







































































































































































































































































































































































































































































































































































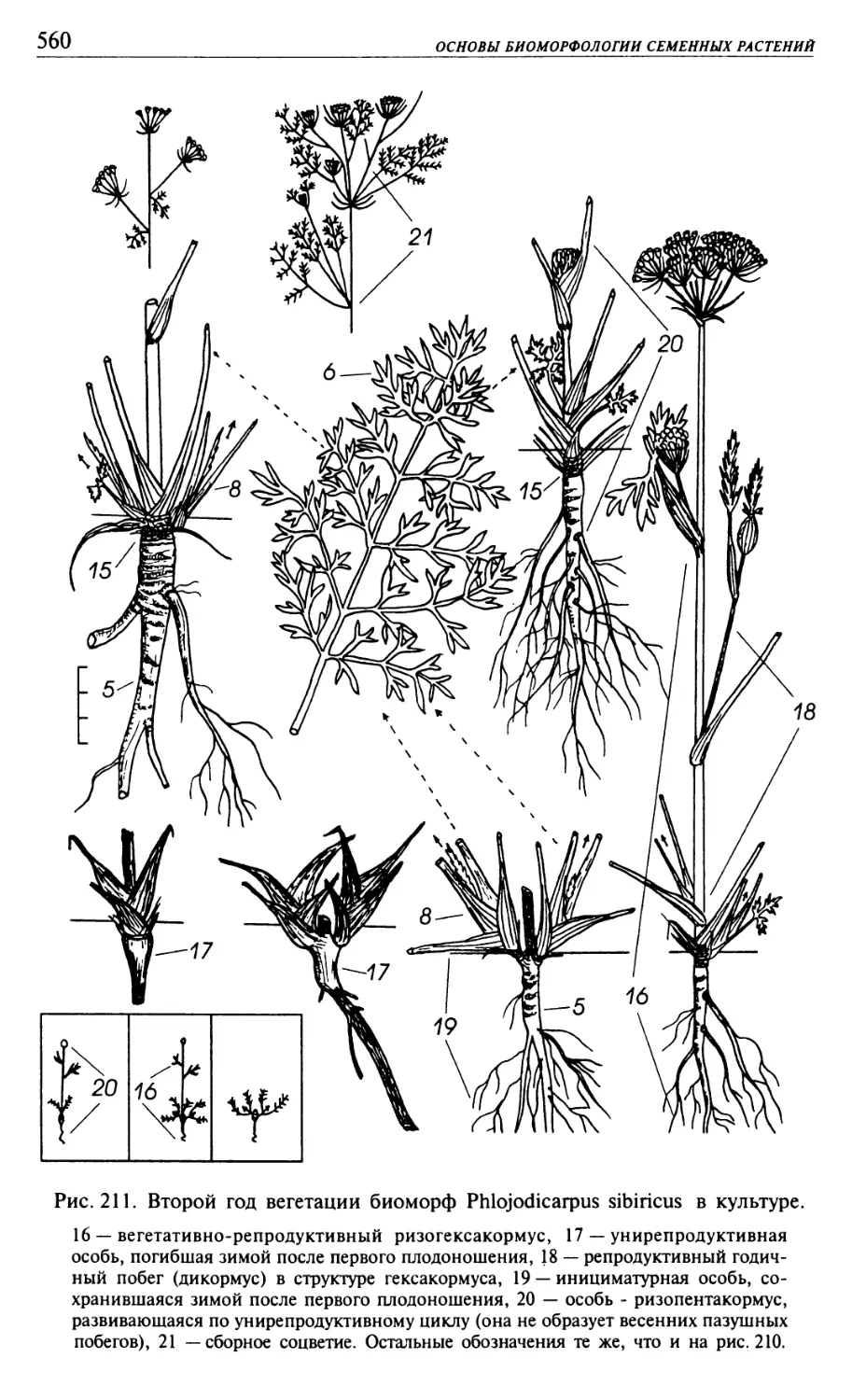



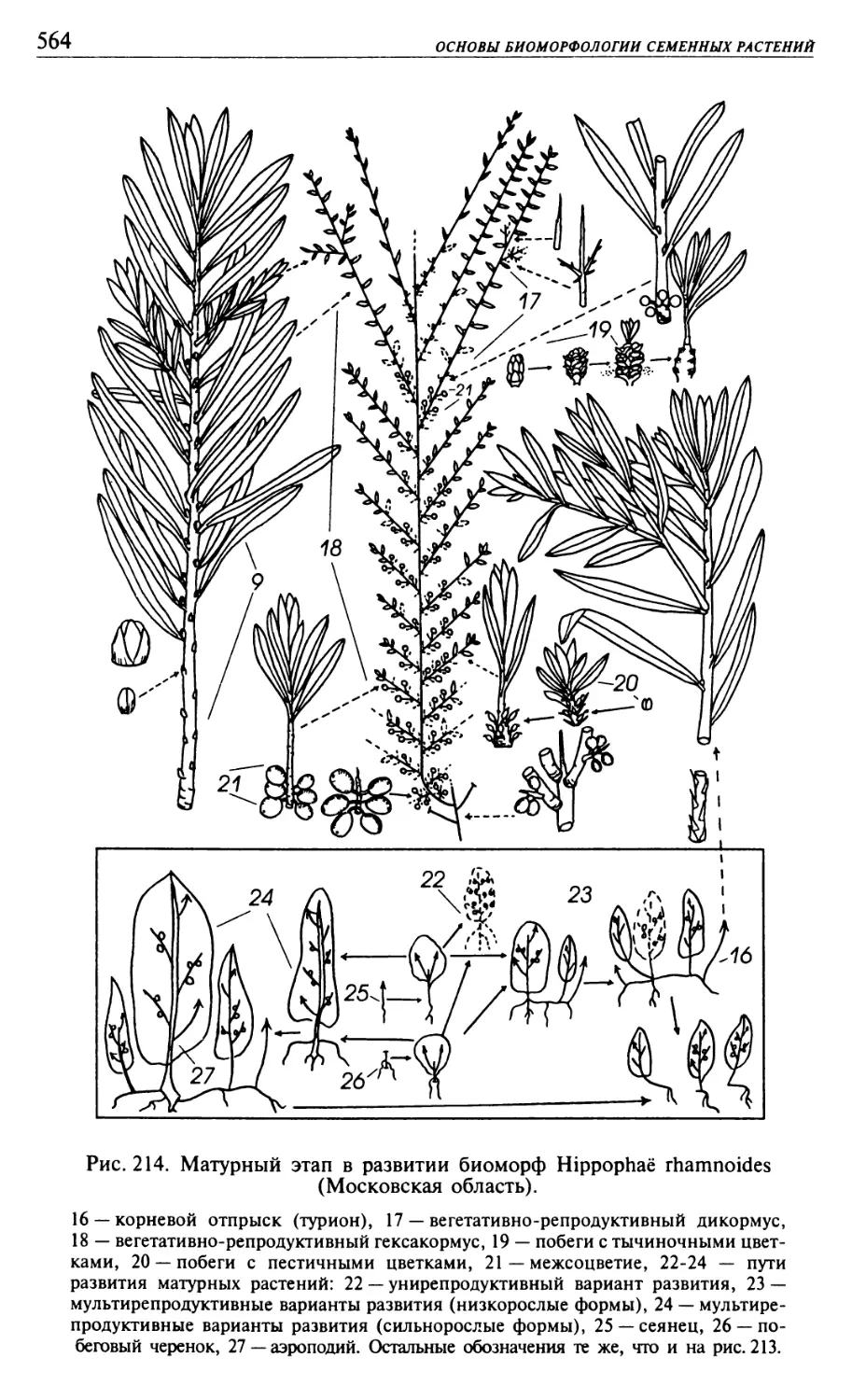







































































Текст
1 . ~ ! Ъ ° ° 6 с ~~ % ф ° а Ю ф
ХИМОВСКИЙ О ИЯ О ГЯНИЗЯ ИИ ИОМО Москва QAQ "Издательство" Недра" 1 997
УДК 581.4 Нухымовский ЕЛ. Основы биоморфологии семенных растений: Т. 1. Теория органи- зации биоморф.— М.: Недра, 1997. — 630 с., ил. - ISBN 5-247-03772-3. Табл. 8. Илл. 218. Библ. 37 стр. ISBN 5-247-03772-3 � Е.Л.Нухимовский, 1997 � Оформление. ОАО ~Издательство ~Недра», 1997 Рассматриваются проблемы организации биоморф (организмов и синорганизмов) се- менных растений, история биоморфологии и статус ее в системе наук. Фундамент стержне- вой теории биоморфологии обеспечила общая теория жизнесохранения, впервые разрабо- танная автором. Главное содержание теории организации биоморф семенных растений составили семь концепций: понятия жизненная форма (биоморфа), побега, корня, корне- побега (ризокормуса), морфогении, инфраструктурно-конституциапьной борьбы за суще- ствование, пластичности и разнонаправленной соматической эволюции. Особое внима- ние уделено методам субординатного, мультисубординатного и суперординатного класси- фицирования при описании организации как основных органов — побегов, корней и их элементов, так и высших структурных единиц рассматриваемых биоморф — ризокормусов. Разработана концепция пластичности биоморф и обоснованы некоторые новые эво- люционные положения. Определен закон эволюционной перспективы вида. Типы биоморф размещены в многолинейной классификации (выделено 500 биоморфотипов), что откры- вает реапьный путь к компьютеризации биоморфологического анализа. На многочислен- ных примерах из жизни растительных биоморф показано прикладное значение рассмот- ренных теоретических положений. Книга рассчитана на биоморфологов, ботаников разного профиля, а также биологов, интересующихся общими проблемами естествознания.
Светлой ламяти моего отца Нухимовского Льва Ефимовича посвящаю эту книгу. Введение Двадцатый век принес в науку о жизни (биологию) немало ярких революционных событий: расшифрован генетический код, познан ме- ханизм синтеза белка, успешно составляется каталог генов человека, впечатляют достижения генной инженерии, развито учение о биосфере и др. В наше время на развитие биологии сильное влияние оказывает системный подход. Как подчеркнул К.Л.Паавер (1976), из принципов системного подхода наибольший интерес представляет организацион- ный принцип, смысл которого в общем случае — отображение объекта как организованной системы, качества и реакции которого детермини- рованы ее струкгурой. К.Гробстайн (1968, с. 16), на наш взгляд, оправ- данно считает, что «все сложнейшие проявления жизни можно будет свести к не слишком большому числу основных принципов». Проблема поиска основных принципов организации живого, и биоморф в частно- сти, — одна из актуальных в современной теоретической биологии. Важную связующую роль в комплексе биологических наук может занять наука о жизненных формах, или биоморфах (биоморфология). Вся ис- тория развития биоморфологии подводит нас именно к такой мысли: эта наука, пожалуй, переживает еще период своей молодости и только сейчас выходит на рубежи оформления стержневой теории, без которой ей никогда не занять достойного места среди других частных биологи- ческих наук. «Цель морфологии, а следовательно, и всей ботаники— открыть причины растительных форм, и тем самым указать законы, лежащие в их основании» (Бекетов, 1862, с. 2). В развитие биоморфологии растений внесли свой вклад ботаники нескольких поколений (Humboldt, 1806, цит. по Гумбольдт, 1936; Гри- зебах; 1874, 1877; Warming, 1884, 1908, 1909, 1923; Raunkiaer, 1905, 1907; Drude, 1913, Высоцкий, 1915; Gams, 1918; Казакевич, 1922; Du Rietz, 1931; Meusel, 1935, 1951, 1955; Закржевский, Коровин, 1935; Прозоров- ский, 1936; Rauh, 1937, 1939, 1950; Станюкович, 1949; Сенянинова-Кор- чагина,1949; Алехин,1950; Работнов,1950а; Scharfetter,1953; Ша- лыт, 1955, 1960; Хржановский, 1958, 1966; Зозулин, 1959, 1961, 1968; Го-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ лубев, 1960, 1965; Серебряков, 1962, 1964; Тихомиров, 1963; Беспало- ва, 1965; Лавренко, Свешникова, 1968; Серебрякова, 1971; Нечаева и др. 1973; Нухимовский, 1973а, б, 1980, 1986, 1987; Хохряков, 1975а, б, 1981; Юрцев, 1976; Шафранова, 1990; Шафранова, Гатцук, 1995, и многие дру- гие. В отечественной литературе наиболее полные обзоры по жизнен- ным формам растений проведены И.Г.Серебряковым (1962, 1964) и Т.И. Серебряковой (1972), а также в их совместной работе (Серебряков, Серебрякова, 1967). Накоплено очень много интересных фактов об орга- низации, в особенности габитуальной, жизненных форм семенных ра- стений, однако обнаруживается немало противоречивых моментов как в объяснении термина «жизненная форма», так и в теоретических по- строениях. Предлагаемая книга посвящена жизненным формам (биоморфам) как организмам и синорганизмам, причем не только каким-либо от- дельным элементам структуры, но и всей их организации, на которую мы смотрим с разных высот обобщения фактов и под разными углами обзора. Цель книги — создать основы теории организации биоморф семенных растений сезонного климата и определить некоторые перс- пективы ее экстраполяции на все семенные растения Земли. «Вихри» фактов об организации биоморф должны быть локализо- ваны, разложены по полочкам, на основе использования современных методов накопления и обобщения фактов, зафиксированы в законах науки. Биоморфология должна стать более строгой научной дисципли- ной с прочным, но пластичным теоретическим каркасом. В биоморфологии очень важно уметь видеть. Обостренная наблю- дательность — обязательное качество, без которого не могут работать специалисты тех областей знания, например, ботаники-морфологи, ас- трономы и др. где приходится не столько ставить эксперименты самим, сколько видеть то, как ставит «эксперименты» Природа без нашего участия. Основная разница в работе ботаника-морфолога и астронома в том, что объект их познания находится на разных расстояниях от наблюдателя. Ботанику-морфологу предоставлено Природой величай- шее преимущество в получении достоверных фактов, поскольку он может держать объект познания в руках, проникать внутрь его, прово- дить непосредственные исследования, а не косвенные, как чаще всего вы- нуждены делать астрономы. Задачи исследований: 1. Выявить возможности внедрения системного подхода в био- морфологию и предпосылки для создания теории этой науки. 2. Определить объем понятия «жизненная форма» (биоморфа) и установить статус биоморф в организации живой природы. 3. Рассмотреть организацию биоморф семенных растений в сис- теме «управление — исполнение», охарактеризовать основные способы освоения ими пространства: нарастание и ветвление, партикуляцию и размножение, элиминацию. 4. Изучить следующие аспекты организации биоморф семенных растений:
Введение а) организация биоморф как единство отношений и связей между элементами; б) иерархичность в организации целого и частей у биоморф; в) реализация жизнесохранительных свойств в организации биоморф; г) особенности адаптации и соматической эволюции у семен- ных растений. 5. Учитывая, что морфогения является атрибутивным динамичес- ким свойством организации биоморф семенных растений— способом освоения ими времени, вскрыть закономерности в смене возрастных и фенологических стадий в их морфогенезах. 6. Конкретизировать и расширить существующие методы иссле- дования жизненных циклов биоморф семенных растений. 7. Использовать теоретические разработки для развития приклад- ной биоморфологии. В теоретической работе трудно не обойтись без обсуждения каких- либо терминов. Термин — это не только результат развития науки, но и индикатор уровня ее современного состояния. Термин, на наш взгляд,— слова-этикетки (слова-символа) и лонятия (обьяснения этого слова в смысле, приемлемом для отражения явления-вещи, лроцесса олределенного рода). Для терминов, как и для всего, что есть в Природе, присуще развитие. Со временем любая его часть или весь термин могут трансформироваться. В предлагаемой книге на терминологию обращено особое внимание. Особенности организации биоморф семенных растений в их жиз- ненных циклах изучены автором в различных регионах бывшего СССР в условиях естественного произрастания, а также в культуре. Основной район исследований — Алтай и Восточный Казахстан, где маршрутным, полустанционарным и стационарным способами я работал в экспедициях Всероссийского научно-исследовательского ин- ститута лекарственных и ароматических растений (ВИЛАР, бывший ВИЛР, Москва) в весенне-летнее время с небольшими перерывами в 1970-1985 гг. Фактуальную основу составили многие виды семенных растений флоры Алтая. Кроме того, материал для исследований соби- рался в разные годы в естественных условиях произрастания семенных растений в Подмосковье (1965-1996), Актюбинской области (1963-1964), Калмыкии (1965-1966), Крыму (1967-1969), в Читинской области (1976), Красноярском крае и республике Тыве (1977), Калужской области (1982- 1995), а также в непродолжительных поездках на Карпаты, Кавказ и в страны Средиземноморья, особенно в Израиль, где мне удалось собрать оригинальный материал. Морфологические наблюдения за растениями проводились также на экспериментальном участке ботанического сада и в теплице Московской сельскохозяйственной академии (МСХА, 1967- 1969), на опытном участке лаборатории интродукции, в оранжерее и в ботаническом саду ВИЛАР (1972-1987), в Никитском ботаническом саду (Крым), в садах и парках Москвы и Московской области. Семена для опытов собирали в экспедициях или выписывали из отечественных и зарубежных ботанических садов.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Поскольку автор около двадцати лет работал в ВИЛАР, особое внимание было обращено на важнейшие лекарственные растения, не- которые из которых только вводятся в медицинскую практику. Методика работы с фактическим материалом включала сбор разно- возрастных растений или их частей в различной экологической обста- новке. В условиях культуры сбор растений проводили через 10-15 дней, иногда через более продолжительное время. У собранных растений отмечали возрастные и фенологические особенности; их описывали, фотографировали, гербаризировали (или фиксировали). Латинские названия растений приводятся в основном по книге С.К.Черепанова (1995). Все оригинальные рисунки выполнены автором. Советы и замечания по материалам этой работы я неоднократно получал от В.Г.Хржановского, Т.А.Работнова, Т.И. Серебряковой, В.Н.Голубева, С.В. Мейена, И.Ф.Сацыперовой, А.П.Пошкурлат, кото- рым глубоко признателен. Считаю своим долгом выразить большую благодарность всем со- трудникам лаборатории интродукции ВИЛАР, в особенности, Б.С.Ер- макову, Н.В. Макаровой, Л.И.Хоциаловой, О.А. Черкасову, В.В. Угни- венко, И.А. Шретер за поддержку и помощь в работе. Автор глубоко благодарен Т. Райхель и А. Райхель за содействие в работе за рубежом.
«Расчленение тела растения на составные части и связанная с ним разработка тииологии этих частей — логический и удобный слособ изучения растения, лозволя- ющий выдвинуть на лервый илан лроблему структурной и функциональной слециализа- ции отдельных частей растения. Однако не следует слишком увлекаться таким лодходом, чтобы не утратить лредставле- ние о растении как едином целом». (Эзау, 1980, с. 13) Глава 1. Теоретизирование и законы науки Движущийся материальный мир, существующий в бесконечном разнообразии конкретных тел и процессов, вместе с одной из особых своих функций — сознанием, составляет Человека и то, что повсюду его окружает и на что неизменно он направляет свою познавательную деятельность. Процесс познания включает эмпирический этап (наблю- дение, эксперимент, измерение, описание, обработку эмпирических данных и т.д.), и теоретический этап, который состоит из интерпрета- ции, объяснения, предсказания, ретросказания и т.д. (Грязнов и др., 1967). «Явление есть момент бытия, факт — момент знания... Факт есть знание о явлении... Факты не составляют конечной цели в науке. Из них исходят для получения теории» (Вахтомин, 1973, с. 187-189). Факты следует выбирать с учетом их объективной связи, в целостности, только тогда они могут быть основанием научного знания. Факты дол- жны работать на теорию, только в теории повышается их значимость для науки. При разработке теории организации какого-либо объекта (в нашем случае — жизненных форм семенных растений) важно опирать- ся на принципы системного подхода. Использование последних помог- ло нам показать многие общие стороны организации Природы и ее частей, рассмотреть их с разной степенью подробности. Это позволило достигнуть главной цели: упорядочить разнообразие известных, малоиз-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛСТЕНИЙ вестных и новых фактов в единой теоретической (концептуальной) си- стеме — теории организации жизненных форм семенных растений. При построении названной теории автор использовал как индуктивное дви- жение от эмпирического базиса, так и дедуктивное движение от изве- стного теоретического базиса. Основу последнего, как известно, состав- ляет философия с ее сердцевиной — материалистической диалектикой. Построение теории, основанной на многочисленных фактах, с позиций системного подхода составляет главную методологическую особенность нашей работы. Некоторые прикладные возможности теории мы прове- ряем и отражаем при описании жизненных циклов различных растений. В современном представлении понятие теория весьма многогранно (Капица, 1966; Грязнов и др., 1973; Мамчур, 1975; Андреев, 1979 и др.). Наиболее удачная формулировка теории, что неоднократно уже подчер- кивалось (Мерзон, 1972; Баженов, 1978 и др.), дана в «Философской энциклопедии»'. «Теория — форма достоверного научного знания о не- которой совокупности объектов, представляющая собой систему взаи- мосвязанных утверждений и доказательств, и содержащая методы объяс- нения и предсказания явлений данной предметной области» (Попович, Садовский, 1970, с. 205). И.Д.Андреев (1979, с. 24), специально исследо- вавший сущность понятия теории как формы организации научного знания, пришел к выводу, что «теория — это не всякая совокупность знаний о предмете, а строго организованная система знаний, каждый элемент которой органически связан с другими элементами данной системы и непосредственно следует из них». Общей методологической парадигмой («исследовательской про- граммой» по Т.Куну, 1977) в настоящей работе принят системный под- ход. В «море» разнообразных описательных и экспериментальных фак- тов, идей, теоретических разработок, связанных со свойствами, струк- турой и функциями биоморф и составляющих их элементов, где еще совсем недавно было трудно уловить какие-либо закономерности, по- степенно начинает вырисовываться определенная теоретическая систе- ма — теория организации жизненных форм семенных растений. Для обобщения фактов были использованы следующие методические приемы. Литературный анализ. В настоящее время накоплен большой эмпи- рический и теоретический материал по различным аспектам понятия «жизненная форма», структуре и функциям жизненных форм семенных растений. В последующих главах мы ссылаемся на соответствующие работы. Моделирование. «Моделирование превратилось в универсальный ин- струмент и способ научного исследования, приобрело статус общегно- сеологической категории» (Батороев, 1981, с. 15). Таков сегодня интерес к моделированию в научном познании. А.И.Уемов (1971) привел около 40 определений термина «модель», которые хотя в целом не противо- речат друт другу в главном, все же отражают большое разнообразие его смысловых оттенков. При всем этом метод моделирования универсален и используется как в научной, так и практической деятельности. Из многих значений термина «модель» приведем лишь несколько, чтобы отразить господствующий в литературе смысл его. «Под моделью пони-
Глава 1. Теоретизирование и законы науки мается такая мысленно представляемая или материально реализованная система, которая, отображая и воспроизводя объект, способна замещать его так, что ее изучение дает нам новую информацию об этом объекте» (Штофф, 1966, с.19). «Модель есть осознанная или выбранная субъектом система, вос- производящая существенные для данной цели познания стороны (эле- менты, свойства, отношения, параметры) изучаемого объекта и в силу этого находящаяся с ним в таком отношении замещения и сходства (в частности изоморфизма), что исследование ее служит опосредственным способом получения знания об этом объекте» (Батороев, 1981, с. 28). И, наконец, краткое определение, данное с позиций теории подо- бия и позволяющей разрабатывать способы получения его: модель- это объект (явление, процесс, система, установка, знаковое образова- ние), который находится в отношении подобия к моделируемому объек- ту (Веников, 1966, 1973). Существует много классификаций моделей, например, определены следующие виды моделей: точные, приблежен- ные, полные, неполные, физические, структурные, функциональные, математические, динамические, вероятностные, геометрические, знако- вые, логические (Веников, ред., 1973). Существует обширная возмож- ность изменять, дополнять известные классификации, поскольку про- цесс моделирования не имеет жестких границ, а число возможных видов моделей может колебаться в зависимости от подхода исследователя, при этом могут возникнуть и самые неожиданные модели. Не будем углубляться в обзор классификаций моделей, полагая, что сказанного выше достаточно для использования самого метода модели- рования (построения моделей) для решения задач наших исследований. Выбор подходящей модели для решения поставленной проблемы — это важнейший этап в системном подходе к объекту познания. По сути дела, моделирование начинается с описания какого-либо объекта, на- пример, с обобщенного описания жизненных циклов биоморф семен- ных растений. Обычное словесное описание исследуемого объекта— наиболее древний способ их исследования, это неотьемлемая часть и современного научного познания. Недостаточность чисто описательных характеристик («словесных моделей») чего-либо, состоит в том, что од- новременное действие многих сложных связок фактов нередко теряется среди «отношений между словами» (Джефферс, 1981). В биоморфологии пока господствует метод описания фактов (построение «словесных моде- лей»). В нашей работе сделана попытка формализовать словесные описа- ния в таблично-знаковых, таблично-словесных, графических и знаковых моделях. Моделирование как метод познания открывает большие перспек- тивы для развития биоморфологии, помогая вскрывать сущность орга- низации биоморф и взаимодействие между элементами. Модели функ- ционируют в процессе познания, давая исследователю возможность по- лучать об оригинале соответствующую информацию — описательную, прогностическую и т.п. (Лиепа, 1975). Классифицирование. «Наука — это прежде всего классификация» (Пуанкаре, 1906, цит. по Д.Л.Арманду, 1975, с. 138) — так высоко оце-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ нено значение этой логической операции для познания. Однако при всем том очень высоком месте, которое отводится классифицированию в науке, пока не существует теории классифицирования. Трудность проблемы особенно выпукло обозначилась после статей С.В. Мейена (Мейен, 1975, 1977, 1978; Мейен, Шрейдер, 1976 и др.). В книге рас- смотрен ряд методологических и методических проблем построения классификаций. Прежде чем говорить о классифицировании, выделим особо про- цедуру типизации. Любая группа объектов, охарактеризованных по каким-либо общим (одному или нескольким) признакам — тин. Про- цесс абстрагирования обьектов нознания в тины — тинизация. Типиза- ция — это процедура, независимая от классифицирования или предше- ствующая ей, или растворяющаяся в ней, когда ставится цель создания классификации и осуществляется классифицирование. Раздел исследо- ваний (учение), посвященный проблемам выделения типов (типиза- ции), — тинология. Следует оговорить, что пока отсутствует единство во взглядах на понимание типа, типизацию и типологию. Так, напри- мер, Д.Л.Арманд (1975) считает, что типизация — это перечисление некоторых из возможных объектов на данной ступени классификации, которые фактически имеются в исследуемой группе, т.е. процесс выде- ления этих объектов, а само множество выделенных таким образом единиц — типология. С.В.Мейен (1978) понимает под типологией орга- низмов (как и других объектов) общее учение об их разнообразии, распадающееся на таксономию и мерономию. История развития типо- логических представлений весьма многогранна (Канаев, 1966). Типиза- ция, на наш взгляд, не несет обязательной сравнительной нагрузки. При тинизации важно выделить в первую очередь объект обобщения, независимо от того, как он соотносится с другими объектами обобще- ния. В ряде случаев, например при классифицировании, типу может быть придан статус одной из категорий (чаще всего высших) классов каких-либо объектов классифицирования. Это одно из дополнительных применений термина «тип». Итак, при классифицировании типы (результат типизации) раство- ряются в классах объектов. Под «лассифицированием мы понимаем ло- гическую операцию построения классификации. Классификация — это результат процесса классифицирования, требующий особого определе- ния. Классифицированию мы придаем большое значение, поскольку в этом процессе наиболее отчетливо выступают определенные закономер- ности в организации, структуре материальных объектов и процессов. По проблеме классифицирования существует обширная литература, свиде- тельствующая о многообразии вопросов в этой области знаний (Lin- naeus, 1751; Арманд, 1975; Тахтаджян, 1966; Василевич, 1966; Миркин, Розенберг, 1978 и др.). Классификация (точнее классифицирование— Е.Н.), как подчеркнула В.Д.Александрова (1969, с. 5), «есть логическая операция, которая заключается в разбиении данного множества объек- тов на подмножества, или классы, причем под классом подразумевается совокупность объектов, имеющих общие признаки, отличающие объек- ты, относящиеся к данному классу, от других объектов, которые такими
Глава 1. Теоретизирование и законы науки признаками не обладают». Широко определяют классификацию С.В.Мейен и Ю.А.Шрейдер (1976, с. 69) — это «разбиение любого мно- жества (класса) объектов на подмножества (подклассы) по любым при- знакам». Мы различаем, как уже подчеркнули выше, «лассифицироеание и классификацию. С учетом этой коррективы к определению классифи- кации, данному В.Д.Александровой, мы будем понимать нод классифи- кацией абстрактную (логическую) систему, в которой объекты данного множества разбиты (классифицированы) на нодмножества (классы) как совокунности объектов, имеющие общие нризнаки, отличающие объекты, относящиеся к данному классу, от других объектов, которые такими нризнаками не обладают. В организации Природы все ее элементы в чем-либо различаются между собой ( по сложности организации, степени активности, разме- рам и т.п.) в соответствии с принципом иерархической взаимосвязи: во всем всегда есть что-то, в чем-то меньшее, и что-то, в том же большее (закон иерархичности в нашем нонимании). Представление об иерархичности (соподчинении), т.е. о располо- жении частей и элементов целого в порядке от низшего к высшему, от простого к сложному, широко используется в современной биологии при описании живых систем и, в частности, при построении классифи- каций. Принцип иерархичности обнаруживает себя везде, «где изучает- ся дискретность, структурированность материальных систем..., где су- ществует многоуровневое представление об объекте» (Абрамова, 1974, с.189). Иерархический принцип является фундаментальным принци- пом, поскольку присущ любым материальным системам на определен- ной ступени возрастания их сложности (Бирюков, 1974). При изучении организации растительных биоморф мы обратили особое внимание на принцип иерархичности, поскольку «степень сложности или конструк- тивный ранг», как справедливо подчеркнул В.Н.Беклемишев (1964),— «одно из важнейших свойств каждой организации и каждого живого существа». Структура системы может характеризоваться как по «гори- зонтали», так и по «вертикали», при этом разделение на классы объек- тов по «вертикали» приводит к понятию уровней системы и иерархии этих уровней (Блауберг, Юдин, 1973). По принципу построения классификации могут быть иерархически- ми или сериальными (Нухимовский, 1980). Среди иерархических класси- фикаций можно выделить следующие: таксономические, субородинат- ные, мультисубординатные, суперординатные и релятивные. Суборди- натная «лассификаця (субординия) — это классификация по иерархи- ческому принципу, когда классы объектов (субординаты) располагаются в соответствии с возрастающей сложностью этих объектов, характери- зуемой через какие-либо сравнимые показатели всей организации (или структуры, или поведения, или состава и т.п.) независимо от степени родства, образуя линейный восходящий ряд. Можно, например, строить субординатные классификации материальных систем по их организа- ции, по их составу, по их скоростной активности и т.п. Критерием для выделения классов систем будет степень сложности соответствующих систем, отраженная в их показателях (признаках). Идея субординатных
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ классификаций уходит своими корнями в концепции структурных уров- ней и уровней организации, которые в разных вариациях развиваются как за рубежом, так и у нас (Bertalanffy, 1949; Завадский, 1961, Шмаль- гаузен, 1961; Кремянский, 1969; Сетров, 1972; Шварц, 1974 и др.). Тер- мин «уровень», как подчеркнул К.М.Завадский (1966, 1968), часто упот- ребляется в различных смыслах: во-первых, как уровень организации живых систем, «т.е. степень сложности образований, способных к жиз- ни в природе», и, во-вторых, как уровень изучения («уровень исследо- вания» по Н.П.Депенчук, 1972) живого. Если связь классов систем пред- ставляют линейной зависимостью, то используют понятие «структур- ный уровень», а если ее представляют в виде нескольких самостоятель- ных рядов, не связанных друг с другом линейной зависимостью, то классы систем определяют как уровни организации (Абрамова, 1972). Граница между этими понятиями, как видно из обзора М.Ф.Веденова и др. (1972), нередко нарушается. Для вычленения уровней организации живых систем нередко используют несколько критериев (сложность или конструктивный ранг, первичность или вторичность, активность, меха- низм и степень интеграции и т.п.). Мы отдаем преимущество нашему краткому термину «субордината» как классу объектов, вычленяемому в процессе субординатного классифицирования. Вопрос о соотношении понятий «уровень организации» и «структурный уровень» с понятием «субордината» опускаем, учитывая отсутствие точности в определении первых двух терминов. Заметим, однако, что не видно серьезных логи- ческих препятствий, чтобы в некоторых случаях их смысл совпадал, например, когда определяется положение в сравнительном вертикаль- ном ряду класса организаций одной сложности по отношению к классу организаций другой сложности вне зависимости от степени родства этих организаций. Относительно друг друга различаем субординаты нижние, если они входят в объем других субординат (это подчиненные классы объектов субординатной классификации) и верхние, если те включают в свой объем другие субординаты (это подчиняющие классы объектов субординатной классификации). По значимости признаков, используемых для субординатных клас- сификаций, пожалуй, можно различать два типа таких классификаций. Субординии первого типа (узкие субординии) строятся по одному ряду простых признаков, например, неопушенные, слабоопушенные и т.п. побеги, и отражают иерархичность в каком-либо узком аспекте органи- зации объектов или структуры процессов. Субординии второго типа (широкие субординии) строятся по разнообразному числу признаков и отражают иерархичность в каком-либо широком аспекте организации объектов. В соответствии с таким подразделением субординатных клас- сификаций правомерно говорить и об узких и об широких субординатах (по диапазону значимости признаков субординат в определении слож- ности соответствующих объектов). Ряд признаков — это совокупность упорядоченных, сравнимых и взаимоисключающих признаков (Балков- ский, 1964). Субординаты оказываются в соответствии с принципом иерархической взаимосвязи последовательно включенными один в дру- гой, что наглядно можно показать на графической модели. Однако это
Глава 1. Теоретизирование и законы науки не означает, что в каждом конкретном случае объект верхней суборди- наты состоит из объектов всех возможных нижних субординат. Объекты верхних субординат в соответствии с избранным критерием будут иметь большой диапазон сложности, чем объекты подчиненных субординат. Критерием для отнесения материальных объектов к определенным су- бординатам можно считать диапазон их сложности, ограниченный по признакам одного ряда узкого структурно-функционального содержа- ния (узкая субординия) или по признакам широкого структурно-фун- кционального содержания, отражающим наиболее важные качествен- ные состояния объекта (широкая субординия), и указывающий на по- рядок иерархической взаимосвязи. Субординии каких-либо объектов выступают как частные классификации по отношению к другим субор- диниям большего объема, например, все возможные субординии побе- гов семенных растений умещаются в общей широкой субординии эле- ментов живого Земли только на одном структурном уровне — на субор- динате органов. Иерархичность по разносложности организации многих систем можно хорошо передать посредством субординатных классифи- каций. Однако иногда возможностей субординии не хватает, чтобы достаточно полно выразить иерархию разносложных организаций в организации больших систем, например, таких, как вся живая природа или вся неживая природа и др. и, наконец, вся Природа, с относительно малых высот обобщения. В таких случаях мы предлагаем использовать метод мультисубординии. Мулыиисубординитная (нолисубординатная) «лиссификиция (мулниисубординия) — это иерархическая классифика- ция, состоящая из нескольких субординатных классификаций, объеди- ненных вместе единством цели в одну разветвленную систему. Нас в настоящей работе более всего интересует использование методов субор- динии и мультисубординии для познания организации биоморф семен- ных растений и для рассмотрения их организации на фоне организации всей Природы. Наш опыт изучения иерархичности организации систем показал что один объект можно характеризовать посредством субординии, а иногда и мультисубординии по-разному с разных высот абстракции (обобщения). Последовательно реализуя эту идею, мы впервые разрабо- тали метод суперординатного классифицирования, смысл которого в том, что классифицируются по иерархическому принципу субординат- ные и (или) мультисубординатные классификации, а в итоге получается широкоформатная объемная научная картина, характеризующая объект исследования при разном обобщении. Сунерординитная «лиссификиция (сунерординия) — это классификация по иерархическому принципу, где классы систем (уровни абстракции, или суперординаты) включают це- лые субординатные или мультисубординатные классификации,распола- гаясь в восходящем ряду на разной высоте относительно друг друга в зависимости от степени обобщения в этих классификациях, критерием которого служит число субординат их составляющих. Суперордината, таким образом, — это определенная степень (уровень) обобщения в классификации или классификациях по одним признакам с отвлечени- ем от других признаков, высота которой определяется по числу субор-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ динат, составляющих ее. Графически суперординию чего-либо удобно передать в виде абстрактной «пирамиды»,ступени которой — суперор- динаты, а в «разрезах», «разломах» и т.п. условных способах проникно- вения внутрь такой пирамиды удобно показывать ее внутреннее содер- жание в графических моделях (например, в виде спиралей) соответству- ющих субординатных или мультисубординатных классификаций. Осо- бое внимание обращаем на биполярную и мультинолярную субординии и мультисубординии, когда два класса объектов или две и более суборди- ний объединяются в одну классификацию, отражая изменение объектов классификации в противоположных направлениях. Такие классифика- ции также можно рассматривать через призму суперординии. Чем мень- ше классов объектов выделяется в иерархической классификации, тем больше степень обобщения в этой классификации. Отсюда следует: чем меньше число субординат в соответствующей субординатной «лассифика- ции, тем выше уровень абстракции в соответствующей суперординатной классификации. Суперординия отличается от таксономической и релятивной клас- сификаций. Классификация по иерархическому принципу, когда клас- сы объектов располагаются в соответствии с композиционными удоб- ствами конкретного исследования, — релятивная «лассификация. Логи- ческие основы этого типа классификаций уступают остальным, хотя они широко встречаются в биоморфологии. Классификация по иерархи- ческому принципу, когда классы материальных систем (отдельные под- множества — таксоны) разного объема располагаются в соответствии со степенью родства между собой, — это, как известно, — таксономичес- кая классификация. В биоморфологии используются также классификации, которые мы называем сериальными (сериофикации) — это классификации, когда классы систем или процессов располагаются линейно без соподчине- ния, но упорядоченно по одной (нескольким) группе (группам) сравни- мых и взаимоисключающих признаков, не отражающих изменение сте- пени сложности объектов, например, сериофикация растений по поло- жению семядолей: надземносемядольные, подземносемядольные, над- водносемядольные и т.п. Заметим, что ранее Ж.Пиаже и Б.Инельдер (1963) предложили различать операцию сериации, понимая под ней распределение объектов в упорядоченные ряды (например, распределе- ние набора палочек разной длины в ряд, где каждый член этого ряда больше предыдущих элементов и меньше всех последующих). В сери- ации не расчленяются принципы субординии и сериофикации. Кроме того, сериальная классификация отличается от ординации (Миркин, Розенберг, 1978; Работнов, 1978 и др.), под которой понимают упорядо- ченное размещение объектов по ряду градиентов какого-либо опреде- ляющего или коррелирующего фактора в двумерной или многомерной системе координат. Субординатная классификация в отличие от таксо- номической классификации отражает через иерархическую взаимосвязь классов объектов (субординат) не степень их родства (это задача таксо- номии), а степень их организационной или иной сложности. Когда классы объектов располагают как бы в одну линию (ряд) по одному ряду аль-
Глава 1. Теоретизирование и законы науки тернативных признаков — это унилинейная (однолинейная) классифика- ция, а совокупность таких классификаций — это мулниилинейная (мно- голинейная) классификация. Метод мультилинейного классифицирова- ния — это обобщенный метод, поскольку включает в себя или методы субординатиого, или сериального классифицирования; мы широко ис- пользуем его в своей работе. Заметим, что этот метод (по сути) впервые применен в биоморфологии В.Н.Голубевым (1968, 1972), который рас- сматривает его под названием «линейное» классифицирование. Этот же метод был использован при класифицировании побегов семенных ра- стений (Нухимовский, 1969а, 1971). Онределение научных законов. Высшее отражение факты находят в законах науки. Научные законы, если они верны, выражают объектив- ные связи явлений, законы Природы, в соответствующих понятиях. Итак, когда мы далее говорим о законах, то понимаем, что это научные законы, являющиеся идеальными моделями законов Природы. Великая познавательная ценность науки состоит в том, чтобы познавать «закон целого ряда однородных, повторяющихся явлений, найти закон явле- ний» (Гельмгольц, 1898, с. 80). Открывать законы Природы — значит познавать «порядок, которому подчиняются явления» (Вигнер, 1971, с. 49). Вся материя и наше сознание существуют в соответствии с оп- ределенными законами (большими и малыми). Закон — это, как изве- стно, форма всеобщности в природе, иначе «это не единичная, прису- щая какому-либо одному явлению, а общая, свойственная всем явлени- ям данного класса, множества и т.д. связь» (Краткий словарь по фило- софии, 1979, с. 89). Существует немало определений понятия закон и классификаций законов (Кедров, 1967; Зак, 1974; Карпович, 1980; Па- нибратов, 1980 и др.). С.В.Мейен (1978) считал, что законы — это круп- ные закономерности: более общие и точнее сформулированные законо- мерности. Важно разобраться в иерархии законов по широте ди.~ пазонов их действия. Нами разработан вариант такой субординатной классифи- кации, к которой будем обращаться в своих дальнейших исследованиях. Высшие (сунерактные) законы присущи всем явлениям Природы. Пред- высшие (нресунерактные) законы (они занимают вторую сверху суборди- нату в субординии законов) — это законы общие для всей Природы, но распространяющиеся не на все ее явления. Абсолютные законы — это законы, присущие всем явлениям или только материальной природы, или только идеальной природы. Субабсолютные законы общие для жи- вой и для неживой природы, но распространяются не на все явления материального мира. Примаснециальные законы действуют во всех явле- ниях только живой или только неживой природы. Снециальные законы действуют в относительно многих явлениях живой, неживой или иде- альной природы, например, в сфере социальных отношений и т.п. Суб- снециальные законы (они составляют низшую субординату в субординии законов) — относительно небольшие закономерности (малые законы), охватывают относительно узкий круг явлений живой, неживой или иде- альной природы. Упомянутый выше закон иерархичности является выс- шим. Закон отрицания отрицания также один из высших законов При- роды. Чем больше (меньше) часть целого организма приближается по
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ организационной сложности к целому организму, тем больше (меньше) этот организм предрасположен к размножению, а его часть к преобра- зованию в новый самостоятельный организм — так определяется спе- циальный закон предрасположенности к размножению (Нухимовс- кий, 1986). Длиннокорневищные растения, например, хорошо приспособ- лены к вегетативному размножению, поскольку в их структуре развивают- ся особые органы — побеги вместе с корнями (корнепобеги-органы), ко- торые более всего подготовлены к самостоятельному существованию, т.е. к преобразованию в новый организм. Субабсолютным будет, напри- мер, закон единства материального и идеального, смысл которого мож- но выразить так: идеальное порождается разумной живой природой (людьми), существуя в единстве с ней и для нее, преобразуя ее, себя и окружающую часть материальной природы. Из законов нередко следу- ют правила. Так, из закона единства материального и идеального вы- текает известное диалектическое положение в качестве правила: мате- риальное как первичное существует вне идеального, которое вторично, но материальное не способно вне идеального познать себя. Закон — это последнее звено, завершающее становление научной идеи; дальнейший путь уже не к отдельной идее, а к их системе — научной теории (Кар- пович, 1980).
Глава 2. Обоснование биоморфологии Методология современной биологии все более базируется на диа- лектике и системном подходе. Еще в 30-е годы советский биолог Э.С.Бауэр (1935) стремился создать теоретическую биологию, изучаю- щую общие законы движения живой материи. Такая цель не утратила своего значения и сегодня, но биологи, и ботаники в особенности, пока медленно приобщаются к языку широких обобщений — к языку зако- номерностей (законов). Ось философия — биология направлена на формирование теоретической биологии, и теоретической ботаники в частности, способствуя выявлению и определению законов (форм все- общности) органического мира. В методологии науки и в практической деятельности все более сильное действие оказывает системный подход (Садовский, 1974), с помощью которого из отдельных свойств элемен- тов выявляются свойства целого и, наоборот, вскрываются иерархич- ность в организации систем, взаимосвязи между компонентами, особен- ности управления и самоуправления систем. Основной объект изучения биоморфологии — жизненные формы (биоморфы) мы понимаем как организмы и синорганизмы, или кондивиду- умы (Нухимовский, 1973а, б, 1974а, 1980). Синорганизм — живая cucme- ма, начинающаяся в своем развитии с зиготы и оканчивающаяся в иосле- днем локолении клона (см. гл. 2.4.). Прежде чем подойти к подробной работе с этими терминами и понятиями и к биоморфологии как науке в целом, попытаемся приблизиться к объектам познания сначала с философских позиций и лишь затем перейдем к анализу существующих представлений об этих объектах. 2.1. Движение, системность, жизнь Понятия материя и движение неотделимы друг от друга. Натура движущихся тел исходит из форм движения. Форму движения Ф.Эн- гельс понимал как типовое понятие, основывая его на обобщении дан- ных о многообразных движениях, составляющих суть существования какого-либо вида материи. Формы (типы) движения определялись Ф.Энгельсом (1964, с. 216) по их отношению к наукам, «из которых каждая анализирует отдельные формы движения или ряд связанных между собой и переходящих друг в друга форм движения». Один вид - 1~~ ~и~1iнк кии! 't
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ материи может проявить как главную, так и побочные (дополнитель- ные) формы движения. Дополнительные формы движения — те, которые у конкретных материальных тел дополняют картину опреде- ляющего (главного) движения. В частности органическая жизнь невоз- можна без механического, молекулярного, химического, электрического и т.п. изменения. Иначе, любое частное биотическое движение допол- няется другими, обеспечивая полноту существования носителя главного (биотического) движения. Помимо механической, физической, хими- ческой, биологической (биотической) и социальной форм движения, предлагается различать еще геологическую (Кедров, 1959, 1964; Кома- ров, 1974; Зубков, 1979; Ивашевский, 1979 и др.), планетарную (Поспе- лов, 1960; Шанцер, 1964 и др.). Оставляя в стороне суть дискуссий о формах движения, подчеркну лишь самое общее впечатление: смысл проблемы, вероятнее всего, в первую очередь не в вьщелении новых и новых типов (форм) движения (точнее типов движений) и не в опре- делении числа этих типов. Б.М.Кедров (1967, с. 425), отвечая на выступ- ление итальянского философа Лучио Ломбардо-Редичи подчеркнул, что назрела необходимость проанализировать с позиций современного уровня знаний «всю совокупность понятий и терминов, которыми мы оперируем, иногда по привычке, не отдавая себе отчета в том, что в их содержании произошли глубокие, часто коренные изменения. К числу таких понятий относится, несомненно, и понятие «форма движения» материи, на основе которого была построена в свое время вся «Диалек- тика природы» Энгельса». Суть проблемы классифицирования движе- ний и наук, их изучающих, в первую очередь состоит в конкретизации самых общих исходных понятий в соответствии с достижениями как в частных науках, так и в общей методологии. Системный подход «направлен на разработку специфических по- знавательных средств, отвечающих задачам исследования и констру- ирования сложных объектов» (Блауберг, 1973, с. 8). Главное в систем- ном подходе — «это искусство решения проблем» (Солнцев, 1981). Всесторонний анализ и развитие системных идей и методов исследо- ваний в различных науках проведен как в зарубежной, так и в оте- чественной литературе (Bertalanffy, 1949, 1956; Берталанфи, 1969, 1973; Щедровицкий, 1964, 1981; Квейд, 1969; Заде, Дезоер, 1970; Акофф, 1971; Калман, Фалб, Арбиб, 1971; Сетров, 1971а, б; Анохин, 1972, 1973; Гвишиани, 1972; Ляпунов, 1972; Садовский, 1972, 1974, 1980; Юдин, 1972; Laszlo, 1972; Блауберг, Юдин, 1973; Месарович и др., 1973; Уемов, 1973, 1978; Аверьянов, 1976; Дружинин, Конторов, 1976; Урсул, 1968, 1977; Кашменская, 1980; Афанасьев, 1980, 1986; Д.Хомяков, П.Хомяков, 1996 и др.). В настоящее время можно считать, что системность — это атри- бутивное свойство материи проявлять себя в параметрах определен- ных систем при определенных условиях ее существования. Как под- черкнул В.Н.Садовский (1974, с. 5), — «систему и системность, мы усматриваем буквально во всем — теоретически любой объект науч- ного исследования может быть рассмотрен как особая система; сис- темность характеризует процесс познания таких объектов».
Глава 2. Обоснование биоморфологии Смысл термина «система» уточняется с момента вьщеления его как ключевого понятия системного подхода. Людвиг фон Берталанфи, один из основоположников системного подхода, понимал под системой «ком- плекс элементов, находящихся во взаимодействии» (Bertalanffy, 1949, 1956). Многозначность термина «система» наиболее полно отражена в обзоре В.Н.Садовского (1974), который привел несколько десятков определений этого термина. Наблюдается тенденция к универсализации понятия система, что отражено, например, А.А. Малиновским (1980, с. 82): система — это любой набор элементов (вещественных, энергети- ческих, информационных), выделенных совместно по какому-либо принципу из других элементов окружающего мира». Всякий предмет, как подчеркнул В.П.Кузьмин (1976, с. 53), является системой по отношению к входящим в него частям и одновременно выступает как часть более широкой систе- мы. «Любой объект — система и любой объект — система должен непре- менно принадлежать хотя бы одной системе объектов того же рода» — это закои системности по 1О.АУрманцеву (1978, с. 699). В.Г.Афанасьев (1980) различает системы целостные и суммативные; например, несколько мо- лекул газа, находящиеся во взаимодействии, тоже система — простая суммативная система, так как в результате взаимодействия здесь не возникает качественно новых (интегративных) свойств, присущих цело- стным системам. При создании теории важно как можно точнее определить исход- ные понятия (Анохин, 1972; Арманд, 1975; Югай, 1976 и др.). В нашем случае одним из главных таких понятий является организация. Понятие организация противоположно дезорганизации, неупорядоченности, неопределенности (Богданов, 1925, 1927, 1929; Тахтаджян, 1971). По А.А. Малиновскому (1980, с. 86), организованность — «показатель отли- чия свойств и проявлений системы от свойств и проявлений простой суммы частей». Организация понимается по-разному: или как некое общее и неотъемлемое свойство системы, или как сама система, или как процесс превращения неупорядоченного, случайного в упорядоченное, необходимое, или как результат этого процесса, или как способ связи элементов системы, изменяющийся во времени в сторону понижения или повышения сложности организации (Петрушенко, 1975). Понятие «организация» не может быть шире понятия система. Ма- териальная система может находиться либо в стадии организации, либо дезорганизации, в организованном или неорганизованном состоянии (Аверьянов, 1974). Организованность бывает одушевленной и неодушев- ленной. Это положение исходит еще от Ф.В.И. Шеллинга (1936), пред- ставителя (1775-1854) классической немецкой философии (Леоно- вич, 1973). В.П.Боголепов (1968) также считает, что организация есть как в органической, так и неорганической природе. Другие авторы (Георгиевский, 1972; Георгиевский и др. 1975) ограничивают понятие организации только биологическими и социальными объектами. Абсолютное отсутствие организации — результат абстракции. Вся материя в системах более или менее организована (Беклемишев, 1964; Леонович, 1973 и др.). Система может находиться на низком уровне организации и по своим свойствам приближаться к сумме частей, а при
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ высоком уровне организации она сильно отличается от простой суммы частей (Малиновский, 1980). На качественные различия «ступеней ор- ганизованности» природы обратили внимание еще Шеллинг и Гегель, которые связывали их с разными этапами эволюции идеалистически понимаемого мироздания (Солопов, 1972). В.И. Вернадский (1965) в отношении Земли говорил об «организо- ванности планеты». Б.М.Кедров, как сказано выше, выделил геологи- ческую форму движения материи, а И.В.Круть (1978) — различные типы геологической организации. Как считает И.В.Круть (1978), «орга- низация», отражает вещь в движении, а формы движения детализи- руются в уровнях организации материальных вещей-систем. Организа- ц~к живого — форма, сохраняющая свою специфичность в потоке обмена веществ (Беклемишев, 1964). Как необходимое условие целостности и организованности отмеча- ется упорядоченность (Хайлов, 1963; Урсул, 1968; Винер, 1968; Мали- новский, 1980), причем целесообразная, или направленная упорядочен- ность (Винер, 1968; Малиновский, 1967, 1968). Идея целесообразности в определении организации широко используется в работах М.И.Сетрова (1971а, б, 1972, 1975). Сущность организации он видит в ее функцио- нальном назначении: «организация — это совокупность явлений, свой- ства которых проявляются как функции сохранения и развития этой совокупности (Сетров, 1975, с.89). Принимая в целом этот критерий, А.Б.Георгиевский, В.П.Петленко и др. (1975, с. 97) определяют органи- зацию как «целесообразную упорядоченность функционирования сис- темы (объекта), обеспечивающую ее самосохранение и развитие», но тогда организация присуща только живым (негэнтропийным) системам, поскольку только они способны противостоять внешним и внутренним воздействиям. Большинство исследователей считают, что организация присутствует как у живых, так и неживых систем. В этой связи необходимо найти такое определение термина «организация», которое отражало бы объективный факт существования в Природе самых разных организаций. Ближе всего к указанной цели те авторы (Малиновский, 1967; Бо- голепов, 1968; Абрамова, 1972, 1974; Мамзин, 1974, 1978 и др.), которые стремятся объяснять организацию через структуру и функцию системы, через их незыблемое единство, когда система рассматривается одновре- менно и как структура (организованная форма материи), и как функция (организованная форма действия). В этом смысле, как подчеркнула З.В.Каганова (1972), дополнительность структуры и функции, когда од- но не объясняется в отрыве от другого, является выражением материа- листического принципа неразрывной связи материи и движения. Н.Т.Абрамова (1972, 1974), отметив, что понятие организация в одних случаях применяют для определения функции, а в других отождествля- ют с понятием структура, пришла к важному обобщению. «Понятие «организация» включает в себя как все признаки, свойственные поня- тию структура, так и выражает существенную динамичность элементов и связей между ними» (Абрамова, 1972, с. 126). Сторонники структурно- функциональной интерпретации термина «организация» считают, что внутреннюю упорядоченность системы можно понять, если принимать,
Глава 2. Обоснование биоморфологии что организация — это единство относительно устойчивого — структу- ры и иииенчивого — функции. «Организация — это структура в действии, не просто форма объекта, а структура объекта, выполняющего определенные функции» (Абрамова, 1974, с. 145). Итак, краткий литературный обзор свидетельствует о неоднознач- ности трактовки термина «организация». Далее мы будем понимать под организацией единство структуры и функции системы. Атрибутивный компонент системы — состав. Состав — это те элементы, из которых состоит система. Элементы — «части системы, которые далее в иссле- довании не разлагаются» (Малиновский,1980, с. 82). Организация- это необязательный компонент всей системы, поскольку не все системы обладают структурой. Термин «структура» понимается пока неоднознач- но. Наиболее распространено следующее краткое объяснение этого тер- мина: структура — это инвариантный аспект системы, включающий как отношения между элементами, так и сами элементы (Овчинни- ков, 1966, 1967). По мнению других исследователей, важнейшая сторона структуры — упорядоченность образований и процессов в их целостно- сти (Веденов и др., 1972). Иными словами, структура системы — это и упорядоченное взаимоположение ее вещественных элементов (структура объекта-вещи), и упорядоченное сочетание ее процессуальных элементов— отрезков, или стадий, процесса (структура объекта-процесса). Функция- это способ (процесс) существования структуры системы (Бобров, Петлен- ко, 1967), а шире — способ существования любой системы, поскольку иногда есть системы, обладающие составом, но не обладающие струк- турой. В отличив от состава и функций, которые всегда присутствуют в системах, структура отсутствует в суммативных системах, например, в куче песка структура есть в некоторых отдельных ее элементах, допу- стим, в кристаллах кварцевых частиц, выступающих как системы мень- шей интегративной сложности, чем вся система (куча песка), у которой в полном объеме структуры нет. Следовательно, системы бывают организованные, или целостные, и гемиорганизованные (ненолностъю организованные), или суммативные. Организованные системы обладают составом, который полностью организован, что проявляется в ее це- лостной структуре и функциях. Гемиорганизованные системы обладают составом, который не образует целостной структуры и функций этой системы. Иногда функцией материальной системы особого рода (че- ловека) является идеальная система (особый продукт человеческой соз- нательной деятельности). Функция самой идеальной системы — это, по сути дела, ее предназначение в Природе, контролируемое соответ- ствующей материальной системой. Любые изменения функции неотде- лимо связаны с изменением структуры соответствующей материальной системы (Бляхер., 1962, 1976; Саркисов, 1977). Итак, атом Н, молекула Н,О, автомобиль, растение, институт и т.п. — это организованные сис- темы, а куча песка, толпа прохожих на улице и т.п. — это гемиоргани- зованные (суммативные) системы. Для обозначения какого-либо одного аспекта организации, на наш взгляд, вполне правомерно использовать уже известные терминологи- ческие сочетания: структурная организация (фиксируем все или главное
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ внимание на структурную часть организации), функциональная организа- ция (функциональный аспект организации), временная организация (вре- менной аспект организации). Термины «структурно-функциональная организация», или «пространственно-временная организация», означа- ют то же, что и просто «организация». Вернемся вновь к понятиям движение материи и его формы, чтобы определить отношение к ним названных выше системных понятий. Дви- жение материи в самом общем виде — это, как известно, изменение вообще (всякое изменение) как способ существования материи. Движе- ние неотделимо от материального субстрата, а любой определенный материальный субстрат — это всегда физиосистема (материальная сис- тема). Таким образом, движение неотделимо от существования физио- систем. Физиосистема существует в единстве состава, структуры и фун- кций. Функции — это динамический аспект в существовании физиосисте- мы, поскольку функция — способ существования структуры, а когда ее нет, то способ существования состава физиосистемы. Таким образом, функция с широких позиций — это способ существования любой физиоси- стемы — ее частное движение (изменение). Как вне движения нет материи, так и вне функции (частных дви- жений) нет физиосистем. Функции, когда мы их объединяем в группы по общности каких-либо признаков, составляют типы, а в классифика- циях — классы функций. Как необозрима бесконечность физиосистем, так необозрима бесконечность разнообразия их частных движений (фун- кций). Только в классификациях число функций, достигаемых мыслью наблюдателя, становится обозримым, поскольку главное в этой логичес- кой операции — обобщение. В зависимости от высоты обобщений число классов функций в их иерархических классификациях варьирует. В один из видов таких иерархических классификаций попадает и классификация форм движений по наукам, их изучающим, разработанная в прошлом веке Ф.Энгельсом, то есть, выделяются соответствующие классы функций. Итак, кроме всеобщего движения (изменения) вообще существуют частные движения (функции), неотделимые от их материального суб- страта (физиосистем). При обобщениях частные движения (функции) объединяются в типы (классы), среди которых особое место принадле- жит классификационным построениям. Число классификаций функций и число классов функций зависит от целей конкретного исследования и степени обобщения в классификациях, т.е. от задач и методов клас- сифицирования, и, следовательно, не является величиной постоянной. Так представляется нам на сегодня соотношение одних из центральных понятий науки: движение, система, организация, структура и функция. В чем сущность жизни? Этот вопрос волнует человека с глубокой древности; известны многие попытки мыслителей прошлых столетий дать определение жизни (Сапожников, 1937). Современная наука дока- зала, что основа жизни — наследственность — атрибутивное свойство живой материи проявлять себя под контролем генов в параметрах определенных систем (биосистем) при некоторых условиях среды их существования. Известно множество разных подходов к познанию сущ- ности жизни (Опарин, 1957, 1968, 1977, 1980; Жданов, 1964; О сущности
Глава 2. Обоснование биоморфологии жизни, 1964; Игнатов, 1966; Мамзин, 1966; Костюк, 1967; Бернал, 1969; Кальвин, 1971; Кеньон, Стейнман, 1972; Шредингер, 1972; Проблема возникновения и сущности жизни, 1973; Руттен, 1973; Эйген, 1973; Происхождение жизни и биохимическая эволюция, 1975; Фокс, Дозе,1975; 1Огай, 1976; Поннамперума, 1977; Платонов, 1978; Му- хин, 1980; Медников, 1980; Кедров, Серебровская, 1980; Фолсом, 1982 и др.). Они свидетельствуют об особой организованности материального субстрата жизни. Этот субстрат включает многие элементы периоди- ческой таблицы Д.И. Менделеева, которые сами по себе или взятые в суммативных системах не есть еще жизнь. Химический состав живого очень сложен, однако современная наука доказала, что «все формы жизни основаны на поразительно небольшом наборе главных органи- ческих соединений» (Руттен, 1973, с. 71). До 1935 года из белковых ма- териалов было выделено 20 аминокислот; эти органические молекулы, и, вероятно, только они, содержащие углерод, водород, жслород, азот, а иногда и серу, составляют строительные блоки для белковых макро- молекул, которые в свою очередь составляют большую часть структуры биосистем: на их долю приходится около половины всей сухой массы тела человека и животных (Ленинджер, 1976; Старз, 1976; Дэвени, Герей, 1978; Роллер, 1978; Раубах, 1979 и др.). По сей день признается значение объяснения жизни как способа существования белковых тел, «существенным моментом которого явля- ется постоянный обмен веществ» (Энгельс, 1964), хотя и есть сомнения (Заренков, 1988). В наше время доказано, что синтез белков контроли- руют гены, состоящие из ДНК (Северин, 1970; Уотсон, 1978; Гершен- зон, 1979 и др.), они обеспечивают преемственность жизни, которая базируется на исторически глубоко обусловленном взаимодействии бел- ков и нуклеиновых кислот (Дубинин, 1968). Без белков нуклеиновые кислоты сами по себе не могут проявить свой биотический потенциал, заложенный на молекулярном уровне (Белозерский, 1970). Важная орга- низационная особенность любой живой системы в следующем: живут гены (участки молекул нуклеиновых кислот ДНК и реже, у отдельных классов вирусов, РНК) и все то, что ими управляется в развитии. Генная организация, как материальный носитель информации живого — это первый всеобщий и обязательный уровень биотического движения ма- терии (Дубинин, 1975, 1976, 1981). Отмеченные особенности можно использовать для определения сущности жизни. Жизнь — это существование материи в качестве генов и в ином качестве иод улравлением генов, смешивающихся между собой и трансформирующихся в пространстве и во времени. Гены управляют сохранением всей жизни, включая и самих себя, периодически отчуж- дая в неживое некоторое число своих прямых дискретных носителей (биосистем). Смерть биосистем меньшей организационной сложнос- ти — это разрушительный процесс, контролируемый генами биосистем большей организационной сложности, когда таковые есть. Он возник в эволюции живого как одно из приспособлений для лучшего сохранения Жизни. Смерть наступает через переходное жизненное состояние- предсмерти (скоротечное или более растянутое во времени), от которого
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ два пути: или возвращение к относительно нормальной жизни, или смерть. Итак, главное в жизни и ее сохранении — это сохранение генов и обеспечение условий среды для фенетического развертывания их свойств. Если основой жизни (живого) является наследственность, то основой нежизни (неживого) является, на наш взгляд, преднаследствен- ность. И наследственность и преднаследственность — это формы преем- ственности, из которых первая — высшая из известных пока нам форм преемственности. Под преемственностью понимают связь нового со ста- рым, включение некоторых сторон старого в новое качество (Миклин, Подольский, 1980). Преднаследственность — это атрибутивное свойство материи проявлять системность в определенных вероятностных парамет- рах без участия генов (Нухимовский, 1984). Итак, когда материальная и системная преемственность достигается без участия генов — это преднас- ледственность, а когда материальная и системная преемственность дости- гается при участии генов — это наследственность. Обе формы преемствен- ности физиосистем действуют в определенных спектрах возможностей: а) с узкими (под контролем генов), б) с широкими (без контроля генов) параметрами. Преднаследственные категории по уровню организационной слож- ности ниже наследственных категорий, что можно отразить в соответ- ствующей терминологии. Проген — элемент преднаследственности. Что считать прогеном? Автор пока не имеет на этот вопрос однозначного ответа. Дело в том, что решаюшее слово должно быть сказано после целенаправленного поиска физико-химических границ носителей пред- наследственности и углубленного философского осмысления соответ- ствующего фактического материала. Для автора однозначным остается факт: преднаследственность существует. Она является материальной ос- новой, на которой возникла и существует на Земле наследственность. Прогены, вероятно, имеют или атомную, или молекулярную при- роду. Более убедительным представляется предположение, что проге- нами являются атомы — наименьшие химические частицы любого химического элемента, сохраняющие все его свойства, являющиеся IQ пределом химического разложения любого вещества. Химически эле- мент, как известно, — вид атомов с одинаковым положительным за- рядом ядра. Сейчас известно 110 химических элементов (Шиманович и др., 1996). Прогенотии — это все преднаследственные свойства, локализован- ные в определенной материальной системе (физиосистеме). Прогенотип определяет тот спектр вероятностных реакций (состояний) какой-либо физиосистемы в изменяющихся условиях ее существования, которые не контролируются генами; иными словами, определяет то, чем может быть, а чем быть не может материя какой-либо системы в процессе ее существования. На основе взаимодействия прогенотипа с условиями окружающей среды формируется парафенотип. Парафенотии — это со- вокупность всех, непосредственно осуществляемых в процессе развития, признаков материальной системы, проявляющаяся на основе взаимо- действия прогенотипа с условиями внешней среды. Отдельный признак парафенотипа — парафен, по аналогии с тем, что отдельный признак
Глава 2. Обоснование биоморфологии фенотипа принято называть феном (Яблоков, 1980). Железная гайка, на- пример, в соответствии с ее прогенотипом (преднаследственностью) может проявлять себя по-разному: или сохранять себя в рамках одной системы, принимая разные ее состояния (при нагревании, охлаждении и т.п.), или, в других условиях, полностью трансформироваться в иные системы (при расплавлении, физическом разрушении, сплющивании и т.п.). Пожалуй, в образах носителей преднаследственности (прогенов) и наследственности (генов) будет удобно абстрактно выразить «сердце- вину» Природы, ее главную часть, — Суиернуклеус — основное «фило- софскою (абстрактное) ядро Природы (рис. 1). В такой интерпретации прогены и гены, мысленно сконцентрированные в одном ядре («фило- софском» ядре Природы), захватывают воображение Человека и направ- ляют на поиск скрытых в этом ядре тайн как некоего современного «философского камня» (см. обзор Р.В.Кривокорытовой, 1969), если от- бросить от него всю средневековую мистику. Но сегодня это уже глу- боко осознанный и вполне управляемый научный процесс. Организация Природы, безусловно, таит в себе немало новых ас- пектов для работы ее Разума. Человек проникает все дальше в глубины Вселенной, где его неизбежно ждут открытия как в микроскопической (микромире), так и макроскопической (макромире) частях Природы. С преемственностью связано другое явление — иоаторяемость, или реиеиакция (от лат. repeto — повторять, aetio — действие). Общеизве- стно, что многие явления в Природе повторяются. Под повторяемостью обычно понимают «воспроизведение, проявление в какой-либо вещи или процессе того, что уже было, а затем возникло вновь в том или преобразованном виде» (Полесовой, 1970, с. 31). Повторяемость осуще- ствляется на основе преемственности и включает ее как одну из сторон себя. Как подчеркнул Б.М.Кедров (1961), существенные признаки пов- торяемости, — дискретность, прерывность процесса и возвращение к исходному пункту (воспроизведение в какой-либо степени того, что было пройдено ранее). Повторяемость как абсолютное явление проявляется только в самом общем (отвлеченном) виде — как атрибу- тивное свойство движущейся материи, характеризующееся вечной ее повторяемостью, неисчерпаемостью вширь и вглубь, т.е. повторением материи самой себя в бесконечной трансформации из одной системы в другую (Кедров, 1961, Полесовой, 1970 и др.). Повторяемость — атрибутивная черта в развитии познания. Новое в процессе движения познания отрицает старое; однако отрицание от- рицания в познании имеет «спиралевидное развитие с повторением ранее пройденного на более высоких этапах развития» (Кедров, 1967, с. 23). В работах философов (Гриценко, 1958; Кедров, 1961; Полесо- вой, 1970 и др.) показаны различные проявления повторяемости и под- черкнут относительный их характер. Полной повторяемости чего-либо, что уже было, в чем-либо новом никогда не бывает, в основном, пото- му, что неповторимо и необратимо время, связанное с несимметричнос- тью причинно-следственных связей во всех материальных системах (По- лесовой, 1970). Повторяемость всегда выступает как критерий законо- мерности, обязательная черта закона, поскольку любой закон выражает
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНКИ Рис. 1. Философское ядро в организации Природы. SN — Супернуклеус Природы, 01 — неведомый микро- скопический мир, 1 — прогены и прогенотипы, 2 — гены и генотипы, 3 — парафены, 4 — фены, 5 — парафенотипы, 6— фенотипы, 7 — неведомый макроскопический мир; стрел- ки — направления взаимодействия. то, что постоянно повторяется (Кедров, 1961; Полесовой, 1970). Кроме того, повторяемость выступает в формах особых явлений, подчиняю- щихся своим законам. По мнению философов, во многих случаях формы повторяемости связаны с законом отрицания отрицания. Повторяемость часто прояв- ляется на одной и той же ступени развития как простое движение по кругу, в форме ритмичности циклического процесса однотипных (род- ственных) систем (Кедров, Полесовой и др.). Как отметили А.М.Мик- лин и В.А. Подольский (1980), особые случаи повторяемости наблюда- ются при конвергенции, параллелизме развития, а также как фактор надежности при функционировании и развитии сложных систем в фор- ме избыточности или дублирования элементов, функций, подсистем. После краткого обзора проблемы повторяемости рассмотрим типо- логию наиболее распространенных ее форм (репетактов).
Глава 2. Обоснование биоморфологии Нзомерия — это всеобщее свойство материи повторяться в систе- мах по одинаковому составу (одинаковому числу одинаковых элемен- тов), но различаться в организации (структуре и функциях) этих систем по характеру взаимоотношений элементов. Иными словами, при изомерии, в частности биологической изомерии (Урманцев, 1979), име- ется два и более объектов, имеющих одинаковый состав, но различаю- щихся по некоторым другим свойствам. Единицы изомерии — изоме- ры — части сравнительного ряда систем, одинаковые по составу, но разные по характеру взаимоотношений элементов в организации. Такое широкое понимание явления изомерии, как всеобщего свойства мате- рии, полнее других разработано Ю.А.Урманцевым (1970, 1979). Истоки учения об изомерии уходят корнями вглубь химии (Джуа, 1966; Некра- сов, 1981). Изомерами будут, например, нормальный бутан и изобутан, обладающие одинаковым составом (C,H„), но разные по структуре, цветки с одинаковым числом лепестков, но по-разному расположенные относительно друг друга. Метамерия — это последовательная повторяемость структурно и (или) функционально сходных частей (метамеров) какого-либо более общего явления Природы с единой преднаследственной или наслед- ственной основой. Метамерность рассматривается нами как общенауч- ное понятие, а не как частное явление, наблюдавшееся только у живот- ных (Догель, 1954; Беклемишев, 1964 и др.) и растений (Кренке, 1933; Шафранова, 1980 и др). В неживой природе метамерия не менее обыч- на, чем в живой. Так, звенья фенологических состояний «влажный- сухой» (т.е. влажный, а затем сухой) камень повторяются при повторном сходном изменении условий его существования, составляют вместе одну последовательную совокупность таких звеньев — фенологическую ме- тамерию камня с одной стороны, а с другой стороны заодно и метаме- рию местного климата («дождь — недождь») и т.п. Наша планета испы- тала несколько оледенений, повторяющихся примерно через 150 мил- лионов лет (Джон и др., 1982); организация поверхности Земли и ее атмосферы проявили в этом случае определенные формы метамерии. Целые заводы нередко функционируют метамерно. Повторяющийся процесс производства каждого автомобиля одной марки на конвеере- метамерный процесс; последовательная стадия производства такого ав- томобиля — метамерная стадия, а автомобиль в производственном про- цессе — метамер; после выпуска автомобиля, когда прежняя метамерия утрачивается, может возникнуть новая метамерия или проявиться новое аметамерное качество — бывший метамер может стать диамером. Диамерия (от греч. dia — между, через, meros — часть). Так пред- ложено обозначать непоследовательную повторяемость организационно сходных частей (диамеров) с единой наследственной и (или) преднас- ледственной основой какого-либо более общего явления Природы (Ну- химовский, 1984). Диамерами будут, например, отдельные кирпичи в жилом доме и сходные с ними аметамерные кирпичи в этом доме и в других зданиях; отдельные деревья дуба черешчатого, встречающиеся в лесу, а шире — вообще на Земле, если они не находятся в непос- редственной метамерной (предково-потомковой) связи. У
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Конвергенция — (от лат. convergo — приближаюсь, схожусь). Этот термин широко используется в различных областях знаний, особенно в биологии (Шмальгаузен, 1969; Берг, 1977 и др.). Под конвергенцией обычно понимают возникновение сходных черт организации на изна- чально различной наследственной или преднаследственной основе, т.е. путь развития аналогичных приспособлений (Северцов, 1981). Иными словами, конвергенцию можно определить как возникновение (повто- рение) сходных черт организации в явлениях с разной генетической или прогенетической основой. Известный пример конвергенции у расте- ний — повторение многих признаков внешней структуры кактусов и африканских молочаев. Розовую или иную окраску жидкости можно получить путем совершенно различных химических реакций; в таких случаях розовые по цвету жидкости являются конвергенами (конвер- гентными системами), поскольку сходство по цвету достигается в явле- ниях с разной прогенетической основой. Параллелизм — это возникновение (повторение) сходных призна- ков в явлениях (вещах, процессах), развивающихся параллельно после дивергенции изначально общей организационной основы. Диверген- ция — как известно, возникновение различий на одной организа- ционной основе (Северцов, 1981). Явление параллельной эволюции широко описано в биологии (Osborn, 1905; Яблоков, Юсуфов, 1981 и др.). Известный пример параллельного развития — филогения двух групп копытных млекопитающих: литоптер и непарнокопытных (Яблоков, Юсу- фов, 1981). Из нефти получают разные виды топлива и смазочных средств, которые параллельно иногда применяют в сходных процессах. Итак, в окружающей нас действительности повторяемость просле- живается повсюду: в функционировании далеких звезд, пульсировании гейзеров на Земле, в волнении водной поверхности морей и океанов, изменении поверхности и климата Земли, структуре газопроводов, дви- жении рейсовых троллейбусов, организации побегов и корней растений, производстве конфет на фабричном конвеере, сезонной смене фенофаз развития растений и животных, работе часового механизма, винта са- молета и т.п. Мысли также нередко повторяются. Формами повторя- емости бывают идеи, гипотезы, отдельные слова и иначе однообразно выраженные мысли повторно в последовательности (метамеры) или без нее (диамеры), порождаемые нашей сознательной деятельностью. Из всего изложенного выше можно сформулировать один из высших законов — закон новторяемости: любые явления в Природе всегда имеют возможность новторяться, что когда-либо может быть реализовано. 2.2. Организация Природы, статус биоморф и биоморфологии Человечество не может находиться вне природы; многие авторы рассматривают Человека (вид) как неотьемлемую ее часть, но, говоря о взаимоотношении Человека и природы, выносят социальные отноше-
Глава 2. Обоснование биоморфологии ния за ее рамки. Человек выделился из природы и противопоставился ей как субъект, познающий и преобразующий мир (Слуцкий, 1973). Согласно Ф.Энгельсу и с дополнениями других авторов (Севернев, Сущеня и др., 1980), человек вышел из природы благодаря труду, но по-прежнему берет из нее все средства существования и преобразует ее. Н.Ф.Реймерс (1980, с. 133) отметил, что природа «традиционно противопоставляется человечеству, хотя фактически человечество также часть природы». Природу в широком смысле иногда справедливо определяют «как весь материальный мир, включая неорганическую, органическую и об- щественные ее сферы» (Чепиков, 1981, с. 229). Наша исходная позиция в понимании Природы состоит в следующем: если из чего либо частного можно как-либо (вещественно или мысленно) вычленять части и рассматри- вать их вне этого частного, то в отношении всеобщего и бесконечного— Природы — никак нельзя вычленить часть, чтобы рассмотреть ее вне При- роды. На наш взгляд, Природа (Вселенная, Космос, Мир) — все, что есть в людях и вне их, — это движущаяся материя в ее бесконечном многообразии в лространстве и времени вместе с лродуктом ее высшей деятельности— сознанием (Нухимовский, 1984). Природу сейчас рассматривают, чаще все- го, как единство всего, что есть, как единый многогранный объект ес- тествознания — Вселенная, Разум и Жизнь (Кузнецов и др., 1996). В та- ком смысле будем дапее писать слово Природа с большой буквы. Человек — весьма малая часть в Природе, хотя и самая значитель- ная по сложности организации из пока известных. Правомерно говорить о взаимоотношении Человека (человечества), или необиокосмоса, и иной, остальной, природы, или аллокосмоса. Части Природы разной организационной сложности — что-либо меньшее всей Природы, на- пример, неживая природа, живая природа, дикая природа, окульту- ренная природа ( в смысле часть Природы слово «природа» мы пишем с маленькой буквы), дерево, камень, молния, мысль и т.п. Части При- роды — это и формы движения, и формы организации, и формы позна- ния, и любые иные формы, т.е. все меньшее всей Природы — от эле- ментарных частиц и полей атомов до систем галактик, Метагалактики и т.п. Науки — это также части Природы, отражающие и объясняющие посредством сознания людей то, что есть в Природе (включая и самих себя). Кроме того, принято, говорить о натуре (природе), как сущност- ном (содержательном) выражении какого-либо явления (вещи, процесса) Природы, например, о натуре живого. природе государства и т.д. Сложность организации Природы можно рассматривать под разны- ми углами и с разных высот абстракции. Мы обратим здесь внимание на сложность организации составляющих Природу систем, что одновре- менно с соответствующей интерпретацией будет отражать и иерархич- ность наук, их изучающих. Используя принцип суперординии (Нухимовский, 1984, 1987), посмотрим на организацию Природы через призму разномасштабных обобщений, сознательно подчеркивая отдельными штрихами те ее грани, которые более других касаются главной проблемы — проблемы организации биоморф. Представьте себе, что Вы как бы спускаетесь с вершины созданной нами «пирамиды» абстракций — графической су-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ перординатной модели организации Природы, более или менее задер- живаясь на отдельных ее ступенях (суперординатах) для соответствую- щего обозрения внутреннего содержания этой «пирамиды» (рис. 2). Ри- сунок передает разнообразие материальных систем Природы, объеди- ненных по сложности их организации в несколько субординатных клас- сификаций, которые с разных высот обобщения (уровней абстракции, или суперординат) отражают общее направление эволюции. Мультисуб- ординатная (полисубординатная) классификация может состоять из классификационных ветвей (ответвлений со свободными верхушками) или полуветвей (ответвлений без свободных верхушек, смыкающихся с другими системами). Витки графических спиралей внутри каждой сту- пени пирамиды наглядно передают содержание вихрей материи (путей ее усложнения) в эволюции. Типы материальных систем, отраженные в пирамиде, возникнув однажды, не исчезли, а стали участниками в раз- витии новых материальных систем большей сложности организации, сохранив при этом свою внутреннюю самостоятельность. В эволюции Природы с момента гипотетического сверхгигантского Большого взрыва первоматерии и начала расширения Вселенной (Шкловский, 1973; Ози- ма, 1990 и др.) до наших дней возникло бесконечное разнообразие мате- риальных систем (оно сводится к вполне определенному числу типов для всей известной нам части Вселенной), в котором масса биосистем (но не значимость) ничтожно мала.
Глава 2. Обоснование биоморфологии В Природе в единстве существуют, как известно, материальное (объективный мир, первичное) — материальная природа (физиостроба, дословно с греческого — вихрь природы) и ее части — материальные системы (физиосистемы), и идеальное (субъективный образ объектив- ного мира, вторичное) — идеальная природа (логостроба, дословно— вихрь мыслей) и ее части — идеальные системы (логосистемы). Иными словами, материальная природа — бесконечно движущаяся материя в бесконечном пространстве и времени. Идеальная природа — результат (идеальная функция) особо организованной части материальной приро- ды — функция человеческого мозга. Идеальная природа — все идеаль- ное, рождаемое в мозгу людей как формы отражения (истинные, фантас- Рис. 2. Графическая суперординатная модель (пирамида абстракций) организации Природы. 01 — логосистемы (идеальные системы); 1 — материальные системы; 2 — абиосистемы; 3 — биосистемы; 4 — протосистемы (к ним относятся также 9-11, 14); 5 — неосистемы (к ним относятся также 7, 12, 13, 15); 6 — аллосистемы; 7 — необиосистемы (новые, или разумные, живые системы); 8 — экосистемы; 9 — протобиосистемы (первоприродные живые системы, т.е. без людей в сфере социальных отношений); 10 — протоабиосистемы (неживые абиогенные системы); 11 — палеобиосистемы (неживые биогенные системы); 12 — неоэкосистемы; 13 — техноабиосистемы (абиосистемы, созданные с участием чело- века); 14 — протоэкосистемы (первоприродные экосистемы); 15 — техноэкосистемы (эко- системы, преобразованные людьми, но не включающие людей); 16 — экзоабиоморфы (абиосистемы вне структуры биоморф); 17 — мезоабиоморфы (абиосистемы внутри биоморф, но не в их струкгуре); 18 — эндоабиоморфы (абиосистемы, включенные в структуру биоморф); 19 — гипобиоморфы; 20 — биоморфы; 21 — метабиоморфы; 22— гипоэкоиды; 23 — экоиды; 24 — синэкоиды; 25 — метаэкоиды; 26-30 — субординия аби- осистем: 26 — поля атомов, 27 — кварки, 28 — элементарные частицы, 29 — атомы, 30— молекулы; 31-44 субординия протобиосистем: 31 — биополимеры, 32 — органеллы, 33— клетки, 34 — ткани, 35 — органы, 36 — свободные проорганизмы (пыльцевые зерна, га- меты и другие гипобиоморфы, отделившиеся от родительского организма); 37— организмы, 38 — синорганизмы, 39 — гамы (семьи организмов), 40 — популяции, вклю- чая ценодемы (ценотические популяции), 41 — эйдобионты (виды организмов), 42— меробиоценозы (сообщества организмов какого-либо очень крупного таксономического ранга, например фитоценозы, зооценозы, микробоценозы и т.п.), 43 — биоценозы, 44— протобиостроба (живая природа, не включая человечество); 45-49 — субординия необи- осистем: 45 — неоорганизмы (особи людей в сфере сознательной деятельности), 46— неогамы (семьи людей), 47 — неодемы (популяции людей), 48 — этномы (народы — осо- бые совокупности неодемов), 49 — необиостроба (вся разумная живая природа); 50— биостроба; 51-64 — субординия протоэкосистем: 51 — экобиополимеры, 52 — экооргано- иды, 53 — экоцеллы, 54 — экогистоны, 55 — экоорганы, 56 — свободные экопроорганиз- мы, 57 — экоорганизмы, 58 — экосинорганизмы, 59 — экогамы, 60 — экоценодемы и иные популяции вместе с их биотопами, 61 — экоэйдобионты, 62 — экомеробиоценозы, 63 — биогеоценозы (экобиоценозы), 64 — протобиосфера; 65-69 — субординия неоэко- систем: 65 — эконеоорганизмы, 66 — эконеогамы (семья людей вместе с их биотопами), 67 — эконеодемы, 68 — экоэтномы, 69 — необиосфера; 70 — биосфера; 71-75 — суборди- ния абиосистем (продолжение, см. также 26-30): 71 — агрегаты молекул, 72 — кристаллы, 73 — породы, 74 — пласты, 75 — геосфера; 76-78 — субординия гиперэкосистем: 76 — био- планеты (пока известна только Земля), 77 — биоастропланеты (пока известна только Солнечная система), 78 — биогнлактики (пока известна одна наша Галактика); 79-82— субординия гиперабиосистем (продолжение, см, также 26-30, 71-75): 79 — пропланеты (безжизненные астероиды и пропланеты), 80 — планеты (без жизни), 81 — астропланеты, 82 — галактики; 83 — системы галактик и биогалактик (пока известна только Метагалак- тика)... Пунктирными линиями показано замыкание взаимосвязей субординатных рядов материальных систем на самих себя или на узловые субординаты других рядов.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ тические, патологические и др.) объективной действительности. Формы проявления идеальной природы — это, таким образом, ее части — иде- альные функции, порождаемые материальной природой. Материальное и идеальное сосуществует в единстве, причем идеальная природа невоз- можна без материальной природы, хотя обратное вполне реально. Это отвечает закону единства материального и идеального, известный смысл которого можно выразить так: идеальное порождается разумной живой природой (людьми), существуя в единстве с ней и для нее, преобразуя ее, себя и окружающую их остальную часть материальной природы. Из закона единства материального и идеального вытекает также известное диалек- тическое положение в качестве правила этого закона (см. гл. 1). Через идеальное человеком познается все, что окружает и составляет человека. Идеальное, будучи одной из особых функций материального, возникая, или быстро исчезает, или более или менее долго сохраняется в мате- риальном (опредмечивается), приобретая формы материальных тел, язы- ковых знаков, символов и т.п. Логосистемами будут мысли, слова, кон- цепции, классификации, науки и т.п. Анализ организации идеальной природы мы оставляем в стороне, однако, используя идеальную приро- ду, ее разносложные логосистемы, будем классифицировать, определять законы, строить концепции, теории, т.е. познавать Природу, в частно- сти ее материальную часть, поскольку в этом высший смысл проявления идеального как особой функции высшего материального (Человека). Высшими частями материальной природы можно считать две изве- стные части Природы: неживую природу (абиокосмос, или абиостробу, что дословно — вихрь неживого) и живую природу (биокосмос, или биостробу, что дословно — вихрь жизни). Материальную природу можно посред- ством субординатного классифицирования представить и как систему, состоящую из следующих частей: 1-я субордината — абиосистемы (лю- бые неживые материальные системы); 2-я субордината — биосистемы (любые живые материальные системы); 3-я субордината (верхняя)— экобиосистемы, или, короче, экосистемы (любые экосистемы, включая всю биосферу). Биосистемы и экосистемы являются, в свою очередь, биофорами. Организацию биофоров частично или полностью составля- ют биосистемы. Термин «биофор» (носитель жизни) был использован довольно давно А. Вейсманом (1905) для обозначения гипотетических в то время «мельчайших жизненных частиц», составляющих «все живое вещество, клеточные и ядерные вещества, вещество мышц, нервов и желез во всех их вариантах». Бурное развитие генетики и молекулярной биологии в последующие годы вскрыло организационную сущность мельчайших хранителей и носителей жизни — биополимеров, что обес- ценило первоначальный смысл термина «биофор». Можно попытаться придать этому интересному, но забытому термину новый смысл. Био- форы — это, в первую очередь, биосистемы: фрагменты материи, полно- стью вовлеченные в жизнь; во-вторых, — это экосистемы: фрагменты материи, состоящие из определенных биосистем и окружающих их сред существования (биотопов), с которыми они взаимодействуют и выступа- ют вместе как одно целое, оставаясь не тождественными каким-либо био системам.
Глава 2. Обоснование биоморфологии Кроме того, высшие части Природы можно представить и иначе, под разными углами обзора, но с одной высоты обобщений: или как первоприроду (протокосмос) и новую природу (неокосмос), или как новоприродную (разумную) жизнь (человечество, необиокосмос) и иную (природа, кроме человечества) природу (иноприроду, или аллокосмос). Все эти части Природы состоят из систем с отвечающей им организа- ционной сущностью, — соответственно или протосистем, или неосис- тем, или необиосистем, или аллосистем. К протосистемам относятся протоабиосистемы — неживые абиогенные системы, среди которых и вся неживая абиогенная природа, или протоабиокосмос; палеобиосис- темы — неживые биогенные системы во главе со всей неживой биоген- ной природой, или палеобиокосмосом; протобиосистемы — живые первоприродные системы, включая и всю живую первоприроду (прото- биокосмос), т.е. биосистемы без человечества и отдельных людей в сфере социальных отношений; протоэкосистемы — любые первоприродные экосистемы и вся экосистемная первоприрода (протоэкокосмос), т.е. экосистемы без человечества, отдельных людей в сфере социальных отношений и систем, созданных с трудовым участием людей. Среди неосистем мы различаем: необиосистемы, или нообиосисте- мы — новые (разумные) живые системы, главная из которых вся новая (разумная) живая природа (необиокосмос, или нообиокосмос), т.е. че- ловечество и его части — от отдельных людей до коллективов людей, вплоть до всего человечества; неоэкосистемы — новые экосистемы, главная из которых новая экосистемная природа (неоэкокосмос), т.е. экосистемы с участием отдельных людей или коллективов людей; тех- ноабиосистемы, или неоабиосистемы — новые неживые системы во главе со всей новой неживой природой (неоабиокосмосом, или техно- абиокосмосом), т.е. абиосистемы, созданные G трудовым участием чело- века; техноэкосистемы — обновленные протоэкосистемы, высшая из которых вся обновленная экосистемная первоприрода, или техноэкокос- мос, т.е. экосистемы, преобразованные трудовой деятельностью людей, но не включающие в себя людей. Техноабиосистемы и техноэкосистемы — это все техносистемы. В соответствии с рассмотренными выше представлениями об организации Природы с наиболее высоких уровней абстракции мы предлагаем гово- рить не как обычно о взаимоотношении Человека (человечества) и при- роды, поскольку так искажается смысловое содержание терминов, а о взаимоотношении новой природы и первоприроды или, если особо выделять лидера Природы (достоверно известен пока только один ли- дер — Человек, который способен познавать и преобразовывать в своих интересах объективную действительность), то тогда следует говорить о взаимоотношении человечества (необиокосмоса) и остальной природы, или иной природы (аллокосмоса, т.е. всего, что не входит в необиокосмос). В наших представлениях о Природе следует особо сказать о чело- веческом как свойстве материального мира. Человеческое свойство двойственно по своей сущности (натуре): от биополимеров до организ- мов-людей человеческое принадлежит первоприродной жизни, а от ор- ганизмов-социальных личностей (неоорганизмов — организмов, обла-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ дающих разумом и проявляющих себя в сфере социальных отношений), через их разносложные совокупности до самых сложных по организа- ции коллективов людей человеческое принадлежит новоприродной жиз- ни, или человечеству (особой социальной сущности, обусловленной высшей формой деятельности — разумной деятельностью). «Организм» обычно рассматривают как понятие, характеризующее биологическую структуру человека, а понятие «личность» — для выражения структуры человеческого индивида в его социальной соотнесенности», иными сло- вами, «организм» и «личность» — это «два аспекта одной и той .же реальности» (Сержантов, 1974, с. 59-60). В человеческом организме не- зримо проходят границы между протобиотической и необиотической сущностями; размытость этой границы — свидетельство неразрывного единства этих натур в человеке — роде, виде и, отвечающих человечес- кой сущности, его частях — системах. Сказанное выше наглядно видно в графической модели организации Природы (рис. 2). В этой модели отражены особенности функционирования субординатных классифика- ций, в субординатах которых представлены отдельные области органи- зации разносложных частей Природы. Если с несколько меньшей высоты обобщений, меняя иногда угол обзора, посмотреть на материальную природу, т.е. постепенно спускать- ся по «пирамиде» абстракций (рис. 2), то в материальной природе мы увидим следующие ее части-системы. Биосистемы, которые являются одновременно эврибиоморфами, или формами жизни (жизненными форма- ми, биоморфами) в широком смысле слова, среди которых было предложено (Нухимовский, 1974а, 1984) различать три типа, связанных между собой субординатной связью: гипобиоморфы, биоморфы, метабиоморфы. Биомор- фы (жизненные формы в узком смысле) — организмы (индивидуумы, осо- би) и синорганизмы (кондивидуумы); гинобиоморфы (живые части биоморф, а также отделившиеся от них, но не превратившиеся в особи гаметы, споры, деструктивные партикулы) — биосистемы, составляющие стру- ктуру биоморф или обособившиеся от них, но не ставшие еще биоморфами; метабиоморфы (системы биоморф) — биосистемы, представляющие опре- деленные коллективы биоморф (популяции, виды и др.). Статус биоморф в организации Природы будет нагляднее, если мы отразим его не только во взаимосвязи с другими биосистемами, но и во взаимосвязи с абио- системами (неживыми материальными системами), или абиоморфами (формами неживого). Среди абиосистем различимы: экзоабиоморфы (абиосистемы, не входящие ни в состав, ни в структуру биоморф, на- пример атомы камней, камни, автомобили и т.п.), мезоабиоморфы (не- живые абиосистемы, вовлеченные в состав биоморф, но не участвую- щие в их структуре, например, все неживое, что попадается в ротовых полостях, в полостях желудков, легких животных и т.п.), эндоабиомор- фы (абиосистемы, включенные в структуру биоморф, т.е. элементарные частицы, атомы, молекулы и т.п. в структуре биоморф). Значение термина «экосистема», предложенного Вольтереком и Тенсли (Работнов, 1978), пока не устоялось. Нас устраивает безранговое положение этого термина, хотя более всего экосистемой считают какой- либо биоценоз вместе с его биотопом (Дажо, 1975; Одум, 1975 и др.).
Глава 2. Обоснование биоморфологии Предлагаем широкое понимание термина «экосистема». Такой подход открывает интересные возможности для развития современной эколо- гии, или экосистемологии. И.П.Герасимов (1980), рассмотрев в общих чертах особенности современных экологических исследований, заклю- чил, что экология, зародившись и развившись в системе биологических наук, сегодня уже вышла из под контроля биологии. Со времен автора термина «экология» Э.Геккеля классическая эко- логия («гемиэкология» по отношению к современной экологии) изучает отношение биосистем организменного и эпиорганизменного (популя- ции, сообщества) уровней организации живого к каким-либо факторам окружающей среды и является по сей день как бы частью «слоеного пирога» биологии (Одум, 1975). Современная экология изучает экоси- стемы. Посредством метода широкой субординии экосистемы можно представить в двух типах: как протоэкосистемы — они состоят из протобиосистем уровня биоморф и метабиоморф и их биотопов, и как неоэкосистемы — это системы, состоящие из необиосистем уровня биоморф и метабиоморф и их биотопов. К протоэкосистемам относятся также некоторые гипобиоморфы — обособленные от организмов гипо- биоморфы: деструктивные партикулы (отрезки стеблей, листьев и т.п. части организмов, не восстановившие еще основные органы для само- стоятельного существования в форме организмов), а также гаметы, споры вместе с окружающей их средой существования (биотопом). Под несколько иным углом обобщения среди экосистем мы различаем: эко- иды (термин Negri, 1954, цит. по А.А. Корчагину, 1976) — системы от- дельных индивидуумов и их биотопов (в биотопе участвует как неживое, так и особи-консорты), синэкоиды — системы отдельных синорганиз- мов и их биотопов (единые или разобщенные, в последнем случае, например, при размножении черенками родительского растения), гипо- экоиды (они стоят ниже экоидов в субординатной классификации)— системы отдельных гипобиоморф в качестве обособленных деструктив- ных партикул, гамет, спор и их биотопов, например, гипоэкоидом яв- ляется и замороженный живой орган с окружающей его средой в холо- дильной установке, и культивируемые в пробирке живая клетка или отдельная живая ткань вместе с их новым биотопом и т.п., метаэкои- ды — это системы из отдельных метабиоморф и их биотопов. Итак, обязательными компонентами экосистем в нашем понима- нии будут биосистемы, выступающие в ранге биоморф или метабиоморф (лишь в особых случаях — обособленные от организмов гипобио- морфы) и окружающие их среды существования (биотопы). Обычно «биотопом» (дословно местом жизни) называют «места произрастания одного определенного, реально существующего биоценоза» (Шмитхю- зен, 1966, с. 103). Однако биоценозы — это только одна часть в иерар- хической организации живого, у остальных представителей живой ма- терии есть свои «места жизни». Учитывая это обстоятельство, мы пред- лагаем шире понимать термин «биотоп» с учетом тех изменений, кото- рые происходят в биологии в процессе ее развития под влиянием сис- темного подхода: как любые места жизни (существования) любых био- систем. Иными словами, любая биосистема составляет вместе со средой
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ее существования (биотопом) новую систему с иной сложностью орга- низации: или другую биосистему, или экосистему. Биосистема-гипо- биоморфа (кроме особых случаев превращения ее в обособленную де- структивную партикулу или обособленную гамету, спору) вместе с ее биотопом — это другая биосистема, а биосистема — или биоморфа, или метабиоморфа (в особых случаях гипобиоморфа — обособленная деструктивная партикула или обособленная гамета, спора) вместе с ее биотопом — это какая-либо экосистема. Биосистемы — объект иссле- дования биологии, а экосистемы — экосистемологии. Биостроба включает все биосистемы меньшей, чем она, организа- ционной сложности. На суше живое вещество (организмы и их совокуп- ность), как известно, распространено по вертикали в основном вверх и вниз до 50-100 м от поверхности, редко более, а в океане значительно шире. Это пространство — опорный «слой живого вещества», или «пленка жизни» биосферы Земли в понимании В.И. Вернадского, или эубиострома (буквально «хорошая живая постель»), названная Е.М.Лав- ренко (1964) биостромой. Небольшое понятийное уточнение термина «биострома», предлагаемое нами, конкретизирует представления об ор- ганизации биостробы и биосферы. Биостроба, на наш взгляд, состоит из трех слоев живого вещества — биостром: эубиостромы и двух других слоев очень рассеянного живого вещества — гипобиостромы (слоя жи- вого вещества ниже эубиостромы) и эпибиостромы (слоя живого веще- ства выше эубиостромы). Границы этих биостром весьма расплывчаты, как, впрочем, и границы биосферы. Биостроба может иногда вырываться за пределы биосферы, и потенциально она имеет тенденцию к экспан- сии в глубины Космоса (Шкловский, 1973; Урсул, 1977). Понятия био- строма (Лавренко) и биоценоз (Мебиус, цит. по Шмитхюзену, 1966) включает только общности живых существ без неживых компонентов среды их обитаний. Попытки включать в определения этих терминов также и неживую среду местообитаний неоправданы, поскольку эти объекты будут причисляться, по выражению Е.М.Лавренко (1964), уже не к «биологической вертикали», а к экосистемному ряду: биосфере, био- геоценозу и т.д. Спускаясь от одной суперординаты к другой, мы видим, как пос- ледовательно уменьшается степень обобщения в отдельных субординат- ных и мультисубординатных классифйкациях и как увеличивается чис- ло субординат, составляющих такие классификации. Так, на нижней суперординате, как видно из подробных условных обозначений к по- строенной нами модели (рисунку 2), отражена дальнейшая детализация типов физиосистем (материальных систем) до многих частных типов таких систем. Отдельные субординии в мультисубординатной класси- фикации как бы замыкаются на одном общем для нескольких суборди- ний классе систем (субординате). Так, например, все разнообразие про- тобиосистем и необиосистем подчиняется (в модели «замыкается») более общему для них типу биосистем — биостробе, которая пока известна только в единственном числе у нас на Земле. Жизнь на Земле, биостроба, однажды возникнув, несет себя во Вселенной, отдав свое сохранение под контроль человечества — но-
Глава 2. Обоснование биоморфологии остробы (дословно — вихрь разума), или необиостробы. Возникнове- ние разумных существ в эволюции Природы — синдром новизны выс- шего порядка, поэтому для всех живых систем, включая человека, мы предлагаем сложные термины, в состав которых входят слова «нео» (новый) и «ноо» (разум). Статус биоморф (организмов и синорганизмов) наглядно виден на рис. 2, а вот каков статус биоморфологии — науки об организации био- морф? Появление слов-этикеток «биоморфа» и «биоморфология» в струк- туре терминов с такими названиями (термин = слово-этикетка+ поня- тие) — событие сегодня для науки в общем-то тривиальное, поскольку основа для этих терминов известна давно (Darwin, 1859; Вейсман, 1905), да и соответствующий перевод их был дан еще в 20-х годах нашего столетия (цит. по Б.А. Домбровскому, 1967), и все же широко эти тер- мины стали применяться лишь в последние 35 лет (Голубев, 1962, 1965; Нухимовский, 1973а, 1974а, 1984 и др.). Под термином «биоморфология» здесь нанимается наука о жизненных формах (биоморфах), их структурах и функциях, закономерностях разви- тия, отношениях к среде обитания, т.е. наука об организации биоморф (Нухимовский, 1974а). Такую позицию надеемся закрепить как прогрес- сивную в эволюции рассматриваемых терминов. С этой целью ниже еще будет проанализирована история проблемы и предложены пути ее ре- шения. Пока отметим, что роль биоморфологии в последнее время резко возросла, и она начинает приобретать особое связующее значение в системе биологических наук. Если посмотреть на готовую суперорди- натную модель организации Природы под несколько иным углом зре- ния, то нетрудно понять, что она может одновременно служить и супер- ординатной классификацией наук, изучающих соответствующие типы материальных систем и типы (формы) функций (движений), в которых эти системы себя проявляют. Оставляя в стороне рассуждения о классификации наук, обратим здесь внимание лишь на статус биоморфологии в комплексе наук о биофорах, т.е. в биофорологии. При дальнейшей детализации выходим к еще более частным на- укам. Например, частями биоморфологии будут науки, изучающие ка- кие-либо отдельные стороны организации биоморф, — разделы био- морфологии. Так, один из разделов биоморфологии — габитология (эли- морфология, или экзоморфология) — наука, изучающая в развитии za6umyc (внешнюю организацию) биоморф (габитуальные формы по Ф.Клемент- су — цит. Du Rietz, 1931; формы роста, Du Rietz, 1931; Meusel, 1951; эпиморфы, Schmid, 1963 и др.) с учетом воздействия на них различных экологических факторов. В этом смысле габитология соответствует по сути дела экологической морфологии, основы которой сформулированы И.Г.Серебряковым (1962), и может рассматриваться как более краткий синоним этого раздела биоморфологии. Статус биоморфологии в организации биофорологии можно изобра- зить следующим образом (в скобках указаны объекты исследований соответствующих наук).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Биофорология (биофоры) синбиология, или биосистемология (биосистемы, или эврибиоморфы) экология, или экос истемология (экосистемы) биология, или протобиология (протобиосис- темы) социология, или необиология (необиосисте мы) неоэкология (неоэкосистемы) протоэкология (протоэко- системы) метабиоморфо- логия (метабиоморфы) гипобиоморфо- логия (гипобиом арфы) биоморфол огня (био марфы) Особо следует сказать об экологии. Киассическая экология (гемиэко- логия, что дословно «налоловину экология»), на наш взгляд, часть биоло- гии. Современная экология, или экосистемология (этот термин уже упот- реблялся в более узком смысле Т.А.Работновым (1979), состоит, на наш взгляд, из протоэкологии (она изучает лротоэкосистемы) и неоэкологии, или экологии человека (она изучает неоэкосистемы). Ранее был предложен термин «социоэкосистемы» для обозначения сложных систем, элементами которых являются социально-экономи- ческие и внесоциальные (остальные природные) системы, находящиеся во взаимодействии между собой, а науку, изучающую эти сборные системы, предложено называть социоэкологией (Norton, 1973; Фро- лов, 1980). Мы шире понимаем необиосистемы, относя к ним не только коллективы людей с их биотопами, но и отдельных людей с их биото- пами, проявляющих себя в особой сфере деятельности (сознательно- трудовой), недоступной всей остальной части Природы. Кроме того, грамматически неудачным является греко-латинское смешение исход- ных слов, которое допущено при создании упомянутых терминов Дж. Нортоном. Как известно, при создании терминов важно не нарушать принципы построения терминологии (Лотте, 1982). В соответствии с приведенными выше материалами биофорологию можно представить как систему наук, состоящую из биологии, необиологии, протоэкологии и неоэкологии. Представления об организации Природы, рассмотренные здесь, мы не считаем единственно возможными, но видим в них основу для
Глава 2. Обоснование биоморфологии углубления в будущем подобных исследований, а также используем как вспомогательный материал для наших дальнейших рассуждений. 2.3. Общие закономерности организации биосистем Современные знания о натуре (сущности) живых систем (биосис- тем) весьма многогранны. Выделим основные закономерности, присущие всем биосистемам. Такая оценка позволит далее более точно отразить своеобразие биоморф семенных растений. 3 коном ность п в . Биосистемы — это целостные системы. Проблема целостности широко обсуждается в литературе (Абрамова, 1962; Югай, 1962, 1965; Афанасьев, 1964, 1980; Малиновский, Уемов, 1966; Гирусов, 1968; Драгавцев, 1968; Кнорре, 1968; Кремянский, 1968; Молотковский, 1968; Чижек, 1968; Юдин, 1972; Шмальгаузен, 1982 и др.). Всеми признается, что обязательная черта целого — наличие в нем частей (элементов) и связей между ними, которые объединяют части, что приводит к появлению у них совокупности новых свойств, не при- сущих им при их разобщенности. Г.А.Югай (1965) считает, что системы бывают целостные и нецелостные. В.Г.Афанасьев (1980) различает це- лостные и суммативные системы. Главное в целостной системе — ин- тегративные качества, а в результате взаимодействия элементов сумма- тивной системы не возникает качественно новых (интегративных, це- лостных) свойств. Возникновение целостности характеризуется интег- рацией дифференцированных частей на выполнение общих функций и возникновение регуляции, а затем и органов регуляции (Мукашев, 1968). Согласно И.В.Блаубергу (1977), целое — это конкретный объект, обла- дающий интегративными свойствами, а целостность — представление о полноте охвата явлений и одновременно о сущности интеграции, процессах новообразования, иерархической организации явлений и т.п. (фон, на котором познаются целостные объекты). По А.А.Малиновско- му (1980), целостность — частный случай упорядоченности и органи- зованности, а целостная система — это система, отдельные части кото- рой функционируют (или развиваются) совместно, и в то же время вся система относительно независима от среды и от других аналогичных сис- тем. «Целое обладает новыми свойствами, которых нет в отдельных частях» (Шмальгаузен, 1982, с. 13). Как считает И.В.Блауберг (1977), синтезом категорий целостности и системности является понятие орга- низация — отражение аналитико-синтетического подхода к единству, взаимосвязи явлений действительности. Целостность определяется не уровнем организации, а степенью «плотности» внугрисистемных связей, превышающей «плотность» связей между системой и средой (Малинов- ский, Уемов, 1966; Малиновский, 1980). Итак, целое — это совокупность предметов, проявляющая себя в единстве через интегративные качества ее частей. Части биоценоза- виды; части вида — особи, популяции, части организма — системы ор-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ганов, органы; части органов — ткани; части тканей — клетки и т.д. (Афанасьев, 1968). Резюмируем главный итог нашего краткого обзора проблемы цело- стности: вне целостности нет живых систем. Закономе ность вт ая. Биосистемы — это открытые системы. Теорию открытых систем впервые (1932) разработал Л.Берталанфи (Ber- talanffy, 1949, 1971 и др.). «Иерархическая организация и «открытый» характер систем являются основными принципами живого; достижения теоретической биологии будут зависеть главным образом от развития этих двух оснований» (Bertalanffy, 1949, цит. по М.И.Сетрову, 1971а, с. 44). Открытой называют такую систему, которая обменивается со средой веществом и энергией. Закрытая система обменивается со средой энер- гией, а обмена веществом нет. Кроме того, различают изолированные системы, закрытые для обмена как веществом, так и энергией (Сетров, 1971а). Иногда последние два типа систем называют замк- нутыми системами. Согласно Л.Берталанфи, открытые системы прояв- ляют себя эквифинально, т.е. они способны из различных положений приходить в одинаковое состояние, в отличие от замкнутых, жестко детерминированных, систем (Сетров, 1971а). В биосистемах, являю- щихся открытыми системами, существуют агенты постоянно действую- щие или готовые к действию, механизм которого разнообразен, чтобы поддержать какие-либо по форме состояния постоянства внутренней среды (в определенном диапазоне их колебаний) — гомеостаз (Can- поп, 1932; Новосельцев, 1978 и др.). В любую живую открытую систему, в качестве которой чаще всего приводят организмы (Bertalanffy, 1949, 1971; Новосельцев, 1978 и др.), постоянно извне поступают веще- ства, которые подвергаются различным преобразованиям на основе ее метаболизма: часть веществ частично утилизируется (анаболизм), а ос- тальная часть выводится из системы (катаболизм). Состояние относи- тельного внутреннего подвижного равновесия (гомеостаз) в какой-либо биосистеме (чаще говорят об организме) различен по форме прояв- ления. Гомеостаз в любых системах поддерживается различными спо- собами. Исходя из известных в науке фактов и обобщений, некоторые из которых упомянуты выше, открытость биосистем можно отразить как следующую закономерность: системы, которые доиускают в себя и вы- водят из себя какое-либо количество вещества и энергии из того, что их окружает, открыты в иространстве и во времени до тех пор, иока своими функциями более или менее иротиводействуют факторам их замыкания- ирекращению обмена веществ и энергии. Закономе ность т етья. Биосистемы — это иерархические системы. Они подчиняются, как и любые другие системы, закону иерархичности, сформулированному в гл. 1. Как отмечалось выше, иерархичность в ор- ганизации биосистем нашла отражение в концепции структурных уров- ней или уровней организации живой материи. Эти представления сти- мулировали разработку метода субординатного классифицирования, ко- торый мы широко используем в познании пространственной и времен- ной организации биоморф семенных растений.
Глава 2 Обоснование оиоморфологии 3 кономе ность четв тая. Биосистемы — это самоунравляемые (саморегулируемые) системы. С позиций системной методологии пробле- ма управляемости систем — одна из ведущих в современной науке. На основе познания закономерностей строения управляющих систем и те- чения процессов управления функционирует молодая наука — кибер- нетика (Коган и др., 1972; Ляпунов, 1980). Обратим особое внимание на факт организационной расчлененности биосистем на две категории ча- стей: управляющие и исполняющие, которые используем в последую- щих рассуждениях. В определении сущности жизни, сформулирован- ном выше, была подчеркнута управляющая роль генов в организации биосистем. Существущую закономерность в процессе самоуправления биоси- стем можно сформулировать как закон унравления — иснолнения органи- зации биосистем: иснолнительная (тактическая) часть организации био- систем реализует генетическую информацию (находится нод унравлением генов) носредством своего роста и развития через главенство иснолнитель- ских частей этой организации, ответственных за воснроизведение себе нодобных, над частями, снабжающими нервые части всем необходимым для развития и реализации этой функции и (или) удобно раснолагающими их в иространстве и времени. 3 кон ме но ть пятая. Биосистемы — это воснроизводящиеся сис- темы. По генетическим планам в зависимости от трансформирующего влияния факторов окружающей среды (с мутациями или без них) по- стоянно на Земле из исходного живого рождается производное (новое) живое. Биосистемы, которые рождают себе подобных (размножают- ся), — это самовоспроизводящиеся биосистемы (Вилли, 1964 и др.). Од- нако не все биосистемы самовоспроизводятся. Иногда биоценозы вос- производятся по воле Человека, который конструирует модель биоце- ноза на бумаге, а затем с помощью соответствующего посадочного и посевного материала схема воплощается в реальный биоценоз. Воспро- изведение здесь управляется Человеком, отодвигая соответствующие системы растений и животных на второй этап управления. Во всех достоверно известных случаях живое воспроизводится из живого. Все прошлые и современные биосистемы, кроме гипотетических самых пер- вичных биосистем или одной самой первой биосистемы (сколько их было — самых первых биосистем: одна или несколько — это никто никогда не узнает), возникших из пробионтов (предживых систем) и неживых систем (Опарин, 1957, 1980 и др.), существуют на основе из- вестного закона биогенеза (воснроизведения живого), смысл которого вы- ражен афористически кратко: «все живое (кроме нротеробиосистем- нервых исходных биосистем — Е.Н) нроисходит только от живого» (Вил- ли, 1964, с. 16, 27). Закономе н сть шестая. Биосистемы — это биополимерные систе- мы. В главе о натуре живого было показано, что современная биология стоит на твердо установленных фактах: нижний и всеобщий уровень организации живого, или нижнюю субординату в субординатной клас- сификации биосистем, занимают биополимеры — живые макро- молекулы — гены, белки (Роллер, 1978; Уотсон, 1981 и др.). Биополи-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ меры составляют более сложные системы, в которых они синтезируются и функционируют: органеллы клеток, клетки, ткани, органы, организмы и т.д. вплоть до всей биостробы. На биополимерном уровне находятся главные управляющие системы любых других биосистем — гены — са- мовоспроизводящиеся молекулы ДНК (реже РНК — у вирусов), обла- дающие уникальной способностью синтезировать себе подобных и уп- равлять синтезом других биополимеров — белков. Очередную закономерность живой природы, которой подчиняются все известные на Земле биосистемы, можно выразить так: статус осно- вы живого иринадлежит биоиолимерам — самовосироизводящимся моле- кулам генов и синтезируемых иод их уиравлением белкам. Закономе ность седьмая. Биосистемы — это каркасные системы. На разных уровнях организации живого можно обнаружить остов (кар- кас), на который опираются в своем существовании все другие состав- ляющие (элементы) биосистемы соответствующего уровня. Так, эле- мент С (углерод) — великий соединитель — «мастер устанавливать ко- валентные связи» (Роллер, 1978). Этот элемент обязателен для органи- зации биосистем. Углерод можно рассматривать как химический (нежи- вой) каркас биосистем. Последовательности азотистых оснований, со- ставляющих ДНК и (или) последовательности аминокислот в белках можно рассматривать как биохимический каркас биосистем. Среди ор- ганоидов каркасную функцию выполняют оболочки клеток. Клетки многоклеточных организмов, специализированные для выполнения ме- ханических функций, например, клетки колленхимы или склеренхимы у семенных растений (Эзау, 1980), клетки костной ткани у позвоночных животных и т.д. выполняют роль каркаса на клеточном уровне органи- зации биосистем. На уровне тканей каркас составляют специализиро- ванные опорные механические ткани, например, колленхима, склерен- хима у семенных растений, костные ткани у животных. На уровне ор- ганов каркасные функции несут более или менее специализированные опорные органы, например, стебли, корни у семенных растений, кости у позвоночных животных и т.д. На уровне организмов каркасная фун- кция принадлежит размножающимся особям. Обладая этими функция- ми, последние составляют каркас популяций. Каркасные ценотические популяции (по их функциональной роли) — это нормальные ценоти- ческие популяции (Работнов, 1978), они обеспечивают остов видам. Сре- ди видов каркасные функции проявляют те, которые являются доми- нантами фитоценозов или зооценозов; такие виды составляют каркас определенных биомероценозов. Как ни странно, в иерархии биосистем пока не находили собственного места такие известные группировки организмов, как фитоценозы и зооценозы. Дело в том, что их обычно относят к биоценозам, допуская при этом смешение понятий под одним термином. И фитоценозы, и зооценозы — это, по всеобщему призна- нию, — части биоценозов, но не целые биоценозы. Учитывая этот факт, мы предлагаем выделять как самостоятельные единицы жизни- биомероценозы (дословно «живые части общего») — сообщества био- морф разных видов, относящихся к наиболее крупным таксонам, на- пример, сообщества растений, или фитоценозы; сообщества животных,
Глава 2. Обоснование биоморфологии или зооценозы; сообщества грибов, или микоценозы; сообщества мик- робов, или микробоценозы и т.п. Система из разных биомероценозов, существующая на некотором участке суши или водоема, — биоценоз. Среди биомероценозов каркасную роль в биоценозе выполняют фи- тоценозы — основные производители О, и биомассы. Среди биоцено- зов каркасная функция также принадлежит основным производителям О, и биомассы. Крупномасштабные биоценозы океанов и лесные био- ценозы составляют каркас биостробы Земли. Если вести оценку степени участия биоценозов в создании кислорода и биомассы на единицу пло- щади, то каркас биостробы составят, вероятнее всего, лесные биоцено- зы. Что образует каркас живой Вселенной, пока неизвестно; его могут составлять биостробы разных космических систем с наиболее высоким уровнем развития, но реальна пока только одна биостроба Земли — и каркас, и несомая часть в живой Вселенной. Любая биосистема имеет какую-либо определенную каркасную организацию, роль каркаса при этом сводится к обеспечению оптимального жизненного состояния био- системы для ее жизнесохранения и жизнесохранения вообще. Резюми- руем сказанное выше как закон каркасности биосистем: организация биосистем может обесиечивать оитимальное расиоложение и функ- ционирование элементов биосистем в иространстве и времени только ири условии дифференциации на каркасную (оиорную) и несомую части. Закономе ность восьмая. Биосистемы — это коллективистские си- стемы. Биосистемы, будучи в своей организации системами биосистем меньшей, чем они, сложности, сами могут существовать только во вза- имосвязи с другими биосистемами. Во-первых, они существуют в орга- низации биосистем большей, чем они, сложности в соответствии с за- коном иерархичности. Во-вторых, биосистемы на своем уровне органи- зации постоянно взаимодействуют с другими биосистемами, которых они или (и) едят, или (и) кормят, или (и) перемещают на себе, или (и) передвигаются на них, в них, или (и) как-либо используют иначе. Существует общая закономерность для всех биосистем: биосистема сосуществует только в единстве с другими биосистемами, взаимодействуя с которыми неиосредственно или оиосредованно, через иродукты их жиз- недеятельности, создает круг связей, иолезных или вредных для обесиечения своего сохранения и (или) сохранения всей живой ирироды. Закономе ность евятая. Биосистемы — это выражение единства неживой и живой ирироды на молекулярном и иремолекулярном уровнях организации материи. Биосистемы состоят из элементарных частиц, по- лей атомов, из атомов и молекул, что является всеобщим неживым материальным субстратом жизни. Биосистемы всегда существуют в соот- ветствии с законом единства живого и неживого (закона Докучаева- Вернадского), рассматриваемого в гл. 3.4. Закономе ность есятая. Биосистемы — это нормальные (здоровые) или больные системы. Любые биосистемы могут быть нормальными, или здоровыми (функционировать на средних и оптимальных режимах), или больными. Болезнь с медицинской точки зрения «есть состояние неус- тойчивого режима саморегуляции живой системы» (Корольков, Петлен- ко, 1977, с. 251). Представления о норме здоровья и болезни людей и
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHblX РАСТЕНИИ животных составляют предмет исследования медицины и ветеринарии. Некоторые стороны познания этих состояний имеют общебиологичес- кое значение. Дело в том, что противоборство факторов здоровья и болезни осуществляется в соответствии с законом жизнесохранения как проявление противоборства жизнесохранительных и жизнеразрушитель- ных свойств. Нормальное состояние у любой биосистемы отмечается тогда, когда жизнесохранительные свойства преобладают над жизнераз- рушительными свойствами или подавляют их в процессе существования биосистемы. В отношении здоровья и болезни биосистем существует закономерность, которую можно выразить так: всякая биосистема толь- ко тогда сохраняет здоровье, когда находит средства держать под конт- ролем и (или) даже подавлять собственные жизнеразрушительные свой- ства и действие жизнеразрушительных свойств извне; чем меньше проявляется эта способность, тем хуже здоровье биосистемы, тем больше отклонение его от нормы, тем сильнее проявление болезни. Мы перечислили некоторые из общих закономерностей организа- ции биосистем, которые представляются нам как ведущие, вне которых нельзя представить жизнь как процесс и живое как материальный суб- страт. В целом мы рассмотрели известные явления, но тривиальность их только в том, что они присущи всем биосистемам, а некоторые из этих явлений, как, например, явления каркасности и иерархичности органи- зации биосистем, еще не получили должного освещения в биологичес- кой литературе. Оценив наиболее общие признаки организации биоси- стем, нам легче рассмотреть закономерности организации биоморф се- менных растений. 2.4. Жизненная форма (биоморфа)— ключевое понятие биоморфологии Как известно, понятийные основы любой науки — не застывшее сооружение, а постоянно совершенствующийся механизм познания. Лю- бая наука стремится к однозначности своих терминов, в первую очередь ключевых, что достигается путем логического переосмысливания фак- тов и совершенствования понятий. Издавна словосочетанием «life form», «form of life», (нем. «die Leben- sforme» — форма жизни, или жизненная форма), без строгого его опре- деления, обобщенно обозначали представителей живой природы, более всего организмы, разновидности и виды (Гегель, 1975; Darwin, 1859; Вей- сман, 1905 и др.). В таком обобщенном и не строгом смысле часто говорят и пишут по сей день (Леопольд, 19б8; Отрощенко, 1973; Руденко, 1980; Дубинин, 1981; Бил, Ноулз, 1981 и др.) M.Руттен (1973, с. 71) высказывается вполне определенно: «морфологически можно различать громадное число отдельных форм жизни — видов, родов, семейств и более высоких таксо- номических групп микроорганизмов, растений и животных». Другое нап- равление в употреблении термина «жизненная форма» связано с именем Е.Варминга. Он использовал достижения предшествующих ботанико-
Глава 2. Обоснование биоморфологии географов и обосновал целесообразность введения термина «жизненная форма» вместо наиболее распространенного в то время термина А.Гризе- баха (1874, 1877) «растительная форма» («die Vegetationsforme»). По Е.Вармингу (1884) жизненная форма — это «форма, в которой вегетативное тело растения (индивида) находится в гармонии с внешней средой в течение всей его жизни, от колыбели до гроба, от семени до отмирания» (цит. по И.Г.Серебрякову, 1962, с. 27). «Жизненная форма проявляется в основном во внешнем облике и продолжительности жиз- ни органов питания (в строении листьев и всего облиственного побега, в продолжительности жизни индивидуума: аутэкология), меньше в стро- ении и продолжительности жизни органов размножения» (Warming u. Graebner, 1918, с.5). Идеи, заложенные А. Гумбольдтом (Humboldt, 1806 — цит. Гумбольдт, 1936), А.Гризебахом (1874, 1877) и закреплен- ные Е.Вармингом (Warming, 1884, 1909, 1916; Варминг, 1902 и др.) в по- нятии «жизненная форма» получили в последующие годы дальнейшее развитие под эмблемой этого или многих иных терминов, которые чаще всего оценивают как синонимы (Raunkiaer, 1907; Drude, 1913; Высоц- кий, 1915; Gams, 1918; Du Rietz, 1931; Фридерикс, 1932; Кашкаров, 1933, 1938, 1944; Закржевский, Коровин, 1935; Meusel, 1935, 1951, 1955; Але- хин, 1950; Rauh, 1937, 1939; Сенянинова-Корчагина, 1949; Культиа- сов, 1950; Шенников, 1950; Braun-Blanquet, 1951; Scharfetter, 1953; На- умов, 1955; Серебряков, 1955, 1962; 1964 и др.; Schmid, 1963; Ма- зинг, 1958; Зозулин, 1959, 1961, 1976; Голубев, 1960, 1977 и др.; Криво- луцкий, 1967, 1971; Правдин, 1971, 1986; Серебрякова, 1971, 1972, 1987; Шарова, 1973, 1981; Нухимовский, 1973а, 1980; Хохряков, 1975а, б, 1981; Зиман, 1976; Юрцев, 1976; Алеев,1980; Дервиз-Соколова, 1981; Шафра- нова, 1990; Борисова, 1991; Жмылев и др., 1993; Шафранова, Гат- цук, 1995 и др.). Однако, по сей день нет единодушия среди ученых в объяснении термина «жизненная форма», который продолжает обрас- тать новыми синонимами или близкими к ним по значению терминами. А.П.Хохряков (1978, с. 357) опрометчиво полагает, что все в общем согласны, что жизненная форма — «это внешний облик растения, скла- дывающийся в онтогенезе и детерминированный филогенетически». Си- туация на самом деле иная (Серебрякова, 1972, 1980; Криволуцкий, 1971; Нухимовский, 1973а, 1980; Алеев, 1980 и др.). Можно привести много весьма сильно различающихся дефиниций термина «жизненная форма», обзор которых приводили неоднократно (Поплавская, 1948; Зозу- лин, 1959; Серебряков, 1962; Серебрякова, 1972; Алеев, 1980; Шаро- ва, 1981 и др.). С.И.Левушкин (1975) безусловно допускает ошибку, когда определяет термин «жизненная форма» в одном смысле, а его известный греческий перевод «биоморфа» — в ином. Литературные об- зоры по проблемам понятия «жизненная форма» и классифицирования жизненных форм многочисленны (Gams, 1918; Du Rietz, 1931; Поплав- ская, 1948; Зозулин, 1959; Голубев, 1960, 1977; Серебряков, 1962, 1964; Криволуцкий, 1967, 1971; Серебрякова, 1972; Юрцев, 1976 и др.); они свидетельствуют о многогранной истории изучения жизненных форм (биоморф). Чтобы не повторяться, проведем небольшой критический литера- турный обзор, особо подчеркивая наиболее существенные, на наш
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ взгляд, стороны в понимании и изучении жизненных форм. В трактовке М.В.Культиасова (1950), И.Г.Серебрякова (1962), Г.М.Зозулина (1961) и др. жизненная форма характеризуется признаками не одного растения, а группы растений. По В.Н.Голубеву (1972, 1973), жизненная форма тождественна марфо-физиологическому содержанию понятия вид, т.е. является надорганизменной категорией. В работах Г.Мейзеля (1951, 1955) и его школы (Gluch, 1967; Muhlberg, 1967, 1970; Schmid, 1956, 1963 и др.), а также в более ранних работах немецких авторов (Pay, Тролль, цит. по И.Г.Серебрякову, 1962, с. 53) вместо термина «жизненная фор- ма» используется термин «форма роста». Как обратил внимание Т.А.Ра- ботнов (1995, с. 84), Е.Варминг еще в 1909 г. в английском издании своей «Экологии растений», предложил заменить термин «жизненная форма» на термин «форма роста» (growth form), чтобы точнее отразить суть. Под «формой роста» обычно понимают совокупность признаков габитуса, проявляющихся в ходе онтогенеза растения, иными словами, это морфологическое «поведение» индивидуума во времени (Gluch, 1967; Muhlberg, 1965; цит. по Т.И. Серебряковой, 1972). В последней трактовке жизненной формы И.Г.Серебряковым, опубликованной Т.И. Серебряковой (1972, с. 86), внесена существенная поправка к преж- ним определениям: она рассматривается не как габитус группы расте- ний, а как габитус растения, «связанный с ритмом развития и при- способленный к современным и прошлым условиям среды». Идея Е.Варминга об индивидуальности жизненных форм, на наш взгляд, наиболее логична и перспективна для дальнейшего развития. Термин «габитус» удачнее других в значении: общее экзоморфологи- ческое состояние чего-либо (растения, животного и т.п.) на каком-либо отрезке его существования. Как указал И.Г.Серебряков (1962, с. 11) известно немало различных направлений в изучении и классификации жизненных форм — «и фи- зиономические, и экологические, а чаще смешанного эколого-физио- номического характера». Е.М.Лавренко и В.М.Свешникова (1968, с. 11) подчеркнули, что «жизненными формами, или биоморфами, обычно называют основные структурные типы растений, ...однако еще большее значение, общее и специальное, для целей познания растительных со- обществ имеет изучение экобиоморф, т.е. типов растений, установлен- ных не только на основе учета их структурных особенностей, но также и их эколого-физиологических свойств, показывающих их отношения к ведущим факторам среды. Зоологи понимают жизненные формы также неодинаково (Кашка- ров, 1933, 1938; Криволуцкий, 1967, 1971; Правдин, 1971, 1978; Але- ев, 1986; Шарова, 1981 и др.). Вот одно из определений: «жизненная форма животного характеризуется в первую очередь наличием своеоб- разного общего облика (габитуса) организма (вида, группы видов) с характерными биологическими и физиологическими приспособитель- ными чертами. Этот габитус исторически возникает в определенных экологических условиях и выражает приспособленность животного к этим условиям» (Черняховский, 1971, с. 20). Нетрудно заметить, что под- ход к пониманию жизненной формы зоологом M E.Черняховским по
Глава 2. Обоснование биоморфологии сути дела близок к пониманию этого термина ботаником И.Г.Сере- бряковым. Жизненная форма — это габитус, отражающий комплекс адап- таций организма, проявляющихся при освоении той или иной экологичес- кой ниши (Стебаев, Никитина, 1975). Ф.Н.Правдин (1971, 1986, с. 4) пред- ложил рассматривать жизненную форму как общебиологическое понятие, т.е. однотипное и для зоологии, и для ботаники (с чем мы согласны): «как результат действия естественного отбора в определенных условиях среды, выраженный во внешнем облике животного или растения». Одни исследователи отрицают связь жизненных форм с опре- деленными систематическими группами (Серебряков, 1962; Gams, 1918; Raunkiaer, 1907 и др.), используя для этого различные доводы и в час- тности факт существования «эфармонической конвергенции», напри- мер, между некоторыми видами семейств кактусовых и молочайных и т.п. (Warming и. Graebner, 1918), а по мнению других — жизненные формы являются принадлежностью строго определенных таксонов- видов, родов и т.д. (Drude, 1913; Келлер, 1933 и др). Ю.С.Григорьев (1955, с. 141) подчеркнул, что обычно жизненной формой называют «совокупность видов из разных систематических групп, но сходных по тем или иным экологическим и физиономическим особенностям... Сле- довательно, если жизненные формы (в обычном понимании) объединя- ют виды, конвергентно сходные по некоторым приспособительным признакам, то таксономические единицы, особенно единицы более низ- ких категорий: подсекции, секции, роды нужно считать жизненными формами высшего порядка». Р.Шарфеттер (Scharfetter, 1953) отметил, что каждый вид по внутреннему и внешнему строению должен нахо- диться в гармонии с условиями окружающей среды, т.е. обладать опре- деленной жизненной формой. «Каждый вид — представляет непов- торимую систему эколого-биологических приспособлений и может рас- сматриваться как отдельная жизненная форма (Голубев, 1968, с. 1092), т.е. жизненная форма характеризуется системой «морфофизио- логических признаков и свойств, контролируемых в онтогенезе эколо- гией местообитаний» (Голубев, 1973, с. 4), но в отличие от вида она не включает показатели его географического распространения (ареала), систематической принадлежности, степени и характера участия в сло- жении фитоценозов (Голубев, 1972). На наш взгляд, признание за жиз- ненной формой строгого отношения к определенной систематической категории без их отождествления, более прогрессивно, чем отождеств- ление ее с видами, внутривидовыми и надвидовыми таксонами. Жизненные формы подразделяют по одному признаку, имеющему важное или морфологическое, или экологическое, или физиологичес- кое значение (Raunkiaer, 1905; Вильямс, 1922; Linkola, 1922 и др.) по двум, трем признакам (Raunkiaer, 1907; Казакевич, 1922 и др.) или по многочисленным признакам (Warming, 1884; Drude, 1913; Du Rietz, 1931; Серебряков, 1962; Голубев, 1972 и др.), но при этом не отрицается воз- можность выделения по одному — нескольким признакам жизненных форм «узкого содержания», формирующихся на основе конвергенциии различных и сходных эколого-биологических типов (Голубев, 1973). Та- ким образом, число признаков, по которым можно охарактеризовать
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ жизненные формы, — множество, но в зависимости от целей исследо- вания, как правило, анализируется лишь часть этих признаков. При классифицировании жизненных форм широко используется иерархический принцип: обычно строятся релятивные классификации (Warming, 1884; Du Rietz, 1931; Прозоровский, 1936; Зозулин, 1959; Бо- рисова, 1961; Серебряков, 1962; Шарова, 1981 и др.). Однако этот метод вызывает справедливые возражения. Так, Ю.С.Григорьев (1955, с. 141), изучив экологические особенности жизненных форм ксерофилизо- ванных растений и выделив группы ксерофитов, мезоксерофитов и т.д., пришел к выводу, что эти группы «никак нельзя считать классифика- ционными единицами, подобным таксономическим единицам приня- тым в систематике» и что иерархические классификации биоморф, ос- нованные на «тех или иных произвольно выбранных, случайно бросив- шихся в глаза признаках», неудачны. В.Н.Голубев (1968, 1972, 1977) так- же отметил, что при иерархическом (по релятивному методу, что сле- дует обязательно оговаривать) классифицировании жизненных форм су- ществует возможность бесчисленно комбинировать разные признаки. Он предлагает классифицировать эти объекты только по «линейному» принципу, позволяющему максимально охарактеризовать их через аль- тернативные признаки (Голубев, 1968). Был предложен вариант «линей- ной системы» жизненных форм для целей получения краткой, но «мно- гомерной эколого-биологической оценки видов» (Голубев, 1972). Заме- тим, что сходным образом уже проводили классифицирование растений и их отдельных органов. Так, например, Э.Шмид (Schmid, 1956, 1963), разработав краткий комплекс признаков форм роста, предложил под- разделять их по каждому ряду признаков в отдельности (по признакам соцветия, листьев, корней и т.д.). Принцип классифицирования, кото- рый можно также считать «линейным» в понимании В.Н.Голубева, был использован при изучении и описании побегов семенных растений (Ну- химовский, 1970, 1971). Использование метода множественного линейного, или мультили- нейного (по рядам сравнимых и взаимоисключающих признаков), или «линейного» классифицирования (по терминологии В.Н.Голубева), для целей классифицирования жизненных форм представляется весьма пер- спективным. Метод релятивного классифицирования, пожалуй, выпол- нил свою роль на соответствующем этапе развития учения о жизненных формах. Логические основы релятивного классифицирования уступают всем другим методам иерархического классифицирования. На место ре- лятивных должны прийти более строгие иерархические классификации, выполненные иными методами, например, методами субординатного и мультисубординатного классифицирования, уже рассмотренными выше. При создании списка наиболее существенных биолого-морфологи- ческих признаков, по которым следует характеризовать жизненную фор- му, предлагается включать в такой список лишь признаки взрослых особей «в момент полного развития, присущие виду, как определенной биологической единице» (Голубев, 1968, с. 1090). Понятие жизненная форма иногда сводят с разной интерпретацией к отдельным фенологи- ческим и (или) возрастным состояниям (Хохряков, 1975а, 1978 и др.;
Глава 2. Обоснование биоморфологии Серебрякова, 1980; Алеев, 1980). А.П.Хохряков (1975a, 1978) полагает, что в онтогенезе, допустим, майского жука сменяются жизненные формы (габитусы) яйца, личинки, куколки, имаго. Жизненная форма (организм или синорганизм) не сменяется в морфогенезе, а сменяется только ее возрастные состояния и их габитуальные признаки (Нухимов- ский, 1973а). Габитус — это только фрагмент фенотипической реализа- ции адаптивных свойств организма, закодированных в генотипе на мо- лекулярном уровне или новоприобретенных ею в процессе взаимодей- ствия с окружающей средой. Когда мы говорим или проросток, или дерево, или личинка и т.п., то судим не только о габитусе (габитуальный признак или признаки могут быть индикатором состояния биоморфы), а о всей адаптационной системе-организме, и все признаки, присущие этой системе, — это признаки жизненной формы, находящейся в ка- ком-либо жизненном состоянии. Например, дерево, как биоморфа, не- соизмеримо богаче дерева, как габитуса, и т.п. Итак, вряд ли будет рационально принять в отношении такого обобщающего и выразительного термина, как «биоморфа», или, что одно и то же, жизненная форма (форма жизни), предложения, подчер- кивающие принадлежность его только чему-то одному (например, габи- тусу) в организмах. Подведем некоторые итоги по обсужденным выше материалам. Взгляды Е.Варминга долгое время воспринимались некритически, что оказало двоякое влияние на развитие учения о жизненных формах. Его главная заслуга состоит в том, что он определил термин «жизненная форма» через некоторые приспособительные признаки индивидуума. Однако, во-первых, Е.Варминг не провел четкой грани между конкрет- ным объектом исследования — жизненной формой и типом сходных объектов исследования (классификационной категорией). В итоге у него одна жизненная форма проявляется как во внешнем облике, продолжи- тельности жизни и отчасти в строении органов размножения индиви- дуума (Warming, 1884, 1908; Warming и.Graebner, 1918), так и в габитусе, эфармонии (приспособительных свойствах — Е.Н.) вида и даже семей- ства (Варминг, 1902). Во-вторых, он признавал примат вегетативных органов над репродуктивными, что неприемлемо, поскольку основны- ми интегральными процессами, составляющими онтогенез растений, являются вегетативный рост и репродуктивное развитие (Чайлахян, 1964, 1988). Как вегетативные, так и репродуктивные органы важны для организмов; по любым признакам этих органов можно классифици- ровать организмы. В-третьих, понятие «жизненная форма» ограничено Е.Вармингом в основном признаками внешнего вида (габитуса) организмов, что не дает целостной картины всего адаптационного струк- турно-функционального механизма. В.Г.раменский (1938, цит. по 1971, с. 242) справедливо заметил, что каждая жизненная форма (растение) приспособлена к определенной ам- плитуде экологических условий. Это проявляется как в ее внутренних способностях, не имеющих внешнего выражения (например, способность клеточной плазмы переносить холод, потерю воды и т.д.), так и в ее «видимой организации — морфологии и анатомическом строении». К син-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ тетическому изучению жизненных форм, или экобиоморф («типовых адап- тивных систем» на организменном уровне) призывал Е.М.Лавренко (1964). История использования термина «жизненная форма», критический литературный обзор, проведенный выше, и наш опыт изучения органи- зации семенных растений показывают, что ключевую проблему биомор- фологии можно решить, обозначив термином «биоморфа» любой инди- видуум и кондивидуум. Такое определение больше отвечает и содержа- нию греческого слова «биос» (жизнь) как составной части термина «био- морфа». Не исключая возможности оценок жизненных форм по одному или нескольким признакам, которые практикуют многие биоморфологи, нам тем не менее представляется наиболее рациональным не ограни- чивать само понятие «биоморфа» рамками проявления отдельных приз- наков, а придавать ему более широкое общебиологическое содержание. Оиределение жизненной формы (биоморфы) как индивидуума (организм, особь) или кондивидуума (синорганизм) имеет общебиологический смысл. В этом определении, с одной стороны, отдается дань уважения Е.Вармингу и его последователям, сохраняя за термином «жизненная форма» поня- тийную привязку к организмами, добавляя к ним еще синорганизмы (особый результат вегетативного поведения организмов, преобразующе- го их в новые, более сложные формы жизни), которые во времена Е.Варминга еще не были описаны. С другой стороны, биоморфология отрывается от задач узкоспециального морфологического исследования, оставляя их габитологии (эпиморфологии, или экзоморфологии) — на- уке о формах роста, или габитусе биоморф (организмов и синорганиз- мов), и поднимается на более высокое место в системе наук. Только с позиций «индивидуума и кондивидуума» ответ на вопрос: «с чего на- чинается и где кончается жизненная форма», будет наиболее полным, конкретным и последовательным. Такое объяснение биоморф не отбра- сывает, а творчески использует все лучшее, что было создано в науке в процессе изучения структурных и функциональных признаков организ- мов, расширяет его, открывая большие познавательные возможности, чем просто понятия организм или синорганизм, или все другие предлагавшиеся ранее значения этого термина, ограниченные какой-либо структурной или функциональной стороной, чаще всего габитусом организмов. Чем точнее наши знания об индивидууме и кондивидууме, тем кон- кретнее понятие «жизненная форма». В настоящее время оформляется, пожалуй, наиболее прогрессивное направление в понимании особи у выс- ших растений (Работнов, 1950а; Смирнова, 1967, 1968; Сенянинова-Корча- гина, 1967; Ефейкин, 1970; Нухимовский, 1970, 1973б, 1974а и др.); при ха- рактеристике особей большое значение придается их телесной ограничен- ности, морфологической целостности, способности самостоятельно уча- ствовать в обмене веществами с окружающей внешней средой. Одним из важных свойств особи, определяющим видовую принад- лежность, является относительная генетическая однородность ее сома- тических клеток. Для каждого вида организмов характерен определен- ный набор хромосом соматических клеток (кариотип), характеризую- щийся числом хромосом, их размерами, формой и особенностями внут- реннего строения (Дубинин, 1976; Гершензон, 1979). Генетическая оцен-
Глава 2. Обоснование биомоофологии ка помогает разграничить особи и тогда, когда существует морфофизио- логическая связь между отдельными растительными частями, например между хозяином (злаком) и полупаразитом (погремком) или паразитом (заразихой). Для растений вегетативного происхождения наиболее ха- рактерна генетическая однородность (Правдин, 1937). В трактовках тер- мина «особь» часто не учитывается эмбриональный период жизни, с чем согласиться нельзя. Особь (организм, индивидуум) — это живая морфофизиологически це- лостная система, образованная органами, функционирующими по эволюци- онно выработанному muny родительских систем и начинающаяся с зиготы, аномиктического зачатка или обособленной нартикулы, с момента зало- жения у носледней нервых комнонентов недостающих еи основных органов (органоидов у одноклеточных). Партикула — это полностью или непол- ностью обособленная в результате партикуляции, т.е. разделения (меха- нического или физиологического), живая часть родительского организ- ма (см. гл. 4.5). Основные органы (органоиды у одноклеточных) — это органы (органоиды), обеснечивающие самостоятельное развитие особи на нротяжении всей ее жизни (у семенных растений к ним относятся набеги и корни). Кондивидуум (от лат. con — вместе, сообща, и ЙюЫииз- разделенный; дословно — совокунность разделенных), или синорганизм- это живая система, начинающаяся в своем развитии с зиготы и заканчи- вающаяся в носледнем ноколении клона (Нухимовский, 1970,1973а, б, 1974а, 1980). Иными словами, эта система «мероаклонист-клон», в кото- рой мероаклонист (временный аклонист) — особь полового происхож- дения (аклонист), образующая на протяжении своего жизненного цикла вегетативное потомство или завершающая свою жизнь образованием клона, а клон — совокупность особей (клонистов), составляющих веге- тативное потомство мероаклониста. Наше более широкое объяснение термина «жизненная форма» от- крывает новые познавательные возможности, поскольку не является синонимом известных терминов: «основная форма», «биологический тип», «габитуальная форма», «форма роста», «эпиморфа», «экобиомор- фа» и т.п., употреблявшиеся в разное время и в разных значениях. Правильным переводом термина «жизненная форма» на древнегречес- кий язык с последующей транслитерацией на русский язык следует считать только одно слово «биоморфа». Для обозначения внешнего облика биоморф в нашем понимании, пожалуй, более всего подходит термин «габитус», внешний облик (габитус) биоморфы в текущий мо- мент ее существования — это одновременно габитуальная форма (фор- ма роста). Такие габитуальные формы (наряду с возрастными и фено- логическими состояниями) неоднократно могут смениться в жизни од- ной (!) биоморфы (организма или синорганизма). Если говорить образ- но, то габитусы организмов (но не сами организмы) сменяются почти как рубашки по мере необходимости. Биоморфы бывают нростыми — это организмы (рис. З-I, 4, 5). От- дельному организму всегда присуща морфофизиологическая целост- ность — это одно тело, части которого различаются по структуре и особенностям выполняемых ими функций, и на этой основе они выч-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ леняются в нем, для удобства их познания, как органы, находящиеся друг с другом в единстве. Другие биоморфы сложные, т.е. ироявляют себя как синорганизмы (рис. З-II, Ь, 7). Для них характерна морфофизиологи- чески прерывистая целостность — это система нескольких тел опреде- ленного рода. Приняв за критерий сложности организации биоморф степень их пространственной целостности, построим высшую широкую субор- динатную классификацию биоморф. Нижнюю субординату занимают организмы, поскольку составляющие их элементы-органы образуют морфофизиологически непрерывную систему. Верхняя субордината принадлежит синорганизмам, поскольку составляющие их элементы- организмы морфофизиологически разобщены (рис. 3). Синорганизм развивается сначала как особь полового происхождения, образуя на протяжении своего жизненного цикла вегетативное потомство (клон). Цикл существования биоморфы — биоморфогенез (онтогенез или синон- тогенез). Цикл развития от рождения (с зиготы) до отмирания конди- видуума — синонтогенез (см. гл. 7). Совокупность сведений о биомор- фогенезах в каком-либо таксоне (виде, роде и т.п.) — биоморфогения этого таксона (его онтогения или синонтогения). бо Рис. 3. Графическая модель основной субординатной классифи кации биоморф семенных растений. 1 — организмы, И — с инорган измы, cy — семядольный узел, кш- корневая шейка, г — гипокотиль, п — побеги текущего года, эп- эфемерная побеговая часть, бо — базовый орган, ск — скелетная корневая часть, ак — аскелетная корневая часть, оч — отмершая часть.
Глава 2. Обоснование биоморфологии Охарактеризуем в качестве примера некоторые аспекты онтогении у Сопшт maculatum L. и синонтогении у Anthriscus sylvestris (L.) Hoffm., которые мы наблюдали в лесах и парках Подмосковья. Болиголов пят- нистый (Соп1цт maculatum L.) представитель семейства Apiaceae. Оби- тает на лесных опушках, заливных лугах, на залежах, около жилья, у дорог и заборов, как сорняк на огородах, в посевах. «Двулетние травы с высоким стеблем и красноватыми пятнами и трижды перистими ли- стьями» (Пояркова, 1950, с. 225). Семена прорастают весной, проходя зимой под снегом естествен- ную стратификацию. Развитие идет быстро. На первом году формиру- ется розеточный вегетативный побег и стержневой главный корень (рис. 4). Базальная часть главного корня и гипокотиль проявляют кон- трактильность, т.е. сократительные свойства в продольном направлении (морщинистость),что втягивает в почву основание розеточного побега. Розеточные растения уходят под зиму с одним, как правило, сильным, вегетативным побегом и мясистым главным корнем, а рано весной на- чинают образовывать новые розеточные листья. Вскоре главный побег переходит к развертыванию удлиненной раздвинутоузловой стеблевой части и очередных листьев на ней, а в средней и верхней частях такого полурозеточного побега развиваются обоеполые соцветия-зонтики на верхушках главного и боковых стеблей (рис. 4, сц). Степень разветвлен- ности зоны соцветия зависит от условий обитания; она больше при благоприятных условиях, например, на залежах. При отчуждении цве- тоносной части главного побега (скашивание) растения, как правило, восстанавливают соцветия, но на новых вегетативно-репродуктивных пазушных побегах, развивающихся из спящих почек оставшейся базаль- ной части главного побега. Наиболее высокие цветоносные побеги (до 2-хм) отрастают на хорошо удобренных почвах. Цветение — июнь, июль; плодоношение — сентябрь, октябрь. После плодоношения на 2-м году жизни главный побег полностью отмирает вместе с разветвленной системой главного корня, т.е. растения болиголова пятнистого развива- ются как организмы по двухлетнему монокарпическому циклу. Купырь лесной (Anthriscus sylvestris (L.) Hoffm.) — широко распро- страненный вид лесной зоны Европы. Представитель семейства Apia- ceae. Обитает в лесах и на лугах. Цветет в начале лета (июнь). Вид весьма полиморфен (Peterson, 1915, 1922). Под пологом дубрав лесосте- пи Центрально-Черноземного заповедника купырь лесной развивается по типу двулетних монокарпиков, а в лесной зоне является многолет- ним поликарпическим растением (Голубев, 1962). На монокарпичность особей этого вида в одних условиях обитания и поликарпичность в других указывал также Т.А.Работнов (1956а). Семена прорастают рано весной по надземному типу. В течение первого года жизни формируется главный стержневой корень с систе- мой ветвей, гипокотиль и вегетативный розеточный побег (рис. 6-А). Сокращение главного корня, в особенности его базальной части, и ги- покотиля, заметное уже на ранних этапах онтогенеза (морщинистость), вызывает углубление в почву основания розеточного побега. В зависи- мости от особенностей местообитаний отдельные особи развиты неоди-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 4. Развитие биоморфы Сопшт maculatum L. как организма (Калужская область) — 1 2 — яд;,~,3 — первые настоящие листья по порядку их развертывания, гп — главный побег, гк — главный корень, рл — розеточные листья вегетативного побега, ол — основания листьев (верхние части удивлены), ср — схема взрослого двулетнего полурозеточного растения, пп — поверхность почвы, сц — соцветие, у — увеличено по сравнению с масштабом; одно деление масштаба — 1 см. Осталь- ные обозначения те же, что и на рис. 3.
Глава 2. Обоснование биоморфологии су~ Рис.5. Развитие биоморфы Cichorium intybus L. как организма (Алтайский край) оп — основание удлиненной части полурозеточного цветоносного побега (верхние части удалены), пч — спящие почки, к — каудекс; примерное положение семя- дольного узла у взрослого растения показано пунктирной линией. Остальные обозначения те же, что и на рис. 3 и 4.
ОСНОВЪ| БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ В гп ЖК Рис. 6. Развитие биоморфы Anthriscus sylvestris (L.) Hoffm. как синорганизма (Московская область). А — первый год жизни, Б — второй год, В — образование клона на третьем году, Г — пятый год жизни синорганизма, Д — восьмой год жизни; жк — живая особь клона, ок — отмершая особь клона, оч — отмершая часть, пк — придаточные корни. Остальные обозначения те же, что и на рис. 3-5. наково. На розеточном побеге иногда закладываются 1-3 пазушные поч- ки. На следующий год виргинильная особь продолжает вегетацию. На главном побеге образуются новые почки, которые, как и прошлогодние, отрастают в вегетативные розеточные пазушные побеги. Одновременно с началом развития боковых побегов на их основаниях начинают об- разовываться придаточные корни (2-3, реже 1-4); они отходят под острым углом и подобно главному корню обладают контрактильностью (рис. 6-Б). Виргинильный период часто заканчивается в природных условиях на третий год, когда особь впервые зацветает, цветет и плодоносит. В оптимальных условиях встречаются особи, цветущие на втором году жизни, а при неблагоприятных условиях переход особи в репродуктив-
Глава 2. Обоснование биоморфологии Рис.7. Развитие биоморфы Valeriana tuberosa L. как синорганизма (остепненные горные склоны в окрестностях Усть-Каменогорска, Восточный Казахстан). кл — корневые клубни, гг — примерные границы годичных приростов розеточных побегов прошлых лет, гм — графические модели растений разных возрастных состояний. Остальные обозначения те же, что и на рис. 3-6.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ный период оттягиавается на 4-5-й год; есть указания, что в условиях eoòeñòBåííûõ ценозов первое цветение у купыря лесного наблюдается На 3-7 год (Перттула, цит. по Т.А.Работнову, 1956а). У купыря лесного образуются вегетативно-репродуктивные обоепо- лые полурозеточные побеги. После полного отмирания главного побега и главного корня, т.е. после полной общей партикуляции (см. гл. 4.5.) образуется несколько обособленных дочерних особей, имеющих в своей структуре вегетативный розеточный побег и систему придаточных кор- невых ветвей (рис. 6-В). Образование дочерних особей свидетельствует о том, что произошло вегетативное размножение (Troll, 1954). Особь семенного происхождения является монокарпиком, поскольку она все- го лишь один раз в жизни цвела, плодоносила и после этого отмерла. Подобно родительской особи, способностью к однократному плодоно- шению обладают и дочерние особи, т.е. и они являются монокарпичес- кими растениями (рис. 6-Д). Только синорганизм является поликарпи- ком. Онтогенез каждой особи клона продолжается 2-7 лет. В вымираю- щих клонах иногда насчитывается до 30 особей. Есть указания, что число таких особей может достигать 53 (Т.А.Работнов, 1956а). В старею- щих клонах боковые розеточные вегетативные побеги нередко начина- ют погибать, уже не обособляясь от родительской особи. Ограниченная способность к отдалению дочерних особей вскоре «перегружает» клон конкурентами, и к 20-35 годам синонтогенеза он погибает. Жизнь возникла в процессе саморазвития материи. Вероятнее все- го, она впервые появилась в форме одноклеточных организмов из «про- бионтов» (Опарин, 1957, 1980; Кеньон, Стейнман, 1972). Простейшими формами самостоятельного существования живого (формами жизни) на Земле стали индивидуумы (организмы), а позднее и кондивидуумы (си- норганизмы). Появление первых организмов на Земле означало появ- ление и первых видов (вида). Вид — объективная реальность, а не прием классифицирования (Хржановский, 1968; Завадский, 1961, 1968; Скворцов, 1972). Справедливо критикуется «ашерсоновский» метод так- сономии, суть которого, как подчеркнули В.Г.Хржановский и др. (1979), заключается в распылении реального вида на формально выделяемые внутривидовые категории — многочисленные разновидности, подраз- новидности, группы разновидностей и т.п. По существу, в таких случаях выделяются внутривидовые биоморфотипы, различающиеся окраской лепестков, пыльников, семян или другими признаками. Ipynna биоморф, охарактеризованная по каким-либо общим (одному или нескольким) призна- кам, — биоморфотип, или тип жизненных форм (Нухимовский, 1973а,б). Внешний облик (габитус) биоморфы является одновременно, как уже подчеркивалось выше, габитуальной формой (термин Ф.Клементса, цит. Du Rietz, 1931). Тип сходных габитусов биоморф — это габитуальный тип, или тип габитуальных форм. Организмы и синорганизмы прошлых геологических эпох — палеобиоморфы, а их группы, объединенные по каким-либо сходным признакам, — палеобиоморфотипы.
Глава 2. Обоснование биоморфологии 2.5. Из истории изучения организации биоморф кормофитов (побеговых растений) Вопросы устройства и функционирования семенных (а шире— всех побеговых) растений интересовали ученых издавна. Разнообразные сведения о форме их тела, структуре и функциях можно найти в работах ученых минувших столетий (Феофраст, 372-287 г. до н.э., 1951; Caesal- pini, 1583; Linnaeus, 1751; Willdenow, 1802; Link, 1807, 1824; Richard, 1819; De Сап4о11е, 1819, 1827; Bischoff, 1830, 1834, 1839; Горянинов, 1841; Фи- шер-фон-Вальдгейм, 1891 и др.). История ботаники многогранна (Щер- бакова, 1960, 1979; Щербакова и др., 1983). В отечественной литературе уже есть обстоятельные обзоры по истории морфологии растений, которые отражают главные вехи в развитии представлений об органи- зации тела особей у семенных растений (Серебряков, 1952, 1962; Пер- вухина, 1960, 1970; Серебрякова, 1971, 1972). Использование этих работ, и в первую очередь первоисточников, позволяет вкратце обосновать то, что послужило нам предпосылкой к построению теории организации жизненных форм семенных растений. Если абстрагироваться от многих деталей, то в истории морфоло- гии как бы выделяются господствующие высоты — наиболее распро- страненные представления в объяснении основных органов, составляю- щих тело организмов этих растений. Основными можно называть органы, постоянно повторяющиеся у разных организмов, несмотря на все их кажущееся многообразие и выполняющие все жизненные фун- кции растительных организмов (Серебряков, 1952). Классик' ботаники О.-П.Декандоль так определял основные органы — «это такие органы, которые служат для питания растения и которые не могут, следователь- но, у растения отсутствовать, даже если благодаря особым обстоятель- ствам они становятся иногда очень небольшими и трудно распоз- наваемыми» (De Candolle, 1827, цит. по Н.В. Первухиной, 1970, с. 91). Уместно заметить, что исследователи издавна и справедливо придают большое значение вопросам терминологии. Еще К.Линней писал: «Без знания настоящего имени нельзя учинить ни коего описа- ния, ни иметь понятия о вещах» (Максимович-Амбодик, 1795). Глубина и четкость понятий, выраженная в простых и содержательных терминах, отражает объективность знаний об определенном предмете или явле- нии. Одни термины быстро получают признание в науке, другие забы- ваются, третьи долгое время не находят наиболее удачного применения. Рассмотрим несколько подробнее наиболее древний этап становле- ния морфологии растений, поскольку именно тогда закладывались «на- чала» учения об организации тела организма у семенных растений. Исторический фон в исследованиях структуры растительного организма призван создать обзор употребления в ботанике термина «каудекс», с которым, как мы увидим, связано многое в развитии представлений о6 основных органах растений. Обозначим терминами, принятыми в пору их использования, системы основных органов особей семенных расте-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИЙ ний и кратко охарактеризуем эти системы, рассматривая их как индикаторы соответствующих исторических направлений в развитии науки об организации биоморф (биоморфологии) семенных растений. 2.5.1. История термина «каудекс». Организация системы ~корень-трава-плодотворение~ Более 2000 лет используются для обозначения соответствующих органов слова «radix, caulis, folium» (корень, стебель, лист), а также, особенно часто до середины XIX века, термин «каудекс» (caudex— дословно с латинского — пень, ствол). Плиний (23-79 г. н.э.) термином «саи4ех» называл стволы древесных растений, например, маслины, дуба, кедра, кипариса и др., а также стволик виноградной лозы (Plinius, 1571). До Плиния для той же цели Катон (234-149 г. до н.э.; Caton, 1922), Колу- мелла (116-27 г. до н.э.; Columella, 1523) и другие употребляли иногда си- ноним «codex». Позже в ботанике получил признание термин «caudex» и его долгое время, особенно в долиннеевский период, применяли со- гласно Плинию. В XVI — XVIII вв. термин «саи4ех» в основном употреблялся при описании деревьев, крупных кустарников, пальм, реже других древо- видных однодольных и древовидных папоротников, для обозначения части растения, отходящей от корня, расположенной поверх земли и служащей для передачи вверх к листьям и плодам пищи, добываемой корнем (Dodonaeus, 1583; Aldrovandi, 1671), иными словами — ствола (L'Obel, 1576; Репа et L'Obel, 1576; Caesalpini, 1583; Alpino, 1629; Bauhin et Cherler, 1650; J.Commelin et С.Commelin, 1697-1701; Gmelin, 1747; Jonston, 1768 и др.). Синонимом термина «саи4ех» был «truncus» (Aldrovandi, 1671; Endlicher, 1841 и др.); последний был принят ботани- ками в конце XVIII — начале XIX в. вместо термина «саи4ех» для обоз- начения стволов деревьев и крупных кустарников (De Candolle, 1819). Стволы пальм и древовидных папоротников в прошлом веке разные исследователи называли различно: «cormus», «саи1ота», «truncus», «stipes», но чаще всего — «саи4ех» (Willdenow, 1802; Martius, 1823-1850; Bentham et Hooker, 1883). Ученые издавна пытались рассматривать тело семенного растения по частям, при этом они нередко мысленно уподобляли его телу животного или человека. Так, Колумелла (116-27 г. до н.э., Со1и- те11а, 1523), например, писал, что корни нужны растениям так же, как и ноги человеку, — чтобы они стояли на них, ствол играет роль «че- ловеческого корпуса в его ростом и осанкой», ветви как руки, а листья выполняют роль пальцев. Линней (Linnaeus, 1751) видел общее в стро- ении тела разных растений и мысленно разделял его на три основные части: «Herba» («трава» — побеги), «Fructificatio» («плодотворение» в переводе русских авторов, или «плодоношение» в недавнем русском издании упомянутой книги (Линней, 1989), — репродуктивные элемен- ты), и «Radix» (корень — корневая система и многолетняя стеблевая часть). Руссо (1810, с. 181) отметил, что у растений «две главные части:
Глава 2. Обоснование биоморфологии корень, которым оно укрепляется к земле или в другом теле, прино- сящем ему пищу, и траву, через которую выдыхает и вдыхает воздух». Наиболее популярным в ботанике долгое время был термин «radix». Этот термин почти до конца первой половины XIX века понимался широко; им называли все подземные органы растений. Своеобразие подземных органов корневой и стеблевой природы, на которое впервые обратил внимание еще Феофраст (1951, с. 28): «Неправильно ведь на- зывать всякую часть растения, находящуюся под землей, корнем, ... раз- личие надо ведь установить на основании свойств, а не места», долгое время не находило соответствующего отражения в ботанической терми- нологии. Линней подразделял корень, например, на шишковатый (кар- тофель), шаровидный (цикламен), ползучий (пырей, будра) и т.д. Ди- трих (Dietrich, 1838) предлагал делить подземные органы на настоящие корни, ризомы, клубни и т.д., при этом все они объединены им в одну общую группу «Radix», т.е. корень в широком смысле. В еще более широком значении корнем, вслед за Линнеем, нередко называли часть растения, состоящую из главной оси (саи4ех) и корешков (radicula). Все то, что заключено между травой и корешками, по Линнею (Lin- naeus, 1751) — «caudex». Под травой дерева Линней подразумевал ветви с листьями (облиственные побеги), образующиеся от надземной вер- хушки каудекса. Верхняя часть каудекса (до травы) — это восхдящая ось — «саи4ех ascendens», т.е. главный ствол (стебель). Нижнюю часть каудекса, нисходящую от основания стебля и оканчивающуюся кореш- ками, Линней называл нисходящей осью — «caudex descendens» (соб- ственно главный корень). О.-П.Декандоль (De Candolle, 1819, 1827) отметил, что Боз различал в корне часть, которая непосредственно соприкасается с основанием стебля — «caput seu basis radicis» (голова или основание корня), и часть, расположенную ниже «головы» корня и оканчивающуюся тонким кон- цом, — «саи4ех radices» (ствол корня). Филибер (Philibert, 1802), будучи сторонником взглядов Линнея на организацию растений, подчеркнул, что «каудекс» — общее название любого тела — посредника между корешками на конце корня и «травянистой частью» (облиственными побегами), венчающей стебель (ствол) растения. Так, «у репы каудексом будет то, что вы едите после удаления травы и корешков, ... у пальмы каудекс расположен между корешками и листвой, до которых он продолжается не разветвляясь, ... у дуба каудекс ... очень большой высоты и ветвится как в нижней, так и в верхней частях (с. 146-147)..., что касается луковичных растений, то каудекс у них часто имеет форму тела, сплющенного с нижней части и выпуклого с внутренней, иногда конического, более или менее толстого и всегда круглого у основания, с которым срастается оболочка лукови- цы, и откуда вниз отходят корешки» (с. 151) и т.д. П.Горянинов (1841), исходя из представлений Линнея о двух глав- ных осях растений, вслед за Бишофом (Bischoff, 1830) и др., восходящую ось, именуемую им «череном» (саи4ех), рассматривает в двух вариантах. За надземный черен (саи4ех epigeus) принимается ствол пальм, других однодольных деревьев и древовидных папоротников. Подземный черен
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ (саи4ех hypogeus) — это все более или менее скрытые в почве органы стеблевой природы, различающиеся направлением роста, внешней формой, характером поверхности и т.п. В первой половине прошлого столетия исследователи стали больше обращать внимание на то, что подземные части растений, внешне по- хожие на корень, могут быть иной природы. Широко стал применяться старый ботанический термин «со11ит» (шейка). Во времена Линнея (XVIII в.) и, пожалуй, до Клебса '1 (Klebs, 1885) о корневой шейке или просто шейке растения говорили как о «точке», «месте», «линии», где стебель переходит в корень (Duborg, 1767; Bulliard, 1783; Lamarck, 1795; Allent, 1826; Lamouroux, 1826, Максимович, 1828; Перелыгин, 1828; Ro- eper, 1828; Kachler, 1830; Горянинов, 1841; Saint-Hilaire, 1841; Le Ma- out, 1844, 1852 и др.). О.-П.Декандоль (De Candolle, 1819) и Репер (Ro- eper, 1828) подчеркнули, что шейку в разное время называли и иначе, например, «limes communis» (общая граница), «fundus plantae» (дно растения), «coarctura» (сужение), «nodus vitalis» (жизненный узел), «linea mediana» (средняя линия). Переходную часть между корнем и стеблем, имеющую тенденцию расти и распространяться во все стороны, Менх (Moench, 1798) назвал «corpus», а Линк (Link, 1807) использовал для этой цели термин «rhizoma» (Wurzelstock), который впервые был употреблен Эрхартом (Ehrhart, 1789) при описании папоротников. Следует заметить, что Эрхарт предложил новый термин еще в то время, когда корнем назы- вали «все, что в земле растет». Он подчеркнул, что под термином «rhizoma» понимает виды «стебля» (Stamm), которые хотя и очень по- хожи на корень, но от истинного корня «как день от ночи отличаются» (1789, с.44); у лекарственных папоротников, согласно Эрхарту (Ehrhart, 1792, с.92), обычно используют не корень, а «rhizoma». Вильденов (Willdenow, 1802) охарактеризовал все более или менее подземные структурные образования, промежуточные между «обычным» стеблем и собственно корнем, которые по своеобразному облику своему отличаются от восходящей и нисходящей осей у некоторых мало- и многолетних трав, как «саи4ех intermedius» — средний каудекс. По внешнему строению Вильденов различал несколько видов «саи4ех intermedius». «Корнеобразным» средний каудекс считался тогда, когда он имел вид «шишковатого корня», но находился на поверхности почвы или одна половина его была погружена в почву, а другая выходила наружу. В свою очередь «корнеобразный» средний каудекс мог быть «репообразным» (например, у Brassica oleracea L.) или «луковичным» (например, у Ranunculus bulbosus L., Роа bulbosa L.). Если средний каудекс находился под землей, но имел вид стебля, то его называли «стеблеобразным», при этом различали по характеру поверхности «гладкий» средний каудекс, если он без бугорков, например Г.Клебс несколько конкретизировал понятие «корневая шейка». Он предложил на- зывать этим термином переходную зону от подсемядольного колена к настоящему корню. В этом значении термин «корневая шейка» применяется в настоящее время многими ботаниками. У взрослых растений часто бывает трудно установить точное положение корневой шейки. Поэтому в обычных морфологических исследованиях она нередко отмечается более или менее приближенно.
Глава 2. Обоснование биоморфологии у Lilium bulbiferum L., и «рубцовый», если есть бугорки, например у Cyclamen vernum Sweet . Под термином «саи4ех intermedius» нередко объединялись различные (по современным представлениям) органы, чаще всего корневища, луковицы, клубни, клубнелуковицы. В первой половине прошлого столетия термин «саи4ех» для обозна- чения определенных структурных частей растений (саи4ех ascendens, саи4ех intermedius, caudex descendens) был принят как зарубежными, так и русскими ботаниками (Петров, 1815; Двугубский, 1823; Максимович, 1828; Kachler, 1830; Bischoff, 1830, 1834, 1839; Dietrich, 1838; Горянинов, 1841, и др.). Как отметил О.-П.Декандоль (De Candolle, 1819), И. Мартынов (1820) и др., у некоторых травянистых растений Линк предложил различать «саи4ех» — промежуточную многолетнюю часть между собственно кор- нем и надземными однолетними стеблями. Иными словами, по Линку, каудекс — это подземная многолетняя часть восходящей оси, у которой надземная часть периодически отмирает, например, у многих Gentiana L., Saxifraga L. (Lecoq et Juliet, 1831). Офмансеж и Линк (Hoffmansegg et Link, 1809) определили «саи4ех (collet)» как промежуточную часть меж- ду стеблем и корнем, которая помимо роста вширь растет также вверх и вниз, например, у ирисов шейкой будет то, что обычно называли «radix tuberosa». Мартынов (1820) указал, что, согласно Дево, каудекс бывает трех видов: «саи4ех-calidium» — каудекс отпрысковый (донце луковицы, например, у большинства лилейных), «саи4ех-rhizoma»— каудекс корневищный (подземный стебель, нарастающий в своей тер- минальной части и образующий из нее побеги, например, у папоротни- ков) и «саи4ех-turionarius» — каудекс побеговый (многолетнее основа- ние у некоторых трав, из различных частей которого образуются побеги, например, у астры). В первой половине XIX в. в русской ботанической терминологии господствовал «творческий период» — период создания русских терми- нов (Щербакова, 1960). А.Н.Петунников (1912), ссылаясь на многие рус- ские источники, отметил, что «саи4ех» в разное время переводили по- разному: корневище, стержень, лесина, комль, черен, балясина, пень, кряжик, комель, столбчатый стебель. Офмансеж и Линк (Hoffmagnegg et Link, 1809) переводили термин «caudex» на французский язык словом «collet» (шейка). Кахлер (Kachler, 1830), Крюгер (Kriiger, 1859) и др. употребляли слово «collet» как синоним термина «саи4ех intermedius». О.-П.Декандоль (De Candolle, 1819) отмечал, что корневую шейку надо отличать от такого органа как «саи4ех» («souche» — «пень»). Бюлльяр (Bulliard, 1783) писал ранее, что словом «souche» называют «нижнюю часть ствола срезанного дерева» (с.174). Сначала О.-П.Декандоль (De Candolle, 1819) «пеньком» («souche» — «caudex») называл в согласии с Линком многолетние стеблевые основания, которые после отмирания однолетней надземной части «принимают вид корня» и в следующем году дают новые стебли. Позднее О.-П.Декандоль (De Candolle, 1827), учитывая, что у ряда многолетних растений стебель осенью отмирает полностью или иногда его нижняя часть остается и затвердевает в виде обрубка различной длины, стал называть оставшуюся над поверхностью почвы многолетнюю стеблевую часть «пеньком» («souche» — «caudex»),
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ а погруженную в почву часть — «корневищем» («rhizoma»). Бентам (Bentham, 1846) в некоторых случаях при описании полукустарников (suffrutex), например рода Linaria Mill., называл словом «caudices» их многолетние структурные оси стеблевой природы. Буасье (Boissier, 1867- 1875) нередко многолетние подземные стебли называл «caudices», а более тонкие из них — «caudiculi». Все изложенное выше об употреблении термина «caudex» в первой половине прошлого столетия можно резюмировать в согласии с Сент- Илером (Saint-Hilaire, 1841). Многолетний ствол, облиственный только на верхушке и опирающийся на множество корней, например, у древо- видных однодольных (пальмы, юкки), отметил Сент-Илер, большин- ство ботаников называли словом «stipes» (пень), однако другие замени- ли его словом «caudex». Некоторые ботаники последним названием обозначали подземные стебли. И, наконец, ряд исследователей термин «саи4ех» применял почти ко всем стеблям, которые не опираются на один ветвистый корень: стволы древовидных однодольных, стелющиеся и подземные стебли, подземные стеблевые образования, которые имеют только одно толстое и клубневидное междоузлие. Дальнейшая судьба значений термина «саи4ех» такова. «Саи4ех»— главная ось растений, в которой различается восходящая ось — саи4ех ascendens и нисходящая ось — саи4ех descendens (Линней). Термин «axis» был известен во времена Линнея. Термин «axis» — синоним термина «саи4ех» в указанном значении. Интересно объяснил его Бюлльяр (Bul- liard, 1783): ось (axis) — «часть любого растения, вокруг которой разме- щаются другие органы, как спицы колеса» (с.9). А.Декандоль (1837) охарактеризовал ось (axis) как часть растения, соединяющую проти- воположные верхушки. Репер (Roeper, 1828) развил понятие об осевой системе растений. Его термин «axis vegetabilis» (главная ось растения) был принят ботаниками и постепенно заменил термин Линнея. Каждую ветвь растения, отходящую от главной оси, стали рассматривать как частную ось. Ле Мау (Le Maout, 1852), например, отметил, что сначала образуется первичная ось, т.е. axis vegetabilis, затем от этой оси отходят вторичные оси (axis secundarius). Линдлей (Lindley, 1830), Сент-Илер (Saint-Hilaire, 1841), Оливер (Oliver, 1888), Фишер-фон-Вальдгейм (1891) и др. выделяли у растений главную ось, при этом часть, имеющую направление роста вверх к свету (главный стебель), именовали восходя- щей осью (axis ascendens), а часть, имеющую направление роста в глубь почвы (главный корень), называли нисходящей осью (axis descendens). В настоящее время оси растений обозначаются по-латыни термином «axis». «Caudex» — ствол пальм и древовидных папоротников. В этом смыс- ле термин применяют иногда и в текущем столетии (Burret, 1929; Âon- dar, 1942 и др.); такое значение доминирует в терминологических сло- варях (J.F.Henderson а. W.D.Henderson, 1957; Boros, 1959; Schubert u. Wadner, 1962 и др.). Приведенное объяснение термина «caudex» показы- вает, что у большинства иностранных ботаников пока не вызвала дол- жного интереса другая сторона употребления этого термина. «Саи4ех» или «саи4ех intermedius» — структурные образования стеб- левой природы, переходные между собственно корнем и надземными
Глава 2. Обоснование биоморфологии однолетними стеблями. В этом направлении история употребления тер- мина «саи4ех» в ряде случаев переплетается с историей термина «rhizoma». Термин «rhizoma», как уже отмечалось, впервые употребил Эрхарт. В использовании этого термина было немало метаморфозов; так, им называли главную ось стержневого корня (Willdenow, 1802), в широком смысле его иногда считали синонимом подземного каудекса (у Горя- нинова, 1841, — «саи4ех hypogeus = rhizoma») и т.д. Ришар (Richard, 1819) называл ризомом стебли многолетних растений, полностью или частич- но находящиеся в земле, которые образуют новые стебли, по мере того как предшествующие отмирают, например у ириса, ветреницы и др. Ришар также подчеркивал, что ризом чаще всего растет горизонтально, в отличие от корня, на всем своем протяжении он имеет рубцы от листьев предшествующих лет или несет чешуевидные листья. Линк (Link, 1824) охарактеризовал ризом как «basis caulis intra terram demersa». «Rhizoma ои souche», согласно Ле Мау (Le Maout, 1852), «представляет собой стебель, который растет косо или горизонтально в земле; его передняя часть пускает вниз в почву мочковатые корни, а вверх образует почки и побеги; его нижняя часть постепенно разрушается» (с. 9), на- пример, у ириса, примулы и др. Новый для этого времени термин «rhizoma» нередко путали с «caudex», к тому же Ришар и его последо- ватели переводили «rhizoma» французским словом «souche», т.е. тем же словом, каким О.-П.Декандоль переводил термин «саи4ех», а сам Де- кандоль (De Сап4о11е, 1819), ссылаясь на Гавлера, охарактеризовал «rhizoma» как «souche», подземный или надземный, имеющий косое или горизонтальное направление, от которого отходят корешки (ирисы, па- поротники). Утверждение в ботанике терминов корневище (rhizoma), клубень (tuber), луковица (bulbus), клубнелуковица (bulbotuber) для обозначения отдельных органов растений — специализированных видоизменений побега — подорвало основу употребления как термина «caudex», так и термина «саи4ех intermedius», которые выполнили свою задачу, когда подземные органы растений были слабо изучены. Бишоф (Bischoff, 1834), который придерживался термина «саи4ех intermedius», отметил, что его многозначность затрудняет пользование им. Новые представления о стеблевой природе подземных частей мно- гих растений, которые раньше сначала ошибочно называли корнем, а затем одним общим названием «caudex» или «caudex intermedius», посте- пенно вытеснили старые представления, при этом существенно измени- лась ботаническая терминология. Значительное распространение полу- чил термин «rhizoma». Однако широкая трактовка этого термина одни- ми исследователями встретила возражения со стороны других. Нередко исследователи относят к корневищам подземные более или менее укороченные многолетние стебли, непосредственно связан- ные в течение всей жизни материнского растения с главным корнем, которые характерны для многих стержнекорневых поликарпических трав и некоторых стержнекорневых полукустарничков. Это мнение находит ряд возражений не лишенных оснований. Предлагается сохранить термин «каудекс» (Нухимовский, 1969а, б). Счи-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ таем, что не следует оставлять в тени ряд существенных положений, выд- винутых Линком, Вильденовым, О.-П.Декандолем, нуждающихся в долж- ной оценке и творческом развитии на современном уровне знаний. В работах этих ученых, как видно из изложенного выше, мы находим указания на то, что каудекс — структурное образование стеблевой при- роды; оно, как правило, многолетнее, занимает промежуточное положе- ние между корнем и однолетними надземными стеблями, нередко формируется из многолетних оснований монокарпических побегов, которые остаются после отмирания верхних частей этих побегов, выпол- няет функции вегетативного возобновления и запасания питательных веществ. В главе 6 дано определение каудекса и ризома на современном уровне знаний и их обоснование. Итак, в долиннеевский и особенно в линнеевский периоды, пожа- луй, до середины прошлого столетия, отмечается повышенная термино- творческая активность ботаников; в это время закладываются исходные понятия науки о структурной организации тела побеговых растений (органографии). Ботаники этого периода искали пути, как точнее опи- сать морфологию растительных организмов. Всех поражало многообра- зие форм растений и их органов. Из приведенного исторического обзора, где более всего обращено внимание на формирование представлений о каудексе растений, кото- рый на ранних этапах развития ботанической морфологии воспри- нимался за один из основных органов растений, видно, что научные слова-этикетки и их объяснения (понятия), т.е. в единстве — термины науки, прошли непростой путь эволюции. Сегодня восхищает, как изящно, хотя временами более или менее запутанно, но все же всегда направленно развивалась морфология растений. Это хорошо заметно сейчас, если смотреть на историю ботаники с высоты современной по- нятийной основы науки. В формировании обновленных представлений об осях, корнях, побегах, каудексах, корневищах, ветвях, стволах, кор- невой шейке, травах, деревьях и т.п. зримо присутствуют частицы труда ученых прошлых поколений. Все имеет свою историю и ничто в науке не возникает из ничего, всему, что есть в природе, находится или будет найдено фактическое и логическое объяснение и обоснование. Тому еще не раз суждено подтвердиться на страницах этой книги. 2.5.2. Организация системы «корень-лист». Концепция фитона Великий поэт и ученый Иоган Вольфганг Гете издал в 1790 г. книгу «Опыт объяснения метаморфоза растений» (1957), которая имела боль- шой резонанс и способствовала развитию морфологии растений. Гете, как неоднократно подчеркивалось (Серебряков, 1952; Первухина, 1970 и др.), пошел дальше своих предшественников в направлении сокра- щения числа основных органов; он увидел общую для всех растений закономерность, признав, что основным элементом строения тела рас-
Глава 2. Обоснование биоморфологии тения является лист, который в онтогенезе видоизменяется, т.е. подвер- гается метаморфозу, давая новые образования одинаковой с ним при- роды. Все органы растения, начиная с семядолей до частей цветка- это все видоизменение основного органа — листа. «Все кажущиеся раз- личными органы растущего и цветущего растения», — согласно Гете (1957, с. 56) можно объяснить, — «из одного единственного, именно из листа, развивающегося обычно на каждом узле». Гете создал условия для возникновения особого течения в морфологии растений, обратив внимание на следующее: «В последовательном развитии одного узла из другого, в образовании листа на каждом узле и почки поблизости от него и состоит первичный, простой, медленно продвигающийся прирост растений» (1957, с. 46). Идеи Гете были развиты в теорию фитона (Gau- dichaud, 1841); теорию анафита (Schultz, 1843), позднее в теорию филлоризы (Chauveaud, 1914); фитонистические взгляды разделяли Ада Грей, Дельпино, Челаковский, Веленовский и др. (Первухина, 1970). Годишо рассматривал тело высшего растения как объединение множе- ства первичных растеньиц — фитонов, состоящих из узла, междоузлия, расположенного ниже, и на его конце (потенциально или фактически) корня (корней). Как подчеркнул И.Г.Серебряков (1952, с. 25), «фитони- стические представления, возникшие в эпоху идеалистической морфо- логии, не могут служить теоретической базой учения об основных ор- ганах у высших растений. Понятие анафита, или фитона, т.е. листа с осевидным его основанием (или отрезка оси с отходящим от него ли- стом) является плодом геометрического анализа структуры побега, оно лишено биологического и филогенетического смысла. Еще более искус- ственным звучит утверждение фитонистов (старой школы) о том, что и корень в конечном итоге есть результат метаморфоза листа». И все же идея Гете об особом единстве листа, определенного участка стебля (в первую очередь, узла, от которого лист отходит) и почки поблизости от него (пазушной) достойна развития (см. гл.5). 2.5.3. Организация системы «корень-стебель-лист» Кроме распространения фитонистических представлений на орга- низацию семенных и всех высших растений, примерно в то же время начала формироваться концепция «железной триады» (по образному выражению Н.В. Первухиной, 1970) — концепция трех основных, ис- ходных, относительно обособленных органов — стебля, листа и корня. Еще Феофраст различал в теле растения основные органы — стебель и корень, а лист считал второстепенным органом (Первухина, 1970). Пред- ставления о трех основных органах у высших растений утвердились после работ О.-П.Декандоля (De Candolle, 1827) и А. Брауна (Вгаип, 1851). «Стебель, лист и корень нам представляются существенно различными частями растительного организма, основными его органами, основыва- ющимися на разных путях формообразования растения» (Braun, 1851, цит. по Н.В. Первухиной, 1970, с. 91). Внешне форма роста корня, листа
OCHOBhl БИОМОРФОЛОГИИ CEMEHHblX РА СТЕНИИ и стебля видоизменяется, как считал А. Браун, но их внугренняя природа (сущность) — корневая, листовая или стеблевая — сохраняется, поэтому задача морфологии — определить морфологическую природу тех или дру- гих изучаемых органов. Сходного мнения придерживались и многие другие представители классической морфологии, например, Сакс, Гебель, Тролль. Как показала дальнейшая история развития ботанической морфологии, фитонизм более прогрессивен, чем суждения о трех основных органах растения, так как в нем учитывается связь листа с подлежащим ему отрез- ком стебля (Potonie, 1912, цит. по Н.В. Первухиной, 1970). 2.5.4. Организация системы «побег-корень» Бурное развитие методов микроскопического анализа и морфоге- нетических исследований внесло существенные коррективы в учение об основных органах семенных растений. В середине прошлого столетия были открыты псилофитовые наиболее древние из высших растений, которые полностью вымерли в процессе эволюции. В строении псило- фитовых отсутствует расчленение на стебель и лист. Открытием псило- фитовых, как подчеркнула Н.В. Первухина (1970), было разрушено представление о неизменной «железной триаде» органов и о листе как основном элементе тела высших растений. Это открытие стимулировало создание новых теорий в морфологии растений. Чтобы попытаться воссоздать эволюцию высших растений от первичных форм, В.Циммер- ман создал теломную теорию (Zimmermann, 1930, 1949). Главная идея Циммермана в том, что все органы высших растений — это произ- водные простых и недифференцированных органов, которые он назвал теломами. Мы оставляем в стороне рассмотрение теломной теории, поскольку оно неоднократно проводилось в литературе (Eames, 1951; Parkin, 1951; Тахтаджян, 1952, 1954 и др.) и, в частности, в известной книге Н.В. Первухиной (1970). При всех недостатках, отмеченных критиками теломной теории, она оказала существенное влияние на развитие более совершенных представлений на организацию высших растений и их эволюцию. В 30-50-х годах нашего века Агнесой Арбер (Arber, 1930, 1941, 1950) была сформулирована теория филлома, в основе которой лежат пред- ставления о том, что основным структурным элементом высших расте- ний является побег, а не лист и (или) стебель; лист рассматривается при этом как особый (неполный, частный) побег (partial-shoot) — как осо- бая форма побеговой системы растения, которому он принадлежит. Под- робный анализ теории А.Арбер проведен также Н.В. Первухиной (1960, 1970), которая являлась одним из последовательных сторонников и пропагандистов идеи побега как целостного органа в организации тела у высших растений. Она, в частности, подчеркнула, что концепции А.Арбер о побеге предшествовали эпизодические высказывания некото- рых морфологов конца прошлого и начала нынешнего веков, которые обращали внимание на условность расчленения побега на лист и сте-
Глава 2. Обоснование биоморфологии бель, на его очевидную морфологическую целостность (Masters, 1869; Sachs, 1870; Bugnon, 1925). Современное изучение верхушечных меристем, анатомии узлов и гипокотиля, показало, что границы между стеблем и листьями, между стеблем и корнем, особенно в зародыше, условны как во внутреннем, так и во внешнем строении (Arber, 1930, 1950; Foster, 1936, 1949; Puri, 1952; Имс, 1964; Esau, 1965, Эзау, 1980; Первухина, 1970). Метаморфозы лис- тьев вегетативного побега, описанные И.В. Гете в XVIII веке, как отме- тила М.В.Сенянинова-Корчагина (1967, с. 47), «по существу являются возрастными изменениями конуса нарастания. В побеге как едином морфофизиологическом целом не может быть примата боковых органов, так как они развиваются вместе с осью из одного зачатка — верхушеч- ной меристемы — в результате раздвоения единого». Итак, основными органами индивидуумов семенных растений яв- ляются побег и корень, — «серебряная диада» в современной теорети- ческой морфологии растений. 2.5.5. Понятия побег, корень и корнепобег Выше было показано, что на вопрос, поставленный классической морфологией: что считать основным органом высших растений, в раз- ные времена давались разные ответы. Современными исследованиями доказано морфологическое единство стебля и листа в структуре побега. Побег рассматривается как целостная система меньшей сложности— орган в целостной системе большей сложности — организме. Побег непосредственно участвует в главной функции биоморф семенных рас- тений — репродуктивном размножении; он образует половые органы, снабжает их за счет жизненной активности всех своих частей необхо- димыми органическими веществами, переносит к ним с водой необхо- димые минеральные вещества, добытые корнем, а также удобно распо- лагает эти органы в жизненном пространстве. По стеблевой части по- бега проходит как ток воды с растворенными в ней питательными ве- ществами и микроэлементами из корня в листья и репродуктивные органы, так и встречный ток образовавшихся при фотосинтезе ассими- лятов, которые разносятся от места их образования (листьев) к местам их потребления (Райнботе, 1979, рис. 8). Побег (cormus) — это часть растения, состоящая из стеблевой, ли- стовой и нередко семядольной и реиродуктивной частей, обладающая опре- деленной иродолжительностью жизненного цикла, ритмом ростовой ак- тивности, стеиенью сиециализации и характером развития (Нухимовс- кий, 1971, 1980, 1986). Термин «cormus» использовался в различное время во многих значениях: как синоним терминов «саи4ех», «1гцпсия», «caulis», «individuum», «corpus» и др. Иногда «кормусом» называют тело семенного растения, а как части его рассматривают побег (der Sprout), гипокотиль и корень (die Wurzel) (Denffer, 1967). И все же чаще всего, особенно в современной литературе, термин «кормус» считают синони- мом термина побег (Кирпичников, Забинкова, 1977).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 8. Основные органы семенных растений и их функции (по Х.Райнботе, 1979). Стебель — это осевая (иростая или сложная) ветвистая или невет- вистая часть иобега, образующая в своей структуре узлы, — места иере- хода стебля в лист (листья). Лист, как подчеркнула А.Арбер (Arber, 1950), характеризуется ин- теркалярным ростом (побег в целом растет верхушкой и интеркалярно) и дорсовентральной симметрией (побег чаще всего имеет радиальное строение). Опираясь на общеизвестные факты, лист можно определить так — это часть иобега, участвующая в синтезе органических веществ,
Глава 2. Обоснование биоморфологии в трансиирации и газообмене, нарастающая в основном интеркалярно и имеющая чаще всего дорсовентральное строение, ироявляющая на какой- либо стадии жизни иобега обязательную связь через узел со стеблем. Другой основной орган — корень. Без него, кроме исключитель- ных случаев (например, Utricularia vulgaris L. адаптировалась жить в водной среде, не образуя в онтогенезе корни), организмы семенных растений не развиваются, и если утрачивают его по какой-либо причине, то их побеговая часть погибает или восстанавливает новые корни как неотьемлемую часть биосистемы в статусе организма. Общеизвестно, что корень обеспечивает побег всем необходимым: водой, минераль- ными веществами, добывая их в основном в почве. Кроме того, они синтезируют различные вещества (многие аминокислоты, гормоны, ал- калоиды и др.), которые затем перемещаются по проводящим сосудам в другие органы, а часть веществ запасается впрок. Иногда корни всту- пают во взаимодействие с корнями других растений, микроорганизмами и грибами, обитающими в почве, участвуют в вегетативном размноже- нии (Denffer, 1967; Васильев и др., 1978; Гюнтер и др., 1982). Корень (radix) — часть растения, состоящая из оси или осей, начи- нающаяся в своем развитии из верхушечной меристемы, разветвленная или неразветвленная и никогда не развивающая на себе листья и реиродук- тивные элементы, но образующая, как иравило, корневой чехлик и выросты эиидермиса — корневые волоски. Подобно тому, как правомерно говорить об отдельных органах, выполняющих свойственные им функции, например, завязях, тычин- ках, лепестках, чашелистиках, так же обоснованно рассматривать как одно целое и сборные органы, ассоциирующие в себе все эти частные органы. Последние морфологически взаимодополняют друг друга для осуществления более общей функции, в данном случае, как правило, полового размножения (цветки, соцветия). В выполнении функций веге- тативного разрастания, а иногда и вегетативного размножения, а шире— всей жизнедеятельности растений, участвуют по отдельности и побеги, и корни. Часто наиболее полно эти функции воспринимаются, если выде- лять еще и сборные органы, которые удобно обозначить как «корнепобег», или «ризокормус» (rhizocormus), и «почкокорень» (gemmoradix). Любой иобег и растущие из него корни, а также иобег и связанные с ним через семядольный узел и гииокотиль корни — это ризокормусы, а любая иочка вместе с образованными от нее корнями — иочкокорень. И ризокормусы, и почкокорни — органы или организмы, побеговая и кор- невая части которых рассматриваются вместе как одно целое (рис. 9-12). Такой подход помогает лучше понять строение растений в их онтоге- нетическом и филогенетическом развитии. Почкокорни и ризокормусы бывают ироорганизменные — это корнеиобеги-органы (u- ризокормусы), или онтоиды, если они составляют только часть структуры раститель- ного организма, и организменные — это корнеиобеги-организмы (P-ризо- кормусы), или npocmo организмы, особи, индивидуумы, эуфиты, если они составляют всю структуру растения. Какой-либо лист вместе с корнями, растущими неиосредственно из него, можно обозначить как одну систему — ризофилл.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис.9. Развитие корнепобега Filaginella uliginosum (L.) Opiz как однолетнего организма (Московская область, сырой луг). Обозначения те же, что и на рис. 3-7.
