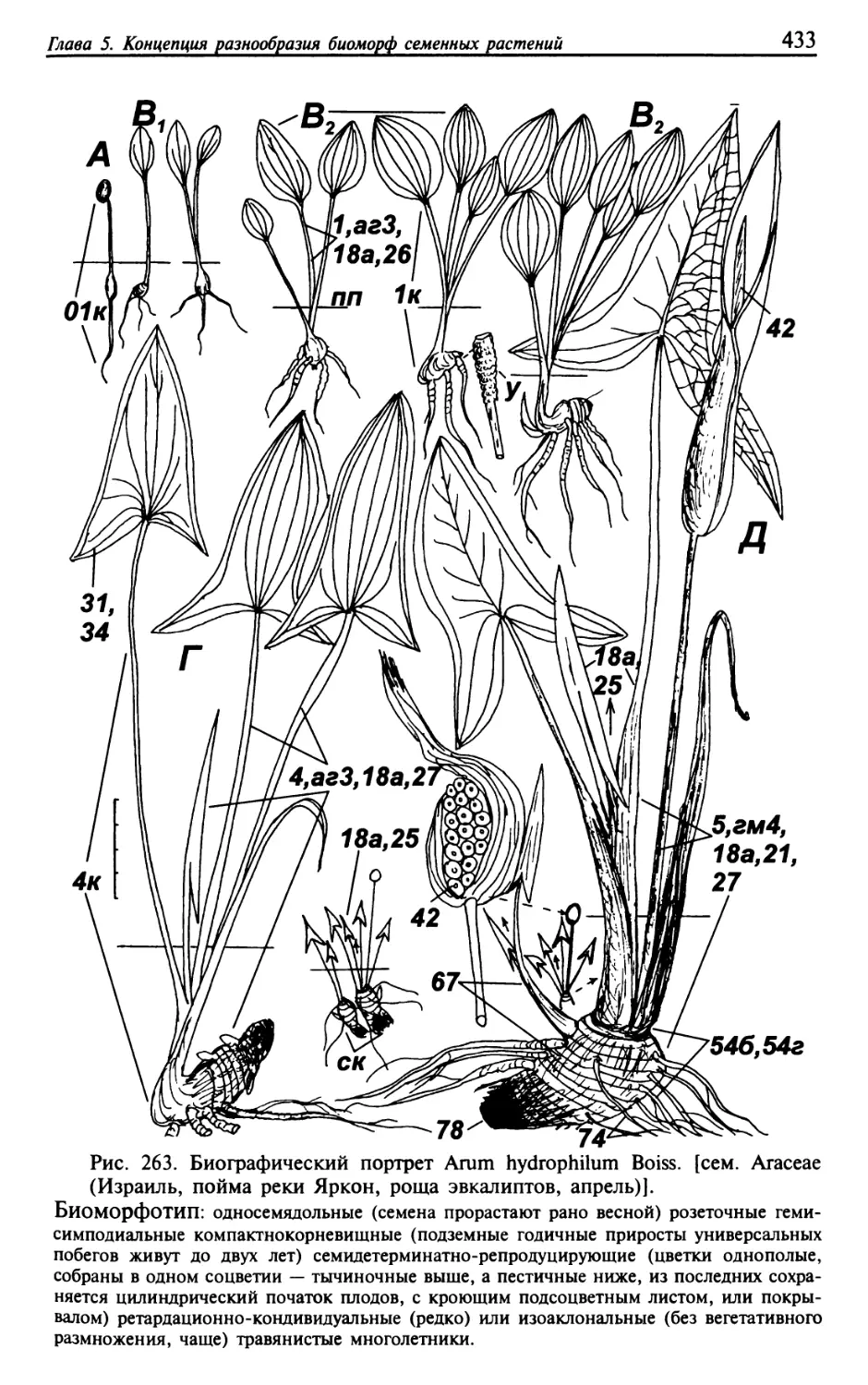


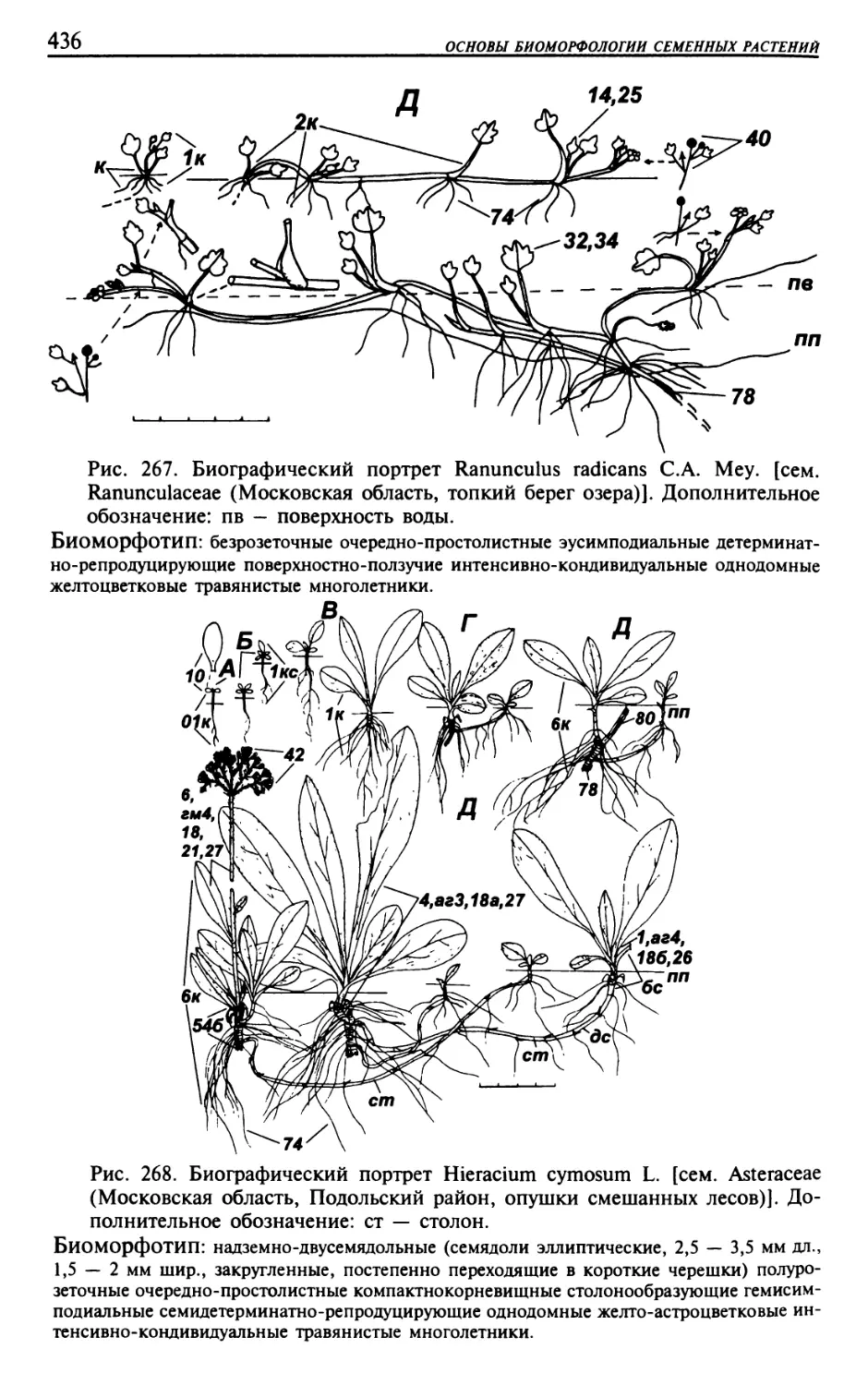







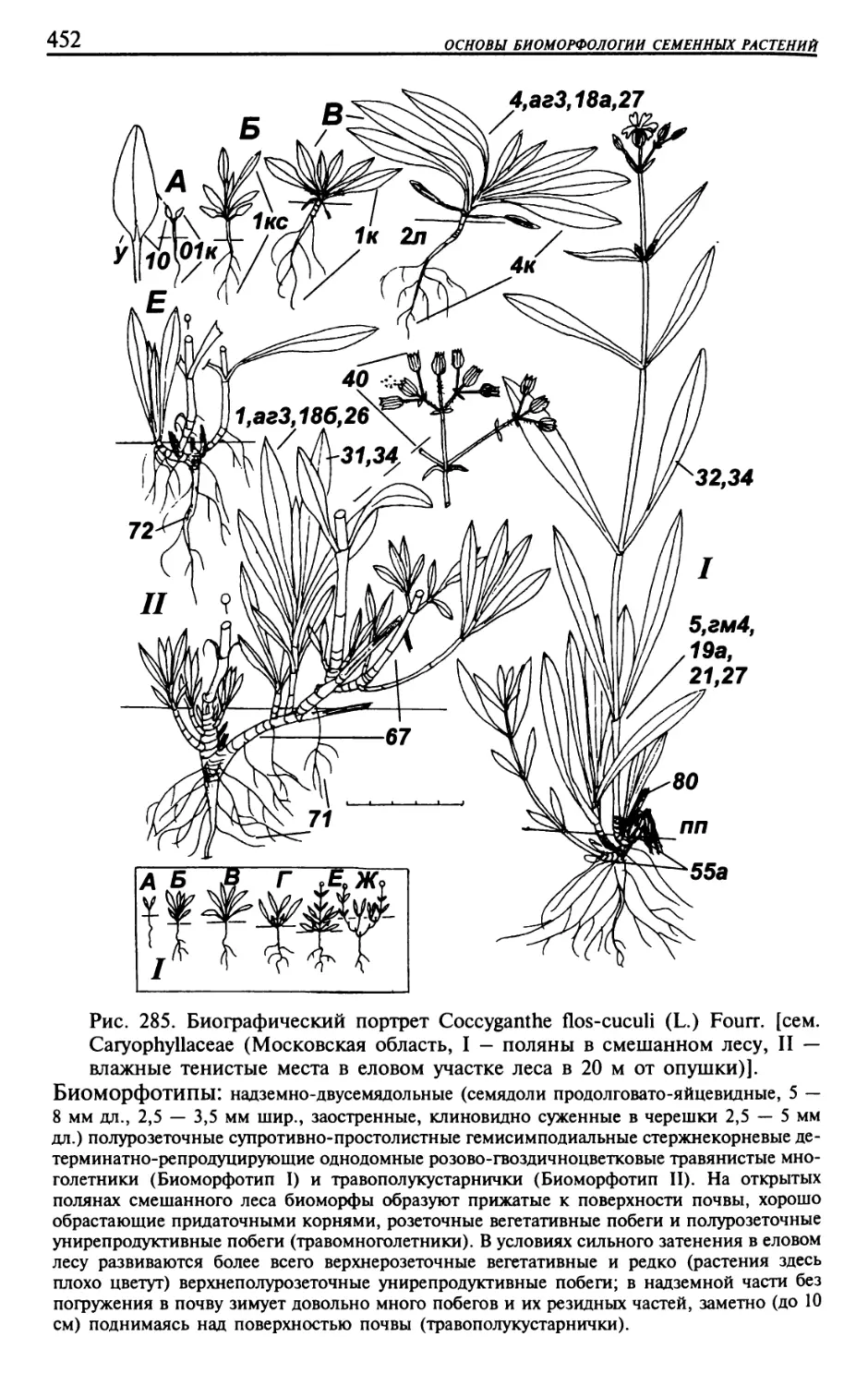

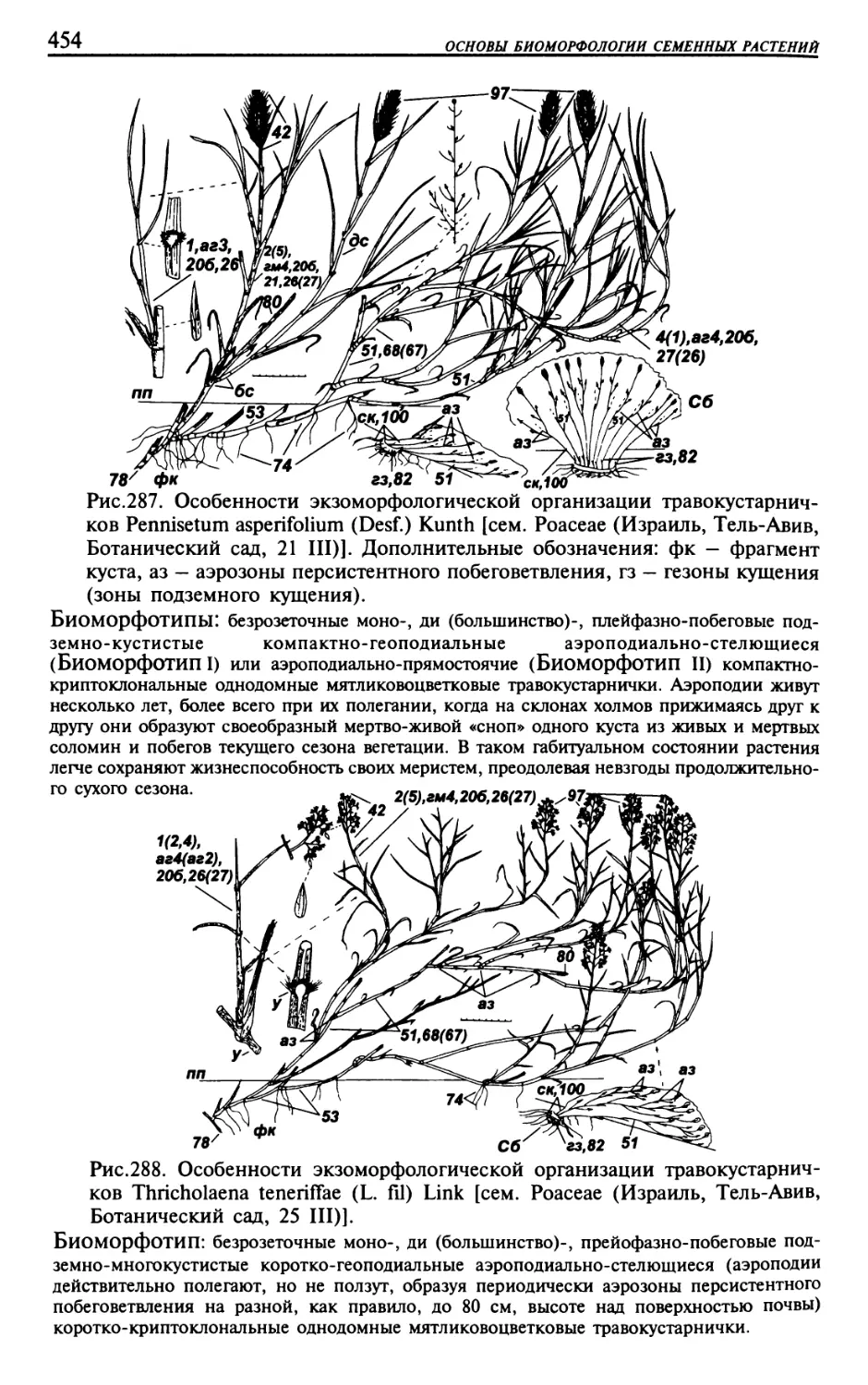
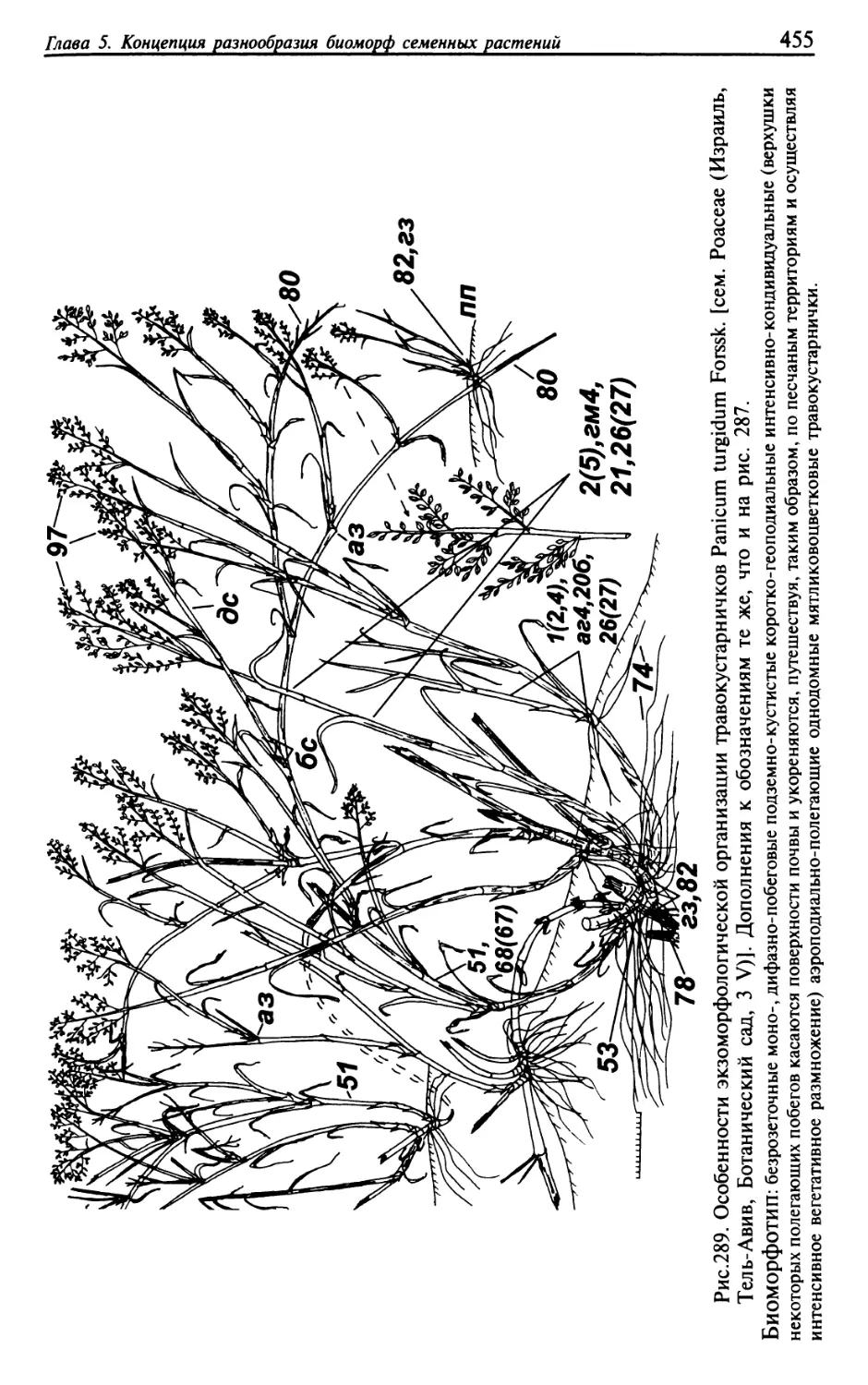

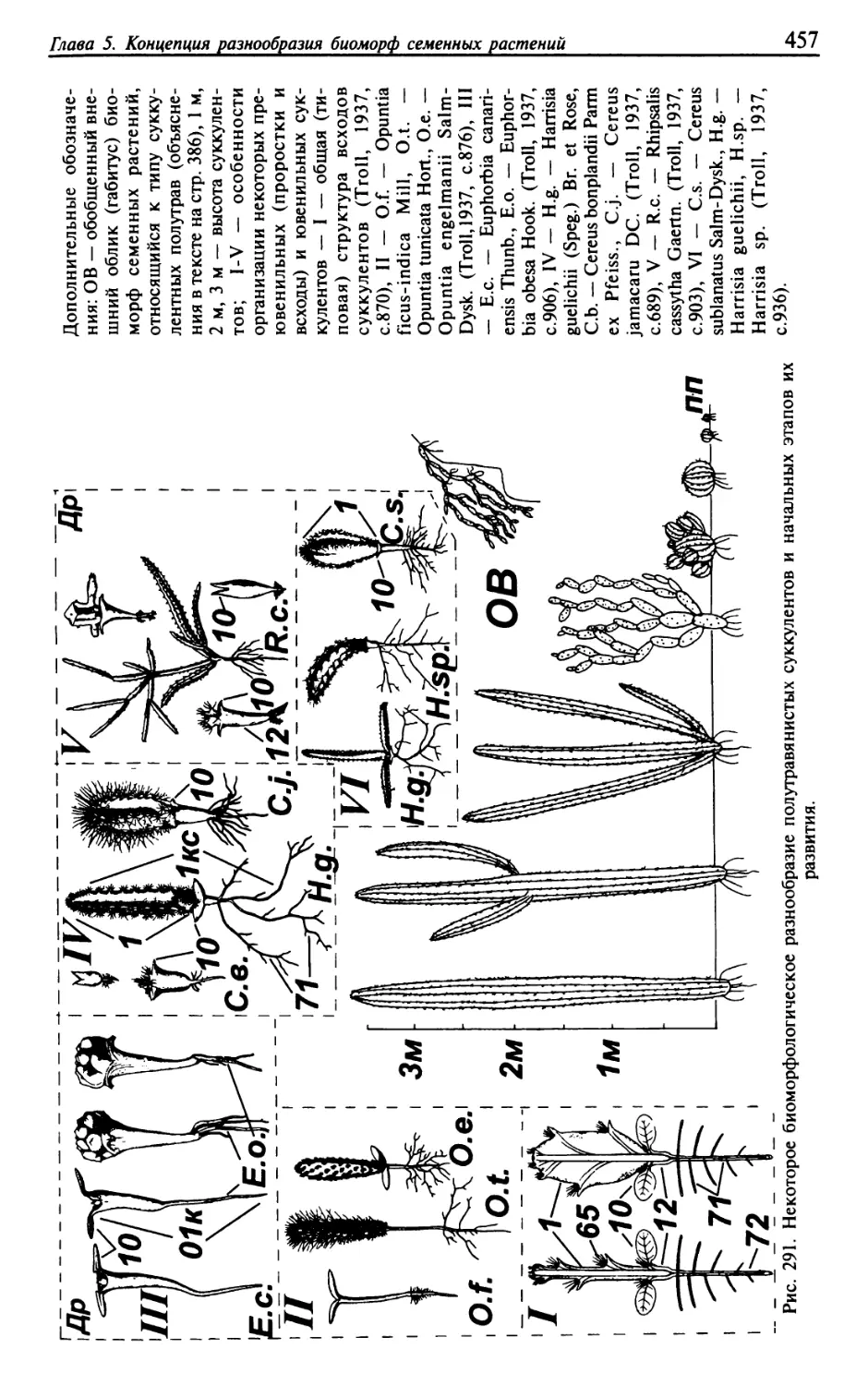

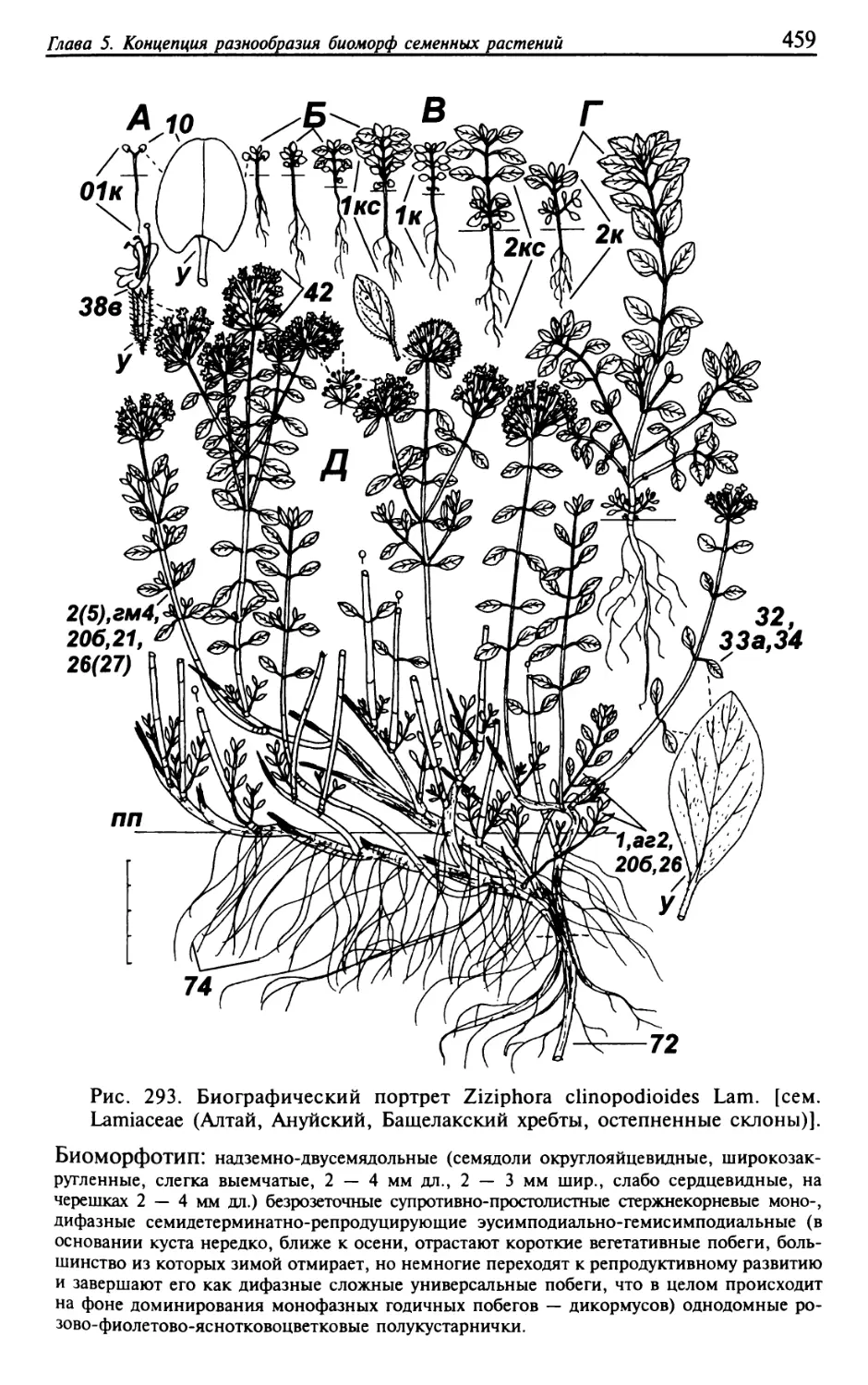
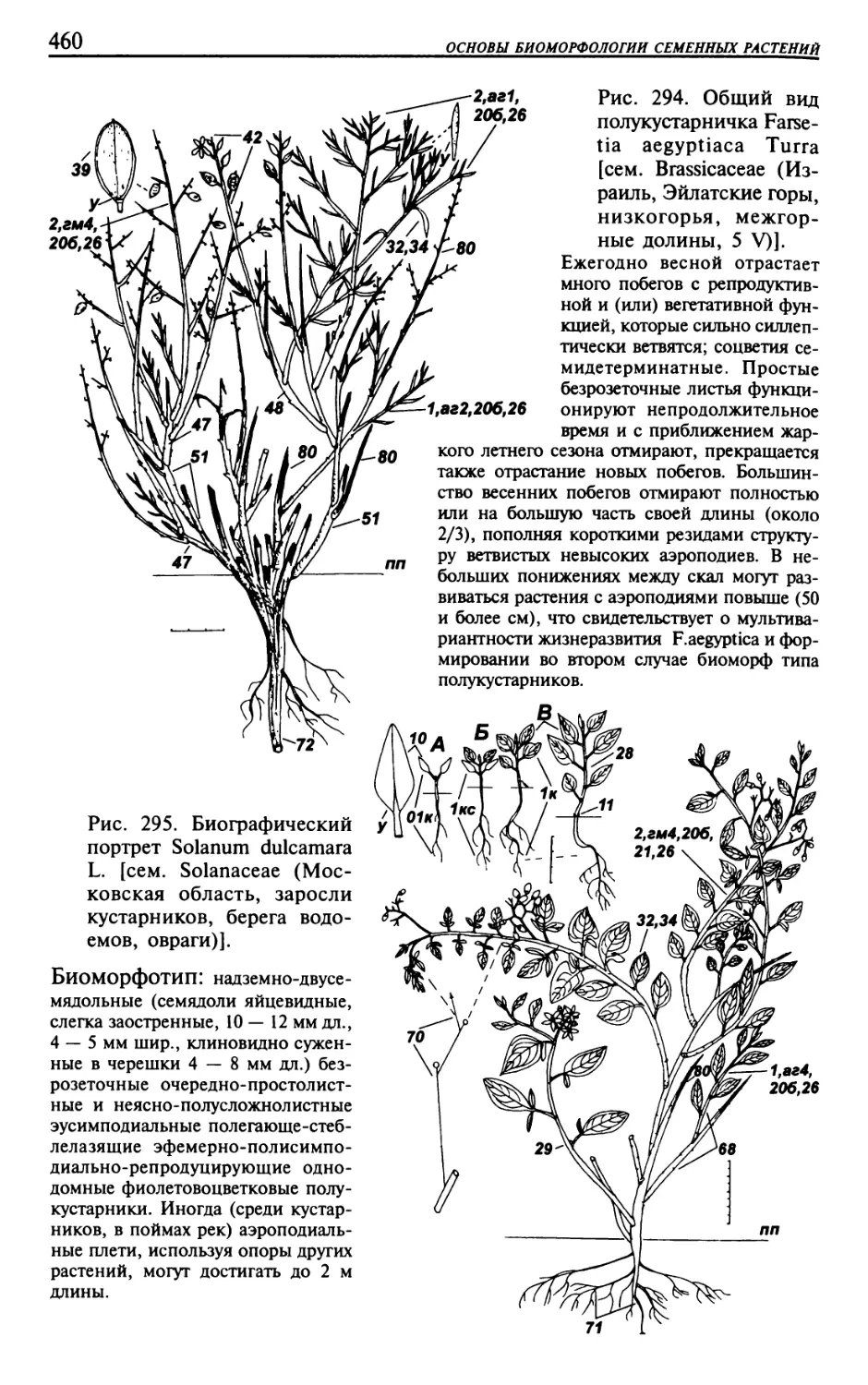
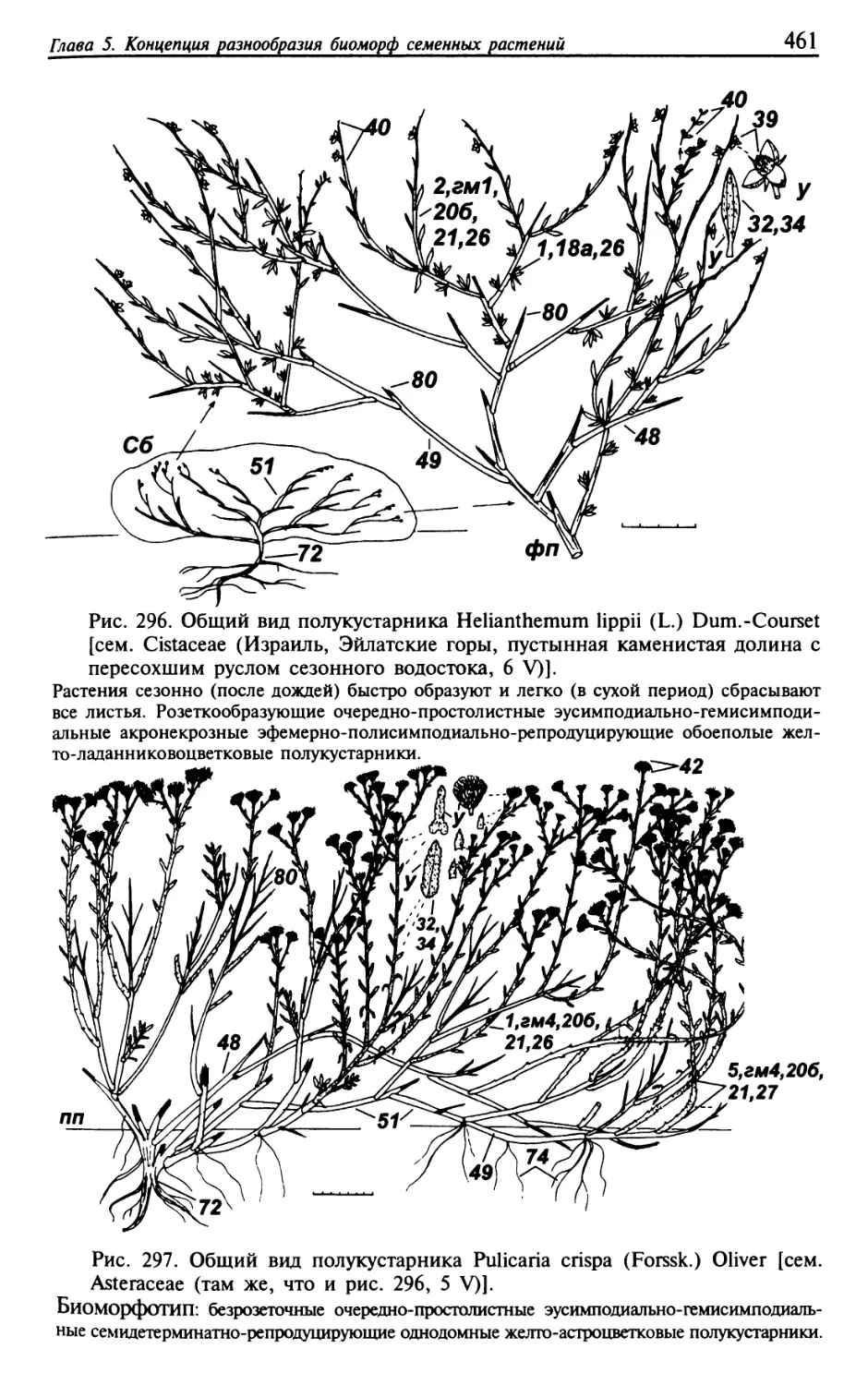


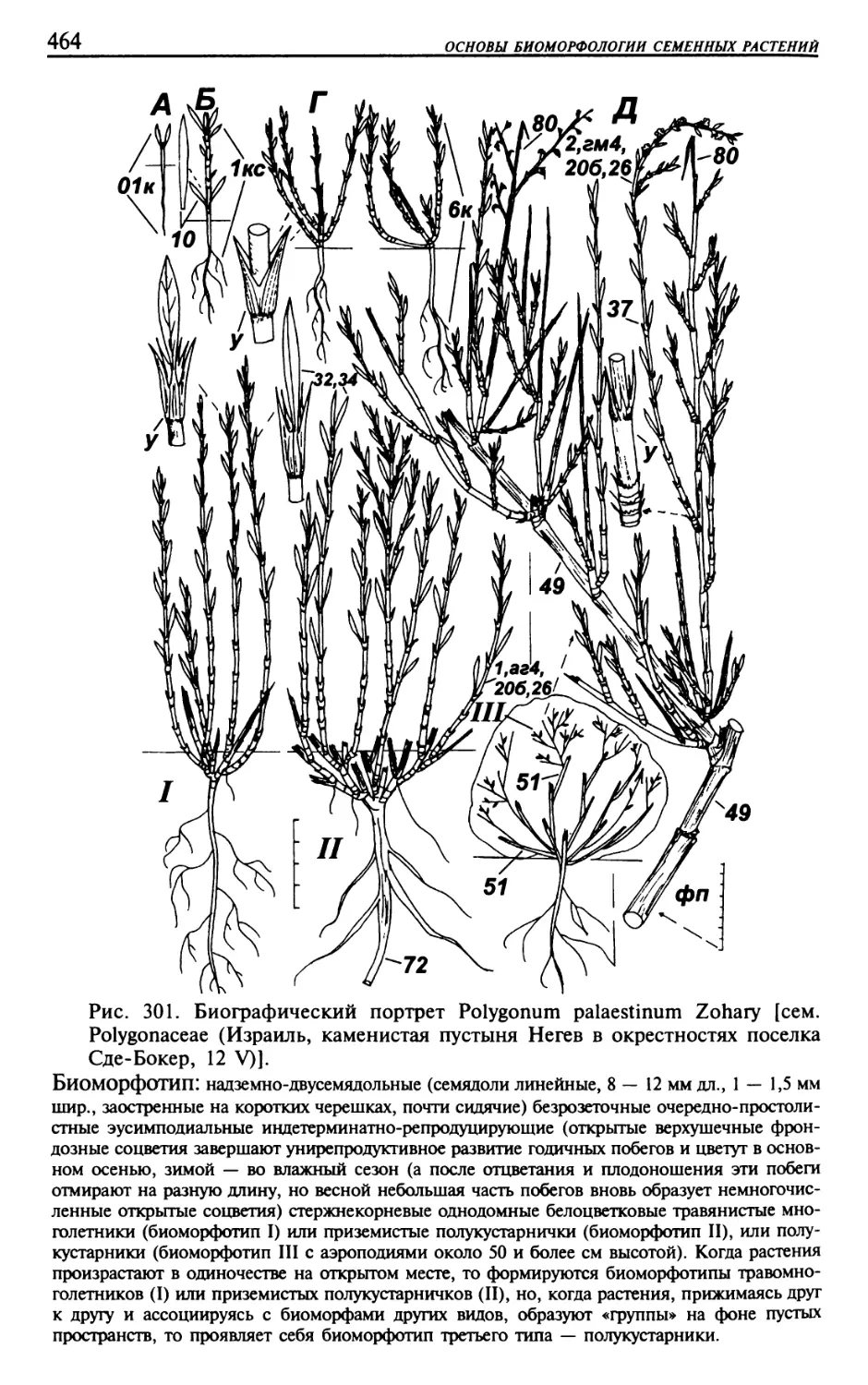


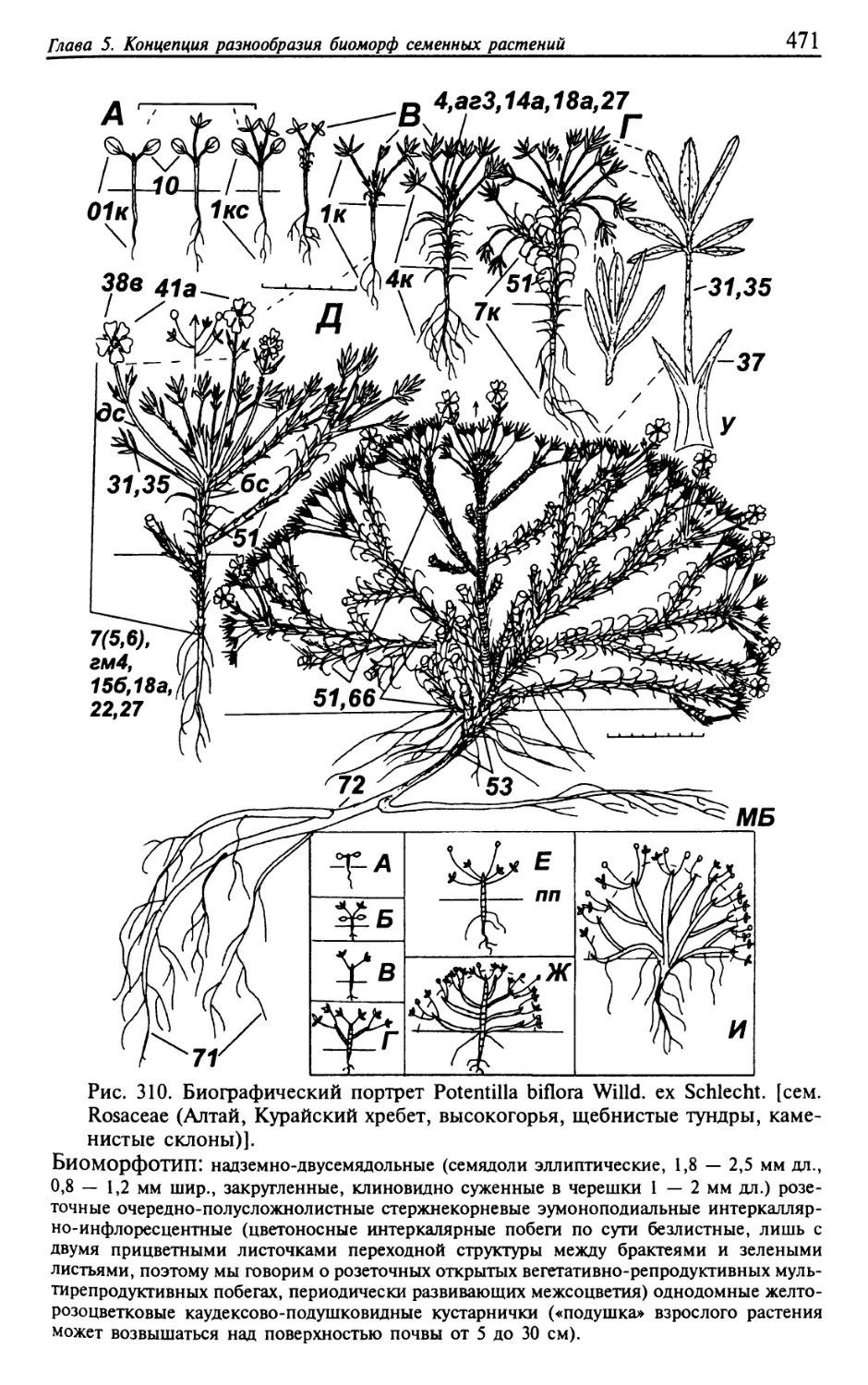
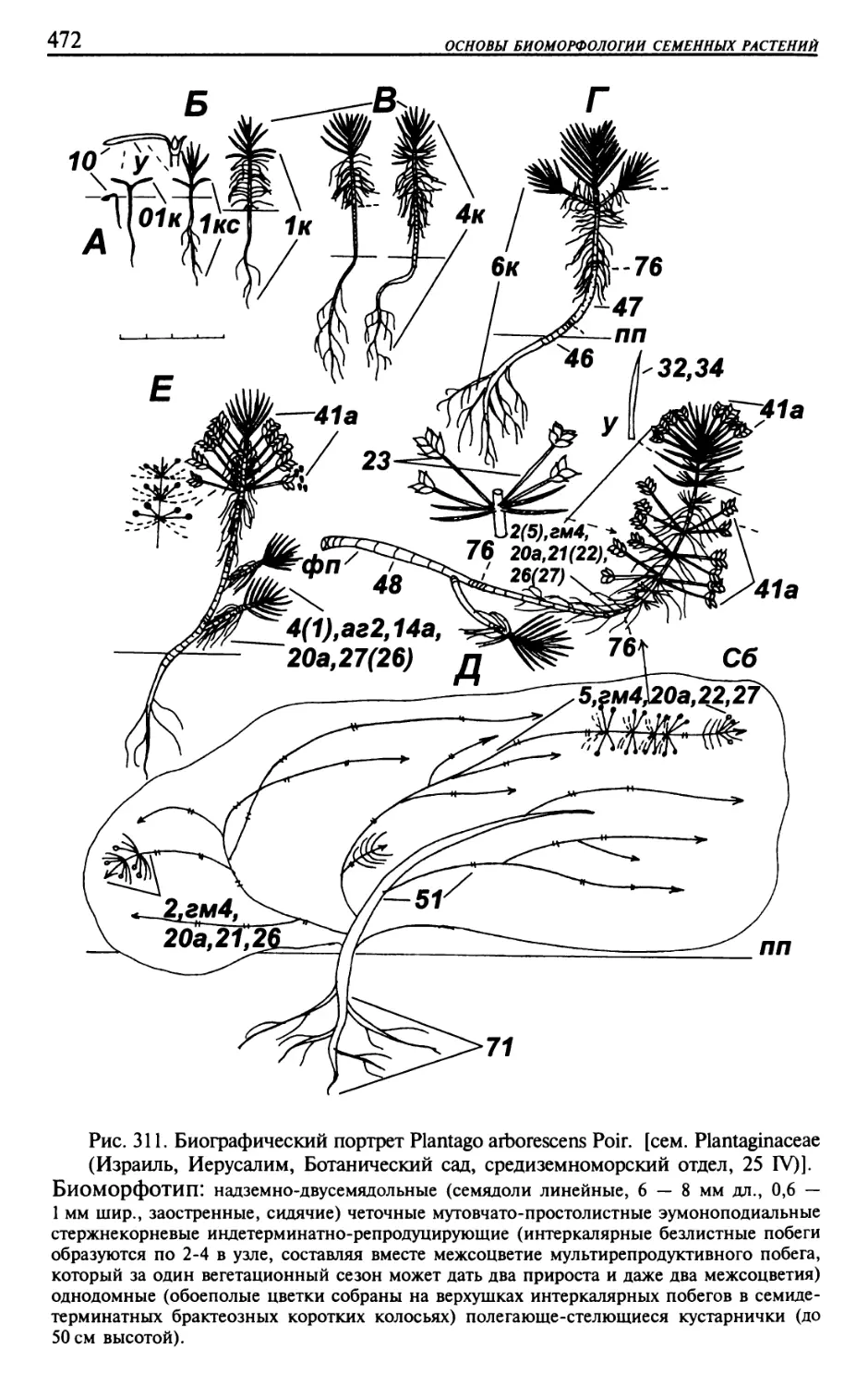

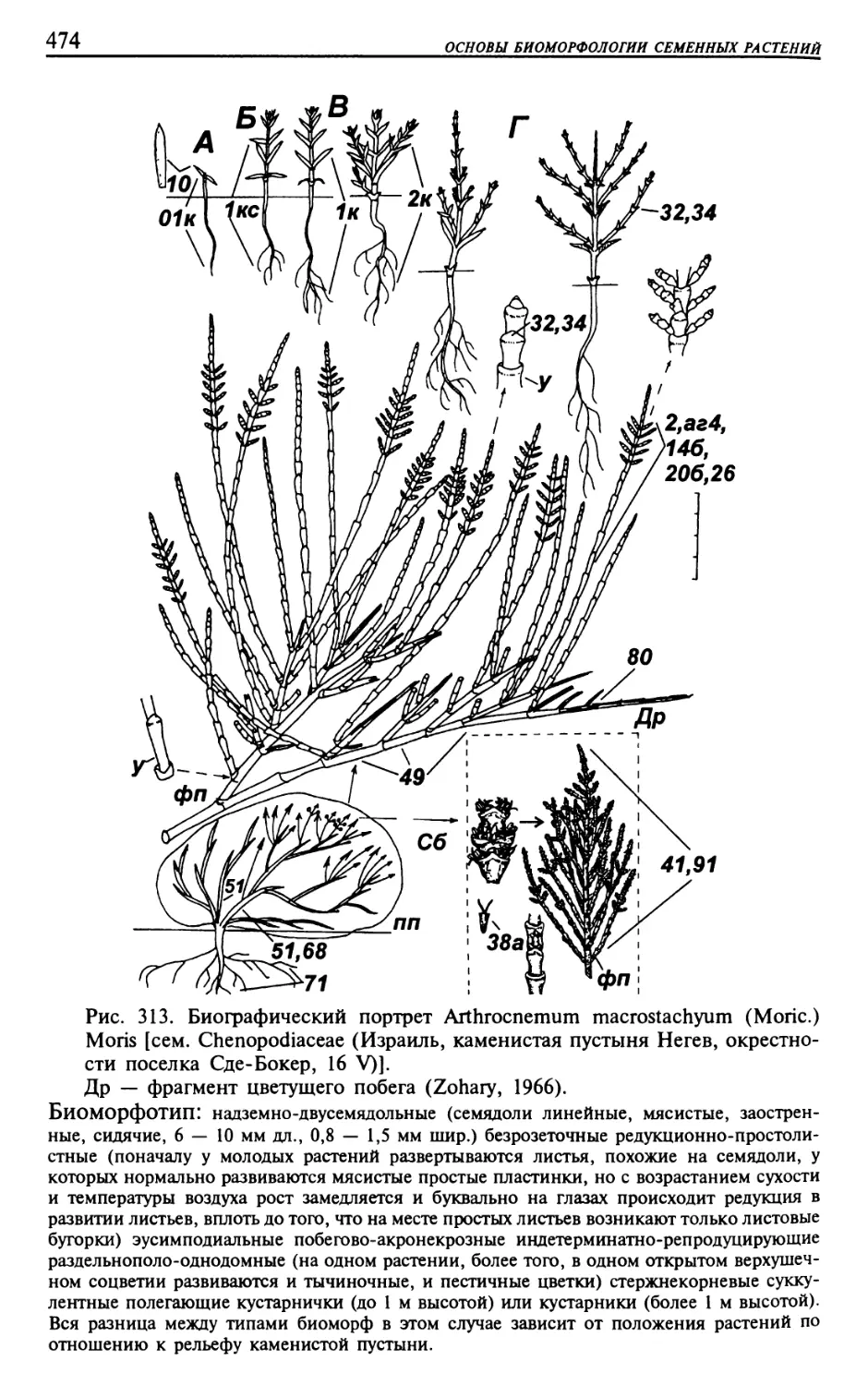
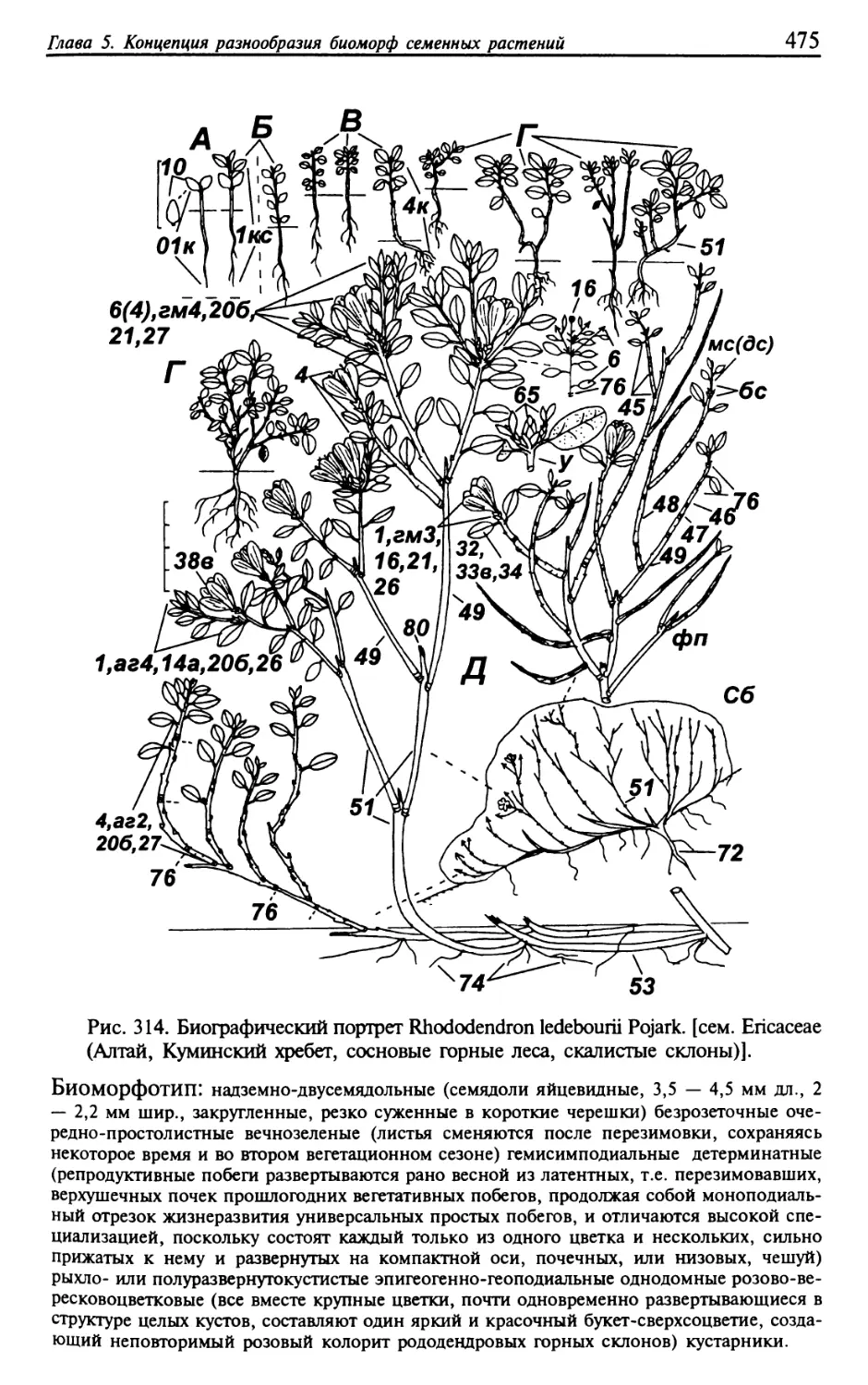


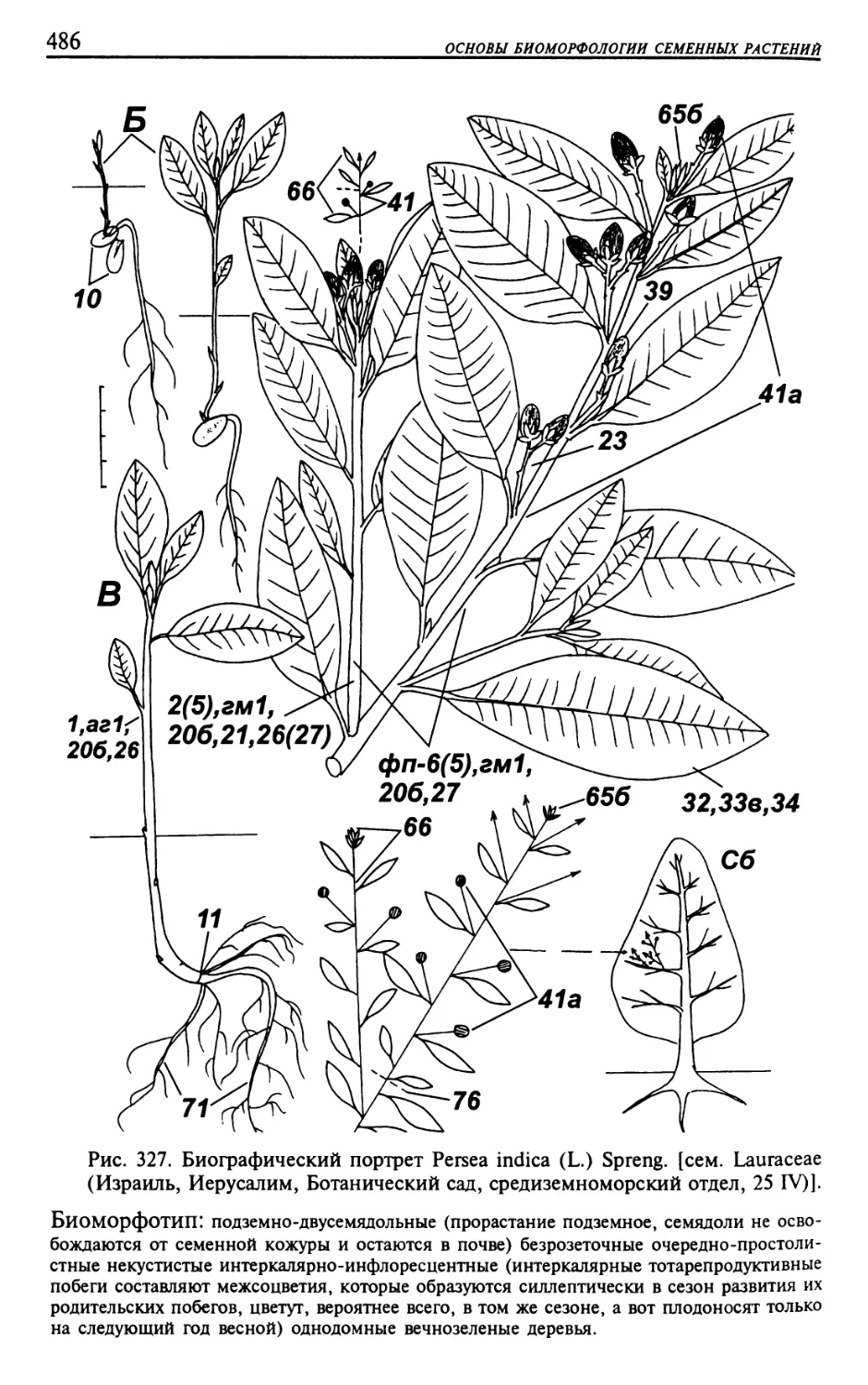
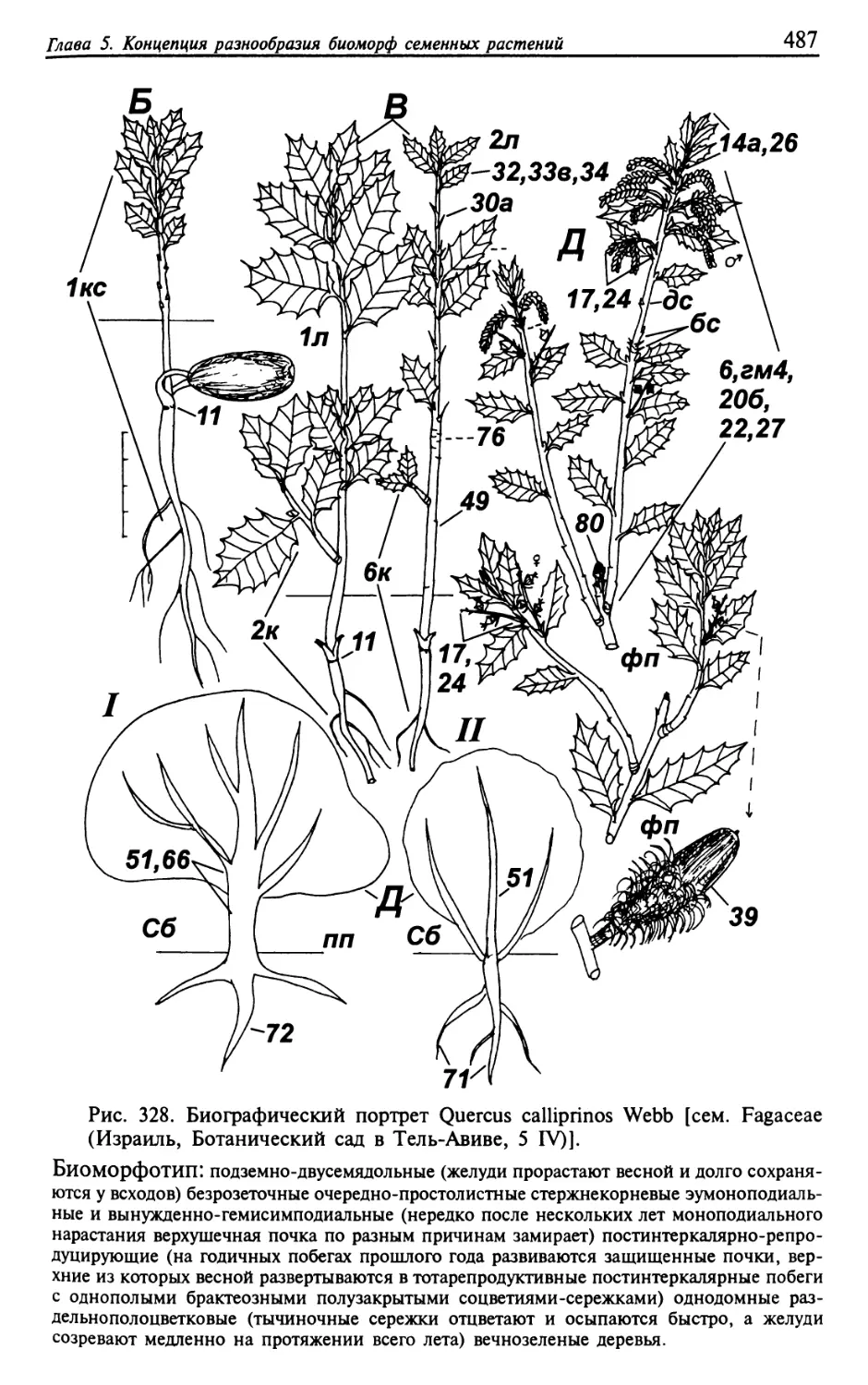


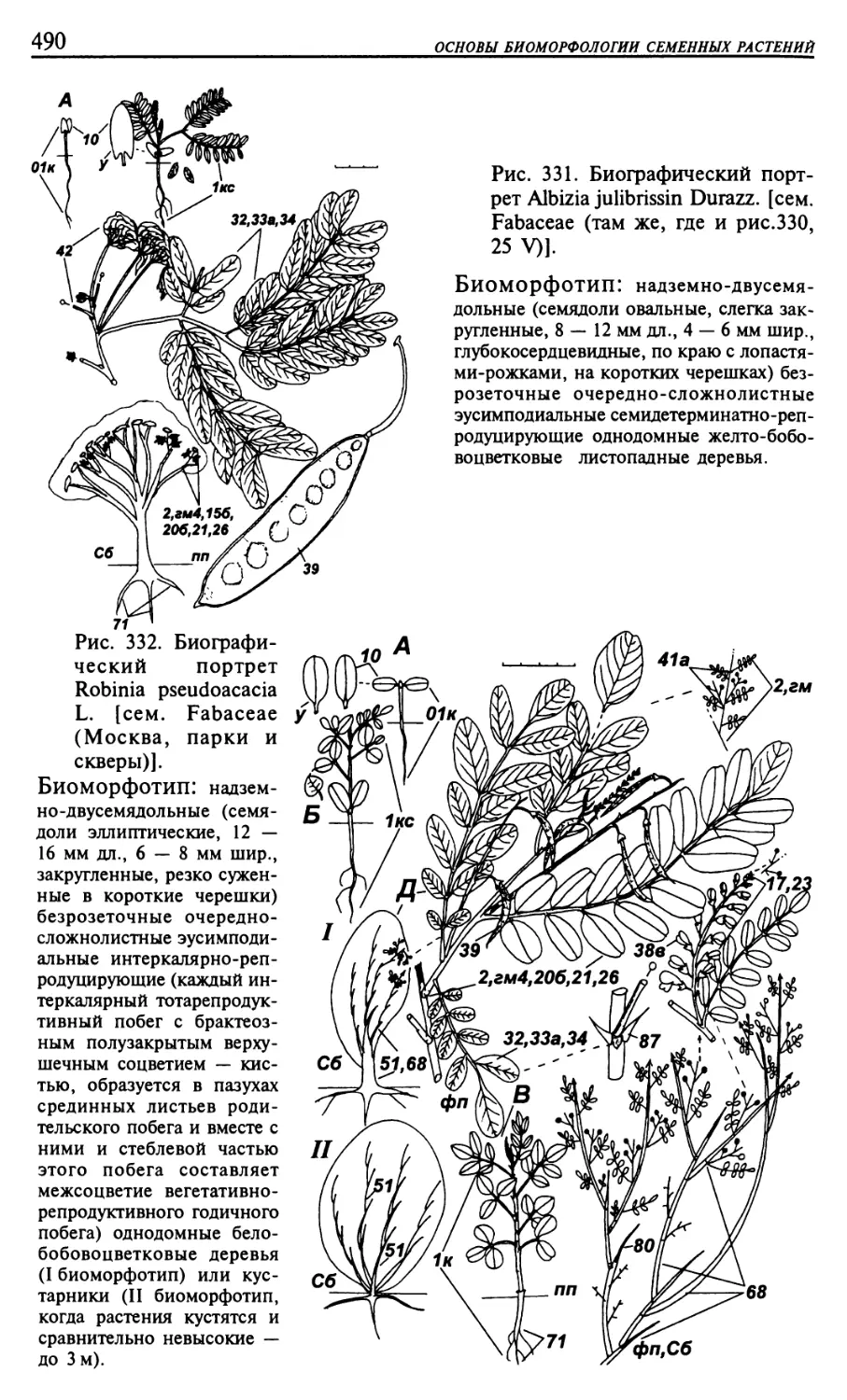








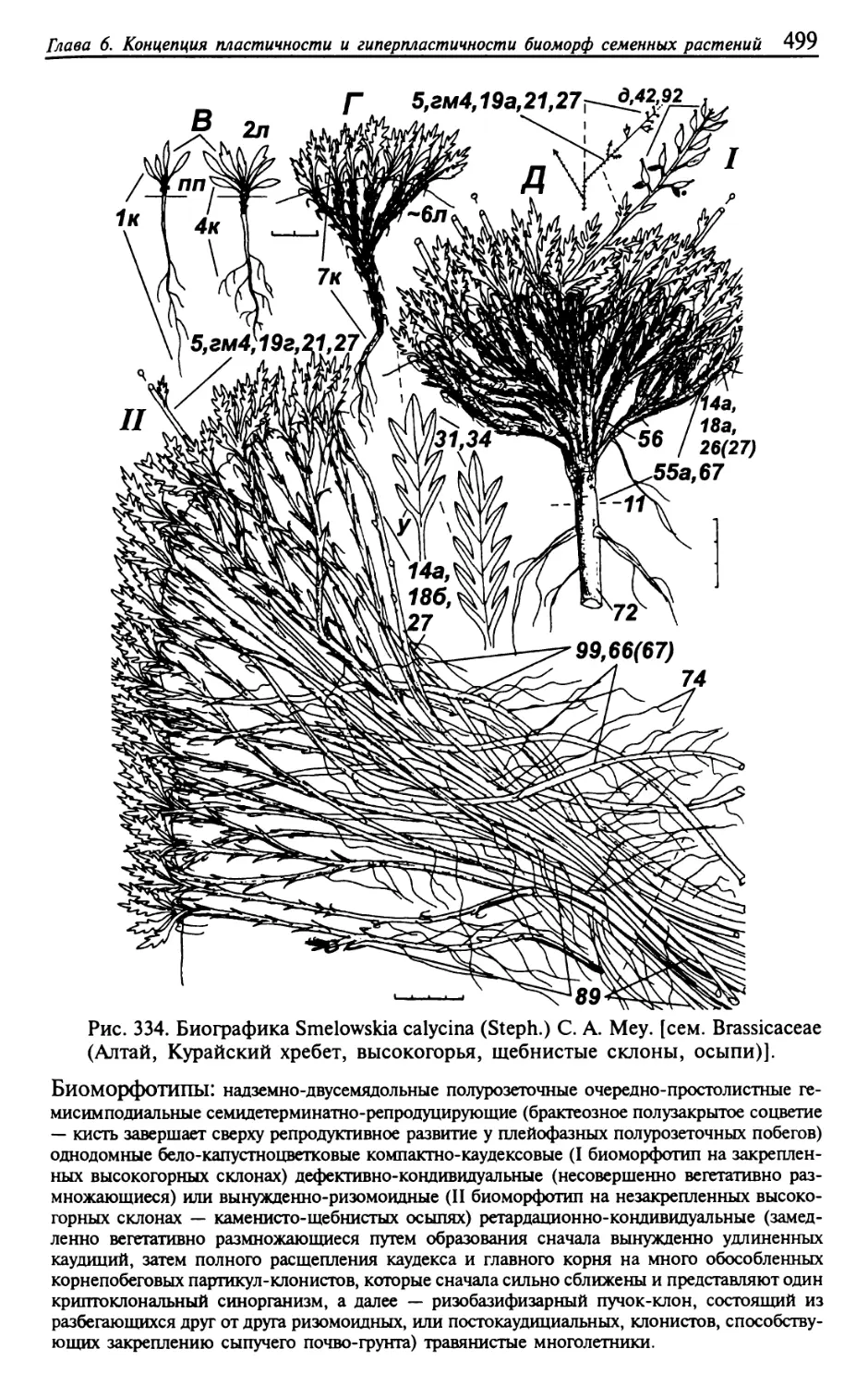







































































































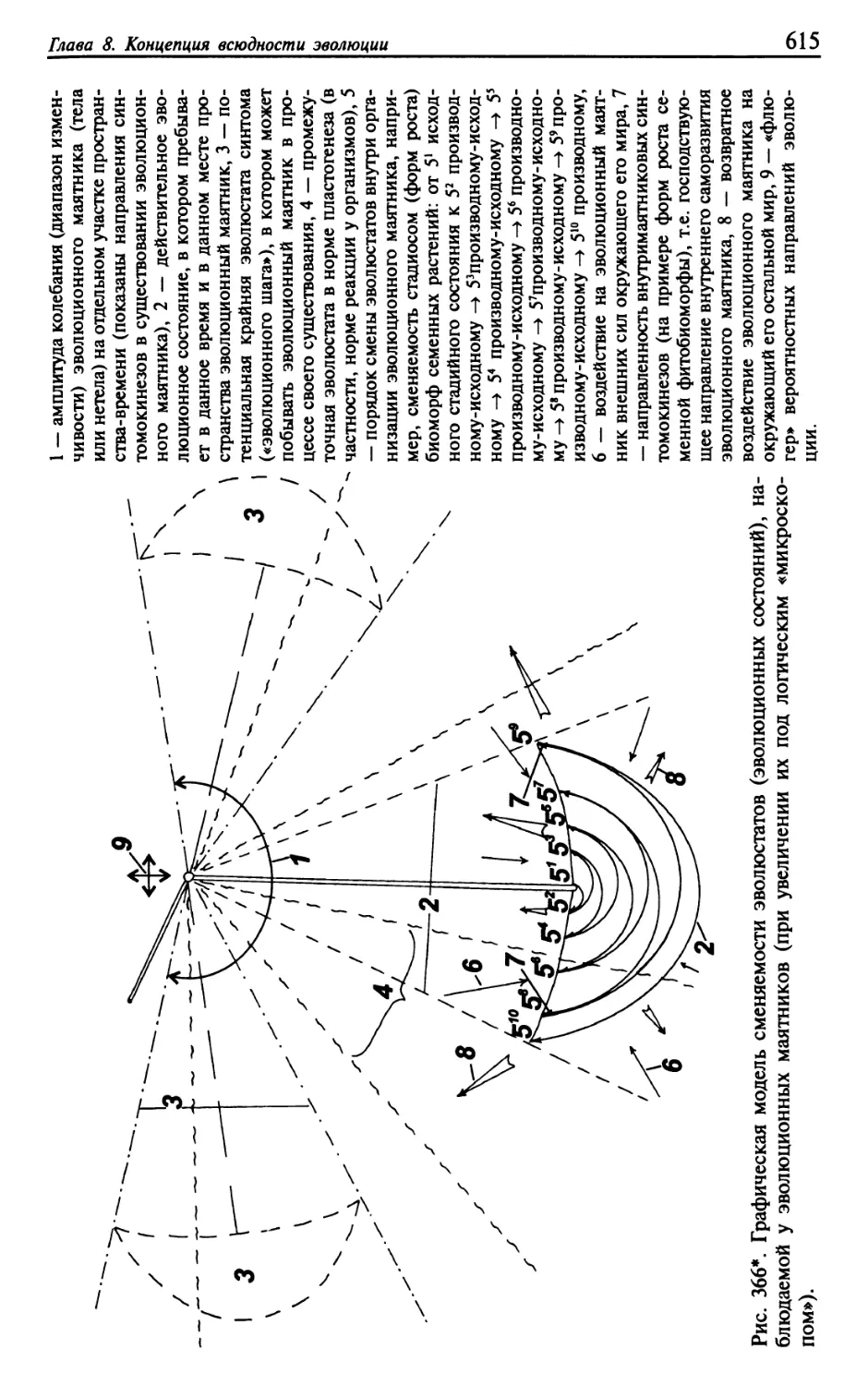

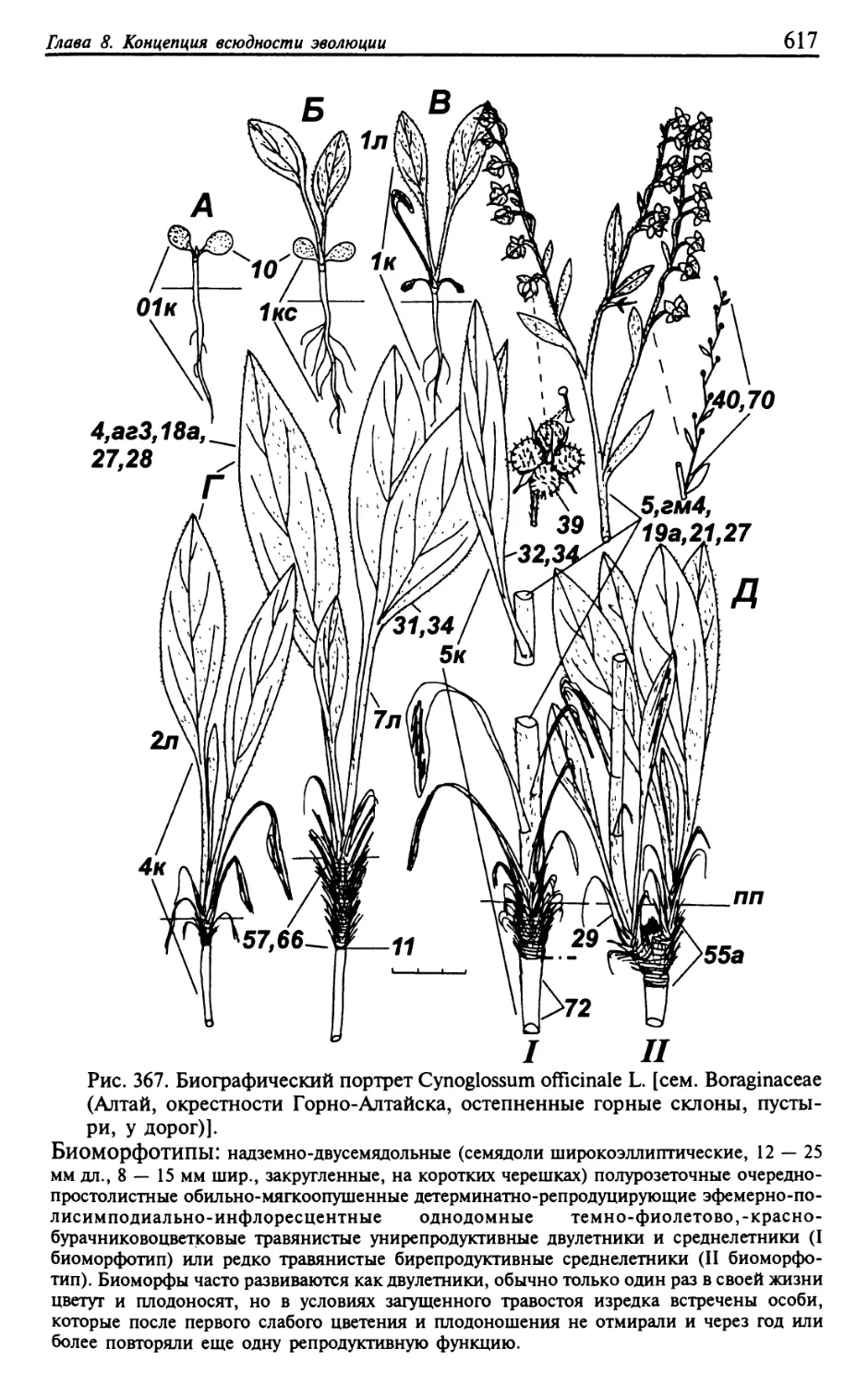
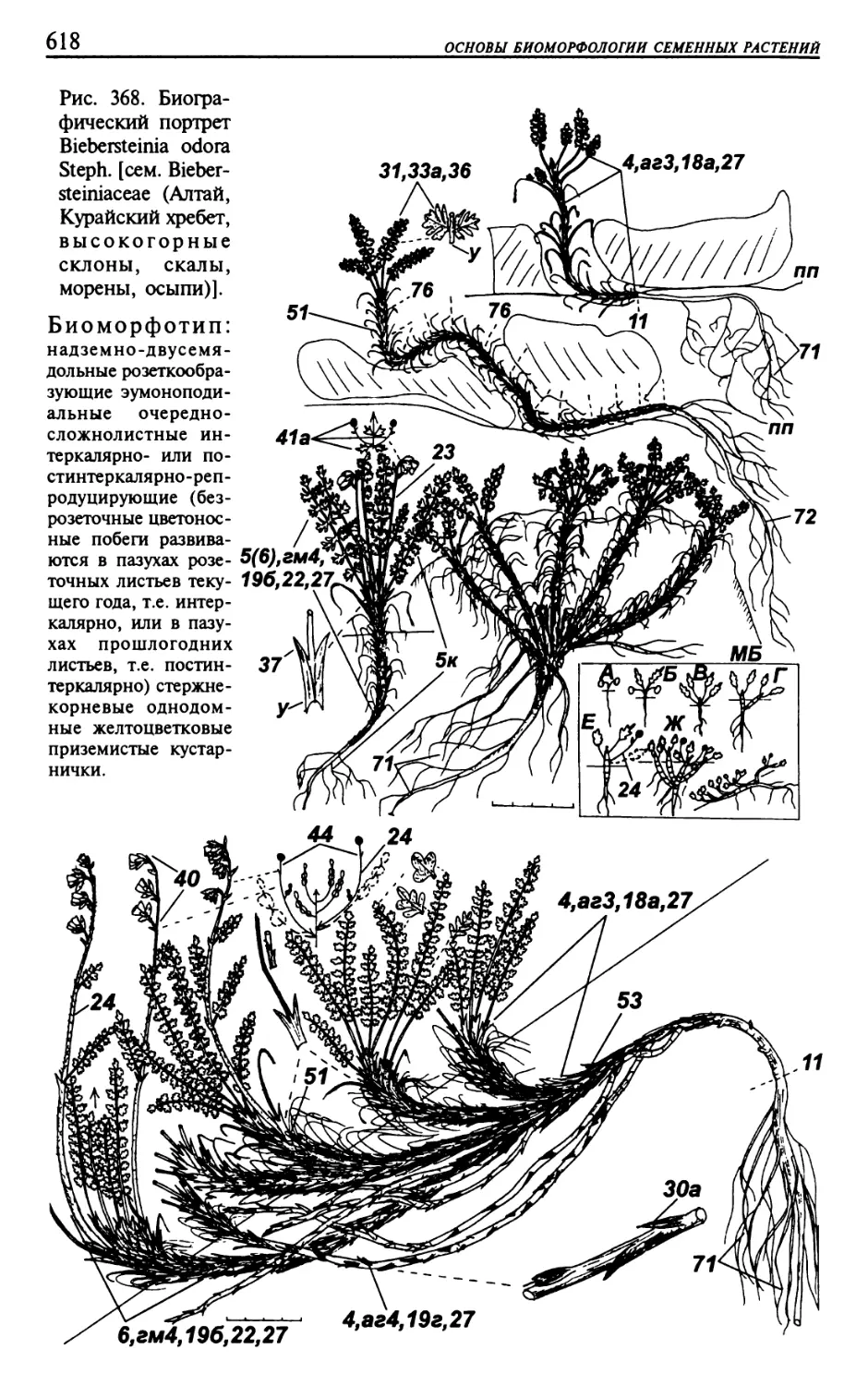
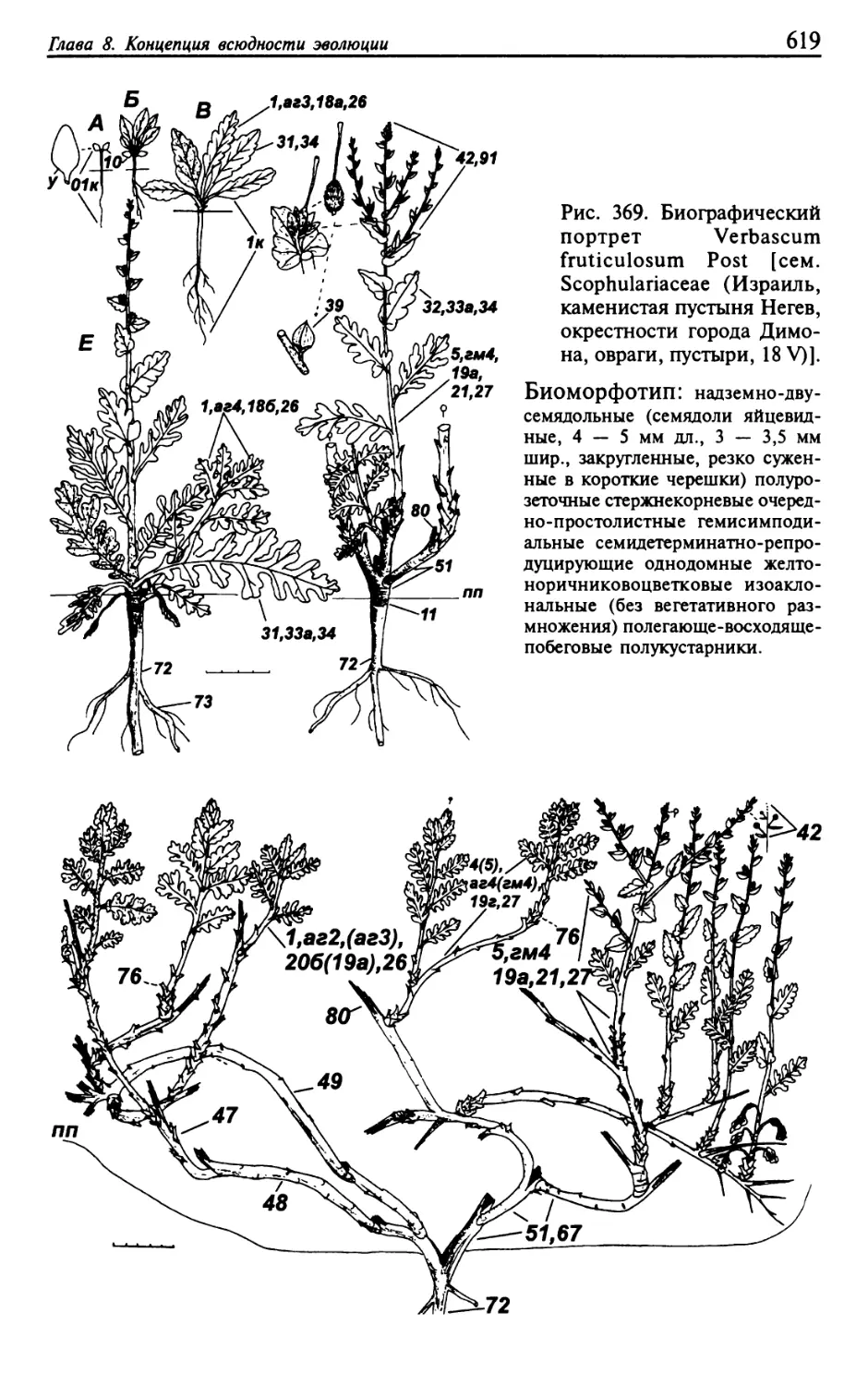

















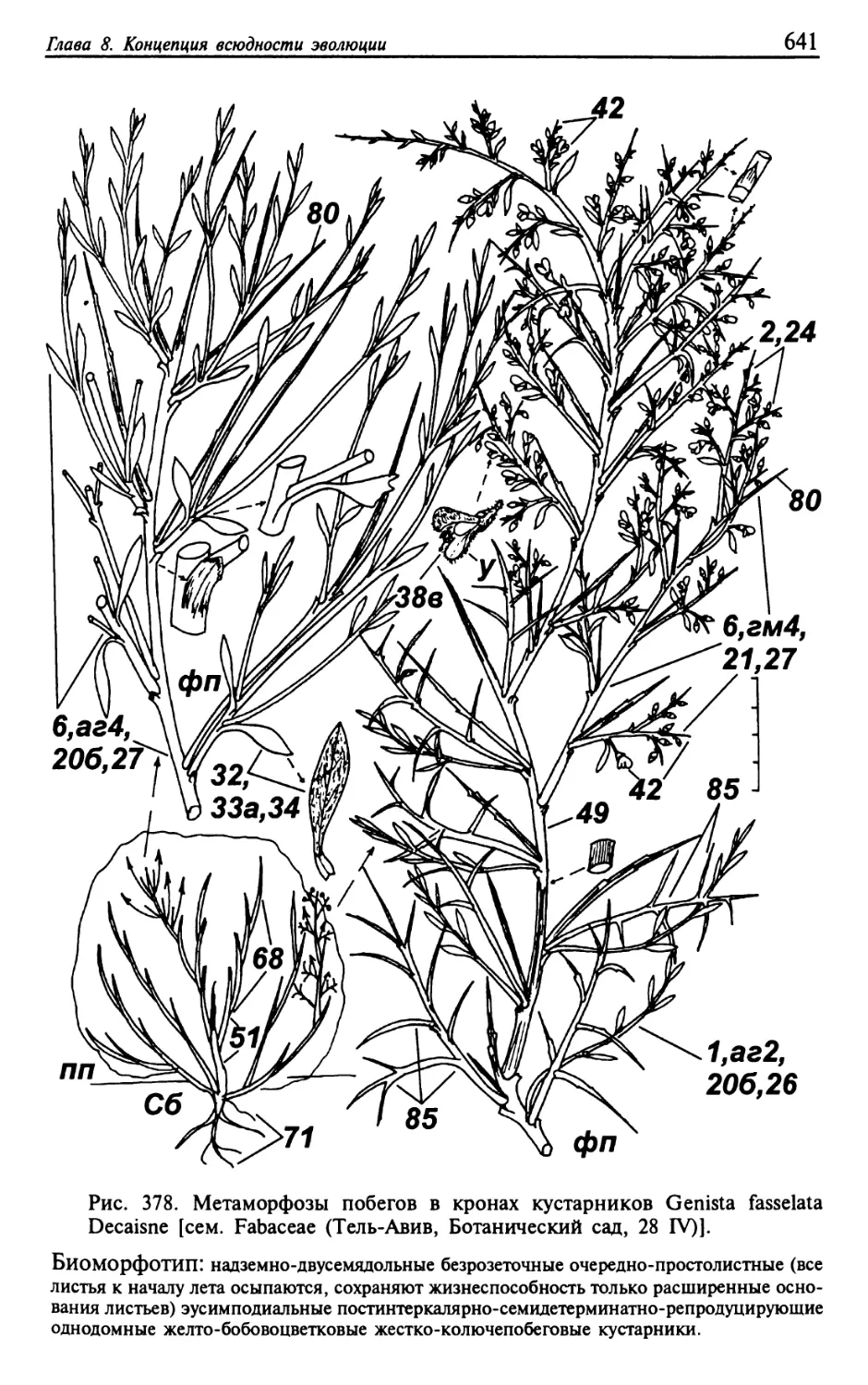
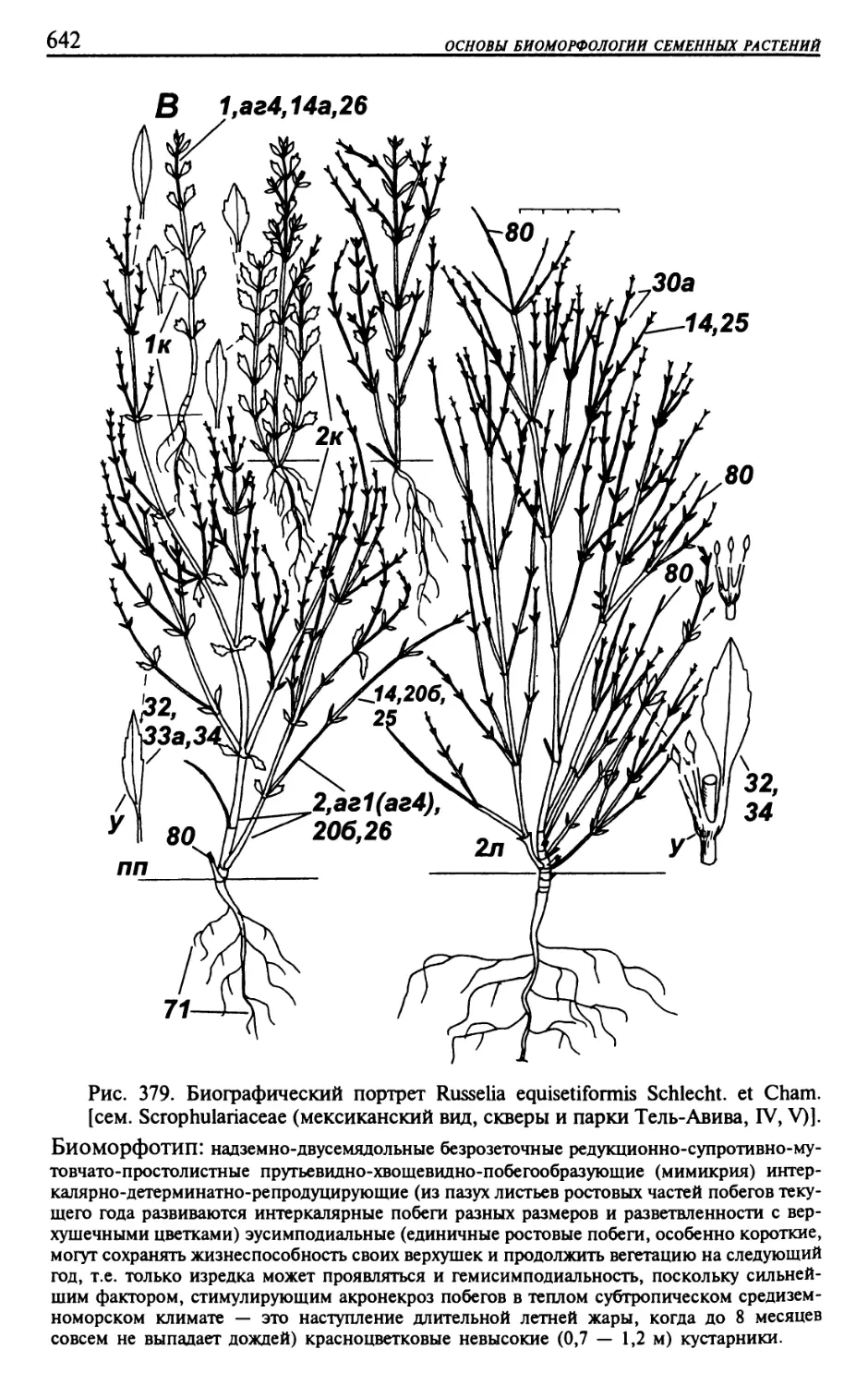
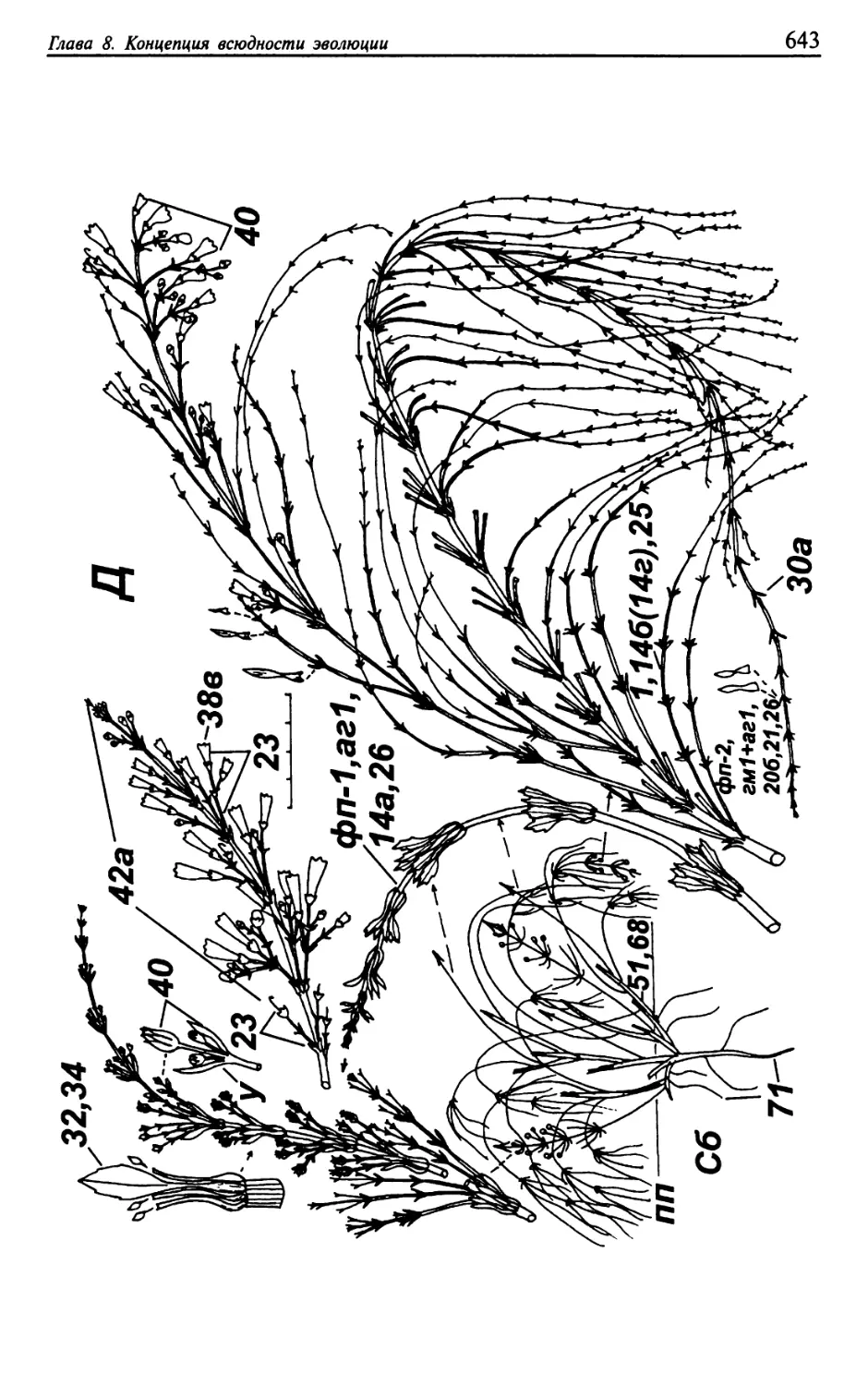
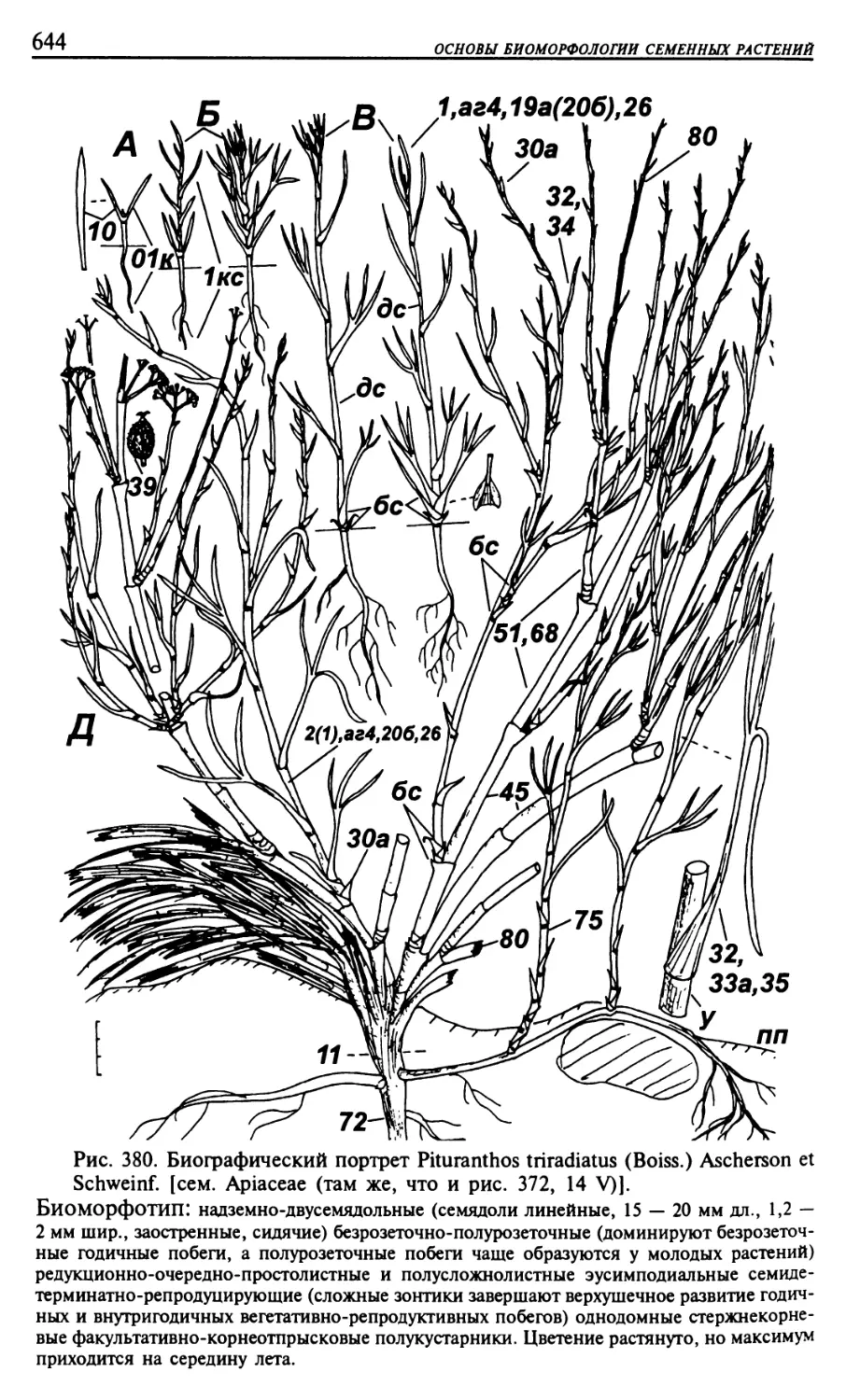




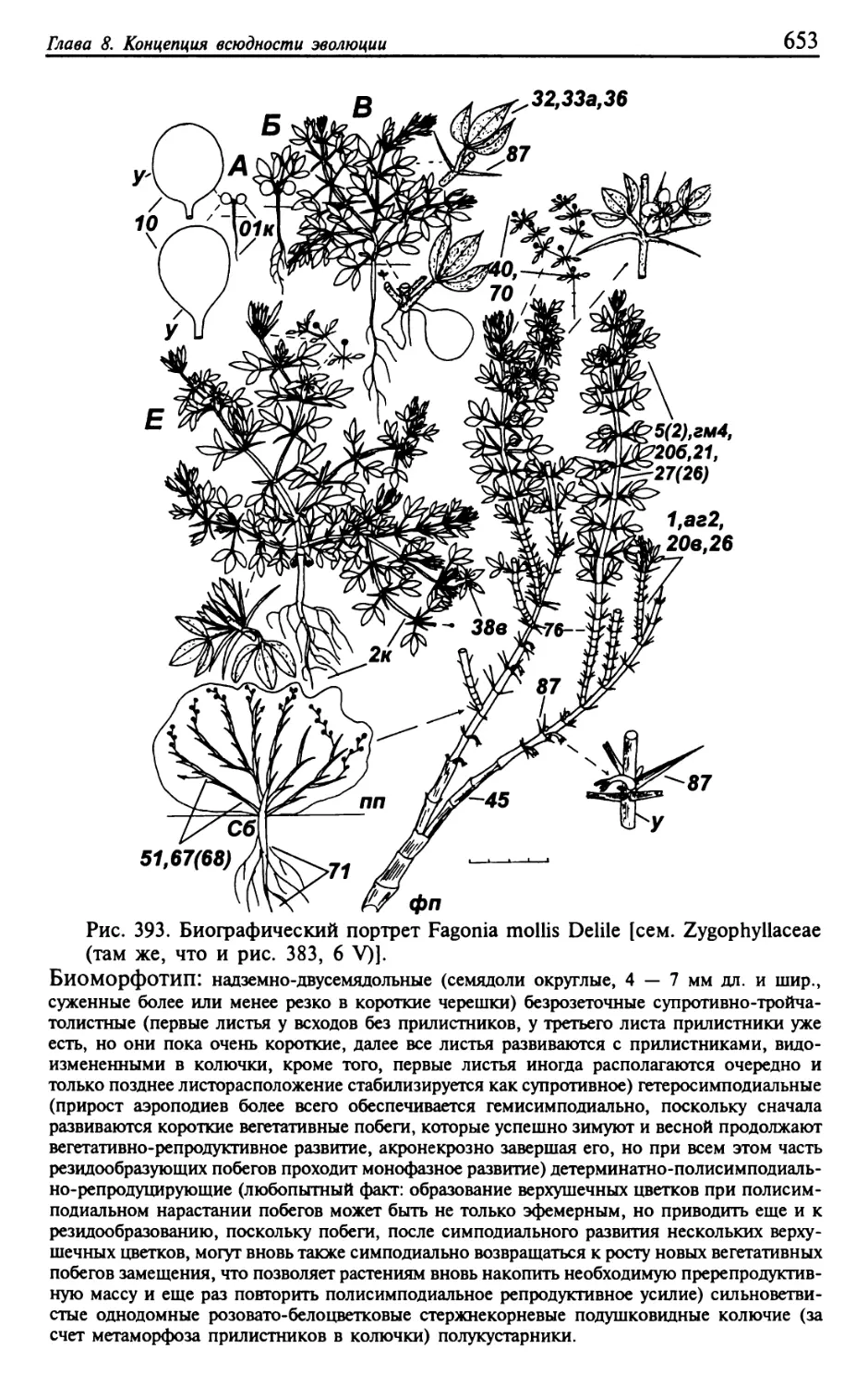

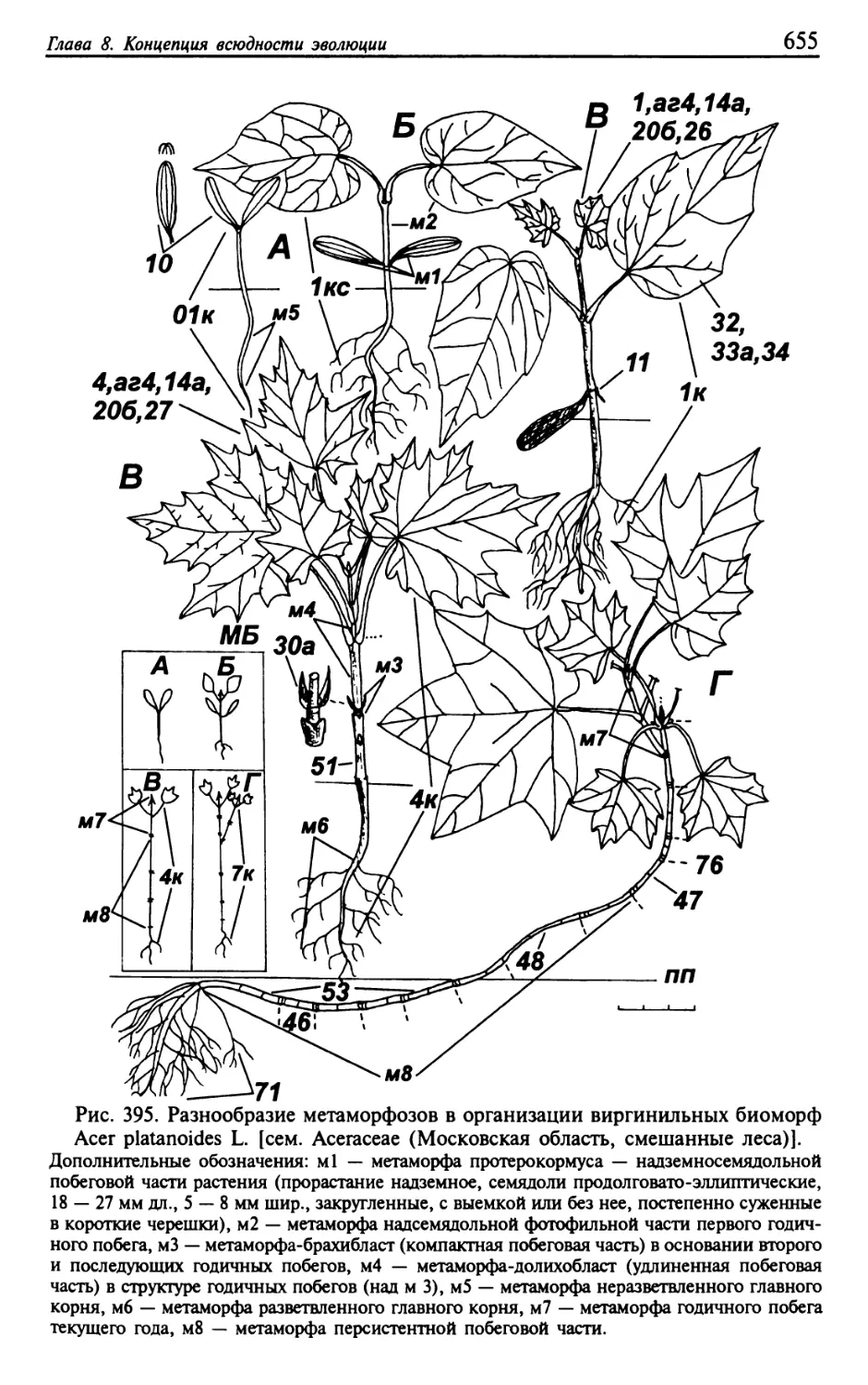
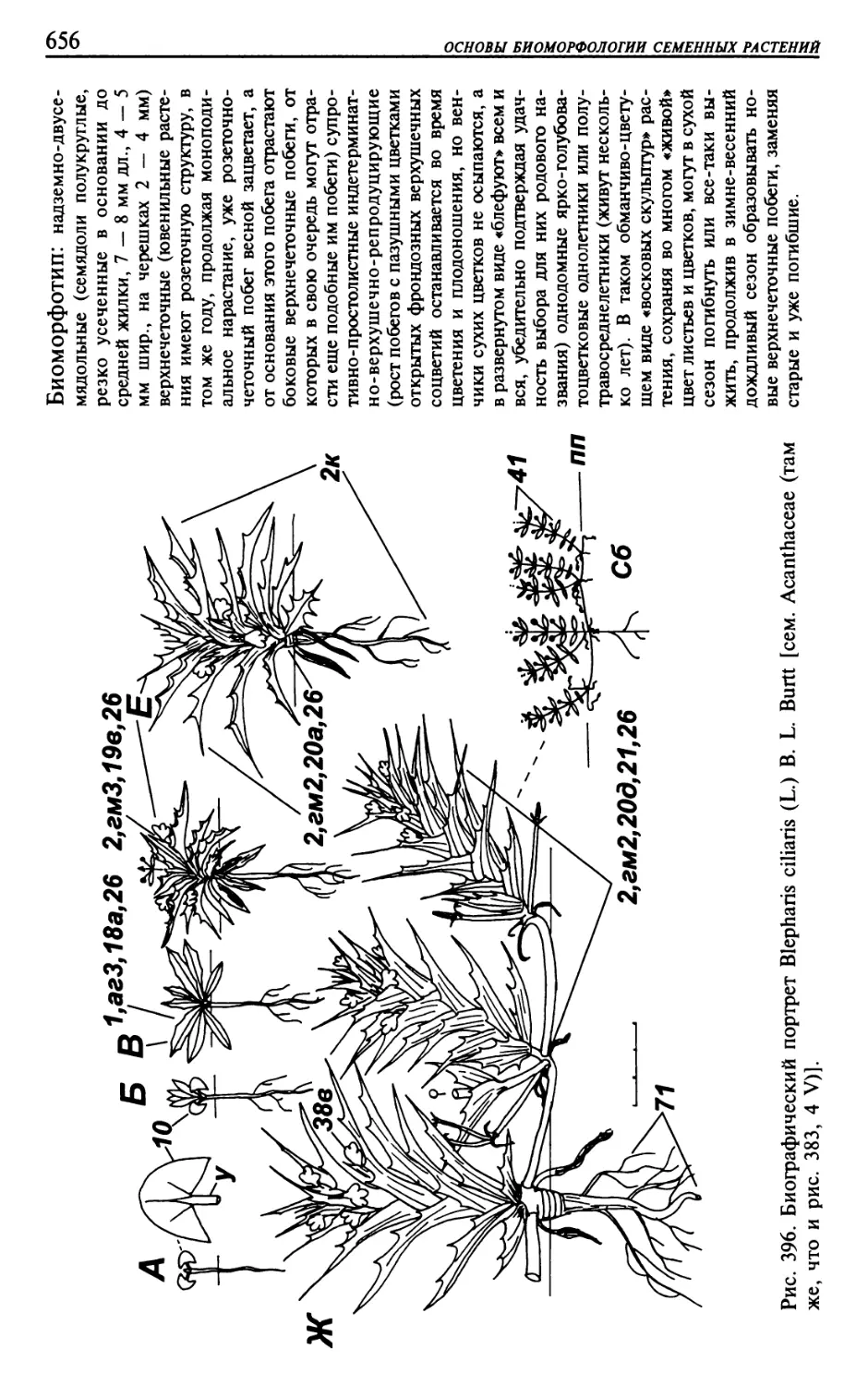
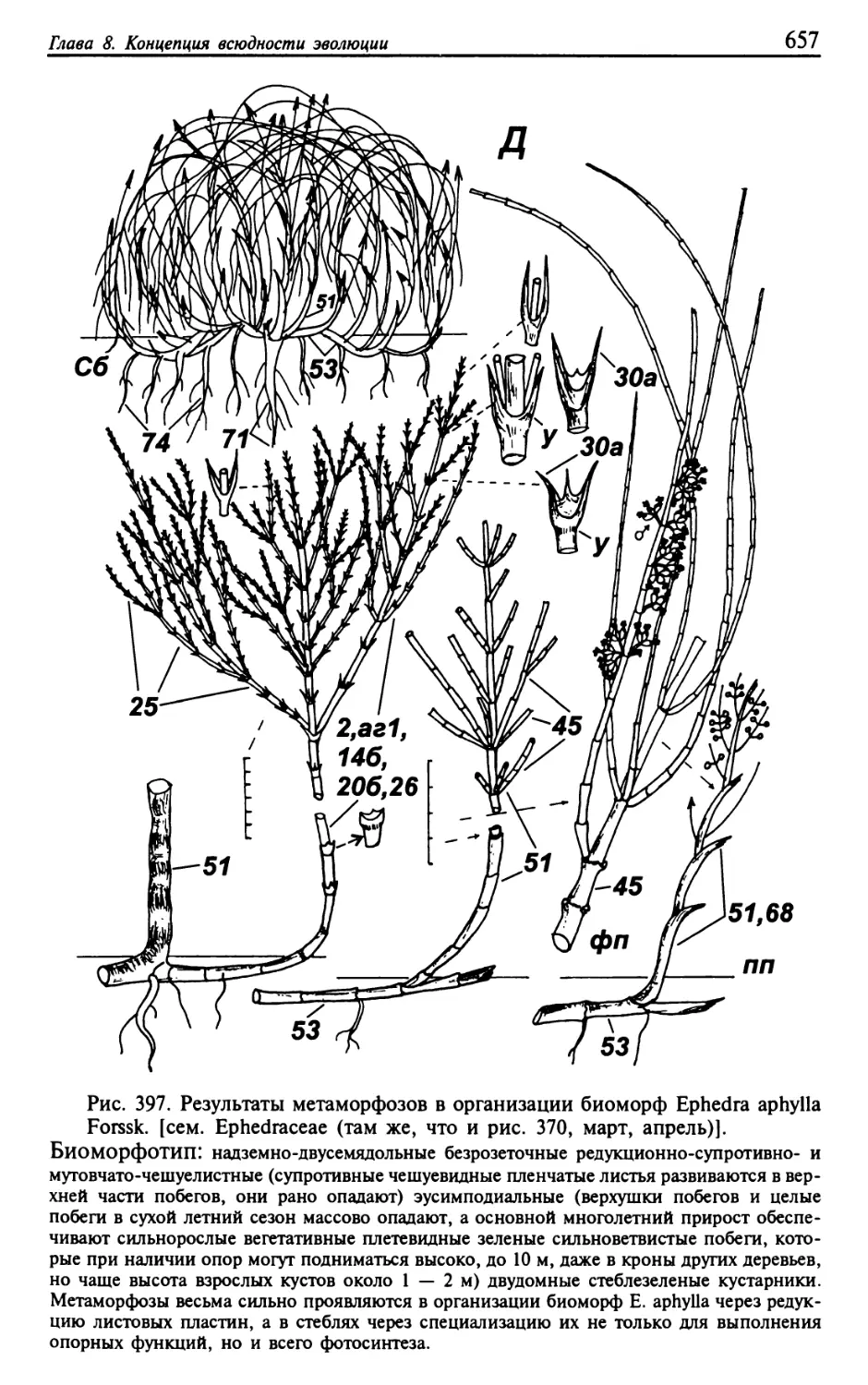
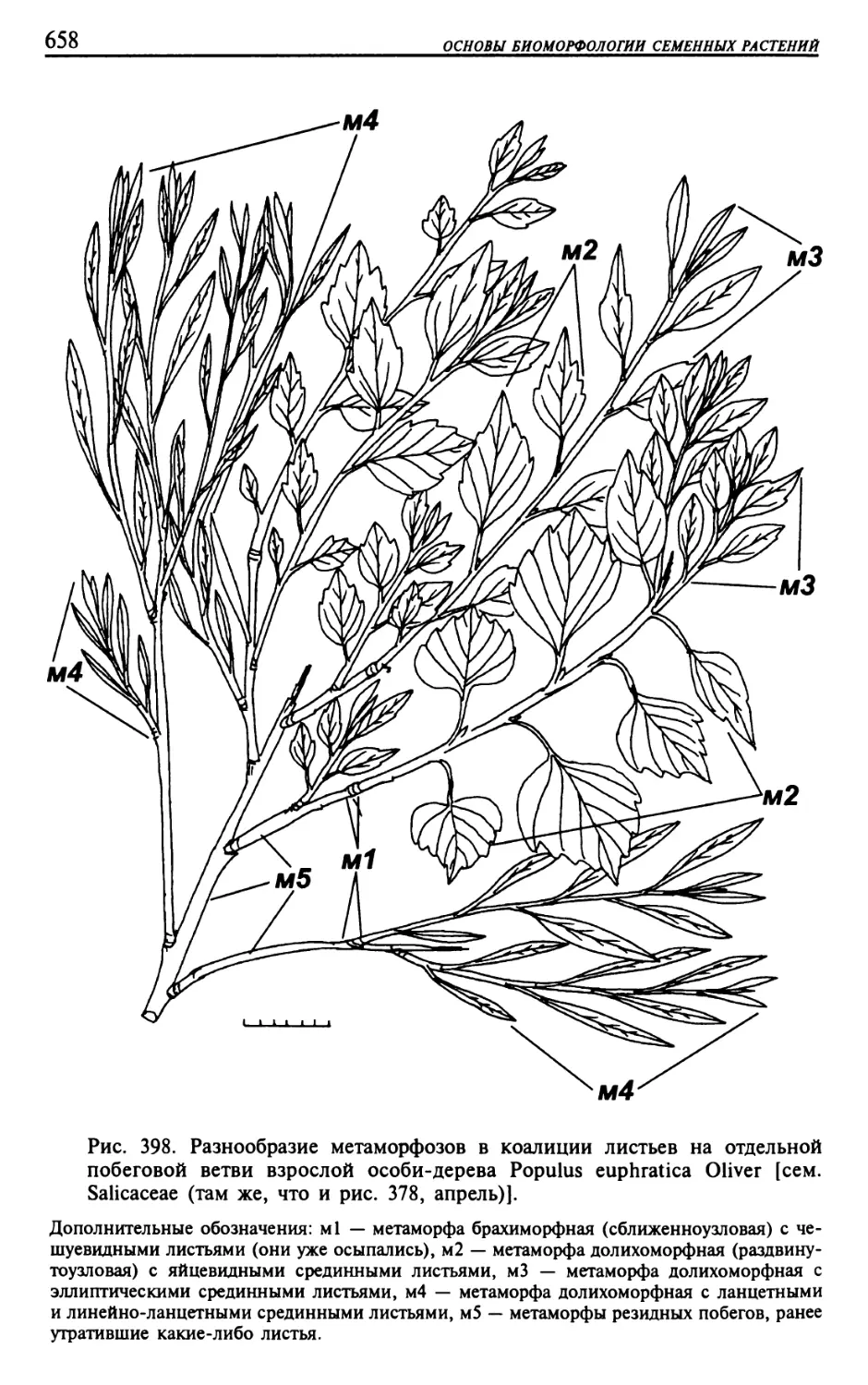



































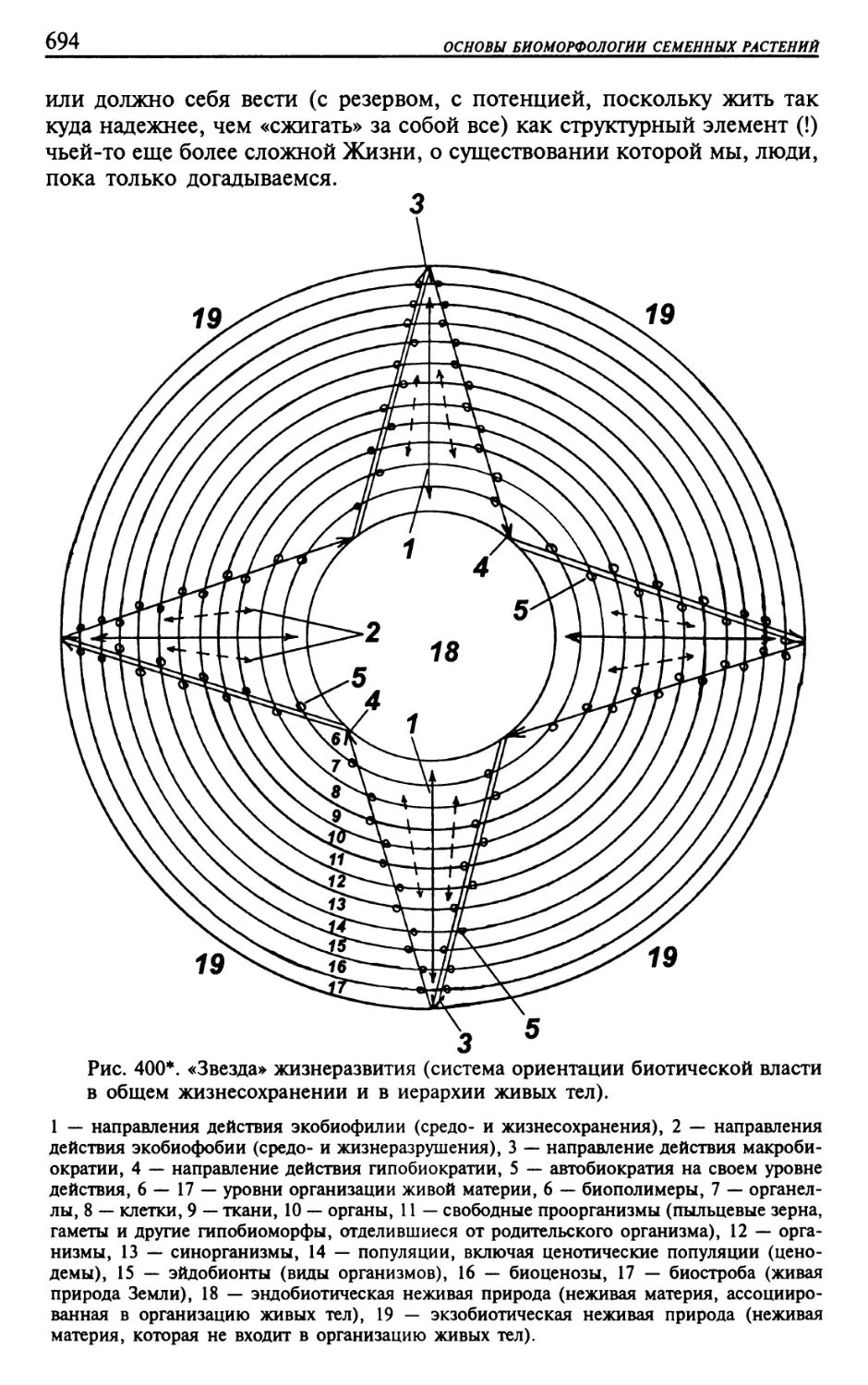












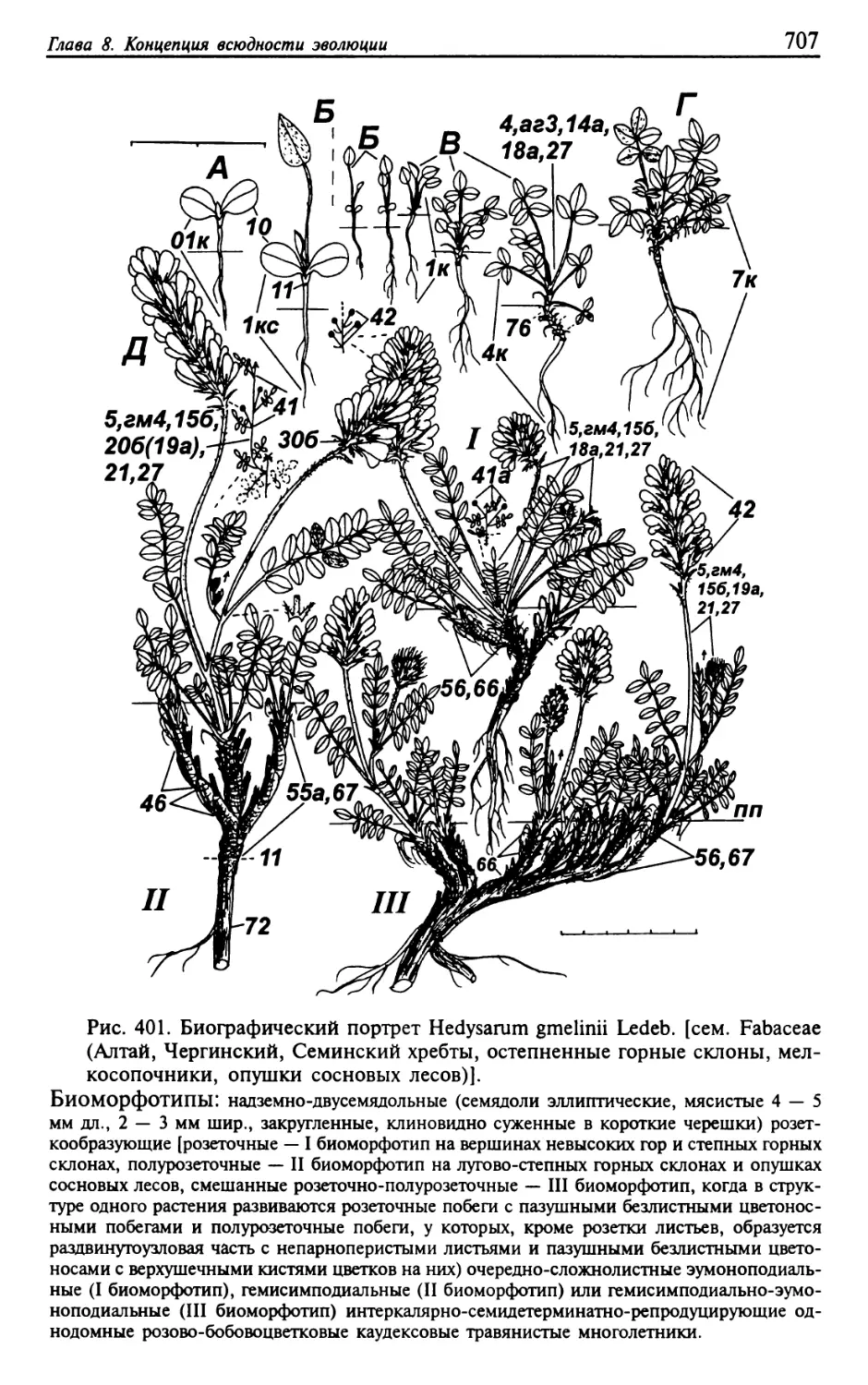
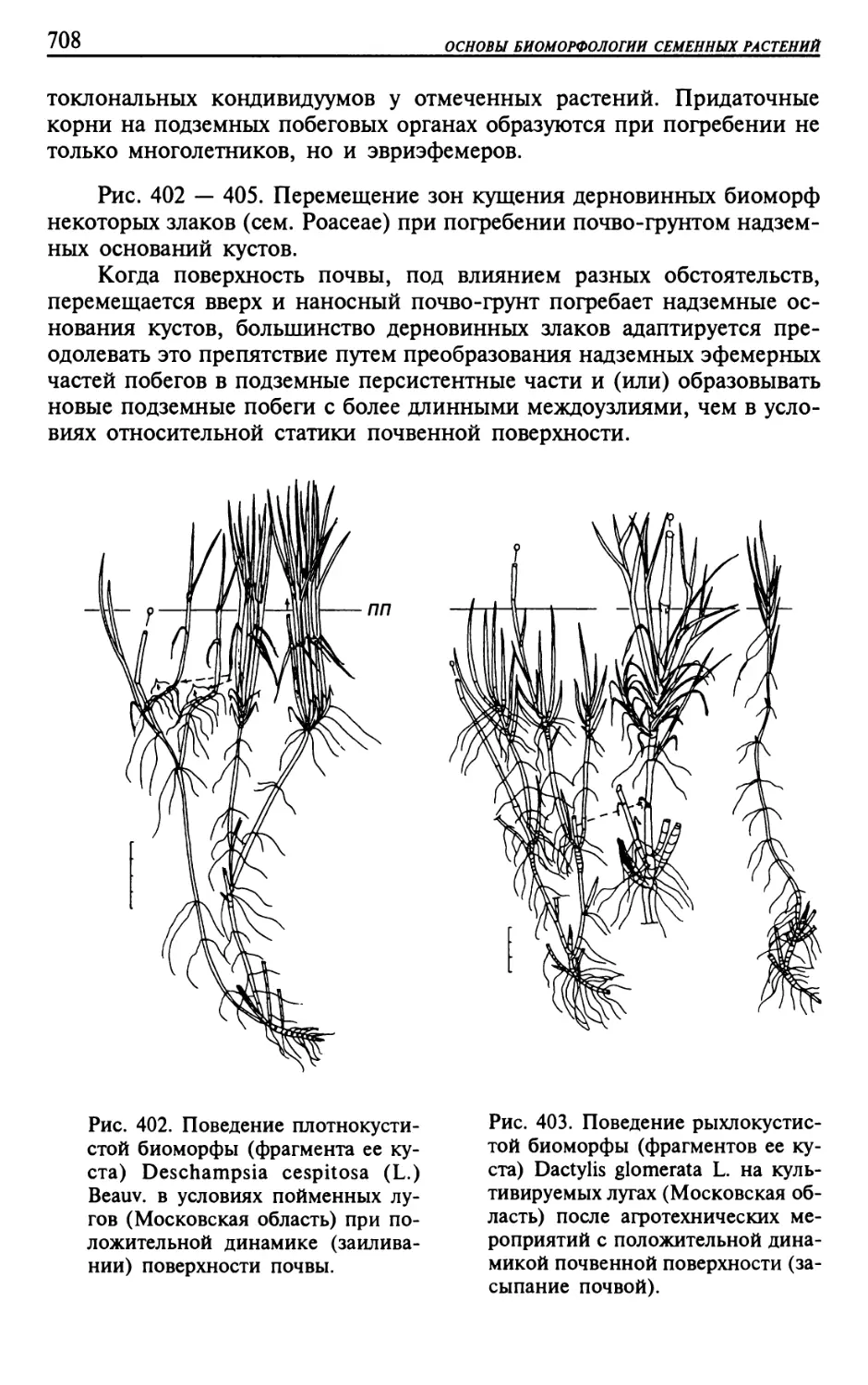





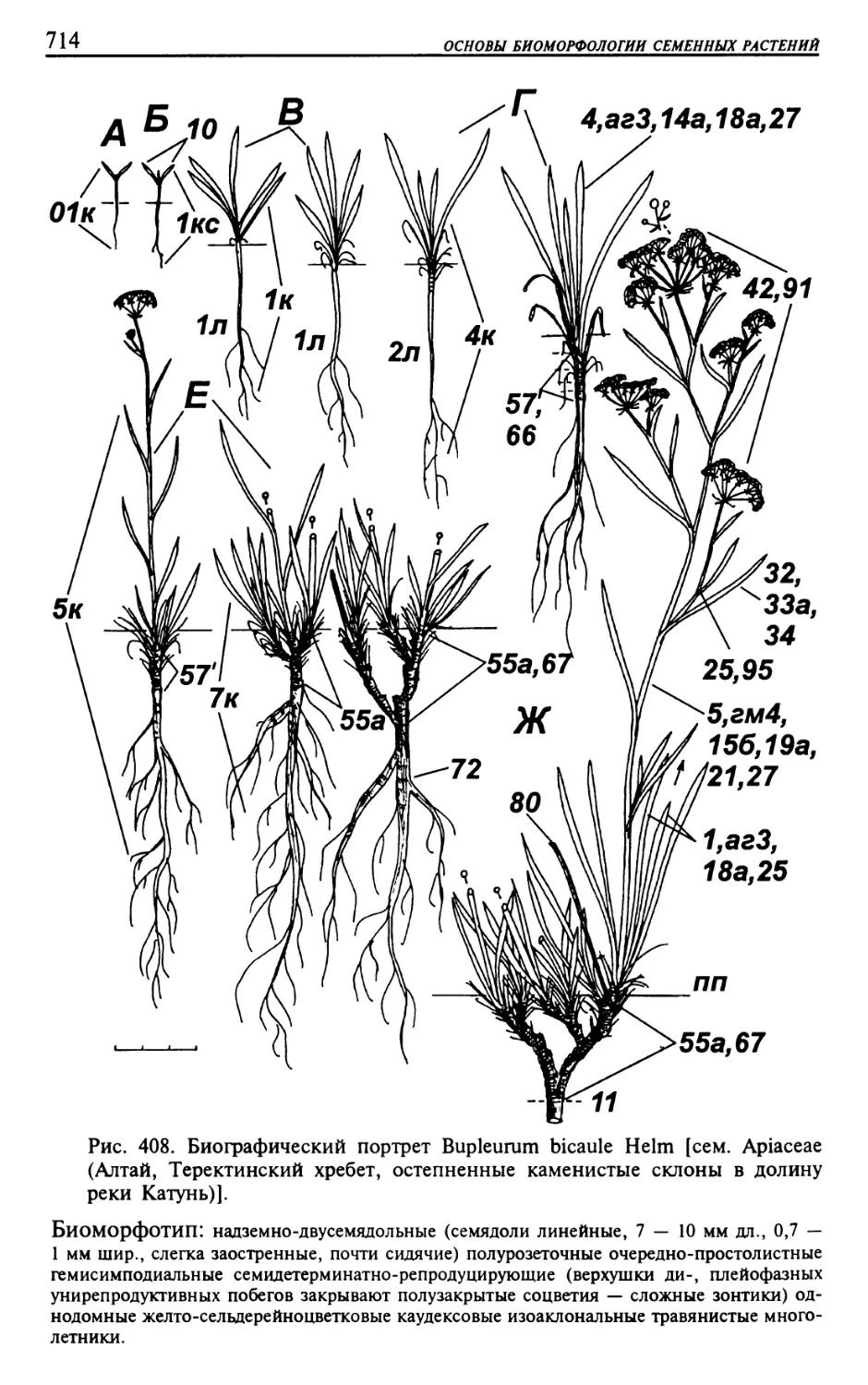
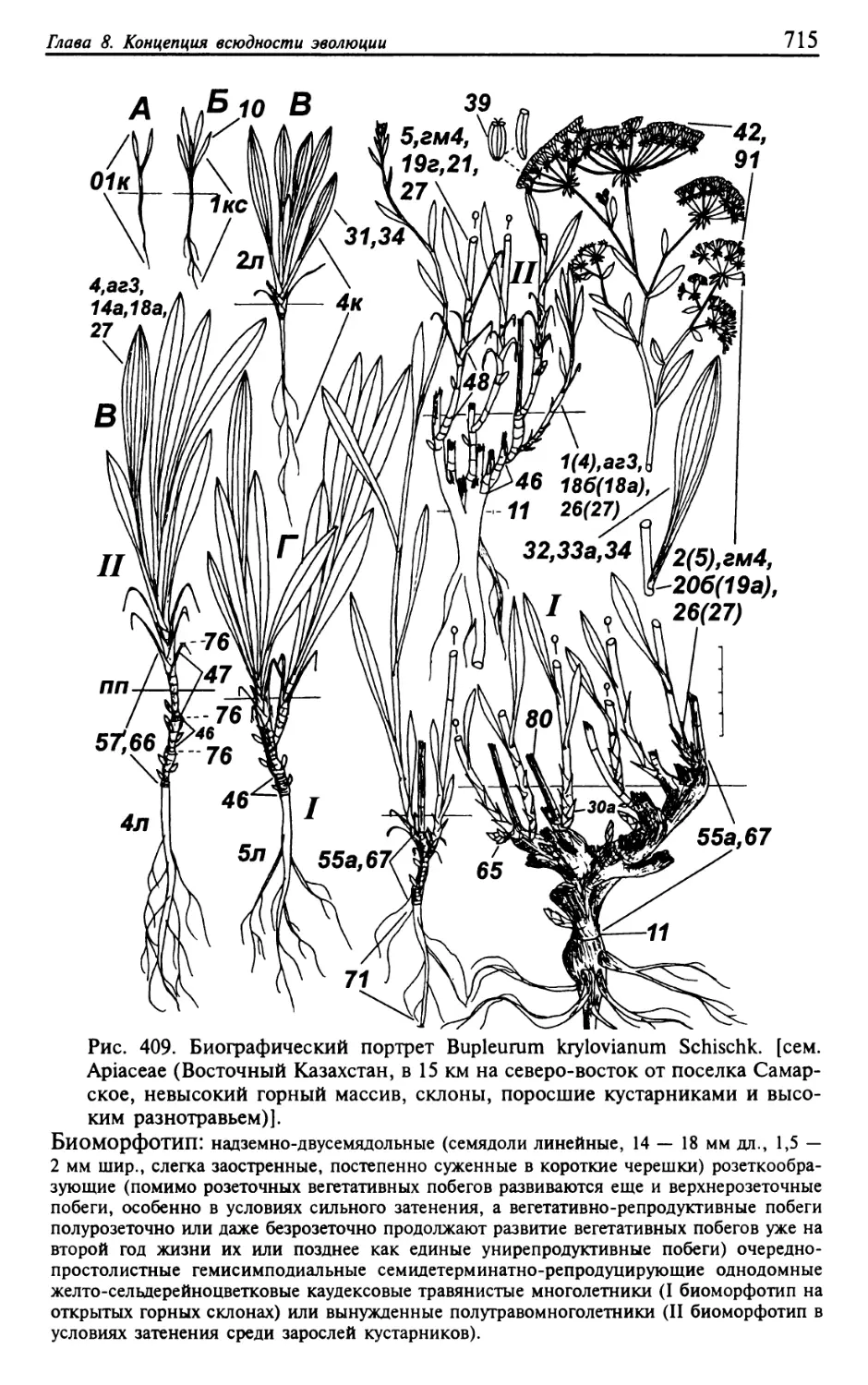
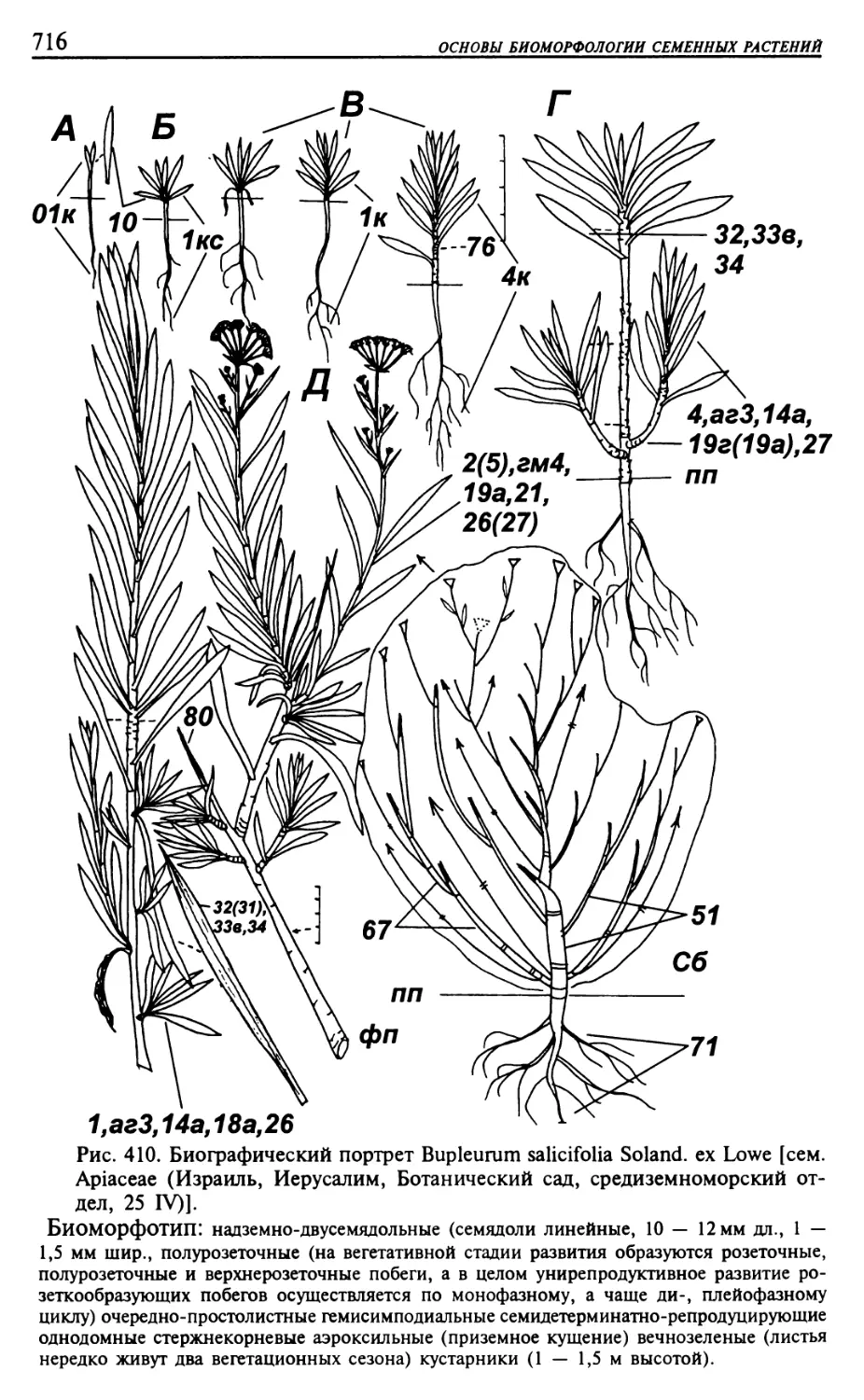

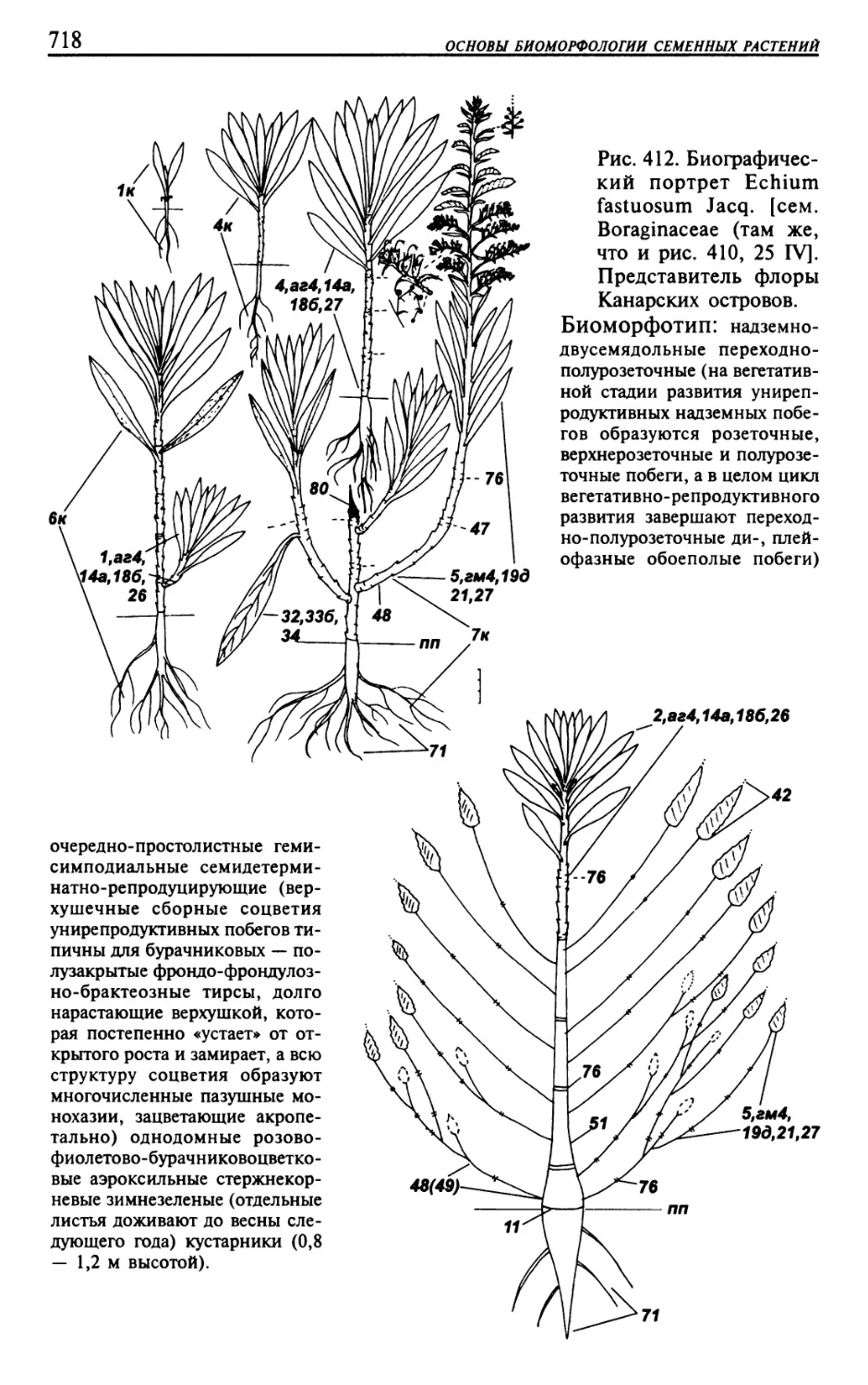











































































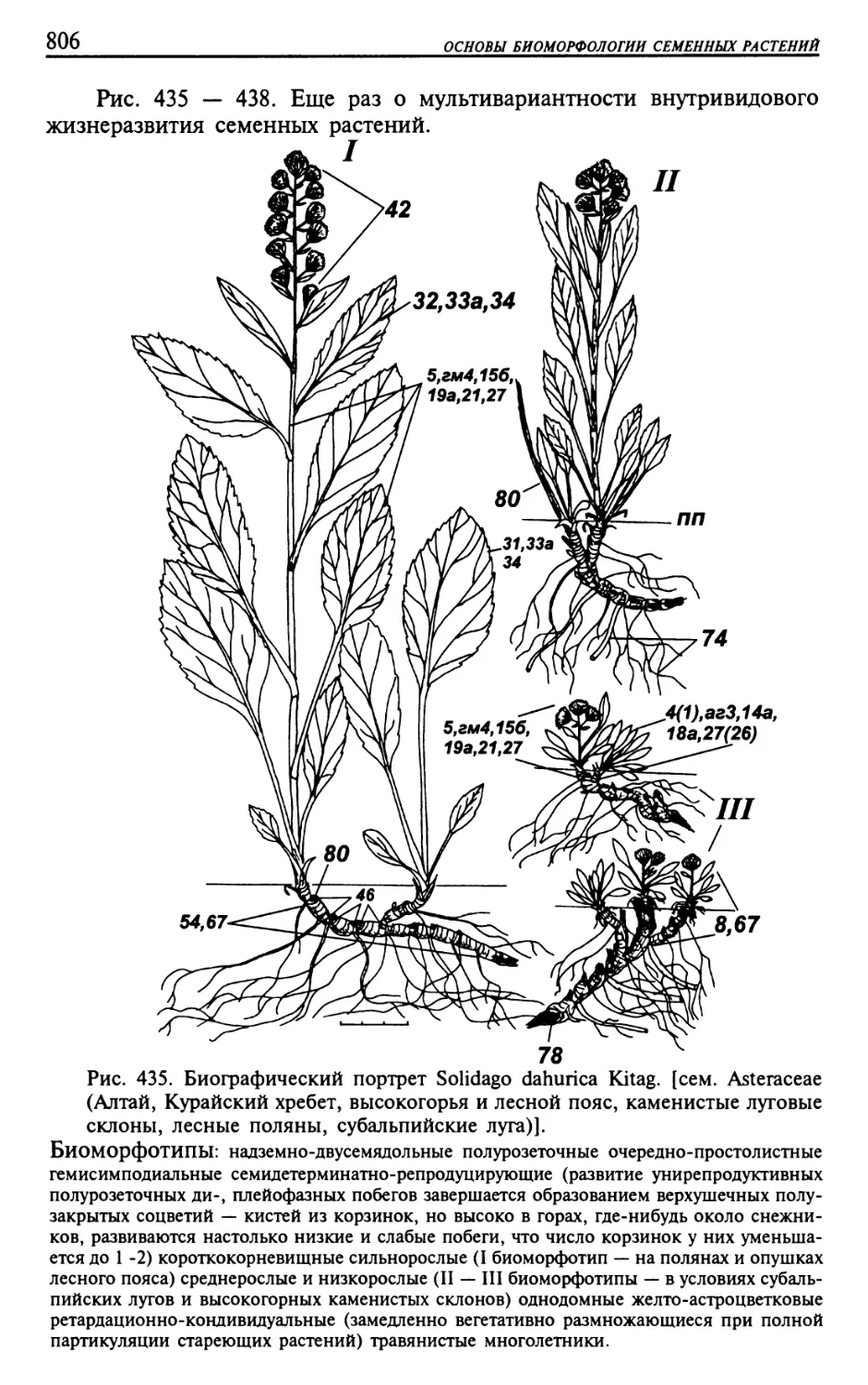
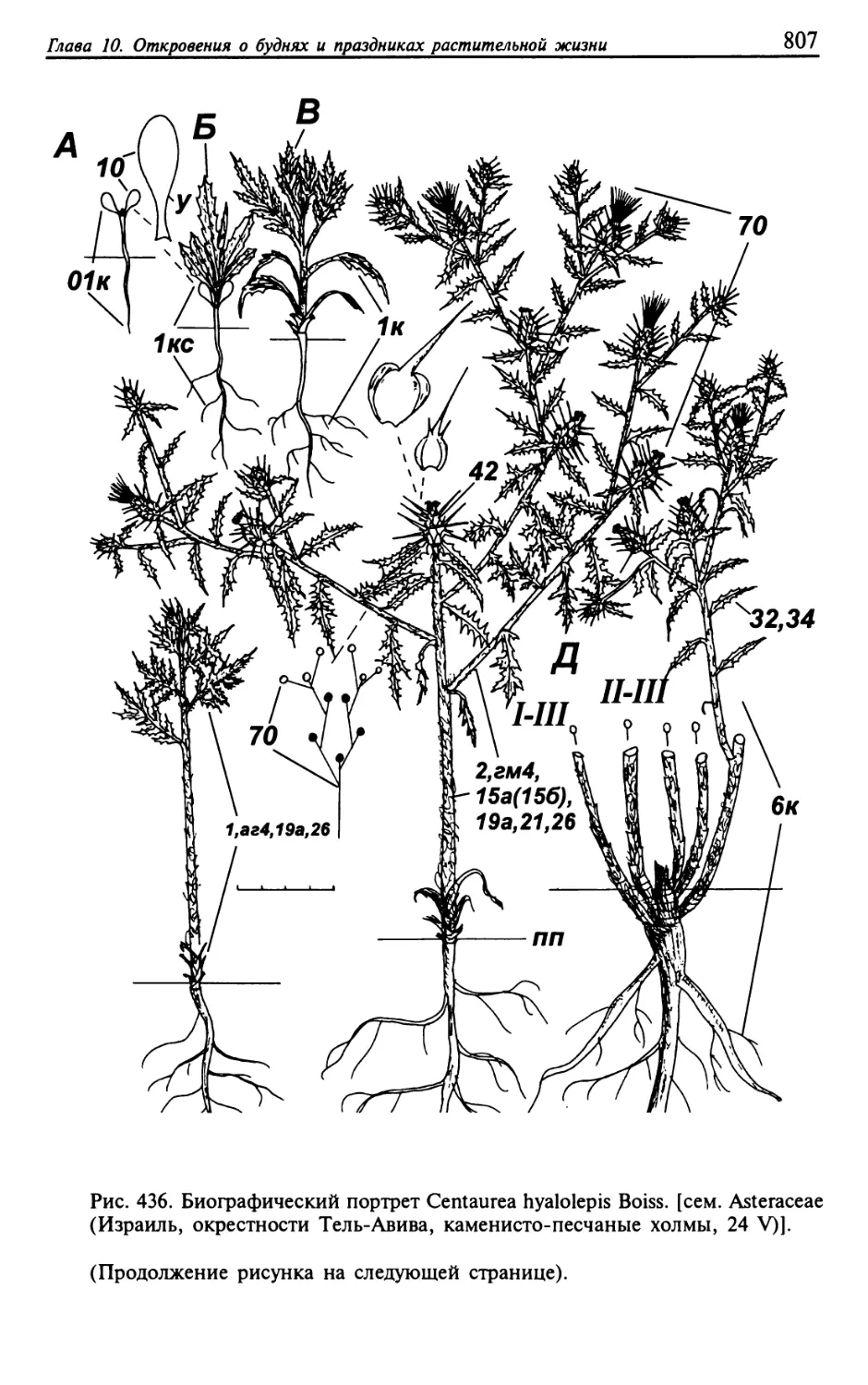

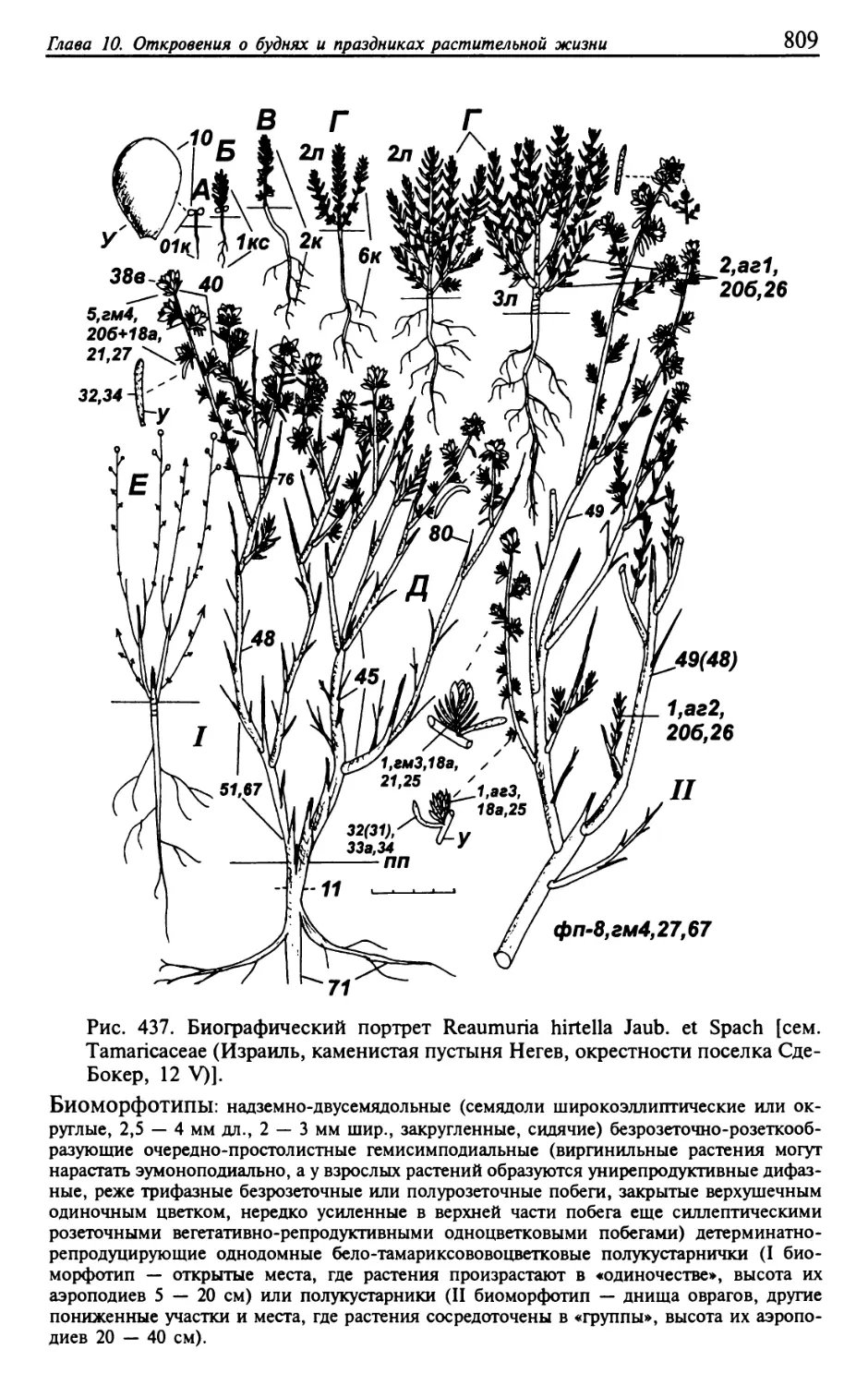
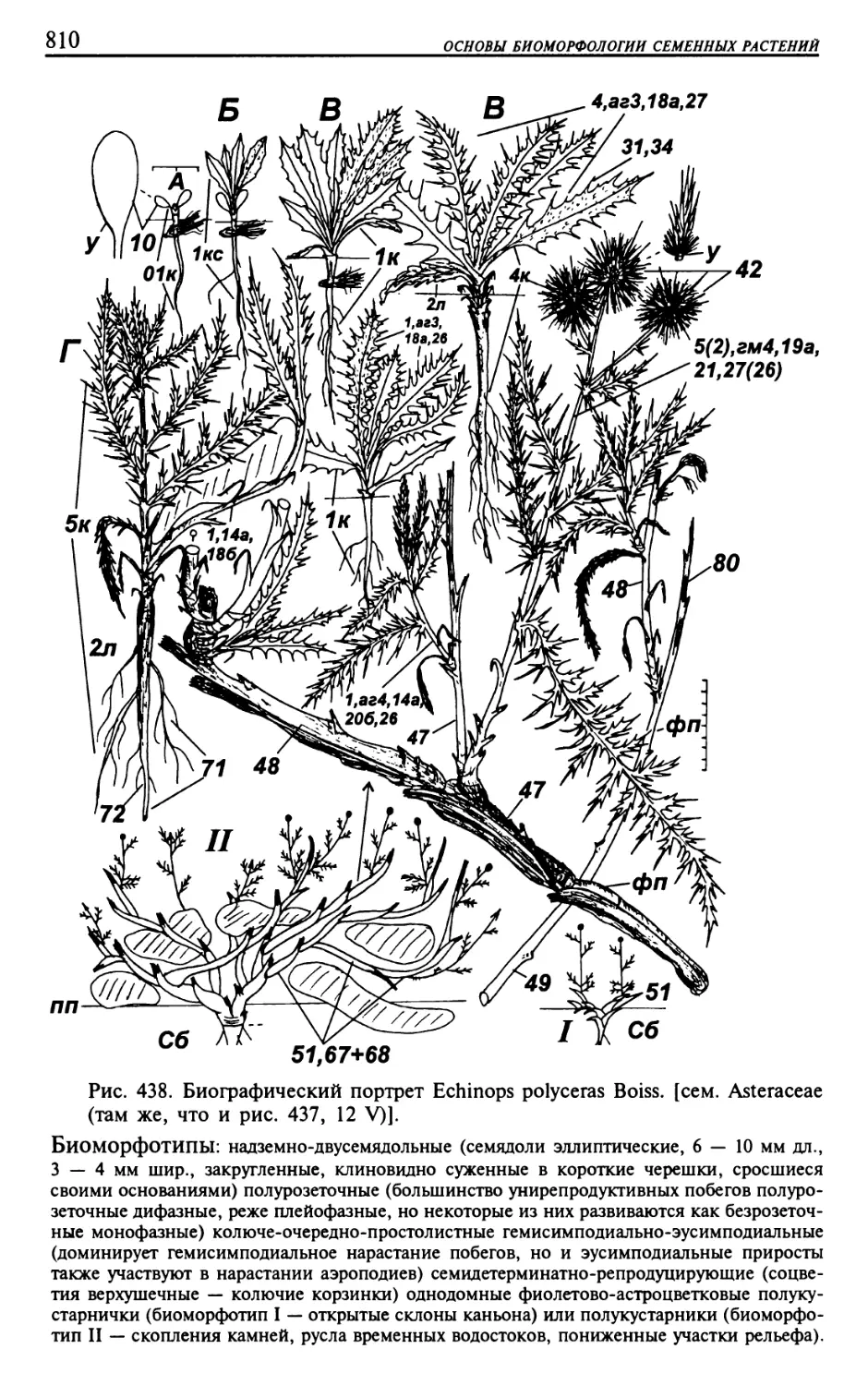






















































