























































































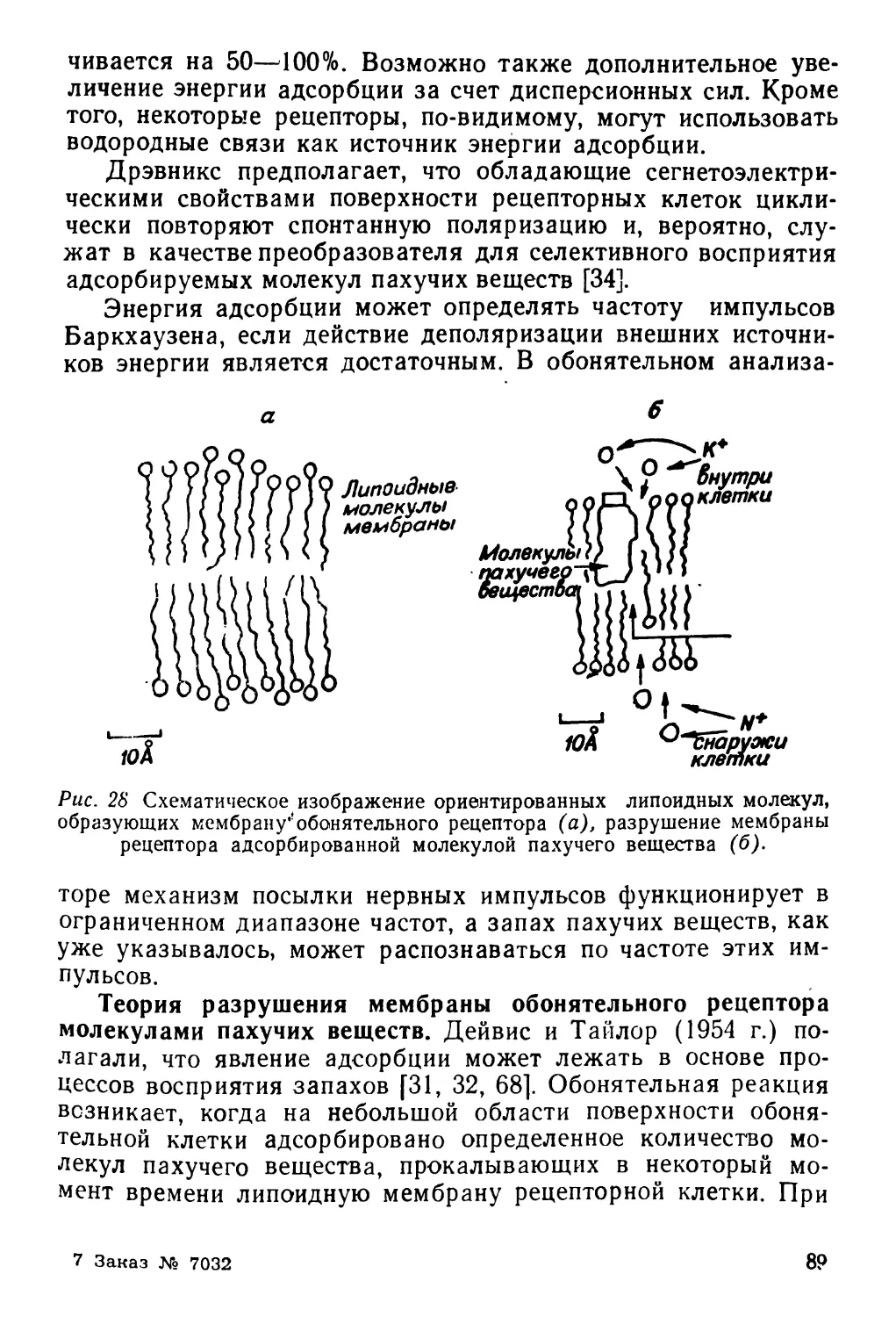






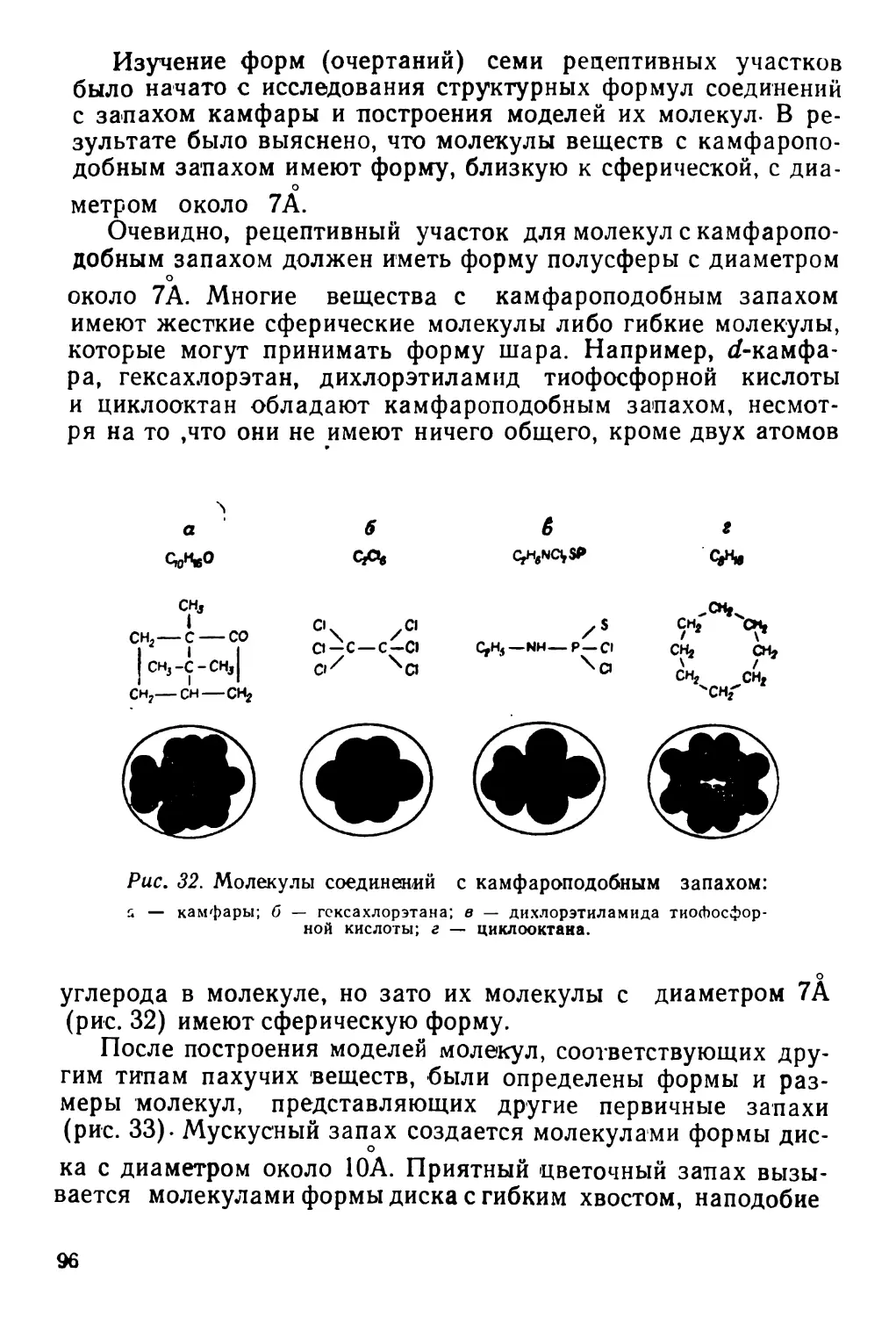































Текст
АКАДЕМИЯ НАУК СССР
СИБИРСКОЕ ОТДЕЛЕНИЕ
ИНСТИТУТ АВТОМАТИКИ И ЭЛЕКТРОМЕТРИИ
Р. М. МАЗИТОВА,
В. Н. ОХОТСКАЯ, Б. И. ПУЧКИН
ОБОНЯНИЕ
И ЕГО МОДЕЛИРОВАНИЕ
ИЗДАТЕЛЬСТВО <НАУКА>
СИБИРСКОЕ ОТДЕЛЕНИЕ
НОВОСИБИРСК
1965
В настоящей работе предпринята попытка систематизировать
данные об исследовании обонятельных анализаторов живых орга-
низмов и выявить возможность объективного измерения количе-
ственных и качественных характеристик запаха. Кратко рассмотре-
ны строение и физиология органов обоняния. Приведены некоторые
свойства пахучих веществ, соотношения запаха и молекулярной
структуры. Описаны существующие теории обоняния, среди которых
особое внимание уделено адсорбционным теориям, дающим возмож-
ность объективного «измерения» запаха и, следовательно, модели-
рования обонятельного анализатора. Рассмотрены осуществленные
модели обонятельного анализатора и намечены возможные направ-
ления подобных исследований.
Книга, построенная на материалах важнейших работ в данной
области, опубликованных за последние 25 лет в Советском Союзе
и за рубежом, предназначена для «научных и инженерно-технических
работников, специализирующихся в области создания новых средств
измерительной техники, и может представить интерес для более ши-
рокого круга читателей, в том числе и биологов.
Ответственный редактор
чл.-корр. АН СССР К. Б. КАРАНДЕЕВ
3-3-13
1355-66
ВВЕДЕНИЕ
Органы чувств живых организмов, служащие для восприя-
тия информации из окружающей среды, обладают рядом до-
стоинств, в том числе высокой чувствительностью, нередко пре-
восходящей чувствительность технических средств. Поэтому
вполне понятно стремление специалистов в области измери-
тельной техники использовать 'сведения о 'процессах сбора
и переработки информации в живых организмах при создании
новых, более совершенных приборов. В связи с этим большое
внимание уделяется детальному исследованию того или иного
анализатора, накоплению достоверных количественных дан-
ных о его структуре, функциях и т. д. Подобные исследова-
ния в настоящее время становятся все более многочисленны-
ми. Значительная их часть относится к изучению обонятель-
ного анализатора. И это не случайно.
Прежде всего до настоящего времени нельзя считать изу-
ченным механизм возбуждения обонятельных рецепторов па-
хучими веществами. Мало изучена взаимосвязь запаха веще-
ства с его физическими и химическими 'свойствами, а сами за-
пахи не имеют количественной оценки, носят предметный
характер (например* фруктовый, цветочный). Недостаточны
наши знания и о процессах передачи и переработки инфор-
мации в ббонятельном анализаторе, обеспечивающих необы-
чайно широкий диапазон ощущаемых запахов и их интенсив-
ностей.
Все эти и многие другие вопросы, связанные с функциони-
рованием обонятельного анализатора, представляют несом-
ненный интерес как для познания процесса обоняния, так
и для использования полученных знаний в практической дея-
тельности людей, например, при создании новых устройств
газового анализа.
3
Накопленные к настоящему времени сведения об обоня-
тельных анализаторах позволили построить первые модели,
частично воспроизводящие функции органов обоняния. Однако
решение этой задачи находится в начальной стадии. Тем по-
нятнее интерес, проявляемый учеными разных специальностей
к изучению обонятелвного анализатора и процесса обоняния.
Авторы выражают свою искреннюю благодарность чл.-
корр. АН СССР К. Б. Карандееву, д-рам мед. наук Я. Д. Фин-
кинштейну и М. Я. Субботину, а также д-ру техн, наук
М. П. Цапенко, взявшим на себя труд орочесть и обсудить
рукопись.
ГЛАВА I
КРАТКИЕ СВЕДЕНИЯ
О МОРФОЛОГИИ И ФИЗИОЛОГИИ
ОБОНЯТЕЛЬНОГО АНАЛИЗАТОРА
Некоторое (представление о восприятии и различении за-
пахов живыми организмами, необходимое для (понимания
дальнейшего материала, возможно только при достаточном
знании строения и физиологии брганов обоняния. Для этого
ниже кратко рассматривается строение органов обоняния поз-
_ воночных (на примере млекопитающих, человека и рыб) и на-
секомых.
1.1. СТРОЕНИЕ ОБОНЯТЕЛЬНОГО АНАЛИЗАТОРА
МЛЕКОПИТАЮЩИХ И ЧЕЛОВЕКА
Восприятие пахучего вещества .в организме осуществляет-
ся участком слизистой (обонятельный эпителий), находящим-
ся в области верхних носовых ходов у человека [2, 4, 43] и за-
нимающим площадь 5 см2 (рис. 1). Обонятельный эпителий
отличается желтоватым цветом от эпителия, выстилающего
остальную часть носовой полости, и содержит обонятельные
рецепторы, опорные и другие клетки (рис. 2). Поверхность
обонятельного эпителия покрыта тонкой пленкой жидкости-
секрета, выделяемой железами Боумена.
Обонятельные клетки имеют форму веретена или бокала
с двумя отростками: периферическим и центральным. Перифе-
рические отростки клеток доходят до поверхности слизистой
5
и заканчиваются булавовидными утолщениями, несущими не-
сколько «ресничек (6—8 у человека, 10—14 у кролика) диа-
метром 0,1 мк и длиной от 1 до 2 мк. Центральные отростки
отходят от клеток в виде тонких нитей (волокон) диаметром
0,2 мк, которые на пути к обонятельным луковицам головного
мозга объединяются в мно-
гочисленные пучки, содер-
жащие от нескольких сотен
до нескольких тысяч нерв-
ных волокон обонятельного
нерва (filae olfactoria) и
проникают через решет-
чатую кость в полость че-
репа. Эти волокна, перепле-
таясь между собой, образу-
ют поверхность обонятель-
ных луковиц. Из внеш-
него слоя луковицы волок-
Puc. 1. Анатомия органа обоняния
человека.
1 -- обонятельная луковица; 2 — обо-
нятельная область.
на входят в клубочки (рис. 3), в которых образуются
контакту аксонов обонятельных рецепторов с дендритами бо-
лее глубоко расположенных митральных и пучковых клеток.
Пучковые клетки служат для передачи информации в свя-
занную с другой ноздрей луковицу. Митральные 1И пучковые
клетки являются вторичными обонятельными нейронами.
Последний слой луковицы содержит
зернистые клетки. Современные мето-
ды исследования не позволяют окон-
чательно решить вопрос о природе и
функциях ' клеток-зерен. Существует
предположение, что они служат для
задержки сигналов, получаемых от
Рис. 2. Микроскопический срез обонятельного
эпителия млекопитающих.
1 — тригеминальный нерв; 2 — волокна обонятельно-
го нерва; 3 — обонятельные рецепторы; 4 — опор-
ные клетки; 5 — окончания тригеминального нерва;
6 — секрет; 7 — обонятельные волоски.
6
неодновременно возбуждаемых рецепторов. Это необходимо
для синхронизации передачи информации от митральных
клеток в обонятельный мозг 1421. Взаимосвязь упомянутых
клеток можно проследить на следующем примере. 1900 клу-
бочков кролика являются своеобразными коммутаторами,
каждый из которых соединяет 26000 входных рецепторных
волокон с 24 митральными и 68 пучковыми клетками [421.
Обонятельные
рецепторы
Волоски Волокна
500000000 50000000
Клубочки
1900
____________-___f
Первичные нейроны\_
24
Пучковые
клетка-
Митральные
клетки
45000
к мозгу
Рис. 3. Схематическое изображение обонятельной системы человека.
Обонятельная луковица кролика связывает 50 000 000 обоня-
тельные рецепторов с 45000—60000 выходных митральных
клеток, аксоны которых образуют обонятельный тракт. На
рис. 3 дано схематическое изображение обонятельной луко-
вицы. Волокна обонятельного тракта подходят к обонятель-
ным центрам второго порядка. Из этих центров берут на-
чало третьи обонятельные нейроны, заканчивающиеся в выс-
шем обонятельном центре.
Для сравнения систем передачи информации в органах
чувств человека заметим, что количество нервных волокон
в обонятельном нерве 100-106, в зрительном нерве 1 • 10б, в слу-
ховом нерве 0,08* 106. Эти данные показывают, какое огромное
количество сообщений способен различать обонятельный ана-
лизатор. * > »
В эпителии, выстилающем носовую полость, найдены окон-
чания тройничного нерва (болевые, тактильные, холодовые,
тепловые, вкусовые)'. Возбуждение окончаний тройничного
нерва может привести к изменению «размеров обонятельной
щели, т. е. изменить доступ воздуха к обонятельному эпи-
телию.
7
1.2. СТРОЕНИЕ ОРГАНА ОБОНЯНИЯ РЫБ
Обонятельный орган рыб представлен двумя складчатыми
мешками, сообщающимися с внешней средой двумя отверсти-
ями на -верхней стенке черепа [41. У некоторых видов рыб обо-
нятельные мешки отсутствуют и обонятельный нерв может
просто оканчиваться в области небольшого пигментного пят-
нышка кожи. В (рецепторном слое органа обоняния описаны
опорные (эпителиальные)
и чувствительные (обоня-
тельные) клетки. Опор-
ные клетки рецепторно-
го слоя карася снабжены
ресничками, у многих из
них замечены следы се-
крета. По форме обоня-
тельные клетки разделя-
ются на три типа: ните-
образные, палочкообраз-
ные, то есть клетки с
периферическим отрост-
ком, имеющим форму уз-
кого цилиндра или тон-
кой извивающейся нити,
и реже попадающиеся
колбочкообразные, у кото-
рых периферический от-
росток резко конусовидно
расширен (рис. 4). Пери-
ферические отростки обо-
нятельных клеток закан-
чиваются нерезко выра-
женным темноокрашен-
ным утолщением — обо-
нятельной булавой, несу-
щей на своей свободной,
Рис. 4. Микроскопический срез
обонятельного эпителия рыбы.
обращенной в полость носа поверхности кисточку чув-
ствительных волосков. От основания обонятельных клеток
отходят обонятельные отростки, которые затем, переходя из
рецепторного слоя в подлежащую соединительную ткань, об-
разуют парные безмякотные обонятельные -нервы. Последние
проникают в черепную коробку, входят ib обонятельные луко-
вицы переднего мозга, где, распадаясь, образуют ряд харак-
8
торных клубочков, <в которых осуществляется контакт (си-
напс) между обонятельными клетками и нейронами луковицы.
В обонятельных луковицах хорошо выражены митральные
клетки, клетки-зерна и особые так называемые эпендиматоид-
ные митральные клетки, которые располагаются в составе
желудочковой эпендимы обонятельных луковиц. Кроме того,
обонятельные луковицы рыб содержат группы сравнительно
небольших звездчатых ганглиозных клеток.
1.3. ОРГАНЫ ОБОНЯНИЯ НАСЕКОМЫХ
Обонятельные органы насекомых—сенсиллы (рис. 5) рас-
положены на отдельных сегментах антенн, усиков [11, 33].
Строение плакоидной (sensilla placoida) и конической (sen-
silla basiconica) сенсилл представлено на рис. 6 и 7. Перифе-
рическая кожистая мембрана плакоидной сенсиллы имеет вид
пластинки толщиной менее одного мик-
рона. Клеточные элементы этой сенсил-
лы состоят из обволакивающих клеток и
Рис. 7. Базикони-
ческая сенсилла?
1 — конусовидная мем-
брана; 2 — обонятельная
палочка; 3 — чувстви-
тельная нервная клетка;
4 — трихогенная клетка.
Рис. 6. Плакоид-
ная сенсилла.
1 — рецепторная пла-
стинка; 2 — обоня-
тельная палочка; 3 —
чувствительная нерв-
ная клетка. 4 —
трихогенная клетка.
Рис. 5. Органы
обоняния насе-
комых»
Д — антенна рабочей
пчелы (видно распре-
деление поровых пла-
стинок); /В — детали
части членика ан-
тенны.
9
биполярных чувствительных нервных клеток (обонятельных
рецепторов), периферические отростки которых, направляясь
к поверхности «мембраны, заканчиваются в вакуоле. Централь-
ные отростки чувствительных клеток объединяются -в центро-
стремительные нервы.
Базиконическая сенсилла отличается от плакоидной глав-
ным образом тем, что ее мембрана имеет конусовидную фор-
му. Количество биполярных чувствительных клеток непостоян-
но. Их 'периферические отростки направлены к основанию ко-
нуса и лежат в жидкости вакуоли.
Таким образом, в сенсиллах насекомых нервные окончания
чувствительных клеток отделены от внешней среды (обоня-
тельного стимула) мембраной (кожицей) толщиной менее
1 мк, состоящей из внутреннего хитино-протеинового слоя
и наружной эпитикулы (прозрачная оболочка). В отличие от
соседних кожистых поверхностей хитино-протеиновый слой
только частично уплотнен, а эпитикула содержит небольшое
количество липоидов. Поэтому можно предположить относи-
тельно легкую проницаемость мембраны. Поверхность мем-
браны сухая, не покрыта ни слизью, ни секретом.
У разных видов насекомых число обонятельных сенсилл
различно. Так, например, у человеческой вши — 9—10, у пче-
линого трутня — 30 000, двукрылые насекомые; питающиеся
навозом, имеют в среднем 820, а насекомые, питающиеся цве-
точной пищей и ориентирующиеся в значительной мере зре-
нием, имеют 494 сенсиллы ит. д. У самцов насекомых, отыски-
вающих самку по запаху, антенны сильнее разветвляются
и имеют гораздо большую поверхность, чем у самок, напри-
мер, у бабочек сзтупний.
Интересны результаты экспериментальных исследований
зависимости обоняния от целости антенны. Известно, что обо-
нятельные сенсиллы пчелы находятся на восьми средних
и концевых члениках (сегментах) и отсутствуют на четырех
первых. Полная ампутация обонятельных члеников одной ан-
тенны почти не отражается на способности пчелы отличать од-
ни запахи от других. Ампутация обеих антенн уничтожает обо-
няние полностью, но достаточно оставить один обонятельный
членик из шестнадцати или даже половину членика, чтобы не-
которая способность обонять сохранилась.
Насекомые способны своими обонятельными органами
ощущать запахи пахучих веществ в крайне малой концен-
трации.
10
1.4. ЭЛЕКТРОФИЗИОЛОГИЯ ОРГАНА ОБОНЯНИЯ
Электрофизиологические исследования позволяют исполь-
зовать для анализа работы обонятельного анализатора харак-
теристики электрических проявлений активности нервных кле-
ток. Известно, что (поверхностная 'мембрана нервных клеток
электрически поляризована [6]. В среде, окружающей клетку,
более 90% составляют положительные ионы натрия и отрица-
тельные ионы хлора. В растворе, находящемся внутри клетки,
основную часть образуют положительные ионы калия, а отри-
цательные представлены достаточно крупными органическими
частицами, которые не могут диффундировать сквозь мембра-
ну клетки. Поэтому концентрация ионов натрия снаружи
примерно в 10 раз выше, чем внутри клетки; концентрация же
ионов калия, напротив, внутри клетки в 30 раз выше, чем сна-
ружи. Хотя проницаемость мембраны для всех ионов невели-
ка, тем не менее она неодинакова для разных ионов; ионы ка-
лия и хлора проходят сквозь эту мембрану гораздо легче, чем
ионы натрия и крупные органические ионы. В результате воз-
никает разность потенциалов, достигающая 60—90л«в, причем
внутреннее содержимое клетки оказывается заряженным от-
рицательно по отношению к внешней среде. Для поддержания
этих различий в концентрации ионов нервная клетка, по-види-
мому, располагает своего рода «насосом», который выкачива-
ет ионы натрия через мембрану клетки наружу с такой же ско-
ростью, с какой они проникают в клетку в направлении элек-
трохимического градиента.
Возможно, (работа этого «насоса» связана с обменом ионов
натрия на ионы калия.
Электрохимические процессы, лежащие в основе нервного
импульса, или, как его называют, потенциала действия, в тече-
ние последних 15 лет удалось в значительной мере выяснить.
Разность потенциалов между внутренней и наружной поверх-
ностями мембраны определяется главным образом различной
проницаемостью мембраны для ионов натрия и калия. Особен-
ность мембраны нервного волокна состоит в том, что степень
ее проницаемости зависит, в свою очередь, от разности потен-
циалов между ее внутренней и наружной поверхностями.
А. Ходжкин и А. Хаксли установили, что искусственное пони-
жение этой разности потенциалов немедленно вызывает повы-
шение проницаемости мембраны для ионов натрия. Когда
ионы натрия проникают сквозь мембрану, они вызывают ло-
кальное погашение части избыточного -отрицательного заряда
внутри клетки, что .приводит к дальнейшему уменьшению раз-
ности потенциалов [6]. Таким же образом в ответ на адекват-
ное раздражение в рецепторных клетках немедленно происхо-
дит местное изменение мембранного потенциала, дающее на-
чало нервнрму импульсу, 'который распространяется по всему
волокну.
Эл ектроф из иологические исследов ан ия органов обоняв и я
проводили многие ученые. Изучалась электрическая ак-
тивность обонятельного эпителия, нервов, луковиц и обоня-
тельного центра головного мозга [3, 14—19, 40, 52, 56, 62, 64,
66, 70—73, 78, 80, 82].
Исследование электрической активности обонятельного
эпителия дает прямую информацию о его реакции, так как при
этом имеет место непосредственный контакт электрода
с рецепторными клетками. Однако этот метод исследова-
ния неизбежно влечет за собой в большей или меньшей
степени нарушение естественного состояния исследуемого
органа.
Электрическая активность обонятельной луковицы (мит-
ральных клеток) не дает точной информации о реакции рецеп-
торов под воздействием пахучих веществ вследствие конвер-
генции нервных волокон обонятельных рецепторов к постси-
наптичеоким митральным клеткам. Так как нервное волокно
служит проводником импульсов, об ответе обонятельного ре-
цептора можно судить по результатам изменения частоты им-
пульсов в его аксоне.
Механизм проведения сигналов от обонятельного эпителия
к луковице может быть выяснен только при одновременной ре-
гистрации их электрической активности.
В ответ на раздражение пахучим веществом в эпителии
появляются медленные — «1монофазные»1 потенциалы с кру^
тым подъемом и медленным опадом (рис. 8), на падающую
часть которых при сильном раздражении часто накладывают-
ся ритмические колебания. Амплитуда, длительность и форма
медленного потенциала зависят от концентрации пахучего ве-
щества, продолжительности раздражения, объема вдуваемой
пахучей -смеси, повторности раздражений при данной глубине
погружения электродов в обонятельный эпителий.
1 Термины «монофазный» и упомянутые ниже «двухфазный», «трех-
фазный» потенциалы взяты из электрофизиологии (см. рис. 8, 9 и 10) и не
соответствуют общепринятой в электротехнике терминологии.
12
Медленный потенциал возрастает по амплитуде с увеличе-
нием концентрации пахучего вещества, а его длительность уве-
личивается с продолжительностью раздражения при неизмен-
ной концентрации [3, 62].
Увеличение концентрации пахучего вещества и увеличение
объема вдуваемой пахучей смеси в широких пределах по воз-
действию на величину медленного потенциала адекватны.
1100 м кв
1сек
Рис. 8. Медленный, потенциал обонятельного эпи-
телия. Горизонтальные линии под кривыми показы-
вают продолжительность предъявления стимула
(амилацетата).
Повторные раздражения приводят к постепенному умень-
шению амплитуды медленного потенциала и впоследствии
к его исчезновению. Полное восстановление медленного по-
тенциала наступает после некоторой паузы.
Форма медленного потенциала изменяется с изменением
типа запаха [62]. Этот результат был получен при исследова-
нии обонятельного эпителия лягушки. А. Л. Бызов и Г. И. Фле-
рова установили, что наиболее четкие и постоянные различия
в ответах выявляются в условиях кратковременных повторных
вдуваний пахучего воздуха. Ими была сделана попытка клас-
сифицировать запахи по форме ответов [3]. Однако некоторы-
ми исследователями [70] установлено, что раздражение одним
и тем же запахом вызывает различные формы медленных по-
тенциалов (рис. 9)1.
1 Аналогичные изменения медленного потенциала были записаны
с усиков (антенн) личинок шелкопряда [56]. *
По мере погружения микроэлектрода «в обонятельный эпи-
телий амплитуда медленного потенциала уменьшается сначала
незначительно (иногда даже увеличивается), и лишь при даль-
нейшем погружении наблюдается ее быстрое уменьшение.
В большинстве случаев при глубине погружения микроэлек-
трода на 250—300 мк ответ полностью исчезает [3].
В обонятельном нерве потенциал действия имеет форму
затухающих колебаний. Частота этих колебаний составляет
в начале возбуждения 7—48 гц и уменьшается к концу до 5—
10 гц. Скорость про-
ведения сигналов
вдоль нерва состав-
ляет 24—27 см!сек.
На эту реакцию, за-
Рис. 9. Медленный .потенциал и вызван-
ные волны.
а — трехфазный медленный потенциал обоня- ’
тельного эпителия; б — электрическая актив-
ность обонятельной луковицы; в — электри-
ческая активность другой, при пересечении ее
обонятельного нерва.
писанную с нервных
волокон, оказывают
влияние вид стиму-
лирующего пахуче-
го вещества, его кон-
центрация и ско-
рость потока паху-
чей смеси в обоня-
тельной щели 178].