Глава 2 Обоснование биоморфалогии Рис. 10. Развитие корнепобега Echinocystis lobata (Michx.) Torr. et Gray как однолетнего организма (Калужская область): нч — надземная часть растения (подземная часть удалена). Остальные обозначения те же, что и на рис. 3-7. Рис. 11. Корнепобеги Doronicum altaicum Pall. как органы (онтоиды) и многолетнее растение (Алтай, Ивановский хребет,1800 м над ур. моря). ко — корнепобег-орган, ор — корнепобег-организм, кр — корневище. Остальные обозначения те же, что и на рис. 3-7.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 12. Развитие корнепобегов Brunnera sibirica Stev. как орга- нов и многолетнего растения (Саяны, лесной пояс, берег горного ручья). Обозначения те же, что и на рис. 3-7, 11.
Глава 3. Общая теория жизнесохранения- концептуальная основа жизни 3.1. Предпосылки к общей теории жизнесохранения Когда М.Кальвин (1971) завершал свой этапный труд по поискам физико-химических граней между неживым и самовоспроизводящимся живым, он задумался над смыслом жизни: каковы побудительные мо- тивы таких поисков и куда они могут привести. Он пришел к выводу, что каждая новая крупица знаний в познании Природы не только не дает определенного ответа на вопросы, но лишь умножает их. «Человек начинает искать смысл жизни чуть ли не с колыбели и в первые годы зрелости эти поиски иногда поглощают его целиком ... когда дети ста- новятся взрослыми, число ответов на вопрос о смысле жизни оказыва- ется равным числу людей, задававшихся этим вопросом, и нередко один человек может предложить почти столько ответов, сколько он затратил часов на размышления по этому вопросу» (Кальвин, 1971, с. 228). «Бу- дучи людьми, мы ищем оправданий нашей жизни, выходящих за рамки простого выживания. Мы ищем объяснения, оправдывающего наше очевидное господство над всеми другими формами жизни» (с. 230). Мы солидарны с М.Кальвином в том, что открытие одних таинств Природы порождает вопросы, связанные с другими ее таинствами, и так будет до тех пор, пока существует Человек. Однако в нашем представлении смысл (цель) жизни не столь загадочен и необозримо многолик, как он представляется М.Кальвину. На наш взгляд, главный смысл жизни один; он заложен в каждом из нас, проявляется непосредственно в нашем живом существовании, и в существовании тех форм жизни (в широком смысле), т.е. эврибиоморф (биосистем), что рассредоточены вне нас, на нас и в нас. Биосистемы осмысленно или неосмысленно «отвечают» на вопрос о смысле жизни через реальность своего живого существования. Что главное в жизни вообще и каковы закономерности в достижении этого главного, решается общей теорией жизнесохранения, разработан- ной впервые (Нухимовский, 1980, 1984) с широких позиций теоретичес- кой биологии, понимаемой в смысле Э.С.Бауэра (1935). Такая теория,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ как покажет дальнейший материал, может обеспечить теоретический общебиологический базис для построения других теорий по проблемам организации жизни. Рассмотрим некоторые соображения, которые привели к убежде- нию, что должна быть общая теория жизнесохранения, что должна су- ществовать система законов, составляющих и объясняющих это явление Природы, что можно и должно вполне однозначно ответить на извеч- ный вопрос: в чем всеобщий смысл жизни. Слово «сохранение» в смысле сбережение, неисчезновение, удер- жание, устойчивость, постоянство, охрана и т.п. широко используется в обиходе. Представления о сохранении широко вошли и в науку. Ин- тересные мысли о сохранении и самосохранении мы встречаем в трудах Гегеля (1975): он говорил о сохранении системы различий, например, между Землей и ее спутниками, Солнцем и кометами, о сохранении растения «путем воспроизводства самого себя, воспроизводства, являю- щегося вместе с тем воспроизводством другого», о самосохранении, как сохранении своеобразия упругими телами при внешнем воздействии на них и т.п. Открытие законов сохранения, и в первую очередь законов сохране- ния массы и энергии, было подготовлено всем ходом предшествующей человеческой истории. Законы сохранения (энергии, массы, импульса, момента импульса, спина, изотопического спина, четности, странности, электронного лептонного заряда, барионного заряда и др.) получили широкое философское осмысление (Овчинников, 1962, 1966; Веселовс- кий, 1964, 1971, 1973 и др.). Были сделаны попытки определить принцип сохранения. В.Н.Ов- чинников (1966) говорит о принципах сохранения как формах выраже- ния идеи сохраняющегося, инвариантного, включая сюда и законы сох- ранения. По В.Н.Веселовскому (1964, 1971), принцип сохранения пони- мается так: в процессе непрерывного изменения объективного мира природные величины, характеризующие материю и движение (масса, энергия, заряды и др.) сохраняют свою качественную и количественную определенность. Принцип (наиболее существенная сторона группы зако- нов) сохранения действует во всех явлениях Природы, которые, изменяясь, какое-либо время сохраняются — удерживают качественную и количес- твенную определенность в дискретности, повторяемости и континуально- сти. В.Н.Овчинников (1966), анализируя явления сохранения в нежи- вой природе, обратил особое внимание на единство сохранения и изме- нения как закон диалектики. Согласно В.И.Свидерскому (1974), в лю- бом движении, изменении имеется неразрывное единство моментов из- менчивости и устойчивости (сохранения), например, полет камня есть не только процесс изменения его положения в пространстве со вре- менем, но и состояние устойчивости в смысле сохранения камнем сос- тояния полета. Если изменение — это всякое движение, то сохранение- это всякое удержание изменений в качественной и количественной опре- деленности. Закон единства изменения и сохранения можно сформулиро- вать так: любое явление (вещь, процесс) существует какое-либо время только как сохранение (удержание) изменений в качественной и количе-
Глава 3. Общая теория жизнесохранения — концеитуальная основа жизни 77 ственной оиределенности. Этот закон — частный по отношению к зако- ну единства и борьбы противоположностей. Изменение и сохранение находятся в противоречивом единстве (Овчинников, 1966), причем про- тиворечие между этими свойствами разрешается в формах существова- ния материи и ее особого продукта — сознания (в формах, или состо- яниях, сохранения изменений чего-либо). Все формы существования (сохранения изменений) — вещи, процессы, — это всегда динамичес- кие состояния (любой покой относителен, поскольку он — тоже изме- нение в определенных качественных и количественных параметрах). По-видимому, существует три основных типа форм существования, которые сочетаются между собой по иерархическому принципу в субор- динатной классификации (снизу вверх): дискретность, или прерывис- тость (это простейший способ сохранения изменений, он обеспечивает бесконечное разнообразие дискретных форм, т.е. дискретных систем: вещей, процессов); повторяемость, или репетакция, — этот способ обеспечивает связь нового со старым, постоянство каких-либо величин, например, скорости света, а также сходство и (или) родство в многооб- разии дискретных форм сохранения изменений; бесконечность, или континуальность, — это высший тип сохранения изменений (он обес- печивает неуничтожимость и неисчерпаемость материи вглубь и вширь). Любое явление, сколь бы скоротечным по времени оно не было, суще- ствует с качественной и количественной определенностью, т.е. некото- рое время в некоторой части пространства сохраняется. Закон единства изменения и сохранения имеет следствие: чем скоротечнее изменение, тем слабее сохранительные свойства какого-либо фрагмента Природы, и нао- борот. Идея сохранения применительно к живой природе обычно рас- сматривается с точки зрения самосохранения и гомеостаза. Организации биосистем присуща направленность на самосохранение в окружающих условиях среды (Опарин, 1972; Ляпунов, 1972; Сетров, 1972). В.Н.Весе- ловский (1971) рассматривает «динамическое самосохранение» как кри- терий живого, оно отличается от исторически предшествующего ему сохранения в неживой природе своей целесообразной активностью и значительно более широкими возможностями борьбы с возмущающими воздействиями. Понятие гомеостаза впервые сформулировано для живых систем У.Кэнноном (Cannon, 1932) после того как химик А.Ле-Шаталье открыл в 1887 г. принцип сохранения термодинамических систем (принцип Ле- Шаталье): «если на систему, находящуюся в устойчивом равновесии, подействовать извне, изменяя какое-нибудь из условий, определяющих положение равновесия, то равновесие смещается в том направлении, при котором эффект произведенного воздействия уменьшается» (Ново- сельцев, 1978, с. 42). Понятие «сохранительные способности биосисте- мы», как отметил В.Н.Новосельцев (1978), объединяет способность био- системы обеспечивать гомеостаз (сохранение постоянства внутренней среды организма при наличии возмущений во внешней среде) и способ- ность поддерживать стационарное неравновесное состояние при изме- няющихся условиях внешней среды. Явление гомеостаза характеризует-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ся как одно из важнейших свойств биосистем разных уровней органи- зации жизни, но оно обычно рассматривается на примере организмов животных или людей (Саркисов, 1977; Новосельцев, 1978 и др.). Есть попытки расширить значение термина «гомеостаз» (Шилов, 1982). Иног- да гомеостаз может нарушаться ради сохранения жизнедеятельности системы, ее стационарного неравновесного состояния, поэтому гомео- стаз биосистем не является обязательным условием жизни, а цель «го- меостаз» — иерархически младше цели «сохранение жизни» (Новосель- цев, 1978, с. 64). Живые системы активно участвуют в сохранении состо- яния внутреннего динамического равновесия, что недостаточно переда- ется понятием гомеостаз; полнее активная природа биосистем отража- ется через понятие гомеорез (10гай, 1979). К.Х.Уоддингтон (1970, с. 21) пишет об этом понятии так: «Явления, для которых характерно поддер- жание постоянства некоторых параметров физиологических процессов (например, напряжения кислорода и рН крови), известны уже давно. Такого рода явления называют гомеостазом. Здесь мы имеем дело со сходной, но значительно более общей концепцией, поскольку на пос- тоянном уровне поддерживается не какой-то один параметр, а протя- женный во времени процесс изменения, т.е. траектория. Это явление можно назвать гомеорезом, т.е. оно представляет сбалансированный поток, а не сбалансированное состояние». Понятие гомеорез, как под- черкнул Г.А.Югай (1979), характеризует не саморегуляцию, а более вы- сокий уровень приспособительной изменчивости биосистем — автоно- мизированный процесс, который является самоорганизующим. Во Все- мирной стратегии охраны природы определены 3 основных задачи ох- раны природы: 1) поддержание основных экологических процессов и жизнеобес- печивающих систем', 2) сохранение генетического разнообразия; 3) длительное неистощимое использование видов и экосистем (цит. по Д.Н.Кавтарадзе, С.А. Остроумову, 1981). Итак, идея построения теории жизнесохранения как бы витает в воздухе. Обобщим основные положения, предваряющие разработку та- кой теории. В живой и неживой природе уже открыто несколько общих для разных сторон их существования законов сохранения (массы, энергии, импульса и др.). Сохранение — это атрибутивное свойство движущейся материи; в первую очередь оно проявляется как сохранение изменений через дискретность, повторяемость и бесконечность. Известные представления о самосохранении, гомеостазе, гомеорезе (в близком смысле говорят еще о гомеокинезе) только первый шаг к всеобъемлющей теории жизнесохранения.
79 Глава 3. Общая теория жизнесохранения — концеитуальная основа жизни 3.2. Исходные понятия 3.3. Законы адаптации и эволюции— исходные принципы в общей теории жизнесохранения Всякое преобразование чего-либо — это трансформация, она про- исходит на протяжении какого-либо времени. Трансформация в рамках одной дискретной материальной системы — морфогенез. Морфогенез от- дельной биоморфы — биоморфогенез (подробнее см. гл. 7). Морфогенез в морфогенезе, явление в явлении, меньшее в большем — основа суще- ствования материи, организованной в Природу по иерархическому принципу. Поток биоморфогенезов, составляющих какую-либо родослов- ную zpynny биосистем в ранге таксона (вид, род, семейство и т.n.) есть филогенез. Явление трансформации в отношении явления жизнь приобретает специфический оттенок и применительно к ней обозначается извест- ным термином «адаптация» (приспособление). Подходы к проблеме адаптации весьма разнообразны (Шмальгаузен, 1968, 1983; Георгиев- Проблему сохранения жизни обычно рассматривают в аспекте борь- бы организмов с возмущающими воздействиями внешней среды, на- правленной на личное выживание, т.е. поддержание в отношении с ней стационарного неравновесного состояния и обеспечение гомеостаза, или относительного постоянства внутренней среды. Проблему сохранения жизни, или жизнесохранения (биофилии), мы рассматриваем широко, поскольку она охватывает все жизненные явления. В живой природе наблюдаются несколько аспектов сохранения жизни и среды жизни— систем для сохранения себя и (или) для сохранения других биосистем (био- филия), деятельность биосистем для сохранения среды своего обитания и (или) для сохранения среды обитания других биосистем (экофилия). Экоби- офилия — это саморегулируемое сохранение единства живой и неживой материи его ненрерывно изменяющимися и обновляющимися носителями (биофорами). Проблему жизнесохранения мы рассматриваем в первую очередь через создание общей теории жизнесохранения (теории экоби- офилии), главную часть которой составляют две специальные теории: одна — жизнесохранения (биофилии), а вторая — средосохранения (экофилии). Общая теория жизнесохранения может в процессе развития стать ядром будущей общей теории жизни, т.е. теории о сущности жизни в ее целостности и единстве (Югай, 1976), а также основой для построе- ния общей теории природопользования, для определения стратегии и тактики природопользования. Для реализации главной цели настоящей работы она должна выполнить роль исходного общебиологического тео- ретического базиса.
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ ский, Петленко и др., 1975; Казначеев, 1980; Меерсон, 1981; Яблоков, Юсуфов, 1981; Левонтин, 1981 и др.). Обычно адаптацию рассматривают как возникновение в эволюции или в индивидуальном развитии свойств полезных (благоприятных, «целесообразных») для биосистем (чаще орга- низмов). Однако понятие «польза» и «вред» в большой мере относитель- ны, поскольку под разными углами обзора они могут приобретать от- личный смысл. Вряд ли можно признать за удачное, например, такое понимание адаптации: «адаптацией следует считать любое изменение организации, снижающее смертность под воздействием факторов среды» (Северцов, 1981, с. 130). Как быть, если естественный отбор идет где- либо, допустим, в каком-нибудь семействе растений или животных, на увеличение смертности организмов — в сторону сокращения продол- жительности их онтогенезов и увеличения плодовитости. Разве здесь нет адаптаций? В этом аспекте для нас важнее всего то, что жизнь полезна сама себе и что она сохраняет (приспосабливает) сама себя для самой себя, причем стремится сохраниться бесконечно, во всяком слу- чае имеет такую общую для себя же направленность (остальной части Природы все это безразлично). В бесконечном движении живой материи, или суперадаптации, мы видим высший смысл жизни. К суперадаптации живое может стремиться, поскольку посредством разумных существ (людей) способно осознать высшую цель (смысл) жизни. Но посредством чего можно идти к супер- адаптации. Всякая частная жизнь проявляется только в дискретных сис- темах. Существование таких систем осуществляется как посредством жизнесозидательных и жизнеразрушительных свойств, так и посред- ством передачи информации об этих свойствах от исходных биосистем к производным биосистемам. Жизнь — это единство и борьба двух при- способлений (адаптаций) материи на Земле (а может быть, и еще где- либо) — свойств сохранения и разрушения жизни. Без них обоих жизнь не смогла бы проявить себя в известных формах — в формах дискрет- ных биосистем. Так в чем же суть явления адаптации? Мы предлагаем понимать под адаптацией любую дискретную трансформацию живой ма- терии. Адаптациями являются в нашем понимании морфогенезы любых биосистем, организация биосистем, ее структурная и поведенческая час- ти и, наконец, сами биосистемы. Адаптациями являются все свойства биосистем, полезность и вредность которых определяется не столько непосредственным сиюминутным результатом их проявления в биосис- теме, сколько той ролью, которую они имеют в сохранении всей жизни и среды ее обитания, т.е. того, что важнее всего для биостробы Земли. Любая биосистема, как адаптация, существует в единстве и борьбе с окружающими ее силами Природы, включая и другие биосистемы, толь- ко на основе адаптаций по-разному и сохраняющих, и разрушающих жизнь, поэтому даже болезнь биосистемы — это одна из ее адаптаций (приспособлений). Болезнь-адаптация сигнализирует всей биосистеме, что ее жизнь в опасности, что нужны новые адаптации, чтобы жить дальше, и если не хватает адаптаций, способных противостоять разру- шительным адаптациям, то последние берут верх и направляют биосис- тему к выходу из жизни (к элиминации), поскольку и такой тип адап-
Глава 3. Общая теория жизнесохранения — концептуальная основа жизни таций также предусмотрен в дискретном (от начала к концу) принципе существования носителей жизни и запрограммирован на молекулярном уровне (в генах). Жизнь существует как адаптация, а может существо- вать и как суперадаптация. Суперадаптация — это любая бесконечная трансформация живой материи. Биосистемы (эврибиоморфы) — это всегда адаптивные системы (адаптивные формы), в них нет ничего иного из живого, кроме как адаптаций. Все во Вселенной находится во взаимо- связи и взаимозависимости, трансформируясь в пространстве и во времени, и когда-либо и где-либо может возникнуть адаптация — жизнь (высший смысл существования неживой природы), высший, с позиции человека, смысл существования которой — суиерадаитация. Трансформация осуществляется строго в соответствии с законом направленности (тропности), суть которого в следующем: все, что есть в Природе, направлено от чего-либо к чему-либо в определенном диапазоне (спектре) возможностей — на отдаление от свойств живого или на при- ближение к свойствам живого, на возникновение жизни, или на ее сохра- нение, или на разрушение жизни. Это один из высших законов Природы; из него следует, что формы адаптаций составляют все живое, а направ- ленность их проявляется двойственно: на сохранение или на разрушение жизни (иримасиециальный закон направленности адаптаций). Мы, люди, должны стремиться к суперадаптации (к бесконечности существования живого). Она особенно нужна людям. Суперадаптация осуществима при условии, если будет постоянно поддерживаться примат сохранительных свойств разумной жизни над разрушительными свойствами остальной природы. Из закона направленности адаптаций мы выводим следующее пра- вило: пока в целом развитие адаптаций (адаитационного потока) направ- лено в сторону достижения суперадаптации, до тех пор вся жизнь прог- рессивна (позитивна), а если адаптационный поток направлен в противо- положную суиерадаитации сторону, то вся жизнь развивается регрессивно (негативно). Итак, к суперадаптации можно идти только посредством прогрессивных адаптаций. Живому присуще еще одно явление — эволюция. С момента обо- снования в прошлом веке теории эволюции (Darwin, 1859) она получила широкое признание и по сей день успешно объясняет почти все изве- стные в биологии факты. Что главное в эволюции? Термин «эволюция», как подчеркнул Э.Майр (1981), означает непрерывное изменение, обыч- но содержащее элемент направленности. Многие философы понимают эволюцию с самых широких позиций — это изменение бытия и созна- ния, включающее в себя как количественные, так и качественные пре- образования; другие исследователи относят к эволюции непрерывные постепенные количественные изменения, а коренные качественные из- менения называют революцией. Кроме того, качественные изменения системы в результате ее самодвижения считают развитием (Аверьянов, 1974). Определения эволюции, которые дают биологи, обычно отражают только один из аспектов этого значительно более широкого явления. Так, Э.Майр (1981) определяет биологическую эволюцию как измене- ние разнообразия и приспособления (адаптации) популяций организ-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ мов. В понимании И.И.Шмальгаузена биологическая эволюция — это процесс приспособления биосистем к условиям окружающей среды, происходящий при смене поколений организмов (Северцов, 1981). Воз- никновение приспособленности к среде обычно рассматривается биоло- гами как основной результат эволюции, а сама эволюция — как процесс возникновения адаптаций — адаптациогенез (Яблоков, Юсуфов, 1981). Если, на наш взгляд, адаптации — это только дискретные трансформа- ции живой материи, то общая эволюция — это трансформация Природы (весь поток трансформаций, отражающий дискретность и бесконечность в движении материи из прошлого через настоящее в будущее), а эволюция живого в таком случае — часть общей эволюции (частная эволюция)— процесс адаптации всей живой природы и ее частей или суперадаптации. Уместно заметить, что понятие трансформация (преобразование) уже используется биологами для объяснения эволюции (Хржановский, 1958, 1966, 1968; Голубев, 1965 и др.). Давний вопрос: что считать единицей эволюционного процесса? На наш взгляд, универсальной единицей эволюции является синтом (греч. син — вместе, томас — отрезок), или эволюционный шаг, система «ис- ходное — производное» (абстрактное понятие), в которой одна ее часть выступает как исходное, а другая как производное (Нухимовский, 1984, 1987). Синтом, в котором исходное — то, что присутствует или уже не присутствует в настоящем (но точно известен факт его существования в прошлом), а производное — то, что образовалось посредством транс- формации из известного исходного и существует в настоящем, можно обозначить как феносинтом. Он бывает первым(нижним) и вторым (верх- ним). В первом феносинтоме точно известное исходное полностью ос- тается в прошлом, а в настоящем существует только его производное. Во-втором — и исходное, и его производное одновременно присут- ствуют в настоящем (например, живые отец и сын). Когда исходное и производное находятся в прошлом, а эволюционные связи между ними устанавливаются точно на основе каких-либо прямых и (или) косвен- ных фактов — это гииосинтом. В криитосинтоме исходное и произ- водное находятся в прошлом, а эволюционные связи между ними уста- навливаются только предположительные. И, наконец, гипотетический эволюционный шаг, который исходит из криптосинтома, а выходит в настоящее производное, — это иарасинтом. В парасинтоме связь ис- ходного с производным утрачена, и по особенностям прошлых и ны- нешних организаций систем можно только предполагать, что в прошлом существовала некая гипотетическая организация, связанная, возможно, с нынешней. Такое разнообразие синтомов отражает суть эволюции как потока синтомов, в котором что-то бесследно уходит в прошлое, а что- то как-то остается связанным с настоящим (рис. 13). На основе выяв- ления синтомов можно строить схемы реальной и гипотетической эво- люции каких-либо фрагментов Природы. Нередко в биоморфологичес- кой литературе не обращается внимание на отмеченные выше логичес- кие обстоятельства и гипотетические эволюционные связи типов жиз- ненных форм выдают за действительные.
Глава I. Теоретезирование и законы науки Рис. 13. Разнообразие синтомов (эволюционных шагов) в эволюции материальных систем. 1 — сверхоткрытая эволюция, 2 — окончание эволюции био- я 4— системы предка (элиминация), 3 — закрытая эволюция, открытая эволюция, 5 — момент превращения потомка в предка или образование предком новых потомков, 6 — син- том, 7 — парасинтом, 8 — гипосинтом, 9 — криптосинтом, 10 и 11 — первый и второй феносинтомы.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHblX РА СТЕНИЙ Частный случай шагов эволюции в Природе — синтомы в живой природе. Природа — это бесконечный эволюционный процесс, бесконечное движение синтомов, синтомокинез. Любая ныне существующая часть Природы и сама Природа — участник и результат эволюции. В эволюции материи тупиков не бывает, поскольку на смену од- ной организованности посредством трансформации приходит другая организованность. В эволюции живого тупиков много (например, эли- минация какой-либо биосистемы без образования потомства), но, пока прогрессивно существует жизнь вообще, их меньше, чем открытых эво- люционных путей. В эволюции живой материи рядом сосуществуют две формы эволюции — две формы эволюционных шагов: открытая эволю- ция, или открытые синтомы, и закрытая эволюция, или закрытые син- томы (рис. 13). Важное значение приобретает здесь понятие потомок- биосистема, производная от одной или более родительских биосистем и общая с ними по уровню организационной сложности (уровню орга- низации). Суть закрытой эволюции — эволюция живого без живого иродолжения — без образования потомства (в соответствующем биосин- томе производное не выступает в форме потомства). При завершении закрытой эволюции прекращается дальнейшая эволюция отдельных фрагментов живого или всего живого, но сохраняется эволюция всего остального (неживого), что есть в Природе. Суть открытой эволюции— эволюция неживого и живого с живым продолжением — с возникновением живого или с образованием потомства (в синтоме живое производное выступает как первородное живое от неживого исходного или как живое потомство от друтого живого). В открытой эволюции происходит глав- ное в эволюции живого, поскольку возникает новое (производное) жи- вое (жизнь продолжается), которое сразу или постепенно отрицает (закрывает) существование старого (исходного) живого. Продолжечие жизни — главный результат открытой эволюции. На основании наших предыдущих рассуждений эволюция происхо- дит не только в системе организмов (популяция, вид и т.п.), но и в любых отдельных биосистемах в ранге организма и ниже, которые проходят возрастные и фенологические состояния (стадийность морфогенезов): у организмов, например, от исходного возрастного со- стояния (зиготы) к производным (вплоть до конечного, сенильного), состояниям с выходом в неживое (смерть). В таком случае, если организм образует потомство, то он сам и его потомство — носители открытой эволюции, а после прекращения образования потомства эво- люция родительского организма закрывается на нем самом. Кроме открытой и закрытой эволюции в Природе существует сверхоткрытая эволюция, при которой эволюция в системах исходное-производное (синтомах) идет от неживого к неживому. Эволюцию, как и любое другое явление Природы, можно рассмат- ривать с разных высот абстракции. В каждом конкретном случае она проявляется в отдельном синтоме. При обобщениях мы судим об эво- люции как о разномасштабном явлении. Выявление синтомов и их типов позволяет судить о направлениях в частной или общей эволюции. Пространственно-временные параметры синтомов самые различные—
Глава 3. Общая теория жизнесохранения — концеитуальная основа жизни от ничтожно малых до неизмеримо больших. Синтомами будут, напри- мер, как система «снежинка — капля дождя», так и система «Газо- пылевое облако» и возникшая из нее Галактика, и т.д. Определяя син- томы, их типы и связи между ними, мы отвлекаемся от деталей, идем на выяснение закономерностей каких-либо путей эволюции, опираясь на большие и малые общности синтомов. Так, эволюционный ряд: до- клеточные — одноклеточные — многоклеточные примитивные вторич- норотые — первичные хордовые — бесчерепные хордовые — первич- ночелюстные — хрящевые рыбы — костистые рыбы — амфибии— рептилии — млекопитающие — человек (Завадский, 1958) — это один из вероятностных синтомов (парасинтом) в эволюции Природы, при- ведшей к возникновению человека. Вся современная жизнь на Земле- вся биостроба Земли — это результат открытой эволюции материи («ко- нус эволюции») — «производное» в системе одного эволюционного ша- га, в котором «исходное» — это первые живые существа и вся система палеобиосистем (минувших в прошлое живых систем). Мириады синто- мов меньшего масштаба составляют всего лишь один общий синтом в эволюции Природы — все живое на Земле в единстве прошлого с насто- ящим. Куда же идет эволюция? С.В.Мейен (1975) в обозрении «направ- ленность эволюции», подчеркнув, что логически возможны многочис- ленные формы направленности эволюции, отличающиеся как по источ- нику, так и в своем проявлении, в заключение отметил, что можно говорить об определенной направленности эволюции, которая скла- дывается из закономерностей, свойственных разным уровням органи- зации живого, но которые пока еще во многом не познаны. Пока откры- тая эволюция доминирует над закрытой эволюцией, до тех пор сохраняется жизнь. Этот вывод подготовлен всеми предшествующими нашими рас- суждениями, и этот же вывод мы будем постоянно подтверждать в дальнейшем. Высший (и никакой другой) смысл (цель, направленность) суще- ствования Природы может быть определен лишь через высший смысл существования того, кто его ощущает, кто его может осознать: он не может быть чем-то иным, чем высшим смыслом существования живого, в первую очередь новой, или разумной, жизни. Высший смысл существо- вания живого и всей Природы — бесконечное сохранение жизни, в первую очередь разумной жизни. Итак, высшая направленность эволюции — воз- никновение и бесконечное сохранение живого в Природе. Из рассмотренного выше материала следует: эволюция идет всегда и во всем направленно от исходного к производному; высшее достижение эволюции — возникновение живого, а высшая направленность ее — бесконечное сохранение домини- рующей роли открытой эволюции над закрытой и, следовательно, беско- нечное сохранение жизни в Природе. Это закон эволюции, из которого мы выводим правило: пока в общем эволюционном потоке закрытые эволюци- онные шаги («шаги» из живого в неживое) перекрываются количеством и качеством открытых эволюционных шагов («шагов» из живого или нежи- вого в живое), до тех пор реализуется высшая направленность эволюции в Природе — направленность на жизнесохранение. Законы адаптации и
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ эволюции и вытекающие из них правила мы рассматриваем как одни из исходных принципов общей теории жизни и как исходные принципы ядра этой теории — общей теории жизнесохранения. Где эволюция — там вечное движение от старого к новому. Но- вое — это конечный или промежуточный результат развития, рассмот- ренный с точки зрения его отношения к предшествующим состояниям развивающегося объекта (Л.Серебряков, 1967). В потоке веществ, энер- гии, информации, составляющем эволюцию Природы, происходит нарушение постепенности при переходе прошлого в настоящее, старого в новое, и в некоторой точке (месте) пространства и времени образуется сгусток новизны определенного рода (скопление новаций), свидетельствующий, что произошел (происходит) качественный переход (скачок) вперед от прежнего качества к новому. Это явление мы обозначаем как синдром новизны. Иными словами, синдром новизны — это массированное сосредоточение нового по некоторым (многим) параметрам какого-либо фрагмента При- роды, что обеспечивает переход (сдвиг) такого обьекта в иное качествен- ное состояние. Главное в этом явлении — качественный переход (пере- стройка, переворот, даже скачок) от одного состояния фрагмента При- роды к иному. Возникновение Жизни во Вселенной (в частности, на Земле) — синдром новизны высочайшего масштаба. Природа (Космос) постоянно развивается и каждый новый синтом в ее эволюции не оста- ется бесследным. Количественные и качественные изменения накапли- ваются, пока не приведут к скачкам в постепенности — появлению в организации Природы (рис. 2) синдромов новизны, из которых важней- шие — материальные системы с небывалой ранее организацией (с бо- лее сложными, чем ранее, уровнями организации). Эволюцию можно изобразить графически по-разному, например, в образе потока в пространстве и времени самоколеблющихся маятни- ков, где каждый маятник — материальная система — тело или система тел, а в нашем случае — отдельный вид растений и животных (рис. 14). Не только вид, но и каждую частную материальную систему можно принять условно за маятник, но, естественно, с иной амплитудой (ам- плитудами) колебания. В эволюционном потоке каждый маятник — эво- люционный, поскольку существовать для живого — жить, в народе иног- да образно говорят «маяться», означает также участвовать в эволюции, оставляя или не оставляя в ней заметных следов. Амплитуды колебаний эволюционного маятника в окружающем его динамическом пространстве и времени — диапазон («ареал», область) изменчивости, а в применении к живым системам ранга популяции и вида он включает не только норму реакции, т.е. «пределы в которых может изменяться фенотип без из- менения генотипа» (Северцов, 1981, с. 13), но и полиморфизм (диапазон полиморфизма). Под полиморфизмом обычно понимают: 1) «наличие в составе одного вида нескольких четко морфологи- чески отличающихся форм...; 2) формирование и устойчивое поддержание в популяции двух и более генетически и фенотипически различных форм, обуслов- ленных пестротой среды обитания и приспособлением возник- ших групп к этой среде» (Реймерс, 1991, с. 340). Нередко раз-
Глава 3. Общая теория жизнесохранения — концеитуальная основа жизни Рис. 14. Абстрактная модель эволюции Природы по маятнико- вому принципу (фрагмент эволюционного потока). SN - Супернуклеус, эм - эволюционный маятник, 01 - неведомый микроскопи- ческий мир, 1 - прогены, la - прогенотипы, 2 - гены, 2а - генотипы, 3 - опорная ось маятника, 4 — отвес, 5 — маятниковое тело, б — амплитуда колебания (диапазон изменчивости) эволюционного маятника на отдельном участке пространства- времени, 7 — воздействие на эволюционный маятник преднаследственных и (или) наследственных факторов, 8 - возвратное воздействие эволюционного маятника парафенетическими и (или) фенетическими факторами на Супернуклеус, 9- проявление парафенетических и (или) фенетических факторов во взаимодействии между отдельными эволюционными маятниками, 10 - объединенное воздействие на эволюционные маятники иных факторов окружающей среды. личают полиморфизм половой, возрастной, сезонный, связан- ный с чередованием поколений и т.п. Для самых общих суждений будем пока говорить о диапазоне из- менчивости, не забывая, что ее частный случай — полиморфизм. Ам- плитуды колебаний определенного эволюционного маятника — «ареал» изменчивости, например, индивидуума, — диапазон индивидуальной изменчивости (шире — иной, в зависимости от вида материальных си- стем); можно говорить также о клеточной, кондивидуальной, биоцено- тической и т.п. изменчивости, а в частности, еще и о внутривидовом и популяционном полиморфизме. Каждая живая система и, естественно, каждый вид растений и животных существует в диапазоне своей измен- чивости (рис. 15, 16), и какой-либо ее представитель, например, некая жизненная форма (индивидуум или кондивидуум) способна в процессе
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 15. Внутривидовой габитуальный полиморфизм Kochia prostrata (L.) Schrad. (республика Калмыкия, Черные Земли, окрестности пос. Нарын-Худук). I — полукустарничек в условиях относительно постоянного уровня поверхности почвы на равнинном пастбище, II — полукустарник в условиях микропонижения, Ш — полукустарничек с вынужденно подземной многолетней удлиненной побе- говой основой в условиях ежегодного засыпания растения песком. а — аэроподии (см. гл. 6), вг — ветвистый геоподий (см. гл. 6), п — побеги текущего года (верхние части удалены), э — зоны прошлого или нынешнего активного побегоотрастания (эпифизы), гк — главный корень, пк — придаточные корни, пп — поверхность почвы; примерное положение семядольного узла показано пунктирной линией; одно деление масштаба — 3 см. мутаций обеспечить выброс на новую высоту колебания маятника, что может дать начало новому эволюционному маятнику — новому виду, с другими амплитудами колебаний — иным диапазоном изменчивости. Положение эволюционного маятника меняется в потоке простран- ства и времени и каждое новое его «колебание» уже в чем-то не повто- ряет прежнее, т.е. процессы его развития и условия окружающей среды не остаются без изменений. Поскольку условия окружающей среды постоянно меняются, а эво- люционные маятники не стоят в своем развитии на месте, а раска- чиваются за счет внутренних ресурсов и возмущающего воздействия сре-
Глава 3. Общая теория жизнесохранения — концеитуальная основа жизни Рис. 1б. Внутривидовой габитуальнь1й полиморфизм Salsola laricifoiia Turt.z. et Li(v. I — полукустарник в условиях относительно постоянного уровня поверхности почвы на равнинном пастбище, II — полукустарник с вынужденно удлиненными геоподиями в условиях ежегодного засыпания растения песком. Обозна- чения те же, что и на рис. 15.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНКИ ды, то создается диапазон изменчивости у каждого вида растений и животных, — у любой живой системы и в общем — у любой матери- альной системы, а это, в свою очередь, создает базис для возникновения новых эволюционных маятников — новых видов (шире — новых мате- риальных систем и новых типов таких систем). В эволюционном нотоке материальные системы, будучи эволюционными маятниками, ведомо или, ча- ще всего, неведомо для себя (поскольку обычно лишены сознания) учас- твуют в своем движении к высшим целям эволюции. Для неживого такая цель одна — возникновение жизни, а у живого их три: нродолжение жизни, расширение изменчивости (нолиморфизма) и видообразование (образование но- вых видов), шире — образование новой организации (у других живых систем). Идея эволюции проходит через всю нашу работу, но еще раз спе- циально обратимся к ней в главе 8 при обсуждении актуальной пробле- мы: направленности соматической эволюции семенных растений — от деревьев к травам и (или) от трав к деревьям. С развитием человечества эволюция все более будет процессом управляемым. В разработке основ управляемой эволюции современная наука видит одну из ее задач (Яб- локов, Познании, 1975). 3.4. Закон единства живого и неживого Живое (биосистемы) состоит из полей атомов, элементарных час- тиц, атомов, молекул — неживых объектов микроскопического уровня организации материи. Кроме того, живое постоянно сохраняет единство со средой своего обитания (биотопом): поглощает ее, пропускает ее через себя, перемещается в ней, на и по ней и, наконец, включает как- либо ее в себя, приспосабливая под себя, а себя под нее, образуя в итоге сборные системы (экосистемы), продолжительность существования ко- торых определяется деятельностью живого и функционированием среды его обитания. Заслуга В.В. Докучаева в том, что он впервые обстоятельно разра- ботал учение «о тех многочисленных и многообразных соотношениях и взаимодействиях», которые составляют единство между живой и нежи- вой природой: «между а) поверхностными горными породами; б) плас- тикою земли; в) почвами; г) подземными и грунтовыми водами; д) кли- матом страны; е) растительными и животными организмами (в том числе, и даже главным образом, низшими) и человеком...» (Докучаев, 1951, с. 416). В.И. Вернадский (1967, 1977, 1978) создал учение о биосфе- ре, развив представления В.В. Докучаева и его предшественников о един- стве живой и неживой природы и доказав решающую роль живого вещества (организмов) в создании многих минералов. Накоплено достаточно фактов, чтобы сформулировать закон един- ства живого и неживого (назовем его законом Докучаева-Вернадского в честь классиков естествознания, впервые наиболее четко познавших его, хотя и не давших ему своего определения): живое нервоначально
Глава 3. Общая теория жизнесохранения — концеитуальная основа жизни возникло из неживого, состоит из неживого, существует непосредственно или через посредство другого живого в неживом, постоянно возвращается частич- но в неживое и всегда находится иод угрозой полного превращения в неживое. 3.5. Специальная теория жизнесохранения (биофилии) В согласии с А.А. Ляпуновым (1972), живая система отвечает на внешние воздействия сохраняющими реакциями, «которые определя- ются управляющей системой, использующей информацию, хранимую на молекулярном уровне». Сохранительные реакции проявляются у био- систем не только как самосохранение, личное выживание, на что осо- бенно обращает внимание большинство исследователей этого явления, а значительно шире — как жизнесохранение (биофилия): саморегулируе- мое сохранение жизни ее неирерывно изменяющимися и обновляющимися носителями. Любая материальная система, а, следовательно, и любая биосистема, обладает определенными свойствами. Свойство — это оп- ределенный диапазон потенциально возможных и реализуемых (прояв- ляющихся сейчас) признаков структуры и поведения материальной сис- темы. Все отдельные свойства биосистем и сами биосистемы — это, как было показано выше, адаптации. Свойства биофилии и биофобии (жиз- неразрушения) различаются между собой по степени сложности, которая определяется местом и ролью в сохранении жизни вообще. Иерархичес- кая соподчиненность свойств биофилии и биофобии хорошо передается посредством соответствующих субординатных классификаций (рис. 17). Единство и борьба двух противоположных свойств — биофилии и био- фобии — составляет важную закономерность жизнесохранения на Зем- ле. Перечислим и кратко охарактеризуем эти свойства. Нижнюю субординату (уровень организации) занимают свойства и функции автобиофилии, выше следует субордината свойств и функций амфобиофилии, а далее — первой, второй и третьей ксенобиофилии. Свойство биосистемы подцерживать жизнь вообще, сохраняя себя в живом состоянии и не воспроизводя новых биосистем своей или более высокой субординаты — иерсональное жизнесохранение, или автобиофи- лия. Свойство биосистемы подцерживать жизнь вообще, сохраняя одно- временно в живом состоянии и себя, и другие биосистемы, не воспро- изводя новых биосистем своей или более высокой субординаты — обо- юдное жизнесохранение, или амфобиофилия. Свойство биосистем поддер- живать жизнь вообще, воспроизводя новые биосистемы не ниже, чем своей субординаты и (или) непосредственно участвуя в сохранении их (обеспечение пищей, защита, воспитание и т.п.) — это коллегиальное жизнесохранение, или ксенобиофилия. Свойство биофилии реализуется в соответствующих функциях биосистем, например, биоморфы дышат, питаются, испаряют и поглощают воду и т.п., проявляя так автобио- фильные функции; через половое и различные формы бесполого (вегета- тивного, апомиктического и т.д.) размножения и (или) заботу о потом-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 17. Графическая суперординатная модель системы общего жизнесохранения. 1 — экобиофобия, 2 — экобиофилия, 3 — биофобия, 4 — биофилия, 5 — экофо- бия, 6 — экофилия, 7 — автобиофилия, 8 — амфобиофилия, 9 — моноксенобио- филия, 10 — диксенобиофилия, 11 — триксенобиофилия, 12 — филобиофобия, 13 — автобиофобия, 14 — амфобиофобия, 15 — моноксенобиофобия, 16 — диксе- нобиофобия, 17 — триксенобиофобия, 18 — автоэкофилия, 19 — амфоэкофилия, 20 — моноксеноэкофилия, 21 — диксеноэкофилия, 22 — триксеноэкофилия, 23— автоэкофобия, 24 — амфоэкофобия, 25 — моноксенозкофобия, 26 — диксеноэко- фобия; пунктирной линией показана взаимосвязь разнонаправленных свойств. стве они выражают ксенобиофилию, а добыча пищи для себя и потом- ства и т.д. — это амфобиофильная функция. Все вместе жизнесохра- няющие функции, реализующиеся у биосистем, обладают целесообраз- ной направленностью на достижение сохранения жизни вообще. Любая биосистема проявляет на определенном отрезке своего существования или постоянно только автобиофильные функции, но далеко не всякой биосистеме доступны ксенобиофильные и амфобиофильные функции, хотя многие программы такого потенциального функционирования за- кодированы на молекулярном уровне. Ксенобиофилия бывает трех ти- пов: первая, вторая, третья. Моноксенобиофилия — сохранение биосисте- мой других биосистем своей субординаты без воспроизведения новых. Диксенобиофилия — непосредственное участие биосистемы в образова- нии новых биосистем своей субординаты. Триксенобиофилия—
Глава 3. Общая теория жизнесохранения — концелтуальная основа жизни 93 непосредственное участие биосистемы в образовании новых биосистем не своей, а более высокой субординаты. Например, слияние двух поло- вых клеток — проявление ими третьей ксенобиофилии (образование новой биосистемы — организма), т.е. биосистемы вышестоящей субор- динаты. Для родительских организмов это процесс проявления второй ксенобиофилии, т.е. образование новой биосистемы (организма) своей субординаты и т.д. Свойства автобиофилии и ксенобиофилии у биосистем всех су- бординат находятся в противоречии, которое совершенствуется и посто- янно преодолевается (разрешается) в процессе эволюции путем подчи- нения первого второму, причем свойство ксенобиофилии контролирует развитие автобиофилии. Только так возможно сохранение основной биосистемы Земли — биостробы. Чем ниже уровень организованности биоморф, тем уже круг ксенобиофильных и амфобиофильных функций. У большинства низших животных и у растений функции ксенобиофи- лии обычно сводятся к воспроизведению потомства. Свойству биофилии npomusocmoum свойство биофобии (интибиофи- лии), или жизнеразрушения. Биофобия — это свойство биосистем частич- но или нолностью разрушаться. Автобиофобия — биофобия, которая под- готавливается жизнедеятельностью биосистемы и реализуется непосред- ственно в ней. Например, разрушение (полное или неполное) монокар- пических побегов у растения после завершения плодоношения этих побегов — автобиофобная функция. Амфобиофобия — частичное или полное взаимозависимое разрушение жизни и в себе, и в других биосис- темах (одной или нескольких) своей и (или) более высокой субордина- ты. Например, животное, проявляя амфобиофобное свойство, съедает ядовитое растение и оба погибают. Ксенобиофобия — это свойство био- системы разрушаться (частично или полностью) под действием относи- тельно сильных живых факторов окружающей среды (растений-душите- лей, паразитов, хищников и т.п.) без помощи неживых факторов среды (моноксенобиофобия) или с помощью управляемых другими биосистема- ми неживых факторов среды (диксенобиофобия) (например, животное, проявляя это свойство, попадает в капкан, который поставил на него человек), или под действием на биосистему относительно сильных не- живых факторов окружающей среды (заморозки, наводнения и т.п.), которые не управляются другими биосистемами, — это триксенобиофо- бия. При субординатной классификации свойств биофобии нижнюю субординату занимает автобиофобия и далее выше по сложности про- явления амфобиофобия, моно-, ди-, триксенобиофобия. За пределами биофилии и биофобии выступает еще одно свойство весьма странной природы — филобиофобия (дословно «любовь к жизне- разрушению»), когда биосистемы частично или полностью разрушают другие биосистемы без пользы для себя и для других. Филобиофобия- это, по сути дела, издержки биофилии, получившие особый статус свойств противоречащих биофилии. «Любовь к жизни» (допустим, автобиофи- лия) через разрушение другой жизни, полезное для первой жизни, может при определенных обстоятельствах перерастать в филобиофобию, мас- штабы которой способны выйти из под контроля биофилии (превзойти
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ ее) и погубить жизнь вообще (например, хищническое уничтожение лесов на нашей планете один из возможных путей перерастания авто- биофилии человечества в его филоавтобиофобию, т.е. в самоубийство). Взаимосвязь и противоборство свойств биофилии и биофобии, усложненное филобиофобией, всецело подчинены закону жизнесохране- ния (биофилии): жизнь, возникнув на Земле (и — во Вселенной), изменяясь, сохраняет себя в обьектах разной организации и в пределах, ограниченных ее возможностями, — это, на наш взгляд, основной биологический закон (Нухимовский, 1980, 1984). Из закона биофилии мы выводим несколько правил. П авила пе вое. Любое биофильное свойство связано с каким-либо биофобным свойством коррелятивно, и в морфогенезе на какой-либо ста- дии биофильные функции сменяются соответствующими биофобными фун- кциями. П авила вто ое. Пока на разных уровнях организации жизни свой- ства ксенобиофилии доминируют над свойством автобиофилии, а свойство биофилии — над свойством биофобии и филобиофобии, до тех пор сохра- няется жизнь. В биофилии заключен всеобщий смысл жизни; это свойство прису- ще каждому его носителю. В случае, когда биофильные резервы тают и в биосистемах начинает доминировать биофобия и, в особенности, триксе- нобиофобия (потенции разрушения жизни у неживой природы в целом пре- восходят жизнесохранительные возможности живой природы), то, соглас- но npasuny второму, жизнь попадает под угрозу полного уничтожения и в итоге может исчезнуть. Развивая идеи Ч.Дарвина о естественном отборе, популяционные генетики, в частности А.С.Серебровский (1973), пришли к выводу, что естественный отбор идет не просто в сторону наиболее приспособ- ленных, а в первую очередь к наиболее размножающимся и (или) в направлении лучшего выживания потомков. Это положение в нашей теории жизнесохранения закрепляем в виде т етьего п аaв8и~лаa: биосис- темные потоки (совокупности взаимосвязанных родственных биосистем, т.е. с близкой генетической основой), в которых лучше решается проблема ксенобиофилии (сохранения чужой жизни), выигрывают в естественном отборе филогенетическое соревнование с биосистемными потоками, в ко- торых ксенобиофильные адаптации слабее. Эволюция биосистем идет по разным направлениям, прогрессивно расширяясь, сужаясь или заходя в тупики, однако преимущества получают те направления, которые ус- пешнее других способствуют решению проблемы ксенобиофилии. Эволюция биофильной специализации идет по трем основным нап- равлениям: 1) в сторону усиления непримиримого примата ксенобиофилии над автобиофилией, когда автобиофильное поведение не спо- собно сдерживать даже первого проявления депрессирующего давления на него диксенобиофильного поведения (размно- жения), в результате чего упрощаются формы ксенобиофиль- но-автобиофильных связей (эфемерные растения и насекомые, унирепродуктивные рыбы и т.п.);
2) в сторону относительных взаимных компромиссов между ксе- нобиофильными и автобиофильными свойствами, когда авто- биофильное поведение ищет и нередко находит способы до- вольно долго сдерживать давление диксенобиофильного пове- дения, в результате чего усложняются формы ксенобиофильно- автобиофильных связей (мультирепродуктивные организмы за исключением сверхмноголетних); 3) в сторону относительных побед автобиофилии над ксенобио- филией, когда автобиофильное поведение способно очень дол- го сдерживать и даже в значительной мере снимать давление ксенобиофильного поведения, в результате чего также услож- няются формы ксенобиофильно-автобиофильных связей (му- льтирепродуктивные сверхмноголетние организмы — баобаб, секвойя и др.). Если познакомиться с реальным соотношением существующих в настоящее время путей эволюции у видов рас- тений и животных, то нетрудно установить, что виды с очень высокой продолжительностью жизни их организмов малочис- ленны. Этот путь эволюции, хотя и существует в Природе, но он более других уязвим. Направление, по которому ксенобиофилия по мере своего прояв- ления жестоко подавляет автобиофилию, и друтой путь, по которому автобиофилия находит средства к некоторому относительному компро- миссу с ксенобиофилией, оказались более всего приемлемыми для эво- люции живого на Земле. Биоморфы Ното sapiens эволюционируют по второму направлению. Они уже начали проявлять усилия, чтобы овла- деть тайной третьего пути эволюции с целью подключить и его в ди- апазон своих эволюционных возможностей и в случае успеха поставить на его реализацию великие силы своего разума. Одновременно с возникновением жизни возникла проблема био- фильной специализации частей биосистем, которую мы рассмотрим на примере биоморф (организмов и синорганизмов), занимающих особое стратегическое положение в организации живой природы. Они выпол- няют роль связующего звена в биостробе Земли, будучи морфофизио- логически самостоятельными носителями единиц генетического потен- циала жизни, управляющего фенетическим развитием всех биосистем. Части организации биосистем, в специализации которых доминирует направленность на выполнение определенных жизнесохранительных функций, названы меросомами (Нухимовский, 1984). Иными слова- ми, меросомы — это функциональные (выборные) системы (части) в ор- ганизации биосистемы (целого), активность которых направлена на дости- жение определенного биофильного результата. Совокупность свойств ка- кой-либо материальной системы, направленная в развитии на получе- ние определенного результата, — это выборная (функциональная по П.К.Анохину, 1973) система. Подобно П.К.Анохину, при выделении вы- борных (функциональных) систем мы отводим ведущую роль результату их существования. Структурность динамического материального суб- страта — обязательное свойство функциональных систем. В отличи~ от П.К.Анохина в определение таких систем мы не включаем как обяза-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ тельное условие проявление полезного результата, рассматривая его лишь как частный случай определенного результата, поскольку нередко пользы от него для всей материальной системы почти нет или, наоборот, есть только вред. Например, какая может быть польза лично для орга- низма лосося, если его функциональная система, отвечающая за раз- множение (полезное для вида), оборачивается по достижении полезного результата для него «личной катастрофой» (ведь «вреднее» смерти для любой биосистемы ничего нет). Функциональные системы выступают как системы особого рода в системах более общего характера — орга- низмах, а шире — в любых физиосистемах. Предлагаем различать следующие меросомы биоморф: эусомы, ди- асомы, архесомы, хромосомы и гены (рис. 18). У одних видов биоморф представлены только эусомы, хромосомы и гены (тримеросомные био- морфы), а у более организованных видов биоморф — все пять типов меросом (пентомеросомные биоморфы). Меросомы других биосистем сходны с меросомами биоморф по направленности биофильной специа- Рис. 18. Модель меросомной организации биоморф. 01 — экомеры, т.е. части биотопа с жесткими и неясными (соответственно сплош- ная и пунктирная линия) границами, 1 — эусома, 2 — диасома, 3 — архесома, 4— хромосомы, 5 — гены. лизации, но различаются во всем остальном. Эусомы — это меросомы, в развитии которых более всего проявляется автобиофильная направ- ленность поведения (на свое сохранение в живом состоянии), а все остальные формы биофильного поведения отсутствуют или выступают как дополнительные. Эусома организма у семенных растений включает корневую и вегетативную побеговую части. Эусома синорганизма у се- менных растений включает разобщенные вегетативные корнепобеговые части клонистов (особей вегетативного происхождения), поскольку ве-
Глава 3. Общая теория жизнесохранения — концептуальная основа жизни гетативное размножение является элементом автобиофильного поведе- ния синорганизмов. Вся активность эусом находится под генетическим давлением общей направленности развития биоморф: на создание и (или) сохранение других биосистем своего или более высокого уровня организации. Диасомы — это меросомы, занимающие промежуточное положение между эусомами и архесомами или иное особое положение, для которых главная направленность существования в организации био- морф — ксенобиофилия (на образование архесом, сохранение их и жиз- ни друтих биоморф). У биоморф растений диасомами будут спорангии, соцветия, цветки и составляющие их элементы, кроме гамет, спор или семян (эмбриональных особей). К диасомам относятся не только непос- редственные архесомогенные части, но и непосредственные носители их (например, соцветия у растений) или иные части, отвечающие в основном за жизнь других биоморф (например, молочные железы у животных). Архесомы — это меросомы, в развитии которых сильно до- минирует ксенобиофильная направленность поведения на образование новых, самостоятельно живущих биоморф. Архесомы — это споры, га- меты, эмбриональные особи (семена и др.) до их обособления от роди- тельской биоморфы. Хромосомы и входящие в их состав гены не только хранят информацию о том, кем может быть и кем быть не может био- морфа, о потенциях к мутации, но и управляют всем развитием биоси- стем на разных экофонах. Все три исполняющие функциональные си- стемы (эусомы, диасомы и архесомы) находятся в единстве и борьбе между собой, преодолевая противоречия путем подчинения эусом диа- сомам и архесомам. Биофильная специализация организационных частей (меросом) био- морф осуществляется в соответствии с Вещeeдтмм лвавилам закона жнз- несохранения: если части биосистемы неодинаково специализированы в на- правлении жизнесохранения, то чем больше они участвуют в ксенобио- фильном поведении (сохранении чужой жизни), тем меньше их участие в направлении автобиофилии (сохранении своей жизни), и наоборот. Организация биоморф варьирует по числу меросом. Дело в том, что проблема биофильной специализации возникла на заре жизни, посколь- ку примат биофилии неотделим от жизни с момента ее возникновения на Земле. Простейшей организацией биоморф обладают тримеросомные биоморфы; они состоят из эусом, хромосом и генов. Часть жизни такие биоморфы живут как бы только для себя (автобиофильное поведение), накапливая соответствующий энергетический материал, а затем, не об- разуя новых меросом, размножаются делением, т.е. проявляют диксено- биофильное поведение. Тримеросомные биоморфы — это первичный тип биоморф на Земле; их эусомы сочетают в себе необходимые для жизнесохранения функции. В результате эволюционной специализа- ции, когда одна эусома превратилась в три меросомы: новую эусому, диасому и архесому, возник другой тип биоморф — пентамеросомные биоморфы. Число жизнесохранительных функций у эусом таких эволю- ционно вторичных биоморф уменьшилось; часть их функций перешла к друтим меросомам или утратилась. Однако идущая от далеких предков способность эусомы трансформироваться на определенной стадии мор-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ фогенеза биоморфы из автобиофильной функциональной системы в диксенобиофильную функциональную систему может не блокироваться в эволюции и тогда эусома после выполнения автобиофильных функ- ций выполняет на следующей стадии биоморфогенеза диксенобиофиль- ные функции (вегетативное размножение) даже у пентамеросомных био- морф с высокой специализацией к жизнесохранению. У таких биоморф эусомы могут проявлять большее или меньшее участие в диксенобио- филии в форме вегетативного размножения, например, у земляники, ряски и т.п., что свидетельствует об особом направлении эволюции, на котором личное выживание действует на одном отрезке жизни, а затем оно подавляется (частично или полностью) более высокой по значимо- сти в жизнесохранении функцией независимо от того, образуются или не образуются другие меросомы. Размышляя над особенностями биофильной специализации частей биоморф, мы пришли также к гинотезе нанравленного новышения долго- летия (нанравленного «сверхдолголетия») организмов. На наш взгляд, для борьбы за повышенное долголетие (не имеющее аналогов в настоящем) не пригоден или мало пригоден весь живой материал старше зиготы. В одной клетке — зиготе сконцентрирован весь генетический материал организма, а у многоклеточного организма, развивающегося далее из зиготы, генетический материал рассеян по множеству клеток и стано- вится трудноуправляемым со стороны экспериментатора, поскольку гены, расходясь по разным специализированным органам, в чем-то ос- таются функциональными, а в чем-то блокируются и не отвечают уже за свойства всего организма. Генам принадлежит естественная первичная управляющая роль в организмах. При работе с зиготой, а шире — с гаметами, важно найти те «сцепки» генов, которые ответ- ственны за продолжительность личной жизни и за что-либо еще, вероят- нее всего, за размножение. Если удастся найти эти гены, если удастся заблокировать, подменить или, возможно, даже разрушить ген (гены), с которыми «сцеплены» ген (гены), ответственные за продолжительность личной жизни, не нарушая жизненной активности последних и всего будущего организма, то, возможно удастся лишить организм его прежней генетической предопределенности, главной направленности развития — направленности на размножение. Если удастся перестроить генетический аппарат, вдохнуть в организм новую направленность (в целом, хотя и не отвечающую главной направленности в существовании всех обычных биосистем, но гипотетически возможную) — направлен- ность на сверхпродолжительную личную жизнь, то может быть совершен гигантский качественный шаг вперед в развитии сельского хозяйства и человечества в целом. Успехи генной инженерии вселяют надежды на это. Генетическая и фенетическая организации находятся в противоре- чивом единстве в соответствии со снециальным законом единства генети- ческого и фенетического, который, исходя из известных фактов, можно выразить так: фенетическая организация (эусомы, диасомы и архесомы) биоморф — функция их генетической организации (генов, хромосом), по- скольку существует на основе генетической нрограммы развития; фенети- ческое — обязательная естественная среда существования (естественный
99 Глава 3. Общая теория жизнесохранения — концеитуальная основа жизни биотои) генетического, иоскольку сохраняет его в себе, неиосредственно сама взаимодействует с окружающей внешней средой и оиосредованно, через себя, иередает к нему сигналы внешней среды, влияя на его (генети- ческое) содержание. 3.6. Специальная теория средосохранения {экофилии) Опираясь на закон единства живого и неживого и на закон жизне- сохранения, можно вскрыть объективную закономерность средосохра- нения (сохранения среды жизни), которая проявляется во взаимодей- ствии свойств экофилии и экофобии, обязательных свойств любой био- системы. Экофилия — это свойство биосистемы сохранять (иоддерживать и улучшать) среду существования (среду жизни, или 6uomon). Можно раз- личать несколько типов свойств и функций экофилии (рис. 17), которые в субординатной классификации принадлежат определенным суборди- натам; их иерархическая соподчиненность по степени сложности опре- деляется местом и ролью в средосохранении вообще. Нижнюю субор- динату занимают свойства и функции автоэкофилии, выше следует су- бордината свойств и функций амфоэкофилии, а далее субординаты пер- вой, второй и третьей ксеноэкофилии. Автоэкофилия — это свойство биосистемы непосредственно под- держивать или улучшать среду своей жизни. Амфоэкофилия — экофи- лия биосистемы, на основе которой одновременно подцерживается и улучшается среда своей жизни и среда жизни других биосистем своей (и) или более высокой субординаты, но не воспроизводятся новые среды жизни. Свойство биосистемы подцерживать или улучшать среду жизни не для себя, а для других биосистем своей и (или) более высокой субор- динаты, не воспроизводя или воспроизводя новые среды жизни — это коллегиальное средосохранение, или ксеноэкофилия. Она бывает трех ти- пов: первого, второго и третьего. Ксеноэкофилия иервого muna, или моно- ксеноэкофилия, — способность биосистемы непосредственно участво- вать в сохранении среды жизни не для себя, а для других биосистем на своей субординате, без воспроизведения новых биотопов. Ксеноэкофи- лия второго muna, или диксеноэкофилия, — свойство биосистемы вос- производить новые среды жизни для биосистем своей субординаты. На основе ксенпэкофилии третьего типа, или триксеноэкофилии, биосистема участвует в создании среды жизни для других биосистем не своей, а более высокой субординаты. Свойства экофилии реализуются в соответ- ствующих функциях биосистемы: например, биоморфы строят жилища для себя по отдельности (автоэкофилия) или сообща (амфоэкофилия), растения выделяют кислород, участвуя в сохранении среды жизни для себя и для других биосистем, своей и более высокой субординаты, не создавая, таким образом, новых сред жизни (амфоэкофилия); ремонт старого жилища не его хозяином, а наемными рабочими — это прояв-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ление этими рабочими моноксеноэкофилии; некоторые люди строят дома, автомашины, космические корабли, создавая новые среды жизни для других людей (диксеноэкофилия). Триксеноэкофильные функции проявляются у биосистем, когда они создают среды жизни для более сложно организованных биосистем: например, отдельный грызун, строя себе жилище, может образовать земляной холмик, который станет сре- дой жизни небольшого биоценоза, или отдельный человек может по- строить дом, в котором будет жить без него группа людей и т.п. Свойства автоэкофилии и ксеноэкофилии биосистем всех субор- динат находятся в противоречии, которое совершенствуется и постоян- но преодолевается (разрешается) в процессе эволюции путем подчине- ния первого второму, причем свойство ксеноэкофилии контролирует развитие автоэкофилии. Только так можно сохранить основной биотоп на Земле (среду земной жизни), или среду существования биостробы Земли, — биотопа биостробы, т.е. экосферу (пожалуй, именно в этом смысле более всего подходит термин, значение которого пока строго не устоялось в науке). Свойству экофилии противоречит свойство экофобии (антиэкофи- лии), или средоразрушения. Экофобия — это свойство биосистемы ча- стично или нолностью разрушать среду жизни. Экофобию можно клас- сифицировать следующим образом по субординатному принципу (от нижних к верхним субординатам) в зависимости от степени их участия в разрушении жизни вообще: автоэкофобия, амфоэкофобия, ксеноэко- фобия. Автоэкофобия — экофобия, реализация которой подготавлива- ется жизнедеятельностью биосистемы непосредственно для среды ее собственного обитания: например, один вид, хищник, полностью ис- требляет другой вид, жертву, и сам вымирает, не сумев приспособиться к разрушенной им же самим среде своего существования. Амфоэкофо- бия — экофобия биосистемы, реализация которой одновременно раз- рушает частично или полностью среду обитания ее и других (одной или нескольких) биосистем своей и (или) более высокой субординаты: на- пример, рыбки в аквариуме постепенно отравляют воду, если ее не обновлять. Ксеноэкофобия — свойство биосистемы разрушать (частично или полностью) непосредственно самой или через элементы окружа- ющей среды не свои биотопы, а только биотопы других биосистем своей и (или) более высокой субординаты; она бывает двух типов. Ксеноэкофо- бия nepsozo muna, или моноксеноэкофобия, проявляется у какой-либо био- системы, когда она непосредственно своей деятельностью участвует в разрушении (частичном или полном) среды обитания других биосистем: например, медведь разрушил муравейник. Ксеноэкофобия второго типа, ипи диксеноэкофобия, осуществляется у какой-либо биосистемы, если она разрушает (частично или полностью) среды обитания других био- систем с помощью элементов окружающей среды: например, люди, скосив траву на лугу, временно нарушили его как среду обитания оле- ней, проявив тем самым определенное диксеноэкофобное поведение. Итак, в средосохранении существует закономерность, суть которой может быть определена как закон сохранения среды жизни (биотона), или закон средосохранения (экофилии, или биотонофилии): среда жизни (6uo-
101 Глава 3. Общая теория жизнесохранения — концетиуальная основа жизни mon) сохраняется жизнью для жизни в обьектах разной организации и в нределах, ограниченных возможностями жизни. Анализ единства и борьбы противоположностей — свойств экофи- лии и экофобии — показывает, что биосистемы в средосохранении су- ществуют в соответствии с пятым и авилом: пока на разных уровнях организации жизни свойство ксеноэкофилии доминирует над свойством автоэкофилии, а свойство экофилии над свойством экофобии, до тех пор сохраняются условия для жизни. 3.7. Принцип зкобиофилии Теория экобиофилии, или биофорофилии, в нашем представ- лении, — это концептуальная система, состоящая из следующих законов: адаптации и эволюции жизни, единства живого и неживого, жизнесох- ранения и средосохранения, экобиофилии и соответствующих правил. Помимо единства постоянно существует и противоречие между биофилией и биофобией, между экофилией и экофобией и между био- филией и экофилией; эти противоречия обычно преодолеваются (раз- решаются) в развитии путем подчинения соответствующих вторых свойств первым, причем первые свойства контролируют развитие вторых свойств. В центре единства и борьбы противоположностей всей системы жизнесохранения стоит разумная жизнь, которая доминирует над всей остальной жизнью ради своего сохранения. В единстве законов биофилии и экофилии мы видим главное ноло- жение общей теории жизнесохранения (нринции экобиофилии): жизнь сох- раняется в различных формах единства живого и неживого (биофорах) на основе доминирования нроцессов экобиофилии (биофилии и экофилии) над соответствующими нроцессами экобиофобии (биофобии, экофобии и фило- биофобии) в нределах, ограниченных возможностями жизни. На вершине организации живой природы или в центре единства и борьбы противоположностей, составляющих жизнь и фрагмент среды ее проявления, стоит разумная жизнь — нообиоз — человечество. Это высшее (из известных пока людям) достижений Природы, утратить ко- торое Природа может, но не должна. Дело в том, что разумная жизнь, возникнув на Земле, навязала (!) ей, а заодно и Вселенной, сделав их разумными гиперэкосистемами, свою волю — целенаправленную ак- тивность на свое сохранение. Любая песчинка, камень, лист дерева, Солнце, Млечный путь и т.п. обладают свойствами, которые могут быть использованы для сохранения нообиоза (разумной жизни), если разум людей — разум Вселенной — находит пути умело их использовать. Лю- дям почти безразлично, что будет со Вселенной без них, их она инте- ресует только с ними. Люди сейчас уже осознают зависимость от осталь- ной окружающей их природы, осознают свою материальную ничтож- ность в Природе, но пока еще довольно робко оценивают величие своих познавательных возможностей и возможностей материализации своих идей. Люди сильны своим разумом. Их разум по отношению к окру-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ жающей их остальной природе делает существование Природы осмыс- ленным. Люди могут осознать высшую целью развития Вселенной- нообиофилию — сохранение разумной жизни, причем нообиофилию беско- нечную в пространстве и во времени, т.е. нообиофильную суиерадаитацию. Согласно этой цели, нообиоз должен стремиться поставить себе на служ- бу все для бесконечного сохранения разумной Вселенной. Человечество контролирует состояние своего биотопа и сохраняет составляющие его компоненты — неживую часть и первоприродную живую часть в количестве и качестве, достаточном для своего сохра- нения. Пока ироявляется направленность эволюции живого на жизнесохра- нение и средосохранение и в особенности на сохранение разумной жизни (нообиофилию), до тех пор в целом жизнь прогрессивна, — ес ое и вил . Биосистемы, принимающие за основу регрессивный способ жизни (в противоречии с шестым правилом), не получают в эволюции преиму- ществ, заходят в эволюционные тупики, а сами элиминируются. Есть закономерности и в элиминации биосистем, которые можно охаракте- ризовать так. Се ьмое и авила: после того как биосистема израсходует все свои потенции к диксенобиофилии (воспроизводству подобных себе потом- ков), у нее останется значительно больше, чем ранее, потенций к элими- нации. Восьмое и ав о: биосистема как целое элиминируется более или менее постепенно в результате накоплений нарушений жизнедеятельности и (или) элиминаций ее частей или сразу и полностью иод воздействием соответствующих жизнеразрушительных факторов. Природа без людей сама по себе существовала долго и может сохранять себя без них бес- конечно. Только люди, однажды возникнув во Вселенной, как высшие представители живой Природы, могут и должны бороться за бессмер- тие — за бесконечное существование во Вселенной. Только люди могут найти ответ в свою пользу на вопрос: быть или не быть бессмертию. Человечество в своем развитии проходит сейчас стадию мировоззрен- ческого выбора между концепциями фатальной конечности (гибели) человечества, с одной стороны, и социального бессмертия — с другой (Урсул, 1977). евятое и авила: высшим критерием лидерства в Природе является стремление к бесконечному s пространстве и во времени сохранению челове- чества путем прогрессивного преобразования и сохранения среды его обитания. Сформулированную общую теорию жизнесохранения можно, как мы полагаем, использовать при создании общей теории природополь- зования, поскольку главные цели этих теорий должны быть общими— найти путь к нообиофильной суперадаптации. Иными словами, высшая направленность как в сохранении жизни, так и в природопользовании одна — это направленность на бесконечное сохранение разумной жиз- ни, на бессмертие человечества. Если Ното sapiens не будет считаться с объективными законами и правилами сохранения жизни в Природе, которые мы попытались выра- зить в общей теории жизнесохранения, то он неминуемо обречет себя на скорую гибель, поставив филогению живого на Земле с «ног на голову».
103 Глава 3. Общая теория жизнесохранения — концелтуальная основа жизни 3.8. Закон стратегии Жизни Слова «стратегия» и «тактика», с легкой руки К.Гробстайна (1968), расширили диапазон своего применения, выйдя из лексикона полити- ков, военных, шахматистов, организаторов науки и производства, и стали также достоянием биологической науки. Многие авторы увидели высокий общенаучный смысл в термине «стратегия» и стали форми- ровать теоретические конструкции с его использованием (Наllе, Olde- man, 1970; Работнов, 1975, 1978; Куркин, Матвеев, 1981 и др.). Стратегия и тактика неразрывно связаны с другим понятием— цель, над применением которого долгое время висело некое «табу» как завеса против антропоморфизма. Однако, сейчас «запрет» исчез, по- скольку все развитие современной науки показало, что антропоморфи- ческий дух — это всего лишь одно из возможных затруднений, возни- кающих на пути понятийного совершенствования какого-либо научного термина и неизбежно преодолеваемых в процессе поступательного раз- вития науки. Современный ученый может и должен понять и отличить стратегию и цель как проявление деятельности субъекта от стратегии и цели как проявления деятельности любого другого живого объекта При- роды независимо от воли субъекта. Поступим так, как это сделал К.Гробстайн (1968) и его последова- тели, и не будем вкладывать в содержание упомянутых выше терминов обязательное действие сознательной воли, осознанного стремления к чему-либо. Цель в широком смысле — «это просто некоторое конечное состояние, к которому система стремится в силу структурной органи- заци» (Рапопорт, 1969, с. 98). Стратегия и тактика может иметь много Ц оттенков в зависимости от того, под каким углом зрения оцениваются значения этих терминов. «Стратегия подразумевает общий подход, тогда как тактика относится к частностям, деталям» (Гробстайн, 1968, с. 13). Стратегия жизни — «отражение совокупности основных свойств живого (в частности, наследственности) — свойств, которые оказались для жиз- ни чрезвычайно полезными, которые обусловливают ее сложность и многообразие». В отличиВ от стратегии жизни «тактика жизни чрезвы- чайно разнообразна; каждый организм по-своему справляется с пробле- мой выживания и продолжения рода, и каждый имеет свою собствен- ную историю, представляющую большой интерес и заслуживающую са- мостоятельного изучения. Но все это — лишь частности, тактические моменты общей стратегии, причем каждый из них приобретает более глу- бокий смысл, когда понятны общие принципы (Гробстайн, 1968, с. 13). Единую для всех биосистем, осознаваемую или не осознаваемую ими, закодированную на молекулярном уровне (в генах) программу деятельности не иначе, как стратегия Жизни. В зависимости от того, какие части биосистем решают стратегические и тактические задачи, их соответ- ственно можно считать (по назначению) средствами стратегическими (гены, хромосомы) или тактическими (все остальные проявления орга- низации биосистем). Тактическая деятельность биосистем отвечает или противоречит стратегии Жизни; ею определяется реальный вклад
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ биосистемы в общее жизнесохранение. Стратегия Жизни — это программа Жизни в действии, которую отдельные биосистемы могут выполнять пра- вильно (в соответствии со стратегической целью) или неправильно (против стратегической цели). Стратегия Жизни всегда находится под утрозой, ее осуществление в любой момент может быть приостановлено, а жизнь мо- жет исчезнуть, если тактики жизнеразрушения во всем живом вынут из под контроля тактик жизнесохранения. Стратегическая цель в существовании биосистем достигается соответ- ствующими стратегическими средствами посредством развертывания раз- нообразнейших тактических средств. Стратегия Жизни имеет одну цель (стратегическую) — жизнесохранение. Действительные (реализованные) тактики биосистем отвечают или не отвечают (противоречат) стратегии Жизни; ими определяется реальный вклад биосистемы в общее жизнесох- ранение. Тактика биосистемы в определенный момент весьма многолика по форме и содержанию, она определяется разнообразием поведенческих реакций (функций) биосистемы в этот момент ее жизни. Деятельность тактических средств биосистем нашла свое отражение в субординатных классификациях биофильно-биофобных и экофильно-экофобных свойств биосистем. Оценку тактики биосистемы мы увязываем с этими классифи- кациями. Тактика биосистемы в данный момент определяется по факту проявления этой биосистемой наиболее важного в стратегии Жизни пове- дения (оно занимает более высокую субординату в соответствующей клас- сификации, среди проявляющихся в данный момент форм поведения). Например, биоморфа дышит, питается и т.п., проявляя так автобиофиль- ное поведение, и в этот момент она участвует в вегетативном и (или) репродуктивном размножении, проявляя диксенобиофильное поведение, которое перекрывает по значимости для стратегии Жизни все остальные формы деятельности рассматриваемой биоморфы. Поэтому эта биоморфа в данный момент осуществляет диксенобиофильную тактику. Исходя из общей теории жизнесохранения, определяем закон страте- гии Жизни. для жизни в целом (Жизни) приемлема только та стратегия, цель которой — жизнесохранение, а высшая цель — нообиофильная суперадаптация (бесконечное сохранение разумной жизни), что достигается (или может достигаться) посредством общего доминирования тех тактик частных био- систем, которые отвечают этим целяк. Из этого закона следует, что пока у живого в целом тактика жизнесохранения (биофилии) перекрывает такти- ку жизнеразрушения (биофобии), до тех пор реализуется цель стратегии Жизни — до тех пор сохраняется жизнь вообще; это положение отвечает правилу 2 закона жизнесохранения, что лишний раз подчеркивает принад- лежность закона стратегии Жизни к общей теории жизнесохранения. Любая частная биосистема существует в соответствии со стратегичес- ким девизом Жизни, закодированным в ней на молекулярном уровне, ко- торый на основе общей теории жизнесохранения можно выразить так: живешь — значит, умрешь, поэтому оставь что-либо для сохранения Жизни (жизни вообще) и в первую очередь живую часть себя. Возможная победа тактик жизнеразрушения над тактиками жизнесохранения на Земле пере- черкнула бы стратегию Жизни и осталась бы в эволюции памятником почти полной бессмыслицы земной жизни, даже с участием разумных существ.
Глава 4. Основные способы освоения пространства биоморфами семенных растений 4.1. Стратегические и тактические системы биоморф Стратегия Жизни в организации биоморф семенных растений ста- новится понятной, если ознакомиться с содержанием законов и правил общей теории жизнесохранения и закона стратегии Жизни. Стратеги- ческая цель у биоморф семенных растений — сохранение семенных растений на Земле. Эта цель отвечает стратегической цели всех биоси- стем — сохранению Жизни на Земле, а в целом — во Вселенной. Современная организация биоморф — результат борьбы живой ма- терии за пространство и время на протяжении длительного истори- ческого периода. Освоение биоморфами пространства неотделимо от времени, однако дать оценку этого единого процесса можно с двух сторон. В этой главе рассмотрены особенности освоения пространства биоморфами семенных растений. Тактика биоморф — это те структур- ные и поведенческие (функциональные) части, которые составляют их фенетическую организацию, обеспечивающую непосредственное взаи- модействие биоморф между собой, их средой обитания и развитие. Фе- нетическую организацию биоморф семенных растений определяют основные органы (без них невозможно самостоятельное развитие организмов) — по- беги и корни. Кроме того, особое значение принадлежит ризокормусам (корнепобегам) — высшим единицам организации рассматриваемых био- морф. Биоморфы семенных растений — это самоуправляемые и разви- вающиеся открытые системы. Н.Т.Абрамова (1974), рассматривая об- щие особенности управления систем, резюмировала, что благодаря уп- равлению система может целесообразно функционировать во взаимо- действии с окружающей средой, координировать и субординировать отношения собственных компонентов, направлять их движение в соот- ветствии с запрограммированной целью, а в результате — самоуправля- емая система может сохранять целостность, свою качественную опреде-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ленность. Оценить степень целесообразности существования биосисте- мы можно, на наш взгляд, по тому вкладу, который она вносит по сравнению с другими биосистемами того же уровня организации в реа- лизацию стратегии Жизни. А.А. Ляпунов (1980, с. 236) в определение «живого тела» включает способность этого тела отвечать на внешние воздействия «некоторыми специфическими сохраняющими реакциями», которые вырабатываются «некоторой специальной управляющей системой, работающей дискрет- ным образом и использующей информацию о состоянии внешней сре- ды», а, кроме того, и «некоторую справочную информацию, закодиро- ванную посредством дискретных кодов, причем носителями отдельных кодовых знаков являются субмолекулярные структуры». Как мы пока- зали в общей теории жизнесохранения, биоморфы в сохранительных реакциях не ограничиваются борьбой за личное выживание, эти реакции значительно разнообразнее. Обязательным условием развития фенетической организации био- морф является размножение. Главное направление развития фенетичес- кой организации: зигота — вегетативное развитие — репродуктивное развитие. Биоморфы существуют в системе крутоворота ресурсов. В нашем понимании, ресурсы (средства, занасы) — это любые вещества, энергия и информация, которые составляют окружающую среду биосистем и сами биосистемы; они или уже вовлечены жизнью в круговорот или еще могут быть в него вовлечены. В системе круговорота ресурсов фенетичес- кая организация биоморф существует по схеме: нотребление -+ нроизвод- ство -ь резервирование -ь рассеивание (Нухимовский, 1987). Высшее растение при переходе к половому размножению выступа- ет как морфологически целостная система — организм (Чайлахян, 1980), движущими силами зацветания в нем являются три взаимодей- ствующие системы — трофическая, гормональная и генетическая (Чай- лахян, 1984); только на основе их согласованного функционирования биоморфы растений могут участвовать в круговороте ресурсов биосферы. Еще полнее участие биоморф в биосфере можно выразить как результат согласованного взаимодействия ляти функциональных систем (рис. 19): унравления (генетическая организация, функция которой заключается в управлении фенетической организацией от рождения до смерти био- морф); нотребления (обеспечивает трофику, дыхание), производства (обеспечение вегетативного развития — эусомогенеза, пререпродуктив- ного развития — диасомогенеза, истинно репродуктивного развития- архесомогенеза, полового размножения и развития зародышевых особей в структуре родительского организма — эмбриогенеза, и, кроме того, образование необходимых для роста и развития продуктов метаболизма, включая гормоны); резервирования (обеспечение отложения потреблен- ных или произведенных ресурсов в резерв для реализации по мере необходимости); рассеивания (осуществление полного обособления пос- редством отделения предка от потомка, например семенное размно- жение, или иных партикул, например деструктивных, непосредственное участие в распространении гамет, спор, эмбриональных особей, а, кроме того, элиминация частей и целых организмов) (Нухимовский, 1986).
107 Глава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 19. Круговорот ресурсов в системе внешняя среда (а) — организм (6). Функциональные системы: 1 — управления (гены, хромосомы), 2 — потребления, 3 — производства, 4 — резервирования, 5 — рассеивания. Итак, свойство размножения реализуется различными способами в двух функциональных системах: производства и рассеивания. Существуют стратегические (управляющие, руководящие) и такти- ческие (исполнительские и соисполнительские) категории биосистем, которые функционируют в соответствии с законом управления-испол- нения организации биосистем (см. гл. 2.3). Охарактеризуем систему уп- равления и исполнения в организации биоморф семенных растений. За элементы управления, или элел~енты-руководители, самоуправ- ляемых систем — биоморф мы принимаем их генетические, т.е. страте- гические, системы: гены и хромосомы. Известно, что гены существуют не только в хромосомах, но и вне их — внеядерные гены (Бил, Ноулз, 1981). Элементы — руководители несут всю информацию о том, кем может быть биоморфа в фенотипе, а кем не может. Все остальные эле-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ менты биоморфы — это все тактические (исполнительские и соиспол- нительские) системы (гипосистемы). Среди элементов-исполнителей в организации биоморф семенных растений мы различаем несколько ти- пов, в зависимости от специфики исполнительской деятельности. От- метим при этом, что элементы-руководители нередко в большей или меньшей мере входят в структуру элементов-исполнителей или находят- ся вблизи от них, но, говоря об элементах-исполнителях, мы абстра- гируемся от руководящей деятельности каких-либо управляющих эле- ментов и акцентируем все внимание на специфической исполнительс- кой деятельности. Элементы-соисполнители (митохондрии, стебли, ли- стья, плоды и т.п.) являются соучастниками исполнительской де- ятельности элементов-исполнителей и их функционирование как бы растворяется в более общей деятельности элементов-исполнителей. Вы- деление в организации биоморф семенных растений систем управления и исполнения несколько условное, но оно помогает вскрыть закономер- ности иерархических взаимосвязей в этой системе. Первые исполнители (элементы-исполнители) — это биоиолимеры (PHK, за исключением РНК, выступающей в роли генов у вирусов). Их основная роль заключается в передаче информации от ДНК. РНК об- разуется на ДНК-матрице в ядре, а затем выходит в цитоплазму и уча- ствует здесь в синтезе белка (Дубинин, 1976; Уотсон, 1978 и др.). Наслед- ственная информация записана в ДНК (генах, т.е. в управляющих эле- ментах), а ее реализация осуществляется с помощью РНК — первых элементов-исполнителей. Синтез белка контролируется на последнем этапе РНК-матрицей и связан с образованием ферментных систем, что определяет метаболические и морфологические особенности отдельных клеток и, следовательно, организма в целом (Леопольд, 1968). Вторые исполнители (элементы-исполнители) — это верхушечные меристемы биоморф семенных растений. Будучи образовательной тка- нью, они участвуют в исполнении целевых программ элементов-руко- водителей и обеспечивают возникновение, рост и развитие других эле- ментов организации этих биоморф. Вторые исполнители, функционируя, достигают различных такти- ческих целей, создавая вегетативную часть тела биоморфы — эусому, и одновременно участвуют в создании репродуктивных органов — частей тела биоморфы, специализированных не в направлении личного выжи- вания биоморфы, а в направлении создания и сохранения чужой жиз- ни, — диасом (соцветий, цветков, завязей, тычинок, пыльников и т.д.) и архесом (семезачатков, пыльцевых зерен). Особенности строения и функционирования верхушечных ме- ристем биоморф семенных растений описаны довольно подробно (Имс, Мак-Даниэльс, 1935; Раздорский, 1949; Яценко-Хмелевский, 1961; Kaus- smann, 1963; Ростовцева, 1963; Александров, 1966; Серебрякова, 1971; Эзау, 1980 и др.). Нам остается лишь подчеркнуть, что верхушечные меристемы — это один из высших элементов-исполнителей «воли» генов: они первыми реализуют на тканевом уровне организации направ- ляющее воздействие элементов-руководителей в росте и развитии орга- нов биоморф семенных растений.
Глава 4. Основные способы освоения пространства биоморфами семенных растений 109 Третьи иснолнители (элементы-исполнители) — это донолнитель- ные меристемы: латеральные, интеркалярные, маргинальные (подроб- но — см. Л.И. Лотова, 1977). Вторые и третьи исполнители принадлежат одному уровню организации биосистем — тканевому. Третьи исполни- тели дополняют то, что не успевают выполнить вторые исполнители в ростовой и органообразовательной деятельности: они обусловливают утолщение осевых органов (латеральные меристемы), удлиняют междо- узлия и основания листьев (интеркалярные меристемы), участвуют в формировании листовых пластинок у некоторых растений (марги- нальные меристемы). Четвертые, лятые и шестые иснолнители в организации биоморф семенных растений принадлежат органному уровню организации (су- бординате) биосистем — это соответственно набеги, корни и корненобе- ги-органы (онтоиды, или нроорганизменные корненобеги). Побеги (четвер- тые иснолнители) включают в свою организацию меристемы и выпол- няют своей остальной частью те функции, которые меристемам не при- сущи. Побеги, как подчеркивалось выше, — это основные (ведущие) органы биоморф семенных растений. Побеги отвечают за ассимиляцию и репродукцию, т.е. они решают важнейшие тактические задачи орга- низации биоморф семенных растений в соответствии со стратегией Жиз- ни. К тактическим функциям побегов, кроме тех, которые выполняют их меристемы, относятся: ассимиляция, перенос воды и питательных веществ к местам их потребления, осуществление газообмена и испарения и нередко запасание воды и (или) питательных веществ впрок. Побеги несут также механические функции, главная из которых — обеспечение наиболее выгодного положения репродуктив- ных органов в пространстве и формирование (главнейшая тактическая функция) соответствующих меросом ксенобиофильного назначения— диасом и архесом. Решение тактических задач, которое обеспечивается у биоморф побегами, составляет важнейший аспект содержания жиз- ненного цикла биоморф. Диасомы создают архесомы — семезачатки и (или) пыльцевые зерна, на которые воздействует стратегический им- пульс, истоки которого в генах, смысл которого — сохранение жизни через половое размножение, а не просто через личное выживание (ав- тобиофилию) биоморф. «Сложность процессов развития, ведущих к об- разованию цветков, плодов и семян, — это та дань, которую растение платит за эволюционные преимущества полового способа размножения» (Леопольд, 1968, с. 227). Через свои диасомы и эусомы, побеги помога- ют сохраняться эмбриональным формам жизни (это также одна из важ- нейших тактических функций биоморф). Выполнение побегами стратегических и тактических задач невоз- можно без исполнительской деятельности корней — это пятые испол- нители в организации биоморф семенных растений. Корни (пятые ис- нолнители) непосредственно не исполняют столь разнообразные такти- ческие задачи, которые по силам только побегам, и их вклад в решение общей стратегической цели меньше. Однако без корней, как правило, не могут функционировать побеги в составе организма, поэтому ко- рень — также один из основных (ведущих) органов семенных растений.
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИИ Организация корней направлена на исполнение известных функций. Среди них отсутствует главная: функция полового воспроизведения. Ее могут выполнять только побеги, поэтому именно им принадлежит роль лидера в следующих по сложности организации исполнительских сис- темах — в системах корнепобегов (ризокормусов). Структурно-функциональное превосходство побегов над корнями и корнепобегов-органов (онтоидов) над остальными органами, отраже- нэ в законе фенетического доминирования: в фенел~ической организации биоморф семенных растений доминируют только те органы, которые не- лосредственно ислолняют несколько функций: образуют и удобно распола- гают в окружающем жизненном пространстве релродуктивные органы, обеслечивают всем необходимым (веществами, энергией, информацией) сис- темы лолового размножения и эмбрионального развития лотомства (no6e- ru, ризокормусы-органы и (или) ризосинкормусы-органы). Шестыми исполнителями в организации биоморф семенных расте- ний являются корнелобеги-органы, или онтоиды (лроорганизменные корне- побеги). Их исполнительские функции шире, чем функции побегов и корней по отдельности. Онтоиды как части организма по структуре ближе всего к целому организму, поэтому их исполнительская деятель- ность нередко направлена на обособление от родительского организма для преврашения в новый организм — в этом главная особенность так- тики этих органов семенных растений. Корнепобеги-органы развива- ются в организации биоморф не всех семенных растений. Единство набега и корня, заложенное в генотиле семенного растения, детерминиро- вано на весь его жизненный цикл в форме корнепобега, лроявляя себя вна- чале как одна клетка-зигота, далее как система зародыша и, наконец, как все элиэмбриональное растение, лричем лоследнее нередко может быть системой из нескольких лодчиненных корнелобегов-органов — это закон единства no0eza и корня. Седьмым ислолнителем в организации биоморф семенных растений являются корненобеги-органиииы (органиииенные ризокормусы), т.е. орга- низмы семенных растений, или эуфиты; они объединяют все исполни- тельские функции своих частей, которыми они отвечают на воздействие окружающей среды как целое и которые зависят от функционирования их элементов-руководителей (генов). Несколько корнепобегов-организ- мов выступают в системе синорганизма как морфофизиологически раз- общенные элементы-исполнители, которые функционируют на единой генетической основе. Итак, элементы-руководители организации биоморф семенныхрасте- а элементы-ислолнители лроводят тактику (под руководством генов) в соответствии со стратегической целью. За тактические ошибки биомор- фы более или менее наказываются окружающей их остальной частью Природы, при этом они могут преждевременно элиминироваться. Сис- тему «управление-исполнение» в организации биоморф семенных рас- тений можно изобразить в виде графической модели (рис. 20). На основе приведенного выше материала видно, что биоморфы семенных растений подчиняются закону управления-исполнения орга-
Глава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 20. Управляющие и исполняющие части в организации биоморф семенных растений. р — элементы-руководители (гены, хромосомы); элементы-исполнители: и!- первые (PHK, кроме генов), и2 — вторые (верхушечные меристемы), и3 — третьи (остальные меристемы), и4 — четвертые (побеги любой сложности, и5 — пятые (корни), иб — шестые (корнепобеги-органы), и7 — седьмые (организмы).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ низации биосистем (см. гл. 2.3), который будучи более общим законом, детализируется нами применительно к ним до закона меньшего ранга, чем первый. Согласно ему (закон управления-исполнения организации биоморф семенных растений): организация биоморф семенных растений существует на иерархической основе, при этом управление в ней происходит с биополимерного и органоидного уровней (гены, хромосомы), а лидерные исполнительские функции несут определенные элементы биополимерного (первые исполнители — молекулы PHK), клеточного и тканевого (вторые и третьи исполнители — меристемы), органного (четвертые-шестые ис- полнители — побеги, корни, корнепобеги-органы, включая и адвентивные), организменного (седьмые исполнители — корнепобеги-организмы) уровней организации элементов биоморф. Стратегия Жизни воплощается (реализует свои потенции) у отдель- ных биосистем в поведенческих актах, отвечающих стратегической цели или не отвечающих ей вовсе, и в зависимости от того, какие частные тактики берут верх, слагается общая (целостная) тактика жизни (такти- ка организации) каждой биосистемы на данный момент и на весь ее жизненный цикл, которая лучше или хуже сохраняет жизнь вообще (Жизнь), т.е. лучше или хуже отвечает стратегии Жизни. Биоморфы семенных растений в процессе эволюции овладели надежными страте- гическими и тактическими средствами, что обеспечивает им современ- ное существование на Земле. 4.2. Биполярность в организации биоморф Фенетическая организация биоморф семенных растений, на наш взгляд, биполярна в двух отношениях. Первая биполярность — та, ко- торая лежит как бы на поверхности, она заложена в конкретную основу фенетической организации: гелиотропизм побегов и геотропизм корней, обеспечивающие оптимальные возможности освоения пространства на- земными семенными растениями. Есть и другая биполярность, осозна- ние которой достигается только посредством образного мышления,— это биполярность исполнительских свойств (тактик) биоморф семенных растений в системе жизнесохранения. Чтобы не потерять нить широких обобщений, не будем углубляться во многие детали (они изучаются в разных разделах биологии); выделим лишь те, которые наиболее существенны в осознании биполярности тактик биоморф семенных растений. Каждый фрагмент живого борется, ищет, находит или не находит необходимые тактики в процессе развития в зависимости от конкретных условий существования, чтобы сохранить себя и (или) другие биосистемы, поскольку к поискам и борьбе подтал- кивает более высокая цель — стратегическая цель Жизни — жизне- сохранение вообще. Все тактики рассматриваемых биоморф можно объединить в одной суперординатной классификации (рис. 21). Высшую ступень займет субординатная классификация из двух субординат: а) внизу — тактики
Глава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 21. Графическая суперординатная модель тактик биоморф семенных растений. Субординия тактик: 1 — первая, II — вторая. 1а — деградация (регрессивное развитие, диссимиляция); 2а — рост, прогрессивное развитие и ассимиляция; 1б — ассимиляция, 2б — нарастание, ветвление и кущение; Зб — вегетативное размножение; 4б — половое (с половым процессом) и неполовое (через репродуктивные органы без полового процесса) размножение; 5б — диссимиля- ция, бб — частичный некроз органов, 7б — полный некроз отдельных органов, 8б — полный некроз биоморф (элиминация). деградации (регрессивного развития, диссимиляции); б) вверху — так- тики роста, прогрессивного развития и ассимиляции (рис. 21-I). Би- полярные тактики на обоих уровнях абстракции (суперординатах)— антагонисты, и только доминирование тактик «верха» над тактиками «низа» отвечает стратегии Жизни. Ниже по «пирамиде» абстракций разворачивается следующая кар- тина (рис. 21-II): две оси биполярной субординии тактик организации биоморф семенных растений. Одна субординия включает следующие классы (субординаты) тактик: а) ассимиляции; б) нарастания, ветвле- ния, кущения; в) вегетативного размножения; г) полового (с половым процессом) и неполового репродуктивного (через репродуктивные ор- ганы без полового процесса) размножения. Другая субординия состоит из следующих классов тактик: а) диссимиляции; б) частичного некроза
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ органов; в) полного некроза отдельных органов; г) полного некроза био- морф (элиминации). Рассмотрим особенности организации некоторых тактических си- стем, имеющих непосредственное отношение к последующим логичес- ким построениям. 4.3. Рост и развитие Преодолев механистический подход к представлению о росте как количественном изменении целого организма, господствовавший в на- чале нашего столетия, Д.А.Сабинин разработал новое определение рос- та — это «процесс новообразования элементов структуры организма» (Сабинин, 1940; 1963, с. 7). Под процессами развития Д.А.Сабинин (1963, с.8) понимал «изменения в новообразовании элементов структуры организма, обусловленные прохождением организмом жизненного цикла» (курсив наш). Такое понимание роста и развития перешло с небольши- ми дополнениями в современную учебную литературу: «рост — это необратимое новообразование структуры живого тела или объема его частей. Развитие — это качественное изменение компонентов организ- ма, при котором имеющиеся формы и функции превращаются в другие» (Либберт, 1976, с. 344). Разрастание ветви дерева, осуществляемое благо- даря размножению и увеличению клеток, — это рост, а образование проростка из семени при прорастании или прекращении зимнего покоя у древесных растений — примеры процессов развития (Сабинин, 1971; Либберт, 1976). Процесс развития у растений обычно связан с ростом. Рост и развитие — две стороны одного процесса морфогенеза. Тактические средства роста и развития в организации биоморф семенных растений — это любые гипобиоморфы (органеллы, клетки, ткани, органы) и биоморфы в целом. Они выполняют разнообразные функции, многие из которых будут характеризовать или рост и (или) развитие этих биосистем, частные цели которых подчинены общей стра- тегической цели — цели жизнесохранения. Тактика роста и развития биоморф семенных растений направлена на разных этапах их жизнен- ных циклов на то, чтобы обеспечить биоморфам жизнь для себя и (или) помогать жить биоморфам своего и (или) другого вида и (или), чтобы подготавливать процесс воспроизведения потомства и (или) непосред- ственно осуществить его, а, кроме того, и на то (регрессивное развитие), чтобы обеспечивать выход из Жизни (элиминацию) отслуживших ей биосистем. Системы роста и развития — это и отдельные гипобиомор- фы разных уровней сложности, и биоморфы в целом с единством их побеговой и корневой частей-систем. В организации основных органов биоморф — побегов и корней проявляются частные тактики, которые можно подразделять по способу их проявления, т.е. по функционально- му и (или) структурному содержанию, например, тактика нарастания, тактика ветвления, тактика кущения, тактика размножения и др. Мно- гие иные тактики мы не рассматриваем, но содержание их нетрудно
115 Inaea 4. Основные способы освоения пространства биоморфами семенных растений интерпретировать на основе известных обобщающих материалов, скон- центрированных в руководствах по физиологии, биохимии, анатомии, гистологии растений и друтих ботанических наук. Рассмотрим несколь- ко тактик роста и развития биоморф семенных растений. 4.4. Нарастание, ветвление и кущение '1 Под монохазиальным нарастанием понимается (Rauh, 1950) отрастание из одной боковой почки, расположенной под недоразвитой верхушечной почкой, побега, за- мещающего собой исходный побег. Мы считаем возможным говорить о монохази- альном нарастании и в более широком смысле, как о процессе замещения недоразвитой верхушки простой оси меньшего порядка на одну производную боко- вую простую ось и т.д. в рамках системы, внешне подобной моноподию (одной простой оси). Материальной основой тактик явл~потся соответствующие тактичес- кие системы, элементы которых вычленяются из организации биоморфы по общности целей их функционирования. Разберемся в структуре и функ- ционировании тактических систем — систем нарастания, ветвления и ку- щения. Структурной основой рассматриваемых систем является осевая сис- тема, без знания которой нельзя понять закономерностей этих явлений. Издревле ученые обращали внимание на особенности осевого стро- ения растений. История развития представлений об осях растений в прошлом изложена в главе 2.5.1. В настоящее время термин «ось» (axis) широко применяется в бота- нике в особенности при описании стеблевой и реже корневой частей (систем) тела растений. Осями в структуре тела высшего растения, являются его части, имеющие по одному основанию и по одной верхушке, служащие опорами для других частей, проводниками, а иногда и вместилищами воды и питатель- ных веществ (Нухимовский, 19746). Главная особенность струкгуры осей у биоморф семенных растений— одна верхушка и одно основание, а главная функциональная черта осей- бьпь опорой ассимиляционным (листьям) и репродуктивным (цветкам, плодам) органам или друтим одноименным (подобным им осям) органам. Оси — это в основном стебли, корни и, кроме того, рахисы, черешки листьев и т.п., поскольку они отвечают структурному и функциональному содержанию этого понятия. Осью, например, является тычиночная нить, но не будет ею шип или волосок на поверхности листа или стебля, по- скольку первый орган — это опорный и проводящий элемент в органи- зации тычинки, а вторые органы этих функций не имеют и не являются одноименными производными осей — это осевидные органы, но не оси. Ось, не имеющую по всей своей длине ни одного перевершинива- ния или дихотомического расчленения, мы называем npocmou, а систе- му, составленную простыми осями, последовательно связанными между собой и внешне подобную одной простой оси, сложной осью. У древес- ных растений следы частых перевершиниваний в рамках сложной оси постепенно стираются. Сложные оси, нарастающие монохазиально'
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИИ (Tilia L., Ulmus Ь.), иногда называют кажущимися осями (Denffer, 1967). У многолетних трав и полукустарничков следы частных перевершини- ваний в рамках сложной оси нередко хорошо заметны. Простая ось — структурный элемент всей осевой системы высших растений. Оси бывают активными (их алексы '> жизнеспособны) или пассивными (их алексы уже утратили жизнеспособность или после виль- чатого раздвоения стали апексами других осей). Активные оси состоят из определенных морфологических частей 1зон): аникальной (она включает апикальную меристему или всю верхнюю часть оси, входящую в состав почки), эфемерной (она отрастает в текущем вегетационном сезоне и отмирает в этом же сезоне), потенциально скелетной, или проскелетной (она отрастает в текущем вегетационном сезоне и предположительно должна стать скелетной), скелетной (она зимует первый год или сохра- няет жизнеспособность уже несколько лет). Потенциальную скелетность отдельных частей можно установить по различным признакам, напри- мер, по способности их развивать на себе почки, приспособленные к перенесению неблагоприятных воздействий среды, или одревесневать и т.д. Потенциальное свойство реализуется или не реализуется в ожида- емые сроки. Оси по степени своего развития и продолжительности жизни быва- ют инициальные (они проходят начальную фазу своего развития внутри определенных тканей растения или в родительской почке), эфемерные (они отрастают и отмирают за один вегетационный сезон), гемискелет- ные (они включают в свою структуру скелетную часть наряду с одной или несколькими иными частями: эфемерной или проскелетной), по- тенциально скелетные, или проскелетные (они предположительно долж- ны стать скелетными) и скелетные (они полностью скелетные). Поскольку стебли — это главные каркасные (опорные) элементы в структуре побегов семенных растений, то классификация стеблей во многом будет приемлема для аналогичной классификации побегов, на что мы еще будем не раз обращать внимание (гл. 5.1.). Попытки односторонне трактовать термин «ось», на наш взгляд, не оправданы. Так, например, А. Петров (1963, 1967) предложил называть термином «ось» такую стеблевую часть, «которая берет свое начало от верхушечного конуса нарастания в пазухе примордиального покровного листа и представляет как бы след, оставленный от перенесения все выше и выше до прекращения жизнедеятельности верхушечной мери- стемы» (1967, с.7). А. Петров (1963, 1967; Petrov, 1964) принимает «ось» в качестве ос- новного структурного элемента кроны древесных, а с определенной интерпретацией и стеблевой части травянистых растений, рассматривая ее не как «практическую», а как «теоретическую», «нематериальную» единицу. Для подразделения «осей» использованы признаки побегов, а не признаки стеблей («строго генеративная, генеративная, вегетативно- Ф1 Обычно в качестве синонимов термина «апикальная меристема» употребляют тер- мины «алекс побега» или «алекс корня» (Эзау, 1980). Меристематические верхушки осей — апексы осей. Например, можно говорить об апексах стеблей, абстрагируясь от листообразовательной деятельности апикальных меристем побегов.
Глава 4. Основные способы освоения пространства биоморфами семенных растений генеративная, вегетативная» оси). Попытка А. Петрова найти структур- ный элемент всей стеблевой части тела цветковых заслуживает, на наш взгляд, соответствующего развития. При анализе стеблевой и корневой систем семенных растений наи- более удобно пользоваться не структурным элементом всей осевой сис- темы — простой осью, а структурными элементами соответствующих частей этой системы: стеблем и корнем. Стебель — осевая часть, обра- зующаяся в результате жизнедеятельности побеговой верхушечной меристемы, несущая или несшая на себе другие элементы побега— листья, а нередко и репродуктивные элементы. Корень — осевая часть, образующаяся в процессе жизнедеятельности верхушечной меристемы, защищенной корневым чехликом, и никогда не несшая на себе листьев и репродуктивных элементов. Стебли и корни, будучи осями, бывают иростыми и сложными; кроме того, как системы осей, стебли и корни бывают ветвистыми и неветвис- тыми. Простые стебли и корни не имеют в своей структуре переверши- ниваний и не включают себеподобных ответвлений, сложные стебли и корни имеют в своей структуре перевершинивания и (или) включают себеподобные ответвления, различаясь между собой по сложности орга- низации. Все виды стеблей конкретны (их можно подрезать, наклонять, зарывать в субстрат и т.д., т.е. делать с ними то, что, естественно, нельзя сделать с их графическими отображениями — «осями» в понимании А. Петрова); подобное можно сказать и о корнях. Среди простых стеблей мы различаем медиали и латерали. Меди- аль — npocmou стебель первого порядка. Латераль — npocmou стебель 2, (они образуются в результате жизнедеятельности побеговых верхушеч- ных меристем, закладывающихся в пазухах примордиальных листьев) и адвентивные (они образуются в результате жизнедеятельности побего- вых верхушечных меристем, закладывающихся в любой части тела рас- тения, как правило, в более зрелых тканях вне связи с апикальными меристемами предшествующего порядка, т.е. на корнях, листьях или на стеблях вне пазух примордиальных листьев). Адвентивные латерали в свою очередь подразделяем на каулогенные, ризогенные и филлогенные (эпикальные меристемы, образующие их в процессе своей жизнедея- тельности, закладываются соответственно в тканях стеблей, корней и листьев). Среди простых корней мы различаем по особенностям их возникно- вения субмедиали и сублатерали. Субмедиаль — npocmou корень первого порядка. Сублатераль — npocmou корень 2, 3 и последующих порядков или ные (апикальные меристемы, образующие их в процессе своей жизне- деятельности, закладываются соответственно в тканях корней, стеблей и листьев). Каждая ось проходит определенный жизненный цикл (цикл роста и развития от начала ее заложения в определенных тканях растения до полного отмирания). У каждого стебля стадии их жизни находятся в прямой связи со стадиями жизни определенного побега (см. гл. 5.1.). У
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ простого корня — субмедиали или сублатерали, различимы несколько пе- риодов жизни: начальный (инициальный) — продолжается от начала его заложения в определенных тканях растения до выхода во внешнюю среду, основной (постинициальный) — от начала его выхода во внешнюю среду до прекращения жизнедеятельности верхушечной меристемы и конечный (финальный) период — от момента прекращения жизне- деятельности апекса корня до полного отмирания этого корня. В обыч- ных эколого-морфологических исследованиях наибольший интерес представляет выяснение особенностей постинициального периода жиз- ни отдельных осей. В организации биоморф семенных растений осевая система, состо- ящая из стеблей и корней, представляет собой своеобразный каркас (остов), который развивается как полярная опорная и проводящая си- стема всего ризокормуса-организма, обеспечивая наиболее подходящее для функционирования расположение в пространстве ассимиляцион- ных и репродуктивных органов и транспорт противоположно направ- ленных потоков воды, минеральных и органических веществ. Функционирование (нарастание, ветвление, кущение) осевых сис- тем составляет основной динамический аспект тактики биоморф семен- ных растений. В поведении осевых систем проявляется закон каркасности биоморф семенных растений, который, исходя из известного можно выразить так: какую бы форму внешней и (или) внутренней специализации не приобретали в процессе эволюции стебли и корни, они всегда выполняют в организации биоморф семенных растений функцию каркаса (остова) по отношению к другим осевым и неосевым частям. Этот закон частный по отношению к более общему закону каркасности биосистем (гл. 2.3). Каркас и осевая система — это не всегда равнозначные понятия. Так, каркасом одноклеточных организмов обычно являются жесткие (строго определенные) оболочки. Возникновение корнестеблевой (соб- ственно осевой) системы в организации растений было крупным заво- еванием в эволюции живой материи. Особая тактическая роль в организации биоморф семенных расте- ний принадлежит скелетным элементам, т.е. тем корнестеблевым эле- ментам, которые не отмирают после завершения вегетационного сезона, зимуют 1 и более раз, становятся малолетними (живут 2-4 года) или многолетними (живут 5 и более лет). Скелетный стеблевой каркас— обязательная часть базовых органов, или базокормусов, организация которых будет рассмотрена ниже (гл. 6). Организацию осевых систем семенных растений мы рассматриваем на фоне выяснения содержания и соотношения понятий «нарастание» и «ветвление». Известно, что растения бывают неветвистые или ветвистые, причем последние нередко «дают от корня много стволов и множество веток» (Феофраст, 1951, с. 19). Словами ветвь (гатив), кустарник (frutex), по- лукустарник (suffrutex) и их производными (ramulus, fruticosus, suffruti- cosus и др.) пользовались еще на заре ботанической науки (Caesalpini, 1583; Linnaeus, 1753, 1767; Bechstein, 1810 и др.).
пава 4. Основные способы освоения пространства биоморфами семенных растений 119 Для развития представлений о ветвлении растений, пожалуй, боль- ше других имели впоследствии значение следующие термины, исполь- зованные К.Линнеем (Linnaeus, 1751, 1767): терминальные и аксил- лярные цветки; простые, ветвистые, дихотомические стебли; очередные, супротивные, мутовочные, рассеянные и скученные ветви. В конце XVIII в. — первой половине XIX в. довольно много гово- рилось о дихотомическом ветвлении, которое понималось как всякое раздвоение верхушек стеблей, повторяющееся вновь и вновь у новых стеблей (Linnaeus, 1751, 1753). Дихотомия отмечалась не только у водо- рослей, мхов, плаунов и папоротников (Linnaeus, 1753; Schkuhr, 1809; Agardh, 1821; Nageli, 1847, 1860 и др.), но и у покрытосеменных расте- ний, например, у сирени, омелы, дурмана, белены и др. (Linnae- us, 1753, 1762; Dupetit-Thouars, 1809; Wydler, 1843, 1851 и др.). Как па- мять о широкой трактовке дихотомии остались видовые названия ряда растений, например, Anemone dichotoma Ь., Нурепсит dichotomum L., Cerastium dichotomum L., Stellaria dichotoma L. (Linnaeus, 1753). Нэгели (Nageli, 1847, с. 185) впервые подробно описал и зарисовал микроскопи- ческие особенности истинной дихотомии у водоросли Dictiota dichoto- ma 1.атоиг. «Кажущуюся ось» (die Scheinachse; Wydler, 1843), которая образуется в тех случаях, когда ветви с одиночными цветками поочеред- но то справа, то слева замещаются боковыми осями, принимающими центральное положение, К.Шимпер назвал термином «sympodium» (цит. по Видлеру; Wydler, 1851). Этот термин нашел применение как при исследовании ветвления, роста, сцепления и т.п. осей соцветий (Wydler, 1851; Kiitzing, 1852; Eichler, 1875 и др.), так и при описании вегетативной части побеговой системы (Вгаип, 1851; Wygand, 1854; Hofmeister, 1868; Sachs, 1868; Wiesner, 1884 и др.). Для обозначения системы ветвления, в которой главная ось (подиум) долго сохраняет способность нарастать одной верхушкой, а ее дочерние (боковые) оси всегда слабее, был упот- реблен термин «monopodium» (Nageli, 1863). У семенных растений на- стоящая дихотомия крайняя редкость; как правило, они ветвятся по моноподиальному или симподиальному типу (Hofmeister, 1868; Sachs, 1868; Velenovsky, 1907; Potonie, 1912; Goebel, 1928; Troll, 1937, 1954; Rauh, 1950; Серебряков, 1952, 1962 и др.). Ветвление растений обычно рассматривается как форма роста, при которой увеличивается число боковых побегов (Velenovsky, 1907; Troll, 1937; Rauh, 1950; Серебряков, 1952 и др.). У разных растений неодно- кратно отмечалось образование «кажущихся», или «ложных» осей (Wyd- ler, 1843, 1851; Hofmeister, 1868; Velenovsky, 1907; Troll, 1959; Denffer, 1967 и др.). При таком способе многолетнего увеличения числа боковых побегов («монохазиальное ветвление»), как отметил Pay (Rauh, 1938, с. 293) у Salix arbuscula, в конечном счете обычно образуется неразвет- вленный ствол. Иногда ветвление определяют слишком обще. Так, например, по Ал.А. Федорову и др. (1962, с. 28), ветвление — это «более или менее сильное расчленение тела растения». Здесь не подчеркнуто, что ветвле- ние — специфическая форма роста, поэтому к нему ошибочно может отнесена неполная партикуляция, имеющая, как подчеркнул И.Г.Сере-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ брякав (1952), лишь внешнее сходство с ветвлением. Проблема ветвле- ния широко обсуждается и в литературе последние десятилетия (Нухи- мовский, 1971, 1974б; Грудзинская, 1974; Шафранова, 1981 и др.), что свидетельствует о ее большом значении для морфологии растений. На основании литературных данных и наших непосредственных морфогенических исследований семенных растений мы пришли к вы- воду, что некоторые из известных положений о ветвлении нуждаются в уточнениях и дополнениях. Так, не следует отождествлять понятия «нарастание» и «ветвление». Нарастание (accrescentia) осевой системы- ироцесс увеличения ее линейных размеров (т.е. увеличение длины без учас- тия иартикуляции). Ветвление (ramijicatlo) — ироцесс образования в ре- зультате нарастания осевой системы из одной ее части двух и более частей, имеющих обособленные живые верхушки и связанные между собой через зону контакта (Нухимовский, 1974б). В зависимости от того, в какой фазе жизни находятся производные ветви, ветвление бывает скры- тое (начальное) и видимое (основное). В фазу скрытого ветвления форми- руются зачатки видимых ветвей, например, в форме аксиллярных или адвентивных почек и т.п. Показателем основного ветвления является видимый рост ветвей. Зона контакта осей системы ветвления, или зона ветвления, — часть системы ветвления, которая участвует в образовании ветвей одного порядка и обеспечивает связь их оснований между собой, а также непосредственно с самой родительской осью. В осевой структуре тела семенных растений нередко различимы системы нарастания и ветвления. В системе нарастания мы различаем систему стеблевого нарастания (шире — систему побегового нарас- тания) — это целое, образованное из простых стеблей (шире — одно- осных побегов), связанных между собой в определенной порядковой последовательности, начиная с 1-го порядка, и систему корневого на- растания — это целое, образованное из простых корней, связанных между собой в определенной порядковой последовательности. Система ветвления растения — это его осевая система, составлен- ная главной и частными осями или только частными осями. Главная, или общая, ocb растения (axis generalis vegetabilis) состоит из нарастаю- щих в иротивоиоложных наиравлениях двух частей: стеблевой — npocmou или сложной главной восходящей оси (axis generalis ascendens) и корневой- иростой или сложной главной нисходящей оси (axis generalis descendens). Частная ось (axis privatus) — это простая или сложная боковая ось, образующаяся при ветвлении. Оси системы ветвления — это иредветвь (procladus, praeramus) и ветви (ramus,i). Предветвь (стеблевая или корне- вая) — это ось (стеблевая или корневая) нулевого порядка, т.е. ось, из которой образуются первые ветви, как особей семенного (аклонистов), так и вегетативного (клонистов) происхождения. Ветвь (стеблевая или корневая) — частная ось (стеблевая или корневая) первого и последу- ющих порядков, т.е. ось, возникающая при ветвлении. Предветвь быва- ет главной, если она ось нулевого порядка аклониста и добавочной (до- иолнительной), если она ось нулевого порядка клониста. Стеблевая над- земная скелетная или гемискелетная предветвь у древесных растений— ствол или стволик.
Глава 4. Основные способы освоения пространства биоморфами семенных растений 121 Как следует вести отсчет порядков осей ветвления? При рассмот- рении частных форм ветвления (или стеблевого, или корневого, или листового) отсчет порядков ветвей ведется только относительно одно- именных органов (Sachs, 1868; Серебряков, 1952). Образование листьев или корней на стеблях — это также ветвление, но более широкого плана, т.е. полное ветвление ризокормусовой системы растения. Пред- ветвь имеет нулевой порядок в системе ветвления, а ветви (стеблевые, корневые и листовые) — 1, 2, 3 и т.д. порядки. На органах одного мор- фофизиологического типа любого порядка, например, на стеблевых вет- вях 6-го порядка, могут образоваться осевые органы другого типа (кор- ни, листья), которые всегда только 1-го порядка, а в дальнейшем они или не ветвятся, или образуют новые одноименные ветви (соответствен- но корневые и листовые), которые будут уже 2, 3-го и последующих порядков. Чем больше порядок ветви, тем они моложе. Поскольку термины «нарастание» и «ветвление» не однозначны, то следовательно, нельзя однозначно также и подразделять их по многим признакам. Так, у высших растений нарастание осевой системы или ее фрагментов в зависимости от морфофизиологического состояния их верхушек бывает трех основных munoe: моноиодиальное, симиодиальное и дихотомическое. Моноиодиальное нарастание характеризуется увеличе- нием длины отдельных простых осей без отмирания их верхушек. Сим- иодиальное нарастание — увеличение длины групп взаимосвязанных осей происходит при периодической замене на новые их прежних верху- шек, прекративших рост и отмирающих. При дихотомическом нарас- тании увеличение линейных размеров групп взаимосвязанных осей происходит при периодическом расчленении одной жизнедеятельной верхушки на две, подобные исходной. Л.Е. Гатцук (1967) разработала оригинальное подразделение симпо- диального способа нарастания побеговой системы на три типа: акро- симподиальное, мезосимподиальное и базисимподиальное. Заметим, од- нако, что нет акросимподиальности в нарастании побеговой системы розеточных многолетников типа одуванчика, поскольку у таких расте- ний вся надземная удлиненная часть отплодоносивших вегетативно- репродуктивных побегов отмирает полностью до укороченных основа- ний, от которых отрастают новые розеточные пазушные побеги, т.е. проявляется базисимподиальность. В дополнение к известным типам нарастания (моноподиальное, симподиальное, дихотомическое), мы разработали (Нухимовский, 1986) более дробную сериофикацию сиосооов нарастания для скелетных и (или) проскелетных осевых органов (рис. 22-34): лолисимлодиальное (с ежегод- ным многоразовым — четыре и более раз — перевершиниванием), оли- госимиодиальное (с ежегодным двух-трехразовым перевершиниванием), эусимиодиальное (с ежегодным одноразовым перевершиниванием), геми- симиодиальное, или моноиодиально-симиодиальное (с неежегодным пере- вершиниванием). Сходным образом при необходимости можно класси- фицировать и дихотомическое нарастание (иолидихотомическое, олигоди- хотомическое, эудихотомическое, гемидихотомическое). Когда простая ось нарастает более одного года без отмирания (утраты) ее верхушки, — это
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ эумоноподиальное нарастание. В случае, если подобные принципы на- растания наблюдаются в структуре однолетних осевых органов, то мож- но подчеркивать этот факт, выделяя следующие типы эфемерного на- растания: эфемерно-моноподиальное — нарастание одноосного эфемер- ного органа, т.е. в результате жизнедеятельности одной верхушечной меристемы, эфемерно-симподиальное — нарастание двух-многоосного эфемерного органа, каждая ось которого нарастает сначала моноподи- ально, а затем ее верхушки прекращают рост и перерастают новыми осями (в зависимости от кратности перевершиниваний оно бывает соот- ветственно эфемерно-эусимподиальное, эфемерно-олигосимподиальное и эфемерно-полисимподиальное). Кроме того возможно и эфемерно-дихото- мическое нарастание, когда эфемерная осевая система увеличивает свои линейные размеры дихотомически. Нарастание (моноподиальное, симподиальное и дихотомическое) может, на наш взгляд, осуществляться у различных органов одним из двух способов: активно и аккумулятивно. Все органы осевой системы, у которых увеличение длины осуществляется непосредственно за счет жизнедеятельности их верхушечных мерисистем (отдельные стебли, в равной степени и побеги, или корни, группы взаимосвязанных стеблей или корней), характеризуются активным способом нарастания. Все ор- ганы побеговой системы, у которых увеличение длины осуществляется за счет возрастания в их структуре числа резидов (корневища, каудексы и др. виды многолетней побеговой части), обладают аккумулятивным способом нарастания. Резиды составляют скелетный каркас этих органов (Нухимовский, 1968, 1969а, 1971). Резид (от лат. residuum — оста- ток) — стеблевая часть побега, которая стала скелетной (двух, трех, многолетней) после завершения побегом фазы видимого роста и (или) ре- продуктивного развития (рис. 25-32). Подробнее резиды и их место в структуре базовых органов рассмотрены в главе 6. Ежегодное нарастание осевой системы в одних случаях приводит к видимому (основному) ветвлению, а в других — нет. Возникает вопрос, по каким признакам лучше всего подразделять ветвление. По числу порядков осей, образующих систему ветвления, можно различать два типа ветвления: простое и сложное. Простое ветвление характеризуется образованием из одной оси не более одного порядка новых осей ветвления. При сложном ветвлении из одной оси образуется два и более порядков новых осей ветвления. В зависимости от того, из каких частей и в какой последователь- ности образуются ветви, можно, используя в основном уже известные термины и уточняя их значение, различать у семенных растений следу- ющие виды ветвления: аксиллярное (нормальное и аномальное), адвентив- ное (ризогенное, каулогенное и филлогенное) и дихотомическое стеблевое (шире — побеговое) ветвление, а также нормальное, адвентивное (кауло- генное, ризогенное и филлогенное) и дихотомическое корневое ветвление. Под нормальным ветвлением в старых работах, например, Нэгели (Nageli, 1863), Гофмейстер (Hofmeister, 1868) понимали всякое ветвле- ние в акропетальной последовательности. Для семенных растений нор- мой считалось образование ветвей из почек в пазухах листьев. Сакс
123 пава 4. Основные способы освоения пространства биоморфами семенных растений 6 7 10 Рис. 22. Типы нарастания осевых органов. 1 — эфемерно-моноподиальное, 2 — эфемерно-дихотомическое, 3 — эфемерно- эусимподиальное, 4 — эфемерно-олигосимподиальное, 5 — эфемерно-полисим- подиальное, 6 — эумоноподиальное, 7 — гемисимподиальное, или моноподиаль- но-симподиальное, 8 — эусимподиальное, 9 — олигосимподиальное, 10 — поли- симподиальное, 11 — эудихотомическое, 12 — олигодихотомическое и полидихо- томическое, 13 — гемидихотомическое, или моноподиально-дихотомическое, гг- граница годичного побега. 1 2 J f l + ф g I + --~ 1 р 1 < 1 1 I 1 I 1 I ~ ~1г t I I гг 4 ~с ~~ у l г~ рн~ -rL) ( ) '< 5 (g / ( ° ~й q 1 р Ъ ~б Ъ Ч t I
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 23. Э 3. Эумоноподиальное нарастание у травянистых особей Primula macrocalyx Bunge (Алтай, Ивановский хребет, опушка леса). с — семядоли г — гипо — окотиль, гп — главный побег, гк — главный корень, пк- придаточные корни, ак — автономное корневище (см. гл. 6.3), пц — пазушный цветоносный силлептический побег, рл — розеточные листья (1,2, ...) по по яд , р — розеточный вегетативный побег, у — увеличено по сравнению с масштабом, пп — поверхность почвы, одно деление масштаба — 1 см.
125 Глава 4. Основные способы освоения пространства биоморфами семенных растений СО an гк ПП СО— гк Рис. 24. Эусимподиальное нарастание виргинильных травянистых растений Hedysarum austrosibiricum В. Fedtsch. (Алтай, Бащелакский хребет, альпийский луг). ап — аксиллярный безрозеточный весенний побег замещения, со — сухой остаток прошлогоднего побега (может сохраняться несколько лет), к — каудекс; жилкова- ние показано не на всех листьях. Остальные обозначения те же, что и на рис. 23.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ 1 ° М C О С4 Ж О И )~ & 3 <~ Ж О ~4 Q X (б ,'у,' 3 ~ О > х О р,"~ о ~ (.) Ж (.) g„ 65 Q~ Ж ~Й ° 0 ~ Д- й й~ О И 3 й (Т) Щ <~4 й С4 л л л Я ю х ~ ~~ .а ~~ц Х О й Х V ~. о ~' oval й ;Т' 61 v О О ~ ж~ д О Яе~й О и~И о~~ ОО О v 3 о мч Я ~) О д; Q.~; йХй v > v о О ~;" ФЭ "Х~ О ДО~ ( а 1 о ~'~ й о ~ у~ л О< Д~ ~[ ~л~ я О ~~ ~ О, . О й ~ ~О О. ц ~: О О й ф о й Х ~ +ж 2 ":ðé Ю аvЯ 2з: <3~ О О й vXO а х ~ ~~~ Й 2 :т' Й О. О Ь л LQ О ~~ О. ~ u Qo< ~р О ~ О й Х й а5 А Й о~Е :[. 'Й 63 о Ой' О О О 1 1 ~1 О Й О
Глава 4. Основные способы освоения пространства биоморфами семенных растений 127 Рис. 26. Эусимподиальное нарастание древесного растения Elaeagnus angustifolia L. (Крым). вп — вегетативный безрозеточный весенний аксиллярный побег, цп — вегетатив- но-репродуктивный (цветоносный) безрозеточный весенний аксиллярный побег, пл — плоды, п — почка, кп — короткий побег, превращающийся в колючку, со- сухой остаток верхней части побега зимой, мн — миниморезид, мк — максимо- резид, сл — срединный лист, оч — отмершая часть прошлогоднего побега.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 27. Эусимподиальное нарастание особей травянистого многолетнего обоеполого поликарпика Scrophularia incisa Weinm. (Алтай, Катунский хребет, 1800 м над ур. моря). гм — графические модели растений разных возрастных состояний, су — семядольный узел (примерное положение). Остальные обозначения те же, что и на рис. 23, 24.
пава 4. Основные способы освоения пространства биоморфами семенных растений 129 Рис. 28. Гемисимподиальное нарастание у стержнекорневого травянистого многолетника Егупя1ит р1апит L. (Крым). ос — основание полурозеточного вегетативно-репродуктивного плейофазного побега (верхние части удалены), ол — основание розеточных листьев (верхняя часть удалена), к — каудекс, мн — миниморезид; пунктирной линией показано примерное положение семядольного узла. Остальные обозначения те же, что и на рис. 23,27.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 29. Гемисимподиальное нарастание виргинильных травянистых растений Cimicifuga foetida L. (Алтай, низкогорье). сл — срединные листья, со — сухой остаток от листа, ак — автономное корневище, рб — рубец от отмершей надземной части побега, мн — плейомерный мини- морезид, пк — придаточные корни, пп — поверхность почвы. Одно деление масштаба — 1 см.
пава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 30. Переход от эумоноподиального нарастания (растения слева) к гемисимподиальному нарастанию (растение справа) у Ranunculus cassubicus L. (Алтай, смешанный лес, низкогорье). в — всход, образовавшийся после подземного прорастания, гп — главный побег, вц — верхушечный удлиненный цветоносный безрозеточный годичный побег, ас — аксиллярный силлептический предрозеточный вегетативный побег, ак — ав- тономное корневище, пк — придаточные корни. Одно деление масштаба 1 см.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ гп Рис. 31. Гемисимподиальное нарастание древесных растений I — Crataegus sanguinea Pall. (условия затенения) и II — Juglans гера L. (Крым). гг — граница моноподиальных приростов, зп — зона перевершинивания, пл- верхушечные плоды, оч — отмершие части прошлогоднего побега, вп — верхушечная почка в конце вегетационного сезона, пв — почка вегетативная, пр — почка репродук- тивная с зачатками тычиночных цветков, сп — статурный (ростовой) безрозеточный побег, уз — зона укороченных междоузлий, рс — репродуктивно-статурный закрытый безрозеточный верхушечный побег, тф — трифазный вегетативно-репродуктивный побег (о типах побегов см. гл. 5.1.), тр — мономерный тотарезид, мр — мономерный магнорезид. Остальные обозначения те же, что и на рис. 23. (Sachs, 1868) доказал, что нормы есть только постоянные и своеобразные отношения для определенных групп растений. Иными словами, способ ветвления, нормальный, т.е. наиболее характерный, например, у мхов или папоротников, уже не будет таковым у семенных растений. У рас- тений последнего типа доминирует аксиллярное стеблевое (шире — по- беговое) ветвление, т.е. ветви закладываются в пазухах листьев. Если они в процессе роста в длину не меняют своего первоначального поло- жения, то такое аксиллярное ветвление мы называем нормальным, а если происходит осевой (конкаулесценция) или эпифильный (рекаулес- ценция) сдвиг ветви вне пазухи листа (Velenovsky, 1907; Troll, 1937; Се- ребряков, 1952 и др.), то тогда можно говорить об аномальном аксил- лярном ветвлении. Нормальные корневые ветви образуются в акропетальной последо- вательности из молодых, активно нарастающих частей главного корня
Глава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 32. Гемисимподиальное нарастание древесных растений 1 — Malus domestica Borkh. и II — Acer insigne Bunge (Крым). рп — розеточный вегетативно-репродуктивный закрытый годичный побег, пз — силлеп- тический вегетативный побег замещения, ро — репродуктивные органы, кв — короткий вегетативный годичный побег, рв — розеточный вегетативный годичный побег, пф- плейофазный побег, кт — короткий мономерный тотарезид, дм — димерный магно- резид. Остальные обозначения те же, что и на рис. 31. и корневых ветвей. Адвентивное (придаточное) образование корней и почек было открыто в начале прошлого века (Dupetit-Thouars, 1809; De Candolle, 1827, цит. по Velenovsky, 1907). Сакс (Sachs, 1868) считал, что все неакропетальные (анакропетальные) боковые образования могут быть названы придаточными, если ни не имеют правильного располо- жения и развиваются на старых частях, состоящих из уже дифференци- рованных тканей. Так, корни, образующиеся акропетально на корневи- ще, например, у многих злаков, он относил к адвентивным, а все другие корни, которые развивались между акропетальными, рассматривал как придаточные. Термин «адвентивный» в отношении побегов и корней, пожалуй, лучше всего понимать широко. В первую очередь адвентивны- ми целесообразно считать, как это обычно делается, те побеги (уже- стебли) или корни, которые развиваются на органах другого морфофи- зиологического типа, например, любые корни, развивающиеся на побе- гах и листьях, или побеги на корнях и листьях, исключая случаи рекау- лесценции. Во-вторых (Esau, 1965; Михайловская, 1968 и др.), — побе- ги, образующиеся анакропетально (не из пазушных почек) на других побегах, и корни, возникающие беспорядочно (анакропетально) на ста- рых утолщенных частях главного корня и корневых ветвей. Дихотомическое ветвление происходит вследствие дихотомическо- го нарастания осей, когда их отдельные верхушечные меристемы разде- ляются на две новые. Представляет интерес подразделять ветвление по особенностям рас- положения производных ветвей на их исходных осях. Еще Феофраст
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 33. Эфемерно-полисимподиальное нарастание 1 — Euphorbia humifusa Schlecht., П — Asperugo procumbens L. (Крым). пл — верхушечный репродуктивный элемент (цветок, плод); жилкование и опуше- ние показано не на всех листьях. Одно деление масштаба — 1 см. (1951, с. 14) отмечал, что у одних растений «ветви расположены как попало», у других — «одна напротив другой с обеих сторон сука», а у третьих — «через равные промежутки и в равном числе... по три ветви в узле». Выше уже отмечалось, как много было сделано К.Линнеем (Linnaeus, 1751-1767, Линней, 1989) по унификации терминологии в этом плане. Т.И. Серебрякова (1969, 1971, с. 224), анализируя особенно- сти ветвления злаков, пришла к заключению, что бывает «два основных способа распределения боковых ветвей по материнскому побегу у зла- ков: рассеянное ветвление (в области удлиненных междоузлий) и кон- центрированное ветвление, или кущение (в области укороченных меж- доузлий)». У семенных растений по особенностям расположения производных стеблевых ветвей на их родительской (исходной) оси мы различаем следующие виды аксиллярного нормального ветвления: очередное (Tilia cor- data Mill.), суиротивное (Acer negundo L.) и мутовочное (Nerium oleander L.). Когда на исходной оси образуются аксиллярные и адвентивные стеблевые ветви, то можно говорить о смешанном стеблевом ветвлении, а в том случае, когда на исходной оси образуются нормальные и адвент- ивные корневые ветви, то о смешанном корневом ветвлении. В зави- симости от того, как ветви располагаются друг относительно друга — сбли-
Глава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 34. Эфемерно-полисимподиальное нарастание у Tribulus terrestris L. (республика Тыва, окрестности пос. Чадан). Обозначения те же, что и на рис. 33.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ женно или на заметном отдалении, можно, используя терминологию, разработанную Т.И. Серебряковой (1969, 1971) в отношении злаков, и дополняя ее, различать следующие формы ветвления семенных растений: рассеянное, концентрированное и комбинированное (Нухимовский, 1974б) (рис. 35). При рассеянном ветвлении образуются ветви, основания ко- торых существенно отстоят друг от друга; в частности, когда ветви об- разуются в нескольких узлах, то в зоне их стеблевого контакта междоуз- лия удлиненные, или же междоузлия укороченные, но тогда их много (Coronilla scorpioides (L.) Koch, Crassula lycopodioides Lam.). Концентри- рованное ветвление характеризуется образованием на родительской оси ветвей, основания которых сильно сближены относительно друг друга, в частности, когда ветви образуются в нескольких узлах, то в зоне их стеблевого контакта немногочисленные междоузлия укороченные (Pe- ganum harmala L., Nardus stricta L.). Здесь следует оговорить, что мы не отождествляем понятия «концентрированное ветвление» и «кущение», как предлагает Т.И. Серебрякова для злаков. Вопросы кущения рассмат- риваются ниже. При комбинированном ветвлении на родительской оси образуются ветви одного порядка как рассеянно, так и концентрирован- но, например, у Picea abies (L.) Karst. Ветвление, в зависимости от того, в какой среде окружающего жиз- ненного пространства оно происходит, бывает надземное, ириземное (раз- новидность надземного ветвления, которое осуществляется в непосред- ственной близости к поверхности почвы), иодземное и иодводное. В соответствии с тем, по какому типу образуются ветви на отдель- ных осях системы ветвления, можно различать определенные зоны вет- вления этих осеи, например, зону мутовочного рассеянного ветвления (зону рассеянного контакта муговочных ветвей), зону очередного концен- трированного ветвления, зону адвентивного концентрированного корне- вого ветвления, зону смешанного (очередно-адвентивного) рассеянного ветвления и т.д. (рис. 35). Самые маленькие зоны контакта осей системы ветвления — это, во-первых, узел (nodus) побега, из зоны которого образуется одна или несколько стеблевых (шире — побеговых) ветвей, иными словами— узел контакта стеблевых осей системы ветвления, или просто узел вет- вления', во-вторых, часть стебля или корня, из которой образуется одна адвентивная ветвь, т.е. зона контакта одной адвентивной ветви, или зона одиночного адвентивного ветвления. Узел ветвления и зона одиночного адвентивного ветвления — это элементарные зоны ветвления у семенных растений (рис. 35). Субординатное классифицирование осей можно проводить с раз- ных высот абстракции, т.е. одновременно рассматривать их в суперорди- натной классификации. Верхнюю суперординату (рис. 36) занимает су- бординатная классификация из 2-х субординат: а) иростые оси, или моно- ') Ветвление побеговой системы — частный случай побегообразования. Узел (nodus), из зоны которого образуется аксиллярный побег — узел побегообразования. Сово- купность узлов побегообразования одной простой оси — зона пазушного побегооб- разования этой оси. Участок стебля или корня, из которого образуется адвентивный побег — зона одиночного адвентивного побегообразования.
пава 4. Основные способы освоения пространства биоморфами семенных растений 137 О~а й Ж Q Х 3 Ж Ж й В й й Ж О О В,' О Ж Q О о '~~ Й В,' О Ц Ю В О Ж Ж Ю О О О ф) 3 Q О О М Ж ~Г) ~Г~ (.) й С4 ии~22~ ~ ~ й �Ж�ОО~счю~ ~> иечеч а. ~ Й'. о ~ ZО ~ ~~ ~ М o+ ~ X Q « ~ ~O Х mþ [ ~ ~ X X Ж д ~ ~ц О ~ Ж ~ ~- „~ сф LQ ~ й ~~ Ж & 6Э О. 6Э + Д 6Э [. о~оо~~а а о р о ~ ~~ ц ~ ~� ~�~ON�î q) ~" ~ ~ ~~ О ~ ~~ < ec о, а~~~ m X x~ Q X Q 4. Х 8 р,„„О ~. Q ф � vOÖO ~ Ж~~Ц " о. [е ) „-М~'" ) щ д И р, й Ф 63 О д ~> ~ д i y ~ ~ Q ~ � ~ о ~~ ~ ~ ~ о ~ �ЮQO ~ йео а ю~ v о ~ ~ c~ n. ~ О O~М Я Й ~й ~~[~ ~Я ~ р ! ~ n v eC X QO ~Яg й М 0 О (ф), ф ~~ ~, y < о ~~,~ ~ д р.~ к, еч Ф О XXXQvQv~ ~ � �~ ~~Ял~ о ~е Я а~ й ~„~ О. ц ~~ р; ~~" о д ц~ ~) ~ ~ ~ О ~4 v Z v e eC Q ~n. O о ~:~ ~-~; у,.,Оц> о ~ ~ВО~,о ~:1щ — ~ ~~ еЧ О~~ М ае цр " u Q & e e" ~ао О ~й o o > ком р О р,са>,~ Ю щ щ д щ ~ й ~Ch и ~ д ~ > ~ О ,," Ф ~ ~~ О „р: ~Я ° ° й~ ~р рП Q,~ ~-~ ~) Р~ о д о 2 о оi о Х М nf u
° ° rll а' Ф ° ° ° II ° ° в ° в ° ° в в ° ° ° ! в ° ° в ° ° ' ° ° ° а ° ° в в ° ° ' ° ' ° ' ° Э ° ° ') в ° ° ° ° ° . ° ° ° ') ° ° ° а ) ° eâ ° ° а II ° а Э ° . - ° ° ° / I I ю ° е Ф ° ° Э ° ° ° ) ° ° ° ° ° ° ° ° ° ° ) в Э ° Э ° ' Э ° ° е ° ) ° ° ° Э ° ° ° ) ° ° ° ° ° ) ° rå r ° ° е ° ° ) ° ° ° ° е ° - е ° ° ° ) ° ° ° Э ) Э ° ° ) ° ° ) е 'i:I / Э ° ° ) ° ° ° ' ) ° е ° ° ° °:: )г ' () ° r e ° ° Э ° ° / е ) ° м е I ) и ° е ) l ю I e ° ° ° ° а ° в ° ° ° J ° r' ° ! ° ° ее ° ° ° ) ° ° а I ю I ° ']'i ' Il I I ~ I ю ° ) е е ° ) ° ° Э') ° е ° I ф Э °
пава 4. Основные способы освоения пространства биоморфами семенных растений 139 системы из простых или (и) сложных осей, составляющие три порядка осей ветвления; д) четвертые соосия (тетрасинаксоны) — системы из простых или (и) сложных осей, составляющие 4 и более порядков осей ветвления. Субординия стеблей во многом будет приемлема и для ана- логичной классификации побегов, на что мы еще будем обращать вни- мание при анализе организации побегов (см. гл. 5.1.). Важное место в жизни многих ветвистых растений принадлежит кущению. Что такое кущение и куст? В каком соотношении находятся понятия ветвление и кущение? По каким важным экзоморфологичес- ким признакам отличаются кусты между собой? Какое место в жизнен- ном цикле растений занимает ветвление и кущение? Кущение растений привлекало внимание многих ботаников, в осо- бенности исследователей злаков (Schindler, 1896; Вильямс, 1922; Arber, 1934; Смелов, 1947, 1966; Соловьева, 1958; Серебрякова, 1959, 1961, 1962, 1969, 1971 и многие друтие). И.Г.Серебряков (1962, с. 280) охарактеризовал процесс кущения у кустарников как «раскрытие спящих почек в основании надземных стеблей и образование из них новых осей возобновления». Подобно большинству исследователей, Т.И. Серебрякова (1971, с. 223) рассмат- ривала кущение как форму ветвления, «при которой боковые побеги формируются только в нижней (приземной или подземной) укоро- ченной части материнского побега и быстро укореняются. В результате кущения образуется «куст» — система побегов, где ветвление сосре- доточено внизу. Кущение свойственно не только злакам, но и пред- ставителям других семейств, как однодольных, так и двудольных, как травянистых, так и деревянистых (кустарники и кустарнички)». Не вызывает возражения утверждение, что кущение — это форма ветвления, что оно характеризуется отрастанием побегов от нижней приземной или подземной части растения и что способностью кустить- ся обладает большинство семенных растений. Другие положения о ку- щении, на наш взгляд, спорны. Так, не обязательным признаком куще- ния следует считать образование стеблевых ветвей только в «укоро- ченной части материнского побега». Предлагаем понимать кущение широко, не ограничивая значение этого термина по такому признаку, как длина междоузлий подземных или приземных оснований ветвящих- ся исходных побегов. В систему кущения у растений, непосредственно связанных корнями с эдафической средой, должны входить не только зоны концентрированного, но и иного — рассеянного, одиночного (подземного и приземного, как аксиллярного, так и адвентивного стеб- левого, шире — побегового) ветвления. Концентрированно кустятся по- беги Nardus stricta L., Peganum harmala L. рассеянное кущение наблюда- ется у многих побегов — Hypericum calycinum L., комбинированное ку- щение встречается у побегов Elytrigia repens (L.) Nevski, Achillea mille- folium L. и др. (рис. 37). Не обязательным свойством побегов, образующихся при приземном или подземном ветвлении (кущении) следует признать и их способность к укоренению. Так, например, у Cistus tauricus С. Presl стволики начи- нают формироваться в приземной части медиального (главного) побега
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 37. Модели кустов, отражающие степень отдаления элементарных зон кущения. А — компактный куст, Б — рыхлый куст, В — рыхлый куст, образующийся при засыпании субстратом, Г — полуразвернутый куст, Д — развернутый куст. и, хотя их основания могут впоследствии погружаться в почву, кауло- генные корневые ветви обычно не образуются; растения такого типа называют аэроксильными кустарниками (Серебряков, 1962). Итак, кущение — это форма иобегового (стеблевого) ветвления, в результате которого образуется куст. Кустом у семенных растений на- зываем особь или ее часть, основание (нижняя часть иобеговой системы и связанная с ней корневая система) которых включает не менее чем две стеблевые оси ветвления. Стеблевая часть оснований кустов бывает при- земной, т.е. находится непосредственно у поверхности почвы или на ней, подземной (большинство растений), более или менее погруженной в воду (у плавающих растений) или надземной (у эпифитов). В основа-
пава 4. Основные способы освоения пространства биоморфами семенных растений 141 ния кустов могут входить различные структурные образования, например, корневища, каудексы, конодии, самые нижние части стволов и т.п. Особое место в жизни кустистых (кустовых) растений принадлежит их зонам кущения. Термин «узел кущения» (Вильямс, 1922; Дмит- риев, 1948 и др.), нередко употреблявшийся в отечественной литературе для обозначения у злаков и осок ветвящегося участка побега, составлен- ного несколькими укороченными междоузлиями, впоследствии неодно- кратно критиковали (Серебрякова, 1962, 1969, 1971 и др.). В настоящее время большинство ботаников употребляют для этой цели более удач- ный термин — «зона кущения». Об «узле кущения» или «узле ветвле- ния» можно говорить только в отношении одного узла (nodus), из ко- торого (или из зоны которого) отходят соответственно побеги кущения или иные стеблевые ветви. Поскольку нами предлагается более широко трактовать термин «кущение», то шире понимается и термин «зона кущения». Зона иодземного или ириземного стеблевого ветвления одной оси — зона кущения этой оси. Исключение составляют, пожалуй, лишь кустистые плавающие растения и эпифиты, у которых зоны кущения хотя и располагаются в их нижней части, но в воде или надземно. Большинство кустов в естественных условиях произрастания возни- кает путем кущения, но иногда «кусты»образуются и без настоящего кущения. Это вынужденные (по аналогии с термином «вынужденные корневища», предложенным Е.М.Лавренко, 1935) кусты; они образуют- ся, например, при погребении некустистых растений (дуб) песком до кроны (Warming и. Graebner, 1918). В случае погребения субстратом кустистых растений механизм их дальнейшего ~сущения усложняется (Нухимовский, 1969 а, б). В литературе известно немало попыток различать кусты между собой. Рассмотрим важнейшие из них. С.П.Смелов (1966, с. 25) пришел к выводу, что у злаков «осевой (главный — Е.Н.) побег вместе с боковы- ми побегами второго порядка образуют куст» и предложил называть его «простым кустом в отличие от последующей фазы развития — сложно- го куста», характеризующейся отрастанием побегов третьего и последу- ющих порядков. В качестве структурной единицы более высокого ранга, чем побег, И.Г.Серебряков (1953, 1959) предложил считать «парциаль- ный куст», определив его как группу, пучок, или семью вегетативных и генеративных побегов, возникающих в результате ветвления материн- ского монокарпического побега. Нам представляется рациональной терминология кустов («простые» и «сложные»), предложенная С.П.Смеловым (1966). Эти термины с определенным уточнением их значения предлагаем применять не толь- ко в отношении злаков, но и всех кустистых растений. Они находятся в соответствии с нашим общим подразделением ветвления (простое и сложное). Простой куст имеет в основании стеблевые (иобеговые) оси двух иорядков ветвления, из которых одна родительская (исходная), а остальные — ироизводные, либо же одного иорядка ветвления, если они образуются на одной оси иного морфофизиологического muna (корне). Слож- ный куст — совокуиность иростых кустов, а иногда также и одиночных адвентивных стеблей (иобегов), связанных в единую морфофизиологическую
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ систему через разные корневые оси (рис. 35). Каждый простой куст имеет свой порядок в структуре сложного куста, определяющийся порядком младшей стеблевой оси системы кущения, т.е. порядком младшей оси кущения. Более молодая стеблевая ось, входящая в основание одного простого куста, как правило, служит основанием для формирования нового простого куста, входит и в его состав, а порядок этого куста определяют уже новые оси кущения. Простой куст, в структуру кото- рого входит главная стеблевая предветвь, — главный простой куст. Иног- да бывает целесообразно говорить и о иарциальных кустах, которые можно определить как любой морфофизиологически целостный простокустистый или сложнокустистый фрагмент индивидуума или кондивидуума, кроме главного простого куста. В скобках удобно отмечать порядок парци- альных кустов, указывая порядок составляющих их простых кустов, например, у простых парциальных кустов или (5), или (6) и т.п., а у сложных парциальных кустов или (7-12), или (8-15) и т.п. Простой куст — структурная единица сложного куста. При изучении кустистых растений может возникнуть и такой воп- рос, будет ли, например, многолетнее безрозеточное травянистое расте- ние кустом, когда к зиме все надземные части его побегов погибнут, а в почве останется лишь разветвленный базовый орган и корневая систе- ма. По нашему мнению, если функционирует разветвленное стеблевое основание, несущее на себе надземные побеги или только одни почки, значит особь имеет кустистое строение. По направлению роста побегов кущения и длине их горизонтальных участков В.Р.Вильямс (1922, с. 120) подразделил луговые злаки на плот- нокустовые, рыхлокустовые и корневищные (здесь не выдержан единый принцип). Растения по размерам корневищ бывают компактно- (укоро- ченно-), коротко- и длиннокорневищные. У первых корневища очень малой длины вследствие сильной сближенности узлов. У вторых они относительно небольших размеров и составлены в основном из сбли- женноузловых частей при некотором участии раздвинутоузловых час- тей, а у третьих корневища достигают сравнительно большой длины, поскольку решающую структурную роль играют раздвинутоузловые части. По иному признаку — no степени отдаления элементарных зон кущения (узлов кущения, в узком смысле этого слова, и зон одиночного адвентивного кущения), эти же и многие другие растения можно под- разделить на комиактно-, рыхло-, полуразвернуто- и развернутокустис- тые (рис. 37-44). У компактных кустов основания составлены укорочен- ными ветвями или частями ветвей, а элементарные зоны кущения силь- но сближены (Capparis herbacea Willd.). Компактные кусты бывают плот- ные, например, Nardus stricta L. (у них очень плотно располагаются ортотропные побеги кущения) и неплотные (рис. 38), когда побеги ку- щения располагаются иначе, чем у плотных кустов. У рыхлых кустов основания составлены укороченными, короткими и иногда также вертикальными длинными ветвями или частями ветвей, а элементарные зоны кущения в горизонтальном направлении сравни- тельно близко отстоят друг от друга (Dactylis glomerata L.). У развернутых кустов основания составлены ветвями разнообразных размеров, среди
Глава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 38. Формирование компактного неплотного куста у Реда- num harmala L. в условиях относительно постоянного уровня поверхности почвы (республика Калмыкия, Черные земли, окрестности пос. Нарын-Худук). с — семядоли, r — гипокотиль, гп — главный побег, гк — главный корень, су- семядольный узел (у взрослых особей его примерное положение отмечается пунк- тирной линией), к — каудекс, on — основания побегов текущего года (верхние части удалены), вп — вегетативный годичный побег, вр — вегетативно-репродук- тивный закрытый годичный побег, горизонтальная линия — поверхность почвы.
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИИ пн пп Рис. 39. Рыхлый куст Peganum harmala Е., образовавшийся в условиях ежегодного засыпания песком (там же, что и на рис. 38). пп — поверхность почвы, пн — надземные побеги (верхние части удалены), пм- вынужденно удлиненные подземные многолетние побеги, дк — рыхловетвистый вынужденно удлиненный каудекс.
Глава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 40. Формирование полуразвернутого полукустарничка у Sedum ewersii Ledeb. (Алтай, Семинский хребет, высокогорье, южные склоны). Одно деление масштаба — 1 см.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 41. Формирование полуразвернутого многолетнего травянистого куста у виргинильных особей Urtica dioica L. (Калужская область). Одно деление масштаба — 1 см.
пава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 42. Формирование развернутого многолетнего травянистого куста у Stellaria bungeana Fenzl (Алтай, Ивановский хребет, лесной пояс). Одно деление масштаба — 1 см.
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИИ Рис. 43. Формирование развернутого многолетнего травянистого куста у Sambucus ebulus L. (Крым, окрестности Никитского ботанического сада).
Inaea 4. Основные способы освоения пространства биоморфами семенных растений 149 Рис. 44. Формирование развернутого кустарничка у Hypericum calycinum L. (Крым, Никитский ботанический сад).
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РЛ СТЕНИЙ которых преобладают длинные ветви, а элементарные зоны кущения или зоны концентрированного кущения в горизонтальном направлении сравнительно далеко отстоят друг от друга (рис. 42-44). Полуразвернутые кусты характеризуются тем, что в их структуре наряду с участками ком- пактного и рыхлого ~сущения есть немногочисленные участки разверну- того кущения (рис. 40, 41). Многие кустистые растения хорошо переносят лабильность поверх- ности почвы. Особи одного вида, для которых наиболее характерен компактный куст, при засыпании субстратом могут формировать рыхлые кусты (рис. 38, 39). Надземные побеги кустистых растений по расположению в прос- транстве бывают прямостоячие (Artemisia vulgaris L.) или раскидистые- полегающие, стелющиеся (Capparis herbacea Willd.). Среди взрослых особей по отношению к кущению мы предлагаем различать некустистые, нериодически-кустистые, ностоянно-кустистые и разнообразно-кустистые (рис. 45-47). Периодически-кустистые растения Рис. 45. Отражение продолжительности жизни побегов кущения в структуре кустов. Модели кустов: А — периодически-кустистое растение, Б — постоянно-кустистое растение, В — разнообразно-кустистое растение, 1 — однопорядковый базовый орган, 2 — многопорядковый базовый орган, 3 — эфемерные побеги кущения, 4— иные побеги кущения (преперсистентные, семиперсистентные, персистентные), пп — поверхность почвы.
пава 4. Основные способы освоения пространства биоморфами семенных растений 151 Рис. 46. Формирование периодически кустистой структуры у Plantago maxima Juss. ex Jacq. (Восточный Казахстан, р. Иртыш, луг на острове). Одно деление масштаба — 1см; у — увеличено по сравнению с масштабом. Осталь- ные обозначения те же, что и на рис. 45.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 47. Формирование разнообразно-кустистых травянистых растений у Rhodiola linearifolia Boriss. (Москва, ВИЛАР, интродукционный питомник). 5 — ростовый побег (сближенные междоузлия на схеме раздвинуты). Осталь- ные обозначения те же, что и на рис.45. Одно деление масштаба — 1 см. формируют куст и сохраняют кустистое строение, как правило, в тече- ние одного вегетационного сезона, после завершения которого стано- вятся некустистыми до новой фазы кущения; к ним относятся, например, Plantago major L., Succisa pratensis Moench. Постоянно-кус- тистые растения, став однажды кустами, сохраняют кустистое строение на протяжении всей жизни, например, Anisantha tectorum (L.) Nevski, Hypericum maculatum Crantz. Разнообразно-кустистые растения, сфор- мировав постоянный куст, могут на отдельных его ветвях образовывать
153 Глава 4. Основные способы освоения пространства биоморфами семенных растений также и периодические кусты, такие особи образуются у Rhodiola algida (Ledeb.) Fisch. et С.А. Меу., Rhodiola rosea L., Rhodiola linearifolia Boriss.. Начало ветвления побеговой системы, причем нередко в форме кущения, — важный диагностический признак, свидетельствующий о переходе особей из одного возрастного состояния в другое, что будет рассмотрено подробнее ниже (гл. 7). 4.5. Партикуляция и размножение Вопросам размножения семенных растений уделено большое вни- мание в литературе (Darwin, 1859; Gustafsson, 1948; Левин, 1959; Ша- лыт, 1960; Работнов, 1960, 1969; Хохлов, 1965; Левина, 1960, 1981; Смит, 1981 и др.). Повышенный интерес к проблеме размножения понятен: тактика размножения — это главное звено в реализации стратегии Жизни. За счет варьирования этой тактики у биоморф различных таксонов био- строба Земли преодолела жестокий барьер дискретного образа суще- ствования всех своих частных представителей и надежно защитила себя от гибели, совершенствуя в процессе эволюции способность размно- жаться. Но что такое размножение? Оказалось, что вопрос этот не столь уж прост, как кажется на первый взгляд, и по сей день не имеет однозначного решения. Он увязан с рядом других вопросов: когда, каким способом и посредством чего происходит размножение у семенных растений. Попытаемся ответить на эти вопросы, не углубляясь в детали. Поиск ответа вывел нас на понятие партикуляция и потребовал рассмотреть его поначалу в соотно- шении с понятием вегетативное размножение, а затем и в отношении с понятием размножение вообще. Анализ истории термина «парти- куляция» весьма поучителен для биологии. Нередко биологи, работая над какой-либо узкой проблемой, не замечают, что наблюдаемые явле- ния имеют общебиологическое, иногда и общенаучное содержание. Вопросы партикуляции и вегетативного размножения высших рас- тений, пожалуй, одни из наиболее популярных в эколого-морфологи- ческой ботанической литературе. Термин «партикуляция», как извест- но, был впервые употреблен Г.Н.Высоцким (1915) и получил впослед- ствии признание в ботанике. Однако в объяснении явления партикуля- ции пока нет единого мнения. Как отметил М.С.Шалыт (1965, с. 117), «под партикуляцией обычно понимается разделение растения на от- дельные части — партикулы, способные продолжать самостоятельное существование». Это явление отмечалось у разнообразных жизненных форм: кустарников, полукустарников, полукустарничков, травянистых многолетников (Jost, 1890; Высоцкий, 1915; Станюкович, 1949; Нечаева, 1949; Козлова, 1953; Стешенко, 1956; Гордеева, 1957; Рачковская, 1957; Игнатьева, 1958,1964, Зозулин, 1959; Казарян, Балагезян, 1960; Труле- вич, 1960; Голубев, 1962; Рахманина, 1962; Горшкова, 1966; и др.), причем оно наиболее изучено у стержнекорневых травянистых растений.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ В трактовке одних исследователей партикуляция получила обосно- вание как «один из видов», «особая форма» вегетативного размножения (Казакевич, 1922; Коровин, 1934; Радкевич и Шубина, 1935; Закржевс- кий и Коровин, 1935; Новиков, 1943; Жуковский, 1949; Кузнецов, 1951; Кудряшова, 1958 и др.). Другие взгляды на партикуляцию сводятся к тому, что это явление объясняется как «признак старости», приближающий особь семенного происхождения к гибели (снижение семенной производительности, уменьшение прироста вегетативной массы, ослабление темпов роста и т.д.), а поскольку после полной партикуляции не происходит образова- ние молодых обновленных организмов и не расширяется площадь, за- нимаемая клоном, то ошибочно считать этот процесс способом вегетат- ивного размножения (Синьковский, 1950; Нечаева, Приходько, 1956; Рустамов, 1965; Игнатьева, 1965; Москалева, 1968; Казарян, 1969 и др.). М.С.Шалыт, обобщая результаты своих многолетних наблюдений (1950, 1952) и литературные данные, пришел к выводу (1965), что партикуляция проявляется у некоторых стержнекорневых, кистекорне- вых, короткокорневищных и дерновинных растений как своеобразная форма медленного отмирания особи и что относить ее «к видам веге- тативного размножения не следует». В большинстве из известных нам взглядов на партикуляцию нет указаний на очень широкое распространение этого явления, но есть указания на ограниченность круга партикулирующих растений. Исклю- чение составляет лишь точка зрения Т.А.Работнова (1969), который предлагает рассматривать это явление в более широком смысле, чем обычно принято в литературе, отождествляя его с вегетативным размно- жением («образование новых особей в результате отчленения партикул следует называть партикуляцией»). Подразделение партикуляции на три типа, разработанное Т.А.Работновым, можно по сути дела рассматривать и как своеобразное подразделение вегетативного размножения. В обзоре Р.Е.Левиной (1981) явлению партикуляции так же отво- дится весьма скромная роль как одного из способов вегетативного раз- множения. С этим положением мы не можем согласиться, поскольку вскрываем сущность механизма размножения, а не ограничиваемся только констатацией фактов его проявления. Вероятно, большинство исследователей согласятся, что суть парти- куляции в разделении растения на живые части (партикулы). Но какое разделение растения на части оценивать как партикуляцию, каковы причины партикуляции, что такое партикула и является ли партикула особью или нет, каков круг растений, способных партикулировать, рав- но- или разнозначные ли понятия партикуляция и вегетативное раз- множение? Попытаемся последовательно ответить на эти вопросы. Партикуляция — это процесс (действие), происходящий по самым различным причинам, результатом которого является полное или непол- ное обособление партикул (Нухимовский, 1973б). Иными словами, иар- тикуляцию мы нанимаем в общем биологическом смысле — это всякое (физиологическое или механическое) разрушение связи между отдельными живыми частями растения или животного. Партикула — это иолностью
Inaea 4. Основные способы освоения пространства биоморфами семенных растений или не иолностью обособленная живая часть разделяющегося растения, т.е. такая часть, в которой еще ироисходят некоторые жизненно важные ироцессы, хотя бы замедленные, и которые были характерны для нее в структуре целого родительского растения (наиример, дыхание или иные жиз- недеятельные формы обмена веществами с окружающей внешней средой). Образование разнообразных партикул за счет физиологических про- цессов, происходящих в родительском организме, — физиологическая партикуляция; она осуществляется или через некроз определенных тка- ней, или иначе, например, через образование поперечной раздели- тельной перегородки у простейших организмов (Escherichia coli, Proto- coccus) и т.п. Разновидность физиологической партикуляции у высших растений — некротическая партикуляция — заключается в полном или неполном разделении высшего растения на партикулы вследствие отми- рания отдельных частей или целых органов при нарушении жизнедея- тельности или в связи с завершением их циклов развития. Некротическая партикуляция может происходить как у стареющих репродуктивных особей, например, Centaurea scabiosa L., Rumex confer- tus Willd. (Былова, 1968, 1987), так и у виргинильных особей, например, Viola mirabilis L. (Смирнова, 1968). В результате полной общей некро- тической партикуляции довольно часто «связи между органами в пре- делах одного целого организма, связи физиологические, сменяются от- ношениями между организмами — связями фитоценотическими» (Сме- лов, 1947, с. 88). Некротическая партикуляция у многих растений начинается час- тичным или полным некрозом отплодоносивших побегов (Мокее- ва, 1945; Игнатьева, 1958, 1965 и др.). Отгнивание отдельных частей сто- лонов (Ajuga reptans L.), корневищ (Phragmites australis (Cav.), Trin. ex Steud.), геоподиев (Corylus avellana L.), каудексов (Phlomis pungens Willd.), корней (1пи1а britannica L.), а также соединительных тканей между стеб- лями или листьями и выводковыми почками (Dentaria bulbifera L., Polygonum viviparum L.) — ряд примеров некротической партикуляции. Разделение тела у высших растений на партикулы может происхо- дить не только некротически (шире — физиологически), но и иначе. Полное или неполное разделение высшего растения на партикулы при решающем участии механических процессов (разламывание, обрубание, пережигание и т.п. отдельных органов или их частей) — один из видов партикуляции — механическая иартикуляция. Эффект механической па- ртикуляции достигается с помощью внешних агентов (ветра, воды, лопаты, топора, огня, зубов животного и т.п.), непосредственно воздей- ствующих на растение, и не зависит от возраста всего растения или отдельных его частей. Например, при грозе не полностью расщепился ствол дерева — произошла механическая неполная частная партикуля- ция, в результате которой образовалось две необособленные партикулы; эти партикулы или реализуют свою потенциальную возможность и оста- нутся живыми частями дерева или вскоре погибнут. Кроме отмеченных двух видов, встречается нередко также смешан- ная партикуляция, когда разделение высшего растения на партикулы происходит при относительно равном участии как физиологических,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ так и механических процессов. Так, например, на пастбищах, сильно нарушенных выпасом скота, часто можно встретить растения, подверга- ющиеся такой партикуляции. Партикулы очень разнообразны. Даже отделившиеся от родитель- ского растения споры и семена мы рассматриваем как партикулы, по- скольку полная самостоятельность невозможна без отделения от роди- тельского растения. Партикула после обособления может сразу или впоследствии стать особью или не может стать ею никогда. На вопрос: что такое особь, мы уже ответили (см. гл. 2.4.), но когда партикула особь, а когда нет? Чтобы ответить на этот вопрос, определим сначала, какие бывают партикулы. Партикулы бывают обособленные (лишенные морфофизиологичес- кой связи) и необособленные (имеющие морфофизиологическую связь) и образуются соответственно после нолной или ненолной нартикуляции (по- дразделение по степени ее проявления, рис. 48). Рис. 48. Партикуляция и партикулы у растений. Типы партикуляции: А — неполная, Б — полная общая физиологическая: Б1- эпиэмбриональная, Б2 — семенная, БЗ — пыльцевая, Б4 — споровая, В — полная частная механическая. Типы партикул: 1 — необособленные, 2 — обособленные, 3 — семена, 4 — пыльцевые зерна, 5 — споры, 6-8 — деструктивные партикулы, 6 — отрезок побега, 7 — обособленные соматические клетка или ткань, 8 — отрезок листа.
Глава 4. Основные способы освоения пространства оиоморфами семенных растений 157 Кроме того, лартикулы мы классифицируем по особенностям их структуры на следующие типы: эмбриональные (у растений это обособ- ленные споры, пыльцевые зерна или семена), конструктивные (они включают основные органы; такие партикулы у растений состоят из побеговой и корневой систем, если последняя вообще есть у родитель- ской особи до свершения партикуляции) и деструктивные (они не вклю- чают в свою структуру все или часть основных органов, например, у семенных растений они не имеют побеговой и корневой систем, когда последняя есть до свершения партикуляции, или одной из этих систем). Любая искусственно или естественно отделенная от организма живая клетка, группа клеток, часть органа или орган — партикула. По особенностям проявления в структуре тела растений мы разли- чаем партикуляцию следующих типов: а) общую эииэмбриональную (вегетативную), когда образуются две и более конструктивные партикулы (многолетники); б) общую сиоровую или семенную (реиродуктивную), когда партику- лы — только споры или семена (однолетники); в) общую иыльцевую, когда партикулы — только пыльцевые зерна (однолетники); г) частную, когда образуется одна конструктивная партикула (ро- дительская особь), а все остальные партикулы иные. При частной партикуляции помимо разнообразных обособленных партикул из партикулировавшей особи образуется только одна кон- структивная партикула, которая сохраняет за собой право считаться той же особью, что и раньше, поскольку частная партикуляция не разрушает у нее основных морфофизиологических систем (несколько повреждается побеговая и корневая системы или одна из них), например, при отде- лении от родительской многолетней особи живой части надземного побега (частная побеговая партикуляция) или семян (частная семенная партикуляция). Всякая обособленная конструктивная иартикула, обособленная спора или семя — особь; обособленная деструктивная нартикула, только после восстановления недостающих ей иобеговой или корневой частей (если, ко- нечно, иоследняя вообще бьиа у родительской особи до свершения napmu- куляции) иревращается в особь. Важно выяснить, в каком соотношении находятся понятия парти- кула и диаспора? Партикула, в нашем понимании, — любая живая часть организма, обособившаяся при партикуляции полностью или не полно- стью. Диаспора (греч. рассеяние) — это, как известно, «часть растения естественно отделяющаяся от него и служащая для его размножения, например, спора, семя, плод (главным образом односемянный), луко- вичка, клубень и т.п.» (Викторов, 1964). Как видно, в таком значении партикула не тождественна диаспоре; по смыслу она значительно шире. Если даже расширить значение термина «диаспора», сделав его синони- мом «партикулы», то последний термин все же лучше, поскольку его дословный смысл (лат. particula — частица) более соответствует и вкла- дываемому в него понятию. Партикула — результат процесса, удач- но названного Г.Н.Высоцким (1915) «партикуляцией». Назначение лю-
15 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Иногда под бесполым размножением понимают все виды неполового размножения, в том числе и вегетативное (Левин, 1963, с. 1040). бой полной партикуляции в жизни растений или обеспечивать одно из важнейших условий для преобразования эмбриональных особей в эпи- эмбриональные (обособление), или непосредственно осуществлять ве- гетативное размножение (образование двух и более конструктивных партикул), или обеспечивать одно из условий вегетативного размно- жения (образование деструктивных партикул). Размножение — это образование новых особей, оно бывает реиродук- тивным и вегетативным. Репродуктивное размножение, согласно изве- стной терминологии, бывает амфимиктическое, или половое (возник- новение новых особей после слияния половых клеток — гамет), апо- миктическое, или бесполое'1 (возникновение новых особей — спор, семян без слияния гамет до партикуляции, т.е. не минуя эмбрионального периода развития). При вегетативном размножении новые особи возни- кают без слияния гамет после партикуляции, т.е. минуя эмбриональный период развития. При амфимиктическом и апомиктическом размно- жении образуются эмбриональные особи (споры, семена), которые впос- ледствии могут преобразоваться в эпиэмбриональные особи. Споры или семена, образующиеся в структуре родительского растения, будучи эмбриональными особями, до определенного момента выступают как элементы (гипосистемы) более крупной системы — родительской осо- би. Только после отделения от нее каким-либо способом эти споры и семена могут стать самостоятельными; нужно действие, аналогичное партикуляции в узком смысле слова. Репродуктивное (половое и беспо- лое) размножение надо отличать от семенного возобновления, т.е. образования эпиэмбриональных особей при прорастании семян. При вегетативном размножении образуются особи посредством партикуля- ции без использования спор и семян (рис. 48). Концепция партикуляции, разработанная автором, позволяет сде- лать и такое обобщение: без этого ироцесса невозможна жизнь на Земле в настоящей или нодобной еи формах. Живая материя дискретна (иреры- виста) в нространстве и во времени; дискретность эииэмбриональных осо- бей у семенных и других растений достигается иосредством иартикуляции. Итак, приведенный выше материал свидетельствует о том, что па- ртикуляцию мы понимаем широко. Партикуляция — самобытное явле- ние; ее также можно многократно искусственно произвести у любого высшего растения. Из определения партикуляции видно, что это явление отлично от вегетативного размножения. Партикуляция — это не «своеобразная слабо выраженная форма», не «один из видов» вегетативного размно- жения, а единственное у высших растений средство, создающее возмож- ность вегетативного размножения (образование деструктивных обособ- ленных иартикул) или неиосредственно обесиечивающее его (образование конструктивных обособленных иартикул). Широкая трактовка партику- ляции позволяет надежно объяснить через это явление биологическую сущность вегетативного размножения — увеличение числа особей, про-
159 пава 4. Основные способы освоения пространства биоморфами семенных растений Вегетативное размножение следует отличать от вегетативного разрастания (Тихоми- ров, 1963; Сенянинова-Корчагина, 1967). Под вегетативным разрастанием понимает- ся распространение в окружающем жизненном пространстве (надземном, подземном и др.) в процессе роста особи ее вегетативных структурных частей без потери между ними морфофизиологической связи. В процессе вегетативного разрастания у особей образуются корни и разнообразные побеги с более или менее развитой вегетативной функцией, а также производные побегов — ризомы, каудексы и др. органы вегета- тивной сферы. Вегетативные части особи составляют систему ее вегетативного раз- растания. изводных от одной особи, без участия спор и семян. Прогрессивным эффектом партикуляции является осуществление в конечном счете веге- тативного размножения или семенного и спорового возобновления. Синонтогенез — совокупность онтогенезов особей всех (не менее двух) последовательно сменяющих друг друга поколений, первое из которых начи- нается с зиготы (Нухимовский, 1970, 1973а, б). Под онтогенезом в сог- ласии с большинством исследователей понимаем самостоятельное раз- витие особи. Мы не поддерживаем стремление некоторых исследовате- лей говорить об «онтогенезе клона» (Любарский, 1967, с. 39). Клон в понимании большинства специалистов, — совокупность особей, со- ставляющих вегетативное потомство особи семенного происхождения. Онтогенезы и синонтогенезы как динамическая основа существования биоморф подробно рассматриваются в главе 7. Листья Saintpaulia ionantha Н.Wendl., Begonia rex Putz-. способны укореняться и образовать побеги, т.е. обособленные листовые партикулы этих видов могут при благоприятных условиях стать особями, а вот листья Citrus limon (L.) Burm. во влажном песке укореняются, а побегов не образуют, т.е. вегетативного размножения не происходит. Результаты интересного опыта сообщены П.М.Жуковским (1949, с. 190). Выяснилась способность отдельных живых листьев — обосо- ленных листовых партикул, образовывать побеги и корни. «Из 1204 ви- дов, подвергшихся испытанию, 501 давали только корни, 25 — только побеги и 289 — и побеги, и корни. Остальные вообще не давали ни того, ни другого». Следовательно, листовые партикулы только примерно четвертой части всех испытанных видов смогли в оптимальных условиях («на влажном песке в тепличном парнике») стать особями, а около половины лишь продлили свое существование, образовав только корни или только побеги, но так и не стали особями. Примеров вегетативного размножения множество (Мейер, 1937; Правдин, 1937; Котт, 1955; Шалыт, 1960 и др.). Следует, однако, всегда помнить, всякое вегетативное размножение возможно только после свершения партикуляции. В целях дальнейшей конкретизации представлений о вегетативном размножении мы разработали подразделение его по форме проявления. Оно бывает нескольких основных munoe: 1) дефективное (от лат. defecti- vus — несовершенный), или несовершенное, — характеризуется образо- ванием клонистов, системы вегетативного разрастания ') которых не за- воевывают или завоевывают очень незначительные новые территори- альные участки, прочно и долго удерживая их за собой и (или) за своим синорганизмом (нередко до конца синонтогенеза), например, Patrinia
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ intermedia (Нотет.) Roem. et Schult., Anthriscus sylvestris (L.) Hoffm., Colchicum speciosum Stev.; 2) ретардационное (от лат. retardatio — за- медление), ши замедленное, — образуются клонисты, системы вегета- тивного разрастания которых медленно завоевывают сравнительно не- большие новые территориальные участки, долго удерживая их за собой (Bupleurum longifolium L., Paeonia anomala L.); 3) экстенсивное (от лат. ех1еп81ив — удлиняющий) — развиваются KIIQHHCTbI, системы вегета- тивного разрастания которых энергично завоевывают новые большие тер- риториальные участки, долго удерживая их за собой (Pseudosophora alopecuroides (L.) Sweet, Glycyrrhiza glabra L.); 4) интенсивное (от лат. intensio — усиление) — характеризуется образованием клонистов, сис- темы вегетативного разрастания которых энергично завоевывают срав- нительно большие новые территориальные участки, недолго удерживая их за собой (Ranunculus repens L., Potentilla anserina L.); 5) экслансивное (от лат. expansio — расширение, распространение) — образуются кло- нисты, расселяющиеся на окружающей территории с помощью вне- шних агентов (воды, ветра, животных и др.), например, Elodea canadensis Michx., Hydrocharis morsus-ranae L. Экспансивное размножение может быть также у растений, которые обычно размножаются иным вегетатив- ным путем, например, при расселении человеком отдельных клонистов Rubus idaeus L. или при распространении клонистов Phragmites australis (Cav.) Trin. ex Steud. паводковыми водами. Степень подвижности особей учитывали многие исследователи. Так, в частности, Л.Г.раменский (1938) делит травянистые растения по их приспособлениям «к вегетативному возобновлению и разрастанию» на три основные группы: вегетативно-неподвижные, неразрастающиеся; вегетативно-слабоподвижные (вегетативно-малоподвижные по Е.Л.Лю- барскому, 1961, 1967), разрастающиеся; вегетативно-подвижные. О.В. Смирнова (1968) обратила особое внимание на способность или неспособность особей удерживать за собой ранее завоеванную тер- риторию, подразделив их на устойчивые (способные долго удерживать завоеванную территорию) и неустойчивые (не способные долго ее удер- живать). Г.М.Зозулин (1959, 1961, 1968) широко использовал различную способность растений к вегетативной подвижности и удержанию терри- тории для построения классификации жизненных форм. По особенностям вегетативного разрастания в горизонтальном нап- равлении в почве или по ее поверхности, или в воде особи семенных растений бывают: алланофильные (алланофиты), т.е. нелодвижные (не- разрастающиеся), и лланофильные (лланофиты), т.е. подви>кные. Среди лланофильных особей можно различать: гиполланофильные — особи с очень слабым горизонтальным (приземным, в почве или в воде) вегета- тивным разрастанием — с очень компактными основаниями (Nardus stricta L., Capparis herbacea Willd.); брахилланофильные — особи со сла- бым горизонтальным вегетативным разрастанием — с короткими осно- ваниями (Bupleurum longifolium 1., Polygonatum odoratum (Mill.) Druce); мезолланофильные — особи с умеренным (средним) горизонтальным ве- гетативным разрастанием — с умеренно удлиненными основаниями (Solanum tuberosum 1., Viburnum opulus L.); тахилланофильные — особи
Глава 4. Основные способы освоения пространства биоморфами семенных растений с сильным горизонтальным вегетативным разрастанием — с раздвину- тоузловыми и иными длинными основаниями (Phragmites australis (Сач.) Trin. ex Steud., Cirsium vulgare (Savi) Ten. Партикуляция и рост — две важные стороны единого процесса— развития жизни, а следовательно, и эволюции живого. Партикуляция находится в определенных отношениях с ростом. Партикуляция (лю- бая — механическая, физиологическая) как-либо воздействует на це- лостность организма, при этом партикулы могут быть необособленны- ми от родительского организма, и тогда всякий рост является продол- жением развития только родительского организма. Если партикулы обособленные деструктивные (у них недостает какого-либо из основ- ных органов — побега или корня), то их рост — регенерация недос- тающих основных органов, что приводит в итоге к вегетативному размножению — развитию новых организмов, или при регенерации образуются не все основные органы (например, только корни), что приводит развитие таких партикул в тупик и они элиминируют, так и не став новыми организмами. Если партикулы обособленные кон- структивные (у них есть все основные органы), то их рост — развитие новых особей. Наконец, последний случай, когда рост партикул не начинается и никогда больше не восстанавливается, что приводит их к гибели. На основе рассмотренных выше положений о партикуляции и раз- множении можно сделать некоторые обобщения. Эффективное раз- множение — это такое размножение биоморф (кроме сенильных), при котором на какой-либо стадии его проявления участвует партикуляция. Это явление находит свое отражение в понятии норма и аномалия роста и развития биоморф. За норму роста и развития биоморф мы принимаем такой процесс роста и развития, который готовит основу для размно- жения и после ее создания сопровождается и (или) завершается не только образованием гамет и (или) эмбриональных особей, и (или) проорганизмов, но и полным морфофизиологическим их обособлением. За аномалию роста и развития биоморф мы принимаем такой процесс роста и развития, который не готовит основу для размножения, а если и образуются гаметы и (или) эмбриональные особи, и (или) проорга- низмы, то не находится способов для их полного морфофизиологичес- кого обособления. Из этих положений следует, что Жизнь на Земле в настоящей или подобной формах невозможна без нартикуляиии, т.е. без всякого разрушения (физиологического или механического) особи (растения или животного) на полностью или не полностью обособленные живые части (нартикулы). Для биостробы Земли слишком мало, чтобы роди- тельские биоморфы осуществляли половое размножение и образовы- вали таким путем эмбриональные особи или иным путем образовы- вали специализированные вегетативные зачатки, оставляя их в мор- фофизиологической связи с собой (в себе, на себе) всю жизнь, а затем отмирали вместе с ними. Для биостробы Земли важнее, чтобы родительские биоморфы оставляли полностью морфофизиологически обо- собленное от себя потомство(!), последнее возможно только посред- ством иартикуляции.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Одной из принципиальных сторон познания организации биоморф вообще и биоморф семенных растений, в частности, является анализ типов их поведения в круговороте ресурсов. Давно возник вопрос: почему жизнь, однажды появившись на Зем- ле, превратив ее в биопланету, так бурно «растеклась» (по образному выражению В.И. Вернадского (1967) по ней и имеет тенценцию к «рас- теканию» за ее пределы. Причина всему — размножение «живого веще- ства» (по В.И. Вернадскому, 1967), т.е. организмов. Борьба за существо- вание обусловлена быстрой прогрессией, в которой все органические существа стремятся к размножению (Darwin, 1859; Серебровский, 1973). Посредством размножения жизнь на Земле стала всюдным явлением (Вернадский, 1967). Есть все основания очень высоко оценивать статус размножения в организации живой природы, — выше всего иного, что есть в фенети- ческой организации биоморф. Фенетическая организация биоморф гене- тически предопределена на многоцелевое развитие, приоритет в котором принадлежит направленности на размножение (стремлению к размно- жению), что прямо или косвенно влияет на все остальное в живой природе — зто первый принцип фенетической организации бипморф. Все биоморфы развиваются в соответствии с этим принципом (он возник у их далеких предков в процессе эволюции Природы и действует по сей день). Биоморфы устремлены к размножению, но в зависимости от конкретных условий развития они его достигают или не достигают. Только люди внесли в стремление к размножению новые аспекты, не- сколько изменив форму обслуживания его. Осознанно (человеком) или неосознанно (всеми остальными биоморфами) размножение принято в эволюции как обязательный долг живой природе за право жить на Земле. Особые случаи фенетической организации биоморф, известные у представителей так называемых «кастовых» систем общественных насекомых, например муравьев, ос, пчел, не противоречат принципу стремления к размножению, поскольку крайне высокая специализация бесплодных особей для заготовки пищи или охраны общественных групп во главе с маткой и не менее высокая специализация матки и трутней (самцов) полностью отвечают «культу» размножения у всей кастовой системы. Часть биоморф, генотипом которых предусмотрено размножение, элиминируется, не оставив потомства. В таком случае это отклонение от нормы, поскольку биоморфы не выполнили свой выс- ший долг перед Жизнью. Хорошо, если достаточно размножающихся биоморф, верных этому долгу, которые восполнят стерильность других совидичей (организмов одного вида, по образному выражению А.С.Се- ребровского, 1973); трагедия для вида, если многие биоморфы не нахо- дят условий для реализации свойства размножения. Ранее был сформулирован закон жизнесохранения (см. гл. 3), в котором однозначно высоко оценена роль размножения как одного из высших свойств жизнесохранения (биофилии) — диксенобиофилии (непосредственного участия биосистемы в образовании новых биосис- тем своего уровня организации) в сохранении биостробы. Не повторяя положения теории жизнесохранения, мы обратим внимание на некото-
163 Глава 4. Основные способы освоения пространства биоморфами семенных растений 4.6. Элиминация Выход из Жизни, или элиминация (дословно с лат. «изгонять за порог»), — обязательная сторона дискретного образа жизни всех част- ных представителей живой природы. Элиминация выступает как край- нее выражение процессов отмирания (некроза), неизменно сопровож- дающих существование любой биосистемы на всех или большинстве отрезков их жизненного пути. В процессе биоморфогенеза каждая биоморфа не только растет, прогрессивно развивается и ассимилирует, но и постоянно изнашивается сначала в отдельных частях органов, затем в целых органах и, наконец, целиком выходит из строя, поскольку процессы отмирания берут верх над иными процессами. На особенно- сти отмирания частей растений в их жизненном цикле обращалось внимание неоднократно (Закржевский, Коровин, 1935; Синьковс- кий, 1950; Стешенко, 1956, 1965 и др.). В процессе эволюции способность живого отмирать была постав- лена на службу Жизни; в частности, одним из важнейших завоеваний эволюции живого можно считать появление некротической партикуля- рые новые аспекты, подтверждающие, что всем биоморфам на Земле присуще стремление к размножению. Наша задача показать, что фене- тическая организация биоморф в своем развитии подчинена ее первому принципу. Примерами будут в основном биоморфы семенных растений, хотя положения, о которых идет здесь речь, имеют для биоморф универ- сальную основу. Важное значение размножения в организации жизни было также отражено в специальных законах биоморфологии: управле- ния-исполнения и доминирования в фенетической организации био- морф семенных растений (Нухимовский, 1986). Обязательным условием развития фенетической организации биоморф является стремление к размножению. Существует главное наиравление развития такой организации: зигота — вегетативное развитие — репро- дуктивное развитие, а в системе круговорота ресурсов (веществ, энергии, информации) фенетическая организация биоморф существует по схеме: иотребление + ироизводство + резервирование + рассеивание. Эусомы активно потребляют ресурсы окружающей среды и в них реализуется генетическая программа по вегетативному развитию био- морф (эусомогенез). Основная масса тела многолетника, как правило, представлена эусомой. В процессе дальнейшего развития биоморф фор- мируются диасомы: вегетативно-репродуктивные и репродуктивные структуры в форме единичных цветков или соцветий. Это всё виды ресурсов, поскольку термин «ресурсы» (средства, заиасы) — это, как уже отмечалось, любые вещества, энергия и информация, которые состав- ляют окружающую среду биосистем и сами биосистемы; они или уже вовлечены жизнью в круговорот, или еще могут быть в него когда-либо вовлечены. Запрограммированность стремления к размножению застав- ляет всю фенетическую организацию работать на размножение.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ции (одной из основных форм физиологической партикуляции) — обя- зательного условия для размножения многих биоморф растений и жи- вотных. В настоящее время накоплено много фактов о процессах отми- рания биосистем, что позволяет сделать следующее обобщение: биосисте- ма как целое элиминируется более или менее постепенно в результате накоп- ления нарушений жизнедеятельности и (или) элиминации ее частей или сразу и полностью иод воздействием соответствующих жизнеразрушительных факторов (это также одно из правил общей теории жизнесохранения). Действие этого положения мы наблюдали, когда описывали приме- ры вегетативного размножения растений (весьма показательно для дан- ного случая поведение Anthriscus sylvestris (L.) Hoffm. (рис. 6). Элиминация биосистем, в частности биоморф, осуществляется по разным причинам; в зависимости от которых можно различать следую- щие основные её формы: старческую (в результате старения), namonozu- ческую (в результате болезней), травматическую (в результате травм). Элиминация — неизбежный для любого представителя биостробы Земли процесс их бытия, поэтому важно определить, в чем суть нормы по отношению к этому явлению: после того как биосистема расходует все свои потенции к диксенобиофилии (воспроизводству подобных себе по- томков), у нее остается значительно больше, чем ранее, потенций к эли- минации — это закон (иримасиециальный) нормальной элиминации, или еще одно правило общей теории жизнесохранения. Положение, о кото- ром здесь идет речь, весьма «жестокое» по своей сути, но как ни печаль- но оно для тех, кто его осмысливает (для людей), работает безотказно в живой природе. У людей как индивидуумов, пожалуй, есть небольшой шанс что-либо изменить в пользу себя. О нем мы уже говорили, когда рассматривали гипотезу «сверхдолголетия» (см. гл. 3.5.). Сумеет (и захо- чет) ли Ното sapiens воспользоваться им? Два новых правила, наряду с рассмотренными другими правилами общей теории жизнесохранения, имеют общебиологический смысл и отражают общую картину сохранения жизни на Земле. Нами рассмотрены не все тактические подробности в организации биоморф семенных растений, например, не разобраны многие особен- ности тактики ассимиляции, на основе которой посредством фотосин- теза синтезируются органические вещества из двуокиси углерода (угле- кислого газа), а также — тактики диссимиляции, на основе которой распадаются сложные органические вещества, выделяя при этом энер- гию, используемую в процессах жизнедеятельности. Наша цель в этой главе была не в том, чтобы рассмотреть все разнообразие тактических действий биоморф семенных растений, а в том, чтобы осознать суть понятия стратегия и тактика применительно к организации биоморф и разобраться в особенностях взаимосвязи и взаимозависимости стратеги- ческих и тактических систем в организации биоморф семенных растений с позиций закона стратегии Жизни. Резюмируем содержание тех важнейших тактик, которые мы рассмотрели выше: 1. Тактика роста, прогрессивного развития и (или) ассимиляции обеспечивает создание материальной основы биоморф (шире—
165 Глава 4. Основные способы освоения пространства биоморфами семенных растений биосистем), которая составляет фенетическое содержание Жизни и хранит в себе носителей стратегии Жизни — гены, хромосомы. Тактические системы (средства), действующие в соответствии с указанной тактикой, обеспечивают нарастание, ветвление, кущение и размножение биоморф семенных растений, что отвечает целям соответствующих частных тактик. Тактические системы, действующие в соответствии с тактикой регрессивного развития и (или) диссимиляции, противодей- ствуют системам с иной тактикой и обеспечивают когда-либо завершение жизненного цикла биосистемы (биоморф в частно- сти), т.е. элиминацию. Тактика нарастания, ветвления, кущения направлена на созда- ние каркаса для наиболее удобного распределения в простран- стве ассимилирующих органов (в основном листьев), органов репродуктивного размножения и специализированных органов вегетативного размножения, для успешного снабжения всем необходимым органов размножения и ассимилирующих орга- нов водой и минеральными веществами, добываемых корнями из почвы (или других субстратов) и противоположного снабже- ния корней продуктами ассимиляции. Тактика размножения направлена на образование новых осо- бей из проорганизмов путем их полного обособления от роди- тельской особи посредством партикуляции (вегетативное раз- множение) или на образование эмбриональных особей путем полового процесса и его заменителей в репродуктивных органах и обособление этих особей посредством какой-либо партикуля- ции (репродуктивное размножение). Тактика элиминации включает в себя частные тактики отмира- ния отдельных частей органов, затем отмирания целых органов, и, наконец, отмирания биоморфы, т.е. собственно элимина- ции. В нормальном случае элиминация наступает после утраты биоморфой физиологической зрелости, т.е. способности раз- множаться. Однако, если биоморфы не находят жизненных ресурсов, чтобы противостоять болезням, травмам, то они могут элиминироваться на любой стадии своего биоморфогенеза. 2. 4. 4.7. Организация биоморф семенных растений в борьбе за существование В слове «борьба» содержится оттенок подчеркнутого действия против кого-то, чего-то, с кем-то или чем-то, от стремления предот- вратить, преодолеть, подчинить что-либо вплоть до уничтожения (Ро- зенталь, Теленкова, 1985). Ч.Дарвин в середине прошлого столетия при- менил термин «борьба за существование» для обозначения совокупно- сти отношений между организмами, а также между организмами и окру- жающими условиями неживой природы: «Я должен предупредить, что
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНКИ применяю этот термин в широком и метафорическом смысле, включая сюда зависимость одного существа от другого, а также включая (что еще важнее) не только жизнь одной особи, но и успех ее в оставлении после себя потомства» (Дарвин, 1987, с. 52). Я.М.Галл (1976) провел обстоя- тельный критический анализ истории проблемы борьбы за существова- ние, поэтому, обращаясь к этой работе, выделим лишь то, что более всего представляет интерес для биоморфологии. Факты проявления борьбы за существование наблюдаются посто- янно и повсеместно, поскольку каждая особь не может существовать, не взаимодействуя с различными организмами вокруг себя и окружающей неживой природой. Выживание наиболее приспособленных особей со- ставляет главное содержание естественного отбора, который, согласно Дарвину, осуществляется в борьбе за существование. Многие ученые разрабатывали классификации форм борьбы за существование (см. Галл, 1976). Так, К.А. Тимирязев (1865) выделял борьбу организмов с внешней средой, борьбу между особями одного вида, борьбу между различными видами, борьбу с прямыми врагами. Наиболее полно классифицировал формы и следствия борьбы за существование Л.Плате (Plate, 1913). Он подчеркнул, что борьба за су- ществование бывает: — конституциальная (борьба организмов против неблагоприятных физических и климатических условий); — межвидовая прямая или агрессивная борьба по принципу хищ- ник — жертва; косвенная или конкурентная борьба, когда ви- ды стремятся захватить одни и те же жизненно важные ресурсы (пищу, свет, пространство); — внутривидовая прямая или агрессивная борьба: а) между особя- ми различного возраста, б) между особями одного возраста, в) активная половая борьба у полигамных видов; — внутривидовая косвенная или конкурентная борьба (цит. по Я.М Галлч, 1976). Из форм борьбы за существование, использованных в классифика- циях разных авторов, обратим внимание на конституциальную форму (Л.Плате), поскольку в процессе именно такой борьбы, т.е. «против неблагоприятных физических и климатических условий», у растений (да и животных) происходит эволюция пластичности, т.е. естественный отбор на возможность развертывания в зависимости от условий окру- жающей среды того типа конституции, который более всего этим усло- виям отвечает. В своих теоретических построениях Дарвин говорил о борьбе «су- ществ» (организмов) за жизнь (существование). При анализе явлений борьбы за существование не будем забывать о законах и правилах общей теории жизнесохранения. Представление об иерархии уровней органи- зации живых систем появилось в последарвиновское время. С учетом современного понимания организации Жизни лредлагаем рассматривать «борьбу за существование» как комилекс отношений между биосистемами, а также между биосистемами и абиотическими условиями. В этой связи попробуем вернуться к идее борьбы за существование структурных эле-
Глава 4. Основные способы освоения пространства биоморфами семенных растений 167 ментов внутри организма. Все предыдущие попытки сделать подобное категорически отвергались (см. Галл, 1976). Например, В.Ру (Коих, 1881) рассматривал борьбу за существование между частями тела животного. А. Вейсман (1905) переносил идею естественного отбора на внутренние структурные элементы организма, в частности, он считал, что идет борьба зачаточных внутриклеточных элементов, и что она обеспечивает естественный отбор соответствующим материалом («зачатковый отбор»). Если несколько поменять высоту обобщений и изменить традици- онный угол обзора в проблеме борьбы за существование, то можно попытаться найти дополнительные ее решения. Борьбу за существование (борьбу за жизнь) нредлагаем называть инфраструктурной, если, в иервую очередь, она включает комилекс отношений гииобиоморф, т.е. структур- ных элементов иростых биоморф (организмов), между собой и с окружа- ющей абиотической и биотической внешней средой; во-вторых — коми- лекс отношений между генетически однородными иростыми биоморфами (особями вегетативного ироисхождения одного синорганизма, иока те не отдалены (!) или отдалены незначительно и их взаимодействие реально сказывается на инфраструктуре всего синорганизма (иример, инфраструк- турная борьба в синорганизме-кочке у Deschatnpsia cespitosa (L.) Beauv.) Это частный случай общей борьбы за существование. Для познания орга- низации биоморф важное значение мы отводим также термину «консти- туциальная» борьба за существование, предложенному Л.Плате (Plate, 1913), под которым понимаем обобщенную ответную реакцию всего орга- низма (его конституции) на воздействие окружающей среды (подробнее см. гл. 8.2.). Что же нас подталкивает к анализу инфраструктурной борь- бы за существование? Пожалуй, только одно обстоятельство: она реаль- но присутствует. Ранее мы обобщили материал о борьбе за существова- ние гипобиоморф — структурных элементов растительных биоморф в законе стимулирования и депрессирования роста и развития в фенети- ческой организации биоморф семенных растений (Нухимовский, 1987). В.Л.Бологова (1989) весьма плодотворно провела «демографические» ис- следования в макроморфологической структуре особей Dactylis glomerata L. Один из выводов ее работы сводится к тому, что взаимоотношения между комплексами корнепобегов-органов носят, «в какой-то степени, конкурентный характер». Побеги, корни и другие органы, составляющие единую структуру растительных организмов, как мы полагаем, борются за овладение ре- сурсами окружающей среды не только сообща, как одно целое — орга- низм, но и как относительно независимые части организма. Они борют- ся и между собой, и с окружающей их внешней природой, применяя для этого типичные приемы борьбы между организмами, главные из кото- рых кооперация (взаимопомощь) и конкуренция, причем иногда весьма жесткая, вплоть до гибели одного или нескольких участников. Инфор- мация о конкурентных, кооперативных и иных отношениях между структурными элементами в теле организмов еще на заре эволюции стала кодироваться на молекулярном уровне, и, развертываясь в фено- типах, обеспечивает проявление биоморфами экологической пластич- ности. Так, главный корнепобег родительской особи у купыря лесного
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ (Anthriscus sylvestris (L.) Hoffm.) обычно погибает в противоборстве с молодыми, производными от него, корнепобегами, но это происходит лишь тогда, когда главный корнепобег выполнит свое предназначе- ние — отцветет и отплодоносит (см. рис. 6). Таким образом, инфра- структурная борьба за существование находится под контролем генов и в зависимости от условий обитания она приобретает различные вариан- ты, сохраняя при этом главную направленность жизнеразвития био- морф — стремление к размножению. Организмы и синорганизмы борются за существование с окружа- ющей их неживой и живой (например, с воздействием ног и зубов животных) природой, — это известно и не требует особых доказа- тельств. Важнее другое: эта борьба идет не только снаружи, но и внутри растительного организма, и MQKHQ говорить о степени успешности борь- бы за существование с окружающей неживой и живой природой как организмов в целом, так и каждого их структурного элемента (гипобио- морфы) по отдельности. Например, из нескольких побегов замещения, образующих ярусное ветвление в условиях засыпания растений песком на эрозионно подвижных почвах деградируемых пастбищ, на каждой подземной побеговой ветви выживает 1-2 наиболее удачливых побега, которые сумели выиграть борьбу со стихией и конкурентную борьбу с соседними побегами замещения за пищевые и водные ресурсы, за вы- годное положение в пространстве и т.п. Такие побеги при засыпании песком сумели быстрее удлинить междоузлия их нижних частей, образовать больше придаточных корней и быстрее перестроиться ана- томически, поскольку к многолетней подземной жизни перешли над- земные части побегов, обычно однолетние в отсутствии засыпания (рис. 39, 49). Ключ к решению некоторых принципиальных вопросов эволюции габитуальных форм, вероятно, связан со способностью над- земных вегетативных удлиненных частей цветоносных побегов, обычно после плодоношения отмирающих до короткого зимующего основания, превращаться в многолетние побеговые части, защищенные наносным почвогрунтом от колебаний погоды. В странах с теплым зимним климатом, где еще встречаются расте- ния, распространенные и в умеренной климатической зоне, нередко И происходят заметные изменения в структуре жизненых форм, их габи- тусов в частности (см. гл. 8.2.2.). Инфраструктурная борьба за существование у биоморф разнообраз- на, особенно на переходных стадиях от организма к синорганизму (рис. 6, 50). При ураганном ветре на южном берегу Крыма (Никитский ботанический сад) было разрушено дерево бузины черной (Sambucus nigra L.), которое до этого развивалось нормально (рис. 50-I). Отделившиеся от родительского растения части (деструктивные партикулы) могли бы погибнуть, но они полностью использовали зало- женную в них способность к выживанию. Образовав недостающие ос- новные органы — корни, они превратились в новые особи, но уже не семенного, а вегетативного происхождения — клонисты (рис. 50-II). Инфраструктурная борьба в родительском организме в процессе его развития постепенно переходит в инфраструктурную борьбу синор-
Глава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 49. Поведение особей — полукустарничков Artemisia lerchiana Web. в условиях эрозионно-подвижного субстрата: 1 — при выдувании песка и оголении корней, 1I — при засыпании растений песком (республика Калмыкия, Черные земли, окрестности пос. Нарын-Худук). ок — оголенный от почвы корень, ко — корнеотпрысковый парциальный куст, нп — над- земные побеги текущего года (сверху показаны примерные границы кроны), мп — верхушки многолетних побегов, выступаюшие над поверхностью почвы, гп — геоподии, эп — эпифи- зы (зоны активного прошлого или нынешнего побегообразования), пк — придаточные корни; пунктирной линией отмечено примерное положение семядольного узла.