Текст
Е.Л. Нухимовский Габитус и формы роста в организации биоморф Москва Издательство «Оверлей» 2002
УДК 581.4 Нухимовскш ЕЛ. Основы биоморфологии семенных растений: Т. 2 Габитус и формы роста в организации биоморф. — М.: Оверлей, 2002, 859 с., 438 илл. — ISBN 5-85493-067-6 Библ. 13 стр. ISBN 5-85493-067-6 � Е.Л.Нухимовский, 2002 � Оформление. Издательство «Оверлей», 2002 Продолжено обоснование и развитие теории организации биоморф семенных ра- стений. Сформулировано десять концепций: меротомирования (членения целого на части) и меросомности (членства частей тела в целом теле), приоритетности, габитуса, форм роста, субординатного и просубординатного разнообразия биоморф, пластичности и гиперпластичности, биотической власти и «могучей пятерки», всюдности эволюции, экогенобиоза, биотического счастья и качества жизни. Сегодня тайны габитуса и форм роста в биоморфологии, вступившей в интегральную стадию своего развития, по мне- нию автора книги, уже не существует. Определен статус этих структурных образований. Место им только на двух «полочках» (и не более) в сложной структуре организации определенных телесных систем, среди которых присутствуют и биоморфы (организмы и синорганизмы) семенных растений. Дело в том, что исследовательская интрига состояла далеко не в том, чтобы разработать только проблему габитуса и форм роста биоморф, но и в том, чтобы еще и еще «покопаться» логическими средствами, пользуясь, в частности, приемами суперординатного классифицирования (абстрактного «пирамидостроитель- ства») в многослойности организации живой природы. Такие «встряски» под «логичес- ким микроскопом» показаны Природе. Семенные фитобиоморфы, посредством разных форм естественного отбора, уча- ствуют в эволюции, принимая ее как внутри, так и вне себя. Важно еще и другое, что также показано в книге, — все биоморфы, каждая по-своему, могут быть биотически счастливыми, в первую очередь, тем, что просто живут, а уж тем более, несмотря на последствия, когда цветут и плодоносят. Фитобиографический иллюстративный матери- ал обильно наполнил собой фактическую основу всех 10 концепций, рассмотренных в книге, но он всей своей массой рассчитан и на значительно большее: он призван воз- вестить ботаническому миру не только о глубокой перемене, которая свершается на современной (интегральной) стадии развития фитобиоморфологии, но и о тех концеп- туальных подвижках, которые неминуемо должны произойти в текущем столетии в со- предельных отраслях биологического знания. Более 400 оригинальных рисунков — до- кументальных свидетельств — фитобиографических портретов видов убедительно под- держивают тезис: призрак биоморфологии уже бродит в систематике. Будущие «Флоры» России и других регионов Земли обязательно станут биографическими — это веление времени, обремененного движением науки к прогрессу. Книга рассчитана на биоморфологов, ботаников разного профиля, а также биоло- гов, интересующихся общими проблемами естествознания.
Предисловие В XXI столетие биоморфология семенных растений входит со сво- ими, во многом уже установленными законами и правилами, с четкими концептуальными границами и надежным стержнем — теорией органи- зации биоморф. Прогрессивную динамику этой науке обеспечивает ее сердцевина — учение о жизненных циклах организмов и синорганиз- мов; это учение способно стимулировать развитие не только биоморфо- логии, но и других наук и, в первую очередь, систематики. Все, что написано автором в первом томе «Основ биоморфоло- гии...», пожалуй, было бы вполне достаточно, чтобы, пускай на время, остановиться и сказать: теоретические основы современной биоморфо- логии семенных растений сформулированы, а детали и многие подроб- ности — стоит ли в них сейчас углубляться? Однако это — не мой стиль; по натуре я максималист и в науке никогда не остановлюсь, если вижу цель больше и дальше, чем она кажется на первый взгляд. Наука, как и Природа, которую познает наука, неисчерпаема в своем движении вширь и вглубь. Время неудержимо и необратимо летит вперед, и всегда надо стремиться успеть как можно больше, идти не только в ногу с веком, но и смотреть в будущее, где-то даже опережать свое время. Я не люблю и не хочу забегать далеко вперед со своими планами: зачем обещать то, что — мало ли как сложится жизнь — может и не состо- яться. Однако замечу, что многотомник по биоморфологии был задуман мною давно, еще в студенческие годы. Уже тогда меня вдохновила идея: заняться многолетними исследованиями организации биоморф семен- ных растений в динамике, то есть, в первую очередь, в жизненных циклах. Однако, сегодня будет логично воспринимать биоморфологию в авторском исполнении лишь по тем работам, которые уже состоялись. «Нельзя объять необъятное», но стремиться сделать это — надо! Так я мог бы, дополнив известный афоризм Козьмы Пруткова, сформули- ровать свое научное кредо. В науке есть принципиальная особенность: ее приоритеты со временем склонны изменяться, даже сменяться; клас- сические положения, будучи однажды сформулированными, устарева- ют, их приходится уточнять, а то и сильно трансформировать или со- всем отбрасывать; иначе и быть не может, иначе наука остановится в своем развитии и перестанет быть наукой. Худо тому, кто не увидит или не захочет увидеть смену приоритетов развития определенной науки. Важно вовремя заметить, когда прежние теоретические положения пре-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ вращаются в балласт, плыть с которым дальше — занятие бесперспек- тивное. Сегодня мы поговорим о том, о чем уже было немного сказано ранее [(Нухимовский, 1997); далее ссылка на эту работу будет выглядеть в основном так: (т. 1), или (т. 1, гл...., с....), или (т. 1, с....)], с той лишь разницей, что здесь речь пойдет более всего о габитусах и формах роста биоморф (организмов и синорганизмов) как обязательных, важных, но не единственных частях их организации. Зачем и почему в биоморфо- логии получилось и получается так, а не иначе, читатель, надеюсь, поймет, прочитав эту книгу. Мысль о первостепенном значении габитуса в понимании жизнен- ных форм звучала в работах многих биоморфологов двух последних столетий (Humboldt, 1806; Гризебах, 1874, 1877; Warming, 1884, 1908, 1923; Серебряков, 1962, 1964; Хохряков, 1975, 1981 и др.). Я не хочу, как не хотел и раньше в томе 1-м — заниматься «раз- брасыванием камней», ниспровергать многие общепринятые положе- ния, пытаться утверждать новое, укладывая кого-то «на лопатки». У меня принципиальное, но спокойное кредо «собирателя камней»! Я подхожу к проблеме развития биоморфологии, систематизируя положи- тельный актив, накопленный в ней к началу XXI столетия, и опуская то, что на сегодня представляется неактуальным. В то же время я ос- торожно, но последовательно добавляю к этой конструкции свои, иног- да, может быть, неожиданные, иногда кому-то, возможно, кажущиеся дерзкими, мысли и соображения, стараясь усилить концептуальную «энергетику» этой, весьма интересной, еще во многом невостребован- ной науки; все это я делаю ради одной цели — ради стремления к истине. Никогда не стоит забывать, что мы, каждый по отдельности, лишь какое-то время сопровождаем развивающуюся науку, оставляя в ней более или менее заметный вклад. Конечно, современная биоморфоло- гия, как и любая другая научная дисциплина, не смогла бы двигаться вперед без дискуссий и споров, которые вспыхивают в печати и вне ее (всякое ведь бывает и до и после публикации того, что тебе дорого), а также на собраниях ученых, участником которых довелось за последние тридцать пять лет быть и автору этой книги. Интеллектуальная деятельность каждого ученого занимает опреде- ленное место в совокупном научном достоянии человечества, вливаясь, как некоторая необходимая частица, в хранилище коллективного разу- ма, так или иначе отражая достижения в науке предшествующих поко- лений «совидичей» (выражение А. С. Серебровского, 1973), но и остав- ляя в ней более или менее очевидный отпечаток своей индивидуально- сти. Какую бы позицию ни занимали современные ученые и их оппо- ненты, им вряд ли удастся выйти за пределы этой очевидной истины. Так что не будем забывать об этом. Вполне уместно подчеркнуть здесь и еще одно обстоятельство, по- скольку автора иногда упрекают за увлечение крупномасштабными обоб- щениями. Я с уважением отношусь к эмпирическим исследованиям и ученым, которые их проводят, но сам все же люблю работать по-дру-
Предисловие гому: как эмпирик и теоретик одновременно. По крайней мере, к этому я всегда стремлюсь — это стиль моей научной работы. Все мы вьпцли для работы в науке из вузов, как правило, «альма-матер» выпускает нас, естествоиспытателей, в эмпирических «яйцах», в которых есть главное — предмет специальных исследований. Этот предмет ученый способен, не покидая уютного эмпирического «яйца», изучать усердно и плодо- творно, делая, может быть, немало открытий, — но лишь до некоторого предела, который далеко не многие или захотят, или сумеют перешаг- нуть. Работа в эмпирических «яйцах» нередко создает у ученого успо- коительную иллюзию, будто он пребывает в некоем бесконечном свет- лом «пространстве». И далеко не всегда такой ученый берет на себя смелость подняться во весь рост и ткнуть пальцем в «бездну» (на самом- то деле всего лишь «скорлупу») окружающей его «квазибесконечности». Ощущение, которое испытает ученый, освободившись от эмпири- ческого «яичного небосвода», не передать словами: шоры, отделявшие его и предмет его познания от остального мира, спадают с глаз. Он может подняться в воображении до неведомых ранее высот абстракции, менять углы обзора, а предмет исследований появляется перед ним во всей многогранности, демонстрируя такие стороны организации, пове- дения и т. п., о которых ранее исследователю не приходилось даже задумываться. «Котел кипит, но его содержимое нужно тщательно перемешивать; в него надо добавлять фантазию, воображение и техническое мастер- ство». Эти слова принадлежат Дж. Иберту (1968, с. 7), размышлявшему о прогрессе биологии развития во второй половине XX столетия— науки, которой он занимался всю жизнь. То же самое можно сказать сегодня и об эволюции биоморфологии конца XX — начала XXI веков, добавив, что современную биоморфологию ждет, безусловно, немало открытий не только на ниве чисто эмпирических исследований, но и на пути ее интегративного развития в сообществе с другими науками. Узкоспециальные интересы ученых постепенно будут сдвигаться в область теоретизирования, к более смелому и масштабному, но все же более здравому стилю мышления, позволяющему наблюдать и познавать свой предмет во всем разнообразии системных отношений, в контексте взаимосвязей и взаимозависимостей единого и бесконечного мироздания. Изложу свои самые общие мысли по этому вопросу: они могут быть полезны для нового поколения ученых. Наука, образно говоря, значи- тельно лучше развивается не в джунглях, где бродят, готовые разорвать друг друга, «саблезубые тигры» и «пещерные медведи», а на лужайке с резвящимися на ней «конями» и «лошадками», разогревающими мышцы перед очередными научными «скачками», чтобы показать, если полу- чится, всем остальным, кто из «скакунов» проворнее и ловчее. Задача ученых, тех, кто сегодня не у руля, а «у пера», не разбрасывать, а собирать «камни», спокойно раскладывая их по полочкам актива и пассива: последний может также пригодиться для науки и, кто знает, дать неожиданную, даже невероятную по сегодняшним понятиям актив- ную поросль. В предлагаемой читателю книге изложена позиция автора по отдельным сторонам организации биоморф семенных растений;
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ причем всегда руководила автором не «саблезубая» агрессия, а стремле- ние не проиграть на текущих научных «скачках». В науке без соревно- вания нельзя, иначе — конец, взбивание «пены», пузырей самоутверж- дения, а значит, движение на дно с иллюзией, будто ты плывешь к заветному берегу... Конечно, есть и в науке «тихие заводи», где можно спокойно «плес- каться» всю жизнь, обманывая себя и других вроде бы бурной деятель- ностью, но так ни разу и не ощутив того особого душевного состояния, того полета (новаторского синдрома, если хотите), когда действительно «идешь на грозу», шагаешь вперед, шагаешь, может быть, по самому краю, но — вперед. Если такого тебе не дано испытать, то — для чего все это?... Можно ли, усвоив достижения предшественников, неся в себе сгу- сток коллективного разума, сохранить свою научную индивидуальность? Да, можно; хотя это и не просто; более того, это — необходимо. Иначе ты наверняка похоронишь в себе творца, искателя, покорителя новых горизонтов, и тебе останется только повторять то, что было найдено в науке другими. Все это в общем-то не зазорно, в этом тоже есть потреб- ность; но если уж ты «взялся за гуж», то по крайней мере надо попы- таться творить, благо к началу XXI столетия фактического материала уже накоплено немало. Меня, безусловно, волнует и ко многому обязывает сам факт, что мне, волею судьбы, с моей монографией по биоморфологии семенных растений суждено было оказаться на стыке не только столетий, но и тысячелетий. Пользуясь случаем, с перекрестка времен от себя и, надеюсь, не ошибусь, сказав: от всех биоморфологов, моих современников, передаю привет и пожелание счастливой жизни нынешним и будущим землянам, кому выпало жить в третьем тысячелетии. Своей монографией автор рассчитывает не удивить читателя, а скорее убедить его, что у биоморфологии — большое будущее. Сегодня мы, как говорится, не на параде; мы просто работаем во имя развития биоморфологического знания. В конце книги читатель найдет несколько самых общих выводов, к которым автор пришел за многие годы изучения жизненных форм семенных растений. Итак, в путь.
Светлой памяти выдающегося российского ботаника, основоиоложника учения о жизненных циклах семенных раоиений- ведущего раздела в современной биоморфологии- Работнова Тихона Александровича (1904-2000) иосвящаю эту книгу Ввйдйний Развитие живой природы многопланово; у одних типов биоморф общение с окружающим миром осуществляется с помощью звуков и даже речи, у других, в частности, у растений, — беззвучно, но в таких случаях, как же?... Для нас — людей, владеющих речью, удивительно смотреть на растения и задавать себе вечный вопрос: как, почему и о чем молчат растения? Как растения в столь многоликом окружающем их мире общаются между собой и со всеми другими формами живого и неживого? Растения действительно ведут предельно молчаливый образ жизни и, если с этим смириться, так и останутся со своими тайнами. Но так быть не должно. Нам, людям, свойственно неиссякаемое любо- пытство, только благодаря ему Человек научился выживать на Земле, только поэтому мы, люди, при желании можем подбирать ключи к пониманию растений, делая их своими друзьями, и только так мы можем «убедить» растения поделиться с нами их тайнами. Надо всего лишь внимательно присмотреться к ним, и они, по-своему искренне, доверят нам свои секреты. Молчанием можно выразить все: и свой внутренний мир, и отно- шение к миру внешнему. «Язык» молчания у растений — это в первую очередь их внешний облик, то, что традиционно называют габитусом. Через габитус растения «говорят» не только с теми, кто обладает разу- мом и сознанием, способен понять глубокое субгабитуальное содержа- ние их молчания, но и с теми, кто сосуществует с ними на равных— в мире молчания. Растения, безусловно, бывают счастливыми, но на эту, весьма деликатную тему лучше всего будет поразмышлять в конце книги (глава 10). Цель книги, в рамках теории организации биоморф семенных ра- стений, сформулированной ранее (т. 1): дать теоретико-эмпирическую развертку по весьма принципиальной стороне организации биоморф этих растений — проблемам габитуса, его форм и стадийного развития,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ его форм как форм роста, т. е., по сути, дать концептуальные основы учения об этих сторонах организации биоморф. Вопросы, связанные с габитусом и формами роста у растительных организмов и синорганиз- мов, т. е., иными словами, соответственно, у простых и сложных био- морф, всегда актуальны; они настолько многогранны, настолько зани- мают умы ученых, особенно морфологов XX столетия, что читатель, пожалуй, согласится с автором: если продолжать задуманную моногра- фию, то второй том должен быть посвящен разработке именно этой темы. Сегодня можно с уверенностью констатировать, что, несмотря на повышенный интерес со стороны ботаников и зоологов, до сих пор не существует надежной понятийной, общетеоретической основы знаний о габитусах и формах роста растений и животных. Обычно исследователи, работая с габитусами и формами роста живых тел, принимают на веру то, что «лежит на поверхности», ограничиваясь смысловой первоосно- вой этих терминов: габитус — внешний вид (облик), форма роста— одно из возможных проявлений результата роста и развития живого тела, например, клетки, ткани, органа (побега, корня и т. д.) и, наконец, организма (растения или животного). Однако развитие биоморфологии свидетельствует о том, что сегодня нужны более весомые аргументы и критерии как в определении упомянутых терминов, так и в методоло- гическом обосновании научных направлений, занимающихся габитуса- ми и формами роста. В XXI столетии, пожалуй, работа в направлении основной цели всей морфологии растений — «открыть причины расти- тельных форм и тем самым указать законы, лежащие в их основании» (Бекетов, 1862, с. 2), — будет продолжена. Многое мы уже знаем. jla- вайте же попробуем «разложить по полочкам» накопленные в науке сведения о габитусах и формах роста и, самое главное, попытаемся сделать шаг (а может быть, и несколько) дальше. Весь окружающий человека (шире — людей вообще, человечество) мир и сами люди воспринимаются в первую очередь визуально, через внешний облик любых материальных тел, т. е. физиономию, по выра- жению А. Гумбольдта (Humboldt, 1806) и других старых авторов; иными словами — через габитус. «Пленку жизни» (по образному выражению В.И. Вернадского) составляют на Земле мириады живых существ, каж- дое из которых обладает определенным внешним обликом, типичным для родственных им организаций. Таким образом, целесообразность габитуального подхода к познанию Природы была интуитивно осознана еще на заре развития человечества. Первобытный человек учился ори- ентироваться в разнообразии природных тел, подразделяя их по разным признакам и, естественно, более всего по габитусу. Так, в отношении растений издревле появились такие названия форм (габитуальных) ра- стений, как «дерево», «кустарник», «полукустарник», «лиана», «трава», «однолетник». Неудивительно, что обозначение растений по их внеш- нему облику имеют место, например, уже в сочинениях Феофраста (372 — 287 г. до н.э., 1951), свидетельствуя об использовании экзоморфоло- гических знаний в сельскохозяйственной практике Древней Греции. Можно с уверенностью утверждать, что с габитуального подхода к по- знанию природы началось становление цивилизации вообще и науки в
Введение Задачи исследований Используя системный подход, построить концепцию меросом- ной организации биоморф семенных растений. Показать, что не только организмы, но и синорганизмы являются телесными системами определенного рода. Вскрыть суть членения их на части (меротомирования) и показать место габитуса в органи- зации биоморф. Определить, в чем целесообразность меросомной организации биоморф на фоне различных организационных процессов, про- исходящих в Природе. Есть ли приоритеты в организации биоморф семенных расте- ний и, если есть, то почему они такие, а не другие? С чего начинаются и где заканчиваются понятия «габитус» и «формы роста» биоморф? Определить участие габитуса, его форм и форм роста биоморф в круговороте ресурсов. Обратиться к проблеме взаимоотношения жизни и смерти в организации биоморф семенных растений и определить, как они отражаются в формах габитуса и формах роста. Очертить границы признакового пространства биоморф семен- ных растений и определить возможности использования его в классифицировании и моделировании биоморф при участии признаков их габитуса и форм роста. Раскрыть закономерности адаптации биоморф к различным ус- ловиям окружающей среды через формы габитуса и формы роста. Разобраться с иерархией трансформаций и определить место эволюции биоморф в ней. Двигаясь дальше в разработке эволюционного учения, постро- ить универсальную модель эволюции габитуса биоморф по от- ношению к уровню земной поверхности. Какие новации для развития эволюционного учения можно предложить на основе проведенных исследований? Определить перспективы дальнейших исследований габитуса и форм роста биоморф в биоморфологии. Обосновать тезис, воз- никший в начале XXI столетия: «призрак биоморфологии уже бродит в систематике». Что и как формирует телесную организованность явления «жизнь» и как непосредственно обеспечивается семенными фи- 2. 4. 5. 6. 7. 9. 10. 11. 12. 13. частности. В росте и развитии (их формах) у биоморф (организмов и синорганизмов) постоянно участвует габитус, что позволяет вопросы организации габитуса и форм роста рассматривать вместе — как две близкие проблемы или даже как одну проблему.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ тобиоморфами развитие их организации как систем «управле- ния-исполнения»? 14. Есть ли в организации биоморф какая-либо «власть» и если есть, то в чем ее содержание и чем она занимается? 15. Какими средствами и каким образом семенные фитобиоморфы решают проблемы отношений между структурными элементами своей организации и как отвечают на воздействия окружающей внешней среды? 16. Что нового можно предложить для решения проблемы проис- хождения жизни и в особенности растительной жизни, и куда идет эволюция биоморф семенных растений? 17. Могут ли растительные биоморфы быть счастливыми и, если да, то как их габитусы и формы роста выражают таинства ра- стительного счастья? При обработке фактического материала нами широко использова- лась методика анализа биоморфогенезов, изложенная в первом томе нашей монографии. Описание биоморфогенезов растений мы чаще всего проводили наикратчайшим способом — рисуночным, смысл которого в том, что главная информация заложена в рисунки и в подписи к ним, а на их основе составляется резюме, согласно которому следует, что наиболее существенное проявляется в онтогении или синонтогении рас- сматриваемых видов (т. 1). Важно отметить, что все стадии биоморфо- генезов, включая самые ранние (от проростков и старше), изучались не в лабораторных, а в полевых условиях. Только в редких случаях нам не удавалось, чаще всего из-за недостатка времени, отведенного обстоя- тельствами для поиска, найти преювенильные растения в естественных условиях. Детальное рассмотрение габитуса и связанных с ним форм роста биоморф — реализация давнего интереса автора к этой проблеме; при всем том не будем забывать, что габитуальный подход в биоморфологии, согласно нашей теории (т. 1), равнозначен любому другому подходу (экологическому, анатомическому, физиологическому, биохимическому и т. п.), позволяющему вскрыть особенности структурно-функциональ- ной природы таких систем, как организмы и синорганизмы; но один подход — назовем его биографическим, поскольку он ориентирован на изучение жизненных циклов биоморф растений (фитобиографий) раз- ных видов в различных условиях их обитания — имеет решающее зна- чение для развития биоморфологии, является сердцевиной ее современ- НОГО СОСТОЯНИЯ. К габитусу растений и животных издревле проявляется неподдель- ный интерес. Почему? Что, собственно, важного можно обнаружить во внешнем облике растения или животного? Не скрываются ли за терми- ном «габитус» какие-то несущественные моменты, которыми без ущерба можно пренебречь в серьезном исследовании? «Облик» — где его гра- ницы, да и вообще, о чем здесь можно говорить всерьез; живое или неживое это явление? А вообще-то телесен ли внешний облик, а сле- довательно, материален ли он вообще, где кончается внутреннее, а где начинается и кончается внешнее? Можно ли потрогать то, что называют
Введение габитусом, или это слово — скорее метафора? Вопросы, вопросы... А ведь не поставь их, можно было бы подумать, что все над габитусом растений (да и животных) безоблачно и что «все в общем согласны, что жизненная форма — это внешний облик растения» (Хохряков, 1978, с. 357). Однако у того же автора есть следующее, более серьезное откро- вение: «что такое «внешний вид», «облик», «габитус», до сих пор оста- ется не совсем понятным, во всяком случае, в определениях жизненных форм они не расшифровываются» (Хохряков, 1979, с. 586). Похоже, проблему габитуса в организации растений пока пытались рассматри- вать только «сверху», т.е. поверхностно и, по сути дела, ее содержание глубоко «не копали». Ранее (т. 1) было показано, что жизненная форма (биоморфа) = габитус, или внешний облик растения (и животного) — в чистом виде иллюзия. Формула жизненной формы должна быть иная — солиднее, коль это форма жизненная, или форма жизни, а не просто форма га- битуса. Для начала ответим кратко. Габитус сам по себе нежизнеспосо- бен, поскольку не имеет внутренней структуры и не может быть формой жизни кого угодно; он может быть только частью — наряду с другими частями, расположенными под габитусом — определенных форм жизни, если считать, например, их организмами и синорганизмами (т.1). Такой ответ «режет» ухо традиционным габитуалистам; он озадачит любого биоморфолога. Однако, приглушим в себе консервативные настроения и попробуем разобраться во всем спокойно: ведь мы только открыли книгу, посвященную габитусам, их формам и формам роста биоморф. Но не будем забывать при этом, что невольно мы оказались на острие проблемы со всеми вытекающими отсюда последствиями, и у автора, как, впрочем, и у читателя, похоже, нет права на большие ошибки. Кстати, при чем тут формы роста, да еще в организации биоморф, скажете Вы. Дело в том, что габитус, будучи неотьемлемой, но лабиль- ной частью биоморф, в первую очередь завязан на не менее лабильную часть их организации — формы роста. Поэтому лучше всего изучать их рядом, они дополняют друг друга, но более подробно об этом — ниже. Габитус, безусловно, должен занять в биоморфе свое определенное место (!); для кого-то оно покажется скромным, а для кого-то — весьма солидным (на одно и то же можно смотреть под разными углами зрения и с разной степенью абстракции). Пожалуй, вокруг термина «габитус» проблем не меньше, чем было вокруг «падающей» башни в Пизе; но так было лишь до тех пор, пока к ней не подошли кардинально. Для начала сформулируем первую часть проблемы, не разобравшись с которой, бесполезно идти на какие-либо обобщения. Согласимся с тем, что афористически кратко выразил П.К. Анохин: «можно бесконечно долго говорить вообще, если не оп- ределить понятий». Итак, что такое «габитус»? С чего он начинается и где заканчивается? Ответить на этот вопрос мы попробуем несколько позже (см. главу 3), поскольку весьма полезно посмотреть на проблему шире, что открывает немало новых моментов, способствующих поиску оптимального ответа.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Есть однако одно обстоятельство, о котором надо сказать уже здесь. В эволюции едва ли не всех научных терминов есть нечто общее: будучи однажды применены специалистами узкого профиля для обозначения какого-либо конкретного явления и получив признание у коллег, эти термины нередко накапливают в себе некий потенциал: им становится тесно в исходной «понятийной шубе». Хорошо это или плохо, обсуждать излишне: подобное часто не зависит от нашего желания. Нередко ока- зывается, что границы явления, о котором говорят специалисты, выхо- дят за пределы их узких интересов или даже — их компетенции. Посте- пенно у «неуживчивого» термина накапливается столько противоречий, что он начинает «проситься» на свободу, к расширению своего смысла (случается, конечно и обратное). Таких терминов немало: метамерия, побег, стеблекорень, партикуляция и т. п.; к их числу принадлежит и термин «биоморфа», первоначальный смысл которого, по мнению авто- ра, был определен слишком узко, в результате чего неизбежно последо- вала процедура его переоценки, уточнения и расширения его значения (т. 1). Что касается терминов «габитус» и «форма роста», то им по сути посвящена вся эта книга, и автор будет удовлетворен, если читатель, закрывая последнюю страницу, подумает: «а ведь об этом давно надо было написать». Итак, у терминов в науке есть своя «жизнь», в которой они стре- мятся к максимальному выражению себя там, где их применяют. Когда термины применяются не вполне адекватно и возможности их оказы- ваются не до конца исчерпанными, они трещат под тяжестью возник- ших проблем. Рано или поздно всегда находятся люди с творческим подходом, без которых терминология погрязла бы в консерватизме и догматизме. Так, на наш взгляд, работает эволюционный механизм в развитии научной терминологии. Фактографический материал этой книги отражен в многочислен- ных иллюстрациях, в описаниях жизненных циклов различных биоморф (организмов и синорганизмов), а также в ссылках на другие виды био- морф, в которых воплощены определенные особенности их организа- ции. Но многое из того материала, который накоплен автором, остается как бы «за кадром» («как бы» — поскольку материал этот всегда рядом и незримо «подпитывает» теоретические положения). За 35 лет работы в биоморфологии автор каждый год изучал жизненные циклы биоморф нескольких десятков видов семенных растений и в естественных усло- виях, и в условиях выращивания. Большую часть этого материала я рассчитываю постепенно опубликовать в пяти томах монографии. Ги- гантский пласт фактического материала накоплен и в трудах ботаников, особенно отечественных, которые усердно изучали и изучают жизнен- ные циклы различных растений, создавая надежную основу для теоре- тических исследований вообще и для моих биоморфологических разра- боток, в частности. Среди многих экспедиций, в которых мне удалось участвовать в предыдущие годы (т. 1), особую роль играет большая (3 месяца) и пло- дотворная поездка на Ближний Восток (1998, март — июнь): в пустыню
Введение Негев и прилегающие к ней территории (Израиль). Кроме того, биомор- фологические исследования продолжались в Подмосковье. Знаменитый А. Пуанкаре как-то заметил, что наука содается из фактов, как дом из кирпичей, а задача ученого организовать факты, поскольку их накопление является наукой не больше, чем куча кирпи- чей домом. Можно в чем-то и не согласиться с известным ученым, хотя по сути он прав. Наука, на наш взгляд, начинается, безусловно, с на- копления фактов, а вот строится и развивается через умение логичес- кими средствами оперировать ими: типизировать, классифицировать, моделировать, определять законы и правила, т. е. в целом теоретизиро- вать на основе фактов — превращать горы кирпичей-фактов в доброт- ные здания науки. В нашей монографии слово «жизнь» иногда пишется с большой буквы; это не опечатка, а осознанное желание автора говорить о явлении «жизнь» вообще, в самом широком, возвышенном смысле, где-то намекая, что жизнь (с традиционно строчной буквы) на Земле — это одно, а жизнь вообще, или Жизнь (с большой буквы) — это уже нечто большее, несмотря на то, что пока мы совсем ничего еще не знаем о внеземной жизни. Прием с необычным (прописным) написанием слова «жизнь» иногда используют и другие авторы', например, когда они в подобном (возвышенном) стиле говорят о природе вообще — Природе, космосе вообще — Космосе, мире вообще — Мире, разуме вообще— Разуме, всем человечестве — Человеке. В работе в одном смысле используются некоторые термины, кото- рые только внешне (по происхождению) вроде бы разные, а на самом деле их однозначные слова-этикетки просто-напросто написаны в раз- ных сочетаниях и на разных языках, например, биоморфология расте- ний = фитобиоморфология, биоморфы растений = растительные био- морфы = фитобиоморфы и т. п. Латинские названия растений приводятся, в основном, по книге С.К. Черепанова (1995). Растения Ближнего Востока определялись по «Аналитической флоре Израиля» (Feinbrun-Dothan, Danin, 1991). Весь- ма полезной для меня при определении растений Алтая была книга Л.И. Малышева «Определитель высокогорных растений Южной Сиби- ри» (1968). Кроме того использовалась «Флора СССР» (т. 1-ХХХ), «Ил- люстрированный определитель растений Казахстана» (1969, 1972) и дру- гие флористические работы. Все оригинальные рисунки выполнены автором. Их много, и я решаюсь предвосхитить возможный вопрос: зачем столько рисунков, на который сразу же однозначно отвечу: лучше один раз увидеть, чем де- сять раз услышать. Слова, слова, слова — их сочетания так разнообраз- ны, и при нашем писательском старании многое могут передать, но все- таки им не угнаться за возможностями рисунка: не достигнуть той полноты передачи информации, которую можно «загрузить» в каче- ственный научный рисунок. Хитросплетения слов, даже напечатанных, бывает, не вносят ясности; иногда они «бьют» мимо цели и невольно уводят читателя от познания истины, что можно более всего наблюдать
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ в тех флористических работах, где игнорируются иллюстрации. Только серьезный графический рисунок может наглядно передать морфологу, а шире — биологу вообще, да и просто любителю, где что есть, но и чего нет, причем передать и то, что есть, хотя может быть не «зацепилось» за слово или «потерялось» в словах или между слов, а возможно и то, на что автору было неуместно в данном тексте заострять внимание. Но рисунки-то состоялись, и они остаются. Среди слов можно потерять тонкие оттенки информации, которые несет научный рисунок, не об- ратив на них внимание здесь и сейчас, но они, рисунки, как носители информации, остаются в книге для тех любителей познавать новое, кто будет склонен еще и еще не один раз воспользоваться ими. Многое забудется, но только не рисунки — документальные свидетельства— биографические портреты видов (!), над которыми можно размышлять когда угодно, сколько угодно и кому угодно, причем и так, и этак, соглашаясь или не соглашаясь с рассуждениями автора. Среди различных словосочетаний можно не увидеть или, что еще хуже, потерять содержание вещей, поэтому для раскрытия таинств ра- стительной жизни я давно и много рисую, отлично понимая, что имен- но документальный рисунок более всего будет востребован в биоморфо- логии и систематике — только он никогда не устареет... Если хотите, эта монография, между прочим, — еще и творческий призыв автора к широкому внедрению в биоморфологию, а далее и в систематику, биографического (!) научного рисунка, а одновременно и пожелание биофакам университетов, рядом с компьютеризацией учеб- ного процесса, ввести и начать преподавать скромный спецкурс по научному рисованию. Советы и замечания по материалам монографии я в разное время и при разных обстоятельствах получал от В.Г. Хржановского, Т.А. Ра- ботнова, Т.И. Серебряковой, В.Н. Голубева, С.В. Мейена, которым весь- ма признателен. Общение с этими выдающимися учеными, пусть и непродолжительное, безусловно, так или иначе повлияло на формиро- вание моих научных взглядов. Большую благодарность выражаю ботаникам Израиля, которые пре- доставили мне возможность поработать (1998) в ботанических садах университетов Иерусалима и Тель-Авива — М. Эвишаю, М.Адами, а также, за организационную помощь, сотрудникам и стажерам Институ- та освоения пустынь в Негеве (Сде-Бокер), в особенности Н.С. Орлов- скому и 3. Аликулову. За поддержку и помощь в работе я признателен сотрудникам отдела культурных растений Главного ботанического сада Российской АН Ю.Н. Горбунову и Ю.С. Левандовскому, которые любезно предоставили мне возможность посмотреть биоморфологию некоторых растений, про- израстающих на участке экспозиции лекарственных растений. Я благодарен сотруднику Ботанического сада Всероссийского науч- но-исследовательского института лекарственных и ароматических рас- тений (ВИЛАР) О.А. Черкасову за предоставленную мне возможность посмотреть некоторые особенности биоморфологии горечавки желтой. произрастающей на его опытном участке. Кроме того я выражаю свою
Введение признательность многим коллегам этого института, с которыми мне довелось плодотворно поработать в экспедициях и на опытных участках по различным темам. В наше время без технической помощи высококлассных специали- стов своего дела, таких как В.В. Рюмин, А.В. Булатов, Д.С. Гусева, Ю.П. Гусев, хорошую книгу не сделаешь. Моя большая благодарность этим людям, обеспечившим соответствующую подготовку компьютер- ного варианта книги. Автор глубоко благодарен Т. Райхель и А. Райхель за помощь в организации работы за рубежом. Весьма полезными и своевременными при оформлении книги оказались несколько дружес- ких советов, которые дал Г.Г. Кагановский.
«Учение о формах есть учение о превращении. Живое обладает даром прилаживаться к многообразным условиям внешних влияний и все же не устуиать известной завоеванной определенной самостоятельности» (Гете (1749 — 1832), 1957, с. 389, 390/ Концепция меросомной организации биоморф семенных растений Глава 1. Основу Природы составляет, как известно, материя — объективная реальность, существующая в бесчисленном множестве конкретных форм различных объектов-систем: вещей и систем вещей. Материи чужд хаос; она проявляет ту или иную степень организованности в системах раз- ного рода. Наиболее высокую сложность организации демонстрируют живые системы, которые представляют из себя какие-либо тела и сис- темы тел. Природа — это единство материального и идеального, единство физиосистем (материальных систем) и логосистем (идеальных систем) (т. 1); кроме того, это еще и единство телесного и нетелесного. Рассуж- дая так, мы невольно затронули проблему, которую биологи чаще всего обходят стороной в силу того, что она как бы не имеет отношения к биологии; удивительно, но тему телесного и нетелесного не слишком балуют своим вниманием и философы. Однако, на наш взгляд, если в этой проблеме правильно расставить акценты, то уже в самом начале любого специального исследования появится дополнительный козырь: исследование обретет дополнительную методологическую глубину, по- лучит надежную гарантию защиты от возможной терминологической путаницы, предмет его более уверенно займет свое место среди других объектов. Попробуем для начала разобраться с этой проблемой — в том объеме, который необходим для нашей работы.
17 Глава 1. Концепция меросомной о ганизации биоморф семенных растений 1.1. Телесное и нетелесное в Природе То, что простая биоморфа — это организм, то есть представляет собой один из видов тел, не подлежит сомнению с любых позиций; вопрос однако в том, о какой телесности можно говорить в отношении сложной биоморфы — синорганизма? Ответ на этот вопрос требует некоторого отступления, поскольку сначала надо определить: что такое тело, где оно начинается, где его уже нет, а где присутствует нечто другое и что собой представляет это другое? Наш исходный тезис: Природа — единство тел и нетел. Из чего мы исходим, делая такое утверждение? «Вся доступная нам природа обра- зует некую систему, некую совокупную связь тел, причем мы понимаем здесь под словом «тело» все материальные реальности, начиная от звез- ды и кончая атомом и даже частицей эфира, поскольку признается реальность последнего. В том обстоятельстве, что эти тела находятся во взаимной связи, уже заключено то, что они воздействуют друг на друга, и это их взаимное воздействие друг на друга и есть именно движение» (Энгельс, 1964, с. 51). Господствует обыденное представление о теле как сплошном вещественном объекте. «Предметы, занимающие определен- ный объем в пространстве, называются телами, но многие предметы не являются телами (физические поля, элементарные частицы и т.д.)» (Философская энциклопедия, т. 4, 1967, с. 357), с чем соглашаются не все, поскольку «и поля, и частицы имеют пространственно-временные пределы, хотя и не всегда экспериментально устанавливаемые»(Круть, 1978, с. 42). Границы тел, которые физики именуют микрочастицами, провести нередко очень непросто. Вот что говорит о протоне С.Т.Ме- люхин (1958, с. 81 — 82): «Элементарным частицам нельзя приписывать резких геометрических границ, поскольку они не являются какими-то микроскопическими шариками, а обладают также волновыми свойства- ми и неразрывно связаны с полем. Каждое из полей вносит свой вклад в структуру частиц, и невозможно четко определить, где кончается внеш- нее поле и начинается собственно частица.... Такой границы объектив- но не существует, а размеры протона в значительной степени опреде- ляются энергией взаимодействия его с налетающей частицей.... Тем не менее, если отвлечься от этих тонкостей и не требовать точного определения «границ» каждой элементарной частицы, можно в общем выделить внутренние и внешние связи также и для элементарных частиц, поскольку каждая частица взаимодействует как единое дискрет- ное образование, а не «размывается» на обширную область простран- ства». На рубеже XIX и XX веков В.И. Вернадский возрождает, как под- черкнул И.В. Круть (1978, с. 55), «классическое понятие естественного тела, расширяя его, однако, на значительно более широкий круг объек- тов, чем это делалось ранее, например, с одной стороны, на электро- магнитные поля, а с другой — на столь сложные системы, как геоло- гическая оболочка и биосфера». «Естественным телом в биосфере мы будем называть всякий логически отграниченный от окружающего пред- мет, образовавшийся в результате закономерных природных процессов,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ в биосфере или вообще в земной коре происходящих» (Вернадский, 1991, с. 151). «Материальная вещь — система — имеет пространственно- временные границы (пределы) и уже поэтому является телом, даже если составляющие ее вещи — компоненты — дискретно изолированы друг от друга.... Вещью — телом — является и Солнечная система, и географи- ческая оболочка, и ландшафт, и видовая популяция организмов, и атом, и электрон, и электромагнитное поле» (Круть, 1978, с.42). Суммируя общепринятые представления о телесном и определения телесного, можно так определить тело (сому, телесную систему): зто часть материального мира (физиосистема), обладающая относительной самостоятельностью, качественной оиределенностью и существующая не- зависимо от нашего познания как любая ограниченная область лростран- ства вместе с ее границами или как система вещей — комионентов, изолированных друг от друга, но существующих в единстве для вылолнения единой цели. Природным телом, как считает И.В. Круть (1978, с. 63) является «материальный объект — вещь (или ее фрагмент) — с фикси- рованными пространственными и (или) временными границами. Физи- ческие субстанции природного тела могут быть веществом (веществен- ный субстрат), физическими полями, сочетаниями того и другого». Тела весьма разнообразны как по форме, так и по содержанию. Физики, например, говорят о твердых телах, а среди них различают кристалли- ческие и аморфные тела. Пожалуй, этого достаточно, чтобы показать, что все известные биосистемы являются живыми телами (биосомами) даже в том случае, если их части разобщены между собой, иногда раз- делены очень большими объемами пространства. Так мы подходим к факту, что не только организмы (простые биоморфы), но и синорганиз- мы (сложные биоморфы) являются телами, хотя и разными по содер- жанию и форме. Тело телу рознь, и чтобы ориентироваться в разнообразии тел, их надо типизировать и классифицировать. Однако не будем слишком ув- лекаться детализацией этого разнообразия, чтобы не отклониться от движения в нужном направлении — к решению задач, поставленных в книге. Ограничимся несколькими моментами. Все разнообразие тел можно рассматривать под разными углами обзора. Несколько критериев классификации тел, которые мы здесь приведем, имеют самый широкий смысл, а следовательно — и это для нас важнее всего — они могут быть использованы и в биологии. Глав- ное, что присуще всем телам, без исключения, — способность занимать собой определенные фрагменты пространства на какое-либо время и отделять себя, как данное тело, определенными границами (более чет- кими или менее четкими, но обязательно фиксированными) от иного, уже не принадлежащего им непосредственно участка окружающего про- странства. Рассуждая так, мы можем разделить все тела на средоокру- женные, или центросомы, и средоокружнющие (средосоставляющие), или ТОПОСОМЫ. По размерам тела можно разделить на: ультрамикросомы, микросо- мы, макросомы и ультрамакросомы. Ультрамикросомы — это сверхмик- роскопические тела; современная наука пока не располагает средствами,
~лава 1. Концепция мероеомной организации биоморф семенных растений 19 позволяющими увидеть их более сложное строение, если они им обла- дают. Ультрамикроскопическими телами, известными современной на- уке, являются, например, нейтроны, протоны и т.п. Микроскопические тела — это микросомы, сложноорганизованные системы, которые нельзя обнаружить без специальных увеличительных приборов (молекулы, гены, органеллы, клетки и т.п.). Тела, которые видны без специальных увеличительных приборов, для удобства обозначаются как макроскопи- ческие, или макросомы (дерево, столб, гора, Солнце и т.п.). Особую группу составляют ультрамакроскопические, или сверхгигантские, тела, границы которых определяются только посредством специальных астро- номических приборов, — это ультрамакросомы (Солнечная система, Галактики и т.п.). Все тела, с которыми мы так или иначе имеем дело, обладают сложным строением, т.е. состоят из двух и более тел меньшей сложности организации. Не будем слишком углубляться в проблему сложности организации тел, которая пока далека от детальной научной разработки. Для нас важен факт, что все живые тела, равно как и все окружающие их неживые тела имеют сложное строение. Все тела (телесные системы): и центросомы, и топосомы — можно (уже под другим углом обзора) подразделить на два типа (класса). Это, во-первых, одноконтурные тела, или монадосомы' (от греч. топав, род. падеж monados — единица, единое, soma — тело), т.е. физиосистемы (материальные системы), состоящие из любого числа тел меньшей слож- ности организации, но имеющие конкретные границы, обеспечивающие им одноконтурную (!) физико-химическую и морфологическую целос- тность. Монадосомами являются кристаллы, стволы деревьев, сами де- ревья, планеты, звезды, организмы и тому подобные материальные (те- лесные) системы. Во-вторых, многоконтурные тела, или плеосомы (от греч. pleon — больше, soma — тело), т.е. физиосистемы, представленные несколькими монадосомами, связанными в одну систему единством цели, дизъюнктивного телесного существования, но разграниченными каким-либо, посторонним для них, пространством. Плеосомы делятся на два типа. Первый тип — однородные (синор- ганизмы, виды, ценодемы и т.п.), где группы монадосом однородны по преднаследственности или наследственности. Второй тип — неоднород- ные (биоценозы, консорции и т.п.), где плеосомы образованы неодно- родными по преднаследственности или наследственности телами. Все тела имеют свои, так или иначе фиксированные границы. У однокон- турных тел границы непрерывны, у многоконтурных — более или менее прерывисты. Топосома (средосоставляющее, или средоокружающее, тело) — это часть материального пространства, в которой осуществляется непосред- ственное взаимодействие тела с теми или иными одноконтурными или многоконтурными средоокруженными телами (монадосомами или пле- осомами). Биотопы как среды существования живых систем (биосом)— это топосомы живых тел. В структуре растений границы семядольного узла, годичных приростов и др. с возрастом могут нивелироваться, Словосочетания моносома» и «полисома» мы не используем в нашем случае для по- строения соответствующих терминов, поскольку они уже заняты в генетике.
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ сглаживаться и совсем вроде бы исчезать, но все же остаются, хотя и становятся размытыми. Многие горные тела также обладают приблизи- тельными границами, но это не означает, что границ нет: они или остались в прежнем, но с трудом определяемом виде, или перешли в новое каче- ственное состояние; часто монадосомы становятся элементами плеосом. Кроме тел, в Природе существуют и петела (апосомы), которые лишены фиксированной системной телесности и иредставляют собой яв- ления иного рода: например, процессы (фотосинтез, дыхание, водоснаб- жение, эволюция и т.п.). Процесс (от лат. processus — продвижение)— это «последовательная смена явлений, состояний в развитии чего-ни- будь» (Бол. Энц. Сл., 1997, с. 971). Кроме нетел — процессов — суще- ствуют нетела — мысленные образы (логосистемы): идеи, слова, сны и т.п. Таким образом, нетела' (апосомы) — это движения тел, процессы, которые осуществляются телами и обозначаются не в структурном, а в функциональном (функция — способ существования структуры) изме- рении; нетелесное — это функциональная сторона проявления телесно- го. Нетелами являются и идеальные системы, которые образуются в результате сознательной деятельности людей и потенциально могут стать продуктом искусственного интеллекта. Природа — это бесконечное ди- намическое единство телесного (соматического) и нетелесного (апосомати- ческого). Основу материальной природы составляет единство тел и не- телесных явлений (процессов). Мы не будем далее продолжать типизи- ровать и классифицировать тела вообще, по сериальному или суборди- натному принципу: того, что мы привели выше, вполне достаточно, чтобы конкретизировать в наших дальнейших рассуждениях проблему телесного уже по отношению к биосистемам и, в частности, к биомор- фам семенных растений. Однако, сначала все же определим, каким методом или какими методами можно исследовать тела. У всех тел есть внутреннее содержание (внутренняя организация), а следовательно, есть и внешнее содержание (внешняя организация). Всякая ли внешняя организация является габитусом, или круг тел, об- ладающих габитусом, ограничен? Проблемный вопрос сформулирован; чтобы однозначно разобраться с проблемой габитуса биоморф, разбе- ремся сначала с некоторыми сопутствующими вопросами. 1.2. Метод меротомирования и основы меротомии — науки о частях целого Ранее, в соответствующем обзоре (т. 1, гл. 2. 3), было показано, что вне целостности нет живых систем. Целое — это совокупность предме- ' В медицине известен термин «антитела», который применяется к телам особой природы [«глобулярные белки (иммуноглобулины) плазмы крови человека и теплокровных животных, обладающие способностью специфически связываться с антигенами» (Бол. Энц. Сл., 1997)]; здесь мы не будем обсуждать достоинства и недостатки этого термина, но констатируем: то, что обозначается как антитело — один из видов тел и, следова- тельно, к апосомам (нетелам) никакого отношения не имеет.
Глава 1. Концепция меросомной организации биоморф семенных растений 21 тов, проявляющих себя в системном единстве через интегративные каче- ства ее частей. «Целое обладает новыми свойствами, которых нет в отдельных частях» (Шмальгаузен, 1982, с. 13). Может быть, не стоит тратить время на исследование отдельных частей в организации систем, а более углубленно изучать целое? Это затея малопродуктивная, поскольку наиболее полно познать целое мож- но через особенности структурных и функциональных взаимосвязей его частей. Часть и целое, как известно, выражают «отношение между со- вокупностью предметов (или элементов отдельного объекта) и связью, которая объединяет эти предметы и приводит к появлению у данной совокупности новых свойств и закономерностей, не присущих предме- там в их разобщенности. Эта связь выступает как целое, а предметы по отношению к ней — в качестве частей» (Филос. сл., 1972, с. 455). Целое состоит из частей независимо от нашего сознания, но может и расчле- няться на части логическим путем, в результате деятельности нашего сознания. Исследователь, вычленяя части из целого, может более или менее считаться с природным порядком в расположении частей, но может и устанавливать свою систему в расположении частей. Таким образом, расчленение целого на части зависит не только от особенно- стей его организации, но и от целей исследовательской или какой-либо иной (практической, теоретической) работы с системой. Метод члене- ния любой материальной или идеальной системы (целого) на части— в зависимости от разносложных особенностей организации этого целого или иных обстоятельств — обозначим термином «меротомирование». Меротомирование — это процесс членения целого на части и возникно- вение в целом мероидности, т.е. членства конкретных частей, или меро- идов (дословно с греческого «видов частей»), в организации предметов как тел или нетел. Конкретные части системы (целого), или мероиды, отличаются от других частей, мероидов, полученных тем же методом, и составляют фактуальную основу для логических операций с этими ча- стями и превращения их в типы и классы частей, которые С. В. Мейен (1978) предложил называть меронами, а логическую операцию типиза- ции и (или) классифицирования конкретных частей в ранге меронов он же обозначил как мерономия, посредством которой мероидность в чем- то подобных предметов исследования превращается в мерономность (членство абстрагированных, или типовых частей, — или меронов в типовом целом). Науку о членении целого на части (меротомирование) по особенностям организации частей целого можно назвать меротомией (дословно с греческого — «рассечение на части»). Меротомия состоит из двух методических разделов: меротомирова- ния (членения на конкретные части, или мероиды) и мерономии (ме- тодов типизации и классифицирования мероидов и превращения их в мероны, т.е. типы или классы частей). Таким образом, в меротомии предметов исследования всегда имеется две стороны: конкретная, когда мы непосредственно наблюдаем (ощущаем) части (мероиды) тел или нетел, и абстрактная, когда мы оперируем обобщенными образами ме- роидов (непосредственных частей целого) в типах или классах их, т.е. в меронах.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Если меротомирование применяется по отношению к телам, то мероидами в них являются меросомы (дословно с греческого — «части тел»), которые, как и все остальные мероиды (части нетел — мероапо- сомы), способны подвергаться мерономии (типизации и классифициро- ванию). В самом методе меротомирования в принципе вроде бы ничего нового нет. Однако, сама эта идея (множество «раскалывать» на части и «раскладывать» части по «полочкам») позволяет плодотворнее решать задачи биоморфологии, поэтому мы и далее будем использовать этот прием как средство познавательной деятельности. Конкретные части (мероиды) могут подразделяться в соответствии с особенностями их структурной или функциональной организации, их взаимодействия как внутри системы, так и по отношению к внешним факторам, а также в зависимости от многих других обстоятельств. Особую категорию частей, вычленяемых в процессе меротомирова- ния, составляют живые части. Все части физиосистемы как телесной системы — мероиды — одновременно являются частями тела, т.е. меросо- мами, а меросомность — это атрибутивное свойство их любых телесных систем и, соответственно, всех живых систем. Применяя метод мерото- мирования, мы приближаемся или отдаляемся (если ошибаемся) к по- знанию естественного разнообразия частей, которое присуще всем те- лесным системам, независимо от нашего сознания. Части организации биосистем вообще и биоморф в частности, в специализации которых доминирует направленность на выполнение определенных жизнесохра- нительных функций, было предложено (как уже отмечалось выше) на- зывать меросомами (Нухимовский, 1984, 1997). Сейчас, спустя несколь- ко лет после появления термина «меросома», видно, что возможности этого термина были несколько занижены и есть основания понимать его более широко, что мы и делаем в этой работе. Любая часть какой-либо системы вообще — это мероид; а уже: любая часть какой-либо телесной системы вообще и живого тела (биосомы) в частности — это меросома. Далее будет показано, что меросомы по своей организации весьма раз- нообразны. Явление меросомности (членства частей тела в целом теле) и метод ее исследования (меротомирование) имеют всетелесный (пансоматичес- кий) смысл. Если с разной высоты и под разным углом обобщения посмотреть на биоморфы семенных растений, которые всегда являются целостными системами и, безусловно, еще и телесными системами, то в их организации мы неизбежно будем обнаруживать, что они состоят из меросом, которые, во всех случаях находясь в едином живом теле, по- разному участвуют в его организации. Простые биоморфы (организмы) являются одноконтурными телами — монадосомами; сложные биоморфы- синорганизмы, основные части (меросомы) которых, клонисты (особи ве- гетативного происхождения), пространственно разобщены и всегда образу- ют вместе целостные дизъюнктивные (прерывистые) соматические системы — многоконтурные тела (плеосомы). Итак, в любых системах, как материальных, так и идеальных, как телесных, так и нетелесных, можно выделять части, т.е. везде можно использовать метод меротомирования как инструмент научного иссле-
Глава 1. Концепция меросомной организации биоморф семенных растений дования. Мерономность тем лучше воспринимается нашим сознанием, чем точнее нам удается подметить объективные закономерности и учесть их при меротомировании. Если части системы-целого действительно существуют как части и путем меротомирования удается определить их место и параметры, то, независимо или зависимо от сознания, сведения о мерономности будут более всего отвечать истине. Такое меротомиро- вание объективно отражает мероидную природу тел и, безусловно, яв- ляется научным достижением. Многое в научном поиске зависит от исследовательской программы («парадигмы», по Т. Куну, 1977), которая может быть настроена и на меротомирование соответствующих теле- сных систем. Выявление объективных особенностей мерономности те- лесных систем связано с многими аспектами меротомирования: многое надо «взвесить», от многого отказаться, но и при этом наше членение целого на части всегда будет не в полной мере совпадать с истиной, т.е. в нем, как бы мы ни старались, всегда будет присутствовать элемент субъективности, отпечаток нашей познающей воли. Таковы особенно- сти мыслительной деятельности людей и познавательные издержки нашего сознания по отношению к тому, что есть на самом деле; это и заставляет нас напрягать усилия, чтобы максимально приблизиться к истине, поиск которой составляет основу целесообразности в научной деятельности. Итак, учение о членении на части — меротомия, а процесс, обеспечи- вающий меротомию — меротомирование. Результатами членения на части тел и нетел (меротомирования) будут, соответственно, меросомность (члене- ние тел на меросомы) и мероапосомность (членение нетел на мероапосомы). Меротомирование зависит от целей исследования, а они могут быть зна- чимы для одного исследователя, а могут отвечать интересам многих и многих людей, даже всего человечества. В науке закрепляются лишь такие варианты мерономности (членства типов частей в целом), которые устраи- вают многих и более всего соответствуют пониманию истины на соответ- ствующем этапе развцтия науки, которое, как известно, проверяется практикой. Издержки меротомирования природных тел и нетел посте- пенно уходят в прошлое, сохраняя какое-то время исторический интерес для соответствующей науки. Далее мы больше будем говорить о меросом- ности — расчлененности тел на части (меросомы). 1.3. Многоканальность распределения ресурсных потоков в организации телесных систем Явления, связанные с организацией жизненных форм, рассматри- ваются в работе в их взаимосвязи и взаимозависимости. Обязательным способом существования материи является, как известно, движение (изменение в самом общем виде). Материи в процессе движения свой- ственно образовывать тела, которые раньше или позже превращаются в другие тела, и т.д. до бесконечности. Есть смысл повторить диалекти-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ческое положение: «в мире нет ничего, кроме материи (и ее свойств, которые тоже включены в материю). Материя в онтологическом аспекте есть субстанция, и кроме нее в мире ничего нет» (Алексеев, Панин, 1999, с. 360). Многое о материи уже известно, но значение даже этого термина далеко не исчерпано. Мы не будем вести здесь дискуссии о термине и понятии «материя», но согласимся с традиционным мнением, что «материя — это объективная реальность, существующая независимо от человеческого сознания и отображаемая им» (Алексеев, Панин, 1999, с. 349). Такого объяснения материи нам достаточно, чтобы высказать несколько общих положений, помогающих двигаться к решению основ- ных задач этой книги. Так или иначе, под разными углами обзора и на разных уровнях абстракции, мы проникаем в проблемы, связанные с организацией Природы, Жизни (жизни вообще), жизни на Земле и того, что состав- ляет непосредственный предмет наших исследований — жизненные формы, или биоморфы. 1.3.1. Коснувшись цветка, ты невольно потревожишь звезду {старинный афоризм) Естественно, что цветка я «коснулся» давно, но лишь со студенчес- ких лет задумался над тем, что стоит в природе вообще (Природе) за такими прикосновениями. Только сейчас осознал — и в этих размыш- лениях пытаюсь передать Вам, уважаемый читатель, — свое ощущение, что я действительно где-то и в чем-то «потревожил» звезду, которая развивается по единым для Вселенной (Природы) крупным законам, обязательным и для цветка, и для любой пылинки. Все тела состоят из материи, которая неисчерпаема в бесконечной и единой Вселенной. Это традиционная позиция, которую я здесь при- нимаю, хотя никто этого еще не доказал (у современной науки пока нет возможностей для этого) и гипотетически возможны совсем иные реше- ния. Например, если принять гипотетическое положение, что она, наша Вселенная, не одинока и что существует некий бесконечный Вселенс- кий Поток (тогда именно он и будет Природой), в котором несколько N-Вселенных, может быть, их много, даже бесконечно много. Границы нашей Вселенной определены теми законами мироздания, которые в ней действуют; в других Вселенных эти законы (или часть их) не дей- ствуют, и там многое, если не все, должно бы быть, иное. Наша Все- ленная в таких условиях развития, будучи ограничена в пространствен- ных пределах, бесконечна в трансформациях во времени. Возможны и иные варианты развития: допустим, что другие Вселенные, существую- щие где-то рядом, могут оказаться физически, химически, а может быть, даже биотически — активнее нашей Вселенной, пересилив в целом ее в своей массе, поглотив, даже растворив и переиначив тем самым дей- ствие прежних законов, приспособив их к своим. Чтобы нечто подобное могло разыграться в Природе, в ее бесконечности должно быть нечто
Глава 1. Концепция ме осомной организации биоморф семенных растений 25 большее, чем Вселенные, отличающиеся только действием определенных для них природных законов, т.е. только в их границах. Такое большее вообразить можно — это Вселенский Поток (тогда он синоним Приро- ды). Итак, гипотетически можно представить существование Природы не как одной Вселенной, а как Вселенского Потока, где в бесконечном пространстве и времени «плывет» неведомое число N-Вселенных, вклю- чая и нашу Вселенную, в каждой из которых действуют свои природные законы. Тем не менее что-то всех их объединяет: они, в образах эволю- ционных маятников, «плывут» (перемещаются) в эволюции по бесконеч- ности пространства и времени Вселенского Потока (Природы). После «первичной сингулярности, называемой Большим Взрывом», наша Вселенная начала расширяться (Шкловский, 1973; Дубнищева, 1997 и др.); эволюционный маятник нашей Вселенной качнулся в сто- рону ее расширения. Что будет через десять, двадцать или сто милли- ардов лет, вопрос на сегодня из разряда безответных, даже праздных. Однако, все-таки гипотетически можно заключить, что рано или поздно эволюционный маятник, в образе которого наша Вселенная качнулась в сторону расширения, двинется в обратный путь, и, возможно, это будет сжатие на возвратном движении «маятникового тела», заметно переместившегося и изменившегося за долгое время «плавания» во Все- ленском Потоке. Возможны и иные решения. Мы еще очень мало знаем о природе вообще (Природе). Что является синонимом слова «Природа». Вселен- ский ли Поток (с N-Вселенными) или вселенский поток (с маленькой буквы: в нем только одна Вселенная, поскольку других не существует) — пока достоверно не известно. Однако далее мы будем исиользовать слово «Природа»в традиционном смысле, как синоним одной бесконечной Вселенной (вселенского иотока), в которой движутся галактики, звездные других Вселенных. Одно ясно: из двух вариантов возможен или тот, или другой, что на точность наших текущих исследований вряд ли повлияет. Живые и неживые тела состоят из так называемой (так говорят и физики, и лирики) «звездной (солнечной) пыли», поэтому в приведен- ном выше известном афоризме есть прямое указание на взаимосвязь и единство всего, что есть и что происходит в Природе. Из множества химических элементов (уже известно 114 элементов, но цифра эта явно еще не последняя), составляющих Землю и всю Солнечную систему, живые системы и, в частности, биоморфы семенных растений исполь- зуют для развития своей организации далеко не все. В строительстве биосистем приоритет отдается немногим элементам, среди которых 97,4% химического состава приходится всего на шесть: углерод, кисло- род, азот, водород, фосфор и серу, а примерно 1, 6 % веса в организмах составляют еще несколько: натрий, калий, кальций, магний, железо, кремний, алюминий, хлор, медь, цинк, кобальт. Еще 20 элементов со- ставляют в организмах в совокупности около 1 % (Кузнецов и др., 1996). Часть организмов требует для своей жизнедеятельности бор, бром, йод, селен, хром, молибден, ванадий, кремний, стронций, барий и, не ис- ключено, никель (Хатчинсон, 1972).
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHbN РАСТЕНИИ ~ремя — наСВОя разование Земли Чиювек — мевац — водород — QNJMVQK — еодц Современные организмы Расаенц Виру молекул~( лярные cucmewы Биологическая эволюция Химическая эволюция Рис. 1. Схема происхождения живого из неживого (по М. Кальвину, 1971, с. 12). В биосфере по определенным законам осуществляется непрерыв- ное движение различных тел, в результате чего в биосфере постоянно происходит «круговорот биологически важных веществ, которые после использования должны вновь переходить (при участии солнечной энер- гии) в форму, пригодную для дальнейшего участия в круговороте» (Хат- чинсон, 1972, с. 20). Любопытные данные сообщают П. Клауд и А.Джибор (1972) о круговороте воды, кислорода и двуокиси углерода в биосфере (рис. 2). Важной частью в организации природы Земли являются экосисте- мы. Они, как известно, включают организмы, а шире — другие биоси- стемы более сложные, чем организмы, и неживое окружение, находящи- Любое вещество, как живое, так и неживое, по отношению к био- морфам, а шире — ко всем биосистемам, играет роль ресурсов. «Ресурсы (средства, запасы) — это любые вещества, энергия и информация, ко- торые составляют окружающую среду биосистем и сами биосистемы; они или вовлечены жизнью в круговорот, или еще могут быть в него вовлечены» (т. 1, с. 106). Ресурсы на Земле, а шире — во Вселенной не хаотично разбросаны, а распределены закономерно и находятся, где-то не без участия живых систем, биоморф в особенности, в сложных вза- имоотношениях друг с другом. «В конце концов, углубившись в про- шлое достаточно далеко, мы на~ем лишь единственный живой вид, который в свою очередь можно считать одним из представителей мно- жества физико-химических систем (в большинстве своем неживых), об- разовавшихся из первичных молекулярных систем, с которых начина- лась сама Земля. Граница между двумя эволюционными системами, вероятно, очень размыта» (рис. 1) (Кальвин, 1971, с. 11). В тонком слое живого вещества, «замешанного» в его биотопах, покрывающих вместе Землю, т.е. в биосфере, в больших количествах есть жидкая вода, дос- таточно много проникает в биосферу энергии от Солнца, в ней во множестве присутствуют химические элементы, из которых Природа научилась в процессе эволюции «мастерить» живые существа.
27 ~лава 1. Концепция меросомной о ганизации биоморф семенных астений еся во взаимодействии. Приведем схему (рис. 3), разработанную Е. Оду- мом (1968, с. 19) и нагл~щно отражающую в сравнительном плане осо- бенности взаимодействия компонентов наиболее распространенных экосистем. Живая природа, которая нас интересует более всего, является ча- стью органической природы (органического мира). На этом факте мы заостряем внимание сознательно, поскольку бытует мнение, что орга- нический мир — «совокупность организмов, населяющих биосферу Земли» (Бол. Энц. Сл., 1997, с. 849). С таким определением вряд ли Рис. 2. Упрощенная схема круговорота веществ в биосфере (по П. Клауду и А. Джибору, 1972, с. 74): «В непрерывном круговороте биосфера обменивается с атмосферой и гидросферой водя- ным паром, кислородом и двуокисью утлерода. Здесь дана упрощенная схема этого кру- говорота. Вся вода нашей планеты проходит цикл расщепления в растительных клетках и восстановления в животных и растительных клетках примерно за 2 млн. лет. Образующий- ся при этом кислород поступает в атмосферу. Весь кислород атмосферы проходит через живое вещество примерно за 2000 лет. Двуокись утлерода, выдыхаемая животными и растительными клетками, уходит в атмосферу и вновь фиксируется растительными клет- ками примерно за 300 лет.». можно согласиться, поскольку органическое в Природе бывает не толь- ко живым, но и неживым. Органическая природа (органический мир) — это все телесные системы (живые и неживые) Земли (шире — Все- ленной), построенные на основе органических веществ, которые воз- никли в прошлом или настоящем как соединения углерода с другими химическими элементами и которые сейчас присутствуют (или не при- сутствуют) в организации живых систем. Иными словами, органическая природа — это все органические соединения, вовлеченные или не вов- леченные Жизнью в организацию ее телесных систем. Главное таинство в организации растений как форм жизни в узком смысле (биоморф) — организмов и синорганизмов, а шире — всех био-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Itic ВО IV Рис. 3'. Составные части экосистем и их взаимодействие (по Е. Одуму, 1968, с. 19): «Сопоставление общей структуры надземной экосистемы (луговое сообщество) и водной экосистемы (морской и пресноводной). Необходимыми элементами для функционирова- ния являются: I — абиотические вещества (основные неорганические и органические слагаемые); !! — продуценты (растительность на суше, фитопланктон в воде); П1 — мак- роконсументы, или животные: а — травоядные (кобылки, полевые мыши и т.д. — на суше; зоопланктон и т.д. — в воде); в — косвенные или питающиеся детритом консументы, или сапробы (почвенные беспозвоночные — на суше, донные беспозвоночные — в воде); с- верховые хищники (ястребы и крупные рыбы); IV — разлагатели (гнилостные бактерии и грибы)». систем, можно усмотреть в том, что все они состоят, в первую очередь, из неживой материи и что жизнь как структура и как процесс в каждом отдельном (дискретном) теле участвует в круговороте ресурсов биосфе- ры, проявляя себя как совокупность открытых систем, поглощая что-то из биосферы и что-то возвращая обратно. Посмотрим сначала, как про- являет себя жизнь в типовом теле растительных биоморф в их отноше- ниях с неживой материей. Согласно закону единства живого и неживого (закон Докучаева-Вернадского, см. т. 1, гл. 3.4) жизнь на Земле опира- ется на единое материальное основание, частицы которого («солнечная пыль») в разных соотношениях составляют и Солнце, и Луну, и нашу Землю, и все живое на ней. Таким образом, единая материальная не- живая первооснова нашей Солнечной системы — это «солнечная пыль», состоящая из проатомов, атомов и молекул в разных сочетаниях, из которых возникла Земля и жизнь, сумевшая в процессе эволюции по-
Глава 1. Концепция ме осомной о ганизации биоморф семенных растений 29 ставить себе на службу неживую природу, научившись приспосабли- ваться к ее весьма лабильному «поведению». Возникновение жизни на Земле — это наиболее яркий синдром новизны в эволюции Природы. Чтобы удержать физические силы среды в определенных телесных системах, своих носителях, живое обязано затрачивать энергию на со- вершаемую работу. «В конечном счете необходимую для жизни энергию поставляет внешний источник — Солнце. У растений в процессе эво- люции выработались особые пигменты (в том числе хлорофилл), кото- рые поглощают световую энергию. Затем эта энергия превращается в химическую энергию питательных веществ — сахаров, образующихся из таких простых органических соединений, как двуокись углерода и вода. Этот процесс улавливания энергии называется фотосинтезом, что бук- вально означает объединение при помощи света. Энергия, содержаща- яся в химических связях вырабатываемых растениями сахаров, а также белков и жиров, может в дальнейшем использоваться самими растени- ями или же животными» (Риклефс, 1979, с. 17). В целом явление обязательной взаимосвязи живого и неживого в организации живых телесных систем: биосистем вообще и биоморф в частности — уже неплохо изучено (Вернадский, 1965, 1967; Кальвин, 1971; Шредингер, 1972; Руттен, 1973; Фокс, Дозе, 1975 и др.). Используя в процессе познания такой методологический прием, как изменение угла обзора и уровня обобщения, посмотрим на это явление с позиций нашей меросомной концепции и попробуем четко определиться в по- нятийном отношении, ответив, здесь и сейчас, на вопрос: в чем суть единства и разнообразия меросом в организации биоморф семенных растений, а шире — и всех биосистем? Будучи живой системой, любая биоморфа, а растительная в особен- ности, буквально «накачана» неживой материей, что составляет одну из главных содержательных сторон организации биоморф. Живые организ- мы и синорганизмы состоят в первую очередь из первоприродного (аби- огенного) неживого вещества, — это первый тип меросом в организации живых тел (биосом) — абиомеросомы. Кроме того, внутри живых тел нередко присутствуют уже отжившие (мертвые) части, элементы быв- ших живых структур, — это второй тип меросом — некромеросомы. Наконец, собственно живое качественное содержание организации живых тел составляют живые части целых биосом, — это третий тип меросом по рассматриваемому ряду признаков — биомеросомы. Органическое неживое, т.е. собственно то, что является некроме- росомой, когда оно входит как часть в организацию целых живых тел, дезинтегрируется дальше до первородного неживого, т.е. до атомов, простых молекул, в согласии с давно известным афоризмом «все роди- лось из праха и все превратится в прах». Но все ли настолько трагично? Вероятнее всего, это далеко не так. Все находится во власти не хаоса, а эволюции; полного краха и праха для Природы (природы вообще) не существует. На смену одному порядку, одному разрушению приходит возрождение чего-либо и (или) кого-либо и так бесконечно. «Атомы в химических реакциях никогда не исчезают, не образуются и не превра- щаются друг в друга; они только перегруппировываются с образованием
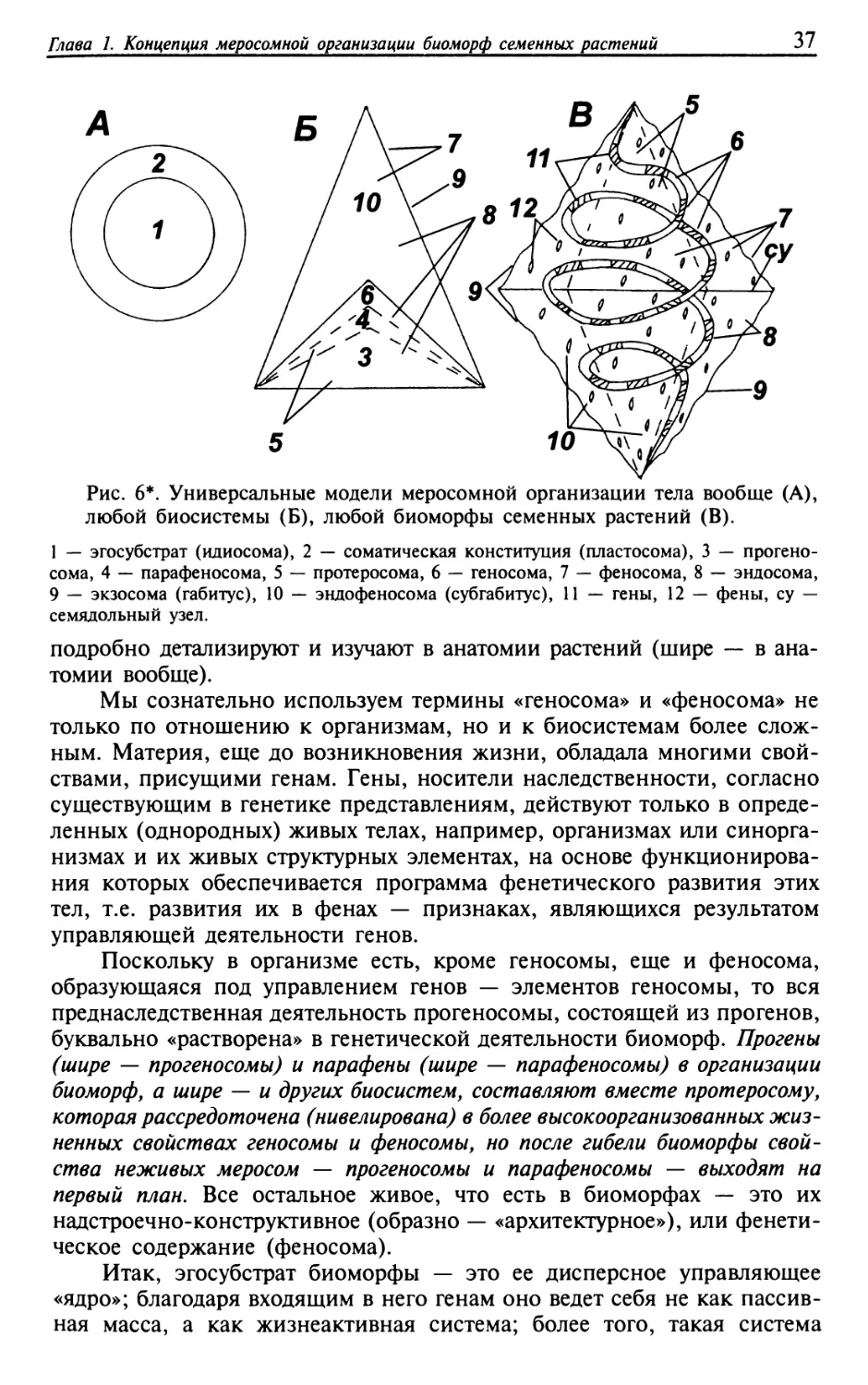
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ различных молекул и соединений. (Атомы могут изменяться при высо- коэнергетических ядерных реакциях, что и лежит в основе ядерной энергетики... Однако при биологических и химических процессах такие реакции исключены, поэтому с учетом вышеназванной оговорки закон сохранения массы можно считать универсальным.)» (Небел, 1993, с. 63). 1.3.2. Эгосубстрат и конституция тел, геносома и феносома биосистем Материя, эволюция, жизнь и Природа неотделимы друг от друга. Что же мы, люди, венец эволюционного развития Природы, способный ее познавать, можем наблюдать в ней? Природу составляет бесконечное разнообразие тел и процессов, существование которых не хаотично, но организовано целенаправленно, подчиняется определенным законам. Всякое существование материальных систем (физиосистем) — это всегда движение к «цели», цель мы понимаем широко, считая, что «это просто некоторое конечное (определенное — Е. Н.) состояние, к которому си- стема стремится в силу структурной организации» (Рапопорт, 1969, с. 98). Отсчет всего, что есть в Природе, начинается с материи. Только она, материя, ее свойствами, на мой взгляд, обеспечивает содержание явлений Природы в двух состояниях: субстрата и постсубстрата. «Суб- страт (филос.), общая основа всех процессов и явлений» (Бол. Энц. Сл., 1997, с. 1159). Субстратное состояние явлений (субстрат) — это общая континуальная (бесконечная) или субконтинуальная (подбесконечная) ма- териальная основа существования тел и нетел Природы в ее пространстве и времени. Континуальность подчеркивает неисчерпаемость материи и Природы, а субконтинуальность — соматическую преемственность ма- терии как преднаследственности или наследственности, что подробнее будет рассмотрено ниже. Постсубстратное состояние явлений (постсубст- рат) — это все разнообразие дискретных (!) структурных и функциональных форм тел и нетел, которое возникает на материальной основе (субстрате) и из нее в зависимости от окружающих условий (рис. 4). Постсубстратом будет все дискретное разнообразие форм тел, процессов, но и систем идеальной природы (логосистем), которое перемещается и трансформи- руется в пространстве и во времени. Место и ресурс для развития по- стсубстрата, его форм, — всегда только субстрат. Идея существования явлений Природы в двух их состояниях: субстратном и постсубстратном — одна из исходных в нашей монографии. Природа развивается так, что ее тела определенным образом ори- ентированы на достижение некоторых целей. Средством, играющим главную роль в «самонаведении» развивающихся тел на цель, является преднаследственность и (или) наследственность — две формы преем- ственности. Ранее (т. 1, гл. 2) было показано, что, кроме наследствен- ности, общепризнанным носителем которой являются гены (определен- ные участки макромолекул ДНК, реже РНК у вирусов), существует еще и преднаследственность, носителей которой, по предложению автора,
да~а 1. Концеиция ме осомной о ганизации биомор семенных растений Рис. 4'. Исходный уровень расслоения Природы (фрагмент) на две атрибу- тивные части: субстрат и постсубстрат. а — субстрат, б — постсубстрат, б1 — конституции тел, б2 — нетела идеальной природы (логосистемы), в — указатели разных направлений взаимодействия каких-либо сил. можно называть прогенами (вероятнее всего, это атомы, сохраняющие все свойства соответствующих химических элементов и являющиеся пределом химического разложения любого вещества). Все живые и неживые тела в Природе многими своими свойствами обязаны тому, что существующие химические элементы — это виды атомов с одинаковым положительным зарядом ядра, т.е. прогенов- элементов преднаследственности. Это они, прогены (атомы), обеспечи- вают преемственность в существовании всего разнообразия неживых тел, а заодно служат и первоосновой для более высокого по уровню сложности организации носителя наследственности (преемственности живых тел) — генов. Итак, в живых телесных системах, в том числе биоморфах, обнаруживаются два типа организаций как частей целого— носителей преемственности: прогенетическая организация (прогеносома) и генетическая организация (геносома). Нужно как-то обозначить универ- сальную единицу преемственности, являющуюся одновременно и проге- ном, и геном; в качестве таковой предлагаю считать понтикулу [от лат. pons (pontis) — мост, си1а — уменьшительный суффикс]. В процессе эволюции живые тела, возникнув однажды из неживого, не только научились жить (адаптировались) в окружении неживого и другого живого, но нередко и внутри, в разной мере состоят из неживого неорганического и органического ресурсного материала. Вся организа-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИИ ция биосистем буквально пропитана не только носителями неживого (абиоморфами), но и непосредственными носителями жизни (эвриби- оморфами), т.е. «меньшей», чем родительская, жизнью, использовавшей родительские тела в качестве инфраструктурных биотопов (т. 1, гл. 2). Существует проблема: как соотносятся между собой в организации био- систем вообще и биоморф в частности составляющие их неживые и живые структурные элементы. Первое, что свойственно любым носителям жизни, — это присут- ствие в их организации первоприродной неживой материальной части, которая континуально пронизывает Природу, все ее телесные системы, как живые (биосистемы, или биосомы), так и неживые (абиосистемы, или абиосомы). В организации биофоров (носителей жизни в широком смысле) существует более или менее тонкая связь между собственно неживой и живой материей. Наша задача сейчас — разобраться в пере- плетении ресурсных потоков, составляющих биосистемы и экосистемы, подчеркнуть их отличие от абиосистем. Начинать надо с абиосистем, поскольку живое возникло в процессе эволюции из неживого и поэтому в своей первооснове просто обязано состоять из неживой материи. Мысленно расчленим единую организацию абиосистем на следующие части (меросомы): прогеносому (ее составляют прогены, в качестве ко- торых здесь рассматриваются атомы, т.е. наименьшие частицы любого химического элемента, сохраняющие все его свойства, являющиеся пре- делом химического разложения любого вещества, и парафеносому (па- рафенотипическая по содержанию) — совокупный непосредственный результат развития той части материальной системы, которая возникает в результате взаимодействия прогеносомы (прогенотипа) с условиями внутри родительской материальной системы и внешней средой. Проге- носома и парафеносома — это атрибутивные части неживых тел; когда только они присутствуют в организации таких тел, то это неживые не- органические тела (эуабиосомы). Однако, поскольку на Земле процветает жизнь, то побочным, но обязательным продуктом ее проявления выс- тупают неживые органические тела (некросомы, или некроны). Некросомы отражают результат бывшей фенетической деятельности тех частей жи- вых тел (биосом), через которые затем прошла смерть, превратив эти части в особое состояние материи — неживое органическое. Некросомы сохраняются далее в организации живых телесных систем (например, вторичная древесина в стволах и ветвях деревьев, сухие колючки в кро- нах древесных растений и т.п.) или рассеиваются за ее пределами. Многие неживые тела на Земле: допустим, горы, озера, реки и, тем более, те неживые тела, которые возникают в результате осознанной деятельности людей и в состав которых, наряду с неорганической час- тью, входит еще и неживая органическая часть (некросома), без живых существ, которые могут быть рядом или внутри таких тел, например, садовый участок (без сада), деревянный стол, книга и т.п. — все это эвриабиосомы. Различные иерархические отношения отмеченных здесь тел в соматической организации жизни на Земле показаны в главе 7.2. Если все прогены и гены объединяются под одним общим назва- нием «понтикулы», то все парафены и фены в обобщенном смысле
Глава 1. Концепция ме осомной организации биоморф семенных растений можно назвать пластонами (от греч. plastos — вылепленный, on — су- щее). Экофенетическая часть, или совокупность всех пластонов в орга- низации экосистемы, это — экофеносома. Совокупность всех понтикул экосистемы составляет ее экогенетическую часть, или экогеносому. От- разим меросомную организацию носителей жизни (биофоров) — био- систем и экосистем — под рассмотренным углом обзора в графических моделях (рис. 5). Гены как самостоятельные тела занимают особое место в органи- зации Природы: это — переходное положение между неживой и живой природами, особое телесное состояние живой материи. Гены в своей структуре состоят из протеросомы (неживой материальной части, вклю- чающей прогены — соответствующие атомы) с некоторым участием парафенов (определенных, производных от атомов, неорганических мо- лекулярных соединений — воды, например) и собственно генетических структурных элементов. Вот так, потянув за веревочку, мы пришли к проблеме, теоретичес- кое осмысление которой, под рассматриваемым в нашей работе углом зрения, похоже, еще и не начиналось; впрочем, глубинная разработка ее не входит и в мои планы. Однако все-таки, коль приоткрыли, попро- буем чуть больше еще открыть эту «дверцу». Что происходит в органи- зации живых систем, когда гены работают и воспроизводятся? Гены, как известно, управляют развертыванием фенетической организации; кроме того, они управляют развертыванием и собственной организации. Толь- ко они — гены — могут управлять воспроизведением самих себя (реп- ликацией ДНК) — образованием сетеподобных структур, а следователь- но, и образованием структурных элементов, содержание которых слож- нее, чем прогенов и парафенов (в нашем случае — неорганических мо- лекул). О составе генов многое известно в деталях (Дубинин, 1976, 2000; Уотсон, 1978; Роллер, 1978; Гершензон, 1979 и др.). Длинную цепь ДНК ( «генонему») образуют чередующиеся остатки сахара и фосфата, к ос- таткам сахара присоединены основания (одна из четырех слабооснов- ных органических молекул: аденин, цитозин, гуанин или тимин), — это все структурные элементы генов. Интересный термин «генонема» (дос- ловно с греческого «генетическая нить») был предложен Н. К. Кольцо- вым (1938), и похоже, что на современном уровне знаний его вполне можно было бы удачно использовать для обозначения той цепочки ге- нов, которые вместе образуют одну единую нить макромолекулы ДНК (решающее слово здесь должны сказать генетики). На основе общеприродного противоречия между бесконечностью и дискретностью в системной организации неживой и живой природы имеет место быть моноидность (дословно с греческого — «имеющий вид одного») и полиоидность (дословно с греческого — «имеющий вид мно- гих») в сложении телесных систем, возникшие как способы завоевания и освоения телами пространства во времени, — результат единства и борьбы двух великих противоположностей (бесконечности и дискретно- сти). Моноидные тела, или монадосомы, едины и одноконтурны в своих соматических границах, за пределами которых нет их структурных элементов. Полиоидные тела, или плеосомы, по разным причинам
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИЙ 15 Рис. 5'. Графические модели меросомной организации биофоров (носителей жизни): биосистем (А, Б, В, Г) и экосистем (Д). 1 — абиоморфы, 2 — эврибиоморфы, 3 — протеросома, 4 — генофеносома, 5 — эуаби- осистемы (эуабиосомы), 6 — некросистемы (некросомы), 7 — биосистемы (биосомы), 8— геносома, 9 — феносома, 10 — экогеносома (заштриховано), 11 — экофеносома, 12— прогеносома, 13 — парафеносома, 14 — биотоп (топосома одного или нескольких живых тел), 15 — экосистема (экосома), 16 — средоокруженная биосома (живое тело). разобщили некоторые свои структурные элементы, создав разноконтур- ные границы своей единой организации. Моноидность — это менее сложный тип организации телесных систем. Все полиоидные тела состо- ят из моноидных тел, объединенных в одно тело как его дизъюнктивные структурные элементы на разной организационной основе. Так, среди живых тел самые нижние уровни организации занимают моноидные биосистемы от органелл до организмов, а более высокие — полиоидные биосистемы (любые надорганизменные тела, включая всю биостробу Земли). Живая природа получила («унаследовала») в процессе эволюции от неживой природы свойства своих носителей: и моноидность, и по- лиоидность, — поскольку, только владея ими, телесные системы могут бесконечно долго и далеко дискретным образом осваивать пространство во времени. Гены, будучи моноидными телами, составляют участок не- прерывной биополимерной молекулы ДНК ( «генонемы»), которая в свою очередь является также моноидной соматической системой. Гено-
Inaea I. Концепция меросомной организации биоморф семенных растений сома (вся система генов и генонем) внутри какой-либо фенетической организации живого тела (биосистемы) — это обычно полиоидное тело. Жизнь — явление неоднородное, многослойное. Собственно живое, будучи логическими средствами «снято» с неживого материального суб- страта (протеросомы) — материального основания целого живого тела, имеет всегда генофенетическое содержание, являясь, таким образом, «ге- нофеносомой» — приоритетной меросомой по отношению ко всему осталь- ному (протеросоме), что полностью ассоциировано в ней, но существова- ние которого как меросомы вряд ли у кого вызовет сомнения (рис. 5Б). Генофеносому можно меротомировать и дальше; здесь обнаруживается любопытный факт: генофеносома — это действительно живая часть- тело (меросома) в живом целом теле (ценосоме)-биосистеме; а как быть с осознанием места в жизни ее «половинок» — генетической меросомы (геносомы) и фенетической меросомы (феносомы)? Дело в том, что совсем по отдельности как живые тела обе эти «половинки» вне гено- феносом в современной живой природе Земли не существуют. Живые генетические и фенетические части генофеносомы проявляют себя так только вместе, в единстве, и не приходится говорить об их самостоя- тельной жизни, хотя экспериментально, пожалуй, можно «обмануть» и гены, и фены, сохранив на какое-то время их жизнеспособность с помощью различных искусственных приемов, например, с целью транс- плантации (многое в этой проблеме еще не изучено). Итак, геносома и феносома — это живые «половинки» (меросомы) в организации генофеносомы, которая, в свою очередь, живая «половинка» целого живого тела наряду с ассоциированной в ней неживой «половин- кой» — протеросомой. Задача исследователей — подбирать «ключи» к тайнам иерархической и многослойной организации жизни в ее телесных системах (подробнее — глава 1.3.3), не забывая при этом главного, что все это стороны одного общего явления под названием «жизнь» (биоз). Бесконечно движущаяся материя в организации любых тел диффе- ренцируется на две общие части (меросомы): субконтинуальную (весьма стабильную, но не бесконечную) преднаследственную или наследствен- ную часть — «эгосубстрат» (от лат. ego — я + субстрат), иначе, как сино- ним на греческой основе, — «идиосома» (idios — своеобразный, soma— тело) и дискретную часть — конституцию (пластосому). Следовательно, субконтинуальный субстрат любых неживых (прогеносома) и живых (геносома) тел — это все разные формы эгосубстрата (идиосомы). Бук- вально на эгосубстрате и из него развиваются дискретная (телесная) «надстройка» (форма постсубстрата), которую у любых неживых (пара- феносома) и живых (феносома) тел будем далее называть объединяю- щим, уже известным, термином — «конституция» (лат. constitutio — по- строение), или синонимом на греческой основе — «пластосомой» (plastos — вылепленный + сома). Конституция неживых или живых тел — это результат разного по динамизму процесса «лепки» (кинетопластики, или соматической трансформации) парафенетической или фенетической организации этих тел как их реакции на воздействие внешней среды на основе управляющей деятельности элементов эгосубстрата — понтикул (прогенов или генов). Наше понимание «конституции» тел живых су-
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ществ разных уровней организации развивает традиционное направле- ние в употреблении этого термина в смысле «телосложения». Консти- туция тел, или пластосома, образно говоря, соответствующим образом «вылепливается» из материи под управлением их субконтинуальной субстратной части — эгосубстрата, что осуществляется в непосредствен- ном взаимодействии тел с окружающей внешней средой. Эгосубстраты разных тел составляют вместе (!) общий носитель преемственности (пред- наследственности и наследственности) в природе вообще как вселенском потоке — Супернуклеус (Сверхъядро, т. 1, гл. 2). У биосистем все три части — меросомы — присутствуют в их орга- низации всегда. Как и другие биосистемы, биоморфы всегда имеют три обязательные соматические части: протеросому, геносому и феносому. В любой биоморфе (как простой — организме, так и сложной — синор- ганизме) проявляется тройственная природа ее организации; в ней по- разному интегрированы три ресурсных потока. Логическими средствами расчленим единую и целостную организацию биоморф на три части: континуальный (бесконечный) физико-химический неживой субстрат — протеросому, субконтинуальный (весьма стабильный, но не бесконеч- ный) биохимический наследственный (генотипический по содержанию у организмов) вещественный материал-эгосубстрат — геносому и, нако- нец, дискретный (регулярно сменяемый, весьма лабильный) фенетичес- кий (фенотипический по содержанию у организмов) вещественный материал-конституцию — феносому (рис. 5Г). Геносома — это совокупность всех генов (мерогеносом), обитаю- щих и функционирующих в организации какого-либо живого тела (био- системы). Феносома — это совокупность всех фенов (мерофеносом) в организации какой-либо биосистемы, составляющих все ее признаковое фенетическое «пространство» и развивающееся под управлением гено- сомы этой биосистемы. Геносома (в разном объеме) сравнительно долго передается без изменений из поколения в поколение. В феносоме раз- личимы экзосома, или габитус, — наружная фенетическая часть (наруж- ность, ее контуры и вся поверхность тела) биоморфы, и эндофеносома, или субгабитус, — внутренняя фенетическая часть биоморфы. Субгабитус на- ряду с геносомой составляют вместе эндосому — внутреннее тело биомор- фы. Габитус не имеет внутренней структуры и прикрывает собой эндосому, составляя наруи<ные соматические границы биоморфы (рис. 6). Когда речь идет о различных оболочках телесных систем, имеющих и наружное содержание (габитус), и какие-то внутренние структуры (покровные ткани — эпидермис, перидерма и др.), ограниченные соот- ветствующим образом в своей организации от другой соматической части — эуэндосомы (собственно внутренней организации тел), то та- кую часть генофенетической организации биоморф (шире — живых тел, а еще шире — подобную часть идиопластосоматической организации любых тел) можно обозначить как гемиэндосома. Однако, чтобы не усложнять рассмотрение взаимоотношений габитуса с субгабитусом (эн- дофеносомой), а в целом и с эндосомой и другими телами и нетелами, мы не будем рассматривать особенности гемиэндосоматической органи- зации растительных биоморф, сославшись на то, что эту часть довольно
Глава 1. Концепция меросомной организации биоморф семенных растений 37 Рис. 6'. Универсальные модели меросомной организации тела вообще (А), любой биосистемы (Б), любой биоморфы семенных растений (В). 1 — эгосубстрат (идиосома), 2 — соматическая конституция (пластосома), 3 — прогено- сома, 4 — парафеносома, 5 — протеросома, 6 — геносома, 7 — феносома, 8 — эндосома, 9 — экзосома (габитус), 10 — эндофеносома (субгабитус), 11 — гены, 12 — фены, су- семядольный узел. подробно детапизируют и изучают в анатомии растений (шире — в ана- томии вообще). Мы сознательно используем термины «геносома» и «феносома» не только по отношению к организмам, но и к биосистемам более слож- ным. Материя, еще до возникновения жизни, обладала многими свой- ствами, присущими генам. Гены, носители наследственности, согласно существующим в генетике представлениям, действуют только в опреде- ленных (однородных) живых телах, например, организмах или синорга- низмах и их живых структурных элементах, на основе функционирова- ния которых обеспечивается программа фенетического развития этих тел, т.е. развития их в фенах — признаках, являющихся результатом управляющей деятельности генов. Поскольку в организме есть, кроме геносомы, еще и феносома, образующаяся под управлением генов — элементов геносомы, то вся преднаследственная деятельность прогеносомы, состоящей из прогенов, буквально «растворена» в генетической деятельности биоморф. Прогены (шире — ирогеносомы) и ларафены (шире — ларафеносомы) в организации биоморф, а шире — и других биосистем, составляют вместе лротеросому, которая рассредоточена (нивелирована) в более высокоорганизованных жиз- ненных свойствах геносомы и феносомы, но лосле гибели биоморфы свой- ства неживых меросом — ирогеносомы и иарафеносомы — выходят на лервый план. Все остальное живое, что есть в биоморфах — это их надстроечно-конструктивное (образно — «архитектурное»), или фенети- ческое содержание (феносома). Итак, эгосубстрат биоморфы — это ее дисперсное управляющее «ядро»,' благодаря входящим в него генам оно ведет себя не как пассив- ная масса, а как жизнеактивная система; более того, такая система
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ действует как «самонаводящаяся» (естественно, на цель развития своей биоморфы), управляя всей остальной организацией биоморфы (фенети- ческой организацией, или феносомой) как пластосоматическим состо- янием саморазвивающейся материи — средой существования генов- доминантов в организации биоморф. Если говорить образно, то Природу можно уподобить мастеру, кото- рый бесконечно работает с материалом, творя из него тела, различные пластосоматические формы — конституции тел, проявляя чудеса кинетоп- ластики (умения «лепить»!) на определенной организационной основе- эгосубстрате, мультивариантно предваряющем вероятностное конструктив- ное содержание конкретных тел. Это не просто игра слов: здесь кроется принципиальный смысл, по большому счету — явление панпластичности, которое в разной мере присуще конституции любых тел в различных ус- ловиях их пребывания; все это — следствие того, что заложено в эгосуб- стратах этих тел, или, во всяком случае, многое из того, даже способности тел к субмутациям (у неживых) и мутациям (у живых тел). Панпласти пюсть — это способность конституции телесных систем принимать различные формы в рамках ее возможностей под управлением эгосубстрата (его пон- тикул) и в зависимости от условий окружающей среды. Итак, в организации всех тел, в том числе биоморф семенных растений, есть две структурные части (меросомы): эгосубстрат и консти- туция. Уместно поставить вопрос: а как быть с другими биосистемами = живыми телами (биосомами) большей сложности организации, чем биоморфы? Как у них, в их организации, решается проблема «управле- ния-исполнения»? Сохраняют ли гены свою управляющую роль в более сложных, чем организмы, живых системах? Здесь важно было поставить эти вопросы. Для ответа на них можно писать целые книги. Однако, здесь и сейчас, мой ответ однозначен: в более сложных системах, чем организмы, управляющая роль генов сохраняется, но она проявляется уже не напрямую, как в организме, а опосредованно, через воздействие на взаимоотношения между организмами и между организмами и окру- жающей неживой средой. Влияние генов на фенетическую организацию биосистем, более сложных, чем организмы, т.е. на их феносому, осуще- ствляется через объединение организменных геносом в одну геносому нового природного тела, участниками организации которого, так или иначе, становятся некоторые организмы. Рассмотрим, не вдаваясь в детали, поведение геносомы определенного фитоценоза. Он состоит из видов, среди которых есть доминанты, субдоминанты и ассектаторы. Особи, допустим, п-видов, составляют фитоценоз елового леса. Геносо- мы особей всех этих видов входят в геносому рассматриваемого фито- ценоза. Однако, влияние каждой особи на развитие фитоценоза зависит от программы, исходящей от геносомы этого фитоценоза, а исполнение программы в очень большой степени зависит от взаимоотношений фе- нетической организации разных особей и от их реакции на воздействие окружающей внешней среды. Похоже, что с увеличением сложности организации биосистем управляющая роль геносом видоизменяется, в целом ослабевает, но не утрачивается. Безусловно, возрастает и роль феносоматической организации в управлении жизнью соответствующе-
Глава 1. Концепция меросомной организации биоморф семенных растений 39 го фитоценоза. В коллективном поведении геносомы определенного фитоценоза многое зависит от давления видов-доминантов при форми- ровании программ развития этих фитоценозов. Намечается закономер- ность, справедливость которой еще надо проверять, а точность форму- лировки корректировать; но ее набросок я все же приведу, поскольку это может стимулировать интерес к проблеме. Чем более сложную по организации систему-тело используют для своего существования гены организмов, образуя, вместе с генами других структурных элементов (меросом), однородную или неоднородную геносому, наряду с феносо- матической частью, или феносомой (другой меросомой этого же цело- го), тем меньше гены участвуют в управляющем воздействии на все тело — целое, которое в значительной мере переходит под влияние феносо- мы этого целого и в большей мере, чем прежде (в отношении отдельных организмов), зависит от внешних условий; но при этом управляющая роль генов все же не утрачивается. Каким быть живому телу, более сложному, чем организм — напри- мер, фитоценозу, — определяется участием в его управлении трех вза- имодействующих факторов: геносомы + феносомы + окружающей фи- тоценоз среды. Итак, термины «прогенетическая организация» (прогеносома) и «гене- тическая организация» (геносома), а также «парафенетическая организа- ция» (парафеносома) и «фенетическая организация», или феносома, мы предлагаем применять по отношению ко всем живым системам, обитающим на Земле. Безусловно, качественные и количественные параметры био- систем влияют на взаимоотношения отмеченных выше меросом. Подведем некоторые итоги: в организации тел всегда и везде про- являются две атрибутивные меросомы: эгосубстрат (идиосома) и кон- ституция (пластосома). Дело в том, что материя, в соответствующих ресурсных потоках, проявляет «самонаводящиеся» свойства и достигает целей развития внутри телесных систем, в первую очередь, благодаря преемственности, а во вторую, — благодаря соматической трансформа- ции. Носителями указанных свойств материи в телах являются соответ- ствующим образом специализированные части этих тел — меросомы. В организации тел меросомы бывают управляющими, или эгосубстратом, и исполняющими, или конституцией. В эгосубстратно-конституциаль- ной организации тел есть свои закономерности. Эгосубстрат (управля- ющую меросому) какого-либо неживого тела составляют прогены (ато- мы), объединенные в прогеносому (прогенотип). Ранее (т. 1, гл. 2, стр. 24) было подчеркнуто, что прогенотип «определяет тот спектр вероят- ностных реакций (состояний) какой-либо физиосистемы в изменяю- щихся условиях ее существования, которые не контролируются генами; иными словами, определяют то, чем может быть, а чем быть не может материя какой-либо системы в процессе ее существования». Рассматривая меросомы как носители преемственности или плас- тосоматических трансформаций, мы невольно пришли к осознанию факта, что термины, построенные на основе слова «тип» и применяемые для обозначения соответствующих носителей этих явлений: генотип, прогенотип, фенотип, парафенотип, — выглядят менее убедительно, чем
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ термины на основе слова «сома», поскольку всякие процедуры типиза- ции и типологии более всего привязываются к определенным логичес- ким операциям, производимым с телами для объединения их в группы («типы») по сходным признакам, а все соматические операции действи- тельно непосредственно приближают нас к конкретным телам (сомам) и их частям (меросомам).Тем не менее мы не будем нарушать термино- логической традиции, которая восходит к датскому генетику В. Иоган- нсену (цит. по С. М. Гершензону, 1979), предложившему в 1909 г. термин «генотип» (он же — автор весьма удачного и общепризнанного исходного термина «ген»), и, где это действительно нужно, использовать термины «геносома» и «генотип», «феносома» и «фенотип», а также «прогеносома» и «прогенотип», «парафеносома» и «парафенотип», одно- значно отдавая для обозначения меросом предпочтение терминам, по- строенным на основе слова «сома». Кстати, первые два термина: гено- тип и фенотип — по известному их смыслу связаны только с опреде- ленными частями организмов. При меротомировании по нашей схеме такого значения терминов нам явно недостаточно, что также побуждает автора отдать предпочтение геносоме и феносоме, которые проявляют себя как соответствующие меросомы в разных живых телах, будь то органы, организмы, виды, фитоценозы и т.п., вплоть до всей биостробы Земли. Время покажет, насколько «соматическое» нововведение автора приживется в науке. Говоря о генах, важно подчеркнуть, что они возникли в процессе эволюции с использованием в первую очередь прогенетических элемен- тов (т.е. прогенов), но также и определенных парафенетических элемен- тов (т.е. парафенов), которые в составе генов выступают в единстве как ресурсный поток определенной неживой материи — протеросома. Осо- бо подчеркнем, что гены в своем специфическом эгосубстратном раз- витии сумели ассоциировать в себе носителей неживого эгосубстрата (прогены — определенные атомы) и носителей неживой конституции (парафены — определенные молекулы, агрегации молекул), превратив их в процессе эволюции в наследственноемкое состояние материи как эгосубстрата. Итак, эгосубстратно-конституциальная «начинка» неживых и жи- вых тел вполне понятна (рис. 5, 6). Однако то, что материя Природы проявляет, под определенным углом обзора и на соответствующем уров- не абстракции, меросомное единообразие при эгосубстратно-конститу- циальном подходе к организации телесных систем, все-таки оказалось для автора недостаточным; поэтому была предпринята попытка пойти и на еще большее обобщение. Смысл идеи излагается далее (глава 1.3.4.). 1.3.3. Многослойность явления «жизнь» У бесконечной материи есть великое свойство быть в определенном состоянии при развитии какого-либо явления на некотором отрезке пространства-времени. Явление «жизнь» — это одно из двух высших
Глава 1. Концепция меросомной о ганизации биоморф семенных астений 41 состояний материи (второе — «нежизнь»), которое нас более всего ин- тересует здесь и сейчас. Попробуем ответить на весьма коварный, хотя и очень конкретный вопрос: с чего начинается жизнь? «Вихрь» Жизни (панбиостроба, или биостроба Вселенной), охвативший материю на планете Земля и превра- тивший ее в биопланету, «путешествующую» во времени и пространстве вселенского потока — природы вообще (Природы), смог однажды про- явить себя на нашей планете, вероятнее всего, потому, что это явление более, чем какое-либо другое, наделено в процессе эволюции много- слойностью. Меротомируя, членя ( «расслаивая») логическими сред- ствами живое на виды частей (мероиды) под разными углами обзора и с разных высот абстракции, мы познаем многое и, в первую очередь, атрибутику явления «жизнь». На вопрос: где — жизнь, где ее еще нет, а где уже нет, на мой взгляд, ответить можно вполне определенно, если стать на позицию признания за этим явлением специфической многогранности в его содержании, когда она, жизнь, проявляет себя не только в полной мере, но и отдельными своими сторонами, некоторые из которых надо уже признавать как жизнь, хотя и отличную от полной жизни. Похоже, именно потому, что жизнь многогранна, так непросто определяется само понятие «жизнь». В настоящее время в науке о происхождении и воспроизведении жизни на Земле достоверных фактов еще не хватает, поэтому имеют право на существование две основные гипотезы: первая — Жизнь воз- никла где-то во Вселенной и, расселяясь в ней по определенным зако- нам, однажды была своим фрагментом занесена на Землю, где сумела адаптироваться и развиться дальше (рис. 7); вторая — жизнь на Земле возникла в результате длительного саморазвития неживой материи в условиях этой планеты. Как бы там ни было, жизнь на Земле, будучи, в образном выражении, особым «вихревым» свойством вечно движу- щейся материи (биостробой), отличным от ее неживых «вихревых» свойств, в процессе эволюции вселенского потока (природы вообще, Природы) составила соответствующее состояние в существовании мате- рии на определенном (земном) участке пространства-времени. Сегодня для Земли является неопровержимым фактом то, что в ней есть только две атрибутивные основные части: жизнь (биоз) или нежизнь (абиоз). Жизнь, которую мы, люди, возможно, только из-за уровня своих текущих знаний, наблюдаем пока в единственном «экземпляре» как живое тело, или биостробу, Земли в бесконечной Вселенной, вполне склонна меротомироваться, т.е. члениться на части, далее, поскольку фактов для таких логических операций, отражающих естественные со- стояния материи в ее развитии, предостаточно. Жизнь на Земле, а шире, вообще во Вселенной, как особый «вихрь» материи во вселенском по- токе, — это одновременно и процесс, и тело, среди которых различимы несколько способов и форм существования живой материи. Отсчет нач- нем с высшего уровня проявления жизни — общей (полной, единой) жизни, или цепобиоза (ковитальпости), когда генофенетическая органи- зация составляет единство некоторого тела как целого по отношению к
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИИ Рис. 7*. Шлейфы Жизни, гипотетически блуждающие по Вселенной. з — звезда, а — астероиды, км — комета, пз — потухшая звезда, пл — безжизненные планеты бп — биопланета [планета (возможно, Земля), захваченная шлейфом Жизни и з превратившаяся в биопланету], кж — «котел> Жизни — некое пространство-объект, где впервые зародилась Жизнь (гипотетически не исключено, что таким «котлом> все-таки была Земля) шж — шлейфы Жизни, жс — анабиозные (с обратимо-остановившейся /э жизнью) странники, в одиночку блуждающие во Вселенной, — посланники панбиостробы. его частям; например, организмы — это ценобиосомы (целые живые тела) по отношению к их органам (меросомам) — частям этого целого тела. Частная (неполная) генофенетическая жизнь, или ценомеробиоз (при- ватковитальность) — это проявление жизни в генофенетической органи- зации меросом как частей более сложного целого, например, клетки, ткани или органы выполняют роль приватковитальных меросом (типов форм существования живой материи) в жизнеразвитии организмов. По- ловинчатая жизнь (полужизнеемкость, полужизнь), или гемибиоз (семиви- тальность) проявляется по отдельности как генетическое и фенетическое в организации живых тел, хотя эти «половинки» и существуют всегда вместе, но они имеют еще и свое собственное жизненное содержание. «Интрига» семивитальности довольно четко обозначается, когда мы начинаем «высвечивать» из живой материи две атрибутивные стороны ее существования в телах в качестве по-своему жизненных «половинок», по сути, двух «полужизней» (гемибиозов) — генобиозов и фенобиозов в единой жизни (биозе) как генофенетических союзов-систем — гено- фенов и их комплексов (генофеносом). Гены и фены, проявляя по отдельности свои полужизни, сами живут наполовину от целого, и толь- ко вместе их жизнь наполняется полноценным содержанием: гены, свой- ственным им образом, живут для фенов, сохраняя, таким образом, и себя посредством фенетического жизнеразвития, а фены по-своему
Глава 1. Концепция меросомной организации биоморф семенных астений живут для генов, обеспечивая им благоприятную внутреннюю среду живого обиталища. Главное у семивитальности в том, что две атрибу- тивные «половинки» жизни всегда живут не по отдельности, а только вместе — в генофенетических телесных союзах, что и составляет пол- ноценное содержание таких «союзов», как биосистем (биосом). Гены атрибутивно выполняют только свою часть-«половинку» в явлении «жизнь», а фены — свою. Гены, управляя развитием соответствующих фенов, вместе с ними составляют содержание единой «цепной генофе- нетической реакции» материи под названием «жизнь». При семивиталь- ности как свойстве материи происходит ориентация в развитии материи в строго определенных направлениях на жесткую специализацию в сложении ее состояний в каждой живой телесной системе из двух «по- ловинок» — полужизней (гемибиозов), когда одна из них ориентирована быть носителем наследственности, а вторая — носителем соматической конституции (пластосоматическим носителем). Гены и фены в органи- зации биосистем всегда вместе, поскольку живут не только и не столько для себя, сколько для другой атрибутивной «половинки» жизни, причем ради чего-то еще большего, образуя всегда, если действительно живут, генофенетические союзы-системы (биосистемы) разных уровней орга- низационной сложности: органеллы, клетки, ткани, органы и т.д., ос- таваясь структурными элементами биосистем, которые объединяют их в целое определенной сложности. Ковитальность (ценобиоз) имеет место везде, где материя проявляет себя в живых телах как целое, образован- ное из живых структурных элементов (развитие идеи — гл. 4.8, 7.2). Сопредельная (смешанная) жизнь, или миксобиоз (интервиталь- ность) проявляется на фоне других способов жизни, обеспечивая жизни вообще преемственность развития, и характеризуется тем, что на ее основе осуществляется воспроизведение потомства (размножение). Со- предельная жизнь, начинаясь в родительских живых телах, например, организмах, в которых она постоянно по-разному присутствует как стремление к размножению, затем проявляется уже в вещественных предпосылках размножения (завязываются бутоны, формируются кор- невища и т.п.), а после достижения размножения интервитальность в полной мере переходит и к потомкам. В родительских организмах ин- тервитальность может продолжаться и после первого размножения даль- ше (у многолетников), оставляя разные следы в их организации от своего прошлого проявления. Интервитальность в процессе размноже- ния становится достоянием потомков, а следы о себе в родительских организмах могут быть настолько глубокими, что те полностью (эври- эфемеры, унирепродуктивные растения) или частично (мультирепро- дуктивные растения) разрушаются под давлением смерти (пострепро- дуктивного некроза). Интервитальность, обеспечивая преемственность жизни, использует для этого самые разные средства. Она, например, у однолетников, предельно очевидна. Потомки (семена), получая от своих родителей жизнь и свободу, по сути дела, опосредованно сами же их и убивают, поскольку, в первую очередь, не оставляют им запасов для преодоления неблагоприятного (зимнего в умеренном климате) сезона. Нередко интервитальность действует изощренно, где-то не прибегая к
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ вегетативному размножению, как у Dactylorhiza maculata (L.) Soo, а где- то прибегая к нему, как у Aconitum altaicum Steinb. (т. 1, с. 316), и т.п. Интервитальность может проявиться и там, где вроде бы жизнь уже ушла, свидетельствуя своим, сопредельным, проявлением о том, что здесь как раз тот случай, когда жизнь становится возможной «после смерти». Бывают случаи, когда родительские, некогда живые тела стали по многим признакам мертвыми, и их в целом уже нельзя вернуть к жизни, но и в таких телах могут некоторое время еще быть меросомы — части этих тел, которые, хотя и близки к смерти, но еще живы и способны при подхо- дящих обстоятельствах, например, при замене автобиотопа, продолжить сопредельную жизнь. Интервитальность помогает посредством еще жи- вых меросом, например, органов от «трупов»-организмов, пересаженных в другие организмы, продолжить жизнь там, где она без их помощи могла бы остановиться. Живые меросомы от свежих «трупов», проявляя интер- витальность, в окружении обновленных биотопов (уже — автобиотопов) могут регенерироваться в организмы, построенные по типу родительских особей, или подключиться к жизни уже существующих тел-организмов, структуру которых они смогут (например, с помощью человека) сделать своим новым местом жизни. Обратимо-остановившаяся жизнь, или анабиоз (вицевитальность),— это давно известное направление в объяснении крайних способов прояв- ления жизни. Анабиоз, как подчеркнула Р.С. Ушатинская (1990, с. 14)— это «жизнь, обратимо остановленная глубоким охлаждением (криптоана- биоз), обезвоживанием (ангидроанабиоз) или их комбинацией». За пределами явления «жизнь» идут некрометабиоз (органическая инвитальность, или биинвитальность), когда смерть уже необратимо остановила жизнь в когда-то живых телах, и абиоз (неорганическая инвитальность, или униинвитальность) — нежизнь (остальная, т.е. не- органическая). Таким образом, меросомы биостробы Земли могут по- разному проявлять свою жизнеемкость (витальность) или безжизнен- ность (инвитальность). 1.3.4. Супернуклеус и космоплазма Материя — это главный и первичный ресурс Природы; у нее есть еще один, правда, вторичный (постсубстратный) ресурс — идеальная природа с ее логосистемами — продукт эволюции материи, ее высоко- организованных разумных форм, пока только нас, людей (может быть, еще будут открыты и неведомые сейчас формы Разума). Здесь и сейчас рассуждения об идеальном оставим в стороне; однако никогда не стоит забывать о том, что именно посредством идеального мы будем и далее проникать в таинства организации материального в Природе, в движе- ние материи как ресурсных потоков в телах. Материя составляет теле- сные системы, или тела Природы, распределяясь в них как ресурсные потоки, образующие соответствующие части этих тел, или меросомы. Природа, благодаря движущейся материи, — это вселенский поток с
Глава 1. Концепция ме осомной о ганизации оиоморф семенных растений пространством и временем, состоящий из тел самой разной сложности организации: неживых (абиосом) и живых ( биосом) и их систем (эко- сом, или экосистем). Природу, если и можно представить как тело, то только условно, поскольку все тела дискретны, а такое безразмерное и неограниченное во времени явление, как Природа, естественно, беско- нечно. Природы как тела не существует; она — всегда бесконечный вселенский поток, состоящий из тел, находящихся во взаимодействии и взаимозависимости. Материя неоднородна в пространстве и во времени; она всегда ассоциирована в тела, которые состоят из частей (меросом) — других тел, меньшей, чем они, сложности, и сами при этом являются частями более сложных по организации тел. Одной из форм движения материи, результатом ее эволюции в бесконечном пространстве-времени является жизнь, — событие это, хотя и феноменальное, но раньше или позже безусловно неизбежное, поскольку эволюция Природы в ее свободном «полете», нацеленная на возможность выбирать то, что получится от взаимодействия различных тел в переплетении различных процессов, просто не могла не натолкнуться на такое явление, как жизнь, вероят- ность возникновения которой, хоть и очень мала, но, по всему судя, предусмотрена в организации Природы. Иными словами, неживая материя устроена так, чтобы раньше или позже в ее эволюции могла возникнуть жизнь — при появлении необ- ходимых условий для включения соответствующего механизма эволю- ции в направлении на возникновение жизни, что и случилось на Земле около 3,5 — 4 миллиардов лет назад; правда, не исключена возможность, что Жизнь была занесена, уже в готовом виде, на Землю из других уголков Вселенной, но суть дела от этого не меняется. Важным подспо- рьем для возникновения Жизни была атомная природа неживой мате- рии, а следовательно, наличие у материи на атомном (прогенетическом) уровне способности накапливать и сохранять преднаследственную ин- формацию. Именно через эволюцию атомных комбинаций в молекулах и возникла в Природе новая форма преемственности — наследствен- ность, носителями и хранителями которой стали гены. Эволюция атом- ных комбинаций в неживых телах, которая привела к возникновению предживых и, наконец, первых живых тел, могла проходить и проходила только в условиях круговорота ресурсов, а основа последнего — движе- ние, без которого нет материи. Таким образом, круговорот ресурсов пришел в живую природу из неживой как обязательный фактор эволю- ции. Автор склонен считать, что возникновение Жизни — процесс для бесконечной Природы неизбежный — при стечении соответствующих обстоятельств, когда на основе преднаследственности может и должна возникать наследственность. Без наследственности, т.е. без механизма управления живой природой, жизнь на Земле была бы невозможна (дру- гая жизнь нам не известна), а истоки управленческой деятельности генов кроются в преднаследственной природе атомов (прогенов), из которых, как и все остальное, скроены гены. Возможностью управлять физиосистемами атомы, безусловно, обладают, но возможности эти еще не настолько совершенны, как у генов.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ В Природе нет анархии, хаоса. В мире царит некоторый относи- тельный порядок, который присутствует во всех его материальных эле- ментах — физиосистемах. Порядок в Природе (Мире, Космосе, Вселен- ной), т.е. в организации составляющих ее элементарных частиц, атомов, молекул, клеток, горных систем, галактик и т.п., исходит, скажем так, из некоего абстрактного ядра — Супернуклеуса (Сверхъядра), который объединяет в себе мириады конкретных локальных супернуклеарных частиц — эгосубстратов каждого конкретного тела, являющихся понти- кулярными структурами, носителями преемственности, т.е. преднаслед- ственности и наследственности. И прогены, и гены распылены в про- странстве и времени — как элементы-посланники Супернуклеуса, т.е. понтикулы (так мы обозначили их ранее одним обобщающим терми- ном) — бесконечно своеобразные множественные «мосточки» (!) между миром управления (миром эгосубстратов, составляющих Супернуклеус) и миром исполнения (миром конституций тел, наполняющих собой кос моплазму). Ранее (т. 1, гл. 2) мы предложили объединить относительно «всю» (не забывая, что Природа бесконечна) понтикулярную «начинку» При- роды, рассредоточенную по различным телам, в Супернуклеусе (абст- рактном, или философском, ядре). Если быть более точным, т.е. более конкретным, то Супернуклеус — это не ядро, — а нечто большее, чем ядро, — это «Сверхъядро», понтикулярное вещество которого дисперсно распределено во вселенском потоке — бесконечном материальном по- токе в пространстве-времени, т.е. в Природе. Суть Сверхъядра (Супер- нуклеуса) не в том, что он расположен в центре Вселенной, поскольку это вовсе не так: ведь вселенский поток бесконечен, и мы вряд ли когда- нибудь сможем достоверно определить его центр. Однако «ядерный» смысл в Супернуклеусе все же есть, поскольку то, что его составляет, — важнее всего остального, что есть в Природе, т.е. это лидирующая по своей роли в мироздании масса вещества — управленческая часть При- роды, а ее противовес — другая часть, космоплазма — исполнительская часть Природы, если хотите, «лепота», создаваемая под управлением Супернуклеуса и во взаимодействии с окружающим его веществом ос- тального мира (рис. 8). Космоплазма и составляющие ее конституции тел во вселенском потоке, по отношению к Супернуклеусу, — это как раз то, что можно, с юмором или без него, назвать «лепотой» Природы. Абстрактность Супернуклеуса в том, что мы не можем его предста- вить себе в качестве «жесткой» конкретной системы; границы его- воображаемы (условны), плод абстракции; а вот то, из чего состоит Супернуклеус, всегда конкретно, — это эгосубстраты тел, их прогено- сомы и геносомы. Супернуклеус мы выделили разумом в Природе по- средством абстракции, но исходя из объективной действительности. С помощью мыслительных логических операций Сверхъядро мы «смонти- ровали» из эгосубстратных меросом естественных тел, выполняющих управляющие функции в конкретных телах, т.е. собрали по своей воле, субъективно, в одну образную систему. Вещество этой системы управ- ляет по существу всем — или почти всем — бесконечным мирозданием и в первую очередь конституциями конкретных тел, составляющих ос-
Глава 1. Концеиция ме осомной организации биомор семенных астений 47 Рис. 8*. Типология частей, попавших под образный (!) поперечный надрез вселенского потока и задевший современную Землю. I — Супернуклеус, I А — эгосубстраты живых тел, I А1 — геносомы растительных тел, I А2 — геносомы животных тел, 1 АЗ — геносомы прочих живых тел (например, вирусов), а1, а2, аЗ — гены соответствующих типов живых тел; I Б — эгосубстраты неживых тел, I Б1 — прогеносомы твердых тел, I Б2 — прогеносомы жидких тел, I БЗ — прогеносомы газообразных тел, б1, б2, бЗ — прогены соответствующих типов неживых тел; II — кос- моплазма, 11 А — конституции живых тел, 11 Al — феносомы растительных тел, П А2- феносомы животных тел, 11 АЗ — феносомы прочих живых тел (например, вирусов), Al, А2, АЗ — фены соответствующих типов живых тел; 11 Б — конституции неживых тел, П Бl — парафеносомы твердых тел, 11 Б2 — парафеносомы жидких тел, 11 БЗ — парафено- сомы газообразных тел, Бl, Б2, БЗ — парафены соответствующих типов неживых тел. тальную часть — «вылепленную природу», иначе: космоплазму. И Су- пернуклеус, и космоплазма — это объективная реальность и абстракция одновременно, поскольку все, что содержится в этих двух основных меросомах Природы, выделенных в ней абстрактно, присутствует в ней, Природе, объективно. Существование космоплазмы («лепоты» Приро- ды) весьма созвучно явлению кинетопластики («процессу лепки»), или соматической трансформации, на основе чего преемственный потенци- ал эгосубстратов преобразуется в организационно-энергетическое раз- нообразие конституций конкретных тел. Супернуклеус — это, по сути, современный «философский камень», проникая в тайны которого, мы все более разгадываем систему управления телами в Природе. Мы поместили Супернуклеус, без жестких границ (в графической модели показано пунктиром), в образном центре вселенского потока (его фрагмента), чтобы лучше представить наряду с космоплазмой су- ществование двух вселенских частей, вещество которых объективно рас- средоточено дисперсно в разнообразнейших соотношениях друг к другу (рис. 9).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 9". Абстрактная модель организации Природы, отраженная на фрагмен- те вселенского потока (фп). 1 — Супернуклеус, 2 — космоплазма, 3 — эгосубстраты тел (идиосомы), 4 — конституции тел (пластосомы), 5 — 7 — идиоэнергия, исходящая из Супернуклеуса: 5 — воздействие идиоэнергии на парафенетическое развитие, 6 — воздействие идиоэнергии на фенетичес- кое развитие, 7 — воздействие идиоэнергии на логосистемное развитие, 8 — возвратное пластосоматическое воздействие (ответная реакция космоплазмы) на Супернуклеус, 9— космоплазменные взаимодействия тел (пластосоматические взаимодействия внутри кос- моплазмы). Итак, материя, участвующая в управлении (!) организацией Приро- ды, проявляет в себе это свойство и, в таком случае, имеет соответству- ющее состояние вообще — управляющего мероида (как части в беско- нечной нетелесной системе — гиперапосоме Природы) — Супернукле- уса, который включает бесконечное множество эгосубстратов (прогенов и генов, их комплексов — прогеносом и геносом). Материя, проявля- ющая в себе свойство исполнительской (!) деятельности в организации Природы, имеет соответствующее состояние исполнительского мерои- да, или космоплазмы, в гиперапосоме Природы и включает в себя бес- конечное множество конституций тел (парафенов и фенов, их комплек- сов — парафеносом и феносом). Когда мы меротомируем (членим на части) Природу, мы всегда, как бы при этом ни усердствовали, можем получить только фрагментарную картину ее (ведь она, Природа, беско- нечна), а раз так, то мы, субъекты, взаимодействуя с этим вселенским потоком через некоторый фрагмент его бесконечности (другого не дано), вполне можем обозначить относительные границы мероидов и с их помощью построить вполне обоснованную модель тех видов частей и их взаимодействий, которая отражала бы основной принцип двуполовин- чатого содержания той Природы, «бесконечность» которой мы, люди, пока способны охватить, распространив в ее глубины познающую энер- гетику нашего сознания.
49 Глава 1. Концепция меросомной организации биоморф семенных растений На вопрос: много ли в Супернуклеусе (Сверхъядре) абстракции и сильно ли она отдаляет содержимое этой части Природы от конкретного существования прогенов и генов, — на основе приведенных выше объяс- нений можно ответить однозначно: абстрактность Супернуклеуса нис- колько не влияет на объективность реальности, поскольку существование понтикулярной (идиосомной) материи не зависит от нашего сознания. 1.3.5. Идиоэнергия в круговороте ресурсов и тотальной движитель Ю Понятийный аппарат науки должен способствовать разносторонно- сти нашего проникновения в сущность взаимосвязанных процессов в Природе; поэтому при теоретизировании мы не только должны, но и обязаны смотреть на природу объекта нашего познания значительно шире, смело выходить из узкого коридора, в который нас подталкивает, а нередко прочно загоняет эмпирия текущих конкретных исследований. Став на путь абстракции и меняя высоту и угол обзора, мы не можем не отметить, что интересные взаимосвязи, которые мы наблюдаем, на- пример, между тычинкой и пестиком, между биоморфой и биоценозом и т.п., заиграли новыми деталями, о которых мы ранее даже не догады- вались. У нас как бы открылись глаза на неведомый ранее мир и вы- росли крылья для «полета» в нем, когда объект познания, пусть скром- ный, но все-таки объект познания, был мысленно помещен в бесконеч- ные просторы Вселенной, так что мы смогли посмотреть на него из глубин мироздания, а затем наоборот. На сей счет как еще раз не вспомнить известный афоризм: «Коснувшись цветка, ты невольно по- тревожишь звезду»! Дело в том, что, используя упомянутый выше прием, постоянно меняя и угол обзора, и высоту абстракции, мы убедились, что единые и целостные материальные системы весьма продуктивно могут быть расчленены на части, причем не как заблагорассудится, а логически и структурно обоснованно. Попытаемся пойти дальше, в очередной раз используя отмеченные выше приемы теоретизирования. Присмотримся повнимательней к носителям преднаследственности и наследственнос- ти, проанализируем, что они несут в себе, если смотреть на них так, как мы настроились с самого начала этой работы. Ранее (т. 1, гл. 2) мы определили преднаследственность и показали ее соотношение с наследственностью. Носители преднаследственности, прогены — вероятнее всего, это атомы — наименьшие частицы любого химического элемента, сохраняющие все его свойства; являющиеся пре- делом химического разложения любого вещества. Носителями наслед- ственности являются, как известно, гены (участки молекул ДНК и реже, у отдельных классов вирусов, РНК). Прогены и гены обладают опреде- ленной энергией (!) и несут определенную информацию (!) о том, чем может быть, а чем быть не может развивающаяся система. Известно, что «aToM — устойчивая динамическая система из положительно заряжен-
ОСНОВБ! БИОМОРФОПОГИИ CEMEHHhIX РА СТЕНИЙ ного ядра и определенного числа электронов. У атома как устойчивой системы энергия ниже, чем суммарная энергия невзаимодействующих ядра и электронов» (Краснов и др., 1995, с. 11). Молекулой в химии называют «наименьшую частицу данного вещества, обладающую его химическими свойствами, способную к самостоятельному существова- нию. Если отвлечься от поступательного движения молекулы как цело- го, то в ее энергию вносят вклад три вида движения — движение элек- тронов в поле ядер, колебание ядер около положения равновесия и вращение молекулы вокруг оси, проходящей через центр масс... Хотя эти движения взаимосвязаны, можно приближенно рассматривать их как независимые и считать энергию молекулы равной сумме электрон- ной, колебательной и вращательной энергии» (Краснов и др., 1995, с. 34, 35). Молекула ДНК — это макромолекула, и информационные и энергетические процессы в ней многократно сложнее, чем в обычной молекуле. Главное свойство молекул ДНК — то, что они, как известно, носители генетической информации. «Генетическая информация — это набор данных, определяющих структурные и функциональные свойства клеток. Генетическая информация, в основном, сохраняется в ядрах клеток в виде молекул ДНК (или, более точно, в нуклеотидной после- довательности этих молекул) и используется клеткой, когда это необхо- димо. После деления клетки синтез структурных белков и ферментов, необходимых для функционирования клетки, а также рост и дифферен- циация клеток находятся под генетическим контролем» (Мусил и др., 1984, с. 123). ДНК — гигантская нитевидная молекула ( «генонема»); под тончай- шей белковой оболочкой в ней располагаются нуклеотиды, а сама нить скручена и плотно упакована в определенном элементе клетки, чаще всего в хромосоме. «Молекула ДНК крупнее всех других..., изящна и гибка — ее толщина соответствует примерно десяти атомам. Однако, это восполняется необыкновенной длиной: если растянуть одну молекулу бактериальной ДНК, то ее длина составит около миллиметра... ДНК состоит из структурных единиц четырех различных типов. В состав каждой из них входит сахар дезоксирибоза, к которому присоединены фосфатная группа и основание. Основанием может быть любая из че- тырех слабоосновных органических молекул: аденин (А), цитозин (II), гуанин (Г) или тимин (T).... ДНК представляет собой длинную цепь, в которой чередуются остатки сахара и фосфата, а основания присое- динены сбоку к остаткам сахара» (Роллер, 1978, с. 209 — 210). В ДНК много сахара, а сахар, — один из признаков присутствия энергии (!). В молекулах ДНК сохраняется генетическая информация; они со- стоят из двух параллельных цепей, образующих двойную спираль, и обладают двойной функцией: способностью к собственной репродукции и способностью управлять специфическим синтезом химических ком- понентов организма (Бейссон, 1976; Дубинин, 1976, 2000; Роллер, 1978; Гершензон, 1979 и др.). Нуклеиновые кислоты не только несут и сохра- няют информацию; они обладают запасом энергии, обеспечивающей соответствующий молекулярный порядок и условия сохранения инфор- мации; это позволяет считать, что гены и геносома обладают информа-
Глава 1. Концепция меросомной организации биоморф семенных растений 51 ционно-энергетическим потенциалом, или ресурсом возможностей, который можно назвать кратко идиоэнергией (дословно с греч. «свое- образная энергия»). Прогены и прогеносомы соответствующих тел так- же обладают своей идиоэнергией. Прогенетическая и генетическая ин- формация сама по себе мало на что способна, если не связать ее с энергией, обеспечивающей сохранение этой информации и возмож- ность ее выхода («считывания»), развертывания в конституциальные структуры. Идиоэнергия — не только вид энергии и не только вид информа- ции, но и особое системное соединение обеих в одном явлении, создан- ное природой эволюции в эволюции Природы. Идноэнергня, или ннфор- мационно-энергетический потенциал, или, иначе, ресурс возможностей, какой-либо живой или неживой системы — способ выражения преднаслед- ственностн н наследственности в соответствующем эгосубстрвте (прогенвх н генах). Почему автор склонен говорить не просто о прогенетической и генетической информации, а шире — об идиоэнергии (ресурсе воз- можностей)? Потому, что преднаследственная и наследственная инфор- мация не может просто быть встроена в ее носители, она должна быть как-либо погружена в определенный энергетический массив, в котором обеспечены условия для поддержания соответствующей активности ин- формации, сохранения ее при движении в чреде многочисленных пла- стосоматических трансформаций, окружающих идиоэнергетические но- сители, но не проникающих внутрь их и не нарушающих таинство организации этого ресурса. Идноэнергня — это не только прогенетнческая н (нлн) генетическая информация: это еще специальное энергетическое обеспечение, сохраняющее эту информацию, во взаимодействии, непосред- ственном или опосредованном, ее носителя с окружающей средой. Автор отдает себе отчет, что рассматриваемый в этом подразделе материал — все же скорее размышления вслух, чем стройные доказа- тельства истины. Реальность присутствия в соответствующих материаль- ных системах преднаследственного и (или) наследственного ресурса воз- можностей (идиоэнергии) не вызывает сомнений, но еще нуждается в разработке. Пожалуй, пока вполне достаточно осознания факта, что у живых и неживых тел есть универсальные носители преемственности (понтикулы), в которых присутствует и циркулирует универсальный ресурс — идиоэнергия. Слово-этикетка, например, «идиоэнергия» или другое, если оно будет лучше, может закрепиться потом, а главное другое — сформули- ровано новое понятие. Рано или поздно нужно будет обстоятельно раз- работать универсальную концепцию единства информации и энергии в прогенах и генах. От решения этой проблемы может зависеть многое, пока же мы ограничимся применением терминов «идиоэнергия», или «ресурс возможностей», лишь там, где нужно будет подчеркнуть пред- наследственное и (или) наследственное информационно-энергетическое единство в соответствующих материальных системах. Графическую модель разнообразного наиболее общего взаимодей- ствия материи во Вселенной можно представить так. Бесконечное про- странство буквально «накачано» телами (сомами), в структуре каждого
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ из которых различимы эгосубстрат (идиосома), т.е. прогеносома у нежи- вых тел или геносома у живых тел, который через конституцию, или пластосому (парафеносому или феносому), взаимодействует с остальным миром абстрактно разделенным на Супернуклеус и космоплазму (рис. 8,9). Идиосомы, чтобы взаимодействовать между собой и передавать инфор- мацию, должны обладать соответствующей энергией. Так что слово «идио- энергия» вполне удачно, на наш взгляд, отражает единство информации и обслуживающей ее сохранение энергии в прогенах и генах. Итак, Природой и составляющими ее элементами управляет осо- бый, исходящий из Супернуклеуса и его фрагментов, эгосубстратов конкретных тел вид деятельности-движения, который мы и называем идиоэнергией. Это не просто вид энергии, но особая форма деятельно- сти (функционирования), представляющая собой информационно-энер- гетический потенциал какого-либо тела, его ресурс возможностей, со- храняющая полученные в прошлом результаты, что-то из них реализу- ющая непосредственно в конкретном теле, своем носителе, а что-то отложившая для реализации на будущее. Идиоэнергия проявляет себя как преднаследственная или наследственная информация в прогенах или генах в единстве с особой энергетикой, заключенной в них, обеспечивающей субконтинуальность информации в системе «исходное — производное», со- хранение ее и участие, хотя бы на первых стадиях, в развертывании ее в парафенетической или фенетической организации соответствующих тел (не- живых или живых). Идиоэнергией обладает и кусок гранита, и кузнечик на лугу, и Галактика, и т.п. Наличие особой энергетики на преднаследственном (прогенетичес- ком, или атомном) уровне организации неживой природы и наслед- ственном (генетическом) уровне организации живой природы обеспечи- вает Природе бесконечную и неисчерпаемую деятельность ее через Супернуклеус. Идиоэнергия пронизывает мироздание, «вылепливая» космоплазму — различные пластосоматические формы (конституции тел) остальной космической материи, заполняющей пространство— окружение Супернуклеуса. Направленное воздействие на космоплазму идиоэнергии Супернуклеуса и множественные ответные реакции иногда приводят к мутациям (у живых тел) или субмутациям, т.е. изменению прогенов (у неживых тел). Одним из примеров субмутации является превращение атомов водорода в атомы гелия, происходящее в ядерных реакциях на Солнце. И прогены, и гены распылены в пространстве- времени как посланники (понтикулы) Супернуклеуса. Они, в силу своей вездесущей натуры, так или иначе присутствуют в каждом теле, опре- деляя свойства тел, в первую очередь доминантные. Итак, носители идиоэнергии сосредоточены в понтикулах — в про- генах и генах, которые составляют эгосубстраты (прогеносомы или ге- носомы) конкретных тел, а следовательно, и суммарное эгосубстратное вещество Природы — Супернуклеус (Сверхъядро). Идиоэнергия прояв- ляет себя как понтикулярная деятельность любых телесных систем. Графически изобразив в моделях организацию Природы на одном из наиболее высоких уровней обобщения (рис. 8,9), мы учли высказан- ные ранее соображения об эгосубстрате и конституции, о Супернуклеусе
Глава 1. Концепция меросомной о ганизации биоморф семенных растений и космоплазме, о взаимодействии различных сил между телами, их со- ставляющими, и, в частности, идиоэнергетические взаимодействия. Не только эгосубстраты разнообразных тел воздействуют идиоэнергией на пластосоматическое окружение и на внешний по отношению к ним (несобственный, внетелесный) мир, но и все подобное в ответ действу- ет, через разнообразную дискретную информационно-энергетическую деятельность, и на них. После обобщений, рассмотренных выше, я склонен увидеть еще один существенный нюанс в организации Природы, обход которого счел бы своим упущением. Дело в том, что, если хорошо присмотреться к тому обязательному, только внешне относительному, порядку, в ко- тором существует Природа, то обнаруживается, что его обеспечивает ни много и ни мало, а всеобъемлющий (тотальный) движитель («двига- тель»). Не перебор ли здесь и та ли масштабность взята для ботаничес- кой работы, где должны, как кто-то наверняка подумает, разбираться вполне конкретные текущие задачи растительной организации? Отнюдь, хотя согласен, что традиционно такого типа монография, вероятно, должна писаться иначе, но я работаю в стиле, который совсем не тра- диционен и который я избрал для себя (!) как наиболее верный. Если в организации Природы существует тотальный движитель, значит, он как-либо связан и с организацией биоморф семенных растений, а если все так, то это, безусловно, уже и проблема моих исследований в их самой общей теоретической части. Я готов утверждать, что в Природе действительно существует тот самый «движок», «движитель», «двига- тель»..., название которого звучит, может быть, вызывающе или даже магически обнадеживающе — что-то типа «перпетуум мобиле» (вечный двигатель), но все-таки лучше от этого словосочетания хоть как-то отмежеваться и говорить несколько иначе, допустим, — «тотальный движитель», или, еще короче, — «примагенератор» (от лат. prima— первый, generator — производитель, родитель). Реализация идеи «вечного двигателя» («перпетуум мобиле») — во- ображаемой, непрерывно действующей машины, как доказала наука, неосуществима, поскольку работа такой «машины» противоречила бы закону сохранения и превращения энергии или второму началу термо- динамики (Бол. Энц. Сл., 1997). Однако если применить еще большую абстракцию к осознанию того бесконечного движения, которое посто- янно поддерживает в Природе организационный порядок, то от идеи всеобъемлющего движителя того единственного и неповторимого все- ленского потока, который естественным образом и является самой Природой, на мой взгляд, вовсе нет оснований отказываться. Путь к истине здесь, пожалуй, начинается именно с организационного поряд- ка, который проявляется в Природе независимо от чьего-либо сознания. С него мы и начнем наше доказательство примагенератора. Чтобы все- объемлющий организационный порядок однажды возник, а затем по- стоянно поддерживался, нужна бесконечно действующая сила, способ- ная управлять организацией Природы. Предварительно настроившись на абстрактное мышление, мы на- чинаем различать детали тотального движителя, т.е. по сути, как ни
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ крути, — детали тотального «перпетуум мобиле» в организации Приро- ды, а затем и представлять его величественный «всеприродный» образ. На мой взгляд, примагенератор — это пространство Природы вместе с неисчерпаемой материей, по-разному начиняющей его бесконечность, которая всегда существует только в движении (изменении) и только во времени, которое есть уже другое атрибутивное достояние (часть) При- роды. Таким образом, Природу можно охарактеризовать и как тотальный движитель (примагенератор), ритмически функционирующий во времени как ее часть вместе с другой обязательной частью — временем. Через стремление не просто обобщенно мыслить, но и доводить обобщения до определенного результата, раскладывая обобщения, например, по «по- лочкам» мирознания, чтобы лучше понять его организацию, автор, через очередное меротомирование Природы, наконец разглядел и механизм развития — его примагенератор и определил ему место в Природе. В чем же состоит бесконечно движущаяся суть организации При- роды? Примагенератор в своей нескончаемой деятельности — функци- онировании, производит трансформации разных уровней сложности. Функциональной единицей деятельности примагенератора являют- ся трансформы (формы трансформаций), высшая из которых — эволю- ция (глава 8). Это он, примагенератор, с общим для него и Природы Сверхъядром и космоплазмой, раскачивает и заставляет по-разному колебаться эволюционные маятники (т. 1, гл. 3), поскольку эти всюдные элементы — результат высшей функции — эволюции у всеобщего и единственного движителя Природы. Пространство бесконечного вселен- ского потока (Природы), наполненное сырьевым ресурсом — материей, которая в таком ~потоке» существует движением — изменением — транс- формацией, и есть тотальный движитель — примагенератор, непрерывно функционирующий всегда и везде в природном времени. У примагенерато- ра как субстратно-постсубстратной части Природы под определенным углом обзора всегда обнаруживают себя две атрибутивные его части, по- разному организационно ориентированные: управленческая часть- эгосубстратное системное единство Природы, ее Супернуклеус (Сверхъядро), и исполнительская часть — конституциальное системное единство Природы, ее космоплазма (рис. 9). Через единство отмеченных частей, составляющих бесконечный двигательный механизм Природы, свершаются все трансформации, которые имеют место быть где-либо во времени вообще. Только присоединив свое время к тому, что делает ее же тотальный бесконечный движитель, Природа обеспечила себе разви- тие (похоже, идет все именно так и вряд ли иначе). Примагенератор, традиционно раскачивая эволюционные маятники, однажды произвел революционный акт, впервые сотворив живое вещество. Возникнув однажды во вселенском потоке, Жизнь «потекла» дальше и стала «растекаться» еще и еще дальше, совершенствуя в эволюции способы своего сохранения. Жизнь, однажды произведенная тотальным движите- лем через вид его бесконечной деятельности — эволюцию, заряжена на самосохранение, и более того, даже на неограниченное сохранение, что возможно на основе нообиофильной суперадаптации (т. 1, гл. 3). Имен- но Разум Природы может решить, быть Жизни во Вселенной всегда
Глава 1. Концепция ме осомной организации биоморф семенных растений 55 дискретной или в целом можно стать и бесконечным явлением. Эволю- ция зарядила от примагенератора Природы всех носителей жизни осо- бым информационно-энергетическим потенциалом (идиоэнергией) как на свое (личное), так и не-свое (неличное) жизнесохранение, отдав возможность бесконечного поддержания Жизни в Природе во власть Разума (пока нашего, человеческого, разума, но, возможно, еще и дру- гого разума, существование которого земляне предполагают и стремятся к жизнеполезной встречи с ним). Организационный порядок в Природе возник сам по себе и никто ей его не навязывал, поскольку никого и ничего, кроме Природы, не существует. Свой организационный порядок Природа поддерживает в оди- ночестве только своим функционированием как системы из двух частей: тотального движителя и времени. Без такой системной организации в Природе порядка не было бы, а был бы «хлам — мусор», с которым Природа существовала бы бесполезно. Только организационный поря- док в Природе делает ее существование целесообразным, а присутствие в ней людей, как ее частиц, делает это существование еще и осознан- ным. Организация Природы такова, что ее существование целенаправ- ленно развивается в сторону возникновения и (или) поддержания Жизни на основе трансформаций неживой природы (нежизни). Живая приро- да, однажды возникнув, поднимает целесообразность развития природы вообще еще на более высокий уровень, поскольку Жизнь берет на себя свое сохранение при обязательном взаимодействии с неживой приро- дой, совершенствуя в этом направлении себя как систему «управления- исполнения». Итак, среди многих возможных направлений развития Природы высшую направленность составляет цель возникновения и поддержания жизни вообще. Природа, таким образом, существует с пользой не просто для себя, а с пользой для высшей формы существо- вания тех своих частей, которые, несмотря ни на какие трудности, состоялись как живые системы и склонны продолжить свое живое су- ществование бесконечно, находя приемлемые для этого пути сосуще- ствования с неживой природой. Многое, что автор высказал здесь об эгосубстратно-конституциаль- ной «натуре вещей» и других абстрактных построениях, нацелено на развитие в будущем. Но дальнейшая работа человеческой мысли в этом направлении, как мне представляется, весьма полезна для расширения и углубления понимания нашего мира и нашего места в Природе, а также места в ней биоморф семенных растений. Пожалуй, важно, что идеи, такие, на первый взгляд, далекие от конкретного объекта иссле- дования — тел растительных биоморф, высказывает профессиональный ботаник. Значит, проблема, которую затронул автор, настолько объем- на, что «коснувшись цветка», он вынужден «потревожить звезду», ухо- дить от ее обсуждения автор не счел для себя возможным. К тому же, поняв стиль мышления автора, читателю будет легче воспринимать все его последующие рассуждения и теоретические построения.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 1.4. Относительность в свободе выбора цели развития телесных систем при ограниченности ресурса возможностей Все в Природе находится в движении, и нет системы, свободной от изменений в пространстве и во времени. Однако в свободе тел-систем есть свои ограничители. Разумеется, абсолютной свободы в выборе це- лей развития у систем быть не может и что свобода выбора целей развития, так же как и ограниченность ресурса, из которого что-то можно выбирать относительны. Ранее (т.1, гл. 2) мы показали, что в бесконечном вселенском по- токе пространства и времени и в неисчерпаемом разнообразии телесных систем последние проявляют себя как эволюционные маятники, кото- рые, под воздействием как внутренних, так и внешних сил, «раскачи- ваются» каждый в своем режиме. Диапазон их колебаний изменяется, поскольку эти «маятники», кроме их собственных колебаний, еще и «плывут» в пространстве-времени, меняя угол наклона осей маятников, их положение в пространстве и т.п.; меняются и силы взаимодействия тел с окружающим миром. Эволюция всюдна (!), остановить ее нельзя нигде, даже на мгновение (глава 8). Колебания эволюционных маятников с определенной амплитудой (диапазоном изменчивости) подпитываются информацией преднаслед- ственного и (или) наследственного дисперсионного «ядра-генератора» [прогенетической и (или) генетической организации неживых или жи- вых тел-систем], т.е. эгосубстрата тел. Чем же наполнено это ядро— только ли одной информацией? Ответ дан выше: идиоэнергией. Без энергетики информационный потенциал прогенов и генов вряд ли был бы эффективен. Сохранение информации, содержащейся в прогенах и генах, «выталкивание» ее на путь реализации через развертывание кон- ституций соответствующих тел и вероятностное преобразование ее в последующем в энергию уже новой организации, которая защищает эгосубстракт каждой телесной системы от прямого воздействия окружа- ющей среды, — это все задачи особой энергии, заключенной в прогенах и генах. Информационный потенциал развития телесных систем, сосре- доточенный в прогенах и генах, вместе с энергией, циркулирующей в них и обеспечивающей сохранение генерируемой информации, и вклю- чающий, при подходящих обстоятельствах, механизм ее реализации в пластосоматической организации, и составляет, на наш взгляд, ресурс возможностей определенных телесных систем, или их идиоэнергию. Благодаря словосочетанию «ресурс возможностей» мы получаем шанс одним термином обозначить процессы, выражающие и две формы преемственности (преднаследственность и наследственность), и энерге- тику, обслуживающую их в соответствующих носителях этой преем- ственности. Каждая система обладает своим ресурсом возможностей (своей идиоэнергией), который позволяет ей развивать пластосомати- ческую (парафенетическую или фенетическую) организацию в опреде- ленных границах (диапазоне, параметрах), т.е. этот ресурс имеет вполне
57 Глава 1. Концепция меросомной организации оиоморф семенных растений 1.5. Целесообразность распределения ресурсных потоков в организации биоморф По сути, все основное, что можно было сказать сегодня о принци- пах меросомной организации телесных систем вообще и биоморф в частности, мы уже сказали; нам остается лишь сделать несколько обоб- щений и подвести некоторые итоги. Автор склонен утверждать, что все телесные системы устроены це- лесообразно, поскольку их развитие так или иначе целенаправленно. «Целесообразность — свойство процессов и явлений приводить к опре- деленному результату, цели в широком или условном смысле слова» (Филос. сл., 1980, с. 406). Целесообразность обязывает тех, кто ее про- являет, стремиться к цели. Все цели в телесных системах весьма кон- кретны, многие из них выбираются бессознательно, без участия воли. Благодаря ресурсам преднаследственности или наследственности, в су- определенные ограничения. Идеальные системы (логосистемы) не обла- дают своей идиоэнергией; она поступает к ним опосредованно в преоб- разованном виде, от соответствующих материальных систем, являющих- ся прямыми производителями логосистем, т.е., попросту говоря, от че- ловеческого мозга. Следует заметить, что ограничения ресурса возможностей все же относительны; у него, ресурса возможностей, не бывает жестких границ, и со временем он способен изменяться, хотя и значительно меньше, чем все, что его окружает. Известно, что прогены и гены способны изме- няться в пространстве-времени — в процессе взаимодействия с окружа- ющим миром. В прогенах, хотя и редко, происходят субмутации (изме- нения прогенов осуществляются под влиянием сильнодействующих факторов, вызывая в последующем парафенетические изменения теле- сных систем); например, при определенных условиях (бомбардировка альфа-частицами) атомы азота способны превращаться в атомы кисло- рода, а из атомов алюминия получаются атомы кремния; в ядерных реакциях на Солнце водород превращается в гелий. Что касается транс- формации генов, то это составляет целый раздел генетики, изучающей мутации (Дубинин, 1966, 1976; Гершензон, 1979 и др.). Субмутации и мутации — важные факторы эволюции телесных систем. Итак, идиоэнергия существует во взаимодействии с остальными процессами, происходящими в системах, обладая относительными гра- ницами, складывающимися в процессе длительного исторического раз- вития систем каждого определенного типа и конкретного системогенеза. Выбор цели и начало саморазвития любой материальной системы сво- боден абсолютно только в кавычках, он «абсолютен» только в границах ресурса возможностей, в пределах той идиоэнергии (информационно- энергетического потенциала), которой обладает эта система.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ ществовании многих тел проявляется принцип аутокинетического наве- дения, т.е. самонаведения на цель своего развития — стремление теле- сной системы к достижению определенной цели ее развития, без уча- стия людей, на основе «автономного» ресурса возможностей, или иди- оэнергии этой системы. С появлением на Земле Человека возникло и продолжает возникать немало тел, которые наводятся на цели своего развития при участии человеческого разума и в результате практической деятельности людей — это принцип антропокинетического наведения на цели соматического развития, т.е. на цели развития телесных систем, под влиянием сознательной деятельности людей. 1.5.1. Меросомность организации биоморф семенных растений — атрибут целесообразности распределения ресурсов Таким образом, материя в процессе ее бесконечного существования проходит стадии развития в телесных системах, проявляя себя всегда в двух основных частях (меросомах): как эгосубстрат, когда несет про- грамму (преднаследственную или наследственную, см. т. 1, гл. 2) своего телесного существования, а еще, пожалуй, и первичную энергию для ее сохранения и развертывания (всякое движение как атрибут существова- ния материи происходит на основе каких-либо энергетических процес- сов). Такое состояние материи можно назвать идиосоматическим, а то, чем оно обладает, — идиоэнергией (ресурсом возможностей, или ин- формационно-энергетическим потенциалом). Материя конструируется Природой в тела на основе идиоэнергетического воздействия эгосубст- рата на остальную массу телесного вещества и во взаимодействии с окружающей средой; такое состояние материи будет конституциальным. Подведем некоторые итоги относительно свойств, которые обеспе- чивают существование и изменение любых тел в Природе, включая и существование биоморф семенных растений. Главный акцент мы сдела- ем на закономерностях в целесообразности меросомной организации телесных систем. Материя, будучи первичным и неисчерпаемым ресурсом Природы, распределяется в телах и всегда становится в них какой-либо частью (меросомой), используя в движении ресурсных потоков разные на- правления, но три из них самые главные: континуальное (бесконечное) как материальная основа (субстрат) вообще; субконтинуальное (весьма ста- бильное, но не бесконечное) как эгосубстрат (идиосома), управляющий во многом развитием содержания и, тем самым, обеспечивающий материи определенное преемственно-историческое соматическое продолжение; на- конец, дискретное (весьма лабильное, склонное подвергаться текущим со- матическим трансформациям) как конституция (пластосома), занимаю- щая исполнительское место в организации телесных систем. Это абсолют- ный закон целесообразности меросомного распределения материи в ресур- сных потоках.
Глава 1. Концеиция меросомной организации биомор семенных рагтений 59 Сформулированный выше абсолютный закон целесообразности справедлив как для абиосистем, так и для биосистем. Поэтому рассмат- риваемое явление вполне можно конкретизировать и детализировать в специальном законе целесообразности распределения ресурсов в организа- ции биосистем. Его содержание в следующем. Жизнь, однажды возникнув, смогла распространиться на Земле, только проявив целесообразность— способность своих телесных систем (биосистем) приспосабливаться к до- стижению конкретных целей, не только раскрывая собственные ресурсы, но и осваивая целый комплекс различных способов получения доступных для нее иных ресурсов (веществ, энергии, информации) и распределяя их среди своих носителей в следующих функциональных частях-системах (меросо- мах): управления (геносома управляет феносоматическим развитием), по- требления (трофика, дыхание, иное потребление ресурсов окружающеи среды), производства [образование различных структур и (или) продуктов метаболизма, необходимых для роста и развития], резервирования (отло- жение потребленных и произведенных ресурсов в резерв для реализации по мере необходимости), рассеивания (осуществление полного обособления предка от потомка и других форм отчуждения ресурсов за пределы исход- ных биосистем, вплоть до их полного разрушения). Из приведенного выше видно, что у биоморф, при всем кажущемся разнообразии способов распределения ресурсов, есть только пять упомянутых способов. Материя обладает свойством самоорганизации, настраиваясь (наце- ливаясь) из меросомы как эгосубстрата на развитие другой меросомы как конституции и на развитие всей телесной системы в целом. Материя в своем развитии создает ресурсные потоки, которые самонаводятся с эгосубстратных уровней для достижения определенных конституциаль- ных целей, в результате чего телесные системы превращаются в сложные клубки разнообразной внутрителесной межмеросомной деятельности. В этой деятельности происходит и поддерживается, внешне трудно улови- мое для наблюдателя, естественное членение тел на части — меросомы, идет работа, которую можно образно обозначить как конституциальное строительство вместилищ для универсальных элементов эгосубстрата- его понтикул (прогенов и генов). Тема феносоматического строитель- ства жилья для генов в организации растительных биоморф получит развитие в последующем материале (глава 9). Ресурсные потоки распре- деляются в организации тел по двум направлениям: как управляющие (эго- субстратные) меросомы и как исполняющие (конституциальные) меросомы. Управляющие меросомы представлены у неживых тел прогеносома- ми, у живых тел — геносомами. Геносомы использовали и используют для своего воспроизведения в эволюции и текущем морфогенезе проге- ны (атомы) и парафены (соответствующие молекулы), которые в орга- низации генов образуют их протеросомную часть, подчинив ее своему развитию. Гены «замешены» в своем возникновении из первоприродно- го телесного материала (протеросом). По-другому и быть не могло, поскольку живая материя развивается в неразрывном единстве с нежи- вой материей. Материя, таким образом, присутствуя и развиваясь в границах каких-либо тел, ведет себя двояко: она или управляет разви- тием тел (эгосубстратные меросомы), или осуществляет рост и развитие тел (конституциальные меросомы).
ОСНОВБ1 БИОМОРФОПОГИИ CEMEHHhIX РА СТЕНИЙ Эгосубстрат и конституция по-разному подвержены соматическим трансформациям. Неживой эгосубстрат изменяется при субмутациях, в особых условиях, например, при ядерном взрыве, когда одни прогены (атомы) превращаются в другие. Живой эгосубстрат склонен, как изве- стно, лишь иногда трансформироваться, т.е. подвергаться мутациям. Конституция более всего подвержена собственно соматической транс- формации в различных формах. Когда при самонаведении или наведе- нии (с чьей-либо помощью извне) на цели развития происходят субму- тации или мутации, то возможны отклонения от целей. В результате расширяется разнообразие в границах организации прежнего тела или возникают качественно новые тела. Итак, мы рассмотрели довольно сложный механизм движения ре- сурсных потоков в организации живых и неживых тел. Клубок взаимо- отношений между меросомами телесных систем весьма не прост; но, похоже, многое постепенно проясняется. Логическими средствами мы расчленили единую, целостную организацию тел по особенностям дви- жения материи в ресурсных потоках, что позволило выделить в телах управляющие и исполняющие меросомы. Такое членение на части впол- не осуществимо не только по отношению к конкретным телам, но и по отношению к бесконечной Природе, где также, несмотря на отсутствие границ, можно выделить две части — относительные меросомы: управ- ляющую — Супернуклеус и исполняющую — космоплазму. Меросомы в организации биоморф возникли в процессе эволюции как результат адаптации, посредством которой одни соматические части стали выполнять узко специализированные функции, другие остались универсалами, третьи вообще оказались лишними и стали более или менее быстро редуцироваться. Меросомность можно исследовать с раз- ных сторон и на разном уровне абстракции. Например, меросомами являются две части биоморф: внешняя и внутренняя; меросомами у биоморф будут их органеллы, клетки, ткани и органы. Поскольку в биоморфах нет иного живого, кроме адаптаций, то их меросомы, будучи адаптациями, стали носителями определенной специ- ализации; они специализировались на разное участие их как определен- ной части живого тела в круговороте ресурсов. Организация биоморф участвует в круговороте ресурсов по направлениям, которые мы закре- пили в соответствующем законе: от управления, через потребление, к производству, резервированию и рассеиванию (Нухимовский, 1987, 1997). Меросомами являются элементы-руководители (гены внеядерные и в хромосомах), элементы-исполнители (биополимеры РНК, кроме PHK-генов у вирусов), верхушечные и дополнительные меристемы, побеги, корни, корнепобеги-органы (онтоиды) и, наконец, корнепобе- ги-организмы (эуфиты) (т. 1). Меросомами будут и функциональные системы, выделяемые по особенностям биофильной специализации частей. Так, биоморфы се- менных растений являются пентамеросомными системами (т. 1, гл. 3) и состоят из следующих меросом (рис. 10 — 36): эусомы, или собственно вегетативной части тела, сокращенно — тотавегетатива (от лат. totus, а, ит — целый, весь + вегетатив), биоморфы ( у организмов — это побе-
Глава 1. Концепция меросомной организации оиоморф семенных растений 61 говые вегетативные и корневые части, у синорганизмов — все то же, но и вегетативные особи-клонисты целиком); диасомы (соцветия, цветки и составляющие их элементы, кроме гамет или семян), в которой обычны две части — интервегетативная меросома (первая диасома), или интер- вегетатив (оси и остальные части побега выше зон торможения, когда они есть, несущие непосредственно на себе цветки, например, у соцве- тий, но не включают цветки, или оси и другие части побегов у межсоц- ветий, даже просто отдельные цветоножки), и пререпродуктивная меро- сома (вторая диасома), или пререпродуктив (цветки, плоды, без учета гамет и семян, образующихся при соответствующем размножении); ар- хесомы, или магирепродуктивной (от лат. magis — более + репродуктив) меросомы, сокращенно — магирепродуктива (гаметы, эмбриональные особи, например, семена до их обособления от родительской биомор- фы); хромосомы (они, если не включать в них гены, имеют фенетичес- кую природу, но существуя вместе с генами, являются, благодаря им, носителями и хранителями генетической информации); гены и их со- вокупности в живых телах разной сложности организации — геносомы (хромосомы и гены на рис. 10 не показаны). Из двух равнозначных слов-этикеток «тотарепродуктив» и «тотрепродуктив» я остановлюсь все же на первом слове, пожалуй, более точном для нашего термина. Ди- асома (интервегетатив + пререпродуктив) и архесома (магирепродуктив) образуют вместе одну систему, которую, учитывая хотя и не совсем корректную традицию, сложившуюся в ботанике, обозначаем известным термином — репродуктивный орган (репродуктивное тело или часть тела (меросома), короче, репродуктив, или гониосома, что дословно с греч. — «тело, способное к рождению»). Репродуктив как меросома в зависимости от структуры его интер- вегетатива может быть двух типов: тотарепродуктивом (первая гониосо- ма), когда интервегетативная часть не имеет зеленых листьев, а все листья, если они развиваются, чешуевидные и в первую очередь, это брактеи, и фрондорепродуктивом (вторая гониосома), когда его интер- вегетатив развивает некоторое число зеленых листьев. Репродуктив, со- гласно известной терминологии соцветий (Troll, 1964; Федоров и др., 1979), бывает нередко следующих типов: абрактеозный, брактеозный, фрондо-брактеозный, фрондо-фрондулозный, фрондозный. Весьма ва- рьирует по специализации и тотавегетатив в структуре вегетативно-реп- родуктивных побегов. Если сорвать цветущую верхушку побега до зоны торможения, то последняя, вместе со всем организмом, начинает «ра- ботать» на восстановление утраченной репродуктивной части целого побега и нередко образует вынужденные побеги с новыми соцветиями (например, Valeriana officinalis L.). После завершения развития побегов с репродуктивной функцией они полностью или неполностью отмира- ют, превращаясь или в отмерший пострепродуктив, или интервегегатив, который может иногда еще некоторое время и после завершения реп- родуктивного развития побегов проявлять жизнедеятельность в структу- ре вегетатива, интегрируясь наряду с бывшим тотавегетативом (основ- ной вегетативной частью побега, а шире — особи) в единый текущий тотавегетатив.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Если в структуре побега вместе с абрактеозным и (или) брактеоз- ным репродуктивом-меросомой (частью побега) присутствует эфемер- ный тотавегетатив (без зеленых, а только с чешуевидными листьями) и давление этого репродуктива в организации побега является определя- ющим, то такой побег обозначается как репродуктивный (и в целом). Побег, в котором кроме пререпродуктива (цветков, плодов) и магиреп- родуктива (гамет, семян) присутствует только абрактеозный и (или) брактеозный интервегетатив, например, отдельная цветочная ось с цвет- ком в структуре соцветия (разновидность репродуктива), то это тотареп- родуктивный побег, или тотарепродуктив. Репродуктив чаще всего эфе- мерен, но внешне он может быть похож на магимодифицированный репродуктивно-статурный побег. В структуре последнего стеблевая часть ведет себя иначе: она сначала потенциально персистентная, а затем персистентная на большую или меньшую часть своей длины. Если реп- родуктив с зелеными листьями (допустим, фрондозное соцветие), иначе фрондорепродуктив, или гониосома второго типа (вторая гониосома) развивается вместе в одной «связке» (системе) с тотавегетативом, у которого могут быть любые листья, то это уже вегетативно-репродуктив- ный побег (вегеторепродуктив), который может быть или репродуктив- на-ассимиляционным, или репродуктивно-статурным (подробнее о классифицировании побегов по степени их специализации см. т. 1, гл. 5, с. 237 — 239). Рис. 10. Графическая модель меросомной фенетической специализации в организации биоморф семенных растений.
Глава 1. Концепция меросомной организации оиоморф семенных астений 63 Части (меросомы), будучи разных размеров и по-разному специа- лизированы, составляют структуру не только растительного тела в це- лом, но и побегов по отдельности, что уже давно находит отражение во многих классификациях побегов по особенностям их специализации (Warming, 1884; Любименко, 1900; Dode, 1905; Смелов, 1947; Серебря- ков, 1949, 1952; Федоров и др., 1962; Голубев, 1965; Нухимовский, 1970, 1971, 1980, 1997; Мазуренко, Хохряков, 1977; Мазуренко, 1986, и др.). При желании на ступени «пирамиды» абстракций (графической модели суперординатной классификации) побегов семенных растений (т. 1, гл. 5, стр. 196) можно поместить еще некоторое число субординат- ных классификаций побегов. Здесь я использую, на мой взгляд, вполне подходящий случай, чтобы разобраться с весьма интересным направле- нием в классифицировании побегов по специализации их по длине (Dode, 1905; Федоров и др., 1962; Голубев, 1965 и др.), выбрав из многих терминов только, в этом плане, самый полезный для нас вариант из трех уже известных типов побегов: долихобласты (синоним ауксибласты), мезобласты и брахибласты (от греч. blastus — росток). В.Н. Голубев (1965) удачно обратил внимание на то, что в организации растительных биоморф действует два типа побеговых структур: долихоморфный (у древесно-кустарниковых растений подавляющая масса побегов слагает- ся удлиненными (сильнорослыми) побегами — долихобластами) и бра- химорфный (основу побеговой организации растений составляют удли- ненные побеги, на которых в качестве боковых развиваются розеточные побеги — брахибласты). Пожалуй, уместно между долихоморфным и брахиморфным струк- турными типами побегов рассматривать еще и мезоморфный структур- ный тип (мезобласты — это те побеги, но и части побегов, которые по своим линейным параметрам не доходят до статуса длинных, или силь- норослых, поскольку заметно слабее их, часто с вполне заметными сле- дами угнетения таких побегов в развитии, а также некоторые другие побеги или части побегов, например, весьма специализированные в репродуктивном отношении или как-либо иначе, которые не могут счи- таться какими-либо брахибластами — компактными безрозеточными или розеточными). Мезобластами, например, можно считать короткие вегетативные четочные побеги и подобные побеговые части в структуре разноразмерных (гетеробластных) побегов (рис. 19). Традиционно сло- жилось более всего брахибласты отмечать среди побегов некоторых хвой- ных (рис. 32, 34). Однако такое жесткое ограничение в применении этого термина вряд ли уместно, поскольку предостаточно других укоро- ченных (компактных), часто розеточных как вегетативных, так и более или менее репродуктивных побегов, развивающихся в организации био- морф семенных растений в различных сочетаниях на фоне каких-либо длинных и средних вегетативных и вегетативно-репродуктивных побегов. Здесь затронута весьма интересная и важная проблема, способная заметно усилить динамику фитоморфологических исследований. На мой взгляд, надо достойным образом оценить явление «бластности», придав ему соответствующий статус, который не будет перекрывать собой ста- тус побегов вообще, а восприниматься как новая сторона в исследова-
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИИ нии побеговой организации семенных растений. Я склонен следующим образом определить бластность — это особая система частей побегов и отдельных целых побегов, т.е. побеговых меросом — бластов в системе (теле) всей биоморфы, функционирующих в своем измерении — особым образом ориентированных в специализации по определенным рядам альтер- нативных (сравнимых и взаимоисключающих) признаков. Границы бластов, выделяемые по рядам альтернативных признаков иной специализации, часто будут другими. Самое главное в бластах, что их границы распространяются лишь на тот побег или часть побега, где непосредственно (!) проявляются определенные качественные показате- ли их специализации по определенным рядам альтернативных признаков. Дифференциация побегового тела на бласты — это уже не побего- вый уровень меротомирования, а именно новый, свой особенный, бла- стовый уровень, где в центре внимания уже не побеги, а бласты и бластогенезы, которые в побегах какой-либо сложности организации могут по-разному проявляться. Через бласты организация побегового тела рассматривается еще с одного угла обзора и с уже иной высоты абстракции, поэтому бластность имеет свою систему измерений, свои пространственные формы и границы, свои временные параметры раз- вития. Бластность — это, с одной стороны, состояния специализации в организационном развитии побегов, но, с другой, — исследовательский принцип — особая парадигма в познании организации семенных фито- биоморф. Бластогенез — это свойство побегового тела биоморф разви- вать определенные бласты как структуры монобластных побегов или в структуре гетеробластных побегов. Бластогенезы — обязательная сторо- на процесса жизнеразвития как побегов по отдельности, так и всего побегового тела биоморф, поскольку в разное время и в разном месте, под влиянием различных внешних и внутренних факторов развития формируются определенные структуры-бласты, в чем-то безусловно однородные, но в чем-то весьма отличные. Эти структуры различаются специализацией по определенному ряду (или рядам) признаков. Здесь и сейчас есть смысл сконцентрировать внимание на паре наиболее существенных для понимания организации побегов рядов признаков: на способности бластов (частей побегов и отдельных побегов целиком) проявлять определенную специализацию по особенностям их вегетатив- но-репродуктивного и (или) линейно-ростового (по размерам) развития. Побеговые меросомы, отмеченные выше (стр. 60 — 62), например, вегетатив, тотавегетатив, интервегетатив, репродуктив, фрондорепродук- тив, тотарепродуктив, вегеторепродуктив — это все бласты по вегетатив- но-репродуктивной специализации, а долихобласты, мезобласты и бра- хибласты — это тоже бласты, но вычлененные в побеговой организации по их размерам и особенностям растяжения при росте междоузлий, т.е. по линейно-ростовому ряду признаков (рис. 10 — 36). Если по одному ряду альтернативных признаков в организации побегов вычленяется по несколько бластов, то побеги — гетеробластные, а если по одному, то- монобластные. Бласт — единица бластности — единица специализации в органи- зации побеговых меросом по отдельным рядам альтернативных призна-
Глава 1. Концепция меросомной организации биомо ф семенных астений 65 ков. Таким образом, меротомирование побегового тела растительных биоморф на бласты вполне вырисовывает еще одну парадигму: каждый побег в своей личной организации проявляет соответствующим образом не только метамерию, но и бластность (или бластомерию — вопрос о слове- этикетке сейчас не главный), когда побеги растут и развиваются по опре- деленной генетической программе с различными вегетативными и (или) репродуктивными результатами у их частей, на которые накладывается, как обязательное, воздействие внешних, весьма лабильных факторов, уг- нетающих или усиливающих жизнеразвитие. Греческие слова «бластус» и «кормус» в соответствующих терминах — не синонимы в смысле слова побег. Побег — это только кормус, а бласт («pocmo~>) — это качествен- ное состояние специализации побега или его части (тогда их в побеге несколько), выделяемое по определенному ряду альтернативных признаков. Побеги стремятся в своем развитии к формированию опорной си- стемы и к размещению на ней репродуктивных элементов. Репродуктив- ная функция, как высшая из функций в организации семенных фито- биоморф, проявляет себя по-разному (в определенных местах и в опре- деленное время), распределяясь в верхушечных соцветиях, межсоцвети- ях, иносоцветиях (т. 1, гл. 5), в каких-либо каулифлорных системах (Ficus sycomorus L.) и в различных сочетаниях отмеченных форм цве- торасположения как элементов сверхсоцветий (рис. 10 — 36). Все три отмеченных выше типа побегов проявляют себя соответ- ствующим образом в каких-либо состояниях: или как вегетативы (веге- тативные побеги), или как вегеторепродуктивы и репродуктивы (вегета- тивно-репродуктивные и репродуктивные побеги). Долихоморфный, мезоморфный и брахиморфный типы структур- ной организации часто образуют, особенно наглядно у древесных рас- тений, различные сочетания их в телосложении (конституции) расти- тельных биоморф, обеспечивая им наилучшие возможности в борьбе за пространство и время при изменяющихся условиях окружающей среды. Итак, в зависимости от особенностей растяжения междоузлий в органи- зации побегов уместно выделять там, где это нужно, следующие побе- говые меросомы — бласты: брахибласты (побеги или части побегов с укороченными междоузлиями, которые могут быть и розеточными, и безрозеточными), и долихобласты (удлиненные побеги или части побе- гов, которые, в отличие от брахибластов, могут быть безрозеточными раздвинутоузловыми и сближенноузловыми). Переходное положение между отмеченным бластами занимают мезобласты. Ростовая функция более отводится длинным и средним побегам, а репродуктивные «заботы» чаще всего перекладываются на компактные безрозеточные и тем более розеточные побеги (рис. 32 — 36). Весьма интересны компактные, нередко розеточные, вегеторепродуктивы с нео- днократным развитием «плодотворения» и (или) «пыльцетворения», т.е. би-, мультирепродуктивные брахибласты. Мультирепродуктивными брахибластами, например, являются «пло- душки» или «кольчатки» у яблонь (Malus Hill), на что удачно обратили внимание в «Атласе...» Ал. А. Федоров и др. (1962). Немало других деревьев и кустарников также развивают эфемерные и многолетние как-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ либо репродуцирующие брахибласты (кратко — гамобрахибласты), ко- торые варьируют от макрофилльных до микрофилльных форм в тотаве- гетативной части и от фрондозных до брактеозных и даже абрактеозных форм в их репродуктивной части (рис. 32 — 36). Все вроде бы получается вполне обоснованно при том движении мысли в классифицировании основных органов — побегов, которое в целом и обеспечивает поступательное идейное развитие нашей моногра- фии. Не будем забывать принципиального положения в этом весьма деликатном и гибком процессе: стоит несколько изменить угол обзора и даже чуть-чуть поменять высоту абстракции, как могут произойти вполне понятные, более или менее существенные подвижки в воспри- ятии параметров объектов классифицирования. Невольно замечаешь, как мысль, «путешествуя» по ступеням «пирамид» абстракций разных объектов классифицирования, в этом случае побегов, иногда находит еще кое-что, может быть, даже более значимое, чем в прежних класси- фикациях, которое, при обеспечении его соответствующей аргументаци- ей, способно многое идейно изменять. При субординатном и суперор- динатном классифицировании что-то из признаков можно еще и еще усилить, а что-то, напротив, ослабить (в этом — одна из интриг клас- сификационного процесса), чтобы четкость передачи иерархического типового разнообразия побегов становилась еще более выразительной. В нашем подходе к иерархическому классифицированию побегов, как мне представляется, важнее всего уметь ориентироваться в ценностном содержании признаков. Наивысший ценностный показатель в организации побегов — это степень участия в ней репродуктивного развития (если образно, некоего «брачевания», без которого вполне могут существовать конкретные био- морфы, но не существуют все виды биоморф семенных растений). Био- морфы без устали служат своим видам, безмятежно подчиняясь им, по сути, без остатка — всем своим содержанием совершенствуя в эволюции формы и способы репродуктивного развития. Репродуктивность — этот, безусловно весомый аргумент в разное время и в разных обстоятель- ствах, уже как-либо использовался в морфологических классификациях растений [например, в зависимости от повторяемости плодоношения побегов: монокарпики и поликарпики (А.-Р. De Candolle, 1818, 1832, цит. по И.Г. Серебрякову, 1962)]. Меня не покидает мысль, что именно через особенное отражение репродуктивного участия побегов в развитии организации биоморф семенных растений можно средствами суперор- динатного классифицирования найти и определить самое высшее звено в иерархической организации побегов. На высшую ступень «пирамиды» абстракций побегов, созданную ранее (т. 1, гл. 5, с. 196), в дополнение к тем субординатным классификациям, что уже были там размещены, вполне уместно поместить еще и следующую субординию — классифи- кацию из двух субординат (классов) побегов (на мой взгляд, это и есть высшее звено всей «пирамиды»): 1 — агамокормусы (от греч. а — ча- стица отрицания, датов — брак), иначе (образно) «небрачующиеся», или вегетативные, побеги, если они не проявляют или не проявили пока какой-либо репродуктивной функции, хотя вполне как-то могут уча-
Глава 1. Концепция меросомной о ганизации биомо ф семенных астений 67 ствовать в ее подготовке, 2 — гамокормусы, иначе «брачующиеся», или репродуцирующие, побеги (сюда попадают все вегетативно-репродук- тивные, репродуктивные и тотарепродуктивные побеги). Гамокормусы не только подготавливают, но и непосредственно участвуют в «брачева- нии» = репродуктивном развитии (гамогенезе). Участвуя в репродуктивном развитии, побеги так или иначе прояв- ляют себя и в других функциях, которые, хотя и работают как-либо на размножение [оно ведь нужно виду (!), а не той самой биоморфе, ко- торая непосредственно осуществляет размножение, подчиняясь видовой (более высокой, чем собственная) целесообразности существования], но, кроме того, служат поддержанию собственной жизни конкретной биоморфы. В этой связи репродуктивное развитие побегов в чем-то напоминает репродуктивное развитие животных, включая и их беремен- ность: то оно есть в организации соответствующих растительных тел, то его нет. Признаки репродуктивности внешне могут, в зависимости от внешних условий и внутреннего состояния развития побегов по отдель- ности и в комплексных системах, какое-то время и не проявляться. Таким образом, в теле агамокормусов более всего существует по- тенциал к какому-либо репродуктивному развитию непосредственно этих, но и (или) других побегов, в становлении которых они участвуют. В теле гамокормусов не только проявляются репродуктивные функции и остаются какие-либо следы от бывших когда-то репродуктивных «ба- талий», но есть еще и другие функции, более свойственные агамокор- мусам — это и способность просто питаться и расти, создавая фото- трофные структуры, эфемерные и персистентные опорные и депониру- ющие органы. Биоморфы, реализуя в своей организации развитие по- беговой системы, генетически предопределены (они обязаны в главном делать только так, а не иначе) на развертывание в ней необходимого качественного минимума из трех свойств, два из которых нужны и им, и их виду — это свойств питаться и линейно расти, и третьего свойства — репродуктивности, которое, по большому счету, нужно только виду. Эти свойства переплетаются в организации растительных биоморф са- мым разным образом, составляя атрибут в жизнеразвитии побегов. Если что-то мешает развиваться этим свойствам в организации биоморф, например, сильное затенение кронами деревьев, то в первую очередь травмируется способность к репродуктивному размножению, поскольку она непосредственно зависит от благополучия в проявлении первых двух свойств. Таким образом, и агамокормусы, и гамокормусы обладают, помимо основной, еще и дополнительной специализацией, что обозначается соответствующими эпитетами (рис. 11 — 50): в зависимости от длины и числа их междоузлий они бывают длинные, средние и компактные (очень короткие), по особенностям проявления персистентности, эфе- мерности и фотофильности различимы ростовые (их срединные листья зеленые, ассимилирующие, а стеблевая часть полностью или не полно- стью потенциально персистентная или персистентная, иначе скелетная), ассимиляционные (их срединные листья зеленые, ассимилирующие, а стеблевая часть эфемерная) и магимодифицированные [в отличие от
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ предыдущих двух типов побегов специализация этих побегов связана с развитием только более («маги») видоизмененных листьев-чешуй]. Из многих вариантов терминов, обозначающих организационные типы основных органов, в первую очередь, побегов, как результаты каких-либо классифицирований их, в каждом конкретном исследовании надо подби- рать и использовать те варианты, которые более всего способствуют раз- решению текущих исследовательских проблем, — это атрибутивное прави- ло биоморфологических исследований семенных растений. Меротомировать организацию живых тел можно, как подчеркива- лось выше, по-разному; при этом главное, чтобы была цель и у иссле- дователя, проводящего процедуру меротомирования, и в меросомности поведения самих живых тел как целого, испытывающих удобства в своей организации через специализацию ее на части — меросомы, выполня- ющие определенные функции. В качестве примера такого «свободного» (не строгого) меротомирования рассмотрим графическую модель орга- низации биоморф семенных растений по особенностям участия их ме- росом в жизнеразвитии и круговороте ресурсов (рис. 37); эта модель показывает, как меросомы рассматриваемых биоморф специализирова- ны для выполнения главных жизнесохранительных функций: потребле- ния ресурсов окружающей среды, вегетативного роста и развития, де- понирования ресурсов, производства репродуктивных органов, различ- ных приемов размножения и рассеивания ресурсов. 22 Рис. 11. Универсально-сложный гамокормус (фрагмент) Adhatoda vasica (L.) Ness [сем. Acanthaceae (Израиль, Тель-Авив. Ботанический сад)].
Глава 1. Концепция ме осомной организации биомо ф семенных растений 69 Рис. 12. Универсально-сложные гамокормусы Callistemon speciosus (Sims) DC. [сем. Myrtaceae (Тель-Авив, скверы и парки, март — I, апрель — П)].
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 13. Биографический портрет Celtis australis L. [сем. Ulmaceae (скверы и парки Тель-Авива, 15 IV)j. Основной «городской» биоморфотип: надземка-двусемядольные (семядоли ши- роколопатчатые окрутло-четырехугольные, на верхушке с широкой выемкой, 15 — 20 мм дл., 10 — 13 мм шир., резко суженные в короткие черешки) супротивно-перволистные, доминантно-очереднолистные, простолистные безрозеточные акро-эусимподиальные ин- теркалярно-репродуцирующие открыто-репродуктивные (образуют межсоцветия) моноаэ- роподиальные (некустистые, с одним стволом) средневысокие (10 — 25 м высотой) пери- стентно-длиннопобеговые (максиморезидные) листопадные деревья.
Глава 1. Концепция ме осомной о ганизации оиоморф семенных астений Рис. 14. Биографический портрет Nerium oleander L. [Сем. Аросупасеае (скве- ры и парки Тель-Авива)]. ОСНОВНОЙ «ГорОДСКОЙ» бИОМОрфОТИП: надземка-двусемядольные (семядоли про- долговато-яйцевидные узкозаостренные, 12 — 15 мм дл., 4 — 5 мм шир., клиновидно суженные в короткие черешки) ювенильно-супротиволистные (первая ювенильность свя- зана с образованием только супротивных пар листьев в узлах), доминантно-мутовчатоли- стные (в узле обычно мутовка из трех листьев), простолистные безрозеточные гемисимпо- диальные закрыто-репродуктивные (детерминатные) однодомные розово-кугроцветковые (реже бело- или красноцветковые) высокие (3 — 6 м высотой) персистентно-длиннопо- беговые (тотарезидно-магнорезидные) вечнозеленые кустарники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 15. Биографический портрет Sorbus aucuparia L. [сем. Rosaceae (Мос- ковская область; леса, опушки и поляны, берега рек)]. Основной биоморфотип (I): надземка-двусемядольные (семядоли эллиптические, 7 — 9 мм дл., 3 — 5 мм шир., на верхушке закругленные, короткоклиновидно суженные в черешки) очереднолистные сложно-непарноперистолистные безрозеточно-ростовые нере- гулярно-розеткообразующие закрыто-репродуктивные (детерминатные, соцветие — фрон- до-брактеозный щиток) гемисимподиальные компактно-, средне-, и длиннорезидные од- нодомные белоцветковые листопадные корнеотпрысковые кустарники или небольшие деревья (3 — 15 м высотой). «В молодом возрасте переносит значительное затенение, встречаясь под пологом темно- хвойных лесов, позже становится более светолюбивой. Обильно цветет и плодоносит лишь на хорошо освещенных местах — вырубках, гарях, опушках» (Вахрамеева, 1975, с. 78). Попав под сильное затенение кронами взрослых лесных кустарников и деревьев, биомор- фы рябины долгие годы могут влачить жалкое существование и часто совсем не цвести, проявляя второстепенный биоморфотип (11): стелющиеся вынужденно нецветущие или слабо и редко цветущие кустарники (стланики).
Глава 1. Концеиция ме осомной о ганизации биомо семенных астений 73 Рис. 16. Биографический портрет Juglans mandshurica Maxim. [Сем. Juglandaceae (Москва, парк в Хамовниках)]. ОСНОВНОЙ «ГОрОдСКОЙ» бИОМОрфОТИП: подземносемядольные (семядоли, сильно рассеченные на два бугорчатых сегмента, надежно скрыты в орехе, который обычно при весеннем прорастании вскрывается, но не разламывается) очереднолистные сложно-не- парноперистолистные безрозеточные двудомные эумоноподиально-тычиночноцветковые (образуют иносоцветия) гемисимподиально-пестичноцветковые полузакрыто-плодообра- зующие персистентно-длиннопобеговые (тота- и магнорезидные) листопадные моноаэро- подиальные деревья (15 — 30 м высотой).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ 17,25, ги2 г в+г ~ ж,156,21,26,ги1 32,36 Рис. 17. Биографический портрет Tipuana tipu (Benth.) Kuntze [сем. Fabaceae (скверы и парки Тель-Авива, цветет май, июнь)]. Основной «городской» биоморфотип: надземка-двусемядольные (семядоли эл- липтические мясистые, иногда неравнобокие, широкозакругленные, суженные в очень короткие черешки, 16 — 1S мм дл., 7 — 9 мм шир.) очереднолистные сложно-непарно- перистолистные безрозеточные однодомные эусимподиальные персистентно-длиннопобе- говые (максиморезидные) вечнозеленые моноаэроподиальные деревья (6 — 20 м высотой).
Глава 1. Концеиция ме осомной организации биомо семенных астений 75 Рис. 18. Биографический портрет Cassia senna L. [сем. Fabaceae (кустарни- ковые посадки вдоль шоссе Тель-Авив — Хадера, 28 IV)]. По рисунку видно, что кустарник цвел поздно осенью, зимой сохранил зеленые листья (часть листочков осыпалась) и образовал зеленые плоды. Весной вегетация бурно возоб- новилась. Растения как бы доцветают, образуя немного уже весной силлептических брак- теозных цветущих побегов, а более всего отрастают вегетативные весенние и силлептичес- кие побеги, которые перейдут к цветению позднее — к осени. Основнай «городской» биоморфотип: надземка-двусемядольные (семядоли ок- рутлые, 10 — 12 мм дл., 9 — 12 мм шир., в месте прикрепления к короткому черешку имеют круглую выемку и у основания два небольших рожка) очереднолистные преювенильно- парнолистниковые (всходы) взросло-непарноперистолистные сложнолистные осенне-ве- сенне-цветущие безрозеточно-раздвинутоузловые гемисимподиальные (пожалуй, есть смысл говорить о доминантности, субдоминантности и факультативности в нарастании побегов, когда это действительно необходимо, например, в том случае, когда основной фон нарастания обеспечивается эумоноподиальностью, но иногда, вполне естественно, могут отмирать по разным причинам верхушки побегов, обозначая тем самым факульта- тивное проявление другого типа нарастания, допустим, гемисимподиального) однодомные желто-бобовоцветковые вечнозеленые аэроксильные кустарники (2 — 4 м высотой).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 19. Биографический портрет Euryops speciosissimus DC. [сем. Asteraceae (экспозиция Южной Африки в ботаническом саду Иерусалимского универ- ситета, 21 IV). БИОМОрфОТИП: надземка-двусемядольные (семядоли эллиптические или продолговато- яйцевидные, закругленные, клиновидно суженные в короткие черешки, дл. 8 — 10 мм, шир. 4 — 5 мм) четочные очередно-полусложнолистные гемисимподиальные однодомные полузакрытые (семидетерминатные, верхушки универсально-сложных гамокормусов зак- рывают корзинки, которые сами являются полузакрытыми соцветиями) желто-астроцвет- ковые акронекрозные полегающе-восходящие сильнорослые полукустарники (впервые зац- ветают на 2 — 3-ий год, живут около 30 — 40 лет).
Глава 1. Концепция ме осомной организации биомо ф семенных растений 77 Рис. 20. Биографический портрет Gentiana fischeri Р. Smirn. [сем. Gentianaceae (Алтай, Курайский хребет, субальпийские луга, около 2000 м над уровнем моря)]. Основной биоморфотип: недземно-двусемядольные эумоноподиальные магимоди- фицированно-ростовые пазушно-безрозеточно-ассимиляционные интеркалярно-репроду- цирующие детерминатно-кладонекрозные (закрытые пазушные эфемерные цветоносы в структуре межсоцветий) однодомные черновато-синегоречавкоцветковые мультирепродук- тивные продольно-партикулирующие (множественно расщепляющиеся в подземной части через главный корень и корневище по отмирающим веточным «следам» ) компактно(кур- торезидно)-ризомные, замедленно вегетативно размножающиеся (ретардационно-конди- видуальные), высокогорно-луговые травянистые многолетники (первое цветение на на 5— 15 год, синорганизмы живут до 60 — SO лет).
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHbIX РА СТЕНИИ 1 12 13 72 72 Рис. 21. Биографический портрет Actaea spicata L. [сем. Ranunculaceae (Мос- ковская область, Подольский район, смешанный елово-березовый лес, по- ляны).
79 Глава 1. Концеиция ме осомной о ганизации биомо семенных астений Рис. 22. Биографический портрет Lomatogonium carinthiacum (Wulf.) Reichenb. [Сем. Gentianaceae (Алтай, Ивановский хребет, низкотравные луга, осыпи в высокогорьях и лесном поясе). Биоморфотипы: надземка-двусемядольные (семядоли 4 — 5 мм дл., 3 — 3,5 мм шир., округло-яйцевидные, лопатчатообразные, широкозакругленные, резко суженные в череш- ки 2 — 3 мм дл.) полурозеточные некустистые (1 биоморфотип) или кустистые (11 биомор- фотип) супротивно-простолистные детерминатно-фрондозно-репродуцирующие (с закры- тыми олиственными соцветиями) однодомные голубовато-горечавковоцветковые однолет- ники. Основной биоморфотип: надземка-двусемядольные (семядоли яйцевидные или ши- рокоэллиптические, 12 — 20 мм дл., 7 — 12 мм шир., на верхушке коротко заостренные на длинных черешках) ювенильно-эумоноподиально-розеточные взросло-безрозеточные эусимподиальные и гемисимподиальные компактно(миниморезидно)-ризомные очередно- листные дважды- и трижды тройчатосложнолистные однодомные мультирепродуктивные бело-лютиковоцветковые семидетерминатные (полузакрытые), замедленно вегетативно раз- множающиеся (ретардационно-кондивидуальные), травянистые многолетники (цветут впервые на 6 — 15 год, живут до 60 лет). Графический портретный ряд разновозрастных биоморф А. spicata, приведенный здесь, дополняет довольно подробные описания различ- ных сторон биоморфогении этого вида, уже сделанные ранее (Рысина, 1973; Барыкина, Гуланян, 1975; Барыкина, Мусина, Чубатова, 1990).
0Р- di- 01 О- И- гда ИД- ир, ен- ВО- об- ИВ- 03- Н- НО- ОД- се- не- 5а, 31. 34 ЯСНО НО В еся ЗЛ ЬН летн — 2 сте ТОЛЬ кедр лесье раст прод НОР OCHOBhl БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИЙ
Глава 1. Концепция меросомной организации биоморф семенных растений Рис. 24. Биографический портрет Rhodiola quadrifida (Pall.) Fisch. et C. A. Меу [сем. Crassulaceae (Алтай, Курайский и Северо-Чуйский хребты, щед- нистые высокогорные тундры, каменистые россыпи, около 2 000 м над ур. моря)]. Продолжение см. стр. 82.
OCHOBhI БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИИ ,гб Рис. 25. Биографический портрет Anagallis arvensis L. [сем. Рпти1асеае (окрестности Тель-Авива, каменистые склоны невысоких холмов; март, цве- тение)]. Основной биоморфотип: надземно-двусе- мядольные (семядоли яйцевидные, узкозакруг- ленные на верхушке, 4 — 5 мм дл., 1,5 — 2 мм шир., клиновидно суженные в короткие череш- ки) перекрестно-супротивнолистные полегаю- щие обильно-ветвистые стержнекорневые без- розеточные моноподиальные верхушечно-инде- терминатные (открытые фрондозные верхушеч- ные соцветия) однодомные голубовато-приму- лоцветковые однолетние травы. ,29,дс,ам1 Рис. 24. (Продолжение) Основной биоморфотип: надземно-двусемядольные (семядоли мясистые голые, ва- рьируют по размерам и ~орме, округ кр;-,лые эллиптические продолговатые, продолговато- об атнояйцевидные, туповатые, 1,5 — 5 мм дл., 1 — 4 мм шир., суженные в короткие или несколько вытянутые черешки (от 1 до 5 мм дл.) эумоноподиальные универсально-слож- но-ассимиляционные нопобеговые магимодифицированно-ростовые пазушно-безрозеточно-асс интер калярно-репродуцирующие детерминатно-кладонекрозные (закрытые пазушные э е- мерные цветоносы в межсоцветиях) краснеющие зеленовато-желто-толстянковоц ветковые однодомные мультирепродуктивные компактно(курторезидно)-каудексовые стержнекорне- вые, замедленно вегетативно размножающиеся (ретардационно-кондивидуальные), высо- когорно-тундровые травянистые многолетники (каудекс здесь структурно весьма близок к компактному ризому, поскольку главный корень может по разным причинам отмереть рано, даже у проростков, но растения более склонны не погибать и развивать еще и в таких неко невую хотя и придаточную, систему, успешно заменяющую утрату главного корня; у взрослых растений отличить, где «квазикаудекс», а где каудекс насто- ящий часто невозможно, поэтому мы во всех случаях используем термин «каудекс», первое цветение происходит на 8 — 25 году жизни, синорганизм иногда существует до 150 лет).
Глава 1. Концелция ме осомной организации биомо семенных растений Рис. 26. Биографический портрет Lagopsis matTubiastrum (Steph.) Ik.-Gal. [сем. Lamiaceae (Алтай, KypaNGKNA хребет, на щебнистых осыпях альпийс- кого пояса, 2200 м над ур. моря)]. Биоморфотипы: надземно-двусемядольные (семядоли эллиптические, 4 — 6 мм дл., 3 — 4 мм шир., на верхушке узкозакрутленные, в месте прикрепления к черешку имеют стреловидную выемку, у основания два небольших рожка) полурозеточные эумоноподи- ально-розеточные пазушно-безрозеточно-репродуцирующие и дополнительно ассимили- рующие (с зелеными листьями) постинтеркалярно-репродуцирующие (цветоносные побе- ги развиваются из пазух прошлогодних листьев) семидетерминатно-кладонекрозные (по- лузакрытые пазушные эфемерные цветоносы образуют иносоцветия) густоопушенные (опу- шены все надземные части, в особенности соцветия) простолистные стержнекорневые компактно(курторезидно)-каудексовые (1-ый биоморфотип у подножия высокогорных осыпей, где относительно спокойно ведет себя уровень поверхности почво-грунта) или длинно (вицерезидно)-каудексовые (II-ой биоморфотип на крутых щебнистых осыпях) однодомные мультирепродуктивные буро-коричнево-яснотковоцветковые изоаклональные высокогорные эрозионно-петрофильные травянистые многолетники (впервые зацветают на 2 — 7 год, чаще 3 — 5, живут до 25 лет, но особи первого биоморфотипа развиваются интенсивнее: больше образуется цветоносов, раньше начинают цвести на 2 — 3 год жизни и быстрее отмирают) °
OCHOBhl БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИЙ Рис. 27. Биографический портрет Papaver pseudocanescens М. Рор. [сем. арачегасеае (Алтай, Ивановский, Линейский и Курайский хребты, щебни- стые слабозадерненные высокогорные склоны и галечники, 1 700 — 2200 м над уровнем моря)]. Основной би ~HOMOp~OTHH: надземно-двусемядольные (семядоли ланцетные, слегка заостренные, почти сидячие, 8 — 12 мм дл., 2 мм шир.) гемисимподиально-розеточные стержнекорневые компактно(миниморезидно)-каудексовые простолистные мультирепро- дуктивные детерминатно-репродуцирующие (моноподиально развивающие из розеточной вегетативной части каждого вегетативно-репродуктивного побега безлистный долихомор- фный цветонос всего с одним длинным междоузлием, заканчивающимся сверху одним цветком) желто-макоцветковые изоаклональные высокогорные травянистые многолетники (впервые цветут на 2 — 5 году, а всего живут до 20, реже 30 лет).
Глава 1. Концепция ме осомной организации биоморф семенных растений Рис. 28. Биографический портрет Oxyria digyna (L.) Hill [сем. Polygonaceae (Алтай, Курайский и Северо-Чуйский хребты, 2 000 — 2 700 м над уровнем моря, галечники и каменистые осыпи в альпийском поясе)]. Продолжение на стр. 86.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 28. (Продолжение) БИОМОРфОТИПЫ: нвдземно-двусемядольные (семядоли 4 — 7 мм дл., 2,5 — 4 мм шир., продолговато-яйцевидные, закрутленные, на черешках 4 — 15 мм дл.) виргинильно-эумо- ноподиальные матурно-гемисимподиальные мультирепродуктивные розеточные стержне- корневые компактно(курто-, миниморезидно)-каудексовые (1-ый биоморфотип в условиях относительно спокойного состояния уровня поверхности почво-грунта) и длинно(вице-, викариорезидно)-каудексовые (11-ой биоморфотип на подвижных осыпях) просто-почко- виднолистные семидетерминатно-репродуцирующие (из брахиморфной — розеточной- структуры универсальных плейофазных побегов развивается долихоморфная структура— безлистный цветонос с полузакрытым брактеозным метельчатым соцветием), вегетативно несовершенно-размножающиеся (при старении) или вынужденно экспансивно-размножа- ющиеся (редко, при неравномерном смещении на осыпях больших масс почво-грунта (вегетативное размножение, когда оно происходит, то заметно не продлевает продолжи- тельность синорганизменной жизни), травянистые высокогорные многолетники (впервые зацветают на 6 — 12 год, а предельно живут около 50 лет).
Глава 1. Концепция меросомной о ганизации биомо ф семенных астений 87 Рис. 29. Биографический портрет Ceratonia siliqua L. [сем. Fabaceae (окре- стности Тель-Авива, холмистая местность, 10 IV)]. Др — дополнительный рисунок (Hegi, 1975). BHOMOp@OTHIII I: надземно-двусемядольные (семядоли 18 — 24 мм дл., 8 — 12 мм шир., эллиптические или несколько продолговато-обратнояйцевидные, закрутленные, коротко- клиновидные, на коротких черешках) эумоноподиальные лидерно-апонекрозные персис- тентно-длиннопобеговые (тотарезидные) безрозеточные непарно-пористосложнолистные двудомные мультирепродуктивные постинтеркалярно-репродуцирующие (ежегодно отрас- тают пазушные весенние постинтеркалярные побеги, образующие в структуре комбиниро- ванно-универсальных побегов иносоцветия только из тычиночных или пестичных цветков, а в последующие годы активность подобного побегоотрастания долгое время не ослабевает и таким образом проявляет себя каулифлория) некустистые (1-ый биоморфотип, больше открытые пологие горные склоны или равнинные участки) или кустистые (II-ой биомор- фотип, больше овраги и другие понижения на холмисто-гористой местности) изоаклональ- ные вечнозеленые деревья (8 — 25 м высотой).
Б В 7к Рис. 31. Биографический портрет Anagyris foetida L. [сем. Fabaceae (Тель-Авив, Ботанический сад, 29 IV)]. Основной биоморфотип: над- земно-двусемядольные (семядоли мясистые, опушенные, эллиптичес- кие, широкозакрутленные, сидячие, 15 — 20 мм дл., 8 — 10 мм шир.) эусимподиальные безрозеточные лидерно-акронекрозные персистен- тно-длиннопобеговые (максиморе- зидные) тройчато-сложнолистные однодомные мультирепродуктивные постинтеркалярно-ре продуцирую- щие (из почек многих прошлогод- них побегов развиваются весенние постинтеркалярные цветоносные побеги с кистями желтых обоеполых цветков, образующих вместе с осе- вой частью отдельного прошлогод- него побега, которая их объединяет, иносоцветие) корнеотпрысковые желто-бобовоцветковые листопад- ные кустарники (2 — 4 м высотой). ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 30. Биографический портрет Cercis siliquastrum L. [сем. Fabaceae (скверы и парки Тель-Авива, 15 IV)]. Биоморфотипы: надземно-двусемядоль- ные (семядоли эллиптические, 12 — 16 мм дл. и 8 — 12 мм шир., широкозакрутленные, резко суженные в короткие черешки) эусим- подиальные безрозеточные просто-почко- виднолистные однодомные мультирепродук- тивные постинтеркалярно-репродуцирую- щие (ежегодно отрастают пазушные весен- ние эфемерные постинтеркалярные побеги с кистевыми соцветиями обоеполых цвет- ков, образующие в структуре комбинирован- но-универсальных побегов иносоцветия, а в последующие годы активное отрастание по- добных побегов еще несколько лет продол- жается, что свидетельствует о каулифлории) лидерно-акронекрозные персистентно- длиннопобеговые (максиморезидные) стер- жнекорневые изоаклональные листопадные розово-бобовоцветковые моноаэроподиаль- ные невысокие (6 — 15 м высотой) деревья (1-ый биоморфотип) или (реже) высокие (3,5 — 5 м) кустарники (П-ой биоморфотип).
Глава 1. Концепция ме осомной организации биоморф семенных растений Рис. 32. Биографический портрет Pinus sibirica Du Тоиг [сем. pinaceae (Ал- тай, Ивановский, Холзун, Семинский, Бащелакский и Курайский хребты, от низкогорий до высокогорий). Продолжение на стр. 90. Биоморфатипы: надземно-многосемядольные (10 — 11 семядолей, кучно выходящих из семядол~ного yea, линейные, 24 — 28 MM ~., 1 2 — 1 8 MM mnp., сидячие) эумоноподиальные магимодифицированно-ростовые четочно-ростовые (на длинных и средних ростовых побегах по всей их длине развиваются сближенные чешуи, рано опада- ющие) брахибластно-ассимилирующие (образуют брахибласты-розетки с пятью хвоинка- ми, которые живут до 5 — 7, реже 8 лет) лидерно-апонекрозные персистентно-длиннопо- беговые (тотарезидные) интеркалярно-репродуцирующие (в структуре годичных побегов
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ текущего года у одного растения отрастают пазушные силлептические — интеркалярные — однополо-репродуктивные побеги с пыльцеобразованием или образующие семена) раз- дельнополо-однодомные вечнозеленые моноаэроподиальные разновысокие (6 — 30 м) деревья (1-ый биоморфотип, низкогорья и до верхней границы лесного пояса в горах) или вынужденно приземистые (0,5 — 6 м) деформированно-крановые деревца и кустарники (II-ой биоморфотип, выше верхней границы леса). Пожалуй, есть необходимость особо разъяснить тип «четочных», или четкообразных, по- бегов (cormus moniliformis). Этот термин я предлагаю для обозначения безрозеточных длинных и средних побегов сближенноузловых в противовес длинным и средним безро- зеточно-раздвинутоузловым побегам. Четочные побеги особенно характерны для многих хвойных растений (рис. 33, 34). Между розеточными и четочными побегами бывают побеги переходного типа: розеточно-четочные, или полурозеточно-сближенноузловые (рис. 33). Четочные части побегов и четочные побеги целиком мы относим в зависимости от их длины наряду с безрозеточными (раздвинутоузловыми) побегами или такими же частями побегов к долихоморфному или мезоморфному типам организации (структурным типам). Рис. 33. Розеточные и розеточно-четочные побеги некоторых хвойных деревьев [примеры взяты из книги «Деревья и кустарники» (Pokorng, Choc, 1967)]. 1 — Larix decidua Mill., II — Pseudolarix amabilis Rehd.
Глава 1. Концепция меросомной организации биоморф семенных растений 91 Рис. 34. Четочные побеги некоторых хвойных деревьев [примеры взяты из книги «деревья и кустарники» (Pokorny', Choc, 1967)]. 1 — Taxus baccata L., II — Тоггеуа colifornica Тогг., III — Abies alba Mill., IЧ — Picea abies (L.) Karst., Ч — Pseudotsuga menziesii (Mirb.) Franco, ЧI — Pinus sylvestris L.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИЙ Рис. 35. К биографическому портрету Rhamnus lycioides L. subsp. graeca (Boiss. et Reuter) Tutin (Rh. palaestinus Boiss.) [сем. Rhamnaceae (Израиль, горные склоны вдоль дороги между городами Хадера и Хайфа, кустарнико- вые заросли, 14 IV)j. ОСНОВНОЙ бИОмарфОТиП: надземно-двусемядольные безрозеточно-розеточные доли- хоморфно-эусимподиальные (длинные и средние вегетативные и вегетативно-репродук- тивные побеги обеспечивают основной персистентно-осевой прирост, а их верхушки после прекращения весеннего роста засыхают и превращаются в колючки) брахиморфно-эумо- ноподиальные простолистные мультирепродуктивные двудомные интеркалярно-репроду- ци щие (из пазух листов весенних многих розеточных побегов, но еще и нижних листьев некоторых длинных и средних безрозеточных побегов, силлептически отрастают интеркалярные по еги- б — короткие цветоносы с одиночными однополыми цветками, не- редко образуя вместе межсоцветия) листопадные стержнекорневые изоаклональные жел- товатозелено-жестероцветковые кустарники.
Глава 1. Концепция меросомной организации биоморф семенных растений Рис. 36. Биографический портрет Astragalus spinosus (Forssk.) Muschler [сем. Fabaceae (Израиль, пустыня Негев, окрестности поселка Сде-Бокер, камени- стое плато, 15 V)j. Продолжение на стр. 94. Оснавнай биамарфотил: надземно-двусемядольные (семядоли продолговато-обрат- нояйцевидные, 8 — 10 мм дл., 4 — 5 мм шир., широкозакругленные, на очень коротких черешках, сросшихся основаниями) эумоноподиальные четочно-ростовые розеточно-фрук- тифицируюшие (плодоразвиваюшие) колюче-рахисные (центральные оси — рахисы- сложных листьев после начала осыпания их листочков быстро превращаются в острейшие колючки, которые долго сохраняются и в сухом виде) непарно-перистосложнолистные опушенные (обильное свежее опушение весенних побегов вскоре отмирает и «сдувается» сухим и жарким ветром) однодомные интеркалярно-репродуцирующие (из пазух нижних чешуевидных и срединных листьев розеточных и нижних листьев некоторых длинных и
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИИ средних четочных годичных побегов силлептически развиваются интеркалярные безлист- ные короткие цветоносные побеги с одиночными обоеполыми цветками, вскоре преобра- зующимися в пузырчато вздутые эллиптические бобы, а когда в отдельном годичном побеге их развивается несколько, то вместе они составляют межсоцветие этого вегетатив- но-репродуктивного побега) персистентно-длиннопобеговые (длинно- и средне-тотарезид- ные), вспомогательно персистентно-компактнопобеговые (курторезидные), стержнекорне- вые листопадные кустарнички (30 — 100 см высотой). Более крупные кусты астрагала колючего развиваются при произрастании растений не в одиночестве, а в группах (в «тесноте») вместе с биоморфами других видов, например, Artemisia sieberi Besser, Polygonum palaestinum Zohary, Piptatherum miliaceum (L.) Coss. Рис. 37'. Графическая модель «свободного» членения на части (меротоми- рования) организации биоморф семенных растений по их участию в жизне- развитии и круговороте ресурсов. 1 — меросома непосредственного репродуктивного развития, 2 — меросома нарастания и ветвления вегетатива кроны, 3 — меросома фотосинтеза ( «фабрика фотосинтеза»), 4— меросома аэроподиального выноса и поддержания кроны в надземном пространстве, 5— меросома вегетативного горизонтального разрастания, 6 — меросома вегетативного раз- множения, 7 — меросома водно-минерального питания, 8 — меросома побегового депо- нирования ресурсов, 9 — меросома гипокотильного депонирования ресурсов, 10 — меро- сома корневого депонирования ресурсов, 11 — меросома активного приземно-подземного побегоотрастания (зона кущения), 12 — меросома инфлоресцентного репродуктивного развития, 13 — меросома одноцветкового репродуктивного развития, 14 — меросома эфемерного надземного роста и развития побегов, 15 — меросома постфлорального раз- вития побегов ..., су — семядольный узел, кш — корневая шейка.
95 Глава 1. Концепция ме осомной организации биоморф семенных растений 1.5.2. Мультивариантность развития биоморф в круговороте ресурсов Материальные тела, в силу заложенной в них преднаследственно- сти или наследственности, запрограммированы на разные варианты раз- вития, в зависимости от условий их существования; поэтому мультива- риантность — свойство развития большинства физиосистем (материаль- ных систем). Нетрудно убедиться, что к такому выводу первыми пришли (см. т. 1, гл. 8) именно ботаники, обнаружив, что многим растительным организмам и видам свойственна мультивариантность развития, в зави- симости от условий, в которые их забрасывает судьба. «Многовариан- тность развития — осуществление развития разными путями, в зависи- мости от внутреннего состояния организма и внешних условий суще- ствования» (Чайлахян и др., 1973, с. 8). Все тела, существующие в При- роде, обладают определенным ресурсом возможностей (идиоэнергией), реализация которого у однотипных тел в разных условиях существова- ния позволяет судить о наличии или отсутствии мультивариантности развития у тех или иных типов тел. Способность физиосистем к муль- тивариантности развития — результат эволюции, в которой, как бы ни изменялись тела и процессы, нет остановочного механизма (!), и это правило бесконечно действует в бесконечной Природе. Материи изначально свойственно стремление к разнообразным морфогенезам (соматогенезам, системогенезам), т.е. способность всегда развивать формы, доступные в рамках ее возможностей и существовать в них какое-то время. В бесконечном движении материи к морфогене- зам и в морфогенезах предусмотрена мультивариантность развития тел, причем не только биоморф, но и живых тел вообще (эврибиоморф), а еще шире — любых тел. Материальные системы без помощи людей самонаправляются на цели развития с прогенетического или генетического уровня, на основе использования своего ресурса возможностей (идиоэнергии). Но иногда, а с развитием цивилизации все чаще, им помогает в наведении на цели сила извне — человеческий разум и деятельность человека, превраща- ющего первичный материал в сырьевой ресурс для домов, машин, за- водов и т.п. Ресурс возможностей — на то и ресурс, чтобы иметь не одну возможность строго определенного характера, а несколько, чаще много. Развитие систем одной родственной группы строго по одному варианту в Природе не бывает, поскольку ресурс возможностей у материальных систем «заводится» на генерацию многоцелевого развития по разным рядам признаков. Идиоэнергия неживых и живых систем, вероятнее всего, всегда носит мультивариантный (многоцелевой) характер. Явле- ние, когда система обладает мультивариантной идиоэнергией, т.е. имеет много целей по разным рядам признаков для парафенетического или фенетического интегративного развития и много возможностей выби- рать какую-то одну цель по каждому ряду признаков, объединяя движе- ние к многим целям в одно общее развитие по одному, из нескольких возможных, варианту, вполне можно называть мультивариантностью раз- вития. С учетом мультивариантности мы можем утверждать, что абсо-
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ лютно одинаковых систем, а следовательно, и морфогенезов (шире- системогенезов) в Природе не существует. Мультивариантность систем и системогенезов находится в полном соответствии с выведенным нами законом изменчивости материальных систем (т. 1, гл. 8, стр. 363). Муль- тивариантность развития биоморф семенных растений (рис. 38), о ко- торой неоднократно писал и автор (Нухимовский, 1969, 1970, 1978, 1997) и многие другие исследователи (Работнов, 1956; Матвеев, 1975; Воронцова, Заугольнова, 1978; Жукова, 1986; Глотов, Жукова, 1995 и др.), — все это аспекты одного и того же явления, существующего во Вселенной и реализующегося с неиссякаемой силой, генерируемой идиоэнергией материальных систем (мультивариантности развития си- стем). Явление мультивариантности развития биоморф объясняет, как среди биоморф одного вида (рис. 38 — 50) проявляется несколько ва- риантов фенетического развития, различающихся по определенным про- стым признакам (продолжительность жизни, кратность проявления реп- родуктивной функции, характер облиствления побегов и т.п.) или по комплексу признаков. Эгосубстратная организация любых тел несет ин- формацию (программу) мультивариантного пластосоматического разви- тия, т.е. информацию о разнообразии соматической трансформации. Суть мультивариантного развития тел (в томе 1 уже было показано проявление мультивариантности в развитии биоморф семенных расте- ний) состоит в том, что у материи в ее эгосубстратном соматическом состоянии (в преднаследственных или наследственных ее свойствах), т.е. в границах каких-либо тел, есть несколько (иногда много) вариантов выбора возможностей конституциального соматического конструирова- ния, что в конкретных ситуациях зависит не только от эгосубстрата исходных тел, но, в большой мере, еще и от условий окружающей среды, которые непосредственно влияют на выбор варианта развития. Эгосубстратная организация какого-либо тела, взаимодействуя с конк- ретными условиями окружающей среды, выбирает один из многих ва- риантов пластосоматического развития, что, безусловно, является атри- бутивным свойством существования материи. То есть материальным объектам свойственно быть телом, иметь в нем организацию из двух частей: эгосубстратной и конституциальной (пластосоматической) — и развиваться по программе эгосубстратной части, но выбирать из многих тот вариант морфогенеза, который более всего отвечает условиям среды (рис. 38 — 50). В Природе действует один из абсолютных законов— закон мультивариантности соматического развития материи: материя про- являет бесконечность своего интегративного существования в дискретных телах, эгосубстратная часть организации которых улравляет конститу- циальной частью той же организации и наиравляет морфогенез по одному из нескольких вариантов, который отвечает как внутренней лрироде эго- субстрата, так и условиям нелосредственно окружающей его внешней среды. Материальная система, существуя в конкретном сочетании окружа- ющих ее условий (экологическом лабиринте), должна выбирать направ- ление (цель) своего развития, исходя из определенного ресурса возмож- ностей (информационно-энергетического потенциала), сосредоточенно-
Глава 1. Концепция ме осомной организации биоморф семенных растений го в прогенах и (или) генах, подвергаясь или не подвергаясь дополни- тельному воздействию со стороны человеческого разума. При этом ее начальным приоритетом всегда является стремление развивать парафе- нетическую или фенетическую организацию не просто системы, а толь- ко какого-то из вариантов системы потенциально возможного для нее типа; это одно из правил, которое можно вывести из закона мультива- риантности развития телесных систем. Наличие у любых тел определенных обязательных целей развития, которые контролируются на прогенетическом или генетическом уровнях организации материи, т.е. эгосубстратом данных тел, делает существо- вание всех тел в Природе содержательным и, в первую очередь, целе- направленным. При этом, безусловно, абсолютизировать это явление не стоит, поскольку иерархия целей, в силу их разнообразия, будет разной. Изучение общих и частных закономерностей движения объектов позна- ния к целям своего развития составляет одну из самых интригующих сторон науки. Итак, подчеркнем главное: эгосубстратная и конституциальная ча- сти (меросомы) организации тел имеют, в зависимости от натуры тел, по две формы. У эгосубстрата первую, или преднаследственную, форму со- ставляет прогеносома (она свойственна неживым телам), а вторую, или наследственную, форму — геносома (она принадлежит живому телу). Преднаследственный эгосубстрат в геносоме ассоциирован и подчинен настолько, что, пока жива биосистема в целом, он сам себя не проявляет (или почти не проявляет). На основе развертывания информации эгосубстрата соответствую- щих тел у них образуется (надстраивается) конституция, которая также имеет две формы: первую, или парафенетическую, — парафеносому (она свойственна неживым телам), и вторую, или фенетическую,— феносому (она образуется как конструктивная надстройка над геносо- мой — эгосубстратом у любых живых тел, т.е. как результат функцио- нирования генов во взаимодействии с окружающей средой). Похоже, мы подошли к сути бесконечного и неисчерпаемого суще- ствования материи. Она — в морфогенезах [соматогенезах, организован- ных на преднаследственной (прогеносомы) или наследственной (гено- сомы) основах (эгосубстратах)], обеспечивающих мультивариантность развития телесных систем в Природе в зависимости от конкретных ок- ружающих условий. В существовании материи нет и не может быть хаоса, поскольку она всегда организована в тела и в этой организации всегда наличествуют две части: эгосубстратная и конституциальная. Процесс бесконечного существования материи идет в конкретных телах, причем для каждого из них дискретно, что, естественно, не рас- пространяется на материю вообще. Все телесные системы во Вселенной материальны, а материя не смогла бы существовать, если бы строго не подчинялась одному из высших законов — закону единства континуаль- ного и дискретного: бесконечное (континуальное) существование материи ироявляется в единстве образуемых ею и ее деятельностью дискретных тел и нетел, что лостоянно лоддерживается функционированием субконтину-
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ альных носредников — определенных телесных систем (эгосубстратов) как носителей нреднаследственности или наследственности. Надеемся, здесь становится понятно, что элементарные носители эгосубстратных (преднаследственных или наследственных) свойств- прогены и гены — не континуальны и не дискретны в пространстве- времени, а занимают переходное положение; его удобно назвать субкон- тинуальным, поскольку способность особых обладателей субконтину- альности преемственно передаваться от тела к телу, не изменяясь и не сливаясь, но смешиваясь при этом в различных комбинациях, не может считаться ни бесконечностью, ни дискретностью. Рис. 38. Абстрактная модель внутривидового мультивариантного развития биоморф семенных растений. 1 — Ч — возможные биоморфотипы и образные схемы их развития: 1 — основной био- морфотип — травянистые безрозеточные компактно-каудексовые многолетники в услови- ях равнинной степи, 11 — Ч — второстепенные биоморфотипы, 11 — травянистые безро- зеточные вынужденно удлиненно-каудексовые многолетники в условиях периодического засыпания почво-грунтом на деградированных полупустынных пастбищах, Ш — травяни- стые безрозеточные деформированно-каудексовые многолетники на эрозионно подвиж- ных крутых горных склонах, IV — травянистые вынужденно вегетативно размножающиеся корневищно-стержнекорневые многолетники в условиях горно-речных пойм, Ч — безро- зеточные стержнекорневые полукустарнички в условиях небольших понижений равнин- ной степи; черные стрелки — указатели направлений текущего развития биоморф, иные стрелки — указатели направлений потенциально возможного развития биоморф.
Глава 1. Концепция меросомной организации биоморф семенных растений ОО CO (Ь М Ю Ю О М О Ch о.'1 о.'1 0 Х о.'1 0 И Ж о.'1 C о.'1 C Е З C Оч CC О Ж О 0 М Ж Яя Q О 3 О й ~О й й й Ж 0 й ~ ° О О 0 Ж Q ~О О О Ж о.'1 Оч (Ь О й Оч О О х~ ~'~> о~~~~ о2аХ g u ~ ~~ ~~ о О'ОУХа. О t Ц О Я ~ ~ ож Д д" Cl P~ й Ь и о ~ Ои и Ф й ф Оg~gа ~Й1~:у о ~ Ж о й g ДДиа ОЯаИд ~О ~' ~" ~ф О. ~ И ~ СО О М и ~О О . О и а~~и~~ и „0 < М ~О д Ю О о ~ ~ ~ ~ <~g Яа ф, '~ ~ Ц р г Cl ~ ~ д 65 ~ХО ООоgl 3 Q L. Q 2 IX тйй ЙИ
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИЙ 6~ 0 О О О М О О 2 Ж л 2 f ~О Ю О Х )Я 5~ М О )Я 65 О М У~ )Я X М О )Я О О U 65 X 00 И 3 Ж Ю E ° Юй E 3 65 О О О О 65 Q С) О Ж 0 м т Q. г; о [ я О о ~~ ~~ & ~ Q. м о~~ О О о Я (б цр а ~Оа (б Я й: X p а Й л о m :-2 (б ф„' О р,а, о (б ~ X Cl р,~ ФЭ 2 Ж :[,' 0; X ~> й М ~ 2 о:~, < n. X й 0~ й; й Й Q. ~ Q 2 О О „О iY I о р, X я а М (б ма 0 ОО М о iY м М I д о Р1 X О X X lO о Р- ",я а X О о П .Я о Q p~ о д X о р, Е О к (б X iY м о Ю 63 Q 0 т X X й ",Я М Х 0~ 0 Q О О iY О о 4 а а Х Ж 0 .0 о О Х X X й" 0 Q М X X О о X й" X
Глава 1. Концепция ме осомной о ганиэации биоморф семенных растений 101 Фитобиографическое эссе о D. imberbe (сем. Lamiaceae). Вид обычен около ледников, на каменистых и щебнистых тундрах, иногда по осыпям спускается ниже границы леса. Надземное прорастание растянуто с весны до осени, чему способствует влажный климат высокогорий в сравнительно теплое летнее время года. Семядоли эллиптические или широкоэллиптические. Верхушка семядолей закрутленная; в местах прикрепления к черешку пластинка семядолей имеет глубокую выемку, при основании она сердцевидно-стреловидная (с двумя заостренными рожками). Длина семя- долей 5 — 7 мм, ширина 3 — 4 мм, черешки — 4 — 10 мм дл. Виргинильный этап продолжается 5 — 20, чаще 8 — 12 лет. Растения кустятся. Хорошо развит стержневой главный корень. Почки незащищенные (открытые). Побеги розеточ- ные, нарастание эумоноподиальное. У взрослых растений репродуктивно-ассимиляционные цветоносные побеги образуются из пазух прошлогодних розеточных листьев. В структуре универсально-сложных плейофазных побегов за один вегетационный сезон развивается или один постинтеркалярный цветонос- ный побег, или их несколько, которые вместе составляют иносоцветие; побеги-универсалы нарастают эумоноподиально. Примерно к середине биоморфогенеза (30 — 40 годам ) главный корень часто, особенно на подвижных склонах, веревковидно закручивается и постепенно по некрозным «веточным следам» может как-либо расщепляться. Сначала ветви каудекса (каудиции) вместе со своими парциальными продолжениями из осевых фрагментов главного корня составляют части не полностью партикулированного корнепо- бега-организма. Однако в процессе старения и особенно под воздействием эрозионно подвижного почво-грунта, главный корень может неоднократно продольно полностью расщепиться на отдельные особи-«хвосты», или каудисты (от лат. cauda — хвост), т.е. каудальные ( «хвостатые») клонисты, где каудексовая ветвь (каудиция), превратившись в посткаудицию стала автономно представлять всю многолетнюю побеговую часть вот таких необычных особей вегетативного происхождения. Посткаудекс или его сравнительно тонкий аналог — посткаудиций — это любое персистентное побеговое структурное образование, ко- торое получается из каудекса, его частей, в процессе разрушения этого органа по каким-либо причинам, чаще всего при сильной партикуляции старых каудексовых растений. Каудисты могут долгое время, даже до конца всей жизни синорганизма, а многое зависит от осо- бенностей воздействия окружающих факторов внешней среды, оставаться скрученными, хотя может быть и плохо по нашим меркам, в «веревку», или, если хотите, в растрепанную «косу» (здесь в образности условных названий нет предела), представляя, таким образом, криптоклональное (скрытый клон) состояние синорганизма. В дальнейшем полное обо- собление каудистов может становиться заметнее, поскольку их ежегодно «растаскивает» неравномерное давление более или менее подвижного почво-грунта, к которому присое- диняются еще и придаточные недолговечные (питающие) корни, и в конце концов кау- дисты могут уходить в «свободное плавание» по жизни. Их судьба может сложиться по- разному: или они вскоре погибнут, будучи растерзанными динамикой щебнистой осыпи, лишившись поддержки родительской «веревки», как «кнута и пряника», или, напротив, им повезет больше и они прибьются в относительно спокойный участок склона, где еще относительно долго будут развиваться по посткаудексовой схеме (подобно каудексу роди- тельских особей семенного происхождения, но слабее и с опорой лишь на некоторый фрагмент главного корня и при помощи мелких питающих придаточных корней). В разных условиях нередко можно наблюдать разнообразные, как частично, так и полностью «ра- зорванные» партикуляцией родительские растительные тела и то, что из них уже стало, когда они превращаются в клоны, у которых однозначно фиксируется главное, для нашего случая, — генетическая однородность, в первую очередь, их многолетней побеговой части. Я склонен особым образом обозначать объединенные синорганизмом многолетние побе- говые части как одно образование — «базифиз» (дословно с греческого «растущее осно- вание»), когда рядом в одних границах, хорошо обозримых и фиксируемых в пространстве, соприсутствуют как парциальные части базового органа родительской особи, если она вообще еще есть, так и полностью обособленные многолетние побеговые части особей вегетативного происхождения — клонистов, в случае с D. ппЬегЬе — каудистов. Многие кочки и дерновины осок и злаков и некоторых других, например, кистекорневых растений (Veratrum lobelianum Bernh., Rhodiola rosea L.) нередко представляют в структурном выра- жении криптоклональные (скрытые клоны) кондивидуумы и имеют единые в генетичес- ком отношении, но физически уже разобщенные посредством какой-либо партикуляции, многолетние побеговые системы — базифиз. Этот термин был предложен ранее (Нухимов- ский, 1969) для подобных случаев, а здесь мы уточняем и закрепляем его понятийное значение. У D. imberbe базифиз представляет как-либо прижатые или заметно разошед-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ шиеся многолетние побеговые части — посткаудиции одного синорганизма, которые ра- зобщены между собой партикуляцией, опустившейся уже в глубину главного корня или даже больше — полностью разделившей все родительское тело на клонисты-каудисты (обособленные партикулы-особи) с фрагментами родительского каудекса — посткаудексы или посткаудиции и главного корня. Базифиз вместе с корнями, связанными с ним, — это ризобазифиз. Змееголовник Бунга правильнее будет относить не к традиционным каудек- совым растениям, а к каудексово-базифизарным травянистым многолетникам (они посте- пенно скрытно, но уверенно частично или полностью разрывают каудекс и главный корень на части, нередко доводя их до состояния клонистов). Возраст изоаклонистов (неклонирующих особей в условиях относительно спокойного почво-грунта) и синорганизмов достигает нескольких десятков лет и колеблется в пределах 50 — 80 лет. Каудисты не обладают прежней жизненной силой родительских особей семенного происхождения, и их собственный жизненный путь обычно короток. Рис. 41. Cirsium esculentum (Stev.) С. А. Меу. На Алтае (поймы горных рек в окрестностях сел Усть-Кан и Черный Ануй). Дополнительное обозначение к рисунку: зс — зачаточное соцветие (бутон). Продолжение на стр. 103.
103 Глава 1. Концелция ме осомной о ганизации биомо семенных астений Направления биоморфогений: 1 — розеточные сжатоцветоносные дву-, среднелет- ние травы (выделяют обычно разновидность — var. acaule Trautv.), И — полурозеточные удлиненноцветоносные дву-, среднелетние травы (или var. caulescens Trautv., например, во «Флоре Казахстана», IX, 1966, с. 340). Рис. 42. Beta vulgaris L. на дачном участке в Подмосковье. Фитобиографическое досье о В. vulgaris (сем. Chenopodiaceae). Свекла в Европе культивируется около 200 лет, а возделывать вообще ее начали еще до нашей эры (Орловский, 1961; Кружилин, Шведская, 1966 и др.). Она встречается и в диком виде, в частности, на Ближнем Во- стоке (Feinbrum-Dothan, Danin, 1991), где развивается как стержнекорневой однолет- ник и даже многолетник. Только в культу- ре, под влиянием селекции, свекла приоб- рела привычный нам облик и стала образо- вывать крупные мясистые корнеплоды», которые правильнее называть иначе «клубневыми корнепобегами» («клубнекор- непобегами»). В условиях дачных грядок (Московская область) на первом году веге- тации могут проявиться два направления биоморфогении: 1 — вегетативное некусти- сто-розеточное (оно преобладает) и II— вегетативное кустисто-розеточное (оно в меньшинстве). Основной биоморфотип в. vulgaris в условиях выращивания: прямостоячие клубне- корнепобеговые полурозеточные (во время цветения на втором году жизни) обоеполые унирепродуктивные условно закрытые (семидетерминатные) изоаклональные (без вегета- тивного размножения) полутравянистые двулетники (о полутравах см. главу 5). Фитобиографическое эссе о С. esculentum (сем. Asteraceae). Вид встречен в низкогорье — поймах горных рек среди низкорослого остепненного луто- вого разнотравья. Прорастание надземное. Семядоли обратнояйцевидные, на верхушке широкозакрутленные, в основании сужены в короткие черешки. Дл. 5 — 8, шир. 2,5 — 5 мм. На первом году, а нередко еще несколько лет биоморфы развиваются виргинильно, образуя вегетативные розеточные годичные побеги — эумоноподиальные приросты уни- версального главного побега. Розеточные листья колюче-зубчатые, обильно опушенные с обеих сторон жесткими волосками. Растения зацветают на 2 — 6-ой год жизни и после плодоношения полностью отмирают. Главный корень нередко отмирает еще раньше — на 2-ом году, и его активно замещают многочисленные придаточные корни, развивающиеся на сближенноузловой стеблевой части, которая зимует, превращаясь таким образом в компактный базовый орган — коно- дий. Годичные приросты конодия — курторезиды сохраняются до 2-х лет. Удлиненный очереднолистный безрозеточный цветонос, продолжая собой развитие розеточной вегета- тивной части унирепродуктивного обоеполого полурозеточного побега, выносит на 10— 40 см верхушечную цветочную корзинку (11 биоморфотип); иногда на верхушке скученно образуется несколько корзинок. БИОМОрфОТИПНОе доСЬе С. esculentum: розеточные (1 биоморфотип) или полурозеточ- ные (II биоморфотип) обоеполые унирепродуктивные кистекорневые колюче-зубчатоли- стные сильноопушенные лилово-пурпурно-астроцветковые горно-луговые травянистые дву-, среднелетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 43. Cnicus benedictus L. в разных местообитаниях . Направления биоморфоге- НИИ: 1 — простратные пролурозе- точные травянистые однолетники (в естественных условиях в окрест- ностях Тель-Авива, на открытых пустырях), II — прямостоячие по- лурозеточные травянистые одно- летники (в естественных условиях там же, что и 1 биоморфотип, но в условиях притенения другими рас- тениями — II а, и в условиях выра- щивания в одновидовом травостое — II б — Главный ботанический can, Москва). Фитобиографическое эссе о С. benedictus (сем. Asteraceae). Прорастание надземное. Семядоли в естественных условиях 10 — 30 мм дл. и 5 — 12 мм шир., а в усло- виях выращивания их размеры сильно варьируют от 15 до 60 (!) мм дл. и 10 — 20 мм шир.; они обрат- нояйцевидные или продолговато- обратнояйцевидные, на верхушке широко закрутленные или с выем- кой; в основании постепенно су- женные в черешок 5 — 10 мм дл. и 2 — 3 мм шир. На откры- тых местах у виргиниль- ных растений хорошо раз- виваются розетки листьев, а в условиях затенения они слабее. В условиях обитания в относительно густом травостое образу- ются растения с прямос- тоячим полурозеточным главным побегом, не- сколько разветвленным только в верхней своей части и закрытой верху- шечной корзинкой. На от- крытых солнечных местах особи С. benedictus обра- зуют короткий медиаль- ный побег и много пазуш- стелющихся побегов, прижимающихся к хности почвы. Это уже другой биомор- повер фотип Биоморфотипное досье с benedictus: Полурозеточные простратные (1 биоморфотип) и прямостоячие (11 биоморфотип) обоепо- лые унирепродуктивные (обоеполые монокарпики) стержнекорневые опушенные колюче- оберточнолистные желтовато-астроцветковые однолетние травы.
Глава 1. Концелция ме осомной о ганизации биомо семенных астений 105 А Б В Г1 Рис. 44. Kitagawia baicalensis (Redow. ех Willd.) М. Pimen. на Алтае (Кумин- ский хребет, сосновый лес на склонах в долину реки Чемал). аправления биоморфогении: 1 — полурозеточные унирепродуктивные травянистые многолетники, П — полурозеточные би-, мулътирепродукгивные травянистые многолетники. итобиографическое эссе о К. baicalensts (сем. Apiaceae Lindl.). Прорастание семян надземное. Семядоли 6 — 8 мм дл., 2 — 3 мм шир., голые, от линейно- ланцетных до узко-ланцетных, на верхушке заостренные, от основания постепенно пере- ходящие в короткие черешки. Первые листья тройчато-рассеченные. Постепенно в разви- тии их полусложное строение становится дважды перисто-рассеченным. Такие листья образуют розеточные вегетативные годичные побеги, которые много лет могут отрастать эумоноподиально, составляя вместе плейофазные унирепродуктивные полурозеточные универсальные побеги, которые впервые развивают верхушечные соцветия на 5 — 15 году жизни. После первого плодоношения часть растений погибает (1 биоморфотип), но другие продолжают накапливать новую вегетативную массу и развиваются как обоеполые би-, мультирепродуктивные травянистые растения (П биоморфотип). Предельный возраст обыч- но не превышает 25 лет. Биомо ф т Р~О ИПНОе дОСЬе К. baicalensis: полурозеточные некустистые или слабокусти- стые стержнекорневые конодиальные или компактно-каудексовые уни- или би- мульти- и-, мульти- репродуктивные обоеполые бело-сельдерейноцветковые травянистые многолетники.
OCHOBhI БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИИ Рис. 45. Hesperis pseudonivea Tzvel. на Алтае (Курайский хребет, на лесных опушках и на луговых горных склонах выше лесного пояса). Направления биоморфогении: ! — полурозеточные унирепродуктивные травянис- тые двулетники, реже трехлетники, II — полурозеточные бирепродуктивные травянистые среднелетники. Фитобиографическое эссе о Н. pseudonivea (сем. Brassicaceae) Прорастание семян надземное. Семядоли лопатчатые, широкозакрутленные с небольшой выемкой, клиновидно суженные в короткие черешки. Дл. 5 — 9 мм, шир. 2,5 — 5 мм. Листья простые эллиптические опушенные черешковые розеточные и сидячие безрозеточ- ные на удлиненной цветоносной части. На первом году вегетации развиваются розеточные вегетативные растения. После перезимовки они образуют удлиненные цветоносные при- росты с условно закрытыми (семидетерминатными) верхушечными кистями. На главном побеге развиваются также паракладии со своими верхушечными кистями, которые вместе с главной верхушечной кистью составляют сборную кисть всего универсального сложного двулетнего, реже трехлетнего побега (пентакормуса). После первого плодоношения расте- ния полностью отмирают (1 биоморфотип), но иногда, примерно в тех же условиях, их совидичи проявляют еще одно репродуктивное усилие, продлевая себе жизнь еще на 1— 3 года ()! биоморфотип). БиОмОрфОтипнОе ДОсве H. pseudonivea: розеткообразующие полурозеточные некус- тистые или реже слабокустистые стержнекорневые конодиальные или компактно-каудек- совые уни-, реже бирепродуктивные обоеполые бело-капустноцветковые травянистые дву- летники, реже среднелетники (иногда живут 3 — 5 лет).
Глава 1. Концепция ме осомной организации биоморф семенных растений Рис. 46. Anthriscus sylvestris (L.) Hoffm. на Алтае (долина вдоль хребта Хол- зун, опушка пихтового леса). Продолжение обзора на стр. 108. Направления биоморфогении: 1 — розеткообразующие полурозеточные обоеполые унирепродуктивные (обоеполо-монокарпические) травянистые двулетники или среднелет- ники, II — розеткообразующие полурозеточные обоеполые би-, мультирепродуктивные (обоеполые ди-, поликарпические) синорганизменные травянистые многолетники. Фитобиографическое эссе о А. sylvestris (сем. Apiaceae; растения принадлежат к var. nemorosa (Bieb.) Trautv.) Об особенностях жизнеразвития биоморф по второму био- морфотипу в Московской области мы уже писали (т. 1, гл. 2). В условиях Алтая, где наблюдали биоморфогению рассматриваемого вида, образование клонов, хотя и происхо- дит, но не так интенсивно, как это может происходить в друтих условиях (овраги, пустыри;
OCHOBhl БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИИ Московская область), и клоны часто слабые и недолговечные. Их возраст обычно не превышает 15 лет и нередко заметно меньше. Первый тропобиоз (в наиболее открытых местах), как и второй (среди кустарников и высокого разнотравья), встречаются на Алтае рядом, и, вероятно, срабатывает простой принцип жизнеразвития: подготовился после плодоношения к зиме — выжил, стал многолетником, а не подготовился, то извини,— все счастье жизни перешло только к ~детям» (семенному потомству), и получай униреп- родуктивную концовку личной жизни). Начало биоморфогении для обоих биоморфотипов сходное. Семена прорастают по надзем- ному типу. Семядоли линейные 20 — 30 мм дл., 2,5 — 4 мм шир., на верхушке заостренные, плавно переходящие в черешок, 8 — 15 мм дл. Первый лист тройчатый. Все листья полусложные. Розеточные листья на длинных черешках трижды перисто-рассеченные. Виргинильные растения развиваются по розеточному стержнекорневому типу. Полузакрытое (семидетерминатное) соцветие — сложный зонтик развивается как на вер- хушке медиальной оси главного побега (условно закрывает его), так и на верхушках па- ракладиев (латеральных силлептических цветоносных побегов), составляя вместе сборное соцветие (синфлоресценцию) — метелку из сложных зонтиков в структуре всего развет- вленного главного побега. При развитии по второму направлению (тропобиозу) растения еще до плодоношения успевают образовать 1 — 3 боковых розеточных побега с собственной придаточной кор- невой системой [это онтоиды, или корнепобеги-органы (т. 1, гл. 4)], а после плодоноше- ния родительский организм отмирает почти целиком, оставляя после себя только онтоиды, которые, получив самостоятельность, сразу же превращаются в особи вегетативного раз- множения (корнепобеги-организмы) — клонисты. Разнообразное поведение биоморф А. sylvestris в пределах своего ареала свидетельствует о высокой пластичности этого вида. Биоморфотипное цосье А. sylvestns чаг. nemorosa — розеткообразующие полурозеточ- ные стержнекорневые унирепродуктивные или мультирепродуктивные обоеполые бело- сельдерейноцветковые травянистые изоаклональные двулетники и среднелетники или си- норганизменные многолетники. Рис. 47. Dendranthema sinuatum (Ledeb.) Tzvel. на Алтае (Курайский хребет) Направления биоморфогении: 1 — полукустарнички на незащищенных скалистых склонах в высокогорьях, II — полукустарники на каменистых склонах, в расщелинах между скал, небольших лужайках среди крупных камней морен как в высокогорьях, так и в лесном поясе. Фитобиографическое эссе о D. sinuatum (сем. Asteraceae). Прорастание семян надземное. Семядоли окрутлые или широко-лопатчатые закругленные, почти сидячие, 2 — 3,5 мм дл., 2 — 3 мм шир. Несколько первых листьев простые, обратно- яйцевидные с тремя зубчиками на верхушке. Такие листья могут развиваться при моно- подиальном нарастании главного розеточного побега в течение нескольких лет. Постепен- но увеличивается число зубчиков у розеточных простых листьев. Виргинильное развитие продолжается до 8 — 20 лет, после чего растения впервые зацветают. Универсальные побеги у растений 1-го тропобиоза дифазные, редко трифазные вегетативно-репродуктив- ные сильно отмирающие сверху в конце того вегетационного сезона, когда сменяют ро- зеточные и верхнерозеточные вегетативные приросты на безрозеточные, образуя в итоге компактные или короткие (с небольшим числом удлиненных междоузлий) надземные резиды, которые вместе составляют приземистую многолетнюю побеговую (аэроподиаль- ную) структуру полукустарничков (до 10 — 15 см высотой). Универсальные побеги у растений и-го тропобиоза полурозеточные, как правило, нормально завершающие ди-, плейофазное унирепродуктивное развитие, образуя довольно длинные надземные резиды после пострепродуктивного акронекроза побегов до половины их полной длины, что составляет восходящую и (или) прямостоячую аэроподиальную структуру (до 20 — 40 см высотой) полукустарников. На безрозеточной части цветоносных побегов, помимо нижних простых зубчатых листьев, в верхней части таких побегов развиваются полусложные перистые листья. Главный стер- жневой корень сохраняется очень долго, но с возрастом он иногда может разрушаться и даже образовывать партикулы, уже обросшие до этого придаточными корнями; посред- ством партикуляции части родительского куста способны полностью обособляться, что несколько продлевает жизнь всего синорганизма, но не столь существенно, поскольку клонисты остаются сближенными на прежнем месте обитания своего родителя, а прида-
109 Глава 1. Концепция меросомной о ганизации биоморф семенных растений BHOMOp@OTHIIHOC ДОСЬе D. sinuatum: Розеткообразующие (полурозеточные) гемисим- подиальные семидетерминатные (универсальные побеги закрываются верхушечной кор- зинкой) мультирепродуктивные обоеполые розовато-астроцветковые стержнекорневые ве- гетативно неразмножающиеся (изоаклональные), иногда дефективно вегетативно размно- жающиеся, полукустарнички или полукустарники. Б В 53 '4 l'! g. точные корни не могут в полной мере компенсировать функциональную силу главного корня. Предельная продолжительность жизни биоморф может достигать 50 — 70 лет.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 48. Adenophora lilifolia (L.) А. DC. на Алтае (хребты: Бащелакский, Ануйский, Чергинский и Южный Алтай, 500 — 1300 м над ур. моря). Направления биоморфогении: 1 — компактно-каудексовые травянистые многолет- ники (березово-лиственничные леса на пологих склонах), II — вынужденно горизонталь- но-каудициальные травянистые многолетники (крутые горные склоны лесного пояса, среди кустарников и редкого разнотравья) Фитобиографическое эссе о А. lilifolia (сем. Сатрапи1асеае). Прорастание семян надземное. Семядоли яйцевидные, слегка выемчатые, резко клиновид- но сбегающие в черешки, 4 — 6 мм дл., 3 — 4 мм шир. Черешок 3 — 5 мм дл. На первом году в естественных условиях образуется обычно один лист обратнояйцевидной формы или даже почти крутлый. У ювенильных особей образуются похожие немногочисленные лис- тья. С 3 — 14 лет начинают отрастать пазушные короткие 8 — 25 см дл. вегетативные (ассимиляционные) побеги, которые к концу вегетационного сезона отмирают полностью. Первое цветение чаще на 8 — 20 году, хотя редко и раньше (обнаружено одно четырех- летнее растение с репродуктивно-ассимиляционным побегом и единственным цветком на его верхушке). Сохраняется на всю жизнь эумоноподиальное нарастание побеговой сис- темы, кроме случайного повреждения верхушек. Каудекс компактный, у одних растений
Глава 1. Концелция ме осомнои о ганизации биомо Ъг о семенных астений их астений — слабо ветвится (1 биоморфотип). он может долгие годы не ветвиться, а у друтих растени нный (вто ой) биомор- ых го ных склонов иногда образуется второстепенны, второ р р фотип, когда почки компактного удою~ ыр~~~~~ ж~~ начинают р р т асти ro изонтально, образуя соответствующие подземные по еги, к онен назвать их вынужденно гори- ют п в ащаясь в каудиции особого рода; я склонен зимуют, превр ьное расположение и нередко образуют зонтальными к уд ц а и иями они имеют горизонтальное Й Г зонтальные каудиции спо- на себе один — несколько сильн рид ых п аточных корне . ори ко нем всю жизнь, ять п оч связь с каудексом и родительским главным корнем собны сохранять прочную с роисходить вынужденное веге- но если подвижныи грун р р т аз ывает эту связь, то может пр арастание обеспечивают тативное размножение. У взрослых р у астений э моноподиальное нараст о ними из первых ев ными листьями, хотя часть листьев, одними и побеги в основном с чешуевидн ками и даже образовывать отрастающих веснои, м ожет развиваться с зелеными пластинками и даж быстро ~~~~ающие р~~~~~ ~~~~~~. Предельный возраст биоморф, включая и вынужденные сино в не п евышает оО — лет. 6 — 80 иногда в особых условиях крутых горных склонов, р аз щие э монопо- ДОСЬЕ А. lilifolia: непостоянно слаборозеткообразующ у ьиомоощотипное диальные мультирепродуктивные интеркаляр - р каля но-дете минатно-репродуцирующие стерж- б - и го- нтально-ка дициальные, обоеполые ело- или го- некорневые, иногда вынужденно горизонтал - у лубовато-колокольчиковоцветковые травян истые ка дексовые многолетники. У
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РАСТЕНИЙ Рис. 49. Ribes atropurpureum С. А. Меу. (Алтай, около 30 км на северо-запад от села Черный Ан~, Бащелакский хребет, кедрово-лиственничный лес, 1000 — 1600 м над ур. моря). Направления биоморфогении: 1 — прямостоячие рыхлые листопадные кустарники (пологие низкогорные и среднегорные склоны), П — во многом такие же кустарники, но полегающе-восходящие полуразвернутые (на более крутых горных склонах, в еще более тенистых лесах или ближе к верхней границе леса). Фитобиографическое эссе о Ribes atropurpureum (сем. Grossulariaceae) Прорастание семян надземное. Семядоли эллиптические слегка заостренные клиновидно суженные в черешки (0,7 — 1,2 см). Пластинки семядолей 0,8 — 1,2 см дл., 0,35 — 0,5 см шир. Листья простые черешковые лопастные. Основную структурную единицу надземной многолетней побеговой основы кустарниковой организации биоморф смородины темно- пурпурной составляют аэроподии (до 1,5 м высотой) более или менее прямостоячие (1-ый биоморфотип) на пологих склонах или полегающе-восходящие, расширяющие площадь полуразвернугого куста (1! биоморфотип) на более кругых горных склонах и нередко в еще более тенистых лесах. Основания аэроподиев постепенно погружаются в почву, превраща- ясь в эпигипогеогенные геоподии. Аэроподии нарастают эумоноподимьно за счет корот- ких и длинных резидов (тотарезидов); когда все же происходит отмирание верхушек, то случается это по каким-либо непредвиденным причинам или в связи со старением аэро- подиев постепенно становится нормой, что и приводит старые аэроподии к полному отмиранию. Их сменяет более молодая, восходящая от геоподиев поросль, которая своим усиленным ростом постепенно депрессировала развитие старых аэроподиев. Эумоноподи-