Величина реакции
не зависит от отно-
воздуха при ее измене-
сительной влажности окружающего воздуха при ее измене-
нии от 0 до 99% и температуре 20°С и в малой степени за-
висит от изменения состава воздуха, используемого для при-
готовления пахучей смеси. С повышением температуры
скорость появления обонятельной реакции увеличивается.
Обонятельная реакция с увеличением температуры в диапазо-
не от 10 до 35°С несколько уменьшается, а в более узких пре-
делах изменения температуры (от 20 до 30°С) остается неиз-
менной.
При стимулировании обонятельного эпителия водным ра-
створом пахучего вещества отмечалось, что обонятельные ре-
цепторы нечувствительны к изменению степени ионизации, ве-
личины pH раствора на несколько единиц и изменениям осмо-
тического давления в диапазоне ±20—25% [78].
Иссл'едовался единичный обонятельный рецептор черепахи.
При стимуляции амиловым и н-бутиловым спиртами, гвоздич-
ным маслом и др. были зарегистрированы импульсные разря.
ды (спайки) одного рецептора [66], положительные по отноше-
14
нию к медленному потенциалу, «монофазные» или «двухфаз-
ные». Амплитуда импульсов составляла 0,5—2 мз, а их
длительность 3—4 мсек. Потенциал действия (колебания) по-
являлся в обонятельных луковицах через 700 мсек после воз-
никновения медленного Й^тенциала в обонятельном эпителии.
Отмечено, что форма и частоты колебаний идентичны колеба-
ниям в обонятельном нерве (рис. 10). Очевидно, митральные
а 1500мкв
б \20мкв
/*^~*>Ч*^***^.............>лг-.. , ..„«im—
У . .. Lllliliii ]50»кв
1сек '
Рис. 10. Медленный потенциал обонятельного эпи-
телия (а), изменение потенциала обонятельного
нерва (б), колебания в обонятельной луковице (в).
клетки обонятельных луковиц получают возбуждение от обо-
нятельного нерва. Частоты колебаний в луковице изменяются
от 7—18 гц в начале до 5—10 гц к концу стимуляции1. Было
также отмечено, что колебания в луковице часто продолжа-
лись дольше, чем в обонятельном нерве, что может быть вы-
звано циркуляцией активности (межклеточными взаимодей-
ствиями) как в самой луковице, так и между луковицей и выс-
шими обонятельными центрами мозга.
После пересечения обонятельного нерва, подходящего к од-
ной из луковиц, в обеих обонятельных луковицах обнаружива-
ются потенциалы действия, являющиеся, вероятно, следствием
активности пучковых клеток.
В литературе описан интересный эксперимент по раздра-
жению обонятельного эпителия некоторыми пахучими вещест-
вами с одновременной регистрацией разного типа ответов
в обонятельном эпителии, нерве и луковице [70]. Различные
пахучие вещества вызывают о/г-ответы (реакции или измене-
ния электрической активности, возникающие с началом сти-
муляции), o/f-ответы (реакции, появляющиеся после оконча-
ния стимуляции) и on—off-ответы в обонятельном эпителии.
1 Подобное предположение согласуется с результатами исследований
на кроликах, приведенными Эдрианом в [15, 16].
15
нерве и луковице. На рис. 11 даны on- и о//-ответы обонятель-
ного эпителия при стимулировании амилацетатом. Изме-
нение типа ответа можно наблюдать и при изменении
интенсивности запаха. Так, например, раздражение малыми
концентрациями эфира вызывае^ о/г-ответ, средними —
on—off-ответ обонятель-
ного эпителия лягушки и
жабы и* большими — off-
ответв эпителии лягушки
(рис. 12).
Повторные раздраже-
ния большими концентра-
циями эфира изменяли
Рис. //.on-и о//-ответы обоня-
тельного эпителия при стиму-
лировании амилацетатом.
а — кривые, записанные с помо-
щью макроэлсктрода; б — кривые,
записанные с помощью микроэлек-
трода.
величину оп-ответов обонятельного эпителия без измене-
ния off-ответов; on-ответы в луковицах в таких услови-
ях имели ту же амплитуду. Можно предположить, что о/г-от-
i/i6 /—-------------- ---------—
Рис. 12. Появление off- ответа обонятельного
эпителия (медленного потенциала) при увеличении
концентрации эфира.
ice*
вет обонятельного эпителия (медленный потенциал) не имеет
никакого отношения к виду on-ответа в луковице. Более того,
off-ответ в луковицах может быть часто и без off-ответа в обо-
16
нятелыном эпителии (рис. 13). До сих пор off-ответ был полу-
чен только у амфибий и рыб. Три типа ответов можно отнести
к трем типам обонятельных клеток у этих животных [70].
Как уже указывалось, на падающую часть характеристи-
ки медленного потенциала накладываются периодические
колебания (рис. 14). Они имеют более высокую час-
тоту, чем колебания в
нерве и луковице, и
возникают с некото- а -------------------------——
рым запаздыванием по
отношению к ним.
Можно полагать, что
колебания потенциала
в обонятельном эпите-
лии непосредственно не
связаны с началом
электрической актив-
ности обонятельного
нерва и луковиц и, сле-
довательно, с обоняни-
ем 171].
Эдриан, изучая обо-
нятельный орган кро-
лика, предположил
возможность разделе-
ния рецепторов на че-
тыре группы в зависи-
Рис. 13. Ответы на стимуляцию эфирам:
а — медленный потенциал обонятельного эпи-
телия; б — колебания в обонятельном нерве;
в — колебания в луковице.
мости от их чувствительности к пахучим веществам с раз-
личной молекулярной структурой: ароматическим соединени-
ям, эфирам жирных кислот, терпенам, сернистым соединениям.
Различение запахов определяется чувствительностью отдель-
ных групп рецепторов, связанных с митральными клетками.
Обнаружено, что более легкие молекулы в малых концентра-
циях будут возбуждать в основном переднюю (anterior) часть
обонятельного эпителия, в то время как тяжелые масла оказы-
вают большее влияние на заднюю (posterior) часть эпителия.
Было показано, что различные участки обонятельного
эпителия проектируются (отображаются) на определенные
области обонятельной луковицы. ’ Так, передняя и дор-
сальная (спинная, верхняя) части обонятельного эпителия
проектируются в передней части луковицы, задняя и вен-
тральная (брюшная, нижняя) — в задней части луковицы
[14—19, 43].
2 Заказ № 7032
17
Таким образом, электрофизиологические исследования по-
зволили установить некоторую связь между электрической ак-
тивностью обонятельного эпителия, нерва и луковицы при сти-
муляции пахучими веществами, определить зависимость ответа
обонятельного эпителия, нерва и луковицы от типа пахучего
вещества и его концентрации. Получен ряд доказательств спе-
цифичности рецепторов и связи различных участков обоня-
тельного эпителия с определенными областями обонятельной
луковицы.
1100 мкв
I 20 мкв
150 мкв
Рис. 14. Электрическая активность обонятельного
эпителия (а и б) нерва (в), луковицы (г). Кри-
вые «а» и «б» записаны усилителями с большой и
малой постоянными времени.
150 мкв
*- —<
1свк
Механизм восприятия запахов и возбуждения обонятель-
ных рецепторов на основании электрофизиологических иссле-
дований определить не удалось.
1.5. НЕКОТОРЫЕ ОСОБЕННОСТИ ОБОНЯТЕЛЬНОГО АНАЛИЗАТОРА
Адаптация и утомление. Длительное воздействие
некоторого запаха может привести более или менее быстро
к неспособности ощущать этот запах, не ослабляя чувстви-
тельности к другим запахам. Это явление, называемое обо-
нятельной адаптацией, может вызываться процессами в обо-
нятельном эпителии, обонятельной луковице и обонятельных
центрах [2, 46,47]. Это можно заметить также на кривых
электрической активности обонятельного эпителия как умень-
шение величины медленного потенциал^ с увеличением вре-
мени предъявления стимула или как уменьшение частоты
импульсов в аксонах постсинаптических нервных клеток при
18
тех же условиях. Электрофизиологические исследования ор-
ганов обоняния (насекомых показали, что явление адаптации
характерно и для них.
Аносмия — дефект органов обоняния, состоящий в не-
способности ощущать или различать некоторые группы па-
хучих веществ [2, 46, 47].
Паралич — ряд веществ (например, формальдегид) па-
рализует чувство обоняния ко всем запахам в равной степе-
ни [2, 46, 47].
Подавление одного запаха другим (маски-
ровка). Анализ обонятельных ощущений возможен только
в случае примерно одинаковой интенсивности запахов ве-
ществ, образующих смесь. Если же интенсивность запаха
одного из веществ больше интенсивности запала другого, то
более сильный запах подавляет слабый, так что запах смеси
почти ничем не отличается от запаха того вещества, которое
пахнет сильнее. Например, метил-ионон, имеющий очень
сильный запах, ведущий к быстрому утомлению обоняния,
подавляет запахи других веществ [2,46—48].
Слияние запахов. Запах смеси приобретает в боль-
шинстве случаев новые качества, не свойственные ее ком-
понентам. Слиться могут не только близкие по своему харак-
теру запахи, но и весьма различные. Так, смешение анетола
и капроновой кислоты, анетола и скатола, бензилацетата
и цитраля, цитраля и пинена приводит к образованию в каж-
дом случае совершенно нового своеобразного запаха смеси.
В настоящее время обнаружено множество пар пахучих
веществ, запахи которых взаимно компенсируют друг друга.
Трудность полной компенсации заключается в необходимости
точно подобрать количества этих веществ. К тому же най-
денные количественные соотношения не вполне одинаковы
для разных людей. Хорошую компенсацию дают следующие
пары: кедровое дерево и каучук, каучук и воск, воск и толу-
анский бальзам, йодоформ и перуанский бальзам, мускус
и горький миндаль, циклогексанол и цинеол, скатол и кума-
рин, этилмеркаптан и эвкалипт и др. [2, 46. 48, 55].
Приведенное в этой главе описание строения обонятель-
ного анализатора и результатов электрофизиологических
исследований хотя и не раскрывает существа механизма вос-
приятия запахов, передачи и преобразования информации,
по используется ниже для анализа различных теорий обоня-
ния и моделей обонятельного анализатора.
2*
ГЛАВА II
ЧУВСТВИТЕЛЬНОСТЬ ОРГАНОВ ОБОНЯНИЯ
Чувствительность органов обоняния характеризуется аб-
солютным, количественным разностным и качественным раз-
личительным порогами [2].
Абсолютный порог — минимальная концентрация пахуче-
го вещества Спор, вызывающая обонятельное ощущение. Ко-
личественный разностный порог определяется минимальной
обнаруживаемой разностью концентраций данного вещества.
Качественный различительный порог — минимальная при-
месь одного вещества к другому, способная вызывать изме-
нение характера ощущения.
При исследований органов обоняния сопоставляются по-
роговые концентрации разньих пахучих веществ, оценивается
чувствительность одного и того же субъекта к определенным
запахам при различных условиях, а также разных субъектов
при одинаковых условиях.
2.1. АБСОЛЮТНЫЙ ПОРОГ ОБОНЯНИЯ
Многие живые организмы имеют необычайно высокую
обонятельную чувствительность —низкие абсолютные поро-
ги. Так, некоторые рыбы ощущают пахучее вещество при
концентрации его 1011 мг/л. Интересно, что миграция рыб,
в частности лосося, определяется выбором течений, имеющих
характерные запахи растений, омываемых водой у истоков
рек в местах нереста [64]. Примерами пахучих веществ с низ-
кими пороговыми концентрациями для человека могут слу-
жить скатол—4-10-7 мг/л, этилмеркаптан—4,4 -10 s мг/л,
тринитробутилтолуол — 5-10 9 мг!л.
20
Данные о 'величине пороговой концентрации важны для
сравнительной оценки интенсивности запаха различных ве-
ществ. В табл. 1 приведены пороговые концентрации (абсо-
лютные пороги) пахучих веществ для человека [2], обладаю-
щих ольфактивным действием1.
В табл. 2 приведены концентрации для веществ, облада-
ющих ольфактивным и тригеминальным действиями2.
По величине абсолютного порога и0, выраженного числом
молекул пахучего вещества во вдыхаемом воздухе при поро-
говой концентрации, и количеству рецепторов в обонятель-
ном эпителии определена чувствительность отдельных обоня-
тельных рецепторов человека [81]. В указанной работе было
принято, что обонятельное ощущение создается не порого-
вым количеством молекул п0 во вдыхаемом воздухе, а числом
молекул пахучего вещества и, адсорбированных на обоня-
тельном эпителии.
При этом
« = «0 • Л f> ' f3, (1)
гдеД — коэффициент, определяющий часть вдыхаемого но-
сом воздуха, проходящего через обонятельную щель;
/г — коэффициент, определяющий часть молекул пахучего
вещества, остающихся в воздухе при достижении ими обо-
нятельной щели; /г<1, так как часть молекул адсорбирует-
ся слизистой, выстилающей носовую полость;
/з— коэффициент, определяющий часть молекул, адсор-
бированных на обонятельном эпителии.
Зная п и число рецепторов в эпителии, можно вычислить
среднее число молекул, приходящееся на рецепторную клет-
ку. Статистический анализ позволяет определить минималь-
нее число молекул п,\, на которое реагирует клетка.
Значение fi было определено из геометрических размеров
носовой полости. Найдено, что при нормальном дыхании
через обонятельную щель протекает 5—10% Общего количе-
ства воздуха, проходящего через носовую полость. При
более высокой скорости потока эта величина достигает
20%. Очевидно, fi может быть принято равным от 0,05—0,1
до 0,2.
1 Вещества ольфактивного действия возбуждают только обонятель-
ные рецепторы.
2 Веществ, возбуждающих обонятельный и тройничный /нервы.
21
Таблица 1
Название вещества Молекулярный вес Пороговая концентрация
X 106 моль/л X ю3 мг)л молекул! л
Бензол 78,1 0,068 5,28 4 • IO***
Толуол 92,1 0,022 2,0 13,4 • 1016
Ксилол 106,1 0,0075 0,8 4,6 • 10^
Псевдокумол 120,1 0,0017 0,2 1,0 • 1015
Дурол 134,1 0,00067 0,09 4,0 • 10*
Амиловый спирт 88,1 0,011 1,0 6,8 • 10*
Изоамиловый спирт . . 88,1 0,0011 0,1 6,8 • 101*
Гептиловый спирт . . . с . . 116,2 0,0086 1,0 5,2 • 10*
Терпинеол 154,5 1,17 180 6,8 • 10*
Цитраль 152,2 0,00066 0,1 4,0 . 10*
Валериановая кислота . . , . . 102,1 0,000098 0,01 6,0 • 10*
Капроновая кислота 116,1 0,0'ЮЗЗ 0,04 2,0 • 10*
Гептиловая кислота 130,1 0,0023 0,3 1,4 • 10*
Каприловая кислота 144,2 0,00035 0,05 2,0 . 10*
Нониловая кислота 158,2 0,000126 0,02 8,0 • 10*
Каприновая кислота ч . . е . . 172,2 0,00029 0,05 17,6 • 10*
Лауриновая кислота 200,2 0,0005 0,1 3,0 • 10*
Изоамилацетат .... 130,1 0,69 900 4,2 • 10*
Кумарин ...... 146,1 0,000068 0,01 4,0 • 10*
Гваякол 124,1 0,03 3,8 2,0 • 10*1
Ванилин 152,1 0,0 000 033 0,0 005 2,0 • 10*’
Анилин : 93,1 0,00049 0,046 3,0 • 10*
Гелиотропин . . * . . 150,1 0,00033 0,05 2,0 • 10*
о-Толуидин ... 107,1 0,00027 0,009 16,4 • 10*
м-Толуидин 107,1 0,00024 0,026 14,8 • 10*
п-Толуидин 107,1 0,00013 0,014 7,8 • 10*
Ксилидин 121,2 0,00019 0,023 11,6 • 10*
Кумидин 135,2 0,000059 0,08 3,6 . 10*
Метиловый эфир антраниловой кислоты - . . • в • 139,1 0,000042 0,006 2,6 • 10*
Тринитробутил — то- луол 283,13 0,000000017 0,000 005 10,8 • 10»
Скатол , . 131,1 0,000003 0,0 004 18,4 . 10*
22
Таблица 2
Название вещества Молекулярный вес Пороговая концентрация
X 106 МОЛЬ / Л X 1о3 мг/л молекул! л
Бром . 79,96 0,42 33,4 2,6 • 1017
Фосфористый водород 34,0 5,9 200,0 3,6 • 1018
Сероводород 34,1 0,235 8,0 14,6 • 101«
Метиловый спирт. . . . 32,0 18,7 600 17,4 • 1018
Этиловый спирт . - . . 46,1 5,42 252 3,2 Щ18
Пропиловый спирт . . . 60,1 ‘ 0,084 5,0 5,0 Ют
Изопропиловый спирт 60,1 0,664 40 4,0 • 1017
НчБуТИЛОВЫЙ спирт 74,1 0,0135 1,0 8,2 • 101>
Изобутиловый спирт . . 74,1 0,0135 1,0 8,2 10i>
Фенол 94,1 0,0425 4,0 2,6 • 1018
Хлорфенол 128,5 0,000034 0,0434 18,0 • 10*2
Уксусный альдегид . . 44,0 0,016 0,7 9,4 • 10»-
Ацетон 58,1 0,069 4,0 4,2 • 1018
Камфара 152,2 0,033 5,0 8,0 • 10**
Муравьиная кислота . . 46,0 0,54 25,0 3,2 • 1017
Пропионовая кислота . . 74,1 0,000675 0,05 4,2 • 1014
н-Масляна я кислота . . 88,1 0,0000113 0,001 7,0 • 101-
Метилацетат 74,0 0,027 2,0 16,4 • lol-
Эфир 74,1 0,0135 1,0 в.2 • 101-
Хлороформ 119,4 0,25 30,0 15,2 • 1018
Бромоформ . . . - . 252,9 0,008 0,2 4,8 • 101*
йодоформ 393,8 0,000154 0,056 8,6 • 10»з
Нитробензол 123,1 0,331 41,0 2,0 . 1017
Пиридин ? 79,1 0,000505 0,04 3,0 • 101*
Этилмеркаптан .... 62,1 0,0000007 0,000044 3,8 • Юн
Диэтилдисульфид . . s 122,2 0,00246 0,3 1,48 • IO*г»
Однако могут иметь место индивидуальные отклонения or
приведенных величин, обусловленные различиями в конфи-
гурации полости носа1.
Для определения величины f2, предполагалось, что моле-
кулы, сталкивающиеся со слизистой мембраной или обоня-
тельным эпителием, оставались «а них. Количество пахучих
молекул в воздухе после адсорбции на слизистом эпителии
носовой полости до достижения ими обонятельной щели мо-
жет быть подсчитано для момента времени t на основа-
нии [81]:
Рт2 t
а=аое~ № , (2)
1 Заметные изменения просвета обонятельной щели могут возникать
вследствие изменений кровенаполнения раковин или выделения слизи.
23
где d—ширина обонятельной щели;
D— коэффициент диффузии пахучего вещества;
а0—общее количество молекул пахучего вещества.
Принимая d=l мм, 0 = 0,08, имеем
а = aQe 80* .
(3)
Если скорость вдыхаемого воздуха определяется расходом
К см3/сек, то время переноса молекул от входа в носовую
полость до обонятельного эпителия составляет примерно
2
t= -у сек, Подстановкой значения t в выражение (3)
получено
’ -80 • 2
f^e V
«О 7- (4)
При нормальном дыхании скорость потока V составляет
250 см3/сек (через каждую ноздрю) и, следовательно,
/2=е-0.64_0>5
(5)
Пользуясь выражением (3), можно получить число неад-
сорбированных молекул в обонятельной области (1—/з), от-
куда
/з=1- е-^- (6)
При нормальном дыхании f3 г= 0,5. Подстановка значений
fi, f‘2 и /з в уравнение (1) дает 2% молекул пахучих веществ,
адсорбирующихся на обонятельном эпителии при нормаль-
ном дыхании. Остальные 98% могут адсорбироваться в сли-
зистой другой части полости носа' или пройдут сквозь обоня-
тельную щель, не соприкасаясь с обонятельным эпителием.
Значения f2 и /з подсчитаны в предположении, что молекула
пахучего вещества, попадая на поверхность слизистой или
обонятельный эпителий, задерживается на ней достаточно
длительное время (порядка секунды или более). При нор-
мальной скорости потока /з увеличивается с увеличением ад-
сорбции, f2— уменьшается. Поэтому количество молекул, до-
стигших эпителия, в некоторых пределах почти не зависит
от интенсивности адсорбции.
24
Нам кажется, что значения /2 и /3 в действительности не
могут быть одинаковыми, так как способность адсорбировать
молекулы пахучего вещества слизистой, выстилающей носо-
вую полость, и обонятельным эпителием различна.
На основании изложенного выше в [81] были определены
пороги возбуждения отдельного рецептора. Известно, что
пороговое значение (по) для меркаптана составляет 109 мо-
лекул. Так как только 2% молекул адсорбируется в обоня-
тельном эпителии, то п = 2-107. Адсорбированные молекулы
могут распределяться по всем обонятельным рецепторам, чис-
ло которых составляет (4—5) 107. Следовательно, на один ре-
цептор в среднем приходится V2 молекулы. Предположив,
что для возбуждения только одного рецептора необходимо
п{ или большее число молекул, легко определить вероятность
возбуждения одного рецептора. А для того, чтобы вызвать
ощущение, очевидно, должна быть возбуждена по крайней
мере одна клетка. Если выбрать слишком большим, веро-
ятность возбуждения только одного рецептора уменьшается.
Статистический анализ показывает, что не должно быть
больше 8 молекул. Вполне вероятно, что пх может быть рав-
но 1. Медленность диффузии в слизистой оболочке не позво-
ляет концентрировать молекулы на специфических рецепто-
рах, особенно чувствительных к данному запаху. Кроме то-
го, молекулы могут быть адсорбированы -на неэффективных
участках (не все клетки обонятельного эпителия, например
опорные клетки, эффективны для восприятия запаха). Все
это приводит к уменьшению значения В той же работе
[81] получено, что для возникновения обонятельного ощуще-
ния необходимо не менее 40 молекул или одновременное
возбуждение 40 рецепторов, каждый из которых реагирует
на одну молекулу.
Интересный ‘способ выражения обонятельных порогов
термодинамической активностью Аоб предложен в [54]:
4 — ^об
‘ 06 Ро ’ (7)
где Роб — парциальное давление паров пахучего вещества,
соответствующее пороговой концентрации;
Ро — давление пара чистой жидкости при 37°С.
Пороговые значения для некоторых пахучих веществ, сог-
ласно [58], даны в табл. 3.
25
Таблица 3
Название вещества Концентра- ция, мг\л Роб атм. . Ю 7 PQ атм (37°С)
Амилацетат 0,039 90 0,013 1 . 10-з
Масляная кислота Искусственный 0,009 18 0,003 1 • 10 з
мускус .... Валериановая кис- 0,00004 0,029 0,0000013 2 . 10 з
лота 0,029 50 0,001 5 . 10 -з
Амиловый спирт 0,225 500 0,007 7 . 10-з
Нитробензол . . 0,146 300 0,0007 40 • Ю-з
Метилсалицилат . 0,10 150 0,00026 40 • 10 з
Табл. 3, показывает, что порог, выраженный Лоб, характе-
ризуется большим постоянством, чем выраженный весом на
единицу объема. В определенных гомологических рядах хи-
мически нейтральных гибких молекул для любого данного
числа атомов углерода Лоб примерно постоянно.
Чистота пахучих веществ, используемых в экспе-
риментах, имеет большое значение, так как некоторые из них
можно обнаружить по запаху даже при величине Аоб около
10-6 [58]. Требования к чистоте особенно важны при иссле-
довании веществ, имеющих очень слабый запах. Для многих
органических соединений чистота 99% обычно считается до-
статочно высокой. Тогда если Лоб примеси составляет 10-6,
а основного вещества 10-4, то оба запаха будут одинаковой
интенсивности. Наиболее высокая степень очистки для опре-
деленных классов соединений допускает 0,01% примеси. По-
видимому, запахи различных химических веществ надо оце-
нивать с некоторой осторожностью. Так, например, резуль-
таты экспериментов с нормальным рядом парафинов позво-
лили утверждать, что пентан имеет неясный запах, а высшие
члены ряда, особенно октан, имеют сильный запах керосина
[48]. Результаты других исследований находятся в полном
противоречии с этим утверждением [58]. В действительности
октан имеет очень слабый запах, мало отличающийся от за-
паха пентана. Этот факт объясняется тем, чго запах кероси-
на или бензина может быть связан с различными типами
колец углеводородов, которые лишь с трудом выделяются
из н-парафинов. Если парафины получены не из керосина,
а из чистого исходного соединения, то они совсем не обла-
дают запахом. Запах ацетилена определяется главным обра-
зом запахом таких примесей, как фосфин.
26
Изложенное позволяет заключить, что запах данного хи-
мического соединения зависит от его пороговой концентра-
ции, методов синтеза этого вещества и исходных материалов
и, следовательно, различных примесей.
2.2. ИЗМЕРЕНИЕ ОБОНЯТЕЛЬНЫХ ПОРОГОВ
Пороговое значение определяется не только интенсивно-
стью запаха и временем его, воздействия, но и скоростью,
с которой пахучая смесь проходит через носовую полость.
Наибольшая чувствительность наблюдается при возрастании
концентрации пахучего вещества с наибольшей скоростью.
Введение пахучего вещества с недостаточной скоростью, на-
пример, если его концентрация увеличивается от 0 до 5 по-
роговых значений в течение одной минуты, не дает никакого
ощущения.
При скорости потока более 4 л/мин обонятельный порог
пахучего вещества стабилизируется
При постоянстве скорости
потока (250 с.м3/сек) зави-
симость порога от времени
воздействия представлена на
рис. 16 и 17 [81]. Если запах'
действует в течение корот-
кого промежутка времени,
порог практически постоя-
нен, а при длительном воз-
действии запаха — пропор-
ционален времени воздей- .
ствия. Для очень малых
длительностей действия за-
паха порог возрастает. Это происходит потому, что объем во-
влекаемого воздуха становится весьма малым; следовательно,
большее значение приобретает объем полости носа, которая
должна быть заполнена прежде, чем пахучее вещество достиг-
нет обонятельного эпителия. Введение поправки на эту поте-
рю отражено штриховой линией на рис. 16. Кривые, приве-
денные на рис. 16 и 17, получены при введении воздуха пу-
тем вдувания [81]. При определении порогов обоняния было
предложено воздействовать запахом в течение одной секун-
ды с последующим десятисекундным перерывом. Соблюдение
(рис. 15).
Арб
ю-Ч
12-
ю-
в-
6
4 ——г-
0 2
Рис. 15. Зависимость обо-
нятельного порога (Аоб)
от скорости потока паху-
чей смеси в обонятель-
ной полости.
-1—I—I—I—I—I---1-Г“*
6 в ю 12 М
Скорость потока, л/мин
27
Рис. 16. Зависимость обонятельного порога от
времени воздействия пахучего вещества.
Рис. 17. Зависимость интенсивности стимула от времени воз-
действия пахучего вещества:
а — для вторичного бутилмеркаптана; б — для метаксилена.
этих условий сохраняет способность обнаруживать запахи на
неограниченное время 158].
Продолжительность ощущения запаха определяется в ос-
новном качеством пахучего вещества и в меньшей степени
зависит от его концентрации.
2.3. РАЗЛИЧИТЕЛЬНЫЕ ПОРОГИ
Вопрос о минимально различаемых концентрациях паху-
чих веществ и о различии пахучих веществ по запаху изучен
недостаточно [42]. Можно предположить, что обонятельные
ощущения в среднем диапазоне интенсивности стимула С
подчиняются закону Вебера-Фехнера:
= const. (8)
о
Как указывалось в начале главы, разностный порог АС—
это минимальное изменение концентрации пахучего вещест-
ва, ощущаемое органами обоняния, по сравнению с первона-
чальной концентрацией Со:
АС-С-Со. (9)
Если часть .информации, идущей к мозгу, зависит от из-
менения концентрации пахучего вещества в воздухе, мозг
будет обучаться распознавать изменение концентрации при
условии, что эти изменения определимы, т. е. если они превы-
шают случайные флуктуации в системе.
Закон, определяющий восприятие заметной разницы, дол-
жен быть статистическим, потому что процесс столкновения
молекул с рецепторами является случайным и подчиняется
статистическому закону, согласно которому среднее квадра-
тичное значение отклонения от среднего числа пахучих мо-
лекул и, попадающих на рецепторы, равно ]/7Г. Для того, что-
бы была воспринята заметная разница, она должна быть
пропорциональна \п. Таким образом, различительный порог
можно выразить через добавочное число молекул Ап, при
котором становится возможным выделить изменение концен-
трации пахучих веществ из флуктуаций столкновений моле-
кул, определяемых средним числом этих молекул. Следова-
тельно, должно быть
An = kV п . ‘
29
Коэффициент пропорциональности k определяет надежность
выделения сигнала из флуктуаций. Соотношение (10) может
быть дано в единицах концентрации пахучих веществ.
2.4. СТАТИСТИЧЕСКОЕ ОБНАРУЖЕНИЕ• ЗАПАХОВ
Порог обонятельной чувствительности не следует считать
точно фиксированным параметром. В действительности он
флуктуирует вблизи некоторого среднего значения. Пред-
ставление о статистическом характере порогов можно полу-
чить, если даже пренебречь флуктуациями в нервных кана-
лах и считать, что порог обонятельной чувствительности пол-
ностью определяется флуктуациями числа адсорбированных
пахучих молекул на обонятельном эпителии [5,42]. При ис-
следовании разностных порогов может определяться нали-
чие (или отсутствие) обонятельного стимула со средней ин-
тенсивностью С на фоне со средней интенсивностью Сф.