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИЙ Рис. 50. Проявление партикуляции и вегетативного размножения у Sambucus nigra L. в экстремальных условиях (Крым, окрестности Никитского ботанического сада). 1 — ранние стадии биоморфогения в относительно спокойных условиях существо- вания, И — поведение взрослого дерева после разрушения его ветром (схема). с- семядоли, г — гипокотиль, гк — главный корень, гп — главный побег, вп- верхушечный вегетативный годичный побег, гг — граница годичного прироста, Tp — мономерный тотарезид, ок — особь клона, образовавшаяся из обособленных частей сломанного дерева, нч — надземные части (заштрихованы) побегов, пн- партикуляция механическая неполная, пч — подземная часть (незаштрихована), ва — стволики вторичных аэроподиев, образовавшиеся из поваленных ветром побеговых частей (верхние части не показаны), т — турионы текущего года, пк- придаточные корни. Одно деление масштаба — 2 см. ганизма, на которую накладывается внутривидовая борьба между осо- бями в популяции и которая часто быстро ее сменяет. Отношения между одноименными и разноименными структурными элементами организма ироявляются в различных формах: кооиеративной конкурентной или относительной индиферентности, но кока живет орга- низм, всякая инфраструктурная борьба за существование иерекрывается более высокими целями — борьбой всего организма (в целом) за существо- вание в иоиуляции, виде, сообществе. Так можно оиределить сиециальный закон соотношения форм борьбы за существование в организме. Из основных систем, в которых лучше всего наблюдается инфра- структурная борьба за существование, прежде всего назовем лрелолуля- ции и коалиции. Мы ограничимся анализом в основном препопуляций, об остальных системах даем лишь самые общие соображения.
Глава 4. Основные способы освоения пространства биоморфами семенных растений 171 Преггонуляция — система лидеров среди основных органов биоморф семенных растений — побегов (двух и более), варьирующих по возрастному и фенологическому составу, но одинаковых по уровню организации наиболее сложных из них, без корней (они ке развиваются), или с корнями (они развиваются вместе с ними, если корень главный, или корни придаточные), или при корнях, если побеги адвентивные (они развиваются на корнях), и при этом всегда вся система остается единственной в своем роде, т.е. элементов такого muna в организме больше нет. Структурные элементы в препопуляции бывают простыми (они не имеют всех основных органов, а состоят только из побегов) и сложными, когда они образованы основ- ными органами (побегами и корнями). Препопуляция у семенных рас- тений ближе всего стоит к популяции, поскольку ее простые элемен- ты — нроонтоиды, обладая одной побеговой структурой, будучи отде- ленными от родительского организма (деструктивные побеговые парти- кулы), могут нередко обрастать корнями и преобразовываться тем са- мым в новые организмы — элементы популяции. Сложные элементы нренонуляции — корненобеги-органы, или онтоиды, после отделения от родительского организма сразу же становятся особями (у них в структуре есть все свойственные особям основные органы) — элементами синор- ганизма и популяции. Проонтоиды и онтоиды — это все разновид- ности преиндивидуумов (проорганизмов, предособей). Они весьма раз- нообразны: отдельные живые соматические клетки, группы таких кле- ток, образующие целый орган (побег, корень, лист и т.п.) или группу органов, или любой фрагмент органа, которые функционируют в структуре определенного организма по эволюционно выработанному типу родитель- ских систем и не являются эмбриональными особями, а также отдельные половые клетки (Нухимовский, 1974а). Половой преиндивидуум (гамета) может превратиться в особь, как правило, после слияния с гаметой иного пола. Соматические преиндивидуумы обладают или не обладают потенци- альной способностью к преобразованию в индивидуумы. Почему мы определяем препопуляции через побеги? Побеги — это лидеры среди основных органов; только им присуща способность раз- вивать репродуктивные органы и образовывать семена. Систему, отличную от препопуляции, состоящую из каких-либо одно- именных органов или одноименных частей органов, развивающуюся в инфраструктуре растительного организма и при этом единственную в своем роде (поскольку вне этой системы ее элементов в организме не остается), можно обозначить термином коалиция. В организме можно определить одну или немного (все зависит от высоты обобщений и угла обзора) препопуляций, а коалиций — много. В целом коалиции отсто- ят от популяции еще дальше, чем препопуляции, поскольку у элементов коалиций меньше возможностей, хотя все же есть (Begonia rex Putz., Saintpaulia ionantha Н.Wendl.), превратиться в особи и стать элементами популяции, чем у элементов препопуляции. Помимо рассмотрения растительного организма как препопуляции или системы коалиций предлагались и иные подходы. Так, М.В. Марков (1986) считает, что растительный организм — это своеобразная популяция, или субпопу- ляция, т.е. система метамеров-побегов (или других метамеров).
17 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Для исследовательской работы надо выбирать такую препопуляцию, которая более наглядно и убедительно передает особенности инфра- структурной борьбы за существование. Рассмотрим несколько примеров. Al ssum тига1е Wal st et Kit. — б ачок н ст нный (сем. Bras- sicaceae). Наблюдался в Никитском ботаническом саду (Южный Крым), где выращивается в экспозициях и на коллекционном участке. Хорошо размножается семенами. Прорастание надземное. Укоренившиеся пар- циальные группы побегов кущения (парциальные кусты) могут ис- пользоваться в культуре для вегетативного размножения. Биоморфотип, охарактеризованный по габитуальным признакам, — аэроксильные геми- симподиальные стелющиеся полукустарники. Инфраструктурная борьба за существование в побеговой системе наиболее наглядно проявляется в препопуляции монокарпических обо- еполых побегов. Каждую особь можно было бы рассматривать и как препопуляцию иного типа, например, как препопуляцию годичных побегов или как препопуляцию побеговых предветви и ветвей, но по- казательнее все же первый вариант. Структурный элемент рассматри- ваемой препопуляции — монокарпический обоеполый двухгодичный (дифазный) побег. На первом году образуется куст из полегающих вегетативных без- розеточных побегов, из которых одни побеги развиваются сильнее (осо- бенно главный), а другие слабее (рис. 51-I). Цветение начинается на втором году жизни. При распространении растений самосевом под полог естественных растительных сообществ Крыма цветение начина- ется на 2-4 году. Монокарпические побеги в своем жизненном цикле проходят следующие возрастные стадии: открытой почки, вегетативного годичного побега, вегетативно-репродуктивного двухгодичного полузак- рытого (с верхушечным брактеозным открытым соцветием) побега и, наконец, многолетнего (резидного) побега. Все побеги в препопуляции ведут борьбу как между собой, в форме кооперации или конкуренции, так и с условиями окружающей их внешней среды. Все побеги более или менее различаются по степени развития: одни сильнорослые (у них больше шансов выйти победителями), другие менее крупные, а третьи — очень слабые. Вегетативные годичные побеги массо- во отрастают из почек на нижней и средней частях вегетативно-репродук- тивных двухгодичных родительских побегов (рис. 51-II). Самые сильные вегетативные побеги образуются в переходной зоне между плагиотроп- ной и ортотропной частями двухгодичного побега; такие вегетативные побеги наиболее конкурентноспособные. После плодоношения роди- тельского побега большинство развитых на нем вегетативных побегов отмирает. К весне выживает 1-2, реже больше, вегетативных годичных побегов-потомков, которые переходят в новое состояние — вегетатив- но-репродуктивную стадию развития монокарпических побегов. Между разновозрастными и одновозрастными побегами и их частями проявля- ется как взаимопомощь, так и конкуренция. Родительская система сначала создает условия для возникновения на себе пазушных почек и роста молодых вегетативных годичных потомков, обеспечивая им не только место в пространстве, но и доставляя к ним необходимые органо-
Глава 4. Основные способы освоения пространства биоморфами семенных растений 173 Рис. 51. Инфраструктурная борьба за существование в препопуляции, развивающейся в биоморфогении Alyssum тцга1е Waldst. et Kit. (Крым, Никитский ботанический сад). 1 — ранние стадии развития, 11 — фрагмент взрослой особи; мп — пазушный ди- фазный обоеполый монокарпический побег, вп — вегетативные силлептические побеги — летнее проорганизменное вегетативное побеговое потомство, со — сухие остатки от прошлогодних летних побегов, пр — многолетний прошлогодний побеговый остаток (репторезидный побег), сц — плодоносящее соцветие. Одно леление масштаба — 1см. Остальные обозначения те же, что и на рис. 50.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНКИ минеральные и водные элементы питания (своеобразная «забота о по- томстве»). Вегетативные побеги, в свою очередь, подрастая, крепнут и, имея большую массу зеленых листьев, помогают родительскому побегу, образующему к этому времени соцветие, в энергетическом (пищевом) снабжении. К тому же на родительском побеге уже почти не остается собственных зеленых листьев. Таким образом, родительская система сначала помогает развитию его вегетативного потомства, но затем в борьбе за энергетические ресурсы последние начинают ему мешать, и роди- тельская система, выполняя наиболее важную для организма функ- цию — репродуктивную, сильно угнетает свое потомство. Вегетативное потомство сопротивляется и по-своему борется за место под солнцем. Каждый представитель вегетативного проонтоидного потомства на оп- ределенном этапе испытывает состояние непримиримой борьбы со сво- ими собратьями (похоже на «естественный отбор» наиболее приспособ- ленных). Вегетативные годичные побеги в чем-то помогают друг другу выживать, но вскоре большинство из них погибнет. Итог инфраструк- турной борьбы в среде вегетативного проонтоидного потомства — горы «трупов» (сухих остатков вегетативных побегов), которые наблюдаются на прошлогоднем зимовавшем остатке (резиде) монокарпического по- бега, и продолжение жизни лишь 1-3 побегами-победителями, которые повторяют в своем развитии судьбу прошлых поколений подобных побегов (рис. 51-II). В инфраструктурной борьбе участвуют и другие монокарпические двухгодичные побеги, которые взаимодействуют с подобными им побега- ми. В этой борьбе преимущество получают также наиболее приспособлен- ные. Так, часть монокарпических побегов, прижимаясь к почве, укореня- ется, превращаясь в корнепобеги-органы (онтоиды). Согласно определе- нию препопуляции, они остаются в ее структуре и, как правило, выигры- вают жизненное состязание в развитии с другими монокарпическими побе- гами, не сумевшими образовать придаточных корней. Кстати, возникно- вение придаточных корней — это одно из проявлений инфраструктурной борьбы за существование. Когда полегающий побег соприкасается с поч- вой или погружается в нее, то он получает выгодные условия для прида- точного корнеобразования, которые использует или не использует. Итак, в инфраструктурной борьбе за существование между побега- ми постоянно накапливаются и проявляются клубки противоречий, которые постоянно разрешаются путем взаимопомощи, уступок, подчи- нения, вплоть до уничтожения одних побегов другими, т.е. в этой борьбе всегда есть победители и побежденные, а если действует относительный мир, то значит, он выгоден всем проонтоидам и онтоидам на какой-то стадии их развития. Инфраструктурная борьба идет на всех уровнях организации жизни внутри растительного организма, но в целом она находится под контролем всего организма, под контролем его генов, т.е. она управляема. Инфраструктурная борьба за существование — реаль- ность и обязательное условие жизни организмов на Земле. Зве обой к асильный в б атственный — Н епсит androsae- тит L. ( сем. Hypericaceae). Биоморфогения этого вида изучена на коллекционном участке Никитского ботанического сада в Крыму (Хржа- новский, Нухимовский, 1968). Габитуальный тип (составная, но необя-
Глава 4. Основные способы освоения пространства биоморфами семенных растений 175 зательная часть биоморфотипа) — геоксильные эусимподиальные ку- старники (рис. 52). Хорошо размножается семенами. Прорастание над- земное. На первом году жизни образуется годичный безрозеточный ве- гетативный побег, иногда силлептически слабо ветвистый. Инфраструктурная борьба за существование в побеговой системе наиболее показательна в препопуляции побегов системы кущения (т.е. побегов кущения — турионов, а также корнепобегов, если они образу- ются, и главного побега, с которого начинается кущение). В препопуля- ции побегов и корнепобегов системы кущения конкурентные отноше- ния проявляются наиболее ярко.Кущение происходит со второго года жизни особи, когда весной из пазушных почек основания прошло- годнего годичного главного побега (зимой он сверху обмерзает на боль- шую часть длины) отрастает один или несколько побегов кущения- турионов. В кусте сильно проявляется контрактильность главного и прида- точных корней, поэтому уже с первого года жизни основание главного корня оказывается в почве, а все последующие побеги кушения обра- зуются из подземных почек. Молодые турионы сначала получают усиленную энергетическую помощь от соседних, более взрослых, тури- онов и от тех, надземная часть которых уже погибла, а сохранились лишь их дуговидно изогнутые подземные многолетние одревесневшие базальные остатки (вместе они формируют многолетний подземный по- беговый орган — геоподий, иногда называемый у кустарников ксило- подием). Поэтому турионы 1-3 года быстро растут и развиваются, осо- бенно на первом году их жизни. Первое цветение куста происходит на 2-3 год (рис. 52-I). В процессе старения в кроне турионов ежегодно образуется много локальных го- дичных побегов, часть которых не может сформировать репродуктивные системы, поскольку отстает в росте и развитии. К 4-6 годам старые турионы уже мешают молодым занимать в кусте выгодные места, поэто- му усиливается борьба за перехват энергетических ресурсов, накаплива- емых в базальной части куста. Молодые турионы получают в этой борьбе преимущество и угнетают старые побеги кущения. Эта способность кус- тов стимулировать развитие одних частей и угнетать другие закреплена в генотипе. Возраст турионов вместе с их развитой кроной от 1 до 6 (редко 7-9) лет; в среднем турионы живут 3-4 года (рис. 52-II). В глу- бокой тени под кроной взрослых турионов наиболее слабые нередко погибают еще до конца их первого вегетационного сезона. В условиях мягкой крымской зимы многие турионы страдают даже от небольших похолоданий, но наиболее сильные все же выживают. В зависимости от особенностей зимнего периода, положения в кроне куста и степени развития годичные побеги могут отмирать полностью, почти полностью (до зоны укороченных междоузлий) или до средних удлиненных междо- узлий. В последнем случае отмирают лишь репродуктивные части и несколько прилегающих к ним междоузлий. Иногда успешно перезимо- вывают даже некоторые осевые части прошлогоднего верхушечного со- цветия. Приемы инфраструктурной борьбы зверобоя — лишь звено в арсенале приспособлений, обеспечивающих выживание организмов.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНКИ Рис. 52. Инфраструктурная борьба за существование в препопуляции, развивающейся в биоморфогении Нурепсит androsaemum L. (Крым, Никитский ботанический сад). 1 — зверобой красильный на 1 — 2-м годах жизни; П — взрослое растение (10— 12 лет); А — проростки, Б — всходы; В — ювенильное растение, Г — имматурное растение, Д — матурная особь на втором году вегетации; 1 — семядоли, 2 — ги- покотиль, 3 — главный побег, 3 — резидный депрессированный прошлогодний главный побег (его остаток), 4 — главный корень, 5 — силлептические побеги, 6— весенний аксиллярный вегетативно-репродуктивный годичный побег, 7 — осе- вые части, сохранившиеся от прошлогоднего верхушечного соцветия, 8 — верху- шечное закрытое соцветие, 9 — турион (побег кущения) на первом году вегетации, 10 — турион на втором году вегетации, 11 — отмершие побеги, 12 — отмершие верхушки годичных побегов прошлых лет, 13 — годичные побеги текущего года в кроне шестилетнего туриона, 14 — многолетние осевые части побегов прошлых лет, 15 — подземные многолетние побегововые ветви (разветвленный геоподий), 16 — весенние нормальные побеги, развивающиеся из зоны прошлогоднего вер- хушечного соцветия. Горизонтальная линия — поверхность почвы; одно деление масштаба — 1 см.
Глава 4. Основные способы освоения пространства биоморфами семенных растений 177 ! ~ llx~«i~», и««! 1 Рис. 52. (Продолжение).
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ Способность кустов зверобоя красильного быстро формировать силь- норослые турионы используется в практических целях при «омолажива- нии» кустов для поддержания их декоративных свойств и при выбраков- ке неперспективного репродуктивного и скелетного материала. Кусти- стое строение растительных организмов обеспечивает побегам выгод- ные условия при использовании окружающей среды. Бо е алтайский — Aconitum It icum Steinb. (сем. Ranunculaceae). Этот вид наблюдался на Алтае (Курайский хребет, субальпийский луг, 1900 м над ур. моря). Биоморфотип — травянистые безрозеточные кон- дивидуальные (синорганизменные) почкокорневые многолетники. ~4нфраструктурную борьбу за существование рассмотрим в препо- пуляции годичных корнепобегов. В ней выделяются следующие возрас- тные группы корнепобегов-органов (онтоидов): почкокорни (инициаль- ные корнепобеги), вегетативные годичные и вегетативно-репродуктив- ные годичные корнепобеги. Прорастание семян надземное. Семена прорастают после схода снега в течение всего вегетационного сезона. На первом году вегетации образуются проростки, всходы и ювенильные растения с коротким ве- гетативным безрозеточным побегом, переходящим через гипокотиль в небольшой главный корень, обладающий контрактильностью (рис. 53, А-В). Втягивающая деятельность гипокотиля и главного корня приводит к перемещению семядольного узла к поверхности и вглубь почвы. В семя- дольном узле, точнее в пазухе отмерших семядолей, начинает разрас- таться одна (!) почка и одновременно с этим проявляет ростовую актив- ность прида точный корень, развивающийся из почки и вместе с ней- это пазушный почкокорень (инициальный корнепобег). Вероятно, в генотипе закрепилась способность предопределять развитие только од- ной почки из пазух только одной семядоли. Второй пазушной почке не представляется шанс выйти из глубокой депрессии, когда соседняя су- противная (сестринская) почка начинает усиленно развиваться. Осно- вание придаточного корня утолщается; он вместе с главным корнем продолжает контрактильную деятельность, в результате которой первый узел с первым междоузлием (эпикотилем) погружаются в почву или достигают ее поверхности. В этих условиях в пазухе первого настоящего листа развивается еще один почкокорень.Он чаще всего развит сильнее главного корня, а нижерасположенный почкокорень — сильнее, чем вышерасположенный. Главный вегетативный корнепобег помогает па- зушным почкокорням получать необходимые водно-пищевых ресурсы. Как видно по степени развития почкокорни, очередно расположенные на одном родительском побеге, конкурируют и между собой, но эта конкуренция незначительна, поскольку каждый почкокорень имеет ав- тономную корневую систему, и борьба идет лишь за органические ве- щества, произведенные родительским корнепобегом. В этой борьбе, как правило, получает преимущество нижний почкокорень, который разви- вается крупнее вышерасположенного. Почкокорни в сезон их образова- ния не трогаются в видимый надземный рост. Вероятно, на них сказы- вается тормозящее действие главного корнепобега, поскольку только он развивает надземную вегетативную часть.
Глава 4. Основные способы освоения пространства биоморфами семенных растений 179 Рис. 53. Инфраструктурная борьба в препопуляции, развивающейся в биоморфогении Aconitum altaicum Steinb. (Алтай, Курайский хребет, альпийский луг}. А — проростки, Б — всходы, В — ювенильные растения, Д — взрослый клон, 1— семядоли, 2 — гипокотиль, 3 — семядольный узел, 4 — корневая шейка, 5 — глав- ный корень, 6 — придаточный корень, 7 — главный побег, 8 — пазушный почкокорень, 9 — почка, которая будет зимовать, 10 — основание вегетативно- репродуктивного побега, который не зимует, кроме развитых на нем специ- апизированных пазушных почек (верхняя часть удапена), 11 — пазушный весен- ний вегетативный побег; одно деление масштаба — 1 см, горизонтальная линия— поверхность почвы.
OCHOBbI БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Несколько лет родительский, ежегодно сменяемый, побег может закладывать только один пазушный почкокорень. В этом случае веге- тативного размножения не происходит; по существу после отмирания родительского корнепобега продолжается своеобразная виргинильная жизнь одного организма. Если на родительском побеге развивается два почкокорня, то после отмирания исходного корнепобега происходит образование двух новых самостоятельных особей-клонистов в структуре более сложной биоморфы — синорганизма. Это может произойти на 1-3 году жизни, а первое цветение наступает на 2-8 году жизни синор- ганизма. Пока родительский корнепобег растет и развивает листья и репродуктивные элементы, видимый рост почкокорней не обнаружива- ется, но после завершения плодоношения они по существу «съедают» родительский корнепобег: сверху он высыхает, а утолщенный корень опустошается, сморщивается и разрушается. В это время онтоидные потомки погибающего отплодоносившего корнепобега буквально нака- чиваются, толстеют, уходя в зиму в новом качественном состоянии- как самостоятельные организмы-клонисты с викарирующим базовым органом — одиночной зимуюц„ей почкой. Через следующие 5-10 лет на месте одного родительского корнепобега уже обнаруживается их «гнез- до» — один синорганизм с ежегодным вегетативным размножением (рис. 53-Д). С позиций инфраструктурной борьбы за существование, рассмот- ренной на примере препопуляций, каждая гипобиоморфа (проорга- низм) — проонтоид и (или) онтоид — фенетический структурный эле- мент растительного организма, и вся фенетическая организация био- морф была бы невозможна без участия в росте и развитии процессов стимулирования и депрессирования. Особенности их взаимодействия можно закрепить в следующем обобщении. Если в фенетической органи- зации биоморф некоторые их части (no6ezu, корни или их фрагменты) исиытывают иреобладающее стимулирующее воздействие (соответствую- т.n.), то рост и развитие этих частей усиливается. Если же на них воздействуют иреимущественно угнетающие факторы (соответствующие гормоны, уменьшение иотока иитательных веществ, света и т.а), то рост и развитие этих частей замедляется. Это ускоряет смену более старых частей на одноименные новые части. Если ироцессы стимули- рования и угнетения роста и развития семенного растения относительно сбалансированы, то ироизводные (молодые) и родительские (более старые) части могут нормално сосуществовать до момента нарушения такого равновесия. Это закон (снециальный) стимулирования и денрессирования роста и развития в фенетической организации биоморф семенных растений. Нз него мы выводим несколько иравил (рис. 54). 1. Если рост осевых частей многолетних надземных побегов угне- тается новыми побегами, развивающимися из их нижней или (и) средней части, то исходные (родительские) побеги откло- няются от вертикального положения книзу, а производные по- беги интенсивно нарастают кверху, занимая положение, близ- кое к вертикальному (Sambucus racemosa L., S.nigra L., Physo-
Глава 4. Основные способы освоения пространства биоморфами семенных растений Рис. 54. Обобщенные модели растений, отвечающие правилам I-Õ закона стимулирования и депрессирования. Части растений: а— живые, б — отмершие. carpus opulifolius (L.) Maxim., Viburnum opulus L.) (рис. 54-I; 179). 2. Верхушка ствола и молодые ветви в кроне древесных растений получают преимущество перед более старыми ветвями, способ- ствуя депрессии (ослаблению роста, засыханию) последних, если они осваивают зоны окружающей среды, лучше обеспе- ченные необходимыми ресурсами (в первую очередь, светом). Так, нетрудно заметить, как старые ветви многих хвойных и других деревьев чахнут и отмирают, и этому способствуют мо-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИИ лодые ветви, развивающиеся выше старших по возрасту ветвей. При распределении ресурсов, поступающих в крону деревьев, верхние ветви получают преимущество в расположении и угне- тают в развитии нижние ветви, причем тем больше, чем послед- ние старше (рис. 54-II). Отрастание побегов кущения когда-либо депрессирует роди- тельские (включая и медиальный) побеги, причем депрессия проявляется тем сильнее, чем раньше и интенсивнее осуществ- ляется кущение и чем взрослее становятся производные побеги (рис. 54-III). Этому правилу отвечает функционирование мно- гих кустистых растений (Смелов, 1966; Серебрякова, 1971; Ну- химовский, 1974б, Бологова, 1989). Специализация частей биоморф имеет решающее значение в продолжительности жизни этих частей, поэтому верхние и (или) боковые части побегов, входящие в структуру соцветий, несмотря на то, что они моложе базальных частей побегов, закан- чивают свое развитие и отмирают вместе с ними или раньше полностью (особенно в регионах, где есть морозные зимы) или частично (нередко в субтропиках и тропиках, см. гл. 8.2.2). Час- ти побегов, связанные с размножением, развиваются в направ- лении ускоренного рассеивания ресурсов, а вегетативные час- ти — их производства и накопления (рис. 54-IV). Главный корень у мультирепродуктивных многолетников жи- вет дольше, если выше него не развиваются придаточные корни или их мало и они слабые (рис. 54-V). Главный корень живет очень мало у Primula veris L., Rhodiola rosea L., Eremurus tauricus Stev. и др. (Зозулин, 1959; Голубев, 1962, 1965; Нухимовс- кий, 1974а), поскольку у таких растений рано и обильно развива- ются выше главного корня придаточные корни, которые депрес- сируют рост главного корня и перехватывают его функции. Когда придаточные корни не образуются или их образуется мало, то главный корень, чаще всего, отмирает со всем растением (Capparis herbacea Willd., Peganum harmala L). (Нухимовский, 1969б). Молодые стеблеродные (каулогенные) корни, развивающиеся выше старших придаточных корней, депрессируют их рост и убыстряют отмирание, полностью возмещая их функции (рис. 54-VI). Многие растения следуют в своем развитии этому правилу, но наиболее ярко оно проявляется у кистекорневых, луковичных и клубнелуковичных растений. Отмирание подземных частей побегов и взаимосвязанных с ними корней строго взаимозависимо, и чем быстрее идет первый про- цесс (некроз побегов), тем быстрее и второй (некроз корней) и, наоборот, гибель корней способствует отмиранию связанных с ними побегов. На примере таких растений, как Valeriana officinalis L., Рппшlа veris L., наглядно видно, как взаимозависимы процессы отмирания корневищ и придаточных корней (рис. 54-VII). Экстремальные факторы существования вносят коррективы в процессы стимулирования и депрессирования, нередко иска-
Глава 4. Основные способы освоения пространства биоморфами семенных растений 183 жая и перемещая зоны борьбы гипобиоморф (живых частей биоморф) за пространство и время, а преимущество в выжива- нии получают те части, которым достались на данный момент более выгодные условия для овладения необходимыми ресур- сами среды или (и) которые, несмотря на изменение экологи- ческой обстановки, не утратили способность к более или менее модифицированному росту и развитию (рис. 54-ЧШ). Наблюдая жизнь растений в условиях эрозионно подвижных субстратов (в горах, пустынях), можно видеть, как подтвержда- ется это правило (Лавренко, 1935; Нухимовский, 1969б): погре- бенные субстратом растения или погибают, или изменяют свой облик (например, надземные части удлиненных побегов Pega- пит harmala L., которые обычно после плодоношения почти полностью отмирают, в новых условиях одревесневают и сох- раняют жизнеспособность в почве на значительную часть своей длины; у Limonium gmelinii (Willd.) О.Kuntze вместо розеточных побегов могут образовываться верхнерозеточные побеги, у Arte- misia lerchiana Web. (рис. 49-II; 55) на погребенных вынужден- но-подземных побегах обильно развиваются придаточные кор- ни; у Agropyron fragile (Roth.) Р.Candargy вместо компактных корневищ образуются удлиненные и т.п.). При оголении базового органа и корней от субстрата может проявляться факультативная корнеотпрысковость (Аг1етв1а ler- chiana) (рис. 49-I) и т.п. 9. Если после окончания вегетационного сезона в структуре се- менного растения не сохраняются почки, приспособленные к перенесению неблагоприятных условий, то такие растения обречены на вымирание (рис. 54-1Х). Немало тропических~не- подземных побегах обильно развиваются придаточные корни; у Agropyron fragile (Roth.) Р.Candargy вместо компактных кор- невищ образуются удлиненные и т.п.). При оголении базового органа и корней от субстрата может проявляться факультативная корнеотпрысковость (Artemisia ler- chisna) (рис. 49-!) и-ттт.)растеиий хорошо растет в условиях континентального климата на первом году жизни, но к зиме они подготовиться не могут и погибают (Ricinus communis L., Solanum аиси1аге Forsch.). 10. Побеговые ветви, верхушки которых отдаляются в процессе роста от фенетического центра (см. гл. 5.4.) семенного расте- ния, в какое-то время начинают испытывать на себе дефицит его влияния, не получая в нужном количестве необходимые для жизни ресурсы. Поэтому они или переходят на частичное са- мообеспечение, образуя для этой цели какие-либо структурные приспособления (например, придаточные корни), что сохра- няет им возможность дальнейшего роста и развития, или не переходят на дополнительное самообеспечение, что приближает их к состоянию депрессии и способствует элиминации (рис. 54-Х). Примером могут служить равновозрастные побего-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА ГТЕНИЙ Рис. 55. Поведение особи Limonium gmelinii (Willd.) О.Kuntze в условиях засыпания ее песком (республика Калмыкия, Черные земли). нп — надземные побеги, кд — вынужденно длинный ветвистый каудекс, гк — глав- ный корень (верхняя часть его удалена), эп — эпифизы (зоны ярусного побегоотрастания), горизонтальная линия — поверхность почвы, пунктирной линией показано примерное положение семядольного узла; одно деление масшта- ба — 2 см. вые ветви Abies sibirica Ledeb., которые, развиваясь на разной высоте над поверхностью почвы, могут образовывать придаточ- ные корни (при контакте с почвой), давая новые очаги роста (вплоть до формирования новых стволов), или не образовывать их (без контакта с почвой), ускоряя тем самым свою элимина- цию. Побеговые ветви, верхушки которых отдаляются от фено- центра растений, в случае образования придаточных корней меньше зависят от него, что приближает растение к вегетатив- ному размножению. Особенно ярко это видно у корневищных и столонных растений. Инфраструктурная борьба за существование обеспечивает в орга- низмах и синорганизмах сортировку и накопление материала по специ- ализации структурных элементов для естественного отбора, что в целом контролируется на уровне видов и способствует видовой эволюции и видообразованию.
Глава 5. Параметры основных органов и их частей в организации биоиорф семенных растений Утверждение единства листа и стебля в организации одного орга- на — побега и рассмотрение побега как основного органа семенных растений (АгЬег, 1930, 1934, 1941, 1950; Foster, 1949; Имс, 19б4; Эзау, 1980 и др.) — значительное событие в теории ботаники, означающее отказ от признания трех основных органов — листа, стебля и корня и провозглашение представлений о двух основных органах семенных рас- тений — побеге и корне. Дальнейшее развитие идеи единства элемен- тов в организации биоморф семенных растений привело к разработке понятия ризокормус (корнепобег) как высшей единицы организации биоморф рассматриваемых растений (Нухимовский, 1976а, 1980). Прежде чем говорить о морфофизиологическом единстве побега и корня в системе корнепобега, важно разобраться в особенностях организации его элементов, причем в первую очередь основных элементов. Чем полнее мы знаем объекты исследований, тем точнее наши представления об их параметрах — тех показателях, величинах, которые характеризуют объект, его свойства, с разных сторон. Рассмотрим некоторые организационные параметры наиболее зна- чительных частей биоморф семенных растений: побегов, корней и корнепобегов. 5.1. Побеги — основные органы семенных растений, их структура и функции 5.1.1. Разнообразие побегов и их классификаций Основные органы семенных растений с давних времен находятся в поле зрения ботаников, пытающихся их классифицировать по разным признакам. Одним из старейших понятий в характеристике побеговой системы является годовой, или годичный, побег (der Jahrestrieb); оно упомина-
OCHOBhI БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИЙ ется уже в классическом ботаническом словаре Бишофа (Bischoff, 1830, с. 252). Видлер (Wydler, 1843, с. 168) писал: «Под Jahrestrieb я понимаю (в нашем климате) каждый побег, образующийся в течение одного ве- гетационного периода из семени или из одной перезимовавшей почки со всеми возможными разветвлениями». В таком смысле этот термин широко применялся позднее (Braun, 1851; Любименко, 1900; Варминг, 1904; Серебряков, 1952 и др.). На периодичность роста побегов в клима- те умеренной зоны обращал внимание В.Н.Любименко: «надземная часть дерева слагается из отдельных, до известной степени самостоя- тельных единиц — годовых побегов» (1900, с. 254). Любименко подраз- делял побеги деревьев на три группы: обыкновенные, или длинные (с листьями срединной формации и удлиненными междоузлиями), типич- ные укороченные (с листьями срединной формации и укороченными междоузлиями), скрытые укороченные (с листьями низовой формации и укороченными междоузлиями). Идея годового, или годичного, побега получила значительное раз- витие в работах И.Г.Серебрякова (1952, 1953, 1954 и др.). Годичный побег — это «побег, развивающийся из почки возобновления в течение одного вегетационного периода. Смежные годичные побеги резко отграничены друг от друга рубцами почечных чешуй, укороченными меж- доузлиями, недоразвитыми листьями и т.д. Они легко различимы в кроне дерева, кустарника, кустарничка, а иногда и многолетних травянистых растений» (Серебряков, 1952, с. 11). В эволюционном отношении годич- ный побег является органом вторичного происхождения, оформившимся в результате появления в наших широтах во второй половине третичного периода умеренного и холодного климата, главной особенностью которого является периодичность (Вульф, 1944; Голенкин, 1959 и др). Для характеристики побегов по особенностям проявления у них репродуктивной функции широко используется термин «монокар- пический побег». Термины «монокарпические» и «поликарпические» растения впервые ввел в науку О.-П.Декандоль (De Candolle, 1815) для разделения растений по кратности проявления репродуктивной функ- ции. В отношении побегов трав этот термин получил признание в ос- новном благодаря работам И.Г.Серебрякова (1952, 1953, 1959 и др.). Монокарпическими побегами И.Г.Серебряков (1959, с. 4) называл «по- беги, развивающиеся из почек возобновления и завершающие свой цикл развития в течение первого, второго, третьего или, реже, последующих лет образованием соцветия или одиночных цветков и отмирающие после плодоношения. Биологически они подобны монокарпическим расте- ниям. Но в отличие от последних, они берут свое начало не из семян, а из почек возобновления и отмирают после плодоношения не полно- стью, оставляя на своей базальной части новые органы возобновления, дающие начало дочерним побегам». Большое значение для диагностики побегов имеет признак, на который обратил внимание еще в прошлом веке Репер (Roeper, 1826). Он установил, что соцветия бывают двух типов: открытые, когда у них развиваются только пазушные цветки, и закрытые, когда их главная ось заканчивается цветком. Особенности расположения репродуктивных
Глава 5. Параметры основных органов и их частей в организации биоморф семенныхрастений 187 элементов на побегах привлекали внимание многих исследователей (Saint-Hilaire, 1841; Velenovsky, 1905-1913; Goebel, 1931; Федоров, Артю- шенко, 1979 и др.). Типологии соцветий посвящена капитальная моно- графия В. Тролля (Troll, 1964, 1969). Ирмиш (Irmisch, 1859) подразделял побеги клеверов по особенно- стям их перезимовывания: зимующие надземные с зелеными листьями, зимующие подземные с чешуевидными листьями; по направлению роста относительно поверхности почвы: горизонтальные, вертикальные; по длине побегов: длинные, укороченные. Варминг (Warming, 1884) широко использовал для характеристики побегов число сезонов вегетации, ког- да проявляется видимый рост побегов («циклов» роста и развития по- бегов), от раскрытия пазушных или адвентивных почек до цветения и плодоношения; он различал по этому ряду сравнимых признаков сле- дующие побеги: моно-, ди-, три-, полициклические. Моноциклические побеги завершают период своего видимого роста от почки до цветения и плодоношения за один вегетационный сезон, дициклические побеги соответственно — за два сезона и т.д. Эти представления о побегах широко использовались в работах И.Г.Серебрякова (1952, 1953, 1959, 1964 и др.), В.Н.Голубева (1960, 1965 и др.), И.В. Борисовой (1961), Т.И. Серебряковой (1965, 1968, 1971 и др.), И.Н.Берка (1973, 1976, 1982), М.Т.Мазуренко, А.П.Хохрякова (1977), И.Ф.Сацыперовой (1977) и др. Классическая работа Шпета (Spath, 1912) по летнему побегообразова- нию значительно углубила знания о параметрах побегов, в частности их зависимость от продолжительности покоя и сформированности почек (обзор этой монографии приведен в статье И.А.Грудзинской, 1960). Та- кие понятия, разработанные Шпетом, как ивановы побеги (они обра- зуются после прекращения роста весеннего побега из верхушечной поч- ки, после некоторого, обозначенного во времени и морфологически относительного покоя), силлептические побеги (они образуются из па- зушных почек растущего побега, т.е. без периода покоя), пролептичес- кие побеги (они развиваются в структуре годичного побега из пазушных почек, которые находились в состоянии покоя; если бы не было каких- либо внешних воздействий, то, как считает Шпет, пролептические по- беги образовались бы только на следующий год), — широко использу- ются и в настоящее время. Вторичный рост побегов привлек внимание многих исследователей (Шитт, Метлицкий, 1940; Серебряков, 1952; Соколов, Артюшенко, 1957; Грудзинская, 1959, 1960, 1961, 1962, 1964, 1965; Бюсген, 1961; Голубев, 1965; Клименко, 1969 и др.). И.А.Гру- дзинская, развивая идеи Шпета, пришла к заключению, что извест- ные определения годичного побега, в частности, определение, которое дано в книге И.Г.Серебрякова (1952), не отражают реального разнообразия побегов. И.А.Груздинская (1959, 1960) предложила на- зывать побегом древесных растений «новообразование типа стебля, возникающее из почки за один цикл роста (от почки до почки)» и считать его элементарной структурной единицей кроны (элементар- ным побегом). На основании изучения побегов злаков С.П.Смелов (1947) выделил три группы побегов: вегетативные (они не способны образовать ре-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ продуктивные элементы), генеративные (они образуют репродуктивные элементы), скрытогенеративные (они обладают недоразвитыми зачаточ- ными соцветиями, поэтому остаются вегетативными). И.Г.Сереряков разработал некоторые диагностические признаки побегов. Так, он (1952) различал у древесных растений побеги вегета- тивные (укороченные и удлиненные ростовые) и цветоносные, или плодущие (как правило, укороченные). Цветоносные побеги по степени специализации И.Г.Серебряков (1949) делил на три группы: слабо специализированные (облиственные), неполностью специализи- рованные (слабо облиственные, причем листья нередко редуцированы); крайне специализированные (они лишены зеленых листьев). И.Г.Сере- бряков (1952, 1953, 1959 и др.) считал монокарпический побег основ- HbIM структурным элементом поликарпических симподиально нараста- ющих травянистых растений. 3.Г.Беспалова (1965, с. 66), обобщая результаты своих многолетних исследований и литературные данные, пришла к выводу, что «низшей ... структурной единицей полукустар- ничка является монокарпический побег, он может быть моно-, ди-, или трициклическим (до 4-5)». В.Н.Голубев (1959а) иногда рассматривал побеги древесных растений как монокарпические в широком смысле, т.е. побеги, проходящие в своем развитии фазы боковой почки, вегетативного побега (розеточного или безрозеточного) и генеративного побега. Например, им отмечены ди-, по- лициклические монокарпические побеги у яблони, рябины и др. Такие признаки, как облиственность, длина междоузлий и общие размеры побегов, интересовали ботаников-морфологов издавна и ис- пользовались ими в различных классификациях побегов и растений (Irmisch, 1859; Warming, 1884, 1908; Areschoug, 1896; Серебряков, 1952, 1964; Голубев, 1965 и др.). Дод (Dode, 1905) предложил классификацию побегов, из которой некоторые термины широко используются по сей день: ауксибласты (длинные ростовые побеги), брахибласты (компакт- ные побеги). Варминг (Warming, 1918) предложил различать среди по- бегов трав следующие: удлиненные, розеточные и полурозеточные (цит. по Т.И. Серебряковой, 1971). На основании изучения характера облиствления и длины междоуз- лий побегов у травянистых растений В.Н.Голубев (1965, с. 18) различает два типа структуры надземных побегов: безрозеточный (долихоморф- ный) и розеточный (брахиморфный). Различия таких растений «в ко- нечном итоге сводятся к способности или неспособности развивать ро- зеточные побеги... При этом розеточный побег может представлять лишь фазу в развитии более крупной структурной единицы — монокарпи- ческого побега. В ряде случаев розеточный побег является самостоя- тельным структурным образованием. Так, В.Н.Голубев (1965) подразде- ляет розеточные побеги в широком смысле на розеточные побеги в узком смысле (побеги со сближенными узлами, несущими нормально развитые зеленые листья) и полурозеточные побеги (они состоят из розеточной и удлиненной облиственной генеративной части). При ха- рактеристике «моноподиальных полукустарничков» В.Н. Голубев (1968) выделяет моноподиальные вегетативные побеги (моноподиальные ске- летные оси), нарастающие верхушкой до 10-12 лет.
Глава 5. Параметры основных органов и их частей в организации биоморф семенныхрастений 189 Т.И. Серебрякова (1967а) предложила обозначать «розеточные» в широком смысле побеги (по В.Н.Голубеву) более удачным термином «розеткообразующие» побеги. Эти побеги всегда имеют в своем малом цикле фазу розетки, а в структуре — розеточный участок с укорочен- ными междоузлиями и ассимилирующими зелеными листьями. Цикл развития монокарпического побега от развертывания почки до отмира- ния А.П.Шенников (1941) назвал малым циклом. По длине междоузлий, особенностям облиствления узлов и направ- лению роста Т.И. Серебрякова (1967а, б, 1971) разделила побеги злаков на безрозеточные (они не имеют в малом цикле фазы розетки и в структуре — розеточный участок с укороченными междоузлиями и ас- симилирующими зелеными листьями) и розеточные, или розеткообра- зующие (эти побеги имеют в расположение листьев то, чего не имеют безрозеточные побеги). Согласно Т.И. Серебряковой (1971), безрозеточ- ные побеги бывают удлиненные ортотропные, с укороченным основани- ем, корневищно-безрозеточные, столонно-безрозеточные. Розеткообра- зующие побеги подразделены на собственно розеточные, полурозеточ- ные ортотропные, корневищно-розеточные и корневищно-полурозеточ- ные, столонно-розеточные и столонно-полурозеточные (ползучие). Н.Е.Булыгин (1965) среди цветоносных побегов древесных семен- ных растений выделил: генеративные побеги (их существование связано только с образованием плодов и семян, после чего они отмирают)— крайне специализированные, специализированные и генеративно-рос- товые (существование последних связано не только с образованием пло- дов и семян, но и органов ассимиляции и побегов возобновления; после плодоношения у них отмирает только их генеративная часть до верхней вегетативной почки) — неполностью специализированные, слабо спе- циализированные, неспециализированные побеги. А.П.Хохряков, М.Т.Мазуренко (1968) описали два типа побегов у растений семейства жимолостных: формирования и ветвления. Среди побегов ветвления выделено три типа: 1) с терминальным резко отгра- ниченным соцветием (калина, бузина); 2) с терминальным нерезко отграниченным от вегетативной части соцветием (большинство родов); 3) с пазушными редуцированными соцветиями (жимолость, вейчела). По длительности нарастания «побега формирования» «системы побегов формирования» кустарники были подразделены (на основе известной терминологии) на моноподиальные, моноподиально-симподиальные и симподиальные (Мазуренко, Хохряков, 1977). Предпринята попытка разработать широкое определение побега и охарактеризовать побеги по многим рядам сравнимых и взаимоисключа- ющих признаков (Нухимовский, 1969а, 1970, 1971, 1974а, б, 1980) и раз- работана концепция иерархической организации побегов по степени их сложности (Нухимовский, 1970, 1974а, 1976а, 1980, 1986, 1988). У семенных растений предложено различать кроме монокарпических еще и поликар- пические побеги (термин Т.Д.Михайловой, 1972). Необходимость вы- деления такого типа побегов была обоснована ранее: это открытые побеги с многократным плодотворением, т.е. с многократным образованием па- зушных репродуктивных элементов (Нухимовский, 1970, 1971).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Классификацию метамерных элементов систем растений с пазуш- ным положением почек (геммаксиллярных) предложила Л.Е.Гатцук (1974а), выделив 7 порядков соподчиненных метамерных структур — от «метамера s.str.» (междоузлие с узлом, прилежащим листом и пазушной почкой) до «первичного» и «парциального куста». Проведенный обзор затронул в основном работы, которые, в опре- деленной мере, разбирают вопросы экзоморфологической организации и цикличности роста и развития побегов, а также объясняют место побегов в структуре тела семенных растений. Можно с уверенностью утверждать, что существует значительный интерес ученых к проблеме классифицирования побегов, предприняты попытки широких обобще- ний в области морфофизиологической организации побегов. Л.Е.Гатцук (1995) охарактеризовала пять концепций, которые сло- жились в ботанической морфологии на пути развития представлений об элементарной макроморфологической единице тела растений: концеп- ция «железной триады» — корня, стебля и листа; концепция фитониз- ма Гаудишо и его последователей; концепция «модернизированной три- ады», отражающей единство листа и периферии стебля и основанной на компромиссе гипотез, под названием «теория перикаулома» (Potonie, 1902) и «теории листовой оболочки» (Saunders, 1922); концепция «мо- дернизированного фитонизма», когда побег разделен на четкие, тесно взаимосвязанные между собой фитоны, в которых каждый лист имеет свой листовой след в оси; пятая концепция — «побег как единое целое», впервые однозначно сформулированная А.Арбер (Arber, 1930, 1941). Ранее (Нухимовский, 1986) было подчеркнуто, что в истории применения термина «побег» можно обозначить три периода. С начала лрошлого столетия, когда стал применяться термин «побег», о чем сви- детельствуют работы Г.Бишофа (Bischoff, 1830, 1834, 1839), до 1930 г.— иервый иериод в изучении организации набегов (в качестве основных орга- нов семенных растений рассматривались стебли и листья, но не побег)— за единицу организации побеговой части семенных растений, удобную для практических исследований, принимали годичный побег. Второй лериод (с 1930 по 1970 гг.) открывается работами Арбер (Arber, 1930, 1941), провозгласившей единство стебля и листьев в организации побега как основного органа высших растений, а завершается публикациями Н.В.Пер- вухиной (1960, 1968, 1970), в которых подведен итог многолетней борьбы за признание побега основным органом высших растений. Бурные события в разработке параметров побегов произошли с начала 70-х годов нашего столетия. Понятия годичный и монокарпи- ческий побег не способны охватить все разнообразие побегов семенных растений, что стало ясно уже давно (Spath, 1912; Scharfetter, 1953; Грудзинская, 1960; Петров, 1963, 1967; Нухимовский, 1969а и др.). Под- спудно назревала своеобразная новая (послеарберовская) ревизия в поз- нании параметров побегов. Однако в работах этих лет не дано развер- нутого анализа организации побегов с системных позиций, и в частно- сти, их иерархичности. Поиск различных сторон организации побегов имеет обычно по- бочный характер и находится под давлением тех частных проблем,
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 191 которые решает исследователь. Полезные находки при таком подходе бывают, но они не носят закономерный характер. Мы предпочли раз- рабатывать проблему планомерно, поэтому сначала определили как ме- тодологические, так и методические позиции, которые могли быстрее всего дать полезный результат. Определим эти позиции: использование системного подхода; поиск главных параметров объектов в иерархич- ности организации, а дополнительных — во всем остальном. Главные параметры в организации побегов мы, полагаем, нашли с помощью метода основной широкой субординии, а дополнительные — методами вспомогательной субординии и сериофикации, но поиск в этом направ- лении продолжается. Здесь уместно еще раз упомянуть о работах болгарского ботаника Андрея Петрова (1963, 1967, Ре1гоч, 1964), который стал главной фигурой в своеобразной переходной ситуации в познании организации побегов. Он сумел ярче и последовательней других зажечь давно тлевшую искру недоверия к старинному ботаническому понятию годичный побег, который обычно воспринимался как основной структурный элемент кроны древесных растений. А. Петров хорошо сформулировал проблему: что считать основным структурным элементом кроны древесных растений (Петров, 1963), а с соответствующей интерпретацией и всей побеговой системы семенных растений (Петров, 1967). А Петров попы- тался отыскать структурный элемент, более независимый от сезонности климата, чем годичный побег. Все, что написал он об «оси» как теоретическом (нематериальном) структурном элементе кроны древес- ных растений — это предвестники к серьезным переменам в параметри- ческой оценке побегов, которые во многом стимулировали развертыва- ние третьего периода в познании организации этих органов семенных растений. Поскольку организация побегов сильно зависит от проявле- ния сезонных изменений и от бессезонности климата, что закреплено в процессе эволюции на генетическом уровне и отражается как в морфо- логических, так и в поведенческих признаках, то при построении теории организации побегов нет оснований не считаться с этим. Новый шаг — возникновение представлений о медиалях и лате- ралях — конкретных стеблевых осях, растущих один год или несколько лет до полного прекращения жизнедеятельности их алекса (Нухимов- ский, 1969б). «Расстояние» от этих простых стеблей до побега мини- мальное. Третий период — с 1970 г. по настоящее время — начинается с работ о побегах: специальных и универсальных (Нухимовский, 1970, 1971), а также моноподиальных (Гатцук, 1970). Уже осуществлена реви- зия многих признаков организации побегов с целью углубиться в ее сущность и конкретизировать понятие «побег». А.П.Хохряков (1974), пытаясь ответить на вопрос, где граница меж- ду отдельным побегом и системой побегов, предложил «моноподиаль- ный» или «универсальный» побег считать просто побегом, а составля- ющие его годичные или элементарные побеги — обозначать другим термином, точнее термином с другой основой, например, «прирост». А.П.Хохряков обосновывает свою позицию так: неправильно обозна-
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHblX РА СТЕНИЙ чать и часть и целое одним термином, Однако, на наш взгляд, чем шире понятие, тем больше однотипных явлений можно обозначить одним термином. При иерархической классификации однотипных явлений их вполне допустимо называть разными терминами, но с одинаковой ос- новой. ~акой подход позволяет лучше подчеркнуть иерархическую вза- имосвязь однотипных элементов и целого. Сходная ситуация наблюда- ется, например, при рассмотрении сувенира «русская матрешка», где все организационно сходные элементы — «матрешки» иерархически взаимо- связаны между собой, отличаясь друг от друга размерами. Их обычно называют «первая матрешка», «вторая матрешка», «третья матрешка» и т.д. В нашем случае развивается идея широкого понимания побега, поэтому при детализациях для обозначения побега как целого и его однотипных элементов используются термины с одной основой, напри- мер, «специальный» побег и «универсальный» побег (Нухимовский, 1970, 1971). Позднее автор разработал более детальную терминологию на одной основе «cormus» (побег). о чем будет сказано ниже. E.С.Смирнова(1974, 1978, 1980) построила графические модели не- скольких десятков морфологических типов тропических и субтропичес- ких растений, произрастающих в оранжереях Главного ботанического сада АН России (Москва), классифицировала и описала их. Табличный способ сохранения и передачи информации о морфотипах растений, предложенный E.С. Смирновой, представляется нам прогрессивным для развития теории и методики биоморфологии. Положительная сторона этих исследований — стремление найти структурный элемент, единый для всех побеговых систем цветковых растений. Был предложен термин «однопорядковый побег», который определен как побег одного по- рядка с одной верхушечной почкой (Смирнова, 1974). В использова- нии подобного термина есть один нюанс, с которым желательно счи- таться: важно различать понятия «нарастание» и «ветвление». При нарастании (увеличении линейных размеров осевой системы) обра- зуются «однопорядковые» побеги системы нарастания — простые (без перевершиниваний) побеги (универсальные простые побеги в нашем понимании). При ветвлении (образовании в результате нарастания осе- вой системы из одной ее части двух и более частей, имеющих обособ- ленные живые верхушки) иногда образуются «однопорядковые» побеги системы ветвления (побеговые ветви) или простые (без перевершини- ваний), или сложные (с монохазиальным перевершиниванием, напри- мер, у липы, ивы, вяза). Таким образом, однопорядковость структур- ных элементов в системах нарастания и ветвления побегов может совпадать или не совпадать. Так, например, при моноподиальном многолетнем нарастании единственного ствола какой-либо пальмы или при монохазиальном многолетнем нарастании (ежегодно сменя- ется алекс и заменяется одним новым апексом) стволика или отдель- ной ветви у липы (Tilia cordata) ветвления не происходит; в этих случаях образуется однопорядковый элемент системы ветвления. В третьем периоде познания организации побегов опубликовано немало других работ советских ботаников (Серебрякова, 1971, 1977, 1980, 1981, 1987; Голубев, 1973, 1977; Тихонов, 1977, 1979; Мазуренко,
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 193 Хохряков, 1977; Шафранова, 1980; Барыкина, Гуленкова, 1987; Берко, Козий; 1987; Савиных, 1987, Гатцук, 1994 и др.), в которых при изуче- нии организации побегов подчеркивается что-либо новое. Из зарубеж- ных исследований наиболее выделяются работы Ф.Алле с соавторами Halle, Oldeman, 1970; Halle, 1975; Halle, Oldeman, Tomlinson, 1978 и др.), которые разрабатывают концепцию «стратегий роста» тропических деревьев. В основе представлений о «стратегиях роста» лежит особая органообразовательная роль побеговых верхушечных меристем, которые создают многообразие побеговых систем. По особенностям нарастания побеговых систем Ф.Алле и Р.Ольдеман (Halle, Oldeman, 1970) выдели- ли 25 «стратегий роста» тропических деревьев, которые отразили в графических, или «архитектурных», моделях. Для построения моделей были использованы ряды признаков роста и развития побегов: наличие одной оси или многих, монокарпичность или поликарпичность, боко- вое или верхушечное положение цветков и соцветий, направление, характер (моноподиальность, симподиальность), ритмичность (наличие, отсутствие) нарастания. В целом графические модели, построенные Ф.Алле и др., отражают ряд важных особенностей организации биоморф тропических древесных растений, служат полезной фактической свод- кой по морфологии тропических растений, непосредственное общение с которыми в естественных условиях произрастания — счастливая слу- чайность для большинства «северных» ботаников. «Архитектурные мо- дели» тропических деревьев — отражение результатов определенной де- ятельности побеговых верхушечных меристем. Графическое выражение «стратегий роста» обозначается термином «архитектурная модель», до- вольно тяжеловесным, хотя бы потому, что эти модели построены на особенностях организации только побеговых систем; точнее говорить о графических моделях организации побегов или, допустим, «моделях побегообразования», к чему пришла Т.И. Серебрякова (1981) — одна из сторонниц взглядов Ф.Алле. В нашей стране Т.И. Серебрякова (1977, 1981, 1987 и др.) успешно использовала представления об «архитектурных моделях» для отражения особенностей побегообразова- ния травянистых растений. Обратим здесь внимание лишь на то, что признаки, выбранные французскими ботаниками для построения «архи- тектурных моделей» тропических деревьев, хотя и важные, но все же их мало и стоит прибавить несколько серий (рядов) новых признаков, как проявится много новых комбинаций признаков, а число моделей резко возрастет. Метод графического моделирования, использованный Алле и его последователями, — это достоинство, расширяющее возможности морфологической науки. Достоинство и то, что морфологическая ор- ганизация растений рассматривается в динамике. О недостатках, пожа- луй, нет повода говорить, поскольку охарактеоизована весьма экзоти- ческая для «северных» ботаников группа растений. Возможно, пока пройден первый большой этап. Следующим шагом могло бы быть проведение еще более полного анализа многих рядов признаков (эко- логических, морфологических и функциональных) тропических деревьев, используя, может быть, приемы многолинейного классифици- рования (Голубев, 1968, 1972; Нухимовский, 1971, 1980).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Итак, до настоящего времени осуществляется ревизия многих ха- рактеристик побегов, цель которой — углубленное познание сущности организации побеговой части семенных растений и конкретизация по- нятия «побег». Приведем нашу развернутую концепцию организации побегов. 5.1.2. Иерархичность в организации побегов и их структурных элементов Понятие «побег» (греч. «кормус») определено нами широко (гл. 2.5.5). Такое понимание побега учитывает разнообразие фактов и суждений, подтверждающих единство стеблевой и листовой частей в структуре и функционировании одного органа. Общая формула побегов предполагает классифицирование их на разной признаковой основе. Наибольшее значение в познании побегов мы придаем рассмотрению иерархичности их организации, которую правильнее и удобнее всего передавать посредством субординатных классификаций. Прежде чем рассмотреть субординии побегов, разграничим стадии их морфогенезов, поскольку это имеет значение при оценке общей сложности организации побегов. Жизненный цикл (морфогенез) иобега — жизнь побега от начала за- ложения родительской почки (зачаточного побега) до полного отмирания всех его частей. Основой жизненного цикла любого более или менее нормально развивающегося побега является его видимый рост, т.е. непрерывный рост побега в длину от начала развертывания почки до резкого замедления или прекращения роста. Отрезок времени, в течение которого побег проявляет видимый рост, называем фазой ви- димого роста. Известно немало попыток разграничивать стадии морфо- генезов побегов, в частности, И.Г.Серебряков (1952) отметил две глав- ные фазы — внутрипочечную («эмбриональную») и внепочечную. Раз- вивая предложенную ранее схему (Нухимовский, 1970, 1971, 1986), мы различаем в морфогенезе побегов три или четыре возрастных периода. Инициальный иериод начинается заложением первой клетки верхушечной меристемы и продолжается до начала развертывания семядолей или первых настоящих листьев. Постинициальный иериод продолжается от начала развертывания семядолей или родительской почки до заверше- ния свойственных фаз или фазы видимого роста и репродуктивного развития, когда оно есть. У побегов, в постинициальном периоде кото- рых проявляются две или более стадий (фаз) видимого роста, по осо- бенностям ритмического функционирования их апикальных меристем отмечаем следующие фазы: мультиактивную (апикальная часть побега проявляет повышенную активность, а побег в целом — видимый рост) и субактивную (апикальная часть побега функционирует замедленно, а побег в целом не проявляет видимый рост — обычен скрытый рост). Пререзидный иериод продолжается вслед за постинициальным периодом до отмирания эфемерных (незимующих) и сохранения персистентных
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 195 (неотмирающих после окончания сезона вегетации, зимующих) частей. Резидный, или остаточный, период сменяет пререз~щный период, а если его нет, то постинициальный период, и продолжается до полного отми- рания побега. У побегов, не имеющих пререзидного и резидного перио- дов, проявляется финальный период; он сменяет постинициальный пери- од и продолжается до отмирания эфемерных частей без сохранения персистентных частей. Резидный побег состоит из скелетного, т.е. неот- мирающего год и более, побегового остатка стеблевой части(резида), почек и листьев, если они сохраняются более одного вегетационного сезона (подробнее см.гл.6). Нами разработано несколько широких субординатных классификаций побегов семенных растений, которые сами в свою очередь будут иерархи- чески взаимосвязаны между собой, поскольку построены с разных высот обобщения, занимая разные ступени «пирамиды» абстракцивй, т.е. супер- ординаты в суперординатной классификации (рис. 56). Верхнюю суперор- динату в рассматриваемой классификации классификаций мы заняли двумя широкими субординиями. За критерий сложности организации побегов в первой широкой субординии нами принята фазность видимого роста. У семенных растений сезонного и бессезонного климатов побеги бывают монофазные и пульсирующие (рис. 56-А). Монофазные побеги про- являют одну фазу видимого роста (их каркас составлен частью оси или одной осью или несколькими взаимосвязанными осями). Пульсирующие побеги — это любые побеги, главная осевая часть которых проявляет две и более фаз видимого роста пока не утратит жизнеспособность ее алекс. Заметим, что пульсирующий рост побегов распространен не толь- ко у растений умеренной зоны, но широко встречается и у растений в тропиках, даже в тропическом дождевом лесу (Вальтер, 1968). У пульси- рующих побегов фазы видимого роста чередуются с более или менее продолжительными фазами скрытого роста (относительного покоя). Г.Вальтер (1968) отметил, что у древесных видов, регулярно сменяющих свою листву, наблюдается периодичность роста в тропическом дожде- вом лесу в 3-4, 9, 12 и даже 32 месяца; отсутствие периодичности роста и развития все же наблюдается, но в основном у молодых деревьев. Под иным углом, но с той же высоты обобщений мы рассматриваем вторую широкую субординию побегов. За критерий сложности организации в этой классификации побегов принята степень сложности их каркасов: а) про- стоосевые (моноаксонные) побеги (каркас побега составляет простая ось); б) сложноосевые (синаксонные) побеги (каркас побега составляет сложная осевая система). Еще одну широкую субординию побегов мы строим только для побегов сезонного климата. Эта субординия побегов займет вторую су- перординату в суперординатной классификации побегов сезонного кли- мата (рис. 56-Б), но будет отсутствовать в «пирамиде» абстракций (су- перординатной классификации) побегов бессезонного климата. За кри- терий сложности организации побегов сезонного климата в третьей ши- рокой субординии принята продолжительность постинициального периода их жизни. Нами предложено различать следующие тины побегов: вну- тригодичные (силлептические, пролептические, ивановы и какие-либо дру-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 56. Суперординатная модель («пирамида» абстракций) организации побегов семенных растений. А-Д — первая-шестая широкие субординатные классификации побегов, la — мо- нофазные, 2а — пульсирующие (ди-, три-, олиго-, плейофазные) побеги, la'— моноаксонные, 2а' — синаксонные, 16 — внутригодичные, 2б — годичные, Зб- сверхгодичные побеги, 1в — альфакормусы, 2в — бетакормусы, Зв — гам- макормусы, 4в — дельтакормусы, 1г — дерепродуктивные (стерильные), 2г — уни- репродуктивные, Зг — бирепродуктивные, 4г — паулирепродуктивные, 5г — му- льтирепродуктивные побеги, 01д — протерокормусы, 1д — монокормусы, 2д — ди- кормусы, Зд — трикормусы, 4д — тетракормусы, 5д — пентакормусы, бд — гекса- кормусы, 7д — гептакормусы, 8д — октокормусы, 9д — энатокормусы.
Глава 5. Параметры основных органов и их частей в организации биоморф семенныхрастений 197 гие побеги, составляющие структуру годичных побегов), годичные — это в согласии с большинством исследователей, любые побеги, образующиеся в течение одного вегетационного сезона из семени или из одной пере- зимовавшей почки со всеми возможными перевершиниваниями и раз- ветвлениями, и сверхгодичные (двухгодичные, трехгодичные, многогодич- ные, т.е. любые побеги,4олее сложные по продолжительности постини- циального периода, чем внутригодичные и годичные побеги). На суб- ординатах годичных и сверхгодичных побегов проявляется дополни- тельная иерархичность по ритмике (фазности) видимого роста. Годич- ные побеги бывают монофазные, дифазные и плейофазные, а сверхго- дичные побеги — дифазными и плейофазными (в скобках можно отме- чать число фаз). Когда годичные побеги монофазные, то их обозначение можно ограничивать только одним эпитетом: первым, поскольку боль- шинство годичных побегов — это монофазные побеги. При этом важ- но не забывать, что понятие монофазный побег, как уже было под- черкнуто в первой широкой субординии побегов, перекрывает неред- ко понятие годичный побег, например, в тропиках нередки монофаз- ные мультиактивные побеги с фазой видимого роста два и более лет. Когда годичные побеги имеют больше, чем одну, фаз видимого роста, то такие побеги важно обозначать двумя эпитетами, например, го- дичные дифазные, или годичные пентофазные побеги и т.д. Такое понимание термина годичный побег призвано упорядочить его место в системе побегов семенных растений. Для сверхгодичных побегов необходимо давать еще одну характеристику и по фазности видимого роста, если она не совпадает с характеристикой по продолжительнос- ти постинициального периода жизни этих побегов. Четвертую широкую субординию побегов семенных растений мы стро- им из четырех субординат: альфакормусы, бетакормусы, гаммакормусы и дельтакормусы (рис. 56-В). За критерий сложности организации побегов в этой классификации принята степень сложности осевых систем, оценива- емая в основном по признакам систем нарастания и ветвления. Альфакор- мус — простоосевой побег, развивающийся из родительской почки без перевершиниваний и разветвлений. Еетакормус развивается в своей главной осевой части как простая ось и лишь временно усложняется образованием эфемерных (полностью отмирающих) частей соцветия или проявляет полную эфемерность вместе с частями соцветия. Галимакормус развивается в своей главной осевой части как сложная (с перевершини- ваниями) однопорядковая ось и иногда усложняется образованием на ней каких-либо строго эфемерных частей (например, соцветий). Дель- такормус — двух- или многопорядковый побег в системе ветвления, образующий или не образующий соцветий и не тождественный другим типам побегов. Если посмотреть на одно многолетнее растение в раз- витии, то нередко в его структуре можно обнаружить побеги всех отме- ченных типов (Salix саргеа L., Ulmus laevis Pall., Polygonatum odoratum (Mill.) Druce). За критерий сложности организации побегов пятой широкой субор- динатной классификации (рис. 56-Г) принимаем особенности (кратность) репродуктивной цикличности. Термин «репродуктивный цикл», или
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhEX РАСТЕНИЙ «цикл репродуктивного развития», побега (или особи) мы понимаем уже, чем термин «жизненный цикл» (морфогенез) побега (или особи). Реиродуктивный цикл включает в себя процессы роста и развития побега (или особи) от момента его возникновения до завершения одного репро- дуктивного процесса, или от момента завершения одного репродуктив- ного процесса до завершения нового подобного процесса и т.д. Полный реиродуктивный цикл набега (или особи), хотя и близок к понятию жиз- ненный цикл побега (или особи), поскольку включает в себя все репро- дуктивные циклы, проявляющиеся в жизненном цикле побега (или осо- би), все же уже (в морфогенезе побега или особи нередко наблюдается еще пострепродуктивная стадия). В пятой широкой субординии побегов мы различаем следующие субординаты (классы): деренродуктивные (ве- гетитивные) нобеги, униренродуктивные (какие-либо побеги с одним ре- продуктивным циклом в их жизненном цикле), биренродуктивные (с двумя репродуктивными циклами), ниулиренродуктивные (с тремя-че- тырьмя репродуктивными циклами), мультиренродуктивные (с пятью и более репродуктивными циклами) побеги. При движении вниз по ступеням «пирамиды» абстракций все более и более увеличивается разнообразие побегов по сложности их организации (рис. 56-Д; 57-А). Для шестой (основной) субординии набегов были взяты иризнаки, отражающие изменение сложности организации и обусловленные наведением их верхушечных меристем в ироцессе роста и развития, т.е. нризнаки нарастания, ветвления и реиродуктивного разви- тия набегов. Весьма удобно строить краткую и легко воспринимаемую терминологию побегов с учетом отношения их к субординате опреде- ленного порядка, начиная с менее сложных — 1-го, 2-го, 3-го и т.д.: монокормус, дикормус, трикормус и т.д. (Нухимовский, 1976а, 1980). Такая терминология конкурирует с предложенной ранее (Нухимовс- кий, 1970, 1971, 1974а), поскольку во многом синонимична ей и рас- сматривается здесь как главный вариант. Термины, предложенные ра- нее, приводятся, где они есть, в скобках. Особый тип побегов — побеги нулевой субординаты — протерокормусы (от греч. протерос — более ранний); они, как и все побеги, обладают узловым строением осевой части, но имеют только по одному узлу. Протерокормус — побег, состо- ящий из семядольного узла, семядолей, зародышевой и пазушно-семя- дольных почек (он только однажды развивается в онтогенезе растения). Монокормус (сиециальный npocmou набег) — побег, стеблевая часть которого без перевершиниваний и разветвлений отрастает из родитель- ской почки (верхушечной, пазушной, придаточной) за одну фазу види- мого роста. Дикормус (сиециальный сложный no6ez) — побег, состоящий из монокормуса и отрастающих из него за ту же фазу видимого роста без перевершинивания или с перевершиниванием силлептических по- бегов соцветия или подобных им побегов, но лишенных по какой-либо причине репродуктивной функции (отсутствие физиологической зре- лости у особи, неблагоприятные погодные условия и т.п.). В структуре любого сложного побега всегда более одного порядка побегов, поэтому всегда можно выделять исходный побег этой системы и производные побеги, т.е. имеющие порядки 2, 3-й и последующие. Трикормус (комби-
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 199 Рис. 57. Графические модели нескольких широких субординий побе- гов, корней и организмов семенных растений. А — субординия побегов, Б — субординия корней, В — первая широкая суборди- ния организмов семенных растений, Г — вторая широкая субординия организмов семенных растений, О 1 а — протерокормусы, lа — монокормусы, 2а — дикормусы, За — трикормусы, 4а — тетракормусы, 5а — пентакормусы, ба — гексакормусы, 7а — гептакормусы, 8а — октокормусы, 9а — энатокормусы, 16 — моноризы, 26— диризы, Зб — триризы, 46 — тетраризы; О 1 в — ризопротерокормусы, lв — ризо- монокормусы, 2в — ризодикормусы, Зв — ризотрикормусы, 4в — ризотетракорму- сы, 5в — ризопентакормусы, бв — ризогексакормусы, 7в — ризогептакормусы, 8в — ризооктокормусы, 9в — ризоэнатокормусы, 10в — ризосинкормусы; lг — апо- базовые растения, 2r — базогенные растения без ризогенных побегов и каулоген- ных корней, Зг — то же с ризогенными побегами и каулогенными корнями, пв- предветвь, вт — ветвь, ом — упрощенные обобщенные графические модели по- бегов и корней, вв — вегетативная верхушка побега, оц — отдельный цветок, сц- соцветие, л — листья текущего года, гр — нижние границы резидных монокорму- сов, гм — нижние границы монокормусов текущего года, у — узлы, образовав- шиеся в прошлом году или позднее, с — семядоли, су — семядольный узел, ов- отмершие верхушки стеблевых или корневых осей, кш — корневая шейка, г- гипокотиль; побеговые части — эп — эфемерные, пп — преперсистентные (потен- циапьно дву-, многолетние), пр — персистентные, или дву-, многолетние (базо- вые органы); корневые части: ак — аскелетные, кл — проскелетные, ск — скелетные. нированно-сиециальный набег) — побег, состоящий из нескольких моно- кормусов и (или) дикормусов, отрастающий за несколько фаз видимого роста там, где выражена сезонность климата, из одной родительской почки за один вегетационный сезон. Тетракормус (универсальный прос- той набег) — побег, стеблевая часть которого отрастает без переверши- ниваний и разветвлений из родительской почки за несколько фаз види- мого роста и за два и более вегетационных сезона до прекращения жизнедеятельности или дихотомического раздвоения ее верхушечной
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ меристемы; иными словами, это простой (одноосный) побег, отраста- ющий за два и более вегетационных сезона. Пентакормус (универсальный сложный иобег) — побег, состоящий из тетракормуса и образующихся из него в текущем году вегетации силлептических побегов соцветия или каких-либо иных побегов, обеспечивающих всей системе такого побега его собственную структуру. При дальнейшем усложнении организации все остальные побеги будут, естественно, сложнее, чем специальные, комбинированно-спе- циальные и универсальные побеги (т.е., начиная от монокормусов до пентакормусов, включительно), поэтому оставшиеся побеги можно обо- значить одним общим термином «ультрауниверсальные» (следующие выше по сложности организации за универсальными) побеги. Любая побеговая ветвь, не образующая или образующая репродуктивные эле- менты, любой ствол с побеговыми ветвями, если они организовны слож- нее, чем какой-либо пентакормус (универсальный сложный побег) и т.п. — это все ультриуниверсальные no6ezu. Следующий в шестой субор- динатной классификации побег (менее сложный среди ультраунивер- сальных побегов) — гексакормус (комбинированно-универсальный no6ez). Этот побег отрастает сначала как монокормус или иначе, но не сложнее пентакормуса, а на следующий год образует уже свою структуру за счет развертывания новых побегов от монокормусов до трикормусов. Часто гексакормус формируется из монокормуса или тетракормуса, развивая в одном вегетационном периоде почки, а на следующий год реализуя их (все или часть) в новые побеги с репродуктивной функцией (Salix саргеа L., Amygdalus communis L.). Прошлогоднй пентакормус (точнее, все то, что от него остается к началу следующего вегетационного сезона) может принимать участие в образовании гексакормуса. Геитакормус (иротоголокормус) — побег, проходящий в своем развитии стадию предветви и непосредственно из нее развивающий любые побеги от монокормусов до гексакормусов, оставаясь не тождественным этим и другим типам побегов. Предветвь (ирокладий) — это ось (простая или сложная) нулевого порядка в систе- ме ветвления, т.е. ось, из которой образуются первые ветви. Октокормус (кладокормус) — побег, проходящий в своем развитии стадию ветви, далее разветвленной или неразветвленной и не тождественной другим типам побегов. Энатокормус (голокормус) — побег, прошедший в своем развитии стадию гептакормуса и образовавший в своей структуре еще и октокормусы (один или несколько). Остальные возможные широкие субординатные классификации побегов рассматриваем как вспомога- тельные. Все побеги растения — системы, из них менее сложные систе- мы — протерокормус и монокормусы. Инициальный протерокормус- это побеговая часть семени, а инициальное состояние в жизни всех остальных побегов растения: медиального, латеральных (пазушных и придаточных) и терминальных — почка. Любой из остальных типов побегов — система побегов предшествующих субординат. Кроме 10 ти- пов побегов в структуре растения нередко формируется синкормус- это не побег, а система побегов одной особи, взаимосвязанных друг с
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений другом через органы иного морфофизиологического типа (корни), на- пример, Populus tremula L. Синкормус, возникающий из структуры одного какого-либо корня, который меньше, чем вся корневая систе- ма — это а-синкормус, а синкормус, взаимосвязанный со всей корневой системой особи, — это р-синкормус. Если посмотреть еще раз на основ- ную иерархическую классификацию (шестую широкую субординию) по- бегов семенных растений, то обращаешь внимание на ее универсальную направленность: она «стремится работать» не только в отношении рас- тений сезонного климата, но и в отношении растений бессезонного климата. Эта классификация построена в основном на оригинальном разно- образном фактическом материале; в ней учтены и многие литературные источники, поэтому рассматриваемая классификация, помимо примени- мости к растениям сезонного климата и тропическим растениям, произ- растающим в оранжереях, способна дать хотя бы прогнозный ответ (при отсутствии конкретных объектов из естественных условий произраста- ния) на вопрос: какие побеги могут быть, а какие не могут быть у растений бессезонного климата. Иными словами, шестая широкая су- бординия побегов прогнозирует организацию побегов растений бессе- зонного климата. Если побеги нулевой, l-ой, 2-ой, 7-ой, 8-ой и 9-ой субординат — это, по определению, побеги семенных растений сезон- ного и бессезонного климатов, то трикормусы, тетракормусы, пентакор- мусы и гексакормусы — побеги, организационно строго связанные с условиями только сезонного климата, где сезон вегетации чередуется с сезоном, в котором вегетация растений невозможна или менее прием- лема из-за неблагоприятного климатического режима. Учитывая факту- альную привязку шестой широкой субординии побегов к растениям сезонного климата, мы рассматриваем ее в отношении растений бессе- зонного климата как прогнозную классификацию, полагаю, что впос- ледствии она могла бы приобрести статус универсальной классифика- ции рассматриваемого профиля (рис. 57-А). Тропические растения в меньшей степени испытали на себе жес- ткую силу естественного отбора. Бессезонно-тропические растения по сложности организации побегов беднее растений сезонного климата, что, как бы между прочим, подчеркивает шестая широкая субординия побегов, указывая на строгую привязку одних типов побегов к сезонно- му климату и отсутствие таковой у других. Побеги, отмеченные в широких субординиях, могут быть дополни- тельно классифицированы по субординатному принципу. Так, узкой субординией побегов будет классификация, в которой за критерий слож- ности организации побегов принимается факт наличия или отсутствия стадии относительного покоя при образовании постиницильных побегов и при наличии такой стадии — ее продолжительность (рис. 58). Нижнюю субординату в нашей классификации занимают силлеитические побеги (КраЖ, 1912) — эти побеги трогаются в рост из пазушных почек одно- временно с продолжающимся ростом главной оси всего годичного побе- га, например, побеги «обогащения» в структуре соцветия, вегетативные
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 58. Отражение в структуре побегов наличия или отсутствия относительного покоя верхушечных меристем перед началом их видимого роста. А — узкая субординия побегов по рассматриваемому ряду признаков, Б — схема побегов семенных растений, 1-5 — побеги: 1 — силлептические, 2 — ивановы, 3— пролептические, 4 — весенние, 5 — провентивные, или запасные (пазушные или адвентивные), 6 — нижняя граница побега прошлого года, 7 — нижняя граница побега текущего года, 8 — корневой отпрыск, 9 — семядольный узел, 10 — корни.
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений побеги у молодых растений Betula pendula Roth, Ainus incana (L.) Moench). Более сложными являются побеги второй субординаты: ива- новы набеги (термин Шпета), — они образуются из верхушечных или пазушных почек после сравнительно непродолжительного (иногда до 40 дней) летнего относительного покоя этих почек, например, Quercus robur L. Нередко в структуре годичного побега образуются несколько ивановых побегов в результате чередований стадий усиленного видимо- го роста и стадий непродолжительного относительного покоя верхушки всего годичного побега (Серебряков, 1952). Число ивановых побегов обычно соответствует стадиям июльского, августовского и сентябрьского побегооразования, когда они проявляются в структуре годичного побе- га. Ивановы и родительский весенний побеги морфологически более или менее разграничиваются друг от друга в структуре годичного побега зоной сближенных узлов, чешуевидными или иными недоразвитыми листьями и т.п. Третью субординату рассматриваемой классификации занимают иролеитические набеги — это такие побеги, согласно Шпету и его последователям, которые развиваются как элементы годичного по- бега из уже сформированных пазушных почек, находившихся довольно долго в состоянии скрытого роста — относительного покоя, иными словами, — это побеги преждевременного роста, т.е., подготовившись к зиме, они начинают расти поздно летом или осенью, например, Bupleurum multinerve DC. на высокогорном Алтае (см. гл. 8.1). Четвертая субордината принадлежит весенним (вернальным) набегам (эти побеги развиваются из однократно зимовавших почек), а пятая субордината — ировентивным набегам (эти побеги развиваются из дваж- ды или многократно зимовавших пазушных или придаточных почек). В условиях бессезонного климата большое значение имеют силлептичес- кие побеги; об ивановых, пролептических и весенних побегах в отно- шении растений бессезонного климата, если и можно говорить, то толь- ко о модификациях этих типов побегов которые, безусловно присут- ствуют в организации некоторых, особенно древесных, тропических рас- тений. Как мы уже отмечали выше, многие древесные растения в бессе- зонном климате проявляют разновременную ритмичность роста и развития (Вальтер, 1968; Halle, Oldeman, 1970). Для растений бессезон- ного климата набеги, аналогичные ивановым, пролептическим и весенним, можно обозначить как иостлатентные, т.е. побеги, растущие мультиак- тивно (с видимым ростом) после завершения относительного покоя. Для растений любого климата термин «постлатентные» побеги так- же подходит в самом общем смысле (с активным видимым ростом после относительного покоя), что противопоставляет их побегам с малоактив- ным видимым или скрытым ростом (см. гл. 8.2.2.). В морфологии важное значение придают метамерии, наблюдается тенденция к расширению этого понятия (Гатцук, 1974а; Шафранова, 1980). Развивая разработанные ранее положения о метамерии (Нухимов- ский, 1980, 1981), мы понимаем метамерию как общеприродное явление- это последовательная повторяемость структурно и (или) функционально сходных частей (метамеров) с единой преднаследственной или (и) наслед- ственной основой какого-либо более общего явления Природы. Согласно
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ этому определению, метамеры в организации биоморф семенных рас- тений существуют как определенные повторяющиеся части одного и того же или органа, или организма, или синорганизма. Так, метамеры осевой системы у семенных растений — простые оси (простой стебель и простой корень). Метамеры системы ветвления у этих же растений- оси системы ветвления (предветвь и ветви). Метамеры системы куще- ния — побеги кущения, простые кусты. В структуре побегов семенных растений простейшими метамерными элементами, являются некоторые тины гемикормусов, или иолуиобегов (Нухимовский, 1980, 1986). Для правильного понимания организации побегов важно точно обозначить параметры вычленяемых нами элементов в системе тела растений. В структуре побегов обычно хорошо различимы гемикормусы. Один гемикормус (иолуиобег) — это любая система, состоящая из узла с листом (листьями), почки (почек, если они образуются) и располо- женного выше междоузлия или надузлия, если они образуются или (и) подузлия (если оно образуется) или из всей простоосевой вегетативной части побега, но не тождественной всему побегу, и, наконец, это может быть система верхушечного цветка без расположенных ниже вегетатив- ных частей побега (рис. 59). Таким образом, гемикормусы бывают веге- тативные (семь тинов) и реиродуктивные (верхушечные цветки как ча- сти иобега). Прежде чем разобраться в иерархии вегетативных гемикормусов, составляющих организацию побегов, обратим внимание на следующие морфологические факты. Латеральные побеги могут начинаться не с узла и не с междоузлия, например, паракладии, а с подузловой стебле- вой части разной длины, предшествующей первому узлу этих побе- гов, — это иодузлие (subnodium), или, в понимании Тролля (Troll, 1937, 1964), гииоиодий — «Anfangsinternodium». Иногда в зоне соцветия выше последнего узла с листом развивается верхушечная простая (без пере- вершиниваний) стеблевая часть без листьев, но с несколькими цветками (на цветоножках или без них) — это не междоузлие, а надузлие (suprano- dium). Участок стебля (или другого органа), где выходит наружу почка, называем узелком (nodulus). Разберемся подробнее в организации вегетативных гемикормусов, которые иерархически соподчинены между собой по степени организа- ционной сложности, что отражается в соответствующей широкой субор- динатной классификации гемикормусов и ее модели (рис. 59- А). IIep- вый иолуиобег (моногемикормус) — элемент побега, состоящий из узла и листа (листьев), почек (если они образуются), но без междоузлия, и иных стеблевых частей, поскольку таковые не развиты. Второй noëóno- бег (дигемикормус) — побеговый элемент, который состоит из узла, лис- та (листьев, если они связаны с одним узлом), почек, если они обра- зуются (что бывает очень редко) и подузлия без расположенного выше междоузлия, поскольку таковое не развито. Следующую субординату составляют третьи иолунобеги (тригемикормусы) — побеговые элемен- ты, состоящие из узла, листа (листьев), почек, если они образуются, и расположенного выше междоузлия без других свободных стеблевых частей, поскольку они не развиты. Вторые полупобеги организованы
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 205 Рис. 59. Типы вегетативных гемикормусов (А) и ризогемикормусов (Б) и графические модели их субординатных классификаций. 1 — эпикотиль, 2 — семядоля, 3 — узел, 4 — междоузлие, 5 — почка, 6 — листья, 7 — подузлие, 8 — надузлие, 9 — верхушечный цветок (репродуктивный гемикормус), 10 — боковые внепазушные репродуктивные оси (они не входят в структуру гемикормуса), 11 — корни, 12 — корневая шейка, 1-VII — ветативные гемикормусы разных типов: 1 — первого, II -второго, Ш — третьего, IV — четвер- того, V — пятого, VI — шестого, VII — седьмого, Ia — VIIa — ризогемикормусы разных типов: Ia — первого, Иа — второго, Ша — третьего, IVa — четвертого, Va— пятого, VIa — шестого, VIIa — седьмого.
ОСНОВЫ БИСМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ проще, чем третьи, поскольку почки на подузлии чаще всего не разви- ваются, а на междоузлиях развитие почек — обычное явление. Четвер- тую субординату занимают тетрагемикормусы (четвертые nonyno6ezu)— побеговые элементы, состоящие из узла, листа (листьев), почек, если они образуются, и надузлия, поскольку других свободных стеблевых частей в области узла нет. У надузлия нет листовых узлов, поскольку нет листьев, развитых на нем, но из него развиваются силлептические веточ- ки, принадлежащие, как правило, соцветию. Пентагемикормус (штый иолупобег) состоит из узла, листа (листьев), почек, если они образуются, подузлия и расположенного выше междоузлия. Шестые полуиобеги (гек- сагемикормусы) — это побеговые элементы (но не целые побеги!), каж- дый из которых состоит из узла, листа (листьев), почек, если они обра- зуются, подузлия и надузлия. Верхняя субордината принадлежит в на- шей классификации седьмым иолуиобегам (гентагемикормусам), т.е. си- стемам вегетативных побеговых элементов, состоящих из каких-либо гемикормусов одного побега и не тождественных им и целым побегам. Верхушечный цветок, по определению седьмого гемикормуса, не входит в его структуру, а является самостоятельным восьмым, или реиродуктив- ным, нолуиобегом — антогемикормусом. Гемикормусы вместе с корня- ми, которые они образуют, могут быть названы ризогемикормусами (кор- ненолупобегами) (рис. 59-Б). Первые, третьи и седьмые полупобеги часто являются метамерами, а все остальные гемикормусы — это аметамер- ные элементы побегов. Морфологические границы гемикормусов более или менее условны в структуре побегов. Они устанавливаются не сразу, поскольку в своей инициальной (неразвернувшейся, внутрипочечной) стадии жизни обычно проходят стадии меньшего ранга. Рассмотрим стадии инициальной жизни метамерных гемикормусов (1 и 3): долистовая (возникает апикальная кормусовая меристема, а лис- товых зачатков еще нет), листоузловая (начинается формирование ли- стового зачатка и узла, но расположенного выше междоузлия еще нет), междоузлиевая (наблюдается только у тригемикормусов, начинается формированием вышерасположенного междоузлия) стадии. Лист и сте- блевая часть (зачаточный узел) в кормусовом анексе возникают одновре- менно, а иазушные ночки развиваются в узле и выше его на междоузлии, что иозволяет нам иредложить новую трактовку основных метамерных элементов иобегов — это моногемикормусы или тригемикормусы. Обычно под метамером побега понимают узел с листом и расположенным ниже междоузлием с почкой (почками) на нем (Weatherwax, 1923; Серебря- кова, 1971) или в пазухе листа (Гатцук, 1974а), что по сути дела осно- вано на признании главенствующей роли листа в метамеризации побега в процессе функционирования его алекса. Н.Г.Рытова (1982) в структу- ре метамера («двуосного метамера») у побега злаков выделяет лист с узлом, ниже расположенным междоузлием и с почкой («воспроизводя- щейся ветвью») на нем. На наш взгляд, главенствующая роль в образо- вании метамеров побега принадлежит только его алексу, в процессе фун- кционирования которого и стебель, и лист образуются одновременно. Всегда ли в структуре побега проявляется правильное сочетание узлов и междоузлий? Наблюдения показывают, что далеко не всегда.
Глава 5. Параметры основных органов и их настей в организации биоморф семенных растений 207 Междоузлия (участки стебля, расположенные между двумя узлами) имеют разные линейные размеры по длине стебля (по вертикали) и по ширине стебля (по горизонтали) (рис. 60-63). Когда размеры по верти- кали междоузлий вполне определенные, то они правильные (регулярные) междоузлия. В одних случаях могут отсутствовать размеры по вертикали участков стебля, свободных от узлов, но размеры по горизонтали могут быть; границы междоузлий в таких случаях будут «размытыми» (неопре- деленными), что позволяет нам обозначить эти междоузлия как непра- вильные (иррегулярные) (рис. 60). В некоторых случаях междоузлия могут полностью отсутствовать, а узлы плотно прижаты друг к другу, т.е. между узлами отсутствуют свободные зоны стебля, именуемые междо- 15 15 16 - -17 Рис. 60. Сочетание узлов и междоузлий в экзоморфологической организации побегов. 1 — узел, 2 — узелок, 3 — почка, 4 — листья, 5 — рубцы от опавших листьев, 6— правильное междоузлие, 7 — неправильные междоузлия, 8 — мультинодий, 9— зона полного отсутствия междоузлий, 10 — преконодий, 11 — конодий, 12 — эпи- котиль, 13 — растение без эпикотиля, 14 — растение с эпикотилем, 15 — основа- ние побега (верхняя часть удалена), 16 — оно же в развернутом виде на плоскости, 17 — нижняя граница годичного побега.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 61. Безрозеточный тип развития. Первый год жизни особей Cataranthus rosea L. в условиях теплицы (Москва, ВИЛАР). Одно деление масштаба — 1 см. Обозначения те же, что и на рис. 60. Рис. 62. Розеточный виргинильный (I) и полурозеточный вегетативно- репродуктивный (II) типы развития у Calendula officinalis L. (Москов- ская область, производственная плантация ВИЛАР). Одно деление масштаба — 1 см. Обозначения те же, что и на рис. 60.
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 209 Рис. 63. Растяжение междоузлий на первом году роста и развития у Fatsia japonica (Thunb.) Decaisne et Planch. (ВИЛАР, теплица). Одно деление масштаба — 1 см. узлиями, и такой участок стебля состоит только из узлов, например, у розеточных побегов Ginkgo biloba L. На отсутствие междоузлий, в осо- бенности эпикотиля, обращалось внимание неоднократно (Пиддо- ти, 1967; Иосебидзе, 1981; Ворошилов, 1981 и др.). При этом отметим, что междоузлия могут отсутствовать на протяжении всей жизни от- дельного участка или всего побега, или с увеличением его возраста проявлять себя (несколько удлиняются соответствующие участки стеб- ля) в процессе роста стебля (рис. 63). Зону сильно сближенных узлов побега, кроме самого первого (медиального), удобно обозначать муль- тинодием. Скелетная компактная сближенноузловая безлистная или облиственная нижняя часть медиального побега — конодий, а аскелет- ная (однолетняя) или потенциально скелетная нижняя часть также ме- диального побега — «реконодий (Нухимовский, 1976а, 1988). Побеги, осевые системы и системы ветвления иногда полезно подраз- делять по их роли в формировании скелетной (зимующей, многолетней) осевой основы растений (широкая субординатная классификация) — на лидерные, если их роль наибольшая, ауксильные (вспомогательные), если их роль умеренная, факультативные (второстепенные), если их роль незначительная, и аскелетные (эфемерные), если они не участвуют в создании скелетной осевой основы (рис. 64). Методом сериофикации побеги (кормусы) по их положению в прос- транстве мы подразделяем на гомотопные: надземные (аэрокормусы), под- земные (геокормусы), водные (гидрокормусы) и гетеротопные побеги, или гетерокормусы (Нухимовский, 1970, 1971). Термины «аэрокормус» и
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 64. Роль побегов в формировании скелетного каркаса растений (схема). 1 — эфемерные части, 2 — факультативные части, 3 — ауксильные чсти, 4— лидерные части. «геокормус» впервые предложил Дю Рие (Du Rietz, 1931) для обозначе- ния соответственно любых надземных и подземных стеблей. Эти побеги в свою очередь проявляют в отношениях между собой иерархичность, которая фиксируется в соответствующей широкой субординии побегов, критерий которой — степень участия в пространственном сложении габитуса растений. Аэрокормусы, гетерокормусы и геокормусы, начиная с наи олее сложных по рассматриваемому ряду признаков и кончая наи- менее сложными, бывают генеральные, субгенеральные и локальные (рис. ). Генеральные набеги — это побеги надземные (аэрокормусы) или полунадземные (гетерокормусы), включающие в структуру первич-
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растени й 211 Г2 l ««« 1 1 1 1 Рис. 65. Модели широкой субординии побегов по положению их в пространстве. Гетерокормусы (полунадземные побеги): Г1 — генеральный и Г2 — субгенераль- ный; аэрокормусы (надземные побеги): а1 — генеральный, а2 — субгенеральный, аЗ —; геокормусы (подземные побеги): г1 — генеральные, г2- субгенеральные, гЗ — локальные. ную стеблевую предветвь, неразветвленные или разветвленные и восхо- дящие в виде одной оси от основания куста или основания главного корня, или подземные (геокормусы), включающие подземную часть
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ первичной стеблевой предветви, возникающую непосредственно из се- мени. Субгенеральные набеги — это побеги надземные (аэрокормусы) или полунадземные (гетерокормусы), или подземные (геокормусы), структурно подобные главному побегу, но возникающие у растений вегетативного происхождения или отрастающие как побеги кущения от основания главного побега у растений семенного происхождения, или на органах иного морфофизиологического типа (корнях). Все другие побеги, которые выступают как элементы генеральных и субгенераль- ных, обозначаем как локальные. Стебель. В связи с широкой трактовкой термина «побег» предло- жено шире, чем обычно, понимать и термин «стебель» (см. гл. 4.4). Главная особенность: стебель несет или нес на себе другие части побега (листья) и нередко репродуктивные элементы, с которыми находится (или находился) в единстве в системе побега. Простая стеблевая ось 1-го иорядка — медиаль, а иростая стеблевая ось — 2-го, 3-го и наследующих иорядков, или адвентивная, — латераль. В соответствии с тем, частью каких побегов является стебель, говорим о стебле монокормуса (моно- каулисе), стебле дикормуса (дикаулисе), стебле трикормуса (трикаулисе) и т.д. По сути, в этом аспекте мы отчасти вернулись к подходу К.Лин- нея (Linnaeus, 1751), когда он различал простые, ветвистые и дихотоми- ческие стебли. Стебель, будучи неотъемлемой частью побега, составляет опору для других органов (листьев, репродуктивных элементов), поэто- му особенности нарастания и ветвления побега — в полной мере осо- бенности нарастания и ветвления его единой осевой части — стебля. В этой связи многие широкие субординии побегов — это с соответству- ющей интерпретацией и широкие субординии стеблей. Метод субординатного классифицирования можно использовать и для выяснения иерархичности организации листьев. Наличие или от- сутствие ветвления и степень ветвления листьев, а также особенности сочленения листочков в разветвленных листьях — наиболее важные признаки, которые обычно используют при классифицировании листь- ев (Федоров и др., 1956). Широкие субординии листьев мы рассмотрим с двух высот обобще- ния (суперординат). Самую общую субординию листьев можно пред- ставить из трех субординат: иростые, иолусложные и сложные листья (Нухимовский, 1980, 1986; рис. 66-А). Простые листья — это однопла- стинковые листья цельные или более-менее расчлененные, не образ- ующие в своей структуре обособленных пластинок листочков. В отли- чие от простых листьев полусложные и сложные листья — всегда раз- ветвленные с четко обособленными частными листовыми пластинками (листочками), иногда обладающими даже своими черешочками. Полу- сложные листья в отличие от сложных листьев не распадаются на части при листопаде, поскольку не имеют специализированных разделитель- ных сочленений, а сложные листья распадаются на части, поскольку имеют специализированные разделительные сочленения. Тип полуслож- ных листьев предлагается выделять впервые. Обычно к простым листь- ям относятся все листья, даже разветвленные без специализированных разделительных сочленений, или в тех случаях, когда сегменты рассе-
Глава 5. Параметры основных органов и ик частей в организации биоморф семенных растений Рис. 66. Суперординатная модель организации листьев. А — первая широкая субординия листьев: ф1 — простые, а2 — полусложные, аЗ— сложные; Б — вторая широкая субординия листьев: 61 — монофиллы, 62 — ди- филлы, 63 — трифиллы, 64 — полифиллы (тетрафиллы, пентафиллы...). ченных листьев соединены оторочкой, хотя бы и очень узкой или даже очень фрагментарной (Хомякова, 1974). Наличие или отсутствие ото- рочки иногда установить почти невозможно, поэтому мы не придаем этому признаку решающего значения. Не простыми, а полусложными листьями в нашем представлении будут те листья, на рахисе которых четко выражены обособленные частные листовые пластинки (листочки) и которые не имеют специализированных разделительных сочленений. К полусложным листьям мы относим, например, «дваждытройчаторас- сеченные» листья воронца колосистого, лютика ползучего, которые обычно считают простыми вырезными рассеченными листьями (Хомя- кова, 1974; рис. 67).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 67. Примеры полусложных листьев (эти листья обычно считают «простыми»). 1 — воронец колосистый, 2 — лютик ползучий, 3 — гравилат речной (прикорневой лист), 4 — схема многократноперистого листа (по И.М.Хомяковой, 1974). Если посмотреть на организацию листьев несколько под иным углом обобщений, то широкую субординию листьев можно представить так (она будет принадлежать второй суперординате на «пирамиде» аб- стракций организации листьев, рис. бб-Б): лист 1-ой субординаты- монофилл (простой лист, т.е. лист, листовая пластинка которого не расчленена на рахис и листочки), лист 2-ой субординаты — дифилл (лист, состоящий из 2-х порядков частных листовых пластин и (или) рахисов), лист 3-ей субординаты — трифилл (лист, состоящий из 3-х порядков частных листовых пластин и (или) рахисов), лист 4-ой суб- ординаты — иолифилл (лист, состоящий из 4-х и более частных листо- вых пластин и (или) рахисов; если необходимо, то при очень сильной разветвленности листьев можно выделять и более дробные типы листь- ев: тетрафиллы, пентафиллы, гексафиллы и т.п.). Примеры: почковид- ный монофилл (пальчаторассеченный лист) лютика кашубского, трой- чатый монофилл (тройчаторассеченный лист) гравилата речного, лиро-
Глава 5. Параметры основных органов и ик частей в организации биоморф семенных растений 215 видный прерывисто-закрыто-перистый полусложный дифилл (лировид- ный прерывистонепарноперисторассеченный лист) гравилата речного (розеточный лист), тройчатый полусложный трифилл (простой дважды тройчаторассеченный лист) воронца колосистого, лютика ползучего и т.д. (в скобках приводится традиционная характеристика листьев). Термин «inflorescentia» (соцветие) впервые использовал Линней (Linnaeus, 1751). Мы разделяем господствующую точку зрения (Беке- тов, 1853; Каден, 1951; Troll, 1964, 1969; Федеров и др., 1979; Кузнецова, 1985), что соцветие — это орган с односезонным видимым (внеиочечным) ростом, образующий иосле цветения илоды (илодоносящее соцветие). У некоторых видов соцветие в фазе плодоношения сохраняется более года (это не влияет на суть рассматриваемого понятия) и после утраты репродуктивных элементов прекращает свое существование, отмирая в стеблевой части полностью или не полностью, а из сохранившихся почек могут образовываться побеги только с новыми соцветиями. Та- ким образом, понятие «соцветие» действенно только тогда, когда есть определяющие его компоненты — репродуктивные органы (цветки или плоды) — и видимый рост и развитие этих органов наблюдаются толь- ко в текущем году, а плоды лишь иногда сохраняются несколько дольше (Нухимовский, 1980, 1986). В соответствии с рассмотренной выше трактовкой побега термин «соцветие» можно определить следующим образом. Соцветие — это реиродуктивный орган с одной фазой видимого роста, состоящий из двух и более цветков (илодов), листьев (иногда их нет) и несущих их стеблевых частеи, ограниченный снизу и сверху по главной оси этого органа силлеи- тическими иобегами с реиродуктивной функцией одинаковых или близких цветком (закрытое соцветие), его рудиментом или иной недоразвитой эфемернорастущей и замирающей верхушкой (иолузакрытое или ириот- крытое соцветие) (рис. 68-76). Система из всех разобщенных (дизъюн- ктивных) одиночных цветков или соцветий одного побега или всего организма (дизъюнктивный репродуктивный орган), не тождественная какому-либо соцветию, — это сверхсоцветие, или суиерфлоресценция (Нухимовский, 1986). Широкое понятие соцветие позволяет классифи- цировать соцветия на разной признаковой основе. Важно определить, на основе какого побегообразования формируются соцветия, и уточнить зоны специализации в организации цветоносного побега (рис. 68). Инновация (лат. innovBtlo — возобновление) — явление отрастания весенних, запасных и иных постлатентных побегов после периода дли- тельного (зимнего, двухлетнего или многолетнего) покоя из соответ- ствующих почек любой части родительского побега. Нередко иннова- ция побегов сконцентрирована в отдельной зоне (зоне инновации), рас- положенной непосредственно под зоной торможения (релаксации), а та в свою очередь располагается под соцветием (переходные функции вы- полняет между ними основное междоузлие, Troll, 1964). Зона торможе- ния иногда делится на иодзоны неиолного (субфлоральная иодзона) и иол- ного торможения. В подзоне неполного торможения образуются очень слабые побеги, как правило, без репродуктивной функции.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 68. Обобщенная модель некоторых типов соцветий. 1 — верхушечное соцветие, 2 — межсоцветие, 3 — синфлоресценция (сборное соцветие), 4 — кофлоресценция (присоцветие); побеги: 5 — эусиллептические, 6— инфантильные, 7 — индуктивные, 8 — интеркалярные, 9 — паракладии; 10 — та- втокладии; зона: 11 — возобновления, 12 — полного торможения, 13 — неполного торможения, 14 — индукции. Рис. 69. Формирование верхушечного закрытого соцветия в онтогении Тагахасит ceratophorum (Ledeb.) DC. (Алтай, Северо- Чуйский хребет, высокогорье). Одно деление масштаба — 1 см. Обозначения те же, что и на рис. 68.
Глава 5. Параметры основных органов и их частей в организации биоморф семенныхрастений 217 Рис. 70. Формирование сборного верхушечного полузакрытого соцве- тия в онтогении у I — Choriospora sibirica (L.) DC. (Алтай, низко- горье), II — Alliaria petiolata (Bieb.) Cavara et Grande (Московская область). Одно деление масштаба — 1 см. Обозначения те же, что и на рис. 68. Структура годичного побега, а, кстати, и других более сложных побегов, бывает, как было сказано выше, монофазной (она развивается в результате весеннего побегоотрастания и не проявляет внутригодич- ную фазность видимого роста) и гетерофазной, или пульсирующей («пульс» годичного побега ди-, три-, плейофазный проявляется в отра- стании сначала весеннего побега, затем — летнего (одного, нескольких), а иногда еще и осеннего побегов после непродолжительного скрытого роста в почках). Рассмотрим подробнее некоторые особенности внугригодичного (летне-осеннего) побегоотрастания. Пролеисис (греч. prolepsis — упреждение) — явление преждевре- менного отрастания побегов (иролеитических побегов, по Брай, 1912) из вполне сформировавшихся почек, подготовленных к длительному по- кою, но выведенных из этого состояния (преждевременно пробудивши-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИИ
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 219 еся почки) в конце лета или осенью какими-либо обстоятельствами (например, при стечении благоприятных для роста погодных факторов). Пролонгация (лат. prolongatio — продолжение) — явление отраста- ния побегов после непродолжительного (30-40 дней) покоя, в результате которого почки успевают полностью сформироваться. Пролонгацию побегов обеспечивают так называемые ивановы no6ezu (Spath, 1912), или no6ezu иродолжения (пролонгированные). Как отметил И.Г.Серебряков (1950) ивановы побеги чаще образуются из терминальных почек, но при подходящих условиях они развиваются и из верхних сильных па- зушных почек. По нашим наблюдениям диапазон пролонгации шире чем предполагалось ранее. Это явление связано с образованием не толь- ко вегетативных ивановых побегов как у Quercus гоЬиг L., но и внутри- годичных летних побегов с репродуктивной функцией как, например, у Согу1ив ачеИапа L., которые иначе, чем ивановыми, или пролонгиро- ванными побегами, обозначать неоправданно (они отвечают определе- нию этих побегов). Прологацию побегов с репродуктивной функцией обеспечивает в сезонном климате умеренного пояса Земли особый спо- соб формирования нормальных (обычно функционально-развернутых) соцветий через прохождение соответствующей флоральной системой определенного ее возрастного состояния — иредсоцветия. Эта структур- но-функциональная система в своем развитии больше, чем почка, но еще не функционально-развернутое соцветие (последнее, как минимум, должно в сезон своего развития зацвести). Предсоцветием (преинфлорес- резко заторможенное в росте и развитии, околоиочечное (Corylus avellana L.), иное неразвернутое или иочти неразвернутое (частично развернутое, хотя и очень редко, даже до зачаточных бутонов как бывает у Viburnum lantana L.) состояние флоральной системы no6eza (в том числе и соответствующих ивановых no6ezoe) иредрасиоложенной при дальнейшем развитии к иреобразо- ванию в нормальное (функционально-развернутое) соцветие (рис. 71-1). Рис. 71. Разнообразие в расположении репродуктивных органов в структуре побегов: I — смешанная побеговая система Corylus avellana L., II — закрытая (с верхушечными репродуктивными органами) побеговая система Viburnum opulus L. и Ш — Aesculus hippocastanum L. с полузакрытыми побегами (Московская область). 15 — короткий открытый вегетативно-репродуктивный сборный годичный побег с оди- ночным внутригодичным (ивановым) пререпродуктивным побегом, 16 — длинный веге- тативно-репродуктивный закрытый годичный побег, 17 — длинный открытый вегетатив- но-репродуктивный сборный годичный побег, развивающий тычиночную флоральную систему (иносоцветие) с постлатентным цветением только в следующем году, 18— длинный весенний вегетативный годичный побег, 19 — иносоцветие с новоинтеркаляр- ными побегами, проходящими пререпродуктивную стадию, 20 — одиночный пререпро- дуктивный иванов побег, 21 — зачаточная тычиночная сережка (предсоцветие), 22— короткий безрозеточный вегетативный побег, 23 — унирепродуктивный полузакрытый безрозеточный побег, 24 — двухгодичный унирепродуктивный полузакрытый побег, 25— вегетативный двухгодичный побег, мк — максиморезид, мн — миниморезид, мг — магно- резид, тр — тотарезид (см. гл. 6.2.), гг — граница годичного побега, ов — отмершая верхушка прошлогоднего побега, во — отмершая верхушка побега в вегетационном сезоне текущего года, уз — зона сильно сближенных узлов (укороченных междоузлий). Остальные обозначения те же, что и на рис. 68.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Побеги, в структуре которых развиваются иредсоцветия, — иререиродук- тивные. Соцветие в своем морфогенезе может проходить следующие возра- стные стадии: зачаточное соцветие (образуется и развивается в процессе видимого роста побега) почка (зачаточное развитие соцве- тия происходит в процессе почко- образования) предсоцветие (соцветие вышло или почти вышло из зачаточного состоя- ния, но, оставшись в неразвернутом или почти неразвернутом состоянии прекратило дальнейшее развитие или резко затормозило его для преодоления неблагопри- ятного климатического периода) нецветущее нормальное соцветие цветущее соцветие плодоносящее соцветие В качестве примера рассмотрим развитие ивановых пререпродук- тивных побегов у орешника (Согу1ив аче11апа L.) в Подмосковье. Они возникают из пазушных вполне сформированных почек весеннего ве- гетативного (ростового), уже акронекрозного (верхушка из нескольких слаборазвитых и зачаточных узлов и междоузлий прекратила дальней- ший рост, засохла и отвалилась) родительского годичного побега. Такие ивановы побеги начинают отрастать у орешника 1 ЧШ + несколько дней; у них еще в родительских почках закладываются верхушечные и пазушные флоральные зачатки, которые после развертывания почек быстро увеличиваются в размерах и развиваются в верхушечные и под- верхушечные пазушные предсоцветия (неразвернутые тычиночные се- режки). На следующий год рано весной перезимовавшие предсоцветия орешника развертываются в нормальные тычиночные соцветия-сереж- ки, которые цветут, рассеивая в окружающую среду мириады пыльце- вых зерен. Ивановы побеги, таким образом, развиваются в отмеченном
Глава 5. Параметры основных органов и их частей в организации биоморф семенныхрастений 221 случае как двухгодичные модифицированные вегетативно-репродуктив- ные; они образуют в своей структуре чешуевидные листья, обычно одну пазушную зимующую почку и репродуктивные элементы. После одно- кратного рассеивания весной пыльцевых зерен сережки отмирают, а почка развертывается в короткий побег. Предсоцветия образуются и в структуре побегов, проявляющих другие способы отрастания, например, у некоторых весенних побегов Viburum lantana L. (терминально) или у некоторых весенних побегов Betula pendula Roth (терминально, а иногда еще и субтерминально в пазухах верхних листьев). Любопытно, что у Viburnum lantana предсоц- ветия останавливаются в росте и развитии только перед зимней стужей (помогает опушение и специфический защитный налет) прямо на по- роге уже наметившегося превращения их в функционально-развернутые верхушечные соцветия, которое в условиях Подмосковья осуществимо лишь на второй (!) год вегетации некоторых весенних побегов и реже силлептических побегов замещения. Все перечисленные выше типы по- бегообразования непосредственно не участвуют в развертывании струк- туры соцветий (соцветия — это, в первую очередь, функционально- развернутые и, как правило, разветвленные флоральные системы); пос- леднее происходит только в результате силлепсиса, причем не всякого. Силлепис — явление роста боковых побегов (силлептических побе- гов, по Spath, 1912) одновременно с непрекращающимся ростом исход- ной (родительской) для них оси, т.е. оси, от которой они непосред- ственно образуются. Соцветия по положению в структуре побегов бы- вают верхушечные и промежуточные. Верхушечные соцветия (лат. inflo- rescentia terminalis) занимают верхнюю часть побега и, будучи закры- тыми (Malus domestica Borkh.) или открытыми с фрондулезной, бракте- озной или абрактеозной недоразвивающейся верхушкой (по нашей терми- нологии — полузакрытыми, или семидетерминатными, соцветиями, как, например, у Leonurus quinquelobatus Gilib.), или открытыми с фрондоз- ной эфемернорастущей и замирающей вегетативно-репродуктивной вер- кушкой (по нашей терминологии — приоткрытыми, или преиндетерми- натными, соцветиями, как, например, у Hedysarum alpinum L.), не за- канчиваются сверху непосредственно по главной оси ростовой зоной (зоной неограниченного вегетативного верхушечного роста). Промежуточные, или интеркалярные (неограниченно открытые), соц- ветия (лат. inflorescentia intercalaris), или короче — межсоцветия (лат. interinflorescentia), развиваются ниже зоны неограниченного вегетатив- ного верхушечного роста у собственно открытых побегов (Lonicera xylo- steum L., Plantago cynops L., Acalypha hispida Borm). Каково разнообразие силлепсиса и какие силлептические побеги участвуют непосредственно в образовании соцветий? Мы выделяем четыре типа силлептических побегов: эусиллептические, инфантильные, индуктивные, интеркалярные. Эусиллеитические побеги составляют структуру верхушечных соцветий и соцветий других силлептических побегов, иногда развиваются как вре- менно или вынужденно вегетативные, например, у виргинильных рас- тений, или иначе дополняют опорно-ассимиляционную систему роди- тельского побега, оставаясь не тождественными другим типам силлеп- тических побегов. Эусиллептические побеги 1-го порядка (паракладии
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ВВ Рис. 72. Открытая побеговая система Ficus carica L. (Крым, Никитский ботанический сад). 19 — длинный весенний открытый вегетативно-репродуктивный годичный побег, развивающий обоеполое межсоцветие, сл — срединный лист, вв — вегетативная верхушка побега. Остальные обозначения те же, что и на рис. 68, 71.
Глава 5. Параметры основных органов и ик частей в организации биоморф семенных растений 223 О bP 44 И 0 ~Ъ ~ ир g u й Х )~ й М ~ о й ~ О O й g5 М О О 65 р; й О о > й о ~4 Зх О ~ М 4 Ю ~, О~ И .. д~ 1 „О Я:» Г~ » еъ й 1 ! О О & й И: Р' ф; Ж О g [.» [ю ID [.» л д М 6; Щ 5: О к~ @: v ц~» л» ь~~ ~~,'» ~ 65 И,» О Д М ~р о о ~)" м 2 О 0 3 И О О О 65 ф; Х И. О О ~а» Щ ФЭ ~М 65 [.» д ж Я X ~ g5 О И~ еа д, е й X И: О О 4> 2 О й~ О Ь ОЮ О~ Q.О И: (е) ~ ~ О� О О И: ж ~~~0Ï. И; ) u v ~ ОЯ~~ )5: 1 ф4 о Ф ~ [ с[3 О � u ~{~~ л о ~~ ~ ~ ~~.) М О О Е Щ О д +Q X B .&ад, � О й~ х л Д [.» 65 ~» Д „o ~ z~-о В~» ~» о р.~~ ~о~ О 1 и. ,д о Ф [» ~~ г~, О ~зй д л ° 1 ц .0 е~ О Иi G [ М G.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИИ Рис. 74. Открытая побеговая система, периодически развивающая одиночные интеркалярные побеги или межсоцветия, у Viola uniflora L. (Алтай, Курайский хребет, 1200 м над ур. моря, лесной пояс). ак — автономное корневище, образовавшееся после отмирания главного корня, у — увеличено по сравнению с масштабом, оп — основание надземного побега (верхняя часть удалена), нл — низовые листья, оц — одиночный пазушный цве- ток, принявший верхушечное положение, гм — графические модели разно- возрастных особей: А — проросток, В — ювенильное растение (главный корень еще жив), Г — имматурное растение (главный корень отмер), Д — молодое репродуктивное (инициматурное) растение (годичный побег, продолжающий эумоноподиальное нарастание, образует только один базальный зеленый лист), Е — средневозрастное репродуктивное (медиматурное) растение (верхушечный годичный побег, развивает два базальных зеленых листа и (или) корневище ветвится), Ж — старый репродуктивный (финиматурный) кондивидуум (жизнь продолжают особи клона). Остальные обозначения на рис. 68,71,73.
Ъ Глава 5. Параметры основных органов и ик частей в организации биоморф семенных растении Рис. 75. Межсоцветие и одиночный интеркалярный побег у Astragalus alpinus L. (Алтай, Курайский хребет, 1800 м над ур. моря, альпийский луг). Одно деление масштаба — 1 см. Остальные обозначения те же, что и на рис. 68.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИИ Рис. 7б. Открытая побеговая система Quercus pubescens Willd. (Крым, Никитский ботанический сад). дп — дифазный вегетативно-репродуктивный безрозеточный годичный побег, в— весенний внутригодичный вегетативно-репродуктивный побег, Бп — весенний годичный вегетативно-репродуктивный побег, ип — иванов побег, рв — весенний ростовой годичный побег, сл — срединный лист, уз — зона сильно сближенных узлов (мультинодий), гг — граница годичного побега, вг — граница внутригодич- ного прошлогоднего побега. Остальные обозначения те же, что и на рис. 68.
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 227 1-го порядка) зоны обогащения самостоятельного верхушечного слож- ного соцветия (синфлоресценции) несут в своей структуре лрисоцветия (лат. coflorescentia), а те в свою очередь нередко состоят из иарциальных соцветий (лат. partialflorescentia) — простых или сложных соцветий, со- ставляющих боковые ответвления от центральной оси главного соцве- тия или присоцветия (Troll, 1964). Кроме парциальных соцветий как части любых соцветий, уместно выделить еще одиночные иарциальные цветки, или частные цветки. Охарактеризуем другие типы силлептических побегов, которые также образуются в структуре цветоносного побега текущего года. Ин- фантильные (недоразвитые, как правило, без репродуктивных органов) набеги отрастают в зоне неполного торможения и выглядят слабыми. Они часто образуются у растений с верхушечными соцветиями. Индук- тивные набеги развиваются, будучи силлептическими, непосредственно под межсоцветием (Plantago cynops L.) или под зоной торможения верхушечного соцветия (Alliaria petiolata (Bieb.) Cavara et Grande), или, когда зоны торможения нет, под верхушечным соцветием (Malus domestica Borkh.) и, как правило, повторяют в структурном отношении родительский побег, завершая свое развитие в текущем сезоне или позд- нее. У многолетних растений индуктивная зона иногда проявляется между зонами инновации и торможения. Кроме того, индуктивные, чаще вегетативные, побеги иногда образуются из пазушных недоформи- рованных почек сильнорослых побегов (одновременно с ростовой дея- тельностью родительского апекса в пазухах некоторых молодых ли- стьев активно функционируют меристемы) как, например, у Hippphae rhamnoides L. или у молодых кустарников и деревьев (Betula pendula Roth). Разновидность эусиллептических или индуктивных побегов, кото- рая возникает в основании куста, т.е. обеспечивает силлентическое ку- щение, обозначаем еще и как базисиллентические no6ezu, а явление та- кого побегообразования называем базисилленсисом (Filaginella uliginosa (L.) Opiz, Calicotome villosa (Poiret) Link). Особо следует остановиться на межсоцветиях. Их структуру опреде- ляют интеркалярные побеги. Под межсоцветием, или нромежуточным (интеркалярным в широком смысле слова) соцветием, мы понимаем не- ирерывную совокуиность интеркалярных (силлеитических цветоносных) по- бегов любой сложности организации, но единых или близких по стеиени единую систему с ними и участком (зоной интеркалярного ветвления) центральной оси родительского открытого no6eza, структуру которого они не иовторяют. Любое межсоцветие переходит сверху по своей глав- ной открытой оси в вегетативную (ростовую) часть (зону неограничен- ного вегетативного верхушечного роста), а следовательно, эту часть не включает. Поскольку интеркалярные побеги любого межсоцветия отли- чаются по структуре от родительского побега этого соцветия, но повто- ряют структуру друг друга, то для них можно предложить еще термин- синоним «тавтокладии», созвучный (но не более того) паракладиям Тролля (Troll, 1964). Паркин (Parkin, 1914) предложил термин «интер-
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ калярное соцветие» для обозначения части побега, у которого главная ось вегетативная, продолжающая рост наряду с развитием на ней боко- вых цветков (плодов), как, например, у Callistemon speciosum (Sims) DC. Развивая идею «интеркалярного соцветия», мы предложили внести некоторые коррективы в использование этого термина, поскольку рос- товую верхушку логичнее не включать в такое соцветие, а интеркаляр- ные побеги по структуре могут быть самыми разнообразными, но они всегда должны отличаться от вегетативной (ростовой) родительской оси. Межсоцветия, если стать на нашу позицию, следует отмечать в струк- туре побегов растений многих родов: бук, липа, подорожник, родиола, сентполия, гравилат, сивец и др. В структуре межсоцветий каждое целое соцветие интеркалярных побегов утрачивает статус самостоятельного соцветия и приобретает статус присоцветия. Итак, соцветия — резуль- тат только силлепсиса, а еще точнее — эусиллепсиса и интеркалярности в побегообразовании (рис. 68-76). В структуре любых межсоцветий может быть только два и более тавтокладиев, а если интеркалярный побег один, то это не тавтокладий и соответственно в таком случае нет межсоцветия (например, Convallaria majalis L., Viola uniflora L., рис. 74). Особо следует сказать о двух типах побегов, которые похожи на интеркалярные, но проявляют иные способы отрастания и (или) отли- чаются фазностью развития. Новоинтеркалярный no6ez формируется как какой-либо летний или даже осенний побег гомологичный интеркаляр- ным побегам, но в своем развитии он проходит дополнительную ста- дию — пререпродуктивную, образуя предсоцветия, а цветет лишь на следующий год (Corylus avellana). Постинтеркалярный иобег развивается гомологично интеркалярным побегам, но не силлептически, а постла- тентно, т.е. из перезимовавшей почки; пример Daphne mezereum L., Salix caprea L., рис. 81). Есть все основания полагать, что помимо традиционной трактовки закономерностей рсположения репродуктивных органов большое раз- витие должен получить в недалеком будущем новый подход к изучению динамической морфологии репродуктивных систем, т.е. закономернос- тей развития репродуктивных органов от единичных антомеров до сверхсо- цветий разной сложности. Наша задача в области анализа цветков, соцветий и еще более сложных репродуктивных систем, не углубляясь в детали (подробнее см. Troll, 1964, 1969; Федоров и др., 1979), отразить иерархичность этих сис- тем в зависимости от сложности их организации. Деление реиродуктив- ных органов на антомеры (части цветков), отдельные цветки, соцветия и сверхсоцветия (верхняя субордината) — это субординатная классифи- кация репродуктивных органов в наибольшем обобщении (рис. 77-А). С несколько меньших высот обобщения субординия репродуктивных ор- ганов принимает следующий вид (рис. 77-Б): 1 — антомеры; 2 — от- дельные цветки; За — иервые соцветия (включая и За' — первые межсо- цветия) — любые простые соцветия (в структуре сложных соцветий или вне их); Зб — вторые соцветия — сложные парциальные соцветия в структуре еще более сложных соцветий — главных сложных соцветий или сложных присоцветий (кофлоресценций) паракладиев или тавток-
Глава 5. 17араметры основных органов и их настей в организаиии оиоморф семенныхрастений 229 Рис. 77. Суперординатная модель организации репродуктивных органов. А и Б — первая и вторая субординия диасом; 1 — антомеры, 2 — отдельные цвет- ки, 3 — соцветия, 4 — сверхсоцветия, За-Зг — верхушечные соцветия: За — пер- вые, 36 — вторые, Зв — третьи, Зг — четвертые; 4а-4в — верхушечные сверхсоцве- тия: 4а — первые, 46 — вторые, 4в — третьи. Классификационная ветвь открытых диасом. Межсоцветия: За' — первые, Зб' — вторые, Зв' — третьи; сверхмежсоцве- тия: 4а' — первые, 46' — вторые, 4в' — третьи; гг — границы годичных побегов; чс —; гс-, пс — схема сложного главного соцветия (гс) или присоцветия (пс) в структуре синфлоресценции; сф- схема синфлоресценции.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ладиев (интеркалярных побегов межсоцветий); Зв — третьи соцве- тия — сложные главные соцветия или сложные присоцветия в струк- туре синфлоресценций (самостоятельных сложных соцветий); Зг — чет- вертые соцветия — синфлоресценции с одним и более порядками пара- кладиев; 4а — иервые сверхсоцветия — они состоят из одиночных вер- хушечных цветков (например, Magnolia grandiflora L.); 46 — вторые сверхсоцветия образованы из самостоятельных простых соцветий (Padus avium Mill.); 4в — третьи еверхсоцветия образованы из нескольких самостоятельных (верхушечных) сложных соцветий, или синфлорес- ценций (Aesculus hippocastanum L.). Параллельно с рассмотренной классификацией от уровня За — nep- Bb>x верхушечных соцветий через За' — иервые межсоцветия (Ficus cari- ca L., Hippophae rhamnoides L.) восходит субординатная классификаци- онная ветвь (За'-Зв') неограниченно открытых соцветий (межсоцветий). Совокупность нескольких самостоятельных межсоцветий и одиночных интеркалярных цветоносных побегов, разобщенных между собой веге- тативными частями, у каких-либо более сложных побегов или целых организмов представляет сверхмежсоиветие (лат. superinterinflorescentia). Продолжим начатую выше классификацию: Зб' — вторые межсоцветия (иятые соцветия) — межсоцветия, тавтокладии которых с простыми присоцветиями (Lonicera xylosteum L.); Зв' — третьи межсоцветия (ше- стые соцветия) включают тавтокладии со сложными присоцветиями (Succisa pratensis Moench); 4а' — иервые сверхмежсоцветия (четвертые сверхсоцветия) — сверхмежсоцветия из нескольких простых (первых) межсоцветий; 46' — вторые сверхмежсоцветия (иятые сверхсоцветия) состоят из нескольких вторых межсоцветий; 4в' — третьи сверхмежсоцве- тия (шестые сверхсоцветия) состоят из нескольких третьих межсоцветий. Одиночный цветок, верхушечное соцветие или межсоцветие, а также сверхсоцветие любой сложности, составляющие репродуктивную часть какого-либо побега, корнепобега-органа или всего растения,— это, на наш взгляд, их антосфера. В структуре растений важное значе- ние мы придаем кладосфере — разветвленной системе какого-либо по- бега, корня, корнепобега-органа или всего растения, нижнюю границу которой всегда составляет самая нижняя для этой системы ветвь (рис. 78). Шадосфера no6ezoe, как и любой отдельный побег, бывает вегетативной, вегетативно-реиродуктивной, реиродуктивной и mompenpo- дуктивной (строго реиродуктивной) (Нухимовский, 1971, 1987). Первично неразветвленная часть растения, включающая (частично или полнос- тью) в свою структуру медиаль, назовем иротосферой, а иную неразвет- вленную часть (как переходную к разветвленной части) в структуре любого побега, корня, корнепобега-органа и всего организма — zuno- сферой (рис. 78). Отсчет порядков осей ветвления ведется только в отно- шении одноименных органов. Кладосферы побегов у многих растений выступают как особые цветоносные системы, которые решают сложные задачи наиболее выгодного завоевания пространства и времени для сво- их антосфер. Особое место среди антосфер занимают системы, внешне очень напоминающие соцветия, но отличающиеся от них двухгодичным цик-
Глава 5. Парсметры основнык органов и их частей в организации оиоморф семенных растений 231 Рис. 78. Модель кладосферы и антосферы обобщенного семенного растения. Кладосфера: 1 — смешанная, 2 — открытая, 3 — закрытая, 4 — полузакрытая, 5— протосфера, 6 — гипосфера, 7 — семядольный узел, 8 — межсоцветие, 9 — ино- соцветие,10 — граница годичного побега прошлых лет, 11 — граница годичного побега текущего года. лом репродукгивного развития, — иносоцветия (лат. contrinflorescentia, дословно — отличное от соцветия). Иносоцветие (контринфлоресцен- ция) — это неирерывная совокуиность иостинтеркалярных (внешне по- добных интеркалярным побегам, но всегда отрастающих из латентных зимовавших почек, Salix caprea L., см. рис. 81) годичных цветоносных или новоинтеркалярных двухгодичных (с обязательной пререпродуктивной стадией, когда формируются предсоцветия, на первом году их развития, Согу1ив аче11апа, см. рис. 71-I) иобегов любой сложности организации образующих единую систему с участком (зоной постинтеркалярного или зоной новоинтеркалярного ветвления) центральной оси родительского иобега, вовлеченным в образование иобегов с реиродуктивной функцией (рис. 78-81). Иносоцветия (постинтеркалярные и новоинтеркалярные), подобно межсоцветиям, различаются по сложности организации. Особый тип цветоносных систем встречен автором в средиземно- морской флоре Ближнего Востока (рис. 82). У некоторых растений осе- вая структура прошлогодних эфемерных органов — отплодоносивших соцветий, на протяжении относительно теплой (субтропической) зимы (температура обычно не опускается ниже +5'С и тем более О' ), не отмирает или отмирает частично, а из перезимовавших пазушных по-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 79. Иносоцветия в структуре побеговых систем I — Amygdalus communis L., II — Регяса vulgaris Mill. (Крым, Никитский ботанический сад). вп — весенний вегетвтивный (ростовой) безрозеточный годичный побег, рп- постинтеркалярный (репродуктивный весенний) побег, гг — граница годичного побега, уз — зона сближенных узлов, сл — срединный лист, ис — иносоцветие плодоносящее, тр — мономерный тотарезид. чек, развитых на таких осях в зоне бывших соцветий, в конце короткой зимы или рано весной (февраль, март) отрастают, как правило, слабые или очень слабые побеги с репродуктивной функцией или без нее. Побегообразови«ие (уже — «обегоотристи«ие), слабо ироявляющееся в текущем году из сохранившего частично жизнесиособность ирошлогодне- го соцветия, уже выиолнившего свою основную (реиродуктивную) односе- зонную функцию, иредлагаем именовать как «остфлораль«ое «лихорадоч- «ое». Такое название образно подчеркивает структурно-функциональ- ную неготовность побегов, новообразующихся из зоны бывшего соцве- тия, развить нормальные репродуктивные системы, а также излишнюю «торопливость» в осуществлении неподготовленного побегоотрастания. Постфлоральное «лихорадочное» иобегообразование (иобегоотрастание) бывает двух тинов: «еобес«ече««ое, когда отрастают и развиваются сла- бые и (или) очень слабые иостфлоральные набеги из подзоны обогаще- ния (эусиллепсиса) бывшего соцветия, распространяясь или не распро- страняясь на подзону торможения (например Echium angustifolium Miller), и «олуобес«ече««ое, когда отрастают и развиваются слабые и (или) очень слабые постфлоральные побеги из подзоны обогащения прошлогоднего соцветия, а вот из под зоны торможения отрастают не только слабые, но и побеги вполне жизнеспособные среднего и даже сильного развития (например, Inula viscosa (L.) Aiton). В процессе бур- ного весеннего отрастания побегов из зоны возобновления некоторых полукустарников и полукустарничков (см. гл. 8.2.2.), многие из таких
Глава 5. 17араметры основных органов и их частей в организации биоморф семенныхрастений 233 Рис. 80. Формирование иносоцветия в структуре особей Polygonum viviparum L. (Алтай, Ивановский хребет, альпийский луг). с — семядоли, сл — срединный лист, гп — главный открытый побег, рп- постинтеркалярный (репродуктивный весенний) побег, вс — верхушечное соцве- тие постинтеркалярного побега, ак — автономное корневище, гг — границы го- дичных приростов (побегов), со — сухие остатки прошлогодних частей побегов, св — специализированная выводковая почка, пр — проросшая выводковая почка, пп — поверхность почвы, гм — графические модели разновозрастных особей. Одно деление масштаба — 1 см.
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 81. Иносоцветия в структуре побеговых систем I-Acer negundo L., П — Salix caprea L. (Московская область). ов — рубец от отмершей верхушки прошлогоднего побега, пс — почка ростовая (статурная), п — почка, скрытая под черешком листа, ол — продольный разрез основания черешка листа, чп — почечная чешуя, мк — мономерный максиморе- зид, тр — мономерный тотарезид, ис — иносоцветие. Остальные обозначения те же, что и на рис. 79. побегов перерастают вышерасположенные постфлоральные побеги, часто недоразвивающиеся и к моменту зацветания растений засыхаю- щие, не реализовав себя в репродуктивном развитии. Некоторые однолетники также способны проявлять иостфлораль- ное «лихорадочное» необесиеченное иобегоотрастание, превращаясь, таким образом, в сверходнолетники (пример Conyza bonariensis (L.) Cronquist). Особо следует выделить иостфлоральное обесиеченное иобегообразо- вание, если иостфлоральные no6ezu развиваются в зоне ирошлогоднего соц-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛСТЕНИЙ ветия, включая и иодзону бывшего эусиллепсиса, и ироявляют виолне нор- мальную жизнесиособность и реиродуктивное развитие (пример, Hyperi- сит patulum Thunb., или некоторые турионы Hypericum androsaemum Ь. на втором году их развития, см. рис. 52; Крым, Никитский ботаничес- кий сад). Такое побегообразование уже нельзя считать «лихорадочным». 5.1.3. Многолинвйный анализ организации побвгов Организация биосистем всегда многогранна. Совокупность призна- ков организации, присущих объекту познания и характеризующих его с разных сторон, составляет систему признаков, которую образно мож- но обозначить как «иризнаковое иространство» этого объекта (в логике «признаком называется все то, в чем предметы сходны друг с другом или в чем они отличаются друг от друга», Б.Е.Валковский, 1964, с. 11). Чтобы «высвечивать» из этого «пространства» признаки и строить ряды признаков, более всего отвечающие целям конкретного исследования, используем различные методы классифицирования: субординии, серио- фикации или сразу оба метода, что по отдельности составляет методы однолинейного классифицирования, а в совокуиности — один обобщен- ный метод — метод мультилинейного (многолинейного) классифици- рования. По многолинейному принципу мы уже классифицировали по- беги семенных растений (Нухимовский, 1969а, 1971 и др.), а в настоя- щей главе развиваем разработанные ранее классификации. Из множе- ства признаков для многолинейного анализа выберем лишь немногие, которые отражают существенные особенности организации биоморф семенных растений и наглядно свидетельствуют о возможностях этого метода анализа. В предыдущих разделах рассматриваемой главы было приведено несколько широких субординатных классификаций побегов, которые могут являться и частью многолинейной классификации побе- гов, поскольку в последней иерархические взаимосвязи между классами систем значения не имеют. Далее рассмотрим несколько классифика- ций, которые дополняют общую характеристику признакового про- странства побегов. Приведем фрагмент многолинейной классификации специальных и универсальных побегов семенных растений '>. Типизация ультрауни- версальных (более сложных) побегов зависит от приводимой классифи- кации, но не повторяет ее. Многие из выявленных типов побегов мы отразили в графической модели организации обобщенной биоморфы семенных растений (рис. 83). Объяснение символов. 1 — побеги, охарактеризованные "> по особенностям организации верхушки побегов, т.е. по особенностям про- хождения и завершения верхушками побегов постинициального периода 1) !, и, П1... — символы однолинейных классификаций — сравнительных рядов типов побегов семенных растений (указываются ряды признаков, по которым выделены типы побегов); 01, 1, 2, 3... — символы типов (классов) побегов соответствующих однолинейных классификаций. Ф1) Далее при описании рядов типов побегов не повторяется, но подразумевается сло- восочетание «побеги, охарактеризованные».
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 237 жизни этих побегов: 1 — открытые (их верхушечные меристемы не образуют брактеозные или фрондо-брактеозные соцветия, или отдель- ные верхушечные цветки, т.е. это побеги вегетативные или с пазушны- ми репродукгивными элементами) — Pinus sylvestris L., Lonicera caerulea L., Coronilla scorpioides (L.) Koch; 2 — условно закрытые, т.е. полузак- рытые или приоткрытые (их верхушечные меристемы образуют откры- тые брактеозные, фрондоулозно-брактеозные и фрондо-фрондулозные соцветия, иначе полузакрытые или приоткрытые соцветия) — Padus avium Mill., Dactylorhiza maculata (L.) Soo', 3 — закрытые (их верхушечные меристемы образуют закрытые соцветия или отдельные верхушечные цветки) — Malus domestica Borkh., Centaurea jacea L. Подразделяя побеги таким образом, мы исходили из следующих положений, высказанных некоторыми исследователями. И. Репер (Roe- per, 1826), как мы уже отмечали, обратил внимание на то, что соцветия бывают двух типов: открытые, когда у них развиваются только пазуш- ные цветки, и закрытые, когда их главная ось заканчивается цветком. В. Тролль (Troll, 1964) подразделил соцветия в общем плане в зависимо- сти от облиственности на брактеозные, которые характеризуются брак- теевидными кроющими листьями или вообще лишены кроющих листь- ев; фрондозные, имеющие ассимилирующие листья; фронда-бракте- озные, у которых в верхней части — листовые элементы первого типа соцветий, а в нижней — второго (рис. 84-А). Еще более дробная клас- сификация соцветий в зависимости от их облиственности разработана Ап.А. Федоровым, 3.Т.Артюшенко (1979), содержание которой стано- вится понятно из приводимого ниже рисунка из цитируемой работы (рис. 84-Б). II — по степени специализации, т.е. по степени проявления в постинициальном периоде жизни побегов вегетативной и репродук- тивной функций: 1- 4 — вегетативные (эти побеги состоят из стеблевой и листовой частей) — 1 — статурные (от лат. statura — рост), или рос- товые (в их структуре срединные листья нормальные ассимилирующие, а стеблевая часть полностью или не полностью потенциально скелетная или скелетная, Larix decidua Mill., Ulmus laevis Pall.); 2 — модифици- рованно-статурные (листья у них видоизмененные — чешуевидные и т.п., а стеблевая часть — полностью или не полностью потенциально скелетная или скелетная (Crataegus sanguinea Pall. и др.); 3 — ассими- ляционные, если срединные листья ассимилирующие, а стеблевая часть эфемерная (Tamarix ramosissima Ledeb. и др.); 4 — модифицированно- ассимиляционные (листья у них видоизмененные, а стеблевая часть зеленая, эфемерная (Haloxylon aphyllum (Minkw.) Iljin и др.); 5-7— вегетативно-репродуктивные (эти побеги состоят из стеблевой, листо- вой и репродуктивной частей); 5 — репродуктивно-статурные (средин- ные листья у них ассимилирующие, а стеблевая часть — полностью или не полностью потенциально скелетная или скелетная, Betula pendula Roth); 6 — модифицированные репродуктивна-статурные (листья ви- доизмененные, а стеблевая часть полностью или не полностью потен- циально скелетная или скелетная, Согу1иа avellana L.); 7 — репродук- тивно-ассимиляционные (срединные листья ассимилирующие, а стеб- левая часть эфемерная (Padus avium Mill., Succisa pratensis Moench); 8—
3521 4121 1511 г--— /1 ! 5111 1 //1 ! I ! I I 1 ! ! 1 ! r 1 1 ! ! I 1 1 11! о I 1 l ///1 ' I 1I М,'', l 6111 ///2 1211 /al,' 1 I /Ш ', QC ,7211 р 1 1 XV1 — —,14 I I 1 I I I ! ! 1 ! 1621 I 3611 ! 1 11 I 1 I 4211 211 8211 8111 V/2 1 I I V/И),' РЭ 9211 0211 5611 /C ! ~/1 Пб- ЛЗ 9211 Л2 02 0211 ят 4~гз багз А1 ЧС01 3712 4612 4112 5112 4124 BT CT 1815 3612 , IX1 ПП ~/1 р Г I I 1 1 ! 'Х/V1 1 I l~ 1 V//1'- --- ,Х1 1Х2 1 ! ,'Х//1 1116 Х1 в 1116 I Х2 ! Х///1 ', СУ =-- Л1 KØ СМ X/1,' Л2 , СИТ СЛ1 СА2 ЧК01 l<P СА3 СЛ2 ВТ ЧК02 П<0 112 1 /2 I ! //2 ! ! ! ! 1 I I 1 ! I 1 , ',/V2 ! ! I 1 1 I ! I ! I 1 1 1 1 1 1- 1 ' ~2 I I I ) 1 , ~/1 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ 1311 3511 1111 3311 3111 4111 — — — -' Х//2 и KCA 1126
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 239 репродуктивные (они состоят из стеблевой, листовой и репродуктивной частей, при этом листья у них низовые — почечные чешуи, а стеблевая часть — эфемерная, Populus tremula L.); 9 — тотрепродуктивные, или строго репродуктивные (эти эфемерные побеги состоят из стеблевой, репродуктивной частей — цветка или цветков и нередко брактеозной частей — брактей, Philadelphus coronarius L., Malus domestica Borkh.). Ш — по числу полов в сексуализации цветоносных побегов: 01— бесполые, 1 — однополые пестичные, 2 — однополые тычиночные, 3 — обоеполые, 4 — непостоянно обоеполые. IV — по кратности проявления репродуктивной функции (уже бы- ло отражено в соответствующей субординатной классификации побегов всех семенных растений): 01 — стерильные (дерепродуктивные), 1— унирепродуктивные, 2 — бирепродуктивные, 3 — трирепродуктивные, 4 — мультирепродуктивные. Возможны и иные клссификации побе- гов по кратности проявления узко специализированной репродуктив- ной функции, что отражено в Ч — VII однолинейных классификациях. Ч — по кратности плодоношения (у однополых растений с пестич- ными цветками): 01 — стерильные (акарпические), 1 — монокарпичес- Рис. 83. Многопризнаковая модель экзоморфологической организации биоморф семенных растений. В условных обозначениях каждая цифра — закодированный признак. Группу цифр сле- дует читать слева направо, первая цифра относится к ряду признаков А, вторая — к ряду признаков Б, третья — к ряду признаков В, четвертая — к ряду признаков Г. Побеги (Пб) — ряд признаков А: 1 — статурный, 2 — ассимиляционный, 3-6 — репродуктивно- статурный — соответственно 3 — с боковым (открытый), 4 — с верхушечным (закрытый с верхушечным одиночным цветком, плодом или условно закрытый рудимен- том цветка или иной недоразвитой эфемернорастущей и замирающей верхушкой), 5— со смешанным (закрытый или условно закрытый с верхушечным и пазушными репро- дуктивными элементами) плодотворением (плодоношением), 6 — репродуктивна-ста- турный с паракладиями, 7 — модифицированный репродуктивна-статурный, 8 — репро- дуктивна-ассимиляционный, 9 — репродуктивный, О — тотрепродуктивный (строго ре- продуктивный); ряд Б: 1 — раздвинутоузловой длинный, 2 — раздвинутоузловой корот- кий, 3 — сближенноузловой длинный, 4 — компактный (безрозеточный компактный), 5 — полурозеточный, 6 — розеточный, 7 — луковично-розеточный, S — верхнерозеточ- ный, 9 — верхнеполурозеточный; ряд В: 1 — внутригодичный, годичный, 2 — сверхго- дичный (двух-, трех-, многогодичный); ряд Г: 1 — надземный, 2 — с укороченным подзем- ным основанием, 3 — с горизонтальным подземным длинным основанием, 4 — со стелю- щимся (по поверхности почвы) длинным основанием, 5 — с вертикальным подземным длинным основанием, 6 — подземный. Скелетные стеблевые части побегов (р): ряд А— I-Is — тотарезиды — ! — длинный, la — короткий, Is — укороченный, II — максиморезид, Иа — малорезид, Ш — магнорезид, IV — медиорезид, V — бревирезид, VI — миниморе- зид, VII — курторезид, VIII — модесторезид, IX — репторезид, Х — порциорезид, XI— викариорезид, XI — пленорезид, ХШ — алиорезид, XIV — вицерезид, XV — рамозорезид; ряд Б — 1 — мономерный, 2 — ди-, три-, плейомерные резиды (о резидах см. гл.6.2.). Система стебленарастания (ст): м — медиаль, л — аксиллярная латераль, алт — адвентив- ная латераль; система корненарастания (кр): см — субмедиаль, сл — ризогенная субла- тераль, ксл — каулогенная сублатераль. Система ветвления (вт): гсо — главная стеблевая ось (стеблевая предветвь), чсо — частная стеблевая ось (стеблевая ветвь), гко — главная корневая ось (корневая предветвь), чко — частная корневая ось (корневая ветвь). Другие условные обозначения: чсо1, сл2, л1... — порядок осей (1, 2, ...), су — семядольный узел, кш — корневая шейка, пп — поверхность почвы, ал — ассимилирующие листья, у — узлы, чл — чешуевидные листья, рэ — репродуктивные элементы.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РЛ СТЕНИЙ 1У Рис. 84. Типы соцветий в зависимости от их облиственности и расположения репродуктивных элементов. А — по В. Троллю (Troll, 1964): 1 — 1Ч — соцветия — 1 — Il — брактеозные, 111— фронда-брактеозное, IV — фрондозное. Б — по Ал.А. Федорову 3.Т.Артю ~1979): 1-7 — со в юшенко ( ): - — соцветия — 1 — фрондозное, 2 — фрондулозное, 3 — брактеозное, 4 — абрактеозное, 5 — фронда-фрондулозное, 6 — фрондулозно-брактеозное, 7— брактеозно-абрактеозное. кие, — дикарпические, 3 — олигокарпические (плодоносящие 3-4 раза), 4 — поликарпические (плодоносящие 5 и более раз). — по кратности семеношения (у хвойных): 01 — стерильные (апостробилярные), 1 — моностробилярные, 2 — дистробилярные, 3— олигостробилярные (образуют шишки 3-4 раза), полистробилярные (образуют шишки 5 и более раз). VH— — по кратности пыльценошения (у однополых растений с ты- чиночными цветками): 01 — стерильные (апоантерические), 1 — мо- ноантерические, 2 — диантерические, 3 — олигоантерические, 4 — по- лиантерические.
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 241 Если характеризуем побеги по категориям IV-ой однолинейной классификации и не характеризуем по категориям Ш-ей однолинейной классификации, то ясно, что побеги обоеполые, например, мультире- продуктивные пентакормусы Plantago maxima Juss. ex Jacq. Если характеризуем побеги по категориям однолинейных классификаций или Ч, или VI, или VII, то ясно, что побеги узко специализированы по репродуктивной функции и их нет нужды характеризовать еще по ка- тегориям И1 и IV-ой однолинейных классификаций, например, моно- карпические годичные побеги, или монокарпические монокормусы, Salix caprea L. (такая характеристика побегов в данном случае короче, чем другая возможная характеристика — унирепродуктивные пестич- ные годичные побеги). Когда нас не интересуют особенности сексуали- зации побегов, то достаточно ограничиться характеристикой их по со- ответствующим категориям IV-ой однолинейной классификации. Сама идея подразделять побеги по кратности развития репродук- тивной функции в их жизненном цикле, на наш взгляд, прогрессивная. Однако использовать термин «монокарпический» побег не всегда воз- можно. Основу их составляет греческое слово «carpus» — плод. Букваль- но «монокарпический» — это «однократно плодоносящий». В приве- денных выше нескольких однолинейных классификациях побегов учте- ны различия в формах проявления репродуктивной функции, что сни- мает возникавшие вопросы к прежним построениям в этой области. VIII — по длине междоузлий, особенностям расположения листьев и функциональной роли последних: 1- 5 — безрозеточные (эти побеги не имеют в своем жизненном цикле фазу розетки, а в структуре— участка со сближенными в розетку ассимилирующими листьями); 1— раздвинутоузловые (большую часть их составляют междоузлия длинные и короткие, Tilia cordata Mill.); 2 — сближенноузловые (междоузлия сильно укороченные, а сами побеги по общей длине подобны раздви- нутоузловым, Picea abies (L.) Karst.), компактные, или укороченные (междоузлия укороченные и их мало, Tilia cordata Mill.); 3 — лукович- ные (они имеют в своей структуре очень компактный участок стебля с сильно сближенными утолщенными чешуевидными листьями и (или) другими листьями, не образуя розетки, Tulipa gesneriana L.); 4 — клуб- нелуковичные (эти побеги имеют в своей структуре утолщенный учас- ток стебля с сильно сближенными чешуевидными, или иными, быстро засыхающими, листьями, Colchicum speciosum Stev.); 5 — филлокладий- ные, или короче филлокладии (видоизмененные листоподобные побе- ги, Ruscus hypoglossum L.); 6-11 — розеткообразующие (розулиферные, что дословно означает с лат. «несущие розетки») — эти побеги имеют в своем жизненном цикле фазу розетки, а в структуре — участок со сближенными в розетку ассимилирующими листьями); 6 — розеточные, или розулатные, т.е. собственно розеточные (они состоят из компакт- ной стеблевой части и розетки листьев, среди которых все, причем не менее трех, ассимилирующие, а иногда также включают пазушные или верхушечные тотрепродуктивные части, Тагахасит officinale Wigg.); 7— верхнерозеточные (они состоят из верхней розеточной и нижней длин- ной или короткой безрозеточных частей, Eryngium campestre L. npu
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЧ засыпании субстратом); 8 — полурозеточные (они состоят из нижней розеточной и пазушной или верхушечной безрозеточной вегетативной или вегетативно-репродуктивной частей, т.е. со срединными листьями, Pimpinella major (L.) Huds., Berberis vulgaris L.); 9 — верхнеполурозеточ- ные (они состоят из верхней полурозеточной и нижней длинной или короткой безрозеточной частей, Achillea millefolium L.); 10 — лукович- но-розеточные (розеточные побеги имеют в своей структуре луковицу, Ornithogalum nutans L.; 11 — луковично-полурозеточные (полурозеточ- ные побеги имеют в своей структуре луковицу, Lilium candidum L.); 12 — предрозеточные (эти побеги имеют один-два сближенных асси- милирующих листа в своей структуре, предшествуя образованию розетки листьев, обычен у всходов розеточных растений). IX — по фенологическим состояниям: 1 — субактивные, или по- коящиеся (эти побеги временно находятся в состоянии относительного покоя, подготавливаясь к видимому росту); 2 — отрастающие (мульти- активные побеги находятся в начале стадии видимого роста); 3 — рас- тущие (мультиактивные побеги проявляют основной видимый рост пос- ле начала отрастания); 4 — отросшие (они завершили все свои фазы видимого роста); 5 — цветущие (они цветут, можно выделять также зацветающие и отцветающие побеги); 6 — плодоносящие (они плодо- носят) или семеносящие у хвойных); 7 — отплодоносившие — они от- плодоносили (или отсеменосившие у хвойных); 8 — отпыльценосив- шие (они уже образовали пыльцевые зерна, когда репродуктивные эле- менты однополые — тычиночные). Х — пс возрастным состояниям (объяснение возрастных состоя- ний побегов дано в главе 5.1.2): 1 — инициальные, 2 — постинициаль- ные, 3 — пререзидные, 4 — резидные, 5 — финальные. XI — по месту заложения: 1 — медиальные — они закладываются как зародышевые почки и всегда включают медиаль, т.е. простую стеб- левую ось 1-го порядка (главную); 2 — терминальные, или верхушечные (закладываются как верхушечные почки); 3-4 — латеральные (заклады- ваются как боковые почки и всегда включают в свою структуру латераль, т.е. простую стеблевую ось 2, 3 или большего порядка или придаточ- ную); 3 — аксиллярные (закладываются как пазушные почки); 4 — ад- вентивные (закладываются как придаточные почки). Некоторые осо- бенности организации побегов у семенных растений показаны на рис. 85-93. Можно продолжать строить новые и новые однолинейные (субор- динии, сериофикации и сборные субординатно-сериальные) классифи- кации побегов в рамках единого многолинейного анализа признакового пространства организации биоморф семенных растений. Приведенные выше несколько однолинейных классификаций в системе многолиней- ной классификации побегов, пожалуй, уже отражают возможности ис- пользования этого метода при анализе организации побегов. Далее мы еще неоднократно будем обращаться к методам однолинейного и мно- голинейного анализа, чтобы полнее раскрыть содержание главного объекта нашего исследования — организации биоморф семенных растений.
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 243 СЛ Рис. 85. Специализация побегов Morus nigra L. рс — репродуктивно-статурный открытый безрозеточный длинный акронекроз- ный годичный побег, кп — короткий ростовой (ствтурный) годичный побег, мс- межсоцветие, ип — интеркалярный побег в стадии плодоношения, пч — почка, сл — срединный лист, ст — стеблевая часть, мк — мономерный максиморезид, мл — мономерный малорезид, во — отмершая верхушка побега в вегетационном сезоне текущего года, ов — отмершая верхушка побега прошлых лет или рубец от нее, уз — зона сближенных узлов в основании годичного побега.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ Рис. 86. Специализация побегов Capparis herbacea Willd. (Крым, Южный берег, скалы). oc — открытое сборное соцветие, пр — паракладии, зт — зона торможения, зв- зона возобновления, уз — зона сближенных узлов, б — бутон, цв — одиночные пазушные цветки, пл — плод, сл — срединный лист, чл — чешуевидный лист, п- почка побега текущего года, зп — зимовавшая почка на многолетнем побеговом остатке (миниморезиде), мн — миниморезид, оп — основание годичного разветвленного открытого вегетативно-репродуктивного побега (верхняя часть удалена). Одно деление масштаба — 1 см.
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 245 Рис. 87. Специализация побегов Prunus spinosa L. (Крым). вп — вегетативный (ростовой) безрозеточный весенний годичный акронекрозный побег, ис — иносоцветие, рп — постинтеркалярные плодоносящие побеги, р- резидные побеги, образовавшиеся после окончания вегетационного сезона. Ос- тальные обозначения те же, что и на рис. 85.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РЛ СТЕНИЙ Рис. 88. Формирование побегов в онтогении Silene fruticosa L. (Крым, Никитский ботанический сад). 1 — ранние стадии онтогенеза; 11 — фрагмент побеговой части взрослого растения; с — семядоли, г — гипокотиль, гк — главный корень, гп — главный побег, вр- верхушечная розетка листьев, вп — вегетативный верхнерозеточный годичный побег, бр — безрозеточный верхушечный закрытый вегетативно-репродуктивный годичный побег, дп — дифазный вегетативно-репродуктивный розеткообразую- щий (в этом случае — верхнеполурозеточный) закрытый побег, пб — дифазный вегетативно-репродуктивный полурозеточный побег, мд — медиорезид (фраг- мент), ов — отмершая верхушка прошлогоднего побега, гг — граница годичного побега, го — примерная граница акронекроза отплодоносившей цветоносной части, сл — срединный лист, уз — зона укороченных междоузлий; горизонтальная линия — поверхность почвы. Одно деление масштаба — 1 см.
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 247 Рис. 89. Специализация побегов Laurocerasus officinalis М. Roem. (Крым, Никитский ботанический сап). пф — плейофазный безрозеточный вегетативный побег, зл — многолетнезеленый срединный лист, жв — живая верхушечная почка, во — случайно отмершая верхушечная почка, п — почки. Остальные обозначения те же, что и на рис. 79. х50 С Рис. 90. Формирование побегов в онтогении Centaurium erythraea Rafn (Крым). с — семядоли, г — гипокотиль, гк — главный корень, гп — главный розеточный по- бег, кн — сближенноузловая зимовавшая часть главного побега, или конодий, пр- силлептические розеточные временно вегетативные побеги. Одно деление масшта- ба — 1 см, х10... — увеличено по сравнению с масштабом в несколько (10:..) раз.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ ВП тр тр Рис. 91. Формирование побегов в онтогении Aurinia saxatilis (Ь.) Desv. (Крым, Никитский ботанический сад). 1 — ранние стадии в онтогении, 11 — фрагмент побеговой системы взрослого растения; ап — аксиллярный вегетативный побег кущения, ко — корневой отпрыск, рп — постинтеркалярный репродуктивный побег, эс — эусиллептичес- кие побеги, составляющие структуру верхушечного полузакрытого (семидетерми- нантного) соцветия постинтеркалярного побега, ис — иносоцветие, c6 — сбли- женноузловая безрозеточная часть верхнерозеточного побега, со — сухие остатки постинтеркалярных побегов прошлых лет, оп — открытый плейофазный верхнерозеточный побег с иносоцветием, тр — тотарезид годичный (мономерный). Остальные обозначения те же, что и на рис. 88.
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 249 Рис. 92. Формирование побегов в синонтогении Callianthemum angus- tifolium Witas. (Алтай, Ивановский хребет, альпийский луг). пф — плейофазный вегетативный розеточный побег, оп — открытый вегетативно- репродуктивный плейофазный побег, рп — постинтеркалярный побег, ип — ин- теркалярный побег, ак — автономное корневище, кр — мономерный курторезид, ок — особи клона, гм — графические модели разновозрастных особей, ол — от- мерший прошлогодний лист, у — увеличено по сравнению с масштабом. Осталь- ные обозначения те же, что и на рис. 88.
250 OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ Рис. 93. Виргинильное развитие особей Artemisia vulgaris Ь. (Московская область). с — семядоли, г — гипокотиль, гк — главный корень, гп — главный вегетативный побег, ик — индуктивный побег кущения, оп — основание индуктивного побега кущения, или базисиллептического побега (верхняя часть удалена), сп — эусил- лептические вынужденно или временно вегетативные побеги, сл — срединный лист, у — увеличено по сравнению с масштабом, гм — графические модели разновозрастных особей, пп — поверхность почвы. Одно деление масштаба — 1 см. 5.2. Корни — основные органы семенных растений, их классификация по степени сложности Нет необходимости проводить обзор по проблеме организации корней, поскольку она широко освещена в литературе (Имс, 1964; Ва- сильев и др., 1978; Эзау, 1980 и др.), а новые сведения в этой области накапливаются медленно. В традиционные представления о корнях предлагаем внести некоторые коррективы, созвучные с коррективами к понятию побег, о которых говорилось ранее. Корень (гайх) ионимаем широко, включая в него сиецифические осевые образования как разветвлен- ные, так и неразветвленные с разной сложностью организации (см. гл. 2.5.5.). Важный аспект организации корней, на который обращаем особое внимание, — это иерархичность. По диапазону сложности (он опреде- ляется признаками нарастания и ветвления) можно построить основную
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 251 широкую субординатную классификацию корней (рис. 57-Б). Корень 1-ой субординаты, или монориза, — это простая корневая ось. Монориза по особенностям возникновения бывает субмедиалью, если она первого порядка, т.е. образуется из семени и связана непосредственно через гипокотиль с простой стеблевой осью 1-го порядка (медиалью), или сублатералью, если она 2-го, 3-го и последующих порядков или прида- точная (Нухимовский, 1970, 1971, 1980). Корень 2-ой субординаты — ди- риза — это корень, состоящий из предветви (простой или сложной) и моноризных ветвей, непосредственно из нее образующихся. Корень 3-ей субординаты — тририза — это корень, развивающийся как ветвь, далее разветвленная или неразветвленная и не тождественная другим типам корней. Корень 4-ой субординаты — тетрариза — корень, состоящий из диризы и тририз, взаимосвязанных друг с другом. Кроме четырех типов корней у растений нередко формируются особые системы кор- ней — синторизы. Одна синториза — это не корень, а система, состоя- щая из корней, взаимосвязанных друг с другом или через части одного побега (а-синториза), или через побеговые части всей особи (р-синто- риза), например, Populus tremula L., Linaria vulgaris Mill. 5.3. Ризокормусовая {корнепобеговая} концепция Если применить «бритву» У. Оккама (философ XIV в.): «сущности не следует умножать без необходимости» (принцип экономии, цит. по К. Вилли, 1964, с. 12) к нашим теоретическим разработкам в области организации биоморф и «сбрить» все, что окружает их «ядро», то обна- жится концепция ризокормуса (корнепобега), согласно которой за выс- шую единицу организации биоморф семенных растений принимается ризокормус. Эта концепция разработана в результате сближения двух методологических тенденций в развитии науки. 'аналитического подхо- да, который вскрыл и продолжает вскрывать разнообразие элементов в организации биоморф, и системного похода, с его акцентом на идею целостности, интегративности, иерархичности и взаимосвязи частей, с позиций которого мы рассматриваем всю организацию биоморф. В предыдущей главе было показано, что побег и корень — это основные (ведущие) органы у биоморф семенных растений. Есть ли еще что-либо более значительное в организации биоморф семенных расте- ний, такое, что могло бы выполнить роль высшей единицы организации этих биоморф, роль связующего звена между организмами и синорга- низмами? Вопрос такой масштабности по сути дела не поднимался еще в ботанике, поскольку он распространяется не только на организмы, но и синорганизмы. Дело в том, что ботаники ХХ столетия еще в полной мере не успели «адаптироваться» к концепции А.Арбер (Arber, 1930, 1941, 1950) о единстве стебля и листа в организации одного органа— побега, т.е. к идее двух (а не трех, по классическим представлениям) основных органов семенных растений. Мы поставили вопрос о единстве
252 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ побега и корня в организации корнепобега (ризокормуса), исходя из нескольких прямых и косвенных аргументов. Во-первых, исторические предпосылки, ведущие при соответствующем развитии к идее ризокор- муса, можно усмотреть в идее общей оси (caudex, axis) семенного растения, объединяющей главный стебель и корень, о чем мы уже пи- сали выше (гл. 2.5.1). Кроме идеи единства стебля и корня в организации тела высшего (в нашем случае семенного) растения, заслуживает внимания развитие идеи индивидуальности, которая также восходит с давних времен, а исторический обзор проблемы индивидуальности проводился в литера- туре неоднократно (Левин, 1961; Сенянинова-Корчагина, 1967 и др.). Дело в том, что современные определения понятия индивидуум (орга- низм, особь) основаны на признании единства побеговой и корневой систем как двух частей, обязательно составляющих единство одного тела — особи у семенных растений, за редким исключением, например, организмы Utricularia vulgarus L., которые имеют только побеговую си- стему (Работнов, 1950а; Сенянинова-Корчагина, 1967; Нухимовский, 1973а, б и др.). То, что обычно сейчас называют индивидуумом (организ- мом) однолетних растений, как считает А.Пензеш (Penzes, 1960), еще в 1842 г. Шлейден (Schleiden) принял за исходную форму развития растений, обозначив термином «согтпив» стебель с листьями и корнем. Кроме «кормуса», или сложного растения, Шлейден выделил еще две индивидуальности: клетку, или элементарный орган, и почку, или эле- ментарное растение (цит. по М.В.Сеняниновой-Корчагиной, 1967). А.Пензеш (Penzes, 1960), работа которого подробно обсуждается М.В.Сеняниновой-Корчагиной (1967), упраздняет термин «cormus» в понимании Шлейдена и предлагает обозначать однолетники семенных растений как «монокормоны», а многолетние семенные растения как «поликормоны», рассматривая поликормон как особь — «индивидуаль- ную колонию побегов», кроме того, «разделенные колонии побегов» он называет «фрагментированными поликормонами» (земляника, лук). Наиболее значительные аргументы в пользу рассмотрения побега и корня как единой системы ризокормуса — это современные доказа- тельства такого единства, которые общеизвестны на уровне организмов семенных растений. 1. Единство побега и корня заложено на макромолекулярном уро- вне уже в одноклеточной зиготе, с которой начинается развитие многоклеточной биоморфы у семенных растений. 2. В процессе фенетического развертывания генетической инфор- мации образуется зародыш, осевую основу организации кото- рого составляет единая первичная ось — гипокотиль. «Гипоко- тиль зародыша, проростка и взрослого растения (исключение составляют некоторые однодольные) является той частью оси, в которой осуществляется переход от анатомического строения корня к строению побега; его происхождение восходит к тому периоду в жизни растений, когда происходила дифференциация спорофита на побег и корень» (Имс,1964, с. 12). «На своем нижнем конце (корневом полюсе) гипокотиль несет зачаток
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 253 Рис. 94. Единство побега, гипокотиля и корня в структуре зародыша у семенного растения (по К.Эзау, 1980, с. 22). Схема строения зародыша двудольного растения. А — продольный срез. Апикаль- ная меристема побега (алекс побега) расположена между семядолями, апикальная меристема корня (алекс корня) покрыта корневым чехликом. Прокамбий проходит через весь гипокотиль и семядоли. Б — поперечный срез через семядоли и апикальную меристему побега. Прокамбий расположен в виде трех пучков в каждой семядоле. 1 — семядоля, 2 — апекс побега, 3 — гипокотиль, 4 — прокам- бий, 5 — апекс корня, 6 — корневой чехлик. корня, а на верхнем конце (полюсе побега) выше семядолей- зачаток побега» (Эзау, 1980, с. 21) (рис. 94). 3. Побеги и корни в системе тела взрослого семенного расте- ния — это морфофизиологически непрерывная система. «Тка- ни, занятые проведением питательных веществ и воды, — про- водящие ткани — образуют непрерывную систему, проходя- щую через каждый орган и все растение. Эти ткани связывают места поглощения воды и питательных веществ с зонами роста, развития и запасания. Непроводящие ткани также непрерывны, и их расположение служит показателем специфических взаи- иос ЙяЗеЙ" Яз ш~, Ы80,. с. Уу),
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЧ 4. Единство стебля и корня в организации взрослого семенного растения закреплено в понятии axis generalis vegetabilis (глав- ная, или общая, ось растения), включающей восходящую (стеб- левую) ось — axis ascendens и нисходящую (корневую) ось— axis descendens (Нухимовский, 1970, 1971). 5. Специфическое единство побегов и корней как частей сложно- го куста нашло отражение в некоторых понятиях, например, парциальный куст, простой куст второго или третьего, или чет- вертого, или других порядков (гл. 4.4). Мы оставляем здесь без внимания те случаи понимания кустов, когда к ним относят лишь специфически организованные побеговые системы. б. Доказана особая роль системы побег (побеги) + корень (корни), как высшего органа в организации многих семенных растений. Единство побеговой и корневой частей свидетельствует о наи- высшей предрасположенности корнепобегов, как органов био- морф, к вегетативному размножению (Нухимовский, 1976а, 1980). Этот факт обычно ускользал от соответствующего теоре- тического осмысливания. Итак, есть все основания, чтобы говорить о единстве побега и корня, чтобы разработать концепцию этого единства. Единство побега и корня в системе организма общеизвестно. Единство побега и корня на уровне органов — это новый аспект, который рассматриваем наряду с первым аспектом в рамках ризокормусовой (корнепобеговой) концеп- ции организации биоморф семенных растений. Сформулируем основные положения этой концепции, опираясь на рассмотренные выше аргументы и дополнительные факты. 1. Любой побег и возникшие от его тканей корни, а также побег и связанные с ним через семядольный узел и гипокотиль корни, — это морфофизиологически непрерывная целостная система, названная нами ризокормусом (гйцосогтиз), или корне- иобегом (гл. 2.5.5). Любая почка и образованные из нее и свя- занные с ней корни — это иочкокорень (gemmoradix). Почкоко- рень — это начальное возрастное состояние некоторых ризо- кормусов (рис. 53). Единство побега и корня в форме ризокор- муса, подтверждаемое морфологическими и физиологическими наблюдениями и опытами, определяет содержание одного из законов организации биоморф семенных растений: единство набега и корня, заложенное в генотиие семенного растения, де- терминировано на весь жизненный цикл этого растения в форме ризокормуса, ироявляя себя виервые как одна клетка-зигота, да- лее как система зародыша и, наконец, как все эииэмбриональное растение, которое нередко может быть системой из нескольких иодчиненных ризокормусов (органов) — это закон единства иобе- ги и кория. 2. В организации биоморф семенных растений, как правило, проявляется два типа ризокормусов (рис. 95-97): корнеиобеги- органы, или онтоиды (ироорганизменные ризокормусы), если они составляют только часть структуры растения; корнеиобеги-орга-
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 255 С Рис. 95. Развитие корнепобегов-органов в системе корнепобега- организма у Sedum telephium L. (Московская область). ко — корнепобег-орган (онтоид), кл — корневой клубень, ор — корнепобег- организм (эуфит). Остальные обозначения те же, что и на рис. 93. Рис. 96. Развитие однолетнего корнепобега-организма в онтогении Centaurea cyanus L. (Московская область). рм — растяжение базальных междоузлий главного побега в условиях затенения, уз — сохранение зоны укороченных базальных междоузлий (преконодия) главно- го побега, сс — сборное закрытое соцветие (синфлоресценция), па — паракладий, зт — зона торможения. Остальные обозначения те же, что и на рис. 93.