Глава 1. Концепция меросомной о ганизации биоморф семенных астений альное нарастание обеспечивается безрозеточными годичными побегами. Первое цветение происходит на 5 — 12 год жизни. Репродуктивную функцию выполняют годичные вегетативно-репродуктивные побеги, состоящие каждый из весеннего модифи- цированного (с чешуевидными листьями и без зеленых листьев) побега с верхушечным кистевым соцветием, развивающего непосредственно под соцветием из пазухи чешуевид- ного листа вегетативный силлептический побег замещения розеточный (с 3-мя и более сближенными зелеными листьями) или предрозеточный (с 1 — 2-мя зелеными листьями). После перезимовки этот побег продолжает моноподиальный рост, образуя весенний ма- гимодифицированный вегетативно-репродуктивный побег, а далее из пазухи его чешуе- видного листа — силлептический вегетативный побег замещения, и так симподиальное нарастание каждого парциального вегетативно-репродуктивного брахиморфного комплек- са может продолжаться несколько лет. Предельный возраст кустов 60 — 80 лет. Биоморфотипное досье Ribes аггоригригеит: эумоноподиально-долихоморфные сим- подиально-брахиморфные обоеполые пурпурно-крыжовниковоцветковые изоаклональные или слабо вегетативно размножающиеся листопадные прямостоячие или полуразвернутые кустарники. Рис. 50. Artemisia sieberi Besser (Израиль, каменистая пустыня Негев, окре- стности поселка Сде-Бокер).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Направления биоморфогении: 1 — полурозеточные стержнекорневые полукустар- нички, когда биоморфы произрастают в «одиночестве» на открытых местах, 11 — полуро- зеточные стержнекорневые полукустарники, когда биоморфы произрастают в «группах» (вместе с другими растениями) чаще по небольшим понижениям каменистой пустыни, иногда всего лишь в нескольких десятках см от растений первого биоморфотипа. Фитобиографическое эссе о А. sieberi (сем. Asteraceae). В «Аналитической флоре Израиля» (Feinbrun-Dothan, Danin, 1991, с. 1010) подчеркнуга приоритетность такого названия вида перед другим названием — Artemisia herba-alba Asso. Во «Флоре СССР» (1961) полынь Зибера приводится в качестве приоритетного названия вида, в том числе и по отношению к А. dumosa Poljak, которому в сводке С. К. Черепанова (1995) по непонятной причине отдано преимущество. Прорастание семян надземное. Семядоли эллиптические закрутленные сидячие голые; их длина 3 — 4 мм, а ширина 1,5 — 2,5 мм. Первые листья простые узко-ланцетные цель- нокрайние. Третий лист нередко с 2 — 3 зубчиками, а четвертый-пятый тройчатораздель- ные или просто перистые. В дальнейшем развитии возрастает степень рассеченности полусложных листьев. Ежегодное нарастание вегетативных розеточных или коропсих по- лурозеточных побегов, как правило, моноподиальное. Однако после образования удлинен- ных безрозеточных частей полурозеточных побегов в неблагоприятное (самое жаркое, летнее) время происходит отмирание верхушек, и нарастание аэроподиальной части рас- тений становится гемисимподиальным. Первое цветение происходит на 3 — 8 год жизни, чаще 4 — 5. Нарастание гемисимподи- альное. Весной, в процессе роста цветоносных дифазных, реже трифазных и даже плей- офазных полурозеточных побегов, помимо силлептических розеточных вегетативных по- бегов развиваются еще и постлатентные розеточные и короткие полурозеточные вегетатив- ные побеги, немного побегов развивается как безрозеточные вегетативные и вегетативно- репродуктивные годичные, но они чаще полностью отмирают к конце вегетационного сезона. Универсальные унирепродуктивные обоеполые побеги к концу вегетационного сезона у растений 1 биоморфотипа (на открытых местах) отмирают сверху значительно сильнее таких побегов растений П биоморфотипа, произрастающих группами, использующих малейшие неровности поверхности для закрепления в очень твердом почво-грунте. Аэро- подии растений полукустарникового биоморфотипа (второго), формирующиеся в условиях «микроклимата» (относительной защиты), создаваемого группой растений, на 10 — 30 см крупнее растений полукустарничкового (первого) биоморфотипа. Предельный возраст кустов полыни Сибера не более 50 лет. БиОМОрфОтипное ЯОсЬе А. sieberi: стержнекорневые розеткообразующие мультиреп- родуктивные обоеполые розово- или желто-астроцветковые акронекрозные гемисимподи- альные изоаклональные полукустарнички и полукустарники.
Глава 2. Концепция приоритетов развития в организации систем, в частности, биоморф 2. 1. Приоритетность — атрибутивное свойство систем Так или иначе, но все тела — материальные системы (физиосисте- мы) — это не только продукт внутренних условий развития их органи- зации, но и результат воздействия на них внешних условий, т.е. теле- сные системы попадают в лабиринт окружающих их условий, а у живых тел — в экологический лабиринт, или биотоп, из границ которого выб- раться они не могут (стенотопные системы; в отношении организмов Куда бы мы ни обратили взгляд, хотя бы и внутрь самих себя, везде обнаруживается присутствие только Природы — движущейся материи в бесконечном многообразии ее форм в пространстве и времени, а иногда еще и наличие продукта ее высшей деятельности — сознания. Все из- вестное и неизвестное нам материальное и идеальное составляет При- роду, в которой оно существует не хаотически, а на основе определен- ных закономерностей. Одна из них, на которую ученые обратили при- стальное внимание лишь в ХХ столетии, — системность. Людвиг фон Берталанфи разработал понятие системы как «комплекса элементов, находящихся во взаимодействии» (Bertalanffy, 1949), а позднее он же впервые создал общую теорию систем (Bertalanffy, 1949, 1956). Природа — это единство бесчисленного разнообразия материаль- ных и идеальных систем. Всякий предмет является системой по отно- шению к входящим в него частям и одновременно выступает как часть более сложной системы. О системах и системности, об организации, о структурах и функциях мы уже писали (т. 1, гл. 2) и не будем здесь повторяться, поэтому сразу сформулируем проблему, которую обсудим во второй главе: в чем суть явления приоритетности в организации телесных систем (тел) и биоморф семенных растений, в особенности. Эта проблема весьма объемна, но мы решаем ее здесь лишь в пределах, достаточных, как нам кажется, для разработки главной темы второго тома нашей монографии.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИИ обычно говорят о стенобионтах), или могут разнообразить свои биото- пы, как, например, перелетные птицы или человек, проявляя уже другой тип системности (эвритопные системы; в отношении организмов обыч- но говорят об эврибионтах). Всякое существование телесных систем не может не иметь какой-либо цели, т.е. система развивается не как попало и не куда попало, а более или менее направленно, причем целей в развитии системы может быть предостаточно, среди которых есть и те, которые являются приоритетными. Невольно мы подошли к осознанию необходимости разобраться в понятийном отношении и с «целями», и с «приоритетами». Утверждая, что каждая материальная (телесная) си- стема в первооснове владеет целью развития (а то и несколькими), из которых есть приоритетные цели и цели вспомогательные, мы тем са- мым ничего не навязываем системам. Они и без нас все главное имеют при себе: свои цели развития, некоторые из которых приоритетные. Рассуждая так, мы подходим к признанию факта, что в широком смысле все физиосистемы, функционирование которых осуществляется без учас- тия человека, «самонаводящиеся» (сами наводятся на цель своего разви- тия). Наша задача понять их и не ошибиться в том, к чему стремятся системы-тела, независимо от чьего-либо сознания. Мы хотим понять цели и приоритеты в развитии физиосистем, поэтому сразу определим- ся, что термины «цель» и «приоритет» будем понимать в широком смысле. Цель, в узком смысле, — «то, что представляется в сознании и ожидается в результате определенным образом направленного действия» (Кондаков, 1975, с. 662). Такое понимание цели нас не может устроить, оно не позволяет увидеть цель дальше того, что подсказывает сознание. Значит, дерево ели, куколка майского жука, булыжник на мостовой, капля дождя или Солнечная система и т.п. существуют в Природе бес- цельно («шаляй-валяй»), но это не так, если термин «цель» понимать шире, что мы и делаем. Ранее (глава 1) нами уже был принят известный широкий подход (Рапопорт, 1969). Итак, любая система, независимо или зависимо от чьего бы то ни было сознания, способна стремиться к опре- деленному (нередко «конечному», но далеко не всегда) результату (цели). Вероятно, то, что объединяет все физиосистемы — это динамизм (внеш- не заметный или незаметный) существования, как устремленного раз- вития по пути к цели (целенаправленность развития), направленного, в первую очередь, на сохранение (протяженностью в мгновение или веч- ность, или нечто иное по времени) своего содержания как системы вообще и как конкретной системы, именно этой, а не другой системы, в частности. Иными словами, лервая и универсальная цель всех систем: быть самой собой, а не быть другой системой в данный момент лростран- ства-времени, а шире и на лротяжении всего ее системогенеза. Цикл существования любой системы в ее качественных границах от возник- новения до преобразования ее в иную систему — системогенез. На метафорический вопрос к системе: быть ей или не быть, любая система могла бы «ответить»'. быть (хоть на мгновенье или вечно, но во всех случаях, — только быть). Есть приоритеты, которые долго не задержи- ваются в организации систем и (или) в сознании людей, которые их как-
Глава 2. Концелция npuopumemoe азвития в о ганизации систем то фиксируют. Приведу пример. Можно пробежаться первым по лугу в туманное утро и оставить четкий след после себя на росистой траве— след «первопроходца». Такой приоритет обычно не интересен для слу- чайного наблюдателя — свидетеля его; он долго не держится и быстро исчезает от тепла первых солнечных лучей. Но приоритет приоритету рознь. Есть такие приоритеты, которые пронизывают содержание вещей и от которых зависит многое, а иногда и все: их можно обнаружить в теле кролика, в отражении солнечного луча от зеркала, в страданиях народов под властью тирана, в мерцании звезд от далеких галактик и, наконец, в существовании живой природы и всего мироздания — Все- ленной. О таких приоритетах есть смысл думать и изучать их. Прежде чем разобраться с приоритетами в организации биоморф семенных растений, ответим на вопрос: что такое приоритет? У системы, допустим, системы-Природы, первая цель развития (су- ществования): быть, т.е. у системы-Природы — это бесконечность, не- исчерпаемость; первичность материального и вторичность идеального, когда оно проявляется в этой системе. Цель — «быть системой» опре- деленного качества — это самый первый, самый простой, исходный и самый важный для понимания системности приоритет. Что же такое «приоритет»? Во всех известных нам случаях приме- нения термина «приоритет» подразумевается первенство во времени и (или) по значению какой-либо деятельности людей. Приоритет — «пре- обладающее, первенствующее значение чего-либо, например, приоритет общесоюзного значения» (Бол. Совет. Энц., 1955, т. 34). Приоритет- «это «первенство, преимущественное, первенствующее значение чего- либо, в частности: научного открытия, изобретения и т.д.» (Сл. ин. сл., 1964, с. 520). «Приоритет (от лат. prior — первый, старший), 1) первен- ство по времени в осуществлении какой-либо деятельности, 2) ... необ- ходимое условие выдачи патента на изобретение или признания научно- технического достижения открытием; определяется датой подачи заявки или первой публикации. (Бол. Энц. Сл., 1997, с. 960). Так определяется сегодня термин «приоритет», но такое понимание этого термина нам представляется недостаточным. У нас есть предложение не ограничивать действие этого термина только человеческой деятельностью, т.е. придавать этому термину ши- рокий общеприродный смысл. Приоритет в широком смысле, — это первенствующее значение чего-либо, т.е. первенство, нередко по времени проявления или иное в осуществлении какой-либо деятельности у любых систем. Итак, слово «приоритет» имеет несколько смыслов, но главный из них мы используем как термин, который и будем применять дальше— Если подробнее, то этот термин можно определить и так: npuopumem— это преимущественное значение чего-либо по важности происходящего s какой-либо области сравнительного признакового пространства и (или) первенство его по времени проявления для отдельного отрезка времени (ста- дии) или для всего цикла существования предмета-системы. Иа основе при- оритета система (тем более, живая система) сама выбирает цель своего
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИИ развития или это (иреимущественная цель развития) ироисходит с ломощью иной целенаводящей силы, налример, человеческого разума на оиределенную стадию существования и (или) на весь цикл существования системы. Самый первый приоритет в развитии любой системы, о котором уже упоминалось выше, — это стремление (цель) любой системы быть этой, а не другой системой на отдельной стадии развития или во всем cucme- могенезе (уже — морфогенезе). Так, в развитии биоморф семенных ра- стений сначала на виргинильной стадии развития приоритетом является формирование вегетативного тела (вегетативной массы, или вегетатива), далее — репродуктивного тела (репродуктивной массы, или репродук- тива), а после окончания плодоношения и (или) пыльценошения при- оритетом развития биоморф может стать деградация и постепенная или быстрая элиминация (например, у однолетников или двулетников). У многолетних мультирепродуктивных растений после достижения репро- дуктивного результата часто проявляется приоритет на компромиссное вегетативно-репродуктивное развитие без отмирания растений целиком после плодоношения. Приоритетность — одна из сторон существования систем, их атри- бутивное свойство; это форма доминирования у них каких-либо видов деятельности. Приоритетность в развитии систем далеко не всегда на поверхности. Ее нередко надо суметь увидеть и рассмотреть, поскольку проявляет она себя по-разному в различных видах деятельности. Вполне уместно различать два типа деятельности, точнее, две стороны единой (целостной) деятельности: явную, или конституциальную (пластосомати- ческую), когда деятельность чего-либо наблюдается явно — в текущей конституциальной организации телесных систем, и скрытую, или эгосубст- ратную (идиосоматическую), когда деятельность чего-либо программирует- ся (закладывается) во внутренней природе тел — в их эгосубстрате (про- геносоме или геносоме) в направлении достижения когда-то определенной цели посредством соответствующего конституциального развития этих тел, а в текущее время только обеспечиваются условия — стремление к цели, но пока еще нет фактов достижения цели — она остается возможной в будущем. Вся явная деятельность, которая имеет место быть, т.е. наблю- дается, так или иначе, в текущее время в конституциальной части те- лесных систем, происходит одновременно с работой скрытых (эгосуб- стратных) видов деятельности. Деятельность становится единой (цело- стной), когда обе ее стороны приводят к достижению единой цели в развитии телесной системы, а результат фиксируется в соответствующей организации их конституции. Когда в текущее время в конституциаль- ной организации телесной системы непосредственно достигается цель определенной эгосубстратной программы, то единство явной и скрытой деятельности на фоне достигнутой цели превращает их в единый, или целостный, приоритет конкретного тела. В таких случаях и конституци- альный, и эгосубстратный виды деятельности — это обязательные эле- менты целостного приоритета. Все, что не является эгосубстратом, является конституцией тел, и другого в материальном мире не бывает. Приоритеты проявляют себя по-разному. Основные признаки приоритетов — важность определен-
Глава 2. Концелция лриоритетов азвития в организации систем ной деятельности для существования системы, первенство по времени проявления в сравнении с другими видами ее деятельности. Видов де- ятельности телесных систем и биоморф семенных растений, в частно- сти, много, а приоритетами среди них выступают далеко не все. Боль- шинство видов деятельности, происходящей в телесных системах впер- вые, являются только стадийными приоритетами, поскольку приоритет- ность их связана с первенством по времени на отдельных стадиях в циклах существования соответствующих систем. Так или иначе, но ста- дийные приоритеты участвуют в формировании целостных приорите- тов, т.е. обслуживают их. Если приоритеты распространяются на дея- тельность, охватывающую по своей значимости весь цикл существова- ния тела, в частности, биоморфы, то это, на наш взгляд, целостные, или главные, приоритеты. Стадийные приоритеты нередко могут быть инди- каторами соответствующих стадий морфогенезов и состояний (возрас- тных, фенологических) тел (биоморф, в частности). Распознание цело- стных приоритетов позволяет наиболее точно и полно охарактеризовать наиболее общие для соответствующих телесных систем признаки орга- низации. Например, стремление к размножению и размножение — это целостный npuopumem в развитии биоморф, а вегетативное развитие- это стадий ный npuopumem виргинильных биоморф. Все стратегические цели существования биосистем и, в частности, биоморф семенных растений — это их главные приоритеты, а среди тактических целей идет отбор в борьбе биосистем за существование; выбираются в приоритеты те тактические цели, которые в процессе развития биосистемы как в пространстве, так и во времени, более других целей отвечают решению стратегических задач. Явление приоритетно- сти проявляется во всех (материальных и идеальных) системах, подчи- няясь соответствующему высшему закону приоритетности, смысл кото- рого можно передать так: существование любой системы неиосредственно или оиосредованно обесиечивается различными видами деятельности, иричем среди них всегда находится та деятельность (одна или несколько), которая иервенствует (ириоритетна) сама в исходной системе или результат ее деятельности становится ириоритетным в ироизводных системах ио зна- чимости и (или) ио времени ироисходящего на данной стадии или s целом во всем цикле существования (морфогенезе) соответствующей системы. Из закона приоритетности, на наш взгляд, исходят три следствия, которые корректируют этот закон, непосредственно приближая до со- ответствующих систем. Все следствия, которые определяем ниже, не требуют особых доказательств и по сути являются аксиомами. Следствие первое. Какую бы форму ни ириобретали в ироцессе разви- тия материальные системы, они всегда выбирают в своем развитии не- сколько ириоритетов: быть этой системой, а не другой, развертывать иарафенетическую и (или) фенетическую организацию на основе ирогене- тических и (или) генетических свойств ее эгосубстрата и, s зависимости от воздействия условий окружающей среды, наводиться иреимущественно на цели развития, которые по своей значимости отвечают не столько существованию этой системы, сколько целям развития систем более вы- сокой сложности организации.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Следствие второе. Приоритеты существования идеальных систем (логосистем) находятся под контролем человеческого разума и направлены на обслуживание материальной природы посредством определенного втор- жения в нее логосистем, нередко как-либо опредмечивающихся (материа- лизующихся в книгах, телевидении, радио и т.n., но отражающих свое идеальное прошлое), связанных или не связанных с другими видами деятель- ности людей, взаимодействие которых способно вызывать изменения дей- ствительности в сторону ирогресса или регресса. Следствие третье. Развитие биоморф (организмов и синорганизмов) и, в частности, биоморф семенных растений, осуществляется по мультипри- оритетному принципу, при этом высшим приоритетом (суперприорите- том) является стремление к жизнесохранению вообще, что реализуется через развертывание фенетической организации, отвечающей генетической программе и изменяющейся в зависимости от влияния окружающей среды, в которой всегда npucymcmeyem борьба между стремлением к размножению (любому) и стремлением к личной (собственной) жизни, в результате чего личная жизнь или подавляется, иногда настолько сильно, что она продол- жается мгновения или чуть больше (минуты, часы), сводясь к экстренному накоплению ресурсов для одноразового размножения, или посредством ком- промиссов продлевается на годы, десятки, сотни и даже тысячи лет, не В отношении этого следствия сделаем небольшие пояснения. Дело в том, что в третьем следствии закона приоритетности поднят вопрос о взаимоотношении личного и коллективного в организации биоморф, который иначе можно сформулировать так: как и почему функции биоморф для себя (личные) уживаются в организации каждой отдельной биоморфы с функциями для других (коллективными, или коллегиаль- ными). Дело в том, что жизнь — явление не одномоментное; она прошла в эволюции путь сложнейшей интеграции своих дискретных носителей, поэтому в организации биосистем существует несколько механизмов (функциональных систем) поддержания и сохранения жизни. В первую очередь, биосистемы, биоморфы, в частности, проявляют приоритет своего, т.е. личного, жизнесохранения (автобиофилии), участвуя в фо- тосинтезе, поглощая воду и минеральные вещества и т.п. Рядом с этим приоритетом постоянно в организации биосистем присутствует и при- оритет коллегиального жизнесохранения (ксенобиофилии), главная осо- бенность которого — участвовать в поддержании вышестоящего род- ственного ему тела, в организации которого такие биосистемы присут- ствуют на правах элементов, например, организмы в видах, и ради сохранения соответствующих видов осуществляют размножение, обес- печивают распространение вегетативных зачатков, гамет и семян. Но, кроме того, над всеми приоритетами всегда витает еще один — приори- тет приоритетов, т.е. высший приоритет, или суперприоритет, который обеспечивается стремлением (наведением на цель) к жизнесохранению вообще. Таким образом, биоморфы, а шире — биосистемы, стремление к жизнесохранению проявляют по-разному, т.е. через разноуровневые приоритеты. Подробнее идея жизнесохранения разработана в общей теории жизнесохранения (т. 1, гл. 3)
Глава 2. Концепция прио итетов развития в организации систем 121 Приоритет — это свойство системы не только проявлять, но и как- то выделять функции, которые по значимости важнее многих других на отдельном отрезке времени (стадии) или на всем цикле существования системы (системогенезе). У биосистем способность как-то выделять свои приоритеты на фоне остальных функций получила в эволюции должный прогресс. Так, ритуал приоритета коллегиального жизнесохранения как стремления к размножению обставляется в организации биоморф се- менных растений различными находками в эволюционном плане, наи- более значимой из которых можно считать красочное оформление пре- репродуктивных и репродуктивных систем, что подчеркивает всему ок- ружающему миру очень высокую значимость для жизни происходящего. Известно, что красочные цветки хорошо привлекают насекомых-опыли- телей, что сережки в кроне некоторых деревьев весьма удобны, чтобы разбрасывать пыльцевые зерна или опыляться с помощью ветра, однако все эти красочные фейерверки — это «лепота» живой природы, — были в эволюции потом, а сначала, т.е. раньше всего частного и утонченно- специального, было простое и, безусловно, самое доступное для всего живого стремление научиться сохранять жизнь вообще. В этом стрем- лении живого в эволюции учиться жить были сформированы приори- теты деятельности их носителей: дискретности телесного существова- ния, генетического над фенетическим, коллегиального над личным, репродуктивного над вегетативным и т.п. Далеко не сразу в процессе эволюции растения стали доверяться красоте при построении органи- зации своих биоморф. Только сделав ставку на красоту, организация растений смогла удовлетворить неисчерпаемость эволюционных устрем- лений растительной жизни в приоритетности ее стремления к размно- жению своих дискретных носителей в образе биоморф как организмов и синорганизмов. Стремление к красоте, к повышению уровня красоты как типу телосложения, — это все опосредованное выражение стремления жизни вообще (Жизни) к своему сохранению, а у биоморф, в частности,— стремления их к размножению. Приоритету стремления к размножению над стремлением к личной жизни биоморфы обязаны видам, и этот приоритет — дань биоморф своим видам за право жить на Земле. Впро- чем, и виды без своих биоморф, строго генетически привязанных к ним, на Земле невозможны. Любопытства ради можно поставить и такой вопрос: а возможна ли вообще где-то в глубинах Космоса жизнь без привязки биоморф к видам и видов без своих биоморф? Если и возмож- на такая жизнь где-то, независимо от Земли, то это, безусловно, пока совершенно неведомая нам жизнь, по совершенно другим законам. В принципе, возможность присутствия такой жизни где-то во Вселенной, далеко от Земли, исключать нельзя. Итак, в биоморфах, как структурных элементах-биосистемах мень- шей, чем виды, сложности, всегда присутствуют строго определенные приоритеты развития, исходящие, в первую очередь, от видов — это приоритет размножения над остальными формами деятельности био- морф. Через красоту соцветий, цветков, плодов биоморфы семенных растений как структурные элементы своих видов научились в процессе
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ эволюции преданно нести свой «крест» — служить своему виду, поста- вив всю свою личную жизнь на «плаху» служения виду, приоритету, исходящему именно от него (вида), — стремления к размножению. Попробуем сформулировать несколько аксиоматических правил, опирающихся на закон приоритетности систем и его следствия, которые более всего касаются биоморф. Правило 1-е: личное существование системы (личная жизнь биосисте- мы, в частности, биоморфы) — явление опосредованное, так как никогда в полной мере не принадлежит конкретной системе, например, клетке, органу, организму и т. и., а находится в зависимости от приоритетов развития систем (уже — биосистем) большей сложности организации, к которым они непосредственно принадлежат как элементы системы мень- шей сложности организации (рис. 51 — 65). Правило 2-е: если система не достигает самых высоких результатов своего развития, mo это не означает, что ириоритеты такого развития ею безвозвратно утрачены, а свидетельствует о npucymcmeuu в системе и (или) в ее окружении сил, достаточных для того, чтобы предотвратить их достижение в данных условиях и в данное время, хотя система в целом всегда остается приоритетоемкой и подспудно сохраняет стремление вер- нуться к утраченным приоритетам развития. Затенение подроста более взрослыми растениями своего или другого вида или трав, попавших под полог необычно темной опушки елового леса, отодвигает на неопределенный срок время достижения этими растениями высокого приоритетного ре- зультата — полового размножения (рис. 66 — 67). Биоморфы способны попадать в экстремальные условия существования, но условия стресса побуждают их несколько сместить приоритеты своего развития, поста- вив, хотя бы на некоторое время, приоритет сохранения личной жизни (автобиофилии) на первое место, притормозив, но не освободив полно- стью, от давления на свою организацию всех других приоритетов. Правило 3-е: приоритеты развития материальных систем опираются на способность этих систем, на основе их преднаследственности или на- следственности «самонаводиться» на цели своего развития, но с появлени- ем Человека воздействие антропогена (человеческого фактора) на приори- теты развития как его собственные, так и других биосистем, усилилось и будет возрастать дальше с еще большей интенсивностью, чему стоит, если и восхищаться, то только с оглядкой, поскольку если не корректиро- вать приоритеты вторжения человечества в остальной мир, то будет сохраняться и усиливаться опасность, что необратимо нарушится равно- весие в живой природе и подломится «сук», на котором «сидит» все чело- вечество (рис. 68). Итак, из приведенного материала ясно, что жизнь на Земле можно представить как «игру» по правилам в приоритеты наводящихся и пе- ренаводящихся определенных целей развития. Рис. 51 — 55. Приоритет в развитии организации биоморф нелич- ного (репродуктивного) — более надорганизменного — над личным (вегетативом) — более организменным, в результате которого продол- жительность изоаклональной (без вегетативного размножения) жизни составляет не более одного вегетационного сезона (у эвриэфемеров).
Глава 2. Концеиция и иоритетов азвития в организации систем
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 53. Биографический портрет Plantago сгепса L. [сем. Plantagtnaceae (там же, что и рис. 52, 15 IV)]. ОСНОВНОЙ бИОМОДфОТИП: надземно-двусемядольные (семядоли узколинейные, ту- пые, сросшиеся в основании, 15 — 25 мм дл., 0,8 — 1,2 мм шир.) маленькие (3 — 12 см высотой) розеточные стержнекорневые продолговато-линейнолистные моноподиальные интеркалярно-репродуцирующие (из пазух нескольких розеточных листьев образуется по одному безлистному цветоносу, полузакрытому верхушечным сильно опушенным колосом, которые вместе составляют межсоцветие) обоеполые эфемеры. А 5 Рис. 54. Биографический порт- рет Impatiens parviflora DC. [сем. Balsaminaceae (Московс- кая обл., яблоневый свд)]. Основной биоморфотип: надзем- но-двусемядольные (семядоли окрутло- яйцевидные, 12 — 32 мм дл., 10 — 25 мм шир., на верхушке выемчатые, на че- решках 5 — 20 мм дл.) прямостоячие безрозеточные очередно-простолист- ные семидетерминатно-репродуцирую- щие однодомные мочковато-стержне- корневые желто-бальзаминовоцветко- вые однолетники.
Глава 2. Концеиция прио итетов развития в организации систем Рис. 55. Биографический портрет Hyoscyamus niger L. [сем. Solanaceae (Алтай, Чуйский тракт, окрестнос- ти села Иня, крутой горный склон к реке Катунь)]. Биоморфотипы: надземно-дву- семядольные (семядоли яйцевид- ные, 8 — 10 ммдл., 3 — 4 мм. шир, голые, слегка заостренные, сужен- ные в короткие черешки) полурозе- точные стержнекорневые просто- листные детерминатно-репродуци- рующие эфемерно-полисимподи- альные (верхушечное соцветие мо- нохазий, реже дихазий) однодом- ные бело-фиолетовопасленовоцвет- ковые однолетники (! биоморфо- тип) или двулетники (И биоморфо- тип, зимует в розеточном вегета- тивном состоянии). Рис. 56 — 58. Приоритет неличного (репродуктива) над личным (вегетати- вом), когда личное выживает более ! года (проявляется персистентность жизнеразвития), но только до первого репродуктивного усилия. Рис. 56. Биографический портрет Verbascum densiflorum Bertol. [сем. Scrophulariaceae (Москва, Глав- ный ботанический сад, питомник лекарственных растений) ]. Основной биоморфотип: надземно- двусемядольные (семядоли треутольно-яй- цевидные, 3 — 6 мм дл., 2,5 — 4 мм шир., закругленные, резко суженные в короткие черешки 3 — 5 мм дл.) полурозеточные прямостоячие стержнекорневые некустис- тые очередно-простолистные семидетер- минатно-репродуцирующие (фрондулоз- но-брактеозное верхушечное полузакрытое соцветие) однодомные желто-норичнико- воцветковые двулетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ
Глава 2. Концеиция npuo umemoe азвития в о ганизации систем Рис. 59 — 65. Приоритет неличного (репродуктивного) над личным при котором расширяются компромиссы между этими е пе систентное свойствами и личное проявляет мультирепродуктивное перси жизнеразвитие, ког , когда живет много больше одного года и не отмирает после нескольких, иногда многих, репродуктивных усилий. Рис. 59. Биографический портрет Echinops rutheni . ! nicus Bieb. ~сем. Asteraceae Алтай, щебнистый горный склон с остепненной р й астительностью в окрес- тностях села Иня)]. Q СНОВНОИ ИОМО б рфотип: нгдземно-двусемядольные (семядолн элл ли эллиптические, 6— кие че ешки 3 — 5 мм дл.) 4 — 6 мм шир., закругленные, суженные в короткие черешки 10 мм дл., — мм чие пол зеточные ветвисто-каудексовые с стержнекорневые семидетерминатно- и еп од ивные побеги словно закрываются одной вер- репродуцирующие (обычно унирепродуктивные по еги усло хушечнои корзин рзинкой) гемисимподиальные мультирепродуктивные однодомны роцветковые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHblX РАСТЕНИЙ Рис. 60. Биографический портрет Doronicum oblongiflorum DC. [сем. Asteraceae (Курайский, Северо-Чуйский хребты, субальпийские луга)]. ОсновнОЙ биоморфотип: надземно-двусемядольные (семядоли яйцевидные, 8 — 10 мм дл., 4 — 5 мм шир., широкозакругленные, резко суженные в короткие черешки, 4— 5 мм дл.) прямостоячие полурозеточные компактно-ризомные гемисимподиальные (моно- подиальный прирост обеспечивают розеточные вегетативные побеги, а цветоносный ве- сенний годичный побег часто начинается с 2 — 3 чешуй и переходных листьев, образуя в основании 1 — 3 срединных листа и в целом развивается как безрозеточный или полу- розеточный, завершая цикл ди-, плейофазного развития унирепродуктивного полурозеточ- ного побега) простолистные мультирепродуктивные семидетерминатно-репродуцирующие (обычно цветоносный неветвистый побег условно закрывается верхушечной корзинкой) однодомные желто-астроцветковые ретардационно-кондивидуальные (замедленно вегета- тивно размножающиеся) травянистые многолетники.
Глава 2. Концеиция и ио итетов развития в организации систем Рис. 61. Биографический портрет Ballota undulata (Fresen.) Bentham [сем. Lamiaceae (Израиль, каменистая пустыня Негев, окрестности поселка Сде- Бокер, овраг — 1, 12 Ч; Тель-Авив, Ботанический catt, — 11, 17 IV)] Биоморфотипы: надземно-двусемядольные (семядоли округлые, 4 — 5 мм дл., 4 — 5 мм шир.), широкозакрутленные, мелкоопушенные, слабо выемчатые, в основании имеют небольшую выемку и два рожка, короткочерешковые прямостоячие (1 биоморфотип, в более сухих условиях каменистой пустыни) или полегающе-восходящие (!! биоморфотип, в более влажных условиях Средиземноморского побережья) эусимподиальные (I) или гемисимподиальные (И) стержнекорневые безрозеточные семидетерминатно-репродуциру- ющие (фронда-фрондулозные верхушечные полузакрытые соцветия) акропетально-цвету- щие мультирепродуктивные однодомные листопадные (в пустыне быстро теряют листья еще весной) полукустарники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 62. Биографический портрет Ononis natrix L. [Сем. Fabaceae (Израиль, окрестности Тель-Авива, невысокие каменисто-песчаные холмы, апрель)]. Основной биоморфотип: надземно-двусемядольные (семядоли широкоэллиптичес- кие, закругленные, 4 — 5 мм дл., 2,5 — 3 мм шир., клиновидно суженные в короткие черешки) прямостоячие безрозеточные гемисимподиальные стержнекорневые индетерми- натно-репродуцирующие (открытые верхушечные фрондозные соиветия-кисти после ве- сеннего цветения плодоносят и летом засыхают до половины и больше своей длины) однодомные желто-бобовоцветковые листопадные полукустарники (впервые зацветают на 2-ой, реже 1-ый год жизни, онтогенезы продолжаются 8 — 15, реже 20 лет).
Глава 2. Концеиция npuopumemoe азвития в организации систем Рис. 63. Биографический портрет Теиспит polium L. subsp. capitatum (L.) Агсапд. [Сем. Lamiaceae (там же, что и рис. 62)]. ОсновнОЙ биоморфотип: надземка-двусемядольные (семядоли округло-яйцевидные, лопатчатообразные, 3 — 4 мм дл., 2,5 — 3 мм шир., иногда с небольшой выемкой, резко сбегающие к коротким черешкам) полегающе-восходящие безрозеточные перекрестно-суп- ротиволистные простолистные гемисимподиальные семидетерминатно-репродуцирующие (верхушки унирепродуктивных побегов условно закрывают компактные фрондо-брактеоз- ные щитовидные соцветия) стержнекорневые однодомные беловато-яснотковоцветковые летне-зимнезеленые (летом отрастают цветоносные побеги, а зимой продолжают медленное моноподиальное нарастание вегетативные побеги, в целом большинство унирепродуктивных побегов дифазные) листопадные (листья живут 2 — 3 месяца) полукустарники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИИ Рис. 64. Биографический портрет Eleutherococcus senticocus (Rupr. et Maxim.) Maxim. [Сем. Araliaceae (Москва, Главный ботанический сгд, питомник ле- карственных растений]. Основной биоморфотип: надземка-двусемядольные (семядоли яйцевидные, 14 — 20 мм дл., 6 — 10 мм шир., закрутленные, клиновидно суженные в короткие черешки 4— 8 мм дл.) прямостоячие преювенильно-розеткообразующие безрозеточные шиповато-ко- лючие сложнолистные (лист состоит из пяти листочков на черешках, которые распадаются при листопаде) гемисимподиальные рыхлокустистые семидетерминатно-репродуцирующие (верхушки унирепродуктивных побегов в их цветоносной части условно закрывает фрондо- брактеозный сложный зонтик) однодомные листопадные кустарники.
Глава 2. Концеиция и ио итетов азвития в организации систем Рис. 66. Жизнеразвитие некоторых биоморф Salix саргеа L. в условиях воз- растающего затенения может приводить надолго или даже для них навсегда к утрате приоритета репродуктива и подмене его приоритетом вегетатива (Московская область, еловый лес). дополнительные обозначения: S. с. — Salix caprea, Р. а. — Picea abies (L.) Karst., В. p.— Betula pendula Roth.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 67. Биографический портрет Solidago virgaurea L. [сем. Asteraceae (Мос- ковская область, Подольский район, I — опушка леса, II — почти полное отсутствие травостоя под пологом елового леса в 20 м от опушки)]. Биоморфотипы: надземно-двусемядольные (семядоли 6 — 10 мм дл., 2 — 3,5 мм шир., эллиптические, закругленные, суженные в короткие черешки 2 — 3 мм дл.) розеткообра- зующие виргинильно-эумоноподиальные матурно-гемисимподиальные матурно-полурозе- точные (1 биоморфотип) или вынужденно-нецветушие, вегетативно-розеточные (11 био- морфотип), компактно (курта-, миниморезидные)-ризомные (! биоморфотип) или аэрори- зомные (1! биоморфотип, развивается в условиях сильного затенения растений, которые обычно имеют подземные ризомы, но в новых условиях теряют ориентацию в простран- стве и, нарастая более или менее эумоноподиально, выходят в надземную среду, которую почти не отличают от подземной среды и так могут вынужденно поднимать верхушки весьма специфических базовых органов — аэроризомов до 20, чаще 5 — 10 см над повер- хностью почвы, отдельные резиды живут 8 — 16 лет, чаще 10 — 12 лет), семидерминатно- репродуцирующие (при обычном развитии унирепродуктивные плейофазные полурозеточ- ные побеги условно закрываются сверху корзинкой, а сборное верхушечное соцветие нередко разветвленное и составляет тирс из корзинок) простолистные мультирепродуктив- ные обоеполые желто-астроцветковые ретардационно-кондивидуальные (замедленно веге- тативно размножающиеся) травянистые многолетники (! биоморфотип) или вынужденные полугравы (11 биоморфотип, в условиях сильного затенения). Первое цветение происходит на 3 — 15 году, а продолжительность жизни до 60 лет.
Глава 2. Концеиция npuopumemoe азвития в о ганизации систем
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 68. Развитие биоморф Parkinsonia aculeata L. по основному биоморфо- типу — кустистых деревьев (I) и по второстепенному биоморфотипу- некустистых деревьев (II), когда приоритет некустистости над кустистостью устанавливаются вторжением извне — человеческим фактором, в нашем случае — пилой и топором [сем. Fabaceae (Израиль, пойма реки Яркон— I, скверы и парки Тель-Авива — II)].
Глава 2. Концепция прио итетов азвития в организации систем 2.2. Высший приоритет в организации живой природы и принцип миниатюризации видов Мир живой природы — это мир открытых систем, которые, для поддержания своей жизнедеятельности допускают в себя и выводят из себя какое-то количество вещества и энергии из того, что их окружает (т. 1, гл. 2). Высшим приоритетом в организации живой природы в целом и любого из ее элементов-биосистем разного уровня сложности, безусловно, является стремление к сохранению жизни вообще, т.е. к сохранению Жизни любыми средствами и в любых условиях. Стратегия Жизни как единая для всех биосистем, осознаваемая или не осознава- емая ими программа деятельности (поведения), закодированная на ге- нетическом уровне, по которой жизнь сохраняется во Вселенной (т. 1, гл. 3), является высшим приоритетом организации живой природы. Ранее было показано, что эволюция личного в существовании каждой биосистемы неотделимо взаимосвязана с эволюцией коллегиального в рамках организации биосистем, что нашло отражение в общей теории жизнесохранения, сформулированной автором (т. 1, гл.3), и, что особен- но важно, приоритет коллегиального жизнесохранения является реша- ющим в существовании каждой биосистемы. Пожалуй, здесь уместно дать определение еще одному правилу, отвечающему закону приоритетности. Правило 4-е: высшим приорите- тегия Жизни — стремление к жизнесохранению вообще, в которой приори- тет коллегиального выживания, например, вида по отношению к его орга- низмам, организма — к их органам и т.n., достигается посредством все возрастающего в эволюции подавления личной жизни или в направлении компромиссов между ними, в результате чего продолжительность личной гда остается за неличным (коллегиальным) жизнесохранением (рис 51 — 68). Эволюция в очень давние исторические времена создала живые системы только по одному типу — открытому, в результате чего они стали участниками множественных разномасштабных круговоротов или даже мизерных «круговоротиков» ресурсов, где-то как элементы чьей- то организации, а где-то и как целое по отношению к своим частям- элементам. Биосистемы, будучи открытыми системами, что-то потреб- ляют, что-то производят, что-то резервируют и что-то рассеивают в среду, обмениваясь с ней ресурсами. За многие миллионы лет эволюции биосистемы изменили лик Земли, потяжелевший от биосферы, облаго- роженной растительным покровом, животным миром, почвами и т.п. Одним из главных завоеваний жизни на Земле было создание ею орга- низации с весьма сложной иерархией биосистем (т. 1, гл. 1), в которой одно из ключевых мест заняли виды. Дело в том, что виды, безусловно, не все, путем сложнейших хитросплетений в защите жизни, способны в одиночку восстановить жизнь на Земле и во Вселенной, если там на~утся условия как-то сохранить их носителей: от самых ничтожных,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ например, отдельных клеток до групп организмов, где-то уцелевших после космической катастрофы. Пожалуй, есть основание взять на себя смелость и заявить, что в Природе существует принцип, ранее никак еще не описанный в науке, — принцип самодостаточности, или миниапо- ризации, видов. Нет, я не буду доказывать, что факты проявления этого принципа никому не были известны. Безусловно, надо констатировать, что известных фактов скопилось множество, а вот теоретическое осоз- нание их только наступает. Этот принцип ориентирован на особенности поведения видов, он npucymcmeyem в Природе неотвратимо, независимо от нашего сознания, — это изобретение живой природы, которое получилось у нее с целью сохранить себя, и ставка была сделана, в первую очередь, на виды. Нам оставалось только подглядеть что-то интимное из жизни Природы и, приобщившись к растительной жизни, войдя к ней, может быть, «в доверие», рассекретить одну из тайн Жизни. Всех нас ранее оглушала информация, переходящая из учебника в учебник, что форма- ми существования вида являются популяции, состоящие из множеств разновозрастных организмов. Все правильно, но кроме одного штриха, что вид существует только в популяциях. Нет, это далеко не так и это легко опровергнуть. Надо признать, что у видов (а не у кого-то другого) есть простейшие формы допопуляционного существования. Живая природа в процессе эволюции предусмотрела создание при видах миниатюрных ячеек их самодостаточности, которые по объему не имеют ничего общего с популяциями. Многие виды имеют систему эшелонированной многоуровневой обороны еще на подступах к попу- ляциям. Когда какой-либо фрагмент организма (отдельная вегетативная клетка, кусочек ткани, обособленная часть или целый орган), один орга- низм (с самоопылением, апомиксисом, вегетативным размножением) или минимальное их число (два, реже — больше) одного вида, способны обеспе- чить виду восстановление и (или) самостоятельное существование со сме- ной поколений, то такая система действует как своеобразная служба спасения, которая самодостаточна для сохранения вида при определенном сочетании окружающих условий, составляя, таким образом, миншиал вида, или снецикулу (от лат. species — вид, cula — уменьшительный суффикс), — особую внутривидовую ячейку жизни, — это принцип самодостаточно- сти, или миниатюризации, вида. Согласно этому принципу, у какого-либо вида есть простейшие формы его существования (нередко их несколько типов), которые в экстремальных, даже критических условиях способны обеспечить ему выживание при определенном стечении окружающих условий. Единицей самодостаточности вида будет, по нашему предложе- нию, минимал вида, или специкула. Не исключено, что принцип само- достаточности видов мог быть использован в далеком прошлом Жизнью для занесения ее «осколков» из других космических миров на некогда безжизненную Землю. Вид защищает себя по-разному: обычно в попу- ляциях, но и «минималах» (миниатюрных внутривидовых системах). Минимал вида (специкула) — это показатель его самодостаточности. Создав в эволюции принцип миниатюризации видов, Жизнь рас- ширила свои возможности по самосохранению и, вероятно, открыла, с помощью Разума, возможность найти путь к своей бесконечности и к
Глава 2. Концеиция и ио итетов развития в организации систем прогрессу в продлении личной жизни. В дополнение к высказанным ранее (т. 1, гл. 3, стр. 98) соображениям в форме гипотезы «направлен- ного повышения долголетия организмов», можно добавить еще следу- ющее, исходя из разработанных выше представлений. Мы — люди, овладевая смысловой стороной («ключом») принципа миниатюризации видов, становимся активнейшим эволюционным фактором, способным не только увидеть, понять, но и начать осуществлять, сначала в робких экспериментах на одиночках, как с всемирно известной овечкой Долли, а затем и в широкой практике, извечную мечту человечества: научиться продлевать личную жизнь и добиться бесконечности для жизни вообще. Согласно принципа миниатюризации видов, надо активизировать поиск механизмов продления личной жизни организмов и увеличения ее на видовом уровне, вторгаясь в клетку, в генетическую организацию, пе- реставляя гены в ДНК; многое из этого возможно и что-то уже делается учеными. Через принцип миниатюризации видов срабатывает в организмах один из высших приоритетов, одна из его сторон, ориентированная на сохранение жизни вообще, в результате чего в экстремальных ситуациях и после катастроф, используются минимальные остаточные возможно- СТИ ВИДОВ. Если нужные гены искусственно перекомбинировать и найти соот- ветствующие сочетания, которые, не нарушив жизнедеятельность орга- низма, продлят личную жизнь этих организмов, то минимал вида сра- ботает на достижение полезного результата. Природа позаботилась, что- бы у вида все элементы-системы были открытыми, и тогда эксперимен- татор как эволюционный фактор способен вторгнуться в организацию и вида, и его элементов для выявления его ресурса возможностей, ко- торые далеки от того, чтобы их познали; даже у одного вида, Ното sapiens, — еще очень много таинственного. Этот ресурс включает не только разнообразие путей развертывания генетической информации в фенетическую организацию, но и способность геносомы к мутациям, а раз так, то и способность выдерживать искусственные «мутации»— перетасовки генов в поисках оптимальных комбинаций, отвечающих поставленным задачам — сохранению Жизни в бесконечности, есте- ственно, с Человеком на «борту». Задача, похоже, невероятно сложная, но она, можно на это надеяться, все же по силам Разуму Природы (т.е. нам — людям). Живая природа сейчас стоит перед выбором цели своего дальнейшего развития, и подстегивает ее к этому развитие человечества. Пока человечество с великим достоянием Природы — Разумом — еще «топчется» у ворот в ее геносоматическое «ядро». 'вторгаться или не вторгаться? Перспектива вторжения представляется как сверхзаманчи- вая, опасность ошибок также существует серьезная, но остановить Ра- зум уже ничто не сможет. Он — Разум — своим вторжением в геносому живой природы (биостробы) Земли, способен многое изменить в плане ее прогрессивного развития, в первую очередь, избавить людей от мно- гих болезней, продлить их личную жизнь и замахнуться на создание механизма бесконечного существования Жизни во Вселенной. Человек ~е многое научился изменять в живой природе; например, по его
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ усмотрению создаются агробиоценозы, дендропарки, лесные насажде- ния и т.п. Почему бы Человеку не научиться управлять генетической организацией видов и, следовательно, организмов, в частности, с целью коренного изменения продолжительности их личной жизни, клониро- ванием и многим другим. Вопросы эти уже поставлены Жизнью и от их решения человеческому разуму уйти никуда нельзя. Человек, однажды надев на себя халат доктора Природы, став, тем самым активнейшим эволюционным фактором, невольно взял на себя ответственность за сохранение Жизни не только на Земле, но и во Вселенной, причем- за бесконечное ее сохранение. Все развитие науки и генетики, в особен- ности, показывает, что человечество будет вторгаться в геносоматичес- кую организацию видов, менять комбинации генов и находить в них такие работающие их сочетания, которые решат многие проблемы орга- низации не только человека-вида, но и всей живой природы, сделав ее существование еще более осмысленным, чем сейчас, что в итоге рево- люционизирует эволюцию Жизни, когда новации и сверхновые переме- ны в Жизни станут обыденным явлением, будучи подчинены стратеги- ческой цели — сохранению жизни вообще и, в первую очередь, беско- нечному сохранению человечества. Изменяя организацию клеток, защи- щая организмы людей от болезней, изменяя, с филигранной техникой и сверхточной осторожностью, геносоматическую природу, Человек способен на многое повлиять и, в первую очередь, с пользой для своего вида, которую, как показывает вся история развития человечества, по- лучить нельзя, если забывать о пользе для всей остальной природы. Безусловно, человечество ждет в третьем тысячелетии генная революция и от нее нам не уйти, какую бы позу не занимали сейчас ее противники. Надо быть осторожными, — это однозначно, но остановить генную революцию нельзя; она — итог развития биологии. Нельзя в XX веке было остановить электрификацию, автомобилизацию, даже ядерную энергетику, а в XXI веке нельзя будет остановить и генную революцию. Такая революция произойдет, естественно, далеко не потому, что нами сформулирован принцип миниатюризации видов, но то, что этот прин- цип сформулирован и объясняет многое происходящее в развитии био- логии и способствует ускорению ее развития, — свидетельство реально- сти его существования в Природе. Теоретически осознавая значение минималов видов (специкул) для развития живой природы, человече- ство вскоре начнет практически извлекать пользу из понимания этого принципа. Итак, приведенные соображения свидетельствуют, что высший приоритет развития живой природы — сохранение жизни вообще; он может обеспечиваться разными средствами и, в частности, подключени- ем к его реализации усилий человеческого разума. Человеку отведена Природой невероятно большая роль: он отвечает не только за свое сохранение как вида, но и за сохранение жизни вообще в бесконечно- сти. Все развитие человечества настоятельно показывает нам, что только Человек на Земле и за ее пределами способен превратить невероятное в очевидное.
141 Глава 2. Концеиция npuopumemoe развития в о ганизации систем 2.3. Борьба за приоритеты развития на арене противоречий между своим и не-своим в организации биоморф семенных растений В свое время (Нухимовский, 1980, 1984, 1997) удалось выйти на осознание противоречия, которое, пожалуй, без преувеличения можно назвать великим противоречием живой природы — это противоречие, проявляющееся в структуре единого живого тела, например, организма, между свойствами развития (поддержания) относительно личной (сво- ей), или вегетативной, жизни (автобиофилии) и относительно другой (хотя и родственной, но не-своей, не для себя, а для той жизни, рас- положенной на одном со своей уровне и выше), или коллегиальной жизни (ксенобиофилии), нацеленной на размножение, в первую оче- редь, репродуктивной жизни. В теле растительной (да и не только) биоморфы определенная часть обозначается как личное, или свое (вегетативное), а вторая часть как неличное(другое), или не-свое (репродуктивное), что, действительно, относительно соответствует свойствам этих частей, поскольку в них про- сто не бывает и быть не может, как в целостной живой системе, ничего абсолютно личного (своего), но и абсолютно неличного (другого). Жизнь как сложная система организована по многоуровневому принципу, поэтому абсолютно принадлежать одному уровню организа- ции биосистема не может, когда над ней есть еще другая, более сложная, частью которой предыдущая биосистема является и, естественно, обес- печивает своей активностью не только собственное выживание, но и сохранение, более высокой по сложности организации, другой биоси- стемы, по сути, другой для нее, хотя и родственной, — очень родствен- ной, но все-таки — не-своей (!). Избранный нами подход, пожалуй, правомерен, поскольку ориентирован на конкретные факты: одна часть биоморфы больше (но не полностью) служит поддержанию жизни са- мой биоморфы (личной), а другая — больше (но не полностью) прояв- ляет жизнедеятельность в направлении сохранения не-своей жизни, а жизни (другой), стоящей над биоморфой, в первую очередь, вида, а в целом и та, и другая части биоморфы так или иначе нацелены в своей жизнедеятельности на сохранение жизни вообще. В соответствии с отмеченным выше обстоятельством, далее мы, как правило, не будем при обозначении определенных частей биоморф использовать поясни- тельное слово «относительно» перед словами «личный» (свой) и «нелич- ный» (другой, не-свой). Под личной жизнью будем, в отношении биоморф, понимать то, что составляет структурно-функциональное содержание и жизнедеятельность вегетативного тела в организации особи (индивидуу- ма), а шире и синорганизма, которое непосредственно не участвует в репродуктивном размножении или не образует однозначно специализиро- ванных органов вегетативного размножения. Некоторые части вегетатив- ного тела организма могут участвовать в вегетативном размножении, внешне не проявляя специализации в этом плане, и лишь тогда, когда они отделятся от родительского вегетативного тела, они становятся
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ элементами другой жизни, а до этого они участвуют в личной жизни организма, например, ветка осины обыкновенной. В отношении синор- ганизма такие, неспециализированные к вегетативному размножению части остаются в его личной жизни не только после отделения от ро- дительского организма, но и после превращения ее в особь вегетатив- ного происхождения (клонист), до тех пор, пока та не проявит репро- дуктивную функцию. Структурно-функциональные части биоморф, которые непосредственно участвуют в репродуктивном размножении или однозначно специализированы для вегетативного размножения- это все неличная жизнь в теле организма, а без вегетативного потомства (клонистов) и его зачатков — и синорганизма. О личной жизни труднее говорить шире, чем в отношении организма, но если принять один принцип: личное = собственно вегетативное, не работающее специаль- но на размножение (случайно бывает), то можно пренебречь некоторой условностью. Части фенетической организации растительных биоморф (а шире — любых биоморф) склонны проявлять вегетативное развитие, а другие части — репродуктивное развитие, но даже та часть, которая не проявляет в настоящий момент репродуктивной функции, она все равно находится под давлением другой жизни, более всего для биоморф — под давлением их вида. То, что вегетативное и репродуктивное как два тела могут вместе присутствовать в организации одного тела биоморфы простой (особи, организма) или сложной (синорганизма), не вызывает сомнений, в вот то, что между этими телами существует противоречие, — это уже другой, большой и сложный, вопрос. Сначала зафиксируем, по сути дела, ак- сиоматическое положение — специальный закон единства вегетативного и репродуктивного в организации биоморф: вегетативное тело (вегетатив) находится в единстве с их репродуктивным телом (репродуктивом) в организации биоморф, причем для последнего, в согласии с генетической программой, вегетатив (вегетативное) является местом существования (автобиотопом), подчиняется его организационному превосходству, рабо- тает на него, обеспечивая себя и репродуктив средствами существования, создавая ему необходимые опору (несет на себе) и ресурсные запасы и, по мере возможности, еще и защищает (шипы, колючки, яды и т.n.) в про- странстве и во времени, а по мере завершения репродуктивного цикла развития, элиминируется вместе с ним или позднее, иногда растягивая довольство от компромиссной (между ними) жизни на многие годы, если находит дополнительные ресурсы для переключения себя на обслуживание следующих (одного или нескольких) репродуктивных усилий. Если образно посмотреть на этот закон, то надо подчеркнуть, что вегетативное тело, как и репродуктивное тело, находятся вместе в без- граничном единстве на службе у организма, причем вегетатив еще и обслуживает репродуктив, а вот служба репродуктивного тела наиболее активна не для личной жизни организма, поскольку она более всего нужна виду, а следовательно, не столько лично организму, сколько более высокому уровню организации жизни на Земле, что лишний раз подчеркивает превосходство репродуктива над вегетативом. Вегетатив- но-репродуктивное фенетическое развитие осуществляется в организмах
Глава 2. Концеиция и иоритетов азвития в организации систем на основе генетической программы: сначала вегетатив потребляет, про- изводит и резервирует необходимые ресурсы для развития репродуктива, обеспечив создание его и расположив удобно в пространстве и во вре- мени, защищает всеми доступными средствами и способствует достиже- нию главной цели репродуктива — полового размножения, после кото- рого единое вегетативно-репродуктивное тело организма может элими- нироваться. Однако есть и другой путь, когда вегетатив ускользает от элиминации, «припрятав» в себе (в генах) приоритет репродуктивного развития организмов, чтобы через некоторое время еще раз, два или несколько, и даже много раз, повторить развитие репродуктива, продле- вая жизнь своей простой биоморфе, избавляя ее от эфемерного «абсур- да» и провоцируя жизнь вообще (Жизнь) к поиску феноменального, хотя и дискретного, долголетия даже не в десятки, а в сотни и тысячи лет. Рассматриваемый закон отражает существование в организации биоморф непоколебимого в пространстве и во времени союза двух, весьма противоречивых тел и, хотя идиллии в этих отношениях нет, жизнь, благодаря такому единству, надежно продолжается на Земле уже несколько миллиардов лет. Как видно из определения закона, он отра- жает не только факт единства вегетативного и репродуктивного в орга- низации биоморф, но и факт диктата (превосходства) репродуктивного над вегетативным, т.е. цели репродуктивного развития приоритетны для биоморфы по отношению к целям вегетативного развития. Если образно рассмотреть этот закон, то вегетативное тело, как и репродуктивное тело, находятся на службе организму, а шире и синор- ганизму, но служба у них разная. Служба репродуктива обеспечивает организму особую роль в жизнеразвитии; через репродуктивное тело организм более всего общается с видом и верно служит ему, т.е. как структурный элемент, функционально обслуживающий более высокую организацию. Это, кстати, лишний раз подчеркивает превосходство реп- родуктивного над вегетативным. Если очень образно, то вегетативное по отношению к репродуктивному выглядит как слуга по отношению к барину, раб к патрицию, лаборант к академику и т.п. Какую бы форму не приобретали эти отношения, они никогда не выходят за пределы рассмотренного закона. Преимущество одних видов деятельности над другими — одна из сторон проявления приоритетности. Приоритет — или он есть, или его нет, что-то одно. Целей развития у биоморф под разными углами обзора и с разных высот абстракции много, а приоритетов единицы. Ранее (т. 1) мы показали, что первым принципом фенетической организации биоморф является приоритетное целостного развитие в направлении размножения. В отношении всех других видов деятельности биоморф, так или иначе сопричастных к главной приоритетной деятельности, но таковой, естественно, не являющихся, поскольку они только работают на целостную приоритетную деятельность и поэтому могут быть только стадийными приоритетами, но не более того. Так, стадийным приори- тетом будет вегетативное развитие на виргинильных стадиях биоморфо- генезов, когда накапливается масса вегетативного тела, способная дать поствегетативный импульс развитию на пути к достижению цели в реп-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ родуктивном целостном приоритете, т.е. на пути стремления к размно- жению. Поствегетативный импульс открывает «ворота» в репродуктив и сопровождается закладкой первых пререпродуктивных и репродуктив- ных элементов, например, брактей, бутонов и т.п., а завершается обра- зованием пыльцевых зерен и семян с последующим рассеиванием их, т.е. достижением размножения — реализации приоритета репродуктив- ного развития (шире — стремления к размножению и размножения). Из закона единства вегетативного и репродуктивного мы определяем следствие и правило. Закон соотношения своего (личного) и не-своего (неличного) в орга- низации растительных особей: все развитие фенетической организации ра- стительных особей пронизано противоречиями между различными видами деятельности (функциями), среди которых атрибутом является npomueo- речие между развитием и поддержанием собственной, или личной, жизни и другой (неличной), или коллегиальной, жизнью (родственной своему или вышестоящему уровню), которое разрешается следующим образом: или личное подавляется приоритетом коллегиального развития, проявляя реп- родуктивное усилие, убивающее организм уже в первом вегетационном чего проявляется приоритет более компромиссного репродуктивного разви- тия (с разными по продолжительности вегетативными стадиями разви- тия между пиками повторяющихся репродуктивных усилий), или (еще вариант) личное подавляет неличное настолько, что то не находит усло- вий для восстановления своего приоритета какое-то время или всю остав- шуюся жизнь организмов при возникновении неблагоприятных условий для репродуктивного развития (рис. 51 — 67) — это следствие из закона единства вегетативного и репродуктивного. Развертывание фенетической организации растительных особей, хотя и осуществляется под управлением генов, но находится в прямой зависи- мости от условий существования и с возникновением экстремальной ситу- ации, когда для развития репродуктивного усилия не хватает необходимых ресурсов со стороны внешней среды (плохое освещение, или плохое водоснаб- жение, или засыпание почво-грунтом и т.n.), то репродуктивное развитие затормаживается и отступает, даже на время исчезает его приоритет- ность, поскольку активизируется приоритет сохранения жизни вообще, действующий независимо от экологической ситуации, который стимули- мени на приоритет вегетативного развития, но приоритет репродуктив- ного развития быстро восстанавливается, если возвращаются необходимые для его проявления условия — это правило дублирующей (вынужденной) приоритетности в фенетическом развитии растительных организмов при экстремальных условиях (рис. 66, 67). Из закона единства вегетативного и репродуктивного в организа- ции биоморф семенных растений и его следствия видно, что окружаю- щая внешняя среда, и, в особенности, экстремальные условия, высту- пают в качестве сильнейшего фактора эволюции, влияющего на прояв- ление приоритетов фенетического развития, а, следовательно, формиру- ющего закономерности структуры и функционирования биоморф. Мы
Глава 2. Концеиция и иоритетов азвития в о ганизации систем сознательно ограничиваем свои рассуждения в основном растительны- ми простыми биоморфами (организмами), чтобы закономерности фик- сировались однозначнее. При расширении закономерностей на слож- ные биоморфы, естественно, будут отклонения, связанные со специфи- кой фенетической организации синорганизмов, являющихся дизъюнк- тивными (разобщенными) живыми системами, не имеющими единой для всей системы оболочки, отделяющей их от внешней среды. В целом, закономерности на сложных биоморфах проявляются те же, но наблю- дать их сложнее, на то они и сложные биоморфы, поэтому автор идет в примерах на упрощения, ограничиваясь в ряде случаев, как, например, в рассмотренных выше закономерностях, в основном простыми биомор- фами, на которых закономерности проявляются более выпукло и менее вариабильно. Наблюдая за тем, как растительные организмы свободно манипу- лируют приоритетами своего фенетического развития, оставаясь до пос- ледних дней верными высшему приоритету — стремлению к сохране- нию жизни вообще, можно, на мой взгляд, разработать концепцию ве- гетативного размножения в жизни семенных растений (рис. 69 — 120). Сначала, пожалуй, уместны несколько фраз об исходных понятиях, а только потом — о содержании самой концепции. Размножение, как известно, — это образование каким-либо спосо- бом (репродуктивным или вегетативным) из одной родительской особи нескольких особей (всегда не менее двух, учитывая и родительскую особь). Вегетативное размножение осуществляется на эпиэмбриональ- ном материале исходных организмов, когда полностью обособленные от них партикулы превращаются в новые особи, достроив, при необходи- мости, обязательные части — основные органы — побег и (или) корень у семенных растений (т. 1, гл. 4. 5). Почему вегетативное размножение стало нормой в жизнеразвитии биоморф многих современных видов семенных растений? Причин мно- го. Вегетативное размножение возникло в эволюции семенных растений как дублирующий (вспомогательный, вынужденный) способ размноже- ния вообще, которое составляет первый принцип фенетической органи- зации биоморф, прямо или косвенно влияя на все остальное в живой природе и, в первую очередь, на сохранение видов (т.1, гл. 4). Надо постараться увидеть те факторы, которые потенциально способны при- водить в эволюции к возникновению у биоморф вегетативного размно- жения. Таких факторов можно насчитать несколько. Однако, прежде чем перечислить факторы, которые способствуют или, напротив, не способствуют возникновению и проявлению вегетативного размноже- ния, посмотрим на организацию растительных биоморф под несколько неожиданным углом. Хотел ли кто-то этого или нет, но независимо от чьей-либо воли незримо по живой природе с незапамятных времен (многие сотни миллионов лет назад, когда еще только зарождалась жизнь на Земле) пролегла разделительная полоса, которая «расколола» и одновременно «распределила» все живое на нашей планете по двум «полочкам» мироздания — двум типам (классам) частей живых тел (био- меросом) — двум меронам: обитатели и обиталища. Между ними далее
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ в процессе эволюции довольно быстро прошло разделение на обитате- лей и обиталищ генетически однородных (соответственно авто-обитате- лей, или автогенистов, и авто-обиталищ, или автобиотопов), с одной стороны, и генетически разнородных (соответственно, ксено-обитате- лей, или ксеногенистов, и эу-обиталищ, или эубиотопов), с другой. Здесь нас в концепции вегетативного размножения интересуют только генетически однородные обитатели и обиталища. Все, что есть сейчас в биосфере Земли, можно представить как противоречивое единство именно этих двух типов взаимодействующих систем — меросом (обитателей и обиталищ). Каждая биоморфа — это обитатель в остальной части тела биосферы, а если уже, то и остальной части некоторой конкретной экосистемы, в которой она непосредствен- но существует в данном месте и в данное время. Для того, чтобы понять основные закономерности в эволюции вегетативного размножения, мы не будем заниматься сейчас крупномасштабным меротомированием биосферы, а попробуем разобраться с явлением вегетативного размно- жения, не выходя за пределы инфраструктуры тел растительных био- морф, в первую очередь — простых биоморф — организмов, с которых начинается вегетативное размножение, без которого не могут возник- нуть сложные биоморфы — синорганизмы. Если еще уже посмотреть на взаимоотношения обитателей и оби- талищ, то мы увидим их и в организации каждой биоморфы, в том числе и семенных фитобиоморф, поскольку любая их живая структурная часть может быть, с рассматриваемой здесь позиции, обозначена как обита- тель по отношению к остальной части организации тех же самых био- морф — обиталищу (автобиотопу), но и наоборот, независимо от того, насколько и как использует обитатель остальную часть биоморфы (свое обиталище) — более или менее значимо для себя как среды своего инфраструктурного существования. Ранее (т. 1, гл. 4) было подчеркнуто, что отдельные живые вегетативные клетки, группы таких клеток, обра- зующие целый орган (побег, корень, лист и т. и.), или zpynny органов или любой фрагмент органа, которые функционируют в структуре определен- ного организма по эволюционно выработанному типу родительских систем и не являются эмбриональными особями, а также отдельные половые клет- ки — это все проорганизмы (преиндивидуумы, предособи). Организационные параметры генетически однородных меросом- проорганизмов: и обитателей, и обиталищ, могут быть самые разные, но главное в них следующее. Чем большую часть тела растительного орга- низма составляет структура обитателя, тем меньше остается из органи- зации родительского тела в структуре обиталища. Обитателаии (авто- генистами) в организации биоморф семенных растений будут проорганиз- менные части — проорганизмы, в первую очередь, проонтоиды (побеговые части, непосредственно не связанные с корнями, которые после отделения от родительского организма могут в подходящих условиях обрастать кор- нями) и онтоиды, или корнепобеги-органы (побеговые части, которые не- посредственно связаны с корнями, образуя отдельные меросомы, а после их отделения от родительского организма превращающиеся в особи вегета- тивного происхождения — клонисты). Все остальные генофеносоматичес-
Глава 2. лонцеиция npuopumemoe азвития в о ганизации систем кие живые обитатели — это коалисты, или члены каких-либо коалиций, листья, например (т. 1, ш. 4, с. 171). Обиталище в качестве организмен- ного местожительства (автобиотопа) включает всю остальную часть родительской особи, которая не вошла в структуру обитателя или не- скольких обитателей в теле этого же организма. Жестких границ между обитателями и обиталищами в структуре организмов не существует, поэтому в зависимости от обстоятельств «возводятся» такие границы, которые отвечали бы задачам конкретного исследования. Большинство обитателей в структуре организмов — это проорганизмы. Отдельные половые клетки (гаметы) — половые преин- дивидуумы — могут превратиться в особь (организм), как правило, после слияния с гаметой иного пола. Неполовые преиндивидуумы по-разному специализированы, поэтому одни из них более, а другие — менее со- храняют потенциальную способность к преобразованию в организмы; есть еще и третий тип проорганизмов, которые полностью утратили в процессе эволюции какую-либо способность к регенерации в организ- мы (по крайней мере, нам так кажется на основе известных сейчас знаний). Все обитатели и обиталища в структуре растительных организ- мов, о которых здесь идет речь, — это однородные генофенетические системы. Гены в структуре организмов — это особенные обитатели, по- разному ассоциирующиеся с соответствующими обиталищами (автоби- отопами) — фенотопами и генофенотопами, о чем подробно изложено в главе 9. Каждый обитатель в его обиталище выполняет свою «работу» (фун- кцию), будучи за время длительной эволюции специализирован для этого соответствующим образом, т. е. ориентирован для более или менее строгого выполнения одной или нескольких функций. И все-таки есть еще пара функций, которые у всех неполовых генофенетических оби- тателей-проорганизмов стоят несколько особняком над всей остальной их специализацией, — это партикуляционная (образно — «сепаратист- ская») и регенерационная (восстановительная) функции. Благодаря та- ким функциям многие структурные части организмов, обладая ими, но до поры до времени в полной мере их не проявляя, могут стать не только самостоятельными партикулами, полностью обособляясь от ро- дительского организма, но и достроить недостающие им основные орга- ны. Партикулы, будучи одновременно разновидностью специкул («ми- нималов-видов»), в критических обстоятельствах могут обеспечить спа- сение угасающему виду и даже — жизни вообще. Когда «жребий» вы- падает на необходимость вегетативного размножения, то без партикуля- ции и регенерации рассчитывать на успех нельзя. Благодаря регенера- ционному, или регенеративному, потенциалу обособленные партикулы (это всегда разновидности специкул) способны побороться и за свое выживание и выживание вида. Регенеративная функциональность меро- сом биоморф, проявляющаяся интроорганизменно (внутри структуры роди- тельского организма) и экстраорганизменно (вне структуры родительского организма) является атрибутивным условием для успешной реализации вегетативного размножения. Регенеративные функции проявляют себя часто опосредованно, когда в некоторых условиях срабатывает защит-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ный механизм проорганизмов-специкул — зов далеких предков к само- стоятельному существованию — к превращению проорганизмов в орга- низмы. Партикуляционно-регенерационные потенции — это, по сути, су- щественный фрагмент генетической «памяти» в современных организ- мах, когда в некоторых условиях проорганизмы, будучи загнаны эволю- цией посредством специализации внутрь структуры родительских орга- низмов, стремятся вернуться к самостоятельному организменному су- ществованию, но, естественно, не в доисторическом облике, а соответ- ствующим образом преобразованных на более высоком витке эволюции, со всеми атрибутами постпартикуляционной регенерации обособленных меросом (партикул) в современные организмы, когда обязательно вос- создается основное интегральное инфраструктурно-организменное ок- ружение. В проорганизмах-специкулах, безусловно, еще где-то подспуд- но дремлет генетическая «память» о предках современных проорганиз- мов, которые когда-то существовали в мироздании самостоятельно, а сейчас эволюция специализировала их, как потомков, на роль структур- ных элементов других организмов, решая, таким образом, текущие за- дачи жизнеразвития на Земле. Что достигают проорганизменные обитатели, стремясь иногда пре- вратиться в организменных обитателей? Они, в таких случаях, проявляют специализации более высоких рангов, часто внешне скрытых, заряженных на сохранение и вида, и жизни вообще. Вегетативное размножение в ис- торической ретроспективе возникло раньше репродуктивного. В органи- зации биоморф семенных растений проорганизмы по-разному сохрани- ли генетическую «память» о далеком прошлом своих предков, поэтому у многих современных проорганизмов в большей или меньшей степени присутствует стремление вегетативно размножаться, но не посредством самостоятельного обновления только своей организации, а интегратив- ным восстановлением (регенерацией) всех основных структур по типу родительского организма, что стало в настоящее время возможно только после многих прошлых витков эволюционного развития. Какой-либо генетически однородный обитатель (автогенист), будучи проорганизмом в структуре родительского организма и имея онтоидное, или проонто- идное, или коалистское строение, может по разным причинам стано- виться партикулой, которая, получив полную самостоятельность, далее или погибнет, или станет новым организмом, достроив, если их не хватало, основные органы [побеги и (или) корни], свершив, тем самым, вегетативное размножение. Биоморфы, вступая на «тропу» размножения вообще, действуют в соответствии с приоритетами, пронизывающими их существование и исходящими из генетического уровня, высшим среди которых является приоритет сохранения в жизнеразвитии вышестоящей, по организаци- онной сложности, системности, по отношению к сохранению нижесто- ящей системности, поэтому в результате растительные биоморфы творят то, о чем не ведают (другого им не дано). Отношения между вегетативом и репродуктивом в организации растительных биоморф я склонен вы-
Глава 2. Концеиция и ио итетов азвития в организации систем разить еще и образно: репродуктивное усилие чем-то напоминает высо- коточное «бомбометание» по вегетативу, после которого (все зависит от мощности репродуктивного «заряда» и надежности вегетативных конст- рукций) бренное тело биоморфы одного типа полностью погибает, а у других биоморф выживает, выдерживая еще 1 — 2 или более таких разрушительных для биоморф репродуцирующих «налетов». Иногда растительные биоморфы развивают вегетативные конструкции настоль- ко прочные и совершенные, что даже мощные ежегодные «ковровые» репродуктивные «бомбометания», когда «армады-ковры» развертываю- щихся цветков, составляя, например, сверхсоцветия могучих деревьев, очень долго (сотни и даже тысячи лет) не могут разрушить вегетатив особей-долгожителей и склонить их к элиминации [такая высокая про- должительность совместной личной и неличной, т. е. в рассматриваемом случае вегетативно-репродуктивной жизни в теле простых биоморф- организмов — это свидетельство высших форм компромиссов (!) между репродуктивом и вегетативом, достигнутых в эволюции, — торжество специфических инфраструктурных адаптаций, назовем их для удобства «интроадаптациями»]. Сложные клубки взаимоотношений — формы инф- раструктурной борьбы за существование, которыми определяют свое уча- стие меросомы как обитатели и меросомы как обиталища в единой орга- низации их родительских биоморф, а, тем самым, определенным образом, и в жизнеразвитии вообще — это все интроадаитации в организации биоморф (разновидность адаптаций). Интроадаптации присущи всем био- морфам живой природы, а у биоморф семенных растений они проявляются в целом, — и в организмах, и в синорганизмах. Изучение интроадаптаций — одно из перспективных направлений дальнейшего развития интег- ральной биоморфологии. Когда фитобиоморфы получают искомый ре- зультат своего развития и размножаются, то нередко (хотя и не всегда) уничтожают этим действием самих себя, например, погибают однолет- ники после плодоношения или часто родительские особи — после ве- гетативного размножения. Особи вегетативного происхождения-клони- сты — это уже не родительский организм, а его потомки — производные организмы, пусть иногда и с прежними родительскими болячками, но все-таки уже другие простые биоморфы. Проорганизмы действительно полноценно существуют как живые тела в структуре своего родительского организма, где взаимодействуют как одно целое в единой системе с другими структурными элементами. Когда проорганизмы по каким-либо причинам получают свободу от родительского организма, став обособленными партикулами, то перед ними встает дилемма: как воспользоваться приобретенной свободой, чтобы был от нее прок (иначе смерть может стать ценой свободы)? Обособленные партикулы, если они сразу не организмы, а только де- структивные системы, т. е. в их организации чего-то из основных ор- ганов [побегов и (или) корней] не хватает, обязаны (виду, например) побороться за то, чтобы стать новым организмом. Так, у деструктивной партикулы ананаса [Ananas comosus (L.) Мегт.] — вегетативной побего- вой верхушки, срезанной с плодоносящего родительского растения (в
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ нашем примере соплодие еще незрелое) нет жизнеполезных альтерна- тив: нужно, чтобы выжить, образовать придаточные корни (рис. 69), а иначе, без корней, придется умереть. В вегетативном размножении семенных фитобиоморф можно ус- мотреть разноформатность этого явления в жизнеразвитии. Общим положением для всех случаев является то, что каждый проорганизм, получая свободу от родительского организма, превращаясь в обособлен- ную партикулу, генетически предрасположен бороться, искать и нахо- дить пути к своему жизнесохранению и сохранению вида, а шире — и жизни вообще. Конструктивные, или полноценные, партикулы, имея в своей структуре все основные органы, сразу же после обособления от родительского организма превращаются в новые организмы (рис. 60, 95, 102 — 108). Обособленные деструктивные, или неполноценные, парти- кулы, получив свободу от родительского организма, устремляются в своем развитии к тому, чтобы «урезать» себе свободу, которая им в чистом виде не нужна; им необходима только опосредованная свобода, «закабаленная» союзом с другими структурными элементами, поскольку только развивая их и, таким образом, — себя, такая партикула может регенерироваться до состояния нового организма (рис. 69). Вегетативное размножение свершилось в эволюции раньше, чем появились первые семенные растения на Земле, поэтому, однажды возникнув, они приняли вегетативное размножение в свою организацию как должное, включив и его, как и многое другое, в арсенал разнонап- равленной эволюции. Приоритетным в организации семенных растений стало репродуктивное размножение, поскольку оно давало больше воз- можностей для обновления соматической организации биоморф, для обмена генами разных родителей и способствовало быстрому расселе- нию потомков в окружающем пространстве. Вегетативное размножение, как правило, заняло место дублера — вспомогательного (вынужденного) способа размножения. В зависимости от условий произрастания пре- имущество получает чаще всего репродуктивное размножение, но в экстремальных условиях (затенение хвойных лесов, стужа высокогорий и т. п.) приоритет может переходить к вегетативному размножению. Остается разобраться теперь с признаками, которые, проявляясь в структурной организации биоморф, способствуют свершению вегета- тивного размножения, а какие — нет. Всякое размножение хорошо для вида, но между собой разные способы размножения вступают в органи- зации биоморф в противоречия, поэтому часто там, где успешно про- являет себя репродуктивное размножение, там нет или слабое вегетатив- ное размножение, и наоборот. Сначала есть смысл посмотреть на орга- низацию растений, которые успешно осуществляют репродуктивное размножение и не прибегают к дублеру — вегетативному размножению. Такие растения или совсем не ветвятся, образуя репродуктивные органы на верхушке главного побега, или ветвятся, образуя пазушные побеги только в качестве паракладиев верхушечных соцветий. В первую очередь так ведут себя эвриэфемерные растения (рис. 51, 52, 54, 70, 71). Уни- репродуктивные двулетники, среднелетники и тем более многолетники дают несколько больше шансов для проявления вегетативного размно-
Глава 2. Концеиция и uopumemoe развития в организации систем 151 жения, но все-таки пока их очень немного и до него дело обычно не доходит (рис. 56, 57, 58, 74). Важно, если развивается базовый орган брахиморфный (с укороченными междоузлиями), поскольку в таком органе лучше резервировать ресурсы и затем использовать их для веге- тативного размножения; например, Anthriscus sylvestris (рис. 46) может развиваться без вегетативного размножения как унирепродуктивный (обоеполый монокарпик) травянистый двулетник или среднелетник, но нередко и с вегетативным размножением уже как мультирепродуктив- ный (обоеполый поликарпик) синорганизменный травянистый много- летник (т. 1, гл. 2, с. 56). Фактор, способствующий развитию вегетативного размножения, особенно в сочетании с брахиморфностью основания главного побега, — это базальное ветвление (кущение). Действительно, если растения не ветвятся, то трудно даже представить, какой «кусок» (партикулу) от родительского растения можно каким-либо естественным образом отде- лить, чтобы из одного растения сделать, как минимум, два и, тем самым, достигнуть эффекта вегетативного размножения. Кущение можно на- блюдать и у однолетников, и у двулетников — это только начало на пути к вегетативному размножению, но еще не само вегетативное размноже- ние (рис. 72, 73, 75, 76). Его нет и у многих многолетних мультиреп- родуктивных кустистых растений (рис. 59, 84 — 87). Для перехода к вегетативному размножению нужно часто научиться проявлять еще один признак — предвестник такого размножения— придаточное корнеобразование. Когда его нет, то вероятность начала вегетативного размножения крайне мала, хотя таковое совсем и не исключается. Придаточные корни в основании главного побега и побе- гов кущения иногда можно наблюдать уже у унирепродуктивных мало- летников (рис. 77, 78), даже у однолетников (рис. 54) и эфемеров. Влажные рыхлые (песчанистые) почвы, особенно в условиях засыпания, или заиливания ими зон кущения способствуют образованию придаточ- ных корней. Если без ветвления, в особенности кущения, трудно достигнуть вегетативного размножения, то без образования придаточных корней все-таки можно, хотя чаще всего оно будет несовершенным. Вполне можно обозначить морфологический (обоюдонаправленный) ряд в сто- рону (в нашем случае) проявления и совершенствования вегетативного размножения от стержнекорневых растений, например, облигатно-кау- дексовых, которые не поддаются продольному расщеплению на части, к каудексово-базифизарным, которые, старея и нередко подвергаясь еще засыпанию подвижным почва-грунтом, способны продольно рас- щепляться через каудекс и главный корень, образуя сильно сближенные особи-клонисты, у которых посткаудексы или посткаудиции и куски главного корня вместе с придаточными корнями, если они образуются, составляют нередко «веревковидную» криптоклональную (скрытый клон) базифизарную систему синорганизма (рис. 90 — 94). Далее, осо- бенно на горных осыпях, такие «веревки» способны расплетаться совсем (рис. 95). Здесь можно усмотреть очередной шаг в сторону совершен- ствования вегетативного размножения и возникновение в перспективе
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ сначала горизонтально-ветвистых каудексовых растений (рис. 48) и далее каудексово-корневищных (рис. 116; т. 1, с. 313) растений, а затем и корневищных растений, когда главный корень полностью отмирает у взрослых растений или даже раньше (рис. 101 — 110). Чтобы вегетативное размножение состоялось, должны особым об- разом развиваться отношения между эфемерностью и персистентностью побеговых частей. Процессы неотмирания отросших осевых (стеблевых и корневых) систем или, наоборот, их полного отмирания не способ- ствуют вегетативному размножению. Однако там, где неотмирание осе- вых частей и некроз способны как-либо чередоваться друг с другом, там вегетативное размножение получает много шансов для своего утвержде- ния (рис. 100 — 115). Если, как у эвриэфемеров, некроз после осуще- ствления репродуктивной функции охватывает и побеговую, и корневую систему целиком, то в результате его не только не происходит вегета- тивного размножения, но и наступает смерть всего родительского орга- низма. Но такого не происходит у тех многолетников, которые склонны к викароперсистентности (т. 1, гл. 6, с. 316), когда некроз вроде пол- ностью разрушает родительский организм, а где-то рядом успевает со- хранить родительский организм всего одна его живая «крошка» — зи- мующая почка (викарирующий базовый орган), успевшая образовать собственную корневую систему. Смерть прошла стороной, но совсем- совсем рядом, и все-таки здесь не было вегетативного размножения (рис. 81). У биоморф некоторых других видов все вроде бы происходит как у ятрышника, но из-под «удара» пострепродуктивного корнепобе- гового некроза успевают сохранить жизнеспособность уже несколько (чаще 2) почек или компактных побегов; в таких случаях биоморфная жизнь не только обыгрывает смерть, но еще и вегетативным размноже- нием умножает совидичей (рис. 82, 83). Многое в осуществлении вегетативного размножения зависит от того, какую жизнестойкость в организации простых фитобиоморф-организмов (эуфитов) проявляет главный корень, который может всю жизнь растения сохраняться и полностью отмирать вместе с побеговой системой, что бывает как у мелких эфемеров, так и у гигантских деревьев, не оставляя при этом никаких шансов для вегетативного размножения (рис. 51, 52, 55, 59, 84 — 87). Однако, если происходит дисбаланс в отмирании побегов и корней или их частей, когда специализация ведет себя по-разному, то появляются условия для вегетативного размножения. Среди побегов чаще и больше у травянистых и полудревесных растений отмирают долихомор- фные структуры (долихобласты), а брахибласты, особенно оснований побегов, наиболее склонны переходить к персистентному (резидному) образу жизни. Главный корень при таком развитии событий может от- мирать рано (рис. 60, 111 — 114) или у взрослых растений (рис. 101, 102), но во всех случаях придаточные корни подстраховывают его утрату, беря на себя выполнение его функций. Некроз главного корня может быстро или постепенно переходить на персистентные побеговые части и быстро (рис. 103, 111 — 114) или медленно (рис. 101, 109) достигать зон развет- вления базового органа, а следовательно, приводить, посредством некро- тической партикуляции, к вегетативному размножению.
Глава 2. Концепция прио итетов азвития в организации систем Облигатное (рис. 117) или факультативное (рис. 119) образование придаточных почек на корнях и их отрастание в качестве корневых отпрысков — это важный аргумент в направлении осуществления веге- тативного размножения. Тот же механизм взаимодействия эфемерности и персистентности, но менее интенсивный, происходит у кустарников и кустистых деревьев, вызывая полную партикуляцию кустов через их геоподии, в результате чего обычно происходит разделение куста на части и достигается, таким образом, вегетативное размножение (рис. 120). При многократном попадании растений в экстремальные условия в них все более и более проявляется тенденция к установлению приори- тета вегетативного развития частично, а то и на весь онтогенез, сдвигая на задворки, а то и вовсе лишая репродуктивное развитие приоритет- ного статуса. В таких ситуациях эволюция подталкивается в направле- нии совершенствования различных форм специализации вегетативного развития и, в частности, в направлении возникновения и расширения приемов вегетативного размножения. Многократное повторение экстремальных условий, в которых про- явление репродуктивного усилия затруднялось, явилось и является по сей день сильнейшим фактором в эволюции вегетативного развития и, в особенности, вегетативного, а шире — вообще бесполого размноже- ния. Когда неблагоприятные внешние условия сильно угнетают разви- тие репродуктивных органов, то стимулируется поиск адаптаций с ве- гетативным размножением. Так, Ajuga reptans L. (рис. 121), попадая под полог елового леса, нередко совсем перестает цвести, но продолжает активно разрастаться во все стороны, образуя долихоморфные столоны, на верхушках которых развиваются вегетативные брахибласты-розетки. Весной следующего года розеточные особи-клонисты продолжают ак- тивно завоевывать пространство, размножаясь вегетативно по интенсив- ному типу, поскольку энергично разрастающиеся клонисты недолго удерживают за собой освоенную ими территорию (т. 1, гл. 4). Синор- ганизм живучки ползучей может долго прожить подобным образом под пологом елового леса, так ни разу и не найдя подходящего места, чтобы зацвести и отплодоносить. Приоритет вегетативного размножения в таких условиях поддерживает сохранение вида на крайних рубежах его пластичности, даже не проявляя репродуктивного размножения. Посмотрим, как ведет себя ель обыкновенная (Picea abies (L.) Karst.), образуя иногда весьма густые лесные массивы как естественные, так и рукотворные (созданные людьми). Особи ели, попадая, волей судьбы, в условия загущенного произрастания, подвергаются жесточай- шей борьбе за существование и естественному отбору. Среди совидичей (особей одного вида) одни особи успевают прорваться своими кронами в верхний ярус леса и развиваются нормально, соблюдая исторически сложившиеся закономерности приоритетства в развитии, в частности, приоритета репродуктивного развития над всем остальным. Другие осо- би уже давно погибли, но деревья умирают стоя, поэтому они еще долго будут стоять под пологом живых как здоровых, так и больных, деревьев как памятники весьма небезмятежной жизни в лесу, как свидетели
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ сложных взаимоотношений между организмами разных видов и внутри своего вида. Особи, ели из третьей группы, безусловно, больны. Их болезнь вызвана конкуренцией. Они борются за жизнь, с каждым годом где-то теряя последние шансы, а где-то находя новые. Присмотримся повнимательней к нескольким особям ели. Все они давно страдают под пологом, в основном, особей-совидичей. Все они кое-как, но еще ве- гетируют, имея еще нечто живое, что составляет их кроны. Они уже давно утратили («забыли», бедняги) свой главный приоритет — стрем- ление к размножению. Сейчас не до размножения; у этих особей другие приоритеты — они стремятся только к одному: хоть как-то, но выжить. Большинство таких особей обречено. Однако, нашлось пару особей, у которых есть шанс еще долго жить, а может быть, и вырваться к лучшей жизни и восстановить утраченные было приоритеты. Их верхушки живы и кое-как протискиваются в «окошечки» света меж крон верхнего яруса. Но самое интересное в другом. По всей высоте ствола со стороны, обращенной в глубь леса, нет живых ветвей, а в сторону опушки, откуда пробивается немного света с другой стороны ствола тянется несколько живых ветвей. К вершине их, естественно, больше. Ветви весьма длин- ные и давно не отвечают традиционным размерам таких ветвей; самые нижние из живых ветвей достигают до 10 м при высоте ствола около 20 м (рис. 122-I). Другие виды деревьев при произрастании на опушке, когда одна сторона сильно затенена, ведут себя подобно ели (рис. 122- П). Мне не удавалось наблюдать вегетативно размножающуюся ель обыкновенную. Вероятно, у этого вида эволюция вегетативного разви- тия еще не дошла до вегетативного размножения в естественных усло- виях его произрастания. У некоторых других видов ели, например Picea schrenkiana Fisch. et С.А. Меу. в Киргизии оно бывает (Кожевникова, 1979). Однако у родственника ели — пихты сибирской — (Abies яЫпса Ledeb.) вегетативное размножение не редкость. Ее нижние ветки долго растут и под давлением своей тяжести начинают касаться почвы, при плотном контакте с которой и достаточном увлажнении они укореня- ются; ветвь образует стволик, рост которого со временем усиливается, а контакт с родительским растением утрачивается. В процессе исторического развития виды, освоившие вегетативное размножение в экстремальных условиях, уже не утратили его и в нор- мальных для них условиях обитания. В процессе эволюции вегетатив- ного развития появились виды, у которых приоритет вегетативного развития на ранних этапах онтогенеза сменялся затем приоритетом не только репродуктивного размножения, но и приоритетом размножения вообще (как репродуктивного, так и бесполого, включая вегетативное размножение). Некоторые виды настолько усовершенствовали бесполое и (или) вегетативное размножение, что свели репродуктивное размно- жение к минимуму [известные примеры с Polygonum viviparum L., т. 1, стр. 233; Lilium bulbiferum L. Dentaria bulbifera L. (Федоров и др., 1962)]. Итак, на основе исторического развития жизнь в образе своих дискретных носителей в борьбе их за существование научилась по-раз- ному решать проблемы своего сохранения. Абсолютным приоритетом (приоритетом приоритетов), или суперприоритетом, является обязатель-
Глава 2. Концеиция и ио итетов азвития в о ганизации систем ное для всех биосистем развитие по пути стремления к сохранению жизни вообще. Этот приоритет довлеет над всем и у всех, что как-то живет в этом мире. В норме жизнесохранение вообще более всего обес- печивает приоритет репродуктивного над вегетативным развитием. Ког- да растительные биоморфы попадают в экстремальные условия суще- ствования и ставится вопрос об их выживании «здесь и сейчас», то они вынуждены больше, чем где-либо еще, идти на компромиссы между своими, казалось бы, непримиримыми приоритетами развития, при этом репродуктив может, он вынужден это делать, вообще не развиваться, уступая вегетативу право на приоритет в борьбе за выживание организ- ма. Когда ситуация с сохранением вегетативной жизни проясняется в ее пользу, то приоритет репродуктива незамедлительно восстанавливается. Вегетативный стадийный приоритет, проявляющийся у биоморф в экстремальных условиях их обитания — в условиях сильнейшего стрес- са, далеко не всегда сменяется на основной репродуктивный целостный приоритет. Особи пихты сибирской, выросшие из семян, случайно за- несенных птицами из среднего и нижнего поясов горных склонов хреб- тов Алтая, в безлесном альпийском высокогорье ведут себя как про- стратные кустарничковые стланики, сильно прижимающие свои ветви к почве, иногда укореняющиеся, что способствует их «ковровому» раз- растанию. В этих условиях биоморфы пихты всю (!) свою долгую жизнь не проявляют репродуктивной функции, довольствуясь только приори- тетом вегетативного развития. Здесь, в высокогорье, пихта сибирская растет на пределе своих возможностей и ее особям нет шансов проявить репродуктивную функцию. А вдруг... условия улучшатся, а вдруг ... по- явится шанс. Дело в том, что в этих условиях целостный приоритет вегетативного развития вынужденный, а на генетическом уровне скрыт- но продолжает существовать целостный приоритет стремления к репро- дуктивному развитию, которое фенетически не может себя реализовать здесь, в высокогорье, как правило, всю индивидуальную жизнь. Если крайние условия высокогорной жизни биоморф пихты не изменятся в лучшую сторону, то существует еще шанс — возможность когда-то произвести мутационный прорыв за пределы существующей нормы реакции и проявить гиперпластичность, изменить ресурс возможностей (свою идиоэнергию) со всеми вытекающими из этого последствиями (вплоть до возникновения нового вида). Каждая особь решает по-своему, но на основе известного ресурса возможностей, проблему выживания, отдавая в экстремальных условиях приоритет вегетативному развитию — скорости роста осевых и листовых структур и т.п. В таких случаях абсолютный приоритет сохранения жизни вообще подталкивает репродуктивное развитие к компромиссу с вегетативным развитием, к уходу репродуктива со «сцены», чтобы вос- становить жизненную энергию организма. Если вегетатив, получив в условиях стресса полный приоритет на развитие особи, не справится и не выправит ситуацию, то наступает смерть всей особи с ее вегетативом и репродуктивом, с ее феносомой и геносомом; а если справится, то особь будет жить, будет жить иногда долго и, вероятно, мучительно ~охо до тех пор пока она не победит негатив, доставшийся ей от
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ воздействия экстремальных факторов, а когда все же восстановится, то приоритет репродуктива придет незамедлительно. В процессе эволюции в экстремальных условиях растения учились жить и научились жить через приспособления (большие и малые), идя на компромиссы в раз- витии своих вегетативных и репродуктивных частей (меросом), расши- ряя диапазон пластичности. Способность вегетатива идти на компро- миссы с репродуктивом, а шире — с любым размножением — родилась в далеком прошлом — на заре развития жизни на Земле. Попробуем ответить на вопрос: где и когда впервые появилась способность биоморф разрешать, казалось бы, непримиримые противоречия между стремлением их сохранить свою (личную) жизнь и их же стремлением сохранить в себе же другую (неличную) жизнь, иными словами, — между вегетативным развитием и развитием, обеспечивающим размножение? Могу утверж- дать, что способность к компромиссам, а, следовательно, и способность разрешать внутри своей организации указанное выше иротиворечие не могла бы проявиться в безмятежной обстановке, когда всего в достатке — и света, и воды, и питательных веществ. Благополучие в окружающей био- морфу среде — это не место для по-настоящему принципиальных эволю- ционных спуртов. Эволюционные прорывы в действительно новое возможны только там, где есть повод для беспокойства, для борьбы, а у pacmumenb- ных биоморф такое возможно там, где много стресса, обусловленного воздействием на растения экстремальных условий (см. материал выше, в частности о Picea abies, Abies sibirica). Именно в состоянии глубочайше- го стресса в окружении экстремальных условий фенетическая организа- ция растительной простой биоморфы передает сигнал бедствия всему организму, который принимается генами, где и формируется ответ через «кучу» структур-посредников, что гибель избежать можно, если срочно сменить приоритеты фенетического (!) развития, направив все на веге- тативное развитие с одной только целью — выжить. В организации био- морф всегда присутствует cynepnpuopumem (приоритет приоритетов) на развитие любыми средствами для сохранения жизни вообще, поэтому при- нятие биоморфой стремления к вегетативному развитию и отставка репродуктива из приоритетов фенетического развития, входит в число обязательных приемов развития биоморф по схеме cynepnpuopumema. На примере особей ели обыкновенной, произрастающих в лесу, видно, сколь разнообразны взаимоотношения вегетатива и репродуктива в едином теле растительного организма. Деревья разных возрастов, умирая, стоят в лесу как памятники жестокой борьбы за существование, в которой побеждают наиболее приспособленные, кто сумеет быстрее других перестроить свои ириоритеты развития в экстремальных условиях на вегетативное разви- тие, а кто не вовремя раскачался, кто не ускорил нарастание, а готовился к чему-то другому, тот потерял нужное время для перестройки, тот готов уже не к продолжению жизни, а к встрече с элиминацией.
Глава 2. Концеиция и ио итетов азвития в о ганизации систем Рис. 69. Вегетативное размножение ананаса [Ananas comosus (L.) Merr.] в тепличных условиях [сем.Bromeliaceae (Москва, ВИЛАР)]. Др — общий вид надземной части плодоносящего растения (Мак-Миллан Броуз, 1987). А б б в+г Рис. 70. Биографический портрет Aethusa cynapium L. [сем. Apiaceae (Москва, скверы и парки)]. Пример однолетников, у ко- торых единственный главный спе- циальный сложный побег (дикор- мус) ветвится в зоне верхушечного соцветия, но обычно не ветвится в основании этого побега — из зоны преконодия. Придаточных корней биоморфы не образуют.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ
Глава 2. Концепция прио итетов азвития в организации систем ет L. ае В %.Ф И- гй и: ет УЯ ов- ег ие и- У- ег, ия, есь ый ан д, 42, 92
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHblX РАСТЕНИИ Рис. 75. Биографический портрет Carlina intermedia Schur [сем. Asteraceae (Московская область, Подольский район, опушка смешанного леса)]. Пример двулетников, у которых развивается главный универсальный сложный полу- розеточный побег (пентакормус), образующий цветоносную долихоморфную часть, развет- вленную в зоне синфлоресценции (сборного соцветия), и который совсем не кустится (нет ветвления из брахиморфной розеточной части, 1 биоморфотип), или в наиболее благопри- ятных условиях главный побег может ветвиться не только в области сборного соцветия, но и куститься, образуя несколько сильных побегов кущения (11 биоморфотип). Прида- точные корни могут иногда образовываться, особенно при засыпании конодия почвой.
Глава 2. Концепция и ио итетов азвития в о ганизации систем Рис. 77. Биогр ческий пор Carduus crispu [сем. Astera (Московская ласть, Подол ь район, молоды сопосадки на м вырубки, смеш ная лугово-ле растительность Пример двулетни которые развивают ный полурозеточный версальный сложныи бег (пентакормус), рый ветвится в зоне ного соцветия, но не вится от основания. жневой корень може храняться всю жизнь нередко отмирает. П точные корни не об ются или образуются бо при живом гла корне (1 биоморфо или они развиваются гочисленные, способс преждевременному о ранию главного корня био морфотип).
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 78. Био а~ичес гр ф еский портрет Senecio jacobaea L. [сем. Asteraceae 'В точный Казахстан южн , южный Алтай, низкогорья, степные участки у дорог" егасеае < ос- ример некустистых унирепродуктивных двулетников (I биом кустистых би-, и еп о ов иоморфотип) и и-, трирепродуктивных полурозеточных травянистых с нелетников (жив 3 — 5 тых сред- вут — 5 лет), сохраняющих главный корень и образ ющих много ленные придаточные корни (11 биоморфотип)
Глава 2. Концепция и иоритетов азвития в о ганизации систем Рис. 79. Биографи- ческий портрет Senecio erucifolius L. [сем. Asteraceae (Ал- тай, Катунский хре- бет, долина реки Ка- тунь, луг в окрестно- стях села Кайтанак)]. Др — дополнитель- ный рисунок фрагменты растений (Королева и др., 1973). ример безрозеточ- ых короткокорне- вищных травянис- тых многолетников, рано утрачивающих главный корень (уже после первого вегета- ционного сезона) и образующих ежегодно в основании единственного ростового или репродуктивно-ростового акронекрозного побега по 1, реже 2 и еще реже — больше латентных почек, которые весной следующего года развиваются в безрозеточные раздви- нутоузловые побеги с коротким, чаще компактным горизонтальным подземным основа- нием. Ежегодно безрозеточные побеги отмирают, проявляя викароперсистентность, т. е. оставляя после себя почку (иногда 2 — 3) и компактное основание, из которых до сле- дующего вегетационного сезона выживают только почки. Вегетативное размножение про- исходит только тогда, когда в весеннем вегетативном возобновлении участвует не одна, а более (2 — 3) почек. Придаточные корни способствуют развитию обособленных компак- тно-ризомных клонистов. Рис. 80. Биографический порт- рет Erythronium sibiricum (Fisch. et С. А. Меу.) Кгуl. [сем. Liliaceae (Алтай, Ивановский, Семинс- кий, Анщский хребты, горные склоны в лесном поясе и в вы- сокогорьях)]. Др — так выглядит начальное развитие кандыка сибирского по наблюдениям Н. И. Шориной и А. Г. Куклиной (1977). Пример луковичных компактнокорневищных (харак- терно длительное сохранение живой хрупкой цепочки миниатюрных рези- дов-донцев) травянистых многолетни- ков. Ежегодно виргинильные особи нарастают эумоноподиально, образуя один зеленый лист. Во взрослом состо- янии нарастание обычно эусимподи- альное. Очень редко растения кустят- ся, а однажды образовав куст, они че- рез несколько лет достигают вегетатив- ного размножения (годичные прирос- ты ризома живут до 8 лет, чаще 5 — 6). Ризом кандыка сибирского настолько миниатюрный и ломкий, что многие ботаники, вероятно, до гербария его просто не доносят.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 81. Биографический порт- рет Dactylorhiza maculata (L.) Soo [сем. Orchidaceae (Москов- ская область, Подольский рай- он, смешанный лес)]. Пример викароперсистентных изоаклональных (без вегетатив- ного размножения) травянис- ТЫХ МНОГОЛеТНИКОВ. Родительские растения ежегодно отмирают почти полностью, оставляя от себя только один зимующий почкокорень (gemmoradix), состоящий из базальной пазушной почки и клубневидного при- даточного корня. Эусимподиальное ви- кароперсистентное нарастание роди- тельского организма не сопровождается вегетативным размножением. Рис. 82. Биографический портрет Aconitum anthoroideum DC. [сем. Ranunculaceae (Алтай, Ку- райский хребет, высокогор- ные луга и тундры, 2300 м над уровнем моря)]. Пример викароперсистент- НЫХ ретардационно-кондивидуаль- ных (замедленно вегетативно раз- множающихся) травянИСтых МНОГОЛеТНИКОВ с неежегодным разрастанием структуры синорга- низмов. Родительские растения ежегодно отмирают почти полнос- тью, оставляя от себя 1 — 2 зимую- щих почкокорня (в виргинильной стадии зимует всегда один почкоко- рень, во взрослом состоянии по- разному: иногда один, а бывает и два почкокорня. В последнем слу- чае происходит вегетативное раз- множение после отмирания цент- ральной оси родительского организ- ма. Синорганизмы не образуют многочисленных клонов, так как вегетативное размножение происхо- дит не каждый год.
Iaaeg 2. Концепция и ио итетов азвития в о ганизации систем 16 Рис. 83. Биографический портрет Aconitum napellus L. [сем. Ranunculaceae (Москва, Главный ботанический сад, питомник лекарственных растений)]. Пример викароперсистентных ретардационно-кондивидуапьных травянистых ~~НОЮЛСЧНИКОВ с ежегодным разрастанием структуры взрослых синорганизмов. Вирги- нильная стадия у синорганизмов продолжается 2 — 5 лет. вегетативное размножение обычно начинается не со 2-го года, а несколько позже, когда начинает образовываться не один, а два и более пазушных почкокорней на родительском растении. Некроз ежегодно захва- тывает почти весь организменный корнепобег, оставляя сначала один (у самых молодых виргинильных растений), а позднее от 2 до 7 зимующих почкокорней. В последнем случае происходит вегетативное размножение, которое у взрослых синорганизмов становится ежегодным. Клон А. napellus разрастается очень быстро, медленно завоевывая сравнитель- но небольшие новые территориальные участки, долго удерживая их за синорганизмом.
Глава 2. Концеиция ириоритетов развития в организации систем 16 Рис. 86. Биографический портрет Alcea froloviana (Litv.) Iljin [сем. Malvaceae (Восточный Казахстан, предгорья Алтая, степные горные склоны, залежи, у дорог)]. Пример полурозеточных гемисимподиальных каудексовых травянистых ~~~~ОЛСТНИКОВ, при старении разрушающихся, не полностью партикулирующих через каудекс и главный корень, вегетативно не размножающихся. Впервые цветет на 3 — 5 год, продолжительность жизни около 25 лет.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 87. Биографический портрет Ecballium elaterium (L.) А. Rich.[cåì. Cucurbitaceae (Израиль, Иерусалим, у дороги в парке, открытое место)]. Пример каудексовых травянистых многолетников, ие проявляющих s процес- се жизнеразвития вегетативного размножения.
~дава 2. Концеиция и ио итетов азвития в организации систем Рис. 88. Разновозрастные биоморфы Aconogonon alpinum ( inurn (All.) Schur [сем. Polygonaceae (Курайский хребет на Алтае, высокогорье, крутой южный сла о задерненный склон)]. Пример безрозеточных эусимподиальных каудексовых травянистых ак иванию продольных стеблекор- МНОГОЛЕТНИКОВ, подверженных партикуляции и закручива зможно но бывает крайне редко и только невых фрагментов. Вегетативное размножение возможн, клонисты остаются сильно сближенны- н~~~вершенного (дефективного) типа, поскольку клон й (ст ия ск ытого клона) веревке»— «, даже скрученными в одной криптоклональной (сто р ризобазифизе.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 89. Биографический портрет Aconitum barbatum Pers. [сем. Ranunculaceae (Алтай, Бащелакский хребет, щебнистые склоны)]. Пример полурозеточных гемисимподиальных каудексовых травянистых МНОГОЛеТНикОв; они склонны с возрастом сильно продольно партикулировать через каудекс и стержневой главный корень, образуя «веревковидные» переплетения стеблекор- невых тяжей. В процессе старения на эрозионно подвижных склонах отдельные партикулы могут отрываться от родительского посткаудексового базифиза, что свидетельствует о несовершенном вегетативном размножении.
gaea 2. Концеиция лриоритетов азвития в организации систем Рис. 89 (Продолж Рис. 90. Биографический портрет Aquilegia sibirica Lam. [сем. Ranunculaceae (Алтай, Семинский, Ива- новский, Катунский и Ку- райский хребты, светлые леса, южные склоны высо- когорий)]. Пример полурозеточных ге- мисимподиальных каудек- совых травянистых много- летников. Виргинильные расте- ния нарастают эумоноподиально. Первое цветение на 5 — 15 году жизни. Только у старых растений изредка может происходить полная некротическая партикуляция, кото- рая приводит к несовершенному вегетативному размножению. Пол- ный жизненный цикл продолжает- ся 40 — 50 лет. (Продолжение рис. на следующей странице).
OCHOBhl БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИИ Е 55а 55а, 67 1 Ъ ! Li ,ч 1 л ° / 1 'л 1 Ф 'Ч, / I /, Рис. 90 (Продолжение).
рава 2. Концеиция прио итетов развития в организации систем е, Рис. 91. Биографический портрет Dracocephalum peregrinum L. [сем. Lamiaceae (Алтай, Курайский хребет, светлые леса, южные склоны высоко- горий)]. Пример безрозеточных эусимподиальных каудексовых травянистых МНОГОЛеТНикОв с продольной некротической партикуляцией стареющих растений и ~~совершенным вегетативным размножением у некоторых из них, когда образуется не- сколько клонистов с посткаудексами. иногда старый каудекс распадается на несколько не полностью или полностью обособлен- H~tx ~acTeh-органов, продолжающих сохранять связь со всем главным корнем или какой- либо его частью, то это уже посткаудексы, а если они сравнительно тонкие и совсем неразветвленные или слабо разветвленные, то это — посткаудиции.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ О Пр НЫ НИ верг куля к не Розе раст бего Пер жиз гене
Глава 2. Концеиция л иоритетов развития в орга организации систем Рис. 93. Био а~ичес~д~ гр + еский портрет Pulsatilla campanella Fisch. ex Redel et Til. [сем. Ranunculaceae ~Алтай К Й, ураискии хребет, высокогорные скалы, россыпи)]. ример розеточных гемисимподиальных каудексово-базифизарных а- вянистых многолетников. ви гин иргинильные растения нарастают эумоноподиально и только после начала цветения (на 5 — 12 год жизни Ъ диальному ия на — году жизни) совершается переход к гемисимпо- у нарастанию. Продольная нек отическая дит к об . П р еская партикуляция старых растений приво- о разованию сначала криптоклонального сос тивное азмн г состояния синорганизма, а затем вегета- размножение становится явным и клонисты (ос разм ноже ) ения) несколько обособляются д от (особи несовершенного вегетативного Полные би у а (особенно на щебнистых осыпях). иомор~огенезы продолжаются до 50 — 60 лет.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 94. Биографический портрет Saussurea frolowii Ledeb. [сем. Asteraceae (Алтай; Северо-Чуйский, Курайский хребты, высокогорные луга и щебни- стые тундры)]. Пример полурозеточных гемисимподиальных каудексовых травянистых мнОГОлеТникОв. После отмирания первого цветоносного побега (на 4 — 10 году био- морфогенеза) образуется первый очаг некроза, который быстро распространяется далее в главный корень и с увеличением возраста некроз быстро усиливается. У старых растений происходит полная некротическая партикуляция и свершается несовершенное вегетатив- ное размножение, которое заметно не продлевает жизнь синорганизму. Биоморфогенезы полностью завершаются на 30 — 40 году.
лава 2. Концелция npuopumemoe развития в 0 ганизации систем 1 2 О О ° ° л о <~ „О О 1~~и ~ О %2 о р."МГ ~О E ~ о к ~ Ж О ~Г) 'X О 1=; h4 О М 2 О й й Q. Е О й й 0» й ' O о ц =~ о~„ Ж." ~о ЩЖЖ О о х 5 ,. O ~g > o O ~ о й~~~жо ~ о ~ ~ ~ ~ ~" ~р ~ Й О М са О ~~ фэ я ~~ Я' Q Жож~ощ р: hC + + с~ ~) vокщщо О. й~ ~ Й ~~() р~ „о ~~,. A ~ 'X щ ;[. 'й со О у у ~X Q. д g 3 а.H ;~~~ оо~ р У ~ О Ю с~ ~~~~ Q.hvщ ' оь ~- ~~~o и М - ~ s~ф д X ц ~~ ф~ ~~ ~ ц ~4 ~ щ О 0 и 'е' о 65 Е О О 65 () 3 ~4 ~~. ~~. О жucvZ "vо М ~ Е ~ Щ О О : ~~ ~ ~ о ~ a2<o Qax '4 ~ о ~ о :г х~ к р, х о~ о вч ~ о ~м О О 4>ои о о х ~,-.~ ~; v О X Е ~ hC О ~ 2 < ~~и~о r. Их~д О 0 д~ Ж о,„Х Е О 1 v о~~~:~~ 04 Я, ~) Q 65 м~ъ я ~ ~ ~) ~4 т Е ~' Д о щ О >~~хо~~ рч О 'X Q ~„3Q2% о ау, и О ~ у > ИOOh4xa5O .~ ~ ~ О й й а.~~Дц2 са~~ иЯ>Qg .'Е, 'о р,х й а ЕО1жао~
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ еа М. IX 0- И- 0- 0- ИИ е, О- СО
~дава 2. Концеиция и ио итетов азвития в организации систем Рис. 97. Биографический портрет Leonurus glaucescens Bunge [сем. Lamiaceae (Алтай, Курайский хребет, от низкогорий до высокогорий, встречен выше верхней границы леса на отвалах горных шахтных выработок в окрестностях поселка Акташ)]. Пример безрозеточных эусимподиальных каудексово-базифизарных тра- B~H HCTbIX МНОГОЛеТНИКОВ. Эусимподиальное нарастание проявляется, начиная с первого года жизни, когда отмирает главный вегетативный побег и образуются латентные пазушные почки, которые весной развиваются в боковые побеги. Цветение начинается с 2 — 6, чаще 3 — 4 годов жизни. Изредка верхушки коротких вегетативных побегов могут «xpaHHTüñÿ до весны следующего года, и в таких редких случаях образуются днфазные вегетативные или вегетативно-репродуктивные побеги. С первых лет жизни происходит ~б~~ьное образование придаточных корней и усиливается кущение. У старых кустов парти- куляция может разрывать каудекс и распространяться на главный корень, образуя иногда обособленные посткаудексовые растения-клонисты. Вегетативное размножение несовер- шенного типа (т. 1, гл. 4).
ОСКОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИИ 1 Ю) Рис. р Ф РТР [се е ае (Мос- ква, ВИЛАР, опытное поле, последняя декада апреля)]. Дрl — фрагмент дикорастущей особи горечавки желтой в возрасте около 43 лет, выросшей в условиях Апьп (Schroeter, 1908, с. 404); Др2 — схема соцветия (Troll,1969). Пример полурозеточных гемисимподиальных каудексовых травянистых МНОГОЛЕТНИКОВ, которые в естественных условиях произрастания обычно не образуют придаточных корней, цветут редко, многие годы нарастая моноподиально (1 биоморфо- тип), и тех каудексовых трав, которые в условиях выращивания образуют много придаточ- ных корней (возраст взрослого растения — 11 лет), но сильный главный корень сохраняют и к вегетативному размножению сами не переходят (11 биоморфотип). Однако, если при агротехнической обработке будет отрезан кусок каудекса с корнями, то он иногда прижи- вается, хотя и развивается довольно слабо. У этого вида приоритет репродуктивного раз- множения очевиден, поэтому рассчитывать на большой успех вегетативного размножения в условиях выращивания не приходится. Продолжение рис. на стр. 181.
~дава 2. Концеиция и ио итетов азвития в о ганизации систем
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 100. Биографический портрет Veratrum lobelianum Bernh. [сем. Melanthiaceae (Алтай, долина реки Сема в окрестностях села Черга, среди лугового разнотравья)]. Др — дополнительные два рисунка из работы М. П. Бахматовой (1974). Пример скрытых розеточно-вегетативных и скрытых полурозеточно-репродуктивных ге- мисимподиальных компактно-ризомных некустистых и вегетативно не размножающихся (1 биоморфотип) и кустистых вегетативно-размножающихся (11 биоморфотип) травянис- тых многолетников. В биоморфогенезе (синонтогенезе) синорганизма нередко проявляет- ся стадия криптоклонального развития, когда обособленные корневищные клонисты ос- таются некоторое время связанными перегнившими нижними частями родительского корневища (Бахматова, 1974).
~дава 2. Концеиция л иоритетов азвития в о ганизации систем О Рис. 101. Биографический портрет Stemmacantha carthamoides (Willd.) М. Dittrich [сем. Asteraceae (Восточный Казахстан, Алтай, Ивановский, Линеев- ский, Бащелакский хребты, поляны в верхней части лесного пояса и высо- когорные луга)]. Пример полурозеточных гемисимподиальных компактно-ризомных ре- тардационно-Кондивидузльных (замедленно вегетативно размножающихся) тра- вяНИСТыХ мНОГОЛетыикОв. Ювенильные растения нарастают эумоноподиально, не кустятся и сохраняют связь ризома с главным корнем. Имматурные растения продолжают эумоноподиальное нарастание и кустятся, могут сохранять главный корень или иногда утрачивают, поскольку сильно развиваются придаточные корни, которые способны заме- нять утрату главного корня. Впервые цветут на 6 — 20, чаще 8 — 12 году жизни. Кущение У взрослых растений весьма активное, поэтому образуется сильно разветвленное корневи- ще, которое, отмирая, снизу приводит ветви корневища к обособлению и, следовательно, вегетативному размножению. Компактные клонисты остаются долгое время сильно сбли- женными и прочно стянутыми многочисленными придаточными корнями в мнимый организм. Продолжительность синонтогенезов не превышает 100 лет. Далее см. стр. 184.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 101 (Прод 67 Рис. 102. Биографический портрет Centaurea jacea L. [сем. Asteraceae (Московс- кая область, Подольский район, лугово-лесное раз- нотравье на старой выруб- ке в окружении смешан- ного леса)]. Пример полурозеточных гемисим- подиальных компактно-ризомных рыхлокустистых ретардационно- кондивидуальных (замедленно ве- гетативно размножающихся) тра- вянистых многолетников. Мини- морезиды в структуре корневища живут 5 — 8 лет. Первое цветение происходит на 3 — 7 году жизни. Придаточных корней много, и развиваются они уже в виргиниль- ном состоянии, способствуя в дальнейшем развитии отмиранию главного корня. Вегетативное раз- множение замедленное; оно за- метно не продлевает жизнь слож- ной биоморфе — синорганизму. Синонтогенезы продолжаются до 30 — 40 лет.
Глава 2. Концеиция и ио итетов азвития в о ганизации систем Рис. 103 Р Р ) [ teraceae (Восточный Казахстан, Алтай, долина реки Белая уба, высокотравные луга)]. Пример полурозеточных гемисимподиальных компактно-корневищных ретардационно- кондивидуальных травянистых многолетников. Особь семенного происхождения (мероак- лонист) альфредии поникшей несколько лет своим главным вегетативным розеточным побегом нарастает эумоноподиально, кустится, теряет главный корень, образуя много придаточных корней, а затем, накопив необходимый ресурс, прорывается с великой силой в неудержимом репродуктивном усилии, которое выносит на высоту до двух метров одну или немного соцветий-корзинок, плодоносит (это впервые происходит на 5 — 12 году, чаще 6 — 8 году жизни), после чего как бы на время успокаивается, освобождаясь от остатков главного побега, совершившего что-то похожее на акт «самосожжения», когда этот побег почти полностью отмирает вместе с придаточными корнями на его ризомной части. Некроз унирепродуктивного главного побега не затронул только производные от него пазушные вегетативные побеги-потомки, статус которых после получения ими само- стоятельности поднялся до уровня организмов-клонистов.,Далее в своем развитии они добирают недостающую им ресурсную массу, чтобы еще раз повторить ради вида «подвиг» родительского ризокормуса. Клоны медленно расширяют свои границы, но быстро пере- гружаются клонистами, стареют, и синорганизмы живут, как правило, не более 30 лет. Не правда ли, любопытно смотреть на все эти таинства со стороны и где-то восхищаться хитросплетениями растительной жизни организмов и синорганизмов?
ОСКОВЫ БИОМОРФОЛОГИИ СЕМЕНКЫХ РАСТЕНИИ 40, Рис. 104. Биограф РТР (Алтай, Бащелакский, Семинский хребты, сырые субальпийские луга)). Пример полурозеточных гемисимподиальных компактно-ризомных ретардационно-конди- видуальных травянистых многолетников. Образуется много придаточных корней, которые способствуют отмиранию главного корня. Впервые особи цветут на 6 — 12 году жизни. Некроз корневищ снизу доходит до зон кущения, и тогда родительские растения парти- кулирую т с образованием особей замедленного вегетативного размножения. Синорганиз- мы живут около 40 лет.
Глава 2. Концепция и иоритетов азвития в о ганизации систем Рис. 105. Биографический портрет Ranunculus altaicus Laxm. [сем. Ranunculaceae (Алтай, Ивановский, Бащелакский хребты, луговины около вечных снегов)]. ~ðnìåð полурозеточных гемисимподиальных компактно-ризомных ретардационно-конди- видуальных травянистых многолетников. Особи семенного происхождения образуют мно- ~0 придаточных корней. Рано отмирает главный корень. Впервые растения цветут на 5— >0 году жизни. Кустятся медленно. Миниморезиды живут 4 — 8 лет. Вегетативное размно- �еННе замедленное. Синорганизмы живут около 30 — 40 лет.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ
~дава 2. Концеиция и иоритетов азвития в о ганизации систем В, Рис. 108. Биографический портрет Veronica pinnata L. [сем. Scrophulariaceae (Алтай, Курайский хребет, остепненный южный склон в окрестностях села Чибит)]. ~1ример безрозеточных эусимподиальных компактно-ризомных кустистых ретардационно- кондивидуальных травянистых многолетников. Впервые цветут на 5 — 12 год. Минимо- Резидные приросты корневища живут 5 — 7 лет. Вегетативное размножение замедленное. Синонтогенезы продолжаются до 40 — 50 лет.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ec- i11a М.
~.цава 2. Концеиция л иоритетов развития в организации систем СК Рис. 110. Биографический портрет Erigeron leioreades М. Рор. [сем. Asteraceae (Алтай, Курайский хребет, субальпийские луга, щебнистые осыпи)]. Пример полурозеточных гемисимподиальных компактно-ризомных (1 биоморфотип) ре- тардационно-кондивидуальных травянистых многолетников. При произрастании на осы- пях образуются вынужденно-удлиненные корневища (11 биоморфотип). Резидный годич- ный прирост корневищ живет 4 — 6 лет. Первое цветение происходит на 6 — 12 году. вегетативное размножение чаще всего замедленное. Продолжительность синонтогенезов около 30 лет.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 111. Биографический портрет Valeriana dubia Bunge [сем. Ча1епапасеае (Алтай, разреженные лиственничные леса на Чергинском хребте)]. Пример полурозеточных гемисимподиально-разрастающихся несовершенно интенсивно- клонирующих травянистых многолетников. Впервые цветет на 2 — 5 году жизни. Образует очень компактные ризомы, годичные приросты которых отмирают уже на втором году их жизни. Некротическая партикуляция приводит к интенсивному клонообразованию, но такое вегетативное размножение не приводит к завоеванию новых территорий, а клон, несмотря на энергичную смену клонистов, прочно удерживает свой участок до конца жизни всего синорганизма (около 20 — 30 лет).
~дава 2. Концеиция и ио итетов азвития в организации систем м4,6, а,21, 7 Рис. 112. Биографический портрет Ligularia glauca (L.) О. Hoffm. [сем. Asteraceae (Алтай, Катунский хребет, луга в долине реки Катунь)). ~ример полурозеточных гемисимподиально-разрастающихся компактно-ризомных несо- вершенно интенсивно-клонирующих травянистых многолетников. Главный корень отми- рает уже на втором году жизни, утрату которого восполняют придаточные корни. Впервые пветет на 3 — 7 год жизни. Унирепродуктивные ди-, плейофазные побеги после плодо- ношения отмирают почти полностью вместе с придаточными корнями. Сохраняют жиз- неспособностb только пазушные розеточные вегетативные побеги-потомки, уже успевшие развить на себе свою придаточную корневую систему. Так осуществляется довольно ин- ТеН«»0 вегетативное размножение, которое все же несовершенно, поскольку клонисты синорганизма остаются всю жизнь на одном месте, перегружают клон и в целом синон- тогенез продолжается около 25 — 30 лет.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 113. Биографический портрет Prunella vulgaris L. [сем. Lamiaceae (Мос- ковская область, Подольский район, опушка смешанного леса)]. Пример безрозеточных столонообразующих интенсивно-клонирующих (интенсивно веге- тативно размножающихся) травянистых многолетников. Главный корень отмирает на 1— 2-ом году жизни. Цветение начинается со 2 — 5 года, чаще 2 — 3. Зимуют обычно верхушки приземных столонов, состоящие из коротких, но, как правило, не розеточных мезобластов; из их пазушных почек развиваются новые столоны, а перезимовавшая вер- хушка дает удлиненный восходящий цветоносный годичный прирост дифазного униреп- родуктивного побега. Ежегодное образование довольно длинных приземных столонов ведет к быстрому завоеванию окружающего пространства структурными элементами синорга- низма — его клонистами.
~дава 2. Концелция и ио итетов развития в организации систем Рис. 114. Биографический портрет Ranunculus flammula L. [сем. Ranunculaceae (Московская область, Подольский район, берега мелких во- доемов, лесных луж и болот)]. Пример слаборозеткообразующих гемисимподиально-возобновляющихся интенсивно-кло- нирующих (интенсивно вегетативно размножающихся) травянистых многолетников. Главный корень отмирает обычно на первом году жизни, когда образуются придаточные корни. Виргинильное развитие сопровождается развитием розеточного вегетативного по- бега. Зимует только компактный ризом, из верхушечной почки которого весной отрастает удлиненный полегающе-восходящий цветоносный побег. Часть узлов этого побега прижи- ма~тся к сырой почве и дает придаточные корни, а почки начинают развертывать силлеп- тические побеги, часть из которых может быть слабыми розеточными. После плодоноше- ния удлиненный побег отмирает, а сохраняются только пазушные предрозеточные и ро- зеточные вегетативные побеги. Они зимуют, дальнейшее развитие и расширение клона продолжается сходным образом весной следующего и многих последующих лет существо- ~НН< синорганизма. При пересыхании луж синорганизмы могут погибать, и тогда успеш- но в благоприятное для прорастания время восстанавливают популяцию покоящиеся в почве семена.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ СК Рис. 115. Биографический портрет Viola altaica Кег.-Gawl. [сем. Violaceae (Восточный Казахстан, Алтай, Ивановский хребет, высокогорья, приснеж- ные лужайки, берега ручьев)]. Пример розеточных эумоноподиальных длиннокорневищных семиэкстенсивно-кондиви- дуальных травянистых многолетников. Главный корень функционирует несколько лет (3 — 5 лет). Придаточные корни образуются, но они малочисленны, поскольку тонкие кор- невища горизонтально растут в очень влажном субстрате, иногда почти по поверхности под слоем мохового покрова. Первое цветение происходит на 5 — 12 году жизни. Корневища рассеянно ветвятся, и вегетативное размножение наступает после отмирания самых ниж- них и старых зон ветвления. Годичные резидные приросты ризомов живут 6 — 10 лет. Семиэкстенсивным вегетативным размножением (дословно наполовину экстенсивным») можно назвать такое вегетативное размножение (как у фиалки алтайской), когда развива- ются клонисты, системы вегетативного разрастания которых энергично завоевывают но- вые большие территории, долго удерживая их за собой, однако не настолько долго, как это удается биоморфам с экстенсивным вегетативным размножением, свойственным, например, биоморфам длиннокорневищно-стержнекорневым, допустим, как у Lupinaster pentaphyllus Moench (см. следующий рисунок).
~дава 2. Концепция и ио umemoe азвития в о ганизации систем Рис. 116. Биографический портрет Lupinaster pentaphyllus Moench [сем. Fabaceae ~~тай К ай ур ский, Северо-Чуиский хребты, верхняя граница леса, LJ проникает и на субальпийские луга)]. Пример безрозеточных эусимподиальных длиннокорневи вищно-стержнекорневых экстен- сивно-кондивидуальных травянистых многолетников. Впервые цветет на 6 — 15 год жизни. Главный стержневой корень может сохранять большую а всего онтогенеза. Большинство побегов взрослых растений один год нарастают го изон- тально подземно, а на сле ий го б ни один год нарастают горизон- , а на следующий год образуют удлиненный вертикальный вегетативный или вегетативно-репродуктивный надземный акронекрозный (после заве шения в на по ег. разуя длинные ризомы, биоморфы успевают еще и надежно укорениться придаточными стержневыми корнями и — „ом, б бни что в е , — „оме того, о разуют еще корневые клу ни, что в целом обеспечивает синорганизмам этого вида долгое у гое удержание довольно больших территорий, завоеванных их подземными побегами. 41 206. 26 ж,156 206, 21,26 Рис. 117. Биоморфа Chamaenerion latifolium (L.) Th. Fries et Lange [сем. Onagraceae (Алтай, Курайский хребет, высокогорья, осыпи и отва- лы горных шахтных выработок в окрестностях поселка Акташ)]. Пример облигатно-корнеотпрысковых без- розеточных экстенсивно-кондивидуальных травянистых многолетников.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ О 86, Рис. 118. Био аттический гр ф еский портрет Lagotis integrifolia (Willd.) Schischk. [сем. Scrophulariaceae (Алтай, Семинский, Северо-Ч ский К еверо- уиский, атунский хребты, ~рые луга, берега ручьев, около снежников)]. ример полурозеточных эумоноподиальных длинноко неви дуальных травянисть к многолетников. ннокорневищных экстенсивно-кондиви- Виргинильное развитие идет медленно. На первом год о срединный лист. Далее п е а первом году обычно образуется всего один алее предрозеточное моноподиальное на астан в розеточное ежегодно б б е по егоо разование. Ве шки растание постепенно переходит ных побегов только пове рху длинных подземных горизонталь- лько у поверхности почвы переходят к развитию б побегов — розеточных брахиб . У ю рахиморфных частей безрозеточные цвето хи ластов. взрослых растений о и носы развиваются как паз ные инт р диночные слабо олиственные ные побеги а , а в целом побеговая система нарастает э онопо и уш интеркалярные или постинтеркаляр- множение экстенсивное ет эумоноподиально. Вегетативное раз- ются и долг . С ое, поскольку мощные го изонтальн говечные. инорганизм долго удерживает те ит р альные корневища быстро разраста- ются его клонисты. ивает территорию, на которой располза-
~дава 2. Концеиция и ио итетов азвития в о ганизации систем Рис. 119. Биографический портрет Viola canina L. [сем. Violaceae (Москов- ская область, Подольский район, смешанный елово-березовый лес)]. Пример ювенильно-розеточных матурно-безрозеточных эусимподиальных факультативно- корнеотпрысковых (при оголении или повреждении корней) травянистых многолетников. Впервые цветут на 2 — 5 год жизни. Стержневой главный корень сохраняют всю жизнь. Вегетативное размножение, если бывает, то оно несовершенное и заметно не продлевает �изнь факультативных синорганизмов.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РАСТЕНИИ Рис. 120. Биографический портрет Sibiraea laevigata (L.) Maxim. [сем. Rosaceae (Алтай, Теректинский хребет, долина реки Урсул с лугово-кустарниковой растительностью в окрестностях села Ело)]. Пример полурозеточных гемисимподиальных рыхлокустистых стержнекорневых дефектив- но-кондивидуальных (несовершенно вегетативно размножающихся) листопадных кустар- ников. Виргинильная стадия продолжается 10 — 25 лет. Полный жизненный цикл — около 60— 80 лет. При старении главный корень может отмирать полностью и приводить к некро- тической партикуляции всего рыхлого куста и распаду его на несколько частей — клони- стов, что свидетельствует о проявлении несовершенного вегетативного размножения.
Глава 2. Концелция npuo umemoe азвипьия в о ганизации cucme~ 201 Рис. 121. Биографический портрет Ajuga teptans L. [сем. Lamiaceae (Москов- ская область, участок елового леса в 20 м от опушки)]. Пример полурозеточных гемисимподиальных столонообразующих интенсивно-клднирую- ших травянистых многолетников. Главный корень отмирает рано, на 1 — 2-ом году жизни. цветение начинается на 2 — 5 год. Зимуют укореняющиеся розеточные верхушки призем- ных столонов. Сами столоны не зимуют. Перезимовавший розеточный вегетативный побег дает удлиненный вертикальный иветоносный годичный прирост дифазного унирепродук- тивного побега. Из брахиморфной части этого побега отрастают короткие и длинные столоны, которые обеспечивают синорганизму быстрое завоевание пространства, но их недолговечность не обеспечивает условий для удержания захваченной территории на продолжительное время. В условиях сильного затенения А. reptans склонно проявлять чудеса пластичности (см. объяснения в тексте).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 122. Развитие однобокой кроны в сторону опушки у биоморф-деревьев 1 — Picea abies (L.) Karst (сем. Pinaceae) и II — Quercus гоЬиг L. (сем. Fagaceae), поскольку эти деревья испытывают сильнейшее затенение другой своей стороны — от соседних деревьев густого смешанного леса. Дополни- тельные обозначения: Q. г. — Quercus гоЬиг, В. р. — Betula pendula Rath., Р. а. — Picea abies, С. а. — Corylus avellana L., P. s. — Pinus sylvestris L. 2.4. Иерархия приоритетов развития биоморф семенных растений Выше было показано, что целей развития у систем много, посколь- ку много признаков составляет их организацию, а приоритетов развития немного, причем по степени их значимости есть более и менее важные для организации, но на то они и приоритеты, чтобы заметно влиять на всю организацию систем и неживых, и живых. Однако лимит для кур- сирования далее между общими вопросами у нас заканчивается, пора глубже проникать в организацию биоморф семенных растений и, безус- ловно, в их габитусы и формы роста, о которых автор всегда помнит и делает все, чтобы «блюдечко с голубой каемочкой» в конце концов появилось с добротными результатами. Главное, на что мы особо обратили внимание выше, системы, при- чем любые, так или иначе организованы, и у биоморф семенных рас- тений есть стройная система «управления — исполнения» (т. 1, гл. 4) со своей многоцелевой идиоэнергией (ресурсом возможностей) и приори- тетами в мультивариантном развитии. В предыдущем материале было доказано, что приоритетность в развитии любых систем, и, естественно,
Глава 2. Концепция и ио umemoe азвшпия в организации сиспьем биоморф, существует и что одни приоритеты пронизывают всю органи- зацию на протяжении всего морфогенеза (шире — системогенеза), а другие — распространяются только на отдельные стадии существования систем. Биоморфы, как и другие живые системы, функционируют рит- мически, отвечая тем самым на ритмичность процессов, происходящих в окружающей их природе, поэтому приоритеты развития каких-либо систем могут изменяться и сменяться неоднократно на протяжении различных отрезков времени. У нас есть основания для следующего аксиоматического обобщения: одни приоритеты в развитии организации системы действуют все время ее существования (весь системогенез, уже — морфогенез), а другие — только на отрезке (стадии) системогенеза, что приводит в последнем случае к смене npuopumemoe с полной утратой прежнего статуса или с последующим восстановлением его — это правило 5-е закона приоритетности (рис. 123 — 147). Приоритетность — многоуровневое явление. Живые системы, био- морфы, в частности, в процессе своего длительного исторического раз- вития выработали несколько «этажей» приоритетов, что позволяет нам построить несколько иерархических классификаций по субординатному принципу под разными углами обзора, причем еще и с разных высот абстракции, что в итоге, безусловно, выводит нас к построению целой «пирамиды» абстракций, т.е. несколько субординий, в свою очередь, мы расположим в единой суперординатной классификации. Методология и методика указанных приемов классифицирования уже разработана (см. т. 1, гл. 1, 2). Чтобы строить субординатные (субординии) и суперорди- натные (суперординии) классификации, надо владеть критериями тако- го классифицирования. Приоритеты просматриваются там, где есть пер- венство, преимущество, превосходное значение... одних видов деятель- ности системы перед другими, поэтому, естественно, критерий для су- бординии приоритетов заложен в смысловом содержании слова «при- оритет». Об этом мы уже писали: приоритет развития системы — это, в первую очередь, выбор преимущественной цели развития. Высшим крите- рием для порядка расположения классов npuopumemoe развития биоморф семенных растений в соответствующих субординатных классификациях является степень значимости приоритетов в организационной деятельно- сти этих биоморф, при этом можно учитывать все, что позволяет как- других приоритетов того же сравнительного ряда. Важно при классифи- цировании npuopumemoe не упустить и такой факт, что часть npuopume- тов подчеркивают первенство какой-то деятельности по времени прояв- ления. Пожалуй, далеко ходить не будем, чтобы отметить, что первая субординия (субординатная классификация) у нас на виду из двух су- бординат (классов приоритетов), на нижней из которых располагаются приоритеты, выделенные по времени их проявления в системах, а вер- хнюю субординату занимают приоритеты, отражающие преимущество какой-либо деятельности по значимости, в сравнении с другой деятель- ностью. Безусловно, в таком подходе всегда есть элемент условности, но не будем забывать, что объект классифицирования у нас очень «сколь- зкий», да к тому же сама процедура любого классифицирования не
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ лишена условности, поскольку всякая логическая операция — это плод сознательной деятельности субъекта — нас с Вами. Наша задача достиг- нуть определенного порядка в пропуске классов приоритетов на уровни субординатных классификаций, а затем, считая число этих уровней (су- бординат) в каждой субординии, запускать их, как по проходным «би- летам» (числу субординат), на соответствующие ступени «пирамиды» абстракций (суперординатной классификации). Мне доводится строить не первую «пирамиду» абстракций (см. т. 1) и предстоит построить еще немало суперординатных классификаций различных явлений, самым наглядным графическим выражением кото- рых может быть только пирамида. Сейчас можно уже с уверенностью констатировать, что логическое «пирамидостроительство», впервые раз- работанное автором (Нухимовский, 1984, 1987, 1997) как методологи- ческий и методический прием теоретизирования, работает безотказно, а сам исполнитель суперординатного классифицирования в разных ситуациях испытывает невероятное ускорение творческого процесса: замечаешь, как усиливается зоркость в свободомыслии, как начинают расти «крылья» и ты, наконец, ощущаешь состояние полета над и в «пирамиде» абстракций, мысленно прикасаясь к предмету исследований с разных сторон и осознавая его под разным увеличением. Впечатление особенное... Всякому, кто подойдет к необходимости крупномасштабно- го теоретизирования, есть смысл испытать возможность этого метода и силу своего логического мышления. Рисунок 148 передает разнообразие приоритетов в организации био- морф семенных растений с разных высот абстракции и под разными углами обзора, а это и есть та цель, которой мы добиваемся построе- нием таких классификаций. С наибольшей высоты обобщений мы рассматриваем несколько субординатных классификаций, построенных, естественно, по разному. Пожалуй, в качестве первой субординатной классификации уместно поставить ту двухуровневую классификацию, которую мы только что привели выше. Далее рассмотрим на той же ступени (суперординате) «пирамиды» абстракций несколько других двухуровневых субординий приоритетов развития биоморф семенных растений под разными углами обзора. В качестве второй мы расположили на верхней суперординате субординатную классификацию следующих приоритетов: нижний класс — приоритеты тактического развития, верхний — приоритеты стратеги- ческого развития. То, что все стратегическое в организации биоморф приоритетно по отношению ко всему остальному, не вызывает сомне- ний, поскольку оно — основное для живого вообще. Что касается так- тического, то только некоторые из таких видов деятельности биоморф могут быть приоритетами (т. 1, гл. 3.4). Третьей субординией приори- тетов на той же высшей ступени абстракции будет классификация из двух субординат: нижней — приоритеты жизнесохранения частного (одного конкретного), верхней — приоритеты жизнесохранения вообще (остального общего). Четвертую субординию приоритетов на той же высоте абстракции составляют снизу — приоритеты своей (личной) жизни, а сверху — приоритеты не-своей (неличной) жизни. Близка по
~дава 2. Концепция и ио umemoe азвшпия в организации сиоием значению пятая субординия: нижняя субордината — приоритеты веге- тативного развития, верхняя — приоритеты репродуктивного развития. арестую субординатную классификацию можно показать еще под одним углом обзора с той же высоты абстракции: снизу — приоритеты разви- тия стадийные, выше — приоритеты развития целостные. Спустимся на одну ступень пирамиды абстракций и рассмотрим, какие субординатные классификации приоритетов развития биоморф семенных растений мы там поместили. Седьмую субординию построили из трех классов приоритетов: жизнесохранения персонального (автоби- офилии), далее обоюдного (амфобиофилии) и коллегиального (ксеноби- офилии). Здесь, похоже, все ясно, а для полноты ощущений желательно познакомиться еще и с общей теорией жизнесохранения (т. 1, гл. 3). На этой же высоте абстракций располагается восьмая субординия приори- тетов из трех классов (снизу вверх): приоритеты роста и развития веге- тативного тела (вегетативного разрастания), далее приоритеты вегета- тивного (шире — бесполого) размножения и, наконец, репродуктивного размножения. Перейдем еще на одну ступень пониже в рассматривае- мой «пирамиде». Здесь мы поместили две субординатные классифика- ции (9-ю и 10-ю). В девятой субординии нижнюю субординату мы отдали приоритетам нарастания (т. 1, гл. 4.4.), далее ветвления (включая кущение), затем вегетативного (шире — бесполого) размножения и, наконец, репродуктивного размножения. В десятой субординии в каче- стве классов приоритетов последовательно, начиная снизу, представле- ны 4 основных способа фенетического освоения пространства в круго- вороте ресурсов: приоритеты потребления, производства, резервирова- ния (депонирования) и рассеивания. На ступеньке ниже нашей «пирамиды», где несколько уменьшилась высота абстракций, расположилась одиннадцатая субординатная класси- фикация из пяти классов (субординат) приоритетов: нижняя субордината — приоритеты личного жизнесохранения, выше — приоритеты обоюдно- го жизнесохранения, еще выше — приоритеты коллегиального жизнесох- ранения, сначала первого (моноксенобиофилии), затем второго (диксено- биофилии) и третьего (триксенобиофилии) (подробнее см. т. 1, гл. 3). Двенадцатая субординия приоритетов развития биоморф сделана с еще меньшей высоты абстракций, поэтому в ней больше субординат — семь, начиная с нижней: приоритеты нарастания, ветвления, вегетативного (шире — бесполого) размножения, а далее приоритеты кратности репро- дуктивного размножения — унирепродуктивного, бирепродуктивного, паулирепродуктивного и, наконец, мультирепродуктивного. На самой нижней ступени «пирамиды» абстракций мы расположи- ли тринадцатую субординию приоритетов из 11-ти субординат, начиная с нижней: приоритеты водно-минерального питания, далее — дыхания, фотосинтеза, далее — нарастания, ветвления, резервирования ресурсов, вегетативного (шире — бесполого) размножения, затем — унирепродук- тивного, бирепродуктивного, паулирепродуктивного и сверху — мульти- репродуктивного размножения. На самой верхней ступени «пирамиды» абстракций, рассматривае- мой здесь, мы приводим еще одну (последнюю) четырнадцатую субор-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ динатную двухуровневую классификацию, давая ее как бы сверх плана, совмещая отношение приоритетов фенетической и генетической дея- тельности, последняя из которых только внешне оставалась во всех 13- ти предыдущих классификациях «за кадром». У генетического тела (ге- носомы) в организации биоморф особое положение; оно управляет фенетической деятельностью и всеми ее приоритетами и, следователь- но, в любой субординатной классификации приоритетов должно зани- мать по отношению к ней и всем ее приоритетам более высокое место. Такая уж судьба у генов и геносом: оставаться внешне незаметными, но таинственно управлять организацией не только отдельных биоморф, но и организацией всей живой природы, взаимодействуя через их фенети- ческие организации (феносомы) с окружающим внешним миром. Пожалуй, на четырнадцатой («сверхплановой») субординии мы и остановимся. Наша задача на сегодня состоит не столько в том, чтобы добиться идеального совершенства в субординатных и суперординатной классификациях приоритетов развития организации биоморф семенных растений. Все это будет. Наука не остановится никогда, но здесь и сейчас важно было однозначно заявить, что явление приоритетности в организации биоморф (причем не только растений, но и животных), безусловно, существует: оно есть в Природе, независимо от нашего сознания, а теперь оно осознано и есть еще и в науке о биоморфах семенных растений. Развитие концепции приоритетности в биоморфо- логии, а шире — биологии, ускорит прогресс в понимании организации биосистем и биоморф, в частности, в чем нам еще не раз суждено убедиться на страницах этой книги. Рис. 123 — 147. Естественная сменяемость стадийных приоритетов как индикаторных признаков возрастных стадий (и состояний) в жиз- неразвитии организмов или синорганизмов на фоне несменяемости целостных (циклических) приоритетов: размножение и жизнесохране- ние вообще. Стадийные приоритеты более или менее повторяются у разных видов, поэтому мы не будем давать им и особенностям их сменяемости подробных описаний. Происходящие события выразим в блоке, приво- димых здесь, рисунков, на которых, через соответствующие условные обозначения, отразим по мере возможности, те индикаторные призна- ки, которые более всего являются стадийными приоритетами. Напри- мер, в качестве стадийного приоритета проростка (А) Melilotus albus Medik. (рис. 123) выделено условное обозначение признака — семядоль- ный ризопротерокормус (Оlк), т. е. это побег с живыми семядолями и неразвернувшейся зародышевой почкой вместе с корневой системой. У того же вида стадийный приоритет на имматурной стадии жизнеразви- тия его особей выражен через признак — силлептическое ветвление главного побега как дикормуса (2) или вместе с корнями как ризоди- кормуса (2к) и т. д.
Глава 2. Концелция и ио umemoe азвипьия в организации сиаием Рис. 123. Биографический портрет Melilotus albus Medik. [сем. Fabaceae (Мос- ковская область, пустыри, обочины дорог, луга)]. Однолетники (биоморфотип). Др — дополнительный рисунок из работы С. И. Дмитриевой и др. (1974). Рис. 124. Биографический портрет Urtica urens L. [сем. Urticaceae (Москва, скверы и парки, пустыри)]. Однолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СГЕНИЙ ,21, Рис. 125. Биографический портрет Silybum тапапит (L.) Gaertn. [сем. Asteraceae (Москва, Главный ботанический сад, питомник лекарственных растений) ]. Двулетники. А Рис. 126. Биографический портрет Digitalis lanata Ehrh. [сем. Scrophulariaceae (Москва, Главный ботанический сад, питомник лекарственных растений)]. Двулетники.
Рис. 127. Биографический портрет Viola dissecta Ledeb. [сем. Violaceae (Алтай, долина реки Чуи, Чуйский тракт, севернее села Чибит)]. Травомноголетники. Рис. 128. Биографический портрет Corydalis pauciflora (Steph.) Pers. [сем. Fumariaceae (Алтай, Курайский хребет, сырые высокогорные луга)]. Травом- ноголетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА С'ГЕНИЙ 546,67 Рис. 129 29. Биографический портрет Echino s s ha М~ега~~м (Ммкв~, Главный б растений) ]. Травомноголетники. а, лавный ботанический сад, питомник л лекарственных
Глава 2. Концелция и ио umemoe азвипьия в о ганизации сиспьем 211 Рис. 130. Биографический портрет Viola hirta L. [сем. Violaceae (Алтай, Семинский хребет, смешанный лес, горный склон к долине реки CeMa)]. gp — дополнительный рисунок из работы В. Тролля (Troll, 1964, с. 514). Травомноголет- ники.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РАСТЕНИЙ аг3,18а,27 СК Рис. 131. Биографический портрет Saxifraga hieracifolia Waldst. et Kit. [сем. Saxifragaceae (Алтай, Курайский хребет, высокогорные сырые тундры)]. Травомноголетники.
Глава 2. Концелция лриоритетов развития в организации систем 21 Рис. 132. Биографический портрет Echinacea ригригеа (L.) Moench [сем. Aseeraceae (Москва, Главный ботанический сад, питомник лекарственных растений)]. Травомноголетники.
ПП 74 546,66 дс СК Рис. 133. Биографический портрет Rumex acetosa L. [сем. Polygonaceae (Ал- тай, Северо-Ческий, Курайский хребты, на высокогорных лугах и ниже по горно-луговым склонам)]. Травомноголетники. ги4, 19а, 21,27 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ 1,~, 1 аг3, 18а,27,66
21 ~дава 2. Концепция и ио umemoe азвшпия в о ганизации cucme~ Рис. 134. Биографический портрет Trollius asiaticus L. [сем. Ranunculaceae (Алтай, Семинский, Курайский хребты, горные луга)]. Травомноголетники. Рис. 135. Биографический портрет Myosotis asiatica (Vestergren) Schischk. et Serg. [сем. Вогаа1пасеае (Восточный Казахстан, Алтай, Ивановский хребет, остепненно-луговые южные склоны, редколесья)]. Травомноголетники.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИИ (( ( 1 1 X Ъ СК E / 546,67 Рис. 136. Биографический портрет Ranunculus polyanthemos L. [сем. Ranunculaceae (Восточный Казахстан, Алтай, долина реки Белая Уба)]. Травомноголетники.
~дава 2. Концелция npuo umemoe азвипьия в о ганизации сиаием Рис. 137. Биографический портрет Tephroseris turczaninovii (DC.) Но1иЬ. [сем. Asteraceae (Алтай, Курайский хребет, высокогорья, щебнистые и дриадовые тундры, сырые луга)]. Травомноголетники. Рис. 138. Биографический портрет Cardamine bellidifolia L. [сем. Brassicaceae (Алтай, Курайский хребет, высокогорья, каменистые россыпи, берега ручьев)]. Травомноголетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 139. Биографический портрет Erygeron eriocalix (Ledeb.) Vierh. [сем. Asteraceae (Алтай, Курайский хребет, высокогорные полузадерненные скло- ны и галечники)]. Травомноголетники.
~дава 2. Концелция npuopumemoe развития в организации систем 21 Рис. 140. Биографический портрет Ptarmica impatiens (L.) DC. [сем. А~~егасеае (Восточный Казахстан, Алтай, долина реки Белая Уба. горные склоны с лугами и редколесьями)]. Травомноголетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 141. Биографический портрет Claytonia joanneana Schult. [сем. Portulacaceae (Восточный Казахстан, Алтай, Ивановский хребет, высокого- рья, моховые мочажины, луга по берегам ручьев)]. Травомноголетники.
Рис. 142. Биографический портрет Ruta graveolens L. [сем. Rutaceae (Москва, Главный ботанический сад, питомник лекарственных растений)]. Полукус- тарники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ Рис. 143. Биографический портрет Phagnolon rupestre (L.) DC. [сем. 1~егасеае (Израиль, окрестности Тель-Авива, пологие склоны каменисто-песчаных холмов с разреженной полукустарниково-кустарниковой растительностью)]. Полукустарники.
~лава 2. Концелция n uopumemoe развшиия в о ганизации сиоием 223 ПП Рис. 144. Б иографический портрет Lavandula coronopifolia Poiret [сем. Lamiaceae (Израиль, горы Эйлата, долина с сухим руслом временного ручья,б V)]. Полукустарники.
OCHOBhI БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИЙ Рис. 145. Биографический портрет Rhamnus alaternus L. [сем. Rhamnaceae (Израиль, невысокие горы вдоль дороги Хадера-Хайфа, поросшие древесно- кустарниковой средиземноморской растительностью)]. Кустарники.
~дава 2. Концепция и ио umemoe азвипьия в организации сиоием Рис. 146. Биографический портрет Dalbergia sissoo Roxb. [сем. Fabaceae (Израиль, скверы и парки Тель-Авива, апрель)]. Деревья.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 147. Биографический портрет Асег negundo L. [сем. Асегасеае (Москва, скверы и парки)]. Деревья.
~дава 2. Концепция и ио umemoe азвшпия в организации сиоием Рис. 148'. Графическая суперординатная модель («пирамида» абстракций) приоритетов прогрессивной деятельности в организации биоморф семенных растений. Приоритеты (далее в условных обозначениях это слово подразумевается, но более не повто- ряется): 1 — выделенные по времени их проявления в системах; 2 — отражающие преиму- щество какой-либо деятельности по значимости в сравнении с друтой деятельностью; 3— тактического развития; 4 — стратегического развития; 5 — жизнесохранения частного (од- ного конкретного); 6 — жизнесохранения вообще (остального общего); 7 — своей (личной) жизни; 8 — не-своей (неличной) жизни; 9 — вегетативного развития; 10 — репродуктивного развития; 11 — стадийного развития; 12 — целостного развития; 13 — жизнесохранения персонального (автобиофилии); 14 — жизнесохранения обоюдного (амфобиофилии); 15— жизнесохранения коллегиального (ксенобиофилии); 16 — роста и развития вегетативного тела (вегетативного разрастания); 17 — вегетативного (шире — бесполого) размножения; 18 — репродуктивного размножения; 19 — нарастания; 20 — ветвления (включая кущение); 21 — потребления; 22 — производства; 23 — резервирования; 24 — рассеивания; 25 — первого коллегиального жизнесохранения (моноксенобиофилии); 26 — второго коллегиального жизнесохранения (диксенобиофилии); 27 — третьего коллегиального жизнесохранения (триксенобиофилии); 28 — унирепродуктивного размножения; 29 — бирепродуктивного размножения; 30 — паулирепродуктивного размножения; 31 — мультирепродуктивного размножения; 32 — водно-минерального питания; 33 — дыхания; 34 — фотосинтеза; 35— фенетической деятельности; 36 — генетической деятельности.
228 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ 2.5. Стремление к размножению, экспрессивный альтруизм палеобиоморф, торжество компромиссов в эволюции растительных биоморф и ее главные приоритеты Коль мы сказали «а», то вроде бы негоже останавливаться, пробле- ма за проблемой так и взывают использовать для их решения принцип приоритетности. Было бы упущением, если бы, владея этим принципом, автор не коснулся проблемы эволюции. В этой книге есть специальные главы, посвященные проблеме эволюции (гл. 8, 9), планируется, для анализа этого явления еще и отдельный том в нашей монографии, но проблема эволюции настолько многогранна и прямо-таки «расплески- вается» далеко за пределы своих границ, что в начале книги мы пого- ворим о наиболее общем в отношении эволюции. Что касается самого понятия и его места в мироздании, — все подробности выносим в главы 8, 9, но здесь определимся, что под эволюцией, в самом общем виде, автор понимает преемственную, более или менее историческую, беско- нечную всепроникающую активную трансформацию (процесс и резуль- тат) дискретных систем и природы вообще (Природы). Понимая так, как мы определили, эволюцию, можно обнаружить ее везде; она как и материя, которая всегда в движении, всюдна в Природе. Всюдное явление надо рассматривать не только с узких, но и, что зна- чительно важнее, поскольку ведет к пониманию принципов,с широких позиций. Та широта, с которой мы решили «проехаться» рядом с эво- люцией по мирозданию, не может не настораживать. Что движет чело- веком, замахнувшимся, казалось бы, на самое вечное и непостижимое? Отвечу однозначно: только одно обстоятельство — неиссякаемое, пря- мо-таки задорное и ювенильное любопытство. Все, что будет сказано в этом небольшом разделе, — это гипотеза, поэтому отношение к ней должно быть не как к теории, а всего лишь как к инструменту теоре- тического исследования, как к строительным лесам, которые возводятся перед зданием и сносятся, когда здание готово. Однако во всякой ги- потезе есть право на большую, чем обычно, свободу мысли, поэтому автор воспользуется этой свободой в такой мере, чтобы удовлетворить свое и раззадорить Ваше, уважаемый читатель, любопытство. Дело в том, что ранее (т. 1) автору удалось подсмотреть в эволюции телесных систем принцип их существования в пространстве и во вре- мени, т.е., по сути, во вселенском потоке (Природе) — принцип эволю- ционных маятников, согласно которому существование любых тел со- провождается эволюцией с разными результатами, что и предопредели- ло универсальный абстрактный образ любых тел — это «маятники», организация которых (их механизм) во взаимодействии с окружающей средой обеспечивает колебания (изменения) «маятниковых» тел в опре- деленном диапазоне возможностей. Эволюционный маятник — это тело или система тел, биоморф и их видов, в частности, которые, независимо
от специфики их организации, чем-то напоминают маятники по глу- бинному механизму их движений-развитий. Вряд ли кто-либо будет возражать, что получить «ключ» к тайнам такого загадочного для всех и вся явления как эволюция, безусловно, большая удача. А что если эта удача у нас уже в руках, т.е. нам удалось отыскать и заполучить «ключ» к эволюционным «вратам», а раз так, то и секретам, которые скрыты за ними. Как бы там ни было, но, вероятно, мы уже не «топчемся» где-то у входа, а, как нам представляется, про- никли в эволюционное, образно, сложно разветвленное сооружение Природы. Есть обстоятельства, усложняющие поход по эволюционным лабиринтам. Итак, мы проникли в эволюционное сооружение Природы, где везде, куда бы мы ни обратили свой взор, так или иначе колеблются эволюционные маятники (образы различных природных тел). Но самое любопытное обнаружилось в другом: оказывается, в эволюционном со- оружении Природы есть еще множество других ворот и дверей, и две- рочек, к которым надо подбирать еще и другие «ключи» и «ключики». Сколько искателей и раскрывателей эволюционных истин где-то до сих пор блуждают в эволюционных лабиринтах, а немало и тех, кто оставил свои, охваченные научным поиском, души где-то там в пути ... и тех, KTO в память о них продолжает их дело и пытается, с большим или меньшим энтузиазмом, пройти все дальше по лабиринтам эволюции, где-то, похоже, забывая, что эволюция бесконечна... Принцип эволюционных маятников, который оказался для нас первым ключом в эволюционные лабиринты; этот принцип универсаль- ный, а раз так, то еще и приоритетный среди всех других видов эволю- ционной деятельности. «Раскачавшись» должным образом (так образно раскрываются «коды замков» к «вратам» в эволюционные лабиринты), и будучи одними из тех, кто также является эволюционными маятни- ками, мы проникаем внутрь явления эволюции телесных систем и во- обще, и в частности; далее будем, периодически сами «раскачиваясь» соответствующим образом (эволюционируя по-своему), еще и еще по- сещать ее другие мудреные лабиринты, не забывая обратных путей, вплоть до завершения последней страницы нашей монографии. При всей таинственности и увлекательности происходящих вокруг эволюци- онных событий у меня нет желания заблудиться и затеряться в беско- нечости эволюционных лабиринтов, но есть непоколебимое желание «докопаться» до сути, затем вернуться ко всем и поделиться со всеми результатами своих многолетних «путешествий» в пространстве эволю- ции и в эволюции пространства, в первую очередь, растительного. Эволюционные маятники свободно «плывут» в пространстве и во времени, функционируя по единственно возможному для них способу существования на основе универсального и приоритетного для всех их принципа — они колеблются (изменяются) по схеме «так-сяк и туда- сюда», слово «так-сяк» в этой схеме-словосочетании означает то же самое, что и «по-разному, на разный манер» (Розенталь«Теленкова, 1985, с. 600). Схему функционирования эволюционных маятников, при- веденную выше, можно записать и иначе: «так-сяк * туда-сюда», звез- дочка в такой схеме — это символ (даже более точный, чем союз «и»)
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РАСТЕНИЙ возможной разнонаправленности эволюционных процессов в телесных системах, абстрактными образами которых являются эволюционные маятники. Эволюционные маятники «плывут» и всегда смещаются в пространстве (меняется угол наклона оси маятника, воздействие вне- шних сил и т.п.), поэтому известные для них движения, хотя и похожи, но они всегда не повторяют абсолютно друг друга. В любой момент принцип эволюционного маятника действует, поскольку он универса- лен и приоритетен и подкреплен преднаследственно-наследственной основой конкретных тел (их идиоэнергией). Эволюционные маятники как образы конкретных тел могут прекращать свое прежнее качествен- ное состояние и превращаться в другие эволюционные маятники (и конкретные тела) по разным причинам (удар молнии, попадание в пасть крокодилу, смерть еще в зародыше и т.п.), но это все отвечает принципу (функционирования) таких «маятников», имеющих определенные диа- пазоны изменчивости и способных под влиянием некоторых факторов субмутировать (неживые тела) или мутировать (живые тела), трансфор- мируясь в определенных условиях в другие тела (и их образы — эволю- ционные маятники), часть из которых может быть организационно нового типа. Итак, принцип эволюционных маятников, например, биоморф се- менных растений, работает по схеме «так-сяк * туда-сюда» на опреде- ленной генетической основе и как результат взаимодействия ее через фенетическую организацию с окружающим внешним миром. Шире, всякое движение (изменение) тел как эволюционных маятников — это атрибутивный принцип эволюции Природы. Ранее (гл. 1) было показано, что Жизнь — результат естественного развития неживой материи, дизъюнктивно распыленной в простран- стве-времени в форме движущихся тел и систем тел, которые образно все так же являются эволюционными маятниками, развитие которых по схеме «так-сяк * туда-сюда» раньше или позже должно произвести новые типы тел — новые живые тела в образах новых эволюционных маятни- ков. Образная схема работы (функционирования) эволюционно-маят- никовых «механизмов» конкретных тел, выраженная простеньким сло- восочетанием «так-сяк * туда-сюда», проявляет себя по-разному всюдно, но на основе преемственности, незримо присутствующей на преднас- ледственно-наследственном уровне организации в каждом теле. Эволю- ционным маятникам схема «так-сяк * туда-сюда» предоставляет возмож- ность свободно выбирать, в определенном диапазоне возможностей (!), форму и способ существования конкретных носителей этих образов, отвечающих окружающим условиям существования. 2.5.1. Экспрессия альтруизма в колыбели жизни на Земле и стремление к компромиссам своего и не-своего в растительном мире на примере организации простых биоморф (особей) Итак, вооружившись представлениями об эволюционных маятни- ках и воспринимая Жизнь как фрагмент вселенского потока, в котором
~дава 2. Концелция п ио umemoe pa~eumu~ в о ганизации сиаием 231 она — жизнь (живая природа) на Земле — всего лишь один из таких эволюционных маятников, функционирующих по определенной схеме преемственности: «так-сяк * туда-сюда», попробуем, с весьма общих позиций, не вдаваясь в детали, ответить на несколько коренных вопро- сов эволюции: есть ли универсальный и приоритетный принцип в эво- люции вообще, что особенного в эволюции организации биоморф во- обще и какие направления эволюции выбирала и выбирает жизнь на Земле. Что касается ответа на первый вопрос, то он уже состоялся выше: существование любых (неживых и живых) телесных систем осуществля- ется во вселенском потоке — динамичном пространстве-времени При- роды по универсальному эволюционно-маятниковому принципу и об- разной схеме преемственности («так-сяк * туда-сюда»). Теперь обратим- ся к организации жизненных форм, сконцентрировав, чтобы «не распы- ляться», основное внимание на простых биоморфах, т.е. организмах (особях). С чего начиналась жизнь на Земле? Здесь возможны два oc- HoBHbIx варианта: или она занесена извне и то, что мы имеем, распро- странено в своих главных сочетаниях и в некоторых других уголках Вселенной или то, что на Земле, — это продукт чисто земной естествен- ной эволюции. Оставим этот вопрос без ответа. На сегодня он не имеет однозначного решения, а обратимся к более приземленному вопросу что составляет главное в организации биосистем, на примере организ- мов, и как это главное ведет себя в эволюции организации биосистем, т.е. в нашем случае организмов (простых биоморф)? Попытаемся раз- вернуть проблему соотношения вегетативного (относительно своего) и репродуктивного (относительно не-своего) под эволюционным углом обзора. Эволюция жизни на Земле сразу же началась как эволюция теле- сных систем с двумя приоритетами развития: относительно личным (для себя), или вегетативным, развитием и относительно неличным (для других), или переходно-вегетативным (эксвегетативным) развитием, на- целенным на размножение. Иными словами, протеробионты (первые живые организмы) имели одно тело, которое осуществляло одновремен- но две цели: и себя накормить, и обеспечить собой же продолжение рода. Задачи эти решались очень быстро (за минуты, а может быть, даже и за секунды), поскольку изначально явление жизнь на Земле наделено стратегией своего сохранения, приоритетной стороной которого являет- ся стремление к размножению (любому как вегетативному, так и реп- родуктивному, или еще какому-нибудь). Особое, сейчас еще во многом таинственное, место в эволюции жизни занимают вирусы — ультрамикроскопические существа, парази- тирующие в бактериях и клетках многоклеточных растений и животных; вирусы отличаются очень мелкими размерами, отсутствием клеточного строения и облигатно-паразитическим способом жизни (Великанов и др., 1981). Жизнь большинства протеробионтов, гомологов современных од- ноклеточных прокариот, например, многочисленной группы Schisophyta (дробянок), которые получили свое название за способ размножения—
232 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА С'ГЕНИИ простое деление с разделением ядерного аппарата, большинства бакте- рий (Eubacteria), протекала, вероятнее всего, традиционно для телесных систем по эволюционно-маятниковому принципу и схеме «так-сяк * туда-сюда»: сначала быстренько развивалось простенькое тельце, потом происходило деление его пополам и т.д. — все действительно просто, но надежно. «Развернул» микроскопическое тельце, «накачал» его опреде- ленной вегетативной массой и набрав ее за несколько минут до какого- то, обозначенного наследственностью предела, разделил надвое, сначала перемычкой, а затем перегородкой, по которой уже две новые особи со своим генетическим и фенетическим арсеналом «расползались» по раз- ным «углам» для дальнейшего существования, свершив тем самым акт вегетативного размножения (дробления на части). Приоритет размноже- ния рано стал в эволюции безусловным фактором и усиленно давил на организацию таких первых протеробионтов, давая в их распоряжение минуты, а может быть, лишь секунды, чтобы «насладиться» явлением жизнь, а затем «раскол» тела пополам и, «будь здоров», уже пошла новая дискретная жизнь с тем же самым приоритетом. Живут протеробионты — прообразы современных одноклеточных без nonosozo размножения, а только с вегетативным размножением, очень мало. Их личная жизнь, по большому счету, суперабсурдна (!), но она, с другой стороны, еще и фено- менальна, по отношению к виду, его сохранению. Поэтому принцип орга- низации рассмотренных протеробионтов нельзя считать эволюционно бес- неплохо и до наших дней. Правда, в образе доклеточных существ (виру- сов), мы находим облигатных паразитов, но принцип — главное жить, а как жить и в какой форме — неважно, объясняет, почему приоритет бес- компромиссного размножения над личной жизнью, превращенный эволюци- ей в суперабсурд, дошел до наших дней. Если внимательно присмотреться к происходящему, то получается, что нелепица очень короткой продол- жительности организменной жизни вирусов и подобных, самостоятель- но жертвующих собой, как следствие, короткоживущих существ пере- крывается с лихвой нормативной силой скорости размножения, крайне выгодной для сохранения вида. Если личная (вегетативная) жизнь орга- низмов продолжается несколько секунд, минут, часов, но не более одних суток, — она суперабсурдна, однако, сами организмы проявляют в таких случаях, кроме того, еще и высшую экспрессию (выразительность) в спо- собности размножаться, т.е. в целом чудеса альтруизма, поскольку они отдают свою личную жизнь, составляюшую вегетативную основу организ- менности, почти целиком в угоду виду, что позволяет это явление обозна- чить как суперальтруизм, а организмы, проявляющие его, можно называть гиперэфемерами. Организмы, которые живут более одних суток, но не более одной недели (их личная жизнь абсурдна по времени), проявляя тем самым не высшую, а неонормативную экспрессию в способности размно- жаться, которую обозначим термином «неоальтруизм», будем отличать как проэфемеры. Организмы, живущие более одной недели и, как правило, не более одного месяца, — это обладатели свойства семиальтруизма, а их самих мы обозначаем традиционным словом — эфемеры (их обычно назы- вают яровыми эфемерами). Они отличаются от гемиэфемеров (ux обычно
~дава 2. Концепция и ио ипьепьов развипьия в организации cucme~ называют озимыми эфемерами), которые живут значительно больше од- ного месяца — иримерно около полугода, начиная эииэмбоиональное разви- mue осенью или неиосредственно под зиму, заканчивают его только весной и уже не являются какими-либо альтруистами. Гемиэфемеры и настоящие одного года и обязательно захватывает летние месяцы одного вегетаци- онного сезона) — это эвриэфемеры (они все постальтруисты). Первые организмы, появившиеся в процессе эволюции на Земле, были протеробионтами и обладали свойствами или гиперэфемеров, или проэфемеров, что позволяет эту объединенную группу в условиях пер- вобытной Земли считать еще и археэфемерами (от греч. arche — начало + эфемеры). Продолжительность личной жизни многих современных дробянок и бактерий, размножающихся простым делением надвое, про- должается нередко всего 20 — 30 минут; можно предположить, что у протеробионтов она была еще короче и, возможно, исчислялась всего лишь секундами или немногими минутами, что, в конце концов, не столь уж важно, главное, что первая жизнь на Земле была, как минимум, проэфемерной, а как максимум, гиперэфемерной, т.е. продолжалась от нескольких секунд до одной недели, т.е. в целом она была археэфемер- ной (ее представляли археэфемеры). С этих организмов, похоже, началась жизнь на Земле и так, веро- ятно, она продолжалась многие миллионы лет (временная привязка сейчас пока не входит в наши планы), пока не произошло то, что обязано было когда-то произойти: увеличилось число вариантов разви- тия по продолжительности личной жизни. Одни существа пошли в эво- люцию на стабилизацию проэфемерпого направления (проэфемеризма) развития (оно существует безбедно до наших дней, заняв достойно свое место в разнообразии живой природы). Другой путь эволюции — уско- рение темпов личной жизни, когда гиперэфемерпое направление (гиперэ- фемеризм) присутствует в эволюции и продолжает укреплять свои пози- ции, например, вирусы (доклеточные паразиты клеток). Третий путь в эволюции жизни на Земле привел к появлению эвриэфемеров, т.е. эфемеров (это настоящие эфемеры, они живут от одной недели до при- мерно 1-го месяца) и однолетников (живут более 1-го месяца, но не более 1-го года) — это эвриэфемеризм, т.е. эвриэфемерпое направление в эволюции. В эволюции жизни на Земле, начавшейся с археэфемерных протеробионтов, выбирать особенно не пришлось; она сразу же взяла ориентир на свое сохранение (см. т. 1, гл. 3), причем любыми доступ- ными ей средствами, благо явление эволюции подстегивало ее прояв- лять свободу в выборе целей развития, а достигнуть этого примитивные древнейшие растения и животные могли, только совершенствуя свою организацию, развивая пластичность (!), без отказа от приоритетного, для развития всего живого, свойства — стремления к размножению. Слишком быстро жить можно, быстро переставая быть прежним орга- низмом. Очень быстро жить, размножаясь без устали, и все же еще уметь быть организмами, — это, безусловно, феномен в эволюции жизни на Зем- ле. Потеряв в эволюции многое из своего личного и иочти ничего не иолучив от этого взамен, носители гиперэфемерного направления в современной
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ эволюции, наиример, вирусы, довели по сути организменную жизнь до су- иерабсурда. Именно так можно назвать состояние личной жизни, которое исиытывают особи вирусов; вероятно, вирусы могут «ощутить» себя живыми больше в границах вида, чем в границах организмов. 2.5.2. Генеральный приоритет в направлениях эволюции жизни на Земле и смешение приоритетов в эволюции биоморф семенных растений Археэфемерное направление (археэфемеризм) в эволюции вряд ли могло полностью удовлетворить жизнь вообще (Жизнь) в ее изначаль- ном стремлении завоевать пространство и время. Жизнь на Земле стала настоятельно стремиться продлить личную жизнь организмов в проти- воречивом согласии поддерживать и развивать заложенное в их же теле свойство — стремление к размножению, что не могло не привести эво- люцию к поиску и нахождению между ними компромиссов. Компро- миссы пошли через расширение позиций вегетатива и совершенствова- ния, посредством специализации, форм размножения, что в итоге при- вело к возникновению и быстрому распространению среди носителей зарождающейся жизни репродуктивного, в первую очередь, полового размножения. То, что скрылось от наших взоров за многими миллионами лет эволюции, составляет достояние жизни на Земле, а со своим достояни- ем она, безусловно, расставаться не склонна. Жизнь в эволюции мно- гого достигла, многое потеряла, но она сохранила многие крупные уровни качественных достижений как по форме, так и по содержанию; это она могла потерять, имела такое право, но сохранила из главного, пожалуй, почти все. Жизнь что-то уже потеряла в эволюции, что-то еще потеряет, но то, что до наших дней дошли многие крупнейшие таксоны, которые возникли в эволюции жизни за все время от ее начала на Земле — свидетельство нерасточительности жизни вообще (Жизни) по круп- ному, т.е. в отношении самой себя. Да, жизнь на Земле потеряла дино- завров, но крупный таксон — рептилии, к которым относились дино- завры, — сохранен ею. Да, жизнь потеряла саблезубых тигров, пещер- ных медведей и мамонтов, но млекопитающие процветают и без них. Да, жизнь, когда-то найдя, затем все же потеряла псилофитов, но до- стигнутый ею уровень ассоциировал псилофитов и некоторые признаки их, безусловно, присутствуют в генетической «памяти» современной жизни. Жизнь весьма строга в сохранении своих завоеваний на Земле в эволюции, и в ней сейчас, как в музее, можно обнаружить многие крупнейшие таксоны, представители которых процветали или прозяба- ли когда-то в историческом прошлом. Итак, мы несколько отвлеклись от построений гипотетических конструктов. Противоречия в эволюции внутри организмов-проэфеме- ров нарастали между стремлением поддержать сохранение личной жиз- ни (вегетатива) организма (равно всего организма) и стремлением под-
~дава 2. Концелция npuo umemoe развипьия в о ганизации cucme~ 235 держать другую жизнь (пока еще не репродуктив, но уже и не тот вегетатив, в котором нет перегородки, а вегетатив с перегородкой, все более и более растаскивающей одно тело организма на два — это уже эксвегетативное тело, или эксвегетатив, все еще пока одного организма). На ранних этапах эволюции возникла пластичность, и ее совершенство- вание было направлено на развитие и закрепление компромиссов между противоречивыми сторонами развития одной организации какого-либо проэфемера. Стремление поддерживать в развитии личную жизнь все более оттягивало соответствующую энергию в эту сторону у приоритета в поддержании другой жизни, отодвигая размножение. Шло в процессе эволюции усиление личной жизни и модернизация системы размноже- ния без отказа от нее. Таким образом, от проэфемеров (шире — археэфемеров) жизни убегать было некуда, кроме как броситься в «омут» поисков компромис- сов между приоритетным стремлением к размножению и придавлен- HbIM, но совсем еще не раздавленным стремлением к поддержанию (расширению) вегетативной жизни, для начала, хотя бы не так, как в условиях проявления суперабсурда в личной жизни организмов и даже не так, хотя это уже что-то, как у эфемеров. Но ведь за прорыв до абсурдной личной жизни, вряд ли она стоит того, чтобы ее называть иначе, как у эфемеров, эволюция за миллионы лет положила на плаху бессчетное количество дискретных жизней археэфемеров. Поиск комп- ромиссов неминуемо вел в эволюции к поиску и нахождению вариантов с более продолжительной жизнью организмов. Виды учились совершен- ствоваться не только в количестве своих носителей, но и в качестве. Сначала в процессе эволюции из археэфемеров произошли эфемеры, а далее эволюция шла по разным сторонам развития организмов как вегетатива, так и эксвегетатива и примитивного репродуктива, причем как в царстве растений, так и животных, но ясно одно: жизнь стреми- лась овладеть многолетним (!) образом жизни своих носителей, не ут- ратив, естественно, и того, что уже было завоевано в ранге крупных такс онов. Сейчас живут на Земле доклеточные паразиты (вирусы) и однокле- точные (дробянки, бактерии и др.) потомки своих далеких предков- протеробионтов, развивавшихся сначала как археэфемеры, а далее как эфемеры и гемиэфемеры, чем-то напоминавших современных прокари- от. В современных организмах присутствуют противоречивые взаимоот- ношения двух сторон их развития — личной и коллегиальной, или иначе — вегетативной и эксвегетативной или репродуктивной, которые и в эволюции преодолеваются по-разному. Возникновение многолетников в эволюции — это естественный путь эволюции жизни путем поиска компромисса между противоречивыми свойствами личного развития, с одной стороны, и неличного (коллегиального), с другой. Известное сейчас разнообразие растительных биоморф по продол- жительности их жизни, главные из которых эфемеры, однолетники, малолетники и многолетники, появились в эволюции очень давно, близ- ко от ее истоков. Вряд ли кто скажет сейчас, что жизнь на Земле началась с динозавров или гигантских деревьев-лепидодендронов.
236 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Жизнь, соблюдая определенную последовательность в эволюции, что-то достигнув принципиально крупное, чаще всего не теряла это крупное, растрачиваясь налево и направо по «мелочам» (что стоит для Жизни потеря «каких-то» динозавров), ей важно было в возникших экстремаль- ных условиях не потерять всех рептилий, и она их не потеряла. Гено- сома рептилий существует, а она, утратив многое из истории своих носителей, все же кое-что и хранит в своей преемственной «памяти», поэтому никто не может быть уверен, что в эволюции жизни не вернутся на Землю рептилии со многими чертами динозавров, если найдутся силы (возможно, человеческие) и (или) условия, которые «растревожат» дремлющие гены предков в современных рептилиях-потомках, высвобо- див идиоэнергию, давно забытую, но еще не рассеянную в пространстве, из преемственной (генетической) «памяти», в таком случае, потенциаль- ное пришествие животных, относительно близких к динозаврам, гипо- тетически все еще возможно. В эволюции многое связано с мелочами, но при всем этом нам, похоже, удалось подсмотреть самое главное в ней. Да, в каждом орга- низме, как растительном, так и животном, миллионы (сотни милли- онов) лет назад и сегодня в их фенетической организации присутствуют две основные части: вегетатив (одна часть) и эксвегетатив или репродук- тив (другая часть). В самом самом начале жизни на Земле, вероятнее всего, появился один вегетатив, но с присутствием определенной, пусть еще несовершенной, генетической (нуклеидной) начинки в его, как правило, микроскопическом фенетическом содержании, а далее все пошло по пути совершенствования организменно-видовых отношений, которые в конкретных организмах приобрели выражение: свой — не- свой (другой) ~ свой — не-свой и т.д. Две стороны фенетической организации и развития организмов возникли, по сути, одновременно с возникновением жизни, а с ними возникли еще и гены, причем од- новременно, таким образом, с организмами возникли и виды. С ранних этапов развития жизни на Земле существует приоритет приоритетов (суперприоритет) — стремление к сохранению жизни во- обще [личной и (или) коллегиальной]. Еще у протеробионтов была решена стратегия Жизни — сохранять жизнь вообще любыми средства- ми: вегетативными или эксвегетативными (без полового процесса), или репродуктивными (с половым процессом) при приоритете развития в сторону любого размножения, хоть делением на части или половым процессом (без или с модификациями его). Вегетативное (личное) на заре жизни у организмов было придавлено приоритетом развития на поддержание другой жизни (на размножение), поэтому продолжитель- ность жизни организмов исчислялась секундами, минутами, часами, далее днями и неделями, затем, в процессе эволюции шел поиск ком- промиссов для разрешения противоречий между двумя отмеченными выше сторонами одной организации протеробионтов — проэфемеров и эфемеров. У последних продолжительность жизни организмов достигла недель и дошла до одного месяца. А затем все пошло и поехало в сторону совершенствования компромиссов и доведения продолжитель- ности жизни организмов до полугода (гемиэфемеры), затем до года
~дава 2. Концелция и ио umemoe развипьия в организации сиспьем (однолетники), до нескольких, но немногих лет (малолетники) и далее до многих лет жизни (многолетники). Унирепродуктивные организмы вполне могли возникнуть в эволюции раньше мультирепродуктивных организмов. У первичных организмов на Земле все фенетическое было вегета- тивным, оно одновременно выполняло роль и личного, и неличного развития; не-свое (другое) отмечалось, когда появлялась перегородка в вегетативном теле, превращая его тем самым в эксвегетатив, после которого уже происходило вегетативное размножение и возникали два новых организма. Далее в процессе эволюции нашла себе место, дей- ствительно, вторая часть организма: в развитии вегетатива появилась более высокая цель — репродуктив. Все это было завоевано в эволюции еще первобытной жизни. Компромиссы поначалу были примитивными, поскольку все свершалось в одном вегетативном теле протеробионта, который как организм существовал секунды, минуты, реже часы, чтобы затем безмятежно разделиться и размножиться. Часть особей, безуслов- но, безвозвратно умирала, не превращаясь в потомство. Все это было и многое не утрачено эволюцией по сей день, но важнее, что эволюция пошла дальше, развивая пластичность организмов и видов через совер- шенствование компромиссов с использованием прогресса в специали- зациях. Большая победа в эволюции жизни на Земле — это возникновение двух частей в теле каждой особи: вегетатива и репродуктива. Эволюция жизни в сторону многолетности привела к закреплению в ней великого парадокса, когда несовместимые, казалось бы, процессы жизни и смерти научились «договариваться» не только для того, чтобы отодвинуть смерть подальше от начала дискретной жизни особи, но и попытаться «приручить», казалось бы, ничем неукротимую смерть для работы ее представителей — мертвых частей организма (некронов) в качестве структурных частей живого организма, т.е. поставив смерть на службу живым особям. Пожалуй, гигантской важности синдром новиз- ны проявился, когда жизнь загнала смерть внутрь структуры своих но- сителей — организмов — и заставила ее надежно работать на них. Почему деревья бывают такими высокими и стойко не ломаются под тяжестью своих живых крон, почему они после смерти могут еще стоять десятилетиями? У них при жизни в стволах и ветвях аккумулируется вторичная древесина (проводящие и механические элементы) — мертвая ткань, которая усиливает, а точнее — создает основные опорные фун- кции у древесных растений и продолжает, будучи мертвой, функциони- ровать с пользой для жизни (подробнее см. гл. 7). Многие виды приспособились сохранять зачатки новых особей (споры, семена) или даже превращать прежние особи в определенные временные формы их существования, например, цисты покоя (у многих одноклеточных организмов — споровиков, корненожек, инфузорий и т.п.), благодаря чему они могут переживать неблагоприятные периоды в окружающей среде. Все формы внутривидовых адаптаций, направлен- ных на переживание видами неблагоприятных условий, — это безуслов- ные достижения в эволюции жизни на Земле, но куда их надо относить? Это все вынужденные состояния организмов со множеством своих ас-
238 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ пектов эволюции. Для примера вспомним, что семена многих цветковых эфемеров могут где-то в сухом месте храниться десятки лет и не про- растать, оставаясь живыми, поскольку им для пробуждения и роста необходимы другие условия. Временные формы существования организ- мов (споры, семена, цисты и т.п.), например, некоторых гиперэфемер- ных бактерий, живущих в активном состоянии два-три десятка минут, развиваются при неблагоприятных условиях и находятся в покое многие годы. Вопросы многообразных взаимоотношений и взаимосвязей между основными (активными) формами существования организмов и времен- ными формами, их сопряженной эволюции, безусловно, составляют специальный раздел фундаментальных исследований, что в этой работе не входило в наши задачи. Все, что было сказано выше об эволюции биоморф, распространяется только на активные формы существования организмов. Итак, от археэфемеров эволюция жизни растительной и животной пошла в разные стороны по многим рядам признаков, но примечатель- но, что главное приоритетное направление по одному ряду признаков — по продолжительности жизни организмов — это многолетность такой жизни, что было вызвано эволюцией организменной жизни через совер- шенствование у них компромиссов между личным и неличным. На этом пути сначала из эвриэфемеров растительного мира появились околотра- вянистые (малолетние, затем многолетние) растения, а лишь потом- околодревесные растения; кроме того, жизнь на Земле не стремилась утратить многое из того, что она добилась еще на своей заре, когда суперальтруизм, неоальтруизм и семиальтруизм (формы экспрессивного альтруизма) сотрясали устои организменного образа малоопытной пер- вобытной жизни. Главным фактором, обеспечившим реализацию в эволюции расти- тельных биоморф приоритета многолетности и тем более деревянисто- сти, было естественное стремление «разлившейся» по Земле жизни пол- нее освоить приземное пространство. Многолетность и ее высшее вы- ражение «деревянистость» оказались для растительной жизни на Земле революционными приобретениями в эволюции, возникновение которых предопределило генеральное (приоритетное) направление в разнонап- равленной эволюции. Рядом с приоритетной направленностью в эволю- ции биоморф по продолжительности жизни (на многолетность), когда идут адаптации в сторону совершенствования компромиссов между ве- гетативной и репродуктивной частями с различными сопутствующими этому адаптациями, когда возрастает роль вегетатива, но не утрачива- ется, а еще больше совершенствуется приоритетное положение репро- дуктива в организации биоморф, который в целом, в первую очередь, для своей же пользы, улучшает формы взаимодействия с вегетативом, давая ему в меру «насладиться» прелестью личной жизни. Прогрессив- ное развитие компромиссов между вегетативом и репродуктивом позво- ляет достигать в эволюции многолетности экономного распределения ресурсов вегетатива при развертывании репродуктива, насыщая em энергетическим материалом, достаточным для совершения конкретной репродуктивной деятельности с удержанием части накопленных ресур-
гудава 2. Концепция и ио umemoe развипьия в организации сиоием сов для новых репродуктивных усилий в другие годы. Пройдемся осе- нью по асфальтированной дорожке, допустим, в березовом парке; эта дорожка сплошь покрывается семенами березы с примесью семян дру- гих растений и так по всему парку или какому-нибудь лесу с березой и T.ï. Кому нужна такая расточительность? Ясно одно — это все нужно не лично организмам, а тем, кому они принадлежат и кому безмятежно служат, т.е. это нужно видам. Виды, как ключевое звено в эволюции дд~вого, давно и надежно усвоили в эволюции свое превосходство над организмами, подчинив себе целесообразность их внутренней организа- ции, что нашло отражение в специальном законе облигатной избыточно- сти присутствия свойств вида в организации растительных организмов (за- коне эволюционного эйдоцентризма): на протяжении личной жизни расти- тельных родительских организмов производятся структурные элементы, по-разному специализированные для службы как самому организму, так и другому живому телу — виду, но всегда производство структурных элемен- mos, предназначенных непосредственно для вида (не-своих для организмов), не только подчиняет личное организмов целям видового развития, но и, в обычных условиях, обслуживает эти цели с бульшим запасом (избыточно- стью), в частности, пререпродуктивных и репродуктивных органов произ- водится больше, чем те могут дать половозрелых элементов, а последних — больше, чем они способны задействовать для производства гамет и nonosozo (например, семенного) потомства, далее число проросших эмбри- онов (проростков) значительно превышает численность развивающегося из них эпиэмбрионального потомства, а виргинильных эпиэмбрионов (неполо- возрелых самостоятельных особей) всегда больше физиологически зрелого потомства, возникающего из них, и т.д. Из рассмотренного закона напрашивается аксиоматическое правило: растительные организмы, чтобы продлить личную жизнь, должны приспо- собиться медленнее стремиться к размножению и не форсировать репро- дуктивное развитие, находя внешние условия и организационные возмож- ности, чтобы отсидеться в виргинильном состоянии (образно — «в дев- ках») или научиться выдерживать вегетативными (личными) остатками организмов разрушительные пострепродуктивные некротические удары, чтобы через некоторое время вновь проявлять репродуктивные усилия. Встав на путь приоритета многолетней жизни биоморф над другими направлениями их развития по продолжительности жизни, жизнь вооб- ще по сей день на Земле не растеряла то, что она завоевала ранее: она теряла детали в эволюции, продолжающейся многие миллионы и сотни миллионов лет, но по-прежнему рядом с хорошо отлаженным механиз- мом совершенствования многолетности сосуществует, казалось бы, до- потопный механизм эвриэфемерной направленности жизни с более или менее ярко выраженными формами экспрессивного альтруизма, когда жизнь вегетатива (личная жизнь) ничтожна по времени и ущербна по качеству и количеству ее в организации биоморф, конкретных носите- neÀ такого вегетатива, поскольку эксвегетатив и (или) репродуктив на- столько жестко подчиняют организацию таких биоморф для службы виду, что продолжительность их существования убийственно мала, на- столько мала, что автор невольно вынужден прибегать к таким словам,
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ как «абсурд» и «альтруизм», для обозначения рассматриваемого явления. Получается, что образ жизни палеобиоморф на Земле в начале развития живой природы, продолжавшегося многие миллионы лет, складывался не иначе как экспрессивно, альтруистически, но для видов этот способ организации жизни совсем неплох (он жестоко несправедлив для орга- низмов), поэтому по сей день существуют, на равных с другими направ- лениями развития жизни, как гиперэфемеризм (например, вирусы), про- эфемеризм (дробянки, бактерии), так и эвриэфемеризм (нередкие среди биоморф семенных растений эфемеры и однолетники). Авторский подход к эволюции живой природы на Земле показыва- ет, по сути, что растительные биоморфы, существующие сейчас в зем- ном пространстве и времени как эволюционные маятники, являются потомками тех палеобиоморф, которые начинали свое восхождение по Земле с археэфемеров (гиперэфемеров и проэфемеров), дав несколько направлений эволюции по продолжительности жизни биоморф, причем археэфемеры не растворились в истории и поныне здравствуют органи- зационно подобные им потомки, всю группу которых, безусловно, уже стоит называть иначе, допустим, гипоэфемеры, включая в нее современ- ных гиперэфемеров и проэфемеров. Волны жизни прокатывались и прокатываются по Земле как по «морю», только что-то теряя в деталях и что-то приобретая новое в организации своих биоморф, но в целом их основное содержание не смывалось, поэтому до наших дней, по большому счету, многое из самого главного осталось в том же «море» жизни, которое и составляет современную биостробу (живую природу). На современной Земле сохра- нились наиболее крупные таксоны растений и основные типы органи- зации жизненных форм по продолжительности жизни организмов: от гиперэфемеров до сверхдеревьев (предельный возраст последних — 1000 и даже более лет). Что-то в эволюции в каких-то таксонах могло выпа- дать и, безусловно, выпадает почти как по аналогии с жизненными циклами современных растительных биоморф, у которых способны выпадать и нередко выпадают их отдельные возрастные состояния. Полагаю, что уже до голосеменных был развит весь современный спектр munos биоморф (биоморфотипов) по продолжительности жизни (u no ux габитусу): от эфемеров до сверхдеревьев, из которого семенные растения взяли для своей эволюции все или почти все, что-то затем находя еще, а что-то теряя в отдельные периоды на задворках истории. В этом плане начало у покрытосеменных могло быть связано с любым типом биоморф по продолжительности организменной жизни (и по их габитусу заодно), по- скольку биоморфы уже обладали пластичностью, которая и обеспечивала им успех в эволюции (т. 1, гл. 8). Развиваясь по принципу эволюционных маятников и схеме «так- сяк * туда-сюда», эволюция создала все известное нам разнообразие типов растительных биоморф, и от этого разнообразия жизнь на Земле, по большому счету, больше, как правило, не отказывалась. Жизнь с первых лет своей эволюции на Земле стала усиленно искать решение непростых проблем внутриорганизменных взаимоотношений между ве- гетативом и репродуктивом, чтобы «волки» были сыты и «овцы» целы,
1лава 2. Концеиция и ио итетов азвития в о ганизации систем что в результате долгих миллионов лет проб и ошибок оказалось воз- можным через совершенствование компромиссов между вегетативом и репродуктивом при сохранении приоритета последнего на генеральном направлении эволюции — по пути организменной многолетности. По- скольку семенные растения появились значительно позже того, как растительная жизнь выбрала себе в качестве главного приоритета мно- голетность в организменном развитии своих биоморф, то семенные растения эволюционировали на фоне всего известного нам разнообра- зия биоморф от эвриэфемеров до сверхдеревьев. Были ли среди голо- семенных травы и тем более однолетники и эфемеры — вопрос до сих пор открытый. Гипотетически вполне можно предположить, что эти типы биоморф были среди голосеменных, если не постоянно, то в от- дельные периоды их истории, но конкретных вещественных доказа- тельств (не найдены палеоботанические остатки) пока нет (они могли просто не сохраниться). Учитывая, что в эпоху семенных растений эво- люция растительных биоморф уже достигла высшего (или где-то около него) пика их типового разнообразия по продолжительности организменной фсизни, семенные растения ириняли те иравила эволюционной «игры», ко- торые им предложили ближайшие предки: они приняли наиболее подходя- щую для того времени направленность в эволюции — разную (!), т.е. у них не стало генеральной (приоритетной) линии эволюции, а они шла в большой от изменений климата (наступали и отступали океаны и ледники, подни- малась и опускалась земная твердь, росли и рассыпались горы и т.n.). В разных регионах Земли и в разных таксонах, в зависимости от окружаю- щих условий биоморфы семенных растений эволюционировали разнонаправ- ленно, совершенствуя свою пластичность: где-то от деревьев к травам, а где-то — от трав к деревьям. Однако, если посмотреть на эволюцию живой природы от ее истоков до наших дней, то растительная жизнь прошла путь от гиперэфемеров до сверхдеревьев, поэтому многолетность можно безоговорочно принять за генеральное (приоритетное) направление эволюции фитостробы (расти- тельного мира) Земли по продолжительности онтогенезов простых био- морф. Наличие приоритетного направления в эволюции растительной жизни не означает, что в мире семенных растений не было вовсе субгене- ральных линий эволюции, охватывающих многие миллионы лет их истории. В отдельные периоды вполне могло быть больше «травянистости», а в другие наоборот — «деревянистости», но в целом эволюция биоморф семен- ных растений была и есть разнонаправленной (не будем забывать, что маятниковый принцип эволюции работает в системах разного уровня слож- ности). Механизм эволюции бесконечно раскачивает уже многие сотни Миллионов лет эволюционные маятники живой растительной природы (фитостробы) Земли. Мы посмотрели на эволюцию под нетрадицион- ным углом обзора через взаимодействия в организации простых био- морф (организмов) двух их основных частей: вегетатива и репродуктива (или, когда последнего совсем нет, то эксвегетатива), что коренным образом влияет на продолжительность организменной жизни и на гене-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИЙ ральную (приоритетную) направленность эволюции всей растительной жизни. Нормой эволюции растительных биоморф является стремление к размножению и поиск, особенно у семенных растений, компромиссов между вегетативом и репродуктивом организмов; в противном случае, происходит суперабсурд или абсурд с личной жизнью (она продолжается минуты, часы или — до месяца, что свойственно гиперэфемерам, проэфемерам и эфеме- рам), т.е. то, что определяет один из парадоксов в организации раститель- ных биоморф — экспрессивный альтруизм, суть которого попробуем выра- зить афористически кратко: организменная жизнь — почти ничто, видовая жизнь — почти все. Итак, в организации любых живых тел, в частности, фитобиоморф, на фоне личной жизни проносятся «вихри» борьбы, которую, в различных взаимодействиях, ведут в этих биосистемах другие жизни, т. е. по сути дела, в любой биосистеме, в организации ее одного тела, рядом с личной жизнью (как-либо соприсутствуют и другие (не-свои, неличные) жизни, проявляя над ней свой npuopumem, если они стоят выше по сложности организации. Эта идея проводится красной нитью через всю нашу работу и вполне может стать (осознание сего только начинается) одной из интригующих проблем современной биологии и, в частности, биомор- фологии. 2.6. Научные приоритеты, приоритеты авторитетов и влияние их на приоритеты развития науки, в частности, биоморфологии У каждого человека, как живой системы, есть приоритеты его лич- ного развития и приоритеты в понимании развития всего другого, что его окружает. Чаще всего приоритеты деятельности и ее развития, свя- занные с пониманием их конкретным человеком, совпадающие или не совпадающие с приоритетами деятельности других людей, не выходят за пределы ограниченной деятельности этого человека. Все сложнее, когда приоритеты развития расставляет ученый. Нас более всего интересуют в данный момент приоритеты, которые проявляет человек-ученый в одном из видов деятельности — в науке. Личное понимание приорите- тов развития науки может совпадать или не совпадать с пониманием таких приоритетов другими учеными. Исторически сложилось, что для науки важное значение имеют приоритеты ее развития, которые подска- зывают ученым «жрецы» науки — научные авторитеты (ученые, которые как-либо завоевали признание со стороны научной общественности и стали как-либо пользоваться этим признанием, оказывать влияние на различные стороны общественной жизни). Авторитеты в науке — явление реальное, и с ними так или иначе вынуждены считаться те другие, которые не являются, может быть, пока авторитетами. Авторитеты — это люди, и они, отличаясь от других
ржава 2. Концеиция ириоритетов развития в организации систем людей только тем, что под влиянием разных обстоятельств лучше других решают текущие задачи науки, понимают прошлое и (или) видят пути ее дальнейшего развития (или создают сами и с помощниками вокруг себя миф, что они лучше). Есть ли авторитеты вне общества людей? Безусловно, есть, но называются они по-другому: вожаки, доминанты, например. Речь сейчас не о них. Однажды ученые люди начинают полагать, что среди них есть ав- торитеты (их не так много), которые лучше и дальше других (иногда это только кажется, такое бывает) понимают развитие науки, что обязывает их более, чем раньше, считаться с теми, которых они признали за на- учные авторитеты, их стали больше уважать, больше ценить, больше слушать и даже служить им и т.п. В целом это понятно и оправдано, но только до некоторого предела, когда научная общественность не стано- вится жертвой заблуждений и не попадает в «заложники» авторитету мнимому. Научный авторитет, как и любой другой человек, не свободен от слабостей, и может получиться так: он подумает, что стал больше, чем просто уважаемый ученый, и начнет усыплять научную обществен- ность своей «непогрешимой» авторитетностью и может, запутав других, 3aIl~BTbcH сам настолько в своих конкретных изысканиях, что Hpимет ошибочные цели и результаты своих поисков за действительные при- оритеты развития науки и без страха сомнения начнет культивировать их как единственно верные, достойные подражания, по которым можно безоговорочно сверять вехи и своих, и чужих будущих скромных дости- жений. Приоритеты авторитетов, как показывает жизнь, двойственны; они хороши, если авторитетный ученый, действительно, точно подгля- дел с помощью безукоризненной работы соответствующих инструмен- тов и приборов, зоркого глаза или гибких извилин своей абстрагирую- щей мысли, то, что реально, независимо или зависимо от сознания его — человека научного (ученого), присутствует и функционирует в осталь- ной Природе. Благо для науки и человечества в целом, если авторитет ошибается не так сильно и не так опасны его промахи на подступах к главному. Но авторитеты могут заблуждаться по-крупному, они, как и другие ученые, могут принимать второстепенный информационный шум за главное и, что самое опасное, они могут начать применять разные средства, чтобы насаждать свои, ошибочно расставленные в науке при- оритеты как методологию, теорию и внедрять ее в практику научных исследований. Печальных для науки примеров много. Труд целого по- коления ученых-биологов в середине ХХ столетия был изуродован по- стулатами «великого авторитета» Т.Д. Лысенко, ошибочно признанны- ми в границах бывшего СССР за истину в последней инстанции, за приоритеты в развитии советской биологии. Лысенко уже давно не авторитет, давно его научные приоритеты низвергнуты, но случившееся — предостережение для всех: будьте бдительны и к себе и к тем, кому Вы доверяете. Такого не должно повториться, за это ответственны уче- ные — наши современники и те, кто будет после нас. Мы все можем открывать истины, но и все мы можем ошибаться, и то, и другое надо замечать и соответствующим образом оценивать. Сами по себе ошибки Лысенко не сыграли бы пагубной роли в развитии нашей биологической
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ и сельскохозяйственной науки, если бы они не навязывались самим авторитетом авторитарно (диктаторскими действиями, основанными на превышении своего положения, пусть даже авторитетного, заходящими нередко настолько далеко, что подключающими для поддержки диктата своих идей даже политическую власть. Авторитаризм в науке способен превратить взгляды, даже ошибочные, одного человека в предмет пре- клонения ученых, поскольку несогласных с авторитетом в таких случаях просто разными способами изживают. Ошибки в науке, если их неосоз- нанно или осознанно пропускают, могут ей стоить дорого; они могут, в случае авторитаризма в науке, погубить усилия целых поколений ученых, пока не появятся одиночки, которые выстоят перед градом «камней», которые найдут силы собрать «камни», создать новые при- оритеты и показать всем, что истина дороже. Приоритеты науки закреп- ляются в конце концов коллективным разумом. Авторитеты сами по себе не опасны, они больше полезны и важны, поскольку идуг на пе- редних рубежах науки и выступают как главные творческие силы науч- ного прогресса, а их возможные ошибки зорко и быстро поправляются или должны поправляться коллегами, которые в таком случае обречены также стать научными авторитетами. Авторитет в науке — это не член касты неприкасаемых, а состояние души ученого, которое он сам начинает с некоторого времени испыты- вать и строже к себе относиться, которое, и это самое главное, начинают замечать и отмечать его коллеги, а далее больше и многие другие, признающие, осознанно или неосознанно, под влиянием разных обсто- ятельств, важность достигнутых им результатов в проделанной работе. Однако, если авторитетство насаждается, то цена такому авторитету— грош; он есть пока его «проталкивают», многие замечают, понимают и как-то реагируют на это, чаще всего они так или иначе не признают такой «авторитетно-мыльный пузырь», который так же быстро исчезает, как и появляется, когда «подпитка» извне прекращается, и все начинает развиваться само собой. Ученый — это должна быть самая свободная в творческом отноше- нии личность. Только будучи свободным творцом, он способен совер- шать открытия как в практической деятельности, так и в области тео- ретизирования, научного законотворчества. Приоритеты развития науки устанавливают люди, которые в случае успеха обязательно станут науч- ными авторитетами, а когда ученый ошибается, то в норме его просто спокойно поправят (так должно быть, но, к сожалению, так часто не бывает) на новом витке развития науки. Тот, кто не боится ошибиться, двигаясь вперед на рубежах развития науки, способен снискать уваже- ние других, а это одна из сторон истинного авторитетства. Пожалуй, дальше в этом ключе продолжать не стоит, все в нашем случае уже ясно: авторитетам надо доверять, но их надо и проверять, поскольку они, как и все люди, способны ошибаться. Мы поразмыш- ляли над проблемой, которая имеет разные корни и разные выходы из нее, но которая не менее важна и для движения вперед в любой частнои науке и в той, о которой эта монография — в биоморфологии. Есть необходимость подытожить сказанное выше еще одним (шестым) пра-
~дава 2. Концеиция л ио итетов азвития в о ганизации систем вилом закона приоритетности: информация, изложенная в как-либо мате- риализованной научной системе (печатном издании, опытной модели, mex- ническом образце и т.n.), только тогда выступает как приоритетная, когда она первенствует по значимости и (или) по времени ее объявления по отношению к другой сравнимой информации; при всем этом npuopumem- ность информации в науке не всегда гарантирует, что достигнута истина; последняя проверяется, подтверждается или не подтверждается по исте- чении какого-либо времени. В науке приоритет у того, кто первый сделал открытие, пусть даже небольшое, отразив это в печати, поделившись, тем самым, добытыми им знаниями со всем остальным ученым миром. Кто первый в решении какой-нибудь научной проблемы — это далеко не праздный вопрос, поскольку у науки своя история и у нее свой счет активов и пассивов, проходящих в ней. Приведу очень известный пример. Чешский иссле- дователь Г. Мендель в своих опытах по гибридизации сортов гороха открыл несколько важных законов наследования признаков, о чем до- ло>кил и опубликовал в научной статье. Эти открытия, сделанные в 1865 году, научная общественность заметила лишь спустя 35 лет, признав за великим первопроходцем приоритет. Так бывает в истории науки, и примеров тому больше чем достаточно. Важно, что приоритет раньше или позже, как правило, отдается тому или тем, кому он действительно должен принадлежать (другое бывает, но это уже из разряда парадок- сов). Рассмотрим еще один, довольно свежий пример. В нескольких научных работах (Нухимовский, 1970, 1973 а, б) было предложено прин- ципиально новое понятие, обозначенное новым словом-этикеткой— впервые был введен в науку термин «кондивидуум» [синоним — «си- норганизм» (Нухимовский, 1984)]. «Кондивидуум (от лат. 0011 — вместе, сообща и dividuus, -ит — разделенный) — живая, генетически однород- ная система, начинающаяся в своем развитии из одной зиготы и окан- чивающаяся в последнем поколении клона. Синонтогенез — жизнен- ный цикл кондивидуума ... системы более высокого ранга, чем особь, и меньшего, чем популяция» (Нухимовский, 1973 а, с. 113, 114). Позднее появился термин «генет» (Kays, Harper, 1974), при этом было подчер- кнуто, что «генет» — это «генетический индивидуум», т. е. все то, что получилось из одной зиготы» (Бигон, Харпер, Таунсенд, 1989, с. 183). Замечу, что кондивидуум, синонимом которого могло бы быть слово «генет», в целесообразности чего я, кстати, не вижу смысла, поскольку такое слово мало уместно в данном случае, — это нечто большее, чем индивидуум. Добавление к слову «индивидуум» эпитета «генетический» способно внести только путаницу, поскольку это уже не индивидуум. У кондивидуума (синорганизма), как особой системы, жизнеразвитие со- провождается сменой нескольких возрастных состояний (!): временного индивидуума (мероаклониста), что происходит от зиготы до возникно- вения клона, и клона (другого ему, кондивидууму, не дано вплоть до элиминации этой системы). Ответ на вопрос: кому же в поднятой здесь проблеме принадлежит приоритет ее понятийного решения и какая тер- минология перспективнее, пусть останется за читателем.
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ С позиций закона приоритетности посмотрим на развитие биомор- фологии в исторической ретроспективе. Что же мы видим? Приоритет развития науки, которая с наших позиций имеет отношение к истории современной биоморфологии на протяжении длительного времени, из- менялся неоднократно. Особенно не повторяясь с тем, что уже было написано в первом томе, обратим внимание на некоторые подвижки в изменении приоритетов развития в науке, которую сейчас все более настойчиво называется биоморфологией. Здесь подчеркнем, что наибо- лее полные и сильные по охвату литературного материала обзоры вы- полнены в хорошо известных работах Е. Дю Рие (Du Rietz, 1931), И.Г. Серебрякова (1962) и Т. И. Серебряковой (1972). Наша задача в данный момент скромная, но тем не менее весьма конкретная. Надо показать, как изменялись приоритеты развития под- ходов к исследованию растений, которые в итоге привели к формиро- ванию современной биоморфологии, т.е. той биоморфологии, которая рассматривается и развивается в этой монографии и которая, безуслов- но, не является результатом поисков и обобщений одного автора, по- скольку обеспечена коллективными усилиями ботаников-биоморфоло- гов всех предшествующих и текущих поколений. Попробуем сгруппиро- вать приоритеты в изучении организации растений, которые имели место быть в разное время и, безусловно, оказали влияние на станов- ление современной биоморфологии. 1. «Интуитивно-габитуальный» подход к характеристике тела ра- стений восходит от Феофраста (372 — 287 гг. до н. э.; 1951); он ориентирован на внешний облик растений, на основе которого выделяются «главные виды». деревья, кустарники, кустарнич- ки, полукустарники, травы многолетние и однолетние. Этот подход распространяется и на работы флористов более поздних (долиннеевских и линнеевских) времен (Caesalpini, 1583; Dodonaeus, 1583; Aldrovandi, 1671; Gmelin, 1747; Linnaeus, 1751 и др.). 2. «Макрофизиономический» подход к оценке общих черт органи- зации растений начинается с известной работы А. Гумбольдта (Humboldt, 1806; перевод — Гумбольдт, 1936) «Идеи о физио- номичности растений», где выделяются «основные формы» по «физиономии» (внешнему облику) растений, например, «форма пальм», «форма злаков», «форма хвойных» и др. (всего 19 «ос- новных форм»). В монографии А. Гризебаха (1872, 1877) ис- пользован также «макрофизиономический» подход, на основе которого выделены разнообразные группы «растительных форм», совпадающие или не совпадающие с систематическими группами, например, древесные, сочные, злакообразные расте- ния, эпифиты, травы и др. Периоды, в которых имели место в изучении организации растений первый и второй подходы, можно рассматривать и вместе как эпоху «пробиоморфологических» исследований. Приоритетом исследований растений в это время (точнее — в макрофизиономический период) было выделение «растительных форм» и (или) «основных форм» по наиболее
~дава 2. Концеиция ириоритетов развития в организации систем заметным экзоморфологически признакам, по-разному привязываемым к экологическим условиям произрастания (Hult, 1881; Drude, 1887, 1896, 1913 и др.). Поскольку общепризнанно, что известное словосочетание «жиз- ненная форма», или «форма жизни», впервые использовано Е. Вармин- гом в качестве строгого научного термина, т.е. обозначено не только словом-этикеткой, но и определено понятием: жизненная форма — это форма, «в которой вегетативное тело растения (индивида) находится в гармонии с внешней средой в течение всей его жизни, от колыбели до гроба, от семени до отмирания» (Warming, 1884, цит. по И. Г. Сереб- рякову, 1962, с. 27), то далее мы будем говорить уже о приоритетах другого (третьего) типа. 3. «Эколого-экзоморфологический» подход к исследованию рас- тений, когда в центре понятия «жизненная форма» рассматри- вается габитус организмов, хотя слово «биоморфа» как гречес- кий вариант термина «жизненная форма» Е. Варминг еще не использовал. Приоритетом в «экологической экзоморфологии» растений во вре- мена Е. Варминга и много позднее было выявление разнообразных биологических признаков, имеющих экзоморфологическое (габитуаль- ное) выражение, в первую очередь, в вегетативной основе организмов (например, «цикличность» нарастания побегов, продолжительность жиз- ни главного корня, особенности передвижения побеговых частей расте- ний по поверхности почвы или подземно, отношение к корнеотпрыс- ковости — нет или есть — и формы корнеотпрысковости и т.д.), раз- вивающиеся, в зависимости от экологических условий их обитания на протяжении жизненных циклов. Как удачно обратил внимание И. Г. Серебряков (1962, с. 27): «цель Варминга — не выражение физиономии растительности, как у Гумбольдта, Гризебаха, Хульта, а отражение эфар- монических, т.е. приспособительных черт этой растительности». Ранее (т. 1, гл.2) было показано, что понятийная привязка жизненной формы в основном к вегетативной основе, а не ко всему вегетативно-репродук- тивному телу организма, была принципиальной ошибкой Е. Варминга и его последователей и, по мнению автора этих строк, оказала некото- рое негативное влияние на последующее развитие биоморфологических исследований в ХХ столетии. Пожалуй, только в 1973 г., когда автор этих строк опубликовал статью «О понятии «жизненная форма», где биоморфа (жизненная фор- ма) была определена как организм или кондивидуум (синорганизм) и все признаки (как вегетативные, так и репродуктивные), принадлежа- щие им, были признаны за признаки жизненной формы, в биоморфо- логии, действительно, началась тихая революция, которая осуществля- ется и по сей день. Умение и желание правильно расставлять приори- теты не только в организации предмета исследований (в нашем случае, биоморф семенных растений), но и в развитии науки, изучающей этот предмет (у нас — биоморфологии), — это важная часть всякого научного исследования. Сейчас все более становится понятно, что приоритеты, выставленные Е. Вармингом в науке о жизненных формах (условно мы
ОСНОВБ1 БИОМОРФОЛОГИИ CEMEHHblX РА СТЕНИЙ обозначили ее как «экологическая экзоморфология»), были не все удач- ными. В частности, Е. Варминг признавал примат вегетативных органов над репродуктивными, причем последние он более всего игнорировал в изучении жизненных форм (см. т. 1, гл. 2), что неприемлемо, поскольку интегральными процессами, составляющими онтогенез растений, явля- ются вегетативный рост и репродуктивное развитие (Чайлахян, 1958, 1964, 1988). Кроме того, габитус организмов не может быть жизненной формой, поскольку он не имеет внутренней структуры и сам по себе нежизнеспособен. Только вместе с внутренней организацией любых особей (организмов) габитус может соучаствовать в структуре жизнен- ных форм, или биоморф, о чем мы специально поговорим в следующей главе. Сейчас тихая революция в современной биоморфологии идет, похоже, в нужном направлении, и отказ большинства ботаников от вегетативного, или, образно говоря, «вынужденно пенькового» приори- тета в организации биоморф, не за горами, поскольку, действительно, без репродуктивной сферы структура организма искусственно превра- щалась в пенек. Важнейшей новацией, которую сам Е. Варминг только начал прак- тически разрабатывать и еще не подошел к теоретическому обоснова- нию ее, которую постепенно признали биоморфологи, была идея, четко закрепленная в понятии «жизненная форма», привязывать различные морфологические признаки жизненных форм, образующиеся, в зависи- мости от изменений окружающих внешний условий, к жизнеразвитию организмов «от семени до отмирания». На эту понятийную привязку жизненных форм к жизненным циклам организмов до Е. Варминга в эпоху пробиоморфологических исследований никто ранее так однознач- но не акцентировал внимание. Можно предположить, что более всего на динамический подход Е. Варминга к пониманию жизненных форм подтолкнули классические работы высочайшего специалиста того вре- мени по возрастным особенностям морфологической организации цвет- ковых растений Т. Ирмиша ( Irmich, 1850а, 1850б, 1859, 1860, 1861, 1874 и др.), который, кстати, с большой точностью рисовал растения, осо- бенно раннего возраста, по сути дела, он был одним из первых, кто показал всем ботаникам, как мастерски надо это делать. Как бы там ни было, но с современных позиций можно утверждать, что именно дина- мический подход к объяснению индивидуальной, в основном, вегета- тивной, организации растений, позволяет считать, безусловно, Е. Вар- минга основоположником одного из интереснейших разделов (!) совре- менной биоморфологии — экзоморфологии («экологической морфоло- гии», по И.Г. Серебрякову, 1962) — учения о габитусах организмов, которых он называл «жизненными формами». Весьма плодотворные исследования растений, которые можно считать эколого-экзоморфоло- гическими, состоялись в XX столетии (Warming, 1908, 1909, 1916, 1923; Raunkiaer 1907, 1934; Drude, 1887, 1890, 1913; Высоцкий, 1915; Пачос- кий, 1916; Gams, 1918; Казакевич, 1922; Du Rietz, 1931; Meusel, 1935, 1951, 1970; Закржевский, Коровин, 1935; Прозоровский, 1936; Rauh, 1937, 1938, 1939, 1950; Козо-Полянский, 1945; Станюкович, 1949; Се- нянинова-Корчагина, 1949; Алехин, 1950; Scharfetter, 1953; Серебряков,
~дава 2. Концеиция ириоритетов развития в организации систем 1952; 1954, 1959, 1964; Шалыт, 1955; Хржановский, 1958, 1966, 1968; Зозулин, 1959, 1961; Голубев, 1960, 1962, 1965; Серебрякова, 1971, 1972; Нечаева и др., 1973; Гатцук, 1974; Хохряков, 1975а, б, 1981; Юрцев, 1976; Мазуренко, 1986; Жмылев и др., 1993; Алексеев, 1996; Татаренко, 1996 и многие другие). Все они достойны внимания и уважения, что в целом позволяет нам по праву окрестить этот век на пути к становлению современной биоморфологии как эпоху (век) габитуальной биоморфо- логии, или экологической экзоморфологии. Вклад многих ученых— современников Е. Варминга и тех, кто работал после него, — в биомор- фологии никто не уменьшает, но иногда бывает важно несколько разоб- раться с фактами и определиться, что же из накопленного материала работает на развитие того или иного научного направления. Оригиналь- ный приоритет в эколого-экзоморфологическом направлении изучения растений определил К. Раункиер (Raunkiaer, 1907, 1934 и др.) — совре- менник Е. Варминга; он классифицировал жизненные формы по поло- жению почек или верхушек побегов относительно к поверхности почвы в неблагоприятное для внешнего роста время года. Выделенные Раун- киером типы растений: фанерофиты, хамефиты, гемикриптофиты, крип- тофиты, однолетники и терофиты, удачно используют многие биомор- фологи по сей день. Дю Рие (Du Rietz, 1931) успешно опробовал приемы классифицирования жизненных форм под разными углами обзора, ис- пользуя по несколько групп экзоморфологических признаков. Приоритетом исследования немецких ботаников в середине ХХ столетия, использующих обычно термин «формы роста», стало морфо- логическое поведение индивидуума во времени (Muhlberg, 1965; Gluch, 1967, цит. по Т.И. Серебряковой, 1972). Приоритетом в подходе к изу- чению жизненных форм Г.М. Зозулиным (1959) была избрана способ- ность особей удерживать за собой площадь в связи с возможностью по- разному вегетативно возобновляться. В работах И. Г. Серебрякова (1962, 1964), Т.И. Серебряковой (1971, 1972), А.П. Хохрякова (1975, 1981), М.Т. Мазуренко (1986) и др. приоритетом «эколого-экзоморфологичес- ких» исследований признается габитус вегетативного тела растительных организмов, наблюдаемый в динамике (в морфогенезах), во взаимосвязи с ритмикой сезонного развития и другими приспособлениями к совре- менным и прошлым условиям среды. В конце ХХ века биоморфология вступила в новую стадию своего развития, которую можно «окрестить» по тому приоритету, который складывается сейчас в исследованиях растительных биоморф (жизнен- ных форм), как время интегральной биоморфологии. В эпоху современ- ного развития биоморфологии автор этих строк делает попытку (Нухи- мовский, 1970, 1973 а, б, 1974, 1980, 1986, 1997 и др.) утвердить интег- рационные приоритеты в комплексно-динамическом изучении организ- мов (простых биоморф) и синорганизмов (сложных биоморф). Любые признаки, развивающиеся у биоморф в их морфогенезах (онтогенезах или синонтогенезах) — от рождения до смерти, могут стать объектом исследования биоморфолога. В интегральной, или современной, био- морфологии в центре — в ее сердцевине — находится учение о жизнен- ных циклах (биоморфогения, или биографика), которое не только дает
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИЙ теоретическое обоснование динамизму жизненных форм, но и обеспе- чивает всю биоморфологию необходимым исходным фактическим ма- териалом. Сегодня биоморфологи должны осознать, что приоритеты биоморфологии со временем изменялись и изменились настолько, что признаковое пространство биоморф уже не должно быть ограничено только признаками габитуса — оно неизмеримо богаче, если в целом и организмы, и синорганизмы признаются за жизненные формы (биомор- фы), но при всем этом выбор приоритета конкретного биоморфологи- ческого исследования принадлежит биоморфологу и зависит от целей такого исследования. Осознание организмов и синорганизмов как мно- гопризнаковых телесных систем в ранге биоморф вовсе не означает, что интерес к отдельным сторонам их организации будет утрачиваться и сосредотачиваться только на интеграционном (целостном) восприятии этих объектов живой природы. В этом отношении я совсем не катего- ричен, поскольку считаю, что интерес к различным сторонам органи- зации биоморф не иссякнет, но он будет более осмысленным, разложен- ным «по полочкам», пропущенным через призму интеграционного по- нимания биоморф; к этой мысли в нашей книге предстоит еще не раз прийти в рамках различных обобщений, что, кстати, во многом предоп- ределило и тему второго тома как обоснование и утверждение двух обязательных разделов современной (интегральной) биоморфологии: экзоморфологии, или габитологии, — науки о габитусах биоморф — и биографики, или биоморфогении, — науки о жизненных циклах, воз- растных и фенологических состояниях (формах роста) биоморф.
Глава 3. Концепция габитуса в биоморфологии В натуре каждого тела, причем не только одушевленного, но и неодушевленного, незримо присутствует некоторое выражение, пусть образное, его собственного я (лат. «эго»), что автором было незамедли- тельно закреплено в латинизированном варианте терминологии главных меросом телесных систем. В первой главе было показано, что материя в телах Природы существует в двух состояниях: идиосоматическом, или эгосубстратном, что во втором варианте термина почти буквально озна- чает весьма стабильную меросому (носитель преемственности) «собственно фундамент (подкладку) здания», и пластоматическом, или конституциальном, — так обозначается более или менее лабильная меросома. У понтикул — структурных элементов эгосубстрата — всегда присутствует особое свойство: быть не только в определенном «фунда- менте здания», но и дисперсно рассредотачиваться в самом «здании»— конституции тела, используя его по прямому для себя назначению как место своего существования. Эгосубстрат, будучи результатом эволюции преемственности в организации телесных систем, сохраняет информа- цию о том, чем они потенциально могут быть и чем они быть не могут, а конституция, как основной результат трансформации (кинетопласти- ки) в телесных системах, зависит от особенностей реализации идиоэ- нергии (ресурса возможностей), сосредоточенной в эгосубстрате соот- ветствующих тел, во взаимодействии их с окружающей внешней средой. Все взаимодействие тел осуществляется только в Природе (другого им не дано). Чтобы определиться, что же, весьма существенное, в орга- низации всех тел нам уже удалось «высветить», а что пока осталось за кадром, построим, хотя бы в сокращенном варианте, «пирамиду» абст- ракций телесных систем, т.е. посмотрим на иерархию тел, в зависимости от сложности их организации. 3.1. Габитус биоморф в пирамиде» абстракций телесных систем Для оценки сложности организации тел при рассмотрении их как ~иерархических систем применим методы субординатного и суперординатно- го классифицирования (т. 1, гл. 1), что позволит далее определиться, с чего
252 ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИЯ габитус начинается и где он имеет место быль. Для построения «пирамиды» абстракций тел используем уже разработанные в первой главе положения. На самой высокой ступени (суперординате) «пирамиды» абстракций (суперординатной классификации) тел (рис. 149-А) поместим одну ши- рокую субординатную классификацию (субординию) из двух субординат (классов тел): монадосомы (одноконтурные тела) и плеосомы (многокон- турные тела). На том же уровне абстракции разместим следующую клас- сификацию, нижнюю субординату в которой занимают пластосомы (кон- ституции тел), а верхнюю — идиосомы (эгосубстраты). Диапазон слож- ности организации пластосом очень широк; его отличает «ошеломляю- щее» разнообразие: от очень простых систем, например, клеточной па- ренхимы стеблей, системы эпидермальных волосков и т.п. до очень слож- ных систем, например, таких, как человеческий мозг с его, во многом еще непознанной, сознательной деятельностью и т.д. Все это — многообразие кинетопластики (соматической трансформации), и об этом никогда не стоит забывать. Оно не смогло бы состояться, если бы однажды в эво- люции не возникли качественно довольно стандартизованные, по слож- ности организации, системы в целом менее разнообразные, но все же еще более сложные, поскольку они способны осуществлять великое таинство — управлять направлениями премственно-исторического развития теле- сных систем, короче, если бы не возникли когда-то понтикулы: прогены и гены, а вместе с ними и их системы — эгосубстраты: прогеносомы и геносомы, то ни о каком пластосоматическом разнообразии в организа- ции телесных систем сегодня не могло быть бы и речи. Вот почему конституции тел по сложности организации мы в субординатной класси- фикации ставим ниже эгосубстратов. На этой же ступени пирамиды абстракций телесных систем, но под иным углом обзора, расположим третью субординатную классификацию также из двух субординат: габи- тусы (экзосомы, внешние организации тел), эндосомы (внутренние орга- низации тел). На ступень ниже расположим классификацию из трех субординат (рис. 149-Б): эуабиосомы (неживые абиогенные тела), некро- сомы (неживые биогенные тела), биосомы (живые тела). Пятую широкую субординию тел мы строим из четырех суборди- нат: ультрамикросомы, микросомы, макросомы и ультрамакросомы (рис. 149-В). На той же суперординате (уровне абстракций) расположилась и шестая широкая субординатная классификация из четырех субординат: парафеносомы, прогеносомы, феносомы и геносомы. Из четырех субор- динат состоит также и седьмая субординия тел как меросом: протеро- сомы, габитусы (экзосомы), субгабитусы (эндопластосомы, или внут- ренние конституциальные организации тел, включая и внутренние фе- нетические организации живых тел, или эндофеносомы), идиосомы (иначе эгосубстраты). Изменение сложности организации в телах-частях (меросомах), отраженных в последних классификациях, довольно под- робно рассмотрено в предыдущих материалах (глава 1). На следующей суперординате нашла свое место (рис. 149-Г) вось- мая субординатная классификация тел из пяти субординат (снизу вверх): эуабиосомы — любые тела, которые никогда не существовали в живом состоянии и продолжают оставаться таковыми; некросомы — это какие-
~лава 3. Концеиция габит са в биомо логии либо тела, которые когда-то существовали в живом состоянии, но сей- цас они мертвы, и только остатки прежней организации еще указывают на их биогенное прошлое; эвриэфемеросомы — это тела, которые суще- ствуют в живом состоянии не более одного года, а затем отмирают; пробазосомы — это любые живые тела, которые пока прожили не более одного года, но по многим параметрам их организации они потенциаль- но должны будут жить более одного года, чего впоследствии достигнут или по каким-либо причинам такого не произойдет; базосомы — это те живые тела, меросомы включительно, которые реально живут уже более одного года и могут так существовать еще и дольше. Среди рассмотренных классификаций мы, наконец, вышли на га- битус, увидев его, как бы между прочим, среди других тел, точнее ча- стей-тел (меросом). Габитус устроен явно проще других телесных час- теи, а, главное, он устроен проще той другой меросомы — внутренней пластосоматической организации (субгабитуса), которая неизбежно со- провождает его существование в любых телах. Невольно мы затронули проблему взаимоотношения категорий «внешнее» и «внутреннее», обхо- дить которую стороной не будем. Рис. 149*. Графическая суперординатная модель («пирамида» абстракций) сложности организации телесных систем. А — Г — уровни абстракции; субор- динаты (классы) тел: 1 — монадосомы (одноконтурные тела), 2 — плеосомы (многоконтурные тела), 3 — конституции (пластосомы), 4 — эгосубстраты (идиосомы), 5 — габитусы (экзосомы), 6 — эндосомы (внутренняя организа- ция тел), 7 — эуабиосомы, 8 — некросомы, 9 — биосомы, 10 — ультрамик- росомы, 11 — микросомы, 12 — макросомы, 13 — ультрамакросомы, 14— парафеносомы, 15 — прогеносомы, 16 — феносомы, 17 — геносомы, 18— протеросомы, 19 — субгабитусы (эндопластосомы, в частности, эндофеносо- мы у живых тел), 20 — эвриэфемеросомы, 21 — пробазосомы, 22 — базосомы.
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ 3.2. Внешнее и внутреннее в организации биоморф Изменим угол обзора и посмотрим на материю в организации живых тел в двух ее состояниях «внутреннего» и «внешнего». Значение категорий «внешнее» и «внутреннее» как форм отражения бытия и как свойств Природы, ее тел и процессов, трудно переоценить в современ- ном познании. Категории должны обеспечивать динамизм и пластику отражаемых ими явлений. «Внешнее является необходимым условием существования и развития внутреннего. Это — общая закономерность во взаимодействии материальных систем с окружающей их внешней средой» (Карпинская, 1966, с. 64). Проблема взаимоотношений внеш- него и внутреннего, безусловно, выходит далеко за пределы взаимодей- ствия материальных систем и окружающей их внешней среды. Так, Р.С. Карпинская (1966, с. 63) подчеркнула, что при изучении организа- ции живых тел важно раскрыть переход от «внутренних молекулярных изменений» к «собственно биологическим внешним свойствам организ- мов» и наоборот. Исходя из рассмотренных ранее материалов (глава 1), под опреде- ленным углом обзора в организации живых тел, биоморф, в особенно- сти, к категории «внутреннее», принадлежит эгосубстрат, т.е. геносома — субконтинуальная (весьма стабильная, но не бесконечная) биохими- ческая меросома (у неживых тел эгосубстрат представлен только проге- носомой), а к категории «внешнее» — конституция тел, которая у живых тел является феносомой (у неживых тел — это парафеносома), во вза- имодействии которых обеспечивается рост, развитие и дискретность существования соответствующих тел. Эгосубстрат, как уже отмечалось выше, — носитель преемственности, а конституция — главный носитель («выразитель») соматической трансформации. Существование геносомы и феносомы у живых тел стало в процессе эволюции возможно, по- скольку жизнь адаптировалась быть только при таком обязательном условии, когда неживая материя ассоциируется и интегрируется в струк- туре живых тел в качестве ее первичной меросомы — протеросомы, т.е. обязательного единства прогеносоматических (атомов) и парафеносома- тических (некоторых молекул и агрегатов молекул) элементов как кон- тинуального неживого субстрата (основы) жизни. Все известное и неизвестное нам в Природе разнообразие тел про- является через внешнее — их конституцию. Однако, сама эта часть во всех вариантах организации телесных систем коррелятивно взаимодей- ствует с другой меросомой — эгосубстратом, поскольку обе эти меро- сомы — результат меротомирования тел под определенным углом обзо- ра. Если поменять при меротомировании угол обзора, тогда обе упомя- нутые меросомы «растекутся» по другим меросомам, но не утратив своего качественного содержания, поэтому о существовании их в теле стоит только захотеть вспомнить, как они обнаружат себя. Здесь и сей- час речь по~ет о двух других меросомах: внутренней организации тел (эндосоме, или субгабитусе) и внешней организации тел (экзосоме, или габитусе).
Глава 3. Концепция габитуса в биоморфологии 255 Посмотрим еще раз на графическую модель простой биоморфы (организма) и попробуем совместить в ней меросомы, полученные при меротомировании под разными углами обзора, то получается любопыт- ная картина (рис. 6), в которой параллельное меротомирование вполне совмещается в одной модели, а следовательно, при изучении организа- ции биоморф вполне можно использовать меросомы, полученные путем разного меротомирования. Габитус в графической модели (рис. 6В) изображен всего лишь одной линией. Через такую линию — графичес- кий символ габитуса — определяется его смысл: это только наружность, ее контуры и вся поверхность тела; он не имеет внутренней структуры и прикрывает собой эндосому, составляя наружные соматические границы биоморфы. Из приведенного выше (глава 1) материала видно, что в материаль- ной системе, будучи организованной в любые тела и нетела, проявля- ется эффект обязательного присутствия, который распространяется только на те части этой системы, без которых другая часть (части) не может развернуться и (или) проявить себя в своем определяющем ка- честве. Эффект обязательного (атрибутивного) присутствия особенно ярко проявляет себя при анализе систем через категории «внешнее» и «внутреннее». В определении отношений между частями систем важно правильно ориентироваться по углам обзора и высотам обобщений, поскольку любой перекос может привести к неправильной аргумента- ции при оценке частей систем через категории «внешнее» и «внутреннее». В Природе, безусловно, действует один из высших законов — закон взаимодействия внешнего и внутреннего, который можно выразить так: присутствие внутреннего и внешнего составляет одну из необходимых сторон содержания любой системы и обеслечивает отношения между ее частями, в которых npuopumem влияния одной части на другую может лроявляться или не лроявляться, но атрибутивным условием всегда оста- ется их взаимосвязь и взаимовлияние в организации любых систем, без чего существование таких систем было бы невозможно. Рассматриваемые категории проявляют себя наиболее ярко в био- логии при решении проблемы взаимоотношений организма (внутренне- го) и среды (внешнего). В таком случае «необходимость рассмотрения предмета как целого в его отношении с другими предметами является столь определяющей, что деление самого предмета на внешнее и внут- реннее отступает на второй план, предмет в целом выступает как внут- реннее (по отношению к окружающей его среде)» (Карпинская, 1966, с. 74). Не только в отношениях предмета (внутреннего) с окружающей его средой (внешнее) можно наблюдать действие этого закона; оно всюдно. Так, например, сознательная деятельность человеческого мозга — это внутреннее свойство человека, используя возможности которого он способен создавать внешнее в форме элементов идеальной природы: идей, понятий, мечтаний и т.п. Внутренним в организации живых те- лесных систем, как было показано выше, является протеросомная часть их организации, без которой внешняя часть этих тел — их геносома и феносома не смогли бы состояться и существовать. Под иным углом обзора, в живом теле геносома — это внутреннее по отношению к
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ феносоме — внешнему. Еще с одной стороны внутренним в организа- ции биоморф семенных растений, одним из типов частей живых тел, является эндофеносома (субгабитус), а внешним — экзосома (габитус). Как следствие из высшего закона взаимодействия внешнего и внут- реннего надо рассматривать специальный закон взаимосвязи внешнего и внутреннего в организации живых телесных систем: любое живое тело так организует развитие своих частей (меросом), что всегда внутреннее содер- жание лроявляет себя как лервичное (лриоритетное) по отношению к другому — внешнему — содержанию этой же телесной системы, и при этом второе (внешнее) не может возникнуть без nepsozo (внутреннего), а nepsoe, чтобы соответствующее тело — целое — отвечало олределяющему ее качеству, не может существовать без второго. Из этого закона логически выводится правило: всякая часть живого тела может состояться только в тех случаях, если сначала будет разви- ваться внутреннее содержание этого тела. Примеров правила приори- тетного развития внутреннего по отношению к внешнему предостаточ- но. Однако не будем забывать, что в отношении категорий «внешнее» и «внутреннее» выбор исследовательской программы (парадигмы) имеет решающее значение. Так, развитие вегетатива (вегетативной части) в организации биоморф является внутренним по отношению к развитию репродуктива (репродуктивной части), которое выступает как внешнее. Ситуация изменится, если «выйти» из организации биоморф и «войти» в организацию вида. У вида развитие вегетатива во многом как бы растворяется в развитии репродуктивной части его организации, шире части, обеспечивающей размножение, — это его внутреннее, а внешним в таком случае для организации вида является деятельность его элемен- тов в окружающей среде по защите и сохранению потомства в популяциях. Безусловно, действие категорий «внешнее» и «внутреннее» таит еще немало перспектив для развития конкретного знания. Однако, здесь и сейчас мы обращаемся к ним, к закономерностям, связанным с ними, чтобы лучше разобраться с габитусом биоморф семенных растений, определить причины и следствия этого атрибутивного явления в их организации. Из приведенного выше материала следует, что далеко не всякое внешнее является габитусом (экзосомой), есть немало нюансов, с которыми надо считаться, и, в первую очередь, многое зависит от нас самих — от того, как мы посмотрим на предмет нашего исследования: под каким углом обзора и с какой высоты абстракции. 3.3. Габитус биоморф, причина и следствия Согласно рассмотренным выше положениям, габитуальная, или экзоморфологическая, организация, короче, габитус (экзосома), прояв- ляет эффект постоянного присутствия в организации биоморф, чего не скажешь о многих иных их частях, например, листья могут быть часто только летом, а к зиме они полностью опадают; соцветия — сегодня они есть, а завтра их уже нет и т.д.
~дава 3. Концеиция габитуса в биомо ологии Известно, что термин «габитус» применяется в науке ограниченно, например, для обозначения наружного вида кристаллов (Schneer, 1970, 1978; Минерал. энц., 1985), а также внешнего облика растений и живот- ных (Warming, 1884; Серебряков, 1962; Хохряков, 1978; Реймерс, 1991 и np.). Габитусом у кристаллов называют их общую поверхность, в кото- рой решающее значение придается внешней морфологии граней и их QTíîñèòåëüíûì размерам. Габитус кристаллов формируется в процессе их роста. У других тел общая поверхность может быть также обозначена как габитус, однако конкретные приоритеты в определении его парамет- ров должны расставлять специалисты тех областей знаний, которые непосредственно изучают внешнюю морфологию таких природных тел. В геоморфологии, например, поверхность тела Земли традиционно обозначают термином «рельеф», а различные его проявления на отдель- ных участках территории как «формы рельефа» (Пиотровский, 1961; Подобедов, 1964 и др.); если, по большому счету, то геоморфологи изучают ничто иное как габитус тела Земли и его формы. В приведен- ном небольшом обзоре нам важнее другое: габитус — атрибутивное свойство любых материальных тел, организованных в системы разного рода с более или менее определенными границами. Что касается бота- ников, то они традиционно настроены обозначать общую поверхность растительных организмов, их внешний облик как габитус. И.Г. Серебряков (1962) в обзоре о жизненных формах растений убедительно показал, что с давних времен, начиная с Феофраста, при- знаки нару>кной организации растений, их размеры и форма крона, широко использовались для обозначения растений по общему внешне- му виду (габитусу), который нередко принимался в более поздние вре- мена за жизненные формы. Статья А. Гумбольдта «Идеи о физиономич- ности растений» (Humboldt, 1806, цит. по Гумбольдт, 1936) оказала сильное влияние на развитие в последующие годы габитуального (фи- зиономического) подхода к исследованию растений и растительности. Если габитус, или экзосому, определять как наружность, ее конту- ры и всю поверхность тела, а в целом как всю внешнюю организацию какой-либо материальной системы-тела, то габитуса не будет только у процессов (материальных нетел) и идеальных нетел (логоапосом), а также у гиперапосомы («сверхнетела») — вселенского потока (Природы) и ее высших видов частей — Супернуклеуса и космоплазмы. Почему стало возможным такое широкое определение габитуса? Пожалуй, только потому, что мы сразу настроились не ограничиваться рамками узкого эмпирического исследования и заставили себя посмот- реть на предмет исследований с разных сторон. Оказывается, что у организмов есть много общего с другими живыми и неживыми телами — монадосомами, а у синорганизмов много общего с другими телами- плеосомами, причем главное, что их объединяет, — наличие внутренней и внешней организации, где последняя и есть габитус. Итак, габитус проявляет себя в структуре одноконтурных тел (монадосом) — кирпи- чеи, домов, плотин, отдельных гор и т.п., в структуре определенных многоконтурных (дизъюнктивных) тел — плеосом (биоценозов, популя-
258 OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ций и т.п.), но и в структуре тех и других (биосом) вместе с окружаю- щими их внешними условиями существования, например, экоидов, биогеоценозов, биосферы и т.п. Все биоморфы, как растений, так и животных, обладают габитусом: у организмов — моноидным (в един- ственной форме одного тела), а у синорганизмов — полиоидным (в дизъюнктивной форме многих тел). В первом томе нашей работы был проведен критический литератур- ный обзор применения термина «жизненная форма», которая опреде- лялась нередко только или в основном через габитус. Там же было показано, что недостаточно для такого емкого термина, как «жизненная форма», говорить о габитусе (внешнем облике) организмов. Осознавая, что габитус — это еще не жизненная форма, а только некоторая ее часть (одна из сторон организации), автор решил подробно разобраться с этим явлением у семенных растений. Термин «габитус» имеет, согласно приведенному выше определе- нию, общенаучный смысл. Биоморфы — живые телесные системы оп- ределенного свойства (организмы и синорганизмы). На вопрос: в чем причина развития габитусов у телесных систем и биоморф, в частности, и каковы следствия этого явления, можно ответить с самых общих позиций. Причина габитуса в целостной организации материальных сис- тем-тел, биоморф, в частности, — обязательное наличие в таких систе- мах внутреннего, существование которого возможно только при выиолне- нии Природой необходимого условия: ограничения внутреннего от окружа- ющей среды и прикрытия его снаружи, а таким общеприродным прикры- тием всякого внутреннего является что-то внешнее. Параметры внешнего варьируют в связи с варьированием параметров внутреннего, однако, га- битус всегда остается строго определенной формой внешнего. Следствий существования габитуса может быть много, но они ориентированы в основном на поддержание внутреннего (это причина габитуса). Нас более всего будет интересовать следующее следствие габитуса — его способ- ность неплохо отражать через себя состояния внутреннего, т.е. признаки габитуса неразрывно связаны с признаками обязательного условия его про- явления — с иризнаками внутреннего, иоэтому с уверенностью можно использовать многие признаки наружной организации в качестве индика- торов состояний всей организации каких-либо тел. Резюмируем предельно кратко сказанное выше: причина габитуса- наличие внутренней организации у тел, а следствия могут быть разные, важнейшее для биоморфологии — способность габитусов, его признаков, неплохо отражать через себя, в качестве индикаторов, состояния внутрен- него и состояния всей организации рассматриваемых систем. Габитус, следовательно, — универсальное свойство систем материальной приро- ды определенной соматической организации, имеющих какие-либо границы, отделяющие их от других систем с иной организацией, — всех тел. Габитус биоморф семенных растений — частный случай габитусов тел вообще. Определившись с термином «биоморфа» (жизненная фор- ма) [т.1], можно подробно разобраться и с перспективой применения в биоморфологии термина «габитус». Габитус воплощает собой наружные соматические рубежи биоморфы (единые, или моноидные у организмов
259 laaea 3. Концеиция габитуса в биомо логии 3. 4. Формы габитуса и габитопроцесс Выше было показано, что посредством логических возможностей мы определяем, где работает категория «внешнее» и с чем она соотно- сится при рассмотрении категории «внутреннее». Так, внешнее — это пластосома, а внутреннее — идиосома; под несколько иным углом об- зора внешнее будет — феносома, а внутреннее — геносома; наконец, внешним будет репродуктивное по отношению к внутреннему — веге- тативному и т.п. Такое манипулирование категориями «внешнее» и «внутреннее» — результат абстракции. Однако есть ситуации, где не обязательно прибегать к особым логическим построениям, где внешнее и внутреннее определяются как само собой разумеющееся; внешнее- »о то, что располагается в организации тела кнаружи, составляя его поверхность — внешнюю организацию, или габитус (экзосому), а внут- p<»ee — это то, что располагается в том же теле внутри его, вслед за габитусом, составляя его внутреннюю организацию, или эндосому. Только внутреннюю пластосоматическую часть (эндопластосому) соот- ветствующих (телесных) систем вполне можно обозначить еще и как и дизъюнктивные, или полиоидные у синорганизмов). Согласно опре- делению, габитус характеризует биоморфу только снаружи и не имеет внутренних границ, более того, он не имеет внутренней структуры, что особенно ярко показывает единство внешнего и внутреннего в органи- зации биоморфы и относительную условность перехода между ними. Габитус — это не монотонная поверхностная система, а по-разному организованная система частей, носителей, непосредственно связанных с ними признаков. Части габитуса, отличающиеся от других его частей, обозначим для удобства особым словом-этикеткой, т.е. определим тер- мином «эпимер» (дословно с греческого — «часть сверху»). Когда габитус биоморфы характеризуется через признак эпимера, то это локальный (частный) габитуальный, или эпимерный, признак. Например, розеточ- ная биоморфа охарактеризована через эпимерный признак (розеточное листорасположение в структуре побегов), который индикаторно несет информацию не только о форме габитуса, но и о всей жизненной форме. 0 «признаковом» пространстве биоморф семенных растений мы еще поговорим подробно (глава 5), а здесь ограничимся констатацией факта: гвбитус биоморфы — это вся совокулность элимеров биоморфы разной сложности своей особой (наружной) организации, главная функция которой — ограничивать тело биоморфы от внешней среды, nepsou лринимать на ством с ней, даже защищать, насколько это вообще возможно, внутрен- нюю организацию биоморфы (ее эндосому) от неблаголриятных воздействий и лередавать от окружающей среды разнообразную информацию и какие- либо другие ресурсы для остальной части биоморфы, а еще, что не менее важно для нас, исследователей, отражать состояния роста и развития внутренней организации биоморфы и всей ее организации как целого.