Мгновенные значения интенсивности фона все время меняют-
ся вокруг среднего значения Сф, вероятность их отклонения
от которого определяется законом Пуассона. Мгновенные
значения воздействующего стимула также флуктуируют, но
вокруг среднего значения С, подчиняясь тому же закону.
Пусть критерием обнаружения различительного порога слу-
жит уровень интенсивности Сь Это означает, что интенсив-
ность запаха, превышающая эту величину, воспринимается
наблюдателем как изменение интенсивности Независимо от
того, вызывается ли это действием флуктуирующего фона
или сигнала.
Увеличение интенсивности стимула при неизменном фоне
и уровне обнаружения увеличивает отношение сигнал—шум»
что приводит к росту вероятности правильного суждения
о присутствии сигнала.' Если построить зависимость вероят-
ности правильного суждения о наличии сигнала от интенсив-
ности обонятельного стимула С, то получается кривая, ана-
логичная психофизической кривой зависимости вероятности
обнаружения стимула от его интенсивности (рис. 18).
При определении порогов необходимо учитывать не толь-
ко флуктуации количества адсорбированных молекул, но
и флуктуации в нервных каналах.
Теория обнаружения сигналов (стимулов) на примере
зрительного анализатора, учитывающая не только флуктуа-
30
ции воспринимаемой информации, но и флуктуации в нерв-
ной системе, была разработана в [74, 75]. На рис. 19 пока-
зано распределение значений нервной активности г на уров-
не, где действует механизм обнаружения сигналов (стиму-
лов). Заданной интенсивности сигналов соответствует
определенное распределение возможных значений нервной
Рис. 18. Зависимость вероят-
ности обнаружения стимула
от его интенсивности.
Рис. 19. Модель обнаружения сигнала
в шумах нервной системы.
активности. Возникновение разброса z обусловлено наличи-
ем шумов в нервном канале. Если бы их не было, то опреде-
ленные интенсивности стимулов соответствовали бы фикси-
рованным значениям нервной активности и наоборот. Таким
образом, кривая распределения вероятных значений нервной
активности в ответ на стимул определенной интенсивности
выражает смесь сигнала с шумом (сш) [74, 75]. Левая кри-
вая на графике выражает распределение вероятности шумов
(ш) в отсутствие сигналов.
Предполагается, что эти распределения нормальны и име-
ют одинаковое стандартное отклонение. Вводится понятие
порогового критерия z0. Когда значение z превосходит эту
величину, оно оценивается как сигнал. Чем больше г, тем
более вероятно, что оно действительно вызвано обонятель-
ным стимулом. Можно выяснить, что определяет величину
порогового критерия. Для удобства введены следующие обо-
значения:
Лш(4)— условная вероятность правильного ответа
О принятии сигнала. Эта вероятность определяется площадью
Лш под кривой распределения справа от ze;
31
ложного реше-
деле 1не пода-
Рш(— )—условная вероятность правильного ответа об-
отсутствии сигнала;
Рсш(~) = 1 “Л:ш(л-) — условная вероятность ложного реше-
ния об отсутствии сигнала, когда он на самом деле подавался.
рш( = 1 —Рш(—)—условная вероятность
ния о принятии сигнала, когда он на самом
вался.
Как уже указывалось, Рсш ( + ) зависит от
стимула. С увеличением последней кривая
(сш) сдвигается вправо, что приводит к увеличению вероят-
ности ^сш(4г), зависящей также от уровня обнаружения z0.’
Кроме того,
интенсивности
распределения
от z0 зависит
рш
Рис. 20. Зависимость вероятности пра-
вильного обнаружения сигнала и ве-
роятности принять шум за сигнал от
выбора критерия обнаружения и отно-
шения сигнал — шум.
(+), т. е. вероятность того, что
шум будет принят за сиг-
нал. Обе эти зависимости
показаны на одном графике
(см. рис. 19).
Значение zo = O соответст-
вует случаю, когда уровень
обнаружения совпадает со
средним значением 2Ш. В
этом случае вероятность то-
го, что шум будет принят за
сигнал, равна половине пло-
щади, ограниченной кри-
вой, т. е. 0,5. Если z0 стре-
мится к — со , то, очевид-
но, Рсй1 ( + ) = Рт ( + ) = 1,
т. е. все принимается за
сигнал. Если z0 стремится к
т. е. все принимается за
шум. Zq должно быть вы-
брано так, чтобы обеспе-
чить наибольшее отношение
. Выбранное таки-м образом значение г0 называют оп-
тимальным критерием. Его <мож»но определить с помощью
графиков >р‘ис. 20. Для этого надо найти точку на кривой, где
наклон равен величине w, определяющейся равенством [63]
* сш
Уш -У,„+
Кщс-г К сш-
(И)
32
где РСш и Ли —априорные вероятности возникновения и от-
сутствия сигнала; их сумма, естественно, равна единице;
V — численные значения четырех перечисленных выше
условных вероятностей. Увеличение Рст или УСшь должно
привести к уменьшению w.
Такой же результат дает уменьшение . Как следует
из рис. 20, уменьшение наклона кривых дает такой оптималь-
ный критерий, при котором наиболее выгодно допустить
большее количество ложных решений о наличии сигнала для
достижения относительно высокого числа правильных ре-
шений о наличии сигнала в присутствии шума.
ГЛАВА III
ПАХУЧИЕ ВЕЩЕСТВА И ИХ СВОЙСТВА
3.1. ФИЗИЧЕСКИЕ СВОЙСТВА
Для возникновения обонятельных ощущений необходимо,
чтобы пахучие вещества вошли в соприкосновение с обоня-
тельными рецепторами в виде паров или, что бывает реже,
в виде мельчайших капелек или пылинок в окружающем воз-
духе. Кроме того, у млекопитающих пахучие вещества, преж-
де чем вступить в контакт с обонятельными клетками, долж-
ны проникнуть через тонкий слой (толщиной 2мк) жидкости,
покрывающей липоидную оболочку-мембрану рецептора.
Известно, что большая группа пахучих веществ (аммиак,
хлор, формалин) возбуждает, кроме окончаний обонятельного
нерва, болевые и тактильные окончания тройничного нерва.
Некоторые вещества действуют на его холодовые (ментол),
тепловые (спирты), а также на вкусовые окончания. Это об-
стоятельство дало повод делить пахучие вещества на возбуж-
дающие только обонятельный нерв (ольфактпвные) и веще-
ства, действующие также на тройничный нерв (общего дей-
ствия).
Ниже рассмотрены свойства пахучих веществ. Некоторые
из них, возможно, определяют механизм восприятия и разли-
чения запахов.
Летучесть. Пахучие вещества должны хотя бы в ми-
нимальной степени обладать способностью испаряться, т. е.
иметь определенную летучесть.
Парциальное давление паров пахучих веществ находится
в некотором соотношении с интенсивностью запаха. Многие
.шахучие вещества, парциальное давление паров которых ве-
34
лико, как, например, эфир, хлороформ, керосин, камфара,
нафталин и др., имеют сравнительно сильный запах, а веще-
ства с низким парциальным давлением паров — парафиновое
масло, глицерин, железо, медь и др.— не обладают им. Одна-
ко прямой зависимости между парциальным давлением и ин-
тенсивностью запаха разных веществ не существует.
Замечено, что органы обоняния неспособны возбуждаться
молекулами с молекулярным весом меньше 17 (аммиак)
и свыше 300 (терпены). Вещества с малыми молекулярными
весами, как, например, водород, кислород, углерод и их сое-
динения, подобные воде, угольной кислоте, метану, практи-
чески не имеют запаха [2, 48]. Исключением являются гало-
гены и озон, которые обладают высокой химической актив-
ностью и, возможно, действуют на окончания тройничного
(нерва.
Интенсивность запаха определяется молекулярным весом
(более тяжелые вещества имеют более сильный запах). Воз-
можно, этот вывод справедлив только для молекул с очень
простой структурой, с двумя или тремя атомами [691. Подоб-
ное предположение не является совершенно точным, ибо сле-
дует помнить об адаптации органов обоняния к таким веще-
ствам, как вода, угольная кислота, которые, как кислород
и азот, постоянно воздействуют на периферическую часть
анализатора. С другой стороны, с увеличением молекулярно-
го веса пахучих веществ уменьшается их летучесть и, сле-
довательно, интенсивность запаха.
Растворимость. Как уже указывалось, обонятельный
эпителий всегда влажен, и обонятельные клетки имеют обо-
лочку, содержащую липоиды. Поэтому многие авторы счита-
ют, что интенсивность запаха определяется растворимостью
пахучих веществ в воде и липоидах [2, 48, 49]. Действительно,
средние члены многих гомологических рядов1, растворимые
в воде и липоидах, обладают сильным запахом, в то же время
низшие члены, растворимые в воде, но не растворимые в ли-
поидах, и высшие, растворимые в липоидах и не растворимые
в воде, имеют слабый запах или не пахнут совсем. Например,
метанол, этанол, не растворимые в липоидах, пахнут слабо;
бутанол, растворимый в воде и липоидах, пахнет сильно.
Диффузия. В естественных условиях различение запа-
ха существенно зависит от диффузии молекул пахучих ве-
1 Каждый последующий член гомологического ряда отличается от
предыдущего на группу СНг. Члены одного гомологического ряда обла-
дают сходными химическими свойствами.
3* 35
ществ в воздухе. Первыми воспринимаются вещества с боль-
шей скоростью диффузии. В табл. 4 приведены значения ско-
ростей диффузии для некоторых пахучих веществ. _ , . Разбавление. Степень Таблицам - naanяилгния и чистотя пя-
Название вещества Скорость диффузии, см\сек хучих веществ влияют на качество запаха. Некоторые используемые в парфюме- рии вещества в чистом виде имеют неприятные запахи, например, индол, присут- ствующий в жасмине, а так- же наиболее важные фикса- торы — цибет и серая амб- ра. Запах разбавленного
Бензол Толуол Ксилол : Псевдокумол Эвгенол Камфара Серный эфир Уксусноэтиловый эфир 0,53 0,53 0,67 0,67 1.3 41 4,4 10,0
раствора может быть силь-
нее концентрированного [48]. Некоторые химические соедине-
ния в одной концентрации, приманивающие насекомых, могут
быть отпугивающими в другой. Если же вещество в малой кон-
центрации отпугивает насекомых, той с увеличением концент-
рации его действие не изменяется. Способность некоторых ве-
ществ приманивать увеличивается с увеличением их концент-
рации до некоторого максимума, затем уменьшается и при
определенных концентрациях переходит в отпугивание. Даже
для химически чистой приманки можно найти концентрацию,
при которой она становится отпугивающей. Примером может
служить реакция домашней мухи на пары изовалерианового
альдегида. Так, концентрация 2-Ю-6 моль!л не вызывает
поведенческой реакции, 1,2-10“ моль!л—максимально при-
манивает, концентрация от 2 10 до 2,8- 10 ~5 моль!л и при-
манивает и отпугивает. При дальнейшем увеличении концент-
рации отпугивающие свойства увеличиваются и будут наи-
большими при концентрации 6-10-5 моль!л.
Адсорбция. Явление адсорбции играет большую роль
в процессе обоняния, так как обонятельный эпителий, покры-
тый секретом, и омывающий его воздух, могут быть рассмот-
рены как гетерогенная система. Поэтому изучение специфич-
ности адсорбции пахучих веществ,’ обусловленной влиянием
типа адсорбента, имеет важное значение для понимания
механизма обоняния. Благодаря адсорбции происходит по-
вышение концентрации пахучих веществ в обонятельном
эпителии, часто находящихся в воздухе в ничтожных
количествах.
36
Адсорбция 'происходит под влиянием молекулярных сил
поверхности адсорбента и ведет к уменьшению свободной по-
верхностной энергии. Молекулы адсорбата, приближаясь из
объема газа или раствора к поверхности раздела фаз, испы-
тывают притяжение со стороны этой поверхности. Таким об-
разом, поверхность адсорбента покрывается тонким слоем
молекул адсорбата. При физической адсорбции молекулы ад-
сорбата сохраняют свою индивидуальность. Адсорбционные
силы при этом имеют ту же природу, что и межмолекулярное
взаимодействие в газах, жидкостях и твердых телах.
Адсорбционные силы притяжения подразделяются на дис-
персионные, вызываемые согласованием движения электро-
нов в сближающихся частицах, и на электростатические, свя-
занные с наличием в молекулах адсорбата постоянных элект-
рических диполей, квадруполей или вообще мультиполей.
В случае неполярного адсорбента проявляются в основном
дисперсионные силы притяжения.
У полярных адсорбентов на поверхности существует элек-
тростатическое поле, взаимодействующее с диполями и муль-
типолями молекул адсорбата, что усиливает адсорбцию,
в особенности когда на поверхности расположены преимуще-
ственно ионы одного знака или сходным образом ориентиро-
ванные диполи.
Молекулы пахучих веществ могут адсорбироваться непо-
средственно на чистой поверхности твердых тел или через
слой конденсированной влаги.
Известно, что все газы и пары в той или иной степени
адсорбируются на поверхности твердых тел. В этом смысле
неадсорбирующихся газов нет. Адсорбируемость данного ве-
щества зависит от условий, в которых происходит адсорбция.
Были исследованы адсорбирующие свойства поверхностей
твердых тел при воздействии на них потока пахучих веществ
в течение 5 мин с последующим определением длительности
существования запаха 148]. Результаты исследования сведены
в табл. 5.
Наиболее полные сведения о природе адсорбционных сил
и механизме адсорбции можно получить, зная величины энер-
гий адсорбции, определяемых как теплоты адсорбции. Теп-
лота адсорбции вообще является важной термодинамической
характеристикой и представляет непосредственную меру из-
менения полной энергии в адсорбционной системе.
Теплота адсорбции зависит от количества адсорбирован-
ных молекул, которое, в свою очередь, определяется количе-
37
Таблица 5
Пахучие вещества Адсорбирующие поверхности Длительность существова- ния запаха
Пиридин Алюминий, стекло, золо- то, свинец, никель, се- ребро Медь, цинк, фарфор Не более нескольких I секунд
» Несколько минут
Железо, сталь От 0,5 до 1 ч
Валериановая Медь, золото, железо, Не более нескольких
кислота свинец, никель, фар- фор, сталь, цинк секунд
То же Железо, серебро, олово Несколько минут
» Стекло 0,5 ч
Скатол Фарфор Запах отсутствует
» Стекло 1,5 ч
» Металлы От 1 до 20 дней
Ионон Стекло, фарфор, серебро Не более нескольких секунд
» Свинец, никель, железо, медь, алюминий От 1 до 4 дней
ством молекул, ударяющихся о поверхность адсорбента
[46, 47]. Число молекул, ударяющихся в секунду о поверх-
ность площадью 1 сл!2, согласно молекулярно-кинетической
теории газов, равно
NP
П~У 2-MRT ’
где Р — давление, дн!см2;
N — число Авогадро;
М — молекулярный вес;
R — газовая постоянная;
Т — абсолютная температура.
Если давление газа выразить в миллиметрах ртутного
столба, то уравнение после замены постоянных числовыми
значениями примет вид
л=3,52 • 1022 , Р-. ^ .
VMT (13)
Молекулы пахучего вещества можно считать не взаимодей-
ствующими друг с другом и подчиняющимися закону Даль-
38
тона. Молекула газа, попадая на поверхность, остается на
ней или упруго отражается. В большинстве случаев она оста-
ется на поверхности на период, зависящий от природы участ-
ка поверхности, с которой она сталкивается, и других факто-
ров: кинетической энергии молекулы (ее собственной, а не
средней), температуры поверхности, природы молекулы ит. д.
Если молекулы остаются на поверхности на некоторое время
(даже очень короткое), средняя концентрация в поверхност-
ном слое будет выше, чем в окружающем воздухе, т. е. будет
иметь место адсорбция пахучих молекул.
В момент адсорбции происходит обмен энергией менаду
молекулами и поверхностью. Для полного теплообмена меж-
ду ними требуется время порядка стократного периода коле-
бания молекул поверхности. Это используется для подсчета
времени т, в течение которого адсорбированная молекула
остается на твердой поверхности
Q
~=\eRr , (14)
где —период колебания адсорбированных молекул пер-
пендикулярно к поверхности;
Q — теплота адсорбции.
Подсчитано значение т, соответствующее различным теп-
лотам адсорбции на твердых поверхностях при комнатной
температуре [27]. Эти данные приведены в табл. 6.
Теплота адсорбции газов и паров составляет от 1—5
(малые молекулы) до 10—20 (большие молекулы) ккал!моль,
а Для аргона, азота, кислорода приблизительно равна
3,5—4 ккал!моль.
При покрытии молекулами адсорбата части поверхности
теплота адсорбции уменьшается до половинного значе-
ния [461.
Число молекул о, адсорбированное на единице поверхно-
сти, равно числу молекул п (12), умноженному на время т,
в течение которого они остаются на ней, т. е.
в=пх. (15)
Таким образом, можно подсчитать приблизительно число
адсорбированных молекул и величину поверхности, покры-
тую молекулами, зная их геометрические размеры. При ком-
39
личество
Таблица 6
Теплота адсор- бции, кка.1\моль сек
3,5 4 10 15 20 4 • IO " 4 • 10-1» 3,2 • 10—« 1,8 • 10--’ 1,0 • 10-2
натной температуре некоторая часть любой поверхности по-
крыта такими газами, как аргон, азот, кислород и т. д. Ко-
-----------------------]х 1м0ЛекуЛ уменьшается из-за отра-
жения от поверхности, полярности,
молекул, молекулярной несиммет-
ричности и поверхностного за-
грязнения. Энергия адсорбции за-
висит от природы и строения моле-
кул адсорбата и их ориентации у
поверхности, а также от природы
и структуры адсорбирующей по-
верхности. Например, при адсорб-
ции в целом неполярных молекул
бензола и н-гексана на поверхности
несущей функциональные группы или ионы (гидратирован
ном силикагеле), теплота адсорбции бензола выше теплоть
адсорбции н-гексана [9]. В этом случае отчетливо проявляется
специфическое взаимодействие л-электронной системы моле[
кулы бензола с гидроксильными группами поверхности силиь
кагеля. При адсорбции этих веществ на неполярной поверх!
ности графитированной сажи взаимодействие их молекул
с адсорбентом определяется только проявлением дисперсион-
ных сил. В соответствии с этим теплоты адсорбции н-гексан^,
в молекуле которого содержится большое число атомов водо-
рода, выше теплот адсорбции бензола.
Адсорбция, кроме теплоты, может характеризоваться из-
менением величины поверхностного натяжения и появлением
контактного потенциала на границе раздела двух сред. Более
подробно эти явления будут рассмотрены ниже.
3.2. СООТНОШЕНИЕ МЕЖДУ ЗАПАХОМ
И МОЛЕКУЛЯРНОЙ СТРУКТУРОЙ ПАХУЧИХ ВЕЩЕСТВ
Вопрос о соотношении химической структуры и запаха
имеет огромное значение для теории обоняния. Многие иссле-
дователи пытались найти определенную и исчерпывающую
связь между химическим строение^м и запахом веществ. Од-
нако запахи некоторых веществ нельзя объяснить с точки
зрения химического строения.
Для более полного представления о связи между химиче-
ским строением и запахом веществ можно все пахучие веще-
ства разделить на три класса [2, 48, 69]:
40
1) вещества, обладающие сходной структурой и имеющие
подобные запахи;
2) вещества, обладающие сходной структурой, но разны-
ми запахами;
3) вещества с различной структурой, но с подобными за-
пахами.
К первому классу веществ относятся, например, бензол
и его гомологи (толуол, ксилол), имеющие похожие специ-
фические сильные запахи. Примером веществ второго клас-
са могут служить некоторые цис- и транс-изомеры1 II, отлича-
ющиеся только пространственным положением групп у двой-
ной связи. Вещества третьего класса иллюстрируются такими
совершенно отличными по химическому строению, но похо-
жими по запаху соединениями, как мускус-макроцикличе-
ский кетон и амбровый мускус — производная бензола.
Из 30 элементов, которые находят в природе в несвязан-
ном состоянии, только семь (при нормальном атмосферном
давлении и температуре) пахнут. Это фтор, хлор, бром, йод,
кислород (в виде озона), фосфор и мышьяк. Интересно, что
все эти вещества обладают только двумя типами запахов:
раздражающим запахом галогенов и чесночным запахом
(фосфор и мышьяк). Озон по запаху напоминает и чеснок
и хлор. Оказывается, что простые молекулы, состоящие из
одинаковых атомов, могут иметь только два запаха — «гало-
геновый» и «чесночный». Некоторые авторы предлагали даже
считать эти запахи первичными. Из семи перечисленных эле-
ментов четыре — галогены — находятся в седьмой группе,
фосфор и мышьяк в пятой и кислород в шестой группе пери-
одической системы. Возможно, существует какая-то связь
между запахом элемента и его способностью функциониро-
вать с высокой валентностью.
Из неорганических соединений пахучими являются глав-
ным образом те, в состав которых входят неметаллические
элементы. Например, H2S, НгБе, CSC12, CS2, H2S2, NH3, все
окислы азота и т. д.
1 Изомеры — вещества, имеющие одинаковый качественный и коли-
чественный состав, одинаковый молекулярный вес, но различное распо-
ложение атомов в молекуле. Цис- и транс-термины, используемые для
обозначения определенных пространственных изомеров:
R с—Н
II II
R2—с—н н—С—R2
ifuc-изомер транс-изомер
41
Соединения фосфора, мышьяка, сурьмы, висмута часто
обладают чесночным запахом.
Большинство пахучих веществ относится к органическим
соединениям и состоит в основном из углерода, водорода,
кислорода, азота, серы.
Существует предположение, что запах химических соеди-
нений определяется наличием в них некоторых групп или
элементов (всего около 18), названных осмофорами. Но на-
личия осмофоров недостаточно для существования запаха.
Запах обусловливается и структурой, в которую входят ос-
мофоры. Эти структуры называются осмогенными, т. е. не-
сущими запах. Введение в осмоген осмофоров в некоторых
случаях усиливает запах, а в некоторых изменяет его или
даже уничтожает совсем. По признаку уменьшения влияния
на запах осмофоры располагаются в следующем порядке:
фосфор, мышьяк, сера, селен, хлор, бром, кислород, азот,
карбонил, карбоксил, эфирная группа, лактон, амин, имин,
лактам и т. д. Двойные связи, несомненно, увеличивают запах,
и, следовательно, бензольное кольцо можно рассматривать
как осмофор. Многие авторы даже считали, что ненасыщен-
ные валентности являются главной причиной запаха химиче-
ских веществ [48, 49].
Большое значение имеет и тип соединения осмофорной
группы с основной структурой пахучей молекулы. Многим
пахучим веществам присуща гетерополярная (ионная) связь
осмофора с осмогеном. Существует мнение, что запах такой
молекулы создается за счет ее поляризации, порождаемой
гетерополярной связью.
Очень интересны факты различия запахов у изомеров,
имеющих незначительные отличия в химическом строении.
Например, децилвалеролактан
I I
сн3снсн, снсо
I
(СН,)9СН3
не пахнет, а изомерный ему 3,7-диметилоктилвалеролактон
i 0 i
СНзСНСН, снсо
СН2СН2СН(СНз)(СН,,)зСН(СНз)2
имеет отчетливый запах.
42
В гетероциклических1 соединениях влияние на запах ока-
зывает положение заместителя (а, 0 или у) относительно ге-
тероатома. Так, например, а-пропилпиридин
N-CH
СН3СН2СН2- С
\\
сн
нс=сн
обладает неопределенным цветочным запахом, а упРопил'
пиридин—явным запахом фиалки.
HC = N
// \\
СН3СН2СН2- с сн
нс=сн
Наиболее удивительны различия в запахах изомеров
в ароматическом ряду. У некоторых ароматических соедине-
ний изменение положения заместителя полностью изменяет
запах2. Например, ортопроизводные бензиловых эфиров об-
ладают приятным цветочным запахом, а парапроизводные
не пахнут совсем. У производных фенилгорчмчного масла, на-
оборот, введение заместителя в параположение придает мас-
лу сильный приятный аромат, а в мета- и ортоположения не
изменяет запах. Для веществ с тремя заместителями в бен-
1 Гетероциклические соединения содержат в кольце один или не-
сколько неутлеродных атомов (N, S, О и т. д.).
2 В бензольном ядре возможны 3 различных относительных положе-
ния двух заместителей:
ортоположение, мет ап сложение, пара поло-
жение.
43
зольном ядре было замечено, что расположение 1, 3, 4 харак-
терно для многих хорошо известных пахучих веществ: вани-
лина, гелиотропина, эвгенола. Поэтому расположение 1, 3, 4
было названо «идеальным расположением». Но наличия
этого расположения еще недостаточно для получения запаха,
в частности, 1, 3, 4-производные метоксиацетофенона почти
не пахнут.
В каждом отдельном случае, кроме места введения замес-
тителя, имеет значение еще и тип и количество осмофорных
групп. Так, для фенилгорчичного масла с двумя осмофорами:
хлором и метильной группой, влияние на запах оказывает
ориентация групп (пара:положение дает анисовый, мета —
едкий, орто—главным образом цветочный запахи). Тип
запаха зависит больше от положения, чем от характера за-
мещающих групп. Однако хлор все же сильнее влияет на
запах, чем метильная группа.
Подобные закономерности свойственны большому количе-
ству пахучих веществ.
Различие в запахах встречается даже у таких мало отли-
чающихся по строению соединений, как оптические изомеры1.
Предполагалось, что оно появляется только в том случае,
если молекулы, составляющие рецепторы, также оптически
активны. В одной из работ [85] сделан вывод, что оптические
изомеры различаются не по качеству, а только по интенсив-
ности запаха. Причем, как было установлено многочисленны-
ми опытами, интенсивность запахов располагается в следую-
щем порядке: левовращающий имеет более сильный запах,
чем мезоформа2, которая пахнет сильнее правовращающего
изомера. Это различие исчезает при тщательной очистке
веществ.
Неодинаковые запахи цис- и транс-изомеров, имеющих
лишь пространственное отличие в строении, объясняются зна-
чительными различиями в их физических и химических свой-
ствах. Цис- и транс-формы стероидов часто оказывают со-
вершенно разное физиологическое действие на организм.
Обычно ф/с-форма более интенсивна по запаху. Из различия
запахов изомеров можно сделать вывод, что обонятельные
1 Оптические изомеры —d и I — энантиомеры — право- и левовра-
щающий— отклоняют в правую или левую сторону поляризованный луч
света.
2 Мезоформа — вещество, молекулы которого содержат 2 асиммет-
ричных атома углерода, взаимно компенсирующих оптическую активность
друг друга, и потому оптически неактивное.
44
органы очень чувствительны даже к незначительным изме-
нениям в строении молекулы.
Молекула может быть пластичной, гибкой, как у алифа-
тических углеводородов, или жесткой, как у циклических
и полициклических углеводородов. При переходе от первых
к последним замечено значительное увеличение интенсивности
запаха. В процессе изучения влияния жесткости молекул на
их запахи наблюдалось, что форма легких молекул играет бо-
лее важную роль, чем тяжелых [69]. Например, разница в ин-
тенсивности запахов цис- и транс-бутена-2 (4 атома углеро-
да) заметна, а у молекул, содержащих, по крайней мере,
10 атомов углерода, незначительна.
Интенсивность запаха молекул некоторых веществ опре-
деляется наличием двойных связей и их положением внутри
молекулы. Бициклические вещества с двойной связью в а-,
0-, у-положениях имеют почти одинаковые запахи, различа-
ющиеся лишь по интенсивности. Вещество, имеющее двойные
связи вдали от гидроксильной группы (у-положение), дает
более слабый запах, чем вещества, у которых двойные связи
расположены в а-и p-положениях по отношению к гидро-
ксильной группе. При хроматографировании на окиси алюми-
ния у-продукт адсорбируется хуже, чем а- и р-продукты.
Следовательно, можно допустить некоторую связь между ин-
тенсивностью запаха вещества и его адсорбируемостью.
Запах некоторых веществ зависит от отношения числа ос-
мофорных групп к количеству групп СН2, входящих в осмо-
ген. Столл [69] определил эту зависимость для макроцикличе-
ского мускуса, кольцо которого состоит из 12—17 групп СН2,
вводя в его молекулу различное количество осмофорных
\ /
групп (СО =, С , —СН2-СН3). Если в качестве осмофорной
/ \
группы взяты СО, зависимость окажется следующей: при
00: СН2 от 0 : 12 до 0 : 17 вещества не пахнут, от 1:10 до
1 : 11 вещества имеют запах мяты и камфары, от 1 : 14 до 1 : 16
запах мускуса, от 2 : 14 до 2 : 16 запах исчезает. Отсюда следу-
ет, что запах частично может быть функцией отношения веса
осмофорной группы к молекулярному весу.
Группы, смежные с осмофором, также влияют на интен-
сивность запаха. Включение метильной группы в положение,
смежное с двойной связью, вызывает почти полное исчезно-
вение запаха.
45
Очень важно установить, как изменяется запах молекулы,
имеющей несколько осмофорных групп. Отмечено, что гера-
ниол, имеющий одну гидроксильную группу, сильно пахнет.