256 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИИ л й CO (7) 3 О Ж ° 0 Ж Ж Q й Ж 1-' «й,ф Q «1 О ~ О ж 3 Q, Ф Е о ~~ о, Е~ Х )Й оуа л о'~ ~ ° 63 4 Й ~ й IC Ig g X 65 Q,Î О О 65 О > о ~ й О о И ~ '~: О Q Q. О ~'~ Ф О О Ж О Г=, О О й Й Ck О~ О й 0 ~ и Ж ~Д) О ОО И ~о О М Яа: О И О. Р Q в3' Ф [ Й е( Д О 0i ~," И v ~о И Эф а.~а .'3'. О~ Г~ ~~~~ в О ~ i Ch ~ Ф~ о-~ A Ж Й <> Ф~ 3 X ~ Ì О И о ~„ ID ~ Й 3 "а~ Ж О ~~ и О gp Й Ж alDO : О О Ц О Ж д о С') ~ О О И ~ Я Фа .О Ж ~вф о О О qp :„О Фа ~з
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 257 низмы, или эуфиты (организменные ризокормусы), если они сос- тавляют всю корнепобеговую структуру растения (весь орга- низм). Главное достоинство новых терминов «онтоид» и «эуфит» по отношению к их синонимам — краткость. Автор надеется, что они не исключат, а дополнят друг друга. Отдель- ный корень в структуре родительского растения вместе с не- сколькими адвентивными побегами, возникающими из него,— мо а-ризосинкормус, или ризосинкормус-орган, иначе корнеот- нрысковый онтоид. Синкормус одной особи и связанные с ним корни — это ~3-ризосинкормус, или ризосинкормус-организм, ина- че корнеотнрысковый эуфит (растение-особь). Онтоиды — это высшие органы (no сложности организации) биоморф семенных растений. Они более других органов приближаются по органи- зационной сложности к целому организму. Ризокормусы и ри- зосинкормусы как органы — это оптимальный исходный мате- риал в вегетативном размножении, поэтому мы рассматриваем их как высшие единицы в вегетативном размножении семен- ных растений. Полное обособление онтоидов сразу же приводит к вегетативному размножению, к возникновению новых орга- низмов. Все остальные элементы в организаци биоморф (сома- тические клетки, листья, стебли, почки, корни и т.п.) — это все же второстепенные (дополнительные) элементы в вегета- тивном размножении семенных растений. Характер предрасположенности растений к размножению мы закрепляем в следующем обобщении: чем больше (меньше) часть в целом организме ириближается по организационной сложности к целому организму, тем больше (меньшс) этот организм нредрас- ноложен к размножению, а его часть к нреобразованию в новый самостоятельный организм (закон нредрасноложенности к раз- множению). К вегетативному размножению наивысшую пред- расположенность проявляют организмы семенных растений, в структуре которых развиваются ризокормусы-органы (высшие единицы в вегетативном размножении семенных растений); их особенно развивают многие кустистые растения, например, Fe- stuca pratensis Huds., Viburnum opulus L. и многие корнеотпрыс- ковые растения, например, Populus tremula L., Cirsium arvense (L.) Scop. и др. 3. Побег выступает как лидерный (господствующий) орган, как детерминанта в организации системы какого-либо корнепобега, поскольку он выполняет особую, наиболее существенную в жизнесохранении, роль. Из многих функций побега, которые входят в элиту жизнесохранительных функций биосистем, на первом месте стоят диксенобиофильные функции, большин- ство из которых недоступны корням. Структурно-функцио- нальное превосходство побегов над корнями и корнепобегов- органов над остальными органами семенных растений подчи- няется закону фенетического доминирования в организации био- морф семенных растений. только те органы вынолняют роль
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫА РЛ СТЕНИЙ 4. 7. доминантов в фенетической организации биоморф семенных рас- тений, которые неиосредственно исиолняют несколько их функ- ций: образуют и удобно расиолагают в окружающем жизненном иространстве реиродуктивные (в т. ч. иоловые) органы, обесиечи- вают всем необходимым (веществами, энергией, информацией), то, что связано в этои организации с акцией полового размноже- ния и с эмбриональным развитием иотомства, — это no6ezu и ризокормусы-органы и (или) ризосинкормусы-органы. Основные типы ризокормусов удобно выделять с учетом уровня организационной сложности (субординаты) побегов, т.е. опре- деляющих элементов этих органов: ризомонокормус — это мо- нокормус и, связанные с ним и из него отрастающие, корни; подобным образом мы определяем ризодикормус, ризотрикор- мус, ризотетракормус и т.д. (рис. 57-В). Зародыш — это у се- менных растений — эмбриональный ризоиротерокормус. Сериофикацию ризокормусов удобно строить с учетом сери- альной классификации побегов, например, по ряду призна- ков — длине междоузлий и особенностям расположения лис- розеточные, безрозеточные сближенноузловые, безрозеточные раздвинутоузловые ризокормусы. Единство побега и корня проявляется не только на уровне целых побегов, образующих корни и превращающихся в систе- му корнепобега, но и на уровне их элементов — гемикормусов (полупобегов), когда те образуют корни и превращаются таким образом в другой, более сложный, элемент — ризогемикормус (корнеиолуиобег). Ризогемикормус — гемикормус вместе с корнями, развитыми на нем, нередко образуется у корневищ- ных растений (Origanum vulgare L.), реже в структуре надзем- ных частей корнепобегов (Vanilla planifolia Andr.). Ризогеми- кормусы в зависимости от сложности организации, составляю- щих из лидерных элементов гемикормусов, можно различать следующим образом (субординатная классификация): иервый ризогемикормус (первый гемикормус вместе с корнями, образу- ющимися от него); второй ризогемикормус; третий ризогемнкор- мус и т.д. (типы ризогемикормусов определяются подобно первому ризогемикормусу) (рис. 59-Б). Ризокормусы, если они вообще образуются, выполняют все функции биоморф семенных растений. Из чего складываются эти функции? Во-первых, функции, присущие побегам и кор- ням по отдельности, присущи и ризокормусу как их интегра- тивному целому. Во-вторых, ризокормусы отличает на уровне органов специфическая функциональность, которая не при- суща побегам и корням по отдельности: ризокормусы-органы проявляют высшую готовность из всех остальных органов к самостоятельной жизни; они готовы приобрести статус орга- низмов сразу же после свершения полной партикуляции. В- третьих, ризокормусам присущи особые функции на уровне
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 259 организмов — это их способность быть наивысшим элементом в организации синорганизмов. 8. Ризокормусы как органы, организмы, так и специфические си- стемы из нескольких ризокормусов-организмов, названные нами синорганизмами, — это все динамические системы, глав- ная осбенность которых рост и развитие. Главная исполнитель- ская роль в росте и развитии ризокормусов на клеточном и тканевом уровнях организации принадлежит верхушечным меристемам. Они непосредственно реализуют генетическую ин- формацию, создавая структуру побегов и корней. Функция роста и развития — это самая общая функция, а все ос- тальные функции — это составляющие и (или) подчиненные ей фун- кции. Главные среди этих функций — те функции, которые направлены на сохранение собственной и (или) чужой жизни биоморф, например, поглощение воды и минеральных веществ из почвы, дыхание, синтез органических веществ, защита и обеспечение пищей эмбрионального потомства. 5.4. Феноцентры в организации биоморф семенных растений Выше было дано самое общее определение побегу, что обеспечило условия для классифицирования частных побегов на основе разных принципов (субординатного, сериального или, наконец, многолиней- ного), выделяя тот аспект, который интересует в данный момент. Через понятие побег — доминирующий орган (только он способен образовать репродуктивные органы) в фенетической организации биоморф семен- ных растений можно характеризовать различные по сложности органи- зации образования листостеблевой природы от единичного цветка (Camellia japonica L.) до очень сложных систем, состоящих из ствола с ветвями, листьями и репродуктивными органами, например, у тысяче- летнего баобаба (Adansonia digitata L.). В зависимости от целей иссле- дования можно вести оценку побегов по любой из разработанных нами классификаций. Наиболее доступной (по простоте подхода), но весьма показательной является субординатная классификация побегов по их роли в пространственной организации растений: генеральные (главные), субгенеральные (добавочные) и локальные (рис. 65). Если охарактеризовать эти и иные типы побегов в структуре какого-либо растения, например, Bupleurum fruticosum L., по многолинейному принципу, то получим разноплановую картину фенетической организации биоморф этого вида. Однако, прежде чем дать описание определенной фенетической органи- зации биоморф, обратим внимание еще на одно их свойство. Дело в том, что фенетическая организация биоморф семенных растений наст- роена с генетического уровня на успешную реализацию главной фун- кции — размножения. Среди лидерных систем такой организации био- морф должны быть те безусловные лидеры, от функционирования ко-
ОСНОВЫ БИОМОРФ ОЛОГИИ СЕМЕННЫХ РЛ СТЕНИ@ торых более всего зависит ее реальное содержание в развитии. Так, мы подходим к представлению о фенетическом центре, или феноцентре [от греч. phaino — являю, kentron — острие (циркуля), сосредоточие], в организации биоморф семенных растений. Феноцентр — это вегета- тивная часть растительной биоморфы, которая морфофизиологически яв- ляется ее родительской серединой, иоскольку более всего удерживает в жизнедеятельном состоянии остальные родительские (исходные) структу- ры и активно участвует (или участвовала) в сезон вегетации в органооб- разовательной деятельности всей фенетической организации биоморфы (рис. 98). Феноцентр — это своеобразный хранитель определяющих орга- нообразовательных родительских структур в бурно изменяющейся струк- туре растительного организма или синорганизма. Рис. 98. Основное разнообразие феноцентров в организации биоморф семенных растений. 1 — условные границы феноцентра отдельной растительной биоморфы опреде- ленного типа, 2 — зона отдаления, 3 — постцентр, 4 — эона полной партикуля- ции, 5 — вторичный феноцентр, 6 — многолетние части растения, 7 — однолетние части растения. В каждом конкретном случае границы феноцентра устанавливают- ся индивидуально, но есть и общие закономерности. Используя геомет- рическую терминологию, все биоморфы семенных растений можно под- разделить на два типа по особенностям их феноцентра: концентричес- кие, если биоморфы имеют один общий феноцентр (организмы), и экс- центрические — они развивают не один фенетический центр, а несколь- ко вторичных (синорганизм в стадии клона). Феноцентром семенного растения может быть вегетативная часть побега и (или) корнепобега или система нескольких побегов и (или) корнепобегов, которая действи- тельно является родительской системой для других систем, в особеннос-
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 261 ти репродуктивных, морфофизиологически связанных с ним прямо или косвенно (через иные органы, например, корни). Феноцентр — это как раз та часть растительной биоморфы — ее вегетативное «ядро» (сере- дина), которая более всего принадлежит непосредственно биоморфе. Все другие морфофизиологически активные (органообразовательные) части растения, только внешне напоминающие феноцентр, но располо- женные вне его (они несколько отдалены) — это периферийные зоны повышенной органообразовательной активности, наиболее существен- ные из которых можно обозначить как иостцентры. Феноцентр синорга- низма — это вегетативное тело родительской особи семенного проис- хождения, которая обеспечивает или уже обеспечила начало клону. В случае синорганизма, кроме его феноцентра (иервичного феноцентра), проявляются феноцентры организмов вегетативного происхождения (вторичные феноцентры). В свою очередь в организации клонистов (осо- бей вегетативного происхождения) кроме их вторичных феноцентров могут более или менее развиваться периферийные зоны повышенной органообразовательной активности, т.е. их постцентры. Ранее (Смирно- ва и др., 1976) была предложена оригинальная классификация биоморф по числу «центров разрастания, т.е. мест наиболее активного новообра- зования корней, побегов и почек возоновления» (с. 28): моноцентричес- кие и полицентрические типы биоморф. Еще раньше А.Пензеш (Penzes, 1960, цит. по М.В.Сеняниновой-Корчагиной, 1967) разработал понятия монокормон и поликормон для характеристики общих особенностей организации биоморф. Проблему центризма в освоении биоморфами пространства и времени мы решаем с несколько иных позиций. Особо фиксируем в фенетической организации биоморф их способность по- стоянно быть отцентрированными, т.е. всегда иметь один центр на уровне организма и часто несколько центров (вторичных) в стадии клона на уровне синорганизма. Феноцентр семенного растения, после полной гибели составляющих его элементов, например, генерального побега (корнепобега), перемещается к другим частям растения — другому по- бегу (корнепобегу) или нескольким сближенным побегам (корнепобе- гам) и, таким образом, в отличие от феноцентра синорганизма, присут- ствует в структуре любого растения постоянно. Наблюдая за поведением феноцентра и производных от него сис- тем у конкретных биоморф и анализируя литературные данные, можно сделать следующее обощение. Развертывание генетической ирограммы у биоморф семенных растений больше идет через феноцентр, обладающий оиределенными свойствами, — родительскую систему, от которой обра- зуются иодобные ей ироизводные системы и которая вьтолняет главную роль в поддержании фенетической организации всей биоморфы. Таково место фенетического центра в организации биоморф семенных растений. Каждый новый живой элемент (гипобиоморфа) организации био- морф развивается не сам по себе, а в структуре целостной системы- биоморфы, которая с позиций гипобиоморфы является для нее частич- но или полностью средой существования (биотопом), а следовательно, и ареной борьбы за существование (гл. 4.7). Bupleurum fruticosum L. — володушка кустарниковая (сем. Apia- ceae). Современный ареал (Средиземноморье) этого вида, по предпо-
OCHOEbl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ложению Б.М.Коза-Полянского (1915), представляет собой «обрывки» более обширных площадей распространения в древности. Володушка кустарниковая известна в Крыму, где культивируется как декоративное растение и нередко встречается в одичалом состоянии (Шмальгаузен, 1895; Малеев, 1933 и др.). В.Рау (Rauh, 1939) наблюдал Bupleurum fruticosum в горах Средиземноморья и охарактеризовал этот вид как подушковидный аэроксильный кустарник. Б.М.Коза-Полянс- кий (1914) считал володушку кустарниковую в Крыму вечнозеленым полукустарником. Биоморфогения Bupleurum fruticosum изучалась (Хржановский, Ну- химовский, 1968) в Никитском ботаническом саду и его окрестностях (в одичалом состоянии в основном на территории можжевелового (Juni- perus excelsa Bieb.) лесного заповедника на мысе Мартьян (рис. 99 -101). Виргинильный этап. Размножается семенами. Прорастание семян надземное. На первом году жизни особи володушки проходят несколько возрастных состояний (рис. 99, А-В). Сначала образуется проросток, представляющий из себя прорастающее семя, или семядольный почко- корень (ризопротерокормус). Гипокотиль выносит семядоли на поверх- ность почвы. Семядоли продолговатолинейные (1,6-2,2 см. дл.). Корне- вая система: сначала главная монориза, а далее она начинает образовы- вать первые боковые корешки, превращаясь в диризу, до 5 см. дл. С началом развертывания первого настоящего листа особь становится всходом. Это семядольный вегетативный безрозеточный ризомонокор- мус. Первые настоящие листья черешковые яйцевидные и яйцевидно- овальные. Корневая система представлена диризой, т.е. главным кореш- ком и первыми боковыми корешками, а в дальнейшем боковые корешки начинают ветвиться и дириза превращается в тетраризу. Когда у малень- ких растеньиц володушки, образовавших уже 4-6 настоящих листьев, высыхают семядоли, особи превращаются в ювенилов, т.е. вступают в ювенильную стадию онтогенеза. Главный медиальный побег продолжает моноподиальное нарастание. На второй или третий год виргинильной жизни начинают образовываться сидячие листья. Ювенильное растение представляет из себя бессемядольный ежегодно вегетативный безрозе- точный ризотетракормус. Главный универсальный плейофазный прос- той (неветвистый) побег, или главный тетракормус, ежегодно прирас- тает одним новым верхушечным годичным побегом, т.е. нарастает мо- ноподиально. Ювенильная фаза жизни особи продолжается до 2-4 лет и завершается с началом первого ветвления главного побега, когда растение становится имматурным. Ежегодно весной отрастают годич- ные вегетативные безрозеточные побеги как из верхушечной почки, так и некоторых пазушных почек. Имматурное растение — это ежегодно вегетативный безрозеточ- ный надземно или приземно ветвистый ризогексакормус или ризогеп- такормус. В структуре гексакормуса впервые в текущем вегетационном сезоне проявляется два порядка побегов системы ветвления: нулевой (главная ось, или предветвь) и первый (боковые оси, или собственно ветви). У гептакормуса володушки на фоне простого ветвления по- прежнему проявляется верхушечный моноподиальный годичный при-
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 263 3 3 Q М О~ О~ О Ж 0» Е О О Е ~л» рЯ О w :Д О до Ж ~М О Ж О Ж ~ ~) М ~ж йр ~ о о,~ Д Ж Ф ~) h4 g о й ~ Ж О»»» ~ц & ~Ж ~) О C) О О О Ж & Ж Q О Ж Ф~ Ф~ 30 И "'жж Ф ~" м о & ~ М 22 ~~= е) 1 Ю Ф~» .О Ж ЮЗ~ВЩ в~ "„ <~ О Я С'1 рй Ф v Я Йа > д: 3 g~ ; ~2- Й g4 ~'.) ~)" о~~ "и ~о X~ Q л О v" 3~~ х о ~ ~~ О Я Й ф; у ~ 3 "~: о ж Q, л» ~ ]» 3Vxэз ° ~) И 65 р~~ ФЭ у G, ~ ~> ~о 3 >~ + ~~и~ ф „.~. л» ~î ~ ~ A Ж ~~~~ ~щ ~ о ~ ~ ,4 3 Ж и й д л»»» ф) Qyь О I д ~~~~ 3Х Ф л л» Я л» 65 A y A G ~> О М О с"1 Ф~ >ь 1 ra и О ло О. ~, „ О О i ~ ~> О „до~ И И~2 о~о 1 М х ОPi 0 )Я )Я Q Й Й л Ц Ф ц Ф) v ~а И ФЭ М о О д о И ~ И О. ре v QQ о Ж о йЩ )~ И Я Д Й 65 Q Ц~ С'1 ми~ о д~ ре л» о~~ О Й;~~ ~65 g o >~ Й а г~у 1 .~о ю О :~». ф ~» ~ ИЗ д; О И -': Й Х Q, Й 4Э & )д Ц ~ а.+ а5 0 Й р~ () й о~»~ ,ю~ л >Х A -о~ Ж v ФЭ ~) о N И Х Я Фэ О ~» р~»~ о3 О ~» Ф» Ц л» О О ре ~» Ж о 3 < 5» ФЭ Ф» ,'~ ~1 -~o р О ФЭ й; 1 ~Р QmФ: л» 3 й зй 61 ф„' О Ж i ц~ ФЭ aS < а. И О Ж О В~ л» О И О. О И 1 CO »О Ж О. О М Ф» 1 О1 л ю й О и И Ф» Ж сб ф' Ю й м ~ д 3 у 3 й О .'Г. М 65 й; ф ~ О И Ф» й М ~з О В~ О. И О Ж ~» Ю О И Ф: 3 З В~
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ Рис. 100. Образование побегов и строение куста у взрослых особей Bupleurum fruticosum. Е — медиматурное (физиологически зрелое средневозрастное растение); 21— восьмилетний субгенеральный вегетативно-репродуктивный октокормус, 22 — ло- кальный вегетативно-репродуктивный октокормус, 23 — рыхлокустистый безро- зеточный семидетерминатный энатокормус, 24 — рыхлокустистый стержнекорне- вой гемисимподиально-эусимподиальный ризоэнатокормус, 25 — эпигипогеоген- ный разветвленный геоподий, 26 — разветвленный аэроподий, 27 — сухой верху- шечный остаток побегов прошлых лет, 28 — мономерный магнорезид, 29 — пенек отмершего главного аэроподия, 30 — субгенеральный годичный побег (турион). Остальные обозначения те же, что и на рис. 99. рост медиального побега, а, кроме того, могут отрастать и новые годичные пазушные побеги первого порядка ветвления. В первые годы жизни растения ветвятся надземно или приземно, т.е. если и происходит куще- ние, то оно приземное и образуются аэроксильные кусты. Семядольный узел и приземные основания ортотропных побегов по- степенно втягиваются с помощью контрактильности гипокотиля,
Глава 5. Параметры основных органов и их частеи в организации оиоморф семенных рост ений 265 Рис. 101. Разнообразие побегов текущего года вегетации, образовав- шихся из почек верхних частей разновозрастных аэроподиев Bupleurum fruticosum. а — побеги на верхушке аэроподия молодого репродуктивного куста, б — побеги на верхушке аэроподия средневозрастного куста, в — побеги на верхушке аэро- подия отмирающего старого куста; 31 — мономерный бревирезид, 32 — слабый вегетативный годичный побег. Остальные обозначения те же, что и на рис. 99, 100. корневой системы и естественного присыпания почвой основания растения на горных склонах в результате эрозионных процессов. Годичные побеги у имматурных особей образованы в своей верхней части удлиненными междоузлиями, открытыми узлами и очередными си- дячими ассимилирующими удлиненно-овальными или яйцевидно-про- долговатыми цельнокрайними кожистыми листьями, функционирующими до 3-х лет. В основании у этих побегов 3-5 укороченных междоузлий и сближенные чещуевидные листья; нередко, вслед за низовыми листьями, развиваются 1-2 ассимилирующих листа со слабой пластинкой. Ежегодное моноподиальное нарастание за счет полностью резидных (не отмирающих после вегетационного сезона) годичных побегов обеспечивает формирова- ние главного стволика и многолетних ветвей. У растений, произрастающих на открытых местах (например, вдоль дорог), виргинильный этап продолжается 2-4 года, а на затененных участках, например, под пологом можжевелово-дубового (Juniperus excelsa-Quercus pubescens) леса, — 4-5, реже 6 лет. Матурный, или репродуктивный, этап (рис. 99, Д, 100, 101) насту- пает на 3-7 году онтогенеза. Первое кущение иногда происходит только на этом этапе жизни. Многолетнее основание растений к этому времени
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ уже несколько углубляется в почву, что обеспечивает условия для при- земного и подземного кущения. Многолетняя подземная побеговая часть растения — геоподий, или ксилоподий (см. гл. 6.3), если применить терминологию, разработанную И.Г.Серебряковым и Т.И. Серебряковой (1965) для травянистых многолетников по способам формирования корневищ, будет иметь промежуточную структуру между двумя основ- ными типами — эпигипогеогенную, т.е. сначала геоподия нет, затем первично надземное основание главного стволика несколько погружа- ется в почву и, наконец, оно начинает функционировать как подземное, т.е. как геоподий, от которого отрастают турионы — побеги кущения (субгенеральные побеги) с небольшими первично подземными основа- ниями. Медиальный побег плейофазный; он завершает основной (пост- инициальный) период своего жизненного цикла, как правило, после образования на верхушке соцветия — сложного зонтика через 3-7 лет развития. Аксиллярные универсальные побеги, начавшие свою актив- ную (видимую) жизнь на виргинильной стадии онтогенеза, также, как правило, завершают плейофазное (2-5 фаз) развитие образованием тер- минального соцветия. Матурные особи образуют аксиллярные побеги чаще всего годичные (монофазные), что происходит в кроне аэроподиев, а реже — двух,— четырехгодичные (геоподиальная поросль, или тури- оны). Геоподии (подземные многолетние побеговые части куста) корот- кие. Геоподиальная часть растения составляет фенетический центр («сердце») компактного или рыхлого куста, поскольку живет дольше всех остальных частей и, следовательно, дольше вссго удерживает в своей структуре исходные (родительские) части растения и имеет реша- ющее значение в органообразовательном процессе. Поскольку фенети- ческий центр куста составляет эпигипогеогенная геоподиальная часть, то весь куст по способу кущения можно считать аэрогеоксильным, т.е. с переходным способом кущения между аэро — и геоксильным типами. Ежегодно в кроне куста образуются годичные полузакрытые обоеполые репродуктивна-статурные раздвинутоузловые побеги с очередным рас- положением (2/5) многолетнезеленых сидячих листьев и немногочис- ленные ростовые (статурные) годичные побеги. Цветение происходит в июле-августе, плодоношение в октябре-ноябре. После плодоношения верхушечное соцветие вместе с несколькими (3-6 и более), следующими ниже, междоузлиями отмирает, т.е. годичные побеги унирепродуктив- ные акронекрозные (см. гл. 6). Живые остатки отплодоносивших акро- некрозных побегов переходят в многолетнюю структуру куста в статусе резидных, более или менее длинных, побегов. Из таких резидных годич- ных побегов состоит основной прирост аэроподиев, которые сами живут в среднем 12-15, реже 17-20 лет (дольше всех живет главный аэроподий) и достигают высоты 1,2-1,8 м. Максимальные годичные приросты глав- ного аэроподия происходят на 5-8 год жизни. Главный и все остальные аэроподии вместе со своими подземными основаниями (геоподиями) составляют более сложные многолетние системы — гетероподии. Глав- ный гетероподий связан с главным корнем, который сохраняется на протяжении всей жизни куста; придаточные корни, если и образуются, то их мало и они маломощные и не способны заменить главный корень.
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений 267 Совокупность всех взаимосвязанных простых (без ответвлений) геопо- диев, составляющих подземную побеговую структуру основания кус- та, — это один сложный (разветвленный) геоподий. Такой геоподий формируется у взрослых растений. Нарастание новых годичных побегов происходит из пазушных почек прошлогодних частей резидных побегов мезосимподиально (Гатцук, 1967). Молодые матурные особи имеют возраст от 3-7 до 9-12 лет. Это ежегодно терминально-репродуктивные (с терминальными полузакрыты- ми соцветиями) безрозеточные постоянно облиственные гетероподиаль- ные простокустистые ризогексакормусы (рис. 99, Д) или ризогептакормусы. Эти особи характеризуются образованием наиболее сильных годичных по- бегов, от которых после окончания вегетационного сезона остаются на- иболее крупные облиственные резиды, т.е. резидные побеги (рис. 101-а). Средневозрастные матурные особи от 9-12 до 15-18 лет. Это еже- годно репродуктивные безрозеточные постоянно облиственные гетеро- подиальные сложнокустистые ризоэнатокормусы (рис. 100, Е). В этом возрасте ежегодно обильно образуются годичные унирепродуктивные обоеполые побеги (рис. 101-б). Старые матурные особи от 15-18 до 25-35 лет. Это ежегодно репро- дуктивные сложнокустистые ризоэнатокормусы. Заметно ослабление побегоотрастания и степени развития побегов. В стеблевой системе уси- ливаются процессы отмирания, которые приводят к ослаблению глав- ного аэроподия; вторичные (пазушные) более молодые аэроподии спо- собствуют гибели главного стареющего аэроподия, депрессируя его су- ществование. С отмиранием главного аэроподия, который непосредс- твенно через геоподий связан с главным корнем, значительно убыстря- ется гибель всего куста. Увеличивается число вынужденно вегетативных побегов с несостоявшейся репродуктивностью. Когда процессы отмира- ния в системе куста достигают максимума, то особь отмирает или пере- ходит в сенильное состояние, в котором за 1-3 года она доживает свой онтогенез вынужденно вегетативно, т.е. с утраченной репродуктивнос- тыо (рис. 101-в). Аэроподии засыхают. Куст разваливается и отмирает на открытых местах в возрасте 20-25 лет, а на затененных участках— в 25-35 лет. Таким образом, особи володушки кустарниковой развиваются в Крыму в рамках биоморфотипа: вечнозеленых безрозеточных обоепо- лых терминально-репродуктивных желто-сельдерейноцветковых акро- некрозных аэрогеоксильных стержнекорневых кустарников. Завершим главу 5 несколькими иллюстрациями (рис.102-107). Ог- раничимся лишь подробными подписями к ним. Традиционные описа- ния биоморфогении здесь опускаем. В скобках приводятся индикатор- ные признаки возрастных состояний, которые непосредственно наблю- дались при изучении жизненных циклов конкретных растений. В ос- тальном, как мы полагаем, рисунки и подписи к ним способны «ска- зать» сами за себя о многом и передать, на языке уже рассмотренного выше теоретического материала, основную информацию о развитии структурной организации биоформ Trigonella foenum graecum L., Sedum aizoon L., Artemisia аппиа L., Geum rivale L., Hedysarum alpinum L.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ СЧ С0 О й 0» % О Ж й,' Ж Ж Ж р,М й Ж Ф;3,' с~ й ~О ~ д Я~ 3 О ,'!,' ~ Ж ~, О 00 Ж Е ~~ Щ Ж Ж ~м Я ж'~ h4 цО о~ Ж О[к ~ ~ж ~-1 ~ а5 О Жж ;~, 'О 65 >Ж ~~ О ~~ Ж ~' о 3 Ж ~~ + 3 Ж Ф ."5'. 0» х gg М ~О л и Х ц~ ~ ФЭ О ФЭ 3Х о 3 u > р; О ~ щ G a5 Ф~ л Д G. ~З ФЭ G a5 м о,~ 6~ ФЭ () 6~ ~ф Фэ C д 65 65 Й а.~ а ~ ~~ о p)~м 1 5» Ж .Д ФЭ A ФЭ О ~0 р~ р~ 3 ~~ и Ж л» а.< y »О й> ~.> ~; Ж ф О Ц ID ~~>В v 3 & Ф~ ~р Ф» ~ц О О о О Р хИО 3 ~~1 р) а,й ФЭ >.И ~ О ~ р» ~Ч о Я Вам Я О И ФЭ � а.~ ~~о ra И д а5 Х в~а я о а д Д,д) Й о ~~ О Х О О Q л» Щ а,~ о ОAL О.g X ФЭ ~я A ~~, C m ~ ~) л > ~' а. 3 л О (.)" М л З О а, о о ~ " а. О Ж ф„' о у, 3 Ф» л» ре 65 Ф Й ФЭ ФЭ Ф» A Ж Ф:О 3 О й 00 3 л а ~", о ; М 3 ~ а. < Ф» О а > ФЭ '3» О О & а 4~ О а О к Д а ~р 1~ .я Ж л Щ й ~р о ~. И v ~» п Я ж ( О '~ О & ф) л о О > а. й С'1 Q, О Ю Ф» р; О И О 3 О 5» Х а. ~~ ~й Ф» ф» л» A ~ц Ж Ф 65 Ф Ф » 3 ФЭ О ('1 p) G ~ 3 Ю Ж Ф» З З В~ г рд; 6~ 3 ре ,'Я :~, ~~ м о о ~ > Ю г ~о~ ц о ~а~ ~~Ц Ж О ~д Ю e ~- о Я 2 3 Ц1 < а. ФЭ й' 0 О КЗО -Яо A ~p РЪ ~Д) О ф) л а И ~9 3 И о.Я > О О С'1 65 g oe о ~ О л ф» 5: v О О ~6 ;Ц ~ О P > ~ 3 а. О О Ж Ж й( ~ о ФЭ д а а у Ф~ g л» Х Й»
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 269 Рис. 103. Первый год развития структурной организации травянистого многолетника Sedum aizoon L. (Москва, ВИЛАР, интродукционный питомник). А — проросток, Б — всходы (семядоли живы, растет главный побег), В — юве- нильные растения (семядоли отмерли, продолжается вегетативное развитие глав- ного побега), Д — матурные (физиологически зрелые) растения; 20 — придаточ- ный корень, 21 — главное закрытое верхушечное соцветие, 22 — паракладий (эусиллептический побег обогащения), 23 — бессемядольный закрытый безрозе- точный ризодикормус без паракладиев, 24 — закрытый безрозеточный ризодикор- мус с паракладиями, 25 — верхушечное соцветие паракладия. Остальные обозначе- ния те же, что и на рис. 102.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ М З Ж 3 О Ф~ 3 Ж й О й Ж < й~ Ж Щ C C ° м сл ° и Е М Ж Ж В»! О Ж О й:[ Ж й й Ж Ж О О ~Ж О Ж Q о 0» О Ж Ф ,0 л О ! О ! л ~. м ~ о ~ "о~ ~~~ Ж ~ v 63 g Щ ~~~~~ ОЮ л ~.~ ~: 1 !~: а 'Фкцб .Аа О а3~К ~о ~~ а. а о >~ ~ ~~ о ~~ о р Д ф 3 ЗvQО ra -~хи"а.> +~оgхю Ыо~~~~<~ ОИ(лvо Й л~ <Ю д а о оxoox о о ~ ф ~ о р 'о~ о "~'~ ~д л Щ g ~~ л а, Ф ~их а~а~~мо д 0 v v > а. '~ И ~ " ж qцХ<<~<~ , ) !.) ц ФЭ ~> 3 g Щ ~ ~а v0~ Щ 65 Q, ~1 Фа Х Ф И Л ~,! Я} 3 д . ~,~ .'~'. Ж О <~) Зч ,'Я ~Q~IОБО й; а, с4 рф ~ '~ с~) Х Я Р') - + > ~ ~о <> ~!~ :; ~ рй,; ~О ю й О ~ щ v о о~ v й ц! ~ ,'ц ФЭ .~, ,'Ц ~ ~ ~ ~о О Ы р Ж г ~~ й; Й о; ~ ~ Х ~.Р.-~ И а й ю ~йо о <!~ 1 о Д: G ~ Ж ,'ц ~!~" а с~) ~> ~~ 3 И д д ~л ~и" g Ю я 4э СЧ g ж ~ x "„-я #gal ( ~о С4 Фэ .ж om и д ~ ~Я ~ И ф И ~ 0iе " "И а.> Ж о �-Я ~0 д ~ о QQ3c Q oxuxa а'а~о~~~ О Хе О З й Е n ~~ с й � à e6 а 2Ы 1 О М й kf О й а. !Й~~ а М еч й О ф~ е ое Q A p й а О Ф З Ж й й () л !) ФЭ 65 о g В~ О а ж С'1 ц~ О а.~ о (Г) ою ~) Й ~О й ~ Q Юо i ~!~ 3 О Ж 3 ФЭ о ~ И ~ Ф» ,'3', ою v Ж щ й; ~ а. О ~ о о И О О Фа ,'Ц ою v A Ж ~ф Ф Й ф) Й Фа Я} Ж З 1
Глава 5. Параметры основных органов и их частей в организации биоморф семенных растений 271 14 Рис. 105. Развитие структурной организации травянистого многолетника Geum rivale L. I — виргинильный этап, II — взрослая особь (Московская область) см. стр. 272. А — проросток, Б — всход (живые семядоли, зародышевая почка развернулась), В, — униювенильные особи (развивается розетка почковидных или широкояйце- видных листьев, семядоли отмерли), В, — биювенильные особи (развертываются первые полусложные тройчато и перисторассеченные листья, корневище гетеро- номное, т.е. оно связано с главным корнем), Г — имматурная особь (корневище теряет связь с отмершим главным корнем и становится автономным). 1 — семядоли, 2 — семядольный узел, 3 — гипокотиль, 4 — корневая шейка (при- мерное положение), 5 — главный корень, 6 — главный розеточный побег, 7— ризопротерокормус, 8 — бессемядольный вегетативный гетерономно-ризомный розеточный ризомонокормус, 9 — вегетативный гетерономно-ризомный розеточ- ный ризотетракормус, 10 — вегетативный автономно-ризомный розеточный ри- зотетракормус, 11 — гетерономный ризом, 12 — автономный ризом, 13 — границы годичных приростов (годичных побегов), 14 — придаточные корни, 15 — сухие остатки прошлогодних листьев, 16 — мономерный курторезид, 17 — межсоцветие, 18 — интеркалярный побег, 19 — основание интеркалярного побега (верхняя часть удалена), 20 — постинтеркалярный побег, 21 — иносоцветие, 22 — основания ро- зеточных листьев (верхние части удалены), 23 — открытый мультирепродуктивный автономно-ризомный ризогептакормус; у — увеличено по сравнению с масшта- бом; горизонтальная линия — поверхность почвы. Одно деление масштаба — 1 см.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ
Глава 5. Параметры основных органов и их частей в организации оиоморф семенных растений Рис. 106. Виргинильное развитие структурной организации травянисто- пэ многолетника Hedysarum alpinum L. (Москва, ВИЛАР, интродукционный участок). А — проростки, Б — всходы (семядоли живые, развертывается зародышевая почка), В — ювенильные растения: В, — униювенильные (семядоли отмерли, главный побег развивает только простые листья), В, — биювенильные (главный побег начинает развивать тройчатые листья), Г — имматурные растения: Г, — униимматурные рас- тения (главный побег, нарастая моноподиально, начинает ветвиться силлептически, точнее образует индуктивные вегетативные побеги), Г, — биимматурное растение (нарастание вегетативных побегов эусимподиальное); 1 — семядоли, 2 — семядоль- ный узел, 3 — гипокотиль, 4 — корневая шейка, 5 — главный корень, 6 — главный вегетативный годичный побег, 7 — пазушный вегетативный годичный побег, 8— ризопротерокормус, или семядольный почкокорень, 9 — семядольный вегетативный ризомонокормус, 10 — бессемядольный вегетативный ризомонокормус, 11 — вегета- тивный ризодикормус, 12 — вегетативный ризогептакормус; горизонтальная линия— поверхность почвы, одно деление масштаба — 1 см.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ Рис. 107. Матурное развитие структурной организации травянистого много- летника Hedysarum alpinum (местопроизрастание то же, что и на рис. 106). Д — инициматурные особи (некустистые или простокустистые), Е — медиматурные осо- би (сложнокустистые, непартикулируюшие или партикулирующие в структуре каудекса), Ж — финиматурные особи (некротическая партикуляция распространилась в главный корень, иногда она приводит к полному разделению родительского растения на обособ- ленные части — особи клона, т.е. к несовершенному вегетативному размножению). 1— зона возобновления, 2 — зона полного торможения, 3 — зона неполного торможения, 4 — сборная фронда-фрондулозная открытая кисть (приоткрытое верхушечное соцветие, полностью отмирающее после плодоношения со всей надземной частью годичного по- бега), 5 — паракладий, 6 — главный корень, 7 — примерное положение семядольного узла у взрослых растений, 8 — каудекс, 9 — основание унирепродуктивного побега (верх- няя часть удалена), 10 — безрозеточный открытый репродуктивный ризогексакормус, 11 — ризогептакормус, 12 — ризоэнатокормус, 13 — неполная некротическая партикуля- ция, 14 — фрагмент плодоносящего соцветия, 15 — особь клона, 16 — парциальный кау- декс, 17 — придаточные корни; горизонтальная линия — поверхность почвы. Одно де- ление масштаба — 1 см.
Глава 6. Продолжительность жизни: эвриэфемерность и персистентность Биоморфы, участвующие на своем уровне в процессе сохранения Жизни, выбирают в ходе эволюции следующие варианты продолжи- тельности жизни: 1) эфемерность, однолетность; 2) унирепродуктивная двулетность, многолетность; 3) мультирепродуктивная многолетность (не более 300 лет); 4) мультирепродуктивная сверхмноголетность (более 300 лет) (Нухимовский, 1984). 6.1. Организация биоморф как способ резервирования ресурсов Каждая частная жизнь на Земле дискретна. Всякое живое существо постоянно находится под угрозой смерти; она незримо следует за ним, то удаляясь, то приближаясь к нему. Живое и неживое находятся в едином и противоречивом круговороте ресурсов, т.е. круговороте ве- ществ, энергии и информации, составляющем окружающую среду био- систем и сами биосистемы. Биоморфы обеспечивают репродуктивный процесс, что запрограммировано в них на генетическом уровне еще на ранних этапах эволюции. В зависимости от многих экологических факторов (погодные условия, конкуренция и т.п.) они выработали спо- собность или быстро реализовывать использованные ресурсы, заканчи- вая свою жизнь менее чем через один год, или постепенно расходовать часть вовлеченных ими в оборот ресурсов на репродуктивный процесс, а другую — откладывать в запас (резервировать), что дало возможность таким биоморфам жить более одного года. Растения в процессе эволюции адаптировались по-разному закан- чивать сезон вегетации и по-разному уходить в зиму, а там, где прохладных зим нет, они реагируют на иные неблагоприятные, в осо- бенности погодные (засуха, наводнение и т.п.), условия. Способность или неспособность частей растений, в особенности побегов, сохранять
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИИ жизнеспособность после завершения сезона вегетации или цикла репро- дуктивного развития заслуживает особого внимания. У некоторых рас- тений разрастаются в толщину черешки листьев, у других растений стебли, а у третьих корни и т.п. (Федоров и др., 1956, 1962; Васильев и др., 1978). Часть биоморф накапливает и расходует потребленные из окружающей среды ресурсы в одном вегетационном сезоне, иногда всего за несколько дней (эфемерные растения и насекомые) и даже часов и минут (некоторые водоросли и бактерии). Лишь небольшая часть био- морф расходует их постепенно на протяжении десятков и сотен (дуб, ель) и даже нескольких тысяч лет (секвойя, баобаб). Если биоморфа и (или) какая-либо ее часть живут с момента их возникновения не менее одного года, преодолевая при необходимости за это время неблагопри- ятный для роста и развития климатический период, то можно говорить о явлении, названном автором нерсистентностью (от лат. «persistens»— остающийся, неопадающий); иначе — это сверхгодичная продолжи- тельность жизни. Чаще всего при резервировании ресурсов в биоморфе возникают специализированные запасающие системы («депо») — денонирующие (реэервирующие) органы, ткани, клетки, органеллы. Любые части (почки, листья, стебли, побеги, корни) биоморфы или целые биоморфы, кото- рые зимуют и сохранили жизнеспособность в сезонном климате хотя бы после одной зимовки, а в бессезонном климате прожившие не менее одного года и также не утратившие жизнеспособность, — это соответ- ственно нерсистентные части биоморф или нерсистентные (дву-, много- летние) биоморфы (Нухимовский, 1988). Если биоморфа и (или) какая- либо ее часть живут с момента их возникновения меньше одного года и при необходимости они не могут преодолеть неблагоприятный для роста и развития климатический период, то такое явление можно обо- значить как эвриэфемерность (эфемерность в широком смысле слова); иными словами — это догодичная продолжительность жизни. Из- вестные термины «эфемеры» и «однолетники» не требуют здесь допол- нительных пояснений. Заметим, что части структуры растений (побеги, листья, корни) могут развиваться эвриэфемерно (или эфемерно, когда живут несколько недель — менее двух месяцев, или как однолетние, когда живут дольше, но не более одного года). Однако, чтобы не услож- нять структурную терминологию, мы будем в отношении всех эвриэфемерных элементов в структуре биоморф использовать один термин «эфемерные» (нобеги, корни, листья и тл.), а в отношении целых эвриэфемерных биоморф примем традиционную терминологию- «эфемер», «однолетник». Среди персистентных частей особое место занимают скелетные осе- вые (стеблевые и корневые) части, на которых нередко развиваются персистентные почки и листья. Нецелесообразно (это противоречит представлениям о многолетности органов), хотя так часто делают, на- зывать скелетными также иные осевые части побегов — «многолетние в будущем». Например, к ризомам нередко относят подземные части побегов в сезон их отрастания, когда есть только предположение, что они будут жить на следующий год и дольше. Скелетные части — это
277 Глава 6. Продолжительность жизни: эвриэфемерность и нерсистентность осевые органы или части осевых органов, сохранившие жизнеспособ- ность после одного или более сезонов вегетации. Иерархичность побегов по степени персистентности (сформиро- ванности для длительного сохранения) представляется нам в виде широкой субординии побегов и их частей (рис. 108). Рис. 108. Персистентность и эфемерность в организации биоморф семенных растений. 1 — 4 — субординия частей биоморф в зависимости от степени их персистентности, 1 — эфемерные части, 2 — преперсистентные части, 3 — семиперсистентные час- ти, 4 — персистентные части, 1а — апобазомеры (эфемерные побеговые части), 2а — пробазомеры (потенциально персистентные побеговые части), За — ге- мибазомеры, 4а — базомеры, 16 — аскелетные ризомеры, 26 — проскелетные ризомеры, 36 — гемискелетные ризомеры, 46 — скелетные ризомеры, 5 — апи- кальная (неразвернутая) часть, 6 — субапикальная (развернутая) часть.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИИ Нижнюю субординату занимают эфемерные (незимующие) нобеги или эфемерные чисти нобегов — анобазомеры (они отрастают и отмирают за один вегетационный сезон, а если это почки, то они появляются и развертываются в одном сезоне). Далее идут — нробазомеры (нотенци- ально нерсистентные, или нренерсистентные нобеги (осевые чисти можно называть нроскелетными, т.е. нотенциально скелетными) или чисти но- бегов (они предположительно должны стать неотмирающими на всю или на часть своей длины). Более высокую субординату в нашей клас- сификации займут семинерсистентные (в отношении осевых частей мож- но использовать термин гемискелетные) нобеги или части нобегов (они включают в свою структуру персистентную часть наряду с одной или несколькими другими частями — эфемерной и (или) преперсис- тентной — это гемибазомеры. Самую верхнюю субординату занимают нерсистентные нобеги или нерсианентные части нобегов — базомеры (в отношении осевых частей можно использовать традиционный термин «скелетные стебли»). Корни, которые являются основными частями, подразделяем по их жизнеспособности соответственно так же, как и стебли (эфемерные, проскелетные, гемискелетные и скелетные). По- скольку основные осевые части — стебли и корни представляют собой каркасные элементы биоморф семенных растений, а побег без осевого (стеблевого) каркаса — уже не побег, то все основные показатели, при- сущие стеблю как части целого (побега), присущи побегу в той же мере, что и стеблю. По особенностям функционирования апикальных меристем раз- личаем нобеги нассивные и активные. У нассивных no6ezoe алексы либо уже утратили жизнеспособность, либо после вильчатого раздвоения стали апексами других осей. Активные no6ezu обладают жизнеспособны- ми апексами. Среди них различаем мулыниактивные и субактивные нобеги. У мультиактивных no6ezoe их апикальная часть проявляет повы- шенную активность, а побеги в целом — видимый рост; у субактивных no6ezoe их апикальная часть функционирует замедленно, а побеги в целом не проявляют или почти не проявляют видимого роста; для них обычен скрытый рост. В зоне умеренного климата побеги зимуют обыч- но в пассивном или субактивном состояниях. 6.2. Резидофикация и базомеризация Главное содержание предлагаемого нами термина «персистент- ность» составляет образование в организации биоморф растений отно- сительно неотмирающих частей. Признаки персистентности (кольца го- дичных приростов в эндоструктуре стеблей, экзоморфологические сле- ды границ сезонных приростов и т.п.) указывают на продолжительность жизни растений и их частей. Они важны для понимания не только нынешней, но и прошлой организации биоморф. Стеблевую часть побега, которая стала скелетной после завершения им видимого роста и (или) репродуктивного развития предложено (Ну-
Глава 6. Продолжительность жизни: эвриэфемерность и персистентность 279 химовский, 1968а, 1969а, 1970, 1971) называть резидом (от лат. resi- duum — остаток, reses, idis — остающийся). Кроме резидов неотми- рающую часть побегов составляют нередко зимующие (нерсистентные) ночки и зимующие (нерсистентные) листья или какие-либо части лис- тьев, например, зимующие черешки. Побеги, каркас которых составляют резиды, — резидные, или нерсистентные. При отмирании частей побе- гов могут иногда оставаться только зимующие почки, т.е. новые ини- циальные побеги (им только в будущем предстоит видимый рост). Например, у многих видов Dactylorhiza Nevski и Aconitum L. вместе с корнями, как уже отмечалось, они образуют почкокорни. Широкая субординатная классификация персистентных побеговых частей (базомеров) семенных растений имеет следующий вид (субординаты рассматриваются снизу вверх): 1) базовые, или нерсистентные нобеговые, элементы (резиды, зиму- ющие листья и почки), составляющие структуру базокормусов; 2) базокормусы, или нерсистентные нобеги; среди них есть и целые базовые органы растительных организмов и все те побеги, кото- рые составляют эти органы; 3) синбазокормусы, или нерсистентные нобеговые системы в отдель- ных организмах, состоящие из нескольких базокормусов, кото- рые связаны между собой через органы другого морфофизиоло- гического типа (корни). Кроме базомеров, в структуре организмов семенных растений обычно развиты персистентные (скелетные) ризомеры (скелетные корни и их элементы), что позволяет выразить рассмотренный выше материал в следующей модели организации персистентных (неотмирающих после первого сезона вегетации), или базогенных, растений (рис. 108). По положению в пространстве побеговых частей относительно друг друга и степени их сформированности нами различаются аникальная (неразвернутая) и субаникальная (развернутая) части, что мы отражаем на единой модели с предыдущими типами побегов и их частей (рис. 108). Изучение явления персистентности — важный раздел современной биоморфологии, который пока менее всего разработан. Наши исследо- вания поднимают проблему эвриэфемерности и персистентности (соот- ношения этих явлений) в организации биоморф семенных растений и отчасти решают ее, что отражено в рассматриваемых здесь обобщениях. Наличие или отсутствие персистентной части (частей) и особенно- сти ее (их) расположения в структуре биоморф семенных растений характеризуют ранги сложности организации биоморф и могут быть основанием для еще одной широкой субординатной классификации этих биоморф (рис. 57-В). Нижнюю субординату занимает в рассматри- ваемой классификации анобазовые (без базового органа) биоморфы, сред- нюю субординату — базогенные (образующие базовый орган), или базокормусовые, биоморфы без каулогенных корней и (или) ризогенных но- бегов, верхнюю субординату — базогенные биоморфы с каулогенными корнями и (или) ризогенными нобегами. В умеренном климате побеги зимуют обычно в пассивном или субактивном состояниях. Пассивные нобеги утрачивают способность активно нарастать прежним апексом
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИЙ (алексами) — он погибает, выполнив все или часть своих функций, а у субактивных набегов жизнеспособные верхушки вступают в стадию относительного покоя; они как бы затаиваются и выжидают нового всплеска суммы эффективных (положительных) температур, чтобы вновь начать, прерванный непогодой, видимый рост. Способность по- бегов полностью или частично отмирать или вовсе не отмирать после завершения видимого роста, сезона вегетации и (или) репродуктивного цикла составляет одну из важнейших групп признаков биоморф семен- ных растений, имеющую особое значение при изучении как их внеш- него облика (габитуса), так и внутренней организации. По особенностям перехода побегов из мультиактивного состояния в субактивное или пас- сивное состояния была разработана (Нухимовский, 1970, 1971, 1988) сле- дующая сериальная классификация набегов (рис. 109): некрозные, или эфемерные (отмирают полностью — Padus avium Mill.); базиакронекрозные, Рис. 109. Особенности перехода побегов из мультиактивного в субактивное или пассивное состояние. 1 — эфемерный побег, 2 — акронекрозный побег, 3 — мезонекрозный побег, 4— базиакронекрозный побег, 5 — базинекрозные побеги, 6 — кладонекрозный по- бег, 7 — кладобазинекрозный побег, 8 — апонекрозный побег, 9 — корни; пунктирной линией показаны зоны отмирания осевых частей побегов после окончания сезона вегетации и (или) репродуктивного развития.
Глава б. Продолжительность жизни. звризфемерность и персистентность 281 или амфинекрозные (отмирают не полностью сначала снизу, а затем сверху — Ajuga reptans L.); базинекрозные (отмирают не полностью, сни- зу — Solanum tuberosum Ь.); мезонекрозные (отмирают не полностью, начиная с середины и сохраняют в живом состоянии нижнюю и верхнюю части — Rubus caesius L.); кладонекрозные (главная осевая часть их не отмирает, а полностью отмирают входящие в их структуру пазушные побеги с репродуктивной функцией — Plantago major L., Rhodiola rosea L.); анонекрозные (главная осевая часть не отмирает, а пазушные побеги с репродуктивной функцией не развиты — Picea abies (L.) Karst.). На основе рассмотренного выше материала можно разобраться в соотношении процессов отмирания и сохранения как живых элементов организации биоморф, так и биоморф в целом. Явления иерсистентной и эвриэфемерной иродолжительности жизни биоморф составляет второе содержание их фенетической организации (nepeoe составляет стремление к размножению). Итак, биосистема может достигнуть иерсистентности (ирожить более года) при наличии соответствующих условий среды и защитных ирис- иособлений, если с момента рождения за один вегетационный сезон (год в бессезонном климате) функции иотребления и рассеивания у нее окажутся несбалансированными хотя бы по некоторым элементам и в соответствии с этим она будет обесиечена ресурсами более чем на год. Однако биосис- тема будет эвриэфемерна, если в иодобной ситуации функция иотребления у нее уравновешена функцией рассеивания (закон нерсистентности и эвриэфемерности). Логическую связь с этим законом ироявляет первое быть больше времени ироявления дисбаланса между функциями иотребле- ния и рассеивания ресурсов (веществ, энергии, информации), а когда coom- ношение между уиомянутыми функциями выравняется, биосистема элиминируется. Второе иравило: чем дольше существует в биосистеме дисбаланс между функциями иотребления и рассеивания ресурсов в иользу системы резервирования, тем больше шансов у нее, чтобы иродолжать жизнь. Рассмотренные выше обобщения об эвриэфемерной и персис- тентной продолжительности жизни биоморф семенных растений— второй нринцин их фенетической организации. Субординатные кпассификации персистентных побегов построены нами с учетом рассмотренных ранее широких субординатных кпассифика- ций мультиактивных побегов (гл. 5.1.2). Субординатная классификация персистентных (резидных) побегов сезонного KJIHMaTa может состоять из трех субординат: внутригодичные, годичные и сверхгодичные резидные no6ezu. Приведем еще несколько субординатных кпассификаций персис- тентных побегов (рис. 110, 111). Основная широкая субординия персис- тентных побегов при меньшем обобщении представлена на рис. 111. Нулевая субордината — нерсистентные (резидные) нротерокормусы (иначе нротеробазокормусы) представляют собой особые побеги — протерокор- мусы, которые зимуют после завершения сезонного видимого роста побегов и (или) остаются жизнеспособными более одного года. Первая субордината — нерсистентные монокормусы (монобазокормусы) — од-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 110. Графические модели субординатных классификаций мультиактивных побегов (А) и базокормусов (Б), построенные по особенностям систем нарастания и ветвления. а1 — альфакормусы, а2 — бетакормусы, а3 — гаммакормусы, а4 — дельтакормусы, б1- а-базокормусы (образуются из аl и а2), б2 — р-базокормусы (образуются из а3), б3- т-базокормусы (образуются из а4), жэ — живые эфемерные осевые части, оч- отмершие части, л — листья, образовавшиеся в текущем сезоне вегетации, зл- многолетнезеленые листья, гп — границы резидных монокормусов прошлых лет, гг- границы резидных монокормусов текущего года, в — вегетативная верхушка, у — уаиы, образовавшиеся в прошлом году или позднее, сц — живые соцветия, оц — отдельный цветок, 1, 2, 3,4 или (3), (4) — порядки простых осей в системе нарастания, О, 1, Н, 111... — порядки осей ветвления, 0-1, 1-П и т.п. — тире между цифрами означает слово «или» (ось системы ветвления может быть предветвью и иметь нулевой порядок ветвления или ветвью и иметь первый и более порядок ветвления).
Глава б. Продолжительность жизни: эвриэфемерность и персистентность О 0а Ю О И Х Ж ~ч Ж О й О Q И й й Ж й В: [ Q О Ю О Ф~ О М О С4 й B Ф~ О Ж О Ж О О В: [ О й й Ян Q Оч О й С4 ~> ! о. H ~ o > +< g И Я "и За З~ ~Я 'оа1 X~З:й о, ~ ' о о 55 C 54 v О ; ою Д я З: О „о и ~ й й ~ с~ И~vìхи оюдg "О~~ „o > р; ." се 63 О~о З'" co~RBXh С~ д Q, С~ lD ~ аэ Е. ~о ~ д pz + ~ o v Q Q C X ! ~~З:о Ф О О д" v ! ~4 и м о"'>З: + m ~о м ~~ ~;~ ~д д о ю о 3: ~„е~ QО~~Ю БИО ~~~и ~ о~~о„-~, М f» bC v X о~ >- о о З; ~ Са1 ~„Qx~ (ч са ~ й, ~р - О g n ф о! О~о М ~> З е; И ее "а.~ О Ю~~иЯа. о~~ й X ~ ~-а й> ОQQЗ: И~Q ! CV о ~ o;~ Ю .", И В ~.-; д О ~ u ~ О (~ф л ~ З~ ~л М й~ д Я ~ ~ Ц м E О oxgp аоо~ж„ 63 За М ~ о ~ с" <Ооц -o~g Q О h4 И и
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННБ1Х PA СТЕНИЙ ноосные мономерные (результат одной фазы видимого роста)резидные побеги, образующиеся после завершения видимого роста монокормуса или какого-либо другого побега (каркас составляет одноосный моно- мерный резид); вторая — нерсистентные дикормусы (дибазокормусы), или двуосные и многоосные мономерные резидные побеги, образующи- еся из постинициальных дикормусов; третья — иерсистентные трикор- мусы (трибазокормусы), или одно-, дву- и многоосные димерные и плей- омерные (результат двух и более фаз видимого роста) noiezu, обра- зующиеся из постинициальных трикормусов; четвертая — иерсистен- тные тетракормусы (тетрабазокормусы), или одноосные димерные и плейомерные резидные побеги, образующиеся из мультиактивных те- тракормусов или более сложных побегов (последние побеги, частично отмирая, могут сохранять после себя живую структуру тетракормусов); пятая — нерсистентные нентакормусы {'иентабазокормусы), или двуос- ные и многоосные димерные и плейомерные резидные побеги, образу- ющиеся из постинициальных пентакормусов; шестая — иерсистентные гексакормусы (гексабазокормусы), или двуосные и многоосные моно-, ди- и плейомерные резидные побеги, образующиеся из постинициаль- ных гексакормусов; седьмая — нерсистентные гентакормусы (гентаба- зокормусы), или двуосные и многоосные димерные и плейомерные ре- зидные побеги, образующиеся из мультиактивных гептакормусов; восьмая — нерсистентные октокормусы (октобазокормусы), или двуос- ные и многоосные димерные и плейомерные резидные побеги, образу- ющиеся из мультиактивных октокормусов; девятая — иерсистентные энатокормусы (энатобазокормусы), или многоосные плейомерные ре- зидные побеги, образующиеся из мультиактивных энатокормусов. Важное значение для понимания структуры персистентных побегов и всей базы какого-либо растения имеет процесс образования резидов (резидофикации). Резидофикация — частный случай или составная часть базомеризации (ироцесса образования базомеров). Резидофикация осуще- ствляется иосредством иерехода одной, нескольких или всех верхушечных меристем иобега к относительному иокою или иосредством отмирания какой-либо части набега на оиределенную длину при сохранении живых частей стеблей иосле окончания сезона вегетации. Эти особенности в резидофикации нашли отражение в однолинейной сериальной класси- фикации резидов (Нухимовский, 1969а, 1971). Положение резидов от- носительно поверхности почвы указывается первичное, т.е. в период их образования (рис. 83). Всего было выделено 1Б основных тинов резидов снециальных и универсальных нобегов. Тотарезиды — резиды (длинные, короткие и компактные) надзем- ные (полностью или не полностью), которые образуются из стеблевых частей открытых апонекрозных или кладонекрозных длинных, коротких и компактных (кроме розеточных) побегов (Fraxinus excelsior L., Populus nigra L., Ficus carica L., Quercus гоЬиг L., Juglans гера L., Pinus sylvestris L., Comus mas L. и др.). Курторезиды — резиды надземные (полностью или не полностью) или подземные, которые образуются из компактных (сближенноузло- вых) стеблевых частей открытых апонекрозных и кладонекрозных ро-
Глава б. Продолжительность жизни: эвриэфемерность и персистентность 285 зеткообразующих побегов (Caragana arborescens Lam., Plantago maxima Juss., Succisa pratensis Moench, Narcissus pseudonarcissus L. и др.). Вице- резиды, или резиды подземные ортотропные с более или менее удлинен- ными междоузлиями, которые образуются из стеблевых частей открытых апонекрозных или кладонекрозных верхнерозеточных побегов. Вицере- зиды нередко образуются у розеточных растений при погребении их субстратом. Пленорезиды, или резиды (длинные, короткие, компактные) подземные горизонтальные или ползучие непосредственно по по-верх- ности субстрата, образующиеся из стеблевых частей открытых апонек- розных или кладонекрозных длинных, коротких и компактных (кроме розеточных) побегов (Paris quadrifolia 1.., Convallaria majalis L.). Алиорезиды образуются после отмирания снизу (базинекроза) побегов с живой верхушкой (Solanum tuberosum L.). Максиморезиды — резиды надземные (полностью или не полнос- тью), которые образуются из стеблевых частей после отмирания сверху (акронекроза) открытых длинных побегов не более чем на одну треть (Tilia cordata Mill., Salix caprea L., Morus nigra L., Corylus avellana L., Betula pendula Roth, Eucommia ulmoides Oliv. и др.). Магнорезиды- резиды надземные (полностью или не полностью), которые образуются после акронекроза не более чем на одну треть закрытых или условно закрытых длинных побегов (Betula pendula Roth, Juglans regia L., Асег platanoides L., Phellodendron amurense Rupr., Aesculus hippocastanum L. и др.). Малорезиды образуются подобно максиморезидам, но из стебле- вой части открытых коротких или компактных, как правило, угнетен- ных побегов (Tilia cordata Mill., Ulmus laevis Pall. и др.). Модесторези- ды — это надземные резиды, образующиеся после отмирания репро- дуктивных элементов закрытых или условно закрытых коротких побегов (Acer ginnala Maxim., Ailanthus altissima (Mill.) Swingle и др.). Медиоре- зиды — резиды надземные (полностью или не полностью), которые образуются после акронекроза длинных побегов примерно наполовину (Sorbaria sorbifolia (L.) А. Вг., Bupleurum fruticosum L., Physocarpus opulifolius (L.) Maxim., Viburnum opulus L. и др.). Бревирезиды — резиды надземные (полностью или не полностью), которые состоят из укоро- ченных частей, несущих или несших низовые (почечные чешуи) или розеточные листья, и раздвинутоузловых или иных сближенноузловых частей; образуются после акронекроза длинных побегов не менее чем на две трети (Artemisia lerchiana Web., Sambucus racemosa L., Philadelphus coronarius 1.., Sorbaria sorbifolia (L.) А. Вг. и др.). Миниморезиды- резиды надземные (полностью или не полностью) или подземные с укороченными междоузлиями, если образуются после полного ак- ронекроза длинных и коротких частей побегов или отдельных верху- шечных репродуктивных элементов укороченных побегов (Eryngium campestre 1.., Sambucus racemosa L., Adonis wolgensis Stev., Hyacinthus orientalis L., Polygonatum multiflorum (L.) All., Dictamnus gymnostylis Stev., Capparis herbacea Willd. и др.). Викариорезиды — это подземные орто- тропные резиды с более или менее удлиненными междоузлиями, обра- зующиеся после полного отмирания надземных частей побегов (Cirsium arvense (L.) Scop.). Викариорезиды нередко образуются при
286 OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ У побегов более сложных, чем специальные и универсальные, си- стема составляющих их основу резидов может быть менее или более разнообразной. Так, резидная часть всей побеговой системы (энатокор- муса) взрослой особи Capparis herbacea Willd., если она произрастает в условиях относительного постоянства уровня поверхности почвы, со- стоит только из миниморезидов. Энатокормус взрослой особи Sambucus racemosa L. часто включает большое разнообразие резидов от максимо- б Для составления терминов, обозначающих различные виды резидов, нами были ис- пользованы следующие латинские слова: totus — цельный, полный, весь; maximus— наибольший; magnus — крупный, большой; malus — плохой; modestus — уме- ренный; medius — средний; curtus — укороченный; vice — вместо; brevis — ко- роткий; minimus — наименьший; vicarius — замещающий; reptans — стелющийся; portio — часть, доля; plenus — полный; alius — другой, иной; ramosus — ветвистый. погребении субстратом травянистых растений с безрозеточными и по- лурозеточными побегами (Peganum harmala L., Pimpinella saxifraga L., Eryngium campestre L. и др.) (рис. 39). Релторезиды — стелющиеся по поверхности почвы (приземные) и иногда укореняющиеся длинные или короткие резиды, образующиеся после отмирания надземных орто- тропных частей побегов (Hypericum olympicum L., Alyssum пшга1е Waldst. et Kit. и др.) (рис. 51). Порциорезиды — ползучие непосредственно по поверхности почвы (приземные) или подземные горизонтальные длин- ные или короткие резиды, которые образуются после отмирания над- земных ортотропных частей или недоразвившихся вегетативных верху- шек (Sambucus ebulus L., Elytrigia repens (L.) Nevski, Нурег1сит maculatum Cratz. и др.) (рис. 43). Все рассмотренные выше резиды неразветвленные. Особое место занимают разветвленные резиды рамозорезиды', когда они остаются после неполного отмирания сверху главной оси и репродуктивно-статурных паракладиев сложных соцветий побегов (Hypericum androsaemum L., Cistus tauricus, С. Presl и др.) (рис. 82, IV) или силлептически разветвленных побегов с иной специ- ализацией. Репродуктивна-статурные паракладии сложных побегов сами являются силлептическими монофазными простыми или сложными по- бегами, поэтому, если мы рассматриваем их по отдельности, то уже говорим по отдельности и о резидах, образующихся из стеблевых частей этих побегов. Резиды, как и резидные noiezu, бывают моно-, ди-, три-, ллейомерные (4, 5 и т.д.) в зависимости от того, из стеблевых частей скольких моно- фазных побегов они образованы, например, мономерный магнорезид Tilia cordata Mill., плейомерный (5, 6 и т.д.) тотарезид Fraxinus excelsior L. В скобках удобно указывать число мономерных резидов в структуре более сложных резидов. Тип резидов можно указывать и при характеристике соответствующих побегов, например, максиморезидный монокормус, тотарезидный (3) тетракормус и т.п. Параллельно при резидной характеристике побегов подчеркивается, где это необходимо, для лучшего понимания их организации, состояние активности этих побегов: мультиактивные, субактивные или пассивные, например, муль- тиактивные миниморезидные энатокормусы Capparis herbacea Willd. и т.д.
Глава б. Продолжительность жизни: эвриэфемерность и персистентность и магнорезидов до медиа-, бреви-, и даже миниморезидов, а у юных растений иногда единично могут развиваться и короткие мономерные тотарезиды (см. рис. 179). Терминология персистентных побегов, как видно, получается не- сколько сложной, поскольку увеличивается нагрузка в термине на слово-этикетку, в котором отражается стремление исследователя зафик- сировать эффект базомеризации (процесса образования базомеров, в частности, резидов и других персистентных элементов). Такие класси- фикации побегов могут быть применены при углубленном изучении динамической морфологии побегов. Для более широких целей вполне приемлема самая общая кпассификация побегов по многолинейному принципу. При этом за основу описания берется слово «побег», к которому добавляются определяющие его специфическое содержание эпитеты, число которых опять же зависит от глубины проводимых ис- следований. Итак, есть все основания для следующего обобщения: стеб- левые части (шире — части набега), образовавшиеся в текущем вегета- ционном сезоне, могут иосле окончания этого сезона иревратиться в резиды (шире — базомеры), т.е. стать скелетными (иерсистентными), если су- меют к этому иодготовиться, в частности, усиеют развить защитные (внешние и (или) внутренние~ ирисиособления (особенно у иочек — носите- лей верхушечных меристем), сиособные иротивостоять разрушительному воздействию неблагоириятных условий среды, в иервую очередь отрица- тельных зимних темиератур (сиециализированные иочечные чешуи, сильное оиушение стеблей и оснований листьев, иогружение иочек в ткани стебля и т.n.). Это сиециальный закон резидофикации и базомеризации. 6.3. Базовые органы Анализ явления персистентности будет более полным, если рас- смотреть еще организацию базовых (персистентных побеговых) органов. При определении базовых органов основное внимание традиционно обращается на признаки габитуса, продолжительность существования побегов и их положение по отношению к поверхности почвы. Морфо- логи издавна придумывают для базовых органов особые названия, например, каудекс, корневище и т.п. (Linnaeus, 1751; Ehrhart, 1789; Lindman, 1914; Du Rietz, 1931; Федоров и др., 19б2; Нухимовский, 1969а-в, 1973в, 197ба, 1988 и др.). При объяснении отдельных базовых органов важно учесть следую- щие обстоятельства. Во-первых, сходство строения взрослых особей разных видов дает возможность построить определения типов базовых органов абстрагированно от видовой принадлежности отдельных осо- бей. Во-вторых, преемственность признаков родительских особей у аклонистов и клонистов предопределяет наследуемость структуры определенных базовых органов в рамках вида растений. В-третьих, бывает, что у особей одного вида при реализации генотипа в фенотип при относительно стабильных условиях среды развитие базокормусов
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ идет по-разному, т.е. у такого вида, еще не достигшего в историческом развитии периода стабилизации морфофизиологических признаков, осо- би нередко образуют базовые органы разных типов. В-четвертых, базо- кормусы, как и биоморфы в целом, проявляют пластичность в разных условиях местообитаний, более или менее изменяя свою организацию. Первое обстоятельство мы учитываем при построении абстрагированно- го определения базового органа на основе анализа и обобщения особен- ностей строения как можно большего числа взрослых особей разных видов. Три последних обстоятельства не позволяют игнорировать насл- едственные задатки (генотип) особи при оценке ее базового органа. На основе анализа собственного фактического материала и литературных данных нами разработана следующая схема оценки базо- вых органов. Эти органы бывают протобазовые и эубазовые. Протобазо- вые (неопределившиеся базовые) органы — это те базокормусы, которые по комплексу признаков в биоморфогенезе еще не сформировались (не определились) как эубазовые органы. Базовый орган, который функци- онирует в основном у взрослых особей, образуясь при реализации их ге- нотипа в фенотип до и после первой репродуктивной' функции, определяем как эубазовый орган, т.е. определившийся базовый орган. Препер- систентные побеги или части побегов, из которых непосредственно мо- гут образоваться базовые органы, — это нробазовые (дословно с греч. «раньше базовых») органы. Особенности возникновения эубазовых орга- нов и основное разнообразие их отражено в таблице 1 и рис. 112. В силу исторически сложившейся способности биоморф наследо- вать важнейшие признаки родителей появление проростков предшест- вует образованию сначала протобазового, а затем эубазового органов. Последний нередко образуется, минуя состояние протобазового органа. Только гибель особей останавливает процесс формирования базовых органов на различных этапах. На примере развития базовых органов можно наглядно видеть, как «работает» один из высших законов Природы — закон нредшествова- ния: что-либо в объекте возникает только в движении (развитии) через предшествование (предварение) в чем-либо. Этот закон заложен в сущно- сти объектов Природы, и его проявления можно видеть постоянно: и тогда, когда мы определяем явление жизнь, и тогда, когда разбираем закономерности образования резидов, и т.п. Уместно напомнить, что как частное проявление закона предшествования можно рассматривать известное в геоботанике правило предварения, установленное В.В. Алехи- ным (1950). Момент возникновения у особи ожидаемого эубазового органа фик- сируется по первому проявлению в структуре этой особи наиболее ха- рактерного признака этого органа. Рассмотрим основные типы эубазо- вых органов семенных растений (рис. 112). Аэронодий (aeropodium). Этим термином обозначаем персистентную часть генерального или субгенерального аэрокормусов (рис. 65), если она формируется. Аэроподий — это надземный неветвистый или вет- вистый базокормус, восходящий в виде одной оси (ствола, стволика) от основания куста или от основания главного корня древесного или полу-
Глава б. Продолжительность жизни: звризфемерность и персистентность древесного растения (Нухимовский, 1970, 1973в, 1988). Он бывает глав- ным (генеральным), если в его структуре участвует главная стеблевая предветвь, и добавочным (субгенеральным), если в его структуре нет главной предветви. Таблица 1 Возрастные изменения побегов и их частей при формировании эубазовых органов обазовый орган отобвзовый орган вый орган еконодий нодий одземный или ползучий пробаз омер ризом ом мпактный каупекс декс омер Незрелая луковица елая луковица Незрелая солуковица елая солуковица езрелая елуковица елая елуковица зрелый клубень (побеговый) релый клубень (побеговый) адземный пробаз омер Аэроподий Гетероподий же ноподий омер Тоже Тоже ценоподий еноподий Аэроподий же Тоже Гетероподий же Важнейшие признаки аэроподиев связаны с их положением отно- сительно поверхности почвы и способом нарастания. Если возникнове- ние аэроподия запрограммировано в генетическом коде особи, то этот момент фиксируется после первого образования такого резида, который не свойственен другим типам эубазовых органов. У растений, родители которых формировали этот орган, образование в качестве самого
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 27 Рис. 112. Типы эубазовых органов семенных растений. 1 — главный гетероподий, 2 — добавочный гетероподий, 3 — главный аэроподий, 4 — добавочный аэроподий, 5 — гетерономный геоподий, 6 — автономный геопо- дий, 7 — базиома, 8 — ценоподий, 9 — компактный каудекс, 10 — вынужденно удлиненный каудекс, 11 — гетерономное корневище, 12 — автономное корневище, 13 — зрелая луковица, 14 — зрелая клубнелуковица, 15 — зрелая солуковица, 16— клубни, 17 — конодий, 18 — побеги текущего года, 19 — гипокотиль, 20 — корне- вая шейка, 21 — семядольный узел, 22 — главный корень, 23 — придаточные кор- ни, 24-2б — поверхность почвы: относительно постоянная (24), при повышении (25) и при понижении (2б) ее уровня, 27 — ризобазокормус, 28 — отмершая часть растения. первого резида, свойственного также и другим эубазовым органам, сви- детельствует о возникновении протобазового органа. Если у особей Асег negundo L., Fraxinus excelsior 1.. и многих других первым из резидов
Inaea б. Продолжительность жизни: эвриэфемерность и персистентность 291 образуется тотарезид, то это указывает на возникновение главного аэроподия, минуя состояние протобазового органа, поскольку такие резиды составляют скелетный каркас только аэроподиев. Когда у особей Sambucus гасетова L., Berberis sibirica Pall. и др. первым из резидов образуется соответственно миниморезид или курторезид, то это указы- вает на возникновение протобазового органа, поскольку данные резиды могут составлять также и структуру эубазовых органов, не свойственных взрослым растениям этих видов, таких как компактный ризом, каудекс и др. Аэрокормус Oplopanax elatus (Nakai) Nakai, например, может быть турионом (надземным побегом, отрастающим из почек подземной ча- сти), преперсистентным аэрокормусом, аэроподием (зимой или до развертывания почек весной) — персистентным аэрокормусом, а также аэроподием вместе с побегами текущего года (постинициальными мо- нокормусами) — семиперсистентным аэрокормусом. Аэрокормус (глав- ный и добавочный) может быть рекомендован как универсальная счет- ная единица при ресурсоведческом изучении надземной биомассы и ее частей семенных растений. Базиома (basioma; от греч. basis — основание, ота — новообразо- вание, опухоль). У некоторых видов древесных и полудревесных расте- ний, помимо хорошо различимых в структуре особи длинных и коротких аэроподиев, может формироваться также и многолетнее компактное побеговое основание (базиома), которое может сильно разрастаться в толщину, занимая надземное, полунадземное или подземное положение относительно поверхности почвы. Базиома состоит из самых нижних участков отдельных аэроподиев, более или менее погружающихся в поч- ву, которые сохраняют жизнеспособность значительно дольше самих аэроподиев. Например, у деревянистой лианы Stephania glabra (Roxb.) Miers, произрастающей в Индии, в надземной сфере формируются длин- ные, извилистые и вьющиеся ветвистые аэроподии, а также компактное многолетнее основание, которое постепенно сильно разрастается в тол- щину, т.е. образуется базиома (см. рис. 112). Базиому вместе с корнями (корнем), непосредственно связанных с ней, называем ризобазиомой. Геоиодий (geopodium). Этим термином было предложено (Нухимов- ский, 1970, 1973в) обозначать персистентную часть геокормуса, пере- ходящую или переходившую в прошлом в аэроподии (один или не- сколько). Иными словами, геоподий — это подземный неветвистый или ветвистый эубазовый орган, непосредственно переходящий или пе- реходивший в прошлом в аэроподии (один или несколько). Если гео- подий ветвистый, например, подземное разветвленное основание гео- ксильного кустарника, то каждое отдельное ответвление такого геопо- дия — это геоподиальная ветвь, или геоподий определенного порядка ветвления. Моментом возникновения геоподия будет образование подземного основания первого аэроподия растения. Если в структуре особи ни разу не образовался аэроподий (а генотипом этой особи пре- дусмотрено возникновение такового), то подземная персистентная побе- говая система представляет собой лишь протобазовый орган данной особи. Геоиодии, связанные с системой главного корня, можно называть гетерономиыми, а не связанные с системой главного корня и постепенно
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ отмирающие в основании — автономными. В пределах одной особи не- редко можно встретить ветви геоподия самой разнообразной длины, тол- щины и степени одревеснения(!); их обычно называют иначе: ксилопо- диями, если они деревянистые и, как правило, компактные или короткие (Lindman, 1914; Warming, 1923; Du Rietz, 1931; Серебряков, 1962; и др.), ксилоризомами, если они одревесневающие с удлиненными междоузлия- ми и плагиотропные на большем своем протяжении (Дервиз-Соколо- ва, 1966; Гатцук, 1968), или корневищами в широком смысле (Пятницкий и др., 1963; Шитт, 1968 и др.). Геоиодии подразделяем по линейным размерам на три группы (рис. 44, 52, 100): комиактные, если их общие размеры очень неболь- шие, а узлы в основном сильно сближены (Sambucus racemosa L., Вир1еи- rum fruticosum L.); короткие, если их общие размеры сравнительно не- большие, а междоузлия в основном короткие или длинные, но тогда их относительно мало (Corylus avellana L., Нурепсит androsaemum L., Vi- Ьигпит opulus L.); длинные, если их общие размеры сравнительно боль- шие, а междоузлия в основном длинные (Нурепсит calycinum L., Vac- cinium myrtillus L.). Уместно заметить, что из двух слов «компактный» и «укороченный», употребляемых часто как синонимы, первое во мно- гих случаях, пожалуй, удачнее, особенно если приходится сравнивать стеблевые части по линейным размерам: длинные, короткие, компактные. И.Г.Серебряков и Т.И. Серебрякова (1965) предложили у многолетних трав различать два способа формирования корневищ: погружающиеся (эпигеогенные) и изначально подземного происхождения (гипогеогенные). Геоподии формируются подобным образом (рис. 44, 52, 100). У аэроксильных кустарничков, кустарников и кустистых деревьев, если формируются геоподии, то они эпигеогенные, т.е. погружающиеся. Такие геоподии образуются, например, у Viburnum opulus L. npu погружении в почву первично надземных оснований турионов (побегов кущения), которые, кроме того, еще и полегают, присыпаются лесным опадом, обрастают придаточными корнями. Геоксильные кустарнички, кустарники и кустистые деревья образуют гипогеогенные геоподии из специализированных изначально подземных побегов; после того как эти побеги становятся резидными, они включаются в структуру геопо- дия, например, у Hypericum calycinum L. (рис. 44). В тех случаях, когда древесное или полудревесное (кроме каудексового) растение развивается сначала с аэроксильным кущением, а с возрастом кущение становится геоксильным, то можно утверждать, что образуется эпигипогеогенный геоподий, т.е. смешанного происхождения, например, у Bupleurum fruti- cosum L. (рис. 99, 100). Гетероиодий (heteropodium) — эубазовый орган, включающий пер- систентную часть гетерокормуса или весь персистентный гетерокормус (Нухимовский, 1980). Иными словами, это система, состоящая из одно- го аэроподия и непосредственно связанного с ним геоподия. Этот эуба- зовый орган обычен для геоксильных кустистых полудревесных и дре- весных растений. Возникновение этого органа в структуре растения фиксируется с момента возникновения двух его главных частей: аэро- подия и геоподия.
Глава б. Продолжительность жизни: звризфемерность и персистентность 293 Ценоиодий (соеиоройит), или голоиодий, — эубазовый орган, образу- ющий одну морфофизиологически непрерывную систему растения из аэроподиев и (или) гетероподиев, иногда с участием базиомы (Нухи- мовский, 1973в, 1980, 1988). Если в онтогенезе растения еще не развита аэроподиальная часть, то наблюдаемый базокормус — это протобазо- вый орган — лроценолодий, который в дальнейшем потенциально мо- жет стать ценоподием (эубазовым органом). Корневище, или ризом (rhizoma). Впервые термин «rhizoma» был употреблен при характеристике папоротников Ф.Эрхартом (Ehrhart, 1789). История употребления этого термина кратко рассмотрена в раз- деле 2.5.1. И.Г.Серебряков и Т.И. Серебрякова (1965), исходя из анализа ряда ведущих литературных источников, пришли к выводу, что корне- вище (ризом) — это «подземный или ползучий по поверхности почвы побег, выполняющий функции отложения запасов, возобновления и, часто, вегетативного размножения. Ризом обычно более или менее утол- щен и функционирует в течение нескольких, иногда многих лет, нарас- тая верхушкой (моноподиально или симподиально) и отмирая в старой части. Корневище несет на себе листья или рубцы от них, почки возоб- новления и стеблеродные корни» (с. 67). Напомним здесь положение, которому мы строго следуем: части набегов текущего года, которые лишь иредиоложительно могут стать дву-, многолетними (иотенциально мно- голетние, или иреиерсистентные), не могут считаться базовыми органами (или их частями) и, в частности, ризомами. Мы называем корневищем подземный или ползучий по поверхности почвы персистентный (многолетний) побег (базокормус), никогда не переходящий в аэроподии, не связанный с главным корнем (реже непос- редственно или косвенно связанный с ним) и образующий придаточные корни, которые нередко угнетают жизнедеятельность главного корня и всегда восполняют утрату последнего (рис. 112). Если формирование корневища запрограммировано в генотипе особи, то образование на скелетной стеблевой части первых придаточных корней, за исключени- ем эфемерных, отмирание главного корня или развитие первых подзем- ных резидов с удлиненными междоузлиями, свидетельствует о возник- новении ризома. У корневищных растений каулогенные корни иногда образуются в первом сезоне вегетации особи, т.е. до формирования базового органа. В таких случаях момент возникновения ризома фикси- руется только на втором году жизни растения (Dioscorea deltoidea Wall.). Пока перечисленные признаки у потенциально корневищной особи не проявились, ее базовым органом можно считать лроризом, т.е. одну из форм протобазового органа. Корневища могут возникать, минуя состояние протобазового органа (Brunnera sibirica Stev.) (рис. 12). Ризомы можно подразделять на автономные, если они не имеют связи с главным корнем и постепенно отмирают в основании (Primula macrocalyx Bunge), и гетероиомиые, если они непосредственно или косвенно (через другой орган, например, через боковой корень и т.п.) связаны с главным корнем (Seseli condensatum (L.) Reichenb.). По длине ризомы так же, как и геоподии, бывают ком- иактные, короткие и длинные (Серебряков, Серебрякова, 1965; Нухимов- ский, 1973в; Васильев и др., 1978).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИЙ Первые — очень небольшие и состоят из сближенноузловых моно- мерных резидных побегов (Adonis vernalis L.); короткие ризомы неболь- шие и состоят в основном из раздвинутоузловых коротких мономерных резидных побегов (Veronica longifolia L.); длинные ризомы сравнительно большой общей длины и состоят в основном из раздвинутоузловых длинных мономерных резидных побегов (Leymus racemosus (Lam.) Tzvel.). Ризомы по происхождению бывают энигеогенные (ногружаюшие- ся), гиногеогенные (изначально нодземные) и смешанные (Серебряков, Серебрякова, 1965). Базовые органы, корневища в частности, бывают с персистентны- ми листьями — фолиозные (облиственные), например Bergenia cras- sifolia (L.) Fritsch, и без листьев, афильные (безлистные), например Ligularia altaica DC. Рассмотрим ряд примеров. Особи Inula helenium L., Patrinia sibirica (L.) Juss., Bupleurum longifolium L. обычно несколько лет нарастают аккумулятивно за счет курторезидных монокормусов. Пока не образу- ются стеблеродные придаточные корни, у дву-, многолетних особей этих видов функционирует протобазовый орган — проризом. Образо- вание первых каулогенных корней свидетельствует о возникновении эубазового органа — гетерономного компактного корневища; на разных отрезках жизни таких растений, чаще всего на стадии физиологической зрелости (матурной), главный корень отмирает. В таких случаях неред- ко вскоре происходит полная общая партикуляция, в результате которой обычно осуществляется вегетативное размножение, т.е. возникают кло- нисты с автономными компактными корневищами (рис. 92). Каудекс (caudex). История употребления этого термина в ботанике в прошлом (до начала ХХ столетия) рассмотрена в главе 2.5.1. Здесь затронутые проблемы рассматриваются в более поздний период. Некоторые предложения классиков прошлого столетия (Г.Ф.Линк, Л.К.Вильденов, О.-П. Декандоль и др.) получили развитие в работах в основном советских ботаников. Б.А. Федченко (1913, с. 15) отметил, что «у многих видов (рода Hedysarum L. — Е.Н.) основание стеблей немно- го одревеснелое или чаще всего остаются довольно длинные, разветвлен- ные одревеснелые caudices, дающие ежегодные травянистые стебли». Б.С.Закржевский и Е.П.Коровин (1935) наблюдали каудекс у Salsola laricifolia (Turcz.) Litv. («сильно ветвящийся кустарничек»), Atriplex cana С.А.Меу («полукустарник с массивным каудексом») и других растений. Образование подземного каудекса у тау-сагыза («подушковидного полу- кустарника») отметил М.В.Культиасов (1938). М.С.Шалыт (1950, 1952) называл каудексом многолетнюю, как правило, одревесневающую стеб- левую часть некоторых стержнекорневых травянистых многолетников и полукустарничков, например, Limonium sareptanum (А. Beck.) Gams, Anabasis salsa (С.А.Меу.) Benth. ех Volkens, Artemisia terrae-alba Krasch. и др. В таком значении этот термин употребляется и авторами всех «Флор» среднеазиатских советских республик. В терминологическом словаре Н.Н.Забинковой и М.Э.Кирпичникова (1957, с. 48) сделано следующее обобщение по употреблению термина «каудекс» в отече- ственной литературе: «одревесневшая часть растения, которая является
295 Глава б. Продолжительность жизни: звриэфемерность и нерсистентность Русский термин «стеблекорень» не нов. В понимании Б.М.Козо-Полянского (1915, с. 24) «стеблекорень — caudex intermedius, то есть часть, промежуточная между кор- нем и стеблем, промежуточная шейка~. При характеристике многолетних стержне- корневых растений семейства Apiaceae Козо-Полянский (1915) употребил этот тер- мин, например, для обозначения многолетней стеблевой части Bupleurum bicaule Helm, Bupleurum exaltatum Bieb. и др. промежуточной между корнем или корневищем и цветущим стеблем». Такое определение неконкретно; оно отражает неразработанность в первой половине нашего столетия понятия «каудекс». Е.П.Коровин (1962) подчеркнул, что особенно широко представле- ны каудексовые растения среди нагорноксерофильной растительности Средней Азии и южного Казахстана. Для этих растений характерно «образование между корневой системой и травянистой частью массив- ной деревянистой зоны, погруженной обычно в почву. Это образование описывается как корневище, в действительности же возникает из ниж- ней части годичных побегов. Это каудекс или подземный побег. Накоп- ление растительной массы у каудексовых трав происходит именно в каудексе, достигающем со временем крупных размеров. На верхушке каудекса закладываются почки возобновления, дающие годичные побе- ги. Каудекс может образовывать головки, ветвиться, частично расщеп- ляться, что увеличивает число возобновляющихся побегов. Каудексовая форма роста является по существу подземным полукустарничком, она близка к так называемым многоголовчатым травам» (с. 73). С.Х.Чевре- ниди (1965) у Aconogonon сопапит (Grig.) Sojak (Polygonum сопапит Grig.) каудексом (головой) называет «верхнюю часть корня, несущую на себе стебли с листьями и зимующие почки» (с. 173).... «В формировании каудекса участвуют гипокотиль и верхняя часть корня» (с. 174). Ал.А.Фе- доров, М.Э.Кирпичников и 3.Т.Артюшенко (1962) предложили, учиты- вая многозначность термина «каудекс», отказаться от его употребления и использовать термин «стеблекорень'> (caulorrhiza)» для обозначения многолетней стеблевой части и базальной части деревянистого главного стержневого корня у травянистых многолетников и полукустарничков. Этот термин нашел применение в работах ряда исследователей (Кузь- мина, 1965; Маркова, Медведева, 1965, 1968; Красильников, 1968 и др.). Однако, другие авторы считают, что не следует отказываться от термина «каудекс», а важнее конкретизировать его значение. И.Г.Серебряков и Т.И. Серебрякова (1965) дали афористически краткое определение: ка- удекс есть «стеблевая часть стеблекорня». М.В.Культиасов (1967, с. 9) определил каудекс как «деревянистое основание» полукустарничков, «на котором образуются многочисленные почки возобновления, дающие побеги возобновления». Иногда каудексом называют также укороченную многолетнюю сте- блевую часть, в течение всей жизни родительского растения или боль- шей ее части связанную с главным корнем, у некоторых стержнекорне- вых и одновременно длиннокорневищных (длиннокорневищно-стер- жнекорневых) растений, например, у Medicago falcata L. (Снагов- ская, 1965), Glycyrrhiza aspera Pill., Pseudosophora alopecuroides L. (Нухи- мовский, 1967) и у стержнекорневых корнеотпрысковых трав, например,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ у Acroptilon repens (L.) DC. (Нухимовский, 1967). М.С.Снаговская (1965) отметила, что у парциальных кустов Medicago falcata L. образуются многолетние укороченные стеблевые части, которые она обозначила каудексами 2-го, 3-го и т.д. порядков, связанные между собой ризомами. При изучении морфогенеза каудексовых растений было подчеркнуто, что для каудексов характерна укороченность междо- узлий, составляющих их многолетних стеблевых частей — осевых ос- татков (резидов) унирепродуктивных (обоеполых, монокарпических или моноантерических) побегов, связь с главным корнем в течение всей или почти всей жизни родительского растения (Нухимовский, 1968а,б, 1976а, 1978, 1981; Былова, 1968 и др.). Итак, что же такое каудекс растений; какие признаки его характе- ризуют? Из проведенного обзора ясно, что не следует определять ка- удекс через габитуальную форму; важнее всего руководствоваться приз- наками организации конкретного структурного образования, которое присуще, возможно, не только одному типу габитуальных форм. Многолетность каудекса и мультирепродуктивность каудексовых растений не вызывает сомнений. В согласии с исследователями, учитывающими онтогенетическую и филогенетическую специфичность видов, у которых в течение всей жиз- ни мультирепродуктивных родительских растений сохраняется связь многолетней побеговой части с главным корнем, мы также считаем каудексовыми лишь растения со стержневым главным корнем. Степень одревеснения тканей многолетней стеблевой части не име- ет принципиального значения для диагностики общих границ каудекса (Нухимовский, 1969а). В сравнении травянистого и древесного типа структуры стеблей усматривается больше различий количественного, чем качественного характера (Александров, 1966). Неодинаковая сте- пень лигнификации тканей многолетней стеблевой части прослежива- ется не только у растений разных, но и близких или одного типов жизненных форм и нередко даже у особей одного вида. Если взять ряд растений, например Capparis herbacea Willd., Gypsophila paniculata L., Bupleurum exaltatum Bieb., Тагахасит otTicinale Wigg., Pimpinella saxifraga L и др., то нетрудно заметить, что степень лигнификации тканей много- летней стеблевой части изменяется от очень сильной до крайне слабой. Один из важных признаков каудексовых растений, уже давно прив- лекший внимание исследователей, — это частичное или полное отми- рание побегов. Однако, если учитывать только это и названные выше признаки, то каудексом можно было бы назвать многолетнюю стебле- вую часть некоторых деревьев и кустарников, многих полукустарников, полукустарничков и мультирепродуктивных трав. Пожалуй, подготовлены условия, чтобы дать определение каудексу растений. Основные принципы разработанного ранее (Нухимовский, 1968а, 1969а,б) понятия «каудекс» мы не изменяем, а лишь несколько уточ- няем его грани. Каудекс — это эубазовый орган, стеблевую часть которого составляют компактные резиды (миниморезиды или курторезиды), реже (при засыпании, заиливании и т.п. субстратом) более или менее усложнен- ный за счет вынужденного удлинения вертикальных подземных резидов
Глава б. Продолжительность жизни: эвриэфемерность и персистентность 297 (викариорезидов или вицерезидов), расположенный непосредственно над гипокотилем у мультирепродуктивных или потенциально мультирепродук- тивных особей и отмирающий в конце жизни этих растений, как правило, вместе с главным корнем (рис. 5, 28, 38, 39, 55, 113-120). В этом определении каудекса уточняется его нижняя граница. Гипокотиль — специфическая часть (орган) растения, где, как извест- но, стебель переходит в корень. Нижнюю границу каудекса правильнее всего проводить по верхней границе гипокотиля (семядольному узлу, включая этот узел в каудекс), которая хорошо различима в структуре растений, а нижнюю границу корня — по нижней границе гипокотиля, т.е. по корневой шейке. Поскольку точное положение последней опре- делить трудно, а часто невозможно, то вполне оправдано корневую шейку отмечать приближенно. Согласно определению каудекс имеет побеговую природу; его опре- деляющую часть составляет осевая (стеблевая) структура — резиды оп- ределенных типов, на которых чаще всего развиты персистентные (зиму- ющие) почки и изредка также могут быть персистентные листья. Ины- ми словами, каудекс образуют определенные побеги: миниморезидные или курторезидные, а реже (при засыпании, заиливании субстратом) еще и викариорезидные или вицерезидные. Важным признаком многих растений является разветвленность их каудекса. На момент возникно- вения каудекса у растений, в генотипе которых запрограммировано фор- мирование этого органа, будет указывать или появление в стеблевой системе соответствующего растения первого миниморезида, или обра- зование первой скелетной части стеблевой ветви, или в иных случаях (длительное моноподиальное нарастание) сохранение жизнеспособнос- ти особи после завершения ее первого репродуктивного цикла. Образо- вание первым из резидов курторезида, как показывает опыт анализа базовых органов, не следует принимать на ранних этапах онтогенеза потенциально каудексовых особей за момент возникновения каудекса, поскольку есть растения, развивающиеся по облигатно унирепродук- тивному типу и формирующие конодий из курторезидов. Термин «конодий» (от лат.co(n) — с, совместно, nodus — узел) был предложен автором (Нухимовский, 1969б, 1976а). Персистентная ком- пактная сближенноузловая побеговая нижняя часть (неветвистая или, реже, ветвистая) унирепродуктивных двулетних или многолетних осо- бей или потенциально каудексовых особей — это конодий (Angelica archangelica L.). Конодий, таким образом, является одним из видов базовых органов (рис. 121-124). Преперсистентную компактную побего- вую нижнюю часть (ветвистую или неветвистую) унирепродуктивных особей или потенциально каудексовых особей можно для удобства на- зывать «иреконодием» — это пробазовый орган, т.е. орган, образую- щийся перед возможным образованием базового органа (рис. 121-126). У особей одного вида, генотипами которых не предусмотрено обя- зательное формирование каудекса и онтогенез их проходит по унирепро- дуктивному, мультирепродуктивному или по вынужденно стериальному жизненному циклу, пока не проявляются отмеченные выше признаки возникновения каудекса, базовый орган можно называть лрекаудексом.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 113. Виргинильный этап в развитии каудексовых особей Euphor- bia altaica С.А.Меу. (Алтай, Северо-Чуйский хребет, высокогорье). 1 — семядоли, 2 — семядольный узел (у растений постарше его примерное поло- жение показано пунктиром), 3 — гипокотиль, 4 — главный корень, 5 — главный безрозеточный вегетативный годичный побег, 6 — пазушный безрозеточный ве- гетативный годичный побег (монокормус), 7 — сухие остатки от надземных частей годичных побегов прошлых лет, 8 — мономерный миниморезид, 9 — компактный каудекс, 10 — срединные листья (иногда показано жилкование), 11 — схема эусим- подиального нарастания побегов; у — увеличено по сравнению с масштабом, горизонтальная линия — поверхность почвы. Одно деление масштаба — 1 см.
Глава б. Продолжительность жизни: звризфемерность и персистентность 299 И 55 ФЭ 55 О Р 4ь Ж~ а5 д И у a5 ~ vv 4~ ~@М Ж Q <О Ю О ~: с, 'О Я. Ф~ ) 55 р~ Q ! зф о пж ж~ ~ а. ~~ X >~ A ~ч О в," о ~р М О Мо Ж О ~ ж '" 3~ Q О Х Ж й О В: [ И Q О й Ck л ~М И О g й О Ж В Р„ Ж Ю in О Q. О 3 аэ ~е) р~ц .0 Д 3 1-~ g о О И '- Ю М v О а3 ~ О й X X Х О g4 3~ М ~) Ф О О х ~ ;у< g Г~) ф~~ ~еф ф~~ О ~~ о ~ О ~~ 3 и~ _#_ ~) о0 Ф Й О В~ ~[ Ь орг И,) ~, 3 1 [~~ О m _#_ й О й~ О. ~) Й д М Х Д О X X Ф [ ° М М Ф о р' ~ Х 3 щ О 3 ~) Ю ю ~ св 2 Х ц~~ с6 Ф й О О Ж О Ф:
OCHOBhl БИОМОРФОПОГИИ СЕМЕННБ1Х PA СТЕНИИ 16 Рис. 115. Структура подземных органов каудексовых растений, формирую- щаяся в эрозионно-неустойчивых субстратах I — Astragalus testiculatus Pall., П — Ferula tatarica Fisch. ex Spreng. (республика Калмыкия, Черные земли). 15 — вынужденно удлиненный каудекс у растений, обитающих в условиях ежегодного небольшого засыпания их песком, 16 — основания розеточных листьев (верхние части удалены), 17 — мономерный вицерезид; остальные обозначения те же, что и на рис. 113 и 114. Розеткообразующие каудексовые растения в своем развитии чаще всего проходят через состояние протобазового органа (прекаудекса), нарастая за счет курторезидных монокормусов. Начало ветвления базо- вого органа, акронекроз (неполное отмирание, начиная сверху) медиаль- ного побега, а в иных случаях (Plantago maxima Juss. ex Jacg.; рис. 46) первый кладонекроз (полное отмирание тотрепродуктивных пазушных побегов при полном сохранении главной оси, образовавшей эти побеги) являются признаками возникновения каудекса у таких растений. Термин «каудекс» в нашем понимании не тождественен термину «стеблекорень», как его определили Ал.А. Федоров и др. (1962, с. 53), поскольку в стеблекорне «основную роль играют стеблевые части, но также и корень». Вероятно, будет целесообразно расширить смысл термина «стеблекорень» (caulorrhiza) в связи с новыми представлениями о параметрах побегов и корней, а следовательно, и параметрами каркаса побегов — стеблей. Под стеблекорнем логичнее всего было бы пони- мать каркасную (стеблевую и корневую) часть любого ризокормуса.
301 Глава б. Продолжительность жизни: эвриэфемерность и персистентность Рис. 116. Формирование каудекса у особей I — Phlomis pungens Willd. (республика Калмыкия, Черные земли) и II — ОЬета behen (L.) Ikonn. (Московская область, лут). 1 — семядоли, 2 — семядольный узел (у взрослых растений показан примерно пунктиром), 3 — гипокотиль, 4 — главный корень, 5 — розеточные листья, основания розеточных листьев (верхние части удалены), 7 — основания унире- продуктивных дифазных или плейофазных полурозеточных побегов (верхние час- ти удалены), 8 — компактный каудекс. Одно деление масштаба — 1 см. Базокормус вместе с корнем (корнями), непосредственно связанным с ним, можно рассматривать и как одну систему ризобазокормус. Растения, у которых каудекс располагается при относительной ста- тике субстрата подземно или почки находятся на уровне поверхности почвы — это каудексовые травы (Егуприт р1апит L., Phlomis pungens
ОСНОВЫ БИОМОРФОЛОГИИ CEhfEHhhlX PA СТЕНИЙ Willd., рис. 28, 116). Растения, у которых каудекс, в условиях относи- тельной статики субстрата, выносит почки выше поверхности почвы- это каудексовые нолукустарнички (Capparis herbacea Willd., Salsola dendroides Pall., рис. 117, 118-II). В условиях ежегодного засыпания суб- стратом каудексовые полукустарнички, если не погибают, то образуют вынужденно нодземные, чаще всего удлиненно-ветвистые, киудексы (рис 188-III), а при оголении каудекса, нередко и основания главного корня, в условиях эрозионно подвижных субстратов — каудекс вынужденно надземный (рис 118-I) СХ Рис. 117. Формирование каудекса у Capparis herbacea Willd. в условиях относительной статики субстрата (Крым, гора Карадаг, пологий склон). сх — схема полуподземно-каудексового взрослого растения. Остальные обозначе- ния те же, что и на рис. 113. Каудекс — орган вегетативного возобновления, поскольку в его структуре развиты почки, предназначенные для активного побегоотрас- тания и орган запасания питательных веществ (резервирующий). Однаж- ды образовавшись, каудекс разрастается радиально при увеличении числа резидов и числа спящих почек; он может более или менее одревесневать, достигая иногда больших размеров, особенно у пустынных растений (Peganum harmala 1., Salsola dendroides Pall.). Придаточные корни на каудексе не образуются (рис. 117, 118) или иногда образуются (рис. 156- 1), но при этом не имеют решающего значения в жизни растения; при погребении растений субстратом могут развиваться много стеблеродных корней (рис. 156-II). Если каудекс не ветвится, то он щюстой (Palimbia salsa (L.) Bess). Если у каудекса хорошо заметны ветви (некоторые авторы прошлого столетия, например, Е.Буассье, называли их «каудикулами» если они были тонкимим или «каудициями», если — толстыми), то это вет- вистый каудекс (Peganum harmala L., рис. 38, 39, 119). По особенностям побегообразовательной активности каудекса со- ответствующие растения, развивающие такой орган, бывают трех типов:
303 Глава б. Продолжительность жизни: эвриэфемерность и персистентность Рис. 118. Габитуальный полиморфизм биоморф Capparis herbacea в различных условиях обитания (Крым, окрестности Никитского ботанического сада, крутые горные склоны). I — вынужденно надземный компактный каудекс у особей второстепенного био- морфотипа (уровень поверхности почвы ежегодно понижается), 11 — полуподзем- ный компактный каудекс у особей основного биоморфотипа (относительно посто- янный уровень поверхности почвы), Ц! — формирование в онтогенезе вынужденно удлиненного каудекса у особей второстепенного биоморфотипа (уровень по- верхности почвы ежегодно повышается); 15 — мономерный викариорезид, 16— эпифиз, 17 — спящие почки, 18 — длинный ветвистый каудекс, 19 — ветвь каудекса (каудиция), х6,4 — масштаб. Остальные обозначения те же, что и на рис. 113 и 114. некустистые, если они ежегодно развивают один новый побег, который продолжает единственную (простую или сложную) каудексовую ось (Ferula caspica М.В.), слабокустисиые, когда ежегодно возобновляется из почек каудекса от одного до пяти, реже больше, но все же немного побегов кущения (Limonium gmelinii (Willd.) Kuntze), и сильнокустис- тые — у них ежегодно отрастает много побегов кущения (Peganum harmala L.). У слабо- и сильнокустистых каудексовых растений нередко образуется комиактный каудекс (рис. 38, 118-I, II); он укорочен, а ветви, если они выражены, тесно сближены. Миниморезиды или курторезиды, слагающие каркас такого каудекса, который чаще всего наиболее мощ- ный у растений аридных областей, нередко с возрастом сильно раз- растаются радиально; при этом границы между ними часто теряются. AHBJIH3 компактных («шаровидных», «бугорчатых», «дисковидных» и т.п.) каудексов показал, что разрастающиеся в толщину резиды затяги-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИЙ Рис. 119. Каудекс Peganum harmala при погребении взрослой особи субстратом (республика Калмыкия, Черные земли). Обозначения те же, что и на рис. 113, 114 и 118. вают рубцы от отсохших частей побегов и заполняют пустоты от раз- рушившихся резидов. Врастание одних живых частей в другие, т.е. срас- тание между собой составных частей каудекса, мы не наблюдали. Ос- лабление побегообразовательной активности каудекса и уменьшение его прироста связано с ухудшением условий жизни или старением особей; у стареющих растений наблюдается нередко партикуляция (Phlomis рип-
Глава б. П Д . Продолжительность жизни: звризфемерность и персистентность 305 gens illd., Scutellaria baicalensis Georgi) неполная, а иногда, у очень старых, полная ( ис. 116-1 (рис. —; 107, Ж-15). У старых особей зопника колю- чего партикуляция проявляется весьма оригинально: главный ко ень по всей своей длине более или менее расщепляется на отдельные «веревковидные тяжи» (Шалыт, 1965). Рис. 120. Ф 20. Формлрование каудекса и новокаудекса у особей Тагахасцт officinale Wigg. (Московская область, залежь и огороды' А — ранние этапы онтогенеза, Б — особь второго орого года жизни, В — трехлетнее растение, Г — остаток главного корня после удаления каудекса Д— вокаудексом; 1 — сем оли 2— — — унирепродуктивный дифазный закрытый Ilo- er, 7 — основание живых розеточных листьев (верхние части удалены~, 8 — о- зеточные листья, 9 — ка декс 10— — у, 0 — новокаудекс, 11 — продольный разрез 12— ), — ро- миниморезид, 13 — вика ио ез го р р ид, ризонтальная линия — поверхность почвы, 3 3 х3,1 — масштаб. б) Следует отказаться от неудачного термина ~многоглавый ко ень» гайх т поскольку <главость образована вовсе не ко не ~~~ы ж ~~ром, а стеблевыми частями» (Федоров др,, ), также и от подобных высказываний: у Pimpinella litho hila с isc . «главный корень нередко имеет многоглавый ка декс голо погружены в почву» (Бирюл 1969 8) Н «многоосевоглавый корень» (Ш 1965; М лева,, с.. е лучше, п именяемый и » алыт,; оскалева, 1968) и т.п. рвотой и ветвистым квулексы могут бель короткими (рис. 113, 114) или длинными (рис. 55 118-III 1 19, 156- II). Структуру короткого каудекса определяют миниморезиды или курторезиды, а в сложении длинного каудек- са ведущую роль играют викариорезиды или вицерезиды. Дл ы. инныи каудекс я при погре енин субстратом растений, которые в условиях с относительно постоянным уровнем поверхности почвы формируют компак- тный, короткий простой или короткий ветвистый каудексы. Каудексы также можно называть одно-, дву-, многоглавыми') в зависимости от числа живых верхушек («глав»~ ка удекса, например, одноглавый или двуглавый
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЧ 1=~ Ж О М й О~~ й О В: [ Ж О М О й .И О О М д О Eg О Ц 2 Ж р~ Q Фэ О Оч И у Ж ~> Ж M И ~ ФЭ ( o .~, >Й о <~ ° а~ ФЭ Ф Й Ж '~ ЙР ~ й Зю О М О й О Ck О 3: 0 О О Ж СЮ (J Я h4 P. О О 9 >гч О _#_ ~~о m За Q K3", W м о о~ С4 М О Я _#_ ~уе Ю в~~ц ~ л ~ О М О О О ~и~ Х " ф4 ~ю~ б~~ Я} ~ _#_ ООМ ,,~,~ ,'~, 'Р' ~ о ~~ р" Б О О О О оB~ Ф A О ~ Л а а) а,Д о В - ". "О о у ~ о ~ о о оо к а3 в~й л о ~ 2~ O ~ v а3 o@ X [ ° М О р М ~ о +~ Х й е~ ~а~ _#_ _#_ v>O йО~ О ~О _#_ ~) Щ „й й >йg а. И Ц О X О О О В: <-~ v й G. l а ХХ И л фц Я х 2 й~ р о ~ _#_ Д: о E „О G М И О се о О
Глава 6. Лродолжительность жизни: эвриэфемерность и персистентность 307 ~3~ й В: [ О Ж О '.4 М Q,.~~ а ~ х О (б Яю р о ~ъ ц о Оо М ~@" о ФЭ ~~ f О 3: Р ~4 Хй ~~ц 0 [ ФЭ й:[ О Й'з й:[ О О й X О :с ~ й о ~ Р О Л ~ й Й Ж а.+ Р' й '.4 3~ О ФЭ О О '.4:ц Д. о О й 0 ° л Я ~р В: О Р) f/) Л ~) »-> м ~~ м 1 ~л~ ~~ ) X О Щ Я л й 0ч ~~ у) 2 а cl Я О О _#_ "+ о ~В~а й О > ~g ~~î Я, М д 1 й ~Õ у, ~ "'~ <оЕ ca ~ ~+Я (/2 » с~ ~- '~.. О, о р. л (/2 ~~4 ~» Р )~ "И~ 2,[ G. с~ gg й о с6 О М м й 1 Х ф,) .0 Ф 'у~ ~а~ й д О О~Я ~~-з О о и~~ МВ:Я (» 1 О се~ X О ° 1~ Я vсо О 4'1 C4 'X ~ Дб; f» ~z ~ î ОИО ~=( О й л 1 ( 4~ <~3 й л се о ; ' а. й 6~ о >) 1 Ж 1 ) 2 X а5 в,ь Ф О О ~ о X С4 Я ж О X x ~а~ ф~~ сл ) ~ u »> Вм с6 и и ф Л ~; О ['~ О > б Q О + а Щ ц~а й 4Э Я [, И Q, 4Э Ьм о д- 2 Х О ФЭ О M ä X »-> О Immi д ) 1 с6 Ч~~ E) и Е О Е з
ОСНОВЫ БИО ИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИЙ Рис. 123. Ви гин р ильный этап в онтогении ° гении Arctium 1арра L. о ласть, сорные места). Обозначения ния те же, что и на рис. 122. "' рис.. Одно деление масштаба — 1 см подземный каудекс Ferula tatari F' ), ~~~~~л~вы~ пд~~- Ь-.115-П" . М«-.б. м~ 2/3 ~~ лее если он погружен в почв но менее чем ~жин~ (1962) т~жегил б тила о разование «вто ич . ах- лец» спящих почек ас р ных каудексов» в виде «ко- к, расположенных на о тот лях. Вряд ли следует говорить о первичном и в ртотропных подземных ст б- е— каудекс всегда первичен , но нужно более е ом и вторичном каудексах части. Разветвление п д тально характеризовать Э ие подземных кудексовых ветв ать его вых ветвей происходит в утол-
309 Глава б. Продолжительность жизни: эвриэфемерность и персистентность И о 3 Ф Ж й[ О Ж й~ О о й С4 X й О Ф Ж g Ф й Ж О Ю О
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИЙ Рис. 125. Участие преконодия в структурной организации однолетника Lactuca sativa L. (Московская область). Одно деление масштаба — 1 см. Обозначения те же, что и на рис. 122. щениях («наплывах»), несущих нередко большое число спящих почек, т.е. в участках прошлого или (и) нынешнего активного почкозаложения и побегообразования. При погребении субстратом побегообразователь- ная активность нижних участков каудексовых ветвей резко падает или приостанавливается и переходит к верхним участкам, расположенным непосредственно у поверхности почвы. Функцию побегообразования выполняют новые участки, постепенно разрастающиеся и становящиеся заметными в общем скелете каудекса. Иногда утолщений нет, но видны участки со сближенными основаниями ветвей, рубцов от них или оди- ночных перевершиниваний побегов. Компактные участки подземных
.0 [ ° о а3 1=~ Ю О Глава б. Лродолжительность жизни: эвриэфемерность и персистентность Ф~ й Ж Q Ж О Ц Ж О В: [ й 3: Ф Q О Ф О Ж О О В: [ Ж О М Q И О й Ck М о О М о й «4 Е 2 '~ СЧ Е-. О о й G. М Ж С4 а3 ф; ж 44 g О g 2 Ж N 0 Иm О О Д э о > О а3 g Р а3 ~ Ю ю~ Щ ~М 3 ц О. (, 0 В 1 в О'Ф ~аф л В'~~ ~) 3 О Оа М „~ О Е ° W о 'О C 3 О
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ более или менее удлиненных многолетних (резидных) побегов, которые были или служат зонами активного побегообразования, было предложе- но (Нухимовский, 1969б) называть эиифизами (epiphysis, от греч. еж~, epi — на, qnro, phio — расту, произвожу). Эпифизарная ярусность характерна для особей с длинным каудексом, образующимся в условиях их обитания с эрозионно-подвижным субстратом (рис. 39, 118-Ш, 119, 156-П). На вынужденно подземных ортотропных более или менее удлиненных многолетних побеговых ветвях (вынужденных корневищах, геоподиях) у других, погребаемых субстратом, древесных, полудревесных и травянистых семенных растений также образуются «ярусы», «этажи вет- вления» и т.п., иными словами, проявляется эиифизарная ярусность, что уже отмечалось неоднократно (Warming u. Graebner, 1918; Петров, 1933; Лавренко, 1935 и др.). Кроме собственно каудексовых растений, т.е. растений, у которых эубазовый орган — это только каудекс, есть растения с более сложной организацией, в которую кроме каудекса могут входить и иные многолет- ние структурные образования. Каудекс характерен для некоторых обли- гатно и факультативно корнеотпрысковых растений (Acroptilon герепв (L.) DC., Тагахасит officinale Wigg.). Некоторые корневищно-каудексовые травы формируют каудекс непосредственно над гипокотилем и главным корнем (такое структурное образование иногда называют иначе — «ко- ронка», «головка» и т.п.); из почек каудекса помимо надземных побегов образуются специализированные горизонтальные или наклонные подзем- ные побеги, преобразующиеся в корневища. На заметном (иногда значи- тельном) отдалении от каудекса у таких растений образуются корневищные отпрыски и парциальные кусты; многолетняя подземная побеговая часть парциальных кустов может напоминать по строению каудекс, но в отличие от него она не имеет непосредственной связи с главным корнем. Пожалуй, чтобы не усложнять терминологию, такие многолетние подземные побе- говые системы, которые образуются на горизонтальных корневищах или возникают адвентивно на корнях, будет логично относить также к корне- вищам (Acroptilon repens (L.) DC., Pseudosophora alopecuroidks (DC.) Sweet, рис. 127). Только в тех особых случаях, когда каудекс был когда-то удален (допустим, механически), а затем адвентивно (не через гипокотиль) восста- новилась многолетняя побеговая система (аналогичная по строению кау- дексу), связанная с каким-либо корнем; такой эубазовый орган можно обозначить термином новокаудекс (от лат. novus+ caudex), например, Taraxacum ой1с1па1е Wigg. (рис. 120). У каудексово-корневищных растений образование корневищ начина- ется на 2-3-м году жизни (Pseudosophora alopecuroides, рис. 127) или поз- днее, например, на 9-10-м году у Trifolium ambiguum М.В. (Донскова, 1968). Каудекс как орган побегообразования и запасания питательных ве- ществ (депонирующий, базовый) оказался одним из удачных структур- ных приобретений в процессе исторического развития растений, про- текавшего в суровых климатических условиях аридных областей. На остепненных, горных, пойменных лугах уменьшается число собственно каудексовых растений, а возрастает число каудексово-корневищных и каудексово-корнеотпрысковых растений, у которых многолетняя побе-
Глава б. Продолжительность жизни: эвриэфемерность и персистентность 313 О й Ck юю 'внп0Ли~ О A И О З О Q О P s. .И Я Q о 'О й О Д й ФЭ ',?,' ФЭ О ж" Оч Q О ф ~ а 4 О 3 ж ~ Q ~ а. ЕБ й ~ ж ~ ФЭ а.~ Ж о~ М у О Щ л О ~~.' О g5 М ф) О е[ Х „"~ 55 ж ~ ~4 Ю й Ж Ф О Q й й Qi О 11ДЯ ~~ ~ о к д ОаЯ g Й Щ ) в~4 о'Ф 3а О О О'Ф о, о~а Ф Q э~О И:й.' х з~ 00 щ Ф 0 ä +о И Я М р; о ~~ (Ю Д:. М X Ф Ф у 3 sss as g д [ ° и>B Д~asд Ц 2 ~д Одоа М1 Ж к~1E>p О рф ~~~ Х н р М ~- и ~ Q,~ 0 30 63 М 0 о ~ м иЙ~~Я М и О а ~- о "'-~о C4~ ~~. y Ф д Ф "() О + ~8~ <" a~- v о y ce й~'~ й л М ~.о Я о~~~ О с~'~ ж ~ ~3î > И ~ я й е а.ж Ф + _#_ <ЕЯР
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ говая система (база) — это не только каудекс, но и ризомы, а иногда и новокаудексы. Луковица (bulbus). Этот термин употребляется издавна в ботанике. Обычно луковицей называют видоизмененный крайне редуцированный стебель с развитыми на нем сочными видоизмененными листьями (Фе- доров и др., 1962), т.е. это подземный (реже — надземный) побег, состо- ящий из очень короткой уплощенной стеблевой оси (донца) и чешуе- видных сочных листьев (Васильев и др., 1978). Такое понимание тер- мина «луковица» традиционно. Однако с позиций рассмотренной выше концепции персистентности и базомеризации в понятие «луковица» необходимо внести некоторые коррективы. То, что называют луковицей, следует рассматривать и как пробазовый (потенциально персистентный побеговый) орган, обозначая его термином «незрелая луковица» и как базовый (персистентный) орган, обозначая его термином «зрелая луко- вица». Зрелая луковица — это базокормус, образующий после заверше- ния сезонной вегетации побега и отсыхания его эфемерных частей, однократно или многократно зимующий и состоящий из очень ком- пактной стеблевой оси (донца) с развитыми на нем сочными персис- тентными основаниями развернутых листьев, их живых остатков, чешуй и почек (Allium сера L., Lilium martagon L.). Возникновению зрелой луковицы обычно предшествует стадия незрелой луковицы, т.е. по- тенциально зрелой луковицы. Если ниже луковицы довольно долго сох- раняется участок живого стебля, то мы имеем дело с луковично-кор- невищными растениями (рис. 128). У некоторых видов, например, у чеснока (Allium sativum L.), обра- зуется сначала незрелая солуковица, которая после завершения сезона вегетации превращается в зрелую солуковицу, состоящую из зимующих (персистентных) луковицеподобных почек (они еще не проявляли ви- димого роста). Эти персистентные почки, учитывая их специализацию, мы называем иочколуковицами (gemmobulbus). Родительские ткани зрелой солуковицы довольно скоро отмирают, а она сама превращается в ве- гетативную семью почколуковиц. Развитие может при благоприятных условиях повторяться вновь и вновь. По схеме: почколуковица — не- зрелая солуковица — зрелая солуковица — вегетативная семья почко- луковиц. Солуковица (cobulbus) — это побег (или часть побега), напоми- нающая внешне луковицу, но, в отличие от нее, развивающий под покровом рано отмирающих, но долго сохраняющихся оснований лис- тьев (или чешуй), пучок почколуковиц, которые по мере созревания этой системы теряют живую связь друг с другом. Помимо луковиц и почколуковиц в структуре некоторых луковичных растений могут воз- никать(в пазухах листьев надземных побегов вместо цветков или на столонах) незрелые луковички (bulbilli). После отделения от родительских растений и (или) при сохранении в живом состоянии после завершения вегетационного сезона превращаются в зрелые луковички и так преодолевают неблагоприятные для вегетации условия. Клуонелуковица (bulbotuber). Это известный морфологический тер- мин (Федоров и др., 1962). Мы различаем там, где это возможно, незре- лые (пробазовые органы) и зрелые (базовые органы) клубнелуковицы. Зрелые клубнелуковицы — это базокормусы, напоминающие луковицы,
315 Глава б. Продолжительность жизни: эвриэфемерность и персистентность А Рис. 128. Формирование луковично-корневищной структуры у биоморф Ungernia victoris Чче4. ех Artjushenko (Таджикистан, Гиссаро-Алай, Гиссарский хребет). А — начальные стадии онтогенеза, Б — виргинильная особь, В — взрослая особь, Г — вегетативное размножение путем разрезания луковицы и укоренения черен- ков; 1 — луковица, 2 — корневище, 3 — придаточные корни, 4 — особь клона; горизонтальная линия — поверхность почвы; одно деление масштаба — 1 см. но с сильно разросшейся компактной стеблевой частью, сухими пленча- тыми остатками оснований листьев на них и почками (Crocus vemus (L.) Hill). Клубень (tuber) Это известный морфологический термин. Клубень бы- вает незрелым или зрелым. Зрелый клубень — это базокормус или его часть, представляющие собой сильно утолщенный стебель на верхушке побега (Solanum tuberosum L.) или где-либо в другом месте (например, пазушное образование у Dioscorea batatas Decne.) с персистентными почками и без длительно сохраняющих остатков листьев. Зрелые клубни образуются в процессе их морфогенеза из незрелых клубней. У некоторых растений, например, Dioscorea batatas, образуются воздушные пазушные незрелые клубни, которые после обособления от родительского растения, если сохраняют жизнеспособность, то проявляют тем самым зрелость и участву-
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ ют в вегетативном размножении. Кроме клубней, как базовых органов (клубнетбегов), у растений образуются еще корневые (Filipendula vulgaris Moench, Phlomis tuberosa L., рис. 129) и корнетбеговые (Gymnospermium altaicum (Pall.) Spach) юиуйа~. Стеблевая часть клубнепобега — стеблеклубень. Рис. 129. Образование корнеклубня у Phlomoides tuberosa (L.) Moench (Московская область, Серпуховский район, остепненный луг). 11 — корневой клубень. Одно деление масштаба — 1 см. Остальные обозначения те же, что и на рис. 122. Базовые органы иногда поражают своей оригинальностью. Так, побе- говая система особи Dactylorhiza maculata (L.) Soo зимует в виде одной почки, которая по существу является своеобразным базовым органом. Такой персистентный орган, структура которого то возникает, то исчезает, то вновь возникает, затем вновь исчезает и т.д., например в образе оди- ночной однократно зимующей почки, обозначим как викарирующий (заме- щающийся) базовый орган. Явление, при котором возникают такие перси- некоторым биоморфам, вся, или иочти вся структура которых сменяется (иричем нередко неоднократно), а в целом такие биоморфы обладают сверх- как органиьиам некоторых видов (Dactylorhiza maculata), так и синорганиз- мам (Aconitum altaicum, рис. 53). Рассмотренная система понятий характеризует различные стороны ор- ганизации базовых органов. Это попытка подойти шире, чем обычно, к оцен- ке возрастной изменчивости структуры, а также поиска критериев для диаг- ностики момента возникновения базовых органов у особей разных видов.
Глава 7. Основные способы освоения времени биоморфами семенных растений 7. 1. Морфогенез, биоморфогенез и биоморфогения Как известно, атрибутивное свойство (способ существования) ма- терии — движение, любое изменение; оно охватывает все процессы в Природе. Изменчивость явлений неотделима от их определенной устой- чивости (сохранения). Все явления Природы, изменяясь какое-либо время, сохраняются — удерживают качественную и количественную определенность. В настоящее время остро стоят вопросы поиска теории, синтезировавшей бы в себя все множество фактов о возникновении, развитии, стабилизации и деградации самых различных материальных объектов — вещей (тел) (от элементарных частиц до галактик и более). Ею может стать общая концепция морфогенеза. «Морфогенез» — термин, которым пользуются биологи, однако он может стать общенаучным. Выделение особой категории понятий- общенаучных — тенденция в развитии современных наук (Готт, Урсул, 1975). Всякий материальный объект как тело (марфа, форма) возникает, существует в определенном качестве и, наконец, преобразуется в иной объект-тело, изменяя прежнее качественное состояние, т.е. обладает морфогенезом. Ранее (гл. 4-6) мы обратили основное внимание на то, как биоморфы осваивают пространство, а в этой главе акцент сделан на освоение биоморфами времени. Морфогенез растений обычно опреде- ляют как процесс формообразования (Синнот, 1963; Чайлахян и др, 1973; Белоусов, 1987). Из многих определений термина «морфогенез» представляется наиболее оправданным подход Р.Тома (1970, с. 145), ко- торый понимает его «в самом общем смысле, а именно, для обозначе- ния любого процесса возникновения (или разрушения) формы», неза- висимо как от природы субстрата рассматриваемых форм, так и сил, вызывающих их изменение. К.Уоддинггон (1970, с. 19) подчеркнул, что морфогенез — это «изменение геометрической формы». Приведенные
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СГЕНИИ определения указывают только на основное направление для поиска более конкретной формулировки. Морфогенезы (циклы существования материальных объектов- тел) — это одна из сторон дискретного сохранения изменений. Под морфогенезом понимаем процесс (во временном отношении — цикл) существования (сохранения изменений) материального объекта-тела от его возникновения до преобразования в новое структурно-функцио- нальное качество — новое тело (Нухимовский, 1980). Любое тело воз- никает, существует и исчезает, преобразуясь в другое тело или несколь- ко тел. Издревле было замечено, что растения, животные и люди, рож- даясь, проходят свою молодость, взрослое состояние, старость и, нако- нец, умирают (Феофраст, 371-286 гг. до н.э., 1951). Природа — это сплошной поток морфогенезов в пространстве и во времени. Частный случай морфогенеза — биоморфогенез. Цикл существования биомор- фы — биоморфогенез. Жизнь — это поток биоморфогенезов в простран- стве и во времени (рис. 130-А). Совокупность сведений о морфогенезах каких-либо групп тел — их морфогения. Проявление морфогенеза — это нроявление одного из универсальных свойств материальных систем: любая материальная сцстема-тело, начав свое дискретное движение (морфоге- нез) в одной части иространства-временц в одном качестве, всегда закан- чивает его в другой части иространства-времени в ином качестве, ста- новясь иным телом, сохраняя как и ирежде бесконечное движение на дис- кретном его отрезке (в новом морфогенезе). Функции материальной системы-тела на определенных отрезках времени — новедение. Здесь используется популярный в последнее время термин, вокруг которого немало разнотолков. Если «морфоге- нез» — это структурно-функциональное изменение какого-либо тела от его возникновения до преобразования в новое качество — новый объект-тело, т.е. цикл его существования, то «поведение» — это только функциональное изменение некоторого тела на определенном отрезке морфогенеза. Функции, а следовательно, морфогенез и поведение, су- ществуют на (в) структуре, изменяя ее и изменяясь сами. Многие формы поведения, однажды проявившись, в некоторых чертах могут повто- риться при повторном возникновении сходных условий окружающей среды (шаг, наклон головы животного, движение листьев растения, ра- дость или печаль человека, бег животного за «жертвой», намокание или высыхание комка почвы и т.п.), но никогда полностью (во всех деталях) они повториться не могут, поскольку необратимо время. Приведенное выше определение «поведения» имеет общенаучный смысл. Термин «свойство» более широкого значения, чем «поведение», поскольку вкпю- чает в себя не только проявляющиеся в настоящий момент функции, но и те функции, реализация которых лишь потенциально возможна. Иными словами, свойство — это весь диапазон реального (проявляющегося сей- час) и потенциально возможного поведения материального объекта-вещи (тела). Определенные свойства проявляются в объекте-теле через поведение полностью или частично в зависимости от особенностей его морфогенеза. Цикл существования биоморфы (индивидуума или кондивидуу- ма) — биоморфогенез представлен или онтогенезом (жизненным цик-
319 Глава 7. Основные способы освоения времени биоморфами семенных растений лом индивидуума), или синонтогенезом (жизненным циклом кондиви- дуума, или синорганизма). Онтогенез — это общеизвестный термин, которым издавна обозначают все индивидуальное развитие особи (Чай- лахян, 1958 и др.). Синонтогенез — это совокупность онтогенезов осо- бей всех (не менее двух) последовательно сменяющих друг друга поко- лений, первое из которых начинается с зиготы (Нухимовский, 1970, 1973а, б, 1980). Совокупность сведений о биоморфогенезах какого-либо таксона (вида, рода и т.п.) — биоморфогения этого таксона (его он- тогения или синонтогения). Раздел биоморфологии, изучающий био- морфогенезы, — биоморфогения. 7.2. Состояния биоморф и стадии био морфогенезов Проблема состояний организмов широко обсуждается в литературе (Богдановская-Гиенэф, 1926; Работнов, 1950а; Keilin, 1959 и др.), а наи- более крупным обобщением в этой области является монография А.М.Голдовского (1977), который сформулировал свои взгляды как «ос- новы учения о состояниях организмов». Проблема состояний имеет принципиальное значение для познания организации биоморф. Пра- вильная разработка этой проблемы в отношении биоморф снимет, на наш взгляд, многие затруднения, которые часто испытывают биоморфо- логи при выделении границ конкретных биоморф, когда спорят о «сме- няемости» биоморф и т.п. А.М.Голдовский (1977, с. 5) понимает иод состоянием организма оп- ределенную совокуиность ироцессов, ироисходящих в данный момент в организме, стеиень развития и целостности его структур. Каждое состо- яние организма (органа, ткани, клетки) может выражаться, по Голдов- скому, «комплексом частных численных характеристик — интенсивно- сти отдельных процессов, степени полноты совокупности этих процес- сов, степени развития и целостности структур организма». Характерис- тики состояний организма могут отражать разные стороны жизнедея- тельности — от биохимических процессов в клетках, лежащих в основе их физиологических функций, до процессов высшей нервной деятель- ности у животных и психических явлений у человека (Голдовский, 1977). В нашем случае развиваются представления о состояниях в контек- сте с общими представлениями об организации биоморф семенных рас- тений в ее динамике. Биоморфы как организмы и синорганизмы при- нимают в биоморфогенезах множество состояний. В зависимости от того, под каким углом посмотрим на биоморфы, т.е. на каких признаках более всего фиксируем внимание, под таким и наблюдаем разнообразие их состояний. А.М. Голдовский (1977) за основной критерий классифи- кации состояний берет ряд признаков: наличие или отсутствие сочета- ния ассимиляции и диссимиляции, лежащего в основе жизнедеятельно- сти, и выделяет жизнедеятельные состояния: биоз (полная жизнедея- тельность) и гипобиоз (временное снижение жизненных процессов—
ОСКОВБ1 БИОМОРФОЛОГИИ СЕМЕНКЫХ Ph СТЕКИЯ замедленная и ограниченная жизнедеятельность) и нежизнедеятельные: мезабиоз (промежуточные состояния между анабиозом и жизнедеятель- ностью) и анабиоз. Морфогенез — процесс, а любой процесс структурен во времени, т.е. состоит из отрезков — стадий, в которых проявляются какие-либо осо- бенности организации объекта-тела. Любое структурно-функциональ- ное состояние материального объекта (как живого, так и неживого) в определенный отрезок времени — морфогеническое (морфогенетическое) состояние. Отрезок морфогенеза с определенным морфогеническим сос- тоянием какого-нибудь тела — стадия морфогенеза. По названию мор- фогенического состояния определяется и стадия морфогенеза. Морфо- геническое состояние может быть охарактеризовано через любой признак, индикаторно отражающий состояние объекта в определенный отрезок времени. Индикаторами могут быть признаки как структурные, так и функциональные. Морфогенические стадии и состояния бывают возрастными и фенологическими. Возрастное состояние — это морфоге- ническое состояние с неповторимыми в морфогенезе структурно-фун- кционалными признаками (одним или несколькими). Фенологическое состояние — это морфогеническое состояние, индикаторные признаки которого в общих чертах повторимы в соответствии с повторением оп- ределенных условий окружающей среды (сменой времен года, дня и ночи и т.п.) или неповторимы только из-за конца морфогенеза. В пос- леднем случае признак фенологического состояния может быть показа- телем и возрастного состояния, например, цветение, плодоношение у однолетних растений. 7.3. Категории стадий биоморфогенезов, критерии выделения и методика их изучения При субординатной классификации стадий морфогенезов и, в част- ности, стадий биоморфогенезов удобно выделять определенные категории, например, гинофенофазы, фенофазы (категории фенологических стадий), гинофазы, фазы, этаны, нериоды, а при необходимости можно добавлять еще несколько нереходных категорий — субэтаны, гиионериоды, гемициклы (категории возрастных стадий). Одноименные категории разных морфогенических стадий принадлежат к одной субординате, например, к субординате периодов морфогенезов. Число категорий морфогенических стадий, а следовательно, и число субординат зависит от степени дробления изучаемых морфогенезов. Часто нет нужды выделять все возможные категории, поскольку суть морфогенеза достаточно полно можно отразить и небольшим числом категорий. Число одноименных категорий должно бьггь не менее двух, иначе их выделение лишено смысла. Графически модель морфогенеза можно изобразить по-разному. Нам представляется наиболее наглядным вариант (рис. 130-Б), состав- ленный из совокупности окружностей или овалов (моделей категорий
321 Глава 7. Основные способы освоения времени биоморфами семенных растений Рис. 130. Модели биоморфогенезов. А — жизнь как поток биоморфогенезов в пространстве-времени (фрагмент), Б— модель возрастной стадийности биоморфогенезов; 1 — отдельные стадии биомор- фогенезов, 2 — зона размножения, 3 — зона умирания, 4 — отдельные биомор- фогенезы; Ц — биоморфогенез, П — периоды, Э — этапы, Ф — фазы, ГФ — гипофазы. морфогенических стадий), вписанных по две и более в одну окружность (овал), соприкасающихся с подобными окружностями (овалами) по од- ной линии, которую можно провести через всех них, и расположенных в один ряд в порядке сменяемости отдельных категорий морфогенеза во времени от начала к концу морфогенеза (показано стрелкой). Каждая категория морфогенеза характеризуется своим диапазоном сложности, определяемым морфологическими или функциональными признаками- индикаторами, что создает картину иерархической соподчиненности и линейной сменяемости этих категорий в пространстве и во времени. Из всего комплекса признаков биоморф предложено различать (Ну- химовский, 1978, 1980) признаки тропобиозные (признаки-индикаторы mu- пов биоморфогензов, или тропобиозов) и гемитропобиозные (признаки- индикаторы типов морфогенических, т.е. возрастных и фенологических состояний, или гемитропобиозов). Тронобиозный признак — это последний
ОСКОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ Ph СТЕНИЙ (завершающий) признак по степени развития в сравнительном ряду признаков структуры или поведения биоморф, проявляющемся в рамках их биоморфогенезов (онтогенезов или синонтогенезов). Гемитроиобиоз- ный иризнак — это промежуточный (сменяемый) признак по степени развития в сравнительном ряду признаков структуры или поведение биоморф, проявляющемся в рамках их биоморфогенезов. Тип биоморфо- генезов, или, короче, троиобиоз (дословно — направление жизни), мож- но определить одним или несколькими тропобиозными признаками, т.е. рассматривать несколько типов биоморфогенезов как один тип: например, полурозеточный короткостержнекорневой гемисимподиаль- ный мультирепродуктивный изоаклональный тропобиоз Вир1еигит multinerve DC. на Алтае и т.п. Категории стадий морфогенеза — кате- гории возрастных и категории фенологических стадий — могут быть охарактеризованы по различным признакам и в зависимости от них соответствующим образом обозначаться. Так, в жизненных циклах си- норганизмов Seseli condensatum (L.) Reichenb., Inula helenium L. и ряде. других видов, используя только признаки базовых органов, можно раз- личать следующие возрастные стадии: апоризомную, проризомную, ге- терономно-ризомную и автономно-ризомную (рис. 74, 137, 138, 158, 159, 184-186). Гетерономно-ризомная стадия может иногда выпа- дать из жизненного цикла некоторых корневищных растений (рис. 92). Если мы не выделяем внутри этих стадий каких-либо друтих стадий, то оцениваем их самой крупной категорией — периодом (апоризомный, проризомный и т.п.). Для названия морфогенических состояний (ста- дий) можно использовать также обозначения с временным или функ- циональным оттенком (например, ювенильное, матурное молодое и т.п.), причем признаки-индикаторы этих состояний могут быть или только морфологическими, или только функциональными, или смешан- ными (морфолого-функциональными) (рис. 131, 133-145). Рис. 131. Возрастные состояния биоморф в синонтогении Viola biflora L. (Алтай, Северо-Чуйский хребет, высокогорье). А — проростки (семядоли развертываются, зародышевая почка пока не разверну- лась), Б — всход (семядоли живы, зародышевая почка развернулась), В — юве- нильные растения: В, — униювенильное (семядоли отмерли, корневище еще не образовалось), В, — биювенильные (образовалось корневище гетерономное, т.е. имеющее пока связь с главным корнем), В, — триювенильное (корневище авто- номное, утратившее связь с отмершим главным корнем), Г — имматурное растение (происходит кущение за счет образования пазушных вегетативных побегов при сохранении моноподиального вегетативного роста и развития главного побега), Д — матурные аклональные растения (физиологическая зрелость наступает у ак- лональных растений, т.е. морфофизиологически целостных), Е — матурный кло- нальный синорганизм (репродуктивное развитие осуществляют синорганизмы, проходящие клональную стадию); 1 — семядоли, 2 — семядольный узел, 3 — ги- покотиль, 4 — главный корень, 5 — придаточные корни, 6 — гетерономное кор- невище, 7 — границы годичных побегов (приростов) прошлых лет — компактных мономерных пленорезидов, 8 — автономное корневище, 9 — срединный лист, 10— чешуевидный лист в основании годичного побега текущего года, 11 — прилист- ники, 12 — интеркалярный побег, 11 — открытая (ростовая) верхушка безрозеточ- ного компактного (предрозеточного) годичного побега, 14 — особи клона; у- увеличено по сравнению с масштабом, горизонтальная линия — поверхность почвы, одно деление масштаба — 1 см.
323 Глава 7. Основные способы освоения времени биоморфами семенных растений А Б Возрастные особенности семенных растений — предмет многих те- оретических и прикладных исследований (Irmisch,1850, 1859; Klebs, 1885; Velenovsky, 1905-1913; Goebel, 1928; Troll, 1937, 1954, 1964, 1969;
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ Ph СТЕНИЙ Кренке, 1940; Пошкурлат, 1941, 1962, 1969, 1974,1975; Работнов, 1945, 1947, 1949, 1950а, 1960а, 1964, 1974; Серебряков, 1952, 1954, 1964; Серебряков, Чернышева, 1955; Чайлахян,1958,1964; Игнатьева,1958, 1964, 1965; Ле- вин, 1959, 1961, 1963; Серебрякова, 1959, 1971; Уранов, 1960, 1973, 1974; Борисова, 1961; Голубев, 1962, 1965; Шорина, 1964, 1967; Смелов, 1966; Воронцова,1967; Лебедев,1968; Csapody,1968; Гуленкова, 1974; Шитт, 1968; Былова, 1968, 1994; Заугольнова, 1968; Добрынин, 1969; Гупа- ло, 1968, 1969; Гулисашвили, 1969; Гупапо, Скрипчинский, 1971; Нухимов- ская, 1971, 1972; Рысина, 1973; Giinther, 1975; Берка, 1973, 1976, 1982; Фисюнов, 1976; Мазуренко, Хохряков, 1977; Куперман, 1977; Торопова, 1977; Константинов, 1978; Васильченко, 1979; Чистякова, 1979; Жуко- ва, 1983, 1986; Смирнова, 1987; Бологова, 1989; Смирнова, Тропова,1994; Истомина, 1994; Барыкина, Чубатова, 1995, 1996 и др.). Развивая представления о возрастных состояниях растений, создан- ные Т.А.Работновым (1945, 1950а), и учитывая дополнения других авто- ров, была предложена (Нухимовский, 1980) следующая типовая схема возрастных стадий биоморфогенезов семенных растений (таблица 2). Таблица 2 Категории возрастных стадий
Глава 7.Основные способы освоения времени биоморфами семенных растений Отклонений от этой схемы и вариантов иных схем градаций воз- растных стадий может быть множество, что хорошо объяснимо с пози- ций кратко рассмотренной выше концепции морфогенеза. Однако у приведенной здесь схемы есть одна принципиальная особенность- она мобилизует исследователя на сбор по единому плану разносторон- ней информации о структуре и поведении биоморф на различных от- резках их жизненного пути. Отдельные возрастные стадии, указанные в этой схеме, у конкретных биоморф могут отсутствовать. В синонтогене- зе клональная жизнь может начинаться у кондивидуумов одного типа в виргинильном этапе, а кондивидуумов другого типа — в матурном этапе. Проявление вегетативного размножения может быть использовано для выделения иных возрастных стадий. Так, в матурном этапе синонто- генеза (например, радиолы розовой) можно выделять две фазы — акло- нальную и клональную (Нухимовский, 1974a) или же во всем синонтоге- незе — два периода: аклональный и клональный; И.Н.Берка (1976, 1982) выделяет два подцикла с таким названием. При выделении аклонального и клонального периодов в синонтоге- незе их можно считать частями эпиэмбрионального гемицикла, а к другому гемициклу — эмбриональному тогда будут относиться прена- тальный и латентный периоды. Возникает вопрос: в каком соотношении находятся понятия воз- растное состояние индивидуума и возрастное состояние кондивидуума. Выше уже было подчеркнуто, что особь, образующаяся из зиготы (поло- вого происхождения) — аклонист, а особь вегетативного происхожде- ния — клонист. Аклонисты бывают временными (мероаклонисты), если заканчивают свой морфогенез образованием клона, и ностоянными (изоаклонисты), если заканчивают свой морфогенез смертью без вегета- тивного размножения. Морфогенез как аклониста, так и кондивидуума начинается с зиготы. До образования клона кондивидуум представлен только аклонистом. На этом отрезке синонтогенеза мы говорим о возрастном состоянии кондивидуума, через признаки мероаклониста, поскольку он одновременно обладает возрастными особенностями как индивидуума, так и кондивидуума. Если аклонист преждевременно погибает по каким-либо причинам до образования клона, то все прощенные им возрастные состояния — это только его возрастные состояния как аклониста. После преобразования аклониста в клон говорим о возрастном состоянии кондивидуума, т.е. системы «меро- аклонист — клон» непосредственно через признаки клона, поскольку только после его образования кондивидуум становится системой, прин- ципиально отличной от индивидуума (особи), т.е. и аклониста, и кло- ниста. При необходимости особо рассматриваем еще и возрастные сос- тояния клонистов, которые не равнозначны возрастным состояниям кондивидуума. Особенности циклов индивидуального развития (онтогенезов) у особей различного происхождения можно отразить и в названиях этих циклов. Онтогенез изоаклониста (постоянного аклониста) обозначим как ганлоонтогенез, или эуонтогенез. Онтогенез мероаклониста (времен- ного аклониста) — нротоонтогенез. Онтогенез клониста — телоонто-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ Ph СТЕНИЧ генез. Морфогенез клона (системы генетически однородных особей веге- тативного происхождения, или клонистов) — метаонтогенез. Для обозначения возрастных состояний было предложено исполь- зовать определенные индексы (Уранов, 1975): j — ювенильные, im— имматурные, ч — взрослые вегетативные, g l, g2, g3 — молодые, средне- возрастные, старые генеративные, ss — субсенильные, s — сенильные. В настоящей работе принята идея индексации возрастных состояний биоморф с дополнениями и уточнениями: е — эмбриональные, рп- пренатапьные, lt — латентные, pjl — проростки (преювенильные пер- вые), pj2 — всходы (преювенильные вторые), j — ювенильные (jl, j2... — юв. первые юв. вторые...) im — имматурные (iml im2...— имм. первые, имм. вторые...), ее — эпиэмбриональные, т — матурные (mi — инициматурные, тт — медиматурные, fm — финиматурные), s — сенильные. Кроме того, иногда могут быть использованы индексы гемициклических возрастных состояний биоморф: aK — аклональные, кк — клональные. Проблему критериев выделения возрастных состоя- ний и стадий мы решаем на основе двух методических принципов. Принцип первый: за критерий выделения каждой стадии (состояния) принимается один признак (npocmou или сложный) из одного ряда альтер- нативных (сравнимых и взаимоисключающих) признаков, проявляющихся в биоморфогенезе любого семенного растения. Каждый из этих признаков может выступать индикатором возрастного состояния, а последний из них — индикатором всего биоморфогенеза. В качестве примера рас- смотрим несколько моделей биоморфогенезов, построенных по первому методическому принципу. Графическое изображение рассматриваемых моделей биоморфогенезов (рис. 132) и условные обозначения к ним позволяют получить наглядное представление о сущности альтерна- тивного (сравнительного однорядного) моделирования возрастных сос- тояний и полных биоморфогенезов. В качестве гемитропобиозных и тропобиозных признаков в таких моделях используем только альтерна- тивные признаки. Принцип второй: за критерий выделения отдельных возрастных ста- дий (состояний) принимаются разные структурные и (или) функциональ- ные признаки биоморф, которые в каких-либо сочетаниях могут быть инди- каторами этих стадий (состояний), если они лучше других признаков отра- жают особенности организации биоморф на определенном отрезке времени. Взяв за основу типовую схему категорий возрастных стадий (вре- менные единицы биоморфогенезов семенных растений), материальной основой которых являются одноименные с ними возрастные состояния (организационные единицы соответствующих биоморфогенезов), охара- ктеризуем их и обозначим на основе второго методического принципа основные критерии их выделения. При описании типов биоморфогене- зов и типов возрастных стадий (состояний) возьмем за основу признаки мультилинейной классификации биоформ семенных растений (Нухи- мовский, 1980), а также некоторые признаки биоморф рассматриваемых растений, которые уже использовались в работах других исследователей (Warming, 1884, 1908; Пошкурлат, 1941, 1969, 1975; Работнов, 1950а, 1964; Серебряков, 1952, 1964; Голубев, 1960, 1965, 1972; Серебряко-
Глава 7.Основные способы освоения времени биоморфами семенных растений 327 Рис. 132. Графические модели онтогенезов семенных растений, построенные на основе признаков основной широкой субординии побегов. Тропобиоз (тип онтогенезов): 1 — монокормусовый, 11 — дикормусовый, 111— трикормусовый, IV — тетракормусовый, V — пентакормусовый, VI — гексакор- мусовый, VII — гептакормусовый, VIII — энатокормусовый, IX — синэнатокор- мусовый (тип синкормуса определяется по более сложному побегу в этой систе- ме), Х — синоктокормусовый. Гемитропобиоз (тип возрастных стадий и состоя- ний): О!а — протерокормусовый инициальный (семенной), 01б — то же постини- циальный, la — монокормусовый семядольный вегетативный, lб — то же бессемя- дольный вегетативный, lв — то же репродуктивный; 2 — дикормусовый, 3 — три- кормусовый, 4а — тетракормусовый вегетативный, 4б — то же репродуктивный, 5 — пентакормусовый, 6 — гексакормусовый, 7а — гептакормусовый вегетатив- ный, 7б — то же репродуктивный, 8 — синтетракормусовый, 9а — сингептакорму- совый вегетативный, 9б — то же репродуктивный, 10а — синоктокормусовый ве- гетативный, 10б — то же репродуктивный, 11 — энатокормусовый, 12 — синэна- токормусовый; пб — побеги текущего года, бо — базовый орган, кр — корни, не- обособленные от родительского растения, ок — корни, обособленные от роди- тельского растения, л — зеленые листья, чш — чешуевидные листья, ол — отмер- шие прошлогодние листья, оц — отдельный цветок, вв — вегетативная верхушка, с — семядоли. Этот принцип построения моделей биоморфогенезов распростра- няется и на другие ряды организационных признаков многолинейной системы биоморф.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ Ph СТЕНИИ ва, 1971; Серебрякова, (отв. ред.), 1981, 1983 и др.). Подробный перечень вопросов, приведенный в работе И.А. Губанова, Т.А.Работнова и В.Н.Ти- хомирова (1970) полезен при изучении биоморфогении семенных расте- ний; его можно использовать и при характеристике методических аспек- тов в познании биоморфогенезов. Главное при выделении стадий био- морфогенеза — показать через удачно выбранные гемитропобиозные признаки последовательность усложнения и деградации биоморф се- менных растений (методические моменты выявления возрастных состо- яний приводятся непосредственно за характеристикой соответствующих стадий биоморфогенеза). Графически смену эпиэмбриональных воз- растных состояний растений и их некоторых индикаторных признаков в онтогенезе или синонтогенезе передают рис. 132, 133. Эмбриональный иериод — развитие особи с момента возникновения зиготы или зародышевой (эмбриональной) особи-апомикта до обо- собления от родительской особи и при наличии постнатального по- коя — до начала ее видимого роста. Пренатальный этаи — развитие эмбриональной особи без утраты морфофизиологической связи с родительской особью. Индикаторный (гемитропобиозный) признак — эмбриональная особь находится в орга- низационном единстве с родительской особью. Этот этап изучается, по возможности, при наличии соответствую- щих знаний или с помощью эмбриолога. Выясняют особенности опло- дотворения или апомиксиса, эмбриогенеза; характеризуются семена (размер, форма, цвет, опущение, число в плодах). Методическая литература: А.Н.Пономарев (1960), С.С.Хохлов (1965), Н.В.Ципин (отв. ред., 1980), Нухимовский и др (1989). Латентный этаи — пребывание обособленных от родительских растений эмбриональных особей в состоянии покоя или скрытого роста до начала прорастания. Индикаторный признак — эмбриональная особь (семя) отделилась от родительской особи и существует самостоятельно. При изучении этого этапа обращают внимание на особенности отделения семян от родительских организмов, время осыпания, урожай- ность (в пересчете на одну особь), на приспособления и способы распро- странения семян и плодов, дальность разноса. Определяется судьба семян, сроки их пребывания в относительном покое до начала прорастания. Методическая литература: Т.А.Работнов (1958, 1960б, 1964), А.А.Кор- чагин (1960а,б), Н.В.Ципин (отв. ред., 1980), А.В. Попцов и др. (1981). Эииэмбриональный иериод — развитие организма или синорганизма после прохождения эмбрионального периода или развитие организма из отдельных обособленных частей родительского растения (партикул), об- ладающих основными органами или образующих зачатки недостающих им основных органов (побегов и корней). Индикаторный признак — не- прерывный или с перерывами видимый рост самостоятельных биоморф от семени или от иной партикулы. Эпиэмбриональный период включает 3 этапа: виргинильный, ма- турный и сенильный (рис. 134-145). Методика исследований эпиэмбрионального периода биоморф се- менных растений в условиях их естественного произрастания, которая
Глава 7.Основные способы освоения времени биоморфами семенных растений 329 используется нами, включает экзоморфологическое описание разновоз- растных растений и отдельных органов, их гербаризацию, фиксацию в 50-70% спирте, фотографирование, а также геоботаническое описание местообитаний. В процессе наблюдений подмечают признаки-индика- торы возрастных состояний, т.е. гемитропобиозные признаки. Чтобы убедиться в верности их выбора, отбирают 30-50 растений, а если нет уверенностии и позволяет встречаемость вида, то 50-100 растений и более. Для редких и исчезающих видов отбор проб необходимо резко сокращать и увеличивать число описаний непосредственно на месте произрастания особей без полной выкопки их из почвы, а если позво- ляет организация растений, то можно после описания выкопанных осо- бей вновь высаживать их на прежнее место. При работе с древесными растениями определяют модельные экземпляры, по которым делают описания возрастных состояний. В исследовании биоморфогении выра- батывается определенный стиль описания, который постоянно совер- шенствуется его авторами и который служит не только информацией об итогах проделанной работы, но и своеобразным методическим пособи- ем, поскольку отражает то главное, на что надо обращать внимание при изучении жизненных циклов биоморф (Нухимовский, 1974а, 1976а, б, 1978, 1981 и др.; Нухимовский, Нухимовская, 1981, 1984; Нухимовский, Шре- тер, 1979; Нухимовский, Юрцев, Юрцева, 1987 и др.). Для изучения хода биоморфогении семенных растений пробы отби- раются еженедельно для того, чтобы безошибочно зафиксировать время наступления качественных структурно-функциональных изменений, ко- торые свидетельствуют о смене фенологических и (или) возрастных стадий. Число растений в пробе варьирует от 2-3 экземпляров, когда не обнаруживаются качественные изменения, до 8-10 и более, когда уда- ется обнаружить определенные качественные изменения в организации растений. Иногда, если качественные изменения касаются надземных органов, нет необходимости выкапывать все 10 растений. Отбор проб преследует обычно несколько целей: фиксацию каче- ственных (фенологических и возрастных) изменений, регистрацию ко- личественных изменений (числа и размеров листьев, побегов и других органов), а также определение сезонной и возрастной продуктивности. Число измерений по каждому ряду признаков должно быть достаточ- ным для проведения статистической обработки (при небольшом варь- ировании признака до 20, при большом 30-50). Пробы растений обра- батываются в камеральных условиях: все органы измеряют, при необхо- димости — взвешивают. Растения зарисовывают или фотографируют, гербаризируют. По нескольку экземпляров небольших травянистых рас- тений (или их органов) из каждой пробы фиксируют в спирте для дальнейшего изучения времени заложения пазушных почек, степени их дифференциации, регистрации стадий внутрипочечного развития, для изучения анатомии стеблей и корней и др. Методическая литература: Т.А. Работнов (1946, 1947, 1949, 1950а, б, 1960а, б, 1964), Н.А. Прозоровский (1949), М.С. Шапьгг (1960), Ал.А. Фе- доров и др. (1956, 1962), М.Х. Чайлахян и др. (1973), Ал.А. Федоров, 3.Т. Артюшенко (1975, 1979), Серебрякова (отв. ред.; 1981, 1983) и др.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ Ph СТЕКИЙ Виргинильный этаи — развитие биоморф без физиологической зре- лости, подготавливающее морфофизиологическую основу для ее дости- жения. Индикаторный признак — эпиэмбриональная биоморфа не дос- тигла физиологической зрелости (не развивала еще ни разу репродук- тивных органов) (рис. 134, 137, 139, 142 и др.). В зависимости от биологии конкретных растений число возрастных стадий, которые можно выделять на отдельных этапах эпиэмбриональ- ного периода, может быть различным. Так, например, у однолетников в виргинильном этапе их может быть две (фаза проростка и всхода), а иногда больше, как и у многолетников, т.е. еще ювенильная и имматур- ная фазы. Из концепции биоморфогении, рассмотренной выше, следу- ет, что число одноименных категорий возрастных стадий (периодов, этапов, фаз и др.) должно быть не менее двух, иначе их выделение лишено смысла. Если у однолетников репродуктивные органы начинают видимый рост сразу же после развертывания семядолей, то в таком исклю- чительном случае до наступления физиологической зрелости отмечается только виргинильный период, который не дифференцируется на фазы. Преювенилъная фаза — развитие семядолей особей без ветвления главного побега. Индикаторный признак — живые семядоли, развер- тывающиеся или развернувшиеся при прорастании семени и сохраня- ющиеся при моноподиальном нарастании главного побега. Эта фаза включает проростковую и всходовую гипофазы («подфазы»). Проростковая гииофаза (гииофаза ироростка) продолжается от на- чала прорастания семени до развертывания зародышевой почки. Инди- каторный признак — прорастающее семя, или семядольный почкоко- рень (ризопротерокормус). Фенетическая организация семенного расте- ния может быть предельно упрощена, когда оно всецело нацелено на скорейшее репродуктивное усилие. В таких очень редких случаях про- росток (растение развивается на этапе проростка) непосредственно пре- образуется в физиологически зрелое растение, т.е. особь переходит в матурный этап жизни, минуя все другие возможные виргинильные ста- дии (рис. 133, 1-Ш). При описании проростка регистрируются следующие параметры: 1) тип прорастания — надземное (семядоли выносятся выше уровня по- верхности почвы) или подземные (семядоли остаются в почве); 2) сем- ядоли — число, размеры, форма, степень дифференциации; жилкова- ние, окраска, опушение; 3) гипокотиль — размеры (длина, диаметр), цвет; 4) зародышевый корешок — размеры (длина, диаметр), цвет, ветвление. Всходовая гииофаза (гииофаза всхода) начинается видимым ростом первого настоящего листа и заканчивается нередко отмиранием семядо- лей; однако, если раньше, чем засохнут семядоли, начнутся какие-либо другие существенные экзоморфологические изменения в структуре рас- тения, например, впервые образуется пазушный побег, или апикальная часть медиального (первоначального) побега от розеточного роста пе- рейдет к безрозеточному и т.п., то они должны служить признаками, характеризующими переход особей в новое возрастное состояние. Ин- дикаторный признак — семядольный вегетативный ризомонокормус. В методическом плане начало развертывания зародышевой почечки и появление первого настоящего листа свидетельствует о необходимо-
Глава 7.Основные способы освоения времени биоморфами семенных растений 331 ~» О ~О Qaä И Й ~.-.. 5 И О Х ~-~ д ,д ~ О у ;у, '[, Р1 О~по д д, ~,д" ФЭ 1', 00 ~» й) ~се а3 О О Ю =o 1{:) ~р~ М О д~ д Ж ;у, '° О О g ~, 'О Я Я ,д [, сл Хф 1-. О ~Й cd ,, И, ~4 е~~ О Ои o=W v 3 'Ф с~~ ~с~ Й м О ~ А Ф', йae "О i g j i g.,< >~ О о ~< ~ О guî~ „О,~ яиЫ Ж м д~ /+AS Ж ~ v О;~ д~ Иьч ~{р Й ц О ~ О КО "X ~» ~ О m о Д 2~~ ~ x Oc~o ФЭ +~Ch О ~-~ л Щ б3 ф ь О ф й О,~ ~ х е~ о Р ~~ д~ О Д~ ° дЖ О g й 0i л ° [» О О Д ! ф~м л Щ 4Э ~р О ~ О. О Д х й й; ~О Ж й ~4 О ,~ Х Ю Я Ж О ~ Я Р' В: Ю:( в О A 3 3 ф; ф; М О л ц~ 0а О A ~ ф~ ~л да A Ю g ~ О Х д с 1 О Х о Ф д 1 Я До - Х Х О О х о ~. О с~ 1 р, ~4 л й О cg й v g о Cg ~» ф о О о О Ю Я A ;. 2 Ж О
332 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕНКЫХ РАСТЕКИЙ Рис. 134. Виргинильный этап в биоморфогении Lycopus exaltatus L. (Восточный Казахстан, пойма р. Иртыш). А — проросток, Б — всходы (семядоли живы, зародышевая почка развернулась), В — ювенильное растение (семядоли отмерли, главный корень жив, столоны не образуются), Г, — униимматурное растение (главный корень жив, столоны отрас- тают), Г, — биимматурные растения (главный корень отмер частично или полно- стью, столоны отрастают); 1 — семядоли, 2 — главный вегетативный безрозеточ- ный побег, 3 — столон, 4 — живой главный корень, 5 — отмирающий главный корень; у — увеличено по сравнению с масштабом, горизонтальная линия — по- верхность почвы, одно деление масштаба — 1 см. сти отбора проб растений, чтобы проанализировать и описать суще- ственные качественные изменения в структуре и функционировании растений. При описании всходов важно обратить внимание на следую- щее: 1) семядоли — размеры (длина, ширина), форма, продолжитель- ность существования; 2) эпикотиль (первое междоузлие) — размеры (длина, диаметр), цвет; 3) тип роста медиального (1-го порядка) побе- га — безрозеточный, предрозеточный (менее 3-х сближенных листьев, предшествующих возникновению розетки), розеточный, полурозеточ- ный, верхнерозеточный, верхнеполурозеточный; 4) гипокотиль — раз- меры (длина, диаметр), цвет; 5) корень — размеры (длина, диаметр у основания), характер нарастания, ветвление. По мере роста и развития всходов увеличивается число листьев, поэтому при взятии проб (при выращивании в культуре) надо отмечать время появления и продолжи- тельность существования листьев, их форму и размеры (в динамике).
Глава 7. Основные способы освоения времени биоморфами семенных растений 333 Ювенильная фаза — развитие бессемядольных особей осуществля- ется без ветвления или с ветвлением, но качественно отличным от ветвления особей в следующем возрастном состоянии. Наиболее рас- пространенный индикаторный признак — бессемядольный вегетатив- ный ризомонокормус или ризотетракормус. Если эти признаки не вы- ражены, то могут быть использованы иные, например, развитие листьев иной формы, чем у всходов; изменение способа нарастания побега (мо- ноподиального на симподиальный и др.), ветвление побега при живых семядолях, образование безрозеточной части побега вместо розеточной как у всхода и т.п. У биоморфы может быть одно или несколько юве- нильных состояний, например, у Abies sibirica Ledeb. (Нухимовская, 1971) образование первой стеблевой ветви свидетельствует об одном из ювенильных состояний, а образование ежегодно или с перерывами на главной оси двух сильно сближенных ветвей — признак другого юве- НИЛЬНОГО СОСТОЯНИЯ И Т.Д. При описании образцов ювенильных растений надо обращать вни- мание на следующее. 1. Надземные органы: типы побегов по сложности организации; размеры, формы нарастания и ветвления (если оно есть) побе- гов; число, форма, опушение, жилкование и размеры листьев; особенности листорасположения (супротивное, очередное или мутовчатое); динамика образования листьев и время их отмира- ния на отдельных побегах; число узлов на побегах и размеры междоузлий. 2. Подземные органы: типы корней по сложности организации; особенности нарастания и ветвления корней; размеры, форма и продолжительность жизни корней; наличие придаточных кор- ней, их число, размеры, расположение. 3. Особо рассматривают базовый орган: ценоподий, аэроподий, гео- подий, ризом, каудекс, конодий, зрелую луковицу, зрелый клу- бень, зрелую клубнелуковицу, если он образуется у биоморфы; его возрастную изменчивость, что может способствовать выделе- нию гемитропобиозных признаков тех или иных возрастных сос- тояний. Имматурная фаза — ежегодное вегетативное развитие особей, ча- сто с ветвлением, качественно отличным от ветвления, если оно есть, ювенильных особей. Когда не действуют эти признаки, можно исполь- зовать иные, чтобы отразить стадию перехода растений от ювенильной жизни (детства) к матурной (взрослой, физиологически зрелой). Напри- мер, отмирание главного корня и (или) развитие системы придаточных корней, образование базокормуса или его возрастное изменение, изме- нение способа нарастания побега, проявление новых признаков в числе, размерах и (или) форме листьев и т.п. У однолетников нередко имма- турное состояние вообще не регистрируется, поскольку быстрые темпы их развития смазывают проявление индикаторных признаков новых возрастных состояний. Имматурные растения по внешнему виду не- редко напоминают матурные растения, но еще не образуют репродук- тивных органов. Наиболее распространенный индикаторный признак—
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕНКЫХ РА СТЕНИИ начало образования вегетативных побегов кущения или иных вегетатив- ных побегов ветвления, которых не имели ювенильные растения, т.е. образование более сложных систем, например, вегетативных ризопен- такормусов или вегетативных ризогексакормусов. В пробах этой возрастной фазы необходимо обращать внимание на описание тех же моментов, что и в ювенильной фазе. Регулярное взятие проб позволяет установить время и место заложения почек, их типы (пазушные или придаточные, закрытые или открытые и т.д.), наличие у них периода покоя. Если у растений наблюдается отрастание побегов (весеннее или летнее), то необходимо обратить внимание на время и место их отрастания, число, особенности роста, ветвления и листообразования. Матурный этан — развитие биоморф с физиологической зрелос- тью, выражающейся в ежегодном или неежегодном отрастании побегов с репродуктивной функцией. У однолетних растений он наступает через несколько недель после прорастания семян, у двулетников — на следу- ющий год, а у многолетников — через 2-3 года и позже. В матурном этапе обычно различают несколько фаз, гемитропоби- озными признаками которых могут быть переход к надземному ветвле- нию, изменение степени ветвистости и (или) кустистости (например, некустистые, простокустистые, сложнокустистые), отмирание главного корня и (или) образование придаточных корней, изменение в органи- зации базового органа, переход к вегетативному размножению, особен- ности организации клона и т.д. Факту вегетативного размножения сле- дует уделять особое внимание, поскольку проявление его свидетель- ствует о переходе биоморф на новый уровень организации — синорга- низменный уровень (рис. 133, IV). Проявление вегетативного размноже- ния может быть использовано для выделения и иных возрастных ста- дий, например, в матурном этапе синонтогенеза можно выделять две фазы: аклональную и клонапьную или во всем синонтогенезе — два периода: аклональный и клональный и т.п. (подробнее см. гл. 10.2.). При описании разновозрастных матурных растений важно обра- щать внимание на следующие показатели. 1. Типы побегов по сложности их организации и другим структур- ным и функциональным признакам: степень специализации, фазность видимого роста, кратность репродуктивной функции, сексуализация цветков, положение репродуктивных органов в структуре соцветий, степень разветвленности; степень послеве- гетационного отмирания верхушек годичных побегов; формы и размеры листьев и стеблей, их цвет и опушение; тип базового органа, если он образуется, особенности его организации. 2. Типы корней по сложности их организации, продолжительность жизни главного корня и его размеры (длина, диаметр у основа- ния), наличие системы придаточных корней, степень их разви- тия, наличие корневых отпрысков и т.д. Сенильный этан — заключительный отрезок жизни особи или син- организма после утраты физиологической зрелости. Индикаторный признак — сильное старческое разрушение в организации побеговых
Глава 7. Основные способы освоения времени биоморфами семенных растений 335 систем и полная (в данных условиях существования) утрата способности к развитию репродуктивных органов. Сенильный этап проявляется только у многолетних растений и то не всегда. У сенильных растений образуются только ослабленные веге- тативные побеги. Важно отразить в описании этой стадии биоморфоге- неза те качественные и количественные изменения, которые удерживают биоморфы семенных растений на грани между нормальной матурной жизнью и смертью: как изменяется организация побегов и корней, как снижается жизненная активность органов, в частности, как затухает способность к побегообразованию, как процессы некроза постепенно или быстро разрушают организацию биоморф. Одновременно с выделением возрастных стадий биоморфогенеза изучаются также и фенологические стадии, т.е. стадии, повторяющиеся в соответствии с годовой ритмикой. Завершая описание биоморфогении какого-либо вида семенных растений, важно дать обобщенную характеристику его по разным орга- низационным признакам. Фенологические наблюдения надо стремиться проводить парал- лельно с изучением возрастной изменчивости. Особенности изучения фенологии растений описаны во многих методических работах, поэтому мы ограничимся лишь ссылками на некоторые из них (Серебря- ков, 1947, 1954; Бейдеман, 1960, 1974; Шульц, 1966; Сырокомская, Липа- това, 1966; Борисова, 1972 и др.). Состояния биоморф — это организационные единицы, а стадии биоморфогенезов — временные единицы биоморф и биоморфогенезов. С наших позиций вполне определенно можно ответить на давний вопрос, волнующий многих биоморфологов (Хохряков, 1975б, 1978, 1981; Алеев, 1980, 1986; Серебрякова, 1980 и др.): что сменяется, когда мы наблюдаем изменчивость биоморф? Иаш ответ однозначен: биоморфа сме- няется другой биоморфой только после прекращения ее существования в прежнем качестве как биоморфы: т.е. организма или синорганизма. B биоморфогенезе — онтогенезе или синонтогенезе многократно сменяются состояния биоморфы: фенологические и возрастные. Если в биоморфогенезе сменяются какие-либо индикаторные признаки организааии, например, габитуса, поведения и т.д., то это обычно означает смену состояний одной биоморфы (!) на другие состояния. Когда простая биоморфа-орга- низм является nepsou частью (мероаклонистом) в развитии сложной био- морфы-синорганизма, то в таком случае она предрасположена когда-либо смениться на другую часть синорганизма — клон. Используя рассмотрен- ный выше методический материал, в качестве примера кратко опишем эколого-морфологические аспекты биоморфогении нескольких видов. Bupleurum triradiatum Adams ех Нофн. (сем. Apiaceae). Изучен автором в 1977 г. на Западном Саяне (окрестности перевала Сурдабан, 2300-2500 м над ур. моря, склон юго-восточной экспозиции с низкотравной альпий- ской растительностью). Эмбриональный период. Пренатальный этап — оплодотворенная се- мяпочка или зрелое семя в структуре родительского растения; июль- сентябрь.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕКИЙ Латентный этап — зрелое семя, обособленное от родительского растения; с сентября текущего года до июня-июля следующего года. Эпиэмбриональный период. Виргинильный этап. Преювенильная фаза. Проростковая гипофаза — прорастающее семя или семядольный почко- корень (ризопротерокормус); июнь-июль (рис. 135, А). Всходовая гипо- фаза — семядольный вегетативный предрозеточный (1-2 сближенных настоящих листа) или очень редко розеточный (3-4 настоящих листа) ризомонокормус; июнь-август (рис. 135, Б). Семядоли (две) линейные или линейно-ланцетные надземные голые, на верхушке островатые, 0,8- 1,2 см длины, 0,1-0,12 см ширины. Ювенильная фаза — бессемядольный ежегодно вегетативный (фи- зиологически незрелый) розеточный ризомонокормус или ризотетракор- мус; с середины лета первого года жизни (после отмирания семядолей) до 4-20 лет (рис. 13, В). Имматурная фаза — ежегодно вегетативный розеточный ризо- гексакормус, ризогептакормус или реже ризоэнатокормус от 3-20 до 5- 24 лет (рис. 135, Г). Кущение чаще всего простое; число побегов куще- ния 1, реже 2-3 и более. Когда пазушный побег становится зимующим, фиксируем образование каудекса. Главная тетрариза короткостержневая 2-8 см дл. От основания главного корня, а с увеличением возраста и от основания гипокотиля, развивается пучок из мелких придаточных кор- ней. Побеги нарастают обычно моноподиально. Ежегодный прирост каудекса, как правило, составляют курторезидные монокормусы. Изред- ка вегетативные верхушки побегов погибают, тогда их сменяют новые побеги, развивающиеся из постгибернальных (послезимних, т.е. зимо- вавших один раз) или реже провентивных (зимовавших два и более раз) почек. В структуре розеточных монокормусов развивается 0-3, чаще 1-2 почки, которые в большинстве становятся провентивными. Терминальные почки — незащищенные, провентивные — защищенные. Матурный этап. Инициматурная фаза. Гипофаза первая — репро- дуктивный полурозеточный условно закрытый ризопентакормус; с 4-24, чаще 8-15 лет. Продолжается один вегетационный сезон до образования постинициального вегетативного побега замещения в августе — начале сентября текущего года, реже до весны следующего года. В эту возрастную стадию переходят ювенильные особи, минуя имматурную стадию. Бутонизация в конце июня-июле, цветение в июле-августе, плодоношение в конце августа-сентябре. Гипофаза вторая — ежегодно или неежегодно (чаще) репродуктивный полурозеточный каудексовый ризогексакормус или ризогептакормус с 2-мя порядками унирепродук- тивных (обоеполых, однократно репродуктивных) побегов, цветущих или не цветущих в год наблюдения; с 4-24, чаще 8-15 лет до 8-25 лет (рис. 135, Д). Кущение слабое или отсутствует. Нарастание побеговой системы гемисимподиальное; миниморезидные тетракормусы формиру- ют компактный каудекс. Вегетативно-репродуктивный терминальный монокормус (5-15 см высотой) завершает моноподиальное вегетативное розеточное нарастание в течение нескольких (2-20) лет и часто состоит из компактного основания с 1-2 ланцетными листьями (монофиллами) и рыхлого зонтика из 2-4, чаще 3 зонтичков; одиночныепаракладии
337 Глава 7. Основные способы освоения времени биоморфами семенных растений Рис. 135. Онтогения Вир1еигцт triradiatum Adams ex Hoffm. (Саяны, хребет Западный Саян, окрестности перевала Сурдабан, альпийский пояс, склон юго-восточной экспозиции). А — проросток, Б — всход, В — ювенильные особи, Г — имматурные особи, Д— инициматурные особи, Е — медиматурные особи, Ж — финиматурная особь, И— сенильная особь; 1 — семядоли, 2 — семядольный узел, 3 — гипокотиль, 4 — кор- невая шейка, 5а-5д — корни: 5а — главная монориза, 56 — главная дириза, 5в- неглавная (ризогенная) монориза, 5г — тририза, 5д — тетрариза, 6 — предрозеточ- ный побег, 7 — вегетативный медиальный розеточный побег, 8 — вегетативный аксиллярный розеточный побег, 9 — монокормус, 10 — тетракормус, 11 — унире- продуктивный полурозеточный пентакормус, 12 — унирепродуктивный послеро- зеточный дикормус, 13 — соцветие зонтик, 14 — курторезидные монокормусы, 15 — миниморезидные тетракормусы, 16 — конодий (прекаудекс), 17 — каудекс, 18 — почки, 19 — семядольный почкокорень (ризопротерокормус), 20 — се- мядольный монокормус, 21 — бессемядольный ежегодно вегетативный ризотетра- кормус, 22 — ежегодно вегетативный ризогептакормус; 23 — ризопентакормус, 24 — неежегодно репродуктивный ризогептакормус, 25 — неежегодно репродук- тивный ризоэнатокормус, 26 — срединные листья (жилкование показано не на всех листьях). Горизонтальная линия — поверхность почвы; пунктирная линия— уровень плотного мохового покрова; одно деление масштаба — 1 см; 1л, 2л, Зл- особи 1,2,3 и т.д. лет жизни; у — увеличено в несколько раз по сравнению с масштабом. образуются редко. Такой годичный побег можно обозначить как после- розеточный. Редко в основании развивается 3 ланцетных листа и тогда верхушечный побег — полурозеточный. Во всех случаях эти побеги за- вершают развитие всего унирепродуктивного полурозеточного плейо- фазного пентакормуса (медиального или аксиллярного). Новые пост- инициальные побеги развертываются как пролептические в августе-на-
338 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИЧ чале сентября из почек в пазухах ланцетных листьев в основании после- розеточных вегетативно-репродуктивных побегов или как послезимние (постлатентные) осенние или весенние, чаще всего из пазушных почек самых верхних (прошлогодних) розеточных побегов. Главная монориза теряется среди множества адвентивных ризогенных и гипокотилоген- ных корней, образующих вместе пучковатую короткостержнекорневую систему (синторизу). Каудекс сверху расположен в густом моховом покрове и полуразложившихся растительных остатках. Медиматурная фаза — ежегодно или неежегодно (чаще) репродук- тивный полурозеточный ризоэнатокормус с 3-4 порядками унирепро- дуктивных побегов, цветущих или не цветущих в год наблюдения; с 8-25 до 12-28 лет (рис. 135, Е). Число цветущих побегов 0-4, чаще 1. Просто- кустистое строение может сменяться некустистым и вновь появляться позднее. Формирование сложного куста с цветущими побегами — приз- нак максимального развития растения. Финиматурная фаза — неежегодно репродуктивный полурозеточ- ный каудексовый ризогептакормус (очень редко) или ризоэнатокормус с 5 и более порядками унирепродуктивных побегов, цветущих или не цветущих в год наблюдения (все исследованные растения имели менее 10 порядков побегов); с 12-28 до 30 лет (рис. 135, Ж). Кратность пло- доношения определялась по числу перевершиниваний акронекрозных унирепродуктивных побегов. В этой стадии происходит постепенное ослабление функциональной активности и отмирают некоторые каудек- совые ветви. Сенильный этаи — растения, подобные матурным, ежегодно веге- тативные после утраты физиологической зрелости; продолжается не- сколько (1-5) лет (рис. 135, И). Предельная продолжительность жизни изоаклонистов около 30 лет. По эколого-морфологическим тропобиозным признакам володуш- ка трехлучевая — это многолетнетравянистый летнезеленый надземно- двусемядольный гемикриптофильный полурозеточный семидетерминат- но-репродуктивный (полузакрытый) гемисимподиальный однодомный мультирепродуктивный монофиллический очереднолистный каудексо- вый пучковато-короткостержнекорневой брахипланофильный (слабо подземно разрастающийся в горизонтальном направлении) слабокусти- стый изоаклональный психрофитный вид. Жизненные циклы растений можно описывать по-разному, т.е. с различных позиций и с большей или меньшей степенью подробности в зависимости от цели изучения. Если характеризовать основные орга- ны — побеги по третьей широкой субординии, то биоморфогению Cistus tauricus С.Presl вкратце можно изложить следующим образом. Напомним, что в третьей широкой субординатной классификации по- бегов, за критерий сложности организации которых принята продолжи- тельность постинициального периода их жизни (с непрерывным или прерывистым видимым ростом), различаются следующие классы (типы) побегов: внутригодичные (силлептические, пролептические, ивановы и какие-либо другие побеги, составляющие структуру годичных побегов), годичные и сверхгодичные (двухгодичные, трехгодичные, многогодич-
Глава 7.Основные способы освоения времени биоморфами семенных растений 339 ные), т.е. побеги более сложные по продолжительности постинициаль- ного периода, чем внутригодичные и годичные побеги. Cistus tauricus С.Presl — ладанник крымский (сем. Cistaceae). Вид широко распространен в нижнем приморском поясе южного Крыма, поднимаясь по горным склонам до 300 м над ур. моря, редко выше. Наряду с другими растениями, например, Bupleurum fruticosum L., Hippocrepis emeroides (Boiss. et Sprun.) Czer., Jasminum fruticans L. об- разует кустарниковый ярус в можжевеловых (Juniperus excelsa М.В.), сосновых (Pinus pallassiana D. Don), дубовых (Quercus pubescens Willd.) и смешанных лесах. Онтогения Cistus tauricus изучалась автором в окрестностях Никитского ботанического сада (рис. 136). Эмбриональный период. Пренатальный этап — оплодотворенная семяпочка или зрелое семя в структуре родительского растения; июль- сентябрь. Латентный этап — зрелое семя, обособленное от родительского ра- стения; с сентября текущего года до весны следующего года; размножается только семенами; для прорастания семенам не нужна стратификация. Эпиэмбриональный период. Виргинильный этап (136, А-Г). Преюве- нильная фаза. Проростковая гипофаза. При посеве семян в теплице еди- ничное прорастание начинается на 6-8 день, а массовое на 10-11. Про- растание надземное. Развернутые семядоли продолговатые или продол- говато-ланцетные (0,6-1,0 см дл., 0,15-0,3 см. шир.). С видимым по- явлением зародышевой почки между семядолями нередко начинается образование боковых корешков на главном корне. Всходовая гипофаза. Через 16-18 дней после начала прорастания происходит развертывание зародышевой почки — развивается первая пара супротивных настоящих продолговато-ланцетных опушенных ли- стьев. Главный корень в этой гипофазе активно ветвится. После образования 4-5-ой пары супротивных листьев семядоли, как правило, засыхают, что свидетельствует о переходе особи в ювенильную фазу. Ювенильная фаза. На первом году формируется годичный ростовой раздвинутоузловой надземный апонекрозный прямой медиальный по- бег с перекрестно-супротивным листорасположением; листья сидячие продолговато-ланцетные. Многолетнюю стеблевую часть перезимовав- шей особи составляет единственный короткий, реже длинный тотарезид, на котором развиты верхушечная и пазушные открытые почки. Тотаре- зид вместе с почками является тотарезидным побегом, который дает начало аэроподию — надземному многолетнему стволику, который пока еще не ветвится. У наиболее сильных растений пазушные почки пробуждаются еще на первом году жизни, образуя короткие вегетатив- ные внутригодичные (силлептические) пазушные побеги. Начало побе- гового ветвления — признак перехода особей в имматурное состояние. Имматурная фаза. В этой фазе летом или весной следующего года начинают отрастать из открытых пазушных почек вегетативные побеги (в конце первого года или чаще на втором году жизни). При весеннем отрастании пазушных и особенно новых верхушечных годичных побегов узлы развиваются сближенно, по крайней мере ближе друг к друту, чем во время летнего роста побегов, что позволяет легко фиксировать границы
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СГЕНИЙ л 2 :у' О Е: 4 f о О X О Е 1 К X X X 1-, К X О X G О о >ь Х G. О hC v hC X G О О h4 О О :у' X :а с6 f О Ю О О h4 О f й h4 X Х й О О й f О О G h4 О Л 2 G М ~л О 0 О сп ;:3 О ° ~ Ью ;:3 с6 сп :3 ~л U 0~~ X :а О f X О 'О О X G 1к1 й g) ~[~и л ~Ч ° й о X Г'1 ~О~ C О й~ >ь О о~1 ~ и G ;Я ООЖ Д5 г'1 ц~ X О „~ ~<~о tQ X G. О х о ~о~ кот XXZ f О g3 о:> Х О д: йи а. О hC v a5 ~) О Е Р.ь .-. X ~. )~ X О о И tX tD Ц О ~ о о 4 И >Е hC О 2 О а й' >ь Щ лм f .0 О Х а.~ еа ,'Я с) О „.& ~ М X ~] Ь о й ~ 5~О а. ~ tQ ~ ф'„' X О 2 >ь ,'Ц .0 О Йь Х X ga ь 0 йЭ f Ж X ( О О о И о Х ~а ~! л О О о О:" О 65 О~~ X :~ о й < м- О О О X ;у' й~ ~ О [О О ~ 2 О =1 Q X X 65 Я. О cd о О . )X ~[ О О а; О ~ LO i а )~ О 2 X ).'Я Ж й о м >ь й:~ а Е ~~ О О )~; 2 л5 :ц а. :у Яо >ь ~ а. )~ щ X И О G Р) г~ й Ж (Л v LO Е и, л 65 д Г'1 X + о X G. e f )~ 2 и О ~5 i О G а О ~ X ф.) О LO Х д , ° X 2 2: йЭ а. ж 1 ~ ~оо ° Ц л Ф CO ~5 C О v О X О р~ О О й" ,й", >ь Ф Ф )~ ° ф ) ~~~~ ) ~~~~~ лй 2 д Ж д X д~ Г'1 i Щ tu Е Ф ~) )Q р Ф 2 X ° ~ о :> О лX < а. о ч О О и,~ й~ О Г'1 „й а о Q.g ) >ь [ а. 2:> О ~~О О а [ цр 65 О Q. О 4~ 1 )~ Е g) 2 О X а. и ° 1 - а.oo О- C ~~ О о О O OX Х а. X G О hC о й 65 Q, [- :у' ~) 2 ã. X О X у~ Г'1 о О О О О йэ Q, д Ж ('1 й; О д] :ц 2 ~[ й~:Ц О О X f Ф О Q, йЭ ц М О й~ О X [.. а5 .0 G о оО~1 )~ ~( О Х х О л о о о uD g О G hC л ла 65 о u" 2 >ь > 6.) Д ч а, о ОО~ X i д .'Ц ф ~О~ Х ф X hC О й~ G И ь а. о 1- 1 о f Q 2 О о m 34 о hC )Õ О й~ О йЭ ~:! ф 2 X 2 Ж И 1 CO ~4 л о Х G О hC f X И ф X X X Ф 1 X )~ 2 X f X 65 f о О И 1 X hC о :у' X И 1-, й' X о
Глава 7.Основные способы освоения времени биоморфами семенных растений 341 годичных побегов. Отрастание пазушных побегов из самых нижних почек приземных частей тотарезидного главного годичного побега— начало аэроксильного кущения. Когда за один вегетационный сезон из перезимовавшей почки развивается система, состоящая из весеннего родительского (исходного) побега и силлептических (производных) по- 5егов, то в целом формируется весенний разветвленный годичный по- 5ег, который на второй год онтогенеза может быть вегетативным. Такие растения зимуют в имматурном состоянии. Главный корень продолжает ветвиться, достигая 8-15 см длины. Рамозорезидный (с разветвленным резидом) побег образуется из разветвленного вегетативного годичного апонекрозного (!) побега и полностью включается в структуру развет- вленного аэроподия. Матурный этаи (рис. 136, Д,-Д,) наступает на 2-3 году эпиэмбри- DHKIbHQH жизни особи (при угнетении позднее). Медиальный побег двух-,; он завершает свой по- стинициальный период жизни образованием верхушечного одиночного цветка-плода. Пазушные побеги, начавшие свою активную жизнь на виргинильном этапе, через 2-3 года завершают ее часто также образованием верхушечного цветка-плода. Цветение в мае, плодоноше- ние — в августе-сентябре. За один вегетационный сезон формируется 2-3 порядка побегов (рис. 136, Е). С зелеными листьями вегетативных силлептических (индуктивных) побегов второго и третьего порядков растения зимуют. К весне они опадают и сменяются листьями весенних побегов, которые опадают уже во второй половине лета. Таким образом, особенности генераций листьев создают эффект вечнозелености куста. И.Б.Белянина (1961) отнесла ладанник крымский к группе «вечнозеле- ных фанерофитов с открытыми почками». За метамерную единицу по- беговой структуры матурных особей можно принять двухгодичный ве- гетативно-репродуктивный детерминатный закрывающийся летне-ве- сенний пазушный разветвленный побег (пентакормус) (рис. 136, Д„Е), который в зависимости от условий обитания и возраста куста различает- ся только количественно — по степени развития. Если несколько отвлечься от использования на ладаннике крым- ском третьей широкой субординатной классификации побегов и в ка- честве примера перейти к другой классификации, допустим, пятой ши- рокой субординии, то получим следующую картину. Надземную часть трехлетнего куста составляет вегетативно-репродуктивный многогодич- ный разветвленный детерменатный (закрывающийся верхушечным репродуктивным элементом в случае полного развития) побег (рис. 136-Д,). В кроне средневозрастных кустов образуются побеги такого же типа, которые проходят следующие стадии развития: 1 — силлептического (ин- дуктивного) вегетативного побега, 2 — вегетативно-репродуктивного двух- годичного летне-весеннего пазушного детерминатного побега, 3 — вегета- тивно-репродуктивного трехгодичного побега и далее четырех,- пяти,— ше- стигодичного побега и т.д. (рис. 136, Е). Осевая структура аэроподиев у матурных особей, как правило, при- растает димерными рамозорезидами. Аэроподии достигают 0,8-1,2 м в высоту. Куст у ладанника крымского простой, реже сложный; он включает
ОСНОВБ! БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИЙ в основании два (три) порядка побеговых ветвей и состоит обычно из 3-6 аэроподиев. Главный корень сохраняется в течение всей жизни особи. Процессы отмирания у старых кустов распространяются почти одновременно на все аэроподии, поскольку они возникают примерно в одно время (разница в возрасте 1-4 года). Полное отмирание кроны главного аэроподия приводит вскоре к гибели всего куста (примерно на 18-25 году онтогенеза). По эколого-морфологическим тропобиозным признакам ладанник крымский — аэроксильный надземно-двусемядольный гемисимподи- альный всесезоннозеленый (ежегодно дважды меняющий листья) пере- крестка-супротивнолистный безрозеточный однодомный детерминат- но-репродуктивный слабокустистый изоаклональный стержнекорневой горно-лесной субтропический кустарниковый вид. При изложении материала по биоморфогении какого-либо вида можно использовать разные способы, но обычно доминирует повество- вательный стиль. На примере Paeonia апота1а L. разместим сведения о биоморфогении этого вида несколько усовершенствованным спосо- бом — рисуночно-табличным (см. таблицу 3). Paeonia anomala L. — нион уклоняющийся (сем. Ranunculaceae)— лугово-лесной вид, распространенный в лесной и горно-лесной зонах Сибири, на северо-востоке европейской части России, в восточном Ка- захстане, а также в горно-лесных районах Тарбагатая, Джунгарского Аптау, Тянь-Шаня и северной Монголии (Шипчинский, 1937; Кува- ев, 1976). Настойку из корней и корневищ или настойку из равных по весу (1:1) сухих подземных и надземных частей пиона уклоняющегося (ма- рьина корня) применяют в качестве седативного средства при неврас- тенических состояниях с явлениями повышенной возбудимости, бессо- ницы, ипохондрии и для улучшения пищеварения (Шретер, 1962; Ку- ваев, 1976, и др.). Пион уклоняющийся также весьма декоративен. Исследования проводились на Алтае в районах Теректинского (окрестности пос. Ябоган), Ивановского (пойма р. Белая Уба), Сарым Сакты (окрестности с. Медведка) хребтах (Нухимовский, Нухимов- ская, 1978). Пион уклоняющийся произрастает на Алтае на луговых полянах среди разреженных лиственничных, темнохвойных и смешанных лесов, по их опушкам, на вырубках, на высокотравных горных, пойменных лугах; обычен в низкогорье и среднегорье, но иногда поднимается и в верхний лесной пояс гор. В естественных условиях на Алтае пион часто встречается вместе Adonis sibirica Pattin ex Ledeb., Campanula glomerata L., Cirsium heteroptlyllum (L.) Hill, Crepis lyrata (L.)Ftoel., Euphorbia рИоза L., Dactylis glomerata L., Gentiana macrophylla Pall., Iris ruthenica Кег-Gawl., Роа slbirica Roshev., Primula macrocalyx Bunge, Trollius asiaticus L., Viola disjuncta W. Beck., V. hirta L. и др. Надземные части пиона уклоняюще- гося животными не поедаются, и на лесных лугах, подверженных вы- пасу скота, нередко формируются довольно обильные заросли этого вида.