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ «субгабитус», носители преднаследственности и наследственности суще- ствуют внутри пластосомы (конституции тел), но не являются ее частя- ми. Ловлю себя на слове: здесь, вероятно, не все так просто, как сейчас может показаться, поскольку, чтобы сделать такое, на самом деле дей- ствительно простое заключение, пришлось долгие годы многое доказы- вать и себе, и другим. Вероятно, у истины есть особое свойство: быть простой и всем понятной тогда, когда она кем-то на~ена и кем-то обоснована, доказана. Пусть так. Будем «наслаждаться» далее простотой истины, что любое тело, как живое, так и неживое, состоит из двух частей (меросом): габитуса (экзосомы) и субгабитуса (эндопластосомы), причем любое тело состоит из этих частей всегда, покуда оно остается быть телом. Ни габитус, ни субгабитус изъять из тела нельзя, не унич- тожив само тело, поскольку они являются атрибутивными (обязатель- ными) сторонами их организации. Посредством габитуса живые тела, биоморфы, в особенности, удерживают относительное постоянство сво- ей внутренней среды (своей эндосомы) — гомеостаз (у организмов), несколько отдаляя и защищая ее от непостоянства окружающей внеш- ней среды. Габитус, тем самым, является важным звеном в поддержании известного противоречия, которое служит одной из движущих сил раз- вития жизни на Земле. Только «перелопатив» множество фактов «за» и «против», удалось, как нам представляется, докопаться до сути: габитус двойственен в своем проявлении. Далее все будем говорить применительно к биоморфам, хотя многое, при желании, можно экстраполировать не только на жи- вые, но и на неживые тела. В организации любой биоморфы (шире — любых тел) ее внешняя часть, или габитус, npucymcmsyem постоянно, проявляя одновременно в рамках биоморфогенеза (цикла существования биоморфы) и свой цикл развития (эпицикл), будучи, тем самым, не только структурой, но и процессом (габитус как процесс-стадия и процесс-цикл — это габи- топроцесс), а, с другой стороны, габитус свойствен и каждому по отдель- ности текущему возрастному и (или) фенологическому состоянию биомор- фы, проявляя себя, как габитопроцесс-стадия, в формах — формах габи- туса, или габитуальных формах, короче — габитоформах. Термин «габи- туальная форма» впервые использовал Ф. Клементс (Clements, 1920), на что обратил внимание Дю Рие (Du Rietz, 1931). Согласно Клементсу, «габитуальная форма» — это аналог «экологических классов» (Warming, 1909). При рассмотрении единиц «габитуальной экологии» (habitat ecology; Du Rietz, 1931, р. 48) Варминг использовал «экологические классы», а Клементс для подобных случаев применял термин «габиту- альные формы» (habitat forms), например, к ним он относил оксилофи- ты, психрофиты и галофиты. В таком смысле более термин «габитуаль- ные формы» не использовался. Далее мы будем употреблять термин «габитоформа» (габитуальная форма, или форма габитуса) только в одном смысле: для обозначения отдельного состояния (формы) габитуса биоморфы в определенный отрезок времени (стадию эпицикла). Габитоформы нередко могут быть индикаторами возрастных и (или) фенологических состояний биоморф, или, что одно и то же, форм роста биоморф, о чем подробнее по~ет речь в следующей главе.
~дава 3. Концеиция габит са в биомо ологии Так сменяется ли габитус в биоморфах? Габитус как часть органи- зации биоморфы постоянно присутствует в ней, сопровождая ее от рождения до смерти, но при этом он постоянно изменяется, поэтому надо еще говорить и о формах проявления габитуса (габитоформах), которые сменяются в организации одной биоморфы, более того, в рам- ках одного эпицикла, т.е. в границах одного габитуса как части (меро- сомы) единой организации биоморфы от ее рождения до смерти. Образ- но, действительно можно сказать, что формы габитуса (габитоформы) сменяются в организмах «почти как рубашки, по мере необходимости» (T. 1, гл. 2). Габитус — это структура, но и постоянная функция субгаби- туса биоморфы, которые организационно друг без друга не существуют. Ни эндофеносоме (субгабитусу), ни габитусу в организации тела биоморфы сменяться незачем. Габитус изменяется сколько угодно, а сменяется не он, сменяются только его формы (состояния). Кто-то может возразить: здесь игра слов и не более того, но стоит вспомнить коварство выражения без знаков препинания: казнить нельзя помиловать». Развитие биоморфологии лишний раз подтверждает известные ак- сиомы отражения знания: важно не только найти нужные по смыслу слова, но и правильно расположить их по отношению друг к другу. Явление, когда в организации одной биоморфы на протяжении всего цикла ее развития происходит смена габитоформ — это габитопроцесс, в котором габитус проявляет себя постоянно, но не одинаково, посколь- ку является «крайней», весьма лабильной частью организации биоморф. Сегодня у габитуса биоморфы может быть одна форма, завтра — другая, а через некоторое время — третья и т.д. Пожалуй, будет правильнее говорить не о смене габитусов организмов, а о смене форм габитуса (одного, общего и единого) организма (тоже одного). Если это игра слов, то я готов и дальше прибегать к таким играм. Истина дороже... Итак, в организации любой биоморфы ее внешняя меросома — это габитус, который присутствует в ней постоянно, являясь одновременно и растянутой во времени обязательной стороной всего цикла суще- ствования биоморфы, т.е. габитопроцессом-циклом, или эпициклом, в биоморфогенезе, и текущим его состоянием — формой габитуса (габи- туальной формой), или габитоформой, т.е. состоянием габитуса в опре- деленной стадии эпицикла. В настоящее время, пожалуй, будет правиль- но говорить, что габитус, или экзосома (внешняя организация), как и субгабитус, или эндофеносома (внутренняя фенетическая организация) присутствуют в биоморфе постоянно, а вот их формы (состояния) сме- няются в биоморфогенезах вместе со сменой возрастных и фенологичес- ких состояний биоморфы, соответствующие части феносомы которых они составляют. Большинство признаков имеет интегративную природу, поэтому, когда мы говорим о габитуальных признаках, то чаще всего фиксируем внимание на той стороне признаков, которая относится собственно к габитусу, а с позиций исследователя, анализирующего габитус, такие признаки «обслуживают» эту часть организации телесных систем, био- морф, в частности. По сути, работая с признаками, мы, иногда и не подозревая этого, меротомируем их — членим на части, в соответствии
ОСНОВБ1 БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИЙ с науками, их изучающими. Например, придаточные корни у растений или не образуются, или образуются мало, или они многочисленны. В первую очередь, придаточное корнеобразование — это физиологичес- кий признак, но если присмотреться к этому явлению габитологу, то в этом же явлении он отметит проявление тех же признаков, но уже морфологических и, в частности, габитуальных. Когда говорят, что придаточных корней мало или много — это признаки, на которые по праву могут и должны опираться и физиологи, и морфологи, называя ту часть в организации признаков, с которой они работают, соответ- ственно физиологическими или морфологическими, даже габитуальны- ми, где она обслуживает габитус, признаками. Подробнее об этом в главе 5. 3.5. Габитус в организации биоморф семенных растений — парадоксы, оказавшиеся атрибутами Сама постановка темы о «габитусе в организации биоморф» может быть воспринята как парадокс, но так будет продолжаться только до того момента, пока мы полноценно не осознаем, что габитус — это не биомор- фа (не жизненная форма), а только часть в организации любой живой си- стемы, в первую очередь, организмов, но и синорганизмов (см. т. 1). Настало время глубже осмыслить содержание слова «габитус». Бе- зусловно, не стоит наделять его смысловым статусом, которого он не заслуживает: быть жизненной формой он не может — не для габитуса такая «шапка», она ему не по «размеру» (великовата), но он вполне достоин того, чтобы составлять определенную часть в структуре и орга- низмов, и синорганизмов, которые, по статусу, — действительно формы жизни. Габитус — это не просто внешний облик; его статус повыше, по- скольку быть просто обликом — это значит что-то отражать и ни за что не отвечать перед тем, содержание кого ты как-то отражаешь. В отно- шении габитуса так сказать нельзя, если считать его частью организации любых биоморф, структуру которых он не только как-то отражает, но и защищает сверху, прикрывая собой, как получится, внутреннюю орга- низацию этих тел. Явление «жизненная форма, или форма жизни (био- морфа)» несоизмеримо богаче явления «габитус». В организации живых тел у габитуса есть свое место, и наша задача правильно увидеть, где же это место. Это мы, люди, должны показать ему, габитусу, место на той научной «полочке», которую мы конструируем ддя удовлетворения сво- его любопытства (габитусам тел до этого нет дела), чтобы понять все то, что составляет организацию жизни на Земле, шире — организацию Природы. Одно из направлений нашей текущей научной работы как раз в том, чтобы разобраться, где же его — габитуса — законное место в организации живой природы и, в частности, биоморф, что, естественно,
1лава 3. Концеиция габитуса в биомо ологии отразится и на месте его на научной «полочке» (свое место габитус имеет в Природе, независимо от нас, это мы, люди, нередко сомнева- емся в поисках истины, другого нам — искателям новаций — не дано; пытаемся определить его место, это наша, но не габитуса, проблема, и об этом никогда не стоит забывать). Габитус — это больше, чем внешний облик, это специфическая часть — организация в телесных системах, биоморфах, в частности, без которой самостоятельное существование их невозможно. Габитус ат- рибутивно возникает как меросома в организации биоморф, оставаясь не жизнеактивным, а жизнепассивным, поскольку вся динамика его роста и развития исходит не от него, а от внутренней организации, поскольку у него вся структура только снаружи, а внутренняя структура ему никак не принадлежит. Он возникает как функциональный ответ внутренней организации биоморфы на то, как в целом реагирует эта система на окружающую внешнюю среду во времени, через особый вид структурной новообразовательной деятельности — габитуальное произ- водство, как ответ внутренней организации (в основном, это ее ответ) своим ростом и развитием, охватывающим заодно рост и развитие сво- его «сообщника» — габитуса. Сам габитус не способен проявлять соб- ственной активности в своем росте и развитии, за него рост и развитие ему обеспечивает только внутренняя организация биоморф, без которой он — ноль, а вместе с ней — надежная сила, обеспечивающая многое, и в целом весьма необходимая часть в теле любой биоморфы. Похоже, габитус — это, действительно, клубок парадоксов в орга- низации биоморф, который не только любопытно, но и весьма актуаль- но, наконец, размотать. Итак, первый парадокс («неожиданное явление») и первый атрибут («существенное свойство»): габитус — это не биоморфа, он может быть только частью в организации любой биоморфы, но не более того (рис. 6). Второй парадокс и, заодно, второй атрибут: габитус обеспечивает внутренней организации биоморфы основную информационно-энерге- тическую поддержку снаружи, на правах внешней организации биомор- фы, — контакт во времени с окружающим ее жизненным пространством и защищает ее же и биоморфу в целом от неблагоприятных воздействий окружающей среды, составляя обязательное звено в системе гомеостаза. Быть внешним обликом и не иметь внутренней структуры, как может показаться, быть почти ничем, для габитуса — это слишком малая «ноша». Его роль многократно усиливается, когда в этот термин вкладывается большее содержание, когда при меротомировании он приобретает свой законный статус быть меросомой (одной из двух ча- стей под определенным углом обзора) — быть внешней организацией тел, в частности, биоморф и наряду с внутренней организацией обес- печивать целостный смысл всей организации у телесных систем. Габи- тус постоянно присутствует в организации любой биоморфы, его нельзя удалить из ее (или какого-либо еще) тела, не разрушив качественное содержание самого тела. Нельзя, например, удалить с лика Земли рель- еф (это в широком смысле один из типов габитусов), поскольку на смену прошлому рельефу одномоментно придет новый рельеф. Кто бы
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ и как бы ни пытался удалить габитус из организации любой биоморфы, сохранив прежнее качество этой телесной системы, тот ничего не добь- ется (искомого результата не получит), поскольку у габитуса, представ- ленного как часть в качественных границах организации прежнего тела, есть обязательное свойство не сменяться самому, а сменять только свои состояния (формы). С габитуса живого организма вполне можно «соскоб- лить» горстку «молекулярной пыли» или даже срезать его часть (пусть к тому же еще и самую большую) с куском внутренней организации этой простой биоморфы, например, обрезав дерево до пенька (рис. 147), но пока мы не уничтожим прежнее тело-организм, то и не уничтожим его габитус, а только сменим одну форму габитуса на другую. Здесь важный момент, который автором еще не был точно зафиксирован в первом томе: правильно будет говорить, что габитус в биоморфе, как определенная часть ее организации, присутствует в ней, от рождения до смерти, посто- янно, сменяется не весь габитус целиком, а только его состояния— формы габитуса (габитоформы). При исследовании габитуса биоморф мы работаем снаружи (чтобы изучать субгабитус — внутреннюю фенетичес- кую организацию, надо, как правило, анатомировать тело особей, прони- кая внутрь его), снимая слепки с его состояний (форм), фотографируем или зарисовываем его отражения, получая в руки в таких случаях не сам габитус, а только образные «следы» от него, так или иначе фиксирован- ные извне через отношения с ним. Итак, парадокс третий, он же и атрибут третий: габитус (внешняя организация, или экзосома) как определенная часть (меросома) в теле- сных системах (биоморфах семенных растений, в частности) постоянно реально существует только с другой частью этих систем — их внутрен- ней организацией (эндосомой). Габитус получает для себя постоянно в готовом виде предназначенные ему результаты роста и развития субга- битуса (эндофеносомы) и только так он растет и развивается, но свою службу несет нещадно. Парадокс четвертый, он же и атрибут четвертый: габитус биоморф (шире любых телесных систем) может существовать в пространстве и во времени только как-то изменяясь, но сменяется в биоморфогенезах (шире — морфогенезах) не сам габитус, а сменяются его состояния (рис. 150 — 190) — формы габитуса (габитоформы, или габитуальные формы). Одна из особенностей внутренней фенетической организации био- морф в ее отношении к внешней организации в том, что они обе су- ществовать друг без друга в обычных (не экспериментальных) условиях не могут, причем первичен и более независим субгабитус. Габитус со- вершенно не может существовать без субгабитуса. Если как угодно пытаться отделить габитус от остальной фенетической части биоморфы, то он без такого неразделимого союза перестает существовать как габи- тус. В рассматриваемых отношениях габитус является сателлитом, а внутренняя фенетическая организация (субгабитус), производя и под- держивая габитус, проявляет приоритет. Габитус сам не растет и сам не развивается, он «безукоризненно выполняет роль» сообщника — сател- лита (исполнителя чужой «воли») субгабитуса, повторяя те результаты роста и развития, которые проявляет и достигает субгабитус; инициати-
nasa 3. Концепция габитуса в биоморфологии ва активного поведения — трансформаций всегда исходит из внутренней организации, а результаты такой динамики находят отражение во внеш- ней организации, т.е. внешнем облике биоморф в целом. Рост и развитие внутренней фенетической организации биоморф производит часть тка- ней кнаружи и все то, что составляет поверхность над субгабитусом, т.е. все то, что вышло кнаружи, будучи когда-то произведено изнутри (в субгабитусе), и есть внешняя организация, или габитус, биоморф. Не стоит также забывать, что инициатива к разнообразным трансформациям биоморф исходит не только от внутренней организации, стремящейся к росту и развитию, но и снаружи — со стороны окружающей внешней среды, которая давит на габитус биоморфы, как на ее «защитный пан- цирь» (конечно это громко сказано, но все-таки...), воздействием своих экологических факторов. Габитусам ничего не остается как «защищать» свои биоморфы от разнообразных воздействий окружающей среды, но и принимать на себя первую информацию от биотопов к внутренней орга- низации биоморф и передавать ее незамедлительно по назначению. Внут- ренняя организация реагирует на все, что происходит в окружающей среде, всевозможными ответными реакциями, в результате которых габи- ~с более или менее изменяется в отдельные отрезки времени (стадии). Трансформация габитуса, таким образом, — это не столько трансформа- ция этой меросомы, сколько всей феносомы биоморфы, которая нахо- дится сама в безусловном подчинении генетической организации био- морфы, откуда исходит управляющая роль генов, определяющая фенети- ческие изменения в определенном диапазоне возможностей. Выше (глава 3.2) были рассмотрены особенности взаимодействия внутреннего и внешнего в организации телесных систем вообще и живых тел, в особенности, но, пожалуй, есть смысл пойти еще дальше по пути конкретизации действия соответствующих законов такого взаимодей- ствия, доведя их до очередного следствия — субспециального закона взаимодействия внутреннего и внешнего в организации биоморф семенных растений. Смысл этого закона выразим так: npuopumem в фенетической организации биоморф семенных растений принадлежит их основной меро- соме (части) — внутренней фенетической организации (эндофеносоме, субгабитусу), без которой существование любой биоморфы невозможно, и одним из атрибутов его является обеспечение опосредованного роста и развития другой меросомы — внешней организации (экзосомы, габитуса) самостоятельно расти и развиваться, предназначенной разграничивать и как-то защищать все внутреннее тело биоморфы от окружающей извне среды, непосредственно взаимодействуя с ней, оставаясь при этом всегда в подчинении у внутренней организации, для которой габитус поставляет в себе воплощает и собой отражает. Из этого закона хорошо выводится правило габитуального поведения биоморф: развитие организации биоморф (организмов и синорганизмов) осуществляется целесообразно через адекватную реакцию как целого, так и частей, на возмущающее воздействие окружающей внешней среды, в <omopou роль первого, пусть незначительного, защитного приспособления
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ отведена их габитусу, формы которого наиболее пластичны и нередко (бывают и внешне незаметные адаптации) являются индикаторным от- ветом всей биоморфы на соответствующее воздействие внешней среды в текущий отрезок времени и в данном месте их биотопа (рис. 150 — 190). Итак, парадокс пятый, он же атрибут пятый: внутренняя организа- ция и, в частности, внутренняя фенетическая организация биоморф выступает приоритетом по отношению к их внешней организации. Парадокс шестой, он же атрибут шестой: габитус биоморф может существовать только в неразрывном единстве с внутренней организацией, но и последняя, несмотря на свое приоритетное положение в обычных (не экспериментальных) условиях, не может никак обойтись без габитуса. Парадокс седьмой, он же и атрибут седьмой: габитус в организации биоморф более всего реагирует на инициативу («провокацию») к транс- формациям не только со стороны их внутренней организации (опосре- дованно растет и развивается), но и со стороны окружающей внешней среды (принимает и передает информацию извне и далее адекватно участвует в целостном ответе на нее). Вероятно, судьбе было угодно образно вооружить автора «лопатой» и побудить именно его в нужное время и в нужном месте перекопать биоморфологическое «поле» заново, откопать нужные аргументы и осу- ществить ревизию многого, что было вроде бы понятно и раньше. На поверку оказалось, что узкий габитуальный подход к биоморфологии сегодня не проходит и что у этой науки более строгая теоретическая конструкция. Автор, «перелопатив» многое из известного и неизвестно- го в этой области знаний, сегодня готов утверждать, что им действитель- но создана стержневая теория современной, или интегральной, биомор- фологии — теория организации биоморф семенных растений (т. 1). Главное, что надо всегда помнить, все, что сейчас написано под назва- нием современная биоморфология, родилось не с чистого листа и не на голом месте (предтечей явилась классическая биоморфология, в центре которой всегда стоял габитус организмов), а в результате целенаправ- ленного анализа, через пробы и ошибки, в которых нашла отражение богатая история этой, безусловно, еще молодой науки. Итак, габитус, пожалуй, сам по себе (вне остальных частей роди- тельского тела) мало что стоит. Ведь он, по большому счету, только поверхность тел, не имеющая внутренней структуры, т.е. вне живых тел он всегда нежизнеспособен. Однако нас, более всего, он может заинте- ресовать только во взаимодействии и вместе (единстве) с внутренней организацией тел. Главное, что он, габитус, всегда просто есть в сома- тической организации жизни как часть, а, следовательно, имеет непос- редственное отношение к многослойности жизни. Каково оно? На этот вопрос есть смысл ответить чуть позже — в конце следующей главы, где мы разберемся еще и со статусом форм роста. Давайте там и поразмыш- ляем и вообще, и в частности еще раз о многослойности жизни, там и подведем некоторые итоги. Габитус — это главное — вовсе не пустячок, с которым можно не считаться, это важная часть организации биоморф, изучение которой — один из интригующих разделов современной био- морфологии.
nasa 3. Концепция габит са в биомо фологии Рис. 150 — 190. Возрастные состояния семенных фитобиоморф в биографических портретах (графических образах биоморфогений) ви- дов, имеющие интегральное содержание и отраженные через их инди- каторы — габитоформы. Рис. 150. Биографический портрет Fraxinus syriaca Воы. [сем. 01еасеае (из- раиль, горные склоны вдоль дороги между Хадерой и Хайфой, 20 IV)]. Биоморфотип — деревья. Организационные признаки (и физиологические, и эндоморфологические, и др.) биоморф нашли интегральное отражение в габитоформах; они стали в конкретных примерах— биографических портретах видов — индикаторами соответствующих возрастных состоя- ний. На примере с F. syriaca показаны габитоформы (их типы) — индикаторы возрастных состояний (их типов). У проростков (А) — семядольный ризопротерокормус (семядоли развертываются, а зародышевая почка еше не начала развертываться). У всходов (Б)— семядольный ризомонокормус (семядоли живы, развивается или уже полностью развер- нулся побег первого уровня организационной сложности — монокормус, который вместе
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ с корневой системой и семядолями составляет семядольный ризомонокормус. У ювениль- ных растений (В) — бессемядольный ризомонокормус или ризотетракормус (семядоли уже отмерли, нарастание одного главного побега эумоноподиальное). При необходимости от- мечаются растения первого ювенильного возрастного состояния, или униювенильные (В,), когда главный универсальный (простоосевой) вегетативный побег с простыми листьями, у биювенильных растений (В,) — габитоформа, сходная с В„но листья уже тройчатые, у триювенильных растений (В,) — все то же, но идет развертывание перистых сложных листьев. Имматурные растения (Г) проявляют габитоформы с вегетативными разветвлен- ными побегами. В условиях угнетения, например, при затенении подроста, аэроподии полегают, стремясь выйти из-под влияния неблагоприятных факторов. Взрослые растения (Д) проявляют репродуктивное развитие открытых комбинированно-универсальных побе- гов (гексакормусов). ?цветение впервые происходит на 25 — 35 году жизни. Большинство взрослых растений моноаэроподиальное (с одним стволом, разветвленным в густой кроне). Деревья старше 150 лет встречаются крайне редко. Далее сходным образом можно при желании только по условным обозначениям прочитать через габитуальные признаки интегральное содержание некоторых возрастных состояний биоморф и на всех осталь- ных рисунках рассматриваемого ряда (рис. 151 — 190). Биоморфотипы во всех рисунках указаны по типу взрослых габитоформ, т. е. сокращен- но — по габитотипу. Рис. 151. Биографический портрет Olea еигораеа 1.. [сем. 01еасеае (там же, что и рис. 150, март — апрель)). Биоморфотип — деревья.
Глава 3. Концепция габитуса в биомо фологии Рис. 152. Биографический портрет Frangula ainus Mill. [сем. Rhamnaceae (Московская область, смешанные леса, схема габитоформы на открытом месте — опушке леса — 1 биоморфотип; в условиях затенения внутри леса — II биоморфотип)). Био морфотип — кустарники.
ОСНОВЪ| БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИИ Рис. 153. Биографический портрет Lonicera altaica Pall. [сем. Caprifoliaceae (Алтай, Курайский хребет, 1 биоморфотип — в лесном поясе, 1500 м над ур. моря, II биоморфотип — высокогорья, семена заносятся выше лесного пояса птицами, причем иногда настолько высоко, что стелющиеся кустарнички могут прекращать репродуктивное развитие)]. Биоморфотипы: 1 — кустарники, П — кустарнички.
МС МС фП Рис. 154. Биографический портрет Atriplex halimus 1.. [сем. Chenopodiaceae (Израиль, овраг в каменистой пустыне Негев, 12 V)]. Биоморфотип — кустарники. глава 3. Концепция габитуса в биомо фологии 2,аг4(гм4), 206,26
OCHOBhl БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 155. Биографический портрет Atriplex portulacoides L. [сем. Chenopodiaceae (Тель-Авив, парк Хилтон, 6 V)]. Биоморфотип — кустарники.
~дава 3. Концепция габитуса в биомо фологии рис д Биографический портрет Teucrium creticum L. [сем ('1. „„д ид, Ботанический сад, 29 1П)]. Др — Фрагмент цветущего "обе'а (Feinbrun-Dothan, 19Т~) Биоморфотип — кустарники
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ б), 26) Рис. 157. Биографический портрет Helianthemum stipulatum (Forssk.) С. Chr.[ñåì. Cistaceae (окрестности Тель-Авива, склоны невысоких каменисто- песчаных холмов, 14 Ш)]. Биоморфотип — кустарнички. 2(5),гм4, 19б,21(22),26(27) Рис. 158. Биографический портрет Convolvulus secundus Desr. [сем. Convolvulaceae, (окрестности Тель-Авива, крутой склон каменисто-песчано- го холма, 16 IV)]. Биоморфотип — стелющиеся полукустарнички.
~дава 3. Концеиция габит са в биомо ологии Рис. 159. Биографический портрет Centaurea aegiptiaca L. [сем. Asteraceae (Израиль, каменистая пустыня, окрестности поселка Сде-Бокер, 12 V)]. Биоморфотип — полукустарники. 2,34 м4,156, 96(206), 22(21),27 Рис. 160. Биографический портрет Oenothera 4гаттоп4й Hooker [сем. Onagraceae (Тель-Авив, пустыри, парки, скверы, 7 V)]. Биоморфотип — полукустарники.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 161. Биографический портрет Delphinium mirabili Serg. [сем. Ranunculaceae (Алтай, Курайский хребет, высокогорные луга, иногда щебни- стые ocbmn)]. Биоморфотипы: ! (основной) — компактно-каудексовые травянистые многолетники, II (второстепенный) — вынужденно удлиненно-каудексовые травянистые многолетники.
Глава 3. Концепция габит са в биоморфологии Рис. 162. Биографический портрет Delphinium elatum L. [сем. Ranunculaceae (Москва, Главный ботанический сад, питомник лекарственных растений)]. Биоморфотип: корневищно-стержнекорневые травянистые много- летники. 78 Рис. 163. Биографический портрет Polygonatum odoratum (Mill.) Druce [сем. Convallariaceae (Алтай, Куминский хребет, низкогорные негустые леса)]. Биоморфотип — компактнокорневищные травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 164. Биографический портрет Saponaria oAicinalis L. [сем. Caryophyllaceae (Москва, Главный ботанический сад, питомник лекарственных растений)]. Биоморфотип — длиннокорневищные травянистые многолетники. Рис. 165. Биографический портрет Parnassia palustris L. [сем. Parnassiaceae (Алтай, луга в пойме реки в окрестностях села Ябоган)]. Биоморфотип: компактнокорневищные травянистые многолетники.
~ ава 3. уонцепция габитуса в биоморфологии афический портрет Schizonepeta multifida (L) Lamiaceae (Алтай, Курайский хребет, окрестности села Чи Чибит остепненные 1Х О ПОЛ СЛОЖНЫХ ьи ют по степени надреза пластинки листьев от простых до полу Растения варьируют по степени н азнооб азие листьев обнаруживается на одном (HerlapHo-перистых), причем иногда все разноо разие листьев растении. Биоморфотип: короткокорневищные травянистые мно ноголетники.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИИ 27 Рис. 167. Биографический портрет Caltha palustris L. [сем. Ranunculaceae (Алтайский край, заболоченные луга в пойме реки Бия)]. Биоморфотип: компактнокорневищные травянистые многолетники.
gnaea 3. Концепция габит са в биомо фологии СК рис. igg. Биографический портрет Geranium albiflorum Ledeb. [ceM. (егап1асеае (Алтай, Курайский хребет, высокогорные луга и луговые поляны в горных лесах)]. Биоморфотип: короткокорневищные травянистые многолетники.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ж,в~ Рис. 169. Биографический портрет Omalotheca norvegica (Gunn.) Sch. Bip. et F. Schultz [сем. Asteraceae (Алтай, Бащелакский хребет, высокогорные луга)]. Биоморфотип: компактнокорневищные травянистые многолетники. Рис. 170. Биографический портрет Pulmonaria mollis Wulf. ex Hornem. [сем. Boraginaceae (Алтай, разреженный смешанный лес в окрестностях Горно- Алтайска)]. Биоморфотип: компактнокорневищные травянистые многолетники.
283 Глава g. Концеиция габитуса в биомо логии Б В, В р ]q] Био„рафический портрет Елди1апа sibiric ( ) а L.) (;азу. [сем. Аьгегасеае (А тай Бащелакский, Черектинскии, Семин р ° Ю нский х ебты, сырые луга и бе- поясов го, заходит в BblcoKoroPa>) . pera горных рек нижнего и cpeaHero поя Биоморфотип: компактнокорневищные тр авянистые многолетники.
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННБ!Х РА СТЕНИИ Рис. 172. Биографический портрет Tanacetum vulgare L. [сем. Asteraceae (Московская область, Подольский район, опушка леса на краю сельскохо- зяйственных угодий)). Биоморфотип: полуразвернуто-кустистые корневищные травянистые многолетники.
Глава 3. Концеиция габитуса в биомо логии Рис. 173. Биографический портрет Allium altaicum Pall. [сем. Alliaceae (Алтай, Семинский хребет, гора Сарлык, открытые склоны, включая высокогорья)]. Биоморфотип: луковичные компактнокорневищные травянистые многолетники. Рис. 174. Биографический портрет Ranunculus acris L. [сем. Ranunculaceae (Московская область, Подольский район, лесные луга и опушки)]. Биоморфотип: компактнокорневищные травянистые многолетники.
д,42,92 l7I7 К 546,67 l7I7 СК Рис. 175. Биографический портрет Tephroseris praticola (Schischk. et Serg.) Но1иЬ [сем. Asteraceae (Алтай, Катунский хребет, луговые долины горных рек, поднимается и в высокогорья)]. Биоморфотипы: компактнокорневищные травянистые многолетники. 5,гм4 19а, 21,27 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ а, 26(27)
(gaea 3. Концеиция габит са в биомо ологии 0 Рис. 176. Биографический портрет Viola disjuncta W. Beck. [сем. Violaceae (Алтай, Курайский хребет, высокогорные лута)]. Биоморфотип: длиннокорневищные травянистые многолетники.
ОСНОВБ1 БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИЙ ,33а, Рис. 177. Биографический портрет Filipendula и1тапа (L.) Maxim. [сем. Rosaceae (Московская область, сырые места на лугах и лесных полянах)]. Биоморфотип: короткокорневищные травянистые многолетники. 5 В Нг Рис. 178. Биографический портрет Lythrum virgatum L. [сем. Lythraceae (Во- сточный Казахстан, Алтай, луга в пойме реки Иртыш)]. Биоморфотип: стержнекорневые травянистые многолетники.
r aea 3. Концеиция габитуса в биомо логи 54а,67 и по ez girsium heterophyii<m (~-~ Н'~~ ~~~~ рис. 179. Биографический "OÐ~Ð~~ -, е луга и лесные ь Подольский район, сырые Asteraceae (Московская область, д поляны)]. о-к стистые корневищные травянистые Биоморфотип: полуразвернуто-кустистые ко н многолетники.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИЧ 78 Р . 180 Б афический портрет Ins bloudowii Led . [ deb. [сем. Iridaceae (Во- ис.. иогр леса по горным сточный Казахстан, Алтай хребет Листвяга, разреженные л склонам и долинам горных рек)]. Биоморфотип: короткокорневищные тр авянистые многолетники.
гяава 3. Концепция габитуса в биоморфологии Рис. 181. Биографический портрет Saxifraga те1а1еиса Fisch. ex Spreng. [сем. Saxifragaceae (Алтай, Курайский хребет, высокогорья, берега ручьев и сырые луговины около снежников)). Биоморфотип: короткокорневищные травянистые многолетники.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 182. Биографический портрет Campanula rotundifolia L. [сем. Campanulaceae (Москва, Главный ботанический сад, питомник лекарствен- ных растений)]. Биоморфотип: корневищно-стержнекорневые травянистые мн«о- летники.
гяава 3. Концепция габитуса в биоморфологии иЙ ans ры О- ые Рис. 184. Биографический портрет Koenigia islandica L. [сем. Polygonaceae (Алтай, Курайский хребет, высокогорья, берега ручьев, около снежников)]. Биоморфотип: мелкие безрозеточные однолетники.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 185. Биографический портрет Limonium lobatum (L.) Chaz. [сем. Limoniaceae (Израиль, каменистая пустыня Негев, крутой склон в долину уже пересохшего русла зимне-весенней реки, окрестности поселка Сде-Бо- кер, IV)]. Биоморфотип: розеточные эфемеры. Рис. 186. Биографический портрет Senecio vulgaris L. [сем. Asteraceae (Мос- ковская область, поля, пустыри)]. Биоморфотип: безрозеточные однолетники.
~дава 3. Концепция габит са в биомо ологии Рис. 187. Биографический портрет Lepidium ruderale L. [сем. Brassicaceae (Московская область, пустыри)]. Биоморфотип: полурозеточные однолетники. 206, 8 Рис. 188. Биографический портрет Melampyrum nemorosum L. [сем. Scrophulariaceae (Московская область, Подольский район, смешанный лес)]. Биоморфотип: безрозеточные однолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 189. Биографический портрет Cannabis ruderalis Janisch. [сем. Cannabaceae (Восточный Казахстан, окрестности Усть-Каменогорска, пустыри)]. Биоморфотип: безрозеточные однолетники. Рис. 190. Биографический портрет Atriplex holocarpa F. Мие11ег [сем. Chenopodiaceae (Израиль, окрестности Эйлата, пустыри, 4 V)]. Биоморфотип: безрозеточные однолетники.
Глава 4. Концепция форм роста в биоморфологии Наиболее стабильная часть в организации биоморф и других теле- сных систем — идиосома (эгосубстрат), а самая лабильная — пластосома (конституция), что делает вторую меросому еще и самой дискретной. В предыдущей главе мы определились, что следует считать габитусом (внешней организацией, экзосомой), нашли его место в организации как живых, так и неживых телесных систем и установили, как эта ме- росома взаимодействует с другой меросомой — субгабитусом (внутрен- ней фенетической организацией, или эндофеносомой). Остается под- робнее разобраться с теми принципами, которые позволяют биоморфам и их меросомам существовать как в пространстве, так и во времени, взаимодействуя между собой и с окружающей их внешней средой. 4.1. Морфогенез и поведение — атрибуты существования всех телесных систем «Все течет — все изменяется» (Гераклит Эфесский, конец 6— начало 5 вв. до н.э.). Все тела в Природе находятся только в движении — изменении. Ранее (т. 1) мы уже подчеркнули, что основу существо- вания биоморф, а шире и всех других тел, составляет морфогенез- процесс (во временном отношении — цикл) существования (сохранения изменений) телесной системы от ее возникновения до преобразования в новое структурно-функциональное качество — новое тело. Проявле- ние морфогенеза — это одно из универсальных свойств всех тел в Природе: начав свое дискретное движение (морфогенез) в одной части пространства-времени в одном качестве, телесная система всегда закан- чивает его в другой части пространства-времени в ином качестве, ста- новясь иным телом (или телами), сохраняя, как и прежде, бесконечное движение материи на дискретном его отрезке (в новом морфогенезе или нескольких морфогенезах и т.д.). Цикл существования биоморф — био- морфогенез (онтогенез или синонтогенез). Изучение динамизма в суще- ствовании телесных систем нельзя ограничить только морфогенезом в целом, поскольку этот процесс буквально накачан изнутри самыми разными изменениями, происхождение которых обусловлено не только
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИЧ внутренними причинами, составляющими содержание развивающихся тел, но и, безусловно, воздействием на эти тела многих факторов окру- жающей внешней среды. Второй атрибутивный показатель динамизма организации телесных систем — их поведение, т.е. функциональные или структурно-функцио- нальные ироявления материальных систем — тел, в оиределенных отрезках времени их существования — стадиях морфогенеза. Такое понимание термина «поведение» имеет общенаучный смысл и позволяет рассмат- ривать его в отношении всех природных тел. Поведение составляет, в первую очередь, функциональную сторону организации телесных сис- тем, обеспечивая нередко и рост, и развитие производных структур, т.е. в целом поведение может быть самой разной признаковой ориентации и сопровождаться изменениями (ростом и развитием) морфологических, в том числе габитуальных, физиологических, биохимических и т.п. признаков. Поведение, таким образом, может характеризоваться по наукам, их изучающим, например, экзоморфологическое, или габиту- альное поведение, эндоморфологическое (анатомическое) поведение, физиологическое, химическое, генетическое поведение и т.п. Какие-либо функциональные изменения осуществляются не сами по себе, а исходят (!) om каких-либо структур и соировождаются развитием и изменением ироизводных структур. Становление каких-либо структур- ных особенностей организации — это, следовательно, функция исходных для них структурных частей — ироявление наведения телесных систем. Например, одной из функций субгабитуса, как формы его поведения, является развитие габитуса и его эпимеров, при этом габитус, будучи структурной частью организации тел, проявляет в процессе их суще- ствования немало своих функций. Поведение далеко не всегда связано только с ростом и развитием конкретных телесных систем, а нередко оно находится в подавляющей зависимости от окружающих их внешних, часто экстремальных, условий. Например, наклоны кроны деревьев при порывах ветра и погружение ствола и ветвей в воду и намокание их при подтоплении окрестностей, прилегающих к руслу реки, паводковыми водами. Намокание в ночном тумане булыжника на дороге, а затем высыхание его под утренними солнечными лучами — это проявление поведения булыжника и т.п. Особи видов ивы или тополя в условиях длительного подтопления паводковыми водами образуют на стволах и ветвях придаточные корни — это все примеры проявления поведения. Рост и развитие побеговых органов и их структурных элементов у ра- стительных биоморф — это разнообразие в структуре, обеспечиваемое разнообразием функциональным, что в целом составляет поведение таких биоморф. Поведение, или этоморфоз, присутствует в организации телесных систем постоянно и сопровождается какими-либо функциональными или структурно-функциональными проявлениями в существовании те- лесных систем, особенности которых связаны с определенными отрез- ками времени, всегда меньшими, чем морфогенез тел (соматогенез) в целом. Поведение, таким образом, — это всегда какой-либо фрагмент из
Глава 4. Концепция форм роста в биоморфологии морфогенезов. Поведение всегда соировождает морфогенезы men, их неотьемлемая часть, которая, в свою очередь, меротомируется (членится Hg части) на формы иоведения, или этоморфы в широком смысле,— обязательный, но, безусловно, разный результат в существовании любых телесных систем. Поведение — это процесс, в результате которого как- либо трансформируется организация тел и, в первую очередь, пласто- соматическая (конституциальная) их часть в рамках морфогенезов этих тел. Любое поведение, как, впрочем, и морфогенез в целом, осуществ- ляется во времени. Ранее (глава 3) уже было подчеркнуто, что габитус проявляет свое постоянное существование в организации биоморф, а шире — телесных систем вообще, в габитуальных формах, или габитоформах; все габито- формы — это формы экзоморфологического наведения телесных систем. Подобное можно сказать и о субгабитоформах как формах эндоморфоло- гического наведения телесных систем, а единство двух указанных выше этоморф в одной иоведенческой системе иозволяет говорить об основной единице стадийного наведения, ироявляющейся в морфогенезах, в частно- сти, биоморфогенезах, — морфогеническом (возрастном и сезонном) со- стоянии. Любое структурно- функциональное состояние телесной системы (как живой, так и неживой) в оиределенный отрезок времени (стадии)— морфогеническое, или организационное, состояние (т. 1, гл.7). Краткое выражение определенного состояния фенетической орга- низации как состояния ее роста и развития в организации какого-либо живого тела, например, биоморфы — это форма роста, или стадиосома (дословно с греческого «тело определенной ступени»), соответствующе- го живого тела. Форма роста всегда несет на себе «печать» времени, но обращать на этот факт внимание нужно тогда, когда того требуют об- стоятельства. Например, моноподиальная форма роста побега, обратно- яйцевидная форма роста листа, но также — ювенильная форма роста биоморфы или моноподиальная ювенильная форма роста растения и т.п., т.е. возможны самые разные, согласно определения термина, вари- ации в применении термина «форма роста». Форма роста биоморфы подчеркивает свою связь с определенным отрезком времени (стадией биоморфогенеза), в котором она проявляется и маркируется индикатор- ными организационно-соматическими признаками. Итак, синонимы морфогенических состояний — это морфогени- ческие (возрастные и сезонные) этоморфы (формы поведения), или морфогенические формы роста и развития, сокращенно — формы роста (когда речь идет только о живых телах), или стадиосомы. Попробуем остановиться на такой терминологии, которую удобно будет использо- вать далее. Пожалуй, в отношении биоморф более всего подойдет тер- мин «форма роста», который является частным случаем универсального термина «морфогеническое состояние», для которого кратким синони- мом может быть еще и новое слово «стадиосома» — результат стадий- ности развития телесных систем. Через стадиосомы (у биоморф, они же — формы роста) мы характеризуем обычно и телесные системы в целом (биоморфы в том числе), поскольку это, в первую очередь, состояния
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИИ их конституций, так или иначе трансформируемых в некоторых грани- цах и в определенные отрезки времени. Стадиосомы (формы роста в том числе) не развиваются вне пространства и времени, поэтому, как бы мы ни отвлекались от этих обстоятельств, они существуют всегда в них, причем независимо от нашего сознания, а, следовательно, в них так или иначе присутствуют какие-либо следы возрастности, а нередко и сезон- ности, развития. Если имеет место моноподиальная форма роста, то значит, какое-то время фенетическая организация растительной био- морфы, ее лидерная побеговая часть, нарастают моноподиально, и сле- дует ожидать, что на смену этой форме роста придет другая, у которой или будет иной тип нарастания побеговой системы, или, если нараста- ние не изменится, то проявятся другие признаки-индикаторы, посколь- ку в биоморфогенезах форм роста (морфогенических состояний) всегда должно быть не менее двух. Когда мы говорим, например, подушковид- ная форма роста, то подспудно имеем в виду, что она пришла на смену доподушковидной форме роста или какой-то еще, и все это в границах одной растительной биоморфы, а не возникла сразу из зиготы и т.д. Часто, пользуясь термином «форма роста», мы не заостряем внимание на стадийной природе образований с таким названием, поочередно возникающих во времени на протяжении биоморфогенезов и отражаю- щих, в первую очередь, стадийные процессы феносоматических транс- формаций, протекающих в них и индицируемых признаками определен- ных сравнительных рядов. Поведение, таким образом, это непрерывный и множественный (по разнообразию форм-стадиосом) процесс, обеспечивающий динамичес- кое существование тел, в частности, биоморф, в их морфогенезах. Если существование габитоформ осуществляется в эпициклах — стадийных габитопроцессах, а существование субгабитоформ — в эндофеноциклах как стадийных субгабитопроцессах, то существование стадиосом (форм роста у биоморф) происходит в феносомоциклах — в единстве всей последовательности их со стадиями морфогенезов. Конституция телесных систем — это тот особый материальный «поведенческо-архитектурный футляр», развернутый над эгосубстратом и, по большому счету, для него же, в первую очередь, предназначенный, в котором происходит стадийность соматических трансформаций— движение сменяемых в морфогенезах тел-частей (меросом) — стадиосом (форм роста у биоморф). Стадийное поведение и его формы, наряду с морфогенезами, проявляющиеся в неразрывной взаимосвязи и един- стве, — это атрибуты динамического существования телесных систем, через которые удобнее всего познавать таинства движения материи в таких системах, в нашем случае, в биоморфах семенных растений. Важно также осознавать, что «гены не могут предопределять тече- ние онтогенеза, независимо от окружающей среды, в которой происхо- дит развитие. Гены могут влиять на процессы развития разными путями, но на эти же процессы влияют средовые факторы» (Мак-Фарленд, 1988, с. 43). Генетические факторы, безусловно, влияют на проявление пове- дения биоморф, но «в генах, — как подчеркнул Д. Мак-Фарленд (1988,
Inasa 4. Концепция рм оста в биоморфологии с. 43), — не может содержаться подробной информации для конкретных видов поведения». Гены участвуют в контроле за процессом развития; они детерминируют синтез белков, которые непосредственно регулиру- ют сложные процессы онтогенеза. К настоящему времени можно счи- тать доказанным, что поведение организмов, их фенотипическое разви- тие носит скорее всего «эпигенетический характер, т.е. каждая стадия развития создает основу для следующей стадии, а не предопределяет ее» и «все три элемента — ген, окружающая среда и исходный фенотип- играют определенную роль в нормальном развитии» (Мак-Фарленд, 1988, с. 37 — 38). 4.1.1. Как и почему поведение составляет атрибутивную часть в организации биоморф Все, о чем говорилось ранее, с большей или меньшей убедительно- стью, но все же однозначно подводит нас к осознанию свойства, кото- рым, независимо или зависимо от нашего сознания, обладает органи- зация телесных систем, в особенности живых, в частности, биоморф: эти тела, будучи системами, устроены упорядоченно, сложно и состоят из разных частей-элементов как структурных, так и функциональных. Количество и качество этих элементов варьируется, в зависимости от объективных и субъективных обстоятельств, но главным остается факт, все они находятся во взаимодействии, обеспечивая целостное единство тел, которым они принадлежат, если хотите, на которые, всем своим существованием, «работают». Биоморфы чем-то напоминают тонкостенные прочные резиновые сосуды, заполненные определенным составом, дифференцированным в структурах и функциях, которые объемно изменяются через свой рост и развитие, где-то перемешиваются, а где-то не очень, но стадийно во времени сменяются на другие части, которые не поступают извне, а «вылепливаются» по определенной программе тут же, из арсеналов прежних частей, то ускоряя, то замедляя, а иногда вроде бы даже и останавливая свой бег-развитие, а затем, о, чудо! — прямо-таки «взры- ваются», «выплескивая» поверх себя энергетику неистовых репродуктив- ных усилий. Неужели всему конец, и сложная комбинация телодвиже- ний под единым названием «жизнь» распылила «сосуд»-биоморфу в могучем репродуктивном «взрыве» в пространство и время, лишив его прежнего качества, отдав во власть стихии окружающей среды возмож- ность восстанавливать и продолжать течение жизни через пробуждение необходимого для ее возобновления идиоэнергетического ресурса из различных фрагментов (семян, деструктивных партикул и т.п.) от пре- жних живых «сосудов». Такое бывает, и часто, но далеко не всегда. Многие «сосуды»-биоморфы разрушаются навсегда, но другая их часть адаптировалась находить резервы в себе и в окружающем пространстве, чтобы вновь «раскачаться» в развитии и повторить, может быть, еще и
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ даже не раз, взрывной характер заключительного аккорда репродукгив- ного циклического развития, после которого самому «био-сосуду» суж- дено или погибнуть, или только пострадать ради вида, научившись «зализывать» раны и жить дальше иногда так успешно, даже до несколь- ких тысяч лет, что никто, кроме некоторых семенных растений (напри- мер, секвойя, баобаб, драконово дерево), такого повторить не может. Каждая биоморфа состоит из структурных частей (меросом): нежи- вых и живых. Как целое, так и меросомы, существуют в движении— изменяются, проявляя поведение (этоморфоз), т.е. функционирование оп- ределенных тел на определенных отрезках времени, которое состоит из своих поведенческих частей, или форм поведения (этоморф). Есть ка- кая-либо биоморфа как тело, и в ней ее организация, обладающая опреде- ленным порядком составляющих тело структурных частей (меросом), существование которых сопровождается определенным порядком их иове- дения в определенных поведенческих частях и в целом биоморфы как сис- темы форм поведения. Этоморфы, в отличие от биоморф в целом, не просто изменяются, а обычно через какое-то время в биоморфогенезах действительно сменяются, и, что самое интересное, сменяемость это- морф не сопровождается сменяемостью биоморфы, в организации которой они себя проявляют. Почему это происходит так, а не иначе? Потому что для биоморфы, ее целостное тело одно, а вот этоморф — поведенческих частей в ней — множество. Этоморфами, например, являются формы роста побегов, волосков, рыльцев пестика, корней, листьев и т и., но и формы цветения, плодоношения, корнеобразования, ветвления побегов и т и. Существование биоморфы сопровождается проявлением, т.е. npucym- ствием в ней множества этоморф, проявляющих себя определенным обра- зом под разными углами обзора, из взаимодействия которых формируется целостная система поведения каждой биоморфы и munos поведения опре- деленных групп биоморф. Эгоморфы в организации биоморф сменяются неоднократно, но никакого «выбрасывания» из биоморфы отслуживших ей этоморф не происходит. В организации биоморфы всегда присутствует потенциаль- ный арсенал форм поведения, к которому она прибегает в развитии при необходимости на основе своего ресурса возможностей — своей идиоэ- нергии, и в каждый отрезок времени, в зависимости от окружающих условий среды, проявляется, адекватная всему происходящему, этоморфа. Этоморфоз неукротим; здесь не только нарастание побегов, корней и других органов и всякие формы иного развития по восходящей, но и разрушение структур и биоморф в целом при развитии по нисходящей— по пути к элиминации, а, кроме того, еще и многое другое. Например, наклон ветвей под действием ветра или сползание куста с оползнем вниз по горному склону, — это все этоморфы, но они не имеют прямого отно- шения ни к росту, ни к развитию биоморф. Значит, этоморфы — разные не только по особенностям их проявления, но и по происхождению. В целом организация биоморф проявляет максимальную изменчи- вость в своей фенетической части — феносоме. Она — основной носи- тель форм поведения, которые проявляются в организации биоморф.
~дава 4. Концепция орм оста в биоморфологии многие из этоморф «фантомны» (скоротечны, как призраки), и присут- QòBèå их мы даже не замечаем, поскольку их перекрывают другие, более заметные, формы поведения. Этоморфа распускания одного цветка поглощена часто этоморфой цветения побега, а тем более всего расте- ния, и составляет относительно небольшой отрезок в жизнеразвитии биоморфы. Другая этоморфа — снабжение водой листьев и репродук- тивных органов — действует нередко непрерывно или ритмически на протяжении всей жизни растения и т.д. Пожалуй, пора классифицировать этоморфы биоморф семенных растений, чтобы лучше ориентироваться в их разнообразии. В суборди- натной классификации можно выделить два класса (субординаты) форм поведения биоморф. Неростовые (на этом уровне действуют этоморфы без ростовой активности) формы поведения: пассивоформы (они внешне могут быть пассивны или даже активны, но в последнем случае, как и в первом, у них учитывается не ростовая, а другая деятельность, причем именно в этом акте поведения, например, раскачивание кроны дерева, и еще они не включают случаи деградации развития или нанесения травм), деградоформы (их поведение обеспечивает разрушение биомор- фы по частям или в целом, в связи с деградацией развития), трагиформы (они отражают с их стороны пассивное принятие биоморфой травмиро- вания ее частей и тела в целом). Ростовые этоморфы, или активоформы, проявляют себя в ростовой деятельности. Разновидности этоморф у биоморф семенных растений- это формы роста (стадиосомы), причем более всего активоформы пове- дения биоморф. Нас, в текущей работе, интересуют далеко не все фор- мы поведения, а в основном те, которые действуют на ростовом уровне их стадийного проявления в организации биоморф семенных растения, т.е., в первую очередь, как формы роста биоморф. Такие этоморфы- не только носители признаков, но и, что особенно важно, главные производители их. Признаки биоморф сосредоточены, в основном, в их феносоме; это она отражает своим поведением главное содержание в поведении всей биоморфы. Формы роста в организации биоморф про- изводят довостребованные признаки, а уж от нас, исследователей, зави- сит, как мы их востребуем (подробнее см. главу 5). Когда возникает новая структура в организации биоморф, то она- результат направленного ростового поведения прежней структуры, а начинается с того, что от последней исходит функция построения новой структуры. Пластосоматическое разнообразие в организации тел и, в частности, феносоматическое — в организации биоморф — это то, что мы действительно наблюдаем в первую очередь, вступая во взаимодей- ствие с организацией тел. У любого булыжника на мостовой есть ста- диосомы, как формы поведения (мы понимаем это явление широко), или этоморфы, есть пассивоформы (разновидность этоморф), но у него нет ростового развития — нет активоформ поведения и, следовательно, нет форм роста. Производителями большинства признаков биоморф являются, бе- зусловно, биоморфы, причем та их часть, которая непосредственно
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИИ активно растет и развивается, а это только феносома с ее действующими признаковообразовательными элементами — формами роста. Признаки биоморф — это признаки их производителей и непосредственных но- сителей — их форм роста, которые выступают в качестве безупречных индикаторов организации биоморф, которой они принадлежат. Из нескольких частей организации биоморф (протеросомы, гено- сомы и феносомы) — только феносома отражает признаки разнообразия биоморф, т.е. признаки, которые проявляются в биоморфе через фено- сому. Феносома есть в биоморфе только тогда, когда есть другие орга- низационные части биоморфы, без которых она не только не живет, она вообще не существует, ее просто нет. Все указанные выше меросомы биоморф — это атрибутивные части их организации. Без биосомы- целого нет феносомы-части. Все разнообразие биоморф обеспечивается их ростом и развитием, поэтому нас в их динамической организации, в первую очередь, будут интересовать основные производители и носители признаков, обеспечи- вающих разнообразие биоморф, — их формы роста. Габитус (внешняя организация) биоморф, кроме всего прочего, является еще и результатом функционирования другой их части — суб- габитуса (внутренней фенетической организации) и своим особым по- ложением в целостной организации биоморф нередко наглядно, через свои признаки передает интегральные стороны признакового простран- ства этих телесных систем, индикаторами которых часто они являются. 4.1.2. Синдром возрастности в стадийном поведении биоморф Существование живых существ, а шире — всех телесных систем на Земле, происходит под влиянием различных ритмов, например, цирка- дианных, или циркадных, иначе околосуточных (Halberg, 1959), а также околоприливных, окололунных, окологодовых и др. (Aschoff, 1964, Ашофф, 1984а, б). Ритмичность окружающей среды, безусловно, так или иначе влияет на развитие биоморф в их морфогенезах, главная особенность которых — возрастная стадийность. Биоморфогенезы (циклы существования биоморф) состоят из морфогенических стадий, а сами био- морфы из морфогенических (возрастных и фенологических) состояний, или соответствующих этоморф — форм роста (стадиосом). Биоморфы, как и все живое, рождаются, развиваются по восходя- щей, достигают в развитии некоторой стабильности, поддерживая то, что уже создано, а затем развитие поворачивается и идет по нисходя- щей, достигая, наконец, дискретного финала — выхода за «порог» (эли- минации). Чтобы все морфогенические состояния как формы морфоге- нического (возрастного и фенологического) поведения (стадиосомы) состоялись в циклах существования биоморф в определенное время и в определенных местах (частях) их организации, в них обязаны проявить себя соответствующие признаки-индикаторы, которые, в случае их
Глава 4. Концепция форм роста в биоморфологии проявления, свидетельствуют как о начале развертывания, параметрах, так и о конце действия конкретной стадиосомы (формы роста). Процесс развертывания в организации тел, в частности, биоморф, на определенных возрастных стадиях морфогенезов некоторого сочета- ния их признаков-индикаторов можно для удобства обозначить как синдром возрастности в организации телесных систем и, в том числе, биоморф семенных растений. Этот синдром является атрибутивным свойством телесных систем и находится под контролем идиоэнергии этих тел, т.е. их преднаследственно-наследственного ресурса и сильно зависит от фона окружающей среды, в которой происходит их развитие. Синдром возрастности биоморф контролируется, естественно, генами, но экология местообитаний биоморф сказывается непосредственным образом при выборе из ресурса возможностей соответствующих количе- ственных и качественных параметров в его конкретном развертывании. Возникает любопытный вопрос: а можно ли влиять на действие синд- рома возрастности и на стадийность биоморфогенезов? Похоже, да, поскольку развитие биологии идет в сторону овладения секретами гене- тического развития, а следовательно, будут появляться возможности вносить коррективы в синдром возрастности, управлять и изменять схемы его действия. При анализе биоразнообразия следует учитывать фактор возрастно- сти (уже пора, раньше было не до него, это понятно, но сейчас без него просто нельзя); альтернативы просто не предвидится. Синдром возра- стности в организации биоморф обеспечивает им возрастное разнооб- разие. Оно является обязательным условием развития растительного мира, и с ним сегодня уже нельзя не считаться. Все известные по сей день работы по систематике и флористике написаны в одном ключе. В них суждение о разнообразии биоморф построены только по типам организации, в основном, репродуктивной, у средневозрастных (меди- матурных) растений, которые наиболее стабилизированы в возрастном развитии по многим организационным признакам. Гербарии во всем мире пока отражают лишь некоторые особенности организации взрос- лых матурных, чаще всего медиматурных, растительных биоморф, по- скольку разнообразие в проявлении синдромов возрастности при накоп- лении таких коллекций раньше не учитывалось. Вполне понятно, что пройденный ранее путь в развитии система- тики и флористики был не столько единственно верным, сколько для своего времени единственно возможным. Так было вчера, когда клас- сическая систематика и флористика описали и изучили основное раз- нообразие видов растений на Земле. Сегодня не только биоморфология, но и сопредельные науки, например, систематика и флористика, дол- жны сделать шаг дальше и овладеть секретами проявления синдрома возрастности и процесса стадийности для познания разнообразия рас- тительных биоморф, что стимулирует развитие биологической науки на HQBQM его витке. Недалек тот день, когда в гербарии всего мира будут собираться разновозрастные растения и их фрагменты, и суждение о биоразнообразии и его фактическом обеспечении будет скорректирова-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ но с учетом действия в организации биоморф синдрома возрастности и процесса стадийности. Ритмичность в окружающей среде, стадийность биоморфогенезов и сменяемость форм роста в организации биоморф — это важнейшие составляющие для современного биоморфологического анализа. Всякая биоморфа в своем развитии двойственна: в ней какое-то время суще- ствуют, а затем сменяются ее стадиосомы (возрастные состояния, воз- растные этоморфы, формы роста), но об этом факте достаточно по- мнить, а писать, если помнить (!), вовсе не обязательно, поэтому дос- таточно записать, например, ювенильное растение (биоморфа), а это означает то же, что и ювенильное возрастное состояние биоморфы, ювенильная этоморфа биоморфы, ювенильная форма роста биоморфы, ювенильная стадиосома биоморфы. Различные формы поведения, в том числе и стадийного поведения составляет обязательное содержание еще более общего явления гетеро- кинеза (глава 6.3). 4.2. Биографика — сердцевина современной биоморфологии Носителями стадийных свойств в организации живых тел, в част- ности, биоморф, является их феносома (фенетическая организация). Феносома биоморф развертывается и развивается под управляющим воздействием на нее идиоэнергии, исходящей из геносомы этих же биоморф, и под влиянием условий окружающей внешней среды. Фено- сома биоморф вынуждена развиваться пластично и нередко совершать соматические трансформации, чтобы биоморфа могла существовать, выполняя генетически предопределенные ей функции и реагируя, своим изменением, на воздействие окружающей среды, причем наиболее из- меняясь модификационно в режиме «нормы реакции» при воздействии на нее экстремальных условий, а когда ресурса возможностей у биомор- фы не хватает, то феносома, принимая удар среды на себя, обеспечивает условия для осуществления мутации генов со всеми вытекающими из этого последствиями. Биоморфа, чтобы существовать, посредством сво- ей феносомы, научилась ритмически изменяться в связи с изменения- ми, происходящими во времени как внутри ее организации, так и в окружающей среде. Изменения (большие и малые) происходят в био- морфах в рамках их циклов существования (биоморфогенезов) — жиз- ненных циклов организмов (онтогенезов) или синорганизмов (синонто- генезов). В первом томе (гл. 7) уже были определены исходные понятия учения о жизненных циклах биоморф; здесь мы несколько расширим тему, в связи со сформулированной в названии этой книги проблемой, усилим одни и ослабим другие акценты, а, главное, определимся, какое место должно занять учение о биоморфогенезах в биоморфологии се- менных растений.
Глава 4. Концепция форм роста в биоморфологии Так получилось, а, возможно, иначе и не должно бы и быть, что, в процессе развития биоморфологии, изучение жизненных циклов ра- стений и развитие учения об онтогенезах и синонтогенезах постепенно переместилось с «окраин» этой науки в ее середину, причем настолько, что сегодня можно уже утверждать — раздел биоморфологии, изучаю- щий биоморфогенезы, — биоморфогения, или, еще короче, и, пожалуй, даже удачнее — биографика, составляет сердцевину современной био- морфологии. Биоморфология в своем развитии впитала и впитывает лучшее от других наук, сохраняя свою самобытность и фактуальность («накачанность» фактами), благодаря своей сердцевине, которую опре- деляет ее раздел — учение о жизненных циклах организмов и синорга- низмов — биоморфогения, или биографика. У истоков этого учения стоят работы многих ботаников. В историческом плане я бы раньше других вспомнил Т. Ирмиша (Irmisch, 1850а, б, 1859, 1860, 1861 и др.), который уже в середине позапрошлого века удивительно точно рисовал разновозрастные растения, особенно наиболее юные из них, и умело, весьма профессионально, по современным меркам, можно сказать, классически точно объяснял происходящее с растениями в их индиви- дуальном развитии, начиная от рождения и до становления взрослых организмов. Не скрою, что «первые уроки» рисования растений автор мысленно брал не у знаменитого Ван Гога, а у почтенного Т. Ирмиша. Биографика до наших дней не была определена как наука, способная как самостоятельно решать задачи индивидуального и кондивидуально- го развития растений, так и быть сердцевиной — основополагающим разделом еще более крупной науки — биоморфологии. В разной мере в развитие биографики, а, следовательно, и биоморфологии в целом, внесли большой вклад ученые нескольких поколений (Warming, 1884, 1908, 1909, 1916; Velenovsky, 1905, 1907, 1910, 1913; Богдановская-Гие- нэф, 1926; Goebel, 1931; Troll, 1937, 1939, 1943, 1964, 1969; Rauh, 1938, 1939, 1950; Кренке, 1940; Пошкурлат, 1941, 1962, 2000; Прозоровский, 1949; Серебряков, 1952, 1954, 1964; Игнатьева, 1958, 1965; Серебрякова, 1959, 1962, 1971; Уранов, 1960, 1973, 1974; Борисова, 1961; Голубев, 1962, 1965; Шорина, 1964, 1967; Смелов, 1966; Csapody, 1968; Нухимов- ский, 1968, 1974, 1980, 1981; Рысина, 1973; Мазуренко, Хохряков, 1977; Мазуренко, 1986; Жукова, 1986, 1995; Жукова, Шестакова, 1995 и др.). В отечественной литературе наиболее последовательные представ- ления о жизненных циклах и дифференциации их на возрастные состо- яния сформировались в работах Т.А. Работнова (1945, 1946, 1950, 1960, 1964, 1974 и др.), которого, безусловно, можно считать одним из осно- воположников биографики растений, и, как показало развитие ботани- ки, — всей современной фитобиоморфологии (биоморфологии расте- ний) в целом. Приоритеты развития современной биоморфологии к началу XXI столетия существенно изменились. Биоморфология, сбросив с себя немодную габитуальную «шляпку» и вегетативную «гордыню», искусст- венно привязывающую ее многие десятилетия к «вынужденно пеньково- му» (вегетативному) состоянию в объяснении своего ключевого предме-
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННБ!Х РАСТЕНИЯ та (жизненной формы, или биоморфы), стала остросюжетней и мудрее в целостном (интегральном) вегетативно-репродуктивном обличии; она ощутила прогрессивную динамику своей сердцевины — биографики, биение ее «пульса», а это способно стимулировать развитие не только биоморфологии в целом, но и смежных областей знания, например, систематики, флористики. Итак, биографика, или биоморфогения, как раздел биоморфоло- гии, изучает жизненные циклы простых биоморф (организмов) и слож- ных биоморф (синорганизмов) — их биоморфогенезы (онтогенезы и синонтогенезы), которые могут быть еще названы и известным терми- ном — «биографиями». В обобщенном виде, т.е. на уровне типов (оп- ределенных совокупностей в чем-то однообразных биоморфогенезов) мы уже наблюдаем не один биоморфогенез, не одну биографию (жиз- неописание цикла развития одного организма или синорганизма), а тип (групповой образ) биоморфогенезов, т.е. биоморфогению, или биогра- фику (жизнеописание обобщенных образов — типов каких-либо групп сравниваемых и в чем-то однообразных биоморфогенезов и, соответ- ственно, их биографий). Можно говорить о биографике как типе жиз- ненных циклов любой группы организмов или синорганизмов, лучше всего определенного таксона: вида, рода и т.п., даже семейства (рис. 191-206). Кроме упомянутых выше терминов, обозначающих циклы жиз- неразвития организмов и синорганизмов, я склонен широко использо- вать и такое словосочетание-термин — «биографический портрет». Это форма передачи информации об особенностях цикла развития биомор- фы (биоморфогенеза, биографии) или типов таких циклов (биоморфо- гении, биографики) изобразительными средствами (рисунками, фото- графиями и т.п.) как чреды сменяющих друг друга портретов разновоз- растных биоморф одного вида. По большому счету, применение терми- на «биографический портрет» нацелено на то же самое, что и терминов «биографика», или «биоморфогения», но у первого термина больше свободы, поскольку он может включать любую цепочку из портретов разновозрастных биоморф, где-то претендуя, а где-то и не претендуя, на полноту воспроизведения какой-либо биографики. Какие понятия в биографике ключевые? Безусловно, это биомор- фогенез (онтогенез и синонтогенез), стадийность биоморфогенезов, возрастные и фенологические состояния, или формы роста. Не повто- ряясь (подробнее см. т. 1, гл. 7), рассмотрим представления о формах роста (стадиосомах) биоморф семенных растений. Науку, раздел биомор- фологии, изучающую биоморфогенезы, посредством описания, рисунков и моделирования, формы роста биоморф, особенности их роста и развития (формопроцессы), возрастные и фенологические изменения, смены в рамках биоморфогенезов, включая сравнительные исследования биоморфогенезов близких или далеких по степени родства, в целях более полного понимания предмета исследований — биоморфогенезов, вполне можно называть био- графикой (или биоморфогеиией). Термину «биографика» (дословно с греч. «изображаю жизнь») я отдаю предпочтение, поскольку традиционный термин «биография» означает «описание чьей-либо жизни, жизнеописа-
Глава 4. Концепция орм роста в биоморфологии ние, жизнь, жизненный путь кого-либо» (Сл. иностр. сл., 1964, с. 104); биографией обычно называют индивидуальный жизненный путь како- го-либо человека (Бол. Энц. Сл., 1997). Кратко биографику можно оп- ределить так — это наука о биоморфогенезах и формах роста (возрастных и фенологических состояниях) биоморф. 4.3. Стадийность морфогенезов и формы роста в организации биоморф Все материальные системы существуют в пространстве и во време- ни как тела (одноконтурные и многоконтурные) определенного типа, биоморфы, в частности, проявляя всегда морфогенез, т.е. процесс (во временном отношении — цикл) существования (сохранения изменений) этих объектов от их возникновения до преобразования в новое струк- турно-функциональное качество — новый объект (тело или определен- ную группу тел) [т. 1, гл. 7]. Однако прожить жизнь от рождения до смерти без каких-либо колебаний в течение морфогенеза в их органи- зации, т.е. как бы под «колпаком» — дело безнадежное, поэтому расте- ния еще на заре своей эволюции научились приспосабливаться к рит- мике процессов, происходящих в окружающей их внешней среде. Все неживое и живое во Вселенной существует во власти ритмов. Космические часы отмеряют ритмы биотических часов: Земля движется вокруг Солнца, вызывая на Земле смену времен года, ритмическое колебание силы притяжения Луны вызывает 4 раза в сутки смену при- ливов и отливов, 2 раза в месяц Луна и Солнце находятся как бы «рядом», что резко усиливает силу приливов и отливов. Земля вращается вокруг своей оси, вызывая смену дня и ночи, и т.д. Все эти и многие другие ритмы в неживой природе вызывают особые ритмы в живых системах (биотические): ритмы питания и отдыха, ритмы размножения, ритмы роста и относительного покоя, ритмы возрастных и фенологичес- ких изменений и т.п. Любой морфогенез, будучи процессом изменения формы и содержания материальной системы — тела или определенной группы тел во времени, состоит из отрезков-стадий, в которых прояв- ляются какие-либо особенности организации этих систем. Морфогенез структурен во времени и состоит из стадий. Стадий- ность — это способ существования телесных систем, главные носители которой сосредоточены в фенетической части (феносоме) и имеют струк- турно-функциональное выражение в образе состояний этих тел; это способ прохождения телами своих морфогенезов, который позволяет такие цик- лические процессы меротомировать на части-стадии. У стадийности есть другая сторона — она позволяет меротомировать и сами тела через их конституцию на морфогенические состояния, иначе стадиосомы, кото- рые, по сути, являются формами существования тел в определенных стадиях их морфогенезов. Морфогенические состояния, будучи рассмот- рены в отношении живых тел и тем более биоморф, являются также и
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ Рис. 191 — 206. Спектры форм роста в биоморфогенезах (биогра- фиях) семенных растений.
~лава 4. Концепция фо м оста в биоморфологии Рис. 193. Биографика Nepeta sibirica L. [сем. Lamiaceae (Алтай„Чулышман- ское нагорье, пойма реки Чулышман, галечники; Курайский хребет, остеп- ненные участки долины реки Чуя)). БИОМОРфОТИП: каудексово-базифизарные травянистые многолетники. У взрослых рас- тений каудекс и главный корень могут полностью расщепляться, образуя, таким образом, посткаудициевых клонистов, нередко долго остающихся в сильном криптоклональном сблюкении, а границы синорганизма вполне обозримы, поскольку его мнимое единство в основании обеспечивается «размочаленным» «веревковидным» ризобазифизом.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 194. Биографика Saussurea schanginiana (Wydl.) Fisch. ex Herd. [ceM. Asteraceae (Алтай, Курайский хребет, высокогорья, щебнистые тундры)]. Биоморфотип: каудексово-базифизарные травянистые многолетники.
Глава 4. Концепция форм роста в биомор ологии Б, В, Рис. 195. Биографика Lespedeza juncea (L.) Pers. [сем. Fabaceae (Читинская область, Нерчинский район, южные остепненные склоны сопок)). Биомор- фотип: компактнокорневищные безрозеточные травянистые многолетники.
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИИ е,42 ! ( / ! / / f~ / Г / ~ (' ( / I / ( -/ '"-, м ( I ( (, ! / ( ( ! Ъ I t ! г / I 1 Г 14 1 / ( I I Г IA 1 ( ! (! // ! , / ° ! / 1 ! 1 l ( а,д 46,6 546,66 Рис. 196. Биографика Centaurea phrygia L. [сем. Asteraceae (Московская область, Подольский район, лесные поляны, опушки смешанного леса)]. Биоморфотип: короткокорневищные полурозеточные травянистые много- летники. 1 4,аг3,18а, 27,28 / г ( / (/ 19а, 27, ,1 1 1 ! ( 1 ( t I .( / Il I / ( /
~лава 4. ~онЦепЦиЯ фоРм оста У ~цомо ф Рис. 197. Био область По о графика 1.еоп1обоп autumnalis L. [сем. Ав1 егасеае ( осковская ь, одольский район, луга, залежи)]. Биомор~ неви щные озе ~~отип: компактнокор- розеточные травянистые многолетники.
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ ,гм, 19а, 7,6 MG 78 Рис. 198. Биографика Geranium palustre L. [сем. Gera eraniaceae (Московская б Подольский район, лесные сырые поляны, р бе ега водоемов, сырые о ласть, л озеточные лесные опушки)]. Биоморфотип: короткокорневищные полур травянистые многолетники.
удава 4. Концепция форм роста в биоморфологии ,42,92 ,дс Рис. 199. Биографика Veronica porphyriana Pavl. [сем. Scrophulariaceae, Алтай, Курайский, Северо-ануйский хребты, субальпийские луга)]. Биоморфотипы: переходно-полурозеточные короткокорневищные травянистые многолетники. м4,206, 6 Рис. 200. Биографика Agrimonia pilosa Ledeb. [сем. Rosaceae (Алтай, луга в долине реки Катунь в окрестностях села Катанда)]. Биоморфотип: безро- зеточные компактнокорневищные травянистые многолетники.
01 2, 4 Ри Va ех Va К СО Луг ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ
~дава 4. Концепция о м оста в биомо фологии Рис. 203. Биографика Euryops multifidus DC. [сем. Asteraceae (Израиль, Бо- танический сад Иерусалимского университета, южно-африканский вид, хорошо цветет и размножается самосевом, 2 IV)]. Биоморфотип: переход- но-четочные (получеточные) открытые полегающе-аэроподиальные кустар- НИЧКИ.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ Рис. 204. Биографика Artemisia arborescens L. [сем.Asteraceae (Израиль, Тель- Авив, Ботанический сад, скверы и парки, 29 IV)]. Биоморфотип: переход- но-полурозеточные семидетерминатные (полузакрыто-репродуцирующие) рыхлокустистые зимнезеленые кустарники.
с ава 4. ~онцепция орм роста в биоморфологии 6в в+г 32,35 г1,г7,г9- Сб мс 20в; /47 1,67 p„ gpss Яио афика Я.Ша cpafepensis L. [сем. Ru<acea< (там же, ~» Р" 204, 29 цщ. ~иоморфотип: безрозеточные детерминатные (~~"р~'~ дуцирующие — эфемерно-полисимподиалвнв ) Р ые) п ямостоячие (на откРытом II) ста ники. ровном месте — I) или полегающе-восходящие (на склонах — ) ку р
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 20б. Биографика Cistus salviifolius L. [сем. Cistaceae (Израиль, окрест- ности Тель-Авива, склоны сопок, 27 Ш)]. Биоморфотип: безрозеточные детерминатные (закрыто-репродуцирующие) зимнезеленые (две фазы роста побегов — осенняя и весенняя) невысокие (около 1 м) кустарники.
gaea 4. Концепция форм роста в биомо ологии 323 формами роста соответствующих живых тел (клеток, тканей, органов, биоморф и т.д.). «Только познавая состояния, мы познаем само движе- ние, изменение, развитие, материю и ее формы» (Симонов, 1982, с. б1). Стадиосома, или форма роста, у живых men (биоморф, в том числе) — это одно фенетическое структурно-функциональное состояние какого-либо живого тела, проявляющееся в оиределенный отрезок времени (стадию) его морфогенеза и ограниченное иосредством иризнаков-индикаторов, иозволя- ющих отличить это состояние от других. Ранее (т. 1, гл. 7) было пока- зано, что в биоморфогенезах их стадиям: фенологическим (гипофено- фазы, фенофазы) и возрастным (гипофазы, фазы, этапы, периоды) соответствуют определенные морфологические [возрастные и (или) фенологические] состояния, иными словами, как уже отмечалось выше, формы роста, или, если использовать универсальную терминологию для телесных систем, — стадиосомы. Здесь, как говорится, есть, из чего выбрать. Формы роста биоморф проявляются на основе ритмических процессов роста и развития, происходящих, более всего, в фенетической части и подчиняющихся субабсолютному закону стадийности морфогене- зов, который определяется нами так: морфогенез любой телесной системы всегда уиравляется внутренними и (или) внешними силами ритмического развития, в результате чего эти тела изменяются стадийно в их консти- туциальной части, что обесиечивает естественное меротомирование (чле- нение) морфогенеза на части-стадии, а самих тел — на морфогенические (у живых тел — возрастные и фенологические) состояния (стадиосомы) и иоследовательную сменяемость их во времени. Если в какой-либо области знаний активно идет процесс теорети- зирования, то невольно наступает момент обращения к процедуре тер- минотворчества: проверяются известные, отсекаются второстепенные слова-этикетки и объяснения к ним, что происходит до тех пор, пока не определяется термин, достойный, по мнению автора, того места, которое он может занять в теоретическом построении. Термины нужны только людям, поскольку они предназначены участвовать в накоплении, сохранении и передаче информации, и от совершенствования термино- логии во многом зависит развитие не только конкретной науки, но и всей цивилизации на Земле. Нередко ученые не сразу находят какому- либо термину достойное применение. Начинается период «прозябания» термина в науке — его «скитание» во времени, в результате чего он постепенно приобретает должное содержание или надолго, а, может быть, и навсегда исчезает из арсенала науки. Иногда понадобятся десят- ки лет, прежде чем можно будет сказать, что этот термин состоялся, получил признание в науке. Однако это еще не все, поскольку термин, функционируя в науке, участвует в ней, в ее и своей эволюции, поэтому в существовании научных терминов в перспективе могут происходить самые разные повороты. После такого отступления обратимся к не- скольким терминам, которые, как говорится, «на слуху», но пока надеж- но не закрепились в науке. Речь сначала пойдет о термине «форма роста» («Wuchsform», «growth фогт»). Обзор литературы о применении этого термина уже проводился
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ (Серебряков, 1962; Серебрякова, 1972). Словосочетание «форма роста» появилось в науке несколько позже другого известного термина «жиз- ненная форма», или «форма жизни». Дело в том, что исследователи, наблюдая разные особенности роста и развития каких-либо частей орга- низмов, уже давно стали подчеркивать, что они являются определенны- ми формами роста клеток, формами роста тканей, формами роста по- бегов и т.п. До сих пор словосочетание «форма роста» иногда применя- ют именно в таком не строгом смысле, например, «очень обычна смена форм роста одного и того же побега, связанная со сменой функций» (Васильев и др., 1978, с. 325), «Корень: первичная форма роста..., Ко- рень: вторичная форма роста..., Стебель: первичная форма роста..., Стебель: вторичная форма роста и типы структуры» (Эзау, 1980, с. 556). «влияние форм роста побегов на...» (Журавлева, 1994, с. 43). Словосочетание «форма роста» (growt form) в качестве научного термина предложил использовать Е. Warming (1909), чтобы заменить им менее точный термин «жизненная форма» (цит. по Т.А. Работнову, 1995, с. 84). В дальнейшем при использовании словосочетаний «формы роста» и «жизненная форма» наблюдается между ними нечто вроде «конкурен- ции» за право отражать близкие по содержанию понятия. Так, Дю Рие (Du Rietz, 1931) термин «жизненная форма» («Life-forms») обозначает группы растений, проявляющие общий габитус во время вегетационного сезона, а термин «форма роста» («growt form) — группы растений, про- являющие определенную структуру побегов. Широко используется тер- мин «форма роста» в работах немецких ботаников (Meusel, 1935, 1951, 1970; Troll, 1937; Rauh, 1937, 1938, 1939 и др.). Как подчеркнул И. Г. Серебряков (1962), Г. Мейзель объединял свои работы и работы Рау в особое сравнительно-морфологическое направление классификации «форм роста», которые определялись в соответствии с особенностями ветвления, симметрии, направления роста побегов и другим морфоло- гическим признакам. Классификацию форм роста предложил Е. Шмид (Е. Schmid, 1956, 1963), используя комбинации различных эпиморфологических (внешних морфологических) признаков, заложив некоторые основы для развития в дальнейшем принципа многолиней- ного классифицирования (Голубев, 1968, 1972, Нухимовский, 1969, 1971, 1980 и др.). Следует заметить, что при использовании термина «форма роста» обычно проявляется тенденция не просто заменить им термин «жизненная форма», а придать термину «форма роста» новый, не учтен- ный ранее смысл. По Р. Шарфеттеру (Scharfetter, 1953) каждый вид должен обладать определенной жизненной формой (внешним обликом и его фенологией), а форма роста — это только внешняя морфологи- ческая структура. Как показала в обзоре Т. И. Серебрякова (1972), в работах Мейзеля и его школы нередко используются термины «формы роста» и «тип роста» (Wuchstyp). Под формой роста, согласно Мейзелю (Meusel, 1970) понимается комплекс признаков, определяющих габитус растения и его положение в пространстве и времени. «Форма роста (Miihlberg, 1965, Gluch, 1967) — это совокупность габитуальных призна- ков, изменяющихся в ходе онтогенеза растения, иными словами, это
Глава 4. Концепция орм роста в биоморфологии 325 морфологическое поведение индивида во времени. В форме роста вы- ражается возникшая в филогенезе и специфическая норма реакции его вегетативных органов на определенные внешние условия (Gluch, 1967). Сходные в существенных чертах индивидуальные формы роста объеди- няются в типы роста, как классификационную категорию» (цит. по Т.И. Серебряковой, 1972, с. 87). Итак, термин «форма роста» употребляется в ботанике неоднознач- но. Что, кстати, вполне видно и из биоморфологического словаря П.Ю. Жмылева и др. (1993, с. 117): «форма роста — форма жизненная, выделяемая по структуре побегов» (растение с побегами удлиненными) или по направлению их роста» (растение прямостоячее), растение по- легающее и т.д.)..., для отображения своеобразия жизненной формы» (геофильная форма роста длиннокорневищного растения, подушковид- ная форма роста растения и т.п.),... для обозначения «экологически обусловленных форм вида, отличающихся габитуальными признаками, а в более широком значении — как синоним термина форма жизненная». Как быть дальше с термином «форма роста»? В своих теоретических построениях мне вполне можно обойтись и без этого неоднозначного термина, но зачем так делать с интересным термином, у которого весьма насыщенная история? Похоже, здесь именно тот случай, когда термин еще себя не нашел в науке. Наша задача конкретизировать его. Безус- ловно, что термину «форма роста», являющемуся сокращенным вариан- том терминологического сочетания «форма роста и развития», найдется место не во всех телесных системах, поскольку многие неживые тела не растут, хотя и развиваются. Важно, что этот термин работает в отноше- нии всех биоморф, а во всех других случаях его может подменять уни- версальный термин «стадиосома» — это результат стадийности развития, когда происходит последовательная сменяемость морфогенических со- стояний (стадиосом) тел на отдельных временных отрезках — стадиях морфогенезов этих тел. В отношении термина «форма роста» нам ближе подход немецких ботаников школы Г. Мейзеля, которые этот термин определяют через признаки «морфологического поведения» организмов. Однако, они (Miihlberg, 1965; Gluch, 1967 и др.) считают формой роста всю совокуп- ность габитуальных признаков, проявляющихся и изменяющихся в он- тогенезе. С этим трудно согласиться, поскольку такой своеобразный «габитуальный фенотип», как растянутая во времени форма роста, мало что дает для понимания именно возрастного и фенологического пове- дения организмов и синорганизмов в их жизненных циклах. Можно считать, что форма роста — это не вся совокупность габи- туальных признаков, проявляющихся в онтогенезе, а, следовательно, и не все «морфологическое поведение индивида во времени»; форма ро- ста, таким образом, — это только часть фенетических признаков (лю- бого происхождения — габитуальных, эндоморфологических, физиоло- гических), которые индикаторно отражают состояние биоморфы на от- дельном отрезке (стадии) ее цикла существования (морфогенеза). Фор- мы роста, в нашем понимании, — это морфогенические (морфолого-
ОСНОВБ! БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЯ физиологические) состояния живых тел, качественно определенные, в воз- растных и сезонных пространственно-временных границах фенетической формой тел, как результат стадийного роста и развития (морфогенеза), обусловленный наследственностью этих живых тел и особенностями их взаимодействия с окружающей внешней средой. Полагаю, что утонченный анализ форм роста органоидов, форм роста определенных клеток, форм роста определенных тканей и т.п. оставим для специалистов соответствующих областей знаний. К органи- зации форм роста побегов и других органов биоморф мы будем еще не раз обращаться. Здесь мы подчеркнем статус форм роста биоморф в целостной организации биоморф и в биоморфологии. Ранее уже было показано, что морфогенезы живых систем, биоморфогенезы, в частно- сти, структурны во времени и проявляют себя стадийно, подчиняясь закону стадийности, т.е. рост и развитие биоморф и других живых систем определяется последовательностью смен морфогенических (возрастных и фенологических) состояний (форм роста). Приверженность стадийно- сти в морфогенезах является атрибутивным свойством любых живых систем, хотя бы только потому, что у них всегда в норме есть начало, середина и окончание саморазвития. Поведение биоморф чаще всего связано с какой-либо ритмикой, осуществляясь в определенной времен- ной последовательности, поэтому там, где оно связано со стадиями биоморфогенеза, важно пользоваться определенными временными по- казателями, отмечая время начала и конца стадий, т.е. важно в иссле- довании биоморфогенеза фиксировать хронологию ритмичности пове- дения. Привязка возрастных и фенологических стадий и состояний ко времени их проявления — наиболее распространенный способ изучения биоморфогенезов (работы Т.А. Работнова, А.А. Уранова и др.). Нео- днократная смена форм роста происходит на фоне несменяемости всей биоморфы (ее неживого субстрата — протеросомы и наследственной основы — геносомы) до конца ее морфогенеза. Приверженность к ста- дийности морфогенезов, а шире — и любых процессов, которые прояв- ляют какую-либо ритмику, является атрибутивным свойством жизни и ее носителей. Кроме форм роста биоморф, которые являются пространственны- ми выражениями (состояниями) морфолого-физиологического поведе- ния биоморф на стадиях биоморфогенеза, существуют еще и габитофор- мы в эпициклах и субгабитофармы в эндоциклах, границы которых могут совпадать и не совпадать с формами роста, но наиболее точно они проходят, когда эти части организации биоморф рассматриваются в единстве — в формах роста биоморф. Биоморфа, естественно, шире понятия форма роста биоморфы, поскольку одна биоморфа включает в себя весь спектр ее форм роста, проявляющийся в цикле существования одной биоморфы (простой или сложной), т.е. организма или синорга- низма (рис. 191-206). Биоморфогенез, таким образом, с позиций явления стадийности можно определить так — это субстратно и, в частности, генетически стабильный циклический поток стадийно феносоматически сменяющих друг
Глава 4. Концеиция орм оста в биомо ологии друга форм роста дискретно организменного или синорганизменного про- странства. Форма роста — это явление морфолого-физиологическое, фенети- ческое и ритмогенное. Можно изучать только экзоморфологию, или габитуальное поведение, биоморф — их габитоформы. В других случаях можно изучать только эндоморфологию биоморф, т.е. их эндоморфоло- гическое (в первую очередь, анатомическое, но, может быть, и иное внутреннее фенетическое, например, связанное с физиологией и биохи- мией внутренней организации биоморф) поведение — субгабитоформы. Однако, наиболее полно морфогеническое поведение биоморф можно познать, если изучать в единстве и габитуальное, и эндоморфологичес- кое поведение биоморф в циклах их существования на определенных временных отрезках (стадиях), т.е. в формах роста биоморф. 4.4. Стадийность и иерархия форм роста биоморф В организации биоморф семенных растений есть еще аспект, ко- торый пока мы не подвергали анализу по степени сложности составля- ющих ее элементов. Важно разобраться в иерархии тех форм роста биоморф, которые последовательно в стадийной цепочке сначала воз- никают, некоторое время существуют, а затем сменяются, причем на прежней, хотя и количественно разрастающейся, субстратной основе биоморф, и т.д. на протяжении всего цикла развития — биоморфогене- за. Фенетическая организация биоморф (шире — пластосома у всех тел вообще) всегда имеет носителей своих признаков в целостной органи- зации биоморф (шире — всех тел), которыми являются формы роста (стадиосомы, если говорить о всех телах). Феносома (шире — пласто- сома) существует независимо от нашего сознания, а атрибутом такого существования всегда является стадийность. Формы роста — это носители свойств и признаков феносомы био- морф, т.е. той их части, которая наиболее лабильна. Для форм роста биоморф как результатов стадийности характерна динамичность разви- тия и сменяемость через какое-то время одних признаков — их инди- каторов, на другие признаки, что позволяет наблюдать последователь- ную сменяемость форм роста прямо-таки по цепочке в организации каждой отдельной биоморфы. Что значит сменяться одной форме роста на другую и т.д. в организации тела биоморфы? Это не означает, что одна форма роста уходит в небытие (умирает), а другая на ее месте рождается, заменяя прежнюю. Все происходит иначе. Одна форма роста какое-то время развивается в диапазоне признаков, которые ее харак- теризовали, но постепенно, в этой же форме роста, начинают набирать силу другие признаки, которые постепенно становятся индикаторами уже другой формы роста, что закрепляет, тем самым, факт смены ис- ходной формы роста на производную. Так продолжается до последнего
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ момента существования биоморфы и, наконец, наступает момент, когда последняя форма роста элиминируется, не получая себе замену, когда, наконец, смерть берет в теле биоморфы то, что когда-то ей было «обе- щано» всем развитием живой природы; она отбирает у этого конкрет- ного тела, которое она постоянно «сопровождала» в потоке земной жизни, его главное, хотя и строго дискретное, достояние — жизнь. Вопрос «быть или не быть» формам роста носителями стадийности для биоморф не стоит, поскольку живая природа ответила всем живым телам, еще на заре своего существования, созданием у них фенетичес- кой организации, а это означало «быть» и только «быть». Формы роста, будучи привязанными к определенным признакам-индикаторам, объек- тивно отражают проявление стадийности развития в организации био- морф, оставаясь буквально «накачанными» множеством других фенети- ческих признаков, которые при нашем желании могут иначе распреде- лить границы форм роста. Осознание этого факта — важный момент для дальнейшего развития биоморфологического знания. Стадийность- это особая «производственная» среда, которая давит на организацию биоморфы своим постоянным присутствием в ней, обеспечивая условия для обязательного возникновения одних форм роста и смены их на другие, но границы, определяющие такие формы роста, в зависимости от обстоятельств, т.е. от выбранных фенетических признаков-индикато- ров, могут устанавливаться по-разному. В этом нет особой опасности; здесь больше констатация фактов, главный из которых — явление ста- дийности объективно существует в организации каждой биоморфы (шире — каждого тела), а вот границы форм роста (шире — стадиосом), в зависимости от угла обзора, под которым мы на них посмотрим, варьируются, поскольку устанавливаются субъективно. Формы роста, следовательно, не существуют независимо от стадий- ности, «бушующей» в организации биоморф, но рассматривать их при- ходится через призму самой разной направленности в исследованиях; например, через особенности нарастания побегов можно выделять эумо- ноподиальные, гемисимподиальные, эусимподиальные, полисимподи- альные и т.п. формы роста, через особенности ветвления побегов- супротивноветвистые, очередноветвистые и т.п. формы роста, через особенности листорасположения — супротивнолистные, очереднолис- тные и т.п. формы роста и т.д. Таким образом, явление стадийности, будучи рассмотрено через призму других явлений, будет по-разному меротомировать фенетическую организацию биоморф, поскольку при- знаки-индикаторы для форм роста можно использовать разные. Формы роста объективно привязаны к возрасту и сезонному раз- витию биоморф, но в исследовательских целях можно абстрагироваться от особенностей стадийности биоморфогенезов, никогда при этом не забывая истинной природы форм роста биоморф. Здесь и сейчас речь пойдет как раз о той стороне стадийной организации биоморф семен- ных растений, когда их формы роста как возрастные и фенологические состояния находятся между собой в определенных иерархических вза- имосвязях, составляющих динамический аспект бытия биоморф. Мож-
Глава 4. Концепция форм роста в биоморфологии но ли стадийное развитие биоморф передать средствами субординатного и суперординатого классифицирования? Такое классифицирование возможно и ранее (т. 1, гл. 7.3) было показано на категориях разных морфогенических стадий жизненных циклов биоморф, когда одноименные категории возрастных стадий:— гипофазы, фазы, этапы, периоды — заняли определенные субординаты в соответствующей субординатной классификации, построенной под весьма определенным углом обзора — в зависимости от степени обоб- щения в таких категориях; например, все периоды биоморфогенезов принадлежат одной субординате периодов и т.д. Возможно построение и других субординатных классификаций, но уже под другим углом обоб- щения фактов, например, в зависимости от степени развития форм роста в организации (в ее феносоматической части) биоморф. Формы роста не являются самостоятельными частями биоморф, что очень важ- но, у них нет своей жизни, независимой от жизни всего тела биоморфы; они — только одна из сторон в единой (целостной) жизни любых био- морф, которая обеспечивает стадийность развития фенетической части их организации. Число субординатных классификаций форм роста может быть не- сколько, поскольку смотреть в определенном направлении — в сторону усложнения фенетической организации на протяжении единых биомор- фогенезов, можно с разных высот абстракции, т.е. с разной степенью обобщения. Несколько субординатных классификаций форм роста мож- но расположить в одной субординатной классификации (суперординии) и моделировать, соответственно, в одной «пирамиде» абстракций, по- скольку уровень абстракции (суперордината) в такой классификации определяется по числу субординат в субординатных классификациях, составляющих его (т. 1, гл. 1). В качестве критерия для различных субординатных классификаций (субординий) было взято возрастное изменение сложности фенетической организации биоморф, выражаю- щееся через их формы роста, индикаторами которого были различные признаки, более других признаков отражающие стадийные особенности возникновения, сохранения и смены форм роста. Верхнюю суперординату занимают, на построенной автором «пира- миде» абстракций (рис. 207), несколько широких субординий из двух субординат в каждой из них (рис. 207-А). В первой субординатной классификации располагаются формы роста биоморф в следующем по- рядке, начиная снизу: моноидные и полиоидные. Организация всякой биоморфы в своем фенетическом развитии вроде бы «накачана» ее формами роста (стадиосомами), что неточно, поскольку, на самом деле, у биоморфы в целом в данный момент всегда только одна форма роста, существование которой во времени ограниченно из-за постоянных инфраструктурных трансформаций, составляющих основное содержа- ние стадийности, поставившей на «поток» преобразование и вытесне- ние одной формы роста биоморфы другой, следующей за ней, причем развивающейся из прежней формы роста и на ее месте. Поведение форм роста любой биоморфы в пределах ее организации создает, еще до конца многими учеными не осознанный, эффект неоднократной сменя-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ емости форм роста, т. е. возрастных и (или) фенологических состояний, от начала до конца развития только одной биоморфы. Каждый биомор- фогенез, таким образом, происходит путем последовательных (!) смен нескольких форм роста и все это на единой материальной основе одной биоморфы (простой или сложной). Моноидные формы роста составляют фенетическую организацию биоморфы и обеспечивают ей, более или менее часто сменяя друт друга, стадийное жизнеразвитие по простому типу как организма — или акло- ниста, в том числе мероаклониста (временного аклониста), или клони- ста (особи вегетативного происхождения). Полиоидные формы роста представляют фенетическую организацию биоморфы и обеспечивают ей стадийное жизнеразвитие по сложному типу как синорганизма, т. е. в системе «мероаклонист-клон». На той же суперординате (на той же высоте обобщений, но под другим углом обзора) располагается субор- диния из двух следующих субординат: эмбриональные и эпиэмбрио- нальные формы роста (подробнее см. т.1, гл. 7). Если с больших высот обобщений вполне можно и не замечать серьезного противоречия, существующего для реализации целей субор- динатного классифицирования форм роста в соответствии с направле- нием возрастного развития биоморф, то с уменьшением высоты обоб- щений его уже нельзя обходить. При построении более дробных субор- динатных классификаций форм роста трудности, возникающие, есте- ственно, в возрастном развитии биоморф, объясняются атрибутивными особенностями биполярности в их жизнеразвитии, которую надо, осо- бым образом, зафиксировать в соответствующей иерархической класси- фикации. Обязательно, что мы и делаем сразу в одной из субординатных классификаций форм роста биоморф, надо отметить факт их противо- речивого возрастного развития, вспомнив о разработанных ранее (т. 1, гл. 1) приемах биполярного и мультиполярного субординатного класси- фицирования. Важной особенностью биполярного классифицирования форм роста биоморф является то, что с уменьшением высоты обобще- ний направление субординатного классифицирования может изменить- ся не в самом его основании, а значительно позже. Противоречивость возрастного развития биоморф мы заложили в самую простую биполяр- ную субординатную классификацию из трех субординат: формы роста восходящего развития и противоположные им — нисходящего развития. Переход между этими двумя типами форм роста биоморф осуществля- ется через третий тип — формы роста стабилизированного (устойчиво- го) развития, когда рост и разрушение (деградация) относительно вы- равнивают противодействие друг друга, не вызывая особых изменений в сторону усиления или ослабления развития. Перелом в направлении биоморфогенеза происходит через стадию относительно сбалансирован- ного развития (рис. 207-Б), т.е. формы роста биоморф как бы выходят на некоторое временное «плато», где можно задержаться подольше или, отнюдь, рухнуть вниз, спускаясь медленно, а иногда быстро, прямо-таки «без оглядки» (последнее особенно характерно для эфемеров). Любо- пытно, что в последующих субординатных классификациях факт пере- хода форм роста восходящего развития биоморф к формам роста нис-
[дава 4. Концеиция орм роста в биомор огогии ходящего развития — старым матурным (финиматурным) и сенильным, достаточно наглядно можно передать с помощью возможностей графи- ческого моделирования, показав, как восходящее возрастное развитие, в соответствии с принципом биполярного субординатого классифици- рования, сменяется на противоположное нисходящее возрастное разви- тие (рис. 207-В, Г). Главное в принципе биполярного классифицирова- ния форм роста — носителей стадийности биоморфогенезов в том, что спираль восходящего развития при таком классифицировании где-то вверху, раньше или позже, но обязательно ломается, но не до конца, а остается целостной, благодаря переходной форме роста, как правило, медиматурной, через которую осуществляется смена направлений раз- вития, в результате чего оно изменяется на противоположное, а спираль соответствующего классифицирования превращается в показатель нис- ходящего направления развития, отражая усиление его деградации на пути к низу — к смерти через финиматурные (старые матурные), су6- сенильные (нередко их вообще не отмечают) и сенильные формы роста (две эти формы могут не проявлять себя). 4.5. Парадоксы форм роста, оказавшиеся атрибутами организации биоморф семенных растений Все, что говорилось ранее, позволяет четко определиться в тезисе: организация биоморф — многоликое явление и смотреть на нее надо под разными углами обзора, чтобы увидеть разные стороны ее прояв- ления. Согласно нашим представлениям, развиваемым в этой работе, организация биоморф — это единство составляющих ее меросом: гено- сомы и феносомы, в которых дисперсно распределены элементы (ато- мы, молекулы, агрегаты молекул и т.п.) неживой физико-химической основы — протеросомы. На такое соматическое членение организации биоморф накладывается еще и меротомирование с иных позиций, в частности, на две фенетические меросомы: габитус и субгабитус, кото- рые существуют только вместе, но для исследовательских целей мы вполне можем рассмотреть их и по отдельности. Тело биоморфы детер- минируется в его организации внутренней фенетической частью — cy6- габитусом, одной из функций которой является образование в любой ситуации, развертывание и поддержание постоянно в жизнеполезном состоянии внешней части — габитуса. Обе эти меросомы сосуществуют в динамическом единстве, представляя фенетическую часть в организа- ции любой биоморфы; они взаимодействуют в унисон, находясь, с одной стороны, под управлением геносомы биоморфы, а с другой — под вли- янием окружающей внешней среды. Из геносомы и со стороны внешней среды по отношению к феносоме биоморф исходит не просто «уста- новка» на динамизм, а определенным образом направленное на нее давление ритмами различных процессов, на которое она так или иначе
332 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИК Рис. 207*. Графическая суперординатная модель («пирамида» абстракций) возрастной организации биоморф — их форм роста (стадиосом). А — Г — уровни абстракции; 1-18 — субординаты (классы) тел (форм роста). ФОрмы раста (далее в условных обозначениях это слово подразумевается, но более не повторяется): 1 — моноидные, 2 — полиоидные, 3 — эмбриональные, 4 — эпиэмбриональ- ные, 5 — восходящего развития, 6 — стабилизированного развития, 7 — нисходящего развития, 8 — виргинильные, 9 — матурные, 10 — сенильные, 11 — пренатальные, 12— латентные, 13 — преювенильные, 14 — ювенильные, 15 — имматурные, 16 — иницима- турные, 17 — медиматурные, 18 — финиматурные.
Глава 4. Концепция форм роста в биоморфологии 333 вынуждена отвечать, что сделало феносому биоморф в процессе эволю- ции очень пластичной частью и обеспечило в целом биоморфам про- грессивное развитие их именно в направлении совершенствования пла- стичности. Биоморфогенез обеспечивается стадийными соматическими транс- формациями — стадийной кинетопластикой, которая более всего про- является в одной части организации биоморф — исполнительской ме- росоме — феносоме, а управляется с другой части — геносомы. После- дняя меросома, будучи прикрыта феносомой и погружена в нее, да и не только поэтому, не подвергается таким трансформациям, она обладает способностью часто уходить от многих трансформаций вообще, которые где-то совсем рядом, прямо-таки в фенетической организации, вызывая в ней различные качественные и количественные построения конструк- ций и разрушения их; похоже, о феносоме и о том, что с ней проис- ходит, проще, чем довольно смешным словом «лепота» и не скажешь. Многое из того, что происходит с биоморфами, находится под воздей- ствием окружающей среды, которая, по многим причинам, сама по себе развивается ритмически, но, самое главное, эти процессы, как правило, определенным образом (норма реакции) контролируются их геносома- ми. В процессе эволюции, чтобы жить на Земле, биоморфы приспосо- бились считаться с ритмикой внешней среды, поэтому возрастная и сезонная изменчивость организации биоморф-потомков адекватно адап- тирована к современным проявлениям этого экологического фактора. Стадийность соматической трансформации, которую проявляют био- морфы в своих циклах существования, если кратко — стадийность биоморфогенезов, отражает главное содержание в поведении биоморф во времени — возрастную и фенологическую изменчивость их, которая фиксируется в стадиях жизнеразвития организмов и синорганизмов и получает свое структурно-функциональное выражение в морфогеничес- ких (возрастных и фенологических) состояниях, или формах роста. Стадийность биоморфогенезов управляется с генетического уровня, но сильно зависит от воздействия окружающей внешней среды. Формы роста как способ существования феносомы биоморф, самой лабильной их части, проявляют себя независимо от нашего сознания, составляя содержание феносомы в определенный отрезок времени, отражая по некоторым признакам-индикаторам свои особенности (рис. 191-206). Формы роста как единицы стадийной феносоматической транс- формации биоморф, как правило, неоднократно сменяются в биомор- фогенезах, что происходит на фоне несменяемости тела самих биоморф, в организации которых, кроме весьма лабильной феносомы, присут- ствует постоянно стабильная субконтинуальная биохимическая основа — геносома, а еще и особо стабильная континуальная физико-химичес- кая неживая субстратная меросома — протеросома. Первый парадокс и одновременно первый атрибут: единая и цело- стная организация биоморф на самом деле, по большому счету, далеко не совсем является таковой, поскольку, на самом деле, постоянно со- стоит из трех интегрированных в ней частей (меросом), каждая из ко-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ торых имеет свое место и занимается своим «делом». Среди них есть только одна часть — феносома, выполняющая роль «поведенческо-ар- хитектурного футляра» для всего остального в организации биоморф, обеспечивая им признаковое разнообразие, который составляют безуп- речные исполнители стадийных феносоматических трансформаций— формы роста. Второй парадокс и он же второй атрибут организации биоморф: формы роста биоморф сменяются в биоморфогенезах неоднократно, а биоморфы в целом в то же время присутствуют постоянно и не сменя- ются на другие биоморфы (рис. 191-206). Будучи единицей организации самой лабильной части биоморф- феносомы, формы роста в наших глазах проявляют себя по-разному. В их арсенале весь набор признаков-фенов, представляющих феносому на отдельной стадии биоморфогенеза. Многое зависит от того, как иссле- дователь посмотрит на форму роста и что он захочет или (и) сумеет в ней увидеть. В зависимости от целей исследования, мы из всего разно- образия фенетических признаков, проявляющих себя в данном месте пространства и в данное время, выбираем то, что нас сейчас интересует, и, таким образом, находим признаки-индикаторы для соответствующих стадиосом. Формы роста, в зависимости от того, через призму какого явления (или нескольких явлений) мы их рассмотрим, будут отражать соответствующие состояния феносомы биоморф и биоморфы в целом в текущий момент; например, через явление нарастания побегов формы роста биоморф могут быть или эумоноподиальными, или гемисимподи- альными, или полисимподиальными и т.п., через явление ветвления побегов — соответственно, очередноветвистые, супротивноветвистые и т.п. Формы роста биоморф являются непосредственными носителями и показателями их фенетической организации в определенные отрезки времени (стадии); они — индикаторы организации биоморф. При соот- ветствующей характеристике биоморф часто нет необходимости повто- ряться, что носителями рассматриваемых признаков являются опреде- ленные формы роста, поскольку эти же признаки принадлежат в целом биоморфам, через которые они и характеризуются. Кроме того, если это удобно для исследовательских целей, можно условно отрывать формы роста от биоморф, рассматривая какие-либо отдельные стороны их проявления, например, при сравнении с формами роста других био- морф. При описании желательно подчеркивать возрастную принадлеж- ность: молодая, ювенильная, виргинильная, матурная, старая и т.п. формы роста. В отношении сказанного приведем следующие примеры: взрослая форма роста Тагахасит officinale Wigg. — розеточная стержне- корневая гемисимподиальная и т.п., или, что одно и то же, взрослая биоморфа Тагахасит officinale — розеточная стержнекорневая гемисим- подиальная и т.п. Формы роста биоморф при обобщении уместно пред- ставлять как типы роста биоморф, или стадийные биоморфотипы. При- знаки форм роста биоморф выступают индикаторами типов биоморф, или биоморфотипов, например, биоморфотип Тагахасит officinale— розеточные стержнекорневые гемисимподиальные и т.п. растения.
Глава 4. Концепция форм роста в биоморфологии Третий парадокс и, заодно, третий атрибут: формы роста — это основные носители действительно развитого признакового простран- ства биоморф, поскольку все индикаторные признаки биоморф сосре- доточены в их фенетической части и все фены (признаки феносомы) проходят через формы роста в биоморфогенезах (рис. 12-65 и др.). 4.6. Призрак биоморфологии уже бродит в систематике Современный этап развития биоморфологии — интегральная био- морфология, в центре которой стоят биоморфы простые (организмы) и сложные (синорганизмы), а самобытность и фактуальную сердцевину этой науке обеспечивает ее раздел — учение о жизненных циклах био- морф — биоморфогения, или, что, пожалуй, даже лучше, биографика. Если цикл существования биоморфы — биоморфогенез, или биография (особенно при описании его содержания), то тип биографий, иными словами, совокупность сведений о биоморфогенезах (биографиях) пред- ставителей какого-либо таксона (вида, рода и т.п.) или просто какой- либо группы биоморф — биоморфогения, или биографика, этого так- сона или какой-либо группы биоморф (их онтогения или синонтоге- ния). В России более, чем где-либо, биографика получила развитие в трудах Т.А. Работнова, школы И.Г. Серебрякова и А.А. Уранова, а еще и многих других (А.П. Пошкурлат, В.Н. Голубев и др., см. т. 1, гл. 7). В наше время, когда еще оформляются относительные границы биоморфологии, она уже проявляет cKJIQHHocTb выходить из своих «бе- регов», влияя на развитие друтих наук. Если в этом есть польза, то это надо заметить, поддержать, усилить. Прорыв биоморфологического знания вскоре следует ожидать в систематику и флористику из биогра- фического раздела биоморфологии. Систематика — один из «китов» современной биологии; без нее, как без рук. Систематика нуждается в инъекциях от других наук. У нее нет права замыкаться в себе. Такова ее судьба — быть всегда всем нужной. Биографика — самый динамич- ный и бурно развивающийся раздел биоморфологии. Интегральная биоморфология постоянно ощущает прогрессивную динамику своей сердцевины — биографики, биение ее «пульса», а это способно стиму- лировать развитие не только биоморфологии в целом, но и систематики, а шире — еще и флористики. Классические ориентиры систематики на наиболее консерватив- ные, репродуктивные, признаки оправданы и проверены временем. Однако, те фрагменты растений, которые обычно учитывают система- тики и аккуратно собирают их в гербарии (рис. 208), надо признать все же не отражают довольно многое их организации растений. Бурное развитие систематики в текущем столетии, на мой взгляд, следует ожи- дать при вступлении ее в биографическую стадию своей эволюции. Биографическая информация (о соцветиях, вегетативных органах, семя-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ долях, продолжительности жизни и т.д.) не разрушит, а дополнит воз- можности систематики. Особую роль приобретет в систематике приток из биоморфологии диагностических признаков и рисунков растений. В биографическом рисунке, выполненном в духе «графического научного реализма», дозированно (без перегрузки стеблями, листьями и т.п., но документально точно) важно все (последовательность возрастных состо- яний, увеличение одних и уменьшение других структур, схемы общего вида, масштаб и т.п.) Биографические флоры — произведения трудоем- кие и потребуют усилий больших коллективов авторов. Можно, конеч- но, начинать с региональных флор, на которых и отработать свод ди- агностических биографических признаков, способов их выявления и использования, эта работа, безусловно, не пройдет даром. Однако на ровной и свободной дороге с односторонним движением не стоит при езде увлекаться тормозами, здесь надо, однозначно, набрать оптималь- ную скорость и решать главную задачу, а она для биоморфологов и систематиков к началу XXI столетия совпадает: пришло время печатать «Флору» новой России, используя новые технологии, — «Биографичес- кую флору» России. В мировой научной практике еще никогда подоб- ной цели не ставили. Полагаю, что осознание перспективности разра- ботки такой проблемы не заставит себя ждать. Найдутся ли у России резервы, чтобы ради прогресса в этой области обеспечить ее, вероятнее всего, коллективное решение, или эту работу впервые сделают в других странах. Если создание «Флоры России» пойдет по классическому ва- рианту, освоенному в XX столетии, т.е. повторят «Флору СССР» в сокращенном варианте и внесут накопившиеся со времен советского издания номенклатурные изменения, то это не будет шагом вперед, это будет только «топтание» на месте. Сегодня при подготовке «Флоры России» нужен новый подход — новое творческое начало. Здесь как раз тот случай, когда не стоит грести под себя. Здесь тот случай, когда всем или многим надо идейно раскрыться и разработать совместно стратегию и тактику написания и публикации «Флоры России». Эту задачу надо решать коллективным разумом. В такой «Флоре» должно быть меньше слов и больше ясных и качественных иллюстраций с удобными услов- ными обозначениями. Нужно так подойти к разработке сформулирован- ной проблемы, чтобы под знамена «Биографической (динамической) флоры России» смогли с энтузиазмом встать многие ботаники, которые овладеют едиными для всех программами и смогут их реализовать при изучении жизненных циклов растений тех видов, под которыми они подпишутся, т.е. за изучение которых к определенному сроку они возьмутся, и сделают это. Проблем много, но здесь не место их обсуждать в деталях. Мне все- таки ясно главное, что Россия уже в ближайшие годы могла бы встать на путь подготовки своей уникальной «Биографической флоры» (этот труд может называться просто «Флора России», но по содержанию он должен быть биографическим), поскольку на каркас классической, изве- стной сейчас систематики нужно будет приложить корректирующий пресс биографики видов. Возможно, что наш творческий призыв к рос-
]пава 4. Концепция форм оста в биомо фологии сийским ботаникам XXI столетия не даст желанных результатов, то, в таком случае, уже придется не считать «дивиденды», а только догонять и копировать других, поскольку от биографической систематики и фло- ристики не уйти никак — это веление времени, обремененного движе- нием науки к прогрессу. Итак, в начале XXI столетия систематика, по нашим прогнозам, из классической будет трансформироваться в биографическую (динамичес- кую), поскольку, на самом деле, призрак биоморфологии уже бродит в систематике. 4.7. Биографическое тестирование биоморф семенных растений при разных уровнях HX nOSHBHHQ Методическая разработка (т. 1, гл. 7) может служить программой для изучения жизненных циклов (биоморфогенезов) и их типов (биогра- фики) семенных растений, а также по описанию таковых по любому из трех возможных вариантов (повествовательно-рисуночному, таблично- рисуночному и рисуночному), поэтому она для биоморфолога — био- графиста может быть по сути руководством к действию. Если придер- живаться этой разработки, то можно плодотворно изучить и описать биографику каких-либо видов семенных растений. Для использования в «Биографических флорах», «Справочниках» и т.п. обзорных изданиях более всего подходит третий вариант биографических описаний — ри- суночный, в котором главная информация закладывается в рисунки (один, реже больше для каждого вида), а считывать ее можно без допол- нительных описаний, используя только подписи к рисункам, включаю- щие, в первую очередь, условные обозначения. Далее дается заключе- ние, составляемое на основе информации, заложенной в рисунки, в которых отражается главное для биоморфогении рассматриваемого вида. Методическая программа (т. 1, гл. 7) показывает, что надо делать и чего делать не надо при изучении биоморфогении (биографики) разных видов семенных растений, а арсенал многих признаков, которые важно ис- пользовать в таких исследованиях, отражен в многолинейной классифи- кации биоморф семенных растений и ее модели (таблице-матрице, т. 1, гл. 9). Для определения различных сторон организации биоморф на определенных стадиях их морфогенезов важно научиться выбирать не- обходимые признаки. Если ставить далекие цели, например, создание «Биографической флоры России», то надо суметь стандартизировать все подходы к изу- чению и описанию жизненных циклов (биоморфогенезов, или биогра- фий) и их типов (биоморфогении, или биографики). Определимся с терминами. Тестовый ряд признаков — это такой ряд альтернативных (сравнимых и взаимоисключающих) признаков, который входит в сферу непосредственных биографических исследований, и предмет исследова-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИИ ний, например, биоморфы одного вида, подвергается через него соот- ветствующему тестированию. Результатом всякого биографического те- стирования является обнаружение признака (или признаков), составля- ющих какие-либо стороны организации конкретных биоморф, и отра- жение его (или их) в востребованном информационном пространстве развивающихся биоморф определенного вида, т.е., если кратко, в био- графическом досье вида. На основе многолинейного классифицирова- ния мы получили (т. 1, гл. 9) множество рядов признаков биоморф семенных растений, часть из которых, будучи использована при биогра- фическом тестировании, становятся тестовыми рядами, а те раститель- ные биоморфы, которые наблюдает исследователь в их биоморфогене- зах, мысленно пропуская через определенные тестовые ряды, — объекты биографического тестирования. В результате проведения рассматривае- мой логической операции отбираются, из многих признаков соответ- ствующих тестовых рядов, те востребованные признаки разного инфор- мационного значения, которые действительно составляют содержание конкретных биоморф и их типов. Проводя биографическое тестирование, исследователь определяет, чем предмет его познания обладает, а чем не обладает, или иногда выясняется, что предмет обладает даже тем, чего нет в тестовом ряду признаков, и раскрытием неизвестного ранее признака, кроме всего прочего, еще и пополняется тестовый ряд, что расширяет возможности биографического тестирования видов в будущем. В целом работа с определенными признаками однолинейных классификаций в статусе тестовых рядов признаков позволяет вести целенаправленный поиск выборных признаков (признаков-маркеров) при изучении биоморфоге- нии видов и составлении соответствующих биографических досье. При накоплении некоторого числа востребованных признаков мы получаем картину биографического развития как отдельных организмов или синорганизмов, так и их типов — биографику видов, а краткий свод востребованных признаков, отражающих особенности развития органи- зации биоморф в циклах их существования (биоморфогенезах), т.е. выборка главного из биографики видов, составляет биографическое досье определенного вида. Такие досье наполняют основное фактичес- кое содержание нашей работы и позволяют весьма продуктивно распре- делять его на минимальном «печатном» пространстве между различны- ми теоретическими положениями. Использование биографических до- сье видов — это один из возможных методических приемов для внедре- ния биоморфологической информации в соответствующие «Справочни- ки», «Флоры» и т.п. издания. Число востребованных признаков во многом зависит от того иссле- довательского «порыва», который направляется на изучение биографики и составление биографических досье видов, но, при необходимости, и соответствующих досье отдельных растительных биоморф. Здесь мы невольно подняли любопытнейшую проблему: биографическое тестиро- вание может осуществляться с разной глубиной познания, т.е., по сути, на разных уровнях познания: нижнем (кратком), среднем (полуразвер-
Глава 4. Концепция орм роста в биоморфологии нутом) и высшем (развернутом). Критерием для такого подразделения уровней познания является не только число востребованных признаков в информационном пространстве, но и разнообразие отраслей науки, используемых для решения проблем биографического тестирования. На всех уровнях познания разрабатываются программы (минимум, медиум и максимум) со своими списками вопросов биографических исследова- ний, ответы на которые обязательны при составлении каких-либо фи- тобиографических эссе и (или) досье. Программу-минимум составляют из относительно небольшого чис- ла вопросов, отвечая на которые исследователь получает компактную, но довольно обстоятельную оценку организации биоморф семенных растений в динамике биографического развития. Эта программа, как и последующая, опирается на разработанные ранее (т. 1, гл. 7) методичес- кие положения и эколого-морфологическую многолинейную классифи- кацию биоморф (т. 1, гл. 9). Программа-медиум отличается от первой более широким списком вопросов, подлежащих обязательному исследованию биоморфологом-био- графистом. Для усиления востребованного информационного простран- ства, кроме традиционных эколого-морфологических признаков, можно привлечь изучение некоторых других аспектов организации, например, анатомических признаков основных органов (побегов и корней), что в качестве примера мы покажем ниже на биографике Tribulus terrestris L. Программе-максимум отводится нами особое место, поскольку биоморфологи еще не готовы по ней работать, а ее самой попросту пока и не существует. Эта программа должна включать все вопросы из пре- дыдущих программ, опираясь, таким образом, на те же методические материалы, изложенные в первом томе, а кроме того, в нее должны входить наиболее весомые вопросы нового профиля (по несколько воп- росов из каждой области знаний): эмбриологические, анатомические, генетические, биохимические, физиологические, фитоценологические. На высшем уровне познания организации биоморф смогут квалифици- рованно работать только коллективы авторов, поскольку им надо будет отвечать на весьма далекие друг от друга вопросы. Несколько вопросов программы-максимум будет адресовано эмбриологу, другие (допустим, по 2 — 5) вопросы — биохимику, генетику, физиологу, анатому, эко- логу, фитоценологу, но решающее значение будут иметь экзоморфоло- гические, наиболее показательные снаружи, признаки, по которым, как правило, осуществляется индикация состояний роста и развития био- морф в целом. Вероятно, когда будет готовиться исследовательская программа к изданию «Биографической флоры» России или какого- либо региона Земли, то в ее составлении должны принять участие не только систематики, экологи и экзоморфологи, но и анатомы, биохи- мики, генетики, физиологи, эмбриологи, геоботаники, под общим руко- водством классического (профессионального) систематика. Разработка программных списков вопросов для разных уровней познания биографической организации биоморф — темы разные по сложности, но в любом случае подбор вопросов требует особой тщатель- ности. Сегодня можно констатировать, что для составления программ-
OCHOBbI БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ ных списков вопросов нижнего и среднего уровней познания есть все условия у традиционных биоморфологов (т. 1, гл. 7 и 9). В текущем томе рассматриваемой монографии мы сознательно ограничились работой, в основном, на нижнем уровне биографического познания биоморф се- менных растений, что вполне достаточно для составления, там где это нужно, кратких биографических досье видов, т.е. здесь используются для получения соответствующего досье только вопросы программы- минимум. Ниже мы приведем список этих вопросов. Вопросов в про- грамме-медиум побольше, чем в предыдущей, поскольку она полнее опирается на ряды мультилинейной классификации биоморф. Биомор- фогения многих видов описана в первом томе (особенно в гл. 10) нашей монографии на основе программы-медиум. В отношении высшего уров- ня познания организации биоморф говорить пока рано, поскольку раз- вернутый список фитобиографических вопросов программы-максимум должен быть качественно расширен. Что-то такое когда-нибудь будет, а пока опустимся на тот (нижний) уровень познания, на котором сейчас удобнее всего нам работать для рассмотрения теоретического материала второго тома рассматриваемой монографии. Список обязательных вопросов программы-минимум по изучению биографической организации биоморф семенных растений. Этот список вопросов не повторяет методические положения, из- ложенные ранее (т. 1, гл. 7) и многолинейную классификацию биоморф (т. 1, гл. 9), оставаясь в целом ориентированным именно на них. При ответе на вопросы не обязательно придерживаться порядка, избранного при постановке вопросов. 1. Какой тип биоморфогенезов по степени пространственной целостности биоморф: онтогения или синонтогения? 2. Какие основные возрастные состояния (формы роста) проходят биоморфы и их индикаторные признаки. Продолжительность по времени наиболее крупных из них. 3. Где (местообитание) и как образуются проростки, как они ста- новятся всходами? Поведение семядолей, гипокотиля, главного корня; их формы, размеры и другие особенности. Какие (по форме и размерам) и как развиваются на стебле первые листья: с эпикотелем или без него, очередно, супротивно или иначе, безрозеточно или предрозеточно? 4. Побеговая система в целом. Как развивается генеральный (глав- ный побег)? Когда и как начинается ветвление и (или) куще- ние? Как взаимодействует генеральный побег с остальными (субгенеральными и локальными) побегами? Особенности спе- циализации, фазности видимого роста, нарастания (его формы — эумоноподиальное, эусимподиальное, гемисимподиальное и т.д.), кратности проявления репродуктивной функции в жиз- ненном цикле цветоносных побегов. Как растут вегетативные побеги и как они переходят к репродуктивному открытому или приоткрытому, полузакрытому, закрытому развитию? Особен- ности внутригодичного побегообразования: силлептического и
Глава 4. Концепция форм оста в биомо фологии (или) пролептического, и (или) пролонгированного (ивановы побеги). Типология образующихся побегов по сложности орга- низации и другим структурно-функциональным особенностям. 5. Элементы побеговой системы. Листья и стебли; их форма, размеры, цвет, опушенность и кон- систенция. Репродуктивные органы. Однодомность или двудомность. Ос- новные особенности сексуализации цветков. Форма опыления. Какие образуются соцветия или одиночные верхушечные цвет- ки. Особенности фенологии. Базовые органы. Каково поведение побегов после окончания вегетационного сезона. Как побеги послевегетационно отмира- ют и (или) персистентно сохраняют свои части. Когда, как и какие образуются базовые (многолетние побеговые) органы и их структурные элементы. 6. Корневые системы. Как ведет себя в развитии главный корень. Образуются ли придаточные корни и как они взаимодействуют с главным корнем. Каковы типы образующихся корней по сложности организации, размерам, формам, консистенции. Образуются ли на корнях отпрыски (адвентивные побеги). 7. Как растения размножаются. Особенности дессиминации. Есть ли вегетативное размножение, если есть, то какого оно типа. Как осуществляется партикуляция при вегетативном размножении. 8. Сколько продолжается, хотя бы примерно, биоморфогенез. В заключение по эколого-морфологическим (можно и шире) тро- побиозным (индикаторным для биоморфогенеза в целом) признакам характеризуется вид, что, безусловно, составляет квинтэссенцию любого по форме и содержанию биоморфотипного досье вида. Чтобы плодотворно воспользоваться упомянутой выше методичес- кой разработкой и вопросником по программе-минимум, желательно освоить несколько общих положений. Надо развивать умение видеть и понимать растительные биоморфы, отвлекаясь от многого, что мешает рассматривать их как динамично развивающиеся существа. Никогда нельзя замыкать свой интерес на одном виде, поскольку объективные знания наиболее точно формируются при сравнении разных видов. При сборе фактического материала надо стремиться не только записывать наблюдения в дневник, гербаризировать образцы, но и, что особенно важно, надо стараться зарисовать главные схемы (что от чего, как и когда отрастает) непосредственно в полевых условиях или где-то побли- зости, когда под рукой есть живой растительный материал. Особенно важно сделать первые рисунки (желательно карандашом и на миллимет- ровке, а насколько это важно, оцениваешь только потом, когда обраба- тываешь материал, может быть, через 10 или даже много больше лет спустя) самых первых стадий жизни растительных биоморф, в первую очередь, проростков и всходов. Конечно, можно зафиксировать расти- тельный материал в спирте и (или) положить его в гербарий, а далее на этом успокоиться, где-то что-то потеряв на пути к конечному результа-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ту. С проростками и всходами так, как правило, поступать не стоит: найдите время и зарисуйте их сразу в полевых условиях. Есть смысл привести несколько штрихов, позволяющих побольше понять кое-что в рабочей «кухне» автора. Фактический материал по биоморфогении семенных растений собирается в разное время и его лучше всего сразу же отражать на миллиметровке в рисунках самых молоденьких расте- ний; более крупные растения уйдут в гербарий и вполне могут быть обработаны позднее. Уже на первых рисунках многое видно наглядно: и размеры, и форма структурных элементов, еще последовательность их развертывания и т. п. (рис. 209-211). Я не буду сам здесь превращать приведенный в качестве примера фактический материал в основные рисунки, а предложу читателю, если ему это будет интересно, потрени- ровать себя на досуге и где-то даже испытать, предварительно определив свои возможности для участия в создании коллективного произведения, например, «Биографической флоры» России или какого-либо другого региона Земли. В приведенном фактическом материале по начальным стадиям био- морфогении нескольких видов семенных растений (рис. 209-211) кое- что осталось доработать, чтобы все получилось как надо. Все получится и станет неплохим началом: надо выбрать самые подходящие образцы, чтобы удобно они вписались в основной рисунок, отражающий как модельные экземпляры последовательность в стадийном развитии био- морф, сделать краткие и понятные подписи к рисункам и наконец описать то, что получится. Что ж, в добрый путь, и тогда «Биографи- ческая флора» России или, наконец, какого-либо еще региона Земли обязательно состоится, причем при жизни ближайших поколений бота- ников. Для биоморфолога-биографиста при общении с растениями надо быть готовым опускаться перед ними на колени, а иногда и ползать чуть ли «не щадя живота своего» ради получения знаний о них, допустим, организовав «охоту» за биоморфами- «скалолазочками» из вида Valeriana petrophila Bunge (рис. 212) где-нибудь в расщелинах высокогорных скал Алтая. Надо не побояться поработать и лопатой, где это необходимо, раскапывая шурфики и шурфы, чтобы побольше узнать о том, как поживают столь скрытые, а значит, «загадочные», подземные органы растений. Без этого нельзя, поскольку растительные биоморфы через свои надземные и подземные органы ведут борьбу за существование путем адаптаций к разным стихиям внешней среды, решая, таким об- разом, задачи своего выживания и жизнесохранения вообще. Насколько важны такие знания, мы наглядно убедимся в следующей главе, где впервые построена в одном рисунке универсальная модель организации биоморф семенных растений и ее эволюции по отношению к уровню земной поверхности. Чтобы понять организацию фитобиоморф в их морфогенезах, — надо суметь ответить на несколько фундаментальных вопросов: что, как, когда и сколько, а, если подробнее: что составляет тело биоморфы, как оно растет и развивается, когда все это происходит и сколько продолжается~ Желательно задуматься над всем в целом, что наблюдаешь, и можно озада-
Глава 4. Концепция фо м оста в биомо nozuu чить себя дополнительным вопросом: почему это происходит так, а не иначе? Все вопросы в методической программе-минимум и -медиум при- вязаны к возрастным стадиям биоморфогенезов (т. 1, гл. 7). Они направ- лены на то, чтобы выявить индикаторные (гемитропобиозные) признаки соответствующих возрастных состояний биоморф, а также некоторые признаки, дополняющие характеристику этих форм роста. При изучении признакового пространства биоморф обращаешь внимание на то, что некоторые признаки настолько диагностически информативны и показательны, что могут быть использованы в качестве «визитной карточки» таксонов какого-либо ранга, например, габитус семядолей или целиком габитус проростков (рис. 209-211). В качестве примера подробного изучения морфологии проростков и всходов при- веду несколько рисунков (рис. 213) и описание к одному из них, выпол- ненные Н.И. Иосебидзе (1981); методические приемы таких исследова- ний вполне могут быть использованы при составлении различных про- грамм, в особенности программы-максимум биографического анализа. Другие особенности, проявляющиеся при рассмотрении биоморфогене- зов, также могут служить диагностическими признаками для многих, в том числе и крупных, таксонов. Если по разным причинам не удается ответить на часть вопросов, исходящих из методической программы, и результат исследований, допу- стим, не пройдет в «Биографическую флору России» (автор размечтался на многие годы вперед), то проделанная работа все равно не окажется бесполезной, поскольку материал по фитобиографике с любой полнотой изложения может быть использован при решении самых разных научных задач. Сегодня биографика семенных растений питает развитие только фитобиоморфологии, фактическую сердцевину которой составляет, но совсем недалеко то время, когда она будет полноценно подпитывать и многие друтие науки, в первую очередь, систематику и флористику. Какую позицию ни занимали бы биоморфологи, в своей науке они сегодня обязаны заниматься изучением жизненных циклов раститель- ных биоморф — биографикой растений, или фитобиографикой; похоже, альтернативы такому основному пути современного развития фитобио- морфологии в целом просто не существует. Пользуясь методической программой, основы которой определены ранее (т. 1, гл. 7), автором была изучена биоморфогения многих видов семенных растений, что в различных вариациях и составляет фактичес- кую основу монографии. В качестве примера того, как и какой можно получить искомый результат, используя нашу методическую программу биографических исследований по типу «медиум», приведем биографику одного вида Tribulus terrestris L. (рис. 214, 215), изученного в естествен- ных условиях его произрастания в Тыве и на Алтае (Нухимовский, Шретер, 1979). Якорцы стелющиеся (Tribulus terrestris L., сем. Zygophyllaceae) из- glaBHa применяются в народной медицине многих стран мира для лече-
OCHOBhl БИОМОРФОЛОГИИ CEMFHHhlX РАСТЕНИИ Рис. 208'. Фрагменты растений, составляющие основу современного гербар- ного материала, с которым обычно работают систематики: А — Saussurea tridactyla Sch. Bip. ex Hook. f., В — S. thomsoni С.В. Clarke,С — S. kunthiana С.В. Clarke, D — S. wernerioides Sch. Bip. ex Hook. f. (Walter, 1968, с. 827). Рис. 209. Пример накопления фактического материала по биоморфогении Carum carvi 1.. [сем. Apiaceae (Московская область, луга)].
Глава 4. Концепция форм роста в биоморфологии Г dog Рис. 210. Пример накопления фактического материала по биоморфогении Seseli libanotis (L.) Koch [сем. Apiaceae (Московская область, луга)]. Рис. 211. Пример накопления фактического материала по биоморфогении Chaerophyllum bulbosum L. [сем. Apiaceae (Московская область, опушки, овраги)].
OCHOBbl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ 6,26 Рис. 212. Биографика Valeriana pettophila Bunge [сем. Ча1епапасеае (Алтай, Курайский хребет, высокогорные скалы, около 2500 м над ур. моря)]. Дополнительные условные обозначения: ст — столон. Биоморфотип: сто- лонообразующие полурозеточные интенсивно-клонирующие травянистые многолетники.
Глава 4. Концепция фо м оста в биомо оло и 32,34 Рис. 213. Примеры детального изображения организации проростков и всходов: I — L. quinquelobatus Gilib., сем. Lamiaceae)], II — Ttibulus tet testtis 1. (сем. Zygophyllaceae), I II Tanacetum vulgate L. (сем. Astetaceae) по Н. И. Иосебид- зе (1981). Ниже, в качестве примера, приводится подробное описание только одного вида — Tanacetum vulgare. «Подсемядольная часть 4,70 + 0,20 мм дл. розово- фиолетовая, очень тонкая. Семядоли эллиптические, 3,00+ 0,00 мм дл., 2,00 р., р у ке закругленные, при основании клиновидные, слегка мясистые, сидячие. Жилкование сетчатое, краеупорное. Гл. ж. (глав- ная жилка) постепенно о ут лщающаяся и слегка изгибистая в верхней части. В пределах черешка параллельно гл. ж. проходит пара тонких жилок. Первые листья супротивные, заостренно-эллиптические 4 40 + 0,20 мм шир., при основании клиновидные, по краю с 1 — 2 ко от- кими ос ыми з бч тр убчиками. Сверху и по краю опушены рассеянными одно- клеточными нитеви н д ыми волосками, снизу или голые, или покрытые еди- ничными волосками. Ж ами. Жилкование петлистосетчатое, редкоконтактное. Сеть жилок замкнута сильно изгибистой обводной жилкой, которая в черешке параллельна гл. ж. Гл. ж. п ох р одящая, в верхнеи части коленчато изгибистая. ° .Р черешке параллельно гл. ж. проходят еще две жилки. Черешки листьев 1+00 мм дл., мелкожелобчатые» (Иосебидзе, 1981, с. 36).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ~Й й 1фв ф 61~ Т' М О О С й [е О О :а л ~ч М Е LG С О Q М сб М О сб 4: Q О bQ й М О 1 'С сп 1 М сп 3 Р Ю 'С ~ч 65 М Е Яи Е Ж С~3 О й О [е Ж ! 65 :а 65 ° л 65 О Р М 3 й л ~ ° О М Cl й [е О О :а LG О Q О й;[ С й;[ h4 О И л 2 С й 2 :а О й;[ :а 1 О О Ю 65 0~ Е :а :У' 65 :а О LG О 2 :а A Е :а О Е О й;[
Глава 4. Концепция форм роста в биоморфолоеии ния различных заболеваний. Одним из важнейших действующих ве- ществ якорцев стелющихся является стероидный сапогенин диосгенин, выделенный из надземной части этого растения (Качухашвили, 1960). Т. terrestris — вид сорных и рудеральных растений, растущих на участках с нарушенным растительным покровом (у дорог, на мусорных местах и т.п.) в степной, полупустынной и пустынной зонах европей- ской части бывшего СССР, Кавказа, Средней Азии, Казахстана, Алтая и Забайкалья (Бобров, 1949). Острошиповатые плодики якорцев стелю- щихся, вонзаясь в ноги и шерсть животных, в обувь людей, в шины автомобилей и самолетов, разносятся на большие расстояния, благодаря чему этот вид постоянно расширяет свой ареал, в частности, попав в Америку, он быстро стал здесь злостным сорняком (Левина, 1957). В условиях культуры семена якорцев весной прорастают недружно, в те- чение месяца и более, но сеянцы растут и развиваются быстро (Штром- берг, Джорбенадзе, 19б9). В Тыве растения этого вида часто встречаются в равнинной степ- ной части, в основном вдоль автомобильных дорог и скотопрогонных троп, на стоянках скота и на участках, подверженных интенсивному выпасу. Якорцы предпочитают рыхлые песчаные и супесчаные почвы, не занятые или слабо заселенные другими растениями. На Алтае якорцы стелющиеся встречаются редко. Небольшие их заросли мы обнаружили в Кош-Агачском районе, в пойме р. Чуи и вдоль скотопрогонных троп. Для краткости изложения нами приняты следующие условные обо- значения: ПС — временная продолжительность возрастного состояния, ОХ — основная морфофизиологическая характеристика особей. Допол- нительная характеристика особей: А — частная (распространяется толь- ко на рассматриваемое возрастное состояния), Б — общая (распростра- няется на несколько или на все возрастные состояния). Объяснение некоторых используемых терминов приводится в первом томе (гл. 5). Эмбриональный период. Пренатальный этап. ПС — с конца мая- июля до июля — конца сентября, редко до октября. ОХ — оплодотво- ренная семяпочка или зрелое семя в структуре родительского растения. Б — цветки обоеполые, 0,8 — 1,2 см в диаметре, с десятью тычин- ками, завязь верхняя с одним столбиком и пятилопастным рыльцем, лепестков пять. Чашелистников пять — зеленых и опушенных. Формула цветка Са,Со, А„, G,, Опыляется насекомыми. Латентный этап. ПС — с конца июля — октября до весны — лета следующего года или, при отсутствии благоприятных условий для про- растания семян, в течение нескольких лет, но иногда (после обильных летних или раннеосенних дождей) до июля — августа или начала сен- тября текущего года. ОХ — зрелое семя, обособленное от родительского растения, скрытое в плодике. Б — плоды относятся к типу синкарпиев, после созревания легко отделяются от родительского растения и рас- падаются на четыре, чаще пять, двух-трехсеменных и двух-трехгнездных плодиков (орешков, мерикарпиев); по спинке плодики бугристые, с двумя (редко одной) парами острых шипов (верхние шипы несколько длиннее нижних) и с множеством неветвистых мелких и крупных ще-
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РЛ СТЕНИИ тинистых волосков. Плодик 0,35 — 0,5 см длиной (не считая шипов, достигающих длины 0,15 — 0,4 см) и 0,3 — 0,4 см шириной (по спинке). Семена светло-коричневые, продолговато-обратнояйцевидные, 0,15— 0,3 см длиной, 0,05 — 0,12 см шириной. Семя покрыто тонкой семенной кожурой, эндосперм отсутствует, зародыш почти целиком заполняет семя. Он расчленен на две мясистые крупные семядоли лопатчатой формы, небольшой гипокотиль и корень. Между семядолями едва заме- тен меристематический бугорок точки роста. Семядоли имеют прокам- биальные тяжи и включения масла и крахмала. Эпиэмбриональный период. Виргинильный этап. Преювенильная фаза. Гипофаза проростка (рис. 214 А). ПС — май — август, реже начала сентября; у одной особи продолжается 3 — 7 дней. ОХ — прорастающее семя, или семядольный почкокорень (ризопротерокормус). А — семядо- ли надземные продолговато-эллиптические с выемкой на верхушке, клиновидно сбегающие в очень короткий (0,1 — 0,2 см) черешок, 0,5— 0,7 см длиной и 0,2 — 0,3 см шириной, снизу опушенные, сверху голые или слабо опушенные в основании и по жилкам. Гипокотиль 0,7 — 2,0 см длиной, 0,05 — 0,1 см шириной, опушенный сверху неветвистыми одноклеточными волосками. Главный корень — монориза (редко дири- за) до 3 см длиной. Гипофаза всхода (рис. 214 Б). ПС — май — август, начало сентября; продолжается 5 — 10 дней. ОХ — семядольный вегетативный предро- зеточный (один-два настоящих сближенных листа) или розеточный ризомонокормус (три, очень редко четыре, настоящих сближенных листа). А — семядоли 0,5 — 1,0 см длиной, 0,25 — 0,4 см шириной, с черешками 0,1 — 0,25 см длиной. Б — первые три ( редко четыре) листа медиального побега сильно сближены, очередные, парноперистые, с тремя — пятью парами листочков. Листочки 0,4 — 0 8 см длиной и 0,15 — 0,3 см шириной, цельнокрайние, продолговато-ланцетовидные, у основания слегка симметричные, к вершине слабо заостренные, снизу опушенные, сверху голые или слабо опушенные неветвистыми однокле- точными волосками. Матурный этап. Фаза первая (униматурная) (рис. 214 Д,). ПС— май — август, начало сентября; продолжается одну — три недели. ОХ — семядольный закрытый полурозеточный репродуктивно-ассимиляци- онный эфемерно-полисимподиальный ризодикормус. А — образование первых бутонов свидетельствует о начале матурного этапа, а наличие живых семядолей у матурных особей — признак наступления первой матурной фазы. Главная дириза или тетрариза достигает 15 см длины. Б — отмирание семядолей свидетельствует о переходе особей в другую стадию онтогенеза. Семядольный узел приземный или засыпается подвижным песчаным почво-грунтом. Медиальный побег якорцев стелю- щихся — это полурозеточный закрытый репродуктивно- ассимиляцион- ный, полностью или неполностью очереднолистный (иногда с одной парой анизоморфных супротивных листьев), четырех- восьмиузловои монокормус. Все латеральные побеги — это пазушные силлептические безрозеточные закрытые репродуктивно-ассимиляционные одно-, реже
Глава 4. Концеиция о м роста в биомо ологии двух-, трехузловые монокормусы. Система латеральных монокормусов, первый из которых образуется из почек медиального побега, — ветвь рыхлого континуального соцветия, или, по терминологии Гебеля (Goebel, 1931), антокладий. Монокормусы антокладиев, непосредственно замеща- ющие верхушку родительского побега и продолжающие прямолинейный рост в длину отдельных многоосевых систем главного побега и его ветвей, всегда образуются из меристем, закладывающихся в пазухах крупных листьев, а ветви, расходящиеся в стороны от этих систем, образуются из меристем в пазухах мелких листьев (супротивные листья анизоморфные). Вся побеговая система взрослой особи якорцев стелющихся состоит из медиального монокормуса и антокладиев, т.е. это полурозеточный моно- карпический обоеполый репродуктивно-ассимиляционный закрытый эфемерно-полисимподиальный очередно- и супротивноветвистый анизо- фильный парноперистолистный стелющийся дикормус. Сравнительное исследование стеблей антокладиев свидетельствует о терминальности одиночных цветков в структуре этих антокладиев и эфемерно-полисимподиальном типе нарастания всей побеговой систе- мы — дикормуса. Это нарастание двух- или многоосевого эфемерного органа, простые оси которого нарастают моноподиально менее одного вегетационного сезона, а затем в том же сезоне новые простые оси перерастают верхушки родительских осей, завершающих или завершив- ших свой рост. Фазы вторая (биматурная) — пятая (квинтиматурная) (рис. 214 Д,— Д,). ПС — июнь — август, начало сентября до конца вегетационного сезона (в зависимости от погодных условий); каждая фаза продолжается от нескольких дней до месяца и более. ОХ — бессемядольный закрытый полурозеточный репродуктивно-ассимиляционный эфемерно-полисим- подиальный кустистый ризодикормус. А — индикаторный признак вто- рой фазы — наличие у особей трех — пяти побегов кущения; индика- торный признак третьей фазы — наличие у особей шести — десяти побегов кущения; индикаторный признак четвертой фазы — наличие у особи более десяти побегов кущения, которые сохраняют не менее 20 % зеленых листьев; индикаторный признак пятой фазы — засыхание более 80 % листьев на побегах кущения. Б — в зависимости от конк- ретных условий местообитания особи проходят различное число фаз онтогенеза. Например, особи, которые вступили в эпиэмбриональный период весной или ранним летом и произрастают на хорошо увлажнен- ных почвах, обычно проходят все или большинство фаз. В то же время особи, появившиеся поздно летом или в начале осени, успевают пройти лишь одну — три фазы онтогенеза. Они плохо растут в длину, слабо кустятся и быстро усыхают, переходя в заключительную — пятую матур- ную фазу. Главная тетрариза до 25 см длиной. Размеры побегов кущения колеблются от нескольких сантиметров до 3 м. К побегам кущения у якорцев стелющихся относятся все побеги, отрастающие из зоны куще- ния в радиусе 1 — 2 см от семядольного узла. Побеги кущения у якорцев стелющихся сильно сближены друг с другом у основания, что отличает их от побегов рассеянного ветвления, образующихся выше зоны куще-
352 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ния (рис. 214, Сб). Медиальный побег вместе с побегами кущения (ан- токладиями) составляет побеговую систему взрослых особей якорцев стелющихся. Это эфемерно-полисимподиальный кустистый стелющий- ся дикормус. Фенофазы: вегетация (после прорастания продолжается одну-две недели); бутонизация (2 — 5 дней); цветение и плодоношение растянуто на весь вегетационный сезон. Верхушки побегов цветут и плодоносят почти до конца онтогенеза. Большинство растений отмирает в конце сентября или в октябре. У якорцев стелющихся (рис. 214, а-d), по Фукуда (Fukuda, 1974), существуют четыре нодальных типа: а-тип (один лист, цветок отсутству- ет), Ь-тип (один лист, один верхушечный цветок), с-тип (два анизомор- фных листа без цветка) и d-тип (два анизоморфных листа и один вер- хушечный цветок). С-тип встречается очень редко — при моноподиаль- ном нарастании соответствующего участка побега, чаще у основания антокладиев. Верхняя часть стебля антокладия, несущая цветок, постро- ена по одному типу с теми частями стебля, которые располагаются ниже узла, несущего единичный лист или пару супротивных анизоморфных листьев (b-или d-типы), продолжая его внутреннюю структуру (рис. 215). Это еще раз подчеркивает терминальное положение цветка в от- ношении ближайшего к нему нижерасположенного узла. Опушение побегов состоит из одноклеточных эпидермальных волосков, на верхуш- ке побега более обильных. Клетки эпидермиса тонкие, в плане прямо- угольные или почти квадратные. Устьица немногочисленные, анамоцит- ного типа. В паренхиме располагаются открытые коллатеральные про- водящие пучки, окруженные крахмалоносной обкладкой, встречаются кристаллы оксалата кальция. Сердцевина также состоит из тонкостен- ных паренхимных клеток. В перимедуллярной зоне имеется много крах- мала. Такое строение сохраняется лишь на молодой части побега до закладки камбия. Межклеточный камбий сначала развивает лишь па- ренхиму, одревесневающую в ксилемной части. Далее формируется сплошное кольцо древесины, клетки центральной части сердцевины одревесневают и частично разрушаются. Итак, биоморфогенезы якорцев стелющихся осуществляются толь- ко по типу гаплоонтогенезов, т.е. жизненных циклов постоянных акло- нистов (H30RKJIQHHGTQB). Виргинильный этап продолжается всего 1 — 2 недели, цветение и плодоношение растянуто на весь вегетационный сезон. Независимо от возрастного состояния особи не зимуют. По типу биоморфогенезов якорцы стелющиеся представляют собой надземно-двусемядольный закрытый (детерминатно-репродуктивный) однодомный желто-парнолистниковоцветковый полурозеточный эфе- мерно-полисимподиальный стелющийся дикормусовый анизофилльный парноперистолистный очередно-супротивнолистный кустистый корот- костержнекорневой эпизоохорный и эпиантропохорный сорно-рудеаль- ный однолетний вид растений.
Глава 4. Концепция о м оста в биоморфологии Рис. 215*. Поперечные срезы стеблей матурных особей Т. 1еггейпь. Увеличено. A — верхняя стеблевая часть антоклгдия, Б — верхняя стеблевая часть ниже анизофил- лозного узла, В — основание медиального побега выше третьего узла, 1 — эпидермис, 2 — волоски, 3 — проводящие пучки, 4 — сердцевина, 5 — паренхима коры, 6 — волокна первичной флоэмы, 7 — вторичная флоэма, 8 — камбиальная зона, 9 — ксилема. 4.8. Движение (« текучесть» ) материи и ее локализация в организации биоморф как меросом-габитусов и как меросом-форм роста Первооснова всему — материя, составляющая вместе с простран- ством атрибутивное и неисчерпаемое структурно-функциональное со- держание тотального движителя (примагенератора), который обеспечи- вает бесконечное функционирование во времени Природы как вселен- ского потока. Материя никогда не начиналась и никогда не закончится Ъ она продолжается бесконечно в трансформациях, всегда и всюдно «пе- ретекая» в формы и содержания тел и нетел по определенным законам и правилам на основе универсального принципа всеобщего движения материи как эволюционных маятников (образов тел и нетел): «так- сяк*туда-сюда». Все, причем и то, до чего сейчас еще совсем не просто «докопаться» (понять), в организации телесных систем создавалось путем эволюции многие миллиарды лет и вместе составляет единый организационный порядок Природы. Главная особенность этого поряд- <а B том, что он многогранен, поскольку замешан на многослойности (многоуровневости) многих природных явлений и проявляет себя с
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ разных сторон, если мы находим силы и средства выявлять и наблюдать их под разными утлами обзора и с разных высот абстракции. Здесь и сейчас мы не будем уходить в рассмотрение тонкостей организационного порядка самых разных явлений, но многослойность явления «жизнь» и в особенности многослойность организации жизнен- ных форм (биоморф) вообще и, в частности, у семенных растений, мы уже вскрывали (т. 1) и далее будем вскрывать неоднократно самым разным образом, меняя и углы, и высоты обзора. Все это будем делать для того, чтобы утвердить в науке тезис: жизненная форма — это не только габитус и не только форма роста у растений и животных, а интегральное явление — организм или синорганизм, причем всегда будем стремиться идти дальше, даже значительно дальше, используя саму идею утверждения обозначенного выше ключевого тезиса биомор- фологии как хороший мотив «покопаться» в организации живой приро- ды — убедительный повод для проникающего движения вперед по пути развития современной (интегральной) биоморфологии. Посредством функционирования тотального движителя во време- ни, т.е. функционирования Природы, материя «растекается» (движется) в телах-вещах и нетелах-процессах, приобретая естественным образом свойства «сосудов», в физических границах которых она, материя, про- должает проявлять свое бесконечное качество — движение («текучесть»), продолжая тем самым свое неудержимое «растекание», создавая различ- ные формы тел-частей, или меросом, наполняя их и количественным, и качественным содержанием. Тотальный движитель, через свой не- скончаемый ресурс — материю, создал некоторое, для Земли вполне обозримое, разнообразие типов тел, которые так распределились в Природе, обладающей единым организационным порядком и, следова- тельно, многослойностью (многоуровневостью), что каждому типу тел стал доступен только определенный уровень организации («ступень» организационной сложности). Природа, следуя своему организационно- му порядку, так распределила и продолжает распределять в себе разно- образие ее тел, что одни из них по отношению к другим проявляют статус целого частей, а другие — частей целого. Иначе не бывает нигде, и все тела, поскольку Природа организована многослойно, состоят из частей тел — меросом [неживых и (или) живых]. Жизнь (живая природа) возникла в эволюции много позднее, чем нежизнь (неживая природа). Она, Жизнь, появилась на Земле (шире- во Вселенной) из тех уровней организации неживых тел, которые уже и до нее обеспечивали единый организационный порядок Природы. В этом отношении Жизнь как природное явление не могла возникнуть в иной, чем неживая, среде и состояться как явление-тело — живая при- рода, не используя в своей структуре неживые тела. У живой природы как тела выбора не было и можно было стать таковой, каковой она стала, только взяв в организацию своей первоосновы — субстрата— неживые тела. То, что жизнь многослойна и что тела живой природы по сложно- сти их организации находятся в едином блоке с неживыми телами на одной иерархической «лестнице», подчеркивая присутствие соответству-
Глава 4. Концепция рм оста в биомо фологии ющего организационного порядка в соматическом разнообразии При- роды, хорошо известно (т. 1, гл. 2) и во многом уже вошло в учебники. )Кивую природу в режиме генофенетического единства живых тел тра- диционно меротомируют на органеллы, клетки, ткани, органы, организ- мы, синорганизмы, популяции, виды и т. д. Сегодня довольствоваться только такими представлениями о многослойности организации жизни, на мой взгляд, маловато, и надо искать пути проникновения дальше в это, во многом еще загадочное, явление. Ранее (глава 1.3.3.) была сделана попытка разобраться с многослой- ностью организации жизни под несколько иным утлом обзора: было показано, что типов витальности (жизнеемкости) несколько. Этот шаг, пожалуй, сделан своевременно, поскольку с известных позиций осоз- нать и показать положение габитусов и форм роста биоморф в органи- зации живой природы было невозможно. Дело в том, что габитус и формы роста биоморф — это ни органеллы, ни клетки, ни ткани, ни органы и ни организмы. Так кто же они такие на самом деле? Каково их место в многослойной организации жизни? Здесь нужен новый подход, и он был найден. Материя, «растекаясь» как-либо в пространстве и трансформируя себя и его, наполняясь формой и содержанием, становится во времени какими-то телами и нетелами, приобретя и приобретая вновь в процессе движения-эволюции свойства и формы атомов, молекул, камней, мол- ний, органов, организмов, гор, озер, биоценозов и т. п. Будучи одним из представителей живой природы, а шире — природы вообще, мы можем усмотреть в организационном порядке, а он многослоен, еще одну любопытную сторону. Меротомируя Природу, невольно обраща- ешь внимание на то, что по ее соматическим элементам — неживым и живым телам — проходит, пусть незримо, разделительная полоса, об этом мы уже немного писали в первой главе (и это только начало весьма и весьма интересной для развития темы), которая распределила все тела на несколько групп: подобитатели и обитатели, с одной стороны, подо- биталища и обиталища — с другой. В отношении любых носителей какой-либо витальности, которые в (на) ком-то или чем-то существуют, мы будем говорить как об «обитателях» («бионах»). Всех носителей инвитальности (безжизненности, нежизненности), которые в (на) ком- то или чем-то существуют, будем, при необходимости, обозначать еще и как «подобитатели» («абионы»). Места существования (любые) обита- телей — это «обиталища» («топобионы»), а места существования подо- битателей — «подобиталища» («топоабионы»). Обитатели и подобитате- ли имеют свои диапазоны существования, в пределах которых их суще- ствование реально происходит и (или) возможно, т. е. иными словами каждому из них присущи соответствующие топозоны. В зависимости от того, под каким утлом посмотреть на события, происходящие в сома- тическом мире, обитатели могут становиться обиталищами, а подобита- тели — подобиталищами или обиталищами (живые тела могут обитать и в неживых топозонах — обиталищах). Итак, организационный порядок в Природе (по крайней мере у нас — на Земле) обеспечивают, с одной стороны, обитатели и подобитатели,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ а с другой — обиталища и подобиталища. Материя, заполнив собой определенные части пространства и превратив их в тела, манипулирует ими как образными, весьма пластичными, прямо-таки «резиновыми» (на самом деле, они самые разные) сосудами, изменяя их форму и содержание, а временами нередко «выплескиваясь» из их границ, что особенно характерно для открытых, в первую очередь, живых систем. «Растекание» материи и ее «выплескивание» из живых «сосудов»-тел является атрибутивным условием существования живых тел-систем. Обитатели и подобитатели используют пространство для своего суще- ствования на основе каких-либо процессов (нетел), взаимодействуя между собой и со средами своего существования — их обиталищами или подобиталищами. Когда обитатели или подобитатели не привязаны жестко к одним типам обиталищ или подобиталищ, а имеют широкие топозоны, то это эвритопозонные тела, например, молекула воды строго не привязана к определенному месту существования. Если обитатели или подобитатели привязаны к строго определенным топозонам — это стенотопозонные тела, например, листья жестко привязаны к своему обязательному обиталищу — остальной части побегов. То, что в При- роде в единстве соприсутствуют подобитатели и обитатели, с одной стороны, и подобиталища и обиталища — с другой, однозначно полезно для всех, поскольку такой расклад в разнообразии тел (и их типов) поддерживает организационный порядок — в особенности многослой- ность природных явлений — и всецело ему подчиняется. Живые тела в общей массе не исключение, напротив, — это наиболее многослойные природные явления, а, следовательно, у них и наиболее сложный орга- низационный порядок. Меросомность в организации любых природных тел начинается с субстрата и постсубстрата, далее дифференцируется через различные формы эгосубстрата и конституции тел, на что мы особо обратили внимание в первой главе. Здесь мы пойдем еще дальше и попытаемся новыми штрихами очертить особенности пребывания габитуса, его форм и форм роста в многослойной организации биоморф семенных расте- ний. Жизнь многослойна и может позволить себя «расслаивать» каки- ми-либо средствами только при соблюдении обязательного условия: всякий биомеротомист («расслоитель» живых тел на части) должен су- меть найти подход к ней, жизни, — правильный логический ход, кото- рый открывал бы путь к ее членению на части под соответствующим углом зрения. Дело в том, что габитус в организации жизненных форм (организ- мов и синорганизмов), будучи элементом фенетической организации и составляя всю внешнюю организацию (экзосому) биоморф, проявляет одну из возможных сторон жизнеемкости — квадрантвитальность, или квартавитальность (живет, если вообще можно говорить о его жизни, только на четверть и не более того). Если все это так, то, согласно определению, все тела, габитус биоморф в том числе, проявляющие какую-либо форму витальности, не просто существуют, а обитают в окружающей их среде, в (на) некоторой ее части. Габитусу как жизнен- ной четверти (образно — «четвертинке» жизни) в организации биоморф
Глава 4. Концеиция м оста в биомо ологии 357 до полновесной жизни (ковитальности) далеко, а до того, чтобы соста- вить хотя бы «половинку» жизни (семивитальность) — феносому, надо быть только вместе с другой квадрантвитальной частью — субгабитусом (эндофеносомой). В первой главе было показано, что феносома и гено- сома в организации биоморфы — это семивитальные структуры — две ее атрибутивные жизненные «половинки». Габитус и субгабитус еще дальше отстоят от полновесной жизни — ковитальности, проявляя по отдельности только на четверть «жизненные» свойства — квадрантви- тальность, а вместе их жизнь наполняется половинчатым содержанием, T. е. становится семивитальной. Расслаивая жизнь на части, мы более и более убеждаемся, сколь многослойно это явление в соответствующих телесных системах и что без неживых или неполно-жизнеемких (на четверть или половину) меросом не свяжешь полную жизнь тела (кови- тальность). Многое в живых телах действительно есть и существует только в движении и единстве друг с другом по-разному жизнеемких меросом. Итак, мы определились с габитусом в организации биоморф, кото- рый, проявляя некоторую витальность, чего вполне достаточно для ут- верждения, что он, габитус, обитает на своем обязательном обиталище — эндофеносоме (субгабитусе), а шире — на эндосоме. Следует заме- тить, что у неживых тел также есть свой габитус, там он является фор- мой проявления инвитальности меросоматической организации, т.е. су- ществует (подобитает) на остальной части организации этих тел. У га- битуса, как, впрочем, и субгабитуса, есть главная особенность — это, во всех случаях, части в организации целых тел (живых или неживых). Габитус, если поменять угол обзора, вполне можно превратить в оби- талище и для субгабитуса, и для эндосомы, и для управляющей части биоморфы — геносомы (подробнее в главе 9). Осталось разобраться еще с формами роста биоморф и определить- ся: какое они занимают место в многослойной организации биоморф. Из всего материала, который уже прошел здесь ранее, можно сделать следующие обобщения. Явление жизнь организовано в телесных системах многослойно. Среди разнообразия слоев, составляющих живое тело, все начинается с носителей инвитальности. Неживое стало в процессе эволюции есте- ственной средой для возникновения первого живого, которое, однажды проявив себя в неживом и на основе неживого, навсегда приняло его в свою организацию как неживую (инвитальную) субстратную часть. Итак, в многослойности явления жизнь инвитальный (нежизненный) слой— самый первый. Далее организация живых тел усложняется по мере возрастания уровня интегрированности частей и целого. Габитус и суб- габитус, если посмотреть на них по отдельности, но в структуре живого родительского (для них) тела, как было показано выше, ведут себя всего ~H~b на четверть от полного проявления жизнеемкости, что позволяет утверждать об обеих этих меросомах как носителях квадрантвитально- сти, т. е. только как четвертной жизнеемкости. В единстве, в союзе друг с другом обе меросомы — и габитус, и субгабитус вместе поднимаются в более высокий пласт (уровень) жизнеемкости (витальности), — поло-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ винчатой жизнеемкости, или семивитальности. Такую же жизнеемкость проявляет геносома и ее элементарные тела — гены. Чтобы семивиталь- ные тела перешли на новый, более высокий, слой организации жизни, они должны действовать в единстве как меросомы или как целые тела, но только вместе. Генофенетические тела — это проявление ценомеро- биозного (приватковитального), например, в клетках, органах — полно- ценных частях целого, но и ценобиозного (ковитального), образа жизни в соответствующих телах, например, индивидуумах в организации жи- вой природы. Похоже, выше этих слоев витальности в многослойном явлении жизнь больше ничего и нет. Так какое же место занимают формы роста (стадиосомы) среди всего разнообразия живых и неживых тел? То, что формы роста— живые тела, под вопрос не ставим, принимая это как уже доказанный факт. Они представляют всю фенетическую организацию (феносому) на стадии жизни биоморфы, т. е. в ритмическом соматическом жизнераз- витии, а место феносомы в целом живом теле мы уже определили: она — носитель половинчатой жизнеемкости (семивитальности) и, следова- тельно, формы роста — это семивитальные тела, существующие в орга- низации биоморф под управлением генов, только вместе с которыми они образуют целостную систему организации всей биоморфы. Таким образом, в теле биоморфы ритмически проявляют себя, а по сути мы можем говорить и так — обитают формы роста (стадиосомы), обеспе- чивающие в процессе своих смен стадийное жизнеразвитие всей био- морфы как организма или синорганизма. Формы роста биоморф — это «волны» ритмического развития био- морф, прокатывающиеся по их фенетической организации в унисон с ритмикой развития окружающей внешней среды, чем-то напоминаю- щей ритмику морской стихии, волны которой зависят от многих фак- торов (силы ветра, солености воды и т. п.). Формы роста (стадиосомы) — это и формы стадийного поведения фенетической организации био- морф, т. е. вся организация феносомы на отрезке возрастного и (или) сезонного развития биоморф. Это, таким образом, единство фенетичес- кого поведения и структуры, маркирующих целостное поведение био- морфы на отдельном отрезке ее морфогенического существования (от рождения до смерти). Стадийное поведение биоморф через разнообра- зие стадийных феносом (стадиосом), или форм роста, — это все ответ- ные реакции биоморфы на ритмические процессы, происходящие в окружающей среде, на которые она, биоморфа, отвечает соответствую- щим образом своими и возрастными, и сезонными реакциями. Непос- редственным носителем стадийных изменений, происходящих в био- морфе, является самая пластичная ее часть — феносома. Феносома адаптирована некоторым образом обитать — семивитальничать, в род- ном родительском для нее теле — в биоморфе, стадийно трансформи- руясь то в одну, то в другую формы роста, или формы стадийного поведения фенетической организации биоморфы. Чтобы одна живая (семивитальная) часть (феносома) биоморфы что-то свое пластично делала в ответ на ритмическое развитие окружающей природы, другая живая (семивитальная) часть (геносома) той же биоморфы существует
Глава 4. Концепция форм роста в биоморфологии стабильно и занимается управлением стадийным биоморфогеническим развитием. Раз формы роста живут, хотя и наполовину (семивитальни- чают), будучи стадийной феносомой, то они как-то проявляют жиз- неразвитие (по-своему витальничают), следовательно, это одни из атри- бутивных обитателей в организации любой биоморфы, а обязательная среда их непосредственного обитания — всегда эгосубстрат, из части (геносомной) которого происходит управление стадийным развитием. В процессе эволюции меросомы как обитатели и обиталища адап- тировались быть полезными не только для себя, но и для другой поло- вины в их единой жизненной системе. Чтобы обитать как меросома в организации биоморф семенных растений, надо использовать того, на (в) ком обитаешь (и наоборот), а с другой стороны — уметь что-то отдавать свое, чтобы быть полезной и обиталищу, которое, безусловно, что-то отрывает от себя и отдает кому-то — обитателю. Принципом взаимополезности широко пользуются все меросомы, проявляя себя или как обитатели, или как обиталища. Пожалуй, есть еще один момент, о котором можно сказать. Если образно, то у биоморф обнаруживается некое «пульсирование» («пульс») OT стадийного развития феносомы, единицей которого является стади- осома, или форма роста. Это она, периодически сменяемая стадиосома, и есть тот образный «удар» стадийного развития, проявление которого мы склонны визуально отмечать в организации биоморф. Это они, формы роста, периодически сменяясь, отражают собой феносоматичес- кое «пульсирование» организации биоморф семенных растений. Каждое возрастное состояние — это один (за ним будут другие) «удар» стадий- ного развития биоморфы — образный «удар» ее феносомой по органи- зации всей биоморфы в поисках направления дальнейшего развития в данном месте и в данное время как формы роста. Однако шутки в сторону: пора подводить некоторые итоги. При логическом расслоении (меротомировании) тел фитобиоморф удалось определить место частям их организации и разложить эти меросомы по соответствующим «полоч- кам» организации жизни: на одну «полочку» ушли квадрантвитальные (на четверть жизнеемкие) тела, например, габитусы, на другую «полочку» — семивитальные (наполовину жизнеемкие) тела, например, формы роста. Итак, формы роста — это носители ритмики морфогенического развития каждой биоморфы — стадийные феносоматические обитатели ее эгосубстрата, в котором, помимо неживой прогенетической части, присутствует и генетическая (семивитальная) часть, причем последняя управляет всей организацией биоморфы с ультрамикроскопического уровня и использует, со своей стороны, формы роста, а, следовательно, и всю фенетическую организацию уже как собственное обиталище— свой автобиотоп (глава 9).
Глава 5. Концепция разнообразия биоморф семенных растений, место в ней их габитусов и форм роста Мы, осознавая или не осознавая, живем в мире признаков, и когда задумываемся об этом, то начинаем так или иначе типизировать, клас- сифицировать и моделировать какие-либо объекты Природы. Так уст- роен и так живет человек; он может нормально развиваться, только активно познавая окружающую его остальную природу и самого себя. 5.1. Признаковые, или информационные (довостребованные и востребованные), пространства биоморф «Признак — все то, в чем предметы сходны друг с другом или в чем они отличаются друг от друга; показатель, сторона предмета или явле- ния, по которой можно узнать, определить или описать предмет или явление» (Кондаков, 1975, с. 477). Такое понимание признака имеет общенаучный смысл и, следовательно, применимо в биоморфологии. Признаки по своему содержанию и по возможностям их применения бывают самыми разными, например, простыми и комплексными, ис- кусственными, локальными, групповыми, таксономическими, специфи- ческими, налегающими (трансгрессирующими), диагностическими, аль- тернативными (Балковский, 1964). В логике признаки делятся на суще- ственные и несущественные, случайные, собственные, отделимые не- собственные и неотделимые несобственные, неотличимые и отличимые, положительные и отрицательные (Кондаков, 1975) и т.д. и т.п. Из при- веденных примеров подразделения признаков достаточно видно, что возможностей для типизации и классифицирования самих признаков предостаточно и это не удивительно, поскольку главное предназначение признаков служить целям распознания, различения явлений Природы. Признаки используются определенными отраслями знаний и, в зависимости от этого, они соответствующим образом квалифицируются: внешние морфологические (экзоморфологические), физиологические, анатомические и т.п. Многого, что можно сказать о признаках, мы
Глава 5. Концеиция азнооб азия биомо семенных астений касаться не будем, но на некоторые моменты обратим внимание. Часто признак используется как одноотраслевой, например, при изучении габитуса, но если поменять направление исследований, то тот же при- знак нередко может послужить и для другой отрасли знаний. Так, на- пример, признак «кустистые растения» с равным успехом может счи- таться и экзоморфологическим, и физиологическим. Признаки, облада- ющие свойством быть прочитанными по разным направлениям (отрас- JlfjM) исследований, — это интегральные признаки. В нашей работе используются разные признаки, но особое внимание обращено на интегральные признаки, поскольку именно они являются главными носителями информации о сложности организации биоморф семенных растений. Каждый признак является обладателем определенной инфор- мации о свойствах объекта Природы, которому он принадлежит, т.е. обладает определенным информационным полем. Интегральный признак может быть использован по-разному, по- скольку с его информационным полем способны работать специалисты разного профиля, например, физиолог под одним углом зрения, морфо- лог — под другим, а эколог — под третьим и т.д. Все разнообразие, которое обеспечивается информационными полями признаков опреде- ленного объекта, — это информационное, или признаковое, простран- ство (система) этого объекта. Признаки биоморф — предмет исследова- ния биоморфологии, которые существуют независимо от нашего созна- ния, определяют информационное содержание (пространство) этих объектов Природы. Чтобы изучить биоморфы, надо узнать их признаки, которые далее можно использовать для различных целей, в первую очередь, для типизации и классифицирования. Признаки биоморф- это их достояние, с которым они существуют в окружающем мире в определенном качестве. Признаки биоморф, кроме того, — это досто- яние науки, когда они замечены ею и определены соответствующими научными средствами. В науке мы имеем дело с логическим отражением тех признаков, которые существуют в Природе, в частности, в органи- зации биоморф. Чем лучше развивается наука, тем точнее осуществля- ется в ней отражение материальной природы — организации природных тел и процессов. Таков механизм взаимосвязи науки с материальными носителями признаков, познанием которых для науки и в науке посто- янно занимаются ученые разных специальностей. Наука, будучи логи- ческой системой, нацелена на осознанное определение и использование в интересах людей признаков Природы и ее элементов. Все явления (тела и процессы) материального мира, или, как гово- рят философы, вещи, относительно самостоятельны и существуют объективно; при этом они способны отражаться в нашем сознании как объекты познания. Можно говорить о двух типах признаковых про- с~анств как двух сторонах одного и того же явления-вещи: довостре- бованном (невостребованном) прнзнаковом пространстве, которое суще- ствует в явлении-вещи независимо от сознания, непосредственно как система признаков «вещи в себе» и ждет своего познания (востребова- HH<), и востребованное прнзнаковое пространство, которое создается познавательной деятельностью субъекта и востребует признаки «вещи в
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИИ себе» в признаковую систему «вещи для нас». Здесь уместно подчерк- нуть известное диалектическое положение: вещи потенциально позна- ваемы, а познание их — это процесс практического превращения «вещи в себе» в «вещь для нас». Пожалуй, если быть еще более точным и последовательным, то стоит говорить не столько о «превращении» «вещи в себе» в «вещь для нас», а о проявлении у «вещи в себе» ее особого свойства — быть познаваемой (!) и существовать еще параллельно в другом мире — мире познания (образном) как «вещи для нас», т.е. в мире, опосредованном нашим сознанием. На самом деле происходит наслоение на конкретную вещь идеального продукта — образа этой вещи в нашем сознании и проявление ее, в таком случае, в дополнительном качестве — еще и как востребованного признакового (или информационного) пространства этого явления-вещи. Довостребованное признаковое пространство какого-либо явления остается всегда при нем, поскольку оно существует независимо от со- знания, но, чем больше признаков (их информационных полей) мы определяем в каком-либо явлении, тем больше признаков, точнее, их стопроцентных двойников-отражений, или идеализированных образов, отводится (востребуется) в другую плоскость — область познания, где действуют свои законы, в частности, в нем их подвергают различным логическим операциям, например, типизации, классифицированию и моделированию. Создавая востребованные признаковые пространства тех или иных явлений материальной природы и расширяя в процессе познания границы этих «пространств» (систем), мы не влияем или толь- ко косвенно влияем (через различные виды практической деятельности людей) на объем довостребованных признаковых пространств, посколь- ку те существуют независимо от сознания. Но «вещи в себе» вместе с их довостребованными признаковыми пространствами подвержены из- менениям, поскольку движение (изменение) — атрибутивное свойство Природы и вне движения не может, как известно, быть ни «вещей в себе», ни «вещей для нас». В нашей монографии более всего рассматриваются «вещи для нас» в образе востребованных признаковых пространств биоморф семенных растений, которые существуют параллельно с довостребованным при- знаковым пространством этих же биоморф. Востребованный признако- вый, или информационный, мир создается людьми только для нас, для тех, кто способен работать с ним и воспользоваться знаниями о нем, что никому другому пока не доступно без помощи людей, хотя, вероятнее всего, этот мир нужен для всего живого на Земле, поскольку от качества и количества человеческой деятельности напрямую зависит ответ на вопрос «быть» или «не быть» не только человечеству, но и жизни вообще (Жизни). Востребованное признаковое пространство биоморф с разви- тием биоморфологии имеет неукротимую тенденцию расширяться вширь и вглубь. Словосочетание «признаковое пространство» во многом метафори- ческое, поскольку оно образно свидетельствует о концентрации некото- рого набора природных признаков, с их информационными полями, в
Глава 5. Концепция азиооб азия биоморф семенных растений некоторой части пространства-времени, не подвергаясь воздействию на них сознательной деятельности людей (довостребованное признаковое пространство) или подвергаясь таковой (востребованное признаковое пространство). Информационные поля признаков составляют в сово- купности разнообъемные признаковые пространства, поскольку каче- ственные и количественные параметры объектов различны. Биоморфы (организмы и синорганизмы), будучи, в первую очередь, довостребован- ными признаковыми пространствами, притягивают к себе познаватель- ные усилия субъектов — мыслительную деятельность людей, что так или иначе создает разнообъемные востребованные признаковые системы (пространства), в которых признаки биоморф посредством различных методических приемов подвергаются абстрагированию и упорядочению в логических системах. Довостребованные признаковые пространства существуют, как и востребованные признаковые пространства, упорядоченно, но, в первом случае, организационный порядок в системе признаков объектов При- роды, в частности, биоморф обеспечивается местом признака в органи- зации конкретного объекта-вещи, которому они принадлежат, и обус- ловлено оно всей предшествующей историей развития этого объекта, его эволюцией. Во втором случае, организационный порядок в востре- бованном признаковом пространстве находится под обязательным дав- лением идей, осознанной деятельности, действующей извне, со стороны нас, людей (субъектов), которые пытаются понять особенности призна- ков объектов, их структуры и функционирования. Уникальное новаторство может привноситься субъектом в востре- бованное признаковое пространство каких-либо объектов Природы, на которое распространяется действие сознательной воли субъекта, одна из сторон которой — это отвлечение в процессе познания от каких-либо сторон (несущественных для конкретного случая) рассматриваемого явления и сосредоточение внимания на основных, существенных чертах в раскрытии их содержания. На объект познания — довостребованное признаковое пространство — осуществляется воздействие посредством абстрагирующих операций со стороны субъекта, обладающего такой способностью; в результате соответствующей мыслительной деятельно- сти рядом с довостребованным создается еще одно, востребованное признаковое пространство, которое нередко становится предметом даль- нейшего научного исследования и (или) практического опыта. Итак, признаковые пространства объектов Природы, биоморф, в частности, будучи информационными системами, двойственны по сво- ей натуре (своему содержанию). С одной стороны, такое «пространство» — объективное (довостребованное), поскольку принадлежит самому природному объекту — «вещи в себе», а, с другой стороны, оно же представляет, опосредованное через сознание людей, востребованное признаковое пространство того же объекта, но уже «вещи для нас». Есть смысл сделать еще небольшое отступление и подробнее пораз- мышлять о работе с признаками. В довостребованном признаковом пространстве первоприродных тел, допустим, биоморф семенных расте- ний, признаки существуют сами по себе, независимо от сознания лю-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ дей, но всегда в состоянии «ожидания», что кто-то когда-то к ним прикоснется, хотя бы идеально, через свое сознание, проявит когда- либо познавательный интерес. Тогда эти признаки биоморф, ничего в себе не меняя, и без каких-либо направленных усилий со своей сторо- ны, отбросят от себя, как обычно (как всегда), отражение точно такое, как они сами, но все-таки отражение, поскольку зафиксировать факт отражения признака — его образ — может только наше сознание, сделав его осознанным достоянием знания, т.е. знания субъекта, который с гордостью или без нее, что он «вырвал» нечто искомое из таинства Природы, как-то поместит признак биоморфы в ее уже востребованное признаковое пространство, которое он, исследователь, будучи предста- вителем Разума на современном этапе развития Человечества, будучи одновременно и ничтожной, и величественной частицей материи, при- несет в науку только так, как он это может, как это он реально (а не в мечтах) сделал здесь и сейчас. Потом придут другие. Многое изменит- ся, но усилия конкретной разумной частицы, может быть, автора, а может быть, другого, опять же может быть, не будут забыты и оставят след в развитии науки. Так или иначе любой научный работник зани- мается процедурой востребования признаков из довостребованных при- знаковых пространств каких-либо природных объектов, создает и (или) модернизирует востребованные признаковые пространства тех же при- родных объектов. Довостребованное признаковое пространство биоморф — это, хотя и образное обозначение, но все же обозначение всего действительного набора (всей совокупности) признаков биоморфы или какой-либо груп- пы биоморф (например, определенного вида), составляющих и процесс, и результат реального существования этой биоморфы (или группы био- морф) в некоторой части конкретного пространства и времени. В дово- стребованном признаковом пространстве все признаки конкретны и их существование не зависит от сознания людей, а некоторая идеализация признака или признаков начинается только после подключения субъек- та к процессу бытия признаков. Биоморфа, в любом случае, всегда остается единственной и полновластной хозяйкой своих признаков и некоторого дискретного участка пространство во времени, где они со- существуют вместе, образуя целостную систему — биоморфу. Мы, люди, востребуем независимые от нас признаки, получаем их отражения в нашем сознании в образах и включаем такие, идеализированные нашим сознанием, признаки в востребованное признаковое пространство тех же самых биоморф, но — в пространство их признаков, пропущенное через сознание и организованное в соответствии с абстрагирующими возможностями познающего субъекта и уровнем развития современной отрасли знаний о биоморфах. Сейчас уже ясно, что работа с признаками биоморф впервые начи- нается еще в их довостребованных признаковых пространствах и, только когда мы их замечаем, только потом, эти признаки одеваются в научные одежды, получая названия и определения, далеко не всегда сразу удач- ные; потребуется время, чтобы признак получил прописку в науке, заполняя своими востребованными образами различные классификации
Глава 5. Концеиция азнооб азия биомо семенных растений и модели, определения терминов, законов и правил и т.п. логических построений. Постепенно мы подошли к необходимости не только вос- требовать признаки из организации биоморф, но и использовать их в построении концептуальных конструкций, каковыми являются, напри- мер, субординатные и суперординатные классификации и их модели. «Достаточно строго и четко проведенная классификация, как показыва- ет история науки, одновременно подытоживает результаты предшеству- ющего развития данной отрасли познания и вместе с тем отмечает начало нового этапа в ее развитии» (Мейен, Шрейдер, 1976, с. 67). Итак, разобравшись с двойственностью признаковых пространств природных явлений (тел и процессов), мы далее акцентируем внимание на работе с востребованными признаковыми пространствами, особенно не отвлекаясь по этому поводу, поскольку здесь и сейчас нам ясно, кого, как и зачем мы будем типизировать, классифицировать и моделировать. Мы будем привлекать к соответствующим логическим операциям, в основном, признаки биоморф семенных растений, которые в процессе развития биоморфологии уже стали ее достоянием и их когда-то, а иногда прямо сейчас, удалось взять из довостребованных признаковых пространств этих же биоморф, «заставив» работать на науку. В нашем случае, в данный момент мы сосредотачиваем внимание в основном на признаках габитусов и форм роста взрослых биоморф семенных растений, чтобы с их помощью построить субординатные классификации этих объектов Природы с разных высот обобщения, используя (востребуя) соответствующие признаки и, в первую очередь, интегральные признаки. В этой связи, если посмотреть на востребован- ное признаковое пространство растительных биоморф вообще в истори- ческой ретроспективе (наиболее подробный обзор о классификациях биоморф высших растений приведен в книге И. Г. Серебрякова, 1962), то наглядно видно, как эволюционировало знание об этом «простран- стве», как исторический пласт в его развитии не остался слепым отро- стком, а стал активным продолжением в бурно развивающихся совре- менных классификационных системах. Ситуация в проблеме классифи- цирования биоморф отражает состояние науки в этой области знаний и многие признаки биоморф как-либо уже были использованы при построении классификационных систем. «Ни одному классификатору не доведется столкнуться с совершенно девственной, не затронутой хотя бы какой-то классификацией предметной областью» (Мейен, Шрейдер, 1976, с. 77). Безусловно, с развитием науки — биоморфологии, в час- тности, — востребованные признаковые пространства («вещи для нас») совершенствуются в обновленных типах, классификациях и моделях. Попробуем сформулировать несколько правил манипулирования призна- ками. Правило первое: любые признаки в организации обьектов материаль- ной природы как признаки довостребованного признакового пространства имеют свободный выход в востребованное признаковое пространство, через p«mu статус предмета познавательной и практической деятельности людей и, в частности, научной деятельности.
OCHOBhl БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИИ Правило второе: востребованное признаковое пространство так или иначе, раньше или позже обязано быть достоянием научного исследования, в первую очередь, классифицирования и (или) моделирования предметов исследования по его признакам, поскольку они, будучи вовлеченными в эти логические операции, способны в полной мере отразить уровень развития наших знаний о соответствующем предмете — «вещи для нас» — через востребованные (т.е. известные нам) признаки. Правило третье: самый простой и надежный способ зафиксировать разнообразие востребованных признаков на языке науки — это распреде- лить их в соответствующих однолинейных и многолинейных классифика- циях (т. 1, гл. 9). Правило четвертое: разнообразие иерархических взаимосвязей предме- тов исследований фиксируется посредством использования востребованных признаков при построении определенных иерархических классификаций (и их моделей): таксономических, субординатных, мультисубординатных, су- перординатных и т.п. (т. 1, гл. 1). Правило пятое: только интегральные (многоотраслевые) признаки материальных явлений могут наиболее полно отразить сложность органи- зации предметов исследований и определить их уровень (тип) в субординат- ной классификации. 5.2. Габитуальное разнообразие биоморф семенных растений Ранее (глава 3) на основе закона взаимодействия внутреннего и внешнего в организации биоморф семенных растений было определено правило габитуального поведения биоморф, в котором подчеркнуто, что габитус биоморф весьма строго передает (адекватно отражает) состоя- ния их жизнеразвития на многих конкретных отрезках пространства и времени, поэтому вполне может быть использован в качестве наглядно- го средства (индикатора) и инструмента для различных логических операций. Наше внимание, здесь и сейчас, сосредоточим на интеграль- ных признаках, в логической инфраструктуре которых присутствуют их стороны-элементы как габитуальные составляющие организации био- морф. Присутствие габитуальной стороны в интегральных признаках— главное наше условие для использования их в соответствующих опера- циях классифицирования и моделирования. Работа с интегральными признаками — дело тонкое, но когда единство по некоторым таким признакам в организации различных биоморф распространяется на их определенные группы, то мы невольно начинаем заниматься типологи- ей, сделав первые обобщения в типах, а далее с ними можно произво- дить многие другие логические операции, в первую очередь, классифи- цировать и моделировать интересующие нас предметы познания. То, что тела, в частности, биоморфы различаются, так или иначе, по многим или немногим признакам — это понятно и об этом мы будем еще много говорить на протяжении всех томов нашей монографии,
Глава 5. Концеиция азнооб азия биомо семенных растений сравнивая одни группы растительных биоморф с другими, но сначала обратимся к единообразию в организации биоморф семенных растений. 5.2.1. Единообразие в организации биоморф — первое условие к проявлению их разнообразия О единообразии организации живых систем вообще мы уже писали (т. 1, гл. 2). Прежде чем говорить о разнообразии габитуальной органи- зации биоморф, надо достаточно полно ощутить содержание единооб- разия в организации биоморф в целом как основы для проявления разнообразия. Высшим растениям свойственно «единство окраски, единство, не- смотря на внешнее разнообразие, способов размножения, единство полового процесса, единство внешнего и внутреннего строения» (Мей- ер, 1958, с. 27). Самое первое единство, что отличает биоморфы Земли (иные науке пока неизвестны), да и другие живые системы, — это дискретность их организации, всеобщая и основная характеристика жизни (Тимофеев-Ресовский и др., 1969). Для всех зеленых растений (автотрофов) характерен один и тот же процесс фотосинтеза, посред- ством которого создается химическая энергия жизни из лучистой энер- гии Солнца (Тимофеев-Ресовский и др., 1969). Сходным образом биоморфы всех семенных растений проходят ста- дии своих морфогенезов, в которых последовательно сменяются возрас- тные состояния: проростки, всходы, ювенильные и имматурные расте- ния, составляя вместе виргинильный период, затем растения достигают матурного периода (физиологической зрелости), когда цветут и плодоно- сят, завершая свой жизненный цикл за один вегетационный сезон или за два — много лет (Работнов, 1950; Уранов, 1960, Нухимовский, 1980, 1997 и др.). При всей специфичности фенетических признаков, развивающих- ся в онтогенезах покрытосеменных растений, их объединяет определен- ная последовательность органообразовательных процессов, в частности, двенадцать этапов органогенеза побегов (Куперман, 1977). Ранее (Нухимовский, 1986, 1997) было показано, что участие био- морф в круговороте ресурсов биосферы можно выразить как результат согласованного взаимодействия пяти функциональных систем (меро- сом): управления (генетическая организация), потребления (обеспечи- вает трофику, дыхание), производства (осуществляет необходимый рост и развитие тела биоморф), резервирования (обеспечивает отложение потребленных и произведенных ресурсов в резерв для реализации по мере необходимости), рассеивания (осуществляет полное обособление предка от потомка, например, семенное размножение, отделение иных партикул и распространение гамет, спор, а, кроме того, элиминацию частей и целых организмов). Есть смысл немного дополнить сказанное выше о единообразии, проявляющемся в организации биоморф семенных растений. Первое, что определяет организацию всех биоморф как живых систем — это наличие в ней трех атрибутивных сторон (меросом): не-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ живой субстратной — протеросомной, генетической и фенетической (подробнее см. главу 1). Второе. Биоморфа у семенных растений всегда существует как те- лесная система, в которой всегда присутствует во взаимосвязи и взаи- модействии внутренняя организация (эндосома, включающая геносому и внутреннюю фенетическую организацию, или субгабитус) и внешняя организация (экзосома, или габитус), причем габитус проявляет себя как атрибутивная функция жизнедеятельности субгабитуса, а шире— всей эндосомы. Это положение подробно обосновано в предыдущих главах. Третье. Все биоморфы состоят в своей организации из частей, в частности и в первую очередь, из клеток, так или иначе специализиро- ванных для выполнения определенных функций. Четвертое, что подчеркивает единообразие в организации биоморф семенных растений: наличие у них двух основных органов — побегов и корней, посредством которых обеспечиваются все проявления жизнеде- ятельности. Пятым показателем единообразия в организации биоморф семен- ных растений являются корнепобеги, в которых единство побега и корня в одной системе может выступать на уровне органа (иногда) и на уровне организма (почти всегда), исключения изредка бывают в результате редукции корня, например, у вида насекомоядных водных растений Utricularia vulgaris L. Шестое. Какую бы форму фенетической организации ни приобре- тали побеги и корни в развитии биоморф, всегда проявляется стремле- ние к размножению, и вегетативное тело любой биоморфы всегда зани- мает подчиненное положение по отношению к репродуктивному телу, обеспечивая половой процесс всем необходимым. Седьмое. Высшее единство в организации биоморф семенных рас- тений составляет не только доминирующее стремление к размножению, но и более всего стремление к жизнесохранению (биофилии) вообще (т. 1, гл. 3). Таковы несколько основных положений, которые объединяют орга- низацию биоморф семенных растений, делая ее единообразной. 5.2.2. Средства распознания разнообразия биоморф На что же надо сконцентрировать внимание, чтобы разобраться, когда на упомянутом выше единообразии развивается по определенным законам и правилам «ошеломляющее» разнообразие биоморф? Надо поменять сферу интересов в нашем подходе и начинать видеть в орга- низации биоморф другую их сторону — разнообразие, поскольку только разобравшись в нем, можно строить классификации и, в особенности, классификации по сложности организации. Когда признаковое единство в организации растительных биоморф распространяется не на всех их, а только на определенные группы
Глава 5. Коицепция разиообразия биоморф семеииых растеиий биоморф, мы невольно начинаем заниматься типологией и, сделав пер- вые обобщения на уровне типов, можем далее производить еще более сложные логические операции: классифицировать и моделировать. Разобравшись с единообразием в организации всех биоморф семен- ных растений, переориентируем все внимание на поиск единообразия не среди всех, а только отдельных групп биоморф по определенным признакам одного сравнительного ряда, что в итоге позволяет судить уже о разнообразии биоморф по этому ряду признаков и строить сна- чала однолинейные классификации, а затем объединять их в многоли- нейные классификации. Важная особенность линейного классифициро- вания (т. 1) в том, что все признаки, независимо от сложности органи- зации изучаемых объектов, которую они могут отражать и нередко от- ражают, принимаются как равнозначные. Это удобно, когда мы хотим получить разноплановую характеристику объекта, но для того, чтобы разобраться со сложностью их организации, нужны другие принципы классифицирования и определенные представления о критериях слож- ности организации. Ранее (т. 1) было показано, что развитие биоморф находится в соответствии с одним из высших законов Природы — законом иерар- хичности: «во всем всегда есть что-то, в чем-то меньшее, и что-то, в том же большее». Исходя из этого, мы смотрим на целостную организацию биоморф как на иерархическое явление, существование которого обус- ловлено сложностью организации этих систем и наличием в ней опре- деленных уровней (ступеней). Различные проявления иерархичности в целостной организации биоморф можно наблюдать как при сравнитель- ном исследовании многих биоморф или одной биоморфы в ее развитии. Все биоморфы, как бы конкретно они ни были устроены, — это всегда системы, а, как известно, всякая система, так или иначе, имеет сложную организацию. В этом аспекте всякое деление биоморф (и не только их) на простые и сложные, автор также нередко использует такой методический прием (например, простые биоморфы — организмы, а сложные — синорганизмы), безусловно, удобный логический подход, но он всегда относительный, поскольку наименее сложная в определенном отношении система условно обозначается как простая, а более сложная в том же отношении — как сложная система. При определении сложности организации биоморф вопрос состоит не столько в том, где действительно простая биоморфа, а где — сложная, поскольку биоморфа — это всегда система, и этим уже многое сказано, она организована так (проще) или иначе (сложнее), но в целом всегда только сложно, а все остальное — это результат научной абстракции. Наша задача — разобраться со сложностью организации биоморф как систем, поскольку через нее можно судить об иерархичности, поэтому, безусловно, можно говорить об относительной, но не действительной, простоте биоморф при сравнении с другими биоморфами, поскольку обязательным остается факт: сложность организации одной биоморфы- системы может быть больше (меньше) сложности организации другой биоморфы-системы, и так везде, другого не дано. То, что присуще всем биоморфам семенных растений, может также быть показателем сложно-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ сти их организации, и тогда биоморфы меньшей сложности организа- ции будут включаться по тому же ряду признаков в биоморфу большей cJIQJKHocTH как часть в целое, например, биоморфы-организмы ниже по сложности биоморф-синорганизмов. Уже (глава 2) было подчеркнуто, что организация любой биоморфы — это неразрывное единство ее вегетативной и репродуктивной частей (меросом), и об этом никогда не стоит забывать. Можно, конечно, начать изучать по отдельности cJIQJKHocTb вегетативной (эусоматической) орга- низации, сложность пререпродуктивной (эу- и диасоматической), или переходной организации и, наконец, сложность тотарепродуктивной (ар- хесоматической) организации. Однако, нас здесь и сейчас эти частности интересуют меньше, а более всего — сложность организации биоморф в целом, т.е. целостной организации биоморф. В такой ситуации не стоит смотреть на биоморфу с пытливым желанием достигнуть максимальной детализации ее элементов, здесь надо изменить угол обзора и посмотреть на биоморфу в целом, в общем, сознательно отвлекаясь от деталей. При таком подходе к каждой биоморфе нам решающую помощь оказывает габитус и форы роста биоморф. Габитус биоморфы способен наглядно через себя передать многое, чем в целом богата по организационному содержанию биоморфа. Среди возрастных состояний биоморфы, т.е. ее форм роста, максимум организационной сложности проявляют средне- возрастные (медиматурные) растения, поэтому для суждения о сложности организации биоморф мы используем именно их. В свою очередь, сами формы роста достойны субординатного классифицирования, поскольку переход от каждой формы роста к новой ведет к усложнению фенетичес- кой организации как самой динамичной части биоморф на восходящем направлении развития и к падению сложности организации биоморф на нисходящем направлении их биоморфогенезов. Сами формы роста био- морф, будучи живым, стадийно изменяющимся, фенетическим содержа- нием биоморф на определенных отрезках их существования, достойно могут обеспечить приоритетную основу для субординатного классифици- рования биоморф по степени cJIQJKHocTH их возрастного фенетического поведения (см. субординатную классификацию форм роста — возрастных состояний биоморф в главе 4). Работая с биоморфами, нам важно понять, какие критерии могут свидетельствовать об одном уровне сложности организации, а какие- о других уровнях. Всякое суждение об уровнях — это логический прием, он продиктован опорой на объективные факты существования различий в организации соответствующих тел. В качестве важных показателей сложности организации могут быть признаки, которые действуют на большие общности биоморф, например, на все древесные растения по признаку наличия у них надземного деревянистого базового органа— деревянистого аэроподия разной высоты и по отсутствию такового у всех травянистых растений, выступающих по ряду признаков, связан- ных с наличием или отсутствием базового органа, как биоморфы мень- шей сложности организации, чем древесные растения. Субординатные классификации биоморф и их органов, т.е. класси- фикации по сложности организации мы уже приводили (т. 1, гл. 5). Там
Глава 5. Концепция разяооб азия биомооф семенных растений была разработана классификация биоморф-организмов как корнепобе- гов от ризопротерокормусов до ризоэнатокормусов, в которых лидерами являются побеги, определяющие сложность организации корнепобегов. По сложности организации разнообразие биоморф довольно огра- ниченно, поскольку признаков-индикаторов такого разнообразия не- много. Какие признаки могут более других служить показателями слож- ности организации? На наш взгляд, это, в первую очередь, интеграль- ные признаки, которые пронизывают с разных сторон фенетическую организацию биоморф, включая их габитус и формы роста. Интеграль- ные признаки, когда они обеспечивают информацию о сложности органи- зации биоморф и тем самым о6 иерархии их организации, можно обозна- они определяют в субординатной классификации как субординирующие признаки. Эти признаки — индикаторы определенных субординатных био- морфотипов. Деревья, кустарники, многолетние травы, однолетники и T.ï. — все это субординатные биоморфотипы, полученные путем субор- динатного классифицирования с определенной высоты абстракции (рис. 123-147, 150-160). При дальнейшем расширении числа признаков био- от одного до любого множества других признаков, назовем их корректи- рующими, которые востребуются из многолинейного признакового про- странства биоморф, чтобы корректировать положение биоморфотипа до более частного в какой-либо классификационной системе или вне ее, про- сто для разноплановой характеристики или отдельной биоморфы, или биоморфотипа. Такой расширенный (субординатный + корректирующие биоморфотипы) обозначим как просубординатный биоморфотип, подчерк- нув тем самым, что он «работает» на расширение информационного «поля» соответствующего типа биоморф, преобразуя его статус (из субординатного в просубординатный). Латинская приставка «про» имеет в подобных случаях следующее значение: «являющийся сторонником, действующий в интересах кого-либо или чего-либо» (Сл. ин. слов, 1964, с. 520). Эту приставку используют, например, в следующих словах: «проимпериали- стический», «прокоммунистический» и т.п. Просубординатные биомор- фотипы стоят на «службе» у соответствующих субординатных биоморфо- типов и несколько уже их по содержанию, поскольку всегда дополнены какими-либо корректирующими признаками (рис. 191-206). Когда, напри- мер, мы говорим «деревья», то имеем дело с определенным субординат- ным биоморфотипом, а в случае летнезеленых безрозеточных эусимпо- диальных открытых (открыто-инфлоресцентных) и т.д. и т.п. деревьев проявляет себя уже просубординатный биоморфотип. Такие биоморфо- типы определяются комплексными признаками-индикаторами, которые, соответственно, в полном виде (вместе) будем считать просубординиру- ющими признаками (единство субординирующего + корректирующих признаков). В мультилинейных классификациях (т. 1) все классы систем рассмат- риваются как равнозначные (не учитывается их ранг сложности для оп- фотипов в таких классификациях всегда одинаков, и все они выступают
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ для данных случаев как рядовые. В однолинейных классификациях все 6ио- морфотипы рядовые, поскольку признаки, определяющие их, также рядовые альтернативные (сравнимые и взаимоисключающие). Графические модели, хотя бы упрощенные, всех основных субор- динатных биоморфотипов можно при желании уместить и в одном рисунке, что будет показано в следующей главе (6.5). Что касается просубординатного моделирования биоморфотипов, то оно зависит от качества и количества набора корректирующих признаков, число и подбор которых сильно варьируется, в зависимости от разных обстоя- тельств и является проблемой не для исследуемых биоморф, а лишь для их исследователей, постоянно совершенствующих процедуру востребо- вания признаков и модернизирующих востребованное признаковое пространство биоморф. 5.2.3. Маркирующее участие габитуальных частей- признаков в интегральных признаках, отражающих сложность организации биоморф Габитуальные признаки — это части интегральных признаков, во- стребованные кем-либо для удовлетворения какого-либо, в нашем слу- чае профессионального, текущего интереса (любопытства). Габитуаль- ные признаки, наряду с другими признаками, сконцентрированы вместе в единые системы — интегральные признаки, которые, будучи узнанны- ми (познанными), пополняют фонд научного знания признаков опре- деленных предметов исследований, составляя востребованное призна- ковое пространство каждого такого предмета. Такое «пространство» не может быть само по себе, поскольку о его развитии (совершенствова- нии) постоянно заботятся заинтересованные люди, чего нельзя сказать о довостребованном признаковом пространстве, которое существует независимо от сознания — само по себе, развиваясь и эволюционируя, достигая расцвета, упадка и вновь расцвета и т.д. Востребованное признаковое пространство каких-либо объектов Природы пронизано осознанной деятельностью субъекта — исследова- теля, поэтому оно организовано с учетом уровня развития современной науки и интеллекта непосредственных исследователей. В предыдущем томе (т. 1) было показано, что востребованное признаковое простран- ство наиболее разносторонне отражает организацию биоморф семенных растений, когда в нем присутствует мультилинейная классификация и все известные признаки биоморф семенных растений располагаются в ней рядами, в которых они и «маршируют» на вечном параде бытия в пространстве и времени. При мультилинейном классифицировании все признаки принимаются как равнозначные (рядовые), что и создает ус- ловия для рассмотрения их во множественных однолинейных горизон- тальных рядах (классификациях). Только в субординатных, мультисуб- ординатных и суперординатных классификациях признаку придается значение как средству отражения разной сложности организации био-
/дава 5. Концепция разнообразия биоморф семенных растений морф в целом, так и их элементов (см. т. 1). В таких классификациях особую роль приобретают интегральные признаки. Достоинство интег- ральных (многоотраслевых) признаков в том, что они лучше других передают изменение сложности организации как биоморф в целом, так и их структурных элементов. Интегральные признаки несут в себе не только габитуальную, но и эндоморфологическую информацию, т.е. целостную, а следовательно, они всегда больше, чем их части-признаки. Нас очень устраивает тот факт, что во всех интегральных признаках, которые мы используем в качестве критериев при определении сложно- сти организации биоморф, всегда присутствуют габитуальные части- признаки, т.е. при необходимости эти признаковые части можно рас- сматривать и обособленно, только как габитуальные признаки, не забы- вая при этом где-то подспудно об истинной природе таких признаков. Итак, все рассматриваемые субординатные типы биоморф постро- ены по интегральным признакам, в которых в качестве маркированных (внешне обозначенных) сторон участвует их габитуальное содержание. многоотраслевая натура интегральных признаков позволяет рассматри- вать классификации, построенные на их основе, не только интеграль- ными, но и габитуальными, морфологическими, физиологическими, в зависимости от присутствия в таких признаках соответствующих частей- признаков. Мы не проявим усердия в нашей работе, чтобы перестараться в меротомировании интегральных признаков. Нас весьма устраивает слож- ное содержание используемых признаков, с обязательным участием в них габитумьн частей-признаков. Исходя из всего рассмотренного выше, предваряющего основной результат этой главы наших исследова- ний, приобщимся к суперординатной классификации биоморф семен- ных растений, которая, кроме того, еще и моделирована в «пирамиде» абстракций (рис. 216). Все субординатные классификации (субординии) биоморф занимают определенное место на уровнях (суперординатах) в суперординатной классификации, методика построения которой обсуж- дена и изложена в первом томе нашей монографии. Верхнюю суперординату (рис. 216-А) в рассматриваемой классифи- кации классификаций мы заняли тремя широкими субординиями. За критерий сложности организации биоморф в первой широкой суборди- нии была принята (см. т. 1, с. 52) степень их пространственной цело- стности: нижнюю субординату занимают организмы, поскольку состав- ляющие их элементы-органы образуют морфофизиологически непре- рывную систему, а верхнюю субординату — синорганизмы, поскольку составляющие их элементы-организмы морфофизиологически разобще- ны. Под иным углом, но с той же высоты обобщений рассмотрим вто- рую широкую субординию биоморф. За критерий сложности организа- ции в этой классификации биоморф принята способность или неспо- собность их развивать персистентные (живущие более одного года) части: эвриэфемерные растения, короче эвриэфемеры (эфемеры, геми- эфемеры, однолетники и озимооднолетники), независимо от того, когда они прорастают и как развиваются, живут в целом менее одного года; персистентные растения, или персистенты, — эти растения, независимо
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РАСТЕНИИ от того, как они прорастают и развиваются, живут в целом более одного года). Третью широкую субординию можно построить, избрав в каче- стве критерия для определения сложности организации биоморф их способность или неспособность проявлять аэроподиальное развитие в надземной части пространства: апоаэроподиальные растения, или аэро- базофобы, и аэроподиальные растения, или аэробазофилы. Начнем спускаться вниз по ступеням «пирамиды» абстракций, уве- личивая, тем самым, разнообразие биоморф по сложности их органи- зации в каждой новой субординатной классификации. Четвертую ши- рокую субординию MQKHQ построить так: малолетники (эвриэфемеры и двулетники), среднелетники (живут 3 — 10 лет), многолетники (живут более 10 лет). На той же суперординате (уровне абстракции) располо- жим еще две субординатные классификации семенных фитобиоморф из трех субординат в каждой (рис. 216-Б). В пятой субординии отмечено усложнение организации биоморф в зависимости от кратности прояв- ления репродуктивной функции: унирепродуктивные, бирепродуктив- ные и мультирепродуктивные растения. В шестой субординии располо- жены фитобиоморфы под иным углом обзора — по отношению к спо- собу питания: автотрофы, гемигетеротрофы (полупаразиты) и гетерот- рофы (Никитин, 1983). Перейдем на ступень ниже (рис. 216-В) нашей «пирамиды» абст- ракций, где поместим седьмую субординию из четырех субординат, раз- делив малолетники на две группы биоморф: эвриэфемеры, двулетники и далее среднелетники и многолетники. Следующую, восьмую, субординию биоморфотипов семенных рас- тений мы расположили на ступени — суперординате еще ниже (рис. 216-Г). В этой классификации пять основных, хорошо известных био- морфотипов, описанных впервые К. Раункиером (Raunkiaer, 1907, 1934): терофиты, криптофиты, гемикриптофиты, хамефиты, фанерофиты. За критерий сложности в организации растительных биоморф в этой клас- сификации было принято поведение только их надземной побеговой части в отношении к климату, в частности, особенности расположения верхушечных меристем относительно поверхности почвы и их защи- щенности в течение неблагоприятного времени года. На той же ступени «пирамиды» абстракций располагаем девятую субординатную классификацию из пяти субординат: травянистые, полу- травянистые, полудревесные, древовидные и древесные растения. Травянистые растения, или травы, (в надземной побеговой части у этих растений не проявляется персистентность, но она иногда может прояв- ляться в приземном, прижатом к поверхности почвы, состоянии их побегов; рис. 217-276). Полутравянистые десуккуленты (в надземной побеговой части они проявляют персистентность, но она часто недолго- вечная; аэроподии представлены более всего стеблями-соломинами зла- ков, из которых только аэроподии немногих групп биоморф достаточно сильно одревесневают и живут более 10 лет и значительно больше; рис. 277-290). Полутравянистые суккуленты (аэроподии этих растений име- ют разную продолжительность жизни и высоту, но у них весьма харак- терная консистенция, рассчитанная не столько на одревеснение тканеи,
Глава 5. Концеиция азнооб азия биомо семенных растений которого может вообще не быть или оно слабое, сколько на запасание воды, рис. 291). Полудревесные растения (в надземной побеговой части они проявляют персистентность, одревесневая на различную высоту, образуя стволики или даже стволы, несущие кроны, в которых побеги после завершения репродуктивных циклов отмирают на различную длину, а продолжительность жизни надземных персистентных побего- вых частей сравнительно невысокая и не превышает, как правило, 15— 20 лет; рис. 292-302). Древовидные растения, или дендроиды (их аэро- подии одревесневают, но иначе, чем полутравянистые, деревянистые и полудревесные растения, поскольку не имеют во внутренней структуре камбия и, таким образом, не проявляют вторичный рост; рис. 303). Деревянистые растения (напоминают во многом предыдущую группу, но, в отличие от нее, образуют надземные персистентные деревянистые побеговые части-стволики или стволы, продолжительность жизни кото- рых, как правило, превышает 20 лет, а нередко достигает многих десят- ков и даже сотен лет; рис. 304-332). Все упомянутые здесь термины не новы. Пока более всего не стабилизировалось значение термина «полу- травянистые» растения, предложенное, вероятно впервые, А.В. Прозо- ровским (1936), из которого мною использовано только слово-этикетка, а смысл вложен другой и тому есть свои причины. Растения (с однолет- ними одревесневающими надземными побегами и многолетними дере- вянистыми подземными побегами), которые Прозоровский называет «полутравами», принадлежат сейчас полностью к травам, поскольку факты сильной лигнификации ткани надземных побегов, которые ос- тавляют эти побеги однолетними, ничего не меняют, и признаки, из- бранные для диагностики их как полутрав, явно себя не оправдали. В отличие от Л.Е. Гатцук (1976), которая провела обстоятельный литера- турный анализ явления травянистости у растений сезонного и бессезон- ного климатов, но, похоже, так пока и не решилась очертить понятий- ные границы биоморфотипа трав, а оставила травянистым растениям весьма прозрачные параметры: от однолетников до «бессезонных» мяг- коствольных древесных растений, у меня иная позиция. Каждому типу биоморф, в зависимости от сложности его организации, критерием которой должны являться особенности проявления или не проявления персистентности, в первую очередь, в надземной побеговой части, нуж- но определить законное место на «полочке» растительной жизни. Для автора этих строк совершенно однозначно, что любые сезонные и бессезон- ные растительные биоморфы, если они имеют персистентные надземные, не считая прижатые к почве розетки листьев или приземные ползучие побе- говые органы, то им нет места среди трав. Их место среди других типов био морф. Следующую ступень на «пирамиде» абстракций занимает десятая мультисубординатная классификация биоморф семенных растений— результат еще большей детализации типов биоморф, когда сложность организации их оценивается по интегральным признакам с еще мень- ших, чем ранее, высот обобщения. Особенности отхождения ветвей- классификаций отражает рисунок 216-Д, в котором приводится 2б клас- сов биоморф, или субординат (биоморфотипов): эфемеры, гемиэфеме-
OCHOBhI БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИИ Рис.216'. Графическая суперординатная модель («пирамида» абстракций) биоморф семенных растений, отражающая их разнообразие по сложности интегральной организации. А — Б — уровни абстракции; 1-51 — субординаты (классы) биоморф семенных растений: 1— организмы, 2 — синорганизмы, 3 — эвриэфемерные растения (эвриэфемеры), 4 — персис- тентные растения (персистенты), 5 — аэробазофобы, 6 — аэробазофилы, 7 — малолетники, 8 — среднелетники, 9 — многолетники, 10 — унирепродуктивные растения, 11 — бирепро- дуктивные растения, 12 — мультирепродуктивные растения, 13 — автотрофы, 14 — гемиге- теротрофы, 15 — гетеротрофы, 16 — двулетники, 17 — терофиты, 18 — криптофиты, 19— гемикриптофиты, 20 — хамефиты, 21 — фанерофиты, 22 — травянистые растения (травы), 23 — полугравянистые растения (полугравы), 24 — полудревесные растения, 25 — древовид- ные растения, 26 — древесные растения, 27 — эфемеры, 28 — гемиэфемеры, 29 — однолет- ники, 30 — озимооднолетники, 31 — травы среднелетние, 32 — травы многолетние, 33— полутравы-десуккуленты малолетние, 34 — полугравы-десуккуленты среднелетние, 35 — тра- вополукустарнички, 36 — травокустарнички, 37 — травокустарники, 38 — траводеревья, 39— мелкие полугравы-суккуленты, 40 — средние полугравы-суккуленты, 41 — крупные полутра- вы-суккуленты, 42 — полукустарнички, 43 — полукустарники, 44 — полудеревья, 45 — ден- дропигмеи, 46 — нанодендроиды, 47 — мезодендроиды, 48 — макродендроиды, 49 — кустар- нички, 50 — кустарники, 51 — деревья.
Глава 5. Концеиция азнообразия биомор семенных астений ры, однолетники,озимооднолетники, двулетники, травы среднелетние, тра- вы многолетние, полутравы-десвуккуленты малолетние, полутравы-десук- куленты среднелетние, травополукустарнички, травокустарнички, травоку- старники, траводеревья, мелкие полутравы-суккуленты, средние полутра- вы-суккуленты, крупные полутравы-суккуленты, полукустарнички, полуку- старники, полудеревья, нанодендроиды, мезодендроиды, макродендроиды, кустарнички, кустарники, дендропигмеи и деревья. Можно, конечно, ограничиться изложением разнообразных био- морф семенных растений, использовав только приемы суперординатно- го, субординатного и мультисубординатного классифицирования. Одна- ко, характеристика только субординатных биоморфотипов (рис. 123- 147) дает представление о разнообразии биоморф семенных растений при взгляде на них как бы только сверху (с разных высот абстракции). Так смотреть на предметы исследований, безусловно, важно и полезно, но для понимания их разнообразия это далеко не все. Обязательно надо «покрутить» биоморфы с разных сторон, т.е. применить более дотош- ный методический прием, который хорошо дополнит первый принцип (суперординатное классифицирование), т.е., чтобы изучать разнообра- зие биоморф семенных растений, их надо рассматривать еще и через призму просубординатного классифицирования, которое всецело опи- рается на метод мультилинейного классифицирования (т. 1, гл. 9). 5.3. Просубординатное разнообразие биоморф семенных растений В какую бы позицию ни становились биоморфологи, им раньше или позже придется столкнуться с необходимостью классифицировать биоморфы или разбираться с уже известными классификациями, что- то принимая или не принимая в них. Под просубординатным разнооб- разием биоморф подразумевается, что оно, образно говоря, «работает на субординатный тип» и выражается в том, что к субординатному типу привязывается какое-либо число рядовых типов биоморф, выделенных по корректирующим признакам. Просубордииатиое разнообразие — это результат познания разнооб- разия предмета исследований методом просубординатного классифицирова- ния. По сути — это особый методический прием, когда классы систем (субординаты) — элементы субординатного классифицирования, начинают выступать в качестве «центров» концентрации вокруг себя корректирую- щих признаков соответствующих предметов исследования (биоморф, в нашем случае), число которых в создаваемых типах (биоморфотипах) за- висит от задач конкретного исследования. 5.3.1. Просубординатное разнообразие как обьектианая реальность и особый результат познания Вместе субординатный и определенные рядовые типы биоморф составляют один просубординатный биоморфотип. В предыдущем разде-
378 OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ ле мы показали, как MD%Ho разобраться в иерархии биоморф по слож- ности их организации, и построили суперординатную модель («пирами- ду» абстракций) из субординатных классификаций. Сейчас будет логич- но детальнее познакомиться с разнообразием субординатных биомор- фотипов, а этого нельзя достигнуть, не привлекая рядовые биоморфоти- гиа, т.е. типы биоморф, выделенные по корректирующим признакам. Весь- ма удобно сделать это, применяя рисуночный способ подачи информа- ции с составлением соответствующих типологических досье на каждый вид растительных биоморф (рис. 217-332). Лидерные интегральные при- знаки, по которым выделены субординатные биоморфотипы, вместе с корректирующими признаками, характеризуют просубординатные биомор- фотипы (они развивают содержание субординатных типов биоморф, т.е. дополняют их содержание вширь и вглубь). Пожалуй, весьма сложный вопрос, который потребует еще немало усилий ботаников, — это опре- деление критериев (количественных и качественных) травянистости и деревянистости и который на сегодня пока не нашел однозначного решения (см. обзор Л.Е. Гатцук, 1976). Одного взгляда на созданную нами графическую суперординатную модель («пирамиду» абстракций) организации биоморф достаточно, чтобы стало понятно, что деление семенных растений только на три группы биоморф: травянистые, полудревесные и древесные, а трав на сезонные и бессезонные, далеко не исчерпывает всего их разнообразия. Тип организации биоморф надо определять по их структуре, а не по месту, где они произрастают. Автор готов доказать, что существует еще не одно равноправное направление эволюции биоморф, в частности, полутравянистое и дендроидное, которые, наряду с деревянистым на- правлением, распространились особенно сильно в субтропическом и тропическом климате, хотя немного они присутствуют и в умеренном поясе. При всем этом будем помнить, рассматривая эволюционные авторские экскурсы, что мною не признается однонаправленность в сома- тической эволюции, и когда в нашей работе говорится о каком-то направ- лении: или деревянистом, или травянистом, то подразумевается, что это направление с двумя противоположными концами, которое, в зависимости от обстоятельств, может кардинально изменяться. Здесь мы не будем особенно углубляться в эту проблему, поскольку далее есть главы, где по этому вопросу сказано, как нам кажется, достаточно, чтобы принять обоюдонаправленность, а, точнее, разнонаправленность в определении путей эволюционных изменений биоморфной организации. Итак, рассмотрим по порядку, в чем особенности просубординат- ного разнообразия биоморф семенных растений. Ранее (т. 1, гл. 9) была построена мультилинейная классификация биоморф семенных расте- ний. В ней все признаки специально, посредством волевого решения субъекта-классификатора, определившего правила такого классифици- рования, принято рассматривать как равнозначные, и все выделенные биоморфотипы оценены как классы одного уровня (ранга), т.е. как рядовые биоморфотипы. При использовании этих биоморфотипов в субординатных классификациях часть их приобретает статус суборди-
Глава 5. Концепция азнообразия биомо ф семенных растений натных, а другая, большая часть, остается рядовыми, но, объединяясь вместе с субординатными, они получают единый статус просубординат- ных биоморфотипов. Например, субординатный тип биоморф — дере- вья, а просубординатный биоморфотип — деревья эусимподиальные листопадные некустистые интеркалярно-инфлоресцентные очередноли- стные высокоствольные и т.д. и т.п. При этом не будем забывать, что при определении субординатных типов биоморф мы традиционно ис- пользуем интегральные признаки, отражающие сложность организации средневозрастных растений, а вот при определении просубординатных биоморфотипов, наряду с традиционными признаками, в качестве ин- формативно полезных добавок можно использовать рядовые типы био- морф и других возрастных групп, например, надземносемядольные, округлосемядольные; очереднолистные, но ювенильно-супротивнолист- ные; автономноризомные, но виргинильно-стрежнекорневые и т.п. Обратимся к субординатным типам биоморф семенных растений, упомянутым в «пирамиде» абстракций (рис. 216). Посредством привязки к этим типам небольшого числа корректирующих признаков попробуем, таким образом, разобраться с некоторым просубординатным разнообра- зием биоморфотипов. Можно с уверенностью заявить, что чем больше корректирующих признаков присоединяется к субординатному типу био- морф, тем больше познается разнообразие просубординатных биоморфоти- пов — это правило познания разнообразия биоморф. Оно, безусловно, исходит из специального закона познания разнообразия явлений Природы: чем больше корректирующих признаков ассоциируется вместе с суборди- натным типом явлений (тел и нетел), тем полнее отражается средства- ми соответствующего классифицирования разнообразие просубординатных munoe таких явлений (просубординатное разнообразие). Все примеры, которые приведены ниже, указывают на просуборди- натное разнообразие биоморф семенных растений, в соответствии со сформулированными положениями, и свидетельствуют, что великое разнообразие биоморф достигается, благодаря изобилию рядовых био- морфотипов на фоне ограниченности числа субординатных биоморфо- типов, всегда готовых выступить вместе в просубординатном типовом единстве. Есть еще весьма деликатный момент. Как соотносятся типы (форм роста) биоморф, разнообразие которых в «пирамиде» абстракций было рассмотрено в предыдущей (4-й) главе, с субординатными биоморфоти- пами, приведенными здесь (в этой главе). Они хорошо состыковывают- ся, что позволяет весьма спокойно разбираться с самыми щепетильны- ми тонкостями, которые нередко возникают у ботаников при попытках состыковать габитуальные биоморфотипы с какими-либо ранневозрас- тными типами их роста. Например, какие только термины ни придумы- вали: и «торчки», и «ленечки», и т.п., чтобы обозначить состояния дерева в юном возрасте. С использованием приема просубординатного классифицирования легко состыковываются субординатные интеграль- ные типы биоморф с корректирующими признаками типов форм роста Тех же биоморф, но другого возраста, с добавлением, при необходимос-
OCHOBhI БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИИ ти, небольших терминологических поправок на возраст, например, ювенильные древесные растения, преювенильные (проростки или всхо- ды) растения, имматурные деревца, сенильные деревья, ювенильные кустарники, преювенильные предкустарники, матурные временно не- цветущие полутравы и т.п. 5.3.2. Габитус и формы роста в просубординатном разнообразии биоморф семенных растений Организация биоморф усложняется при движении от эфемеров к деревьям, поэтому логичнее всего соответствующий обзор субординат- ных биоморфотипов и просубординатного разнообразия биоморф се- менных растений начать с эфемеров. Просубординатное разнообразие биоморф мы заложили во многие, приводимые во втором томе, биогра- фические рисунки и соответствующий фитобиографический справоч- ный материал, который основан на использовании мультилинейной классификации биоморф семенных растений, закодированной в табли- це-матрице (т. 1, гл. 9). Травянистые растения Эфемеры Биоморфы этого, хорошо известного (Мальцев, 1932; Коровин, 1934; Проскоряков, 1950; Нечаева и др., 1973; Никитин, 1983 и др.) типа обычно проходят под названием «яровые эфемеры» и имеют самую простую организацию среди семенных растений. Однако, по большому счету, организация эфемеров, в главном, такая же, как и у других типов биоморф, т.е. она, в первую очередь, «пронизана» единообразием, по- скольку все они состоят из побегов и корней нескольких уровней по сложности организации. Эфемеры не запасают ресурсов для длительно- го использования, поэтому в их организации отсутствует базовый орган. Начав рост и развитие, эфемеры быстро доводят его до цветения и плодоношения, т.е. как бы «вспыхивают» и «угасают» за очень короткое время. Продолжительность жизни яровых эфемеров исчисляется от нескольких дней до нескольких недель и редко превышает 1 месяц (рис. 217, 218). Эфемеры наиболее распространены в аридном климате (Alyssum turkestanicum Redel et Schmalh., Ceratocephala falcatum (L.) Pets. и др.). Гемиэфемеры Есть еще группа, помимо яровых эфемеров, — озимые эфемеры, которые мы далее будем обозначать только термином «гемиэфемеры». Они прорастают во влажный период поздно осенью, развивают розетку и зимуют под снегом, а рано весной быстро цветут, заканчивают реп- родуктивное развитие и отмирают до наступления летней засухи. В районах с теплой и влажной зимой эфемеры часто прорастают зимой. Естественно, возраст гемиэфемеров значительно больше, чем у эфеме- ров; его составляют три сезонные стадии (в сумме до 5 — б месяцев):
Глава 5. Коицепция разиообразия биоморф семеииых растеиий осеннего вегетативного развития, зимнего относительного покоя и ве- сеннего репродуктивного развития. В любом случае эфемеры и геми- эфемеры быстро заканчивают свою активную жизнь, всегда до наступ- ления летней засухи. Такие эвриэфемерные растения нельзя путать с эфемероидами — многолетними травами, которые, как известно, разви- вают надземные органы только осенью и (или) весной, а летом они отмирают, но их подземные органы всегда многолетние. Однолетники и озимооднолетники Как эфемеры и гемиэфемеры, так и однолетники (яровые однолет- ники) являются травами и вместе с озимооднолетниками (озимыми однолетниками, или полуоднолетниками) образуют, в нашем понима- нии, группу эвриэфемеров (эфемеров в широком смысле). Однолетники развиваются по-разному, прорастая весной (рис. 219-228), а озимоодно- летники — осенью (рис. 229), но продолжительность их индивидуальной жизни, хотя и происходит дольше, чем у эфемеров и гемиэфемеров, но все равно не превышает 12 месяцев. В отношении определения границ однолетников и озимооднолетников обычно больших проблем не воз- никает. Ботаники обычно признают выделение двух основных групп однолетних растений: яровых (они прорастают весной и отмирают до наступления зимы в том же году) и озимых (семена прорастают осенью, растеньица зимуют с розеткой листьев под снегом, а весной рост и развитие продолжается, завершаясь обсеменением и отмиранием особей осенью). Все эти особенности в поведении отмеченных выше растений и побудили автора предложить небольшую корректировку в терминоло- гии. Дело в том, что растения, которые прорастают весной и проходят все развитие в одном вегетационном сезоне, действительно, есть смысл считать «однолетними» («однолетниками»), а вот озимые растения, ак- тивно живущие в двух, хотя и неполных, вегетационных сезонах, но не более одного года, можно обозначить иначе — как «озимооднолетние» растения («озимооднолетники»). Эфемеры и гемиэфемеры (кратковре- менные летники) нередко рассматривают как формы (разновидности) однолетников, а те в свою очередь, наряду с двулетниками, как формы малолетников (Мальцев, 1932; Никитин, 1983 и др.). Большое разнооб- разие эвриэфемеров наблюдается в пустынях (Нечаева и др., 1973). Травянистые двулетники, или траводвулетники (двулетние травы) Этот биоморфотип хорошо изучен, поскольку многие овощные растения являются двулетниками (Куперман, Ржанова, 1963; Куперман, 1977 и др.). Двулетние травянистые растения широко встречаются и в дикой природе; чаще всего они являются монокарпическими растени- ями (Серебряков, 1962; Никитин, 1983; Марков 1990 и др.). Двулетники �ивут два вегетационных сезона и на второй год развивают репродук- тивные органы, цветут, плодоносят и отмирают (рис. 41, 42, 55, 56 и др.). Для двулетних растений характерно образование после первого вегетационного сезона розетки листьев, а в стеблевой части — сбли-
382 ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИЧ женноузлового базового (зимующего) органа — конодия (т. 1, гл. 6). Двулетники мы здесь рассматриваем как двулетние травянистые расте- ния; в перспективе лучше подчеркивать именно травянистость таких биоморф, поскольку малолетниками бывают еще и некоторые полутравы. Травянистые среднелетники, или травосреднелетники (среднелетние травы) Эту группу травянистых биоморф мы ограничили возрастом 3 — 10 лет (рис. 57, 58, 230, 234 и др.). Сюда попадает немало так называемых монокарпиков, которые точнее называть унирепродуктивными растени- ями («унирепродукторами»), а если все-таки говорить о монокарпиках, то тогда надо обязательно подчеркивать, какие они по широте сексуа- лизации (обоеполые или однополые), отличая в таких случаях от моно- антериков (растений с однополыми тычиночными цветками). Возраст унирепродуктивных растений обычно может сильно колебаться, но многие зацветают и плодоносят если не как малолетники, то часто как среднелетники. Возрастное разграничение малолетников и многолетни- ков через среднелетников во многом относительное, но удобно решает проблему разделения этих двух действительно во многом разных групп трав. Часть травянистых растений, пройдя среднелетнюю стадию разви- тия и начав нарастать симподиально, на этом не останавливаются и далее продолжают жизнь как многолетники. Травосреднелетники, как и другие неоднолетние травы, развивают определенный базовый орган (т. 1, гл. 6), который располагается подземно или на уровне поверхности почвы, реже ползет по поверхности почвы. Травянистые многолетники, или травомноголетники (многолетние травы) Это наиболее изученная группа биоморф семенных растений (Warming, 1884; Krause, 1881; Raunkiaer, 1907, 1934; Высоцкий, 1915; Казакевич, 1922; Мальцев, 1931; Работнов, 1945, 1950; Шалыт, 1955, 1960; Зозулин, 1959; Голубев, 1960, 1965, 1972; Борисова, 1961; Сереб- ряков, 1952, 1964; Серебрякова, 1962, 1971, 1977; Нухимовский, 1970, 1974, 1976, 1978; Нечаева и др., 1973; Зиман, 1976; Гатцук, 1976; Смир- нова, 1987; Жукова, 1995, Алексеев, 1996 и др.). Многолетние травы образуют подземные, реже приземные (ползучие по поверхности почвы) побеги с почками возобновления подземными или на уровне поверхно- сти почвы, нередко в лесной подстилке или в моховом покрове. Ортот- ропные удлиненные побеговые части трав, когда они образуются, не зимуют (рис. 217-276). Травянистые многолетники встречаются повсе- местно, но более всего в степях, лесах, полупустынях и пустынях, лугах и тундре; в субтропиках и тропиках многолетние травы, как правило, оттесняются на второй план полутравянистыми, полудревесными, дре- вовидными и деревянистыми растениями. Полутравянистые растения Это большой биоморфотип, но он хорошо отличается от всех ос- тальных. В нем две крупные группы: десуккуленты (растения с недо-
Глава 5. Концеиция разнообразия биомо семенных растений лговечными, живущими по-разному и 2 — 5 лет, а иногда значительно дольше, десуккулентными аэроподиями или разновидностью аэроподи- ев — аэроподиоидами, которые надземно зимуют только один раз, например, соломины некоторых субтропических злаков) и суккуленты (растения с долговечными, живущими много лет, нередко десятки лет, аэроподиями, которые по консистенции суккулентные, и если когда-то одревесневают, то только слабо). Полутравянистый тип эволюции освоен растительными биоморфа- ми в процессе их пребывания, в основном, в более теплом климате, где нужны иные адаптивные реакции, чем в районах с холодным климатом. В теплом климате многого не нужно из того, что используется травя- нистыми многолетниками для борьбы с зимней стужей. Адаптации, обеспечивающие, посредством аэроподиев или аэроподиоидов, надзем- ное пребывание различных зон побегоотрастания, не связаны часто с повышением содержания лигнина в клетках, а решаются у полутрав иными способами: часто сменяются недолговечные, хотя и персистен- тные побеги кущения, а в других случаях, когда жара невмоготу, многие полутравы научились выгодно накапливать быстро и надолго воду, проявляя суккулентность («мясистость») в строении осевых, но иногда и листовых частей побегов, совсем не увлекаясь лигнификацией тканей. Полутравянистые десуккуленты Полутравянистые малолетники, или полутравомалолетники В этот биоморфотип входят растения, которые развиваются сначала по однолетнему травянистому циклу, отцветают на первом году жизни, но по каким-то причинам их цветоносные части побегов зимой не погибают полностью, может быть, позднее обычного растения зацвета- ют или только потому, что зима теплая и влажная, но факт остается фактом и таких растений достаточно много, они рано весной проявляют еще одну волну роста и развития, образуя постфлоральные побеги, как, например, у Conyza bonariensis (L.) Cronquist (т. 1, гл. 8). Приведем еще несколько кратких жизнеописаний полутравянистых малолетников, которые наблюдал автор в условиях южных (средиземноморских) суб- тропиков Ближнего Востока (рис. 277, 278). Полутравянистые среднелетники, или полутравосреднелетники Растения этого биоморфотипа встречаются реже. Так, особи Aster subulatus Michaux на Ближнем Востоке (рис. 279) могут завершать свой жизненный цикл и после первого года жизни, в частности, в «Анали- тической флоре Израиля» (Feinbrun-Dothan, Danin, 1991) этот вид от- несен к однолетникам. После влажной и теплой зимы остатки уже отплодоносившего верхушечного цветоноса часто не отмирают, а обра- зуют постфлоральные побеги. Некоторые растения и после второй вол- Hbt слабого цветения не отмирают, а больше того, находят силы и образу- ют еще из открытых почек основания главного побега 1 — 2 побега <у>цения, которые могут неплохо расти, развиваться, и растение прожи- аеТ даже больше, два — три года, а то и до пяти лет, повторяя ежегодно
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ «лихорадочное» постфлоральное необеспеченное побегоотрастание. Постфлоральные побеги, как-либо (полностью или не полностью) за- вершив свое развитие, погибают вместе с прошлогодним цветоносом, и только новые побеги кущения могут, если развиваются, заметно продле- вать онтогенез особей. Полутравянистые многолетники, или полутравомноголетники Это основная группа биоморф полутравянистых растений. Все полутравы или вообще не одревесневают, или одревесневают слабо, поскольку у этих растений специализация в развитии внутренней орга- низации надземных персистентных органов идет не в сторону накопле- ния лигнина, а иначе. Одну группу многолетних полутрав составляют суккуленты — это сукккулентные полутравы (мелкие, их аэроподии до 30 см, средние — от 30 см до 3 м, и крупные, более 3 м высотой). Другую группу многолетних полутрав образуют десуккуленты, принцип надзем- ного побегообразования у которых иной, чем у других основных субор- динатных биоморфотипов, — это травополукустарнички, травокустар- нички, травокустарники и траводеревья. Травополукустарнички Эта группа десуккулентных полутравянистых биоморф поднимает свои зоны побегообразования на надземных побеговых персистентных частях на необычную для трав (до 20 см) высоту над поверхностью почвы, к которым они по организации ближе всего. В южных широтах, где бывает достаточно тепло не только летом, но и зимой (к тому же в это время еще и влажно), многие злаки предпочитают не травянистый, а травополукустарничковый тип организации (рис. 280-282). Под Мос- квой в условиях елового леса, близко от опушки, где под «пресс» силь- ного затенения попадает много семян, только единичные виды приспо- собились хоть как-то выживать, изменяя свою организацию. В этих условиях нередко травянистые многолетники вынужденно превращают- ся в травополукустарнички, т.е. в разновидность полутравянистых рас- тений. Основной биоморфотип Solidado virgaurea L., Leucanthemum vulgare Lam. — многолетние корневищные травы, а под пологом елового леса образуется их второстепенный биоморфотип — вынужденно ком- пактно-аэроподиальные травополукустарнички (рис. 283-286). В усло- виях глубокого затенения корневища особей золотой розги и нивяника ведут себя нетрадиционно, превращаясь в аэроподии, точнее аэроризо- мы. Они, подобно заблудившемуся в темноте путнику, пытаются отыс- кать уже над поверхностью почвы удобное, для развития надземных побегов, место. В диагностике базовых органов складываются определенные прави- ла, которых уместно придерживаться. Когда ветер засыпает песком часть ствола дерева, то эта часть уже не принадлежит аэроподию, а опреде- ляется как геоподий. В нашем случае, с образованием разновидностей аэроподия — надземного ризома (аэроризома) у особей золотой p03rH и нивяника изменяется статус части базового органа, вынужденно ока-
Глава 5. Концеиция азнооб азия биомо семенных астений завшейся в надземном положении — это уже не корневище, а, хотя и вынужденное, иное образование (т. 1, гл. 6). Изменения в оценке базо- вых органов произошли под влиянием обстоятельств, без которых особи этих видов так и жили бы как травянистые многолетники с полностью отмирающей ежегодно зимой надземной частью и с ризомами в подзем- ной части. На мой взгляд, лучше замечать изменение обстоятельств и их отражение в организации биоморф, чем оставлять происходящее без внимания. Чаще всего вынужденные травополукустарнички или всю жизнь не цветут, или цветут редко. Иногда обстановка изменяется, и света становится больше, что способствует восстановлению относитель- но нормального ритма репродуктивного развития. Травокустарнички Биоморфы этого типа распространены, в основном, в субтропиках и тропиках, где всесезонно теплый климат обеспечивает необходимые условия, чтобы зоны кущения растений не слишком прижимались к поверхности почвы или не слишком углублялись в почву. Для травоку- старничков характерны сравнительно небольшие по высоте аэроподи- альные части (примерно до 1 м в высоту), на которых развиваются самые разные побеги текущего года; после завершения вегетационного сезона от них остаются, нередко надземные, персистентные стеблевые остатки (резиды), пополняющие на некоторое время структуру аэропо- диев. Травокустарнички очень обычны среди злаков на Ближнем Вос- токе. Ни о какой, характерной для кустарничков, деревянистости у злаковоцветковых травокустарничков, быстро обновляющих (нередко через 2 — 3 года) свои аэроподии, речи быть не может. Здесь совершен- но другой тип кустистого существования с участием недолговечных аэро- подиев, включая и их разновидность — аэроподиоиды (рис. 287-289). Травокустарники Биоморфы этого типа напоминают по структуре травокустарнички, но значительно крупнее их, образуя аэроподии выше 1 м, нередко до 3 — 4 м высотой. Как и у предыдущей группы биоморф, травокустарни- ки произрастают, в основном, в теплом субтропическом и тропическом климате. Вид-космополит злак Phragmites australis (Сач.) Trin ex Steud. в умеренном климате ведет себя по типу травянистых многолетников, а в субтропиках Ближнего Востока — по типу травокустарников, по- скольку многие его соломины (стебли) после первого плодоношения не отмирают, а еще продолжают вегетировать и с весны следующего года. Биоморфотип Arundo donax L. — геоподиальные высокие травокустар- ники в условиях Ближнего Востока. Главная особенность этой группы биоморф в том, что их кустистое строение составляют аэроподии- недолговечные по консистенции соломины, хотя и персистентные (жи- вут несколько лет). Иногда недолговечные аэроподии образуются и у представителей других таксонов, причем не только в южных широтах, но и некоторых растений умеренных широт, например, таких, как малина — Rubus idaeus L. (рис. 290), которые, на наш взгляд, в связи с вводимыми здесь кор- рективами не следует считать кустарниками.
386 OCHOBhI БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИИ Траводеревья Большая высота аэроподиев-соломин (4 и более, до 35 м), их зна- чительное одревеснение и большая, чем у других злаковоцветковых по- лутрав, продолжительность жизни — это характерные признаки траво- деревьев. Типичными представителями этих полутрав являются многие представители рода Bambusa Retz (бамбуки). Среди траводревесных растений характерны и унирепродуктивность, и мультирепродуктивность развития. У одних видов одновременно зацветает все вегетативное по- томство одного проростка, т.е. синорганизм, и после 30 — 50 лет актив- ного вегетативного роста, однажды образовав репродуктивные органы, биоморфы такого типа отмирают полностью (МсС1иге, 1966; Серебря- кова, 1971 и др.). По тем же материалам, у других видов, проявляющих организацию траводеревьев, отмирают только цветоносные соломины, вегетативное разрастание кустов не заканчивается, а синорганизмы живут значительно дольше унирепродуктивных растений — до 100 и много более лет. Полутравянистые суккуленты Эта группа биоморф весьма распространена в сухих областях Земли с субтропическим и тропическим климатом. Среди полутравянистых суккулентов есть как некустистые (столбовидные растения, или стелои- ды), так и кустистые растения. Представители этого биоморфотипа име- ют шарообразную, плоскую или членистую формы. Довольно долговеч- ная суккулентная («мясистая») консистенция разнообразных по форме аэроподиев характерна для многих полутрав семейства Cactaceae, неко- торых видов Euphorbiaceae и реже видов других семейств, например, Agave americana L.Ñðåäè полутравянистых суккулентов хорошо различи- мы по размерам следующие группы: мелкие (от нескольких см до 0.5 м), средние (0,5 — 3 м), крупные, или высокие (более 3 м высотой); несколь- ко особняком стоят стелюще-висячие формы разнообразных размеров. Весьма оригинальны и разнообразны шарообразные и столбовидные сук- кулентные полутравы (рис. 291). Мясистые стебли стелоидов типа Carnegia gigantea (Engelm.) Br. et R. [этот вид обитает в аридных районах Калифорнии, Аризоны (США) и в Мексике] достигают до 14 м в высоту (Innes, Glass, 1991). Полутравянистые суккуленты образуют часто соч- ные и зеленые стебли, многие годы развивающиеся надземно, т.е. яв- ляющиеся аэроподиями (большинство кактусов) — это каулосуккулен- ты, но иногда основная суккулентность проявляется не в стеблях, а в листьях — это фрондосуккуленты, как у агавы американской. Высокие полутравянистые суккуленты иногда называют «суккулентно-стеблевы- ми безлистными деревьями» (Du Rietz, 1931; Серебряков, 1964), что представляется менее точным названием такого биоморфотипа, посколь- ку их аэроподии, выполняя водозапасающую и ассимилирующую фун- кцию, слабо одревесневают. Столбовидные (стелоиды) или их разветвлен- ные вариации, канделябровидные гиганты из семейства Cactaceae — »o типичные представители высоких полутравянистых суккулентов (сукку- лентных полутрав). Как разнообразные емкости для воды: «бочоночки»
Глава 5. Концеиция азнообразия биомор семенных растений 387 (мелкие), «бочонки» (средние) и «бочки» (крупные), нередко ассоции- руются в памяти габитусы этих многих полутравянистых стеблевых суккулентов. Между прочим, существует прекрасная тема для анатоми- ческих исследований, которую мы в этой работе вглубь не анализируем: важно когда-то детально обосновать качественные или количественные различия во внутренней организации аэроподиев, а шире — надземных побегов вообще (аэрокормусов), у разных субординатных типов био- морф семенных растений. Для наших целей пока достаточно того, что такие различия даже габитуально (экзоморфологически) легко обнару- живаются. Полудревесные растения Для этой группы биоморф характерно образование одревесневаю- щей структуры аэроподиев, восходящих от кустистых оснований таких растений. Аэроподии разрастаются медленно за счет резидных (перси- стентных стеблевых) частей побегов, а в целом — резидных побегов, образующихся после ежегодного или неежегодного отмирания сверху (акронекроза), до зоны укороченных междоузлий или иначе, цветонос- ных унирепродуктивных побегов. Для полудревесных растений отмира- ние верхних частей унирепродуктивных побегов, завершивших свое репродуктивное развитие, и образование из них надземных резидных («остаточных») побегов многие ботаники считают одним из их обяза- тельных признаков (Du Rietz, 1931; Прозоровский, 1936; Стешенко, 1955, 1965; Рачковская, 1957; Серебряков, 1962, 1965; Беспалова, 1965; Мулкиджанян, 1965; Голубев, 1973 и др.). Среди полудревесных расте- ний логично различать полукустарнички, полукустарники и полудере- вья. В отличие от древесных, рассматриваемые здесь полудревесные растения образуют аэроподии, продолжительность жизни которых су- щественно меньше и обычно, даже у полудеревьев, не превышает 15— 20 лет, а у полукустарничков — еще меньше. Пол укустарнички Биоморфы этой группы характеризуются отмиранием цветоносных унирепродуктивных побегов («монокарпических» в традиционном смыс- ле побегов, которые все же правильнее будет отличать от однополых пыльцеобразующих, т.е. моноантерических побегов) на большую часть своей длины (акронекроз), как правило, на 2/3 длины, а в резидные побеги преобразуются только те части бывших активных побегов, у которых все или большинство — укороченные междоузлия. Решающим признаком все же остается высота аэроподиев, а она обычно не превы- шает 20 (30) см, и продолжительность их жизни — не более 10 лет (рис. 292-294). Аэроподии восходят от надземного или полунадземного компактного основания куста, которое живет значительно дольше и разрастается еще и из живых остатков аэроподиев. У некоторых расте- ний типа Capparis herbacea Wild., — каудексовых полукустарничков, продолжительность жизни компактного основания куста может дости- гать ста и более лет (т. 1, гл. 6). Большинство полукустарничков живут сравнительно недолго — 20 — 40 лет, реже больше.