Если же в его молекулу добавить еще одну гидроксильную
группу, то образуется непахнущий гликоль 169]. Все гликоли
<не пахнут, независимо от того, тяжелые они или легкие.
Этилметилкетон с одной кетонной группой обладает сильным
запахом. Диацетил с двумя кетонными группами пахнет
меньше, но, если в его молекуле одну группу СО заменить
на СН2, получится вещество с гораздо более сильным за-
пахом. При введении двух кетонных групп в молекулу цикло-
гексадеканона мускусный запах исчезает.
Наличие двух осмофорных групп СО и СНз в тяже-
лой и легкой молекулах не влияет на запах. Введение в мо-
лекулу пахучего вещества второй гидроксильной группы
уничтожает запах во всех случаях. Введение в молекулу вто-
рой кетонной групшы усиливает запах малых молекул,
(С2Н5СОСНз — этилметилкетон), но устраняет у больших
(С]6Н3оО — циклогексадеканон).
количество атомов углерода >
Рис. 21. Зависимость обонятельного
порога (Лоб) от числа атомов углеро-
да в молекуле пахучего вещества.
1 — н-спирты; 2 — н-парафуны; 3-1-
хлорпарафины.
Замещение одной гидро-
ксильной группы на кетон-
ную увеличивает интенсив-
ность запаха. Эпоксидные
группы (—СН—СН—) уси-
\ /
О
ливают запах больших мо-
лекул. Добавление двойных
связей в молекулу пахучего
вещества увеличивает ин-
тенсивность запаха, но при
сочетании этих связей с дру-
гими осмофорами — умень-
шает. Двойные связи осо-
бенно влияют на запах ма-
лых молекул, но на запах
больших молекул это влия-
ние незначительно. Если
классифицировать осмофо-
ры по признаку ослабле-
ния интенсивности запаха
в больших молекулах при
введении этих осмофоров, то
они располагаются в следу-
46
ющем порядке: гидроксильная, кетонная, альдегидная, эфир-
ная, эпоксидная группы и двойная связь. Замечено, что этот
порядок распространяется на силу адсорбции этих веществ
на окиси алюминия при хроматографическом анализе-
Как полагают, интенсивность запаха, а следовательно, по-
роговые концентрации определяются размерами молекул па-
хучего вещества. Зависимость обонятельного порога Лоб
(термодинамическая активность) от числа атомов углерода
в молекуле пахучего вещества представлена на рис. 21. Эта
зависимость показывает, что оптимальные характеристики
возбуждения (наиболее низкий порог) имеют молекулы
с 4—5 атомами углерода 158].
Из изложенного выше нельзя сделать заключение о том,
какие химические и физические свойства дают возможность
тому или иному веществу возбуждать обонятельные клетки,
хотя очевидна некоторая связь между адсорбируемостью па-
хучего вещества, формой и размерами его молекул и запахом.
Не определен также критерий для составления научно обос-
нованной классификации запахов. Возможно, недостаточ-
ность наших знаний зависит от того, что до сих пор разделе-
ние различных веществ на пахучие и непахучие, а также
стремление разными способами группировать запахи дела-
лось на основании ощущений человека. Такого рода оценка
является в достаточной мере субъективной, так как обоня-
тельный анализатор человека не в состоянии дифференциро-
вать все запахи, существующие в природе.
Стремление установить соотношение между запахом и хи-
мической структурой побудило многих исследователей зани-
маться детальным изучением этого вопроса.
Так, в последние годы [20, 21, 22] стало возможным гово-
рить о более тесной связи качества запахов с формой и раз-
мерами молекул пахучих веществ, это позволило не толь-
ко рационально классифицировать запахи, но и предсказы-
вать запахи синтезированных веществ (подробное изложение
этого вопроса дано в главе V).
ГЛАВА IV
ОСНОВНЫЕ КЛАССИФИКАЦИИ ЗАПАХОВ
И ТЕОРИИ ОБОНЯНИЯ1
Процесс восприятия запаха можно разделить на следую-
щие стадии:
1) воздействие пахучих веществ на обонятельный эпите-
лий и возбуждение первых нейронов обонятельного анализа-
тора;
2) передача возбуждения по обонятельным нервам в мозг
с помощью электрических импульсов;
3) переработка информации в центральной нервной
системе.
В большей части теорий авторы пытаются объяснить
процесс обоняния и механизм возбуждения обонятельных
рецепторов, связывая физические и химические свойства
пахучих веществ с их запахами. Существуют противоречивые
мнения о том, необходим ли контакт молекул пахучего ве-
щества с обонятельным эпителием или последний может
возбуждаться под воздействием волновых процессов, опреде-
ляемых молекулярными колебаниями; если контакт пахучих
молекул с обонятельными рецепторами необходим, то явля-
ется ли процесс возбуждения химическим или физическим.
Многочисленные теории обоняния могут быть разделены
на три группы:
1) теории, постулирующие возбуждение обонятельной
области при контакте молекул пахучего вещества с обоня-
1 Термин «теория» используется нами постольку, поскольку он при-
меняется авторами цитированных работ, хотя, по существу, следовало бы
говорить о гипотезах.
48
тельным эпителием, процесс возбуждения предполагается
химическим;
2) то же самое, что в п. 1, но процесс возбуждения пред-
полагается физическим (возможно, колебательное возбуж-
дение) ;
3) волновые теории, не требующие непосредственного
контакта молекулы пахучего вещества с обонятельным эпи-
телием.
Основной трудностью в оценке этих теорий является не-
возможность количественно охарактеризовать возбудитель--
запах, так как наше представление о нем является чисто
предметным. В некоторых случаях мы пытаемся группиро-
вать запахи родственных веществ или предметов, говоря
о цветочном, фруктовом и других запахах.
4.1. КЛАССИФИКАЦИЯ ЗАПАХОВ ПО СУБЪЕКТИВНОМУ ПРИЗНАКУ
Разные системы классификации основаны на некоторых
элементах сходства специфических запахов, позволяющих
группировать их. При составлении классификации прихо-
дится учитывать, что обонянием мы различаем в каждом
отдельном предмете только один нерасчлененный признак,
в то время как глаз дает восемь категорий признаков: цвет,
форму, величину, удаление, объемность, направление, покой,
движение [48].
С точки зрения эмоциональных впечатлений, вызываемых
запахами, можно говорить о приятных, безразличных, непри-
ятных, отвратительных и тошнотворных запахах.
Вероятно, самая старая классификация запахов может
быть отнесена к 1756 г. В ней запахи были разделены на
семь классов: 1) ароматические; 2) бальзамические; 3) ам-
бромускусные; 4) каприловые; 5) чесночные; 6) дурманя-
щие; 7) смердящие [2, 48].
Приведем еще одну систему классификации, предложен-
ную Цваардемакером в 1895 г. и уточненную в 1914 г. В ней
все запахи пахучих веществ разделены на 9 классов: 1) эфир-
ные; 2) ароматические (камфароподобные, пряные, анисо-
вые, лимонные, миндальные); 3) бальзамические (цветочные,
лилейные, ванильные); 4) амбромускусные; 5) чесночные
(луковичные, мышьяковистые, галоидные); 6) пригорелые
(жженое кофе, поджаренный хлеб); 7) каприловые; 8) про-
тивные (наркотические запахи, запах клопов); 9) тошно-
4 Заказ № 7032
49
ЩБетоиный
Пряный
Гчилостный
__—is фруктовый
А Пригорелый
----Смолистый
Рис. 22. Призма Хеннинга.
творные (запах падали, трупные запахи, скатол и т. д.)
[2, 48]- Большим недостатком этой классификации следует
считать некоторую произвольность в отношении отнесения
тех или иных веществ к тому или иному классу. Та-
кая произвольность существует и в других системах класси-
фикации.
Хеннинг (1924 г.) пытался графически изобразить все
обонятельные ощущения в виде призмы, на углах которой
обозначены шесть основных запахов в зависимости от поло-
жения осмофорных групп в молекулах (рис. 22): 1) цветоч-
ный; 2) фруктовый; 3|) гнило-
стный; 4) пряный; 5) пригоре-
лый; 6) смолистый [2, 48]. Все
запахи, которые не могут быть
прямо отнесены к одному из
перечисленных шести классов,
должны занять в этой призме
положение на ребрах, гранях
или внутри в зависимости от
того, с каким или со скольки-
ми классами обнаруживается
у них сходство.
Рассмотренные выше классификации субъективны. С их
помощью нельзя предсказать запахи вновь синтезированных
веществ. Схема обонятельных ощущений Хеннинга построена
по аналогии со схемами цветовых или вкусовых ощущений.
Если в отношении цвета можно твердо говорить о трех ос-
новных цветах, а в отношении вкуса — о четырех основных
ощущениях (горький, соленый, кислый, сладкий), то выде-
лить основные обонятельные ощущения можно только пред-
положительно. Между обонятельными ощущениями нет не-
прерывного перехода, который позволил бы расположить
их в определенные ряды.
В 1927 г. Крокер и Хендерсон постулировали существова-
ние четырех основных типов запахов — ароматного, кислого,
горелого, каприлового, воспринимаемых четырьмя видами
обонятельных рецепторов [48, 49], и возможность получения
любого запаха из этих основных, интенсивность которых из
меняется от 0 (отсутствие запаха) до 8 (очень сильный за-
пах). Таким образом, с помощью условных характеристик от
0000 до 8888 можно определять различные запахи, причем
первая цифра соответствует интенсивности ароматного запаха,
вторая — кислого, третья — горелого и четвертая — каприло-
50
вого. Например, запах розы может быть представлен харак-
теристикой 6423, указывающей, что интенсивность ароматно-
го запаха 6, кислого — 4, горелого — 2 и каприлового — 3.
Этот метод оценки запаха также субъективен.
4.2. КОЛЕБАТЕЛЬНЫЕ ТЕОРИИ
И КЛАССИФИКАЦИЯ ЗАПАХОВ
Структурные особенности молекул химических соедине-
ний, в том числе и молекул пахучих веществ, находят свое
отражение в спектрах поглощения и излучения этими веще-
ствами ультрафиолетовых, инфракрасных лучей и в комбина-
ционных спектрах рассеяния света. Это послужило основани-
ем для попыток создания теорий обоняния и построения
систем классификации запахов, основанных на физических
свойствах. веществ.
Спектры атомов и молекул определяются свойствами
электронных оболочек, колебаниями атомных ядер в моле-
кулах и вращением молекул, а также воздействием массы
и структуры атомных ядер на положение энергетических
уровней. Кроме того, спектры зависят от взаимодействия ато-
мов и молекул с окружающей средой.
Молекулярные спектры очень сложны, так как возможны
различные электронные переходы в молекулах (электронные
спектры), колебательные переходы с изменением колебатель-
ных состояний ядер атомов, входящих в состав молекулы
(колебательный спектр), и изменения вращательных состоя-
ний молекулы (вращательный спектр). Эти спектры располо-
жены в различных областях длин волн. Колебательно-враща-
тельные спектры расположены в ближней инфракрасной
части спектра от 0,2 до 40 мк (в волновых числах1 * от 8-103
до 250 см~л ). Вращательные спектры расположены в более
далекой инфракрасной части спектра до 1,5 мм (т. е. от 250
до 6 см~х). Электронные спектры, усложняющиеся колеба-
тельной и вращательной структурами, представляют собой
систему характерных полос, располагающихся от ультрафио-
летовой до ближней инфракрасной области. Каждая полоса
поглощения соответствуег возбуждению колебательного со-
стояния молекулы. Число возможных колебаний молекулы
1 Волновое число см~х
к
где X — длина волны в микронах.
4
51
определяется ее структурой. Если в молекуле содержится п
атомов, то число возможных колебаний не превышает (Зм—6).
Частота колебаний определяется массой частиц и силой свя-
зи, удерживающей частицы вместе. Чем больше масса, тем
ниже ее частота; чем больше силы связи, тем выше частота.
Иногда часть молекулы может колебаться почти независимо
от остальной, поэтому определенные группы атомов, при-
надлежащие к основному скелету, -имеют собственные часто-
ты. Однако среди всех (Зп—6) существует много других,
менее локализованных молекулярных колебаний, определяе-
мых растяжением (сжатием), изгибом и скручиванием ядер-
ного скелета. Они обычно имеют частоты, соответствующие
длинам волн в диапазоне 6—50 мк. Амплитуды различных
колебаний могут определяться из спектра поглощения. Зона
поглощения, связанная с обычным колебанием молекул,
может не проявиться с равной яркостью в инфракрасном
спектре поглощения вследствие специфической зависимости
колебаний от распределения электрических зарядов.
В положении теплового равновесия все энергетические
уровни населяются в соответствии с функцией энергетиче-
ского распределения Больцмана. Необходимо учитывать не
только основную частоту колебания, но и обертоны и комби-
национные уровни.
Массу частиц, конфигурацию и силы связи пары d- и I-
оптичеаких энантиометров мож)но считать тождественными.
Оптические изомеры обладают некоторым рядом колебатель-
ных энергетических уровней. Различие в частотах колебания
между d- и Z-оптическими изомерами, с одной стороны, и ме-
зоформой — с другой, не будет очень большим.
Свыше 90 лет назад Оугл высказал предположение, «что
волновая теория запаха... возможно, станет единственно пра-
вильной» [77]. Это высказывание является прототипом целой
серии подобных теорий, выдвинутых в последующие годы.
Однако она, как и все теории этого класса, не объясняет
явления усталости органов обоняния и изменения качества
запаха при разбавлении.
Хейникс (1919 г.) считал, что характер запаха обусловлен
спектром поглощения ультрафиолетовых лучей разными
пахучими веществами. Им была предложена соответствую-
щая классификация запахов (табл. 7) [2, 48]. Данные этой
таблицы хорошо не проверены.
Другие авторы предполагали возможность резонанса
колебаний валентных электронов молекул пахучего вещества
52
с колебаниями электронов чувствительных нервных оконча-
ний, а также возможность более определенной связи запаха
с частотами колебаний молекул пахучих веществ в некото-
ром диапазоне. Причиной молекулярных колебаний счита-
лась ненасыщенность химических соединений [48].
Таблица 7
Название вещества
Спектр поглощения, мк
Галоидопроизводные разных соеди- нений 0,350—0,330
Триметиламин, сероуглерод и ве- щества с гнилостным запахом . 0,330—0,300
Вещества с противными запахами . 0,300—0,280
Вещества с пригорелым запахом 0,280—0,260
Вещества с пряным запахом. . 0,260—0,240
Ванилин, гераниол 0,240—0,220
Эфирные запахи, запах фруктов . 0,220—0,200
Дайсон (1938 г.) утверждал, что молекула пахучего веще-
ства должна иметь колебания в диапазоне 3—7 мк инфра-
красной области спектра [35]. Предполагалось, что молекулы
должны обладать энергетическим уровнем в этом диапазоне
частот, соответствующем основным колебаниям молекул.
В табл. 8 представлена спектральная характеристика некото-
рых групп пахучих веществ.
Диапазон 3—7 мк, возможно, был выбран потому, что не-
которые молекулы с сильными запахами содержат структур-
ные группы, имеющие основные частоты колебаний, соответ-
ствующие этому диапазону S—Н-группа—4 мк, С—О-груп-
па—5,88 мк, С—С-группа—6 мк).
Однако для некоторых из упомянутых выше структурных
групп имеются характерные частоты колебаний и в других
частях спектра. Встречаются вещества, совершенно лишен-
ные запаха, но обладающие спектром, характерным для па-
хучих веществ, например, глицерин (3,47; 3,38; 3,11 мк). Тео-
рия Дайсона, подобно множеству более ранних и более позд-
них теорий, основанных на молекулярных колебаниях, не
объясняет различия запахов определенных изомеров.
Теория о селективном инфракрасном излучении органов
обоняния с длиной волны 8—14 мк была предложена и про-
S3
верена экспериментально (1949 г.) на пчелах Беком и Майл-
сом [26]. Предполагалось, что длина волны излучаемых коле-
баний определяется типом и размером воспринимающего
органа. Согласно этой теории, чувствительный орган или
рецептор должен различать охлаждение под воздействием се-
лективного поглощения инфракрасного излучения пахучими
веществами. Охлаждение воспринимающего органа или ре-
цептора должно вызывать Та нервный блица 8 импульс. Эта теория была опровергнута следую- щими экспериментами [77]. Мед, спектр погло-
Классы веществ Спектр поглощения, мк
Имины Альдегиды Кетоны Сложные эфиры, терпены . . Хлорангидриды Ацетилен Галоидопроизводные аромати- ческих углеводородов . . Меркаптан и вещества, обла- дающие сложным запахом . Эфиры Высшие алифатические угле- водороды Ароматические углеводороды Гетероциклические соединения и аммиачные основания . . . Пиридин и пиррол 8—14 мк, вводился в нос. щения которого в ин- 6,25 фракрасном диапазоне 5,88 характеризуется дли- ной волны 8—14 мк, 5’2б нельзя заменить дру- 4J6 гими веществами с по- добным спектром по- 4»35 глощения. Пчелы и че- 400 ловек легко отличают 3j0 запах этих веществ от запаха меда. Метил- 3,57 меркаптан, 1помещенный 3,33 в тонкую полиэтилено- 313 вую трубку, пропуска- 3,03 ющую инфракрасные лучи с длиной волны Ощущение запаха отсутствовало,
хотя меркаптан имеет интенсивную полосу поглощения с дли-
ной волны около 10 мк и мог иметь, согласно этой теории,
запах меда. В тепловом равновесии ощущение запаха остает-
ся неизменным, хотя процесс излучения должен был бы от-
сутствовать.
Эксперименты с пчелами повторил Джонстон. Он пред-
положил, что селективность привлечения определяется дей-
ствием приманки, излучающей инфракрасные лучи [44]. Вы-
двинутая в этой работе теория заключается в том, что инфра-
красное излучение 1меда с длиной волны 10 мк резонирует
с антенной пчел. Теория инфракрасного излучения была под-
держана в 1957 г. Народным. Он использовал результаты
исследований [59], в которых показано, что самка мотылька
Сатурний излучает сигнал с длиной волны отЗ до 11 мк, аво-
54
лоски на антенне самца, имеющие длину от 40 до 80 мк,
для приема этого сигнала изменяются примерно на 4 мк.
Если считать, что самка является радиопередатчиком фик-
сированных частот от 100 до 27,3 мгмггц, означает ли это, что
самец мотылька имеет настраивающуюся антенну в качестве
рецептора для обнаружения самки? Поведение пчел в раз-
личных условиях наводит на мысль, что восприятие запаха
может быть связано с явлением резонанса.
Между антенной и грудной клеткой насекомого существу-
ет электростатическое поле, разность потенциалов между ни-
ми составляет 2 в. Структура антенны мотылька имеет сход-
ство с магнетроном и обладает несколькими сотнями полос-
тей и волосков, резонирующих с длиною волны излучения
10 мк.
Казалось бы, запах является инфракрасным излучением-
Однако если излучение инфракрасного спектрометра напра-
вить на обонятельный эпителий и пройти весь инфракрасный
спектр, то резонанс в обонятельном органе не обнаружива-
ется ни при какой длине волны. По-видимому, необходим
контакт пахучего вещества с поверхностью обонятельного
эпителия.
Оттосон сумел покрыть носовую полость очень тонкой
пленкой пластического материала, пропускающего большие
дозы инфракрасного излучения в диапазоне 3—14 мк. В обо-
нятельный эпителий через пленку вводился капиллярный
микроэлектрод. При воздействии пахучих веществ реакция
воспринимающего органа отсутствовала и вновь появилась
после снятия пленки. Этот эксперимент опровергает теорию
инфракрасного излучения или поглощения и другие колеба-
тельные теории, так как использованные пахучие вещества,
бутанол и амилацетат, имеют интенсивное поглощение в ши-
роком диапазоне длин волн, для которых пленка не являет-
ся преградой [77].
Тем не менее, дальнейшие исследования в области коле-
бательных теорий обоняния были проведены Райтом с целью
установления соотношения между запахом и спектром паху-
чих веществ [83—85].
Было отмечено, что частоты молекулярных колебаний, вы-
бранные Дайсоном, характеризуют функциональные группы
молекул (ОН, СН, СО и т. д.). Поэтому, если запах может
быть соотнесен с частотами этих групп, то в равной степени
запах можно связать с этими функциональными группами.
С другой стороны, появление линий в инфракрасном спектре
55
свыше 10 мк определяется общей конфигурацией молекулы,
так что соотношение aainaixa с колебаниями молекулы будет
более приемлемым при более низких частотах. Кроме того,
формула Планка показывает, что при температуре тела наи-
большую вероятность имеют нормальные колебания с дли-
ной волны, значительно большей 10 мк [36].
Райт предполагал, что для возбуждения органа обоняния
молекула пахучего вещества передает энергию колебания.
При этом создается нервный импульс.
Другой механизм восприятия запахов предполагает, что
при окислении молекулы вещества должен излучаться квант
энергии. Если допустить это, то эпителий должен флуоресци-
ровать или фосфоресцировать [77].
Райт показал, что большое число веществ с запахом нит-
робензола имеет инфракрасный спектр в диапазоне 40--
67 мк. Предсказание на основании спектральной характери-
стики нитробензольного запаха бутиронитрила подтверди-
лось экспериментально.
Слабым местом этой гипотезы является то, что все 60 сме-
сей, названных Райтом как имеющие запах нитробензола,
были производными бензола или тиофена, и характер их
спектра является просто свойством ядерных скелетов этих
молекул. Существует большое число различных веществ,
имеющих частоту в этом же диапазоне и отличных по запаху
от нитробензола.
H2S (3,79; 3,83; 7,75 мк) и D2S (5; 5,28; 10,55 мк) не ле-
жат в рассмотренном Райтом диапазоне (свыше 10 мк). но
обладают сильным одинаковым запахом. Идентичность запаха,
возможно, является следствием обмена молекул между H2S
и D2S в ольфакторной области. Такое объяснение, конечно,
нельзя считать удовлетворительным. Вероятно, такая гипо-
теза основана на произвольном выборе экспериментальных
данных, подтверждающих выбор частотного диапазона мо-
лекулярных колебаний.
Измерение инфракрасного спектра поглощения большого
числа репеллентов для насекомых (москитов) должно было
иллюстрировать основу их действия—наличие полосы погло-
щения инфракрасного излучения с длиной волны 25 мк. Од-
нако полученные Райтом данные для шести репеллентов недо-
статочно убедительны: в трех случаях полоса слаба, а водном
отсутствует совсем (21,76 мк). В то же время многие другие
вещества, удовлетворяющие этому условию, не являются спе-
56
цифическими репеллентами для насекомых (пропилацетат,
циклогексан, пропиловый спирт).
Интересна попытка Райта [83] в 1961 г. косвенно прове-
рить гипотезу о связи запаха с низкочастотными колеба-
ниями органических молекул. Он проводил испытания с нит-
робензолом (54,64; 24,51 мк) и метилсалицилатом (54,05;
37,73; 28,09; 23,87 мк). Если приведенные спектральные ха-
рактеристики этих пахучих веществ правильны и полны, то их
запахи должны быть в основном подобными. Запах метилса-
лицилата в отличие от запаха нитробензола изменяется от
миндалеподобного до запаха озимых, что может быть связа-
но с дополнительными линиями в спектре. Отсюда следует,
что небольшое количество метилсалицилата, добавленное
в нитробензол, может легче ощущаться, чем в обратном слу-
чае. После экспериментов на 12 субъектах были определены
пороговые концентрации пахучих веществ и пороговые кон-
центрации каждого пахучего вещества при постоянной кон-
центрации второго. Результаты экспериментов показали, что
запахом озимых в десять раз сильнее маскируется миндаль-
ный запах нитробензола, чем наоборот, т. е. они довольно
отчетливо согласуются с предположением, сделанным на ос-
нове гипотезы. Несмотря на очевидные недостатки гипотезы
Райта, в 1963 г. вновь было сделано допущение о том, что
низкочастотные колебания молекулы объясняют физическую
природу запаха [84]. Эта гипотеза объясняет продолжитель-
ные неудачи химиков в поисках связи между запахом и хи-
мическим составом, так как в отличие от химических теорий
она рассматривает молекулы прежде всего как механическую
систему материальных точек и упругих сил. Она объясняет
также высокую информационную емкость обонятельного ап-
парата позвоночных, чувствительных к значительному коли-
честву частот колебаний постулированием множества типов
рецепторов, каждый из которых настроен на ограниченный
диапазон частот. При этом вещество, воздействующее на
один и только один тип рецептора, будет иметь первичный
запах. Эти рецепторы могут создавать нервные импульсы,
когда пахучие молекулы имеют колебания соответствующей
частоты, обычно низкой и трудно контролируемой (инфра-
красный диапазон поглощения находится вне диапазона из-
мерения большинства спектрометров). Это чисто технически
препятствует проверке гипотезы прямым путем, когда уста-
навливается связь отдельных запахов с отдельными комбина-
циями низкочастотных колебаний. Наличие приборов не об-
легчает эту задачу из-за сложности ощущения запахов даже
качественно простых групп пахучих веществ, например, нату-
рального и искусственного мускуса или многих веществ
с камфароподобным запахом.
Чувствительность обонятельного ощущения насекомых
выше чувствительности позвоночных. Возможно, в обонятель-
ном анализаторе насекомых используется тот же самый ос-
новной механизм, но, видимо, намного упрощенный. Мозг
насекомых не может справляться с большим потоком инфор-
мации, да и нет необходимости в высокой информационной
емкости обонятельного аппарата. Все, что требуется от обо-
нятельного анализатора для качественной дискриминации
запахов — это простой ответ «да» или «нет» на вопрос о при-
сутствии определенного запаха пахучего вещества. Соответ-
ствующая реакция поведения следует автоматически.
Возможно, для этой цели достаточно сенсорного (чувст-
вительного) аппарата, настроенного на узкий диапазон
частот [84].
Некоторый интерес представляет работа Народного (1957),
посвященная «векторному анализу» запахов [60]. Им было
постулировано: 1) каждый запах может быть изображен век-
тором в трехмерном пространстве в прямоугольной системе
координат; 2) добавление нового запаха аналогично вектор-
ному сложению, в результате которого появляется «результи-
рующий» запах; 3) вычитание запахов (маскировка, нейтра-
лизация) аналогично вычитанию векторов, в результате чего
запах может исчезнуть (например, запахи кумарина и рези-
ны практически нейтрализуются, исчезает резиновый запах
или при недостаточной компенсации появляется кумариновый
или ванилиновый); 4) интенсивность запаха определяется ре-
зультирующим вектором запаха и кратчайшим расстоянием
между линией действия этого вектора и воспринимающим
органом.
Для получения математических соотношений в качестве
единичного вектора может быть принят вектор, соответствую-
щий запаху ванилина при следующих условиях: воздух, насы-
щенный ванилиновыми и водяными парами при 20°С и нор-
мальном атмосферном давлении, протекает со скоростью
5 л/ыин через «нормальный» нос. В качестве «нормального»
носа может быть принят обонятельный аппарат насекомого,
дающий определенный электрический ответ на заданный раз-
дражитель. Векторный анализ позволяет объяснить многие
явления в обонянии, например компенсацию запахов.
58
Каротиноидная теория. В последние годы (1961,
1962) большое внимание было привлечено к каротиноидной
теории обоняния [28, 29, 57]. Как уже указывалось, запах
любого пахучего вещества может быть связан до некоторой
степени с низкочастотными колебаниями молекул, определя-
ющими поглощение в инфракрасной области спектра (свыше
16,67 мк). Однако до сих пор неизвестен механизм восприя-
тия и рецепторы энергии колебаний молекул пахучих ве-
ществ. Предполагалось, что обонятельный эпителий должен
иметь на своей поверхности молекулы, способные взаимодей-
ствовать с пахучим веществом. Если взаимодействие пахуче-
го вещества и рецепторов в пределах эпителиальных клеток
отсутствует, то можно считать, что молекулярные «рецепто-
ры», воспринимающие запах молекулы, будут локализоваться
в секрете на поверхности эпителия.
Была сделана попытка исследовать природу соединений
этого промежуточного пространства. Известно, что поверх-
ность обонятельного эпителия имеет желтоватую окраску,
а поверхность дыхательного эпителия не окрашена. Известно
также, что желтые участки содержат каротиноиды. Это под-
тверждается проведенными на животных (коровах) экспери-
ментами, показавшими, что обонятельный эпителий содержит
свободные каротиноиды, свободный ватамин А, а также, свя-
занные с протеинами каротиноиды.
Авторами теории предполагалось, что протеиносвязываю-
шие каротиноиды обонятельного эпителия являются рецептора-
ми энергии молекул пахучего вещества, проникающего в носо-
вую полость. Механизм восприятия энергии не выяснен.
Известна способность каротиноидов существовать в много-
численных цис-и транс-формах, а также то, что только дей-
ствие света на них вызывает появление геометрической изо-
меризации. Можно предположить, что обмен энергией моле-
кулы пахучего вещества с обонятельным каротиноидом вле-
чет за собой подобные изменения молекулы.
Гипотеза подтверждается тем, что у людей и у животных,
находившихся на бескаротиноидной диете, ослабляется обоня-
ние, в большинстве случаев обоняние восстанавливается пол-
ностью или частично (в 50 случаях из 56 по данным наблюде-
ний) три лечении витамином А.
Эта гипотеза встретила серьезные возражения Моултона,
который утверждает, что если каротиноиды действительно яв-
ляются обонятельными рецепторами, то они должны находить-
ся обязательно в обонятельном эпителии позвоночных [57].