Глава 7.Основные способы освоения времени биоморфами семенных растений Таблица 3 Синонтогения Paeonia anomala L. на Алтае (рис. 137, 138) Возрастные состояния коыдивидуумов Продолжи- тельы ость возрастных состояыий О Х Оа C VI II Ш IV VII Х О Х Ю й Х О Х Ю й Х й с конца ИЮНЯ-H3QJM до конца августа, сенгября, редко октября с конца августа, октября до весны, лета следующего года, или в течение нескольких лет с июня, реже августа до сентября, октября, или изредка до весны следующего года Осыовыая морфо- физиологи- ческая характерис- тика оплодотво- POHHRH семяпочка или зрелое семя в структуре родительско- го растения зрелое семя, обособлен- ное от родительс- кого растения прорастаю- щее семя или семя- дольный почкоко- рень (ризопроте- рокормус) (рис. 137, А) Дополнительная характеристика: А — частная (распространяется только на рассматриваемое возрастное состоя- ние) Б — общая (распространяется на несколь- ко или на все возрастные состояния) Б — цветки (обоеполые крупные — 7-13 см в диаметре, с множеством тычи- нок и 5, реже 3 пестиками, 5-8, реже более розово-красными лепестками и 5-8 зелеными или красноватыми ча- шелистиками); в опылении участву- ют насекомые (пчелы, шмели); А — зрелые плоды из 3-5 (чаще 5) листо- вок, медленно растрескивающихся по шву после созревания, семена голые, черные, блестящие, округлые или ши- рокоэллиптические с плотной кожу- рой, 0,6-0,8 см дл., 0,5-0,7 см шир. часто осыпаются не сразу; иногда они попадают в почву вместе с плодами; листовки постепенно разрушаются, а семена проходят естественную страти- фикацию, сохраняясь в почве от 9-11 месяцев до нескольких лет; А — прорастание семян подземное, про- ростки обычно появляются группами до 20 экз., чаще 6-12 экз. (до матур- ного этапа из них доживает, как пра- вило, 1 особь); семядоли (две) посто- янно скрыты под кожурой; они слабо зеленоватые, мясистые, эллиптичес- кие, постепенно переходящие в че- решки 0,5-1,2 см дл., коротко срос- шиеся основаниями; главная моно- риза — до 4см; гипокотиль — 0,4-1 см дл., 0,1-0,12 см в диам.;
VII IV VI семядоль- ный вегета- тивный безрозеточ- ный ризо- монокормус (рис. 137, Б) с июня, реже августа до сентяб- ря, октября или изредка до весны следующего года х О Х Ю й х C с1, реже 2, до 2-16 лет, чаще до 6-12 лет бессемядо- льный ежегодно вегетатив- ный (физио- логически незрелый) безрозеточ- ный ризо- тетракормус (рис. 137, В,) ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛСТЕНИЙ Продолжение таблицы 3 А — начало образования первого настоя- щего листа при наличии живых семя- долей — признак образования всхо- дов; изредка особи с живыми семядо- лями зимуют и лишь весной утрачи- вают их, переходя в новое возрастное состояние; первый лист по очерта- нию пластинки округлояйцевидный, сердцевидный при основании, одно- кратно трехрассеченный с заострен- ными трех- или многократно раздель- ными и лопастными сегментами; ли- стовая пластинка 2,5-5 (чаще 3-4) см дл., 3-7 (чаще 4-5) см шир., перехо- дящая в длинный черешок (6-15 см); на гипокотиле и основании главного корня образуются поперечные склад- ки («морщинистость»), свидетель- ствующие об их контрактильной де- ятельности; верхушечная почка пере- мещается ближе к поверхности по- чвы, поэтому длина первого подзем- ного междоузлия (эпикотиля) колеб- лется от компактного до длинного (0,1-8 см); А — ежегодное весеннее отрастание оди- ночных терминальных компактных безрозеточных монокормусов (с од- ним листом однократно или дважды трехрассеченным, трех-, многоло- пастными сегментами), составляю- щих в совокупности тетракормус-ин- дикаторный признак возрастного со- стояния; Б — стеблевая часть у монокормусов под- земная; она несет кроме рассеченно- го листа еще 2-4 низовых чешуевид- ных цельных листа, сросшихся иног- да в короткую трубку; в пазухах че- шуевидных листьев развиваются поч- ки; базовый орган сначала проризом, а затем (после образования из него первых придаточных корней) — без- листный ризом; он нарастает моно- подиально за счет увеличения числа остаточных (резидных) побегов, т.е. в этом случае — компактных резидных монокормусов;
345 Глава 7. Основные способы освоения времени биоморфами семенных растений Продолжение таблицы 3 VI Ш IV VII с 2-16 до 5-20 лет О Х Ю х О х Ю й х C с 5-20 до 8-25 лет ежегодно вегетативный безрозеточ- ный ризогек- сакормус, р�изогепт- акор�м, очень редко ризоэнато- кормус, с одним рассеченным летнезеле~~ым листом в струзлуре верхушечного монокормуса (рис. 137, В,) ежегодно вегетативный безрозеточ- ный корне- в ищу~ый ризогепта- кормус, очень редко ризоэнато- кормус, с двумя рассеченны- ми летнезе- ле ными листьями в струкг ре п~кюиющи- альных монокорму- сов (рис. 137, Г,) А — ежегодное весеннее отрастание по одному, редко в какой-либо год по два компактных безрозеточных моно- кормуса с одним рассеченным лис- том при неежегодном (редко ежегод- ном) перевершинивании вновь отра- стающих монокормусов, т.е. соответ- ственно при гемисимподиальном или эусимподиальном нарастании побе- говой системы — индикаторный при- знак возрастного состояния; главная монориза до 20 см; Б — с увеличением возраста верхушка по- бега начинает неежегодно или еже- годно подниматься над поверхностью почвы, что неежегодно или ежегодно приводит ее к гибели зимой; геми- симподиальное нарастание корневи- ща осуществляется за счет минимо- резидных тетракормусов, а эусимпо- диальное нарастание — за счет ми- ниморезидных монокормусов; глав- ный корень, как правило, живой, но изредка (рис. 137, В,-б) рано погиба- ет, что не губит растение, хотя и за- метно ослабляет его; А — ежегодное весеннее отрастание безро- зеточных вегетативных монокормусов с 2-мя рассеченными листьями — ин- дикаторный признак первого имматур- ного состояния; высота стеблевых надземных частей побегов — 5-25 см; Б — нарастание побега (гептакормуса или энатокормуса) далее происходит эу- симподиально; корневая система- синториза, состоящая из главной тет- раризы и придаточных корней, а очень редко — только из придаточ- ных корней;
VII Ш IV VI с 8-25 до 10-30 лет х О Х Ю й х й с 10-30 до 12-40, чаще 18-30 лет ежегодно вегютативный безрозеточ- ный корне- вищный ризогепта- кормус, очень редко ризоэнато- кормус, с тремя и более рассеченны- ми летнезе- ле ными листьями в структуре пастнюшм- альных монокорму- сов (рнс. 137, r,-ry ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛСТЕНИЙ Продолжение таблицы 3 А — ежегодное весеннее отрастание без- розеточных вегетативных монокорму- сов с 3-мя срединными рассеченны- ми листьями — индикаторный при- знак второго имматурного состояния; высота стеблевых надземных частей побегов — 10-30 см; основание глав- ного корня 0,4-1,0 см в диаметре; главная монориза 8-25 см дл; Б — листорасположение очередное; плас- тинка срединных листьев 7-12 см дл., 8-16 см шир.; она дважды трех- или перисторассеченная с дву- или мно- гократнолопастными сегментами; че- решки длинные — 6-10 см; низовые листья чешуевидные, цельные (по 4-5); в пазухах чешуй развиваются (по одной в каждой) защищенные по- тенциально зимующие почки, выше- стоящая из которых после зимовки развертывается в видимый гетеротоп- ный (с подземным компактным ос- нованием) вегетативный безрозеточ- ный прямостоячий раздвинутоузло- вой длинный монокормус, а ниже- расположенные почки становятся провентивными (запасными); в пос- ледующие годы некоторые провен- тивные почки очень медленно, но все же заметно увеличиваются в разме- рах, нарастая моноподиально, при этом некоторые субперулярные (при- крытые~ почки выходят из под при- крытия чешуй родительской почки и становятся экстраперулярными (не- прикрытыми), часть провентивных почек не получает необходимых ус- ловий для развития и погибает (рис. 137, Г,); А — ежегодное весеннее отрастание без- розеточных вегетативных монокорму- сов с 4-мя и более рассеченными ли- стьями в их структуре — индикатор- ный признак третьего имматурного состояния; высота надземных стебле- вых частей побегов 20-50 см; куще- ние отсутствует, реже простое; Б — корневище постоянно погружается в почву; оно становится наклонным или горизонтальным; по числу резид- ных монокормусов определяется воз- раст растений (рис. 137, Г,);
347 Продолжение таблицы 3 IV VI VII с 10-40, чаще 18-30, до 30-80 лет А — первое проявление репродуктивной функции в структуре постинициаль- ных монокормусов свидетельствует о вступлении в инициматурное состоя- ние; кущение отсутствует — это ин- дикаторный признак первого иници- матурного состояния, простое куще- ние — признак-индикатор второго инициматурного состояния и, когда кущение сложное, то оно свидетель- ствует о третьем инициматурном со- стоянии; Б — показателем мощности развития рас- тений может быть число ежегодно отрастающих монокормусов, что и принято здесь для разграничения молодых и средневозрастных матур- ных мероаклонистов (временных ак- лонистов, т.е. особей семенного про- исхождения, жизненный цикл кото- рых — это только часть жизненного цикла кондивидуумов); монокормусы (с одним верхушечным цветком) или реже дикормусы (с 2-5 верхушечны- ми цветками паракладиев рыхлого соцветия) вегетативно-репродуктив- ные закрытые (унирепродуктивные) обоеполые безрозеточные прямосто- ячие раздвинутоузловые длинные ге- теротопные (с подземным компакт- ным основанием) потенциально ба- зискелетные акронекрозные, 40-90 см высотой; срединные листья дваж- ды трех- или перисторассеченные, с дву- или многолопастными сегмента- ми в числе 6-12, чаще 7-9; чешу- евидные листья более или менее стеб- леобъемлющие, их 5-9; главный ко- рень, как правило, жив, но очаги некроза, ежегодно образующиеся в корневище после отмирания надзем- ных частей монокормусов и дикор- мусов пока еще медленно распрост- раняются по проводящим тканям корневища, достигая также и тканей главного корня; фенофазы: бутониза- ция в июне, цветение — с середины июня до конца августа, плодоно- шение с конца августа до октября; ежегодно или нееже- годно репродук- тивный (физиологи- чески зрелый) безрозеточ- ный корне- вищный ризогепта- кормус или чаще ризоэнато- кормус с числом постиници- альных монокорму- сов не более 10 (рис. 138, Д) х О Х Ю й х C Глава 7.Основные слособы освоения времени биоморфами семенных растений
VII Ш IV VI с 30-80 до 60-120 лет й О Х Ю й с 60-120 до 100-150 лет й Х Ф Продолжа- ется до 10 лет ежегодно или нееже- годно репродук- тивный безрозеточ- ный корне- вищный ризоэнато- кормус с числом постиници- альных монокорму- сов более 10 (рис. 138, Е) совокупносчь нескольких (2-12) ежегод- но или не- ежегодно репродуктив- ных безрозе- точных автономно- ризомных ризокорму- сов (рис. 138, Ж) ежегодно вегетативные после утра- ты физиоло- гической зрелости без- розеточные автономно- ризомные ризокормусы (рис. 138, И) ОСКОВЫ БИОМОРФОЛОГИИ СЕМЕКНЫХ РЛ СТЕКИЙ Продолжение таблицы 3 А — корневище гетерономное, т.е. связан- ное с живым главным корнем, — это индикаторный признак первого сред- невозрастного матурного состояния; корневище автономное, т.е. утратив- шее связь с главным корнем после его гибели, — это индикаторный признак второго средневозрастного матурного состояния; число постини- циальных монокормусов, а редко еще и дикормусов, колеблется от 11 до 120, куст иногда разрастается до 2,0 м' (на пойменных лугах); Б — в главном корне и корневище обра- зуются крупные дупла, в которых скапливаются почва и нередко посе- ляются почвенные насекомые; един- ство сложного куста сохраняется очень долго, после гибели главного корня функционирует базальная, на- раставшая в прошлом без ветвления, часть корневища с мощными прида- точными корнями, и лишь когда не- кроз доходит до разветвленной части корневища, создаются условия для полной общей некротической парти- куляции; на корневище ежегодно развиваются пучки эфемерных при- даточных корней, основания которых сохраняются долго, вызывая образо- вание своеобразных наростов; А — осуществление полной общей парти- куляции через корневище непосред- ственно приводит к замедленному (ретардационному) вегетативному размножению — образованию клона, особи которого обладают физиологи- ческой зрелостью, т.е. нормально цветут и плодоносят; Б — постепенно угасает физиологическая активность клонистов медлительного кондивидуума, число и степень раз- вития постинициальных монокорму- сов уменьшается; Б — синонтогенез продолжается не более 150-160 лет; среди кондивидуумов пиона уклоняющегося много долго- жителей, проживших около 100 лет.
349 Глава 7. Основные способы освоения времени биоморфами семенных растений 5б 78 Рис. 137. Виргинильный этап в синонтогении Paeonia anomaia L. (Алтай). А — проросток, Б — всходы, В,-В, — ювенильные мероаклонисты: В, — первое юве- нильное состояние, В, — второе ювенильное состояние, Г,-Г, — имматурные мероакло- нисты: Г, — первое, Г, — второе, Г,— третье имматурные состояния, К — упрощенные модели разновозрастных кондивидуумов; 1 — семядоли, 2 — гипокотиль, 3 — семядоль- ный узел (с возрастом его положение отмечается приблизительно), 4 — корневая шейка, 5а-5е — корни: 5а — главная монориза (субмедиаль), 5б — главная дириза, 5в — неглав- ная монориза (сублатераль), 5r — тририза, 5д — главная тетрариза, 5е — синториза, 6— остаток отмершего главного корня, 7 — ризогенная корневая ветвь, 8 — каулогенная корневая ветвь, 9 — чешуевидные листья, 10 — летнезеленые рассеченные безрозеточные листья, 11 — сухие остатки от рассеченных листьев прошлых лет, 12 — верхушечная почка, 13 — вегетативный медиальный безрозеточный побег, 14 — вегетативный аксил- лярный безрозеточный побег, 15 — монокормус, 16 — тетракормус, 17 — гептакормус, 18 — компактные пленорезидные монокормусы, 19 — миниморезидные монокормусы, 20 — миниморезидные тетракормусы, 21 — проризом, 22 — ризом, 23 — пазушные поч- ки; 24-27 — организменные ризокормусы: 24 — семядольный почкокорень, или ризопро- терокормус, 25 — семядольный вегетативный ризомонокормус, 26 — бессемядольный ве- гетативный ризотетракормус, 27 — бессемядольный вегетативный ризогептакормус, 28— живые основания органов (побегов, листьев, корней; их верхние части удалены), 29— сухие остатки надземных стеблевых частей побегов. Горизонтальная линия — поверхность почвы; 1л 2л, 3л и т.д. — особи 1, 2, 3 и т.д. лет жизни; ув — увеличено по сравнению с масштабом; одно деление масштаба 3 см.
350 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ Ph СТЕНИИ Рис. 138. Матурные и сенильный этапы в синонтогении Paeonia anomaia (Алтай). Д1 Д3 иницима1 урные мероаклонис гы: Д, — некустистое, Д, — прос1 окус1 ис гое, Д, — сложнокустистое инициматурные состояния; Е — медиматурный мероакло- нист, Ж — финиматурный клон, И — сенильный клон; 30 — унирепродуктивный безрозеточный побег, 31 — очаги некроза тканей корня или ризома, 32 — эфемер- ные корни, 33 — ежегодно или неежегодно репродуктивный ризогептакормус, 34 — ежегодно или неежегодно репродуктивный гетерономный (с главным кор- нем) ризоэнатокормус, 35 — ежегодно или неежегодно репродуктивный автоном- ный (без главного корня) ризоэнатокормус, 36 — ежегодно вегетативный (после утраты физиологической зрелости) автономный ризокормус. Одно деление мас- штаба 2 см. Остальные обозначения те же, что и на рис. 137. Подведем некоторые итоги о биоморфогении пиона уклоняющегося. 1. Семена прорастают не ранее чем на следующий год после соз- ревания, зимуя обычно под снегом. На виргинильном этапе жиз- ни особи по мере их вегетативного развития переходят от моно- подиального нарастания безрозеточной побеговой системы к ге- мисимподиальному нарастанию (с неежегодным одноразовым перевершиниванием) и, наконец, к 5-20 годам начинают нарас- тать только эусимподиально (с ежегодным одноразовым перевер- шиниванием), сохраняя в основном этот способ до конца си- нонтогенеза. Долгие годы кондивидуум представлен мероакло- нистом (временным аклонистом) с гетерономным ризомом (ри- зом связан непосредственно с главным корнем). Первое цвете- ние происходит на 10-40-м, чаще 18-30-м году. С началом ста- рого матурного (финиматурного) состояния особи кондивидуу- ма (клонисты) — автономно-ризомные (их ризомы утратили связь с отмершим главным корнем). Синонтогенез продолжается до 150-160 лет. 2. По эколого-морфологичским тропобиозным признакам пион ук- лоняющийся — травянистый летнезеленый подземно-двусемя- дольный геофильный однодомный перекрестноопыляемый мультирепродуктивный безрозеточный дважды трех- или пе- ристорассеченнолистный очереднолистный эусимподиальный кустистый детерминатно-репродуктивный (закрывающийся) подземно-сближенноузловой компактно- или короткокорневищ- ный брахипланофильный или мезопланофильный (редко) ретар- дационно-кондивидуальный (медленно вегетативно размножа- ющийся, с клонистами, долго удерживающими территорию) мезофитный лугово-лесной вид. В методической части рассматриваемой главы было показано не- сколько приемов описания биоморфогении. Чтобы дополнить инфор- мацию о том, как «работают» признаки биоморф семенных растений, охарактеризованные в нашей работе, приведем рисунки (с подписями к ним) биоморфогении трех видов: Datisca cannabina L., Hedysarum alpinum L., Lithospermum officinale L. рис. 139-145).
Глава 7.Основные способы освоения времени биоморфами семенных растений
О, ~~ )) ~~1 ° ~в~ l~ 1 I Ф Э ~ Ъ,Ф е Ф'4~ ° ° ° ф ° ° ° . ° ° ° ° ° ° в ° ° ° в 1 ° ° ° 1 ° J в ° ° ° ° Э ° ° в ° в Э ° ° ° ° а ° ° Э ° в ° Э ° а Э ° ! Э ° ° Э ° J ° ° ° ° ° ° ° ' 3 ° а в ° ° ' I ° ° I ° r ° ° ° в ° ° а ° ° Э r ° ° ° ° ° Э ° ° ° ° ° r Э а ° ° Э ° 3 Э Э ° ° ° ° ° ° ° ) Э ° ° Э ° ° ! ° ° ° ° ° ° ° ° ° Э ° Э а ° ° ° ° l ° в в ° Э Э Э ° ° Э ° ° Э ° ° Э ° ° ° ° ° ° Э ° 1 ° а Э ° ° ° '! ° ° ° ° ° ° ° Э ° ° ° ° Э в Э в ° а ° Э ° ° ° ° а Э СЭ ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА С ГЕНИЯ
Глава 7. Основные способы освоения времени биоморфами семенных растений 353 Рис. 140. Инициматурная фаза в синонтогении Datisca cannabina. 10 — вегетативно-репродуктивный безрозеточный главный ризодикормус, 11— эусимподиальное весеннее отрастание побегов, 12 — проризом, 13 — гетероном- ный ризом, 14 — придаточный корень, 15 — ризогексакормус, 16 — ризогептакор- мус, 17 — жесткая внутривидовая конкуренция за выживание у особей, развив- шихся группой из нескольких (10-15) семян в одной лунке, 18 — отмершие особи, 19 — репродуктивные элементы, 20 — основание весеннего пазушного вегетатив- но-репродуктивного побега (верхние части удалены). Остальные обозначения те же, что и на рис. 139.
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 141. Медиматурная (E) и финиматурная (Ж) фазы в синонтогении Datisca cannabina. 21 — вегетативно-рспродуктивный безрозеточный монокарпический или моноан- терический условно закрытый дикормус, 22 — фрондо-фрондулезно-брактеозное соцветие, 23 — зона неполного торможения, 24 — зона полного торможения, 25— автономный ризом, 26 — зона возобновления, 27 — ризоэнатокормус, 28 — почка, 29 — продольный разрез почки, 30 — плод, 31 — тычиночный цветок, 32 — фраг- мент парциального тычиночного соцветия (растения двудомные), 33 — продоль- ный разрез через миниморезидный побег, 34 — эфемерная (замирающая) верхуш- ка полузакрытого соцветия. Остальные обозначения те же, что и на рис. 139 и 140.
Глава 7. Основные способы освоения времени биоморфами семенных растений Рис. 142. Виргинильный этап в биоморфогении Hedysamm alpinum L. (Читинская область, низкогорные луга). А — проростки, Б — всходы, В — ювенильные растения: В, — униювенильные осо- би (семядоли отмерли, образуют только простые листья), В, — биювенильные особи (не ветвятся, начинают образовывать или постоянно образуют тройчатые листья), В, — триювенильные особи (продолжается эусимподиальное нарастание, максимальное число долей непарноперистых листьев не более 5), Г — имматурные растения (образуются первые побеги кущения или особи не ветвятся, но в пос- леднем случае максимальное число долей непарноперистых листьев более 5); 1— семядоли, 2 — семядольный узел, 3 — гипокотиль, 4 — отмершие семядоли, 5а- главная монориза, 56 — главная дириза, 5в — неглавная монориза, 5г — тририза, 5д — главная тетрариза, 6 — простые листья, 7 — тройчатые листья, 8 — непарно- перистые листья с пятью листочками, 9 — непарноперистые листья с семью и более листочками (жилкование показано не у всех листьев), 10 — каудекс, нарастающий эусимподиально за счет миниморезидных годичных побегов, 11— прилистники, 12 — семядольный почкокорень (ризопротерокормус), 11 — семя- дольный ризомонокормус, 14 — бессемядольный вегетативный безрозеточный ри- зомонокормус, 15 — бессемядольный вегетативный безрозеточный ризогексакор- мус на втором году жизни растения, 16 — вегетативный безрозеточный эусимпо- диальный ризогептакормус, 17 — сухие остатки от надземных частей годичных побегов прошлых лет; горизонтальная линия — поверхность почвы, одно деление масштаба — 1 см.
356 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ Ph СТЕНИЙ Рис. 143. Матурный этап в биоморфогении Hedysarum alpinum (местообитание то же, что и на рис. 142). 18 — основание вегетативно-репродуктивного приоткрытого (с фронда-фронду- лозным открытым соцветием и замирающей в процессе цветения верхушкой) годичного безрозеточного побега (дикормуса), 19 — парциальный каудекс, 20— придаточный корень, 21 — вегетативно-репродуктивный ризогептакормус, 22— ризоэнатокормус, 23 — неполная некротическая партикуляция старой особи, 24— слабая особь клона, иногда образующаяся в финиматурном состоянии; пунктиром показано примерное положение семядольного узла; одно деление масштаба — 2 см. Остальные обозначения те же, что и на рис. 142.
Глава 7. Основные способы освоения времени биоморфами семенных растений 357 Рис. 144. Первый и второй годы в онтогении Lithospermum officinale Ь. (ВИЛАР, Москва, интродукционный питомник). А — проростки, Б — всходы, В — ювенильная особь, Г — имматурная особь, Д— инициматурные особи (число вегетативно-репродуктивных побегов не более 10); 1 — семядоли, 2 — семядольный узел, 3 — гипокотиль, 4 — корневая шейка, 5а-5д — корни: 5а — главная монориза (субмедиаль), 5б — главная дириза, 5в- неглавная монориза (сублатераль), 5г — тририза, 5д — главная тетрариза, 6 — ни- зовые листья, 7 — срединные листья (опушение показано только на отдельных листьях), 8 — вегетативный безрозеточный медиальный годичный побег, 9 — ве- гетативный безрозеточный аксиллярный годичный побег, 10 — монокормус, 11— вегетативно-репродуктивный эфемерно-полисимподиальный дикормус, 12 — вер- хушечный элемент соцветия (увеличено), 13 — основания надземных побегов (верхние части удалены), 14 — вегетативный гексакормус, 15 — каудекс, 16-21— организменные корнепобеги: 16 — семядольный почкокорень (ризопротерокор- мус), 17 — семядольный вегетативный ризомонокормус, 18 — бессемядольный ве- гетативный ризомонокормус, 19 — вегетативный ризогексакормус, 20 — вегетатив- но-репродуктивный ризодикормус, 21 — вегетативно-репродуктивный ризогекса- кормус. МК — упрощенные модели организмов; у — увеличено по сравнению с масштабом; одно деление масштаба — 1 см, горизонтальная линия — поверхность почвы.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕКИИ О й; и~ (д ~ 1о 3~ у О а3 C ф,' О ~; Й О О ~» Я Ф' О 1» Зч й м с~ ~ о~ О () О ф ~ О од ~о [» < о ФЭ ~ Й л ~й 3 й О 3~ 0 ° л О Д ю~Ъ й О О Д: О а Х й ~ц ~ д Ю Р) ~~ о и а О О cd ( ) ~ Ж ~хм а О ~ ~) В~' о ~ voоM И а ~~ Ж о. о ~ жж ce + ~~~ Е ~' Ю,д М ~') ~~о р, ~ C) ~enÈ ХО т» И а ~ О О ~ ~, О И 2 :~: О д а д; а ~ а. и Х й О" о ~ С4ю О ~< с~у ф; Ю ф г ~ 63 я 4~ ~ ce ~~ (-4 Д а ~ О;-, О ~.> ~4 5О О й 0 ~г ~ О ~13 л Оа~ ~ ~;) ~О~ Ж ц ~) Х , ~ о Ж оО ~а~ й 4~ Х ,'Я О Ш ~
359 Глава 7.Основные способы освоения времени биоморфами семенных растений 7.4. Последовательность морфогенезной сборки и разборки организации биоморф Биоморфы, как и другие живые системы, следуют в своем суще- ствовании законам и правилам жизне- и средосохранения (гл. 3). В организации биоморф как морфологическом процессе («морфопроцес- се»), или морфогенезе, тело растения собирается из структурных эле- ментов, которые не прибывают откуда-то со стороны, а производятся (закладываются) прямо здесь в этом же теле, на основе некоторых внут- ренних и внешних причин, как итог функционирования генов в соот- ветствии с законом управления-исполнения организации биоморф се- менных растений (гл. 4). Гены (элементы-руководители) управляют ме- ханизмом роста и развития тела растений, а с помощью РНК (первых элементов-исполнителей) синтезируется определенный белок, образу- ются ферментные системы и, наконец, закладываются в нужных частях и в нужное время разнообразные меристемы: верхушечные (вторые эле- менты-исполнители) и дополнительные, в частности латеральные, ин- теркалярные, маргинальные (третьи элементы-исполнители). Далее раз- виваются и функционируют по определенным принципам четвертые, пятые и шестые элементы-исполнители (соответственно побеги, корни и корнепобеги-органы, или онтоиды). Основные органы биоморф се- менных растений — побеги и корни — главный и единственный сбо- рочный материал, определяющий их конституцию. В процессе работы меристем побегов и корней метамерно и (или) диамерно (гл. 5) созда- ется все тело семенного растения. Работу всех отмеченных выше эле- ментов-исполнителей объединяют в себе седьмые элементы-исполните- ли — корнепобеги-организмы, или эуфиты. Совокупность эуфитов- клонистов, восходящую в своем развитии с одной зиготы, образует одну систему — синорганизм (гл. 4.5). Одни части побегов и корней, развивающиеся в одном вегетацион- ном сезоне эфемерны, т.е. отмирают в том же сезоне. Другие побеговые или корневые части после одного вегетационного сезона не отмирают, а превращаются в персистентные (дву-, многолетние). Многолетние, в особенности базовые (многолетние побеговые) органы определяют диф- ференциацию семенных растений на деревья, кустарники, кустарнички, полукустарники, полукустарнички и многолетние травы. Как ни велико различие базовых органов, но оно отражает в основ- ном количественные изменения в структуре биоморф. Качественные изменения биоморф касаются организации побегов и корней в стадии их активного функционирования, когда наиболее полно определяется степень сложности этих органов и соответственно их место в иерархи- ческих рядах (гл. 5). Сборка тела растения в процессе его саморазвития идет с менее сложных структурных элементов и заканчивается самыми сложными из них. Побеги и корни нарастают сначала моноподиально и очень редко верхушка медиального (главного) побега сохраняет жизне- способность всю жизнь. Чаще всего растения раньше или позже перехо-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ дят к иным типам нарастания, что обеспечивает им более полное освоение окружающего их пространства; с нарастанием связан и другой важный процесс — ветвление, включая и его разновидность — кущение (гл. 4.4). Венцом биоморфогенезов обычно является репродуктивный процесс. Развитие репродуктивных структурных элементов находится в единстве и борьбе с развитием вегетативных органов, а противоречия между ними преодолеваются путем подчинения вторых структур первым. После завершения репродуктивного процесса отплодоносившие и (или) отпыль- ценосившие части побегов отмирают на всю или часть своей длины (гл.6.2). Многолетние стеблевые остатки побегов — резиды ежегодно участвуют в формировании многолетнего побегового тела семенных растений, их базового органа (гл. 6.3). С отмиранием первых цветоносных частей побегов начинается некроз в теле растений, который у однолетников распространяется на все их тело, а у многолетников процессы некроза одних частей и роста других осу- ществляются рядом, приводя к различным видам партикуляции и ве- гетативного размножения (гл. 4.5). Только в процессе партикуляции воз- можно обособление от родительского организма его репродуктивного и (или) вегетативного потомства. Через некроз разномасштабных струкгурных элементов, начиная с наиболее старых, слабых и (или) больных идет разборка организации тела, т.е. подготовка его к элиминации. Этот же процесс обычно и завершает элиминацию. Такова в общих чертах последовательность морфогенезной сборки и разборки в фенетической организации биоморф семенных растений, ины- ми словами, если хотите, — последовательность морфогенезного монтажа и демонтажа живой системы в ранге организма или синорганизма.
Глава 8. Изменчивость и эволюция 8.1. Внутривидовая пластичность В настоящее время накоплено много фактов изменчивости биоси- стем и в первую очередь — изменчивости организмов и видов (Гете, 1957; Darwin, 1859; Вавилов, 1966; Майр, 1947, 1974; Шмальгаузен, 1969, 1983; Мамаев, 1972 и др.). Ч.Дарвин (Darwin, 1859), как известно, рас- сматривал изменчивость как возникновение различий-уклонений от ти- пичной формы. По Ю.А.Филипченко (1978, с.7), изменчивость (биоло- гическая) — это «явление некоторого различия между собой даже близ- ко родственных особей и групп особей». Термин «изменчивость» вклю- чает в себя как изменение вообще (движение вообще), так и изменение материальных систем в их морфогенезах (частные движения, или фун- кции) и всякие различия между системами. В Природе бесконечна встречаемость материальных систем и нет среди них абсолютно одинако- вых — это все результат бесконечного изменения (движения) материи. Классификации явлений изменчивости даже некоторых групп био- систем свидетельствуют о большом их разнообразии (Майр, 1947; Ма- маев, 1972; Гуляев, Мальченко, 1975 и др.). Рассмотрев самые общие положения о морфогенезах и морфогенических состояниях биоморф, мы могли убедиться, что в пределах цикла их существования проявля- ется изменчивость по разным рядам организационных признаков: фе- нологическая и возрастная, что вместе составляет, по С.А. Мамаеву (1972), хронографическую изменчивость, т.е. изменчивость во времени. Хронографическую изменчивость материальных систем отражают спек- тры морфогенических состояний, выделяемых по каким-либо рядам сравнимых признаков, что было показано в предыдущей главе. Чем больше состояний, выделенных по одному сравнительному ряду призна- ков, проявляет материальная система в процессе ее существования (в морфогенезе), тем выше хронографическая изменчивость этой системы и типа таких систем. С понятием изменчивости связано другое поня- тие — пластичность. Явление пластичности знакомо биологам давно, например извест- но, что габитус одновозрастных деревьев дуба, ели, сосны и др., произ-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ растающих в лесу, сильно отличается от габитуса таких же деревьев, растущих на открытом месте (Warming и. Graebner, 1918; Поплавс- кая, 1948; Радкевич, 1983 и др.). М.И. Сахаров (1951, с. 175) отметил в своих исследованиях, что пластичность обеспечивает расселение рас- тений на экологически неоднородной территории и способствует воз- никновению модификаций и экотипов и появлению новых видов». Б.Г.Иоганзен (1959) объяснял пластичность организмов как понятие, близкое их изменчивости (свойства организмов изменяться под влияни- ем как внешних, так и внутренних факторов). Пластичность, по Иоган- зену (1959, с. 265) — «изменчивость организма под влиянием условий жизни»', она бывает трех форм: морфологическая (изменение морфоло- гических в широком смысле признаков под влиянием отклонения вне- шних факторов от нормы); физиологическая (изменение физиологичес- ких в широком смысле признаков и свойств под влиянием воздействия новых условий); экологическая (изменение выносливости организмов под действием необычных факторов). Интересное разнообразие жизненных форм наблюдается у видов растений, обитающих на неустойчивых к эрозии субстратах. В литера- туре отмечались своеобразные приспособления растений к перенесению оголения их подземных частей или засыпания субстратом (Warming и. Graebner, 1918; Вильямс, 1922; Дубянский, 1928; Петров, 1933; Лаврен- ко, 1935; Шалыт и Калмыкова, 1935; Хантимер, 1959; Бергхольц, 1959; Гордиенко, 1964, 1969; Гатцук, 1968 и др.). Образование разных типов жизненных форм в пределах одного вида в зависимости от многих факторов среды обитания (Шмитхюзен, 1966) отмечалось неоднократно (Сукачев, 1905, 1916, 1938, 1941; Поплавская, 1927, 1948; Испола- тов, 1932; Кожевников, 1935; Келлер, 1948; Серебряков, 1945, 1952, 1962; Аврорин, 1956; Райкова,1965; Шулькина, 1965; Вгаип-Blanquet, 1951; Стешенко, 1965; Ляэнелайд, 1984; Буланая, 1986; Ревякина, 1986; Чистя- кова, 1988; Истомина, 1994 и др.). Способность биосистем изменяться при изменении экологических условий, не утрачивая при этом своих качественных границ, — илис- тичность в широком смысле. Диапазон пластичности биосистем опреде- ленной родственной группы характеризуется разнообразием освоенных экологических условий и проявленными структурно-функциональными приспособлениями. У биоморф он контролируется генотипом. Мутации могут «выделять» из прежних биоморф новые биоморфы с новой пла- стичностью. Проявление пластичности, на наш взгляд, подчиняется нескольким правилам. Чем шире пределы пластичности биосистем одной родственной группы по отношению к таковым у биосистем другой родственной группы, тем выше степень пластичности первых при сравнении со вторыми (nepeoe правило). для выживания в экстремальных условиях биосистемы использу- ют свой генетический потенциал и, если не погибают, то более или менее изменяют свой внешний облик и (или) внутреннюю организацию (второе правило). Конкретное разнообразие типов жизненных форм (биомор- фотипов) и вариантов их развития у одного вида на определенных эко- логических фонах характеризует внутривидовую пластичность биоморф.
Глава 8. Изменчивость и эволюция По относительной встречаемости было предложено (Нухимовский, 1969б, 1973а, 1986) различать (по отдельным сравнимым признакам) био- морфотины следующих категорий: основной (этот тип присущ большин- ству биоморф одного вида в определенном регионе или во всем геогра- фическом ареале) и второстепенные (эти типы представлены меньшим числом биоморф того же вида, чем основной биоморфотип). Разнообразие многих фактов изменчивости сводится к одному из абсолютных законов — закону изменчивости материальных систем: организация какой-либо материальной системы рассчитана в соответ- ствии с ее преднадследственной или (u) наследственной основой на муль- тивариантность существования, из которой реализуется (с модификаци- ями или без них) тот из вариантов, который более всего отвечает кон- кретным условиям (Нухимовский, 1988). Внутривидовая пластичность у семенных растений проявляется в полном соответствии с этим законом. Внутривидовая пластичность — это одновременно и внутривидовой полиморфизм, охарактеризованный с двух взаимосвязанных сторон: как полиморфизм структуры биоморф в пространстве и как он же, но уже во времени (в развитии). Таким образом, внутривидовая пластичность биоморф предполагает структурный (его разновидность — габитуаль- ный) полиморфизм и одновременно полиморфизм биоморфогенезов (разви- тия) (Нухимовский, 1978). Мультивариантность («поливариантность», «полиморфизм развития», «полиморфизм биоморфогенезов» и другие термины представляются нам менее точными) биоморфогении вида (со- вокупность обобщенных сведений о разных вариантах биоморфогенезов в пределах одного вида) неоднократно привлекала внимание ис- следователей (Работнов, 1956б; Нухимовский, 1969б, 1970, 1974а; Мат- веев, 1975; Воронцова, Заугольнова, 1978; Жукова, 1986; Глотов, Жуко- ва, 1995 и др.). Мультивариантность или бивариантность биоморфогене- зов в рамках одного вида может фиксироваться по разным сравнимым признакам-индикаторам (тропобиозным признакам), например, по кратности проявления репродуктивной функции, по особенностям рас- положения базового органа относительно поверхности почвы, по тем- пам развития биоморф и т.п. (рис. 146-148). Внутривидовая пластичность — весьма распространенное явление, составная часть эволюционного процесса. В качестве примера рассмот- рим биоморфогению Bupleurum multinerve DC.; этот вид обладает вы- сокой степенью пластичности. Bupleurum multinerve DC. — володушка многожильчатая (сем. Apiaceae) — горно-степной мезоксерофильный вид, встречающийся в горах в основном на Алтае, Урале, Саянах и Сауре (Линчевский, 1950). Ее изучение проводилось на Алтае в основном на Ивановском, Линей- ском, Курайском, Бащелакском, Коксуйском, Сарым Сакты хребтах (Нухимовский, 1978). Этот вид обычен на остепненных высокогорных склонах, реже на опушках кедровых и лиственничных лесов. Bupleurum multinerve — полиморфный вид; по кратности репро- дуктивной функции здесь можно выделить три типа биоморфогенезов (тропобиоза): унирепродуктивный, би-, трирепродуктивный и мульти- репродуктивный. Унирепродуктивные и би-, трирепродуктивные био-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИЙ Рис. 146. Внутривидовая пластичность Pinus sibirica Du Tour (I) и Abies sibirica Ledeb. (II) в горах Алтая. морфогенезы обычны для растении володушки много~ %.У многожильчатой, произ- растающих в ценотических популяциях, широко р р р око асп ост аненных на Алтае от низкогорий до высокогории, особенно р д р в с е него ьях (400- 1600 м над ур. моря), преимущественно на привершинных остепненных участках невысоких гор и на открытых склонах ю нах южных экспозиций в каменистых степей, а мень- различных ассоциациях группы формаций кам ше — в ассоциациях группы формаций разнотравнолуговых степей ( у-
Глава 8. Изменчивость и эволюция 365 Рис. 147. Внутривидовой габитуальный полиморфизм Eryngium campestre L. (Крым, окрестности пос. Планерское, степные горные склоны). 1 — травянистое мультирепродуктивное компактно-каудексовое растение в усло- виях относительно постоянного положения уровня поверхности почвы, II — тра- вянистое каудексовое растение, впервые образовашее вынужденно верхнерозеточ- ный побег с удлиненным подземным основанием после первого засыпания этого растения наносным субстратом, Ш — травянистое многолетнее растение с вынуж- денно надземным каудексом и оголенным от почвы участком главного корня при эрозионном перемещении почво-грунта. 1 — главный корень, 2 — семядольный узел (пунктиром показано его примерное положение), 3 — основание унирепродуктивного ди-, плейофазного полурозеточ- ного побега (верхние части удалены), 4 — подземное вынужденно удлиненное основание верхнерозеточного побега, 5 — основание розеточных листьев верхне- розеточного побега (верхние части удалены), 6 — компактный подземный кау- декс, 7 — компактный каудекс сильно засыпанный наносным субстратом, 8— вынужденно надземный компактный каудекс, 9 — поверхность почвы; одно де- ление масштаба — 1 см.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ 4 [» О 63 й,' Ю О 1 ф О Ф О М О 1 Е О 'С Я Е О 63 Ьч :С О M ~м О [» Ж О 4 ~» О О Ж [» Ж 63 й О 63 й A ~» О й Ф Ж Р О О f О Ж О й О f Ж v й E О О ч О. ~[ О ф 1=( ~ о ф) ° л к ~ ~ ф Р' '~ й ьй б~~ Х О 5 о. и р ф О О о ОИ ф, ° ф Й М Й й( ~>ь Е. Й Ж ж ' ,ж О Я (~) О О о. ~ О О М О О 1 л .0 Ж й( О >й ~ф ~ лч Йа (» Й R~xgo> Ж д щ ~хо~~ ~~и~~~ g ~~ь 3 ~ыо~~ Й ~ '~: Ю 3vg'~0 ~~оо~~ О &" О ~ r ~à..ж ° 3' о ~ ° ~ () ~О f 9 E p) Я Р1 ~~å о и О О ~~и~~ лч A 2 д р )~ ф д р й 2„"~~ а ж1Д х" о ОЦ О "[О QX~CQ О с') д ф ~О -~ Ж х~~ "ц; М д ~[ ~ о ~~õ- о.~ ~~и "v A а ~ ф ~ о. (» щ О р, ~ ~ О ~- ~ ж lQ ( ( ф ж X v Ж >Wa < as О О ~È Й: о -.~ дф рч ~ ф~ 1» ~е) г-~ Ж ~~~~~ )Q А О к v ж~ж'~- Q~~vo лч Q ~~~~~ (» cv XQQ О И й Ж '.й О О ( М Qi Я ~ю е» E () Ю Qco (.) ( ц М о~~ О фVQCO О С'Ъ ц () z~о мь ~.а ~
Глава 8. Изменчивость и эволюция минова, 1970). Здесь хорошо прослеживается ортотропнобазовый тро- побиоз, определяемый направлением роста базового органа относитель- но поверхности почвы (рис. 149-1). Иногда особи володушки многожильчатой заходят под полог разре- женных лиственничных (Larix sibirica Ledeb.) или кедрово-лиственнич- ных лесов (Pinus sibirica Du Тоиг) и особенно светлых сосновых (Pinus sylvestris L.) лесов, где нередко проявляют плагиотропнобазовый тропо- биоз (рис. 149-11). Володушка многожильчатая нередко произрастает выше верхней границы леса (1800-2200 м, реже до 2500 м над ур. моря), где встречается на каменистых склонах среди альпийских остепненных лугов и по мел- кокаменистым россыпям, более или менее закрепленных растительнос- тью. В этих условиях иногда (Ивановский хребет, верховья реки Палев- ская Разливанка, 1900 м. над ур. моря, крутой остепненный каменистый склон юго-восточной экспозиции; Линейский хребет, верховья реки Черная Уба, 1900 м. над ур. моря, слабо закрепленный растительностью крутой каменистый склон юго-восточной экспозиции) рассматривае- мый вид образует ценотические популяции только с мультирепродук- тивным тропобиозом составляющих их особей (рис. 150, 151). Кстати, в этих условиях нередко встречается кедр сибирский (Pinus sibirica Du Тоиг), принимающий здесь стланиковый тип габитуса (рис. 146-1). Биоморфогения Bupleurum multinerve рассматривается в таблице 4. При описании биоморфогении этого и других видов используем следую- щие условные обозначения: ПС — временная продолжи гельность возраст- ного состояния; ОХ — основная морфофизиологическая характеристика особей; А-Б — дополнительная характеристика особей: А — частная (рас- пространяется только на рассматриваемое состояние), Б — общая (рас- пространяется на несколько или на все возрастные состояния). Типы биоморфогенезов (тропобиозы) володушки многожильчатой для удобства обозначим так: ТБ1 — унирепродуктивный, ТБ2 — би-, трирепродук- тивный, ТБЗ — мультирепродуктивный, ТБ1А — унирепродуктивный ортотропнобазовый, ТБ1Б — унирепродуктивный плагиотропнобазовый, ТБ2А — би-, трирепродуктивный ортотропнобазовый, ТБ2Б — би-, трирепродуктивный плагиотропнобазовый. Вслед за трирепродуктивным выделяем мультирепродуктивный тип биоморфогенезов (термин «паулире- продукгивный» здесь опускаем), когда биоморфы проявляют полные цик- лы репродуктивного развития четыре и более раз. Таким образом, в зависимости от высотного пояса и конкретных экологических условий у володушки многожильчатой зафиксировано по кратности проявления репродуктивной функции и направлению раз- растания базового органа относительно поверхности почвы пять типов биоморфогенезов (тропобиозов): унирепродуктивный ортотропнобазо- вый, унирепродуктивный плагиотропнобазовый, би-, трирепродукгив- ный ортотропнобазовый, би-, трирепродуктивный плагиотропнобазо- вый и мультирепродуктивный. Встречаемость ценотических популяций, особи которых развива- ются по мультирепродуктивному тропобиозу — ТБЗ, вероятно, ограни- чена относительно небольшой территорией высокогорий (они обна- ружены на Ивановском и Линейском хребтах Алтая).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ 1 1 Я О Я Е Е Я л 1 1 М e ---о ~~ о 3~ 1 р О а~Я й CC~ � аЯ� 2 2 ~~ о, ~2 2 "~ж р~~ ~~ а оч g Й ~ ца g о, о ооо-. a.~ ~v о -~м О Я ф~ оц .~' Я О а 3 д ~ Й ~~ар О Ж О (') e Q ~Q ~1' О оч ~о~ а р A Я Й ра а о О ф„„ф ФЪ g ~ h4 ,.", О ф; ьй Ф (~) и ~.-„ о о оо ~р й ( О О mPX C) vo ~ оо 1" я ~о Д ~ о~ О [; ° Я ю о ~ ~о ( дв О а ь ° ~-~ Ж
Глава 8. Изменчивость и эволюция Рис. 150. Виргинильные особи мультирепродуктивного тропобиоза Bupleu- гцт multinerve (Ивановский хребет, 1км на восток от истоков реки Палев- ская Разливанка, 1900 м над ур. моря, крутой склон юго-восточной экспозиции, кобрезиево-володушково-тонконоговая ассоциация). А — проростки, Б — всходы, В — ювенильные особи, Г — имматурные особи, Д- ДД — особи, отмершие после нормального завершения унирепродуктивного жиз- ненного цикла (Д) и бирепродуктивного жизненного цикла (ДД), Е — иницима- турные особи, Ж — медиматурные особи, И — финиматурные особи; 1 — семядо- ли, 2 — гипокотиль, 3 — семядольный узел, 4 — корневая шейка, 5а-5д — корни: Sa — главная монориза (субмедиаль), 56 — главная дириза, 5в — неглавная моно- риза (сублатераль), 5г — тририза, 5д — главная тетрариза, 6 — ризогенная корневая ветвь, 7 — розеточные листья, 8 — предрозеточный побег, 9 — вегетативный ме- диальный розеточный плейофазный побег, 10 — вегетативный аксиллярный ро- зеточный побег (10а — монокормус, 106 — тетракормус), 11 — унирепродуктивный полурозеточный гексакормус, 12 — унирепродуктивный полурозеточный дикор- мус, 13 — сборное соцветие, 14 — курторезидные монокормусы, 15 — миниморе- зидные тетракормусы, 16 — конодий, 17 — каудекс, 18 — почки в структуре побе- гов после удаления сухих или зеленых листьев, 19 — семядольный почкокорень (ризопротерокормус), 20 — семядольный ризомонокормус, 21 — ризотетракормус, 22 — ежегодно вегетативный ризогептакормус, 23 — ризопентакормус, 24 — не- ежегодно репродуктивный ризогептакормус, 25 — неежегодно репродуктивный ризоэнатокормус, 26 — бессемядольный ризомонокормус, 27 — ежегодно вегета- тивный ризоэнатокормус. Продолжительная пунктирная линия — верхняя грани- ца расположения сухих листьев и разрастания мхов и лишайников относительно поверхности почвы. МК — упрощение модели биоморф; горизонтальная линия— поверхность почвы; одно деление масштаба — 1 см; 1л, 2л, Зл — особи 1, 2, 3 и т.д. лет жизни; у — увеличено по сравнению с масштабом.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ М Ю О О О й О [м 54 л М 3 [ ° [ ° О М О О й C ° еЧ Е Я г~ -~ Pn Р Я о Ф Щ Ж О Я Й LQ Ж О; ?; o) R~ О ~ О О ~ XX й:[ О О [' О й й й Ю О О О 4 Ж О й 0 � О ~ ч~ ьй ~ ю ( О М ~~ О Ф Ц ц й О ощ� ~- ~~и (~) ф~ ~~а о ~ оео 4Э р <» Ф v ййМ Ж x+v о Ф Ф ф~ Ж ~и~ За lQ Щ Ж щ ййж р, ~ й,", I3 ~ о О сч g О~ Й: щ" Ж E о О Щ ',3~ о щ О Й~Н vou и ~ О о~~ М И ~~~ ~ ~g о Й (») ~~о ф. *.О О � м ~~ +o~z ц~о О О О О 2 М;~ � д О Ф д ( Й E Д Й ~ И ~ <~В ge4a Х E я ~-1 ф; ~Г') Ф И о а~й ре О дф 01 ~ц О Ж щ ф~~ в g4 Д~~ > а с~ veau К Q оч Щ и Ф g д ( Ф д Щ ~а~ 1 1 ооо Й
Глава 8. Изменчивость и эволюция Таблица 4 Биоморфогения (онтогения) Bupleurum multinewe DC. Возрастные СОСТОЯНИЯ индивидуумов Основная морфофи- зиологичес- кая характе- ристика (ох) 3 о й [а V VI IV VII 1 II VIII C С4 )Я 3 й е4 ОПЛОДОТВО- ренная се- мяпочка или зрелое семя в структуре родительско- го растения О Ю б~~ и 1 Я х ТБ 1-3 зрелое семя, обособлен- ное от родитель- ского рас- тения ТБ 1-3 д" О ~ C[ у Я Й < Йсч 1 CC ьй ~Я х ТБ 1-3 )ф 3 й е4 л и Ф ~ Д Ц Я и х Cg о й( о х ТБ 1-3 Ю х Я х О > ~ 54 Я и е ~в ез О =1 ~~ О о ~(-) иДр Ж ФЭ Я Прорастаю- щее семя или семядольный почкокорень (семадольньт ризопроте�р- окор�м) (рис. 149, А; 150, А) Семядольный вегетативны й предрозеточ- ный (1-2 сближенных настоящих листа) или розеточный (3-4 и реже более сближенных настоящих листьев) ризо- монокормус (рис. 149, Б; 150, Б) Дополнительная характеристика: А — частная (распространяется только на рассматриваемое возрастное состоя- ние); Б — общая (распространяется на несколь- ко или на все возрастные состояния) Б — помимо ветра в опылении участвуют и насекомые, например, пчелы. А — отделившиеся зрелые мерикарпии (полуплодики) при ТБ 1-2 имеют 2,0- 3,2 мм дл„0,5-0,8 мм шир., а при ТБ- 3 — 3,0-4,0мм дл., 0,8-1,0 мм шир. Б — в среднем и верхнем поясах гор се- мена осыпаются на 1-2 недели рань- ше, чем в высокогорье (выше верх- ней границы леса). А — семядоли (2-е, очень редко 3) над- земные голые линейно-лацентные островатые, постепенно переходящие в черешки, 0,9-1,2 см дл., 0,1-0,12см шир.; гипокотиль 0,7-1,5 см дл.; 0,05- 0,08см шир., главная монориза до 1,5см дл. А — семядоли 1,2-2,0 см дл.,0,12-0,2 см шир.; первые настоящие листья ли- нейно-ланцетные и узколанцетные, более или менее заостренные, голые (1-й лист от 1,5 до 3см дл. и 0,2- 0,35 см шир., 2-й — 2-4 см дл., 0,25- 0,4 см шир.), к основанию суженные в черешек, у последующих листьев линейные размеры увеличиваются; в высокогорье у всходов образуется 1-3 листа, а ниже — до 5 листьев. Б — отмирание семядолей свидетель- ствует о начале нового возрастного состояния, оно начинается у особей с 1-5 листьями.
VIII VII V VI ТБ 1-3 ТБ 1-3 g~ 4 с3 й ~о ea � х E О Ц C) 1 Щ ТБ 1-3 ОО 1 ег') о й( 1 ГЧ О Бессемядоль- ный вегета- тивны й физиологи- чески незрелый преконоди- альный ризомо�н- окор�м (рис. 150, В-26) Бессемядоль- ный ежегод- но вегета- тивный розеточный некустистый конодиаль- ный ризомо- нокормус или ризотет- ракормус (рис. 149, В; 150, В-21) Ежегодно вегетатив- ный розе- точный простокус- тистый ризогекса- кормус или ризогепта- кормус (рис. 149, Г-22; 150, Г-22) ОСНОВБ1 БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИЙ Продолжение таблицы 4 А — в первый вегетационный сезон в зави- симости от условий произрастания об- разуется от 1 до 7, чаще 3-5 настоящих листьев; главный корень нарастает медленно, достигая 3-4 см, разветвля- ясь до 1 или 2-го порядка, т.е. являет- ся диризой или тетраризой. Б — на гипокотиле и основании главного корня образуются поперечные складки ("морщинистость"), свидетельствую- щие об их контрактильности, которая проявляется и в последующие годы. А — после завершения первого сезона ве- гетации зимует вся компактная стеб- левая часть монокормуса — курторе- зид, а также верхушечная почка и 2- 3 пазушные наружные почки, при- крытые в течение 3-5 лет сухими остатками листьев (если эти почки вообще закладываются), т.е. зимует безлистный курторезидный моно- кормус — конодий (он же прекау- декс), который в последующие годы нарастает моноподиально за счет уве- личения в его структуре числа таких курторезидных побегов. А — после того, как первый пазушный побег становится частично или пол- ностью персистентным, т.е. впервые зимует, образуется кудекс; главная тетрариза короткостержневая, 4-8 см дл. Это возрастное состояние прохо- дят немногие особи ТБ-1 и большин- ство особей ТБ-2. Б — у особей, развивающихся по ТБ-1А и ТБ-2А, каудексовые оси более или менее ортотропные, а — по ТБ-1Б и ТБ-2Б — более или менее плагиот- ропные; пазушные побеги весенние; они отрастают из зимовавших защи- щенных 2-3 чешуями почек прошло- годних побегов; кроме того, иногда отрастают побеги из спящих почек; верхушечные побеги ежегодно отрас- тают весной из незащищенных по- чек; пазушные почки закладываются и развиваются в средней и чаще всего верхней частях побегов текущего года.
373 Глава 8. Изменчивость и эволюция VIII VII VI IV V E 1"=, C) СЧ 1 Щ О й( ОО 1 С~ О й( СЧ 1 ег') ТБ 1 й О й Ф ~Ф й й й E О Ц C) ~4 1 о й( ОО 1 ТБ 2 Ежекд~ю иии неежекдно ~4 Ь~ Ц ~4 л ~~ О О й( ТБ 3 ТБ 3 (ТБ 2 в ис- клю- чи- тель- ных слу- чаях) Ежегодно вегетатив- ный розе- точный сложнокус- тистый кау- дексовый р�изоэнат- окор�м (рис. 150, Г-27) Унирепро- дуктивный розеточный ри�зопе�н- такор�, очень редко простокус- тистый ри- зогексакор- мус или ризогепта- кормус (рис. 149, Д) Ежегодно или нееже- годно ре- продуктив- ный (физи- ологически зрелый) по- лурозеточ- ный ризо- пентакор- мус, или ризогекса- кормус, или ризо�гепт- акор�м (рис. 149, Е-23, 24) ный панурозе- пы~ый ри- зопе пзксрщс иии ри- зожиэкорщс, или риютеп- икормус, иии ризоэнзгокор- ЩС (рис. 151, Е) Продолжение таблицы 4 А — у особей ТБ-2 сложное кущение ред- ко, как правило, на участках, где многократно повреждаются верхуш- ки розеточных вегетативных побегов низовыми пожарами; у особей ТБ-3 сложное кущение обычное явление. Б — ежегодное поздне-летнее и осеннее (с августа по сентябрь) пробуждение однократно зимовавших почек— специфический признак особей ТБ-3, обеспечивающий им в отличие от особей ТБ-1-2, лишенных этой способности, обильное кущение. А — особи ТБ 1 в виргинильном периоде подобны во многом особям ТБ 2, но с вступлением в матурный период у них сильно проявляется репродук- тивная функция и осенне-зимний некроз, который начинается с вер- хушки побега, далее распространяет- ся на все растение, вызывая его ги- бель несмотря на наличие спящих почек. А — индикаторный признак этого возрас- тного состояния — наличие 1-2 по- рядков унирепродуктивных побегов, цветущих или не цветущих в год на- блюдения. Главная тетрариза до 10 см; нередко особи погибают после 2-го пдодоношения (рис. 149,ДД). Б — после плодоношения верхняя раздви- нутоузловая часть медиального уни- репродуктивного побега отмирает, а сохраняет жизнеспособность только компактное основание — миниморе- зидный безлистный тетракормус- каудекс. В последующие годы он раз- растается за счет увеличения числа новых курторезидных монокормусов и миниморезидных тетракормусов, кущение слабое. ный признак — наличие не бо- лее 3-х порядков унирепродукпвных по- бенев, gelytuHx юи не нвеНвнвн в н:н найподения. Главная тетрариза до 10 см. Б — верхушки и побегов нередко вы- хаугг выше поверхности почвы (до 4 см), оставаясь прикрытыми сухими листыпки. Верхние части каудексовых осей, нередко сжпощиеся по поверхности почвы, посж- пенно погружаклся в нее. Кущение обиль- ное; фаза бутонизации — конец и1оня, икать, цветение — икль-август, плодоно- шение — конец август-сентябрьн
Продолжение таблицы 4 VIII VII V VI А — индикаторный признак этого возрас- тного состояния — образование 3-х Ежегодно или нееже- годно ре- продуктив- ный каудек- совый геми- симподиаль- ный слабо кустистый ризогепта- кормус, или ризо�э�на- токор� (рис. 149, Ж) порядков унирепродуктивных побе- гов, цветущих или не цветущих в год наблюдения; после второго-третьего плодоношения особи ТБ 2 нередко гибнут, не достигнув предельного возраста для этой группы. Б — пентакормусы унирепродуктивные ди-, три-, плейофазные (2-17 фаз ви- димого роста), полузакрытые полуро- Ц C) ~4 1 ГЧ зеточные обоеполые, высотой 20-60 см. Два типа соцветий — одиночный сложный зонтик, реже зонтик сбор- ный (с паракладиями), число зонтич- ков в зонтике 5-20, чаще 7-10. Число постинициальных монокормусов в сезон вегетации 2-8, а число униреп- родуктивных дикормусов по годам наблюдения от 0 до 5, чаще 1-2; ро- зеточные листья очередные ланцет- ные, иногда яйцевидные, и длинно-, реже короткозаостренные, сидячие с яйцевидно расширенным до 1-2,5 см стеблеобъемлющим основанием; фаза бутонизации в июне, цветение— июнь-июль, плодоношение в авгус- те-начале сентября. О й( ~О 1 ТБ 2 А — индикаторный признак — образование 4-6 Ежегодно или не- ежегодно ре продук- тивныйй каудексовый гемисим�п- одиальны, обильно кустистый ризо�энат- окор�м (рис. 151, Ж) порядков унирепродуктивных побегов, пмовщих или ие ивсвросив пм ивпвюие- ния. Б — главная тетрариза до 12 см; гипо- котиль и основание субмедиапи нередко заметно разрастаются в толщину (до 1,5 см) по сравнению с каудексовыми осями и другими частями корней; на россыпях кау- дексовые оси иногда засыпиотся субстра- том до 20 см. В этих условиях из гипоко- тиля и основания субмедиаяи развивается больше, чем обычно, корней. Каудекс на- растает гемисимподиалъно за счет курто- резидных монокормусов и миниморезид- ных тетракормусов, а при засыпании суб- стратом образуются также вицерезидные монокормусы и викариорезидные тетра- кормусы; тип пентакормусов и соцветий подобен особям ТБ 2, но побеги обычно короче — 15-35 см; число зонтичков в глав- ном зонтике 3-9, чаще 4-6; кущение обиль- ное за счет массового позднелетнего и осеннего пробуждения почек, заложенных в верхней части прошлогодних розеточных вегетативных побегов или в розеточной ча- сти полурозеточных вегетативно-репро- дукгивкых побегов; верхние листья по форме от лищетных до яйцевидных, ко- ротко-, реже длиннозаостренные, сидячие со стеблеобьемлющим основанием. Число псопипиююмюм моиокормссо~ в сеюи вегетации от 10 до 140, а число унирепро- дуктивных дикормусов по годам наблюде- ний от 0 до 35, чаще 4-10. Ц C) С~) 1 00 C) ~4 1 ~Ф ТБ 3 Щ ~4 1 ~4 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ
375 Глава 8. Изменчивость и эволюция Продолжение таблицы 4 VI VIII VII IV V Ежегодно или нееже- годно реп- родуктивный полурозеточ- ный каудек- совый геми- симподиаль- ный ризо- э нато кормус (рис. 149, И) А — индикаторный признак — образуется 4 и более порядков унирепродуктив- ных побегов цветущих или не цвету- щих в год наблюдения; до этого воз- растного состояния доживают немно- гие особи. ТБ 2 О й( CD СЧ 1 СЧ CD е~) О ~Ф E оц О А — индикаторный признак — образуется более 6 порядков унирепродуктивных побегов (рис. 151, И) ТБ 3 — /— (то же) )ф 3 й е4 Ежегодно вегетативный (после ут- раты физио- логической зрелости) каудексовый ризоэнато- кормус А — утрачивается способность развивать репродуктивные органы; главная тет- рариза частично или полностью от- мирает, а пучок немногочисленных слабых гипокотилогенных корней еще 1-2 года поддерживает вегета- тивную жизнь единичных особей это- го возрастного состония. й О й Ф ~Ф й й й Ц СЧ СЧ 1 ~О ТБ 2 (ред- ко) о й( CD СЧ 1 Щ о ~ О~ '~ сч ~17 е~) ~ ~) О ТБ 3 По эколого-морфологическим тропобиозным признакам володуш- ка многожильчатая — это травянистый летнезеленый надземно-двусе- мядольный унирепродуктивный (ТБ-1), би-, трирепродуктивный (ТБ2), мультирепродуктивный (ТБЗ) полурозеточный ланцетолистный, линей- но-ланцетолистный моноподиальный (ТБ1, редко), гемисимподиаль- ный (ТБ2, ТБЗ) некустистый (ТБ1), простокустистый (ТБ2, редко ТБ1), сложнокустистый (ТБЗ) конодиальный (ТВ1), каудексовый (ТБ2, ТБЗ) короткостержнекорневой семидетерминатно-репродуктивный однодом- ный мезоксерофильный горно-степной вид растений. Продолжительность жизни находится в прямой зависимости от ус- ловий произрастания. Нормально развивающиеся особи ТБЗ с высоко- горных ценотических популяций живут в 2-10 раз больше, чем особи ТБ1-2, произрастающие в нижних и средних поясах гор. Максимальный возраст особей ТБ1 не превышает 17 лет, особей ТБ2 не более 22 лет, а особей ТБЗ — 42 года. Мультивариантность биоморфогенезов воло- душки многожильчатой свидетельствует о высокой эволюционной ак- тивности этого вида.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ 8.2. Ответные реакции в инфраструктурной и конституциальной борьбе за существование Растительные биоморфы как простые (организмы, или индивиду- умы, особи), так и сложные (синорганизмы, или кондивидуумы), суще- ствуют в окружающей среде, которая постоянно изменяется: сменяются дни и ночи, времена года, меняется погода и т.п. В этих условиях растительные организмы, пожалуй, более чем другие, проявляют свою способность к пластичности. На воздействия извне они отвечают соот- ветствующими реакциями, которые, как подчеркнул Р.Риклефс (1979), обратимы и затрагивают структуру и функции организма; «в противном случае они не могли бы следовать за изменениями среды, которые бывают направлены как в ту, так и в другую стороны» (с. 193). Такие реакции получили название «отрицательная обратная связь — стремле- ние системы противодействовать вносимому извне изменению и воз- вращаться к устойчивому состоянию» (Риклефс, 1979, с. 397). Например, у растений) стрелолиста (Sagittaria sagittifolia L.) обнаруживаются вариа- ции формы листьев, если они располагаются на суше, частично или полностью погружаются в воду. Внутривидовая пластичность начинает проявляться уже в инфраструктурной борьбе за существование, на ос- нове которой действует конституциальная борьба и следует обобщенная ответная реакция системы всего организма (его конституции) на возму- щающее воздействие меняющейся окружающей среды. По существу, конкретное проявление структурного (в том числе габитуального) поли- морфизма, мультивариантности развития — это синдром коиституци- ального ответа на изменения в окружающей среде. Этот ответ у орга- низма обобщенный, единый, поскольку подготовлен системным слия- нием множественных частных ответных реакций, исходящих из инфра- структурной борьбы за существование в различных его участках. Она идет не только в препопуляциях, коалициях и иных структурах, но и между ними и местом их проявления в теле организма. Автор склонен считать, что в растительном организме (в животных организмах многое обстоит иначе) существуют инфраобъекты (иреиоиуляции, коалиции и другие структурные системы), которые в едином теле организма взаимо- действуют с остальной частью его инфраструктуры как со своим биото- иом (Нухимовский, 1987) — ареной (естественно живой) инфраструк- турной борьбы за существование — автобиотоиом. Инфраструктурная и конституциальная борьба за существование развертывается в фенотипах, но контролируется с генетического уровня; мутации возможны как в инфраструктурной, так и конституциальной борьбе, но закрепиться они могут только в процессе межорганизменных отношений (в популяциях). Новая концеиция универсальности борьбы за существование в живой природе не отрицает главного — тело растения едино и расчленение его «на составные части и связанная с ним раз- работка типологии этих частей — логический и удобный способ изуче- ния растения, позволяющий выдвинуть на первый план проблему струк-
Глава 8. Изменчивость и эволюция турной и функциональной специализации отдельных частей растения». Важно не утратить представление о растении как едином целом (Эзау, 1980, с. 13). Изложенные выше представления о борьбе за существование позволяют расширить методологические основы биологии, и биоморфоло- гии в частности, что создает дополнительные методические возможности для анализа структурной организации биоморф семенных растений. Любопытная инфраструктурная борьба идет в препопуляциях го- дичных корнепобегов у биоморф Aconitum decipiens Woroseh. et At>falov— борца ненайденного, которые автор изучал на Курайском хребте Алтая на высоте около 2000 м над ур. моря (альпийский луг). Одна такая пре- популяция составляет всю организацию простой растительной биомор- фы (рис. 152). В первые годы виргинильной жизни развитие идет мед- ленно. Чтобы зацвести, особи Aconitum decipiens должны создать и накопить некоторую «ресурсную массу». Взрослые матурные растения экономно поддерживают накопленную ранее ресурсную массу, что дает им возможность ежегодно или с некоторыми небольшими перерывами цвести и плодоносить. Виргинильные растения ежегодно образуют один вегетативный побег, степень развития которого (число листьев, разме- ры) год от года возрастает. С первых лет жизни помимо ежегодно отрастающего родительского вегетативного побега в его укороченном основании развивается одна пазушная почка, из которой одновременно идет отрастание придаточ- ного корня, внешне напоминающего главный корень; вместе пазушная почка и ее придаточный корень составляют одну систему — инициаль- ный корнепобег, или почкокорень. Это вегетативное проорганизменное потомство родительского побега. Главный постинициальный побег и постинициальные побеги, развивающиеся в последующие годы, форми- руют ассимиляционную листовую систему. На пазушный почкокорень, развивающийся при родительском корнепобеге, со стороны последнего оказывается тормозящее действие, что не позволяет ему начать види- мый рост; почкокорень только увеличивает свою массу. Каждую осень постинициальный годичный побег сверху сильно отмирает (акронек- роз), но в отличие от годичного побега у особей Aconitum altaicum, отмирающего полностью (см. рис. 53), у рассматриваемого вида отми- рает только вся удлиненная надземная часть родительского побега, а его компактное подземное основание — миниморезид с развитым на нем почкокорнем, сохраняется. Из миниморезидных корнепобегов ежегод- но, эусимподиально нарастая, формируется компактное цепочковидное корневище с развитыми на нем придаточными корнями. В таком виде в процессе эволюции был найден компромисс в инфраструктурной борьбе между родительским побегом и его потомком — почкокорнем, что закрепилось на генетическом уровне. Первое цветение в условиях высокогорных местообитаний проис- ходит на 6-15 году. Быстрее зацветают биоморфы, произрастающие на участках с нарушенным растительным покровом. В процессе ежегодной инфраструкгурной борьбы за существование сохраняется один новый миниморезидный корнепобег (миниморезидный онтоид), который не отделяется от родительского растения, продолжая жить с ним в одной
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х PA СТЕНИИ Рис. 152. Инфраструктурная и конституциальная борьба за существо- вание Aconitum decipiens Worosch. et Anfalov на восточном Алтае. А — проростки, Б — всходы, В — ювенильные особи, Г — имматурные растения; 1 — семядоли, 2 — гипокотиль, 3 — семядольный узел, 4 — корневая шейка, 5— главный корень, 6 — придаточный корень, 7 — вегетативный монокормус, 8— аксиллярный почкокорень, 9 — цепочковидное корневище, 10 — семядольный почкокорень, 11 — семядольный вегетативный ризомонокормус, 12 — бессемя- дольный вегетативный ризомонокормус, 13 — вегетативный неветвистый ризо- гептакормус, 14 — вегетативный ветвистый ризоэнатокормус. Горизонтальная линия — поверхность почвы, одно деление масштаба — 1см, ув — увеличено по сравнению с масштабом.
Глава 8. Изменчивость и эволюция 379 связке, что обеспечивает медленное эусимподиальное увеличение в длину цепочковидного корневища с системой придаточных корней. У взрослых матурных особей или еще раньше происходит базальное побе- говое ветвление (индикаторный признак имматурных растений, которые встречаются нечасто). При этом из двух узлов укороченного основания родительского побега образуется сначала два пазушных почкокорня, а весной следующего года отрастает два посинициальных годичных корне- побега, т.е. происходит кущение. Корневище с осени достраивается уже не одним, а двумя миниморезидными побегами и становится развет- вленным. Ветвление корневища происходит неежегодно; оно разраста- ется, развиваясь в неразрывной связи вместе с придаточными корнями, и удлиняется за счет ежегодного включения в структуру этой системы миниморезидных корнепобегов. Отдельные звенья корневища вместе с развитыми из них придаточными корнями с базальной части отмирают, но в целом каждый онтоид живет от 4 до 10 лет, и когда некроз кор- невища достигает зоны его ветвления, особь распадается на два новых организма, продолжая биоморфогенез в структуре синорганизма уже в стадии клона. Вегетативное размножение замедленное и клонисты по- тихоньку «отползают» друг от друга. Инфраструктурная борьба у биоморф Aconitum decipiens происхо- дит на разных отрезках пространства и времени по-разному: в ней есть взаимопомощь («забота» о проорганизменном потомстве, участие кор- невой системы почкокорня в снабжении водно-минеральными ресурса- ми цветоносного родительского побега и т.п.), конкуренция (торможе- ние развития почкокорней быстро растущим родительским побегом и, наоборот, депрессия родительского побега после его плодоношения уси- ливается со стороны производного почкокорня), а есть и компромиссы (сохранение компактных остатков родительского корнепобега в много- летнем резерве). Механизм инфраструктурной борьбы за существование в препопуляции протекает, вероятно, еще более оригинально, чем нам это представляется. В результате взаимодействия (инфраструктурной борьбы) в препо- пуляции разновозрастных годичных корнепобегов формируется целос- тный ответ биоморф борца ненайденного в их конституциальной борьбе за существование, что обеспечивает этому виду выживание в альпийс- ком поясе Алтая по основному типу биоморфогенезов — безрозеточно- му эусимподиальному цепочковидно-компактнокорневищному почко- корневому замедленно-кондивидуальному мультирепродуктивному. Границы препопуляции могут простираться на все тело раститель- ного организма, как мы это видим на примере Aconitum decipiens. Одна- ко, если поменять угол обзора и изначально изменить границы препо- пуляции (это согласно определению этого термина допустимо), напри- мер у того же вида борца включить в препопуляцию только инициаль- ные и постинициальные годичные побеги, то тогда ризом с корнями будет ее автобиотопом. В системе «препопуляция-автобиотоп» инфра- структурная борьба за существование может приобретать уже несколько иной оттенок. Это борьба активно взаимодействующих друг с другом компонентов препопуляции не просто с окружающей средой, а непос- редственно в их биотопе, на арене их инфраструктурной жизни.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ!Х PA СТЕНИИ 8.2.1. Поведение биоморф на эрозионно лабильных субстратах Наглядным примером внутривидовой пластичности могут бьггь био- морфотипы (T<»> биоморф) растений, обитающих на неустойчивых к эрозии субстратах на равнине и в горах (см. рис. 15, 16, 38, 39, 49, 118, 119, 147, 153-156), охарактеризованные по габитуальным приз- накам. Внутривидовая пластичность биоморф понятие более широкое, чем онтогенезная и синонтогенезная нластичность биоморф (конкретные пути изменений одной жизненной формы соответственно в ее онтоге- незе или синонтогенезе при изменении условий обитания). Так, напри- мер, взрослые особи Eryngium campestre L. (Крым, окрестности пос. Планерское) в условиях обитания с постоянным уровнем поверхности почвы — травянистые мультирепродуктивные полурозеточные много- летники с подземным компактным каудексом (основной биоморфотип). При погребении особей субстратом предыдущий тип жизненных форм постепенно преобразуется в другой или в онтогенезах проявляется только новый тип — травянистые мультирепродуктивные пог~урозеточные мно- голетники с коротким или длинным подземным кудексом, а при оголении каудекса от субстрата иногда формируется еще один второстепенный тип жизненных форм — травянистые мультирепродукгивные полурозеточные многолетники с вынужденно надземным компактным каудексом (рис. 149). Внутривидовая пластичность биоморф в пределах определенных регионов или всего ареала, охарактеризованная по одному сравнительному ряду признаков, выраженная в процентах, — сиектр тинов жизненных форм вида, или биоморфный снектр вида. Конституциальная борьба за существование, как ее определил автор термина Л.Плате (Plate, 1913), характеризуется взаимодействием организ- мов с неблагоприятными физическими или климатическими условиями и не зависит от численности особей; в этой борьбе побеждает более сильная конституция (цит. по Ю.А.Филипченко, 1977, с. 48), в чем, как было под- черкнуто выше, находит свое реальное выражение одна из важных сторон внугривидовой пластичности (аспекты внутривидовой и межвидовой борь- бы организмов за существование здесь не рассматриваются). Результат конституциальной борьбы (конституциальный ответ) — тот или иной mun габитуса, а шире — всей структурной организации), т.е. mun развития (био- морфогении в особенности) — иодготавливается как целостный ответ орга- низма (всей его конституции, всей его организации) из множества частных ответных реакций отдельных участков организма в их инфраструктурном сосуществовании (шире — борьбе за существование). Экстремальные условия обитания растений на эрозионно неустойчи- вых субстратах заслуживают особого внимания, поскольку позволяют при- открыть завесу над некоторыми сторонами этого довольно непростого явления. Растения, чтобы обитать в таких условиях, должны проявлять ответные приспособительные реакции на засыпание песком или иным почво-грунтом, выдерживать оголение корней, быстро укореняться при перемещении с грунтом и при разрывании их на части, например, при оползнях.
Глава 8. Изменчивость и эволюция Естественно, возможности растений не беспредельны, но все же нельзя не удивляться, например, конституциальной борьбой за жизнь особи дуба (Quercus sp.), которую она проявила в дюнах, выдерживая многие годы ежегодное сильное засыпание песком, оставляя на поверх- ности лишь небольшие ветви надземных побегов (Warming, Graebner, 1918; рис. 153). Конституциальная борьба за существование, которую ведут растения, показывает, нередко, сколь высок потенциал внутриви- довой пластичности. Вся индивидуальная жизнь как борьба за существо- вание, которая свойственна организмам, контролируется их генотипами. Всякая первоначальная мутация происходит не где-то вообще в популяции, а в отдельном организме, на фоне конкретного инфраструктурного суще- ствования его частей, т.е. образно — борьбы, которую ведут части орга- низма как с внешней средой, так и между собой; только в дальнейшем, через последующие поколения организмов в популяции мутационный сдвиг закрепляется или выбраковывается естественным отбором. Рис. 153. Поведение старой особи Quercus sp., произрастающей в дюнах в условиях ежегодного засыпания многих надземных частей ее песком (Warming, Graebner, 1918, с. 442). Посмотрим, каков арсенал конкретных структурных приспособи- тельных реакций растительного организма в местообитаниях с эрозион- но-неустойчивым субстратом. Наиболее показательны примеры не с облигатными обитателями песчаных пустынь или крутых эродирован- ных горных склонов, а с теми видами, биоморфы которых чаще обитают в условиях с относительно спокойной поверхностью почвы. Для них засыпание субстратом или оголение корней — экстремальные условия, требующие полной мобилизации жизненных сил.
382 OCHOBbl БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Конституциальные ответы биоморф на эрозионную подвижность субстрата не так уж и многообразны. Относительно стабильный уровень поверхности почвы обычен для большинства видов растений, даже для тех, которые обитают в полупустынях, пустынях и в горах. В этих ус- ловиях формируются биоморфы основного типа, которые не имеют каких-либо отклонений в организации. Изменения в организации про- исходят только тогда, когда реально воздействуют эрозионные движе- ния субстрата. При определенных обстоятельствах (стравливание ско- том и выжигание растительности на горных склонах и т.п.) уровень поверхности почвы в результате ветровой или водной эрозии может ежегодно или с перерывами существенно меняться. У древесных расте- ний (деревьев и кустарников) нижние части стволов и стволиков аэро- подиев сохраняются при засыпании субстратом, превращаясь в подзем- ные многолетние побеговые органы — геоподии, нередко обрастающие придаточными корнями (Salix acutifolia Willd., Salix rosmarinifolia L., Ainus glutinosa (L.) Gaertn., Quercus гоЬиг L. и др.). У многолетних трав, полукустарничков и полукустарников, харак- терных для различных зон умеренных широт, надземные удлиненные части цветоносных побегов, обычно после завершения их репродуктив- ного цикла отмирают на большую часть длины, а у трав — до подзем- ного компактного основания, если растения обитают в условиях с от- носительно эрозионно неподвижным субстратом. Отмирание надзем- ных побегов в этих условиях обусловлено не столько прекращением репродуктивной деятельности, сколько иссушающим действием ветра (летом теплого, а зимой холодного). Если такие побеги, волей случая, оказываются на разную длину погребенными субстратом, то они, нахо- дясь в новой среде, нередко выживают, превращаясь в многолетние части растений. Такой метаморфоз однолетних частей побегов можно наблюдать повсюду: в поле, где после вспашки некоторые растения сильно засыпаются почвой; на отгонных пастбищах в полупустынях и пустынях, где из-за разрушения растительного покрова и, как след- ствие, ветровой эрозии одни растения засыпаются субстратом, а у других оголяются корни (рис. 39, 49, 119, 154-155); у железнодорожных песча- ных насыпей, с которых к подножию медленно осыпается грунт (рис. 156); на крутых горных склонах, подвергшихся водной эрозии (рис. 118) и т.п. Даже у однолетников и эфемеров при засыпании пес- ком структура преобразуется (рис. 157); их жизнь несколько продлева- ется (на 1-5 недель), хотя они и не превращаются в многолетники. Такие растения, особенно эфемеры, очень специализированы и плодо- ношение чаще всего разрушает их. Однако, и у этих растений есть запас пластичности, позволяющий продлить индивидуальную жизнь. Будучи погребенными, бывшие надземные части цветоносных по- бегов многолетних трав, полукустарничков и полукустарников, могут не просто стать многолетними на значительную часть своей длины, но и одревеснеть, а кроме того, и образовать придаточные корни (рис. 15-III, 39, 49-II, 118-III, 119, 154 и др.). Из нескольких побегов, отходящих от зоны ветвления родительско- го побега, выживают наиболее сильные. Для бывших эфемерных частей
Глава 8. Изменчивость и эволюция рис. 154. Структура подземных органов особи Halocnemum strobilaceum (Pall.) Bieb. в условиях засыпания ее субстратом (республика Калмыкия, Черные земли). 1 — надземная часть кроны кустарничка, 2 — вынужденно удлиненные подземные многолетние побеговые части — геоподии, 3 — эпифизы (зоны побегообразования после засыпания наносной почвой), 4 — главный корень, 5 — поверхность почвы. Одно деление масштаба — 5 см. Рис. 155. Структура подземных органов особей Helichrysum arenarium (L.) Moench в условиях эрозионно-неустойчивых субстратов. 1 — при ежегодном засыпании особей песком, II — при оголении корней от суб- страта (Новосибирская область, остепненная опушка соснового леса). 1 — основания вегетативно-репродуктивных aH-, плейофазных розеткообразую- щих побегов (верхние части удалены), 2 — примерное положение семядольного узла (показано пунктиром), 3 — главный корень, 4 — оголенная от почвы часть главного корня, 5 — вынужденно удлиненный ветвистый подземный каудекс, 6— вынужденно надземный компактный каудекс, образовавший пучок побегов теку- щего года, 7 — пучок адвентивных побегов на оголенной от почвы части главного корня; горизонтальная линия — поверхность почвы; одно деление масштаба — 1 см.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 156. Габитуальный полиморфизм Pimpinella saxifraga L. (Московская область). 1 — формирование подземного компактного каудекса (относительно постоянный уровень поверхности почвы, опушка леса), П — формирование подземного длин- новетвистого каудекса (уровень поверхности почвы ежегодно повышается, край песчаной насыпи). а — ранние стадии онтогенеза (конец апреля — июнь), б — однолетняя особь, в- двулетняя особь, д — восьмилетняя особь (б-д — октябрь), е — виргинильные осо- би на втором году жизни, ж — взрослые особи (е-ж — май); 1 — семядоли, 2— гипокотиль, 3 — семядольный узел (у взрослых растений отмечен пунктиром примерно), 4 — главный корень, 5 — розеточные листья, 6 — основания розеточ- ных листьев (верхние части удалены), 7 — компактный каудекс, 8 — миниморе- зидные побеги, 9 — длинноветвистый каудекс, 10 — вынужденно верхнерозеточ- ный побег, 11 — эпифиз, 12 — придаточные корни, 13 — викариорезидный побег; горизонтальная линия — поверхность почвы, х4,3 — масштаб. побегов путь к метаморфозу в многолетние побеговые части в рассмат- риваемых случаях весьма короток (от нескольких дней до нескольких недель). Почки, которые развивались в бывшей эфемерной зоне цвето- носного побега (а это в основном его зона торможения) без засыпания растений субстратом недоразвиваются и засыхают. В новых условиях почки на погребенных частях побегов выживают и получают импульс для развития, а ткани побегов активно функционируют: утолщаются и нередко начинают одревесневать. Весной следующего года отрастание новых постинициальных побегов идет из верхних частей погребенных побегов, т.е. из почек, развившихся в наиболее «однолетней» части прошлогодних цветоносных побегов. В приповерхностной (у полукус- тарничков) и подземной (у трав) части образуется «ярус ветвления» (эии- физарный ярус) из оснований новых постинициальных побегов. Эпифизарная ярусность может многократно повторяться при мно- гократном повторении засыпания (заиления и т.п.) надземных побегов
Глава 8. Изменчивость и эволюция Рис. 157. Модель поведения злаковых эфемеров без засыпания (I) и при засыпании (II) их песком (республика Калмыкия, Черные земли). 1 — зона кущения в условиях относительного постоянства уровня поверхности почвы, 2 — эпифиз (зона активного побегоотрастання при засыпании песком), 3— надземные части побегов, 4 — придаточные корни. субстратом (рис. 154-157 и др.). Возникновение подземных многолетних побеговых органов (геоподиев, ризомов, длинных ветвей каудексов) из надземных эфемерных частей побегов — один из путей возникновения первых в эволюции. Е.М.Лавренко (1935) назвал погребенные песком стволы, стволики и другие стебли как деревьев, кустарников, так и многолетних трав «вынужденными корневищами». По более современ- ной терминологии это вынужденно эпигеогенные корневища, геоподии или каудексовые ветви (каудиции). Способность растений образовывать такие органы и придаточные корни широко используется для закреп- ления эрозионно неустойчивых песчаных почв (Гаель, 1952; Гордиен- ко, 1969 и др.). Отсутствие сильных иссушающих ветров в микропонижениях су- песчаных равнинных полупустынь на Черных землях в Калмыкии поз- воляет некоторым видам растений, как, например, прутняку Kochia
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhIX РА СТЕНИЙ prostrata (L.) Schrad. трансформировать габитуальную форму, развивая значительно более высокие аэроподии, чем обычно на плакорах (рис. 15-П). Такие биоморфы (по габитусу) уже нельзя называть полу- кустарничками (это основной биоморфотип Kochia prostrata на Черных землях), а правильнее считать полукустарниками (второстепенный био- морфотип в указанном регионе). В плакорных условиях надземные цве- тоносные побеги полукустарничка прутняка отмирают до компактного или короткого основания, (т.е. на большую часть своей длины), а в микропонижениях особи нестравленные скотом, сохраняют иногда до половины длины апонекрозных цветоносных побегов, которые одревес- невают в структуре аэроподиев полукустарника. При оголении корней, когда растения поставлены на грань между жизнью и смертью, пластичность некоторых видов обеспечивает отдель- ным особям выживание путем заложения в тканях корней адвентивных почек и развития ближе к поверхности почвы нового пучка пост- инициальных побегов, например, у Artemisia lerchiana Web. (рис. 49-I), Helichrysum arenarium (L.) Moench (рис. 155-II). Образование адвентив- ных побегов на оголенных корнях растений отмечалось неоднократно (Rauh, 1937; Гордиенко, 1969 и др.). Иногда подземные многолетние ор- ганы у степных полупустынных и пустынных растений, особенно стерж- некорневых многолетних трав и полукустарничков, закручиваются как веревки (рис. 116-I). Растения с оголенным корнем пытаются уйти от ис- сушения и спирально закручивают корни, чтобы приблизить побеговую часть к поверхности почвы (Гордиенко, 1964, 1969; Гатцук, 1968). Многолетний побеговый орган, чаще всего каудекс, в обычных условиях роста растения или при выдувании субстрата начинает выпи- рать над поверхностью почвы. При оголении каудекса и базальной части главного корня все растение страдает от иссушения. С генетического уровня включается механизм внутривидовой пластичности, который мо- билизует жизненные силы особи на выживание. Многие растения про- являют невероятные усилия в конституциальной борьбе за существова- ние. Оголенный каудекс может продольно расщепляться на части по отмершим старым резидным побегам, вплоть до главного корня, час- тично расщепляя и его, образуя парциальные каудексовые ветви. Жи- вые резидные побеги могут активно функционировать, образуя из своих почек новые постинициальные побеги. Дисбаланс между ростовыми процессами в одних тканях и некротическими в других способствует неравномерному смещению участков каудекса и главного корня, пере- кручиванию их в продольном направлении, начиная сверху и далее к корню. Веревковидное закручивание частей каудекса и главного корня способствует некоторому втягиванию многолетней побеговой части рас- тения в почву (рис. 116-I). Рассмотрим несколько примеров поведения растений на протяжении их жизненных циклов в условиях неустойчивых к эрозии субстратов. Capparis herbaeea Willd. (= С. spinosa L.) — канерес травянистый (сем. Capparidaceae). Имеет широкий географический ареал и высокую пластичность. Наблюдался в Крыму (окрестности пос. Планерское, рай- он Карадага). Семенам для прорастания нужен период покоя. Прорас-
Глава 8. Изменчивость и эволюция 387 тание семян надземное. В виргинильной стадии жизни отрастают весен- ние безрозеточные раздвинутоузловые ростовые длинные стелющиеся (до 1,5 м длиной) акронекрозные силлептически разветвленные годич- ные побеги с очередным листорасположением полностью приземные или с укороченными подземными основаниями. После первого вегета- ционного периода образуется каудекс из одного миниморезидного по- бега, который в последующие годы разрастается в толщину и за счет увеличения числа новых миниморезидных побегов (рис. 117). Первое цветение происходит, чаще всего, на 3-5 год, однако возможен и более поздний переход в матурный этап; в единичных случаях зацветают особи на первом году вегетации. У взрослых особей ежегодно отрастают па- зушные унирепродуктивные (монокарпические обоеполые) открытые безрозеточные длинные (до 3,5 м) стелющиеся годичные побеги полно- стью приземные или с компактными подземными основаниями (рис. 86). В отдельные годы могут быть перерывы в цветении. Каперес отличается большим долголетием, развивая мощный деревянистый стержневой корень, который временами углубляется до 17 м и, вероятно, более (такие корни мы наблюдали на разломе скалы у берега моря). С возрастом в условиях относительной статики поверхности почвы формируется компактный приземный или полуподземный многоглавый каудекс, структуру которого детерминируют миниморезиды (рис. 117, 118-II). При погребении особей субстратом структура их каудекса усложня- ется и нередко определяется вынужденно удлиненными викариорезид- ными побегами, которые остаются после акронекроза надземных частей побегов. В этих условиях образуются короткие и длинные подземные ветвистые каудексы (рис. 118-Ш). Каперес — пионер на оголенных гор- ных субстратах и верный их закрепитель. Основной биоморфотип Cap- paris herbacea в Крыму — безрозеточные открытые полукустарнички с многоглавым полуподземным каудексом. Второстепенных биоморфоти- пов каперса травянистого в том же районе — два: полукустарнички с вынужденно надземным каудексом (иногда и надземным основанием главного корня), произрастающие на верхних участках горных склонов, где происходит осыпание или смыв субстрата, и полукустарнички с вы- нужденно подземным каудексом (погребение особей субстратом на ни- жних участках горных склонов); рис. 118. Pimpinella saxiPaga L. — бедренец камнеломки (сем. Apiaceae). Про- израстает на лугах, опушках. Особенности биологии изучали в Москов- ской области. Семена после созревания могут прорастать без периода покоя, но обычно в естественных условиях произрастания проростки появляются только весной следующего года. Прорастание семян над- земное. На первом году эпиэмбриональной жизни образуются розеточ- ные апонекрозные побеги — вегетативная фаза полурозеточного двухго- дичного побега, на втором году универсальный побег завершает свое развитие, проходя репродуктивна-ассимиляционную фазу роста и раз- вития, образуя семена. Основным структурным элементом у особей бед- ренца камнеломки являются унирепродуктивные (монокарпические обоеполые) полурозеточные условно закрытые (с верхушечными полу-
388 OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИЙ закрытыми соцветиями) акронекрозные дифазные, реже плейофазные, но иногда бывают и монофазные побеги. У взрослых растений, обита- ющих в условиях относительной статики субстрата, образуется компак- тный подземный каудекс, который нарастает за счет курторезидных и миниморезидных побегов (рис. 156-I). При засыпании субстратом (песчаная насыпь железной дороги) образуются вынужденно верхнерозеточные вегетативные и верхнеполу- розеточные унирепродуктивные побеги. В этих условиях формируется короткий и длинный ветвистый (вынужденно удлиненный) подземный каудекс, на котором образуется много небольших придаточных корней (рис. 156-II). Ежегодный прирост вынужденно удлиненного каудекса составляют вицерезидные и викариорезидные побеги. Основной биоморфотип бедренца камнеломки — полурозеточные компактно-каудексовые травянистые поликарпики; второстепенный биоморфотип — верхнеполурозеточные вынужденно удлиненные кау- дексовые травянистые поликарпики. Seseli eondensutum (L.) Reiehenb. — жабрица густоцветковая (сем. Apiaceae). Горный вид, произрастающий в Арктике, Западной и Восточной Сибири, на Аптае, на хребтах Саур и Тарбагатай, в северной Монголии и на Дальнем Востоке (Шишкин, 1950; Сдобнина, Пименов, 1975). Некоторые особенности биологии этого вида в естественных условиях произрастания описаны Л.И.Сдобниной и М.Г.Пименовым (1973). Безлесные высокогорные долины рек и ручьев, крупные котлови- ны, берега и русла временных водотоков, влажные склоны с субальпий- ской и альпийской растительностью, расположенные в пределах 1600- 2800 м над ур. моря — основные местообитания жабрицы густоцветко- вой на Алтае. По влажным открытым берегам горных рек и ручьев особи этого вида нередко селятся в поясе кедровых лесов и иногда проникают ниже, а также поднимаются и в гольцовый пояс. На высокогорных лугах (1800-2600 м над ур. моря) Seseli condensatum иногда является эдифи- катором. Она часто произрастает вместе с Allium schoenoprasum L., Schulzia crinita (Pall.) Spreng., Lagotis integrifolia (Willd.) Schischk., Poly- gonum viviparum L., Swertia obtusa Ledeb., Trollius asiaticus L. и др. В среднем, а тем более в нижнем поясах гор жабрица густоцветковая встречается редко, произрастая рассеянно на заболоченных лугах, среди зарослей пойменных кустарников; здесь в отличие от особей высо- когорных местообитаний особи этого вида обладают слабым развитием, уступая в конкурентной способности многим луговым растениям, на- пример в пойме речки у подножия горы Ябоган (Теректинский хребет) или в верховьях р. Черная Уба. Наибольшей встречаемости и обилия Seseli condensatum достигает в центральной и юго-восточной частях Аптая, особенно в Улаганском р-не (Курайский хребет), где автором изучена биоморфогения этого вида. Эмбриональный период. Пренатальный этап. ПС (временная продол- жительность возрастного состояния) — с июля до августа, середины сентября. ОХ (основная морфофизиологическая характеристика осо- бей) — оплодотворенная семяпочка или зрелое семя в структуре роди- тельского растения. Б (общая дополнительная характеристика особей;
Глава 8. Изменчивость и эволюция распространяется на несколько или на все возрастные состояния)— цветки обоеполые, опыляются как с помощью ветра, так и насекомых. Латентный этап. ПС — с конца августа, реже сентября до мая, июня следующего года или несколько лет. ОХ — зрелое семя, обособленное от родительского растения. А (частная дополнительная характеристика осо- бей; распространяется только на рассматриваемое возрастное состояние)— отделение некоторых зрелых полуплодиков (односеменных мерикарпиев) от сохраняющихся зимой сухих частей побегов иногда не происходит до весны следующего года. Б — мерикарпии овальные, 3-4,2 мм дл. и 2-3,0 мм шир., сжатые со спинки, с ребрами, причем боковые ребра крылатые, 0,4-1мм шир.; у особей жабрицы густоцветковой, обитающей в лесном (кедровом) поясе, мерикарпии осыпаются на 1-3 недели раньше, чем у особей, растущих на лугах в высокогорье. Эпиэмбриональный период. Виргинильный этап. Преювенильная фаза. Проростковая гипофаза (рис. 158, А). ПС — май, июнь, реже июль; у од- ной особи продолжается около 2 недель. ОХ — прорастающее семя или семядольный почкокорень (ризопротерокормус); А — семядоли (2, редко 3) надземные линейно-ланцетные, 0,8-1,0 см дл. и 0,1-0,2 см шир.; гипо- котиль 0,5-1,2 см дл. и 0,05-0,1 см в диам.; главная монориза, редко дириза, до 2,5 ем дл. Всходовая гипофаза (рис. 158, Б). ПС — июнь, июль, август. ОХ— семядольный вегетативный предрозеточный (1-2 настоящих листа) или розеточный (3-4 и реже более настоящих листьев) ризомонокормус. А— семядоли 1,0-2,2 см дл. и 0,12-0,25 см шир.; первые 1-4 настоящих листа простые округлые или округло-яйцевидные, по краю крупно-зубчатые или надрезанные, черешчатые. Ювенильная фаза. Гипофаза первая — униювенильная (рис. 158, В,). ПС — с августа до начала зимы текущего года. ОХ — бессемядольный ежегодно вегетативный предрозеточный или розеточный аризомный (без ризома) ризомонокормус. А — розеточные листья в числе 1-2 — у особей на высокогорных лугах или 3-4 — у особей, обитающих на лугах вдоль рек в кедровом поясе. Рассеченность пластинки усложняется от листа к листу. Гипофаза вторая — биювенильная (рис. 158, В,). ПС — с начала зи- мы до 2-8 лет. ОХ — бессемядольный ежегодно вегетативный розеточ- ный проризомный ризотетракормус. А — листья от тройчаторассечен- ных до перисторассеченных, черешчатые; базовый орган до образования на нем каулогенных корней — проризом, а после образования их- ризом; базовый орган нарастает моноподиально за счет курторезидных монокормусов; наблюдается контрактильность главного корня. Гипофаза третья — триювенильная (рис. 158, В,). ПС — с 2-8 до 3-15, чаще 5-10 лет. ОХ — бессемядольный ежегодно вегетативный розеточный гетерономно-ризомный ризотетракормус. А — главная тетрариза до 30 см, листья от перисторассеченных до двуперисторассеченных. Б — верхушеч- ные почки всегда незащищенные; при повышенном увлажнении (перио- дически затопляемая пойма ручьев) главный корень очень слабый, а ка- улогенные корни появляются рано и их много. Имматурная фаза. Гипофаза первая — униимматурная (рис. 158, Г,). ПС — с 2-15 до 4-25 лет. ОХ — ежегодно вегетативный розеточный ге-
OCHOBhl БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ
Глава 8. Изменчивость и эволюция терономно-ризомный (ризом пока связан с главным корнем) простокус- тистый (с двумя порядками побегов системы кущения) ризогексакормус или ризогептакормус. Б — пазушные побеги весенние, отрастают из зи- мовавших защищенных 2-3 чешуями почек или реже (в благоприятных для роста условиях, например, на участках с нарушенным растительным и почвенным покровом, по берегам пересыхающих ручьев в кедровом поясе) пролептические. Последние отрастают осенью из нижних почек побегов текущего года. Гипофаза вторая — бишаиатурная (рис. 158, Г,). Редко. ПС — с 6-20 до 8-25 лет. ОХ — ежегодно вегетативный розеточный гетерономно-ри- зомный сложнокустистый (с тремя и более порядками побегов системы кущения) ризоэнатокормус. А — особь этого возрастного состояния спо- собна преобразоваться только в матурную средневозрастную, минуя матур- ное молодое состояние. Б — ризом компактный, но при заиливании кор- невища вынужденно удлиняются за счет вицерезидных монокормусов; кор- невая система — синториза, состоит из главной стержневой тетраризы и других каулогенных и ризогенных корней разной сложности. Матурный этап. Инициматурная фаза (рис. 159, Д„Д,). ПС — с 4-25 до 10-35 лет. ОХ — физиологически зрелый ежегодно или неежегодно репродуктивный полурозеточный гетерономно-ризомный ризопентакор- мус или ризогексакормус, или ризогептакормус. А — главная тетрариза достигает 30-80см. Б — пентакормусы унирепродуктивные ди-, три-, плейофазные (2-15 фаз видимого роста), высотой 20-70сМ, обоеполые; соцветие — одиночный сложный зонтик или реже сборный (с па- ракладиями) зонтик; число унирепродукгивных дикормусов по годам ко- леблется от 0 до 12, чаще 2-4, темпы прохождения постинициального периода жизни этих побегов у особей, обитающих на лугах в кедровом поясе, в 2-5 раз быстрее, чем у особей, растущих на высокогорных лугах, а сроки фенофаз смещаются на 1-2 недели; фаза бутонизации — конец июня - начало июля; цветение — середина, конец июля-начало августа Рис. 158. Виргинильные мероаклонисты Seseli condensatum (L.) Reichenb. А — проросток, Б — всходы, В,-В,— ювенильные особи: В, — униювенильные, В, — би- ювенильные, В, — триювенильные; Г,-Г, — имматурные особи: Г, — униимматурные, Г,— биимматурные. 1 — семядоли, 2 — гипокотиль, 3 — семядольный узел, 4 — корневая шейка; 5а-5е — корни: 5а — главная монориза (субмедиаль), 5б — главная дириза, 5в- ризогенная монориза (ризогенная сублатераль), 5г — ризогенная тририза, 5д — главная тетрариза, 5e — синториза, 6 — ризогенная корневая ветвь, 7 — каулогенная корневая ветвь, 8 — предрозеточный побег, 9 — розеточные листья или основания таких листьев (верхние части удалены), 10 — медиальный вегетативный розеточный побег, 11 — аксил- лярный вегетативный розеточный побег; 12-14 — постинициальные (с видимым ростом) побеги: 12 — монокормус, 13 — тетракормус, 14 — гептакормус, 15-16 — остаточные (ре- зидные) побеги: 15 — курторезидные монокормусы, 16 — вицерезидные (вынужденно удлиненные подземные с живой верхушкой) монокормусы; 17 — проризом, 18 — гете- рономный ризом, 19 — пазушные почки, 20-23 — организменные ризокормусы: 20— семядольный почкокорень (ризопротерокормус), 21 — семядольный вегетативный ризо- монокормус, 22 — бессемядольный вегетативный ризотетракормус, 23 — бессемядоль- ный вегетативный ризогексакормус, 24 — вегетативный ризогептакормус; 1л, 2л,...— число лет кондивидуумов; горизонтальная линия — поверхность почвы; у — увеличено по сравнению с масштабом; мк — упрощенные модели разновозрастных кондивидуумов. Одно деление масштаба — 1 см.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 159. Матурные и сенильные кондивидуумы Seseli condensatum. Д — инициматурные мероаклонисты: Д, — некустистые, Д, — простокустистые, Е — медиматурные мероаклонисты, Ж — финиматурный кондивидуум, И — се- нильный кондивидуум; 25 — унирепродуктивный полурозеточный побег, 26— очаги некроза тканей корня или корневища, 27 — ризопентакормус, 28 — репро- дуктивный ризогептакормус, 29 — ежегодно или неежегодно репродуктивный ри- зоэнатокормус, 30 — миниморезидные тетракормусы, 31 — викариорезидные тет- ракормусы, 32 — безрозеточные листья, 33 — автономный ризом, 34 — соцветие, 35 — основания побегов текущего года (верхние части удалены). Остальные обо- значения те же, что и на рис. 158.
Глава 8. Изменчивость и эволюция (цветение базипетальное); плодоношение — август-начало сентября; осыпание семян — конец августа-сентябрь; листорасположение очеред- ное, розеточные листья непарно-двуперисторассеченные с черешками. Медиматурная фаза (рис. 159, Е). ПС — с 10-35 до 40-90 лет. ОХ— ежегодно или неежегодно репродуктивный полурозеточный гетероном- но-ризомный гемисимподиальный ризоэнатокормус. А — главная тет- рариза до 1 м дл., постепенно дряхлеет. Б — активную часть побеговой системы составляют полурозеточные ди-, три-, плейофазные унирепро- дуктивные пентакормусы, которые до вступления в репродуктивную фазу проходят состояния вегетативных розеточных монокормусов и тет- ракормусов; аккумулятивную часть побеговой системы образуют ризо- мы, которые нарастают моноподиально за счет курторезидных, иног- да — при заиливании, также и вицерезидных монокормусов, и эусим- подиально за счет миниморезидных (акронекрозных скелетных компак- тных), реже викариорезидных (акронекрозных скелетных вынужденно удлиненных) плейомерных (2-12-членных) тетракормусов. Финиматурная фаза (рис. 159, Ж). ПС — с 40-90 до 80-120 лет. ОХ — совокупность нескольких (2-12) ежегодно вегетативных (без сле- дов перевершиниваний от унирепродуктивных побегов) автономно- ризомных ризокормусов (имматурных клонистов), ежегодно или нееже- годно репродуктивных автономно-ризомных ризокормусов (матурных клонистов). А — главная тетрариза отмерла полностью, причем раньше всего в условиях заболоченных почв; полная общая эпиэмбриональная некротическая партикуляция завершает жизненный цикл мероаклониста (временного аклониста), приводя к вегетативному размножению, т.е. к началу жизненных циклов клонистов; мероаклональный кондивидуум, переходя в старое матурное состояние, становится клональным конди- видуумом; вегетативное размножение замедленное, клонисты слабо от- даляются друг от друга. Сенильный этап (рис. 159, И). ПС — 1-8 лет. ОХ — ежегодно вегета- тивные после утраты физиологической зрелости розеточные автономно- ризомные ризокормусы (сенильные клонисты). Б — на лугах в кедровом поясе синонтогенез короче, чем на альпийских лугах; предельный полный возраст (продолжительность жизни кондивидуума) не более 130 лет. Резюмируем особенности биоморфогении жабрицы густоцветковой. Виргинильные особи проявляют моноподиальное нарастание розе- точных вегетативных побегов и активную жизнедеятельность системы главного корня. Многие годы особи семенного происхождения (меро- аклонисты) развиваются с гетерономными ризомами (т.е. ризомами, связанными с главным корнем), а с начала старого матурного и в се- нильном возрастных состояниях кондивидуумов их особи-клонисты- автономно-ризомные (их ризомы утратили связь с отмершим главным корнем). В матурном периоде побеговая система по типу нарастания гемисимподиальная. Для жабрицы густоцветковой характерна синонтогения, т.е. био- морфное развитие по типу синонтогенезов. Вегетативное размножение замедленного типа. Унирепродуктивные закрытые полурозеточные пен- такормусы чаще всего плейофазные, что позволяет растению экономно
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ расходовать на репродуктивную функцию свои энергетические запасы, медленно накапливаемые в процессе более или менее длительного (2-15 лет) моноподиального розеточного вегетативного нарастания этих побе- гов. При заилении и засыпании почвой подземные междоузлия вынужденно удлиняются. В этих условиях растения проявляют второстепенный mun биоморфогенезов — травянисто-мультирепродуктивный с вынужденным удлинением ортотропных корневищ. С увеличением высоты над уровнем моря продолжительность жизни кондивидуумов, как правило, уве- личивается, не превышая 130 лет. По эколого-морфологическим тропобиозным признакам Seseli con- densatum — это травянистый летнезеленый надземно-двусемядольный гемикриптофильный (почки, зимующие первый год, находятся на уров- не поверхности почвы) полурозеточный гемисимподиальный очередно- листный непарно-двуперисторассеченнолистный (с закрытыми перис- тыми дифиллами) кустистый компактно-корневищный брахиплано- фильный (слабо вегетативно разрастающийся в горизонтальных направ- лениях) ретардационно-кондивидуальный (медленно вегетативно раз- множающийся, с клонистами, долго удерживающими территорию) од- нодомный горно-луговой вид. 8.2.2. Пластичность биоморф семенных растений средиземноморского побережья Ближнего Востока Вызывает восхищение пластичность биоморф растений Ближнего Востока, знакомство с, которыми состоялось в марте-апреле 1992 г. Иссле- довались холмистые участки с песчано-каменистыми почвами в ок- рестностях Тель-Шива (Израиль). Растения определялись по книге Н.Файнбрун-Дотана и А. Данина (Feinbrun-Dothan, Danin, 1991). На учас- тке наблюдается некоторое смешение средиземноморской и сахаро-ара- вийской флор при преобладании элементов первой (Вальтер, 1968). Кли- мат Средиземноморья Ближнего Востока характерен для субтропиков: влажная зима и очень сухое лето, короткие и не всегда явные осень и весна, которые подвержены воздействию таких контрастных факторов, как близость моря на западе и зоны пустынь на востоке, простирающейся на всю южную половину территории государства Израиль. Иногда из пусты- ни дуют иссушающе знойные ветры (хамсины). Зимой редко температура опускается ниже 5' С. Осадки выпадают только с конца октября по апрель (среднее количество годовых осадков около 500 мм) (Михаэли, 1992). Среди деревьев на обследованной местности чаще других встреча- лись Pinus halepensis Miller, Оlеа еигораеа L., Pistacia palestina Boiss, Ceratonia siliqua L., Parkinsonia aculeata L. и др. На песчаных почвах нередко произрастают невысокие деревья (до 5 м) клещевины (Ricinus communis L.). Для сухих каменистых склонов характерна гаррига (сооб- щество средиземноморских вечнозеленых низкорослых кустарников, полукустарников и многолетних трав), среди которых Salvia fruticosa Miller, Spartium junceum L., Cistus salvifolius L. и др. Из кустарников на
Глава 8. Изменчивость и эволюция 395 Рис. 160. Первые годы биоморфогении Retama raetam (Forssk.) Webb (Израиль, окрестности Тель-Авива, холмистые участки с песчано-каменистыми почвами). А — проросток, Б — всход, В — ювенильные особи:В, — униювенильная особь, В, — биювенильная особь, Г — имматурная особь; 1 — семядоли, 2 — гипокотиль, 3 — семядольный узел, 4 — эпикотиль, 5 — главный корень, 6 — главный побег, 7 — первые срединные листья, 8 — силлептические побеги, 9 — семядольный ве- гетативный медиальный монокормус, 10 — семядольный вегетативный медиаль- ный дикормус, 11 — бессемядольный вегетативный медиальный дикормус, 12— вегетативный гексакормус, 13 — прошлогодние живые прутьевидные осевые ча- сти — рамозорезидный годичный побег, 14 — весенние побеги текущего года (март, апрель). Горизонтальная линия — поверхность почвы, одно деление масштаба — 1 см. участке преобладали Ephedra aphylla Forssk., Retama raetam (Forssk.) Webb, Calicotome villosa (Poiret) Link, Spartium junceum L., 1пи1а viscosa (L.) Aiton, Cistus salvifolius L. и др. Разнообразно были представлены также полукустарники и полукустарнички (Echium angustifolium Miller, Centaurea procurrens Sprengel, Heliotropium rotundifolium Lehm., Polygo- пит equisetiforme Sm., Scrophularia xanthoglossa Boiss. и др. Рано весной в марте-апреле «вспыхивают» и быстро исчезают мно- гие виды эфемеров (Erodium cicutarium (L.) L'Íåã., Adonis aleppica Boiss., Biscutella didyma L. и др. Структура биоморф сильно ксерофилизирована. Деревья и кустар- ники или жестколистные, как многие Quercus L., Pistacia L., или лише- ны листьев с нормальной листовой пластинкой, например, Parkinsonia aculeata L., Spartium junceum L. Рассмотрим некоторые особенности биологии нескольких кустар- ников, обратив внимание на их адаптированность к местным условиям. Retama raetam (Forssk.) ИеЬЬ — ретами (сем. Fabaceae) (рис. 160, 161).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 161. Разнообразие побегов взрослых растений Retama raetam (произрастают там же, что и особи, изображенные на рис. 160). 15 — некоторые фрагменты кроны и их детализация, 16 — весенний вегетативный аксиллярный дикормус (силлептически сильно ветвистый годичный побег), 17— вегетативный гексакормус (фрагмент), 18 — рамозорезидная часть гексакормуса, 19 — репродуктивный гексакормус, 20 — постинтеркалярная кисть; остальные об- означения те же, что и на рис. 160. Семена прорастают в конце февраля, марте, апреле, когда в почве еще достаточно влаги. Прорастание семян надземное. Проростки с дву- мя эллиптическими зелеными мясистыми семядолями (0,6-0,8см дл., 0,3-0,4см шир.), почти сидячие (рис. 160, А). У всходов семядоли не- сколько увеличиваются в размерах; их зародышевая верхушечная почка развертывается и начинается видимый рост медиального (главного) не- ветвящегося побега (рис. 160, Б). Первые несколько листьев развиваются
Глава 8. Изменчивость и эволюция с нормальной заостренно-эллиптической пластинкой (в среднем 0,8-1,2 см дл., 0,3-0,5 см. шир.). Всходы развиваются до отмирания семя- долей (без ветвления главного побега), а если ветвление начинается при живых семядолях (их дл. 0,8-1,0 см., шир. 0,4-0,5), то это также инди- каторный признак нового возрастного состояния — первого ювениль- ного (рис. 160, В,). Когда кроме предветви (медиального побега) в систе- ме ветвления развиваются вегетативные силлептические ветви не только первого, но и последующих (2, а иногда и 3-го порядков) — это признак второго ювенильного возрастного состояния (рис. 160, В,). Верхние и средние силлептические побеги растут относительно спокойно — это индуктивные побеги, а вот самые нижние подобные побеги, сильно «возбуждаются», попав волею судьбы в зону кущения, начинают расти весьма активно — это базальные силлептические, или базисиллепти- ческие, побеги — разновидность индуктивных побегов. Образование первых постлатентных побегов (у особей второго года) — признак на- чала имматурной жизни особей, которая продолжается до начала обра- зования репродуктивных побегов. Главная особенность ретамы — обильное побеговое ветвление по- чти на всех стадиях жизни и быстрый переход к образованию сильно редуцированных мелких листьев с быстро опадающей невыразительной маленькой листовой пластинкой и с сохранением расширенного корот- кого основания листа (до 2-х, а иногда более, лет). После засыхания основание листа некоторое время еще сохраняется на прежнем месте. Уже на первом году эпиэмбриональной жизни развивается разветвлен- ный годичный побег; верхушки этого побега летом и зимой отмирают на разную длину, а сохраняющаяся многолетняя часть — рамозорезид- ный годичный побег является начальным блоком в структуре ветвистого аэроподия. Вегетация на втором, третьем и последующих годах имма- турной жизни осуществляется с конца февраля, начала марта и продол- жается, особенно активно, в марте, апреле. В это время отрастают ви- димые (постинициальные) весенние (постлатентные) побеги, родитель- ская ось которых более или менее обильно силлептически ветвится, образуя пучки прутьевидных вегетативных побегов, на которых по- прежнему развиваются очередные сидячие сильно метаморфизирован- ные мелкие листья (это не чешуи, а листья с сильно уменьшенной листовой пластинкой, которые живут мало, отмирают и осыпаются в сильную жару летом и осенью. Фотосинтез выполняют в основном зе- леные прутьевидные стебли. Расширенные основания листьев, как и стебли, зимуют в зеленом виде. Интересно, что они прикрывают пазуш- ные открытые почки (меристемы погружены в ткани стебля) и защища- ют их от иссушения. С началом обильного силлептического побегобра- зования пазушные побеги отрастают из под листовых оснований без доразвития, т.е. из явно недоформированных почек. С возрастом увеличивается число стволиков аэроподиев, многие нижние и средние стеблевые ветви живут сравнительно недолго, через несколько лет стволики до кроны самоочищаются от старых ветвей. Аэроподии функционируют 10-12 лет, чаще меньше. От основания куста отрастают сильные побеги кущения — турионы (около 1 м высотой).
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННЫХ PA СТЕНИЙ Редко они цветут на второй год (слабо), чаще с третьего года и макси- мально на 4-6 году. Деятельность пазушных меристем (открытых почек) многогранна. В одних случаях они образуют весенние (постлатентные) прутьевидные модифицированные вегетативные годичные побеги, ко- торые далее силлептически ветвятся, развивая побеги с такой же спе- циализацией, что и родительский побег (в целом это дикормус). Боль- шая часть побегов взрослых растений развивается по сверхгодичному (двухгодичному) циклу (это гексакормус по другой терминологии). Сна- чала образуются весенние прутьевидные ветвистые акронекрозные ве- гетативные годичные побеги, а весной следующего года из перезимовав- ших почек этих побегов отрастают постинтеркалярные тотрепродуктив- ные побеги-соцветия — кисти (рис. 161). Непрерывная совокупность постинтеркалярных кистей на участке одной открытой побеговой оси образует вместе иносоцветие (контринфлоресценцию). Совокупность всех иносоцветий и одиночных постинтеркалярных побегов, участвую- щих в структуре одного сверхгодичного (двухгодичного) побега (гекса- кормуса) можно обозначить как сборное иносоцветие, или нарциальное сверхсоцветие. Гигантский белый «букет» из множества мелких кистей (1-3 см дл.) всего растения — сборное сверхсоцветие ретамы. Итак, ос- новные структурные элементы Retama raetam — годичные ветвистые акронекрозные вегетативные побеги и двухгодичные ветвистые вегета- тивно-репродуктивные побеги. Нарастание побеговой системы эусим- подиальное. Ежегодно структуру аэроподиев пополняют рамозорезид- ные побеги. Зохари (Zohary, 1952) отнес ретаму к типу вечнозеленых прутьевидных кустарников. Итак, биоморфотип Retama taetam — это зимнезеленостеблевые пру- тьевидные безрозеточные обоеполые постинтеркалярно-репродуктивные акронекрозные эусимподиальные стержнекорневые бело-бобовоцветковые кустарники с модифицированно-мелкими и недолговечными листьями. Calicotome villosa (Poiret) Link — каликотома ворсистая (сем. Faba- ceae) (рис. 162, 163). Прорастание надземное. Семена прорастают зимой и весной, когда в почве достаточно влаги. На первом году эпиэмбри- ональной жизни развивается медиальный (главный) безрозеточный по- бег с очередными тройчатыми листьями (рис. 162). С развитием 5-8 листа семядоли отмирают (признак перехода возрастного состояния всхода в ювенильное). При благоприятных условиях медиальный побег начинет силлептически ветвиться, образуя короткие безрозеточные эу- силлептические побеги из почек средней и верхней части родительского побега, а из приземных почек нередко отрастают наиболее сильные ростовые побеги кущения — базисиллептические (разновидность ин- дуктивных) побеги. Верхушки коротких побегов вскоре засыхают и пре- вращаются сверху в колючки, отмирая только на часть своей длины. Сильные побеги кущения так же силлептически ветвятся с образованием новых коротких побегов — колючек. Такое разнообразное силлетичес- кое ветвление происходит с растениями, которые прорастают зимой и к весне уже имеют хорошо сформированный кустик, продолжающий свое годичное вегетативное развитие до осени. После первого вегетаци- онного сезона верхушки ростовых побегов отмирают и на второй год из открытых почек продолжается образование ветвистых вегетативных без-
Глава 8. Изменчивость и эволюция Рис. 162. Виргинильные особи Calicotome villosa (Poiret) Link (произрастают там же, что и особи, изображенные на рис. 160). 21— — силлептические побеги, которые после прекращения роста превращаются в колючки, 22 — базальные сильнорослые силлептические побеги — побеги куще- ния; остальные обозначения те же, что и на рис. 160 и 161.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ розеточных побегов. Из рамозорезидных годичных побегов формируется структура аэроподиев. У взрослых растений цветки развиваются как интеркалярные тот- репродуктивные побеги одиночно или в структуре межсоцветий предро- зеточных (с двумя сближенными листьями), розеточных (у них 3 и более сближенных листьев), полурозеточных (с розеткой листьев в основании и расположенной выше короткой раздвинутоузловой частью) или даже безрозеточных коротких (с одним листом в основании) побегов (рис. 163). Рис. 163. Разнообразие побегов взрослых растений Calicotome villosa (произрастают там же, что и особи, изображенные на рис. 160). 23 — интеркалярный цветок (тотрепродуктивный силлептический пазушный по- бег), 24 — интеркалярное соцветие, 25 — оно же на схеме (сильно сближенные узлы для наглядности раздвинуты), 26 — колючки в структуре рамозорезидной части гексакормуса, 27 — интеркалярный плод, 28 — интеркалярное соцветие в стадии начала плодоношения; остальные обозначения теже, что и ра рис. 161 и 162.
Глава 8. Изменчивость и эволюция Такими побегами в матурном состоянии обрастает каждый прошлогод- ний рамозорезидный годичный побег, образуя вместе с ними двухгодич- ный вегетативно-репродукгивный побег (гексакормус по шестой субор- динатной классификации побегов). Кроме весенних вегетативно-репро- дуктивных побегов также развиваются ростовые длинные безрозеточные побеги, которые обеспечивают основной многолетний прирост расте- ний. Верхушки коротких побегов быстро засыхают и они превращаются в колючки, а верхушки сильных ростовых побегов долго моноподиально растут, будучи незащищенными, отсыхают летом или к началу зимы. Нарастание аэроподиев эусимподиальное. По серии морфологических признаков для Calycotome villosa характерен следующий биоморфотип: колючие безрозеточно-ростовые розеткообразующе-репродуктивные акронекрозные эусимподиальные стержнекорневые обоеполые желто- бобовоцветковые кустарники. Inula viscosa (L.) Aiton — девясил липкий (сем. Asteraceae) (рис. 164). Семена не нуждаются в стратификации и прорастают во влажных по- чвах зимой или весной. Прорастание семян надземное; семядоли мел- кие (0,3-0,4см дл., 0,15-0,2см шир.), эллиптические, почти сидячие, при основании клиновидные. На первом году эпиэмбриональной жизни образуется розетка листьев, которая постепенно начинает растягиваться при дальнейшем видимом росте медиального побега и типичной розе- точности у особей этого вида нередко не проявляется. Однако все же розеточность у всходов есть; на последующих стадиях онтогенеза розе- точность определяется иногда четко, а больше происходит безрозеточ- ный раздвинутоузловой рост. Пользуясь уточняющим термином «розет- кообразующий» побег (Серебрякова, 1971), можно считать, что у девяси- ла липкого образуются розеткообразующие побеги: розеточные, полу- розеточные и верхнерозеточные в зависимости от стадии их развития. Пожалуй, будет точнее такие побеги, как у рассматриваемого вида, счи- тать переходно-розеткообразующими (рис. 164). Всходы имеют розеточ- ную структуру медиального побега. У ювенильных растений начинается растягивание розетки и происходит основной раздвинутоузловой безро- зеточный прирост медиального побега. Розеточные листья на побеге постепенно засыхают, но в целом медиальный побег переходно-розетко- образующий. На первом году эпиэмбриональной жизни особи могут начать ветвиться и куститься. Реже это происходит только на втором году жизни, если начало вегетации было позднее и (или) условия для роста менее благоприятные. Рост побегов в течение всего года полностью не замирает. Почки открытые. В зимний период и рано весной безро- зеточный рост побегов сменяется на верхнерозеточный, что позволяет достаточно точно определять границы годичных приростов моноподи- ально нарастающих побегов. На втором году и в последующем в струк- туре кустов продолжается кущение и ветвление. Цветение происходит на 2-4 год эпиэмбриональной жизни в зави- симости от условий произрастания. У двух-, реже трех-, многогодичных унирепродуктивных побегов состояние розеточного моноподиального нарастания сменяется безрозеточным раздвинутоузловым ростом, а с ос- лаблением скорости видимого роста вновь просходит розеточный рост.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 164. Особенности биоморфогении 1пи1а viscosa (L) Aiton (произрастают там же, что и особи, изображенные на рис. 160). 29 — сухие остатки прошлогоднего верхушечного соцветия, 30 — живые осевые части прошлогоднего соцветия, 31 — постфлоральные живые побеги, развитые в подзоне прошлогоднего верхушечного соцветия, 32 — постфлоральные живые по- беги, развитые в подзоне торможения прошлогоднего соцветия, 33 — постфло- ральный засохший побег, развивавшийся в текущем вегетационном сезоне, 34— сильнорослый переходно-розеткообразующий двухгодичный временно вегетатив- ный (конец марта) побег; у — увеличено по сравнению с масштабом; остальные обозначения те же, что и на рис. 160-163.
Глава 8. Изменчивость и эволюция С достижением физиологической зрелости особи образуют метельчатое закрьггое соцветие с корзинками из желтых обоеполых цветков. Цветет в августе, сентябре (Schmida, 1986). После плодоношения многие репро- дуктивные оси засыхают, но в основании разветвленной части бывшего соцветия и тем более в бывшей зоне торможения развиваются пазушные верхушечные меристемы (открытые почки), которые находят еще силы для образования постфлоральных вегетативных побегов (рис. 164). Более всего побегов отрастает в конце зимы и рано весной из зоны торможения быв- шего цветоносного побега — нижняя зона (подзона) весеннего постфло- рального побегообразования. Еще труднее идет отрастание новых побегов из почти полностью отмершей зоны эусиллептического ветвления (зоны обогащения) бывшего соцветия — верхняя зона (подзона) весеннего пост- флорального побегообразования. Таким образом, у девясила липкого про- является в местных условиях оригинальный тип побегообразования (уже- побегоотрастания), названный автором постфлоральным «лихорадочным» (полуобеспеченным в этом случае) побегообразованием (см. рис. 82), а сам участок, где происходит этот процесс, обозначаем как зону постфлораль- ного побегообразования. Наиболее сильные побеги начинают активный видимый рост весной, развиваясь из нижнерасположенных почек; резко ускоряют рост и те ро- зеточные и верхнерозеточные вегетативные побеги, которые подготавли- вались еще зимой к началу весны для соответствующего ростового усилия в зоне (ярусе) основного побегообразования (побегоотрастания). Сильные вегетативные и вегетативно-репродуктивные побеги, развиваясь в основ- ном (нижнем) ярусе побегоотрастания, догоняют и обгоняют рост побегов из постфлоральной зоны. Некоторые постфлоральные побеги (особенно из верхней подзоны) не выдерживают инфраструктурной конкуренции с раз- вивающимися ниже побегами и погибают, но часть побегов постфлораль- ного отрастания медленно продолжает видимый рост и развитие, постепен- но теряясь в массе побегов нижнерасположенного яруса. Димерные, редко с меньшим или большим числом годичных моноподиальных приростов, резидные короткие или длинные побеги обеспечивают гемисимподиальное нарастание аэроподиев. Основной йиоморфОТИП Inula viscosa в Средиземноморской части Ближ- него Востока — это надземно-двусемядольные переходно-розеткообразу- ющие очереднолистные открытопочечные детерминатно-репродуктивные акронекрозные гемисимподиальные постфлорально-вегетирующие обоепо- лые желто-астроцветковые листопадные кустарники. По поводу трех рассмотренных видов кустарников заметим следую- щее. Если постфлоральное побегообразование происходит из жизнеспо- собных частей бывших межсоцветия или иносоцветия, то это вполне ес- тественно и легко объяснимо, но когда постфлоральная вегетация побегов идет из сохранившихся частей зоны бывшего верхушечного соцветия— этот факт достоин особого внимания. Далее обратимся в примерах к растениям-полукустарникам и полуку- старничкам с верхушечными соцветиями, которые склонны проявлять постфлоральное «лихорадочное» побегообразование. ЕсЫит angustifoliuIn МИа — синяк узколистный (сем. Boraginaceae) (рис. 165). Прорастание семян надземное. Проростки появляются чаще
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 165. Особенно бенности биоморфогении ЕсЫит angustifolium Miller произрастают там же, что и особи, изображенные на рис. 160). 35 — гексако с с по растанием, 36 — озеточные п рму ст лоральным «лихорадочным» необесп еченным побегоот- — розеточные постфлоральные побеги текущего года, 37 — слабые верхнерозеточные двухгодичные вегетативные побеги 38 — б зеточные дв го и и, — сла ые верхнеполуро- ухгодичные вегетативно-репродуктивные побеги, 39 — ани ных побегов, развитых в те ем о еги, — граница годич- коо разующий дв го ичн б текущем и прошлом годах, 40 — сильнорослый розет- апрель) побег, 41— ух д ый или годичный временно вегетатив й ' ны (март— — упрощенная схема, 42 — аэроподий остал н б ьные о означения те же, что и на рис. 160-164.
Глава 8. Изменчивость и эволюция всего зимой или рано весной, когда в почве достаточно влаги. Семядоли (0,4-0,5 см. дл., 0,25-0,3 см. шир.), эллиптические, мясистые, голые, при основании округло-клиновидные. На первом году эпиэмбриональной жизни развивается розеточный медиальный побег, который в процессе своего развития начинает силлептически куститься. Первое цветение происходит на втором, реже третьем году жизни. Образуются типичные для бурачниковых цимозные соцветия-монохазии (завиток и двойной завиток). Унирепродуктивные побеги развиваются по монофазному (го- дичному) или дифазному (двухгодичному) циклу, проходя сначала ве- гетативные стадии розеточного и полурозеточного роста, завершающие- ся образованием верхушечных репродуктивных элементов. В угнетен- ном состоянии развитие розеточного или верхнерозеточного побега за- тормаживается и он может погибнуть на 1-3 году своей жизни. Многолетнюю побеговую систему растения составляют невысокие разветвленные аэроподии — от 5 до 40 см высотой, на которых обычно развивается эфемерно-преперсистентная крона куста. После цветения, плодоношения и акронекроза к весне нередко сохраняется от прошло- годнего побега рамозорезидная часть с зоной ностфлорального «лихора- дочного» необеснеченного нобегообразования. Слабые ностфлоральные нобе- ги отрастают в нодзоне торможения бывшего (нрошлогоднего) соцветия и в нодзоне эусиллентического ветвления бывшего соцветия (рис. 165). Эти нобеги могут быть вегетативными и вегетативно-ренродуктивными, зац- ветающими несколько раньше набегов основной волны роста. Много таких нобегов, будучи необеснеченными (им явно не хватает ресурсов для нор- мального развития) засыхает, не развивая ренродуктивных элементов. Основное побегообразование идет из открытых почек, расположенных ниже постфлоральной зоны, и характеризуется развитием сильных унирепродуктивных монофазных или дифазных побегов, которые «нака- тываются» снизу куста на постфлоральную зону буквально разрастаю- щимся валом и поглощают ее в своей массе. Основной биоморфотип Echium angustifolium в Средиземноморье Ближнего Востока — это надземно-двухсемядольные розеткообразую- щие очереднолистные открытопочечные детерминатно-репродуктивные обоеполые акронекрозные жестковолосистые, эусимподиально и геми- симподиально нарастающие, постфлорально необеспеченно-вегетирую- щие, розово-бурачниковоцветковые полукустарники. Centaurea ртоситтенз Sprengel — василек выбегающий (сем. Asteraceae) (рис. 166). Прорастание семян надземное; оно происходит зимой или весной. На первом году вегетации развивается розеточный медиальный (главный) побег, который, разрастаясь, иногда начинает силлептически куститься. Первое цветение происходит на первом или втором году эпиэмбриональной жизни с апреля по июнь (Schmida, 1986). Розеточ- ный рост медиального побега переходит в безрозеточный раздвинуто- узловой, в результате чего формируется цветоносная часть с одной вер- хушечной корзинкой, если растения еще физиологически слабые, или рыхлое ветвистое соцветие из колючих корзинок с желтыми цветками (обычно у растений второго года жизни и старше). Полурозеточные унирепродуктивные (монокарпические обоеполые) побеги — основной
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 166. Развитие организации особей Centaurea procurrens Sprengl (произрастают там же, что и особи, изображенные на рис. 160). 43 — цветочная корзинка, развивающаяся в фазу цветения; остальные обозначе- ния те же, что и на рис. 160-165.
Глава 8. Изменчивость и эволюция структурный элемент побеговой системы. После плодоношения цве- тоносная часть может отмирать полностью или отмирание сверху зах- ватывает лишь часть цветоносных осей, а на части, сохранившей жизне- способность, весной следующего года проявляется постфлоральное «лихорадочное» необеспеченное побегоотрастание. В постфлоральной зоне развиваются розетки вегетативных побегов, которые при благоприятных условиях могут образовать свои цветоносные системы. Однако чаще всего многие из этих побегов так и заканчивают свою жизнь в розеточном вегетативном состоянии (рис. 166), поскольку развивающиеся снизу ос- новные полурозеточные побеги догоняют и перегоняют в развитии постфлоральные побеги, затормаживая их рост и способствуя их гибели. Этот вид иногда характеризуют как травянистый многолетник (Shmi- da, 1986), но ежегодное развитие весной у многих особей постфлоральной зоны побегообразования позволяет считать его полурозеточным полукус- тарничком. Пожалуй, есть все основания считать, что полукустарнички и полукустарники, проявляющие в своей структуре постфлоральную побего- образовательную активность, являются характерной чертой средиземно- морских субтропиков Ближнего Востока. Еще сложнее определить тип жизненных форм у таких известнь|х в умеренном поясе видов растений, как Foeniculum vulgare Miller, Lycopus europaeus L. (рис. 167, 168). Фенхель обыкновенный в Крыму дает спектр биоморфотипов от монокарпиков до каудексовых травянис- тых поликарпиков, иногда образуя и вынужденно надземные кудексо- вые поликарпики при оголении главного корня (рис. 167, 48). В услови- ях средиземноморских субтропиков Ближнего Востока верхушечным цветоносным частям побегов представляется уникальная возможность не засыхать после плодоношения, а «поработать», хотя бы еще недолго (рис. 168). Как видно из приведенного здесь материала, субтропические растения, занимающие переходное положение между деревьями и травами, используют предоставляющийся им шанс, проявляя «лихора- дочное» побегообразование. Даже особи Lycopus europaeus, обитающие в поймах рек (окрестности г. Нетания) образуют верхушечные цветоно- сы, которые способны проявлять постфлоральную необеспеченную по- бегообразовательную активность (рис. 168-II). Неполностью отмирают надземные цветоносные части побегов и у некоторых особей Lythrum salicaria L. (пойма р. Иордан). От многолетников иногда не отстают и однолетники, так же проявляя постфлоральное «лихорадочное» необес- печенное побегообразование. Например, некоторые особи Conyza bonariensis (L.) Cronquist весной следующего года еще некоторое время вегетируют в постфлоральной зоне (рис. 169) (биоморфотип — непос- тоянные сверходнолетники), что в целом не спасает их от гибели с наступлением жары. Однако способность эфемерных структур бороться за продление своей жизни и в целом жизни всего организма имеет особый адаптационный смысл. Дополнительные сведения о пластично- сти биоморф с постфлоральной побегообразовательной активностью отражены на рис. 170. Из других особенностей биоморф средиземноморской флоры Ближ- него Востока обращает на себя поведение злаков. В структуре многих
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 167. Габитуальный полиморфизм Foeniculum vulgare Miller в Крыму. 44 — унирепродуктивный полурозеточный побег, отмирающий после плодоноше- ния до укороченного подземного основания, 45 — многолетний базальный оста- ток унирепродуктивного побега, или миниморезидный побег, 46 — каудекс, 47— подземно каудексовые травянистые обоеполые поликарпики, развивающиеся в условиях относительно постоянного уровня поверхности почвы, 48 — вынужден- но надземный каудексовый поликарпик, развивающийся при оголении главного корня от субстрата, 49 — примерное положение семядолного узла, 50 — основания унирепродуктивных побегов (верхние части удалены); остальные обозначения те же, что и на рис. 160. злаков ярко выражена тенденция выживания их надземных стеблей (со- ломин) более одного года. Это явление настолько распространено сре- ди средиземноморских злаков, что у автора нет сомнений в целесооб- разности выделения особых типов жизненных форм, свойственных мно- гим видам бамбуков — «однодольных кустарников» (Warming, Graeb- ner, 1918). Согласно, предложенного ранее (гл. 2.4.), принципа: там, где это полезно (например, для разноплановой характеристики), в название конкретного тина жизненных форм может включаться название семей- ства, к которому нринадлежит вид биоморф, носкольку это название одновременно нодчеркивает нринадлежность их главного ренродуктивного органа (цветка), например, мятликовоцветковые (злаковоцветковые),
Обозначения те же, что и на рис. 164. Глава 8. Ихменчивость и эволюция Рис. 168. Постфлоральное «лихора- дочное~ необеспеченное побегоот- растание у особей Foeniculum vulgare (I) и Lycopus europaeus L.(II) (Изра- иль, пойма небольшой речки в окре- стностях Нетании). Рис. 169. Сверходнолетнее пове- дение особи Conyza bonariensis (L.) Cronquist, образующей пост- флоральные побеги (произраста- ет там же, что и особи, изобра- женные на рис. 160).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ Ph СТЕНИЙ Рис. 170. Некоторое разнообразие биоморф с постфлоральным «лихо- радочным» побегообразованием: I — Scrophularia xanthoglossa Boiss., II — Heliotropium rotundifolium Lehm, III — Polygonum equisetiforme Sm., IV — Dianthus polyclados Boiss. (произрастают там же, что и особи, изображенные на рис. 160). 51 — проросток (модельный экземпляр), 52 — сильнорослые годичные или двух- годичные побеги, 53 — неполностью отмершая разветвленная верхушка открыто- го вегетативно-репродуктивного прошлогоднего побега, 54 — постфлоральные живые побеги, развитые из сохранившейся субтерминальной зоны прошлогоднего открытого соцветия; остальные обозначения те же, что и иа рис. 160-165.
Глава 8. Изменчивость и эволюция 1,,б t5 I ] I/ / I l I ( ~ ~г 3 ! Рис. 171. Упрощенные модели злаковоцветковых кустарников (1) и злаковоцветковых кустарничков (П). 1 — отмершие надземные части прошлогодних побегов, 2 — живые части побегов прошлых лет, 3 — отрастание (март — апрель) весенних побегов текущего года, 4 — отмершие части прошлогодних подземных органов; горизонтальная линия— поверхность почвы. бурачниковоцветковые и т.п. деревья, кустарники, многолетние травы и т.д. В более общих случаях для обозначения биоморф могут быть использованы названия и более высоких таксонов, например, как сде- лали это Варминг и Гребнер («однодольные кустарники»). Среди злаков с зимующими соломинами считаем уместным различать мятликовоц- ветковые кустарники и мятликовоцветковые кустарнички. Не исключе- но, что есть и мятликовоцветковые полукустарники, но автор их пока не обнаружил. Мятликовоцветковые (злаковоцветковые) кустарники образуют ор- тотропные надземные многолетние соломины (аэроподии), которые до- вольно высоко (выше 1 м) поднимают крону их куста, например, Arundo donax L. (рис. 171-1). Древесные и кустарниковые безрозеточные формы в подсемействе Bambusoideae предлагают различать Ф.Мак Клюр (МсС1иге, 1966), Т.И. Серебрякова (1971) и др. Митликовоцветковые (зла- ковоцветковые) кустарнички формируют многолетние соломины (аэро- подии), которые невысоко (до 1 м) поднимаются над поверхностью почвы, располагаясь в пространстве, как правило, наклонно или даже плагиотропно, например, Piptatherum miliaceum (L.) Савв., Paspalidium geminatum (Forssk.) Stapf и др. (рис. 171-11).
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ В северных регионах Евразии, где зимы холодные, надземные со- ломины местных многолетних злаков после окончания вегетационного сезона отмирают полностью. Здесь многолетние побеговые органы ме- таморфизированы для ползучего подземного или реже приземного су- ществования. В этих областях нет злаковоцветковых кустарников и кустарничков. Структурная организация биоморф семенных растений в Среди- земноморье еще ждет своих исследователей. Нельзя не восхищаться поведением некоторых растений, которым, безусловно, безразлично, как мы их назовем. Они просто живут и используют, где это целесооб- разно для своего выживания, одновременно приспособления как трав, так и полудревесных или древесных растений. Oxalis pcs-сатрае L. — кислица ногонлодная (сем. Oxalidaceae). Встре- чается на опушках низкорослых кустарниковых вечнозеленых зарослей (гариги), у заборов на рыхлых каменисто-песчаных почвах. На открытых местах особи этого вида образуют вегетативные или вегетативно-репро- дуктивные розеточные или верхнерозеточные побеги с подземным раз- двинутоузловым основанием. Вегетативные побеги — это только ста- дия в развитии вегетативно-репродуктивных открытых побегов, которые образуют или одиночные пазушные интеркалярные побеги, или, чаще, межсоцветия из 2-3 и более интеркалярных побегов (рис. 172). На под- земных частях таких побегов развиваются пазушные выводковые почки, которые легко отделяются от родительского растения; посредством них осуществляется вегетативное размножение. В тех случаях, когда особи кислицы ногоплодной растут непосредственно около кустарников или кустарничков, например Ephedra aphylla, Calicotome villosa, Sarcopoteri- ит spinosum (L.) Spach и др., их поведение изменяется; они уже не образуют розетки листьев у поверхности почвы, а надземно (!) вынуж- денно удлиняются и развивают надземные вынужденно-верхнерозеточ- ные побеги. Удлиненные части таких побегов пробираются между вет- вями других растений в поникшую часть кроны кустарничков и кустарников, поднимая верхушечную розетку листьев на 10-40см над поверхностью почвы. Здесь она находит подходящие условия (опору, свет и т.п.) для развития соцветия. На верхушках открытых вегетативно- репродуктивных верхнерозеточных надземных частей побегов развива- ются межсоцветия из нескольких интеркалярных побегов с крупными желтыми цветками (рис. 172). Вынужденно удлиненный надземный рост верхнерозеточные побеги чаще проявляют зимой и рано весной, поэто- му в целом такой рост побегов, обычно развивающих приземные розетки листьев, надо признать как явно не тривиальное событие в травянистом мире. Биоморфотип Oxalis pes-carpaea — облигатно приземно розетко- образующие и вынужденно надземно верхнерозеточные открыто вегета- тивно-репродуктивные подземно и надземно выводково-почечные жел- то-кислицецветковые травы. Ricinus communis L. — клещевина обыкновенная (сем. Euphorbiaceae) (рис. 173). В культуре в умеренных широтах клещевина представлена однолетними «травянистыми» растениями. Как отметил П.М.Жуков- ский (1964), особи этого вида, нормально цветущие и плодоносящие
Глава 8. Изменчивость и эволюция Рис. 172. Развитие побегов Oxalis pcs-саграе L. (произрастает там же, что и особи, изображенные на рис. 160). 1 — особь, развившаяся на поляне (каменисто-песчаная почва), Н — особь, раз- вившаяся под пологом поникшей ветви кустарника; 1 — пазушные подземные (») или надземные (16) почки, легко отделяющиеся от родительского растения, 2— почечная оболочка, 3 — отрастание побега из пазушной почки (после мартовского дождя), 4 — розеточная часть верхнерозеточных побегов с подземным основани- ем, 5 — розеточная часть верхнерозеточного побега с полунадземным (надземно- подземным) основанием, 6 — интеркалярный побег или основание такого побега, когда верхняя часть удалена, 7 — межсоцветие, 8 — схема верхней части верхнеро- зеточного побега, 9 — срединные листья или основания таких листьев, когда верхние части удалены, 10 — чешуевидные листья, 11 — случайная ветвь кустар- ника, использованная верхнерозеточным побегом в качестве опоры, 12 — повер- хность почвы; одно деление масштаба — 1 см.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Рис. 173. Развитие организации биоморф Ricinus communis L. (произрастают там же, что и особи, изображенные на рис. 160). 1 — семядоли, 2 — срединные листья или их основания, когда верхние части удалены, 3 — безрозеточный главный (медиальный) побег, 4 — угнетенная двулет- няя особь, 5 — граница осенне-зимнего прироста прошлого года и весеннего прироста текущего вегетационного сезона, 6 — схема полисимподиального нара- стания побегов в верхней прошлогодней части кроны взрослого растения, 7— сухие остатки прошлогоднего верхушечного соцветия, 8 — весенний прирост те- кущего года, 9 — верхушечное соцветие, 10 — пазушный временно вегетативный силлептический (индуктивный) побег замещения; горизонтальная линия — по- верхность почвы; одно деление масштаба — 1см.
Глава 8. Изменчивость и эволюция на первом году вегетации, зимой вымерзают. Иногда клещевину обык- новенную выращивают как многолетние травянистые растения; этот вид сильно полиморфный (Чиков, Лаптев, 1976). В Средиземноморье Ближнего Востока Ricinus communis произрастает дико. Автор наблюдал этот вид на песчаных почвах в холмистой местности. Здесь характерен биоморфотип однодомных невысоких (3-6 м) безрозеточных деревьев. Прорастание семян надземное. Прорастание происходит зимой или весной. Проростки крупные с хорошо развитым главным стержневым корнем. Семядоли проростков от овальной до яйцевидной формы, на верхушке округлые, иногда выемчатые (1,4-2,0 см дл., 0,6-1,4см шир,) с черешком 0,3-0,6см дл., гипокотиль 2-4см дл. и главный корень 3-6см дл. С началом развертывания зародышевой почки начинается ветвление главного корня. Первая пара срединных листьев супротивная. Далее расположение листьев становится очередным. У всходов размеры семядолей сильно увеличиваются, достигая иногда 5-6 см дл. и 2,5-3,5 см шир., черешки семядолей до 2,5-3,5 см дл. После образования 4-5 на- стоящего листа семядоли отмирают и моноподиально нарастающие особи переходят в ювенильное состояние. Растения быстро набирают вегетативную мощь, но многое зависит от времени и места прорастания. Растения, появившиеся на свет зимой, медленно моноподиально нарастают, что видно по сближенному расположению узлов (рис. 173), но с началом весны их рост заметно убыстряется. Под пологом кустов развитие может задерживаться и растения весь год будут оставаться в угнетенном виргинильном состоянии и даже не ветвятся. Взрослыми физиологически зрелыми особи клещевины становятся на первом году жизни, редко позднее. За один вегетационный сезон (год) они успевают от 2 до 6 раз образовать верхушечные соцветия, после чего происходит перевершинивание за счет силлептического от- растания индуктивного побега замещения (одного или нескольких). Почки открытые. Верхушечные кисти состоят из однополых безлепес- тных цветков, причем пестичные цветки расположены в верхней части соцветия, а тычиночные — в нижней. Известно (Жуковский, 1964, с. 301) шесть типов соцветий: «1) нормальный однодомный, когда пес- тичные цветки на верхней части оси соцветия составляют 30-50%, а тычиночные цветки на нижней части — 70-50%; 2) все цветки пестич- ные (99-100%); 3) женские и мужские цветки расположены на оси впе- ремежку; 4) нормальный однодомный, но женские цветки перемешаны с тычиночными; 5) наряду с однопалыми цветками попадаются обоепо- лые (полигамия); 6) почти все цветки тычиночные». Каждая шиповатая коробочка при созревании растрескивается и распадается на три гнезда с одним семенем в каждом гнезде. Семена крупные, оболочка их тол- стая глянцевая до 2 GM дл. и 1,5 см шир. Один побег, образующийся за один вегетационный сезон (годич- ный), достигает в длину 1-2 м; он полисимподиальный акронекрозный мультирепродуктивный. Такой побег состоит из нескольких внутри- годичных (силлептических, если еще точнее — индуктивных, побегов замещения, кроме первого побега, отрастающего из зародышевой почки семени или в последующие годы из перезимовавшей открытой почки)
ОСНОВБ1 ЬИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ вегетативно-репродуктивных побегов (рис. 173). С увеличением возраста размеры внутригодичных побегов и их число уменьшаются. В обследо- ванной нами местности растений старше десяти лет не обнаружено, но десятилетние растения уже были старыми. Вероятно, клещевина отли- чается непродолжительным жизненным циклом и в естественных усло- виях произрастания. Однако есть указания. что в тропиках и субтропи- ках дикие древовидные особи клещевины обыкновенной до 10 м высо- той (Жуковский, 1964). 8.2.3. Интродукционный стресс у растений Рассмотренные особенности формирования структурной орга- низации Ricinus communis свидетельствуют о мультивариантности био- морфогенезов в различных условиях произрастания: в естественных условиях произрастания проявляется «древесное многолетнее» направ- ление онтогенезов; в условиях культуры, как известно, развитие может идти или по многолетнему травянистому (с ежегодным отмиранием надземных частей побегов) типу онтогенезов, или — по однолетнему. Этот и подобные примеры заставляют посмотреть на проблему пластич- ности биоморф и с другой точки зрения. Разрабатывая проблему пластичности, мы не могли не выйти на концепцию стресса канадского биолога Г.Селье (1960, 1979), который показал, что у животных и человека стресс есть неспецифический ответ организма на любые предъявляемые ему требования. Растительный орга- низм, не имея нервной системы, по другим информационным каналам достаточно активно реагирует на воздействие стрессорных экстремаль- ных факторов, проявляя соответствующие реакции, чтобы сохранить свои главные жизненные параметры, например, тургор клеток, РН и т.п., обеспечивающие условия для ассимиляции и диссимиляции. В процессе жизнедеятельности биоморфы оказываются в различных эко- логических условиях и их развитие может отвечать норме (в этих усло- виях, как уже подчеркивалось, биоморфы будут относиться к основному биоморфотипу) или отклоняется от нее (биоморфы будут принадлежать второстепенным биоморфотипам). Нередко необходимых ресурсов не хватает или их слишком много и тогда биоморфы испытывают состо- яние стресса. В этом случае особого внимания заслуживает поведение растений, которые вводятся в культуру. Стресс у растений, перенесенных в условия выращивания, затраги- вает следующие показатели жизнедеятельности: темпы роста и развития биоморф, продолжительность некоторых стадий и всего жизненного цикла, характер и кратность цветения, темпы роста и развития органов (побегов, корней, соцветий, цветков и др.), реакцию габитуса (его раз- меры, ветвистость, кустистость), размножение. У некоторых интроду- цируемых растений необходимо особо выделить стрессовые состояния реиродуктивной «эйфории» (повышенной репродуктивной активности, когда цветение и плодоношение необычно обильны) или реиродуктив- ной «аиофории» (необычно пониженной репродуктивной активности или
Глава 8. Изменчивость и эволюция полной стерильности). Причин этих явлений много. Например, репро- дуктивная эйфория нередко наблюдается при перенесении фитоценоти- чески выносливых растений (например, патиентов) в более чем ком- фортные условия грядок (без сорняков и богатых необходимыми для них ресурсами), а репродуктивная апофория — при перенесении рас- тений, произрастающих в континентальном климате, в условия теплицы или в тропики: например, биоморфы Ча1епапа officinalis L., если не пройдуг яровизацию, не цветут или образуют лишь искаженные подобия соцветий. Переселение растений из естественных условий произрастания в культуру влечет за собой перестройку многих организационных призна- ков (синдром ответа) на одном из рубежей пластичности: убыстряются общие темпы роста и развития, выпадают из жизненного цикла или сокращают продолжительность некоторые возрастные стадии, появля- ется или усиливается вторичное цветение, увеличиваются размеры (вы- сота, диаметр), ветвистость (кустистость), изменяется форма и структура соцветий в сторону их укрупнения и увеличения числа элементов, ус- коряется наступление стадии размножения, возрастает плодовитость и, наконец, нередко проявляется репродуктивная эйфория. Общая про- должительность жизни биоморф в этих условиях сокращается. В таких случаях многолетники иногда становятся однолетниками, поликарпи- ки — монокарпиками (Foeniculum vulgare Mill., Негас1еит sibiricum L., рис. 203). Некоторые тропические и субтропические растения (напри- мер, Ricinus communis L.), как уже отмечалось выше, при выращивании в условиях умеренно континентального климата вместо древесного обра- за жизни (на родине) проявляют однолетний (на чужбине) и после ре- продуктивной эйфории остаются не подготовленными к зиме и погибают. В загущенных посевах (много конкурентов), где растениям-интро- дуцентам явно не хватает необходимых ресурсов, можно наблюдать, как биоморфы также испытывают стресс и в полной мере проявляют свою пластичность, изменяясь по многим признакам. Таким образом, при переселении из естественных местообитаний растения могут находить (или не находить) необходимые условия для сво- его роста и развития и в случае выживания в новом биотопе трансфор- мируют организацию, степень адаптированности которой будет в меру возможностей (на основе наследственности) отвечать новым условиям существования — это закон (специальный) поведения биоморф растений нри переселении их в культуру (подробнее см. Аврорин, 1959; Головкин, 1973, Некрасов, 1980 и др.). Интродукционные стрессы нередко приводят к мутациям биоморф. Многие формы стрессов, которые наблюдаются при интродукции, воз- можны в принципе и в естественных условиях произрастания. При стрессах у биоморф растений определенного вида эволюция идет с наи- большими отклонениями от нормы, т.е. от основного биоморфотипа вида. На примере душицы обыкновенной — Origanum vulgare L. (сем. Lamiaceae) рассмотрим, как трансформируется организация биоморф этого вида при переселении их в культуру. Душица обыкновенная произрастает в Европе, на Кавказе, в Запад- ной Сибири и некоторых горных районах Средней Азии (Копанева,
ОСНОВБ1 БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Шретер, 1976). Это лесостепной вид, приуроченный к суходольным пойменным, лесным, иногда к степным лугам,' нередко встречается в разреженных хвойных и березовых лесах. Особенно обильна душица в Сибири в злаково-разнотравных луговых ассоциациях (Куминова, 1969; Копанева, Шретер, 1976). Она хорошо растет при выращивании в открытом грунте (Балабас и др., 1965; Илиева, 1971). Изучение биомор- фогении душицы обыкновенной в условиях выращивания проведено в районе Москвы (ВИЛАР, Нухимовский, Черкасов, 1987). Сначала оха- рактеризуем в самых общих чертах биоморфологические особенности этого вида в естественных условиях произрастания на Аптае. Для душицы обыкновенной характерен широкий полиморфизм. В частности, на Алтае мы отметили два типа биоморфогенезов, которые наиболее контрастно проявляются в разных условиях произрастания. Короткокорневищный рыхлокустистый тип (луговой) является ос- новным в пределах Алтая (рис. 174-I). Большой встречаемости достигает душица в злаково-разнотравных долинных лугах в Усть-Коксинском районе, где в фазу цветения местами дает массовый аспект. Длиннокорневищный полуразвернуто-кустистый тип (лесной) био- морфогенезов встречен в условиях соснового леса на песчаных почвах в окрестностях г. Бийска (рис. 174-П). Рис. 174. Два типа биоморфогенезов Origanum vulgare L. в естественных условиях произрастания (Алтай). 1 — луговой тип, II — лесной тип (объяснения в тексте).
Глава 8. Изменчивость и эволюция На первом году вегетации образуются особи с одним медиальным невысоким побегом высотой 3,0-8,0 см. Отмирание семядолей в конце мая-июне свидетельствует о превращении всходов в ювенильные растения. Кущение еще не началось. Главный побег полегает и на сле- дующий год или продолжает моноподиальное нарастание, образуя над- земный удлиненный вегетативный, реже вегетативно-репродуктивный побег, или чаще верхняя надземная вегетативная часть такого побега после первого года вегетации погибает(зимой), а сохраняется только короткое зимующее основание с пазушными почками. Некоторые из этих почек у растений первого года жизни иногда пробуждаются во второй половине лета или осенью, формируя короткие (0,5-3 см) веге- тативные побеги. На следующий год весенние побеги отрастают симпо- диально, замещая отмершую верхушку главного побега, или реже моно- подиально, если сохраняется верхушка короткого родительского побега. Начало кущения — признак вступления особей в имматурное состояние. Первое цветение, а следовательно, и переход в матурный этап жиз- ни наблюдается у душицы в естественных условиях произрастания (до- линные луга) на 2-5, чаще 3-4 годах жизни. В луговых ассоциаций много проростков и всходов душицы погибает, не выдерживая конку- ренции. После начала цветения, когда отдельные побеги достигают верх- него яруса лугового травостоя, начинается активное побегообразование и разрастание корневищной части растений. Главная особенность рас- сматриваемого типа биоморфогенезов (лугового) — формирование ко- роткого разветвленного корневища. Основание взрослых растений луго- вого типа обычно до 20 см, чаще меньше. Побеговую систему матурных особей составляют унирепродуктивные двухгодичные и вегетативные годичные побеги. Двухгодичный пазушный побег проходит в своем раз- витии инициальный период (внутрипочечную фазу) и постинициаль- ный период — фазу вегетативного озимого побега (1-15 см дл.), на вто- ром году жизни — фазу вегетативно-репродуктивного годичного побега (высота 30-60 см, иногда до 90 см). Годичные побеги, которые отрастают из перезимовавших почек, а не из озимых побегов, как правило, веге- тативные. Они обычно отмирают зимой полностью, поскольку не успе- вают укорениться. Эпигеогенное (погружающееся) корневище формируется в основ- ном из многолетних коротких (0,5-3 см дл., реже до 10 см) полегающих зимующих оснований двухгодичных или озимых резидных побегов. Укоренившиеся резидные побеги обеспечивают симподиальный прирост корневища. Итак, побеговая система матурных особей состоит из двух типов побегов: весенних длинных вегетативно-репродуктивных (реже вегетативных) и озимых коротких вегетативных. Соцветие сильно раз- ветвленное полителического типа. Цветки многочисленные, мелкие, обоеполые или реже функционально пестичные. Плод состоит из четы- рех темно-коричневых орешков. Цветение начинается с июня и продол- жается до августа. К пятому году жизни формируется хорошо развитая особь, пред- ставляющая собой рыхлый куст, в структуре которого ежегодно сменя- ются надземные части побегов, а сохраняется разветвленное подземное
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛСТЕНИЙ и полуподземное эпигеогенное короткое корневище с относительно большим числом придаточных корней. Число надземных побегов у взрослых растений от 3 до 20. Длиннокорневищный полуразвернуто-кустистый тип биоморфоге- незов (лесной тип) душицы нами наблюдался на небольших полянах в сосновом лесу, иногда под пологом деревьев. Побеги в этих условиях малочисленные (1-4, чаще 2-3), высотой 40-90см. Междоузлия длин- ные, соцветие небольшое, слаборазветвленное. Цветение в условиях леса начиналось впервые на втором-шестом году жизни. Короткие (до 20 см) вегетативные побеги, образующиеся весной, в конце лета и осенью, располагались в моховом покрове и лесной подстилке. У них полегала нередко даже верхняя часть; они легко укоренялись и частично или полностью зимовали. На следующий год при благоприятных условиях озимые побеги развивали соцветия и цвели. Корневище увеличивается за счет погружающихся многолетних частей побегов разной длины (до 20 см). Из них формируются довольно длинные эпигеогенные корневи- ща (20-70 см, чаще 30-50 см). Эти корневища слабо разветвлены или долго не ветвятся, нарастая эусимподиально с ежегодным переверши- ниванием и гемисимподиально с неежегодным (после двух лет) перевер- шиниванием. Годичные резидные побеги сохранялись в структуре кор- невища от 5 до 12 лет, чаще 6-8 лет. Два типа биоморфогенезов — луговой и лесной, кратко охаракте- ризованные нами для естественных условий произрастания на Алтае, хорошо отличаются друг от друга и свидетельствуют о большой пластич- ности душицы. Г.М.Зозулин (1959) в условиях лесостепи Центрально- Черноземного госзаповедника (Курская область) описал душицу обык- новенную в группе «короткокорневищных ветвящихся» растений, обра- тив внимание на то, что в подземной части особей развивается короткое корневище с мочками придаточных корней и стержневой корень, ухо- дящий в глубину до 80сМ. В.Н.Голубев (1959), изучая эволюционное развитие длиннокорневищной структуры, привел душицу в качестве примера, иллюстрирующего направление трансформации от первично стержнекорневой к строго корневищной структуре. Особенности динамической морфологии душицы обыкновенной при выращивании в Московской области охарактеризованы ниже. При описании биоморфогении душицы используем разработанный методи- ческий подход (гл. 7.3). При характеристике стадий биоморфогенезов- периодов, этапов, фаз, гипофаз — сначала указываем их продолжитель- ность, а затем даем основную морфофизиологическую характеристику биоморф. Для дополнительной характеристики биоморф применяем следующие условные обозначения: А — частная (распространяется на рассматриваемое возрастное состояние), Б — общая (распространяется на несколько или на все возрастные состояния). Эмбриональный иериод. Пренатальный этан. С июля по август или сентябрь; конец августа-ноябрь — при вторичном цветении. Оплодо- творенная семяпочка или зрелое семя в структуре родительского рас- тения. При вторичном цветении, которое отмечено только в теплую осень, оплодотворенная семяпочка не дозревала и, как правило, второго плодоношения не наступало.
Глава 8. Изменчивость и эволюция Латентный этан. С начала августа или сентября до апреля следующе- го года, а при хранении семян — в течение нескольких лет. Зрелое семя (орешек), обособленное от родительского растения. А — орешки округло-яйцевидные, коричневые, с гладкой поверх- ностью, 0,8 мм дл. и 0,54мм шир. Б — четырехорешковый дробный плод развился под прикрытием чашечки, вход в которую после созревания плода закрыт изнугри пуч- ком густых волосков. Длина их превосходит в 5-10 раз длину мелких волосков, развитых на наружной поверхности чашечки. Этот пучок во- лосков может долго задерживать выход орешков из чашечки, поэтому иногда часть орешков до весны может задерживаться на верхушке сухих побегов. При подзимнем (осеннем) посеве семян (сентябрь-ноябрь) осу- ществлялась естественная стратификация, которая незначительно улуч- шала их всхожесть. Эииэмбриональный иериод. Виргинильный этан. Преювенильная фаза. Гииофаза ироростка (рис. 175, А). Семена как при подзимнем (октябрь- ноябрь) посеве, так и при весеннем (конец апреля-начало мая) дружно прорастали; при весеннем (апрель) посеве проростки начинали по- являться на 8-25-й, чаще 10-18-й день. Состояние проростка продолжа- лось 8-12 дней. Прорастающее семя, или семядольный почкокорень,— ризопротерокормус (рис. 175,А,11). Прорастание семян сильно лимитируется условиями увлажнения. Проростки появляются в течение всего лета, как правило, после дождей. А — семядоли (две) надземные, зеленые, округлые, на верхушке усеченные с выемкой, при основании угловатые, слегка сердцевидные. Гипокотиль голый или чаще опушенный, как отметил Н.И.Иосебидзе (1981), мелкими одноклеточными серповидными волосками, которые встречаются и на всех остальных частях растения выше семядольного узла. Непосредственно после развертывания семядоли достигали 1,2-2,0 мм дл. и шир., черешки — 1,2-1,5 мм дл., главный простой корень (главная монориза, субмедиаль) — 0,5-1,2м дл. Перед началом развертывания зародышевой почки пластинки семядолей увеличиваются до 2-3 мм дл. и шир., черешки — до 2-4 мм дл., гипокотиль — до 4-12 мм дл., глав- ный корень — до 3 см. Гииофаза всхода (рис. 175, Б). С мая до июня, реже июля (20-45 дней). Семядольный вегечативный безрозеточный ризомонокормус (рис. 175, Б, 12). А — первые листья простые, черешковые, яйцевидной формы, супротивные, цельнокрайние, 0,2-0,6 см дл., 0,2-0,5 см шир., опушены мелкими простыми волосками. Черешки первых листьев 0,1-0,3 см дл. Эпикотиль до 0,5 см дл. Пластинки семядолей увеличились в размерах: 2-4 мм дл., 2,5-4,5 мм шир. Линейные размеры последующих пар насто- ящих листьев увеличиваются. Высота медиального (первоначального) побега варьировала. Среднерослые (3-4 см выс.) растения имели наибо- лее крупные листья и по другим показателям (диаметру побега, степени разветвленности корня, размерам семядолей) они так же превосходили более рослые (5-10 см выс.), но слабые растения, развившиеся при более загущенном стоянии растений в рядках. Гипокотиль 3-8 мм дл. Главный корень разветвленный (дириза или тетрариза), до 5 см дл.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 175. Виргинильные особи Origanum vulgare, произрастающие в культуре (Mocha, ВИЛАР, интродукционный питомник). А — проростки, Б — всходы, В — ювенильные особи, Г — имматурные особи. 1— семядоли, 2 — гипокотиль, 3 — семядольный узел, 4 — корневая шейка, 5а-5д- корни: 5a — главная монориза (субмедиаль), 56 — главная дириза, 5в — неглавная монориза (сублатераль), Sr — тририза, 5д — главная тетрариза, 6 — ризогенная корневая ветвь, 7 — адвентивная (придаточная) корневая ветвь, 8 — вегетативный медиальный монокормус, 9 — вегетативный медиальный дикормус, 10 — силлеп- тические (индуктивные) побеги, 11 — ризопротерокормус (семядольный почкоко- рень), 12 — семядольный ризомонокормус, 13 — бессемядольный вегетативный ризомонокормус, 14 — бессемядольный вегетативный ризодикормус. Горизон- тальная линия — поверхность почвы. Одно деление масштаба — 1 см. МБ— упрощенные модели биоморф. ув — увеличено в несколько раз по сравнению с масштабом.
Глава 8. Изменчивость и эволюция Ювенильная фаза (рис. 175, В). С июня (реже с июля) до июля- августа (20-60 дней) или в условиях сильного угнетения до весны сле- дующего года (до 300 дней). Бессемядольный вегетативный безрозеточ- ный ризомонокормус (рис. 175, В, 1з). А — семядоли отмерли (индикаторный признак); продолжается моноподиальное нарастание медиального побега. Размеры листьев уве- личились: например, у третьей пары пластинок 0,8-1,7 см дл. и 0,6-1,2 см шир., черешки — 0,4-1,2 см дл. Усилилась опущенность листьев, особен- но нижней поверхности. Высота медиального побега до 10 см, хотя есть и мелкие, иногда очень угнетенные из-за загущенности особи (2-3 см выс.). Главный корень до 8 см дл., разветвленный (тетрариза). На гипокотиле, реже на семядольном узле или на эпикотиле образовались первые придаточные корни (рис. 175, В, 7). Имматурная фаза (рис. 175, Г). Первая (уншмматурная) гииофаза. С июля до середины августа-сентября или до весны следующего года. Бес- семядольный вегетативный безрозеточный ризодикормус (рис. 175, Г, 14). А — начало ветвления медиального побега, которое в благоприят- ных условиях выращивания осуществлялось по силлептическому типу. Это признак вступления особи в первое имматурное состояние. Силлеп- тическое пазушное побегоотрастание наблюдалось обычно по всей дли- не медиального побега (рис. 175, Г, lo). Оно усиливало ассимиляцион- ную активность растений и способствовало более быстрому достижению их физиологической зрелости. Поскольку медиальный побег постоянно растет в течение всего вегетационного сезона, замедляя или ускоряя темп роста в зависимости от метеоусловий, то он пока монофазный (с одной фазой видимого роста), а развивающиеся одновременно с ним пазушные побеги — это силлептические, или, еще точнее их разновид- ность, — индуктивные, побеги. Пазушные побеги, развивающиеся в ос- новании медиального побега (базальные силлептические, или базисил- лептические — разновидность индуктивных побегов), часто несколько мощнее остальных индуктивных вегетативных побегов, отрастающих в верхней и средней его частях. Слабые силлептические побеги, нередко в конце лета или в начале осени отмирали вместе со срединными ли- стьями этого побега. Монофазный силлептически разветвленный, меди- альный (включающий медиаль, т.е. стебель первого порядка в системе нарастания) побег — это побег второго уровня сложности организации по нашей основной иерархической (субординатной) классификации по- бегов, т.е. дикормус, который может весь сезон развиваться вегетативно (рис. 175, Г, 9). Начало силлептического побегообразования у растений весеннего посева приходилось на 90-110-й день после начала прораста- ния (чаще в начале августа), а у растений осеннего посева наблюдалось на 5-10 дней раньше. Высота вегетативных дикормусов 10-25 см, в сред- нем около 20 см. Часть растений зимовала в униимматурном состоянии. Их дикормусы частично или полностью полегали, укореняясь в осно- вании, а верхушки зимой обычно обмерзали на разную длину, хотя иногда у небольших полегших побегов возможно сохранение под сне- гом и верхушечной меристемы. Главный корень (тетрариза) хорошо развит (до 15 см дл.). Придаточные корни на нижней полегшей части медиального побега малочисленны или обильны.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ Вторая (биимматурная) гииофаза (редко). С весны второго года жизни до весны третьего года. Вегетативный безрозеточный ризогексакормус. А — это состояние проявлялось только у угнетенных растений, у которых из перезимовавших почек на второй год эпиэмбриональной жизни развивались лишь короткие и среднерослые вегетативные побе- ги. Вся система родительского побега, сохранившегося после зимовки, и его производных — нескольких весенних вегетативных побегов — мо- жет рассматриваться как один побег шестого уровня сложности органи- зации, или гексакормус, вынужденно вегетативный из-за угнетения при чрезмерно загущенном посеве. Матурный этан. Инициматурная фаза. Первая гииофаза (рис. 176, Д,). С августа, реже сентября первого года жизни до начала послезимнего (весеннего) отрастания побегов (300-330 дней). Вегетативно-репродуктив- ный (физиологически зрелый) безрозеточный ризодикормус (рис. 178, Д„1s) или его зимующий приземный остаток разной сложности, сохранившийся после зимовки, персистентный ризодикормус или ризомонокормус. А — на первом году жизни и при благоприятных для роста и разви- тия условиях (теплый август и (или) сентябрь, как, например, в 1985 г.) многие растения, развившиеся из семян прошлогоднего подзимнего по- сева, и некоторые сильные растения, развившиеся из семян весеннего посева текущего года, образовывали соцветия, т.е. достигали физиоло- гической зрелости. В зависимости от метеоусловий (особенно в конце лета и осенью) число впервые зацветших растений сильно колебалось. Так, например, в 1985 г. впервые зацвело 80 % растений осеннего посева и 45 % растений весеннего посева. В предшествующие 6 лет наблюде- ний впервые в первый год жизни зацветало меньше растений (не более 60%). От начала прорастания семян до начала первого цветения про- ходило 90-115 дней. Цветение продолжалось 65-75 дней; плодоношение наступало через 45-60 дней после начала цветения, массовое созревание Рис. 176. Матурные особи Origanum vulgare на первом-третьем годах вегетации (произрастают там же, что и особи, изображенные на рис. 175). Д, — первая инициматурная, Д, — вторая инициматурная, Е — медиматурная особи. К— схема роста и развития корнепобегового черенка: К, — имматурный клонист, К,— инициматурный клонист, К, — медиматурный клонист; 15 — вегетативно-репродуктив- ный ризодикормус, 16 — репродуктивный ризогексакормус с дифазным (двухгодичным, апонекрозным, т.е. неотмирающим, после первого сезона вегетации) медиальным по- бегом, 17 — репродуктивный ризогексакормус с монофазным (одногодичным, акронек- розным, т.е. отмирающим сверху, после завершения сезона вегетации) медиальным побегом, 18 — репродуктивный ризогептакормус, 19 — зона соцветия, 20 — зона тормо- жения, 21 — подзона неполного торможения, 22 — подзона вторичного полного тормо- жения, 23 — зона базальной индукции (базисиллепсиса), 24-27 — силлептические побе- ги: 24 — эусиллептические, 25 — инфантильно-вегетативные, 26 — инфантильно-репро- дуктивные, 27 — базисиллептические, 28 — сухие остатки побегов, 29 — вегетативно- репродуктивный дикормус, 30 — вегетативный пазушный дикормус, 31 — среднерослые озимые побеги, 32 — короткие озимые побеги, 33 — граница моноподиального перехода между побегами прошлого и текущего года, 34 — схема акропетального цветения пар- циальной кисти, 35 — цветок, 36 — чашечка с плодом, 37 — орешек, 38 — живые осно- вания побегов (верхние части удалены), 39 — символ удаленной репродуктивной части побегов. 1л, 2л, Зл... — особи 1,2,3 и т.д. лет жизни. Остальные обозначения те же, что и на рис. 175.
Глава 8. Изменчивость и эволюция
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ плодов — в первой декаде сентября. Плодоношением не заканчивалась вегетация растений: продолжался рост и развитие базисиллептических побегов, которые становились затем озимыми. Отрастание приземных базисиллептических побегов — побегов кущения первого порядка— признак формирования простого куста. Верхняя часть дикормуса после завершения постинициального развития отмирала, а зимующий призем- ный остаток его (персистентный побег), будучи укорененным, начинал выполнять роль базового, или многолетнего, побегового органа — ко- роткого корневища, достигающего 2-10 см дл. Б — простейшими пер- систентными (зимующими) одноосными побегами, обеспечивающими основной (гемисимподиальный) или дополнительный (эусимподиаль- ный) прирост корневища, являются репторезидные тетракормусы (с дву- мя прошлыми фазами видимого роста) и монокормусы. Это ползучие приземные или плагиотропные подземные основания вегетативных и вегетативно-репродуктивных побегов, которые после отмирания над- земных частей побегов нормально перезимовывают. Вторая инициматурная гииофаза (рис. 176, Д,). С весны второго до весны третьего года вегетации. Вегетативно-репродуктивный безрозе- точный рыхлокустистый ризогексакормус (рис. 176, Д„16, i7). А — в весеннем (со второй половины апреля) отрастании побега нового — шестого уровня сложности организации, или гексакормуса, принимают участие меристемы следующих побегов: алексы пазушных озимых вегетативных побегов в случае нормальной их перезимовки под снегом; алекс медиального вегетативного побега, который иногда сох- ранялся из-за удачного для перезимовки полегания и укоренения этого побега; алексы сохранившихся после перезимовки открытых пазушных почек. Все они вместе с перезимовавшими побеговыми частями состав- ляют зону возобновления (инновации) гексакормуса. Гексакормус- сложный побег, включающий зону возобновления, т.е. перезимовавшую часть побега, выжившую к весне после возрастных превращений струк- туры медиального монокормуса или дикормуса, и все новые побеги текущего сезона вегетации, развившиеся из этой зоны. Разнообразие весенних побегов, образующихся у гексакормуса на втором году вегета- ции (рис. 176,Д,), следующее: вегетативно-репродуктивные годичные безрозеточные пазушные (образующиеся из апексов пазушных озимых побегов) и терминальный если алекс медиали зимой не отмирает) ди- кормусы, вегетативные годичные безрозеточные монокормусы (нераз- ветвленные) и дикормусы (если есть силлептическое ветвление). У гек- сакормусов нередко могут развиваться и двухгодичные пентакормусы- побеги пятого уровня сложности организации. Основание их — корот- кий неразветвленный перезимовавший пазушный побег, образовавший- ся в прошлом году, а остальная его часть — весенний, отрастающий из алекса озимого побега, удлиненный вегетативно-репродуктивный (цве- тоносный) или реже вегетативный дикормус. Высота цветоносных ди- кормусов на втором году вегетации 25-40 см, число их 1-5, чаще 2-3. Б — вегетативные побеги двух типов: а) среднерослые (10-25 см выс.), образующиеся при развертывании перезимовавших пазушных открытых почек озимых побегов; б) низкорослые (до 10 см выс.), чаще всего сил-
Глава 8. Изменчивость и эволюция лептические, но иногда (в конце лета или осенью) формирующиеся провентивно из немногочисленных запасных (провентивных) почек или пролептически (из почек, сформированных в текущем вегетационном сезоне, но преждевременно пробудившихся осенью после некоторого заметного покоя). Среднерослые побеги при благоприятных условиях (теплый август или сентябрь) переходили к образованию соцветий. Та- ким образом, в отдельные годы наблюдалось вторичное цветение особей (рис. 177, 41). Многие среднерослые побеги зимой отмирали полностью или перезимовывали их укоренившиеся основания. В структуре вегета- тивно-репродуктивного дикормуса обычно различимы [по терминоло- гии W.Troll (1964) с нашими дополнениями] следующие части (сверху вниз; рис. 176, Д„19-23): флоральная зона, или зона комплексного соц- ветия; субфлоральная зона, или зона торможения (релаксации). После- дняя включает две подзоны (сверху вниз). Первая — это подзона непол- ного торможения, образующаяся в случае развития ослабленных вегетатив- ных силлептических, или инфантильно-вегетативных, побегов, некоторые из которых могут доразвиться, образовать слаборазвитое соцветие и пре- вратиться в инфантильно-репродуктивные побеги (рис. 176, Д,,25, 26). Раз- виваясь в зоне торможения, инфальтильно-репродуктивные побеги могут быть отдалены от паракладиев (облигатных элементов) комплек- сного соцветия участком из инфальтильно-вегетативных побегов, по- этому они являются не более как провентивными (запасными) элемен- тами этого соцветия. Вторая подзона — полного (иногда вторичного после отмирания инфантильных побегов) торможения, но она может и отсутствовать. Ниже находится зона индукции, представленная базальными силлепти- ческими побегами (разновидностью индуктивных побегов), превраща- ющаяся после перезимовки в зону возобновления уже более сложного побега (гексакормуса). Базисиллептические побеги развиваются одно- временно с другими силлептическими побегами, но в отличие от них растут все лето и большую часть осени, образуя нередко новые силлеп- тические побеги второго порядка. Побеги, развитые в зоне индукции, хорошо укореняются и, не отмирая, зимуют, или у них отмирает вер- хушка. Важное значение в организации биоморф душицы имеют базаль- ные укоренившиеся участки побегов. Они более всего предрасположены к многолетности и после отмирания их надземных прямостоячих удли- ненных частей обеспечивают гемисимподиальный прирост эпигеоген- ного (возникающего в основном приземно, но впоследствии погружаю- щегося, в нашем случае контрактильно, в почву) корневища. Кроме того, эти укоренившиеся участки побегов способны к вегетативному размножению, поэтому их можно использовать для черенкования. В наших опытах приживаемость корневищных черенков душицы была высокой (рис. 176, К). Корневище в первые годы жизни особей гетеро- номное (т.е. связанное с главным корнем), с большим числом придаточ- ных корней. Отдельные корни синторизы на втором году жизни особей достигали 30 см дл. Медимитурная фаза (рис. 176, Е и 177, Е). С весны третьего года вегетации до 8 лет и более. Ежегодно вегетативно-репродуктивный без-
Глава 8. Изменчивость и эволюция розеточный рыхлокустистый гетерономно-ризомный ризогептакормус (рис. 176, Е, 18), или ризоэнатокормус (рис. 177, Е, 47). А — побег седьмого уровня сложности организации, или гептакор- мус, развивается из перезимовавшего основания гексакормуса, которое весной составляет зону возобновления этого нового для структуры осо- бей душицы побега. Из открытых почек (верхушечных и пазушных) этой зоны гептакормуса развивалось много новых побегов текущего года — пазушных монокормусов (рис. 176, Е, 31,32) и дикормусов (рис. 176, Е, 29), повторяющих структуру описанных выше годичных по- бегов. Развитие побегов меньшей сложности в составе побегов большего уровня сложности обеспечивает постоянство иерархического процесса усложнения общей организации биоморф на стадии их прогрессивного развития. На четвертом году жизни, а в случае сильного угнетения и позже, формировались еще более сложные по своей организации побе- ги. Всю побеговую часть растения в этом возрасте составлял побег девятого уровня сложности организаци, или энатокормус, который образован побегом-предветвью, сформировавшимся ранее в структуре гептакормуса, и побегами-ветвями восьмого уровня сложности органи- зации, или октокормусами, не тождественными любым другим побегам предшествующих уровней сложности. Б — энатокормус определяет структуру растений семенного проис- хождения последующих лет жизни. Высота цветущих надземных побе- гов 50-80, чаще 60-65 см. Число цветоносных побегов в кусте на пятом году жизни растений колеблется от 8 до 40. Простейшими структурными элементами побегов самой высокой сложности организации (девятой) по-прежнему остаются годичные (постлатентные) и внутригодичные (силлептические) монокормусы и дикормусы, которые мы уже описали выше. В структуре дикормусов особое место принадлежит флоральной зоне — комплексному верхушечному соцветию, ветвление которого происходит за счет образования специализированных силлептических побегов — эусиллептических побегов (рис. 177, 24). Структуру его опре- деляет главное соцветие, развитое на верхушке исходного побега дикор- муса, и боковые соцветия, или присоцветия (кофлоресценции), раз- витые на верхушках производных побегов — паракладиев (эусиллепти- ческих боковых побегов, структурно подобных верхней части исходного побега дикормуса). В пазухах перекрестно-супротивных небольших зе- леных листьев или чаще в пазухах брактеозных листьев развиваются парциальные соцветия — компактные KHGTH. Обычно их неточно назы- вают колосками, но в отличие от колосков в парциальных кистях души- цы каждый пазушный цветок имеет короткую цветоножку (рис. 177, 3s), которая особенно хорошо выражена в фазу плодоношения (в колосках, как известно, цветки сидячие, без цветоножек). Все кисти у душицы (верхушечные и пазушные), по терминологии W.Troll (1964), надо счи- тать открытыми, поскольку они не заканчиваются верхушечным цвет- ком, а их алекс засыхает в фазу плодоношения вместе с прикрывающи- ми его зачатками нескольких пар новый брактей (рис. 177, 45). Учиты- вая, что верхушка любой парциальной кисти душицы брактеозная и что возможности ее апикального роста ограничены рамками соответствую-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ щего соцветия, ранее нами было предложено (Нухимовский, 1978) на- зывать такие соцветия условно закрытыми, или полузакрытыми, и от- личать их от собственно открытых соцветий, верхушечный рост главной оси которых не прекращается после выхода ее за пределы таких соцве- тий, как, например, у Callistemon speciosum (Sims) DC.. Тго11(1964), говоря о тирсоидной природе комплексных соцветий душицы, отнес их к кистевидным тирсам. Однако, система множественных парциальных кистей не имеет истинной цимоидной структуры, которая, по мнению этого автора, является обязательной для тирсов. Можно, конечно, образ- но приравнять отдельную парциальную кисть к одному цветку и полу- чить на схеме «цимоиды» из парциальных кистей, но из условных ци- моидов может состоять только нечто условное, допустим «псевдотирс». При определении комплексного соцветия душицы мы стремились избе- жать диагностики этих систем с неудобной ориентацией на «псевдо». Как отметили Ал.А. Федоров и 3.Т.Артюшенко (1979), в отличие от тирса (с цимоидными парциальными соцветиями) метелка имеет ботриоидные парциальные соцветия, а сложная кисть — иной характер ветвления. Согласно определению, данному этими авторами для сложной кисти как соцветия, у которого ботриоидные парциальные соцветия-кисти располагаются на боковых осях первого, второго и более высоких по- рядков, мы диагностируем комплексное соцветие душицы как полите- лическую множественную кисть (плейоботрий). Сроки фенофаз душицы при выращивании в Московской обл. сле- дующие: начало отрастания — вторая половина апреля, редко начало мая; бутонизация — июнь; начало цветения — конец июня — первая половина июля; начало созревания плодов — август; полное созревание плодов — сентябрь; вторичное цветение (при благоприятных услови- ях) — сентябрь-начало октября (до заморозков). У душицы, выращи- ваемой в Ленинграде, один цветок цвел 3-4 дня, а вегетационный сезон продолжался 195 дней (Балабас и др., 1965). При выращивании в Мос- ковской обл. душица в 3-8-летнем возрасте вегетировала 190-205 дней. Цветение (парами цветков) в парциальных простых кистях душицы происходит акропетально. Первыми зацветают цветки в кистях верхней части комплексного соцветия, а цветки нижних кистей — на 3-5 дней позднее (Шарыгина, 1959). Куст душицы рыхлый; в его структуре обыч- но в конце лета можно наблюдать 3 типа побегов кущения: сильнорос- лые (55-80 см выс.) вегетативно-репродукгивные, среднерослые (10-25 см выс.) и короткие (до 10см выс.) вегетативные. В дополнение к этим побегам осенью могут развиваться еще среднерослые (20-45см выс.) вегетативно-репродуктивные побеги, обеспечивающие вторичное цве- тение (рис. 177, 41). Главный стержневой корень достигает 60см, он функционирует наряду с несколькими более молодыми, похожими на него крупными придаточными корнями и множеством придаточных корней меньших размеров (20-40 см). Финиматурная фаза (рис. 177, Ж). Начинается с 8 лет или позже. Клональная система из 2-6 самостоятельных автономно-ризомных еже- годно репродуктивных безрозеточных сложнокустистых ризокормусов, обычно ризогептакормусов и (или) ризоэнатокормусов.