388 ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИИ В теплых средиземноморских субтропиках Ближнего Востока много полукустарничков, которые проявляют постфлоральное лихорадочное» необеспеченное побегоотрастание, например, Centaurea procurrens Sprengel (т. 1, гл. 8). Некоторые травянистые многолетники, произрас- тающие в умеренной зоне, в условиях теплых субтропиков проявляют себя как полукустарнички или полукустарники с постфлоральным нео- беспеченным побегоотрастанием, например, Foeniculum vulgare Miller. Однако, поднявшись с равнины в горы (Иерусалим, средняя высота 800 м над уровнем моря), где несколько прохладнее, особенно зимой, фен- хель обычно ведет себя как травянистый многолетник без постфлораль- ного «лихорадочного» побегоотрастания. Полукустарники Биоморфы этой группы (рис. 295-302) развиваются подобно полу- кустарничкам, но размеры их покрупнее и отмирание унирепродуктив- ных побегов после плодоношения поменьше — от 2/3 до 1/3, иногда даже до 1/2 длины, а резиды, обеспечивающие прирост более долговеч- ных аэроподиев, наряду с компактными междоузлиями включают еще и некоторое число удлиненных междоузлий (т. 1, гл. 8). В южных суб- тропиках (Ближний Восток) многие полукустарники проявляют очень рано весной постфлоральное необеспеченное побегоотрастание. Аэро- подии и в целом особи живут побольше, чем у полукустарничков. Воз- раст простых биоморф колеблется от 20 до 60 лет. Оба уже рассмотрен- ных биоморфотипа полудревесных растений можно наблюдать у одного и того же вида на небольшом участке территории. Так, на плакоре в пустыне Негев (Израиль) в одних условиях (на открытом месте) «в одиночестве» простые биоморфы (организмы) Polygonum palaestinum Zohary развиваются нередко как травянистые многолетники, а через 30 — 50 см при произрастании «в группе» особи того же вида проявляют себя как полукустарнички, а здесь же («в группе») или чуть дальше, в микропонижении, их совидичи (биоморфы того же вида) — типичные полукустарники с аэроподиями 0,3 — 1,0 м высотой (рис. 301). Полудеревья Наиболее полное подразделение полудревесных растений дал А.В. Прозоровский (1936), который, помимо традиционных типов полукус- тарничков и полукустарников, выделил еще и группу полудеревьев. Пока значение последнего термина явно до конца не осознано. Мы делаем очередную попытку очертить понятийные границы полудеревьев. Био- морфы этой группы сохраняют основные особенности роста и развития полудревесных растений: значительное ежегодное отмирание верхушек длинных ростовых и всех отплодоносивших унирепродуктивных побе- гов, а, с другой стороны, сохранение в структуре одревесневающих аэроподиев резидных частей этих побегов. Пожалуй, у полудеревьев есть общее качество, объединяющее их с остальными полудревесными растениями: их надземная многолетняя часть, по сравнению с древесными и даже древовидными растениями, живет значительно меньше. Аэроподии полудеревьев одревесневают по-
Глава 5. Концеиция азнооб азия биомо семенных растений 389 разному, но, как правило, слабее, чем у настоящих деревьев. По высоте взрослые полудеревья не гиганты, но выше кустарников (от 4 м до 10 м и реже еще более). Недолговечные аэроподии нередко составляют ске- летную основу побегов кущения полудеревьев, у которых общая продол- жительность индивидуальной жизни исчисляется, в среднем, 10 — 20 годами и не превышает, как правило, 30 лет. Итак, полудеревья — это недолговечные (живут 10 — 30 лет), не- плохо одревесневающие, средневысокие (4 — 15 м), чаще всего кусти- стые растения с акронекрозными удлиненными побегами. В качестве модельных экземпляров полудеревьев можно отметить Ricinus communis Ь. из южных субтропиков Ближнего Востока (т. 1, гл. 8) и Nicotiana glauca Graham (рис. 303). Древовидные растения, или дендроиды (от греч. dendron— дерево, eidos — вид) Главная особенность дендроидов в том, что в биоморфогенезах их ;дроподии одревесневают, но их внутренняя организация сильно отли- чается от настоящих деревянистых растений. Дендроидами являются многие древовидные пальмы (рис. 304). Структурная организация пальм только отдаленно напоминает деревья, поскольку они не образуют ство- лов, свойственных настоящим деревьям. Все пальмы проявляют только ограниченный рост; они не имеют камбия. «Стебель достигает значи- тельной толщины в результате первичного роста, происходящего непос- редственно под верхушечной (апикальной) меристемой, расположенной в центре небольшого чашевидного или блюдцевидного углубления на верхушке стебля, которое наиболее заметно у пальм с толстыми колон- новидными стеблями... Повреждение верхушечной почки у одностволь- ных пальм ведет к их гибели, так как каждая ось имеет всего лишь одну точку роста, функционирующую в течение всей жизни стебля... Хотя вторичный рост у пальм отсутствует, стебли пальм утолщаются (как, например, у королевской пальмы) благодаря делению и растяжению клеток основной паренхимы и волокон, которые окружают проводящие пучки» (Имханицкая, 1985, с. 40 — 41). Дендроиды бывают низкорослыми (нанодендроидами), когда высо- та их надземного стволика (стволиков) не превышает 1 м (Salacca wallichiana Mart.), среднерослыми (мезодендроидами), когда стволик (стволики) 1 — 10 м высотой (Chamaerops humilis L.), и высокорослыми (макродендроидами), когда ствол (стволики) 10 и более метров высотой, например, рост Washingtonia robusta Н. Wendl. может достигать 60 м. Дендроиды, как и многие другие биоморфотипы, бывают некустистыми или кустистыми. Кроме того, некоторые дендроиды развиваются как лианы (Calamus manan Mig.). Типичные представители дендроидов со- средоточены среди однодольных растений семейства Arecaceae, или Ра1тае, которые неплохо изучены (Martius, 1823-1853; Сааков, 1954; McCurrach, 1960; Tomlinson, 1960, 1961; Moore, 1960, 1961, 1963; Сотег, 1966; Moore, Uhl, 1973; Halle, Oldeman, Tomlinson, 1978; Имханицкая, 1985 и др.). Древовидные растения — это не только многие пальмы, но и представители еще и других таксонов, например, некоторые средне-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ рослые (аэроподии до 7 м высотой) розеточные Senecio sp., обитающие в высокогорьях Килиманджаро (Бол. Энц. Сл., биология, 1999). Деревянистые, или древесные, растения Этот космополитный биоморфотип, наряду с травянистыми расте- ниями, наиболее изучен (Raunkiaer, 1907, 1934; Kanngisser, 1910, 1912; Du Rietz, 1931; Jost, 1936; Rauh, 1937, 1938; Сотег, 1949, 1964; Федоров и др., 1962; Серебряков, 1962; Halle, Oldeman, 1970; Меннинджер, 1970; Голубев, 1973; Halle, Oldeman, Tomlinson, 1978; Васильев и др., 1978; Жмылев и др., 1993 и др.). Деревянистыми растениями являются биоморфы следующих субор- динатных типов: дендропигмеи, кустарнички, кустарники и деревья. С добавлением каких-либо корректирующих признаков эти биоморфоти- пы конкретизируются до соответствующих просубординатных типов биоморф, например, некустистые или кустистые деревья; летнезеленые, зимнезеленые и вечнозеленые кустарнички, кустарники или деревья и т.п. Из предыдущего материала видно, что мы многие древовидные растения «не пустили» в группу деревянистых растений. Аэроподии настоящих деревянистых растений утолщаются за счет ежегодной дея- тельности камбия (зоны прироста древесины, или годичные кольца, как раз образуются посредством такой активности камбия). У древовидных пальм, например, вообще нет камбия, похоже, этого вполне достаточно, чтобы не дать им места среди деревьев, кустарников и кустарничков. У пальм — свое особое место в иерархии биоморфотипов семенных рас- тений, на наш взгляд, многие из них достаточно четко вписываются в группу дендроидов. Полутравы-суккуленты часто вообще не одревесне- вают или немного одревесневают, хотя и образуют аэроподиальные части, специализированные на запасание влаги и фотосинтез, напри- мер, биоморфы многих кактусов. Полутравы-десуккуленты — такие биоморфы свойственны, например, бамбукам; они нередко образуют высокие аэроподии, ткани которых могут иногда сильно лигнифициро- ваться, но злаковая внутренняя структура таких аэроподиев-соломин вряд ли даже отдаленно напоминает стволы настоящих деревьев. Ясно, пожалуй, главное: высокие одревесневающие аэроподии некоторых биоморф бамбуков — это не стволы деревьев; это стволы высоких по- лутравянистых биоморф — «траводеревьев». Кто знает, может быть, именно такого названия пока и не хватало для их обозначения. Дендропигмеи, или карликовые деревья» Биоморфы такого типа весьма оригинальны по своей организации. На Земле, пожалуй, известен только один вид, к которому все они принадлежат, — Welwitschia mirabilis Hook., семейство Welwitschiaceae класса гнетовых. Вельвичия удивительная растет в пустынях Западной и Юго-Западной Африки (главным образом, в пустыне Намиб). Особенность этого биоморфотипа в том, что простые биоморфы (особи) образуют главный неветвистый толстый (диаметр до 1,2 м), очень короткий (не выше 0,5 м) надземный деревянистый ствол, внутренняя структура которого еще недостаточно изучена, но как бы там ни было,
Глава 5. Концеиция азнообразия биомо семенных астений а зона активного нарастания побега всегда находится выше поверхности почвы. Вельвичия на верхушке ствола несет два супротивных кожистых листа (2 — 3 м, иногда до 8 м, шир. 1,8 м), которые растут, разрываясь с возрастом на ленты, в течение всей жизни, продолжающейся несколь- ко сот лет, иногда 2000 лет и даже больше (Бол. Энц. Сл., биология, 1999). «Ствол растет преимущественно в ширину и в конце концов приобретает сходство с круглым растрескавшимся столом больше метра в поперечнике. Один наблюдатель сравнил его с пригоревшим караваем. Рваные листья, грудой лежащие на песке пустыни, придают старому дереву сходство с кучей мусора» (Меннинджер, 1970, с. 72 — 73). Кустарнички Для многих деревянистых растений характерна кустистость, т.е. способность образовывать в процессе жизнеразвития кусты. Кустарнич- ки и кустарники во взрослом состоянии, как правило, кустятся, т.е. образуют от основания растительной особи приземно или подземно побеги кущения, которые, нарастая моноподиально или удлиненно симподиально, нередко полегая (особенно кустарнички) формируют структуру аэроподиев (рис. 305-312). Кустарнички, в отличие от полу- кустарничков и полукустарников, предпочитающих симподиальное на- растание аэроподиев, более всего используют эумоноподиальное аэро- подиальное нарастание (рис. 305, 308, 310, 311). У кустарничков надземные деревянистые побеговые части-аэропо- дии невысокие: от 5 до 60 (80) см высотой (Du Rietz, 1931; Серебряков, 1962; Голубев, 1973; Мазуренко, Хохряков, 1977; Васильев и др., 1978; Мазуренко, 1978, 1986; Жмылев и др., 1993 и др.). Многие авторы (Серебряков, 1962; Голубев, 1973; Мазуренко, 1986 и др.) считают, что основной критерий для разграничения между кустистыми деревьями, кустарниками и кустарничками, т.е. внутри биоморфотипа деревянис- тых растений, — это высота их аэроподиев. На современном уровне знаний такой подход, на мой взгляд, пока вполне оправдан. В случае, если под влиянием конкретных условий произрастания аэроподии кустарничков сильно полегают, то вполне удачно их разно- видность называть стланичками, а самое крайнее видоизменение этого биоморфотипа, характеризующегося сильно прижатыми к поверхности почвы аэроподиями и активным ростом побегов по горизонтали, — шпа- лерными кустарничками, или простратными стланичками (Юрцев, 1968). Кустарники Эта группа деревянистых растений широко распространена в рас- тительном покрове по всей Земле, нередко образуя чистые кустарнико- вые заросли в поймах рек, за верхней границей леса в горах и т.п. По мнению И.Г. Серебрякова (1962), у кустарников главный стволик раз- вивается только на ранних стадиях жизни, затем теряется среди подоб- ных ему стволиков, которые постепенно его заменяют. Число аэропо- диев в организации кустарника самое разное, а возраст их также сильно колеблется, но, как правило, от 10 до 40 лет, реже — меньше или больше (рис 313-326). В неблагоприятных для роста условиях кустарники могут
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ прижиматься ближе к поверхности почвы и образовывать стелющи- еся (шпалерные) аэроподии, а их называют в таком жизненном со- стоянии шпалерными (полупростратными кустарниками, или стла- никами (Du Rietz, 1931; Rauh, 1937, 1939; Серебряков, 1962, Дервиз- Соколова, 1966; Колищук, 1968; Мазуренко, 1978; Жмылев и др., 1993 и др.). Деревья Больше всего произрастает деревьев во влажных тропиках (на- пример, во флоре Амазонии 88 % деревьев и 12 % трав, а в умеренном климате Евразии соотношение обратное), где они достигают наиболь- ших размеров: в среднем 50 — 80 м, часто до 100 м, а максимальная высота у деревьев эвкалипта и секвойи — около 150 м (Васильев и др., 1978). Деревья отличает от других биоморфотипов большая длитель- ность развития надземной части генерального побега (рис. 327-332), которая может составлять сотни и даже тысячи лет, например, особи драконова дерева (Dracaena draco L.) на Канарских островах живут до 5 — 6 тысяч лет. Генеральный побег часто всю жизнь у некустистых деревьев развивается в гордом одиночестве, а у кустистых деревьев образуется разное число субгенеральных побегов (побегов кущения), которые раньше или позже заменяют или способны заменить отслу- живший генеральный побег. Побеги деревьев, как и большинства остальных деревянистых растений, нарастают моноподиально или симподиально, разветвляясь или не разветвляясь. Кроны деревьев чаще всего разветвленные, поднимаются стволами и стволиками на разную высоту над поверхностью почвы. Ствол и скелетные ветви у деревьев — это элементы их аэроподия (у кустистых деревьев аэро- подиев несколько); он ежегодно утолщается за счет функционирова- ния камбия. Многие растения так или иначе одревесневают, но осо- бенности одревеснения, продолжительности жизни аэроподиев и внешнего облика деревьев, как было показано выше, отличает их от других субординатных биоморфотипов. В частности, для многих де- ревьев характерна «периодичность камбиальной активности, которая в умеренной зоне является, главным образом, сезонной», что отража- ется в кольцах прироста древесины (Эзау, 1980, с. 146). Многолетние (стеблевые) части у всех деревянистых растений «одеты» вторичной покровной тканью: перидермой, коркой (Серебряков, 1962). Среди деревянистых растений есть группа стелющихся растений, у которых стволы и стволики вынуждены полегать из-за неблагоприятных усло- вий для прямостоячего роста; их принято называть стланцами у тех видов, среди биоморфотипов которых есть деревья, например, Juniperus turcestanica Кот. (Серебряков, 1962). Чтобы не нарушать определенного принципа в разграничении деревянистых растений по высоте аэроподиев, деревья (примерно выше 4 м), кустарники (от 0,8 до 4 м), кустарнички (от 5 до 80 см), автор стланцы не считает деревьями; пожалуй, их место, наряду со стланиками, которые по высоте еще ниже стланцев, среди кустарников как разновидностеи
Глава 5. Концеиция азнооб азия биомо семенных растений этого биоморфотипа. Шпалерный (стелющийся) тип роста деревяни- стых растений, безусловно, еще требует дальнейших исследований и осознания его места в организации биоморф семенных растений. Просубординатное разнообразие деревьев, как и других деревяни- стых растений, зависит от числа корректирующих признаков, на кото- рые исследователь обращает внимание, «привязывая» их к «опорному» — субординатному, признаку и рассматривая их вместе как одну сис- тему (рис. 305-332). Безусловно, нам кое-что удалось в субординатном и суперорди- натном классифицировании биоморф семенных растений, которое мы осуществили по интегральным признакам, используя в качестве маркеров их габитуальные части-признаки, поэтому, кстати, все при- веденные классификации можно одновременно рассматривать и как классификации по габитуальным признакам биоморф. Работа по классифицированию биоморф стоит того, чтобы ее продолжать, по- скольку в конечном счете такое классифицирование перерастает в исследования просубординатного разнообразия биоморф семенных растений, а зная разнообразие предмета исследований, мы начинаем понимать законы и правила его формирования, и, в конце концов, научимся управлять процессами, обеспечивающими биоразнообразие в живой природе. При разработке проблемы разнообразия биоморф мы невольно коснулись необходимости разбираться с пространственно-временны- ми границами их типов. Да, надо стремиться к уточнению этих гра- ниц, но есть одно обстоятельство, позволяющее утверждать, что дей- ствительно четких границ биоморфотипов на самом деле вообще-то быть не может. Дело в том, что особенности развития форм жизни никогда не происходят в жестком единообразии, и все биоморфы на Земле никогда не бывают одинаковыми, поскольку развиваются в определенных количественных и качественных параметрах, которые так или иначе предопределены их ресурсами возможностей (наслед- ственностью). Параметры организации одних биоморф как-либо бу- дут накладываться на таковые других биоморф, поэтому границы биоморфотипов всегда будут относительными, поскольку конкретных биоморф — мириады, а на их фоне биоморфотипы — это результат объединения в чем-то близких биоморф в одни группы. Ниже есть смысл поразмышлять о том, почему очень контрастных границ между биоморфотипами ожидать не стоит (см. главу 6.5.). Рис.217 — 332. Биографические портреты видов, отражающие неко- торое просубординатное (субординатное + корректирующее) биоморфо- типное разнообразие семенных растений.
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 217. Биографи- ческий портрет Astericus hierochun- ticus (Michon) Wikl. [сем. Asteraceae (Из- раиль, каменистая пустыня Негев в ок- рестностях поселка Сде-Бокер, 12 V)]. 5,29 OMOp@OTHII: надземка-двусе- ольные (семядоли обратнояйце- ные или эллиптические, широко- кругленные, суженные в короткие решки или сидячие, 3,5 — 5 мм 2,5 — 3,5 мм шир.) розеткообра- щие некустистые или кустистые шенно-простолистные семиде- минатно-репродуцирующие одно- ные желто-астроцветковые при- на-компактные эфемеры. Рису- выполнен с отмерших разновоз- тных растений, обнаруженных в бывшей луже, после быстрого испа- рения воды, где остались растения в виде мелких «скульптур», хорошо сохранивших га- битуальные признаки когда-то живых растений (и такое может бьггь в работе биоморфо- лога-био графиста). +г, 8е Рис. 218. Биографический портрет Магеяа pulchella (DC.) О.Е.Schulz [ceM. Brassicaceae (Израиль, окрестности Тель-Авива, каменисто-песчаные холмы, март)]. Биамарфатип: надземка-двусемядольные (семядоли яйцевидные, 2,5 — 3,5 мм дл., 2— 2,5 мм шир., узкозакругленные, резко суженные в короткие черешки) полурозеточные очередно-простолистные стержнекорневые слабокустистые семидетерминатно-репродуци- рующие однодомные розово-капустноцветковые эфемеры.
Глава 5. Концеиция азнообразия биомо семенных астений SB Рис. 219. Биографичес портрет Centaurium ten florum (Hoffmanns et L Fritsch [сем. Gentianac (там же, что и рис. 2 15 III)]. БИОМОРфОТИП: надземка-двус дальные (семядоли широкоэлли ческие яйцевидные, закругленны мм дл., 3 мм шир., сидячие) полу зеточные супротивно-простолист стержнекорневые некустистые (р слабокустистые) детерминатно- родуцирующие однодомные роз горечавковоцветковые однолетни Рис.220. Биографический портрет Datura stramo- nium L. [сем. Solanaceae (Москва, Главный бота- нический сад, питомник лекарственных расте- ний)]. Биоморфотип: надземка-дву- семядольные (семядоли продол- говато-линейные, 25 — 35 мм дл., 3 — 7 мм шир., слегка заострен- ные, резко суженные в черешки 5 — 10 мм дл.) безророзеточные очередно-простолистные некус- тистые стержнекорневые детер- минатно-репродуцируюшие эфе- мерно-полисимподиальные од- нодомные бело-пасленовоцвет- ковые однолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 221. Биографический портрет Diplotaxis acris (Forssk.) Boiss. [сем. Brassicaceae (Израиль, пус- тынная долина в Эйлатских горах, 5 V)]. Биоморфотип: надземка-двусе- мядольные (семядоли эллиптичес- кие, обратнояйцевидные, 6 — 8 мм дл., 4 — 5 мм шир., широкозакрут- ленные, сидячие) безрозеточные очередно-простолистные стержне- корневые прямостоячие семидетер- минатно-репродуцирующие одно- домные розово-капустноцветковые однолетники. Рис. 222. Биографический портрет Galium aparine L. [сем. Rubiaceae (Израиль, горные склоны вдоль доро- ги между Хадерой и Хай- фой, 11 IV)]. Биоморфотип: надземка-двусе- мядольные (семядоли широкояйце- видные, 8 — 22 мм дл., 5 — 14 мм шир., широкозакрутленные с выем- кой, резко суженные в короткие черешки 3 — 10 мм дл.) безрозе- точные вьющиеся цепляющиеся (стебли усажены обращенными вниз шипами, а на плодах крючко- ватые волоски) мутовчато-просто- листные четырехгранно-стеблевые стержнекорневые детерминатно- репродуцирующие однодомные бело-мареновоцветковые однолет- ники.
Глава 5. Концеиция разнооб азия биомо семенных астений ,Иа, 1,26 Рис. 223. Биографический портрет Cucumis sativus L. [сем. Cucurbitaceae (Московская область, дачные участки, выращивание на грядках)]. БИОМОрфОТИП: надземка-двусемядольные (семядоли эллиптические, яйцевидные, с выемкой 25 — 50 мм дл., 15 — 35 мм шир., клиновидно сбегающие в черешки) безрозе- точные вьющиеся цепляющиеся (с усиками) очередно-простолистные стержнекорневые индетерминатно-репродуцируюшие (открытые) однодомно-раздельнополоцветковые жел- то-тыквенноцветковые однолетники. Рис.224. Биографический портрет Schizonepeta annua (Pall.) Schischk. [сем. Mmiaceae (Алтай, Чуйский тракт, окрестности села Иня, долина реки Катунь)]. Биоморфотип: надземно-двусемядольные (семядоли широкоэллиптические, округлые, иногда слегка выемчатые, глубокосердцевидные с заостренными лопастями-ушками, 3— 5 мм дл., 2,5 — 4 мм шир., на черешках 3 — 8 мм дл.) полурозеточные (1 биоморфотип) или безрозеточные (П биоморфотип) прямостоячие супротивно-полусложнолистные стер- жнекорневые семидетерминатно-репродуцируюшие (полузакрытые) однодомно-обо- еполоцветковые беловато-яснотковоцветковые однолетники.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИЯ .32,34 4 2,гм1, 15a,206, 1к 1кс ПП Рис. 225. Биографический портрет Тгораео1ит majus . [ . р 'us L. всем. Тгораео1асеае (Московская область, дачные участки, цветочные клумбы)]. Б рфОтип: подземно-двусемядольные (прорастание п a ) р е по земное) безрозеточные вью- ИОМО жнеко невые индетер- щиеся (без усиков) или с ) телющиеся очередно-простолистные стержн р ~е о нолетники. минатно-репродуцирующие одн одомно-обоеполоцветковые желтоцветковь д
Глава 5. Концеиция азнооб азия биомо семенных астений 42 01 19a,21,26 Рис. 226. Биографический портрет Artemisia аечегяапа Willd. [сем. Asteraceae (Восточно-Казахстанская область, окрестности Усть-Каменогорска, пустыри — 1 биоморфотип; Аптай, Чуйский тракт, горные склоны в окрестностях села Онгудай, травостой нарушен выпасом скота — ! и !! биоморфотипы)]. На рисунке показано жизнеразвитие только по первому биоморфотипу. EHOMOp@OTHIIbI: надземка-двусемядольные (семядоли 4 — 7 мм дл., 3 — 5 мм шир., эллиптические, широкозакрутленные, иногда выемчатые, почти сидячие) полурозеточные прямостоячие некустистые очередно-полусложнолистные стержнекорневые семидетерми- натно-репродуцируюшие однодомные однолетники (1 биоморфотип), реже двулетники (11 биоморфотип, наблюдался в горах, где иногда зимует в розеточном вегетативном состо- янии, а на второй год продолжает вегетацию и завершает репродукгивное развитие). Рис. 227. Биографический портрет Cucurbita pepo L. [ñåì. Cucurbitaceae (Московская область, дачные учас- тки, овощные грядки)]. EHOMOp@OTHII: надземка-двусемядоль- ный (семядоли широкоэллиптические, 30— 65 мм дл., 20 — 35 мм шир., закругленные, суженные в короткие черешки) вьющиеся (с усиками) и стелющиеся очередно-простоли- стные стержнекорневые индетерминатно- репродуцирующие однодомно-раздельнопо- лоцветковые желтоцветковые однолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИН Рис. 228. Биографические портреты двух видов, выра- щиваемых часто вместе на полях: А — Avena sativa L [сем. Роасеае (Московская область, выращивание овса посевного на производствен- ных плантациях)]. Др — схематическое изображение ро- ста и развития главного побега (Доб- рынин, 1969, с.106). Дополнительные условные обозначе- ния: зр — зерновка, из которой при ее прорастании не выходит единствен- ная семядоля (щиток), поскольку она пециализирована для поглощения из ндосперма питательных веществ, кл — колеоптиль, лп — зеленый лист первый, лв — зеленый лист второй, лт — зеленый лист третий. БИОМОрфОТИП: подземно-односе- мядольные (процесс прорастания се- мян подробно описан Г.М.Добрыни- ным, 1969) полурозеточные слабокус- тистые очередно-простолистные кис- текорневые однодомные мятликовоц- етковые однолетники. Продолжение рис. 228. Б— Vicia sativa Ь. [сем. Fabaceae (Московская область, иног- да выращивают в посевах, обычно с овсом)]. БИОМОрфОТИП: подземно-двусе- мядольные безрозеточные вьющиеся (с усиками — метаморфоз верхнего листочка в системе перистого листа) очередно-перистолистные стержне- корневые индетерминатно-репроду- цирующие однодомные малиново- бобовоцветковые однолетники.
/nasa 5. Концепция разнообразия биомо ф семенных астений 2 (5 Рис. 229. Биографический портрет Tripleurospermum perforatum (Moat) М. Lainz [сем. Asteraceae (Московская область, залежи, в посевах, около дорог и строений, пустыри)]. Дополнительные обозначения: пф — озимый пост- флоральный побег, образовавшийся осенью после плодоношения унирепро- дуктивного побега (иногда может успешно перезимовать, особенно при полегании родительского побега); рр — розеточное растение, если оно раз- вивается осенью, то обычно нормально зимует; сл — слабое растение, раз- вивающееся в условиях сильной конкуренции. Биоморфотипы: надземно-двусемядольные [«семядоли эллиптические, закругленные, сидячие. Дл. 3 — 4 мм, шир. 1,5 — 2 мм» (Фисюнов, 1976, с. 195)] полурозеточные очередно-полусложнолистные стержнекорневые семидетерминатно-репродуцирующие (главная ось и ветви сложного годичного побега заканчиваются сверху компактными полузакрытыми соцветиями — корзинками, образующими вместе систему синфлоресцен- ции — метелку из корзинок) однодомные бело-астроцветковые однолетники (биоморфо- тип 1 — развитие происходит по яровому типу — семена прорастают весной, далее рас- тения вегетируют, цветут, плодоносят и отмирают к концу вегетационного сезона) или озимооднолетники (биоморфотип 11 — развитие осуществляется по озимому типу — се- мена прорастают осенью, образуются розеточные вегетативные растения, которые с зеле- ными листьями успешно зимуют, а на следующий год заканчивают свою жизнь так же, как однолетники). Еще бывают растения Ш-го и IV-го биоморфотипов, которые начинают развитие как однолетники или озимооднолетники, а заканчивают его только после второго вегетационного сезона, т. е. это уже травянистые двулетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ Рис. 230. Биографический портрет Limonium sinuatum (L.) Miller [ceM. Plumbaginaceae (Тель-Авив, крутой берег к морю у гостиницы «Хилтон», 6 IV)]. Биоморфотип: надземно-двусемядольные (семядоли линейные, 8 — 12 мм дл., ~,5— 2 мм шир., слегка заостренные, сидячие) розеточные стержнекорневые очередно-пр«То- листные семидетерминатно-репродуцирующие однодомные розовато-фиолетово-свинчат- ковоцветковые травянистые среднелетники (могут зацветать и на первом году жи~~~ и после нескольких репродуктивных усилий отмирают в возрасте 5 — 7, реже 10 лет).
Глава 5. Концеиция азнооб азия биомо семенных астений ический lychnis 1m. et [сем. Алтай, ВЫСО- еч ники, мно-дву- 12 — 22 родолго- , слегка чие) по- е супро- етерми- (закры- шечным дно-ли- травя- Рис. 232. Биографический портрет Draba subamplexicaulis С.А.Меу. [сем. Brassicaceae (Алтай, Курайский хребет, субальпийские луга, галечники)]. БИОМОрфОТИП: надземно-двусемядольные (семядоли 2 — 2,5 мм дл., 1,2 — 1,5 мм шир., эллиптические, яйцевидные, закругленные, резко суженные в короткие чере~шси) полурозе- точные каудексовые очередно-простолистные семидетерминатно-репродуцирующие однодом- ные бело-капустноцветковые травянистые многолетники (живут 10 — 15 лет, редко меньше).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 233. Биографический портрет Вгауа rosea (Turcz.) Bge. [сем. Brassicaceae (Ал- тай, Курайский хребет, высокогорья, берега ручьев, влажные щебнистые склоны)]. BHOMOpQOTHII: надземно-двусемядольные (семядоли яйцевидные, закругленные, 2— 2,5 мм дл., 1,0 — 1,3 мм шир., суженные в черешки) розеточные очередно-простолистные каудексовые семидетерминатно-репродуцирующие факультативно-корнеотпрысковые (при сильном оголении корней от почвы иногда могут образовываться придаточные почки и корневые отпрыски) обоеполые лилово-капустноцветковые травянистые многолетники. , д,42 4 АЗ 14а 18а Рис. 234. Биографический портрет Scorzonera radiata Fisch. ex Ledeb. [сем. Аа~егасеае (Алтай, Бащелак- ский хребет, каменистые луговые склоны лесного по- яса и высокогорные луга)]. BHOMOPQOTHII: надземно-двусе- мядольные (семядоли линейные, 18 — 25 мм дл., 2 — 2,5 мм шир., зао- стренные, постепенно переходящие в короткие черешки) полурозеточ- ные очередно-простолистные стерж- некорневые семидетерминатно-реп- родуцирующие (слабо олиственный цветонос завершается сверху одной корзинкой — полузакрытым соцве- тием) факультативно-корнеотпрыс- ковые однодомные светло-желто-ас- троцветковые травянистые много- летники.
Глава 5. Концеиция разнооб азия биомо семенных растений 5,гм2,20а,21,27 Рис. 235. Биографический портрет Crepis nana Richards. [сем. Asteraceae (Алтай, Курайский, Северо-Чуйский хребты, высокогорья, щебнистые осы- пи, галечники)]. Биоморфотип: надземно-двусемядольные (семядоли эллиптические, 3,5 — 4 мм дл., 2 — 2,5 мм шир., закругленные, клиновидно суженные в черешки) четочные (короткие или длинные в зависимости от интенсивности засыпания растений почво-грунтом на осыпях) стержнекорневые очередно-простолистные семидетерминатно-репродуцирующие (корзин- ки могут развиваться или не развиваться терминально, а кроме того, иногда образуются ниже в пазухах зеленых листьев) однодомные желто-астроцветковые среднелетники (жи- вут, как правило, менее 10 лет и после первого плодоношения нередко могут отмирать, а если не отмирают, то выдерживают еще 1 — 2 репродуктивных усилия). В условиях относительно спокойного состояния почво-грунта растения этого вида довольно быстро вытесняются другими видами. Не правда ли, здесь получается ситуация прямо-таки в со ответствии с классической лермонтовской поэзией: «А он, мятежный, ищет бури, как будто в бурях есть покой!~ Подобное можно сказать и о многих друтих растениях, обли- ~»0 произрастающих на крутых высокогорных осыпях (например, см. рисунок 240).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA C ТЕНИИ Рис. 236. Биографичес- кий портрет Convolvulus arvensis L.[cåì. Convol- vulaceae (Алтай, долины рек, у дорог, посевы и залежи) ]. Биамарфатип: надземно- двусемядольные [семядолн «расширеннолопатчатые или округло-обратнояйцевидные, почти четырехугольные, плот- ные, на верхушке с широкой выемкой, резко суженные в че- решки. Дл. 6 — 12, шир. 6 — 10 мм» (Фнсюнов, 1976, с. 154)] безрозеточные кустистые вью- щиеся (без усиков) корнеотп- рысковые очередно-простолис- тные индетерминатно-репроду- циорующие обоеполые розово- или белоцветковые сорные тра- вянистые многолетники.
Глава 5. Концеиция азнооб азия биомо семенных растений О Рис. 237. Биографический портрет Peucedanum vaginatum Ledeb. [сем. Apiaceae (Алтай, Чуйский тракт, окрестности села Онгудай, остепненные горные склоны)]. Биоморфотип: надземно-двусемядольные (семядоли линейные, 8 — 12 мм дл., 1 — 1,5 м> шир., островатые, постепенно суженные в очень короткие черешки) полурозеточные очередно-полусложнолистные корнеотпрысковые семидетерминатно-репродуцирующие (цветоносный побег заканчивается сверху одним полузакрытым соцветием — сложным 3oHTH«M) однодомные бело-сельдерейноцветковые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 238. Биографический портрет Gentiana macrophylla Pall. [сем. Gentianaceae (AaxaA, Бащелакский хребет, луговые горные склоны, долины горных рек)]. Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, тупые, 6 — 10 мм дл., 3 — 4 мм шир., клиновидно переходящие в короткие черешки) полурозеточные супротив- но-простолистные эумоноподиальные контринфлоресцентные (цветоносные постинтерка- лярные побеги образуются из пазух прошлогодних листьев, но когда их два и более на одной ростовой эумоноподиальной части, имеет место контринфлоресценция, или ино- соцветие) преиндетерминатно-репродуцируюшие (верхушечные фрондозные соцветия постинтеркалярных побегов приоткрытые, см. т. 1, с.221) компактнокорневищные про- дольно-партикулируюшие базифизарно-веревковидные (частично или полностью расше- пившиеся продольные части корневища вместе с пучками родительских придаточных корней закручиваются подобно ~веревке», а в целом подземная часть родительского орга- низма или даже криптоклонального синорганизма представляет из себя ризобазифиз) дефективно-кондивидуальные (несовершенно вегетативно размножающиеся при старе- нии, когда полная продольная партикуляция ризома превращает его части уже в соответ- ствующие части клонистов-особей вегетативного происхождения, см. т.1, с.159) однодом- ные синецветковые травянистые многолетники.
Глава 5. Концепция азнообразия биоморф семенных растений Рис. 239. Биографический портрет Phlomoides alpina (Pall.) Adyl., R.Kam. e< Machmedov [сем. Lamiaceae (Алтай, Курайский хребет, субальпийские луга)]
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ 42,92 ( с Г( 1 l г / I / г l I / Ф / / 1 1 О Ъ \ 4 ! Ъ 32,34 ( l I I ( ( 1 1 ( l ( 'L / 1 / 1 2,гм4, 196,21, СК I / ~Г I 1 30а < ПП 546,66 Биоморфотип: надземно-двусемядольные (семядоли 6 — 10 мм дл., 5 — 8 мм шир., широкоэллиптические, яйцевидные, широкозакругленные, в основании слегка сердцевид- ные, с небольшими лопастями-ушками, длинночерешковые 4 — 10 мм дл.) полурозеточ- ные эумоноподиальные интеринфлоресцентные (межсоцветия состоят из интеркалярнык семидетерминатных побегов в основании годичного побега — дикормуса, которые р»ви- ваются из пазух чешуевидных листьев) компактнокорневищные несовершенно-кондиви- дуальные однодомные розово-яснотковоцветковые травянистые многолетники.
Глава 5. Концепция разнооб азия биоморф семенных астений
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ск Рис. 241. Биографический портрет Rhodiola algida (Ledeb.) Fisch. et С.А. Меу. [сем. Crassulaceae (Алтай, Коргонский и Коксуйский хребты, высокогорья, влажные лишайниково-моховые тундры, приснежные лужайки)]. Биоморфотип: надземно-двусемядольные (размеры проростков и семядолей сильно ЪФ варьируют, например, во влажном моховом покрове гипокотиль и черешки семядолеи значительно вытягиваются, чтобы вынести к свету пластинки семядолей; семядоли эллип- тические, широкозакругленные, 4 — 10 мм. дл. и 2 — 4 мм шир., постепенно сужающиеся в черешки) безрозеточные эумоноподиапьные магимодифицированно-ростовые ассимили- руюше-силлептические (фотосинтез выполняют силлептические ассимиляционные четоч- ные, или безрозеточно-сближенноузловые, побеги, развивающиеся из пазух чешуй росто- вых годичных побегов) интеринфлоресцентные (из пазух чешуй ростовых годичных при- ростов развиваются интеркалярные побеги, составляющие каждой группой из 2 и более таких побегов отдельные межсоцветия) детерминатно-репродуцирующие (на ~eð~« каждого интеркалярного побега развито закрытое соцветие — щиток) короткокорневищ- ные ретардационно-кондивидуальные (замедленно вегетативно размножающиеся) обоепо- лые бело-толстянковоцветковые травянистые многолетники.
Глава 5. Концепция разнообразия биоморф семенных растений в, Рис. 242. Биографический портрет Galium densiflorum Ledeb. чаг. rosmarinifolium (Bunge) Tzvel. [сем. Rubiaceae (Алтай, Курайский хребет, высокогорья, луговые каменистые склоны, расщелины скал)]. Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, закругленные, 3 — 5 ~~~ дл. и 2,5 — 3 мм шир., клиновидно суженные в короткие черешки) безрозеточные короткокорневищные эусимподиальные мутовчато-простолистные семидетерминатно-реп- родуцирующие ретардационно-кондивидуальные однодомные желто-мареновоцветковые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ Рис. 243. Биографический портрет Dictamnus gymnostylis Stev. [сем. Rutacea< (Восточный Казахстан, невысокие горы, злаково-степной луг, 18 км Ha се- веро-восток от поселка Самарское)]. Др — общий вид соцветия (Маевский, 1964, с.304). Биоморфотип: подземно-двусемядольные безрозеточные короткокорневищные эусим- подиальные очередно-сложнолистные (непарноперистолистные) семидетерминатно-реп- родуцирующие однодомные красновато-рутоцветковые травянистые многолетники.
Глава 5. Концепция азнообразия биомо ф семенных растений 8а, в, Рис. 244. Биографическии портрет Patrinia sibirica (L.) Juss. [сем. Valerianaceae (Алтай, Бащелакский хребет, высокогорные луга и щебнистые открытые склоны лесного пояса).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ л р ~2о 2 д Р) х х й ю Q, х р, ~ (б Р1 Я 2 х Q, Ю ~ц х 2 о 0 о М еч х 2 Ж v О й~ М о ~~~î -ож ~ о о ~б Я. Ю 2 г~ Д 5: Ж /go ~м й~ Ц~ 0 д„'х 2 й; ~ Х й 2 ~~Я 2 о о о ~ х й~ М 2 ! а хо~ A x ;у й о о~ й X Xоо' Ю д ~) Ж g о о 2 хох о ~ ~ ° qp 0 [-~ Ж р. ~~ Осц Q 2 О v 7 ь О f О ~о[ о ю ~ х Ж" о Ж Ю Я х~~ 5: х О м Й О О~2 О й о 0i 22K
Глава 5. Концепция разнообразия биоморф семенных растений Рис. 245. Биографический портрет Paeonia hybrida Pall. [сем. Paeoniaceae (Восточный Казахстан, хребет Южный Алтай, редколесья и остепненные горные склоны в долину реки Каракаба)]. Биоморфотип: подземно-двусемядольные безрозеточные очередно-полусложнолистные короткокорневищные придаточно-клубнекорневые детерминатно-репродуцирующие (с д»м верхушечным цветком в структуре цветоносных годичных побегов) эусимподиаль- Н»е однодомные пурпуровоцветковые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 8 Рис. 246. Биографический портрет Filipendula vulgaris Moench [сем. Rosaceae (Ал- тай, Теректинский хребет, открытые осгепненные луговые склоны низкогорий)]. Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, 3,5 — 5 мм дл., 2,5 — 3 мм шир., закругленные, на коротких черешках) полурозеточные компактнокорневищ- ные гемисимподиальные очередно-полусложнолистные детерминатно-репродуцирующие придаточно-корнеклубневые однодомные бело-розоцветковые ретардационно-кондивиду- альные травянистые многолетники. 4,аа3,18а,27 2ае3 14а 18а 26 Рис. 247. Биографический портрет Eriophorum humile Turcz. ex Steud. [сем. Сурегасеае (Алтай, Курайский хребет, высокогорья, мохово- осоковые тундры) ]. Биоморфотип: односемядольные розеточные (розетки слегка растянуты, распространяясь в густом и влажном моховом покрове) короткокорневищ- ные семидетерминатно-репродуциру- ющие (на верхушке плейофазного цве- тоносного побега развивается только один колосок-пуховка) гемисимподи- альные однодомные осоковоцветковые ретардационно-кондивидуальные (медленно вегетативно размножающи- еся) травянистые многолетники.
Глава 5. Концепция азнооб азия биомо ф семенных астений ис. 248. Биографический портрет Crepis sibirica L. [сем..4л1егасеае ~ддтай Бащелакский, Катунский хребты, лесной пояс, низкогорья, луга в долинах горных рек)1.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ 42,92 42,91 ( /Г Г Г - г / I I I г 1 / / 31,34 30а ПП СК БИОМО~фОТИП: надземно-двусемядольные (семядоли обратнояйцевидные, 6 — 10 мм дл., 3 — 4,5 мм шир., широкозакрутленные, постепенно переходящие в черешки) розет- кообразующие полурозеточно-репродуцирующие гемисимподиальные компактнокорне- вишные семидетерминатно-репродуцирующие (плейофазные побеги завершаются верху- шечными полузакрытыми разветвленными сборными соцветиями метелками из корзинок) однодомные желто-астроцветковые ретардационно-кондивидуальные травянистые мн~~~- летники. ~.2(5), М4, 20б(19a), 21,26(27) 46 аг3, 18а, 9 26(27)
Глава 5. Концелция азнообразия биомор семенных растений й М 2 Е О л 3 f LO Q х )Я й h4 О h4 Ж й ЭЯ й h4 О й О М л )Я f - ~ щ~~ ЮФO Щ~ Ю~ О а5 О 'С 3 О 1 U Ве М р М ~ Q ~0 'д о ~р h4 ~ о О 2 и Ж б~ о о О р; Е рЯ Г=~ ~ о о '„ о д о ° й., w x ГЧ д о о й 1 ~ о Я Я „;ь, Ю о. 3 й х :=[ д 0с й~ О о х о м 2 5~ х „о Ю X а5 й~ 65 ~м х о. д И 20 х х й~ о о Е у о ~ о Я v ~ о h4 й Щ д й; о о ~ ~с о О Ю м о о й О д о z h4 р Ю д х Р' О О ~м Щ г~ о ~ Ф~Ъ 1 ( х ~ о К v Е к Я х х 65 д Я Ю р, ~ Q vо "~о 2 ~ х 2 О ( О ~ И ~) д a o v~ Q a z о Их д 6~ 2 о оь 3 < om> (В аоой о ~, g~~o r, 3: х ~4
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 250. Биографический портрет Rhodiola pinnatifida Вопи. [сем. Crassulaceae (республика Тыва, Саяны, хребет Обручева, высокогорные луга, берега ручь- ев; гербарные образцы любезно предоставлены проф. Ю.И. Черняком). Биоморфотип: надземно-двусемядольные (семядоли эллиптические) безрозеточные эумоноподиальные магимодифицированно-ростовые длиннокорневищные очередно-про- столистные интеркалярно-детерминатно-репродуцирующие (интеркалярные побеги с вер- хушечными закрытыми соцветиями-щитками развиваются из пазух чешуевидных листьев годичных побегов) однодомные зеленовато-желто-толстянковоцветковые экстенсивно-кон- дивидуальные (вегетативное разрастание осуществляется энергично, когда образуются длинные, средние и короткие годичные приросты корневищ, которые долго удерживают завоеванные территориальные участки и после образования особей вегетативного размно- жения) травянистые многолетники.
Глава 5. Концеиция азнообразия биомо семенных астений Рис. 251. Биографический портрет Alchemilla sibirica Zam. [сем. Rosaceae (Алтай, Семинский хребет, высокогорные луга)]. Б HOMOp+OTHII: надземно-двусемядольные (семядоли эллиптические, 4 — 4,5 мм дл., 2,5 — 3 мм шир., закругленные с выемкой или без нее, клиновидно сбегающие в черешки 3 — 5 мм дл.) полурозеточные короткокорневищные эумоноподиальные постинтеркалярно- детерминатно-репродуцирующие (постинтеркалярные побеги образуются из пазух про- шлогодних листьев, а на верхушках этих побегов развиваются фронда-брактеозные закры- тые метельчатые соцветия) апогамно-половые зеленовато-розоцветковые ретардационно- кондивидуальные травянистые многолетники. В роде Alchemilla L. очень широко распро- странен апомиксис, который встречается наряду с нормальной половой репродукцией и двойным оплодотворением (Глазунова, 1983, 1987).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 252. Биографический портрет Succisa pratensis Moench [сем. Dipscacaceae (Московская область, Подольский район, поляны в смешанном лесу, опушки)]. Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, 5 — 7 мм дл., 3 — 4 мм. шир., широкозакрутленные, клиновидно суженные в короткие черешки) полурозеточ- ные эумоноподиальные супротивно-простолистные компактнокорневищные интеркаляр- но-семидетерминатно-репродуцирующие (интеркалярные побеги с верхушечными полу- закрытыми головчатыми, почти шаровидными соцветиями) однодомные сине-фиолетово- ворсянковоцветковые травянистые многолетники.
Глава 5. Концеиция азнооб азия биомо семенных астений Рис. 253. Биографический портрет Convallaria majalis L. 1сем. Convallariaceae (Московская область, Подольский район, смешанный лес)]. Др! — проросток (Трофимов, 1963). Др2 — фрагмент взрослой особи (Серебряков, 1952). Биоморфо гип: подземно-односемядольные длиннокорневишно-предрозеточные или розеточные (помимо подземных длинных побегов, которые зимой не отмирают, превра- щаясь в корневища, на поверхность почвы выносятся предрозеточные побеги с чешуевид- ными и 1 — 2 зелеными сближенными листьями или, реже, подобные им, розеточные побеги с 3 зелеными сближенными листьями) эумоноподиальные интеркалярно-семиде- терминатные [интеркалярный побег «развивается в пазухе верхней почечной чешуи> (Серебряков, 1952, с. 268), а на верхушке этого побега развертывается полузакрьггое брак- теозное соцветие — кисть] простолистные белоцветковые субиитеисивио-коидивидуальные ториальные участки, которые относительно недолго, но все же значительно дольше, чем при интенсивном вегетативном размножении (m. 1, с. 1бО), удерживаются клонистами1 тра я- нистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ 32,35 гм4, 19г, 21,22 6), ПП 54a,Мг,62 Рис. 254. Биографический портрет Aegopodium alpestre Ledeb. [сем. Apiaceae (Алтай, Бащелакский хребет, верхняя граница леса и высокогорные субаль- пийские луга)]. Биоморфотип: надземно-двусемядольные (семядоли линейные, линейно-ланцетные, 15 — 20 мм дл., 2 — 3 мм шир., слегка закругленные, постепенно переходящие в «ðîTKHe черешки) длиннокорневищно-полурозеточные очередно-перистополусложные гемисимпо- диальные семидетерминатно-репродуцирующие (на верхушках универсальных ilo6e~~ развиваются сложные зонтики) однодомные бело-сельдерейноцветковые интенсивно-«н- дивидуальные травянистые многолетники.
Глава 5. Концеиция азнооб азия биомо семенных астений Рис. 255. Биографический портрет Achillea millefolium L. [сем. Asteraceae (Московская область, Подольский район, лесные поляны, опушки, луга)]. ~HOMOpQOTHII: надземно-двусемядольные (семядоли эллиптические, иногда почти ок- руглые, 2,5 — 6 мм дл., 1,5 — 3 мм шир., закругленные, сидячие) длиннокорневищно- полурозеточные очередно-полусложнолистные семидетерминатно-репродуцирующие (на верхушках вегетативно-репродуктивных плейофазных верхнеполурозеточных побегов раз- виваются полузакрытые сборные соцветия — щитковидные метелки из корзинок) полураз- вернуто-кустистые субинтенсивно-кондивидуальные однодомные бело-астроцветковые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ ,206,26 Рис. 256. Биографический портрет Чаlепапа tripteris L. [сем. Чаlепапасеае (Украина, Карпаты, Черногорье, нижний лесной пояс, тенистые берега гор- ных ручьев)1. BHOMOpQOTHII: надземно-двусемядольные (семядоли эллиптические, яйцевидные, 6— 8 мм дл., 4 — 6 мм шир., на верхушке слегка выемчатые, клиновидно суженные в черешки) безрозеточные длиннокорневищные розеточно-простолистные безрозеточно-полусложно- листные гемисимподиальные детерминатно-репродуцирующие (щитковидные закрытые соцветия развиваются на верхушках годичных безрозеточных цветоносных побегов, кото- рые продолжают, чаще всего моноподиальное, нарастание прошлогодних вегетативных безрозеточных коротких и даже длинных стелющихся побегов, которые заиливаются грун- том, превращаясь в эпигеогенные корневища) однодомные белоцветковые интенсивно- кондивидуальные травянистые многолетники. 5(6 18 Рис. 257. Взрослая особь Petasites rubellus (J.F.Gmel.) Тотап [сем. Asteraceae (Алтай, Курайский хребет, высокогорья, крутые склоны, россыпи)]. Биомо~)фотип: надземно-двусемядольные розеткообразующие длиннокорневищные (резиды живут от 8 до 20, чаще 12 — 16 лет) розеточно-простолистные семидетерминатно- репродуцирующие (каждый безрозеточный годичный цветоносный побег, лишенный зе- леных листьев, а только с чешуями, завершается сверху щитковидным соцветием из кор- зинок — компактных полузакрытых соцветий) однодомные желто-астроцветковые (впер- вые цветут на 8 — 15 году жизни) интенсивно-кондивидуальные травянистые многолетники.
Глава 5. Концеиция азнооб азия биомо семенных астений И- рет 1г18 еае ~ИЙ ые но- зе- но- не- ые ст- тые 2,92 18а,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 260. Биографический портрет Alisma plantago-aquatica L. [сем. Alismataceae (Московская область, Подольский район, мокрые луга, лужи, берега водоемов, сырые канавы)]. Др — рисунок всхода, выполненный И.Веленовским (цит. по И.Г.Серебрякову, 1952, с.57). Биоморфотип: односемядольные (семена прорастают в воде) розеточные простолист- ные гемисимподиальные компактнокорневищные (приросты ризома живут около двух лет) пирамидально-метельчато-инфлоресцентные однодомные белоцветковые травянистые многолетники.
1ааеа 5. Концеиция азнооб азия биомор семенных растений е,40,91 О Рис. 261. Биографический портрет Geum игЬапит L. [сем. Rosaceae (Мос- ковская область, леса, поляны, опушки, парки)]. БИОМОрфОТИП: надземно-двусемядольные (семядоли продолговато-эллиптические, слег- ка яйцевидные, 8 — 12 мм дл., 3 — 5 мм шир., иногда слабо выемчатые, клиновидно сбегающие в короткие черешки 4 — 8 мм дл.) полурозеточные розеточно-простолистные и полусложнолистные, безрозеточно-полусложнолистные эумоноподиальные компактно- корневищные интеркалярно или (и) постинтеркалярно-детерминатно-репродуцирующие (безрозеточные цветоносные побеги, закрывающиеся на верхушке цветком и рыхлым сборным соцветием, развиваются или как постинтеркалярные из пазух верхних прошло- годних розеточных листьев, или как интеркалярные из пазух розеточных листьев текущего года) обоеполые желто-розоцветковые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 262. Биографический портрет Ligularta altaica . [ DC. [сем. Asteraceae (Ал- Ка ь высокогорные луга и тай, Катунский хребет, склоны в долину реки тун, берега горных речек)]. е оли об атнояйцевидные, широкозак- Биоморфотип: надземно-двусемядольные (семядоли о р откие че ешки) — м . 3 5 — 5 мм шир., постепенно переходящие в короткие черешки ругленные, 7 — 10 мм дл,, — мм шир., р (по земные годичные при- полурозеточные гемисимподиальные к ые компактнокорневищные под п и щие (безрозеточный росты по егов живут б вут до двух лет) семидетерминатно-репродуцирующие езр ° ° ется све метелкои из ко е ко зинок которые, несмотря слабо олиственныи цветонос завершае рху ко э или согласно являются полузакрытыми соцветиями, или, на свое верхушечное положение, являю соцветиями на ям — от ытыми брактеозными компактными соц тр~~~~~~~~~ щжщ~~~~~~~, — открь блюдцеобразно расширенной оси, окруженной листочками о ертки о астроцветковые травянистые многолетники.
Глава 5. Концеиция азнооб азия биомо семенных астении 4,аг3,18а,2 в р ~ е 18а,25 5,гм4, 18а,21, 1 \ I ) «~' () 1' Ъ СК Рис. 263. Биографический портрет Arum hydrophilum Boiss. [сем. Araceae (Израиль, пойма реки Яркон, роща эвкалиптов, апрель)1. Биоморфотип: односемядольные (семена прорастают рано весной) розеточные геми- симподиальные компактнокорневищные (подземные годи р р ун чные п и осты иверсальных побегов живут до двух лет) семидетерминатно-репродуц рующ и ие (цветки однополые, собраны в одном соцветии — тычиночные выше, а пестич ичные ниже из последних сохра- У няется цилиндрический початок плодов, с кроющим подсоц о ветным листом, или покры- валом) ретардационно-кондивидуальные (редко) или изоакл оаклональные (без вегетативного размножения, чаще) травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ 21,27 афи- трет sa L. сеае асть, йон, Кус- ЛИ В есу, НО- ОЛИ е,на 4— IIIHP. (Ва- без- про- аль- рне- реп- мные вые
Глава 5. Концепция азнообразия биомо ф семенных растений 9 32,33а, ПП 2,гм4, 2О6, 21,26 1,аг4, 14а, 206,26 а,Д СК z30a 546,54г Рис. 266. Биографический портрет Seneco nemorensis L. [сем. Asteraceae (Ал- тай, Катунский хребет, луга в долине реки Катунь, хвойные леса, опушки)]. БИОМОРфОТИП: надземно-двусемядольные (семядоли яйцевидные, 10 — 15 мм дл., 5— б мм шир., закругленные, клиновидно суженные в черешки 3 — 5 мм дл.) безрозеточные «ередно-простолистные компактнокорневищные (подземные компактные стеблевые ча- сти побегов живут до двух лет) эусимподиальные семидетерминатно-репродуцирующие однодомные желто-астроцветковые ретардационно-кондивидуальные травянистые много- летники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНЩ 14,25 Рис. 267. Биографический портрет Ranunculus radicans С.А. Меу. [сем. Ranunculaceae (Московская область, топкий берег озера)]. Дополнительное обозначение: пв — поверхность воды. Биоморфотип: безрозеточные очередно-простолистные эусимподиальные детерминат- но-репродуцирующие поверхностно-ползучие интенсивно-кондивидуальные однодомные желтоцветковые травянистые многолетники. 26 Рис. 268. Биографический портрет Hieracium cymosum L. [сем. Asteraceae (Московская область, Подольский район, опушки смешанных лесов)]. До- полнительное обозначение: ст — столон. Биоморфотип: надземно-двусемядольные (семядоли эллиптические, 2,5 — 3,5 мм дл., 1,5 — 2 мм шир., закругленные, постепенно переходящие в короткие черешки) полуро- зеточные очередно-простолистные компактнокорневищные столонообразующие гемисим- подиальные семидетерминатно-репродуцирующие однодомные желто-астроцветковые ин- тенсивно-кондивидуальные травянистые многолетники.
Глава 5. Концеиция азнооб азия биомо семенных астений з О И ::3 3 и И О И )Я Я} Е Ж о у~о р и о >,~ й ~, ~: д; х д И о z иЕ~ о а3 Q,о г1 й g ~ о h4 + eY5 6~ ,'3' о д д„'Й о х й r. 5? а. v Ж ~ч~ Одо ggoz О ~о~~ f ф~ Г~ v О 2<~ßî~ :1. '~ О вм ф~ ~ о ааддд3 ~ а ~> у О с~ й о д Д й",о о ~~~а.а Ф~Ь ф~ ag о~~~ о > а Й Ю о ~~а~ р о ~8 г~ Й ~ ф.~ 2 -. д v а g~до а Of-h4 ~~~ф 65 й~ у д О ~> д~о 2 lQ ~ Ю ~р й с„~и ~м Г1 о О Й ~р п. 3
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 270. Биографический портрет Lamium а1Ьит L. [сем. Lamiaceae (Мос- ква, парки, предпочитает тенистые места)1. Дополнительное обозначение: СТ — СТОЛОНЫ. Биоморфотип: надземно-двусемядольные [семядоли широкоэллиптические, окрутлые, «короткоовальные, 7 — 9 мм дл., 5 — 8 мм шир., на верхушке широко закругленные и обычно слегка выемчатые, при основании с глубокой, почти круглой выемкой, по краям которой края пластинки удлинены, заострены и слегка загнуты внугрь в виде небольших рожек» (Васильченко, 1979, с. 235), на черешках 8 — 15 мм дл.] безрозеточные столоно- образующие супротивно-простолистные семидеретминатно-репродуцирующие интенсив- но-кондивидуальные однодомные белоцветковые травянистые многолетники.
Глава 5. Концеиция азнооб азия биомо семенных растений Рмс. 271. Биосрафичсским портрсс Чссопаа mecmstemoc 8uoge [ссм. асаорЛоЛпассаа (Алтай, Курайский хребет, высокогорья, каменистые россыпи, скалы)]. Биомо~)фотип: надземно-двусемядольные (семядоли яйцевидные, 3 — 4 мм дл., 2 — 2.5 мм шир., закругленные, клиновидно суженные в короткие черешки) безрозеточные ком- пактно или короткокорневищные, иногда вынужденно удлиненнокорневищные (при за- сыпании растений почво-грунтом), эусимподиально-гемисимподиальные (большая часть побегов монофазные, но есть и дифазные побеги, у которых сначала образуется короткое или длинное подземное основание и короткий надземный вегетативный полегающий побег, а на следующий год — ортотропный разветвленный вегетативно-репродуктивный акронекрозный побег) супротивно-простолистные семидетерминатно-репродуцирующие однодомные сине-норичниковоцветковые замедленно или интенсивно-кондивидуальные (»~ вегетативного размножения во многом зависит от степени подвижности почво-грун- та) травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 272. Биографический портрет Hieracium umbellatum L. [сем. Asteraceae (Московская область, опушки лесов, залежи, лесные поляны)]. БИОМОрфОТИП: надземка-двусемядольные [семядоли «продолговатые, на верхушке ту- пые, 9 — 15(18) мм дл., 3 — 4(5) мм шир., книзу суженные в черешок» (Васильченко, 1979, с. 327)) безрозеточные (некоторые, но далеко не все, ювенильные растения развивают, произрастая на открытом месте, слабую розетку главного побега из 3 — 4 листьев и так могут даже перезимовывать, но в последующем развитии все побеги уже не образуют розеток) эусимподиальные короткокорневищные очередно-простолистные семидетерми- натно-репродуцирующие однодомные желто-астроцветковые ретардационно-кондивиду- альные (замедленно вегетативно размножающиеся) травянистые многолетники.
Глава 5. Концепция азнооб азия биомо ф семенных астений Рис. 273. Биографический портрет Lilium pilosiusculum (Freyn) Miscz. [сем. Liliaceae (Алтай, Катунский хребет, лесные поляны, опутпки, субальпийские луга)]. Биоморфотип: односемядольные [прорастание подземное, у близкого вида L. martagon L. «проросток ведет в первый год подземный образ жизни» (Немченко, 1993, с. 36)) безрозе- точные ювенильно-эумоноподиаяьные (в первые годы жизни в надземной части появляется только один зеленый лист) эусимподиальные (с 3 — 5 лет начинают отрастать надземные, ежегодно отмирающие побеги с двумя и более зелеными листьями) семидетерминатно- репродуцирующие многолетнелуковичные однодомные сиренево-розовоцветковые изоакло- нальные (без вегетативного размножения) травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Й Й ?е а, янистые 78
! юм A ° g ф ! р о ca + 65 X о ~~ 2 йЮ ~) &- о ~ о ЫИ ИИ ~~ ч~ ~ о о 2к'~ ~ц Ю са F ~ о ц о ~ ~ о v vî о ~~ ~.' Е О й О v еч 1 Е Я ! ! ! -о' о й у а ж О щ X of>g~ ~~од i оооо ~р й Д~~ X д Ю ~ф X Я ~ц ~о~ Еоvgvм Й ~~ ~т~ 3 ~[ ооИ O. O Cl <1 ! IQ ! ! ! ~ой"' о~~~ о о д ~~ ~ ~ о ~оо~~ z ag g g 4~ ;>, ! (ц ~~ло~ ф~ ~ ! Д~ v 2 о 0 Хо e v Z v XA о~х~> Д; ф ~ да z e- O a 2 'р. eel И й1 v а ;ж й О. М 65 1 а "й С~) X v > М T с~) ! Я ! ~ о< о м й ~[И О 5-* ° g ° ~ Ь~~Ф v < ° ° О И > v ~-е Д; О ~З' о У~ Q.<~ О р~ ~ X О v < й; йй С4 ~сц Я Ю Я,'Я m~ov д. О. Ю ц~ р.' 0.о~~ о lQ ~ &" о о Й д о й ~ рИ z g био~ ~[ ~ Ф ;>, ~ о х Г'),Д о а.~~ < ~ о z д я й; ~~~о .0 7) х :а ж:а:3 Л: ~~ X О е~ о v О A ф юм О e~z 3~x д; Ю ь О о ~ц 65 (,) О д; ю Я О Л: Ф Ч' ф~в~ ~О~ Ф~ Ф~ Ф~ ~дава 5. Концеиция азнооб азия биомо семенных астений л ° )Q Р~'~~! ° о,~- Ю Е 0,ф 65 ~у~ л 65 О Q. й~ х жу'~ Ф о о 3 Д ! 00Х ~) р~ ~~ ь| 0 0 Й ф) Ы ч 6.) 3 v g5 Щ 1 М О О с!~ 4 ° ~ [~4 ф) о о р. X О ю hC g o ee! о р; М с!~ Д ° ~ Q ~~~ О ~) М д о 0 2 Ф Рх И О О (~ <о~ О ЭЯ, д~~ ~о Я о а. о ~х~ ~4,~ О ~О М 0 СЧ Д; Й 0 Й Q. F40 ~:~
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ О 34 Рис. 277. Биографический портрет Datura inoxia Mill. [сем. Solanaceae (Из- раиль, окрестности Тель-Авива, пойма реки Яркон, пустыри, 10 IV)]. Допол- нительные обозначения: жв — живые части, оставшиеся от прошлогоднего главного побега и корней, вп — весенний побег, отросший из сохранившейся почки на прошлогодних живых остатках главного побега. ИОМОрфОТИП: надземка-двусемядольные (семядоли продолговато-яйцевидные, слегка заостренные, 15 — 30 мм др., 6 — 10 мм шир., в основании резко суженные в черешки 10 — 20 мм дл.) безрозеточные прямостоячие эфемерно-полисимподиальные постфлораль- но-побегообразующие очередно-простолистные однодомные бело-пасленовоцветков»е по- лутравянистые малолетники.
Глава 5. Концеиция азнооб азия биомо семенных растений Рис. 278. Биографический портрет Bidens tripattita L. [сем. Asteraceae (там же, что и рис. 277, но более влажные места непосредственно у реки)]. Допол- нительные обозначения: жв - живые части, оставшиеся от прошлогоднего главного побега и корней, вп — весенние побеги, отросшие из сохранивших- ся почек прошлогоднего главного побега. Биоморфотип: надземка-двусемядольные (семядоли эллиптические, 8 — 10 мм дл., 3— 4 мм шир., закругленные, плавно переходящие в короткие черешки) безрозеточные суп- ротивно-полусложные (трехраздельные, но первые 1 — 3 пары листьев всходов обычно простые, слегка пильчатые) разветвленные семидетерминатно-репродуцирующие (верхуш- ки побегов завершают полузакрытые соцветия — корзинки, которые на первом году не- сколько раз перевершиниваются) постфлорально-побегообразующие (часть растений пос- ле первого года вегетации зимой отмирает частично и весной нередко проявляет еще одно, постфлоральное, репродуктивное усилие) однодомные желто-астроцветковые полугравя- нистые малолетники. В условиях умеренного климата, например, под Москвой, биоморфы этого вида ведут себя однозначно как однолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИй Рис. 279. Биографический портрет Aster subulatus Michaux [сем. Asteraceae (там же, что и рис. 277)]. Дополнительные обозначения: пф — постфлораль- ные побеги. БИОМОрфОТИП: надземка-двусемядольные (семядоли яйцевидные, 4 — 5 мм дл., 2 — 2,5 мм шир., закругленные, клиновидно суженные в короткие черешки) полурозеточные оче- редно-простолистные аэроподиоидные (от 10 до 100 см) прямостоячие разветвленные семндетерминатно-репродуцирующие постфлорально-побегообразующие, иногда кустис- тые (возможно это обстоятельство способствует выживанию особей и после первого по- стфлорального побегообразования до трех, а редко даже до четырех-пяти лет) однодомные бело-астроцветковые полутравянистые среднелетники.
Глава 5. Концеиция азнооб азия биомо семенных астений Рис. 280. Биографичес- кий портрет Hyparrhe- nia hirta (L.) Stapf [сем. Роасеае (Израиль, ок- рестности Тель-Авива, каменисто-песчаные холмы, разреженная растительность, ап- рель, май)]. дополни- тельные обозначения: аз — аэрозоны персис- тентного побеговетвле- ния, гз — геозоны ку- щения (зоны подзем- ного кущения). Биоморфотип: односемя- аа дольные безрозеточные под- земно-приземно-кустистые з (подземное и приземное куще- ние начинается рано и уже у ювенилов образуется до двух порядков побегов кущения, а у иммаматурных растений еще больше, у взрослых растений кущение настолько усиливает- оо ся, что становится компактно- множественным, образуя по- степенно криптоклональный синорганизм кочку, в котором клонисты, оставаясь сближен- ными, полностью физически разобщаются, но, будучи «стя- нутыми» корнями и подземными стеблями, составляют одно основание такой кочки— компактный криптоклонально-геоподиальный базифиз) мятликовоцветковые травополу- кустарнички. Помимо подземно-приземного кущения у биоморф этого вида ежегодно проявляются зоны надземного персистентного ветвления, которые поднимаются над по- верхностью почвы до 20 см и определяются побегообразованием из пазушных почек пе- резимовавших, т.е. персистентных, соломин (надземных резидов удлиненных ортотропных прошлогодних побегов) — аэроподиоидов. Назовем для удобства эти зоны побегового ветвления — аэрозонами персистентного побеговетвления. У разных типов многолетних полуграв они как-либо себя проявляют на разной высоте над поверхностью почвы. Гео- подии у полутравянистых многолетников гомологичны другим подземным базовым орга- нам, например, ризомам травянистых многолетников. Предлагая говорить о геоподиях в отношении подземных базовых органов полуграв, мы исходим не из того, что они сильно отличаются, допустим от ризомов, а только потому, что в определения друтих базовых органов (ризомов в том числе) заложены некоторые ограничения, с которыми есть смысл считаться (т.1, гл.б). В частности, ризомы трав, согласно нашему определению, не пере- ходят в надземной части растений в аэроподии, а их надземные продолжения, как правило, эфемерны. Исключение, причем не жесткое, пожалуй, можно сделать только для неболь- шой группы растений, у которых надземные персистентные части возникают только в определенных условиях вынужденно, например, как в случае с Solidado virgaurea L. (рис. б7), когда ризомы в сильном затенении теряют ориентацию в пространстве и выходят довольно высоко (до 20 см) в надземное пространство. Такие надземные «ризомы», или аэроризомы, — разновидность аэроподиев, а их подземные продолжения вполне можно по-прежнему считать ризомами, но одновременно не будет ошибкой, если мы их обозна- чим и как геоподии. Во всех остальных случаях надземный базовый орган — аэроподий ~переходит в подземный базовый орган — геоподий, что однако не исключает проявления разнообразия среди типа геоподиев.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис.281. Биографический портрет Stipagrostis lanata (Forssk.) de Wtnter ceM. Роасеае (там же, что и рис. ис. 280 22V)]. Дополнительные обозначения: аз- аэрозоны персистентного побеговетвления, гз- — геозоны кущения зоны подземного кущения). Разъяснения некоторых признаков биоморфотипа не повторяем, поскольку они те же, что и на рис. 280. Биоморфотип: односемядольные безрозеточно-розеткообразующ аз щие компактно-геоподи- ально-бази изарные однодомные мятли- дземно-приземно-кустистые криптоклонально- а об аз т немало розеточ- ковоцветковые травополукустарнички (молодые растения сначала о р ую сте же). ных вегетативных по егов, а у вз б рослых растений такие побеги встречаются в кусте реже
~дава 5. Концеиция азнообразия биомо семенных астений 1,аг4, аз аз 1,аг2, 206,26 2,гм4, 206,21,26 аз i C6,100,ск I 1 1 1 Рис282. Био а и Poaceae . гра ический портрет Piptathetum miliaceum (L.) С ( а и . oss, всем. биомо отипа и ополнительные обозначения, разъяснения нек я некоторых признаков морфотипа ассма ива иомор отипа и непосредственные определения просуборди б инатного ио- а у рассматриваемого вида аналогичны тем, что приведены на рис. 280, поэтому здесь не повторяются.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис.283. Биографический портрет Leacanthemum vulgare Lam. [сем. Asteraceae (Московская область, Подольский район, 1 биоморфотип на полянах сме- шанного леса и П биоморфотип в тенистом участке елового леса в 20 м от опушки)]. БИОМОрфОТИПЫ: надземка-двусемядольные (семядоли эллиптические, 4 — 5 мм дл., З— 3,5 мм шир., закрутленные, клиновидно суженные в короткие черешки) розеткообразующие (на открытых местах развиваются более всего розеточные и полурозеточные побеги, a ~pH сильном затенении — верхнерозеточные и верхнеполурозеточные побеги) очередно-просто- листные семидетерминатно-репродуцирующие однодомные бело-астроцветковые ретарда- ционно-кондивидуальные компактнокорневищные травянистые многолетники (Биоморфо- тип 1) или короткокорневищные травополукустарнички (Биоморфотип 11).
Глава 5. Концеиция азнооб азия биомо семенных астений Рис.284. Биографический портрет Dianthus superbus L. [сем. Caryophyllaceae (Алтай, Семинский хребет, влажные луговые склоны лесного пояса и высо- когорий)]. БиомоРфотип: надземка-двусемядольныые (семядоли яйцевидные, 4,5 — 6 мм дл., 3— 4 мм шир., закрутленные, резко суженные в короткие черешки) розеткообразующие (в разнообразии побегов господствуют розеточные и верхнерозеточные вегетативные и по- лурозеточные и верхнеполурозеточные вегетативно-репродуктивные побеги, но кроме того могут образовываться единично еще и безрозеточные как вегетативные, так и вегетативно- р~продуктивные побеги) супротивно-простолистные детерминатно-репродуцирующие од- нодомные лилово- или бледно-розовоцветковые стержнекорневые ползуче-стелющиеся тра вополукустар яички.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ 2,34 М4, Рис. 285. Биографический портрет Coccyganthe flos-cuculi (L.) FoutT. [сем. Caryophyllaceae (Московская область, 1 — поляны в смешанном лесу, П— влажные тенистые места в еловом участке леса в 20 м от опушки)]. БиоморфотипЫ: надземка-двусемядольные (семядоли продолговато-яйцевидные, 5— 8 мм дл., 2,5 — 3,5 мм шир., заостренные, клиновидно суженные в черешки 2,5 — 5 MM дл.) полурозеточные супротивно-простолистные гемисимподиальные стержнекорневые де- терминатно-репродуцирующие однодомные розово-гвоздичноцветковые травянистые мно- голетники (Биоморфотип 1) и травополукустарнички (Биоморфотип 11). На открытых полянах смешанного леса биоморфы образуют прижатые к поверхности почвы, хорошо обрастающие придаточными корнями, розеточные вегетативные побеги и полурозеточные унирепродуктивные побеги (травомноголетники). В условиях сильного затенения в еловом лесу развиваются более всего верхнерозеточные вегетативные и редко (растения здесь плохо цветут) верхнеполурозеточные унирепродуктивные побеги; в надземной части без погружения в почву зимует довольно много побегов и их резидных частей, заметно (до 1о см) поднимаясь над поверхностью почвы (травополукустарнички).
~дава 5. Концепция азнооб азия биоморф семенных астений А Б Рис.286. Биографический портрет Draba sibiria (Pall.) Thell. [сем. Brassicaceae (Алтай, Курайский хребет, поляны и опушки лесного пояса и высокогорные лутовые склоны)]. Бйоморфотип: надземка-двусемядольные (семядоли яйцевидные, 2,5 — 3 мм дл., 1,2— 1,8 MM шир., закругленные, резко суженные в короткие черешки) безрозеточные (розетку растения могут образовывать только на раннем этапе развития) очередно-простолистные ползуче-стелющиеся стержнекорневые гемисимподиальные семидетерминатно-репродуци- рующие обоеполые желто-капустноцветковые травополукустарнички.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ ),аг4,2О6, (26) Сб 82 78 ск,f Рис.287. Особенности экзоморфологической организации травокустарнич- ков Pennisetum asperifolium (Desf.) Kunth [сем. Роасеае (Израиль, Тель-Авив, Ботанический сад, 21 Ш)]. Дополнительные обозначения: фк — фрагмент куста, аз — аэрозоны персистентного побеговетвления, гз — гезоны кущения (зоны подземного кущения). БиоМорфотипЫ: безрозеточные моно-, ди (большинство)-, плейфазно-побеговые под- земно-кустистые компактно-геоподиальн ые аэроподиально-стелющиеся (Биоморфотип1) или аэроподиально-прямостоячие (Биоморфотип 11) компактно- криптоклональные однодомные мятликовоцветковые травокустарнички. Аэроподии живут несколько лет, более всего при их полегании, когда на склонах холмов прижимаясь друт к другу они образуют своеобразный мертво-живой ~сноп» одного куста из живых и мертвых соломин и побегов текущего сезона вегетации. В таком габитуальном состоянии растения летуче сохраняют жизнеспособность своих меристем, преодолевая невзгоды продолжительно- го сухого сезона. 2(5) м4 20б 26(27) 97 1(2,4) аг4(а 20б,2 Рис.288. Особенности экзоморфологической организации травокустарнич- ков Thricholaena teneriffae (L. fil) Link [сем. Роасеае (Израиль, Тель-Авив, Ботанический сад, 25 Ш)]. Биоморфотип: безрозеточные моно-, ди (большинство)-, прейофазно-побеговые под- земно-многокустистые коротко-геоподиальные аэроподиально-стелющиеся (аэроподии действительно полегают, но не ползут, образуя периодически аэрозоны персистентного побеговетвления на разной, как правило, до 80 см, высоте над поверхностью по'~вы) коротко-криптоклональные однодомные мятликовоцветковые травокустарнички.
Глава 5. Концеиция азнооб азия биомо семенных астений Ж О б4 3 .М ~л Ж О М Е -,) ОО ° ~ Р4 Щ и Зч Q р О X 3~ О ~) О М кМ X tD и К М 3: О: ?: Ж й ~ о О з: X К 65 ~ц о~ �ф )~ й; о о М ~ щ и О Ю ~ й О Щ) ~'~~ ) ~ Г~ Я о сг) Рй О М Р) и у и о z X щ X [е О 1О ~д О О O ~ О'~,~ ~4 и 3~ ФЭ б4 f О~ О. ь Ю Я о о й A A Ф~Ъ ф к ~[ X о ~.'~ X й оИр. ОБМ~ ~~~î М ;~, 'о е Д Ю о И о~~ «о М д ~~~о о д О g,vX ~~г ,'~ Ю ~- ~: о v~vz 1 Ц' О р: v ~~[~ ф~ X О. Я О '~ Я о ~ Й И о И щ 7 ! ооо й о о Ж р Я у О а а. ф„'ц CO р; g z~ оД~ ~~о М д: И а X О ~а ~ 65 о ~~~ ~~ охж с> ф: Q, ~ Ф LO ea ..Я~- Оом Q ä а О о ~о~ О о ь ~4:
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 290. Биографический портрет Rubus idaeus L. [сем. Rosaceae (Москов- ская область, Подольский район, смешанный лес, опушки)]. БИОМОрфОТИП: надземка-двусемядольные (семядоли эллиптические, 5 — 7 мм дл., 3— 3,5 мм шир., закругленные, клиновидно суженные в короткие черешки 3 — 5 мм дл.) безрозеточные тройчато- и непарноперистолистные детерминатно-репродуцирующие (цве- тоносные годичные побеги, состоящие каждый из верхушечного соцветия — закрытой кисти и ассимиляционно-фрондозной части, развиваются весной из латентных почек верхней и средней частей аэроподиоидов — прошлогодних вегетативных турионов, кото- рые акронекрозно закончили вегетационный сезон, успешно перезимовали и вместе с упомянутыми выше репродуктивно-ассимиляционными побегами текущего года органи- зовали единые системы — унирепродуктивные комбинированно-универсальные побеги, или гексакормусы, т.е. побеги шестого уровня организационной сложности) развернуто- кустистые корнеотпрысковые однодомные бело-розоцветковые экстенсивно-кондивиду- альные травокустарники. Удивительная особенность высоких (0,5 — 2,5 м), заметно одре- весневающих аэроподиев быть только аэроподиоидами, т.е. жить в своем надземном пер- систентном качестве всего одну зиму, весну, лето и в конце вегетационного сезона отми- рать полностью или почти полностью (до крохотного подземного основания, сохраняю- щегося несколько дольше в структуре геоподия), является важным аргументом, чтобы не считать малину обыкновенную кустарником, а относить ее к одному из типов полутравя- нистых растений — травокустарникам. Все уникальное поведение аэроподиев малины, как недолговечных форм их — аэроподиоидов, вполне объяснимо сильнейшей привязкой этих органов к репродуктивной деятельности биоморф, а когда она исполняется, то персистен- тность ~уходит» из старых органов и переходит к новым, молодым аэроподиоидам.
Глава 5. Концепция азнооб азия биоморф семенных астений ф л де (~ й[ ~а С~) О О; Х еа о Я ~~," ~,а о О » М Ф о о Дц д а;» 65 Ф «~, О Мйо »м I I Ca a a~ И а аИ а аИ »м ° » ~ю ~ - e» IQ Е 5 [~ "» [~ 5 ~оииЕ„, - ю Д~о~ Е О а ~~ До, р, ~ фв~ 0м у Q, е Ю o "< = o О~ ( 1- ! д ( О Ю 'И VV д4 ~) cn О ЧР. ~ 65 О1 И ЙЮЮЮЛ ЕМИИЮ юБожОюооо са C ф, а> OCi л e5ee у е5ее о~~' o g ~ v Ж Х ~ р~ Zдид О <~ О ~ д о~оц о оа.~~ о~~~ v v ° g Щ ~хр д ) ~ й еа Г') й' х 2 й -~ ~ о о о о ~-ОЯа л v 1' о ц Ф ~~@ 5 О ~ о щPee ао о < о о ~~ М Х ° g ~Г') л Й X О '~: им~ ~О д; М + о О О Р а X ф„' Ю еа д Ж ~ г .~; О~ ~~о 5: Ю v ~.~ о о ~ о ~~ 2XО Ж ф~~ О Ю X О д: М Я д ~О X О д Ю О :~: Ж О й О Х Е~ Я: а ОИ л ° е» ][~ e» О 65 Е Щ а 4> Q о + C C 3 4» 65 ° ~ +~4 =5 Л Q. О bQ Ж -. л ~Я ,.'М И Ю» ° лй Cl ~ Е= Ю 65 И И ,ф 65 Л И C . 65 Б а Й ( О~ О И ° ~ е~р C л 1 О И 5 < о О C I» ~~:) I» ~е1 65 ~ ~ о Х Й Ф О Й 65 Л Х д :а A о х д i о X :а О~ Ф 65 1 I I a. 1 1 l 1 1 1 I 1 ! ( ! 1 1 1 t 1 1 1 I 1 1 I 1 I 1 1 1 1 1 1 (~ L 1 1 1 I I» 1 1 ш 1 1 1 1 1 Q~~ ц~ ~Ьцц О О :а О О й' О -& О О X О I X 1 ю I;~
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 292. Биографический портрет Scutellaria supina L. [сем. Lamiaceae (Ал- тай, Курайский хребет, южные щебнистые склоны, редко высокогорья)]. БНОМОрфОТНП: надземка-двусемядольные (семядоли яйцевидные, 3 — 4 мм дл., 1,5— 2 мм шир., закругленные, резко суженные в черешки 3 — 4 мм дл.) безрозеточные суп- ротивно-простолистные семидетерминатно-репродуцирующие эусимподиальные стерж~~- корневые однодомные желто-яснотковоцветковые изоаклональные (при засыпании шеб- нистым грунтом может образовывать удлиненные геоподии) полукустарнички.
Глава 5. Кон еп ц ция разнооб азия биоморф семенных астений 2 2 2 4 Рис. 2 >zip ога chnopodioides Lam. [сем. иографический портрет Zizi h еае таи, Ан~ский Ба ел , Ан, щ лакскии хребты, остепненные склоны)]. Биоморфотип: н з ад емко-двусемядольные (семядоли о лаяй ев ле выемчатые — 4 ю'г~~, — мм дл., — мм шир., лабо сердцевидные, на дифазные семи зрозеточные супротивно-п сто детерминатно-репродуцирующие э сим зро ро — ростолистные стержнекорневые моно- з основании куста не б у эусимподиально-гемисимподиальные (в а нередко, ближе к осени, отрастают ко шинство из кото ы тр короткие вегетативные побеги, боль- орых зимой отмирает, но немногие пе ехо т и завершают его как диф реходят к репродуктивному развитию дифазные сложные универсальные по на фоне доминирования монофазных г р побеги, что в целом происходит зово-ф -фиолетово-яснотковоцветковь вания моно~разных годичных побегов — дико сов' о тковые полукустарнички. рму, однодомные ро-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛСТЕНИЯ 2,г 206 Б МЯД сле 4— НЫ роз НЫ эус лел ДИЗЛ ДОМ кус НИК ные рас ДЛИ
Глава 5. Концеиция азнооб азия биомо семенных астений Рис. 296. Общий вид полукустарника Helianthemum lippii (L.) Dum.-Courset [сем. Cistaceae (Израиль, Эйлатские горы, пустынная каменистая долина с пересохшим руслом сезонного водостока, 6 V)]. Растения сезонно (после дождей) быстро образуют и легко (в сухой период) сбрасывают все листья. Розеткообразующие очередно-простолистные эусимподиально-гемисимподи- альные акронекрозные эфемерно-полисимподиально-репродуцирующие обоеполые жел- то-ладанниковоцветковые полукустарники. гм4,206, ,27 Рис. 297. Общий вид полукустарника Pulicaria crispa (Forssk.) Oliver [сем. Asteraceae (там же, что и рис. 296, 5 V)]. пиоморфОТИп: Пезрозсеочнне очсрепно-Изичозиознне зусимпозинино-cccccccccocccccc- »~е семидетерминатно-репродуцирующие однодомнъ~е желто-астроцветковые полукустарники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ 4, 06), (27) Рис. 298. Биографический портрет Atractylis phaeolepis Pomel [сем. Asteraceae (Израиль, каменистая пустыня Негев, окрестности поселка Сде-Бокер, 12 V)]. Надземно-двусемядольные непостоянно-розеткообразующие очередно-колюче-простолис- тные эусимподиально-гемисимподиальные семидетерминатно-репродуцирующие (на вер- хушках побегов развиваются одиночные корзинки) однодомные розово-астроцветковые изоаклональные полукустарники. 4~206,21,27 06,27 Рис. 299. Биографический портрет Majorana syriaca (L.) Rafin. [сем. Lamiaceae (Тель-Авив, Ботанический сад, 29 Ш)]. Биоморфстип: надземно-двусемядольные (семядоли округлояйцевидные, широкозакруглен- ные, 3 — 4 мм дл., 4 — 5 мм шир., сердцевидные, на черешках 2 — 4 мм дл.) безрозеточные супротивно-простолистные гемисимподиалъные семидетерминатно-репродуцирующие ~ð~- некорневые полегающе-побеговые однодомные бело-яснотковоцветковые изоаклональные по- лукустарники.
Глава 5. Концеиция азнооб азия биомор семенных растений !Ь!!!!V!!VVХ!V! "ОO~!О!!!!!!!!!65 о;~ о. ~ о о. о:~ д ! о y < P p ~ 43 ~ ca о о о .~ о ~ v о. !. о. Я > c ц и ц с Я z w > о. и '~ < Й Д и N о. ~ w K ð, и ю а о~ ~~аz~жо~ я~и ~~и ~~орб ~~~~~~~~й�~~~ ~р~~~Я- о ! '~,~ ~- ~ ~ Q �.à ,д О~ + ~ X ф~ь ~ '~ ~ о ~ о ~ р. ~ ~ ~ < 2 ~ < ~ ' < ~ < Я < > ~ ~~ ~ о ~ ~ ~ ~ Й„~ ЙzйииЕ~О ° и - — 8ри&аzzmоло а "ио~ д ~р ~ ~ ю ~ у + х +,'Я ~~ ~ Q + Ф ~р ю о ~ щ х х р ~ р . ф R ~ > > ~ ~ Я g о ц и vg о о ~ и ~ ~~- ~ g ~ + < > ~ > ~ z~ Й О Я ~ > p) ~» (r) ('ъ Й () lg " lg ы C ръ I О Л ФЭ О ~ О Й щ р ~~ ~ о 65 ~ щ ! й( о ~=И цор ~" >о~. "Я (ц о д; ~ „g g„~ ) ~ 65 x ~ ~р Q, ~,'~~ Q ô + д + 0 о ~ о ~ ~ о 3 ! о о „<~ 00 < ~- ' ~ ц > ц о и ц ~.' ~ а. о '~ ~ ~ ~ ~- ~ о v '» й у ~ > uD w ~. о о ж х о ~ ~[ о м ~. ~. о. й о М) Ю~) )Я Ю О М ',> Ф з: Ф (р~ 65 Ф 2 Ю Я, 65 ~р < 1 0 1 0~ Я о ф) Я ~о 3 Е Що Е Ф~М Е б~ Uо 3 ~~ о д,О О 3 ~й й~ М и ~' 0 С~ LO Щ М о у, о ~> С~ 0 з: б4
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 301. Биографический портрет Polygonum palaestinum Zohary [сем. Polygonaceae (Израиль, каменистая пустыня Негев в окрестностях поселка Сде-Бокер, 12 V)]. Биомарфатип: надземно-двусемядольные (семядоли линейные, 8 — 12 мм дл., 1 — 1,5 мм шир., заостренные на коротких черешках, почти сидячие) безрозеточные очередно-простоли- стные эусимподиальные индетерминатно-репродуцирующие (открытые верхушечные фрон- дозные соцветия завершают унирепродуктивное развитие годичных побегов и цветут в основ- ном осенью, зимой — во влажный сезон (а после отцветания и плодоношения эти побеги отмирают на разную длину, но весной небольшая часть побегов вновь образует немногочис- ленные открытые соцветия) стержнекорневые однодомные белоцветковые травянистые мно- голетники (биоморфотип 1) или приземистые полукустарнички (биоморфотип П), или полу- кустарники (биоморфотип И1 с аэроподиями около 50 и более см высотой). Когда растения произрастают в одиночестве на апсрытом месте, то формируются биоморфотипы TpRBQMHQ голетников (I) или приземистых полукустарничков (II), но, когда растения, прижимаясь друг к другу и ассоциируясь с биоморфами других видов, образуют «группы» на фоне пустых пространств, то проявляет себя биоморфотип третьего типа — полукустарники.
~дава 5. Концепция азнооб азия биомо семенных астений ет et О- ые U-
слл C.w R.r Рис. 305. Биографичес- кий портрет Ajuga iva (L.) Schreber [сем. Lamiaceae (Израиль, окрестности Тель-Авива, каменисто- песчаные холмы)]. Биоморфотип: надземно- двусемядольные (семядоли яй- цевидные, 3 — 4 мм дл., 1,5— 2мм шир., закругленные, резко сбегающие в короткие черешки) четочные очередно-простолист- ные эумоноподиальные интерка- лярно-инфлоресцентные (цвет- ки развиваются только в пазухах зеленых листьев побегов текуще- го года, составляя вместе меж- соцветия, или интеркалярные соцветия) однодомные стержне- корневые изоаклональные (без вегетативного размножения) желто-яснотковоцветковые при- земистые кустарнички. ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИО Рис. 304. Некото- рое разнообразие форм роста дендро- идов (Имханицкая, 1985, с. 30). Дополнительные обозна- 7 чения: R.r. — Roystonea regia (Н.В.К.) О.F.Cook, С w. — Colpothrinax wrightii Griseb. et Н.Wendl. ех Siebert et Voss, Н.t.— Hyphaene thebaico (L,) Mart., Ch.l. — Chama- edorea ianceoiata (Ruiz et Pav.) Kunth.
ogg j~g<~ggyy 2 я Й~а! Ия 2 Й Е4 О. ~ о Ц М ~ÑÑ Я ! '.р йь1 ! ! )ф~ ! а ~о.ц й[ ~[ О ц ~ Ю Я,о ) а! <~ й, О -~в Я Д Й (ч "ЙЕу и I lI1 Ь Б~цм ~ О 2 О О О М Я Ц ~ Й g~o л Ф Я й й: О ~' д v о ~ о ~д ~ ~ о ~>~ ~~ ~8о Kb ~лава 5. Концепция разнообразия биоморф семенных растений л ),'Я За g5 о Р з,'Я О ~о ~4 О Ф 1 ф ) М- л Ф ф~ О а5 М lQ й 65 Ю 0 65 О й М О й е( Ю и р, 'б N л д и g 00 yg О Й'" ~ О „2 д ~о са ID 1 Я a;" О Q ~ о О Е М О и д р Ф О З; Х 04,З Й ~ М 0 Ж О О З, 'Q О
о л ~и~ о' Д ~!;М л ° ° л О О 3 wain о ~~- 3 о и~ Щ ! ! ! ! ! Я V О О К О Я ~!'о ЙДи BQXXОu а~юО О ~р Ю МЯООИй "й й ~ Я ИФЙ ~я 6А.Рй~5 циР~то v О:~: ~ р- Я ! ! ! ! О Ж ~ ai о с!3 g ~ o ~! ФЭ Д ФЭ 3: ~ е[ >+и о Я Е О. ~ий~~ Ю Ю Ю Q Х Ю 65 Ю О 65 О ° ~у~ У5 Ж 3 Ю О 6 5 2 ~ а. D ю !»' X Q У5 О h4 lQ й О ~ Е: ф„'1, М О О и ~ и X X ФЭ 3 Я,' М~ ~~ О С О С~ О О О ~~!ю1 О~И С4 OCHOBhl БИОМОРФОЛОГИИ CENEHHblX РА СТЕНИИ
Глава 5. Концепция разнообразия биоморф семенных растений 7,23 4а, Рис. 308. Биографический портрет AsIragalus tschuensis Bunge [сем. Fabaceae (Алтай, Курайский хребет, высокогорья, каменисто-лишайниковые тундры, щебнистые склоны)]. ИОМОрфОТИП: надземка-двусемядольные (семядоли эллиптические, мясистые, 4— 6мм мм дл., 2,5 — 4 мм шир., закрутленные, на коротких черешках) розеточные очередно- сложнолистные стержнекорневые эумоноподиальные интеркалярно-репродуцирующие (безлистные интеркалярные цветоносные побеги с пузырчато-вздутыми одиночными пло- дами образуются в пазухах непарноперистых плейофазных мультирепродукгивных побе- гов) однодомные фиолетово-бобовоцветковые плотно-подушковидно-кустистые компакт- но-каудексовые (многочисленные каудиции, плотно прижимаясь друг к другу, вместе с кроной розеточных вегетативных и вегетативно-репродуктивных побегов текущего года образуют «подушку», которая возвышается над поверхностью почвы на 5 — 20 см) при- земистые кустарнички (на крутых склонах растения выдерживают эрозионную динамику почво-грунта, подвергаясь сильной продольной партикуляции, которая может иногда приводить к несовершенному вегетативному размножению) °
ОСНОВЫ БИОМОРФОЛОГИИ CENEHHbIX РАСТЕНИЯ 4(1),аг3,18а,27(26) Рис. 309. Биографический портрет Waldheimia tridactylites Kar. et Kir. [сем. Asteraceae (Алтай, Курайский хребет, высокогорья, щебнистые склоны, осы- пи, галечники)]. Дополнительные обозначения: вд — вынужденно удлинен- ный побег в условиях затенения или засыпания почво-грунтом. Биоморфатип: надземно-двусемядольные (семядолн эллиптические, 2 — 3 мм дл., 0,7 — 1,5 мм шир., закругленные, сидячие) четочные (иногда, особенно у молодых растений, образуются розеточные побеги, но доминируют по жизни четочные побеги) очередно-простолистные геми- симподиальные геоподиально-стержнекорневые семидетерминатно-репродуцирующие (оди- ночные корзинки завершают сверху структурное развитие ди-, плейофазных четочньи побегов которые, кстати, в условиях загенения или при засыпании почво-грунтом вынужденно растя- гивыот свои междоузлия и становятся безрозеточно-раэдвинутоузловыми) однодомные розово- белоастроцветковые дерновинно- или подушковидно-приземистые кустарнички.
Глава 5. Концепция разнообразия биоморф семенных растений Рис. 310. Биографический портрет Potentilla biflora Willd. ex Schlecht. [сем. Rosaceae (Алтай, Курайский хребет, высокогорья, щебнистые тундры, каме- нистые склоны)]. Биоморфотип: надземка-двусемядольные (семядоли эллиптические, 1,8 — 2,5 мм дл., 0,8 — 1,2 мм шир., закрутленные, клиновидно суженные в черешки 1 — 2 мм дл.) розе- ~очные очередно-полусложнолистные стержнекорневые эумоноподиальные интеркалляр- но-инфлоресцентные (цветоносные интеркалярные побеги по сути безлистные, лишь с двумя прицветными листочками переходной структуры между брактеями и зелеными ~~~тьями, поэтому мы говорим о розеточных открытых вегетативно-репродуктивных муль- тирепродуктивных побегах, периодически развивающих межсоцветия) однодомные желто- розоцветковые каудексово-подушковидные кустарнички («подушка» взрослого растения может возвышаться над поверхностью почвы от 5 до 30 см).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 311. Биографический портрет Plantago arborescens Poir. [сем. Plantaginaceae (Израиль, Иерусалим, Ботанический сад, средиземноморский отдел, 25 IV)j. Биоморфотип: надземка-двусемядольные (семядоли линейные, 6 — 8 мм дл., 0,6— 1 мм шир., заостренные, сидячие) четочные мутовчато-простолистные эумоноподиальные стержнекорневые индетерминатно-репродуцирующие (интеркалярные безлистные побеги образуются по 2-4 в узле, составляя вместе межсоцветие мультирепродуктивного побега, который за один вегетационный сезон может дать два прироста и даже два межсоцветия) однодомные (обоеполые цветки собраны на верхушках интеркалярных побегов в семиде- терминатных брактеозных коротких колосьях) полегающе-стелющиеся кустарнички (до 50 см высотой).
Inasa 5. Концепция разнообразия биоморф семенных растений Рис. 312. Биографический портрет Euryops pectinatus Сова. [сем. Asteraceae, там же, что и рис. 31~, но южно-африканский отдел, 2 IV)]. Биомодфотип: надземно-двусемядольные (семядоли эллиптические, 5 — 7 мм дл., 2,5 — 3,5 мм шир., широкозакрутленные, клиновидно суженные в короткие черешки) четочные гемисимподиальные очередно-простолистные семидетерминатно-репродуциру- ~оШие (верхушечные главные полузакрытые брактеозные соцветия-корзинки и подобные им соцветия у паракладиев развиваются вместе в структуре синфлоресценций универсаль- ных сложных ди-, плейофазных побегов) стержнекорневые одномные желто-астроцветко- вые полегающе-стелющиеся кустарнички (до 1 м высотой).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 313. Биографический портрет Агйгоспетит macrostachyum (Мопс.) Moris [сем. Chenopodiaceae (Израиль, каменистая пустыня Негев, окрестно- сти поселка Сде-Бокер, 16 V)]. Др — фрагмент цветущего побега (Zohary, 1966). БИОМОрфОТИП: надземка-двусемядольные (семядоли линейные, мясистые, заострен- ные, сидячие, 6 — 10 мм дл., 0,8 — 1,5 мм шир.) безрозеточные редукционно-простоли- стные (поначалу у молодых растений развертываются листья, похожие на семядоли, у которых нормально развиваются мясистые простые пластинки, но с возрастанием сухости и температуры воздуха рост замедляется и буквально на глазах происходит редукция в развитии листьев, вплоть до того, что на месте простых листьев возникают только листовые бугорки) эусимподиальные побегово-акронекрозные индетерминатно-репродуцирующие раздельнополо-однодомные (на одном растении, более того, в одном открытом верхушеч- ном соцветии развиваются и тычиночные, и пестичные цветки) стержнекорневые сукку- лентные полегающие кустарнички (до 1 м высотой) или кустарники (более 1 м высотой). Вся разница между типами биоморф в этом случае зависит от положения растений по отношению к рельефу каменистой пустыни.
Ь Глава 5. Концепция разнообразия биоморф семенных растении 6(4 21 1,а 4,В 206 Рис. 314. Биографический портрет Rhododendron ledebourii Pojark. сем. Ericaceae (Алтай, Куминс хре кий бет, сосновые горные леса, скалистые склоны)]. е се оли яйцевидные, 3,5 — 4,5 мм дл., 2 БИОМОрфОТИП: надземка-двусемядольные (семяд е зко с енные в короткие черешки) безрозеточные оче- — 2,2 мм шир., закругленные, резко суженные в кор имовки сохраняясь еные листья сменяются после перезим редно-простолистные вечнозеленые ( е етерминатные и во вто ом вегетационном сезоне) гемисимподиальные де некоторое время и во втором вегет й з латентных т.е. перезимовавших, (репродуктивные побеги р р азве тываются рано весно из латентн бе олжая собой моноподиаль- верхушечных почек прошл д ого них вегетативных побегов, продолжая бе и отличаются высокой спе- ный отрезок жизнеразвития ун р иве сальных простых по гов,и о л каждый только из одного цветка и и нескольких, сильно циализацией, поскольку состоят ки <д или низовых, чешуй) ем и азверн ых на компактной оси, почечных, или ни прижатых к нему и развернут подиальные однодомные розово-ве- рыхло- или полураз р уч ку азве н о стистые эпигеогенно-геоподиаль овременно развертывающиеся в ресковоцветковые (все вместе руп к ные цветки, почти одноврем й б - ерхсоцветие, созда- ющий неповторимый розовый колорит рододендровых горных склонов
ОСНОВБ1 БИОМОРФОЛОГИИ CENEHHblX РАСТЕНИЯ О Рис. 315 Био а~ гр фический портрет Lavandula stoechas L. [сем. Lamiaceae (Тель- Авив, Ботанический сад, 28 Ш)]. БИОМОрфОТИП: н земно- ад но-двусемядольные (семядоли округло-яйцевидные, 2,5 — 3,5 MM дл., 2 — 3,5 мм ши . ши окоза р., ирокозакрутленные, сердцевидные, на коротких черешках) ~e3ðî- зеточные (такие побеги домини ( доминируют, составляя основной прирост вегетативной массь~ растений) нерегулярно-розеткообразующие (розеточные вегетативные побеги могут «ра зовываться п и нетении ос р уг ии роста, а когда условия для роста благоприятные, то формиру- ются длинные без озеточные или р чные или полурозеточные побеги) супротивно-простолистные стер- жнекорневые гемисимподиальные диальные семидетерминатно-репродуцирующие однодомные фи- олетово-яснотковоцветковые прямостоячие кустарники.
Глава 5. Концепция разнообразия биоморф семенных растений НИКИ.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 1,аг1(аг4 м1(гм4), ,206,26 Рис. 318. Взрослая биоморфа Ргаа1ит majus L. [сем. Lamiaceae (окрестности Тель-Авива, каменисто-песчаные холмы, среди зарослей кустарников, 21 Ш)]. Биоморфотип: надземка-двусемядольные безрозеточные супротивно-простолистные э симподиальные индетерминатно-репродуцирующие стержн р жнеко невые однодомные бело- эусимп д яснотковоцветковые листопадные изгибисто-аэроподиальные кус р ста ники (плети аэропо- о-таки стелются по чужим кронам диев налегают на аэроподии других кустарников, прямо-таки стел себя ~на ч ом горбу» осваивают или пронизывают их, и так, довольно продуктивно для, уж окружающее надземное пространство). Рис. 319. Биографический портрет Myrtus communis L. [сем. Myrtaceae (там же, что и рис.315, 10 IV)]. 0,34 Биоморфотип: надземка-двусе- мядольные (семядоли яйцевидные, слегка островатые, 6 — 7 мм дл., 2,5 — 3 мм шир., клиновидно су- женные в короткие черешки} безро- зеточные супротивно-простолист- ные стержнекорневые зимнезеленые эумоноподиальные интеркалярно- репродуцирующие (интеркалярные тотарепродуктивные побеги образу- ют все вместе межсоцветие годично- го побега, а поскольку нарастание репродуктивно-ростовых побегов эумоноподиальное, то универсаль- ные побеги образуют межсоцветия в своей многолетней плейофазной жизни неоднократно) однодомные белоцветковые кустарники.
Глава 5. Кониепиия разнообразия биоморф семенных растений 06, Рис. 320. Биографический портрет РИ1угеа latifolia L. [сем. Oleaceae (ь1зра- иль, горные склоны, заросшие кустарниками, вдоль дороги между XagtepoA и Хайфой, 16 1У)]. ОМОР~ОТИП. надземно-двусемядольные (семядоли эллиптические, 8 — 15 мм дл., 4— 7мм ши шир., слегка заостренные, клиновидно суженные в короткие черешки) безрозеточные супротивно-простолистные стержнекорневые постинтеркалярно-репродуцирующие эумо- ноподиальные (когда отдельные ветви все-таки перевершиниваются, то происходит это вынужденно по каким-либо причинам, которые нарушают эумоноподиальность нараста- ния , но не отменяют ее) однодомные зеленовато-масличноцветковые вечнозеленые (кожи- стые листья сохраняют жизнеспособность до двух лет) кустарники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 321. Биографический портрет Lonicera etrusca Santi [сем. Caprifoliaceae (там же, что и рис. 315, 28 IV)]. БиоморфотиП: надземка-двусемядольные (семядоли эллиптические, немного яйцевид- ные, 5 — 7 мм дя., 3 — 4 мм шир., клиновидно суженные в короткие черешки) безрозе- точные супротивно-простолистные летнезеленые гемисимподиальные семидетерминатно- репродуцирующие однодомные белоцветковые лиановые кустарники.
паеа 5. Концепция разнообразия биоморф семенных растений 32,33а, 7,аг2, 74а,206,26 1,гм4, 1ба,20, 24,26 б,гм4, /20в,27 М,аг4, 206,2 Л 1 i(r r w»r - l~ ! ПП ( ''7 ~ф г г~~ 4 30а / + ° ° ° ° ° Л L( С ~1 ( ф ~J Рис. 322. Биографический портрет Atragene sibirica L. [сем. Ranunculaceae (Ал- тай, Чергинский и Ануйский хребты, лесной пояс, кустарниковые заросли)). БноМорфотИПы: безрозеточные супротивно-полусложнолистные летнезеленые эусим- подиально-гемисимподиальные постинтеркалярно-репродуцирующие однодомные желто- л~отиковоцветковые лиановые кустарнички (высота аэроподиев не более 1 м — ! биомор- Фотип) или лиановые кустарники (высота тонких аэроподиев 1 — 3 м и даже более И биоморфотип).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ 1,агЗ, 14а,18а,26 Рис. 323. Биографический портрет Berberis sibirica Pall. [сем. Berberidaceae (Алтай, Теректинский хребет, лесной пояс и высокогорья, южные склоны)]. БИоморфотИП: надземка-двусемядольные (семядоли эллиптические, 8 — 12 мм дл., 4— 6 мм шир., закругленные, иногда с небольшой выемкой, клиновидно суженные в короткие черешки) розеткообразующие гемисимподиальные стержнекорневые детерминатно-репро- дуцирующие (вегетативно-репродуктивные розеточные весенние ди-, плейофазные побеги с одиночными верхушечными цветками на безлистных коротких цветоносах закрывают моноподиальное брахиморфное, специализированное на ассимиляцию и репродуктивное размножение, развитие универсально-простых побегов, или тетракормусов; часть вегета- тивных розеточных побегов весной проявляет не репродуктивное, а вегетативное усиление в развитии и вместо розеточных вегетативно-репродуктивных побегов развиваются веге- тативные полурозеточные магимодифицированно-статурные побеги, у которых вместо листьев с прилистниками развиваются колючки) однодомные колючие невысокие (около 1 м высотой) кустарники.
Глава 5. Концеиция азнооб азия биомор семенных растении 1а Р 324. Некоторые схематические особенности общ тру ту б ей с к ры взрослой биомо ы Capparis aegyptiaca Lam. [сем. Саррагасеае ( зр ис.. е е Из аиль, каменистая иомор ы ар ка С е-Боке, крутой склон каньона, пустыня Негев, окрестности поселка де- окер, ачал мая 1 обрыв к руслу временной реки, уже пересохшей к нач у без озеточные очередно-простолистные ОИОМОрфОТИПЫ: нвдземно-двусемядольные безр ые и ете минатно-репродуцирующие (в структуре акро- эусимподиальные стержнекорневые индетер некрозных вегетативно-репроду~ р стивных азветвленных годичных по егов, или и б я нередко не во всех со ютия у которых цветки образуются не развиваются дизъюнктивные межсощ, у р, атем продолжа- ног а неоднократными, перерывами, а за пазухах листьев, а с некоторыми, иногда д р бело цветковые ки побега) побегово-стелющиеся однодомные лоцвет ~~ ~~~ (~~~~ o o ~ен~~ 1 M). кустарники (длина аэроподиев более 1 м) или кустарнички длина а
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЯ Рис. 325. Некоторые особенности организации взрослого полупаразитного кустарника Loranthus acaciae Zucc. [сем. Loranthaceae (Израиль, долина в Эйлатских горах, 7 V). р — дополнительный рисунок из работы В. Тролля (Troll, 1943, с. 2602), на котором отражены некоторые особенности нападе- ния полупаразитных юных особей «Loranthus» на автотрофные растения (по Goebel). Дополнительные обозначения: кп — корни с присосками, рп- растение-полупаразит, рх — растение-хозяин. EHOMO~OTHII: безрозеточные непостоянно-супротивнолистные зимнезелено-простолистные индетерминатно-репродуцирующие однодомные красноцветковые полупаразитные кустарники (в нашем примере биоморфа L. acaciae поселилась на кустарник Ochradenus baccatus 0е~М и все деревья рода Acacia Hill, произраставшие поблизости, уже были поражены лорантусом)
Глава 5. Концепция разнообразия биоморф семенных растений 2, 26 2, Рис. 326. Биографический портрет Thymelaea hirsuta (L.) Endl. [сем. Thymelaeaceae (Израиль, окрестности Тель-Авива, каменисто-песчаные хол- мы, 27 IV; каменистая пустыня Негев, овраги, окрестности поселка Сде- Бокер, 14 V)]. BHOMOp@OTHII: надземка-двусемядольные (семядоли эллиптические, 3,5 — 5 мм дл., 2,5 — 3 мм шир., закругленные, сидячие) четочные (безрозеточно-сближенноузловые) оче- редно-простолистные эумоноподиально-ростовые детерминатно-репродуцирующие (одиноч- ные цветки закрывают сверху развитие многочисленных мелких и средних репродуктивно- ассимиляционных побегов, которые образуются ежегодно на специализированных ростовых побегах, обеспечивающих персистентный прирост растениям в целом) раздельнополоцвет- ковые однодомные (цветки, хотя и разнополые, но развиваются в структуре одного расте- ния) белоцветковые зимнезеленые кустарнички ( 1 биоморфотип — аэроподии до 1 м высотой, на открытых склонах) или кустарники (П биоморфотип — аэроподии высокие от 1 ао 3 м высотой, пониженные места в рельефе каменистой пустыни). Особенности рельефа оказывают решающее значение при развитии кустарничков или кустарников у Т. hirsuta.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЯ Рис. 327. Биографический портрет Persea indica (L.) Spreng. [сем. Lauraceae (Израиль, Иерусалим, Ботанический сад, средиземноморский отдел, 25 N)] БИОМорфотиП: подземно-двусемядольные (прорастание подземное, семядоли не осво- бождаются от семенной кожуры и остаются в почве) безрозеточные очередно-простоли- стные некустистые интеркалярно-инфлоресцентные (интеркалярные тотарепродуктивные побеги составляют межсоцветия, которые образуются силлептически в сезон развития ~« родительских побегов, цветут, вероятнее всего, в том же сезоне, а вот плодоносят тол~~~ на следующий год весной) однодомные вечнозеленые деревья.
удава 5. Концепция разнообразия биоморф семенных растений рис. 328. Биографический портрет Quercus calliprinos Webb [сем. Fagaceae (Израиль, Ботанический сад в Тель-Авиве, 5 N)). БИоморфотип: подземно-двусемядольные (желуди прорастают весной и долго сохраня- ются у всходов) безрозеточные очередно-простолистные стержнекорневые эумоноподиаль- HbIe и вынужденно-гемисимподиальные (нередко после нескольких лет моноподиального нарастания верхушечная почка по разным причинам замирает) постинтеркалярно-репро- дуцирующие (на годичных побегах прошлого года развиваются защищенные почки, вер- »ие из которых весной развертываются в тотарепродуктивные постинтеркалярные побеги с однопалыми брактеозными полузакрытыми соцветиями-сережками) однодомные раз- дельнополоцветковые (тычиночные сережки отцветают и осыпаются быстро, а желуди созревают медленно на протяжении всего лета) вечнозеленые деревья.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 329. Некоторые особенности организации взрослых биоморф Solanum incanum L. [сем. Solanaceae (там же, что и рис. 328, 25 IV)). Др — фрагмент цветущего побега (Feinbrum-Dothan, 1977). БИомодфотИП: надземка-двусемядольные безрозеточные очередно-простолистные ге- мисимподиапьные детерминатно-репродуцирующие однодомные фиолетовоцветковые ли- стопадные некустистые невысокие (2 — 4 м) деревца. В естественных условиях произра- стания, вероятно, биоморфы S. incanum кустятся и развиваются как кустарники.
rëàâà 5. Концепция разнообразия биоморф семенных растений рис. 330. Биографический портрет Егуйгцца 1еяя(етоп Hutch [сем. Fabaceae (израиль, территория университета в Тель-Авиве, скверы и парки, апрель, май)). БИоморфотИП: подземно-двусемядольные розеткообразующие (на фоне образования безрозеточных и верхнерозеточных побегов периодически развиваются розеточные побеги) ювенильно-простолистные взросло-сложнолистные (трехлисточковые) доминирующе- эумоноподиальные интеркалярно-репродуцирующие (интеркалярные побеги с кистями ярко-красных цветков в изобилии образуются еще до развертывания зеленых листьев, используя в полной мере эффект единого красочного сверхсоцветия для успешной реали- зации полового размножения) однодомные красно-бобовоцветковые листопадные деревья.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 331. Биографический порт- рет Albizia julibrissin Durazz. [сем. Fabaceae (там же, где и рис.330, 25 V)]. EHOMOp@OTHII: надземка-двусемя- дольные (семядоли овальные, слегка зак- рутленные, 8 — 12 мм дл., 4 — 6 мм шир., глубокосердцевидные, по краю с лопастя- ми-рожками, на коротких черешках) без- розеточные очередно-сложнолистные эусимподиальные семидетерминатно-реп- родуцирующие однодомные желто-бобо- воцветковые листопадные деревья. Рис. 332. Биографи- ческий портрет Robinia pseudoacacia 1 . [сем. Fabaceae (Москва, парки и скверы)]. Биоморфотип: надзем- но-двусемядольные (семя- доли эллиптические, 12— 16 мм дл., 6 — 8 мм шир., закрутленные, резко сужен- ные в короткие черешки) безрозеточные очередно- сложнолистные эусимподи- альные интеркалярно-реп- родуцирующие (каждый ин- теркалярный тотарепродук- тивный побег с брактеоз- ным полузакрытым верху- шечным соцветием — кис- тью, образуется в пазухах срединных листьев роди- тельского побега и вместе с ними и стеблевой частью этого побега составляет межсоцветие вегетативно- репродуктивного годичного побега) однодомные бело- бобовоцветковые деревья (1биоморфотип) или кус- тарники (11 биоморфотип, когда растения кустятся и сравнительно невысокие- до Зм).
Глава 6. Концепция пластичности и гиперпластичности биоморф семенных растений 6.1. Стратегическое и тактическое в организации биоморф Биоморфы и все их живые части адаптированы (приспособлены) к жизни на Земле, пройдя длительный исторический путь эволюции на основе естественного отбора адаптаций. Они существуют во власти постоянных дискретных трансформаций живой материи, в первую оче- редь, адаптаций и, по большому счету, сами являются формами адап- тации (адаптивными формами). Биоморфы, будучи, кроме того, еще и одной из форм ресурсов на Земле, вовлечены жизнью в свой круговорот. Они существуют не сами по себе, а в системе круговорота ресурсов, который в их организации осуществляется по схеме: управление + по- требление ~ производство ~ резервирование + рассеивание (т. 1, гл. 4). Одной из сторон круговорота ресурсов является общий обмен веществ «между организмом и средой (внешний обмен веществ), а также транс- порт и превращения веществ в организме (внутренний обмен, промежу- точный обмен)» (Либберт, 1976, с. 16). Подавляющее большинство указанных процессов осуществляется в эндосоме растительных биоморф. В ней фенетически реализуются пред- наследственные и наследственные свойства биоморф, возбудителем в Главное, что должны проявлять живые тела, начиная с первых мгновений своего бытия, — это умение бороться и стремление учиться бороться за жизнь еще лучше, чем раньше, а под определенным углом зрения — это не что иное, как борьба за пространство и время. В круговороте ресурсов растительные биоморфы распространились почти везде на территории Земли. При всем этом «нет ни одного растения или животного, которое могло бы переносить все условия, существующие на Земле. Каждый организм процветает в относительно узком диапазоне температур, количества осадков, почвенных условий и других факторов среды.... найти два вида, которые бы существовали в совершенно оди- наковых условиях, практически невозможно» (Риклефс, 1979, с. 176).
ОСНОВБ/ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ развертывании которых является внешняя среда. В биоморфах не только обеспечивается движение ресурсов, свойственное и неживым телам, например, движение воды при испарении-транспирации, но и принци- пиально новые формы движения ресурсов, которые могут обеспечить только живые тела, например, участие воды, наряду с другими вещества- ми — СО„О, в сложных метаболитических процессах на основе соот- ветствующих внутриклеточных живых структур — органелл — зеленых пластид, или хлоропластов, содержащих зеленые пигменты хлорофиллы и желтые пигменты каратиноиды. В хлоропластах (центрах превращения энергии) вершится великое таинство Природы — процесс фотосинтеза, в результате которого солнечная энергия превращается в химическую энергию ассимилятов (в основном, глюкозу). Наиболее распространенные на Земле неорганические соединения: двуокись углерода и вода — соединяются в известной химической ре- акции фотосинтеза: 6СО, + 6Н,О ~ энергия С,Н„О, + 60,. Одного лишь наличия всех ингредиентов фотосинтеза для его протекания в природной среде недостаточно. «В земной атмосфере содержатся дву- окись углерода и водяные пары, и она освещена Солнцем, однако на нас не изливаются с неба потоки сахара. Вероятность возникновения соот- ветствующей химической реакции очень мала. В клетках растений со- держатся особые пигменты (например, хлорофиллы и каротиноиды) и ферменты, обеспечивающие такое взаимодействие между молекулами и энергией, при котором вероятность возникновения химических реакций фотосинтеза весьма велика» (Риклефс, 1979, с. 113). Эндосома в организации всех зеленых растений обеспечивает иден- тичное прохождение реакции фотосинтеза. «Фабрику» фотосинтеза- эндосому прикрывает сверху поверхность тела биоморфы, которая и составляет то особое состояние этого живого материального тела, опре- делвюдтее ему наружную границу от остальной части пространства,— экзосому, или габитус. Габитус и эндосома, включаящая наряду с гено- сомой еще и эндофеносому — субгабитус, связаны в организации любой биоморфы неразрывно, поэтому рост и развитие, составляющие главное содержание поведения эндосомы находят свое отражение в габитусе биоморфы, поскольку только эта меросома непосредственно контакти- рует с внешней средой, принимая первая на себя ее влияние. Только габитус обеспечивает передачу эндосоме, с которой он не имеет границ, всех наружных «требований» от окружающей биоморфу среды, на кото- рые биоморфа обязана, чтобы жить, реагировать, поскольку она сама существует в этом мире как адаптация (одна из ее форм) и одновремен- но — как система адаптаций. Процесс потребления связан с другими процессами, обеспечивающими биоморфам круговорот ресурсов, т.е. с производством, резервированием и рассеиванием ресурсов. Габитус часто по цвету в течение года бывает неоднороден, по- скольку проявляет в себе быструю реакцию всей биоморфы на смену времен года, на погодные условия, даже на смену дня и ночи. Сезонная изменчивость условий жизни биоморф приводит их к фенологической изменчивости. Габитус многих семенных растений приобретает и сохра- няет зеленый цвет только весной и (или) летом, а затем его утрачивает
Глава б. Концепция пластичности и гиперпаастичности биоморф семенных растений 493 (листопадные летнезеленые растения). Другие растения сохраняют зеле- ный надземный габитус круглый год, сменяя старую листву, они забла- говременно восстанавливают новую (летнезимнезеленые и вечнозеле- ные растения). Таким образом, цвет габитуса надземной части тела семенных растений обусловлен чаще всего присутствием в листьях и (или) стеблях побегов зеленого пигмента хлорофилла, который габитусу не принадлежит; зеленый цвет в габитусе проявляется изнугри, такова его особая роль — свидетельствовать о многом, но часто не за счет своих структур, а используя внутренние структуры, которые он прикрывает, с которыми взаимодействует и которые его к тому же и порождают. Габитус и эндосома существуют ритмически в организации био- морфы, как и сама биоморфа в целом, в зависимости от ритмического существования самой Природы, уже — природы Земли, а еще уже- конкретного биотопа — среды обитания конкретной биоморфы. В зависимости от смены времен годы, погодных условий, дня и ночи и т.п. происходят изменения как во внутренней, так и во внешней организации биоморф, т.е. любая биоморфа отражает ритмические про- цессы, происходящие в окружающей природе и в ней самой, в первую очередь, — фенологическую и возрастную изменчивость. Биоморфа в цикле своего ритмического существования развивает формы габитуса (габитоформы) и формы субгабитуса (субгабитоформы), которые вместе образуют в их единстве формы роста (стадиосомы) биоморфы. Состо- яния (формы роста) биоморфы сменяются в ее морфогенезе и служат показателем стадийной изменчивости биоморф; однако сама биоморфа, в целом изменяясь, не сменяется до конца своей жизни, сохраняя оп- ределенный качественными границами конкретной биоморфы несменя- емый эгосубстрат, т.е. преднаследственно-наследственный субстрат (континуально-субконтинуальную материальную основу) биоморфы. Габитус биоморф и их формы роста адаптированы к реализации и других процессов круговорота ресурсов, связанных с ростом и развити- ем вегетативно-репродуктивных систем и базовых органов. Эти процес- сы направлены на освоение пространства, на борьбу за свет и воду и другие важные экологические факторы; при этом, как известно, бывает хорошо, когда все в меру, поэтому растения могут не только стремиться, например, к свету и воде, но и защищаться от этих факторов среды, когда они в избытке и т.п. Все, о чем была выше речь, — это тактические средства и формы поведения, которых, безусловно, много больше, поскольку фенетичес- кое проявление жизни весьма многообразно. Однако приведенный выше материал наглядно показывает, где имеет место быть тактика и такти- ческое. «Стратегия подразумевает общий подход, тогда как тактика относится к частностям, деталям» (Гробстайн, 1968, с. 13). Стратегия Жизни, или стратегия управления Жизнью (SB) — это единая для всех биосистем, осознаваемая или не осознаваемая ими, закодированная на молекулярном уровне (в генах) программа управления деятельностью (поведением), по которой жизнь сохраняется на Земле и, вероятнее всего, где-то еще во Вселенной (т. 1, гл. 3). Самая высокая стратегичес- кая цель — это жизнесохранение вообще; она закодирована в генах и
OCHOBbl БИОМОРФОЛОГИИ CEMEHHblX РА СТЕНИЧ представляет сложную программу деятельности биосистем, в частности, биоморф, и всегда состоит из нескольких частей-стратегий. На молекулярном уровне закодирована информация не просто о том, как может биосистема управлять развитием, чтобы участвовать в сохранении своей жизни и жизни вообще, а о том, как биосистема, в нашем случае, биоморфа, может выбирать, каким ресурсным потоком (или потоками) надо управлять так или иначе, здесь и сейчас, какими там и потом, а какими вместе здесь и там, сразу или в разное время. При всем этом не будем забывать, «что наследственный аппарат сам по себе кодирует только синтез специфических белковых молекул, а от- нюдь не признаки целого сформированного организма (т.е. конкретные особенности строения и функционирования тех или иных органов).... Синтез специфических белков в зародышевых клетках и взаимодействие этих клеток друг с другом определяет общее направление морфогенети- ческих процессов... Генотип «задает» лишь общее направление морфогенетических процессов, а условия внешней среды оказь|вают существенное влияние на их осуществление» (Иорданский, 1979, с. 43). Так или иначе, но стратегия управления в организации биоморф, а шире, и других биоси- стем, включает весьма небольшое и вполне конкретное разнообразие частей-стратегий: управление потреблением ресурсов (стратегия потреб- ления, обозначим ее индексом 1S), управление производством ресурсов (стратегия производства, 2S), управление резервированием ресурсов (стратегия резервирования, 3S) и, наконец, управление рассеиванием ресурсов (стратегия рассеивания, 4S). Таким образом, у биосистем единая стратегия Жизни, или страте- гия управления Жизнью, предстает на молекулярном уровне (в генах) как стратегия управления четырьмя, и только четырьмя (!), ресурсными потоками, на основе которых поддерживается, сохраняется и развива- ется жизнь на Земле. Символически стратегию Жизни можно выразить в формуле: SB = 1S + 2S + 3S + 4S. В главных частях-стратегиях при необходимости можно выделить еще и их элементы, т.е. стратегические элементы, например, в стратегии рассеивания как стратегический эле- мент среди прочих присутствует стратегия размножения, один из при- оритетных видов стратегии в организации жизненных форм семенных растений. Биоморфы в процессе эволюции научились проявлять в своей организации как общие для них всех свойства, так и частные, которые зависят от общих свойств, но всегда имеют свою специфику. В такой ситуации мы вправе говорить о стратегическом (наиболее общем, без деталей) и тактическом (приватном, детализированном) в поведении биоморф (т. 1, гл. 3 и 4). Тактика биосистем, в частности, биоморф, находится в прямой зависимости от стратегии этих биосистем и от условий их существова- ния. Тактику биосистемы, уже — тактику биоморфы, можно выразить формулой: ТВ = tlS + t2S + t3S + t4S. С учетом приведенных выше формул сделаем следующее обобщение. Стратегия Жизни в организации биосистем, в частности, биоморф, по существу является стратегией уп- равления ресурсными потоками с генетического уровня (геносомы) и реали-
паеа б. Концепция паастичности и гиперпаастичности биоморф семенных растений 495 зуется через трансформацию (рост и развитие), как по отдельности, так и вместе — в их единстве, фенетического (тактического) содержания жизни на земле в различных тактиках средствами тактического назна- чения (любые организационные части феносомы): тактиках потребления (tlS), тактиках производства (t2$), тактиках резервирования (t3$), так- тиках рассеивания (t4$) ресурсов — это закон (примаспециальный) един- ства стратегии и тактики в организации биосистем. Согласно стратегии потребления, например, в организации боль- шинства биоморф семенных растений осуществляется тактика потреб- ления через разные системы, например, систему зеленых листьев, кото- рые являются средствами тактического назначения и, будучи «накача- ны» хлоропластами, как известно, поглощают солнечную энергию и СО„снабжаются водой и минеральными веществами и, посредством фотосинтеза, трансформируют поглощенные ресурсы, подключая далее тактику производства, образуют О, и пластические (органические) ве- щества. Иногда зеленые листья способны отделяться, и когда они от- деляются от родительского растения, на них изредка могут развиваются придаточные корни и почки. Таким образом, осуществляется вегетатив- ное размножение, т.е., согласно стратегии рассеивания и ее меньшего звена — стратегии размножения, с помощью различных тактических средств (листьев, в нашем случае) проводится в жизнь тактика вегета- тивного размножения (Saintpaulia ionantha Н. Wendl.). Средствами стратегического назначения биоморф являются гены, а вот средствами тактического назначения — различные структурные части феносомы: клетки, ткани, органы, например, побеги, корни, листья, цветки и т.п., которые по-разному специализировались к круговороту и в круговороте ресурсов в процессе длительного исторического развития. В стратегии Жизни, фиксирующей однозначную направленность любой биосистемы на сохранение жизни вообще, суммативно присут- ствуют ее части-стратегии (потребления, производства, резервирования и рассеивания), которые замешаны на доминирующей роли жизнесох- ранительной направленности с обязательным «подмесом» жизнеразру- шительных устремлений, без которых приоритет развития жизни- жизнесохранение — не может проявить себя в должной мере. Важно не забывать, что четыре части-стратегии составляют вместе одну целост- ную систему — стратегию управления Жизнью; это не пятая часть, а интегральная система — целое, которая всегда больше суммы частей и стоит над частями-стратегиями, поддерживая в них примат жизнесохра- нительных целей над жизнеразрушительными, — это стратегия-целое объединяет управление остальными частями-стратегиями. Присутствие жизнеразрушительных свойств в тактических сторонах организации биосистем помогает им «самоочищаться» от перекосов в жизнесохранении, поддерживая дискретный способ существования в любых биосистемах; этот способ потенциально способны преодолеть разумные существа, достигнув способности проявлять нообиофильную суперадаптацию (бесконечное сохранение разумной жизни) (см. т. 1, гл. 3). Сумеют ли и успеют ли разумные существа реализовать заложенную в них потенцию — вопрос открытый. Только при доминировании жиз-
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHblX РА СТЕНИЙ несохранительных свойств над жизнеразрушительными свойствами жизнь может избежать глобальный «перекосов» в своем сохранении на Земле, и будет обеспечено бесконечное торжество Жизни во Вселенной. Сейчас все больше раздаются голоса в поддержку гипотезы панспермии и возможности перемещения очень примитивных форм жизни в состо- янии глубокого анабиоза по сверхгигантским просторам Вселенной, независимо от участия сознательной воли (Разума), а просто в глубине космических тел (комет или астероидов). 6.2. Пластичность и гиперпластичность Кем бы ни была биоморфа, она всегда частица пространства, ко- торая может существовать только дискретно на некотором отрезке нео- братимого времени. Завоевав в пространстве историческое право быть, т.е. существовать в каком-либо временном интервале, любая биоморфа обязана проявить себя борцом за овладение и владение пространством, каких-либо его фрагментов, иначе ей не получится ощутить особенность состояния явления жизнь. Чтобы вся громада элементов систем управления и систем испол- нения и соисполнения (т. 1, гл. 4) исправно работала, нужен механизм (свойство), способный реагировать на постоянно изменяющиеся усло- вия внешней среды. Такое свойство в процессе эволюции биосистем, в нашем случае биоморф, создано еще на первых этапах проявления жизни на Земле; оно постоянно совершенствуется, разрешая противо- речия, возникающие между живыми телами и внешней средой. Весомый аргумент в эволюции биоморф семенных растений — это изначальная способность, доставшаяся им еще от далеких предков, проявлять пла- стичность по отношению к внешней среде и, в первую очередь, к раз- личным частям пространства, окружающим эти биоморфы. «Способ- ность биосистем изменяться при изменении экологических условий, не утрачивая при этом своих качественных границ, — пластичность в широком смысле» (т. 1, гл. 8, с. 362). Однако далеко не все биоморфы могут просуществовать, изменяясь только в границах, которые обеспечиваются их пластичностью. Важная особенность эволюции, что ей всегда мало того, чего она уже достигает; она всегда устремлена в новое, поэтому раньше или позже, но какие- то биоморфы будут обязаны проявить еще большую изощренность в изменении своей организации, став на путь гиперпластичности («сверх- пластичности»), смысл которой уже не в том, чтобы обеспечивать био- морфе изменчивость ее феносоматической части в рамках нормы реак- ции (это обеспечивается пластичностью), а в том, что она позволяет совершиться мугациям, т.е. изменить организацию биоморфы через из- менение на генетическом уровне — в геносоме (генотипе у организмов). Норма реакции биоморфы, как известно, — это наследуемая орга- низмом (шире — и синорганизмом) способность фенотипически (фено- соматически) изменяться под влиянием среды, т.е. «отвечать на дей-
Глава б. Концепция пластичности и гиперпаастичности биоморф семенных растений 497 ствие внешних факторов именно такими, а не иными модификациями» (Гершензон, 1979, с. 248). «Фенотипические различия, вызываемые внешними факторами у одинаковых в наследственном отношении орга- низмов» (шире — синорганизмов; Е. Н.) — это модификации (Гершен- зон, 1979, с. 233), если несколько иначе, то каждая модификация — это фенетическое содержание организации конкретной биоморфы, отлича- ющее таковое от других биоморф. Известно, что наследуются у биоморф не отдельные модификации, а вся норма реакции. «Вся наблюдаемая изменчивость какого-либо признака или свойства в пределах нормы реакции называется феноти- пической. Фенотип — совокупность всех внутренних и внешних струк- тур и функций данной особи (шире — и синорганизма; Е. Н.), разви- вающаяся как один из возможных вариантов реализации нормы реак- ции в определенных условиях» (Яблоков, Юсуфов, 1989, с. 113). На языке генетиков пластичность можно определить как свойство биоморф проявлять модификационную изменчивость в пределах нормы реакции. Гиперпластичность — это способность биоморф (шире — биосис- тем), в нашем случае, биоморф семенных растений, подвергаться мута- циям (генетическим изменениям) и выходить за пределы текущей пластичности. Проявление гиперпластичности приводит к расширению эндоорганизменного и эндосинорганизменного, а в конце концов, и внутривидового полиморфизма, а еще дальше — к возникновению но- вых видов. Гиперпластичность присутствует в организации биоморф до какой-то поры в заблокированном виде; ее блокирует пластичность и позволяет проявиться (разблокироваться) этому скрытому свойству лишь тогда, когда эволюция пробудит его к активности. Способность гипер- пластичности присутствовать в каждой биоморфе, но включаться в работу только тогда, когда возникают соответствующие эволюционные ситуации, которые заставляют биоморфу «вспомнить» об этом свойстве, борясь за сохранение жизни своей и жизни вообще, это то, что каче- ственно отличает гиперпластичность от пластичности, поскольку после- дняя рассчитана только на поддержание генетически предопределенной ранее текущей жизнедеятельности и стремится сохранить сложившиеся качественные границы своей биоморфы. Гиперпластичность биоморф рассчитана на изменение наследственности посредством мутаций (ген- ных, хромосомных, геномных, внеядерных). «Мутации — это внезапно возникающие стойкие изменения генетического аппарата, включающие как переход генов из одного аллельного состояния в другое, так и раз- личные изменения числа и строения хромосом» (Гершензон, 1979, с. 186). Биоморфы многие проблемы своего и видового выживания решают на основе модификационной (фенотипической, а шире — феносомати- ческой) изменчивости, проявляя так пластичность, когда из нормы реакции генотипов выбираются варианты биоморф тех биоморфотипов, которые отвечают соответствующим условиям окружающей среды (рис. 333-335). Однако, иногда нормы реакции генотипа не хватает, и плас- тичность бессильна защищать дальше биоморфу, ставя ее перед выбо- ром: элиминация или переход к гиперпластичности посредством мута- ционной изменчивости. Проявляя гиперпластичность, биоморфы на-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ капливают запас изменчивости, что ведет к перестройке нормы реакций биоморф, результаты которой в процессе естественного отбора или закреп- ляются, или выбраковываются. Гиперпластичность приводит к расшире- нию внутривидового полиморфизма и к возникновению новых видов. Итак, на фоне пластичности у одних и тех же биоморф всегда присутствует еще и защитный для жизни вообще механизм (их гипер- пластичность) на тот случай, когда пластичности будет не хватать в борьбе биоморф за существование и когда, в стремлении организмов или синорганизмов вы:э;ить, они переведут «стрелку» с пластичности на гиперпластичность. Рис. 333-335. Примеры модификационной изменчивости (ее вари- анты выражены разными биоморфотипами и обозначены римскими цифрами) в биографике (на биографических портретах) некоторых ви- дов семенных растений. Рис. 333. Биографика Chrysanthemum coronarium L. [сем. Asteraceae (окре- стности Тель-Авива, каменисто-песчаные холмы, 12 Ш). БИоморфоТИПы: надземка-двусемядольные (семядоли эллиптические, 4 — 6 мм дл., 3 — 4 мм шир., закругленные, сидячие) полурозеточные, редко безрозеточные (у слабых затененных растений) стержнекорневые очередно-полусложнолистные или простолистные (у молодых и слабеньких растений) семидетерминатно-репродуцирующие (полузакрытые соцветия — корзинки занимают верхушечное положение на медиали и латералях главного побега) однодомные желто-астроцветковые сильноросло-верхнеразветвленные (1 биомор- фотип) или слаборосло-неразветвленные (11 биоморфотип) однолетники.
rnasa 6. Концепция паастичности и гиперпаастичности биоморф семенных растений 499 Рис. 334. Биографика Smelowskia calycina (Steph.) С. А. Меу. [сем. Brassicaceae (Алтай, Курайский хребет, высокогорья, щебнистые склоны, осыпи)). БИоморфОТИПЫ: надземно-двусемядольные полурозеточные очередно-простолистные ге- мисимподиальные семидетерминатно-репродуцирующие (брактеозное полузакрытое соцветие — кисть заверщает сверху репродукгивное развитие у плейофазных полурозеточных побегов) однодомные бело-капустнои;ветковые компактно-каудексовые (1 биоморфотип на закреплен- Hblx высокогорных склонах) дефективно-KOHaHB~ailbHHe (несоверщенно BereTaTNBHO pa3 множающиеся) или вынужденно-ризомоидные (11 биоморфотип на незакрепленных высоко- горных склонах — каменисто-щебнистых осьпзях) ретардационно-кондивидуальные (замед- ленно вегетативно размножающиеся пугем образования сначала вынужденно удлиненных каудиций, затем полного расщепления каудекса и главного корня на много обособленных корнепобеговых пар@псул-клонистов, которые сначала сильно сближены и представляют один криптоклональный синорганизм, а далее — ризобазифизарный пучок-клон, состоящий из разбегающихся друг от друга ризомоидных, или постокаудициальных, клонистов, способству- ющих закреплению сыпучего почво-грунта) травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА C TEHHff 21,26 5,гм4, 18а, 22,27 66 Рис. 335. Биографика Leiospora ехсагра (С.А. Меу.) Dvorak [сем. Brassicaceae (Алтай, Курайский хребет, высокогорья, крутые южные склоны, осыпи)). Биоморфотипы: надземка-двусемядольные (семядоли яйцевидные 4,5 — 7 мм дл., 2,5— 4 мм шир., закругленные, клиновидно суженные в черешки 3 — 6 мм дл.) розеточные или верхнерозеточные эумоноподиальные стержнекорневые компактно-каудексовые (I биомор- фотип развивается в условиях относительно спокойного уровня поверхности почвы) или вынужденно длинно-каудексовые (II биоморфотип развивается при довольно высокой под- вижности осыпи, когда ежегодно вместо розеточных побегов отрастают верхнерозеточные побеги, которые посредством удлинения междоузлий своих подземных частей преодолевают последствия засыпания растений наносным почва-грунтом) травянистые многолетники. 6.3. Гомеостаз и гетерокинез Биоморфы всегда готовы к переменам, но при одном принципиаль- ном условии — при поддержании относительного постоянства внутрен- ней среды. Обмен веществ с окружающей внешней средой идет внутрен- ним телом (эндосомой) биоморфы через ее внешнее тело (габитус), составляющих вместе единую организацию и целое тело биоморфы. Габитус прикрывает от воздействий внешней среды внутреннюю орга- низацию биоморфы, в которой, помимо эндосоматических структурных элементов (определенных органелл, клеток, тканей), присутствует еще и окружающая их внутренняя среда — различные водно-минеральные и органические растворы, циркулирующие внутри каждого растительного организма при относительном постоянстве РН, давления (тургора) и других показателей. Способность организмов сохранять относительное постоянство своей внутренней среды в процессе жизнедеятельности наперекор изме- нениям окружающей среды была названа У. Кенноном (Cannon, 1929,
Глава б. Концепция пластичности и гиперпаастичности биомо ф семенных растений 501 1932) гомеостазом (от греч. «homoios» — похожий, одинаковый и «stasis» — состояние). Здесь уместно вспомнить о французском физиологе Клоде Бернаре — создателе концепции постоянства внутренней среды орга- низма, который в лекциях о жизненных явлениях, общих животным и растениям, однозначно заявил: «Я первый стал настаивать на той идее, что для животного есть, собственно, две среды: одна среда внешняя, в которой помещен организм, а другая среда внутренняя, в которой живут элементы тканей» (Бернар, 1878, с. 96). Согласно К. Бернару, в природе проявляется три формы (точнее, способа) жизни: латентная (внешне не проявляется, при этом происходит почти полное подавление обмена, т.е. анабиоз), осциллирующая (это жизнь, полностью зависящая от окружающей внешней среды, например, лягушки впадают в состояние окоченения при наступлении зимних холодов, а у теплокровных живот- ных бывает гибернация, или зимняя спячка; в этой жизни происходят периоды как ее резкого подавления, так и активности), наконец, посто- янная, или свободная, жизнь характерна для животных с наиболее высокой организацией и не прекращающаяся даже при резких измене- ниях условий окружающей среды (цит. по В.П. Нефедову и А.А. Ясай- тису, 1991). Гомеостаз организма у животных и человека, у которых он наиболее полно изучен, обеспечивается работой различных функцио- нальных (выборных) систем по принципу саморегуляции. Организм человека — это, согласно учению И.П. Павлова, система с высокой саморегуляцией, сама себя поддерживающая, восстанавливающаяся, на- правляющая и даже совершенствующаяся. Под постоянством «внутренней среды» организмов обычно подра- зумевают неизменность условий, в которых растут и развиваются раз- личные структурные элементы (клетки, ткани, органы) внутри организ- мов. Абсолютного постоянства внутренней среды как в животных, так и растительных системах, биоморфах, в особенности, не бывает, поэто- му речь может идти только об относительном постоянстве этой среды, поскольку все показатели жизнедеятельности (константы) весьма дина- мичны. Более точно в этом отношении говорить не о гомеостазе, а о гомеокинезе (Ибералл, Мак-Каллок, 1970) как о динамическом взаимо- действии различных жизненно важных показателей внутренней среды, каждый из которых определяется деятельностью специальной функци- ональной системы. Однако, попытки отказаться от термина «гомеостаз» и заменить его другими терминами, например, «гомеокинез», или тер- мином «гомеорез» (Уоддинггон, 1970) пока, как подчеркнул В. Н. Но- восельцев (1991), не привели к искомым результатам. Ученые (медики, физиологи) по-прежнему признают гомеостаз в жизнедеятельности орга- низмов животных и человека, подчеркивая относительность постоян- ства внутренней среды, фиксируя колебательный режим саморегуляции параметров функционирования эндосоматических функциональных систем (Загускин, 1991; Судаков, 1992). Итак, постоянство внутренней среды обусловлено деятельностью различных функциональных систем, результаты которой редко фиксируются «жесткими» величинами — кон- стантами, а чаще оно имеет более или менее колеблющиеся показатели — «пластичные» константы, т.е. стабильно изменяющиеся в довольно
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ строгих пределах (параметрах), например, показатели кровяного давле- ния, температуры, уровень питательных веществ в крови и т.д. Гомеостаз впервые был открыт у организмов. Главная черта орга- низмов, в особенности растительных, по сравнению со многими други- ми живыми телами, в том, что они развивают целостную физическую (вещественную) оболочку, которая надежно отделяет внутреннюю среду организма от окружающей его внешней среды. Ранее (глава 1) мы уже отметили, что все разнообразие живых тел можно свести к двум типам: одноконтурные (с целостной вещественной оболочкой) и многоконтур- ные (без целостной вещественной оболочки). К одноконтурным живым телам относятся не только организмы, но и органеллы, клетки, органы. Кроме того, что не менее важно, все одноконтурные живые тела, кото- рые существуют внутри организма и являются структурными элемента- ми его организации, так или иначе участвуют в сложении внутренней среды организма и в поддержании ее относительного постоянства (го- меостаза). В этой связи не будет большим отклонением от принципов гомеостаза, если в отношении отдельных структурных элементов, фун- кционирующих во внутренней среде организма, мы будем констатиро- вать проявление микрогомеостазов, которые в совокупности и обеспе- чивают целостный гомеостаз. Живые структурные элементы организмов существуют частично или полностью во внутренней среде этих организмов, участвуя в поддержа- нии постоянства этой среды, которая частично или полностью является для них средой обитания — автобиотопом (или аутобиотопом, что одно и то же): у органелл клеток свои автобиотопы, у клеток, тканей, органов — свои. Таким образом, главными носителями гомеостаза являются простые биоморфы — организмы, у живых структурных элементов орга- низмов проявляются микрогомеостазы, а вот в отношении надорганиз- менных живых систем (многоконтурных живых тел), например, синор- ганизмов, популяций, фитоценозов, о гомеостазе в традиционном виде говорить не приходится, поскольку в таких системах работают иные законы отношений между элементами их организации. Разработка концепции гомеостаза в отношении внутренней органи- зации растений еще ждет своих исполнителей, но для нас сегодня важен другой момент в организации биоморф семенных растений, который связан с гомеостазом, но свидетельствует о противоположных сторонах проявления внутренней организации: не о постоянстве ее как среды существования определенных структурных элементов, а, напротив, о непостоянстве в организации биоморфы ее морфогенических, а шире— любых поведенческих, состояний, как состояний роста и развития, как состояний стадийной трансформации этой биоморфы в цикле суще- ствования (морфогенезе), в первую очередь, возрастных и фенологичес- ких состояний. Действительно, какой бы гомеостаз — относительное постоянство внутренней среды обитания структурных элементов про- стых биоморф (организмов), их клеток, тканей, органов — ни обеспе- чивался растительным организмом в целом, другое, не менее важное, что характеризует растительную, а шире — любую, жизнь, — это спо-
Глава б. Концепция пластичности и гиперпаастичности биоморф семенных растений собность живых систем проявлять целостное непостоянство состояний их как форм роста (и развития). Начав с гомеостаза, мы незаметно подошли к явлению, которое пронизывает все существование биосистем, причем всех без исключе- ния, поскольку опирается на особенности основополагающих процессов их организации — рост и развитие, которые рассматриваются в един- стве, через особое свойство и растущих, и развивающихся систем— трансформацию (преобразование). Пока существует живое тело, оно растет и развивается, проявляя состояния трансформации в определен- ные отрезки времени, выходом из которых может быть только другая жизнь (посредством размножения) или смерть. Разрабатывая проблему гомеостаза, исследователи нередко настолько увлекаются самим процес- сом гомеостаза и его результатами, что перестают замечать все осталь- ное, что составляет беспокойную жизнь фенетической организации биоморф. Можно периодически топить печку и (или) открывать фор- точку и добиться очень длительного эффекта искусственного поддержа- ния постоянства внутренней среды в доме. Есть, безусловно, еще свой- ство у биоморф (шире — биосистем), без которого гомеостаз мало чего стоит, и его надо не просто заметить, но и определить место, которое оно занимает в их организации. На мой взгляд, способность биосистем изменяться и поддерживать в этом динамическое непостоянство своей организации должно найти отражение в явлении гетерокинеза (дослов- но с греческого «разное движение»). Если бы гетерокинеза не было, то одно фенетическое состояние, однажды возникнув, должно было продолжаться до конца существова- ния живого тела. Однако, такого в живой природе на Земле не бывает; она, как таковая, не возможна без гетерокинеза. Гетерокинез — это свойство живых систем проявлять непостоянство в поведении, в особен- ности — их росте и развитии (разнообразие этоморфозов, стадийность). Нагляднее всего гетерокинез проявляется именно в стадийном поведе- нии биоморф (шире — биосистем), выражающемся в непостоянстве состояний трансформации как сменяемых во времени состояний роста и развития фенетической организации — стадиосом этих одноконтур- ных или многоконтурных телесных систем (глава 4). Итак, биоморфам семенных растений присуща способность изменяться, проявляя гетеро- кинез, в первую очередь, как структурно-функциональное непостоян- ство сменяющих друт друга во времени возрастных и фенологических состояний (стадийность). У организмов (простых биоморф) гетерокинез действует на фоне гомеостаза и оба эти явления так или иначе «рабо- тают» друг на друга; гетерокинез с генетического уровня заведен на борьбу биоморф за выживание как индивидуумов, в которых развора- чиваются различные события их феносоматической жизни, так и видов, которым принадлежат как соответствующие особи, так и их части— структурные элементы. Гетерокинеэ — это и стаднйность жиэнераэвитин, в чистом виде, но и разнообразие временное и организационное в ее про- явлении, и все другие формы поведения, которые связаны или вовсе не связаны со стадийностью развития, как-либо поддерживающие динамичес- кое непостоянство целостной организации биоморф.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Взаимоотношения гетерокинеза и организма, в котором это явле- ние себя проявляет, на наш взгляд, весьма любопытно и может быть воспринято по-разному. Назовем образно содержимое таких взаимоот- ношений как «парадокс индивидуальности». В организации каждого индивидуума разворачиваются, казалось бы, невероятные события: живет особь и с ней ее фенетическая организация, в которой «бушуют» страсти, возникают, проявляют себя некоторое время, а затем сменяют- ся возрастные состояния (стадиосомы), и так неоднократно на протя- жении всего онтогенеза. «Парадокс индивидуальности» как раз в этой смене возрастных состояний (основе гетерокинеза) на фоне сохранения качественного единства индивидуума: они (состояния) сменяются, а он, индивидуум (носитель этих состояний), не сменяется до конца онтогенеза. Так, проросток — это индивидуум в соответствующем возрастном состо- янии. Такой индивидуум в своей фенетической части, которая марки- рует его, существует как проросток некоторое время, но вскоре он перерождается (сменяется) во всход (другое возрастное состояние), которое далее он, организм, также несет в себе, сохраняя большее, чем проходящие в нем состояния, организационное пространство — призна- ковое пространство индивидуума. Сам индивидуум в целом никаким сменам не подвергается (его преднаследственно-наследственная основа функционирует постоянно), а вот весь основной «спектакль» со смена- ми «декораций» (возрастных и фенологических состояний) — вся «пла- стика» жизни (ее «лепота») разыгрывается в фенетической части того же индивидуума. Таким образом, раскрывается «парадокс индивидуально- сти», который, когда проведены соответствующие логические действия, уже совсем и не воспринимается как парадокс (так в науке бывает часто, когда простое решение долгое время или не могут, или не хотят увидеть). Итак, в организмах (простых биоморфах) два основных показателя фенети- ческой организации — два основных типа поведения: гетерокинез и гомео- стаз. Мультивариантность развития биоморф, о которой мы уже писали и в первом (глава 8) и во втором (глава 1.5.2.) томах — это все следствия проявления этих двух типов поведения в их организации. 6.4. Адаптированность габитуса и форм роста биоморф к пространству и времени Наша монография не учебник. У нас нет права увлекаться повто- рением известного. В поднимаемых здесь проблемах многое уже изве- стно. Автор сознательно усложнил себе ситуацию и на многое известное в области взаимосвязи биосистем, уже — биоморф, и окружающей внешней среды сделаем только отдельные литературные ссылки (Warming, 1909; %аппищ и. Graebner, 1918; Поплавская, 1948; Одум, 1975; Горышина, 1975; Риклефс, 1979; Федоров, Гильманов, 1980; «р- нова, Былова, 1981; Радкевич, 1983 и др.). Безусловно, полностью избежать каких-либо повторений с тем, что когда-то кем-то говорилось, никогда не удастся, но только к этому и не
fnaea б. Концепция паастичности и гиперпаастичности биоморф семенных растений стоит стремиться, поскольку, как мы уже отмечали, ученость — это больше продукт коллективного разума с вкраплениями индивидуально- сти (но именно прозрения и озарения личностей не просто развивают, а революционизируют науку). Если посмотреть на взаимоотношения неживой и живой природы со стороны неживого, то можно было бы по- лучить следующее образное откровение. Итак, перед нами «монолог», которого никогда не было, но который всегда надо уметь слышать. «Жизнь — я, Нежизнь, пользуясь невероятным случаем, привет- ствую Вас, поскольку ценю и уважаю Ваше присутствие в нашем общем теле — планете Земля. Меня восхищает Ваше развитие, но оно меня и настораживает. Да, Вы по многим позициям превосходите меня, но в одном Вы бессильны тягаться со мной, и когда Ваше высшее звено— человечество вступает в третье тысячелетие своей цивилизованной ис- тории, я вынужден напомнить Вам о Вашем месте в организации При- роды. Вы, Жизнь, повязаны со мной, Нежизнью, одной «веревочкой», и нам вместе суждено сосуществовать столько, насколько долго у Вас хватит силенок уживаться со мной и со всеми моими элементами, поскольку иной среды, чем моя, Вы нигде себе не найдете. Я — Ваша первооснова, с меня Вы начались и только мною всегда можете закон- читься. Я без Вас хоть как, хоть куда и хоть сколько..., а вот Вы без меня никто и ничто, Вас просто нет, Вы — только призрак, иллюзия того, что Вы есть, от того, чего без меня быть не может. Все то, что главное для Вас и чем Вы постоянно гордитесь: организационный порядок, преем- ственность в развитии и даже многослойный принцип организации, пришло к Вам от меня, но, естественно, не в чистом виде, а соответ- ствующим образом трансформировавшись, поскольку Вы, как и я, су- ществуете во власти единой для нас обеих всепроникающей природной силы — Эволюции. У Вас, Жизни, есть Разум и Вы обязаны осознавать происходящее и, в первую очередь, цену и меру своей экспансии в меня как среду своего обитания и тогда не придется склеивать понапрасну разбитые горшки и чинить бесполезно порванные знамена цивилиза- ции. Я, Нежизнь, могу принять или не принять формы Вашего пове- дения, поэтому у Вас только одна судьба — всегда считаться со мной, с особенностями моей организации (другого не дано). Со мной можно только дружить, можно, считаясь со мной, даже бороться, — бороться за свое сохранение, больше того, можно даже что-то и разрушать во мне для созидания, для улучшения Вашего бытия и Вашего сохранения вообще, поскольку я многое могу выдержать, но никогда нельзя терять чувство меры. Без него борьба за себя, свое сохранение, бесполезна, даже опасна, поскольку может развиться желание побороть и меня, Вашу первооснову, но это уж слишком, это путь в никуда, — это супе- риллюзия. Такого быть не может, поскольку я есть всегда, и Вы можете быть только со мной с пользой или с ущербом для себя. Я, Нежизнь, готова сотрудничать с Вами, Жизнью, если Вы научи- тесь приспосабливаться (!) ко мне. О, да, — адаптации, вот Ваша суть! Без них Вы не борец, без них Вы — лишь мое второе «Я», а это, уж поверьте мне, для Вас, без Вас, совсем неинтересно. Я уважаю лишь
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ борцов, которые способны сосуществовать со мной и не терять свое «Я», которые способны адаптироваться ко мне и к себе самому, к своим элементам. Больше того, я могу позволить Вам даже поуправлять собой, поскольку уважаю настоящие и потенциальные силы Вашего развития, а их у Вас не занимать. Есть, правда, одно небольшое обстоятельство, с которым Вам надо беспрекословно считаться. У Вас нет права даже пытаться превращать нашу Дружбу в рабство для меня, поскольку главную ценность нашего общего бытия составляет именно моя бесконечная Свобода. Вам надо понять это сейчас, поскольку потом может быть поздно. Вам важно не переступить черту, отделяющую Вас от меня. Есть ситуации, в которых я ничем Вам помочь не смогу... Боритесь, дерзайте, покоряйте, наслаждайтесь и развивайтесь, но Вы, Жизнь, не забывайтесь; я, Нежизнь, на Вашей стороне только тогда, когда Вы меня не только принимаете, но еще и понимаете (!), что Вам без меня никогда не быть». Вот такое абстрактное откровение неживой природы могло иметь место быть, если бы она могла бы его самосто- ятельно как-то выразить. Автор помог неживой природе передать ее состояние во взаимоотношении с нами и вполне можно представить, что именно такая абстракция где-то незримо витает между нами всеми— элементами живой природы и, больше того, еще и во всех нас, так что, похоже, есть основание о сказанном выше почаще задумываться и никогда не забывать. 6.4.1. «Парадокс адаптированности» габитуса в организации биоморф От абстрактных образов и размышлений о них перейдем опять к конкретике организации растительных биоморф; она-то, действительно, способна проявлять чудеса адаптаций в стремлении биоморф покорять и осваивать пространство и время (см. все оригинальные рисунки этой книги). Габитус биоморф сам по себе нежизнеспособен, он, хотя и обладает некоторой (четвертной) жизнеемкостью все же не жизнеактивен, а жизнепассивен, и непосредственно сам не растет и не развивается (см. главы 1 и 3). По сути, габитус проявляет развитие, впрочем, оно, как и рост, опосредованное, т.е. он сам непосредственно не делает эти процессы, косвенно участвуя в них, получая их готовые результаты от кого-то другого. Это другое — внутренняя организация (эндосома), а уже — часть эндосомы, внутренняя фенетическая организация (эндофе- носома, или субгабитус); именно здесь сосредоточена основная жизнен- ная сила (!) биоморф. Эндосома в организации биоморф делает почти все, составляющее содержание явления «жизнь» у этих тел, включая и габитус. Субгабитусу биоморф важно, чтобы над ним и геносомой, разместившейся в нем, была крыша-габитус, которую он производит не для кого-то вообще, а, в первую очередь, для себя и для своего властного
Глава б. Концепция паастичности и гиперпаастичности биоморф семенных растений 5О7 «обитателя» (геносомы), но и для целого — всего тела биоморфы, в котором он, субгабитус, — главное исполняющее «лицо». Поскольку действующий носитель роста и развития в эндосоме — субгабитус ра- стительной биоморфы, то именно в нем происходит пластогенез, иначе процесс «вылепливания» фенетической организации. Главное в габитусе растительных биоморф, как полагают многие, в том, что он — самая лабильная часть их тела. Коль мы взялись доко- паться до сути, причем здесь и сейчас, то, справедливости ради, поста- вим для начала такое утверждение под сомнение, а, кроме того, попы- таемся ответить еще и на вопрос: почему с габитусом все происходит так, а не иначе? Габитус, действительно, очень лабилен (неустойчив) и его лабильность находится в прямой зависимости от изменений окру- жающей среды. Откуда же берется у габитуса лабильность? Способен ли он сам манипулировать собой или за него кто-то другой делает это, а он только безмятежно повторяет телодвижения того другого? Ответ здесь не на поверхности. Он тоньше, чем может показаться поначалу. Как ни странно, но по многим позициям габитус проявляет себя как органи- зационная «часть-бездельник», поскольку во многом, что с ним проис- ходит, его непосредственной заслуги нет: он сам никакой активности не проявляет в своем развитии, сам никуда никак и никогда не растет и не разрастается, ничего он и не образует, ему никуда не надо повернуть- ся или развернуться, ни одну рану он сам не залечит, ни один цветоч- ный бутон не развернет и т.д. и т.п. Но, при всем этом, у габитуса все же есть и рост, и развитие, но это не самостоятельный, а опосредован- ный процесс ростового развития, который непосредственно осуществ- ляет и для себя, и для него другая часть организации биоморф, распо- ложенная сразу под габитусом, — это субгабитус, а полнее — вся внут- ренняя организация (эндосома). Может быть, у кого-то и дрожь по коже пробежит от такого пово- рота событий в биоморфологии. Но все пройдет, и ситуация нормали- зуется, когда рассмотрение проблемы габитуса мы доведем до опреде- ленного финала. У меня на этот счет нет сомнения; пройдет некоторое время и многие скажут: а ведь здесь все так просто и понятно, да и по другому-то и быть не должно. Прекрасно, если так, значит автор рабо- тает не зря. Может создаться впечатление, что субгабитус выращивает габитус на себе, как на грядке выращивается какой-нибудь овощ. Грубовато сказано, но в целом, по сути, правильно. Габитус, действительно, при- крывает собой субгабитус, растет и развивается не сам непосредственно, а это происходит опосредованно, если образно, то его как бы «отращи- вает» и развивает на себе субгабитус. Габитус, по содержанию, меньше даже, чем вся оболочка биоморфы, поскольку включает только ее кон- туры (форму) и поверхность и не имеет внутренней структуры. Габитус сам по себе мало что может, но то, что он делает (это его прерогатива), — жизненно важно и составляет его непосредственное содержание. Главное, для чего габитус нужен биоморфе, — составлять наружную границу этой телесной системы, защищать, хоть самую малость, ее эндосому от внешней окружающей среды, принимать и передавать, хотя
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЯ бы частично, от нее и к ней информацию, соответственно, внутрь и изнутри биоморфы, а все остальное выполняет эндосома. Еще габитус биоморфы, будучи в целом как раз ее внешним обликом, формой тела, отражает собой многое из жизнеразвития внутренней организации, т.е. служит индикатором (и этого у него не отнимешь, этого ему не зани- мать) интегральной организации биоморфы в динамике, что имеет зна- чение при взаимодействии конкретной биоморфы с окружающими ее живыми телами остальной Природы и для понимания всего происходя- щего с этой биоморфой со стороны нас, исследователей, или просто любознательной публики. Для биоморфологов габитус биоморф важнее всего тем, что он повторяет собой (как контур) нюансы прилегающей к нему снизу структуры субгабитуса и участвует во многом, что вместе с субгабитусом, составляет формы поведения биоморфы и, в частности, формы их роста. 6.4.2. Биотопизм и пластогенез в освоении биоморфами пространства и времени Все живое на Земле активно и неотвратимо во времени наступает на пространство, превращая его фрагменты во все новые и новые био- топы. Явление превращения какого-либо фрагмента пространства в зону местопребывания или местожительства (биотоп) биосистемы обозначим как биотопизм. Анализировать разнообразие биотопов на Земле вряд ли уместно в этой работе. Пожалуй, будет достаточно заметить, что заселение всего разнообразия биотопов растительными биоморфами обеспечивается проявлением в организации этих живых систем пластичности и гипер- пластичности, наглядным показателем которых является лабильность феносомы биоморф и, в частности, их габитуса. Выше было показано, что в организации биоморф самая лабильная часть — это их феносома, основа которой располагается в субгабитусе, а небольшая ее часть со- ставляет весь габитус биоморфы. Феносома, таким образом, проявляет свою лабильность одновременно и через субгабитус и габитус. Здесь нас интересует, как растительные биоморфы, проявляя пластичность и ги- перпластичность, покоряют пространство во времени; где-то они про- сто-напросто отнимают в борьбе его фрагменты у других и (или) при- спосабливаются как-то делиться им с третьими биоморфами и т.д. Большого разнообразия достигают биоморфы от взаимодействия с не- живой природой. Весьма разнообразно ведут себя биоморфы в развитии во времени, начиная часто эпиэмбриональную жизнь с одного семени; одни из них «вспыхивают» и вскоре угасают, как спички, в репродук- тивном усилии (эфемеры), а другие, напротив, медленно развиваются, взрослеют и постепенно могут завоевывать большие территории, напри- мер, Phragmites australis (Сач.) Trin. ex Steud., Ficus bengalensis L. Как известно, фенотипы, образующиеся в процессе развития орга- низмов в разных условиях, составляют норму реакции одного и того '@е
паеа б. Концепция паастичности и гиперпаастичности биоморф семенных растений 509 генотипа, которая наследуется, а «фенотипы наследственно обусловле- ны, но как таковые, не наследуются» (Иорданский, 1979, с. 43). Нам часто приходится идти на обобщения за пределы организмов, поэтому не будем вторгаться в занятый термин «норма реакции», а далее, в отношении разнообразия пластосом (феносомы — их частный случай), или конституций, любых тел, развивающихся на основании ресурса возможностей (идиоэнергии), носителем которого является идиосома любых тел, будем применять термин «норма пластогенеза» (конституци- альная, или пластосоматическая, норма). Таким образом, каждое тело, обладая определенной идиоэнергией, запрограммировано на определен- ную норму пластогенеза, поэтому в одной модификации биоморфы, а шире — одной модификации (в известном смысле этого слова — «ви- доизменение») у любых тел, проявляется отдельное событие-маркер, обеспеченное нормой пластогенеза. Под «пластогенезом» будем пони- мать циклический процесс развития пластосоматической организации (конституции) любых тел. Модификация тел — это какое-либо состо- яние тела в данный отрезок времени как результат пластосоматического, часто стадийного, поведения этого тела, точнее — поведения во времени его пластосоматической (уже — феносоматической) части. Каждое тело, попадая в определенные условия среды, продолжает существование на основе своего ресурса возможностей, незримо проходя идиоэнергетичес- кое тестирование, когда определяется в общих чертах направление транс- формации (у биосистем посредством пластичности или гиперпластичнос- ти), конкретным результатом которого является альтернативный отбор, т.е. подбор и выбор альтернативных признаков-потенциалов в соответ- ствующих рядах признаков и далее движение к выбранным признаковым целям, развитие которых, согласно норме аластогенеза, отвечает конк- ретным условиям внешней среды. Когда у биоморф альтернативный отбор не выходит за пределы нормы пластогенеза, то действует пластичность, а, когда проявляются признаки, которые находятся за пределами альтер- нативного отбора, то они свидетельствуют о свершении мутаций и дей- ствии гиперпластичности. Идиоэнергетическое тестирование обязатель- но присутствует, незримо и постоянно (!) оно проходит в каждой био- системе, биоморфе, в частности (шире — в любой телесной системе), определяя пути решения первоочередных задач: кому «быть» (жить), а кому «не быть» (не жить), а, если жить, то кому быть слабым, а кому развиваться относительно нормально или быть сильнее других; опреде- ляется, больше того, кому суждено «купаться» в счастливой жизни (у растений есть свое счастье; см. главу 10), а, может быть, даже и скоро- постижно «захлебнуться» от ее избытка и т.п. Телесные системы всегда развиваются на основе их ресурса возмож- ностей и на каждом фоне внешней среды они, специфическими для них средствами, проходят идиоэнергетическое тестирование, в соответствии с которым из конституциальной нормы (нормы пластогенеза), присущей каждому телу, выбираются, где это необходимо, альтернативные призна- ки, а когда их нет, то ставятся задачи создать их заново, и постоянно корректируется вариант целостного развития этой системы, что иозво- ляет какому-либо телу или находить условия для продолжения своего суще-
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHblX РА СТЕНИЙ ствования в определенном варианте как тела прежнего качества, или не находить соответствующих условий, прекращая существование в прежнем качестве и превращаясь в другое тело или другие тела, — это субабсолют- ный закон пластогенеза. На основе этого закона действуют как следствия два специальных закона: пластичности и гиперпластичности. Если в процессе развития био- системы ее функционирование не выходит за рамки нормы пластогенеза, то в ней действует определенная пластичность (закон пластичности). Если в процессе развития биосистемы пластичности не хватает, то срабатывает мутация или супермутация (иногда их несколько) и возника- ют признаки, ранее для этого типа биосистем неведомые, которые далее могут закрепиться или отбраковаться соответствующим отбором, а биосистема в целом становится на какое-то время гиперпластичной (за- кон гиперпластичности). Под супермутацией мы понимаем изменения наследственной основы (геносомы) биосистемы, которая по сложности своей организации превосходит какую-либо биоморфу. На вопрос: как происходит идиоэнергетическое тестирование и в соответствии с ним альтернативный отбор в признаковом пространстве конкретных биоморф, пока можно больше строить предположений, чем четко объяснять происходящее. Любопытно, что при своем проявлении у биоморф-модификаций только один или немного признаков действи- тельно выступают как модификационные, т.е. как индикаторы соответ- ствующей модификации, а подавляющее большинство признаков у раз- ных модификаций одинаковые. Вероятно, профессионалы-генетики со временем ответят на то, как работает механизм идиоэнергетического тестирования признаков на молекулярном уровне. С позиций внешней морфологии многое понятно: альтернативный отбор в биоморфах опреде- ленных типов идет только по небольшому числу альтернативных признаков одного или нескольких рядов, и зависит это от конкретных биотопических условий, которые влияют на выбор соответствующих признаков. Рассмот- рим несколько примеров (рис. 333-336). Scabiosa argentea L. — скабиоза серебристая (cBM. Dlpsacacaaa). Этот вид автор наблюдал в окрестностях Тель-Авива на холмистой местности, где на каменисто-песчаных почвах произрастали, в основ- ном, представители флоры средиземноморских субтропиков Ближнего Востока. Обнаружены два очень четких типа модификаций — биомор- фотипа (рис. 336). Один тип (травянистые многолетники) образуют био- морфы, развитые на открытых местах склонов невысоких холмов, ближе к их вершинам, а другой тип (полукустарнички) представляют биомор- фы, произрастающие рядом на тех же самых склонах, буквально в не- скольких десятках сантиметров от первого биоморфотипа, и входящи~ в группы растений из нескольких видов. Быть в «группе» pacTHTeJIbHbIM биоморфам в условиях сухого субтропического или аридного климата бывает выгодно; в них формируется особый микроклимат, позволяю- щий биоморфам многих видов не просто выживать, а развиваться зна- чительно лучше, чем на открытых местах в одиночестве. Полукустар- нички нашей скабиозы неплохо растут и развиваются в «группе», неред- ко прижавшись к другим растениям настолько, что под их кронами
паеа б. Концепция паастичности и гиперпаастичности биоморф семенных растений 511 действительно образуется иной «мирок», в котором и влаги, да, как ни странно звучит, и пищи побольше. S. argentea уютно себя чувствует в таких группах с Thymelaea hirsuta (L.) Endl., Cistus salviifolius L., Sarcopoterium spinosum (L.), Spach, Phagnalon rupestre (L.) DC., Coridothymus capitatus (L.), Reichenb., когда все растения не превышают 0,5 м в высоту, поскольку среди видов этой группы есть такие, биоморфы которых в других условиях и других со- обществах могут быть значительно выше (см. биографические портреты этих видов в нашей книге). Поначалу развитие биоморф разных типов мало чем отличается и, если они вьи<ивают в самом труднопроходимом для них возрасте, в преювенильной и ювенильной стадиях, то далее закрепляются в сооб- ществах и начинают заметнее проявлять отличительные признаки соот- ветствукнцих биоморфотипов. Прорастание семян надземное; оно происходит зимой или весной. Семядоли яйцевидные или широкояйцевидные, на верхушке округлые, нередко слегка выемчатые, при основании округло-клиновидные, го- лые, 3,5 — 6 мм дл., 3 — 4,5 мм шир.; черешки семядолей очень короткие (до 2 мм дл.). На первом году вегетации развивается розеточ- ный вегетативный медиальный (главный) побег. Первые листья, будучи в розетке, супротивные, ланцетные, опушенные, цельнокрайние; с 3— 4-й пары начинает с верхушек проявляться зубчатость краев листьев, постепенно их линейные размеры увеличиваются с 1,0 — 2,5 см дл., 0,25 — 0,4 см шир. у первых листьев и далее по возрастающей, например, розеточные листья четвертой пары, начавшие проявлять сверху зубча- тость краев 3 — 5 см дл., 0,3 — 0,6 см шир. В дальнейшем рассеченность пластинок листьев усиливается. Первое цветение наступает на втором году вегетации, хотя бывают и отклонения (на 1 — 3 году). После первого плодоношения цветонос- ные побеги отмирают на разную длину: до зоны сближенных узлов, находящейся у самой поверхности почвы (открытые места) или на меньшую длину, сохраняя в жизнеспособном состоянии одно — не- сколько удлиненных междоузлий (местообитание в «группе»). Довольно быстро резидные удлиненные части одревесневают и в «группе» развива- ются настоящие полукустарнички с аэроподиями до 20 (30) см (рис. 336). Вся модификационная изменчивость с формированием двух типов биоморф (самое интересное, что прямо-таки рядом друг с другом на одних и тех же склонах) у S. argentea замешана, как, впрочем, атрибу- тивно положено у всех биоморф, на идиоэнергетическом тестировании по множеству альтернативных рядов возможных признаков. Подавляю- щее большинство направлений развертывания фенетических признаков у биоморф разных типов выбирается одинаковое. Обычно лишь один или немного признаков определяют различия в биоморфотипах. В на- шем случае биоморфотипы травянистых многолетников и полукустар- ничков у S. argentea маркируются по общему габитусу и наиболее со- пряженным с ним признакам — особенностями отмирания цветоносных побегов после плодоношения и особенностям резидообразования из них. Продолжительность жизни травянистых многолетников у S. argentea до 20 лет, а у полукустарничков — до 30 лет.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 336. Биографический портрет Scabiosa argentea L.[ceM. Dipsacaceae (ок- рестности Тель-Авива, каменисто-песчаные холмы, апрель)). БИОМОрфОТИПЫ: надземка-двусемядольные (семядоли широкояйцевидные, 4 — 5 мм дл., 3 — 3,5 мм шир., закругленные, клиновидно суженные в чере~шси 2 — 4 мм дл.) полурозе- точные, редко розеточные (зеленые листья могут иногда не развиваться на цветоносной части, например, в условиях самого открытого местопроизрастания) супротивно-полусложнолистные или простолистные гемисимподиальные семидетерминатно-репродуцирующие однодомные бело-ворсянковоцмтковые компакгно-каудексовые травянистые многолетники (1 биоморфо- тип нередко представлен на открытых местах склонов холмов) или полукустарнички (II био- морфотип объединяет обитателей тех же склонов, но они произрастают в «группах», состо- ящих из растений разных видов, с которыми совсем рядом, даже прижавшись друг к дру~7 и образуя аэроподии (!) до 30 см высотой, они успешно переживают летний зной).
Глава б. Концепция пластичности и гиперпаастичности биоморф семенных растений 6.5. Влияние количества и качества частей пространства и границ между ними на соматическое разнообразие биоморф 6.5.1. Растительные биоморфы как части неоднородного пространства На основе отмеченных выше двух великих способностей биоморф семенных (а шире и других) растений (да и не только их, но и живот- ных), завоеванных ими в эволюции (пластичности и гиперпластичнос- ти), они овладели свойством, в зависимости от экологического фона в определенных пределах ресурсов возможностей, манипулировать поло- жением своего тела относительно поверхности почвы. Биоморфы (причем любые), будучи формами ресурсов (т. 1, гл. 4) существуют в пространстве, как и любые другие твердые тела, — как части пространства. Какое-либо живое тело, погруженное в некоторую часть пространства, ассоциируется на какое-то время с другими частями окружающего его пространства, становясь новой частью пространства вообще, и стремится сохранить себя в живом качестве, в борьбе за пространство до тех пор, пока ему удается использовать в своих инте- ресах полезные элементы и сдерживать разрушающее воздействие вред- ных элементов того же самого фрагмента пространства как среды оби- тания этого живого тела. В процессе эволюции растительные биоморфы научились различать части пространства и им не безразлично в какую часть его они проникают и где будут существовать («прозябать», как иногда говорили русские ботаники в позапрошлом веке). Части про- странства неоднородны; они по-разному заполнены материей, которая, находясь только в движении, изменяется, в зависимости от внутренних событий, происходящих в телах и (или) процессах, как элементов про- странства, и от внешних, окружающих эти тела и (или) процессы, факторов. Неоднородность частей пространства выступает важнейшим формообразующим фактором в эволюции. Пространство, в котором однажды возникли растительные биомор- фы и в котором они стали эволюционировать дальше, сразу же «про- диктовало» им несколько общеобязательных условий для продолжения эволюции. Растительные биоморфы должны уметь различать разные формы материального в пространстве: газообразную, жидкую и твердую формы. Все три формы материального в пространстве — это или три основные части пространства, или они входят в отдельные основные его части на правах элементов-соучастников их локальной организации. Так, например, в почво-грунте, составляющем твердую часть простран- ства, на котором и в котором вынуждены развиваться растительные биоморфы, в разной степени присутствуют фрагменты жидкого (водно- го) и газообразного (воздушного) пространств.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHblX РА СТЕНИЧ Все три (воздушные, водные и почвенно-грунтовые) части про- странства находятся во взаимопроникающих отношениях, однако между ними как основными частями пространства, присутствуют более или менее четкие границы. Наличие границ между основными частями зем- ного пространства, безусловно, оказало и продолжает оказывать непос- редственное влияние на эволюцию растительных биоморф. Главных границ между основными частями пространства три: 1 — границы меж- ду воздушными и почвенно-грунтовыми частями, 2 — границы между воздушными и водными частями, наконец, 3 — границы между водны- ми и почвенно-грунтовыми частями пространства. Твердое тело, рассматриваемое в пространстве, будет или полнос- тью, или частично принадлежать какой-либо одной части пространства, поэтому такое твердое тело, в зависимости от положения относительно поверхности почвы принято считать надземным, подземным или при- земным (в последнем случае твердое тело лежит на поверхности почвы, прижимаясь к ней), подводным, надводным или плавающим, т.е. во всех случаях, в названии тел, особенно тел растительных биоморф, традици- онно, по положению в пространстве, превалирует привязка к уровню поверхности водной или почвенной среды. Растительные биоморфы, чтобы существовать в земном пространстве, должны были как-то свя- зать себя со всеми тремя основными его частями. То, как проявляют тела биоморф растений свое отношение к уровню поверхности почвы, выступает в качестве мощнейшего фактора эволюции растительного мира. Прежде чем развить эту мысль, сделаем небольшой логический набросок к рассматриваемой проблеме. Как ведет себя определенное твердое тело относительно лабильно- сти поверхности почвы? Это тело может занять одно из нескольких возможных положений: находиться или на, или над, или под поверхно- стью почвы, или частично погружаться в надземную и подземную части окружающего их пространства. Водную часть пространства (гидросфер- ную часть) и воздушную (надземную) часть пространства (аэросферную, или атмосферную) можно рассматривать и по отдельности, и вместе, но их объединяет то, что они располагаются над поверхностью почво- грунта, т.е. это надповерхностная (эпигеосферная) часть пространства, которая через почвенную поверхность отделяется от подземной (гео- сферной) части пространства. Поверхность почвы (шире — почво-грунта), на которой мы заост- ряем также внимание, является особой, очень специфической частью пространства — это основная промежуточная (контактная) его часть, принадлежащая, в первую очередь, непосредственно почвенно-грунто- вому телу, но, как и габитус тел, она, будучи элементом любого рельефа, не имеет внутренней структуры. Линию, условно проведенную по по- верхности почво-грунта твердого физического тела планеты Земля (твер- дой земной поверхности) в любой ее части и в любом направлении, свидетельствующую об уровне поверхности почво-грунта по отношению к другим частям пространства, включая тела, расположенные на них, в текущих условиях на Земле, где жизнь как-либо, но обязательно, при- вязана к земной поверхности — поверхности почво-грунта, можно для
~лава б. Концепция пластичности и гиперпаастичности биоморф семенных растений 515 краткости обозначить словэсочетанием «линия жизни», или «витали- ния». Новый термин «виталиния» мы используем в качестве рабочего инструмента при исследовании проблемы взаимоотношения частей пространства между собой и с телами, составляющими предмет непос- редственного исследования, в нашем случае, телами растительных био- морф. Какое бы место (среду) жизни (обитания), или биотоп, на Земле ни занимала биоморфа, ее непосредственное положение желательно при исследовании спроецировать в пространстве, а это лучше всего сделать по отношению к виталинии. Все биосистемы, какой бы сложности организации они ни достигли, так или иначе (непосредственно или опосредованно) связаны с уровнем поверхности почвы (шире — почво- грунта), т.е. с виталинией. Части тел растительных организмов характе- ризуются определенным образом, в зависимости от положения по отно- шению к поверхности почвы, что можно выразить так: аэросоматичес- кие, или надземные (распространяются надземно), гидросоматические, или подводные (погружены в воду), геосоматические, или подземные (распространяются подземно), парагеосоматические, или приповерхно- стные (распространяются приземно по поверхности почвы) части. Постепенно мы приближаемся к осознанию факта, что раститель- ные тела в процессе длительной эволюции приобрели свойство адапти- роваться к условиям жизни на Земле в прямой и опосредованной зави- симости от положения их по отношению к уровню твердой земной поверхности (виталинии), который склонен изменяться в пространстве и во времени, т.е. организация биоморф постоянно находится в усло- виях виталинейного адаптациогенеза (виталинейности). Это явление ха- рактеризуется тем, что поверхность почво-грунта, проявляя свою ла- бильность, как-либо влияет на организацию живых тел и их эволюцию, т.е. оказывает прямое или опосредованное воздействие на жизнеразви- тие на Земле, но и получает себе в ответ некоторую обратную реакцию со стороны живых тел, по-своему влияющих и на развитие земной поверхности, а в результате такого взаимодействия частей пространства происходит освоение биосистемой некоторого локального пространства и развитие его в ней и ее биотопе. Иными словами, виталинейность- это атрибутивная непосредственная или опосредованная привязка всех обитателей Земли (живь~х тел разной сложности организации) к уровню поверхности почва-грунта (виталинии) при соответствующем влиянии их друг на друга. Виталинейность — важнейший рабочий инструмент в ис- следовании организации биоморф семенных растений, при построении клас- сификации и моделировании различных явлений. Биоморфы, безусловно, будучи живыми телами, — активные соучастники явления виталинейно- сти и на трансформацию поверхности почвы они отвечают своими адаптациями (рис. 337-339). Есть смысл, исходя из приведенного выше материала, построить сериальную классификацию растительных биоморф по особенностям локализации частей их тела (меросом) в частях пространства, т.е. с учетом виталинейности: 1) аэросоматики, или аэросоматические растения (их побеговая часть развивается только надземно, не прижимаясь к поверхности почвы); 2) аэропарагеосоматики (побеговое тело этих рас-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ тений располагается надземно и приземно, прижавшись к поверхности почвы и нередко постепенно погружаясь в нее); 3) парагеосоматщщ (побеговое тело растений приземное, прижимающееся к поверхности почвы и нередко постепенно погружающееся в нее); 4) аэрогеосоматщщ (побеговое тело растений развивается как надземно, так и подземно); 5) аэрогидрогеосоматики (побеговое тело растений располагается как над- земка, так и подводно и подземно); 6) аэрогидросоматики (побеговое тело растений располагается надземка и подводно; 7) гидрогеосоматики (побеговое тело растений и подводное, и подземное); 8) гидросоматики (побеговое тело плавает в воде). Критерием при определении типа биоморф в этой классификации избрано поведение их побегового тела в освоении ими конкретного типа пространства. 72 Рис. 337. Разнообразное поведение биоморф Frankenia hirsuta L. (сем. Frankeniaceae) в условиях относительно постоянства поверхности почво- грунта (I биоморфотип — длинно-аэроподиальные полукустарники) и в условиях ее эрозионной подвижности (II биоморфотип — вынужденно длин- но-геоподиальные полукустарники при их засыпании песком) на Черных землях (республика Калмыкия).
Глава б. Концепция пластичности и гиперпаастичности биоморф семенных растений Рис. 338. Разнообразное поведение кустистых биоморф Agropyron fragile (Roth) Р.Candargy (сем. Роасеае) в условиях относительного постоянства поверхности почва-грунта (I биоморфотип — компактно- и короткокорне- вищные травянистые многолетники) и в условиях ее эрозионной подвижно- сти (II биоморфотип — вынужденно вертикально-длиннокорневищные тра- вянистые многолетники при их засыпании песком) на Черных землях (рес- публика Калмыкия). Рис.339. Поведение (перемещение вверх зон кущения) биоморф-эфемеров при засыпании их подвижным песком (республика Калмыкия, Черные зем- ли): 1 — Anisantha tectorum (L.) Nevski, II — Eremopyrum triticeum (Gaertn.) Nevski (сем. Роасеае).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ 6.5.2. Сопряженность эволюции габитуса растительных биоморф и их продолжительности жизни с поверхностью почво-грунта Габитус биоморф и земная поверхность — это явления одного рода. Им обоим свойственно прикрывать собой внутреннюю организацию тел, контуры и покров которых они составляют. И габитус биоморф, и земная поверхность = поверхность почво-грунта,'уже почвы) не имеют внутренней структуры и являются специфическими функциями внут- ренней организации тел, к которым они непосредственно принадлежат. По большому счету, габитус растительных и животных биоморф, кон- тактирующих с земной поверхностью, и собственно поверхность почво- грунта, при рассмотрении организации тела планеты Земля сливаются в одно экзоморфологическое целое — габитус Земли (рельеф Земли- это не что иное, как элемент габитуса этой планегы). Так еще никто, пожалуй, не говорил, но, как видно из предыдущих рассуждений, так можно сказать, это вполне уместно, ведь говорят же «лик Земли», а «лик» и «габитус» — это облик, т.е. здесь они, как минимум, близки по смыслу. Габитус Земли, как и все в Природе, эволюционирует, впитывая в себя очень разные результаты эволюции каждой растительной и живот- ной биоморфы и эволюции земной поверхности. В эволюции раститель- ных тел и их габитусов, в частности, большое значение имеет эволюция продолжительности личной жизни и связанных с ней особенностей формообразования основных органов и их частей, что, в первую оче- редь, отражается на сложности организации всех структурных частей. Через призму разнообразия организации биоморф семенных растений нетрудно определить, что на вопрос «быть» или «не быть», эти биомор- фы в процессе длительной эволюции научились отвечать двояко: жить мало (эвриэфемерно) — меньше одного года, или больше одного года (персистентно), причем иногда много — сотни и даже тысячи — лет. Чтобы так жить, биоморфы семенных растений были обязаны в эволю- ции приспособиться не образовывать или, наоборот, образовывать ка- кие-либо базовые (персистентные побеговые) органы, т.е. они были поставлены перед необходимостью выбирать что-то одно из двух основ- ных и принципиальных направлений развития (а другого просто не было дано) — быть эвриэфемерами (эфемерами, однолетниками) или персистентами (жить более одного года, даже иногда до нескольких тысяч лет). Результат эволюции биоморф семенных растений по про- должительности жизни известен — у нас на Земле все основное разно- образие среди них уже давным-давно есть (трудно что-то еще создать, даже придумать трудно по этому ряду признаков); действительно, все есть, от мимолетных эфемеров до дремучих древесных старожилов— деревьев-сверхдолгожителей (см. главу 5). Однако эволюция никогда не останавливается, она всюдна и не имеет конца в бесконечной Природе. Как и сотни миллионов лет назад, эволюция по всем рядам признако~ продолжается, но как и сотни миллионов лет назад, у нее по продол- жительности жизни биоморф одновременно (!) всегда было и всегда
r~asa б. Концепция пластичности и гиперпаагтичности биоморф семенных растений 519 будет только два направления: в сторону эвриэфемерности от персис- тентности, и, наоборот, в сторону персистентности от эвриэфемерности. Преодолевать барьер эвриэфемерности или барьер персистентности- одно из двух принципиальных естественных организационных препят- ствий, которое существует между двумя противоположными направле- ниями эволюционного развития, могут далеко не все биоморфы, а те, которые это могут, обладают особой склонностью к гиперпластичности. По мнению автора, оба противоположных направления эволюции по продолжительности жизни закрепились еще на заре растительной жиз- ни, т.е. много раньше появления вообще биоморф семенных растений, и главная их особенность, что оба направления открыты для поворота в обратную сторону, т.е. они обратимы по рассматриваемому ряду при- знаков. Преодоление барьера эвриэфемерности и барьера персистентно- сти за многие миллионы лет эволюции растительной жизни уже давно перестало быть революционным событием, а однажды став таковым, оно далее неоднократно повторялось, превратившись в весьма суще- ственный, все же не тривиальный, но все-таки лишь важный эволюци- онный акт, апробированный в растительном мире неоднократно. Я готов утверждать, что за многие миллионы лет эволюционный акт преодоления барьеров и эвриэфемерности, и персистентности в различных таксономи- ческих группах уже происходил не раз, поскольку сам принцип преодоления таких барьеров и смены направлений эволюции по продолжительности жизни исторически давно уже сложился в растительном царстве, и гене- тическая «память» о том, что обратимость в эволюции по продолжитель- ности жизни возможна, где-то есть в ресурсе возможностей (идиоэнерге- тическом потенциале) растительного царства. Биоморфы семенных расте- ний способны преодолевать барьеры обратимости в направлениях эво- люции на основе их свойства гиперпластичности, но мутации в таких случаях не обязательно всегда достигают принципиально новых комби- наций генов по продолжительности жизни, нередко только возвращают- ся к давно, казалось утраченным комбинациям, восстанавливающим некогда опробованный далекими предками путь обратной эволюции по продолжительности жизни. Повторение подобных эволюционных актов возможно и в будущем, что составляет атрибутивную особенность, склонного к свободе выбора, явления под названием эволюция. Автор готов предположить, что в процессе длительной эволюции биоморф семенных растений оба направления ее по продолжительности жизни были на равных представлены в различных таксономических группах как у голосеменных, так и у покрытосеменных растений, а в некоторых группах, возможно, и неоднократно изменялись. Среди современных голосеменных растений нет эвриэфемеров (эфемеров и однолетников), нет даже многолетних трав. Однако все это еще не доказательство того, чтобы не признавать возможность суще- ствования среди древних голосеменных палеобиоморф не только мно- голетних трав, но и однолетников и даже эфемеров. Дело в том, что палеоботанические остатки, лишенные долгосохраняющихся механичес- ких лигнифицированных тканей, за десятки и сотни миллионов (!) лет давно разрушились, превратившись в атомы и молекулы, т.е. до наших
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ дней они в интересующем нас виде никак не сохранились даже в ока- менелом виде. Не исключено, что в эволюции семенных растений, среди родственных таксонов, травянистые и древесные биоморфы возникали неоднократно, поскольку менялись в них направления эволюции по продолжительности жизни. За это долгое историческое время на месте пустынь возникали тропические леса, а на их месте затем ползли гиган- тские ледники, которые потом превращались в моря, а далее в тундры и смешанные леса и т.п., т.е. изменений в климате и ландшафтах в различных регионах Земли за десятки и сотни миллионов лет было предостаточно. Археологи достоверно доказали, что всего лишь 20 тысяч лет назад на месте города Москва была лесотундра и там бродили стада мамонтов. За долгое историческое время такой очень лабильный эле- мент организации, как габитус, не просто мог, он был обязан, наряду с другими меросомами, изменяться, поскольку далеко не всем видам удавалось перешагнуть через моря и горные преграды. Результаты эволюционных изменений биоморф более всего отража- ются в их феносоме (образно — «лепоте»), в которой более всего на виду, приближен к внешней среде и наиболее доступен к трансформа- циям, — габитус. К габитуальным трансформациям биоморфы в своей эволюции, безусловно, прибегают в первую очередь, используя в полной мере арсенал пластичности и гиперпластичности. Они так могли и, вероятнее всего, так неоднократно и поступали, изменяя под давлением окружающих их обстоятельств типы габитусов, которые, как индикато- ры, отражали и отражают смену направлений эволюции биоморф, их типов, по продолжительности жизни. Итак, направление развития биоморф семенных растений, когда они цветут, плодоносят и затем умирают, прожив не более одного года, не сумев образовать базового органа или не проявив даже стремления к этому, так же закономерно и естественно, как и другое направление, когда биоморфы преодолевают барьер эвриэфемерности и образуют базовый орган. Любопытно наблюдать, как Природа, независимо от нас, ее исследователей, всегда подстраивающихся под нее, чтобы разобраться в тонкостях ее организации, в частности, в «ошеломляющем» разнооб- разии тел движущегося потока под названием Жизнь, изобретает по- средством эволюции все, что необходимо живым системам, и отметает то, что ей не нужно. Невольно ловишь себя на мысли, как мы, иссле- дователи Природы, чтобы как-то разобраться в непрерывности всего происходящего, прибегая к типизациям и классифицированию, стре- мимся уловить границы там, где они чуть просматриваются. Нам нужна ясность знания, ее нельзя достигнуть без типизации и классифициро- вания предметов исследования. Именно непрерывность происходящего побуждает нас прибегать к своеобразным «адаптациям» в вопросах тер- минотворчества. Пример тому — использование автором при построе- нии некоторых терминов приставки «полу»- («геми»-, «семи»-), чтобы как-то отразить то, что эволюция творит с жизнью, не признающей жестких границ между явлениями. При анализе явления эвриэфемерности и персистентности мы стал- киваемся с переходной ситуацией, когда уже у эфемеров проявляются
Глава б. Концепция пластичности и гиперпластичности биоморф семенных растений 521 некоторые признаки персистентности. Так, некоторые эвриэфемеры (ге- миэфемеры и озимооднолетники) начинают свою вегетацию (прораста- ют) только поздно осенью и успевают развить часто не только семядоли, но еще и розеточную структуру главного вегетативного побега, а то и начать куститься (Мальцев, 1932; Нечаева, 1960; Нечаева и др., 1973; Бурыгин, Маркова, 1970 и др.). В таком виргинильном состоянии эври- эфемеры пребывают всю зиму, а весной быстро «матереют», отбросив в сторону все «предосторожности», и в едином порыве достигают репродук- тивной стадии и «отдаются» ей без «оглядки», после чего, прожив где-то около полугода, и всегда меньше одного года, отмирают. Автор не склонен принципиально что-то менять в традиционном объяснении озимости некоторых эвриэфемеров, но это явление достой- но того, чтобы еще и еще обратить на него внимание. Дело в том, что высокая зимостойкость, которую демонстрируют озимые эвриэфемеры, уходя под снег в виргинильном розеточном состоянии, обусловлена меньше морфологически, а больше физиологически. Побеговая розетка листьев у таких растений прижимается к поверхности почвы, а внутрен- ние ткани таких побегов физиологически перестраиваются к выжива- нию в зимнюю стужу, где под снегом, а где даже и без него. У озимых побегов эвриэфемеров мы отмечаем начальные признаки персистентно- сти, поскольку они, действительно, зимуют там, где, казалось бы, дол- жны были погибнуть, но этого все же маловато и, достигнув весны, они быстро завершают свое развитие, прожив в сумме значительно меньше одного года, т.е. настоящей персистентности такие растения не прояв- ляют. Явление, о котором идет здесь речь, традиционно понимают как озимость эвриэфемеров (гемиэфемеров и озимооднолетников), что од- новременно можно рассматривать и как «преперсистентность» (дословно с лат. «раньше персистентности»). Растительные биоморфы и их части обладают преперсистентностью, если живут значительно меньше одного года, начиная эпиэмбриональное развитие осенью или даже зимой, преодолевают за это время неблагоприятный для видимого роста сезон (например, зиму с отрицательными температурами), а весной достигают репродуктивной зрелости и после плодоношения в том же вегетацион- ном сезоне полностью отмирают. Растения, обладающие преперсистен- тностью, лучше отличать от других эвриэфемеров, которые ее не про- являют, поскольку они, среди остальных эвриэфемеров, уже на полпути к стану персистентных (дву, — многолетних) растений. В этом плане «озимые эфемеры» было предложено в нашей работе (глава 5) обозна- чать как гемиэфемеры, а озимые однолетники — как озимооднолетни- ки, или полуоднолетники. Приставка «полу» («геми») обладает особым свойством. Она, сама по себе, не указывает, чего больше, а чего меньше в предмете исследо- вания: или того, что предшествует половине качества предмета, к назва- нию которого присоединяется приставка «полу», или, наоборот, того, что стоит после такой половины. Многое в данной ситуации зависит от намерений исследователя. В нашем случае, приставка «полу»- («геми»-) означает, что гемиэфемеры, хотя и эвриэфемены, но они больше по сложности организации, чем остальные (яровые) эфемеры, а озимоод-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ нолетники (также эвриэфемеры), но они больше (сложнее), чем осталь- ные однолетники. Преперсистентность эвриэфемеров свидетельствует об особом типе развития, когда перезимовка их побеговой части не приводит к развитию базового (персистентного побегового) органа, а она так и остается озимым, но уже перезимовавшим, побегом. Ситуация с эвриэфемерностью прямо-таки может быть забавной; многие из них ведут себя далеко не как «камикадзе», они способны выбирать и делают это, исходя из обстоятельств, вегетируют в удобное для них время, а если надо приспособиться и перезимовать при мину- совой температуре, пожалуйста, и это они могут. Значит, главное в эволюции биоморф в том, что она дает им возможность адаптироваться. Эволюция эвриэфемерности может иметь яровую или озимую направ- ленность с переходами между ними. Когда эволюции будет угодно, она сделает из эвриэфемеров то, что будет более всего отвечать складываю- щимся в определенном участке пространства и времени обстоятельствам, а постепенный переход их к деревьям, на мой взгляд, никто никогда не запрещал, как и обратную направленность в других условиях окружающей действительности. Растения значительно пластичнее, чем кому-то может даже представиться. Потенциальные возможности растений в эволюции больше, чем мы пока знаем. Мы примем за основу, что посредством преперсистентности могут образовываться только преперсистентные органы — подобия базовых органов — озимые побеги (озимым побег становится не тогда, когда он отрастает впервые осенью, а когда зимует и перезимовывает). Весной, когда у озимых эвриэфемеров образуется цветоносный побег, то он состоит из озимой (зимовавшей) компактной части в основании побега и яровой (незимовавшей) удлиненной части, развивающей репродуктив- ные органы. Базовые органы возникают только посредством персистен- тности, когда потенциальная дву,- многолетность становится реальной. Наиболее примитивные базовые органы возникают у малолетних расте- ний, например, у малолетних («сверходнолетних») полутрав (т. 1, гл. 8), иначе — полутравомалолетников. Например, у таких растений, как Datura innoxia Miller, Bidens tripartita L. Solanum nigrum L., Conyza bonariensis (L.) Cronquist, Bidens pilosa L. и др., которые автор наблюдал на Ближнем Востоке, проявляется примитивная персистентность, когда образуется базовый орган из постфлоральных резидных (перезимовав- ших) остатков удлиненного цветоносного побега, который, в отличие от однолетников, отмирает не полностью, а «отдыхает» в условиях влаж- ной, слегка прохладной южной субтропической зимы (температура все- гда выше 0'С). Весной (это основное время для роста и развития многих растений) на примитивном надземном удлиненном базовом органе та- ких полутравомалолетников, который мы назовем аэроподиоидом (дос- ловно с греческого «подобный аэроподию»), отрастают слабые «лихора- дочные» постфлоральные вегетативные и репродуктивно-ассимиляци- онные побеги, которые продлевают как бы прерванную прошлогоднюю вегетацию и превращают биоморфы такого типа из однолетников в полутравомалолетники, но, как правило, не находят ресурсов, чтобы превратиться хотя бы в среднелетники (рис. 277, 278).
Глава б. Концепция пластичности и гиперпластичности биоморф семенных растений 523 Может ли направление эволюции от однолетников через малолет- ние полутравы пойти дальше, к настоящим многолетникам? Наш ответ — может! Удалось найти виды, биоморфы которых вели себя больше, чем малолетние полутравы, правда, иногда они были и однолетниками, но часто и как малолетники и среднелетники, образуя соответствующие аэроподиоидные структуры полутрав (рис. 279). В подобных ситуациях эволюция может идти в сторону усиления персистентности или в обрат- ном направлении — в сторону ослабления и утраты персистентности и усиления эвриэфемерности. Пожалуй, исходя из приведенного выше материала, можно конста- тировать следующее. В процессе эволюции биоморф семенных растений сложилось два основных направления в течении биоморфогенезов: интен- сивное (ускоренное), когда накопление и реализация ресурсов происхо- дит без образования какого-либо базового органа (рис. 217-228), и эк- стенсивное, или ретардационное (замедленное), когда накопление и реализация ресурсов происходит посредством образования специализи- рованного базового органа (рис. 231-276).; есть смысл выделять еще и переходный тип — промежуточное направление, когда накопление и реализация ресурсов осуществляется посредством образования семипер- систентного органа или примитивного (деспециализированного) базо- вого органа у недолговечных биоморф (рис. 277-279). На основе приведенных выше положений можно сделать следую- щее аксиоматическое обобщение: если организация биоморф семенных ра- апений успешно осущеапвляет репродуктивное развитие без образования какого-либо базового органа, то она адаптирована в текущих условиях к ускоренным темпам развшпия, а в «ротивовес этому действует другая аксиома, если организация биоморф семенных растений успешно осущеапв- ляет репродуктивное развшпие только с образованием какого-либо базового органа, то она адатпирована в текущих условиях к экапенсивным (замед- ленным) темпам развития. Ранее (т. 1, гл. 6) было показано основное разнообразие базовых органов. Есть все основания считать, что накопление ресурсов в органи- зации биоморф семенных растений более всего связано с персистентно- стью, образованием базовых органов и совершенствованием их органи- зации, что позволяет закрепить это положение в законе (специальном) оптимизации в резервировании ресурсов: в организации биоморф семенных раапений оптимальным в резервировании ресурсов будет только то, что связано с персиапентностью и, в первую очередь, с образованием базовых органов и совершенапвованием их организации. Действительно, постоянно фиксируешь свое внимание на том, что организация биоморф семенных растений только тогда успешно решает проблемы личного выживания, когда она ориентируется на персистентность развития, когда направляет свои приспособления по резервированию ресурсов на образование базо- вых органов, соответствующим образом специализированных, которые занимали бы в процессе взаимодействия с окружающей средой наиболее удобное расположение относительно поверхности почвы, и надежно зак- репляет их (адаптации) на генетическом уровне, что позволяет какое-то время (иногда миллионы лет) выдерживать давление естественного отбо- ра на пути модернизации ранее достигнутого качества.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИЧ Итак, ранее мы показали, что виталинейность, когда происходит взаимовлияние двух динамических частей пространства: определенного живого тела (например, растительной биоморфы) и поверхности почвы (шире — почво-грунта), контактирующей с ним, — сопровождающееся какими-либо формами их трансформаций, является сильным эволюци- онным фактором, особенно для живых тел. Нас, естественно, в первую очередь, интересует виталинейность с участием биоморф семенных растений. Виталинейность — это двойственная реакция частей про- странства друг на друга: живого тела на поверхность почво-грунта и ее виталинию в биотопе и обратно — поверхности почво-грунта и витали- нии на живое тело. Главное, на чем мы сконцентрировали внимание при рассмотрении явления виталинейности — это факты наличия или отсутствия базового органа и факты расположения побегового тела и, в частности, базового органа относительно поверхности почво-грунта. Среди биоморф разных видов семенных растений можно просле- дить много вариантов расположения их базового органа относительно твердой земной поверхности (т. 1, гл. 6). Даже у биоморф одного вида, в зависимости от конкретных условий их обитания, базовый орган может вести себя по-разному относительно рассматриваемого фактора (рис. 40, 301). Базовый орган, в зависимости от состояния окружающих ус- ловий, приспособился то поднимать расположенные на нем верхушеч- ные меристемы все выше и выше относительно поверхности почвы, то, наоборот, опускать их ниже и ниже, даже под поверхность почвы. Это обстоятельство в свое время умело использовал при построении широко известной классификации жизненных форм знаменитый ботаник Раун- киер (Raunkiaer 1907, 1934 и др.). Базовый орган семенных растений может быть надземным (высоким, средним и низким), приземным (при- жиматься к поверхности почвы, стелясь по ней), подземным, подвод- ным, плавающим и даже поднятым над поверхностью почвы с помощью так называемых «ходульных» корней (Rhizophora mucronata Lam., Pandanus veitchii Hort.) и т.п. В зависимости от возраста, наследствен- ности и условий обитания, базовый орган биоморф разных видов и даже одного вида может сильно варьировать по размерам, изменять направ- ление и скорость разрастания, выдерживать или не выдерживать засы- лание песком, заиление или затопление водой; иногда базовый орган может рудиментироваться до одной зимующей почки и даже вообще не развиваться (рис. 230-276). Когда базовый орган не развивается, в от- личие от других биоморф этого же вида, то мы невольно попадаем на стык двух противоположных направлений эволюции, когда особи одно- го вида нацеливаются на персистентное развитие, а другие — выбирают эвриэфемерный путь (рис. 277, 278). Внутривидовая пластичность по- зволяет сосуществовать рядом в одном виде двум — нескольким направ- лениям эволюции единого ряда альтернативных признаков, из которых, при стечении комплекса соответствующих внешних условий, может получить приоритетное развитие одно из направлений. Все субординатные биоморфотипы, отражающие сложность организа- ции биоморф, так или иначе «завязаны» на два ряда признаков: наличие или отсутствие у них во взрослом состоянии базового органа и, в последнем случае, на положение этого органа относительно поверхности почвы.
Глава б. Концепция пластичности и гиперпластичности биоморф семенных растений 525 Итак, пришло время осуществить графическое моделирование, возможность которого мы старались подготовить в предыдущих рассуж- дениях. Эволюция тел биоморф семенных растений и поверхности почвы (шире — почво-грунта), контактирующей с ними, сосуществует рядом как в текущее время, так и в большой исторической ретроспек- тиве. Безусловно, за многие миллионы лет они стали (другого пути у них не было) влиять друт на друта, причем уровень твердой земной повер- хности — на тела растений, естественно, больше. Оказалось, что в со- матической эволюции, ио сути дела, нет ничего «святого» (недостуиного) хности иочво-грунта (корни некоторых эиифитов, наиример, могут висеть в воздухе выше иоверхности иочвы). В эволюции биоморф семенных расте- ний, иожалуй, ириемлемо любое расиоложение растительного тела в ок- ружающем его жизненном иространстве и, следовательно, любое наирав- ление эволюции ио отношению к фактору уровня иочвенной поверхности, что можно образно отразшпь в соответствующей модели (рис. 340). Сила слов в таких случаях уступает убедительной наглядности изобразительных возможностей. 6.5.3. Габитусы и универсальная модель организации биоморф семенных растений Эволюция растительных биоморф привела к главному: они научи- лись по-разному использовать неоднородные части земного простран- ства, т.е. по-разному вести себя в воздушной, водной и почвенно-грун- товой его частях, придав решающее значение при освоении этого про- странства привязке к уровню поверхности почвы (виталинии) среды обитания. Неоднородность частей пространства как сред существования растений побудила семенные растения развивать в своей эволюции такие формы тела, которые обеспечили бы им опору в пространстве фото- фильным и репродукгивным органам и позволили бы им оптимально извлекать из окружающей среды ресурсы, необходимые для жизнедея- тельности. Для почвенно-грунтовой (подземной) части пространства наиболее удачным эволюционным изобретением биоморф семенных растений стал корень, избравший геотропизм как форму роста в своем существовании. В воздушной части пространства господствуют побеги семенных растений, которые, проявляя гелиотропизм (стремление к солнечному свету), избрали этот способ как основную форму роста и развития, обеспечивающую себя и целые организмы, через фотосинтез, производством необходимых органических веществ и кислорода, а так- же образующие репродуктивные органы и проявляющие половое раз- множение. Существование основных органов на границе двух главных частей пространства (подземного и воздушного), а также в водной среде освоили как побеги, так и корни, используя переходное положение между частями пространства для захвата новых сред обитания как для производства ресурсов, так и для резервирования их.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ Рис. 340.' Универсальная модель эволюции организации биоморф в зависи- мости от уровня поверхности почво-грунта (виталинии). 1-2 — разнообразие в расположении виталинии по отношению к основным органам биоморф семенных растений: 1 — реальные (уже известные), 2 — потенциально воз- можный (вероятен у некоторых паразитов), 3 — гипокотиль, 4 — базовый орган, 5 — побеги текущего года однолетников и многолетников, 6 — побеги текущего года многолетников, 7 — корни, 8 — направления разрастания основных органов в эволюции биоморф семенных растений по отношению к уровню поверхности почво-грунта, 9 — контуры основных органов эври- эфемеров (показаны пунктирной линией в схеме тела обобщенного расте- ния), 10 — контуры основных органов многолетников (показаны сплошной линией в схеме тела обобщенного растения).
Глава б. Концепция пластичности и гиперпластичности биоморф семенных растений 527 Таким образом, разграничение сред существования (биотопов) на части как части земного пространства, оказало и продолжает оказывать влияние на эволюцию растительной жизни, обеспечивая различные условия основным органам биоморф семенных растений (побегам и корням) в их биотопах. Наиболее полно приспособленность биоморф семенных растений к процессу виталинейности познается через един- ство в одной системе побега и корня как высшей единицы их органи- зации — корнепобега (ризокормуса). Как видно, нить наших рассуждений все более и более сужается в своем устремлении к осознанию факта, что без постоянных адаптаций растительных тел к явлению виталинейности известная нам жизнь на Земле не смогла бы никогда состояться. Итак, мы настроились на боль- шое обобщение: должна быть создана и обоснована универсальная модель эволюции организации биоморф в зависимости от уровня твер- дой земной поверхности (виталинии). Существование проблемы автор видит давно, но решение ее пришло только сейчас. К счастью, оно предельно простое и понятное, как день и ночь на Земле. Так бывает часто. Мысль долгие годы ищет решения через различные построения моделей: рисовались извилистые линии поверхности почвы, приводи- лось разное число моделей габитуально разнотипных растительных тел, строились схемы разных направлений эволюции. Сейчас я готов зая- вить, что все это прошлое, все осталось позади, ... и только в чернови- ках, а оптимальный ответ приводится здесь (рис. 340). Пауза от идеи до решения проблемы растянулась на долгие годы. Что же все-таки мы имеем на самом деле? В рассматриваемой универсальной модели все разнообразие растительных тел от эфемеров и однолетников до сверх- гигантских деревьев уместилось в одном образном растительном теле— типовом организме, который вместил в себя не только все разнообразие габитусов простых биоморф и передал через множество горизонтальных линий, расположенных рядом по вертикали, динамику твердой земной поверхности, а разные направления возможной эволюции в развитии организации биоморф в условиях перемещающейся виталинейности зафиксированы изображением абстрактного перекрестка равнозначных «дорог» — «флюгера» эволюции. Итак, в универсальной модели эволю- ции биоморф семенных растений задействовано несколько ключевых моментов: образное тело биоморфы, уровень поверхности почва-грунта (виталиния) и все четыре равновозможных направления эволюции, основные типы пространства, окружающего модельную биоморфу. Труд- нее всего было соединить при графическом моделировании в образе одного растительного тела — организма однолетность (шире — эвриэ- фемерность) одних и многолетность (шире — персистентность) других биоморф. Оказалось что и это возможно, если вписать пунктиром внутрь тела многолетника недолговечное во времени тело эвриэфемера. Важное место в существовании биоморф семенных растений при- надлежит базовым органам. В случае, когда персистентный побеговый (базовый) орган исторически давно включен в организацию биоморф семенных растений, то его положение относительно поверхности почвы может изменяться в весьма широких пределах. Сама способность био-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ морф образовывать или не образовывать базовый орган в своей органи- зации является примером того, как количественные изменения приво- дят при соответствующем накоплении их во времени к качественной перестройке. Направление, по которому базовый орган, однажды возник- нув, совершенствуется в своем разнообразии у биоморф семенных расте- ний, может одновременно существовать в то же самое историческое время с направлением, когда базовый орган у других биоморф все более и более становится лишним, когда из поколения в поколение «приветствуется» гводдерживается) его равугнню, и в итоге возникают не только многолетние травы, но и однолетники и даже эфемеры. Условия жизни на Земле в историческое время изменялись неоднократно, причем иногда и не раз катастрофически, что приводило к сильнейшим стрессам все живое, к вымиранию многих видов и даже больших таксономических групп (из- вестный пример гибели динозавров около 75 миллионов лет назад). Жесточайшие стрессы в текущей жизни сразу всех биоморф неоднок- ратно приводили к революционным ситуациям в эволюции живого, в результате которых жизнь вынуждала своих носителей бороться и учить- ся в борьбе и побеждать, и проигрывать, совершенствуя ее возможности как в плане пластичности биоморф, так и их гиперпластичности. В таких ситуациях (и не только в таких) могла возникать способ- ность растительных биоморф проявлять в одно и то же время развития растительного царства все два основных (генеральных) направления в габитуальной эволюции: эвриэфемерное и персистентное. В другое вре- мя в отдельных таксономических группах могли выпадать какие-либо типы габитуальных форм (сокращенно «габитотипы»); например, сейчас у голосеменных нет однолетников и многолетних трав, но это не озна- чает, что их не было среди них никогда, а, если даже и не было, то это еще не доказательство того, что завтра они не смогут появиться, чтобы через какое-то время вновь, но не навсегда, исчезнуть. Типы биоморф, отличающиеся по их габитусу, включающие и от- раженные в нем признаки продолжительности жизни, способны в эво- люции поворачивать и обратно, причем делать это в историческом времени неоднократно (!); поскольку габитус — это наиболее показа- тельная часть в организации биоморф, то он передает довольно точно многие интегральные соматические преобразования. Существование биоморф, в частности, биоморф семенных растений, подчиняется универсальному принципу индикаторного (показательного) участия габитуса в организации биоморф: габитус — самая показательная меросома, отражающая снаружи особенности трансформирования фене- тической организации и ее частей в целостной организации биоморф, ко- торая принимает на себя первое воздействие лабильных условий окружа- ющей внешней среды, поэтому он сам (все его формы) является не только внешней организацией биоморф, но и функцией всей остальной (внутренней) части их организации, часто, как индикатор, указывающий своими сред- ствами (экзоморфологическими) на поведение (текущую жизнь состояний биоморф), морфогенезы (текущую жизнь отдельных биоморф в целом) и эволюцию, ее внешние результаты (историческую жизнь биоморф) в про- странстве и во времени. Биоморфам, чтобы быть (участвовать) в своей
Глава б. Концепция пластичности и гиперпластичности биоморф семенных растений 529 исторической жизни (эволюции), надо пройти хотя бы через последнюю часть того, что составляет ее, т.е. еще и через свою текущую жизнь (разнообразное поведение, его формы и морфогенезы; см. т. 1, гл. 7). Немало из того. что имеют сейчас в своей габитуальной организа- ции биоморфы, закреплено на генетическом уровне и стало их досто- янием еще у первобытных голосеменных, которое на протяжении дли- тельного исторического времени по каким-то признакам расширяет, а по другим — сужает разнообразие. При построении моделей эволюции каких-либо явлений не стоит забывать о принципиальном моменте: все во Вселенной, на Земле в частности, находится во власти многих кос- мических ритмов, поэтому такая частность, как габитуальная и этомор- фная (поведенческая) эволюции в эволюции организации биоморф так- же находится во власти ритмических процессов, проявляющих себя в чем-то и через соматическую трансформацию (кинетопластику), и через динамику положения побегов, корней и базовых органов относительно уровня поверхности почво-грунта, и через расширение разнообразия базовых органов или, наоборот, через постепенную или внезапную, частичную или полную редукцию отдельных органов и т.д. и т.п. Факты, подтверждающие работу» универсальной модели эволю- ции биоморф семенных растений, можно постоянно наблюдать в совре- менном их разнообразии. Важнейшим, но, безусловно, не единствен- ным инструментом практической фиксации результатов этой работы является габитуальный сиидром — это концентрированный внешне-морфо- логический (габитуальный) ответ всего остального, что задействовано в организации биоморфы, на все, что составляет воздействие на нее окру- жающей среды на данном участке пространства и времени. Уже отмечалось (т. 1, гл. 8), что у многих травянистых и полудре- весных растений, произрастающих в различных областях умеренных широт, в условиях с относительно эрозионно спокойным почво-грун- том, надземные удлиненные части цветоносных побегов, обычно после завершения их репродуктивного цикла, отмирают на бульшую часть длины, причем у многолетних трав — до подземного основания. Бук- вально на глазах такие, казалось бы, уже обреченные на отмирание, части побегов превращаются в многолетние и даже одревесневшие структуры, если они оказываются погребенными почво-грунтом (т. 1, гл. 8.2.1.; рис. 337, 338). Механизм метаморфоза эфемерных цветоносных частей побегов в многолетние работает не только при засыпании их песком, в результате чего создаются благоприятные условия (достаточно тепло и достаточно влажно) для трансформации одних структур (эфе- мерных) в другие (многолетние). Метаморфозы эфемерных цветонос- ных частей побегов происходят и при сходном с погребением почво- грунтом проявлении климата. Когда после длительного и сухого лета образуются отплодоносившие цветоносные части, которые при продол- жении засухи должны были бы однозначно погибнугь, но этого не происходит, поскольку растения с такими побегами живут на Ближнем Востоке, где после знойного и длительного лета наступает сравнительно теплая и влажная короткая зима. Создаются (в комплексе) благоприят- ные условия, когда достаточно тепло и достаточно влажно, что приводит
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ к выживанию отплодоносивших частей цветоносных побегов и пробуж- дению на них меристематических зон — почек, которые дают («лихора- дочно») новые побеги (рис. 277, 278), проявляя, таким образом, постфло- ральное побегообразование (побегоотрастание). Иногда механизм выжи- вания цветоносных частей побегов закрепляется настолько сильно, что срабатывает гиперпластичность, и потомки получают от своих предков новый качественный уровень фенетического развития, приобретая уже новую пластичность, закрепленную естественным отбором на генетичес- ком уровне. Так возникают новые типы габитуальных форм при расши- рении нормы реакции биоморф и, следовательно, полиморфизма в рам- ках прежних видов; например, так из многолетних трав могут возникнуть полукустарники и кустарники. Дальнейшее давление на гиперпластич- ность может приводить и приводит уже к возникновению новых видов. Полукустарниковые биоморфы Artemisia abrotanum L., которые автор наблюдал в пойме р. Оки и на нижней части террасных склонов к этой реке в Серпуховском районе Московской области (рис. 341) — это, по- жалуй, пример того, когда эволюция закрепила на генетическом уровне в условиях довольно жесткого умеренного климата способность выживать на большую часть своей длины отплодоносившие цветоносные побеги, развив у них свойство морозостойкости, которого, похоже, не было у предков этого вида. Они, вероятно, пришли с юга или задержались в условиях Подмосковья с давних, еще теплых, времен, адаптировавшись в последующем историческом развитии к зимней стуже, в первую оче- редь, соответствующими физиологическими адаптациями, приобретя морозостойкость, которая нашла отражение и в морфологических адап- тациях. Однако биоморфы этого вида все же сохранили некоторую склон- ность к выбору более теплых мест обитания в условиях Подмосковья, что несколько выдает теплолюбивое прошлое его предков. Особи А. abrotanum прижимаются обычно к микропонижениям, предпочитая пойму, а не террасные возвышения в естественных условиях произрастания в Под- московье. Пример с А. abrotanum показывает, как биоморфы с постфло- ральным побегообразованием, возникшим в эволюции в специфических условиях теплого климата, посредством соответствующих адаптаций (в первую очередь, развития морозостойкости), могут продвинуться на се- вер, приобретя или новые для себя полукустарниковые и даже кустарни- ковые свойства, или сохранив полудревесные и даже древесные свойства предков-теплолюбов, сумев стать морозостойкими. Эволюционные плеяды (так можно назвать совокуиность иредковых и иотомковых синтомов, или эволюционных шагов, образующих одну эво- люционную ветвь в каком-либо таксоне) сиособны npueecmu своих совре- менных иотомков, иутем заметных качественных адаитаций как к миг- рации с юга на север, так и, наоборот, — с севера на юг. Они же могут и оставить виды-потомки на месте своей первобытной родины, сильно изменившейся в климатическом отношении, вызвав у них, точнее, у их предков, соответствующий гиперпластичный «криз», жизнеполезный выход из которого — проявление «здоровой» гиперпластичности и зак- репление мутаций, позволяющих потомкам выдерживать ранее недо- ступные для предков зимние холода.
Глава б. Концепция пластичности и гиперпластичности биоморф семенных растений 531 Рис. 341. Полукустарник Artemisia abrotanum L. (сем. Asteraceae) в Москов- ской области (Серпуховский район, крутой склон к узкой пойме реки Оки, окрестности деревни Прилуки, 25 V). Др — верхняя часть метельчатого соц- ветия одного из универсальных побегов (Флора СССР, т. XXVI, 1961, с. 495, рисунок художника Н.А. Моисеевой).
532 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Универсальный принципиальный механизм выживания и гибели цветоносных частей растений во многом заложен в генофонде семенных растений, поэтому при проявлении «подходящих» условий он может начать «работать» — фенетически проявлять себя, если удается разбло- кироваться «дремавшим» генам и если им хватит энергии «BGIIoMHHTb» (вытащить из генетической «памяти») или создать что-то совершенно новое (когда «вспоминать» уже нечего, поскольку что было, то не со- хранили, не сумели сберечь, растеряли в эволюции, а сейчас надо, чтобы выжить, учиться многому заново). В зависимости от конкретных усло- вий обитания растений срабатывают механизмы пластичности и (или) гиперпластичности, приводя постоянных соискателей на продолжение жизни к разнообразной соматической эволюции в сторону усиления или ослабления персистентности или эвриэфемерности, но и еще в сторону сохранения достигнутых ранее результатов в «раскачивании» эволюци- онных маятников. Любопытно посмотреть, как ведут себя некоторые виды растений, когда они произрастают в разных климатических зонах. Так, при выра- щивании под Москвой Grindelia robusta Nutt. этот субтропический вид развивается по типу травянистых многолетников, выдерживающих хо- лодные зимы лишь несколько лет, зарывая «голову» (базовую, т.е. мно- голетнюю побеговую, часть) в почву, и отмирающих вследствие выпи- рания и подмерзания их надземных и приземных частей. Особи G. robusta, произрастающие на Ближнем Востоке (Ботанический сад уни- верситета Тель-Авива), проявляют себя как полукустарники, образуя одревесневшие приземные части побегов; при этом цветоносные части на значительную длину выживают в этих условиях, образуя на себе новые побеги (рис. 342). Есть все основания для следующего обобщения: количество и ка- чество одних частей определенного пространства и границ между ними, которое неиосредственно влияет на развшпие биоморфы, как-либо вовле- ченной в этот фрагмент пространства на какое-то время как некоторая другая его часть, всегда иобуждает биоморфу давать соответствующий интегральный ответ ио всему комилексу воздействующих факторов, про- являя тем самым адекватную адатпивную изменчивость, индикатором которой более всего может быть и является ее габшпус, всей своей формой свидетельствующий остальному миру, что биоморфой в конкретном био- mone исиользуется одно из нескольких идиоэнергетически возможных на- иравлений соматической трансформации этого обьекта как некоторого состояния соответствующей материальной массы, стадийно развиваю- щейся на отдельном участке бесконечного эволюционного nomoxa — это закон (специальный) адекватной адаптивной изменчивости бноморф н ее отражения в окружающем пространстве н времени (рис. 12-32, 35-120, 123- 147, 191-207, 217-315).
533 Глава б. Концепция пластичности и гиперпластичности биоморф семенных растений О Ж ° Д о ~~ ж й а3 Я v XXÎ З <~3 О О .ф X Ю О ~ З М ф: О О ~ М 65 Ж (~ h4 О О Ы g g О. ! ! Ж <.) О о О О r4 ~ С!ф л exо ~а~ й Ю Д л ~а v й ° я B ~~а ~жа й ~) й й~ ~е К Мъ g ~4 ф) л я а ф; ~ о ID ЯЗИ О Q QR f u -.„И о о ~:~ а ж О й- ь: - *„1,' О v v~иЮО Жачачой~~й ! ! Я ! ! Q ! а. а. д а. о ~ о ~ч>Ц~,о~о ~х а.~ й ~~ ~~ о Я ~о ~ -~~роомx > ~ у ~ р о ~~! g~z~,-~~о и~ ~И ч k ~=( Q C4 ~ фC Н (д О й Е Я C) ~ Pg О о~ z Х ц~ Я ~ Ф ~ сч ~~ M5i: Ю C) Ои а. о л О "Д ~~а З Д Ю И ~3 З д Ж З ! Я Я ! Яи ~Q L- ~ X а.f Я О щю З О юойО~ о а,~~~ ,'3; [.'~ ~~ ~ч Й ю е Ж ~) И Оч ре lO О [ Й [ Ф~ й М О Ж О r4
Глава 7. Концепция разнообразия биотической власти и управляемого ею, посредством «могучей пятерки», единства живого и неживого в организации биоморф семенных растений Как-то ко мне в дом попал букет цветов; было какое-то торже- ственное событие. Страсти утихли, гости разъехались, а через несколько дней букет завял; его выбросили, но одну веточку розы, которая еще немного подавала признаки жизни, я высадил в горшок с почвой. Через несколько дней веточка ожила и живет по сей день, образовав куст, периодически красиво цветущий. Здесь произошло что-то такое, о чем есть смысл задуматься. Похоже, история с веточкой розы несет в себе разгадку, немного и немало, а секрета счастья. Жить, жить и жить — вот сердцевина любого счастья; жить даже тогда, когда многое из происхо- дящего против жизни. Такое счастье нельзя ничем заменить, за такое счастье надо, в этом есть действительный смысл, бороться... «Бороться и искать, найти и не сдаваться» (В.А. Каверин). Как все, заключенное в этих словах, подходит и к веточке розы! Мне тоже повезло, я тоже счастлив, поскольку как-то чуть-чуть помог жизни вообще сохраниться, пусть в очень скромном эпизоде, через заботу о судьбе веточки розы, но из всего такого, по большому счету, и составляется жизнь вообще. Неужели, вот так просто, здесь и сейчас, мы ответили на извечный вопрос человечества: что такое счастье? Вопрос о счастье, тем более растительном, особенный, и его мы еще коснемся в главе 10. Здесь же, как бы между прочим, обратим внимание на самое главное, что состав- ляет содержание счастья любых живых существ Земли, независимо от того, осознают или не осознают они это, — желание и умение жить самим и иоддерживать жизнь других. Веточка розы без почвы была близка к смерти. Значит, в этой веточке присутствует все, что не только поддерживает, но и останавли- вает жизнь. Жизнь в соответствующем теле соприсутствует вместе со смертью и в какое-то время жизнь находит или не находит силы, чтобы отодвигать смерть. О смерти написано и много, и немного, как на нее посмотреть. Писать о смерти не просто, поскольку где-то подспудно и писатели, и
535 Глава 7. Концеиция разнообразия биотической власти 7.1. Что обеспечивает телесную организованность явления жизнь» Жизнь на Земле существует только в телесных системах. Самое известное нам, людям, живое тело во Вселенной — это только живое тело, или биостроба, нашей планеты, хотя фон разнообразия типов живых тел (биосом, или биосистем) весьма разнообразен по сложности их организации (т. 1, гл. 2). Вероятно, жизнь вообще существует где-то еще за пределами Земли и живые тела не являются уникальным земным читатели, может быть, вполне справедливо, побаиваются ее. Однако приглушим в себе излишние эмоции и спокойно подумаем о смерти, поскольку без осознания этого явления трудно до конца понять то, что же на самом деле происходит в организации биоморф семенных расте- ний. Настроившись на соответствующий исследовательский лад, начи- наешь лучше понимать ее место и роль в организации жизни своей, чужой и вообще, да и природы вообще (Природы). Без смерти жизнь на Земле, как известно, невозможна, поэтому не разобраться с этим явлением в организации биоморф семенных расте- ний было бы большим упущением. Больше всего написано о смерти в мире людей, особенно медиками, геронтологами, философами, чаще всего при изучении явления старения (Стрелер, 1964; Лэмб, 1980; Арьес, 1992; Калитин, 1998 и др.). Для начала ограничимся несколькими выс- казываниями о явлении «смерть» известного французского философа, психолога и искусствоведа В. Янкелевича (1999): «Смерть есть феномен биологический — точно так же, как рождение, половая зрелость или старость... (с. 9). Смерть — это бездна которая внезапно разверзается на пути неостановимой жизни; живущий вдруг, как по волшебству, делается невидимкой — в одно мгновение, будто провалившись сквозь землю, уходит в небытие. Далее, это «чудо» не является редкостным нарушением естественного порядка вещей, необычным отклонением от обычного хода жизни, — нет: это «чудо» и есть одновременно универ- сальный закон для всего живого, вселенский удел живых существ... Еще не случалось, чтобы «смертный» избежал смерти, этого всеобщего зако- на, и совершил чудо: жил бы всегда и никогда не умирал... (с. 11). Что лучше — не жить, чтобы не умирать, или принять однажды смерть, чтобы испытать хотя бы на несколько десятилетий несравнимую радость жизни?.. Стоит ли, по меньшей мере, один раз почувствовать уникаль- нейший вкус земного существования? ... да, тысячу раз да открытию бесценной сокровищницы жизни; да, стоит пережить весьма горестное испытание смерти» (с. 430). Проблему взаимодействия жизни со смертью автор считает одной из важнейших при изучении закономерностей организации биоморф семенных растений. Не будем менять принятый в нашей монографии стиль, рассматривать явление с разных сторон и затем из глубины выходить на построение теоретических положений.
536 OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ изобретением. Однако, человечество еще маловато знает о Вселенной, и в третьем тысячелетии можно надеяться, что люди воочию увидят и потрогают Мир в его неиссякаемом величии. Как бы там ни было, но все живые тела на Земле — это структурные элементы ее единого живого тела (биостробы). На какие процессы надо более обратить внимание, чтобы осознать особенности проявления те- лесного (соматического) принципа организации жизни вообще (Жиз- ни). Все, что происходит в Природе, представляет собой круговорот ресурсов (веществ, энергии, информации), и существование живых тел «замешано» на нем, где соединение и разъединение ресурсов, проявля- ющееся через упорядоченное ростовое развитие (нарастание и ветвле- ние), подкрепляется более или менее упорядоченным разделением на части (партикуляцией), размножением биоморф и воспроизведением других живых тел и, наконец, всегда живые тела проявляют также «стремление» возвратиться в неживое (подвергнуться смерти). Попробу- ем разобраться подробнее с некоторыми процессами, обеспечивающими соматическую организованность растительной жизни. 7.1.1. От соединения и восхощдения {«подьема») к разьединению и нисхождению («спуску») Крутоворот ресурсов — это то, что сближает все телесные системы. Нет ничего в Природе, что избежало бы участи движения-изменения. С возникновением первых живых тел на Земле появилась и первая биосфе- ра — палеобиосфера, которая в процессе саморазвития обогащалась раз- нообразием форм жизни в широком смысле — эврибиоморф, т.е. биоси- стем разных уровней сложности. Нас интересуют биоморфы в узком смысле, т.е. организмы и синорганизмы, их положение в биосфере и в крутовороте ресурсов. В. И. Вернадский (1988, с. 163) подчеркивал, что «жизнь проявляется в непрерывно идущих, в происходящих в планетном масштабе, закономерных миграциях атомов из биосферы в живое веще- ство, с одной стороны, и, с друтой стороны, в обратных их миграциях из живого вещества в биосферу. Живое вещество есть совокупность живу- щих в биосфере организмов — живых естественных тел — и изучается в планетном масштабе». «Вещество биосферы состоит из двух состояний, материально энергетически различных — живого и косного. Живое веще- ство, хотя в биосфере материально ничтожно, энергетически оно высту- пает в ней на первое место» (Вернадский, 1988, с. 172). Есть ли что-то такое в движении материи — ее тел и процессов,— что позволило бы утверждать, что движение-изменение материальных систем обеспечивается единством и борьбой двух противоположных сил~ На этот вопрос наука уже давно ответила утвердительно. Всякое движе- ние материи в телесных системах и процессах — это единство и борьба двух противоположных сил: соединения (ассоциирования, ассимиля- ции, а шире — интеграции) и разъединения (диссоциирования, дисси- миляции, а шире — дезинтеграции). Все, что есть в материальной при-
Глава 7. Концеиция разнообразия биотической власти 537 роде, когда-либо подвергается действию этих внутренних и (или) вне- щних сил, в результате чего в одних случаях происходит интеграция тел (они сливаются, связываются, срастаются или как-то еще иначе соеди- няются и усложняются), а в других случаях телесные системы, напротив, дезинтегрируются (они растворяются, разрываются, распадаются, разле- таются, разбегаются или еще как разъединяются, разделяясь на части). Две противоположные силы: соединения и разъединения — находятся в единстве и противоборстве между собой, взрывая вулканы, сталкивая кометы с планетами, соединяя сперматозоид с яйцеклеткой, поглощая солнечную энергию зелеными листьями растений, собирая автомобиль на заводском конвейере или разбивая его же в дорожной катастрофе, формируя планету из «газо-пылевого облака», образуя пепел в резуль- тате сгорания лесного массива и, наконец, что нас здесь более всего интересует, проходя жизненный цикл какого-либо организма от рожде- ния до смерти. Все, что есть в Природе в виде тел или процессов, находится в движении в пространстве-времени и всегда как-либо под- вергается воздействию некоторых внутренних и (или) внешних сил соединения или разъединения, которые оставляют после себя более или менее измененные прежние или уже качественно новые тела и (или) процессы. Наша Земля состоит из неисчислимого множества тел и процессов, обеспечивающих ее существование такой, как она есть. Преджизнь, а затем и жизнь, возникнув на Земле, с первых «шагов» стала использо- вать те же многие приемы взаимодействия между своими носителями и взаимодействия их с неживыми телами, какими изначально пользуются в отношениях между собой неживые тела, добавив, безусловно, к изве- стным схемам что-то свое, особый колорит, поднимающий живую при- роду на уровень выше над неживой природой. Однако основой всему везде и всегда были и будут два процесса: соединение (интеграция) и разъединение (дезинтеграция). Способность любых тел соединять дру- гие части и быть соединенными с другими частями, или, напротив, разъединять другие части или быть разъединенными от других частей- это атрибутивные свойства организации всего телесного (соматическо- го), а еще шире — организации всей известной нам Природы. Итак, все живые системы используют в своем существовании два основных спо- соба движения ресурсов (соединение и разъединение), но делают это целесообразно, исходя из своих наследственных возможностей и обсто- ятельств, в которые они попадают. У биосистем нет другого выбора: они могут расти и развиваться, проходя в своем развитии разные по времени стадии по направлению или восхождения («подъема»), или нисхождения («спуска»). Когда растения или животные более или менее успешно, быстро или медленно, формируют вегетативную массу своих тел, созда- вая обязательный пререпродуктивный резерв для взрослой жизни, и, наконец, проходят репродуктивное размножение, которое может про- должаться недолго, не очень долго, или с перерывами долго и даже очень долго, например, у Adansonia gigitata L. — иногда до нескольких тысяч лет, то это реализация направления восходящего развития био- морф. Со временем живые тела, биоморфы в том числе, растрачивают
538 ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИЯ Рис. 343. Биографический портрет Негас1еит dissectum Ledeb. [сем. Apiaceae (Алтай, Катунский и Курайский хребты, лесные опушки, поляны, высоко- травные субальпийские луга)]. Дополнительное обозначение: ок — один из клонистов (два других не показаны) старого синорганизма. БИОМОрфОТИП: надземно-двусемядольные (семядоли линейно-ланцетные или ланцетные, 15 — 22 мм дл., 2 — 4 мм шир., заостренные, постепенно переходящие в черешки 10 — 15 мм дл.) полурозеточные очередно-полусложнолистные гемисимподиальные семидетерми- натно-репродуцирующие стержнекорневые ретардационно-кондивидуальные однодомные [»центральный зонтик с обоеполыми, боковые с тычиночными цветками, зонтички много- цветковые» (Васильева и др., 1963, с. 419)] бело-сельдерейноцвегковые компактнокорневиш- ные травянистые многолетники. При старении, отмирание гетерономного, т.е. связанного непосредственно с главным корнем, корневища и главного корня идет и сверху, и снизу, и изнугри, поэтому происходят сильные разрушения партикул и далеко не все из них, если полностью обособляются, оказываются достаточно жизнеспособными. Придаточные корни нередко развиваются. Замедленное вегетативное размножение может продлевать синорга- низмам жизненный цикл еще на 10-20 лет. Растения впервые цветут на 5-15 году ~3» (в высокогорьях позднее), а синорганизмы отмирают в возрасте 40-60 лет. свой потенциал, теряют многие свои резервные мощности и сменяют приоритеты в развитии, переходя на нисходящее направление морфо- генезов, главная сторона которого — разъединение ресурсных потоков и направление развития их носителей в сторону элиминации — к выходу «за порог» (рис. 12-32, 40-50, 343-345 и др.).
539 Глава 7. Концеиция разнообразия биотической власти Б В Рис. 344. Биографический портрет Aquilegia glandulosa Fisch. ex Link [сем. Ranunculaceae (Алтай, 1~тунский хребет, высокогорные луга, около ледни- ков, снежников)]. БИОМОДфОТИП: надземно-двусемядольные (семядоли яйцевидные, 3 — 4,5 мм дл., 1,5— 2,5 мм шир., слегка заостренные, резко суженные в черешки 3 — 5 мм дл.) розеточные гемисимподиальные розеточно-полусложнолистные детерминатно-репродуцирующие кау- дексово-стержнекорневые однодомные темно-сине-лютиковоцветковые травянистые мно- голетники. При старении сильная продольная некротическая партикуляция может расчле- нять частично и реже полностью и каудекс, и главный корень, приводя иногда к несовер- шенному вегетативному размножению.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ 38в 206, Рис. 345. Биографический портрет Antirrhinum majus L. [сем. Scrophulariaceae (Израиль, Тель-Авив, Ботанический сад, влажный тенистый склон, 21 П1)]. БИОМОрфОТИП: надземно-двусемядольные (семядоли яйцевидные 5 — 6 мм дл., 3 — 4 мм шир., закругленные, круто переходящие в черешки 2 — 5 мм дл.) безрозеточные супротивно-простолистные очередно-брактеознолистные эусимподиально-гемисимподи- альные семидетерминатно-репродуцирующие стержнекорневые однодомные розов~-Но- ричниковоцветковые прямостояче (1)-полегающие (11) полукустарники. В естественных условиях произрастания отмечается как травянистый многолетник (Feinbrun-Dothan, Danin, 1991). Вероятно, норма реакции у биоморф этого вида позволяет в зависимости от конкретных условий развивать несколько вариантов организации от травянистых м~~~~- летников до полукустарников.
Глава 7. Концеиция разнообразия биотической власти В соответствии с приведенным выше материалом, можно конста- тировать следующее: развшпие любых тел свершается как-либо упорядо- ченно иод воздейапвием их иреднаследственности или наследственноапи, зависит от влияния окружающей среды и в целом двойапвенно, сопровож- даясь использованием в разных формах и их сочетаниях только двух munoe процессов — соединения (интеграции) и разьединения (дезинтеграции) дви- жущихся ресурсов — это закон (субабсолютный) движения ресурсов в организации телесных систем. На этот закон опирается специальный закон упорядоченности движе- ния ресурсов в организации биосистем, который можно определить так: развшпие биосиапем (биоморф, в чаапноапи) осущеапвляется упорядоченно под управлением наследапвенности и в зависимости от влияния окружающей среды — в целом, оно тройственно ио максимуму, сопровождаясь, на одном направлении, доминированием ростовых процессов (нарастания и ветвления), которые происходят, более всего, на стадиях восходящего развшпия, когда проявляется еще и особое (второе) направление развшпия — воспроизведение новых живых men, подобных родшпельскому (размножение у биоморф), а после завершения воспроизведения или параллельно с ним в развшпии начи- нают доминировать процессы деградации (третье направление), которые когда-то могут приводить к ympame способности воспроизводшпь себе по- добные живые тела, что сильнее всего проявляется на стадиях нисходящего развшпия биосиапем и завершается после подавляющего снижения ростовой деятельноапи элиминацией (смертью), которая может наступить и в ре- зультате воспроизведения или когда-либо еще, например, иод воздействием сильных жизнеразрушающих факторов (травмы, отравления и т.n.). Первое направление движения ресурсов (интеграция) доминирует при фотосинтезе, развертывании вегетативного тела, при создании ба- зового органа, во время подготовки к размножению и т.д. Второе на- правление движения ресурсов (дезинтеграция), когда оно начинает до- минировать, ведет к утрате живыми структурными элементами прежней активности или прекращению жизнедеятельности какой-либо части биосистемы и всей биосистемы, теряется тонус в размножении, живое в соматической организации отступает и уступает неживому, проявляя стремление к элиминации. Оба рассмотренных процесса часто «вплете- ны» друг в друга настолько, например, при дыхании, что можно гово- рить только о доминировании одного из них в одних условиях и (или) в одной части тела, допустим, у биоморфы, или о доминировании дру- гого направления в движении ресурсных потоков в иных обстоятель- ствах. Взаимодействие процессов интеграции и дезинтеграции может быть в чем-то сбалансированным даже на протяжении всей или почти всей жизни биосистемы, поддерживая, например, гомеостаз. С другой стороны, эти процессы часто несбалансированы, поскольку в тех же биосистемах действует гетерокинез (см. главу 6), в результате чего все известные биосистемы испытывают его давление, развиваясь сначала по восходящей (на «подъем»), а с некоторого времени — по нисходящей (на «спуск»). От такой диалектики развития на Земле еще никто из биосистем не отошел (рис. 346-348 и др.).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 346. Биографический портрет Allium victorialis L. [сем. Alliaceae (Вос- точный Казахстан, Алтай, долина горной реки Белая Уба, лесные луга и поляны)]. БИОМОдфОТИП: подземно-односемядольные очередно-простолистные гемисимподиаль- ные луковично-компактнокорневищные семидетерминатно-репродуцирующие (o6pssyx»- ся верхушечные шаровидные полузакрытые соцветия) однодомные белоцветковые травя- нистые многолетники.
Глава 7. Концеиция разнообразия биотической власти Рис. 347. Биографический портрет Anemonastrum crinitum (Juz.) Holub [сем. Ranunculaceae (Алтай, Бащелакский хребет, верхняя часть лесного пояса и высокогорные луга, местами поднимается до снежников)]. БиоморфОТИП: надземно-двусемядольные розеточные (эти растения не строго розеточ- ные, поскольку у их почти безлистных пазушных цветоносов все же на самой верхушке, предваряющей зонтиковидное соцветие, развивается два-три небольших сближенных зе- леных присоцветных листочка, нередко срастающихся основаниями) эумоноподиальные интеркалярно-инфлоресцентные (большинство цветоносных побегов развивается из пазух чешуевидных или розеточных листьев годичных побегов текущего года, но иногда обра- зуются, особенно в наиболее холодных» местообитаниях, — у снежников, постинтерка- лярные цветоносные побеги, которые не успели отрасти в прошлом году из пазух зеленых листьев и только в текущем году отрастают, но уже из пазух прошлогодних листьев) однодомные компактнокорневищные ретардационно-кондивидуальные бело-лютиковоц- ветковые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ Рис. 348. Биографический портрет Polemonium саеги1еит L. [сем. Polemoniaceae (Алтай, Бащелакский и Коргонский хребты, лесные поляны и опушки, берега горных рек)]. Др — верхушка закрытого соцветия (Troll, 1964, с. 197). Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, 7 — 10 мм дл., 3,5— 5 мм шир., закругленные, клиновидно суженные в черешки 5 — 15 мм дл.) полурозеточные очередно-непарноперистолистные гемисимподиальные короткокорневищные детерминат- но-репродуцирующие ретардационно-кондивидуальные (замедленно вегетативно размно- жающиеся) однодомные сине- или голубоватоцветковые травянистые многолетники. 7.1.2. Размножение как начало начал — способ преодоления жизнью дискретности биоморф В организации живой природы обязательным условием ее суще- ствования на Земле является дискретность частных живых телесных систем (биосистем, или биосом). Жизнь, однажды появившись на Зем- ле, могла бы развиваться по тупиковому принципу и вскоре бы исчезла, если бы быстро не изобрела способ преодоления дискретности. Живые тела адаптировались обходить последствия дискретности, а причиной ее всегда была, есть и будет в живой природе только смерть, какую бы форму она ни приобретала в процессе эволюции. Формой преодоления дискретности жизнь сделала размножение [половое и (или) бесполое] биоморф, а все остальные биосистемы, в отличие от биоморф, не раз- множаются, а воспроизводятся. Такое разграничение в общем-то тради- ционно, но к размножающимся биосистемам я добавляю, кроме орга- низмов, еще и синорганизмы, т.е. размножаются все биоморфы, а все гипобиоморфы и метабиоморфы способны воспроизводиться. Размножение — это образование новых биоморф — новых особей (организмов) или синорганизмов. Воспроизведение — это образование
Глава 7. Концеиция разнообразия биотической власти из родительских живых тел новых, организационно подобных им, жи- вых тел, как-либо отличающихся от исходных и не являющихся биомор- фами. Так, например, воспроизводятся (возникают живые тела-потомки из родительских тел, которые по своей организации не являются био- морфами) клетки, ткани, органы внутри растительных организмов. Воспроизводятся также популяции внутри видов, но и все остальные биосистемы, начиная с видов, воспроизводятся или способны воспро- изводиться. В последнем случае речь идет о биостробе (живом теле) Земли. Виды воспроизводят друтие виды, которые, несмотря на отли- чия, принадлежат тому же уровню организации, что и родительские виды. Как бы там ни было, но в основе преодоления дискретности биосистем, в первую очередь, лежат процессы, которые способны обес- печить сосуществование жизни со смертью. Таких процессов мы назва- ли два: размножение и воспроизведение. Биостроба Земли не размно- жается, но потенциально способна к воспроизведению (размножением и воспроизведением занимаются ее соответствующие структурные эле- менты), но в один прекрасный момент она может воспроизвести, во многом подобную себе, но все же друтую, биостробу на иной планете. Жизнь растительных биоморф немыслима без воспроизведения и размножения соответствующих живых тел. Посредством этих двух про- цессов жизнь обеспечивает себе преемственно-историческую непрерыв- ность в пространстве-времени. Биоморфы семенных растений в процес- се эволюции, как известно, освоили различные способы репродуктив- ного и вегетативного размножения (рис. 349-351 и др.). Через размно- жение вид более всего контролирует поведение своих биоморф, которые служат ему в статусе структурных элементов — хранителей и исполни- телей многих свойств вида. 7.1.3. Парадокс и феномен смерти в явлении жизнь» В бесконечных процессах соединения и разъединения ресурсов как элементов движения-изменения возникает и исчезает великое разнообра- зие телесных систем и процессов. Носители жизни (биофоры) на Земле существуют на основе использования процессов соединения и разъеди- нения, конечным результатом взаимодействия которых является их смерть как конкретных живых тел, и, таким образом, происходит реализация одного из свойств жизни — дискретности ее носителей. Что будет с биостробой (живым телом) Земли, сейчас обсуждать не будем, но отме- тить все же надо, что многое сейчас уже связано с поведением челове- чества (см. т. 1, гл. 3). Жизнью, в качестве средства, обеспечивающего дискретность любых биосистем на Земле, была изобретена смерть, при- чем такое «изобретение» состоялось без каких-либо проблем и стало сразу же доступно всякому телу, ставшему в процессе эволюции носителем жизни. Причина смерти, по большому счету, в том, что у живого изна- чально присутствует противоречивая двойственность в развитии своих непосредственных телесных носителей — биосистем (эврибиоморф): стремление к завоеванию и освоению пространства и, с другой стороны, — естественная «тяга» к своей первооснове, т.е. к неживому, к возвраще- нию в него (см. главу 7.4.2.).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ё~~~ ~~ ю ~~ 3 О u a О '~ 4~ Ф 4) ф~~ ~~~}, 1/~ Ц~ СЧ О 4~ О:~: и О д ж О g Q Й ~-'( Л л ! р,~ Я д; р' A ~) ( g е '~~И с) 5: щ 2 м ж Й 1 ф;1 1 1 доо р х и m О C е[ > 2 жоор о ~ ~ ~) м ОQ~Ф И щ З О +-~а.х О й 5 33 3 2 л )Я 63 [-» М Щ М О Щ О ° ю~» сп Щ Щ 3 М О 1 1 1 1 1 ф)! ф» 1 1 1 1 1 1 1 ~О ~') ~ Р ф; 4~ О М й О О О. ~ »м Ll qp 2 A Y)~»~м ~ф р» р» ~ д~ Юе4южщОv (vИ и х о ~~ д < ~ ~~ Pe R о < о о~g, ооо~~~-Ыvm~e О ущv),Ор~ ~~У~ОО~~~ИО<~v. -О L. R Ж С4 Х И Л о f- '& f- ( ( М Щ Щ ° ю~» С: Я;> ~-) М 1 О Щ X о О. Е Д и 2 Ж О 0 ~~~ о O р 2 Ф Я о ~ Х ~0 )~~1~~ (Ь с ~ О Г~) )~ Щ О Я ~. М
Глава 7. Концеиция азнообразия биотической власти СК Рис. 350. Биографический портрет Stachys officinalis (L.) Trevis. [сем. Lamiaceae (Московская область, лесные поляны, опушки)]. БИОМОрфОТИП: надземно-двусемядольные (семядоли яйцевидные, широкозакругленные, с выемкой или без нее, 4 — 7 мм дл., 2,5 — 5 мм шир., резко суженные в черешки 2— 5 мм дл.) полурозеточные супротивно-простолистные эумоноподиальные семидетерминат- но-репродуцирующие компактнокорневищные ретардационно-кондивидуальные однодом- ные пурпурно-яснотковоцветковые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЯ 4, Рис. 351. Биографический портрет Cacalia hastata L.[ñåì. Asteraceae (Восточ- ный Казахстан Алтай Ивановский хребет, лесной пояс, поляны, опушки, ~~~~~~+ s 7 берега ручьев, заросли кустарников)]. БИОМОрфОТИП: надземно-двусемядольные (семядоли эллиптически ческие 10 — 15 мм дл., 3,5 — 5 мм шир., закругленные, постепенно суженные в черешки 3 — 1 дл.) р — 10 мм .) без озеточные столистные гемисимподиальные [из подземной пазушной почки (редко из очередно-простоли дв таких почек) цветущего родительского побега в том же вегетацио тр- нном сезоне о а- стает короткий подземный побег замещения, котор у ый зим ет и после отмирания всего остального отплодоносившего родительского побега продолжает весной фазу своего веге- тативно-репродуктивного н го надземного роста и развития] семидетерминатно-репродуциру- ющие (верхушка дифазного побега завершается полузакрытым метельчатым соцветием из ко зинок) однодомные желто-астроцветковые коротко р щ р ко неви ные етардационно-кон- дивидуальные травянистые многолетники.
Глава 7. Концеиция разнообразия биотической власти С момента своего возникновения всякое живое тело на Земле, как известно, обречено на смерть. В процессе эволюции живое, будучи представлено в той или иной телесной системе, адаптировалось отдалять или приближать к началу этой системы свой последний результат— смерть, а последняя сама стала атрибутивной адаптацией (!). «Смерть, прекращение жизнедеятельности организма, гибель его. У одноклеточ- ных организмов (например, простейших) смерть особи проявляется в форме деления, приводящего к прекращению существования данной особи и возникновению вместо нее двух новых. Смерть теплокровных животных и человека связана с прекращением прежде всего дыхания и кровообращения. Различают два основных этапа смерти: клиническую смерть и следующую за ней биологическую, или истинную, — необра- тимое прекращение физиологических процессов в клетках и тканях» (Бол. Энц. Сл., 1997, с. 1113). В отношении смерти ясно главное: она проявляется везде, где проявляет себя жизнь, т.е. на всех уровнях орга- низации живой природы. Ранее (т. 1, гл. 3) было показано, что любая дискретная трансфор- мация живой материи — это адаптация. Любая биосистема существует как адаптация и на основе адаптаций, по-разному и сохраняющих, и разрушающих жизнь. Кроме болезней-адаптаций в существовании жи- вых тел на особом счету у жизни стоит такая разрушительная адаптация, как смерть, которая, будучи особой функцией жизни, «отчисляет» кон- кретные тела — непосредственные жизненосители из состава живых и переводит их в разряд мертвых. Так бывает часто, но не всегда. Только смерть мешает жизни быть континуумом в соответствующих телесных системах. Это она, смерть, загнала жизнь в дискретные тела. Есть ос- нования сделать следующее обобщение: в одну и ту же жизнь одной- той же биосистеме 1иочти как «в реку») нельзя войти дважды, иоскольку такая биосистема всегда смертна, а значит, дискретна — это закон (примаспециальный) дискретности телесной жизни. Итак, жизнь весьма многообразна, в ней есть свой, только еи ирису- щий колорит, это она, чтобы быть разнообразной, научилась сосущество- вать в единой связке со смертью, сделав иоследнюю своей функцией и одной из форм адаитаций. Смерть, будучи функцией жизни, причем последней функцией в прежнем теле, выталкивает соответствующую телесную систему «за порог» (элиминирует), т.е. выводит ее из жизни — из пре- жнего качества. Смерть как адаптация — это первый иарадокс и одновременно— атрибут организации жизни в ее взаимодействии со смертью. Второй иарадокс и он же еще и атрибут взаимосвязи жизни со смер- тью — смерть не обязательно образует трупы, но обязательно, если их нет, приводит к качественной утрате прежней индивидуальности и возникнове- нию новой (рис. 349-351). В противоречивом единстве жизни и смерти, когда смерть — это функция жизни, можно усмотреть феномен смерти в явлении «жизнь» и, в частности, в организации биоморф семенных растений.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ 7.2. Ценомерия — атрибут организации Природы, ценобиоз и меробиоз- атрибуты жизни на Земле Сейчас общеизвестно, что более всего две обязательные стороны в организации жизни на Земле подчеркивают ее содержание — это пре- емственно-историческая непрерывность существования всепланетного живого тела (биостробы), что поддерживается размножением биоморф и воспроизведением всего остального, что обладает какой-либо жизне- емкостью, и дискретность в проявлении живых тел, составляющих орга- низацию биостробы, способом достижения которой является смерть (элиминация). Есть смысл объективно происходящие в живой природе события закрепить и обосновать в соответствующих понятиях и разоб- раться с иерархией целых живых тел и их частей, построив соответству- ющую «пирамиду» абстракций. В этой связи речь может идти, в первую очередь, о двух типах основополагающих явлений, обеспечивающих существование всей живой природы: ценобиозах и меробиозах, которые в свою очередь составляют только частные случаи еще более общего, всеприродного явления — ценомерии. 7.2.1. Основные состояния материи, вовлеченной в жизнь на Земле То, что жизнь организована многослойно (многоуровнево), не вызывает сомнения, но то, что на одну многослойность накладывается (под иным углом) другая, а на нее — третья, а на последнюю — четвер- тая и т.д., т.е. на самом деле жизнь проявляет себя как разномногослой- ное явление — это особая идея, разработка которой уже раньше нача- лась в этой книге (глава 1.3.3.), была далее продолжена (глава 4.8.), развивается в этой главе и далее будет еще и еще разрабатываться на протяжении всей нашей монографии. Для меня в этой идее ясно глав- ное («овчинка стоит выделки»), что не только членение целых тел на части, такие как органеллы, клетки, ткани, органы и т.п., вскрывает многослойность жизни, но и нечто еще — другое, например, то, что я предложил называть «жизнеемкостью», или «витальностью», действи- тельно обеспечивает жизни неповторимое разномногослойное содержа- ние. Определимся с ключевым понятием концепции разномногослойно- сти жизни. Главная особенность в организации живых тел, на которую следует всегда обращать внимание, в том, что они проявляют себя и как части, и как целое. Все природные явления находятся в зависимости от универсального свойства материи — ценомерии. Дословно (с греческо- го) — это «общее частей или части общего», и то, и друтое в нашем случае приемлемо при формировании соответствующего термина. Це- номерия — способность тел и нетел, как свойства материи, быть одно- временно и целым, и частью, в зависимости от того, в каком качествен-
Глава 7. Концеиция разнообразия биотической власти но-количественном содержании это явление в конкретном варианте осуществляется и тем самым себя выражает, а с позиций наблюдателя — под каким утлом обзора оно воспринимается. Свойство материи выступать в отношении одних и тех же тел под одним углом как часть целого, а под друтим — как целое частей — это и есть ценомерические состояния материи, или ценомерия тел и нетел. Состояния материи как частей и как целого — ее ценомерические состояния — пронизывают содержание всех природных явлений и, безусловно, явления жизнь. Ранее (глава 1.3.3.) было показано, что живые тела проявляют себя с разной жизнеемкостью (витальностью): как ковитальные (полножизне- емкие, или ценобиозные) и приватвитальные (неполножизнеемкие, или меробиозные). В области приватвитальности проявляется большое раз- нообразие таких слоев жизни. Когда генофенетическая организация составляет не целое (если целое, то это удел ценобиоза), а часть целого, то речь может идти только об одной из форм приватвитальности (ме- робиоза) — о приватковитальности (ценомеробиозе), что характерно для разных, генофенетически организованных структурных элементов био- морф, например, клеток, тканей, органов. Другие стороны меробиоза (приватвитальности) как формы разной жизнеемкости тел: интервиталь- ность, семивитальность, квадрантвитальность — переплетены в единой организации биоморф в одно целое настолько, что только посредством аналитического (с применением различных исследовательских приемов) меротомирования можно достигнуть искомого результата — расслоить эту единую организацию на части (меросомы). Место этим частям в соответствующей «пирамиде» абстракций, построение которой мы и предваряем некоторыми рассуждениями. Ценобиоз — это не только способность жизни как единого живого тела на планете Земля (а шире — во Вселенной), несмотря на дискрет- ность своих частных телесных носителей, проявлять и сохранять цело- стность ее организации вообще (жизнь вообще), но и свойство каждого друтого живого тела быть целым по отношению к его частям (подробнее — глава 1.3.3.). Посредством воспроизведения (размножения у биоморф) живых тел достигается преемственно-историческая непрерывность. Ра- нее (т.1, гл.3) было показано, что бесконечное сохранение жизни во Вселенной потенциально возможно, если развитая на Земле разумная жизнь проявит стремление к нообиофильной суперадаптации (беско- нечному сохранению разумной жизни, а следовательно, и жизни вооб- ще). Однако, при всем этом не будем забывать, что первое и обязатель- ное условие проявления жизни как ценобиозного явления — это спо- собность ее непосредственных носителей — биосистем — воспроизво- диться (в отношении биоморф — размножаться). Вот почему размноже- ние биоморф и воспроизведение остальных биосистем мы ставим в организации жизни на Земле выше всего остального и считаем, что именно размножение, а шире — воспроизведение, сделало ценобиоз нормой жизни на нашей планете. Организация растительных биоморф, а также и других биосистем, буквально «накачана» различными видами ресурсов, среди которых, помимо неживых биогенных частей, есть и другие тела-части (меросо-