59
Однако каротиноиды не обнаружены в обонятельном эпителии
кролика, овцы, крысы и овиньи. Был изучен также состав пиг-
ментов обонятельного эпителия черных и белых крыс и мышей.
Несмотря на утверждение об аносмичности альбиносов,
не найдено существенных различий ни в окраске, ни в коли-
честве, ни в составе этих пигментов у черных и белых крыс
и мышей.
На основе изложенного выше Моултон приходит к выводу,
что каротиноиды обонятельной ткани рогатого скота не имеют
функционального значения [57]. Влияние же витамина А на
восстановление обоняния объясняется зависимостью реакции
на запах от функционального состояния мембраны обоня-
тельного эпителия, имеющего высокую чувствительность к из-
менениям концентрации витамина А в организме.
Моултон, ссылаясь на работу Томсона, считает, что каро-
тиноидной теории обоняния присущи недостатки и противо-
речия, свойственные всем колебательным теориям. Однако не
все замечания Моултона обоснованы [29].
Так, возможно, каротиноиды не были обнаружены в обоня-
тельном эпителии из-за трудности эксперимента. Каротиноиды
являются важными составляющими здоровой ткани, их отсут-
ствие вызывает изменение в обонятельном эпителии. Это под-
тверждает каротиноидную гипотезу обоняния. Большая часть
пигментов присутствует в защитных клетках и железах Боу-
мена. Неизвестно, имеется ли пигмент в значительном количе-
стве в рецепторах. Вероятно, что каротиноиды в секрете боу-
меновых желез, омывающем ворсинки рецепторов, и могут
являться молекулярными «рецепторами», чувствительными
к запаху. Гистохимия обонятельного эпителия недостаточно
изучена, чтобы исключить роль каротиноидов в восприятии
запахов.
Все же основные положения Моултона, связанные с крити-
кой колебательных теорий обоняния, изложенных в [77], авто-
ры каротиноидной теории опровергнуть не смогли.
Заканчивая рассмотрение колебательных теорий, приведем
основные противоречия, содержащиеся в любой из них.
1. Молекулы изотопов имеют одинаковый запах, хотя су-
ществуют различия в частотах колебаний молекул. Типичный
набор таких прототипов: H2S и D2S; СН3ОН, CH3OD и CD3OD;
CH3SH и CH3SD; СНзСНО и CH3CDO и т.д.
2. Основные стереоизомеры имеют различные запахи. Опти-
ческие изомеры (d и /) имеют идентичный набор частот, но
различные запахи.
60
3. Некоторые лары молекул с почти одинаковым набором
частот колебаний имеют существенно различные запахи.
4. В колебательной теории нет удовлетворительного объяс-
нения влиянию малых примесей на запах.
4.3. ХИМИЧЕСКИЕ ТЕОРИИ ОБОНЯНИЯ
Авторы химических теорий связывают процесс обоняния
с некоторыми химическими процессами, определяемыми струк-
турой пахучих веществ и некоторыми другими их свойствами.
Особое значение придается ненасыщенности химических сое-
динений, растворимости в воде и липоидах, наличию остаточ-
ного сродства.
Интенсивность запаха (по Цваардемакеру) зависит от ле-
тучести, распыляем ости, легкости адсорбции на поверхности
водной мембраны и легкости растворения в липоидах.
В 1944 г. Манкриф предложил химическую теорию запа-
хов, которая указывала, что пахучие вещества должны быть
летучими и иметь соответствующую растворимость в протеи-
нах. (Эта растворимость достигает наибольшего значения
в гомологических рядах при С4 или С6). Растворимость свя-
зана со строением молекулы и существует некоторая связь
между запахом и молекулярной структурой (см. 3.2.). Интен-
сивность пахучих веществ может определяться их летучестью.
Как можно объяснить качество ощущаемых запахов сточ-
ки зрения такого рода теорий? Предполагалось наличие не-
большого количества основных запахов (от 4 до 12), воспри-
нимаемых соответствующими рецепторами, покрытыми жиро-
протеиновым веществом различного состава. В таком случае
каждая из групп рецепторов будет возбуждаться тем пахучим
веществом, которое лучше, чем другие, растворяется в протеи-
новом веществе, присущем именно данной группе рецепторов.
Таким образом, несколько рецепторных систем с различными
характеристиками будут вызывать ощущения большого коли-
чества различных запахов.
Позднее, в 1949 г., теория Манкрифа была несколько из-
менена. Согласно новой теории, кроме летучести пахучего
вещества, придавалось значение конфигурации его молекул
и предполагалось наличие соответствующей конфигурации ре-
цептивных участков [48]. Вещества с молекулами фиксирован-
ной формы (насыщенные) могут соответствовать одному
и только одному типу рецептивного участка.
61
Как уже указывалось, ненасыщенные химические соедине-
ния часто обладают сильным запахом, (Возможно потому, что
их молекулы могут существовать в разных непрерывно изме-
няющихся положениях и формах, обусловленных подвижно-
стью л-электронюв, образующих двойную связь. Например,
бензол может существовать в пяти предельных структурах.
Ненасыщенные вещества с постоянно меняющейся формой мо-
лекул, вероятно, могут приспособиться к конфигурации раз-
личных рецептивных участков. Не исключено, что молекулы
мембраны рецепторов тоже могут иметь изменяющиеся
формы.
Если молекула пахучего вещества имеет двойную связь
вблизи полярной группы, то она благодаря большой 'неста-
бильности может легко приспособляться к рецептивному
участку.
Манкриф (1951 г.) предполагает следующий порядок вос-
приятия запахов [48].
1. Пахучее вещество непрерывно испаряется в атмосферу.
2. Некоторое количество молекул пахучего вещества при
вдохе поступает через носовые ходы к обонятельным рецепто-
рам.
3. Молекулы пахучего вещества скапливаются на соответ-
ствующих участках обонятельного рецептора.
4. При скоплении происходит обмен энергией между моле-
кулами пахучего вещества и молекулами мембраны рецептора
(возможно, благодаря уменьшению поверхностного натяже-
ния!) .
5. Обмен энергией вызывает появление электрических им-
пульсов «и передачу их в мозг.
6. В результате указанных явлений возникает ощущение
запаха в высшем обонятельном центре.
Для ощущения запахов по этой теории необходим контакт
пахучей молекулы с обонятельным рецептором, но неизвест-
но, будет ли обоняние результатом поглощения пахучих мо-
лекул тканями, растворения их в липоидах или химической
реакцией между пахучим веществом и рецептором.
62
Для выяснения механизма обоняния Маллинз (1955 г.) рас-
смотрел соотношения между растворимостью некоторых хими-
ческих веществ в липоидах, с одной стороны, и их наркотиче-
ским действием и запахом, с другой [58]. Соотношение между
растворимостью некоторых химических веществ в липоидах
и их наркотическим действием позволило считать термодина-
мическую активность вещества мерой его фармакологической
эффективности (Фегюсон и Бринк). Известно также, что су-
ществует зависимость пороговой активности для наркоза
Днар) длины цепи гомологического ряда для веществ, ко-
торые вводятся в мембрану клетки с различными плотностями
сил сцепления или параметрами растворимости 6 [58]. Плот-
ность сил сцепления молекул (мембраны клетки неодинакова
для различных участков нервной системы: мозг и синапсы име-
ют небольшое значение параметров растворимости, перифери-
ческий нерв — намного большее.
Одним из наиболее удобных методов подхода к механизму
обоняния, по мнению Маллинза, было бы измерение порогов
обоняния, выраженных термодинамическими активностями
Лоб) в пределах разных гомологических' рядов с различной
плотностью сил сцепления, так как подобные измерения поз-
воляют сделать некоторые выводы относительно природы внут-
ренних сил притяжения между молекулами самих мембран
рецепторов.
Измерялись пороги обоняния человека (ДОб) Для н-пара-
финов с низкой плотностью сил сцепления (6 = 7), /-хлорпара-
финов со средней (6 = 8,51) и спиртов с высокими и переменны-
ми значениями плотности сил сцепления (6=9—14,5). Полу-
ченные результаты уже были показаны на рис. 21, где дана
зависимость обонятельного порога (Доб) от числа атомов уг-
лерода в молекуле. Пунктирная линия на том же рисунке изо-
бражает термодинамическую активность, необходимую для
наркоза человека и подопытных животных парафинами и хлор-
парафинами. Из рисунка видно, что малые и большие моле-
кулы имеют сходные пороги наркоза и обоняния и, как уже
указывалось (п. 3.2), молекулы с четырьмя и пятью алифа-
тическими атомами углерода имеют оптимальные характери-
стики возбуждения обоняния.
Маллинз предполагает существование трех типов рецепто-
ров, мембраны которых имеют плотности сил сцепления, соот-
ветствующие 6 = 8, 10, 12.
Нельзя считать, что парафины, хлорпарафины и спирты до-
стигают пороговой концентрации только на рецепторах с соот-
63
ветствующими 'плотностями сил оцепления молекул в мембра-
не, т. е. что парафины достигают некоторой концентрации толь-
ко на рецепторах с плотностью сил сцепления, характерной
для парафинов (6 = 7).
Наибольшая концентрация парафина б|удет все же на ре-
цепторах с 6 = 8. Возможно, она будет достаточной для нар-
котизации некоторых из них. Поэтому удобно выбрать рецеп-
тор с б = 12 для определения того, будет ли происходить
стимуляция 'парафинами и хлориарафинами, а рецептор
с 6 = 8 — спиртами. Тогда возникает предположение, что
адаптация к запаху парафина эквивалентна наркозу у неко-
Таблица 9
Вещества, к которым должны адаптиро- ваться обонятельные клетки (в 10 порого- вых концентрациях) 0 Измерение порогов ве ществ О Измене- ние по- рогов, % Подсчитанное значе- ние мольфракции ада пт. вещества в ре- цепторах (хЮ—4)
г=8 о=10 S=12
Бутан . „ . „ . 7 Бутиловый спирт Бутан 11 0 100 10 1
Бутиловый спирт « 11 7 0 10 100 100
Ацетон . . . ь 10 7 100 20 100 20
10 Бутиловый спирт 11 500 20 100 20
торых рецепторов, имеющих 6=8, а возбуждение чувства обо-
няния требует совместной активности рецепторов с б = 8
и б 12.
Если обонятельный эпителий можно было бы разделить на
группы рецепторов с различной плотностью сил сцепления мо-
лекул мембраны, исследуя то пространство, в котором вещест-
ва с различной плотностью сил сцепления изменяют порог
других веществ с различными б, то можно было бы начертить
схему распределения этих рецепторов.
Были проведены эксперименты для выяснения влияния
адаптации к одному веществу на пороги других (табл. 9) [58].
Так как адаптация к бутану не влияет на порог бутилового
спирта и адаптация к бутиловому спирту не влияет на порог
бутана, можно предположить, что в обонятельном эпителии
имеется по крайней мере два типа рецепторов, отличающихся
64
Рис. 23. Зависимость концентраций паху-
чего вещества хмем с б = 8 на мембранах
рецепторов с б = 8, 10, 12 от времени дей-
ствия стимула. Время дано в произвольных
единицах.
запахов в широком диапазоне интен-
одновременное возбуждение паху-
плотностью сил сцепления пли внутренними силами притяже-
ния молекул в их мембранах. Но действие ацетона на порог
бутилового спирта больше, чем на порог бутана, поэтому мо-
жно предположить и третий тип рецептора.
Малые подпороговые концентрации наркотиков и некото-
рые химические вещества оказывают возбуждающее влияние
на нервные структуры и могут вызывать понижение порога
возбуждения, вследствие чего может произойти самовозбуж-
дение. Например, камфара является таким веществом и, что
весьма интересно, вызывает обонятельное ощущение.
Уменьшение порога возбуждения нервных клеток до само-
возбуждения при достижении в их мембранах определенной
мольфракции химиче-
ского вещества может
быть рассмотрено как
некоторая модель ме-
ханизма обоняния.
Результаты экспери-
ментов по определе-
нию изменения поро-
гов одних веществ под
влиянием адаптации к
другим (см. табл. 9) не
дают возможности по-
строить точную схему
распределения рецеп-
торов с различными
параметрами раствори-
мости.
Система восприятия
сивностей предполагает
чим веществом всех трех типов рецепторов. Тогда, чтобы
обнаружить специфические запахи, центральная нервная
система должна анализировать только разницу во времени
между началом возбуждения различных типов рецепторов.
Задача различения запахов становится проблемой предска-
зания динамики поглощения молекул рецепторами с различ-
ными б, так как молекулы легче всего проникают и возбужда-
ют рецепторы со сходными силами сцепления.
Иллюстрацией предполагаемой системы восприятия запа-
хов может служить рис. 23, на котором показано действие ве-
щества с б = 8 с десятикратной пороговой концентрацией на
обонятельный эпителий. Кривые поглощения приняты линей-
5 Заказ № 7032
65
Табл ица 10
Спирты о Последователь- ность возбуждения рецепторов по о
Метиловый 14,5 12-10-8
Этиловый 13,5 12—10—8
Пропиловый 12,0 12-10-8
Бутиловый п,о 12\» 10/b
Амиловый 10,5 10-12—8
Гексиловый 10,0 ю / 8 1 \12
Гептиловый 9,5 Ю-8-12
Октиловый 9,0 10\ J2 8/12
Дециловый 8,5 8-10-12
атамами углерода,
группа включает
параметров раст-
ными во времени. На этом же рисунке представлены уровни
возбуждения мембран рецепторов в зависимости от концент-
рации вещества: нормальный, самовозбуждения, второй нор-
мальный и уровень наркоза. Результат возбуждения одиноч-
ных рецепторов дан в верхней части рис. 23. Длина цепочки
импульсов зависит от времени изменения концентрации паху-
чего вещества в мембране от одной до двух единиц, соответ-
ствующих уровню самовозбуждения.
Качественные различия
запахов могут быть показа-
ны на примере трех групп
н-спиртов. Известно, что
спирты с 1—3 атомами уг-
лерода имеют сходные за-
пахи, так же как и спирты
с 4—5
Третья
спирты с 6—8 атомами уг-
лерода. В табл. 10 даны
значения
воримости 6 для н-спиртов
и предполагаемая последо-
вательность возбуждения ре-
цепторов для каждого из
спиртов на основании сход-
ства 6 спиртов и 6 рецепто-
ров. С увеличением гомоло-
гического ряда появляются
различия во времени воз-
буждения и в концентрации
пахучего вещества в мем-
бране, обусловленные возрастанием молекулярного объема.
Поэтому спирты первой группы имеют сходные характеристи-
ки возбуждения, тогда как бутиловый спирт, отличающийся
по запаху от первой группы, характеризуется иной последо-
вательностью возбуждения.
Для других химических соединений с увеличением числа
атомов углерода 6 также приближается к 8,5 и уже не может
влиять на характер запаха (рис. 24). Поэтому запах больших
молекул зависит от формы и размеров молекулы, а также
от температуры плавления. Запахи алифатических yi-
леводородов с гибкими молекулами неопределенной формы
и высокими температурами плавления по мере возрастания
6G
стом гомологического ряда: для спир-
тов (1), кислот (2), метилового и этило-
вого эфиров (3), I -хлорпарафинов (4),
парафинов (5).
гомологических рядов становятся трудно различимыми. Мак-
роциклические кетоны с низкими температурами плавления
и жесткими молекулами, легко проникающими в мемб-
раны рецепторов, имеют хорошо различимый сильный запах
мускуса.
Результаты электрофизиологических исследований дают
возможность оценить приемлемость предполагаемой системы
восприятия запахов. Так, исследования электрической актив-
ности обонятельной луковицы [15] показали, что стимуляция
ацетоном (6=10) вызывает появление ряда импульсов боль-
шой амплитуды с об-
щей продолжительностью
0,1 сек, при каждом вдо-
хе. При стимуляции амил-
ацетатом (6=9) возни-
кает совокупность им-
пульсов большой и малой
амплитуды продолжи-
тельностью 0,2 сек, а при
стимуляции пентаном (6=
= 71) — только импульсы
малой амплитуды про-
должительностью 0,3 сек.
Маллинз предлагает сле-
дующее объяснение этих
результатов: ацетон бы-
стрее всего проникает во
все типы рецепторных
клеток, так как он имеет
параметр растворимости средний между 8 и 12. Ответы раз-
ных типов рецепторов будут синхронными, поэтому импуль-
сы с большой амплитудой можно считать суммарным отве-
том. Амилацетат по параметру растворимости ближе к пара-
финам, чем к спиртам. Можно предположить, что он будет
вызывать синхронный ответ рецепторов с 6=12. Пентан име-
ет параметр растворимости, значительно отличающийся от
среднего для всех рецепторов. Очевидно, ответы всех рецеп-
торов на раздражение пентаном будут асинхронными, а сум-
марный ответ может быть представлен в виде длинного ряда
импульсов малой амплитуды.
Вещества с одинаковыми и сходными параметрами раство-
римости вызывают близкие между собой ответы обонятельной
луковицы; например, пиридин и ацетон; ксилол, эвкалипт
5*
67
Рис. 25. Зависимость количества ре-
цептивных участков от их размера.
и амилацетат; октан и пентан и др. Однако электрофизиологи-
ческие исследо1вания обонятельной луковицы могут не дать
тончайших отличительных признаков, которые придают запа-
хам индивидуальность, так как в реакции на некоторое паху-
чее вещество может участвовать от 100 до 1000 волокон с 6 = 8
и одно волокно с 6 = 12. Если рецепторы трех различных типов
распределены «случайно по всему обонятельному эпителию и ес-
ли есть 106 рецепторов каждого типа, то вероятность электро-
физиологического обнаружения данной обонятельной харак-
теристики (ответа) беско-
нечно мала.
Наиболее удовлетвори-
тельной моделью мембраны
клетки Маллинз считает по-
верхностный слой из ци-
линдрических макромолекул
диаметром около 40 А и
длиною 1С0 А (толщина
мембраны). Эти молекулы
расположены в правильный
шестиугольник так, что про-
межуток («пора») между
каждыми тремя молекула-
ми имеет форму треуголь-
ника, в который можно впи-
сать круг диаметром 6 А.
При комнатной температуре
фактические размеры дан-
ного промежутка должны
обладать некоторой диспер-
сией, так что мембрана фак-
тически включает участки
(«поры») различных разме-
ров. На рис. 25 дана зависимость количества участков от их
диаметра и показано положение молекул трех различных
размеров. Сделаны предположения, что данная молекула мо-
жет вызывать наркоз, если она просто занимает достаточное
количество участков. Молекула возбуждает рецептор, если
она по своим размерам полностью соответствует занимаемо-
му участку1.
1 Для простоты рассматриваются молекулы сферической формы.
68
Верхняя кривая рис. 25 дана для малых молекул, средняя —
для средних и нижняя — для больших по сравнению со сред-
ними размерами участка мембраны рецептора. Черная полоса
на -каждой из 'кривых определяет 'количество участков, кото-
рые могут возбуждаться: заштрихованная часть диаграммы
соответствует количеству участков, которые могут быть заня-
ты при наркотизации. Малые и большие молекулы могут воз-
буждать только небольшое количество участков, наибольшее
количество участков возбуждается под действием молекул
средних размеров. Чувствительность мембраны к возбужде-
нию зависит от способности молекул создавать наибольшее
количество нарушений структуры мембраны в возможно более
короткий срок. Количество нарушений, производимое в едини-
цу времени, зависит: 1) от концентрации молекул с оптималь-
ной для проникновения формой; 2) от части молекул с такой
ориентацией относительно поверхности клетки, что их скорость
проникновения близка к максимальной; 3) от изменения струк-
туры мембраны, вызванной изменением сил взаимодействия
между молекулами мембраны под действием молекул паху-
чих веществ.
Пахучие вещества с гибкими молекулами определенных
форм являются значительно менее эффективными возбудите-
лями обоняния, чем вещества с жесткими молекулами. Низ-
кие пороги обоняния последних, по-видимому, являются ре-
зультатом легкости проникновения этих молекул в мембра-
ну, ограничивающую рецептор. Можно считать молекулу крат-
ковременно оказывающей влияние на всю мембрану клетки.
Существует мнение, что многие вещества способны вызы-
вать и возбуждение и наркоз возбудимых структур в зависи-
мости только от концентрации.
Ферментативные теории. Некоторые авторы пред-
полагали, что первичной стадией процесса обоняния является
воздействие молекул пахучего вещества на обонятельный эпи-
телий, в результате чего некоторые метаболические1, вероятно
ферментативные, процессы замедляются или ускоряются [77,
85]. Это может быть как в случае стереоспецифичности фер-
мента только к одному пахучему веществу (одному опреде-
ленному запаху), так и к одной из основных групп пахучих
веществ (первичному запаху). Энергия колебания молекулы
пахучего вещества, возможно, играет при этом случайную роль
в общем процессе энергетического перераспределения.
1 Метаболические процессы — процессы обмена веществ.
69
Если 'восприятие запаха зависело бы от каких-либо фер-
ментативных процессов, то стереоспецифичность ферментов
затрудняла бы объяснение идентичности качества и интен-
сивности запаха оптических антиподов [85]. Сходство запаха
оптических изомеров, несмотря на стереоспецифичность фер-
ментов, возможно при условии высокой и неспецифичной ядо-
витости пахучих веществ, которой они в общем просто не
имеют. Отсюда следует, что процесс обоняния не может, по-
видимому, быть ферментативным преобразованием, управля-
емым молекулой пахучего вещества.
Возникновение обонятельного восприятия, вероятно, зави-
сит скорее от физического, чем от химического взаимодейст-
вия, и ферменты могут быть вовлечены только во вторичный
процесс восстановления реактивности обонятельных клеток
после того, как молекулы пахучих веществ сделают свою рабо-
ту. Это положение кажется достаточно основательным неко-
торым авторам для исключения любой химической или фер-
ментативной теории обонятельного (возбуждения [85].
Создание теорий обоняния определялось не только необ-
ходимостью выяснения природы запаха и механизма его вос-
приятия, но и общепризнанной важностью объективного мето-
да измерения запаха. Такой метод устранил бы влияние физио-
логического и психологического факторов в субъективном
методе измерения запахов. Более того, объективный метод
качественного и количественного измерения, возможно, поз-
волил бы расширить диапазон измеряемых интенсивностей
запаха за счет подпороговых. К сожалению, ни одна из при-
веденных в этой главе теорий не дает возможности подой-
ти к вопросам объективного измерения запахов, выяснения
его природы и механизма восприятия.
4.4. АДСОРБЦИОННЫЕ ТЕОРИИ ОБОНЯНИЯ
При рассмотрении последней группы теорий обоняния, от-
носимых нами к адсорбционным, делается попытка не только
выяснить возможный механизм восприятия качественно раз-
личных запахов, но и приблизиться к возможности их объек-
тивного измерения [31, 32, 34, 37, 46—54].
Адсорбционные теории объясняют важнейшие свойства
обоняния, а именно:
1) большую чувствительность, так как адсорбция являет-
ся процессом концентрации, при котором относительно малое
70
.1
количество пахучих молекул в потоке воздуха может быть се-
лективно собрано обонятельным эпителием;
2) мгновенное ощущение запаха при введении пахучего
вещества и его быстрое исчезновение с устранением пахучего
вещества, так как адсорбция является динамическим про-
цессом.
Природа запаха до сих пор не выяснена, но, вероятно, он
связан с некоторыми физико-химическими свойствами малых
количеств примесей пахучего вещества в воздухе [37]. Тогда за-
дача объективного измерения запаха может быть сведена к оп-
ределению примесей пахучих веществ в воздухе преимущест-
венно методами, использующими преобразование измеряемой
величины в электрическую. Известны попытки создания таких
методов определения концентрации некоторых пахучих ве-
ществ [37]. Даже малое количество примесей пахучих веществ
в воздухе может вызывать относительно большие изменения в
таких явлениях, как: 1) теплота адсорбции на твердых адсор-
бентах; 2) поверхностное натяжение жидкости в контакте с
воздухом; 3) поверхностный потенциал жидкости и твердых
тел; 4) эмиссия электронов или положительных ионов от нагре-
тых поверхностей.
Все указанные выше явления классифицированы как по-
верхностные, т. е. они подвержены влянию характеристик вне-
шнего молекулярного слоя, адсорбированного на поверхности
тела. Эти поверхностные явления положены в основу ряда тео-
рий и могут быть использованы для объективного измерения
запаха.
Теории, основанные на восприятии органами обоняния
теплоты адсорбции. Так как процессы адсорбции экзотер-
мичны, Манкриф высказал предположение о возможности
объяснить механизм восприятия запахов с помощью только
одного процесса адсорбции, полагая выделяющееся тепло
причиной возбуждения обонятельных рецепторов [49, 50]1.
Существование большого количества качественно отличных
запахов и пространственного различия в возбуждении обоня-
тельного эпителия под действием различных пахучих
веществ предполагает селективность адсорбции [14, 19],
т. е. существование отличающихся друг от друга по своему
строению рецептивных участков, селективно адсорбирующих
молекулы пахучих веществ.
1 В то время Манкриф еще .не отрицал возможности растворения
пахучих веществ в липоидах или химической реакции в обонятельных ре-
цепторах.
71
Для выяснения зависимости запаха вещества от его ад-
сорбционных характеристик исследовалась селективность ад-
сорбции различных пахучих веществ на различных адсорбен-
тах (активированном угле, активированном алюминии, силика-
геле. активированной фуллеровой земле, растительном жире)
[49, 50]. Для каждой пары (адсорбент—адсорбат) определя-
лось критическое время контакта, т. е. время, необходимое для
полной адсорбции пахучих молекул. .Его величина составляет
0.04—1,0 сек. и может служить характеристикой этих пар.
Таблица II
Таблица 12
Критическое время * контакта, сек Принятое обозна- чение Название вещества Адсорбционная характеристика
0,059—0,081 0 Амиловый спирт . . 21519
0,082—0,111 1 н-Бутиловыи спирт Четыреххлористый уг 01409
0,112—0,152 2 лерод .... 24899
0,153—0,208 3 Алиллкаприлат . . 21537
Этилацетат . . . . 21449
0,209-0,285 4 Этиллактан . . . . 00419
0,286 -0,390 5 Метилсалицилат . . 01535
Сероуглерод . . . 46999
0.391—0,534 6 Пиридин . . . . . 22668
0,535—0,732 7 Ацетон Масло лимонного сор- 43789
0,733-1,0 8 го ....... 42446
Больше 1 сек 9 Лук Испражнения . . . 03789 00999
В табл. 11 приведены принятые Манкрифом обозначения для
критического времени контакта.
Используя эти обозначения, можно написать критическое
время адсорбции амилового спирта: на активированном угле
(0,117 сек)—2; на силикагеле (0,106 сек) — 1; на активиро-
ванном алюминии (0,338 сек)—5; на активированной фулле-
ровой земле (0,096 сек) —1; на растительном жире (более
1 сек) — 9.
Для характеристики адсорбционных свойств амилового
спирта можно использовать совокупность обозначений крити-
ческого времени контакта на пяти адсорбентах1 (21519), кото-
рая показывает, что амиловый спирт очень легко адсорбирует-
1 Последовательность обозначений та же, что и при перечислении
адсорбентов.
72
ся на силикагеле (1) и активированной фуллеровой земле (1),
легко на активированном угле (2), менее легко на активиро-
ванном алюминии (5) и очень трудно на растительном жи-
ре (9i). В табл. 12 даны адсорбционные характеристики не-
которых пахучих веществ для упомянутых выше пяти адсор-
бентов.
Если процесс обоняния связан с адсорбцией, то, вероятно,
адсорбционные характеристики могут быть и характеристика-
ми запахов. В самом деле, пахучие вещества с различными за-
пахами имеют разные адсорбционные характеристики и, нао-
борот, вещества со схожими запахами имеют близкие адсорб-
ционные характеристики. Так, например, амиловый (21519) и
н-бутиловый (01409) спирты имеют резкие запахи типа си-
вушного масла. Аллилкаприлат (21537) и этилацетат (21449)
имеют похожие фруктовые запахи. Так же было проведено
сравнение адсорбционных характеристик соединений с очень
похожими запахами, но разными структурами. Результаты
этих экспериментов показывают, что адсорбционные характе-
ристики связываются с запахами и до некоторой степени не
зависят от химического строения вещества. Например, а-ионон
(кетон с циклическим строением)
Н3С СН3
С
Н,С СН-СН-СН-СОСНз
I I
н,с ссн3
\ 4
сн
и метил-октин-карбоксилат (эфир с линейной цепочкой)
СН3(СН2)5С=С • СООСНз имеют сильные фиалковые запахи.
Подобие их запахов может быть сопоставлено с адсорбцион-
ными характеристиками а-ионона (11227) и метил-октин-
карбоксилата (21326).
Прямые доказательства адсорбции пахучих молекул внут-
ренней поверхностью носовой полости и особенно обонятель-
ным эпителием были получены в 1955 г. Манкрифом [50].
Быстрая деодоризация1 воздуха, пахнущего бензальдеги-
1 Деодоризация — устранение запаха.
73
дом (лимонным сорго, метил-октин-карбоксилатом, ацетоном
и др.)» в присутствии части обонятельного эпителия овцы, во-
ла и кролика подтверждает существование процесса поглоще-
ния. Разные части головы, даже подбородок, язык и внутрен-
ние ткани носовой полости благодаря сложности структуры
имеют адсорбирующие свойства, сравнимые с характеристика-
ми неорганических адсорбентов. Однако они значительно усту-
пают адсорбирующим свойствам обонятельного эпителия.
В табл. 13 приведены результаты двух экспериментов по део-
доризации воздуха различными адсорбентами.