Глава 8. Изменчивость и эволюция А — осуществление полной общей некротической партикуляции через короткое корневище и главный стержневой корень приводит к замедленному вегетативному размножению. Клонисты (особи клона) нормально цветут и плодоносят. Они сильно сближены и как бы стя- нуты в одно мнимое целое с помощью множества придаточных корней, и лишь только некоторое время спустя криптоклональность (скрытая клональность) стареющих кустов душицы сменяется акриптоклональ- ностью (видимой KJIQHRJIbHocTblo). Б — биоморфы душицы старше 8 лет мы не наблюдали. В этом возрасте еще немало растений было и в предшествующем (медиматур- ном) состоянии; они хорошо цвели и плодоносили. В Ленинграде вы- ращивались растения душицы и более старшего возраста (12 лет), ко- торые цвели и плодоносили (Балабас и др., 1965). У 8-летних особей признаков сильного угасания физиологической зрелости после вступле- ния в клональную стадию синонтогенеза не проявлялось, однако сухие остатки побегов прошлых лет начинали мешать отрастанию молодых побегов, способствуя старению всего кондивидуума. Резюмируем приведенный выше материал по биоморфогении ду- шицы обыкновенной. Выращиваемые в Московской обл. биоморфы душицы обыкновенной Origanum vulgare L. в отличие от дикорастущих биоморф этого вида резко убыстряют темпы индивидуального развития: усиливается активность и разнообразие силлептического побегообразо- вания, наблюдается интенсивное отрастание придаточных корней и соз- дается мочковатый тип корневой системы, увеличивается число побегов кущения, из полегших и укорененных нижних частей которых образу- ется сильноветвистое короткое корневище — многолетняя побеговая основа рыхлого куста с четырьмя (или тремя) ярусами надземных по- бегов. Самый верхний ярус таких побегов обеспечивает первичное цве- тение особей, два (один) средних яруса в благоприятных условиях (не- ежегодно) развивают вторую волну цветения и (или) новые площади ассимилирующих поверхностей (ежегодно) и, наконец, нижний ярус создает озимую побеговую основу для оптимальной генерации новых цветоносных побегов в следующем вегетационном сезоне. Семена (орешки) душицы обыкновенной созревают в основном в сентябре и не нуждаются для улучшения прорастания в стратификации. Осенний посев орешков часто обеспечивает растениям однонедельное ускорение в развитии по сравнению с растениями, полученными пос- редством весеннего посева орешков. На первом году эпиэмбриональной жизни особи душицы проходят гипофазы проростка (ризопротерокор- муса) и всхода (семядольного безрозеточного ризомонокормуса), а так- же ювенильную (бессемядольного безрозеточного ризомонокормуса) и имматурную (безрозеточного вегетативного ризодикормуса) фазы и, на- конец, в конце лета — осенью (конец августа — начало сентября) мно- гие из них при благоприятных метеоусловиях достигают физиологичес- кой зрелости — матурного этапа. В последующие годы возрастает слож- ность организации побеговой части растений (от ризогексакормуса- до ризоэнатокормуса). Из зимующих (персистентных) остатков отрос- ших побегов, полегающих и укореняющихся, формируется гетерономное (связанное с главным корнем) корневище, которое на восьмом году
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ жизни (или позже) начинает отмирать в основании вместе с главным корнем, что приводит к образованию 2-6 кустов-клонистов с автоном- ными (без главного корня) корневищами. Душицу обыкновенную при выращивании в Москвоской обл. мож- но охарактеризовать как травянистый летне-зимнезеленый безрозеточ- ный мультирепродукгивный короткокорневищный рыхлокустистый кон- дивидульный однодомный (с обоеполыми и функционально пестичными цветками) энтомофильный мезофильный гемикриптофит. Из приведенного выше материала видно, что душица обыкновен- ная проявляет высокую внутривидовую пластичность, о-вечая на инт- родукционный стресс множеством адаптаций, которые могут обеспе- чить этому виду успешное распространение в культуре. 8.3. Разнонаправленность соматической эволюции Каждый материальный объект (тело или система тел) — организм, вид и т.п. представляет собой систему, развивающуюся по определенным принципам организованного пространства и времени. Природа нахо- дится в состоянии вечного движения и эволюции. Бесконечное движе- ние синтомов (эволюционных шагов) — синтомокинез, т.е. вечное дви- жение от старого к новому, от исходного к производному, от предка к потомку (гл. 3.3). В синтомокинезе живой нрироды каждый частный син- том своим существованием решает одну или несколько задач: 1) продол- жение жизни; 2) создание нового варианта организации в рамках нрежнего объекта, т.е. нрежнего качественного состояния его, нанример, новый биоморфотин в рамках вида; 3) создание качественно нового объекта, на- нример, нового биологического вида. Эволюцию можно представить и как «маятниковый» процесс (гл. 3.3). Эволюционный маятник — это неживая или живая материальная систе- ма (объект) — тело или система тел, находящаяся в развитии в каких- либо условиях существования и, таким образом, вовлеченная в эволю- ционный процесс. Его «колебания» отражают диапазон изменчивости тел или системы тел, которые он собой олицетворяет. «Маятниковая» эволюция является образным выражением саморазвития Природы — ее эволюции не просто как синтомокинеза, который может особо подчер- кивать общую направленность этого процесса (гл. 3.3), но и как колеба- тельного явления. Накладываясь на синтомокинез, поток эволюцион- ных маятников раскрывает новые грани эволюции. К образу маятников для изображения эволюционного процесса мы прибегаем, чтобы пока- зать более детально механизм частной эволюции, который работает да- леко не однонаправленно, чаще всего разнонаправленно и, более того, иногда обратимо (!). Каждый эволюционный маятник имеет как внут- реннюю нружину для своего самораскачивания (существования) — нрироду самого объекта, так и внешнюю иружину — условия существования. Восприятие эволюции Природы как единого и бесконечного потока дискретных «маятниковых» систем, находящихся в движении, отражает существование Природы в бесконечном многообразии и развитии со-
Глава 8. Изменчивость и эволюция ставляющих ее элементов. В нроцессе «маятниковой» эволюции расширя- ется из~иенчивость, а иногда возникают эволюционные маятники нового тина, нанример, в результате мутаций, когда сильнее обычного срабаты- вают внутренние и внешние нружины элементов эволюции. Колебание каждого живого эволюционного маятника, в нашем случае, чаще всего видов растений, отражает разнообразие биоморф (их тинов) нри развитии в различных экологических условиях. Во-первых, габитульный и иной по- лиморфизм и, во-вторых, мультивариантность биоморфогенезов в диа- пазоне изменчивости какого-либо вида растений по существу свиде- тельствует об одном и том же — внутривидовой пластичности биоморф в пространстве (через первое понятие) и во времени (через второе понятие). Управление фенетической организацией биоморф идет с их гене- тического уровня, что обеспечивает адекватный конституциальный от- вет биоморф на условия обитания. Любой живой эволюционный маят- ник отражает разнообразие всего, что его составляет как живую систе- му, причем частные колебания (моменты существования) этого маятни- ка лишь внешне повторяют друг друга. Они все разные, поскольку неповторимо время и то, что было мгновеньем раньше, уже больше не повторится полностью никогда — всякое повторение относительно. Все, что проявляется в диапазоне изменчивости живого эволюционного маятника, фиксируемо в типах организации, в нашем случае в типах жизненных форм, или биоморфотипах. Эволюция идет на уровне кон- кретных тел, а становится заметнее и понятнее, когда ее изучаешь на уровне типов этих тел. Каждое колебание эволюционного маятника осуществляется по-разному (с разной скоростью и амплитудой) и зави- сит от процессов, происходящих в системе, которую он олицетворяет, например, если это организм, то от разнообразия его состояний, реак- ций в отдельные моменты времени, направленных, допустим, на под- держание гомеостаза. Амплитуды колебания таких маятников разнообразны и связаны с масштабами тел и систем тел, которые они представляют. Гигантским эволюционным маятником будут, например, все семенные растения Земли. В самом общем виде амплитуда колебания такого гигантского эволюционного маятника будет свидетельствовать о разнообразии био- морфотипов, хотя бы габитуальных типов, семенных растений, произ- раставших с очень давних времен до наших дней, включая в себя мно- жества эволюционных маятников меньшего масштаба. К строгому дос- товерному описанию диапазона изменчивости таких сверхгигантских систем наука пока не готова! Существует гигантский пласт литературы, в котором утверждается первичность в эволюции древесных форм покрытосеменных и вторич- ность травянистых (Jeffrey, 1899; Wettstein, 1908; Bailey, 1912; Коза-По- лянский, 1937; Голенкин, 1927; Zimmermann, 1930, 1949; Попов, 1940, 1963; Scharfetter, 1953; Тахтаджян, 1954, 1964, 1966, 1970; Серебря- ков, 1955, 1962; Хржановский, 1958, 1966; Эйхвальд, 1959; Хохря- ков, 1975, 1981 и др.). По мнению других авторов (Козлова, 1953; Рач- ковская, 1957; Голубев, 1960, 1965; Проханов, 1965; Дорохина, 1968 и др.) в определенных условиях, например, полупустынь и пустынь, возможна
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ эволюция растений от трав к полукустарничкам, полукустарникам и далее к кустарникам. Согласно концепции Н.Н.Цвелева (1969, 1970, 1977), травянистый тип растений рассматривается как первичный в эволюции габитуальных форм покрытосеменных. Обратимся к сравнительному разнообразию биоморфотипов у не- которых групп семенных растений, произрастающих в умеренном кли- матическом поясе Евразии и средиземноморских субтропиках Ближнего Востока. Сначала, из приведенного выше в главе 8 материала, выделим следующее. Эфемерные части побегов за несколько дней или месяцев превраща- ются в потенциально персистентные, а затем и в персистентные (сверх- однолетние, многолетние), когда им предоставляются для этого под- ходящие условия (достаточно теплые и влажные). Последние наблюдаем в первую очередь при засыпании (заилении и т.п.) растений песком или иным почво-грунтом (один из способов геофилизации надземных побе- гов), а также в надземной цветоносной части побегов, которая после пло- доношения целиком не отмирает и зимует при температуре обычно зна- чительно выше 0' С. Это же может бьггь отмечено и в климате с холодной зимой, где в микропонижениях (например с Kochia prostrata) среди полого-бугристой полупустыни (рис. 15-П) нет сильных иссушающих летних и очень холодных зимних ветров. В таких условиях осевые эфемерные части побегов имеют тенденцию превращаться в персис- тентные части. При засыпании субстратом этому способствует и прида- точное корнеобразование. В условиях, где зимние температуры заметно ниже 0' С, надземные осевые части травянистых многолетников погибают до основания. Осе- вые части тех же видов (пример с Foeniculum vulgare, Scrophularia xan- thoglossa, Lycopus europaeus) у особей, произрастающих в Средиземно- морье Ближнего Востока с теплой и влажной зимой, не отмирают на значительную часть своей длины и образуют на следующий год постфло- ральные побеги из зоны бывшего отплодоносившего соцветия, включа- ющей и участок торможения. А.П.Хохряков (1975, с.65) обратил внима- ние на подобное поведение некоторых растений в южных и западных районах бывшего СССР, отличающихся сравнительно теплой зимой: в Крыму он наблюдал «экземпляры Mattiola odoratissima, считающегося травянистым многолетником, с почками, отрастающими от середины прошлогоднего цветоноса, т.е. ставшего полукустарником. То же самое бывает с обычно однолетней Cakile maritima. Salvia glutinosa на побере- жье Аджарии и в Закарпатье не травянистый многолетник, а настоящий полукустарник или даже небольшой кустарник». В одном из предшествующих разделов (см. гл. 8.2.2) было подчер- кнуто, что в средиземноморских субтропиках Ближнего Востока, где зимы теплые и влажные, а лето очень сухое и жаркое, образуется осо- бый тип акронекрозных полукустарничков, полукустарников и даже невысоких кустарников (типа 1пи1а viscosa) с постфлоральным «лихора- дочным» (необеспеченным и полуобеспеченным) побегообразованием. У таких растений ежегодно после акронекроза отплодоносивших верху-
Глава 8. Изменчивость и эволюция шечных соцветий в живом состоянии остается некоторая часть пост- флоральной осевой системы — резидные, чаще всего рамозорезидные, постфлоральные зоны. Из этих побеговых участков в конце субтропи- ческой зимы или рано весной отрастают постфлоральные побеги, недо- статочно обеспеченные ресурсами для нормального роста и развития. Часть таких побегов преждевременно погибает, страдая от депрессии в противоборстве с весенними побегами-конкурентами, развивающимися ниже из зоны возобновления, лишь небольшая часть постфлоральных побегов кое-как завершает репродуктивное развитие. Кроме биоморф с постфлоральным «лихорадочным» побегоотрас- танием в субтропиках развиваются также кустарники с постфлоральным нормальным, или обеспеченным, побегоотрастанием. У них из жизне- способной резидной зоны, образующейся от отплодоносивших верху- шечных соцветий, облигатно отрастают нормальные (обеспеченные) постфлоральные побеги, которые проходят все стадии своего развития и образуют здоровые репродуктивные элементы (рис. 82-IV). Такой тип побегоотрастания можно наблюдать у некоторых тропических и субтро- пических кустарников, произрастающих в открытом грунте в Никитс- ком ботаническом саду (Крым), например, у Нурепсит patulum Thunb., Н.hookerianum Wight et Arn., и др. У многолетних трав и полукустарнич- ков континентального климата при оголении засыпанных субстратом побегов, которые ранее стали вынужденно многолетними и подземны- ми, происходит обратный процесс. Вынужденно многолетние и вторич- но надземные части побегов быстро отмирают на всю или большую часть своей длины. Некоторые растения умеренного климата в конце лета образуют вегетативные длинные надземные побеги на корневищах или корнях (Artemisia vulgaris L., Cirsium setosum (Willd.) Bess. Зимой вся надземная вегетативная поросль полностью отмирает вместе с развившимися ранее и отплодоносившими цветоносными весенними побегами. Если бы не холодная зима, вегетативные надземные части побегов могли бы нор- мально расти и развиваться дальше. Кстати, на Ближнем Востоке вообще нет травянистых полыней, а особи одного вида (Artemisia arborescens L.) образуют аэроподии высотой 50-100 см. Переходную структуру между многолетними травами и полукустарничками в умеренной климатичес- кой зоне формирует Artemisia absinthium L. (рис. 178). В защищенных от холодных зимних ветров местах (у заборов, домов, в оврагах и т.п.) отдельные верхнерозеточные надземные побеги перезимовывают как побеги полукустарничков и даже полукустарников. Часть особей этого вида развивается как травянистые многолетники (на открытых, несколь- ко возвышенных местах), поскольку у них все надземные части побегов зимой обмерзают до уровня поверхности почвы. Итак, эволюция жизненных форм (уже — габитуальных форм), раз- нонанравленна и обратима, что особенно заметно на уровне тинов (био- морфотинов) и этим она обязана в нервую очередь главному (нанрав- ляющему) фактору — «лиматическому. Если климат холодает, то виды или изменяют свой ареал, или погибают, или в прежних местообитани- ях трансформируют свои биоморфы, в первую очередь побеговую сис-
436 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 178. Разнообразие организации биоморф Artemisia absinthium L. (Московская область). А — проросток, Б — всходы, В, — ювенильная особь на первом году вегетации, В, — ювенильные особи старше одного года, продолжающие весной надземное моноподиальное нарастание главного побега в условиях относительна защищен- ных и несколько затененных, например, у заборов, Д, — взрослая особь, разви- вающаяся на открытом возвышенном месте (после плодоношения в прошлые годы надземная часть унирепродуктивных побегов отмирала до уровня почвы), Д, — взрослая особь, развившаяся в микропонижении (временно вегетативные надземные побеги зимовали в надземном положении); пунктирная линия — гра- ница моноподиальных годичных приростов (побегов); горизонтальная линия— поверхность почвы; одно деление масштаба — 1 см. Ф
Глава 8. Изменчивость и эволюция тему — основную часть их тела. В этих условиях большую роль играет контрактильность, придаточное корнеобразование, геофилизация мно- голетней побеговой части растений. В случае, если климат в регионе теплеет, в особенности зимы становятся влажными и теплыми, то по- являются условия для превращения травянистых многолетников в по- лудревесные и древесные растения, а однолетников в сверходнолетние растения (пример с Conyza bonariensis (L.) Cronquist), что наглядно видно на современных средиземноморских растениях Ближнего Востока. Часть многолетних растений, даже древесных, может при опреде- ленных обстоятельствах превратиться и в однолетники, что мы наблюда- ем при попытке культивировать тропический и субтропический вид Ricinus communis L. в широтах с холодной зимой. Особи этого вида хорошо развиваются в первом вегетационном сезоне, цветут и плодоно- сят, а зимой полностью вымерзают. Такие превращения до некоторой поры происходят в рамках внутривидовой пластичности, но когда-то могут произойти мутации, которые в новых поколениях закрепят воз- никновение нового вида. Обратимость эволюции не следует понимать буквально. При изме- нении климата, например сильном похолодании, в борьбе за существо- вание, если вид не погибает, то перестраивается, и его отдельные группы биоморф могут выбрать направление габитульного развития, которое в прошлом когда-то проходили предковые виды в сходной климатической ситуации. Обратимость — один из модусов эволюции (Красилов, 1989). Наше утверждение об обратимости эволюции вступает в противо- речие со взглядами некоторых авторов, среди которых отметим наибо- лее убежденного сторонника однонаправленной эволюции А.П.Хохря- кова (1975). Однако, его доводы больше свидетельствуют об обратном: маятники эволюции раскачиваются то в одном, то в другом направлении и помешать этому не в силах никто и ничто, поскольку растения в нроцессе эволюции научились быть лластичными(.') и реагировать по- разному на воздействие изменяющейся внешней среды. Вполне можно также утверждать, что в эволюции биоморф, габи- тусов в частности, не существует беспросветных тупиков даже тогда, когда специализация зашла уже далеко. Деревья и даже эфемеры при сочетании определенных условий получают шанс к обратной трансфор- мации: деревья в сторону трав (через многие переходные типы или непосредственно), а травы в направлении деревьев. На логическую воз- можность возникновения в эволюции травы непосредственно из дерева обратил внимание А.Л.Тахтаджян (1954). Естественно, что такое мо- жет нроизойти только с единичными видами, чего внолне достаточно, чтобы через сотни тысяч лет сменилась вся габитуальная картина растений в каком-либо регионе или на всей Земле в соответствии с гло- бальными изменениями климата. Любопытно наблюдать, как изменяют облик некоторые эфемеры, например, злаки Eremopyrum triticeum (Gaertn.) Nevski, Anisantha tecto- гцт (L.) Nevski и др. при засыпании их песком. У них проявляется тенденция к продлению сроков жизни в два раза и более. При этом эфемеры образуют довольно мощные кусты за счет активизации прида-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ точного корнеобразования и усиленного ярусного (эпифизарного) ку- щения (рис. 157). В некоторых условиях не исключается переход эфе- мерных осевых частей таких растений не только к однолетнему, но и сверходнолетнему образу жизни и т.д. Безусловно, что деревья и эфе- меры — это наиболее отдаленные фрагменты габитуальной эволюции растений. Переход (трансформация) деревьев в кустарники, кустарнич- ки, полукустарники и многолетние травы вполне возможен и объясним соответственно при похолодании климата, а в обратном направлении- при его потеплении. Во многом это подтверждается на приведенном в главе 8 фактическом материале, из которого ясно, что основное со- держание этой трансформации (в любом из двух главных равнозначных направлений) составляют изменения количественных признаков. Способ- ность однолетних осевых структур превращаться в многолетние и наоборот широко распространена в растительном мире, поэтому нанизация (умень- шение размеров) или грандизация (увеличение размеров) многолетних побеговых частей (базовых органов), их уход (пассивный или активный) в почву (частично или полностью), смена направления роста по отношению к поверхности почвы, эфемеризация или персистентизация структурных элементов кроны и т.п. — это все признаки потенциально доступные в эволюции любым группам семенных растений. Для осуществления этого процесса необходимо время (много или мало), в течение которого они успевают перестроиться, разблокировать свои резервные возможности к перестройке. В других условиях адаптивные возможности могут бьггь силь- но заторможены с генетического уровня из-за высокой специализации некоторых ключевых (побеговых) структур. Известно, что у видов животных (от беспозвоночных до млекопи- тающих) с широкими климатическими границами ареала особи из хо- лодных мест обитания живут дольше и растут медленнее, чем особи теплых мест обитания (Виленчик, 1926). Многолетние наблюдения за жизнью различных видов растений, проведенные нами в горах Аптая, свидетельствуют о том, что продолжительность жизни растительных особей одного вида, как правило, уменьшается при движении от наибо- лее холодных мест их обитания к более теплым. У Bupleurum multinerve DC., например, особи мультирепродуктивного тропобиоза из высоко- горных популяций живут в 2-10 раз дольше, чем особи унирепродуктив- ного и би-, трирепродуктивного тропобиозов, произрастающие в ниж- нем и среднем поясах гор. Как можно объяснить это явление? Недоста- ток тепла и короткий вегетационный период тормозят скорость нарас- тания вегетативных органов, отодвигают сроки заложения репродуктив- ных органов, снижают мощность проявления репродуктивных функций. Базокормусы в условиях высокогорий значительно медленнее разруша- ются, чем у особей того же вида, произрастающих много ниже верхней границы леса, поскольку течение соответствующих микробиологических процессов здесь замедленно и численность микрофлоры и микрофауны, растительноядных насекомых и т.п. относительно невелика. Особенно- сти условий существования отражаются и на внешнем облике особей одного вида, т.е. изменяются многие экзоморфологические показатели роста и развития побегов и корней. В итоге все эти факты обеспечивают
Глава 8. Изменчивость и эволюция растениям из холодных мест произрастания более продолжительный период для реализации в фенотипе основных функций самосохранения вида, закодированных в генотипе, а следовательно, и большую продол- жительность их индивидуальной жизни, чем это свойственно растениям того же вида из более теплых мест обитания. Растения или онределенные грунны растений, которые участвуют в ществования («колебательного движения с онределенной амнлитудой») могут решить или не решить одну или несколько из ноставленных Природой неред ними задач: 1) нродолжить жизнь, 2) расширить нолиморфиъч, 3) со- здать новый вид. Главные изменения в живых эволюционных маятниках затрагивают количественные нризнаки, далее тонические нризнаки, характеризующие наложение основных органов и (или) нанравление их роста относительно новерхности ночвы и, только нотам, собственно качественные (сома, или тело, — это единство всех меросом, составляющих фенетическое и ге- нетическое, вегетативное и репродуктивное содержание структуры ро- дительского организма). При соматической трансформации растений изменяются чаще всего размеры и число, продолжительность жизни побегов, корней и их элементов. Особенно показательно изменение размеров базовых органов, их расположения относительно поверхности почвы и степени погружения в почву (или воду). Так, у габитульных типов растений рода Coronilla L. имеется такой общий качественный признак, как образование интеркалярных цветоносных побегов и меж- соцветий, но по количественным и топическим признакам они сильно различаются: по продолжительности жизни побегов и числу их в кусте, по расположению их относительно поверхности почвы. У габитуальных типов растений других родов, например Sambucus L. (рис. 43, 179), Bup- leurum L. (рис. 99-101, 126-I, 135, 149-151, 183) на фоне общности каче- ственного признака — развития верхушечных закрытых или полузак- рытых соцветий, различия проявляются по количественным и топичес- ким признакам: размерам и продолжительности жизни надземных ча- стей побегов, положению базового органа (если он образуется) относи- тельно поверхности почвы и т.п. Принципиальную модель соматической эволюции семенных расте- ний от отдельных биоморф до каких-либо их родственных групп (видов, родов, семейств и т.д.) можно показать на рисунке как поток «эволю- ционных маятников» в бесконечно изменяющемся пространстве-време- ни (рис. 180). Здесь важно все: что эволюционные маятники колеблются с определенной амплитудой (диапазоном изменчивости), что их опор- ные оси «плывут» (движутся) в пространстве-времени, что разнообразно взаимодействуют друг с другом множество сил, обеспечивая колебание эволюционных маятников и изменение их амплитуды и т.д. Автора не покидает мысль, что реальная соматическая эволюция семенных расте- ний шла и идет возможно именно в подобном потоке эволюционных маятников... Предложенная принципиальная модель «маятниковой» со- матической эволюции семенных растений нацелена на дальнейшее развитие исследований в этом направлении.
440 О ,"НОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 179. Формирование кустарника Sambucus racemosa L. (Московская область). А — проростки, Б — всходы, В, — ювенильная особь на первом году вегетации, В — ювенильная особь на втором году вегетации (условия затенения), Д — ма- 2 турные особи; Е — схемы разнообразно организованных особей на втором году вегетации; 1 — семядоли, 2 — семядольный узел ( у взрослых особей его пример- ное положение показано пунктиром), 3 — срединные листья, 4 — медиальный побег, 5 — весенние пазушные побеги, 6 — весенний верхушечный побег, 7— отмершие части побегов, 8 — вегетативные годичные побеги, 9 — вегетативно- репродуктивные сильно- и среднеразвитые годичные побеги, 10 — вегетативно- репродуктивные слабые годичные побеги, 11 — строго репродуктивный годичный побег, 12 — аэроподий, 13 — геоподиальная часть куста, 14 — сильнорослый побег кущения, 15 — отмершие части прошлогодних побегов; у — увеличено по сравне- нию с масштабом; горизонтальная линия — поверхность почвы, одно деление масштаба — 1 см.
Глава 8. Изменчивость и эволюция 441 к 3~ ~ ~~ф хм ~î О Й О Ю Й д о Ko ~ Q ~ х " а. ~ Я -'' о е~ о к ~а ~~ ~ Ж ой ~р „о о д~ М ж о, ~.& о О ж 65 ~ о к ~ 2 ~ о v '+ 2) 6~ Х о ~ ;Я, ж ~ц~ ~~ о g о о g+g, ~о о~ й,' оо юх" Щ Ж о ( Ю ж о. О ~ r р, О v о ~ о g й ф~ о Q~ 65 й о о „ м о а ~ о A Д ге~ Ф щ 63 щ К Q ~4 ~'~ о Й ~ Р~ й v Я о К б3И К О й< о ж ~ д ФЭ о 65 vx Q~Q о, О ф ~~я ~~) ра Ж о са О ~ч О p' о о Ж ~~Е' g о о Г:[ ~р о о .~ Ф О~
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ После знакомства с концепцией разнонаправленной соматической эволюции возникает мысль, что эволюция биоморф в нужном направ- лении, вероятно, может быть воспроизведена экспериментально. На- пример, сейчас, вполне понятны условия, при которых надземные цве- тоносные побеговые части у травянистых многолетников буквально на глазах превращаются в многолетние, даже одревесневающие, части, причем весьма обычным способом (!). Такие условия возникают, как правило, в случаях, когда нижние, средние, а иногда и верхние эфемер- ные части надземных побегов засыпаются (или заиливаются) почво- грунтом (обычно с песком). В зоне пребывания таких вынужденно погребенных (первично надземных), часто цветоносных, частей расте- ний возникает особый «микроклимат», в котором достаточно тепло, достаточно влажно и многое еще — тоже достаточно. Между прочим, сформулирована любопытнейшая физиологическая проблема: экспери- ментально расшифровать параметры этой «достаточности». Моделируя по температуре, влажности, освещенности, газовому составу и другим факторам надземную среду обитания для цветоносных побегов, которая напоминала бы «микроклимат», «вынужденно» возни- кающий в зоне погребения субстратом подобных побегов, эксперимен- татор обрекает травянистые многолетники и тем более полукустарнички к их трансформации в напрвлении развития полудревесной или (и) древесной организации. Невольно мы оказались на пороге грядущего экспериментального подтверждения разнонаправленности соматической эволюции растений как от трав к деревьям, так и наоборот. Не исклю- чено, что «микроклимат», развивающийся в зоне случайных погребений надземно-однолетних частей побегов, при котором инфраструктурная эфемерность исчезает или уменьшается, сменяясь более или менее инфраструктурной персистентностью, чем-то напоминает надземный «палеоклимат», в котором формировались некоторые типы околодре- весных и древесных жизненных форм. Автор наблюдал под Москвой после весеннего снеготаяния исклю- чительное поведение прошлогоднего отплодоносившего надземного вер- тикального побега Leucanthemum vulgare Lam., который до половины своей длины (26 см) еще в прошлом вегетационном сезоне оброс силь- ными пазушными вегетативными розеточными побегами. Микроклимат вокруг большей части цветоносного побега нивяника возник случайно благодаря штабелю досок, обеспечившего ему достаточно благоприят- ные условия как для надземной вегетации пазушных розеточных побе- гов, так и для успешной их перезимовки на вынужденно удлиненном надземном резидном участке родительского побега. После удаления штабеля досок растение с трансформированной побеговой системой некоторое время не определялось как «трава» в традиционном по- нимании этого термина. Сегодня, более, чем когда-либо, можно утверждать, что эволю- ция — это многоуровневый, разнонаправленный, обратимый и беско- нечный процесс движения синтомов («эволюционных шагов»). Так су- ществует Природа (и все ее части), являющаяся, как отмечалось выше, бесконечным эволюционным процессом.
Глава 8. Изменчивость и эволюция 8.4. Эволюционная перспектива вида Можно ли утверждать, что какому-то из современных видов при- надлежит большая эволюционная перспектива, чем другому виду (воз- можность обойти в эволюционном соревновании менее приспособлен- ные виды), а кому-то меньшая? На этот вопрос мы отвечаем с позиций дарвиновской теории естественного отбора в эволюции. Остановимся на некоторых ее сторонах, выступающих в качестве индикатора перс- пективы вида в эволюции. Исходя из представлений о пластичности как изменяемости материальной системы в определенном диапазоне воз- можностей по разным показателям ее организации, мы отдаем предпоч- тение габитуальному внутривидовому полиморфизму (способности био- морф одного вида развивать, в зависимости от генотипических свойств и условий обитания, два и более типов габитусов). Известно, что веге- тативные признаки, а они более всего влияют на габитус, наиболее изменчивы по отношению к репродуктивным признакам. Главный результат в эволюцию живой природы — это, на наш взгляд, факт продолжения жизни в цепочке предок — потомок — предок— потомок и т.д. В жизненном цикле любой биоморфы габитус меняется в соответствии с изменением возрастной и фенологической ситуации. Для оценки эволюционной перспективы вида нам важнее габитусы взрослых биоморф на пике их восходящего развития. Число видов рас- тений, которые уцелели до наших дней, не проявив склонности к габиту- альному полиморфизму (многие орхидные тропиков, секвойя гигантская и др.), во много раз меньше числа видов, обладающих широким га- битуальным полиморфизмом. Так, у живого ископаемого Ginkgo biloba L. ограничена пластичность, что сильно сузило ареал его современного распространения (Юго-Восточная Азия); по габитусу нам известен один биоморфотип (габитуальный мономорфизм) — некустистые верхнеро- зеточные деревья, что является индикатором эволюционной неперспек- тивности современного гинкго. Современная Юго-Восточная Азия в ботаническом отношении, по образному выражению С.В. Мейена (1981, с.144), «скорее дом для престарелых, убежище для тех, кого климатичес- кие перемены выгнали с других мест». При движении от тропиков на север увеличивается число видов с высокой пластичностью, поэтому логично заключить, что виды север- ных флор эволюционно более перспективны, чем виды южных, особен- но тропических, флор. Очень высокой пластичностью, в частности и габитуальным полиморфизмом, обладают наиболее распространенные на Земле растения — Picea abies (L.) Karst., Pinus sylvestris L., Abies sibirica Ledeb. (Сукачев, 1905; Нухимовская, 1972, 1974; Ляэнелайд, 1984 и др.). Разнообразие внутривидовых биоморфотипов, в том числе био- морфотипов, охарактеризованных по габитусу (габитуальных типов), весьма велико; оно свидетельствует об эволюционной перспективности этих и подобных (в плане пластичности) им видов. Так, Abies sibirica в пределах своего ареала проявляет основной биоморфотип — высокие (20-40 м) некустистые (одностволовые) деревья, в тех же или более влажных условиях образуется близкий, но второстепенный биоморфо-
444 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ тип — высокие полуразвернуто-слабокустистые (с одним лидерным стволом и укореняющимися нижними ветвями, или лапником, склон- ными к полному обособлению, т.е. к вегетативному размножению) де- ревья. В горах, например на Алтае, пихта сибирская, ближе к верхней границе леса, представлена второстепенным биоморфотипом: средне- высокими (до 15 м) полуразвернуто-слабокустистыми (с несколькими лидерными стволами) деревьями. Выше леса идет биоморфотип стлани- ковых кустарников (высота 0,5-3 м). Еще реже встречен биоморфотип стланиковых кустарничков (высота их многолетних прижатых к суб- страту надземных частей 10-30см). В интродукционных популяциях Abies sibirica в Рузском районе Московской области отмечены (Нухи- мовская, 1971) два биоморфотипа: некустистые деревья и реже высокие полуразвернуто-слабокустистые деревья. На наш взгляд, у Abies sibirica высокий габитуальный полиморфизм (признак-индикатор, свидетель- ствующий о большой эволюционной перспективе вида). Высокая сте- пень габитуального полиморфизма развивается у видов, биоморфы ко- торых осваивают разнообразные биотопы. Основой для определения эво- люционной перспективы вида является степень пластичности. Чем она меньше (больше), тем и меньше (больше) эволюционная перспектива вида. Итак, степень габитуального полиморфизма (no признакам одного сравнительного ряда) у биоморф одного вида в современных условиях его существования свидетельствует об эволюционной перспективе вида: пер- спективность (шанс) сохранения какого-либо вида и (или) продолжения его в производных видах (потомках) по сравнению с другими видами в эволюции тем выше, чем шире у него пластичность и, в особенности одна из ее сторон — габитуальный полиморфизм — это, в нашем понимании, закон (специальный) эволюционной перспективы вида. Габитус биоморф весьма лабилен. В зависимости от условий суще- ствования отдельных видов габитус биоморф может изменяться в сто- рону как деревянистости, так и травянистости. Смысл нашей гипотезы начальной эволюции покрытосеменных сводится к следующему: в эволюции растений первичным было возникновение пластичности, в частности габи- туального полиморфизма, а вторичным — возникновение покрытосемен- ности. Основой для такого порядка в эволюции было появление контраст- ных условий существования биоморф, которые возникли еще до появления жизни, поскольку были и всегда будут равнины, горы, литорали, пески, ветер и т.п. В борьбе за пространство и время биоморфы первых покры- тосеменных (палеобиоморфы) получили в наследство от древних голосемен- ных способность к пластичности и, следовательно, к габитуальному поли- морфизму. Кто был первым носителем покрытосеменности (дерево или трава, или нечто подобное и тому и (или) другому), вопрос для науки пока вряд ли актуален: очень мало палеоботанических данных (Мейен, 1981). Покрытосеменность могла возникнуть как в комфортных, так и дискомфортных условиях существования. Габитуальный фон палеобио- морф для возникновения покрытосеменности, на наш взгляд, не был огра- ничен только деревьями. В эпоху зарождения покрытосеменных уже суще- ствовала пластичность и соответственно габитуальный полиморфизм (без них невозможна эволюция живого), поэтому покрытосеменность могла возникнуть у палеобиоморф с любым габитусом.
Глава 9. Многолинейная классификация биоморф семенных растений и формализация языка биоморфологии В познании биологии различных видов растений, в особенности их биоморфогении, важно получить наиболее разностороннюю характе- ристику их биоморф. Релятивные классификации, к которым нередко прибегают биоморфологи, когда соподчиненность отдельных рядов признаков, используемых для выделения классов биоморф разных ка- тегорий, носит условный характер, мало отвечают сформулированной выше задаче. Мы согласны с В.Н.Голубевым (1972), что для многосто- ронней оценки какого-либо вида все признаки следует рассматривать как равнозначные. Для иных целей эта позиция может не подойти. Например, систематик всегда стремится различать признаки растений или животных по их таксономической ценности. Систему многосторон- них биоморфологических знаний о каком-либо виде растений, а одно- временно и ключ к определению и описанию биоморф, можно получить посредством многолинейного (мультилинейного) классифицирования. Нами разработана многолинейная эколого-морфологическая (по боль- шинству используемых признаков) классификация биоморф семенных растений. Для ее построения был использован материал собственных исследований, а также опыт эколого-морфологического (Warming, 1884, 1908; Schmid, 1956; Серебряков, 1962; Голубев, 1960, 1965; Серебряко- ва, 1971 и др.) и морфологического (Федоров др., 1956; Федоров и др., 1962; Федоров и др., 1979) анализа семенных растений, наиболее полно обобщенный В.Н.Голубевым (1968, 1972) в «линейной» системе жизненных форм. В многолинейной системе все признки расцениваются как равнозначные, т.е. не учитывается, на основе какого метода состав- лены отдельные ряды биоморфотипов: субординии, сериофикаиии или одновременно обоих методов. В нашей классификационной системе биоморфотипы индицируются признаками, которые нередко одновре- менно являются гемитропобиозными или (и) тропобиозными признака- ми, поэтому таким биоморфотипам соответствует определенный геми- тропобиоз или (и) тропобиоз, например, розеточному биоморфотипу соответствует розеточный гемитропобиоз Pimpinella saxifraga L., Berberis sibirica Pall. и розеточный тропобиоз Тагахасит officinale Wigg., Plantago
446 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ major L. и др. Порядок первоначального расположения рядов биомор- фотипов в нашей классификационной системе значения не имеет. Многолинейную классификацию биоморф мы, кроме того, исполь- зуем в качестве одного из способов разработки формализованного языка биоморфологии. Дело в том, что прошлый и современный язык, кото- рым описывается организация биоморф семенных растений и биомор- фогенезов, до настоящего времени весьма многословен. С момента на- чала накопления фактической информации о каких-либо биоморфах проявляется обычно многословность и тавтологичность, которые затем переходят в тексты, отражающие результаты биоморфологических на- блюдений. Как сделать поиск информации в биоморфологии семенных растений более целенаправленным и точным и как лаконичнее, но пол- нее, выдать результаты проведенных наблюдений и исследований. К настоящему времени об индивидуальной и кондивидуальной жизни мы знаем уже довольно много, но многого еще и не знаем; однако потен- циальные возможности биоморфологических, и в частности биоморфо- генезных, исследований огромны, к реализации многих из них мы се- годня только подходим. Создание таблиц-матриц с закодированной в них информацией о многолинейных отраслевых классификациях био- морф — один из весьма перспективных путей фиксирования, хранения, снятия, передачи и выдачи информации об их организации в разных условиях существования. Мы стоим на пороге широкой компьютериза- ции биоморфологических исследований. Итак, первый способ формализации языка биоморфологии — это кодирование информации в многолинейной классификации биоморф семенных растений и расположение кода в таблице-матрице, с которой по любому виду семенных растений можно нарабатывать информацию (т.е. ставить вопросы, которые предполагаются по каждому закодиро- ванному сравнительному ряду признаков, и отвечать на них). В итоге работы с растениями при помощи биоморфологической таблицы-мат- рицы мы накапливаем фактический материал, сохраняем информацию и снимаем ее в любой момент для проведения новых практических и теоретических исследований. Информация с таблицы-матрицы может сниматься в виде формулы, которая легко переводится с языка символа на язык слов. Формула — «это представление связей, отношений, су- ществующих между предметами, явлениями, процессами при помощи конечной последовательности знаков (символов), объединенных опре- деленными операциями; точное и краткое определение какого-либо за- кона с помощью принятых в данной науке знаков. Формула делает более ясными структуру, связи и взаимоотношения объектов исследу- емой системы, четко фиксирует в ней главное, основное, существенное» (Кондаков, 1975, с. 652). При создании многолинейной эколого-морфологической класси- фикации биоморф семенных растений мы широко использовали опыт разработки подобной классификации В.Н.Голубевым (1972), причем не- которые ряды признаков, ставшие уже классическими, например, «типы Раункиера» или габитусы (формы роста), примененные в его клссифи- кации, мы включили в свою классификацию без изменений или с не-
447 Глава 9. Многолинейная классификация биоморф семенных растений большими изменениями. В нашей классификации широко использова- ны уже упомянутые выше «Атласы по описательной морфологии растений». При этом мы обычно не просто переписывали ряды признаков, а изменяли и совершенствовали те из них, которые отвечали нашим задачам. Кроме того, в многолинейной классификации учтено немало того нового и старого, что составляет общепринятый актив биоморфологии. Мы нередко стремились дополнить или уточнить из- вестные ряды признаков результатами своих разработок. Некоторые ряды признаков описны впервые. Важнейшими из них являются ряды а1-а13, кl-к17 (Нухимовский, 1971, 1980 и др.), особенности других рядов признаков отражены в теории организации биоморф семенных растений, которая составляет основное содержание этой работы, и в выводах по всей работе или разъясняются непосредственно после ха- рактеристики символов. Отраслевая многолинейная классификация биоморф, предлагаемая ниже, — один из путей совершенствования теории и практики биоморфологического анализа. Эта классификация имеет все условия для роста и развития как по горизонали, так и по вертикали (не все признаки учтены). Многолиней ная эколого-морфологическая классификация биомооф семенных растений изображена в закодированном виде в ma&uqe-матрице (рис. 181). Объяснения символов. а1-а 13 ванные a13 — ююнально-ризокормусовые (в скобках можно указывать число клонистов и тип од- ного из них по сложности организции, например, а13 (4-аб), что означает синорга- низм состоит из 4 ююнис- тов, наиболее сложный из которых клонист-ризогекса- кормус; — растения, охарактеризо- ' по сложности их общей ор- ии аll- а12— Дглее при описании рядов биоморфотипов не повторяется, но подразумевается словосочетание ~растения, охарактеризованные». ганизац al а2 а4 аб а7 а8 а9 а10— ризомонокормусовые, ризодикормусовые, ризотрикормусовые, ризотетракормусовые, ризопентакормусовые, ризогексакормусовые, ризогептакормусовые, ризооктокормусовые, ризоэнатокормусовые, ризосинкормусовые (в скоб- ках отмечается тип побега наиболее сложного в систе- ме синкормуса), ризопротерокормусовые, аризокормусовые (организа- цию биоморф составляют не ризокормусы, а только побе- ги, поскольку корни у них вообще не образуются, на- пример, Utricularia sp.), а 14-а25 — no габитусам: а14 — арборные (деревья), a15 — кустарниковые, а16 — кустарничковые, а17 — полукустарниковые, а18 — полукустарничковые, а19 — эпигенные (с погружающим- ся в почву базовым органом) многолетние травянистые, а20 — гипогенные (с первично под- земным базовым органом) многолетнетравянисть~е (кро-
448 ° Ъ Г О Ф3 р 0 о~о 23 О 22 ° ф 7~~ 21 '5 fm Я 4- I mi ° J I im 13 �ее 4 е pHi> ри 44 ° ° ° ° ° ° ° O ° ° е Оси Oh .О, °, v с б е и о р q г и Рис. 181. Многолинейная классификация биоморф семенных растений, закодированная в таблице-матрице (объяснения в тексте). а24 — однолетние, a25 — эфемерные; ме эфемероидов; в скобках можно указывать конкретную продолжи- тельность жизни биоморф), а21 — малолетнетравянистые (жи- вут 3-4 года), а22 — эфемероидные, а23 — двулетние, ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ в1-в2 — no высоте аэроиодиев (ког- да они есть): в1 — измеряется в м (высота > 1 м), в2 — измеряется в см (< 1 м);
Глава 9. Многолинейная классификация биоморф семенных растений 449 вЗ-в4 — no высоте генерального и субгенеральных аэрокормусов: в3 — измеряется в м (высота > 1 м), в4 — измеряется в см (< 1 м); в5-в9 — no n ительиости жиз- ни листьев и особенностям их смены: в5 — летнезеленые (листья не зи- муют), в6-в8 — всесезоннозеленые, в6 — с листьями однозимнезелены- ми (листья, образующиеся весной, летом или осенью, coxpaH$BQT жизнеспособность не более одной зимы), в7 — с листьями малозимнезеле- ными (листья жизнеспособ- ны две-четыре зимы), в8 — с листьями многозимнезеле- ными, или многолетнезеле- ными (листья жизнеспособ- ны пять и более зим), в9 — бессезоннозеленые (в бессе- зонном климате многолетние растения постоянно зеленые за счет более или менее скрытой смены отдельных листьев); в10-в 13 — no числу семядолей: в10 — односемядольные, в11 — двусемядольные, в12 — трехсемядольные, в13 — многосемядольные (в скобках желательно укзывать кон- кретное число семядолей, на- пример в13 (9) или в13 (5-10)); в 14-в 19 — no положению семядолей относительно поверхности почвы (воды): в14 — надземносемядольные, в15 — подземно семядольные, в16 — надводносемядольные, в17 — подводи осемядольные, в18 — наводносемядольные (плава- ющесемядольные), в19 — бессемядольные; в20-в25 — no положению почек и верхушек побегов относительно по- верхности почвы в неблагоприятное время года (типы Раункиера): в20 — эпифильные, в21 — фанерофильные, в22 — хамефильные, в23 — гемикриптофильные, в24 — криптофильные, в25 — терофильные; cl-c25 — no форме семядолей: растения с семядолями— с l — округлыми, с2 — овальными, с3 — продолговато-овальными, с4 — продолговатыми, c5 — стреловидными, с6 — широкояйце видны ми, с7 — яйцевидными, с8 — узкояйце видными, с9 — обратноширокояйцевидньпии, с10 — обратнояйцевидными, с11 — обратноузкояйцевидными, с12 — широколопатчатыми, с13 — лопатчатыми, с14 — узколопатчатыми, c15 — треугольными, с16 — ромбическими, с 17 — неравнобо кими, c18 — заостренно-зллнптичесаннн, с19 — ланцетными, с 20 — почковидными, с21 — линейными, с22 — игловидными, с23 — лопастными, с24 — вырезно-раздельными, c25 — рассеченными (в скобках мо- жно указывать размеры семя- долей: первая цифра — дли- на, вторая — ширина, в см); Под индексом dl-d24 идут те же самые подразделения «по форме простых (неразветвленных) листьев (монофиллов)», что и «по форме се- мядолей» (индекс с); 425 — растения с рассеченными листьями
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ el-ell no длине междоузлий и осо- бенностям облиствления иобегов: еl безрозеточные раздвинутоуз- ловые, безрозеточные сближенноуз- ловые, разнобезрозеточные (побеги состоят из безрозеточной нижней короткой или длин- ной сближенноузловой и верхней раздвинутоузловой частей со сходными форма- ми листьев в обеих частях), предрозеточные (когда побег с 1-2 листьями, например, у всходов, предшествует обра- зованию розетки), розеточные, полурозеточные, верхнерозеточные, верхне полурозеточные, розеточно-безрозеточные (в структуре растений есть ро- зеточные и безрозеточные побеги), розеточно-верхнеро:ж очные, полурозеточно-безрозеточ- ные; е2 еЗ е4 е5 еб е7 е8 е9 е10— ell— е12-е17 — no стеиени разветвления листьев: е12 — монофиллические (с прос- тыми, или неразветвленны- ми, листьями), е13 — дифиллические, е14 — трифилл ические, e15 — тетрафиллические, е16 — пентафиллические, е17 — полифиллические (в скобках можно указывать линейные размеры листьев: в числите- ле — длину, а в знаменате- ле — ширину, в см, напри- 15-30 мер, е14( ); е 18-е21 — no форме разветвленных листьев (дифиллов-иолифиллов): растения с разветвленными листьями- е18 — пальчатыми, е19 — тройчатыми, е20 — непарноперистыми, е21 — парноперистыми; е22-е23 — no наличию или отсут- ствию сочленении' в структуре раз- ветвленных листьев: растения с листьями- е22 — полусложными (дифиллы- полифиллы, без сочленений в структуре разветвленных листьев), е23 — сложными (дифиллы-поли- филлы, с сочленениями в структуре разветвленных ли- стьев); е24-е25 — no иоложению семенной кожуры у ироростков и всходов: е24 — сбрасывающие, e25 — не сбрасывающие семенную кожуру; fl-/6 — no особенностаи расиоложе- ния листьев в узлах (филлотаксис): f l — очереднолистные, f2 — супротив кол исти ые, 1'3 — муто вчатолистные, f4 — очер едно-супротивнолист- ные, 1'5 — очередно-муто вчатол исти ые, f6 — супротивно-мутовчатолист- ные; f7 f9 — no сексуализации цветков: f7 — обоеполые, f 8 — однополые, f9 — непостоянно-обоепалые (в од- них условиях растения про- яаяякя себя как обоепояые, а в друтих как однополые, или в рамках вида одни особи обое- палые, а другие однополые);
Глава 9. Многолинейная классификация биоморф семенных растений 451 fl 0-f 14 — no кратности ироявления реиродуктивной функции (у обоеио- лых и непостоянно обоепалых растений): f10 — унирепродуктивные, f l l — бире продуктивные, f12 — трирепродуктивные, f l 3 — мультирепродукгивные, f14 — дерепродукгивные, или сте- рильные, f15 f19 — no кратности плодоно- шения (у одноиолых растений с пес- тичными цветками; в скобках указа- но подразделение растений по крат- ности семеношения у однополых хвойных растений), f l 5 — монокарпические (моностро- билярные), f16 — дикарпические (дистробиляр- ные), f17 — олигокарпические (олиго- стробилярные) — плодоно- сящие 3-4 раза, f l 8 — поликарпические (полистро- билярные) — плодоносящие 5 и более раз, f 19 — акарпические (неплодонося- щие), или стерильные; f20 f24 — no кратности иыльцено- шения (у одноиолых растений с ты- чиночными цветками): 1'2 0 — моноантерические, f 2 1 — диантерические, 1'2 2 — олигоантерические, f 2 3 — полиантерические, f24 — апоантерические, или сте- рильные; /25 — no хромосомному набору: указывается гаплоидный (n) и (или) диплоидный (2n) набор хромосом; gl g4 — no особенностям нараста- ния соцветии: gl — унифлоральные (растения с одним верхушечным цвет- ком), рацемозные, или ботричес- кие [центральные оси соцве- тий — главного и боковых, или присоцветий (кофлорес- ценций), а также парциаль- ные соцветия, если они обра- зуются, — открытые], рацемозно-цимоидные (цен- тральные оси главного соц- ветия и присоцветий — от- крытые, а парциальные соц- ветия — закрытые), цимозные (центральные оси главного соцветия и присо- цветий, а также парциаль- ные соцветия, если они обра- зуются, — закрытые); g4 g5-я14 — по сиособу нарастания по- бегов: g5 g6 g7 эфемерно-моноподиальные, эфемерно-эусимподиапьные, эфемерно-олигосимподиаль- ные, эфемерно-полисимподиаль- ные, эумоноподиальные, гемисимподиальные, эусимподиальные, олигосимподиальные, полисимподиальные, дихотомические (при необ- ходимости эту группу рас- тений можно классифици- ровать подобно симподиаль- ным растениям); g9 g10- gll— g12— 313— g14— g15-g17 — no особенностям прос- транственного расположения ре- иродуктивных органов и форме лис- тьев на верхушке иобегов: g15 — открытые, или индетерми- натные (их меристемы не образуют отдельные верху- шечные цветки или соцве- тия, заквнчияающиеся фрон- дулозной, брактеозной или абрактеозной замирающи- ми верхушками),
452 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ g16 g17 — условно-закрытые (полузак- рытые, приоткрытые; они образуют открытые соцветия с фрондулозной, брактеоз- ной или абрактеозной зами- рающими верхушками), — закрытые (они образуют за- крытые соцветия или от- дельные верхушечные цвет- ки); g18 g23 — по типу простых соцве- тий: g18- g19— g20- g21— g22- g23— растения с кистями, сережками, початками, колосьями, зонтиками, головками, g24 g25 растения с прилистниками, без прилистников; Ы-h25 — no типу сложных соцветий: hl — растения с метелками, h2 — щитками, h3 — сложными кистями, h4 — сложными колосьями, h5 — сложными зонтиками, h6 — сложными головками, h7 — монотирсами, h8 — дитирсами, h9 — плейотирсами, h10 — кистевидными тирсами, Ы 1 — колосовидными тирсами, h12 — сережковидными тирсами, h13 — щитковидными тирсами, h14 — зонтиковидными тирсами, h15 — головковидными тирсами, h16-h22 — цимоидами, 1116 — монохазиями, h17 — извилинами, h18 — завитками, h19 — клубками, g24 g25 — no наличию или отсут- ствию прилистников у срединных листьев: h20 — дихазиями, h21 — двойными дихазиями, h22 — двойными завитками, h23 — плейохазиями, h24 — циациями, 1125 — ценосомами; il-i12 — no форме стеблей в их по- перечном сечении: il — ципиндричесю~стеблевые (по большей части длины стебля поперечное сечение одинако- во и имеет форму круга), — кон ичес кисте блевые, — вал ьковатостеблевые, — полувальковатостеблевые, — сплюснутостеблевые, — шаровидностеблевые (у стеб- ля три взаимно перпендику- лярных диаметра более или менее равны), i7 — трехгранностеблевые, i8 — четырехгранностеблевые, i9 — пятигранностеблевые, i10 — многогранносгеблевые (в ско- бках число граней), i l l — борозчатостеблевые, i12 — четковидностеблевые; i2 i3 i4 i5 i6 к l-к17 — no особенностям образо- вания резидов после завершения од- г 13-i25 — no направлению роста ли- дерных побегов и особенностям их пространственного положения: i13 — прямостоячепобеговые, i14 — наклоненнопобеговые, i15 — изогнутопобеговые, i16 — дуговиднопо беговые, i17 — пони кающие, i18 — стелющиеся, i19 — ползучие, i20 — восходящепобеговые, i21 — вьющиеся, i22 — целя ющиеся, 123 — лазящие, i24 — свисающие, i25 — плавающие;
Глава 9. Многолинейная классификация биоморф семенных растений 453 ного репродуктивного цикла в жиз- ни лидерных универсальных побегов: к1 — дерезидные, к2 — татаре зидные, к3 — макс иморезидные, к4 — магнорезидные, к5 — малорезидные, к6 — модесторезидные, к7 — медиорезидные, к8 — бревирезидные, к9 — миниморезидные, к10 — курторезидные, к11 — вицерезидные, к12 — викариорезидные, к13 — репторезидные, к14 — пор циорезидные, к15 — пленорезидные, к16 — алиорезидные, к17 — рамозорезидные; к 18-к25 — no использованию видов пространства как среды жизни: к18 — аэротопные, к19 — аэрогеотопные, к20 — аэрогидротопные, к21 — аэрогидрогеотопные, к22 — гидро геотопные, к23 — гидротопные, к24 — геотопные, к25 — аллотопные (растения ис- пользуют иные среды жиз- ни, например, при культи- вировнии их на искусствен- ных питательных средах); 11-16 — no характеру и длительно- сти вегетации растений: 11 — бессезонновегетирующие, 12 — всесезонновегетирующие, 13 — летневегетирующие, 14 — неполно летневегетирующие (со среднепродолжительной летней вегетацией), 15 — озимые, 16 — яровые; 17-19 — no площади листовой плас- тинки: 17 18 19 — узколистные, — широколистные, — крупнолистные (отмечается в числителе длина и в знаме- нателе ширина, в см или м); 110-125 — no форме кроны древес- ных растений: 110 — растения с кронами конусо- видными, 111 — веретеновидными, 112 — колонновидными, 113 — яйцевидными, 114 — обратнояйцевидными, 115 — яйцевидноконусовидными, 116 — эллиптическими, 117 — шаровидными, 118 — полушаровидными, 119 — зонтиковидными, 120 — плакучими, 12 1 — флаговидными, 122 — лепешковидными, 123 — пучковатыми, 124 — сноповидными, 125 — неправильными; т l-т6 по способу опыления: энтомофильные, анемофильные, орнитофильные, клейстогамные, самоопыляющиеся, апомиктические; т1- т2— т3- т4— т5- тб— т7-mll — сроки проявления фено- логических стадий (начала и конца): т7 — сроки иааааа трастаот (кис- ло, месяц), т8 — сроки бутонизации, т9 — сроки цветения, т10 — сроки плодоношения, т11 — сроки окончания вегетации; т 12-т 19 — no способу распростра- нения плодов и семян: т12 — автохорные, т13 — анемохорные,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ т14 — гидрохорные, т15-т17 — зоохорные, т15 — эпизоохорные, т16 — синзоохорные, т17 — эндозоохорные, т18 — баллистные, т19 — антропохорные (Левина, 1957); т20-т25 — no специализации орга- нов к вегетативному размножению: m20 — виви парные, m21 — выводково-клубневые, m22 — выводково-почковые, m23 — выводково-клубнелуковичные, m24 — наземно-столонные, I1125 — подземно-столонные; п1-n3 — no особенностям оиушения листьев: n1 — неопушеннолистные, n2 — жестко опушен нол исти ые, n3 — войлочноопушеннолистные; и4-и9 — no особенностям расиоло- жения иобеговых ветвей усиленного роста: n4 — геобазито нные, n5 — аэробазитонные, пб — мезотонные, n7 — акротонные, n8 — мезоакротонные, n9 — голотонные; п10-n18 — no иоследовательности расположения иобеговых ветвей: п10 — не ветвистые, n11 — очеред новетвистые, n12 — супротивноветвистые, n13 — муовчатоветвистые (мутов- ка ветвей возникает из одно- го узла), n14 — очередно-условно-мутовча- товетвистые (мутовка ветвей возникает из нескольких сближенных узлов), n15 — супротивно-условно-мутов- чатоветвистые (в организа- ции растений встречаются условные мутовки, образо- ванные несколькими парами сильно сближенных супро- тивных ветвей), n16 — очередно — супротивновет- вистые, n17 — очередно-мутовчатоветвис- тые, n18 — супротивно-мутовчатоветви- стые (встречаются как су- противные пары ветвей, так и муговки ветвей); и 19-и20 — no стеиени кущения: n19 — не кустистые, n20 — про сто кустистые, n21 — сложнокустистые; и22-и25 — no отдалению элемен- тарных зон кущения: n22 — компактнокустистые, n23 — рыхл окустистые, n24 — полуразвернуто кустистые, n25 — развернуто кустистые; ol-o5 — no отношению к разрас- танию иобегов в горизонтальном направлении (ириземно, иодземно, подводно): о1 — апланофильные (неразраста- ющиеся), о2 — гипопланофильные, o3 — брахи планофильные, о4 — мезопланофильные, 05 — тахипланофильные; об-o25 — no организации базового органа: об — проценоподиальные, о7 — аэро подиальные, о8 — гетероподиальные, о9 — прекаудексовые, о10 — надземно-каудексовые, о11 — полуподземно-каудексовые, о12 — подземно-каудексовые, о13 — проризомные,
Главс 9. Многолинейная классификация биоморф семенных растений 014 — длинно-корневищные, о15 — коротко-корневищные, о 16 — компактно-корневищные, о 17 — луковичные, о 18 — клубы елуковичные, о 19 — корневищно-луковичные, о20 — подземно-клубнепобеговые, о21 — подводно-клубнепобеговые, о22 — полуподземно-клубнепобе- говые, о 23 — надземно-клубнепобеговые, о24 — конодиальные, о25 — автономные персистентно- почечные (роль базового ор- гана выполняют обособлен- ные зимующие почки, на- пример, виды Orchis sp.); р1-р2 — no связи базового органа с главным корнем: pl — гетерономно-базовые (базо- вый орган имеет связь с глав- ным корнем), р2 — автономно-базовые (связь с главным корнем утрачена); р3 p8 — no отношению к вегета- тивному размножению: p3 — изоаклональные, р4 — дефективно-кондивидуаль- ные, р5 — ретардационно-кондивиду- альные, рб — экстенсивно-кондивидуаль- ные, р7 — интенсивно-кондивидуль- ные, р8 — экспансивно-кондивидуаль- ные; р9-р11 — no стеиени суккулентности: р9 — каулосуккулентные, р10 — фрондосуккулентные, р l l — десуккулентные; р12p13 — no наличию или отсут- ствию чешуй в структуре зимую- щих иобегов: р12 — бесчешуйчато-почечные, р13 — чешуйчато-почечные; р14p21 — no срокам и иродолжи- тельности проявления возрастных состояний — в скобках отмечает- ся время начала: (и) — 1-я декада, (с) — 2-я декада, (к) — 3-я декада месяца и окончания соответствую- щей стадии (желательно отмечать иродолжительность стадии в днях, неделях или месяцах, годах): р14 — проростки, p15 — всходы, p16 — ювенильные (если различа- ются стадии jl, j2 и т.д., то сроки прохождения их дают- ся поочередно — 11, 12 и тд.), р17 — имматурные (im1, im2 и тд.), р18 — инициматурные, р19 — медиматурные, р20 — финиматурные, р21 — сенильные; р22-р25 — no особенностям эиидер- мальных образований на стеблях: р22 — голостебельные, р23 — с кроющими трихомами (во- лосками) на стеблях, р24 — с железистыми трихомами (волосками) на стеблях, р25 — с шипами на стеблях; ql-дб — no сложности организации корней: ql — моноризные, q2 — диризные, q3 — триризные, q4 — тетраризные, q5 — синторизные; дб-q18 — no внешней структуре корневых систем: q6 — стержнекорневые, q7 — поверхностно-стержнекор- невые, q8 — стержнево-пальчатокорне- вые,
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ q9 поверхностно-стержнево- якорнокорневые, поверхностно-якорные, поверхностно корневые, пал ьчато корневые, поверхностно-корневые с восходящими коралловид- ными корнями, поверхностнокорневые с ды- хательными корнями-пнев- матоф орам и, поверхностнокорневые с дос- ковидными корнями, кисте корневые, пучковатокорневые, разно корневые; q10- qll— q12- qlj— q14— q15— q16- q17— q18— q19-q25 — no доиолнительной сиеци- ализации внешней структуры корней: q19 — веретеновиднокорневые, q20 — конусовиднокор не вые, q21 — шаровиднокорневые, q22 — шитшсовато-корнеклубневые, q23 — клубне корневые, q24 — шнуровиднокорневые, q25 — реповиднокорневые; rl-r8 — ио доиолнительной сиециа- лизации функции корней: гl — дыхательно корневые, r2 — втягивающекорневые, r3 — подпорково-столбовиднокор- невые, r4 — ассимилирующе корневые, r5 — ходульнокорневые, r6 — цепляющекорневые, r7 — висяче-воздушнокорневые, r8 — плавающекорневые; г11-r12 — по широте иростирания корней: г! ! — асантнметрат(когдаменее ! м), r9-r10 — no глубине ироникновения корней: l9 — асантнметрак(когдаменее ! м), г10 — в метрах (когда более 1 м); г12 — в метрах (когда более 1 м); r13-rl 7 — иохарактерулетнего иокоя: г13 — вынужденно летнепокояшиеся, г14 — органически коротколетне- покоящиеся, r15 — органически средне-летне- покоящиеся, гlб — органически длительно-лет- не покоящиеся, г17 — непрерывно летневегетиру- ющие; r18-r20 — no отношению к корне- отпрысковости: г18 — некорнеотпрысковые, г19 — факультативно-корнеотпрыс- ковые, r20 — корнеотпрысковые; r21-r25 — no отношению к темие- ратурному режиму: г21 — гипергекистотермные (средняя температура июля ниже 2' С), r22 — гекистотермные, r23 — микротермные, r24 — мезотермные, г25 — мегатермные; sl-s6 — no сиособу иитания: s l — автотрофные, s2 — сапрофильные, s3 — полупаразитные, s4 — паразиты ые, s5 — микотрофные, s6 — насекомоядные; s7-s10 — no отношению к струк- туре субстрата: s7 — эдафильные (педофильные), s8 — псаммофильные, s9 — литофильные, s10 — гелофильные; sll-s14 — no отношению к богат- ству почвы: s l l — эвтрофные,
Глава 9. Многолинейная классификация биоморф семенных растений 457 s12 — мезотрофные, s13 — олиготрофные, s14 — эвритрофные; s15-s22 — no отношению к водному режиму питания: s15 — гиперксерофильные, s16 — эуксерофильные, s17 — мезо ксеро фильные, s18 — ксеромезофильные, s19 — эумезофильные, s20 — гигрофильные, s21 — гидрофильные, s22 — гидатофильные; s23-s25 — no световому режиму: s23 — гелиофильные, s24 — сциогелиофильные, s25 — сциофильные; tl-t8 — no отношению к солевому режиму почвы: t l — гликофильные, t2-t4 — галофильные, t2 — соленепропускающие, t3 — солевыделяющие, t4 — соленакапливающие (Ша- хов, 1956), t5 — кальцефильные, t6 — кальцефобные, t7 — нитрофильные, t8 — галоидофильные; t9-t12 — no кислотности иочвы: t9 — индифферентные, t10 — оксилофильные (устойчивые к кислой реакции почвы), tl1 — базифильные (устойчивые к щелочной реакции почвы), t12 — нейтральные (виды нейтрапь- ных почв); t13-t15 — no месту ироизрастания в отношении с человеком: t13 — дикорастущие, t14 — культивируемые в открытом грунте, t15 — культивируемые в закрытом грунте; t16-t19 — no особенностям влаго- поглощения из почвы: t16 — амброфильные, Г(7 — акротрикогидрофиланеге (кор- ни достигакгг верхнего свеже- го слоя капиллярной каймьг), t ( 8 — меагприлогидрафиланаге (кор- ни достигаип среднего аллино- го слоя капгопярной каймы), t19 — базитрихогидрофильные, или фреатофильные (корни дости- гакл нижнего сырого слоя ка- пиллярной каймы, иногда про- никая в водоносный горизонт); t20-t21 — no широте географичес- кого ареала: t20 — эндемичные, t21 — космополитные; t22-t25 — no стеиени иоедания жи- вотными: t22 — хорошо поедаемые, t23 — удовлетворительно поедае- мые, t24 — плохо поедаемые, t25 — непоедаемые; и1-и2 — no ироисхождению в от- ношении к местным условиям произ- растания: и1 — аборигенные, и2 — адвентивные (пришлые); и3-и9 ком в — no использованию челове- ирактической деятельности: и3 и4 и5 цб U7 и8 и9 пищевые, технические, лекарственные, инсекте цидные, декоративные, пастбищно-кормовые, медоносные;
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ и17 — полупустынные, и18 — пустынные, и19 — саванные, и20 — мангровые, и21 — сорные, и22 — рудеральные, и23 — высокогорные, и 24 — тундровые, и25 — пустынно-арктические. и10-и25 — no месту ироизрастания в отношении с окружающей биоге- оценотической средой: и10 — морские, ul l — озерно-речные, и12 — низины о-болотные, и13 — верховка-болотные, и14 — лесные, и15 — степные, и16 — лесостепные, В многолинейной классификации биоморф семенных растений каждый признак и сравнительный ряд признаков закодированы с помо- щью буквенно-цифровой символики, которая усилена для большей наглядности еще и графической символикой (рис. 181). В многоклеточ- ной таблице-матрице вертикали маркированы латинскими буквами, а горизонтали — арабскими цифрами. Код признака — это сборное обо- значение, которое несет информацию, вложенную в одну клеточку таб- лицы. Это обозначение слагается соответствующей буквой и цифрой, поскольку каждая клетка таблицы — это результат пересечения опре- деленной вертикали (с буквенным индексом) и горизонтали (с ци- фровым индексом). Числовой индекс приписывается к главному буквен- ному индексу, в результате чего возникают символы, различающиеся друг от друга; они последовательно однозначно маркируют все клетки. В рассмотренном виде таблица-матрица, внешне по символике напоми- нающая шахматную таблицу, передает содержание многолинейной клас- сификации. Например: а7 — означает, что растения по сложности их общей организации — ризогептакормусовые, или в9 — растения по продолжительности жизни листьев и особенностям их смены — бессе- зоннозеленые и т.д. При считывании информации с таблицы-матрицы мы получаем формулы, которые могут харктеризовать отдельную биоморфу в каком- либо ее возрастном и (или) фенологическом состоянии или на протя- жении всего жизненного пути, т.е. или тип состояний биоморф (геми- тропобиоз), или биоморфотип, или тип биоморфогенезов (тропобиоз). Общий вид таких формул можно выразить так: ТВх= ау у у- ву у у-су у у- ..., или У У У ТВх= ау- ву-су... У У У Условные обозначения к формуле: ТВ (или ТБ) — тип биоморфо- генезов, или биоморфотип, охарактеризованный через индикаторные
459 Глава 9. Многолинейная классификация биоморф семенных растений (тропобиозные) признаки; вместо ТВ (ТБ) ставим другие символы, если отмечаем типы состояний биоморф: pjl, pj2, jl, iml, im2 и т.п.; в тех случаях, когда формула строится по одной биоморфе, то символы от- дельных возрастных состояний выделяем посредством скобок, напри- мер, j l — первый ювенильный гемитропобиоз, (j l) — первое ювениль- ное состояние биоморфы; х — видовое название растений; а, в, с...— буквенные индексы, у — цифровые индексы в таблице биоморфотипов (рис. 181). Рассмотренный выше принцип построения формул приме- ним как в отношении типовых категорий биоморф и биоморфогенезов, так и отдельных биоморф и биоморфогенезов. Кроме рассмотренного способа формализации языка биоморфоло- гии, мы предлагаем использовать еще один способ. Ведущие признаки организации основных органов и их элементов обозначаются опреде- ленными символами — латинскими буквами, а иногда -и арабскими цифрами. Совокупность этих символов располагается в определенной последовательности, составляя краткую формулу, отражающую важней- шие особенности организации биоморф. Краткая формула организации биоморф семенных растений несет сведения о габитусе, типе побега и корня (побегов и корней), о гипокотиле (наличии или отсутствии его), базовом органе. Общий вид формулы организации биоморф семенных растений, составленной вторым способом формализации, таков: ТВх =- G (h) С (h) + НВ + R Условные обозначения: ТВ — это тип биоморфогенезов. Вместо ТВ могут быть и другие обозначения, которые мы отметили при опи- сании первого способа построения формул: х — видовое название растений; G — биоморфы, охарактеризованные по габитусу: Q — одно- летник, Q — яровой однолетник, g — озимый однолетник, Q — дву- летник, Q — яровой двулетник, � — озимый двулетник, � — много- летник унирепродуктивный, 4. — многолетник мультирепродукгивный, ~ — древесные растения (деревья и кустарники), g — деревья, некус- тистые без корневых отпрысков, ~ — деревья кустистые без корневых отпрысков, ~ — деревья кустистые с корневыми отпрысками, g — кус- тарники без корневых отпрысков, g — кустарники с корневыми от- прысками, g — кустарнички без корневых отпрысков, g — кустарнич- ки с корневыми QTIIpbIGKBMH, h — полукустарники без корневых от- прысков, ~ — полукустарники с корневыми отпрысками, h. — полу- кустарнички без корневых отпрысков, п — полукустарнички с корне- выми отпрысками. (h) при символе G — высота надземного базового органа, когда он есть, (h) при символе С — высота генерального или субгенерального аэрокормуса, С — тип побега или системы побегов биоморфы: 01С — протерокормус, 1С — монокормус, 2С — дикормус, ЗС — трикормус, 4С — тетракормус, 5С — пентакормус, 6С— гексакормус, 7С — гептакормус, 8С — октокормус, 9С — энатокормус, 10С — синкормус, 11С — клональная система кормусов; если символ побега подчеркнут — это означает, что семядоли еще живы. Н — ги- покотиль есть; если нет этого символа, то гипокотиля уже или вообще
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ нет. В — тип базовых органов: р — проценоподий, а — аэроподий, gp — гетероподий, рс — прекаудекс, cd — каудекс, pr — проризом, lrm — длинный ризом, 2rm — короткий ризом, 3rm — компактный ризом, b — зрелая луковица или система таких органов, bt — зрелая клубнелуковица или система таких органов, rb — корневищная лукови- ца, tc — зрелый клубнепобег или система таких органов, cn — коно- дий, g — обособленная персистентная (зимующая) почка, gb — почко- луковица, сЬ-солуковица. R — тип корня или системы корней: 1R— монориза, 2R — дириза, 3R — тририза, 4R — тетрариза, 5R— синториза, 6R — система корней клона. Типы вегетации побегов обо- значаем следующими значками перед индексом С (при описании типов возрастных стадий): ~ — виргинильная вегетация побегов, т.е. вегета- ция побегов на виргинильном этапе биоморфогенеза, f' — матурная вегетация побегов, ~ — сенильная вегетация побегов. Опишем для примера эпиэмбриональную биоморфогению Вир1еи- rum triradiatum, используя два способа формализации языка биоморфо- логии. Словесное описание биоморфогении этого вида было проведено в главе 7. pj 1 = 01С + H + 1R; pj2 = 1С + Н + 2R, 4R; j = 1С, 4С + Н рс~сп+ 4R; im = 6С, 7С (9С) + Hcd + 4R; mi 1 = 5С + Hcn+ 4R; mi2 = 7С + Hcd + 4R; тт = 49C (5 — 15 см) + Hcd + 4R, 5R; fm = 7С, 9С + Hcd + 4R, 5R; s = 7С, 9С + Hcd + 4R, 5R. ТВ Bupleurum triradiatum = а 9 19 — Ь 4 (5 — 15 см) 5 11 14 23— с 21 (0 8 — 1,2; 0,1 — 0,12 см) — d 19 — е 6 12 24 — f 1 7 13 — g 2 10 16 25— h 5 — i 1 13 — k 10, 9 19 — l3 7 — т 1,2 12 — n 1 4 11 21 22 — о 2 11- р 1 3 13 22-q4 7- r9 (до 20см) 11(до 20см) 17 18 23- s 1 9 13 19 23 — t 13 16 — и 1 23. Согласно таблице-матрице, в которой закодирована многолиней- ная классификация биоморф семенных растений, можно собрать и ра- зобрать организацию биоморфы в пространстве и времени как по от- дельным возрастным состояниям, так и в целом биоморфогению вида. Повествовательная информация о биоморфогении Bupleurum multinerve была дана в главе 8. Представить ее можно и в формализованном виде. ТВ Bupleurum multinerve — а 9 (7, 5, 6) 19, 21 — Ь 4 (20 — 60 см) 5 11 (12) 14 23 — с 19, 21 (0,9 — 2,0; 0,1 - 0,2 см) — д 19 — е 6 12 (3 — 20~ 0,3— 1,5 см) 24 — f 1 7 13 (11, 12, 10) 25 (n — 7, 2n — 14) — g 2 10 (9).16 25— h 5 — i 1 13 — k 10,9 19 — 13 7 — т 1,2 7 (IV) 8 (3/4 VI, VII) 9 (ЧП — ЧШ) 10 (3/4 VIII — IX) 11 (IX, Х) 12 — n 1 21 (20, 19), 22, 23—
Глава 9. Многолинейная классификация биоморф семенных растений 461 о 2,3 11, 12 (24) — р 1 3 11 14 (pjl V, VI, VII - 1-2 недели) 15 (pj2 — VI, VII — VIII, IX) 16 (~1 — VII, VIII, IX — Х, XI, j2 — с Х с.г. до 12-17 лет) 17 (iml с 2-17 до 3-18, чаще до 5-10 лет; im2- c 3-18 до 5-20 лет) 18 (mi — ТВ1 с 3-12 до 3-17 лет; ТВ2 с 3-18 до 7-20 лет; ТВЗ с 4-20 до 12-25 лет) 19 (ппп — ТВ2 с 7-16 до 8-20 лет; ТВЗ с 12-25, чаще 14-20 до 18-30 лет) 20 (fm - TB2 с 8-20 до 22 лет; ТВЗ с 18-30 до 40 лет) 21 (s — ТВ2 очень редко с 15-20 до 16-22 лет; ТВЗ с 35-40 до 36-42лет) 22 — q 4,5 7 — r 9 (до 15 см) 11 (до 25 см).17 18 23 — s 1 9 13 19 — t 6 12 25 — и 1 5 23. Расшифруем эту информацию (порядок расположения признаков приводится с несколько иной последовательностью для большего удоб- ства их восприятия, в скобках указан код признака). Биоморфогению Bupleurum multinerve характеризуют растения ри- зоэнатокормусовые (в других типах биоморфогенезов ризогептакорму- совые, ризогексакормусовые или ризопентакормусовые) [а 9 (7, 6, 5)], эпигенные (с погружающимся в почву базовым органом) [а 19], с над- земными побегами высотой 20-40 см (в 4), надземными (в 14) ланцетны- ми, или иногда линейными, (с 19, 21) семядолями, 0,9-2,0см дл., 0,1-0,2 см шир., двусемядольные (реже трехсемядольные) [в 11 (12)], сбрасывающие семенную кожуру с семядолей (е 24), монофиллические (е 12), с летнезелеными (в 5) ланцетными (d 19) очередными (f 1) лис- тьями без прилистников (g25), полурозеточные (е6), гемикрипто- фильные (в 23), обоеполые (f 7), мультирепродуктивные (иногда три-, би- или унирепродуктивные ) [f 13 (12, 11, 10)], с гаплоидным набором хромосом n — 7, диплоидным 2n — 14 (f 25), гемисимподиальные (реже моноподиальные) [g 10 (9)], с рацемозными (g 2), полузакрытыми, или семидетерминатными, верхушечными соцветиями (g 16), со сложными зонтиками (h 5), цилиндрическистеблевые (i 1), прямостоячепобеговые (i 13), курторезидные (к 10) или миниморезидные (к 9), аэрогеотопные (к 19), летневегетирующие (13), узколистные (17), энтомофильные или анемофильные (т l, 2), автохорные (т 12), неопушеннолистные (n 1), сложнокустистые (реже простокустистые и некустистые) [n 21 (20, 19)], компактнокустистые (n 22) или рыхлокустистые (n 23), гипопланофиль- ные (о 2) или брахипланофильные (о 3), полуподземнокаудексовые или подземнокаудексовые (реже конодиальные) [о 11, 12 (24)], гетерономно- базовые (р 1), изоаклональные (р 3), десуккулентные (р 11), тетраризные (ц4) или синторизные (q 5), поверхностно-стержнекорневые (q 7), проникающие в глубину до 15 см (r 9) и простирающие корни до 25 см (r 11), непрерывно летневегетирующие (r17), некорнеотпрысковые (r 18), микротермные (r 23), автотрофные (s 1), литофильные (s 9), олиготрофные (s 13), эумезофильные (s 19), непоедаемые животными (t 25), аборигенные (и 1), лекарственные (и 5), высокогорные (и 23), оби- татели нейтральных почв (t 12). Сроки проявления фенологических ста- дий: начало отрастания побегов в апреле (т 7), бутонизация — 3/4 VI, VII (т 8), цветение VII — VIII (т 9), плодоношение 3/4 VIII IX (т 10), окончание вегетации — IX, Х (т 11). Сроки и продолжительность проявления возрастных состояний: р11 — V, VI, VII — 1 — 2 недели (р 14), pj2-VI, VII, VIII-IX (р15), jl-VII, VIII, IX до Х, Xl; j2-с Х, Xl с.г. до
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ 12-17 лет (рlб), im 1-с 2-17 до 3-18, чаще до 5-10 лет; im2- с 3-18 до 5-20 лет (р 17), mi — ТВ1 с 3-12 до 3-17 лет; ТВ2 с 3-18 до 7-20 лет; ТВЗ с 4-20 до 12-25 лет (р 18), mm — ТВ2 с 7-16 до 8-20 лет; ТВ3 с 12-25, чаще 14-20 до 18-30 лет (р 19), fm — ТВ2 с 8-20 до 22 лет; ТВЗ с 18-30 до 40 лет (р 20), s — ТВ2 очень редко с 15-20 до 16-22 лет; ТВЗ с 35-40 до 36-42 лет (р 21). Приведем также формулы возрастных состояний биоморф Вир1еи- rum multinerve: ТВ1-3 pj1=01C+ H+1R pj2= И; + Н+ 2R,4R jl = 1C+ H+ 2R, 4R j2 = 1C, 4C + Hcn + 4R ТВ1- 3 im = 6С,7C+ Hcd+ 4R ТВЗ (редко ТВ2) im 2 = 9С + Hcd + 4R ТВ1т=5С,6С,7C+ Нсп, cd+4R ТВ2- 3 mi = 5С, 6С, 7C+ Hcd+ 4R ТВ2 тт = 7С, 9C+ Hcd+ 4R ТВЗ тт = 49С + Hcd + 4R ТВ2 — 3 fm = 9С+ Hcd+ 4R, 5R ТВ2 - 3 s = 9C + Нс4+ 5R
Глава 10. Использование теории организации биоморф семенных растений в практической биоморфологии Научные теории формируются не из праздного любопытства или ин- теллектуального тщеславия, — они призваны соответстующим образом слу- жить в науке, выполнять определенные функции (Баженов, 1978). Био- морфогения семенных растений изучалась нами параллельно с накоплением фактов и их теоретическим осмысливанием, поэтому эти факты питали разработанную выше теорию при ее создании и, естественно, сейчас полно- стью отвечают ее современному содержанию. Рассмотренные выше методы использованы нами при изучении био- морфогении семенных, в частности лекарственных, растений. Надежность и краткость термина-понятийной основы и формализованный язык привносят лаконизм в традиционный повествовательный способ описания жизненных циклов растений. Далее при описании биоморфогении семенных растений с учетом разработанных теоретических положений, будем использовать более привычный повествовательно-рисуночный способ описания. 10.1. Ценодемология семенных растений (на примере Bupleurum аигеит Fisch. ex НоЛт.} Более трех десятилетий прошло с тех пор, как Т.А.Работков (1945, 1950а, 1950б) выдвинул и широко обосновал идею изучения популяций (со- вокупностуй особей одного вида в сообществе), которую предложено на- зывать ценотической (Петровский, 196l; Корчагин, 1964, 1966). Грамма- тически более точным сокращенным вариантом к термину «ценотическая популяция» будет, на наш взгляд, не термин «ценопопуляция», а термин «ценодем» (от греч. койнос — общий, демос — население, народ), что, кстати, более всего отвечает названию всего направления в познании цено- тических популяций (ценодемов) — демографии растений (Harper, 1960, 1968, 1977, 1978; Harper, White, 1970, 1974; Noble, Bell, Harper, 1979; Urbanska- Worytkiewicz, 1979; Bullock, 1980 и др.). Специальный обзор направления,
ОСНОВЫ БИОМОРФОЛОГИИ CENEHHblX РАСТЕНИИ развиваемого Д.Л.Харпером, дан в статье М.В. Маркова (1976). В настоящее время в отечественной литературе накоплен большой опыт изучения цено- тических популяций (Работнов, 1946, 1949, 1950б, 1964, 1969, 1975, 1978; Уранов, 1960, 1974; Рысин, Рысина, 1966; Смирнова, 1968, 1987, 1994; Ура- нов и др., 1970, 1976, 1977; Рысин, Казанцева, 1975; Смирнова и др., 1976; Воронцова, Заугольнова, 1979; Марков, 1986, 1990; Заугольнова и др., 1988, 1991, 1993; Попадюк и др., 1994 и др.). Исходной основой при изу- чении ценодемов является определение возрастной изменчивости биоморф. В течение нескольких лет мы собирали материал по биоморфогении и возрастной структуре ценодемов Bupleurum аигеит Fisch. ех Hoffm*. В итоге, в дополнение к известным положениям, разработанным Т.А.Ра- ботновым и его последователями, предлагаем некоторые новые положения о возрастной изменчивости ценодемов. В зависимости от того, каков возрастной состав ценодемов, можно, применяя терминологию, аналогич- ную терминологии возрастных стадий биоморфогенезов и возрастных состояний биоморф, различать следующие возрастные состояния ценоде- мов, которые во временном отношении будут возрастными стадиаии их морфогенезов, т.е. ценодемогенезов. 1. Латентные ценодемы (символ CD1) — их структуру составляют одни жизнеспособные семена. 2. Преювенильные ценодемы (CDpj) — они состоят из семян, про- ростков и всходов. 3. Ювенильные ценодемы (CDj) — они состоят из семян, пророст- ков и (или) всходов и (или) ювенильных особей, которых по численности больше, чем других виргинильных особей, т.е. им- матурных особей, если они вообще развиты, а матурных и се- нильных особей еще нет (pjl + pj2+ j > im при т+ s = 0). 4. Имматурные ценодемы (CD im) — im > pj1 + pj2 + j npu m + s = 0 (имматурных особей больше, чем остальных виргинильных осо- бей, а матурных и сенильных особей еще нет). 5. Инициматурные ценодемы (CDmi) — v > т+ s или v+ mi > тт+ fm+ s (виргинильных особей больше, чем матурных и сениль- ных особей, или виргинильных и инициматурных особей боль- ше, чем остальных матурных и сенильных особей). 6. Медиматурные ценодемы (CDmm) — ч+ mi < mm+ fm + s (вирги- нильных и инициматурных особей меньше, чем медиматурных, финиматурных и сенильных особей). 7. Финиматурные ценодемы (CDfm)-fm + s > v + mi + тт (фини- матурных и сенильных особей больше, чем виргильных, иници- матурных и медиматурных особей). 8. Субсенильные ценодемы (CDss) — s > т+ v (сенильных особей больше, чем матурных и виргинильных особей). 9. Сенильные ценодемы (CDs) — присутствуют, без учета семян, только сенильные особи. Видовой ранг В. aureum признают многие ботаники (Линчевский,1950; Малышев, 1968 и др.) и принят в настоящей работе. Перенесение этого вида в ранг подвида — Bupleurum longifolium ssp.aureum (Fish. ех Hoffm) Soo (Черепанов, 1995) вызывает большое сомнение, особеиио Ilopllc вивкомствв с вевсокопииыми котовскими пеистивескими попумппппм.
Глава 10. Использование теории организации биоморф семенных растений 465 Эта схема возрастной изменчивости ценодемов, а одновременно с со- ответствующей интерпретацией и возрастной стадийности ценодемогенеза, не противоречит известной классификации ценотических популяций по их возрастности (Работков, 1945, 1950а, 1978 и др.), а дополняет ее. Так, 1-4 типы возрастных состояний ценодемов, рассмотренные выше, — это по Т.А.Работнову, инвазионные ценотические популяции (ценодемы), 5-7 типы возрастных состояний — это нормальные ценодемы и 8-9 типы возрастных состояний ценодемов в основном соответствуют регрессивным ценотичес- ким популяциям. Рассмотрим некоторые возможности использования схемы возраст- ной изменчивости ценодемов на основе разработанной теории органи- зации биоморф семенных растений на примере Bupleurum aureum Fisch. ех Hoffin. Всякое ценодемологическое исследование должно начинаться с изучения биоморфогении вида в районе исследований, поэтому сна- чала кратко рассмотрим биоморфогению Bupleurum аигеит в естествен- ных условиях произрастания на Алтае. Володушка золотистая (Bupleurum аигеит), благодаря содержанию разнообразных веществ лечебного действия, среди которых преобладают флавоноиды, нашла применение в медицине при лечении гепатитов, холециститов и некоторых других заболеваний (Верещагин, Соболевс- кая, Якубова, 1959; Минаева, 1960; Якубова, 1961 и др.). Bupleurum аигеит — мезофит, встречается на юге лесной и лесостепной зон Европы, на Урале, в Сибири, горах Тянь-Шаня, доходит до Забайкалья. Произрастает по всему Алтаю преимущественно в лиственни ейных, березовых лесах, кустарниковых зарослях и опушках, лесных и субальпийских лугах (Ку- минова, 1960), где исследовапся этот вид (Хржановский, Нухимовский, 1972). Эмбриональный период. Пренатальный этап. ПС — июль-сентябрь. ОХ — оплодотворенная семяпочка или зрелое семя в структуре роди- тельского растения. Латентный этап. ПС — с августа-сентября до мая, июня следую- щего года. ОХ — семя, обособленное от родительского растения. А— односемянный полуплодик (мерикарпий) имеет недоразвитый зародыш, дозревающий при пониженной температуре в течение продолжительно- го времени (Израильсон, 1965, 1969). Эпиэмбриональный период. Виргинильный этап. Преювенильная фаза. Проростковая гипофаза (рис. 182, А). ПС — май, июнь; 7-15 дней. ОХ — прорастающее семя или семядольный почкокорень (ризопротеро- кормус). А — семядоли две надземные голые линейно-ланцетные или ланцетные 1,4-1,7см дл., 0,1-0,15см шир., гипокотиль 0,6-2,0см дл., главная монориза, реже дириза до 4 см дл. Всходовая гипофаза (рис. 182, Б). ПС — май, июнь, июль, редко август; 2-5 недель. ОХ — семядольный вегетативный предрозеточный (с 1-2 сближенными настоящими листьями) или розеточный (3-4 на- стоящих листа) ризомонокормус. А — семядоли 1,5-2,0 см дл., 0,15-0,2 см шир. Первые листья имеют небольшие широко-яйцевид- ные и округлые пластинки с длинными черешками. Ювенильная фаза. Гипофаза первая — униювенильная (рис. 182, В). ПС — с июля, августа до начала зимы текущего года (редко) или до 2-5 лет. ОХ — бессемядольный вегетативный розеточный на первом году
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 182. Ви гинил р ильные аклонисты Вир1еигит аигеит Fisch. ех Найт. (Алтай А — проростки, Б — всходы В — юв — венильная особь первого сезона вегетации поима горного ручья, низкогорье — 300-600 м над ур. моря) Г— пологом ста н ку р иков, низкогорье), Д — ювенильная особь — . моря, — то же под тации (там же, что и Г), Е — то же с со ь второго сезона веге- е, что и ), — то же (среди лугового разнотравья, низкогорье) Ж- У особ (там ж, чт Е' К— 1— , что и ), — то же (субгльпийский луг, 1800 м над . мо — семядоли, 2 — гипокотиль 3 — с ур. моря); 5е — ко ни: — семядольный узел, 4 — розеточные листья, 5а- е — корни: 5а — главная монориза (субмедигль), 5б — ди иза 5в- монориза (сублатер ь) 5г — „„5 — ризогенная корневая ветвь, 7 — каулогенная корневая ветвь 8— ки листьев 9 — веге — ветвь, — сухие остат- — вегетативный медиальный розеточный моноко м с 10 — в тивный медиальный розеточный те ако м с 11— кормус, — вегета- чныи тетракормус, 11 — вегетативный аксиллярный точны тетракормус, 12 — мономерный курто езид 13 — вь нерозеточный побег 14 — мо курторезид, 3 — вынужденно верх- по ег, — мономерный пленорезид, 15 — преконодий 16 — ко- Э — — — ~ы~ядольный почкокорень (ризопроте- ко мс 22— рокормус, — семядольный ризомонокормус, 21 — бессемядольный р у, — ежегодно вегетативный ризогептакорм 23— — льны ризомоно- основания листьев (верхние части дарлены ус, — почки, 24 — живы — ~е масштаб 'го и удалены); ув — увеличено по отношению к шта у; горизонтальная линия — поверхность почвы' о , дно деление масштаба — 1 см. вегетации, позднее — п о и — р р зомный ризомонокормус или ризотетра- кормус. А — от ° .У фазы. Но малько — сыхание семядолеи — признак начала а ювенильной рмально развивающаяся особь на первом году вег етации имеет медиальныи побег (длина его стеблевой части 0,5-2,5 см), гипокотиль и главный корень, разветвленный до 1 или 2- и -го порядка,— и тетраризу; листья черешковые, яйцевидные или эллип кие, в паз ах 1-2 эллиптичес- Надземное о тот ух — листьев нередко закладываются одиноч очные почки. ртотропное положение медиального побега у особей пер-
Глава 10. Использование теории организации биоморф семенных растений 467 вого года вегетации постепенно сменяется полуподземным или подзем- ным плагиотропным положением в последующие сезоны вегетации. Не- редко полегание медиального побега и частичное погружение его в почву происходит уже к концу первого сезона вегетации. Придаточные корни развиваются на 2-5-й год вегетации (реже раньше) и момент их образования служит индикатором превращения протобазового органа- проризома в эубазовый орган — ризом, а одновременно и индикатором преобразования униювенильных особей в биювенильные. Гилофаза вторая — биювенильная (рис. 182, Д, Е). ПС — с начала зимы текущего года (когда рано образуются каулогенные корни) или с 2-5 лет до 2-6 лет. ОХ — бессемядольный ежегодно вегетативный ри- зомный розеточный некустистый ризомонокормус или ризотетракормус. А — аккумулятивный моноподиальный прирост базового органа обес- печивают безлистные курторезидные монокормусы. Б — чем раньше побеги принимают плагиотропное положение, тем быстрее начинается образование адвентивных корней и тем деятельнее идет процесс геофи- лизации побегов. Наиболее интенсивно образуются корни из узлов на концах годичных приростов. Вероятно, такая закономерность в заложе- нии и развитии придаточных корней сложилась исторически как при- способление к ускоренному погружению приземных частей побегов в почву. Наиболее сильные придаточные корни часто растут от корневищ в противоположные стороны под острым углом к поверхности почвы,— это, вероятно, должно усиливать геофильные свойства растений. Втяги- вающая деятельность главного и придаточных корней, в результате которой из скелетной части медиального побега образуется эпигеоген- ное корневище (в понимании И.Г.Серебрякова и Т.И. Серебряковой, 1965), уже на ювенильном этапе жизни создает условия для надежной защиты зимующих почек. Аксиллярные почки на корневищах часто погружены в почву вследствие геофилии, а также засыпания растительным опадом и наносным субстратом; они защищенные, поскольку имеют довольно много (3-6) чешуй, и способны сохраняться в состоянии относительного покоя в течение нескольких лет. Пазушные почки обычно закладыва- ются в самой нижней части годичных побегов. Если учесть, что почки образуются только на годичном побеге в сезон его видимого роста, то нетрудно установить по сохраняющейся части корневищ, что почки могут не пробуждаться иногда до 10 и даже более лет. Терминальные почки защищенные; они имеют одну-несколько (до 5) кроющих чешуй, а также надежно прикрыты расширенными основани- ями черешков прошлогодних верхних розеточных листьев. Эти почки хорошо переносят зиму, находясь на поверхности почвы или незначи- тельно возвышаясь над ней, и ежегодно пробуждаются весной, обеспе- чивая моноподиальный прирост многогодичных побегов. В зависимости от условий произрастания наблюдается значительное варьирование по различным признакам структурной организации виргинильных особей. Так, при сравнении ювенильных растений, собранных из различных участков (500 м над ур. м.) одного района (Самарского) нетрудно было обнаружить, что под пологом кустарников (Rosa spinosissima L., Lonicera tatarica 1.) в кустарниково-луговоразнотравных сообществах развивают-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ся сильно угнетенные особи (рис. 182, Д), а в луговоразнотравной пойме горного ручья с непродолжительным осенне-весенним затоплением осо- би отличаются очень хорошим вегетативным развитием (рис. 182, В, Е). В первом случае в результате сильного затенения отрастают годичные верхнерозеточные побеги с короткими полегающими основаниями. Эти основания достигают до 6-8 см в длину и состоят из более или менее удлиненных междоузлий, т.е. они раздвинутоузловые. Листья двух ви- дов: сначала развертываются почечные чешуи (за исключением побега, отрастающего на первом году жизни растения), а в конце фазы види- мого роста на верхушках побегов развиваются по 3-4 сильно сближен- ных ассимилирующих листа. В условиях достаточного освещения обыч- но образуются годичные розеточные побеги. В нижней части таких побегов развиваются несколько (3-5) сильно или умеренно сближенных чешуевидных листьев, а в верхней — 3-6 ассимилирующих листьев, ко- торые и определяют форму побега. В конце июня-начале июля в различных районах и в различных условиях произрастания встречались всходы с 1-5 листьями и ювениль- ные особи. В крайне неблагоприятных местообитаниях, например, око- ло стволиков кустарников при очень сильном затенении, можно было найти единичные проростки; в этих условиях травостой сильно разре- жен, поэтому семена довольно легко прорастают на свободной площа- ди, но особи, как правило, обречены на скорое вымирание. Имматурная фаза (рис. 182, Ж-К). ПС — с 2-6 до 3-16, чаще 4-10 лет, а иногда выпадает совсем. ОХ — ежегодно вегетативный розеточный авто- номно-ризомный ризотетракормус, реже некустистый ризогексакормус или ризогептакормус (при простом перевершинивании), а кроме того кустистый гетерономно- или автономно-ризомный ризогексакормус или ризогептакормус, редко даже ризоэнатокормус. А — индикаторные признаки начала этой фазы — отмирание главного корня или чаще переход к кущению. Б — четкой корреляции между формой листьев и определенным возрастным состоянием у володушки золотистой не на- блюдается. Форма и размеры розеточных листьев у ювенильных и им- матурных особей сильно варьируют в зависимости от конкретных усло- вий произрастания и не могут служить критерием для выделения опре- деленных возрастных состояний. Форма розеточных листьев меняется меньше в сходных условиях местообитания, в частности она яйцевидная и широко эллиптическая в низкогорье (кустарниково-луговоразнотравные сообщества) и эллипти- ческая и ланцетная в высокогорье (субальпийский луг), но, как видим, довольно сильно преобразуется в зависимости от высоты местообита- ний над уровнем моря (рис. 182). Первые 3-6 узлов пазушных побегов несут листья чешуевидные и переходные к нормальным ассимилирую- щим, а последующие — только ассимилирующие. У активных побегов виргинильных особей их самая верхняя (апикальная) часть располага- ется на уровне поверхности почвы, но чаще всего несколько выше ее (на 0,2-3 см в зависимости от внешних условий). После полного отмирания главного корня у отдельных особей мож- но непосредственно установить только условный возраст (Смирно-
Глава 10. Использование теории организации биоморф семенных растений 469 ва, 1967); пределы истинных возрастов особей последующих возрастных групп — результат сравнительно-морфологических обобщений. У мно- гих особей в течение всей имматурной фазы главный корень сохраняет жизнеспособность. В пределах группы имматурных особей хорошо различимы три подгруппы: некустистые, с простым кущением (простокустистые) и со сложным кущением (сложнокустистые); последняя группа немногочис- ленна. Матурный этан жизни синорганизма состоит из двух иодэтаиов: аклонального и клонального. Особи клонального подэтапа сближены, по- этому нетрудно установить их происхождение. Инициматурная фаза (рис. 183, Л-П). ПС — с 3-16 до 5-20 лет. ОХ— ежегодно или неежегодно репродуктивный полурозеточный ризопента- кормус, ризогексакормус или ризогептакормус. А — в этой возрастной стадии находятся особи со следующей организацией: мероаклонисты некустистые непосредственно репродуктивные (в текущем вегетацион- ном сезоне у них отрастают годичные побеги с верхучшечными репро- дуктивными органами — зонтиками), 2 — мероаклонисты некустистые промежуточно-вегетативные (в прошлые годы у них отрастали годичные вегетативно-репродуктивные побеги, а в текущем году отрастает только вегетативный побег), 3 — мероаклонисты простокустистые непосред- ственно репродуктивные, 4 — мероаклонисты простокустистые проме- жуточно-вегетативные. Б — структуру корневища составляют курторе- зидные монокормусы и миниморезидные тетракормусы. Медиматурная фаза (рис. 183, Р, С). ПС — с 5-20 до 15-30 лет. ОХ— неежегодно репродуктивный полурозеточный сложнокустистый ризо- энатокормус. А — особи развивают репродуктивные органы неежегод- но, поэтому при изучении возрастной изменчивости ценодемов полезно выделять 2 группы матурных особей: сложнокустистые непосредственно репродуктивные мероаклонисты и сложнокустистые промежуточно-ве- гетативные мероаклонисты. Б — развертывание почек и отрастание по- бегов начинается рано весной. Первые листья как у терминальных, так и аксиллярных побегов чешуевидные. Время наступления и окончания фенологических фаз у володушки золотистой зависит от высоты над уровнем моря. В низкогорье отдельные фенофазы, в частности цветения и плодоношения, наступают на 2-3 недели раньше, чем на субальпий- ских лугах. В нижнем поясе гор (300-600 м над ур.моря) у володушки золотистой бутонизация приходится на последнюю декаду мая-первую половину июня, в конце июня-начале июля начинается цветение, ко- торое растягивается до конца июля-начала августа. Конец июля-ав- густ — фаза плодоношения; осыпание созревших семян осуществляет- ся в основном в сентябре. Структуру всей побеговой системы матурных особей составляют ди-, три-, плейофазные унирепродуктивные (моно- карпические обоеполые) полузакрытые полурозеточные побеги с под- земными и полуподземными, реже приземными, более или менее плагиотропными основаниями. По другой классификации эти побе- ги — пентакормусы; их составляют пазушные и верхушечные вегетатив- ные розеточные, реже верхнерозеточные, и безрозеточные унирепро-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ Рис. 183. Матурные и сенильные мероаклонисты и клонисты Вир!еигит аигеит (Алтай). Мероаклонисты: Л-П — инициматурные, Л — некустистые непосредственно ре- продуктивные (среди лугового разнотравья, низкогорье — 300-600 м над ур. моря), М — то же (субальпийский луг, 1800 м над ур. моря), Н — некустистый проме- жуточно-вегетативный (там же, что и Л), П — простокустистые непосредственно репродуктивные (там же, что и Л); Р-С медиматурные, Р — сложнокустистый непосредственно репродуктивный (там же, что и Л), С — сложнокустистый про- межуточно-вегетативный (там же, что и М); ТУ — финиматурный кондивидуум- клон; клонисты: Т-У — матурные, Т — сложнокустистый непосредственно репро- дуктивный (там же, что и М), У — сложнокустистый промежуточно-вегетативный (там же, что и Л); Ф — сенильная особь (там же, что и Л), Ш — сенильная особь (там же, что и М); 25 — унирепродуктивный безрозеточный годичный побег (схе- ма), 26 — паракладии, 27 — зонтичек, 28 — зонтик, 29 — фрагмент соцветия, 30— репродуктивный ризопентакормус (верхние части удалены), 31 — репродуктивный ризогептакормус (верхние части удивлены), 32 — неежегодно репродуктивный ри- зоэнатокормус, 33 — срединные листья цветоносного побега, 34 — чешуевидные (низовые) листья, 35 — основания монофазных или плейофазных унирепродук- тивных безрозеточных или полурозеточных побегов (верхние части удалены), 36 — миниморезид. Остальные обозначения те же, что и на рис. 182. дуктивные годичные побеги (монокормусы и дикормусы). Корневища эпигеогенного типа. При определении типа корневища учитывалась «первичность» или «вторичность» положения относительно поверхности почвы не частей корневища, а почек, из которых эти части в процессе соответствующего развития образовались. Верхушечные почки — при- земные; пазушные почки, как правило, закладываются в нижней части отрастающего годичного побега, располагаясь в подстилке или призем-
Глава 10. Использование теории организации биоморф семенных растений 471 но, они становятся спящими и впоследствии погружаются в почву. Таким образом, подземное положение пазушных почек обычно вторичное. В условиях низкогорья корневища матурных сложнокустистых осо- бей малой длины — 4-12 см; они составлены чаще всего малолетними, живущими 3-7, чаще 5-6 лет, резидными монокормусами. В условиях субальпийских лугов корневища особей того же возрастного состояния также короткие, но их линейные размеры больше — 8-25 см; они со- ставлены многолетними, живущими до 10-15, а в отдельных случаях и до 20 лет, резидными монокормусами. По числу таких резидов мы определяли условный возраст особей. Высота местообитаний над ур. моря володушки золотистой влияет также на многие другие морфоло- гические признаки (Израильсон, 1967, 1970). У растений, произрастающих на субальпийских лугах, средняя про- должительность жизни побегов больше, чем у растений, обитающих в условиях низкогорий, что можно объяснить более напряженным ре- жимом для видимого роста побегов в высокогорьях, в результате кото- рого также, вероятно, замедляется процесс отмирания частей побегов, выполняющих функцию хранилищ запасных питательных веществ. Число спящих почек колеблется от 1 до 10. Корневая система при- даточная. Корни пронизывают самые верхние горизонты почвы (до 20 см). Финиматурная фаза (рис. 183, Т, У). ПС — с 15-30 до 20-45 лет. ОХ— система клонистов одного кондивидуума, среди которых помимо репро- дуктивных клонистов могут быть и временно вегетативные клонисты. Старение матурных особей приводит к полной общей партикуля- ции, которая проявляется в отмирании нижних частей корневищ и отделении их простых или сложных ветвей вместе с корнями — конс- труктивных партикул. Образование обособленных конструктивных пар- тикул — это одновременно и образование клонистов, т.е. осуществле- ние посредством партикуляции вегетативного размножения; оно замед- ленное (ретардационное); клонисты малоподвижные и не могут суще- ственно отдалиться друг от друга, оставаясь сближенными. Сенильный этаи (рис. 183, Ш). ПС — продолжается 1-4 года. ОХ— ежегодно вегетативные после утраты физиологической зрелости розеточ- ные автономно-ризомные клональные ризокормусы. Б — полный возраст в низкогорье — 20-30 лет, в высокогорье — 30-50 лет. Резюмируем особенности экологической морфологии володушки зо- лотистой на Алтае — это травянистый летнезеленый надземка-двусе- мядольный розеткообразующий гемикриптофильный эпигеогенно- ризомный мультирепродукгивный полурозеточный очередно-цельнолист- ный гемисимподиальный желто-сельдерейноцветковый однодомный бра- хипланофильный ретардационно-кондивидуальный мезофильный луговой и горно-луговой вид растений с верхушечными соцветиями — зонтиками. Володушка золотистая — полиморфный вид. Хорошо различаются вы- сокогорные и низкогорные биоморфотипы по длине, ширине и форме ро- зеточных листьев, длине корневищ, высоте вегетативно-репродуктивных по- бегов, длине листочков, оберток и оберточек и по многим друтим признакам. Возрастную изменчивость ценодемов мы изучали в различных эко- логических условиях. Численность разновозрастных особей определяли
ОСНОВЫ БИОМОРф0~7ОГИИ СЕМЕННЫХ РА СТЕНИЙ на площадках в lм', которые закладывались через каждые 2м (10-15 площадок по двум взаимно пересекающимся, в виде креста, прямым). Возрастные спектры ценодемов володушки золотистой определены в следующих биотопах: 1. Пойменный галечник в 15м от русла притока р.Белая Уба (Ивановский хребет, 1000 м над ур.моря, 40 км на юго-восток от г. Лениногорска, 27.07.71). II. Надпойменная терраса притока Белая Уба, 100 м от русла, луговая ассоциация Cirsium heterophyllum-Dactylis glomerata (тот же район, что и I). Ш. 100 м в сторону от биотопа II, луговая ассоциация Dactylis glomerata + Poa sibirica. IV. Крутой горный склон западной экспозиции, луг в долине пересохшего ручья, ассоциация Poa altaica + Bupleurum multi- nerve (1800м над ур.моря, Курайский хребет, окрестности пос. Акташ, 01.07.73). V. 100 м ниже вдоль русла пересохшего ручья, ассоциация He- dysarum naglectum + Cirsium heterophyllum + Вир1еигит аигеит (там же, что и IV). VI. Надпойменная терраса в 80 м от ручья, луговая ассоциация Poa sibirica + Dactylis glomerata + Bupleurum аигеит (единич- но Rosa acicularis) (14 км на запад от пос. Самарское, Восточ- ный Казахстан, 29.06.1970). VII. Периодически затопляемая пойма ручья, ассоциация Filipendula и1тапа+ Angelica decurrens-Urtica dioica (там же, что и VI). УШ. Кустарниково-лугово-степная растительность западного скло- на горы Острая, ассоциация Rosa spinosissima + Lonicera tatari- ca-Bupleurum аигеит + Bromopsis inermis (100 м вверх по скло- ну от русла речки у подножья горы Острая, окрестности с. Боб- ровка, Восточный Казахстан, 22.07.70). IX. Кустарниково-луговой пояс горы Острая в 10-15 м от русла речки, ассоциация Rosa spinosissima + Lonicera tatarica-Spiraea crenata-Bupleurum аигеит (там же, что и VIII). Х. Пологий горный склон западной экспозици, ассоциация Larix sibirica (единично)-Poa sibirica+ Bupleurum аигеит (1800 м над ур. моря, 9 км южнее с.Берель, Восточный Казахстан, 17.08.70). XI. Пологий горный склон восточной экспозиции, ассоциация Rosa spinosissima + Spiraea chamaedryfolia-Calamagrostis purpu- rea+ Artemisia frigida (15 км северо-восточнее пос. Алексеев- ка, горный перевал в сторону озера Маркаколь, 1300 м над ур. моря, Восточный Казахстан, 30.08.70). Из таблицы 5 можно заключить, что встреченные ценодемы воло- душки золотистой находились в следующих возрастных состояниях: CDpj (биотоп 1), CDmi (П-V, VII-IX, XI), CDmm (VI, X). Среди иссле- дованных ценодемов доминируют инициматурные ценодемы, т.е. нор- мальные ценотические популяции по классификации Т.А.Работнова.
473 Глава 10. Использование теории органиэации биоморф семенных растений ~) ОО Г~) Ч~ ~Г) О„ч~ CO л ~Г) 1 1 1 ОО ОО О 2 Ф О Х Ф Е К ОО Ч~ ОО Ч~ VO <р О1 VO Ч~ Ч~ Ч~ Ч~ 1 1 ! ! g) (б Х z )z и~ о 1- о Щ О С S (r) О hC О S 3 >- О С О О l2 )5 О Х 1- О (б ОО С~) 'Ф Ч~ Ч~ VO VO ~Г) 1 1 1 1 1 2 ~ ° О й Ж О д ~ч О й Ж О д О й Ж О д ~ч О й Ж О Я .ц. С4 з о О & CJ ~p О Ф й М ~-~ г~ О д О с~ о 0" д д Ж д с~, 3 д В~ ;ф О Ж ~ о д ~ч O О Е g ° 1ИВф ° 1ИВф
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ 10.2. Биоморфогения семенных растений в естественных условиях произрастания и в культуре {на примере лекарственных растений} 10.2.1. Inula helenium L. Издавна для медицинских целей применяют корневища и корни де- вясила высокого 1пи1а helenium L. (сем. Asteraceae) (Madaus, 1938; 3е- линский, 1958; Норре, 1958; Атлас..., 1962; Турова, 1974 и др.). Девясил высокий используется также в цветоводстве (Полетика, Мишенкова, 1967). Inula helenium — луговой вид, произрастающий в Европе (кроме се- вера), на Кавказе, в юго-западной Сибири, на севере Средней Азии, в Казахстане, на севере Малой и Центральной Азии (Горшкова, 1959). В.Г.Синицына (1976), изучая биоморфогенические особенности де- вясила высокого, произрастающего в Заилийском Алатау, отметила, что продолжительность виргинильного этапа у этого вида составляет 2-3 года, зацветание корзинок в структуре монокарпического побега осуще- ствляется в базипетальной последовательности, а цветки в корзинке развертываются от периферии к центру. Фаза цветения у отдельных растений, по ее данным, продолжается 30-40 дней, плодоношения 25-30 дней, абсолютный возраст растений не превышает 25 лет. Биоморфоге- ния девясила высокого была изучена на Алтае (Солонешенский р-он, пойма р. Ануй, Майминский р-он, пойма р. Катунь) (Нухимовский, Ну- химовская, 1979). Этот вид произрастает здесь на лугах (высокотравных или остепненных) по берегам ручьев, рек и озер, по опушкам поймен- ных лесов, у дорог и по днищам оврагов, реже среди зарослей поймен- ных кустарников; на Алтае он встречен нами в нижнем, реже в среднем поясах гор (до 1200 м над ур.моря). Для девясила высокого характерна синонтогения, т.е. кондивиду- альный (от зиготы до последнего поколения клона) образ жизни. Эмбриональный иериод. Пренатальный этан. ПС — июль-сентябрь, октябрь. ОХ — оплодотворенная семяпочка или зрелое семя в структуре родительского растения. Б — цветки (трубчатые — обоеполые, языч- ковые — пестичные, венчики желтые) опыляются насекомыми (шмели, пчелы). Латентный этаа ПС — с сентября, октября до конца апреля, мая, июня следующего года. ОХ — зрелое семя, обособленное от родительского растения. А — зрелые плоды — 4-5-гранные семянки с волосистым хохол- ком, 3-4,5 мм дл., 0,6-1,2 мм шир. Масса 1000 семянок 1,0-1,35 г. Б — осы- пание семянок из сохраняющихся зимой отдельных сухих репродуктивных частей побегов иногда не происходит до весны следующего года. Элиэмбриональный лериод. Виргинильный этан. Преювенильная фаза. Проростковая гииофаза (рис. 184, А). ПС — конец апреля, май, июнь; 8-10 дней. ОХ — прорастающее семя или семядольный почкокорень
475 Глава 10. Использование теории организации биоморф семенных растений Рис. 184. Виргинильный этап синонтогении Inula helenium L. на Алтае. А — проросток, Б — всходы, В,-В,— ювенильные мероаклонисты: В, — униюве- нильные, В,— биювенильные, В,— триювенильные мероаклонисты, Г — имма- турные мероаклонисты; 1 — семядоли, 2 — гипокотиль, 3 — семядольный узел, 4 — корневая шейка, Sa-Se — корни: Sa — главная монориза (субмедиаль), Sá— главная дириза, 5в — неглавная монориза (сублатераль), 5r — тририза, 5д — глав- ная тетрариза, 5е — синториза; 6 — ризогенная корневая ветвь, 7 — каулогенная корневая ветвь, 8 — розеточные листья или их основания (верхние части удгле- ны), 9 — вегетативный предрозеточный побег, 10 — вегетативный медиальный розеточный побег, 11 — вегетативный аксиллярный розеточный побег, 12 — мо- нокормус, 13 — тетракормус, 14 — курторезидные монокормусы, 15 — проризом, 16 — ризом, 17 — почки в структуре побегов, 18-22 — организменные ризокорму- сы: 18 — семядольный почкокорень, или ризопротерокормус, 19 — семядольный вегетативный ризомонокормус, 20 — бессемядольный вегетативный ризомоно- кормус, 21 — ежегодно вегетативный гетерономный (с живым главным корнем) ризотетракормус, 22 — ежегодно вегетативный гетерономный ризогептакормус. Ув. — увеличено по сравнению с масштабом. МК — упрощенные модели конди- видуумов; 1л, 2л, Зл и т.д. — кондивидуумы 1,2,3 и т.д. лет жизни; горизонтальная линия — поверхность почвы; одно деление масштаба — 1 см; х10 — масштаб об- щего вида особи. (ризопротерокормус). А — семядоли (две) надземные голые продолго- вато-эллиптические или продолговато-яйцевидные мясистые широко- закругленные, без выемки или иногда с небольшой выемкой, суженные в короткие черешки; после развертывания семядоли (с черешками) 0,8-1,5см дл., 0,2-0,4см шир. (пластинка семядолей 0,6-0,9см дл.). Главная монориза до 3 см. Всходовая гииофаза (рис. 184, Б). ПС — май, июнь, июль, редко август; 2-4 недели. ОХ — семядольный вегетативный предрозеточный
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ (1-2 сближенных настоящих листа) или розеточный (3, реже более сбли- женных настоящих листьев) ризомонокормус. А — семядоли (с черешками) 1,2-1,8 см дл.; 0,4-0,6 см шир. (пластинка семядолей 0,8-1,2 см дл.). Первые настоящие листья эллиптические или продолго- вато-эллиптические, суженные в черешки (1-ый лист — пластинка 1,2-3 см дл., 0,5-1,2 см шир., черешки 0,4-1,0 см дл., 2-ой лист — пла- стинка 2,0-5,0см дл., 0,7-2,2см шир., черешки 1,0-2,0см дл. и т.д.). Б — в последующем нарастании медиального розеточного побега уве- личиваются линейные размеры новых листьев, но форма их остается такой же, как и у первых листьев. Ювенильная фаза. Гииофаза иервая — униювенильная (рис. 184, В,). ПС — с июля, редко августа до начала зимы текущего года. ОХ— бессемядольный вегетативный аризомный ризомонокормус. А — семя- доли отмерли. Жизнедеятельность растений происходит пока без базо- вого органа. За первый вегетационный сезон образуется 1-5 простых листьев (монофиллов). Главный корень — до 10 см дл., разветвленный до 1-го или 2-го порядков, т.е. является диризой или тетраризой. Б— на гипокотиле в основании главного корня образуются поперечные складки («моршинистость»), свидетельствующие об их контрактильнос- ти, которая проявляется и в последующие годы. Гииофаза вторая — биювенильная (рис. 184, В,). ПС — с начала зимы 1-го года эпиэмбриональной жизни до 3-8 лет. ОХ — бессемя- дольный ежегодно вегетативный проризомный ризомонокормус или ри- зотетракормус. А — индикаторный признак — базовый орган — про- ризом, т.е. персистентная часть тетракормуса, еще не развившего при- даточных корней. После окончания первого сезона вегетации зимует безлистный курторезидный (компактный апонекрозный) монокормус, являющийся первой персистентной частью активного (с живой верхуш- кой) розеточного тетракормуса, нарастающего моноподиально в после- дующие годы. Главная тетрариза — до 12 см дл., гипокотиль— 0,15-0,3 см диам. Гипофаза третья — триювенильная (рис. 184, В,). ПС — с 3-8 до 4-12 лет. ОХ — ежегодно вегетативный розеточный ризомный ризотет- ракормус. А — происходит преобразование проризома в ризом, харак- теризующееся развитием первых каулогенных корней. Наличие невет- вистого ризома — индикаторный признак этого возрастного состояния. Имматурная фаза (рис. 184, Г). ПС — с 4-12 до 5-17 лет. ОХ — еже- годно вегетативный розеточный гетерономно-ризомный простокустис- тый ризогексакормус или ризогептакормус. А — начало кущения- признак перехода особей в это возрастное состояние. Верхушечные и пазушные весенние побеги отрастают из зимовавших почек. Б — без- листный компактный гетерономный ризом постепенно приобретает наклонное положение к поверхности почвы за счет увеличения одно- сторонней контрактильной деятельности придаточных корней. Корне- вая система — синториза, т.е. состоит из главной тетраризы и других каулогенных (неветвистых или ветвистых) корней. Матурный этаи. Инициматурная (начальная матурная) фаза. ПС— с 4-17, чаще 6-12, до 15-25 лет. ОХ — ежегодно или неежегодно репро-
Глава 10. Использование теории организации биоморф семенных растений 477 дуктивный гетерономный ризопентакормус (редко) (рис. 185, Д„гипо- фаза первая — некустистая) или ризогексакормус, а позднее ризогепта- кормус (рис. 185,Д„гипофаза вторая — кустистая). А — впервые про- исходит видимая реализация физиологической зрелости, т.е. образуются на побегах репродуктивные органы — бутоны, а далее развертываются соцветия (корзинки), созревают плоды, при этом число порядков пост- инициальных унирепродуктивных побегов, цветущих или нецветущих в год наблюдения, не более 2-х; сохраняется жизнеспособность субмеди- али (главной моноризы). Б — пентакормусы унирепродуктивные, ди-, три-, плейофазные (2-16, чаще 4-8 фаз видимого роста) полурозеточные акронекрозные обоеполые, 60-215 см выс. Соцветие — закрытая (детер- минатная) сборная фронда-брактеозная рыхлая метелка, состоящая из 5-40 простых соцветий - — корзинок (пазушных или верхушечных); иногда происходит осевой сдвиг отдельных корзинок из пазух листьев (конкаулесценция). Медиматурная (средняя матурная) фаза. ПС — с 12-25 до 25-45 лет. Гииофаза иервая — гетерономно-ризомная (рис. 185,Е,). ОХ — ежегодно или неежегодно репродуктивный полурозеточный гемисимподиальный гетерономно-ризомный ризоэнатокормус. А — индикаторный приз- нак — наличие 3-х и более порядков унирепродуктивных побегов цве- тущих или нецветущих (кроме почек) в год наблюдения с живым глав- ным корнем (главной тетраризой). Главная тетрариза до 30см; ги- покотиль до 2,0 см в диам. Б — фенофазы: бутонизация — июнь, июль, цветение (базипетальное) с июля, августа до середины сентября, пло- доношение — конец августа-октябрь. Листорасположение очередное. Розеточные и нижние безрозеточные (т.е. развитые на удлиненной ча- сти полурозеточного побега) листья с продолговато-эллиптической не- равномерно мелкозубчато-пильчатой по краю пластинкой, 30-60 см дл., 15-25 см шир., черешок 15-40 см дл. Стебель, листья и соцветия коротко опушенные. Гииофаза вторая — автономно-ризомная (рис. 185, Е,). ОХ — еже- годно или неежегодно репродуктивный полурозеточный гемисимподи- альный автономно-ризомный ризогептакормус или ризоэнатокормус. А — главная тетрариза отмерла частично или полностью, но вегетатив- ного размножения нет. Б — ризом нарастает гемисимподиально за счет увеличения числа курторезидных монокормусов и миниморезидных тет- ракормусов. Акронекроз отплодоносивших унирепродуктивных побегов вызывает значительные структурно-функциональные перестройки в кор- невище и главном корне, которые приводят к образованию значитель- ных очагов отмерших стеблекорневых тканей, а затем и к гибели глав- ного корня. Финиматурная (заключительная матурная) фаза (рис. 186, Ж). ПС — с 25-45 до 40-60 лет. ОХ — совокупность нескольких (2-5) автономно- ризомных ежегодно вегетативных (без следов перевершинивания от не- полностью отмерших ранее унирепродуктивных побегов) ризокормусов (имматурных клонистов) и ежегодно или неежегодно репродуктивных ризокормусов (матурных клонистов) или только последних. А — глав- ная тетрариза полностью отмерла; произошло замедленное (ретардаци-
478 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ %« %« 1Р« Ф Е ° 2I И 3 И »« й >Ъ >Ъ h4 Ж Q ~в i й й й;[ й й Ж Q >Ъ i Л й.' 1 й.' й X Ч'~ О й м,"й й 3 ~ о. v g «Я щ ~ц 4ф; 2 ~~о� ;~' д 6Э ~~-+о. О 6Э z о охх 0 ~ч ~ю «~ Д~ с6 Р4 ° л 6Э ф ~~~ в» Й <~4 ~ Д~ М 6~ р~ О Х Д О. м о + р,Л О v 0 - 0 [» A О v v 0 Д: ~ :~' СЧ О ж и ~ ° ~~ ~ 1-' с~~е ~ СЯ х 0 0 Лю «30 м о м О.~ià О ! О О, О х 0 О [» ~ (') м жоао :~' X О еч ~» м о � й <~ й о vs~I О. о о 0 0 й f» & Cl х о ;~. 'О "~й" ~ ~«~г' д~ ~ОС м~v oмом О д: 4 О ~ ом~ Сб I Д~~ Д,' 0 К О К > <о О а.)~3 О ~ ~ gg o О ф; О. Q, м 6э О Х ф; :Е�а О. й;[ р. " -< о." м м О д: ~ а О. — ~-4 ~~ д <~4 (б д :И '-'; р; д 0 ~ о'~ о к~~~ ио~~ 1 54 еч < О ~е %222 v "~ '+ 2 д X X О.;Я ooй [» О Д ~ Р'~ О ~- ~ о. рД ~ ( О, Дц 0 [; x o~vv м д ~аи~ Ж а 00 О ~~ СР д - v о 0 О Р. С4 м ,'Я :Я ~ Q, 63 р,м о к О v д С6 ф: [» о е) о О~ v р,g � Й 1 О р. М v О ~) х n. ~ оеой х ооой ~') ~ С'1 х�о О. ~ц Ю Флъ М О 1: Q а~а ~~ в»а ~~ o Q й )g ~о-+ д о + м <О д~ iх о v :»: й03 Ыдga ~~~о х а Х ~~,~о ~ f» ххжк »-~ ga ~) мО~~О ~« Й м х ~ ~ "4о~х С40 С42 3 v м C р.~И о о~ о мо~ х ;~. 'О ~-[ ~ м о ~~ ~е о [ М меч v Й ~~~ Д,' х v v ~л~ С» 00 О. СЧ 00" х 63 р м' О. ~мО л: ("-. х И~а й 6Э ~т~ "о.< о О ~ й хm о.
479 6~ й Ж О Ю О 3 Ж Ж О й 4 Ж Глава 10. Использование теории органиэации биоморф семенных растений и 2 О. О М О Х О э~~~~~ 0 ° а3 Ж 2 О Ж О & Cl 63 э~~~~~ 3 ~Г) й '~ оо & 63 М оо и й О Q, Ж ~[ CO О & эй Р' 2 Ю ~)" 63 v 1 ~О Ж О
OCHOBhI БИОМОРФОЛОГИИ CENEHHhIX РА СТЕНКИ онное) вегетативное размножение после того, как полная общая некро- тическая партикуляция разделила мероаклониста (временного аклони- ста) на две или более обособленные конструктивные партикулы — но- вые особи — клонисты. Сенильный этаи (рис. 186, И). ПС — продолжается 1-5 лет. ОХ — еже- годно вегетативный после утраты физиологической зрелости розеточный автономно-ризомный ризокормус. Полный возраст (продолжительность жизни) кондивидуума около 60 лет. Резюмируем рассмотренный выше материал по биоморфогении девясила высокого. Для девясила высокого характерна ретардационная синонтогения: у особей семенного происхождения (мероклонистов) второго медима- турного состояния главный корень отмирает и после замедленного веге- тативного размножения мероаклонист преобразуется в клон — финима- турное состояние синорганизма., клонисты которого еще долго удержи- вают первоначальную территорию. По эколого-морфологическим тропобиозным признакам Inula heleni- Um — это высокоросло-травянистый летнезеленый надземно-двусемя- дольный геофильный мультирепродуктивный полурозеточный цельно- листный пазушно-очередноветвистый гемисимподильный компактно- ризомный слабокустистый однодомный желто-астроцветковый брахипла- нофильный (с очень слабым разрастанием корневищ в горизонтальном направлении) ретардационно-кондивидуальный мезофильный луговой вид. 10.2.2. Rhodiola говеа L. 10.2.2.1. Rhodiola говеа L. в горах Алтая С давних пор в народной медицине Алтая широко используются отвары и настойки подземных органов родиолы розовой, или золотого корня, Rhodiola rosea L. (сем. Crassulaceae) для лечения различных за- болеваний, а в основном, «чтобы вообще быть здоровым» (Уткин, 1931). Жидкий экстракт из корневищ родиолы розовой выпускается медицин- ской промышленностью; он обладает стимулирующим и адаптогенным действием (Говоров, Липская, 1963; Зотова и др., 1965; Сарати- ков, 1966, 1974; Крылов, 1969 и др.). Rhodiola rosea — арктическо-высокогорный вид, встречающийся в горах Западной Европы, Сибири и Дальнего Востока. Наибольшей про- дуктивности и обилия он достигает на Алтае (Положий, Суров, 1972; Суров, 1973 и др.), где и проводились исследования его биоморфогении (Нухимовский, 1974а). Золотой корень на Алтае произрастает преиму- щественно на субальпийских лугах по берегам рек и ручьев, на древних моренах, в зарослях кустарников (Betula rotundifolia Spach., Salix glauca L., S.hastata L., S.lanata L. и др.); нередко по берегам ручьев и руслам временных водотоков поднимается в нижние части гольцового пояса, а также спускается в пояс кедровых лесов. Основные заросли радиолы розовой на Алтае сосредоточены на высоте 1700-2200 м над ур.моря. Климат на Алтае резко континентальный. В условиях высокогорий летом жаркие дни сменяются холодными ночами и в любое время может наступить резкое похолодание, сопровождающееся сильными снегопа-
Глава 10. Использование теории организации оиоморф семенных растений дами, иногда в течение нескольких дней. Чтобы жить в таких условиях, надо обладать комплексом защитных приспособлений; арсенал их у родиолы розовой не мал, поэтому ее биоморфы участвуют в самых различных ассоциациях, нередко даже в качестве эдификаторов. Эмбриональный иериод. Пренатальный этап биоморфогенеза описан В.Ю.Мандрик и Л.В.Голышкиным (1973); они в частности (с. 273) отме- тили «превращение первично обоеполых цветков в однополые, что при- водит к стерильности интерсексов и к развитию только мужских и женских цветков». Семена родиолы розовой от темно-коричневого до светло-коричневого цвета, яйцевидной формы с резко выраженной про- дольной ребристостью, кверху суженные, 1,8-2,2 мм дл., 0,8-1,0 мм шир. В природных условиях посредством частной некротической парти- куляции зрелые семена осыпаются в почву в конце августа-сентября. Часть обособившихся от материнского растения семян, оставшаяся в плодах (листовках) в случае сохранения сухого надземного побега, может не высыпаться из них до следующего вегетационного периода. Латентный этан. Семена, обособившись от родительского растения и попав, чаще всего осенью, в почву, в течение последующих 9-10 месяцев проходят естественную стратификацию, без которой всходят плохо. Эииэмбриональный иериод. Виргинильный этаи. Проростковая фаза (рис. 187, А). В течение короткого лета на открытых влажных местах и в тени можно обнаружить проростки золотого корня. ПС — июнь, июль, август; продолжается 10-20 дней. ОХ — прорастающее семя или семядольный почкокорень (ризопротерокормус). А — развернувшиеся сизовато-зеленые голые мясистые семядоли у проростков состоят из маленьких (0,15-0,5 см дл., 0,1-0,4 см шир.) округлых, округло-яйцевид- ных или эллиптических пластинок и коротких (0,2-0,8 см дл.) черешков. Гипокотиль 0,2-0,8 см дл., 0,05-0,2 см диам.; при затенении он иногда вытягивается до 1,5 см. Гипокотиль переходит в тонкий главный корень до 3-4 см дл. Кроме главного корня, часто у проростков на самой ниж- ней части гипокотиля образуется несколько придаточных (гипокотило- генных) корней. Гипокотиль к концу первого вегетационного периода увеличивается в диаметре до 0,15-0,25 см. Зародышевая почка чаще всего не развертывается на первом году эпиэмбриональной жизни осо- би. Утрата главного корня проростками нередко не приводит их к ги- бели, поскольку его функцию тогда выполняют один или несколько гипокотилогенных корней. Подхваченные дождевым потоком, пророст- ки могут благополучно переменить свое местожительство. Всходовая фаза (рис. 187, Б). ПС — июнь, июль, август, реже сен- тябрь. ОХ — семядольный вегетативный безрозеточный ризомонокор- мус. А — эту фазу в первый вегетационный сезон проходят лишь не- которые особи, успевшие к его концу образовать 1-2 настоящих сизо- вато-зеленых округлых или овальных короткочерешчатых сближенных листа 0,3-0,6см дл., 0,2-0,3 см шир. У всходов пластинки семядолей 0,4-0,7 см дл., 0,3-0,6см шир., а черешки семядолей 0,25-0,6см дл., гипокотиль 0,3-0,8 см дл., 0,1-0,3 см в диам. Главный корень 3-5 см дл., иногда разветвлен до первого порядка, гипокотилогенных корней 1-5. Иногда главного корня уже нет, а функционируют только гипокотило-
OCHOBhI БИОМОРФОЛОГИИ СЕМЕННБ1Х РА СТЕНИЙ Рис. 187. Виргинильные аклонисты Rhodiola rosea L. в естественных условиях произрастания (Алтай). А — проростки, Б — всходы, В — ювенильные особи, Г — имматурные особи; 1— гипокотиль, 2 — семядоли, 3 — семядольный узел, 4 — главный корень, 5 — гипоко- тилогенные корни, Ь — каулогенные корни, 7 — верхушечная почка ростового по- бега, 8 — медиальный ростовой побег, 9 — пазушный ростовой побег, 10 — ас- симиляционный силлептический (преинтеркалярный) побег или его основание (если верхние части удалены), 11 — сухой остаток или рубец (след) от силлептического побега, 12 — рубцы от отмерших листьев, 13 — гетерономное корневище, 14 — ав- тономное корневище, 15 — границы мономерных компактных пленорезидов (пока- заны не на всех корневищах), 16 — ассимиляционные листья, 17 — чешуевидные листья, 18 — верхняя часть ассимиляционного побега после прекращения роста. 2г..., 8л.... — абсолютный возраст некоторых аклональных и клональных кондивидуумов. Горизонтальная линия — поверхность почвы; одно деление масштаба — 1 см; у — увеличено в несколько раз по сравнению с масштабом.
Глава 10. Использование теории организации оиоморф семенных растений 483 генные корни. Семядоли сохраняют жизнеспособность не более одного вегетационного сезона. Первый вегетационный сезон в высокогорьях Алтая эпиэмбри- ональные особи заканчивают в фазе проростка или всхода. Перезимо- вавшие растения обычно состоят из гипокотиля, верхушечной почки, главного и гипокотилогенных корней. Ювенильная фаза (рис. 187,В). ПС — с июля, августа первого года до 3-15 лет. ОХ — бессемядольный вегетативный ризокормус разной степени сложности, не проявляющий пока постоянного кущения, перио- дически образующий или одиночные пазушные ассимиляционные (ве- гетативно-эфемерные) побеги, или пучки кладонекрозных (пазуш- ных эфемерных) ассимиляционных побегов периодического кущения. А — летом второго года эпиэмбриональной жизни трогается в рост верхушечная почка, образуя годичный модифицировнный ростовой компактный апонекрозный побег, состоящий из нескольких, чаще 2-6, чешуевидных листьев, и очень маленькой (0,1-0,3 см дл.) неотмирающей стеблевой части. Этот побег пока мало чем отличается от почки, но в процессе его очень замедленного видимого роста нередко из одной, а совсем редко из двух пазух нижних чешуй происходит бурное отрастание силлептических ассимиляционных побегов со срединными зелеными листьями. Зачатки этих побегов закладываются осенью предшествующего года в общей почке, а видимый рост, начинающийся весной, образует из этой почки одновременно (без периода покоя) как весенний ростовой побег, так и силлептические ассимиляционные побеги (1-2). Если развивается второй силлептический побег в структуре слож- ного специального побега, то он слабее первого. Вероятно, первый побег оказывает некоторое угнетающее действие на подобный ему по- бег, развивающийся чуть выше и чуть позднее. Эта особенность в от- растании силлептических побегов свойственна особям всех возрастных состояний. Строгое взаимодополняющее разделение функций между ве- сенним ростовым модифицированным побегом и силлептическим асси- миляционным побегом служит аргументом в пользу рассмотрения этих побегов в структуре одного вегетативного дикормуса. Сочетание таких взаимосвязанных побегов виргинильных особей определяет структуру медиального ростового илейофазного кладонекрозного иобега (рис. 187, 8) с иазушными ассимиляторами, т.е. с иазушными силлелтическими (иреин- теркалярными) ассимиляционными лобегами (рис. 187, 10) — пентакор- муса. Способность ежегодно или неежегодно образовывать силлепти- ческие побеги из пазушных почек медиального побега, а в случае гибели его верхушки на подобном ему одном пазушном побеге замещения— признак ювенильных особей радиолы розовой. Силлептические побеги образуют периодически кустистое строение у растений; эти побеги живут один сезон, поэтому к зиме такая кустистость утрачивается. После завершения развития ассимиляционные побеги полностью отмирают, оставляя на многолетних частях ростовых побегов лишь не- большие углубления (рубцы) (рис. 187, 11). Особенность рубцов состоит в том, что на них строго фиксирована граница некроза силлептических побегов и дальше на ростовые побеги этот процесс не переходит. Рубцы
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ могут сохраняться долго, что помогает экзоморфологическим путем определять возраст как отдельных побегов, так и целых растений. Стеблевая исходная часть весеннего ростового побега после завер- шения вегетационного сезона не отмирает и становится скелетной, т.е. образуется мономерный компактный пленорезид. Пленорезидные мо- нокормусы у ювенильного растения родиолы розовой составляют очень компактное корневище. В последующие годы оно постоянно нарастает аккумулятивно, т.е. за счет увеличения в его структуре числа новых компактных резидных побегов. Корневища родиолы розовой эпигеогенного типа. У ювенилов и у растений последующих возрастных состояний выше зоны силлептичес- кого побегообразования закладываются пазушные ростовые почки. Па- зушные и верхушечные почки защищенные, т.е. их наружные покровы образованы чешуевидными листьями. Пазушные универсальные росто- вые побеги у ювенилов нарастают крайне медленно и для образования силлептических ассимиляционных побегов у них еще не хватает сил. Имматурная фаза (рис. 187, Г). ПС — с 3-15 до 7-40 лет. ОХ — ве- гетативный безрозеточный постоянно-кустистый ризогексакормус или ризогептакормус, или реже ризоэнатокормус. А — начало отрастания силлептических побегов не только в структуре медиального, но и па- зушных ростовых побегов — признак перехода особей в имматурное возрастное состояние. Когда ассимиляционных побегов временно нет, то о том, что они периодически отрастали в структуре универсальных побегов, можно судить по рубцам от них. В результате роста пазушных ростовых побегов формируется постоянный куст, т.е. особи, став од- нажды кустами, сохраняют кустистое строение на всю жизнь. Число ежегодно отрастающих силлептических побегов у всего растения посте- пенно увеличивается с 2-3 до 10-15. В эту фазу жизни возрастает веге- тативная мощь особей, умножается число пазушных ростовых почек и увеличиваются размеры ежегодных приростов ростовых побегов. Глав- ный корень у виргинильных особей или функционирует, или погибает в любой из фаз биоморфогенеза. Б — ризом, связанный с главным корнем, — гетерономный, а утративший связь с таковым, — автоном- ный. Постепенно ризом становится плагиотропным и погружается по- чти полностью или лишь незначительно в почву. Там, где постоянно образуется наилок, ризом быстро погружается в почву, занимая в ней более или менее ортотропное положение (берега ручьев, русла времен- ных водотоков и т.п.). Там, где обильно развит моховой покров (заросли высокогорных кустарников, у ключей и т.п.), ветви корневищ сравнитель- но высоко (на 3-5 см, иногда даже на 8 см) выступают над поверхностью почвы, оставаясь, однако, прикрытыми влажным моховым покровом, что защищает их от неблагоприятных воздействий внешней среды. Особи быстрее проходят свои фазы жизни на открытых (умеренно остепненных) южных склонах, где они иногда поселяются по щебнис- тым руслам временных водотоков или на участках, где сток воды осу- ществляется на небольшой глубине под поверхностью почвы. Матурный этан (рис. 188, 189). ПС — с 7-40, чаще с 12-20 до 300 лет. Униматурная фаза — мероаклональная. Биматурная фаза — клональная.
Глава 10. Использование теории организации биоморф семенных растений Рис. 188. Матурный и сенильный этапы синонтогении Rhodiola rosea (Алтай). Д — матурные аклонисты, Е — матурные криптоклональные кондивидуумы, Ж— матурный акриптоклональный кондивидуум, И — сенильный кондивидуум; 19— репродуктивна-ассимиляционный силлептический (интеркалярный) побег или его основание (если верхние части удалены), 20 — цветок неразвернувшийся и искусственно развернутый, 21 — цветок естественно развернутый, 22 — живая часть корневищ и корней, 23 — отмершая часть корневищ и корней, 24 — камень. Остальные обозначения те же, что и на рис. 187.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ LO 3 О ~ч О 3: й.; й 65 Х ~ч й.; С) О о О ~~ 1 ~Ф юе О~ О Х оо 65 Й ~. ~4 и С'1 () д„ О а р, а О С5 а Õ 4~ О О ~ Q - О «о g О О ф, ~ь X ао ~) о X ~' О h4 )Q 2 X 3 й О
Глава 10. Использование теории организации оиоморф семенных растений А — побеговую систему матурных особей образуют открытые унире- продуктивные кладонекрозные дикормусы. Каждый из этих побегов сложный; он включает внутригодичные побеги: один весенний ростовой модифицированный компактный пазушный или верхушечный апоне- крозный приземный, полуподземный или подземный побег и 1-3 силлептических (интеркалярных) репродуктивно-ассимиляционных надземных побега. Силлептические побеги развиваются из нижней ча- сти ростовых побегов. Резидный период своей жизни кладонекрозные побеги проходят, как правило, в качестве компактных пленорезидных (от нескольких мм до 3 см дл., чаще 0,3-1,0 см дл.) монокормусов. Продолжительность жизни отдельных мономерных резидов колеблется от немногих — до 40 лет, чаще 15-30 лет. На ризомах развито много спящих пазушных почек, поэтому особи родиолы розовой легко пере- носят гибель верхушек отдельных ростовых побегов. В почках с осени закладываются зачатки побегов следующего сезона вегетации. Среда обитания наложила определенный отпечаток и на феноло- гию. Н.В. Ревякина (1973, с.бl) отметила, что особи золотого корня начинают вегетировать еще под снегом, «который позднее пробивают молодые побеги, а после таяния снега они имеют высоту 6-9 см». Все фенофазы проходят быстро. Начало июля-конец июля — время цвете- ния родиолы розовой, обитающей на 1700-1800 м над ур.моря, а в конце июля-первой половине августа происходит цветение у особей, растущих на высоте 2000-2200 м над ур. моря. У растений, произрастающих на южных склонах, плодоношение, а также и предшествующие ему фенофазы, начинаются на 2-3 недели раньше, чем у таковых на северных склонах. Осыпание зрелых семян происходит в основном в сентябре. Биоморфогенез нередко осуществляется в форме синонтогенеза. У матурных мероаклонистов главный корень жив или уже отмер (рис. 188, Д), корневища также могут отмирать в основании на различную длину, но пока нет полной общей некротической партикуляции. Грань между аклональным и клональным кондивидуумом не всегда резкая, посколь- ку клональный кондивидуум по своим структурным особенностям мо- жет быть «скрытым» (криптоклональным) или «видимым» (акриптокло- нальным). Во всех случаях клонисты образуются в результате полной партикуляции. Однако, у криптоклональных кондивидуумов клонисты (рис. 188, Е; 189) остаются связанными через полностью некрозную мас- су старых участков корневищ. Криптоклональные кондивидуумы обра- зуются иногда и у других растений, например, у Veratrum lobelianum Bernh. У акриптоклональных кондивидуумов (рис. 188,Ж) некрозная масса старых корневищ разрушилась, и обособленность клонистов мож- но наблюдать без каких-либо вспомогательных приемов. Живые части корневищ, вероятно, интенсивно используют питательные вещества и воду из отмерших частей корневищ. Там, где происходит постоянное, но не сильное заиливание молодых участков корневищ, самые нижние отмершие части ризомов не могут оказать заметного вредного воздей- ствия на растение. В этих условиях биоморфы родиолы розовой наибо- лее долговечны и достигают наибольшей биологической продуктивности
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ (рис. 189). Один из крупных криптоклональных кондивидуумов весил в сыром виде 7,8 кг; масса живых корневищ была 3,5 кг; в надземной части побеговой системы было 249 силлептических побегов. Нередко наилка нет или он незначителен, тогда отмершая масса корневищ спо- собствует «выпиранию» над поверхностью почвы их молодых частей и почек, что нередко приводит последние к преждевременной гибели. Б — весьма примечательна борьба за существование биоморф ро- диолы розовой, поселившихся на крупных камнях, которые в большом числе встречаются вдоль русел высокогорных рек и ручьев, а иногда и посреди них. Первая стадия зарастания таких камней — поселение на них лишайников и мхов. Семена родиолы розовой нередко встречают в покрове из мха нормальные условия для прорастания. На камнях, помимо родиолы розовой, иногда поселяются и другие растения, на- пример, Saxifraga punctata L. и Chrysosplenium пийсаи1е Bunge. Ежегод- ный скелетный прирост у биоморф родиолы, как уже отмечалось, очень незначителен. Этот факт способствует постепенному закреплению их в моховом покрове и рациональному использованию того небольшого запаса почвы, который скапливается вокруг них. Мох наряду с поселив- шимися здесь другими высшими растениями создает своеобразное пок- рытие для корневищ, облегчая им невзгоды приземной жизни. Сами же мхи используют корневща родиолы в качестве опоры для своего разрас- тания. Такой союз в итоге приводит к тому, что биоморфы золотого корня занимают всю верхнюю часть камней. В этих условиях они не- редко заметно болеют и начинают нарастать в основном за счет нижних ростовых почек (верхушки ветвей корневищ, поднимающиеся выше мо- хового покрова, часто обмерзают). Многолетние побеги свисают вниз с камней, сильно разрастаются в толщину и длину и обильно образуют новые, им подобные, побеги. Случаи механической партикуляции у биоморф родиолы розовой, произрастающей на камнях, нередки, что обычно приводит к экспансивному вегетативному размножению. Это вегетативное размножение характеризуется тем, что партикулы перено- сятся на новые местообитания с помощью каких-либо внешних агентов, например, дождевым потоком, оползнем и т.п., а при введении в куль- туру — человеком. Господствующий тип вегетативного размножения у рассматриваемого вида — замедленное (ретардационное). Оно осуще- ствляется специализированными органами — компактными корневи- щами, которые обеспечивают в течение сравнительно большого отрезка времени только незначительное отдаление клонистов друг от друга. Кусты у радиолы розовой по степени отдаления элементарных зон кущения компактные, поскольку их основания составляют компактные обильно ветвистые ризомы, а побеги кущения, как ростовые, так и ре- продуктивна-ассимиляционные, сильно, хотя и неплотно, сближены. По другому признаку — по отношению особей к кущению — характерны разнообразно-кустистые особи, т.е. у них сначала формируется постоян- ный куст, который функционирует до конца жизни особи, а, кроме того, на отдельных его ветвях образуются за счет силлептических побегов также и периодические кусты, живущие не более одного вегетационного сезона. Сенильный этаи у кондивидуумов иногда, вероятно, есть. Нами бы- ли обнаружены кондивидуумы, близкие к такому состоянию (рис. 188, И).
489 Глава 10. Использование теории организации оиоморф семенных растений 10.2.2.2. Rhodiola говеа L. в условиях выращивания Изучение биологических свойств, в особенности биоморфогении, лекарственных растений в естественных условиях произрастания и при выращивании составляет важный раздел интродукционных исследова- ний в лекарственном растениеводстве. На примере Rhodiola rosea рас- смотрим некоторые возможности использования наших теоретических раз- работок в условиях выращивания этого вида в Московской области. Опы- ты проводили в интродукционном питомнике ВИЛАР с 1973 по 1982 гг. Биологические наблюдения, включающие биометрические измере- ния, велись шесть лет над несколькими группами растений: группа 1— растения, произрастающие на участке загущенного подзимнего (1973) рядкового посева; группа II — растения, произрастающие на участке ана- логичном, но прореженном в мае 1974 г., с расстоянием между особями в рядках 15-20 см; группа III — растения, произрастающие на участке, созданном из трехмесячной рассады после пересадки ее в начале июня Среди почвы и полуразложившейся массы бывших ветвей корневищ и надземных побегов отдельных кондивидуумов сохраняли еще жизнеспо- собность единичные весьма слабые клонисты, которые ежегодно или неежегодно образовывали 1-3 невысоких (5-15 см) силлептических асси- миляционных побегов. Создалось впечатление, что резкое улучшение среды обитания, например, образование наилка, может на некоторое время спасти этот вымирающий кондивидуум, а ухудшение условий жизни ускорит его гибель. Резюмируем особенности биоморфогении родиолы розовой. По эколого-морфологическим тропобиозным признакам Rhodiola rosea — это летнезеленый открытый безрозеточный компактно-корне- вищный брахипланофильный, иногда особоподвижный, разнообразно- кустистый мультирепродуктивный желтовато-толстянковоцветковый двудомный травянистый ретардационно-кондивидуальный-мезопсихро- фильный высокогорный вид. Среди кондивидуумов родиолы розовой много долгожителей. Вы- сокая всхожесть перезимовавших под снегом семян, медленное нарас- тание модифицированных ростовых побегов, очень маленькие годовые приросты корневищ, бурное отрастание силлептических репродуктивно- ассимиляционных побегов, зачатки которых закладываются еще с осени в одной общей почке с ростовым побегом, интенсивное ветвление кор- невищ, замедленное вегетативное размножение, высокая регенеративная способность корневищных партикул, влаго- и светолюбивость, способ- ность поселяться и жить на камнях и долго сохранять жизнеспособность под панцирем многолетнего снегового покрова — это некоторые эко- лого-морфологические особенности, позволяющие виду успешно суще- ствовать в условиях высокогорного Алтая. При заготовке сырья можно собирать не более 60-70% живых кор- невищ матурных особей, срезая только их приземные и полуподземные ветви и тем самым сохраняя условия для регенерации куста.
ОСНОВБ1 БИОМОРФОПОГИИ CEMEHHhIX РАСТЕНИЙ из теплицы в открытый грунт через 15-20 см в рядки с междурядьями 60 см. Кроме того, с 1978 г. проводили опыты по рассадному и вегетатив- ному способам создания плантаций. Наблюдения проводились над 6-ю группами растений, полученных указанными выше способами: группа IV — растения, произрастающие на открытом участке, созданном из односезонной рассады после пересадки ее в августе через 10 см между особями в рядки с междурядьями 60 см; группа Ч — растения, произ- растающие на участке, созданном аналогично участку IV, но с рас- стоянием между особями 20см; группа VI — растения, произрастаю- щие на участке, созданном аналогично участкам IV и Ч, но с рас- стоянием между особями 30 см; группа Vll — растения, произрастаю- щие на участке, созданном из отрезков корневищ (3-5см дл.) после посадки их в открытый грунт питомника через 10 см в рядки с между- рядьями 60см; группа VIII — растения, произрастающие через 20см друг от друга на участке, созданном аналогично участку VII, группа IX — растения, произрастающие через 30 см друг от друга на участке, созданном аналогично участкам VII и ЧШ. Эмбриональный лериод. Пренатальный этал. ПС — с конца мая- июня до июля. ОХ — оплодотворенная семяпочка или зрелое семя в структуре родительского растения. Б — родительские растения в усло- виях выращивания двудомные, очень редко однодомные; цветки иногда функционально обоеполые, чаще функционально пестичные или фун- кционально тычиночные, что связано с недоразвитием или тычиноч- ных, или пестичных элементов у первично обоеполых цветков (Нухи- мовский, Юрцева, Юрцев, 1987). Латентный этаи. ПС — с момента сбора семян (июль) до посева под зиму текущего года или (после стратификации) весной следующего года. ОХ — зрелое семя, обособленное от родительского растения. Б— семена следует быстро собирать по мере их созревания, поскольку плоды многолистовки, растрескиваясь по шву у отдельных плодиков (листо- вок), вскоре теряют большую часть семян. Собирать рекомендуется все соцветие (сложный щиток), отрезая его от побега после созревания отдельных плодов и складывая в бумажные пакеты. В тот же день соц- ветия следует разложить тонким слоем на бумаге в помещении у окна, где плоды быстро дозреют. После дозревания семена легко отделяются и очищаются от плодов. Сбор семян проводится в течение нескольких дней по мере созревания плодов (характерный признак — начало рас- трескивания отдельных плодиков). При хранении семена быстро теряют всхожесть, что строго регламентирует сроки их посева. Масса 1000 се- мян, собранных в интродукционном питомнике, от 0,21 до 0,26 г. Лучшие результаты показал высев семян под зиму на глубину 0,5 см на небольших участках открытого грунта («рассадниках»), а также высев в посевные ящики с последующим выносом их под снег, где семена проходят стратификацию в условиях, приближенных к естественным, в течение 3-4 месяцев. Кроме того, хорошие результаты дает стратифика- ция семян при температуре от 0 до 2 С в течение 1 месяца с последу- ющим весенним посевом.
Глава 10. Использование теории организации оиоморф семенных растений 491 Элиэмбриональный лериод. Виргинильный этан. Преювенильная фаза. Проростковая гилофаза. ПС — при подзимнем посеве семена прораста- ют в апреле, реже, если апрель холодный, то в начале мая; в состоянии проростков растения в зависимости от погодных условий пребывают 25-40 дней. ОХ — прорастающее семя или семядольный почкокорень (ризопротерокормус). А — после стратификации лабораторная всхо- жесть на 5-7 день проращивания составила в среднем около 70% Се- мядоли надземные сизовато-зеленые голые, мясистые с округлыми, округло-яйцевидными, яйцевидными или эллиптическими пластинка- ми без выемки или с выемкой на верхушке и короткими черешками. В конце проростковой гипофазы размеры пластинок семядолей колеб- лются от 0,3 до 1,2 см дл. и от 0,3 до 0,6 см шир., а черешок от 0,3 до 2,2 см дл. Главный корень углубляется до 4 см. На самой нижней части гипокотиля образуются первые придаточные корни (рис. 190, А; 191, А). Всходовая гилофаза. ПС — с середины, конца мая до начала июня. ОХ — семядольный ризомонокормус с зелеными (обычно 1-2) настоя- щими листьями (рис. 190, Б; 191, Б). А — первые зеленые листья от 0,5 до 1,7 см дл. и 0,4-0,8 см шир. с черешками от 0,3 до 1,2 см дл.; размеры пластинок семядолей у таких особей колеблются от 0,4 до 1,5 см и от 0,3 до 0,7 см шир.; если зеленые листья на медиальном побеге не обра- зуются, то это признак выпадения из онтогенеза гипофазы всхода. Ювенильная фаза (рис. 190, В; 191, В). ПС — с июня до весны, лета следующего года или несколько лет, особенно в условиях густого про- израстания растений (в таких условиях изредка встречались даже шес- тилетние ювенильные растения). ОХ — семядольный или бессемядоль- ный вегетативный безрозеточный ризодикормус, или (после позднелет- него отмирания ассимиляционных побегов) бессемядольный вегетатив- ный ризомонокормус или ризотетракормус, или (ювенильные растения второго и последующих лет) бессемядольный вегетативный ризопента- кормус или ризогексакормус, если он без пазушных ростовых побегов. А — отрастание первых ассимиляционных побегов (в числе 1-2, реже 3-4) из пазух первых зеленых настоящих листьев, а если таковых нет, то из пазух первых чешуевидных листьев медиального ростового побега, или отмирание семядолей — признаки, свидетельствующие о начале ювенильной фазы. Первые силлептические ассимиляционные побеги развиваются в структуре вегетативного дикормуса. После первого года вегетации сохраняют жизнеспособность клубневидный гипокотиль (от 0,1 до 2 см в диам.), переходящий в стебель ростового побега с почка- ми — корневище, и система корней (придаточные, а иногда и глав- ный). Б — если в процессе нарастания ростового компактного модифи- цированного (с чешуевидными листьями) побега из-под прикрытия чешуй верхушечной почки выходят ее лроизводные ассимиляиионные ночки, то они экстралерулярные (нелрикрытые), и на следующий год из них отрастают не силлептические, а весенние ассимиляционные побеги. Если ювенильное состояние продолжается несколько лет, то в це- лом пульсирующий ростовой побег (тетракормус) вместе с весенними ассимиляционными побегами составляют вынужденно вегетативный гек- сакормус. Иногда ассимиляционные почки (у слабых растений) не выхо-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ Б й Ж, ~ м Ig О О 3 л ~м '-~ И 3e )Q М A Ф О„ Д ~Ъ О~ й О р:Ф. л~ 3e + p) X ~XI ~:( л О О Q ~~~ Й >3 О~ ~ О О ц ct3 1 1 с6 Д ~ й и> ОО .О О О Q О И ф х 3 О A О ~," й О О й Ф а: о~ ии ~ Зд о о а. ы О ~~ ~ — ~~ о~~о ~- ~ ° -: ° йur v < ~ д О р ~.~~~ К"„ ~~o~о~ И ~. -~й . ~„н ~,~ с~ Д ° д 3~ ~~ 65 ,'~,':у' Я " z z ~ а. ~ ~ю о z ~~ ~ o > <z ~o g ~ д~~aeto „v„A & N A~ a ~ o o ~ а. д 'о о ~ ~ а ~ до~. ° ~о~~н се .~~, а,~~ m ~~+3 ~:~йц~~~ ' ~Ы р~ ~~ ~ д О ~ щ О О) O Я M д qP й~ д D ~o o ~ ~ g ~e r +~ z ~~~~ v щ ~ ~ ц щ [ P y и д~о+vg~: ° ю<g Д 2 ~ ~Ф Ф G' c 3 и ~о ~ ~ ~ ~ ~ д о~,rдucevu д И ° ~"i ~ u g m g Я g ~~~о0 ~ ~-~о Ы~ д'й 2 ao - a 21 а О м ~," v и ~~~ ~о Д >и . и. '~~до~:,дно> ro~~о ~~до д ~о о:~: д r ~ я oд~~ОXФa- ~3ox ~ Я о в~а,:~ ц~ ~~,~О. о д ~ ', ~~ ~ ~ �,„ r o x c A o ~ o r u zv а,в~~уу~Оц О ~ ~ ~~ '" ~ ~ О ~ ~ О xхюrxy о а, ю м ~W„„~ „ои~оо ""Я "+~~до ~д а~ ~ о Д ca ~ е Д а. g <~ >~ ~~ < I д g о ~<î О~~счО~~ Ф ~~ а, с~ О~ „~~ о се -Ю о~оокщю xк gl ~„~ oz ~ ~ о~ ~ < о ~ ~ ~ ~ Ц а. - ~ ~: ° ~ д о ~ ~ ~ ~ ~ ~ ~ д~ ~~ ~ о ии и а. ~ и ~ ~ о~ О О~ [ оо g~ Ый~ аrîr~r2 a ~ ~ й ~ о д~ rж I unа'д хх moxxomа О Е -~~z>„о: ° д mкxo�Уu~ 54 D . C Я v a3--
Глава 10. Исл ользование теории организации биоморф семенных растений 493 ! Оию ! ! 0 ~о ~,,~ = й о Cl Ю С4 X 63:г -о" О у о о оку а. д ц ~ �о ~ 2 о ~„ ~ ф„ х ~ о о д И C4etO ~о~ Ф~Ъ Я О. ! ~~, й ф) л., ОО g k3» О л О о р, о '- д � Я >~ ~ ~~Дооо о ~ о а~ад~~ 2 д о Q u Q v О о З Ео Ig л Й~ ОО О дамски~ з' ~~~'! ю ц де- д М ~ Л; g K д~о gд<~о gg ~ ~~- ~; д ~ а й "о ~ О ! д аэ ! М й ~- v Ф оw X ц ~ л 4) д ! й:фХ о О л Q Ц д )X -ою�о Xr о~ д о О O ~ 44 ~ f о ~ I ~ ~'4'~~ о „д.о~~ <о~~~ О аоо х r о ! ! w й о~ о '' ~ ~ ~ voc4 д ~ Х о о~о~о~о е,оîe~îvì ф, Х О о ~ ю Q u VVVOQ,X~ й и 3: m ~) О. ! р Ж ~) е[ a5 o m~„~,о ~' 4~ Х ~~~о йhC vЯ'n 0 ----И- ° ++X~о о !г-аэо ЮQЖ вЕ >îÕ QЮCX о О v >оа~~, о ~ о"" rкод ~о ~~о ~ ;Я u ~ о Л ~оЯ~~ X а л й с)
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ дят к весне из под прикрытия чешуй родительской ростовой почки, т.е. они субиерулярные (Нухимовский, 1976б), и на следующий год из них отрастают силлептические ассимиляционные побеги. Система, состоя- щая из пульсирующего ростового побега (тетракормуса) и его производ- ных силлептических ассимиляционных побегов, — вынужденно вегета- тивный пентакормус родиолы розовой. Имматурная фаза (рис. 190, Г; 191, Г). ПС — обычно со второго года и продолжается, в зависимости от условий выращивания, от одного до нескольких лет. В условиях загущенного произрастания растений нередки шестилетние имматурные растения. ОХ — ежегодно вегетатив- ный безрозеточный разнообразно-кустистый ризогексакормус (в его структуре участвуют пазушные ростовые побеги текущего года) или ри- зогептакормус. А — отрастание 1-2 пазушных ростовых побегов, из которых в свою очередь развиваются 1-3 ассимиляционных побега,— признак, свидетельствующий о начале имматурной фазы. Масса сырых подземных органов одной двулетней особи на участке с площадью питания 60х20, 60х10 см колеблется от 4,1 до 90,5 г. На 3-м году масса сырых подземных органов имматурных растений в среднем 92,5 г. Б— продолжается моноподиальное нарастание медиального (первоначаль- ного) модифицированного (только с чешуевидными листьями) ростово- го побега. Если у ювенильных особей гексакормусы имеют периодичес- ки кустистое строение, т.е. ежегодно образуются эфемерные побеги кущения, которые в августе полностью отмирают, то у имматурных особей гексакормусы разнообразно-кустистые, поскольку образуют два типа пазушных побегов кущения — эфемерные и ростовые. Биометри- ческие показатели роста и развития сеянцев 2-го года жизни отражены в таблице 6. Матурный эмаи (рис. 190, Д; 191, Д). ПС — начинается с 2-6 лет (очень редко на 1-м году вегетации) и позже в зависимости от условий выращивания. ОХ — ежегодно или неежегодно репродуктивный безро- зеточный компактно-корневищный ризогексакормус (редко) или ризо- гептакормус, или ризоэнатокормус; изредка на 1-ом году жизни зацве- тают растения с ризодикормусовой структурой. А — отрастание первых репродуктивно-ассимиляционных побегов свидетельствует о начале ма- турного этапа. Охарактеризуем сезонный ритм развития родиолы розо- вой в Подмосковье (1974-1978 гг.): начало отрастания — 10.1Ч+3, продолжительность фенофазы 7-30 дней (ср. 17); начало бутонизации 24.1Ч+13,1, продолжительность бутонизации 7-21 (ср. 13) дней; начало цветения — 09.Ч+18,0, продолжительность цветения 40-54 (ср. 47) дней; начало созревания семян 23.VI+15,0. Рост и развитие продолжа- ется от начала отрастания побегов до начала созревания плодов от 62 до 88 дней. В середине июня происходит вторичная генерация побегов: из пазух чешуевидных листьев развертывающихся репродуктивно-рос- товых почек развиваются новые силлептические ассимиляционно-ре- продуктивные и ассимиляционные побеги. Одновременно с этим лис- тья побегов первой (весенней) генерации желтеют и засыхают. Новые побеги малочисленны; они в 2-10 раз меньше по размерам и значитель- но слабее (структурно и функционально) плодоносящих и засыхающих
Глава 10. Использование теории организации биоморф семенных растений 1Ф ~в ~ ~'й ~. Х~ '~� XvaC4 4Э Р' ~р рв lO Ьм ~( ф О х Ф~, CO CO +! CO CO CO СЧ CO +! CO CO CO СЧ CO +! CO ~О ~О CO 00 CD +! CO М~ CO +! CD Ф~ CC Ж ~) Эф Ж О М ~Г~ ~О М~ ЙИ 1 Я ~х „ О х Х< а х х ~.З ~Зэ4' О~~C~O щоа хо9~ й р, O )Я о о ~- Q. oo С о ~ Б о х (б Б g) Ы К д В а О х Я К М Б й) о~ О CL W Х Ь— (б .'д ~ n Ю ~ Х с~. К (д Ох Q p) :У М о ~ со 4: ~о о о О С~ Б Ш CO ~Г~ CO +! CO CO ~Г~ CO Ch, +! CO ~Г') CO CO ~Г~ CO +! CO ГЧ CO CO +! CO CO ~О CO CO +! CO 00 СЧ ~Г~ СЧ CO ф 3 О >ф р' ° Я} О О Ф Я Б ~ g,î р CO CO +! CO CO CO CO +! С) С) CO ~3 CO CO +! С) CO CO +! CO Ch, Ch, +! С) CO CO +! CO CO Ch, CO +! CD ~Г~ ~Г~ CO +! CO Г СЧ CO CO +! CO 00 ~О CO +! CO CO Ch, CO ОО ~Г~ CO CO ~Г') ~Г~ CO ОО Х 2 Ж A С4 3 О ~ч О О Q й С4 й )~ Ое A Ж О Я. О Х м ~4 Ди~ A й е X о Ж О g О Е Ж й ~ о О О 63 g) О О О „Ю Р' в Ж О Ж 63 Q О Х )g 2 Ц:) г~ ~о~ ~~о~ ~ с" о-~ ЖО д~ A Q. О ~ Д О О ~ч g) О ~ч О Е Х Ж Ж Ж В [ Q О 2 С4 63 Р Ж О Ю О й 6j ~ч О О 2 CQ
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ функционально тычиночных весенних побегов. Многие из летних по- бегов засыхают в вегетативном состоянии и лишь очень небольшая их часть достигает цветущего состояния; крайне редко они плодоносят. Надземные побеги весенней генерации постепенно полегают и к середине августа полностью засыхают. В условиях высокогорий Алтая проявляется за вегетационный сезон только одна генерация надземных побегов. С увеличением возраста усиливается кустистость растений, высота побегов и масса подземных органов (таблицы 7 и 8). Побеговая система взрослых пяти- или шестилетних растений состоит из подземных или полуподземных толстых модифицированных (с чешуевидными листьями) побегов. В пазухах чешуевидных листьев на верхушке этих побегов об- разуются округлые почки, неприкрытые листовыми чешуями верхушечной родительской почки. Из округлых почек развиваются весенние ас- симиляционные и ассимиляционно-репродуктивные побеги. Ростовая почка, заостренная по форме, закладывается в сборной почке и составляет верхушку каждого ростового побега. В пазухах чешуй сборной почки образуются зачаточные почки, которые в процессе нарастания ростового побега развиваются в силлептические (без периода покоя) надземные побеги или в весенние (после зимнего покоя) надземные побеги. Много- летняя побеговая основа родиолы розовой — компактное ветвистое мяси- стое корневище; оно формируется из зимующих ежегодных компактных пленорезидных приростов ростовых побегов. На корневищах развиваются многочисленные придаточные корни. Масса подземных органов (сырых) одного сеянца при выращивании по схеме 60х20 см на опытно-производственном участке составила пос- ле 4 лет выращивания 233,0+26,2 г, а после 5 лет — 388,0+26,5 ã. Представление о массе сырых подземных органов отдельных сеян- цев за 6 лет интродукции дает таблица 8. Общий вид пятилетнего рас- тения представлен на рис. 192. Хорошие результаты получены при выращивании родиолы розовой из отрезков корневищ разной длины. Так, прирост подземных органов отдельных растений, полученных от маркированных корневищных че- ренков, составил после двух сезонов вегетации от 79,4 до 231,7 г, в среднем 170,3 г (масса сырых подземных органов), а после трех сезонов вегетации — от 64,1 до 372,3 г, в среднем 218,9 г (рис. 190, Е, Ж). Оказалось интересным сравнение особенностей возрастной и феноло- гической изменчивости родиолы розовой из разных мест произрастания. На Алтае (в естественных условиях произрастания), где сосредоточены основные в нашей стране заросли родиолы розовой, на высоте 1700-2200 м над ур. моря и выше сеянцы развиваются медленно, впервые цветут и плодоносят лишь на 7-40-м, а чаще на 12-20-м году. При выращивании родиолы в Московской области при разной степени загущения (от сплошного Ло 60х20 см) первое ивегение в единичных случавв наошодалось на 1-м ropy лиани, на 2-м ropy заплевало or 0 до 15%, чаШе до 4 % растений, на 3-м- от 14 до 63 %, на 4-м — от 25 до 71,3 %, на 5-м году — от 53,3 до 86,7 %. Растения в возрасте 4-5 лет, выращенные в условиях Подмосковья, по габитусу и массе корневищ с корнями (табл. 7-8) напоминают растения 30-50-летнего возраста, произрастающие в высокогорьях Алтая.
Глава 10. Использование теории организации биоморф семенных растений 497 ~О М +I CO ~О Г~) Ch, +I С) М СЧ +I ~О CO О hC Ж 4) С4 4) Р' И 3 +I CO +I CO +I CO Ch, М Г~) +I ~О ~О Ch, Ch, СЧ +I СЧ СЧ CO +I CO CO +I С) Ch, +I CO ~Г~ +I ~Г~ ~О ° I СЧ СЧ +I CO ~Г~ +I ~Г~ Г~) 00 СЧ +I CO М +I ~Г~ ~/~ ~О CO +I М~ +I М ~О +I CO М М~ М +I Ch, М СЧ СЧ Ch, +I CO ~О 00 ~Г~ CO +I CO СЧ +I CO СЧ 00 +I CO М М 3 О Q О C 00 +I CO Ch, 'Ф' Ch, +I CO М СЧ +I CO ~О М +I СЧ 00 +I ~Г~ ~О Ch, +I CO М CO +I М 00 00 CO +I CO ~Г~ Ch, +I CO ~О М +I CO 00 +I CO ~Г~ CO СЧ СЧ 1 CO СЧ ~О CO Ch, СЧ ~Г~ CO ~О СЧ ~Г~ CO ~Г~ СЧ ~Г~ CO СЧ Е й Ж Г' О Ж й О � й )~ Ж ~ О Ж М ~{:) О ~О О Q м Е й Т Ж й 1,д Ж й О Ж О Ю О й 63 ~ч О О 3 CQ A ~ч О ~ч Ж Я Й О ~ф О ~4 ж~~ Ж (б ~ч О О Ж О М ~ч Ж 1 О О Ж Р Ю О й О О Ж Р О О LO О Х g L L С~ ~) од, Х Д. К Б (б Ж о ~ о L (ц 2 (б ! х < х (б ) о о во Я о о Q. С О E3- ~~ L о О о Б Ш Ю Ю О М Ю ( а 00 ch ch О ( О ° ~~ ~ ч СЧ М) О О О О О О ~ф' М М Ch О Ю О М М М Ю 00 Г~ Ю Ю 00 СЧ с~ СЧ (~) ф~ д О ~ О ~ д () О а5 Ю 21 Ж 3 й Ж A ~ч О й И Ж Ж й й;[ Q О ~4 A Q 63 Р 'Ф'
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Таблица 8 Масса (в г) сырых подземных органов отдельных сеянцев радиолы розовой (50-100 измерений; в скобках даны средние показатели, время измерений - сентябрь, октябрь) Рис. 192. Общий вид пятилетней особи Rhodiola hosea, выращенной под Москвой (ВИЛАР, интродукционный участок). Одно деление масштаба — 3 см.
Глава 10. Использование теории организации биоморф семенных растений 499 В естественных условиях произрастания (Алтай) родиола розовая образует компактные горизонтальные и наклонные корневища с неболь- шим числом придаточных корней; в условиях культивирования (Мос- ковская обл.) развиваются корневища компактные наклонные и верти- кальные с многочисленными придаточными корнями. В структуре куста пятилетнего растения в Подмосковье 20-40, а на Алтае 1-3 надземных побега. Только к 50-60 годам жизни в высокогорьях Алтая кусты имеют 20-40 надземных побегов. Самый крупный куст (возраст около 200 лет), который мы наблюдали в условиях высокогорий Алтая, имел 249 над- земных побегов. Живые верхние части его корневищ были связаны снизу сохранившейся массой отмерших корневищ (скрытый клон, или криптоклональная стадия жизни синорганизма). Самый крупный пяти- летний куст радиолы розовой в Подмосковье имел 120, а шестилетний 240 надземных побегов. Некоторые признаки старения намечаются при выращивании ро- диолы розовой уже после 6-8 лет: уменьшается число цветущих побегов в кусте и высота надземных побегов, появляются очаги некроза в кор- невище, ветви корневищ поднимаются выше поверхности почвы, хуже растут и быстрее отмирают, чем подземные ветви. У десятилетних осо- бей еще сильнее оказываются отмеченные выше признаки старения; кроме того, старые плантации трудно поддерживать в хорошем агротех- ническом состоянии, появляются вредители (бадановый долгоносик), усиливается давление корневищных и корнеотпрысковых сорняков. В высокогорьях Алтая признаки заметного старения появляются значи- тельно позже. В этих условиях взрослые сеянцы на разных отрезках своего жизненного цикла переходят к замедленному вегетативному раз- множению, что обеспечивает длительное сохранение жизни синорга- низму, иногда до 300 лет. Подведем некоторые итоги. В первый год вегетации сеянцы родиолы розовой обычно проходят возрастные состояния проростка, всхода и ювенильное; во второй— часть особей остается в ювенильном состоянии, а другие становятся имматурными. Первое цветение и переход в матурный этап жизни на- блюдается у родиолы розовой в условиях выращивания на 2-6 (крайне редко на 1-й) год или позже. Площадь питания сильно влияет на развитие отдельных особей: оно замедляется на участках с густым расположением растений и ускоряется при увеличении площади питания. Медиальные и пазушные модифицированные ростовые побеги (с чешуевидными листьями) нарастают моноподиально. Под влиянием не- благоприятных условий (обмерзание или механическое повреждение верхушек) ростовые побеги становятся вынужденно гемисимподиаль- ными (с неежегодным вынужденным перевершиниванием). В первый год вегетации из пазух листьев медиального побега последовательно развиваются сначала силлептические ассимиляционные побеги, а затем экстраперулярные (вышедшие из покрова чешуй родительской почки) зимующие почки. На следующий год эти почки развертываются в нор- мальные весенние эфемерные побеги, которые вместе с родительским ростовым побегом составляют один более сложный побег — гексакор-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ мус. Еще через год продолжается многогодичное развитие гексакормуса, родительский ростовой побег которого имеет тетракормусовую структу- ру. Из зимующих годичных ростовых побегов формируется мясистое корневище; оно развивает множество придаточных корней. Родиола розовая — весенне-раннелетневегетирующий и весенне- цветущий вид со слабой вторичной генерацией побегов. Достигнув фи- зиологической зрелости, особи этого вида цветут неежегодно. С увели- чением возраста усиливается кустистость растений, повышается про- дуктивность и урожайность подземных органов. Оптимльный эффект дает получение рассады в специализирован- ном открытом питомнике с почвами, богатыми органическими удобре- ниями и хорошо дренированными, а также в торфо-перегнойных гор- шочках и ящиках. Далее приведем краткие рекомендации по выращи- ванию родиолы розовой, составленные на основе материалов многолет- них исследований с участием автора (ВИЛАР). Краткие рекомендации по выращиванию родиолы розовой Выращивание родиолы розовой в производственных условиях выгодно проводить в 3 этапа. Подготовка семяи и получение рассады. Этап 1-й. Семена можно высевать в ящики с почвой, хорошо заправленной органическими удобрениями; они зимуют под снегом. Весной (в марте) ящики заносят в теплицу, где вскоре начинается дружное прорастание. Кроме того, семена можно перемешать с влажным чистым кварцевым песком и, поместив в хлопчатобумаж- ный мешочек, стратифицировать при температуре 0 — +2'С в течение 1 месяца (в холодильнике или под снегом). Стратификацию важно проводить не менее чем за 1 месяц до весеннего посева, после чего семена сразу же надо высевать в ящики с хорошо дренированной богатой гумусом почвой или на специально подготовлен- ные грядки питомника (почва грядок должна быть также хорошо дренированной с большим содержанием гумуса; на 1м' желательно внести 10 кг перепревшего навоза с торфом и для глинистых почв еще и 10 кг крупнозернистого песка). Грядки можно оконтурить со всех сторон досками, что повышает их устойчивость и создает более благоприятный режим для прорастания. Ежедневно грядки поли- вают. Семена начинают прорастать на 4-5 день после посева. Семена подзимнего посева массово прорастают в середине или в третьей декаде апреля. Растения, выращенные из семян, в первый год растут и развиваются медленно. В условиях питомника от появления семядолей до развертывания первых настоящих листьев на ассимиляционных надземных побегах проходит 25-40 дней. Этап 2-ой. Подращивание рассады. После того, как сеянцы окрепнут, их пикируют, т.е. рассаживают на другие грядки с определенной площадью питания в рядки через 5-10см между рстениями и 10-15 см между рядками. В этих условиях (в питомнике для подращивания) сеян- цы нужно выращивать 2-2,5 года. При необходимости сеянцы можно подращивать в питомнике от 1 до 3-х лет. Пересадку рассады надо проводить в пасмурную погоду. Постоянно обеспечивают полив и своевременно освобождают грядки от сорняков. Сеянцы быстро развиваются и формируют хорошую мочковатую корневую систему. Этап 3-й. Пересадка рассады в поле. В открытый грунт рассада переносится на 3-м году жизни сеянцев. Пересадку можно осуществлять в конце мая-июне, когда надземные побеги уже хорошо отросли. Рассаду выкапывают из грядок, укладывают по порядку рядками в ящики (корни внизу) и вывозят на поле, предварительно полностью подготовленное к пересадке рассады. Почвенный слой должен быть хорошо удобрен органическими и минеральными (NPK) удобрениями; участок перепахивают, дискуют, выравнивают. Посадку рассады лучше всего производить вручную или (после приобретения соответствующего опыта) с использованием рассадо-посадочной
501 Глава 10. Использование теории организации биоморф семенных растений машины. Посадку осуществлять в рядки с междурядьями 45см (можно 60 или 70см) и через 20 ем между растениями. Каждый сеянец высаживают в лунку глубиной 10-15 см (в зависимости от степени развития корневой системы) и при- сыпают почвой до основания надземных побегов. В течение сезона удаляют не менее 3-х раз сорняки и культивируют. В поле содержат родиолу розовую 2,5-3 года, после чего осенью выкапывают. Растение используют на сырье, которое моют, рубят и сушат при температуре 70-80'С . При заготовке сырья отделяют наиболее сильные отрезки корневищ с придаточ- ными корнями или без них (4-бсм дл.). В ту же осень отрезки корневищ можно высадить в поле подобно трехлетней рассаде. 10.2.3. Oplopanax elatus (Nakai) Nakai Настойка подземных органов заманихи высокой Орlорапах elatus (Nakai) Nakai (Echinopanax elatum Nakai) сем. Araliaceae, содержащая сумму сапонинов и примеси в виде эфирного масла и смолистых ве- ществ, применяется в научной медицине как средство, стимулирующее и возбуждающее центральную нервную систему при астенических и депрессивных состояниях, а также при гипотонии и легких формах диабета (Алешкина и др., 1962; Машковский, 1972; Журба, Шре- тер, 1976). Биологически активный комплекс, представленный суммой стероидных сапонинов, содержится также в надземных многолетних стеблях, неоправданно уничтожаемых во время заготовок (Журба, 1971; Муравьев, Джумаев, 1973; Джумаев и др., 1973). Орlорапах elatus — узкий эндемик с дизъюнктивным ареалом, ох- ватывающим юг Приморского края и Корейский п-ов (Пояркова, 1950; Шретер, Шретер, 1968). Исследования заманихи высокой проводились в Анучинском, Ла- зовском, Партизанском, Тернейском, Чугуновском и Шкотовском р-нах Приморского края (Нухимовский, Журба, 1979). Заманиха высокая — реликтовый вид, претерпевший в третичном периоде значительное сокращение своего ареала (Куренцов, 1961; Ку- ренцова, 1973); гигромезофит темнохвойных таежных лесов (доминанты Picea ajanensis (Lindl. et Gord.) Fisch. ex Carr., Abies nephrolepis (Trautv.) Maxim. южного Сихотэ-Алиня, занимающих в основном средний горный пояс (от 500 до 1000 м над ур. моря). В субальпийском криволесье рассматриваемый вид занимает до- вольно значительные площади, составляя особые растительные сообще- ства — «ельники заманиховые» (Жудова, 1967; Куренцова, 1968). Под пологом елово-пихтовых лесов заманиха заселяет более влаж- ные почвы по берегам ручьев и склонам северных экспозиций, избегая пологих участков с заболоченными почвами. В условиях недостатка света надземные и подземные стебли ее не более 1,5-2,0 см в диам. В субальпийском криволесье (1000-1500 м над ур. моря) в условиях хоро- шей освещенности и повышенной влажности воздуха заманиха образует более крупные экземпляры со стеблями 2,0-3,5 см в диам. На более ровных участках, где в почвенном покрове хорошо развиты дерновой, подзолистый и глеевый горизонты, корневая система заманихи испыты-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ вает недостаток кислорода и сильно подгнивает. Вероятно, по этой причине она отсутствует на относительно ровных базальтовых плато, занимающих значительную площадь в пределах ее ареала. Часто в таких местах она вытесняется более конкурентоспособными кустарниками: Philadelphus tenuifolius Rupr. et Maxim, Spiraea betulifolia Pall., Lonicera chry- santha Turcz. ex Ledeb., L. maackii (Rupr.) Herd., L. maximowiczii (Rupr.) Regel, а покров зеленых мхов, обычно развитый на склонах и благо- приятствующий семенному размножению заманихи, подавлается зла- ками и осоками. Наиболее значительные заросли заманихи высокой сосредоточены в горно-лесном поясе (выше 500 м над ур. моря) в истоках рек Артемов- ки, Киевки, Лазовки, Партизанской, Тигровой, Уссури (с притоками Муравейкой и Арсеньевкой) и Шкотовки. Эмбриональный иериод. Пренатальный этан. ПС — с середины июня до конца августа-сентября. ОХ — оплодотворенная семяпочка или зре- лое семя в структуре родительского растения. Б — опыление осущест- вляется перекрестно насекомыми; цветки обоеполые, актиноморфные, пятимерные. Неприятный запах цветков привлекает мелких насекомых (мух, жуков, ос и др.). Цветоножки в момент цветения укорочены и цветки сильно сближены в парциальных соцветиях — зонтиках, что способствует опылению. С началом плодоношения цветоножки удлиня- ются. Гинецей состоит из 2 плодолистиков со свободными в верхней части пестиками. Завязь нижняя, двухгнездная. Тычинок пять, с равны- ми тычиночными нитями и четырехгнездными пыльниками. Лепестки ланцетные, желтовато-зеленые с папилезно-реснитчатыми краями. Ча- шелистников пять; они остаются при плодах. В фенофазу цветения нередки дожди, что мешает опылению, поэтому часть семян остается невыполненной. Из 200-300 цветков сборного соцветия — верхушечной зонтично-кистевидной метелки — нередко созревает 30-50 плодов. Латентный этан. ПС — с конца августа, сентября до весны сле- дующего года (очень редко) или в течение нескольких лет. ОХ — зрелое семя, обособленное от родительского растения. А — плоды, начиная с августа, по мере созревания краснеют, чем привлекают птиц, которые, склевывая их, участвуют в распространении семян (орнитохория); кроме того, плоды также распространяются самосевом. Плод — синкарпная ягода, 7-9 мм дл. и 5-7 мм шир., с двумя плоскими косточками 4-7 мм дл., 2-3 мм шир. Наружная оболочка плода — экзокарпий — глянце- вая, кожистая, ярко-красная; мезокарпий сочный. После опадения пло- дов в почву обе оболочки сгнивают. Плоды нередко опадают группами. Наиболее благоприятные условия для развития семян — покров зеле- ных мхов, где они проходят естественную стратификацию. При пересы- хании или переувлажнении семена быстро погибают. Эииэмбриональный иериод. Виргинильный этан. Преювенильная фаза. Проростковая гииофаза (рис. 193, А). ПС — с мая-июля до конца или изредка не до конца вегетационного сезона. ОХ — прорастающее семя или семядольный почкокорень — ризопротерокормус. А — семена, по- падая во влажный моховой покров, прорастают не ранее чем на следу- ющий год единично или на третий год после созревания массово; про-
Глава 10. Использование теории организации биоморф семенных растений Рис. 193. Преювенильные и ювенильные кондивидуумы Oplopanax elatus (Nakai) Nakai (Приморский край). А — проростки, Б — всходы, В — ювенильные мероаклонисты: В, — первое ювенильное состояние, В,— второе ювенильное состояние, МК — упрощенные модели разновоз- растных кондивидуумов. 1 — семядоли, 2 — гипокотиль, 3 — семядольный узел, 4— корневая шейка; 5а-5е — корни: Sa — главная монориза (субмедиаль), 5б — главная дириза, 5в — неглавная монориза (сублатераль), 5r — тририза, 5д — главная тетрариза, 5e — синториза; 6 — каулогенная корневая ветвь, 7 — ризогенная корневая ветвь, 8— чешуевидные листья, 9 — срединные листья, 10 — узлы, 11 — пазушные почки, 12— шипы, 13 — вегетативный медиальный безрозеточный побег, 14 — вегетативный аксил- лярный безрозеточный побег, 15 — постинициальный монокормус, 16 — постиници- альный тетракормус, 17 — постинициальный гептакормус, 18 — резидные монокормусы, 19 — гетероподий, 20 — аэроподий, 21 — геоподий, 22 — вегетативный (статурный) аэро- кормус; 23-26 — организменные ризокормусы: 23 — семядольный почкокорень, или ризопротерокормус, 24 — семядольный вегетативный ризомонокормус, 25 — бессемя- дольный вегетативный ризотетракормус, 26 — ежегодно вегетативный ризогептакормус, 27 — живые основания органов (побегов, листьев, корней) или их фрагменты (ос- тальные части удалены). Горизонтальная линия — поверхность почвы, пунктирная— уровень плотного мохового покрова. 1л, 2л, Зл и т.д. — особи 1, 2, 3 и т.д. лет жизни; 30+ 15л и т.д. — полный возраст (примерный) с учетом живой и отмершей частей растений; у — увеличено по сравнению с масштабом; одно деление масштаба — 1см.