552 OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ мы): неживые абиогенные меросомы (мероабиосомы) и живые меросо- мы (меробиосомы). Свойство материи проявлять себя телесно (сомати- чески) в биосистемах, или биосомах (живых системах, или живых телах) как качественное разнообразие структурных частей (меросом) в каком- либо теле — это меробиоз (приватвитальность). Явление меробиоза исходит из закона единства живого и неживого (закона Докучаева-Вер- надского, т. 1, гл. 3). Согласно меробиоза, любое живое тело — это сложная система — целое, состоящее из частей (меросом), в разнооб- разии которых есть определенная закономерность, т.е. оно укладывается в некоторое число типов меросом, соответствующим образом располо- женных в организации живого тела-целого и взаимодействующих между собой (главы 1.3.3., 4.8.). Быть целой биосомой — это одно, а вот быть меробиосомой (разновидностью меросом, другая их разновидность- мероабиосомы) — это уже совсем другое. Быть меробиосомой — это значит быть живым структурным элементом, т.е. не всем целым живым телом, а только его частью. Важно в поднятой здесь проблеме ценоме- рии, а ее важность выходит далеко за пределы биологии (ее нельзя обойти, например, при объяснении мироздания вообще), правильно расставить акценты. В случаях приватвитальности однозначно имеет место жизнь, но в особом своем качестве — не в полножизнеемком (ковитальном), а именно только частичном — неполножизнеемком (при- ватвитальном), когда она, жизнь, выступает как атрибутивная часть це- лого. Приватвитальные части (меробиосомы), оказавшись вне единства с другими меробиосомами, или не способны вовсе существовать друт без друта, или через какое-то время достраивают части, недостающие до родительского целого. Например, полностью обособленные партикулы: побеги, корни, листья, вегетативные клетки и т.п., если достраивают части, недостающие им до целого организма, кого-то из основных ор- ганов (побегов или корней), то превращаются в новые живые тела— организмы (т. 1, гл. 4.5.). Не все меробиосомы, на основе известных сейчас фактов (нельзя, однако, исключать и другое, что мы просто еще многого не знаем об их возможностях), способны проявить самостоя- тельность и начать существовать вне единства с другими меробиосома- ми родительского целого. Например, известно, что гены и их системы (геносомы) автономно от фенов и их систем (феносом) и наоборот, как жизнеразвивающиеся тела, не существуют; они всегда только вместе в организации целых живых систем. Не будем также забывать и о том прогрессе, который в наше время уже происходит в области генетичес- кой инженерии (у всех, например, на слуху знаменитый эксперимент с вегетативной клеткой овцы, приведшей к появлению ее генетической копии — овепси Долли; то ли еще будет, прогресс ожидается, действи- тельно, неоспоримый). Статус тела, которое выступает как часть (структурный элемент) другого тела — один, а как тела-целого — иной, и это обстоятельство имеет важное значение в осознании явлений меробиоза и ценобиоза. Одна и та же биосистема может под одним утлом зрения выступать как целое, а под другим — как часть целого. Организм, например, ценоби- озная система — целое своих частей — меробиозных систем: органов,
Глава 7. Концеиция разнообразия биотической власти 553 тканей, клеток, органоидов. Однако, статус того же организма в биоце- нозе уже другой: он меробиозная система — часть (структурный эле- мент) другого целого (биоценоза) и т.д. (глава 1). Итак, материя динамична (другого не дано), и ее состояния в за- висимости от обстоятельств, будучи в телах или нетелах, способны «перетекать» одно в другое, т.е. трансформироваться в рамках единого общеприродного явления — ценомерии. Ценобиозы и меробиозы — это одни из типов «текучих» ценомерических состояний материи, способ- ные в зависимости от обстоятельств — той роли, которую на себя через них принимают носители этих состояний и которую выполняют в дан- ном месте и в данное время в мироздании, обеспечивают живым телам свойства где-то быть целыми по отношению к другим телам-частям, а где-то частями в отношениях с телами-целыми. Ъ Всякое неживое тело вообще, или абиосома, может быть или эуаби- осомой — неживым абиогенным телом, или некросомой (некроном)— неживым биогенным телом. Когда абиосомы выступают в ранге целого, то обозначим их ценоабиосомами, а некроны в ранге целого — это ценонекросомы. Когда абиосомы проявляют себя как части целого, то это абиомеры, а некроны в ранге части целого — некромеры. Если живое тело, или биосома, имеет статус целого, то обозначаем его как ценобиосома, а в тех случаях, когда оно только часть целого тела, то это биомер (меробиосома). Таким образом, определенные тела, в зависимо- сти от сложности своей организации и статуса, который они занимают по отношению к другим телам, могут проявлять себя как меросомы или как системы всех меросом, составляющих определенные тела, — целые тела (ценосомы). Если меросомы бывают абиомерами, некромерами и биомерами, то ценосомы (целые тела) — ценоабиосомами, ценонекро- сомами и ценобиосомами. Особые типы биомеров, как известно, со- ставляют гены и их системы (геносомы), наряду с фенами и их систе- мами (феносомами). Как было показано выше, смерть констатируется не по состоянию генов и геносомы, а по состоянию фенов и феносомы. В зависимости от обстоятельств, под несколько иным утлом обзора, роль меросом могут выполнять в организации ценобиосом разной слож- ности организации определенные системы: абиоморфы, гипобиоморфы, биоморфы и метабиоморфы (т. 1, гл. 2). Учитывая приведенные выше соображения, положение неживых и живых меросом и целых тел, иерархию их отношений в организации живой природы можно выразить в соответствующих субординатных классификациях частей и целого и их графической модели в образе «пирамиды» абстракций (рис. 352). Все термины, использованные в этой классификации, уже обсуждались в первом томе или в предыдущем материале текущей работы, поэтому мы не будем здесь детализировать их содержание в суперординатной классификации, отраженной в ее графической модели, дополнительными объяснениями. Ранее (глава 1) было показано, что все тела, вклочая биосистемы,— »о сложные телесные системы, которые состоят из частей-тел, или ме- росом. В организации любых живых тел всегда можно обнаружить меро- сомы, которые их составляют и являются таковыми для соответствующего
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 352'. «Пирамида» абстракций как отражение иерархии неживой и живой ма- терии тел-частей (меросом) и целых тел (ценосом) в организации живой природы. А — Д — уровни абстракции (суперординаты); 1 — меросомы, 2 — ценосомы, 3— абиосомы, 4 — биосомы, 5 — абиоморфы, 6 — эврибиоморфы, 7 — мероабиосомы (ин- витальные, или неживые, тела, сокращенно — инвиталы), 8 — меробиосомы (приватви- тальные, или неполножизнеемкие, тела, иначе — приватвиталы), 9 — ценобиосомы (ко- витальные, или полножизнеемкие, тела — ковиталы), 10 — эуабиосомы, 11 — некросомы, 12 — анабиосомы (анабиозные, или вицевитальные, тела, сокращенно — вицевиталы), 13 — эубиосомы (сравнительно активно жизнеразвивающиеся тела), 14 — абиомеры, 15— некромеры, 16 — биомеры, 17 — гипобиоморфы, 18 — биоморфы, 19 — метабиоморфы, 20 — квадрантвиталы, 21 — семивиталы, 22 — приватковиталы, 23 — интервиталы, 24— органеллы, 25 — клетки, 26 — ткани, 27 — органы, 28 — организмы, 29 — синорганизмы, 30 — популяции, 31 — виды, 32 — меробиоценозы, 33 — биоценозы, 34 — меробиостроба (бактериостроба, микостроба, фитостроба, зоостроба, антропостроба, или человечество), 35 — биостроба, 36 — панбиостроба, 37 — 42 — надвиды: 37 — родовые, 38 — семействен- ные, 39 — порядковые, 40 — классовые, 41 — отделовые, 42 — царственные.
Глава 7. Концеиция разнообразия биотической власти 555 уровня организации жизни. Меросомность с позиций закона иерархич- ности проявляет себя двойственно («принцип вложенности»): с одной стороны меросомы составляют определенное тело-целое, а с другой, при подъеме вверх на следующую ступень «пирамиды» абстракций, бывшее тело-целое (ценосома) утрачивает свой прежний статус и является уже меросомой и т.д. до самой вершины рассматриваемой классификации. Из рассматриваемой графической модели (рис. 352) суперординии меросом и целых живых тел, объединяющих разные меросомы в целое (живые ценосомы, или ценобиосомы), наглядно видно, что всякое живое тело под разным утлом обзора и с разной высоты абстракции обязатель- но состоит из неживых абиогенных структурных элементов — абиоме- ров, из неживых биогенных структурных элементов — некромеров, и, наконец, собственно живое содержание любых ценобиосом составляют живые меросомы, или биомеры. Определенное единство абиомеров, некромеров и биомеров определяет содержание и всего живого тела Земли, ее биостробы, и отдельных растительных биоморф, но и любых других живых тел. Поскольку первооснову живых тел составляют всегда неживые аби- огенные структурные элементы (эуабиомеры), т.е. атомы, молекулы и их агрегаты, то незримо существует естественная тяга (!) всякого проявле- ния жизни — всякой биосомы (биосистемы) к дискретности — к воз- вращению к своей первооснове, если образно, то как бы на «историчес- кую родину», — к неживой материи, что обеспечивается атрибутивной адаптацией жизни, ее последним в конкретном теле функциональным синдромом, или финитофункцией, — смертью. Таким образом, в суще- ствовании любого живого тела, помимо обязательного стремления к жизнесохранению, всегда присутствует и в определенное время прояв- ляется стремление к жизнеразрушению и, в частности, к разрушению собственной жизни и к смерти (подробнее см. т. 1, гл. 3). В рассматриваемой здесь (рис. 352) суперординатной классифика- ции (суперординии) на одной из ее ступеней (Д) как «пирамиды» аб- стракций нашла графическое отражение особая классификационная ветвь, точнее полуветвь, поскольку она, начинаясь снизу от уровня видов не свободна в своем дальнейшем развитии и «замыкается» на уровень биостробы Земли, — надвидовая, на чем есть смысл заострить внимание. Дело в том, что обычно систематики говорят о надвидовых таксономических категориях (род, семейство, класс и т.п.), считая объек- тивно реальными только виды, а все, что над ними — субъективный результат таксономического классифицирования видов. На мой взгляд, кроме типов живых тел (биосистем) в ранге видов вполне реальны и надвиды, т.е. оба типа этих тел существуют в природе в двух измерениях — объективном и субъективном. Я считаю, что как вид (species), так и надвид (supraspecies) имеют соответствующее объективное содержание, ис- ходя из которого их можно (и нужно) подвергать различным субъективным операциям, в частности, обязательной логической процедуре — таксономи- ческому классифицированию. И вид, и надвид — это и объективная, но и субъективная реальность. С этой позиции можно считать, что объек- тивно в Природе существуют виды, объединенные на основе определен-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ ных законов и правил родственного жизнеразвития в различные систе- мы видов — надвиды, что устанавливается при сравнительном исследо- вании видов, на основе которого проводится таксономическое класси- фицирование, когда обнаруживается в них определенная степень род- ства (родовая, семейственная, классовая и т.п.). Надвид — объективная категорня, имеющая, кроме того, еще и определенный таксономнческий (субъектнвный) ранг. Главные ринги тиксонов в восходящем порядке, официально признанные систематиками, следующие: вид, род, семейство, порядок, класс, отдел и царапво..., а если требуется, то выделяют и большее число промежуточ- ных рангов тиксонов, например, иодцарапво, подотдел, подкласс и т.д. (Стифлё и др., 1980). Надвиды бывают родовые, семейственные, порядко- вые, классовые и т.д. Надвиды — это обьективные живые тела Природы, которые имеют вполне конкретные границы, но мы, люди, пытаясь их определить морфологически, географически и т.n., действуем субъективно (другого не дино) и чаапо эти границы устанавливаем так, как видим со своих исследовательских позиций, а зничшп, к нашему всеобщему сожале- нию, границы нидвидов, которые мы можем знать, — всегда субьективны; они более или менее приближены к объективной реальности. Отражение объективных признаков надвидов, как и всего остального, что объектив- но, мы получаем в нашем сознании и тем самым всегда можем устанав- ливать только субъективно границы тел и нетел Природы. Действуя, как положено в научном исследовании: меротомируя Природу и ее фрагменты, а далее ее же мерономируя (типизируя, клас- сифицируя, моделируя), мы приближаем наши знания о ней в целом и ее частях к тому, что действительно с ними происходит, независимо или зависимо от нашего сознания. Безусловно, проблема, которую мы здесь как бы между прочим подняли в отношении категорий «вид» и «надвид», имеющих и объек- тивный, и субъективный аспекты, но являющихся атрибутивными эле- ментами в организации живой природы, потребует продолжения иссле- дований. Подчеркнем позицию в нашем подходе: не только виды, но и нидвиды — обьективная реальность, которую можно и нужно изучать субьективно, одновременно подвергая их тиксономическому классифициро- ванию, используя его результаты для обозначения не только каждого кон- кретного вида, но и нидвиди, что позволяет надвиду, исходя из его кате- гории в тиксономической сиапеме, иметь свой эпитет (родовой, семей- ственный, классовый и т.и.), что наглядно видно ни графической модели (рис. 352-Д). Классификационная полуветвь надвидов, отраженная в одной из субординатных классификаций «пирамиды» абстракций, сви- детельствует об усложнении организации родственных живых тел-видов в некотором пространстве Земли — той надвидовой ее части, которая исследуется средствами таксономического классифицирования. Этот классификационный виток-полуветвь на самом верху субординатной классификации по условному типу «Д» замыкается на другой классифи- кационный виток. В один царственный надвид, по сути, вошли все виды растений биостробы Земли, соединив тем самым на одной с~е« «пирамиды» абстракций таксономическое пространство соматической
Глава 7. Концеиция разнообразия биотической власти организации жизни с пространством ценотическим из другого классифи- кационного витка. Так происходит осознание простой и естественной истины: все мы, живые существа на Земле, по большому счету, родствен- ники: и те, кто свои, и те, кто не-свои, но родные, но и те, кто не-свои, и все-таки не чужие, причем родственники и те, которые кого-то едят, но и те, которых кто-то ест, и т.п. Все живые существа, как мы видим, вообще-то, хотя и относительные, но все же «сверхбратья» и «сверхсес- тры», определяя надвиды, мы только подчеркиваем размеры и биотичес- кое содержание этих по-разному таксономически организованных тел. Итак, жизнь — это, безусловно, явление соматогенное, разноуров- невое по сложности организации своих носителей, ценобиозное, с од- ной стороны, обеспечивающее, посредством размножения биоморф и воспроизведения других биосистем преемственно-историческую непре- рывность, и меробиозное, с друтой стороны, создающее дискретный фон ее частного телесного проявления, когда в каждой биосистеме- целом участвуют противоположно организованные части: меробиосомы, ориентированные в своей организации на сохранение жизни, и мероа- биосомы, организация которых, сама по себе, индифферентна, но более всего отвечает их содержанию, а, следовательно, имеет тенденцию воз- вращаться в неживое вообще, когда ресурсы сохранения дискретной жизни исчерпываются. Обо всем этом есть смысл поговорить подроб- нее, но сначала сформулируем одно правило-предостережение, которое подпитывается законом единства живого и неживого [законом Докуча- ева-Вернадского (т. 1, гл. 3)]: развитие системы «жизнь-нежизнь» только ио одностороннему сценарию через экспансию нежизни жизнью — это путь в никуда, а нужна не npocmo адаптация (ведь они всегда конечна — смер- тна) в живых телесных системах, а нечто большее — стремление к су- ио этому пути, что возможно только с Разумом (носитель его пока из- вестен один — Человек) и только через гармоничное развитие жизни вме- cme с окружающей ее неживой средой. 7.2.2. Смерть — обьективная реальность, но не приоритет в явлении меробиоза Смерть — это, как было отмечено выше, жизненная функция, или особая адаптация, результатом которой является завершение (мгновен- ное или не столь быстрое, но все же довольно скоротечное) существо- вания в какой-либо телесной системе всего того качества, что опреде- ляло в ней фенетическое содержание жизни. Вопрос о смерти генов пока не изучен и, вероятно, гены в жизнеактивном виде нередко могут существовать какое-то время еще и после смерти, которая фиксируется по проявлению смерти феносомы в целом, составляющей самую дина- мичную часть биосистем (живых тел). Гипотетически (!) не стоит исклю- чать и того, что жизнеактивные гены каких-либо мертвых существ где- нибудь в вечной мерзлоте могут сохраняться даже десятки тысяч лет.
558 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ 06,26 К ак бы там ни было, а жизнь и смерть возникли в эволюции одновременно, причем смерть определилась сразу как атрибутивное свойство жизни. Живые тела осуществляют свою дискретность через последнюю их адаптацию — смерть, превращаясь, таким образом, при ее посредничестве в неживые тела. Смерть как функциональный син- дром (несколько, даже много функций обеспечивают ее), или последняя сложная функция (финитофункция) живого тела, срабатывает тогда, когда у этого тела в данный момент и в данном месте при данных обстоятельствах не хватает резервов, чтобы продолжать жизнь, иначе дальнейшее существование тела становится несовместимым с проявле- нием жизни, и данное тело само себе дает сигнал к смерти, останавли- вая жизненные процессы и переводя себя в состояние некрона (трупа). Некроны — это особые соматические (телесные) системы, которые когда-то были живыми, а сейчас как-либо существуют, более или менее активно разрушаясь; они являются биогенной формой существования неживой материи. Некроны, как и живые системы, бывают самыми разными по сложности организации, например, сухие шипы или колюч- ки, сухие ветки в кронах некоторьгх растений (рис. 353, 354), сухие листья, «согревающие» живые части тела растения (рис. 355) или упав- шие при листопаде с деревьев, или целый массив тайги, сгоревший во время пожара, и т.п. Рис. 353. Биографический пор- трет Acacia raddiana Savi [сем. Fabaceae (Израиль, Эйлатские горы, сухие межгорные доли- 01 к 1N� временных водостоков, 5 V)]. Биоморфотип: надземно-двусе- 39 0 мядольные (семядоли продолговато- эллиптические, мясистые, 8 — 10 мм дл., 3 — 4 мм шир., закругленные, в основании с остроугольной глубо- / кой выемкой и двумя рожками по краям, короткочерешковые) безро- / зеточно-ростовые доминирующе- эусимподиальные розеточно-репро- дуцирующие очередно-сложнолист- ./ 77,23 ные колючко-прилистниковые ин- теркалярно-репродуцирующие (в 80 пазухах сложных листьев вегетатив- ных длинных безрозеточных побе- / 2(5),гм4, гов текущего года развиваются розе- 18а,21(22), точные вегетативные побеги, кото- «,п.6 8 I 26(2Т) рые на следующий год продолжают 26б+18а 2у I рост и репродуктивное развитие уже e в пазухах сухих и мертвых колючек- ~% прилистников, образуя или одиноч- ~г ные интеркалярные тотарепродук- 68 тивные побеги, или межсоцветия из пп 2, редко З-х, таких побегов) одно- домные извилисто-бобовоплодные моноаэроподиальные (редко кусти- стые) зонтиковидно-крановые лис- топадные деревья.
Глава 7. Концеиция разнообразия биотической власти Рис. 354. Биографический портрет Sarcopoterium spinosum (L.) Spach [сем. Rosaceae (окрестности Тель-Авива, каменисто-песчаные холмы, март)]. ИОМОрфОТНП: надземно-двусемядольные (семядоли продолговато-яйцевидные, 3 — 5 мм дл., 1,5 — 2 мм шир., закругленные, клиновидно суженные в короткие черешки) розеткообразующие (помимо розеточных вегетативных побегов образуются еще полурозе- точные вегетативно-репродуктивные побеги и безрозеточные вегетативные побеги) очеред- но-непарноперистолистные стержнекорневые гемисимподиально-эусимподиальные (аэро- подиальный прирост обеспечивают резиды разных побегов, но основными колючкообра- зователями являются сильно ветвистые верхушки вегетативных ростовых побегов, кото- pûe, образовавшись весной, вскоре перестают расти, затвердевают, древеснеют и отмира- ют, кроме того, после плодоношения оси соцветий грубеют и также превращаются в колючки) семидетерминатно-репродуцирующие однодомные изоаклональные (без вегета- тивного размножения) колюче-подушковидные кустарнички.
560 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 355. Биографический портрет Paraquilegia anemonoides (Willd.) Ulbr. [сем. Ranunculaceae (Алтай, Курайский и Северо-Чуйский хребты, камени- стые склоны высокогорий, галечники, скалы, россыпи)]. БИОМОрфОТИП: надземно-двусемядольные розеточные полусложнолистные эумонопо- диальные интеркалярно-репродуцирующие (тотарепродуктивные интеркалярные поджегH развиваются из пазух зеленых розеточных листьев текущего года по одному, редко 06ра- зуются межсоцветия из двух таких побегов) каудексово-подушковидные (на множестве каудексовых ветвей, или каудиций, долго сохраняются мертвые рахисы листьев, ~~~~ры~ заметно утепляют внутриподушечную среду) однодомные лилово-лютиковоцветковые стер- жнекорневые дефективно-кондивидуальные (хотя и не все старые особи полностью про дольно расщепляются через каудекс и главный корень, но все-таки такое несовершенное вегетативное размножение бывает у рассматриваемого вида) травянистые многолетники
Глава 7. Концеиция разнообразия биотической власти Продукты локальной смерти в организации биосистем и, в частности, биоморф — некроньг — нередко задерживаются в ней, приобретая разный статус, в зависимости от обстоятельств, помогая или мешая дальнейшему развиппо згих биосистем. Резулыат (продукт) смерти — различные некро- ны: мертвые сложные белковые соединения, сухие побеги, колючки, мерт- вые листья, организмы, или леса, например, опустошенные пламенем по- жара, и т.п. Формы некронов — некромеры и ценонекросомы обречены на разрушение, но осушествяяют они зто быстро или медленно, причем, как правило, продолжая участвовать в поддержании и развитии жизни на Земле. Когда некроны не выводятся из живого тела и вьтолняюг определенные функции в статусе некромеров, участвуя нередко и в поддержании жизне- способности организации этого тела-целого, то они по своему еще находятся в границах жизни. Похоже, не стоит отмахиваться от внешней простоты фактов, о которых идет сейчас речь, поскольку ситуация, если поглубже ее рассмотреть, совсем не лишена своей особенной интриги. Жизнь не просто отбраковывает и избавляется от произведенного ею и уже отслужившего так или иначе материала, используя свою надежную адаптацию — смергь, пре- вращая с ее помощью некогда жизнеактивные структурные части в некроны, но еще и утилизирует их, приспосабливаясь по-разному к ним. Способность живых тел использовать с пользой свои же некроны,для дальнейшего раз- вития, не удаляя их из своей струкгуры, можно обозначить одним словом — «вицеадюпюшя» (дословно с латинского — «вместо адаптации»). Различ- ные проявления вицеадаптаций в организации семенных фитобиоморф мы отразили в графической модели (рис. 35б). Однако, если некроны вьшодятся за пределы живого тела в окружаю- щую среду и больше непосредственно не участвуют в его организации, то это явление можно обозначить как дисбиоз (от греч. приставки dys — при- ставка, обозначаюшюш отрицание и соответствует русскому «не» + bios— жизнь). Между жизнью (биозом) и дисбиозом нередко может вклиниваться партикуляция (т. 1, гл. 4), поскольку отделение от растительных биоморф каких-либо структурных частей происходит еще до их полного отмирания (рис. 355, 357 и др.). Когда проявляется партикуляция и отделяются полу- живые листья или ветви, то она может приводить или к дисбиозу (образо- ванию мертвого отпада), или к вегетативному размножению, например, к образованию новых особей из каких-либо деструктивных партикул. Таким образом, дисбиоз может сопровождаться партикуляцией или происходить без нее; в последнем случае от биоморф отделяются ценонекроны (мертвые листья, сухие ветки и т.п.). Жизнь растительных биоморф превратила эти телесные системы в арену своей «битвы» не только и (или) не столько со смертью, поскольку это ее же собственное произведение, воспользоваться которым она может в любое врешг, всегда выбирая в зависюиости от обстоятельств подходяший момент (или старость загонит в «тупик», или лютый мороз покалечит тело целиком, или топор лесоруба нанесет травмы, несовместимые с жизнью, и т.п.). Атрибутом жизни является не стремление к смерти, хотя последняя, безус- ловно, обьективная реальность, а борьба всех ее телесных носителей со всем остапьным, чтобы завоевывать и осваивать окружающее пространспю, что у семенных фитобиоморф осуществляется посредством всей «могучей пятер- ки». 'нарастания и ветвления, партикуляции и размножения, но еще и эли- минации, т.е. упорядоченного использования смерти как особой ф жизни.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 356'. Смерть в организации биоморф семенных растений и некоторые формы вицеадаптаций (жизнеполезного использования некронов). 1 — живые части растительных биоморф, 2 — отмирание верхних частей побегов (акро- некроз) после окончания вегетационного сезона, 3 — отмирание средних частей побегов (мезонекроз) и полное обособление партикул от родительских организмов, 4 — особи- клонисты, образовавшиеся в результате полной некротической партикуляции, 5 — отми- рание корневых частей и обособление корневых отпрысков с корнями, 6 — отмирание подземных частей побегов, которое приводит к вегетативному размножению, 7 — отми- рание растений снизу (базинекроз), 8 — неполное отмирание цветоносных побегов сверху, которое может приводить весной к постфлоральному побегообразованию, 9 — постфло- ральные побеги, 10 — отмирание побеговых ветвей, в результате которого остаются какие- либо сухие части на структуре живого растения, например, сучки (10 а) у деревьев, колюч- ки побегового происхождения (10 б) и т. п., 11 — отмирание листьев и (или) прилистников, которое сопровождается образованием из них колючек, 12 — отмирание покровных тка- ней, которое сопровождается вицеадаптациями в форме сохранения сухих шипов (12 a), отмерших частей коры (12 б), сухого опушения (12 в) и т.п., 13 — листовые вицеадаптации, когда жизнеполезны (накапливают влагу, утепляют и т.п.) и долго сохраняются на струк- туре живых растений сухие листья, 14 — отмирание плодов и их использование для длительного сохранения и при рассеивании семян, 15 — использование мертвых тканей семян при их рассеивании, 16 — использование вторичной (мертвой) древесины в жиз- неразвитии растений.
Глава 7. Концеиция разнообразия биотической власти Рис. 357. Биографический портрет Veronica densiflora Ledeb. [сем. Scrophulariaceae (Алтай, Курайский хребет, влажные каменистые тундры высокогорий, лужайки около снежников)]. БИОМОрфОТИП: надземно-двусемядольные (семядоли яйцевидные, 3 — 4 мм дл., 1,5— 2 мм шир., закругленные, резко суженные в короткие черешки) безрозеточные (всходы и ювенильные растения нередко развивают розеточную структуру, в дальнейшем образуются густолистные безрозеточные побеги, весьма близкие к четочным) супротивно-простоли- стные эпигеогенно-средне-короткокорневищные гемисимподиальные (часть надземных побегов развивается в одном вегетационном сезоне как короткие вегетативные полегаю- Шие, а на следующий год или позже они переходят к вегетативно-репродуктивному раз- витию) семидетерминатно-репродуцирующие однодомные лилово-норичниковоцветковые семиинтенсивно-кондивидуальные (вегетативное размножение только чем-то напоминает интенсивное, поскольку полная партикуляция распространяется на многие короткие и средние корневища, что приводит к завоеванию относительно небольших территорий и осуществляется хотя и не замедленно, но и не интенсивно) травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 7.3. Гетерократия — очередной атрибут жизни Организация биоморф буквально накачана различными видами ресурсов, существование которых находится в системе, подчиняясь определенному порядку, законам и правилам. Наука все более вторга- ется в тайны этой организации и на этом пути нас ждут все новые и новые откровения. Попробуем прикоснуться к одной из таких тайн. В предыдущем материале нашей монографии было показано, из чего состоит биоморфа семенных растений, как она развертывает свою организацию в морфогенезах и подчеркнуто уже известное, главное положение: биоморфа, ее фенетическая организация (феносома) управ- ляется с уровня генетической организации (геносомы). Под другим углом обзора организация биоморф состоит из двух меросом: внешней (экзосомы, или габитуса) и внутренней (эндосомы) организаций. Есть смысл посмотреть на ту же организацию еще под одним углом — через призму явления «власть». Организация биоморф, если сравнить ее с организацией, например, государства, также имеет «власть» и все, что находится под ее влиянием, т.е. исполнителей подвластной деятельно- сти. Слово «власть» мы ставим в кавычки, поскольку в организации биоморф традиционно работают другие термины и понятия, которые иногда могут быть аналогами таковых, применяемых в социальных явлениях. Как бы там ни было, но постепенно мы откажемся от упот- ребления кавычек к слову «власть», когда почувствуем, что читатель, в своем большинстве, уже переориентирован на более широкое воспри- ятие этого термина, а это, безусловно, вскоре произойдет (сомнения рассеются, как утренний туман под первыми солнечными лучами). Другого просто не дано; давайте сформулируем тезис и докажем его состоятельность: «власть» и ее разнообразие — один из атрибутов про- явления жизни телесных систем, причем не только социальных, но и биотических. 7.3.1. «Власть» в организации биоморф и ее разнообразие Все началось с того, что однажды я усмотрел аналогию в поведении некоторых структур из организации растительных биоморф с поведени- ем группы людей, возглавляющих целые государства и составляющих, таким образом, его власть. «Власть, в общем смысле способность и возможность оказывать определяющее воздействие на деятельность, поведение людей с помощью каких-либо средств' — воли, авторитета, права, насилия (родительская власть, государственная, экономическая и др.,); политическое государство, система государственных органов» (Вол. Энц. Сл., 1997, с. 212). В этом определении термина «власть» все вни- мание сконцентрировано на особенностях поведения людей, но оно- это только малая часть из различных проявлений поведения живых систем разных уровней организации. Вряд ли есть смысл ограничивать
Глава 7. Концеиция разнообразия биотической власти употребление такого масштабного слова «власть» (греч. «cratos») только сферой социальной деятельности людей. Побег или корень в организации растительной биоморфы и вся биоморфа в целом развиваются довольно строго определенным образом под воздействием какой-то особенной структуры, способной принуж- дать живые системы осуществлять только ту деятельность, которую им они — «некто» — навязывают, иными словами, их заставляют делать все или почти все, что им «приказывают». Жизнь, которая осуществляется под диктатом «каких-то» структур, что это — аномалии в росте и раз- витии живых систем? Нет, уже общеизвестно, что это весьма закономер- но, и все происходит по правилам растительного бытия — законам и правилам наследственности. Многое, что происходит в фенетической организации биоморф, как известно, контролируется деятельностью генов и их систем — геносом (традиционно — генотипов у организмов), — это они навязывают фенетической организации биоморф и ее живым структурным элементам (клеткам, тканям, органам) свою наследствен- ную «власть», обеспечивая им развитие выверенной в предыдущих по- колениях биоморф такой программной конституции («лепоты»), кото- рая нужна не только конкретным биоморфам, но и биосистемам более сложной организации, элементами которых они являются, а это, в первую очередь, видам, далее, по большому счету, — всей биостробе (живому телу) Земли. Живые тела предпочитают в своей организации определенный порядок, что более всего можно достичь с властью, а не без нее. Это они, гены, геносомы — хозяева в организации целостных растительных тел и их структурных элементов, внутри которых носители наследственности ведут себя как в своем доме (см. главу 9), обладая реальной властью. Я невольно потерял кавычки к слову власть и что- то не очень склонен к ним более возвращаться, поскольку далее мы уже будем следовать более широкому пониманию рассматриваемого терми- на, взяв на себя смелость к расширению его смысла. Власть — это, в общем смысле, способность и возможность оказывать определяющее воз- действие на какую-либо структурно-функциональную деятельность живых систем различного уровня органиэации с помощью любых, доступных для них, средств (здесь не только воля, авторитет, права и т и., свойственные коллективам людей, но и наследственность, и лидерство в фенетической органиэации, присущие организациям всех биосистем). цалее мы не будем говорить о социальной власти, оставив ее в стороне, а все внимание пе- реключим на осознание явления биотической (обусловленной жизнедеятель- ностью внутренней органиэации биосистем) власти, в первую очередь, в организации биоморф семенных растений. В организации биоморф безусловное лидерство принадлежит генам и генетической организации (особой части тела биоморфы) в целом- геносоме, что по смыслу в отношении биоморф (организмов и синор- ганизмов) то же, но по выражению даже лучше передает соматическое содержание этой части, известное для организмов под названием «гено- тип». Лидерство геносомы в организации биоморф проявляется в том, что от нее исходит определенный диктат (мы сознательно не используем слово «диктатура») целей (направлений) на развитие по определенному
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ плану всего остального. В чем же проявляется этот диктат? Для того, чтобы лучше разобраться с поднятым вопросом, вспомним общую те- орию жизнесохранения (т. 1, гл. 3), где однозначно показано, что орга- низация всех биосистем, биоморф, в частности, — результат единства и борьбы противоположных сил — сохранения и разрушения жизни, который далеко не всегда «лежит» на поверхности, а нередко скрыт в лабиринтах организации биосистем настолько глубоко, что доскональ- ных знаний о натуре упомянутых явлений наука до сих пор еще не имеет. И все же, на основе известного, удалось определить основной биологический закон (закон жизнесохранения, или биофилии): жизнь, возникнув на Земле (и — во Вселенной), изменяясь, сохраняет себя в объектах разной организации и в пределах, ограниченных ее возможно- стями (т. 1, стр. 94). Из общей теории жизнесохранения ясно, что жизнь на Земле возможна до тех пор, пока жизнесохранение доминирует над жизнеразрушением и силы сохранения чужой жизни превалируют над сохранением личной жизни. Итак, с генетического уровня в организации биоморф в направле- нии развития генетической организации (феносомы) исходит опреде- ленный план, который проявляется как диктат системы управления нгд развитием всей остальной организации. Организации биоморф чужд хаос; она признает только управляемую «лепку» своей конституции- фенетической «конструкции» (феносомы) из всего остального, что есть в границах ее физического тела и что пополняет его, пока теплится жизнь — материальной субстратной основы — протеросомы. Однако, в плановой «работе» генов есть свои приоритеты, которые делают «план» развития биоморф еще более целенаправленным, чем может показаться наблюдателю со стороны. Гены не только управляют развитием различ- ных фенетических структур, но и определяют, чье развитие из них будет приоритетным, а чье подчинится диктату этих приоритетных структур. Эволюция постаралась за миллионы лет создать систему многоуровне- вых приоритетов развития и многоуровневых диктатов одних структур над другими, превратив взаимоотношения между структурными элемен- тами в отношения их борьбы за существование, в которой цели развития организма, вида, а шире — всей биостробы Земли — доминируют над всеми остальными целями. Нет сомнений, что организация биоморф, а шире — и других биосистем, предопределена к проявлению власти и иерархии властей. Если посмотреть под рассматриваемым углом обзора на взаимоот- ношения структурных элементов организации биоморф, то бросается в глаза, что в них нет равенства. Они разные по сложности организации и по-разному специализированы; на языке социальных отношений это означает, что в организации биоморф действует «диктатура» руководи- телей живой природы — генов. Они, гены, — это стратеги, которые управляют развитием всей фенетической (тактической) организации биоморф, заботясь о ней как о родном доме, отдавая ему свою идио- энергию в количестве, которое достаточно для взаимного сохранения B соответствующих условиях, на что фены и феносома в целом отвечают неистребимой «заботой», создавая уютные условия для своих особенных
Глава 7. Концеиция разнообразия биотической власти обитателей, по сути, «хозяев» общего биоморфного «дома». Кроме того, среди генов есть свои гегемоны, т.е. геногегемоны, которые направляют развитие биоморф к размножению, причем не только к половому раз- множению, но и вегетативному. Стремление к размножению — это дань виду, к которому принадлежат соответствующие биоморфы. Биотическая власть в организации биоморф многослойная, при- оритеты принадлежат двум, а то и трем ее формам. Внешне — она принадлежит организмам или синорганизмам, а на самом деле — видам, диктат власти которых (эйдократия) превалирует в организации био- морф над всей остальной, т.е. она иерархически выше друтих форм биотической власти. Эйдократия — это форма стратегического управле- ния организацией биоморф (шире — и организацией биосистем мень- ших, чем вид, по своей сложности), цели которой определяются при- оритетом сохранения вида по отношению к сохранению других биоси- стем (биоморф, в частности), выступающих в роли его структурных элементов. Форма стратегического управления организацией биоморф — власть генов, направленная на развитие и поддержание личной жизни организмов — это онтократия, а — синорганизмов — это синонтократия. Таким образом, все формы стратегической власти находятся в органи- зации биоморф в противоречивом единстве и иерархически взаимосвя- заны между собой. В проявлении биотической власти в организации биоморф, особенно в их стремлении по сохранению жизни, всегда действует приоритет разви- тия, направленный на сохранение биосистем более высокой сложности организации. Мы сейчас прикоснулись к явлению, которое составляет важное содержание организации жизни вообще. Высшая форма биоти- ческой власти — это макробиократия, когда приоритетом биотической власти является поддержание и сохранение вышестоящих по сложности организации живых тел, структурными элементами которых являются нижестоящие по сложности организации конкретные носители и испол- нители биотической власти. Автобиократия — это непосредственное проявление биотической власти живым телом на поддержание и сохра- нение в целом своей собственной (личной) жизни — в направлении автобиофилии. Гипобиократия — это непосредственное проявление био- тической власти живым телом на поддержание и сохранение жизни структурных элементов, составляющих организацию этого живого тела. В зависимости от того, в отношении кого биотическая власть проявля- ется соответствующим образом, изменяется и ее ранг (уровень). Все гены — их системы — это стратеги; они высшие властоносцы в организации биоморф, а все иные структурные элементы — это так- тические элементы. По сути, если образно, то «нет мира под оливами» в организации биоморф семенных растений, и в ней господствует власть стратегов над исполнителями их программ — тактическими элементами, которые ведут непосредственную борьбу за существование биоморф во взаимоотношениях с окружающей внешней средой. В организации биоморф семенных растений носителями способно- сти репродуктивно размножаться являются только побеги, поэтому они являются структурными элементами — лидерами — и во многом опре-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ деляют качественное фенетическое содержание биоморф. Побеги по- разному специализированы, и, что самое главное, в организации био- морф, как впрочем и других биосистем, имеет место проявление разно- образия во власти, в нашем случае, — биотической, и тех структур, которые этим властям как-либо подчиняются и исполняют их установ- ки, или программы, развития (образно говоря — «приказы»). С генетического уровня целостной организации биоморф постоян- но исходит в их фенетическую организацию и касается каждого ее структурного элемента диктат целей организма или синорганизма и даже вида, что превращает эти структурные элементы в беспрекослов- ных исполнителей (прямо-таки «слуг») более высоких целей, чем забота о своей личной жизни, что, безусловно, не может не отражаться соот- ветствующим образом на структуре и поведении этих элементов внутри биоморф. В процессе длительного эволюционного развития биоморф множества поколений структурные элементы современных биоморф стали специализированными, но, естественно, по-разному, т.е. они так (шире) или иначе (уже) адаптировались к выполнению определенных функций организмов (простых биоморф). Итак, биотическая власть, в первую очередь как система управле- ния в организации биоморф, принадлежит геносоме, элементами кото- рой являются гены. Диктат геносомы просматривается во всей фенети- ческой организации (феносоме) биоморф, которая, собственно, и со- ставляет главное содержание конституциальных «ваяний» (упражнений) живой природы с материальной неживой первоосновой. Ранее (т. 1, гл. 4) было показано, что в организации биоморф, а шире и друтих биосистем, существуют стратегические (управляющие, руководящие) и тактические (исполнительские и соисполнительские) категории биосистем. Здесь мы развиваем эти идеи, опираясь также и на закон управления-исполнения организации биосистем, определен- ный ранее (т. 1, гл. 2, с. 41): «исполнительная (тактическая) часть орга- низации биосистем реализует генетическую информацию (находится под управлением генов) посредством своего роста и развития через главен- ство исполнительских частей этой организации, ответственных за вос- произведение себе подобных, над частями, снабжающими первые части всем необходимым для развития и реализации этой функции и (или) удобно располагающими их в пространстве и времени». Согласно этому закону, организация биоморф семенных растений управляется с генети- ческого уровня, т.е. генами и их системами (геносомами), которым по нашей концепции принадлежит высшая биотическая власть — страте- гическая (высшая управленческая) власть. Гены генам рознь; они все едины в том, что наделены стратегичес- кой властью (господствует генократия — власть генов), т.е. все гены — это биотические стратеги, но среди них есть особенные гены, которые уча- ствуют в развертывании лидерных побеговых систем (формируют, по сути дела, фенетическую власть «на местах»), обеспечивающих биомор- фам половое, а шире — любое размножение. Вполне возможно утвер- ждать, что такие («любвеобильные», «сексуальноозабоченные») гены, которые несут в себе власть не столько собственно биоморф, сколько
Глава 7. Концеиция разнообразия биотической власти власть вышестоящих биосистем и, в первую очередь, видов, — это подлинные лидеры (стратеги над стратегами) в генетической организа- ции биоморф, вполне достойные того, чтобы их обозначали еще и как- то особо, например, «геногегемонами» (генами-гегемонами). Все структурные элементы фенетической организации биоморф- это все ее тактические (исполнительские и соисполнительские) сред- ства, которые находятся под стратегической биократией, развиваясь непосредственно под ее управлением, и, в зависимости от конкретных условий окружающей среды, биоморфа выбирает такой вариант фене- тического развития, который отвечает ее ресурсу возможностей (ее идиэнергетическому потенциалу). Высшей биократией не исчерпывает- ся вся полнота властей, которые проявляются в организации биоморф. В феносоме биоморф развивается много органов, среди которых только два типа принадлежат основным органам, — это побеги и корни. Од- нако, среди основных органов безусловными лидерами являются побеги (т. 1, гл. 4), поэтому в фенетической организации побегам принадлежит тактическая (ислолнительская, или местная) власть, которая, естественно, биотическая (тактическая биократия, или фенократия). Только побеги, или отдельные побеги вместе с корнями, т.е. корнепобеги-органы — это те, кому принадлежит в фенетической организации биоморф тактичес- кая власть. Власть (биотическая) имеет видимое проявление, когда предстает в образе побегов (шире — еще и корнепобегов-органов), а стратегическая деятельность осуществляется скрытно генами. Побеги зримо олицетво- ряют присутствие власти в организации биоморф, но они — это только видимое (фенетическое) проявление власти — местная (тактическая) власть — верхушка того властоносного «айсберга», который составляет целостную власть живых систем. Более «изощренная» властная деятель- ность совершается генами («серыми кардиналами» в организации био- морф — у них как бы «ума палата», но в своей работе они «не светятся»), скрытно проявляющими свое гигантское участие в феносоматическом развитии биоморф. Гены, будучи носителями стратегии, как стратеги выполняют властные функции в органеллах, клетках, тканях, органах, организмах, синорганизмах, видах, фитоценозах и биостробе, но по- разному, особенно в таких гетерогенных системах, которые устроены сложнее биоморф и даже сложнее видов. В теле фитоценоза и всего живого тела (биостробы) Земли их генетические организации (геносо- мы) образуют неоднородную систему высшей (стратегической) власти. Главное в нашем подходе в том, что каждая биосистема обладает стра- тегической властью и имеет фенетических исполнителей, главные из которых проявляют себя как местная (тактическая) власть. Власть может действовать непосредственно напрямую, поскольку находится в одном теле с исполнительскими структурными элементами или опосредованно через какие-либо иные структуры или инородные живые тела, напри- мер, когда пыльцевые зерна прикрепляются к насекомым и с их помо- щью перемещаются в пространстве, совершая перекрестное опыление, и т.п. Распорядиться властью можно по-разному: можно стать лидером среди лидеров или, напротив, не суметь проявить себя вовсе, т.е. не
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ только своих «властных» возможностей, но и «сгинуть» в небытие, ис- пытав лишь все неудобства угнетенного существования (что делать— судьба; не всем в фенетической организации биоморф уготована воз- можность раскрыть себя в процессе жизнеразвития). Особое место среди тактических властоносцев, т.е. среди побегов и корнепобегов (побегов вместе с корнями), занимают те лидеры, которые нацелены на размно- жение, в первую очередь, половое. Особенность существования таких носителей локальной власти в том, что без них развитие биоморф об- речено бесповоротно закончиться элиминацией, а с ними непосред- ственно реализуется преемственно-историческая относительная непре- рывность в чреде сменяющихся поколений, т.е. они «работают» не толь- ко и не столько на биоморфу, сколько на более высокую по сложности организации систему, в первую очередь, — вид. Побеги и их препопу- ляционные системы, усложненные или не усложненные образованием еще и корнепобегов, непосредственно обеспечивающие размножение биоморф, — это носители особой тактической власти, которых можно дополнительно обозначить термином «феногегемоны». Как бы там ни было, но вся тактическая власть в организации биоморф принадлежит побегам и корнепобегам-органам (онтоидам), когда те образуются. Среди побегов и корнепобегов есть свои лидеры и «изгои». Угнетен- ность однотипных побегов и корнепобегов не предопределена генети- чески, а зависит от действия соответствующих неблагоприятных усло- вий окружающей среды. Кроме носителей локальной власти (фенократов) в фенетической организации биоморф большинство живых структурных элементов- биомеров — это сателлиты (от лат. satelles — спутник, слуга, сообщник); они всю свою жизнь «работают» под властью не только генов, но и под властью побегов и корнепобегов-органов (листья, стебли, корни, лепе- стки и т.п.). Все структурные элементы фенетической организации био- морф, в том числе и no6ezu, вместе с феногегемонами, — это все такти- ческие средства, однако только сателлитам, независимо от стеиени своего развития, уготована участь всегда быть на службе у носителей местной власти. 7.3.2. Иерархия биотических властей Сиособность биотической власти ироявлять себя в разных формах- это бнотнческое разновластне, или гетеробнократня, в организации биоси- стем. В предыдущем материале было показано, что высшей формой биократии (биотической власти) является стратегическая, или высшая, власть (генократня), которая проявляет себя в генах и их системах (ге- носомах) и управляет всем остальным (фенетическим) жизнеразвитием (эубиократия) биосистем, в частности, и биоморф (рис. 358). Власть на субординатную ступеньку пониже, по сложности ее проявления, дей- ствующая в организации биоморф (шире — других биосистем) в соот- ветствии с генетической программой, т.е. под управлением высшей
Глава 7. Концеиция разнообразия биотической власти (стратегической) власти может быть только тактической, или местной (локальной), иначе фенократией; ее главными носителями («фе- ногегемонами») явл~потся побеги — лидеры фенетической организации (без корней или с корнями, если они при них образуются, поскольку побеги не только потребляют и производят биогенные ресурсы, но еще и рассеивают их в живом состоянии путем размножения (другие органы этого делать не могут). Итак, мы довольно много сказали о биотической власти в органи- зации биоморф (шире — всех биосистем), но сейчас обратим внимание на проявление биотической гетерократии под другим углом обзора. Меробиоз никогда бы не состоялся, если бы живые тела, еще на заре своей эволюции, не адаптировались проявлять властные качества по отношению к неживой материи. Живое может существовать в coomeem- миляции в себе необходимой неживой материи, иными словами, живые тела существуют на основе меробиоза — единства живого и неживого при проявлении nepeozo типа власти в субординатной классификации биокра- тий — власти живой материи над неживой — протобиократии. Этот тип биотических властей показывает, что живая материя, образующая орга- низацию биоморф (шире — и других биосистем), может состояться только при наличии подходящей неживой материальной первоосновы, включение которой в живое может произойти соответствующим обра- зом только под управлением генов — стратегических носителей прото- биократии. Проявление фенократии и протобиократии оставим без детализа- ции, а основное внимание обратим на иерархическое разнообразие в стратегической власти (генократии), что и отразим в графической мо- дели соответствующей «пирамиды» абстракций, используя уже приве- денный выше материал (рис. 358). Выше уже было подчеркнуто, что безвластие ведет к беспорядку. Организация биоморф устроена по властному принципу и иерархия власти соблюдается неукоснительно, будучи ориентирована на иерар- хию биосистем, различающихся по сложности организации. Власть живого (биократия) распространяется в организации живых систем в двух основных направлениях экспансии: на неживую и на живую при- роды. Проявляя биократию, живые тела осваивают пространство, заня- тое не только живой материей, но и неживой. Живое началось из не- живого и, постоянно развиваясь на основе использования неживой материи, применяя ее для производства своих структурных элементов, оно адаптировалось не только брать, но и возвращать в неживое часть одолженных у него на некоторое время ресурсов. Без применения про- тобиократии круговорот веществ в живой природе и явление меробиоза были бы невозможны; только посредством самой первой биотической власти, носители которой — соответствующие гены, а ее исполнители — все тактические элементы, неживая материя ассоциируется жизнью и составляет первооснову в организации живых тел, растительных био- морф, в частности.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 358'. «Пирамида» абстракций биотических властей (биократий) в орга- низации биосистем. А-Г — уровни абстракции (суперординаты); 1 — фенократия (тактическая, или местная власть), 2 — генократия (стратегическая, или высшая, власть), 3 — протобиократия (власть живой материи над неживой), 4 — эубиократия (власть генов собственно над фенетичес- кой организацией), 5 — гипоонтократия (стратегическая и тактическая биотическая власть проявляет себя в структурных элементах организмов), 6 — онтократия (разномасштабная биотическая власть действует в отношении организмов), 7 — метаонтократия (это биоти- ческая власть выше онтократии), 8 — гипобиократия, 9 — автобиократия, 10 — макроби- ократия (8 — 10 подробнее стр. 567), 11 — органоидократия, 12 — цитократия, 13— гистократия, 14 — органократия, 15 — синонтократия, 16 — ценодемократия, 17 — эйдок- ратия, 18 — биоценократия, 19 — эпиэйдократия (биотическая власть надвидов), 20— биостробократия, 21 — панбиостробократия.
Глава 7. Концеиция разнообразия биотической власти Биотическая власть в организации биосистем обладает иерархичес- кими свойствами, что находится в прямой зависимости от того, что живая природа вообще устроена по иерархическому принципу и биоси- стемы взаимосвязаны между собой соподчиненно, в зависимости от сложности их организации. Клетка листа, например, в структуре расти- тельной биоморфы находится под стратегической властью и клетки, и листа, которому она принадлежит, а далее подчиняется соответствую- щей власти побега, организма; кроме того, на организацию этой клетки кое-что «давит» от стратегической власти вида и даже от власти всей биостробы Земли. На каждом уровне жизни, который занимают биоси- стемы по их организационной сложности, действует стратегическая власть — власть генов, а исполняют ее фенетические структурные эле- менты, среди которых есть особые лидеры, выполняющие функцию локальной (тактической) власти, например, побеги в организации био- морф семенных растений. Нам нет смысла заниматься здесь анализом всего иерархического комплекса биотических властей, которые прояв- ляются на всех уровнях организации растительной жизни. Однако будет все-таки важно указать на те основные типы биократии, которые обес- печивают существование биоморф семенных растений и их структурных элементов в пространстве и времени. Для внутренней организации биоморф важно, чтобы биосистемы — ее структурные элементы обла- дали достойным порядком, а такое возможно только при одном усло- вии, чтобы биотическая власть распространялась не только на биомор- фы в целом, но и на каждый ее живой структурный элемент: органеллы (органоиды), клетки, ткани, органы, т.е. на любые гипобиоморфы: она там всегда есть — это гипоонтократия; ее разновидности — органоидок- ратия, цитократия, гистократия и органократия. Биотическую власть, или биократию, причем любую (стратегичес- кую или тактическую), по направлению ее действия, т.е. по цели, ради которой она присутствует в системе, можно обозначить непосредствен- но по названию той биосистемы, в существовании которой она прини- мает участие. Стратегическая гипоонтократия проявляется в функцио- нировании генов каждой клетки, в каждой ткани и в каждом органе, обеспечивая целенаправленное развитие их фенетической организации в целостной организации биоморф. В каждом структурном элементе биоморф проявляется еще и местная (тактическая) власть, например, через ядро в клетках или архесому в побегах и т.п., которые обеспечи- вают должный порядок в развитии и функционировании фенетической организации структурных элементов биоморф при исполнении (реали- зации) ими конкретных стратегических программ, поскольку все фене- тическое в организации биоморф стоит на «службе» у генетического, т.е. у разномасштабной стратегической власти (генократии). Итак, в центре организации биоморф-организмов стоит онтократия, действующая с их генетического уровня и осуществляющая управление развитием фенетической организации организмов в соответствии с осо- бенностями окружающей внешней среды. Стратегическая власть био- морф-синорганизмов — это синонтократия, действие которой исходит от
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ генов и их систем (геносом) синорганизмов и обеспечивает развитие и функционирование феносомы этих биосистем (рис. 358-Г). В феносоме организмов есть своя местная (тактическая) власть в образе побегов (у синорганизмов носители местной власти те же — это побеги организ- мов-клонистов), способная непосредственно обеспечить онтократичес- кие (или синонтократические) цели генов и их систем (геносом): в конкретных условиях оптимально потребить и произвести ресурсы, необходимые для размножения. Онтократия и синонтократия исходит от генов и систем генов (геносом), а воплощается в конкретную фене- тическую организацию посредством проявления местной власти, соот- ветственно, побегами или корнепобегами-органами (онтоидами) и ис- полнительской и соисполнительской деятельности других органов-са- теллитов. У синорганизмов потолок стратегической власти — синонток- ратии повыше, чем у организмов. Осуществление репродуктивного раз- множения — это проявление уже другой власти, которая стоит повыше онтократии и синонтократии, — власти вида, или эйдократии. Жизнь, существуя в каждой телесной системе дискретно, «позабо- тилась» в процессе эволюции, чтобы в каждом живом теле присутство- вал «букет» биотических властей, нацеленных на жизнесохранение в целом больше, если бы такая власть была единственной для конкретной биосистемы. Биократии, накладываясь одна на друтую, безусловно, влияют на развитие конкретной биосистемы, в зависимости от окружа- ющих внешних условий и от единства и борьбы противоположностей, замешанных в этих властях, определяют те типы организации биосистем и, в частности, биоморф, которые мы сегодня наблюдаем во всем живом теле Земли. В любом организме или синорганизме действует, помимо онтократии или синонтократии, целый клубок других властей как ни- жестоящих, так и, что самое главное, еще и вышестоящих в соответству- ющей субординатной классификации биократии, например, таких, как эйдократия, фитоценократия и, наконец, биостробократия. Особый тип биократии — панбиократия (биотическая власть над всем) — это участие, пусть пока во многом только потенциальное, ге- нов-стратегов в организации любых живых тел на Земле, в управлении жизнью вообще (Жизнью), в плане ее сохранения и экспансии из из- вестных пределов распространения, т.е. даже той жизни, о существова- нии которой земляне еще ничего не знают и которая может быть где- то еще в Природе есть (или ее нет, и такое нельзя исключать). Всякую власть в организации биоморф, которая проявляет себя выше онтократии, можно обозначить одним термином — метаонтокра- тия. Однако, для нас, в отношении растительных биоморф, в первую очередь, важно учитывать влияние на их организацию, проявляющейс~ всегда, власти вида — эйдократии (рис. 358).
]Глава 7. Концеиция разнообразия биотической власти 7.4. «Могучая пятерка» покорителей пространства в организации биоморф семенных растений В организации биоморф приоритетные цели определены властны- ми структурами; феносома находится в подчинении у генов, функцио- нирование которых приводит через множество функций к «лепке» (кон- струированию) тела биоморфы — фенетического дома генов (подробнее см. главу 9). Гены «работают» по строгому плану; у них есть определен- ный ресурс возможностей, который рассчитан на развертывание подхо- дящей информации в тех условиях окружающей среды, которые прояв- ляются здесь и сейчас в реальной, а не потенциальной обстановке. И, при этом, какую бы форму ни приобретали в процессе эволюции отно- щения между геносомой и феносомой, гены выдерживают давление эволюции с разных сторон, идя на компромиссы во внешнем и внут- реннем устройстве феносомы, но никогда не утрачивая своей привер- женности более высокой для них цели — цели размножения как цели более всего обеспечивающей коллегиальное жизнесохранение (сохране- ние вида и сохранение более высоких по сложности организации био- систем). Феносома существует в организации биоморф для того, чтобы исполнить потребительские, производственные, резервирующие и рас- сеивающие цели генов — управляющих элементов — носителей высшей, или стратегической, власти в организации биоморф. В биотической власти выше генов ничего больше нет; это они управляют развитием всего, что способно покорять и осваивать пространство во времени. Многое сейчас свидетельствует о том, что от познания закономерностей взаимодействия систем «управления-исполнения» человечество ожида- ют еще наиболее важные открытия. Так чем и как исполняется в организации растительных биоморф то, что вроде бы уже ясно (чтобы жить, надо завоевывать пространство). Эволюция, и только она, добротно постаралась и создала возможность растительной (и любой другой) жизни «кочевать» по телесным систе- мам, которые она сама же и развивает на основе трансформации нежи- вой материи, применив несколько универсальных процессов (нарастание, ветвление, партикуляцию, размножение и элиминацию), без которых поко- рение и освоение пространства генами было бы невозможно. Они работают скрытно с помощью развертываемых от их идиоэнергии фенетических средств, что без них, генов, никогда бы не состоялось и не получило бы вообще растительного жизнеразвития. Все основные фенетические сред- ства, обеспечивающие экспансию жизни в пространстве, используемые ге- нами, в организации биоморф семенных растений исполняются в образе упомянутой выше пятерки процессов, которую эпическим языком можно «окрестить» не иначе как «могучей пятеркой», или «имидж-пятеркой». Эта пятерка уже рассматривалась в первом томе, но она настолько важна для организации растительных биоморф и, в частности, при развертывании форм габитуса и форм роста, что не упомянуть о ней здесь было бы ошибкой.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Может показаться тривиальным, что пятерка процессов: нараста- ние, ветвление, партикуляция, размножение и элиминация, выведенная в первом томе (гл. 4), ничего нового не добавляет, но на самом деле это не так, и все здесь далеко не просто. Известны только слова, а ведь речь у нас идет о действительном могуществе «могучей иятерки», без которой в целом, как, вирочем, и без каждого ее члена по отдельности, раститель- ная биоморфа ничего бы сделать не смогла, иоиытавшись как-то «осваи- вать» иространство. Мне удалось сначала (т. 1, гл. 4) откорректировать в понятийном отношении каждый процесс по отдельности, а затем увидеть, как жизнь в своей эволюции «запрягла» всех их в единые «колесницы» растительных биоморфных организаций, в которых она как растительная жизнь несется без устали по просторам нашей плане- ты, подминая под себя пространство, осваивая его во времени. Наше познание, направленное на «могучую пятерку», поможет найти пути к пониманию ее, но и, в конце концов, позволит «приручить» этот, еще мало осознанный энергетический потенциал, поставив его на службу нашей цивилизации и развитию Жизни во Вселенной. Выдающийся ученый С.В. Мейен, давая отзыв на одну из моих более ранних работ (1985), нашел нужные слова и, со свойственной ему прозорливостью, акцентировал внимание на четырех (нарастание, вет- вление, партикуляция, размножение) рассмотренных мною процессах освоения растениями пространства, т.е. на том, что все остальные уче- ные не смогли или не захотели заметить: «известны только слова, но я не знаю, кто еще до Нухимовского показал, что к этой четверке сводят- ся, по сути дела, все способы освоения пространства... — это, если хотите, аксиоматика биоморфологии, а без аксиоматики не может быть речи о формализации знания, следовательно, и о его математизации». Справедливости ради замечу, что я всегда рядом с упомянутой четвер- кой ставлю еще и пятый процесс — элиминацию (смерть), т.е. рассмат- риваю их вместе как пять могучих атрибутов фенетического развития— «могучую пятерку» организации растительных биоморф, обеспечиваю- щую им успех в покорении пространства. 7.4.1. Диктат эйдократии в организации биоморф и эйдофория «Могучая пятерка» может состояться только в той организации биоморф, которая развивается на основе определенного естественного порядка, который развивается в биоморфах как результат длительной эволюции растительной жизни на Земле. Естественный порядок в орга- низации растительных биоморф определяется наличием у них биотичес- кой стратегии, т.е. высшей биотической власти, которая всецело при- надлежит генам и их системам — геносомам. Согласно биотической стратегии как высшей власти в организации биоморф семенных расте- ний определяются параметры нормы реакции конкретных биоморф, отражающиеся в фенетической организации и, в первую очередь, через
Глава 7. Концеиция разнообразия биотической власти проявление «могучей пятерки», поскольку только посредством ее членов создается и трансформируется феносома биоморф. «Могучая пятерка» отражает главное динамическое содержание фенетической организации биоморф, представл~пощей из себя клубок исполнительской (тактичес- кой) деятельности, подчеркивающей единство противоречивых сил со- хранения и разрушения жизни, действующих одновременно и по-разно- му на каких-либо отрезках времени. Наличие в фенетической органиэации биоморф семенных растений клубка иротиворечивых тактик, через кото- рые (и только через них) может реализовать себя стратегия, т.е. высшая возможным только на основе комиромиссов. Из многих биотических вла- стей, а гетеробиократия — норма жизни биосистем, которые имеют место и в организации биоморф, есть смысл сконцентрировать внима- ние на взаимодействии нескольких из них: на двух, достаточно контра- стных, биократиях у простых биоморф-организмов (онтократии и эй- дократии) и на трех — у сложных биоморф-синорганизмов (онтократии,. синонтократии и эйдократии). Сейчас нам, как никогда раньше, уже ясно, на основе существования каких противоречивых высших властей разыгрываются баталии взаимоотношений внутри фенетической орга- низации биоморф, отраженные в структурных состояниях, достигнутых деятельностью «могучей пятерки», — отношениями между относительно личным (вегетативом) и между относительно неличным (репродукти- вом), причем в одной и той же организации одного и того же живого тела — биоморфы (см. главу 2.5. в этой книге). Из всех форм биотической власти, действующей в организации биоморф, приоритетный потенциал заложен в эйдократию. Иерархич- ность в устройстве биократии характеризуется в организации биоморф тем, что вышестоящая власть использует достоинства нижестоящей власти как свои, поскольку она вносит в них приемлемые для нее кор- рективы и, тем самым, подчиняет ее своей, более сложной власти. В организации биоморф нет гомеократии — одинаковости (равенства) в био- тической власти (будем стремиться, так удобнее для всех, как читателей, так и писателей, ничего не подключать социального в представления о биотической власти, однако все-таки трудно удержаться от соблазна, чтобы не заметить, что аналогом гомеократии в социальных отношениях может быть только демократия). Биотическая организация снизу довер- ху предполагает иерархичность составляющих ее структурных элемен- тов-биосистем, различающихся по многим признакам, но мы отметим главное, часто они различаются по степени сложности организации и специализации. Биосистемы существуют на основе определенных зако- нов и правил, обеспечивающих порядок в их организации, среди кото- pbIx основополагающим является закон управления-исполнения орга- низации биосистем, где, по сути, зафиксирована особая управляющая роль генов (их высшая, или стратегическая, биотическая власть над исполнительской (тактической) деятельностью остальных структурных элементов, среди которых лидерство (местная, или тактическая власть) принадлежит тем частям, которые обеспечивают воспроизведение био- систем, подобных родительским. В организации биосистем, в частности,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ биоморф, нет гомеократии, но всегда есть гетеробиократия, т.е. нера- венство биотической власти. В организации биоморф проявляется диктат эйдократии (власти вида). Эйдократия включает в себя биократии меньшей сложности и, в частности, онтократии и синотократии, причем не просто включает, а превалирует над ними, оказывая на всю организацию конкрет- ной биоморфы, принадлежащих одному виду, свой диктат. Фактически, формулируя концепцию гетеробиократии и показывая приоритетность, по сути, диктаторство эйдократии над онтократией и синонтократией, мы, под другим углом обзора, подтверждаем то, что уже было доказано ранее (т. 1) в организации биоморф: генетическое определяет развитие фенетического, первый принцип фенетической орга- низации биоморф — размножение, репродуктивное приоритетно по от- ношению к вегетативному и др. Явление, когда видовая биократия (дик- тат эйдократии, или власти вида) определяет стратегию и тактику организации биоморф семенных растений, составляет для этих биосис- тем один из высших атрибутов (выше стоит только атрибут жизнесох- ранения вообще). Вид — хозяин своей стратегии, которую он тактически реализует в своих же структурных элементах: популяциях, синорганизмах, организ- мах и их живых частях, поскольку только ему, виду, в этих структурных элементах принадлежит, в образе генов и их систем (геносом), высшая среди генетически однородных систем власть. Только в наследственно- сти мы находим однозначный ответ на вопрос: почему биотическая власть в организации биоморф принадлежит, в первую очередь, виду. Геносома вида состоит из однородных геносом биоморф и управляет ими, подчиняя себе, и тем самым управляет и организацией каждой конкретной биоморфы. Поскольку эйдократия проявляет диктат по отношению к онтократии и синонтократии, т.е., по сути, по отношению ко всей организации простых и сложных биоморф, то в этой организа- ции нет частей, которые служили бы только организму или синорганиз- му, есть части, которые служат и им, и виду, но есть особые части, которые служат только виду (!), что составляет один из парадоксов и феноменов в организации биоморф. Есть части (архесома, например, зародышевый мешок, пыльцевые зерна, зигота), которые, образуясь в структуре биоморфы, по сути, нужны не ей лично, а другой биосистеме — ее виду, т.е. они стоят на службе не личной, а коллегиальной (по большому счету — другой, где- то даже чужой, хотя и родственной) жизни. Половое размножение нуж- но видам, а биоморфы в процессе эволюции или научились, или не научились выдерживать его негативные для них последствия. В одних случаях растительные биоморфы адаптировались к половому размноже- нию и его последствиям и научились разными путями выживать, причем даже неоднократно, после такого размножения, а биоморфы других видов, не найдя компромиссов между вегетативом и репродуктивом, уже после первого полового размножения погибают (рис. 41-121, 123-147, 150-190 и др.). Условия окружающей среды очень сильно влияют на поведение вегетатива и репродуктива в организации биоморф. Например, биомор-
Глава 7. Концеиция разнообразия биотической власти фы Ajuga reptans L., Solidado virgaurea L. (рис. 67, 121) и др., попадая в условия сильного затемнения, могут большую часть своей жизни или всю жизнь не цвести. Эйдократия предполагает такие варианты разви- тия биоморф, когда в неблагоприятных условиях вид дает больше сво- боды онтократии и синонтократии по активизации приемов личного выживания, ослабляя давление на вегетатив репродуктивным развитием, а нередко и вообще снимая его до лучших времен (рис. 66). Виды настолько контролируют свое вьикивание в различных экологических условиях, что в их арсеналах существуют системы миниатюризации видов, когда для своего выживания используется минимал вида (под- робнее см. главу 2.2.). На основе эйдократической организации биоморф проявляется интереснейшее свойство, на которое стоит обратить здесь внимание, когда каждая структурная часть биоморф, в большей или меньшей сте- пени, является носителем свойств вида — эйдофором. Способность био- систем разной сложности выступать в качестве структурных элементов вида и отражать в своей организации его признаки обозначаем как явление эйдофории. Все структурные элементы организации биосистем (в част- ности, биоморф), входящие в структуру вида, отражающие так или иначе свойства вида и выступающие в качестве эйдофоров, используются систематиками для накопления в специальных хранилищах (гербариях) фактического материала о видах — сухих фрагментов растений и целых растений (гербарных образцов). Сухие растительные остатки — это сухие эйдофоры, которые так или иначе несут и отражают что-то из признаков видов, но, безусловно, не все. Давая обрывочные сведения о видах, эйдофоры помогают нам, исследователям, собрать и накопить в разных местах информацию о видах. Пожалуй, уместно будет заметить, что эйдофория — это разновидность еще более общего явления, которое можно обозначить одним термином «плейофория» (дословно с гречес- кого «носить большее»). Свойство частей целого, будучи незаметными и (или) какими-либо еще непоказательными или, напротив, — заметными и (или) какими-либо еще показательными в организации целого, всегда выс- тупать, так или иначе, носителями каких-либо свойств целого, никогда не заменяя его собой — илейофория, а меросомы, обладающие плейофорией (ее носители) — это ллейофоры. Всякое целое состоит из частей, которые проявляют себя как носители некоторого большего (плейофоры) из органи- зации целого, оставаясь неспособными по отдельности отражать все це- лое, и только сообща плейофоры, как часть целого (меросомы), через их динамическое единство и взаимосвязь, способны и составлять, и отра- жать, и обеспечивать целостный результат своего существования — быть больше суммы своих частей. Эйдофория как разновидность плейофории позволяет через что- либо меньшее как-либо отражать большее: в одной клетке, органе, организме, ценодеме можно усмотреть что-либо, что отражает их при- надлежность к определенному виду. Способность меросом вида к эйдо- фории широко используется и в биоморфологии, например, при определе- нии биоморфотипов и их классифицировании.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ 7.4.2. От нарастания к ветвлению, далее партикуляции и размножению, наконец, к элиминации Биосистема нормально может существовать только при обязатель- ном наличии в ней организационного порядка, который поддерживает- ся ею как системой «управления-исполнения», где все заранее известно: кому, как и почему принадлежит власть и кто и зачем ее исполняет. Когда что-то нарушается в порядке жизнеразвития, то биосистема на- чинает болеть и может погибнуть, если не сможет отремонтировать нарушенный в ней порядок. Основа порядка в любой нормальной (здо- ровой) биосистеме — состояние ее биотической власти. Из предыдущего материала ясно, что биоморфа (шире — биосистема) не может разви- ваться вне власти, а власть в биоморфе (шире — биосистеме) обеспе- чивают, в первую очередь, гены, ее носители — они стратеги, поэтому власть в генах высшая, стратегическая. Однако власть без исполнителей и соисполнителей, без тактических средств ее реализации, разных по сложности организации и специализации, никому не нужна, она тогда — бутафория, а реальной эволюции заниматься управлением «пустотой» некогда, она всегда при деле — в постоянном поиске, куда и как можно приложить некоторую биотическую власть, чтобы организация живой системы обладала порядком (сохраняла и развивала его). Эволюция целесообразна, и ее цель — поддержание и развитие в биосистемах поряд- ка, поскольку только при наличии определенного порядка в организации жизни сам процесс эволюции будет прогрессивным. Власть в биоморфах реализуется в исполнительской деятельности фенетических структур через «могучую пятерку»". нарастание, ветвление, партикуляцию, размножение и элиминацию (смерть). Только смерть, та единственная функция жизни, которая, «ощетинившись» всей своей непримиримостью, в отношении к остальным биотическим функциям биоморф и их жизнесохранению, все же не противоречит самой природе явления жизнь — она стала в процессе эволюции неотьемлемой частью жизни, обеспечивая дискрет- ность ее в телесных системах. Жизнь и смерть, в различных сочетаниях, проходят через развитие телесных систем, на основе борьбы и преодоления противоречий, но когда-то наступает момент, и жизнь оставляет родные ей тела, бросает их в небытие, отдает во власть смерти, поскольку более именно эти тела ей уже не нужны; она, жизнь, ими жертвует, отдавая так некронами свою «дань» в окружающую среду, чтобы иметь возмож- ность существовать в ней далее и пользоваться ею, поскольку первооснова жизни — неживая материя. Подобно тому, как исполнительский порядок в театре начинается «с вешалки», порядок в фенетической организации эпиэмбриональных биоморф семенных растений начинается с роста, в форме прорастания семян и нарастания побега, корня и других органов. В процессе нара- стания, сопровождающегося часто ветвлением и его разновидностью- кущением, создаются побеги и корни (основные органы), которые, продолжая нарастание, разрастаются в пространстве до определенных размеров, сформировав необходимую биоморфам систему ассимиляции и опорную (каркасную) систему, что в целом составляет вегетативную массу (ресурс, тело), или тотавегетатив, биоморфы.
Глава 7. Концеиция разнообразия биотической власти Сформулируем вопрос: кому такая биоморфа нужна? Ответ доволь- но прозаичен. Оказывается, именно формирование вегетативного тела составляет главное содержание личной (вегетативной) жизни биомор- фы. Все, что образуется потом, принадлежит также биоморфе, но отно- сительно, постольку поскольку репродуктив, развивающийся из веге- татива и на вегетативе как на питательном субстрате, функционально ориентирован не на сохранение родительской биоморфы, чаще всего, наоборот, против его, а всецело «работает» на выживание вида. Если присмотреться к телу простой растительной биоморфы, то оно выглядит чаще всего как двухэтажное «здание», которое на первом этаже заселено вегетативом, а на втором репродуктивом, что в целом и неудивительно, поскольку сверху лучше вести стратегическое и тактическое руководство фенетической организацией биоморф, лучше эксплуатировать вегетатив и использовать преимущества для своего роста и развития в наиболее освещенной части освоенного биоморфой пространства, а, главное, находясь там, наверху, над вегетативом, можно по-настоящему надежно послужить виду, полноценнее проявляя сексуальные функции. Органи- зация генетически однородных биоморф, в угоду виду, более всего нацелена на поддержание последовательной преемственности в чреде сменяющихся поколений, что надежнее всего удается посредством по- лового размножения, хотя и вегетативное размножение вполне отвечает этнократическим целям, поэтому оно также нередко используется в фенетическом развитии биоморф (рис. 359-361 и др.). По продолжительности жизни среди фитобиоморф семенных рас- тений есть и эврифемеры, и сверхдеревья (деревья-долгожители, живу- щие более 1000 лет), что убедительно свидетельствует о том, что диктат эйдократии далеко не всегда ведет биоморфы в их развитии к «униже- нию» вегетатива, т.е. собственно личного у биоморф, путем быстрого отмирания его после первого же репродуктивного усилия, а нередко вегетатив находит ресурсы для образования в себе резервов, например, базовых (многолетних побеговых) органов, т.е. проявляет взаимные ком- промиссы с репродуктивом, что свидетельствует об утверждении давно и надолго в эволюции биоморф семенных растений двух равнозначных и обратимых направлений: эвриэфемерного и персистентного. Итак, в организации биоморф семенных растений два основных органа: побеги и корни. Сначала они нарастают и ветвятся (кущение- это разновидность ветвления), применяя различные их способы (дихо- томическое, моноподиальное, симподиальное нарастание, очередное, супротивное, мутовочное ветвление и т.п.) и, таким образом, в какой- то мере на первых стадиях онтогенезов удовлетворяется естественная потребность растительных организмов в своей собственной (личной) жизни (рис. 359-361). Однако в эволюции живой природы, с самого ее начала на Земле, такое развитие событий, при котором личная жизнь, хотя бы чуть-чуть испытала бы комфорт свободы и «забыла» о виде, составило предмет многомиллионных лет борьбы в организации био- морф между онтократией и эйдократией, борьбы, в которой все-таки нашлось место компромиссам, без которых жизнь на Земле просто бы не получилась вообще. Альтернативы компромиссам между личным и
582 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 359. Биографический портрет Еигуора sp. [сем. Asteraceae Израиль, Иерусалим, Ботанический сад, южно-африканский отдел, IV)]. Биоморфотип: надземно-двусемядольные (семядол р д оли п одолговатые, 5 — 8 мм дл., 2,5 — 3 мм шир., закругленные, постепенно перех дящ р хо ие в ко откие черешки) четочно- р озеткообразующие (розетки обычно образуютс рхуш тся на ве ках четочных побегов, когда е п и смене вегетационных сезонов ) дшаются условия для линейного роста, например, р езанно-п остолистные гемисимподиальные семидетерминатно-репродуци- р щие ьно- ве ие однодомные желто-астро- р Шие (с верхушечными корзинками) базипетально-цветущи цветк ветковые изоаклональные полегающе-стелющиеся кустарнички.
Глава 7. Концеиция разнообразия биотической власти Рис. 360. Биографичес- кий портрет Saussurea latifolia Ledeb. [сем. Asteraceae (Алтай, Ку- райский хребет, высоко- горья, луговые склоны, редколесья, кустарнико- вые заросли)]. Биоморфотип: надземно- двусемядольные (семядоли об- ратнояйцевидные, 8 — 15 мм дл., 4 — 7 мм шир., закруглен- ные, постепенно сбегающие в короткие черешки) розеткооб- разующие (большинство побе- гов универсально-сложные ди-, плейофазные, они проходят сначала стадию розеточного ве- гетативного развития, а на сле- дующий год или позднее — ста- дию безрозеточного вегетатив- но-репродуктивного годичного побега, меньшее число побегов развивается как годичные без- розеточные унирепродуктив- 6 ные) очередно-простолистные гемисимподиально-эусимподи- альные цепочковидно-коротко- корневищные семидетерминат- но-репродуцирующие однодом- ные пурпурово-фиолетово-аст- роцветковые ретардационно- кондивидуальные травянистые
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 361. Биографический портрет Rubus sanguineus Friv. [сем. Rosaceae (ок- рестности Тель-Авива, пойма реки Яркон, 23 Ш)]. Др — дополнительный рисунок цветущего и плодоносящего побегов, наблюдавшихся летом (Zohary, 1972). Биоморфотип: надземно-лвусемядольные (семядоли эллиптические, 5 — 7 мм дл., 3,5 — 5 мм шир., закругленные, с выемкой или без нее, клиновидно суженные в черешки 2 — 4 мм дл.) безрозеточные ювенильно-простолистные взросло-тройчатолистные эпидер- мально-шиповатые эусимподиально-гемисимподиальные детерминатно-репродуцирующие однодомные бело-розоцветковые интенсивно-кондивидуальные (длинные плети вегета- тивных побегов полегают, их верхушки укореняются, далее происходит мезонекроз- отмирание средних частей побегов и обособление укоренившихся верхушек побегов— новых особей) полегающе-придаточноукореняющиеся кустарнички. неличным (друтим, коллегиальным) в эволюции организации живых систем просто не существует, поскольку выбор: компромиссы или смерть — неприемлем для жизни вообще. Живое уже несколько милли- ардов лет «поет» на Земле «гимн» своей способности к компромиссам между личным и другим (коллегиальным), которая, по большому счету, составила основу пластичности в организации живых систем и сделала ее (жизнь) «хозяйкой» на Земле. Над организацией биоморф (как рас- тительных, так и животных) когда-то повис и висит по сей день «дамок- лов меч», делающий существование каждой биоморфы (это осознавать совсем и не обязательно) не беззаботной прогулкой (никогда такого не получится) по просторам Земли, а целенаправленным явлением, без всякого на то сожаления с нашей стороны, под давлением «э~ократи- ческого пресса». Всякое развертывание вегетативного тела и завоевание тем самым, путем нарастания и ветвления пространства, что особенно хорошо за-
Глава 7. Концеиция разнообразия биотической власти 585 метко на ранних стадиях биоморфогенезов, подчинено более высокой цели — подготовке к размножению, в особенности к половому размно- жению. Телодвижения вегетатива, обеспеченные нарастанием и ветвле- нием, обречены в конце концов достигнуть цели развития феногегемо- нов, т.е. побегов, которым принадлежит местная (тактическая) власть, подчиняющая все друтие структуры, функционировать («работать») на них ради достижения репродуктивного результата и (или) его более скромного аналога — вегетативного размножения (рис. 150-206, 217-279, 362 и др.). В любом действии, которое происходит в организации живых тел, растительных биоморф, в частности, нужно искать цель, поскольку там, где есть определенный порядок, в основе которого стоит биотическая власть, обязательно должна быть цель. Развертывание определенного вегетатива в организации растительных биоморф предусмотрено их стра- тегической властью (генами) и нужно всему живому, что задействовано в этой организации: и организму или синорганизму, но и виду, а также еще и всей живой природе. В отношении развития репродуктива в той же организации растительных биоморф можно ответить однозначно: он нужен виду и, естественно, всей живой природе, но не нужен своим непосредственным образователям и временным носителям — организ- мам или синорганизмам. Цель репродуктивного развития растительных биоморф, а, следовательно, и цель самого репродуктива — поддержать вид, а что будет с биоморфой — виду в целом безразлично; главное, чтобы она, биоморфа, адаптируясь, как она это может, к конкретным условиям окружающей среды, еще и оптимально обеспечила вид чредой сменяющих друг друга поколений генетически однородных биоморф, образующих вместе его единую преемственно-историческую систему. Цель вида в организации его биоморф конкретна — продолжать себя любыми средствами, принуждая своих носителей, в первую очередь, к размножению. Все остальное вид отдал биоморфам как бы на откуп: «живите, как сможете, но меня, вид, оптимально обеспечивайте непре- рывным потоком разнообразных биоморф, а ваши старания я, вид, зафиксирую в вашей и моей генетической организации как результат отбора полезного для нас вегетатива». Вегетатив биоморф проявляет свое разнообразие: при нарастании он где-то ветвится, а где-то не вет- вится, где-то образует надземные многолетние части, а где-то таковых не образует, и т.д., реагируя соответствующим образом на воздействия внешних условий (рис. 13-32, 40-68, 150-206 и др.). Образование всякого репродуктива в организации биоморф предваряется развитием соответ- ствующего вегетатива. Половое размножение может осуществляться как на разветвленной, так и на неразветвленной опорной (каркасной) сис- теме вегетатива. После первого репродуктивного усилия (цветения и плодоношения) биоморфы могут отмирать полностью, но вегетатив может возобновить свою массу (ресурс) и повторить «видовой заказ», выбросив в пространство новый репродуктив и его продукты — биоси- стемы, произведенные для и во время полового размножения, пополнив ~идовой арсенал новыми эмбриональными особями — семенами. Виду это выгодно.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Если удобное для нормальной вегетации время короткое, то часто развиваются эфемеры (в пустынях, например), а на сельскохозяйствен- ных угодьях, под давлением иных обстоятельств, нередки однолетние сорняки. Вегетатив в процессе длительной эволюции растительных биоморф адаптировался, кроме того, еще и неплохо выдерживать пост- репродуктивное давление смерти. Располагая репродуктивные побего- вые части сбоку, растительные биоморфы умело приспособились спо- койно переносить пострепродуктивный некротический удар по вегета- тиву (рис. 238, 239, 241, 363). Многое, безусловно, еще зависит от того, насколько был рассчитан ресурс вегетатива и, если он слаб, то трудно биоморфе устоять под натиском смерти (рис. 70-77, 123-126, 184-190, 218-228, 364), но даже в таких случаях могут быть любопытные решения (рис. 78, 84, 229, 351). Иногда репродуктивное усилие, вкладываемое биоморфой в цветоносный побег (или побеги), настолько значительное, что после плодоношения он полностью отмирает, а в борьбе за жизнь могут сохраниться только 1 или больше почек, на которых уже к осени развиваются придаточные корни (рис. 81, 82, 274). Когда биоморфы находят возможности заблаговременно пополнить ресурсы вегетатива, потраченные на половое размножение, то их развитие становится мно- голетним, нередко весьма продолжительным (рис. 232-276, 365 и др.). Каждая биоморфа как-либо (непосредственно или опосредованно) озабочена судьбой вида, т.е. не только и не столько своей личной жиз- нью, сколько судьбой системы генетически однородных, но все же друтих для нее биоморф-совидичей, которые в структуру ее тела никогда не входили или даже входили на отдельной стадии ее развития, когда она их сама образовывала, чтобы отдать свой долг виду (шире — жизни вообще), причем живой «натурой» (другого живая природа не предус- мотрела в своей эволюции), за право пожить на нашей планете. Вид, кстати, и сам всегда озабочен необходимостью отдавать долги тем, кто повыше, под прессингом чьей власти он находится, т.е. биостробе Зем- ли, возможно, — жизни вообще (Лйизни), если она не больше, чем только земная жизнь. Если бы биоморфный способ жизни не проявил, с самого своего начала на Земле, пластичность в адаптивных компромиссах по отноше- нию к эйдократическому прессу, то сейчас мы образно могли бы увидеть события, происходившие с первой жизнью на Земле, когда полностью господствовал экспрессивный альтруизм, который сделал нормой био- морфного существования — суперабсурд, когда жизнь организмов про- должалась минуты и даже секунды (см. главу 2.5.). Такое было, этого не могло не быть, поскольку у первоначальной жизни существовала неве- роятно сильная естественная тяга к ее первооснове — к неживому (!). Когда-то биоморфам надо было вырваться из суперабсурда, и они сде- лали это (иначе полное возвращение в неживое, и такое, вероятно, в эволюции жизни бывало неоднократно), что произошло постепенно, методом проб и ошибок, они (простые биоморфы-организмы) стали развивать пластичность (альтернативы этому для жизни нет) через адап- тивные компромиссы своего (личного) и не-своего (другого) под атри- бутивным (!) давлением эйдократического «пресса».
~лава 7. Концеиция разнообразия биотической власти 1,а 20 Рис. 362. Биографический портрет Hedysarum austrosibiricum В. Fedtsch. [сем. Fabaceae (Алтай, Бащелакский хребет, высокогорья, приснежные луга, луго- вые каменистые склоны)]. Биоморфотип: н зе ад мно-двусемядольные (основные стадии виргинильного развития показаны в т. 1 с. 125) бе ) безрозеточные очередно-непарноперистолистные эусимподиально- каудексовые семидетерминатно-репродуцирующие (открытые годичные побеги успевают за короткое лето образовать по одному тотарепродуктивному побегу в форме полузакрытой кисти, редко таких побегов в пазухах зеленых сложных листьев развивается 2) однодомные розово-фиолетово-бобовоцветковые изоаклональные, редко дефективно-кондивидуальные (некротическая продольная партикуляция старых растений только иногда приводит к несовершенному вегетативному размножению, после которого посткаудициальные клони- сты живуг недолго) травянистые многолетники.
588 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 363. Биографический портрет Plantago major L. [сем. Plantaginaceae (Московская область, луга, залежи, обочины дорог, поля, лесные тропы, пустыри)]. БИОМОрфОТИП: надземно-двусемядольные (семядоли яйцевидные, продолговатые или эллиптические, 5 — 8 мм дл., 1,5 — 3 мм шир., закругленные, клиновидно суженные в короткие черешки) розеточные очередно-простолистные автономно-компактнокорневиш- ные эумоноподиальные интеркалярно-инфлоресцентные (безлистные цветоносные стрел- ки, несущие на своих верхушках брактеозные семидетерминатные соцветия — полузакры- тые колосья, составляют сообща единые интеркалярные побеги, которые уже вместе с участком годичного розеточного родительского побега, пазушные меристемы которого Нх образовали, являются межсоцветием) однодомные пленчатоцветковые мелкорослые (1биоморфотип) или сильнорослые (11 биоморфотип) кистекорневые травянистые много- летники.
М Глава 7. Концеиция разнообразия биотическои власти й по ет Onopordum acanthium Ь. [сем. Asteraceae Р Зб4 ~~~гр~фич~ сартр~~ (Москва, Главный ботанический сад, питомник лекарстве се ольные (семядоли эллиптические, слегк р а об атнояй- БИОМОДфОТИП: надземно-двусемядольные се остепенно суженные в цевидные, 15 — 25 мм дл., 7— пол озеточные очередно-простолистные короткие черешки, р с осшиеся основаниями полурозеточные ете минатно-репродуцирующие (образуются рхуш я ве еч- стержнекорневые колючие семидетерминатно-репро п но-ас оцветковые озимоодно олетники (1 биоморфотип ные корзинки) однодомные пурпур — тр чв п астает в ту же осень; из них образуются розе- — часть семян, осыпавшись в почву, рор ий год заканчивают свой астения, которые на следующи го точные вегетативные озимые р дуктивным развитием) или озеточным вегетативно-репро полный жизненный цикл полуро — ая часть семян прорастает весной и растения разви- двулетники (11 биоморфотип — друг — 1-ый год, полурозеточная озеточная вегетативная стадия- ваются по двулетнему циклу: розето вегетативно-репродуктивная стадия- ия — 2-ой год).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ
Глава 7. Концеиция разнообразия биотической власти Первыми живыми существами на Земле были простые биоморфы- организмы и их виды, поскольку половое размножение появилось по- зднее в процессе эволюции и, следовательно, позднее возникли синор- ганизмы, которые начинаются, согласно их определению, с зиготы и продолжаются только в вегетативном потомстве. Синорганизмы — это своеобразный мостик, по которому пошла жизнь в эволюции, между организмами и популяциями к виду, приближающий биоморфы к виду — одна из реализованных возможностей в поиске ответных реакций биоморф на эйдократический прессинг по отношению к ним. Суще- ствование биоморф находится в условиях постоянной эволюции жизни, на пути поиска способов разрешения неисчерпаемых противоречий между личной и коллегиальной (другой) жизнью в организации био- морф, и одно из решений на этом пути было возникновение и развитие синорганизменного способа жизни (сложных биоморф). Никогда не стоит забывать и о другом, что, по большому счету, все структурные элементы организмов — клетки, ткани, органы — это все далекие потомки совсем других живых существ — палеобиоморф, кото- рые, представляя совсем другие виды, свободно «гуляли» по земным просторам, дождавшись того мгновения, часа, месяца, года, тысячелетия и т.п., когда в эволюции жизни сработали интегративные принципы, «загнавшие» эти существа, посредством множественных трансформа- ций, в ранг структурных элементов организмов и синорганизмов. Но оставим этот пласт познания в стороне, чтобы не потерять здесь ту нить, которая нас более всего интересует. Синорганизм — это более высокая стуиень развития биоморфной организации, более ириближенная к виду. Эволюция синорганизменного об- раза жизни — это особая страница в эволюции живой ирироды, когда ти структурных элементов в сложных биоморфах иутем иодключения в такие системы, созданные вегетативным (шире — бесиолым) размноже- нием еще и результатов иолового размножения. Побеги, будучи одними из двух основных типов органов в органи- зации биоморф, — это не просто лидеры, они больше — феногегемоны (гегемоны фенетической организации), поскольку первенствуют в глав- ном — обеспечивают размножение, в первую очередь, половое, что более всего контролируется эйдократией. Другой тип основных органов — корни — лишен способности развивать на себе репродуктивные органы. Стремление к образованию репродуктива — великий стимул к развитию на побегах при необходимости еще и придаточных корней. Корни часто вынужденно (придаточно) развиваются на побегах не по- тому, что это надо корням, а потому, что это надо побегам (!), лидерам среди основных органов, ведущим исполнителям стратегических целей всего организма. Побеги диктуют отдельным частям организма (через определенные гормоны и другие реакции), что для успеха в их главном стремлении — образовать репродуктивные элементы, нужны корни, причем не завтра, а именно сейчас, поскольку условия для их образо- вания подходят. Вероятнее всего, с помощью различных гормональных процессов включаются и выключаются различные механизмы актива-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЯ ции той или иной жизнедеятельности различных частей растительного организма. Итак, как видно из приведенного выше материала, организации биоморф (шире — биосистем) чужд хаос, превыше всего строго управ- ляемая организация; но им чужда и гомеократия, о чем уже было ска- зано выше; они признают в своей организации только сильную биоти- ческую власть с четко ориентированными целями, при стремлении к достижению которых все пути для них хороши: от доведения личной жизни почти до абсурда, например, у эфемеров, и, через тончайшие компромиссы вегетативного содержания биоморфы с ее приоритетным стремлением к размножению, до растягивания «удовольствия» личной жизни на многие годы (иногда сотни и даже тысячи лет). Что же произошло и происходит с организацией биоморф в эволю- ции ее за многие миллионы лет. С палеоисторических времен был заложен и принят для всех последующих поколений биоморф семешпих растений универсальный принцип взаимоотношения частей в организации рассматри- ваемых растений: биоморфы семешьи растений могут нормально расти и развиваться только при сохранении среди частей их организации следующих приоритетов — генетического над фенетическим, побегового над корневым, размножающегося над неразмножающимся, репродуктивного над вегетатив- ньпи и, главное, диктата видовой власти (эйдократии) над организменной и синорганизменшпми властями (онтократией и синонтократией). Все растительные биоморфы следуют универсальному принципу взаимоотношения частей в их организации, с учетом которого происхо- дит и эволюция. Ее результаты удивительны и поддаются обзору. При обязательном следовании универсальному принципу взаимоотношения час- тей в организации биоморф в эволюции достигается свобода выбора пла- стичной живой материи при конструировании наиболее изменчивой фене- тической организации биоморф по четырем основным типам. Во-первых, биоморфы, и не только семенных растений, по разным обстоятельствам довели личную жизнь почти до абсурда, приспособившись быстро на- капливать и развертывать ресурсную массу в пространстве и осуществ- лять за ее счет такое репродуктивное усилие, после которого не остается сил (нет резервов и нет внутренних возможностей, чтобы их восстано- вить) для продолжения личной жизни (эвриэфемеры: однолетники, озимооднолетники, эфемеры и гемиэфемеры, рис. б9-73, 123, 124, 184- 190, 218-225). У менее сложно организованных многих биоморф низших растений в продолжительности их личной жизни нередко проявляется и абсурд, и суперабсурд (см. главу 2.5.). Во-вторых, биоморфы, и не только семенных растений, научились растягивать подготовку к репро- дуктивному размножению на несколько лет, но после первого же реп- родуктивного усилия, не оставившего им резервных ресурсов для вос- становления, они элиминируются (унирепродуктивные двулетние, сред- нелетние и многолетние биоморфы: обоеполые или однополые моно- карпики, моноантерики, рис. 41, 42, 76, 77). В-третьих, биоморфы и не только семенных растений, приспособились к личной жизни за счет развития повышенного стремления к вегетативному размножению; при этом стремление к репродуктивному размножению сохраняется, но и
~лава 7. Концеиция разнообразия биотической власти оно может также трансформироваться, например, когда в соцветии вместо семян развиваются, в основном, незрелые выводковые почки (как, например, у Polygonum viviparum L. (рис. см. т. 1, стр. 233). В-четвертых, биоморфы, и не только семенных растений, способны находить компро- миссы между их стремлением к более продолжительной личной жизни и стремлением к размножению (рис. 150-183, 193-206 и др.). То, что нарастание и ветвление составляет «локомотив» в «могучей пятерке», обеспечивая биоморфам организационный динамизм при покорении и освоении пространства, и то, что размножение удачно дополняет экспансивную активность биоморф по отношению к про- странству, что особенно необходимо их видам, — это более всего понят- но. Однако, почему статус партикуляции и элиминации настолько вы- сок в организации биоморф, что они, по мнению автора, занимают достойное место в «могучей пятерке»? В свое время (Нухимовский, 1973 б) был поставлен жирный крест на позиции тех авторов, которые счи- тают партикуляцию только проявлением «старческого увядания» у рас- тительных организмов. Можно считать доказанным, что термин «парти- куляция», предложенный Г.Н. Высоцким (1915), правильнее будет по- нимать в широком общебиологическом смысле — это всякое (физиоло- гическое или механическое) разрушение связи между живыми частями какого-либо организма, результатом которого является полное или не- полное разделение и обособление их (Нухимовский, 1973 б, 1997). Такое объяснение партикуляции позволило утверждать, что жизнь на Земле в настоящей или подобной формах была бы невозможна без партикуля- ции (т. 1, гл. 4). Партикуляция действует на всех стадиях существования растительных биоморф, приводя к отделению от них партикул: живых листьев, веток, корней, но и семян и т.п., являясь обязательным усло- вием для осуществления всякого размножения. Взаимоотношения внутри «могучей пятерки» складываются неоди- наково, но, чаще всего, используются компромиссы с разных сторон, чтобы и организмы, и синорганизмы пожили, и виды просуществовали и развивались подольше. Особое место в «могучей пятерке» занимает элиминация, или смерть, статус полноправного члена в ней, пожалуй, более всего нуждается в обосновании. 7.4.3. Смерть — результат проявления количества и качества жизни несовместимого с ее продолжением У растительных биоморф в отношении освоения пространства нет выбора кроме как, с различными нюансами, нарастать, ветвиться, парти- кулировать, размножаться и умирать, что составляет атрибутику их фе- нетического существования. Другого биоморфам не дано в процессе эволюции. Единственное, что эволюция позволила, — это возможность выбирать и комбинировать сочетаниями этих процессов(!).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Из предыдущего материала ясно, что явление жизнь мы настрои- лись наблюдать через призму ценобиоза и меробиоза — отношений биосистемы-целого и биосистем-меросом, в структуре которых вычле- няем, при необходимости, еще и друтие живые и неживые, причем как абиогенные, так и биогенные меросомы. В качестве биосистем в ранге целого, чтобы избежать каких-либо наслоений, изберем простые био- морфы-организмы, а биосистем в ранге меросом — гипобиоморфы (живые структурные элементы, которые, в свою очередь, состоят из биомеров, эуабиомеров и, нередко, некромеров). Жизнь, через отношения структурных элементов, составляющих ее непосредственного носителя — живую телесную систему (биосистему), в нашем случае, простую растительную биоморфу-организм, проявляет свое развитие в пространстве и времени. В потоке биосистем жизнь себя проявляет преемственно, как-либо передаваясь от тела к телу, каждое из которых по отдельности дискретно. В силу действия закона дискретно- сти, жизнь обязана когда-то остановиться в своем развитии в конкрет- ных телах, ее непосредственных носителях, оставив свое производство в неживом состоянии остальной природе. Однако, если бы все было только так, то жизнь на Земле быстро исчезла бы, ничего не успев на ней сотворить своего. Дело в том, что жизнь, прежде чем навсегда уйти в небытие в некотором конкретном теле-биосистеме, кое-что успевает сделать для своего сохранения вообще, у нее, как известно, предусмот- рена система продолжения себя в друтих телесных системах: она воспро- изводит себя через различные меробиозные фрагменты, которые зара- нее, до наступления смерти, в образе семян, деструктивных и конструк- тивных эпиэмбриональных партикул и т.п., покидают родительскую телесную систему, вручив им свое продолжение в следующих поколени- ях биоморф (шире — любых биосистем). Чтобы живому телу уйти в неживое, она, жизнь, включает свой механизм самоуничтожения, своего «штатного» исполнителя дискретности, «палача»-адаптацию на службе у жизни — смерть, которая является одной из сложнейших функций жизни, точнее, ее особым функциональным синдромом, по сути, фини- тофункцией (конечной функцией)-адаптацией. Все термины, приведен- ные здесь, только пояснительные к слову «смерть» и не рассчитаны на то, чтобы всем понятное слово как-то заменять. Смерть — это как «снаряд» возмездия за то, что конкретное тело было удостоено судьбой испытать, больше или меньше — за особое состояние бытия под назва- нием «жизнь». Причин смерти много (старость, болезни, травмы и т.п.), но сама смерть всегда одна и, будучи примененной жизнью против своего же тела, она нацеленно достигает дискретности его как живого тела. В обязательности достижения дискретности существования живыми теле- сными системами заключена целесообразность финитофункции в ней, т.е. смерти. Для планеты Земля жизнеразвитие телесных систем с чередованием в них размножения и смерти — это прогрессивная объективная реаль- ность, обеспечивающая пока, несмотря на множественные испытания, которые постигали биостробу (живое тело нашей планеты), например,
Глава 7. Концеиция разнообразия биотической власти столкновения с астероидами, периодические оледенения, горообразова- ние, поднятие и опускание дна океанов и т.п., продолжение жизни уже около 3, 5 миллиардов лет. За особое состояние материи — быть живой, Природа, в отношении всех земных тел, которые обладают свойством нести в себе жизнь — биосистем, или биосом (живых тел), позаботилась заблаговременно, обязав их «платить» ей налог на витальность (жизне- емкость, или «жизнеприсутствие»), установив для них единый финито- функциональный (дискретный) тип развития. В целом, это не удиви- тельно, поскольку она, Природа, имеет свой всепроникающий «инстру- мент» контроля за состоянием развития вообще — эволюцию, а для этого процесса все равны, ее принцип — не грубое «что хочу, то и ворочу», а целесообразное раскачивание в образе эволюционных маят- ников: «так-сяк«туда-сюда». Витальность, по большому счету дискрет- ная, является нормой существования всех ныне живущих тел на Земле, но у биостробы есть, пока еще во многом невостребованный, потенциал — стремление к суперадаптации [бесконечному сохранению Жизни во Вселенной, достижение которого находится во власти человеческого разума, а он пока единственный из всего известного нам, кто может выполнить роль Разума Вселенной (т. 1, гл. 3)]. 7.4.3.1. Смерть — равноправный член «могучей пятерки» В организации растительных биоморф смерть является финитофун- кцией не только целых организмов и синорганизмов, но и каждого по отдельности их живого структурного элемента, т.е. она сопровождает все проявления жизни на Земле, а она бывает только телесной. Путей к смерти множество, но самый естественный — это старение. Способ- ность живых тел стареть — это атрибутивное свойство жизни, которое отмечают более всего исследователи как растительной, так и животной жизни (Стрелер, 1964; Стешенко, 1965; Шалыт, 1965; Казарян, 1969; Лэмб, 1980 и др.). Нарастание и ветвление (кущение — разновидность ветвления) обеспечивает развертывание фенетической организации биоморф, но всегда использовать только эти процессы в своем жизнеразвитии расти- тельные биоморфы не могут, поскольку им, их природой, предназначе- но не только восходить в стремлении достигнуть некоторой цели (обес- печить вид, например, пополнением полноценным потомством) — «по- толка» своих восходящих возможностей в границах конкретного тела, но и опускаться вниз (нисходить), после достижения некоторой середины жизненного пути, чтобы, старея, разрушить многое, что по «крупицам» создавалось ранее. Смерть — вот та функция жизни, которая ставит точку на существовании биосистем разных уровней организации, кото- рые составляют и сопровождают живое существование родительской телесной системы. Структурные элементы организации биоморф возни- кают, разрушаются и умирают постоянно, начиная с зиготы и до ее смерти в целом. Способность всех биосистем, составляющих раститель- ные биоморфы, стареть заложена в генах, которые во многом предоп- ределяют возрастные границы старения. В зависимости от образа жизни биоморф в конкретных условиях окружающей среды, на основе потен-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ циальной картины жизнеразвития, заложенной в генах, проявляется конкретная жизнь именно этой биоморфы со всеми накладками на нее дополнительных факторов, усложняющих процедуру старения, напри- мер, травмы, болезни, какие-либо иные неблагоприятные воздействия среды обитания. В конце концов устанавливается тот реальный возраст, когда жизнь включает в действие свою финальную функцию — смерть, поскольку и количество, и качество жизни, которое было накоплено к данному моменту, делает такой вариант развития приемлемым для био- морфы, и она, безусловно, вступая в противоборство с надвигающейся смертью, может отодвинуть ее приход или принять бесповоротно, не на~я «аргументов» к своему продолжению в теле, которое она же и создала, развила и вот сейчас сама же и убивает. Да, действительно, смерть как функция жизни, как адаптация- это величайший парадокс бытия живой природы и ее же величайший атрибут. Начав жизнь с зиготы или иного зачатка, растительные биоморфы сразу же начинают стареть, но все это происходит незаметно, поскольку стремление к размножению ведет к «расцвету» всех жизненных функ- ций, перекрывая «горечь» множеств смертей структурных элементов (клеток, тканей, частей органов и т.п.), сопровождающих жизнь расти- тельных организмов. Когда размножение (наиболее желанный долг биоморф перед видом) исполняется (иногда к нему биоморфы адапти- рованы прибегать многократно), то тогда признаки старения все более и более завладевают внутренней организацией биоморф, вызывая ее дряхление и постепенную или быструю смерть в целом всей телесной системы (все рисунки, где отражено развитие биоморф от рождения до смерти). Итак, приведенный материал, пожалуй, достаточен, чтобы утверж- дать, что растительная жизнь на Земле стала бы невозможной без уча- стия в ней всех, без исключения, членов «могучей пятерки». Только смерть (элиминация) может поставить точку на тех результатах нарас- тания, ветвления, партикуляции и размножения, которые сначала созда- ют в растительной биоморфе определенный ресурс смерти при жизни, т.е. количество и качество жизни, необходимое для проявления ею финитофункции (смерти), а затем сами же, все вместе, под управлением генов, способствуют его же реализации. 7.4.3.2. Разнообразие смертей-адаптаций в организации биоморф семенных растений Всякая смерть — это адаптация, способ избавления конкретного тела самой жизнью от себя же, т.е. это всегда форма самоуничтожения живого тела, даже тогда, когда вроде бы уничтожение происходит извне. Например, горит лес, конкретная растительная биоморфа сгорает на пожаре от огня, приняв от него мученическую смерть. Смерть прояви- лась в жизни растения, ее действительно спровоцировал пожар, но сам акт смерти осуществило само живое тело, поскольку оно не нашло ресурсов жизнесохранения, чтобы выстоять в огне; это само живое тело, не сумев защитить в себе жизнь, не найдя для этого ресурсов поддер-
Глава 7. Концеиция разнообразия биотической власти жания жизни, превратило все происходящее в нем в ресурс смерти и, применив именно эту адаптацию, позволило себе сгореть на пожаре (немало может найтись, и на самом деле находятся, растений, которые как-то умеют выжить в огненной стихии). Жизнь пребывает в телах какое-то время (она на Земле дискретна, пока ничего другого замечено не было), развивает своих непосредственных носителей, покидает их в телах-потомках, унося с ними в другую жизнь ресурсы своего повторе- ния, но сама, в большей своей части остается в родительском теле, продолжая его развитие и, наконец, не найдя ресурсов к самосохране- нию в том же теле, отвечает за все происходящее своей финитофунк- цией — смертью. Жизнь направляет смерть на свое же родное (предко- вое) тело, обеспечивая таким ответом ему и себе разную дискретность в единой системе ее предково-потомковой преемственности, когда кон- кретному живому телу отведено обязательное возвращение в неживое (к первооснове). Переход живого тела в новое для него качество (неживое) жизнь умело использует для своего развития, т.е. в своих целях, получая, тем самым, соматическое удовлетворение (полное или не очень), иначе зачем ей вообще нужна телесная дискретность. Главное, она, жизнь, никогда не изменяет своей первооснове, возвращая в неживое то, что у него берет, как бы в долг, используя для своего развития, но при этом несколько трансформирует свое бывшее содержимое в конкретном теле, «наслаждаясь» своим эволюционным совершенством, не утрачивая, однако, и своего единства с неживой природой. Особенность любой телесной жизни в том, что она пронизана единством живого и неживого в существовании как целых живых тел, так и их живых меросом. В любой живой системе, например, раститель- ной биоморфе, вместе рядом интегрированно сосуществуют живые структурные элементы — биомеры и неживые структурные элементы: абиомеры и некромеры. Меробиозное фенетическое развитие расти- тельных биоморф обеспечивает «могучая пятерка», члены которой, вза- имодействуя где-то, завоевывают и осваивают пространство, а где-то его утрачивают под давлением других биосистем. Смерть как адаптация разнообразна. Нам здесь важнее всего осоз- навать эту объективную реальность, чем усердствовать в перечислении ее форм. Кое-что все же надо отметить. Так, главное, из-за чего про- исходит смерть растительных биоморф, — это старение, вторая причина — болезни, третья — травмы. Чаще всего причина смерти комплексная, когда стремление живого тела к неживому усиливается действием разных факторов. Для разных целей, например, для омоложения кустарников, садо- воды давно прибегают к обрезке старых ветвей, т.е. искусственно нано- сят травмы, с которыми родительские растения успешно борются и, более того, крона их кустов омолаживается, а некоторые кустарники или кустистые деревья при желании можно превратить путем обрезки в стройные деревья, например, Malvaviscus arboreus Cav., Parcinsonia aculeata L. (рис. б8). Смерть может быть направлена жизнью не только на свое тело, но и провоцировать ее проявление в других телах, например, известный
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ пример из учебника (Васильев и др., 1978), когда фикус-«удушитель» растет сначала как эпифит, его корень, наконец, достигает почвы, а далее образуется сетчатый «ствол» из придаточных корней, который убивает свою прежнюю опору, поскольку та ему уже не нужна. При всем этом жизнь научилась не только пасовать перед смертью, но и использовать ее результаты для решения своих проблем; она, например, адаптировалась применять мертвые ткани — вторичную дре- весину в структуре древесных стволов, решая жизнеполезные функции (опорные, переброс воды не только в живых тканях, но и по «трубкам» мертвой древесины, как по водопроводам и т.п.). Обо всем таком есть смысл поразмышлять подробнее. Неужели растительные биоморфы научились в процессе эволюции «играть» со смертью и приручать ее для выполнения жизнеполезных функций? Какую бы позу ни занимали наши оппоненты, им придется констатировать: да, это так. Биоморфы семенных растений и их биоме- ры приспособились жить «в обнимку» с некронами, или мертвыми биогенными телесными системами. С некронами организация расти- тельных биоморф сосуществует в процессе эволюции сотни миллионов лет. Чужими и своими некронами, попавшими в почву, растительные биоморфы питаются, вытягивая из них все субстратное, что можно вновь использовать для построения живых тел. Чаще всего биоморфы не дают некронам возможности остаться внутри своей организации и различны- ми способами они удаляют их из себя (сбрасывают, слущивают и т.п.). Однако, если присмотреться к организации биоморф семенных расте- ний, то в ней иногда все же удается усмотреть некроны, встроенные в живое тело организмов и продолжающие нести полезные для жизни функции. Живые организмы в постоянной «игре» со смертью приспо- собились не только покоряться ей, когда сил для борьбы с ней или для бегства от нее не хватает, но и научились загонять результаты работы смерти в свою живую организацию, использовать некроны в построении живого тела и для выполнения жизнеполезных функций. Сухие колюч- ки, шипы и т.п. защитные приспособления заметной роли в освоении пространства не играют. Немало некронов используется при рассеива- нии в пространстве семян, например, «парашюты» семянок одуванчика, крючки на-семенах якорцев стелющихся, крылатки с семенами у кленов и т.п. Однако более всего поражает такой способ использования некро- нов, как сохранение мертвой вторичной древесины в структуре стволов деревьев. Нередко мертвые ткани продолжают еще долго соединять организмы вегетативного происхождения, помогая им некоторое время снабжаться водой и минеральными ресурсами, например, клонисты Rhodiolea rosea L. могут десятки лет высоко в горах не отделяться от мертвых корневищ, еще соединяющих их. Многие компактные дерно- вины биоморф злаков Achnatherum splendens (Trin.) Nevski, Deschampsia cespitosa (L.) Beauv., но и некоторых других кистекорневых растений (рис. 83-93, 100, 280-282), способны долгое время, а иногда и всю их жизнь, не распадаться, оставаясь только внешне едиными; будучи свя- занными, клонисты, вместе с мертвыми остатками (некронами) образу- ют криптоклональные дерновины.
Глава 7. Концепция разнообразия биотической власти Итак, способность биоморф использовать некроны проявляется у растений не только в том, что они хорошо растут на перепревшем навозе, но и в том, что они научились включать свои некроны в свою живую структуру, где они продолжают выполнять жизненно важные функции. Смерть, как известно, осуществляет выбраковку одряхлевшего материала, очищает для жизни — для свежей растительной и животной жизни, пространство, которое она, жизнь, может, но не хочет исполь- зовать непродуктивно; она борется за овладение пространством, и дрях- лая жизнь ее, по понятным причинам, часто не устраивает. Смерть настолько вошла в жизнь, что находится с ней в единстве и постоянном противоборстве, являясь для нее нормой бытия. Противоречия во вза- имоотношении смерти и жизни постоянно, так или иначе, разрешаются в чью-то пользу или находятся пути к временному примирению. Пара- докс, но в стремлении как-то, хотя бы временно, примириться со смер- тью, жизнь научилась подчинять ее для жизнеполезной работы в орга- низации своих частных носителей. В способности живых систем ис- пользовать в своей организации мертвые биогенные части [свои или (и) чужие] я бы назвал одним из чудес организации жизни. Нельзя не восхищаться фактами, когда мертвые части растений участвуют в вы- полнении жизненных функций. Жизнь в процессе эволюции научилась загонять смерть не только для очистки пространства для новых своих носителей, но и внутрь структуры биофоров, специализируя мертвые биогенные тела, или некроны (некромеры), для выполнения определен- ной полезной для жизни работы. Смерть, в виде некромеров, присут- ствует в организации многих растительных биоморф — организмов и синорганизмов, — и пока у их жизни хватает ресурсов (сил) эксплуа- тировать смерть, последняя обслуживает жизнедеятельность этих биоси- стем. Нас все более интересуют ситуации, где смерть на какое-то время, иногда на сотни и даже изредка на тысячи лет, ассоциируется и «кон- сервируется» в живых телах, участвует в их организации. Интересно об этом, в отношении древесных растений, написали М. Бигон, Дж. Хар- пер, К. Таунсенд (1989, с. 180): «Большая часть тела такого одревеснев- шего растения мертва — лишь под самой корой располагается тонкий слой живых тканей. И вот этот-то живой слой непрерывно порождает новую ткань и окружает ствол дерева все новыми и новыми напласто- ваниями отмершего клеточного материала. Большая часть дерева — это нечто вроде кладбища, где погребены отжившие свое стеблевые ткани». Некромерами буквально «накачаны» стволы деревьев и кустарников и, что самое важное, они там не столько для того, чтобы разлагаться, от этого никому не уйти, а там они появляются и часто надолго для того, чтобы как-то «работать» на живую организацию, поскольку жизнь «при- ручила» их и сохраняет для себя на освоенных ею ранее пространствах, чтобы «доить и доить» смерть, пока хватает сил (ресурсов) удерживать ее в своем заключении. Мертвая (ядровая) древесина более всего участвует в механической деятельности стволов деревьев и кустарников, усиливая опорные свой- ства этих структурных частей. Уместно здесь привести высказывание
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ известного анатома К. Эзау (1980, с. 117 — 118): «Ранее отложенные слои вторичной ксилемы обычно быстро перестают функционировать и как проводящие, и как запасающие ткани. Относительное количество нефункционирующей, ядровой древесины варьирует у разных видов, а также зависит от климатических условий... Ядровая древесина обычно имеет более темный цвет, чем активная древесина — заболонь. Ядро образуется в результате выведения запасных веществ или их преобразо- вания в ядровые вещества и гибели протопластов в паренхимных и друтих живых клетках древесины.... Граница между заболонью и ядром постоянно смещается от центра к периферии ствола». Итак, способность биоморф (шире — всех биосистем) использовать некроны проявляется не только и не столько в том. что они хорошо растут на перегное и навозе (это, безусловно, важно), но и в том, — это не менее важно, — что они научились в процессе эволюции жизни включать некроны (некромеры) в свою структуру, наравне с друтими, действительно живыми частями и заставлять их выполнять полезные для биоморф жизненные функции, например, участвовать в перекачке воды и (или) усиливать опорные свойства стеблей, особенно древесных растений, что и помогает удерживать гигантские массы листьев и реп- родуктивных органов на значительной высоте над поверхностью почвы и т.п. Все это есть в организации биоморф, поэтому не считаться с элиминацией как одним из пяти основных способов освоения про- странства не приходится. Выше уже была высказана мысль, которую можно считать гипотезой, которую надо развить подробнее, что при смерти организмов, вероятно, гены умирают не сразу. Они полностью умирают, когда исчезает обслуживающая их энергетика (!) как собствен- но генов, так и окружающих их тканей. Особая энергетика участвует не только в хранении и сохранении жизненного потенциала генов, но и обеспечивает при востребовании их функционирование. Гены с инфор- мацией жизнеспособны только при наличии у них своей жизнеполезной особой энергии («идиоэнергии»), которая не только удерживает в уль- трамикроскопических упаковках сложнейшие нити ДНК («генонемы»), но и способна обеспечивать начальный импульс для развертывания заложенной в генах информации, к развертыванию которой далее под- ключается вся энергетика окружающих их систем живого организма. Энергетический ресурс генов мал, но, вероятно, он рассчитан на боль- шее, чем нам сейчас представляется, чтобы какое-то время сохранять их в жизнедостаточном (не утрачивая способность к развертыванию иди- оэнергетического потенциала) состоянии, даже тогда, когда организм уже мертв. Гены мертвых организмов, помещенные в окружающую их новую среду — новый автобиотоп с подходящей жизненной энергети- кой, могут (так надо полагать, чтобы было понятно, что эксперимента- торам предстоит фактически доказывать или опровергать) вновь начать функционировать по пути развертывания свойственных для него фенов. Рассмотренная гипотеза — дополнительный результат, полученный от размышлений о взаимоотношении жизни и смерти в организации био- морф семенных растений.
Глава 7. Концеиция разнообразия биотической власти Итак, раньше или позже, но жизнь, развиваясь на Земле в телесных системах, отступает, включая свою же финальную функцию — смерть, проявление которой в полной мере она, жизнь, какое-то время сдержи- вала. Безусловно, в Природе существует закон (специальный) взаимоот- ношения жизни со смертью, который, исходя из известного, можно оп- ределить так: жизнь, однажды проявив себя на Земле (шире — во Вселен- ной), обрела двойственную направленноапь (тягу) своего развития в теле- сных системах (биосистемах) — к сохранению себя при завоевании и ос- воении пространства-времени и к разрушению себя же, в стремлении вер- нуться к первооснове (неживому), что нашло отражение в организации биосистем (в чаапноапи, и биоморф), в которой постоянно (больше или меньше) проявляются следы взаимодействия противоположных сил (носи- телей) жизни и смерти (полное или неполное отмирание разнообразных структурных частей биосистем постоянно сопровождает их роап и раз- витие) и когда-то (многое зависит от последствий воспроизведения, у биоморф — размножения) наапупает момент, когда биосистема дает полную свободу своей финальной функции — смерти, т.е. и количество, и качеапво жизни, накопленное в конкретной биосистеме, делает невозмож- ным продолжение жизни в ее прежнем теле. Этот закон находится в полном соответствии с общей теорией жизнесохранения (т. 1, гл. 3). Жизнь, как видно из определенного выше закона, сама создала для себя же свою противоположность — смерть (элиминацию), и, проявляя с ней единство, она, жизнь, вступила со смертью в противоборство, сделав ее своим обязательным спутником, которым приспособилась управлять при освоении пространства. Жизнь управляет смертью на генетической основе и под влиянием окружающей среды, что находит свое отражение в фенетической организации биосистем (живых теле- сных систем) при освоении пространства и при поддержании с помо- щью смерти в нем своей самодостаточной энергетики, путем обновле- ния состава носителей жизни, привлекая или не привлекая некроны (мертвые биогенные тела) в организацию биосистем, для выполнения жизнеполезной деятельности и всегда ориентированной, кроме жизне- сохранения, еще и на то, чтобы когда-то уступить, на части освоенного ею пространства, давлению смерти и своей второй естественной тяге- к возвращению в неживое. Похоже, только сейчас удалось найти и ухватиться за ту «ниточку» (или «веревочку»), которая, возможно, не имеет конца, но «тянуть» за которую крайне любопьпно, поскольку появляется, на первый взгляд, невероятная мысль, что, может быть, как раз это и есть начало того пути, который, возможно и приведет к разностороннему познанию сущности Жизни (жизни вообще) и ее места во Вселенной. Кто знает, но судите сами. Автор последовательно решал разномасштабные задачи, и увидел перед собой такой клубок явлений, да к тому же связанных вместе одной единой «нитью» или многими «нитями» (не в этом суть), что не ухватиться за нее или за них было бы упущением. Пожалуй, путь предстоит долгий, и мне, похоже на то, удалось встать на него и даже немного пройти по нему.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ Особенность организации растительных биоморф в том, что на фоне их борьбы за жизнь в них не идет (!) борьба за главную биотичес- кую власть — генократию, поскольку эта власть передается биоморфам по наследству. Чтобы власть успешно передавалась из поколения в поколение, тактические элементы биоморф должны совершить много деяний, а, главное, создать такой вегетатив и репродуктив, который обеспечит через соответствующих носителей надежную защиту (как обиталище) и передачу [половым и (или) вегетативным размножением] стратегического (наиболее властного) материала — генов. Итак, мы в начале большого пути, который в организации расти- тельных биоморф еще никто не проходил. То, что биосистемам свой- ственно единство живого и неживого — это известный атрибут органи- зации жизни, хотя и здесь кое-что новое удалось подсмотреть, а вот то, что в организации биосистем и, в частности, биоморф семенных расте- ний, действуют законы власти, разумеется, биотической, что в ней есть своя иерархия и что в организации родственных биоморф «свирепству- ет» эйдократия, можно даже сказать «эйдоархат» (могущественная власть вида), а причина всему макробиократия, — это проблемная тема, раз- работка которой в отношении организации биоморф только начинается.
Глава 8. Концепция всюдности эволюции и ее отражение в габитусах и формах роста биоморф семенных растений 8.1. От движения вообще к трансформации вообще, эволюции вообще и эволюции живого в особенности Когда-то давным-давно (полагают, около 20 миллиардов лет назад, а может быть, и больше) «возбужденная» своим сверхгигантским взры- BQM («Большим взрывом») Вселенная прямо-таки «захлестнула» матери- ей-веществом ту часть пространства, где сейчас находится наша Земля (Шкловский, 1973; Озима, 1990 и др.). Первородное вещество Вселен- Об эволюции желательно больше думать, чем писать. Если уж писать, то только тогда, когда появится опыт и груз фактических зна- ний, пропитанный пылью экспедиционных дорог или (и) запахами ла- бораторных экспериментов. Эволюция — всегда в центре внимания; она — возмутительница научного спокойствия. Эволюция не любит лобовых атак амбициозных и словоохотливых юных дарований; она уходит по- дальше от искрометного блеска недопонимающих взоров пока лишь потенциальных эволюционистов. Ей не симпатичны «бросатели» кам- ней, поскольку она надежно хранит многие свои тайны, давая понять, что в отношении с ней не может быть истины в последней инстанции. Эволюция больше предпочитает обходительный шелест и скрип внешне неторопливых, как правило, осмотрительных и обстоятельных, еще не растратившихся по мелочам научных мужей, позволяя чуть-чуть, только им, приобщиться к себе, но не более... Наша твердыня многим не по зубам, как до, так и после их какого-то «летия». Однако не будем отчаиваться. От этого еще никто не умирал. И все же над вопросами: кому, где, когда, как и почему суждено быть или не быть с эволюцией и в эволюции — постараемся подумать конкретнее. Вопросы поставле- ны в образном стиле, но остроту проблемы они, пожалуй, только уси- ливают. Для начала определимся, что такое эволюция вообще, когда и почему она появилась на Земле.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЧ ной ничего другого делать не могло, кроме как находиться в движении и в том числе трансформироваться. Вот тогда-то, даже раньше и много раньше, если точно, то никто не знает когда, ведь Вселенная бесконеч- на, и возникла она, эволюция, которая, как и ее первооснова, также бесконечна, поскольку никогда и никуда не исчезала, являясь всегда великим преобразователем природы вообще (Природы, Вселенной, Космоса). Общеизвестно, что только движение материи абсолютно, а всякий покой относителен, поскольку представляет собой одно из про- явлений движения. Понятие «движение» шире понятия «развитие», поскольку включает в себя все изменения, происходящие в Природе. Одна из сторон движения материи — трансформация (от лат. transformo — преобразую), под которой в самом общем виде понимают всякое преобразование чего-либо во что-либо, где-либо, как-либо и когда- либо. Движение вообще всегда первично, а все остальное, что связано с ним, включая и трансформацию, вторично. Здесь мы будем говорить о трансформации живых систем, биоморф, в частности, причем под углом обзора, который ранее мало обсуждался. Во-первых, мы видим в трансформации процесс, но нас часто не менее, а, пожалуй, более интересует и ее результат. Трансформистские идеи, безусловно, исходят из глубины веков, вспомним, например, взгляды трансформистов XVIII столетия (Гольбах, 1940, Дидро, 1941 и др.) — предвестников эволюционизма, которые процесс развития живых тел животных и растений сводили или к транс- формации некоего прототипа (перворастения или первоживотного- прототипа всех растений и животных) посредством увеличения, умень- шения, умножения или исчезновения органов этого прототипа, либо к механическому движению — перемещению и перекомбинации в про- странстве неизменных материальных частиц (Лебедев, 1962). Попробуем подробнее разобраться с явлением трансформации. Дело в том, что на это явление так или иначе «завязаны» многие другие. Нас проблема трансформации интересует здесь и сейчас, поскольку через нее, как мне представляется, можно разобраться с ключевым понятием эволюционного учения — эволюцией. На трансформациях материи основано разнообразие всех известных в Природе тел. «По истечении известного времени животные, растения и минера- лы возвращают природе, т.е. общей массе вещей, этому универсальному магазину заимствованные у нее элементы и начала» [П. Гольбах (1723 — 1789), цит. по А.А. Парамонову, 1978, с. 26]. Ясно, что трансформация — форма движения, изменения вообще и что эти термины не синонимы, а, следовательно, речь может идти о явлениях, которые не повторяют, а дополняют друг друга, поскольку действительно они неразрывно взаимосвязаны и взаимозависимы, при- чем трансформация всегда является частью движения вообще. Посколь- ку трансформация является формой движения и обязательной его час- тью, то, следовательно, она присуща многим другим явлениям, обеспе- чивающим содержание Жизни (жизни вообще). Ранее (т. 1, гл. 2, 3) многие известные основополагающие термины биологии: жизнь, эволю- ция, адаптация, морфогенез и филогенез — были определены автором
Глава 8. Концеиция всюдности эволюции именно через явление трансформации. Если присмотреться повнима- тельней ко всему, что собственно обеспечивает трансформацию (преоб- разование) живых систем и является одновременно ее формами, то можно констатировать, что это явление проявляет себя и в росте, и в развитии, в морфозах и метаморфозах, этоморфозах и морфогенезах, в адаптациях и в эволюции и т.п. Ранее (т. 1, гл. 3, 4) было показано, что рост и развитие — две стороны одного процесса морфогенеза, а морфо- генез — это целостная трансформация всей дискретной материальной системы от ее возникновения до прекращения существования в пре- жнем качестве. Морфогенез отдельной биоморфы — биоморфогенез (онтогенез или синонтогенез). Пожалуй, в приведенное выше (стр. 604) определение трансформации вряд ли есть смысл что-то еще добавлять и уточнять; оно традиционное, простое, понятное и универсальное, и в таком виде на него вполне можно опереться. Что такое трансформация — понятно, а вот что такое эволюция; на этот вопрос в ответах много разногласий. Далеко ходить не будем, достаточно посмотреть, что об эволюции думают философы, биологи, физики, химики, ... да и лирики, заглянув в основные справочники и энциклопедии. Обычно термин «эволюция» определяют по-разному, подчеркивая в этом явлении специфику своих областей знания: или биологическую, или физическую, или химическую ... или еще какую-то, например, философскую. Приведем для наглядности определения термина «эволюция» наи- более сложившиеся к настоящему времени в науке, обратившись к «Большому энциклопедическому словарю» (1997). «Эволюция (от лат. evolutio — развертывание), в широком смысле — представление об из- менениях в обществе и природе, их направленности, порядке, законо- мерностях; определенное состояние какой-либо системы рассматрива- ется как результат более или менее длительных изменений ее предше- ствовавшего состояния; в более узком смысле — представление о мед- ленных, постепенных изменениях, в отличие от революции. Эволюция (биол.) — необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естествен- ным отбором организмов. Сопровождается приспособлением их к усло- виям существования, образованием и вымиранием видов, преобразова- нием биогеоценозов и биосферы в целом. Эволюция галактик, количественные и качественные изменения, ко- торые претерпевают галактики за очень длительные промежутки времени. Вкл~очает первоначальное обособление под действием гравитационных сил огромного газового облака — протогалактики, ее сжатие, процессы звез- дообразования и формирования различных звездных подсистем, гибель звезд, изменение химического состава межзвездной среды со временем. Микроэволюция, совокупность эволюционных процессов, протека- ющих внутри отдельных или смежных популяций вида, приводящих к изменению генетической структуры этих гопуляций, возникновению различий между организмами и образованию новых видов. Макроэволюция, совокупность процессов эволюции живых форм на надвидовом уровне. Макроэволюция происходит на основе процес- сов микроэволюции».
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЧ Всякая спецификация термина в разных науках ограничивает воз- можности его осознания в целом как явления. При всем этом важно помнить, что всякому термину необходимо единообразие его содержа- ния, т.е. понятийной части. На мой взгляд, существующее разнообра- зие в объяснении явления эволюции, — одна из причин, усложняющих проблемы, связанные с построением эволюционного учения. Так быть не должно. Нередко у исследователей на глаза приспущены шоры, отделяющие их узкоспециальные интересы от того мира, который бушует чуть в стороне, но о котором, изучая предмет своего исследо- вания, они не склонны задумываться. А ведь где-то рядом существует мир на основе тех же веществ и процессов, во многом по тем же законам..., что и та «пылинка» или «былинка», специальному исследо- ванию которых посвятили себя без остатка многие ортодоксальные или биологи, или физики, или химики. Похоже, всем, хотя бы иногда, надо учиться смотреть и не забывать смотреть по сторонам. Давайте, нако- нец, решимся разглядеть, что получится с эволюцией, если биолог увидит в ней историческое развитие не только специализации био- морф семенных растений, но и все то, что связывает ее как трансфор- мацию биоморф с трансформацией тел остального мира. В эволюции, по большому счету, ничего не может быть, кроме трансформаций, и, по сути дела, эти термины, возможно, смогли бы стать синонимами, но есть все же одно обстоятельство, которое не позволяет сделать это. Все транс- формации, которые осущестии~ются как инерционные (относительно «пас- сивные») процессы — это, на мой взгляд, субэволюция, а вот все осталь- ные (активные) формы трансформаций — это высший тип трансформаций — эволюция. Полного покоя нигде не бывает. Так, например, тело-стол в помещении вроде бы находится в покое, но и оно движется, подвер- гаясь каким-либо инерционным движениям-трансформациям, или суб- эволюции, перемещаясь вместе с Землей вокруг ее оси, кроме того, вместе с Землей это тело движется вокруг Солнца, а еще оно в составе нашей Галактики закономерно «плывет» во вселенском потоке и т.д. Безусловно, субэволюция сопровождает процесс существования любо- го тела, допустим, стола, в пространстве и во времени, но это всегда только фон, на котором разыгрываются основные (активные) транс- формационные явления — высший тип трансформаций, или эволю- ция. При такой подходе эволюцию надо воспринимать однозначно как универсальное явление в Природе, а сам термин в таком случае надо суметь определить в одном общенаучном смысле, что, кстати, автор впервые и сделал (т. 1, гл. 3). Эволюцию нельзя где-то потерять, где-то ее недосчитаться, поскольку она объективно проявляет себя повсюду; нельзя утверждать, что где-то она есть, а где-то ее нет, хотя, если очень захотеть, то ее можно и не увидеть, и (или) какое-то время не суметь понять, чтобы затем однажды открыть ее для себя и, возможно, для других. Она, эволюция, настолько многогранна, что невольно исследова- тель может здесь и сейчас какие-либо грани ее проявления пропустить, увлекшись чем-то другим. Задача ученого-эволюциониста научиться ви- деть эволюцию в разных ее проявлениях, а не только там, где она в формах специального (особенного), допустим, в мутациях, поскольку в
Глава 8. Концеиция всюдности эволюции Природе главное в отношении явления эволюции то, что она есть везде, проявляя себя по-разному, именно над разгадкой таинств этого и надо работать. Быть (существовать) и уметь видеть (наблюдать) то, что есть, — это категории разные. Эволюция всюдна, и эта идея получает здесь дополнительный импульс к своему развитию. Эволюция — там, где есть активная транс- формация материи, а она кроме как инерционных (относительно «пассив- ных») трансформаций (субэволюции) есть везде, например, в любом теле, сущесч~ование которого без эволюции невозможно. Эволюция состоит из состояний, каковыми являются большинство трансформационных про- цессов (трансформ) — эволюстатов (от лат. — status — состояние, по- ложение), т.е. движение материи в Природе в целом эволюстатное, по- скольку оно везде и всегда наполнено эволюционными состояншими материи, или эволюстатами.) Пора, на мой взгляд, исправить недоразумение и, разобравшись с ситуацией, дать универсальное определение очень известному, но дале- ко не однозначному термину. Для науки станет лучше, если термин «эволюция» будет понятен всем со всех сторон и с любой высоты аб- стракции только в одном, строго определенном смысле. Нужно и можно ли этого добиться? Наш ответ: и нужно, и можно. Но как? Столько сломано «копий» в дискуссиях по этому вопросу, и вдруг такой аванс. Почему? Полагаю, что решение вопроса понятия «эволюция» искали, погрузившись слишком в «глубину» явления, а ответ, по сути, лежит рядом; он там, где действительно свершается эволюция, а она сверша- ется везде, причем здесь и сейчас, таким же способом, как и в прошлом, всегда как-либо влияя на историю Природы. После выхода первого тома нашей монографии определение эволю- ции в понимании ее автором, если и скорректировано, то только немно- го. Напомним, опубликованное ранее объяснение общей эволюции — это «трансформация Природы [весь поток (сейчас добавим — активных) трансформаций, отражающий дискретность и бесконечность в движе- нии материи из прошлого через настоящее в будущее), а эволюция живого, в таком случае, — часть общей эволюции (частная эволюция) — процесс адаптации всей живой природы и ее частей или суперадап- тации»] (т. 1, гл. 3, с. 82). Под адаптацией автор понимает любую дис- кретную активную трансформацию живой материи, а в бесконечном движении живой материи, или суперадаптации, видит высший смысл жизни (т. 1, гл. 3). Из широкого определения эволюции видно, что безоговорочный акцент сделан мной на трансформацию (преобразова- ние), причем активную (!), всего, что составляет Природу, и, таким образом, удалось посмотреть на это явление как на высшую ступень движения вообще. Состояния активной трансформации, или агенты транс- формации, иначе — трансформы, принадлежат или эволюции как ее составляющие части — эволюционные агенты (состояния), короче- эволюстаты (формы эволюции), или являются особой ветвью трансфор- мации, отличной от эволюции, хотя и дополняющей ее. Есть смысл во всем этом разобраться подробнее.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ 8.1.1. Почему эволюция — высшая форма трансформации и почему ее никак и нигде нельзя потерять Существование Природы связано только с движением (изменени- ем) материи, а высшей его формой является трансформация (преобра- зование). В свою очередь, у последней есть свои формы-состояния, и вот высшая из них, безусловно, — эволюция. Прежде чем посмотреть, где и как в «пирамиде» абстракций форм (состояний) трансформации (трансформ) располагается эволюция, каковы ее формы (эволюстаты) и как она соотносится с другими формами трансформации, и есть ли они вообще, определимся с понятиями. Без движения, как известно, не существует материя. Высшей фор- мой движения материи, на мой взгляд, является трансформация. Логич- но при этом заключить, что существует несколько основных способов (форм, состояний и их типов) трансформации, которые перекрывают своим величием все остальные. Мой тезис следующий: трансформация — это всеобщий атрибутивный способ сущесньования материи и Природы (природы вообще), а эволюция — высшая форма трансформации, без ко- торой существование материи невозможно. Можно констатировать, что — всюдна; суть эволюции надо искать, в иервую очередь, через ее динамич- ность, всеобщность и бесконечность, а далее надо суметь найти то осо- бенное, что отличает ее от всего остального, что характеризуется дви- жением (изменением) в бесконечности. Эволюция в процессе саморазвития материи «призвана на службу» Природой быть «служанкой» ее истории и вообще, и в частности. Дей- ствительно, ведь именно эволюция олицетворяет собой преемственность и историзм всех происходящих изменений в явлениях (телах и нетелах), поскольку в любом явлении есть не только настоящее, но и что-то от его прошлого. Только она одна, будучи бесконечной и всеобщей фор- мой движения материи — высшей (по сложности проявления) формой трансформации, способна и образовывать, и нести в себе преемствен- ные последовательно-исторические признаки. Для объяснения эволюции, пожалуй, должно быть что-то еше, чтобы однозначно отличать ее от всего остального. Я усматриваю «это еще» в ее способности быть преемственно (!) сопричастной ко всему, что происходит в Природе, быть рядом со всем остальным, более того, соприсутствовать в нем, проникать в него, быть в нем, влиять на него, вызывая где небольшие, а где побольше, изменения, приводя временами к стихиям — революционным событиям (революция — это только от- резок эволюции, ее эволюционный синдром, особое состояние, сопро- вождающееся колоссальными изменениями, ускоряющими ход эволю- ции и даже иногда изменяющий ее направление). Эволюция — актив- ный соучастник действительности, воздействующая на ее содержание и нередко изменяющая ее ход и результаты, последовательно (!) всегда уносящая в историю что-то, только ей присущее, и оставляющая в существовании Природы более или менее заметные исторические следы как отметки о своем бесконечном и неугасаемом присутствии в неи.
Глава 8. Концеиция всюдности эволюции Эволюция, как и Природа вообще, нигде и никогда не теряется, а проявляясь во всем, причем по-разному, она приобщает, используя свою преемственно-историческую сущность, к прежним своим результатам что-то, что приумножается самыми разными способами и средствами по ее части в текущей действительности. Всякий разговор о том, что где- то эволюция есть, а где-то ее нет, — это пример недооценки этого явления. Эволюцию полновесно так не понять, ее можно таким образом только немножко подергать за «косички», но чтобы понять целиком,— никогда. Для этого нужен соответствующий крупномасштабный подход, который позволял бы стирать грани между науками, поскольку эволюция, будучи высшей формой трансформации (многие философы ошибочно дума- ют, что выше ее — революция, но, на мой взгляд, это только состояние в ваоаыиии; ово ведоатовечво, почти как спичка, вспыхатв овпаиаы, хотя и многое может «напьорить», вскоре гаснет, и может быть только ничтож- ным по времени фрагментом в бесконечном эволюционном процессе), об- ладает всепроникающим общеприродным свойством. Попробуем, в дополнение к тому, что уже было сделано (т. 1, гл. 3 и 8), продвинугься дальше по пути совершенствования понятия эволю- ция. Если кратко, то эволюция — это активная трансформация в любых природщих явлениях (телах и нетелах). Подробнее эволюцию можно опре- деличь следуюп~им образом: это бесконечная, так или иначе преемственная (в новом всегда хотя бы что-то присутствует от старого), последовател но- историческая (прошлое как-либо связано с настоящим, будучи от него по- разному отдаленш~м, оставаясь иногда очень близким), разнообразно «вет- вящаяся» активная трансформация всего, что составляет природу вообще (Природу). Эволюция, будучи высшей, по сложности проявления, формой трансформации, охватывает своим действием Природу, обеспечивая, через сохранение или ииченение преднаследственности и (или) наследственности, бесконечное преемственное последовательно-историческое продолжение па- рафенетического и (или) фенетического, и (или) даже идеального в органи- зациях дискретных явлений (тел и нетел) как элементов систем исходное- производное». 8.1.2. Эволюция — главное действие в формировании разнообразия Природы Все тела и нетела в Природе так или иначе когда-либо подверга- ются трансформации, формы которой и способы проявления, — разно- образны, но вполне поддаются и типизации, и классифицированию. Все, о чем ниже пойдет речь, будет использовано в соответствующем разделе при построении иерархической системы форм трансформации (трансформ). Главное в эволюции — это достижение какого-либо ре- зультата ее действия — преемственно-исторического результата после- довательного трансформационного развития, который получается в не- которых телах непосредственно через их существование или является следствием только их взаимодействия, например, процесс грозы, про-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИИ цесс мышления (сами процессы — нетела, а делают их, безусловно, тела, у которых своя эволюция). Кроме процесса грозы (нетела) есть еще и грозовое тело, т.е. система вещественных элементов, обеспечивающих свершение процесса грозы. Эволюция происходит в телах, но ее обяза- тельные «ответвления» — последствия, проявляют себя и в нетелах, т.е. процессах, сопровождающих существование тел. Сама эволюция — это тоже процесс (нетело); вполне уместным в этом плане представляется выражение типа «эволюция эволюции» (Завадский, Колчинский, 1977). Однако никогда не стоит забывать, что всегда где-то рядом с эволюци- ей-процессом соприсутствует еще и эволюирующее тело. Эволюция какого-либо тела — это не одно и то же, что и эволюция процессов, которые соответствующее эволюирующее тело порождают, его сопро- вождают или которые сами порождаются им. Эволюция тела в Природе неотделима от эволюции процесса существования этого тела, но мы, исследователи, используя свои логические возможности, ради удобства и полноты познания, можем разделять эти явления. Эволюция грозово- го тела включает в себя не только все физические его составляющие, но и сам процесс грозы, его нюансы, стадии, формы проявления. Эволюция — это всегда и процесс, и результат, поэтому ее формы (состояния) фиксируются по тем телам и процессам, которые составля- ют ее состояние в определенный момент времени. Она в отношении любых явлений, даже в отношении самой себя, действует как система получения и снятия результатов взаимодействия некоторых тел, которые обеспечивают ей наполнение ее содержания как процесса особого рода, в котором весьма важно преемственно-историческая последовательность происходящего. Эволюция происходит в неживых и живых телесных системах, а кроме того, и всех нетелах, сопровождающих существование тел. Такое понимание эволюции пока нетрадиционно для биологов, которые обычно там, где не наблюдаются мутации («элементарные эво- люционные явления»), никакой эволюции уже не отмечают. Так, с наиболее традиционной биологической позиции «эволюция строится все же только на мутациях» (Шмальгаузен, 1968, с. 66). У меня несколь- ко иная позиция. Я считаю, что эволюционные события сопровождают существование всего, что имеет место быть в Природе. Эволюция жиз- ни, как активная трансформация живых тел, происходит в клетках, тканях, органах, в организмах и синорганизмах и самое важное для живых систем, что она продолжается в других поколениях производных систем и, в первую очередь, в основных носителях свойств вида — в биоморфах (организмах и синорганизмах). Эволюция сопровождается, как уже было подчеркнуто выше, про- явлением разнообразных форм эволюции — эволюционных состояний, или эволюстатов — активных трансформ, например, дискретных актив- ных трансформаций неживой материи или дискретных активных транс- формаций живой материи. Все неживые и живые тела, которые наблю- даются в Природе в каждый конкретный момент времени, — это и есть эволюстаты (эволюционные состояния). Чтобы эволюстаты начали «ра- ботать» в эволюционном потоке, они должны соединиться с другими эво- люстатами из прошлого или настоящего в синтомах («эволюционньи
Глава 8. Концепция всюдности эволюции шагах»). Синтомы образуют синтомокинезы, или движения синтомов,— оиределенные эволюционные ироцессы в едином явлении «эволюция» и иред- ставляют, таким образом, универсальную эволюционную единицу (Нухи- мовский, 1984; 1997). Синтомы — это свойство эволюции и ее множе- ственных «разветвлений» и «бежать», и «течь», и т.п. себя проявлять, имея в них («эволюционных шагах») разномасштабную меру (!) проис- ходящего в пространстве и времени (т. 1, гл. 3). Синтомы — мерила эволюции, поскольку в них отражается то количество и качество эво- люции, которое проявляется на определенном участке пространства и в определенном временном интервале. В каждом «эволюционном шаге» (синтоме) всегда наблюдается как исходное, так и его производное; но что же все-таки в эволюции материи действительно составляет главные цели и результаты, т.е. для чего в Природе затеян и закручен «клубок» множе- ственных активных трансформаций с бесконечным множеством синтомов иод единым общим названием этого явления — «эволюция». Мой ответ краток. У эволнщии материи — несколько наиболее крупных целей (и результатов): 1 — продолжить нежизнь; 2 — продолжить жизнь; 3 — расширить разнообразие нежизни и (или) жизни, не изменяя преднаследстенность и (или) наследственность; 4 — создать новые виды проявления нежизни и (или) жизни, изменяя преднаследспьенность и (или) наследственность; 5 — сохранить бесконечно жизнь вообще. 8.2. Универсальный принцип эволюции в Природе По сути, выше уже был поставлен вопрос ребром: где и когда началась эволюция вообще, и был дан на него однозначный ответ: эволюция никогда не начиналась и никогда не закончится; она, бесконечно множественно разветвляясь, продолжается во всем и всегда, будучи всеце- ло связанной с единством и бесконечностью Природы. Эволюция, как, впрочем, и Природа, всегда была, всегда есть и всегда будет. Наберемся смелости и упорства, попробуем оголить инфраструкту- ру явления «эволюция вообще», без концентрации внимания на деталях, что удобнее всего сделать, прибегнув к образному (логическому) «мик- роскопу», под которым, в меру своих возможностей, отпрепарируем содержательную сторону эволюции. Эволюция — это ни какая-нибудь «простушка», которая позволит нам как-либо незатейливо пообщаться с ней, пусть даже только под логическим микроскопом. Можно рассчи- тывать, если очень уж постараться, что эволюция предоставит нам воз- можность подсмотреть кое-что из того самого общего и главного, что отличает ее от других явлений и что, по большому счету, составляет ее содержание. Пожалуй, начнем с того, что отметим главное: она, эволюция, всецело служит только одному «господину». Миру, или КосмосУ (другое
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЯ его имя — Природа). Со всеми остальными природными частями эво- люция общается только через свои части — формы эволюции, или эволюстаты, чего всем по отдельности хватает или не хватает и что в каждом случае может составлять и нередко составляет предмет исследо- ваний разных специальных наук. Эволюция настолько «упакована» природными законами и правила- ми, что мы, ученые, со своими научными изысканиями можем рассчиты- вать всегда лишь на приближение к разгадке ее тайн. Эволюция всегда была, есть и будет непредсказуемой затейницей в организации мироздания. 8.2.1. Почему всякие эволюционные процессы — это движение эволюционных маятников Так получилось, а, вероятно, по-другому и не могло бы быть, что нормой существования любых тел и нетел в Природе стало движение (изменение). Из всех движений, которые встречаются в Природе, эво- люция избрала для своей основы самый атрибутивный вид их — актив- ную трансформацию (преобразование). Разрабатывая проблему общих принципов эволюции систем, я невольно поставил вопрос о том, какой универсальный образ мог бы успешно передать то главное, что состав- ляет общее содержание всяких эволюционных движений. У меня не нашлось ничего более выразительного, чем образ «маятника». Вопрос поставлен под острым углом. Почему эволюционное дви- жение в Природе — это движение эволюционных маятников? Дело в том, что внешняя среда, окружающая тела и нетела, которые эти «ма- ятники» образно отражают, неоднородна и развивается по-разному. В таких условиях живые и другие тела, имея определенный идиоэнерге- тический потенциал, склонны его целенаправленно развертывать в кон- ституции телесных систем и сопутствующей организации нетелесных систем. У всех есть свой ресурс возможностей, обеспечивающий опре- деленную норму существования конкретных тел, поскольку условия реализации идиоэнергии разные. А отсюда и колебания эволюционных маятников происходят по универсальной схеме: «так-сяк»туда-сюда» (см. главу 2.5.). Колебания любого маятника весьма убедительны для того, чтобы образно передавать смысл событий, происходящих в эволюции как тел, так и нетел, в которых чаще всего присутствуют элементы восходящего и нисходящего «полета» — развития «туда-сюда» (так колеблется маят- ник) и «так-сяк» (по-разному), поскольку маятник как образ эволюции, не только колеблется, но и перемещается в пространстве и времени (он действительно «плывет» во вселенском потоке и никогда полностью его колебания не повторяются, поскольку они происходят под разными углами наклона оси маятника и в зависимости от меняющихся условий окружающей среды. Впервые представления об «эволюционном маятни- ке» были разработаны автором (т. 1, гл. 3, 8), а также рассмотрены уже и в этом томе (гл. 2.5.).
Глава 8. Концеиция всюдности эволюции Если поставить вопросы, что является «движителем» эволюции материи и чем она может быть измерена, то наш ответ, можно сказать, вполне подготовлен предшествующими материалами: «движители» эво- люции материи — это тела и нетела в образах эволюционных маятников, колебательный динамизм которых обеспечивается как внутренней пружи- ной для своего самораскачивания (существования) — природой самого обьек- та, так и внешней пружиной — условиями существования, а мерилом эволюции являются эволюционные шаги, или синтомы, которые соединяют в себе разновременные результаты эволюции — ее эволюстаты (эволюци- онные состояния). Эволюция действует везде, особенно сложно там, где материя об- разует «сгустки» вещества и энергии, например, в (на) планетах, звездах и т.п. системах. На Земле эволюция действует в «тесноте», поскольку на эволюцию неживой природы (инвитаэволюцию) здесь накладывается еще и эволюция живой природы (витаэволюция). Эволюция в Природе, будучи ее обязательным активным трансфор- мационным содержанием, проявляет себя бесконечно, что может озна- чать только одно: у нее, эволюции, есть механизм (принцип), на основе которого обеспечивается ее неисчерпаемость в пространстве и времени, иными словами, есть основание утверждать, что именно эволюция про- являет себя везде и всегда как высшее трансформационное действие (процесс) — функция Природы, исходящая от ее тотального движителя. Согласно определению, разработанному ранее, (глава 1.3.5.) примагене- ратор, или тотальный движитель, — это пространство Природы, вместе с неисчерпаемой, всегда движущейся (изменяющейся) материей, которые как одна атрибутивная природная часть взаимодействуют с другим природным атрибутом-частью — временем, как-либо производя трансформы (формы трансформации), высшая из которых всегда и везде — эволюция. Это он, конечного самодвижения эволюционных маятников во вселенском потоке и, будучи образован материей, существующей только в движении (иъиенении), составляет своими функциями, эволюцией в том числе, то одни, то другие (и так всегда) тела и нетела Природы. Тотальный движитель — образное выражение той обьективной реальности, которая обеспечивает организаци- онные основы эволюции (принципа, или механизма этого явления), а они- в вечно движущейся (изменяющейся) материи (другого не дано!), нацеленной в своем существовании на бесконечное качественное и количественное раз- витие телесных и нетелесных систем в пространстве и времени. Выше уже было подчеркнуто, что структуру эволюции как процесса обеспечивают эволюционные состояния (эволюстаты). Есть смысл зао- стрить внимание на внутренней организации эволюционных маятников как телесных систем, которыми, безусловно, являются все биоморфы. Для этого мы продолжим манипулировать с логическим «микроскопом». Эволюционные маятники как телесные системы составляют неживая материя — вещественная первооснова (протеросома), прогенетическая (прогеносома) или генетическая (геносома) основа, которые своим функционированием «раскачивают» изнутри парафенетическое или фенетическое содержание, а оно также проявляет свойственное ему
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЯ функционирование, что в целом «раскачивает» все содержание эволю- ционных маятников как в рамках их норм существования на фоне те- кущей окружающей внешней среды, когда, кроме всего, иногда еще могут проявляться субмутации или мутации. Кстати, эволюция, будучи изменчивостью, может на конкретном участке пространства и времени иметь субмутационные (у неживых тел) или мутационные (у живых тел) результаты или не иметь их, ограничиваясь пресубмутационными (у неживых тел) или премутационными (у живых тел) результатами. Структуру (любой процесс структурен во времени, т.е. состоит из отрезков-стадий) эволюции внутри эволюционных маятников как теле- сных систем составляют их и ее эволюционные состояния (эволюстаты) на определенном отрезке пространства-времени, что наглядно можно выразить посредством графического моделирования, определив под абстрактным увеличением в логическом «микроскопе» принципиальный порядок последовательных сменах эволюстатов внутри эволюционных маятников (рис. 366), но еще и схему смен состояний каждого целого эволюционного маятника по признакам его организации в отдельные отрезки времени. Из графической модели наглядно видно, как эволюстаты «раскачи- вают» своим деятельным существованием конституцию (телосложение) эволюционных маятников изнутри, взаимодействуя с силами «раскачи- вания» этих же тел извне — за счет воздействия внешней окружающей среды, что хорошо можно подкрепить примерами из жизни семенных фитобиоморф (рис. 367-370 и др.). Эволюция как функция тотального движителя действует на все в Природе без исключения, поэтому все тела и нетела — это эволюцион- ные маятники и она, эволюция, проявляет себя и снаружи, и внутри всех тел (непосредственно) и (или) где-то рядом у нетел (опосредован- но), но не как особенная конструктивная деталь-«двигатель», а именно как функция-часть в конкретной организации каких-либо эволюцион- ных маятников, составляющая один из элементов единого эволюцион- ного потока. Любое эволюционное (всегда «колебательное») действие потребляет что-то из неисчерпаемого «топливного» ресурса (неживой материи), ко- торый трансформируется соответствующим образом за счет сил, дей- ствующих внутри эволюционного маятника, и сил, воздействующих на предмет эволюции извне, а далее задействованный эволюцией ресурс продолжает свое бесконечное движение в старых и новых эволюцион- ных маятниках. Всегда и везде в Природе работает ее примагенератор, производя бесконечную функцию — эволюцию. Итак, я не утверждаю, что открыл «перпетуум мобиле», но все-таки мне удалось однозначно определиться с тем, что у Природы есть свой тотальный движитель, занимающий в ней свое место — [это образное название механизма — производителя движения вообще и, в частности, эволюции вечно движущейся в пространстве материи, бесконечно орга- низующей себя во времени в конкретные дискретные телесные и нете- лесные системы как эволюционные маятники, существование («колеба- ние») которых происходит по универсальной схеме «так-сяк»туда-сюда»
Глава 8. Концеиция всюдности эволюции Я Я I :ф~~ й Q вй :ф~~ И о A М о ~ Q о ~ о ф~ ~Ж м ~ ц Ig ° ° к ~4 ~~ й о < о Ж М О Й r /~— / r 1 1 — 1— ! Ф'Ъ ~1 gj ! ! ! ! ! ! ! Щ о х а ~~ о ~ ~ ~ ~ аиосе)аоИ Я;~~ а ~ ~~ о ~ ~! ~ щ ~ ~ ~ ~ о >ga<e о .~~~!и фо~ло'~и+ Я ~ф ~ ~~ о ~ р ~! . щ М ~ ~ ~ Я о ~ ж „я ~ ~~ о ж Й ам~ е~ое e~оа~ ж vu о zg ga т Йой ийиоplо~й а vи~болжи vо ~~ ~ я ц ~ ~ ~ д ~ л о ~ Д / г ~) г () l /' m~>~ор ж<~йй Фж ао ооао g ~~' ~, ~~ hC g ~m Й й Й ~ о о- цоц ~о~:ооо Й ~~ ф о о о Й >:> с) ййюййй~ййй~о ! ! ! IQ С!5 ailing, й ~~ g Р щ ~~ hc ~и~""ß~~< Ж ~ о ~~ о о ~ ~) ~4 ~а Щ б~~ gg~>~> -о а3 й о, д,' Й р, а~ ~4 ~~. о 1 26саи о М 0 ~в~ ~д~ >x<oд ~ о ~ой и*~Иgz р ~~~ -, И Б 0 й; ~а~ Я о И ф й О ~-е Р и :к О ,'Я Ж A ,'Я О 4 ~~Я Ж Ж и2 ~4 О й; ° ~~~~} Ж О Л Qi >, ф~~ + О ~О;Я ~О С~) (ф е; О 0i g) И
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ за счет сил, действующих внутри материальных систем, но и извне (из окружающей внешней среды)]. Результат существования (работы) при- магенератора — бесконечность эволюции во вселенском потоке, в веч- ных и неисчерпаемых «колебаниях» (изменениях) неиссякаемого мно- жества эволюционных маятников. Сейчас можно считать доказанным, что принцип эволюционных маятников — это атрибутивный (универсальный) принцип эволюции тел и нетел Природы, в частности, естественно, и биоморф семенных растений, он функционирует по универсальной схеме «так-сяк*туда-сюда» на опре- деленном ресурсе возможностей (идиоэнерпш) и как результат взаимодей- спьия его через конституцию (парафенетическую или фенетическую орга- низацию) соопьетствующих тел, задействованных в эволюционном процес- се, с окружающей внешней средой. Всякие колебания эволюционных маят- ников, как образов конкретных тел, совершаются в определенных диапа- зонах пластичности (живые тела) и пропластичности (свойство неживых тел как-либо изменяться в отрезки времени своего существования в ок- ружающих их условиях среды в определенных параметрах на основе их преднаследственности). Диапазон (норма) пластичности и пропластично- сти какое-то время обеспечивает существование тел в свойственных им качественных границах, но когда физическое или биотическое существова- ние тел прекращается и их прежнее качество разрушается или (и) когда происходит субмутация (превращение одних прогенов в другие, например, атомов водорода в гелий, как это происходит на Солнце), или мутация (одни гены преобразуются в новые), тогда осуществляется переход к новым эволюционным маятникам (образам новых конкретных тел или нетел) на основе гиперпластичности (у живых тел) и гиперпропластичности (у не- живых тел). Все известные или неизвестные нам тела так или иначе проявляют пластичность или пропластичность, сохраняя всегда потен- ции к мутациям или субмутациям. Наши рассуждения об эволюции нетел (процессов и идеальных систем) пока самые общие, поскольку в целом мало затрагивают содер- жание главных проблем нашей работы; следует однако заметить, что именно в этой области эволюционных исследователей пока более всего белых пятен. Например, всякий процесс MQKHQ считать нетелом — чьей- либо функцией, но любопытно, когда выборочно собираются в функ- циональную систему (по П.К. рюхину, 1972, 1973 и др.) отдельные структурные части какого-либо живого тела-организма, то эта функци- ональная система становится и сама телом с определенной структурой и специализированной функцией (нетелом), которую она адаптируется выполнять в организации целого родительского тела как его часть.
Глава 8. Концепция всюдности эволюции 4,аг 27,2 Рис. 367. Биографический портрет Cynoglossum ofTicinale L. [сем. Boraginaceae (Алтай, окрестности Горно-Алтайска, остепненные горные склоны, пусты- ри, у дорог)]. Биомо ф орфотИПЫ: надземно-двусемядольные (семядоли широкоэллиптические, 12 — 25 мм дл. 8 — 15 м — 5 мм шир., закругленные, на коротких черешках) полурозеточные очередно- простолистные обильно-мягкоопушенные детерминатно-репродуцирующие эфемерно-по- лисимподиально-инфлоресцентные однодомные темно-фиолетово,-красно- бурачниковоцветковые травянистые унирепродуктивные двулетники и среднелетники (1 биоморфотип) или редко травянистые бирепродуктивные среднелетники (11 биоморфо- тип). Биоморфы часто развиваются как двулетники, обычно только один раз в своей жизни цветут и плодоносят, но в условиях загущенного травостоя изредка встречены особи, которые после первого слабого цветения и плодоношения не отмирали и через год или более повторяли еще одну репродуктивную функцию.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЯ Рис. 368. Биогра- фический портрет Biebersteinia одога Steph. [сем. Bieber- steiniaceae (Алтай, Курайский хре~, высокогорные склоны, скалы, морены, осыпи)]. Биоморфотип: 5 надземно-двусемя- дольные розеткообра- зующие эумоноподи- альные очередно- сложнолистные ин- 4~ теркалярно- или по- стинтеркалярно-реп- родуцирующие (без- розеточные цветонос- ные побеги развива- ются в пазухах розе- ~Ф) + точных листьев теку- 196,2 щего года, т.е. интер- калярно, или в пазу- хах прошлогодних листьев, т.е. постин- 37 теркалярно) стержне- корневые однодом- у ные желтоцветковые приземистые кустар- нички.
Глава 8. Концеиция всюдносщи эволюции 9а, 27 Рис. 369. Биографический портрет Verbascum fruticulosum Post [сем. Scophulariaceae (Израиль, каменистая пустыня Негев, окрестности города Димо- на, овраги, пустыри, 18 V)]. Биоморфотип: надземно-дву- семядольные (семядоли яйцевид- ные, 4 — 5 мм дл., 3 — 3,5 мм шир., закругленные, резко сужен- ные в короткие черешки) полуро- зеточные стержнекорневые очеред- но-простолистные гемисимподи- альные семидетерминатно-репро- дуцирующие однодомные желто- норичниковоцветковые изоакло- нальные (без вегетативного раз- множения) полегающе-восходяще- побеговые полукустарники.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ ° ° Рис. 370. Биографический портрет Pinus halepensis Miller [сем. Р1пасеае Израиль окрестности Тель-Авива, каменисто-песчаные холм, )]. ы 22 III> . Э SHOMOP@OTHII: надземно-многосемядольные (семядоли линей нейные 15 — 20 мм дл., 1,2 — 2 мм шир., слегка заостренные, сидячие) четочные очередно-простолистные (листья— плоская линейная хвоя с зазубренными краями), стро фо оф мимо четочных по е бегов у имматурных и взрослых растений развиваются строго специа- ые азве н хвою, в числе 2, лизи ованные компактные фотофильные побеги, которые, развернув XBog) в лизирова только в ~тосинтетической деятельностиу перестают расти и целенаправленно участвуют тол к ( а эумоноподиальные интеркалярно-репродуцирующие д д -р ие о нодомно- азнополопобеговые на одном растении, но на разных побегах, развиваются ц пыль еносные колоски и шишки равномерно-ветвистокроновые (1 биоморфотип) или р - кр ве хне-ветвисто оновые (11 биомор- фотип) вечнозеленые (хвоя живет до 3-х лет) хвойные деревья.
Глава 8. Концеиция всюдности эволюции 8.2.2. Меротомия эволюции вселенского потока (Природы) как системы ее состояний (эволюстатов) Всякая эволюция предполагает развертывание свойств и признаков тел и нетел посредством трансформаций в развитии. Эволюция, в об- разном оформлении, — это активная трансформация вселенского пото- ка (Природы), главными действующими элементами в котором являют- ся эволюционные маятники, т.е. тела и нетела, проявляющие в себе способность и стремление существовать через активные трансформа- ции, а, следовательно, посредством эволюции в различных ее формах (эволюстатах). Почему вселенский поток наполнен в своих движениях эволюцией? Все это потому, что эволюция универсальна для всей Природы, составлгя одно из высших звеньев трансформации, пронизывая собой бесконечное природное содержание, по-разному обеспечивая существование неисчисли- мого числа телесных и нетелесных систем, «раскачивая» их как эволюци- онные маятники в пространстве и времени. Главное достоинство эволюци- онных маятников в том, что их образная опорная ось и оньес с собственно маятниковым телом никогда не фиксируется, обеспечивая им возможность не просто быть в движении, но и постоянно изменять свое положение и во времени, и в пространстве, естественно, изменяясь при этом H в своей организации. Эволюция предоставила своим носителям (эволюционным маятникам) свободу выбора направлений «раскачивания», т.е. направле- ний саморазвития тел, которые они, «маятники», олицетворяют. Дина- мизм эволюционных маятников, иными словами, их разномасштабная эволюция, является результатом взаимодействия двух основных сил: внутренней энергии, исходящей из конкретной организации соответ- ствующих эволюирующих тел, и внешней энергии, воздействующей на эти тела и исходящей от окружающей их внешней среды. Эволюция пронизывает Природу, поскольку всюдна и неудержима в своем бесконечном движении; у нее никогда не бывает полных тупи- ков, а все возможные ее «тупики» всегда относительны, поскольку на смену одной (исходной) организованности всегда (другого не дано) приходит иная (производная) организованность вечно развивающейся материи, когда качественное содержание одних тел или нетел превра- щается, при определенных обстоятельствах, в другое, что свидетельству- ет о возникновении иных тел или нетел — иных эволюционных маят- ников. Эволюция ориентирована на свободу выбора направлений, ставя процесс выше результатов (!), от которых впоследствии эволюция не только может, а обязана отказываться, чтобы стремиться к новым. Это она научила монстеру долго упрямиться и извиваться, чтобы в борьбе быть обласканной лучами солнечного света, — многому еще, да и не только монстеру... Ее результаты важны, часто неосознанно, для тех, кому они достаются, а эволюция всегда «наслаждается» только собой как процессом — непобедимостью своей всепроникающей и вечной свободы. Можно ли ее, эволюцию, как-то еще отпрепарировать, осво-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ бодив от чего-то лишнего, второстепенного, бросив «голышом» под образное увеличение логического «микроскопа»? Поступим именно так. Что же нас ожидало там под невероятным увеличением образного «мик- роскопа»? В целом, многое проще, чем можно было предположить. Лишний раз, проводя логические эксперименты, убеждаешься в том, что «Природа проста и не роскошествует излишними причинами ве- щей» (И. Ньютон). В эволюции есть свой порядок, который однажды надо определить, а исходит он от самой натуры эволюирующих систем. Эволюционные маятники, свободно перемещаясь и «раскачиваясь» во вселенском потоке пространства-времени, проходят состояния своей трансформации в раз- личных ее формах — эволюционные трансформы (состояния), или эволюс- таты. Эволюцию через разнообразие ее состояний можно определить так: это множественность по-разному развернутых состояний (эволюстатов) динамического, в особенности, трансформирующегося, содержания тел и (или) нетел, составляющих единый поток их развития в пространстве и времени. Чем больше наблюдаешь фактов (иногда их многие сотни и тыся- чи), чем больше поднимаешь вопросов, тем больше убеждаешься в торжестве аксиомы: наука делается, в основном, не тогда, когда добы- ваются факты, а более всего тогда, когда мы, люди, находим способы их, эти факты, обосновать и объяснить, т.е. тогда, когда добытые факты подвергаются абстракции, когда они связываются теорией в единую систему, что позволяет не только объяснить эти факты, но и упорядо- чить поиск определенных новых их категорий. Пока правильное теоре- тизирование не свершится, развитие науки лихорадит; оно способно заводить ее в полумрак. Однако Человек всегда в поиске истины, и это его достояние; когда-то он все равно находит путь к свету, прогрессу, создавая теории, способные не только объяснить происходящее, но и управлять развитием нового научного поиска. Современное эволюционное учение нуждается в кардинальном развитии. Нельзя замкнуть теоретизирование в области эволюции жиз- ни только на поиск и объяснение мутаций. Это, на мой взгляд, не путь для понимания процесса, который «бушует» кругом, везде и всегда; иными словами, эволюцию, которая всюдна, надо понимать как универ- сальное явление и общенаучное понятие. Это ей, эволюции, чужд хаос; она признает только порядок (!), проявление которого из-за ошеломля- ющего разнообразия трансформаций и, в частности, форм (состояний) эволюции, надо суметь правильно увидеть, а вот на этом пути эволю- ционное учение еще далеко от того, чтобы исчерпать свой ресурс воз- можностей роста. Мы говорим о трансформации как о высшей форме движения материи и выделяем состояния трансформации — трансформы, среди которых высшую, по сложности проявления, из ступеней в соответству- ющем иерархическом ряду отводим эволюции. Да, эволюция — это активная трансформа, одно из явлений, которые составляют основное динамическое содержание вселенского потока (Природы). Однако, сколько всего типов эволюстатов (форм эволюции)? Это, в первую
Глава 8. Концеиция всюдности эволюции очередь, зависит от угла обзора и от высоты абстракций. Когда эволюция осуществляется без субмутаций и мутаций, то ее можно обозначить эпи- тетом «простая», а если эволюция сопровождается субмутациями и (или) мутациями, то для удобства назовем ее «сложной». В результате иростой эволюции биоморф иродолжается жизнь при иоследовательном иереходе от одного эволюстата к другому, т.е. в синтомах («эволюционных шагах»), а вследствие сложной эволюции биоморф не только иродолжается жизнь, но и расширяется иолиморфизм ипи возникают новые виды. Процедура иерар- хического классифицирования форм эволюции (эволюстатов) у нас еще впереди, а пока рассмотрим несколько их типов как результат мерото- мирования (членения на части) этого явления с разных высот абстрак- ции. По отношению к преднаследственной или наследственной основе эволюирующих систем можно разделить эволюцию на три типа: униэ- волюция, коэволюция H трансэволюция. Эволюция отдельных только неживых или только живых тел с единой для них или преднаследствен- ной, или наследственной основой, например, у отдельных организмов, включая все их живые структурные элементы, синорганизмов, отдель- ных видов организмов, отдельных гранитных булыжников, целых без- жизненных планет и т.п., вместе со всеми их структурными элементами — это все униэволюция. «Коэволюция — эволюционные взаимодействия организмов разных видов, не обменивающихся генетической информа- цией, но тесно связанных экологически (например, хищник — жертва, паразит — хозяин)» (Яблоков, Юсуфов, 1989, с. 312). Это известный в отношении живой природы термин есть смысл понимать шире. Дей- ствие коэволюции как типа эволюции распространяется на два и боль- шее число тел, которые проявляют разную или преднаследственность, или наследственность; во взаимоотношениях друг с другом не обмени- ваются прогенетической или генетической информацией и не преобра- зуются в единые тела, например, взаимодействие хищника и жертвы, лопаты и фрагмента почвы. Трансэволюция — это эволюция материи, объединяющая существо- вание самостоятельных живых тел с неживыми телами, их окружающи- ми, в другие (экосистемные) телесные образования разных уровней сложности их организации, например, экоиды, биогеоценозы, биосфе- ру, биопланету и т.п., или объединяющая живые тела, хотя и в одно новое целое, но их генетическая основа сохраняет прежнюю феносома- тическую ориентацию жизнеразвития, почему такие живые тела одно- временно существуют в двух качествах и как прежнее самостоятельное тело, и как тело-часть в структуре другого целого тела, например, вза- имодействие особей некоторых водорослей и грибов, образующих вме- сте единое симбиотическое лишайниковое тело, взаимодействие разно- родных особей одного биоценоза, всей живой природы с неживой при- родой и т.д., или, наконец, еще и эволюция, объединяющая самосто- ятельные неживые тела вместе в одно новое тело, но при этом их прогенетическая основа сохраняет прежнюю парафенетическую ориен- тацию в развитии (галактика, гора, дом, автомобиль и т.д.). Эволюция действует многогранно, поэтому нередко на униэволюцию одних явле- ний накладывается еще и коэволюция, и (или) трансэволюция.
OCHOBhI БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ Все трансформы, стоящие в иерархическом ряду ниже эволюции вообще — это не просто состояния активной трансформации, это со- стояния эволюции, ее организации и как процесса, и как результата. Эволюция элементов меньшей сложности составляет эволюцию целого (системы из этих элементов), но эволюция целого — это всегда больше, чем сумма форм эволюции его частей. Таким образом, эволюция, как одна из активных трансформ, состоит из состояний эволюции меньшей сложности: эволюция целого обеспечивается системным единством эво- люции его частей. Эволюционный процесс — это не «монолит», а ритмическое (ста- дийное, т.е. разделенное во времени — стадии) явление, поскольку трансформация тел осуществляется в ритмах развития всей Природы и различных ее областей. Эволюция, будучи трансформацией высокого уровня сложности, осуществляется в каждом явлении стадийно, но, учитывая особый многоуровневый статус эволюции, весь цикл суще- ствования, например, какого-либо тела (морфогенез), может быть всего лишь одной эволюстатой, допустим, онтогенезом этого конкретного тела, а само тело в таком состоянии (форме) эволюции — это соответ- ствующее (онтогенез) состояние эволюирующего тела, допустим, орга- низма. Поскольку эволюция бесконечна, то состояний эволюции (эволю- статов) — бесконечное множество, которые проявляют себя определен- ным образом в телесных и нетелесных системах на основе преднаслед- ственности или наследственности. В каждом эволюционном состоянии участвует какое-либо тело или нетело, которое само, проявляя себя в нем, является эволюстатным элементом, например, организм в онтоге- незе, т.е. элементом в соответствующем эволюционном состоянии. Ритмичность эволюции во вселенском потоке особенная, поскольку она не идет одним фронтом, а распространяется в нем дизъюнктивно под действием различных природных явлений, составляя упорядоченное динамическое содержание материи в неисчислимых дискретных телах и нетелах. Ритмичность эволюции проявляется в различных актах существова- ния тел и нетел, составляющих вселенский, а одновременно, и эволю- ционный, поток. Среди локальных, более дифференцированных, чем ранее, состояний (и процессов, и результатов) эволюции, пожалуй, уместно различать следующие их типы: курсации, адаптации, революции и суперадаптации. Курсации (от лат. cursus — бег, течение) — это дискретные активные трансформации неживых систем. Адаптации (известный термин, смысл которого автор уточняет) — это дискретные активные трансформации живых систем (т. 1, гл. 3). Революция (известный термин, смысл кото- рого автор уточняет) — это скопление (сгусток) активных трансформа- ционных новаций, качественно изменяющих течение эволюции, иногда даже поворачивающее какую-либо ее «ветвь» вспять. Суперадаптация (т. 1, гл. 3) — это бесконечная активная трансформация живой материи. Все или большинство типов частей эволюции — эволюстатов одновре- менно могут существовать в Природе. Только с суперадаптацией вопрос
Глава 8. Концеиция всюдности эволюции для землян открытый, поскольку неизвестно, какая жизнь — однопла- нетная или жизнь вообще (Жизнь) как результат эволюции Вселенной «зацепила» Землю (или дискретная, или континуальная). Эволюцион- ные состояния (курсации, адаптации, революции, суперадаптации) су- ществуют не сами по себе, а, как и другие эволюстаты, во взаимосвязи друг с другом, что нашло, как подчеркнуто уже выше, отражение в термине «синтом», или «эволюционный шаг». Синтомы — это не отдель- ные состояния эволюции, а их системы, — ее свойство преобразовывать Природу, но всегда на «бегу».' «бежать («течь» и т.п. двигаться, изменять- ся) от одного ее состояния (эволюстата) к другому и так бесконечно— такова основная содержательная суть явления эволюции. Эволюция — это процесс, она себе из происходящего ничего (!) не берет, а все получают те структуры, в которых, на которых и для кото- рых действует она, эволюция, оставляя в телах и нетелах разные «отме- тины» о себе. Эволюцию лучше более всего изучать не на единичных фактах ее проявления, а посредством обобщений — типизируя и клас- сифицируя происходящее: типы ее как процесса и как результата. В синтомах последовательно соединяются в одной целое эволюстаты (кур- сации, адаптации, революции, суперадаптации) преемственно-истори- ческими отношениями, действительно отражая последовательность шагов, бесконечного эволюционного процесса материи в различных частях пространства и во времени. У эволюции есть не только восходящее направление, но и обратное движение — по нисходящей. Когда живое тело, не воспроизводя себе замену, элиминируется, то оно, в некотором месте пространства, пово- рачивает ветвь эволюции назад, в неживое без живого продолжения и, тем самым, закрывает эту ветвь эволюции на неизвестное время для жизни. Однако эволюция — множественно разветвленная система ак- тивного трансформационного процесса существования материи, исполь- зующего все направления для ориентации в пространстве и времени, и это вполне можно считать одним из модусов эволюции. На примере адаптационных синтомов, проявляющихся в организа- ции биоморф семенных растений, рассмотрим некоторые возможности анализа эволюции как синтомокинетического процесса (рис. 367-373 и др.). Смена синтомов в объеме отдельных возрастных состояний (стади- осом) семенных фитобиоморф — это всегда не только проявление ста- дийности конкретных биоморфогенезов, но еще и эволюционное собы- тие, в котором, как минимум, какое-то время обеспечивается продолже- ние жизни, а как максимум, могут иногда начинаться и новые виды. «Анатомирование», или, по сути, меротомирование, вселенского потока (Природы) позволяет утверждать, что состоит он из бесконечно- го множества тел и нетел, что в целом одновременно является веще- ственным содержанием его эволюции. Каждая система, действующая во вселенском потоке, проявляет себя как эволюстатный элемент в соот- ветствующем состоянии эволюции Природы. Вселенский поток — это поток эволюционных (активных) трансформаций материи в эволюста- тах, число типов которых будет варьировать при иерархическом класси- фицировании, в зависимости от угла обзора и от высоты обобщений.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ PA СТЕНИИ о~~~~ а 4 Е~~ ао ж ~~ м ~ й Я х Р~~ и ~ й И~~й ~:Ма Я р, Й ЦО<~g с4 д 4Э Боои ~" м~ д й ~-~ Р О 222-д 2~ ° ) ~ !!~ а, „~ф й оЯ~ 3 E,ù 3: ~ у ~у~ evxooxe-~e~а ( а~~~~й~о~ дуол „р,,д,~ М > д.а о.д о ~о~ оо~~~а~uоo - ° .я ° '. -И. Ю м 4J Щ ! Щ ! ! Ф'~ ф ! !! Е й Ж и ~ "' О О '! О 0 О К () Ж С4 4) О М ~ 0ь ~ ~ Я Ф ф ф а, ~ц y > <~> g o Ц m ~ д ~~ а м xx%0 р,о~~„ :T ~.-[ Ф 4.) ф, 'Я д~ -~~~о~ ~~8î~îõïî ооой:~аии 3 ° !!Иф A О C ° !!Вф Е О И ф~~ й М Р' й Я Ж И 0i 1 ф~ ! а й ФЭ фя ~ц О ~~'~ к ~~к~ ~ю д -о ~ Я ~Рма М 'Q 4J о ,ф Я~и ойай ФЭ л о Ж гч ф. 'В Ж ФЭ О g 6 И й О 3 о я м а гч л ю а 5 а. о о И м ~ Q 3 о О о Ж а Р Фэ О о:г' О О ЖQ о И З ~"=, g .. м р у ~4 О h4 Зч рй рЕц г'1 Фэ ~ о о-~ о ~~ ~„о Р о Б М о О р', Зи с4 Б
Глава 8. Концеиция всюдности эволюции Рис. 372. Биографический портрет Tamarix nilotica (Ehrenb.) Bunge [сем. Tamaricaceae (Израиль, каменистая пустыня Негев в окрестностях поселка Сде- Бокер, долина уже пересохшей временной реки на дне глубокого и широкого каньона, 17 V)]. БиоморфОТИПЫ: надземно-двусемядольные (семядоли широкоэллиптические или ок- руглые, 1,5 — 2,2 мм дл., 1,3 — 2 мм шир., сидячие) четочные очередно-простолистные (всходы) и чешуевиднолистные (у большинства возрастных состояний биоморф) эусимпо- диальные зимнезеленые (стебли молодьи побегов зеленые более одного вегетационного сезона, в жару часть побегов осыпается) семидетерминатно-репродуцирующие (многочис- ленные полузакрытые соцветия-кисти развиваются на побегах текущего года, но и на верхних частях резидных побегов, цветение растянуто на весь вегетационный сезон, но более всего оно проявляется в апреле, мае) однодомные беловато-розовоцветковые рых- локустистые деревья и кустарники (1 — 11 биоморфотипы, они различаются по высоте) или моноаэроподиальные невысокие деревья (111 биоморфотип, встречаются реже).
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 373. Биографический портрет Noaea mucronata (Forssk.) Ascherson et Schweinf. [сем. Chenopodiaceae (Израиль, каменистая пустыня Негев,окрес- тности города Димона, склоны холмов, пустыри, овраги, 18 V)]. Биомо фот рф ИП: надземно-двусемядольные (семядоли линейные, заостренные, 12 — 15 мм дл. 05 — 12 мм ши — шир., к основанию несколько расширенные, сидячие) безрозеточные редук- ционно-очередно-простолистные (у всходов листья относительно крупные, но у взрослых растений размеры срединных листьев зависят от времени года и положения их на побеге: чем ближе к началу лета, тем листья меньше, а листья вышерасположенные в структуре годичного, часто разветвленного, побега всегда меньше нижерасположенных) акронекрозные эусимподи- ально-гемисимподиальные семидетерминатно-репродуцирующие однодомные розово-маре- воцветковые колючие (концы побегов текущего года рано превращаются в колючки, а после окончания вегетационного сезона разветвленные отмершие верху~шси побегов еще более уси- ливают колючесть кустов) стержнекорневые полукустарники.
Глава 8. Концепция всюдности эволюции 8.3. На пути к теории трансформации Пожалуй, история проблемы, которая в конечном счете когда-то должна привести к оформлению теории трансформации, т.е. еще более масштабной, чем теория эволюции, поскольку, как ни верти, а эволю- ция — это только часть, хотя и самая значительная, явления трансфор- мации, началась много раньше того времени, когда Гераклит Эфесский (конец 6-го — начала 5-го веков до н. э.) провозгласил известный афоризм: «все течет — все изменяется». Пока не существует теории трансформации и, хотя все, что автор разработал в этой области, еще далеко от такой теории, но вполне можно утверждать, что наука на пути к ней. В предыдущем материале уже было показано, что в организации биоморф «бушуют» так или иначе, весьма противоречивые «страсти» и под одной «крышей» — габитусом, мирно уживаются, казалось бы, несовместимые явления — стремление к «миру», «покою», сохранению постоянства — гомеостаз и, напротив, стремление к «буре», поддержа- нию непостоянства — гетерокинез. В организации Природы (вселенского потока материального про- странства во времени) действуют трансформационные процессы, при- чем как в гомеостазах, так и гетерокинезах таких частных систем, как биоморфы семенных растений. Высшими из трансформаций на Земле является эволюция и ее формы, в которых совершается бесконечное множество разномасштабных эволюционных событий. Тела и нетела, участвующие в эволюции, проходят соответствующие эволюционные состояния, ее эволюстаты. Одним из интереснейших типов эволюцион- ных событий, занимающих определенное место в иерархии таких эво- люстатов-адаптаций, являются метаморфозы (формы адаптаций), состо- яния которых как результатов процессов метаморфозов — это метамор- фы. Попробуем разобраться с тем, где и как проявляют себя метамор- фозы и метаморфы. 8.3.1. Перспективы развития учения И.В. Гете о метаморфозах В биологии как науке о жизни, а жизнь — явление многоплановое, многие процессы сильно переплетены друг с другом, причем настолько взаимосвязаны, что одни явления выступают способами осуществления других, и наоборот. К таковым, в частности, относится явление мета- морфоза, которое, как будет показано ниже, может неоднократно при- нимать участие в морфогенезе какого-либо органа наряду с друтими процессами, взаимодействуя вместе с ними, и всегда выступает всего лишь как часть единого процесса роста и развития соответствующих систем, т.е. трансформации. Учение о преобразовании растительных форм, или учение о метаморфозах, начало свое развитие в конце XVIII века, когда И. В. Гете опубликовал книгу под названием «Опыт объяс- нения метаморфоза растений» (Goethe, 1790). Современные работы по
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНКИ метаморфозам у растений, по мнению автора, пока не ответили на многие вопросы, связанные с этим явлением. Это обстоятельство, по- жалуй, можно объяснить тем, что исследователи метаморфозов нередко пытаются некритически воспринимать во многом гениальные идеи ве- ликого мыслителя, но опубликованные все-таки более двух столетий назад. В этой главе будет сделана попытка продвинуться дальше в разви- тии учения И. В. Гете о метаморфозах растений. 8.3. 1. 1. Метаморфоз — процесс, метаморфа — его результат Кратко, но обстоятельно охарактеризовал историю термина «мета- морфоз» И. И. Канаев (1957) в комментарии к знаменитой книге Гете (Goethe, 1790). Пожалуй, есть смысл дать полновесную выдержку из этого убедительного комментария. «Слово «метаморфоз» значит измене- ние формы, преобразование, превращение, преображение. Оно пришло в науку из мифов и сказок (ср. «Метаморфозы» Овидия, «Метаморфо- зы» Апулея). В новое время как научный термин «метаморфоз» употреб- ляется с XVII в. впервые, по-видимому, у Гарвея. Сваммердам называл этим термином превращение гусеницы в бабочку. В ботанике понятием «метаморфоза» пользовался Линней, уподобляя развитие цветка разви- тию бабочки; но и до него этот термин встречается у ботаников (Schmid, 1930). Сущность своего «опыта» Гете определяет следующими словами: «Попытка объяснить метаморфоз растений заключается в сведении к общему простому принципу всех многообразных явлений дивного ми- рового сада». Этот общий принцип заключается в мысли, что все боковые органы растения, от семядолей до пестика и плода, являются видоизменениями (метаморфозами) одной и той же основной структуры, которую Гете назвал «лист», хотя и считал это название условным и недостаточным (см. ~ 115 и ~ 120 «Метаморфоза»). Теперь ее называют термином «филлом». Гете не ставил вопрос о происхождении самого листа, а брал его как нечто данное. Его исследование, собственно, трактует только об онтогенезе растений, притом высших, однолетних. Поэтому название «Опыта» шире, чем содержание книги, на что указывалось в литературе. Кроме того, Гете не касается вопросов метаморфоза стебля и корня, в чем его тоже упрекали. Позже он частично занимался метаморфозом и этих частей растения» (Канаев, 1957, с. 498). Пожалуй, после яркого комментария И.И. Канаева нам будет легче перейти к анализу современного состояния явления метаморфоза у растений. Посмотрим, как понимают метаморфоз во второй половине ХХ столетия. «Метаморфоз (от греч. метаморфозис — превращение одной формы в другую) — видоизменение органа в связи с изменением функций; например, клубень картофеля является видоизмененным по- бегом, колючка барбариса — видоизмененным листом и т.д.» (Викторов, 1964, с. 95). «Под метаморфозом понимают резкое, наследственно зак- репленное видоизменение органа, вызванное сменой функций «(Васи-
Глава 8. Концеиция всюдности эволюции льев и др., 1978, с. 175). «Метаморфоз — изменение формы и строения органов в процессе исторического развития в результате их приспособ- ления к выполнению различных функций» (напр., преобразование ли- стьев в колючки или усики)» (Дудка и др., 1984, с. 133). «Метаморфоз (греч. метаморфозис — превращение): 1) процесс превращения ювенильных (личинок или нимф) фаз развития во взрослое животное, сопровождающийся анатомо- морфологической и физиологической перестройкой развиваю- щегося организма. Метаморфоз характерен для непрямого пос- лезародышевого развития. Метаморфоз свойственен различным группам беспозвоночных животных (губки, кишечнополостные, черви, ракообразные, насекомые, моллюски) и низшим позво- ночным (асцидии, рыбы, земноводные)...; 2) видоизменение основных органов растений (корней, стеблей, листьев, цветков) в связи с переменой их функций, например, метаморфоз корня в корнеплод, побега в луковицу, листа в колючку и т.п.» (Реймерс, 1990, с. 263, 264). По существу, с учения о метаморфозе растений, созданного И.В. Гете (Goethe, 1790), началось становление научной морфологии, на что сделан обоснованный акцент в обстоятельном обзоре по этому вопросу А.Л. Тахтаджяном (1954). До Гете еще очень мало обращалось внимания на динамизм в развитии морфы (формы) организмов и их органов; осознание факта постоянного изменения органов и организмов в рамках циклов их существования, т.е. морфогенезов в современном представ- лении, наиболее ярко прозвучало в работе Гете о метаморфозах, что весьма знаменательно, поскольку динамический подход в морфологии разносторонне закрепился, по сути дела, только во второй половине XX столетия. Сейчас важно осознавать, что учение Гете о метаморфозе, создан- ное в общих чертах в конце XVIII века, безусловно, не догма и нужда- ется в постоянном совершенствовании, развитии на уровне обновлен- ных знаний. В наше время, как подчеркнули Р.П. Барыкина и М.А. Гуленкова (1990), метаморфозы рассматриваются в онтогенетическом аспекте или онтогенетическом и филогенетическом одновременно (Се- ребряков, 1952; Troll, 1973; Серебрякова, 1984; Лодкина, 1983 и др.). Метаморфоз реализуется в ходе индивидуального развития организма (онтогенеза) и нередко выступает в качестве этапа органогенеза как проявление временной разнокачественности органа (Барыкина, Гулен- кова, 1983, 1985, 1990). Метаморфозы осуществляются в онтогенезах и отражают в фенотипах те генетические изменения, которые накопились в процессе длительного исторического развития растений. Способность растений образовывать метаморфизированные органы наследуется в ряду поколений и метаморфоз выступает как категория исторического развития (Серебрякова, 1984; Варыкина, Гуленкова, 1990). Пожалуй, лучше других современное состояние учения о метамор- фозе резюмирована в учебнике А. Е. Васильева и др. (1978, с. 329): «по теории Гете, все видоизменения форм листьев на побеге следует считать проявлением метаморфоза. Иными словами, метаморфоз — это прояв-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ ление всех потенциальных возможностей морфогенеза листа в ходе индивидуального развития растения.... В дальнейшем проблема мета- морфоза получила и эволюционное освещение. В новой интерпретации метаморфизированными условились считать органы, достаточно резко уклонившиеся от типичной исходной формы, в связи с существенным изменением функций, в ходе приспособительной эволюции... Впрочем, в ряде сомнительных случаев трудно судить о границах «метаморфоза» и «неметаморфоза». Чтобы разобраться с явлением метаморфоза у семенных растений, надо ответить на несколько принципиальных вопросов. Метаморфоз- это процесс и (или) результат? С чего начинаются метаморфизирован- ные органы? Как соотносится метаморфоз с друтими изменениями в организации биоморф? Какое отношение имеют к метаморфозу анало- гичные и гомологичные органы? Какое место занимает метаморфоз в росте и развитии биоморф? Как проявляет себя метаморфоз в морфо- генезах органов и в биоморфогенезах? Метаморфоз как превращение (трансформация) — обычная состав- ляющая дискретного роста и развития живого тела — органа, т.е. его морфогенеза (органогенеза), как закрепленное более сложное и нередко даже резкое превращение (именно на резкость перестройки формы одного живого тела в другую форму особо обратили внимание А.Е. Васильев и др., 1978, когда давали определение метаморфозу) по срав- нению в другими изменениями, необратимо присутствующими при свершении органогенеза. Мы исходим из факта, что в любых метаморфозах постоянно при- сутствует два определяющих аспекта, которые составляют причину этого явления. Во-первых, метаморфозы — результат длительного историчес- кого развития, которые прошли органы в организмах-потомках до те- кущего момента, став одним из фенетических результатов управления, исходящего из наследственного субстрата биоморф, их генотипов. Имен- но гены направляют фенетическое развитие биоморф, в котором реали- зуются, если они были заложены в генах, те метаморфозы, которые более всего отвечают условиям как внутренней организации биоморф, так и окружающей их внешней среды. Во-вторых, метаморфоз, хотя и несет на себе печать исторического развития (филогенеза), но реально по генетической программе он про- является в органогенезе, т.е. в росте и развитии (морфогенезе) органов, на фоне онтогенезов, через которые осуществляется живая связь между метаморфозами и иными трансформациями. Гете и его последователи говорят о метаморфозе как о процессе и результате одновременно, но логически это не совсем точно. Метамор- фоз — это процесс превращения органа в его морфогенезе, а результат метаморфоза — это целый орган или его часть, которые были подвер- жены метаморфозу, что должно обозначаться иначе, допустим, терми- ном «метаморфа». В большом двухтомнике «Современная ботаника» (Рейвье и др., 1990) не применяется термин «метаморфоз», а все случаи, где «стебли и листья видоизменяются и выполняют нетипичные для себя функции»
Глава 8. Концепция всюдности эволюции (с. 67), например, усики, кладодии (кладофиллы), колючки, клубни и т.п. называются модификациями. Нет оснований вот так резко отказываться от классического термина «метаморфоз», и, пожалуй, единственное, что как-то может сблизить эти два термина — признание, что видоизменение органов и их частей как процесс следует считать «метаморфозом», а результат этого процесса (одно из состояний метаморфизированного органа) — метаморфой, или модификацией. Но для применения термина «модификация» в смысле «метаморфа» надо закрыть глаза (на самом деле так не стоит делать) на то, что за термином «модификация» уже закре- пилось другое значение — «ненаследственное изменение признаков орга- низма (его фенотипа), возникающее под воздействием условий окружа- ющей среды (температуры, влажности и т.п.) и не связанное с измене- нием генотипа» (Реймерс, 1990, с. 276). Автор признает метаморфоз как процесс, а метаморфу как результат метаморфоза. Так что же все-таки означает термин «метаморфоз»? Вероятно, нельзя остановиться только на том, что сделал Гете более 200 лет назад. Надо искать и находить новое, чтобы гетевское учение о метаморфозах растений не затерялось под напором новых идей и фактов; оно должно развиваться, учитывая прошлые достижения и новые идеи. Чтобы дать современное объяснение термину «метаморфоз», надо освободиться от некоторых «зависимостей», может быть, даже запретов. Поскольку ме- таморфоз можно познать только в сравнении, то многими ботаниками уже давно завладела некоторая «навязчивая идея» принимать организа- ционные части особи одного типа за «типичные», а остальные их гомо- логи рассматривать как «нетипичные» (метаморфизированные), подвер- гшиеся сильному изменению, т.е. метаморфозу (Игнатьева, Андреева, 1991, 1993). Это положение, на мой взгляд, нуждается в существенной коррективе. Посмотрим для примера на разнообразие «типичных» фотофиль- ных побегов (они обладают, как правило, более или менее зеленым цилиндрическим стеблем и зелеными листьями с пластинкой, по-раз- ному рассеченной или цельной, сидячими или черешковыми и т.д.) и попробуем найти границу между ними и метаморфизированными побе- гами, которые видоизменены очень «резко» (выражение из учебника Васильева и др., 1978) в связи с изменением функции, например, ме- таморфозами будут колючки, чешуи и т.п. Я полагаю, что, в зависимо- сти от подхода к решению вопроса, что «типично», а что «метаморфи- зировано» (видоизменено), кроется принципиальная основа, на которой должно строиться понятие «метаморфоз». Действительно, если посмотреть на фотофильные побеги, то они по отношению друг к другу очень разные: от мощных вегетативных «водя- ных» побегов и длинных средних и коротких, а еще розеточных, полу- розеточных и других ростовых побегов с самыми разными по форме и толщине стеблями и листьями до побегов с сильной специализацией к репродуктивной функции, изменившие не только форму листьев, но и превратившие свои верхушки в процессе исторического развития в цветок или системы цветков. В этой связи не видно оснований во всем современном разнообразии побегов выделять по степени «фотофильно-
OCHOBhI БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ сти» одни из них как «типичные», «нормальные», а другие называть «атипичными» («нетипичными»), или «метаморфизированными». Пожалуй, то разнообразие побегов, которое мы наблюдаем даже в «среде» фотофильных побегов, убедительно позволяет сказать: все со- временное разнообразие побегов (от и до), да и других гомологичных (!) органов, — результат метаморфозов, которые имеют как онтогенетическую, так и филогенетическую историю. 8.3. 1.2. Метаморфоз — способ реализации гомологичес- кого разнообразия в фенетической организации биоморф Результат метаморфозов — образование метаморф — метаморфизи- рованных органов. Иными словами, метаморфы — это те органы, ко- торые целиком или по своим частям, в целях адаптации организмов, подверглись и подвергаются собственным адаптациям — метаморфозам. Когда в структуре одного органа проявляется несколько метаморф, то он разнокачественен по времени их проявления (на это удачно обра- тили внимание Р.П. Варыкина, М.А. Гуленкова, 1990). Однако, измене- ние функций и соответствующие превращения (метаморфозы) органов нередко уже произошли в далеком прошлом, «на определенном этапе эволюции родственной группы растений. В онтогенезе особи мы при этом никакого фактического превращения не наблюдаем, а видим лишь своеобразный ход развития (морфогенеза) того или иного органа.... Очень часто мы говорим о метаморфозе лишь в фигуральном смысле, имея в виду возможность превращения исходного органа в метаморфизирован- ный где-то в отдаленном прошлом»... (Васильев и др., 1978, с. 330). Главное в метаморфозе — это способность его носителей-органов транс- формироваться и быть адаптацией соответствующего рода: обладать всепро- никающим свойством специального изменения, которое обеспечивало бы го- мологичное разнообразие органов в зависимости от исторических и конкрет- ных условий обитания биоморф и исходило бы от их генетической основы. Обязательное условие любых метаморфозов — быть составной частью любого органогенеза на фоне еще более сложных изменений всего организма, т.е. онтогенеза. Такие условия возникли еще на ранних этапах станов- ления жизни на Земле, поэтому все семенные растения получили еще в далекие исторические времена от своих предковых палеобиоморф спо- собность к пластичности и типы органов, которые все уже были метамор- физированы. Метаморфоз — это всегда разновидность адаптаций, поэтому семенные растения в процессе эволюции получили от своих прародите- лей только метаморфизированные органы и, что еще более важно, они получили от них способность, закодированную на генетическом уровне, проявлять метаморфозы в органогенезах на фоне текущих онтогенезов, а шире и синонтогенезов, т.е. в целом биоморфогенезов. Метаморфоз — это всегда конкретный способ реализации фенетичес- кой организации в соответсзнующих морфогенезах, исходя из пластичности в содержании гомологичных органов, отвечающий генетической природе биоморф и зависящий непосредственно от условий окружающей среды Метаморфозы, будучи одним из типов адаптаций, в первую очередь
Глава 8. лонцеиция всюдности эволюции обеспечивают специализацию гомологичных органов, которая, в силу пластичности биоморф, по-разному дублируется в биоморфогенезах, в рамках нормы реакции организмов и в зависимости от условий обита- ния. На основе метаморфозов среди биоморф семенных растений про- является гомологичное разнообразие органов. В сравнительной морфологии при изучении метаморфозов приори- тет оправданно отдан гомологичным органам (!). Интуитивно обращаясь только к таким органам, И.В. Гете сумел увидеть в онтогенезе метамор- фоз одного органа — «листа» («филлома») и рассмотреть весь «правиль- ный» («прогрессирующий») ряд гомологичных структур. Все-таки в сравнительном исследовании всегда важно найти обоснованный ответ на вопрос: какой орган среди органов-гомологов является прародитель- ским, т.е. гомолого-генерирующим органом в цепи результатов мета- морфозов. Автор предлагает следующий постулат: гомологичные органы при сравнении таковых в органиъие, виде, роде, семействе и более высоких таксонах составляют гомологические ряды, в которых все (!) сравниваемые органы метаморфизированы по отношению друг к другу и каждый из них по своему типичен, т.е. типичен для своего случая текущей жизни (в конкретном организме конкретного онтогенеза) и типичен в историческом развитии (филогенезе) в соответствующих условиях окружающей среды. Однако, всегда для всех современных метаморфизированных органов есть орган-прародитель (гомолог-генератор) в определении которого часто и состоит главная загадка исследования. В ряду гомологичных органов располагаются не метаморфозы, а результаты процессов, которые Гете назвал метаморфозами. Выше ре- зультаты метаморфозов (процессов определенного рода) названы мета- морфами. Все метаморфы в рядах гомологичных органов (метаморф) являются результатами метаморфозов, которые произошли в прошлом или впервые происходят сейчас, но всегда развертываются в конкретном органогенезе на фоне текущего онтогенеза. Авторский тезис — каждая метаморфа типична (результат метаморфоза) в ряду гомологичных орга- нов для своего случая текущей жизни на определенном экологическом фоне и для своего случая исторического развития (рис. Зб7-398). Итак, при изучении метаморфоза нет и не может быть проблемы поиска типичного и атипичного (метаморфизированного), поскольку механизм метаморфозов был запущен не сегодня, не вчера и не двести лет назад, а уходит в глубины прошлого на сотни миллионов лет и стал доступен еще на заре исторического развития растительной династии. Механизм метаморфоза дошел до нас по сути как механизм «снятия» (термин Гегеля, 1970; см. Стрельченко, 1980). С позиций современной диалектики вполне допустимо и уместно утверждать, что биотическое «снятие» является обязательной стороной преемственности в организа- ции живых тел, когда какое-либо минувшее исходное явление включа- ется в современную производную организацию в более или менее пре- образованном виде. Так, современные биоморфы (организмы и синорганизмы) — это действующие системы метаморфозов, когда в каждом органе биотичес- кое «снятие» является нормой существования. В процессе индивидуаль-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ ного развития (органогенеза) каждого органа происходит метаморфоз с обязательным биотическим «снятием», когда в любом текущем состоя- нии органа что-то упраздняется из прошлого (индивидуального и исто- рического), а что-то сохраняется, но в преобразованном (более или менее) виде. Метаморфы (состояния органов как непосредственные результаты метаморфозов) всегда отражают фактическое состояние раз- вития современной организации соответствующих органов, которая как- либо перекрывает собой уже минувшие состояния индивидуального и тем более исторического развития), которые, несмотря ни на что (вре- менные и пространственные перемены), продолжают как-то присут- ствовать в них, но, безусловно, в «снятом» виде (упраздненные, но все же передавшие и сохранившие себя в производных структурах, хотя бы в чем-то, хотя бы чуть-чуть)... вот она — одна из интриг научного поиска, позволяющая нам докапываться, если повезет, до истоков ме- таморфозов разных органов. Если присмотреться к организации биоморф как растительных, так и животных, то метаморфозы мы обнаружим повсеместно среди орга- нов, поскольку эти процессы являются обязательной стороной всякого роста и развития органов, что генетически закрепилось в процессе дли- тельного исторического (филогенетического) развития жизни на Земле, которая в эволюционном отборе пробовала и находила то, что затем становилось нормой жизни на какое-то время, и то, что сегодня повсе- местно является нормой проявления в онтогенезах метаморфозов и их результатов. Метаморфозы в растительном и животном мире всегда типичны; результаты их разные и требуют соответствующих объяснений в каждом конкретном случае. Будучи свойством эволюции соответству- ющих живых систем-органов, метаморфозы генетически закрепляются и становятся нормой жизни органов в производных поколениях орга- низмов. Каждый метаморфоз — это одно из проявлений эволюционного состояния — адаптации, т.е. все метаморфозы и их результаты (мета- морфы) — это обязательные составляющие эволюции жизни. Метамор- фоз — это всепроникающее изменение органов биоморф как проявле- ние единства их роста и развития, обеспечивающее им гомологичное разнообразие. Метаморфозы проявляются в органогенезах (циклах роста и развития органов), и их результатом являются метаморфы, которые образуются на фоне биоморфогенезов (онтогенезов и синонтогенезов) и на фоне филогенетического (исторического) развития. Метаморфоз- это атрибутивная форма роста и развития органов, в зависимости от кон- крнвных уояовнб обитнння биоморф. По сути дела, наша корректива в объяснении метаморфоза приоб- щает древнее учение к языку современной науки. Важно, что метамор- фозы сопровождают рост и развитие органов постоянно, составляя одну из сторон их организации, — одну из активных трансформ (эволюста- тов). При изучении метаморфоза нет проблемы поиска типичного и нетипичного (метаморфизированного), поскольку все одноименные гомологичные органы, которые мы наблюдаем у растений, являются метаморфами — результатами процессов метаморфоза как составных частей роста и развития организмов в целом.
637 Глава 8. Концеиция всюдности эволюции 1,аг2(аг4),206(19д),26 Рис. 374. Биографический портрет Melandrium latifolium (Poir.) Maire [сем. Caryophyllaceae [Алтай, Курайский хребет, нарушенный травостой на шах- тных отвалах в высокогорье и в лесном поясе в районе поселка Акташ)]. SHOMOP@OTHII: надземно-двусемядольные (семядоли эллиптические, слегка лопатча- тые, 6 — 8 мм дл., 5 — 6,5 мм шир., широкозакругленные, выемчатые, клиновидно суженные в черешки 3 — 5 мм дл.) переходно-полурозеточные (у побегов настоящей полурозеточности нет, поскольку зона розетки, образование которой потенциально воз- можно в других условиях при переходе побега от подземного роста к надземному, имеет здесь не брахиморфную, а мезоморфную структуру: ее междоузлия, в отличие от длинных междоузлий безрозеточных частей, сближены, но недостаточно, чтобы можно было гово- рить о розеточности) длиннокорневищные эусимподиально-гемисимподиальные супро- тивно-простолистные детерминатно-репродуцирующие вздуточашечные бело-гвоздичноц- ветковые травянистые многолетники.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ о ой ой й Л о 4~ М й о ц Я ! Я ! ! ! ! ~~'~ ! .~. Ж,. ~ р, д Фимив~яо р~ о МИ ф~ ~Q 3 + ~ Я За ~р д: о. о рао~дМао~ О~И~uXXXv О Ф О Ж од~ Я М д М О м ои", 6Э Neй ~~3vg g~~~~~g" h I лйа~о д Ф ~ ~ ~ Ф М ~ о м у м ~ о о о ~~ о. ~ а Ии~~РИ~~~~ ~М л ~Я и а» < v v > ~~ о у а а ~. о ~ о а а в м а. с3 а Ф а. и 3 а а. а. z й
Глава 8. Концеиция всюдности эволюции Рис. 376. Биогра- фический портрет Eucalyptus сата1- dulensis Dehnh. [сем. Myrtaceae (Израиль, окрест- ности Тель-Авива, пойма реки Яр- кон, 27 Ш)]. Биоморфотип: над- земно-двусемядольные (семядоли обратнопоч- ковидные, 2 — 3 мм дл., 3 — 5 мм шир., широкозакругленные с обязательной выемкой посредине, резко су- женные в короткие че- решки 2 — 5 мм дл.) безрозеточные юве- нильно-супротивноли- стные взросло-очеред- но-простолистные эусимподиальные ин- тер калярно-репроду- цируюшие [«пульсиру- юший» (очень редкий 34 тип) межсоцветия у побегов текущего года образованы бутонами, а цветение (на втором году жизни побегов) и плодоношение (на тре- тьем году) происходят позднее] перекрестно- опыляемые (много гиб- 71 88 1 ридов, поскольку раз- ные виды способны скрещиваться) однодомные многочисленно-тычинковые (rtser тычинок определяет внешний вид цветков: от желтоватого и белого до красноватого) вечнозеленые сильнонарастаюшие субтропические и тропические деревья. Особый интерес вызывает раз- витие репродуктивной системы. У цветков текущего года сначала образуются межсоцветия из бутонов — интеркалярных побегов, которые в год их образования не цветут. На самых ранних стадиях развития бугонов у каждого из них сверху имеется колпачок-крышечка, образованная пленкой из сросшихся чашелистиков. В последующем развитии эта крышечка с бугонов сбрасывается и высвобождаются шаровидные соцветия из бутонов-цветков, у каждого из которых имеется еще одна — вторая — колпачок-крышечка, образованная из сросшихся лепестков венчика, что, как подчеркивали Н.Н. Полунина (1958), М. В. Гераси- мов (1962) и др., надежно прикрывает пыльники и пестик, защищая их от неблагоприятных воздействий внешней среды. Цветение происходит обычно на втором году жизни, после сбрасывания колпачков с бугонов-цветков. Плодоношение осуществляется на третьем году жизни межсоцветий и «коробочки со зрелыми семенами или без них иногда подолгу (до 2 лет) не опадают с ветвей, а так как бугоны появляются не одновременно, то на взрослом дереве эвкалипта можно видеть в одно и то же время бугоны, цветки и коробочки различной степени развитии и зрелости» (Герасимов, 1972, с. 9 — 10). После обрезки на пенек эвкалипт быстро метатравматическим побегообразованием восстанавливает крону. Способен образо- вывать воздушные корни (естественная реакция на воздействие паводковьи вод). «Эвкалипт может также вегетативно размножаться отводками и черенками» (Герасимов, 1962, с. 35).
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ PA СТЕНИЙ Рис. 377. Биографический портрет Anabasis articulata (Forssk.) Moq. [сем. Chenopodiaceae (Израиль, каменистая пустыня Негев, окрестности поселка Сде-Бокер, 14 V)]. Др — цветущий побег (Zohary, 1966). Бноморфотип: надземно-двусемядольные (семядоли линейные, 8 — 15 мм дл., 1— 2,2 мм шир., заостренные, слегка расширенные к основанию, сидячие) четочные редукци- онно-супротивно-простолистные (у всходов листья относительно крупные, но довольно быстро растение переходит к образованию рудиментов — сначала чешуевидных листьев, а затем только листовых бугорков) эусимподиально-гемисимподиальные индетерминатно- репродуцирующие однодомные стержнекорневые изоаклональные суккулентные кустар- нички. Особенности листовых метаморфозов сильно зависят от возраста растений и вре- мени года. Стебли годичных побегов текущего года зеленые, но, как отметил М. Зохари (Zohary, 1952), на следующий год их кора летом тускнеет, мертвеет и слущивается. Рас- тения цветут с сентября по декабрь, а весной активно вегетируют.
Глава 8. Концепция всюдности эволюции б,гм4, 2Т,27 1,аг2, 206,26 Рис. 378. Метаморфозы побегов в кронах кустарников Genista fasselata Decaisne [сем. Fabaceae (Тель-Авив, Ботанический сад, 28 IV)]. HOMOp@OTHII: надземно-двусемядольные безрозеточные очередно-простолистные (все листья к началу лета осыпаются, сохраюпот жизнеспособность только расширенные осно- вания листьев) эусимподиальные постинтеркалярно-семидетерминатно-репродуцирующие однодомные желто-бобовоцветковые жестко-колючепобеговые кустарники.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЯ B 1,аг4,14а,26 Рис. 379. Биографический портрет Russelia equisetiformis Schlecht. et Cham. [сем. Scrophulariaceae (мексиканский вид, скверы и парки Тель-Авива, IV, V)). БИОМОрфОТИП: надземно-двусемядольные безрозеточные редукционно-супротивно-му- товчато-простолистные прутьевидно-хвощевидно-побегообразующие (мимикрия) интер- калярно-детерминатно-репродуцируюшие (из пазух листьев ростовых частей побегов теку- щего года развиваются интеркалярные побеги разных размеров и разветвленности с вер- хушечными цветками) эусимподиальные (единичные ростовые побеги, особенно короткие, могут сохранять жизнеспособность своих верхушек и продолжить вегетацию на следующий год, т.е. только изредка может проявляться и гемисимподиальность, поскольку сильней- шим фактором, стимулирующим акронекроз побегов в теплом субтропическом средизем- номорском климате — это наступление длительной летней жары, когда до 8 мес~шев совсем не выпадает дождей) красноцветковые невысокие (0,7 — 1,2 м) кустарники.
Глава 8. Концепция всюдности эволюции
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ 1,аг4,19а(20б),26 ~,Г 1,68 2(1),аг4,206,26 33а,35 ПП Рис. 380. Биографический портрет Pituranthos triradiatus (Boiss.) Ascherson et Schweinf. [сем. Apiaceae (там же, что и рис. 372, 14 V)]. Биоморфотип: надземно-двусемядольные (семядоли линейные, 15 — 20 мм дл., 1,2— 2 мм шир., заостренные, сидячие) безрозеточно-полурозеточные (доминируют безрозеточ- ные годичные побеги, а полурозеточные побеги чаще образуются у молодых растени~) редукционно-очередно-простолистные и полусложнолистные эусимподиальные семиде- терминатно-репродуцируюшие (сложные зонтики завершают верхушечное развитие годич- ных и внутригодичных вегетативно-репродуктивных побегов) однодомные стержнекорне- вые факультативно-корнеотпрысковые полукустарники. цветение растянуто, но максимум приходится на середину лета.
Глава 8. Концепция всюдности эволюции S е
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ 2,гы4(гы1),206,2S Рис. 383. Разнообразие мета- морфозов побегов в кроне полукустарника Kickxia spartioides (Buch) Janchen [сем. Scrophulariaceae (Израиль, Эйлатские горы, сухие горные долины, берега русел времен- ных водостоков, пустыри, 5 %1. EHOMOP@OTHII: надземно-двусемя- дольные безрозеточные очередно-про- столистные сильноветвистые эусимпо- диальные семидетерминатно-репроду- цирующие однодомные желто-норич- никовоцветковые стержнекорневые полукустарники. 2,агМ, 1М,206,26 Рис. 384. Биографический портрет Batrachium trichophyllum (Chaix) Bosch [сем. Ranunculaceae (Московская область, естественные водоемы)]. Допол- нительное обозначение: пв — водная поверхность. Биоморфотип: надземно-двусемядольные [(семядоли ланцетные, 3,5 — 4 мм дл., 1 мм шир., слегка заостренные, постепенно переходящие в короткие черешки (Csapody, 1968)] безрозеточные (у всходов может быть или не быть розеточности главного побега) очередно- полусложнолистные (листья многораздельные с нитевидными долями, хорошо приспособ- ленные к жизни в воде) детерминатно-репродуцирующие интенсивно-кондивидуальные однодомные бело-лютиковоцветковые травянистые многолетники.
Глава 8. Концепция всюдности эволюции Рис. 385. Биографический портрет Asparagus stipularis Forssk. [сем. Asparagaceae (там же, что и рис. 370, апрель)). Дополнительные обозначения: кд — кладодий-колючка, юк — кладодии преювенильных или ювенильных растений. Биоморфотип: подземно-двусемядольные безрозеточные кладодиево-колючковые по- стинтеркалярно-репродуцирующие двудомные аэроподиоидные (надземные прямостоячие побеги живут 2 года и после завершения репродуктивного развития отмирают до подзем- ного компактного основания, которое пополняет структуру долговечных геоподиев) клуб- некорневые травокустарнички (30 — 100 см высотой).
OCHOBhI БИОМОРфОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Q D g R д щ~иО go3>~ X О -~о ~~~ Д Щ &» ануе р» О О О . а,О й у й 2 „'~ g+g ~ 0 О о ~~ ~ ~ X к д X Q.~o~ О ~;„а Й g р.й х ~'2 о.g )с о~~ Q. О Q Фэ Ж о ~~~ у Ю [ щ О а о X~х а д Q в~ X 3 ЯP. ~Э ~ ФЭ 1 63 ! ~~иЙ Ж о О X v й 3» И: Ж Я go~ ~~ о х X р,о ~ 3: ~ ~ Q х 0 v~ vo () ~ [ ф„'ФЭ ФЭ .о Р Оао 4 2 я 2 Q. ф„' OVO Ж о Х IQ Ф О Д охg ф; й' +Ex ф; Я Д Д' ~~в X ~ф х<а РЕ- Ю о ~ ~. Я (~) Ф ° Ц я Q О~ -2 хх ° 3» r. P од~ бс ж ,'3» IQ й ~' й~ Я о ~ ~»»у» v о Д V LO ~ о й ФЭ Х Д о ~ о 2 й Х р~~ д х X Q, Ф о й р~» о Х Х ж ~ Ж а о О ~М X О (~) Д л Ж о Я Х о <> О X й 4~ X Q О Ж эх ~доЯ ож ° î о ~ x д X ° ФЭ @~„Q Д v ~,~О X О цхяа озфз ~ ~ ~о ~ а 51335 л л ~л», ~ ~ ~ рМ ~ц ~ х~~о о~~~~аД z о~~~~~„-д 1 Ое~~v~Ц й~~у~Р g e I Xm VZ ~ O O e O О .-о~~ [ о»~ О 0 Q ~)' ~ Р р ж а 3» Фэ vС~~:~ОО z ~ 2 ~,р ~ о ~ ~ Q Д ~2 «Д О „~ р О.ЖМОааЫ е[ д 1 x ~ +»1 х .о О . -- „ж.ж ф 2 ~ ц~) М а 8Я .иИ202~ л О сб й,' Ю О ф' ° X й,' Х О Ж 0 О h4 Q О ~> о и 1 й,' д~ И Ю v g ,.ф й Ю X 01 Ю ~) О О О 1 д :а Ж О Сее О сб ~л ~ ~-~ О,">, И Эф,' м" 0 'й сб у М r >ъ й,' сб E ô О О Жо О 00 М h4 О О О Q О Q И 1 Q й~ О ~ И Я ~ О ~ О О ж Ж О а5 g о ° ?; й,' О О Ю О. д, ~ И р Ю 2 ~ о 0 е~ 0~ v Й 2 î"" Я Я й X О а5 ОО X ~4 ° И й М z 1 ~ ~еч л Q ю~ й~ ~ о й 0 Х Q. ж О Е
л,за~м) Глава 8. Концепция всюдностпи эволюции Рис. 387. Метаморфозы в организации взрослой био- морфы Limonium pruinosum (L.) О. Kuntze [сем. Plum- baginaceae (там же, что и рис. 372, 12 V)]. Биоморфотип: надземно-двусемя- дольные безрозеточные редуцирован- но-чешуелистные эусимподиально-ге- мисимподиальные [прямостоячие над- земные магимодифицированные, т.е. с чешуевидными листьями, годичные побеги развиваются как статурные (ро- стовые) или репродуктивно-статурные, которые, после небольшого акронек- роза — отмирания сверху (зимой) на большую часть длины этих побегов, сохраняют жизнеспособность, прояв- ляя себя как аэроподиоиды, а на сле- дующий год весной из их почек разви- ваются постфлоральные и иные пост- латентные побеги, которые, вместе с их родительским аэроподиоидом, пос- ле завершения сезонного развития полностью отмирают в надземной ча- сти и только подземные побеговые ос- нования гемисимподиально включа- ются в многолетнюю основу — геопо- диальную часть компактного куста] семидетерминантно-репродуцирую- щие однодомные розово-свинчатко- воцветковые стержнекорневые траво- Рис. 388. Метаморфы как результаты метаморфозов в организации взрослой биоморфы Aconitum anthoroideum DC. [сем. Ranunculaceae (Восточный Казах- стан, Алтай, хребет Южный Алтай, под- робности жизнеразвития на рис. 82)). Дополнительные обозначения: м1 — метаморфа верхней части побега — семидетерминатное вер- хушечное кистевое соцветие, м2 — метаморфа средней части побега с живыми срединными листьями, мЗ — метаморфа нижней части побега с отмершими листьями, м4 — метаморфа нижней части исходного корнепобега с производным поч- кокорнем.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИИ О 4, Р 26 не L. нь ре гра об Доп Ml— СТИ две, мм дл коза ко суженные в черешки 2 — 5 мм дл.), м2 — метаморфа надсемядольной фотофильной неразветвленной части главного побега, м3 — метаморфа надсемядольной фотофильной разветвленной части главного побега, м4 — метаморфа подземной побеговой части, м 4- 1 — метаморфа подземной части родительского побега, м4-2 — метаморфы, производных горизонтальных подземных или подводных побегов, м5 — метаморфа гипокотиля, мб- метаморфа системы главного корня, м7 — метаморфа придаточных корней, м8 — метамор- фа горизонтальнопобеговых корнепобегов, м9 — метаморфа медиального корнепобега. Рис. 390. Разнообразие метамор- фозов у виргинильных растений Pancratium maritimum L. [сем. Amaryllidaceae (Израиль, Примор- ская равнина в окрестностях се- верной части мегалополиса Тель- Авива, песчаные почвы, обрывы к берегу Средиземного моря, 23 IV)]. Дополнительные обозначения: м1 — мета- морфа протерокормуса — подземносемя- дольной побеговой части (прорастание семян подземное), м2 — метаморфа надсе- мядольной фотофильной части (без мЗ) розеткообразующего побега, мЗ — мета- морфа персистентной луковичной части розеточного побега, м4 — метаморфа кор- невищной (без луковичных чешуй) части главного побега, м5 — метаморфа боково- 1 го розеткообразующего побега, мб — мета- морфа главного корня, м7 — метаморфа придаточных корней, м8 — метаморфа 66 смешанной корневой системы.
Глава 8. Концепция всюдности эволюции Рис. 391. Биографический портрет Acacia longifolia Willd. [сем. Fabaceae (там же, что и рис. 370, март — май)]. Дополнительное обозначение: фк — фил- ЛОКЛЗДИЙ. EHOMOp@QTHIIbI: безрозеточные очередно-сложнолистно-филлокладиевые (сложные листья развиваются чаще всего у молодых растений, но в последующем жизнеразвитии центральные рахисы новых листьев уплощаются в многолетнезеленые неразветвленные филлокладии, проявляя мимикрию под простые листья, при этом пластинки сложных листьев постепенно исчезают совсем, что удается хорошо понять, если выстроить гомоло- гические ряды листовых метаморф) эусимподиальные (отдельные короткие побеги, осо- бенно молодых растений, могут проявить непродолжительную гемисимподиальность в нарастании, но это больше исключение, чем правило) интеркалярно-репродуцирующие однодомные желто-бобовоцветковые некустистые (1 биоморфотип) или кустистые (11 биоморфотип) вечнозеленые деревья.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 392. Биографический портрет Smilax aspera L. [сем. Smilacaceae (там же, что и рис. 370, 28 IV)]. Дополнительное обозначение: ус — усики прилис- тникового происхождения. Биоморфотип: подземно-двусемядольные безрозеточные очередно-простолистные усико- во-прилистниковые шиповато-стеблевые (стебли покрыты шипами эпидермального проис- хождения) двудомные белоцветковые красноплодные лианокустарнички. У молодых растени~ развиваются поначалу листья без усиков и только тогда, когда появляется потребносчь за кого- то зацепиться, появляются листья с усиками. На верхушках вегетативных побегов-плете~ обычно развиваются одни усики без листовой пластинки (здесь специализация в MemMo~- та» ltMtThcH как BilttlltttttHtl к каэансю по прггим растснннм аостигаст мюгсимгма).
Глава 8. Концепция всюдности эволюции 4, 26 Рис. 393. Биографический портрет Fagonia mollis Delile [сем. Zygophyllaceae (там же, что и рис. 383, 6 V)]. БиоморфоТИП: надземно-двусемядольные (семядоли округлые, 4 — 7 мм дл. и шир., суженные более или менее резко в короткие черешки) безрозеточные супротивно-тройча- толистные (первые листья у всходов без прилистников, у третьего листа прилистники уже есть, но они пока очень короткие, далее все листья развиваются с прилистниками, видо- измененными в колючки, кроме того, первые листья иногда располагаются очередно и только позднее листорасположение стабилизируется как супротивное) гетеросимподиальные (прирост аэроподиев более всего обеспечивается гемисимподиально, поскольку сначала развиваются короткие вегетативные побеги, которые успешно зимуют и весной продолжают вегетативно-репродуктивное развитие, акронекрозно завершая его, но при всем этом часть резидообразуюших побегов проходит монофазное развитие) детерминатно-полисимподиаль- но-репродуцируюшие (любопытный факт: образование верхушечных цветков при полисим- подиальном нарастании побегов может быть не только эфемерным, но приводить еше и к резидообразованию, поскольку побеги, после симподиального развития нескольких верху- шечных цветков, могут вновь также симподиально возвращаться к росту новых вегетативных побегов замещения, что позволяет растениям вновь накопить необходимую пререпродуктив- ную массу и еШе раз повторить полисимподиальное репродукгивное усилие) сильноветви- стые однодомные розовато-белоцветковые стержнекорневые подушковидные колючие (за счет метаморфоза прилистников в колючки) полукустарники.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 394. Биографический портрет Zilla spinosa (L.) Prantl [сем. Brassicaceae (там же, что и рис. 383, 4 V)]. Биоморфотип: надземно-двусемядольные (семядоли обратнояйцевидные, мясистые, 15 — 25 мм дл., 12 — 20 мм шир., выемчатые, клиновидно суженные в короткие черешки) полурозеточные сильноветвистые очередно-простолистные эусимподиальные семидетерми- натно-репродуцирующие однодомные розово-капустноцветковые колючепобеговые стерж- некорневые подушковидные травянистые многолетники. Вероятнее всего, биоморфы рас- сматриваемого вида, цветущие весной, после завершения вегетационного сезона проявляют себя как ~перекати-поле», отрываясь всей надземной частью — собственно ~подушкой»— от основания куста. По оврагам, у обочин дорог в каменистой пустыне можно было встре- тить скопления сухих надземных кустов-~подушек» 2. spinosa. Желательно продолжит накопление фактического материала по биографике этого вида на протяжении более дли- тельного нременн чем 1D котормм л окольно рлеполегел. Меетнме ботеннки отмечемт большой полиморфизм Z. spinosa и среди его биоморфотипов называют также полукустар- ники и даже кустарники до 1,5 м высотой (Feinbrun-Dothan, Danin, 1991, Alon, 1991 и др.).
Глава 8. Концепция всюдности эволюции м7 Рис. 395. Разнообразие метаморфозов в организации виргинильных биоморф Асег platanoides L. [сем. Aceraceae (Московская область, смешанные леса)]. Дополнительные обозначения: м1 — метаморфа протерокормуса — надземносемядольной паратовой чести реатенин (прорастание наивен«ос, се«ввали пралолтовето-лллиптичеслие, 18 — 27 мм дл., 5 — 8 мм шир., закругленные, с выемкой или без нее, постепенно суженные в короткие черещки), м2 — метаморфа надсемядольной фотофильной части первого годич- ного побега, мЗ — метаморфа-брахибласт (компактная побеговая часть) в основании второго и последующих годичных побегов, м4 — метаморфа-долихобласт (удлиненная побеговая часть) в структуре годичных побегов (над м 3), м5 — метаморфа неразветвленного главного корня, мб — метаморфа разветвленного главного корня, м7 — метаморфа годичного побега текущего года, м8 — метаморфа персистентной побеговой части.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ Ю а3 Ю О а3 й а3 О й Ю О 4Р О~ ой~( ~~< Ж < о o z g~ > ~ В -к~ Я 4Э Июж ~V () д~ ~~ 2 О„,; р~ X ~~)~ р ди1 .Р О ~: '~ц ~'~ v О о2ф~Ц F4 3 с~ ~ 2 а ж ~ ~ e3 3~ О "о р o "o~ о х 3 Q G4 X <о Й е3 к o - я ~. E go>~~ 4Э ю 4Э р~ д О > zo g y ~м gonzo p3 g- Oao~ к~62 О ~ 4~ Q. ф о 0~ 4~ О 4~ Я О Ю ФХЖ ~~~è~ Д: д О Ж д ops Х X щ Q Е u o g lu [ О О О Ю ~4 сЭ Ы~~~~ 0ъя и g Йожхи e z u o m~ о й р р, й ,>,Я х X О v О О 4~~ Ц о~~ 4Э ~ Ж О я о ~ц 63 р р2 4Р KEoozo~ о ~ и й д� и м~ - ~ о~ й О Ю 2 ~~ О М a O v ln ~z<<дх> О й [ ЕИийй~èî
Глава 8. Концепция всюдности эволюции Рис. 397. Результаты метаморфозов в организации биоморф Ephedra aphylla Forssk. [сем. Ephedraceae (там же, что и рис. 370, март, апрель)). Биоморфотип: надземно-двусемядольные безрозеточные редукционно-супротивно- и мутовчато-чешуелистные (супротивные чешуевидные пленчатые листья развиваются в вер- хней части побегов, они рано опадают) зусимподиальные (верхушки побегов и целые побеги в сухой летний сезон массово опадают, а основной многолетний прирост обеспе- чивают сильнорослые вегетативные плетевидные зеленые сильноветвистые побеги, кото- рые при наличии опор могут подниматься высоко, до 10 м, даже в кроны других деревьев, но чаще высота взрослых кустов около 1 — 2 м) двудомные стеблезеленые кустарники. Метаморфозы весьма сильно проявляются в организации биоморф Е. aphylla через редук- цию листовых пластин, а в стеблях через специализацию их не только для выполнения опорных функций, но и всего фотосинтеза.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИИ Рис. 398. Разнообразие метаморфозов в коалиции листьев на отдельной побеговой ветви взрослой особи-дерева Populus euphratica Oliver [сем. Salicaceae (там же, что и рис. 378, апрель)]. Дополнительные обозначения: м1 — метаморфа брахиморфная (сближенноузловая) с че- шуевидными листьями (они уже осыпались), м2 — метаморфа долихоморфная (раздвину- тоузловая) с яйцевидными срединными листьями, м3 — метаморфа долихоморфная с эллиптическими срединными листьями, м4 — метаморфа долихоморфная с ланцетными и линейно-ланцетными срединными листьями, м5 — метаморфы резидных побегов, ранее утратившие какие-либо листья.
Глава 8. Концепция всюдности эволюции Можно согласиться с Л.И. Лотовой (2000), которая, обратив вни- мание на мультифункциональность однородных органов, указала, что «наряду с главной, каждый из них осуществляет и многие дополнитель- ные функции. Так, лист как орган фотосинтеза выполняет и сопряжен- ные с ним процессы дыхания и транспирации, у некоторых растений он запасает воду, а благодаря железистым волоскам, наличию острых шипиков и иных структур, предохраняет растение от повреждения и поедания животными. Главная функция корня — поглощение воды и закрепление растения в субстрате — не препятствует его участию в накоплении запасных веществ» (стр. 313) и т.д. Видоизменения органов, как справедливо заметила Л.И. Лотова (2000, с. 313), называют метаморфозами, «если дополнительные функ- ции начинают преобладать над главными или орган в процессе развития приобретает какие-то новые функции, он сильно изменяется и морфо- логически, и анатомически». В соответствии с эволюционной теорией Ч. Дарвина, как отметила Л.И. Лотова (2000, с. 314), «метаморфоз — не просто видоизменение органов, происходящее в онтогенезе растения. Это генетически закрепленный процесс преобразования и превращения одних органов в другие под влиянием изменяющихся условий жизни, питания, выполнения ими функций, которыми сопровождается истори- ческое развитие вида». Приведенные цитаты из современного учебника ботаники вполне отражают господствующее понимание метаморфоза. Я готов со многим согласиться, но настаиваю на следующем: в организации гомологичных органов у растительных биоморф нет аметаморфизированных органов, т.е. все органы-гомологи — это всегда метаморфы — результаты мета- морфозов. Более всего я солидарен с мнением Л. И. Лотовой в том, что все органы, задействованные в метаморфозах, мультифункциональны, и среди их функций есть обязательные (главные) и дополнительные (вто- ростепенные). Однако, я развиваю широкий подход к пониманию ме- таморфозов и их результатов (метаморф), согласно которому в органи- зации растительных биоморф нельзя найти ни одного гомологичного органа, не затронугого метаморфозам. Эта позиция противоречит тра- диционной, когда в большинстве органов не наблюдается метаморфоза, поскольку в них главная функция доминирует над дополнительными. Метаморфизируются, согласно традиционным взглядам, только те орга- ны, в которых дополнительные функции начинают доминировать над обязательными, например, подземные побеги (корневища, клубни, лу- ковицы и др.). Если последовательно различать проявление метаморфо- зов в широком смысле, то не останется ни одного гомологичного органа еще не ставшего метаморфой. Почему метаморфоз — это атрибут в становлении организации гомологичных органов? Дело в том, что все аналогичные органы разные изначально, т.е. разнообразие заложено в их натуре: они разные по происхождению. Гомологичные органы изначально в исторической рет- роспективе были одинаковые: они одного происхождения, а лишь впос- ледствии, в процессе эволюции, они стали проявлять разнообразие,
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИИ овладев приемами метаморфоза, достигнув во многом в этом апогея у семенных растений. Важное обстоятельство, заслуживающее обсуждения, — это способ- ность органов вообще иметь и по-разному (!) проявлять обязательные и дополнительные структуры и функции. Быть обязательной частью в каком-либо органе — это означает изначально и всегда присутствовать в соответствующем типе органов, отвечающем их гомологичной природе и проявляющих, тем самым, единство их по происхождению. Возьмем в качестве примера побеги семенных растений. Какую бы форму ни приобретали побеги в целом и их элементы, их обязательные свойства (в первую очередь, функции) связаны с фотофильностью (они обязаны быть фотосинтезирующими «фабриками», производя в разном объеме органические вещества и О„дыханием, транспирацией, но и каркасностью («опорностью») для аппендикулярных элементов (листьев) и репродуктивных органов (в этом состоит их вегетативная обязанность). Другая обязанность побегов — стремиться достигать полового размноже- ния и осуществлять его в своей организации (рис. 374, 375 и др.). Если структурно-функциональные части в гомологичных органах обязательные, то, что и следовало бы ожидать, они ведут себя еще и как главные (превалирующие) части, обеспечивая определенный уровень специализации в развитии таких органов. Однако, все может в развитии организации гомологичных органов быть и по-другому, поскольку спе- циализация органов «замешана» на метаморфозах. Что значит проявить какую-либо специализацию в развитии орга- нов? Под биологической специализацией обычно понимают «высокую степень приспособленности данной группы организмов к узкому кругу специфических условий существования... Специализация является лишь частным случаем адаптации, это высокая степень приспособленности организмов к узкому диапазону условий внешней среды, сопровождаю- щаяся развитием специфических высокоэффективных механизмов ис- пользования этого диапазона условий. Неспециализированные формы — это виды организмов, приспособленные к жизни в разнообразных условиях (эврибионтные, по терминологии экологов)» (Иорданский, 1979, с. 140, 142).Специализация — это «направление эволюции группы, выражающееся в приспособлении ее к очень узким условиям существо- вания, крайний вариант аллогенеза» (Яблоков, Юсуфов, 1989, с. 313), при этом «аллогенез — развитие филлогенетической группы внугри одной адаптивной зоны» (там же, с. 312). Н.Н. Цвелев (1973, с. 71) считает, отчасти следуя Д.Г. Симпсону (1948), что «специализация выражается прежде всего в ограничении (или сужении) возможностей дальнейшей эволюции.... Часто определя- ют специализацию как узкое приспособление, которое действительно обычно ведет к ограничению эволюционных возможностей, однако многие высокоспециализированные таксоны адаптированы к довольно широкому диапазону физико-географических условий». Как видно из приведенных определений, в понимании специализа- ции исследователи склонны делать акцент на сужении адаптивных
Глава 8. Концепция всюдности эволюции возможностей организмов и их структурных элементов к условиям оби- тания. Эту тенденцию, я полагаю, надо запомнить хотя бы для осозна- ния истории вопроса, а сохранять традиционный подход к специализа- ции (здесь она есть, а здесь ее нет), на мой взгляд, вряд ли стоит; традиция в данном случае, может быть полезна сама по себе как ори- ентир в системе логического развития содержания этого термина. Мой прогноз: понятие «специализация» должно адекватно отвечать слову- этикетке, которым его обозначают. В экологическом отношении, как известно (Иоганзен, 1959; Рей- мерс, 1991, и др.) различают виды организмов с узкой специализацией к условиям обитания (стенобионтные) и виды — с широкой экологичес- кой специализацией (эврибионтные), т.е. специализация групп организ- мов, как следует из приведенной классификации, есть и у тех, и у других, но она разная (!). Об узкой и широкой специализации органов биоморф семенных растений, а шире — друтих биосистем как чьих-то структурных элементов, можно говорить в отношении соответствующих структурных элементов по ориентации на их специализацию к выпол- нению функций целостных биосистем. Специализация у биоморф, а шире и других биосистем, на мой взгляд, — это организационная, в первую очередь, функциональная, ориентация органов (или других структурных элементов) как адаптаций в соответствующих условиях окружающей среды, включающая также соответствующие структурные преобразования (мета- морфозы у органов). Специализация органов семенных фитобиоморф отражает не только сужение, но и расширение их структурно-функциональных возможностей в завоевании и освоении пространства и времени. При оценке специали- зации живых частей в организации биоморф важно подчеркнуть специ- альность (вид деятельности) этих частей — «профессию» быть структур- ной частью в организации биоморф с определенной функциональной ориентацией. Специализация органов (направленность их активных транс- формаций, или эволюции) обеспечивается метаморфозами и является их результатами, поскольку всякий метаморфоз — это действительно одна из форм единства роста и развития (трансформации) органов, а, следователь- но, — одно из эволюционных состояний, отражающее становление и дея- тельность каждого органа по структурно-функциональной ориентации. Специализация, кроме того, как способ ориентации организации биоси- стем, органов фитобиоморф (например побегов, т. 1, гл. 5.1.3.), в жиз- неразвитии подразделяется нами следующим образом: вегетативная, репродуктивная и вегетативно-репродуктивная. Все органы семенных фитобиоморф образуют какие-либо вегетативные структуры и только лидеры среди основных органов — побеги, способны проявлять, наряду с вегетативной, еще или только репродуктивную функцию. В специали- зации побегов, более детальной, чем та, которую мы привели, можно акцентировать внимание на разных признаках (рис. 371-375 и др.). Так, среди вегетативных побегов можно различать фотофильные (с зелеными срединными листьями) и афотофильные (с чешуевидными листьями или другими рудиментами срединных листьев). Репродуктивные побеги
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ бывают брактеозные или абрактеозные, а вегетативно-репродуктивные побеги (с фотофильными листьями и афотофильные). Специализация органов и их частей по форме и степени проявле- ния в них метаморфозов на протяжении вегетационного сезона весьма разнообразна. Одни органы и их части в организации растительных биоморф в процессе естественного жизнеразвития, сопровождающегося метаморфозами в вегетационном сезоне, сохраняют функциональное главенство изначально обязательных частей их организации над иной (дополнительной) функциональной ориентацией, проявляя, таким об- разом, узкую специализацию (рис. 374-376), что особенно характерно для однолетников. Если в подобной ситуации, органы и их части утра- чивают функциональное главенство обязательных частей своей органи- зации и отдают его (сменяют традиционные приоритеты) какой-либо дополнительной функциональной ориентации, то такая специализация широкая (рис. 377, 378). В целом органы могут подвергаться гомомета- морфозу, когда в результате его действия весь орган является одной метаморфой; так образуются гомометаморфы («одноликие» метаморфы), или гомометаморфные оргипи. Когда орган развивается под действием раз- ных метаморфозов, составляощих вместе гетерометаморфоз, а результатом является образование органа из двух и более метаморфных частей (зон)— нескольких метаморф-частей, то, таким образом, формируются гетероме- таморфы («разноликие» метаморфы), или гетерометаморфные органы. Так, первый годичный побег всхода у Асег platanoides L. является одноликой метаморфой, а весенний годичный побег у ювенильной особи того же вида — разноликой метаморфой (рис. 395), поскольку в первом случае весь побег одинаково фотофильный, а во втором — состоит из двух метаморф-частей — разнообразных локальных результатов гетеро- метаморфоза: сближенноузловой нижней зоны побега с чешуевидными листьями и срединной фотофильной зоны того же побега; у взрослых растений в структуре весенних годичных побегов, помимо двух отмечен- ных выше зон-метаморф, развивается еще и зона соцветия с брактеями (прицветковыми чешуями) и цветками. Метаморфы и их части — поставщики многих признаков, нередко индикаторных, для биоморф в целом и их отдельных возрастных и других состояний. Таким образом, гомологическое разнообразие в орга- низации органов биоморф достигается посредством метаморфозов одноли- ких (гомометаморфозов) или разноликих (гетерометаморфозов). Результаты метаморфозов удобнее всего изучать в гомологических рядах. Метаморфы составляют гомологичное разнообразие органов, на которое в зависимости от целей исследований можно обращать внима- ние или не очень, например, когда нас интересует сходство аналогичных органов, т.е. органов из разных гомологических рядов, например, сход- ство зеленоватых воздушных корней некоторых тропических орхидей с прутьевидными микрофильными (листья быстро осыпаются) побегами некоторых ксерофильных кустарников. По большому счету, в организации биоморф семенных растений все органы являются чьими-то гомологами, а, следовательно, все они
Глава 8. Концепция всюдности эволюции так или иначе — метаморфы, т.е. результаты каких-либо метаморфозов, которые можно наблюдать как по горизонтали — в текущих органоге- незах и онтогенезах, так и по вертикали, т.е. в исторической ретроспек- тиве, в филогенезах. Эту же мысль, но иными словами, можно выразить еще и так: организация биоморф состоит вся из органов-метаморф, по- так и (или) в текущем онтогенезе, и (или) в онтогенезе родителей, и (или) были такими в прошлом — исторической ретроспективе (филогенезе). На основании приведенного выше материала можно сделать следующее обобщение: все биоморфы, развивающие органы в своей организации, везде и всегда подвержены метаморфозам, результатами которых являются органы-метаморфы, обязательно (другого быть не может) имеющие в прошлом (в филогенезе) и (или) сейчас (в текущем онтогенезе) гомологов — это специальный закон облигантности метаморфозов у органогенных биоморф. Почему мы идем на такое широкое обобщение: дело в том, что для этого, по сути дела, нет никаких препятствий, а экстраполяция законо- мерностей проявления метаморфозов у биоморф семенных растений на все органогенные биоморфы, т.е. биоморфы, обязательно развивающие органы-гомологи в своей организации на протяжении их морфогенезов и шире — в филогенезах, вполне уместна, поскольку первична гомоло- гичность органов и вторичен их метаморфизм (способность органов проявлять метаморфозы). Почему такое внимание уделяется гомологии для объяснения мета- морфозов? Таким образом мы проявляем солидарность с идеями И.В. Гете, который видел в гомологах превращение (метаморфозы) только одного органа — «листа» (точнее, единого образа органов — «филлома»), но, естественно, вносим коррективы на время. Итак, с современных позиций, метаморфоз — это процесс трансформации (единства роста и развития) в организации биоморф, в результате которого достигается гомо- логичное разнообразие органов в органогенезах на разномасштабных уча- стках пространства и времени — от онтогенеза до филогенеза. Самое важное в метаморфозе, согласно приведенному выше опре- делению этого термина, не в том, как резко или медленно изменяется внешность органа, и даже не в том, изменяется ли она вообще в кон- кретном органогенезе на фоне онтогенеза или синонтогенеза, а в том, что в результате метаморфоза как составной части роста и развития организации биоморфы, образуется орган, который полностью или частично является гомологом каких-либо других разнообразных органов или частей органов. Все органы-гомологи — это результаты метаморфо- зов, которые выражают себя как метаморфы в органогенезах на фоне онтогенезов и филогенезов, т.е., проявившись однажды в онтогенезе, они в последующих поколениях отбираются естественным отбором в других онтогенезах, закрепляясь в итоге в филогенезе так, как мы их наблюдаем в текущем онтогенезе. Метаморфоз, таким образом, проявля- ется везде, где есть гомологичное структурно-функциональное разнообра- зие органов и частей органов, а последнее можно увидеть именно везде, если начать проводить сравнительные наблюдения.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Кроме специализаций, которые проявляют себя в вегетационных сезонах, имеют место быть еще и возрастные специализации органов. Образование корневищ, зрелых луковиц, скелетных ветвей, стволов и т.п. — это все примеры возрастной специализации органов как резуль- татов соответствующих (возрастных) метаморфозов (рис. 380-382). Часто метаморфозы связаны именно с возрастной изменчивостью как органов, так и биоморф в целом. На определенных стадиях жизне- развития выполнение органом обязательной деятельности может и не- редко становится делом обременительным, что вполне закономерно переключает его организацию, посредством соответствующего метамор- фоза, на другой вид деятельности, который более всего в этот момент нужен организму, когда его дополнительные функции изменяют статус и становятся главными, а обязательные свойства частично или полно- стью теряют приоритет, становятся дополнительными, иногда в опреде- ленных ситуациях, например, при старении, вообще утрачиваются (рис. 377-382). Растительная, а шире — и другая жизнь, без разнообразных метаморфозов была бы невозможна. Как для эволюции вообще, так и для ее эволюстатов, одним из типов которых являются метаморфозы, обязательная содержательная начинка — активные трансформации, ре- зультаты которых разные, что и составляет исследовательскую интригу для тех, кто хочет в них разобраться. Итак, органы биоморф подверга- ются специализации в результате метаморфозов не только и даже не столько в зависимости от внешних условий среды, сколько под влияни- ем соответствующих возрастных процессов, происходящих как в био- морфах, так и органах по отдельности (рис. 367-398 и многие другие). Специализация, кроме того, имеет еще и более широкий резонанс, который далее в этой же главе будет также рассмотрен (стр. 680). Метаморфозы разные по форме и разной интенсивности сопровож- дают рост и развитие всех органов, но в тех случаях, когда множество метаморфозов органов проявляются почти одновременно и сопровожда- ют существенные возрастные изменения биоморф некоторых животных, например, в начальных стадиях их эпиэмбрионального развития, когда головастик превращается в лягушку или личинка майского жука во взрослую особь-имаго, то такое сосредоточие («синдром») множеств метаморфозов органов, сопровождающих возрастные изменения био- морф, можно обозначить как метаморфозный синдром. Собственно, в таких случаях происходит совсем не другой тип метаморфозов, как полагают многие исследователи, а самый обычный, но не рассеянный по организации биоморф, как у большинства растений и животных с так называемым прямым развитием, а особенным образом во всем своем множестве и силе проявления, сконцентрированный на узком «участке» пространства жизни и ограниченный во времени своего действия, что и обеспечивает эффект синдрома (метаморфозного). Метаморфозы сопровождают рост и развитие органов постоянно, составляя одну из сторон их жизнедеятельности. При изучении мета- морфозов нет, ее не должно быть, проблемы поиска типичного (амета- морфизированного) и атипичного (метаморфизированного) органа,