Таблица 13
Время дсолоризации, сек
Адсорбент бензальдегид этилацетат
эксперим. 1 эксперим. 11 эксперим. I эксперим. П
Активированный уголь Эксперимент не
(0,6 г, 6—8 меш.). 75 150 проводился
Активированный уголь (0,6 г, 4—12 меш.). . Ткань из подбородка 75 65 НО 122
(1г) 80 90 130 115
Ткань из кончика языка
(1г) Передняя часть носовой 60 75 85 80
выстилки (1г) на рас- стоянии 5 см от входа
в носовую ролость животного 55 45 101 95
То же, на расстоянии
10 см 70 55 105 85
Пигментированный эпи-
телий (1 см2) .... 25 32 32 52
То же 32 27 — —
» 22 40 — —
» 30 28 — —
26 31 — —
При продувании пахучего воздуха через носовые пути голо-
вы убитого животного (пахучий воздух вдувался в одну нозд-
рю и исследовался на выходе из другой) запах сначала не ощу-
щается и появляется в выходном потоке после насыщения обо-
нятельного эпителия молекулами пахучих веществ. Очевидно,
имеет место быстрый и эффективный процесс концентрации па-
хучих молекул на обонятельном эпителии. Поглощающие свой-
74
<лва обонятельного эпителия в основном сохраняются в тече-
ние 24—30 ч после убоя животного. Один и тот же объект (го-
лова) может многократно использоваться в экспериментах
с различными пахучими веществами при промежуточном про-
мывании носовых путей струей чистого воздуха. Оказалось,
что адсорбционные свойства объекта остаются неизменными.
Таким образом, поглощение является, скорее, физическим,
чем биохимическим процессом, т. е. эпителий ведет себя как
физический преобразователь, а не физиологический объект
при адсорбции пахучих веществ.
Результаты экспериментов показали некоторую обрати-
мость процесса поглощения. Так, например, при продувании
чистого воздуха через носовые пути животного спустя некото-
рое время после прекращения вдувания пахучей смеси ясно
ощущался быстро ослабевающий запах последней. В качест-
ве пахучих веществ использовались лук, фенилацетилен, пири-
дин, амилацетат и др. Подобный же эксперимент показал пред-
почтительность поглощения пахучих веществ обонятельным
эпителием, а именно: после поглощения одного пахучего веще-
ства и последующего промывания чистым воздухом носовой
полости до полного исчезновения запаха на выходе возможно
вытеснение ранее адсорбированного пахучего вещества при
пропускании другого пахучего вещества. Такими парами паху-
чих веществ являются метил-октин-карбоксилат и аммиак, аце-
тон и аммиак и др. И обратимость, и предпочтительность ха-
рактеризуют процесс физической адсорбции.
Величина адсорбирующей поверхности обонятельного эпи-
телия намного больше занимаемой им площади. Для овцы при
площади обонятельного эпителия 5 см2 (на каждой стороне)
адсорбирующая поверхность составляет 0,1 м2у если считать
поверхность волосков булавы гладкой [50]. В действительно-
сти адсорбирующая поверхность может быть значительно
большей.
В качестве вероятных процессов возбуждения обонятель-
ных рецепторов Манкрифом были проанализированы мгновен-
ный контакт молекулы пахучего вещества и рецептора, хими-
ческая реакция между ними, растворение молекул пахучего
вещества в тканях рецептора, адсорбция пахучих молекул на
рецепторной поверхности.
При мгновенном контакте молекулы пахучего газа, уда-
ряясь о поверхность рецептора и отражаясь от него, будут
создавать давление, одинаковое как при вдыхании чистого,
так и пахучего воздуха. Следовательно, добавление молекул
75
пахучего вещества в чистый воздух не создает новых условий
для возбуждения обонятельных рецепторов путем мгновенно-
го контакта.
Некоторые авторы химических теорий предполагают воз-
можность существования в обонятельном рецепторе тех фер-
ментов, которые способны вызывать быструю реакцию между
молекулами пахучего вещества и некоторыми веществами в
обонятельной клетке. Продукты реакции в таком случае дол-
жны быть удалены током крови. Однако многие пахучие ве-
щества полностью насыщены и химически нереактивны.
. Растворимость пахучих веществ вряд ли может быть при-
чиной обонятельного ощущения, так как многие вещества с
сильным запахом нерастворимы в жидкостях живого орга-
низма.
Физическая адсорбция может, по-видимому, являться пер-
воначальной ступенью процесса обоняния, что подтверждается
способностью обонятельного эпителия адсорбировать пахучие
вещества. Возможность различения интенсивности запаха, ве-
роятно, может определяться тем, что при равновесии между па-
хучими молекулами, находящимися в воздухе и адсорбирован-
ными на обонятельной поверхности, площадь, покрытая этими
молекулами, пропорциональна их концентрации во вдыхаемом
воздухе. Качественная дискриминация запаха предполагается
зависящей от того, какая зона рецептивной поверхности по-
крывается слоем адсорбированных пахучих молекул. Запол-
нение различных участков этой поверхности определяется
конфигурацией молекул. Среднее время, необходимое для по-
крытия поверхности рецептора молекулами пахучего вещест-
ва, является характеристикой, описывающей запах.
Трудно предположить, каким образом могут ощущаться мо-
лекулы пахучих веществ при очень большом разбавлении, если
отсутствует процесс селективной адсорбции, при котором про-
исходит концентрация молекул пахучего вещества на обоня-
тельном эпителии.
Теория адсорбции была использована Манкрифом для из-
мерения и классификации запахов, так как адсорбция веществ
с различными запахами на ряде неорганических адсорбентов
различна, а вещества со сходными запахами не очень отли-
чаются по своим адсорбционным характеристикам. При этом
сходство двух запахов, определенное сравнением адсорбцион-
ных характеристик, подтверждается методом обонятельной
адаптации и органолептической (субъективной) оценкой
[48, 55].
76
Основываясь на теории адсорбции, Мидлтон (1956 г.) по-
стулировал работу обонятельных рецепторов следующим об-
разом: молекулы пахучего газа адсорбируются на волосках
булавы и на концах нервных волокон, и тепловая энергия, вы-
деляемая в процессе адсорбции, преобразуется рецепторами
в электрический импульс [46, 47]. Если постулируемое спра-
ведливо, оказывается возможным объяснить различные явле-
ния, связанные с обонянием, свойствами нервной системы, ад-
сорбцией газов и строением поверхности обонятельного эпите-
лия. Ниже эти явления объясняются.
1. Адаптация и усталость. Адаптация — харак-
терное свойство клеток мозга, нервных волокон и чувствитель-
ных рецепторов. В последних адаптация может наступать еще
более быстро, чем в нервных волокнах.
Резонно ожидать, что обонятельные рецепторы, подобно ре-
цепторам другого вида, быстро приходят в состояние равнове-
сия с некоторым данным уровнем возбуждения. При условии,
что этот уровень ниже уровня, дающего максимальную обоня-
тельную реакцию, дальнейшее увеличение концентрации па-
хучего вещества вызывает повторное восприятие. Это свойст-
во органов обоняния часто используется парфюмерами при ис-
следовании смеси пахучих веществ. Запах наиболее летучего
пахучего вещества распознается первым, так как оно быстрее
достигает своей максимальной концентрации, при которой
обонятельная поверхность приходит в равновесное состо-
яние и адаптируется к нему, после чего оно больше не ощу-
щается. Затем ощущается другое летучее вещество до наступ-
ления адаптации при достижении им максимальной адсорб-
ции и т. д.
Адаптация — одно из наиболее важных свойств сенсорной
системы. Этот процесс является существенным для распозна-
вания отдельных составляющих в смеси запахов [47].
2. А н о с м и я. Способность обонять «нормальный» диапа-
зон запахов определяется отсутствием каких-либо поврежде-
ний или дефектов в обонятельных рецепторах, нервных волок-
нах и в получающей информацию области мозга. Тот факт, что
некоторые люди не могут воспринимать определенные запахи,
ощущая остальные, подтверждает предположение о том, что
каждый запах возбуждает специализированную часть обоня-
тельного анализатора. Если часть системы повреждена, воз-
буждение рецептора не может восприниматься, в то время как
другие запахи могут еще различаться неповрежденными ча-
стями системы. Исследования Эдриана подтверждают, что раз-
77
личные участки обонятельной области эпителия кролика яв-
ляются специфически чувствительными к различным типам
возбуждающих молекул [14—19].
3. Паралич. Формальдегид парализует восприятие за-
пахов всех типов. Эта способность формальдегида может быть
следствием его реакций1 с реактивным амидом и водородом
сульфгидрильной группы протеиновых молекул, по-видимому
являющихся центрами адсорбции в обонятельной области [47].
Если рецептивные участки имеют свойство селективной адсорб-
ции молекул пахучих веществ, то «выключение» химической
реакцией некоторого количества центров активной адсорбции
уменьшит адсорбцию этих молекул и, следовательно, увели-
чит пороговую концентрацию, требуемую для их ощущения.
4. Особенности обоняния насекомых и жи-
вотных. Насекомое, как уже было отмечено, обладает
весьма высокой чувствительностью обоняния. Обонятельные
волокна насекомых в отличие от обонятельного эпителия жи-
вотных не утоплены во влаге. Таким образом, молекулы паху-
чего вещества адсорбируются на «сухих» концевых органах —
волосках, вызывая обонятельные ощущения при крайне малых
молекулярных концентрациях.
Носовая полость животных всегда насыщена влагой, пре-
пятствующей адсорбции молекул пахучих веществ в активных
центрах поверхности адсорбента. Поэтому ограничивается
возможность адсорбции на этой поверхности молекул тех па-
хучих веществ, теплота адсорбции которых меньше теплоты
адсорбции воды.
Животные имеют различную чувствительность к пахучим
веществам. Например, органы обоняния кролика считают бо-
лее чувствительными, чем человека. Если плотность булав на
единицу поверхности у кролика и человека одинакова, то, оче-
видно, кролик будет иметь значительно большую адсорбиру-
ющую поверхность и большую чувствительность обоняния,
так как он имеет примерно в два раза большее количество
вслосков на каждой булаве, чем человек [47].
5. Соотношение между молекулярной струк-
турой и запахом. Влияние молекулярной структуры
пахучих веществ на их запах (п. 3.2) может быть рассмотрено
с точки зрения теории адсорбции. Известно, что адсорбция зна-
чительно изменяется от участка к участку поверхности. Так,
1 -NH3+HCHO->-N(CHoOH)-
- SH + НСНО -г HS - S—СНо
78
первые молекулы газа, адсорбированные на графите, имеют по-
чти вдвое большую теплоту адсорбции, чем последующие мо-
лекулы [27]. Это показывает, что величина адсорбции в нес-
кольких активных центрах может быть значительно больше,
чем на остальной части поверхности.
Волокна волосков состоят из 18 аминокислот в различных
пропорциях и комбинациях. Вероятно, волоски одной булавы
имеют подобную структуру, а волоски разных булав отлича-
ются по своей структуре. Поэтому различные области обоня-
тельного эпителйя будут эффективно реагировать на различ-
ные пахучие вещества, что согласуется с уже упомянутыми
результатами исследований Эдриана специфической чувст-
вительности различных участков обонятельного эпителия
[14—19].
Поверхность обонятельного эпителия неоднородна, и неко-
торые ее части имеют для определенных молекул большее при-
тяжение (теплоту адсорбции), чем для других. Распознавание
типа запаха будет зависеть от возбуждения обонятельных ре-
цепторов, расположенных на предпочтительных для данных
молекул участках эпителия, и получающей сообщения от них
области обонятельного мозга.
Величина адсорбции молекул на поверхности зависит от
структуры поверхности. Поэтому запахи членов гомологичес-
кого ряда будут различными. С ростом гомологического ряда
парциальное давление паров вещества понижается и часть по-
верхности обонятельного эпителия, покрытая одномолекуляр-
ным слоем, увеличивается. Интенсивность запаха вследствие
этого увеличивается. У высших членов гомологического ряда
скорость молекул уменьшается и, следовательно, уменьшается
скорость диффузии их на поверхность обонятельного эпителия
из окружающего воздуха. Таким образом, с ростом гомологи-
ческого ряда интенсивность запаха вначале будет увеличивать-
ся, а затем уменьшаться.
Запах более летучих веществ (высокое парциальное дав-
ление) быстрее воспринимается, но не устойчив, менее лету-
чих воспринимается медленнее и более устойчив при равной
концентрации пахучих веществ [47].
6. А н а л и з запаха. Если интенсивность двух запахов
приблизительно равна и состав смеси постоянен, вначале вос-
принимается смесь запахов, затем каждый запах в отдельно-
сти: слабый после более сильного, та?; как адаптация уменьша-
ет реакцию на более сильный запах быстрее, чем на слабый.
Это означает, что адаптация является функцией интенсивно-
79
<ти стимула. Отсутствие селективной чувствительности обоня-
тельных рецепторов привело бы к адаптации этих рецепторов
к смеси запахов. Ощущаемое качество запаха не изменялось
бы во времени [47].
7. Интенсивность восприятия запаха. По
аналогии с законом Вебера—Фехнера, применяемым в случае
ощущения тепла, давления, звука и т. д., интенсивность вос-
приятия запаха предполагается логарифмической функцией
интенсивности стимула. В случае восприятия запаха стимул
можно рассмотреть как энергию, передаваемую адсорбиро-
ванными молекулами на поверхность адсорбента. Для любого
газа количество энергии пропорционально числу молекул па-
хучего вещества, адсорбированных в рецептивной области в
единицу времени. Это число адсорбированных молекул прямо
пропорционально парциальному давлению и обратно пропор-
ционально молекулярному весу и температуре. Поскольку те-
плота адсорбции, молекулярный вес и температура постоянны,
интенсивность возбуждения для каждого данного вещества
пропорциональна парциальному давлению вещества.
Реакция нерва на увеличивающееся возбуждение не явля-
ется постоянной. С увеличением интенсивности стимула растет
количество возбужденных одиночных волокон, имеющих более
высокие пороги возбуждения. Распределение специфических
типов рецепторов с различными порогами возбуждения случай-
но. Какая-либо информация относительно закона распределе-
ния вероятностей отсутствует. Предполагается, что это нор-
мальное (гауссово) распределение. В этом случае любые дан-
ные относительно интенсивности возбуждения запахом дадут
прямолинейную зависимость величины ответа в функции от
стимула. Данные по восприятию человеком интенсивности за-
паха анетола в растворе белого масла приведены в [42].
На рис. 26 представлена зависимость интенсивности ответа
от логарифма объемной концентрации анетола. Интенсивность
ответа определена по отношению к ответу на 100%-ный ане-
тол. Предполагается, что объемная концентрация прямо про-
порциональна давлению паров. Реакция домашней мухи на
изовалериановый альдегид показана на рис. 27. Зависимость
величины процентного ответа от молярной концентрации дана
в [33]. В обоих случаях в пределах ошибки эксперимента гра-
фик изображается прямой линией, и поэтому распределение
числа рецепторов одного типа с одинаковыми порогами можно
полагать гауссовым [47]. Теперь ясно, почему закон Вебера—
80
Фехнера справедлив для среднего диапазона концентраций,
но не для предельных их значений.
Попытаемся связать интенсивность восприятия запаха с
параметрами стимула. Известно, что интенсивность восприятия
прямо пропорциональна частоте электрических импульсов,
являющейся функцией возбуждения и пропорциональной ко-
личеству адсорбированного вещества на волосках соответству-
ющих рецепторных клеток. Если число молекул любого газа,
адсорбированных на единице поверхности в единицу времени,
а, число молекул, падающих на единицу поверхности в секун-
ду, п и среднее время, в течение которого каждая молекула
остается на поверхности, т, то
<з = пг, (16)
где а может служить мерой стимула.
Подставляя значения п и г из (13) и (14), получим
а=NproeQIRT I V2*MRT. (17
Эту формулу можно представить в виде
0 = kp, (18)
NtoeQIRT у
где к=у2 ПРИ постоянно” температуре, р — пар-
циальное давление газа. Это означает, что количество адсор-
бированного вещества прямо пропорционально парциальному
Рис. 26. Зависимость интен- Рис. 27. Зависимость величины про-
сивности ответа в процентах центного ответа от молярной коицент-
от логарифма объемной кон- рации изо-валерианового альдегида^
центрацни анетола.
В- 5*10*-
В. 4«КГ5-
I 3>1O'S-
J 2*/0'5-
§ <o-s
\ 8‘Ю*-
1 б’ЛГ6-
5 5'Ю"6-
440^
10 ' 30 50 '^0 90
Процентной ответ
6 Заказ № 7032
давлению газа, энергия адсорбции постоянна (но не одинако-
ва) для всех рассматриваемых видов молекул, и эти молекулы
не настолько уплотнены, чтобы быть помехой для адсорбции
молекул другого вида.
Для 100%-ного одномолекулярного слоя
<т0=^Л ,
(19)
где —в молекулах на единицу поверхности, ps— давление
насыщенных паров. Для некоторого другого значения а, соот-
ветствующего давлению р,
<J=kp,
откуда
Таким образом, если интенсивность возбуждения пропорци-
ональна количеству адсорбированного вещества, то она про-
Таблица 14
Название вещества Пороговая концентрация
zJcm3 молекул/ см3 (пи) молекул / см-
Этиловый спирт Фенол й . . Пиридин .... Ванилин . . . . 5 • Юз 1,2 • Ю-о 4 • 10-е 2 • 10—13 6,5 • 10-1-’ 7,7 • 1016 3,0 • 10й 7,9 • 10» 1,6 • 1017 1,8 • Юн 4,5 • 109 8,5 • 10^
порциональна и парциальному давлению паров- этого вещест-
ва. Следовательно, отношение давления пара во вдыхаемом
воздухе к давлению насыщенного пара также может служить
мерой интенсивности стимула.
8. Обонятельный порог. В табл. 14 представлены
некоторые данные о пороговых концентрациях пахучих ве-
ществ [46, 47].
Если пороговые концентрации паров преобразовать в пар-
циальные давления, можно подсчитать отношение порогового
давления паров пахучих веществ к давлению насыщенных па-
ров. Для определения давления паров при пороговых кон-
82
центрациях используем выражение (13)
/го1О“22)/Л17/3,52 мм рт. ст., (21)
где значения nQ взяты из табл. 14. Полученные результаты
приведены в табл. 15
Из табл. 15 видно, что при пороговой концентрации мини-
мальный стимул соответствует
—10 12 . Для значений р,
Ps
близких к ps (если ps имеет большое абсолютное значе-
ние), слоем молекул одного вещества будут покрыты неспеци-
фичные для него области обонятельного эпителия, в результа-
те чего запах этого вещества может восприниматься как слож-
ный. С уменьшением р воспринимаемый запах будет ощущать-
ся как запах одного вещества (простой запах).
Таблица 15
Название вещества р, мм рт. ст. Ps, мм рт. ст. P'PS
Этиловый спирт . 5,37 10* 88 6,1 • 10-g
Фенол 8,65 • 10 -ю 6,7 • 10-1 1,3 • Ю-о
Пиридин * . . . 1,98 • 10-и 29 6,8 • 10-13
Ванилин ...... 5,19 • 10-is 2 • Ю-з 2,6 • 10-12
Примечание. ps экстраполировано к 320 С. Использо-
вались данные о давлении паров и формула logp -ftlogP-j-Q,
где к и Ci—постоянные, Т— абсолютная температура.
Следует ожидать, что любые типы волосков способны ад-
сорбировать все молекулы, теплота адсорбции которых боль-
ше теплоты адсорбции воды. При передаче к мозгу импульсно-
го сообщения о возбуждении рецепторов качество ощущения
зависит от положения возбужденного рецептора в обонятель-
ной области. Можно ожидать также, что адсорбционные харак-
теристики рецептивных участков в пределах обонятельной об-
ласти не остаются неизменными. Если это так, то для любого
вещества теплота адсорбции Q будет изменяться по всей пло-
щади, на которой молекулы будут адсорбироваться от опти-
мальной до ничтожно малой [47]. Таким образом, обонятель-
ные пороги зависят ст адсорбции веществ на обонятельном
эпителии.
Й*
83
9. Различие между запахами. Если взять смесь
двух пахучих веществ (I и II), то число молекул каждого из
них, адсорбированное на обонятельном эпителии, будет:
а = (qo)i ft Pi .
1 1+ft Pi + ftiPn
— (qo)ii ftiPii
11 1 -f-dj pi -\-bnpn
где
. __ Mi__________________' ___________Mu__________
' “ (<r0), V 2k Mi RT ’ " ~(<y0)u V 2nMnRT '
(22)
(23)
При низких парциальных давлениях, соответствующих порого-
вым концентрациям, эти выражения примут вид
Hi =(<t0)i bi Pi 5
Пп = (о0)ц ^»Рп.
(24)
Отсюда следует, что в этих условиях адсорбция одного ве-
щества не зависит от адсорбции другого. При относительном
увеличении давления вещества 1 или при bit значительно
большем, чем Ьп, величина biPi будет значительно больше
Ьпрп, т. е.
Qi >Qh для T! =(t0)i eQIRT ;
TO
„ _(qo)i ft Pl .
1 1+ft Pi ’
(25)
On =
(gp)ii ftipn
1 + ft />i
(26)
Это значит, что адсорбция вещества I не зависит от ад-
сорбции вещества II, но адсорбция вещества II подавляется
адсорбцией вещества I. Это может иметь место для смеси воз-
духа с парами органического вещества, вытесняющими воз-
дух с поверхности, к которой они имеют доступ вследствие
их значительно большей теплоты адсорбции.
Для паров органических веществ т составляет около 10“s
сек., а для воздуха 1010~ сек. Таким образом, молекулы органи-
ческих веществ остаются на поверхности намного дольше мо-
84
лекул воздуха, быстро вытесняя последние. Если теплота ад-
сорбции Q для вещества I больше, чем для вещества II, то при
возбуждении одного рецептора этими двумя пахучими веще-
ствами значение b соответственно увеличивается для веще-
ства I.
Рассмотрим воздействие двух пахучих веществ I и II на
на два рецептора, отличающихся тем, что наиболее активные
участки рецептора А имеют теплоту адсорбции 15 ккал!моль
для вещества 1 и 10 ккал/моль для вещества II, а рецептор
В имеет теплоту адсорбции 15 ккал/моль для вещества II и
10 ккал/моль для вещества I. Парциальные давления паров
этих веществ малы и примерно одинаковы. Изменение Q от 10
до 15 ккал/моль увеличивает т и соответственно b примерно в
5-104 раз. Поэтому рецептор А будет возбуждаться в основном
веществом!, а рецептор В—молекулами вещества II. При уве-
личении парциального давления паров этих веществ, когда ак-
тивные участки рецептора А для вещества I и рецептора В
для вещества II уже заняты молекулами этих веществ, тепло-
та адсорбции вещества I на рецепторе А будет ниже
15 ккал/моль, но незначительно изменяется для вещества II.
Поэтому каждое вещество уменьшает адсорбцию другого ве-
щества на соответствующем рецепторе и адсорбируется вме-
сто него. Таким образом, вещество II может восприниматься
как вещество 1 и наоборот. Если парциальное давление паров
возрастает, то запахи трудно различать, так как специфичес-
кие свойства рецепторов будут менее выражены.
10. Изменение качества запаха при раз-
бавлении. Явления, описанные выше, лежат в основе изме-
нения качества запаха при изменении концентрации пахучего
вещества. Вблизи пороговых давлений первые молекулы, ад-
сорбированные на определенных типах рецепторных клеток, на
которых они имеют максимальную адсорбцию, будут создавать
специфическое ощущение, соответствующее качественным ха-
рактеристикам рецепторов этого типа. При увеличении пар-
циального давления будут возбуждаться другие рецепторы,
и поэтому должен изменяться общий характер ощущений.
Вещества, которые обладают компенсирующим действием,
будут также изменять качество ощущения при увеличении пар-
циальных давлений.
Качество запаха натурального продукта, являющегося
смесью веществ, должно изменяться с разбавлением, потому
что изменяется общее давление паров. Так как пороговые дав-
ления пара достигаются в некоторой последовательности, то
83
его молекулы могут возбуждать различные рецепторы на их
специфически активных участках.
11. Влияние скорости воздушного потока.
При пороговой концентрации пахучего вещества адсорбирует-
ся достаточное для возбуждения обонятельного рецептора ко-
личество молекул в течение определенного промежутка време-
ни. Так как молекулы пахучего вещества удаляются при ад-
сорбции из газа, то парциальное давление пара пахучего ве-
щества над адсорбированным слоем понижается и может до-
стичь первоначального значения только в процессе диффузии.
Увеличение скорости воздушного потока увеличивает ско-
рость диффузии и уменьшает время, необходимое для дости-
жения в адсорбированном слое пороговой концентрации, не из-
меняя величины последней. При этом увеличивается скорость
испарения влаги с поверхности обонятельного эпителия, дав-
ление ее паров уменьшается, становится ниже давления насы-
щенных паров, что приводит к понижению пороговой концент-
рации пахучего вещества.
Пары воды с небольшой примесью паров пахучего вещест-
ва можно считать смесью двух газов. Если взять в качестве
пахучего вещества ванилин, пороговое давление которого
5-10"15л«л( рт. ст., то количество адсорбируемых молекул ва-
нилина в соответствии с выражением (23) будет
__ (^о)ван * ^ван * Рван
1 4“ ^ванРван“Ь^воды‘ Рводы
но так как
^воды * Рводы _|08
^ван * Рван
ТО
_ __ (^о)ван * ^ван ‘ Рван
иван 1 1 А п
Г i «'воды ‘ Рводы
С увеличением скорости газового потока понижается дав-
ление паров воды, соответственно уменьшается величина
£В0Ды • Рводы и Пороговое давление паров ванилина.
12. Подавление одного запаха другим
(маскировка). Маскировка сильными запахами некото-
рых пахучих веществ (например, метилиононов) возникает
благодаря большому значению теплот адсорбции для многих
типов рецепторов. В результате этого уменьшается количест-
во адсорбированных молекул других пахучих веществ, име-
ющих даже более высокую теплоту адсорбции на специфи-
86
чески активных участках рецепторов. Тем самым подавляет-
ся специфическое качество запаха и изменяется общий запах.
Теория, основанная на изменении поверхностного натя-
жения. Поверхностное натяжение жидкости при адсорб-
ции пахучих веществ заметно уменьшается [37]. Вели-
чина поверхностного натяжения может быть измерена раз-
личными методами без разрушения поверхности жид-
кости. К их числу относится и метод висящей капли. Величи-
на поверхностного натяжения может определяться в этом слу-
чае измерением размеров висящей капли по ее фотографии,
прямым измерением отраженного изображения капли или ко-
личества капель в единицу времени. Под воздействием запаха
поверхностное натяжение изменяется во времени в течение
30—60 мин, достигая некоторого стабильного значения. Раз-
личные пахучие вещества вызывают различные изменения по-
верхностного натяжения определенных жидкостей с «чувстви-
тельным поверхностным натяжением». Однако следует учиты-
вать, что насыщение окружающего воздуха влагой также мо-
жет уменьшать поверхностное натяжение от 460 до350дн/си,
если в качестве подобной жидкости применена ртуть. Поэтому
для объективной оценки запахов необходимо использовать
«водно-стабилизированную ртуть». Поверхностное натяжение
дистиллированной воды под действием эвгенола изменялось
от 71,6 до 48,6 дн!см.
Изменение поверхностного натяжения наблюдается только
в случаях органолептического обнаружения запаха. К сожале-
нию, пока выявлено небольшое количество жидкостей с «чув-
ствительным поверхностным натяжением». Недостаток указан-
ного метода заключается также в трудности преобразования
изменения поверхностного натяжения в электрическую вели-
чину.
Теория, основанная на восприятии органами обоня-
ния контактного потенциала. Помимо изменения поверх-
ностного натяжения, адсорбция пахучих веществ, как
уже указывалось, сопровождается появлением контакт-
ного потенциала, который также может быть исполь-
зован для объективного измерения запахов и для объяснения
возможного механизма восприятия запахов живыми организ-
мами [25, 34]. Контактный потенциал на адсорбирующих по-
верхностях возникает благодаря наличию молекулярных сил
взаимодействия. Обонятельные рецепторы в большинстве слу-
чаев обнаруживают запах при наличии 105—107 молекул паху-
чего вещества в кубическом сантиметре воздуха, различают
87
небольшое отличие в качестве этих молекул и передают инфор-
мацию по нервам в мозг. Однако даже если каждая адсорби-
рованная молекула могла бы участвовать в создании контакт-
ного потенциала, то результирующее изменение потенциала
поверхности составляло бы только доли милливольта. Усиле-
ние этого эффекта может быть достигнуто применением сег-
нетоэлектрических материалов в качестве адсорбентов.
Если молекулы пахучего вещества адсорбируются на мно-
гих участках сегнетоэлектрика, поле поляризации ориентиро-
ванных и индуцированных диполей может привести к спонтан-
ной поляризации сегнетоэлектрика. Вероятность поляризации
в течение данного интервала времени будет пропорциональна
количеству адсорбированных молекул в данной области. Ре-
зультирующий импульс, возникающий в рецепторе, может слу-
жить источником больших сигналов в обонятельном нерве. В
процессе подготовки к новому импульсу рецептор ведет себя
как несегнетоэлектрическое вещество.