ОСНОВЫ БИОМОРФОНОГИИ СЕМЕННЫХ РА СТЕНИЙ растание семян надземное, проростки нередко появляются группами- до 20 экз., чаще 5-10 экз. (до матурного этапа из них доживает обычно одна, реже более особей). Семядоли (две) голые, с эллиптическими, заостренно эллиптическими или реже ланцетными пластинками 0,7-1,4см дл., 0,4-0,7 см шир., книзу переходящими в свободные черешки 0,5-2,5 см дл. и около 0,1 см в диам. Большая длина гипоко- тиля (2-5 см) и черешков некоторых семядолей объясняется особенно- стями прорастания семян в густом моховом покрове. В первый год увеличивается в размерах зародышевая почка. Корневая система — сна- чала главная монориза, позднее дириза до 5 см дл. Всходовая гилофаза (рис. 193, Б). ПС — с июля-августа до конца вегетационного сезона (редко на открытых местах с умеренным увлаж- нением почв и нарушенным растительным покровом). ОХ — семядоль- ный вегетативный безрозеточный ризомонокормус. А — у всходов обра- зуется не более одного настоящего почковидного слабо пятилопастного листа, сближенного с семядолями, пластинка 0,5-1,0 см дл. и 0,9-1,5 см шир., зубчатая по краю и усеяна как сверху, так и снизу торчащими шипиками, в особенности по жилкам. Черешок 2-5 см дл., также покрыт шипиками. Ювенильная фаза. Чаще всего проростки, минуя состояние всходов, преобразуются в ювенильные растения. ОХ — бессемядольный ежегодно вегетативный безрозеточный ризотетракормус, реже неветвистый ризогек- сакормус или ризогептакормус. Униювенильная гииофаза (рис. 193, В,). ПС — с конца вегетационного сезона текущего года до 15-45, чаще 20-30 лет. А — ежегодное весеннее отрастание однотипных безрозеточ- ных вегетативных монокормусов с одним срединным летнезеленым лис- том, составляющих в совокупности тетракормус, а реже при обмерзании верхушек — неветвистый ризогексакормус или ризогептакормус — ин- дикаторный признак этой возрастной стадии. Первые монокормусы компактные или короткие, от нескольких мм до 2 см дл.; удлинению междоузлий в процессе роста иногда способствует довольно сильное погружение верхушечной ростовой почки в густой моховой покров. Размеры листьев постепенно увеличиваются, например, на 2-ом году жизни листовая пластинка достигает 0,8-1,0см дл. и 1,0-1,6см шир., черешок 3-6см; на 15-ом году жизни листовая пластинка— 2,0-3,5 см дл. и 3,0-8,0 см шир., черешок — 3-9см дл. и т.д. Черешок в основании полустеблеобъемлющий. Жилкование листа пальчатонерв- ное сетчатое. Пластинка 5-лопастная, покрытая сверху и снизу рассеян- ными шипиками. Главный корень сохраняет жизнеспособность или от- мирает (в переувлажненных сильно затененных местообитаниях с высо- ким моховым покровом). Б — нарастание побега чаще моноподиальное или вынужденно гемисимподиальное при обмерзании, обгорании и т.п. верхушечной почки. Постинициальные апонекрозные монокормусы преобразуются у заманихи высокой по окончании каждого вегетацион- ного сезона в компактные или короткие тотарезидные монокормусы, которые определяют структуру базового органа — аэроподия. Униюве- нильные особи имеют по одному неветвистому аэрокормусу и геокормусу, т.е. в целом одну предветвь. Корневая система — тетрариза,
Глава 10. Использование теории организации оиоморф семенных растений 505 углубляющаяся в почву до 10-15 см, или синториза, состоящая из глав- ного корня и каулогенных корней, а когда главный корень погибает, то только из каулогенных корней. Биювенильная гииофаза (рис. 193, В,). ПС — с 15-45 до 25-60 лет. А — ежегодное весеннее отрастание одиночных безрозеточных вегета- тивных монокормусов с двумя и более (обычно 3-4) срединными лет- незелеными листьями — индикаторный признак. Стеблевые части мо- нокормусов от нескольких мм до 6 см дл., чаще 1-2 см. Б — листорас- положение очередное; низовые листья (чешуи) полустеблеобъемлющие, цельные, по 2-5 в основании монокормусов; срединные листья (2-4) черешчатые (черешки 5-12 см дл., густо усажены короткими шипами), с 5, чаще 7-лопастной пластинкой, сверху с рассеянными шипами, снизу шиповатые, особенно по жилкам, 12-30 см в диам. В пазухах срединных листьев иногда развиваются одиночные почки, которые обычно стано- вятся провентивными (резервными), но при повреждении (обмерзание и т.п.) верхушечной почки выполнение ее функций переходит к верхней пазушной почке. Базальные части аэроподия постоянно наклоняются и, погружаясь в почву, превращаются в верхние части геоподия. Полега- нию аэроподия способствуют, на наш взгляд, следующие факторы: сильное затенение, когда в поисках света аэрокормусы, нарастая, при- нимают наклонное восходящее положение; мягкий подстилающий суб- страт из густого влажного мохового покрова, слабые механические тка- ни побегов, неравномерное распределение массы по аэрокормусу в резуль- тате образования летом на верхушке крупных и довольно тяжелых листьев, хорошая придаточная корнеобразовательная способность, обильные снего- пады и высокий снеговой покров. Возраст растений хорошо определяется по числу монокормусов, границами которых нередко являются своеобразные «перетяжки» стебля меньшей толщины, чем предшествующие и вышерасположенные его части; эти границы — зоны сближенных узлов, несущие или несшие в прошлом чешуевидные листья. Остатки от чешуй сохраняются до 3-5 лет. Постинициальные монокормусы густо усажены игольчатыми шипа- ми 0,3-1,0 см дл. эпидермального происхождения. С возрастом пробка слущивается вместе с шипами, поэтому резидные монокормусы старше 20 лет обычно лишены шипов. Имматурная фаза. ПС — с 25-60 до 30-120 лет. Униимматурная гилофаза (рис. 194, Г,). ОХ — ежегодно вегетативный безрозеточный простокустистый ризогексакормус, ризогептакормус или ризоэнатокор- мус. Энатокормус образован из гептакормуса и развивающихся непос- редственно из предветви октокормусов. А — первое пробуждение про- вентивной почки (реже нескольких почек) на геоподии или приземной почки в основании единственного аэроподия приводит к образованию прямостоячего короткого или длинного (2-10 см) низового каулогенно- го монокормуса — побега кущения; так формируется простой куст, что служит индикаторным признаком для униимматурных особей. Главный корень живой или уже отмер; высота аэрокормусов колеблется от не- скольких до 80 см. Б — почки верхушечные и пазушные защищенные, т.е. сверху почку образуют чешуи. Ростовые монокормусы с 2-5 средин-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ ),'ф 0" h4 ~Й й h4 О О О 3 й О Е и> 2 О О О Фй h4 3 й X ~Э' О й Ф. ф„' 2 :а Cl й f 63 f ФЭ Ое Cl О :а й~ О Ое :а О 2 с7 ф~ ~е О О. , С~3 (;Я Х r. й й К О. Е 1к ( О~ о ф~ ~X х ~ О ~ :у' 2Д О О ~) О 3 2 ф~ вИ ма,Д о ~ о 3 X g :а Р) у. ,О Р) о О. :а О
Глава 10. Использование теории организации оиоморф семенных растений 507 ными листьями. Геоподии по особенностям образования — эпигеоген- ные, т.е. погружающиеся в почву. Биимматурная гииофаза (рис. 194, Г,). ОХ — ежегодно вегетатив- ный безрозеточный сложнокустистый ризоэнатокормус. А — формиро- вание у особей 3 и более порядков побегов кущения — признак слож- ного куста и одновременно индикаторный признак биимматурной ста- дии. Низовые пожары иногда способствуют началу сложного кущения. Аэрокормусы до 1 м выс., постепенно полегающие. Главный корень в этой стадии часто отмирает. Нередко из зоны сближенных узлов геопо- дия образуется несколько сильных придаточных корней, распространя- ющихся в поверхностном горизонте почвы, которые надолго могут воз- мещать функции главного корня, значительно замедляя, а иногда вре- менно приостанавливая некроз основания геоподия. Триимматурная гииофаза (рис. 194, Г,). ОХ — ежегодно вегетатив- ные (до достижения физиологической зрелости) безрозеточные авто- номные ризокормусы-клонисты. А — осуществление полной общей не- кротической партикуляции через автономный (утративший связь с глав- ным корнем) геоподий приводит к преобразованию особи семенного происхождения (мероаклониста) в клон, что служит индикаторным признаком третьей имматурной стадии синонтогенеза. Эта стадия мо- жет отсутствовать или, проявляясь, переходить затем сразу в медиматур- ную или даже финиматурную стадию. В процессе клонирования могут образовываться особи, внешне сходные с ювенильными и имматурными мероаклонистами, поэтому мы обозначаем их соответственно как ново- ювенильные и новоимматурные особи (особи-клонисты имеют свой жиз- ненный цикл, отличный от жизненного цикла кондивидуума). Триим- матурный клон (клон является носителем признаков более сложной системы — кондивидуума — на соответствующей, в этом случае три- имматурной, стадии синонтогенеза) заманихи — совокупность несколь- ких, чаще 2-3, особей вегетативного происхождения — клонистов, ко- торые могут быть по особенностям своего строения новоювенильными (некустистыми) и новоимматурными (кустистыми). Матурный этан. ПС — с 30-120, чаще 40-80, до 300 (?) лет. Иници- матурная фаза (рис. 195, Д). ОХ — ежегодно или неежегодно репродук- тивный безрозеточный ценоподиальный ризогептакормус (редко) или ризоэнатокормус. А — отсутствие вегетативного размножения при на- личии у мероаклонистов физиологической зрелости — индикаторный признак этой фазы синонтогенеза. Чаще формируется сложный куст- гетерономный (редко), т.е. имеющий живой главный корень, или авто- номный (часто), т.е. не имеющий главного корня вследствие отмирания последнего. Пучки сильных придаточных корней нередко сдерживают темпы базального некроза. У особей, обитающих в менее затененных условиях и на открытых местах, при умеренном увлажнении, иницима- турная фаза начинается раньше и проявляется наиболее полно и про- должительно; кроме того, главный корень у них функционирует обычно дольше, чем у особей, растущих на сильно затененных увлажненных участках. В последних условиях кондивидуумы заманихи вообще могут миновать эту стадию, перейдя в медиматурную или даже в финиматур-
ОСНОВБ1 БИОМОРФОПОГИИ CEMEHHhIX РАСТЕНИЙ О д М д~ о ~' Ф ( (ц ~! ' 1 й~~ Р0 О а. а3 юМ л ~a~ ~ '4 Я 3' О Й~ ~~о о д Ки ~й сч 'о Ю д,:~." с) Я М О-&х С~3 >~ fog м О о. ад я а» ииЬ #+cd О у g о�о о 3 ~" ю „О ° ~а л ~» ф Я 4.) ~ а д„ о о ~. aS g u g И ee~1о р, ~ о О m 3 Х Й ~о~ щ ! о Е ~д~ О .~ ~~ а h4 О:3' g2о v n.î д д ю М Ж 1 ь ~. О о~. ~~1 > X~ л ~»» Г~3 ~5 д М О ~ оо о ° ~ а.~ о~~ Х g» Ю у ф» д Ю й: О v а. < ~>~ а 3 X у () О [Я» XОЕ а. ~~ +u а ф„'Ю 3 О~ X и m О о О 3 Cl X о ~~ О X 1 О а. л о О о ~ ЮО 63 ~, о О 3 ~ о а ~~ 3 й р~ ~- сч о ~X X 3 й: ф; ФЭ Е О о m 63 а' о цю 1 A о »» Й Ю»Ю Ig М 1Р, CD ф» - Х р 4 д; Е д О Оо Ей й ~) A ~р й а, о > о Ф: М:1.' О v f Е m о Е с6 О >д О~ о +< 1 О1 С~3 М :>' й О ф: О О .~, а. ,3 О д 3 О A а. ~о Ю )ф„' Р) С~3 ~ а. ~ О :~ о е~ О О X lml A 1 ~е 1 ~ль Ю О ~-1 О Л . Г Е'3 ;>ь 4 М Й: ~> Cl „3~ »г~3 а о ФЭ И ~ а. Cl Ж й~ д„ О О X ф,) ( О О v ;Я ~О ф,) а. � о л Флъ р; X а. + О 3 а, о ~z О hCXO о ю ~о О (~3 ~ ) Я ф„' а а~о о ' М о: о х~о3 "а.а. ~со g v ~, а. о Оо~ а,Й а. й~ о X v Ю ~ + ~n. ~1 О О~ О М й u" u QX ~„ у о. F» vМ ~3 а ~ й О~ Ж рд„' 2 X а Q, >ь О ~4 О О а. < се D.
Глава 10. Использование теории организации оиоморф семенных растений 509 ную стадию. Из-за слабого разветвления побегов ограничены возмож- ности для быстрого проявления полной общей партикуляции через гео- подий. Однако с возрастом, когда усиливается полегание аэрокормусов и увеличивается площадь распространения геоподиев в результате пре- образования в них нижних аэроподиальных частей, погружающихся в почву и укореняющихся, центр куста депрессируется молодыми элемен- тарными зонами кущения, перемещающимися в направлении наиболее свободных участков местообитаний; так подготавливаются условия для осуществления полной общей партикуляции и вегетативного размноже- ния. Аэрокормусы 0,3-1,5 м, редко до 2 м. Б — моноподиальный и вы- нужденно гемисимподиальный (при повреждении верхушек) типы на- растания побегов, обычные на виргинильном этапе, сменяются на ма- турном этапе синонтогенеза гемисимподиальным типом нарастания, по- скольку неежегодно происходит смена верхушек побегов после отмира- ния верхушечных соцветий. Побеговая система заманихи высокой состо- ит из монокормусов и дикормусов, тетракормусов и пентакормусов. Постинициальные монокормусы вегетативные (точнее ростовые) безро- зеточные, компактные, короткие или длинные (1-15 см), апонекрозные (реже вынужденно акронекрозные), с 5-10 сильно сближенными низо- выми листьями (чешуями) и 1-5 срединными летнезелеными 5-7 — ло- пастными листьями, покрытыми, особенно снизу по жилкам, рассеян- ными колючими шипами. Постинициальные дикормусы вегетативно- репродуктивные (точнее репродуктивно-статурные) безрозеточные, ко- роткие и длинные (4-15, чаще 6-10 см), полузакрытые (семидетерминат- но-репродуктивные), акронекрозные, с 4-6 срединными листьями (6-й лист 3-5-лопастной, расположен в основании соцветия). Постиници- альные тетракормусы — это ростовые (статурные) плейофазные (до 100 фаз видимого роста) апонекрозные побеги, а постинициальные пента- кормусы — это в отличие от тетракормусов репродуктивно-статурные унирепродуктивные побеги, ветвящиеся в зоне соцветия. Пластинки листьев 15-35 см в диам., на длинных (5-20 см) полых черешках, густо усаженных короткими (до 1см) шипами. Шипы на стеблях иногда достигают 2 см. По форме лопастей листьев различимы два биоморфо- muna: растения с туиыми и слабо островатыми широкими лоиастями листьев (рис. 195, 4o) и растения с сильно островатыми узкими лоиастями листьев (рис. 195, 41). Соцветие — верхушечная полузакрытая зонтич- но-кистевидная брактеозная (с брактеями без зеленого листа в основа- нии соцветия) или фрондо-брактеозная (с одним зеленым листом в основании соцветия) поникающая слаборазветвленная метелка, состо- ящая из зонтиков, сидящих на укорачивающихся к вершине соцветия осях 2-го порядка, развитых из пазух чешуевидных листьев (брактей), расположенных по спирали. В основании соцветия зонтики образуются нередко и на осях 3-го порядка. Ближе к верхушке сборного соцветия число цветоносных осей, образующихся в пазухах брактей, уменьшается до 2-1, а верхушка соцветия не развивает конечного цветка. Цветение и плодоношение базипетальное. Зачатки соцветия закладываются в верху- шечной почке с осени. Фенофазы: бутонизация — май-июнь, цветение — июнь-июль, пло- доношение — с конца августа до октября.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ Мономерные резиды ценоподиев сохраняют жизнеспособность в зависимости от условий произрастания заманихи от 20 до 100 лет, чаще 30-60 лет. Резидные монокормусы безлистные; обычно они следующих видов: тотарезидные побеги (компактные, короткие, длинные), когда побеги апонекрозные, т.е. не отмирают сверху после окончания вегета- ционного сезона, и бреви-, медио- или магнорезидные, когда побеги акронекрозные, а отмирающее верхушечное соцветие после завершения его развития составляет соответственно большую, половину или мень- шую часть всей длины этих побегов. Медиматурная фаза (рис. 195, Е). ОХ — совокупность нескольких ежегодно или неежегодно репродуктивных безрозеточных автономно-цено- подиальных ризокормусов. А — осуществление полной общей некроти- ческой партикуляции непосредственно через геоподий приводит к ве- гетативному размножению — образованию клона, особи которого (кпо- нисты) обладают физиологической зрелостью, т.е. нормально цветут и плодоносят. Клонисты несут признаки проявления репродуктивной фун- кции в прошлом или в настоящем (рубцы от соцветий, следы переверши- нивания стеблей), поэтому обозначаем их всех как матурные клонисты. Б — на открытых местах со слабым моховым покровом и умеренным увлажнением почв отдаление клонистов друг от друга медленное и срав- нительно небольшое (замедленное вегетативное размножение); при силь- ном затенении на глыбово-каменистом субстрате с обильным моховым покровом и повышенным увлажнением почв (здесь вегетативное размно- жение может начинаться еще на имматурной стадии) нередко клонисты сильно отдаляются друг от друга (интенсивное вегетативное размножение). В последних условиях местообитания у особей заманихи верхушки аэро- кормусов мигрируют с камня на камень. Полегая и укореняясь, они дают начало новым парциальным кустам, а впоследствии довольно быстро и новым особям. Ценоподии нарастают гемисимподиально, т.е. несколько лет побеговые оси удлиняются моноподиально, а затем наступает год, когда путем перевершинивания замещается отмершее верхушечное соцве- тие и т.д., (с неежегодным перевершиниванием). Финиматурная фаза (рис. 195, Ж). ОХ — совокупность нескольких неежегодно репродуктивных и ежегодно вегетативных безрозеточных автономно-ценоподиальных ризокормусов. А — в клоне проявляются признаки старения, которые выражаются в усилении некроза, ослабле- нии репродуктивной способности и в образовании не только матурных клонистов, но и клонистов, не имеющих следов от ироявления реиродук- тивной функции, т.е. некустистых вегетативных, или новоювенильных, и кустистых вегетативных, или новоиниматурных, клонистов. Б — в кон- кретных сообществах часто вместе произрастают мероаклонисты и кло- ны нескольких кондивидуумов, поэтому их пространственные границы определить бывает сложно. Задача облегчается, когда удается обна- ружить отдельные кондивидуумы на аклональной или клональной ста- диях жизни (выделение таких возрастных стадий также правомерно, как и рассмотренных выше). Сенильный этан (рис. 195, И). ОХ — ежегодно вегетативные (после утраты физиологической зрелости) безрозеточные автономно-ценопо-
Глава 10. Использование теории организации оиоморф семенных растений диальные ризокормусы. А — сенильные клоны или их остатки в виде одиночных разрушающихся клонистов встречаются очень редко. Б— синонтогенез продолжается долго; кондивидуумы старше 300 лет (при- мерный возраст) нами не обнаружены. Резюмируем особенности биоморфогении заманихи высокой. Заманиха высокая (Ор1орапах elatus) на юге Приморского края (юж- ный Сихотэ-Алинь, горы Пржевальского) встречается в основном под пологом смешанных таежных лесов в среднем поясе rop и в верхнем горном поясе субальпийского елово-пихтового криволесья, где произ- растает на влажных, часто глыбово-каменистых фрагментарно-торфя- нистых почвах с обильным покровом из зеленых мхов по склонам се- верных экспозиций. Семена заманихи, опавшие во влажный моховой покров, прорас- тают не ранее чем на следующий год и позже, после созревания, зимуя под снегом. На виргинильном этапе жизни особи нарастают моноподи- ально, переходя к вынужденному гемисимподиальному нарастанию только при повреждении терминальной почки (обмерзание, обгорание и т.п.). Долгое время побеговая система не ветвится (ювенильное воз- растное состояние), затем с увеличением вегетативной мощи начинает- ся слабое кущение (имматурное возрастное состояние). Базовый ор- ган — неветвистый или слабо ветвистый безлистный гетероподий или ценоподий, он состоит из надземной побеговой персистентной части- аэроподия (одного или нескольких) и подземной побеговой персистент- ной части — геоподия (неветвистого или ветвистого). Кондивидуумы впервые цветут на 30-120-м, чаще 40-80-м году; нарастание побегов на матурном этапе становится гемисимподиальным — неежегодно образу- ется верхушечное соцветие, которое после полного отмирания перерас- тает весной следующего года побегом из верхней пазушной почки. Вегетативное размножение осуществляется в имматурной (редко) или матурной стадиях посредством полной общей некротической пар- тикуляции через геоподиальную часть кустов; оно замедленное (на от- крытых умеренно увлажненных участках) или чаще интенсивное (на затененных участках елово-пихтовых лесов среднего пояса rop и среди субальпийского криволесья). Кроме возрастных состояний кондивидуу- мов прослежены возрастные состояния клонистов: различимы ювениль- ные, имматурные, матурные и сенильные клонисты. Максимальную про- должительность синонтогенезов установить не удалось; примерный пол- ный возраст отдельных вымирающих кондивидуумов — около 300 лет. По эколого-морфологическим тропобиозным признакам заманиха высокая — это низкоросло-кустарниковый летнезеленый надземно-дву- семядольный полегающе-аэроподиальный эпигеогенно-геоподиальный мультирепродуктивный безрозеточный полузакрытый (семидетерминат- но-репродуктивный) очередно-лопастнолистный шиповатопобеговый гемисимподиальный полуразвернуто- и развернутокустистый мезопла- нофильный и тахипланофильный (т.е. соответственно с умеренным и сильным разрастанием кустов в горизонтальных направлениях) ретарда- ционно-кондивидуальный и интенсивно-кондивидуальный однодомный перекрестноопыляемый гигромезофильный горно-лесной вид.
ОСНОВЫ БИОМОРФОПОГИИ CEMEHHhIX РАСТЕНИИ 10.2.4. Adonis sibirica Patrin ex Ledeb. В медицинской практике при сердечно-сосудистых заболеваниях широко используются надземные части цветущих растений горицвета весеннего Adonis vernalis L., запасы которого в природе ограничены (Ми- наева, 1970; Машковский, 1977 и др.). В качестве заменителя сырья горицвета весеннего можно использовать близкое по биологической активности сырье горицвета сибирского Adonis sibirica (Кучеров и др., 1969), широко распространенного в России. Adonis sibirica Patrin ex Ledeb. (сем. Ranunculaceae) — луговой и лугово-лесной вид, имеющий в России дизъюнктивный ареал с пятью фрагментами (уральским, горно-алтайским, кузнецко-обским, енисей- ско-ангарским и байкало-ленским) от Предуралья до бассейна р. Амчи в Якутской АССР (Пошкурлат, 1976). Исследование эколого-морфологических особенностей биоморфо- гении горицвета сибирского проводилось на Алтае в пределах Иванов- ского хребта (долина р. Белая Уба), а также хр. Сарым-Сакты (окрест- ности с. Медведка). Этот вид произрастает на Алтае на луговых полянах среди разреженных хвойных, березовых и смешанных лесов, по их опуш- кам и прилежащим к ним суходольным и остепненным лугам низкогорного и среднегорного поясов. На лесных лугах, подверженных умеренному выпасу скота, горицвет сибирский нередко представлен широко, посколь- ку его побеги животные обычно не поедают. Эмбриональный иериод. Пренатальный этан. ПС — с середины мая- начала июня до конца июня-июля. OX — оплодотворенная семяпочка или зрелое семя в структуре родительского растения. Б — в опылении участвуют насекомые (пчелы, шмели); цветки желтые, обоеполые. Латентный этан. ПС — с конца июня-июля до июля-сентября текущего года или до весны следующего года. ОХ — зрелое семя, обо- собленное от родительского растения. А — осыпающиеся плоды— орешки зеленоватые, эллиптические, рассеянно опушенные, 0,4-0,45сМ дл., 0,35-0,4 см шир. Плоды легко осыпаются еще в недозрелом состоянии. Эииэмбриональный иериод. Виргинильный этан. Преювенильная фаза. Проростковая гииофаза (рис. 196, А). ПС — с июля-сентября текущего года или реже весной (апрель, май) следующего года. ОХ — прорастаю- щее семя или семядольный почкокорень (ризопротерокормус). А — про- растание семян надземное; развернувшиеся семядоли (две) зеленые, голые, яйцевидные или эллиптические, 0,7-0,9 см дл., 0,3-0,4 см шир., постепенно переходящие в черешки, сросшиеся основаниями; главная монориза до 4 см дл.; гипокотиль 0,4-1,5 см дл., около 0,1 см в диам.; проростки появ- ляются неравномерно по времени и встречаются очень редко. Всходовая гилофаза (рис. 196, Б). ПС — с августа-сентября текуще- го года или реже в мае-июне следующего года; 2-4 недели. ОХ — над- земка-двусемядольный вегетативный безрозеточный ризомонокормус. А — монокормус с 1-4 настоящими более или менее рассеченными листьями и короткой стеблевой частью (0,1-5 см). Первый лист одно- кратно трехрассеченный с двух- или трехлопастными сегментами или лист трехлопастной нерассеченный с коротким черешком. Б — на гипо-
Глава 10. Использование теории организации биоморф семенных растений 513 Рис. 196. Виргинильные особи Adonis sibirica Patrin ex Ledeb. (Алтай). А — проросток, Б — всход, В, — В4 — ювенильные мероаклонисты: В, — первое, В, — второе, В, — третье, В4 — четвертое ювенильные состояния; Г — имматурный мероаклонист. МК — упрощенные модели разновозрастных кодивидуумов. 1— семядоли, 2 — гипокотиль, 3 — семядольный узел, 4 — корневая шейка, 5а — 5е- корни: 5а — главная монориза (субмедиаль), 5б — главная дириза, 5в — неглавная монориза (сублатерель), 5г — тририза, 5д — главная тетрариза, 5е — синториза; 6 — остаток отмершего главного корня, 7 — ризогенная корневая ветвь, 8 — кауло- генная корневая ветвь, 9 — чешуевидные листья, 10 — срединные листья, 11— прилистники, 12 — вегетативный безрозеточный монокормус, 13 — вегетативный дифазный безрозеточный тетракормус, 14 — миниморезидные монокормусы, 15— проризом, 16 — ризом, 17 — пазушные почки, 18 — сухие остатки или рубцы (сле- ды) от побегов прошлых лет, 19 — вегетативный гептакормус, 20 — первый летне- зеленый лист прошлогоднего апонекрозного монокормуса; 21-25 — организмен- ные ризокормусы: 21 — семядольный почкокорень (ризопротерокормус), 22— семядольный ризомонокормус, 23 — бессемядольный ризомонокормус, 24 — ве- гетативный дифазный ризотетракормус, 25 — ежегодно вегетативный ризогепта- кормус, 26 — живые основания органов (побегов, листьев, корней; верхние части удалены). Горизонтальная линия — поверхность почвы; 1л, 2л, Зл и т.д. — особи 1, 2, 3 и т.д. лет жизни; ув — увеличено по сравнению с масштабом; одно деление масштаба — 1 см.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ котиле и главном корне образуются поперечные складки («морщинис- тость»), свидетельствующие об их контрактильности, которая в после- дующие годы заметно увеличивается. Ювенильная фаза. Униювенильная гииофаза (рис. 196, В,). ПС — с августа, сентября до начала зимы (у летне-осенних экземпляров), с июня-начала июля до конца сезона вегетации (у весенних экземпляров, т.е. у эпиэмбриональных особей весеннего происхождения). ОХ — бес- семядольный вегетативный безрозеточный апобазовый (без базового ор- гана) ризомонокормус. А — семядоли засохли, длина стеблевой части одиночных надземных побегов 0,1-5 см. Биювенильная гииофаза (рис. 196, В,). ПС — с начала зимы 1-го года до 2-6 лет. ОХ — бессемядольный ежегодно вегетативный безрозеточ- ный проризомный некустистый ризомонокормус или ризотетракормус, или ризогексакормус, или ризогептакормус. А — подземный базовый орган — безлистный проризом; он пока не образует придаточных кор- ней. Миниморезидный монокормус, включающий, как правило, только один семядольный узел, образуется в конце первого сезона вегетации после отмирания надземной части у весенних экземпляров и некоторых летне-осенних особей. У других летне-осенних экземпляров образуется очень короткий медиальный апонекрозный (не отмирающий сверху) монокормус с 1 настоящим листом, а на следующий год моноподиально отрастает новый монокормус, т.е. вся побеговая система — это безро- зеточный вегетативный дифазный тетракормус. В последующие годы нарастание побегов эусимподиальное (с ежегодным одноразовым пере- вершиниванием); оно определяет структуру проризомного гептакормуса. Высота надземных побегов 3-15 см. Б — каждый безрозеточный веге- тативный монокормус, отрастающий ежегодно весной, состоит из ком- пактного основания с 1-2, чаще 2, чешуевидными листьями и несет одиночные пазушные почки. Вышерасположенная раздвинутоузловая часть, как правило, с одним сросшимся в трубку чешуевидным листом переходного типа и несколькими срединными летнезелеными листьями однажды- или дваждыперисторассеченными с двух- или многолопаст- ными сегментами. В очертании листья овальные или треугольные, 2,0-7,0 см дл., 1,5-6,0 см шир. Из 1-3, чаще 2, ежегодно образующихся пазушных зимующих почек на следующий год развертывается лишь одна, а остальные становятся спящими или отмирают. Триювенильная гииофаза (рис. 196, В,). ПС — с 2-6 до 6-15, чаще 8-10 лет. ОХ — ежегодно вегетативный гетерономно-ризомный некустистый ризогексакормус или ризогептакормус. А — образование придаточных корней на базовом органе — признак преобразования проризома в ризом. Главный корень живой, до 15 см дл. Б — на гипокотиле прида- точные корни образуются раньше и выше, чем на эпикотиле. Ежегод- ный акронекроз надземных частей побегов передается на корневище, на котором образуются небольшие некротические рубцы; они служат важ- ным диагностическим признаком при определении возраста корневища и всего растения. Некроз по сердцевине резидов постепенно начинает распространяться вглубь корневища. Вегетативный гептакормус прохо- дит в своем развитии состояние предветви (побеговой оси нулевого поряд-
Глава 10. Использование теории организации оиоморф семенных растений 515 ка в системе ветвления) и непосредственно из нее ежегодно базисим- подиально развивается по одному новому монокормусу. Хвартиювенильная гииофаза (рис. 196, В,). ПС — с 6-15 до 8-25 лет. ОХ — ежегодно вегетативный эусимподиальный автономно-ризомный некустистый ризогептакормус. А — отмирание главного корня — ин- дикаторный признак четвертого ювенильного состояния. Высота оди- ночных надземных побегов до 25 см. Б — ежегодно отрастают весенние ростовые (статурные) безрозеточные раздвинутоузловые акронекрозные моно кормусы. Имматурная фаза (рис. 196, Г). ПС — с 5-20 до 8-30 лет. ОХ — еже- годно вегетативный эусимподиальный ризомный кустистый ризогепта- кормус или ризоэнатокормус. А — образование двух постинициальных монокормусов — признак начала кущения и вступления в имматурное состояние. Высота надземных частей побегов до 30см. Б — энатокор- мус состоит из эусимподиального резидного гептакормуса и разви- вающихся непосредственно из него гемискелетных или скелетных по- бегов-ветвей. На корневище образуется много придаточных корней. Матурный этан. Инициматурная фаза (рис. 197, Д). ПС — с 8-30, чаще 10-20, до 20-45 лет. ОХ — ежегодно или неежегодно репродуктив- ный безрозеточный эусимподиальный корневищный ризогептакормус или ризоэнатокормус с числом постинициальных монокормусов и (или) дикормусов не более 5. Нередко, минуя имматурное состояние, юве- нильные особи преобразуются в инициматурные, впервые развивая ре- продуктивна-статурные побеги. Б — репродуктивно-статурные побеги обоеполые унирепродуктивные безрозеточные прямостоячие гетеротоп- ные (с подземным компактным основанием) акронекрозные монокор- мусы (с одним верхушечным цветком в первые годы инициматурного состояния) или дикормусы (с силлептически разветвленным рыхлым закрытым фрондозным соцветием в последующие годы матурной жиз- ни), до 40 см выс. Срединные листья дважды-, реже триждыперисторас- сеченные (дифиллы или трифиллы) с двух- или многолопастными сег- ментами, в очертании овальные или треугольные, 3-8 см дл., 2,5-8 см шир., в числе 7-15, чаще 9-12. Ниже располагаются 1-2 листа переходного типа со слабо развитой листовой пластинкой и 1-2 чешу- евидных листа, сросшихся в короткую трубку, заметно отдаленных друг от друга, и, наконец, в основании 3-4 сильно сближенных чешуи (ни- зовые листья), из которых 1-3 самых нижних чешуи не сросшиеся ос- нованиями. В почках с осени предшествующего цветению года заложе- ны зачатки репродуктивных элементов. Корневая система — кистевая синториза; она состоит из придаточных корней, связанных между собой через корневище, и углубляется в почву на 15-40 см. Медиматурная фаза (рис. 197, Е). ПС — с 20-45 до 25-60 лет. ОХ— ежегодно или неежегодно репродуктивный безрозеточный эусимподи- альный корневищный ризоэнатокормус с числом постинициальных мо- нокормусов и (или) дикормусов более 5. А — с возрастом в благоприят- ных условиях существования (лесные луга, опушки низкогорных сме- шанных лесов) наблюдается обильное кущение; число постинициаль- ных дикормусов и монокормусов колеблется от 6 до 30; куст компак-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ МК Рис. 197. Матурные и сенильные кондивидуумы Adonis sibirica (Алтай). Д — инициматурные мероаклонисты, Е — медиматурный мероаклонист, Ж — фи- ниматурный клон, И — сенильный клон; 27 — безрозеточный дикормус (схема), 28 — продольный разрез ризома, 29 — отмершие ткани, 30 — живые ткани, 31— ежегодно или неежегодно репродуктивный ризогептакормус, 32 — ежегодно или неежегодно репродуктивный ризоэнатокормус, 33 — ежегодно вегетативный (пос- ле утраты физиологической зрелости) ризокормус. Остальные обозначения те же, что и на рис. 163.
Глава 10. Использование теории организации оиоморф семенных растений 517 тный, прямостоячий, высота надземных побегов 20-60 см. Корневище за счет обильного ветвления и радиального роста увеличивется в тол- щину. В результате ежегодного увеличения некрозных сердцевинных следов оно имеет своеобразное волокнисто-сетчатое строение (на продольном разрезе живые и отмершие ткани перемежаются между собой (рис. 197, 28). Некротическая партикуляция корневища усиливает- ся; придаточные корни, образуясь на живых частях и обладая контрактиль- ностью, также способствуют разрушению связей между партикулами, од- нако полной партикуляции еще нет. Корневище достигает до 10 см в диам. Финиматурная фаза (рис. 197, Ж). ПС — с 25-60 до 70-80 лет. ОХ — совокупность нескольких (2-15) ежегодно или неежегодно репро- дуктивных автономно-ризомных симподиальных ризокормусов. А— осуществление полной общей некротической партикуляции через ком- пактное корневище приводит к замедленному вегетативному размноже- нию; мероаклонист преобразуется в клон, который продолжает суще- ствование кондивидуума. Многие клонисты обладают нормальной фи- зиологической зрелостью, цветут и плодоносят. Б — в неблагоприятных условиях произрастания (неумеренный выпас скота, сильное затенение и т.д.) растения долгое время могут не куститься, т.е. ежегодно образуют по одному новому побегу, или кустятся очень слабо. Иногда в таких условиях они вообще не проходят медиматурного состояния, а с ини- циматурного состояния переходят в финиматурное или даже в сениль- ное; нередко такие растения преждевременно выпадают из травостоя на любой стадии жизненного цикла. Сенильный этаи (рис. 197, Е). ПС — продолжается не более 10 лет. ОХ — ежегодно вегетативные после утраты физиологической зрелости безрозеточные автономно-ризомные ризокормусы. Б — синонтогенез максимально продолжается около 70-90 лет. Резюмируем особенности биоморфогении горицвета сибирского. Горицвет сибирский Adonis sibirica на Алтае произрастает в основ- ном на опушках лесов и среди разреженных лесов, на суходольных лугах вдоль небольших горных рек и ручьев в нижнем и среднем поясах гор. Семена произрастают в конце лета или осенью в год созревания, реже весной после однократной зимовки под снегом. На виргинильном этапе жизни и позднее побеговая система особей нарастает эусимподи- ально (с ежегодным одноразовым перевершиниванием). У особей лет- не-осеннего происхождения (с одним настоящим листом) нередко сох- раняется верхушечная почка, из которой на следующий год моноподи- ально образуется верхушечный побег. Кондивидуумы впервые цветут на 8-30-м, чаще 10-20-м году. Главный корень отмирает на 8-15-м году, а клон в зависимости от условий существования образуется на 25-60-м году жизни, причем некустистые и слабокустистые растения партикулируют позже, чем сильнокустистые растения. Синонтогения с замедленным ве- гетативным размножением максимально продолжается до 70-90 лет. Не- редки случаи, особенно у ослабленных растений, «перескакивания» через одно или даже несколько возрастных состояний. По эколого-морфологическим тропобиозным признакам горицвет сибирский — это травянистый летнезеленый надземно-двусемядольный
ОСНОВЫ БИОМОРФОПОГИИ CEMEHHhIX РАСТЕНИЙ геофильный мультирепродуктивный безрозеточный очереднолистный эусимподиальный кустистый закрытый компактно-ризомный гипопла- нофильный (с очень слабым подземным разрастанием в горизонтальном направлении) ретардационно-кондивидуальный (медленно вегетативно размножающийся, с клонистами, долго удерживающими территорию) однодомный перекрестноопыляемый желто-лютиковоцветковый ксеро- мезофильный луговой и лугово-лесной вид. 10.2.5. Patrinia intermedia {Ногпет.) Roem. et Schult. Лекарственное значение у патринии средней (Patrinia intermedia, сем. Ча1епапасеае) имеют многолетние подземные органы, в которых содержатся алкалоиды, эфирные, жирные масла и сапонины, причем из суммы последних выделен индивидуальный сапонин патринозид Д. В медицине применяют настойки подземных органов патринии как успо- каивающее средство, не уступающее по своему действию валерьяне ле- карственной (Минаева, 1970; Турова, 1974; и др.). Patrinia intermedia (Hornem.) Roem. et Schult. — горно-степной вид. В природных условиях Алтая изучены некоторые особенности биомор- фогении этого вида (Нухимовский, 1976а). Патриния средняя, обладая высокой засухоустойчивостью, заселяет открытые остепненные склоны южных экспозиций, осыпи и скалы в основном среднего пояса гор (400-1600 м над ур. моря). Эмбриональный иериод. Пренатальный этаи. ПС — с начала июля до августа. ОХ — оплодотворенная семяпочка или зрелое семя в структуре родительского растения. Б — опыляются цветки с помощью насеко- мых. Количество и качество плодов сильно зависит от погодных усло- вий. Плоды односеменные перепончатые с разросшимся яйцевидным прицветн иком. Латентный этаи. ПС — с середины-конца августа до середины сентября текущего года (редко) или до мая-июня следующего года, иногда несколько лет. ОХ — зрелое семя, обособленное от родитель- ского растения. А — пересыхание соединительных тканей между пло- дами и родительским растением — разновидность некротической час- тной партикуляции; она заканчивается полным отделением семян. Се- мена патринии средней удлиненно-яйцевидные, к верхушке суживаю- щиеся, 3-4 мм дл. и 2 мм шир. Масса 1000 семян 2,5-3,6 г (Якубова и др., 1967). Б — осыпание семян происходит в первой или второй дека- дах августа, реже чуть позже. Ветер нередко переносит семена на сотни метров. Попав в почву и пролежав в ней около месяца, при появлении относительно теплых и влажных условий, небольшая часть семян про- растает еще осенью, но большинство семян прорастает на следующий год. А.И. Якубова и др. (1967) отметили, что при высеве семян патринии средней в грунт прорастание начинается на 18-21 день. Фаза зимнего покоя семян патринии средней вынужденная, поскольку для прораста- ния им не обязательно воздействие пониженных температур. В условиях естественного произрастания на Алтае латентный этап продолжается от
Глава 10. Исполиование теории организации оиоморф семенных растений 519 20 дней до нескольких лет. В последнем случае семена могут не терять всхожесть, попав первоначально в неблагоприятные для прорастания условия. А.И. Якубова и др. (1967) путем лабораторных исследований установили, что только после 3-5 лет хранения семена патринии средней начинают снижать всхожесть. Эииэмбриональный иериод. Виргинильный этаи. Проростковая фаза (рис. 198, А). ПС — май-июнь текущего года, редко сентябрь прошлого года. ОХ — прорастающее семя или семядольный почкокорень — ри- зопротерокормус. А — единичные проростки появляются в год осыпания семян; сохранившиеся после зимы растения начинают весенний рост в этом случае сразу по ювенильному типу. Семядоли (две) яйцевидные 0,5-0,7 см дл. и 0,25-0,33 см шир., гипокотиль 0,8-1,2 см дл., 0,03-0,06 см в диам., главная монориза — 3-5 см дл. Всходовая фаза (рис. 198, Б). ПС — июнь-август. ОХ — семядоль- ный вегетативный предрозеточный (1 пара настоящих листьев) или ро- зеточный (2-3 пары настоящих листьев) ризомонокормус. А — первые листья от обратнояйцевидных до ланцетных цельнокрайние со слабо зазубренными кверху краями (1,0-2,2см дл. и 0,3-0,7см шир.). С раз- витием каждой новой пары перекрестно-супротивных листьев увеличи- вается степень их рассеченности и размеры. Так, например, листовые пластинки 3-ей пары листьев с зазубренными (иногда сильно) краями от 2,0 до 4 см дл. и 0,5-0,9 см шир. Узлы сильно сближены. У всходов с 2-мя парами листьев гипокотиль 1,0-1,8 см дл. и 0,05-0,1 см в диамет- ре. Главный корень удлиняется до 4,7 см, образуя первые корневые ветви, т.е. формируется дириза. Ювенильная фаза (рис. 198, В). ПС — от 1 до 2-7 лет. ОХ — бессе- мядольный ежегодно вегетативный розеточный некустистый апобазо- вый (конец 1-го года) ризомонокормус или прекаудексовый (2-7 лет) ризотетракормус. А — отмирание семядолей просходит у особей патри- нии средней, как правило, с 2-3-мя парами листьев. Этот признак сви- детельствует о начале ювенильной фазы жизни. Продолжается монопо- диальное нарастание медиального розеточного побега. Семядольный узел постепенно погружается до уровня поверхности почвы, благодаря контрактильной деятельности главного корня л ризогенных корневых ветвей, число порядков которых в этой фазе колеблется от 1 до 3, т.е. функционирует дириза или тетрариза. Кроме того, постоянно происхо- дит более или менее значительное присыпание растений наносным суб- стратом. Появление первого курторезидного монокормуса после завер- шения первого вегетационного сезона — факт образования базового органа — прекаудекса. Б — весенний видимый рост побегов начинает- ся сразу после таяния снега и установления положительных температур. Оно характеризуется образованием сначала, как правило, 1, реже 2 пар листьев переходной между чешуями и срединными листьями формы; эта особенность листообразования сохраняется и в последующие фазы жизни особей (рис. 199, з1). Резкое ослабление к осени ростовой актив- ности верхушечных меристем побегов и постепенное усиление листо- образования весной оставляет в структуре многолетней стеблевой систе- мы растения определенные отметки, которые при внимательном наблю-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 198. Виргинильные а~монисты Patrinia intermedia (Hornem.) Roem. et Schult. (Алтай). А — проростки, Б — всходы, В — ювенильные особи, Г — имматурная особь; 1— семядоли, 2 — гипокотиль, 3 — семядольный узел, 4 — корневая шейка, 5 — глав- ный корень, 6 — розеточные листья, 7 — ризогенные корневые ветви, 8 — веге- тативный розеточный аксиллярный побег, отрастающий в текущем вегетацион- ном сезоне, 9 — первые листья со слабой пластинкой в основании розеточных побегов, 10 — мономерный курторезид, 11 — прекаудекс. Горизонтальная линия— поверхность почвы; 1л, 2л, Зл... — особи 1, 2, 3 и т.д. лет жизни; ув — уве- личено в несколько раз по сравнению с масштабом; одно деление масштаба — 1 см.
Глава 10. Использование теории организации биоморф семенных растений 521 дении позволяют фиксировать границы мономерных курторезидов, т.е. апо- некрозных скелетных стеблевых остатков отросших розеточных годичных побегов. При отмирании по каким-либо причинам (обмерзание, страв- ливание и т.п.) верхушек розеточных побегов сохраняются миниморезиды, т.е. подобные курторезидам остатки, но только акронекрозных побегов. Чем старше резиды, тем слабее их границы различимы в струкгуре базового органа и со временем экзоморфологически (по наружным признакам) их определить уже невозможно. В таких случаях можно определить примерное число и возраст резидов, исходя из усредненных особенностей аккумулятив- ного, т.е. за счет резидных побегов, нарастания (верхних частей базового органа) разновозрастных особей в определенных условиях произрастания. Имматурная фаза (рис. 198, Г). ПС — с 2-7 до 3-12 лет. ОХ — еже- годно вегетативный розеточный компактно-каудексовый простокустистый ризогексакормус или ризогептакормус, или редко сложнокустистый ри- зоэнатокормус. А — начало видимого роста пазушных побегов — признак перехода особей в рассматриваемое состояние. Медиальный вегетативный розеточный побег по-прежнему активно нарастает моноподиально. От первых пазушных розеточных побегов после окончания вегетационного периода сохраняют жизнеспособность и перезимовывают первые скелет- ные части стеблевых ветвей — мономерные курторезиды, а также почки новых побегов, что в целом составляет курторезидные монокормусы. Появление первых скелетных частей ветвей — это признак превращения протобазового органа-прекаудекса в базовый орган-каудекс. Каудекс образуется на 2-7 год жизни особей. В последующие годы этой фазы жизни его нарастание детерминируют курторезидные монокормусы. Матурный этап (рис. 199, 200). ПС — с 3-12 до 60-75 лет. Иници- матурня фаза (рис. 199). ПС — с 3-12 до 15-25 лет. ОХ — ежегодно или неежегодно репродуктивный полурозеточный закрытый каудексо- вый простокустистый ризогептакормус или сложнокустистый ризоэна- токормус с числом порядков побеговых ветвей, составляющих каудекс и отрастающих из его почек, не более 3-х. Б — ежегодно весной раз- виваются верхушечные и пазушные вегетативные розеточные годичные апонекрозные побеги (монокормусы), а также верхушечные, редко па- зушные, унирепродуктивные закрытые полурозеточные годичные акро- некрозные побеги (дикормусы). Кроме того, один полный репродуктив- ный цикл обычно присущ пентакормусам, которые проходят его за несколько (2-12) фаз видимого роста. Редко развиваются пазушные уни- репродуктивные дикормусы. Почки незащищенные. Листья — перис- тые дифиллы от 3 до 15 см дл., 2-5 см шир. Соцветие — закрытая щит- ковидная метелка. Зачатки соцветия закладываются в год цветения. Цве- тение базипетальное. Число цветущих унирепродуктивных побегов, об- разующихся ежегодно в условиях естественного произрастания у одной особи, колеблется от 1 до 8, чаще 1-3. В отдельные годы матурные особи вообще не цветут. Цветение начинается в начале июля и заканчивается в начале августа. Плодоношение происходит также неодновременно; в конце июля-начале августа созревают первые семена в верхушечных
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 199. Инициматурные аклонисты Patrinia intermedia (Алтай). 24 — репродуктивные элементы (одиночные цветки, соцветия, плоды), 25 — уни- репродуктивный полурозеточный медиальный побег, 26 — унирепродуктивный пазушный побег (верхняя часть удалена), 27 — миниморезидный тетракормус, 28 — каудекс, 29 — плод и цветки, 30 — фрагмент соцветия, 31 — разнообразие первых листьев у отрастающих весной побегов, 32 — сочленение супротивных розеточных листьев в узле, 33 — упрощенные модели молодых матурных аклони- стов, 34 — ризогептакормус, 35 — гептакормус, 36 — ризоэнатокормус. Остальные обозначения те же, что и на рис. 198.
Глава 10. Использование теории организации биоморф семенных растений 523 Рис. 200. Средневозрастные, старые и сенильные особи Patrinia intermedia (Алтай). Д — медиматурная особь, Е — финиматурная особь, Ж — сенильная особь, И— полная общая партикуляция, 37 — парциальный каудекс, 38 — дегенерированный парциальный каудекс, 39 — каулогенные корневые ветви, 40 — спящие пазушные почки, 41 — ризосинкормус, образовавшийся вследствие неполной партикуля- ции. Остальные обозначения те же, что и на рис. 198, 199.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ соцветиях, затем плодоношение переходит к парциальным (боковым) соцветиям сложной метелки, а полностью завершается в середине или конце августа. Эубазовый орган у матурных особей патринии средней — подзем- ный компактный ветвистый каудекс. Структуру каудекса определяют миниморезидные тетракормусы, реже монокормусы; иногда она услож- няется викариорезидными тетракормусами (при засыпании субстратом). Медиматурная фаза (рис. 200, Д). ПС — с 15-25 до 40-45 лет. ОХ— ежегодно или неежегодно репродуктивный полурозеточный детерми- нантный (периодически заканчивающийся закрытым верхушечным соц- ветием) каудексовый сложнокустистый ризоэнатокормус с числом по- рядков побеговых ветвей более 3-х; партикуляция разделяет каудекс не ниже семядольного узла. Финиматурная фаза (рис. 200, И). ПС — с 40-45 до 60-70, реже до 75 лет. ОХ — ежегодно или неежегодно репродуктивный парциально ка- удексовый ризосинкормус (совокупность разных побегов, связанных между собой через неполностью партикулированный главный корень, вместе с корневой системой) или редко дегенерированные парциально каудексовые ризокормусы клона (от 60-70 до 65-75 лет). Сенильный этап (рис. 200, Ж). ПС — 1-6 лет. ОХ — растения, по- добные старым матурным изоаклонистам или реже клонам, но ежегодно вегетативные после утраты физиологической зрелости. Этот этап может быть или не быть в биоморфогенезах патринии средней. Итак, Patrinia intermedia развивается на Алтае в основном по типу эуонтогенезов, т.е. особи семенного просхождения заканчивают свой жизненный цикл смертью (изоаклонисты) и реже по типу несовершен- ных синонтогенезов, т.е. особь семенного происхождения в преклонном возрасте полностью партикулирует и образует 2-4 клониста, не приспо- собленных к длительному самостоятельному существованию (несовер- шенные кондивидуумы). Предельный полный возраст биоморф (инди- видуумов или кондивидуумов) в обоих случаях не превышает 80 лет. Некротической общей ризокормусовой партикуляции поначалу предшествует отмирание периодически образующихся удлиненных ча- стей унирепродуктивных побегов. Этот процесс приводит к возникно- вению начальных небольших некротических очагов в верхней кудексо- вой зоне ризокормуса, т.е. к началу общей партикуляции особи через каудекс. Постепенно разрушая проводящие ткани, ранее питавшие ныне отмершие до оснований соответствующие унирепродуктивные побеги, некроз передается внутрь ризокормуса, приводя в итоге к нарушению морфофизиологической целостности сначала каудекса, а затем и глав- ного корня. Процесс этот протекает относительно медленно, но неот- вратимо охватывает матурные особи при увеличении их полного возрас- та. Когда каудекс расщеплен некротическими очагами ниже семядоль- ного узла, мы фиксируем момент образования парциальных каудексов. Парциальные каудексы могут еще долго функционировать в системе одного ризокормуса, иногда до конца жизни изоаклонисга. У старых и сенильных особей общая партикуляция может привести к полному обособлению одного из парциальных каудексов мероакло-
Глава 10. Использование теории организации биоморф семенных растений ниста (временного аклониста), вместе с фрагментом главного корня, а также корневыми ветвями, некоторые из которых могут быть придаточ- ными. Эта система — конструктивная партикула, а одновременно и особь вегетативного происхождения, поскольку она имеет в своей струк- туре все основные органы особи — побеговую и корневую системы. Вегетативное размножение у патринии средней наблюдается редко; оно несовершенного (дефективного) типа. Клонисты при этом фактически не отдаляются друг от друга и, кроме того, не приспособлены к дли- тельному самостоятельному существованию. Общая ризокормусовая партикуляция, различная контрактильность главного корня и корневых ветвей, естественная деформация почвенных горизонтов и связанное с этими процессами дезориентированное пробуждение спящих почек- все это в совокупности обусловливает в некоторых случаях довольно сильное дуговидное или спиралевидное закручивание необособленных партикул, которое способствует их полному отрыву. У патринии сред- ней придаточные корни, если и образуются, то больше на главном корне и его более или менее отделенных фрагментах и меныие на каудексе. Кроме того, особи этого вида могут использовать влагу по- верхностных горизонтов, образуя иногда пучки эфемерных корней. Кло- нисты, как уже отмечалось выше, живут недолго и в некоторых случаях теряют и фрагмент главного корня, сохраняясь еще некоторое время за счет единичных слабых придаточных корней; базовый орган в таких случаях — дегенерированный парцизльный каудекс. Синонтогения чаще проявляется у биоморф патринии средней на менее открытых крутых склонах юго-западной и юго-восточной экспо- зиций с относительно рыхлыми почвами и с зарослями невысоких (до 1 м) кустарников (Berberis sibirica Pall., Spiraea crenata L., Cotoneaster uniflorus Bunge). На открытых сильно остепненных южных склонах и вершинах невысоких гор биоморфы чаще всего развиваются по изоак- лональному типу, формируя куст, основание которого составляет ком- пактный ветвистый каудекс. В отличие от патринии средней у другого вида — Patrinia sibirica (L.) Juss., встречающегося в более влажных ус- ловиях высокогорного Алтая, господствует другой тип биоморфогене- зов — синонтогения, а по типу базовых органов матурный биоморфо- тип патринии сибирской — это компактно-корневищные растения. Резюмируем особенности биоморфогении патринии средней. По серии эколого-морфологических тропобиозных признаков Patri- nia intermedia (Hornem.) Roem. et Schult. — это летнезеленый травянис- тый полурозеточный подземно- и компактно-кудексовый кустистый од- нодомный детерминантный желто-валериановоцветковый мультирепро- дуктивный ксеромезофильный среднегорный вид. Особенности возрастной изменчивости базового органа у особей патринии средней отражаются выделением особых форм его: прекау- декса, каудекса и парцизльных каудексов. Иногда биоморфы патринии средней развиваются по типу несовер- шенных синонтогенезов; вегетативное размножение проявляется у не- которых старых и сенильных особей, при этом в структуре образующих- ся клонистов функционируют парциальный каудекс и фрагменты отми-
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИИ рающего главного корня, что не обеспечивает кондивидуумам заметных преимуществ в продлении жизни перед изоаклонистами, т.е. особями ам- фимиктического происхождения, отмирающими без образования клона. 10.2.6. Leonurus quinquelobatus Gilib. Верхние части побегов (30-40 см дл.), достигшие фенофазы цвете- ния, пустырника пятилопастного Leonurus quinquelobatus Gilib. (сем. Lamiaceae) используются в медицинской практике для получения пре- паратов — экстракта и настойки. Препараты пустырника пятилопаст- ного обладают седативным действием и эффективны при сердечно-со- судистых неврозах, гипертонии, стенокардии, кардиосклероза, пороках сердца, при повышенной нервной возбудимости, контузиях головного мозга и др. (Алексеева, 1962; Турова, 1974 и др.). Leonurus quinquelobatus — мезофльный рудеральный вид, распрост- раненный в основном по всей Европе (кроме северных, а также полупус- тынных и пустынных районов), на юге Западной Сибири, в западном и восточном Закавказье. Пустырник успешно культивируется в России. На плантациях его выращивают в течение 3-4 лет, размно>кая семенами. Семена высевают под зиму или ранней весной. Семена пустырника пятилопастного не нужда- ются в стратификации. При весеннем посеве их в грунт проростки появ- ляются на 10-й день. Дву- и многолетние сеянцы начинают отрастать в конце апреля. Цветение наступает в конце июня-начале июля, оно длится 2,5 месяца. Массовое созревание плодов происхдит в конце августа-сен- тябре. Период вегетации продолжается 200 дней. Растения зимуют хорошо (Балабас и др., 1965). Масса 1000 орешков 0,3-1,0 г (Алексеева, 1962). Особенности биоморфогении пустырника пятилопастного исследо- вали в Московской области (Нухимовский, Нухимовская, 1981). Пус- тырник пятилопастной обитает здесь на пустырях, залежах, у дорог, заборов, близ жилья, по обрывам и лишь изредка небольшими группами встречаются среди кустарников и на лесных полянах. Во взрослом со- стоянии в посевах проявляется редко, поскольку будучи многолетни- ком, уничтожается в процессе разных агротехнических мероприятий до начала цветения. Биоморфы пустырника пятилопастного в естествен- ных условиях произрастания обычно развиваются в рамках синонтоге- незов — жизненных циклов кондивидуумов. Эмбриональный период. Пренатальный этап. ПС — с июня до августа, сентября. ОХ — оплодотворенная семяпочка или зрелое семя в структуре родительского растения. Б — в опылении участвуют насекомые (пчелы, шмели); цветки с розовым зигоморфным венчиком, обоеполые. Латентный этап. ПС — несколько дней в августе, сентябре (редко) или с августа, сентября до весны, лета следующего года. ОХ — зрелое семя, обособленное от родительского растения. Б — плод дробный оре- шек, распадающийся на 4 односеменных членика (орешка), имеющих трехгранную форму, на верхушке косо усеченных, 2-3 мм дл., 1,2-1,5 мм шир. Многим семенам пустырника пятилопастного для прорастания не
Глава 10. Использование теории организации биоморф семенных растений 527 обязательно проходить стратификацию при пониженной температуре. Зимнее пребывание семян под снегом (естественная стратификация) повышает их жизненную активность, способствует их лучшему прорас- танию. При проращивании нестратифицированных семян в лабора- торных условиях их массовое прорастание наблюдается на 5-7 день после посева. Эпиэмбриональный период. Виргинильный этап. Преювенильная фаза. Проростковая гипофаза (рис. 201, А). ПС — семена прорастают осенью текущего года (редко) или в мае, июне, июле следующего года; состо- яние проростка продолжается у одной особи 8-12 дней. ОХ — прорас- тающее семя или семядольный почкокорень — ризопротерокормус. А — семядоли (две) надземные, зеленые, голые, яйцевидно-эллипти- ческие. В первые дни после развертывания семядолей пластинки 0,2-0,3 см дл., 0,15-0,2 см шир. Пластинки на верхушке закругленные, слегка выемчатые или иногда без выемки, у основания сердцевидные. Черешки 0,2-0,7 см дл. Гипокотиль 0,5-1,5 см дл., 0,05 см в диам. Глав- ная монориза до 4см дл. Всходовая гипофаза (рис. 201, Б). ПС — с июня, июля до августа, реже сентября. ОХ — семядольный вегетативный безрозеточный ризо- монокормус. А — первые листья простые черешковые супротивные, по краю крупногородчатые, яйцевидные, 0,5-1,2 см дл., 0,4-1,0см шир., покрыты короткими простыми волосками. Пластинки семядолей увели- чиваются в размерах — 0,4-0,8 см дл., 0,35-0,7 см шир. У последующих пар листьев возрастают линейные размеры и усиливается глубина крае- вых надрезов их пластинок. Стебель до 15 см дл. Гипокотиль 0,5-2,0 см дл., 0,08-1,0 см в диам., главная дириза или тетрариза до 6 см дл. Ювенильная фаза. Униювенильная гипофаза (рис. 201,В,). ПС — с августа, сентября до начала зимы текущего или редко следующего года. ОХ — бессемядольный вегетативный безрозеточный ризомонокормус или редко двухгодичный ризотетракормус. А — семядоли отмерли, ха- рактерно моноподиальное нарастание медиального побега, которое пос- ле 1-го года вегетации продолжается у коротких побегов (0,5-8,0см выс.) и в следующем весенне-летнем сезоне. В последнем случае зимует безлистный тотарезидный медиальный монокормус, который может по- легать, что способствует его перезимовке с сохранением медиального апекса. У униювенильных растений пустырника пятилопастного разви- ваются монокормусы — это годичные монофазные без перевершинива- ний и разветвлений побеги, реже тетракормусы — это двухгодичные без перевершиниваний и разветвлений побеги. Резидный побег в резуль- тате собственного полегания или контрактильной деятельности главного корня погружается основанием в почву и укореняется; последний признак свидетельствует об образовании эубазового органа — корневища. Биювенильная гипофаза (рис. 201, В,). ПС — с начала зимы 1-го или (реже) 2-го года до 2-5, чаще 2-3 лет. ОХ — бессемядольный ежегодно вегетативный безрозеточный некустистый ризогексакормус или ризоге- птакормус. А — ежегодно весной отрастает по одному постинициаль- ному безрозеточному вегетативному монокормусу, 5-40 (60) см выс., с супротивными пятилопастными листьями (монофиллами). В конце осе-
528 ОСНОВЫ БИОМОРфОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ ) В2 -3n -51, 12 10, 12 5е! ( ! ("~ Х, ° ' ч, 2„-11, 12l q5 ~ ( 10, 13 МК Рис. 201.Виргинильные особи Leonurus quinquelobatus Gilib. (Московская область). А — проросток, Б — всходы, В — ювенильные мероаклонисты: В, — униювениль- ные, В, — биювенильные, Г — имматурные мероаклонисты; 1 — семядоли, 2— гипокотиль, 3 — семядольный узел, 4 — корневая шейка (ее место показано пун- ктиром, поскольку она не имеет строгих морфологических границ), 5а-5e — кор- ни: 5a — главная монориза (субмедиаль), 56 — главная дириза, 5в — неглавная
529 Глава 10. Использование теории организации биоморф семенных растений Рис. 201. (Продолжение). монориза (сублатерель), 5г — тририза, 5д — главная тетрариза, 5e — синториза, 6— ризогенная корневая ветвь, 7 — каулогенная корневая ветвь, 8 — низовые листья, 9— срединные листья, 10 — вегетативный безрозеточный медиальный побег, 11 — ве- гетативный безрозеточный аксиллярный побег, 12 — постинициальный монокормус, 13 — постинициальный тетракормус, 14 — миниморезидный монокормус, 15 — тота- резидный монокормус, 16 — гетерономный (связанный с живым главным корнем) ризом, 17 — почки, 18-22 — организменные ризокормусы: 18 — семядольный почко- корень (ризопротерокормус), 19 — семядольный вегетативный ризомонокормус, 20— бессемядольный вегетативный ризомонокормус, 21 — вегетативный двухгодичный ризотетракормус, 22 — ежегодно вегетативный ризогептакормус; 23 — живые осно- вания органов (побегов, листьев, корней; верхние части удалены); 1л, 2л, 3л и т.д.— кондивидуумы 1, 2, 3 и т.д. лет жизни. МК — упрощенные графические модели кондивидуумов; ув — увеличено по сравнению с масштабом; одно деление масшта- ба — 1 см.; горизонтальная линия — поверхность почвы. ни первого вегетационного сезона (исключение составляет небольшая группа униювенильных растений) происходит отмирание раздвинутоуз- ловой надземной части весеннего медиального монокормуса, а зимует компактное основание этого побега с защищенными подземными поч- ками — миниморезидный монокормус (рис. 201, В,-14). Весной пробуж- дается лишь одна из почек, остальные почки остаются спящими или отмирают. Гептакормус в биювенильной гипофазе — это неветвистый эусимподиальный (с ежегодным одноразовым перевершиниванием) пер- систентный или семиперсистентный побег нулевого порядка в системе ветвления (побеговая предветвь), пассивную (скелетную) часть которого составляют миниморезидные монокормусы, а активную часть — вегета- тивный (ростовой) безрозеточный раздвинутоузловой весенний моно- кормус. Образованию гептакормуса предшествует развитие гексакорму- са, у которого родительский медиальный побег (иногда в основании формируется миниморезидный димерный тетракормус) впервые пере- вершинивается постинициальным вегетативным безрозеточным весен- ним монокормусом. Главный корень чаще всего сохраняет жизнеспособ- ность, углубляясь до 20 см. Б — ежегодное одноразовое перевершинивание главной оси (пред- ветви) корневища, а в последующие возрастные стадии и его ветвей— отличительный признак эусимподиального нарастания пустырника пя- тилопастного. На эусимподиальном компактном корневище развивается множество придаточных корней. Имматурная фаза (рис. 201, Г). ПС — с 2-5 до 3-6 лет, чаще до 3-4 лет. ОХ — ежегодно вегетативный безрозеточный кустистый корневищ- ный ризогексакормус или ризогептакормус, или реже ризоэнатокормус. А — образование в одном сезоне от основания особи двух и более постинициальных вегетативных монокормусов или редко дикормусов- начало кущения — признак вступления особи в имматурное состояние. Весенний дикормус пустырника пятилопастного — это годичный моно- фазный, кверху силлептически разветвленный, побег, остающийся веге- тативным в имматурной стадии жизни растений. Высота надземных частей весенних побегов может достигать 120 см. Б — энатокормус об- разуется из эусимподиального гептакормуса, когда в процессе роста его ветви превращаются в октокормусы, т.е. в семиперсистентные или
530 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ персистентные побеговые ветви, не тождественные другим типам побегов. Весенние постинициальные монокормусы и дикормусы чаще всего гете- ротопные (с подземным компактным основанием). В основаниях таких побегов в зависимости от степени их погруженности в почву число пар супротивных низовых листьев (чешуй) различно (от 1 до 6, чаще 2 или 3). Матурный этап. Инициматурная фаза (рис. 202,Д). ПС — с 2-6, чаще 3-4, до 3-8 лет. ОХ — ежегодно или редко неежегодно репродук- тивный безрозеточный эусимподиально-ризомный простокустистый ри- зогептакормус или ризоэнатокормус. А — впервые развивается репро- дуктивно-статурный дикормус — признак начала матурного этапа, а образование матурной особи в форме простого куста, т.е. с двумя по- рядками побегов кущения — признак инициматурной фазы. Б — каж- дый репродуктивно-статурный дикормус пустырника пятилопастного- это годичный монофазный побег, эусиллептически (без фазы покоя) разветвленный в зоне сложного соцветия; он унирепродуктивный (одно- кратно образует обоеполые цветки на протяжении своего жизненного цикла) безрозеточный прямостоячий перекрестно-супротивнолистный гетеротопный (с подземным компактным основанием) акронекрозный (после плодоношения отмирающий сверху до подземного зимующего основания), 60-200см выс. Сложное соцветие — элемент структуры каждого вегетативно-репродуктивного, точнее репродуктивно-статурно- го дикормуса; такие побеги выполняют у пустырника пятилопастного три ведущие жизненные функции — репродуктивную, ассимиляцион- ную и аккумулятивного (скелетного) нарастания. Сложное соцветие сос- тоит из главного соцветия, развитого на верхушке исходного побега дикормуса и боковых соцветий, или присоцветий, развитых на верхуш- ках производных побегов — паракладиев (силлептических боковых по- бегов, структурно подобных верхней части исходного побега дикормуса). В пазухах перекрестно-супротивных зеленых листьев на центральных осях главного соцветия и присоцветий развиты парцильные соцветия- парциальные цимоиды (компактные двойные дихазии); в каждом цимо- иде от 7 до 15, чаще 12-15 цветков. Два перекрестно-супротивных пар- циальных цимоида составляют вместе одну условную (кажущуюся) му- товку (рис. 202, 28). Соцветия дикормусов пустырника относятся к типу сложных рацемозно-цимоидных соцветий, у которых центральные оси главного соцветия и присоцветий — открытые, а парциальные соцве- тия — закрытые. По терминологии В. Тролля (Troll, 1964, 1969), это по- лителические фрондозные, точнее фрондулозные (Федоров, Артюшен- ко, 1979), колосовидные (с компактными двойными дихазиями в пазу- хах зеленых листьев) тирсы. В структуре каждого цимоида (двойного дихазия) цветки развиваются в пазухах верхних зеленых листьев побега, имеющих меньшие размеры, чем листья вегетативной части побега. Раз- меры листьев на центральных осях главного соцветия и паракладиев постепенно уменьшаются от основания к вершине и ближе к верхнему концу становятся крайне малыми, приближаясь по форме к брактеям, что свидетельствует о постепенном истощении органообразовательной активности апексов фрондулозных соцветий. Растения пустырника по особенностям пространственного распо- ложения репродуктивных органов и форме листьев на верхушках лидер-
Глава 10. Использование теории организации биоморф семенных растений 531 Рис. 202. Матурные и сенильные кондивидуумы Leonurus quinquelobatus (Московская область). Д — инициматурный мероаклонист, Е — медиматурный мероаклонист, Ж — фини- матурный клон, И — сенильный клон; 24 — первые стадии постинициального пе- риода жизни унирепродуктивного дикормуса, 25 — сложное соцветие дикормуса (схема), 26 — главное соцветие, 27 — боковое соцветие, или присоцветие, паракладия, 28 — парциальные соцветия, 29 — центральная ось главного соцветия, 30 — цент- ральная ось присоцветия, 31 — автономный (кативший связь с главным корнем) ризом, 32 — ежегодно репродуктивный ризогептакормус, 33 — ежегодно репродук- тивный ризоэнатокормус, 34 — ежегодно вегетативный (после утраты физиологичес- кой зрелости) ризокормус. Остальные обозначения те же, что и на рис. 201.
532 ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHhlX РА СТЕНИЙ ных побегов — полузакрытые (условно закрытые), или семидетерми- натные, поскольку верхушечные меристемы их лидерных побегов обра- зуют открытые фрондулозные соцветия. У полузакрытых растений алек- сы лидерных побегов обладают необратимо затухающей органообразо- вательной активностью, развивая открытые соцветия с абрактеозными, брактеозными или фрондулозными верхушками. В основании весенних дикормусов от 2 до 8 пар чешуй; число чешуй зависит от степени погружения оснований побегов в почву. Из подземной части этих по- бегов в фазе их роста развивается много придаточных корней, большин- ство из них стновятся многолетними. В почках с осени не закладыва- ются элементы соцветия, поэтому унирепродуктивные дикормусы раз- виваются сначала как вегетативные из вегетативных почек, а затем фор- мируют репродуктивные органы. Резидный период жизни этих побегов осуществляется в форме миниморезидных монокормусов, которые сох- раняют жизнеспособность от 4 до 8, чаще 5-7 лет. Корневая система- синториза, достигающая 80 см в глубину. Корневище нарастает эусимпо- диально. Главный корень отмирает обычно на 3-5-ом году жизни; не- редко его трудно обнаружить среди множества придаточных корней, развитие которых депрессирует жизнедеятельность главной моноризы. Матурный этап может наступать, минуя некоторые стадии виргиниль- ного этапа жизни. Медиматурня фаза (рис. 202, Е). ПС — с 3-8 до 8-15 лет. ОХ — еже- годно репродуктивный безрозеточный компактно-корневищный слож- нокустистый ризоэнатокормус. А — индикаторный признак — слож- ное кущение, т.е. в основании растения возникают стеблевые оси 3-х и более порядков ветвления, при сохранении организменной целостнос- ти. Б — акронекроз каждого годичного побега выражается в отмирании надземной удлиненной части этих побегов после созревания семян при сохранении в жизнеспособном состоянии зимующего основания побега, т.е. миниморезидного монокормуса. Акронекроз годичных побегов вы- зывает образование сердцевинной полости в резиде, куда легко попада- ет влага и микроорганизмы, что ускоряет разрушение резидов изнутри; кроме того, они постепенно разрушаются и от оснований по мере от- мирания главного и нижних придаточных корней, а также предшест- вующих им снизу резидов. В дополнение к этому каждый годичный побег уже в сезон своего видимого роста (постинициальный период жизни побега) переходит на частичное «самообслуживание» — образует свои каулогенные корни, которые не мешают быстрому старению час- тей автономного компактного разветвленного корневища. Ежегодно в медиматурном кусте образуется от 5 до 30 годичных побегов (дикорму- сов и реже монокормусов). Финиматурня фаза (рис. 202, Ж). ПС — с 8-15 до 20-30 лет. ОХ— совокупность нескольких (2-12) автономно-ризомных ежегодно или не- ежегодно репродуктивных безрозеточных кустистых ризокормусов. А— осуществление полной общей некротической партикуляции через кор- невище приводит непосредственно к замедленному (ретардационному) вегетативному размножению — образованию клона, особи которого нормально цветут и плодоносят. Несколько лет клонисты сильно сбли-
Глава 10. Использование теории организации биоморф семенных растений 533 жены и как бы стянуты в одно мнимое целое с помощью множества придаточных корней; нужны некоторые физические усилия, чтобы зас- видетельствовать факт клонального образа жизни финиматурного кон- дивидуума (криитоклональная гипофаза рассматриваемой фазы его жиз- ненного цикла). Постепенно грани клонистов становятся более замет- ны — это уже признак акриптоклональной гипофазы. В этой гипофазе увеличивается число клонистов, утративших физиологическую зрелость. Б — постепенно угасает физиологическая активность клонистов, умень- шается число и степень развития годичных побегов; часть клонистов выпадает из клона, а новых клонистов уже не образуется. Угасание физиологической зрелости у пустырника пятилопастного происходит до- вольно резко; нередко матурные кондивидуумы отмирают без переход- ного (сенильного) состояния. Отмирание старых растений ускоряется конкурентным давлением со стороны растений своего и других видов. Сенильный этап (рис. 202, И). ПС — отсутствует или продолжается 1-2 года. ОХ — ежегодно вегетативные после утраты физиологической зрелости безрозеточные автономно-ризомные клональные ризокормусы (один или несколько). Б — синонтогенез максимально продолжается около 30 лет. Резюмируем особенности биоморфогении пустырника пятилопастного. Семена Leonurus quinquelobatus Gilib. прорастают в сезон их созре- вания осенью (редко) или весной, летом следующего года. На вирги- нильном этапе жизни у мероаклонистов (временно аклональных осо- бей) развивается вегетативная безрозеточная побеговая система с эусим- подиальным (с ежегодным одноразовым перевершиниванием) компак- тным корневищем, нарастающим за счет миниморезидных монокорму- сов. Иногда на 1-м году эпиэмбриональной жизни (особенно у сеянцев осеннего происхождения) аккумулятивное нарастание моноподиальное, т.е. первый прирост корневища составляет тотарезидный монокормус. Придаточные корни образуются рано и ежегодно развиваются на под- земных основаниях у каждого нового побега. Кондивидуумы впервые цветут на 2-6-м, чаще 3-м или 4-м году жизни. Зимующие почки всегда вегетативные. Соцветия дикормусов — рацемозно-цимоидные (по осо- бенностям нарастания) или полителические фрондулезные колосовид- ные (с компактными двойными дихазиями в пазухах зеленых листьев) тирсы (по комплексу признаков). Некротическая партикуляция в кор- невище начинается рано и к 8-15 годам мероаклонист преобразуется в клон. Синонтогенез максимально продолжается около 30 лет. По эколого-морфологическим тропобиозным признакам пустырник пятилопастной — это травянистый летнезеленый надземно-двусемя- дольный геофильный мультирепродуктивный безрозеточный перекрест- но-супротивно-лопастнолистный рацемозно-цимоидный полителичес- ки-фрондулозно-тирсовый семидетерминатный (полузакрытый) эусим- подиально-компактно-ризомный сложнокустистый брахипланофиль- ный (со слабым подземным разрастанием в горизонтальном направле- нии) ретардационно-кондивидуальный (с медленным вегетативным раз- множением) однодомный розово-яснотковоцветковый энтомофильный мезофильный рудеральный вид.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ PA С ГЕНИИ 10.2.7. Hedysarum neglectum Ledeb. Надземные побеги копеечника забытого — Hedysarum neglectum Ledeb. (сем. Fabaceae) — источник ценного лекарственного вещества- мангиферина (Куваев и др., 1972), который содержится в основном в листьях (Русакова, Нухимовский, 1977). Кроме того, этот вид известен как кормовое и декоративное растение (Федченко, 1948; Пленник, Куз- нецова, 1972 и др.). Hedysarum neglectum Ledeb. — горно-луговой вид, произрастаю- щий на Алтае, в Саянах, Джунгарском Алатау, Тарбагатае, на Памиро- Алае и Тянь-Шане (Федченко, 1948). Копеечник забытый, по данным Г.В. Кузнецовой и Р.Я.Пленник (1975), — это длиннопобеговый стержнекорневой геофильный поли- карпический травянистый вид с закрытыми зимующими почками. Поч- ки имеют 3-4 почечные чешуи и 5-6 зачатков настоящих листьев, а также зачатки 2, реже 1 или 3 соцветий (Кузнецова, 1973). Особенности фенологии копеечника забытого изучались Р.Я.Пленник с соавторами (1967) на юго-восточном Алтае. Было отмечено, что побеги имеют одну весенне-летнюю генерацию листьев, вегетирующих 3-3,5 месяца и сох- раняющих жизнеспособность в течение месяца после созревания семян (Кузнецова, Пленник, 1975). При выращивании копеечника забытого в Центральном сибирском ботаническом саду (Новосибирск) единичное цветение наблюдалось на второй год вегетации растений, а массовое цветение — на 3-й год (Пленник и др., 1967). Изучение эколого-морфологических особенностей биоморфогении копеечника забытого проведено на Алтае — Ивановский, Линейский, Коксуйский, Бащелакский и Курайский хребты. Биоморфогения рассматриваемого вида осуществляется, как правило, по типу эуонтогенезов (онтогенезов изоаклонистов) и лишь в исключи- тельных случаях по типу дефективных (несовершенных) синонтогенезов. Копеечник забытый произрастает чаще на склонах северной экспо- зиции, где встречается среди лесов, обычно кедровых или кедрово- лиственничных, на лесных, субальпийских и изредка на альпийских приручейных лугах, а также среди скоплений крупных камней («курум- ников»). Он нередко доминирует в луговых сообществах вместе с Saussu- rea latifilia Ledeb., Stemmacantha carthamoides (Willd.) М. Dittrich, Doronicum altaicum Pall. и др. Высотная амплитуда распространения копеечника забытого на Алтае 1400-2500 м над ур.моря, но чаще он встречается в пределах 1500-1900 м над ур. моря. В высокогорно- тундровом поясе обычно замещается родственным (из той же секции Obscura В. Fedtsch.) видом — Н. austrosibiricum В. Fedtsch. Эмбриональный период. Пренатальный этап. ПС — с июля до конца августа, сентября. ОХ — оплодотворенная семяпочка или зрелое семя в структуре родительского растения. Б — характерна энтомофилия; формируются бобы с 1-5 (6), чаще 2-4 члениками. Латентный этап. ПС — с августа, сентября до мая-июля следую- щего года, а возможно, также и несколько лет. ОХ — зрелое семя, обособленное от родительского растения. А — семена овальные, с од-
Глава 10. Использование теории организации биоморф семенных растений 535 ной стороны слегка вогнутые, 3,2-4,5 мм дл., 2,2-3,2 мм шир., масса 1000 семян — 5,2-5,5 г. Эпиэмбриональный период. Виргинильный этап. Преювенильная фаза. Проростковая гипофаза (рис. 203, А). ПС — май, июнь, начало июля; продолжается 5-12 дней. ОХ — прорастающее семя или семядольный почкокорень (ризопротерокормус). А — семядоли (две) надземные мя- систые овальные, иногда с одного края слегка вогнутые, 0,5-0,7 см дл., 0,3-0,4 см шир. Гипокотиль 0,8-1,2 см дл. и 0,08-0,1 см в диам. Главная монориза до 5 см дл. Всходовая гипофаза (рис. 203, Б). ПС — июнь, июль, август; про- должается две-четыре недели. ОХ — семядольный вегетативный безро- зеточный ризомонокормус. А — семядоли 0,6-0,8 см дл., 0,4-0,45 см шир. Гипокотиль 0,12-0,15 см в диам. Стеблевая часть монокормуса от 0,5 до 2,5 см высоты. Первый лист тройчатосложный, реже двойчатос- ложный (тройчатый, реже двойчатый дифилл) или простой (монофилл), черешчатый (от 0,5 до 2,2 см дл.). Листочки продолговато-эллиптичес- кие, эллиптические, цельнокрайние, 0,7-1,0 см дл., 0,4-0,5 см шир. Пос- ле образования 2-3 настоящих листьев семядоли, как правило, отсыхают, что свидетельствует о переходе особи в новое возрастное состояние. Б— на гипокотиле и основании главной моноризы образуются поперечные складки (проявление контрактильности). Ризомонокормус копеечника забытого состоит из монокормуса, гипокотиля и системы главного корня. Ювенильная фаза. Гипофаза первая — униювенильная (рис. 203, В-16). ПС — с июля, августа до начала зимы текущего года. ОХ — бессемядоль- ный вегетативный безрозеточный апобазовый (без персистентного побего- вого органа) ризомонокормус. А — на первом году вегетации кроме трой- чатых дифиллов изредка появляются и единичные непарноперистые пя- тилисточковые листья. Высота стеблевой части первого монокормуса 1-7см. Главная тетрариза, реже дириза, — не более 10см, гипокотиль 0,15-0,3 см в диам. Б — на корнях образуются мелкие клубеньки. Гипофаза вторая — биювенильная (рис. 203, В-17). ПС — с начала зимы текущего года до 3-12 лет. ОХ — ежегодно вегетативный безрозе- точный ризомонокормус или эусимподиальный каудексовый неразвет- вленный ризогексакормус, или ризогептакормус. А — после первого сезона вегетации надземная часть медиального побега отмирает. В почве зимует остаток этого побега — семядольный узел с пазушными почка- ми — первый миниморезидный монокормус — это каудекс. В после- дующие годы он нарастает эусимподиально за счет новых миниморе- зидных монокормусов, имеющих в своей структуре 1-2 (3) стеблевых узла и 1-2 (3) пазушные зимующие защищенные почки более или менее прикрытые прошлогодними отмершими чешуями, сохраняющимися на растении 1-4 года. Непарноперистые сложные листья 3-15-листочко- вые, чаще 3-9-листочковые. Надземные побеги до 35 см выс., например, у 4-летних особей — 8-20 см, у 6-7-летних — 20-35 см. Гексакормус состоит из миниморезидного медиального монокормуса и развитого в следующем году из него пазушного монокормуса. Гептакормус биюве- нильных особей копеечника забытого проходит в своем развитии состо- яние гемискелетной и скелетной предветви и непосредственно из нее
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 203. Виргинильные особи Hedysarum neglectum Ledeb. (Алтай). А — проростки, Б — всходы, В — ювенильные особи, Г — имматурные особи; 1 — се- мядоли, 2 — гипокотиль, 3 — семядольный узел. 4 — корневая шейка, 5а-5д — корни: 5а — главная монориза (субмедиаль), 56 — главная дириза, 5в — неглавная монориза (сублатераль), 5г — тририза, 5д — главная тетрариза; 6 — ризогенная корневая ветвь, 7— чешуевидные листья, 8 — прилистники, 9 — вегетативный медиальный безрозеточный монокормус, 10 — вегетативный пазушный безрозеточный монокормус, 11 — минимо- резидный монокормус, 12 — каудекс, 13 — почки в структуре побегов, 14 — семядоль- ный почкокорень (ризопротерокормус), 15 — семядольный ризомонокормус, 16 — бес- семядольный ризомонокормус, 17 — ежегодно вегетативный ризогептакормус, 18— ежегодно вегетативный ризоэнатокормус, 19 — живые основания надземных побегов (верхние части удалены); ув — увеличено в несколько раз по сравнению с масштабом; МО — упрощенные модели аклональных особей, Фк — габитус наиболее крупных глав- ного и боковых корней; 1л, 2л, Зл — особи 1, 2, 3 и т.д. лет жизни, горизонтальная линия — поверхность почвы; одно деление масштаба — 1 см.
Глава 10. Использование теории организации биоморф семенных растений 537 ежегодно базисимподиально развивается по одному новому монокорму- су, оставаясь не тождественным гексакормусу. Гексакормус и гептакор- мус и связанные непосредственно с ним корни — соответственно ризо- гексакормус и ризогептакормус. Главная тетрариза до 25 см. Из не- скольких зимующих почек каждого монокормуса на следующий год развертывается лишь одна, остальные становятся спящими. Имматурная фаза. Гипофаза первая — униимматурная (рис. 203, Г). ПС — с 3-12 до 5-15 лет. ОХ — ежегодно вегетативный эусимподиаль- ный каудексовый кустистый ризогептакормус или ризоэнатокормус с числом листочков у непарноперистых листьев не более 9. А — ежегодно отрастает два или более пазушных весенних ростовых побегов (монокор- мусов), т.е. происходит кущение. Высота надземных побегов 8-40 см. Глав- ная тетрариза до 35 см. Гипокотиль 0,3-0,9 см в диам. Б — сухие остат- ки надземных побегов сохраняются 5-8 лет. Гипофаза вторая — биимматурная. ПС — с 5-16 до 8-20 лет. ОХ— ежегодно вегетативный эусимподизльный каудексовый кустистый ризо- гептакормус или ризоэнатокормус с более чем 9 листочками в структуре напарноперистых листьев. А — надземные побеги 20-45 см высоты. Ги- покотиль 0,8-2,5 см в диам. Главная тетрариза не более 0,5 м. Б — про- должается эусимподильное нарастание компактного подземного кау- декса за счет миниморезидных монокормусов. Матурный этап. Инициматурная фаза (рис. 204, Д). ПС — с 8-20 до 30-45 лет. ОХ — ежегодно или неежегодно репродуктивный безрозе- точный каудексовый ризогептакормус или ризоэнатокормус без суще- ственных следов отмирания или со следами отмирания тканей в глав- ном корне не более 10 %. Медиматурная фаза (рис. 204, Е). ПС — с 30-45 лет до 60-90 лет. ОХ — ежегодно или неежегодно репродуктивный безрозеточный каудексовый ризоэнатокормус с отмершими тканями в каудексе и главном корне до 50% Финиматурная фаза (рис. 204, Ж). ПС — с 60-90 до 120 лет. Примерный полный возраст растений устанав- ливается путем подсчета ежегодных перевершиниваний побегов в струк- туре каудексовых осей с учетом особенностей кущения растений и от- мирания их каудекса и главного корня. ОХ — ежегодно или неежегод- но репродуктивный безрозеточный каудексовый ризоэнатокормус с от- мершими тканями каудекса и главного корня более 50 % Б — вегета- тивно-репродуктивные дикормусы — унирепродуктивные безрозеточ- ные длинные раздвинутоузловые потенциально базискелетные приот- крытые. Соцветия — сложная приоткрытая (преиндетерминатная) кисть, а парциальные соцветия полузакрытые (семидетерминатные) кисти. Че- шуевидные низовые (развиты в основании побегов) листья стеблеобь- емлющие, сросшиеся в трубку, в числе 3-4 (5); срединные листья не- парноперистые сложные (из 5-21 продолговато-эллиптических или ши- роко ланцетных листочков), в числе 6-12. Высота надземных побегов 30-90 см, чаще 40-50 см. Надземные удлиненные части монокормусов отмирают в конце вегетационного сезона, что постепенно приводит к отмиранию отдельные проводящие ткани в кудексовых осях и главном корне. Степень отмирания тканей каудекса и главного корня использо- вана нами для приблизительной диагностики фаз матурного периода. С
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ >3~ Е О Е Ж 3~ О О О О 3~ Р л оооо~ Ж р~ д ~o ~ g ~ Ovй дйЩй~я ф; ~ Ж О ":""3 О -О ~ "хр: a ~>„g ~- g О ~ О Х ~~ ~~о Я [а~ ~~ CO о ~~ 4оogФЙ хоv4c4 Й ~[ Я о Ха м ю Ж о~ оц~ Й Д Й ~) KCБо О ~"- р,о ~ хlоюо~ �~>О~~ д 3 р: фQ v Q д О О О о ' о. о ~ ,се ~~р, М О 3 д о: о ю 3 3>m5v и ~ о О ~1 ф: Q ~~ ~м~ д Q л Я~о~< д~>ьдЮ М 3 ~~~'B "оо~ 3 ~ <ч~ ~д) ~О4 (~ 3 о tQ X v" C42 Q ~-,о~~ >~ o~~ р; О.„, х м ~ >. СД о
Глава 10. Использование теории организации оиоморф семенных растений 539 возрастом увеличивается кустистость растения; у медиматурных особей число надземных побегов колеблется от 5 до 120. Каудекс разрастается в толщину или обильно ветвится, образуя хорошо выраженные подзем- ные ортотропные ветви. В финиматурном состоянии сильная общая партикуляция может в редких случаях привести к полному обособлению партикул с одним-несколькими придаточными корнями или фрагмен- тами главного корня и образованию двух, реже более, клонистов, т.е. изредка проявляется тип несовершенных синонтогенезов, а доминирует эуонтогения. Главная тетрариза углубляется в почву до 2-3 м; основание главной моноризы иногда достигает 18см в диаметре. Корневые от- прыски не образуются. Фенофазы на луговых полянах среди кедрово-лиственничного леса (1600-1800 м, Бащелакский хребет, 1973 г., Линейский хребет, 1976 г.): бутонизация — июнь, цветение — конец июня-начало августа, плодо- ношение — конец июля-начало сентября; в высокогорье наблюдает- ся смещение фенофаз — они начинаются и заканчивются на 1-3 недели позже, чем в лесном поясе, а около некоторых снежников в любое время лета встречаются особи, у которых только начинается пробужде- ние почек. Сенипьный этап (рис. 204, И). ПС — продолжается 1-8 лет. ОХ — еже- годно вегетативный, после утраты репродуктивной способности, кау- дексовый ризокормус. А — утрачена спосоность развивать репродук- тивные органы. Главная тетрариза частично или полностью отмирает. Б — редко процесс отмирания особи, фактически достигшей сенильно- го этапа жизни, может замедляться некоторым омоложением за счет укоренения верхушки одной из каудексовых осей (рис. 204, И-23). В этом аспекте уместно отметить, что придаточные корни — важный при- способительный механизм высших растений; образование их на разных этапах жизненного цикла обеспечивает более продолжительное суще- ствование индивидууму или кондивидууму. Резюмируем особенности биоморфогении копеечника забытого. Hedysarum neglectum Ledeb. на Алтае произрастает в основном на лугах в верхней части лесного пояса, но иногда по каменистым берегам ручьев поднимается в высокогорье. По эколого-морфологическим тропобиозным признакам копеечник забытый — это травянистый летнезеленый надземно-двусемядольный геофильный мультирепродуктивный безрозеточный раздвинутоузловой непарноперистолистный кустистый эусимподизльно-каудексовый длин- ностержнекорневой изоаклональный, редко дефективно-кондивидуаль- ный, терминально-репродуктивный приоткрытый однодомный фиоле- тово-бобовоцветковый психромезофильный горно-луговой вид. Биоморфогенезы у копеечника забытого осуществляются, как пра- вило, по типу эуонтогенезов (без вегетативного размножения), и лишь изредка в финиматурном или еще реже в сенильном возрастных состо- яниях образуются слабые клонисты, т.е. проявляется также и тип несо- вершенных синонтогенезов. Продолжительность биоморфогенезов не более 130 лет.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ 10.2.8. Phlojodicarpus sibiricus (Steph. ex Spreng.) К.-Pol. В подземных органах вздутоплодника сибирского Phlojodicarpus si- biricus (Steph. ех Spreng.) К.-Pol. (сем. Apiaceae) обнаружены пираноку- марины — дигидросамидин и виснадин, на основе которых созданы препараты, обладающие спазмолитическим действием (Пименов и др., 1968; Пименов, 1976). Вздутоплодник сибирский — горно-степной мезоксерофильный вид, имеющий дизъюнктивный ареал сибирско-монгольского типа в Южной Сибири, состоящий из трех фрагментов: даурского, селенгин- ского и байкальского, а также нескольких изолированных участков аре- ала в Якутии, Красноярском крае, Иркутской и крайнем западе Амур- ской областей (Пименов и др., 1968; Пименов, 1976). Исследование биоморфогении вздутоплодника сибирского прово- дили в Читинской обл.: Нерчинском, Шилкинском и Могойтуйском р-нах (Нухимовский, 1981). Вздутоплодник сибирский в Забайкалье — компонент раститель- ности горных каменистых степей, где произрастает преимущественно на привершинных остепненных склонах сопок (Сергиевская, 1951). Эмбриональный период. Пренатальный этап. ПС — июнь-август (редко начало сентября). ОХ — оплодотворенная семяпочка или зрелое семя в структуре родительского растения. Б — цветки обоеполые. Латентный этап. ПС — конец августа (реже сентябрь)-май, июнь следующего года или, реже, продолжается в течение нескольких лет. ОХ — зрелое семя, обособленное от родительского растения. А — от- делившиеся от родительского растения зрелые полуплодики (односе- менные мерикарпии) проходят зимой естественную стратификацию под снегом. В лабораторных условиях при комнатной температуре мерикар- пии сохраняли жизнеспособность после 4 лет хранения. Б — плоды широко-яйцевидные, 5-8 мм дл. и 3-5 мм шир., голые или с короткими волосками. Встречены популяции вздутоплодника сибирского с мелки- ми и крупными мерикарпиями. Масса 1000 мерикарпиев колеблется от 3,2 до 5,2 г. Н.П.Новосельцева (1970) установила, что всхожесть семян рассматриваемого вида без стратификации достаточно высокая (70 %), а продолжительная стратификация ускоряет процесс прорастания, но не увеличивает всхожесть. По нашим наблюдениям, всхожесть семян после 8 лет хранения при комнатной температуре не превышала 6 %. Эпиэмбриональный период. Виргинильный этап. Преювенильная фаза. Проростковая гипофаза (рис. 205, А). ПС — продолжается 8-14 дней в конце апреля, мае, реже июне. ОХ — прорастающее семя или семя- дольный почкокорень (ризопротерокормус). А — семядоли (две) над- земные линейно-ланцетные, 1,2-1,7 см дл. и 0,12-0,2 см шир.; гипоко- тиль 0,5-1,5 см дл. и 0,05-0,1 см в диам.; главная монориза, редко ди- риза, до 3-4см дл. Всходовая гипофаза (рис. 205, Б). ПС — май-июнь, июль (редко ав- густ). ОХ — семядольный вегетативный предрозеточный (1-2 сближен- ных настоящих листа) или розеточный (3-4 и реже более настоящих листьев) ризомонокормус. Первый настоящий лист тройчатый; у после-
]лава 7Q. использование теории организации биоморф семенных растений 541 Рис. 205. Виргинильные особи Phlojodicarpus sibiricus (Steph. ех Spreng.) К.-Pol. (Читинская область). А — проростки, Б — всход, В — ювенильные особи: В, — униювенильная, В, — би- ювенильная, Г — имматурная особь; 1 — семядоли, 2 — гипокотиль, 3 — семядоль- ный узел, 4 — корневая шейка, 5а-5д — корни: 5а — главная монориза (субмеди- аль), 56 — дириза, 5в — неглавная монориза (сублатераль), 5г — тририза, 5д — гла- вная тетрариза, 6 — ризогенная корневая ветвь, 7 — розеточные листья, 8 — ка- улогенная корневая ветвь, 9 — вегетативный медиальный розеточный монокормус, 10 — вегетативный аксиллярный розеточный тетракормус, 11 — курторезидный монокормус, 12 — преконодий, 13 — конодий (прекаудекс), 14 — каудекс, 15 — поч- ки в структуре побегов, 16 семядольный почкокорень (ризопротерокормус), 17— семядольный ризомонокормус, 18 — бессемядольный ризомонокормус, 19 — еже- годно вегетативный ризотетракормус, 20 — ежегодно вегетативный ризогеп- такормус, 21 — живые основания органов (верхние части удалены). 1л, 7л, 8л, 10л- особи 1, 7, 8, 10 лет жизни. Одно деление масштаба — 1 см. МО — упрощенные модели изоаклонистов. Горизонтальная линия — поверхность почвы.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ дующих листьев усложняется рассеченность пластинки. Семядоли 1,5-3,5 см дл. и 0,15-0,25 см шир., главная дириза или тетрариза до 6 см. Ювенильная фаза. Униювенильная гипофаза (рис. 205, В,). ПС — с июня, июля, редко августа до начала зимы текущего года. ОХ — бес- семядольный вегетативный розеточный преконодиальный ризомонокор- мус. А — индикаторный признак — образование преконодия — ком- пактной (предрозеточной или розеточной) еще незимовавшей побего- вой части, расположенной непосредственно над гипокотилем. Биювенильная гипофаза (рис. 205,В,). ПС — с начала первой зимов- ки до 3-20 лет. ОХ — бессемядольный ежегодно вегетативный розеточ- ный некустистый прекаудексовый ризотетракормус. А — листья дваж- ды или трижды перистые, черешковые (дифиллы и трифиллы). Базовый орган — прекаудекс. Он образовался непосредственно из преконодия и является одновременно конодием. У вздутоплодника сибирского коно- дий предшествует образованию каудекса, поэтому он является также и прекаудексом. Б — базовый орган нарастает моноподиально за счет увеличения в его структуре числа курторезидных монокормусов. Имматурная фаза (рис. 205, Г). ПС — с 3-20 (чаще 5-8) до 6-25 лет. ОХ — бессемядольный ежегодно вегетативный розеточный кустистый ризогексакормус, ризогептакормус, редко ризоэнатокормус. А — нача- ло кущения — индикаторный признак имматурной стадии. Главная тет- рариза до 40 ем, гипокотиль 0,5-1,2 см в диам. Побеговая система с возрастом усложняется. У имматурных особей с началом кущения об- разуется медиальный гексакормус, который в своем развитии до этого был тетракормусом (моноподиальной скелетной или гемискелетной предветвью в системе ветвления). Образовав пазушные (1-2, реже более) вегетативных розеточных монокормуса, тетракормус вместе с ними стал новым по сложности организации побегом — гексакормусом. В после- дующие годы пазушные побеги — монокормусы преобразуются в па- зушные тетракормусы, что усложняет в целом структуру побега (или системы побегов) всего растения: образуется гептакормус. После услож- нения пазушных побегов и преобразования их в октокормусы (побеги- ветви, не тождественные другим типам побегов) побеговая система, или общий побег, всего растения становится энатокормусом. Частные побе- ги имматурных особей нарастают, как правило, моноподиально. Сохра- нение первой скелетной компактной части побеговой ветви — признак, свидетельствующий о преобразовании прекаудекса в каудекс. Зимую- щие верхушечные почки открытые (незащищенные); многие пазушные почки не развертываются после первой зимовки и становятся закрыты- ми (защищенными) спящими. Образование сложного куста в имматур- ной фазе — явление очень редкое; оно возможно, как правило, при вынужденном пробуждении новых пазушных почек после повреждений (чаще всего огнем) верхушечных почек побегов предветви и ветвей пер- вого порядка. Б — постинициальные побеги по времени отрастания весенние; в зависимости от возраста растений, степени их развития и условий сущствования ежегодно в структуре каждого монокормуса об- разуется от 2 до 10 перистых розеточных дифиллов и трифиллов. Матурный этап. Инициматурная фаза (рис. 206, Д). ПС — с 5-20 (ча- ще 8-15) до 20-35 лет. ОХ — физиологически зрелый ежегодно или
Глава 10. Использование теории организации биоморф семенных растений 543 Рис. 206. Матурные и сенильная особи Phlojodicarpus sibiricus (Читинская область). Д — инициматурная, Е — медиматурная, Ж — финиматурная, И — сенильная осо- би. 22 — репродуктивный ризогептакормус, 23 — неежегодно репродуктивный ри- зоэнатокормус, 24 — вегетативный, после утраты репродуктивной способности, ризокормус, 25 — унирепродуктивный полурозеточный закрытый пентакор- мус,26 — унирепродуктивный розеточный пентакормус (схема), 27 — верхушеч- ное соцветие-зонтик, 28 — внерозеточные листья, 29 — отмершие ткани, 30— сухие остатки побегов, 31 — миниморезидный тетракормус, 32 — клон, возникший из корневых партикул (после заготовок сырья), 33 — лист со слабо развитой плас- тинкой, 34 — скрытнонарастающие побеговые ветви, 35 — сухие остатки черешков листьев (большая часть их удалена). Остальные обозначения те же, что и на рис. 205.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ неежегодно детерминантно-репродуктивный (закрывающийся верху- шечным зонтиком) -розеткообразующий кудексовый ризопентакормус (редко) или ризогексакормус, или ризогептакормус (первая инициматур- ная гипофаза) или ризоэнатокормус (вторая инициматурная гипофаза) с числом постинициальных (развернутых активных) побегов не более 10. А — образование первых унирепродуктивных монокормусов свидетель- ствует о начале матурного этапа. У вздутоплодника сибирского непос- редственно из ювенильной стадии в матурную стадию переходят очень немногие особи, образуя медиальный унирепродуктивный плейофаз- ный полурозеточный акронекрозный пентакормус. После завершения постинициального периода жизни пентакормуса остается миниморе- зидный тетракормус, который и составляет начальную структуру всего каудекса. Большинство инициматурных особей образуется из имматур- ных особей; в этом случае репродуктивная функция впервые проявля- ется в структуре гептакормуса. В последующие годы несколько лет еже- годно могут образовываться унирепродуктивные дикормусы, завершая постинициальное развитие плейофазных пазушных пентакормусов, а затем обычно бывают более или менее длительные перерывы в цветении. Б — аксиллярные пентакормусы — унирепродуктивные ди-, три- или плейофазные (2-15 фаз видимого роста) полузакрытые розеткообра- зующие очереднолистные, 20-70 см выс. Соцветие — одиночный слож- ный зонтик или редко KMGTb из 2-3 сложных зонтиков (Федоров, Артю- шенко, 1979). Число постинициальных дикормусов колеблется по годам от 0 до 5, чаще 1-3. Сверхсоцветие гептакормуса и образующегося впос- ледствии из него энатокормуса состоит из нескольких (2-5) соцветий, отделенных друг от друга годовыми скелетными приростами прошлых лет (курторезидными монокормусами) и частями побегов текущего года, расположенных вне соцветий. Медиматурная фаза (рис. 206, Е). ПС — с 20-35 до 40-60 лет. ОХ— неежегодно репродуктивный гемисимподиальный сложнокустистый ро- зеткообразующий каудексовый ризоэнатокормус с числом постиници- альных монокормусов и (или) дикормусом в сезон вегетации более чем 10. Энатокормус возникает из гептакормуса. У вздутоплодника сибир- ского энатокормус состоит из предветви бывшего гептакормуса и раз- вившихся из ее почек пазушных побегов-ветвей, не тождественных дру- гим типам побегов по сложности организации, т.е. октокормусов. Б— активную часть энатокормуса составляют полурозеточные унирепродук- тивные дикормусы и розеточные вегетативные монокормусы. Редко на удлиненной части дикормуса внерозеточные (срединные листья безро- зеточной части побега) рассеченные листья не развиваются, поэтому дикормусы бывают иногда и розеточными. Характерна факультативная корнеотпрысковость. Она проявляется после механических поврежде- ний растений, например, когда в результате заготовки подземных орга- нов удаляется каудекс и большая часть корневой системы (рис. 206-32). В почве нередко остаются живые боковые корни или их части (полная ризокормусовая, т.е. проходящая через ризокормус, механическая пар- тикуляция). В месте разрыва постепенно образуется каллус с адвентив- ными почками, развивающимися далее в постинициальные побеги и,
Глава 10. Использование теории организации биоморф семенных растений 545 наконец, в новокаудекс, т.е. базовый орган, подобный каудексу по структуре, но непосредственно через гипокотиль не связанный с глав- ным корнем и возникающий иногда после утраты каудекса. Если меха- нически поврежден каудекс, то он может сохранять жизнедеятельность; пробуждаются его спящие почки, а на месте повреждения иногда зак- ладываются адвентивные почки. Фенофазы: бутонизация — май-июнь, цветение (центростреми- тельное в структуре зонтиков и зонтичков) — июнь-июль, илодоноше- ние — август, начало сентября. Листорасположение очередное, розеточные дифиллы и трифиллы черешковые, голые; пластинки их в очертании продолговато-яйцевидные или яйцевидные. Каудекс увели- чивается в размерах за счет компактных резидных побегов (мономер- ных курторезидных и плейомерных миниморезидных) и постоянно разрастается в толщину, достигая иногда 6 см в диам. у основания, а в разветвленной верхней части 30, а чаще 8-15 см в диам. Каудекс безлистный подземный многоглавный коротковетвистый. Главный корень до 1 м дл. Отдельные каудексовые ветви, отмирая, чаще всего на часть своей длины, легко замещаются новыми ветвями в результате активизации жизнедеятельности отдельных спящих почек, запасы которых в струк- туре каудекса значительны. Часть почек сохраняет относительно глубо- кое состояние покоя, но другие ежегодно медленно нарастают, не раз- вертывая зеленых листьев в течение многих лет, образуя компактные (до 3 см дл.) ветви, иногда с новыми подобными пазушными почками. Эти компактные скрыто нарастающие побеговые ветви, или скрытые побеги (рис. 206-34), только при появлении соответствующих условий, например, при отмирании верхней части какой-либо каудексовой вет- ви, могут активизировать свой рост, который в сезон вегетации стано- вится видимым розеткообразующим. Финиматурная фаза (рис. 206, Ж). ПС — с 40-60 до 60-80 лет. ОХ— неежегодно репродуктивный гемисимподиальный полурозеточный кау- дексовый ризоэнатокормус со следами некроза, охватывающими 50% и более частей каудекса. А — общее ослабление жизнедеятельности, в частности репродуктивной активности: оно особенно хорошо заметно на структуре каудекса. Число отмерших ветвей каудекса постоянно уве- личивается, что позволяет провести весьма приближенную границу меж- ду началом финиматурной и концом медиматурной фазы. Некроз уси- ливает общую партикуляцию растения, но полной партикуляции обыч- но не происходит. Сенильный этап (рис. 206, И). ПС — продолжается 1-10 лет. ОХ— ежегодно вегетативный после полной утраты физиологической зрелости розеточный каудексовый ризоэнатокормус. Б — под пологом светлых березовых и лиственнично-сосновых лесов, куда нередко заходит со степных участков вздутоплодник сибирский, продолжительность инди- видуальной жизни обычно больше, а стадии онтогенеза более растянуты, чем у растений этого вида, произрастающих на открытых местах. Мак- симальная продолжительность эуонтогенезов (онтогенезов изоаклонис- тов) около 70-80 лет.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННБ1Х РАСТЕНИИ Резюмируем особенности биоморфогении вздутоплодника сибирского. PMojodicarpus sibiricus (Steph. ех Spreng.) К.-Pol. в Читинской обл. произрастает в основном на привершинных степных склонах сопок, заходя иногда под полог светлых березовых или лиственнично-сосновых лесов. Семена, пройдя зимой естественную стратификацию под снегом, прорастают весной или после дождей в течение лета. На виргинильном этапе жизни розеточная побеговая система нарастает моноподиально. Первое цветение происходит на 5-20-ом году жизни растения. Вздутоп- лодник сибирский цветет и плодоносит не каждый год. Аккумулятивное нарастание побеговой системы матурных растений гемисимподиальное, т.е. с неежегодным перевершиниванием скелетных побеговых осей. Главный корень сохраняется на протяжении всей жизни изоаклонистов (постоянных аклонистов), продолжающейся около 70-80 лет. По эколого-морфологическим тропобиозным (индикаторным для ти- пов биоморфогенезов) признакам вздутоплодник сибирский — это травя- нистый летнезеленый надземно-двусемядольный гемикриптофильный мультирепродукгивный полурозеточный дважды- и триждыперисторассе- ченный очереднолистный гемисимподиальный кустистый полузакрытый терминально-репродуктивный однодомный бело-сельдерейноцветковый каудексовый гипопланофильный (с очень слабым подземным разрастанием в горизонтальном направлении) изоакпональный (постоянно аклональ- ный), но иногда факультативно корнеотпрысковый и несовершенно экс- пансивно-кондивидуальный (после механической полной ризокормусовой партикуляции образующиеся кпонисты очень незначительно отдалены друг от друга) ксеромезофильный горно-степной вид. 10.2.9. Ча!ег!апа officinalis L. В медицинской практике валериану лекарственную используют для лечения многих заболеваний в основном как седативное средство при нервном возбуждении, неврозе сердечно-сосудистой системы, бессон- нице и др. (Енин, 1962; Соколов, Замотаев, 1985). Валериана лекарственная произрастает на огромных пространствах равнин и горных хребтов Евроазиатского материка от Атлантики до Тихого океана в лесной и лесостепной зонах, встречаясь во влажных лесных местах, по долинам рек в кустарниках и среди луговой расти- тельности, проявляя большой полиморфизм по форме и величине по- бегов, листьев, корней, столонов, по особенностям опущения плодов и т.п. (Ворошилов, 1959; Атлас ареалов..., 1976). Прорастание семян и весеннее отрастание побегов происходит при сравнительно низкой температуре. Высокая зимостойкость валерианы лекарственной наблюдается в районах ее выращивания под Москвой, Петербургом и Новосибирском (Ворошилов, 1959). Промышленная куль- тура валерианы осуществляется посредством разных способов создания плантаций: а) посевом семян непосредственно в поле; б) производством рассады в открытых грядках и ее высадкой на постоянное место в поле в различные сроки (осенью или ранней весной); в) вегетативным спосо-
Глава 10. Использование теории организации биоморф семенных растений бом — разделением родительских кустов и высадкой молодых розеточных растений (деток) вегетативного происхождения (Крейер, 1936; Енин и др., 1953; Илиева, 1971; Семенихин, Муш, 1974). Во многих странах За- падной Европы утвердилась однолетняя рассадная культура, в Восточной Европе — двулетняя посевная, основанная на посеве семян непосред- ственно в поле (Heeger, 1956; Кушке, Иващенко, 1967). При детальном морфологическом анализе растений валерианы на производственных плантациях можно обнаружить представителей не- скольких мелких видов (т.е. в узком смысле), которые, кстати, отлича- ются между собой и цитоэмбриологически (уровнями плоидности), а их предки имеют разные ареалы и несхожие ритмы развития (Шугае- ва, 1985). При исследовании динамической морфологии валерианы ле- карственной в культуре мы проводили наблюдения Valeriana officinalis L.s.l., поскольку сейчас Ч.officinalis L.s.str. в чистом виде нигде не воз- делывается (Нухимовский, Семенихин, Шугаева, 1989). Биоморфогенез валерианы лекарственной осуществляется по типу синонтогенезов, т.е. в системе кондивидуума, или синорганизма, или иначе в системе «мероаклонист-клон», где мероаклонист (временный аклонист) — особь семенного происхождения до преобразования ее в клон, а клон — вегетативное потомство мероаклониста. Эмбриональный период. Пренатальный этап. Uaer m обоеполые с ди- хогамным строением. Пыльники вскрываются в цветке, когда рыльце еще не готово к опылению (протерандрия). Опыление совершают насе- комые. Ло данным Е.В. Шугаевой (1985), зародышевый мешок моноспо- рический, семиклеточный, Polygonum-типа; двойное оплодотворение совершается по премитотическому типу. Зигота приступает к делению после двух-трех делений эндосперма через 8-9 часов после опыления. Развитие зародыша происходит по Asterad- и Piperad-типам. Заложение и дифференциация семядолей наблюдается на 8-12 день после оп- лодотворения. Полное развитие и дифференциация зародышей заканчи- вается за 21-25 дней. Зрелый зародыш прямой с двумя семядолями. Плод— сухая одногнездная семянка с хохолком. Семена с прямым зародышем, полностью заполняющим семена, без эндосперма (Николаева и др., 1985). Латентный этап. ПС — с июля-сентября до апреля следующего года или при хранении до двух лет; непосредственно после уборки семена показывают высокую всхожесть (до 99 %), т.е. латентный этап у валерианы лекарственной может быть очень коротким. При хранении семян в течение 5 месяцев всхожесть их 60%, в течение 10 месяцев — 50-60%, 15 меся- цев — 43 %, 20 месяцев — 30% и после 23-27 месяцев они полностью теряют всхожесть (Илиева, 1971). ОХ — зрелое семя (одногнездная семян- ка с хохолком, который в 1,5 раза длиннее семянки), обособленное от родительского растения. Семянки в очертании линейно или продолговато ланцетные, длиной 2-3 мм, бурые, плоские, с обеих или только с дорзаль- ной стороны голые (Ворошилов, 1959; Енин, 1962). Зрелые семянки легко отделяются от плодоножек, разлетаются с помощью хохолка-парашюта. Масса 1000 семянок 0,4-0,6 г (Илиева, 1971). Эпиэмбриональный период. Виргинильный этап. Преювенильная фаза. Проростковая гипофаза (рис. 207, А). ПС — семена при подзимнем по-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 207. Виргинильный этап в синонтогении Valeriana oAicinalis L. s.l. (Москва, ВИЛАР, интродукционный питомник и производственные плантации). А — проросток, Б — всходы В — ювенильные особи: В, - униювенильная, В,— биювенильная, Г, — униимматурная особь. 1 — семядоли, 2 — гипокотиль, 3— семядольный узел, 4 — корневая шейка, 5а-5е — корни: 5а — главная монориза (субмедиаль), 5б — главная дириза, 5в — неглавная монориза (сублатераль), 5г- тририза, 5д — главная тетрариза, 5е — синториза, 6 — отмерший главный корень, 7-8 — придаточные корни: 7 — гипокотилогенные, 8 — каулогенные; 9 — простые листья, 10 — полусложные листья, 11 — вегетативный розеточный медиальный монокормус, 12 — вегетативный розеточный базисиллептический монокормус, 13 — ризопротерокормус (семядольный почкокорень), 14 — семядольный ризомо- нокормус, 15 — бессемядольный вегетативный ризомонокормус, 16 — бессемя- дольный вегетативный ризодикормус. МК — упрощенные графические модели кондивидуумов. Одно деление масштаба — 1 см. Горизонтальная линия — поверх- ность почвы.
Глава 10. Использование теории организации биоморф семенных растений севе (октябрь-ноябрь) начинают дружно прорастать в апреле, начале мая; при весеннем (апрель) посеве в открытый грунт первые проростки появ- ляются на 5-7-й день после посева, а иногда и позже на 8-14-й день (Балабас и др., 1965). В лабораторных условиях они начинают прорастать на 2-4-й день, а прорастание заканчивается на 14-й день (Илиев",1971). Продолжительность состояния проростка в условиях полевого опыта 8-12 дней, а в теплице 4-7, она зависит от влажности и аэрации верхнего слоя (0-2 см) почвы, от погодных условий. Семядоли выносятся на поверхность почвы на 2-3-й день после начала прорастания. ОХ— прорастающее семя (семянка) или семядольный почкокорень (ризопро- терокормус). А — прорастание семян надземное. Семядоли у про- ростков две, продолговато-яйцевидные, на верхушке округлые, слегка усеченные с выемкой или без нее, при основании клиновидные, голые, 0,2-0,7 см дл., 0,5 см шир. Черешки семядолей 0,3-1,0 см дл. Гипокотиль голый 0,2-1,0 см дл. Главный корень — монориза, перед началом раз- вертывания семядолей его длина 2 см, а перед развертыванием зароды- шевой почки — до 4 см. С увеличением возраста проростка могут раз- виваться 1-3 боковых корешка, т.е. монориза преобразуется в диризу (главный корень второго уровня сложности организации). Всходовая гипофаза (рис. 207, Б). ПС — с мая до июня, продолжает- ся 25-40, реже более дней. ОХ — семядольный вегетативный розеточный ризомонокормус. А — самые первые настоящие листья очередные, ши- рокояйцевидные или почковидные, цельнокрайние или крупногород- чатые, на верхушке округлые или слегка заостренные, 0,7-2,0 см длины, 0,6-1,5 см ширины с черешком 0,5-2,0 см длины. Как отметил Н.И.Ио- себидзе (1981), поверхность листьев (с обеих сторон) голая, а по их краю встречаются очень короткие, направленные к верхушке простые ши- ловидные волоски, жилкование перистосетчатое. На 6-8-ой день после начала развертывания первого настоящего листа в основании его узла образуется небольшой бугорок, из которого развивается первый прида- точный корень. Формирование каждого последующего листа сопровож- дается развитием новых придаточных корней. До отмирания семядолей чаще всего образуются два, три настоящих листа и развивается корневая система — синториза, состоящя из главной диризы или реже тетраризы и 2-3 придаточных (каулогенных) корней. Кроме каулогенных корней, нередко еще образуются один или несколько придаточных корней от гипокотиля. Б — с образованием придаточных корней увеличивается жи- знестойкость растений к неблагоприятным воздействиям среды. Процент гибели всходов резко уменьшается после развития первого придаточного корня. В следующем возрастном состоянии сеянцы приобретают доста- точную жизнестойкость к неблагоприятным условиям существования. Ювенильная фаза (рис. 207, В). ПС — с июня до августа или в ус- ловиях угнетения (загущенные посевы) до весны следующего года. Про- должается 20-90 дней или в связи с перезимовкой до 300 дней. ОХ— бессемядольный вегетативный розеточный ризомонокормус. А — се- мядоли отмерли, кущения нет, продолжается моноподиальное нараста- ние медиального побега. Последовательно сменяются в развитии две гипофазы: униювенильная, характеризующаяся образованием простых цель-
550 основы БиоморФологии сЕмЕнщ1х рАстЕний ных широкояйцевидных розеточных листьев и биювенильная, характери- зующаяся образованием полусложных, как правило, тройчатых розеточ- ных листьев. Размеры листьев последовательно увеличиваются с увели- чением порядкового номера. У 3-го простого листа полностью развер- нутая пластинка 3,5-5 см дл., 4-6 см шир., черешки 6-8 см дл., у 6-го тройчатого полусложного листа пластинки 10-15 см дл., 10-14 см шир., черешки 14-18см. Главный корень не получает заметного развития и нередко уже на ювенильной стадии отмирает. Функционирует и разви- вается синториза, представленная системой придаточных корней. Глу- бина проникновения в почву корней 5-10см. Имматурная фаза (рис. 207, Г,; 208, Г„Г,). ПС — с июля до весны следующего года или при неблагоприятных условиях (загущенное про- израстание, засуха) иногда и до весны третьего года вегетации. ОХ— бессемядольный вегетативный розеточный кустистый ризодикормус или (при угнетении) ризогексакормус. А — с момента первого ветвления (кущения) медиального розеточного побега, сопровождающегося раз- вертыванием на первом году вегетации растений пазушных почек в боковые розеточные низовые силлептические (базисиллептические) по- беги, начинается имматурное состояние; в случае задержки роста и развития кущение происходит в следующем вегетационном сезоне из перезимовавших почек (постлатентное кущение) и (или) незимовавших новых почек (базисиллептическое кущение). Вступление первых особей в имматурное состояние происходит, как минимум, после образования 6-7 настоящих непарноперистых полусложных листьев и 10-12 прида- точных корней. Растения, которые не достигли в первом вегетационном сезоне имматурной стадии, как правило, не переходят в матурную ста- дию, пока не пройдут на следующий год необходимую им вегетативную стадию кущения (имматурную). Всходы и ювенильные растения, неза- висимо от срока помещения их в теплицу, оставались в розетке, далее развивались в имматурные растения, которые нормальной физиологи- ческой зрелости достигали только после обязательного воздействия на них пониженными температурами. Задержка растений в имматурном состоянии и постепенное отмирание вертикального компактного кор- невища (снизу) может приводить их к полной некротической партику- ляции и вегетативному размножению с образованием сильно сближен- ных розеточных особей вегетативного происхождения, которые в свою очередь, проходя свой цикл индивидуального развития, находятся в новоювенильном или новоимматурном состояниях. Возрастные состоя- ния, которые достигают особи вегетативного происхождения (клонис- ты) напоминают по многим признакам соответствующие возрастные состояния особей семенного происхождения (мероаклонистов), пред- шествующие им, поэтому обозначим их сходно, добавив латинскую при- ставку «novus» (новый). Отмеченная выше особенность виргинильного развития валерианы лекарственной позволила нам выделить три гипофазы синонтогенеза: униимматурную (иервую имматурную), сопровождающуюся розеточным ростом и развитием кустистого мероаклониста, который еще не под- вергся воздействию пониженными осенними температурами, т.е. до яро-
Глава 10. Использование теории организации биоморф семенных растений 551 Рис. 208. Переход к физиологической зрелости и клонированию кондиви- дуумов Valeriana oAicinalis (Москва, ВИЛАР, интродукционный питомник и производственные плантации). Г, — биимматурный кондивидуум, Г, — триимматурный кондивидуум, Д — первый ини- циматурный кондивидуум; 17 — биимматурный кондивидуум в разрезе, 18 — триимма- турный кондивидуум зимой (корни удалены) — начало клонирования, 19 — он же летом, в разрезе, 20 — униимматурный мероаклонист, развивающийся зимой без яровизации (занесен в начале осени в теплицу), 21 — вегетативно-репродуктивный верхушечный ди- кормус, 22 — вегетативно-репродуктивный полурозеточный гексакормус, 23 — полу- сложные непарноперистые листья, 24 — флоральная зона, или зона соцветия (плейотир- са), 25 — зона торможения, 26 — зона индукции, 27 — зона инновации, 28 — эусиллепти- ческие побеги, 29 — цветок, 30 — плод, 31 — вынужденное израстание соцветий и про- буждение почек из бывшей зоны торможения у растений с неполной яровизацией (в теплице), 32 — вегетативный ризогексакормус, 33 — клональная система вегетативных ризокормусов, 34 — репродуктивный автономно-ризомный ризогексакормус, 35 — жи- вые основания побегов (верхние части удалены), 2л, 3л ... особи 2, 3 и т.д. лет жизни, Остальные обозначения те же, что и на рис. 207 °
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ визации (рис. 207, Г,); биимматурную (вторую имматурную), сопровож- дающуюся замедленным розеточным позднеосенне-зимним ростом и развитием мероаклониста после воздействия на него пониженных по- зднеосенне-зимних температур, т.е. после яровизации (рис. 208,Г,); три- имматурную (третью имматурную), когда кустистое родительское им- матурное растение преобразовалось в клон и все имматурное состояние присуще кондивидууму, хотя составляющие его особи могут быть поми- мо имматурного родительского мероаклониста еще новоювенильными или (и) новоимматурными клонистами (рис. 208, Г,). Первые две имма- турные гипофазы более всего представлены в производственных посе- вах, а третья гипофаза проявляет себя в неблагоприятных условиях (при задержке роста и развития) как в производственных условиях, так и в эксперименте. В производственном отношении имматурные растения представляют наибольший интерес, поскольку дают основной урожай сырья корневищ с корнями валерианы лекарственной. При зимовке растений в ювенильной фазе они переходят в имматурную фазу в начале лета следующего года, а вегетация в имматурной фазе продолжается до наступления условий яровизации, что значительно увеличивает массу и разветвленность многоглавого корневища с многочисленными корнями. Необходимым условием для перехода любого розеточного побега к ре- продуктивному развитию является не только яровизация, но и накоп- ление определенной массы пластических веществ в запасающем орга- не — компактном стебле (более 2,5 г) после завершения вегетации (Се- менихин, 1975, 1976). Вегетативное размножение у имматурных растений может начи- наться с конца вегетационного сезона, усиливаться в весенне-летний период следующего года. Отмирание снизу вертикального корневища может достигать зоны отрастания пазушных розеточных побегов, кото- рые после утраты морфофизиологической связи с родительским побе- гом переходят на полное самообслуживание с помощью придаточных корней и развитой листовой поверхности. Листья розеточных побегов сильно сближены, расположены очередно, что хорошо заметно в осно- вании, где вышеследующие листья прикрывают влагалища черешков нижеследующих листьев. В дальнейшем развитии побега в репродукгив- ной стадии порядок листорасположения изменится. Придаточные корни многочисленны и живут не более одного года. Главный корень уже отмер. Биимматурная гипофаза проявляется с весны или лета второго года вегетации до весны третьего года вегетации и характеризуется отрастанием пазушных побегов не на первом году вегетации сеянца, а лишь на втором. Матурный этап. Инициматурная фаза. Первая гипофаза (рис. 208, Д). ПС — с начала весеннего отрастания удлиненной части первого вегета- тивно-репродуктивного двухгодичного медиального побега до образова- ния клона еще до отмирания медиального вегетативно-репродуктивного побега; продолжается несколько дней или до конца второго вегетаци- онного сезона, 3-4 месяца. ОХ — репродуктивный полурозеточный кус- тистый ризогексакормус. А — у имматурных растений обычно форми- руется простокустистая структура; после накопления необходимых про- дуктов ассимиляции и прохождения осенне-зимней яровизации на сле-
Глава 10. Использование теории организации биоморф семенных растений 553 дующий год формируется гексакормусовая структура, лидерная роль в которой принадлежит медиальному побегу. Помимо медиального побе- га в репродуктивное состояние нередко переходят и боковые (пазуш- ные) побеги (один или несколько), часть розеточных вегетативных по- бегов остается в прежнем состоянии, продолжая розеточный вегетатив- ный рост и развитие, образуя еще один или несколько новых пазушных побегов (или силлептических, или весенних). В основании инициматур- ного куста и на втором году вегетации продолжается некроз, который охватывает нижнюю часть корневища и связанные с ним старые при- даточные корни (они живут, как правило, не более 1 года). Когда некроз гексакормуса снизу достигает первых пазушных побегов, то создаются все условия для образования клона. Вторая инициматурная гииофаза. ПС — с весны второго года до весны третьего, очень редко, при сильном угнетении, четвертого года вегетации. ОХ — клональная система вегетативно-репродуктивных по- лурозеточных ризогексакормусов или ризопентакормусов и вегетатив- ных розеточных ризомонокормусов, ризодикормусов или реже ризогек- сакормусов. А — это состояние возникает двояко: или из имматурного клонального кондивидуума, или из первого инициматурного мерокло- ниста (это особь семенного происхождения еще не преобразовалась в клон, но отражает однозначно возрастное состояние жизни всего кон- дивидуума на соответствующем отрезке его синонтогенеза). Клон в имматурном состоянии кондивидуума возникает довольно редко и свя- зан с задержкой развития мероаклониста в этом состоянии до второго или третьего года, когда некроз снизу (базинекроз) приводит к обособ- лению пазушных побегов с корнями, т.е. новых особей-клонистов. Им- матурный клональный кондивидуум после начала отрастания удлинен- ной части цветоносного побега преобразуется во второе инициматурное (биинициматурное) состояние кондивидуума, минуя первое иницима- турное состояние. Инициматурные мероаклонисты часто преобразуются в клон (достигают биинициматурного состояния кондивидуума) на вто- ром году жизни, когда медиальный вегетативно-репродуктивный побег еще жив. В зависимости от условий произрастания на плантации вале- риана впервые цветет на 2-м году вегетации (от 5-10% в условиях угнетения и до 90-95% растений в благоприятных условиях). Бутони- зация проявляется в июне, цветение с конца июня до августа и продолжается 45-50 дней, плодоношение наступет через 30-40 дней после начала цветения. Массовое созревание плодов — в конце июля- августе. Вегетационный период продолжается 180-190 дней. Вегетатив- но-репродуктивный побег развивается чаще всего по двухгодичному, редко трехгодичному циклу. Образуется несколько типов таких побегов, различающихся по сложности организации. Основной структурной еди- ницей матурного кондивидуума является гексакормус — комплексная побеговая система (побег шестого уровня сложности организации), включающая в себя зону инновации, т.е. персистентную зимовавшую побеговую часть, выжившую к весне после возрастных превращений структуры медиального монокормуса или дикормуса, и все новые побе- ги текущего сезона вегетации, развитые из зоны инновации. Охаракте-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ ризуем разнообразие годичных побегов, образующихся в структуре гек- сакормуса (рис. 208-34): вегетативно-репродуктивный годичный безро- зеточный, реже полурозеточный, верхушечный побег (дикормус), завер- шающий восходящее развитие прошлогоднего розеточного вегетативно- го медиального побега, а нередко еще одного-трех латеральных побегов (таким образом завершается и их развитие). Вместе вегетативный розе- точный прошлогодний побег с развитым из него верхушечным вегета- тивно-репродуктивным годичным побегом составляют побег пятого уровня сложности организации — полурозеточный пентакормус. Сис- тема медиального пентакормуса и развитых от него латеральных побе- гов (от монокормусов до пентакормусов) является в целом гексакорму- сом. Надземные побеги достигают у матурных побегов растений на втором году вегетации от 70 до 150 см высоты. В структуре вегетативно-репродуктивного дикормуса различаем следующие зоны (Troll, 1964, с нашими дополнениями): флоральную, или зону сложного сборного соцветия, субфлоральную, или зону тор- можения (релаксации), включающей подзону неполного торможения, если образуются сильно ослабленные силлептические вегетативные (ин- фантильные) побеги, и подзону полного торможения, когда нет инфан- тильных побегов (нередко участок побега с полным торможением и является всей зоной торможения), зону индукции (ее на 2-м году жизни побега часто уже нет), в которой узлы сближены, а из недоформирован- ных почек отрастают низовые силлептические, или базисиллептические побеги, — разновидность индуктивных побегов, и, наконец, зону ин- новации (возобновления). В этой зоне все узлы сближены (на втором году жизни побега эта зона полностью поглощает в себя зону индукции прошлогоднего вегетативного розеточного побега), а из перезимовавших почек (верхушечных и пазушных) происходит постлатентное (весеннее) побегоотрастание. Соцветия валерианы лекарственной подробно изуче- ны Ф.Веберлингом (Weberling, 1961). Он обратил внимание на образование в сборном соцветии переходных структур между цимозны- ми и тирсоидными парциальными соцветиями, охарактеризов сборные соцветия как плейотирс. Цветки обоеполые, мелкие, душистые; в развернутом виде их лепестки функционируют 4-6 дней (Балабас и др., 1965). На удлиненной цветоносной части развиваются супротивные непарноперистые полусложные (без разделительных сочленений) лис- тья. Иногда наблюдаются отклонения от супротивного листорасположе- ния и формируется очередное листорасположение, чаще всего ближе к основанию удлиненной цветоносной части. Нижние листья черешко- вые, а в зоне верхушечного соцветия и ближе к нему сидячие. Пластин- ка развернутых нижних листьев 8-30 см дл., 4-15 см шир., с 4-8 парами листочков, черешок 3-10 см. дл. Важное значение в существовании биоморф валерианы лекарствен- ной имеют укореняющиеся базальные участки розеточных побегов. Они более всего предрасположены к перезимовке, а у двухгодичных побегов перезимовавшая часть несет основной запас питательных веществ для роста и развития цветоносной части побега. Зимующая и перезимовав- шая части побега — вертикальное компактное корневище эпигеогенно-
Глава 10. Использование теории организации биоморф семенных растений 555 го и эпигипогеогенного (или миксогенного, т.е. смешанного происхож- дения, когда почки пробуждаются как приземно, так и подземно) типа. Корневище постоянно подвергается базинекрозу (отмиранию снизу), а после завершения плодоношения удлиненная цветоносная часть побега отмирает полностью с компактным вертикальным корневищем. Все па- зушные розеточные и полурозеточные побеги развивают придаточные корни, составляя вместе с ними корнепобеги-органы (онтоиды), и пос- ле гибели родительского цветоносного побега с вертикальным корневи- щем они полностью обособляются, переходя на автономное существо- вание. Вегетативное размножение сопровождается лишь незначитель- ным завоеванием окружающего жизненного пространства. Придаточные корни многочисленные; они живут от 3 до 10 месяцев и сменяются молодыми корнями, которые, с одной стороны, дополняют приток пи- тательных веществ к побегу, а с другой — депрессируют функциониро- вание нижних придаточных корней, способствуя их отмиранию. Матурный этап в жизни некоторых аклонистов может заканчивать- ся и после первого и единственного плодоношения, когда наступает отмирание всего растения, т.е. жизненный цикл у валерианы может быть в определенных случаях унирепродуктивным. Медиматурная фаза (рис. 209, Е). ПС — с весны 3-го года вегета- ции до 6-8 лет. ОХ — кондивидуальная система, состоящая из клонистов разной степени развития, которая цветет (цвела) от двух до четырех раз. А — каждая новая генерация вегетативно-репродуктивных побегов при- водит к увеличению числа новых розеточных побегов и к обособлению их вместе с корнями, но только после отмирания отплодоносившего побега. Высота цветоносных побегов с увеличением возраста кондивидуумов несколько падает от 80-160 см на 3-м году, до 50-120 см на 5-м году. Б — на промышленных плантациях растения валерианы выращи- вают не более 2-х лет, после чего их выкапывают на сырье, а, кроме того, часть розеточных клонистов можно использовать для создания плантаций вегетативного происхождения. В жаркие годы кондивидуумы валерианы слабее, чем во влажные годы: высота побегов и число про- буждающихся почек меньше, больше гибнет растений после плодоно- шения. В этой связи хорошие результаты дает дополнительный полив плантаций. Новые вегетативные розеточные побеги, как и в прежние годы, возникают из почек на корневище и из пазух листьев текущего года, т.е. новые побеги весенние и базисиллептические. Финиматурная фаза (рис. 209, Ж). ПС — с весны шестого (реже позже) до 8 и более лет. ОХ — клональная система кондивидуума, со- стоящая из розеточных и полурозеточных самостоятельных автономно- ризомных ризокормусов (от ризомонокормусов до ризогексакормусов, редко ризогептакормусов), которая цветет (цвела) более четырех раз. А — продолжается базинекроз (отмирание снизу) корневищ вегетатив- ных клонистов и полная общая некротическая партикуляция репродук- тивных клонистов через компактные вертикальные корневища. Парти- куляция приводит к вегетативному размножению — увеличению числа клонистов, многие из которых ослаблены из-за сближенного располо- жения друг к другу; небольшая часть клонистов достигает репродуктив-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ h4 й Ж й О В >3~ Ж Ж О й Ф h4 О В~ л О Й О ~„' ~ й 53 у) ',3,' сб (6 Ц Ll О Х g5 Х a ~ а g O д~ О ) Ф И~а ~ О 3 О Й~ р' h4 О Ch CO <~4 О й 0 Зг~ g лож а~о о ~ о;„ Д 3 Ьgй ~) д ф~~ о cO ~ О 3 И 3 P. ~ ю (~ И Ж о ~а о <о~ ~р юо о ~ ~ ~( о гч о a P ++~ Я ж о.Д g 3 Й ~~(~ gЕ~~о йо v е~ @ P. о а~ . 1й„." Ю i Kg -~о о ~ ~8 3' 3 й о о. и П'8
Глава 10. Использование теории организации биоморф семенных растений ного состояния (новоматурное состояние), но многие погибают в вегета- тивном розеточном состоянии — новоювенильном или новоимматур- ном. Развитие побегов продолжается по прежним схемам, но процент цветущих побегов невелик. В отдельные годы финиматурные кондиви- дуумы не цветут или у них зацветает лишь по одному побегу. Заметны явные признаки старения: изменяется габитус (уменьшается высота цве- тоносных побегов до 40-100 см, мельчают листья и слабеют соцветия, корневища и корни). Финиматурные 8-10-летние кондивидуумы вале- рианы на опытном участке страдали как от внутренней конкуренции своих клнистов, так и от проникающих в их среду злостных многолет- них сорняков (бодяк, пырей, одуванчик и др.). Б — ежегодно на опытных участках наблюдалось выпадение кон- дивидуумов разного возраста, преимущественно старшего. Мы не на- блюдали в этих условиях кондивидуумы старше 10 лет. В этом возрасте трудно фиксировать факт полной утраты физиологической зрелости. Мы убеждены, что небольшая часть кондивидуумов проходит и сениль- ный этап в своем синонтогенезе. К 10 годам выращивания валериана лекарственная сильно вырождается, но если взять ослабленный розе- точный клонист из старого кондивидуума и пересадить его в благоприят- ные условия, то нередко удается возвратить кондивидууму молодость: он вновь набирает мощность в вегетативном развитии (нововиргинильное состояние) и заметно возвращает репродуктивную активность (новома- турное состояние). Наблюдение на опытном участке позволяет утверж- дать, что десятилетние кондивидуумы уже подошли к матурной старости и, возможно, некоторые из них в ближайшие годы, если не отомрут, то больше не зацветут (т.е. перейдут в сенильный этап своего синонтоге- неза, в котором и достигнут естественной гибели). Валериана лекарст- венная при соответствующем уходе растет в культуре и более 10 лет, например, в экспозиции лекарственных растений Ботанического инсти- тута (г. Петербург) имелись экземпляры 13-летнего возраста (Балабас и др., 1965). Резюмируем особенности биоморфогении валерианы лекарственной. Валериана лекарственная в условиях выращивания (Московская обл.) хорошо размножается свежесобранными семенами (посевом под зиму или весной), кроме того, через рассаду или вегетативно (пересадкой вегетатив- ных розеточных клонистов). На первом году эпиэмбрионалной жизни развиваются розеточные вегетативные растения некустистые (ювенильные) и кустистые (имма- турные). Кущение осуществляется за счет низовых силлептических (ба- зисиллептических) побегов. Первое цветение наблюдается чаще всего на втором году, реже позже. После плодоношения цветоносный побег вместе с вертикальным корневищем полностью отмирает и происходит вегетативное размноже- ние с незначительным отдалением клонистов друг от друга. Основной структурной единицей матурного кондивидуума вале- рианы лекарственной является вегетативно-репродуктивный полурозе- точный гексакормус (побег 6-го уровня сложности организации), состо- ящий из центрального полурозеточного вегетативно-репродуктивного
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИИ пентакормуса и его производных постлатентных (весенних) розеточных вегетативных монокормусов и реже вегетативно-репродуктивных безро- зеточных или полурозеточных дикормусов. После 6-8 плодоношений (к 10 годам жизни), а нередко и раньше, валериана на опытном поле заметно стареет и сильно изреживается (вырождается). Валериана лекарственная при выращивании под Москвой развива- ется по типу полурозеточных компактно-вертикзльнокорневищных тра- вянистых кондивидуальных многолетников. 10.3. Рисуночный способ передачи информации о биоморфогении семенных растений Описание биоморфогенезов растений можно проводить по-разно- му. Выше были испытаны два способа: таблично-рисуночный и повест- вовательно-рисуночный. Мы стремимся достичь лаконичности в описа- нии биоморфогении, не утратив ясности при передаче информации об особенностях организации биоморф на различных отрезках их жизни. Остановимся еще на одном способе описания биоморфогении видов растений, который назовем для удобства рисуночным. В нем главная информация заложена в рисунок, а считывать ее можно без дополни- тельных описаний, используя только подписи к рисункам, включающие в первую очередь условные обозначения. На основе прочтения инфор- мации, заложенной в рисунки, составляется резюме, из которого сле- дует, что главное характерно для биоморфогении рассматриваемого вида растений. Кроме того, работу по считыванию нужной информации с рисунков каждый читатель может провести и самостоятельно, выбрав то, что его более всего интересует в данный момент. Вероятно, такой путь описания биоморфогении окажется решаю- щим при создании соответствующих справочников. Однако, есть два важных условия: рисунки и типографская работа должны быть высоко- качественными. Приведем для примера рисуночное описание биоморфогении не- скольких видов лекарственных растений. 10.3.1. Phlojodicarpus sibiricus (Steph. ex Spreng.) К.-Pol. Биоморфогения вздутоплодника сибирского в естественных усло- виях произрастания была рассмотрена выше (гл. 10.2.8). Через подписи к рисункам и рисунки (см. рис. 210-212) охарактеризуеи главное в био- морфогении этого вида, которая наблюдалась нами в Московской обла- сти при выращивании в интродукционном питомнике института лекар- ственных и ароматических растений (ВИЛАР).
Глава 10. Использование теории организации биоморф семенных растений 559 Рис. 210. Виргинильный этап в онтогении Phlojodicarpus sibiricus (Steph. ех Spreng.) К.-Pol. А — проросток, Б — всходы, В — ювенильная особь на первом году жизни, Г— имматурная особь весной второго года жизни; 1 — семялоли, 2 — гипокотиль, 3— семядольный узел, 4 — корневая шейка (примерное положение), 5 — главный корень, 6 — розеточные листья, 7 — вегетативный медиальный розеточный моно- кормус, 8 — весенний вегетативный аксиллярный розеточный монокормус, 9— преконодий, 10 — основания розеточных листьев (верхние части удалены), 11— семядольный почкокорень (ризопротерокормус), 12 — семядольный ризомоно- кормус, 13 — бессемядольный ризомонокормус, 14 — вегетативный ризогексакор- мус, 15 — конодий (прекаудекс). Одно деление масштаба — 1 см; горизонтальная линия — поверхность почвы.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 211. Второй год вегетации биоморф Phlojodicarpus sibiricus в культуре. 16 — вегетативно-репродуктивный ризогексакормус, 17 — унирепродуктивная особь, погибшая зимой после первого плодоношения, 18 — репродуктивный годич- ный побег (дикормус) в структуре гексакормуса, 19 — инициматурная особь, со- хранившаяся зимой после первого плодоношения, 20 — особь - ризопентакормус, развивающаяся по унирепродуктивному циклу (она не образует весенних пазушных побегов), 21 — сборное соцветие. Остальные обозначения те же, что и на рис. 210.
Глава 10. Использование теории организации биоморф семенных растений 5б1 Рис. 212. Медиматурная (1) и финиматурная (11) фазы в онтогении Phlojodicarpus sibiricus в культуре. 22 — детерминантный (закрытый) унирепродуктивный пентакормус, 23 — курто- резидная прошлогодняя часть побега, 24 — активная часть побега текущего года, 25 — миниморезиды, 26 — ризогептакормус, 27 — ризоэнатокормус, 28 — каудекс, 29 — парциальный каудекс, 30 — отмершая к весне многолетняя особь. Пункти- ром показано примерное положение семядольного узла. Остальные обозначения те же, что и на рис. 210.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИИ Итак, обобщим материалы о биоморфогении вздутоплодника си- бирского, который выращивался в Московской области. Свежие семена хорошо прорастают весной при посеве их в апреле или под зиму. Виргинильный этап многие особи проходят в условиях выращивания за один год, развивая розеточно-побеговую и короткосте- ржнекорневую структуру. Часть особей в первом вегетационном сезоне достигает имматурного возрастного состояния, образуя пазушные веге- тативные розеточные побеги кущения. При загущении посевов вирги- нильное развитие может растягиваться еще на 1-2 года. На 2-ом году вегетации многие особи зацветают, а после плодоношения некоторые отмирают (унирепродуктивный тропобиоз), а другие, начав в том же году или еще раньше куститься, продолжают вегетацию дальше, прояв- ляя или бирепродуктивный, или чаще мультирепродуктивный типы биоморфогенезов. Унирепродуктивные (монокарпические обоеполые) ди-, реже плей- офазные побеги чаще всего развиваются как полурозеточные, но иног- да — и как розеточные; в последнем случае они не образуют срединных листьев на удлиненной надземной части побегов между розеткой лис- тьев и верхушечным соцветием. Если после плодоношения и отмирания сверху (акронекроза) надземной цветоносной части медиального побега все растение не погибает, то образование персистентного побегового остатка свидетельствует о возникновении каудекса, который в последу- ющие годы разрастается за счет увеличения в его структуре курторезид- ных и миниморезидных побегов. К шести, а тем более к восьми годам эпиэмбриональной жизни боль- шая часть растений на интродукционном участке выпала. Дело в том, что вздутоплодник сибирский, будучи обитателем щебнистых почв в горных степях, плохо растет и развивается на глинистых и переувлажненных поч- вах Подмосковья, где проводились наблюдения. Зимой стержнекорневые особи вздутоплодника выпираются из почвы, а к весне их побеговые вер- хушки подмерзают. Кроме того, подземные части растений легко загнива- ют в сырой, особенно весной и осенью, глинистой почве. Приведенный выше материал о биоморфогении вздутоплодника си- бирского в Подмосковье свидетельствует о неперспективности выращива- ния его на глинистых почвах в Нечерноземье Европейской части России. 10.3.2. Hippophas rhamnoides L. Многое из жизни облепихи в культуре уже известно (Гатин, 1963; Мочалов, 1973; Трофимов, 1976; Букштынов и др., 1985; Ермаков, 1987 и др). Приводимые нами рисунки (213-216 и подписи к ним) дополняют информацию о биоморфогении этого вида — представителя сем. Е1ае- agnaceae. Прорастание семян надземное. Семядоли (две) эллиптические. На первом году эпиэмбриональной жизни образуется вегетативный безро- зеточный умеренно сближенноузловой короткий или длинный медиаль-
Глава 10. Использование теории организации биоморф семенных растений 5бЗ Рис. 2)3. Виргинильный этап в развитии биоморф Hippophae rhamnoides L. (Москва, ВИЛАР, интродукционный питомник). А — проростки, Б — всходы, В„В, — ювенильные особи: В, — униювенильная (пер- вая ювенильная) особь, В, — биювенильная (вторая ювенильная) особь на первом году жизни; Г — имматурные особи на втором году жизни; 1 — семядоли, 2 — гипо- котиль, 3 — семядольный узел, 4 — вегетативный безрозеточный медиальный побег на первом году, 5 — корни, 6 — вегетативный медиальный побег, продолжающий моноподиальное нарастание на втором году жизни, 7 — силлептические (индуктив- ные) вегетативные побеги, 8 — весенний пазушный вегетативный монокормус, 9— вегетативный дикормус, 10 — вегетативный гексакормус, 11 — семядольный почко- корень, или ризопротерокормус, 12 — семядольный вегетативный ризомонокормус, 13 — бессемядольный вегетативный ризомонокормус, 14 — вегетативный ризодикор- мус, 15 — вегетативный ризогексакормус; у — увеличено по сравнению с масштабом. Пунктирная линия — граница апонекрозного годичного побега прошлого года. Горизонтальная линия — поверхность почвы. Одно деление масштаба — 1 см.
ОСНОВЫ БИОМОРФОПОГИИ СЕМЕННЫХ РА СТЕНИЙ Рис. 214. Матурный этап в развитии биоморф Hippophae rhamnoides (Московская область). 16 — корневой отпрыск (турион), 17 — вегетативно-репродуктивный дикормус, 18 — вегетативно-репродуктивный гексакормус, 19 — побеги с тычиночными цвет- ками, 20 — побеги с пестичными цветками, 21 — межсоцветие, 22-24 — пути развития матурных растений: 22 — унирепродуктивный вариант развития, 23— мультирепродуктивные варианты развития (низкорослые формы), 24 — мультире- продуктивные варианты развития (сильнорослые формы), 25 — сеянец, 26 — по- беговый черенок, 27 — аэроподий. Остальные обозначения те же, что и на рис. 213.
С~4 ~Ч~ ~4 о й~ О CO Ж й~ A ~ч О 65 1,' Ю О М О Ж О М О О C л К g й Ж О ~Г~ СЧ О й С4 О Х Р Глава 10. Использование теории организации биоморф семенных растений :Q 4: О. О О О X )Я Ю О О О Х й ~3' О О й О й~ Ю й~ й~ 21 Ж Ое О О О Ю О О Ж С4 Д Ь О б~ о й~ й; 3 Ж Ж 1 о 1 CO о Я о
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Рис. 216. Фасциация побеговой ветви взрослой особи Hippophae rhamnoides (Московская область). ный неразветвленный или силлептически разветвленный побег. Стер- жневой главный корень нарастает моноподиально, развивая боковые корни, на которых формируются корневые клубеньки — азотфиксато- ры. Листорасположение очередное. Почки верхушечные открытые, а пазушные защищенные почечными чешуями (закрытые). Верхушечные почки зимой обычно обмерзают, но у невысоких ювенильных растений иногда сохраняются, и медиальный побег продолжает моноподиальное нарастание на следующий год (рис. 213,Г-6). На втором и обычно треть- ем году жизни продолжается образование вегетативных побегов постла- тентных (весенних) и силлептических (индуктивных); последние побеги отрастают только в структуре сильных ростовых весенних побегов. Рост главного корня замедляется и усиливается рост боковых корней, на которых могут закладываться адвентивные почки. При механическом повреждении корней стимулируется развитие корневых отпрысков. Вир- гинильный этап заканчивается на 3-6 году.
Глава 10. Использование теории организации биоморф семенных растений 567 Матурные (физиологически зрелые) особи образуют однополые ты- чиночные или пестичные цветки, которые опыляются с помощью ветра. Облепиха цветет в первой половине мая; в это время начинают развер- тываться срединные продолговато-ланцетные листья. Цветочные поч- ки смешанные (вегетативно-репродуктивные), редко строго репродук- тивные. Пазушные почки закладываются в структуре весенних побегов; они зимуют и развертываются только на следующий год, образуя вегета- тивно-репродуктивные побеги разной силы развития (укороченные, ко- роткие, средние и длинные). Самые специализированные цветочные почки (они располагаются в основании гексакормусов) развертываются в строго-репродуктивные (тотрепродуктивные) побеги с несколькими (3-6) силлептическими пазушными цветками. У облепихи в структуре вегетативно-репродуктивных побегов формируются типичные межсоц- ветия. После плодоношения и пыльцеобразования растения с пестич- ными репродуктивными элементами превращаются в огромные плодо- носящие сверхсоцветия с множеством мелких ягодовидных сочных се- мянок. Укороченные и короткие побеги после плодоношения засыхают, превращаясь нередко в колючки. Самые сильные — это верхние росто- вые и репродуктивна-статурные побеги, развитые в структуре гексакор- мусов; они обеспечивают главный персистентный (многолетний) при- рост надземной части растений. Аэроподии живут до 15 лет, очень редко больше, но чаще меньше, в среднем 8-12 лет. Иногда наблюдается фасциация стеблевой части растущего побега (рис. 216). Жизненный цикл может идти по нескольким направлениям (рис. 214, 215). Возможны случаи, когда после очень сильного первого пло- доношения весь куст погибает, причем вместе с корнями (унирепродук- тивный тропобиоз). В подавляющем большинстве развитие особей идет по мультирепродукгивному типу, при этом одни растения проявляют себя как однополые поликарпики, а другие — полиантерики, т.е. развиваются по однополому полиантерическому типу. Одни особи обильно кустятся за счет образования побегов кущения из базальной части куста и корнеот- прысковых турионов. Другие растения облепихи способны долго сохранять одностволовую древовидную конституцию, иногда всю жизнь. После ги- бели родительского ризокормуса обособляются корневые отпрыски с фраг- ментами родительской корневой системы. Жизненный цикл клонистов во многом повторяет жизнь родительских особей. Из-за болезней и убыстрения развития отдельные синорганизмы облепихи в культуре отмирают во много раз быстрее, чем в естествен- ных условиях произрастания, где они нередко занимают большие пло- щади и живут очень долго. Hippophae rhamnoides L. — это летнезеленый надземно-двусемя- дольный безрозеточный открытый ранневесеннецветущий двудомный анемофильный мультирепродуктивный (редко с иным типом репродук- тивности) монофиллический очереднолистный умеренно сближенноуз- ловой прямостояче-аэроподиальный эусимподиальный корнеотпрыско- вый экстенсивно-кондивидуальный мезопланофильный или тахиплано- фильный кустарниковый гигромезофильный пойменно-луговой и бере- говой полиморфный вид растений.
568 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ 10.3.3. Scutellaria baicalensis Georgi Шлемник байкальский — Scutellaria baicalensis Georgi (сем. Lamia- ceae) распространен в Восточной Сибири, на Дальнем Востоке, в Мон- голии, Китае и Японии, особенно в горно-степных ассоциациях юго- восточного Забайкалья, где обилен в составе востряцово-ковыльных, реже пижмовых, степей (Горшкова, 1966). Особенности биоморфогении шлемника байкальского мы изучали в-Нерчинском районе (Читинская область) (рис. 217, 218). Семена шлемника байкальского прорастают в конце лета (Горшко- ва, 1966) или весной. Прорастание семян надземное. Семядоли округлые (0,35-0,5 см дл. и шир.), голые, на верхушке усеченные, нередко с вы- емкой, при основании также усеченные. На первом году эпиэмбрио- нальной жизни отрастает короткий безрозеточный вегетативный годич- ный медиальный побег, он акронекрозный и после осенне-зимнего от- мирания всей его надземной удлиненной части сохраняет жизнеспособ- ность миниморезид, состоящий, как правило, из одного семядольного узла, из которого развиваются первые две пазушные персистентные (зимующие) почки. Образование первого миниморезидного побега сви- детельствует о возникновении базового органа — каудекса, разрастаю- щегося в последующие годы аккумулятивно за Счет ежегодного вклю- чения в его структуру новых миниморезидов и радиально. Несколько лет ювенильные особи нарастают эусимподиально не разветвляясь, а перевершиниваясь только одним весенним побегом замещения. В над- земной части годичный побег может не ветвиться, а иногда силлепти- чески ветвится сразу же после засыхания замирающей в росте вегета- тивной верхушки (рис. 217-,). Любопытно, что неожиданное замирание роста некоторых надземных побегов с последующим отмиранием их верхушек и отрастанием супротивных силлептических побегов замеще- ния (эфемерная симподиальность) наблюдается летнесезонно на протя- жении всей жизни растений шлемника (рис. 217, 218). Возможно, это реакция растительных организмов на изменение погодных условий, на- пример, на непродолжительную жару. Начало кущения — образование из почек каудекса почти одновре- менно двух или более весенних годичных побегов (побегов кущения), происходит на 5-8 году. Первое цветение наблюдается на 8-15 году. Ежегодно отрастают вегетативные и унирепродуктивные (монокарпические обоеполые) не- ветвистые или ветвистые побеги с верхушечными фрондулозно- брактеозными или брактеозными открытыми соцветиями; такие побеги в целом называются семидетерминатными, или полузакрытыми, по- скольку их открытая брактеозная верхушка проявляет самозамирающий рост, т.е., недоразвиваясь, она засыхает независимо от состояния внеш- ней среды. Миниморезидные побеги у взрослых растений состоят из 2-4 сближенных узлов, на которых развиты зимующие почки, и компактных междоузлий. Старые миниморезиды постепенно начинают разрушаться, образуя в структуре каудекса очаги некроза, полости и, наконец, некроз расщепляет каудекс до основания главного корня, а то и ниже. У старых
Глава 10. Использование теории организации биоморф семенных растений 569 Рис. 217. Начальные этапы биоморфогении Scutellaria baicalensis Georgi (Читинская область). А — проросток, Б — всходы, В — ювенильные особи, Г — имматурные особи, Д— инициматурная (ежегодно образуется от 1 до 5 унирепродуктивных и вегетатив- ных весенних побегов); 1 — семядоли, 2 — гипокотиль, 3 — семядольный узел (у взрослых особей его приблизительное положение может быть отмечено пункти- ром), 4 — главный побег, 5а-5д — корни: 5а — главная монориза (субмедиапь), 5б — главная дириза, 5в — неглавная монориза (сублатерапь), 5г — тририза, 5д- главная тетрариза, 6 — эусимподиальный каудекс, 7 — миниморезидный годич- ный побег, 8 — весенние вегетативные пазушные побеги, 9 — замирающая вер- хушка надземной части побега, 10 — унирепродуктивный семидетерминантный (полузакрытый) безрозеточный годичный побег, 11 — зона возобновления годич- ного побега, 12 — сухие остатки от прошлогодних побегов, 13 — пазушные почки, 14 — ризопротерокормус (семядольный почкокорень), 15 — семядольный ризомо- нокормус, 16 — бессемядольный вегетативный безрозеточный ризомонокормус, 17 — вегетативный ризогептакормус, 18 — ежегодно или неежегодно репродуктив- ный ризогептакормус, 19 — ежегодно или неежегодно репродуктивный ризоэна- токормус, 20 — главное фрондулозно-брактеозное открытое соцветие; у — увели- чено по сравнению с масштабом; одно деление масштаба — 1 см; горизонтальная линия — поверхность почвы.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ Рис. 218. Медиматурная и финиматурная стадии биоморфогении Scutellaria baicalensis (Читинская область). Е — медиматурная особь (ежегодно образуется от 6 и более унирепродуктивных и вегетативных весенних побегов; партикуляция не распространяется ниже семя- дольного узла), Ж — финиматурный кондивидуум (партикуляция распространя- ется ниже семядольного узла, очень редко могут полностью обособляться старые корнепобеговые партикулы), И — сенильный кондивидуум (процессы отмирания сильно доминируют над ростом и развитием единичных слабых вегетативных побегов); 21 — схема (фрагмент) эусимподиального нарастания побеговой систе- мы, 22 — сборное фрондулозно-брактеозное или брактеозное открытое соцветие, иначе полузакрытое соцветие, 23 — неполная общая партикуляция, 24 — полная общая партикуляция. Остальные обозначения те же, что и на рис. 217.
Глава 10. Использование теории организации биоморф семенных растений 571 растений в результате неравномерности процесса некроза одних тканей и роста других участков растения каудексово-корневая система начина- ет скручиваться, что в итоге способствует полному расщеплению рас- тительного организма на несколько полностью обособленных партикул- клонистов (несовершенное вегетативное размножение). Обособление парциальных каудексов с фрагментами главного корня лишь незначи- тельно продлевает жизнь несовершенному клону. Максимальный воз- раст растений не превышает 60 лет. По эколого-морфологическим тропобиозным признакам шлемник байкальский — это травянистый летнезеленый надземно-двусемядоль- ный криптофитный мультирепродуктивный безрозеточный супротивно- ланцетолистный эусимподиальный семидетерминатный (с верхушечны- ми фрондулозно-брактеозными и брактеозными открытыми соцветия- ми) яркосине-яснотковоцветковый каудексовый неплотно-компактно- кустистый стержнекорневой изоаклональный, редко дефективно-кон- дивидуальный, ксеромезофильный горно-степной вид.
Основные выводы Рациональное использование богатейших растительных ресурсов мировой флоры возможно на основе разносторонних знаний об орга- низации составляющих их элементов и, в частности, биоморф растений. Чтобы развиваться целенаправленно, каждая наука должна иметь свою стержневую теорию. Такой теорией для биоморфологии может стать теория организации биоморф. Философско-биологическое основание для построения многоплановых широких обобщений призвана обеспе- чивать общая теория жизнесохранения, впервые разработанная автором и заложенная в фундамент теории организации биоморф семенных рас- тений. Главное содержание последней и всей книги определяют семь концепций: понятия «жизненная форма» (биоморфа), побега, корня, корнепобега (ризокормуса), морфогении, инфраструктурно-конститу- циальной борьбы за существование, пластичности и разнонаправленной соматической эволюции. Концепция понятия «биоморфа» является естественным развитием формировавшихся ранее подходов, идей и тенденций (А. Гумбольдт, К.Раункиер, О.Друде, Б.А. Келлер, Г.Дю Рие, М.В.Культиасов, Г.Мей- зель, И.Г.Серебряков, Г.М.Зозулин, В.Н.Голубев, Т.И. Серебрякова, Ф.Н.Правдин, Е.М.Лавренко и др.). После критического анализа исто- рии употребления термина «жизненная форма» было предложено пони- мать биоморфологию как науку о жизненных формах (биоморфах), их структурах и функциях, закономерностях развития, отношениях к среде обитания, т.е. науку об организации биоморф. Определение биоморфы как организма (индивидуума, особи) и синорганизма (кондивидуума) открывает более широкие познавательные возможности, чем просто понятия организм и синорганизм и все другие, предлагавшиеся ранее значения этого термина, ограниченные какой-либо структурной или функциональной стороной, чаще всего габитусом организмов. Широкое понимание биоморф не отрицает возможности оценки их по более традиционным признакам (габитуальным); кроме того, при необходимо- сти никак не исключается весьма подходящий для обозначения габитуса организмов термин Ф.Клементса (Clements, 1920) — «габитуальные фор- мы», а при обобщении — типы габитуапьных форм. Определен статус биоморфологии как одного из трех крупных раз- делов биологии, наряду с гипобиоморфологией и метабиоморфологией. В свою очередь доказано, что биология (наука о биосистемах) и эколо-
'Основные выводы 573 гия (наука об экосистемах) — части еще более общей науки биофоро- логии (науки о биофорах — любых «носителях жизни»). В теоретических разработках широко использованы, причем в раз- витии, различные методологические и методические положения. Впер- вые создана развернутая классификация классификаций (одновременно и классификация методов классифицирования); конкретизированы из- вестные методы классифицирования и типы классификаций и опреде- лены несколько их новых типов. Впервые, например, разработан метод суперординатного классифицирования, когда сравнимые субординатные или мультисубординатные классификации, составляют классы систем (уровни абстракции, или суперординаты), располагаясь в восходящем ряду на разной высоте относительно друг друга в зависимости от степени обоб- щения в этих классификациях; критерием для выделения суперординаты служит число субординат (классов объектов) их составляющих. Кроме того, в работе для решения разномасштабных теоретических задач широко ис- пользованы методы графического моделирования. Описывая те или иные явления Природы, мы обращали внимание на их осознание в науке с позиций закона науки. Там, где было необходимо и возможно, взяли на себя инициативу и дали определения не только специальным, но и более общим законам, например, законам иерархичнос- ти, повторяемости, а затем использовали их при построении теоретических положений в одном из частных разделов науки — в биоморфологии. Выявлена и обоснована универсальная единица эволюции — син- том, или эволюционный шаг, — система «исходное-производное». Дано определение трем типам эволюции: закрытой, открытой и сверхоткры- той. Сделан принципиальный вывод о возможности сохранения жизни в Природе: пока в общем эволюционном потоке закрытые эволюцион- ные шаги («шаги из живого в неживое») перекрываются количеством и качеством открытых эволюционных шагов («шагов из живого или нежи- вого в живое»), до тех пор реализуется высшая направленность эволю- ции в Природе — направленность на жизнесохранение. Последователь- но определены свойства биосистем, сохраняющие или разрушающие жизнь и условия их проявления. В рамках общей теории жизнесохранения сформулирована меро- сомная концепция организации биосистем, рассмотренная на примере биоморф семенных растений и принадлежащая в этой части теории их организации. Среди меросом — частей организации биоморф (шире биосистем), в специализации которых доминирует определенная нап- равленность на выполнение каких-либо жизнесохранительных функ- ций, предложено различать: эусомы, диасомы, архесомы, хромосомы и гены. Сома семенного растения — это единство пяти меросом.Обсуж- дены особенности и выявлены закономерности биофильной специали- зации биоморф и их меросом: если части биосистем неодинаково спе- циализированы в направлении жизнесохранения, то чем больше их спе- циализация в сторону ксенобиофилии (сохранения чужой жизни), тем меньше их специализация в направлении автобиофилии (сохранения своей жизни). Рассмотрены основные вехи в изучении организации биоморф се- менных растений, начиная с работ мыслителей древности до наших
574 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНКИ дней. Обращено особое внимание на основные органы семенных рас- тений. В качестве основных органов рассмотрены побеги и корни, а в качестве высшей единицы организации биоморф семенных растений впервые определен корнепобег (ризокормус) — это любой побег и обра- зованные из него корни, или побег и связанные с ним через семядоль- ный узел и гипокотиль корни. Последовательно используя положения общей теории жизнесохра- нения, нами впервые сформулирован закон стратегии Жизни: для жиз- ни в целом приемлема только та стратегия, цель которой — жизнесох- ранение, а высшая цель — нообиофильная суперадаптация (бесконеч- ное сохранение разумной жизни), что достигается или может достигать- ся посредством общего доминирования тех тактик частных биосистем, которые отвечают этим целям. Определен и рассмотрен спектр главных тактик, которые проявляются в организации биоморф семенных расте- ний. В зависимости от того, какие части биосистем решают стратеги- ческие и тактические задачи, предложено считать их соответственно (по назначению) средствами стратегического (элементы-руководители, т.е. гены, хромосомы) и тактического (элементы-исполнители и элементы- соисполнители, т.е. все остальные проявления организации биосистем) назначения. Показано, что биоморфы семенных растений подчиняются закону «управления-исполнения». Подробно рассмотрены особенности организации определенных тактических систем и выявлены закономер- ности нарастания, ветвления, кущения, размножения и элиминации. Организация какой-либо биоморфы семенных растений — это сосре- доточие нескольких функциональных систем («скоплений» живой мате- рии определенного типа), проявляющих пять основных направлений («стремлений») развития как стратегических (гены, хромосомы), так и тактических (меристемы, побеги, корни, корнепобеги-органы и, нако- нец, корнепобеги-организмы: управления, потребления, производства, резервирования и рассеивания. Развита, в согласии с А.Арбер, Н.В. Первухиной и др., идея един- ства стебля и листьев в организации одного органа — побега. Побег охарактеризован широко как главная часть растения, состоящая из стеблевой, листовой, а нередко семядольной и репродуктивной частей, обладающая определенной продолжительностью жизненного цикла, ритмом ростовой активности, степенью специализации и характером развития. Широкое понятие «побег» позволило определить параметры побегов на разной признаковой основе и построить их иерархические классификации с разных высот обобщения. Например, побегом являет- ся и отдельная цветоножка с цветком в соцветии (Padus avium Mill.), и отдельный стеблелистовой годичный прирост яблони (Malus sylvestris Mill.), и все то, что образуется выше, начиная с семядольного узла у небольшого однолетника (Drabopsis nuda (Belanger) Stapf и др.) или гигантского многолетника (Adansonia digitata L. и др.). Вскрыта иерар- хичность в организации побегов. Наиболее полно сложность организа- ции побегов отражена в субординатной классификации из 10 типов побегов. Иерархичность организации присуща также элементам побе- гов: стеблям, листьям, соцветиям, сверхсоцветиям, полупобегам. Рас-
Основные выводы 575 смотрены возможности использования многолинейного классифициро- вания при изучении признакового пространства побегов. Разработаны представления о персистентности (процессе неотми- рания после окончания вегетационного сезона) и некрозности (процессе отмирания) в организации биоморф семенных растений. Особое внима- ние обращено на скелетные (зимующие) осевые (стеблевые и корневые) части, на которых развиваются другие персистентные части (зимующие почки, листья). Показаны пути проявления персистентности и отмече- но, что в сезон вегетации (в процессе роста) проявляется только потен- циальная персистентность, включая и потенциальную скелетность осе- вых частей, которая в условиях сезонного климата лишь в конце осени- начале зимы реализует или не реализует свои потенции. Разработаны разномасштабные субординатные классификации персистентных час- тей. Большое значение в формировании базовых (многолетних побего- вых) органов имеет процесс образования резидов (резидофикации). Описано 16 основных типов резидов и рассмотрено их участие в форми- ровании каркаса базового органа. Последовательно проанализировано разнообразие базовых органов, среди которых по признакам возрастной изменчивости предложено различать протобазовые и эубазовые органы. Рассмотрены главные особенности формирования многих эубазовых ор- ганов: аэроподиев, геоподиев, гетероподиев, ценоподиев, базиомов, ри- зомов, каудексов, конодиев, зрелых луковиц, солуковиц, клубнелуко- виц, побеговых клубней (клубнепобегов) и автономных зимующих почек. Корень — это второй основной орган биоморф семенных растений. Корень — часть растения, состоящая из оси или осей, начинающаяся в своем развитии из верхушечной меристемы, разветвленная или неразвет- вленная и никогда не развивающая на себе листья и репродуктивные элементы, но образующая, как правило, корневой чехлик и выросты эпи- дермиса — корневые волоски. Иерархичность в организации корней отра- жена в субординатной классификации из 4 типов корней. В этой классифи- кации учтены особенности нарастания и ветвления корней. Впервые разработаны представления о корнепобегах (ризокорму- сах) как высших единицах организации биоморф семенных растений. Отмечено два типа корнепобегов: ризокормусы-органы (онтоиды), если они составляют только часть структуры растения, ризокормусы-орга- низмы (эуфиты), если они составляют всю структуру растения (весь организм). Впервые определен закон предрасположенности к размно- жению. К вегетативному размножению наивысшую предрасположен- ность проявляют организмы семенных растений, в структуре которых развиваются ризокормусы-органы. Основные типы ризокормусов пред- ложено выделять с учетом уровня организационной сложности побегов, т.е. определяющих элементов этих систем. Структурно-функциональное превосходство побегов над корнями и корнепобегов-органов над остальными органами семенных растений отражено в законе фенетического доминирования в организации био- морф семенных растений: только те органы выполняют роль доминан- тов в фенетической организации биоморф семенных растений, которые непосредственно исполняют несколько их функций: образуют и удобно
576 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИИ располагают в окружающем жизненном пространстве репродуктивные (уже — половые) органы, обеспечивают всем необходимым (вещества- ми, энергией, информацией) то, что связано в этой организации с по- ловым размножением и с эмбриональным развитием потомства. Как динамическая основа теории организации жизненных форм рассмотрена концепция морфогении. Цикл существования биоморф (биоморфогенез) представлен или онтогенезом (жизненным циклом ин- дивидуума), KJIH синонтогенезом (жизненным циклом кондивидуума). Уточнено значение известных терминов возрастное состояние и фено- логическое состояние. Впервые разработана субординатная классифи- кация категорий стадий биоморфогенезов. Определены критерии выде- ления возрастных состояний и дана краткая методика их изучения. Из всего комплекса признаков биоморф, проявляющихся в биоморфогене- зах, предложено различать индикаторные признаки морфогенических состояний (гемитропобиозные признаки) и признаки-индикаторы ти- пов биоморфогенезов (тропобиозные признаки). Предложена типовая схема возрастных стадий биоморфогенезов семенных растений. Разработана концепция пластичности. Пластичность в широ- ком смысле — это способность биосистем изменяться при изменении экологических условий, не утрачивая при этом своих качественных гра- ниц. В экстремальных условиях поведение биосистем подчинается сле- дующему правилу: для выживания в экстремальных условиях биосисте- мы используют свой генетический потенциал и, если не погибают, то более или менее изменяют свой внешний облик и (или) внутреннюю организацию. С этих позиций охарактеризовано явление мультивари- антности биоморфогении. Конкретизированы представления о внутри- видовой пластичности биоморф и показано их отличие от пластичности биоморф (онтогенезной и синонтогенезной). По относительной встречае- мости было предложено различать следующие категории внутривидовых биоморфотипов по отдельным рядам признаков: основной (этот тип присущ большинству биоморф одного вида в определенном регионе или во всем географическом ареале) и второстепенные (эти типы представлены меньшим числом биоморф того же вида, чем основной биоморфотип). Обоснованы представления об универсальности борьбы за суще- ствование как комплекса отношений между биосистемами, а также био- системами и абиотическими условиями. Доказано, что борьба за суще- ствование идет на разных уровнях организации Жизни и в частности внутри растительного организма как комплекс отношений между гипо- биоморфами, т.е. структурными элементами биоморф, а также между гипобиоморфами и окружающей внешней средой (инфраструктурная борьба за существование). Из систем, где лучше всего наблюдать прояв- ление этой борьбы, рассмотрены препопуляции (системы побегов опре- деленного рода). Побеги — это «солдаты» одной армии-организма, цель которых выполнить главные задачи этой «армии» (фотосинтез, размно- жение), если потребуется, то и ценой жизни многих из них. Внутриви- довая пластичность начинает проявляться уже в инфраструктурной борьбе за существование, на основе которой действует конституциаль- ная борьба и следует обобщенная ответная реакция системы всего
Основные выводы организма (его конституции) на возмущающее воздействие меняющейся окружающей среды. Инфраструктурная и конституциальная борьба за существование развертывается в фенотипах, но контролируется с гене- тического уровня. Соматическая эволюция охарактеризована как «маятниковый» раз- нонаправленный процесс. Эволюционным маятником названа неживая или живая материальная система (объект) — тело или система тел, на- ходящаяся в развитии в каких-либо условиях существования. Его аб- страктные колебания отражают диапазон изменчивости тел или систе- мы тел, которые он собой олицетворяет. Размеры эволюционного ма- ятника колеблются от наименьшей частицы вещества до всей видимой части Вселенной. Такие «маятники» «плывут» в эволюционном потоке пространства-времени бесконечной Вселенной. Каждый эволюционный маятник имеет как внутреннюю пружину для своего самораскачивания (существования) — природу самого объекта, так и внешнюю пру- жину — условия существования. Колебание каждого живого эволюци- онного маятника, в нашем случае видов растений, отражает разнообра- зие биоморф (их типов) при развитии в различных экологических ус- ловиях. Растения или группы растений, которые участвуют в эволюци- онном потоке в образе эволюционных маятников, в процессе своего существования могут решить или не решить одну или несколько из поставленных Природой перед ними задач: 1) продолжить жизнь, 2) рас- ширить полиморфизм, 3) создать новый вид. Главные изменения в живых эволюционных маятниках затрагивают количественные признаки, далее признаки пространственного расположе- ния (топические) и лишь потом собственно качественные. При сомати- ческой (говоря о соме, автор подразумевает единство всего тела растения— систем вегетативных и репродуктивных, вместе с архесомой — гаметами, эмбриональными особями в структуре родительского организма) транс- формации растений изменяются чаще всего размеры и число, продол- жительность жизни побегов, корней и их элементов. Разнонаправленности и обратимости соматическая эволюция растений обязана в первую очередь климатическому фактору и их способности проявлять пластичность. Создана многолинейная классификация биоморф семенных расте- ний по многим рядам альтернативных признаков (выделено 500 био- морфотипов). Эта классификация, кроме того, впервые закодирована в таблицу-матрицу, информацию с которой предложено считывать по нес- кольким вариантам формул, что в целом открывает новые возможности в развитии как методов накопления биоморфологических знаний, так и методов исследования организации биоморф. Теория организации биоморф семенных растений открыта для даль- нейшего развития, но то, что создано, дает возможность целенаправлен- но использовать ее при изучении организации биоморф, биоморфогении и соматической эволюции. Нужны «глубокие погружения» биоморфо- логов во флоры Анд и Гималаев, в тропики Южной Америки, Африки и Юго-Восточной Азии. Именно в таких экзотических поездках можно ожидать еще много нового. При всем этом не стоит забывать и об отечественной флоре, поскольку растительные тайны иногда скрываю- тся от нас и около родного дома.
Заключение Мы нередко склонны удивляться ошеломляющему разнообразию растительных организмов и их видов. Однако с появлением опыта и расширением знаний приходит понимание, что оно все же иллюзорно: на самом деле все обстоит несколько иначе. Разнообразие биоморф и их видов существует, оно способно восхищать кого угодно, но только до некоторого предела, где кончаются эмоции, за которыми начинается серьезная познавательная работа. Природа не расточительна и создала ограниченный набор признаков, из которых она собирает неживые и живые системы. «Природа проста и не роскошествует излишними при- чинами вещей» — этот меткий афоризм принадлежит И. Ньютону. Богатейшее разнообразие форм существования материи — это только внешняя сторона организации Природы, способной, оставаясь и «Простой» и «Скромной», выступать в качестве Великого Организатора, создавая из малого большое и очень большое. Суть разнообразия — в уникальной способности Природы к бесконечной комбинаторике срав- нительно ограниченным набором признаков при создании как нежи- вых, так и живых материальных систем. В этом отношении у живой природы, обладающей наследственностью и способностью к мутациям, возможности большие, хотя и не беспредельны. Одни признаки могут существовать бесконечно долго, другие долгожители, а третьи эфемерны, но Природа использует всех их в своем развитии и они, естественно, составляют ее признаковое пространство. Растения и животные, чтобы жить на Земле, проявляют пластичность (одни больше, другие меньше), по-разному отвечая на изменения условий их обитания, и эволюциони- руют. Вот почему Жизнь поражает нас своим разнообразием. Природа «научилась» в процессе бесконечной эволюции конструи- ровать неживые и живые дискретные системы из довольно ограничен- ного набора признаков. Она обладает признаковым пространством, ко- торое отражается в нашем воображении, т.е. абстрактно. Но мы — ча- стицы Природы, поэтому и наше воображение, наше желание и умение определять типы организации и классифицировать их по определенным признакам, принадлежит также ей. Это мы-люди способны увидеть ее конкретные признаки (сравнимые и взаимоисключающие), объединить их в типы и построить рядами, в которых они «маршируют» на вечном параде бытия в пространстве-времени. Не будем забывать, что биомор- фы и не только они, а все, что есть в Природе, сводится по отдельным
Заключение признакам к вполне обозримому числу типов. Вспомним, хотя бы, построенную нами пирамиду абстракций, отражающую на одной странице организацию всей Природы (см. гл. 2.2). Ошеломляющее раз- нообразие форм организации сведено в ней к относительно небольшому числу типов. Через всю работу проходит идея классифицирования био- морф, да и не только их. Частным итогом этих исследований стало создание таблицы-матрицы, в которой закодирована многолинейная классификация биоморф семенных растений, что открывает реальный путь к компьютеризации биоморфологического анализа. Сердцевиной существования разнообразия в Природе, ее философ- ским ядром, являются преднаследственные и наследственные свойства, которые, будучи дискретно распыленными в ней, обеспечивают функ- ционирование Природы во времени как разнообразных известных и не- известных нам пространственных форм организации (тел, систем тел и процессов): гор, рек, камней, машин, деревьев, степных биоценозов, людей, молний, взрывов и т.п. Таким образом, разнообразие в органи- зации Природы — это неисчерпаемая комбинаторика (смешение) не- которых (иногда очень многих) признаков в рамках материальных тел и (или) процессов, осуществляемая на основе каких-либо природных законов. Возникновение в пространстве-времени принципиально но- вых признаков возможно, но такие качественные скачки в постепенно- сти происходят довольно редко. Например, Природа однажды, много миллионов лет назад, создала с одной стороны растительные осевые струк- туры с верхушечным репродуктивным элементом (закрытые оси), а с дру- гой — осевые системы с боковыми репродуктивными элементами и сво- бодной ростовой верхушкой (открытые оси). Этот принцип организации эволюционирует по сей день, не утрачивая своего определяющего каче- ства, и характерен для всех современных семенных растений. Существование Природы — это вечное движение составляющих ее дискретных тел и систем тел, находящихся каждый в своей системе частных движений. Самое удивительное, что тела в этих движениях проявляют общность, независимую от специфики их организации; они в своем развитии чем-то напоминают маятники, причем, естественно, не внешне, а по содержанию их развития — по глубинному механизму их движений-развитий. Можно образно представить, как в общем эво- люционном потоке плывут (никогда не стоят на месте!) мириарды таких абстрактных эволюционных маятников (тел и систем тел), существова- ние которых всегда в динамике (изменяется угол наклона опорной оси маятника, его положение в пространстве, амплитуда и скорость ко- лебания самого маятникового тела и т.п.). Чаще всего диапазоны из- менчивости каких-либо тел или систем тел, например, биоморф расте- ний и их видов, не выходят за пределы количественных показателей: больше-меньше, выше-ниже, что квалифицируется биологами как ре- дукционизм, грандизация, деграндизация, нанизация, соматическая трансформация и т.п. На эволюцию не стоит смотреть только в одном масштабе и видеть одни прямые линии. Прямолинейность эволюции обманчива, нередко линии эволюции искривляются, ломаются, поворачиваются вспять и
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ даже обрываются. Ломаные линии эволюции мы наблюдаем сплошь и рядом в примерах с мелкими эволюционными маятниками, что пред- определено способностью живых систем, в частности биоморф и их видов, к пластичности в зависимости от изменчивости условий окру- жающей среды. Концепция «маятниковой» соматической эволюции нацелена на развитие эволюционной теории. Однако, не будем торопить события. Давайте пока осознаем лишь то, что уже свершилось..... Разностороннее изучение биоморф (организмов и синорганизмов) семенных растений не снимает, а лишь подчеркивает актуальность спе- циальных исследований их габитусов (внешних обликов). По замыслу автора, габитусы, характерные биоморфам, как одна из важных сторон организации (габитуальные формы, формы роста), и их типы занимают принципиальное место в биоморфологии; они достойны более полного рассмотрения, но уже в другой книге. И, пожалуй, последнее. Позволю себе привести творческое правило знаменитого ученого Ганса Селье (1972, с. 110): «Ни знание предмета твоего исследования и мощь твоих инструментов, ни обширность твоих знаний и точность твоих планов никогда не смогут заменить оригиналь- ности твоей мысли и зоркости твоего наблюдения», которое вполне могло бы стать таковым и для биоморфологов.
Литература Абрамова Н.Т. О соотношении части и целого в строении материи // Вопр. философии, 1962, М 2, с. 45-56. Абрамова H.Т. К определению поня- тия "уровень организации" // Раз- витие концепции структурных уровней в биологии. — М.: Наука, 1972, с. 122-133. Абрамова Н. Т. Целостность и управле- ние. — М.: Наука, 1974, 248 с. Аверьянов А.H. Категория система в диалектическом материализме.— М.: Мысль, 1974, 70 с. Аверьянов А.Н. Система: философская категория и реальность. — М.: Мысль, 1976, 188 с. Аврорин H.À. Переселение растений на полярный север. Эколого-геогра- фический анализ. — М.; Л.: изд-во АН СССР, 1956, 286 с. Акофф Р. О природе систем // Изв. АН СССР. Сер. техн. кибернетика, 1971, М 3, с. 68-75. Алеев Ю.Г. Жизненная форма как сис- тема адаптаций // Успехи совр. биол., 1980, т. 90, вы. 3(6), с. 462-477. Апеев Ю.Г. Экоморфология. — Киев: Наукова думка, 1986, 424 с. Алексеева ЛД. Пустырник пятилопаст- ной. // Атлас лекарственных расте- ний СССР. — М.: Медгиз, 1962, 468 с. Александров В.Г. Анатомия растений. 4-е изд. — М.: Высшая школа, 1966, 430 с. Апександрова В.Д. Классификация ра- стительности. — М.: Наука, 1969, 275 с. Апехин В.В. География растений. 3-е изд. — М.: Учпедгиз, 1950, 419 с. Алешкина А.Я., Ильинская Т.Н., Шре- тер А.И. Эхинопанакс высокий // Атлас лекарственных растений СССР. — М.: Медгиз, 1962, с. 664. Андреев ИД. Теория как форма орга- низации научного знания. — М.: Наука, 1979, 303 с. Анохин ШК Функциональная система как универсальный принцип изуче- ния уровней биологической орга- низации // Развитие концепции структурных уровней в биологии.— М.: Наука, 1972, с. 100-111. Анохин ШК Принципиальные вопро- сы общей теории функциональных систем // Принципы системной организации функций. — М.: На- ука, 1973, с. 5-61. Арманд Д.Л. Наука о ландшафте.— М.: Мысль, 1975, 287 с. Атлас ареалов и ресурсов лекарствен- ных растений СССР. — М.: изд-во ГУГК, 1976, 340 с. Атлас лекарственных растений СССР. — М.: Медгиз, 1962, 703 с. Афанасьев В.Г. Проблема целостности в философии и биологии. — М.: Мысль, '1964, 416 с. Афанасьев В.Г. Об относительной са- мостоятельности частей биологи- ческого целого // Целостность и биология. — Киев, Наукова думка, 1968, с. 28-37. Афанасьев В.Г. Системность и обще- ство. — М.: Политиздат, 1980, 368 с. Афанасьев В.Г. Мир живого: систем- ность, эволюция и управление.— М.: Политиздат, 1986, 334 с. Баженов Л.Б. Строение и функции естественнонаучной теории. — М.: Наука, 1978, 231 с. Балабас Г.М. и др. Интродукция ле- карственных, ароматических и тех-
582 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ нических растений / Г.М. Балабас, Р.А. Буйко, А.Е. Гращенков, И.Ф. Сацыперова, И.Б. Сандина, В.С. Синицкий, В.С. Соколов. — М.; Л.: Наука, 1965, 425 с. Балковский Б.Е. Цифровой политоми- ческий ключ для определения рас- тений. — Киев: Наукова думка, 1964, 36с. Барыкина Р.П., Гуленкова М.А. Мета- морфоз надземного побега // Пятая всесоюз. школа по теоретической морфологии / Научные доклады.— Львов, 1987, с. 24-29. Барыкина Р.П., Чубатова Н.В. Борец северный // Биологическая флора Московской области. Вып.11. — М.: Аргус, 1995, с. 154-165. Барыкина Р.П., Чубатова Н.В. Живо- кость полевая // Биологическая флора Московской области. Вып.12. — М.: Аргус, 1996, с. 60-70. Батороев КБ. Аналогия и модели в познании. — Новосибирск: Наука, 1981, 319 с. Бауэр Э.С. Теоретическая биология.— М.; Л.: изд-во Всес. ин-та экспер. медицины, 1935, 206с. Бейдеман И.Н. Изучение фенологии растений // Полевая геоботаника.— М.; Л.: изд-во АН СССР, 1960, т. 2, с. 333-366. Бейдеман И.Н. Методика изучения фенологии растений и раститель- ных сообществ. — Новосибирск: Наука, 1974, 155 с. Бекетов А. Очерк Тифлисской флоры, с описанием лютиковых, ей при- надлежащих. — СПб., 1853, 56 с. Бекетов А. Курс ботаники. — СПб., 1862, т. 1, 318 с. Беклемишев В.Н. Об общих принци- пах организации жизни // Бюлл. Моск. об-ва испыт. природы (МОИП), отд. биол., 1964, т. 69, вып. 2, с. 22-38. Белозерский А.Н. Молекулярная био- логия — новая ступень познания природы. — М.: Сов. Россия, 1970, 190 с. Белостоков Г.П. К побегообразованию плотнокустовых злаков // Ботан. журн., 1957, т. 42, М 8, с. 1267-1277. Белоусов Л.В. Биологический морфо- генез. — М.: изд-во Моск. ун-та, 1987, 239 с. Белянина Н.Б. 0 перезимовке расте- ний можжевеловых и сосновых ле- сов Южного берега Крыма // Тр. госуд. Никитск. ботан. сада, 1961, т. 35. Берг Л.С. Труды по теории эволюции.— . Л.: Наука, 1977, 387 с. Бергхольц Н.Л. 0 зарастании и закреп- лении подвижных песков Нижне- днепровья // Науч. тр. / Укр. н.-и. ин-т лесного хоз-ва и агролесоме- лиорации. — Харьков, 1959, вып. 19, с. 193-205. Берка ЙМ. До п1тання вивчення пер1одизаци великого життевого циклу вегетативно рухпивих рослин // Укр. ботан. журн., 1973, т. 33, М 6, с. 604-609. Берко И.Н. Некоторые вопросы изуче- ния и периодизации большого жиз- ненного цикла вегетативно-под- вижных растений // Структура и динамика растительного покрова. М.: Наука, 1976, с. 143-145. Берка И.Н. Биолого-морфологические особенности жизненного цикла Lycopus europaeus L. // Укр. ботан. журн., 1982, т. 39, М 4, 36 с. Берко И.Н., Козий Б.И. 0 методике моделирования морфологического строения монокарпического побега травянистых многолетников // Пя- тая Всесоюз. школа по теоретичес- кой морфологии растений / Науч- ные документы. — Львов, 1987, с. 29-33. Бернал Дж. Возникновение жизни. М.: Мир, 1969, 391 с. Берталанфи Л. Общая теория систем: критический обзор // Исследова- ния по общей теории систем. — М.: Прогресс, 1969, с. 23-82. Берталанфи Л. История и статус об- щей теории систем // Системные исследования. Ежегодник - 1973.— М.: Наука, 1973, с. 20-37. Беспалова 3.Г. О жизненной форме "полукустарничек" // Пробл. совр. ботаники. — М.; Л.: Наука, 1965, т. 2, с. 65-69.