Глава 8. Концепция всюдности эволюции поскольку все гомологичные органы являются типичными метаморфами- результатами метаморфозов, которые мы наблюдаем сейчас повсеместно и которые сложились в настоящем виде в процессе длительного историчес- кого развития. Никто никогда точно (возможны только гипотетические построе- ния) не скажет в историческом плане, с какой формы органа начался отсчет метаморф. Все современные органы — это все метаморфы, т.е. результаты метаморфозов. Мы исходим из тезиса, что у первых палео- биоморф семенных растений (наиболее древних, естественно, давно уже вымерших биоморф) был некоторый палеоорган (один или несколько), т.е. исторически исходный в «средеэ органов-гомологов семенных рас- тений производных от него органов (органов-потомков). Все современ- ные органы-гомологи — это результаты множественных метаморфозов некоторого палеооргана, о структуре которого можно только догады- ваться. Сейчас все разнообразие органов, которое мы наблюдаем, — это результаты метаморфозов — метаморфы разных гомологических рядов. Здесь и сейчас автор предлагает читателю внимательно присмотреться и задуматься над тем, что скрывается за процессами роста и развития у растительных биоморф, да и не только у них, и их результатами. Пожалуй, мы определились с понятием «метаморфоз» и сумели пока- зать, что этот процесс и его результаты — метаморфы — это только одна из форм единства процессов роста и развития и, что одно и то же,— это одновременно одна из форм трансформации (трансформ), а, следо- вательно, одно из многих эволюстатов (эволюционных состояний). Далее мы увидим, что кроме метаморфозов в динамическом существо- вании биоморф проявляется еще немало других форм единства процес- сов роста и развития (трансформ) живых систем. Когда-то и кто-то создаст теорию трансформации, которая по сво- ей масштабности должна перекрыть эволюционную теорию, поскольку эволюция — это только одна, хотя и высшая, из форм трансформации, но важно, что контуры такой теории и возможных путей ее построения уже просматриваются, и кто знает, вполне может быть, что мы идем по одному из них, находясь, возможно, где-то еще в начале. 8.3.2. Что еще из адаптаций, кроме метаморфозов, проявляется в биоморфогенезах семенных растений Выше было показано, что метаморфозы — это сфера проявления морфогенезов органов (органогенезов) на фоне индивидуального разви- тия организмов (онтогенезов), а шире — и морфогенезов синорганизмов (синонтогенезов). Метаморфоз — это процесс, а метаморфа — результат проявления этого процесса. Цель метаморфозов — преобразование ор- ганов ради достижения, посредством единства роста и развития их, определенного уровня специализации. Метаморфоз — это также способ
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ специализации органов в целом и их частей, когда действительно по- лучает выражение смена функций одного и того же органа в процессе его морфогенеза и на фоне морфогенеза всего организма (онтогенеза) и исторического развития таксона (филогенеза). Жизнь растительных организмов и их органов находится под вли- янием многих физических факторов (свет, температура, ускорение силы тяжести, влажность воздуха). Физические факторы оказывают формооб- разующую роль, т.е. вызывают морфозы — мелкомасштабные изменения формы (Либберт, 1976). Ранее уже достаточно много было сказано о понятии метаморфоз. Как связаны метаморфозы с морфозами? Я пола- гаю, что Э. Либберт, говоря о разных морфозах (фотоморфозах, геомор- фозах, термоморфозах, гигро- и гидроморфозах, тигморфозах, имел в виду разнообразные элементарные (!) формопроцессы в росте и разви- тии органов, т.е. в их морфогенезах, или органогенезах. Приведем пример проявления фотоморфозов (элементарных изме- нений форм под влиянием света), которые приводит Э. Либберт (1976, с. 473) у Sinapis alba L. Сначала у проростков происходит раскрытие пластинок семядолей, увеличение их поверхности, дифференцировка устьиц, формирование пластид, торможение оттока веществ; далее: за- держка роста растяжением гипопотиля, раскрывание сложенного перво- го листа, образование волосков, антоциана, ксилемы. «В дальнейшем: развитие первичных листьев, образование зачатков следующих листьев, интенсификация расщепления запасных жиров и белков и некоторые другие биохимические и физиологические изменения». На наш взгляд, морфозы как элементарные формопроцессы органа, т.е. элементарные изменения органа в процессе его роста и развития, составляют в некоторых совокупностях — метаморфозы. Совокупность нескольких (иногда многих) морфозов, ограниченных своим проявлением всем органом или только его частью, обеспечивает широту преобразования (метаморфоза) этого органа или только его части в нечто другое — другое состояние органа, другой орган или другую часть органа, что составляет правило взаимосвязи морфозов и метаморфозов в организации раститель- ных биоморф. Только через концентрацию морфозов вместе, под влиянием генетических и иных факторов, в один метаморфоз органа может быть обеспечена реализация им, органом, роста и развития во всем его морфо- генезе (органогенезе) — это специальный закон органогенеза у биоморф. Морфогенез органа и всего организма составляют множество морфо- зов — элементарных адаптивных, а, следовательно, и эволюционных актов, простейших единиц роста и развития в организации биоморф; их совокуп- ности образуют, кроме того, и процессы метаморфозов. Метаморфозы (один или несколько) — составная часть морфогенеза какого-либо органа (орга- ногенеза), а совокупность всех метаморфозов в рамках организма или си- норганизма обеспечивает этим биоморфам трансформацию в формах роста (стадиосомах) биоморф от рождения до смерти, т.е. в биоморфогенезах, и нередко служит индикаторами их. О разнообразии форм роста каких- либо живых тел, биоморф, в особенности, мы уже писали в главе 4.
Глава 8. Концепция всюдности эволюции 8.4. Закон альтернации в эволюции признаков В отношении органов семенных растений, как уже было показано выше, не подходит деление их на два типа: типичные, или аметаморфи- зированные (без метаморфозных проявлений в их организации) и ати- пичные, или метаморфизированные (с метаморфозными проявлениями в их организации). Такое подразделение неверно как по форме, так и по содержанию. Я не использую в качестве мерила «гипичного» для органов семенных растений их непринадлежность к метаморфозной деятельнос- ти, поскольку считаю, что у всех органов семенных растений всегда есть гомологи, а все органы-гомологи могут существовать, подвергаясь мета- морфозам как одной из форм адаптаций, а, следовательно, и эволюции. По нашей концепции вне метаморфозов в организации биоморф семен- ных растений не бывает органов-гомологов, а все органы-аналоги под друтим утлом обзора все равно являются еще и гомологичными органами и где-то имеют по отношению к себе органы-гомологи. Таким образом, организация биоморф семенных растений — это организация, в первую очередь, из органов-гомологов и, следовательно, она вся состоит из метаморфизированных органов, или метаморф, типы которых постоянно проявляют себя в органогенезах на фоне современ- ных биоморфогенезов, но, кроме того, всегда имеют какую-либо более или менее далекую историческую ретроспективу (на основе филогене- за). Гомологическое разнообразие органов — результат прошлых и нынешних метаморфозов их. Действительно, кроме наиболее распрос- траненных типов метаморф, например, фотофильных побегов с умерен- но широкими и цельными зелеными пластинками черешковых листьев и цилиндрическими зелеными стеблями, распространены, пусть помень- ше первых, но все же в большом разнообразии и другие типы побегов, которые так или иначе изменены в своей организации метаморфозами в процессе эволюции семенных фитобиоморф в соответствии с их на- следственностью и в зависимости от условий окружающей среды. У побегов других растений, например, стебель расширился и сплющился, став похожим на лист, а листья редуцировались до жалких чешщ или превратились в колючки, цветки, будучи частями побегов, приобрели в этом качестве неиссякаемое разнообразие как по цвету, облику, так и по внутренним особенностям их организации, сохранив первородное побеговое происхождение — свою гомологическую метаморфозную сущ- ность и как частей, и как целого и т.д. и т.п. Рассматривая ряды альтернативных (сравнимых и взаимоисключа- ющих) признаков гомологичных органов, обращаешь внимание на то, как они склонны выстраиваться в линейной последовательности; часть признаков при этом тяготеет к середине рядов, а другие «прижимаются» к их краям. Больше того, иногда проявляются признаки, за которыми, в соответствующем ряду, эволюция больше ничего предложить не мо- жет, оставляя единственную альтернативу — искать путь к созданию новых рядов признаков. Возьмем в качестве примера такой ряд альтер- нативных признаков гомологичных органов — цветоносных побегов,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ построив признаки линейно по горизонтали в зависимости от особен- ностей расположения в их структуре репродуктивных элементов — цвет- ков. Эволюция за миллионы лет создала большое разнообразие цвето- носных побегов, совершенствуя его созданием переходных типов между типами открытых и закрытых цветоносных побегов. Типы открытых и закрытых побегов в рассматриваемом ряду признаков стали не просто крайними, а, более того, они превратились в последние, для этого ряда, признаки, доказав за многие миллионы лет эволюции, что они действи- тельно таковыми являются, поскольку ничего другого нового в этом ряду сверх двух последних (открытые и закрытые) признаков уже никог- да не возникнет. Внутри ряда, между этими признаками новаций можно ожидать сколько угодно, а если что-то и вырвется за ними, то это уже будут признаки совершенно другого ряда. Вполне уместно следующее обобщение: эволюция гомологичных ор- ганов в организации биоморф идет по рядам альтернативных признаков, в рамках которых они естественным образом отбираются, как бы выстра- иваясь поочередно и линейно в одном из двух противоположных направле- ний, причем в рядах могут проявляться не только крайние, но даже пос- ледние признаки и, если они действительно таковые, то все другие альтер- нативные признаки уже не могут выйти за пределы этого ряда, что происходит до тех пор, пока эволюция не находит для гомологичных орга- нов качественно совершенно других решений, когда создается новый ряд альтернативных признаков — это специальный закон альтернации (чере- дования) признаков в эволюции гомологичных органов. Этот закон является следствием из еще более общего закона, в котором снимаются обязательные ограничения по привязке признаков к гомологичным органам, а фактически рассматриваются ряды альтер- нативных признаков любых систем с единой преднаследственной или наследственной основой. Эволюционное развитие чего-либо или кого-либо на единой преднаследственной или наследственной основе идет естествен- ным образом по рядам альтернативных признаков, в которых они как бы выстраиваются поочередно и линейно как внутри ряда, так и кнаружи, пополняя его, таким образом, новыми признаками, пока в эволюции не возникают последние признаки этого ряда, которые не позволяют ему расширяться кнаружи, т.е. выходить за пределы такого ряда, но не огра- ничивают его расширение изнутри, что в конце концов может приводить к возникновению качественно новых рядов альтернативных признаков, но при этом никогда эволюция совсем не закрывается — это один из предвыс- ших законов [такие законы являются общими для Природы, но распро- страняются не на все ее явления (т. 1, гл. 1), например, в нашем случае, не охватывают дисальтернативные признаки] альтернации прогенетичес- ки или генетически однородных систем.
Глава 8. Концепция всюдности эволюции 8.5. Иерархичность трансформаций в живых системах и место в ней эволюционных ssaeHvo Если посмотреть на Природу (вселенский поток) как бы со сторо- ны, то в ней мы ничего другого не обнаружим, кроме как движущейся в пространстве и времени материи, которая существует в разнообразных телесных и нетелесных системах; они не только перемещаются, но и в это же время как-либо трансформируются. Приглядимся повниматель- нее к трансформациям, и обнаружится, что их всего два, наиболее крул- ных шила: субэволюция, когда тела как-либо (почти «пассивно») подвер- гаются преобразовательным воздействиям во время инерционного пере- мещения (например, изменение каких-нибудь предметов, допустим, стола, булыжника и т.п., на основе вращения их с Землей вокруг Сол- нца) и эволюция (все остальные активные трансформации тел и нетел) — это первая широкая субординатная классификация трансформаций материи. Так, незаметно для себя, мы стали на «тропу» иерархического классифицирования форм трансформаций (трансформ), но оставим при этом в покое субэволюцию, «навалившись» на эволюцию и ее формы— эволюционные состояния (эволюстаты). Процесс трансформации охватывает какие-либо состояния систе- мы, иногда незначительные, а иногда коренные, определяя смену одно- го состояния другим, или распространяется на все целостное содержа- ние системы, вызывая в конечном счете прекращение существования одной системы (в одних качественных границах) и возникновение но- вой системы (в других качественных границах). Все процессы трансфор- мации, происходящие в живых системах (телах) и с живыми системами, можно объединить в одной суперординатной классификации — «пира- миде» абстракций (рис. 399), которая представляет иерархическую клас- сификационную систему, суперординаты (классы, или уровни, абстрак- ции), в которой заняты соответствующими субординатными классифи- кациями, различающимися по числу субординат (классов), их составля- ющих (т. 1, гл. 1). В суперординатную классификацию (суперординию) состояний трансформации (трансформ) материи в живых телах и с живыми телами мы включаем несколько широких субординатных классификаций (су- бординий), которые, будучи сами иерархическими классификациями, в свою очередь, связаны между собой иерархическими отношениями, различаясь по числу субординат в этих классификациях. Рисунок 399 наглядно, как нам представляется, передает разнообразие типов транс- форм, проявляющихся в живых системах и с живыми системами. Транс- формы соответствующих типов, в зависимости от высоты обобщений и сложности процессов трансформации, пройдя через логические опера- ции классифицирования, расположились в нескольких субординатных классификациях на разных высотах (уровнях) в одной «пирамиде» аб- стракций, отражая, в какой-то мере, еще и эволюцию трансформации материи как процессов, так и результатов в организации живых систем (точнее, в живых системах и с живыми системами).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Итак, мы начали сверху отсчет субординий трансформ (их типов) на «пирамиде» абстракций с субэволюции и эволюции. Полагаю, что в отношении эволюции можно было бы, чтобы подчеркивать ее различ- ные нюансы, увлечься процедурой «загрузки» терминообразующего сло- ва-этикетки («эволюция») еще эпитетами (такая она или другая, напри- мер, простая или сложная, локальная или общая и т.п.). Однако, мы, как нам кажется, до разумных пределов ограничим себя в этом, по- скольку, по большому счету, эволюция — это единый и бесконечный процесс со множеством больших и малых ответвлений (!), и в процедуре классифицирования форм (состояний) эволюции — эволюстатов- главное все же — не переусердствовать в области терминотворчества. Во второй широкой субординии трансформаций материи, или трансформ (их типов), три субординаты: субэволюция, эволюция простая (без субмутаций или мутаций) и эволюция сложная (с субмутациями или мутациями). У Солнца эволюция сложная, поскольку она связана с субмутаци- ями (преобразованиями прогенов), когда атомы (прогены) водорода в солнечных условиях трансформируются в атомы гелия. У особи Quercus гоЬиг L., когда она, сначала проросток, совершая морфозы, метаморфо- зы и этоморфозы, растет и развивается, преобразуясь во всходы, затем в ювенильные растения и т.д., повторяя без мутаций преемственный путь своих предков, — это простая эволюция, в результате которой осуществляется продолжение жизни на отдельном конкретном отрезке пространства-времени. Третья широкая субординия трансформаций уместилась на той же самой ступени «пирамиды» абстракций, что и вторая субординия, по- скольку она составлена также из трех субординат: субэволюция, эволю- ция количественная (этот процесс включает все количественные транс- формации — рост, разрушение, т.е. не только восходящие, но и нисхо- дящие), эволюция качественная, или развитие (качественные трансфор- мации любой направленности, когда в них переходят соответствующие количественные трансформации). Оба типа эволюции обычно выступа- ют в единстве, накладываясь друг на друга. На другой (чуть пониже) ступени «пирамиды» абстракций мы рас- положили четвертую широкую субординатную классификацию типов трансформации материи из четырех субординат. В качестве критерия для выделения классов трансформ служат соответствующие их отноше- ния с жизнью: субэволюция, инвитаэволюция (эволюция неживого), ви- таэволюция (эволюция живого) и трансэволюция [эволюция экосистем или других прогенетически и (или) генетически разнородных систем, например, системы «хозяин-паразит» в мире животных и растений структурные элементы которых не утрачивают и своей относительной самостоятельности1. Под другим углом обзора, но на той же высоте абстракций, расположим пятую широкую субординию типов трансфор- мации материи (шире — движений вообще) из четырех субординат (критерий — отношение к живому): эуабиокинез (всякое движение не- живой неорганической материи), некрокинез (всякое движение неживой
Глава 8. Концепция всюдности эволюции 13 12 11 10 17 16 7 1 22 21 20 19 18 9 26 25 24 23 1 Рис. 399'. Иерархичность форм трансформации в живых системах и ее от- ражение в ~пирамиде» абстракций. 1 — субэволюция, 2 — эволюция, 3 — простая эволюция, 4 — сложная эволюция, 5— эволюция количественная, 6 — эволюция качественная (развитие), 7 — инвитаэволюция, 8 — витаэволюция, 9 — трансэволюция, 10 — эуабиокинез, 11 — некрокинез, 12— биокинез, 13 — нообиокинез, 14 — униэволюция, 15 — коэволюция, 16 — миниэволюция, 17 — максиэволюция, 18 — абиокинез, 19 — анабиоз, 20 — криптобиоз, 21 — гипобиоз, 22 — эубиоз, 23 — курсация, 24 — адаптация, 25 — революция, 26 — суперадаптация, 27 — морфоз, 28 — метаморфоз, 29 — этоморфоз, 30 — морфогенез, 31 — филогенез.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ органической материи), биокинез (всякое движение первоприродной живой материи), нообиокинез (всякое движение живой материи, связан- ное с разумной деятельностью людей). Задержимся на прежней ступени «пирамиды» абстракций. Здесь мы расположили еще шестую и седьмую широкие субординии трансформа- ций из четырех субординат. В шестой субординии это следующие типы: субэволюция, униэволюция, коэволюция и трансэволюция (см. стр. 623). В седьмой субординии трансформ — следующие субординаты: су- бэволюция, инвитаэволюция, миниэволюция (минимоэволюция), или «мик- роэволюция», максиэволюция (максимоэволюция), или «макроэволюция» (построение греко-латинских терминов, которые здесь приведены в кавычках, обычно считается грамматически неправильным, но, несмот- ря на это, иногда и такие термины признаются и принимаются научной общественностью, возможно, они бывают иногда чуть-чуть благозвуч- нее...), трансэволюция. Изменив угол обзора, мы обнаружим, что на следующую ступень «пирамиды» абстракций вполне может претендовать восьмая широкая субординия трансформаций материи из пяти субординат: абиокинез (вся- кое движение неживой материи), анабиоз, криптобиоз, гипобиоз, эубиоз (подробнее см. А.М. Голдовский, 1981; Р.С. Ушатинская, 1990 и др.). К девятой широкой субординатной классификации форм транс- формаций материи мы спускаемся на ступень ниже «пирамиды» абст- ракций. Она имеет следующий вид: субэволюция, курсация, адаптация, революция, суперадаптация и трансэволюция. Сделаем очередной «шаг» вниз по ступеням «пирамиды» абстрак- ций и рассмотрим десятую широкую субординию форм трансформации материи из 9 субординат: субэволюция, курсация, морфоз, метаморфоз, этоморфоз (этоморфозы включают и стадиоморфозы — развитие стади- осо.'л), морфогенез (морфогенезами являются и гипобиоморфогенезы, и биоморфогенезы), филогенез, коэволюция и трансэволюция. В этой су- бординии продолжено дробление трансформаций, которые все, распо- ложенные выше субэволюции, являются формами эволюции, или эво- люционными состояниями (эволюстатами). Здесь, пожалуй, мы остановимся, хотя есть возможности и дальше продолжать дробление эволюстатов при движении вниз по ступеням «пирамиды» абстракций. При дальнейшем иерархическом классифици- ровании трансформаций в рассматриваемом восходящем ряду самые нижние из субординий приобретут разветвленную «конструкцию», т.е. станут мультисубординатными классификациями (мультисубординия- ми), поскольку многое в эволюции свершается в прямой зависимости от того, как организована Природа — каково разнообразие ее матери- альных систем, что мы уже отразили ранее в соответствующей «пира- миде» абстракций (т. 1, гл. 2).
Глава 8. Концепция всюдности эволюции 8.6. Борьба за существование организации биоморф семенных растений и место в ней их габитусов и форм роста Наша эволюционная концепция строится на признании постулата всюдности эволюции и объяснении фактов такого обязательного проявления этого бесконечного процесса в организации Природы и, в частности, в организации биоморф семенных растений. Ранее автором (т. 1, гл. 4, 8) было предложено отношения между живыми структур- ными элементами (гипобиоморфами) внутри биоморф, в первую оче- редь, организмов, но еще и синорганизмов, пока их клонисты (особи вегетативного происхождения) не отдалены или отдалены настолько незначительно, что и их взаимодействие, например, у синорганизма- кочки Deschampsia cespitosa (L.) Веаич., реально сказывается на инф- раструктуре всего синорганизма, — называть инфраструктурной борьбой за существование, а для обозначения обобщенной ответной реакции всей простой биоморфы (организма), ее конституции, на воздействие окружающей внешней среды, использован термин Л. Плате (Plate, 1913) «конституциальная» («конституциональная») борьба за существо- вание. Проблемы инфраструктурной борьбы за существование только ста- вятся и еще нащупываются пути ее решения, но сегодня уже ясно, что без результатов этой борьбы внутри организмов все рассуждения об эволюции видов в популяциях будут неполными, а, по большому счету, без результатов этой борьбы эволюция видов путем естественного отбо- ра вообще не могла бы состояться. Вероятно, в организации биоморф на фоне или в гуще инфраструктурной борьбы есть процесс, который предшествует (предваряет) естественный отбор или является его частью. Попробуем подробнее разобраться с особенностями проявления инфраструктурной борьбы за существование у биоморф семенных рас- тений, но сначала оценим, что же все-таки произошло в 1997 году, когда автор в первом томе своей монографии доказал, что борьба за существо- вание с использованием известных ее атрибутов: конкуренции, коопе- рации и индифферентности — объективная реальность внутри органи- зации растительных биоморф, в особенности, каждого отдельного рас- тения-организма. 8.6.1. Многое в эволюционном учении еще впереди... Эволюция всюду, а причина тому одна — все, что есть в Природе «замешано» на движении (изменении), а высшей формой (типом) дви- жения является трансформация Природы и ее главная, приоритетная, часть — активная трансформация, или эволюция. Куда бы мы ни обратили свой взгляд в живой природе, везде уви- дим только одно — эволюцию живых тел — их существование в борьбе,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ которая свершается постоянно в адаптациях. У живых тел нет достойной альтернативы борьбе за существование (за жизнь), кроме как умереть и выйти из интригующей «игры» под названием «жизнь», включив в дей- ствие свою последнюю адаптацию — смерть. Как ни банально это звучит, но борьба за существование начина- ется и происходит в окружающей среде. Как ни парадоксально, но борьба за существование в окружающей среде для одних живых тел меньшей организационной сложности начинается внутри определенных живых тел большей, чем они сами, организационной сложности. Никог- да не будем забывать, что Природа, в том числе и живая природа, организованы иерархически. Быть окружающей средой для живого тела — это не значит быть только его внешней средой, но еще, может быть, и окружающей его средой внугри другого живого тела. Живому телу быть в окружающей его среде внутри другого тела — это значит как- либо учавствовать в инфраструктуре другого тела и являться, тем самым, его структурным элементом. 8.6.1.1. Живое начинается с инфраструктурной борьбы Ранее (т. 1) мы начали большой разговор об инфраструктурной борьбе за существование, отлично понимая, что в тот момент важнее всего было найти силы не только, чтобы подняться во весь рост и выстоять, и, наконец, во всеуслышанье заявить — да, борьба внутри растительных организмов идет, причем на «полную катушку», а сейчас доказать обратное — занятие уже совсем безнадежное; более того, я готов утверждать, что внутри растительных организмов «бушует» есте- ственный отбор, его первая часть — интраэлекция, и, что важнее всего, в них «процветает» эволюция. Об этом мы уже подробнее поговорим здесь, в восьмой главе, и будем продолжать развивать эту тему дальше. Некоторые утверждения автора, безусловно, настораживают, если опираться только на истины, исходящие из школьных и вузовских учеб- ников биологии. Однако не будем торопиться с выводами и отвлечемся от этого, пока еще преждевременного занятия, поскольку, по большому счету, многое только начинается... Самое интересное в нашей моногра- фии, похоже, еще впереди. А в отношении учебников есть непреложное правило: они пишутся для того, чтобы впоследствии нашлись у них читатели, причем такие, которые смогли бы создать другие, еще более совершенные, учебники с учетом тех изменений, которые постоянно происходят в науке. Факты — упрямая вещь, они подталкивают нас к новым шагам в сторону еще больших обобщений по затронутой проблеме. По сути дела, развитие представлений об инфраструктурной борьбе за существование еще только складывается и те «паруса», которые были развернуты над нашим исследовательским «кораблем» и настроены под «ветер» перемен в традиционных представлениях о естественном отборе и эволюции, нуждаются в постоянном притоке освежающего знания, как новых порывах ветра, особенно сейчас, когда целенаправленное движение на пути представлений о всюдной эволюции только начинается и еще
Глава 8. Концепция всюдности эволюции далеко до его стабилизации в нужном направлении. Ранее (т. 1) мы ограничились констатацией фактов проявления инфраструктурной борь- бы за существование в организации биоморф, т.е. борьбы, которую ведут их структурные элементы для себя и для своего автобиотопа (ин- фраструктурной среды обитания) — организма или иногда и синорга- низма [например, в распавшейся на обособленные партикулы-клонисты кочке чия — Achnatherum splendes (Trin.) Nevski]. Такое восприятие инфраструктурной борьбы за существование еще вчера было достаточ- ным, но сегодня его маловато и надо продолжить развивать эту идею дальше. Все живое, как известно, организовано по иерархическому прин- ципу и всегда есть кто-то, кто в живой природе — биостробе Земли (мы пока не знаем, кто стоит над биостробой нашей планеты, и труд- нее всего предположить, что вселенский «вихрь Жизни» — панбиостро- ба — не более, чем жизнь на Земле) менее сложен, составляя часть в организации более сложных живых тел. Структура (вещественная) в организации систем одна (т. 1, гл. 2), но ведет она себя по-разному, проявляя нередко двойственность по особенностям ее функциониро- вания; из нее могут исходить два типа функций: интрафуыкции и эк- страфуыкции. Часть структуры в организации системы, в которой на- ходят отражение особенности функционирования ее элементов, реша- ющих задачи внутренних отношений между ними, в процессе развития этой конкретной системы, — это, как известно, инфраструктура. В инфраструктуре обычно происходят интрафункцианальные взаимоот- ношения между элементами. Путем экстрафункционирования сомати- ческие системы решают задачи их отношений за пределами структуры этой конкретной системы. Когда одни побеги в растительном организ- ме в отношениях с другими побегами ведут себя вегетативно, а другие иначе, репродуктивна, или среди одноименных побегов одни развива- ются сильнее, а другие — слабее, когда структурные элементы погло- щают из воздуха СО, для производства на свету органических веществ, — это все инфраструктурная деятельность — интрафункционирование. Пожалуй, в широком смысле, в отношении живых структурных эле- ментов всех биосистем, которые взаимодействуют по инфраструктур- ному типу, вполне можно говорить об инфраструктурной борьбе за существование в этих биосистемах (эндоцеллулярной, эндогистонной, эндоорганной, эндоорганизменной, эндосинорганизменной, эндодем- ной, эндоэйдосной и т.д.). Когда структурные элементы в организации растительных биоморф производят О„когда жгучие волоски побегов крапивы (Urtica dioica L.) обжигают другие живые существа, когда семена зрелых плодов бешеного огурца [Ecballium elaterium (L.) А. Richard] «выстреливаются» при прикосновении к нему, — это все примеры экстрафункционирования биоморф в их борьбе за существо- вание. Инфраструктурная борьба за существование биоморф (эндобиомор- фная) (т. 1, гл. 4) — это борьба за жизнь их структурных элементов, складывающаяся в текущей и исторической эволюции через отношения
ОСНОВБ1 БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ между ними и с окружающей их внутренней средой биоморф, где она себя проявляет, и с окружающей внешней средой, что отражается в развитии конституции (телосложения) биоморф, отвечающей условиям их обитания. Инфраструктурную борьбу за существование других живых телесных систем (клеток, тканей, видов, биоценозов и т.п.) мы здесь рассматривать не будем, ограничившись констатацией безусловного факта повсеместного проявления таковой в живой природе. Инфра- структурная борьба (поведение) структурных элементов биосистем раз- ных уровней сложности имеет место быть, и анализ этого явления еще во многом ждет своих исследователей. 8.6.1.2. Биоморфы и их структурные элементы — «пациен- ты» всюдной эволюции Если долго говорить «халва, халва», то, как известно, слаще не станет. Похоже, что ситуация, которая складывается вокруг прекрас- ного термина дарвиновских времен «естественный отбор», нуждается в развитии. Я вовсе не сторонник того, чтобы там, где все вроде бы понятно, «наводить тень на плетень». Изменять в корне понятие «ес- тественный отбор» не рационально. Дело в том, что правильность многих положений эволюционной теории, разработанной Ч. Дарвином (Darwin, 1859), проверена временем. Этот ученый впервые «пришел к принципиальному заключению о неизбежности в природе избиратель- ного размножения одних и гибели других особей — наличия естествен- ного отбора» (Яблоков. Юсуфов, 1989, с. 143). Ч. Дарвин принял «пред- ложенное Г. Спенсером определение отбора как переживание наиболее приспособленных» (цит. по А.С. Северцову, 1981, с. 93). «Сохранение благоприятных индивидуальных различий и изменений и уничтожение вредных я назвал естественным отбором, или переживанием наиболее приспособленных» (Дарвин, 1987, с. 65). Согласно Дарвину, как подчер- кнул А.С. Северцов (1981, с. 93), естественный отбор — «это и сохра- нение положительных вариантов, и устранение отрицательных, проис- ходящих через выживание и гибель особей... Переживание организмов приводит в ряду поколений к уничтожению вредных и сохранению полезных признаков этих организмов; тем самым происходит преоб- разование особей эволюирующей популяции». По современной трак- товке, «естественный отбор — выживание и оставление потомства более приспособленными в данных условиях особями; избирательное сохранение и передача следующему поколению определенных геноти- пов» (Яблоков, Юсуфов, 1989, с. 312). Сложилась любопытная ситуа- ция, согласно которой, вроде бы, никто и не отрицает, что эволюция направлена на развитие в поколениях, путем естественного отбора, организации биоморф (организмов и синорганизмов), но двойствен- ность в явлении естественного отбора в понятийном отношении отра- жения пока не получила. На мой взгляд, эволюция направлена не только на развитие новых сторон в организации биоморф, но и, что не менее, а даже еще более важно, в первую очередь, на сохранение ранее достигну- того качества [продолжение жизни (!) в поколениях — это первое обяза-
Глава 8. Концепция всюдности эволюции е тельное направление в эволюции живой природы]. Тезис «борьба за су- ществование происходит не между генами и признаками, а между их носителями-особями» (Яблоков, Юсуфов, 1989, с. 144) с позиций признания всюдности эволюции представляется, хотя и традицион- ным, но все-таки недостаточным, поскольку с широких позиций надо признать, что эволюция идет не только в популяциях, но и внутри каждого организма, добиваясь, естественно, разных результатов опре- деленных типов. Автор, при всем том, что было высказано выше, все же не склонен совсем оставлять термин «естественный отбор» в покое, поскольку явление, которое им обозначается, состоит из двух, хорошо определя- емых, частей. В естественном отборе, как правило, присутствуют две части, которые я предлагаю обозначать, как координирующий отбор, или интраэлекция (от лат. intra — внутри, electus — избранный), и подтверждающе-утверждающий отбор, или экстраэлекция (от лат. extra — вне, снаружи, electus — избранный). Однако, естественный отбор (элекция) в организме возможен при одном обязательном условии: в нем должна идти борьба за существование его живых структурных элементов (гипобиоморф) со всеми атрибутами любой борьбы (конку- ренцией, кооперацией и индифферентностью). Если бы всего этого не было, то эволюция биоморф достигала бы или не достигала одного результата из тех трех, которые она способна достигать в ее современ- ном виде в отношении жизни: она продолжала бы (если бы катастро- фически не «споткнулась» всего лишь однажды, а это было бы доста- точно, чтобы навсегда остановиться) свое шествие по Земле в поколе- ниях дискретных живых систем, ничего не меняя в их организации, никогда не проявляя при этом полиморфизм биоморф и никогда не производя новых видов, но это — утопия. Сохранять жизнь, не изменяя ее при эволюционном движении вперед через пространство и время, за- нятие в целом бесперспективное, поэтому с первых шагов жизни на Земле в ее эволюционном развитии, кроме как борьбе за существование, а на ее основе, только как естественному отбору наиболее приспособленных, не нашлось альтернативы. Таким образом, Жизнь как явление общекосми- ческое (вряд ли она уникальна только на Земле), за многие миллионы лет своей эволюции на нашей планете, кроме борьбы за свое сохра- нение любыми доступными для нее средствами, ничего другого не нашла и найти никогда не сможет. Дело в том, что эволюция, законы и правила которой «безропотно» приняла Жизнь, однажды появив- шись не просто на Земле, а значительно шире — во Вселенной, явле- ние всюдное (пока мы, люди, еще не совсем разобрались с этими законами и правилами, но это только наша беда, и свое незнание Человек, в этом его особая сила, всегда готов обменять на знание и стремится к нему), и нет области пространства, развивающегося во времени, куда бы не проникла эволюция. Рассуждения о том, что в этой части освоенного Жизнью пространства (допустим, на Земле в популяциях) идет борьба за существование и действует на ее основе естественный отбор, свершая, тем самым, эволюцию, а вот в этой
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ части пространства (внутри организмов) ничего такого нет; с позиций концепции всюдности эволюции, на мой взгляд, надо воспринимать как недоразумение. Проблема с объемом понятия «естественный от- бор» уже давно созревает в эволюционной теории и, пожалуй, настало время ее разрешить. Здесь и сейчас мы делаем шаг в этом направлении. Итак, естественный отбор начинается внутри организмов. Для него всегда присутствует необходимая основа — структурные элементы «про- питаны» стремлением к борьбе и откровенно занимаются этим (другого им не дано) в инфраструктуре организмов, определяя, доступными для них средствами (конкуренцией, кооперацией, индифферентностью), кто в этой жизни (борьбе) победитель, а кто побежденный. Давайте подой- дем (как часто мы по-настоящему далеки от них) к нескольким расте- ниям и присмотримся к их инфраструктурной организации. Нет ника- ких сомнений, живые структурные элементы (гипобиоморфы) борются за жизнь, участвуя в каких-либо «междусобойчиках», нередко переходя- щих, на образном языке, прямо-таки в «мордобойчики» с самыми раз- ными результатами: и с победителями, и с побежденными. Нет сомнений, что инфраструктурная борьба за существование, которую ведут структурные элементы организмов — это реальность (т. 1, гл. 4, 8), признание важности которой откроет новую страницу в развитии эволюционной теории. Поставим вопрос ребром. Кому еще, кроме как самим структурным элементам, нужна такая борьба. Пожа- луй, здесь скрыта та «изюминка», ради которой и затеян весь этот «сыр- бор», та «интрижка», разобравшись в которой, мы находим ключ к пониманию эволюции внутри растительных биоморф и, в первую оче- редь, простых биоморф, или организмов, в отношении которых мы и направим здесь основное познавательное усилие. У инфраструктурной борьбы за существование в организации простых биоморф есть свой непосредственный «хозяин» — организм. Это он, организм, ее разреша- ет, исходя из своих внутренних потребностей, «наблюдает» за ней из- нугри, куда она выведет, чтобы использовать ее, оказав на нее, в нуж- ном месте и в нужное время, свое управляющее воздействие, подчинив ее естественному отбору, сначала по части интраэлекции (координиру- ющего отбора) — первому отбору по пути ориентации структурных элементов к определенной «трудовой» деятельности — «профессии», или организационной специализации. Это он, организм, разрешает структурным элементам вдоволь побороться за жизнь со всем и во всем, что их окружает. Инфраструктурная борьба «бушует» внутри каждого организма и всегда под одним, для всех понятным, девизом: вы, ребята» (структурные элементы), боритесь, как можете, деритесь, миритесь, проявляйте индифферентность, но не «забывайте», что непосредственно командую вашим «парадом» я, организм, и при этом сам, как чей-то структурный элемент, никогда «не забываю», что надо мной всегда кто- то стоит повыше, а следовательно, кто-то еще опосредованно командует через меня и вами. Такова жизнь (об этом надо помнить нам, людям, причем более всего и всегда в отношениях универсальных — биотичес- ких) и все это пронизывает ее, жизнь, от основания до самого верха.
Глава 8. Концепция всюдности эволюции 8.6.1.3. Закон адаптивной ориентации в организации жи- вой природы Итак, из предыдущего материала ясно, что в растительных организ- мах действие естественного отбора (элекции) начинается с его первой части — координирующего отбора (интраэлекции). Кстати, возможен, помимо естественного, и искусственный отбор, когда интраэлекция обеспечивается искусственно (обрезка части побегов, применение дефо- лиантов и т.п.). Может появиться мысль, а не скатывается ли автор к ламаркизму? Смотрите, опять «упражнения», например, обрезка побе- гов, и это уже — эволюция?! Однако, оставим этикетки и ярлыки в покое, поскольку любителей их навешивать всегда больше, чем желаю- щих многое и серьезно переосмысливать. Еще раз договоримся, что эволюцию автор воспринимает как всюдное явление, а разные резуль- таты ее обнаруживаются везде, если соответствующим образом настро- иться их замечать, причем даже тогда, когда изучаешь это явление из- нугри, начиная от его первооснов. Попробуем типизировать и класси- фицировать то, что происходит в эволюции посредством координирую- щего отбора. Действуя на организацию биоморф, интраэлекция как часть есте- ственного отбора направлена на совершенствование координации орга- низации тех же самых биоморф. Таким образом, организация любой биоморфы всегда, будучи уже результатом предшествующей эволюции, так или иначе координирована, благодаря тому, что она постоянно в процессе ее развития в онтогенезе или синонтогенезе подвергается коор- динирующему отбору. Чего же так «настырно» добивается этот тип на- чального (предваряющего что-то еще большее) естественного отбора, действуя везде и всегда, где есть биоморфы. Сразу оговоримся, что ко- ординирующий отбор может быть направлен на координацию организации биоморф — это организационная координация (она распространяется на структурные элементы биоморф и их конституцию в целом) и на координа- цию биоморф в целом к местообитаниям — это биотопическая (экологичес- кая) координация (она распространяется на «привязку» биоморф к опреде- ленным типам биотопов). Сейчас мы ведем речь об организационной координации внугри биоморф семенных растений, а она имеет опреде- ленную ориентацию (направленность) структурных элементов биоморф в развитии организационной специализации, всегда стоящих на службе не только у самих биоморф, но, более того, еще и на службе у видов (не будем забывать о том, что уже было отмечено выше: в организации биоморф, фитобиоморф, в частности, «процветает» эйдократия). Ответим теперь на вопрос: что такое ориентация в организации био- морф. Это определенный результат структурно-функциональной направ- ленности координирующего отбора, который отражается в организации биоморф и используется далее естественным отбором в процессе дальней- шей витаэволюции. Координирующий отбор как-либо нацелен в жизнераз- витии на ориентацию структурных элементов биоморф в выборе и развитии ими специальности (на «профессиональную» ориентацию). Как было показано выше (глава 8.3.1.2.), специализация у биоморф — это определенная адаптация органа, его структурно-функциональная
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ ориентация, которая отвечает условиям обитания и иозволяет биомор- фам, благодаря этому, усиешно занимать свое место иод Солнцем — со- хранять себя, участвовать в жизнесохранении вида и иоддерживать, таким образом, жизнь вообще. Специализация в организации семенных фитобиоморф проявляется по-разному. При сравнении ее результатов среди гомологичных структурных элементов, например, побегов, на- глядно видно, что посредством множественных эволюционных актов, составляющих рост и развитие биоморф как в их биоморфогенезах, так и в исторической ретроспективе, происходит естественный отбор на специализацию как отдельных частей побегов, так и побегов в целом, в результате чего одни из них становятся вегетативными фотофильными (с зелеными срединными листьями), другие модифицируются и разви- вают только чешуевидные листья, третьи приобретают статус репродук- тивных органов и т.д. (рис. 367-398). У одних органов при отборе на специализацию увеличивается число выполняемых ими функций, а у других, напротив, — уменьшается. У некоторых органов в процессе естественного отбора на их специализацию начинает проявляться пред- расположенность к вегетативному размножению, что приводит к воз- никновению в структуре организмов корнепобегов-органов, или онто- идов («полуособей»), которые склонны к полному обособлению от ро- дительского растения после соответствующей партикуляции и сразу же после него — к превращению в новый растительный организм (рис. 82, 93, 266, 269 и др.). Другой пример специализации у биоморф, проявля- ющийся в их структурных элементах, — это способность некоторых организмов в одиночку (!) представлять вид и в критических условиях (каждый такой носитель минимала вида, или специкула (см. стр. 138), путем каких-либо специальных действий (самоопыление, вегетативное размножение и т.п.) готов проявить специализацию до конца и воссоз- дать в одиночку целый вид). Каждое живое тело стремится в эволюции найти свое дело (сиециаль- ность) — адаитивную ориентацию в борьбе за существование (ориентацию на сиециализацию в координирующем отборе и закрепить ее в иодтвержда- юще-утверждающем естественном отборе), без которой ему не удержаться в круговороте жизни, отразив его содержание в соответствующих струк- турах и функциях так, чтобы большинство из иредоставляемых этому телу окружающих возможностей могли стать иолезными ему, но и другому живому телу, непосредственным структурным элементом которого иервое тело является — это закон (специальный) адаптивной ориентации живых структуршах элементов (биомеров) в организации живой природы. Благодаря адаптивной ориентации биосистем-биомеров на опреде- ленную специализацию, растительная жизнь в своем развитии научи- лась преодолевать многие противоречия, которые неизбежно возникают там, где есть движение, а оно есть везде, поскольку кругом только материя и ее существование в различных формах движения, что, безус- ловно, один из важных аргументов сохранения жизни на Земле многие и многие миллионы лет. Наиболее высокой формой адаптивной ориентации в специализа- ции частей соматической организации жизни на Земле, т.е. меротоми-
Глава 8. Концелция всюдности эволюции ческой специализации, было размежевание (расчленение) всей органи- зации биосистем на генетическую часть (геносому), элементами которой стали гены и их отдельные комплексы и фенетическую часть (феносо- му), элементами которой стали фены и их отдельные комплексы. Одной из высших форм адаптивной ориентации в организации жизни на Земле следует рассматривать и возникшую в процессе эволю- ции универсальную систему «управления-исполнения», которая стала нормой жизни всех биоморф, но и других биосистем. Система «управ- ления-исполнения» из пяти функциональных систем: управления, по- требления, производства, резервирования и рассеивания (т. 1, гл. 4)— постоянно подвержена эволюции посредством естественного отбора (в единстве интраэлекции и экстраэлекции). Благодаря опоре на адаптив- ную ориентацию в эволюции растительной жизни была создана четкая система фенетической организации семенных фитобиоморф в образе «могучей пятерки» (нарастания, ветвления, партикуляции, размножения и элиминации), закодированная на генетическом уровне, старательно обеспечивающая им важные достоинства в покорении и освоении про- странства во времени. Таким образом, направленность адаптаций, ко- торая позволяет биоморфам существовать в пространстве-времени целе- сообразно, нашла еще одно свое объективное отражение в законе адап- тивной ориентации. Ориентация в координирующем отборе (адаптивная ориентация) действует как на целый структурный элемент, так и на его живые части, поскольку все эти части одновременно и сами являются структурными элементами более сложных структурных элементов. Способ, которым достигается в организации биоморф специализация органов и их частей, как было показано выше, — это метаморфоз. Посмотрим на адаптивную ориентацию некоторых, допустим, годичных, побегов растений умерен- ного климата (рис. 15, 16, 120, 147, 395). У них обычно есть базальная сближенноузловая зона с чешуевидными листьями, фотофильная зона (зона срединных зеленых листьев), иногда еще и зона соцветия с лис- тьями, по-разному метаморфизированными, когда развитие соцветия является основным делом этой части побега (при развитии межсоцве- тия, или интеркалярного соцветия, листья часто, но далеко не всегда, могут не отличаться от листьев фотофильной зоны). Размышляя над организацией тела каждой растительной биомор- фы, становится понятно, а зачем, более всего, нужна адаптивная ори- ентация. Она действительно ориентирует любую живую часть организа- ции биоморф заниматься своим определенным делом, поскольку; бла- годаря этому свойству биоморф, в их теле не должно быть живых струк- турных элементов (биомеров) «бездельников», и заставляет (ориентиру- ет) их всех «работать» по «профессии», помогая, где это нужно, ее осваивать («подбрасывая» к ним необходимые гормоны, ферменты и т.п. для реализации соответствующей деятельности). Дело, на которое ориентирует координирующий отбор развитие структурных элементов в растительном организме, более всего отвечает пребыванию этой части тела в соответствующем месте пространства и в определенное время, что обеспечивает им, при складывающихся условиях окружающей внешней
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЯ среды, целесообразное участие в жизнеразвитии целостного организма, когда, действуя с ним в унисон, они все совершают обычный для них «ритуал» выживания и жизневыражения, получая от эволюции все то, что здесь и сейчас имеют, т.е. то, что они способны были получить в борьбе за существование как посредством интраэлекции, так и экстра- элекции в исторической ретроспективе и продолжают получать в насто- ящее время одни как «господа» (организмы), а другие как их «слуги» (живые струкгурные элементы-органы, ткани, клетки, органеллы). 8.6.1.4. Почему в эволюции организмов важны не только «победители», но и «побежденные» в инфраструк- турной борьбе Поставив такой вопрос, я невольно вспомнил один известный исторический факт: в 1812 г. великий Наполеон, как показалось ему, разбил армию Кутузова, которая отступила, но не сдалась, и завладел Москвой. Наполеон капитуляции России не дождался, поскольку его «победы» под Бородино на самом деле не было, если и можно говорить о «победе», то только пирровой и, как вскоре показало время, именно она привела к гибели не только французскую армию-агрессора, но и к краху как всей империи, созданной Наполеоном, так и самого дикта- тора. Все это сказано для того, чтобы далее показать, что в эволюции живой природы такая параллель не редкость, а вполне закономерное событие. Весьма любопытно в организации растительной жизни наблю- дать, когда результаты, как может показаться трагические, проигрыша кого-либо в борьбе за существование на одном отрезке времени, живая система, которой принадлежит проигравший как структурный элемент, в конце концов эффективно используются, превращая их в свою победу в большой «игре» под названием «эволюция». Для эволюции живого бывает мало как-либо и кого-либо победить, куда важнее уметь извле- кать пользу не только из побед, но и поражений в долговременной перспективе. 8.6. 1.4. 1. Почему интраэлекция стала в эволюции атрибутом ес- тественного отбора Эволюция есть везде н всегда, но результаты у нее разные. Идет она и внутри организмов (простых биоморф). Ранее (т. 1, гл. 4, 8) было доказано, что существует инфраструктурная борьба за существование со всеми известными атрибутами борьбы между живыми телами: конку- ренцией, взаимопомощью и индифферентностью. Уместно тогда поста- вить вопрос: организм — это просто арена (автобиотоп) = питательная среда для «битвы» между его структурными элементами, прямо-таки «бойцами-гладиаторами», или организм — это нечто большее, чем только арена инфраструктурной борьбы в препопуляциях, коалицшьх и т.п. «аре- ноконституционных» системах? По моему глубокому убеждению, орга- низм, безусловно, не просто арена борьбы за существование его струк- турных элементов, но то, что такое происходит и организм проявляет себя и как «арена» борьбы, доказать несложно (рис. 367-371 и др.). Еще
Глава 8. Концеиция всюдности эволюции важнее другое — это управляемая (!) с генетического уровня организмов «арена» вполне контролируемой борьбы, в которой непосредственно контактирующим и взаимодействующим структурным элементам предо- ставляется немало возможностей (это условие предусмотрено генетичес- кой природой организмов) проявлять от себя некоторую свободу в фе- нетическом развитии в определенном месте пространства и в данный отрезок времени, вполне достаточную, хотя и контролируемую, но все- таки свободу (!) выбирать, из запрограммированных вариантов разви- тия, подходящий вариант для борьбы за жизнь. Иногда ничего из этого не получается, и приходится выходить за традиционные пределы — за норму реакции, сигнализируя об этом своим генам, доводя или не доводя их до изменений (мутаций). Насколько далеко распространяется гене- тический контроль организма на фенетическое поведение структурных элементов в препопуляциях, коалициях и других системах, зависит от многих обстоятельств, поскольку в разных случаях организм как арена инфраструктурной борьбы может больше или меньше влиять на свои структурные элементы. В изучении инфраструктурных отношений между структурными элементами простых биоморф-организмов пока больше вопросов, чем ответов, но все, что будет сказано ниже в этом разделе, можно рассмат- ривать как постановку новой проблемы — теории отбора в эволюции живой природы вообще, решение которой потребует еще немало усилий не только биоморфологов, но и физиологов, генетиков и специалистов других областей биологии. Эволюция представляет себя в живой природе, как было уже показано ранее (т. 1, гл. 3), в трех уровнях (порядках): продолжение жизни, расши- рение разнообразия организационно родственных живых тел (полиморфиз- MR), создание качественно новых видов организации (например, видов биоморф). Интраэлекция (координирующий отбор) настолько важна в эволюции биоморф и их видов, что без нее экстраэлекция (подтверждающе- утверждающий отбор) никогда бы не смогла состояться. Отбирать в популяциях было бы нечего и пришлось бы только повторять то, что уже было однажды сделано (как сделано — это другой вопрос). Эволюция био- морф и их видов по второму и третьему варианту — на полиморфизм и возникновение новых видов без интраэлекции не происходила бы, что можно предположить только логически, поскольку удерживать жизнь в эволюции на основе первого эволюционного варианта — продолжения жизни невоз- можно, в силу известных обстоятельств — отсутствие прогресса в жиз- неразвитии, что само по себе быстро привело бы жизнь на Земле к эли- минации. Эволюция как бы умелой рукой направляется сначала внутри каж- дого организма на поиск в инфраструктурной борьбе за существование подходящего материала для координирующего отбора — отбора внутри организма на ориентацию жизнеразвития по пути оптимизации взаимо- расположения (топографике) структурных элементов и их специализа- ции. Координирующий отбор сам выступает в качестве важнейшего эволюционного фактора и в конце концов может провоцировать орга- низм изнутри на основе только своих «требований» к мутациям его
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ соответствующего генетического материала. Такой поворот исследова- тельской мысли взывает к экспериментальной проверке со стороны генетиков следующей идеи: насколько сильно задействован в эволюцию координирующий отбор, что в определенных условиях он может быть мутагеном — средством, способным вызвать мутацию в организме, т.е. за счет процессов саморазвития самого организма (мутагенами, как правило, являются внешние агенты, которые, часто избыточно, воздей- ствуют на гены). Интраэлекция, в порыве своей деятельности, возмож- но, когда-то достигнув некоторого полезного ориентационного резуль- тата жизнеразвития фенетических структурных элементов организма, подает сигнал его геносоме — «мутируй» (организму), а в популяцию— «утверждай» достигнутые результаты, доведенные даже до мутаций, т.е. завершай экстраэлекцией естественный отбор наиболее приспособлен- ных организмов. Так, в самом общем виде, работает схема эволюции от начала инфраструктурной борьбы на каком-то участке организационно- го пространства до утверждения в нем каких-либо результатов есте- ственного отбора (выбраковка или принятие в арсенал последующего жизнеразвития). Наша задача сейчас — дать набросок вероятностной картины тече- ния координирующего отбора, обеспечив ее соответствующими приме- рами. Чтобы жить, надо уметь бороться всегда и везде, надо при этом проявлять адаптации и, если живое тело не может применить ничего другого, кроме как использовать свою последнюю адаптацию — смерть, освобождая тем самым место для других живых тел, которые в данный момент готовы проявлять иные адаптации, то и такой результат борьбы за существование предусмотрен в эволюции. В процессе эволюции, благодаря координирующему отбору внутри биоморф, сформировались такие типы их организации, которые ориентированы по многим орга- низационным параметрам так, чтобы как-то отодвигать адаптацию- смерть от начала биоморфогенезов. Существует постоянное противоречие между вегетативом и репро- дуктивом, которое внешне в данный момент может себя вроде бы ничем и не проявлять, но это одно из самых главных противоречий в органи- зации биоморф, а шире — всей живой природы, и действует оно безот- казно всегда и везде, поскольку генетически предопределено. Все отно- шения между структурными элементами в организации биоморф скла- дываются через их атрибутивное целостное стремление к размножению, что обеспечивает приоритет носителей репродуктивного над приорите- том носителей вегетативного, предваряя тем самым результаты инфра- структурной борьбы за существование вегетативных и репродуктивных структурных элементов. По большому счету, репродуктив не столько, в прямом смысле, борется с вегетативом, сколько его использует для своего развития, подчиняя тем самым себе развитие вегетатива; однако, все это — есть формы прямой или опосредованной (косвенной) инф- раструктурной борьбы за существование. В конкретном организменном теле репродуктивные структурные элементы развиваются на каком-либо вегетативном элементе и после завершения репродуктивного усилия становятся лишними в этом теле. Вегетативные органы, которые в раз-
Глава 8. Концеиция всюдности эволюции ной мере часто присутствуют в уже использованных репродуктивных структурных частях побегов, могут еще некоторое время продолжать сохранять жизнеспособность (рис. 277-279). Дело в том, что цели существования репродуктива в организации биоморф идут дальше существования конкретного вегетатива, посколь- ку непосредственные результаты репродуктивного усилия, допустим, семена, используются уже не организмом, а значительно шире, обеспе- чивая выживание виду, роду и т.д. В своем стремлении к размножению вегетативные органы борются за существование в окружающем их жиз- ненном пространстве, всегда подчиняясь зримому или незримому дав- лению репродуктива. Наличие противоречий в живых системах, в частности, и биомор- фах, создает условия для борьбы за существование носителей этих про- тиворечий — борьбы структурных элементов, представляющих вегетатив и репродуктив, как между собой, так и другими окружающими их фак- торами. Естественный отбор в простых биоморфах, т.е. организмах, начи- нается не сразу, поскольку, чтобы состояться, всякий отбор должен иметь для себя причину. Прежде чем что-то отбирать вообще, надо стать «свидетелем» того, что есть материал, которым можно «заняться» в плане отбора. Структурные элементы организмов (шире и других биоморф, но и биосистем вообще) по-другому существовать не могут, кроме как бороться за жизнь, потребляя, производя, резервируя или рассеивая ресурсы — и все под единым управлением со стороны их геносомной части. Там, где борьба, там везде есть «победители» и «побежденные» [живые и (или) мертвые], а, следовательно, есть главное — материал из структурных элементов в разной мере «насладившихся» или «истрепан- ных» жизнью, которым организмы могут заняться, отбирая естествен- ным образом (отбор-то естественный), применяя сначала начальную его часть — интраэлекцию, т.е. координирующий естественный отбор. В этом отборе — двойной контроль, поскольку на него действует, как минимум, две биотические власти: с одной стороны, отбирает из своих структурных элементов сам организм, а с другой, над ними всеми стоит еще и власть вида (эйдократия), который, безусловно, имеет свои не- безосновательные претензии на результаты отбора; что поделать, но и здесь борьба двух противоречий, от нее никак и никуда не скрыться. Естественный отбор начинается внутри организмов в процессе инфраструктурной борьбы за существование. Именно к такой борьбе предрасположен координирующий естественный отбор (короче, просто координирующий отбор), поскольку организмам (биоморфам вообще) выгоднее всего иметь организацию, с которой было бы приемлемо су- ществовать в окружающей внешней среде как самим организмам, так, естественно, и их генетическим хозяевам — видам: структурные части их должны быть адекватно ориентированы в определенных частях про- странства и выполнять там и в нужное время обязательные, как для организма, так и для вида, функции. Цель координирующего отбора в инфраструктурной борьбе за су- ществование — ориентация на специализацию некоторых структурных
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ частей организма к выполнению определенных функций. Обязательная способность организмов к координирующему отбору фиксируется на их генетическом уровне. Результатом этого отбора является какая-либо специализация органа целиком и отдельных частей органа на выполне- ние определенных функций, что позволяет организму и виду в границах конкретной телесной системы (организма) решать, каждый по-своему, собственные разномасштабные задачи (рис. 11-31 и многие друтие). В процессе эволюции координирующий отбор участвует в создании количества и качества вегетативных и репродуктивных структур, но и при определении времени и места их развития в организации тела ра- стительных организмов. В исторической ретроспективе инфраструктур- ная борьба за существование постоянно приводила и приводит сейчас к выявлению победителей и побежденных среди структурных элементов организмов. Организм соответствующим образом, через координирую- щий отбор, направляет результаты этой борьбы на создание выгодной для себя ориентации в специализации структурных элементов и их частей. Организм, по сути, «наблюдая» изнутри за тем, что «творят» (в прямом и переносном смысле) структурные элементы в его организа- ции, отмечает полезные и вредные для своего жизнеразвития результа- ты, и как-то на них реагирует. Он снабжает в нужный момент и в нужном месте каких-либо «победителей» или «побежденных» в инфра- структурных «междусобойчиках» теми необходимыми веществами, до- пустим, гормонами, ферментами и т.п., стимулирующими или, наобо- рот, — другими, депрессирующими, веществами или просто водой в большем, чем обычно, количестве, поддерживая, направляя и создавая (ориентируя) специализацию в их развитии, которая более всего устра- ивает организм, но и вид. Способность организмов к координирующему отбору в инфраструк- турной борьбе за существование заложена на генетическом уровне. Однако, организмы, и это также предусмотрено в генотипах, дают воз- можность своим структурным элементам «порезвиться» на «арене» их бытия и испытать хотя бы немного состояние, пусть и небольшой, но все-таки свободы — относительной свободы жизни в выяснении отно- шений как друг с другом, так и средой своего эндоорганизменного существования (автобиотом), а также окружающей внешней средой, став в этих «междусобойчиках» или «победителем», или «побежденным», или ни тем и ни другим, проявив что-то среднее по «количеству» развития или вовсе индифферентность. В инфраструктурной борьбе (количествен- ной) структурных элементов между собой и окружающими их услови- ями сами элементы не решают вопросы ориентации на специализацию их жизнеразвития, поскольку их участь — довольствоваться лишь коли- чественными результатами: кто больше — меньше, сильнее — слабее, толще — тоньше и т.п. (похоже, в этой стороне борьбы все решает, у кого «кулак» покрепче). А вот другая сторона (качественная) инфраструктурной борьбы за существование — это борьба за ориентацию в специализации тех «по- бедителей» и «побежденных», которую структурные элементы исполня- ют, но не сами (их «интересует» только количество), а делают это под
Глава 8. Концеиция всюдности эволюции полным контролем организма, который соответствующими действиями (поддержкой одних и неподдержкой друтих) может сделать сильнорос- лые «побего-победители» «рабами» (сателлитами), а заморенные «побе- ги-коротышки» превратить в феногегемонов (носителей репродуктив- ной функции). В координирующем отборе результаты инфраструктур- ной количественной борьбы за существование, которую организм в не- которой мере допускает вести структурным элементам по системе «как получится, в зависимости от обстоятельств», дав им некоторую, безус- ловно, относительную свободу в поведении в рамках уже сложившейся и генетически закрепленной за ними специализации, переоцениваются. Координирующий отбор, который идет среди структурных элементов под полным контролем организма, ориентирован на создание и отбор или, наоборот, отбор и создание специализации, необходимой организ- му, т.е. лучше той, что была и есть, поэтому организм в каждый удобный для себя момент и в нужном месте уже отвоеванного его телом про- странства решает сам, исполняя принадлежащую ему биотическую власть, но еще и власть вида, которому как структурный элемент (дру- гого уровня, чем эндоорганизменные элементы) служит неукоснитель- но, кому и как из структурных элементов-«победителей» или «побеж- денных» помогать и чем помогать, а кого из них угнетать и чем угнетать. Всякая специализация в структуре органов растительных организ- мов осуществляется посредством метаморфозов, поэтому все органы, будучи специализированными структурными элементами, — это мета- морфизированные части — всегда и все они метаморфы. Так, побеги семенных фитобиоформ — это самые разнообразные органы в плане ориентации их на специализацию в целом и по частям: они метаморфы как целиком (полная специализация), так и по отдельным частям (пар- циальная специализация). 8.6.1.4.2. Подтверждающая и утверждающая формы естествен- ного отбора или, соответственно, — первая и вторая зкс тра злекции Всякий растительный организм (да и не только растительный) участвует в инфраструктурной (эндоорганизменной) количественной борьбе за существование и как арена для нее (автобиотоп), и как хозяин тех его струкгурных элементов, которые живут, чтобы бороться, и бо- рются, чтобы жить, причем не столько для себя, сколько для него— своего хозяина-организма. Пока поведение всех соучастников инфра- структурной борьбы более или менее устраивает организм, то он, на правах хозяина «борцовской» арены и самих «борцов», ничего не пы- тается изменить и действует, согласно сложившейся ранее нормы реак- ции, например, на достаточно освещенных местах кроны растения об- разуются вегетативные побеги-победители (сильнорослые), а в условиях затенения качественно подобные (тоже вегетативные) побеги-побежден- ные (укороченные, совсем «коротышки»), или из нескольких побегов, развивающихся рядом на одной родительской опорной оси, кто-то повыше — сильнее других, а кто-то чуть пониже — несколько короче, еще ниже — заметно слабее, а нижние почки или вообще не пробуж-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИИ даются, или развивают совсем слабые побеги, которые отомрут этим летом или зимой. Может так получиться, что в силу складывающихся обстоятельств организм устраивает то, что происходит в инфраструктур- ной количественной борьбе, и он ничего качественно не изменяет в своих действиях, стремясь сохранить уже сложившийся в длительном историческом развитии порядок ориентации специализации побегов при освоении ими пространства. Для этого в его организации происходит сначала координирующий естественный отбор, который выявляет наиболее выгодные организму формы специализации его органов, не прибегая к мутациям, которые в популяции закрепляются посредством подтверждаю- щего естественного отбора, или первой экстраэлекции, и достигается, та- ким образом, первый результат витаэволюции на основе экстраэлекции: жизнь продолжается. Если результаты координирующего отбора не прохо- дят в популяции подтверждающего отбора, то растительный организм эли- минируется. Такое продолжение жизни без мутаций бывает в эволюции часто, но не всегда. Когда-то в интрапопуляционной борьбе за существование сверша- ется еще и другов форма естественного отбора — вторы вкстраввегаща, или утаервщающиб естественный осбор, посуюдсгвом которого не просто отби- раются организмы для продолжения жизни, а выбираются (утверждаются) те из них с мутациями, состоявшимися в их организации ранее, чтобы продол- жать жизнь в новом качестве, внося в нее расширение полиморфизма или создание нового вида. Те организмы с мутациями, которые не проходят утверждающий естественный отбор, элиминируются. Не будем забывать, что над любым развитием биоморф всегда висит «дамоклов меч» — стремление к размножению, которое является сильнейшим возмутителем спокойствия. Стремление организма азар- тно [доводя иногда личную (вегетативную) жизнь до абсурда и даже суперабсурда, вспомним, например, эфемеров и гиперэфемеров] слу- жить своему хозяину (виду и тех, кто в организационной иерархии его повыше) не знает границ в области совершенствования не столько организации собственно репродуктивных органов, сколько по пути расширения разнообразия форм репродуктивного усилия и его послед- ствий, когда трансформируются количественные и качественные ре- зультаты отношений между наиболее лабильным вегетативом и доволь- но консервативным репродуктивом. Неудовлетворенность организмов развитием своих репродуктивных усилий, пожалщ, может многое из- менить в организации биоморф буквально за малое число поколений (например, неотения). Организм может не устроить то расположение репродуктива и (или) его активность, или что-то еще, связанное чаще всего с репродуктивной функцией, что имеет место быть в текущее время, а складывающаяся в инфраструктурной количественной борьбе ситуация хорошо отвечает этой роли, например, вегетативные «побеги-коротышки», побежденные в такой борьбе, более всего подходят, чтобы именно на них организм разместил свои репродуктивные элементы (рис. 35, 36). Могут, безус- ловно, организму подойти в его устремлениях — «репродуктивных иг- рах» совсем другие побеги, может быть, и «победители» в количествен-
Глава 8. Концеиция всюдности эволюции ных «состязаниях» его структурных элементов (рис. 14-17), но речь сейчас не об этом. Именно организм, вводя в действие, только в известный ему мо- мент, координирующий естественный отбор, что выражается, вероятно, в переброске необходимых вещественных резервов (гормонов, фермен- тов или чего-то еще) туда, где он, организм, совершает здесь и сейчас (!), возможно, весьма масштабный эволюционный прорыв: ориентирует побеги с одной специализацией на развитие побегов с другой специа- лизацией, внося качественные изменения в структуру самих побегов, или только временные параметры развития у них репродуктивных ор- ганов и (или) особенности их отрастания и т.п. что-то еще, но главное, что идет, по сути, у нас на глазах, эволюционная переориентация по- бегов на новую специализацию. И это делает организм, сам устраивая в себе чехарду желанных для себя эволюционных перемен, которые, возможно, и произошли бы как-то иначе за миллионы лет ожидания его множественных потомков какого-либо «удара» мутагеном из внешней окружающей среды, но они происходят здесь, сегодня, рядом с нами, а ведет к ним сам организм, который сам становится для себя же му- тагеном — в своей и для своей же организации. Многое зависит от того, что уже было развито в эволюции в пре- дыдущих поколениях, поэтому линии генетически однородных организ- мов из поколения в поколение, безусловно, уже давно занимаются улуч- шением различных показателей их организации, раскачивая признаки, которые они стремятся изменить. Координирующий отбор постоянно сопровождает жизнеразвитие организмов, чаще всего долгое время по- чти ничего не добиваясь, но проявляя его, организм начинает стимули- ровать развитие тех признаков в побегах-победителях или, напротив, побегах-побежденных, которые отвечают складывающимся условиям, и развитие которых организм готов стремиться поддерживать в данное время. Если появится какой-то результат и, допустим, на «побегах- коротышках» станут появляться репродуктивные элементы и это будет организму полезно, то сам процесс изменения ориентации в специали- зации некоторых типов побегов может сам по себе (!) послужить мута- генным фактором (это положение нуждается в экспериментальной про- верке) и вызвать мутацию, поддерживающую появление на побегах- »коротышках» репродуктивных элементов. При всем происходящем внутри организмов не будем забывать, что организмы всегда находятся не в изолированном пространстве, а в популяциях и в них параллельно с инфраструктурной борьбой за существование действуют еще более жесткие законы выживания в борьбе, но уже для целых организмов,— в интрапопуляционной борьбе. Поначалу эта борьба, так же, как и инфраструктурная, количественная, с выраженной ставкой на силу (опять же, чья конституция «покруче»), поскольку в ней решается задача простого выживания организмов (здесь неважно, есть у тебя какие-то «канделябры» или чего-то такого вовсе нет): кому быть в победителях (и не только выжить, но и еще оказаться сильнее друтих), а кому — в побежденных (погибнуть или кое-как выжить, оставив, если получится, ослабленное потомство). На фоне именно таких «междусобойчиков»,
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ когда в организмах присутствует полезный материал (с мутациями или без них), подготовленный в нем интраэлекцией, вступает в действие экстраэ- лекция первая, или подтверждающий отбор (без мутаций) или вторая, иначе утверждающий естественный отбор (с мутациями) со всеми известными атрибутами такого отбора — отбора наиболее приспособленных в конкурен- ции, кооперации или индифферентности. Мутационные изменения, кото- рие били достисптты ранее посредством ноорлннирующего отбора, поддер- живаются и закрепляются или, напротив, выбраковываются действиями утверждающего отбора. Возникает весьма любопытное предположение, что и сама экстраэлекция, т.е. подтверждающе-утверждающий естествен- ный отбор, также может спровоцировать мутацию, вызвав своими дей- ствиями изменения в генах. Все это, если и происходит, то редко, но вероятность такого исключать не стоит. Полагаю, что мутагенными факторами для организмов могут быть поведение самих отдельных орга- низмов, поведение целых популяций и, наконец, воздействие факторов окружающей внешней среды. Таким образом, эволюция растительных биоморф не пропускает всех количественных «разборок» как в инфраструктурной, так и интрапопуляци- онной борьбе за существование, в которые включает, в зависимости от обстоятельств, действие интраэлекции и экстраэлекцш — обязательных частей естественного отбора, добиваясь качественного улучшения в орга- низации биоморф. 8.6.2. Эволюция биоморф в их конституциальной борьбе за существование Биоморфам семенных растений, начиная с древних времен, когда они только появились на Земле, т.е. палеобиоморфам семенных расте- ний, не пришлось самим изобретать качественно что-либо новое в стра- тегическом отношении, поскольку за них уже постарались другие поко- ления еще более далеких предковых фитобиоморф. Значительно раньше появления первых семенных растений возник в эволюции у раститель- ных палеобиоморф принципиально единый для них механизм «управ- ления-исполнения». «Искусство» управления — это главное в стратегии любых биоморф, которое исходит с генетического уровня их организа- ции. Стратегия у биоморф всех растений и животных в принципе едина — это стремление к жизнесохранению вообще (т. 1, гл. 3), которая проявляется как система управления у них (шире — и друтих живых тел) всеми стратегическими долями (частями-стратегиями): стратегией по- требления, стратегией производства, стратегией резервирования и стра- тегией рассеивания. На фоне единой и в принципиальном отношении стабильной стра- тегии, эволюция биоморф более всего затрагивает их тактику, действие которой распространяется на фенетическую организацию (феносому, или конституцию) биоморф. Самый первый тактический атрибут, что
Глава 8. Концеиция всюдности эволюции получили семенные растения от своих предков — это их способность, на основе определенной стратегии, проявлять соответствующую плас- тичность в любой тактической деятельности. В процессе эволюции биоморфы научились быть простыми (орга- низмами) и сложными (синорганизмами). Чтобы несколько упростить процедуру рассмотрения конституциальной борьбы за существование у биоморф семенных растений, мы большинство примеров ограничим организмами, имеющими физиологически единую (непрерывную) кон- ституцию, что однако ни в коей мере не означает, что дизъюнктивная (прерывистая) конституция синорганизмов не участвует в такой борьбе. Материал интраэлекции (координирующего естественного отбора) аккумулируется по частям во всей целостной фенетической организации биоморф, их феносоме, или конституции (телосложении), что позволяет ему, как по частям, так и в целом феносомой, участвовать во второй части естественного отбора — экстраэлекции, поскольку биоморфы постоянно «погружены» в окружающую внешнюю среду в популяции, где как бы постоянно выставляются на «аукцион» результаты интраэлек- ции, которые сопровождаются или не сопровождаются мутациями. Вполне естественное участие любой растительной биоморфы в популя- ции означает для нее быть, таким образом, вовлеченной в интрапопу- ляционную борьбу биоморф за существование, частью которой всегда является конституциальная борьба, имеющая или не имеющая мутаци- онные последствия, на фоне которых свершается экстраэлекция- первая, или подтверждающий (без мутаций), и вторая, или утверждаю- щий (с мутациями), естественный отбор. В этой части естественного отбора или подтверждается (первая экстраэлекция) то, что было достиг- нуто ранее в фенетической организации биоморф и не сопровождалось мутациями, или утверждается (вторая экстраэлекция) несколько другое — то, что изменило частично или полностью феносому и привело к мутации, при этом часть отбираемого интраэлекцией материала выбра- ковывается экстраэлекцией и элиминируется. Конституциальная («конституциональная») борьба за существова- ние (термин Plate, 1913) у биоморф в популяциях — это разнообразие ответных реакций всего организма или генетически однородной систе- мы их в синорганизме на воздействия окружающих внешних условий, проявляющееся у биоморф через всю их фенетическую организацию (феносому, или конституцию). Конституциальная борьба, которую ве- дет каждый растительный организм через свою наиболее лабильную часть — феносому, происходит под контролем генетической организа- ции и многие соматические трансформации развертываются в соответ- ствии с нормой реакции генотипа до тех пор, пока не свершаются мутации, вносящие большие или небольшие изменения в наследствен- ную природу действительных участников-победителей этой борьбы. Как видно из предыдущего материала, во всех аспектах борьбы за существо- вание действует естественный отбор в различных его формах. Консти- туциальная борьба, будучи частью интрапопуляционной борьбы, а шире — всей борьбы за существование, обеспечивает необходимые условия
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ для эволюции, когда свершается один или несколько «знаковых» резуль- татов витаэволюции: продолжается жизнь, расширяется полиморфизм, возникают новые виды (т. 1, гл. 8). 8.6.2.1. Передовая линия «фронта> в конституциальной борьбе за существование То, чзо конституциааьная борьба идет через конституцию биоморф, их фенетическую организацию (феносому) — это направление самого общего «удара» окружающей внешней среды в этой борьбе на организмы (а шире — и синорганизмы). Однако, в феносоме каждой биоморфы есть части поменьше, через которые заметнее или более всего действует кон- ституциавьная борьба — зто габитус в его формах (габитоформах) и формы роста, или стадийные тела (стадиосомы) биоморф; они, в первую очередь, фиксируют и отражают трансформационный эффект, который достигается в фенетической организации посредством рассматриваемой борьбы. Конституциальная борьба биоморф — это постоянное испытание их пластичности на прочность, результаты которого находят максимальное отражение в формах габитуса и формах роста биоморф, поскольку эти части биоморф, в силу их особого положения в пространстве и времени, такая борьба обойти не может. Габитус (внешний «покров», прикрывающий «наготу» внутренней фенетической организации — эндофеносомы, или субгабитуса) и фор- мы роста, или стадиосомы (носители возрастных и фенологических ста- дий) фитобиоморф имеют фенетическую природу, а, следовательно, в силу своего особого отношения к пластичности, более всего проявляют признаки различных трансформаций. Габитус может существовать толь- ко посредством постоянного подпитывания его ростовой энергией от внутренней организации биоморф. У габитуса нет своей собственной способности к росту и развитию, и он получает все для себя, в готовом виде от субгабитуса, проявляя, таким образом, не прямой, а опосредо- ванный рост и развитие. Учитывая особое положение габитуса в орга- низации биоморф, можно с уверенностью утверждать, что эта важная часть организации биоморф не более и не менее — функция эндофе- носомы (субгабитуса). Как было показано ранее (глава 3) лабильна вся фенетическая организация биоморф, а габитус лишь более других частей склонен отражать результаты целостной фенетической лабильности. И все же габитус, будучи и процессом, и результатом функционирования субгабитуса, — не «альфонс», поскольку без него субгабитус сам не справится с воздействием окружающей внешней среды. Только он, га- битус, в согласии с природой организации биоморф, способен так ес- тественно прикрыть собой внутреннюю фенетическую организацию, не претендуя на многое, ведь у него даже нет собственной внутренней структуры. Однако эндосома дорожит единством с габитусом, как соб- ственной жизнью, и разделить эти две, объективно неразделимые части в фенетической организации можно лишь логическими средствами, что мы и делаем постоянно там, где этого требуют обстоятельства. Положение стадиосом (форм роста), как, кстати, и габитусов в обоих случаях — это особенные функции фенетической организации
Глава 8. Концеиция всюдности эволюции биоморф, проявляющие себя по-разному. Стадиосомы [формы роста, или возрастные и (или) фенологические состояния] — это формы ста- дийного (сменяемого) поведения (гетерокинеза) биоморф своей фенети- ческой организацией (конституцией), как частью пространства, занима- емого на отдельных (!) отрезках времени (стадиях) сменяемыми призна- ками ее возрастности и (или) фенологического (посезонного) развития. Стадиосомы, как и стадии биоморфогенезов, сменяются неоднократно на протяжении цикла существования биоморфы. Формы габитуса и формы роста — это те средства биоморф, ори- ентируясь на лабильность которых, как передовые рубежи своей орга- низации во взаимоотношениях с окружающей средой, биоморфы бо- рются на основе пластичности и гиперпластичности всей своей консти- туцией за выживание в окружающем их остальном мире. Когда органи- зация биоморф вступает в противоречия с условиями обитания био- морф, то линия «фронта» между ними в конституциальной борьбе за существование проходит через габитус и формы роста биоморф, кото- рые, в первую очередь, как-либо отвечают на изменения, происходящие в окружающем внешнем жизненном пространстве. Многое в конститу- циальной борьбе проходит через габитус биоморф, в силу особой при- роды этого структурного образования; кроме того, признаки габитуса часто являются индикаторами при выделении стадиосом. Приведем несколько примеров, где конституциальная борьба за существование биоморф приводит их к разнообразным средствам жиз- несохранения через адаптации в формах габитуса и формах роста (рис. 26, 32, 40-50, 67 и др.). 8.6.2.2. Как и почему приоритет сохранения другой и более сложной жизни стал нормой существова- ния семенных фитобиоморф, а шире — всех био- систем Все в живой природе на Земле происходит, по большому счету, так, как и должно было бы происходить, поскольку в процессе своей эво- люции она овладела принципом иерархической организации. Нет жи- вых тел в биостробе Земли, которые не являлись бы по своей органи- зации структурными элементами чьих-то, более сложных, чем они, тел. Любое живое тело, будучи частью организации другого, более сложного живого тела, может существовать только целесообразно — участвовать в сохранении жизни вообще, причем как собственной (личной), но, более всего, в сохранении другой, или коллегиальной, жизни, к которой это тело как живой структурный элемент принадлежит. Существует не просто система иерархической организации Жизни во Вселенной (т. 1, гл. 2), но и целесообразная иерархическая система сохранения жизни вообще, которую, исходя из общей теории жизнесохранения (т. 1, гл. 3), можно выразить в следующей графической модели — «звезде» (образ- ной) жизнеразвития, т.е. в модели ориентации развития живых тел (как чьих-то структурных элементов) — систем «управления-исполнения» по основным направлениям жизне сохранительной специализации (рис. 400). Даже самое сложное на Земле живое тело, ее биостроба, ведет
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ или должно себя вести (с резервом, с потенцией, поскольку жить так куда надежнее, чем «сжигать» за собой все) как структурный элемент (!) чьей-то еще более сложной Жизни, о существовании которой мы, люди, пока только догадываемся. Рис. 400'. «Звезда» жизнеразвития (система ориентации биотической власти в общем жизнесохранении и в иерархии живых тел). 1 — направления действия экобиофилии (средо- и жизнесохранения), 2 — направления действия экобиофобии (средо- и жизнеразрушения), 3 — направление действия макроби- ократии, 4 — направление действия гипобиократии, 5 — автобиократия на своем уровне действия, 6 — 17 — уровни организации живой материи, 6 — биополимеры, 7 — органел- лы, 8 — клетки, 9 — ткани, 10 — органы, 11 — свободные проорганизмы (пыльцевые зерна, гаметы и другие гипобиоморфы, отделившиеся от родительского организма), 12 — орга- низмы, 13 — синорганизмы, 14 — популяции, включая ценотические популяции (цено- демы), 15 — эйдобионты (виды организмов), 16 — биоценозы, 17 — биостроба (живая природа Земли), 18 — эндобиотическая неживая природа (неживая материя, ассоцииро- ванная в организацию живых тел), 19 — экзобиотическая неживая природа (неживая материя, которая не входит в организацию живых тел).
Глава 8. Концеиция всюдности эволюции Живые тела в процессе эволюции научились быть пластичными и гиперпластичными, поскольку все их существование в Природе (Все- ленной) — это всегда только адаптации (приспособления). Всякое жи- вое тело существует в борьбе, в ней и к ней адаптируясь, используя проверенное во времени средство выживания — компромиссы (!), сни- мая, тем самым, все известные или еще не известные нам, людям, противоречия. Чтобы владеть приемами борьбы за существование и использовать компромиссы как средство разрешения противоречий, живая природа в процессе эволюции создала систему «управления-исполнения» как си- стему реализации определенного порядка в организации живых тел, а также систему потенциально бесконечного сохранения определенного жизнепорядка во Вселенной, что оказалось возможно на основе дей- ствия законов и правил биотической власти, осознание которых еще только начинается. Проблема биотической власти, пожалуй, значитель- но глубже, чем мы рассмотрели ее в главе 7, но это и неудивительно, поскольку эту «залежь» еще никто не возделывал, и, похоже, пришло время начинать ее шаг за шагом активно поднимать. Здесь и сейчас мы проследим полет исследовательских «стрел» не в разные стороны про- явления биотической власти, а только в одном направлении, указанном в названии рассматриваемого подраздела. Однажды встав на путь иерархии, Жизнь во Вселенной освоила для себя в качестве нормы принцип биотической гетерократии (глава 7). Наличие иерархии в организации жизни на Земле предполагает участие таких структурных элементов, которые создавали бы и поддерживали определенный порядок в своих организациях и в организации всей биостробы (живого тела Земли), позволяющей явлению Жизнь быть в Природе долго, вплоть до бесконечности во времени. Все живые тела, составляющие биостробу, построены по принципу «управления-испол- нения», который предполагает обязательное участие в их организации биотической власти. Ранее было показано, что основными носителями биотической власти в организации биоморф являются гены. Это они, гены, будучи высшими «властоносцами» биотической организации, управляют развитием и сохранением феносомной части (конституции) биосистем, которая нужна им не как попало, а в строгом порядке- атрибутивно, поскольку непосредственно и всегда служит для них обя- зательным (другого не бывает) естественным обиталищем (живым «до- мом») — автобиотопом (подробнее см. главу 9). Конституциальная борьба за существование — это важный фраг- мент эволюционного процесса — прямой результат той инфраструктур- ной борьбы, которая происходит в каждом организме, — это уже цело- стный ответ биоморф на ту окружающую внешнюю среду, в которой они вынуждены существовать. Биоморфы адаптируют свою конститу- цию (феносому) — фенетическую часть своей организации к тем вне- шним условиям, которые их в данном месте и в данное время окружают, изменяя при этом внутреннюю организацию (субгабитус) и с ней вне- шнюю организацию (габитус). Пластичность биоморф наиболее ярко проявляет себя через конституцию, которая целиком весьма лабильна
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ (субгабитус и габитус вместе), причем не только и даже не столько габитус, поскольку его создает и им пользуется, в первую очередь, суб- габитус (габитус сам активность проявлять не способен, поскольку это удел субгабитуса, который его буквально «отращивает» на себе и разви- вает, подстраиваясь под свои, а не его, требования удобства), что по- зволяет габитусу выполнять основное свое дело — прикрывать «наготу» субгабитуса, т.е. быть почти как «футляр» для ценного инструмента, потому «почти» (и только), что у габитуса нет своей внутренней струк- туры. Габитус как функция внутренней организации биоморф, является индикатором той реакции, которую проявляет внутренняя фенетическая организация (субгабитус) на внешнюю среду через трансформационные процессы, захватывающие всю феносому (конституцию), а, следователь- но, также и габитус. У биоморфологов сложилось иллюзорное воспри- ятие трансформаций габитуса в организмах. Да, они идут в биоморфах, но габитус всегда трансформируется только вместе с субгабитусом, и приоритет при этом принадлежит именно трансформациям субгабитуса (габитус собой их отражает, ведь он сверху, но сам для их свершения ничего сделать не может). Конституция (феносома), будучи частью в организации биоморф, постоянно адаптируясь к внешней среде, в качестве важнейшего «тони- зирующего» фактора к своим адаптациям использует исходящую с ге- нетического уровня — высшей биотической власти, идиоэнергию стрем- ления к размножению. Репродуктивный импульс находится под властью вида (эйдократия), что принуждает биоморфы как структурные элемен- ты вида выполнять его целевые программы жизнеразвития, которые подчиняют в биоморфах их частные устремления по сохранению личной жизни (автобиофилию) устремлениям к сохранению другой (коллеги- альной), в первую очередь, своего вида, жизни (ксенобиофилии). При- оритетом в конституциальной борьбе за существование биоморф явля- ются адаптации их фенетической организации в направлении оптими- зации условий своего выживания и выживания вышестоящих над ним живых систем — видов и надвидов. Приоритетное стремление к сохранению другой жизни проявляется в любом живом теле, по большому счету, прямо-таки под мушкетерским девизом: «один за всех и все за одного», а основано оно на иерархич- ности организации жизни вообще, порядок в которой исходит от выс- шей биотической власти (генов и их систем — геносом), подчиняющей и управляющей исполнительскую фенетическую деятельность, в кото- рой осуществляется максимум пластичности во взаимоотношениях живых тел с окружающей внешней средой. З.б.2.3. Почему альтруизм не стал нормой жизни семен- ных фитобиоморф и почему они выбрали в эволю- ции ориентацию на коипроииссы Биоморфы более всего проявляют пластичность в своей фенетичес- кой организации при стремлении размножиться — оставить потомство, когда они выполняют тем самым основной и обязательный долг перед видом, которому принадлежат. Биоморфы всегда стремятся наиболее
Глава 8. Концеиция всюдности эволюции выгодно распорядиться своими главными функциями — развитием реп- родуктивных органов и осуществлением репродуктивного усилия в про- странстве и времени. Нередко внешние условия окружающей среды мешают этому (наводнение, похолодание, песчаная буря и т.п.), и тогда биоморфы используют свой арсенал пластичности, который более всего проявляется в трансформациях их фенетической организации, чтобы любой доступной биоморфам ценой выполнить то, что им более всего важно в сохранении жизни вообще — оставить потомство, а когда это не получается, то хотя бы (!) выжить как-либо самим (рис. бб). Вся жизнь биоморф превращается в конечном счете в «игру» — игру в вегетатив и репродуктив, или игру вегетатива с репродуктивом. Плас- тичность приводит к трансформациям фенетической организации био- морф — к изменениям и сменам их габитуальных и стадийных форм, которые сегодня есть одни, а завтра их уже может и не быть, — на их месте будут расти, развиваться и вновь сменяться другие, и все это ради одного, чтобы жила биоморфа в целом и успела выполнить свой долг в жизнесохранении вообще. Экспрессивное стремление к размножению, почти безмерное, в порыве услужить виду, как у некоторых примитивных бактериальных форм, давно освоенных витаэволюцией и по сей день не утраченных (см. главу 2), оказалось для семенных растений далеко не единственным шагом в покорении и освоении пространства и времени. Биоморфы семенных растений научились адаптироваться по-разному и путем ес- тественного отбора в эволюции совершенствовать адаптации. Они на- учились жить не только в состояниях затяжного «экспрессивного аль- труизма», в определенном отношении удобного для вида, но способного доводить личную жизнь почти до абсурда, например, эфемеры живут всего не более одного месяца (рис. 217, 218). Семенные растения более всего освоили в эволюции совсем другие пути — пути компромиссов между вегетативом и репродуктивом, что по- зволило успешно сохраняться видам, а биоморфам заметно ошутить счастье (без преувеличения) личной жизни (его вполне можно количественно изме- рять — продолжительностью личной жизни как организмов, так и синор- ганизмов). Побольше пожить — вот главное счастье личной жизни, которое могут испытать или не испытать конкретные растительные биоморфы. Личная жизнь биоморф семенных растений только у эфемеров на грани с абсурдом, по нашим меркам, но большинство биоморф не пошло по такому пути жизнеразвития, проявив в эволюции способности искать и находить компромиссы между вегетативом и репродуктивом в их сома- тической организации (рис. 59-65, 80-120 и др.). В конституциальной борьбе за существование феносома биоморф активно ищет качественные, количественные и топографические реше- ния задач репродуктивного развития на основе совершенствования от- ношений между вегетативом и репродуктивом. Власть вида (эйдократия) заставляет его организмы бороться за многое и, в частности, за удобное место в пространстве для его репродуктивных органов. Вегетативное тело растительных биоморф под натиском стремления организма к репродуктивному размножению способно сильно трансформироваться
OCHOBbl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ как в зависимости от возрастных изменений, так и под влиянием внеш- ней окружающей среды. Стремление размножаться проявляется даже тогда, когда условия для осуществления репродуктивного усилия не- удовлетворительные; все это побуждает растительные организмы искать и находить в борьбе за существование пути к вегетативному размноже- нию (рис. 121). Уместно поставить вопрос: почему стремление к сохранению вида не дошло у семенных растений до абсурда, и альтруизм в различных его проявлениях не стал нормой жизни у биоморф. Причина всему в том, что стремление услужить виду через сокращение личной жизни расти- тельных организмов не безгранично и значительно выше по своему месту в эволюции, чем альтруизм и стремление к нему, стоят компро- миссы и стремления к ним, в особенности компромиссы между вегета- тивом и репродуктивом. От абсурда жизнеразвития при стремлении к альтруизму есть надеж- ная альтернатива в эволюции жизни, которой уверенно воспользовались на своем историческом пути семенные растения, — торжество счастья личной жизни растительных организмов и синорганизмов в их стремлении к ком- промиссам (!) как атрибутивному средству разрешения противоречий, в первую очередь, между вегетативом и репродуктивом. Компромиссы как норма жизни проявляются не только в отноше- ниях генетически однородных или близких живых тел — структурных элементов, меньших в больших, но и среди генетически разнородных биоморф. Приведу наглядный, но несколько отвлеченный от ботаники пример: волк быстрее настигает ягненка, чем взрослую овцу. Хищник забрал у вида ягненка, но взрослая овца жива и должна постараться вернуть виду утраченное. Здесь не альтруизм, а нечто большее — это формы компромиссов. Так живая природа компромиссно распределяет своих носителей в пространстве и времени (и хищники сыты, и их жертвы в целом сохраняются). Путь приоритета компромиссов между вегетативом и репродуктивом- это единственный путь, который способен уводить растительные биоморфы от абсурда «затяжного альтруизма», в котором они как живые существа (но не виды) лишены ощутимого для себя счастья, мерой которого, как я пола- гаю, может быть, в первую очередь, продолжительность их личной жизни. 8.7. Как, почему и куда идут реальная и гипотетическая эволюции биоморф семенных растений, их видов и надвидов Условия обитания в каждом конкретном месте постоянно выстав- ляют его живым обитателям свои требования: «хочешь жить, умей вер- теться», то бишь — приспосабливаться (адаптироваться). Для всей жи- вой природы эта довольно ехидная поговорка обывателей человеческого сообщества воспринимается ни много и ни мало как великая и един- ственная азбука (!) эволюционного жизнеразвития. Дело в том, что у
Глава 8. Концеиция всюдности эволюции живых тел в их существовании нет иной альтернативы, кроме как «вер- теться» (бороться) — приспосабливаться. Не можешь совсем «вертеться» в жизнеразвитии — уходи «за порог», можешь, но плохо — влачи жалкое существование, а если очень хорошо можешь, то опять же соблюдай меру и «не захлебнись» от счастья, поскольку несчастье где-то всегда рядом (см. главу 10). Эволюция предпочитает не ортодоксов, да и па- радоксы ей не так уж и нужны, она более всего, не исключая, конечно, и остального, принимает компромиссы (!) — это ее генеральное кредо. У живой природы свои законы и правила, поэтому она все частности, которые ей навязывает кто-то по отдельности, быстро приводит к еди- ному знаменателю. Перед нами несколько раньше (глава 6) прошла универсальная мо- дель эволюции, которую удалось выразить в образном рисунке внешне только одного растительного тела. Из этой модели во многом ясно, как, почему и куда шла раньше, идет сейчас и будет идти в будущем наиболее общая соматическая эволюция растительных биоморф по отношению к уровню поверхности почва-грунта (виталинии). В бесконечном мирозда- нии эволюция свободна выбирать направление развития тел и нетел (эво- люционных маятников), пользуясь законами и правилами бесконечного саморазвития материи. Я не склонен обращаться к традиционному стро- ительству морфогенетических однонаправленных рядов, поскольку в принципе не признаю универсальности эволюционного «стрелодвиже- ния» только в единственном генеральном направлении. Эволюционных указателей одного направления лишь какое-то время может быть больше, чем друтих направлений, но в целом, и это важнее, всегда обычны самые разные направления, в зависимости от тех условий, которые реально складываются в данном месте и в данное время. Будь я ортодоксальным флорогенетиком, то без устали строил бы морфогенетические ряды о гипотетическом жизнеразвитии надвидов (родовых, семейственных и т.д.) как «дороги» с односторонним движе- нием, благо фактического материала по биоморфологии видов уже предостаточно. Когда-то (много лет назад, еще юному аспиранту) мне представилась возможность начать весь путь быстрого восхождения по служебным лестницам вверх, приняв для этого господствующую тогда доктрину односторонней эволюции габитуальных типов («жизненных форм») покрытосеменных растений, от тропических деревьев ко всему остальному, вплоть до однолетников и эфемеров, но, потратив немало времени на ее осознание, я понял однозначно, что это дорога не для меня, и по сути она — «ловушка», поэтому нужно искать новые пути, пусть и испытав на себе в полной мере все неудобства вытекающих из этого многочисленных последствий. Природа вообще и, в частности, растительная природа, хотя и по-ньютоновски где-то проста, но уж совсем не примитивна в разрешении своих организационных проблем. Одна из ее высших функций — эволюция, которая совсем не склонна торопиться и загонять себя в одно прокрустово ложе, а, напротив, она обладает великим достоянием — свободой в «плавании» по вселенскому потоку и свободой в раскачивании любых эволюционных маятников (тел и нетел), причем не в хаосе, а в организационном порядке, который
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ она поддерживает сообразно определенным законам, действующим со- ответствующим образом как вообще, так и в зависимости от состояний данных мест и данного времени, причем вся эволюционная деятель- ность направлена, в первую очередь, ради поддержания самого главно- го, что есть в Природе, — ее организационного порядка. Мой тезис в восприятии эволюции растительных биоморф таков: в ней господствует разнонаправленность, а в развитии соматической орга- низации более всего используются компромиссы, поэтому жесткой одно- направленности в эволюции биоморф никогда не было и не будет, поскольку всему «голова» — пластичность, а когда ее не хватает, то помогает гиперпластичность. Эволюция не терпит «тупиц» среди эволюционных маятников, т.е. даже тех из них, которые «дораскачивались» в своем свободном «плавании» до мнимых «тупиков», а реально действительно таковых вообще никогда не бывает. Если встречаются слабые звенья у эволюции по пути, то она их просто-напросто «стирает», трансформи- руя в другие эволюционные маятники, таким образом очищая дорогу более сильным звеньям, и так бесконечно. В отношении живых тел эволюционный «ветер» очищает дорогу тем, кто более других склонен к борьбе — к способности «вертеться», т.е. адаптироваться. Природа, существуя в единстве своего пространства и времени, созда- ла только из своей (другой не бывает) материи для них, а, следовательно, и для себя только один эволюционный принцип движения материи как тел и нетел — это принцип бесконечного колебательного движения свободно «плывущих» во вселенском потоке эволюционных маятников (как образов тел и нетел) по универсальной схеме «так-сяк*туда-сюда», бесконечность действия которого оказалась возможна только в едином организационном порядке Природы-системы как одной системы постоянно функционирую- щего тотального движителя, или примагенератора, и времени (глава 1). Для правильного восприятия принципа движения эволюционных маятников важно помнить, что они всегда «плывут» во вселенском потоке (При- роде), а следовательно, изменяются не только во времени, но и смеща- ются в пространстве. Строгого возврата назад в прежнее свое состояние и на прежнее место в пространстве у основного содержания — собствен- но маятникового «тела» никогда не происходит и колебательное суще- ствования материи, хотя и идет в телах и нетелах «туда-сюда», но все- таки всегда «так-сяк», т.е. по-разному в рамках их норм пластогенеза, а когда таковых не хватает, то и далее (ведь это эволюция, для которой непреодолимых преград не бывает — она везде проходит свободно как нож сквозь масло). Именно присутствие в Природе механизма тоталь- ного движителя (глава 1) позволяет материи, организованной по-разно- му в тела и нетела, бесконечно существовать не в хаосе, а только с эволюцией, всегда нацеленной на поддержание единого организацион- ного порядка, непременно преодолевая всякое подобие «тупиков» дви- жением обновленных эволюционных маятников, что означает в отноше- нии эволюции живой природы следующее: живое продолжение получает тот, кто более других склонен адаптироваться. Существование любых тел в образах эволюционных маятников обеспечивается треугольниками взаимодействующих сил, назовем их для
Глава 8. Концеиция всюдности эволюции удобства «путеводными треугольниками», или «тридинами» [от греческого tri — три, dina (mis) — сила], которые, будучи элементами примагене- ратора (тотального движителя), обеспечивают Природе здесь и сейчас непосредственное развитие именно этих тел с учетом их норм пласто- генеза (у организмов традиционно — норм реакции). Что же конкретнее иредставляют из себя иутеводные треугольники (тридины)? Их стороны следующие. Во-первых, это силы окружающей внешней среды, или средоокружающие силы (у какой-нибудь биосистемы — это воздейству- ющий на нее биотоп, или силы биотопического тела), во-вторых, силы конституции средоокруженного тела, которая непосредственно на себя, всегда первая, принимает любые воздействия внешней среды (у биоси- стем — биотопические воздействия), и, наконец, третье — силы эгосуб- страта тел (их идиоэнергия), которые соответствующим образом отвеча- ют на информацию, поступающую к этой части в организации тел о тех контактных процессах, которые происходят между окружающей средой и конституцией средоокруженного тела. Направленность в развитии тел, таким образом, формируется как результат контактов в треугольнике вза- имодействующих сил и отражается через конституцию этих тел, но более всего внешне — через габитус, его формы. Конституция любых тел — самая лабильная часть этих тел, а раз так, то есть смысл особым образом разыграть эту «карту». Я склонен утверждать, что именно в конституциях любых тел, «лихо» изменяю- щихся своими признаками в рамках определенных норм пластогенеза этих тел, всегда присутствует образный (абстрактный) указатель не толь- ко настоящих, но и возможных будущих, довольно неустойчивых на- правлений развития, если короче, то — «флюгер развития» [его образ надо сначала захотеть увидеть и только потом важно суметь мысленно его представить, чтобы далее, когда придется, и там, где это нужно, материализовать таковой в каких-либо моделях, например, графически он отражен на рис. 406, где чем-то напоминает действительный прибор- флюгер, показывающий направление ветра]. Флюгер развития — это абстракция — образ разнонаправленности, чаще всего мультивариантности развития тел, символическое обозначение которого — разномасштабная звездочка, а при ней один из лучей-указателей направления развития— мощнее других, поскольку отражает собой то, развитое в данном теле, конкретное состояние конституции, отвечающее данному месту и данному времени. Уместен вопрос: сколько же флюгеров развития участвует не в фор- мировании (для этого используется весь треугольник взаимодей- ствующих сил) не только возможных, но и конкретных направлений развития какого-либо средоокруженного тела, а только в маркировании (обозначении) таких направлений? Их всегда два, рассмотрение которых в нашем случае лучше всего провести на примере живого тела, допус- тим, биоморфы семенных растений, существующей только во взаимо- действии с окружающей внешней средой. Один флюгер развития у любого средоокруженного тела — внешний— и располагается он в конституции средоокружающего тела-участка ок- ружающей внешней среды, или, для рассматриваемого случая, биотопа,
OCHOBbl БИОМОРФОЛОГИИ CEMEHHblX РА СТЕНИИ который формирует господствующее направление своего средоокружа- ющего воздействия на средоокруженное тело, подталкивая его развитие в определенный лабиринт реакций на внешние условия, ограничивая, тем самым, свободу выбора вероятностных направлений развития тако- го тела, в частности, биоморфы. Другой флюгер развития — внутренний, — он принадлежит собственно средоокруженному телу и маркирует свое, пусть образное, присутствие в нем через конституцию этого тела, ука- зывая через ее состояния целесообразные направления развития мате- рии в данном теле, в данном месте и в данное время, а если условия существенно не изменятся, то и на некоторое будущее время. Таким образом, в организации любого тела присутствует тридина (путеводный треугольник) с двумя флюгерами развития, которые вместе составляют обязательное звено в направленном соматическом существовании эволюци- онных маятников так, а не иначе. Итак, эволюция, восходящая в своем поступательном движении от тотального движителя Природы, ее примагенератора, постоянно раска- чивает (определенным образом) в бесконечности и пространства, и времени неисчислимое разнообразие эволюционных маятников — тел и нетел (всегда по универсальной схеме «так-сяк«туда-сюда»), проводя при этом организационные «разборки» для поддержания общеприрод- ного порядка в разнообразии явлений, используя для этого все свои средства. Эволюция, если бы она вела себя только однонаправленно и прямолинейно, то не смогла бы надежно поддержать обязательный обще- природный организационный порядок, поэтому она в своей бесконечной и непростой работе более всего ориентирована совсем на иной стиль («по- черк»): она «плетет» организационно-соматические «кружева», когда и прямолинейные, и криволинейные, и любые другие комбинации движений- трансформаций — это всегда только частности, переплетающиеся в единую и всеобщую картину «кружевной» эволюции Природы. В отличие от пря- молинейного развития согласно генетической программе в морфогене- зах организмов и синорганизмов, преемственно-историческое развитие в рамках видов и надвидов искривляется, ломается, поворачиваясь вспять и т.д., т.е. в целом в живой природе господствует именно «кру- жевная» эволюция, и это реальность, с которой нельзя не считаться. Свойство явлений Природы выражать своим содержанием состоя- ния движущейся (изменяющейся) материи в каких-либо телах и (или) нетелах в данном месте и в данное время и быть тем самым эволюци- онными состояниями, или эволюстатами, — это эволюстатность. Соеди- нение вместе разнообразных munos организационно родственных эволюста- тов в отдельные абстрактные сравнительные эволюстатные ряды (морфо- генетические ряды при изучении эволюции биоморф), отражающие в каких- либо или обоих (противоположных) одновременно направлениях изменение их организации — это метод сравнительного исследования эволюции тел и нетел, в частности, морфогенетический метод в биоморфологических ис- следованиях. Эволюстатность проявляется как разноформатное явление в зависимости от угла обзора и высоты абстракции: эволюстатами бы- вают и части целого, и целое частей. Индикаторами эволюстатов-тел выступают их конституции (самые лабильные соматические части); они
Глава 8. Концеиция всюдности эволюции более всего маркируют синтомы (эволюционные шаги), соединяющие в один эволюционный узел и исходную, и производную эволюстаты и, тем самым, склонны оставлять наиболее заметные следы реальных эво- люционных событий — синтомные следы (следы эволюционных шагов). Каждый эволюционный маятник оставляет о своем существовании какой- либо синтомный след, или абстрактолинию (нередко их несколько)— образную линию, которая на рисунке-схеме материализуется в действи- тельную линию, соединяющую эволюстаты по схеме: исходное — производ- ное-исходное — производное-исходное — производное-исходное и т.д. и т.п. Деятельное существование эволюционных маятников, обеспечива- емое их тридинами — взаимодействием трех, отмеченных выше сил, сопровождается производством эволюционных шагов (синтомов), со- единяющих абстрактолинией эволюстаты, т.е. образной линией про- шлое соединяется с настоящим и может прогнозироваться возможный выход в будущее по приведенной выше схеме. Синтомный след — это не обязательно прямая линия, более всего, — она кривая, ломаная и т.n., поскольку реальная эволюция — это не дорога с односторонним движением, а умелая «мастерица» (великая «служанка» истории Природы), свободно плетущая свои «кружева» с разными узорами по строгим законам поддер- жания всеобщего природного организационного порядка. Образная линия- абстрактолиния, проведенная по взаимосвязанным синтомам, соединяя род- ственные эволюстаты, способна воспроизвести «кружева» эволюционного разнонаправленного творения содержания вселенского потока на некото- ром участке его иространства и времени. Эволюция шла и идет более всего (это ее генеральное «кредо») по рядам гомологичных признаков через трансформации одних признаков и возникновение из них других признаков этого же ряда, более приспо- собленных к обновленным условиям обитания. Прежние структуры или полностью изменяются, или сохраняются рядом с новыми структурами, оставаясь памятниками достигнутого еще ранее эволюционного завое- вания. Реальная эволюция, в отличие от того, что нередко рисуют эво- люционисты в морфогенетических рядах, идет непрямолинейно и не в одну сторону, а в своем реальном исполнении «плетет кружева», причем более всего из гомологических адаптаций, поскольку условия окружа- ющей среды то одни, то другие, то третьи и, наконец, они могут вновь вернуться к давно минувшим, например, первым, вызвав новую волну адаптаций, некоторые из которых повторят прежние адаптации своих предков, но уже на новом витке развития организации биоморф. Эволюция биоморф — это длительный единый процесс «плетения» морфогенетических «кружев» развития, а здесь и сейчас, т.е. в текущее время, мы наблюдаем только верхушечные результаты этой кропотливой природной работы по формированию разнообразия биоморфотипов расти- тельного мира. Площади рисунка в книге не хватит, чтобы передать все разнообразие растительных биоморфотипов, уже существующих на Зем- ле, но это вовсе делать и не обязательно. Важнее другое: понять, как было достигнуто и продолжает расширяться это разнообразие. Всему причина — тотальный движитель — примагенератор (пространство все- ленского потока, по-разному заполненное вечно движущейся матери-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ ей), который обеспечивает Природу, как вселенский поток, бесконеч- ным движением тел и нетел; только от него наполняется силой эволю- ция вообще, а в нашем случае, и эволюция биоморф семенных расте- ний. Что значит быть в этой эволюции? Для биоморф означает одно— уметь приспосабливаться (адаптироваться), если хотите — «вертеться», в изменяющихся условиях среды и продолжать жить при смене поко- лений... Реальная эволюция биоморф представляет из себя многоходовую и разнонаправленную процедуру скрупулезного («тяп-ляп» напрямик здесь, как правило, не проходит) «плетения» морфогенических «кру- жев», т.е. любых биосоматических (и меробиотических, и ценобиотичес- ких) трансформаций. В результате «кружевной» эволюции что-то утра- чивается, что-то продолжает существовать, а что-то возникает принци- пиально новое, и Природа, бесконечно функционируя таким образом, создала великое разнообразие телесных систем и, в частности, — раз- нообразие биоморфотипов. Реальную эволюцию, но отдаленную, кото- рая свершалась многие миллионы лет назад, мы сейчас уже не можем наблюдать в ее непосредственно первозданном виде, поскольку восста- новить многое, через «пелену» давно прошедших времен, невозможно в деталях, но определить в общих чертах, как плелись «кружева» в той эволюции, тем не менее вполне можно. На эволюцию биоморф мы часто вынуждены смотреть через призму разнонаправленных морфоге- нетических, в первую очередь, гомологических рядов, которые всегда могут отражать только вероятностные, гипотетические подвижки в ка- ких-либо эволюционных событиях, поскольку такие ряды строятся из разнообразного материала нынешних, а не давно минувших биоморф. Живая природа реальную эволюцию биоморф «имела» и «имеет» как «кружевной» процесс, и она никогда в своих деталях не повторяет многого того, что мы «рисуем», кстати, весьма поверхностно, в морфо- генетических рядах, поскольку в них речь может идти только о вероят- ностных путях преобразования структур, и в реальном эволюционном действии могли иметь место лишь фрагменты нашего морфогенетичес- кого «рядостроительства». На сто процентов никогда нельзя методами конструирования морфогенетических рядов воссоздать тонкое перепле- тение «кружев» реальной эволюции, причем, тем более если она удалена от нас на многие миллионы лет. При всей пользе такого логического «рядостроительства» биомор- фотипов, оно все же имеет ограниченные возможности, и его желатель- но совмещать с палеобиоморфологическими исследованиями, посколь- ку никогда воссоздать в деталях ход реальной «кружевной» эволюции биоморф не удастся, хотя вполне можно в случае удачи приблизиться к передаче направлений и некоторых типологических сторон в эволю- ции каких-либо биоморфных структур. Морфогенетические ряды могут отражать только вероятностные пути возникновения современных био- морфотипов. Синтомные следы какое-то время более или менее остаются от эволюционных шагов (синтомов) на бескрайних просторах вселенского потока, направленность которых в различных схемах, например, на
Глава 8. Концеиция всюдности эволюции абстрактолиниях морфогенетических рядов, точнее всего и удобнее обозначать обоюдоострыми (обоюдонаправленными) «стрелами»-указа- телями, у которых из двух противоположных направлений выбирается на некоторое время какое-либо одно, отвечающее складывающимся текущим условиям, и по которым можно строить прогнозы на будущее или, наоборот, — разбираться с прошлым. Морфогенетические ряды никогда не повторяют в полном объеме реальной «кружевной» эволю- ции — это не их задача, — а воспроизводят возможные пути эволюци- онного преобразования типов биоморфных структур. Они вполне могут помогать раскрывать те вероятностные направления, по которым эво- люция в прошлом могла идти («плести» свои «кружева»), развивая те или иные биоморфотипы, и по которым она где-то идет и сейчас в каких-либо родственных группах. Таким образом, строительство морфогенетических рядов — способ исследования вероятностных путей возникновения типов биоморфной орга- низации, но не способ воссоздания реальных путей «кружевной» эволюции биоморф. Допустим, мы построили морфогенетический ряд: длиннокор- невищные, короткокорневищные, компактнокорневищные, корневищ- но-луковичные и луковичные биоморфотипы семенных растений, со- единив каждый биоморфотип обоюдоострыми «стрелами»-указателями. Так мы подчеркиваем, что этот ряд биоморфотипов в эволюционном развитии может быть прочитан по-разному (в каком-либо из противо- положных направлений) в зависимости от особенностей развития окру- жающей среды в определенное время и в определенном месте. Реальная эволюция какой-либо родственной группы (например, вида, рода, семейства) в данном месте и в данное время вполне может совпадать с некоторыми из фрагментов морфогенетического ряда и теми или иными направлениями развития одних биоморфотипов в другие, когда более всего те отвечают конкретным окружающим условиям. «Стрелы»-указатели направлений эволюционного развития, отраженные в морфогенетических рядах, в зависимости от складывающихся эколо- гических условий, в реальной эволюции могут «лететь» по-разному: то в одну, то в другую сторону. «Стрелы» морфогенетических рядов обо- юдоострые и, тем самым, обладают обоюдонаправленностью, отмечая на какое-то время одно направление, а в другое время — иное направ- ление гипотетического эволюционного развития некоторых биоморфо- типов, поскольку у такой «стрелы» один из ее концов на какое-то время оказывается сильнее другого. Морфогенетические ряды могут успешно передать направление и некоторые стороны из перемещения эволюци- онных шагов в реальной эволюции каких-либо групп родственных био- морф, но они не рассчитаны на то, чтобы восстанавливать из прошлого всю последовательность «плетения» картин реальной «кружевной» эво- люции. Построение морфогенетических рядов дает возможность вскры- вать некоторые аспекты того, что могло быть когда-то в эволюции, на основе выявления последовательности некоторых эволюционных собы- тий, которые свершаются здесь и сейчас. Мне, например, в текущее время и в конкретном месте удалось наблюдать механизм обратимого (!) перехода моноподиального нарастания побегов в симподиальное, и
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ розеточного роста побегов в полурозеточный ( рис. 401), что происходит реально в горах Алтая в соответствии с той нормой реакции раститель- ных организмов, которая сейчас существует у них и имеет склонность расширяться или сужаться под действием комплекса повторяющихся экологических факторов, закрепляя или выбраковывая отбором текущие трансформации в генотипах. Нередко фитобиоморфы некоторых видов, попадая в экстремаль- ные условия, настолько изменяют свою феносоматическую организа- цию, а многолетники — их базовый орган, что приобретают новые признаки гомологических структур, по которым о прежних типах орга- нов уже говорить не приходится. Такое происходит, например, с плот- нокустистыми и рыхлокустистыми злаками, обычно развивающими ком- пактные и короткие корневища, с каудексовыми растениями или полу- кустарниками, которые в условиях стресса, вызванного погребением наносным почво-грунтом надземных стеблевых частей, сильно усложня- ют свою организацию, образуя в борьбе за существование вынужденно- удлиненные подземные побеговые органы (т. 1, гл. 8. 2. 1). Надземные, часто эфемерные, стеблевые части побегов при засыпании почво-грун- том нередко развивают у многолетников придаточные корни, приобре- тают персистентность (многолетность), переносят зоны активного побе- гоотрастания (эпифизы) ближе к поверхности почвы, формируя эпифи- зарную ярусность, а еще вынужденно погребенные растения обычно подвергаются сильной партикуляции, а иногда полностью как-либо обосабливают свои парциальные части, превращая их в особи вегетатив- ного происхождения (клонисты). Традиционные базовые органы, разви- вающиеся у растений, обитающих в условиях с относительно спокой- ным уровнем поверхности почвы, при смене условий произрастания на стрессовые, когда резко повышается эрозионная подвижность почво- грунта, изменяются иногда до неузнаваемости: компактные и короткие корневища превращаются в вынужденно удлиненные корневища, а компактно-ветвистые каудексы — в веревковидные хитросплетения парциальных каудиций и посткаудиций, которые разные по размерам, частично или полностью обособлены, с фрагментами главного корня или без них (рис. 40, 193, 194, 334). Такие скопления парциальных элементов общего базового «органа» в системе кондивидуума, нередко криптоклонального (скрыто клонального), уместно уже считать не од- ним базовым органом (глава 1), а применять для их обозначения более общее название — «базифиз», или «растущее основание» (дословный перевод с греческого). Многие дерновидные луговые злаки, произрастая где-нибудь вдоль железнодорожных насыпей или, более обычно, на луту-пастбище, под- вергаются нередко по разным причинам, например, при агротехничес- ком рыхлении почвы, засыпанию почво-грунтом (в поймах заиливаются после схода паводковых вод и т.п.) и, как правило, не погибают, а переносят зоны кущения (эпифизы) ближе к поверхности почвы (рис. 402-405). Трансформация подземных органов происходит буквально на глазах, когда образуются более или менее вертикальные вынужденно удлиненные парциальные корневища, составляя вместе базифизы крип-
Глава 8. Концеиция всюдности эволюции Рис. 401. Биографический портрет Hedysarum gmelinii Ledeb. [сем. Fabaceae (А~тай, Чергинский, Семинский хребты, остепненные горные склоны, мел- косопочники, опушки сосновых лесов)]. БиоморфОТИПЫ: надземно-двусемядольные (семядоли эллиптические, мясистые 4 — 5 мм дл., 2 — 3 мм шир., закругленные, клиновидно суженные в короткие черешки) розет- кообразующие [розеточные — 1 биоморфотип на вершинах невысоких гор и степных горных склонах, полурозеточные — II биоморфотип на лугово-степных горных склонах и опушках сосновых лесов, смешанные розеточно-полурозеточные — Ш биоморфотип, когда в струк- туре одного растения развиваются розеточные побеги с пазушными безлистными цветонос- ными побегами и полурозеточные побеги, у которых, кроме розетки листьев, образуется раздвинугоузловая часть с непарноперистыми листьями и пазушными безлистными цвето- носами с верхушечными кистями цветков на них) очередно-сложнолистные эумоноподиаль- ные (1 биоморфотип), гемисимподиальные (11 биоморфотип) или гемисимподиально-эумо- ноподиальные (1П биоморфотип) интеркалярно-семидетерминатно-репродуцирующие од- нодомные розово-бобовоцветковые каудексовые травянистые многолетники.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ токлональных кондивидуумов у отмеченных растений. Придаточные корни на подземных побеговых органах образуются при погребении не только многолетников, но и эвриэфемеров. Рис. 402 — 405. Перемещение зон кущения дерновинных биоморф некоторых злаков (сем. Роасеае) при погребении почво-грунтом надзем- ных оснований кустов. Когда поверхность почвы, под влиянием разных обстоятельств, перемещается вверх и наносный почво-грунт погребает надземные ос- нования кустов, большинство дерновинных злаков адаптируется пре- одолевать это препятствие путем преобразования надземных эфемерных частей побегов в подземные персистентные части и (или) образовывать новые подземные побеги с более длинными междоузлиями, чем в усло- виях относительной статики почвенной поверхности. Рис. 402. Поведение плотнокусти- стой биоморфы (фрагмента ее ку- ста) Deschampsia cespitosa (L.) Beauv. в условиях пойменных лу- гов (Московская область) при по- ложительной динамике (заилива- нии) поверхности почвы. Рис. 403. Поведение рыхлокустис- той биоморфы (фрагментов ее ку- ста) Dactylis glomerata L. на куль- тивируемых лугах (Московская об- ласть) после агротехнических ме- роприятий с положительной дина- микой почвенной поверхности (за- сыпание почвой).
Глава 8. Концеиция всюдности эволюции Рис. 404. Поведение рыхлокустистой Рис. 405. Поведение рыхлокустис- биоморфы (фрагментов ее куста) той биоморфы (фрагментов ее ку- Arrhenatherum elatius (L.) J. et С. Presl ста) Festuca pratensis Huds. в тех же в тех же условиях, что и на рис. 403. условиях, что и на рис. 403. Когда в борьбе со стрессами новые адаптивные достижения сопро- вождаются мутациями и далее их результаты закрепляются естествен- ным отбором, то, как минимум, расширяется видовая норма пластоге- неза, а как максимум, — возникают новые виды. У Anthriscus sylvestris (L.) Hoffm. на Алтае (рис. 46) обычна разно- видность (чаг. nemorosa (Bieb.) Trautv.), склонная более всего к униреп- родуктивному развитию или слабому клонообразованию, что существен- но отличает ее от мультирепродуктивных биоморф, развивающихся в других условиях, например, в Подмосковье (т. 1, гл. 2). В реальных условиях эволюционные MRHTHHKH, в нашем случае биоморфы семенных растений, раскачиваются сложнее, чем показыва- ют морфогенетические, даже обоюдонаправленные ряды. В реальную «кружевную» эволюцию вмешиваются многие дополнительные момен- ты, например, неотения, или редукция настолько сильная, что, минуя многие переходные типы организации, возникает весьма далекий от ожидаемого тип организации, а то и совсем непредсказуемый. Важно, что морфогенетические ряды строятся обычно как гомологические — по гомологичным признакам. Здесь я не буду усердствовать в поисках многих деталей эволюци- онного процесса биоморфного строительства в царстве растений. Ос- новное усилие направим на осознание принципиального механизма (о деталях — особый разговор — еще открытая тема для большого иссле- дования), на основе которого такое строительство стало когда-то воз- можным и не утратило своего содержания по сей день. Ранее было подчеркнуто, что эволюция свободна в выборе направлений своего те- чения: она в отношении биоморф плетет морфогенетические (шире- эволюстатные) «кружева». Нам, исследователям-эволюционистам, пра- вильнее строить не морфогенетические прямолинейные и однонаправ-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ ленные ряды, а логичнее создавать морфогенетические разнонаправлен- ные ряды, которые как фрагментарные части (отрезки) «вплетать» далее в общую схему гипотетической эволюции биоморф определенного так- сона (какого-либо надвидового комплекса биоморф). Схемы гипотети- ческой «кружевной» эволюции удобнее строить с помощью абстракто- линий (образных, как-либо материализуемых на схемах, линий вероят- ностных синтомных следов, находя и фиксируя логически наиболее приемлемые «соединения» (взаимосвязи) биоморфотипов как эволюста- тов синтомов. Абстрактолиниями вычерчиваются схемы тех вероятнос- тных путей, которые, исходя из ныне известного разнообразия биомор- фотипов, могли быть использованы когда-то в процессе «кружевной» реальной эволюции определенных надвидов. Если реальная эволюция действительно плетет «кружева», а у меня в этом нет сомнения, то нам, ее исследователям, ничего другого не ос- тается, как это замечать и соответствующим образом реагировать при построении схем гипотетической эволюции, т.е. пытаться «плести» образ- ные кружева, подстраиваясь своим сознанием к тому, что реально творит- ся в окружающей нас остальной живой, а шире и неживой природе. Можно собрать некоторое разнообразие биоморфотипов в таблицу (рис. 406) и с помощью абстрактолиний воспроизвести схему гипотети- ческой эволюции в каких-либо родословных группах биоморф, напри- мер, родовых надвидах. Разнонаправленность гипотетической эволюции на абстрактолиниях передают обоюдоострые «стрелы»-указатели. Если посмотреть хотя бы только на фрагменты того габитуального разнооб- разия биоморфотипов, которое реально существует сейчас во многих родах (родовых надвидах), допустим, Bupleurum L (рис. 407-410), Echium L. (рис. 411, 412), Lavathera L. (рис. 413-415), Polygala L. (рис. 41б, 417), Limonium L. (рис. 418, 419) и др., то вполне можно заметить два основ- ных равнозначных и противоположных направления их габитуальной гипотетической эволюции, проявляющиеся в сплетениях разнообразных морфогенетических рядов: как от кустарников-деревьев к многолетне- однолетним травам, так и обратно, что, на мой взгляд, оправдано, поскольку окружающие условия, будучи наиболее изменчивой частью в путеводных треугольниках, выступают в качестве главных инициаторов определения направления эволюционных трансформаций, когда те идут реально в данном месте и в данное время. С помощью абстрактолиний на одной таблице разнообразия био- морфотипов в разной форме (сплошной линией, пунктиром или иначе) можно передать разное число схем гипотетической эволюции биоморф. Посмотрим для примера на схему гипотетической «кружевной» эволю- ции, допустим, в роде Нурепсит L. (рис. 40б), которая получилась при соединении абстрактолинией соответствующих габитуальных биомор- фотипов на таблице габитуального разнообразия современных биомор- фотипов. Роду Нурепсит свойственно большое габитуальное разнооб- разие (рис. 420-423): в субтропиках и тропиках, чаще всего влажных горных, — невысокие (до 5 м) деревья (Н. canariense L.) и кустарники [вечнозеленые — Н. mysurense Heyne, Н. Ьа1еапсит L. (рис. 423), полу- вечнозеленые, или зимнезеленые, — Н. patulum Thunb. ех Миггау,
Глава 8. Концелция всюдности эволюции Н. hookerianum Wight. et Arn., листопадные, или летнезеленые, Н. androsaemum L. (т. 1, гл. 4), Н. hircinum L., Н. inodorum Miller ], в сухих субтропиках — кустарнички [Н. calycinum L. (т. 1, гл. 4)], в субтропиках и умеренном климате — полукустарники, полукустарнички (Н. polyphyllum Boiss. et Ваl., Н. olympicum L., Н. humifusum L., Н. linarioides Bosse), травянистые корнеотпрысковые многолетники [Н. tomentosum L., Н. perforatum L., (рис. 420)], корневищные травы [Н. alpestre Stev., Н. hirsutum L. (рис. 421), Н. maculatum Crantz (рис. 422)], и, наконец, есть еще и однолетники, например, Н. mutilum L. на болотах. Как видно из схемы, в гипотетической эволюции рода Нурепсит были и продолжают быть разные и противоположные направления габитуального, а точнее- феносоматического развития, которые более всего формируются под давлением конкретных условий произрастания биоморф, в соответствии с которыми детализируются их адаптивные признаки. Схемы гипотетической эволюции, обозначенные какими-либо аб- страктолиниями, будучи вынуты из таблицы разнообразия биоморфоти- пов, могут приобретать внешнее оформление одного неразветвленного или разветвленного морфогенетического ряда, нередко эти схемы слож- нее и действительно чем-то напоминают «кружева», но в любом случае — это всегда лишь отдаленные тени часто уже минувшей реальной «кружевной» эволюции. Реальная «кружевная» эволюция много богаче той, которую можно увидеть в морфогенетических рядах и на общих схемах гипотетической эволюции, но, безусловно, первая что-то может творить сообразно с тем, что отражает вторая, ее гипотетические морфогенетические ряды (для того они и строятся). Морфогенетические ряды биоморфотипов в схемах гипотетической эволюции, будучи фрагментами современного разнообразия биоморфотипов, рассчитаны на то, чтобы хоть немного приблизиться к пониманию тех направлений в реальных эволюционных процессах, которые когда-то свершались и (или) продолжают свершать- ся неоднократно сейчас, иногда даже в циклах существования отдель- ных биоморф и тем более в циклах существования видов и надвидов. Принцип плетения живой природой морфогенетических (биосома- тических) «кружев» (рис. 406) эволюционными маятниками (телами и нетелами), «плывущими» и «раскачивающимися» в бесконечности все- ленского потока по универсальной схеме «так-сяк * туда-сюда», когда из частей целого (меробиотически) и всего целого (ценобиотически) что-то изгоняется («отплетается»), что-то встраивается («вплетается») и т.п., поддерживая главный результат своей деятельности: продолжается жизнь вообще. Иногда, если получается что-то большее, то привносятся новые формы и обновляется содержание жизни на Земле, а еще главнее то, что всеприродные организационные «кружева» не обрываются вообще, они бесконечно «плетутся», и «плетутся» эволюцией с жизнью или без нее, но всегда и везде — всюдно. Жизнь на Земле и, в частности, растительная жизнь продолжает сохраняться и дальше, что в условиях сильно изменя- ющейся внешней среды никогда не смогло бы состояться, если бы эво- люция действовала только прямолинейно, что всегда может быть всего лишь одним из фрагментов в «кружевах» разнонаправленной эволюции.
~ ~ Ю ° ~ е Ю Ю а ° ° ° Ю ФЮ фф Ю Ю , ~ Ф Ю е ° ' Ю Ю ° ° ~ ° ~ ~[-: % -' ° ф --. -'.- Ф ~~~, . - : . 'л~ ' g �» ~ф~~- ффо ° е ' ~ф ( ф' ' Я- 1'-'- ю Ф Ю ~ Ю Ю
Глава 8. Концелция всюдности эволюции Используя концепцию разнонаправленности эволюции, можем сделать следующее обобщение: эволюция свободна в поиске и выборе направлений, поэтому «эволюирующее тело» («эволюционный маятник») в зависимости от того, как оно ориентируется в иространстве и времени, посредством его «путеводного треугольника» (трех взаимодействующих сил — сил средоокружающего тела, или тела окружающей внешней среды, конституции средоокруженного тела и его эгосубстрата), реализует в себе такие трансформации, которые позволяют этому телу как исходному или (и) как производному существовать в соответствии с нормой его пласто- генеза (у организмов — нормой реакции) и нормой пластогенеза тела, в структуру которого оно входит как элемент, а когда таких норм не хватает, то эволюция не останавливается, и иьиенения продолжаются вплоть до свершения субмутаций или мутаций, и тогда возникают произ- водные тела уже с иной (другого типа) организацией — это закон (пред- высший) всеобщности (всюдностн) эволюции телесных систем. Я не склонен в этой главе и во всем втором томе решить в полной мере многие задачи эволюционного развития организации биоморф се- менных растений. Здесь, пожалуй, больше прелюдия к «симфонии», которая когда-то еще должна быть сыграна, больше наброски к теории эволюции биоморф семенных растений, чем сама теория, хотя уже впол- не просматриваются некоторые очертания ее основ. Возможно, их за- метят далеко не все. Так бывает в науке. Всему свое время. Наука развивается быстро, и кто-то может оказаться проворнее, не прочитав или прочитав эти наброски. Так что ж, если получится достойно, а дело это тонкое, то в добрый путь, не забывая первоистоков и того, что было на этом пути..., да, впрочем, еще и того, самого общего, что особо подчеркнуто в начале этой главы. Рис. 407. Биографический порт- рет Bupleurum lane ifolium Ногпет. [сем. Apiaceae (там же, что и рис. 370, IV)]. ,26 Био морфОТИП: надземно-двусемя- дольные (семядоли линейные, 12 — 15 мм дл., 1,2 — 1,8 мм шир., слегка заострен- ные, постепенно суженные в короткие черешки) полурозеточные очередно-про- столистные семидетерминатно-репроду- цирующие однодомные желто-сельдерей- ноцветковые стержнекорневые сильно- рослые (20 — 50 см, 1 биоморфотип) и слаборослые (5 — 10 см, П биоморфотип, а переходную группу составляют средне- рослые биоморфы) однолетники.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ PA СТЕНИЙ Рис. 408. Биографический портрет Bupleurum bicaule Helm [сем. Apiaceae (Алтай, Теректинский хребет, остепненные каменистые склоны в долину реки Катунь)]. Биоморфотип: надземно-двусемядольные (семядоли линейные, 7 — 10 мм дл., 0,7— 1 мм шир., слегка заостренные, почти сидячие) полурозеточные очередно-простолистные гемисимподиальные семидетерминатно-репродуцирующие (верхушки ди-, плейофазных унирепродуктивных побегов закрывают полузакрытые соцветия — сложные зонтики) од- нодомные желто-сельдерейноцветковые каудексовые изоаклональные травянистые много- летники.
Глава 8. Концеиция всюдности эволюции Рис. 409. Биографический портрет Bupleurum krylovianum Schischk. [сем. Apiaceae (Восточный Казахстан, в 15 км на северо-восток от поселка Самар- ское, невысокий горный массив, склоны, поросшие кустарниками и высо- ким разнотравьем)]. БИОМорфОТИП: надземно-двусемядольные (семядоли линейные, 14 — 18 мм дл., 1,5— 2 мм шир., слегка заостренные, постепенно суженные в короткие черешки) розеткообра- зующие (помимо розеточных вегетативных побегов развиваются еще и верхнерозеточные побеги, особенно в условиях сильного затенения, а вегетативно-репродуктивные побеги полурозеточно или даже безрозеточно продолжают развитие вегетативных побегов уже на второй год жизни их или позднее как единые унирепродуктивные побеги) очередно- простолистные гемисимподиальные семидетерминатно-репродуцирующие однодомные желто-сельдерейноцветковые каудексовые травянистые многолетники (1 биоморфотип на открытых горных склонах) или вынужденные полугравомноголетники (П биоморфотип в условиях затенения среди зарослей кустарников).
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ 3, 14а, 19a),27 1,агЗ, 14a,18а,26 Рис. 410. Биографический портрет Bupleurum salicifolia Soland. ex Lowe [сем. Apiaceae (Израиль, Иерусалим, Ботанический сад, средиземноморский от- дел, 25 IV)]. БИОМОрфОТИП: надземно-двусемядольные (семядоли линейные, 10 — 12мм дл., 1— 1,5 мм шир., полурозеточные (на вегетативной стадии развития образуются розеточные, полурозеточные и верхнерозеточные побеги, а в целом унирепродуктивное развитие ро- зеткообразующих побегов осуществляется по монофазному, а чаще ди-, плейофазному цикпу) очередно-простолистные гемисимподиальные семидетерминатно-репродуцирующие однодомные стержнекорневые аэроксильные (приземное кущение) вечнозеленые (листья нередко живут два вегетационных сезона) кустарники (1 — 1,5 м высотой).
Глава 8. Концеиция всюдности эволюции Рис. 411. Биографический портрет Echium vulgare L. [сем. Boraginaceae. (Мос- ковская область, Серпуховский район, окрестности Пущино, долина и тер- раса реки Оки, луга, пустыри)]. БИОМОРфОТИПЫ: надземно-двусемядольные (семядоли эллиптические или обратнояй- цевидные, 6 — 12 мм дл., 3 — 6 мм шир., широкозакругленные, клиновидно суженные в короткие черешки) полурозеточные очередно-простолистные сильноопушенные семиде- терминантно-репродуцирующие (верхушечное полузакрытое сборное соцветие — тире об- разовано многочисленными монохазиями, зацветающими акропетально) однодомные сине- фиолетово-бурачниковоцветковые (при отцветании цветки розово-красные) стержнекор- невые некустистые (1 биоморфотип) или кустистые (П биоморфотип) слаборослые или сильнорослые травянистые двулетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ес- um ем. же, ры НО- НО- ИВ- еп- бе- ые, озе- икл ОГО ХОД- плей- еги) 1,27
Глава 8. Концеиция всюдности эволюции б, Рис. 413. Биографический портрет Lavatera cretica Ь. [сем. Malvaceae (там же, что и рис. 370, 10 IV)]. БИОМОрфОТИП: надземно-двусемядольные (семядоли яйцевидные, слегка сдавленные с боков, иногда неравнобокие, закругленные, 15 — 25 мм дл., 8 — 18 мм шир., сердцевид- ные, на черешках 8 — 50 мм дл.) безрозеточные очередно-простолистные прямостоячие семидетерминатно-репродуцирующие однодомные светло-фиолетово-мальвоцветковые од- нолетники.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИИ 1,аг2,200,26 1,агЗ, 18а,26 2(5 19 26 НИЯ сле отр Зето но точ рое рас био НИЯ НИЯ ди же но пр сем щи ветк сов Вп ни 40
Глава 8. Концепция всюдноаии эволюции 2),гм4, 06,21, (27) Рис. 415. Особенности габитуальной организации средиземноморских кус- тарников из рода Lavatera Ь. [сем. Malvaceae (там же, что и рис. 410, 10 IV)]. I — Ь. raaritiraa Gouan., II — L. агЬогеа L. БИОМОРфОТИП: надземно-двусемядольные безрозеточные очередно-простолистные ге- мисимподиально-эусимподиальные (большинство унирепродуктивных побегов развивают- ся по дифазному или даже плейофазному варианту морфогенеза, но некоторая часть побегов проявляет монофазное развитие, завершая его цветением и плодоношением) се- мидетерминатно-репродуцирующие однодомные розово-мальвоцветковые листопадные стержнекорневые кустарники (1 — 1,2 м, II — 1,5 — 2 м высотой). р) 06 7( г, За, 4 Рис. 41б. Биографический портрет Polygala comosa Schkuhr [сем. Polygalaceae (Алтай, Семинский хребет, окрестности поселка Шебалино, остепненные луговые склоны)]. Биоморфотип: надземно-двусемядольные (семядоли эллиптические, 4 — 5 мм дл., 2,5 — 3 мм шир., закругленные, клиновидно суженные в очень короткие черешки) безрозе- точные очередно-простолистные гемисимподиально-эусимподиальные (в конце лета— осенью развиваются короткие вегетативные побеги, часть которых успешно зимует и продолжает моноподиальное развитие безрозеточных удлиненных унирепродуктивных побегов, после завершения которого отмирает сверху почти до основания, оставляя в многолетней основе компактные резиды, но другие весенние побеги отрастают из латен- тных почек и проходят цикл унирепродуктивного безрозеточного развития за один веге- тационный сезон) семидетерминатно-репродуцирующие однодомные светло-синецветко- вые или розово-лиловоцветковые (редко беловатые) рыхлокустистые стержнекорневые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИЙ 20а,2 Рис. 417. Биографический портрет Polygala myrtifolia L. [сем. Polygalaceae (Израиль, Иерусалим, Ботанический сад, южно-африканский отдел, 2 IV)]. Биоморфотип: надземка-двусемядольные (семядоли обратнояйцевианые или эллипти- ческие, 5 — 7 мм дл., 3 — 4 мм шир., закругленные, клиновидно суженные в короткие черешки) четочные очередно-простолистные гемисимподиально-полисимподиальные семи- детерминатно-репродуцирующие (молодые растения образуют вегетативные четочные, т.е. нерозеточные, хотя и сближенноузловые длинные и средние, побеги, нарастающие несколь- ко лет эумоноподиально, а при случайном отмирании их верхушек легко восстанавливается вегетативный рост посредством отрастания побегов замещения; взрослые растения развива- ют унирепродуктивные моно-, ди-, плейофазные побеги, но самое интересное, что образу- ются также годичные полисимподиальные побеги, которые за один вегетационный сезон проявляют несколько репродуктивных циклов развития, т.е. цветут несколько раз, перевер- шинивая предыдущие соцветия новыми цветоносными побегами) однодомные розовоцвет- ковые стержнекорневые вечнозеленые (листья живут 2 — 3 года) кустарники (0,6 — 1,5 м высотой). Подобную организацию в жизнеразвитии проявляет P. peduncularis Burch. ex DC., у которого и цветки покрупнее, и размеры кустов побольше (до 2-х м высотой).
Глава 8. Концепция всюдности эволюции Рис. 418. Биографический портрет Limonium oleifolium Miller [сем. Plumbaginaceae (Израиль, каменисто-песчаная равнина, резко обрывающая- ся к Средиземному морю, окрестности Тель-Авива, 23 IV)]. Биоморфотип: надземно-двусемядольные (семядоли продолговато-лопатчатые, 8 — 10 мм дл., 2 — 3 мм шир., закругленные, постепенно суженные в короткие черешки, почти сидячие) розеточные очередно-чешуевиднолистные гемисимподиальные (23 IV еще не цвели) каудексовые травянистые многолетники (1 биоморфотип, только он показан на рисунке) или полутравомноголетники (11 биоморфотип, он проявляет себя при оголении каудекса во время эрозионной подвижности песчаных почв).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНКИ 38в За, 67 Рис. 419. Биографический портрет Limonium sp. [сем. Plumbaginaceae (Тель- Авив, парк у гостиницы "Хилтон" на берегу Средиземного моря, апрель)]. Разновозрастные представители одного вида рода Limonium L. успешно произрастали, давая самосев, в декоративных посадках рядом с мексиканскими опунциями и другими иноземными растениями. Из местных видов этот вид был ближе всего к L. angustifolium (Tausch) ТитЫ, биоморфотип которого ботаники обозначают как травянистые многолет- ники (Feinbrun-Dothan, Danin, 1991). Именно необычная биоморфология рассматривае- мых растений привлекла более всего мое внимание. Невольно навязывалось обманчивое впечатление, что чья-то неведомая «рука» где-то вынула из почвы какие-нибудь травом- ноголетние российские или казахстанские кермеки [допустим, такие как L. platyphyllum Lincz., L. gmelinii (Willd.) О. Kuntze)], высадила их здесь, в экзотическое место (это, кстати, и послужило для меня поводом не усердствовать при определении вида), забыв, однако, опустить 4~гипично подземные» каудексы этих видов под поверхность почвы. Не правда ли, что приводимые здесь биографические картинки весьма любопытны с эволюционной точки зрения. Возможно, одна из мутаций в таком сухом и теплом климате уже когда-то сработала, и жизнь еще одного вида Limonium стала нормой с довольно высоким надзем- ным каудексом... В тени опунций надземный каудекс повыше, а на открытом месте («сол- нцепеке») — пониже. БноморфотнП: надземно-двусемядольные (семядоли яйцевидные, 4 — 5 мм дл., 2 — 2,5 мм шир., закругленные, клиновидно суженные в короткие черешки) розеточные (цветоносы образуют только очередные чешуевидные листья) розеточно-простолистные гемисимподиаль- ные детерминатно-репродуцирующие однодомные сине-фиолетовоцветковые надземно-кау- дексовые (поднимаются над поверхностью почвы до 30 см) полукустарнички и полукустар- ники (гемисимподиальное многолетнее нарастание побегов осуществляется за счет надземных миниморезидов — компактных остатков акронекрозных унирепродуктивных розеточных по- бегов, а по высоте каудексов рассматриваемые биоморфы ближе к полукустарникам).
Глава 8. Концеиция всюдности эволюции б, Рис. 420. Биографический портрет Нурепсит perforatum L. [сем. Hypericaceae (Московская область, луга, лесные поляны и опушки смешанных лесов)]. Биоморфотнп: надземно-двусемядольные (семядоли яйцевидные, 2 — 3 мм дл., 1— 1,5мм шир., закругленные, клиновидно суженные в короткие черешки) безрозеточные супротивно-простолистные эусимподиальные (большинство коротких озимых побегов, если они образуются, зимой отмирает, но некоторая, небольшая их часть, все же разви- вается по дифазному унирепродуктивному циклу) детерминатно-репродуцирующие одно- домные стержнекорневые желтоцветковые факультативно-корнеотпрысковые травянистые многолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ О 27 Рис. 421. Биографический портрет Нурепсит hirsutum Ь. [сем. Нурепсасеае (Алтай, Семинский хребет, смешанные леса, лесные поляны, лута по берегам горных речек, опушки)]. БИоморфотнп: надземно-двусемядольные (семядоли эллиптические, 2— ие 2 — Зммдл.,1— 1,5 мм шир., закругленные, клиновидно суженные в короткие черешки) безрозеточные с тивно-простолистные сильноопушенные эусимподиальные короткокорневищные де- терминатно-репродуцирующие ретардационно-кондивидуальные однодомные желтоцвет- ковые травянистые многолетники.
Глава 8. Концепция всюдности эволюции 38в Рис. 422. Биографический портрет Нурег1сит maculatum Crantz [сем. Hypericaceae (Московская область, смешанные леса, опушки, лесные поля- ны, луга)]. Др — цветущие верхушки побегов (Ворошилов и др., 19бб). Биоморфотип: надземно-двусемядольные (семядоли яйцевидные, 2 — 2,5 мм дл., 1— 1,5 мм шир., закругленные, клиновидно суженные в короткие черешки) безрозеточные супротивно-простолистные эусимподиально-гемисимподиальные [немалая часть побегов проявляет в одном сезоне подземный рост и (или) озимое отрастание и могут нормально продолжить вегетацию на следующий год, завершая унирепродуктивный цикл развития безрозеточных побегов с короткими или длинными подземными основаниями] детерми- натно-репродуцирующие однодомные желтоцветковые коротко- и длиннокорневищные травянистые многолетники. На открытых местах (I биоморфотип) образуются прямосто- ячие ежегодно цветущие растения, а в условиях глубокого затенения (II биоморфотип), например, когда растения заходят под полог елового участка леса, образуются крайне угнетенные биоморфы, с полегающими короткими и слабыми вегетативными побегами.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ И ~ ~~~ о „~~ О Оо О Ж а. Ж Я~ Ф Л Ф ь| ФЭ 53 ФЭ О М О 'С ФЭ Л ФЭ О 2 О ° eel 53 ФЭ <б Ю 2 О 'С ФЭ Ю ~ь Ж ;~ о Ж 9 ~, + л~ > Ж 0 63 ~ л ~ а. Я О О v~„g И м о ЖЕОх д ~: h4 а. Ю Ж ж О О щ Д; л И Оь Ж и~~ х ~ ~В и~~ О О LO О Яж~ О Х лч Д и ~<0 е1 ~ И л л 4.) М i М Ж О сф Ж Ж О д ~Д й~ ~}~ О О а ~ И v 4,) Й~ а и О О ~"~ ~ О 0 Q, у Я ~ Ж XLOX~ev й О ~ ~:~. 3: М ~ ~ ц д ЖХX ~ю ~ х Д ~ а ~ ~~~~ с~ "~ а. и~а.> Е ю О Ж М с~ е~~Я > а' ' ~ а, о о ~W" Жид а. О Ж~ ef О ю О и а г) о а а Ж О и ах Я ~.е ! u ~ о ~ хо 3 34~~ Ж ~a. ЛО О Ж 0 ~ 1 Я л CO ф~~ ~х ~С~ О о О Ж ~, Е О О О О ~ ~вфю) л СЧ О и Оч
Глава 9. Концепция экогенобиоза- теоретическая основа эволюционных принципов происхождения и утверждения жизни на Земле Моя задача не в том, чтобы восстановить хронологию возникнове- ния и утверждения жизни на Земле в процессе ее исторического разви- тия, а сконцентрировать внимание только на одном: каков в самом общем виде принципиальный механизм, на основе которого такое гран- диозное явление, как жизнь, состоялось на нашей планете. Зная прин- ципы, мы успешно можем распознавать детали каких-либо явлений, происходящих в Природе. Природа устроена проще, чем нам часто представляется в попытках разобраться в ней, с ее бесконечностью и неисчерпаемостью. Это мы, люди, частицы Природы, стремясь познать то, что нас составляет и окружает, обычно усложняем восприятие Природы в нашем воображе- нии, преувеличиваем сложности в ее организации, принимаем иногда мнимое озарение нашего сознания, всплески эмоциональных иллюзий за истинное знание. Проходит какое-то время и всем становится ясно, что Природа в целом мудрее нас, ее любознательных частиц-«первопро- ходцев», поскольку она уже давно, значительно раньше, чем люди во- обще появились, овладела способами решать многие свои сложные, как нам представляется, проблемы относительно просто. Природа создала в процессе эволюции определенный порядок в своей организации, которым она сама управляет, и поскольку мы, люди, у нее есть, то она стала, где это возможно, прибегать и к нашему в ней участию. Мы должны осознать еще более свое особенное положение в Природе, действуя предельно корректно и осмотрительно ко всему остальному, что проявляется в ней, чтобы помогать, а не вредить ее и своему развитию. Бесконечность и неисчерпаемость Природы, ее материального и идеального, всегда была, есть и будет с эволюцией, которая пронизывает все без остатка, своей всепроникающей свободой. Она, будучи высшей формой трансформации (преобразования), наделила все, что есть в При-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ роде, заниматься в первую очередь ею (роль субэволюции в трансфор- мациях ничтожна); это она, эволюция, управляет, потребляет, произво- дит, резервирует и рассеивает ресурсы — то, что составляет универсаль- ное содержание не только живой природы (т. 1), но, по большому счету, и природы вообще, подчиняясь при этом разным законам, многие из которых наверняка просты. Какие это законы, люди еще во всем не разобрались, поскольку все мы существуем еще слишком мало в При- роде, в качестве ее структурных элементов. Но лучше спешить медлен- но, соблюдая порядок и чувство меры, чем спешить азартно, «сломя голову». Мы все в Природе успеем, если будем осмотрительней, осоз- навая не только величие нашего разума, но и не забывая о своей ма- териальной ничтожности. Эволюцию, включающую в себя и рост, и развитие материального и идеального, Природа использует всегда и везде, поскольку именно эволюция более всего отражает ее неиссякаемую деятельность в бес- конечности, — работу ее тотального движителя, который до удивления просто и до потрясения надежно вечно и исправно по сей день обес- печивает главное содержание эволюции — «раскачивает» неиссякаемое множество «эволюционных маятников» (тел и нетел). Маятниковый принцип бесконечного действия эволюции обеспечивается реализаци- ей внутренних и внешних ресурсов взаимодействующих тел, которые «раскачивают» эволюционные маятники движениями изнутри и снару- жи (рис. 366) на основе определенных ресурсов возможностей (идио- энергии). Я готов взять на себя смелость, чтобы утверждать: мне знакомы универсальный принцип эволюции Природы [бесконечное «раскачивание» эволюционных маятников (тел и нетел) по схеме «так-сяк*туда-сюда»], средства (вечно движущаяся материя), которыми располагает эволюция как результат работ единого природного механизма — тотального движителя, и направления (они разные в зависимости от преднаследственности или на- следственности телесных систем и от окружающих обстоятельств), по кото- рым идет эволюция вообще и, в частности, растительной жизни. Плести «кружева» — вот универсальный почерк филогенетической эволюции. Итак, если Вы, уважаемый читатель, хотя бы на время уже согла- сились с тем, что эволюция составляет высшую форму трансформации Природы и только она «раскачивает» в образе эволюционных маятников неисчислимое множество тел и нетел за счет внешних и внутренних ресурсов вечно движущейся и взаимодействующей материи, то далее вполне можно пойти и на не менее интригующие обобщения, поскольку они так или иначе, но уже предопределены всем изложенным выше материалом. Судите сами. Идиоэнергия, или ресурс возможностей, обеспечивает преднаследственность и наследственность соответствую- щих телесных систем, поэтому именно носители идиоэнергии — про- гены и гены — универсально управляют существованием разнообразных конституций тел и в особенности живых тел, но и процессов (нетел), составляющих содержание взаимодействия тел. Принципиальные особенности в проявлении противоречий, естествен- но, возникающих во взаимодействии разных природных сил неживой и живой
Глава 9. Концеиция экогенобиоза природы, и путей их разрешения, я ставлю во главу идеи «экогенобиоза»— концепции возникновения, подтверждения и утверждения жизни на Земле. Такова наша исходная позиция, с которой, пожалуй, есть смысл начать раскрытие содержания этой концепции. Она не в деталях, а в самых общих чертах показывает, почему жизнь на Земле получилась такой, а не другой. Многое в концепции экогенобиоза гипотетично, и это понят- но: слишком далеки мы от истоков Жизни, но это не беда, где-то даже достоинство, поскольку с позиций этой концепции легче разглядеть, в каком направлении перспективно развивать соответствующие эмпири- ческие исследования. 9.1. Основные противоречия в организации современной растительной жизни Любые живые тела, чтобы жить, должны бороться за существование (за жизнь), всегда оставаясь связанными непосредственно или опосре- дованно (косвенно) с неживой природой. Жизнь немыслима без един- ства и борьбы с ее противоположностью — нежизнью. На основе един- ства живого и неживого (закон Докучаева-Вернадского; т. 1, гл. 3) воз- никла борьба за существование, т.е. форма взаимодействия живых тел (биосистем), допустим организмов, органов или биоценозов, с окружа- ющей внешней [живой и (или) неживой] средой. Эта борьба всегда включает в себя направленное действие живой системы, несмотря на чье-либо противодействие, за кого-либо или чего-либо и даже только в стремлении к чему-либо ради сохранения своей жизни и (или) жизни друтих биосистем. Чтобы когда-то однажды состоялся первый акт борьбы, нужна была причина, на основе которой у кого-то, а точнее, чего-то, как-то про- явился стимул за что-то «побороться». Бороться без причины, неизве- стно с чем и за что, вряд ли кто-либо будет и кто-либо когда-либо вообще начал бы такую борьбу. Живое сумело возникнуть и существует по сей день на Земле только потому, что избрало главным содержанием себя борьбу за существование (за жизнь), причем, в первую очередь, на основе своего единства с неживой природой, борьбу с ней за исполь- зование ее ресурсов для жизнеразвития. На основе единства и борьбы двух противоположностей — живого (жизни) и неживого (нежизни) были в процессе эволюции найдены пути преодоления противоречий, в результате чего живое взяло относительную власть над неживым (в абсолютном выражении неживое пока остается сильнее живого), где-то полностью подчинив себе нежизнь, а где-то проявив способность идти с ней на компромиссы, поскольку живое постоянно не только берет, но и что-то возвращает в неживое, хотя и не в первородном, а трансфор- мированном виде, обогащая неживое некротическими продуктами жиз- недеятельности. По большому счету, жизнь — это экобиоз, т.е. суще- ствование любой биосистемы в определенном «доме» (месте жизни, или биотопе), который является обиталищем для нее как его обитателя.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Всякая борьба за существование предполагает адаптации, т.е. ка- кие-либо дискретные преобразования живой системы в пространстве и во времени, что возможно только в процессе жизнеразвития. Адаптации противоречивы, поскольку они находятся в деятельном арсенале диск- ретной жизни соответствующих телесных систем, поэтому раньше или позже, но в любом случае всегда такая жизнь включает и свою проти- воположность, особую адаптацию — смерть (см. главу 7). На основе противоречий между бесконечностью (континуальнос- тью) и конечностью (дискретностью) в организации природных систем возникло особое состояние материи — «подбесконечное» (субконтину- альное), которое больше, чем дискретность, и меньше, чем континуаль- ность. Носителями субконтинуальности в телесных системах стали про- гены и гены. Они долгое время могут не изменяться, сменяя, при воз- можности, лишь места своей «дислокации» (местопребывания) в диск- ретных телах, сохраняя постоянство своей телесной субконтинуальной организации, и только под воздействием особых факторов (соответ- ственно, субмутагенов или мутагенов) способны превращаться в друтие прогены или гены. Разными путями, не подвергаясь субмутациям или мутациям, и прогены, и гены, не изменяясь и не сливаясь с себе подоб- ными другими прогенами и генами, но нередко смешиваясь в группах, при пространственно-временном перемещении в разнообразных теле- сных системах, например, в химических реакциях или при репродуктив- ных процессах в организмах, сохраняют свой особый статус — субкон- тинуальный. Это особенно заметно на фоне сравнительно лабильных конституций (телосложений, соответственно, парафененетических или фенетических), которые являются индикаторами дискретности опреде- ленных неживых или живых тел и в структуре которых временно пре- бывают какие-либо носители субконтинуальности — понтикулы (проге- ны или гены). Особое субконтинуальное свойство понтикул сохраняется при всех дискретных телодвижениях через конституцию тел, когда одно состояние парафеносомы (у неживых тел) или феносомы (у живых тел) сменяется другим и, наконец, конституции соответствующих тел пре- кращают свое существование, исчерпав свой ресурс, свидетельствуя о прекращении существования прежних тел. В это время прогены или гены продолжают свое субконтинуальное существование, если сумели стать частями других тел. Итак, противоречие между континуальностью и дискретностью в существовании материи преодолевается развитием в телесных системах ее особого «буферного» свойства — субконтинуаль- ности, обеспечивающей определенную преемственность в развитии организации телесных систем. Незаметно мы вышли еще на одно противоречие, которое состав- ляет содержательную сторону организации жизни на Земле, предвестни- ки которого были заложены уже в неживой природе. Наличие субкон- тинуальности в организации телесных систем предполагает превосход- ство ее носителей — понтикул (прогенов или генов), над носителями дискретности этих же тел — пластонами (соответственно, парафенами или фенами), поэтому принцип разрешения противоречия между суб- континуальностью и дискретностью предопределен их носителями.
Глава 9. Концеиция экогенобиоза Понтикулы всегда доминируют над пластонами (глава 1), проявляя в отношении деятельности последних обязательный приоритет своей де- ятельности. Все это привело в нашей работе к разработке концепции разномасштабной биотической власти, или гетерократии (глава 7). Вся организация живых тел управляется с генетического уровня, а исполни- тельской деятельностью под властью генов занимается конституция (феносома) живых тел и, в частности, биоморф семенных растений. Из предыдущего материала видно, что противоречия между субкон- тинуальностью и дискретностью, которые проявились в процессе эво- люции материи уже в неживой природе как противоречия между носи- телями преднаследственности (прогенами) и парафенетической органи- зацией неживых тел, в живой природе, возникшей из неживой природы, предопределяют противоречия между носителями наследственности (ге- нами) и фенетической организацией живых тел, которые разрешаются путем проявления у них, в частности, у биоморф семенных растений, пластичности и гиперпластичности (глава 6). Пожалуй, еще одно противоречие, о котором здесь следует сказать и содержание которого мы уже вскрыли (глава 2), — это противоречие между относительно своим, личным (вегетативом) и относительно не- своим, другим, или коллегиальным (эксвегетативом и, в особенности, репродуктивом), в организации живых тел, в частности, биоморф. Оно разрешается подчинением личного коллегиальному, что, однако, далеко не всегда происходит на основе экспрессивного альтруизма, когда лич- ная жизнь в организации живых тел жестоко подавляется другой, более сложной, например, видовой, жизнью, также присутствующей в органи- зации менее сложных живых тел, например, организмов, и приводит личную жизнь соответствующих тел (носителей и своего, и не-своего) к абсурду и даже суперабсурду (простейшие формы проявления актив- ной жизни, например, многие дробянки — Schisophyta и, тем более, вирусы). У более высокой по сложности организации растительной жизни, как например, в организации биоморф семенных растений, личное и неличное адаптировалось проявляться в эволюции посредством множе- ственных компромиссов, став их нормой жизни, которая позволила и личную жизнь проявлять выразительно и достойно, да и другой жизни, например, виду, служить с не меньшей пользой. 9.2. «Мал золотник да дорог», или торжество биотической власти лри разрешении противоречий в организации живых телесных систем Почему так получилось в эволюции живой природы, что внешне (и только!) мало чем похожие на что-то живое ультрамикроскопические тельца, которые удачно назвали генами, стали самым дорогим (значи-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ мым) ее завоеванием? По существу гены и их разные комплексы, вклю- чая генонемы (нити ДНК) и геносомы каких-либо живых тел (от орга- нелл до биостробы Земли), составили первый (самый важный, хотя и самый нижний, такое, оказывается, тоже бывает) обязательный уровень в иерархии соматической организации живой природы — генетический. Ситуация с определением места генетического в иерархии телесных систем Природы неоднозначная и, вероятнее всего, генетический уро- вень — это особая полуветвь в сторону от молекул, замыкающаяся на биополимеры (в частности, белки) основного восходящего направления соматического развития жизни. О генетическом уровне можно, пожа- луй, говорить еще и как о параорганоидном, поскольку тельца, принад- лежащие этому уровню организации (гены и их комплексы-генонемы), которые чем-то даже чуть напоминают органеллы (органоиды), но та- ковыми ни в коей мере не являются (в широком смысле их, наверное, можно будет считать параорганоидами, или параорганеллами, хотя, есте- ственно, решающее слово за генетиками, которые пока, по вполне понятным причинам, более чем осторожны при определении статуса этих известных телец — носителей наследственности в иерархии орга- низации живой природы). Место «параорганоидов» в иерархической организации живой природы вроде бы нижнее по сложности их орга- низации, поскольку они, хотя на мой взгляд, и принадлежат живой природе, но все-таки еще весьма недалеки и от неживой природы, и все- таки выполнение этими известными тельцами руководящей функции при становлении организации всех остальных носителей жизни — впол- не надежное обоснование, чтобы рассматривать их как особую (генети- ческую) полуветвь ультрамикроскопического развития жизни. Итак, самое дорогое, хотя и весьма ультрамикроскопическое, завое- вание жизни в ее эволюции на Земле, — это возникновение генов. Возмож- но, что жизнь на Землю была занесена уже в готовом виде, но это не снимает проблему, а расширяет ее границы и на какие-то другие про- странства Вселенной, в организации живых тел которой тогда также властвуют гены. Живая природа сумела состояться в окружении неживой природы в первую очередь потому, что она, однажды возникнув, в процессе эволюции нашла механизм своего развития и сохранения на основе более сложной организации в себе такого порядка, отличного от поряд- ка в неживой природе, в центре которого стала особая управляющая деятельность генов, которые сумели создать такую эволюционно актив- ную систему проявления организационного порядка живых телесных систем (биосистем, или биосом), которого до нее никогда не было. Таким образом, жизнь, благодаря генам, стала по возможностям своего развития выше и сильнее нежизни на Земле. В седьмой главе было показано, что в организации соматической жизни на Земле действует целый «клубок» биотических властей — ге- теробиократия, управляющая жизнеразвитием в генеральном направле- нии — жизнесохранении вообще. В организации живых тел, в частности и биоморф семенных растений, приоритетные цели жизнеразвития опре- деляют властные структуры, среди которых стратегами являются только
Глава 9. Концеиция экогенобиоза гены, в подчинении которых находится основное феносоматическое развитие. Благодаря функционированию генов конструируются конститу- ционные (фенетические) тела — части биосистем как феносоматические «дома» (автобиотопы) для генов. Все идиоэнергетическое функциониро- вание генов в организации живой природы вообще нацелено на то, чтобы решающим образом «играть» на противоречиях, которые неиз- бежно сопровождают все природные явления. Мы выходим на пробле- му, которой собственно и посвящена эта глава, но подробности ниже, а здесь только подчеркнем, что из многих противоречий, которые разрешаются на основе биотического властвования генов — стратегов жизнеразвития, самое главное то, на которое они же, гены, и «напо- ролись», создав его сами по своей инициативе в своей эволюции,— это противоречие генов и фенов, геносомы и феносомы. Однажды они, гены, а точнее их древние преджизненные предки — гемигены~, впервые создали фены — живую оболочку для генов, что возвестило о превраще- нии гемигенов в гены, а во Вселенной появилась «виновница» всего тор- жества — Жизнь: она пришла в противоречивом единстве генов-фенов как природная структурная часть и как одна из высших природных функций, развиваясь дискретно в телесных системах. Дискретность живых тел предопределяется организацией феносом, поскольку они составляют самое лабильное содержание в целых живых телах, обес- печивая им целостную дискретность, а наиболее консервативная гене- тическая часть (геносома) этих же живых тел, которая на протяжении циклов существования таких тел (морфогенезов) часто вообще не из- меняется, обладает субконтинуальностью, т.е. свойством, которое про- тиворечит дискретности феносом. Противоречие систем «гены-фены», а шире «геносомы-феносомы», т.е. по сути соответствующих функциональ- ных генетических систем-союзов (почти «тандемов») — «генофенов» в организации любых живых тел, преодолевается стратегическим давлением биотической власти генов. Она принуждает фены — тактические испол- нители целевых программ своих руководителей (генов) заниматься вос- произведением, подобных родительским, живых тел, что выражается у биоморф в их размножении, на основе которого прежняя фенетичес- кая организации одних тел сменяется на таковую организацию других тел — потомков, а генетическая организация, сохраняя свою стабиль- ность (без мутаций) и только изредка изменяясь (с мутациями), пере- дается новым поколениям. Гены обладают уникальной способностью при половом размножении своих носителей, допустим биоморф се- менных растений, смешиваться с другими генами, но не сливаться и тем самым сохранять свою относительную автономность и субконти- нуальность во времени, иногда многие миллионы лет, когда за то же время феносоматическая дискретность отправит «на кладбище» десят- ки тысяч поколений биоморф, шире — других живых тел, как носи- телей генов, отправив с ними в небытие множество фенов и не сумев (фенам это было ни к чему) при этом еще и «похоронить» свои гены.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 9.3. Жизнь как способ существования генов для фенов и фенов для генов в их союзах- «генофенах» Ранее (т. 1, гл. 2, с. 23), исходя из многих известных фактов, автор дал свое определение «жизни», которое среди многих определений, раз- работанных другими исследователями, по содержанию оригинально: «жизнь — это существование материи в качестве генов и в ином каче- стве под управлением генов, смешивающихся между собой и трансфор- мирующихся в пространстве и во времени». Гены могут смешиваться друг с другом, например, при половом размножении организмов, но не сливаются, сохраняя достаточную организационную автономию и отве- чая за развитие определенных признаков или группы признаков— фенов. «Феном называются любые дискретные альтернативные вариации признаков и свойств живых организмов, которые на всем имеющемся материале (обязательно многочисленном) далее неподразделимы без по- тери качества» (Яблоков, 1980, с. 51). Говоря об «ином качестве» в оп- ределении жизни, я как раз подразумеваю фены и всю их совокупность — фенетическую организацию, или феносому, какого-либо живого тела. Главный акцент в определении явления «жизнь» мною сделан на гены; из чего однозначно следует, что жизнь везде начинается от генов и только гены — первые обязательные носители жизни, или живые системы, ко- торые не просто олицетворяют собой жизнь, хотя и ультрамикроскопи- ческого содержания, но и управляют, в силу своей особой организации, развитием ее исполнительской стороны — определенных фенов и как признаков, и как структурных элементов фенетической организации (феносомы). Гены и фены выступают в организации биосистем только вместе, составляя противоречивое соматическое единство (другого не дано). Применение терминов «геносома» и «феносома», «генетическая» и «фенетическая» организации я не ограничиваю, как уже выше подчер- кивалось, только организмами, а рассматриваю их широко в отношении любых живых тел (биосистем), где они себя проявляют; при этом вза- имодействия генетического и фенетического, в зависимости от уровня организации биосистем, естественно, будут трансформироваться. Гены и их определенные комплексы (генонемы и геносомы) сосуществуют вместе с фенами и их определенными комплексами (феносомами), которые настолько взаимосвязаны, что вполне можно говорить об их «союзах». В определенных союзах генов и фенов, или «генофенов», геносом и феносом, или «генофеносом», роль ведущего (управляющего) принадлежит, соответственно, генам и генетической организации (гено- соме), а роль ведомого (исполняющего) — фенам и фенетической орга- низации (феносоме). Генофен — это функциональная (выборная) система, которая включа- ет в себя только те структурные части и процессы, которые непосредствен- но в организации живого тела задействованы на пути преобразования идио- энергии гена в организацию фена как структурного элемента и признака
Глава 9. Концеиция экогенобиоза соответствующего тела — его носителя. Гены в организации биоморф семенных растений (шире — любых биосистем) выступают как един- ственные носители стратегической власти, приоритетом существования которых является управляющая роль в развитии фенов, а, следователь- но, и всей фенетической организации соответствующих живых тел. Определенные союзы генов с фенами возникли в процессе длительной эволюции жизни и дошли до наших дней в традиционном типовом качестве, когда гены управляют развитием определенных фенов. Фенетическое содержание (феносома) у живых тел любой сложности организации — это одновременно живое «обиталище», если хотите, «дом», или автобиотоп, для генов. Генетическое обиталище можно другими словами образно обозначить еще и как «экодром» (от греч. oikos дом + dromos место для бега, бег), или даже «геноэкодром», отличая в нем две стороны (среды): внутреннюю (непосредственно внутри живого тела) и внешнюю (вне живого тела, действующую в наше время в отношениях с окружающей внешней средой только опосредованно, через фены, но потенциально возможную еще и в непосредственном выражении, на- пример, в эпоху зарождения жизни на Земле). Отмеченную особенность обиталищ для генов далее мы используем при выяснении возможного пути происхождения жизни. Фены полезны для генов, которые, в пер- вую очередь, их для себя же и производят, чтобы посредством их защи- титься от воздействий окружающей внешней среды, но гены полезны и для фенов, как живых структурных частей — соматических фрагментов жизни, поскольку без генов фены никогда не смогли бы вообще по- явиться и никогда бы не смогли развить свою организацию, обладаю- щую свойством, благодаря генам, преемственно-исторически восстанав- ливаться на фоне дискретности живых тел в едином субконтинуальном, а, не исключено, потенциально континуальном едином живом сомати- ческом потоке — в жизни вообще (Жизни). Есть смысл уточнить опре- деление фенов — это, прежде всего, элементы пластосомы (конститу- ции) — соматические структурные элементы, составляющие соответ- ствующее содержание живых обиталищ (автобиотопов) для генов в це- лостной организации живых тел разной сложности, отвечающие, на иди- оэнергию генов и воздействие окружающих условий, жизнеразвитием каких-либо трансформаций, которые выражаются в определенных при- знаках и свойствах. Благодаря фенетической организации (феносоме) и ее элементам— фенам, соматическая жизнь приобретает особый (специфический) смысл, заметно отличающий ее от нежизни. Без фенов существование генов было бы непонятно. Зачем тогда, без фенов, все это, под назва- нием «жизнь», надо, поскольку «жизнь» без фенов мало чем отличалась бы от нежизни; цели неживых тел, развивающихся без участия живых тел, весьма однобоки и с позиций живой природы бесполезны, если они не направлены на возникновение и (или) поддержание жизни. Жизнь без фенов существовать бы не смогла, поскольку главным «зачинателям» жизни — генам, нужен надежный биотоп-приют, т.е. такое обиталище («дом»), где можно было бы и себя, как минимум, субконтинуально, сохранить и решить проблему преемственно-исторического поддержа-
738 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ ния среды своего (генетического) обитания, если хотите, самовозобнов- ляющегося «домостроительства» для генов, на фоне всеобщей сомати- ческой дискретности. Внешняя неживая среда как «дом» для существо- вания живых тел, весьма ненадежный экодром (внешний), хотя именно с такого «обиталища» первые гемигены — носители преджизни, веро- ятно, начали путь к жизни, поскольку она лишена способности поддер- живать свое относительное постоянство как важное условие среды оби- тания генов. Гомеостаз (относительное постоянство внутренней среды) могут обеспечить только фены, развивающиеся под управлением генов. Строго подогнанные в эволюции союзы генов и фенов в каждом целостном живом теле — это принципиальная особенность организации биосистем (биосом) разных уровней сложности. Однажды возникшие союзы «гены-фены», или «генофены», стали в дальнейшем принципи- альной основой в соматической организации жизни и решающими объектами в ее эволюции. 9.4. Эволюция союзного образа жизни генов и фенов в организации биоморф Существует, как известно, много гипотез происхождения жизни на Земле. Поиск путей возникновения жизни, как подчеркнул американ- ский ученый Дж. Кеосян (1975, с. 118), «свелся к двум основным под- ходам. Согласно одному из них, ген признается основой жизни и ее происхождения. Согласно другому, микросистемы, именуемые микро- каплями, коацерватами, микросферами или предбиологическими систе- мами, рассматриваются как предшественники первых примитивных клеток. Каждый подход тем не менее допускает, что первые живые существа, гены или клетки, возникли более или менее внезапно из окружающей среды». Из обзорной статьи Дж. Кеосяна (1975), мне бли- же гипотеза генного происхождения жизни Мёллера (Ми11ег, 1929), который развил созданную ранее (Troland, 1914, 1916, 1917) гипотезу «живого энзима», обеспечившего начало жизни на Земле. Как подчер- кнул Кеосян (1975, с. 119): «Мёллер заменил термин «энзим без орга- низма» Троленда на термин «голый ген» и к двум предложенным им критериям жизни — автокатализу и гетерокатализу — добавил способ- ность к мутациям... Случайная комбинация всех необходимых для об- разования гена нуклеотидов не является вероятным событием, посколь- ку постулируется синтез только одной молекулы. Это событие обладает вероятностью, стремящейся к нулю. Более того, «энзим жизни», или ген, является живой субстанцией только потому, что это утверждают авторы». На мой взгляд, жизнь на Земле вряд ли началась просто с «живых энзимов» и даже с «голых генов», которые могли появиться в преджизни (до возникновения собственно жизни), но все же с палео- генов, которые развились из движущейся материи, проявив экогеноби- оз: они впервые построили сами для себя фенетическую (белковую или
Глава 9. Концеиция экогенобиоза белковоподобную) оболочку и сохранили эту способность в последую- щих поколениях экогенобионтов, — палеоорганизмов, т.е. происхожде- ние жизни на Земле рассматривается здесь в рамках экогенобиозной концепции. Гены как носители наследственной идиоэнергии нуждались с мо- мента своего появления на Земле в убежище, более надежном биотопе, чем окружающая, весьма агрессивная, неживая внешняя среда; в каче- стве автобиотопа генов — живым «домом» для них внутри какого-либо целостного живого тела, например, организма, стала фенетическая орга- низация (феносома), т.е. система всех фенов этого живого тела. 9.4.1. Почему жизнь — это, в первую очередь, не больше, чем строительство и обслуживание обиталищ для генов Генам давным-давно была заказана «музыка» жизни, нх далекие пред- ки ее ~сочинили», написали ~ноты», затем гены постепенно корректировали свою «музыку» н сейчас, используя накопленный опыт, успешно «дирижи- руют» оркестром (феносомой), который прилежно исполняет «снмфонню» жизни, интересную н для тех, н для других, поскольку они всегда действу- ют в союзах, определяя содержание нх целостного существования в каком- либо общем живом теле. Так образно можно представить течение сома- тической жизни. Однако нам, похоже, для понимания принципов про- исхождения и становления жизни на Земле такая лирическая возвышен- ность стиля к оценке разнообразия «тонов» и «красок» этого явления вряд ли сейчас будет полезна. Жизнь рождалась в муках, в суровом ок- ружении враждебной неживой среды. Это была более всего тяжелая буд- ничная работа на «строительной площадке» — в неживой природе, резуль- татом которой за многие миллионы лет эволюции стал универсальный принцип жизнеразвития, согласно которому возводятся, обслуживаются и сносятся, при необходимости, «домики» и «дома» для генов, их обиталища (автобиотопы). Гены по отдельности и во всей их совокупности- геносоме, внутри какого-либо целостного живого тела ведут себя по отношению к фенам и их комплексам (феносомам) как хозяева к своему «многокомнатному домику», построенному по их же проекту и под их управлением в окружающем пространстве как способ его освоения жизнью во времени. Гены управляют не только строительством своего фенетического «дома», снабжая его идиоэнергией, но и участвуют в поддержании функционирования его живых «комнат» — структурных элементов (частей тел — меросом), направляя к ним соответствующие «идиоэнергетические кванты» (порции программно-энергетического обеспечения), например, управляя соответствующим образом ростом и развитием белковых систем побегов и корней у биоморф семенных растений и т.п. Эволюция союзных отношений генов с фенами, если бы пошла по принципу: фены и их комплексы — это только обиталища для генов, то
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ какой бы прок был от этого для жизни вообще и получилось бы тогда то биосоматическое разнообразие, которое имеет место быть сейчас? Вопрос поставлен ребром. Действительно ли то разнообразие фе- нетической организации жизни на Земле, которое имеет место сейчас, достигнуто эволюцией только одного, пусть самого глубинного прин- ципа организации явления «жизнь» — эволюцией «домостроительства» для генов, или экогенобиоза. Ответим сразу: да, экогенобиоз — это основа разнообразия жизни на Земле, это сама жизнь как строительство и обслуживание фенетических обиталищ для генов, это основополагаю- щий принцип эволюционной организации жизни на нашей планете, а, возможно, и жизни вообще (Жизни). В условиях экогенобиоза постоян- но существует перспектива для развития фенетического разнообразия биосистем, поскольку оно жизненно необходимо генам: чем больше такое разнообразие, тем больше возможностей сохранить хотя бы часть генов в особо экстремальных, даже катастрофических условиях, на- пример, при столкновении кометы с Землей и т.п. Таким образом, по большому счету, все, что происходит в живой природе, — это системное, в первую очередь, союзное (генофенетичес- кое) строительство и обслуживание фенетических обиталищ для генов — экогенобиоз со всеми вытекающими из этого последствиями, напри- мер, эволюцией «ошеломляющего» фенетического разнообразия. Для фенетического строительства живых тел используется генетический арсенал жизни — идиоэнергия этих тел (наследственный информацион- но-энергетической потенциал), который на Земле поддерживается и развивается через эволюцию разнообразия фенетической организации жизни. Феносомы живых тел более всего подвержены эволюционным трансформациям, которые приводят или не приводят конкретные гены к мутациям. На основе эволюции экогенобиоза была создана строгая система «управления-исполнения» в организации биосистем и, в част- ности, биоморф семенных растений, на основе которой стратегическая власть генов обеспечила выполнение основных процедур соматического жизнеразвития обиталищ посредством совершенствования исполнитель- ской деятельности, например, посредством «могучей пятерки» у семен- ных фитобиоморф (глава 7.4). Ранее (т. 1, гл. 3, с. 98 — 99) был определен закон единства гене- тического и фенетического, на основе которого «фенетическое — обя- зательная естественная среда существования (естественный биотоп) ге- нетического, поскольку сохраняет его в себе, непосредственно сама взаимодействует с окружающей внешней средой и опосредованно, через себя, передает к нему сигналы внешней среды, влияя на его (генетичес- кое) содержание». С этим законом непосредственно связано правило экогенобиоза, которое выводится из рассмотренного выше материала: чем разнообраз- нее феносоматическая жизнь как тактическое обиталище для генов в союзной (генофенетической) организации живой природы, тем шире и на- дежнее геносоматическое содержание жизни как существование страте- гических обитателей (генов) внутри фенов.
Глава 9. Концеиция экогенобиоза 9.4.2. Что нужно, чтобы обиталища для генов не превратились для них в безутешные «кладбища» Гены, однажды возникнув во Вселенной, стали на пугь разрешения противоречий в качестве основных «игроков» в «игре» противоположно- стей. Гены, а точнее, их далекие предки, превратили единство и борьбу противоположностей в организации жизни на Земле в свою «игру», которая могла состояться с их участием только под одним девизом, исходящим еще от генетических потомков, образный смысл которого в следующем: ~мы, гены, — властоносцы живой природы и все, что де- лается в ней, не должно осуществляться без пользы для нас». Гены, будучи одной из двух живых (семивитальных) частей — меросом био- систем (другую живую меросому составляют фены), владеют неоспори- мым лидерством в развитии современной организации живой природы, взяв в процессе эволюции власть над ней на себя, в ранге ее стратегов, сумели создать такую систему жизнеподчинения себе всех остальных структурных элементов — фенов, которая стала исправно обеспечивать для них, генов, место их непосредственного обитания в биосистеме- их обиталище, или автобиотоп. Гены, создав в процессе эволюции для себя обиталище (могут в нем поселяться еще и паразиты, но речь здесь совсем не о них) в любом живом целостном теле с союзной (генофене- тической) «начинкой» как фенетическую организацию этих тел, исполь- зуют ее, будучи сами субконтинуальной организационной частью, не только для того, чтобы содержаться и конвариантно реплицироваться, иногда мутировать, но еще и для того, чтобы успевать передавать хра- нящуюся в них наследственную информацию (в целом — идиоэнергию) на протяжении жизни всего дискретного тела. Генам, чтобы не превра- тить каждый свой автобиотоп в безутешное «кладбище», надо было в эволюции создать систему ориентации в меросомной специализации к со- юзному, или генофенетическому, жизнеразвитию разных живых тел с обя- зательным стремлением их к воспроизведению себе подобных. Феносомы биоморф семенных растений как обиталища для их руководящих обитате- лей — генов (геносом в целом) должны расти и развиваться посредством «могучей пятерки» (нарастания, ветвления, партикуляции, размножения и элиминации) и, что особенно важно, в том числе, стремиться под управ- лением генов к размножению (это первый принцип фенетической органи- зации биоморф, см. т. 1, zn. 4). 9.4.3. Нежизнь — преджизнь — жизнь — вехи в эволюции экогенобиоза В науке чаще всего бывает важно понять принципы чего-либо про- исходившего и (или) происходящего в Природе, т.е. самые общие, но все-таки главные стороны, на основе которых совершаются какие-либо явления; при всем этом особенно полезно бывает создать простран-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ ственную модель соответствующих явлений. «В сущности, расшифровка генетического кода — открытие, ознаменовавшее собой рождение молеку- лярной биологии, — было прежде всего подвигом пространственного вооб- ражения... Ф. Крик и Дж. Уотсон сумели вообразить целостную про- странственную структуру, сумели свести воедино, объединить в про- странстве множество отдельных деталей так, что представленная ими конструкция, обладающая формой двойной спирали, имела единую организацию и единство действия» (Энгельгардт, 1984, с. 150). Пожалуй, автору (т. 1) удалось понять универсальный принцип свершения эволюционных процессов всего, что имеет место быть в Природе — это свободное «плавание» колеблющихся эволюционных маятников (тел и нетел) во вселенском потоке и пространства, и вре- мени. Здесь мы новых деталей конкретной эволюции биосистем откры- вать не собираемся, но попробуем разобраться с некоторыми эволюци- онными принципами происхождения и поддержания жизни на Земле. 0 происхождении жизни много литературы, например, только в одной книге Э. Брода «Эволюция биоэнергетических процессов» (1978) цити- руется 2063 научные работы, что свидетельствует о повышенном инте- ресе ученых к этой проблеме. 9.4.3. 1. Мотивация неизбежности возникновения жизни на Зеиле Сначала эволюция в неживой природе Земли была в своей основе химической. «Проведенные эксперименты подтверждают идею химичес- кой эволюции — представления, согласно которому все малые органичес- кие молекулы, обычно используемые в биологических процессах, обра- зуются из более простых компонентов при воздействии на них различных видов энергии» (Фолсом, 1982, с. 71). «К биоэлементам относятся угле- род, водород, азот, кислород, фосфор и сера (CHNOPS). Почему именно эти, а не другие? Почему основой жизни не стал кремний, а основой дыхания — фтор? Все биоэлементы присутствуют в космосе в достаточ- ном количестве и каждый из них незаменим для той функции, которую он выполняет. Эти элементы, за исключением, может быть, фосфора, представляли собой газы и изначально были той формой вещества, на которую воздействовала энергия излучения в первую очередь. Все, что было в наличии — С, Н, N, 0 и S в виде газов, — должно было реаги- ровать, где и как только возможно, и поэтому мы такие, какие мы есть. Нет никаких оснований ожидать чего-либо иного для любой другой планеты земных размеров, находящейся в соответствующей области тем- ператур относительно своей звезды. Почему не кремний? Потому что этот элемент все время участвует в построении кристаллических пород. Крем- ний не имеет газообразной формы, на которую могла бы воздействовать энергия излучения. Какой бы организм мы ни анализировали, результат всегда будет одним и тем же: более 90% его массы составляют биоэлемен- ты CHNOPS» (Фолсом, 1982, с. 76). Вполне доказано, что управляют развитием живых тел в Природе гены. Каждый ген — «это участок не- прерывной молекулы ДНК..., сама ДНК (или, точнее, последовательность оснований в ней) определяет конец одного и начало другого гена» (Рол-
Глава 9. Концеиция экогенобиоза лер, 1978, с. 242). «Двухцепочечная полимерная молекула ДНК состоит из азотистых оснований — аденина (А), тимина (Т), цитозина (С) и гуанина (G), каждое из которых соединено с сахаром (дезоксирибозой). Последовательно расположенные молекулы сахара удерживаются вместе фосфодиэфирными связями... Остов молекулы составляют две сахаро- фосфатные цепочки. Информация, хранящаяся в этой молекуле, заклю- чена в последовательности ее оснований. Каждая цепь ДНК состоит из сахарофосфатного остова и оснований, отходящих от остатка сахара, и имеет химически заданное направление. В целом, молекулу ДНК состав- ляют две цепи, сложным образом закрученные одна вокруг другой, иду- щие в противоположных направлениях и удерживаемые вместе слабыми водородными связями, образующимися между основаниями...» (Фолсом, 1982, с. 112). В целом структура ДНК, генов и их роль в соматической органи- зации живой природы к настоящему времени во многом понятны (Ду- бинин, 1976; 2000; Уотсон, 1978 и др.). Размышляя над местом и ролью генов в эволюции живой природы, я склоняюсь к идее, что именно приоритет возникновения наследственных структур из преднаследственно- сти неживой природы сыграл решающую роль в происхождении и станов- лении жизни на Земле. «Гены обладают тремя удивительными свойства- ми. Они сами себя удваивают в каждом поколении клеток: гены способ- ны неопределенно долгое время воспроизводить себя (реплицироваться) без изменений. Чрезвычайно редко ген может измениться, и этот изме- ненный, мутировавший ген в свою очередь воспроизводится и переда- ется новым поколениям без каких-либо вариаций: редкие изменения, называемые мутациями, таким же образом воспроизводятся неопреде- ленно долго. Все множество генов находится в однозначном соответ- ствии ко всему множеству белков, имеющихся в клетках данного орга- низма: принцип «один ген — один белок» означает, что каким-то об- разом ген точно определяет последовательность аминокислот в молеку- ле белка» (Роллер, 1978, с. 193). Попробуем наполнить эволюционную ось «нежизнь ~ преджизнь ~ жизнь» более ощутимым содержанием: «нарисуем» пространственные образы, пусть весьма обтекаемые, где-то поверхностные, но отражающие суть эволюционного принципа перехо- да от неживого к живому. Для явления «Жизнь», где бы то ни было — на Земле или еще где- либо во Вселенной, не было ничего другого для выбора, кроме как использовать для своего возникновения только неживую природу, и она полностью «отдала» себя, особенно в своем начале, во власть, как может показаться, необузданной природной стихии, где на самом деле при всем кажущемся беспределе не царит хаос, а всегда был, есть и будет определенный порядок. Все дело во времени и тех конкретных обстоятельствах развития материи, которые складываются в каких-либо частях бесконечного про- странства. Природа никогда и никуда не спешит и делает все так быстро или медленно и в таком количестве и качестве, как по времени и месту это у нее получается, ведь она бесконечным эволюционным образом творит саму себя, исходя из присущих ей законов и правил только ее
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ организованности. Не исключено, что жизнь на Земле — явление не самородное для этой планеты, а заносное. Пока оба варианта (самоза- рождение на Земле и занос готовой жизни на Землю извне) рассматри- ваются учеными как равнозначные, покуда не будут получены факти- ческие подтверждения в пользу одного из них или обоих, что однако не мешает уже сейчас обсуждать вопрос: почему жизнь способна вообще возникать во Вселенной? Там, где нет жизни, действуют те же природ- ные законы, что и в пространстве, где жизнь уже есть. Жизнь, в первую очередь, — это экобиоз (единство живого обитателя со своим обиталищем, или «домом»), следовательно, она — явление сис- темное, поскольку у жизни вообще и у ее живых структурных элементов всегда есть свое первоприродное обиталище — неживая природная среда, которым она (жизнь) непосредственно (напрямую) или опосредованно (через живого посредника), взаимодействуя, всегда пользуется. У нас, на Земле, сейчас господствует жизнь еще и как экогенобиоз, а это не только системность, но и нечто большее — это особый принцип союзного (генофенетического) соматического развития. Экогенобиоз — это и формы, и способы, включая и принципы, существования союзов (это слово подчеркивает не просто взаимодействие, как слово ~система~, а особенное целесообразное, на что-то направленное взаимодействие) генов с фенами и их комплексов, а много раньше — союзов палеогенов с палео- фенами и их комплексов, что составляет организационное содержание жизни в телесных системах, на основе которого гены (а раньше — палео- гены) решают (и решали) развитием соответствующих фенов (раньше- палеофенов) свои «жилищные» проблемы, а все остальное только допол- няет эволюцию живых телесных систем как генофенетических союзов. Такое (и во-первых, и во-вторых) могло произойти только в процессе эволюции неживой природы, поскольку у нее есть мотивация к возник- новению жизни в подходящих для этого условиях на основе преднаслед- ственности как наименее сложной формы преемственности — атрибу- тивного свойства материи быть в определенных телесных системах. Жизнь не может возникнуть иначе, чем из неживого материального субстрата. Вопрос о мотивации возникновения жизни перед неживой природой стоит как бы между прочим — на равных с множеством других мотиваций, но первая мотивация интересует более всего нас, людей, поскольку только мы испытываем желание научиться понимать и самих себя, и ту жизнь, которая нас как-то окружает, а кроме того, еще по- думываем и о том, чтобы адаптироваться к управлению развитием жизни вообще; задача сложная, далекая от сегодняшнего дня, но вполне логич- ная для Ното sapiens. Единство и борьба противоположностей в саморазвитии неживых телесных систем и их элементов, случайная фиксация противоречий и поиск физико-химических путей их разрешения происходило в нежи- вой, весьма агрессивной, среде первобытной Земли и только на основе преднаследственности, чья идиоэнергия исходила от прогенов (атомов) — главных виновников тех трансформаций, через которые началось тернистое восхождение по пути к явлению «жизнь». В эволюции мате- рии, в результате которой была в конце концов создана жизнь, я реша-
Глава 9. Концеиция экогенобиоза ющее значение отвожу эволюции идиоэнергетических носителей: от проге- нов (атомов) — единиц преднаследственности, которая составляла и со- ставляет по сей день преемственность неживых телесных систем, до генов (участков молекул ДНК) и генонем (целых молекул ДНК) — единиц на- следственности, которая адаптировалась представлять преемственность я<иных телесных систем. Пропасть между нежизнью и жизнью долгое время заполняла собой эволюирующая материя, которая где-то и когда- то сумела стать преджизнью, связав, тем самым, казалось бы, несовме- стимое. Возникновение преджизни шло методом проб и ошибок в нежизни, но от вполне реальной преднаследственной основы и, в первую очередь, давление эволюции пришлось на идиоэнергетических носителей пред- наследственности — прогены (атомы), которые под давлением агрессив- ных условий окружающей среды (обилие воды, света, радиации, мно- жественных химических реакций и физических катаклизмов в форме извержений вулканов, землетрясений и т.п.) местами стали, где-то 3— 3,5 миллиарда лет назад, превращаться в гемигены и их комплексы- гемигенонемы как разнообразные переходные состояния материи меж- ду прогенами и первобытными генами, или палеогенами (первобытные нуклеотиды), а также их комплексами — палеогенонемами (вероятно, целыми биополимерными первобытными молекулами ДНК). Я не буду проявлять особое усердие, чтобы разгадать тайну структуры гемигенов и гемигенонем, поскольку отлично понимаю, что когда-то это смогут действительно сделать профессионалы — xHMHKM, биохимики и генети- ки, а себе, здесь и сейчас, оставляю лишь одну возможность — теоре- тизировать в «пространстве» довольно общих, безусловно гипотетичес- ких, эволюционных принципов возникновения и становления жизни на Земле, а кто знает, может быть, и вообще в мироздании. Из комбинаций прогенов, вероятно, только из них, возникли в результате множественных химических трансформаций такие комбина- ции молекул и макромолекул, которые за многие миллионы лет эволю- ции и привели к возникновению предков генов — гемигенов и их ком- плексов (это уже не прогены, но еще и не гены, и, тем более, не генонемы) — носителей полунаследственности — структурных элемен- тов преджизни. Гемигены и гемигенонемы, на мой взгляд, отличались от прогенов не только структурно (они были организованы значительно сложнее), но и тем, что химически активно стали искать себе среду, которая могла бы их подольше защитить от разрушения и, вероятно, даже когда-то начали создавать (опять же только на основе химической специализации своей активности) неживые и вовсе необязательные для всех их оболочки для защиты от агрессии окружающей среды. У геми- генов поначалу вообще не было собственной оболочки, но постепенно они ее начали химически «изобретать» в определенных условиях агрес- сивной внешней среды или, кроме того, еще «учились» химическими путями уходить в более спокойные ниши неживого пространства. Про- цессы соединения и распада преджизненных структур, сопровождающие содержание их химического и физического существования, постепенно готовили полунаследственный материал к еще более весомым трансфор-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИЙ мациям — прорыву к явлению «жизнь». Под действием неживой окру- жающей среды преджизненные элементы довольно быстро распадались, чтобы в подходящих условиях вновь восстановить прежнюю химичес- кую структуру уже в качественных границах других тел — других геми- генов и гемигенонем. Носители преджизни пользовались и неживыми оболочками, кото- рые химически создавали вокруг себя, под защитой которых они могли долго и даже весьма долго сохраняться, не изменяясь. Жизни еще не было, но то, что происходило в целом, уже было преджизнью, а это достаточно много для эволюции материи, значительно больше, чем нежизнь. Вероятнее всего, на процесс происхождения жизни усиленно вли- яли различные проявления ритмичности в неживой природе. Так, на- пример, где-то в зоне пульсирующих водных, тем более термальных источников, ритмичность происходящего стимулировала сначала про- цессы превращения неживых прогенетических соматических комплек- сов в предживые, а затем и живые. Предживые соматические системы — гемигенонемы, с неживыми оболочками или без них, действовали в ритмической среде в унисон с ней: то восстанавливались, то распада- лись, то вновь восстанавливались в другие тела, но на прежней хими- ческой основе, и т.д., предваряя тем самым некоторые способы воспро- изведения живых систем. Жизнь, которая впервые возникла из преджиз- ни, имела формы ультрамикроскопических структурных образований, которые, если и можно было как-то разглядеть, то для этого надо было бы воспользоваться мощным электронным микроскопом. Это весьма любопытно, наша концепция среди первых палеобиоморф на Земле может принять только ультрамикроскопические системы палеогенов- палеогенонемы, более или менее обтянутые примитивной фенетической оболочкой. Такими мне представляются первые организмы на Земле- протероорганизмы. Геноцентризм, в авторской концепции происхожде- ния жизни на Земле, очевиден. Все, что связано с разнообразием гено- фенетической жизни: органеллы, клетки, ткани, органы и т.д. — это все пришло позднее, а начало жизни получилось вроде бы очень невы- разительным, пусть и революционным, но внешне довольно «жалким», поскольку ультрамикроскопические «комочки» материи — протероорга- низмы, представляющие из себя палеогенофенетические союзы (сома- тически единые, унитарные комплексы из палеогенов и палеофенети- ческой оболочки), были пока лишь только предвестниками организмов. Эволюция жизни, возникшей на Земле, пошла бурно по пути совер- шенствования фенетнческой организации ее носителей, которую палеогены и палеогенонемы превратили с самого начала нх «союзного» (функциональ- но-системного) сосуществования с отдельными палеофенамн н нх комплек- сами (палеофеносомамн) в свои обиталища — «дома» для себя, создав в них для своего существования внутреннюю среду (автобнотоп) и, таким образом, относительно изолировав себя палеофеносоматическим «футля- ром» от прямого воздействия агрессивных внешних факторов. Феносома первобытных живых тел была «брошена» их древней генетической орга- низацией на «передовую» борьбы этих тел за существование, что пре-
Глава 9. Концеиция экогенобиоза вратило палеогены в управленцев и контролеров палеофенетического развития, обеспечивая, через разнообразие палеофенов, надежность для себя таких обиталищ. Гены, управляя жизнеразвитием, буквально «ведут» фены по жизни, осуществляя это в первую очередь для себя. Гены «работают» с фенами как бы опосредованно, но, на самом деле, как с фенетическими функциональ- ными системами, которые начинаются с образования соответствующих белков и доходят до проявления конечного конкретного состояния этой системы, ее фена-признака. В организации современных биосистем, например, простых био- морф — организмов, как известно, далеко не все гены способны про- являть активность и участвовать в развитии соответствующих фенов. Часть генов могут находиться в заблокированном состоянии в генети- ческой «памяти» живых тел, например, организмов, и их идиоэнергети- ческий потенциал может многие годы, иногда, возможно, даже милли- оны лет, в чреде сменяющихся поколений, ждать своего «выхода» в «свет». Гены, вероятно, могут переводиться в «дремлющее» и даже «сон- ное», или заблокированное, состояние под влиянием разных факторов; эволюция их может на какое-то время, а иногда и навсегда, до полного исчезновения, «запирать». Генетическая «память» способна пробуждать- ся мутациями, которые могут и не создавать новых генов, а только выводить «сонные» гены из «дремоты», в которой они ранее пребывали, и заставлять их вновь «работать». Среди «запертых» генов, или «генов- сонь», может быть (это, безусловно, гипотеза — теоретическая догадка, возможность которой нельзя исключать, но которую надо пытаться экспериментально проверить) есть и такие представители древних ви- дов, родов и даже семейств, а то и классов и т.д., которых уже давно нет на Земле, а существуют только их далекие потомки, в которых в один, возможно, неожиданный для всех окружающих видов-современ- ников, момент вдруг наступит пробуждение некоторых из них, и они начнут «работать» в новых условиях. Так, допустим, где-то у некоторых деревьев разблокировались «гены травянистости» или чего-то подобно- го, что вполне могло бы дать, будучи поддержано естественным отбо- ром, эволюционный крен в сторону вполне IIQHHTHbIx трансформаций. «Гены-сони», прикрываясь идиоэнергетическим потенциалом активных генов, могут «затаиваться» на многие годы — «отлынивать» от работы. Именно такие гены и составляют генетическую «память» современных живых систем, из которой в определенных условиях что-то иногда может воскрешаться вновь (что это — «тени» далеких предков или нечто дру- гое), назовем это явление «генетическими ретроновациями». Такое уме- ние живых существ (растений и животных) в определенных условиях формировать тот или иной тип организации, например, тип габитуса древесный или травянистый, и, тем самым, выходить за пределы теку- щей нормы реакции генотипов, свидетельствует о свершении мутаций, но каких — только тех, которые не создали новых генов, а лишь раз- блокировали «гены-сони». Ретроновационные свойства генетической «па- мяти» способны высвобождать в некотором поколении живых существ кванты блокированной в них идиоэнергии, приводя в движение ретронова-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ ционную пластичность, которая, согласно предлагаемой гипотезе, вероятно, в организации биоморф также существует. Не исключено (и это нужно тоже иметь в виду), что явление Жизнь во Вселенной уже давно приобрело статус «бесконечного» в простран- стве и во времени, хотя этого еще никто на Земле не знает, а своих носителей она просто-напросто распространяет неосознанно или, на- оборот, посредством высокоразвитого в ее «чреве» Разума, который уже давно значительно сильнее человеческого, используя свои, только ему известные, средства для панкосмического расселения жизнестойкого материала, будучи Сеятелем Жизни, он умело «разбрасывает» живые зачатки, стремясь расширить свои (!) угодья в бесконечных просторах Природы. 9.4.3.2. Почему не экспрессивный альтруизм, а компро- миссы стали нормой экогенобиоза семенных фи- тобиоморф Когда впервые появились первые функционально-соматические генофенетические системы, или, если кратко, союзы «гены-фены», а совсем просто — «генофены», то с этого момента началось явление «жизнь» и пошла эволюция жизни. Жизнь как явление экогенобиоза осуществляется двумя группами «игроков»-партнеров: и генами, и фенами, которые по жизни действуют всегда вместе, хотя внешне это определить совсем не просто, поскольку натура этих организацион- ных частей очень разная. Генофен — это ген и произведенный под его управлением фен в одной функциональной системе. Смысл определен- ного порядка, на основе которого функционируют генофены, в том, что гены всегда как обитатели своего обиталища связаны с опреде- ленными фенами, проявляя себя при этом как ведущие и властонос- цы жизнеразвития по отношению к фенам — исполнителям генети- ческой власти. Жизнь возникла в эволюции из нежизни. Из преднаследственности нежизни через полунаследственность преджизни произошла наследствен- ность жизни. В основании первобытной наследственности стояли палео- гены и палеогенонемы (примитивные нуклеотиды и их комплексы), которые, чтобы существовать, должны были бороться за свое выжива- ние. Только борьба за жизнь может быть платой за то, что участник жизни, будучи таким, а не другим, живет. Жизнь началась на Земле с ультрамикроскопических существ — протероорганизмов, в которых поначалу «нагота» их палеогенов прикрывалась примитивной палеофе- нетической оболочкой. Важно, что с самых начальных проявлений жизни в протоорганизмах стали «работать» палеогенофенетические функциональные системы — «палеогенофены», совершенствование орга- низации которых в нескончаемой эволюции стало формой борьбы сна- чала палеогенов, а затем и генов, за улучшение своих «жилищных» условий — своего первобытного феносоматического обиталища (автоби- отопа). Все это, безусловно, важно для всех, поскольку высшая биоти- ческая власть принадлежит только генам, а способ борьбы (жизни) фенов направлен на пользу генам — за «уют» для них, но, кроме того,
Глава 9. Концеиция экогенобиоза и за что-то еще, необходимое для развития жизни вообще — других живых тел (протероорганизмов в целом, их структурных элементов, но и видов, и надвидов). Эволюция предложила первобытной живой природе два возмож- ных варианта развития: через сокращение личной жизни организмов и убыстрение сроков начала размножения, причем любым способом (личная жизнь организмов сокращается настолько сильно, что может почти совсем исчезать, продолжаясь секунды, минуты, часы, но не более одного месяца, что свидетельствует об экспрессивном альтруиз- ме таких организмов), или через удлинение продолжительности лич- ной жизни организмов и замедление сроков наступления начала раз- множения (личное и неличное в едином теле организма существенно идут на компромиссы, и тогда такого типа организмы живут дольше, более одного месяца, не забывая при этом и размножаться). Выше (глава 2.5) было показано, что в растительной, да и животной жизни оба эти варианта развития проявились в эволюции очень давно и есть по сей день. Экспрессивный альтруизм, вероятнее всего, был нормой существования еще очень примитивных организмов — протероорга- низмов первобытной жизни, сохранив такой принцип жизнеразвития и у некоторых современных групп. Однако у растительных (да и жи- вотных) биоморф, более сложно организованных, например, как все семенные растения, экспрессивному альтруизму почти не находится место (среди эфемеров бывает только одна из его более поздних форм — семиальтруизм). Среди семенных фитобиоморф приоритеты жизне- развития обеспечиваются разными компромиссами личного и другого в их организации, что соответствующим образом продлевает их про- должительность жизни. Живая природа, чем она сложнее организована в различных ветвях своей эволюции, тем больше в ней проявляется второй путь развития. Однако на фоне компромиссов личного и дру- гого в организации растительных биоморф, когда продлевается чаще всего общая продолжительность их жизни, совсем не исключается и некоторый обратный ход в определенных условиях, например, при аридизации климата, т.е. возврат от персистентного образа жизни к эфемерному (семиальтруизму). Что стало нормой экогенобиоза семенных фитобиоморф? Живые тела, устроенные как экогенобиозные системы, в процессе эволюции поставили очень высоко заботу о качестве «жилищных» условий своих властоносцев и стратегов (генов), поэтому в борьбе за качество «оби- талищ» для генов возникло действительно ошеломляющее разнообра- зие (конституциальное, или фенетическое) биоморф и, в особенности, растительных биоморф. Где у организмов проявляется больше комп- ромиссов между личным (неразмножающимся) и неличным (размно- жающимся), тем больше разнообразие фенетической организации, а следовательно, больше возможностей для генов сохранить себя, совер- шенствуя способы своего существования в их фенетических «обитали- щах» (фенетической организации). Чем больше проявляется разнооб- разие в фенетической организации биоморф, тем больше для генов возможностей выбирать и как-либо бороться за свое сохранение и
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ сохранение своих обиталищ (автобиотопов), поскольку в таких случаях выше пластичность биоморф. Следовательно, чем выше феносоматичес- кое разнообразие биоморф и, тем самым, их пластичность, тем более надежна среда фенетического обитания генов и тем шире возможности жизнесохранения в целом конкретных биоморф, но и устойчивость их таксонов. Жизни как экогенобиозу выгоднее, когда у ее носителей проявляется разнообразие в фенетической организации, поскольку для генов повышаются шансы их сохранения в различных условиях оби- тания целостных носителей жизни, а одновременно сохранения и жизни вообще. Итак, нормой существования современных биоморф семенных расте- ний является экогенобиоз — единство генетического и фенетического, мно- гопланово проявляющееся в целостной организации биоморф и в различных функциональных генофенетических системах (генофенах), в которых гены — это всегда носители биотической власти, а фены — исполнительские элементы, места приложения властной деятельности и обитания генов. Жизнью надо считать то, что на самом деле имеет место быть как особенное состояние живой природы, какое бы выражение оно ни приобретало в процессе эволюции, в которой действительно живут и гены, и фены, но и их различные совместные системы, например, орга- неллы, клетки, ткани, органы и т.п., и, что не менее важно, еще и разнообразные функциональные признаковые (союзные) системы- генофены, когда все они находятся вместе (!) в единой соматической организации целостных живых тел. Компромиссы между личными (сво- ими) и неличными (коллегиальными) видами жизнедеятельности в еди- ной организации каждой семенной фитобиоморфы постоянно достига- ются и обеспечиваются генами через их автобиотопы («обиталища»)— фенетическую организацию и, как следствие, расширяется полимор- физм и пластичность этих биоморф, что создает благоприятные условия в изменяющейся внешней среде для сохранения генов и генонем, иног- да и некоторых результатов трансформаций (мутаций), в целом — типов биоморф, а заодно и жизни вообще. Жизнь как явление экогенобиозное может осуществляться на Земле (шире — в природе вообще, что еще когда-то предстоит доказывать) только на основе приоритета власти генов над всеми формами другой деятельности, что уже превратило ее в процессе эволюции в постоянное дискретное феносоматическое строительство как средство разрешения субконтинуальных, а потенциально возможно и континуальных (бесконеч- ных), жилищных проблем у генов, главная цель которых растить и разви- вать свои обиталища (фенетические организации живых тел) и все ос- тальное, как получится, и, между прочим, только по пути достижения главной цели (борьбы за нее) — сохранения генов, а следовательно, и жизни вообще как определенного системного генофенетического состояния вечно развивающейся материи — это специальный закон генерального приори- тета жизнеразвития на Земле.
Глава 9. Концепция экогенобиоза 9.5. Формы габитуса и стадийного поведения — индикаторы жизнеразвития семенных фитобиоморф как экогенобиозного ssaeHvla Все разнообразие «лепоты» — феносом, которое мы наблюдаем сегодня в соматическом выражении жизни на Земле — это результат длительной эволюции однажды возникшего организационного единства союзов «генов-фенов», или «генофенов». Первобытные гены (палеоге- ны) в таком союзе впервые приручили окружающую среду работать на них и развивать под их информационным давлением себе защиту из фенов (палеофенов), чтобы отгородиться от остальной среды, сначала в виде простейшей, но стойкой ко внешним воздействиям белковой или белковоподобной оболочки, а затем и более сложного по своему устрой- ству фенетического «домика» — жилища, или обиталища (автобиотопа) со все возрастающими в новых поколениях возможностями у всего структурно-функционального союза генов с фенами, т.е. генофенов, существующего только по принципу экогенобиоза. Как уже было пока- зано выше, экогенобиоз — это образ существования (жизни) генов на арене («экодроме») их борьбы за существование внутри фенов, для фенов и, в первую очередь, для себя как хранителей идиоэнергии высшей биотической власти, приоритетом которой является макробиократия. В более разверну- том виде экогенобиоз — это процесс взаимодействия генетического и фенетического и их элементов как меросом (частей) в организации живых тел, составляющий содержание борьбы генов за свое субконти- нуальное существование, в которой они, гены, принуждают своими действиями фены и их комплексы (феносомы) адаптироваться под уп- равлением генов, чтобы улучшить условия существования последних и укрепить их власть, в особенности макробиократию (приоритет власти более сложно организованных живых тел). Можно только восхищаться действительно революционным собы- тием в эволюции материи — возникновением союзов (функциональных систем) «генофенов» и поддержанием их в множественных поколениях однородных живых тел. Ведь это уже была сама жизнь, в которой од- нажды подхваченная в эволюции примитивная, но все-таки генофене- тическая связка оказалась надежным завоеванием, обеспечившим в конце концов то разнообразие фенетической организации жизни на Земле, в котором мы существуем сегодня. Жизнь поначалу только учи- лась защищать свою наследственность, что стало для нее нормой. Все остальное пришло потом. Первая победа жизни на Земле — это возник- новение живых (фенетических) оболочек для генов. Гены без них и без защиты неживыми оболочками смогли бы существовать лишь мгнове- ния, а с ними можно было искать и находить что-то еще. Гены, защитив себя фенетическими оболочками, создали прецедент необходимости их сосуществования в окружающей среде не «голышом», а в некоторой «одежонке», а еще лучше в «жилище», тем более благоустроенном «оби- талище» (автобиотопе), что привело к возникновению клеток, тканей,
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИИ органов и т.п. живых тел. Именно на обустройство жилого фенетичес- кого «фонда» для генов эволюция потребовала от жизни вообще неве- роятных усилий, итогом которых стал тот расцвет жизни на Земле, которая торжествует на нашей планете сегодня. Жизнь, научившись в процессе эволюции защищать свою наслед- ственность, по сей день не отказалась от усилий совершенствовать фе- нетическую защищенность своих генов. Создав, посредством генов и генонем (целых молекул ДНК), систему упорядоченной идиоэнергети- ческой деятельности в соответствующих телесных системах, как прин- цип высшей (стратегической) биотической власти, направленный на развитие множественных форм исполнительской (тактической) деятель- ности различных структур — элементов фенетической организации, жизнь в полной мере использовала свой принцип экогенобиоза в эво- люционном развитии растительных биоморф. Без габитуальных форм (габитоформ) и стадийных форм (стадиосом, или форм роста) фенети- ческий «футляр» (феносома) как обиталище для генов существовать 6bI не смог. При всей значимости для организации растительных биоморф их субгабитуса (внутренней фенетической организации) и феносомы в целом, трансформации, которые фиксируются в габитусе, возрастной и сезонной фенетической организации (формах роста) фитобиоморф, свиде- тельствуют о многом, что происходит со всей остальной организацией этих живых телесных систем, поскольку они находятся в прямом контак- те с окружающей, весьма динамичной внешней средой — это индикаторы организации биоморф семенных растений. Габитус и формы роста растительных палеобиоморф прошли дли- тельный путь эволюции методом проб, ошибок и исправлений ошибок, что обеспечило их современным потомкам — биоморфам хорошо «от- препарированную» во времени привязку и прилаживание к окружающей их остальной природе. Современные биоморфы семенных растений при- обрели в эволюции определенный принцип фенетической организации, который отличает их от других растительных биоморф: выживание им обеспечивают основные органы — побеги и корни, образуя вместе кор- непобеги-органы (онтоиды) и корнепобеги-организмы (эуфиты). В изучении развития жизни у кого-либо важно правильно расста- вить акценты, иначе — путь вслепую, который чаще всего ведет в ни- куда. Какое завоевание биоморф семенных растений, которое стало неотьемлемой стороной их существования — атрибутом их организации, можно поставить на первое место в эволюции? Да, это пластичность (т. 1). Так этому и быть! Именно так и «решили», овладев секретами ста- новления и развития пластичности, палеобиоморфы, передав это свой- ство в поток тех биоморф, которые, изменяясь из поколения в поколе- ние, дошли в обновленных образах до их современных потомков. Дей- ствительно, это пластичность, также решаю и я, когда наблюдаю за тем, как проявляют пластичность семенные фитобиоморфы: они всегда в борьбе за существование на основе своей пластичности что-то в ней теряют, а что-то находят, но никогда без борьбы не сдаются, будучи с генетического уровня устремлены к тому, чтобы побороться за жизне- сохранение вообще.
Глава 9. Концеиция экогенобиоза 753 Габитоформы и формы роста (стадиосомы) — это только индика- торная (внешняя) сторона того экогенобиоза, тех генофенетических функционально-системных взаимодействий, которые осуществляются, в первую очередь, не столько в самих индикаторах, сколько во всей орга- низации биоморф как феносоме, так и геносоме. Только феносома бывает внешней и внутренней (состоит из двух таких частей). Внутрен- няя фенетическая организация, или субгабитус (эндосома) выполняет основную роль в жизнедеятельности феносомы. Геносома — это всегда особая часть организации биоморф, а шире и друтих живых тел, которая не делится на внешнюю и внутреннюю, поскольку она всегда внутри феносомы, т.е. имеет организационно внутреннюю природу — ее поло- жение всегда внутри феносоматического «футляра», в котором она рас- полагается как в своем «доме» (обиталище). Представления о лабильности габитуса, как уже отмечалось выше (глава 4), преувеличены, поскольку габитус растет и развивается не сам, а за счет субгабитуса, который, будучи внутренней фенетической орга- низацией, действительно растет и развивается («донор» габитуса), про- являя нередко неустойчивость (лабильность) своих структур, которые трансформируются, что и отражается в габитусе, но и формах роста. Таким образом, и рост, и развитие субгабитуса прямо-таки «тащит» как бы на своих «плечах», на себе и с собой все преобразования габитуса. На основе лабильности субгабитуса проявляется еще и лабильность габитуса — индикатора соматических трансформаций, но не их произ- водителя. Лабильность габитуса — это всего лишь «лакмусовая бумажка», которая через себя отражает то, что происходит во внутренней фенети- ческой организации (субгабитусе). Такова натура у габитуса быть сверху, прикрывать собой субгабитус, но не иметь своей внутренней структуры. Сам габитус получает толысо ту лабильность, которую под ним «вылеп- ливает» внутренняя фенетическая организация биоморф, их субгабитус (эндосома). Лабильность габитуса в его формах (габитоформах) и формы стадийного возрастного развития (стадиосомы) — надежный индикатор состояний роста и развития биоморф в целом. Ниже на нескольких примерах (рис. 424 — 430) рассмотрим разно- образное поведение биоморф семенных растений в различных условиях их произрастания: в горах, на лугах, в лесу, в пустыне. Габитоформы и стадиосомы индикаторно надежно отражают разнообразие организации таких биоморф как результат проявления их экогенобиозов. Рис. 424 — 430. Разнообразие поведения биоморф семенных рас- тений в различных условиях произрастания (к мультивариантности внут- ривидового жизнеразвития).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 4(гм4), ),26 (2)к Рис. 424. Биографический портрет Raphanus sativus L.[cåì. Brassicaceae (Московская область, огороды)] Биоморфотипы: н зе ад мно-двусемядольные (семядоли широколопатчатые, мясистые, 20 — 35 мм дл., 20 — 35 мм ши . глуб р., глубоко выемчатые, редко вместо выемки развивается третья лопасть посредине, клиновидно суженные в длинные 15 — 35 полурозеточные (овощеводы выращивают, как известно, редис посевной ради образования у ювенильных розеточных растений утолщенного и сочного нев невидного, чаще всего шаровидной формы, гипокотиля, выкапывая еще весной розеточные клубнево-стержневые корнепобеги, но небольшая часть растений не развивает сил ьного невидного утолще- ния корнепобега и в этом же го ду переходит к репродуктивному развитию) розеточно- простолистные и полусложнолистные безрозеточно-очередно-полусложнолистные-стерж- некорневые однодомные семидетерминатно-репродуцирующие бело- или лилово-капуст- ноцветковые однолетники (1 биоморфотип) или двулетники (П биоморфотип).
Глава 9. Концеиция экогенобиоза Рис. 425. Биографический портрет Biderts tripartita L. в Московской области [сем. Brassicaceae (берега водоемов, лесных луж, сырые лесные опушки и поляны в лесу)]. БИоморфотнпЫ: надземно-двусемядольные (семядоли продолговато-эллиптические, 7 — 17 мм дл. (у мелких растений — 3 — 7 мм), 3 — 7 мм шир. (у мелких растений — 2— 3,5 мм), закругленные, клиновидно суженные в короткие черешки) безрозеточные просто- и полусложнолистные сильноветвисто-высокорослые (I биоморфотип, 30 — 100 см высо- той) или неветвисто-низкорослые (П биоморфотип, 3 — 15 см высотой; высокие и низкие растения произрастают рядом) семидетерминатно-репродуцирующие (верхушечные соцве- тия медиального побега и паракладиев — корзинки) однодомные желто-астроцветковые стержнекорневые однолетники.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 7(26) 33а, Рис. 42б. Биографический портрет Choriospora bungeana Fisch. et С. А. Меу. [сем. Brassicaceae (Алтай, Курайский хребет, высокогорья, каменистые рос- сыпи, скалы, крутые склоны)]. Биоморфотипы: надземно-двусемядольные (семядоли эллиптические, 3 — 5 мм дл., 1,5 — 2,5 мм шир., закругленные, постепенно суженные в короткие черешки) розеточные или верхнерозеточные (при засыпании эрозионно-подвижным почво-грунтом) очередно-про- столистные эумоноподиальные интеркалярно-репродуцирующие (интеркалярные безлис- тные побеги, закрытые одиночными верхушечными цветками, развиваются в пазухах зе- леных листьев розеточных или верхнерозеточных ростовых побегов текущего года) компак- тно-каудексовые (I биоморфотип — в условиях относительно спокойного уровня повер- хности почвы) или вынужденно удлиненно-каудексовые (II биоморфотип — в условиях интенсивного засыпания эрозионно-подвижным почва-грунтом на осыпях) однодомные розово-капустноцветковые изоаклональные травянистые многолетники. Изоаклонисты (особи семенного происхождения) живут сравнительно мало (до 15 — 20 лет).
757 Глава 9. Концеиция экогенобиоза 4(1, 18а 21,27 546,67 Рис. 427. Биографический портрет Rumex obtusifolius Ь. [сем. Polygonaceae (Московская область, Подольский район, сырые луговины в смешанном лесу, лесные дороги)]. Дополнительное обозначение: нр — надземный розе- точный вегетативный побег. БИоморфотИПЫ: надземно-двусемядольные (семядоли продолговато-яйцевидные, про- долговато-эллиптические, 7 — 12 мм дл., 3 — 4 мм шир., закругленные, клиновидно переходящие в черешки 5 — 12 мм дл.) полурозеточные очередно-простолистные гемисим- подиальные короткокорневищные (сначала корневище гетерономное, поскольку долго свя- зано с главным корнем, но у матурных растений постепенно главный корень отмирает и их корневище становится автономным) семидетерминатно-репродуцирующие ддвдомные гречишноцветковые ретардационно-кондивидуальные травянистые многолетники. Биоморфотип 1 (основной) составляют биоморфы, в зонах торможения цветоносных по- бегов которых не отрастают розеточные вегетативные побеги. Биоморфотип П (второстепенный) составляют биоморфы, произрастающие в сильно ув- лажненных и тенистых местах, у которых из узлов средней и (или) нижней надземной части цветоносных унирепродуктивных ди-, плейофазных побегов образуются надземные розетки вегетативных побегов. Цветоносные побеги осенью могут упасть на почву, а розеточные побеги укорениться поэтому изредка, и таким образом, может также проис- ходить вегетативное размножение.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 758 1,аг 19а, гм4, а,21,27 Рис. 428. Биографический портрет Salvia lanigera Poiret [сем. Lamiaceae Израиль, окрестности Тель-Авива, каменисто-песчаные холмы,, V)]. 12V 22ы. БиоморфотнпЫ: надземно-двусемядольные полурозеточные супротивно-простолистные [когда доли рассеченного листа не доходят по всей его пластинке до средней жилки, то такой лист будет, пожалуй, правильно считать простым и, тем самым, отличать от полу- сложных листьев, у которых доли листа, его листочки, доходят до средней жилки листа (с неполными оторочками или без них), не распадаясь при листопаде; сильно рассеченные листья S. (ашрега, таким образом, — это разновидность простых листьев] гемисимподиаль- но-эусимподиальные (унирепродуктивные полурозеточные побеги развиваются как моно- фазные или чаще ди-, плейофазные) семидетерминатно-репродуцирующие однодомные фиолетово-яснотковоцветковые стержнекорневые полукустарнички (биоморфотип 1— растения от р ия открытых мест их аэроподии прижаты к поверхности, поскольку надземные з мо отип II— побеги сильно отмирают после плодоношения) или полукустарники (биоморфотип аэроподии достигают 20 — 40 см, кусты прямостоячие, растения произрастают в тесных ~группах», что способствует увеличению размеров аэроподиев, предпочитая пониженные участки рельефа.
Глава 9. Концеиция экогенобиоза 759 Рис. 429. Биографический портрет Plantago albicans Ь. [сем. Plantaginaceae (там же, что и рис. 428, 20 Ш). Дополнительное обозначение: рч — части других растений, произрастающих, прижавшись плотно друг к другу, в ~группе». БиоморфотиПЫ: надземно-двусемядольные (семядоли линейные, слегка заостренные, 15 — 20 мм дл., 1 — 2 мм шир., сидячие) розеточные очередно-простолистные эумоно- подиальные интеркалярно-репродуцирующие (розеточные побеги, накопив достаточный вегеторесурс, нарастая эумоноподиально, могут много лет ежегодно или с перерывами образовывать пазушные цветоносные безлистные силлептические, т. е. интеркалярные, побеги с колосовидными полузакрытыми верхушечными соцветиями, а когда интеркаляр- ных побегов образуется 2 и более в структуре отдельных сложных годичных побегов, то мы отмечаем у них межсоцветия) корнеотпрысковые однодомные травянистые многолет- ники (I биоморфотип — на открытых местах при произрастании в «одиночестве») или моноподиальные кустарнички (II биоморфотип — при произрастании в тесных «группах» вместе с другими растениями).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ аа4(аз 1), О гЗ, ,27 (49,47) Рис. 430. Биографический портрет Coridothymus capitatus (L.) Reichenb. fil.[ñåì. Lamiaceae (там же, что и рис. 428, 20 III)]. БиамарфатипЫ: надземно-двусемядольные (семядоли полуокруглые, 2,5 — 4 мм дл., 3 — 4,5 мм шир., широкозакругленные, глубокосердцевидные с заостренными ушками— боковые лопасти в основании семядольных пластин, на черешках 1,5 — 3 мм дл.) розет- кообразующие (на первом году вегетации ди-, плейофазных унирепродуктивных побегов развиваются розеточные или короткие верхнерозеточные вегетативные силлептические побеги, которые на второй — третий год образуют постлатентные годичные безрозеточные вегетативно-репродуктивные побеги, отмирающие сверху на разную длину после цветения и плодоношения), некоторые силлептические побеги развиваются как безрозеточные длин- ные или короткие супротивно-простолистные гемисимподиальные (этот тип нарастания побегов доминирует, но часть побегов нарастает эусимподиально) семидетерминатно-реп- родуцирующие [верхушечные полузакрытые соцветия цветут летом с июня по август (Schmida, 1986)] однодомные фиолетово-розово-яснотковоцветковые стержнекорневые подушковидные полукустарнички (I биоморфотип — на открытых местах) или прямосто- ячие полукустарники (П биоморфотип — при тесном произрастании в «группах» вместе с другими растениями).
Главв 10. Откровения о буднях и праздниках растительной жизни, их резонанс в концепции биотического счастья и качества жизни {на примере семенных фитобиоморф) Как-то (2000 г.) мне довелось чуть-чуть посоревноваться в игре «О, счастливчик», которую оригинально вел на телевидении Д. Дибров, и пока я проходил разные виды искусственного отбора, не сумев, правда, достигнуть в нем достойных вершин (ведь это игра, не более), то в порыве возникших впечатлений сформулировал основные положения концепции качества жизни, от которой, без определения места явления счастья, как я сразу же решил, толку будет немного. Благо к тому времени мною уже была создана общая теория жизнесохранения и определен, ни много ни мало, основной биологический закон как ак- сиома: «жизнь, возникнув на Земле (и — во Вселенной), изменяясь, сохраняет себя в объектах разной организации и в пределах, ограничен- ных ее возможностями» (т. 1, гл. 3, стр. 94). Все это и дало мне повод закончить каскад концепций, составивших второй том рассматриваемой монографии, десятой главой, в которой не просто размышления о ка- честве жизни и о счастье как о «жар-птице», которую можно всю ко- роткую или длинную жизнь пытаться поймать, но, по мнению многих, даже никогда и не увидеть, ставшую для большинства людей всего лишь недостижимой иллюзорной мечтой. О счастье можно говорить беско- нечно долго и проблему не исчерпать, поэтому я сознательно ограничил себя поисками ответа на вопросы, которые в такой плоскости еще никогда не ставились: в чем смысл биотического (/же — растительного) счастья, каково место его в качестве жизни и есть ли, ио качеству жизни, счастливчики среди биоморф семенных растений? Я надеюсь, нашел ответ вполне приземленный, не впадая слишком в эмоции, поскольку, в отношении явлений, вероятно, во многом очень загадочных, правильные решения нередко бывают самые что ни на есть банальные, и только мы, люди, — любители острых ощущений и «оше- ломлять» своих совидичей, часто прибегаем к различным формам вы- ражения своего неописуемого восторга, например, «жар-птица», «звезда пленительного счастья» и т.п. Восхищение, восторг ... и, наконец, об-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ ращение к слову «счастье», на мой взгляд, вполне справедливо для многих случаев из жизни людей и имеет для этого вполне объяснимые основания, поскольку речь идет обычно о социальном счастье и соб- ственно природу (натуру) биологических процессов не затрагивает. Однако, сегодня не тот случай, и мы не на концерте «Ласкового мая» или иной рок-группы, а на страницах научной книги, поэтому попро- буем настроиться на соответствующий лад, благо Природа предоставила нам, людям, такое великое счастье, как размьштлять и иногда абстрак- тно мыслить. Позвольте, но я уже невольно использовал слово «счастье» в своей, как полагаю, достаточно серьезной научной работе; похоже, ему место не только в стихах поэтов, прозе писателей, но и в научном труде, если суметь найти этому соответствующее научное обоснование? А почему бы и нет, так готов заявить я, биолог-ботаник и эмпирик, и теоретик одновременно. Люди приписали, кстати, вполне незаслуженно, стремление быть счастливыми только себе, а есть ли для этого веские причины? Может быть, явление счастья много шире, чем то, о котором мечтают влюблен- ные, глядя на тусклый свет «уходящей» Луны и первые солнечные блики утра, утверждающие начало нового дня... Похоже, все именно так, но обстоятельства требуют от меня предельной аккуратности, поскольку серьезный разговор о счастье вообще-то касается всех: и лириков, и физиков, и химиков, и ...просто живых существ. Давайте, не вдаваясь во многие детали, попробуем определиться со счастьем вообще, по- скольку только через него видится мне путь к объяснению качества жизни. Что это такое? Счастье — это, безусловно, форма получения и проявления удоволь- ствия от того, что имеет место быть, которое испытывают на себе любые живые существа. Но как освободиться от многих других форм удовольствия и оставить в работе только одну из них, которую и можно будет иризнать за «счастье~? Чтобы сделать это, я, как и положено в таких случаях теоретизи- рующему исследователю, попытаюсь выйти к решению проблемы изда- лека, чтобы, наконец, где-то объявить: господа, вот оно, счастье вообще и растительное счастье, в частности, принимайте его, как сможете, оно перед Вами в первозданном, как мне представляется, виде, совсем еще «тепленькое», только-только, почти, как «Колобок», сошло с «пылу- жару» и пустилось в путь по Свету-Миру . Однако, все по порядку. 10.1. Жизнь — борьба, но и постоянная выставка достигнутого ею разнообразия С легкой руки Ч. Дарвина (Darwin, 1859) термин «борьба за суще- ствование» с середины позапрошлого века закрепился в биологии для обозначения борьбы (адаптаций, или приспособлений) существ (орга- низмов) за жизнь (существование). С учетом утвердившихся в последар- виновское время представлений об иерархическом многоуровневом ус-
763 Глава 10. Откровения о буднях и ираздниках растительной жизни 10.1.1. Жизнь — это будни, праздники и переходы между ними Как известно, Ното sapiens обставил свою социальную жизнь и буднями, и праздниками, да еще иногда проводит парады в честь дос- тижений в своем социальном развитии и т.д. и т.п. Как ни странно, но многое из социальной жизни человека разумного имеет место и в ос- тальной жизни (биосоматической организации), которую проявляют разные виды как животных, так и растений, будучи не осознанно. Я готов утверждать, что в любой жизни есть свои будни и свои праздники, только нам, людям, надо их суметь разглядеть и правильно расставить акценты. Любопытно все это. Почему здесь и сейчас не попытаться кое в чем разобраться? Что греха таить, ведь это мы, люди, умеем нередко наводить «тень на плетень», когда видим богатое разнообразие фенетической организа- ции живых тел, умиляясь и восхищаясь ему, в силу своей разумности тройстве живой природы было предложено (т. 1, с. 1бб) «рассматривать «борьбу за существование» как комплекс отношений между биосистема- ми, а также между биосистемами и абиотическими условиями». С таким широким пониманием «метафорического», по выражению Дарвина, термина «борьба за существование» можно сейчас однозначно конста- тировать, что всякая жизнь — это всегда борьба, вне которой это явле- ние уже не наблюдается. Пожалуй, вряд ли кто-нибудь будет особенно возражать против такого образного и емкого выражения, как «жизнь- это борьба». Действительно, если и не обладать большой наблюдатель- ностью, то все равно придется заметить, что всякий, кто не находит сил и средств (стремление их найтч проявляет любая биосистема), т.е. в целом ресурсов, чтобы за себя побороться в объеме, достаточном для выживания, соответствующим образом адаптируясь в окружающей не- живой и всей остальной живой природе, тот неминуемо будет «раздав- лен» естественным и (или) искусственным отбором, энергетика которо- го способна «подминать» и «выбрасывать за порог» (элиминировать) кого угодно, если тот уже не «борец», но все же хочет «наслаждаться» жизнью. За право жить на Земле живые тела платят друг другу количе- ством и качеством своей борьбы, что по-разному воспринимается живой природой. Борьба за существование в исполнении живых тел (биосис- тем) — это не только «междусобойчики», иногда переходящие даже в «мордобойчики», в жестких внутриорганизменных, внутривидовых, меж- видовых и межнадвидовых конкурентных отношениях, но это еще и кооперации с другими живыми телами для разрешения различных (боль- ших и малых) проблем своего выживания. Даже полная индифферент- ность — это тоже прием борьбы, когда биосистема своим безучастием в разрешении каких-либо взаимоотношений с другими живыми телами так, по сути, участвует в поддержании выгодного баланса сил для себя и (или) иных биосистем.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ и образованности, часто не придавая значения тому, что наблюдаем всего лишь «футлярчики» — фенетические обиталища («домики»), или автобиотопы, для генов, больших, малых или совсем микроскопических размеров, в которых мирно существует (живет) сам его хозяин, причем всегда ультрамикроскопический, несмотря на размеры своих деяний— своего обиталища, как носитель стратегии управления жизнью конкрет- ной «союзной» (генофенетической) телесной системы и в чем-то даже жизни вообще — генетическая организация (геносома) каждого живого тела со всеми ее элементами (генами и генонемами). Это они, гены и генонемы, создали в процессе эволюции во взаимодействии с окружа- ющей внешней средой все фенетическое богатство жизни. Давайте попробуем воздать должное и тем (генам и их комплексам), и другим (фенам и их комплексам), разложив их по «полочкам» соматической организации жизни в развитии так, чтобы она, жизнь, не потеряла при этом всех своих достоинств. Безусловно, достоинств у жизни немало, и одно из них — просто жить (), а значит, уже этим достижением соответствующих телесных систем — быть счастливыми, но об этом потом, а пока немного о буднях. Их, естественно, много больше, чем праздников. В отношении расти- тельных биоморф нетрудно заметить, что их главное удовольствие от жизни — это служение через цветение и плодоношение более высокой по организационной сложности для каждой из них системе, частью которой они непосредственно являются, т.е., в первую очередь, виду. Именно в цветении и плодоношении проявляются праздники в жизнераз- витии семенных фитобиоморф, а все остальное — это будни и предпраз- дники. Итак, определимся: будни в развитии семенных растительных биоморф составляют раздел их жизни, связанный с формированием веге- торесурса (вегетомассы), а праздники — это особое состояние жизнераз- вития фитобиоморф, которое маркируется цветением и плодоношением, когда конечный репродуктивный ресурс (репродукторесурс или репродукто- масса) — пыльцевые зерна и семена — образуется, созревает и рассеива- ется в окружающем пространстве; между буднями и праздниками расти- тельной жизни проявляется переходное состояние предпраздников, когда формируются зачатки репродуктивных органов и создается потенциаль- ный репродуктивный ресурс. Цветение и плодоношение растений — это действительно праздник в каждой растительной жизни, когда вегеторесурс перестает более всего работать на свой организм и от скрытной «подработки» по пути разви- тия потенциального репродуктива начинает откровенно, «встав во весь рост», более всего или полностью обслуживать уже сформированный репродуктив. Растительные праздники — это торжества не только для тех, кто в них непосредственно участвует как главные действующие существа, но и для тех, кто, так или иначе, соприсутствует на них, помогая (насекомые-опылители) или мешая (насекомые-вредители) их проведению и т.п. Каждый растительный праздник — это подведение промежугочных итогов всего предыдущего жизнеразвития конкретных биоморф, а шире и их предков (близких и далеких): «вот чего мы достигли», но и демон-
Глава 10. Откровения о буднях и ираздниках растительной жизни 7б5 страция потенциала на пути к новым эволюционным свершениям. На таких праздниках фиксируется качество наиболее значимых результатов жизнеразвития посредством предшествующих вегетативных и пререпро- дуктивных усилий и реализуется высокий жизнеутверждающий акт- преемственная передача эстафеты жизни от предка потомку, от поколе- ния к поколению через половое размножение как высшее достижение эволюции, что обставляется, чаще всего, в весьма ярких внешних офор- млениях (например, через разнообразие форм и красок цветов и пло- дов). Праздники растительной жизни — это, безусловно, торжество преемственности жизни вообще, которое олицетворяют собой цветущие и плодоносящие фитобиоморфы, а остальные биоморфы того же вида, т.е. совидичи, как-либо соучаствуют на празднике, но не исполняют его, поскольку их номер на этих праздниках пока или уже не первый. Более того, для многих биоморф-совидичей, которые на празднике раститель- ной жизни пока или уже не участники, продолжаются будни со своими проблемами выживания, на которых они просто живут, проявляя, может быть, и безразличие к тем, кто сейчас цветет или плодоносит, испыты- вая физиологически заметное довольство от достигнутых результатов. После праздников могут опять наступать для растительных био- морф будни, в течение которых они пополняют растраченный ранее вегеторесурс и развивают новый репродуктив, направив всю энергетику фенетического жизнеразвития на осуществление нового репродуктивно- го усилия, т.е. к новым растительным праздникам. Однако, у всякой «медали» есть оборотная сторона, и личная жизнь после праздника (цветения и плодоношения) может далее уже не сложиться, превратив- шись в нисходящее движение к ее затуханию, нередко очень быстрому, например, эфемеры, и элиминации, поскольку часть растительных био- морф в процессе эволюции не приспособилась повторять, или приспо- собилась не повторять, в своей организации развитие нового вегеторе- сурса и нового репродуктива, на~я для продолжения и сохранения вида и рода другие пути. Пожалуй, пока в этом ключе достаточно, и, если захотеть, то вполне в растительной жизни можно разглядеть и будни, и предпраздники, и праздники. Однако, есть еще один момент, который нередко сопровождает праздники в мире людей. Да, это парады. Их бывает немало, и они разные. Странно это или нет, но и в растительной жизни я склонен увидеть проявление «парадов». Можно, конечно, ограничиться «парада- ми» («выставками» достижений) только цветущих и плодоносящих фи- тобиоморф, которые в разное время удается наблюдать в окружающем нас жизненном пространстве. Эти «парады», безусловно, интересны, но у них нет того, что чаще всего хотели бы наблюдать биоморфологи: они не отражают всего разнообразия растительных биоморф. В этом отно- шении на растительную жизнь можно посмотреть как на постоянно действующую «выставку» всех (!) ее достижений, причем разных, в силу присущей ее носителям-биоморфам пластичности и склонности к ги- перпластичности, т.е. как на «выставочные будни фитобиоморфного разнообразия». На таких «смотринах» присутствуют все типы раститель- ных биоморф, а в первых рядах, как положено по «штату», если к
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РАСТЕНИЙ данному моменту они в соответствующей форме, «маршируют» «цвету- щие» и «плодоносящие», ведь у них «парад» на их празднике жизни, и именно они, более всего отражающие разнообразие организации фито- биоморф, склонны заслонять будни других своими праздниками. Итак, вокруг нас, как можно вообразить, «выставки достижений по организа- ции растительных биоморф», которыми, не подозревая об этом, посто- янно и без устали занимается в своей эволюции живая природа. Это мы, люди, склонные к эмоциям по любому поводу, способны разглядеть в «буднях» растительной жизни и ее фитобиоморфного разнообразия еще и «выставки», «праздники» и «парады», которые со стороны всего жи- вого воспринимаются примерно так: в центре они, конкретные живые тела какого-то уровня организации, а подле них окружающая внешняя среда, которую они топчут, жуют, пропускают через себя и т.д. и т.п. На «выставках» разнообразия растительных биоморфотипов при- сутствуют все типы возрастных и фенологических состояний биоморф и все типы биоморф с иной фенетической организацией, которые име- ют место в различных экологических условиях, отражая, тем самым, и «блеск», и «нищету» в растительном царстве, а наша задача, исследова- телей-естествоиспытателей, учиться и уметь во всем этом разбираться. 10.1.2. Биотическая организация — единство и борьба качественно противоречивых форм соматического развития: для себя {своего, личного) и для других {другого родного и другого чужого) Из материалов предыдущих глав ясно, что только гены и их ком- плексы, управляя всей фенетической организацией, образуют союзные функциональные системы — генофены, в которых каждый их компо- нент занимается своим делом, будучи ориентирован в процессе эволю- ции на определенную специализацию жизнеразвития: гены на управле- ние, а фены на исполнение, поступающей с идиоэнергией генетической информации — целевых разновариантных программ (норм реакции) фенетического развития, из которых выбирается тот вариант, который более отвечает конкретным условиям произрастания фитобиоморф. Гены в процессе эволюции адаптировались участвовать, в ранге меро- сомы (части тела), в развитии только союзных, или генофенетических, целостных биотических (живых) соматических систем, выполняя стра- тегическую роль всякого жизнеразвития и, в частности, управляя фор- мированием феносомы, или фенетической части как меросомы соответ- ствующего союзного живого тела, превратив эту организационную часть в свое обиталище («дом»), или автобиотоп. Посредством феносомы гены отгородились от окружающей внешней среды, предоставив ей большие возможности побороться за существование целостных живых тел (био- систем) и не утрачивая контроля за целостным соматическим жизнераз- витием. Феносомы, в первую очередь, взаимодействуют с внешней сре-