Атенштедт [23, 24] исследовал сегнетоэлектрические свой-
ства тканей живых организмов. Такими свойствами обладают
гожа морской свинки, лягушки, кожица насекомых. Чрезвы-
чайно высокие значения диэлектрической постоянной 8> 106
были обнаружены в тканях мышц и печени собаки. Значение
< 104 было получено и многими другими авторами на тканях
других животных. Недавно исследованные сегнетоэлектрики,
такие как гуанидин, глициносульфат и тиомочевина, имеют
сходные с органическими соединениями структуры. Можно
предположить, что в протеиновых структурах также возника-
ют явления поляризации.
Селективность восприятия может основываться на чувстви-
тельности предполагаемого механизма генерации импульсов
к величине энергии адсорбции порядка 1—2 ккал в диапазоне
5—15 ккал. Ниже определенного уровня энергии генерируе-
мые импульсы будут слишком неясными (редкими)). При пре-
вышении некоторого уровня энергии частота импульсов была
бы слишком большой для передачи их по нерву.
Для обонятельных рецепторов дифференциация запахов
становится возможной, если рецепторы приспосабливаются к
определенным диапазонам энергии адсорбции.
В результате дипольного взаимодействия и взаимодействия
дисперсионного вида энергия адсорбции достигает величины
5—15 ккал. Два полюса диполя на расстоянии г=ЗА взаимо-
действуют с энергией 4,5ятшл. Если же размеры диполей соиз-
меримы с оазделяющим их расстоянием, это значение увели-
88
чивается на 50—100%. Возможно также дополнительное уве-
личение энергии адсорбции за счет дисперсионных сил. Кроме
того, некоторые рецепторы, по-видимому, могут использовать
водородные связи как источник энергии адсорбции.
Дрэвникс предполагает, что обладающие сегнетоэлектри-
ческими свойствами поверхности рецепторных клеток цикли-
чески повторяют спонтанную поляризацию и, вероятно, слу-
жат в качестве преобразователя для селективного восприятия
адсорбируемых молекул пахучих веществ [34}
Энергия адсорбции может определять частоту импульсов
Баркхаузена, если действие деполяризации внешних источни-
ков энергии является достаточным. В обонятельном анализа-
Рис. 28 Схематическое изображение ориентированных липоидных молекул,
образующих мембрану’*обонятельного рецептора (а), разрушение мембраны
рецептора адсорбированной молекулой пахучего вещества (б).
торе механизм посылки нервных импульсов функционирует в
ограниченном диапазоне частот, а запах пахучих веществ, как
уже указывалось, может распознаваться по частоте этих им-
пульсов.
Теория разрушения мембраны обонятельного рецептора
молекулами пахучих веществ. Дейвис и Тайлор (1954 г.) по-
лагали, что явление адсорбции может лежать в основе про-
цессов восприятия запахов [31, 32, 68]. Обонятельная реакция
возникает, когда на небольшой области поверхности обоня-
тельной клетки адсорбировано определенное количество мо-
лекул пахучего вещества, прокалывающих в некоторый мо-
мент времени липоидную мембрану рецепторной клетки. При
7 Заказ № 7032
89
этом концентрация ионов внутри и вне клетки уравновешива-
ется (рис. 28).
Локальное разрушение мембраны зависит от объема и кон-
фигурации молекул. Большие и жесткие молекулы (р-иононы,
ксилол, мускус) легче разрушают мембранную структуру клет-
ки и имеют более сильный запах, чем малые и гибкие молеку-
лы (метиловый спирт, этиловый спирт с большими пороговыми
концентрациями).
Авторы показали, что красные кровяные клетки особенно
удобны как модель для исследования подобного механизма,
так как при разрушении их мембраны — гемолизе гемогло-
бин освобождается с заметным изменением внешнего вида
модели.
В результате экспериментов было найдено, что пахучие
вещества ускоряют вызванный сапонином гемолиз. На рис. 29
показана зависимость скорости гемолиза от концентрации са-
понина с добавлением и без добавления р-ионона. Очевидно,
молекулы пахучего вещества, ослабляя мембрану красных
кровяных клеток, ускоряют процесс проникновения в клетку
сапонина. Степень ускорения гемолиза можно характеризо-
вать величиной А:
1
С — молярная концентрация пахучего вещества;
t и /о — время, в течение которого модель подвергается
гемолизу в присутствии пахучего вещества и без него.
Если существует определенное соотношение между интен-
сивностью запаха пахучего вещества и степенью ускорения
гемолиза, то оно может служить некоторым подтверждением
правильности теории «разрушения» и возбуждение обоня-
тельных рецепторов можно рассматривать как некоторый
механизм, аналогичный гемолизу.
На рис. 30 показана степень ускорения гемолиза некоторы-
ми пахучими веществами в зависимости от величины их поро-
говой концентрации. Вещества, сильно пахнущие, например
ксилольный мускус и р-ионон, являются сильными ускорителя-
ми, вещества с большими пороговыми концентрациями — фе-
90
нол и метанол — слабыми. В ряду изомерных метилкумари-
нов 4-метильное соединение, относительно малоэффективный
ускоритель, имеет запах меньшей силы, чем другие (для неко-
торых людей кажется даже лишенным запаха). При замеще-
нии фтором всех атомов водорода в бутиловом спирте вместо
слабого спиртового возникает более сильный камфароподоб-
ный запах. Таким образом, можно считать установленным, что
с увеличением интенсивности запаха увеличивается воздейст-
вие пахучего вещества на скорость гемолиза.
Оптические изомеры незначительно отличаются друг от
друга по степени ускорения этого процесса и, как было отме-
чено выше, имеют сходные запахи.
2
t .
0-020-
0-018
0-016 -
0-014 -
0-012 -
0-010 -
0-008-
0-006-
0-004-
0-002-
Концентрация сапонина,мкг/мл
Рис. 29. Скорость гемолиза (зави-
симость от времени гемолиза) для
разных концентраций сапонина. В
присутствии 0,0001 М (3-ионона
скорость гемолиза возрастает. Эк-
сперимент проводился при t = 25°С
и pH = 7,2.
1 — сапонин ионон; 2 — сапонин.
может служить гигантский аксон
шие клетки растений, ориентированные
мозможности подобные плазме мембраны.
Рис. 30. Зависимость степени ус-
корения гемолиза пахучими веще-
ствами от величины обонятельного
порога.
Более удобным объектом
для наблюдения, вместо
красных кровяных клеток,
кальмара и некоторые боль-
пленки липоидов, по
91
Величина . обонятельных порогов зависит от энергии ад-
сорбции молекул пахучего вещества на мембране обонятель-
ного рецептора, покрытой секретом (на границе липоид—во-
да), и от тех нарушений мембраны, которые создаются ад-
сорбированными молекулами. Оба фактора можно оценить
количественно, что позволяет определить величину обоня-
тельных порогов различных пахучих веществ. Энергия адсорб-
ции может быть подсчитана по свободной энергии молекул
пахучего вещества при прохождении их из воздуха на грани-
цу липоид — вода. Далее, если критическое количество моле-
кул р пахучего вещества адсорбировано на небольшой пло-
щади а поверхности рецептора, наступает локальное разру-
шение его мембраны. Для некоторых пахучих веществ (0-ионо-
нов), с сильным запахом, т. е. с низкими обонятельными по-
рогами (п) порядка 108 молекул1см3 воздуха, р. может быть
близким к единице. Пахучие вещества с более высокими обо-
нятельными порогами (фенол, этанол) и малыми размерами
молекул создают локальное разрушение мембраны при зна-
чительно большем р.
Рассмотрим зависимость величины обонятельного порога
для различных пахучих веществ от энергии адсорбции и зна-
чения р.
Изотерма Лангмюра для мономолекулярного слоя паху-
чего вещества на обонятельной мембране будет иметь вид:
где х — среднее количество адсорбированных молекул на
квадратный сантиметр поверхности мембраны обо-
нятельного рецептора;
С — концентрация пахучего вещества, молекул/см3;
d — толщина поверхности мембраны (около 10 А);
К — коэффициент адсорбции молекул пахучего вещества
на границе липоид—вода.
Если значение С мало, то выражение (28) принимает вид
x = KCd. (29)
Предположим, что обонятельная мембрана каждой клет-
ки содержит а участков площадью а бш2. Тогда среднее ко-
личество адсорбированных молекул на каждом участке бу-
дет ха.
92
Если на любом из этих участков количество адсорбиро-
ванных молекул р>ах, возможна стимуляция рецептора на
пороговом уровне. Вероятность того, что р молекул адсорби-
ровано на некотором участке, может быть представлена за-
коном распределения Пуассона
Рр=‘г”‘~^- (»)
Тогда количество участков N обонятельного рецептора, со-
держащих р молекул, может быть представлено как
N=aPp=ae~ax . (31)
Для возбуждения рецептора достаточно локальное разруше-
ние мембраны на одном участке, т. е. М=1- При этих услови-
ях
l = . (ах)Р . (32)
р! '
Логарифмируя и подставляя С=п, получим
logn+logA = —J- loga+ logacf+ (33)
p p
Коэффициент К определяется экспериментально; значения
а, а и d могут быть получены из физиологических данных
об обонятельных порогах для сильных и слабых запахов па-
хучих веществ. Так, для 0-ионона значение обонятельного
порога составляет «=1,610* молекул! см3,т. е. logn=8,2; logK=
=8,35; а р = 1, следовательно,
16,55=—log aad. (34)
Для пахучих веществ со слабым запахом принимается р=24,
a (logn-|-logK) приближается к 22. Тогда из выражения (33)
следует
22=—^-r-loga+logad+l . (35)
Совместное решение уравнений (34), (35) дает значения
loga=4,64 и log ad=—21,19.
93
Подставляя эти значения ц (33), получим основное уравнение
для расчета обонятельных порогов
logn+log?C- - И21.19. (36)
Р Р
Рис. 31 позволяет сравнить расчетные и экспериментально
полученные значения обонятельных порогов.
Авторы теории рассмотрели и проанализировали получен-
ные результаты. Так как log а найден равным 4,64, то количе-
ство участков а на каждом рецепторе составляет 44 000;
logad=—21,19,a d=10A,тогда a
64 ’10-16с>и2, т.е.а=64А2.
Поперечное сечение мо-
лекулы р-ионона состав-
ляет 47А2, что соответ-
ствует принятому значе-
нию р=1. Активная по-
верхность каждой обоня-
тельной клетки ad дости-
гает 2,8 • 10~ 0 он2 в отли-
чие от физиологических
данных (10-8—10~6 ом2).
Рис. 31. Сравнение полученных
экспериментально и расчетных
данных об обонятельных поро-
гах.
О — нормальные спирты; X — нор-
мальные углеводороды; ф — орга-
нические соединения.
Однако, как отмечают авторы, только окончания обонятель-
ных волосков имеют доступ к воздуху, вся остальная поверх-
ность обонятельного рецептора утоплена в секрете. Поэто-
му значение активной площади, полученное физиологами,
может быть уменьшено на 1—2 порядка.
Если обозначить через До площадь поперечного сечения
молекулы пахучего вещества, то рД0=а, (37)
т. е. — является линейной функцией размера молекулы и
мерой ее разрушающей силы.
Нам кажется, что рассмотренная теория была бы более
обоснованной, если бы значения обонятельных порогов были
94
взяты с учетом количества молекул, адсорбированных на сли-
зистой носовой полости до достижения ими обонятельной ще-
ли. Кроме того, в теории не уделено внимания вопросам каче-
ственной дискриминации запахов.
Стереохимическая теория обоняния. Для подтвержде-
ния и развития гипотезы Манкрифа необходимо ответить
на два вопроса: 1) что такое «первичный запах»; 2) ка-
кое имеет очертание рецептивный участок для каждого пер-
вичного запаха? С этой целью Эймур рассмотрел характе-
ристики пахучих соединений. Результатом его исследований
было сообщение о семи первичных запахах и создание в 1952 г.
стереохимической теории обоняния [20, 21, 22]. Эта теория
дает подробное описание формы и химических свойств мо-
лекул семи первичных запахов и предполагаемых соответст-
вующих им рецептивных участков. Для выявления первичных
запахов Эймур исследовал 600 пахучих органических со-
единений.
Если гипотеза специфических рецептивных участков вер-
на, первичные запахи должны появляться значительно чаще
сложных, составленных из двух или более первичных запа-
хов. В самом деле, определенные запахи встречаются значи-
тельно чаще других. Например, из 600 рассмотренных
органических соединений более 100 имеют камфароподобный
запах, отнесенный Эймуром к простым, и только 6 имеют за-
пах кедрового дерева.
По всей вероятности, камфароподобный запах является
одним из первичных. Подобным образом были выделены семь
возможных первичных запахов: камфароподобный, мускус-
ный, цветочный, мятный, эфирный (эфироподобный), острый
и гнилостный. Любой известный сложный запах можно соста-
вить смешиванием нескольких первичных запахов.
Семи первичным запахам должны соответствовать семь
различных типов обонятельных рецепторов. Можно предста-
вить рецептивные участки в виде ультрамикроскопических
щелей или углублений различных форм и размеров в мембра-
не рецепторной клетки. Тогда каждая щель (или каждое уг-
лубление) должно принимать молекулу соответствующей кон-
фигурации и размеров. При этом может возникнуть нервный
импульс вследствие дезориентации и деполяризации мембра-
ны рецепторной клетки. Некоторые молекулы могут соответ-
ствовать двум (или более) различным щелям. Вещество, мо-
лекулы которого воспринимаются двумя и более типами рецеп-
торов, имеют сложный запах.
95
Изучение форм (очертаний) семи рецептивных участков
было начато с исследования структурных формул соединений
с запахом камфары и построения моделей их молекул- В ре-
зультате было выяснено, что молекулы веществ с камфаропо-
добным запахом имеют форму, близкую к сферической, с диа-
метром около 7А.
Очевидно, рецептивный участок для молекул с камфаропо-
добным запахом должен иметь форму полусферы с диаметром
около 7А. Многие вещества с камфароподобным запахом
имеют жесткие сферические молекулы либо гибкие молекулы,
которые могут принимать форму шара. Например, d-камфа-
ра, гексахлорэтан, дихлорэтиламид тиофосфорной кислоты
и циклооктан обладают камфароподобным запахом, несмот-
ря на то ,что они не имеют ничего общего, кроме двух атомов
6
5
Ср*
СН,
сн2—с — со
| СН3-С-СНЭ|
сн7—СН—СИг
а /С1
ci—с —C-CI
c»z ХС1
а
ОЛО
Рис. 32. Молекулы соединений с камфароподобным запахом:
а — камфары; б — гсксахлорэтана; в — дихлорэтиламида тиофосфор-
ной кислоты; г — циклооктана.
углерода в молекуле, но зато их молекулы с диаметром 7А
(рис. 32) имеют сферическую форму.
После построения моделей молекул, соответствующих дру-
гим типам пахучих веществ, были определены формы и раз-
меры молекул, представляющих другие первичные запахи
(рис. 33). Мускусный запах создается молекулами формы дис-
ка с диаметром около 10А. Приятный цветочный запах вызы-
вается молекулами формы диска с гибким хвостом, наподобие
96
Камфарный
а
Мускусный
Цветочный
6
Мятный Эфирный
Рис. 33. Модели молекул веществ, создающих первичные
запахи, и соответствующие им рецептивные участки.
9Г
воздушного змея. Свежий мятный запах издается молекула-
ми формы клина с электрически поляризованными группа-
ми атомов, способными образовывать водородную связь вбли-
зи вершины клина. Эфирный запах связывается с молекулами
стержневидной формы или другими тонкими молекулами.
Предполагается, что специфические рецептивные участки
должны иметь форму и величину, соответствующие форме
и величине молекулы (см. рис. 33).
Острый и гнилостный запахи являются исключением из
приведенной схемы соответствия форм и размеров молекул
и рецептивных участков. Молекулы веществ с такими запаха-
ми несут электрические заряды. Класс острых запахов издает-
ся химическими соединениями, молекулы которых вследствие
недостатка электронов имеют положительный заряд и силь-
ное притяжение к электронам; такие молекулы называются
электрофильными. Гнилостные запахи, с другой стороны, вы-
зываются молекулами с избытком электронов — нуклеофиль-
ными. Они испытывают сильное притяжение к ядрам сосед-
них атомов.
Для проверки справедливости стереохимической теории
был проведен ряд экспериментов.
Согласно теории, форма или заряд молекулы определяют
ее запах. Если синтезировать вещество с молекулами опреде-
ленной формы, то оно должно обладать предсказанным за-
ранее запахом, соответствующим форме молекул.
Молекула, состоящая из трех цепочек с одним атомом уг-
лерода, четвертая валентность которого занята атомом водо-
рода
Н СООСНз
С
Н3СООС-СН2-СН2 СН2-СН2-СООСН3
может попасть на рецептивный участок змеевидной формы,
соответствующий цветочному запаху, клинообразной формы—
мятному запаху или одной из своих цепочек на участок
стержневидной формы (эфирный запах). Такая молекула
должна иметь фруктовый запах, составленный из трех пер-
вичных.
При замещении атома водорода сравнительно большой
метильной группой (четвертая связь атома углерода) новая
98
форма молекулы не будет соответствовать рецептивным уча-
сткам змеевидной и клинообразной формы, но одна из ветвей
этой молекулы все же может занять участок стержневидной
формы
СН3 СООСНз
С
Н3СООС-СН2-СН2 СН2-СН2-СООСН3
Вещество с такими молекулами, согласно стереохимической
теории, должно обладать эфирным запахом. Эксперимент
подтвердил правильность предположения фруктового запаха
(виноградоподобного) первого соединения и эфироподобного
запаха второго. Изложенное позволяет понять, почему запах
определенных соединений с бензольным кольцом резко изме-
няется при изменении положения группы атомов.
Была сделана попытка создать сложный запах, сущест-
вующий в природе, смешением первичных запахов. Молеку-
лы вещества с запахом кедрового дерева имеют форму, соот-
ветствующую рецептивным участкам камфароподобных, мус-
кусных, цветочных и мятных запахов. Испытывая различные
комбинации этих четырех первичных запахов, после 86 проб
Джонстон синтезировал запах кедрового дерева. Таким же
образом был синтезирован запах сандалового дерева-
Следующие две группы экспериментов были проведены
для выявления первичных запахов. Молекула, издающая ос-
новной первичный запах, может соответствовать только од-
ному рецептивному участку определенной формы и величины.
Молекулы одинаковой или подобной формы и размеров будут
иметь подобные запахи, молекулы с различными формами
пахнут совершенно различно.
Интересный эксперимент по определению способности пчел
распознавать два запаха был проведен Джонстоном. Один
из двух запахов был назван «правильным» (с ним была свя-
зана подкормка пчел сахарным сиропом), второй — «непра-
вильным» (с ним должен был ассоциироваться «удар» элек-
трическим током). Эта пара запахов могла относиться к
одной первичной группе или разным первичным группам (на-
пример, к цветочной и мятной). Вначале на столик перед
ульем ставили по паре пузырьков с пахучими веществами
и приучали пчел к распознаванию «'правильного» и «непра-
вильного» запаха в каждой паре. Можно предположить, что
99
-после такого обучения пчелы будут в основном стремиться
к «правильному» запаху и будут избегать другого «неправиль-
ного» запаха. Оказалось, что пчелы легко различают разные
первичные запахи (мятный и цветочный) и очень затрудняют-
ся в различении двух запахов в пределах одной первичной
(мятной) группы. Эти эксперименты показали, что обонятель-
ная система пчелы, подобно обонятельной системе человека,
может быть основана на стереохимическом принципе, хотя ор-
ган обоняния пчелы (антенна) отличен от органа обоняния
человека.
Основной целью одного из экспериментов было выяснение
сходства запахов различных химических соединений с почти
одинаковой формой молекул. Испытывали пять соединений,
принадлежавших трем различным химическим классам. Все
они отличаются друг от друга молекулярным строением, но
имеют форму диска и должны обладать мускусным запахом.
Действительно, запахи этих веществ органолептически опре-
делялись как мускусные, и возникали затруднения в различе-
нии по запаху этих пяти совершенно разных химических ве-
ществ.
Все эти эксперименты только косвенно подтверждают сте-
реохимическую теорию. Гестелэнд, исследуя единичные обо-
нятельные нервные клетки лягушки, нашел, что различные
клетки отвечают селективно на различные запахи. Было об-
наружено восемь типов рецепторов. Пять из них восприни-
мают пять первичных запахов, постулированных стереохими-
ческой теорией (камфароподобный, мускусный, эфирный,
острый и гнилостный). Этот эксперимент можно считать
прямым доказательством справедливости стереохимической
теории.
Некоторое эмпирическое соотношение между спектром па-
хучего вещества и запахом не противоречит стереохимической
теории обоняния, так как инфракрасный спектр в диапазоне
частот с волновым числом ниже 1000 смг\ зависит в основ-
ном от общей конфигурации молекулы.
Как же можно объяснить случаи частичной аносмии с точ-
ки зрения стереохимической теории обоняния [21]?
Можно предположить, что рецептивные участки для вос-
приятия некоторых первичных запахов имеют ненормальную
форму. Тогда аносмия некоторых субъектов, например, к ма-
кроциклическим и стероидным мускусам с сохранением чув-
ствительности к нитромускусу возможна, если рецептивные
участки для мускуса имеют меньшую длину (например, 10,5А
100
вместо нормальной длины 11,5А) при их обычной ширине
9А. Рецептивные участки такого размера не допускают ад-
сорбции более длинных стероидных и макроциклических мус-
кусных молекул, но адсорбируют более короткие и более ши-
рокие нитромускусные молекулы.
Может быть и другой случай аносмии, например, к цианис-
той кислоте, которую некоторые лица не способны обонять,
но для которой обычно характерен запах горького миндаля.
Однако запах цианистой кислоты можно считать и острым.
Молекулы цианистой кислоты очень малы, значительно мень-
ше каждого из трех рецептивных участков специфических для
камфароподобного, цветочного и мятного запахов, которые
как было постулировано, составляют запах горького миндаля.
Цианистая кислота обладает большим числом особых
свойств: активно полимеризуется, химически сильно реактив-
на и исключительно ядовита. Свойства этой кислоты можно
рассматривать как причину ее запаха. Индивидуальное не-
постоянство в чувствительности отдельных обонятельных ре-
цепторов различных субъектов к анестезирующему действию
цианистой кислоты объясняет частичную аносмию, так как
рецепторы могут анестезироваться уже первыми молекулами
кислоты. Поэтому различными наблюдателями ощущаются
разные запахи при стимулировании парами цианистой кисло-
ты. Возможно, она имеет сама по себе только едкий запах, но,
соединяясь в носовой полости с некоторыми другими вещест-
вами, создает миндальный запах. Такое объяснение запаха
цианистой кислоты только подтверждает стереохимическую
теорию обоняния.
В данной главе приведены современные адсорбционные
теории, положенные в основу описания возможных механиз-
мов восприятия запахов. Показано, что в процессе обоняния
важную роль играет физическая адсорбция, форма и размеры
молекул. Однако до сих пор трудно сказать, будет ли связано
восприятие запахов с теплотой адсорбции, контактным потен-
циалом или поверхностным натяжением. Возможно также, что
все эти явления оказывают действие на возбуждение обоня-
тельного рецептора.
Большинство авторов подчеркивают селективность адсорб-
ции на рецептивных участках. Однако Дейвис и Тайлор счи-
тают мембраны рецепторов однородными, недифференциро-
ванными. В то же время доказательства, приведенные Эйму-
ром, Манкрифом и др., также достаточно убедительны для под-
тверждения существования различных рецептивных участков.
101
4.5. ИНФОРМАЦИОННАЯ ТЕОРИЯ ОБОНЯНИЯ
'у
Вопросам переработки информации в обонятельном ана-
лизаторе была посвящена статья под названием «Информа-
ционная теория обоняния» [41].
Человек обычно различает несколько тысяч запахов, пар-
фюмер — десятки тысяч. Количество органических соедине-
ний порядка 1 000 000, и все они имеют различные запахи-Для
восприятия такого количества запахов необходима большая
информационная емкость обонятельной системы. Известно,
что каждый нейрон может давать только on- и off-ответы.
Поэтому большая информационная емкость возможна при
объединении нескольких нейронов в группы. Например, от
трех нейронов могут быть получены восемь сигналов (23 = 8).
Если же мы допускаем, что различается по крайней мере
10000 запахов, тогда 2N ^10000, где N — число нейронов
в группе, тогда М=13. Можно предположить, что число мит-
ральных клеток (24), отходящих от каждого клубочка, яв-
ляется этим числом N. Тогда количество возможных запахов
224 ^ 16 000 000.
Известно также, что длительность нервного импульса рав-
на 1 мсек, время восстановления 15—100 мсек,т. е. время им-
пульса составляет 1/50 общего времени восстановления. Моле-
кулярные столкновения с обонятельными рецепторами носят
случайный характер. Чтобы согласовать появление ответов
всех обонятельных рецепторов, необходимо синхронизирую-
щее устройство. Зернистые клетки обонятельных луковиц мог-
ли бы выполнять эти функции. Авторы теории полагают, что
эти клетки получают информацию от рецепторов, задержива-
ют ее на определенное время и затем передают в центральную
нервную систему. Синхронизирующий сигнал может посту-
пать из мозга. Эдриан отметил, что при сильном возбуждении
у кролика импульсы от митральных клеток синхронизируются
с поверхностными волнами.
Опознание запахов возможно с помощью обратной связи
в обонятельной луковице (от митральной клетки к предше-
ствующему нервному уровню) и синапса с двойной связью.
Наличие пахучего вещества определяется цепочкой следую-
щих друг за другом импульсов от группы рецепторов. Под-
пороговая интенсивность запаха или его отсутствие вызывает
появление случайных импульсов. Если к этому синапсу по-
ступает цепочка импульсов от рецепторов и подобная после-
довательность импульсов по цепи обратной связи с задерж-
102
кой на несколько временных интервалов, то возникает на-
дежный ответ митральной клетки. Появление случайных им-
пульсов не вызывает такого ответа, информация в мозг не
передается.
Как уже , указывалось, в п. 1.1, у кролика имеется
50 000 000 первичных нейронов — обонятельных рецепторов,
45 000 — вторичных нейронов — митральных клеток.
Предполагая 24 вида рецепторов, получим для вос-
приятия каждого типа запаха 2 100 000 первичных и 1900
вторичных нейронов. Восприятие интенсивности должно быть
пропорциональным некоторому количеству возбуждаемых
в определенный момент времени нейронов, входящих в число
1900, и интенсивность стимула характеризуется степенью воз-
буждения этих 1900 идентичных групп нейронов. Шкала ин-
тенсивности может быть получена на основании изложенных
выше количественных соотношений групп нейронов и двух
законов: закона нормальных флуктуаций в статистических си-
стемах и логарифмического закона различения, закона Вебе-
ра—Фехнера. Закон, определяющий восприятие уже заметной
разницы в интенсивности, должен быть статистическим, так
как столкновение молекул с рецептором является случайным
процессом. Если наблюдается возбуждение М нейронов, ве-
личина флуктуации составляет Поэтому для восприятия
увеличения интенсивности раздражителя количество возбуж-
денных нейронов должно быть увеличено больше чем на
рХГ“г. е.
N2>Nt+V~N7, (38)
где ЛГ1 — количество возбужденных нейронов при одном
уровне интенсивности;
N2 — количество возбужденных нейтронов при другом
воспринимаемом уровне интенсивности.
Используя выражение (27) применительно ко вторичным
нейронам в луковице (от 1 до 1900 нейронов), получим 30 вос-
принимаемых уровней интенсивностей.
Для первичных нейронов (обонятельных рецепторов), по-
видимому, используется другой принцип функциональной ор-
ганизации, согласно известному закону Вебера—Фёхнера,
который может быть выражен в следующей форме:
-£-=№, (39)
103
где С — концентрация в воспринимаемом диапазоне интен-
сивностей;
Со — пороговая концентрация;
а —величина заметной разницы концентрации;
К—постоянная, равная отношению концентраций при
а=1. Если концентрации С2 и Ci близки по величине, можно
написать
(40)
Отсюда следует, что 2 100 000 (50 000 000:24) первичных
нейронов распределяются на 30 уровней интенсивности, что
диапазон воспринимаемых химических концентраций, т. е. от-
1
Клубочки
Вторичные
1
1,9 х 105
Первичные нейроны
нейроны
Уробни обнару- ’
женин
2100000 1900
Рис, 34. Модель структурной организации соедине-
ния первичных и вторичных обонятельных нейро-
нов.
ношение концентрации, соответствующей максимальному
ощущению, к пороговой составляет примерно 190 000 и что
для получения заметной разницы в обонятельном ощущении
необходимо увеличение концентрации на 52%, т. е. К =1,52.
Отношение числа первичных нейронов к числу вторичных
нейронов также должно изменяться в 190 000 раз. Число
190 000 — динамический диапазон системы.
104
Эти теоретические выводы согласуются с эксперименталь-
ными данными.
Предполагаемая схема восприятия интенсивности приве-
дена на рис. 34.
«Информационная теория запахов» дает возможность
представить себе обонятельный анализатор в виде специали-
зированной вычислительной системы. Однако авторы теории
предполагают существование 24 основных запахов, тогда как
в соответствии со стереохимической теорией, имеющей до-
вольно серьезное экспериментальное подтверждение, более
разумно говорить о семи основных запахах. Поэтому схема
восприятия запахов может быть несколько изменена в части
восприятия качества запаха.
8 Заказ № 7032
ГЛАВА V
МОДЕЛИРОВАНИЕ
ОБОНЯТЕЛЬНОГО АНАЛИЗАТОРА
Моделирование как метод познания имеет широкое при-
менение во многих областях современной науки. В бионике
моделирование используется для изучения биологических
процессов и систем для воспроизведения их на инженерном
уровне.