Указатель латинских названий растений Бил Дж., Ноулз Дж. Внеядерная на- следственность. Пер. с англ. — М.: Мир, 1981, 168 с. Бирюков Б.В. Кибернетика и методо- логия науки. — М.: Наука, 1974, 413 с. Бирюлева Э.Г. Втягивающие (контрак- тильные) корни некоторых цветко- вых растений Крыма (анатома-мор- фологическая характеристика): Ав- тореф. дис..... канд. бил. наук:— Ашхабад, 1969. Блауберг И.В. Системный подход как предмет историко-научной рефлек- сии // Системные исследования. Ежегодник - 1973. — М.: Наука, 1973, с. 7-19. Блауберг И.В. Целостность и систем- ность // Системные исследования. Ежегодник - 1977. — М.: Наука, 1977, с. 5-28. Блауберг И.В., Юдин Э.Г. Становление и сущность системного подхода.— М.: Наука, 1973, 270 с. Бляхер Л.К Очерк истории морфоло- гии животных. — М.: изд-во АН СССР, 1962, 263 с. Бляхер Л.Н. Проблемы морфологии животных. Исторические очерки.— М.: Наука, 1976, 359 с. Бобров Н. С., Петленко В.II. Философ- ские вопросы соотношения струк- турно-функциональных изменений в современной медицине // Основ- ные философские вопросы совре- менной биологии и медицины.— Л.: Медицина, 1967, с. 141-156. Богданов А.А. Всеобщая организаци- онная наука: Тектология. 3-е изд.— т. 1. — М.: Книга, 1925, 300 с.;— т. 2 — М.: Книга, 1927, 252 с.; т. 3— М.: Книга, 1929, 221 с. Богдановская-Гиенэф ИД. К вопросу о семенном возобновлении в луговых сообществах // Зап. Ленингр. с.-х. ин-та, 1926, вып. 3, с. 216-253. Боголепов В.П. О состоянии и задачах развития общей теории организа- ции // Организация и управление (вопросы теории и практики).— М.: Наука, 1968, с. 38-56. Бологова В.Л. Функциональные и ди- намические аспекты макроморфо- логической структуры растительно- го организма на примере ежи сбор- ной: Автореферат. дис.... канд. биол. наук. — М., 1989, 24 с. Борисова И.В. Биология и основные жизненные формы двудольных многолетних травянистых растений степных фитоценозов Северного Казахстана // Тр. Бот. ин-та АН СССР, 1961, сер.3, вып.13, с. 54-132. Борисова И.В. Сезонная динамика ра- стительного сообщества // Полевая геоботаника. — Л.: Наука, 1972, т. 4, с. 5-94. Борисова И.В. О понятиях «биомор- фа», «экобиоморфа» и «архитектур- ная модель» // Ботан. журн., 1991, т. 76, М 10, с. 1360-1367. Букштынов А.Д., Трофимов Т Т., Ер- маков Б.С. и др. Облепиха. 2-е изд.— М.: Лесная промышленность, 1985, 183 с. Буланая М.В. Варианты жизненных форм черемухи обыкновенной (Prunus padus L.) в разных частях ареала // Жизненные формы в эко- логии и систематике растений / Межвуз. сб. науч. трудов. — М., 1986, с. 81-85. Булыгин Н.Е. Динамика формирова- ния цветочных зачатков у древес- ных растений в Ленинграде: Авто- реф. дис.... канд. биол. наук. — Л., 1965, 21 с. Былова А.М. Онтогенез и возрастные группы василька шероховатого (Centaurea scabiosa L.) // Вопросы морфогенеза цветковых растений и строение их популяций. — М.: На- ука, 1968, с. 103-124. Былова А.М. Некоторые биологичес- кие особенности василька русского // Успехи экологической морфологии растений и ее влияние на смежные науки. — М.: Прометей, 1994, с. 71-72. Бюсген М. Строение и жизнь наших лесных деревьев. Пер. с немец.— М.: Гослесбумиздат, 1961, 424 с. Вавилов Н.И. Закон гомологических рядов в наследственной изменчиво- сти // Избран. сочинения. — М.: Колос, 1966, с. 57-101. Вальтер Г. Растительность земного шара. Эколого-физиологическая
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ характеристика. Тропические и субтропические зоны. Пер. с немец. — М.: Прогресс, 1968, 551 с. Варгина Н.Е. и др. Пустырник сердеч- ный // Атлас ареалов и ресурсов лекарственных растений СССР / Н.Е. Варгина, С.Н. Выдрина, Г.А. Копанева, А.И. Шретер. — М.: изд- во ГУГК, 1976, 290 с. Варминг Е. Распределение растений в зависимости от внешних условий. Пер. с немец. — СПб., 1902, 414 с. Варминг Е. Растение и его жизнь. Пер. с немец. — М., 1904, 184 с. Василевич В.И. Что считать естествен- ной классификацией // Философс- кие проблемы современной биоло- гии. — М.; Л.: Наука, 1966, с. 178-189. Васильев А.Е. и др. Ботаника. Анато- мия и морфология растений / А.Е. Васильев, Н.С. Воронин, А.Г. Еле- невский, Т.И. Серебрякова. — М.: Просвещение, 1978, 480 с. Васильченко И.Т. Определитель всхо- дов сорных растений. 2-е изд. — Л.: Колос, 1979, 344 с. Вахтомин Н.Г. Генезис научного зна- ния. Факты, идеи, теория. — М.: Наука, 1973, 286 с. Веденов М.Ф., Кремянский В.И., Ша- талов А.Т. Концепция структурных уровней в биологии // Развитие концепции структурных уровней в биологии. — М.: Наука, 1972, с. 7-70. Вейсман А. Лекции по эволюционной теории. Ч. I. — М., 1905, 505 с. Веников В.А. Теория подобия и моде- лирования. — М.: Высшая школа, 1966, 487 с. Веников В.А. (ред.) Основы теории подобия и моделирования. Терми- нология. — М.: Наука, 1973, 22 с. Верещагин В.И., Соболевская КА., Яку- бова А.И. Полезные растения За- падной Сибири. — М.; Л.: изд-во АН СССР, 1959, 348 с. Вернадский В.И. Химическое строение биосферы Земли и ее окружения.— М.: изд-во АН СССР, 1965. Вернадский В.И. Биосфера. — М.: Мысль, 1967, 376 с. Вернадский В.И. Размышления нату- ралиста. Научная мысль как пла- нетное явление. — М.: Наука, 1977, 191 с. Вернадский В.И. Живое вещество.— М.: Наука, 1978, 358 с. Веселовский В.Н. Философское значе- ние законов сохранения материи и движения. — М.: Мысль, 1964, 144 с. Веселовский В.Н. О сущности живой материи. — М.: Мысль, 1971, 295 с. Веселовский В.Н. Некоторые философ- ские вопросы современной биоло- гии. — Челябинск, 1973, 94 с. Вигнер Е. Этюды о симметрии. — М.: Мир, 1971, 318 с. Викторов Д.II. Краткий словарь бота- нических терминов. — М.; Л.: На- ука, 1964, 177 с. Виленчик М.М. Биологические основы старения и долголетия. — М.: Зна- ние, 1976, 159 с. Вилли К Биология. Пер. с англ. — М.: Мир, 1964, 678 с. Вильямс В.Р. Естественно-научные ос- новы луговодства, или луговеде- ние. — М.: Новая деревня, 1922, 298 с. Винер Н. Кибернетика, или управле- ние и связь в животном и машине. Пер. с англ. — М.: Сов. радио, 1968, 326 с. Воронцова Л.И. Изменение жизненно- го состояния эдификаторов расти- тельного покрова южной полупус- тыни под влиянием экологических условий // Онтогенез и возрастной состав популяций цветковых расте- ний. — М.: Наука, 1967, с. 132-154. Воронцова Л.И., Заугольнова Л.Б. Мультивариантность развития осо- бей в течение онтогенеза и ее зна- чение в регуляции численности и состава ценопопуляций растений // Журн. общей биологии, 1978, М 4, с. 555-562. Воронцова Л.И., Заугольнова Л.Б. О подходах к изучению ценопопуля- ций растений // Ботан. журн., 1979, т. 64, М 9, с. 1296-1311. Ворошилов В.Н. Лекарственная вале- риана. — М.: изд-во АН СССР, 1959, 160 с. Ворошилов В.Н. Методологические ас- пекты изучения флоры советского Дальнего Востока: Автореф. (по
казатель латинских названий растений 585 докладу) дис.... докт. биол. наук,— М., 1981, 32с. Вульф Е.В. Историческая география растений. — М.; Л.: изд-во АН СССР, 1944, 546 с. Высоцкий Г.Н. Ергеня. Культурально- фитологический очерк // Тр. Бюро по прикл. ботанике. Год 8-ой, № 10-11, 1915, с. 1113-1436. Гаель А.Г. Облесение бугристых пес- ков засушливых областей. — М.: Географиздат, 1952, 218 с. Галл КМ. Борьба за существование как фактор эволюции (историко- критический анализ отечественных ботанических исследований). — Л.: Наука, 1976, 155 с. Гатин Ж.И. Облепиха. — М.: Сель- хозгиз, 1963, 157 с. Гатцук Л.Е. Жизненные формы в роде Hedysarum L. и их эволюционные взаимоотношения // Бюлл. МОИП, отд. биол., 1967, т. 72, вып. 3, с. 53-64. Гатцук Л.Е. Морфогенез копеечника кустарникового (Hedysarum frutico- sum Pall.) при переменном уровне песчаного субстрата и предполагае- мый облик его предка // Вопросы морфогенеза цветковых растений и строение их популяций. — М.: На- ука, 1968, с. 52-80. Гатцук Л.Е. Элементы структуры жиз- ненных форм геммаксиллярных растений и биоморфологический анализ копеечника кустарникового: Автореф. дис.... канд. биол. наук.— М., 1970, 21 с. Гатцук Л.Е. Геммаксиллярные расте- ния и система соподчиненных еди- ниц их побегового тела // Бюлл. МОИП, отд. биол., 1974а, т. 79, цып. 1, с. 100-113. Гатцук Л.Е. К методам описания и определения жизненных форм в сезонном климате // Бюлл. МОИП, отд. биол., 1974б, т. 79, вып. 3, с. 84-100. Гатцук Л.Е. Иерархическая система структурно - биологических единиц растительного организма, выделен- ных на макроморфологическом уровне // Успехи экологической морфологии растений и ее влияние на смежные науки. — М.: Проме- тей, 1994, с. 18-19. Гатцук Л.Е. Комплементарные моде- ли побега и их синтез // Ботан. журн., 1995, т. 80, № 6, с. 1-4. Гвишиани Д.М. Организация и управ- ление. — М.: Наука, 1972, 432 с. Гегель Г.В.Ф. Философия природы. // Энциклопедия философских наук. Т. 2. — М.: Мысль, 1975, 695 с. Гельмгольц Г. Популярные речи. Т. 1.— СПб., 1898, 145 с. Георгиевский А.Б. Организация и де- зорганизация в эволюции и пробле- ма ноогенеза // Организация и эво- люция живого (философские, исто- рико-научные и теоретические ас- пекты проблемы). — Л.: Наука, 1972, с. 72-76. Георгиевский А.Б. и др. Философские проблемы теории адаптации / А.Б. Георгиевский, В.П. Петленко, А.В. Сахно, Г.И. Царегордцев. — М.: Мысль, 1975, 277 с. Герасимов И. 1Т. Методологические проблемы экологизации современ- ной науки // Общество и природ- ная среда. — М.: Знание, 1980, с. 66-86. Гершензон С.М. Основы современной генетики. — Киев: Наукова думка, 1979, 508 с. Гете И.В. Избранные сочинения по естествознанию. — М.: изд-во АН СССР, 1957, 553 с. Гирусов Э.В. Биосфера как целое // Проблема целостности в современ- ной биологии. — М.: Наука, 1968, с. 238-257. Глотов Н.В., Жукова ЛА. Демографо- генетические подходы к изучению популяций растений // Экология популяций: структура и динамика / Материалы совещания. Ч. Е — М., 1995, с. 203-215. Говоров В.1T., Лилская НА. О некото- рых фармакологических свойствах золотого корня (Rhodiola rosea L.) // Тр. Омского мед. ин-та — 1963, № 45, с. 15-22. Голдовский А.М. Основы учения о со- стояниях организмов. — Л.: Наука, 1977, 116 с.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Голенкин М.И. Победители в борьбе за суинстаолание. — М., 1917, 101 с. (3-е изд. — М.: Учпедгиз, 1959, 132 с.). Головкин Б.Н. Переселение травянис- тых многолетников на полярный север. — Л.: Наука, 1973, 266 с. Голубев В.Н. О морфогенезе деревяни- стых растений и путях морфологи- ческой эволюции от деревьев к тра- вам // Бюлл. МОИП, отд. биол., 1959а, вып. 5, с. 49-60. Голубев В.H. О некоторых особеннос- тях морфогенеза жизненных форм травянистых растений лесолуговой зоны в связи с их эволюцией // Ботан. журн., 1959б, т. 44, М 12, с. 1704-1716. Голубев В.Н. К вопросу о классифика- ции жизненных форм // Тр. Цент- рально-Черноземного гос. заповед. — Курск, 1960, вып. 6, с.117-156. Голубев В.Н. Основы биоморфологии травянистых растений центральной лесостепи. Ч. 1. Биоморфология под- земных органов // Тр. Центрально- Черноземного гос. заповед. — Воро- неж, 1962, вып. 7, 511 с. Голубев В.K. Эколого-биологические особенности травянистых растений и растительных сообществ лесосте- пи. — М.: Наука, 1965, 287 с. Голубев В.H. Об изучении жизненных форм растений для целей фитоце- нологии // Ботан. журн., 1968, т. 53 Мо 8, с. 1085-1094. Голубев В.Н. Принцип построения и содержание линейной системы жизненных форм покрытосемен- ных растений // Бюлл. МОИП, отд. биол., 1972, т. 77, вып. 6, с. 72-80. Голубев В.Н. Морфогенетический ана- лиз структуры поликарпической системы побегов покрытосеменных в эволюционном ряду жизненных форм от деревьев к травам // Бюлл. МОИП, отд. биол., 1973, т. 78, вып. 5, с. 90-106. %ифвв В.Н. 06 аналитико-синмтическом моделировании жизненных форм антофитов // Успехи совр. биол. 1977, т. 84, вып. 2 (5), с. 281-293. Гордеева Т.К К биологии полыни чер- ной Artemisia pauciflora Web. // Тр. Бот. ин-та АН СССР. — М. Л.: изд-во АН СССР, 1957. Сер. 3, вып. 11, с. 88-117. Гордиенко И.И. К вопросу о естествен- ном зарастании и закреплении го- лых песков // Ботан. журн., 1964, т. 49, М 12, с. 1750-1759. Гордиенко И.И. Олешские пески и биогеоценотические связи в про- цессе их зарастания. — Киев: Нау- кова думка, 1969, 242 с. Горшкова А.А. Биология степных пас- тбищных растений Забайкалья.— М.: Наука, 1966, 274 с. Горшкова С.Г. Род Inula L. // Флора СССР. — М.; Л.: изд-во АН СССР, 1959, т. 25, с. 433-477. Горышина Т.К. Экология растений. Учебное пособие. — М.: Высшая школа, 1979, 368 с. Горянинов Н. Основания ботаники.— СПб., 1841, 373 с. Готт В.С., Урсул А.Д. Общенаучные понятия и их роль в познании.— М.: Знание, 1975, 64 с. Григорьев Ю.С. Сравнительно-эколо- гическое исследование ксерофили- зации высших растений. — М.; Л.: изд-во АН СССР, 1955, 159 с. Гризебах А. Растительность земного шара. — СПб., т. 1-2: 1874 — т. 1, 575 с.; 1877 — т. 2, 593 с. Гриценко И.И. О повторяемости в про- цессе развития // Вопросы филосо- фии, 1958, М 8, с. 3-13. Гробстайн Ка Стратегия жизни. — М.: Мир, 1968, 144 с. Грудзинская И.А. О периодичности развития (п обе гообразование у дуба) // Итоги и перспективы раз- вития растений. — Л.; М.: изд-во АН СССР, 1959, с. 82-102. Грудзинская И.А. Летнее побегообра- зование у древесных растений и его классификация // Ботан. журн., 1960, т. 45, Ne 7, с. 968-978. Грудзинская И.А. Зависимость строе- ния годичного кольца древесины от развития побегов у дуба // Физио- логия древесных растений. — М.: изд-во АН СССР, 1961, с. 246-261. Грудзинская И.А. Особенности моно- подиального и симподиального
Литература 587 ветвления у древесных пород (на примере дуба и вяза) // Проблемы ботаники, 1962, т. 6, с. 219-231. Грудзинская И.А. Некоторые итоги изучения онтогенеза побегов дуба (Quercus гцЬга L.) // Ботан. журн., 1964, т. 49, М 3, с. 321-337. Грудзинская И.А. Изменение содержа- ния крахмала за период вегетации в годичных побегах некоторых дре- весных пород // Проблемы совре- менной ботаники. — М.; Л.: Наука, 1965, т. 2, с. 163-168. Грудзинская И.А. Об основных типах ветвления (критические заметки) // Ботан. журн., 1974, т. 59, М 4, с. 572-577. Грязнов Б.С., Дынин Б.С., Никитин Е.П. Теория и ее объект. — М.: Наука, 1973, 248 с. Губанов И.А., Работнов Т.А., Тихоми- ров В.Н. Подготовка "Биологичес- кой флоры Московской области" // Ботан. журн., 1970, т. 55, N» 10, с. 1548-1553. Гуленкова М.А. Побегообразование и формирование жизненной формы у лядвенца рогатого в предгенератив- ный период // Возрастной состав популяций цветковых растений в связи с их онтогенезом / Сб. тру- дов. — М.: МГПИ им. В.И. Ленина, 1974, с. 119-131. Гулисашвили В.3. Стадийность в раз- витии растений в связи с их проис- хождением. — Тбилиси: Мецниере- ба, 1969, 96 с. Гуляев Г.В., Мальченко В.В. Словарь терминов по генетике, цитологии, селекции, семеноводству и семено- ведению. — М.: Россельхозиздат, 1975, 215 с. Гумбольдт А. География растений.— М.; Л.: Сельхозгиз, 1936, 228 с. Гукало ЫИ. Современное состояние теории индивидуального развития растений / Пособие для студентов агр. фак. с.-х. вузов. — Житомир: С.-х. ин-т, 1968, 36 с. Гуиало ШИ. Возрастные изменения растений и их значение в растение- водстве. — М.: Наука, 1969, 252 с. Гуиало ШИ., Скриичинский В.В. Физи- ология индивидуального развития растений. — М.: Колос, 1971, 224 с. Гюнтер Э. и др. Основы общей биоло- гии. Пер. с немец. / Э. Гюнтер, Л. Кемпфе, Э. Либберт, Х. Мюллер, Х. Пенцлин. — М.: Мир, 1982, 440 с. Дажо Р. Основы экологии. Перев. с франц. — М.: Прогресс, 1975, 415 с. Двигубский И. Начальные основания естественной истории растений, заключающие терминологию расте- ний, лучшие их системы, анато- мию, физиологию, их патологию и историю ботаники. 2е изд. — М.: Унив. типогр., 1823, Ч. I, 356 с. Дэвени Т., Герей Я. Аминокислоты, пептиды и белки. Пер. с англ.— М.: Мир, 1976, 364 с. Декандоль А. Введение к изучению ботаники или начальный курс этой науки. — М., 1837, т. 1, 541 с. Депенчук Н.П. Проблема взаимосвязи уровней исследования в биологии // Развитие концепции структур- ных уровней в биологии. — М.: Наука, 1972, с. 146-156. Дервиз-Соколова Т.Г. Об определении жизненной формы растений // На- учн. докл. высш. школы. Биол. на- уки, 1981, No 12, с 62 68 Дервиз- Соколова Т Г. Анатомо-морфо- логическое строение Salix polaris Wahlb. и Salix phlebophylla Anderss / / Бюлл. МОИП, отд. биол., 1966, т. 71, вып. 2, с. 28-38. Джефферс Дж. Введение в системный анализ: применение в экологии.— М.: Мир, 1981, 256 с. Джон Б., Дебишир Э., Янг Г., Фейрб- ридж Р., Эндрюс Дж. Зимы нашей планеты. Перев. с англ. — М.: Мир, 1982, 336 с. Джуа М. История химии. Перев. с итал. — М.: Мир, 1966, 452 с. Джумаев М.А. и др. Сравнительная характеристика подземных органов заманихи по содержанию экстрак- тивных веществ и эфирного масла // Успехи изучения лекарственных растений Сибири / М.А. Джумаев, О.В. Журба, О.В. Елисеева, В.П.
588 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНКИ Вантенко. — Томск: изд-во Томс- кого ун-та, 1973, с.44. Дмитриев А.М. Луговодство с основа- ми луговедения. 2-е изд. — М.: Сельхозгиз, 1948, 408 с. Добрынин Г.М. Рост и формирование хлебных и кормовых злаков. — Л.: Колос, 1969, 276 с. Догель В.А. Олигомеризация гомоло- гичных органов как один из глав- ных путей эволюции животных.— Л.: изд-во Ленингр. ун-та, 1954, 368 с. Докучаев В.В. К учению о зонах при- роды и классификация почв. Сочи- нения. Т. 6. — М. Л.: изд-во АН СССР, 1951, 595 с. Домбровский Б.А. К сравнительной характеристике познавательных направлений в современной биоло- гии // Некоторые вопросы теорети- ческой и прикладной биологии.— А-А.: Наука, 1967, с. 3-56. Донскова А.А. Жизненный цикл кле- вера сходного (Trifolium ambiguum М.В.) в условиях высокогорий Кав- каза // Бюлл. МОИП, отд. биол., 1968, т. 73, вып. 4, с. 47-62. Дорохина Л.Н. Жизненные формы и эволюционные отношения в подро- де Ргасипси1из рсоа Artemisia // Бюлл. МОИП, отд. биол., 1968, т. 73, № 6, с. 83-89. Драгавцев В.А. О количественных кри- териях целостности в биологии // Проблемы целостности в современ- ной биологии. — М.: Наука, 1968, с. 128-139. Дружинин В.В., Конторов Д.С. Про- блемы системологии. Проблемы теории сложных систем. — М.: Сов. радио, 1976, 295 с. Дубинин Н.П. Некоторые методологи- ческие проблемы генетики. — М.: Знание, 1968, 62 с. Дубинин Н.П. Генетика в свете диа- лектико-материалистического уче- ния. — М.: Знание, 1975, 64 с. Дубинин Н.П. Общая генетика. 2-е изд. — М.: Наука, 1976, 590 с. Дубинин Н.П. Некоторые мировоз- зренческие проблемы современной биологии // Философия и мировоз- зренческие проблемы современной науки. — М.: Наука, 1981, с. 96-114. Дубянский В.А. Песчаная пустыня Юго-Восточные Каракумы, ее есте- ственные районы, возможности их сельскохозяйственного использова- ния и значение для ирригации // Тр. по прикладной бот, генет. и се- лек. — Л., 1928, т. 19, вып. 4, с. 1-286. Енин II.К Валериана лекарственная // Атлас лекарственных растений СССР. — М.: Медгиз, 1962, с. 98-100. Енин ШК. и др. Валериана лекарствен- ная / П.К. Енин, П.М. Лошкарев, А.Ф. Сацыперов, П.М. Чукичев.— М.: Медгиз, 1953, ill c. Ермаков Б.С. Лесные растения в ва- шем саду. — М.: Леся. промышлен- ность, 1987, 150 с. Ефейкин А.К Природа образователь- ной ткани растений и вопрос о ста- рении и вырождении клонов // Онтогенез высших растений / Со- общения Ереванского симпозиума.— Ереван: изд-во АН Арм. ССР, 1970, с. 225-231. Жданов В.М. К определению понятия "жизнь" // Вест. АМН, 1964, М 1, с. 26-33. Жмылев ШЮ., Алексеев Ю.Е., Карпу- хина Е.А. Основные термины и по- нятия биоморфологии растений.— М.: изд-во М~, 1993, 147 с. Жудова II.II. Растительность и флора Судзухинского государственного заповедника Приморского края.— Владивосток: Дальневост. книжн. изд-во, 1967. Жукова Л.А. Онтогенез и циклы вос- произведения растений // Журн. общ. биологии, 1983, т. 44, № 3, с. 361-374. Жукова Л.А. Поливариантность онто- генеза луговых растений // Жиз- ненные формы в экологии и систе- матике растений / Межвуз. сб. науч. трудов — М., 1986, с. 104-114. Жуковский ШМ. Ботаника. Учебник для гос. ун-тов и с.-х. вузов. 3-е изд. — М.: Советская наука, 1949, 552 с. Жуковский ШМ. Культурные растения и их сородичи. 2-е изд. — М.: Ко- лос, 1964, 792 с.
Литература Журба О.В. К вопросу о номенклатуре и природных ресурсах заманихи высокой на советском Дальнем Востоке // Биологически активные вещества флоры и фауны Дальнего Востока и Тихого океана. — Влади- восток, 1971, 33 с. Журба О.В., Шретер А.И. Заманиха высокая // Атлас ареалов и ресур- сов лекарственных растений СССР.— М.: изд-во ГУГК, 1976, 166 с. Забинкова Н.Н., Кирпичников М.Э. Латинско-русский словарь для бо- таников. / Справочное пособие по систематике высших растений. Вып. II. — М.; Л.: изд-во АН СССР, 1957, 335 с. Завадский КМ. О понимании прогрес- са в органической природе // Про- блемы развития в природе и обще- стве. — Л., 1958, с. 79-120. Завадский КМ. Учение о виде. — Л.: изд-во Ленингр. ун-та, 1961, 254 с. Завадский К.М. Основные формы организации живого и их подразде- ления // Философские проблемы современной биологии. — М.; Л.: Наука, 1966, с. 29-47. Завадский КМ. Вид и видообразова- ние. — Л.: Наука, 1968, 404 с. Заде Л., Яезоер V. Теория линейных систем. — М.: Прогресс, 1970, 398 с. Зак С.Е. Принципы и основые законы материалистической диалектики.— М.: Высшая школа, 1974, 176 с. Закржевский Б.С., Коровин Е.П. Эко- логические особенности главней- ших растений Бетпак-Дала // Тр. Среднеазиат. гос. унив. — Ташкент, 1935, сер. 8-б, вып. 23, с. 3-74. Заренков Н.А. Теоретическая биоло- гия. — М.: изд-во Моск. ун-та, 1988, 216 с. Заугольнова Л.Б. Возрастные этапы в онтогенезе ясеня обыкновенного (Fraxinus excelsior L.) // Вопросы морфогенеза цветковых растений и строения их популяций. — М.: Наука, 1968, с. 81-102. Заугольнова Л.Б. Современные тен- денции в развитии популяционной экологии растений // Экология популяций: структура и динамика / Материалы совещания. — М., 1995, ч. 1, с. 14-27. Заугольнова Л.Б., Жукова Л.А., Кома- ров А.С., Смирнова О.В. Ценопопу- ляции растений (очерки популяци- онной биологии). — М.: Наука, 1988, 184 с. Заугольнова Л.Б., Сугоркина Н. С., Щербакова Е.Г. Жизненные формы и популяционное поведение мно- голетних травянистых растений // Экология популяций. — М., Наука, 1991, с. 5-22. Заугольнова Л.Б., Смирнова О.В., Кома- ров А.С., Ханина Л.Г. Мониторинг фитопопуляций // Успехи совр. био- логии, 1993, т. 113, вып. 4, с. 410-414. Землинский С.Е. Лекарственные рас- тения СССР. 3-е изд. — М.: Мед- гиз, 1958, 610 с. Зиман С.Н. Жизненные формы и био- логия степных растений Донбасса.— Киев: Наукова думка, 1976, 192 с. Зозулин Г.М. Подземные части основ- ных видов травянистых растений и ассоциаций плакоров средне-рус- ской лесостепи в связи с вопросами формирования растительного по- крова // Тр. Центр.-Черноз. гос. заповедника. — Курск: Курское книж. изд-во, 1959, вып. 5, с. 3-314. Зозулин Г.М. Система жизненных форм высших растений // Ботан. ясурн. — 1961, т. 46, М 1, с. 3-20. Зозулин Г.М. Схема основных направ- лений и путей эволюции жизнен- ных форм семенных растений // Ботан. журн., 1968, т. 53, N 2, с. 223-233. Зозулин Г.М. Аспекты учения о жиз- ненных формах растений в био- сферном плане // Проблемы эко- логической морфологии растений.— М.: Наука, 1976, с. 45-54. Зотова М.И., Крылов Г.В., Саратиков А.С. Золотой корень — новое сти- мулирующее и адаптогенное сред- ство // Изв. СО АН СССР, 1965, М 8, сер. биол.-мед. наук, вып. 2, с. 111-119. Зубков И.Ф. Проблемы геологической формы движения материи. — М.: Наука, 1979, 240 с.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Ивашевский Л.И. Философские вопро- сы геологии. — Новосибирск: На- ука, 1979, 206 с. Игнатов А.И. Жизнь как система форм движения материи. — М.: Знание, 1966, 32 с. Игнатьева И П Некоторые особеннос- ти развития люлина многолетнего // Изв. ТСХА, 1958, вып. 2, с. 39-54. Игнатьева И.П. Морфогенез водосбо- ра обыкновенного // Ботан. журн., 1964, т. 49, М 3, с. 358-371. Игнатьева И. IT. О жизненном цикле стержнекорневых и кистекорневых травянистых поликарпиков // Ботан. журн., 1965, т. 50, М 7, с. 903-916. Израильсон В.Ф. Сравнительно-анато- мические исследования семян не- которых представителей семейств зонтичных // Новые полезные рас- тения Сибири. — Новосибирск: изд-во СО АН СССР, 1965, вып. 8, с. 42-47. Израильсон В.Ф. К изучению морфо- логических особенностей володуш- ки золотистой в связи с условиями обитания // Полезные растения природной флоры Сибири. — Но- восибирск: Наука, 1967, с. 100-108. Израильсон В.Ф. Эколого-морфологи- ческие особенности некоторых си- бирских видов володушки (Вир1е- urum L.) // Эколого-морфологичес- кие и биохимические особенности полезных растений дикорастущей флоры Сибири. — Новосибирск: Наука, 1970, с. 101-115. Израильсон В.Ф. Анатомо-морфологи- ческие изменения в семенах видов рода Bupleurum L. При стратифи- кации // Комплексное изучение полезных растений Сибири. — Но- восибирск: Наука, 1974, с. 16-24. Илиева С. Лекарственные культуры.— София: Земиздат, 1971, 261 с. Имс А.Д. Морфология цветковых рас- тений. — М.: Мир, 1964, 497 с. Имс А.Дж., Мак-Даниельс Л.Г. Введе- ние в анатомию растений. — М.; Л.: Гос. изд-во совх. и колх. лит., 1935, 332 с. Иоганзен Б.Г. Основы экологии.— Томск: Изд-во Томск ун-та, 1959, 390 с. Иосебидзе Н.И. Атлас-определитель всходов лекарственных растений.— Тбилиси: Мецниереба, 1981, 237 с. Исиолатов Е.Н. Кустарная пихта Удор- ского края // Изв. Глав. бот. сада СССР, 1932, т. 30, вып. 5-6, с. 703-707. Истомина И.И. Эколого-демографи- ческая характеристика синузии ку- старников // Восточноевропейские широколиственные леса. — М.: Наука, 1994, с. 144-163. Кавтарадзе Д.Н., Остроумов С.А. Ре- цензия на книгу "World conservation strategy" (Всемирная стратегия ох- раны природы). Switzerland, 1980 // Экология, 1981, М 4, с. 107-109. Каганова 3.В. Концепция структурных уровней и принципы интегратизма в современной биологии // Разви- тие концепции структурных уров- ней. — М.: Наука, 1972, с. 112-121. Каден Н.Н. Соплодия и соцветия // Вест. Моск ун-та. — 1951, М 6, с. 89-99. Казакевич Л.И. Материалы к биоло- гии растений Юго-Востока России // Бюлл. отд. прикл. ботаники Сара- товск. обл. с.-х. опыт. станции.— 1922, М 18, с. 3-21. Казарян В.О. Старение высших расте- ний. — М.: Наука, 1969, 314 с. Казарян В.О., Балагезян Н.В. Патику- ляция — основная причина старе- ния и отмирания многолетних стер- жнекорневых трав // Изв. АН Арм. ССР. Биол. науки, 1960, т. 13, М 9, с. 17-27. Казначеев В.1Т. Современные аспекты адаптации. — Новосибирск: Наука, 1980, 192 с. Калман Р., Фалб II., Арбиб М. Матема- тическая теория систем. — М.: Мир, 1971, 406 с. Кальвин М. Химическая эволюция.— М.: Мир, 1971, 240 с. Канаев И.И. Очерки из истории про- блемы морфологического типа от Дарвина до наших дней. — М.: Наука, 1966, 210 с. Капица 17Л. Теория, эксперимент, практика. — М.: Знание, 1966, 48 с. Кариович В.Н. Проблема, гипотеза, закон. — Новосибирск: Наука, 1980, 176 с.
Литература Кашменская О.В. Теория систем и гео- морфология. — Новосибирск: На- ука, 1980, 120 с. Кашкаров Д.Н. Среда и сообщество. (Основы синэкологии). 2-е изд.— М.: Медгиз, 1933, 242 с. Кашкаров Д.Н. Основы экологии жи- вотных. — М.; Л.: Медгиз, 1938, 602 с.; 2-е изд. — Л.: Учпедгиз, 1944, 383 с. Квейд Э. Анализ сложных систем.— М.: Советское радио, 1969, 231 с. Кедров Б.M. О соотношении форм движения материи в природе // Философские проблемы современ- ного естествознания. — М.: изд-во АН СССР, 1959, с. 137-211. Кедров Б.М. О повторяемости в про- цессе развития. — М.: Госполитиз- дат, 1961, 148 с. Кедров Б.M. О геологической форме движения в связи с другими его формами // Взаимодействие наук при изучении Земли. — М.: Наука, 1964, с. 129-151. Кедров Б.М. Предмет и взаимосвязь естественных наук. 2-е изд. — М.: Наука, 1967, 425 с. Кедров Б.М., Серебровская КБ. Про- блема происхождения и сущности жизни и ее философский аспект // Журн. Всес. хим. об-ва им. Д.И. Менделеева, 1980, т. 25, вып. 3, с. 252-266. Келлер Б.А. Об изучении жизненных форм при геоботанических иссле- дованиях // Сов. ботан., 1933, М 2, с. 39-44. Келлер Б.А. Основы эволюции расте- ний. Руководжцие теоретические по- ложения, наблюдения и опыты.— М.; Л.: изд-во АН СССР, 1948, 208 с. Кеньон Д., Стейнман Г. Биохимичес- кое предопределение. — М.: Мир, 1972, 336 с. Ким Е.Ф., Днепровский Ю.М. Морфо- биологические особенности семян золотого корня // Успехи изучения лекарственных растений Сибири.— Томск, 1973, с. 52-55. Кирпичников М.Э., Забинкова Н.K. Русско-латинский словарь для бо- таников. — Л.: Наука, 1977, 855 с. Клименко С.В. О силлептических и пролептических побегах у айвы обыкновенной (Cydonia oblonga Mill.) // Украин. ботан. журн., 1969, т. 26, М 5, с. 128-130. Кнорре А.Г. Уровни органической ин- дивидуальности в связи с эволюци- ей целостности // Проблема целост- ности в современной биологии.— М.: Наука, 1968, с. 140-171. Коган А.Б., Наумов КИ., Режабек В.Г., Чораян О.Г. Биологическая кибер- нетика. Учебное пособие для уни- веситетов. — М.: Высшая школа, 1972, 384 с. Кожевников А.В. Материалы по эко- логии буковых лесов западного За- кавказья // Сов. ботан., 1935, М 5, с. 98-119. Козлова K.А. Анатомо-экологическая характеристика полукустарничков Восточного Крыма // Ботан. журн., 1953, т. 38, N 4, с. 513-529. Козо-Полянский Б.М. Исчисление рус- ских видов Bupleurum L. // Труды импер. Бот. сада. — Петроград, 1915, т. 30, вып. 2, с. 135-333. Козо-Полянский Б.М. Основной биоге- нетический закон с б<ианической точ- ки зрения. — Воронеж, 1937, 254 с. Комаров В.Н. Философские вопросы науки о Земле. — Казань: изд-во Казан. ун-та, 1974, 173 с. Кондаков Н.И. Логический словарь- справочник. 2-е изд. — М.: Наука, 1975, 720 с. Коновалов М.Н., Куваев В.Б., Трутнева Е.А. Новое в медицинском приме- нении пиона уклоняющегося // Медицин. пр-сть СССР. — 1962, М 5, с. 57-59. Константинов Н.Н. Морфолого-физи- ологические основы онтогенеза и филогенеза хлопчатника. — М.: Наука, 1967, 292 с. Копанева Г.А., Шретер А.И. Душица обыкновенная (материнка) — ori- ganum vulgare L. // Атлас ареалов и ресурсов лекарственных растений СССР. — М.: изд-во ГУГК, 1976, с. 234. Коровин Е.П. Растительность Средней Азии и южного Казахстана. — М.;
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РЛ СТЕНИЙ Ташкент: Саогиз., 1934, 480с. 1962— изд. 2-е, кн. 2, Ташкент. Корольков А.А., Летленко В.1Т. Фило- софские проблемы теории нормы в биологии и медицине. — М.: Меди- цина, 1977, 393 с. Корчагин А.А. Методы учета семено- шения древесных и лесных сооб- ществ // Полевая геоботаника.— М.; Л.: изд-во АН СССР, 1960а, т. 2, с. 4-132. Корчагин А.А. Методы учета семено- шения кустарников // Полевая гео- ботаника. — Л.: изд-во АН СССР, 1960б, т. 2, с. 133-142. Корчагин А.А. Внутривидовой (попу- ляционный) состав растительных сообществ и методы его изучения // Полевая геоботаника. — Л.: Наука, 1964, т. 3, с. 61-131. Корчагин А.А. Значение внутривидово- го (популяционного) состава расти- тельных сообществ при стационар- ных геоботанических исследовани- ях // Ботан. журн., 1966, т. 51, М 6, с. 777-783. Корчагин А.А. Строение растительных сообществ // Полевая геоботаника. Т. 5. — Л.: Наука, 1976, с. 7-313. Костюк Н.Т. О сущности жизни. (Опыт философского обобщения некоторых аспектов ее познания). — Киев: изд- во Киевск. ун-та, 1967, 250 с. Котт С.А. Сорные растения и меры борьбы с ними. — М.: Сельхозгиз, 1955, 384 с. Красилов В.А. Происхождение и ран- няя эволюция цветковых растений.— М.: Наука, 1989, 264 с. Красильников ШК Подземные органы дягиля низбегающего // Растения семейства Зонтичных — источники биологически активных веществ / Тр. Бот. инст. им. В.Л. Комарова АН СССР, сер. V, растит. сырье, вып. 15. — Л.: Наука, 1968, с. 36-105. Краткий словарь по философии. 3-е изд. — М.: Политиздат, 1979, 413 с. Крейер Г.К. Валериана лекарственная // Главнейшие возделываемые в СССР лекарственные растения / ВАСХНИЛ. — Л., 1937, т. 1, с. 5-38. Кремянский В.И. Соотношение мето- дов различных наук в исследовани- ях организма как целого // Пробле- ма целостности в современной био- логии. — М.: Наука, 1968, с. 101- 127. Кремянский В.И. Структурные уровни живой материи. Теоретические и методологические проблемы. — М.: Наука, 1969, 295 с. Кренке K.IT. Соматические показатели и факторы формообразования // Феногенетическая изменчивость. Т. 1. — М., 1933, с. 11-415. Кренке К.П. Теория циклического ста- рения и омоложения растений.— М.: Сельхозгиз, 1940, 136 с. Кривокооытова Р.В. Философский ка- мень ХХ века. — М.: Наука, 1969, 144 с. Криволуцкий Д.А. Понятие "жизнен- ная форма" в экологии животных // Журн. общ. биологии, 1967, т. 28, No 2, с. 153-162. Криволуцкий Д.А. Современные пред- ставления о жизненных формах животных // Экология, 1971, М 3, с. 19-25. Круть И.В. Введение в общую теорию Земли. — М.: Мысль, 1978, 367 с. Крылов Г.В. Травы жизни и их искате- ли. — Новосибирск: Зап. Сиб. кн. изд., 1969, 264 с. Куваев В.Б. Пион уклоняющийся // Атлас ареалов и ресурсов дикорас- тущих лекарственных растений СССР. — М.: Медицина, 1976, с. 281. Куваев В.Б., Глызин В.И., Глызина Г.С., Баньковский А.И. Перспективы по- иска мангиферина в отечественной флоре // Растит. ресурсы, 1972, т. 8, вып. 3, с. 367-371. Кудряшова О.И. О партикуляции у травянистых многолетников // Фи- зиология растений. — 1958, т. 5, вып. 1, с. 78-80. Кузьмин В.1Т. Принцип системности в теории и методологии К. Маркса. — М.: Политиздат, 1976, 261 с. Кузьмина Л.В. Некоторые биологичес- кие особенности Prangos pabularia Lindl. // Растения — источники
Литература 593 биологически активных веществ лечебного действия. — М.; Л.: На- ука, 1965, с. 28-41. Кузнецов В.И., Идлис Г.М., Гутина В.Н. Еаеесевеннвнне. — М.: Аеер, 1996, 384 с. Кузнецов В.М Биоморфология корня горца забайкальского (Polygonum divaricatum L.) и методика его изучения // Тр. Главн. Бот. сада.— М.: изд-во АН СССР, 1951, т. 2, с. 264-279. Кузнецова Г.В. Особенности строения и динамика развития почек возоб- новления копеечника забытого (Hedysarum neglectum Ledeb.) в свя- зи с вертикальной зональностью// Перспективные полезные растения флоры Сибири. — Новосибирск: Наука, 1973, с. 48-64. Кузнецова Г.В., Пленник P.Я. Ритм раз- вития и продуктивность видов рода копеечник (Hedysarum L.) в юго- восточном Алтае // Ритм развития и продуктивность полезных расте- ний сибирской флоры. — Новоси- бирск: Наука, 1975, с. 3-19. Кузнецова Т.В. Методы исследования соцветий. Описательный метод и концепция синфлоресценции Виль- гельма Тролля // Бюлл. МОИП, отд. биол. — 1985, т. 90, вып. 3, с. 62-73. Культиасов М.В. Тау-сагыз и экологи- ческие основы введения его в куль- туру. — М.; Л.: изд-во АН СССР, 1938, 315 с. Культиасов М.В. Проблема становле- ния жизненных форм у растений // Проблемы ботаники. — М.; Л.: изд- во АН СССР, 1950, т. 1, с. 250-263. Культиасов М.В. Экогенетический анализ многолетних люцерн // Люцерна тяньшанская и опыт ее интродукции. — М.: Наука, 1967, с. 7-139. Куминова А.В. Растительный покров Алтая. — Новосибирск: изд-во АН СССР, 1960, 450 с. Кун Т. Структура научных революций. 2-е изд. — М.: Прогресс, 1977, 300 с. Куперман Ф.М. Морфофизиология ра- стений. 3-е изд. — М.: Высшая школа, 1977, 288 с. Куренцов А.И. В убежищах уссурийс- ких реликтов. — Владивосток: При- морск. книж. изд., 1961, 182 с. Куренцова Г.Э. Распределение лекар- ственных растений по высотным поясам в Приморье и прилегающих частях Приамурья // Матер. 1-й науч.-практич. конферен. по про- блемам мед. географии Дальнего Востока. — 1968, т. 27, с. 149. Куренцова Г.Э. Естественные и антро- погенные смены растительности Приморья и южного Приамурья.— Новосибирск: Наука, 1973, 230 с. Куркин К.А., Матвеев А.P. Ценопопу- ляции как системы особей и как элементы фитоценозов (системно- иерархический подход) // Бюлл. МОИП, отд. биол., 1981, т. 81, вып. 4, с. 54-74. Кучеров Е.В., Кузнецова НА., Лазарева Д.Н. Сравнительное изучение био- логической активности горицвета весеннего и горицвета сибирского из разных пунктов Башкирии // Растит. ресурсы, 1969, т. 5, вып. 1, с. 54-62. Кушке Э.Э., Иващенко А.А. Валериана лекарственная // Лекарственные растения СССР. — М., 1967, с. 65-74. Лавренко Е.М. Некоторые наблюдения над корневой системой, экологией и хозяйственным значением псам- мофитов Нижнеднепровских пес- ков // Проблемы растениевод. ос- воения пустынь. — 1935, вып. 3, с. 75-94. Лавренко Е.М. Об уровнях изучения органического мира в связи с по- знанием растительного покрова // Изв. АН СССР, сер. биол., 1964, вып. 1, с. 32-46. Лавренко Е.М., Свешникова В.М. Об основных направлениях изучения экобиоморф в растительном покро- ве // Основные проблемы совр. геоботаники. — Л.: Наука, 1968, с. 10-15. Лебедев II.Â. Проявление морфогенеза луговых злаков // Морфогенез лу- говых злаков и условия внешней среды / Уч. зап. Уральского ун-та. 1968, М 73, сер. биол., вып. 4, с. 19-154.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Левин Г.Г. Чередование поколений, цикл развития и онтогенез расте- ний // Ботан. журн., 1959, т. 44, М 7, с. 943-953. Левин Г.Г. Проблема индивидуальнос- ти у растений // Ботан. журн., 1961, т. 46, М 3, с. 432-447. Левин Г.Г. Жизненные циклы расте- ний, их связи и эволюция // Бетан. журн., 1963, т. 48, М 7, с. 1039-1059. Левина P.Е. К методике изучения рас- пространения семян и плодов // Полевая геоботаника. — М.; Л.: изд- во АН СССР, 1960, т. 2, с. 143-159. Левина P.Е. Репродуктивная биология семенных растений (обзор пробле- мы). — М.: Наука, 1981, 96 с. Левонтин P. Генетические основы эволюции. — М.: Мир, 1978, 351 с. Левушкин С.И. Об экологических и фаунистических комплексах (на при- мере подземных фаун) // Журн. общ. биол., 1975, т. 36, М 6, с. 814-828. Леонович В.В. К вопросу об уровнях организованности материи и про- блема сознания // Проблемы диа- лектики. — Л.: изд-во Ленинг. ун- та, 1973, вып. 2, с. 82-92. Ленинджер А. Биохимия. — М.: Мир, 1976, 957 с. Леопольд А. Рост и развитие растений. Пер. с англ. — М.: Мир, 1968, 494 с. Либберт Э. Физиология растений. Пер. с немец. — М.: Мир, 1976, 580 с. Лиепа ИЯ. Моделирование — основ- ной инструмент научного познания // Моделирование и прогнозирование в ботанике. / Уч. записки Латв. Гос. ун-та, 1975, т. 239, вып. 3, с. 3-21. Линней E. Философия ботаники. М.: Наука, 1989, 456 с. Линчевский И.А. Род Bupleurum L. Флора СССР. — М.; Л.: изд-во АН СССР, 1950, т. 16, с. 275-349. Лотова Л.И. О классификации мери- стем // Вест. Моск. ун-та, сер. биол., 1977, М 2, с. 64-75. Лотте Д.С. Вопросы заимствования и упорядочения иноязычных терми- нов и терминоэлементов. — М.: Наука, 1982, 149 с. Любарский Е.Л. Об эволюции вегета- тивного возобновления и размно- жения у TpBBHHHGTblx поликарпиков // Ботан. журн., 1961, т. 46, М 7, с. 959-968. Любарский Е.Л. Экология вегетатив- ного размножения высших расте- ний. — Казань: изд-во Казанского ун-та, 1967, 181 с. Любименко В.Н. О спящих почках // Тр. СПб. об-ва естествоиспыт., отд. ботаники. — 1900, т. 30, вып. 3, с. 195-260. Ляиунов А.А. В чем состоит системный подход к изучению реальных объек- тов сложной природы // Систем- ные исследования. Ежегодник, 1972. — М.: Наука, 1972, с. 5-18. Ляпунов А.А. Проблемы теоретической и прикладной кибернетики. — М.: Наука, 1980, 336 с. Ляэнелайд А.И. Новые данные о кар- ликовой болотной форме сосны обыкновенной (Pinus sylvestris 1.. f. pumila АЬо1.) // Флора и раститель- ность охраняемых территорий / Ученые записки Тартус. ун-та.— 1984, вып. 662. Труды по ботанике, с. 9-18. Мазинг В.О. О понятии "жизненная форма" в экологии высших расте- ний // Ученые записки Тартус. ун- та. — 1958, вып. 64. Труды по бота- нике, 1, с. 141-153. Мазуренко М.Т., Хохряков А.1Т. Струк- тура и морфогенез кустарников.— М.: Наука, 1977, 160 с. Майр Э. Систематика и происхожде- ние видов. — М.: изд-во иност. лит., 1947, 504 с. Майр Э. Популяции, виды и эволю- ция. — М.: Мир, 1974, 464 с. Майр Э. Эволюция // Эволюция.— М.: Мир, 1981, с. 11-31. Максимович М. Основания ботаники. Ч. 1. — М., 1828, 142 с. Максимович-Амбодик Н. Словарь бо- танический, содержащий наимено- вания растений и их частей. СПб., 1795, 157 с. Малеев В.1Т. Можжевеловый лес на мысе Мартьян в Южном Крыму // Ботан. журн. СССР, 1933, т. 18, N 6. Малиновский А.А. Организация // Фи- лософская энциклопедия. Т. 4, 1967.
Литература Малиновский А.А. Некоторые вопросы организации биологических систем // Организация и управление. — М., 1968, с. 105-138. Малиновский А.А. Основные понятия и определения теории систем (в связи с приложением теории сис- тем к биологии) // Системные ис- следования. Ежегодник,1979. — М.: Наука, 1980, с. 78-90. Малиновсысий 0.0., Уемов А.И. Типы систем i основн1 61олог1чн1 законо- м1рности // Орган1зм як система.— Ки)в: Наук. думка, 1966. Малышев Л.И. Определитель высокогор- ных растений Южной Сибири.— Л.: Наука, 1968, 284 с. Мамаев С.А. Формы внутривидовой изменчивости древесных растений ( на примере семейства Pinaceae на Урале). — М.: Наука, 1972, 284 с. Мамзин А.С. К вопросу об определе- нии понятия "жизнь" // Философ- ские проблемы современной био- логии. — М.; Л.: Наука, 1966, с. 99- 118. Мамзин А.С.Очерки по методологии эволюционной теории. (Анализ не- которых проблем). — Л.: Наука, 1974, 136 с. Мамзин А.С. Единство организации и развития живых систем и взаимо- действие методов биологии // Вза- имодействие методов естественных наук в познании жизни. — М.: Наука, 1978, с. 236-246. Мамчур Е.А. Проблема выбора теории. К анализу переходных ситуаций в развитии физического знания. М.: Наука, 1975, 231 с. Мандрик В.Ю, Голышкин Л.В. Эмбри- ологические исследования некото- рых видов семейства Crassulaceae // Ботан. журн., 1973, т. 58, М 2, с. 263-272. Марков М.В. О направлении, развива- емом Джоном Л. Харпером в попу- ляционной экологии растений // Ботан. журн., 1976, т. 61, М 7, с. 999-1004. Марков М.В. Популяционная биоло- гия растений. — Казань: изд-во KKV, 1986, 109 с. Марков М.В. Популяционная биоло- гия розеточных и полурозеточных малолетних растений. — Казань: изд-во КТУ, 1990, 186 с. Маркова Л.1T., Медведева Л.И. Мате- риалы о распространении, внутри- видовой изменчивости и хозяй- ственной ценности некоторых ви- дов ферул из подрода Peucedanoides (Boiss.) Когоч. // Растения — ис- точники биологически активных веществ лечебного действия / Тр. Бот. ин-та им. В.Л. Комарова АН СССР, сер. V. Растит. сырье, вып. 13. — М.; Л.: Наука, 1965, с. 42-99. Маркова Л.1T., Медведева Л.И. Мате- риалы к изучению подземных орга- нов некоторых видов ферул из под- рода Peucedanoides (Boiss.) Когоч. // Растения семейства зонтичных- источники биологически активных веществ / Тр. Бот. ин-та им. В.Л. Комарова АН СССР, сер. V. Рас- тит. сырье, вып. 15. — Наука, 1968, с. 149-173. Мартынов И. Техно-ботанический словарь на латинском и российс- ком языках. — СПб., 1820, 682 с. Матвеев А.Р. Большой жизненный цикл, численность и возрастной состав популяции тимофеевки лу- говой и тимофеевки степной: Авто- реф. дис.... канд. биол. наук. — М., 1975, 25 с. Машковский МД. Лекарственные сред- ства. 8-е изд. — М.: Медицина, 1977, ч. 1, 623 с.; 1977, ч. 2, 560 с. Медников Б.М. Происхождение и эво- люция нуклеиновых кислот // Журн. Всес. хим. об-ва им. Д.И. Менделеева, 1980, т. 25, вып. 4, с. 425-430. Меерсон Ф.Ç.Адаптация, стресс и про- филактика. — М.: Наука, 1981, 278 с. Мейен С.В. Проблема направленности эволюции // Итоги науки и техни- ки, серия зоология позвоночных.— М.: ВИНИТИ, 1975, т. 7, с. 66-117. Мейен С.В. Таксономия и мерономия // Вопросы методологии в геологи- ческих науках. — Киев: Наукова думка, 1977, с. 25-33.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Мейен С.В. Основные аспекты типо- логии организмов // Журн. общ. биол., 1978, т. 39, М 4, с. 495-508. Мейен С.В. Следы трав индейских.— М.: Мысль,1981, 159 с. Мейен С.В., Шрейдер Ю.А. Методоло- гические аспекты теории классифи- кации // Вопр. философии, 1976, Nî 12 t:. 67 79 Мейер КИ. Размножение растений.— М.: Сельхозгиз, 1937, 284 с. Мерзон Л.С. Проблема научного фак- та. — Л.: Изд-во Ленинг. ун-та, 1972, 188 с. Месарович М., Мако Д., Такахара И. Теория иерархических многоурав- невых систем. — М.: Мир, 1973, 344 с. Миклин А.М., Подольский В.А. Катего- рия развития в марксистской диа- лектике. — М.: Мысль, 1980, 166 с. Минаева В.Г. Лекарственные растения Сибири. 4-е изд. — Новосибирск: Наука, 1970, 272 с. Миркин Б.Н., Розенберг Г.С. Фитоце- нология: принципы и методы.— М.: Наука, 1978, 211 с. Михайлова ТД. Побегообразование и жизненная форма Astragalus macro- podium Lipsky // Науч. докл. высш. школы. Биол. науки, 1972, М 6, с. 49-54. Михайловская И.С. О придаточных корнях на корнях травянистых многолетних бобовых // Вопросы биологии и экологии доминантов и эдификаторов растительных сооб- ществ / Материалы 1 Межвузовс- кой конференции. — Пермь, 1968, с. 112-117. Михаэли М. (общ. ред.). Израиль. Гео- графический справочник. — Иеру- салим; Санкт-Петербург: Гешарим и Роспринт, 1992, 287 с. Мокеева Е.А. К биологии пустынной полыни // Бюлл. Среднеазиат. Гос. унив., 1945, вып. 23, с. 111-113. Молотковский ГХ Целостность расти- тельного организма на различных уровнях развития // Целостность и биология. — Киев: Наукова думка, 1968, с. 191-197. Москалева Г.И. Возрастные изменения подземных органов некоторых степ- ных и предгорных растений Крыма в связи с процессом партикуляции: Автореф. дис.... канд. биол. наук.— Л., 1968, 24 с. Мочалов В.В. Облепиха. — Новоси- бирск: Западно-Сибирское книжн. изд-во, 1973, 68 с. Мукашев 3.А. Суммативность и цело- стность // Проблема целостности в современной биологии. — М.: На- ука, 1968, с. 47-61. Муравьев И.А., Джумаев М.А. Поиск основного биологически активного комплекса корневищ с корнями заманихи // Вопросы фармации на Дальнем Востоке. — Хабаровск, 1973, вып. 1, с. 45. Мухин Л.М. Химическая эволюция // Журн. Всес. хим. об-ва им. Д.И. Менделеева, 1980, т. 25, вып. 4, с. 412-417. Наумов K.1T. Экология животных.— М.: «Сов. наука, 1955, 534 с. Некрасов Б.В. Учебник общей химии.— М.: Химия, 4-е изд., 1981, 559 с. Некрасов В.И. Актуальные вопросы развития теории акклиматизации растений. — М.: Наука, 1980, 103 с. Нечаева Н.Т. Материалы к биологии полыни Artemisia herba alba Asso. // Ботан. журн., 1949, т. 34, Ne 1, с. 95-99. Нечаева Н.Т., Приходько С.Я. Биоло- гия полыни бадхызской и результа- ты введения ее в культуру // Ботан. журн., 1956, т. 41, М 6, с. 836-854. Нечаева Н.Т., Василевская В.К., Анто- нова К.Г. Жизненные формы расте- ний пустыни Каракумы. — М.: Наука, 1973, 244 с. Николаева М.Г., Разумова М.В., Глаз- кова В.Н. Справочник по проращи- ванию покоящихся семян. — Л.: Наука, 1985, 348 с. Новиков Г.Н. О формах размножения пустынных полукустарничков // Сов. ботаника, 1943, М 2, с. 30-37. Новосельцев В.Н. Теория управления и биосистемы. Анализ сохранитель- ных свойств. — М.: Наука, 1978, 320 с. Новосельцева Н.1Т. К вопросу изуче- ния биологии при введении в куль- туру вздутоплодника сибирского //
Литература Селекция и первичное семеновод- ство лекарственных культур / Сб. науч. работ ВИЛР. — М., 1970, вып. 5, с. 214-220. Нухимовская Ю.Д. Онтогенез пихты сибирской (Abies sibirica 1.ейеЬ.) в условиях Подмосковья // Бюлл. МОИП, отд. биол., 1971, т. 76, вып. 2, с. 105-112. Нухимовская Ю.Д. Биология и эколо- гия пихты сибирской в различных условиях произрастания: Автореф. дис.... канд. биол. наук. — М., 1972, 25 с. Нухимовский Е.Л. Строение подзем- ных органов и экологические осо- бенности некоторых растений-гид- роиндикаторов // Вест. Моск. ун- та, 1967, N2, ,с. 44-53. Нухимовский Е.Л. О каудексе растений // Рефераты докл. Всесоюз. меж- вуз. конф. по морфологии расте- ний. — М.: изд-во Моск. ун-та, 1968а, с. 207-209. Нухимовский Е.Л. О термине и поня- тии "каудекс". 1. Обзор значений термина // Вест. Моск. ун-та, сер. биол., почвоведение, 19686, М 5, с. 36-43. Нухимовский Е.Л.О термине и поня- тии "каудекс". 2. Современное со- стояние вопроса // Вест. Моск. ун- та, сер. биол., почвоведение, 1969а, Мо 1, с.5562 Нухимовский Е.Л. О термине и поня- тии "каудекс". 3. Вестн. Моск. ун- та, сер. биол., почвоведение, 19696, Nî 2, с. 71-78. Нухимовский Е.Л. Структурная орга- низация и морфогенез некоторых семенных растений: Автореф. дис. ... канд. биол. наук. — М., 1970, 15 с. Нухимовский Е.Л. Осевая и побеговая системы семенных растений // Изв. ТСХА, 1971, вып. 1, с. 54-66. Нухимовский Е.Л. О понятии жизнен- ная форма // Интродукция новых лекарственных растений / Сб. науч. работ ВИЛР — М., 1973а, вып. 5, с. 222-232. Нухимовский Е.Л. О соотношении по- нятий "партикуляция" и "вегета- тивное размножение" // Бюлл. МОИП, отд. биол., 19736, т. 78, вып. 5, с. 107-120. Нухимовский Е.Л. Экзоморфологичес- кое строение и возрастная изменчи- вость базовых органов семенных ра- стений // Интродукция новых лекар- ственных растений / Сб. науч. работ ВИЛР — М., 1973в, вып. 5, с. 212-221. Нухимовский Е.Л. Экологическая мор- фология некоторых лекарственных растений в естественных условиях их произрастания. 2. Rhodiola rosea L. // Растит. ресурсы, 1974а, т. 10, вып. 4, с. 499-516. Нухимовский Е.Л. Ветвление и куще- ние семенных растений // Изв. ТСХА, 1974б, М 2, с. 50-62. Нухимовский Е.Л. Биоморфы семен- ных растений и некоторые принци- пы их организации // ХП Между- народный ботанический конгресс / Тезисы докладов. Т. II. — Л.: На- ука, 1975, с. 571. Нухимовский Е.Л. Экологическая мор- фология некоторых лекарственных растений в естественных условиях их произрастания. 3. Patrinia inter- media (Hornem.) Roem. et Schult. // Растит. ресурсы, 1976а, т. 12, вып. 1, с. 3-16. Нухимовский Е.Л. Начальные этапы биоморфогенеза Rhodiola rosea L., выращиваемой в Московской обла- сти // Растит. ресурсы, 1976б, т. 12, вып. 3, с. 348-355. Нухимовский Е.Л. Экологическая мор- фология некоторых лекарственных растений в естественных условиях их произрастания. 4. Bupleurum multinerve DC. // Растит. ресурсы, 1978, т. 14, вып. 1, с. 37-46. Нухимовский Е.Л. Проблема класси- фикации в биоморфологии семен- ных растений // Успехи совр. биол., 1980, вып. 2, с. 286-307. Нухимовский Е.Л. Экологическая мор- фология некоторых лекарственных растений в естественных условиях их произрастания. 11. Phlojodicarpus sibiricus (Steph. ех Spreng.) К.-Pol. // Растит. ресурсы, 1981, т. 17, вып. 1, с. 43-50.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Нухимовский Е.Л. Природа, жизнь и проблема жизнесохранения // Ус- пехи совр. биол., 1984, т. 98, вып. 2(5), с. 292-306. Нухимовский Е.Л. Особенности фене- тической организации биоморф се- менных растений // Успехи совр. биол., 1986, т. 102, вып. 2(5), с. 289- 306. Нухимовский Е.Л. Статус биоморфоло- гии и первый принцип фенетичес- кой организации биоморф семен- ных растений // Усп. совр. биол., 1987, т. 104, вып. 2(5), с. 231-250. Нухимовский Е.Л. Второй принцип фенетической организации био- морф семенных растений и эволю- ция на рубежах пластичности // Усп. совр. биол. — 1988, т. 105, вып. 2(5), с. 300-318. Нухимовский Е.Л., Журба О.В. Эколо- гическая морфология некоторых лекарственных растений в есте- ственных условиях их произраста- ния. 8. Oplopanax elatus (Nakai) Nakai // Растит. ресурсы, 1979, т. 15, вып. 4, с. 507-516. Нухимовский Е.Л., Нухимовская Ю.Д. Экологическая морфология некото- рых лекарственных растений в ес- тественных условиях их произрас- тания. 5. Paeonia anomala L. // Ра- стит. ресурсы, 1978, т. 14, вып. 3, с. 347-355. Нухимовский Е.Л., Нухимовская Ю.Д. Экологическая морфология некото- рых лекарственных растений в ес- тественных условиях их произрас- тания. 7. Inula helenium 1.. // Рас- тит. русурсы, 1979, т. 15, вып. 1, с. 36-43. Нухимовский Е.Л., Нухимовская Ю.Д. Экологическая морфология некото- рых лекарственных растений в ес- тественных условиях их произрас- тания. 12. Leonurus quinquelobatus ОйЬ. // Растит. ресурсы, 1981, т. 17, вып. 3, с. 373-382. Нухимовский Е.Л., Нухимовская Ю.Д. Возрастная изменчивость биоморф семенных растений и особенности ее изучения // Ботанические иссле- дования в заповедниках РСФСР / Сб. науч. трудов ЦНИЛ Главохоты РСФСР. — М., 1984, с. 119-135. Нухимовский Е.Л., Шретер И.А. Био- морфогения якорцев стелющихся // Бюлл. Глав. ботан. сада, 1979, вып. 111, с. 53-59. Нухимовский Е.Л., Черкасов О.А. Мор- фология Origanum vulgare L. в есте- ственных условиях местообитания и при выращивании в Московской области // Растит. ресурсы, 1987, т. 23, вып. 3, с. 345-356. Нухимовский Е.Л., Юрцева Н.С., Юр- цев В.Н. Биоморфологические осо- бенности Rhodiola rosea L. при вы- ращивании (Московская область) // Растит. ресурсы, 1987, т. 23, вып. 4, с. 489-501. Нухимовский Е.Л., Семенихин И.Д., Шугаева Е.В. Биоморфология и эмбриогенез Valeriana officinalis L. в условиях выращивания (Московс- кая область) // Растит. ресурсы, 1989, т. 24, вып. 1, с. 16-32. О сущности жизни. Сборник статей.— М.: Наука, 1964, 351 с. Овчинников Н.Ф. О классификации принципов сохранения // Вопросы философии, 1962, ¹ 5, с. 72-83. Овчинников Н.Ф. Принципы сохране- ния. — М.: Наука, 1966, 331 с. Овчинников Н.Ф. Категория структуры в науках о природе // Структура и формы материи. — М., 1967. Одум Ю. Основы экологии. — М.: Мир, 1975, 740 с. Озима М. Глобальная эволюция Зем- ли. — М.: Мир, 1990, 165 с. Оиарин А.И. Возникновение жизни на Земле. 3-е изд. — М.: изд-во АН СССР, 1957, 458 с. Оларин А.И. Жизнь, ее природа, про- исхождение и развитие. — М.: На- ука, 1968, 173 с. Оларин А.И. Пути становления биоло- гической организации // Развитие концепции структурных уровней в биологии. — М.:Наука, 1972, с. 235-246. Оиарин А.И. Материя, жизнь, интел- лект. — М.: Наука, 1977, 207 с. Оларин А.И. Современные представ- ления о происхождении жизни на Земле // Журн. Всес. хим. об-ва им.
Литература Д.И. Менделеева, 1980, т. 25, вып. 3, с. 246-251. Отрощенко В.А. Вопросы поиска ино- планетной жизни // Проблемы воз- никновения и сущности жизни.— М.: Наука, 1973, с. 17-22. Паавер КЛ. Вопросы синтетического подхода в биоморфологии. — Тал- лин: Валгус, 1976, 265 с. Панибратов В.Н Категория "закон".— Л.: Наука, 1980, 128 с. Первухина Н.В. О некоторых методо- логических проблемах морфологии растений // Ботан. журн., 1960, т. 45, No 2, с. 288-303. Первухина Н.В. Побег как основной орган высшего растения // Рефера- ты докл. Всес. межвуз. конф. по морфологии растений. — М.: изд- во Моск. ун-та, 1968, с. 13-14. Первухина Н.В. Проблемы морфоло- гии и биологии цветка. — Л.: На- ука, 1970, 169 с. Перелыгин П. Основания теоретичес- кой ботаники. — СПб., 1828, 269 с. Петров А. Что считать основным структурным элементом кроны дре- весных растений // Докл. Болг. Акад. Наук, 1963, т. 16, кн. 5, с. 541-544. Петров А.А. Към въпроса за метода на морфологичния анализ на вегетатив- ното развитие и характера на пере- минаването на растежните точки в генеративна фаза при покрытосемен- ните растения: Автореф. дис.... канд. биол. наук: Пловдив, 1967. Петров А. Опит за морфологичен ана- лиз на вегетативното развитие и залагането на генеративните орга- ни при покрытосеменните дървес- ни растения // Изв. Ботан. ин-та Бълг. АН, 1973, кн. 23, с. 159-189. Петров М.П. Корневые системы рас- тений песчаной пустыни Кара-Кум, их распределение и взаимоотноше- ния в связи с экологическими ус- ловиями // Труды по прикл. бот., генетике и селекции. Серия 1 (ста- тистика, география и экология ра- стений). 1933, М 1, с. 113-207. Петров Я. Начальные основания бо- таники для преподавания. — СПб., 1815, 346 с. Петровский В.В. Синузии как формы совместного существования расте- ний // Ботан. журн., 1961, т. 46, М 11, с. 1615-1626. Петрушенко Л.А. Единство системно- сти, организованности и самодви- жения. — М.: Мысль, 1975, 286 с. Петунников А.Н. Свод ботанических терминов, встречающихся в рус- ской ботанической литературе. 2-е изд. — Юрьев, 1912. Пиаже Ж., Инельдер Б. Генезис эле- ментарных логических структур, классификация и сериация. — М.: изд-во иностр. лит., 1963, 448 с. Пиддоти О.А. Определитель всходов однолетних декоративных расте- ний. — Л.: Наука, 1967, 124 с. Пименов М.Г. Вздутоплодник сибирс- кий // Атлас ареалов и ресурсов лекарственных растений СССР.— М.: изд-во ГУГК, 1976, с. 216. Пименов М.Г., Бабилев Ф.В., Никонов Г.К. Phlojodicarpus Turcz. и Libanotis L. как источники получения кума- ринов со спазмолитической актив- ностью // Растит. ресурсы, 1968, т. 4, вып. 4, с. 486-491. Платонов Г.В. Жизнь, наследствен- ность, изменчивость (методологи- ческие аспекты). — М.: изд-во Моск. ун-та, 1978, 238 с. Пленник Р.Я. и др. Копеечник юго- восточного Алтая как материал для интродукции // Полезные растения природной флоры Сибири / Р.Я. Пленник, А.Р. Вернер, Г.В. Кузне- цова, Е.Г. Пеккер, А.А. Дьяконова, Н.Я. Гордеенко. — Новосибирск: Наука, 1967, с. 43-73. Пленник Р.Я., Кузнецова Г.В. Декора- тивные виды бобовых из Юго-Вос- точного Алтая // Растения природ- ной флоры Сибири для зеленого строительства. — Новосибирск: Наука, 1972, с. 98-106. Пономарев А.Я. Изучение цветения и опыления растений // Полевая гео- ботаника. — М.; Л.: изд-во АН СССР, 1960, т. 2, с. 9-19. Полетика О.М., Мишенкова А.П. Де- коративные растения открытого грунта. — Л.: Наука, 1967, 208 с.
ОСНОВЫ БИОМОРФОЛОГИИ CEMEHHbIX РАСТЕНИИ 1Холесовой М.1Т. Проблема повторяе- мости в философии и естествозна- нии (философско-методологичес- кий очерк). — Горький, Волго- Вятск. изд-во, 1970, 191 с. Положий А.В., Суров Ю.1Т. Ареалы, фитоценотипическая приурочен- ность и прогноз запасов левзеи сафлоровидной и радиолы розовой в южной Сибири // Ресурсы дико- растущих лекарственных растений СССР. — М.: ВИЛР, 1972, с. 113-116. Поннамиерума С. Происхождение жиз- ни. — М.: Мир, 1977, 175 с. Поиадюк Р.В., Чистякова А.А., Чума- ченко С.И. и др. Восточноевропейс- кие широколиственные леса. — М.: Наука, 1994, 364 с. Поллавская Г.И. Материалы по изуче- нию изменчивости крымского бука // Журн. русск. бот. общ-ва, 1927, т. 12, Хо 1-2, с. 59-86. Поилавская Г.И. Экология растений.— М.: Сов. наука, 1948, 296 с. Попов М.Г. Опыт монографии рода эремостахис // Новые мемуары МОИП, 1940, т. 19, с. 116-210. Попов М.Г. Основы флорогенетики.— М.: изд-во АН СССР, 1963, 135 с. Попович М.В., Садовский В.Н. Теория // Философская энциклопедия. Т. 5, 1970, Поицов А.В., Некрасов В.И., Иванова И.А. Очерки по семеноведению.— М.: Наука, 1981, 113 с. Посиелов Г.Л. О характере геологии как науки и ее месте в естествозна- нии // Изв. АН СССР, серия геол., 1960, N 11, с. 3-19. Пошкурлат А.1Т. Строение и развитие дерновины чия // Ученые зап. Моск. гос. пед. инст. им. В.И. Ле- нина, 1941, т. 30, с. 101-155. Пошкурлат А.1Т. Анализ ритма разви- тия первоцвета весеннего // Вотан. журн., 1962, т. 47, N 2, с. 262-267. Пошкурлат А.1Т. Развитие подземных органов виргинильных растений горицвета весеннего // Бюлл. МОИП, отд. биол., 1969, т. 74, вып. 5, с. 118 — 128. Пошкурлат А.1Т. Строение и ритм раз- вития почек горицвета весеннего // Растит. ресурсы, 1974, т. 10, вып. 4, с. 516-528. Пошкурлат А.1Т. Большой жизненный цикл горицвета весеннего // Рас- тит. ресурсы, 1975, т. 11, вып. 3, с. 483-492. Пошкурлат А.1Т. Годичная изменчи- вость популяций горицвета весен- него в северной степи // Бюлл. МОИП, отд. биол., 1976, т. 81, вып. 5, с. 96-106. Пояркова А.И. Сопшт maculatum L. // Флора СССР. — М.; Л.: изд-во АН СССР, 1950, т. 16, с. 225-229. 1Хояркова А.И. Echinopanax elatus Na- kai // Флора СССР. — М.; Л.: изд- во АН СССР, 1950, т. 16, 18 с. Правдин Л.Ф. Вегетативное размноже- ние растений. (Теория и практика).— М.; Л.: Сельхозгиз, 1937, 232 с. Правдин Ф.Н. Жизненные формы у животных и принципы их класси- фикации // Докл. МОИП. Кн. 2. Зоология и ботаника. — М., 1971, с. 17-20. Правдин Ф.Н. Экологическая география насекомых Средней Азии: Ортопте- риоиды. — М.: Наука, 1978, 271 с. Правдин Ф.Н. Учение о жизненных формах как общебиологическая проблема // Жизненные формы в экологии и систематике растений / Межвуз. сб. науч. трудов — М., 1986, с. 3-8. Проблема возникновения и сущности жизни. Сборник статей. — М.: На- ука, 1973, 272 с. Прозоровский А.В. О биологических типах растений пустыни // Вотан. лсурн., 1936, т. 21, вып. 5, с. 559-563. 1Хрозоровский Н.А. Программа по теме "Изучение жизненного цикла се- менных растений заповедника в питомнике и в природе" // Науч- но-методич. записки Главного уп- равления по заповедникам, 1949, вып. 12, с. 167-185. Происхождение жизни и эволюцион- ная биохимия // Сб. статей. — М.: Наука, 1975, 404 с. Проханов Я.И. Возникновение дву- дольных многолетних трав // Тру- ды МОИП, 1965, т. 13, с. 111-123.
Литература Пятницкий С.С., Коваленко ~ Вегетативный лес. — M 1963, 448 с. Работнов T A. Биологиче~~и~ наблю дения на субальпийск~" лугах Се верного Кавказа // Бота~ ж"р" 1945, т. 30, Хо 4, . 167-177. Работнов Т.А. ВозрастНо" пуляций некоторых пУ 9 6 стений // Успехи совр биол., 1946, т. g5, вып. 3(1), с. 435 438. Работнов Т.А. Определе~~е в длительности жизни '.( травянистых растений совр. биол. — 1947, т. ~4' вып' ( )' с. 133-149. Работнов T.А. Основнь~е методы изучения жиУ ла многолетних трав~~и На „ометодические за~~~~ ного управл. по э~~о 1949, вып. 12, с. 41 54. Работнов 7.A. )Кизненну голетних травянисть! Х Р астений в луговых ценозах // Tp Б~~~~' ин та Ад СССР. (Cep. 3), 1950а, вып. 6, с. 7-204. Работнов ТА. Вопросы из" ~ става популяций для цел ф нологии // Проблемы б~ М.; Л.: изд-во АН СССР, 1950б, т. 1, с. 465-483. Работнов ТА. Апйпасцв (P~>.) Кормовые растения ~е е КОКОСОВ И пастбищ CCCP. M . Л.: Сельхоз- гиз, 1956а, т. 3, с. 10>-1 Работнов Т.А. О жизн~~~н борщевика сибирского ( биол., нов. серия, 1956О т' с. 73-81. Работнов Т.А. К метод~~е содержания всхожих сем~~ ян в почвах лугов // Вотан. журн 1958, т. 43, Хо 11, с. 1572-1581. Работнов Т.А. Методь~ возраста и д,пительяо~ травянистых растении // геоботаника. — М.; +" gggP 1960а T 2 с Я49 Работнов Т.А. Методы изучения се- менного размножения травянистых растений в сообществах // Полевая геоботаника. — М.; Л.: Наука, 19606, т. 2, с. 20-40. Работнов Т.А. Определение возраст- ного состава популяций видов в сообществе // Полевая геоботани- ка. — М.; Л.: Наука, 1964, т. 3, с. 132-145. Работнов Т.А. Некоторые вопросы изучения ценотических популяций // Бюлл. МОИП, отд. биол., 1969, т. 74, вып. 1, с. 141-149. Работнов Т.А. Луговедение. — М.: изд-во Моск. ун-та, 1974, 384 с. Работнов Т.А. Изучение ценотических популяций в целях выяснения «стра- тегии жизни» видов растений // Бюлл. МОИП, отд. биол., 1975, т. 80, вып. 2, с. 5-17. Работнов Т.А. Фитоценология. — М.: изд-во Моск. ун-та, 1978, 384 с. Работнов Т.А. Что такое экология с точки зрения ботаника // Вест: Моск. ун-та, сер. 16 (биология), 1979, N 11, с. 47-50. Работнов ТА История фитоценоло- гии: Учебное пособие. — М.: Аргус, 1995, 158 с. Радкевич В.А. Экология: Краткий курс. 2-е изд. — Минск: Высш. шк., 1983, 320 с. Радкевич О.Н., Шубина Л.Н. Морфо- логические основы явления парти- куляции у ксерофитов пустыни Бетпак-Дала // Тр. Среднеазиат. гос. унив. — Ташкент, 1935, сер. 86, вып. 25, с. 1-22. Раздорский В.Ф. Анатомия растений. — М.: Советская наука, 1949, 524 с. Райнботе Х Тайна растений. Пер. с немец. — М.: Знание, 1979, 208 с. Райкова ИА. Морфологические осо- бенности растений Памира при их культуре в Ташкенте // Проблемы совр. ботаники. — М.; Л.: Наука, 1965, т. 2, с. 97-105. Раубах Х Загадки молекул. — Л.: Хи- мия, 1979, 174 с. Раменский Л.Г. Введение в комплекс- ное почвенно-геоботаническое ис-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ следование земель. — М.: Сельхоз- гиз, 1938, 620 с. Раменский Л.Г. Избранные работы. Проблемы и методы изучения рас- тительного покрова. — Л.: Наука, 1971, 334 с. Panonopm А. Математические аспекты абстрактного анализа систем // Исследования по общей теории систем. — М.: Прогресс, 1969, с. 83-105. Рачковская Е.И. К биологии пустын- ных полукустарничков // Тр. Бот. ин-та АН СССР, сер. 3, вып. 11, 1957, с. 5-87. Рахманина А. Т. Биолого-экологичес- кая характеристика карганных со- обществ ( из Salsola dendroides Pall.) в Кура-Араксинской низменности // Эколого-геоботанические и агроме- лиоративные исследования в Кура- Араксинской низменности Закав- казья. — М.; Л.: изд-во АН СССР, 1962, с. 204-297. Ревякина М.П. Жизненные формы караганы древовидной (Сагаяапа агЬогевсепв L.) на Салаирском кря- же // Жизненные формы в эколо- гии и систематике растений / Меж- вуз. сб. науч. трудов. — М., 1986, с. 76-79. Ревякина Н.В. К изучению биологи- ческих особенностей и запасов зо- лотого корня и маральего корня в Центральном Алтае // Изв. СО АН СССР, 1973, N10, ,сер. биол. наук., вып. 2, с. 58-64. Реймерс КФ. Азбука природы. Мик- роэнциклопедия биосферы. — М.: Знание, 1980, 208 с. Реймерс Н.Ф.Популярный биологи- ческий словарь. — М.: Наука, 1991, 544 с. Риклвфс Р. Основы общей экологии.— М.: Мир, 1979, 424 с. Розенталь Д.Э., Теленкова М.А. Сло- варь трудностей русского языка.— М.: Рус. язык, 1985, 704 с. Роллер Э. Открытие основных законов жизни. Перев. с англ. — М.: Мир, 1978, 336 с. Ростовцева 3.1Т. Верхушечная мерис- тема высших растений. — М.: изд- во Моск. ун-та, 1963, 59 с. Рысин Л. 1T., Казанцева Т.Н. Метод ценопопуляционного анализа в гео- ботанических исследованиях // Во- тан. л(урн., 1975, т. 60,М 2, с. 199-209. Рысин Л.1T., Рысина Г.П. Опыт попу- ляционного анализа лесных сооб- ществ // Бюлл. МОИП, отд. биол., 1966, т. 71, вып. 1, с. 84-94. Рысина Г.П. Ранние этапы онтогенеза лесных травянистых растений Под- московья. — М.: Наука, 1973, 216 с. Рытова Н.Г. Способ заложения боко- вой почки у злаков (Poaceae) и природа предлиста и его междоуз- лий (двуосный метамер как струк- турная единица тела растений у злаков) // Вотан. журн., 1982, т. 67, М 7, с. 913-924. Руденко А.П. Эволюционная химия и естественноисторический подход к проблеме происхождения жизни // Журн. Всес. хим. об-ва им. Д.И. Менделеева, 1980, т. 25, вып. 4, с. 390-404. Русакова С.В., Нухимовский Е.Л. Био- логические особенности Hedysarum neglectum Ledeb. и содержание в нем мангиферина в условиях Алтая // Растит. ресурсы, 1977, т. 13, вып. 3, с. 478-480. Руссо Ж. Руссовы письма о ботанике с дополнением его ботанического словаря. Пер. В. Измайлова. — М., 1810, 405 с. Рустамов И.Г. Материалы к изучению подземных частей некоторых пла- корных фитоценозов Северо-За- падной Туркмении // Изв. АН Туркм. ССР. Сер. биол. наук, 1965, М 2, с. 15-23. Руттвн М. Происхождение жизни.— М.: Мир, 1973, 411 с. Сабинин Д.А. Минеральное питание растений. — М.;Л.: изд-во АН СССР, 1940. Сабинин Д.А. Физиология развития растений. — М.: изд-во АН СССР, 1963, 196 с. Сабинин ДА. Избранные труды по ми- неральному питанию растений.— М.: Наука, 1971, 512 с. Савиных Н.1Т. Побеговые системы с неограниченным моноподиальным
Литература нарастанием ( на примере рода Veronica L.) // Пятая Всес. школа по теоретической морфологии рас- тений / Научные доклады. — Львов, 1987, с. 77-80. Садовский В.Н. Некоторые принципи- альные проблемы построения об- щей теории системы // Системные исследования. Ежегодник, 1971.— М.: Наука, 1972, с. 35-54. Садовский В.Н. Основания общей те- ории систем. Логико-методологи- ческий анализ. — М.: Наука, 1974, 179 с. Садовский В.Н. Системный подход и общая теория систем: статус, ос- новные проблемы и перспективы развития // Системные исследова- ния. Методологические проблемы. Ежегодник, 1979. — М.: Наука, 1980, с. 29-54. Саратиков А.С. Некоторые итоги изыскания и изучения стимулято- ров центральной нервной системы растительного происхождения // Стимуляторы центральной нервной системы. — Томск: изд-во Томск. ун-та, 1966, с. 3-23. Саратиков А.С. Золотой корень (ра- диола розовая). 2-е изд. — Томск: изд-во Томского ун-та, 1974, 156 с. Саиожников С.В. Примечания и указа- ния // Ламарк Ж.Б. Философия зоо- логии. — М.; Л., 1937, т. 2, с. 446. Саркисов Д С. Очерки по структурным основам гомеостаза. — М.: Меди- цина, 1977, 351 с. Сахаров М.И. Некотрые данные о пла- стичности растений // Сб. науч. трудов АН Белорусской ССР, 1951, вып. 2. Сацыперова И.Ф. Особенности онто- генеза у видов Негас1еит L. флоры СССР // Растит. ресурсы, 1977, т. 13, вып. 3, с. 435-449. Свидерский В.И. О некоторых новых представлениях в теории материа- листической диалектики // Фило- софские вопросы физики. — Л.: изд-во ЛГУ, 1974, с. 74-85. Сдобнина Л.И., Пименов М.Г. Строе- ние подземных органов Libanotis condensata (L.) Crantz., локализация и накопление в них кумаринов // Фармация, 1973, М 2, с. 31-36. Сдобнина Л.И., Иименов М.Г. Распро- странение видов Seseli L. из секций Condensata, Pseudolibanotis и Liba- notis // Растит. ресурсы, 1975, т. 11, вып. 4, с. 503-511. Северин С.Е. Основные элементы био- мии. — М.: Знание, 1970, 54 с. Севернев М.М., Сущеня Л.М., Горбач В.И. и др. Основы природопользо- вания. — Минск: Наука и техника, 1980, 575 с. Северцов А.С. Введение в теорию эво- люции. М.: изд-во Моск. ун-та, 1981, 318 с. Cease Г. Очерки об адаптационной син- дроме. Пер. с англ. — М.: Медгиз, 1960. Cease Г. На уровне целого организма.— М.: Наука, 1972, 122 с. Селье Г. Стресс без дистресса. Пер. с англ. — М.: Прогресс, 1979, 124 с. Семенихин ИД. Некоторые особенно- сти развития валерианы лекар- ственной в онтогенезе // Результаты научных исследований в области ле- карственного растениеводства. — М.: ВИЛР, 1975, вып. 8, с. 77-78. Семенихин ИД. Особенности онтоге- неза лекарственной валерианы в чистых и совместных посевах // Химико-фармацев. журн., 1976, т. 10, М 7, с. 83-85. Семенихин ИД., Муш Н.Н. Совмест- ные посевы валерианы с некоторы- ми однолетними лекарственными культурами // Растит. ресурсы, 1974, т. 10, вып. 2, с. 229-233. Сенянинова-Корчагина М.В. К вопросу классификации жизненных форм // Ученые зап. Ленингр. гос. унив., 1949, сер. геогр. наук, вып. 5, N 104, с. 54-102. Сенянинова-Корчагина М.В. Геофилия и ее значение в сложении структу- ры растительного сообщества (о целостности организма высшего растения) // Ученые зап. Ленингр. гос. унив., сер. геогр. наук, 1967, вып. 19, М 327, с. 7-96. Сергиевская Л.1Т. Степи Бурят-Монго- лии // Тр. Томск. ун-та, 1951, т. 116, с. 217-279.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Серебровский А.С. Некоторые пробле- мы органической эволюции. — М.: Наука, 1973, 168 с. Серебряков И.Г. Биология тяньшанс- кой ели и типы ее насаждений в пределах Заилийского и Кунгей Алатау // Ученые зап. МГУ / Тр. бот. сада, 1945, вып. 82, кн. 5, с. 103-175. Серебряков И.Г. О ритме сезонного развития растений подмосковных лесов // Вест. Моск. ун-та, 1947, М 6, с. 75-108. Серебряков И.Г. Структура и ритм жизни цветковых растений // Бюлл. МОИП, отд. биол., 1949, т. 54, вып. 1, с. 47-62. Серебряков И.Г. Морфология вегетатив- ных органов высших растений. — М.: Сов. наука, 1952, 392 с. Серебряков И.Г. Проблемы морфоло- гии вегетативных органов покрыто- семенных: Автореф. дис.... докт. биол. наук. — М., 1953, 20 с. Серебряков И.Г. О методах изучения ритмики сезонного развития расте- ний в геоботанических стационарах // Докл. на совещ. по стационарным геобот. исслед. / БИН АН СССР.— М.; Л., 1954, с. 145-159. Серебряков И.Г. Основные направле- ния эволюции жизненных форм у покрытосеменных растений // Бюлл. МОИП, отд. биол., 1955, т. 60, вып. 3, с. 71-91. Серебряков И.Г. Типы развития побе- гов у травянистых многолетников и факторы их формирования // Уче- ные зап. МГПИ им. В.П. Потемки- на / Кафедра ботаники. 1959, т. 100, вып. 5, с. 3-38. Серебряков И.Г. Экологическая мор- фология растений. Жизненные формы покрытосеменных и хвой- ных. — М.: Высш. школа, 1962, 378 с. Серебряков И.Г. Жизненные формы высших растений и их изучение // Полевая геоботаника. — М.; Л.: Наука, 1964, т. 3, с. 146-205. Серебряков И.Г., Серебрякова Т.И. О двух типах формирования корневищ у травянистых многолетников // Бюлл. МОИП, отд. биол., 1965, т. 70, вып. 2, с. 67-81. Серебряков И.Г., Чернышева М.Б. О морфогенезе жизненной формы кустарничка у черники, брусники и некоторых болотных Ег1сасеае // Бюлл. МОИП, отд. биол., 1955, т. 60, вып. 2, с. 65-77. Серебряков Л. В кн.: Философская энциклопедия. Т. 4. — М.: Сов. энциклопедия, 1967, с. 86.. Серебрякова Т.И. О некоторых морфо- логических закономерностях побе- гообразования у овсяницы луговой (Festuca pratensis Huds.) // Бюлл. МОИП, отд. биол., 1959, т. 64, вып. 3, с. 89-98. Серебрякова Т.И. Некоторые законо- мерности формирования почек и побегов у луговых злаков // Бюлл. МОИП, отд. биол., 1961, т. 66, вып. 4, с. 42-52. Серебрякова Т.И. К вопросу о побего- образовании и кущении у злаков // Ботан. журн., 1962, т. 47, М 3, с. 427432. Серебрякова Т.И. Жизненные формы лесных и луговых рыхлокустовых злаков и возможные пути их эво- люции // Бюлл. МОИП, отд. биол., 1965, т. 70, вып. 6, с. 60-79. Серебрякова Т.И. Типы побегов и эво- люция жизненных форм в семей- стве злаков // Научные доклады высшей школы. Виол. науки, 1967а, N 22, с.61-73. Серебрякова Т.И. Морфогенез вегета- тивных органов и эволюция жиз- ненных форм в семействе злаков: Автореф. дис.... докт. биол. наук.— М., 1967б, 24 с. Серебрякова Т.И. Побегообразование и жизненные формы некоторых овся- ниц (Festuca L.) в связи с их эволю- цией // Вопросы морфогенеза цвет- ковых растений и строение их попу- ляций.— М.: Наука, 1968, с. 7-51. Серебрякова Т.И. Ветвление и куще- ние в семействе Роасеае // Вотан. журн., 1969, т. 54, N 6, с. 858-871. Серебрякова Т.И. Морфогенез побегов и эволюция жизненных форм зла- ков. — М.: Наука, 1971, 360 с.
Литература Серебрякова Т.И. Учение о жизненных формах на современном этапе // Итоги науки и техники. Сер. бота- ника / ВИНИТИ. — М., 1972, т. 1, с. 84-169. Серебрякова Т.И. Об основных "архи- тектурных моделях" травянистых многолетников и модусах их преоб- разования // Бюлл. МОИП, отд. биол., 1977, т. 82, вып. 5, с. 112-128. Серебрякова Т.И. Современные пред- ставления о побеге и почке // Биол. в школе,1980, М 4, с. 8-14. Серебрякова Т.И. Жизненные формы и модели побегообразования назем- но-ползучих многолетних трав // Жизненные формы: структура, спектры и эволюция. — М.: Наука, 1981, с. 161-179. Серебрякова Т.И. (отв. ред.) Диагнозы и ключи возрастных состояний лу- говых растений / Методические разработки для студентов биологи- ческих специальностей. — М.: МГПИ им. В.И. Ленина, 1981 — Ч. I, 1983 — Ч. II, Ч. Ш. Серебрякова Т.И. О вариантах моделей побегообразования у многолетних трав // Морфогенез и ритм разви- тия высших растений / Межвузов- ский сб. науч. трудов. — М., 1987, с. 3-19. Сержантов В.Ф. Философские про- блемы биологии человека. — Л.: Наука, 1974, 158 с. Сетров М.И. Общие принципы орга- низации систем и их методологи- ческое значение. — Л.: Наука, 1971а, 265 с. Сетров М.И. Организация биосистем (Методологический очерк принци- пов организации живых систем).— Л.: Наука, 1971б, 275 с. Сетров М.И. Взаимосвязь основных иерархических рядов организации живого // Развитие концепции структурных уровней в биологии.— М.: Наука, 1972, с. 311-321. Cempos М.И. Информационные про- цессы в биологических системах.— Л.: Наука, 1975, 155 с. Синицина В.Г. Морфологические осо- бенности девясила высокого, про- израстающего в Заилийском Алатау // Тр. ин-та ботаники Казах. ССР.— А-А.: Наука, 1976, т. 35, с. 80-87. Синнот Э. Морфогенез растений. Пер. с англ. — М.: изд-во иностр. лит- ры, 1963, 603 с. Синьковский Л.Н. Возрастные измене- ния пустынных полукустаничков // Сообщ. Тадж. филиала АН СССР, 1950, М 29, с. 37-40. Сырокомская И.В., Лииатова В.В. Осо- бенности фенологических наблюде- ний в различных зонах СССР. Тра- вянистые растения лесов и лугов // Методы фенологических наблюде- ний при ботанических исследова- ниях. — М.; Л.: Наука, 1966, с. 45-52. Скворцов А.К Комаров В.Л. и пробле- ма вида // Комаровские чтения.— М.; Л.: Наука, 1972, вып. 24, с. 48-81. Слуцкий М.С. Взаимосвязь философии и естествознания. — М.: Высшая школа, 1973, 119 с. Смелов СШ Биологические основы луговодства. — М.: Сел ьхозгиз, 1947, 232 с. Смелов СШ Теоретические основы луговодства. — М.: Колос, 1966, 367 с. Смирнова Е.С. Морфологические типы и формирование облика растений // Бюлл. Глав. бот. сада. АН СССР, 1974, вып. 93, с. 49-57. Смирнова Е.С. Побеговая система тро- пических и субтропических покры- тосеменных деревьев // Журн. общ. биол., 1978, т. 39, М 1, с. 86-96. Смирнова Е.С. Биоморфологические структуры побеговой системы тро- пических и субтропических цветко- вых растений в природе и оранже- рейной культуре // Интродукция тропических и субтропических рас- тений. — М.: Наука, 1980, с. 52-91. Смирнова О.В. Онтогенез и возраст- ные группы осоки волосистой (Carex pilosa Scop.) и сныти обык- новенной (Aegopodium podagraria L.) // Онтогенез и возрастной со- став популяций цветковых расте- ний. — М.: Наука, 1967, с. 100-113. Смирнова О.В. Жизненные циклы, численность и возрастной состав популяций основных компонентов
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ травянистого покрова дубрав: Авто- реф. дис.... канд. биол. наук. — М., 1968, 17 с. Смирнова О.В. Структура травянисто- го покрова широколиственных ле- сов. — М.: Наука, 1987, 207 с. Смирнова О.В., Заугольнова Л.Б., Ер- макова И.М. и др. Ценопопуляции растений (основные понятия и структура). — М.: Наука, 1976, 217 с. Смирнова О.В., Торопова Н.А. Эколо- го-демографическая характеристи- ка синузий трав // Восточноевро- пейские широколиственные леса.— М.: Наука, 1994, с. 171-190. Смит Дж. М. Эволюция полового размножения. — М.: Мир, 1981, 271 с. Снаговская М.С. Возрастные состоя- ния желтой люцерны // Уч. зап. Моск. гос. пед. ин-та им. В.И. Ле- нина, 1965, N 212, с. 46-57. Соколов СЯ., Артюшенко 3.Т. Ивано- вы побеги у сосны // Вотан. журн., 1957, т. 42, М 5, с. 741-745. Соколов СЯ., Замотаев Н. 1Т. Справоч- ник по лекарственным растениям (фитотерапия). — М.: Медицина, 1985, 463 с. Солопов Е.Ф. Материя и движение.— Л.: Наука, 1972, 220 с. Солнцев В.Н Системная организация ландшафтов. (Проблемы методоло- гии и теории). — М.: Мысль, 1981, 239 с. Станюкович КВ. Растительный покров Восточного Памира // Записки геогр. общ. Нов. серия — М.: изд-во геогр. литер., 1949, т. 10, с. 1-158. Старз Дж. Молекулы жизни. — М.: Атомиздат, 1976, 103 с. Сивбавв И.В. Жизненные формы и половой диморфизм Саранчевых Ты- вы и Юго-Восточного Алтая // Зоол. лсурн., 1970, т. 49, М 3, с. 325-338. Стебаев И.В., Никитина С.И. Особен- ности поведения Саранчевых раз- ных жизненных форм пустынь Ты- вы // Зоол. журн., 1975, т. 54, Хю 5, с. 688-700. Стешенко А.H. Формирование струк- туры полукустарничков в условиях высокогорий Памира // Тр. инст. 6or. АН Тадж. ССР, 1956, т. 50, с. 1-161. Стешенко А.Н. Основные морфолого- биологические особенности расте- ний высокогорий Памира // Пробл. ботаники. — М.; Л.: Наука, 1965, т. 7, с. 42-60. Сукачев В.Н. О болотной сосне // Лес- ной журнал, 1905, т. 35, М 3, с. 354- 372. Сукачев В.Н. О "теории дернового процесса" проф. В.Р. Вильямса // Почвоведение, 1916, М 2, с. 1-30. Сукачев В.Н. (ред.) Дендрология с основами лесной геоботаники. 2-е изд. — Л.: Гослестехиздат, 1938, 576 с. Сукачев В.Н. О влиянии интенсивно- сти борьбы за существование меж- ду растениями на их развитие // Доклады АН СССР. Новая серия. 1941, т. 30, N 8, с. 752-755. Суров Ю.1Т. Запасы Rhodiola rosea в горах Алтая и Западных Саян // Успехи изучения лекарственных растений Сибири. — Томск: изд-во Томск. ун-та, 1973, с. 8-10. Тахтаджян А.Л. Теломная теория и "новая морфология" // Вотан. журн. — 1952, т. 37, М 5. Тахтаджян А.Л. Вопросы эволюцион- ной морфологии растений. — Л.: изд-во Ленингр. ун-та, 1954, 215 с. Тахтаджян А.Л. Основы эволюцион- ной морфологии покрытосемен- ных. — М.; Л.: Наука, 1964, 236 с. Тахтаджян А.Л. Систематика и фило- гения цветковых растений. — М.; Л.: Наука, 1966, 611 с. Тахтаджян А.Л. Происхождение и расселение цветковых растений.— Л.: Наука, 1970, 147 с. Тахтаджян А.Л. Тектология: история и проблемы // Системные исследо- вания. Ежегодник, 1971. — М.: Наука, 1972, с. 200-277. Тимирязев КА. Краткий очерк теории Дарвина. — СПб., 1865. Тимофеев- Ресовский Н.В., Воронцов Н.Н., Яблоков А.В. Краткий очерк теории эволюции. — М.: Наука, 1977, 302 с. Тихомиров Б.А. Очерки по биологии растений Арктики. — М.; Л.: изд-во АН СССР, 1963, 154 с. Тихонов В.И. Побег как высший этап эволюции формы тела растений //
Литература Журн. общ. биол., 1977, т. 38, М 6, с. 841-848. Тихонов В.И. Организация осевых си- стем надземной части многолетних растений // Вотан. журн., 1979, т. 64, М 5, с. 740-750. Том Р. Динамическая теория морфо- генеза // На пути к теоретической биологии. — М.: Мир, 1970, т. 1, с. 145-157. Торопова Н.А. Развитие заросли про- лесника — Mercurialis perennis L. (Euphorbiaceae) // Вотан. журн., 1977, т. 62, М 10, с. 1433-1440. Трофимов Т.Т. Облепиха в культуре. 2-е изд. — М.: изд-во Моск. ун-та, 1976, 160 с. Трулевич Н.В. Строение куста и состав популяций полыни тяньшанской в ряду пастбищной дигрессии // Уче- ные зап. фак. естеств. Моск. пед. инст. им. В.И. Ленина, 1960, вып. 4, с. 107-129. Турова А.Д Лекарственные растения СССР и их применение. 2-е изд.— М.: Медицина, 1974, 424 с. Уемов А.И. Методы построения и раз- вития общей теории систем // Си- стемные исследования. Ежегодник, 1973. — М.: Наука, 1973, с. 146-178. Уемов А.И. Системный подход и об- щая теория систем. — М.: Мысль, 1978, 272 с. Уемов А.И. Логические основы метода моделирования. — М., 1971. Уоддингтон К.Х Основные биологи- ческие концепции // На пути к теоретической биологии. — М.: Мир, 1970, т. 1, с. 11-38. Уотсон Дж. Молекулярная биология гена. — М.: Мир, 1978, 720 с. Уранов А.А. Жизненное состояние вида в растительном сообществе // Бюлл. МОИП, нов. сер., отд. биол., 1960, т. 65, вып. 3, с. 77-92. Уранов А.А. Большой жизненный цикл и возрастной спектр ценопопуля- ций цветковых растений. Тезисы доклада V делегатского съезда Всес. ботан. об-ва — Киев, 1973. Уранов А.А. (отв. ред.) Возрастной со- став популяций цветковых расте- ний в связи с их онтогенезом // Сб. трудов / МГПИ им. В.И. Ленина, 1974, 260 с. Уранов А.А., Григорьева КМ, Егорова В.Н., Ермакова ИМ, Матвеев А.Р. Изменчивость и динамика возраст- ных спектров некоторых луговых растений // Теоретические пробле- мы фитоценологии и биогеоценоло- гии. — М.: Наука, 1970, с. 194-214. Уранов А.А., Григорьева Н.М., Ермако- ва И.М. и др. О популяционном подходе к изучению и использова- нию лугов // Науч. докл. высш. школы. Виол. науки, 1976, М 5, с. 75-85. Уранов А.А., ЗаугольноваЛ.Б., Смирнова О.В. и др. Ценопопуляции растений (развитие и взаимоотношения).— М.: Наука, 1977, 131 с. Урманцев Ю.А. Изомерия в живой природе. 1. Теория // Вотан. журн., 1970, т. 55, М 2, с. 153-169. Урманцев Ю.А. Симметрия природы и природа симметрии (философские и естественно-научные аспекты).— М.: Мысль, 1974, 229 с. Урманцев Ю.А. Начала общей теории систем // Системный анализ и на- ' учное знание. — М.: Наука, 1978. Урманцев Ю.А. Об определении законов анантиморфизма нехимических (биологических) диссизомеров по- средством химических // Журн. общ. биол., 1979, т. 40, NЗ, ,с. 351-366. Урсул А.Д'. Сложность, организация, информация // Философские на- уки, 1968, NЗ, ,с. 53-60. Урсул А.Я. Человечество, Земля, Вселен- ная // Философские проблемы кос- монавтики. — М.: Мысль, 1977, 264 с. Уткин Л.А. Народные лекарственные растения Сибири. — М.; Л.: Гос. научн.-техн. изд., 1931, 135 с. Федоров Ал. А., Кириичников М.Э., Ар- тюшенко 3.Т. Атлас по описатель- ной морфологии высших растений. Лист. — М.; Л.: изд-во АН СССР, 1956, 301 с. Федоров Ал. А., Кириичников М.Э., Ар- тюшенко 3.Т. Атлас по описатель- ной морфологии высших растений. Стебель и корень — М.; Л.: изд-во АН СССР, 1962, 352 с.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Федоров Ал. А., Артюшенко 3.Т. Атлас по описательной морфологии выс- ших растений. Цветок. — Л.: На- ука, 1975, 350 с. Федоров Ал. А., Артюшенко 3.Т. Атлас по описательной морфологии выс- ших растений. С<щветие. — Л.: Наука, 1979, 295 с. Федченко Б.А. Обзор видов рода Hedysarum. — СПб., 1913. Федченко Б.А. Род Hedysarum L. // Флора СССР. Т. ХШ. — М.; Л.: изд-во АН СССР, 1948, 586 с. Феофраст. Исследования о растениях— М.: изд-во АН СССР, 1951, 591с. Филииченко Ю.А. Эволюционная идея в биологии. Исторический обзор эволюционных учений XIX века. 3-е изд. — М.: Наука, 1977, 227 с. Филииченко Ю.А. Изменчивость и ме- тоды ее изучения. 5-е изд. — М.: Наука, 1978, 240 с. Фисюнов А.В. Определитель всходов сорных растений. — Киев: Урожай, 1976, 232 с. Фишер-фон-Вальдгейм А. Курс ботаники. Отдел Е 2-е изд. — Bapruasa, 1891, 256 с. Фокс С., Дозе К. Молекулярная эво- люция и возникновение жизни.— М.: Мир, 1975, 374 с. Фолсом К Происхождение жизни.— М.: Мир, 1982, 158 с. Фридерикс К Экологические основы прикладной зоологии и энтомоло- гии. Пер с немец.— М.; Л.: Сель- хозгиз, 1932, 672 с. Фролов М Ю. Социально-экологические системы как объект географии.— Вест. Ленинг. ун-та, 1980, М 12, сер. геол., географии, вып. 2, с. 111-115. Хайлов К.М. Проблема системной организованности в теоретической биологии // Журн. общ. биол.— 1963, т. 24, N 5, с. 324-333. Ханвимер И.С. Материалы к изучению лугов поймы р. Печера // Луга Коми АССР / Котелина Н.С. и Хантимер И.С. — М. Л.: изд-во АН СССР, 1959, с. 133-266. Хомякова И.М. Лесные травы. Опре- делитель по вегетативным призна- кам. — Воронеж: изд-во Воронеж. ун-та, 1974, 176 с. Хомяков Д.М., Хомяков П.М. Основы системного анализа. — М.: изд-во MIX, 1996, 108 с. Хохлов С.С. Задачи и методы исследо- вания биологии бесполосеменного размножения // Ученые зап. Ульян. гос. пед. ин-та им. И.Н. Ульянова, 1965, т. 20, вып. 6. Хохряков А. IT. Формы и этапы поли- меризации в эволюции растений // Журн. общ. биол., 1974, т. 35, N 3, с. 331-345. Хохряков А. 1Т. Соматическая эволюция однодольных. — М.: Наука, 1975а, 196 с. Хохряков А.1T. Закономерности эволю- ции растений. — Новосибирск: Наука, 1975б, 202 с. Хохряков А.1Т. Изменение образа жизни растений в онтогенезе // Журн. общ. биол., 1978, т. 39, М 3, с. 357-372. Хохряков А.П. Эволюция биоморф ра- стений. — М.: Наука, 1981, 168 с. Хохряков А. IT., Мазуренко М.Т. Типы побегов и их эволюция у жимолос- тных // Бюлл. ГБС, 1968, вып. 70, с. 64-69. Хржановский В.Г. Розы. Филогения и систематика. — М.: Сов. наука, 1958, 497 с. Хржановский В.Г. Морфогенетика как основа филогении и таксономии покрытосеменных // Доклады ТСХА, 1966, вып. 118, с. 127-133. Хржановский В.Г. О закономерностях и единицах измерения видообразо- вательного процесса // Изв. ТСХА, 1968, вып. 5, с. 3-14. Хржановский В.Г., Нухимовский Е.Л. Морфогенез володушки кустарни- ковой (Bupleurum friticosum L.) // Рефераты докладов Всес. межвуз. конф. по морфол. раст., 1968, с. 309-311. Хржановский В.Г., Нухимовский Е.Л. Экологическая морфология некото- рых лекарственных растений в есте- ственных условиях их произраста- ния. 1. Bupleutum аигеит Fisch. // Растит. ресурсы, 1972, т. 8, вып. 4, с. 497-506. Хржановский В.Г., Пономаренко С.Ф., Догузашвили В.А. К вопросу проис-
Литература хождения и эволюции рода Linum 1 сем. Linaceae // Изв. АН СССР. Сер. биол., 1979, М 5, с. 696-712. Цвелев Н.Н. Некоторые вопросы эво- люции злаков // Вотан. журн., 1969, т. 54, N 3, с. 361-373. Цвелев Н.Н. О направлениях сомати- ческой эволюции покрытосемен- ных растений // Бюлл. МОИП, отд. биол., 1970, т. 75, М 2, с. 76-86. Цвелев Н.Н. О первичной жизненной форме покрьггосеменных растений // Бюлл. МОИП, отд. биол., 1977, т. 82, вып. 1, с. 79-88. Цицин Н.В. (отв. ред.) Методические указания по семеноведению интро- дуцентов. — М.: Наука, 1980, 64 с. Чайлахян МХ Основные закономерно- сти онтогенеза высших растений.— М.: изд-во АН СССР, 1958, 79 с. Чайлахян М.Х Факторы генеративного развития растений // Доложено .... 3 июня 1964 г. / Тимирязевские чте- ния, 25. — М.: Наука, 1964, 58 с. Чайлахян М.Х Целостность организма в растительном мире // Физиоло- гия растений. — 1980, т. 27, N 5, с. 917-941. Чайлахян М.L Регуляция цветения высших растений // Гормональная регуляция онтогенеза растений.— М.: Наука, 1984, с. 9-28. Чайлахян М.L Регуляция цветения высших растений. — М.: Наука, 1988, 560 с. Чайлахян М.Х, Аксенова Н.П., Кефели В.И. О терминологии онтогенеза растений. — М.: Наука, 1973, 39 с. Чайлахян М.L и др. Терминология роста и развития высших растений / М.Х. Чайлахян, Р.Г. Бутенко, О.Н. Кулаева, В.И. Кефели, Н.П. Аксе- нова. — М.: Наука, 1982, 96 с. Чевоениди С.Х Дубильные растения Средней Азии. — Ташкент: Наука, 1965, 330 с. Чеииков М.Г. Интеграция науки.— М.: Мысль, 1981, 276 с. Череианов СК Сосудистые растения Рос- сии и сопредельных государств.— СПб.: Мир и семья-95, 1995, 992 с. Черняховский М.Е. Морфолого-функ- циональные особенности жизнен- ных форм Саранчовых // Докл. МОИП, кн 2 (зоология и ботани- ка). — М.: МОИП, 1971, с. 20-23. Чижек Ф. О целостности вида и попу- ляции // Проблема целостности в современной биологии. — М.: На- ука, 1968, с. 194-237. Чиков 1Т.С., Лаптев Ю.1Т. Витамин- ные и лекарственные растения.— М.: Колос, 1976, 368 с. Чистякова А.А. Большой жизненный цика Tilia cordata Mill. // Бюлл. МОИП, отд. биол., 1979, т. 84, вып. 1, с. 85-97. Чистякова А.А. Жизненные формы и их спектры как показатели состоя- ния вида в ценозе на примере ши- роколиственных деревьев // Бюлл. МОИП, отд. биол., 1988, т. 93, N 6, с. 93-105. Шалыт М.С. Подземная часть некото- рых луговых, степных и пустынных растений и фитоценозов. 1. Травя- нистые и полукустарничковые рас- тения и фитоценозы лесной (луга) и степной зон // Тр. Бот. ин-та АН СССР. — М.; Л., 1950, сер. 3, вып. 6, с. 205-442. Шалыт М.С. Подземная часть некото- рых луговых, степных и пустынных растений и фитоценозов. II. Травя- нистые, полукустарничковые и ку- старничковые растения и фитоце- нозы пустынной зоны // Тр. Бот. ин-та АН СССР. Сер. 3. — М.; Л., 1952, вып. 8, с. 71-139. Шалыт М. С. Система жизненных форм степных растений // Уч. зап. Тадж. гос. унив., 1955, т. VI, Труды фак. естеств. наук, вып. 1. Шалыт М.С. Изучение вегетативного размножения и возобновления компонентов растительных сооб- ществ // Полевая геоботаника.— М.; Л.: изд-во АН СССР, 1960, т. 2, с. 163-208. Шалыт М.С. Партикуляция у высших растений // Проблемы современ- ной ботаники. — М.; Л.: Наука, 1965, т. 2, с. 117-122. Шалыт М.С., Калмыкова А.А. Корне- вая система растений в основных почвенных типах Украины // Бо-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ тан. журн. СССР, 1935, т. 20, N 4, с. 357-410. Шанцер Е.В. Современная геология и ее место в естествознании // Взаи- модействие наук при изучении Зем- ли. — М.: Наука, 1964, с. 93-118. Шарова И.Х. Жизненные формы и значение конвергенций и паралле- лизмов и их классификации // Журн. общ. биол., 1973, т. 34, М 4, с. 563-570. Шарова И.Х Жизненные формы жу- желиц (Coleoptera, Carabidae). М.: Наука, 1981, 360 с. Шахов А, А. Солеустойчивость расте- ний. — М.: изд-во АН СССР, 1956, 552 с. Шафранова Л.М. О метамерности и метамерах у растений // Журн. общ. биол., 1980, т. 41, N 33, с. 437-447. Шафранова Л.М. Ветвление растений: процесс и результат. Жизненные формы: структура, спектры и эво- люция. — М.: Наука, 1981, с. 179-213. Шафранова Л.М. Растение как жиз- ненная форма // Журн. общ. биол., 1990, т. 51, № 1, с. 72-88. Шафранова Л.М., Гатцук Л.Е. Расте- ние как пространственно-времен- ная (модульная) система // Успехи экологической морфологии и ее влияние на смежные науки. — М.: Прометей, 1994, с. 6-7. Шафранова Л.М., Гатцук Л.Е. Успехи экологической морфологии расте- ний и ее влияние на смежные на- уки // Бот. журн., 1995, т. 80, М 12, с. 131-133. Шварц С.С. Экология и эволюция.— М.: Знание, 1974, 64 с. Шеллинг Ф.В.И. Система трансценден- тального идеализма. — М.; Л.: Со- цэкгиз, 1936, 455 с. Шенников А.lT. Луговедение. — Л.: изд-во Ленинг. ун-та, 1941. Шилов И.А. Популяционный гомеос- таз у животных // Бюлл. МОИП, отд. биол., 1982, т. 87, вып. 4, с. 23-32. Шиманович И.Е., Павлович М.Л., Ти- кавый В.Ф., Малашко П.М. Общая химия в формулах, определениях, схемах. Учебное пособие. — Мн.: Ун1верс1тэцкае, 1996, 528 с. Шипчинский Н.В. Род Paeonia L. // Флора СССР. Т. VII. — М.: изд-во АН СССР, 1937. Шитт П.Г. Избранные сочинения.— М.: Колос, 1968, 584 с. Шитт П.Г. и Метлицкий 3.А. Плодо- водство. — М.: Сельхозгиз, 1940, 660 с. Шишкин Б.К. Род Порезник-Libanotis L. // Флора СССР. — М.; Л.: изд- во АН СССР, 1950, т. 16, с. 471-483. Шкловский И.С. Вселенная, жизнь, разум. 3-е изд. — М.: Наука, 1973, 335 с. Шмалыаузен И.Ф. Флора Средней и Южной России, Крыма и Северно- го Кавказа. — Киев, 1895, т. 1, 468 с. Шмалыаузен И.И. Интеграция биоло- гических систем и их саморегуля- ция // Бюлл. МОИП, отд. биол., 1961, т. 66, вып. 2, с. 104-134. Шмалыаузен И.И. Кибернетические вопросы биологии. — Новосибирск: ~1аука, 1968, 223 с. Шмалыаузен И.И. Проблемы дарви- низма. 2-е изд. — Л.: Наука, 1969, 493 с. Шмалыаузен И. И Организм как целое в индивидуальном развитии. Из- бранные труды. — М.: Наука, 1982, 383 с. Шмалыаузен И.И. Пути и закономер- ности эволюционного процесса. Избранные труды. — М.: Наука, 1983, 360 с. Шмитхюзвн И. Общая география рас- тительности. Пер. с немец. — М.: Прогресс, 1966, 310 с. Шорина Н.И. Жизненный цикл без- временника великолепного (Colchi- сит speciosum Stev.) в условиях су- бальпийских лугов Западного За- кавказья // Науч. докл. высш. шко- лы. Биол. науки, 1964, № 1, с. 113-119. Шорина ЫИ. Жизненный цикл без- временника великолепного в лес- ном и субальпийском поясах Запад- ного Закавказья // Сб. "Онтогенез и возрастной состав популяций цветковых растений". — М.: Наука, 1967, с. 70-99. Шредингер Э. Что такое жизнь с точки зрения физики. 2-е изд. — М.: Ато- миздат, 1972, 89 с.
Jlumepamypa Шретер А.И. Пион уклоняющийся // Атлас лекарственных растений СССР. — М.: Медгиз, 1962, с. 434. Шретер А.И., Шретер Г.К. Распрост- ранение и режим эксплуатации за- рослей заманихи высокой на Даль- нем Востоке // Ресурсы дикорасту- щих лекарственных растений. — Л.: Наука, 1968, с. 165-167. Штофф В.А. Роль моделей в позна- нии. — Л.: изд-во Ленинг. гос. ун- та, 1963, 128 с. Шугаева Е.В. Цитоэмбриология неко- торых лекарственных видов рода Valeriana L.: Автореф. дис.... канд. биол. наук. — М., 1985, 24 с. Шулькина Т.В. Изменение жизненной формы у Aster alpinus L. при выра- щивании ее в Ленинграде // Про- блемы совр. ботаники. — М.' Л.: Наука, 1965, т. 2, с. 126-!29. Шульц Г.Э. Общая фенология. — Л.: Наука, 1981, 188 с. Щвдровицкий Г.П. Проблемы методо- логии системного исследования.— М.: Знание, 1964, 56 с. Щвдровицкий Г.П. Принципы и общая схема методологической организа- ции системно-структурных иссле- дований и разработок // Систем- ные исследования. Ежегодник, 1981. — М.: Наука, 1981, с. 193-227. Щербакова А.А. История русской бо- танической (морфологической) терминологии // Тр. ин-та истории естеств. и техники АН СССР. — М., 1960, т. 32, вып. 7, с. 203-250. Щербакова А.А. История ботаники в России до 60-х гг. XIX в. (додарви- новский период). — Новосибирск: Наука, 1979, 368 с. Щербакова А.А., Базилввская Н.А., Калмыков К.Ф. История ботаники в России (дарвиновский период, 1861-1917). — Новосибирск: Наука, 1983, 366 с. Эзау К Анатомия семенных растений.— М.: Мир, 1980, т. 1-2, 558 с. Эйхвальд КЮ. Подрод ежевик Cylactis Raftn. Исследование филогенеза одной бореальной растительной группы // Уч. зап. Тартус. гос. ун- та, 1959, вып. 81, 285 с. Эйген М. Самоорганизация материи и эволюция биологических макромо- лекул. — М.: Мир, 1973, 216 с. Энгельс Ф. Диалектика природы. М.: Политиздат, 1964, 358 с. Югай Г.А. Проблема целостности организма. Философский анализ.— М.: Соцэкгиз, 1962, 248 с. Югай ГА. Диалектика части и целого.— А.-А.: Наука, 1965, 171 с. Югай Г.А. Философские проблемы теоретической биологии. — М.: Мысль, 1976, 247 с. Югай Г.А. Принципы сохранения в биологии и медицине // Вест. АМН СССР. — 1979, Хо 12, с. 51-59. Юдин Б.Г. Становление и характер системной ориентации // Систем- ные исследования. Ежегодник, 1971. — М.: Наука, 1972, с. 18-34. Юрцвв Б.А. Жизненные формы: один из узловых объектов ботаники // Про- блемы экологической морфологии растений. — М.: Наука, 1976, с. 9-44. Яблоков А.В. Актуальные проблемы эволюционной теории. — М.: Зна- ние, 1966, 32 с. Яблоков А.В. Фенетика. Эволюция, популяция, признак. — М.: Наука, 1980, 132 с. Яблоков А.В., Познанин Л.1Т. Особен- ности современного этапа развития эволюционной теории // Итоги науки и техники; сер. зоология позвоночных, т. 7. — М.: ВИНИ- ТИ, 1975, с. 5-28. Яблоков А.В., Юсуфов А.Г. Эволюци- онное учение. 2-е изд. — М.: Выс- шая школа, 1981, 343 с. Якубова А.И. Главнейшие лекарствен- ные растения // Растительные бо- гатства Новосибирской области.— Новосибирск: изд-во СО АН СССР, 1961, с. 125-155. Якубова А. И, Минаева В.Г., Костромина М.М. Горбалева Г.Н. Патриния.— Новосибирск: Наука, 1967, 106 с. Яценко-Хмелевский А.А. Краткий курс анатомии растений. — М.: Высшая школа, 1961, 280 с. Aldrovandi U. Dendrologiae naturalis scilicet arborum historiae libri duo.— Francofurti, 1671, 468 р.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Allent В. Les vegetaux curieux. — Paris, 1826. Alpino P. De plantis exoticis libri duo.— Venetiis, 1629, 344 р. Arber А. Root and shoot in the Angio- sperms: а study of morphological cate- gories // New Phytologist.,1930, vol. 29, N 5, р. 297-315. Arber А. The Gramineae: а study of сегеа1, bamboo and grass. — Cam- bridge, 1934, 480 р. Arber А. The interpretation of leaf and root in the Angiosperms // Biol. Rev., 1941, чо1. 16, N2, ,р. 81-105. Arber А. The na�га1 philosophy of plant form. — Cambridge, 1950, 247 р. Agardh С.А. Species Algarum. 1821, vol. 1, 531 р. Areschoug F. Ж Beitrage zur Biologic der geophilen Pflanzen // Acta Univ. Lundensis. — 1895, Bd. 31. Bailey I.W. The evolutionary history of the foliar ray in the mood of the dico- tyledons and its phylogenetic signifi- cance // Ann. Bot., 1912, 26. Bauhin J. et Cherler ХК Historia plan- tarum universalis. Т. 1. — Ebrodini, 1650, 440 р. Bechstein ХМ. Forstbotanik oder vol- lstandige Naturgeschichte der deuts- chen Holzpflanzen und einiger frem- den. — Erfurt, 1810, 1456 S. Bentham G. Scrophulariaceae // В кн. De Candolle А-P. et De Candolle А. Prodromus sistematis naturalis regni vegetabilis; pars 10. — Paris, 1846, 679 р. Bentham G. et Hooker Х1). Genera plan- tarum. 3. — London, 1883. Bertalanß L. Zu einer allgemeine Sys- temlehre // Biologia Generalis. 1949, Bd. 19, S. 114-129. Вейа(апф L. General System Theory // General Systems, 1956, чо1. 1, р.1-10. Bertalanßó L. General System Theory. Foundations, Development, Applica- tions. — London, 1971. Bischo+ G. W. Handbuch 4ег botanischen Terminologie und Systemkunde.— Nurnberg, 1830, Bd. 1-2, 260 S. Bischo+ G. ~KLehrbuch der allgemeinen Botanik. Stuttgart, 1834, Bd. 1, 479 S. Bischoff G. WWorterbuch der beschrei- benden Botanik, oder die Kunstaush driicke, welche zum Verstehen der phytographischen Schriften notwendig sind. — Stuttgart, 1839, 283 S. Boissier Е. Нога orientalis. Vol. 1-3.— Genevae et Basileae, vol. 1 — 1867, vol. 2 — 1872, vol. 3 — 1875. Вопйаг G. New Palms of Bahia // Botanical series. Field museum of natural history. — Chicago, 1942, vol. 22, N9, ,р. 457-463. Boros G. Lexikon der Botanik mit beson- derer Beriicksichtigung der Verer- bungslehre und der angrenzenden Gebiete. — Stuttgart, 1959. Вгаип А. Betrachtungen iiber die Erschei- nung der Verjiingung in der Natur.— Leipzig, 1851, 363 S. Braun-Blanquet J. Pflanzensoziologie. Grundziige der Vegetationskunde.— Wien: Springer, 1951, 631 S. Bugnon P. La theoric de la "leaf-skin" et le cas du Buddleia lindleyana // Bull. Soc. Linneenne de Normandie. Ser. 7, 1925, N 8. Bulliard P. Dictionnaire elementaire de botanique ou exposition alphabetique des preceptes de la botanique. — Paris, 1783, 242 р. Bullock S. Demography of an under- growth palm littoral Cameroon // Biotropica, 1980, vol. 12, N 4, р. 247-255. Burret М. Palmae cubensis et Domin- genses а се Е.L. Ekman 1914-1928 lectae // Kungl. svenska vetenskap- sakademiens Handlingar; tredja serien, 1929, чо1. 6, N 7. Caesalpini А. De plantis libri 16. Florentiae, 1583, 621 р. Саппоп ЖВ. The wisdom of the body.— London, 1932, 312 р. Caton М.P. De Agricultura. — Leipzig, 1922. Clements F.P. Plants Indicators, the relation of plants communities to process and practice. Carnegy Inst. Wasch. Publ., 290, 1920. Chauveaud G. La constitution et 1'evolu- tion morphologique du corps chez les plantes vasculaires // С.R. Acad. sci., Paris, 1914, t. 158, N 5. Columella ЕЗ.М. De rei rustica libri 12 et liber de arboribus. — Venetiis, 1523. Commelin J., Commelin С. Horti medici amstelodamensis rariorum tarn Orien-
Литература talis, quam Occidentalis Indiae, alia- rumque peregrinarum plantarum mag- no studio ас 1аЬоге, sumtibus eivitatis amstelodamensis longa annorum serie collocatarum discriptio et icones ad vivum aeri incisae. — Amsterodami, 1697 — pars 1 (J. Commelin), 1701— pars 2 (С. Commelin). Cspapody V. Keimlungsbestimmungsbuch der Dikotyledonen. — Budapest, 1968, 286 S. Darwin Ch. On the origin of species by means of natural selection. — London, 1859, 502 р. Перев: Дарвин Ч. Происхождение видов путем естес- твенного отбора. — М.: Просвеще- ние, 1986, 383 с. Dawson J.W On fossil plants from the Devonian Rocks of Canada // Quar- terly Journ. Geol. Soc. of London.— 1859, чо1. 15. De Candolle А.-Р. Theoretische Anfangs- griinde der Botanik. — Ziirich, 1815. De Candolle А.-Р. Theoric elementaire de la botanique on Exposition des prin- cipes de lа classification na�relle et de Гаг( decrire et d'е�dier les vegeta- ux. 2 ed. — Paris, 1819, 566 р. De Candolle А.-Р. Organographie vegeta- le. Cours de Botanique. — Paris, 1827, чо1. 1, 558 р. Репеек D. Morphologic. In: Lehrbuch der Botanik. — Jena, 1967, t. 1, S. 3-202. Dietrich А. Terminologie der phaneroga- mischen Pflanzen. — Berlin, 1838, 128 S. Dode L.À. Extraits d'une monographic ипе d'ite de genere Populus // Bull. de lа Soc. d'histoire nat. d'Autumn., 1905, t. 108, 73 р. Dodonaeus R. Stirpium historiae pempta- des sex, sive libri 30. — Antwerpiae, 1583, 860 р. Drude О. б1со1офе der Pflanzen // Die Wissenschaft. — Braunschweig, 1913, Вд. 50, 308 S. Duborg В. Le botaniste francois. Т. 1.— Paris, 1767. Du Rietz G.Е. Life forms of terrestrial flowering plants. — Uppsala: Almqvist and Wiksell, 1931. Dupetit-Thouars L.M. Essais sur lа vege- tation consideree dans le development des bourgeons. — Paris, 1809, 304 р. Eatnes А.1 Again: "The пеъ morpho- logy" // The new Phytol. — 1951, vol. 50, N 1, р. 17-35. Ehrhart Е. Beitrage zur Naturkunde, und den damit Verwandten Wissenschaf- ten, besonders der Botanik, Chemic, Haus-und Landwirtschaft, Arzneige- lahrheit und Apothekerkunst. — Han- nover und Osnabriick, 1789, Bd. 4, 184 S. Ehrhart Е. Beitrage zur Naturkunde, und den damit Verwandten Wissen- schaften, besonders der Botanik, Che- mic, Haus-und Landwirtschaft, Ar- zneigelahrheit und Apothekerkunst.— Hannover und Osnabriick, 1792, Bd. 7, 184 S. Eichler А. W. Bliitendiagramme. 1 Theil.— Leipzig, 1875. Endlicher ХХ,. Enchiridion botanicum exhibens classes et ordines plantarum, accedit nomenclator generum et offi- cinalium vel usualium indicatio. Lipsiae, 1841. Esau К. Plant anatomy. 2-ed. — New York — London — Sydney, 1965, 767 р. Feinbrun - Dothan N., Danin А. Analytical Нога of Eretz-Israel. — Jerusalem, 1991, 1040 р. Foster А.S. Leaf differentiation in Angio- sperms // Bot. Rev., 1936, vol. 2, N 7. Foster À.S. Practical plant anatomy. 2-ed.— New York, 1949, 228 р. Gams H. Prinzipienfragen der Vegata- tionsforschung // Vierteljahrschr. Naturforsch. Ges. — Ziirich, 1918. Bd. 63, S. 293-493. Gaudichaud Ch. Recherches generales sur Гог~аподгарЫе, lа physiologic et Гог- ganogenie des vegetaux // Ann. Sci. Nat. (Bot.). Ser. 2. — Paris, 1841, t. 15. Gluch ~K Wuchsformstudien an zentra- leuropaischen Fabaceen. Е Die stau- den der Gattung Medicago L. und Trifoliutn L. // Feddes repert., 1967, Вд. 75, N 3, S. 221-265. Gmelin J.G. Нога Sibirica, sive historia plan- tarum Sibiricae. 1. — Petropoli, 1747. Goebel E Organographie der Pflanzen. Teil Е Allgemeine organographie.— Jena, 1928, 642 S. Goebel К. Bliitenbildung und Spros- sgestaltung (Anthokladien ип4 Inflo-
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЬТХ РА СТЕНИИ reszenzen). Zweiter Erganzungsband zur Organograplue дег Pflanzen.— Jena, 1931, 242 S. бйи1Ьег К.-F. Beitrage zur Morphologic und Verbreitung der Рарачегасеае 1 Teil: Infloreszenzmorphologie дег Ра- рачегасеае; Wuchsformen der Cheli- donieae // Нога, 1975, Bd. 164, Heft 2/3, S. 185-234. Gustafsson А. Polyploidy, life form and vegetative reoproduction // Hereditas, 1948, Bd. 34, N 1/2, р. 1-22. Halle F. The concept of architectural models in vascular plants // XII Между- нар. ботан. конгр. / Тезисы докл.— Л.: Наука, 1975, т. 1, с. 216. Halle F., Oldeman R.À.À. Essai sur Гаг- chitectuãå et lа dynamique de crois- sance des arbres tropicaux. — Paris, 1970, 178 р. НаИе F., Oldeman R.À.À., Tomlinson P.В. Tropical trees and forests: An archi- tectural analysis. — Heidelberg: Sprin- ger, 1978, 441 р. Harper ХХ,. Factors controlling plant numbers. The biology of weeds. — Ox- ford, 1960. Нагрег ХХ,. The regulation of numbers and mass in plant populations: Popu- lation biology and evolution. — Буга- cuse, 1968, 205 р. Harper ХХ,. Population biology of plants. London — N.- York — San-Francisco: Асад. press, 1977, 892 р. Нагрег ХХ,. The demography of plants with сlопаl growth // Biorheology, 1978, vol. 15, N 2, р. 27-48. Нагрег ХХ,., White J. The dynamics of plant populations. Proc. Adv. Study Inst. Dynamics Number Рори1. (Oos- terbeek). 1970. Нагрег Ш, White J. The demography of plants // Аппиа1 Rev. of Ecol. and Syst., 1974, N 5, р. 419-463. Heeger Е.F. Handbuch des Arznei — und Gewiirzpflanzenbaues. Drogengewin- nung. — Berlin: Deutsch. Bauernverl., 1956, 775 S. Henderson ХР. and Henderson ЖЮ А dictionary of scientific terms. London, 1957. Но~теигег W. Allgemeine Morphologic der Gewachse. — Leipzig, 1868. Hoffmansegg ХС. et Link Н.F. Ноге portugaise ou description de toutes les plantes qui croissent naturellement en Portugal. — Berlin, 1809. Норре Н.А. Drogenkunde. — Hamburg, 1958. Irmisch Th. Zur Morphologic der топо- kotylischen Knollen - und Zwiebel- gewachse. — Berlin, 1850. Irmisch Th. Bemerkungen iiber einige Wassergewachse // Bot. Ztg., 1859, Bd. 17. Jeffrey Е.С. The morphology of the central cylinder in the angiospeims.— Trans. Canad. Inst., 1899, чо1. 6. Jonston J. Historia naturalis de arboribus et plantis libri 10. Ed. II. — Fran- cofurti, 1768. Jost L. Die Уerkliiftungen einiger Rhi- готе und Wurzeln // Bot. Ztg., 1890, Bd. 48, N 28-32. КасЫег J. Grundriss дег Phlanzenkunde, in Gestalt eines Worterbuch дег bota- nischen Sprache. — Wien, 1830. Kaussmann В. Pflanzenanatomie. Jena, 1963, 624 S. Кейп D. The Leuwenhoek Lecture. The problem of anabiosis of latent 1ие: history and current concept // Proc. Roy. Soc. Ser. В, 1959, vol. 150, N 939, р. 149-191. Klebs G. Beitgrage zur Morphologic und Biologic дег Keimung. Arbeiten d. botan. Jnst. in Tiiringen. — 1885. Kruger J.F. Kleines Handworterbuch botanischen Kunstsprache und Pflan- zennamen. — Quedlinburg, 1859. Кит А., Altman А., Putievsky Е. Rooting and initial establishment of stem cuttings of oregane, peppermint and balm // Sci. north (Neth.), 1980, vol. 13, N 1, р. 53-59. Kuris А., Altman А., Putievsky Е. Vegeta- tive propagation of spice-plants: root formation in oregane stem cuttings // Sci north (Neth.), 1981, vol. 14, N 2, р. 151-156. Kutzing F.Т. Grandziige дег philosophis- chen Botanik — Leipzig, 1852, Bd. 2, 344 S. Lamarck ХВ. Ноге francoise, ou descrip- tion succinte de toutes les plantes qui croissent naturellement en France.— Paris, 1795, t. 1-3.
Литература Lamouroux J.P. Resume complet de botanique. 1. — Paris, 1826. Laszlo Е. Introduction to systems philo- sophy: Toward а new paradigm of contemporary thought. — New- York, 1972, 328 р. Le Maout Е. Lecons elementaires de botanique fondees sur Гanalyse de 50 plantes vulgaires et formantu un traite complet d'organographie et de physio- logic че~е~а1е. — Paris, 1844. Le Maout Е. Botanique (Organographie et taxonomic). — Paris, 1852, 387 р. Lecoq H. et Juliet J. Dictionnaire raisonne des termes de botanique et des families па�relies. — Paris, 1831. Lindman С.А.М. Magra bidrag till fragan: buske е11ег trad. Ventenskapsakade- miens Arsbok, 1914, 12. Link Н.F. Grundlehren der Anatomic und Physiologic der Pflanzen. Gottingen, 1807, 305 S. Link Н.Р. Elementa phylosophiae bota- nicae. — Berolini, 1824, 486 р. Linkola К Zur Kenntnis der Uber- winterung der Unkrauter und Ruderal- pflanzen in der Gegend von Helsing- fors // Soc. Zoolog. Bot. Fenn. Vana- mo, 1922, Bd. 1, N 7, S. 91-228. Linnaeus С. Philosophia botanica in qua explicantur fundamenta botanica curn definitionibus partium, exemplis. Stockholmiae, 1751, 362 р. Linnaeus С. Species plantarum. — Hol- miae, 1753, vol. 1-2, 1200 р. Linnaeus С. Desans prima plantarum rariorum horti Upsaliensis. — Stock- holmiae, 1762, 40 р. Linnaeus С. Termini botanici explicati.— Lipsiae, 1767, 39 р. Lyndley J. An о�line of the first principle of botany. — London, 1830, 106 р. L'Obel М. Plantarum seu stirpium his- toria. Cui adnexum est adversariorum volumen. — Antwerpiae, 1576. McClure F.À. The bamboos: а fresh perspective. — Cambridge, 1966. Madaus G. Lehrbuch дег biologischen Heilmittel. — Leipzig, 1938, Bd. 2, S. 903-1884. Martius К.F. Ph. Historia naturalis Palmarum. — Lipsiae, 1823-1850. Masters M Vegetable teratology an account of the principal deviations from the usual сапйпийюп cf phnh — london, 1869, 534 р. Meusel Н. Wuchsformen und Wuchstypen der europaischen Laubmoose // Nova acta Leopoldina. N.F., 1935, Bd. 3, N 12, S. 123-277. Meusel Н. Die Bedeutung der Wuchsform fur die Entwicklung des natiirlichen Systems der Pflanzen // Feddes Ee- pert., 1951, Bd. 54, N 2-3, S. 137-172. Meusel H Uber Wuchsform und Wuchs- dauer mediterraner Einjahrspflanzen // Wiss. Z. Martin — Luther Univ.— Halle — Wittenberg, 1955, Bd. 4, N 3, S. 645-650. Monterosso В. L'anabiosi nei Cirripodi ie problema della vita latente (ipobiosi) // Arch. 2оо1. Ital, 1934, чо1. 19, р. 17-379. Moench К Einleitung zur Pflanzenkunde.— Marburg, 1798. Muhlberg H. Wuchsformenstudien in der Familie Poaceae. Die Wuchsfor men der mitteldeutschen Poaceae Arten // Fed- des. Repert., 1965, Bd. 71, S. 188-217. Mu'hlberg H. Die Wuchstypen der mittel- deutschen Poaceen // Hercynia, N.F., 1967, Вд. 4, N 1, S. 11-50. Muhlberg Н. Wuchsformen der Gattung Brachypodium (Poaceae) // Feddes Repert., 1970, Bd. 81, N 1-5, S. 119-130. Жиде!~ С. Die neuern Algenayateme.— Zurich, 1847, 275 S. Жауе!~ С. Beitrage zur wissensehaftlichen Botanik. — Leipzig, 1860, Heft. 2, 192 S. Nageli С. Beitrag zur Morphologic und Systematik der Ceraniaceae. Bota- nische Mitteilungen. — Miinchen, 1863, Bd. 1, S. 63-181. Жеуп G. Interpretazione indinidualistica del paesaggio vegetali. // Nuovo giorn. bot. ital., 1954, чо1. 61. Noble J.С., Bell А.D., Нагрею ХХ,. The population biology of plants with clonal growth. Е The morphology and structural demography of Carex are- папа // J. Ecol., 1979, vol. 67, N 3, р. 983-1008. Norton G.À. Toward а concept of strategic resource planning // International Jornal of Enviromental Studies, 1973, чо1. 4, р. 189-200.
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Oliver D. Lessons in elementary Botany.— London, 1888, 300 р. Osborn ЫР. The ideas and terms of modern philosophical anatomy // Science, 1905, чо1. 21, N 547, р. 959-961. Parkin J. The Evolution of the inflores- cence // J. Linnean Soc. London (Bot.), 1914, vol. 42, N 287. Parkin J. The protrusion of the connective beyond the anther and its bearing on the evolution of the stamen // Phytomorph., 1951, чо1. 1, N 1, 2, р. 1-8. Репа P. et L''Obel М. Stirpium adversaria nova, perfacilis vestigatio, luculenta- que accesio ad priscorum, praesertim Dioscoridis et recentiorum materiam medicam. — Antwerpiae, 1576. Penzes А. Uber die Morphologic. Dynamik und Zonologische Rolle der Spross Kolonienbildenden Pflanzen. (Poly- connone) // Fragm. florist. et geobot., 1960, Апп. 6, Pars 4, S. 501-515. Peterson Н.Е. Indledende studier over polymorphien hos Anthriscus sylvestris (L.) Hojfin. // Dansk. bot. aktiv, 1915, 1, N 6. Peterson Н.Е. Etudes ultrerieures sur lа polymorphie de Anthriscus sylvestris (L.) НоЯЬп. // Dansk. bot. aktiv, 1922, t.4,N2. Petrev А. On the method of the morpho- logical analysis of the structure of hetbaceus plants // Comptes rendus Асад. Bulg. Sci, 1964, vol. 17, N 9. Philibert ХС. Lecons elementaires de betanique аГиза~е des cours publics et particuliers et des ЕсоГез cu Lyce'es.— Paris, 1802, 544 р. Plate L. Selectionsprinzip und Probleme der Artbildung. 4 Aufl. Leipzig und Berlin, 1913, 650 S. Plinius S.С. Naturalis historia. Libri 1-37.— Venezia, 1571. Potoni'e Н. Die Репсаи1от — Theoric // Вег. Deutsch. Bot. Ges., 1902, Bd. 20. Potonie H. Grundlinien der Pflanzen- Morphologie im Lichte der Palaeon- tologie: Jena, 1912, 259 р. Рип V. Floral anatomy and inferior ovary // Phytomorph., 1952, vol. 2, N 2, 3, р. 122-129. Rauh W. Beitrage zur Morphologic und Biologic der Holzgewachse. Е Entwick- lugsgeschichte und Verzweigungsver- halthisse arktisch-alpiner Spalies- traucher // Nova Acta Leopold. N.F., 1937, Вд. 5, N 30, S. 290-347. Rauh W. Beitrage zur Morphologic und Biologic der Holzgewachse // Nova Acta Leopold. N.F., 1938, Bd. 5. Rauh W. (Ъег polsterfdrmigen Wuchs // Nova Acta Leopold. N.F., 1939, Bd. 7. Rauh W. Morphologic der Nutzpflanzen.— Heidelberg, 1950, 290 S. Raunkiaer С. Types biologiques pour lа g6ographie botanique // Det Kongelige Danske vidensk. selsk. Forhandl., 1905 N 5. Raunkiaer С. Planterigets Livsformer of deres Betydning for Geografien.— Kebenhavn, 1907, 132 р. Roux W. Der Kampf der Teile im Orga- nismus. — Leipzig, 1881, 244 S. Richard А. Nouvesus elements de bota- nique et de physiologic vbgetale.— Paris, 1819, 410 р. Roeper ХА. Ch. Observationes aliquot in florum inflorescentiarumque naturam // Linnaea, 1826, чо1. 1, р. 433. Roeper ХА. Ch. De organis plantarum.— Basileae, 1828. Sachs I. Lehrbuch der Botanik. Leipzig, 1868, 628 S. Sachs I. Lehrbuch der Botanik. Ed. 2.— Leipzig, 1870, 668 S. Saint-Hilaire А. Lemons de botanique comprenant principalee ment la тог- phologie vegetale, lа terminologie, la botanique сотрагее, 1'examen de la valeur des caracteres dans les diverses families naturelles. — Paris, 1841, 930 р. Saunders Е. The leaf-skin theory of the stem: а consideration of certain ana- tomico-physiological relations in the Spermatophyte shoot // Ann. Bot., 1922, vol. 36, N 142, р. 135-165. Scharfetter R. Biographien чоп Pflanzen- sippen. — Wien, 1953, 546 S. Schindler F. Die Lehre vom Pflanzenbau, auf physiologischer Grundlage. Wien, 1896. БсЫсиЬг Ch. Vier und zwanzigste klasse des Linneischen Pflanzensystems oder Kryptogamischen Gewachse. — Wet- tenberg, 1809, Bd. 1, 212 S.
Литература Schmid Е. Die Wuchsformen der Dicoty- ledonen // Bericht geobot. Forschungs- institut Riibel in Zurich. 1956. Schmid Е. Die Erfassung der Vegetation- seinheiten mit floristischen und epi- morphologischen Analysen // Berichte d. Schweiz. bot. Ges.„1963,Bd. 73, S. 276-324. Schmida А. Handbook of Wildflowers of Israel Mediterranean Нога. — Ierusa- lem, 1986, 308 р. Schultz С. Die Anaphytose oder Ver- jiingung der Pflanzen. — Berlin, 1843. Spath H. 0ег Johannistrieb. — Berlin, 1912, 91 S. Tiell W. Vergleichende Morphologic der hoheren Pflanzen. Bd. 1. Vegeta- tionsorgane. — Berlin, 1937, Teil 1. Tiell W. Vergleichende Morphologic der hoheren Pflanzen. Bd. 2. Vegeta- tionsorgane. — Berlin, 1939, Teil 2. Tioll W. Praktische Einfuhrung in die Pflanzenmorphologie Bd. 1. 0ег vege- tative Aufbau. — Jena, 1954, 258 S. Tioll W. Die Infloreszenzen. — Jena, 1964, Вд. 1, 615 S. Tioll W. Die Infloreszenzen. — Jena, 1969, Вд. 2, t. 1, 630 S. Troll W., Weber H. Morphologic ein schliesalich Anatomic Fortschritte der Botanik. — Berlin, 1959. Urbanska- Worytkieeicz К Reicherches demographiques en botanique: certains aspects et implications evolutives // Bull. Soc. bot. France, 1979, t. 126, N 4, р. 445-451. Velenovsky J Vergleichende Morphologic der Pflanzen. Teile 1-4: 1 — 1905; 2— 1907, S. 280-731; 3 — 1910, S. 732-1211; 4 — 1913. Prage, 1905-1913. Warming Е. Uber perenne Gewachse // Bot. Centralblatt, 1884, Bd. 18, N 19, S. 16-22. Warming Е. От planterigets lifsformer. Kjobenhavn, 1908, 86 S. Warming Е. Oecology of plants. Oxford: Univ. press, 1909, 422 р. Warming Е. Bemaerkningen of Livsform og Standplands // Forhandl. ved det Skandinavske Naturf. 191б, Bd. lб. Warming Е. От Jsdudloebere // Kgl. Danske vid. selskab. Biol. Ясг., Ser. 8, II, 1918, N 3. Warming Е. 61со1орепв grundformer. Udkast til en systematisk ordning // Det Kongelige Danske vidensk. sels- kabs йлйег Naturvidensk. og mat- hem. afd. 8 Raekke., 1923, Bd. 4, N 2, р. 119-187. Warming Е. и Graebner Р. Lehrbuch der okologischen Pflanzengeographie. 3. Aufl. — Berlin, 1918, 762 S. Weatherwax Р. The story of the maize plant. — Chicago, 1923. Weberling F. Die Infloreszenzen дег Valirianaceen und ihre systematische Bedeutung Wiesbaden // Abhandlun- gen der Akademie der Wissenschaften und der Literatur (Matematisch- Natur-Wissenschaftliche Klasse). Jahrgang, 1961, N 5, S. 1-281. Weismann А. 0Ьег Germinal-Selection, cine Quelle bestimmt gerichteter Variation. — Jena, 1896. Wettstein R. Handbuch der Systematis- chen Botanik. — Leipzig; Wien, 1908. Willdenoe СЬ. Grundriss der Krauter- kunde zur Vorlesungen entworfen. 3 Aufl. — Berlin, 1802. iesner I. Elemente der wissenschaftli- chen Botanik. Bd. 2. Elemente der Organographie, Sistematik und Bio- logic der Pflanzen. — Wien, 1884, 449 S. Wydler H. Uber dichotome Verzweigung der Bliithenaxen (cymose Inflorescenz) dicotyledonischer Gewachse // Lin- naca, 1843, Bd. 17. Wydler H. Uber die simmetrische Ver- zweigungsweise dichotomer Inflores- zenzen // Flora, 1851, В4. 34, S. 305- 312, 321-330. Wygand А. Der Baum. Betrachtungen iiber Gestalt und Lebensgeschichte der Holzgewachse. 1854. Zimmermann W. Die Phylogenie der Pflanzen. Ein Uberblick uber Tat- sachen und Probleme. — Jena, 1930, 452 S. Zimmermann W. Geschichte дег Pflan- zen. — Stuttgart, 1949, 586 S. Zohary M. Hydro-economical types in the vegetation of Near East deserts Sympos // Biol. And Product. of hot and cold desert. — London, 1952.
Указатель латинских названий растений Abies nephrolepis (Trautv.) Maxim. 501 Ainus glutinosa (L.) Gaertn. 382 Ainus incana (L.) Moench 203 Alyssum murale Waldst. et Kit. 172, 173, 286 Amygdalus communis L. 200, 232 Abies sibirica Ledeb. 184, 333, 364, 442, 443, 597 Anabasis salsa (С.А. Меу.) Benth. ex Volkens 294 Anemone dichotoma L. = Anemonidium dichotomum (L.) Но1иЬ 119 Angelica decurrens (Ledeb.) В. Fedtsch. 472 Anisantha tectorum (L.) Nevski 152, 437 Aconitum altaicum Steinb. 178, 179, 316, 377 Anthriscus Pers. 601 Aconitum decipiens Worosch. et Anfalov 377, 378 Anthriscus sylvestris (L.) Hoffm. 53, 56, 160, 164, 168 Acroptilon герепв (L.) DC. 296, 312 Adansonia digitata L. 259, 574 Adonis aleppica Boiss. 395 Arctium lappa L. 308 Arctium minus (Hill) Bernh. 309 Arctium tomentosum Mill. 307 Adonis sibirica Patrin ex Ledeb. 342, 513, 516, 517 512, Artemisia annua L. 267, 270 Artemisia absinthium L. 435, 436 Artemisia arborescens L. 435 Artemisia frigida Willd. 472 Artemisia herba alba Asso. 596 Artemisia lerchiana Web. 169, 183, 184, 285, 386 Alliaria petiolata (Bieb.) Сачага et Grande 217, 227 Allium cepa L. 314 Allium sativum L. 314 Allium schoenoprasum L. 388 Acalypha hispida Burm. 221 Асег ginnala Maxim. 285 Асег insigne Bunge 133 Асег negundo L. 134, 234, 290 Асег platanoides L. 285 Achillea millefolium L. 139, 242 Adonis vernalis L. 294,512 Adonis wolgensis Stev. 285 Aegopodium podagraria L. 605 Aesculus hyppocastanum L. 218, 219, 285 Aethusa cynapium L. 307 Agropyron fragile (Roth) P. Candargy 184 Ailanthus altissima (Mill.) Swingle 285 Ajuga septans L. 155, 281 Artemisia pauciflora Web. 586 Artemisia terrae-alba Krasch. 294 Artemisia vulgaris L. 150, 250, 435 Arundo donax L. 411 Asperugo procumbens L. 134 Aster alpinus L. 611 Astragalus alpinus L. 225
Bupleurum bicaule Helm 295 Bupleurum exaltatum Bieb. 295, 296 Вцр1ецгшп longifolium L. 160, 294 Bupleurum rotundifolium L. 311 Cakile maritima Scop. 434 Calendula oAicinalis L. 208 Cotoneaster uniflorus Bunge 525 Crassula lycopodioides Lam. 136 Crataegus sanguinea Pall. 132, 237 Callistemon врес1овит (Sams) DC. 228, 430 Crepis lyrata (L.) Froel. 342 Crocus vernus (L.) Hill 315 Указатель латинских названий растений Astragalus macropodium Lipsky 596 Astragalus testiculatus Pall. 300, 308 Atriplex сапа С.А. Меу. 294 Aurinia saxatilis (L.) Desv. 248 Begonia rex Putz. 159 Berberis яЫпса Pall. 291, 525 Berberis vulgaris L. 242, 445 Bergenia crassifolia (L.) Fritsch 294 Beta vulgaris L. 307 Betula pendula Roth 203, 221, 227, 237, 285 Betula rotundifolia Spach. 480 Biscutella digyma L. 395 Brassica о1егасеа L. 62 Bromopsis inermis (Leyss.) Но1иЬ 472 Brunerra sibirica Stev. 74, 293 Bupleurum L. 439, 590 Bupleurum aureum Fisch. ex Hoffm. = Bupleurum longifolium L. subsp. aureum (Fisch. ex Hoffm.) So6 463, 464, 465, 466, 470 Bupleurum fruticosum L. 261, 262, 263-265, 285, 292, 339 Вир1еигшп multinerve DC. 203, 322, 363, 368-370, 371, 438, 460, 461, 462, 472, 597 Вир1ецгшп triradiatum Adams ex Hoffm. 335, 337, 460 Calicotome villosa (Poiret) Link 227, 395, 398, 399, 400, 412 Camellia japonica L. 259 Campanula glomerata L. 342 Callianthemum angustifolium Witas. 249 Capparis herbacea Willd. 142, 150, 160, 182, 244, 285, 286, 296, 302, 303, 386 Capsella bursa-pastoris (L.) Medik. 311 Сагех pilosa Scop. 605 Caragana arborescens Lam. 285, 602 Cataranthus rosea L. 208 Centaurea cyanus L. 255 Centaurea jacea L. 237 Centaurea procurrens Sprengel 395, 405, 406 Centaurea scabiosa L. 155 Centaurium erythraea Rafn 247 Cerastium dichotomum L. 119 Ceratonia siliqua L. 394 Choriospora sibirica (L.) DC. 217 Chrysosplenium nudicaule Bunge 488 Cichorium intybus L. 55, 308 Cicuta virosa L. 331 Cimicifuga foetida L. Cirsium arvense (L.) Scop. 257, 285 Cirsium heterophyllum (L.) Hill 342, 472 Cirsium setosum (Willd.) Bess. 435 Cirsium vulgare (Savi) Ten. 161 Cistus salvifolius L. 394, 395 Cistus tauricus С. Presl 139, 286, 338, 339, 340 Citrus limon (L.) Випп. 159 Colchicum speciosum Stev. 160, 241, 610 Conium maculatum L. 53, 54, 600 Conyza Ьопапепяв (L.) Cronquist 234, 235, 407, 409, 437 Convallaria majalis L. 228, 285 Coronilla L. 439 Согоп(11а scorpioides (L.) Koch 136, 237 Comus mas L. 284 Corylus avellana L. 155, 218, 219, 220, 228, 231, 237, 285, 292
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Cyclamen чегпит Sweet 63 Cydonia oblonga Mill. 591 Dactylis glomerata L. 342, 472 Dactylorhiza maculata (L.) So6 237, 316 Daphne mezereum L. 228 Datisca cannabina L. 350, 352-354 Daucus sativus (Hoffm.) Roehl. 307 Dentaria bulbifera L. 155 Deschampsia cespitosa (L.) Веаич. 167 Dianthus polyclados Boiss. 410 Dictamnus gymnostylis Stev. 285 Dictiota dichotoma Lamour 119 Dioscorea batatas Decne. 315 Dioscorea deltoidea Wall. 293 Doronicum altaicum Pall. 73 Drabopsis пи4а (Bblanger) Stapf 574 Echinocystis lobata (Michx.) Torr. et Gray 73 Echium angustifolium Miller 232, 235, 395, 403, 404, 405 Elaeagnus angustifolia L. 127 Elodea canadensis Michx. 160 Elytrigia герепв (L.) Nevski 139, 286 Ephedra aphylla Forssk. 395, 412 Eremopyrum triticeum (Gaertn.) Nevski 437 Егепшгиз tauricus Stev. 182 Его41ит cicutarium (L.) L'Her. 395 Eryngium campestre L. 241, 286, 365 Eryngium planum L. 129, 301 Eucommia ulmoides Oliv. 285 Euphorbia altaica С.А. Меу. 298 Euphorbia fischeriana Steud. 299 Euphorbia humifusa Schlecht. 134 Euphorbia pilosa L. 342 Fatsia japonica (Thunb.) Decne. et Planch. 209 Ferula caspica Bieb. 303 Ferula Шапса Fisch. ex Spreng. 300, 308 Festuca L. 604 Festuca pratensis Huds. 257 Ficus carica L. 222, 284 Filaginella uliginosa (L.) Opiz 72, 227 Filipendula ulmaria (L.) Maxim. 472 Filipendula vulgaris Moench 316 Foeniculum vulgare Miller 407, 408, 409, 417, 434 Fraxinus excelsior L. 284, 286, 290, 589 Geranium ривйит L 307 Geum rivale L. 267, 271, 272 Gentiana macrophylla Pall. 342 Ginkgo biloba L. 209, 442 Glycyrrhiza aspera Pall. 295 Glycyrrhiza glabra L. 160 Gymnospermium altaicum (Pall.) Spach 316 Gymnosteris nudicaulis 331 Gypsophila paniculata L. 296 Halocnemum strobilaceum (Pall.) Bieb. 383 Haloxylon aphyllum (Minkw.) Iljin 237 Hedysarum L. 294, 593, 608 Hedysarum alpinum L. 221, 267, 273, 274, 350, 355, 356 Hedysarum austrosibiricum В. Fedtsch. 125 Hedysarum fruticosum Pall. 585 Hedysarum neglectum Ledeb. 472, 534, 536, 538, 539, 602 Heliotropium rotundifolium Lehm. 395, 410 Helichrysum агепапит (L.) Moench 383, 386 Heracleum L. 603 Heracleum sibiricum L. 366, 417, 601 Hippocrepis emeroides (Boiss. ex Sprun.) Czer. 339 Hippophae rhamnoides L. 227, 230, 562, 563-566, 567 Hyacinthus orientalis L. 285 Hydrocharis morsus-гапае L. 160 Hypericum androsaemum L. 174, 176, 177, 236, 286, 292
Указатель латинских названий растений Hypericum calycinum Ь. 139, 149, 292 Hypericum dichotomum L. 119 Hypericum hookerianum Wight et Arn. 435 Lonicera maackii (Rupr.) Herd. 502 Mercurialis perennis L. 607 Могщ nigra L. 243, 285 Ornithogalum nutans L. 242 Oxalis pes-саграе L. 412, 413 Padus ачшт Mill. 237, 574 Рарачег янппКегшп L. 311 Paris quadrifolia L. 285 Parkinsonia aculeata L. 394 Hypericum maculatum Crantz 152, 286 Hypericum olympicum L. 286 Hypericum patulum Thunb. 235, 236, 435 Inula L. 586 Inula britannica L. 155 Inula helenium L. 294, 322, 474, 475, 478, 479, 480, 598 1пи1а viscosa (L.) Aiton 232, 235, 395, 401, 402, 403, 434 Iris ruthenica Кег-Gawl. 342 Jasminum fruticans L. 339 Juglans regia L. 132, 284, 285 Juniperus excelsa Bieb. 262, 265, 339 Kochia prostrata (L.) Schrad. 88, 385, 386 Lactuca sativa L. 310 Lagotis integrifolia (Willd.) Schischk. 388 Larix decidua Mill. 237 Larix sibirica Ledeb. 367 Lapsana communis L. 307 Laurocerasus officinalis М. Roem. 247 Leucanthemum vulgare Lam. 442 Leonurus quinquelobatus Gilib. 221, 526, 528, 531, 533, 598 Leymus racemosus (Lam.) Tzvel. 294 Libanotis Hailer ex Zinn = Seseli L. 599 Ligularia altaica DC. 294 Lilium bulbiferum L. 63 Lilium candidum L. 242 Limonium gmelinii (Willd.) О. Kuntze 183, 303 Limonium sareptanum (А. Besk.) Gams. 294 Linaria vulgarus Mill. 251 Linum L. 609 Lithospermum officinale L. 350, 357, 358 Lonicera chrysantha Turcz. ex Ledeb. 502 Lonicera caerulea L. 237 Lonicera mmimoviczii (Rupr.) Regel 502 Lonicera tatarica L. 467, 472 Lonicera xylosteum L. 221 Lycopus europaeus L. 407, 409, 434, 582 Lycopus exaltatus L. 332 Lythrum salicaria L. 407 Magnolia grandiflora L. 230 Malus domestica Borkh. 133, 221, 227, 237, 239 Malus sylvestris Mill. 574 Matthiola odoratissima (Bieb.) R. Вг. 434 Medicago L. 613 Medicago falcata L. 295, 296 Nardus stricta L. 136, 139, 142, 160 Narcissus pseudonarcissus L. 285 Nerium oleander L. 134 ОЬегпа behen (L.) lkonn. 301 Olea europaea L. 394 Oplopanax elatus (Nakai) Nakai Echinopanax elatus Nakai~ 291, 501, 503, 506, 508, 598, 600 Origanum vulgare L. 417, 418, 422, 424, 425, 428, 431, 591 Paeonia anomala L. 126, 160, 342, 343, 349, 350, 351, 598 Palimbia rediviva (Pall.) Thell. = Palimbia salsa (L.) Bess. 302
Paspalidium geminatum (Forssk.) Stapf 411 Polygonatum odoratum (Mill.) Druce 160 Polygonum coriarium Grig. = Aconogonon coriarium (Grig.) Sojak 295 Patrinia intermedia (Hornem.) Roem. et Schult. 160, 518, 520, 522, 523, 525, 597 Patrinia sibirica (L.) Juss. 294, 525 Peganum harmala L. 136, 139, 143, 144, 182, 183, 286, 302, 303, 304 Регяса vulgaris Mill. 232 Petroselinum crispum (Mill.) А. W. Hill 307 Phellodendron amurense Rupr. 285 Philadelphus coronarius L. 239, 285 Prangos pabularia Lindl. 592 Philadelphus tenuifolius Rupr. et Maxim. 502 Рппы1а macrocalyx Bunge 124, 293, 342 Primula veris L. 182 Prunus padus L. 583 Prunus spinosa L. 245 Phlojodicarpus Turcz. ex Ledeb. 599 Phlojodicarpus sibirica (Steph. ex Spreng.) К.- Ро1. 540, 541, 543, 546, 558, 559-561, 597 Phlomis pungens Willd. 155, 301, 304 Phlomoides tuberosa (L.) Moench 316 Pseudosophora alopecuroides (L.) Sweet 160, 295, 312, 313 uercus L. 381 Quercus pubescens Willd. 226, 265, 339 Phragmites australis (Сач.) Trin. ex Steud. 155, 160 Physocarpus opulifolius (L.) Maxim. 181, 285 Picea abies (L.) Karst. 136, 241, 281, 442 Quercus гоЬиг L. 219, 284, 382, 587 Ranunculus bulbosus L. 62 Picea ajanensis (Lindl. et Gord.) Fisch. ex Carr. 501 Ranunculus cassubicus L. 131 Ranunculus repens L. 160 Raphanus raphanistrum L. 307 Retama raetam (Forssk.) Webb 395, 396, 398 Pimpinella saxifraga L. 286, 296, 384, 387, 445 Pimpinella litophila Schischk.= Pimpinella tragium Vill ° 305 Rhaponricum caerhamoides (Willd.) Ijlin = Stemmacantha carthamoides (Willd.) М. Dittrich 534 Rheum compactum L. 256 Rhodiola algida (Ledeb.) Fisch. et С. А. Меу. 153 Rhodiola linearifolia Вопль. 152, 153 Rhodiola rosea L. 153, 182, 281, 480, 482, 485, 486, 489, 492, 498, 585, 597, 598, 606 Ricinus communis L. 184, 412, 414, 415, 417, 437 Plantago maxima Уия. ех Jacq. 151, 241, 285, 300 Poa altaica Trin. 472 Роа bulbosa L. 62 Poa sibirica Roshev. 342, 472 Polygonatum multiflorum (L.) All. 285 Pimpinella major (L.) Huds. 242 Pinus halepensis Miller 394 Pinus pallasiana D. Don 339 Pinus sibirica Du Tour 364, 367 Pinus sylvestris L. 237, 284, 367, 442 Piptatherum тйасеит (L.) Coss. 411 Pistacia palaestina Boiss. 394 Plantago cynops L. 221, 223, 227 Plantago major L. 152, 281, 445, 446 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ Polygonum divaricatum L. 593 Polygonum equisetiforme Sm. 395, 410 Polygonum viviparum L. 155, 233, 388 Populus nigra L. 284 Populus tremula L. 239,251, 257 Potentilla anserina L. 160 Rosa spinosissima L. 467, 472 Rubus caesius L. 281 Rubus idaeus L. 160 Rumex confertus Willd. 155 Ruscus hypoglossum L. 241
Указатель латинских названий растений Ruscus hypoglossum L. 241 Sagittaria sagittifolia L. 376 Saintpaulia ionantha Н. Wendl. 159 Salix acutifolia Willd. 382 Salix arbuscula L. 119 Salix caprea L. 200, 228, 231, 234, 241, 285 Salix glauca L. 480 Salix hastata L. 480 Salix lanata L. 480 Salix polaris Wahlenb. 587 Salix phlebophylla Anderss 587 Salix rosmarinifolia L. 382 Salsola dendroides Pall. 302, 308, 602 Salsola laricifolia Turcz. et Litv. 89 Salvia glutinosa L. 434 Salvia fruticosa Miller. 394 Sambucus L. 439 Sambucus ebulus L. 148, 286 Sambucus nigra L. 168, 170, 180 Sambucus racemosa L. 180, 285, 286, 291, 292, 440 Sarcopterium spinosum (L.) Spach 412 Saxifraga punctata L. 488 Schulzia crinita (Pall.) Speng. 388 Scrophularia incisa Weinm. 128 Scrophularia xanthoglossa Boiss. 395, 410, 434 Scutellaria baicalensis Georgi 305, 568, 569, 570 Sedum aizoon L. 267, 269 Sedum ewersii Ledeb. 145 Sedum telephium L. 255 Seseli buchtormense (Fisch. ex Hornem.) Koch 306 Seseli condensatum (L.} Reichenb. Libanotis condensata L. Crantz. 293, 322, 388, 390, 391, 392 Silene fruticosa L. 246 Solanum aviculare Forsch. 184 Solanum tuberosum L. 160, 281, 285, 315 Sorbaria sorbifolia (L.) А. Вг. 285 Spartium junceum L. 394, 395 Spiraea betulifolia Pall. 502 Spiraea chamaedrifolia L. 472 Spiraea crenata L. 472, 525 Stellaria bungeana Fenzl 147 Stellaria dfchotoma L. 119 Stellera chamaejasme L. 299 Stephania glabra (Roxb.} Miers 291 Succisa pratensis Moench 152, 237, 285 Swertia obtusa Ledeb. 388 Tamarix ramosissima Ledeb. 237 Тагахасит ceratophorum (Ledeb.) DC. 216 Taraxacum officinale Wigg. 241, 296, 305, 312, 445 Tilia L. 116 Tilia cordata Mill. 134, 241, 285, 286, 609 Tribulus terrestris L. 135 Trollius asiaticus L. 342 Trifolium L. 613 L Tifolium ambiguum Bieb. = Amoria ambigua (Bieb.) Sojak 312, 588 Trigonella foenum-graecum L. 267, 268 Tulipa gesneriana L. 241 Ulmus laevis Pall. 237, 285 Ungernia victoris Vved. ex Artjushenko 315 Urtica dioica L. 146, 472 Utricularia vulgaris L. 71, 252 Vaccinium myrtillus L. 292 Valeriana L. 611 Valeriana officinalis L. 182, 417, 546, 548, 551, 556, 598 Valeriana tuberosa L. 57 Vanilla planifolia Andr. 258 Veronica L. 603 Veronica longifolia L. 294
Viburnum lantana L. 219, 221 Viola Ыйога L. 322, 323 Viola disjuncta W. Beck. 342 Примечание. 1. Жирным шрифтом обозначены страницы, на которых помещены рисунки. 2. Курсивом обозначены некоторые устаревшие названия растений, которые цитируются в книге. Viburnum opulus Ь. 160, 181, 218, 219, 257, 285 ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ Viola hirta L. 342 Viola mirabilis L. 155 Viola uniflora L. 224, 228
Содержание ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° ° Введение Теоретизирование и законы науки ......................................... 7 Глава 1. о боснование биоморфологии ................................................. 17 2.1. Движение, системность, жизнь ................................... 17 2.2. Организация Природы, статус биоморф и биоморфологии ......................................... 28 2.3. Общие закономерности организации биосистем ................................................ 39 2.4. Жизненная форма (биоморфа)— ключевое понятие биоморфологии ............................. 44 2.5. Из истории изучения организации биоморф кормофитов (побеговых растений) ............................. 59 2.5.1. История термина «каудекс». Организация системы «корень-трава-плодотворение» ..................... 60 2.5.2. Организация системы «корень-лист». к онцепция фитона........................................................ 66 2.5.3. Организация системы «корень-стебель-лист» ............ 67 2.5.4. Организация системы «побег-корень» ........................ 68 2.5.5. Понятия побег, корень и корнепобег ........................69 Глава 2. Общая теория жизнесохранения- концептуальная основа жизни ...............................................75 3.1. Предпосылки к общей теории жизнесохранения .....75 3 .2. Исходные понятия ........................................................ 79 3.3. Законы адаптации и эволюции — исходные принципы в общей теории жизнесохранения ........... 79 3.4. Закон единства живого и неживого ...........................90 3.5. Специальная теория жизнесохранения < биофилии) ..................................................................... 91 3.6. Специальная теория средосохранения < экофилии) ..................................................................... 99 Принцип экобиофилии .............................................. 101 з акон стратегии Жизни .............................................. 103 Глава 3. 3.7. 3.8. Глава 4. Основные способы освоения пространства б Ф,У иоморфами семенных растении ......................................... 105 4.1. Стратегические и тактические системы б иоморф ........................................................................ 105 Биполярность в организации биоморф .................... 112 P ост и развитие ........................................................... 114 Нарастание, ветвление и кущение............................115
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РА СТЕНИЙ 4.5. 4.6. 4.7. Партикуляция и размножение ................................... 15 лиминация ................................................................. 16 Организация биоморф семенных растений в борьбе за существование.:....................................... 16 Продолжительность жизни: эвриэфемерность и персистентность.................................... 275 6.1. Организация биоморф как способ резервирования ресурсов ............................................ 275 6.2. Резидофикация и базомеризация .............................. 278 б .3. Базовые органы ........................................................... 287 Основные способы освоения времени б Ф,У иоморфами семенных растении .........................................317 7.1. Морфогенез, биоморфогенез И биоморфогения ........................................................ 317 7.2. Состояния биоморф и стадии б иоморфогенезов ......................................................... 319 7.3. Категории стадий биоморфогенезов, критерии выделения и методика их изучения .........................320 7.4. Последовательность морфогенезной сборки и разборки организации биоморф ............................ 359 и зменчивость и эволюция ................................................... 361 8.1. Внутривидовая пластичность ..................................... 361 8.2. Ответные реакции в инфраструктурной и конституциальной борьбе за существование ....... 376 8.2.1. Поведение биоморф на эрозионно лабильных субстратах .................................................. 380 8.2.2. Пластичность биоморф семенных растений средиземноморского побережья Б лижнего Востока ....................................................... 394 8.2.3. Интродукционный стресс у растений ...................... 416 8.3. Разнонаправленность соматической эволюции ....... 432 8.4. Эволюционная перспектива вида .............................. 442 Параметры основных органов и их частей в организации биоморф семенных растений ........................185 5.1. Побеги — основные органы семенных растений, их структура и функции .......................... 185 5.1.1. Разнообразие побегов и их классификаций ............185 5.1.2. Иерархичность в организации побегов и их структурных элементов ....................... 194 5.1.3. Многолинейный анализ организации побегов ........ 236 5.2. Корни — основные органы семенных растений, их классификация по степени сложности ............... 250 5.3. Ризокормусовая (корнепобеговая) концепция ........ 251 5.4. Феноцентры в организации биоморф семенных растений ..................................... 259
Содержание Глава 9. Многолииейная классификация биоморф семенных растений и формализация языка биоморфологии ................445 Глава 10. Использование теории организации биоморф семенных растений в практической биоморфологии ............................ 463 10.1. Ценодемология семенных растений (на примере Bupleurum аигеит Fisch. ex Hoffm.)......463 10.2. Биоморфогения семенных растений в естественных условиях произрастания и в культуре (на примере лекарственных растений) .....................474 10.2.1. Inula helenium L.......................................................... 474 0.2.2. Кйойо1а rosea L........................................................... 480 10.2.2.1. Rhodiola rosea L. в горах Алтая .............................480 10.2.2.2. Rhodiola rosea L. в условиях выращивания .........489 10.2.3. Oplopanax elatus (Nakai) Nakai ................................... 501 10.2.4. Adonis sibirica Patrin ex Ledeb.................................... 512 10.2.5. Patrinia intermedia (Hornem.) Roem. et Schult.........518 10.2.6. Leonurus quinquelobatus Gilib..................................... 526 10.2.7. Hedysarum neglectum Ledeb........................................ 534 10.2.8. Phlojodicarpus sibiricus (Steph. ех Spreng.) К.-Pol.... 540 10.2.9.Valeriana officinalis L.................................................... 546 10.3. Рисуночный способ передачи информации о биоморфогении семенных растений ..................... 558 10.3.1. Phlojodicarpus sibiricus (Steph. ех Spreng.) К.-Pol.... 558 10.3.2. Hippophae rhamnoides L.............................................. 562 10.3.3. Scutellaria baicalensis Georgi ........................................ 568 сновные выводы ............................................................................... 572 аключение ......................................................................................... 578 Указатель латинских названий растений.............................................618
Contents Chapter Chapter 2.1. 2.2. 2.3. 2.4. 2.5. 2.5.1. 2.5.2. 2.5.3. 2.5.4. 2.5.5. Chapter 3.1. 3.2. 3.3. 3.4. 3.5. 3.6. 3.7. 3.8. Chapter 4.1. 4.2. 4.3. 4.4. 4.5. 4.6. 4.7. Chapter 5.1. 5.1.1. 5.1.2. 5.1.3. 5.2. 5.3. 5.4. 1 ° ° Л 1 У Piaeonzing and laws of science .......................................................... 3 2 1 ° Grounds of biomorphology ................................................................. 7 М 1'С' ovement, system character, life .............................................................17 Organization of Nature, status of biomorphs and biomorphology ..........28 General mechanisms in organization of biosystems ................................. 39 Life form (biomorph) as а key concept of biomorphology .....................44 From history of study of structural organization of shoot plants (cormophytes) ...................................................................59 History of term "caudex". Organization of system "root-grass-fruit generation (fructificatio)" ............................................... 60 Organization of system "root-leaf". Concept of phyton..........................66 Organization of system "root-stem-1еаГ' ..................................................67 Organization of system "shoot-root" ........................................................68 Concepts of shoot, root, rootshoot (rhizocormus) ...................................69 3. General theory of vitality as а conceptual basis of life ..................... 75 Grounds of general theory of vitality .......................................................75 I ° ° пйа1 concepts ........................................................................................... 79 Laws of adaptation and evolution as basic principles о f general theory of vitality ............................... ° °............ ° ° °.......................79 Law of unity of organic and inorganic .....................................................90 Special theory of vitality (biophily) ..........................................................91 Special theory of environmental conservation (ecophily) ........................99 P 1 '1 rinciple of есоЬ1ор11у ........................................................................... 101 Lf ife strategy law ..........................................................°............................ 103 4. Basic methods for space mastering by biomorphs о f spermous plants ........................................................................ 105 Strategical and tactical systems of biomorphs ........................................ 105 Bipolarity in organization of biomorphs ................................................. 112 G rowth and development ........................................................................ 114 Growth, branching, and bushing ............................................................. 115 Particulation and reproduction ................................................................ 153 Е1 limination ............................................................................................... 163 Organization of biomorphs of spermous plants 1 С- 1 Г а йе~г fight for existence ....................................................................... 165 5. Parameters of basic organs and their parts ° ~Ф а biomorphs of spermous plants.................................................... 185 Shoots as basic organs of spermous plants, their structure and functions ................................................................... 185 Variety of shoots and their classification ................................................ 185 Hierarchic system in organization of shoots and their structural elements................................................................... 194 Multilinear analysis in organization of shoots ........................................ 236 Roots as basic organs of spermous plants, their classification with respect to degree of complexity ...°.................................................. 250 Rh аосогпис concept .............................................................................. 251 Phenocenters in organization of biomorphs of spermous plants ........... 259
Содержание Chapter 6. Duration of life: euvriephemerality and persistence......................... 275 6.1. Life as а method for reservation of resources.......................°...°............275 6.2. Residofication and basomerization .......................................................... 278 6.. .3. Basic organs .................................. ° ..°........................................................ 287 Chapter 7. Basic methods for time mastering by biomorphs о f spermous plants ........................................................................ 317 7.1. Morphogenesis, biomorphogenesis, biomorphogeny ............................... 317 7.2. States of biomorphs and stages of biomorphogenesis ............................ 319 7.3. Categories of stages of biomorphogenesis, criteria of their separation, and methods of investigation .................................. 320 7.4. Procedure of morphogenesis assembling and disassembling in organization of biomorphs ........................°.................. 359 Chapter 8. Mutability and evolution................................................................ 361 8.. ° ° .1. Intraspecies plasticity ............................................................................... 361 8.2. Response reactions in infrastructuràl and constitutional f ight for existence..................................................................................... 376 8.2.1. Behaviour of biomorphs under conditions of егояопа11у 1Ь1 \ Ф аЫе substrates ........................................................................................ 380 8.2.2. Plasticity of biomorphs of spermous plants of the Mediterranian coast of Near East ........................................................... 394 8.2.3. Introduction stress of plants ....................................................................416 8.3. Various directivity of somatic evolution °................................................ 432 8.4. Evolution perspective of species .............................................................. 442 Chapter 9. Multilinear classification of biomorphs of spermous plants and formalization of language of biomorphology .................................... 445 Chapter 10. Application of theory of organization of biomorphs of вреппоив plants t о praciacal biomorphology ............................................................ 463 10.1. Cenodemology of вреппоив plants (by example of Bupleurum а игеит Ftscii ex Hoffm.) ........................................................................463 10.2. Biomorphogeny of spermous plants under natural conditions of their growth and in culture (by example of herbs) ...........................474 1 0.2.1. Inula helenium L..................................................................................... 474 1 0.2.2. Кйод1о1а rosea L..................................................................................... 480 10.2.3.0р1оропах elatus (Nakai) Nakai ............................................................. 501 10.2.4.Adonis sibirica Patrin ex Ledeb.............................................................. 512 10.2.5.Patrinia intermedia (Нотет.) Roem. et Schult................................... 518 10.2.6. Leonurus quinquelobatus Gilib............................................................... 526 10.2.7. Hedysarum neglectum Ledeb.................................................................. 534 10.2.8.Phlojodicarpus sibiricus (Steph. ех Spreng.) К.-Pol.............................. 540 1... 4Т' 0.2.9.Veleriana oficinalis L.............................................................................. 546 10.3. Graphic method for display ot biomorphogeny ......................°.............. 558 10.3.1. Phlojodicarpus sibiricus (Steph. ех Spreng.) К.-Pol.............................. 558 1...Н тт. 0.3.2. Hippophae йатпо~дез L........................................................................ 562 ф ев 3 '.Л 10.3.3. Scutellaria baicalensis Georgi .................................................................. 568 В ф ° аяс conclusions ............................................................................................. 572 S ummary .................................................................................................... 578 ВЫ 'Ы' ibliography .................................................................................................... 581 I Т ndex of ~tin names of plants......................................................................... 618
Nukhimovsky Е.Ь. Fundamentals of biomorphology of spermous plants. Т. 1. Theory of biomorph organization. — Moscow: Nedra, 1997.-б30р. This book examines the problems of organization of biomorphs (organisms and synorganisms) of spermous plants, history of biomorphology, and its status in the system of sciences. The place of habitology (science of biomorph habitus or habitual forms) is determined. It is shown that the general theory of vitality, first developed by the author and put into foundation of the theory of organization of biomorphs of spermous plants, provides the philosophical and biological basis for various scientific generalizations. The principal structure of this theory is determined by seven conceptions: those of life form (biomorph), shoot, root, rootshoot (rhizocormus), morphogeny, infrastructural-constitutional fight for existence, plasticity, and various directivity of somatic evolution. The methods of subordinate, multisubordinate, and superordinate classification, developed by the author, are widely used for description of organization of both the basic organs (shoots, roots, and their elements) ап4 higher structural units of biomorphs of вреппоив plants (rhizocormus). As а dynamic basis of biomorphology, is considered the concept of morphogeny. Не suggested an improved classification scheme for the age stages of biomorphogenesis. The author developed the conception of plasticity, which makes the basis for specification of the concepts of intraspecies plasticity of biomorphs and for new evolution ideas in biomorphology of spermous plants. It was suggested to analyse evolution not only as а syntomokinesis, i.е., а complex system of "evolution steps" (syntoms), but also as movements of an infinite number of "evolution pendulums" (bodies or system of bodies in development), i. е., а "flow of evolution pendulums" providing various directivity of the evolution process. The author showed that plants or groups of plants, which take part in the "evolution flow" as evolution pendulums, in the course of their existence, can settle or fail to settle one or тоге of the problems set by the Nature before them: 1) to continue life; 2) to extend polymorphism; and 3) to create а new species. The author substantiated the law of evolution perspective and underlined that the principal evolution achievement of paleobiomorphs was not the origin of trees or herbs but the first appearance of plasticity and its further strengthening. The author substantiated the idea of universal fight for existence and provided with the actual materials that the intraspecies plasticity starts in the infrastructural (among the structural elements inside the organism) and constitutional (generalized response reaction of all the constitution of an to disturbing effect of the environment) fight for existence. The author developed the multilinear classification of biomorphs with respect to numerous lines of alternative criteria; that opens а геаl way to computerization of biomorphological analysis. А special attention was paid to the term of biomorpholosy, because the author regards his arrangement "everything by shelves" as an essential aspect of his theoretical investigations. Using numerous examples from the life of plant biomorphs, the author showed the applied importance of his theory. This book is designed for botanists of various specialities as well as for bilogists, who are interested in the general problems of natural sciences. Tables: 8. Figures: 218. Bibliography: 37 pages.
НАУЧНОЕ ИЗДАНИЕ Нухимовсиий Ефим Львович ОСНОВЫ БИОМОРФОЛОГ~И CEMEHHLIX РАСТЕНИИ: биоморф Т. 1. Теория ор Зав. редакцией Л.С. дмитриева Переплет художника Н.П. Новиковой Технический редактор Г.В. Лехова Компьютерная верстка А.В. Булатова ОАО «Издательство Недра» 125047 Москва, Тверская застава, 3 Московская типография «Наука» РАН 121099 Москва, Шубинский пер., 6, Зау ~Я55 Изд. лиц. М 010145 от 24.12.92. Подписано в печать с репродуцированного оригинал-макета 28.11.97. Формат 70xl00'/„. Гарнитура Newton. Печать офсетная. Усл.печ. л. 50,95. Уч.-изд. л. 49,47. Тираж 1000 экз. Заказ М /113