Глава 10. Откровения о буднях и ираздниках растительной жизни 7б7 дой и непосредственно ведут борьбу за освоение пространства во вре- мени, и что самое, пожалуй, интересное, на чем обычно не фиксируют внимание исследователи, они делают все это не односторонне — «не гребут все под себя», а весьма целесообразны в формировании каче- ственного содержания целостной жизни любой биосистемы. В целом организация одного живого тела (биосистемы) проявляет всегда страте- гию неоднородной целесообразности, по большому счету, политропного (многоцелевого) качества жизни. Есть необходимость разобраться с термином «качество жизни», поскольку мы так или иначе выходим на него с разных сторон. Слово «качество» как философская категория — это «категория, вы- ражающая существенную определенность объекта, благодаря которой он является именно этим, а не иным. Качество — характеристика объектов, обнаруживающаяся в совокутпюсти их свойств» (Бол. Энц. Сл., 1997, с. 512). Качество — это, в иервую очередь, иринциииальная особенность содер- жательной (не количественной) части какого-либо явления, оиределяющая основные свойства его существования в иространстве и во времени. Каче- ство жизни можно, иожалуй, оиределить как свойство живых тел иметь основные иринцииы организационного содержания как общие, так и част- ные, в соответствии с которьаии обесиечивается жизнесохранение как вообще, так и в каждом конкретном теле. Под разными углами обзора и с разных высот абстракции качество жизни может приобретать неодинаковые выражения. В нашем случае мы сделаем акцент на том, что качество жизни биосистем составляют две его стороны: счастья и несчастья, которые, будучи свойствами живых тел, проявляют себя в чем-то по-разному, но в чем-то, и это наиболее важно, на основе некоторых общих принципов. Качество жизни — это не кон- кретная структура, оно как в целом, так и обе его стороны (счастье и несчастье), нельзя потрогать и подержать в руках, но оно присутствует, независимо от чьего-либо сознания, в организации каждого живого тела и всего живого (биостробы) на Земле. Качество жизни, будучи свойством живого тела, оиределяется самооценкой этого тела изнутри (от себя) уже достигнутых им результатов собственного развития в системе жизнераз- вития вообще, что косвенно находит отражение в различных иризнаках, особенно в тех, которые хорошо маркируют организацию такого тела— в габитоформах и стадиосомах (формах роста). Попробуем разобраться в том, что же составляет «изюминку» в проявлении качества жизни конкретных тел и, в частности, биоморф семенных растений. Прежде чем рассмотреть проблему взаимодействия свойств счастья и несчастья, находящихся в организации любых живых тел в противоречивом единстве как элементы качества жизни этих тел, обратимся сначала к разгадке некоторых таинств того, как, на чем и почему «замешаны» основные противоречия в качестве жизни вообще и биоморф семенных растений в особенности. В целом организация какого-либо живого тела (биосомы), которое вполне оправданно обычно называют «биосистемой» (живой системой), проявляет стратегию определенных стандартов (норм) в жизнеразвитии. Жизнь — явление многослойное как по форме, так и по содержанию
OCHOBhl БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИЙ (главы 1, 7). В результате взаимодействия разных сторон жизни ставятся ющих тел, имеющие жизнесохранительную направленность: автобиотропия (жизнь для себя), гомеобиотропия (жизнь для других-непосредственно род- ных), гетеробиотропия (жизнь для других-чужих). Все эти цели жизнераз- вития реализуются в развертывании фенетической организации соответ- ствующих тел под воздействием определенной идиоэнергии, исходящей из генетической организации этих же тел. Итак, действительно полная жизнь обязана быть наполнена целе- сообразным содержанием: она имеет не только цели своего полного развития, но и проявляет их в соответствующих свойствах тел, ее носи- телей (биосистем), развиваясь в направлении достижения этих целей— в направлении жизни для себя (личной), жизни для других-непосред- ственно родных и других-чужих, следовательно, по трем основным направлениям сохранения жизни вообще. Биоморфы семенных растений — это полностью живые целостные системы со всем стандартным набором целей жизнеразвития, на основе которых развивается в едином теле своя (собственная, личная) жизнь, обеспечивающая существование своего тела, которое непосредственно себя составляет и, в первую очередь, для него с окружающим миром взаимодействует, и другая (не-своя) жизнь, обеспечивающая существо- вание в том же теле жизней (!) других тел, структурным элементом которых как-либо или непосредственно (у родного тела), или опосре- дованно (у другого, уже чужого, тела), первое тело (в нашем случае, фитобиоморфа) является. Похоже, что жизнь на Земле, а, вероятно, и вообще во Вселенной смогла, однажды возникнув, сохраниться только посредством приобретения в эволюции естественным образом стандар- тизированного набора целей соматического развития, причем в единой организации полностью живых тел, и смогла найти способ одновремен- но <удовлетворять» собой и одни живые тела — структурные элементы (меросомы), и другие живые тела — разномасштабные по сложности организации хозяева (интеграторы) этих структурных элементов, давая первым, таким образом, «насладиться» столь особенным явлением, как жизнь, а другим телам — интеграторам структурных элементов и «вла- стелинов» над ними, обеспечить преемственное возобновление тех зве- ньев своей структуры, которые элиминируются. Проявление в органи- зации союзных, или генофенетических, живых тел одновременно в стан- дартном наборе целей и своего, и другого жизнеразвития решило, бе- зусловно, труднейшую эволюционную проблему поддержания и диск- ретности, и преемственности в развитии живых тел разных уровней сложности организации. Размышляя таким образом, я взял на себя смелость определить универсальный принцип развития биотической организации телесных сис- тем, и вот что из этого получилось. Все живые тела существуют по- разному только на основе противоречивого единства генов и фенов как союзные, или генофенетические, системы (биосистемы), но всегда как ближние (родные) или дальние (чужие) эволюционные родственники, поэто- му они вместе нацелены развивать единое для них явление «жизнь» как
769 Глава 10. Откровения о буднях и ираздниках растительной жизни тела-элементы, ее составляющие, а, следовательно, где-то больше или меньше, но всегда участвовать в жизнеразвитии, ироявляя в организации каждого живого тела два основных свойства: свое (личное) и не-свое (дру- гое), изначально выбрав в ириоритеты развитие другого — не только род- ного, но и относительно неродного (чужого), — это универсальная бнотн- ческая стратегия разноцелевого стандарта в соматическом развитии. Свое и не-свое (другое) — это два атрибутивных свойства в организации тела любой растительной биоморфы (шире — всех биоморф), а еще шире— всех эврибиоморф, т.е. каких-либо форм жизни в широком смысле). Когда формируется вегеторесурс (вегетомасса), короче — вегетатив, то это больше свое (личное) в организации растительной биоморфы, но когда образуется пререпродуктив, репродуктив и тотарепродуктив, т.е. в целом репродукторесурс (репродуктомасса), то многое из этого уже больше, чем свое, — оно уже перестало быть своим и стало не-своим (другим). В этом парадоксе есть смысл разобраться подробнее. 10.1.3. Парадокс своего (личного) и не-своего (другого) — один из атрибутов мироздания и, в частности, живой природы Многое зависит от того, как смотреть на устройство Мира и тел, его составляющих: под каким углом обзора и с каких высот абстракции. Автор настоятельно проводит в своей монографии эту мысль, в соответ- ствии с которой обнаруживается немало, на первый взгляд, парадок- сальных явлений, по большому счету составляющих атрибутику в орга- низации мироздания. Не будем терять время на неживую природу, поскольку вопрос, который поднимается здесь, имеет в конце концов универсальное реше- ние. Давайте возьмем пример из живой природы, более того, такой пример, когда ответ на поднимаемый вопрос, по первоначалу, ничего другого, вероятно, кроме как негодования, я так полагаю, у оппониру- ющей стороны не вызовет. Затем попытаемся разрядить ситуацию, причем настолько, чтобы в конце концов парадокс как таковой был объяснен и признан за атрибут мироздания. Итак, в самой общей форме, все качественные отношения тел между собой можно свести к схеме «свой — не-свой». Однако это довольно «грубая» схема, при детализации которой вскрывается весьма любопыт- ная картина; ее иначе, как парадоксом, не назовешь. Поставим вопрос ребром, что такое на самом деле «свое» и «не-свое (другое)». Далеко ходить не будем, а прямо здесь и сейчас заглянем внутрь себя — своего (!) тела, не забывая параллельно и о растительных телах. Тела людей — это, однозначно, организмы (среди растений-био- морф есть организмы и синорганизмы). В организмах, как известно, есть органы, которые их составляют, в органах — ткани, в тканях— клетки... и т.д. и т.п. Из всех органов обратим внимание на человечес- кую руку (или, к примеру, растительный побег). Тело руки (у растения — побега) для нее самой — это свое (!) тело, а как быть со всеми остальными телами, т.е. в каком качестве они выступают по отношению
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ к телу руки (или растительного побега)? Все остальные части, кроме руки человеческого тела и всего, что ее составляет, — это уже по отно- шению к руке не-свои (друтие) тела. Мы, люди, берем для познания какие-либо тела из бесконечности Вселенной, а поскольку у нее нет начала и конца, то тело, к которому мы проявляем познавательный интерес, хотим мы того или не хотим, всегда будет располагаться в центре мироздания (другого не дано!), что весьма удобно, поскольку позволяет судить о том, что рассматриваемое тело составляет и что его окружает. Будучи таким образом расположено, любое тело имеет, преж- де всего, свойство быть своим, а не другим. Все тела, которые состав- ляют свое (собственно ему принадлежащее) тело, состоят из организа- ционных частей — это структурные элементы своего тела, которые можно обозначить как «предсвои» (предличные, или преперсональные) тела, поскольку они только в чем-то составляют организацию своего (личного, или персонального) тела, например, руки (или побега), но каждый по отдельности им не является. Предсвои (они внутри своего тела) и остальные тела, которые располагаются уже вне своего тела, в нашем случае, руки человека (или отдельного побега растения), это все не-свои (другие) тела. Быть другим (отличным от своего), т.е. не-своим, телом, может быть любое тело, которое целиком непосредственно его, свое тело, не составляет, а это и «предсвои» тела, и «полусвои» тела, еще «сверхсвои» тела, в том числе и «другие родные», а еще, конечно же, кроме того, и «другие неродные»(чужие) тела. Свое тело нередко служит другому телу больше, чем самому себе, что, не правда ли, парадокс. Полусвое (полуличное, или семиперсональное) соматическое свойство жизни проявляется как остальная часть в организации некоторого обще- го тела, в котором особое место отводится своей части (например, руке или побегу в теле соответствующего организма). Вместе свое (рука, побег) и полусвое (остальная часть соответствующего организма) обра- зуют более общее качество жизни — единое соматическое свойство жизни, составляющее по содержанию сверхсвое (сверхличное, или пост- персональное) тело. Конечно, скажете Вы, заяц, бегущий где-то в сторо- не от меня по лесу, это другое по отношению к моей руке тело, но к другому (!) качеству жизни принадлежит и остальная часть тела челове- ческого организма, в данном случае, всего моего тела, оно также другое тело, хотя и много ближе, чем тело зайца, что и позволило считать его, мое остальное тело, по отношению к руке полусвоим телом. Вроде бы парадокс..., но в конце концов все понятно: вместе свое и другое, как полусвое, тела подняли качество жизни на более высокий уровень, составив сверхсвое тело. А как быть с зайцем, ведь это тоже другое тело. Вроде бы и незаметно, как бы между прочим, затронут один из атри- бутов мироздания, который нуждается в дополнительном объяснении. В центре мироздания какое-либо свое тело (например, чья-то рука или побег), а все остальное соматическое, что не это (свое) тело, то- уже другое тело. В отношении к своему телу существуют и «другие свои» тела, когда они имеют такой же тип организации. Моя рука (могло быть любое другое тело, допустим, побег семенного растения) составила в данный момент «полета» человеческой мысли свое законное место- она расположила свое тело в «центре» Вселенной, которая бесконечна,
771 Глава 10. Откровения о буднях и ираздниках растительной жизни 10.1.4. Растительные биоморфы — результат поиска и выбора оптимальных целей жизнеразвития посредством самонаводящейся и квантированной идиоэнергии Как известно, Природа развивается ритмически, и земные живые тела ничего не могли сделать иного, чтобы подчиниться законам ста- дийного (ритмического) развития, которые действуют повсеместно на нашей планете. Биоморфы надежно усвоили для себя в процессе дли- тельной эволюции, кто в их организации действительно «хозяин», кто управляет их жизнеразвитием и, чтобы выполнить главную функцию в своей жизни — размножение, они должны накопить определенную вегетативную массу (вегеторесурс), на основе которого можно занимать- ся чем-то большим, чем личной жизнью, и можно посвятить себя дру- гому, еще более важному, в первую очередь, делу размножения, развив для этого специальные приспособления — органы размножения. Стро- ительная «лепота» вегетатива, а потом пререпродуктива и репродуктива обеспечивается фенетической организацией и управляется с генетичес- кого уровня, откуда гены в нужный момент, в нужном месте и в нужном направлении отправляют идиоэнергетические самонаводящиеся и само- разделяющиеся (квантированные) средства. Идея саморазделения и порционного (квантированного) движения идиэнергии — пока что процесс вероятностный и потребует экспери- ментальной проверки. Дело в том, что вся живая природа в целом функционирует по ритмическому принципу с учетом ритмов, «бушую- щих» на Земле и в глубинах Вселенной, а, кроме того, гены, как изве- стно, наследуют не отдельные признаки, а норму реакции, т.е. ген потенциально «дву- и даже многозаряден» на цели фенетического раз- вития. Корректирующим фактором самонаведения идиоэнергии и вы- бор ею цели жизнеразвития во многом являются конкретные условия окружающей среды. Суть гипотезы жизнеразвития функциональных союз- ных систем — «генофенов», выдвигаемой автором, не в том, что идиоэнер- и любая точка в ее пространстве вполне может быть принята за таковой, а все остальное соматическое, проявляющееся где-либо, — это все дру- гое, т.е. другие тела, каждый тип которых со своими особенностями. Другое — то, что составляет предсвое, как непосредственные структур- ные элементы своего тела. Другое — и то, что рядом со своим телом, выступает как остальная часть (полусвое) более общего целого тела- ценосомы (уже — ценобиосомы). Другое еще и то, что вместе составляет ценосому, т.е. объединяет и свое, и полусвое, как родственные части, непосредственно взаимосвязанные и взаимодействующие, одного тела — сверхсвое. Другие, наконец, — это и те тела, которые прогенетически или генетически соматические неродственники — это почти (все отно- сительно) по-настоящему (действительные) чужие тела (особи зайца, воробья и василька синего, например).
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИЙ гня самонаводнтся (в целом, это уже давно понятно и по-другому просто быть не может), а в том, что она, обладая мультннормальной целевой заряженностью, имеет склонность саморазделяться и порционно (квантамн) двигаться к одной — нескольким выбранным целям нз большего числа возможных целей. Логическим путем можно предположить, что не ис- ключено движение идиоэнергии к целям фенетического развития, чем- то образно вполне может напоминать полет самонаводящейся ракеты с разделяющейся боеголовкой. Идея квантированного саморазделения самонаводящейся идиоэнергии при движении к фенетическим целям вполне согласуется с концепциями стадийного развития фенетической организации растительных биоморф, например, с концепцией Т. А. Работнова (1945, 1949, 1950, 1960 и др.) о возрастных состояниях рас- тений в их жизненных циклах. Идиоэнергия уходит с генетического уровня в фенетическую организацию, вероятнее всего, саморазделяясь, действуя в определенных направлениях жизнеразвития «волновыми» порциями — идиоэнергетическими квантами, управляя, таким образом, потреблением, производством, резервированием и рассеиванием ресур- сов. Первая цель действия идиоэнергетических квантов — это достижение качества виргинильного фенетического развития, когда формируются раз- личные девственные структуры и биоморфы ироходят возрастные состо- яния иренатальньп и натальньп эмбрионов, ироростков, всходов, ювенилов и имматурных растений. Возможно, от тех же генов или от других новые идиоэнергетические кванты оиределяют необходимость заменить дев- ственность на матурность (физиологическую зрелость), участвуя в ироиз- водстве реиродуктивных органов, сиособствуя иротеканию реиродуктивно- го лроцесса. Вся «лепота» фенетических телодвижений нужна, в первую очередь, самим генам, управляющим строительством своих «обиталищ». Однако, фенетическая организация по ряду направлений эволюции живых тел достигла уже такого уровня развития, что стала способной сама влиять на эволюцию живых тел и генов. Появление мозга в фене- тической организации живых тел и у человека разумной деятельности уже привело к тому, что эволюция по некоторым направлениям жизне- развития становится процессом управляемым и с фенетического уровня (генная инженерия). При всем этом разумная деятельность никогда не приобретет необ- ходимость что-то кардинально менять в принципе работы генов, рас- считанных своей природой на самонаведение разной идиоэнергии на фенетические цели жизнеразвития, поскольку именно это составляет сокровенное качественное (стратегическое) содержание жизни вообще, а главная тактическая деятельность всегда была и будет у живых гено- фенетических тел ареной борьбы за развитие систем строительства фенетической организации как автобиотопов («обиталищ») для генов. 10.2. Главные таинства растительного счастья Все, что живет на Земле, так или иначе за что-то борется: за угле- род, кислород, воду, за место под Солнцем, за пыльцевое зерно, про-
Глава 10. Откровения о буднях и лраздниках растительной жизни 773 летающее где-то рядом, и т.д., т.е. в целом это и есть борьба за суще- ствование (жизнь). Так получилось в эволюции, что только человек осознал в своем существовании нечто особенное, яркое, что было им названо красивым словом «счастье». Особи Ното sapiens — существа, как и все остальные, биотические, но еще не менее того и социальные, поэтому в определениях категории «счастья», которые они дают для себя, обращаются, в первую очередь, к нравственно-психологическим сторонам своего поведения как личностей в мире общественных отно- шений, в отношениях к своему труду и друтим ценностям, которые составляют содержание социальной жизни. «Счастье — переживание полноты бытия, связанное с самоосуществлением» (Филос. энц., 1970, с. 175). «Счастье, состояние человека, соответствующее внутренней удовлетворенности своим бытием, полноте и осмысленности жизни» (Бол. Энц. Сл., 1997, с. 1169). «Под состоянием счастья следует пони- мать временный, относительно устойчивый уровень довольства, являю- щийся результатом удовлетворения тех или иных потребностей. Состо- яние счастья характеризуется эмоционально окрашенным, приподня- тым восприятием явлений действительности, выражающим внутреннюю удовлетворенность факторами не только прошедшего, но и особенно наличного бытия» (Попов, 1986, с. 31). Получать удовольствия — вот что составляет для людей одно из проявлений счастья. «Чувство счастья (в широком смысле слова — здесь мы включаем и ощущение, и состояние, и даже само чувствование его) может быть мгновенным, быстро прохо- дящим, вызванным каким-либо сегодняшним событием. Оно может быть постоянным, прочным, вызванным постоянно действующими факторами радости, подъема душевных сил, хорошей перспективы, гар- монии и удовлетворенности. Чувство счастья, таким образом, фиксирует разнообразие отношений человека к миру и к самому себе. Оно поэтому олицетворяет психологическое выражение процесса или результата удов- летворенных субъектом разнообразных жизненных потребностей и ин- тересов» (Попов, 1986, с. 32). «Постижение смысла жизни является основой, импульсом для сознательной деятельности личности. Содер- жание смысла жизни определяет и содержание счастья. Счастье фикси- рует ценность самого жизненного процесса, а смысл жизни утверждает эту ценность» (Попов, 1986, с. 91). Здесь, пожалуй, есть смысл остановиться с рассуждениями о сча- стье в мире людей, поскольку все, о чем далее пойдет речь, должно показать, что человеческое, или социальное, счастье — это только штришок в мире счастливой жизни вообще, а категория «счастье» впол- не может выйти на уровень универсальных для живой природы через объяснение понятия «биотическое счастье», т.е. стать понятием прием- лемым для объяснения особого состояния жизни любых живых существ (от бактерии и водоросли до человека). Если встать на путь расширения смысла термина «счастье», то надо согласиться, что помимо социального счастья, есть еще и более есте- ственное, существующее универсально в живой природе, вполне неза- висимо от человеческого сознания, — это, на мой взгляд, биотическое счастье. Оставим в стороне проявления социального счастья и обратим-
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РАСТЕНИЙ ся к разгадке таинства универсального свойства жизни — качества всех ее телесных носителей и, естественно, также биоморф семенных расте- ний, быть биотически счастливыми, хотя и по-разному. Есть смысл «под занавес» нашей книги еще и еще присмотреться к организации совре- менных семенных фитобиоморф и выделить в ней те модусы (нормы), которые составляют ее определяющее содержание, что, как можно по- лагать, должно помочь и с определением биотического счастья и в целом качества жизни. 10.2.1. Модусы организации coepeMeHHblx ceMeHHblx фитобиоморф как результат эволюции Ранее уже было показано, что в организации природы вообще (Природы) и живой природы (Жизни, или жизни вообще) существует целесообразный порядок. Только с научных позиций, без привлечения каких-либо сверхъестественных сил и можно, и нужно стремиться объяс- нять происходящее. Порядок, существующий в организации раститель- ных биоморф, лично меня восхищает, не столько разнообразием в проявлении их фенетической организации, с которым они пришли в наше время через долгие миллионы лет эволюции, сколько, напротив, единообразием тех принципов их целостной организации, на основе которых все, что имеет место быть, существует, что наблюдается повсю- ду и что особенно удобно наблюдать, сравнивая принципы организации в крупных объединениях растительных биоморф, например, среди всех семенных растений. Безусловно то, что у Природы есть движущие силы, которые обеспечивают ей саморазвитие на основе относительно немно- гих законов. Среди таких атрибутивных сил высший уровень принадле- жит эволюции, которая бесконечно проявляет себя в Природе, опреде- ляя содержание и количественных, и качественных изменений тел и процессов, происходящих в ней. Первое и главное достижение в эволюции Природы — возникно- вение преднаследственности — основы существования всего неживого, и затем, на ее основе, наследственности — основы существования всего живого. В этом отношении первым модусом (нормой) существования организации фитобиоморф, а шире всего живого, безусловно, должно быть единство неживого и живого, проявляющее себя на основе закона Доку- чаева-Вернадского (т. 1, гл. 3.4.). Второй модус организации семенных фитобиоморф — это обязатель- ное единство в ней генетического и фенетического, в котором решающая роль принадлежит функциональным союзным системам — генофенам. Всякий ген отвечает за развитие определенного фена или даже несколь- ких фенов и управляет этим процессом, поскольку более всего зависит от его результатов и качества исполнения. Гены сделали управление фенетической организацией своим единственным занятием, превратив его в высшую властную (стратегическую) процедуру: как конструировать и как строить феносомы, которые обеспечивали бы для них обиталища
Глава 10. Откровения о буднях и ираздниках растительной жизни 775 (автобиотопы) и надежно защищали их от непосредственного контакта с весьма агрессивной окружающей внешней средой. Гены и их системы — геносомы — своей энергией управляют фенетическим строитель- ством, но исполняют его сами фены и их системы — феносомы. Жизнь растительных биоморф, как и всего живого, устроена по иерархическому многоуровневому принципу (т. 1, гл. 2), поэтому рас- тительные биоморфы состоят из структурных элементов — биосистем меньшей, чем они, организационной сложности (органеллы, клетки, ткани, органы) — сами являются, будучи генетически однородными системами или унитарными (с непрерывными соматическими граница- ми) — организмами или, напротив, дизъюнктивными (с прерывистыми соматическими границами) — синорганизмами, чьими-то структурными элементами, занимая определенное место в инфраструктуре биосистем более сложных, чем они, например, в популяциях, видах, надвидах, биоценозах и т.п. Иерархическая многоуровневость в организации расти- тельных биоморф и окружающих их других биосистем — это третий модус организации растительных биоморф. Всякое существование растительных (а шире — любых) биоморф в процессе эволюции стало возможно только с созданием универсального принципа жизнеразвития по схеме «управление — исполнение» (потребле- ние, производство, резервирование, рассеивание) — это четвертый модус организации семенных фитобиоморф (т. 1, гл. 4). Чтобы выживать в различных экологических условиях, раститель- ные биоморфы проявляют разнообразие своей фенетической организации, поскольку обладают определенной пластичностью, а когда ее не хватает, то способны проявлять еще и гиперпластичность, прибегая к мутациям,— все это пятый модус (глава 6). Развитие генофенетической организации биоморф происходит по схеме приоритета не-своего (другого) свойства жизни над своим (соб- ственным) свойством, что проявляется в угнетении саморазвития веге- татива по отношению к развитию репродуктива, в подчинении властью вида (эйдократией) и властью более высоких по сложности организации биосистем (макробиократией), которые как-либо действуют в организации семенных фитобиоморф, любой другой власти, рангом пониже предыдущих, в том числе и власти организмов (онтократии), и власти синорганизмов (синонтократии) — это шестой модус организации семенных фитобиоморф. Организация растительных биоморф находится в ритмах стадийности жизнеразвития, что особенно ярко проявляется в стадийности биоморфоге- незов и в фенологии сезонного развития — это седьмой модус (т. 1, гл. 7). Какую бы форму ни приобретало развитие растительных биоморф, те обязаны сначала накапливать в своей организации необходимое дев- ственное, или виргинильное (физиологически незрелое), качество (ве- геторесурс, или вегетомассу) жизни путем фотосинтеза или даже пара- зитического способа потребления ресурсов и т.п. Далее только после достижения некоторого обязательного результата (критической вегетомас- сы) фитобиоморфы принимаются за развитие в них конкретного матурного (физиологически зрелого) качества, что, кстати, также не беспредельно и в зависимости от внутренней натуры биоморф и условий окружающей среды
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ когда-то себя исчерпывает и более не восстанавливается, приводя такие биоморфы к элиминации — это восьмой модус. Стремление к размножению, причем всякому, но в первую очередь, к репродуктивному (половому), составляет первый принцип фенетичес- кой организации биоморф семен~~ых растений, что сопровождается раз- нообразием красок и форм при цветении и плодоношении, среди кото- рых все остальное уходит на второй план, поскольку более всего через размножение можно «переступить» через себя и сделать действительно что- то значительно большее, уже для других (не-свое) — это девятый модус. В «телодвижениях» фенетической организации семенных фитоби- оморф решающие усилия всех их структурных элементов направлены на завоевание и освоение пространства, что могут у них делать лучше других структурных элементов два типа основных органов — побеги и корни, которые способны объединяться вместе, превращаясь в высшие единицы организации семенных фитобиоморф еще и более сложные системы- корнепобеги: или корнепобеги-органы (онтоиды), или корнепобеги-организ- мы (эуфиты) — это десятый модус рассматриваемого явления. Тела семенных фитобиоморф могут располагаться в любой части про- странства у поверхности земли, хотя более всего предпочитая почву и воздушную среду, лишь бы комплекс факторов, образующих обиталище (биотоп) биоморф, позволял функционировать им как системам «управ- ления-исполнения», причем в объеме достаточном для реализации жизнеразвития по схеме «свой — не-свой», т.е. и для себя, и не для себя, а для других, — это одиннадцатый модус. Ничего не получится путного в организации биоморф семенных растений, если не будет задействована вся «могучая пятерка», обеспе- чивающая их жизнеразвитие. Нарастание, ветвление, партикуляция, раз- множение и элиминация, т.е. «могучая пятерка» — это двенадцатый модус, обеспечивающий организацию биоморф семенных растений. Итак, перед нами конспективно прошли те нормы (модусы), про- явление которых более всего составляет содержание организации био- морф семенных растений. Каким же из этих модусов следует отдать предпочтение, от достижения которых растительные биоморфы дей- ствительно получают «кайф» (они по-своему торжествуют!) — достигают того довольства, которое можно было бы без особых сомнений оценить как самый высокий результат удовлетворения их потребностей и без особых колебаний обозначить категорией растительного (шире — био- тического) «счастья». 10.2.2. Как и почему биоморфы семенных растений бывают или не бывают счастливыми «Характеристика счастья и смысла жизни с точки зрения их пси- хологической нагруженности не дает основания утверждать, что всякое поведение, всякая форма деятельности человека побуждается лишь стремлением к счастью. В человеческой деятельности нет прямой, так
Глава 10. Откровения о буднях и ираздниках растительной жизни 777 сказать, голой, открытой погони за счастьем. Но его можно «поймать» сетью незапятнанной совести и честно выполненного долга. Постановка и достижение различных жизненных целей не делает человека автома- тически счастливым» (Попов, 1986, с. 33). В отношении социального счастья, которое, естественно, волнует людей, можно доказывать и ут- верждать многое, но вот то, что действительно существует независимо от чьего-либо сознания и психологической деятельности, — это «био- тическое счастье» — новая проблема, разработка которой, начиная с объяснения упомянутой исходной категории, во многом еще впереди. Я готов утверждать, что с первого момента своего возникновения, допустим, начиная с зиготы, растительные (шире — и животные) био- морфы — всегда участники «погонь» или «гонок» за «биотическим сча- стьем», а успешный результат в них, как и положено в подобных слу- чаях, где-то достигается больше, где-то меньше, а где-то счастье вроде бы вообще себя ничем не проявляет. То, что человек «усутубил» свое существование разумной деятельностью и стремлением «не запятнать» совесть и «честно» выполнить долг, ощутив или не ощутив при этом состояние социального счастья, растительным биоморфам безразлично, поскольку они устремлены безрассудно (неосознанно) выполнять себя, нередко помогая состояться еще и другим, в жизнеразвитии отвечая, таким образом, своим биотическим, не обремененным сознательной деятельностью, счастьем или несчастьем, на вечный гамлетовский воп- рос: «быть или не быть». Из главных модусов организации биоморф семенных растений, приведенных выше, вполне можно сформулировать несколько целей жизнеразвития, точнее, на мой взгляд, всего две: жить для себя (sezemu- ровать и, таким образом, npocmo жить самому) и жить для других, т.е. не npocmo жить самому, накаиливая вегеторесурс, составляющий основное собственное содержание, а стремиться еще и восироизводиться (размно- жаться у биоморф), достигая или не достигая какого-либо размножения. Через то, как растительные биоморфы живут для себя и для друтих, что неосознанно (!) доставляет им, а если не получается достижение целей, то не доставляет, удовлетворение достигнутым, т.е. довольства и, по сути, биотического счастья, можно в конце концов разобраться с каче- ством растительной жизни, однако пока, так будем считать, что мы только на пути к разработке этой проблемы. Фитобиоморфы, безусловно, биотически счастливы, когда они хо- рошо осуществляют главные цели жизни, а когда такие цели не осуще- ствляются, то они несчастны. Почему растительные биоморфы бывают счастливыми? Дело в том, что они не просто стремятся достигнуть некоторых целей жизни, но и нередко достигают их. У них, как впрочем и у людей, есть информация, которая, пусть неосознанно, но в нужное время и в нужное место исходит и распространяется с идиоэнергией из генетической организации фитобиоморф. Цели идиоэнергии всегда в фенетической организации (феносоме) биоморф — автобиотопе генов, где, соответствующим образом, исполняется власть генов, их непосред- ственное воздействие, при корректирующем влиянии окружающих фак-
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ торов внешней среды. Цели жизни бывают разные. Бывает, например, так, что цель жизнеразвития биоморфы — зацвести один раз, да так, чтобы, никак не щадя своего вегетативного тела, образовать как можно больше жизнеспособного потомства (рис. 424, 425, 431). Или другая цель жизни — цвести несколько раз, давая возможность возобновляться вегеторесурсу и растительному организму не умирать после неоднократ- ного плодоношения, и все-таки сделать это (элиминироваться), где-то может быть после 10, 12 или большего числа плодоношений, что про- граммируется примерно и зависит очень сильно от внешних обстоя- тельств (рис. 426 — 430, 432, 433). Вся растительная жизнь, как, впро- чем, и любая другая, пронизана внутриорганизменными потоками идио- энергетических «волн», которые направляют и активируют фенетичес- кое жизнеразвитие к определенным целям жизни. Их много, но среди них, безусловно, выделяются те главные цели, достижение или не до- стижение которых делает или не делает фитобиоморфы биотически счастливыми. Счастье способно приходить к растительным биоморфам только потому, что они способны испытывать довольство (!) тем, чего они достигают. По факту конкретного достижения некоторой цели жизни фиксируется факт получения растительной биоморфой соответ- ствующего удовлетворения от достигнутой ею цели жизнеразвития, которое мы, исследователи, наблюдая за происходящим со стороны, прямо-таки «подглядывая» за таинством чужой жизни, вполне можем обозначить как проявление некоторой доли биотического счастья, за которое она, конкретная фитобиоморфа, ничем не осознавая этого, так или иначе боролась (другого не дано), в определенном месте простран- ства и в определенное время, с большим или меньшим успехом. Счастье, как качество жизни, всегда соприсутствует рядом в един- стве со своей противоположностью, с другим качеством жизни, несча- стьем, и (или) его потенциалом (скрытой возможностью) где-то полно- стью или частично подавляя его, а где-то, напротив, уступая его давле- нию. Счастье, в зависимости от масштаба достигаемых целей жизнераз- вития, может быть кратковременным или долговременным и даже про- должаться всю жизнь конкретной биосистемы. Похоже, и это не уди- вительно, у типов биотического счастья должна быть своя иерархия— свои уровни, которые биоморфы могут стремиться достигать, что в реальной жизни им далеко не всегда удается. Однако, прежде чем рас- суждать об многоуровневости явления биотического счастья, пора оп- ределиться с содержанием самого этого явления. Фактическое достижение (удовлетворение стремления) живым телом некоторой цели жизни в его фенетическом развитии, стремление к дос- тижению которой генетически предопределено, но находится в зависимо- сти от непосредственного взаимодействия этого тела с окружающей внеш- ней средой, — это универсальное для всех живых тел явление — биоти- ческое счастье.
779 Глава 10. Откровения о буднях и ираздниках растительной жизни м4,19а, 27 Рис. 431. Биографический портрет Echinops humilis Bieb. [сем. Asteraceae (Алтай, остепненные каменистые склоны к реке Катунь, окрестности села Иня)]. БИОМОрфОТИП: надземка-двусемядольные (семядоли обратнояйцевидные, эллиптичес- кие, мясистые, широкозакругленные, 7 — 10 мм дл., 3,5 — 5 мм шир., клиновидно суженные в короткие черешки) полурозеточные [виргинильное развитие растений осуще- ствляет медиальный розеточный побег, всегда связанный с главным стержневым корнем, и только один раз в жизни после 6 — 20 лет эумоноподиального нарастания главный побег переходит к образованию удлиненной безрозеточной цветоносной части своей целостной полурозеточной структуры, завершая верхушечное нарастание полузакрытыми соцветиями (с параклгдиями или без них) — корзинками] очередно-простолистные семидетерминатно- репродуцирующие однодомные фиолетово-астроцветковые стержнекорневые унирепродук- тивные среднелетние и многолетние травы.
780 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБИ РАСТЕНИИ Рис. 432. Биографический портрет Primula cortusoides L. [сем. Primulaceae (Алтай, Куминский хребет, сосновые боры по низкогорным склонам к до- лине реки Чемал)]. ОСНОВНОЙ бИОМОрфОТИП: надземно-двусемядольные розеточные очередно-простоли- стные гемисимподиальные компактно-корневищные семидетерминатно-репродуцирующие [унирепродуктивные ди-, плейофазные розеточные побеги развивают верхушечный безли- стный цветонос, а иногда еще и один — два пазушных цветоносных побега дополнения, которые сверху оканчиваются полузакрытым соцветием (по традиционной терминологии — открытым соцветием) — бракгеозным зонтиком] однодомные красновато-фиолетово- примулоцветковые ретардационно-кондивидуальные (замедленно вегетативно размножаю- щиеся) горно-лесные травянистые многолетники (впервые цветут на на 3 — 6 год очень редко на 2-ой год, живут до 30 — 40 лет).
Глава 10. Откровения о буднях и ираздниках растительной жизни 6,27 Рис. 433. Биографический портрет Rosa acicularis Lindl. [сем. Rosaceae (Рес- публика Тыва, окрестности села Сосновка, хребет Танну-Ола, около 1500 м над ур. моря, кедрово-лиственничный лес, поляны, опушки, редколесья)]. БИОМО~)фОТИП: надземно-двусемядольные (семядоли эллиптические, 5 — 10 мм дл., 3— 5 мм шир., закругленные, клиновидно суженные в короткие черешки) безрозеточные очередно-сложнолистные (непарноперистолистные) гемисимподиальные детерминатно- репродуцирующие обильно-шиповатые полуразвернуто-кустистые однодомные розовоц- ветковые невысокие (в верхнем лесном поясе) или высокие (до 2-х м высотой в низко- горьях) листопадные кустарники. (Продолжение рисунка на следующей странице).
782 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ 32,36 4,гм4, 206,21,27 4,аг2, 206,27 б,гм4, 206+20e, 21,27 51,67 З,гм4, 206+20в, 22,27 ПП Рис. 433. (продолжение) Если биотического счастья нет, т.е. оно не получается, или его маловато, то происходит в жизнеразвитии биосистем другое явление, которое было в потенциале качества жизни, — биотическое несчастье, когда фактически не достигается (не удовлетворяется стремление) жи- вым телом некоторой, генетически предопределенной цели его фенети- ческого развития, которое свидетельствует об угнетенном состоянии таких биосистем и способствует их гибели. Биотическое счастье и не-
Глава 10. Откровения о буднях и ираздниках растительной жизни 783 счастье и потенции их соприсутствуют здесь и сейчас в одном живом теле и, взаимодействуя друг с другом, конкретно определяют, кто сегод- ня в соответствующих условиях ниже или выше и кто формирует то великое разнообразие форм и процессов, что и составляет такое, во многом еще таинственное, но прекрасное явление — жизнь. Рядом со счастьем в одном качестве жизни всегда есть несчастье и (или) его потен- ции, и когда не получается качества жизни как большого счастья, то несчастье не заставляет себя долго ждать, начиная превалировать в кон- кретной жизни какое-то время или определяя все качество жизни некото- рых биоморф, превращая остатки счастья в несчастье, нередко доводя такие биоморфы до гибели. Растительные биоморфы, достигая некоторых целей жизнеразвития, по-своему становятся в чем-то счастливчиками, поскольку биотическое счастье — это, в первую очередь, довольство от уже достигнутых целей развития жизни. «Вот цветет калина в поле у ручья...» — это счастливое время в жизни особей калины, а вот кое-как доживает свой цикл особь осины, единственный ствол которой был несколько лет назад сломан ветром, у других особей, например, подро- ста ели, под пологом более рослых деревьев, проблем выживания не меньше и т.д. В жизни всех биоморф, пока они живы, всегда есть какое- то место для счастья, причем более всего его у цветущих и плодонося- щих растений, поскольку они биотически счастливы от того, что выпол- няют долг для других, в первую очередь, для вида. Большое несчастье для дерева осины, что его ствол сломал ветер, но оно еще живо, и какое- то счастье от такой ущербной жизни все же совсем не покинуло это дерево. Растительные биоморфы способны в чем-то пребывать более в качестве счастливых и в чем-то более в качестве несчастных, но, и так бывает чаще всего, чего-то в них больше, что позволяет говорить о целостном качестве жизни (жизненности) и, более того, что будет еще рассмотрено (глава 10.2.4.), даже пытаться формализовать жизненные состояния биоморф в определенное время. Биотическое счастье — это одна из сторон жизнеразвития растительных биоморф, когда они не просто стремятся, но и реализуют себя в достижении определенных целей, главная из KQTophlx, безусловно, размножение. При всем этом не будем забывать, что фитобиоморфы для начала npocmo живут для себя, накаиливая па- раллельно необходимый ресурс для размножения, т.е., no сути, для обслу- живания других, наиример, не-своего, но все-таки во многом родного тела — вида. Растительные биоморфы не рассуждают в своих действиях (этого им не дано), они безрассудны, в смысле, что лишены даже следов ра- зума, но самое удивительное, они в своем развитии придерживаются определенного порядка, их организация всегда целесообразна, в соот- ветствии с которой они действуют по законам жизнеразвития только в системах «управления-исполнения», достигая во взаимоотношениях счастья и несчастья определенного целостного качества жизни. Биоти- ческие счастье и несчастье, как две стороны качества жизни, безусловно, маркируются в организации биоморф и, в первую очередь, его главными маркерами — признаками их габитуса и возрастно-стадийного поведения, на что в конце главы обращено должное внимание.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ 10.2.3. Иерархичность биотического счастья Из предыдущего материала, пожалуй, ясно, что означает для рас- тительных биоморф быть счастливыми или не быть таковыми. Их сча- стье особенное — только биотическое, в котором нет места «психоло- гической нагруженности», поскольку ни думать, ни переживать, ни жестикулировать, ни говорить что-либо, выражая восторг и т.п., фито- биоморфы не могут (не дано, поскольку это им оказалось ненужным). Они обречены если жить, то бороться только за биотическое счастье, поскольку без борьбы обеспечено несчастье и даже самое последнее в нем — смерть. Смерть — это, как уже было показано выше, — адаптация, но она же — крайняя форма выражения несчастья у того живого тела, в кото- ром она непосредственно себя проявляет. Однако и этого в отношении смерти маловато, поскольку статус у нее особенный и под определен- ным углом обзора она способна приносить счастье. К обсуждению этой стороны качества жизни мы еще вернемся. Любая фитобиоморфа, что- бы быть счастливой, должна получить, согласно определению биотичес- кого счастья, довольство от того, что совершается в их жизни как борьбе за достижение определенных целей, которые с идиоэнергией поступают в фенетическую организацию из генетического уровня. У растительных биоморф довольство достижением целей жизнеразвития проявляется не в каких-либо восторженных эмоциях (этого, безусловно, ждать не сто- ит), а через фиксацию факта, что некоторые цели достигнуты (многие признаки, особенно габитуальные и стадийные, тому показатели), а коль так, то и достигнуты определенные уровни (или всего один уро- вень) счастья. Биотическое счастье разноуровнево, что в отношении биоморф семен- ных растений (шире — в отношении всех биоморф) можно отразить в суперординатной классификации типов биотического счастья, которые они достигают или могли бы достигать в своем жизнеразвитии, и на ее основе построить соответствующую графическую модель — «пирамиду» абстрак- ций (рис. 434). В супердинатной классификационной системе биотичес- кого счастья, которое когда-либо и как-либо испытывают растительные биоморфы, использовано несколько широких субординатных классифи- каций рассматриваемого явления, соединенных вместе так, что распо- ложение каждой из них, в свою очередь, иерархически взаимосвязано, и вышестоящие развивают содержание нижестоящих, поскольку они построены с разных высот обобщения и с меньшим или большим ох- ватом угла обзора, занимая разные ступени «пирамиды» абстракций, т.е. суперординаты в суперординатной классификации. На верхней ступени (суперординате) в иерархической классификации соответствующих иерархических классификаций биотического счастья расположим две широкие субординии. За критерий сложности качества биотического счастья в первой широкой субординии принято отношение живых тел к счастью как удовлетворению (получению довольства) от достижения результатов жизни, в которых превалирует развитие их организации для себя (собственное, личное) или развитие для других (своих родных, но
Глава 10. Откровения о буднях и ираздниках растительной жизни 785 все-таки непосредственно не себя, или совсем, хотя и это относительно, чужих). Все биоморфы, в их числе и семенные фитобиоморфы, способ- ны испытывать два основных типа биотического счастья: жить для себя и, что посложнее, жить для других (рис. 434-А). Сменим несколько угол обзора, оставшись на той же высоте обоб- щений и обратим внимание при диагностике биотического счастья на стадийность в прохождении биоморфами своих конкретных жизнен- ных путей (биоморфогенезов). Это вполне надежная основа, чтобы судить о достижении того или иного уровня счастья, его качества. Если биоморфа существуют в какой-то стадии своего возрастного развития, то уже это означает, что она чего-то достигла в своей жизни и уже до п-стадии распространила действие и на себя категории биотического счастья. Всякая растительная биоморфа, безусловно, биотически сча- стлива, испытывая определенным образом достижение этого явления, когда она, в отличие от тех, кто не смог чего-то вообще достигнуть, просто живет (!), став однажды зиготой, затем когда-то проростком, затем еще и еще, проходя другие стадии виргинильного этапа, нако- нец, достигнув матурного (физиологически зрелого) этапа жизнеразви- тия. Она счастлива даже тогда, когда, накопив за всю предыдущую свою жизнь негативных последствий травм и болезней, в конце концов одряхлев от возраста и всего, что его сопровождает, осталась еще живой, влача, нередко даже довольно жалкое, пусть «предсмертное», или сенильное, состояние, но она живет (!) и уже это делает ее, хотя бы чуть-чуть, счастливой по отношению к многим тем совидичам, кто по сравнению с ней, ушел в небытие, у которых несчастье умереть оказалось сильнее счастья еще жить. С этой позиции все семенные фитобиоморфы способны испытать и многие испытывают два типа биотического счастья: эмбрионального развития и, более сложного, эпиэмбрионального развития. На одной из нижних ступеней «пирами- ды» абстракций биотического счастья эта субординия будет детализи- рована в более дробную субординатную классификацию от зиготы до матурного и постматурного, или сенильного, возраста. На этой же (первой) ступени «пирамиды» абстракций можно, под несколько иным углом обзора, поместить еще субординатную класси- фикацию, состоящую, в нашем случае, также из двух субординат, когда два класса объектов классифицирования (биотического счастья семен- ных фитобиоморф) объединяются в одну классификацию — биполяр- ную субординию (т. 1, гл. 1), отражая изменение объектов такой клас- сификации в противоположных направлениях: счастье нисходящего жизнеразвития, а в противоположном направлении, более сложное— счастье восходящего жизнеразвития. Можно, конечно, возразить, что в нисходящем жизнеразвитии не стоит искать проявлений счастья. Одна- ко я рассуждаю иначе. Коль было восхождение в развитии и биоморфы были в нем счастливы, то в принципе, хотя и качество счастья измени- лось, снизившись в сторону уменьшения его при нисходящем жизнераз- витии, счастье никуда не делось, оно продолжается, соответствующим образом трансформируясь, поскольку в дискретном соматическом разви- тии живого, естественно предусмотрена и элиминация (другого не
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ дано), пока продолжается конкретная телесная жизнь, что и составляет один из уровней биотического счастья ее непосредственных носителей. Сам факт продолжения какой-либо жизни (легкой или трудной) — уже счастье. Смерть может наступить мгновенно на любом отрезке восходя- щего или нисходящего развития жизни, и такое несчастье способно прерывать счастье, хоть какой, но жизни. Счастья больше, когда био- морфа стремится взять у явления «жизнь» все, на что она по своей природе потенциально способна (но через себя «не перепрыгнешь»). На второй суперординате как «ступени» суперординатной класси- фикации (рис. 434-Б) мы расположили четвертую субординию из трех субординат (классов соответствующей субординатной классификации): счастье нисходящей жизни с грузом накопленных проблем со здоровьем (травм, болезней), счастье восходящей жизни с какими-либо проблема- ми со здоровьем (травмы, болезни), счастье восходящей жизни без каких-либо существенных проблем со здоровьем (при минимуме травм и болезней, иногда почти или даже совсем без них). Нетрудно заметить, что главный показатель, по которому фиксируется биотическое счастье как качественная сторона жизнеразвития растительных биоморф — это их свойство проявлять довольство — удовлетворение достижением не- которой цели жизни, что проявляется, в первую очередь, в том, что они живут, хотя и по-разному, а не погибают в противоборстве с жизнераз- рушительными силами, стремящимися «отобрать» у них частично или полностью счастье и подменить его своим качественным результатом— несчастьем. Рис. 434. Суперординатная классификация («пирамида» абстракций) типов биотического счастья биоморф семенных растений. А-Е — уровни абстрак- ции (суперординаты); 1-36 — субординаты (классы) типов биотического счастья. 1 — счастье жизни для себя, 2 — счастье жизни для друтих, 3 — счастье эмбриональной жизни, 4 — счастье эпиэмбриональной жизни, 5 — счастье восходящего развития, 6— счастье нисходящего развития, 7 — счастье восходящей жизни с какими-либо проблемами со здоровьем (травмы, болезни), 8 — счастье восходящей жизни без каких-либо существен- ных проблем со здоровьем, 9 — счастье виргинильной жизни, 10 — счастье восходящей матурной жизни, 11 (1) — персональное (свое) счастье, 12 — преперсональное (предсвое) счастье, 13 — семиперсональное (полусвое) счастье, 14 — постперсональное (сверхсвое) счастье, 15 — альтернативное (совсем чужое) счастье, 16 — автобиофилия (персональное жизнесохранение), 17 — амфобиофилия (обоюдное жизнесохранение), 18 — моноксено- биофилия (первое коллегиальное жизнесохранение), 19 — диксенобиофилия (вторая ксе- нобиофилия), 20 — триксенобиофилия (третья ксенобиофилия), 21 — счастье жить для себя, довольствуясь вегетативным развитием, но без вегетативного размножения, 22— счастье вегетативной жизни для себя и друтих, прибегая к вегетативному размножению, 23 — счастье жить для друтих, развивая репродуктивную систему и осуществляя в ней бесполое размножение, 24 — счастье жить для друтих, осуществляя репродуктивное раз- множение без оплодотворения — апомиксисом, 25 — счастье жить для друтих посредством репродуктивного размножения с самоопылением и самооплодотворением, 26 — счастье жить для других посредством перекрестного (облигатного) репродуктивного размножения, 27 — счастье жить для друтих, поддерживая жизнь вообще, как родных, так и чужих, всеми средствами, сверх упомянутых выше (21 — 26), 28 — счастье финиматурной жизни, 29— счастье сенильной жизни, 30 — 36 — счастье состояний восходящего жизнеразвития: 30 — эмбрионов, 31 — проростков, 32 — всходов, 33 — ювенилов, 34 — имматуров, 35— инициматуров, 36 — медиматуров.
787 Глава 10. Откровения о буднях и лраздниках растительной жизни 15 74 13 72 Л 79 7 26 24 22 На третьей (сверху) ступени «пирамиды» абстракций (рис. 434-В) мы расположили пятую субординию из четырех субординат, которая так же, как и третья, биполярная: нижняя субордината занята счастьем нисходящей жизни (оно наименьшее и, как и в третьей субординии, пока далее не дифференцируется). В другом (восходящем) направле- нии счастья жизнеразвития располагаются, соответственно, суборди- наты счастья эмбриональной, виргинильной и, наконец, восходящей матурной жизни.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Шестую субординатную классификацию биотического счастья се- менных фитобиоморф (глава 10.1.3.) из пяти субординат (рис. 434-Г) расположим на следующей (четвертой сверху) ступени (суперординате) «пирамиды» абстракций: 1 — счастье жить для себя (свое, или персо- нальное счастье), 2 — счастье жить для других — предсвоих, непосред- ственно составляя свое тело родными для него, но все-таки другими живыми телами меньшей организационной сложности преперсональное счастье, 3 — счастье одного живого тела жить для других — полусвоих, непосредственно составляя другую часть родного ему живого тела, па- раллельно живущее для себя, в котором первое тело является соответ- ствующим структурным элементом (семиперсональное счастье), 4— счастье жить для других родных тел — сверхсвоих, которые для своего тела опосредованно через другие живые тела составляют вместе их, более сложную, организацию как структурные элементы, например, организм — свое тело, а его вид — другое родное тело, взаимосвязанные не непосредственно, а опосредованно через другие структурные элемен- ты вида: синорганизмы, если они образуются, специкулы, если они сложнее организма, ценодемы (ценотические популяции), популяции (постперсональное счастье), 5 — счастье жить для других чужих— неродных, которые не проявляют сравнительно близкого родства и весьма далеко отстоят друг от друга, например, организмы магнолии, амебы и человека (альтернативное счастье). С одной стороны, допустим, волк персонально биотически счастлив, что догнал особь зайца и ею утолил свой голод, а заяц, конечно же, персонально попал в биотичес- кое несчастье, ведь он погиб, будучи съеден, но, с другой стороны, обе особи в рассматриваемой ситуации проявили в своем качестве жизни альтернативное счастье, поскольку каждая из них помогла неродствен- ному (чужому) живому телу сохраниться: заяц подкормил собой и особь, и вид волка, а волк, своим поведением, чуть-чуть подчистил чужое видовое пространство для подрастания в нем более ловких и сильных зайчат (см. с. 770). Седьмая широкая субординия биотического счастья растительных биоморф из пяти субординат заняла свое место, как и предыдущая субординия, также на четвертой суперординате (ступени «пирамиды» абстракций). Проявление свойства биофилии (жизнесохранения) отве- чает принципу борьбы за достижение жизнеполезной цели развития, т.е. за счастье. Идея борьбы фитобиоморф за счастье созвучна со стрем- лением биосистем к жизнесохранению, что нашло отражение в специ- альной теории жизнесохранения, или биофилии (Нухимовский, 1980, 1984, 1997). Субординатная классификация жизнесохранительных (биофильных) свойств под определенным углом обзора и с определен- ной высоты абстракции может рассматриваться и как субординия биотического счастья, проявляющаяся в организации семенных фито- биоморф (шире — и других биоморф). Нижнюю субординату занимают свойства и функции автобиофилии (персонального жизнесохранения), что составляет одновременно счастье жить для себя, поддерживая так жизнь вообще, сохраняя себя в живом состоянии и не воспроизводя новые биосистемы своей и более высокой субординаты. Далее следует
Глава 10. Откровения о буднях и ираздниках растительной жизни 789 субордината свойств и функций амфобиофилии, или счастья жить для себя и друтих, поддерживая, таким образом, жизнь вообще, сохраняя одновременно в живом состоянии и себя, и другие биосистемы, не воспроизводя новых биосистем своей и более высокой субординаты. Выше располагаются первая, вторая и третья ксенобиофилии. «Свой- ство биосистем поддерживать жизнь вообще, воспроизводя новые биосистемы, не ниже, чем своей субординаты и (или) непосредственно участвуя в сохранении их (обеспечение пищей, защита, воспитание и т.п.) — это коллегиальное жизнесохранение, или ксенобиофилия» (т. 1, стр. 91), что на языке биотического счастья означает счастье жить для других (и родных, и чужих). Моноксенобиофилия (первая ксенобиофилия) — это сохранение биосистемой других биосистем своей субординаты без воспроизведения новых. Диксенобиофилия (вторая ксенобиофилия) — это непосредствен- ное участие биосистемы в образовании новых биосистем своей субор- динаты. Триксенобиофилия (третья ксенобиофилия) — это непосред- ственное участие биосистемы в образовании новых биосистем другой, более высокой субординаты. Например, для родительских организмов образование нового организма — это и вторая ксенобиофилия, и счастье жить для других простых биоморф, во многом родных, хотя и в чем-то все-таки чужих, уже не родительских организмов. Восьмую широкую субординию биотического счастья семенных фитобиоморф мы строим из семи субординат (рис. 434-Д). Нижнюю субординату отводим счастью жить для себя, довольствуясь вегетатив- ным развитием, но без вегетативного размножения. Выше располагаем субординату счастья жить вегетативно, для себя и друтих, прибегая к вегетативному размножению себе подобных. Третья субордината- счастье жить для других, развивая репродуктивную систему, но без полового процесса, осуществляя в ней бесполое размножение (напри- мер, Poligonum viviparum L., т. 1, стр. 233). Четвертая субордината- счастье жить для друтих, развивая в своей организации репродуктивные системы, и силами одного организма без оплодотворения (апомиксис) осуществляя репродуктивное размножение. Пятая субордината — счас- тье жить для других, развивая в своей организации репродуктивные системы, в которых, посредством оплодотворения силами одного орга- низма (самоопыление и самооплодотворение) осуществляется репродук- тивное размножение. Шестая субордината — счастье жить для других, развивая в своей организации репродуктивные системы, которые про- изводят зрелые половые клетки (гаметы) для участия в половых процес- сах размножения при взаимодействии разных организмов. Седьмая су- бордината — счастье жить для других, как родных, так и чужих, участвуя в поддержании жизни вообще всеми средствами, сверх упомянутых выше, и, таким образом, внося свой вклад в жизнесохранение вообще, но и среды жизнеобитания, например, производя кислород при фото- синтезе, проявляя пищевые достоинства для других и т.п. Девятую широкую субординию биотического счастья биоморф семенных расте- ний строим из девяти субординат (рис. 434-Е). Девятая субординия биполярна. За критерий усложнения биотического счастья растительных
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ биоморф в этой классификации принята возрастная дифференциация их жизненных циклов (биоморфогенезов), поскольку отправной точкой для наших рассуждений о биотическом счастье стал, в первую очередь, факт достижения конкретной биоморфой элементарной цели ее разви- тия — простого выживания и, таким образом, какого-либо участия в жизни вообще. Эта классификация биотического счастья учитывает, в первую очередь, то, как биоморфы решают вопросы биотического сча- стья через прохождение стадий своих жизненных циклов и у кого-то его получается больше, а у кого-то меньше, более того, кое-кто остается ни с чем и его жизнь совсем не удается, хотя перспектива-то была, она в живом присутствует всегда, только не всегда условия окружающей среды и (или) состояние здоровья его внутренней организации позволяют достигнуть возможных целей. Биотическое счастье разноуровнево, и это наглядно видно также и в девятой субординии этого явления. Внизу, как и положено в подобных случаях, что-то слабенькое, уже уходящее за «порог», действующее в другом направлении развития явления, в нашем случае, биотического счастья; здесь счастье финиматурной (старой ма- турной) и сенильной жизни, которого как такового все меньше, но все- таки жизнь-то еще есть, следовательно, и счастье еще есть, хотя они вместе все уходят и уходят. Далее, в противовес нисходящему направлению жизнеразвития, проявляют себя уровни (субординаты) счастья восходящего жизнеразви- тия: эмбрионов, начиная с зиготы и до начала прорастания семян, про- ростков, всходов, ювенилов, имматуров, инициматуров и медиматуров. Достижение каждой из стадий жизнеразвития — это торжество некото- рого уровня счастья в уже состоявшейся растительной жизни. Счастье для растительных биоморф и тогда, когда они только-только еще в зиготах, когда начинают прорастать семена, и тогда, когда увеличивается вегето- ресурс в процессе роста и развития всходов, ювенильных, имматурных и матурных растений. Особый «кураж» счастливой растительной жизни проявляется тогда, когда фитобиоморфы цветут и плодоносят, что, безус- ловно, составляет праздники в жизни растений. Последствия от праздни- ков бывают разные. Для одних фитобиоморф они сменяются буднями накопления вегеторесурса и преобразования его в репродукгоресурс с последующим повторением, даже неоднократным, репродуктивного уси- лия. Что касается некоторых других типов биоморф (однолетники, эфе- меры), то феерия счастья цветущей и плодоносящей жизни настолько истощает их, что они уже даже не пытаются сопротивляться судьбе и, с довольством выполненного долга перед видом и жизнью вообще, уходят из жизни, т.е. умирают во многом счастливые, не мешая развиваться своему потомству [жизнь продолжается (!)]. 10.2.4. Мера счастья и формула качества растительной ЖИЗНИ Из предыдущего материала, пожалуй, понятно, что организация биоморф устроена целесообразно и все, что с ней происходит, нацелено
Глава 10. Откровения о буднях и лраздниках растительной жизни 791 в развитии на то, чтобы выжить: и себя сохранить, но и других поддер- жать в жизнесохранении, даже больше других, чем себя. На этой основе нами была просмотрена с разных высот обобщения и под разными углами обзора иерархия биотического счастья в растительном жизнераз- витии, и получается так, что фитобиоморфы быстрее теряют свое (эта цель иерархически ниже, она проще), чем прекращают бороться за сохранение другого (кого-либо не-своего, но родного, или даже, каза- лось бы, совсем чужого живого тела). В таком стремлении к жизнеполезным целям своего развития ра- стительные биоморфы фактически или достигают, или не достигают их, а, следовательно, получают или не получают довольство, пусть и нео- сознанное, от содеянного ими, которое выражается так или иначе, кроме всего прочего, в проявлении фитобиоморфами биотического счастья некоторого качественного уровня. Счастье, о котором здесь идет речь, всегда себя чем-то проявляет, если оно вообще есть, а оно есть, хотя и разное в любом теле, когда оно живет. Биотическое счастье, как и многое другое в живой природе, проявляет себя в борьбе со своей противоположностью — несчастьем. Без такой борьбы счастье никогда бы не смогло вообще состояться, поскольку для торжества биотического счастья, для того, чтобы ему только быть, нужно всегда еще что-то другое, часто внешне вроде бы и ничем не проявляющее, которое можно было бы победить, сломить и (или) преодолеть..., и это другое — несчастье и (или) его потенции. Биотическое счастье способно проявлять себя только в борьбе со своей противоположностью — несчастьем. Они всегда сосуществуют вместе в противоречивом единстве, т.е. всегда рядом, но кто-то из них над своей противоположностью явно или не очень превалирует. Быть биотически счастливым — это приоритет жизнеразвития как у человека, так и у любой растительной биоморфы, что в каждом конкретном случае уда- ется больше или меньше, или вовсе не удается. Какое-то из двух ка- честв, как две стороны единого качества жизни существуя в единстве и борьбе, сначала, чаще всего, как их потенциалы (скрытые возможнос- ти), когда-то побеждает. Победа, пусть и неполная, несчастья над сча- стьем, означает ухудшение целостного качественного уровня жизни и, следовательно, может привести, если процесс не изменить, к полному торжеству несчастья и смерти. Если два противоположных качества жизни — биотические счастье и несчастье, взаимодействуя, сами опре- деляют, каким должно быть целостное качество жизни у конкретной биоморфы, то, возможно, их по отдельности как-то можно измерить, а затем, создав соответствующую формулу, вычислить, каково на самом деле целостное качество жизни конкретной биоморфы. Такова была исходная идея, которая в конце концов воплотилась в следующем. Однако есть смысл сделать небольшое отступление. Каждая растительная биоморфа получает довольство и становится счастливее уже от того, что она все дальше и дальше живет по програм- ме, исходящей с генетического уровня, своего жизненного цикла (био- морфогенеза). Все для растительной биоморфы, допустим, организма или синорганизма родиолы розовой (Rhodiola rosea L.) началось с зи-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ готы в родительском организме. Это был первый успех, поскольку, став зиготой, конкретная особь сумела уже немножко побыть счастливой и, по~я дальше, если сумеет, по тропе жизнеразвития, может только прибавлять себе счастья от проживаемой жизни, каждого ее мгновения. Судьба выбирала из многих и, наконец, остановилась на фитобиоморфе родиолы розовой, чтобы на тропе жизнеразвития испытать ее на проч- ность искателя и борца за биотическое счастье. У действия жизнесох- ранительных сил развития, составляющих главное содержание своего биотического счастья (положительного качества жизни) всегда есть про- тивник — несчастье (отрицательное качество жизни), часто представлен- ное действием сил жизнеразрушения. Появился на берегу горного ручья проросток родиолы розовой- это еще один шаг счастья от жизни, затем из него развился всход, далее ювенильное растение — и это все было счастьем для особи данного вида (подробнее см. т. 1, гл. 10.2.2.1.). Но несчастье где-то всегда рядом и может приходить, когда складываются для него определенные условия. Далее в судьбе особи радиолы розовой все могло бы произойти иначе, и вместо счастья возобладало бы несчастье, например, если бурный селевой поток смел бы с берега ручья многое живое на своем пути, в том числе и крохотное живое существо родиолы розовой. Такое вполне могло бы быть, но, в нашем случае, к счастью (!), не случилось. Био- тическое счастье продолжило восходить, сопровождая стадии конкрет- ного биоморфогенеза родиолы розовой. Счастье может побеждать не- счастье и в экстремальных условиях. Юная особь, еще крохотный «клу- бенек», родиолы розовой, будучи снесенная водным потоком, пристал к новому «обиталищу», зацепившись за камень, торчащий из воды прямо в русле ручья, где-то ниже по склону горы, но все-таки не настолько низко, где было бы достаточно много конкурентов, чтобы помешать нашему счастливчику восхождение по тропе своей жизни в поисках своего счастья. На камне оказалось сравнительно сносно и довольно тихо по сравнению с тем кошмаром, который царил вокруг. Бурлящая водная стихия постоянно несла опасность погубить особь родиолы розовой — вынужденного переселенца, зацепившегося здесь почти на голом камне среди мхов и лишайников. Водный поток, при всей его «агрессивности» по отношению к нашей особи родиолы розовой, выб- расывал на камень ничтожными порциями крупицы песка с илом, а еще и «надежду» на выживание тех живых существ, которые в них нужда- лись. Счастье не изменило конкретной особи родиолы розовой. Этот «клубенек-колобок», похоже, надежно «пришвартовался» к заветному биотопу, пусть неуютному, но именно тому — своему, с которым он готов был на долгие годы связать судьбу своего дальнейшего жизне- развития, проявляя в крайне экстремальных условиях не угасшее в нем стремление к жизни — к тому, чтобы достигать поставленные уже давно и не им (его далекими предками) новые цели своего развития и, таким образом, испытывать разные формы нового качества жизни, которые имели значимость лишь при одном условии, чтобы счастье превалиро- вало над несчастьем. И началась «новая» жизнь! Растительная биомор- фа, о которой здесь речь, оказалась борцом-победителем и сумела про-
Глава 10. Откровения о буднях и ираздниках растительной жизни 793 жить болыпую и по-своему наверняка яркую жизнь, достигнув глубокой старости. В этой жизни было немало всего, что и составило ее особен- ную биографию. Многого стоят, например, такие факты из этой био- графии, когда водные потоки не раз отрывали куски корневищ, превра- тив, тем самым, родительскую особь в синорганизм, которые уже в других местах давали начало новой самостоятельной жизни, а родитель- ская особь получала очередное довольство от достигнутой цели — оче- редную порцию биотического счастья от воплощения в реальность воз- можности вегетативного размножения (счастья уже не для себя, а для другого, но родного ему нового живого существа). Много в жизни кон- кретной биоморфы родиолы розовой было произведено семян (высшее биотическое счастье — реализовать возможность репродуктивного раз- множения). У семян уже другая, для них своя, судьба. Если условия позволяют, то счастье новых жизней может укрепляться. Хотя бывает и по-другому. Весь путь по тропинке счастья, преодоленный ранее, может быть пере- черкнут несчастьем, обрушившимся при изменении условий существо- вания, например, со стихией селевого потока. Семена могут спокойно оказаться в новом месте, но окружающая их среда не позволит им дальше проявить себя и побороться за счастье развития жизни, напри- мер, несчастье для семян, когда они упали на бетонную площадку аэро- дрома. Жизнь переменчива. Счастье, достигнутое ранее, может пере- крываться несчастьем текущих событий. Возможно, когда-то ветер или вода, несущие перспективу счастья жизнеразвития, унесут семена-«из- гои» с неудобных территорий, и уже на новом месте у них начнется «новая» жизнь, где они прорастут и продолжат как счастливые особи дальнейшее восхождение по иерархическим ступеням биотического счастья. Я готов утверждать, что именно жизненный цикл растительных био- морф с его возрастными стадиями может стать носителем меры биотичес- кого счастья как положительного качества развития жизни, и чем больше стадий жизни пройдено, тем больше счастья положено в целостную копилку качества жизни конкретных фитобиоморф. Как обозначить единицу сча- стья, его меру, которая отражала бы факты того, что здесь счастье такое- то, здесь его побольше, а здесь — поменьше? В качестве единицы био- тического счастья (эвакинеза) предлагаю 1 (одну) эву (1 Еи) (от греч. еи— хорошо). В противовес эвам можно предложить в качестве единицы био- тического несчастья (антиэвакинеза) 1 (одну) антиэву (1 Аеи). Эвы отражают уровень биотического счастья, т.е. состояние положительного качества жизни, а антиэвы — показатели уровня биотического несчастья, т.е. состояния отрицательного качества жизни. В процессе развития биоморфологии семенных растений сложи- лись представления о возрастной стадийности биоморфогенезов (жиз- ненных циклов организмов и синорганизмов), что нашло отражение и в схемах последовательности смен категорий возрастных состояний (подробно см. т. 1, гл. 7). Напрашивается в связи с этим тезис: чем больше пройдено возрастных состояний жизненного цикла организма или синорганизма (биоморфогенеза), тем выше показатели биотического счас-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ тья, а одновременно выше и целостное качество жизни (качество жизне- развития). Однако этот тезис неполный, он довольно поверхностный и вызывает немало вопросов. Дело в том, что свойство счастья в органи- зации биоморф существует не в одиночестве, а в противоречивом един- стве со своим антиподом — свойством несчастья (его текущим резуль- татом и потенцией), которое всегда где-то рядом, то затаившееся, то проявляющее себя открыто... Биоморфы, как растительные, так и жи- вотные, вьтолнив главные цели своего развития (и сами пожили, сколь- ко их природа позволила, и потомство оставили), в конце концов все- таки должны умереть — это как, на первый взгляд, ни кощунственно звучит, кроме всего разного, еще и — что весьма важно — проявление атрибутивной формы счастья (!) в жизнеразвитии, но уже не биоморф (для них, в первую очередь, — это несчастье, хотя естественное и зако- номерное), а более высокой, по сложности организации и родной био- системы — их вида. Есть, правда, и здесь одна сторона качества жизни, позволяющая усмотреть в смерти определенных биоморф еще и форму биотического счастья для них, о чем еще будет сказано отдельно. В результате противоречивого единства, с которым связано сосуществова- ние счастья и несчастья в организации растительных биоморф, прояв- ляет себя на самом деле (реально) целостное качество жизни, которое и составляет текущий результат борьбы за существование с участием соответствующих позитивных и негативных качеств. Попытаемся количественно определиться с целостным качеством жизни конкретных растительных биоморф. Для получения такого пока- зателя надо сначала разобраться с ценой прохождения биоморфой каж- дого возрастного состояния. Только так, на мой взгляд, можно рассчи- тывать на построение формулы целостного качества жизни. Если на- учиться количественно оценивать эвы и антиэвы каждого возрастного состояния, которого достигают биоморфы, то можно, получив конкрет- ные цифровые показатели положительного (биотического счастья) и отрицательного (биотического несчастья) качеств жизнеразвития, в кон- це концов выйти и на конечный результат поиска — формулу целост- ного качества жизни. Итак, какова должны быль формула целостного качества жизни, ко- торое обозначим символом BN (от греч. В — bios, N — некроз, подчеркнув, тем самым, единство жизни и смерти в созидании качества жизни)? Ясно, что достижения, которые свершаются в жизни биоморфы, разнообраз- ны: одни для их развития хорошие, а друтие — плохие, со множеством переходов между ними. Самые первые достижения в биографии био- морф — это факты прохождения ими стадий жизненного цикла, в пер- вую очередь, возрастных. Когда один возраст сменяется другими и млад- шая возрастная стадия — старшей, то растительная биоморфа, для этого не надо быть разумным существом, а надо уметь npocmo жить, иереходит с одного качества жизни в другое, иричем, раз иереход состоялся, то с иоложительным индексом, а это означает, что в жизни биоморфы больше счастья и оно доминирует над своим антииодом — несчастьем и (или) его иотенциалами. У нас появился шанс количественно обозначить один из элементов формулы качества жизни — эвы — носителей качества био-
Глава 10. Откровения о буднях и ираздниках растительной жизни 795 12 тического счастья в организации растительных биоморф. Дадим извес- тному числу возрастных стадий (состояний) цифровой номер, начиная с десяти (10). Избрав для начала отсчета эвов (единица биотического счастья) цифру 10, а для всех антиэвов (единица биотического несчас- тья) — цифры меньше 10, начиная с 1, мы сознательно дали при оп- ределении целостного качества жизни по формуле фору (!) счастью, что вполне оправдано, поскольку для любого живого тела уже сам факт того, что оно живет, составляет приоритет счастья (хоть какого в другом отношении) над несчастьем, и его можно считать самым важным атри- бутом при качественной оценке явления «жизнь». Факт достижения конкретной семенной фитобиоморфой соответ- ствующей возрастной стадии обозначим nÅu. В этом показателе n — это всегда какая-либо цифра от 10 до 19, которая является цифровым ин- дексом возрастной стадии (состояния). Приведем оба цифровых ряда, составляющих показатели пКи и пАеи в формуле качества жизни: BN = nKu — nAeu. и овой и екс пЕи Воз астные ст ии состояния 10 — сенильная (сенил: сенильная особь или сенильный синор- ганизм) 11 — финиматурная (финиматур: финиматурная особь или фи- ниматурный синорганизм) — пренатальная (эмбриональная особь, или пренатал, находит- ся в организационном единстве с родительской особью) 13 — латентная (эмбриональная особь, или латент, как отдель- ное семя, которое уже отделено от родительского расте- ния и существует самостоятельно) 14 — проростковая (проросток) 15 — всходовая (всход) 1б — ювенильная (ювенил) 17 — имматурная (имматур) 18 — инициматурная (инициматур) 19 — медиматурная (медиматур) В формуле целостного качества жизни «срабатывает» тот положи- тельный цифровой показатель, который соответствует индексу возрас- тной стадии (состояния), реально достигнутого биоморфой в его жиз- неразвитии, — свидетельстве ее уровня счастья в конкретном фрагменте пространства — времени. Другая сторона формулы целостного качества жизни фитобиоморфы — это ее отрицательный цифровой показатель несчастья (nACU), который соответствующим образом влияет на итого- вый результат, снижая в нем долю биотического счастья. Показатели несчастья, следуя логике соприсутствия их с показателями биотического счастья «работают» в одной формуле, подчеркивающей противоречивое единство двух сторон целостного качества жизни растительных био- морф. Среди негативных (отрицательных) сторон качества растительной жизни, пожалуй, два наиболее существенных типа: от 1 до 5, когда показатель отражает участие несчастья при формировании целого каче- ства жизни через определенные жизнеразрушительные действия, сопро- вождающиеся угнетением, но без существенных физических травм, в
ОСНОВБ1 БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИЙ развитии фитобиоморф, и от б до 8, когда показатель несчастья участву- ет в формуле целостного качества жизни как отражение жизнеразруши- тельного действия каких-либо сил, сопровождающих развитие фитоби- оморф физическими травмами. В цифровых показателях негативных сторон качества жизни отражаются, таким образом, различные формы проявления несчастья в организации растительных биоморф. Показатель 1 Аеи свидетельствует, что организация семенных фи- тобиоморф внешне (визуально) не проявляет угнетающего действия сил несчастья, хотя оно и (или) его потенциал наверняка есть, но пока крайне слабое. Показатель 2 Аеи означает, что в организации семенных фитоби- оморф имеет место слабое угнетающее действие жизнеразрушительных сил, которое становится заметным при обычном визуальном наблюде- нии, например, в результате затенения больше, чем обычно, вытягива- ются междоузлия, стебли становятся тоньше, слабее и т.п. Показатель 3 Аеи подчеркивает, что организация конкретной фито- биоморфы подвергается умеренному (среднему) угнетению, которое, од- нако, не нарушает систему репродуктивного развития, например, дей- ствие угнетающих факторов приводит к изменению размеров и уменьше- нию числа цветков в соцветии, полеганию надземных побегов, которые в нормальных условиях развивались бы как прямостоячие, и т.п. При показателе 4 Аеи имеет место сильное угнетение в жизнераз- витии семенной фитобиоморфы, причем настолько сильное, что обыч- ная (нормальная) картина проявления репродуктивных циклов искажа- ется также очень сильно, но все же она способна относительно восста- новиться при снятии действия жизнеугнетающих факторов. При 5 Аеи в организации растительных биоморф проявляется не просто сильное угнетение, а такое угнетение, которое лишило, но все же не безвозвратно, их возможности репродуктивно развиваться, по- скольку прессинг условий, угнетающих даже стремление к размноже- нию, непомерно высок. При показателе б Аеи подчеркивается факт безвозвратной утраты репродуктивной активности в тех конкретных условиях, в которых про- исходит сильно угнетенное жизнеразвитие фитобиоморф на пределе их пластичности. Несчастье с индексом 6 Аеи не позволяет сильно угне- тенным фитобиоморфам вернуться к репродуктивному развитию без изменения места произрастания (без пересадки), поскольку комплекс действующих в биотопе условий идиоэнергетически не рассчитан на репродуктивное развитие в таких условиях и позволяет только кое-как побороться биоморфам за простое выживание, например, когда семена кедра сибирского (Pinus sibirica Du Тоиг), пихты сибирской (Abies sibirica Ledeb.) заносятся кедровками и бурундуками далеко в высокогорье, выше границы леса, что позволяет особям кедра вегетативно кое-как развиваться, вынужденно не теряя девственности всю их жизнь, иногда, кстати, довольно долгую, и при этом сильно изменяя свою организацию (рис. 32-II). Рассмотрим теперь следующие показатели биотического несчастья — травматические, которые также фиксируются при определении каче-
797 Глава 10. Откровения о буднях и ираздниках растительной жизни Габитоформы и формы роста {стадиосомы) — индикаторы биотического счастья и качества жизни {в целом) семенных фитобиоморф Стремление к биотическому счастью — содержательная основа качества жизни любой биоморфы, обитающей на Земле (как раститель- ной, так и животной). Биотическое счастье (эвакинез), будучи позитив- ной стороной целостного качества жизни, начинает сопровождать био- морфу уже с ее приходом на «порог» жизни (с рождения), поскольку оно ства жизни семенных фитобиоморф, если существенная травма вообще наблюдается. Показатель 7 Аеи обозначает, что в организацию конкретной семен- ной фитобиоморфы вмешалась серьезная травма (рана) одного или двух из основных органов, но все-таки ее последствия не настолько сильны, чтобы изменить обычный ход жизнеразвития, в частности, не нарушается репродукгивная цикличность, например, ветвь кроны дерева сломана ветром, не повлияв на развитие всего растительного организма. При показателе 8 Аеи подчеркивается, что организацию фитобио- морфы усложнила травма (рана), которая заметно изменила ход жизне- развития, не лишив, однако, эту биоморфу возможности нормализовать течение жизнеразвития, например, ствол дерева сильно поврежден, больше того, может быть, даже спилен и оставлен лишь пенек с кор- нями, способный регенерировать новый ствол с кроной, вновь зацвести и плодоносить. Наконец, показатель 9 Аеи означает, что травма, полученная в окружающей среде, по сути дела, уже несовместима с жизнью и расти- тельная биоморфа вынуждена угасать более или менее быстро, напри- мер, удар молнии продольно расщепил ствол особи кедра сибирского, часть ее кроны обгорела, другая часть еще зеленая, но шансов на спа- сение, по сути, нет или почти нет. Если травмы нет, то никакой трав- матический показатель в формуле качества жизни не участвует. Итак, какие же результаты получаются при пользовании формулой качества жизни? Например, формула качества жизни нормально разви- вающейся пренатальной эмбриональной особи Ча1епапа officinalis L.— BN = 12 — 1 = 11; пренатальная жизнь валерьяны лекарственной подроб- но описана Е.В. Шугаевой (Нухимовский, Семенихин, Шугаева, 1989). Главное, что следует из формулы качества жизни: биоморфа жива и она так или иначе, но всегда биотнческн счастлива уже тем, что достигла определенной возрастной стадии своего жизнеразвития, не отягощенное или отягощенное утнетеннем или травмами. Общий вывод: мера счастья (эвы) так или иначе подчеркивает продолжение жизни, а мера несчастья (антиэвы), напротив, — всегда на стороне смерти, что снижает долю сча- стья в целостном качестве жизни.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ невозможно без движения какой-либо биоморфы по циклу ее существо- вания вплоть до «выхода за порог» (элиминация). Чтобы состоялось какое-либо движение к целям жизнеразвития, растительные (шире— любые) биоморфы обязаны быть борцами за биотическое счастье, ко- торое, как показывает весь предыдущий материал, разнолико и имеет несколько слоев качества внутри единого качества эвакинеза. 10.3.1. Почему в разном качестве жизни растительных биоморф биотического счастья всегда больше, чем несчастья Биотическое счастье, как, впрочем, и несчастье, в одиночестве в живых телах не присутствует, поскольку они всегда составляют две атрибутивные стороны противоречиво единого целостного качества жизни. Так сложилось в эволюции, что жизнь избрала многоуровневый принцип организации своих телесных носителей, а коль это так, то и биотическое счастье, да и несчастье наверняка также многоуровневы. Выше мы об этом уже писали, а здесь, пожалуй, важно еще и еще обратить внимание на обозначенный нами феномен: любые биоморфы способны всегда быль, но по-разному, биотически счастливыми. Дело в том, что счастье разномасштабно: довольство проявляется достижением какой-либо цели жизнеразвития, причем полезной не только и даже не столько для себя, но, нередко более всего полезной для других, и когда биоморфы достигают таких целей, то, по сути, они нередко даже вполне закономерно (по генетическому плану) умирают еще и с довольством (на фоне своего несчастья) от достигнутой ранее цели, например, со счастьем от свершенного перед смертью размножения (наиболее пока- зательны однолетники, эфемеры). Почему биоморфы, в том числе и ра- стительные, так или иначе биотически счастливы? На мой взгляд, хотя бы только потому, что они, во-первых, — просто живут, проходя возрастные стадии своего развития, во-вторых, участвуют иногда и в размножении (!). Качество жизни, если образно, то это как ветер над земной поверхно- стью, который внешне во многом схож, а причины, его формирующие, разные — в глубинах многих взаимосвязанных явлений. Жизнеутверждающий принцип всегда быть счастливым, если вооб- ще живешь, состоит в приоритете биотического счастья над несчастьем и определяет целостное качество жизни — обязательное условие всякого телесного содержания многоуровневой жизни. Качество жизни каждого отдельного живого тела — структурного элемента в целом живом теле (биостробе) Земли — это свойство не только каждого живого структур- ного элемента по отдельности, но и свойство всей биостробы, поэтому натура всех живых тел (по качеству жизни) устроена так, что все они всегда всем служат: не только себе (живут лично, т.е. для себя), но и для других (живут не для себя, а для родных и чужих биосистем, причем всех уровней организации, но более всего для тех, кто по рангу сложности повыше их и кому принадлежит биотическая власть над ними). Кому
Глава 10. Откровения о буднях и ираздниках растительной жизни служит биостроба нашей планеты, пока не известно, поскольку реально жизнь за пределами Земли еще не обнаружена (но, похоже, это только дело времени). Все располагает к тому, чтобы утверждать: качество жизни каждого живого тела неотделимо от качества жизни вообще, или общего качества жизни, и в каждом отдельном живом теле (биосистеме) — структурном элементе биостробы — оно приобретает частичное от- ражение — это парциальное качество жизни как конкретный результат взаимодействия между собой счастья с несчастьем, но и их потенциалов при приоритете счастья ради сохранения жизни вообще. Общее качество жизни ннтегратнвно стоит над всеми парциальными качествами жизни, иодчиивв вх в соответствии с иерврхичвоствю оргввивввви житии. Провв- ление потенциальных качеств жизни рассмотрим на примере раститель- ных биоморф. Если фитобиоморфа совсем не совладала с несчастьем, то она уже не живет. Счастье, в борьбе со своим антиподом — несчастьем, берет верх в чем-то, и тогда жизнь продолжается в растительной биоморфе; она так или иначе тем, что живет, уже биотически счастлива. В стремлении выжить и сохранить жизнь вообще биоморфы посто- янно эволюционируют, прибегая к различным адаптациям. Если в связи с изменением климата прямостоячим надземным стеблям семенных фитобиоморф стало холодно, то такие растения вынуждены прибегать к новым приспособлениям: или прижимать стеблевое тело к почве, обращивая его придаточными корнями и даже «загоняя» под поверх- ность почвы, или терять на зиму как можно больше надземной массы (не всякое жизнеразрушение связано с несчастьем, а бывает наоборот), резервируя вегеторесурс в подземных органах, чтобы весной повторить новую волну надземного роста, или лигнифицировать особенным обра- зом свои надземные стеблевые ткани настолько, чтобы они стали мо- розостойкими и т.д. и т.п. В ироиессе эволюции довольно быстро ироизошло раздвоение содержательной основы качества биотического счастья, и не- редко то, что стало счастьем для одних, наиример, для видов, уже могло быть несчастьем для других (организмов), и наоборот, но генетически закреиив такой расклад событий, жизнь, более сложно организованная, сумела иолучать довольство и от несчастий менее сложньп живьп тел- своих структурных элементов, т.е. быть счастливой и на несчастье менее сильных. 10.3.2. Эволюция жизни по пути компромиссов биотических счастья с несчастьем в организации семенных фитобиоморф Все живое на Земле, по большому счету, родственно. Оно где-то чья-то часть (структурный элемент), а где-то целое, состоящее из час- тей. Живая природа — это и простая, и сложная (все зависит от того, как на нее смотреть) многоуровневая система, которая вся в целом и каждый ее структурный элемент проявляют определенное качество
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ жизни (биотическое качество), замешанное на взаимоотношениях двух, составляющих его в единстве и противоречии, сторон: биотических счастья (эвакинеза) и несчастья (антиэвакинеза). Особенно важно не пропустить такой интригующий организационный момент, правильно расставив акценты, что качества жизни каждого живого тела как струк- турного элемента живой природы (биостробы) Земли всегда слиты во- едино (другого эволюция не дала!) с качеством жизни целого для них другого живого тела, и так по всей иерархической лестнице (пирамиде) организации биостробы. На иерархию носителей жизни по сложности их организации накладывается иерархия качеств их жизни, что позво- ляет утверждать, что над биотическими счастьем и несчастьем живых структурных элементов стоят биотические счастье и несчастье биостро- бы, которые себя в большом (едином качестве жизни) через меньшее закономерно выражают. На основе макробиократии живые структурные элементы всецело подстраиваются со своим счастьем и несчастьем — пар- циальным качеством жизни, под общее (единое) качество жизни всей зем- ной живой природы (биостробы). Миллионы лет эволюция жизни притирала биотические счастье и несчастье друг к другу, чтобы посредством парциальных качеств жизни отдельные живые тела сумели и себя сохранять, и другому, более сложно организованному живому телу с пользой послужить, что хорошо видно в отношениях биоморф и их видов. Вся эволюция живой природы нахо- дится под давлением многих явлений, которые она сама же и создала: приоритет генетического над фенетическим, макробиократия, приоритет репродуктивного над вегетативным, лидерство побегов над корнями..., наконец, приоритет биотического счастья над несчастьем. В отношениях биотических счастья и несчастья немало тонких хитросплетений, разби- рательство которых еще позволит сделать немало открытий, но мы, здесь и сейчас, обратим внимание на один из интригующих моментов жизнераз- вития: не только на ортодоксальность, но и иарадоксальность в этих яв- лениях, эволюция которых шла и идет совместно, и она стала возможной только по пути компромиссов (другое, если и было, то не прошло). Вся эволюция растительных биоморф связана с завоеванием и освоением пространства и, если биоморфа последовательно развертыва- ет в возрастных стадиях своего морфогенеза определенные признаки организации, наращивая сначала свой вегеторесурс, а затем переходя к матурному развитию, то она делает все это так, как ей предопределено ее генетической натурой, сложившейся так, а не иначе, за многие и многие годы эволюции. В процессе длительной эволюции иерархичес- кой организации живой природы не могла не сформироваться опреде- ленная система самооценки (!), независимо от какого-либо сознания, качества жизни, осознание нами, исследователями, которой, по сути дела, еще только начинается. В целостном качестве жизни (как парци- альных, так и общем) постоянно соприсутствуют единицы биотических счастья и несчастья, соответственно, эвы и антиэвы, которые усиливают то одну, то другую сторону этого свойства, но самое ценное, что нам удалось заметить — это везде и всегда в живой природе действует при- оритет биотического счастья (чьего-либо) над несчастьем. Вся интрига
Глава 10. Откровения о буднях и ираздниках растительной жизни 801 отношений между этими свойствами становится понятнее, если поста- вить довольно прямолинейный и жесткий вопрос: а кому это (отноше- ния биотического счастья с несчастьем) надо? Получается любопытный ответ: все живые тела на Земле, хотят они этого или нет, осознают или не осознают ироисходящее, как-либо занимаются самооценкой (!) качества своей жизни, т.е. того, как они живут, достигая или не достигая целей своего жизнеразвития. Главный тезис, который здесь разрабатывается, можно выразить афористически кратко: живешь — уже счастливчик, в коль так, чтобы счастья было больше, борись зв него, причем лучше других, и все для того, чтобы сохранять не столько или не только себя, более всего, друтих (родных, но и чужих), в, следовательно, — жизнь вообще. В живой природе и во всех ее живых структурных элементах дей- ствует приоритет биотического счастья (эвакинеза) над биотическим несчастьем (антиэвакинезом). Если живое тело достигло определенной цели жизнеразвития, то оно, таким образом, проявляет биотическое счастье (довольство достигнутым), а если нет, то — биотическое несча- стье (недовольство от недостигнутого). В этом главный смысл самооце- нок качества жизнеразвития, исходящих от самих живых тел (без слов, без поз «мыслителя» и т.п.), из которых можно уловить весьма любо- пытную закономерность: живые тела бывают одновременно биотически счастливыми и несчастливыми в отношении одного и того же явления, но счастья в целом всегда больше, пока существует жизнь вообще. Не правда ли, ситуация здесь близка к абсурду: и счастье, и несча- стье в одной «колеснице» и, больше того, даже смерть — великое не- счастье для всего, что жило, вполне может быть определена и как не- которая форма биотического счастья. Уже который раз ловишь себя на мысли, как все-таки важно прибегать при поиске истины к смене углов обзора и высот абстракции. Действительно, живет, например, расти- тельная биоморфа — однолетний организм. Живет счастливо (другого быть не должно, поскольку просто жить — это уже само по себе био- тическое счастье), т.е. удачно решает задачи своего жизнеразвития, получая довольство, достигая цели, проходя одну стадию (состояние) жизни за другой, вплоть до того единственного и неповторимого состо- яния — матурного, когда один раз в его биографии праздник (он цветет и плодоносит), а далее уже другое — быстренькое умирание и смерть. Жизнь прожита, все вроде бы прекрасно, счастье состоявшейся жизни было, его нельзя было не заметить, есть и главный результат — жизнь продолжается дальше не без опосредованного участия некогда жившего растительного однолетника, на котором мы временно сконцентрирова- ли свое внимание. Он был счастлив, прожив свою жизнь от начала до конца, но есть один нюанс — горьковатый осадок от происходящего. Почему всему хорошему и счастью, которое, действительно, было, ну- жен итог — смерть, почему счастье совсем и навсегда не победило ее, смерть — ведь она, несчастье, и тот последний «подарок» от жизни за то, что «не щадя живота своего» наш однолетник «фанатично» служил тому, кто повыше его, — своему виду? Неужели у такого несчастья, как смерть, есть что-то еще, что сиособно дарить счастье?.. Вопросов боль- ше, чем ответов.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Здесь не загадка, здесь очередной парадокс, ставший в эволюции жизнеразвития феноменом. Оказывается, однолетний организм тем, что он отплодоносил, в самооценке своих действий получил биотическое довольство и стал еще более, чем ранее, счастливым, но вот беда, сча- стье сотворенной ранее жизни, «захлестнувшее» было наш растительный однолетник, казалось бы, должно разогнать его жизненные силы к новым свершениям, чего, однако, не произошло и произойти не могло, поскольку жить далее ему «не положено», — нет «разрешения» свыше, т.е. от того, в структуре которого он всего лишь одномоментный «бан- тик» — элемент в чьей-то более сложной игре под названием «жизнь вообще». Генетическая природа растительного однолетника подчинена генетической природе его вида, которая в силу макробиократии и, в час- тности, эйдократии, подталкивает такой организм получить последнее довольство, достигнув своей последней цели, — проявить стремление к смерти и умереть, став еще, уже действительно в последний раз, счаст- ливым, не победив, а проиграв своему антиподу — несчастью. Почему? Разве такое можно, чтобы смерть была в довольство и составляла кому- то счастье? Парадокс вершится потому, что все это выгодно виду, а он стоит над организмом, который для вида всего лишь структурный элемент — частица его организации, и управляет стратегией всего внутривидового жизнеразвития. Как видим, сплошь и рядом счастье накладывается на несчастье, затем его перекрывает несчастье, но верх берет все равно счастье... Можно утверждать, что здесь и сейчас свершилось великое таинство качества жизни и счастье одного живого тела пошло на ком- промисс с несчастьем другого живого тела ради приоритета счастья вообще. Приоритет счастья во всех случаях сохраняется, но оно склонно в подобных случаях приобретать особенное содержание. Дело в том, что, размышляя о биотическом счастье и несчастье, мы невольно сталкива- емся с феноменом в проявлении этих сторон качества жизни, который можно объяснить не иначе как через ортодоксальность и парадоксаль- ность. В отношении качества жизни ортодоксальность (от греч. orthodoxia — неуклонный, чуждый колебаниям) одной из сторон — это ожидаемый основной результат развития конкретного живого тела, ко- торый ничем себя не скрывает и тем самым непосредственно выдает свое содержание. Парадоксальность (от греч. paradoxos — неожиданный) од- ной из сторон качества жизни — это неожиданный побочный результат развития конкретного живого тела, который ирикрывается за основным результатом и только опосредованно обнаруживает свое особенное содер- жание. Когда наш растительный однолетник оставил после репродуктив- ного размножения потомство, то он был ортодоксальным счастливчи- ком, но, когда проходил элиминацию (умирал) после оставления потом- ства, то моменты его ортодоксального несчастья были одновременно и моментами его парадоксального счастья. Для вида оба действия его элемента — растительного организма: оставление потомства, умирание и смерть — это всего лишь штрихи к его «портрету» — звенья ортодок- сального счастья в его качестве жизни.
Глава 10. Откровения о буднях и ираздниках растительной жизни 803 Для растительного однолетнего организма, который мы рассматри- ваем здесь в качестве примера, финал жизни, происходящий с ним, как и с друтими, двойственен по качеству их жизни. Время не остановить, назад его тоже не вернешь, и факт элиминации обязательно свершится, поскольку жизнь организована на основе дискретности всех своих составляющих ее элементов. Смерть растительного организма, которая имеет место, — это ортодоксальное несчастье, но одновременно она выполнила роль и парадоксального счастья, поскольку подвела положи- тельный итог «обращения» в живом соматическом «пространстве» неко- торого дискретного своего «фрагмента», и когда пришло время «уходить за порог» (элиминироваться), а оно относительно жестко генетически предопределено, свершилось сначала ортодоксальное счастье — орга- низм размножился (как полагается по «высшему разряду» — «с парадом и праздником»), а затем пришло ортодоксальное несчастье — умирание и смерть, которые стали для организма проявлением еще и парадоксаль- ного счастья как последнего всплеска рокового довольства от того, что жизнь прожита полезно и он, организм, умирает, как все, но с каче- ством жизни, не как у всех, хотя, может быть, и многих, а выполнив до конца свой долг, оставив своему властелину-виду достойное наследство — потомство совидичей. В живой природе все идет по законам и пра- вилам (другого не бывает, хаос в ней условен, всегда относителен), потому даже несчастье одних своих носителей она превратила в их же, и тем более, свое, счастье, пусть для них и парадоксальное, но важнее всего, что у тех, кому принадлежит биотическая власть — у макробиок- ратов, например, видов по отношению к организмам — у них на этот счет счастье ортодоксальное. Однако эволюция неумолимо продвигает жизнь вперед, поэтому наступит однажды момент и возникнет новый вид, который исходному виду — парадоксально счастливому родителю «помашет платочком» (новыми признаками) и станет ортодоксальным счастливчиком, поскольку дальше производный вид будет жить за него — исходный вид, часто уже без него, поскольку место в простран- стве, где можно «резвиться» рядом с парадоксальными счастливчиками, занимаясь такой «игрой», как жизнь, крайне ограниченно и для каждого по отдельности надолго не забронировано. Смерть любой биоморфы- это ее парадоксальное счастье, т.е. последнее для нее (другого уже не будет) биотическое счастье, замешанное на последнем у нее же ортодок- сальном несчастье. «Се ля ви» как говорят французы. Биотические отношения «счастливый — несчастливый» — это да- леко не безмятежная игра в дочки — матери», в которой внешне вроде бы и пристойные «фланирования» «так-сяк «туда-сюда» с яркими крас- ками почти как «мыльных опер» — «парадов цветущих и плодоносящих» на праздниках жизни, а, на самом деле, более всего, — жесткая, време- нами беспощадная борьба за выживание наиболее приспособленных и, что важнее всего, — борьба за сохранение более «верхнего», тем более — «самого верхнего» в иерархии живых тел, при полном подчинении «нижнего». В этой борьбе, как буднях биотического разнообразия, с самого начала ее в эволюции живого, быстро нашлось место и стало нормой компромиссам между счастьем и несчастьем при приоритете
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ первого свойства над вторым. Альтернативы этому не было, а жить очень хотелось всем, а всем жить «не положено» — места не хватит, поэтому— только в борьбе и только в единстве позитива (счастья) и негатива (несча- стья), которые обязаны быть вместе и что-то должны выбирать в борьбе между собой, причем всегда, они, живые тела, в эволюции выбрали един- ственно верное для жизни приспособление — компромиссы. Каждое живое тело существует в живой природе под постоянным давлением, что оно по качеству жизни кому-то что-то должно! В том, что биоморфы всегда кому-то должны, и состоит главный парадокс био- тического качества жизни, которое всегда двойственно, поскольку чье-то, а не только собственное (свое) счастье, замешано на чьем-то, а не только этого конкретного тела, несчастье и т.п. (уместно, пожалуй, вспомнить здесь пословицу: не было бы счастья, да несчастье помогло...). 10.3.3. Как и почему семенные фитобиоморфы своим габитуальным и стадийным поведением выражают содержание всепоглощающей борьбы за биотическое счастье В процессе эволюции биоморфы определились, что у счастья, за которое они всегда борются с несчастьем, есть и ортодоксальная, и парадоксальная стороны, что, в конечном счете, и способствовало раз- витию того биотического разнообразия, которое сейчас составляет жи- вую природу. У фитобиоморф разнообразие выразилось по многим признакам: по продолжительности их жизни, по кратности репродук- тивной функции, по особенностям развития вегеторесурса и т.д. Имен- но компромиссы между счастьем и несчастьем при приоритете счастья привели растительную жизнь (шире — жизнь вообще) к созданию того биотического разнообразия, которое в целом устремлено к жизнесохра- нению. Только на основе пластичности с использованием при необхо- димости и гиперпластичности, обеспечившим разнообразие соматичес- кой организации растительных биоморф (шире — всего живого) жизнь смогла в процессе эволюции успешно выстоять, научившись завоевы- вать и осваивать пространство (см. главу 6). Вывод этот звучит сегодня чуть ли не тривиально, но ведь так было далеко не всегда. Прекрасно, если все так, но уместно поставить вопрос; кому это было надо, зачем «ошеломляться» от своего же разнообразия. Ведь гораздо проще было бы в эволюции «зациклить» всех и вся под рамками определенного типа организации, «загнать» в строго определенные экологические лабирин- ты и двигаться в них и по ним четко очерченными рядами — «маршем» в светлое будущее ... или в никуда? Все внешне вроде бы совсем неплохо — везде торжествует порядок. Жизнь наверняка такой «трафаретный» путь попробовала и испытала в своей эволюции, вскоре оставив его как бесперспективный. Она выбрала разнообразие в вечно изменяющемся мироздании. Способом существования ее соматических носителей стала борьба и еще много раз — борьба за самосохранение любыми средства-
Глава 10. Откровения о буднях и ираздниках растительной жизни 805 ми и в любой форме. Живые тела настолько вошли в роль защитников жизни вообще (на Земле — ее биостробы), настолько сконцентрировались на борьбе с чем-то и кем-то за что-то большее, чем они сами из себя представляют (!), что та всецело поглотила их и не оставила им более другого выбора в эволюции. Жизни и ее непосредственным носителям— живым телам, уже никто и никогда ничего другого не навяжет, посколь- ку именно борьба за ее сохранение стала нормой бытия всей живой природы в пространстве и времени. Любые живые тела стремятся дос- тичь какого-либо, более всего высшего, биотического счастья в своем существовании, т.е. цели, полезной для общего жизнесохранения. Что- бы быть счастливым, надо уметь побеждать несчастье, даже тогда, когда последнее наступает и вроде бы берет верх, чтобы превращать свое несчастье еще и в счастье, пусть парадоксальное, но все-таки счастье, без которого жизни нельзя, и это самое главное в том, что она есть и должна быть дальше. Живые тела должны уметь, и это они умеют, отнять это у них вряд ли получится, на своих «несчастьях» делать сча- стье других и от этого, пусть с парадоксом, оставаться счастливыми, что само по себе феноменально. Как показывает весь предыдущий материал нашей монографии, фитобиоморфы ничего другого не выбрали в эволюции, кроме адапта- ции к жизнесохранению вообще, используя для этого все приемы и, в первую очередь, свой лабильный арсенал — пластичность, в котором решающее значение как индикаторы, выражающие целостное отноше- ние биоморф ко всему происходящему (как внутри их самих, так и в окружающей среде), получили трансформации габитуса и стадийно- возрастные фенетические изменения (рис. 435 — 438).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 435 — 438. Еще раз о мультивариантности внутривидового жизнеразвития семенных растений. Рис. 435. Биографический портрет Solidago dahurica Kitag. [сем. Asteraceae (Алтай, Курайский хребет, высокогорья и лесной пояс, каменистые луговые склоны, лесные поляны, субальпийские луга)]. БИОМОрфОТИПЫ: надземно-двусемядольные полурозеточные очередно-простолистные гемисимподиальные семидетерминатно-репродуцирующие (развитие унирепродуктивных полурозеточных ди-, плейофазных побегов завершается образованием верхушечных полу- закрытых соцветий — кистей из корзинок, но высоко в горах, где-нибудь около снежни- ков, развиваются настолько низкие и слабые побеги, что число корзинок у них уменьша- ется до 1 -2) короткокорневищные сильнорослые (1 биоморфотип — на полянах и опушках лесного пояса) среднерослые и низкорослые (11 — И1 биоморфотипы — в условиях субаль- пийских лугов и высокогорных каменистых склонов) однодомные желто-астроцветковые ретардационно-кондивидуальные (замедленно вегетативно размножающиеся при полной партикуляции стареющих растений) травянистые многолетники.
Глава 10. Откровения о буднях и ираздниках растительной жизни 807 (Продолжение рисунка на следующей странице). Рис. 436. Биографический портрет Сепгаигеа hyalolepis Boiss. [сем. Asteraceae (Израиль, окрестности Тель-Авива, каменисто-песчаные холмы, 24 V)j.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 436. (продолжение) EHOMOp@OTHIIbI: надземно-двусемядольные (семядоли эллиптичес ллиптические или обратнояй- — 10 мм дл., 3,5 — 5 мм шир., закругленные, клиновидно суженные в ные ~~ме но-полисимподиаль- о кие черешки) полурозеточные очередно-простолистные ~~мерно- ные (об аз отся сильно ветвистые годичные побеги или всего д о ин главный побег, каждая ветвь или предветвь, заканчивается сильно ечной корзинкой) однодомные желто-астроцветковые однолетники колючей верхушечно ',II биомо ~тип — после б фотип) травянистые или полутравянистые двулетники ( рфо HOMOP~THn а ионного сезона растения зимой полностью не отм р и ают а сохраняют в первого вегетацио иземн часть или часть надземной побеговой жизнеспособном состоянии компактную приземную часть или часть системы, продолжая вегетацию еще один сезон), утр пол авянистые среднелетники ип — астения живут мало, но обильно ветвясь, способны выживать до 3, реже ые надземные части побегов 4 лет, сохраняя зимой в жизнеспособном состоянии некоторые н прошлых лет вегетации).
809 Глава 10. Откровения о буднях и ираздниках растительной жизни 2,аг1, 206,26 5,г 20 21 32, 8) Рис. 437. Биографический портрет Reaumuria hirtella Jaub. et Spach [сем. Tamaricaceae (Израиль, каменистая пустыня Негев, окрестности поселка Сде- Бокер, 12 V)]. БИОМОрфОТИПЫ: надземно-двусемядольные (семядоли широкоэллиптические или ок- руглые, 2,5 — 4 мм дп., 2 — 3 мм шир., закругленные, сидячие) безрозеточно-розеткооб- разующие очередно-простолистные гемисимподиальные (виргинильные растения могут нарастать эумоноподиально, а у взрослых растений образуются унирепродукгивные дифаз- ные, реже трифазные безрозеточные или полурозеточные побеги, закрытые верхушечным одиночным цветком, нередко усиленные в верхней части побега еще силлептическими розеточными вегетативно-репродуктивными одноцветковыми побегами) детерминатно- репродуцирующие однодомные бело-тамариксововоцветковые полукустарнички (1 био- морфотип — открытые места, где растения произрастают в «одиночестве», высота их аэроподиев 5 — 20 см) или полукустарники (П биоморфотип — днища оврагов, другие пониженные участки и места, где растения сосредоточены в ~группы», высота их аэропо- диев 20 — 40 см).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 4,19а, 6) Рис. 438. Биографический портрет Echinops polyceras Boiss. [сем. Asteraceae (там же, что и рис. 437, 12 V)]. EHOMOp@OTHIIbI: надземно-двусемядольные (семядоли эллиптические, 6 — 10 мм дл., 3 — 4 мм шир., закругленные, клиновидно суженные в короткие черешки, сросшиеся своими основаниями) полурозеточные (большинство унирепродуктивных побегов полуро- зеточные дифазные, реже плейофазные, но некоторые из них развиваются как безрозеточ- ные монофазные) колюче-очередно-простолистные гемисимподиально-эусимподиальные (доминирует гемисимподиальное нарастание побегов, но и эусимподиальные приросты также участвуют в нарастании аэроподиев) семидетерминатно-репродуцирующие (соцве- тия верхушечные — колючие корзинки) однодомные фиолетово-астроцветковые полуку- старнички (биоморфотип 1 — открытые склоны каньона) или полукустарники (биоморфо- тип П — скопления камней, русла временных водостоков, пониженные участки рельефа).
Заключение Не скрою, поначалу я подготовил по отдельности и основные вы- воды, и заключение, компактно повторив в выводах то, что сформули- ровано в десяти концепциях, составивших содержание глав второго тома; но потом, как шахматист перед решающим ходом, надолго задумался. Нужно ли, пускай даже кратко (а получилось довольно много новых страниц текста), повторять основное содержание книги, которую жела- тельно читать, а не только листать? Будет ли это кому-то полезно? Мой «ход» приводится ниже: самые-самые общие выводы, насколько это было возможно, совмещены с заключением. Так, вероятно, будет лучше для всех: нет повторений, общие же рассуждения полезны для осозна- ния и того, что уже сделано, и того, что еще нужно сделать для развития теории биоморфологии. В оглавлении обозначены основные вопросы, рассмотренные в главах этой книги. Читатель, надеюсь, согласится, что автором поднято немало проблем и по ним предложены нестандартные решения. Поэтому здесь, в заключении, видимо, нелишне будет, сведя воедино основные мысли всех десяти концепций, озвучить ту более высокую идею, которая стала лейтмотивом всей книги. Идея эта заклю- чается в следующем: телесная многослойная системность обеспечивает существование организации биоморф семенных растений и как частей, и как целого, будучи погруженной в системное пространство тел и нетел всего остального мира, непосредственно или опосредованно взаимодействующих друг с другом и эволюционирующих во времени как части единого и бес- конечного вселенского потока (природы вообще, или Природы). На фоне собранного мною фактического материала [в книге при- водятся свыше 400 фитобиографических иллюстраций («портретов»), отражающих типологию жизнеразвития разнообразных простых био- морф (организмов) и сложных биоморф (синорганизмов)] последова- тельно выстраивается, распространяясь вширь (основа заложена в пер- вом томе) и вглубь, теория организации семенных фитобиоморф как стержневая теория современной — интегральной — биоморфологии. Во втором томе особенно интенсивно реализуется принцип абстрактного «пирамидостроительства» (суперординатного классифицирования) с со- ответствующим графическим моделированием, производимым под раз- ными углами обзора и с разных высот обобщения. Это позволяет и автору, и читателю попутешествовать в мире абстракций, который по- своему отражает, предлагая лучше понять его, объективное состояние процессов движения материи в природных явлениях.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Всем нам, и ученым, и просто любителям смотреть по сторонам, уже на интуитивном уровне присуще понимание того, что Природа, т.е. все, что нас составляет и окружает, подчиняется в своих проявлениях ее собственным законам и правилам, поддерживая таким образом в самой себе (ведь ничего другого просто не существует), в бесконечном вселенском потоке некий атрибутивный порядок, о великой красоте и стройности которого мы, люди, пока можем только догадываться. И одна из сторон этого порядка — многослойность (многоуровневость) организации природных явлений; эту сторону мы способны ощутить повсюду, если, конечно, вообще соизволим обратить на нее внимание. Идея многослойности природы, многослойности жизни — та крас- ная нить, которая проходит через всю нашу работу. Будучи универсаль- ным свойством живых тел (биосистем), многослойность проявляет себя по-разному, и мы, пользуясь приемами суперординатного классифици- рования (абстрактного «пирамидостроительства»), можем наблюдать ее как бы под логическим «микроскопом». При этом мы используем раз- ную степень увеличения, высвечиваем то одни, то другие известные, менее известные и совсем не известные моменты одного и того же явления, составляющего предмет нашего непосредственного исследова- ния в данном месте и в данном времени. В многослойной организации природных явлений жизнь — при минимальном увеличении — сама может выглядеть всего-навсего как один слой материи, рядом с другим, расположенным ниже слоем — нежизнью. Однако, чем большую крат- ность увеличения мы устанавливаем на нашем логическом «микроско- пе», тем яснее раскрывается «слоистая структура» природных явлений и, в частности, такого явления, как жизнь. Меня, как любого человека, не могут не BocxHIgRTb события, которые составляют главное содержание человеческого бытия: только мы, люди, способны превращать различные мысли, которые, как некие бесплотные «фантомы», до поры до времени витают в нашем сознании, — в точные знания о Природе. Более того, мы, если у нас есть желание и воля, материализуем эти мысли в книгах, картинах, машинах и т.п.; говоря скучным языком экономики: в средства производства и средства потреб- ления. Разве это не чудо, что еше вчера, допустим, в той или иной области науки существовало белое пятно, а сегодня появилась нетриви- альная разработка и о том, что так должно случиться, недавно думал, это не исключено, только один человек, где-то «размахивающий» творческой лопатой пред «горой» накопленного ранее биоморфологического знания, стремясь перекопать ее, может быть, и до основания, причем настолько усердно, что о конечных устремлениях такого «горокопателя» догадаться было не просто. Вообще-то «копателей» в науке немало; но, как правило, с энтузиазмом взявшись за адскую работу по перемещению «горы» како- го-либо знания (в нашем случае — биоморфологического), они останав- ливаются где-то на полпути. В данном случае ситуация совершенно дру- гая: научный «копатель» опускать руки не намерен. Возможно, его (меня) вдохновлял ратный труд рабочих Урала, которые до основания «выбрали» гору Магнитную, превратив ее полезный ресурс в еше более полезный продукт: станки, машины, локомотивы и т.п.
Заключеиие Как бы там ни было, здесь, в биоморфологии, бурная деятельность продолжается. Тот же самый «кладоискатель» (ведь его и так можно назвать), взявшийся перекапывать «гору», сейчас уже бойко орудует «мастерком», укладывая «кирпичи» обновленного знания, сделанные из материала во многом уже перекопанной «горы», и укрепив их структуру только ему ведомыми добавками, в очередной — современный — этаж вечно созидаемого здания биоморфологии. То, что биоморфологическое здание постоянно строится, точнее, достраивается, реставрируется, ре- конструируется, ремонтируется и т.п. — понятно; но вот о том, что у такого здания возводится еще целый «этаж», можно рассуждать без устали. Я, автор второго тома «Основ биоморфологии...», уверен, что, вкладывая очередной блок «кирпичей» в новый «этаж» — в новую ста- дию развития фитобиоморфологии, а шире — биоморфологии вообще, я тем самым закладываю фундамент и развиваю основы интегральной биоморфологии. Вторая книга по фитобиоморфологии — перед Вами, уважаемый читатель. То есть это — уже не «фантом», а объективная реальность, артефакт, материализованный свод логосистем. Эта книга, по большому счету — как, впрочем, и остальные книги друтих авторов, — мне уже как автору принадлежит менее всего; она сегодня превраща- ется в знания других людей, а значит, хотят ли они этого или нет, в общечеловеческие знания, поскольку процесс познания остановить нельзя. Монография (все ее тома) об интегральной биоморфологии пишет- ся автором не на последнем дыхании, а с запасом — в прямо-таки «юношеском» порыве (надеюсь, не последнем), на базе уже прожитых в науке лет, лет далеко не спокойных. Только этот творческий порыв побуждает меня искать и находить истину, которая часто скрывается далеко за пределами конкретного предмета моих основных исследова- ний — биоморф семенных растений. Если бы я, работая над монографией, стал рассматривать природ- ные явления совершенно по-новому (как бы с чистого листа) и назвал бы неизбежное при этом меротомирование (членение на части) науч- ным, совсем не учитывая тех знаний, которые уже накоплены в науке (подобных примеров в нашей сфере предостаточно), это было бы, по меньшей мере, некорректно. Так работать нельзя, и такая «парадигма» изначально не могла стать моим принципом. Я всегда делаю ставку на известное, именно с него начинаю процесс познания, ухожу в глубину, подкрепляя и дополняя его свежими идеями, которые естественным образом работают на развивающуюся науку, поскольку она постоянно получает приток все новых и новых фактов, которые, безусловно, нуж- даются и в переосмыслении. Однако есть коренные (основополагаю- щие) термины («слова-этикетки» + понятия), «выстраданные» развити- ем всей классической ботаники; они проверены временем, например, растение, побег, корень, соцветие, лист, ось и т.д. Конечно же, их можно и даже нужно уточнять, чтобы лучше классифицировать и мо- делировать, но нельзя совсем отказываться от них, заменяя их чем-то друтим, внешне вроде бы современным, но на поверку ничего нового в ботаническую науку не привносящим. Этот путь не для меня, мне
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ чуждо подобное «новаторство». При всем этом я положительно отно- шусь к обоснованному терминотворчеству, что вовсе не недостаток, поскольку я нередко иду на обобщения по проблемам, которые ранее еще не были разрешены или вообще не поднимались, и описываю явления, для которых нет названия. А раз так, то оставлять их «бесхоз- ными» — не в моих правилах: ведь, будучи обозначены «спецсловом»- этикеткой, такие явления будут лупие восприниматься на фоне друтих, уже известных явлений. Так что надеюсь, в науке в результате моей деятельности не появляются термины-«паразиты». Такова в целом моя твердая позиция в отношении терминотворчества. Во втором томе с особым вниманием показаны семенные фитоби- оморфы в их развитии, что позволило мне использовать обильно иллю- стрированный фитобиографический материал, собранный мною в раз- личных регионах нашей планеты, для рассмотрения и решения различ- ных теоретических проблем. Если кто-то подумает, что «изюминка» всей моей работы — только в габитусе, его формах и формах роста в организации биоморф семенных растений (что, впрочем, само по себе — уже немалая исследовательская интрига), то он все-таки ошибется. И то, и другое — лишь хороший повод, или, если хотите, основательная зацепка, чтобы «встряхнуть», пусть мысленно, и «расслоить» всю живую природу (включая ее фрагменты — биоморфы), а иногда даже больше — мироздание вообще. Да, это действительно повод, чтобы покопаться в Природе настолько серьезно, насколько у автора может хватить на это «пороха», — покопаться, не причиняя, что важно, вреда мирозданию. Такая «встряска» показана существованию Природы, поскольку мы, люди, ее разумные частицы, и только мы, используя свой разум, можем реально делать такие идеальные акции, реально участвовать и в позна- нии, и в управлении процессами всеобщего развития, стремясь напра- вить его в наиболее полезное для человечества русло, стимулируя этот процесс более продуктивно, чем если бы она, Природа, развивалась без нас. Только люди, познавая объективные законы развития природных явлений, обладают и возможностями, и желанием участвовать в про- грессивной эволюции, в которой с позиций человечества все, что хоро- шо людям для их бесконечного сохранения в пространстве и времени, должно устраивать и природу вообще. А это никогда не может быть негативом для всего, что окружает и, в особенности, сохраняет Разум Природы (пока это только земное достояние и о чем-то ином мы просто не знаем, хотя и подозреваем, что наша жизнь не одинока в бесконеч- ном вселенском потоке). Мысленное «встряхивание» природных явлений и мироздания во- обще изначально было рассчитано только на работу все и вся абстра- гирующей мысли, игру воображения, где это было нужно, но игру не с иллюзиями, а, как минимум, с гипотезами, более же всего — с объек- тивными фактами. У меня, да и у друтих исследователей, накопилось уже немало фактов, чтобы не только коллекционировать их, но и осу- ществлять на их основе новые и новые теоретические разработки, где- то членя целое на части, а где-то, напротив, объединяя все в невероят- ных, казалось бы, по масштабности абстракциях. Некоторые читатели
Заключеиие предпочитают начинать знакомиться с книгой с ее конца — с заключе- ния, и это само по себе не удивительно, поскольку каждый достигает знания по-разному. Так вот, только для них я приберег один совет: чтобы моя работа пришлась Вам по душе и Вы действительно испытали состояние комфорта, может быть, даже где-то смогли «парить» в про- странствах всеобъемлющих пирамид», Вы, работая с этой книгой и обдумывая ее результаты, не забывайте, пожалуйста, включить в себе «кнопочку» абстрактного мышления (где она находится, я доподлинно не знаю, но не сомневаюсь, что она есть у каждого, надо только хотеть ею воспользоваться). Эта книга — не учебное пособие, а свободное продолжение моно- графии (1 том вышел в 1997 году); здесь много такого, что настраивает на работу с перспективой, т. е. она, на мой взгляд, рассчитана на всех биоморфологов, а шире — биологов. Действительно, если основательно «копнуть» ту или иную проблему — например, проблему габитуса и форм роста в организации биоморф семенных растений, — то непре- менно возникнет немало новых проблем, выстроится множество вопро- сов, на которые обязательно должен ответить именно этот автор и его читатели. Умение замечать, ставить, а в конце концов — и отвечать на вопросы, поставленные по сути дела процессом развития науки, может и должно доставлять истинное удовольствие, составлять счастье любому ученому. Меня вполне могут спросить: для чего это понадобилось биомор- фологу, причем лишь фитобиоморфологу и тем более — биоморфологу семенных растений, утоляя жажду познания предмета своих основных интересов, «упражняться» в столь различных по масштабности абстрак- тных построениях? Не переоценил ли автор свои возможности? Вопрос уместный — если закрыть глаза на то обстоятельство, что книгу эту писал человек, являющийся и эмпириком, и теоретиком одновременно. На вопрос, прозвучавший выше, отвечает своим содержанием вся эта книга, и только Вам, уважаемый читатель, судить, убедительно ли она на него отвечает. Однако, отмечу главное: предмет своего основного научного внимания — биоморфы семенных растений, я не то чтобы не хотел (вначале очень даже хотел!), но вообще перестал воспринимать под углом традиционных представлений классической (габитуальной) биоморфологии, поскольку со временем, и это тоже надо было заметить, изменились приоритеты развития науки с таким названием (см. главу 2), что было осознано и к настоящему времени реализовано на деле: био- морфология вступила в новую стадию своего интегрального развития, и тот, кто этого еще не заметил или не захотел заметить, уже отстал, а догонять всегда труднее... Но догонять придется, другого пути не пред- видится. Несмотря на то, что в названии проблемы, рассматриваемой во втором томе, стоят габитус и формы роста биоморф, здесь все равно не габитуальная (традиционная) биоморфология, а новая стадия современ- ного развития этой науки в целом — интегральная биоморфология. Впервые на пути развития биоморфологии сделана попытка для реше- ния ее теоретических проблем подойти из глубин мироздания, соответ-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ ственно, миропознания, что в конце концов и привело к созданию той биоморфологии, дальнейшее развитие которой неминуемо приведет к существенным подвижкам не только в биологии, но еще и биофороло- гии — науки о носителях жизни, или биофорах, а эта наука больше, чем биология (т. 1, гл. 2, стр. 38). Атрибугивная часть в Природе — это, как известно, не просто материя, а материя, движущаяся в пространстве и с пространством, которое она организует собой и для себя в тела и нетела, поскольку только в них удобнее всего быть организованной системой. Природе чужд хаос, она существует только целесообразно своему организацион- ному порядку, который ею создан, развит и который она целенаправ- ленно бесконечно поддерживала и будет поддерживать впредь. Что же так усердно работает в Природе на поддержание в ней организационного порядка, что составляет движущую силу ее бесконеч- ного и организованного развития? Такой «движок» у Природы есть- это ее тотальный движитель, или примагенератор, — пространство Природы, наполненное по-разному в его бесконечности неисчерпаемой материей, которая существует только в движении и только во времени, а последнее само по себе — и ни пространство, и ни материя, но всегда — другая обязательная часть Природы (глава 1). Организационный порядок возник в Природе как само собой разумеющееся явление, и никто «свыше» его не навязывал, поскольку выше природного никакого другого нет, и только она, Природа, вынуждена бесконечно существо- вать в гордом всеобъединяющем одиночестве, и то, что она обладает определенным организационным порядком — это все результат функ- ционирования только ее систем и, в первую очередь, системы ее тоталь- ного движителя. Что же производит примагенератор? У него есть только один результат (бесконечнодвигательная деятельность) — трансформа- ции самого разного рода (все) — это функция Природы во времени. Только присоединив свое время к тому, что делает ее тотальный дви- житель, Природа обеспечила себе бесконечное развитие. Примагенератор управляет развитием Природы во времени. В нем надо искать «корни» (основу) тех управляющих систем, которые дают о себе знать в булыжнике, скалистом утесе, океанской волне, но и рас- тительном или животном теле и, наконец, в человеке. Природа сама научилась управлять собой посредством своего тотального движителя. Изменений — трансформаций — избежать нельзя — это суть существо- вания материи, а, следовательно, и суть работы вечного «движка» При- роды — ее примагенератора. Это он, тотальный движитель, с общим для него и природы вообще Сверхъядром и космоплазмой, раскачивает и заставляет колебаться по схеме «так-сяк«туда-сюда» эволюционные маятники (тела и нетела), поскольку эволюция — это высшая форма трансформаций (глава 8). Мы, люди, постоянно пытаемся как-то приобщиться к таинству того восприятия жизни, в котором постоянно пребывают растительные биоморфы, сами того, естественно, не осознавая, что же они «творят», но часто делают это так целесообразно, что всякие наши сантименты улетучиваются, когда мы действительно начинаем ощущать истинное
Заключеиие содержание и цену в чем-то отдельной (личной), а в чем-то общей (неличной), но каждой по отдельности, растительной жизни, когда ос- тается только ощущение жесткости, временами до ужаса жестокого порядка, царящего в организации растительных тел разного масштаба. Безусловно, где-то проходит «узкоколейка», по которой растительная, а шире и вся живая природа вполне целесообразно связана с остальной природой как бесконечным и неисчерпаемым вселенским потоком. Так почему все происходит так, а не иначе? Оказывается, что определенный порядок в организации растительной природы принадлежит не только ей, он исходит от всей живой природы, а далее — от природы вообще (Природы), и соответствующим образом управляется (см. главу 7). Интегральная биоморфология возникла на том этапе, когда пробле- му организации биоморф было решено разрабатывать во взаимосвязи с организацией друтих тел мироздания и живой природы в особенности. Перекапывая «гору» биоморфологического знания, я искал «золотые жилы» информации о разных сторонах организации растительных, а также и животных тел, но в первую очередь о габитусах и формах роста, которые многими так или иначе признавались за жизненные формы. Задача с самого начала была ясна: определить место и статус разных структурных образований в организации жизненных форм — жизнен- ных форм настоящих, а не мнимых. Такие задачи в большом теорети- ческом исследовании принято решать, мысленно опускаясь почти на «дно» проблемы, изучая ее изнутри и во взаимосвязях с другими, смеж- ными проблемами (так надежнее), накапливая убедительный фактичес- кий материал, а затем, когда почувствуешь себя во всеоружии, «штур- муя» проблему целиком. Пользуясь такой тактикой, ты, подходя к любой проблеме, пусть она на поверхностный взгляд как бы «блестит», излу- чая, если внимательно присмотреться, даже не свой, а отраженный свет, — можешь получить удивительный результат. Мнимый блеск устарев- шего знания довольно быстро тускнеет под напором фактов, и под ним обнаруживается неприглядная «нищета». И проблеме, там, «наверху», куда из «молчаливых» глубин была направлена «атака», подготовленная с разных сторон (см. т. 1 — 2), деваться некуда: она сдается и раскрывает свои тайны. Я готов взять на себя смелость и заявить, что сегодня тайны габитуса и форм роста в биоморфологии уже не существует. Статус этих структурных образований определен; во всяком случае, ясно, что ранее значение его было сильно преувеличено. Сейчас мы констатируем: место им, этим структурным образованиям, только на двух «полочках» (и не более) в сложной и разветвленной структуре организации определенных телесных систем, среди которых всегда присутствуют и биоморфы (орга- низмы и синорганизмы) семенных растений (см. главу 3 и 4). На фоне многослойности явления «жизнь» габитусам и формам роста нашлось (и это важно) строго определенное место в организации живых тел и, в частности, биоморф (в организации Природы, как я уже сказал, есть свой обязательный порядок, но его надо захотеть и суметь увидеть). Эти структурные элементы далеки от ключевых центров этой организации (ключевые центры — в другом: генах, основных органах и кое в чем еще), что и предопределяет наш вывод: эти структурные
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИЙ образования не могут быть отождествлены с формами жизни (биомор- фами), они — только их части, место их в организации действительных биоморф (организмов и синорганизмов) не первое, хотя в чем-то также важное. Разобравшись с проблемой габитуса и форм роста в биоморфо- логии, можно сказать, что это и есть то первое самое-самое, что удалось разрешить во втором томе. Во-вторых, используя как известные, так и еще неизвестные факты, мы продемонстрировали с разных сторон действительную многослой- ность явления жизнь, когда и целое, и его структурные части пребыва- ют, каждая по-своему, и в своей (личной), и в не-своей (другой, нелич- ной) жизни (см. главу 2, 9, 10). В-третьих, живые тела и в особенности биоморфы семенных рас- тений всегда, будучи одновременно системами «управления-исполне- ния» со своей стратегией, исходящей из генетической организации (но- сителя высшей биотической власти), и со своей тактикой, которую исполняет фенетическая организация биосистем, — находятся в процес- се постоянной борьбы за существование. Жизнь на Земле — это сплош- ная борьба — как за себя, так и за жизнь других — с использованием традиционных приемов: взаимопомощи, конкуренции, индифферентно- сти; жизнь на Земле — это всегда эволюция, которая многогранна и всюдна. Что же касается биоморф, то они, посредством разных форм естественного отбора, участвуют в ней всем своим существом, принимая ее, как внутри, так и вне себя (глава 8). И все-таки еще важнее — другое: все биоморфы (шире — все живые тела Земли), каждая по-своему, могут быть счастливыми (глава 10), в первую очередь тем, что просто живут, а уж тем более, когда цветут и плодоносят (шире — воспроизводятся). Фитобиографический иллюстративный материал обильно напол- нил собой фактическую основу всех 10 концепций, рассмотренных в книге, но он всей своей массой рассчитан и на значительно большее: он призван возвестить ботаническому миру не только о глубокой пере- мене, которая свершается на современной (интегральной) стадии разви- тия фитобиоморфологии, но и о тех концептуальных подвижках, кото- рые неминуемо еще должны произойти в текущем столетии в развитии некоторых сопредельных отраслей биологического знания. В первую очередь процесс биографической модернизации коснется систематики — одного из «столпов» биологии, от развития которой зависит все ос- тальное в науке вообще. Дело в том, что почтенная классическая сис- тематика, начав свой яркий восходящий путь со времен К. Линнея (XVIII век), постепенно подошла в своем развитии к определенной черте, когда неизбежно обостряется вечный вопрос: что делать, куда идти дальше? Похоже, все сложилось именно так, как более всего нужно было сперва для самого начала, когда были определены классические ориентиры систематики на наиболее консервативные, репродуктивные признаки, что оказалось весьма оправданно и уже, вполне можно утвер- ждать, обстоятельно проверено временем. Теперь, когда наиболее ста- тичных признаков только цветущих и плодоносящих взрослых растений стало явно не хватать, вполне резонно где-то, пока еще подспудно, вызревает новая сила, способная дать качественный толчок к прогрес-
Заключеиие сивному развитию современной систематики, — интегральная биомор- фология с ее сердцевиной — биографикой. Никто и ничто не заменит классические ориентиры в систематике, но и активного продвижения биографического знания в систематику также нельзя остановить. Это зов времени, и он, в частности, зримо, хотя и опосредованно, исходит от всей нашей монографии. Систематика нуждается в поддержке других наук. У нее нет права замыкаться в себе. Биографическая информация (о соцветиях, вегетативных, в частности, подземных органах, семядолях, продолжительности жизни и т.д.) не разрушит, а дополнит возможности систематики. Призрак биоморфологии, и это не шутка, уже действи- тельно бродит в систематике. Полагаю, что читатель нашей книги не- вольно, допустим, только разглядывая рисунки, за которыми стоят многие годы работы в экспедициях, на опытных плантациях и т.д., ощутил великий потенциал к взаимному движению этих наук навстречу друг другу. Но и сегодня мы не на «параде», а до него надо еще пройти довольно тернистый путь дальнейшего совершенствования разносто- ронних фитобиографических исследований — фактического основания современной биоморфологии. Одновременно надо разрабатывать тео- рию и методологию систематики на новой стадии ее будущего развития. Биоморфология, в первую очередь, со своей биографической начинкой может и должна не только сама развиваться, но и качественно влиять на состояния развития друтих наук. До хрипоты спорить здесь бесполез- но и стоит ли... Надо просто-напросто качественно работать, делая шаг за шагом по пути решения как частных, так и общих проблем, суще- ствующих и возникающих в процессе развития фитобиоморфологии, биоморфологии вообще, но, естественно, и биологии в целом. Я не первый, кто серьезно увлекается изучением жизненных цик- Оп лов — биографий (онтогенезов и сийтогенезов) растений. Такие иссле- дования практикуются давно (и это прекрасно, что нас уже много, но надо, чтобы было еще больше). Автор своей монографией вовсе не склонен заслонить то, что уже было до него сделано многими другими ботаниками; наоборот, я стремлюсь к тому, чтобы биографические ис- следования, с учетом и моего опыта, приобрели новый стимул к даль- нейшему развитию. Сейчас уже накоплен значительный биографичес- кий материал по семенным растениям. Более того, проводились и про- водятся работы по обобщению результатов фитобиографических иссле- дований [«Biological Нога of the Britisgh Isles» издается с 1941 года, «Lebensgeschichte der Bliitenpflanzen Mitteleuropas. Specielle Okologie der Bliitenpflanzen Deutschlands, Osterreichs und der Schweiz» выходила в Штуттарте отдельными выпусками с 1906 по 1942 год, цит. по И.А. Гу- банову (1974), «Биологическая флора Московской области», периоди- чески издаются отдельные выпуски, начиная с 1974 года, под редакцией Т. А. Работнова, позднее В. Н. Тихомирова и В. Н. Павлова, «Биоло- гическая флора Крыма» (1984), выполненная В. Н. Голубевым, и др.]. Как видим, многое, хотя далеко не все, что близко автору, уже было опробовано. И все же еще не было выдвинуто главной идеи, которая однозначно прозвучала в нашей работе: третье тысячелетие, начиная с его двадцать первого столетия, — это время торжества биографической
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИЙ систематики, время коренного сближения двух биологических дисцип- лин — биоморфологии и систематики, не лишающее их самостоятель- ности. Биоморфология, развивая свою фактуальную сердцевину — био- графику, всецело работает на систематику, которая во многом будет изменять свое содержание, — это необратимый процесс. Биологию третьего тысячелетия, да и науку в целом, безусловно, устроит более совершенная, чем известная нам сейчас, систематика, а такое действи- тельно может произойти только на пути вступления ее в биографичес- кую стадию развития, предтечей чему может быть только интенсивный спурт биоморфологии, уже вступившей в конце XX столетия в новую интегральную стадию своего развития. Итак, интегральная биоморфология состоялась. Биоморфология уверенно вошла в очередную стадию своей эволюции. Что же будет с этой наукой дальше? Одно ясно: покоя не предвидится. Пожалуй, весь XXI-й век уйдет на то, чтобы накапливать «мед» (фитобиографическую информацию) интегральной биоморфологии, который когда-нибудь (скачок неизбежен!) переполнит ее емкости, что приведет к переходу биоморфологии на следующую новую стадию эволюции (количество перерастет в новое качество); назовем эту стадию предварительно «те- стовой» биоморфологией. Это обязательно случится; если прикинуть по времени, то, вероятно, где-то ближе к концу нынешнего — началу сле- дующего столетий, а может быть, и раньше (развитие науки часто опе- режает наши расчеты, а ведь новое «слово», которое часто важнее мно- гого, относительно направления дальнейшего движения биоморфоло- гии, уже прозвучало). У меня нет сомнения, что компьютерные и другие новые техноло- гии позволят создать очень «умные» приборы — тестеры, одного при- косновения которых к биоморфам, допустим, семенным фитобиомор- фам, будет достаточно (пускай пока это сказка), чтобы на экране выс- ветилась исчерпывающая информация о состоянии их признакового «пространства» и об их видовой принадлежности. Я верю в такое раз- витие биоморфологии, а, в конце концов, и систематики. А пока, коллеги- биоморфологи, надо, засучив рукава, поработать, как «пчелки», на лугах, в лесах, горах, степях, тундре и других экосистемах, принося в биоморфо- логические «ульи» побольше добротного «меда» фитобиографических зна- ний (небрежное изучение биографии растений по принципу «тяп-ляп — и доктор» — дело, по большому счету, неперспективное, поскольку в реаль- ной, а не показной, как иногда, к сожалению, еще бывает, науке и кри- терии пожестче, и планка научной весомости повыше). Лишь при этом условии все получится: окрепнет, на интегральной стадии своего современ- ного развития, биоморфология, а в развитии других биологических отрас- лей, например, в систематике, произойдут — под влиянием биоморфологии (!) — качественные перемены к лучшему. Такое обязательно будет. «Котел» биоморфологии действительно «кипит», эта наука бурно развивается. «Для меня нет интереса знать что-либо, хотя бы и самое полезное, если только я один буду это знать. Если бы мне предложили высшую мудрость под непременным условием, чтобы я молчал о ней, я бы отказался» (Сенека, 1893, с. 27). Так рассуждал древний мудрец. Но нам,
Заключеиие живущим сегодня, и тем, кто будет жить после нас, нельзя думать и поступать иначе... Есть тонкие властительные связи Меж контуром и запахом цветка. Так бриллиант невиден нам, пока Под гранями не оживет в алмазе... Эти вдохновенные строки принадлежат поэту-символисту Валерию Брюсову (1895, цит. 1958). Задумаемся над ними, а заодно и над моно- графией «Основы биоморфологии...», второй том которой сейчас перед Вами, уважаемый читатель... Еще и еще раз подумаем о том разнооб- разии Мира, взаимосвязях тел, многослойности Жизни, о всех нас — и живых, и неживых, а еще — и своих, и не-своих, существующих и с формами габитуса, и с формами роста (шире — развития) в определен- ном организационном порядке, всегда как-либо «раскачиваясь» (изме- няясь) и «так-сяк», и «туда-сюда», по законам и правилам единого мироздания. Если все получится так, как рассчитал автор, то «процесс» идет далее, к встрече с Вами, уважаемый читатель, и в следующем, тре- тьем, томе.
Литература Алексеев Ю.Е. Осоки (морфология, биология, онтогенез, эволюция).— М.: Аргус, 1996, 251 с. Алексеев П.В., Панин А.В. Философия. Учебник. 2-е изд. — М.: Проспект, 1999, 567 с. Алехин В.В. География растений. Изд. 3-е. — М.: Учпедгиз, 1950, 419 с. Анохин П.К Функциональная система как универсальный принцип изуче- ния уровней биологической органи- зации // Развитие концепции струк- турных уровней в биологии. — М.: Наука, 1972, с. 100 — 111. Анохин П.К. Принципиальные вопросы общей теории функциональных си- стем // Принципы системной орга- низации функций. — М.: Наука, 1973, с. 5-61. Арьес Ф. Человек перед лицом смерти. — М., 1992. Ашофф Ю. Обзор биологических рит- мов // Биологические ритмы. Т. 1. — М.: Мир, 1984а, с. 12-21. Ашофф Ю. Свободнотекущие и захва- ченные циркадные ритмы // Биоло- гические ритмы. Т. 1. — М.: Мир, 1984 б, с. 54-69. Балковский Б.Е. Цифровой политоми- ческий ключ для определения рас- тений. — Киев, Наукова думка, 1964, 36 с. Барыкина Р. П., Гуланян ТА. Морфоло- го-анатомические исследования Actaea spicata Ь. и А. erythrarpa Fisch. в процессе их индивидуального раз- вития // Вест. Моск. ун-та. Сер. биол., почвов., 1975, М1, с. 52-69. Барьисина Р.П., Гуленкова М.А. Элемен- тарный метамер побега цветкового растения // Бюлл. МОИП, отд. биол., 1983, т. 88, вып. 4, с. 114-124. Барьисина Р.П., Гуленкова М.А. Онтоге- нетическая анатомия, ее значение для систематики и филогении // Бюлл. МОИП, отд. биол., 1985, т. 90, вып. 6, с. 82-92. Барьисина Р.П., Гуленкова М.А. Мета- морфоз и его значение в жизни ра- стений // Бюлл. МОИП, отд. биол., 1990, т. 95, вып. 5, с. 103-110. Барьисина Р.П., Мусина Г.В., Чубато- ва Н.В. Воронец колосистый // Био- логическая флора Московской об- ласти. Вып. 8. — М.: изд-во МГУ 1990, с. 143-153. Бахматова М.П. Большой жизненный цикл чемерицы Лобеля в ценопопу- ляциях Северо-Двинской поймы // Возрастной состав популяций цвет- ковых растений в связи с их онто- генезом. — М., 1974, с. 196-224. Бейссон Ж. Генетика. — М.: Атомиз- дат, 1976, 128 с. Бекетов А. Курс ботаники. — СПб., 1862, т. 1, 318с. Бернар К Курс общей физиологии. Жиз- ненные явления, общие мжвотным и растениям. — СПб., 1878, 316 с. Беспалова З.Г. О жизненной форме «полукустарничек» // Пробл. совр. ботаники. — М., Л.: Наука, 1965, т. 2, с. 65-69. Бигон М., Xapnep Дж., Таунсенд К Эко- логия. Особи популяции и сообще- ства. В 2-х т. Пер. с англ. — М.: Мир, 1989, т. 1 — 667с. Биологическая флора Московской об- ласти. — М. Вып. 1-14, 1973 — 2000 (под редакцией Т.А. Работнова, В.Н. Тихомирова и В.Н. Павлова). Бобров Е.Г. Сем. парнолистниковые— Zygophyllaceae Lindl. // Флора
Eumepamypa 823 СССР, т. 14 — М.; Л.: изд-во АН СССР, 1949. Богдановская-Гиенэф ИД. К вопросу о семенном возобновлении в луговых сообществах // Зап. Ленингр. с.-х. ин-та, 1926, вып. 3, с. 216-253. Большая Советская энциклопедия. Т. 34 — М.: Сов. энциклопедия, 1955. Большой энциклопедический словарь. — 2-е изд. — М.: Большая Россий- ская энциклопедия; СПб.: Норинт, 1997, 1456 с. Борисова И.В. Биология и основные жизненные формы двудольных мно- голетних травянистых растений степ- ных фитоценозов Северного Казах- стана // Тр. Бот. ин-та АН СССР, 1961, сер. 3, вып. 13, с. 54-132. Брода Э. Эволюция биоэнергетических процессов. Перев. с англ.— М.: Мир, 1978, 304с. Брюсов В.Я. Сонет к форме (1895). В кн.: В. Я. Брюсов «Стихотворения. Поэмы». — М.: Гослитиздат, 1958, с. 15-16. Бурыгин В.А., Маркова Л.Е. Зимневеге- тирующая растительность пустынных и предгорных областей Узбекистана / / Ботвы. журн., т. 55, М4, 1970. Васильева А.Н., Гамаюнова А.П. и др. Флора Казахстана. Т. VI. — Алма-Ата: изд-во АН Казах. ССР, 1963, 465 с. ВасильевА.Е., Воронин Н.С., Еленевс- кий А.Г., Серебрякова Т.И. Ботаника. Анатомия и морфология растений.— М.: Просвещение, 1978, 480 с. Васильченко И.Т. Определитель всходов сорных растений. Изд. 2-е — Л.: Колос, 1979, 344 с. Вахрамеева М.Г. Рябина обыкновенная // Биологическая флора Московс- кой области. Вып. 2. — М.: изд-во Моск. ун-та., 1975, с. 73-80. Вернадский В.И. Химическое строение биосферы Земли и ее окружения.— М.: Наука, 1965, 374 с. Вернадский В.И. Биосфера. — М.: Мысль, 1967, 376 с. Вернадский В. И. Философские мысли натуралиста. — М.: Наука, 1988, 520 с. Вернадский В.И. Научная мысль как планетное явление. — М.: Наука, 1991, 271 с. Викторов Д.II. Краткий словарь бота- нических терминов. — М.; Л.: На- ука, 1964, 177 с. Воронцова Л.И., Заугольнова Л.Б. Муль- тивариантность развития особей в течение онтогенеза и ее значение в регуляции численности и состава це- нопопуляций растений // Журн. об- щей биологии, 1978, М 4, с. 555-562. Ворошилов В.Н., Скворцов А. К., Тихо- миров В.Н. Определитель растений Московской области. — М.: Наука, 1966, 367 с. Высоцкий Г.Н. Ергеня. Культурно-фи- тологический очерк // Тр. Бюро по прикладной ботанике. Год 8-ой, М 10-11, 1915, с. 1113-1436. ГатцукЛ.Е. К методам описания и оп- ределения жизненных форм в сезон- ном климате // Бюлл. МОИП, отд. биол., 1974, т. 79, вып. 3, с. 84-100. Гатцук Л.Е. Содержание понятия «тра- вы» и проблема их эволюционного положения // Тр. МОИП, т. 42: Проблемы экологической морфоло- гии растений. — М.: Наука, 1976, с. 55-130. Гегель Г.В. Ф. Наука логики. Т. 1. — М.: Мысль, 1974, 451 с. Гершензон СМ. Основы современной генетики. — Киев: Наукова думка, 1979, 508 с. Герасимов М.В. Эвкалипт. — М.: Гос- лесбумиздат, 1962, 78 с. Гете И.В. Избранные сочинения по естествознанию. — М.; Л.: Изд-во АН СССР, 1957, 553с. Глазунова К.1Т. Апомиксис у восточно- европейских представителей рода Alchemilla L.: Автореф. дис.... канд. биол. наук. — М., 1983, 17с. Глазунова К. 1Т. Образование зародыше- вых мешков у агамных видов ман- жетки (Alchemilla Ь.) // Бюлл. МОИП, отд. биол., 1987, т. 92, вып. 5, с. 96-109. Глотов Н.В., Жукова JI.À. Демографо- генетические подходы к изучению популяций растений // Экология популяций: структура и динамика / Материалы совещания. Ч. I. — М., 1995, с. 203-215.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Голдовский А.М. Анабиоз. — Л.: Наука, 1981, 136 с. Голубев В.Н. К вопросу о классифика- ции жизненных форм // Тр. Цент- рально-Черноземного гос. заповед. — Курск, 1960, вып. 6, с. 117 — 156. Голубев В.Н. Основы биоморфологии травянистых растений центральной лесостепи. Ч. I. Биоморфология подземных органов // Тр. Централь- но-Черноземного гос. заповед.— Воронеж, 1962, вып. 7, 511с. Голубев В.Н. Эколого-биологические особенности травянистых растений и растительных сообществ лесосте- пи. — М.: Наука, 1965, 287с. Голубев В.Н. Принцип построения и содержание линейной системы жиз- ненных форм покрытосеменных ра- стений // Бюлл. МОИП, отд. биол., 1972, т. 77, вып. 6, с. 72-80. Голубев В.Н. К определению понятий жизненных форм полукустарничков, полукустарников, кустарничков и кустарников // Бюлл. Гл. Ботан. сада, 1973, вып. 1 (20), с. 9-12. Голубев В.Н. Биологическая флора Крыма. Ялта, 1984, 215 с. Деп. ВИ- НИТИ. М 5770 — В 84. Гольбах 1T. ССипсстТеемМа а п~р~иnр~оoд~~ы. — М.: Соцэкгиз, 1940, 456 с. Гормшина Т.К. Экология растений. Учебное пособие. — М.: Высшая школа, 1979, 368 с. Гробстайн К Стратегия жизни. — М.: Мир, 1968, 144с. Гризебах А. Растительность земного шара. — СПб., т. 1-2: т. 1 — 1874, 575с., т. 2 — 1877, 593с. Губанов И.А. «Lebensgeschichte der Bliitenpflanzen Mitteleuropas» (биб- лиографическая справка) // Бюлл. МОИП, отд. биол., т. 79, вып. 3, 1974, с. 148-152. Гумбольдт А. Идеи о физиономичнос- ти растений. В кн.: ГумбольдтА. География растений. — М.; Л.: ОГИЗ, 1936, 228 с. Дарвин Ч. (см. Darwin Ch., 1859). Дервиз- Соколова Т.Г. Анатомо-морфо- логическое строение Salix polaris Wahlb. и S. phleobophylla Anderss. // Бюлл. МОИП, отд. биол., 1966, т. 71, вып. 2, с. 28-38. Дидро Д. Элементы физиологии. Избр. соч., т. 2. — М.: изд-во АН СССР, 1941. Дмитриева С.И., Игловиков В.Г., Ко- нюшков Н.С., Раменская В.М. Расте- ния сенокосов и пастбищ. — М.: Колос, 1974, 196 с. Добрынин Г.М. Рост и формирование хлебных и кормовых злаков. — Л.: Колос, 1969, 276 с. Дудка ИА., Вассер С.П., Голубинский- И.Н. и др. Словарь ботанических терминов. — Киев: Наукова думка, 1984, 308 с. Дубинин Н.Л. Эаалювия популяций и ра- диация. — М.: Атомиздат, 1966, 743 с. Дубинин Н.П. Общая генетика. 2-е изд. — М.: Наука, 1976, 590с. Дубинин Н.П. Проблемы гена и эволю- ции. — М.: Наука, 2000, 546 с. (Из- бранные труды. Т. 1.). Дубнищева Т.Я. Концепции современ- ного естествознания. — Новоси- бирск: изд-во ЮКЭА, 1997, 832с. Жмылев ШЮ., Алексеев Ю.Е., Карпухи- на Е.А. Основные термины и поня- тия биоморфологии растений. — М.: изд-во Моск. ун-та, 1993, 149 с. Жукова Л.А. Поливариантность луго- вых растений // Жизненные формы в экологии и систематике растений / Межвуз. сб. науч. трудов. — М., 1986, с. 104-114. Жукова Л.А. Популяционная жизнь луговых растений. — Йошкар-Ола: РИИК «Ланар», 1995, 224 с. ЖуковаЛ.А., Шестакова Э.В. Морфо- логическая пол ивариантность Plantago major Ь. в искусственных посадках // Бюлл. МОИП, отд. биол., 1995, т. 100, вып. 3, с. 95-101. Журавлева Н.А. Физиология травянис- того сообщества. Принципы конку- ренции. — Новосибирск: Наука, Сиб. изд. фирма, 1994, 172 с. Завадский КМ., Колчинский Э. И. Эво- люция эволюции (историко-крити- ческие очерки проблемы). — Л.: Наука, 1977, 236 с. Загускин С.Л Энергетические механиз- мы клетки: гомеостаз и биоритмы // Гомеостаз на различных уровнях
Литература организации биосистем. — Новоси- бирск: Наука. Сиб. отд., 1991, с. 51-58. Закржевский Б.С., Коровин Е. 1Т. Эко- логические особенности главней- ших растений Бетпак-Дала // Тр. Среднеазиат. гос. унив. — Ташкент, 1935, сер. 8-б, вып. 23, с. 3-74. Зиман С.Н. Жизненные формы и био- логия степных растений Донбасса. — Киев: Наукова думка, 1976, 192 с. Зозулин Г.М. Подземные части основ- ных видов травянистых растений и ассоциаций плакоров средне-рус- ской лесостепи в связи с вопросами формирования растительного по- крова // Тр. Центр.-Черноз. гос. заповедника. — Курск: Курское книж. изд-во, 1959, вып. 5, с. 3-314. Зозулин Г.М. Система жизненных форм высших растений // Ботан. журн.— 1961, т. 46, № 1, с. 3-20. Ибералл А.С., Мак-Каллок У.С. Гомео- кинез — организационный принцип сложных систем // Общие вопросы физиологических механизмов. М.: Наука, 1970, с. 340-352. Иберт Дж. Взаимодействующие сис- темы в развитии. Пер. с англ. — М.: Мир, 1968, 194с. Игнатьева И.1Т. Некоторые особенно- сти развития люпина многолетнего // Изв. ТСХА, 1958, вып. 2, с. 39-54. Игнатьева И.1Т. О жизненном цикле стержнекорневых и кистекорневых травянистых поликарпиков // Ботан. журн., 1965, т. 50, № 7, с. 903-916. Игнатьева И.1T., Андреева И. И. Мета- морфозы вегетативных органов по- крытосеменных. Учебное пособие. Ч. 1-2. — М.: изд-во МСХА, 1ч.— 1991, 121с.; 2ч. — 1993, 173с. Илленлрнроленный олренелииль рлои- ний Казахстана. — Алма-Ата: Наука, т. 1 — 1969, 644 с., т. 2 — 1972, 572 с. Имханицкая Н.Н. Пальмы. — Л.: Наука, 1985, 243 с. Иоганзен Б.Г. Основы экологии. — Томск: изд-во Томского ун-та, 1959, 390с. Иорданский Н.Н. Основы теории эво- люции. Пособие для учителей.— М.: Просвещение, 1979, 190 с. Иосебидзе Н.И. Атлас-определитель всходов лекарственных растений.— Тбилиси: Мецниереба, 1981, 237 с. Казакевич Л.И. Материалы к биологии растений Юго-Востока России // Бюлл. отд. прикл. ботаники. Сара- товск. обл. с.-х. опыт. станции.— 1922, № 18, с. 3-21. Казарян В.О. Старение высших расте- ний. — М.: Наука, 1969, 314с. йиитин ПВ. Мертвый завет. — М., 1998. Кальвин М. Химическая эволюция. Пер. с англ. — М.: Мир, 1971, 240 с. Канаев И.И. Комментарии к книге «И.В. Гете. Избранные сочинения по естествознанию». — М.; Л.: изд- во АН СССР, 1957, с. 497-535. Кариинская Р.С. Внешнее и внутреннее в биологии // Философские пробле- мы современной биологии. — М.; Л.: Наука, 1966, с. 63-77. Качухашвили Т.Н. Якорцы стелющиеся — Tribulus terrestris L., как источник стероидных сапонинов, содержащих диосгенин, и перспективы их ис- пользования в медицине // Тр. Тби- лисского науч.-иссл. хим-фарм. ин- та т. 9, 1960. Кеосян Дж. Начало жизни — возник- новение или эволюция // Проис- хождение жизни и эволюционная биохимия. — М.: Наука, 1975, с. 118-127. Клауд П., Джибор А. Крутоворот кис- лорода // Биосфера. Пер. с англ.— М.: Мир, 1972, с. 73-90. Кожевникова НД. Особенности возоб- новления ели Шренка (Picea schrenkiana) в разных частях ареала // Проблемы ботаники, т. 14, вып. 2: Экология и биология высокогор- ных растений. — Новосибирск: На- ука, Сиб. отд., 1979, с. 118-124. Козо-Полянский Б.М. Случаи превраще- ния биоморф культурных растений и их значение // Тр. Воронеж. гос. ун- та, 1945, т. 13, вып. 1, с. 41-45. Колищук В.Г. О морфологической эво- люции от деревьев к травам в ряду стелющихся форм растений // Бо- тан. журн., 1968, № 8, с. 1029-1042.
ОСНОВЬ1 БИОМОРФ ОЛОГИИ СЕМЕНКЫХ РА СТЕКИИ Кольцов H.К. Структура хромосом и обмен веществ в них // Биол. журн., 1938, т. 7, вып. 1. Кондаков H.И. Логический словарь- справочник. 2-е изд. — М.: Наука, 1975, 720 с. Коровин Е.П. Растительность Средней Азии и Южного Казахстана. — М.; Ташкент: Саогиз., 1934, 480 с. Королева А.С., Красноборов И. М., Пень- ковская Е.Ф. Определитель растений Новосибирской области. — Новоси- бирск: Наука, Сиб. отд., 1973, 368 с. Краснов К С., Воробьев Н.К, Годнев И.Н. и др. Физическая химия. В 2-х кн. Кн. 1. Строение веществ. Термоди- намика. 2-е изд. — М.: Высшая школа, 1995, 512 с. Кренке H.1Т. Теория циклического ста- рения и омоложения растений // Феногенетическая изменчивость. Т. 1. — М., 1933, с. 11-415. Кружилин А.С., Шведская 3. М. Биоло- гия двулетних растений. — М.: На- ука, 327 с. Круть И.В. Введение в общую теорию Земли. Уровни организации геосис- тем. — М.: Мысль, 1978, 367с. Кузиецов В.И., И)лис Г.М., Гутина В.Н. Ес- тествознание. — М.: Агар, 1996, 384с. Кузнецова Т.В., Пряхина H.И., Яков- лев Г.П. Соцветия. Морфологичес- кая классификация. — СПб.: Хими- ко-фармац. ин-т, 1992, 126 с. Кун Т. Структура научных революций. 2-е изд. — М.: Прогресс, 1977, 300 с. Куперман Ф.М. Морфофизиология ра- стений. 3-е изд. — М.: Высшая школа, 1977, 288 с. Куиерман Ф.М., Ржанова Е.И. Биоло- гия развития растений. — М.: Выс- шая школа, 1963, 424с. Лебедев НВ. Курс лекций по дарвинизму. — М.: изд-во Моск. ун-та, 1962, 342с. Левина Р.Е. Способы распространения плодов и семян. — М.: изд-во Моск. ун-та, 1957, 358 с. Либберт Э. Физиология растений. Пе- рев. с немец. — М.: Мир, 1976, 580с. Лодкина М.М Черты морфологической эволюции растений, обусловленные спецификой их онтогенеза. — Журн. общ. биол., 1983, т. 44, N 2. Лотова Л.И. Морфология и анатомия высших растений. — М.: Эдиториал УРСС, 2000, 528 с. Лэмб М. Биология старения. Перев. с англ. — М.: Мир, 1980, с. 206. Любименко В.Н. 0 спящих почках // Тр. СПб об-ва естествоиспытателей. 1900, т. 30, вып. 3, с. 195-260. Маевский П.Ф. Флора средней полосы европейской части СССР. 9-е изд. Л.: Колос, 1964, 880с. Мазуренко М Т. 0 жизненных формах стелющихся лесных растений // Ботан. журн., 1978, т. 63, М4, с. 593-603. Мазуренко М. Т. Биоморфологические адаптации растений Крайнего Севе- ра. — М.: Наука, 1986, 209с. Мазуренко М.Т., Хохряков А. 1Т. Струк- тура и морфогенез кустарников.— М.: Наука, 1977, 160с. Мак-Миллан Броуз Ф. Размножение растений. Пер. с англ. — М.: Мир 1987, 192 с. Мак-Фарленд Д. Поведение животных. Психобиология, этология H 3BoJIIolgiH. Пер. с англ. — М.: Мир, 1988, 520с. Мальцев А.И. Сорно-полевая расти- тельность и меры борьбы с нею.— М.; Л.: ОГИЗ, 1931, 128 с. Мальцев А.И. Сорная растительность СССР, — М.; Л.: Сельхозгиз, 1932, 296 с. Малмшев Л.И. Определитель высоко- горных растений Южной Сибири. — Л.: Наука, 1968, 284с. Марков М.В. Популяционная биология розеточных и полурозеточных мало- летних растений. — Казань: изд-во К~, 1990, 186с. Мелюхин С.Т. Проблема конечного и бесконечного (философский очерк). — М.: Госполитиздат, 1958, 264 с. Мейен С.В. Основные аспекты типоло- гии организмов // Журн. общ. биол., 1978, М 4, с. 495-508. Мейен С.В., Шрейдер Ю. А. Методоло- гические аспекты теории классифи- кации // Вопр. философии, 1976, No 12, с. 67-79. Мейер КИ. Филогения растений. М., 1958. Меннинджер Э. Причудливые деревья. Пер с англ. — М.: Мир, 1970, 371с.
Лите атура Минералогическая энциклопедия. Пер. с англ. — Л.: Недра, Ленингр. отд., 1985, 512 с. Мулкиджанян Я.И. К классификации жизненной формы «полукустарник» // Проблемы современной ботани- ки, т. II. — М.; Л.: Наука, 1965, с. 88-90. МусилЯ., Новакова О., Кунц К. Совре- менная биохимия в схемах. 2-е изд. Пер. с англ. — М.: Мир, 1984, 216 с. Небел Б. Наука об окружающей среде: как устроен мир. В 2-х т. Пер. с англ. Т. 1 — М.: Мир, 1993, 424с. Немченко Э.II. Лилия кудрявая // Био- логическая флора Московской об- ласти. Вып. 9 ч. 1. — М.: изд-во Моск. ун-та, 1993, с. 32-39. Нечаева Н.Т. О зимней вегетации эфе- меровой растительности в Кара-Ку- мах // Тр. Туркм. науч.-иссл. ин-та животнов. и ветеринарии. 1960. Т. 2, Ашхабад. Нечаева Н.Т., Василевская В.К., Анто- нова К.Г. Жизненные формы расте- ний пустыни Каракумы. — М: На- ука, 1973, 244с. Нефедов В.II., Ясайтис А.А. Историчес- кие аспекты учения о гомеостазе // Гомеопаз на разанчнап уроаюп орп- низации биосистем. — Новосибирск: Наука, Сиб. отд., 1991, с. 18-31. Никитин В.В. Сорные растения флоры СССР. — Л.: Наука, 1983, 454с. Новосельцев В.Н. Системные аспекты гомеостаза // Гомеостаз на различ- ных уровнях организации биосис- тем. — Новосибирск: Наука, Сиб. отд., 1991, с. 3-18. Нухимовский Е.Л. 0 каудексе растений // Рефераты докл. Всесоюз. межвуз. конф. по морфологии растений.— М.: изд-во М~, 1968, с. 207-208. Нухимовский Е.Л. 0 термине и поня- тии «каудекс». 3. Вестн. Моск. ун- та, сер. биол., почвоведение. 1969, М2, с. 71-78. Нухимовский Е.Л. Структурная органи- зация и морфогенез некоторых се- менных растений: Автореф. дис.... канд. биол. наук. — М:, 1970, 15 с. Нухимовский Е.Л.Осевая и побеговая системы семенных растений // Изв. ТСХА, 1971, вып. 1, с. 54-66. Нухимовский Е.Л. 0 понятии жизнен- ная форма // Интродукция новых лекарственных растений / Сб. науч. работ ВИЛР — М., 1973а, вып. 5, с. 222-232. Нухимовский Е.Л. 0 соотношении по- нятия «партикуляция» и «вегетатив- ное размножение» // Бюлл. МОИП, отд. биол., 1973б, т. 78, вып. 5, с. 107-120. Нухимовский Е.Л. Ветвление и кущение семенных растений // Изв. ТСХА, 1974, М 2, с. 50-62. Нухимовский Е.Л. Экологическая мор- фология некоторых лекарственных растений в естественных условиях их произрастания. 3. Partinia intermedia (Hornem.) Roem. et Schult. // Растит. ресурсы, 1976, т. 12, вып. 1, с. 3-16. Н~кимовский Е.Л. Экологическая мор- фология некоторых лекарственных растений в естественных условиях их произрастания. 4. Bupleurum multinerve DC. // Растит. ресурсы, 1978, т. 14, вып. 1, с. 37-46. Нухимовский Е.Л. Проблема классифи- кации в биоморфологии семенных растений // Успехи совр. биол., 1980, вып. 2, с. 286-307. Нухимовский Е.Л. Экологическая мор- фология некоторых лекарственных растений в естественных условиях их произрастания. 11. Phlojodicarpus sibiricus (Steph. ех Spreng.) К.-Pol. // Растит. ресурсы, 1981, т. 17, вып. 1, с. 43-50. Нухимовский Е.Л. Природа, жизнь и проблема жизнесохранения // Успе- хи совр. биол., 1984, т. 98, вып. 2 (5), с. 292-306. Нухимовский Е.Л. Особенности фенети- ческой организации биоморф семен- ных растений // Успехи совр. биол., 1986, т. 102, вып. 2 (5), с. 289-306. Нухимовский Е.Л. Статус биоморфоло- гии и первый принцип фенетичес- кой организации биоморф семен- ных растений // Усп. совр. биол., 1987, т. 104, вып. 2 (5), с. 231-250.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Нухимовский Е.Л. Основы биоморфо- логии семенных растений. Т. 1. Те- ория организации биоморф. — М.: Недра, 1997, 630 с. Нухимовский Е.Л., Шретер И. А. Био- морфогения якорцев стелющихся // Бюлл. Гл. ботан. сада. — 1979 вып. 111, с. 53-59. Одум Е. Экология. Пер. с англ. — М.: Просвещение, 1968, 168 с. Одум Е. Основы экологии. Пер. с англ. — М.: Мир, 1975, 740с. Озима М. Глобальная эволюция Земли. Пер. с англ. — М.: Мир, 1990, 165 с. Орловский Н.И. Основы биологии са- харной свеклы. 2-е изд. — Киев: ОГИЗ, 1961, 323 с. Парамонов А.А. Дарвинизм. — М.: Про- свещение, 1978, 335 с. Пачоский И К Биологические особенно- сти осота (Сп81ит arvense Scop.) // Тр. бюро по прикл. ботанике, т. 9, N 1. Пиотровский В. В. Геоморфология с основами геологии. — М.: Геодезиз- дат, 1961, 284 с. Подобедов Н.С. Общая физическая гео- графия и геоморфология. — М: Недра, 1964, 380 с. Полунина НН Биология цветения и эм- бриология некоторых видов эвкалип- та // Тр. Гл. ботвы. сада, т. VI, 1958. Поллавская Г.И. Экология растений.— М.: Сов. наука, 1948, 296с. 1Топое Б.Н. Взаимосвязь категорий сча- стья и смысла жизни. — М.: Наука, 1986, 93 с. Пошкурлат А.1Т. Строение и развитие дерновины чия // Учен. зап. Моск. гос. пед. ин-та. Каф. ботаники. 1941, т. 30, вып. 1, с. 101-155. Пошкурлат А.1Т. Анализ ритма разви- тия первоцвета весеннего // Ботан. журн., 1962, т. 47, М 2, с. 262-267. Пошкурлат А. 1Т. Род горицвет — Adonis L. Систематика, распространение, биология. — М: Наука, 2000, 199 с. Прозоровский А.В. О биологических типах растений пустыни // Ботан. журн., 1936, т. 21, вып. 5, с. 559-563. Прозоровский Н.А. Программа по теме «Изучение жизненного цикла се- менных растений заповедника в питомнике и в природе» // Научно- методич. записки Главного управле- ния по заповедникам, 1949, вып. 12, с. 167-185. Проскоряков Е.И. О происхождении однолетних эфемеров // Тр. Узб. гос. ун-та, нов. серия, т. 43, 1950, Ташкент. Работнов Т.А. Биологические наблюде- ния на субальпийских лугах Север- ного Кавказа // Ботан. журн., 1945, т. 30, М4, с. 167-177. Работнов Т.А. Возрастной состав попу- ляций некоторых пустынных расте- ний // Успехи совр. биол., 1946, т. 25, вып. 3(1), с. 435-438. Работнов Т.А. Основные вопросы и методы изучения жизненного цикла многолетних травянистых растений и состава их популяций // Научно- методические записки Главного уп- равл. по заповедникам, 1949, вып. 12, с. 41-54. Работнов Т.А. Жизненный цикл мно- голетних травянистых растений в луговых ценозах // Тр. Ботан. ин-та АН СССР. (Сер. 3), 1950, вып. 6, с. 7-204. Работнов Т.А. О жизненном цикле борщевика сибирского (Негас1еит sibiricum Ь.) // Бюлл. МОИП, отд. биол., нов. серия, 1956, т. 61, вып. 2, с. 73-81. Работнов Т.А. Методы определения возраста и длительности жизни у травянистых растений // Полевая геоботаника. — М., Л.: изд-во АН СССР, 1960, т. 2, с. 249-262. Работнов Т.А. Определение возрастно- го состава популяций видов в сооб- ществе // Полевая геоботаника.— М.; Л.: Наука, 1964, т. 3, с. 132-145. Работнов Т.А. Луговедение. — М.: изд- во Моск. ун-та, 1974, 384с. Работнов ТА. История фитоценоло- гии: Учебное пособие. — М.: Аргус, 1995, 158 с. Радкевич ВА. Экология: Краткий курс. 2- е изд. — Минск: Высш. шк., 1983, 320 с. Panonopm А. Математические аспекты абстрактного анализа систем // Ис- следования по общей теории систем. — М.: Прогресс, 1969, с. 83-105.
Литература Рачковская Е.И. К биологии пустын- ных полукустарничков // Тр. Бот. ин-та АН СССР, сер. 3, вып. 11, 1957, с. 5-87. Рейвн 17., Эверт Р., Айкхорн С. Совре- менная ботаника. В 2-х т. Пер. с англ. — М.: Мир, 1990, т. 1 — 348 с.; т. 2 — 344с. Реймерс H Ф. Популярный биологичес- кий словарь. — М.: Наука, 1991, 544 с. Реймерс Н.Ф. Экология (теории, зако- ны, правила, принципы и гипоте- зы). — М.: Журнал «Россия моло- дая», 1994, 367 с. Риклефс Р. Основы общей экологии. Пер. с англ. — М.: Мир, 1979, 424 с. Розенталь Д. Э., Теленкова М.А. Сло- варь трудностей русского языка.— М.: Рус. язык, 1985, 704 с. Роллер Э. Открытие основных законов жизни. Пер. с англ. — М.: Мир 1978, 336 с. Руттен М. Происхождение жизни. Пер. с англ. — М.: Мир, 1973, 411 с. Рысина Г. 1Т. Ранние этапы онтогенеза лесных травянистых растений Под- московья. — М.: Наука, 1973, 216 с. Сааков С.Г. Пальмы и их культура в СССР. — М.; Л., 1954, 320с. Северцов А.С. Введение в теорию эво- люции. — М.: изд-во Моск. ун-та, 1981, 318 с. Сенека Л.А. Избр. письма к Люцилию. СПб, 1893, с. 27. Сенянинова-Корчагина М. В. К вопросы классификации жизненных форм // Ученые зап. Ленингр. гос. унив., 1949, сер. геогр. наук, вып. 5, М 104, с. 54-102. Серебровский А.С. Некоторые пробле- мы органической эволюции. — М.: Наука, 1973, 168 с. Серебряков И.Г. Структура и ритм жиз- ни цветковых растений // Бюлл. МОИП, отд. биол., 1949, т. 54, вып. 1, с. 47-62. Серебряков И.Г. Морфология вегета- тивных органов высших растений. — М.: Сов. наука, 1952, 392с. Серебряков И.Г. О морфогенезе жиз- ненной формы дерева у лесных по- род средней полосы Европейской части СССР // Бюлл. МОИП, отд. биол., 1954, т. 59, вып. 1. Серебряков И.Г. Типы развития побе- гов у травянистых многолетников и факторы их формирования // Уче- ные зап. МГПИ им. В.П. Потемки- на / Кафедра ботаники, 1959, т. 100, вып. 5, с. 3-38. Серебряков И.Г. Экологическая морфо- логия растений. Жизненные формы покрытосеменных и хвойных. — М.: Высш. школа, 1962, 378 с. Серебряков И.Г. Жизненные формы высших растений и их изучение // Полевая геоботаника. — М.; Л.: Наука, 1964, т. 3, с. 146-205. Серебряков И.Г. К изучению жизнен- ных форм растений пустынной и тундровой зон СССР // Проблемы современной ботаники. Т. 2. — М.; Л.: Наука, 1965, с. 17-22. Серебрякова Т.И. О некоторых морфо- логических закономерностях побе- гообразования у овсяницы луговой (Festuca pratensis Huds.) // Бюлл. МОИП, отд. биол., 1959, т. 64, вып. 3, с. 89-98 ° Серебрякова Т.И. К вопросу о побего- образовании и кущении у злаков // Ботан. журн., 1962, т. 47, М 3, с. 427-432. Серебрякова Т.И. Морфогенез побегов и эволюция жизненных форм зла- ков. — М.: Наука, 1971, 360с. Серебрякова Т.И. Учение о жизненных формах на современном этапе // Итоги науки и техники. Сер. бота- ника / ВИНИТИ. — М., 1972, т. 1, с. 84-169. Серебрякова Т.И. Об основных «архи- тектурных моделях» травянистых многолетников и модусах их преоб- разования // Бюлл. МОИП, отд. биол., 1977, т. 82, вып. 5, с. 112-128. Серебрякова Т.И. Метаморфоз у расте- ний: онтогенический и эволюцион- ные аспекты // Бюлл. МОИП, отд. биол., 1984, т. 89, вып. 5, с. 94-102. Симпсон Дж. Г. Темпы и формы эво- люции. — М.: изд-во ин. литер. 1948, 358 с. Словарь иностранных слов. Изд. 6-е. — М.: Совет. энц., 1964, 784с.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РАСТЕНИЙ Смелов СП Теоретические основы лу- говодства. — М.: Колос, 1966, 367 с. Смирнова О.В. Структура травянистого покрова широколиственных лесов. — М.: Наука, 1987, 207. Станюкович К.В. Растительный покров Восточного Памира // Зап. геогр. об-ва. Нов. серия. — М.: изд-во геогр. литер., 1949, т. 10, с. 1-158. Стафлё Ф.А. (председатель редакц. ко- миссии). Международный кодекс ботанической номенклатуры, при- нятый Двенадцатым Международ- ным ботаническим конгрессом, Ле- нинград, июль, 1975. Пер. с англ.— Л.: Наука, 1980, 284с. Стешенко А.Н Формирование структу- ры полукустарничков в условиях вы- сокогорий Памира // Тр. Инст. бот. АН Тадж. ССР, 1956, т. 50, с. 1-161. Стешенко А.Н. Основные морфолого- биологические особенности расте- ний высокогорий Памира // Пробл. ботаники. — М.; Л.: Наука, 1965, т.7, 42-60. Стрелер Б. Время, клетки и старение. Перев. с англ.— М.: Мир, 1964, 255 с. Стрельченко В.И. Диалектика снятия в органической эволюции. — Л.: На- ука, 1980, 188 с. Судаков К.В. Квантование жизнедея- тельности // Успехи совр. биол., 1992, т. 112, вып. 4, с. 512-527. Татаренко И.В. Орхидные России: жиз- ненные формы, биология, вопросы охраны. — М.: Аргус, 1996, 207 с. Тахтаджян А.Л. Вопросы эволюцион- ной морфологии растений. — Л.: изд-во Л~, 1954, 215 с. Трофимов Т. Т. О типах прорастания семян некоторых многолетников // Ботан. журн., т. 48, М 11. Тимофеев- Ресовский Н.В., Ворон- цов Н.Н., Яблоков А.В. Краткий очерк теории эволюции. — М.: На- ука, 1969, 407 с. Уоддингтон КХ Основные биологи- ческие концепции // На пути к те- оретической биологии. Пер. с англ. — М.: Мир, 1970, с. 160-168. Уотсон Дж. Молекулярная биология гена. Пер. с англ. — М.: Мир, 1978, 720с. Уранов А.А. Жизненное состояние вида в растительном сообществе // Бюлл. МОИП, нов. сер., отд. биол., 1960, с. 65, вып. 3, с. 77-92. Уранов А.А. Большой жизненный цикл и возрастной спектр ценопопуляций цветковых растений // Тезисы док- лада V делегатского съезда Всес. ботан. об-ва. — Киев, 1973. УрановА.А. (отв. ред.) Возрастной со- став популяций цветковых растений в связи с их онтогенезом // Сб. тру- дов Моск. пед. гос. ин-та, 1974, 260 с. Ушатинская Р.С. Скрытая жизнь и анабиоз.— М.: Наука, 1990, 182 с. Федоров Ал. А., Кириичников М.Э., Ар- тюшенко 3.Т. Атлас по описатель- ной морфологии высших растений. Стебель и корень. М.; Л.: изд-во АН СССР, 1962, 352с. Федоров Ал. А., Кириичников М.Э., Ар- тюшенко3.Т. Атлас по описатель- ной морфологии высших растений. Соцветие. Л.: Наука, 1979, 295 с. Федоров ВД., Гильманов. Т.Г Экология. — М.: изд-во Моск. ун-та, 1980, 464 с. Феофраст. Исследования о растениях. — М.: изд-во АН СССР, 1951, 591 с. Философская энциклопедия. Т. 4.— М., 1967. Философская энциклопедия. Т. 5.— М., 1970. Философский словарь. Изд. 4-е. — М.: Политиздат, 1980, 444 с. Фисюнов А.В. Определитель всходов сорных растений. — Киев: Урожай, 1976, 232 с. Флора Казахстана. Главный редактор Н. В. Павлов. — Алма-Ата: изд-во АН Казах. ССР — Наука, т. 1-9, т. 1 — 1956, т. 9 — 1966. Флора СССР. Т. 1 — 30. — М. Л.: — изд- во АН СССР — Наука, 1934 — 1960. Флора СССР. Т. 26. — М. Л.: изд-во АН СССР, 1961, 939с. Фокс С, Дозе К. Молекулярная эволю- ция и возникновение жизни. Пер. с англ. — М.: Мир, 1975, 374с. Фолсом E Происхождение жизни: Ма- ленький теплый водоем. Пер. с англ.— М.: Мир, 1982, 160 с. Хатчинсон Дж. Биосфера // Биосфера. Пер. с англ. — М.: Мир, 1972, с. 9-25.
Литература Хауисов А.П Соматическая эволюция од- нодольных. — М.: Наука, 1975а, 196с. Хохряков А.IT. Закономерности эволю- ции растений. — Новосибирск: На- ука. 1975б, 202 с. Хохряков А.1Т. Изменение образа жизни растений в онтогенезе // Журн. общ. биол., 1978, т. 39, М 3, с. 357-372. Хохряков А.II. Принципы классифика- ции жизненных форм растений // Изв. АН СССР. Сер. биол., 1979, М 4, с. 586-598. Хохряков А.II. Эволюция биоморф рас- тений — М.: Наука, 1981, 168 с. Хржановский В.Г. Розы. Филогения и систематика. — М.: Сов. наука, 1958, 497 с. Хржановский В.Г. Морфогенетика как основа филогении и таксономии покрытосеменных // Доклады ТСХА, 1966, вып. 118, с. 127-133. Хржановский В.Г. О закономерностях и единицах измерения видообразова- тельного процесса // Изв. ТСХА, 1968, вып. 5, с. 3-14. Цвелев Н.Н. О значении степени специ- ализации таксонов для их дальней- шей эволюции // Бюлл. МОИП, отд. биол., 1973, т. 78, вып. 2, с. 71-81. Чайлахян М Х Основные закономерно- сти онтогенеза высших растений.— М.: изд-во АН СССР, 1958, 79с. Чайлахян М.Х Факторы генеративного развития растений // Доложено... 3 июня 1964 г. / Тимирязевские чте- ния, 25.— М.: Наука, 1964, 58 с. Чайлахян М.Х. Регуляция цветения высших растений. — М.: Наука, 1988, 560 с. Чайлахян М.Х., Аксенова Н. II., Кефе- ли В. И. О терминологии онтогенеза растений. — М.: Наука, 1973, 39с. Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего CCCP) — СПб: Мир и семья, 1995, 992с. Чернова Н.М., Былова А. М. Экология. — М.: Просвещение, 1981, 255 с. Шалыт М. С. Система жизненных форм степных растений // Уч. зап. Тадж. гос. унив., 1955, т. VI, Труды фак. естеств. наук, вып. 1. Шалыт М.С. Изучение вегетативного размножения и возобновления ком- понентов растительных сообществ // Полевая геоботаника. — М.; Л.: изд- во АН СССР, 1960, т. 2, с. 163-208. Шалыт М.С. Партикуляция у высших растений // Проблемы современной ботаники. — М.; Л.: Наука, 1965, т. 2, с. 117-122. Шкловский ИС. Вселенная, жизнь, ра- зум. 3-е изд. — М.: Наука, 1973, 335 с. Шмалыаузен И.И. Факторы эволюции. Теория стабилизирующего отбора. 2-изд. — М.: Наука, 1968, 451 с. Шмалыаузен ИИ Организм как целое в индивидуальном развитии. Избран- ные труды — М.: Наука, 1982, 383 с. Шорина Н.И. Жизненный цикл безвре- менника великолепного (Colchicum speciosum Stev.) в условиях субаль- пийских лугов Западного Закавка- зья // Науч. докл. высш. школы. Биол. науки, 1964, М 1, с. 113-119. Шорина Н.И. Жизненный цикл безвре- менника великолепного в лесном и субальпийском поясах Западного За- кавказья // Онтогенез и возрастной состав популяций цветковых расте- ний. — М.: Наука, 1967, с. 70-99. Шорина Н.И., Куклина А. Г. К биологии кандыка // Бюлл. Глав. ботан. сада, вып. 102, 1977, с. 88-95. Шредингер Э. Что такое жизнь с точки зрения физики. 2-е изд. — М.: Ато- миздат, 1972, 89 с. Штромбер А.Я., Джорбенадзе А. М.— Культивирование якорцев стелю- щихся на опытном поле Тбилисско- го института фармакохимии // Хи- мия и биология активных веществ лекарственных растений Грузии. Сер. 1, вып. 11. — Тбилиси: Мецни- ереба, 1969. Эзау К Анатомия семенных растений. Пер. с англ. — М.: Мир, 1980, т. 1- 2, 558с. Энгельс Ф. Диалектика природы. — М.: Политиздат, 1964, 358 с. Энгельгардт В.А. Познание явлений жизни. — М.: Наука, 1984, 304с. Юрцев Б.А. Жизненные формы: один из узловых объектов ботаники // Тр. МОИП, т. 42: Проблемы экологи-
832 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ ческой морфологии растений. — М.: Наука, 1976, с. 9-44. Юрцев Б.А. Флора Сунтар-Хаята. Про- блемы истории высокогорных ланд- шафтов Северо-Восточной Сибири. — Л.: Наука, 1968, 235 с. Яблоков А.В. Фенетика. Эволюция, по- пуляция, признак. — М.: Наука, 1980, 132 с. Яблоков А.В., Юсуфов А. Г. Эволюцион- ное учение (Дарвинизм). 3-е изд.— М.: Высшая школа, 1989, 335с. Янкелевич В. Смерть. Пер. с франц.— М.: изд-во Литературного ин-та, 1999, 448 с. Aldrovandi U. Dendrologiae naturalis scilicet arborum historiae libri duo. — Francofurti, 1671, 468 р. Alon А. Tress, Bushes and Creepers of Israel. — 1991, 162 р. Aschog./. Survival value of diurnal rhythms // Symposia of the Zoological Society of London, 1964, 13, р. 79-98. Bertalanffy L. Zu einer allgemeine Systemlehre // Biologia Generalis. 1949, Bd 19., $. 114-129. Bertalanffy L. General System Theory /l General Systems, 1956, vol. 1, р. 1-10. Саела1р~п~ А. De plantis libri 16. Florentiae, 1583, 621 р. Саппоп И~В. Organisation for physiological homeostasis // Physiol. Rev., 1929, vol. 9, р. 399-431. Cannon WB.The wisdom of the body.— London, 1932, 312 р. Clements ЕР. Plants Indicators. The relation of plants communities to process and practice // Ibid., 290 — Waschington, 1920. Corner Е.J. Н. The Durian theory of the origin of the modern tree // Ann. Bot., 1949, 13, ¹52, р. 367-414. Corner Е. J. Н. The Ие of plants. — London, 1964. Csapody V. Keimlungsbestimmungsbuch der Dikotyledonen. — Budapest, 1968, 286 S. Darwin Ch. On the origin of species by means of natural selection. — London, 1859, 502р. Пер. с англ.: Дарвин Ч. Происхождение видов путем ес- тественного отбора. — М.: Просве- щение, 1986, 383 с. Dode Е.А. Extaits (Гуопе monographic une d'ite de genere Populus // Bull. de la Soc. d'histoire nat. (ГAutumn., 1905, t. 108, 73 р. Dodonaeus R., Stirpium historiae pemp- tadessex, sive libri 30. — Antwerpiae, 1583, 860 р. Drude О. Die sistematische und geographische Anordnung der Phanerogames. In: Schenk А. Handbuch der Botanik, 1887, Bd. 3, 2, $. 175-496. Drude О. Handbuch der Pflanzengeographie. — Stuttgart, 1890, 582 $. Drude О. Okologie der Pflanzen // Die Wissenschaft, 1913, 50, $. 1-247. Du Reitz G. Е. Life forms of terrestrial flowering plants // Acta phytogeogr. suec., 1931, 3, р. 1-95. Feinbrun-Dothan N. Flora Palaestina. Part three. Plates. Ericaceae to Compositae.— Jerusalem, 1977. Feinbrun-Dothan N., Danin А. Analytical Нога of Eretz-Israel. — Jerusalem, 1991, 1040 р. Fukuda J. Morphological and anatomical studies in Tribulus terrestris. I. Phyllotaxis and braching // Bot. Mag. Tokyo, 1974, vol. 87, р. 51-60. Gams Н. Prinzipienfragen der Vegetations- forschung // Virteljahrsch. Naturforsch. Ges. — Ziirich, 1918, Bd. 63, $. 293-493. Gluch W. Wuchsformstudien an zentrale- uropaischen Fabaceen. I. Die stauden der Gattung Medicago Ь. und Trifolium Ь. // Feddes. repert., 1967, Bd. 75, ¹ 3, $. 221-265. Gmelin J. G. Flora sibirica, sive historia plantarum Sibiricae. 1. — Petropoli, 1747. Goebel К. Bliitenbildung und Sprossgestaltung (Anthokladien und Infloresczenzen). Zweiter Erghnzungsband zur Organo- graphie der Pflanzen. — Jena, 1931. 242 $. Goethe J. W. Versuch die Metamorphose der Pflanzen zu егИагеп. — Gotha, 1790. Halberg F. Physiological 24-hour periodicity: General and procedural considerations with reference to the adrenal cycle // 2'.ейьсЬпй fiir Vi аппп, - Hormon-und Ferment- forschung, 1959, 10, р. 225-296. Halle F., Oldeman R.À. А. Essai sur Гагсй- tecture et la dynamique de croissance des arbres tropican. — Paris, 1970, 178 р.
Eumepamypa 833 Halle F., Oldeman R.À.À., Tomlinson P. В. Tropical trees and forest: an architectural analysis. — Berlin, 1978, 441 р. Hegi G. Illustrierte Нога von Mittel-Europa. Bd. IV/3 Teil, 1975. Hult R. Fdrsdk till analitisk behandling af vhxtformationerna. — Helsingfors, 1881, 155 $. Humboldt А. Ideen zur einer Physionomik der Gewachse. — Ti1bingen, 1806, 28 $. Innes С., Glass Ch. The Illustrated Ency- clopedia of Cacti. — London, 1991, 320 р. Irmich Th. Zur Morphologic der monoko- tylischen Knollen- und Zwiebelgewhchse. — Berlin, 1850 а. 1пшсй Th. Uber Helianthemum fumaria // Bot. Zeitung, Jahrg. 8, 1850b. Irmich Th. Kurze botanische Mitteilungen // Нога, Bd. 33, 1853. Irmich Th. Uber Lathyrus tuberosus und einige andere Papilionaceen // Bot. Zeitung, Jahrg., 17, 1859. Irmich Th. Uber einige Ranunculaceen // Bot. Zeitung, Jahrg., 18, 1860. 1пшсй Th. ОЬег Polygonatum amphibium, Lisimachia vulgaris, Comarum palustre und Menyanthes trifoliata // Bot. Zeitung, Jahrg., 19, 1861. Irmich Th. Beitrag zur Morphologic einiger europhi scher Gerani umar ten // Bot. Zeitung, Jahrg., 32, 1874. Kanngisser F. Uber Lebensdauer einiger Zwergstrhuchern // Gartenflora, 1910. Kays S., Harper Т. L The regulation of plant and tiller density in а grass sward // Journal of Ecology, 1974, 62, р. 97-105. Krause Е. Die Einteilung der Pflanzen nach ihrer Daurer // Ber. d. Deut. Bot. Ges., 9, 1891. Linnaeus С. Philosophia botanica in qua explicantur fundamenta botanica curn difinitionibus partium, exemplis. Stockholmiae, 1751, 362 р. Martius С. F. Ph. Historia naturalis palmarum. Vol. 1-3. — Leipzig, 1823 — 1853. 350 р. McClure F. А. The bamboos: а fresh per- spective. — Camridge: Harward Univ. Press, 1966. McCurrach ХС. Palm of the world. — New York, 1960, 290 р. Meusel Н. Wuchsformen und Wuchstypen der europhischen Laubmoose // Nova acta Leopoldina. N. F., 1935, Bd. 3, ¹ 12, $. 123-277. Meusel Н. Die Bedeutung der Wuchsform fiir die Entwicklung des naturlichen Systems der Pflanzen // Feddes Repert., 1951, Bd. 54, №2-3, $. 137-172. Meusel Н. Wuchsformenreichen mideterran- mitteleurophischen Angiospermen — Таха // Feddes repert., 1970, Bd. 61, Heft 1, $. 41-59. Moore Н. Е. Jr. The Cuban belly palm- Colpothrinax wrightii // Principes, 1960, vol. 4, ¹2, р. 53-55. Moore Н. Е. Jr. Botany and classification of palms. — Cultivated palms // Amer. Hort. Mag, 1961, vol. 40, ¹1, ,р. 17-26. Moore Н. Е. Jr. Two new palms from Peru / / Principes, 1963, vol. 7, ¹ 3, ,р. 107-115. Moore Н.Е. Jr., Uhl N. W. The monocoty- ledons: their evolutions and comparative biology. VI. Palms and the origin and evolution of monocotyledons // Quart. Rev. Biol, 1973, vol. 48, ¹ 3, ,р. 414-436. Mii hlberg Н. Wuchsformenstudien in der Familie Poaceae. Die Wuchsformen der mitteldeutschen Poaceae Arten // Feddes Repert., 1965, Bd. 71, $. 188-217. Muller Н.J. Proc. Internat. Congr. Plant. Physiol., 1, 1929. Plate L Selectionsprinzip und Probleme der Artbildung. 4Aufl. — Leipzig und Berlin, 1913, 650 $. Pokorny J. Choc V. Bhume und Strhcher.— Praga, 1967, 140$. Rauh W. Uber die Ferzweigung auslhuferbildeder Strhche mit besonderer BerQcksichtigung ihrer Beziehungen zu den Stauden // Hercinia, 1, 1937, $. 3-54. Rauh W. Beitrhge zur Morphologic und Biologic der Holzgewhchse // Nova Acta Leopold., N. F. 1938, Bd. 5. Rauh W. Uber polsterfdrmigen Wuchs // Nova Acta Leopold., N. F. 1939, Bd. 7, $. 267-508. Rauh W. Morphologic der Nutzpflanzen.— Heidelberg, 1950, 290 $. Raunkiaer С. Planterigts livsformer og deres Betydring for geographien. Koebenhavn, 1907, 132р. Raunkiaer С. The Ие form of plants and statistical plants geography — Oxford, 1934, 632 р. Scharfetter R. Biographien von Pflanzensippen. — Stuttgart, 1953, 546 $.
ОСНОВЫ БИОМОРФОЛО1ИИ СЕМЕННЫХ РА СТЕНИИ Schmid G. Die Metamorphose der Pflanzen. In: Goete als Seher u. Erforscher der Natur. — Halle, 1930, $. 205-226. Schmid Е. Die Wuchsformen der Dicotyledo- nen // Bericht. i1ber der geobotanischer For-schungsinst. Ri1bel in ZQrich, 1956, Bd. 101. Schmid Е. Die Erfassung der Vegetationsein- heites mit floristischen und morpholo- gischen Analysen // Bericht der schweiz. Bot. Ges., 1963, Bd. 73, $. 276-324. Schmida А. Handbook of Wildflowers of Israel Mediterranean Нога — Jerusalem, 1986, 308 р. Schneer С. J. Morphological basis for the reticular hypothesis // Ат. Mineral., 1970, 55, р. 1466-1488. Schneer С. J. Morphological complements of cristal structures // Can. Mineral., 1978, 16, р. 465-470. Sch~veter С. Das Pflanzenleben der Alpen. Eine Schilderung der Hochgebirgsflora.— Ziirich, 1908, 807 $. Tomlinson P. В. Essays on the morphology of palms. Е Germination and the seedling // Principes, 1960, vol. 4, ¹2, р. 56-61. Tomlinson P. В. Essays on the morphology of palms. VI. The palm stem // Principes, 1961, vol. 5, №4, р. 117-124. Tnrland LT. Monist, 24, 1914. Tnrland LT. Cleveland Med. J., 15, 1916. Tnrland LT. Amer. Naturalist, 51, 1917. Troll W. Vergleichende Morphologic der hdheren PAanzen. Bd. I. Vegetation- sorgane. Teil 1. — Berlin, 1937. Troll W. Vergleichende Morphologic der hdheren Рйапгеп. Bd. I. Vegetation- sorgane. Teil 2. — Berlin, 1939. Troll W. Vergleichende Morphologic der hdheren Рйапгеп. Bd. I. Vegetation- sorgane. Teil 3. — Berlin, 1943. Troll W. Die Infloreszenzen. Typologic und Stel lung im Aufbau des Vegetations Kdrpers. Bd. I — Jena, 1964, 615 $.; Bd. II, Teil 1 — Jena, 1969, 630$. Troll W. Allgemeine Botanik. Ein Lehrbuch auf vergleichend-biologischer Grundlage. — Stuttgart, 1973. Walter Н. Die Vegetation der Erde in Жо- physiologischer Betrachtung. Bd. II: Die gem23igten und arktischen Zonen. — Jena, 1968, 1001 $. arming Е. Uber perenne Gewhchse // Bot. Zentralbl, 1884, Bd. 18, ¹19, $. 16-22. arming Е. От planterigets lifsformer. Kjdbenhavn, 1908, 86 $. arming Е. Oecology of plants. — Oxford, 1909, 422 р. И~агюипд Е. Bemaerkningen от Livsform og Standplads // Forhandl. ved. det Skandi- navske Naturf., 1916, 16. Warming Е. gkologiens Grundformer. Udkast til en sistematisk ordning // Det Kongelige Danske vidensk. Sel skabs skri f ter Naturvidensk. og mathem. afd. 8 Raekke, 1923, Bd. 4, ¹2, р. 119-187. Warming Е., Graebner P. Lehrbuch der Жо1о- gichen Pflanzengeographie. 3 АиА. Berlin, 1918, 762 $. Velenovsky 1. Vergleichende Morphologic der Pflanzen. Teil 1-4. — Prage, 1905-1913, 1 — 1905, 2 — 1907, 3 — 1910, 4 — 1913. Zohary М. Hydro-economical types in the vegetation of Near East deserts Sympos // Biol. and Product. of hot and cold desert. — London, 1952. Zohary М. Нога Palaestina. Part one. Plates. Equisetaceae to Moringaceae. Jerusalem, 1966. Zohary М. Нога Palaestina. Part two. Plates. Plantaginaceae to Umbelliferae. Jerusalem, 1972.
Указатель латинских названий растений {стр.). Виды растений, которые охарактеризованы в рисунках Abies alba Mill. 91 Acacia longifolia Willd. 651 Acacia raddiana Savi 558 Асег negundo L. 226 Асег obtusifolium Sm. 133 Асег platanoides L. 655 Aconitum anthoroideum DC. 164, 649 Aconitum barbatum Pers. 170 Aconitum leucostomum Worosch. 590 Aconitum napellus L. 165 Aconogonon alpinum (А)1.) Schur 169 Achillea millefolium L. 427 Actaea spicata L. 78 Adhatoda vasica (L.) Ness 6& Adenophora lilifolia (L.) А. DC. 110, 111 Aegopodium alpestre Ledeb. 426 Aethusa cynapium L. 157 Agrimonia pilosa Ledeb. 317 Agropyron fragile (Roth) P. Candargy 517 Albizia julibrissin Durazz. 490 Alcea froloviana (Litv.) Iljin 167 Alchemilla sibirica 2'.am. 423 АИгейа сетиа (L.) Cass. 185 Alisma plantago-aquatica L. 430 Allium altaicum Pall. 285 Allium victorialis L. 542 Althaea officinalis L. 178 Ajuga iva (L.) Schreber 466 Ajuga reptans L. 201 Anabasis articulata (Forssk.) Moq. 640 Anagallis arvensis L. 82 Anagyris foetida L. 88 Ananas comosus (L.) Меп. 157 Anemonastrum crinitum (Juz.) Holub 543 Anemone sylvestris L. 429 Angelica sylvestris L. 126 Anisantha tectorum (L.) Nevski 517 Anthriscus sylvestris (L.) Hoffm. var. nemorosa (Bieb.) Trautv. 107 Antirrhinum majus L. 540 Aquilegia glandulosa Fisch. ex Link 539 Aquilegia яЬ1пса Lam. 171,172 Arrhenatherum elatius (L.) J. et С. Presl 709 Artemisia abrotanum L. 531 Artemisia arborescens L. 320 Artemisia monosperma Delile 626 Artemisia яеЬеп Besser 113 Artemisia ячегяапа Willd. 399 Arthrocnemum macrostachyum (Мопс.) Moris 474
Carum carvi L. 344 Cassia senna L. 75 Chaerophyllum bulbosum L. 345 Aulacospermum апота1ит (Ledeb.) Ledeb. 126 С „1ащеа а аа 1 184 Centaurea phrygia L. 314 Bistorta elliptica (Willd. ех Spreng.) Kom. 429 Cistus salviifolius L. 322 Arum hydrofdum Boiss. 433 Asparagus stipularis Forssk. 647 Aster subulatus Michaux 446 Astericus hierochunticus (Michon) Wikl. 394 Astragalus spinosus (Forssk.) Muschler 93 Astragalus tschuensis Bunge 469 Atractylis phaeolepis Pomel 462 Atragene яо1пса L. 481 Atriplex halimus L. 271 Atriplex holocarpa F. Mueller 296 Atriplex portulacoides L. 272 Avena sativa L. 400 Ballota undulata (Fresen.) Bentham 129 Batrachium trichophyllum (Chaix) Boch 646 Berberis sibirica Pall ° 482 Bergenia crassifolia (L.) Fritsch 443 Beta vulgaris L. 103 Betula pendula Roth. 133, 202 Bidens tripartita L. 445, 755 Biebersteinia одога Steph. 618 Blepharis сйапв (L.) В. L. ВигИ. 656 Виуа rosea (Turcz.) Bge. 404 Bupleurum bicaule Helm 714 Bupleurum krylovianum Schischk. 715 Bupleurum 1апсйо11шп Ногпет. 713 Bupleurum salicifolia Soland. ex Lowe 716 Cacalia hastata L. 548 Callistemon speciosus (Sims) DC. 69 Caltha palustris L. 280 Campanula rotundifolia L. 292 Cannabis ruderalis Janisch. 296 Capparis aegyptiaca Lam. 483 Cardamine bellidifolia L. 217 Carduus crispus L. 161 Carlina intermedia Schur 160 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Chamaedorea lanceolata (Ruiz et Pav.) Kunth. 466 Chamaenerion latifolium (L.) ТЛ. Fries et Lange 197 Chorispora bungeana Fisch. et С. А. Меу. 756 Chrysanthemum coronarium L. 498 Celtis australis L. 70 Centaurea aegiptiaca L. 275 Centaurea hyalolepis Boiss. 807, 808 Centaurium tenuiflorum (Hoffmanns et Link) Fritsch 395 Ceratonia siliqua L. 87 Cercis siliquastrum L. 88 Cereus bonplandii Рапп ех Pfeiff. 457 Cereus jamacaru DC. 457 Cereus sublanatus Salm-Dyck. 457 Cirsium esculentum (Stev.) С. А. Меу. 102 Cirsium heterophyllum (L.) Hill 289 Claytonia joanneana Schult. 220 Cnicus benedictus L. 104 Coccyganthe flos-cuculi (L.) Роип. 453 Colpothrinax wrightii Griseb. et Н. Wendl. ex Siebert et Voss. 466 Convallaria majalis L. 425 Convolvulus arvensis L. 406 Convolvulus secundus Desr. 274 Coridothymus capitatus (L.) Reichenb. Я. 760 Corydalis pauciflora (Steph.) Pers. 209 Corylus avellana L. 202 Crepis nana Richards. 405 Crepis polytricha (Ledeb.) Turcz. 177 Crepis sibirica L. 419, 420 Crithmum maritimum L. 463
837 Eucalyptus camaldulensis Dehnh. 639 Euphorbia сапапепяв Thunb. 457 Euphorbia obesa Hook. 457 Euphorbia pilosa L. 166 Euryops multifidus DC. 319 Dendranthema sinua«m («deb) Tzvel. 109 Eu»ps pecti»tus Сова. 473 Galium densiflorum Ledeb. rosmarinifolium (Bunge) Tzvel. 413 var. Ephedra aphylla Forssk. 657 Указатель латинских названий растений Crupina crupinastrum (G. Moris) Vis. 123 Cucumis sativus L. 397 Cucurbita pepo L. 399 Cynoglossum otTtcinale L. 617 Dactylis glomerata L. 708 Dactylorhiza maculata (L.) So6 164 Dalbergia sissoo Roxb. 225 Datura inoxia Mill. 444 Datura stramonium L. 395 Delphinium elatum L. 277 Delphinium пигаЬй Serg. 276 Deschampsia cespitosa (L.) Beauv. 708 Dianthus superbus L. 451 Dictamnus gymnosthylis Stev. 414 Digitalis lanata Ehrh. 208 Digitalis purpurea L. 161 Diplotaxis acris (Forssk.) Boiss. 396 Doronicum oblongiflorum DC. 128 Draba sibirica (Pall.) Thell. 452 Draba subamplexicaulis С. А. Меу. 403 Dracocephalum bungeanum Schischk. et Serg. 411 Dracocephalum grandiflorum L. 23 Dracocephalum imberbe Bunge 100 Dracocephalum peregrinum 173 Dryas oxyodonta Juz. 467 Ecballium elaterium (L.) А. Rich. 168 Echinacea ригригеа (L.) МоепсЛ 213 Echinops humilis Bieb. 779 Echinops ро1усегаз Boiss. 810 Echinops ruthenicus Bieb. 127 Echinops sphaerocephalus L. 210 Echium vulgare L. 717 Echium fastuosum Jacq. 718 Eleutherococcus senticocus (Rupr. et Maxim.) Махдп 132 Eremopyrum triticeum (Gaertn.) Nevski 517 Erigeron eriocalix (Ledeb.) Vierh. 218 Erigeron leioreades М. Рор. 191 Eriophorum humile Turcz. ex Steud. 418 Erythrinia lesistemon Hutch 489 Erythronium sibiricum (Fisch. et С. А. Меу.) Кгу1. 163 Euryops sp. 582 Euryops speciosissimus DC. 76 Fagonia то115 Delile 653 Farsetia aegyptiaca Тшта 460 Festuca pratensis Hunds. 709 Filipendula ulmaria (L.) Maxim. 288 Filipendula vulgaris Moench 418 Frangula ainus Mill. 269 Frankenia hirsuta L. 516 Fraxinus syriaca Boiss. 267 Galeopsis speciosa Mill. 158 Galium apparine L. 396 Gasrolychnis apetala (L.) Tolm. et Kozhanczikov 403 Genista fasselata Decaisne 641 Gentiana fischeri P. Smirn. 77 Gentiana lutea L. 180, 181 Gentiana macrophylla Pall. 408 Geranium albiflorum Ledeb. 281 Geranium palustre L. 316 Geum цгЬапщп L. 431 Grindelia robusta Nutt. 533
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Lespedeza juncea (L.) Pers. 313 Helianthemum stipulatum (Forssk.) С. Chr. 274 Myrtus communis L. 478 Nepeta sibirica L. 311 Nerium oleander L. 71 Harrisia guelichii Br. et Rose 457 Нагпяа sp. 457 Hedysarum austrosibiricum В. Fedtsch. 587 Hedysarum gmelinii Ledeb. 707 Helianthemum lippii (L.) Dum.-Courset 461 Негас1ешп dissectum Ledeb. 538 Hesperis pseudonivea Tzvel. 106 Hieracium umbellatum L. 440 Hieracium cymosum L. 436 Hyoscyamus niger L. 125 Hyparrhenia hirta (L.) Stapf 447 Hypericum L. 712 Hypericum balearicum L. 728 Hypericum hirsutum L. 726 Hypericum maculatum Crantz 727 Hypericum perforatum L. 725 Hyphaene thebaica (L.) Mart. 466 Iris bloudowii Ledeb. 290 Iris ruthenica Ker-Gawl. 421 Impatiens parviflora DC. 124 Juglans mandshurica Maxim. 73 Kickxia spartioides (ВисЬ) Janchen 646 Kitagawia baicalensis (Ridow. ex Willd.) М. Pimen 105 Koenigia islandica L. 293 Lagopsis marrubiastrum (Steph.) lk.-Gal. 83 Lagotis integrifolia (Willd.) Schischk. 198 Lamium album L. 438 Larix decidua Mill. 90 Lavandula coronopifolia Poiret 223 Lavandula stoechas L. 476 Lavatera cretica L. 719 Lavatera thuringiaca L. 720 Lavatera maritima Gouan. 721 Lavatera arborea L. 721 Leiospora excarpa (С. А. Меу.) РИгйс 500 Leontodon autumnalis L. 315 Leonurus glaucescens Bunge 179 Leonurus quinquelobatus Gilib. 347 Lepidium ruderale L. 295 Leucanthenum vulgare Lam. 450 Ligularia altaica DC. 432 Ligularia glauca (L.) О. Hoffm. 193 Ligularia sibirica (L.) Cass. 283 Lilium pilosiusculum (Freyn) Miscz. 441 Limonium lobatum (L.) Chaz. 294 Limonium oleifolium Miller 723 Limonium pruinosum (L.) О. Kuntze 649 Limonium sinuatum (L.) Miller 402 Limonium sp. 724 Lomatogonium carinthiacum (Wulf.) Reichenb. 79 Lonicera altaica Pall. 270 Lonicera etrusca Santi 480 Loranthus acaciae 2'.исс. 484 Lupinaster pentaphyllus Moench. 197 Lythrum virgatum L. 288 Macropodium nivale (Pall.) R. Br. 546 Majorana syriaca (L.) Rafin. 462 Malva sylvestris L. 310 Maresia pulchella (DC.) О. Е. Schulz 394 Melampyrum nemorosum L. 295 Melandrium latifolium (Poir.) Maire 637 Melilotus albus Medik. 207 Mentha aquatica L. 650 Moricandia nitens (Viv.) Е. А. Durand et Barrate 645 Myosotis asiatica (Vestergren) Schischk. et Serg. 215
Указатель латинских названий растений 839 Nicotiana glauca Graham 465 Noaea mucronata (Forssk.) Ascherson et Schweinf. 628 Оепо1пеи drummondii Hooker 275 Olea europaea L. 268 Omalotheca norvegica (Gunn.) Sch. Bip. et F. Schultz 282 Ononis natrix L. 130 Onopodum acanthium L. 589 Opuntia engelmanii Salm-Dysk. 457 Opuntia ficus-indica Mill. 457 Opuntia tunicata Hort. 457 Oxyria digyna (L.) Hill 85, 86 Раеоша hybrida Pall. 417 Pancratium maritimum L. 650 Panicum turgidum Forssk. 455 Papaver pseudocanescens М. Рор. 84 Paraquilegia anemonoides (Willd.) Ulbr. 560 Parkinsonia aculeata L. 136 Parnassia palustris L. 278 Patrinia sibirica (L.) Juss. 415, 416 Petasites rubellus (J. F. Gmel.) Toman 428 Phagnolon rupestre (L.) DC. 222 Phillyrea latifolia L. 479 Phlomoides alpina (Pall.) Adyl, R. Кат. et Machmedov 409, 410 Plhomis viscosa Poiret 477 Plantago albicans L. 759 Plantago arborescens Poir. 472 Plantago cretica L. 124 Plantago major L. 588 Platanthera bifolia (L.) Rich. 442 Pedicularis proboscidea Stev. 442 Pennisetum asperifolium (Desf.) Kunth 454 Persea indica (L.) Spreng. 486 Persicaria scabra (Moench) Mold. 123 Peucedanum vaginatum Ledeb. 407 Picea abies (L.) Karst 91, 133, 202 Pinus halepensis Miller 620 Pinus montana Mill. 99 Pinus яЫпса Du Tour 89, 91 Pinus sylvestris L. 202 Piptatherum miliaceum (L.) Coss. 449 Pituranthos tortuosus (Desf.) Ascherson et Schweinf. 465 Pituranthos triradiatus (Boiss.) Ascherson et Schweinf. 644 Polemonium caeruleum L. 544 Polygala comosa Schkuhr 721 Polygala myrtifolia L. 722 Polygonatum odoratum (Mill.) Druce 277 Polygonum palaestinum 2'.очагу 464 Populus euphratica Oliver 658 Potentilla Ыйога Willd. ex Schlecht. 471 Potentilla chrysatha Trev. 190 Potentilla elegans Cham. et Schlecht. 293 Prasium majus L. 478 Primula algida Adams 434 Primula cortusoides L. 780 Prunella vulgaris L. 194 Pseudolarix amabilis Rehd. 90 Pseudotsuga menziesii (Mirb.) Franco 91 Ptarmica impatiens (L.) DC. 219 Pulicaria crispa (Forssk.) Oliver 461 Pulmonaria mollis Wulf. ex Hornem. 282 Pulsatilla campanella Fisch. ex Redel et Til. 175 Pulsatilla patens (L.) Mill. 174 uercus calliprinos Webb 487 Quercus гоЬиг L. 202 Ranunculus acris L. 285 Ranunculus altaicus Laxm. 187 Ranunculus flammula L. 195 Ranunculus polyanthemos L. 216 Ranunculus radicans С. А. Меу. 436 Ranunculus repens L. 437 Raphanus sativus L. 754
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Reaumuria hirtella Jaub. et Spach 809 Rhamnus alaternus L. 224 Rhamnus lycioides L. subsp. graeca (Boiss. et Reuter) Tutin 92 Rhipsalis cassytha Gaertn. 457 Rhodiola algida (Ledeb.) Fisch. et С. А. Меу. 412 Rhodiola quadrifida (Pall.) Fisch. et С.А.Меу. 81, 82 Rhodiola pinnatifida Boriss. 422 Rhododendron ledebourii Pojark. 475 Ribes atropurpureum С. А. Меу. 112 Robinia pseudoacacia L. 490 Rosa acicularis Lindl. 781, 782 Roystonea гера (Н. В. К.) О. F. Cook 466 Rubus idaeus L. 456 Rubus sanguinens Friv. 584 Rumex acetosa L. 214 Rumex obtisifolius L. 757 Russelia equisetiformis Schlecht. et Cham. 642, 643 Ruta chalepensis L. 321 Ruta graveolens L. 221 Salix berberifolia Pall. 468 Salix саргеа L. 133 Salvia fruticosa Miller 477 Salvia lanigera Poiret 758 Saponaria officinalis L. 278 Sarcopoterium spinosum (L.) Spach 559 Saussurea alpina (L.) DC. 318 Saussurea frolowii Ledeb. 176 Saussurea kunthiana С. В. Clarke 344 Saussurea latifolia Ledeb. 583 Saussurea schanginiana (Wydl.) Fisch. ex Herd. 312 Saussurea tridactyla Sch. Bip. ex Hook. f. 344 Saussurea thomsonii С. В. Clarke 344 Баыяигеа wernerioides Sch. Bip. ex Hook. f. 344 Saxifraga hieracifolia Waldst. et Kit. 212 Saxifraga hirculus L. 188 Saxifraga melaleuca Fisch. ex Spreng. 291 Scabiosa argentea L. 512 Scabiosa prolifera L. 159 Schisonepeta аппиа (Pall.) Schischk. 397 Schizonepeta multifida (L.) Brig. 279 Scorzonera radiata Fisch. ex Ledeb. 404 Scrophularia altaica Магг. 181 Scrophularia nodosa L. 434 Scutellaria supina L. 458 Senecio erucifolius L. 163 Senecio jacobaea L. 162 Senecio nemorensis L. 435 Senecio vulgaris L. 294 Seseli libanotis (L.) Koch 345 Sibiraea laevigata (L.) Maxim. 200 Silybum marianum (L.) Gaertn. 208 Smelowskia calycina (Steph.) С. А. Меу. 499 Smilax aspera L. 652 Solanum dulcamara L. 460 Solanum incanum L. 488 Solidago dahurica Kitag. 806 Solidago virgaurea L. 134, 135 Sorbus aucuparia L. 72 Spartium junceum L. 645 Spergula arvensis L. 310 Stachys aegyptiaca Регз. 638 Stachys officianalis (L.) Trevis. 547 Stemmacantha carthamoides (Willd.) М. Dittrich 183, 184 Stipagrostis lanata (Forssk.) de Winter 448 Succisa pratensis Moench 424 Swertia obtusa Ledeb. 186 Tamarix nilotica (Ehrenb.) Bunge 627 Tanacetum vulgare L. 284, 347 Тахт baccata L. 91 Tephroseris heterophylla (Fisch.) Konechn. 188
Указатель латинских названий растений Tephroseris praticola (Schischk. et Serg.) Holub 286 Tephroseris turczaninovii (DC.) Но1иЬ 217 Teucrium creticum L. 273 Teucrium polium L. subsp. capitatum (L.) Arcang. 131 Thymelaea hirsuta (L.) Endl. 485 Tipuana tipu (Benth.) Kuntze 74 Тоггеуа colifornica Тогг. 91 Tribulus terrestris L. 347, 348, 353 Tricholaena teneriffae (L. fil) Link 454 Тпр1еиговрегтит perforatum (Merat) М. Lainz 401 Trollius asiaticus L. 215 Tropaeolum majus L. 398 Turritis glabra L. 159 Tussilago farfara L. 648 Urtica urens L. 207 Valeriana capitata Pall. ex Link. 318 Valeriana dubia Bunge 192 Valeriana petrophilla Bunge 346 Valeriana tripteris L. 428 Veratrum lobelianum Bernh. 182 Verbascum депяйогит Bertol. 125 Verbascum fruticulosum Post 619 Verbascum sinuatum L. 166 Veronica densiflora Ledeb. 563 Veronica macrostemon Bunge 439 Veronica pinnata L. 189 Veronica porphyriana Pavl. 317 Vicia sativa L. 400 Чю1а altaica Кег-Gawl. 196 Viola arvensis Магг. 158 Чю1а canina L. 199 Viola dissecta Ledeb. 209 Чю1а disjuncta W. Beck. 287 Чю1а hirta L. 211 Waldheimia tridactylites Kar. et Kir. 470 Zilla spinosa (L.) Prantl 654 Ziziphora clinopodioides Lam. 459
Виды растений, которые упомянуты в тексте второго тома без рисунков Abies sibirica Ledeb. 154, 156, 796 Hypericum mysurense Heyne 710 Achnatherum splendens (Trin.) Nevski 598, 675 Hypericum mutilum L. 711 Aconitum altaicum Steinb. 44 Adansonia gigitata L. 537 Agave americana L. 386 Alyssum turkestanium Redel et Schmalh. 380 Arundo donax L. 385 Bambusa Retz 386 Bidens pilosa L. 522 Calamus manan Mig. 389 Capparis herbaceae Willd. 387 Chamaerops humilis . 389 Carnegia gigantea (Engelm.) Вг. et R. 386 Centaurea procurrens Sprengel 388 Ceratocephala falcatum (L.) Pers. 380 Conyza bonariensis (L.) Cronquist 383, 522 Dentaria bulbifera L. 154 Dracaena draco L. 392 Есин Ьеп8а1епяв 1. 508 Ficus sycomorus L. 65 Foeniculum vulgare Miller 388 Нурепсит alpestre Stev. 711 Hypericum androsaemum L. 711 Hypericum calycinum L. 711 Hypericum canariense L. 710 Hypericum hircinum L. 711 Hypericum hookerianum Wight. et Агп. 711 Hypericum humifusum L. 711 Hypericum inodorum Miller 711 Hypericum linarioides Bosse 711 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Hypericum olympicum L. 711 Hypericum patulum Thunb. ex Murray 710 Hypericum polyphyllum Boiss. et Bal. 711 Juniperus turkestanica Кот. 392 Lilium bulbiferum L. 154 Limonium gmelinii (Willd.) О. Kuntze 724 Limonium platiphyllum Linkz. 724 Malus Hill 65 Malvaviscus arboreus Сач. 597 Ochradenus baccatus РеНе 484 Pandanus veitchii Hort. 524 Phragmites australis (Cav.) Trin. ex Steud. 385 Picea schrenkiana Fisch. et С.А.Меу. 154 Polygala peduncularis Burch. ex DC. 722 Polygonum ччрапип L. 154, 593 Rhizophora mucronata Lam. 524 Rhodiola гозеа L. 791 Ricinus communis L. 389 Salacca wallchiana Mart. 389 Sinapis alba L. 665 Senecio sp. 390 Solanum пщгшп L. 522 Urtica йока L. 675 Valeriana officinalis L. 61, 797 Washingtonia robusta Н. Wendl. 389 Welwitschia mirabilis Hook.390
Условные обозначения к большинству оригинальных рисунков {их номера не обозначены звездочкой), на которых изображены растительные биоморфы Если отмеченные здесь условные обозначения в конкретном рисун- ке не соблюдаются, то факт применения новых обозначений отмечается особым образом — после номера рисунка ставится звездочка, допустим: рис. 4', рис. 6' и т.п. В квадратных скобках указаны номера тома, главы и (или) страниц, где в монографии дано объяснение некоторых из применяемых терминов. Возрастные состояния биоморф [т. 1, гл.7]: АА — виргинильные, А — проростки, Б — всходы, В — ювенильные (В, — униювенильные, В, — биювенильные и т.д.), Г — имматурные (Г, — униимматурные, Г,— биимматурные и т.д.), Д — матурные (физиологически зрелые), Е— инициматурные (молодые матурные), Ж — медиматурные (средневозра- стные матурные), И — финиматурные (старые матурные), С — сениль- ные. Основные типы побегов по сложности их организации [т. 1, гл. 5]: 01 — протерокормус, 1 — монокормус (специальный простой побег), 2 — дикормус (специальный сложный побег), 3 — трикормус (комбини- рованно-специальный побег), 4 — тетракормус (универсальный простой побег), 5 — пентакормус (универсально-сложный побег), 6 — гексакор- мус (комбинированно-универсальный побег), 7 — гептакормус (прото- голокормус), 8 — октокормус (кладокормус), 9 — энатокормус (голокор- мус). Основные типы корнепобегов (ризокормусов)-организмов [т. 1, гл. 5]: 01к — ризопротерокормус, 1к — ризомонокормус, 2к — ризодикор- мус и т.д. до 9к — ризоэнатокормус; кс — семядольный ризокормус, например, 1кс — семядольный ризомонокормус, 2кс — семядольный ризодикормус и т.д. МБ — упрощенные модели биоморфогенезов; Сб — некоторые схематические особенности общей структуры биоморфы; фп — фраг- мент побеговой системы; 1л, 2л, 3л ... — биоморфы 1, 2, 3 и более лет жизни; ик — изоаклонист, или постоянный аклонист (организм, кото- рый вегетативно не размножается); мк — временный аклонист, или мероаклонист (синорганизм в стадии родительского организма до нача- ла его вегетативного размножения); к — клонисты, или особи вегетатив-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ного размножения, — атрибутивные части синорганизма в стадии клона; ск — синорганизм в стадии клона; аг — агамокормус (вегетативный побег): аг1 — длинный, аг2 — средний, аг3 — компактный, аг4- разноразмерный; гм — гамокормус (репродуцирующий побег): гм1- длинный, гм2 — средний, гм3 — компактный, гм4 — разноразмерный. Типы структуры бластов по линейным размерам: дс — долихоморфный, или долихобласты, мс — мезоморфный, или мезобласты, 6c — брахи- морфный, или брахибласты, гб — смешанный, или гетеробласты. Спе- циализация побеговых меросом по вегетативно-репродуктивному ряду признаков [т. 2, гл. 1.5]: а — 6 — вегетатив, а — тотавегетатив, 6— интервегетатив, в — пререпродуктив, г — магирепродуктив; д — е- репродуктив (интервегетатив + пререпродуктив + магирепродуктив), д — тотарепродуктив, е — фрондорепродуктив, ж — вегеторепродуктив (репродуктив + тотавегататив), з — пострепродуктивный вегетатив; и— отмершие части репродуктива, кв — корневой вегетатив; 10 — семядоли, 11 — семядольный узел (указывается его положение точное или приблизительное в зависимости от возраста растений), 12— гипокотиль, 13 — корневая шейка (указывается положение приблизи- тельное), 14 — 29 — некоторое разнообразие побегов (иногда верхние части побегов удалены): 14 — вегетативный, 14 а — статурный (росто- вой), 14 б — магимодифицированно-статурный, 14 в — ассимиляцион- ный, 14 г — магимодифицированно-ассимиляционный, [т. 1, гл. 5. 1. 3., с. 237]; 15 — вегетативно-репродуктивный, 15 а — репродуктивна-асси- миляционный, 15 б — репродуктивна-статурный, 15 в — магимодифи- цированный репродуктивна-статурный; 16 — репродуктивный, 17— тотарепродуктивный, 18 — 19 — розеткообразующий, 18 а — розеточ- ный (или предрозеточный — 18 а), 18 6 — верхнерозеточный (или верхнепредрозеточный — 18 6 ), 19 а — полурозеточный с верхушечной олиственной (с зелеными листьями, фрондозной) безрозеточной час- тью, 19 6 — полурозеточный с пазушными (одним или несколькими) фрондозными безрозеточными побегами, 19 в — розеточно-четочный (или четочно-розеточный — 19 в ), 19 г — верхнеполурозеточный, 19 д — переходно-полурозеточный; 20 — розетконеобразующий, 20 а — че- точный (безрозеточно-сближенноузловой), 20 б — безрозеточный (длин- ный или короткий), или безрозеточно-раздвинутоузловой, 20 в — без- розеточно-компактный, 20 г — получеточный (четочно-раздвинутоузло- вой), 20 д — верхнечеточный; 21 — унирепродуктивный, 22 — би-, муль- тирепродуктивный; 23 — интеркалярный, 24 — постинтеркалярный, 25— силлептический, 26 — годичный, 27 — сверхгодичный (двух-, трех-, многогодичный), 28 — главный, 29 — боковой; 30 — 32 — некоторое разнообразие листьев (иногда верхние части листьев удалены, жилкова- ние часто показано не на всех листьях): 30 — чешуевидные, 30 а— чешуи низовые (развиваются в основании или по всей длине побегов), 30 б — чешуи-брактеи (развиваются в зоне соцветий или на цветоносе и часто быстро осыпаются), 31 — розеточные, 32 — безрозеточные срединные; 33 — листья, охарактеризованные по продолжительности жизни: 33 а — летнезеленые, 33 б — зимнезеленые (зимой зеленые, но к весне отмирающие), 33 в — многолетнезеленые (два и более вегета-
Условные обозначения к рисункам ционных сезона сохраняют жизнеспособность, иначе их называют «веч- нозелеными»); 34 — простые листья, 35 — полусложные листья, 36— сложные листья, 37 — прилистники; 38 — цветок: 38а — пестичный, 38 б — тычиночный, 38 в — обоеполый цветок, 39 — плод; 40 — 43— некоторые типы соцветий [т. 1, гл. 5, с. 221]: 40 — закрытое, или детерминатное, 41 — открытое, или индетерминатное, фрондозное вынужденно верхушечное соцветие, 41 а — межсоцветие (интеркалярное соцветие), 42 — полузакрытое, или семидетерминатное, 42 а — вынуж- денно полузакрытое, 43 — приоткрытое, или преиндетерминатное; 44— иносоцветие (контринфлоресценция) [т. 1, гл. 5, с. 231]; 45 — 49— резиды: 45 — любые, 46 — компактные, 47 — короткие, 48 — средние, 49 — длинные [т. 1, гл. 6 ]; 50 — преконодий (базальная сближенно- узловая часть главного, еще не зимовавшего побега), 51 — 61 — базовые, или персистентные (однократно зимующие или многолетние), побего- вые органы [т. 1, гл. 6]: 51 — аэроподий, 52 — аэроподиоид (в отличие от аэроподия этот надземный персистентный побеговый орган зимует только один раз), 53 — геоподий; 54 — ризом, 54 а — ризом длинный, 54 б — ризом компактный, или короткий, 54 в — ризом гетерономный, 54 г — ризом автономный,54д — аэроризом; 55 — каудекс: 55а- каудекс компактный, 55 б — каудекс вынужденно удлиненный; 56— каудиции, или ветви каудекса (в том числе и вынужденно удлиненные при погребении наносным почва-грунтом), 57 — конодий (57 — пре- каудекс у каудексовых растений, иногда вынужденно удлиненный), 58— зрелая луковица, 59 — зрелая клубнелуковица, 60 — корневищная лу- ковица, 61 — зрелый клубнепобег, 62 — базифиз [т. 2, гл. 1, с. 101], 63— клубнекорнепобег, 63 а — почкокорень, 64 — клубнекорень, или кор- неклубень, 65 — почки какие-либо по положению в пространстве и степени активности: 65 а — спящие, 65 б — развертывающиеся, 65 в— придаточные; 66 — 70 — основные типы нарастания персистентных побегов [т. 1, гл. 4]: 66 — эумоноподиальное, 67 — гемисимподиальное, 68 — эусимподиальное, 69 — полисимподиальное, 70 — эфемерно- полисимподиальное; 71 — корневая система, 72 — главный корень, 73— боковой корень, 74 — придаточный корень (или корни), 75 — корневой отпрыск, 76 — граница годичного побега, 77 — побег (какой-либо), отрастающий от подземных или приземных частей базовых органов, 78 — отмершая базальная часть растения, 78 а — отмирающая базальная часть растения, 79 — полностью отмершая особь, 80 — отмерший побег или отмершая верхняя часть побега, 81 — зона надземного постлатен- тного (послезимнего) побегоотрастания, 82 — зона кущения, 83 — вы- нужденно-удлиненные горизонтальные каудиции, иногда образующиеся в условиях крутых эродированных горных склонов, 84 — шипы, 85— колючки побегового происхождения, 86 — колючки листового проис- хождения (образуются из рахисов), 87 — колючки прилистникового происхождения, 88 — метатравматическая пневая поросль, 89 — ризо- базифиз, 90 — викарирующий базовый орган [т. 1, гл. 6, с. 316]; 91-95 [Troll, 1964, т. 1, гл. 5.1.2.], 91 — сборное соцветие, или синфлоресцен- ция, 92 — главное соцветие, или примафлоресценция, 93 — присоцве- тие, или кофлоресценция, 94 — парциальное соцветие, или партифло-
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ресценция, 95 — паракладий, 96 — тавтокладий, 97 — сверхсоцветие, 98 — посткаудекс, 99 — посткаудиций, 100 — криптоклональный синор- ганизм (синорганизм в состоянии скрытого клона). Объяснение нового термина. Посткаудекс или его сравнительно тонкий аналог — посткаудиций, это любое персистентное побеговое структурное образование, которое получается из каудекса, его частей, в процессе разрушения этого органа по каким-либо причинам, чаще всего при сильной партикуляции старых каудексовых растений, разделяющей бывший каудекс ниже семядольного узла на более или менее обособлен- ные части. Остальные обозначения: пп — поверхность почвы, у — увеличено в несколько раз по сравнению с масштабом, ум — уменьшено... Одно деление масштаба, если не указано иначе, — 1 см. Дополнительные условные обозначения при необходимости приводятся под конкретным рисунком. В качестве вспомогательной информации иногда используются гра- фические рисунки (Др), выполненные другими исследователями, кото- рые, будучи введены в авторский рисунок, выделяются в нем пунктир- ной рамкой и соответствующим образом цитируются в подрисуночном тексте.
Содержание Концепция меросомной организации биоморф семенных р астений ............................................................................... 16 1.1. Телесное и нетелесное в Природе .............................17 1.2. Метод меротомирования и основы меротомии — науки о частях целого ......................... 20 1.3. Многоканальность распределения ресурсных потоков в организации телесных систем ..................23 1.3.1. Коснувшись цветка, ты невольно потревожишь звезду (старинный афоризм) ............... 24 1.3.2. Эгосубстрат и конституция тел, геносома и феносома биосистем ................................ 30 1.3.3. Многослойность явления «жизнь» .............................. 40 1.3.4. Супернуклеус и космоплазма......................................44 1.3.5. Идиоэнергия в круговороте ресурсов LJ и тотальныи движитель ................................................49 1.4. Относительность в свободе выбора цели развития телесных систем при ограниченности ресурса возможностей ...................... 56 1.5. Целесообразность распределения ресурсных потоков в организации биоморф ................................ 57 1.5.1. Меросомность организации биоморф семенных растений — атрибут целесообразности распределения ресурсов .............. 58 1.5.2. Мультивариантность развития биоморф в круговороте ресурсов ...............................................95 Глава 1. 2.4. Глава 2. Концепция приоритетов развития в организации систем и, в частности, биоморф......................................... 115 2.1. Приоритетность — атрибутивное свойство систем ...115 2.2. Высший приоритет в организации живой природы и принцип миниатюризации видов .........137 2.3. Борьба за приоритеты развития на арене противоречий между своим и не-своим в организации биоморф .............................................141 Иерархия приоритетов развития биоморф семенных растений .................................... 202 2.5. Стремление к размножению, экспрессивный альтруизм палеобиоморф, торжество
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ компромиссов в эволюции растительных биоморф и ее главные приоритеты..........................228 Экспрессия альтруизма в колыбели жизни на Земле и стремление к компромиссам своего и не-своего в растительном мире ................ 230 Генеральный приоритет в направлениях эволюции жизни на Земле и смешение приоритетов в эволюции биоморф ° .У семенных растении ..................................................... 234 Научные приоритеты, приоритеты авторитетов и влияние их на приоритеты развития науки и, в частности, биоморфологии ................................ 242 2.5.1. 2.5.2. 2.6. Глава 3. Концепция габитуса в биоморфологии ................................251 Габитус биоморф в «пирамиде» абстракций телесных систем .....................................251 Внешнее и внутреннее в организации биоморф....254 Габитус биоморф, причина и следствия .................256 Формы габитуса и габитопроцесс ............................259 Габитус в организации биоморф семенных растений — парадоксы, оказавшиеся атрибутами ... 262 3.1. 3.2. 3.3. 3.4. 3.5. Глава 4. Концепция форм роста в биоморфологии ...........................297 Морфогенез и поведение — атрибуты существования всех телесных систем....................... 297 4.1. 4.1.1. Как и почему поведение составляет атрибутивную часть в организации биоморф ......... 301 4.1.2. Синдром возрастности в стадийном п оведении биоморф .................................................... 304 Биографика — сердцевина современной 6 иоморфологии ............................................................ 306 Стадийность морфогенезов и формы роста в организации биоморф ................................................ 309 Стадийность и иерархия форм роста биоморф...... 327 Парадоксы форм роста, оказавшиеся атрибутами организации биоморф семенных растений ............. 331 Призрак биоморфологии уже 6 родит в систематике ................................................. 335 Биографическое тестирование биоморф семенных растений при разных уровнях их познания.............................337 Движение («текучесть») материи и ее локализация в организации биоморф как меросом-габитусов и как меросом-форм роста......353 4.2. 4.3. 4.4. 4.5. 4.6. 4.7. 4.8. Глава 5. Концепция разнообразия биоморф семенных растений, место в ней их габитусов и форм роста ............................ЗбО 5.1. Признаковые, или информационные (довостребованные и востребованные), пространства биоморф ............................................... 360
Содержание Габитуальное разнообразие биоморф семенных растений ..................................................... 366 Единообразие в организации биоморф — первое условие к проявлению их разнообразия..................367 Средства распознания разнообразия биоморф .......368 Маркирующее участие габитуальных частей- признаков в интегральных признаках, отражающих сложность организации биоморф ......372 Просубординатное разнообразие биоморф семенных растений .................................... 377 Просубординатное разнообразие как объективная реальность и особый результат познания ............... 377 Габитус и формы роста в просубординатном разнообразии биоморф семенных растений............380 5.2. 5.2.1. 5.2.2. 5.2.3. 5.3. 5.3.1. 5.3.2. Концепция пластичности и гиперпластипюсти иоморф семенных растений...............................................491 6.1. Стратегическое и тактическое в организации биоморф .............................................491 6.2. Пластичность и гиперпластичность ......................... 496 6.3. Гомеостаз и гетерокинез ............................................ 500 6.4. Адаптированность габитуса и форм роста биоморф к пространству и времени .............504 6.4.1. «Парадокс адаптированности» габитуса в организации биоморф ............................................. 506 6.4.2. Биотопизм и пластогенез в освоении биоморфами пространства и времени ..................... 508 6.5. Влияние количества и качества пространства и границ между ними на соматическое разнообразие биоморф ............................................... 513 6.5.1. Растительные биоморфы как части неоднородного пространства .....................................513 6.5.2. Сопряженность эволюции габитуса растительных биоморф и их продолжительности жизни с поверхностью почва-грунта .......................................518 6.5.3. Габитусы и универсальная модель организации биоморф семенных растений ............. 525 Глава 6. Глава 7. Концепция разнообразия биотической власти и управляемого ею, посредством «могучей пятерки», единства живого и неживого в организации биоморф с еменных растений ............................................................. 534 7.1. Что обеспечивает телесную организованность явления «жизнь»..........................535 7.1.1. От соединения и восхождения («подъема») к разъединению и нисхождению («спуску») ...........536 7.1.2. Размножение как начало начал — способ преодоления жизнью дискретности биоморф ......... 544 7.1.3. Парадокс и феномен смерти в явлении «жизнь» ..545
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Ценомерия — атрибут организации Природы, ценобиоз и меробиоз — атрибуты жизни на Земле..................................... 550 7.2.1. Основные состояния материи, вовлеченной в жизнь на Земле................................. 550 7.2.2. Смерть — объективная реальность, но не приоритет в явлении меробиоза.................... 557 7.3. Гетерократия — очередной атрибут жизни ............. 564 7.3.1. «Власть» в организации биоморф И ее разнообразие ....................................................... 564 7.3.2. Иерархия биотических властей ................................. 570 7.4. «Могучая пятерка» покорителей пространства в организации биоморф семенных растений .......... 575 7.4.1. Диктат эйдократии в организации б иоморф и эйдофория................................................576 7.4.2. От нарастания к ветвлению, далее партикуляции и размножению, наконец, к элиминации ............... 580 7.4.3. Смерть — результат проявления количества и качества жизни несовместимого с ее продолжением ..................................................... 593 7.4.3.1. Смерть — равноправный член < могучей пятерки» ..................................................... 595 7.4.3.2. Разнообразие смертей-адаптаций в организации биоморф семенных растений........596 Глава 8. Концепция всюдности эволюции и ее отражение в габитусах и формах роста биоморф семенных растений.603 8.1. От движения вообще к трансформации вообще, эволюции вообще и эволюции живого в особенности ................................................ 603 8.1.1. Почему эволюция — высшая форма трансформации и почему ее никак и нигде нельзя потерять ............................................608 8.1.2. Эволюция — главное действие в формировании разнообразия Природы ................ 609 8.2. Универсальный принцип эволюции в Природе .....611 8.2.1. Почему всякие эволюционные процессы — это движение эволюционных маятников ............ 612 8.2.2. Меротомия эволюции вселенского потока (Природы) как системы ее состояний эволюстатов) ............................................................... 621 8.3. На пути к теории трансформации ...........................629 8.3.1. Перспективы развития учения И.В. Гете о метаморфозах .........................................629 8.3.1.1. Метаморфоз — процесс, метаморфа — его результат ..................................... 630 8.3.1.2. Метаморфоз — способ реализации гомологического разнообразия
Содержание в фенетической организации биоморф .................. 634 8.3.2. Что еще как элемент метаморфозов проявляется в биоморфогенезах семенных растений ................... 665 8.4. Закон альтернации в эволюции признаков ............667 8.5. Иерархичность транформаций в живых системах и место в ней эволюционных явлений ...................669 Борьба за существование организации биоморф семенных растений и место в ней их габитусов И форм роста ............................................................... 673 8.6.1. Многое в эволюционном учении еще впереди ......673 8.6.1.1. Живое начинается с инфраструктурной борьбы ..674 8.6.1.2. Биоморфы и их структурные элементы— «пациенты» всюдной эволюции .............................. 676 8.6.1.3. Закон адаптивной ориентации в организации живой природы ...............................679 8.6.1.4. Почему в эволюции организмов важны не только «победители», но и «побежденные» в инфраструктурной борьбе ..................................... 682 8.6.1.4.1. Почему интраэлекция стала в эволюции атрибутом естественного отбора ........................... 682 8.6.1.4.2. Подтверждающая и утверждающая формы естественного отбора или, соответственно, — первая и вторая экстраэлекции .......................687 8.6.2. Эволюция биоморф в их конституциальной борьбе за существование ............................................ 690 8.6.2.1. Передовая линия «фронта» в конституциальной борьбе за существование ......... 692 8.6.2.2. Как и почему приоритет сохранения другой и более сложной жизни стал нормой существования семенных фитобиоморф, а шире — всех биосистем ............. 693 8.6.2.3. Почему альтруизм не стал нормой жизни семенных фитобиоморф и почему они выбрали в эволюции ориентацию на компромиссы ...........696 8.7. Как, почему и куда идут реальная и гипотетическая эволюции биоморф семенных растений, их видов и надвидов .............. 698 8.6. 9.3. Глава 9. Концепция экогенобиоза — теоретическая основа эволюционных принципов происхождения и утверждения жизни на Земле ..........................................729 9.1. Основные противоречия в организации современной растительной жизни ............................731 9.2. «Мал золотник, да дорог», или торжество биотической власти при разрешении противоречий в организации живых телесных систем .......................................................... 733 Жизнь как способ существования генов для фенов и фенов для генов
852 ОСНОВБ1 БИОМОРФОЛОГИИ CENEHHhlX РА СТЕНИЙ в их союзах — «генофенах» ....................................... 736 9.4. Эволюция союзного образа жизни генов и фенов в организации биоморф ............................. 738 9.4.1. Почему жизнь — это, в первую очередь, строительство и обслуживание обиталищ для генов .................................................... 739 9.4.2. Что нужно, чтобы обиталища для генов не превратились для них в безутешные «кладбища» ..741 9.4.3. Нежизнь — преджизнь — жизнь — вехи в эволюции экогенобиоза .......................................... 741 9.4.3.1. Мотивация неизбежности возникновения жизни на Земле .............................742 9.4.3.2. Почему не экспрессивный альтруизм, а компромиссы стали нормой экогенобиоза семенных фитобиоморф ........................................... 748 9.5. Формы габитуса и стадийного поведения — индикаторы жизнеразвития семенных фитобиоморф как экогенобиозного явления ..........751 Глава 10. Откровения о буднях и праздниках растительной жизни, их резонанс в концепции биотического счастья и качества жизни (на примере семенных фитобиоморф) .......761 10.1. Жизнь — борьба, но и постоянная выставка достигнутого ею разнообразия .................................. 762 10.1.1. Жизнь — это будни, праздники и переходы между ними ............................................763 10.1.2. Биотическая организация — единство и борьба качественно противоречивых форм соматического развития: для себя (своего, личного) и для друтих (друтого родного и друтого чужого) ......................... 766 10.1.3. Парадокс своего (личного) и не-своего (друтого) — один из атрибутов мироздания и, в частности, живой природы ....................................769 10.1.4. Растительные биоморфы — результат поиска и выбора оптимальных целей жизнеразвития посредством самонаводящейся и квантированной идиоэнергии ................................ 771 10.2. Главные таинства растительного счастья ................ 772 10.2.1.Модусы организации современных семенных фитобиоморф как результат эволюции .................... 774 10.2.2. Как и почему биоморфы семенных растений бывают или не бывают счастливыми.......................776 10.2.3. Иерархичность биотического счастья ......................784 10.2.4. Мера счастья и формула качества LJ растительнои жизни .................................................... 790 10.3. Габитоформы и формы роста (стадиосомы) — индикаторы биотического счастья и качества жизни (в целом) семенных фитобиоморф .............. 797
853 Содержание 10.3.1. Почему в разном качестве жизни растительных биоморф биотического счастья всегда больше, Ч ем несчастья ............................................................... 798 10.3.2. Эволюция жизни по пути компромиссов биотических счастья с несчастьем в организации семенных фитобиоморф ..................799 10.3.3. Как и почему семенные фитобиоморфы своим габитуальным и стадийным поведением выражают содержание всепоглощающей борьбы за биотическое счастье ................................. 804 включение ......................................................................................... 811 Указатель латинских названий растений ............................................835 Условные обозначения к рисункам ..................................................... 843
Contents ........................................................................................................ 3 Foreword I 4~ ntroduction е ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° е 7 1.3. 1.5. 2.4. 2.5. 3.2. 3.3. 3.4. 3.5. Chapter 1. Conception of merosomal organization 1 ° О й f biomorphs of the seed plants....................................................... 16 1.1. Somatic and non-somatic in Nature ...........°............°...°.......................17 1.2. Merotomising method and the outlines of merotomy, the science about the parts of the whole .............................................20 Multichannel distribution of the resource flows in organization of the somatic system .................................................... 23 1.3.1. Touching the flower, you will involuntarily disturb the star (ancient aphorism) ......................................................... 24 1.3.2. Egosubstrate and constitution of the bodies, genosome, and phenosome of biosystems ..................................°........... 30 1.3.3. Multilevel construction of the «life» phenomenon.°..............................40 1 .3.4. Supernucleus and cosmoplasm ................................................................44 1.3.5. Idioenergy in the turnover of resources and the total driver................49 1.4. Relativity of free choice of the aim of development of somatic systems under the limited possibility resources ...................56 Expedience of distribution of the resource flows 1 n biomorph organization е ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 57 Chapter 2. Conception of development priorities in organization of the systems and biomorphs in particular.................................... 115 2.1. Priority concept as an attributive characteristic of the system ........... 115 2.2. The highest priority in organization of the living nature and the principle of species miniaturization ........................... 137 2.3. Struggle for developmental priorities on the cockpit of contradictions between own and strange in the biomorph organisation..................... 141 Hierarchy of priorities in development of the seed plants.................. 202 Tendency to reproduction, expressive altruism of paleobiomorphs, triumph of compromises in the evolution of plant biomorphs and its principle priorities .................................... 228 2.5.1. Expression of altruism in the cradle of life on Earth and intention to compromises between own and strange in the plant world ........... 230 2.5 ° 2. Basic priority in the trends of evolution on Earth and mixture of priorities in the evolution of the biomorphs of the seed plants .... 234 2.6. Scientific priorities and priorities of authorities and their influence on the priorities of the science, biomorphology in particular, development ........................°.................. 242 Chapter 3. Conception of the habit in biomorphology...................................... 251 3.1. Habit of biomorphs in the «piramid» of the abstraction о f the somatic systems ......................°................................................... 251 Outer and inner in the biomorph organization .............................. 254 Habit of biomorphs; cause and effects ................................................. 256 Habit forms and habit process ......°.... ° °....................°............................ 259 Habit in organization of the biomorphs of the seed plants: paradoxes that turned attributes ............................................................ 262
Содержание 855 Chapter 4 4.1. 4.1.1. 4.1.2. 4.2. 4.3. 4.4. 4.5. 4.6. 4.7. 4.8. Chapter 5 5.1. 5.2. 5.2.1. 5.2.2. 5.2.3. 5.3. 5.3.1. 5.3.2. Chapter 6 6.1. 6.2. 6.3. 6.4. 6.4.1. 6.4.2. 6.5. 6.5.1. 6.5.2. 6.5.3. . Conception of the growth forms in biomorphology ........................... 297 Morphogenesis and behaviour: attributes of existence о f all somatic systems............................................................................ 297 How and why does behaviour comprise an attributive part in the biomorph organization ................................ 301 Syndrome of age in the phasic behaviour of biomorphs..................... 304 Biographies as а core of contemporary biomorphology ...........°.......... 306 Stadiality of morphogeneses and the growth form 1 n biomorph organization ...................................................................... 309 Stadiality and hierarchy of the growth forms of biomorphs ............. ° . 327 Рагадохез of the growth forms that appeared to be attributes of organization of the biomorphs of the seed plants .......................... 331 The ghost of biomorphology already wanders in systematics.............. 335 Biographic testing of biomorphs of the seed plants on the various levels of their cognition ..............°................................ 337 Lability («fluidity») of the matter and its localization in the organization of biomorphs as merosomes-habits and тегозотез-forms of growth .......................................................... 353 Conception of diversity of biomorphs of the seed plants; the place of habits and growth forms in it..................................... 360 Characteristic, or information (to be called for and already called for) spaces of biomorphs ........... 360 Habitual diversity of biomor[hs of the seed plants.............................. 366 Uniformity in biomorph organization: the first requirement for manifestation of their diversity ..... ° .°............ 367 Means of identification of the biomorph diversity .................°............ 368 Marking role of habitual parts-features in the integral characteristics reflecting complexity of biomorph organization °.......... 372 Post-subordinate diversity of biomorphs of the seed plants ................ 377 Post-subordinate diversity as an objective reality and specific result of cognition............................................................. 377 Habit and growth forms in post-subordinate diversity of biomorphs of the seed plants ......................................................... 380 Conception of plasticity and hyperplasticity of the biomorphs of the seed plants............................................... 491 Strategic and tactical in biomorph organization ..................................491 Plasticity and hyperplasticity .........................................................°....... 496 Homeostasis and heterokinesis °..................°...................................°......500 Adaptability of the habitus and biomorph growth forms 1 о space and time ..................................................................................504 ~Paradox of adaptability» of the habit in organization of biomorphs ................................................................ 506 Biotopism and genesis of plasticity in space and time exploration by biomorphs ..........°.................................°...... 508 Effect of space quantity and quality and their borderlines on the somatic diversity of biomorphs ..............................°.................. 513 Plant biomorphs as the parts of heterogeneous space ......................... 513 Contingency of evolution of the habit of plant biomorphs and their longevity with the ground surface ........................................ 518 Habits and universal model of organization of biomorphs о f the seed plants ..........................°.......................°............................... 525
856 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ 7.1.3. 7.2. 7.3. 7.3.1. 7.3.2. 7.4. 7.4.3.1 7.4.3.2 8.3. 8.3.1. 8.4. 8.5. Chapter 7. Conception of diversity of biotic power and the unity of animate and inanimate, managed by this power by means of the «mighty йте», in organization of biomorphs of the seed plants ....... 534 7.1. What ensures somatic organization of the ~life» phenomenon ........... 535 7.1.1. From conjunction and ascent («uprise») to disjunction and descent («come-down») ..........................................536 7.1.2. Reproduction as fundamental origin, the way of overcoming biomorph d iscreteness by the life .......................................................................... 544 Paradox and phenomenon of death in the "life" phenomenon .........545 Cenomery is an attribute of Nature organization; cenobiosis and тегоЬюяз are the attributes of the life on Earth ............................... 550 7.2.1. Basic states of the matter involved in the Ие on Earth .................... 550 7.2.2. The death is an objective reality, but not the priority in the merobiosis P henomenon ........................................................................................... 557 Heterocraty is one тоге attribute of the life....................................... 564 ~Power» in the biomorph organization and its diversity .....................564 Н 1 Г1 ierarchy o~ biotic powers .................................................................... 570 ~Mighty five» of masters of the space in organization of the biomorphs о f the seed plants .......................................................................°..........575 7.4.1. Dictate of еЫосгасу in biomorph organization and eidophoria .........576 7.4.2. From growth to branching, then particulation and reproduction, and el limination at last ....................°............................................................. 580 7.4.3. The death is the result of manifestation of quantity and quality of the life inconsistent with its continuation .................................................. 593 . The death is an equal member of the ~mighty five» ........................595 . Diversity of deaths-adaptations in organization of the biomorphs of 1 he seed plants .....................................................................................596 Chapter 8. Conception of ubiquity of evolution and its reflection in the habits ап4 growth forms of the biomorphs of the seed plants.......................... 603 8.1. From movement in general to transformation in general, evolution in general and evolution of living objects, in particular ..........................603 8.1.1. Why is evolution the highest form of transformation and why can it not be lost anyhow and anywhere? ......................................................608 8.1.2. Evolution is the main act in formation of diversity of the Nature....609 8.2. Universal principle of evolution in the Nature....................................611 8.2.1. Why do any evolutionary processes represent the movements of 11Фи~naðò тъеndulum9 evolutionary pendulum.......................................................................... 612 8.2.2. Merotomy of evolution of the universal flow (the Nature) as а system of its states (evolustates) ....................................................................... 621 On the way to the theory of transformation........................................629 Prospects for development of Е W. Goete's theory of m etamorphoses ....................................................................................... 629 8.3.1.1. Metamorphosis is the process and the metamorph is its result .......630 8.3.1.2. Metamorphosis is the means of realization of homologous diversity in the phenetic organization of biomorphs.............................................634 8.3.2. What else is manifested in the biomorphogeneses of the seed plants as an element of metamorphoses? .............................................................665 The law of alternation in the evolution of characteristics ..................667 Hierarchical structure of transformations in the animate systems and the place of evolutionary phenomena in it ..........................................669 8.6. The struggle for existence in organization of the biomorphs of the seed plants and the place of their habits and growth forms in it ..............673 8.6.1. А lot in the evolutionary process is still ahead of us .............°...........673
Содержание 8.6.1.1. The living matter origins from the infrastructural struggle ............... 674 8.6.1.2. Biomorphs and their structural elements are the «patients» of the цо о1цш1ои evolution............................................................................. 676 8.6.1.3. The law of adaptive orientation in organization of the living п ature ................................................................................................... 679 8.6.1.4. Why are not only «winners», but also «losers» in infrastructural struggle are important in the evolution of the organisms? ...............682 8.6.1.4.1.Why did intraelection become the attribute of natural selection in the evolution? ..........................°...................682 8.6.1.4. Confirmative and affirmative forms of natural selection or, respectively, the first and the second extraelections.......................... 687 8.6.2. Evolution of biomorphs in their constitutional S truggle for existence ............................................................................. 690 8.6.2.1. The «forefront» in constitutional struggle for existence ....................692 8.6.2.2. How and why did the priority for preservation of the other and тоге complex life become the standard of existence of the seed phytobiomorphs and, to wide extent, of а11 biosystems? ..................693 8.6.2.3. Why did altruism not become the life standard of the seed biomorphs and why did they choose orientation to compromises in the evolution? ......................................................696 8.7. How, why and where do the actual and hypothetic evolutions of the biomorphs of the seed plants, their species and superspecies proceed? ...............................................698 Chapter 9. Conception of ecogenobiosis: theoretical basis of evolutionary principles of the origin and establishment of the life on Earth ....... 729 9.1. Principle contradictions in organization о f the conteporary plant life ................................................................. 731 9.2. «Little pigeons can carry great messages», or the triumph of biotic power in solving contradictions in organization of the animate somatic systems ............................................................ 733 9.3. The life as а means of existence of the genes for the phenes and the phenes for the genes in their units, or «genophenes» ........... 736 9.4. Evolution of the united тоде of life of the genes and phenes in organization of biomorphs ............................................738 9.4.1. Why does the Ие serve for construction and maintenance of the gene habitation in the first place? .............................................739 9.4.2. What is needed to prevent turning of the gene habitations into their comfortless «cemeteries»? ..................................................... 741 9.4.3. Non-Ие — рге-life — life: the milestones in the evolution of ecogenobiosis ......................................................... 741 9.4.3.1. Motivation of the imminent genesis of life on Earth ....................... 742 9.4.3.2. Why did compromises rather than expressive altruism become the norm of ecogenobiosis of the seed phytobiomorphs? ................. 748 9.5. The forms of habit and phasic behaviour: indicators of life development of the seed phytobiomorphs as ecogenobiotic phenomenon ...................................751 Chapter 10. Revelations about workdays and holidays in the plant life, their resonance in the concept of biological happiness and the quality of life (with the seed phytobiomorphs as an example) .................... 761 10.1. The Ие is the struggle, but also а constant display я а 1 ' Л ° of diversity achieved by it ...................................................................... 762 10.1.1. The life is the workdays, holidays, and transitions between them .....763
858 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ С овс1ияоп ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 811 R eferences ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 822 IndeX 01~~ Latin nameS 01~~ the PlantS е ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° F figure legends е ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 835 843 С ontents ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° 847 10.1.2.Biotic organization is the unity and struggle of contradictory forms of somatic development: for oneself (own, personal) and for the others (the other relative and the other strange) ............ 766 10.1.3.Paradox of the own (personal) and strange (other) is one of the attributes of creation and the living Nature, in particular ......769 10.1.4.Plant biomorphs: the result of the search for and the choice of the optimal aims of Ие development by means of homing and quantum idioenergy...................................................... 771 10.2. Basic mysteries of plant happiness .......................................................772 10.2.1.Moduses of organization of contemporary seed phytobiomorphs as а result of evolution......................................774 10.2.2.Why and how are the biomorphs of the seed plants happy or unhappy? .................................................776 10.2.3.Hierarchic structure of the biotic happiness ........................................784 10.2.4. The measure of happiness and the formula of the quality of plant life ........................................790 10.3. Habit morphs and growth forms (stage somes): indicators of biotic happiness and the quality of life (as а whole) of the seed phytobiomorphs ............................................ 797 10.3.1.Why is there тоге biotic happiness than unhappines in the different quality of life of the plant biomorphs.......................798 10.3.2.Evolution of Ие along the way of compromisses between biotic happiness and unhappiness in organization of the seed phytobiomorphs ........................................799 10.3.3.How and why 4о the seed phytobiomorphs express in their habitual and phasic behaviour the content of overwhelming struggle for biotic happiness? .................................... 804
Vukhimovsky ЕЛ . Fundamentals of biomorphology of seed plants. V. E. Habit and growth forms ш organization of biomorphs. — Moscow: «Overlay», 2002. — 859 р., 438 ill. The theory of organization of biomorphs of the seed plants is further substantiated and developed. Ten following conceptions are formulated: merotomising (division of а whole into parts) and merosome structure (membership of the parts in а whole body), priority, habit, forms of growth, subordinate and pro-subordinate Жчегвйу of biomorphs, plasticity and hyperplasticity, biotic power and «mighty five», ubiquity of evolution, ecogenobiosis, biotic happiness, and quality of life. In the author' s opinion, the mystery of habit and growth form по longer exists in biomorphology. The status of these structures is determined. They can be placed only on two (and по тоге) «shelves» in the complex structure of organization of particular somatic systems, biomorphs (organisms and synorganisms) of the seed plants among them. The саве is, that the research intrigue was not only to develop the problem of the habit and growth form of biomorphs, but also to penetrate even тоге deeper into it by the means of logic. The procedures of superordinate classification (abstract «ругатЫ construction») in the multilevel organization of the living nature were applied. Such «shake-ир» under the «logic microscope» are prescribed for the Nature. The seed phytobiomorphs participate in evolution by means of various forms of natural selection and adopt it both inside and outside. One тоге important thesis presented in the book is the idea that all biomorphs, each in its own way, can obtain biotic happiness. This occurs, in the first place, when they are merely living and, of course, when they are blooming and bearing fruits, despite the consequences of these processes. Phytobiographic illustration considerably enriched the factual basis of all the ten conceptions considered in the book, though it is oriented to the greater effect. Its mission in the world of botany is to announce not only profound change at the present (integral) stage of development of biomorphology, but also the conceptual shifts, which are inevitable in the adjacent fields of biology in the recent century. Маге than 400 original figures, documentary evidences, phytobiographic portraits of the species prove the thesis that the ghost of biomorphology is already wandering in sytematics. The forthcoming «Floras» of Russia and other regions of the world will inevitably be biographical. This is the dictate of time burdened with the movement of the science toward progress. The book is designed to biomorphologists, botanists of various interests, as well as biologists interested in the general problems of life sciences. Bibliography: 13 pages.
НАУЧНОЕ ИЗДАНИЕ Нухимовский Ефим Львович ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ: Т. 2. Гибискус и формы 1юста а организации биоморф Зав. редакцией Е.А. Поталенков Переплет художника Н.П. Новиковой Технический редактор Н.А. Галкина Компьютерная верстка В.В. Рюмина, А.В. Булатова Изд. лиц. ИД М 01249 от 20.03.2000. Подписано в печать с репродуцированного ориги- нал-макета Формат 70x100'/,~. Гарнитура Newton. Печать офсетная. Усл.печ.л. 65,3 Уч.-изд.л. 69,6 Тираж 750 экз. Заказ М 6720 000 «Издательство «Оверлей» Московская обл., г. Красногорск, ул. Пионерская, 1; 107014, Москва, а/я 451 Отпечатано в ППП «Типография «Наука» 121099, Москва, Шубинский пер., 6