При моделировании совершается известная схематизация
и абстракция, выделяются и воспроизводятся в модели те
свойства и стороны исходного объекта, прототипа, которые
выступают для нас как существенные на данном этапе по-
знания. Поэтому необходимо выявление аналогии между
«системой объекта» и «системой модели», которые можно
осуществить на различных уровнях (результат, поведение
или функция, структура, материал).
В настоящее время для бионики особенно важно функци-
ональное моделирование. Некоторый специфический подход
к биологическим системам с точки зрения их функциональ-
ной, а не структурной организации назван «относительной
биологией» [65]. Это открывает новые возможности техниче-
ской реализации биологических систем. Появляется возмож-
ность конструирования и изучения близких аналогов реаль-
ных биологических систем с целью обогащения наших
знаний об этих системах и усовершенствования методов
воспроизведения важных биологических процессов в техни-
ческих приложениях.
Рассмотрим существующие функциональные модели пе-
риферической части органа обоняния, в некотором отноше-
нии соответствующие оригиналу и более доступные для изу-
чения, чем обонятельный анализатор.
106
Полезность рассматриваемых ниже Моделей будет опре-
деляться не их абсолютной правильностью или соответствием
обонятельному анализатору, а тем, что они могут дать более
рациональный подход к исследованию обоняния или помочь
в раскрытии истинной природы обоняния. Модель может
обеспечить основу для исследования многих аспектов обо-
няния.
В основу большинства из известных нам моделей обо-
нятельного анализатора были положены адсорбционные
теории.
Изменение контактного (потенциала под воздействием па-
хучих веществ было использовано Древниксом в его модели,
предназначенной для объективного измерения запаха 125].
В качестве адсорбентов были использованы сульфат железа,
борная кислота, альбумин, бихромат калия, хлористый нат-
рий и т. д. Адсорбенты наносились на неподвижные золотые
пластинки. Измерялась величина контактного потенциала при
воздействии разных пахучих веществ. Пары пахучих веществ
использованы при относительно высоком парциальном дав-
лении (100—300 мм рт. ст.), чтобы получить хорошее запол-
нение адсорбирующих поверхностей адсорбатом.
Величина контактного потенциала (определяется матери-
алом адсорбирующей поверхности и типом пахучего веще-
ства) изменялась в следующих пределах: под воздействием
ацетона на всех адсорбентах измеренное значение потенциала
было +360 мв, под действием тиофена на большинстве ад-
сорбентов от —100 до —180 мв. Для тиофена на адсорбиру-
ющей поверхности окиси серебра величина контактного по-
тенциала составляла +400 мв, а на поверхности борной
кислоты —40 мв.
Наибольшие изменения величины контактного потенциала
происходят в течение 30 сек, после соприкосновения с паху-
чим веществом.
Конструктивно описываемая модель состоит из четырех
неподвижных и одной подвижной золотых пластин, помещен-
ных под стеклянный колпак, в который может поступать па-
хучая смесь. Неподвижные пластины покрыты разными ад-
сорбентами, контактные потенциалы которых измеряются при
вращении над ними подвижной пластины, соединенной через
сопротивление на землю. Падение напряжения на сопротив-
лении является мерой величины поверхностного потенциала
и измеряется с помощью осциллографа (рис. 35).
8*
107
Относительное изменение контактного потенциала четы-
рех селективных поверхностей дает возможность судить о ка-
чественной и количественной характеристиках пахучих ве-
ществ. Недостатком этой модели является влияние влаги
и температуры пахучей смеси на результат измерения. Поэ-
тому в модели для создания пахучей смеси используется не
атмосферный воздух, а азот. Модель Древникса не адапти-
руется к пахучим веществам.
Рис. 35. Принципиальная схема мо-
дели Древникса.
А — неподвижные пластины; В — под-
вижная пластина; С — осциллограф; D—
привод.
Рис. 36. Термометрическое уст-
ройство модели Манкрифа (а),
мостовая схема (б).
Прибор для измерения и классификации запахов, разра-
ботанный Манкрифом, основан на использовании теплоты
адсорбции [54]. Чувствительным элементом прибора является
полупроводниковое термосопротивление, поверхность которо-
го покрыта различными адсорбирующими пленками, содер-
жащими активированный уголь, активированный алюминий,
силикагель и т. д. Если пленка не содержит даже высокоак-
тивных адсорбентов, подобных указанным выше, она сама по
себе имеет свойства адсорбента, которые являются достаточ-
ными для определения существования пахучих веществ в ок-
ружающем воздухе. Подобные пленки легче восстанавлива-
ются, освобождаются от пахучих веществ. Вследствие этого
в приборе были использованы пленки из ацетат-бутирата
целлюлозы, поливинилхлорида, вискозной целлюлозы, молоч-
ного казеина, резины, желатина, парафинового воска, аль-
гината натрия и кальция и др. Материал пленки не имеет
108
большого значения, так как процесс восприятия запахов яв-
ляется, по-видимому, физическим. Пленки должны быть
изготовлены из химически нереагирующих, стабильных во
времени и достаточно механически прочных веществ. Неко-
торые пленки обладают способностью селективной адсорбции
определенных пахучих веществ. Таким образом, открывается
возможность распознавания запахов с помощью набора
различных пленок.
Покрытое пленкой полупроводниковое термосопротивле-
ние (Л), находящееся в специальной камере, включалось
в четырехплечий мост (рис. 36). Пахучая смесыпредваритель-
но увлажняется, что соответствует тому увлажнению, которое
имеет место в носовой полости и устраняет чувствительность
к воде и продувается сквозь камеру с термосопротивлением.
При этом наблюдается увеличение температуры адсорбента
и, следовательно, изменение сопротивления полупроводнико-
вого термосопротивления.
Для исключения влияния случайных изменений темпера-
туры окружающей среды в смежное плечо (С) моста вклю-
чается вместо фиксированного сопротивления термосопротив-
ление, не покрытое адсорбентом. Это термосопротивление
монтируется рядом с основным рабочим (Л), и оба находятся
всегда в одинаковых температурных условиях. Для увеличе-
ния чувствительности устройства можно включить в схему
еще два термосопротивления: одно, покрытое адсорбентом,
в плечо D, второе для температурной компенсации в плечо В.
Выходной сигнал моста подается на вход усилителя постов
янногс тока с коэффициентом усиления- 1000.- Усиленный
сигнал измеряется милли- или микроамперметром й Одновре-
менно подается на записывающее устройство. ’ В качестве
пахучих веществ использовали дистиллированную воду, аце-
тон в различных концентрациях, метанол, этанол, н-пропа-
нол, бутанол, аммиак, амиловый спирт, амилацетат, бензол,
хлороформ и многие другие. Схема этого устройства приве-
дена на рис. 37. Показания прибора в миллиамперах при про-
текании пахучей смеси через камеру термосопротивлений со
скоростью 1 л!мин даны в табл. 16.
При обтекании пахучей смесью блока термосопротивле-
ний стрелка выходного прибора отклоняется на несколько се-
кунд до определенного положения, а затем медленно возвра-
щается к своему первоначальному положению (рис/ 38).
Для модели свойственно явление «усталости»., Е.сл.и же пре-
рвать поток пахучего вещества к прибору и заменить его
109
чистым воздухом, стрелка будет возвращаться к исходному
положению почти мгновенно. Для очень тонких пленок
(2-45 мк) явление усталости наступает более быстро. Обоня-
Рис. 37. Принципиальная схема модели.
а — колба с раствором пахучего вещества; б — вход пахучей смеси;
в — увлажнитель; г — перфорированная металлическая трубка; д —
блок термосопротивлений; е — расходомер; ж — устройство для сгла-
живания пульсации; з — воздушный насос.
тельная усталость может начинаться по истечении I сек
с момента возникновения первой реакции прибора на запах.
Более толстые пленки подвергаются действию усталости спу-
стя примерно 0,5 мин после появления первой реакции.
Таблица 16
Пахучее вещество Полихлорвиниловая пленка Пленка из ацетата целлюлозы
Дистиллированная во- 0,02 0,03
да
Ацетон ь . . , „ g Метанол . . . . . 9,3 8,0
4,2 2,2
Этанол , . . . й . 1,8 0,63
н-Пропанол . g . в 1,6 0,52
Изопропанол . . . 2,3 0,41
н-Бутанол . . . . 4 0,52 0,23
Изобутанол » . . . 0,55 0,12
втор-Бутанол . . 9 * 1,2 0,18
трест-Бута нол . 2,3 -0,84
Амиловый спирт . . Аммиак (25%-ный 0,30 0,25
водный раствор) . Триэтиламин (5%-ный 2,0 7,5
водный раствор) . 1,2 —0,12
Амилацетат . . Б • 1,2 0,05
Метилен хлорид . . . 18 11
Бутировая кислота . —2,5 —2,0
Бензол ...... 7,0 0,08
Толуол в 2,8 0,04
110
Результаты экспериментов имеют хорошую повторяемость,
например, при многократном воздействии хлороформом, пов-*
теряющемся через 1 мин, показания выходного прибора были
79, 82, 82, 81, 84 ма, а при воздействии ацетоном — 50, 49, 48,
48, 48 ма. Прибор достаточно чувствителен для восприятия
запахов некоторых цветов (розы, бузины и др.). Показания
прибора определяются концентрацией пахучего вещества
и его качеством (рис. 39). Время, необходимое для достиже-
ния максимального показания выходного прибора, не зави-
Рис. 38. Кривые, полученные при разных концентрациях аце
тона в воде.
сит от концентрации вещества. Время полной адаптации
увеличивается с уменьшением концентрации пахучего веще-
ства (рис. 40).
Из всего, изложенного выше, следует, что использование
адсорбента в непосредственном контакте с термометрическим
устройством любого вида может служить моделью обонятель-
ного эпителия. Сходство модели с оригиналом еще больше
увеличивается, если используемое термометрическое устрой-
ство преобразует тепловые изменения в электрические. Не-
смотря на это, модель остается грубой.
Сравним чувствительность модели с чувствительностью
обонятельного анализатора. Соотношение между энергией
адсорбции и температурными изменениями определяет чув-
ствительность прибора к пахучим веществам. Приближенные
подсчеты показывают, что отклонение в 1 ма выходного при-
бора вызывается адсорбцией 10“9 г адсорбируемого вещест-
ва. Наинизшие пороговые значения меркаптана для живых
организмов составляют 1010 молекул или 10~12 г. Вещества,
подобные ионону и мускусу, ощущаются живыми организма-
111
ми в весьма малых количествах. Однако большая часть запа-
хов воспринимается ими только в больших концентрациях.
Минимальная концентрация ацетона в воде, ощущаемая
в виде запаха, составляет 0,03%. Воздух в непосредственной
близости от этой воды содержит ацетона около 2.10~7 г1см\
По отношению к ацетону прибор чувствительнее носа, но
в отношении других пахучих веществ с очень низкой поро-
Рис. 40. Зависимость показаний выходного
прибора во времени после преобразования
кривых в прямоугольную систему коор-
динат.
а — для 100% ацетона; б — для раствора аце-
тона в воде.
1мко
Рис. 39. Зависимость пока-
заний прибора от концент-
рации пахучих веществ:
а — метиленхлоцида в раство-
ре пропиленгликоля; б — аце-
тона в воде.
говой концентрацией (мускус, ванилин и др.) прибор имеет
более низкую чувствительность. Кривые, полученные с по-
мощью прибора, имеют большое сходство с кривыми электри-
ческой активности обонятельного эпителия лягушки (см.
рис. 8). Кроме этого могут быть отмечены следующие сход-
ные черты прибора с обонятельным анализатором человека:
1) немедленный ответ на .пахучее вещество;
2) ответ сразу же исчезает с удалением запаха;
3) модель достаточно чувствительна, к запахам некото-
рых веществ чувствительнее человеческого носа;
4) необходимо движение воздуха над воспринимающей
поверхностью;
5) прибор «устает» и должен отдыхать перед последую-
щими опытами;
6) модель быстрее адаптируется к сильным запахам, чем
к слабым;
7) модель позволяет отличать запахи различных веществ;
8) модель показывает ограничение реакции на сильные
запахи, когда дальнейшее увеличение стимула не дает от-
вета.
112
Следует упомянуть и о так называемом «электрическом
носе», созданном Розано и Скепсом [38,39]. В этом приборе
в качестве преобразователя используется электролитическая
ячейка, в которой молекулы пахучих веществ адсорбируются
и окисляются на границе раздела жидкость—газ. Таким об-
разом, под воздействием пахучего вещества изменяется ве-
личина электрического тока. Эти изменения можно исполь-
зовать для опознания определенных запахов.
Конструктивно чувствительный элемент прибора оформлен
в виде маленького стакана. Его чувствительность в 100 раз
выше чувствительности человеческого носа к некоторым спир-
там (например, этиловому).
Известна также модель органа обоняния («электронная
ищейка»), работающая на принципе избирательного погло-
щения ультрафиолетового излучения [61]. Специальная ульт-
рафиолетовая лампа сфокусирована на чувствительной труб-
ке детектора. При прохождении газа между лампой и детек-
тором часть ультрафиолетового излучения поглощается.
В устройстве предусмотрена сигнализация, которая показы-
вает достижение определенной концентрации газа.
Модель обладает высокой чувствительностью, сравнимой
с чувствительностью носа собаки. Определяемое количество
пахучего вещества составляет 0,00001%.
Это устройство уже нашло применение на фабриках хи-
мической чистки для обнаружения токсичного газа, перхлор-
этилена. Оно может также обнаруживать запахи бензина,
красок, лаков, аммиака, кислот и т. д. Авторы считают воз-
можности модели неограниченными.
Модели обонятельного анализатора могут найти широкое
применение в химической, пищевой и парфюмерной промыш-
ленности, медицинской диагностике, при дезодоризации кон-
диционированного воздуха, для определения места утечки
в газопроводах и др.
В заключение отметим, что работы последних лет дают
возможность объективной оценки качества и интенсивности
запахов и даже некоторое представление о природе запахов,
хотя до сих пор механизм восприятия запаха окончательно
не выяснен.
Обонятельный эпителий может быть представлен как по-
верхность с миллионами внедренных в нее воспринимающих
элементов-преобразователей, а обонятельный анализатор
в целом как система, предназначенная для пространственно-
временного анализа. Большие аналитические способности
11S
этой системы увеличиваются вследствие совершенства и диф-
ференциации различных типов преобразователей (рецеп-
торов).
Дальнейшие работы, направленные на техническое вос-
произведение обонятельного анализатора, должны основы-
ваться, по-видимому, на серьезных биологических исследова-
ниях, включая биологическое, главным образом функциональ-
ное, моделирование обонятельного эпителия в первую очередь,
сравнительном анализе «системы объекта» с «системой
модели», усовершенствовании модели, а также изучении
физических и химических свойств пахучих веществ, в част-
ности, форм и размеров их молекул и селективной адсорби-
руемости на некоторых адсорбентах.
ЛИТЕРАТУРА
L М- Брейзье. Электрическая активность нервной системы. М., 1955.
2. А. И. Бронштейн. Вкус и обоняние. Изд-во АН СССР, 1950.
3< А. Л. Бызов, Г. И. Флеров а. Электрофизиологическое исследование
обонятельного эпителия лягушки. Биофизика, 1964, т. IX, вып. 2.
4. Я« А. Винников, Л. А. Т и т о в а. Морфология органа обоняния. Мед-
гиз, 1957.
5. В. Д. Глезер, И. И. Ц у к к е р м а и. Информация и зрение. М., Изд-во
АН СССР, .1961.
6. Живая клетка. М., Изд-во иностр, лит., 1962.
7, Н. А. Ж у х о в и ц к и й, А. А. Ш в а р ц м а н. Физическая химия. Метал-
лургиздат, 111963.
,8< Г. К л а у с. Кибернетика и философия. М., Изд-во иностр, лит., 1963.
9. М. В. С е р д о б о в. Теплота адсорбции некоторых веществ с различной
электронной структурой на адсорбентах разной природы. Автореф. канд.
дисс., М., 1964.
40. Л. В< Смирнов. К вопросу о моделировании биологических явлений.
— Сб. «Применение математических методов в биологии», № 2, Л.,
Изд-во ЛГУ, 4963.
11. Б. Н. Шванвич. Курс общей энтомологии. Изд-во «Советская нау-
ка», 1949.
12. В. А. Штоф ф. О роли моделей в познании. Л., Изд-во ЛГУ, 1963.
13. Л. Б. Чичибабин. Основные начала органической химии. М., Гос-
х 1миздат, (1963.
14. Г. D. Adrian. The activity of the mammalian olfactory bulb. EEG Clin
Neurophysiol., 2, 1950.
15. E. D. Adrian. Sensory messages and sensation. Acta Physiol., Scandi-
navica, 29, 1953.
16. E. D. Adrian. Potential oscillation in the olfactory organ. J. Physiol.,
128, 1955.
17. E. D. Adrian. The action of the mammalian olfactory organ. J. Laryn-
golog. otol., 70, 1, 1956.
18. E. D. Adrian. Transmission on information from the olfactory organ.
Проблемы современной физиологии нервной и мышечной систем.
Тбилиси, 1956.
19. Е. D. Adrian. Electrical oscillations recorded from the olfactory
organ. J, Physiol., 136, 2, 1957.
20. J. E. A mo ore. Stereochemical theory of olfaction. Nature, 198,
4877, 1963.
115
21 J. E. A m oo re, B. Fu liman. Stereochemical theory of olfaction-
Nature, 199, 4896, 1963.
22. J. E. A mo ore, J. W. Johnston, M. Rubin. The Stereochemical
Theory of Odor, Scientific American, February, 1964.
23. H. Athenstaedt. Die ferroelektrischen und piezoelektrischen Eigen-
schaften der Organismen. Die Naturwissenschaften, vol. 47 (19), 1960.
24. H. Athenstaedt. Ferroelektrische und piezoelektrische Eigenschaften
biologisch bedeutsamer Stoffe. Die Naturwissenschaften, vol. 48 (13), 1961.
25. A. R. F. lays groundwork for synthetic nose. Chemical and engineering
news, April 3. 1961.
26. L. H. Beck, W. R. Miles. Infrared absorbtion in field studies of
olfaction in honeybees. Proc, of the National Academy of Sciences, 35, 292,
1949.
27. J. H. de Boer. The Dynamical Character of Adsorbtion. Clarendon
Press, 34, 1953.
28. M. H. В г i ggs, R. B. Duncan. Odour receptors. Nature, vol. 191
4795, September 23, 1961.
29. M. H. Bri ggs, R. B. Duncan. A letter to the editor. Nature, vol.
195, 4848, September, 1962.
30. K. Buijs, C. J. H. Schutte, F. Vester. Absens of correlation
between odour and molecular vibration. Nature, vol. 192, 4804, November
25, 1961.
31. S. T. Davies, F. U. T у 1 о r. A model system for olfactory mem-
brane, Nature, vol. 174, 4432, 1954.
32. S. T. Davies, F. U. T у 1 о г. Olfactory thresholds: A test of new
theory. The Perfumery and Essential Oil Rec., 1, 1955.
33. V. G. D e t h i e r. The physiology of olfaction in insects. Ann N. Y.
Acad. Sci., vol. 58, 2, 1954.
34. Andrew Dravnieks. Possible mechanisme of olfaction. Nature, vol
194, 4825, April 21, 1962.
35. G. M. Dyson. Scientific basis of odour. Chemistry and Industry, 57,
647, 1938.
36. G. M. Dyson. A letter to the editor. Nature, 4409, May 1, 1954.
37. S. R. Eaton, S. E. Christian, S. A. Campbell. Surface phenome-
na related to odor measurements. Ann. N. Y. Acad. Sci., vol. 58, 2, 1954.
38. Electric nose sensitive. Science News Letters, vol. 84, 12, 180, 1963.
39. Facts on smell machines aired by scientists. Science News Letters, 30,
1963.
40. M. A. Gerebzoff. Olfaction. J. de Physiol., 45, 1953.
41. T. W. Goodwin. The comparative biochemistry of the carotenoids.
Chapman and Hall, London, 1952.
42. R. M. Hainer, A. E. E m s 1 i e, A. Jacobson. Information theory
of olfaction. Ann. N. Y. Acad. Sci., vol. 58, 2, 1954.
43. Handbook of physiology. Washington, 1959.
44. I. I. Johnston. Infrared loss theory of olfaction. Untenable Physio-
logical Zoology, 26, 266, 1953.
45. N. Lindgren. Bionics. Electronics, 6, 7, 9, 11, 1962.
46. A. W. M i d d 1 e t о n. Smell—the physical sense. Perfumery and Essen-
tial Oil Rec., June, 1956.
47. A. W. M i d d 1 e t о n. Smell—the physical sense. Perfumery and Essen-
tial Oil Rec., July, 1956.
48. R. W. Mon crieff. The chemical senses. Leonard Hill, London, 195L
116
49. R. W. Moncrieff. The characterization of odours. The Journa lot
Physiology, vol. 125, 3, 1954.
50. R. W. M о n с г i e f f. The sorptive properties of the olfactory membrane.
The Journal of Physiology, vol. 130, 3, 1955.
51. R. W. Moncrieff. Olfactory adaptation and odour likeness. The
Journal of Physiology, vol. 133, 2, 1956.
52. R. W. Moncrieff. Depth encephalography a technique for recording
human olfactory responses. Perfumery and Essential Oil Rec., 1956.
53. R. W. Moncrieff. Olfactory adaptation and odor - intensity. The
American Journal of Psychology, vol. LXX, No. 1, March, 1957.
54. R. W. Moncrieff. An instrument for measuring and classifying odo-
urs. Journal of applied physiology, vol. 16, 4, July, 1961.
55. R. W. Moncrieff. Odor compensation. Drug and Cosmetic Industry,
December, 1961.
56. H. Morita, S. Jamas hita. Receptor potentials recorded from
sensilla basiconica on the antenna of the silkworm larvae, bombyx mori.
The Journal of the Experimental Biology, vol. 38, 4, 1961.
57. D. G. M о u 11 о n. A letter to the editor. Nature, vol. 195, 4848, Septem-
ber, 1962.
58. L. S. Mullins. Olfaction. Ann. N. Y. Acad. Sci., vol. 62, October 27,
1955.
59. L. H. Narodny. The resonance of smell. The Perfumery and Essen-
tial Oil Rec., vol. 48, 1, 1957.
60. L. H. Narodny. The vector analysis of odour. The Perfumery and
Essential Oil Rec., 1, January, 1957.
61. New „Electronic Bloodhound" outperforms the real dog. ISA Journal,
vol. 11, 4, April, 1964.
62. D. О ttoson. Analysis of the electrical activity of the olfactory epithe-
lium. Acta Physiol. Scand., 35, Suppl. 122, 1954.
63. W. W. Peterson, I. G. Birdsall, W. C. Fox. The theory of
signal detectability. Trans. IRE, PGIT—4, 171, 1954.
64. C. L. Prosser, F. A. Brown. Comparative animal physiology.
Philadelphia, 1961.
65. R. Rosen. Relational biology and bionics. IEEE, Transactions on Mili-
tary Electronics, vol. MIL—7, April—July, 2—3, 1963.
66. T. Shibuya, S. Shibuya. Olfactory epithelium: Unitary responses
in the tortoise. Science, 140, 3566, 1963.
67. E. Skramlik. Uber die zur minimalen Erregung menschlichen
Geruchs—bzw. Geschmackssinnes notwendigen Molekulmengen. Pflug.
Arch., 249, 702, 1948.
68. Solid/Liquid interface (washing, etc) and cel 1/water interface. Proceedings
of the Second International Congress of Surface Activity, IV, London,
1957.
69. M. Stoll. Facts old and new concerning relationships between molecu-
lar structure and odour. Molecular structure and organoleptic quality, 1957.
70. S. F. Takagi, T. Shibuya. Electrical activity of lower olfactory
nervous system of toad. Electrical activity of single cells, Tokyo, Japan,
1960.
71. S. F. Takagi, T. Shibuya. The electrical activity of the olfactory epithe-
lium studied with micro-and macro-electrodes. The Jap. J. Physiol.,
vol. 10, 4, 1960.
72, S. F. Takagi, T. Shibuya. Potential oscillations in the lower
olfactory pathway of the toad. Nature, 186, 724, 1960.
117
73. S. F. Takagi, T. Shibuya. The potential oscillations observed in
the olfactory epithelium, nerve and bulb of the toad and frog. The Jap. J.
Physiol., vol. 10, 5, 1960.
74. P. W. Tanner, J. A. Swets. The human use of information,. 1.
Signal detection for the case of the signal known exactly. Trans. IRE
PGIT—4, 213, 1954.
75. P. W. Ta n n e r, J. A. Swets. Signal detection for the case of an
unknown signal parameter. Trans. IRE, PGIT--4, 222, 1954.
76. Textbook of physiology and biochemistry. George H. Bell, Norman David-
son, Harald Scarborouch, Robert Garry, Edinburgh, London, 1961.
77. H. W. Thompson. Some comments on theories of smell. Molecular
structure and organoleptic quality, 1957.
78. Don Tucker. Physical variables in the olfactory stimulation processk.
The Journal of General Physiology, vol. 46, 3, January, 1963.
79. Turk Amos. Odorous atmospheric gases and vapors: properties, collec-
tion and analysis. Ann. N. Y. Acad. Sci., vol. 58, 2, 1954.
80. M. Uruha, S. Twase. Olfactory bulb response of rabbit. Science,
vol. 133, 3456, March 24, 1961.
81. Hesel de Vries, Minz Stuiver. Sensory communication. 1961.
82. R. R. Walsh. Single cell spike activity in the olfactory bulb. Amer.
J. Physiol., vol. 186, 2, 1956.
83. R. H. Wright. Odour and chemical constitution. Nature, 4409, May, 1954.
84. R. H. Wright. Odour and molecular vibration. Nature, vol. 190,4781,
June, 1961.
85. R. H. Wright. Odour of optical isomers. Nature, vol. 198, 4882, May
25, 1963.
ОГЛАВЛЕНИЕ
Введение. . • t а . . . ....................., . . . . 3
Глава I. Краткие сведения о морфологии и физиологии обо-
нятельного анализатора .............................. 5
1-1. Строение обонятельного анализатора млекопита-
ющих и человека .................................... 5
1.2 Строение органа обоняния рыб . . ...............8
1.3 Органы обоняния насекомых ... , ... . 9
1.4 Электрофизиология органа обоняния.................11
1.5 Некоторые особенности обонятельного анализатора 18
Глава II. Чувствительность органов обоняния................20
2.1 Абсолютный порог обоняния . ......................20
2.2 Измерение обонятельных порогов...................27
2.3 Различительные пороги............................29
2.4 Статистическое обнаружение запахов...............30
Глава III. Пахучие вещества и их свойства..................34
3.1 Физические свойства...............................34
3.2 Соотношение между запахом и молекулярной струк-
турой пахучих веществ...............................40
Глава IV. Основные классификации запахов и теории обо-
няния....................................................48
4.1 Классификация запахов по субъективному признаку 49
4.2 Колебательные теории и классификация запахов . 51
4.3 Химические теории обоняния..................61
4.4 Адсорбционные теории обоняния................70
4.5 Информационная теория обоняния.............102
Глава V. Моделирование обонятельного анализатора . . . 106
Литература . . , . . . , . в..............................115
Р. М. Мазитова, В. Н. Охотская, Б. И. Пучкин.
ОБОНЯНИЕ
* И ЕГО МОДЕЛИРОВАНИЙ
Редактор Л. В. Шалина
Обложка художника В. И. Селихова
Художественный редактор В. Г. Бурыкин
Технический редактор Т. К. Овчинникова
Корректоры- В. Г. Постникова, Л. И. Коршунова
Сдано в набор 3 июля 1965 г. Подписано в печать 10 декабря 1965 г. MH071S5
Бумага 60X^90/16, 7,5 печ. л., 6,4 уч.-изд. л. Тираж 2300.
Издательство «Наука». Сибирское отделение, Новосибирск, Советская, 20.
Типография изд-ва «Омская правда». Омск-56, проспект Маркса, 39. Заказ 703
Цена 45 коп.
ЗАМЕЧЕННЫЕ ОПЕЧАТКИ
Стра- I ница I Строка Напечатано Следует читать
7 Подпись к рис. 3 . человека кролика
24 12 сверху скорость расход
27 15 сверху скорости расходе
27 рис. 15 скорость расход
31 1 снизу Ze ZQ
36 15 снизу 1,2 • 10 ю 1,2 • 10-ь
36 14 снизу 2.10 2.10-5
52 19 снизу энантиометров энантиомеров
58 6 снизу со скоростью с расходом
71 23 сверху вляние влияние
78 1 снизу (в сноске) —SH + HCHO+HS->S— —сн2 -SH+HCHO+HS+C- —CH2-S+H2O
84 2 снизу 1010- 10-ю
88 21 снизу 10* Е > 10*
88 18 снизу органическими протеиновыми
94 17 сверху 2,8 • Ю-о 2,8 • 10-ю
НО 11 снизу в табл. 16 трест-бутанол трет-бутанол
117 1 сверху The Journa lot The"journal of
118 10 снизу S. Twase S. Iwase
1966-1823
45 коп
АКЛ ДЕ МИЯ
НАУМ
. Лучкин
СССР
СИБИРСКОЕ
О1Д1ПЕНИЕ
1966
I л
1 V
обоняние и его
MoqeAupobciHue