



























































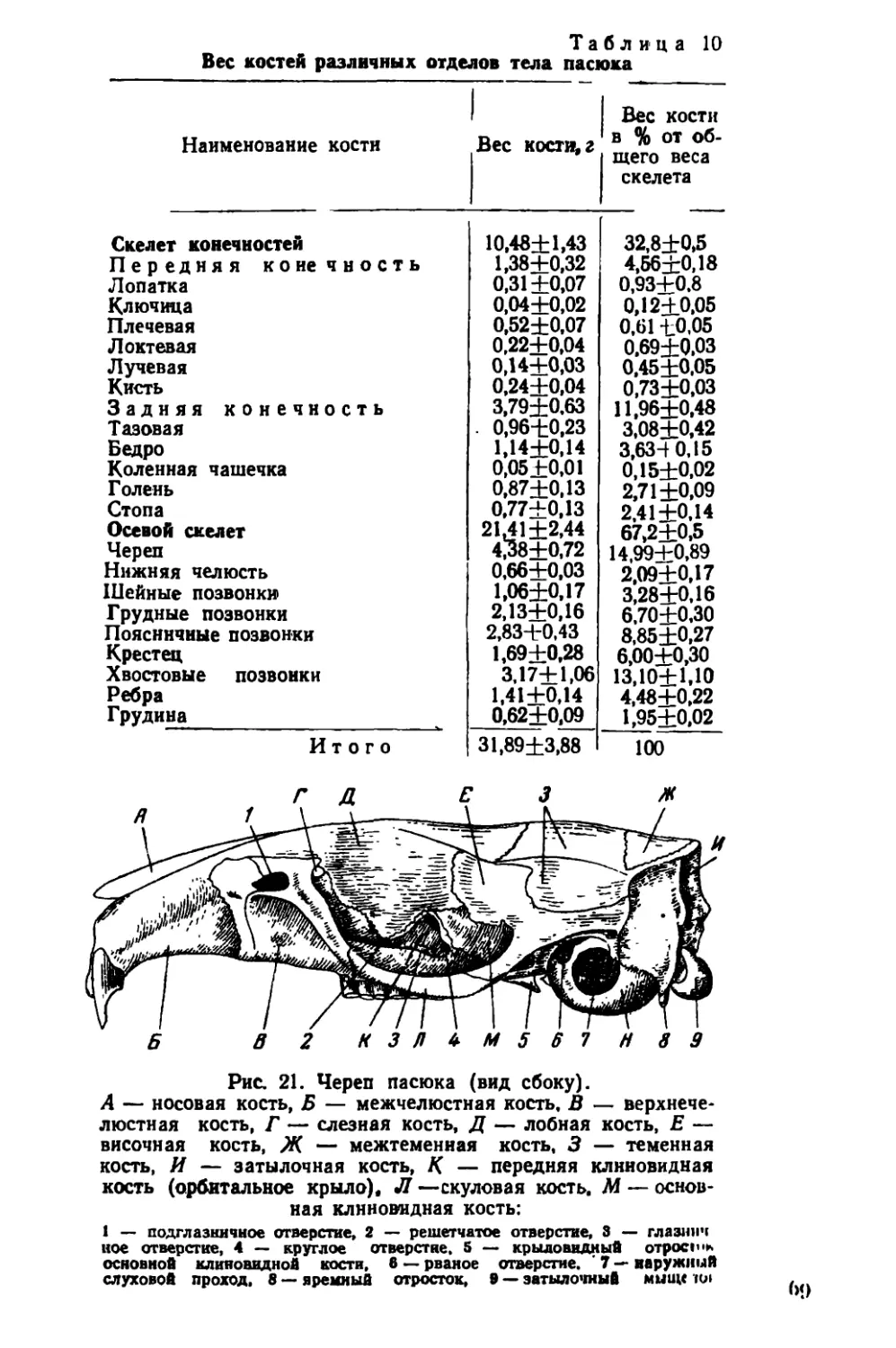


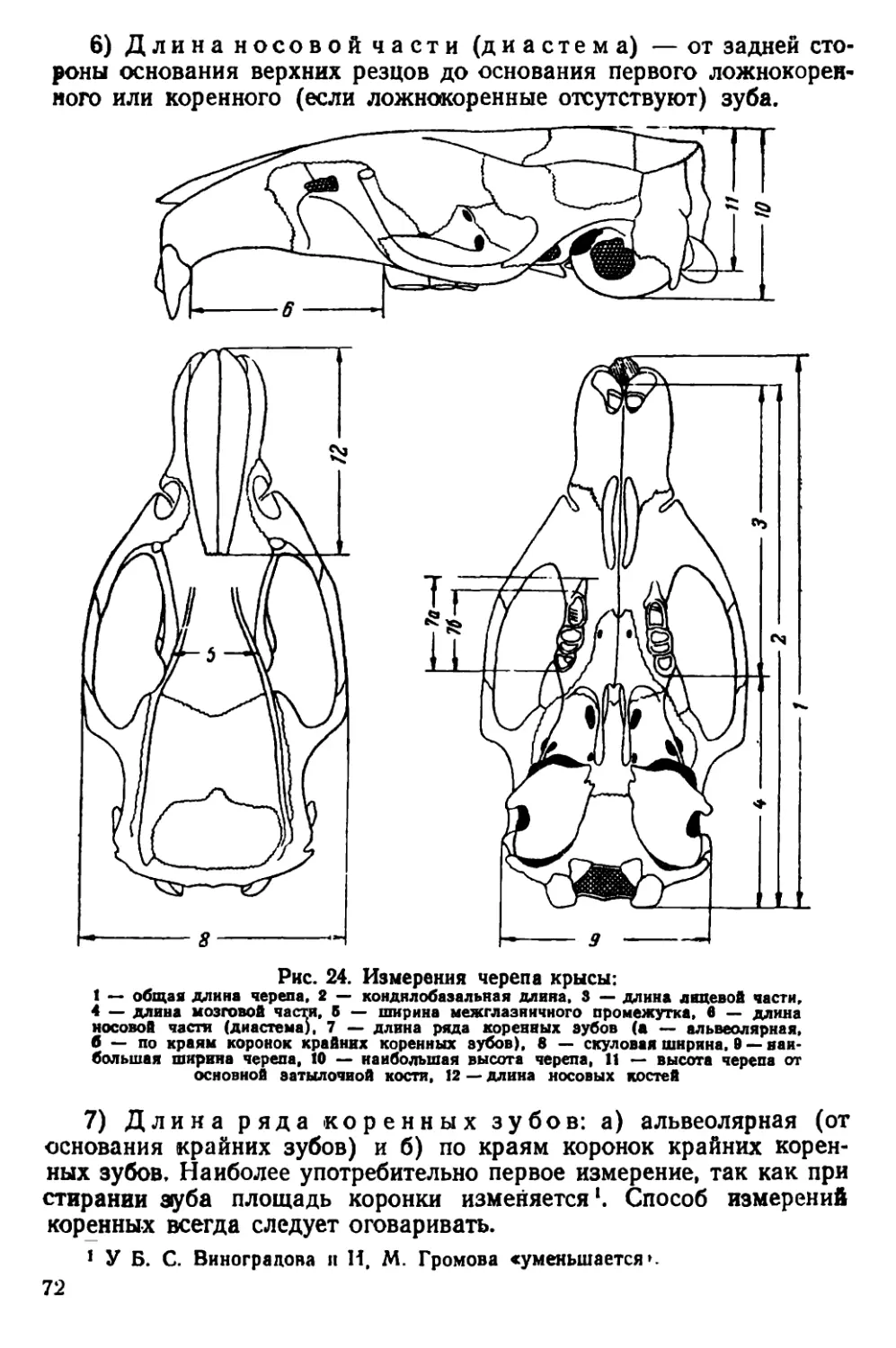
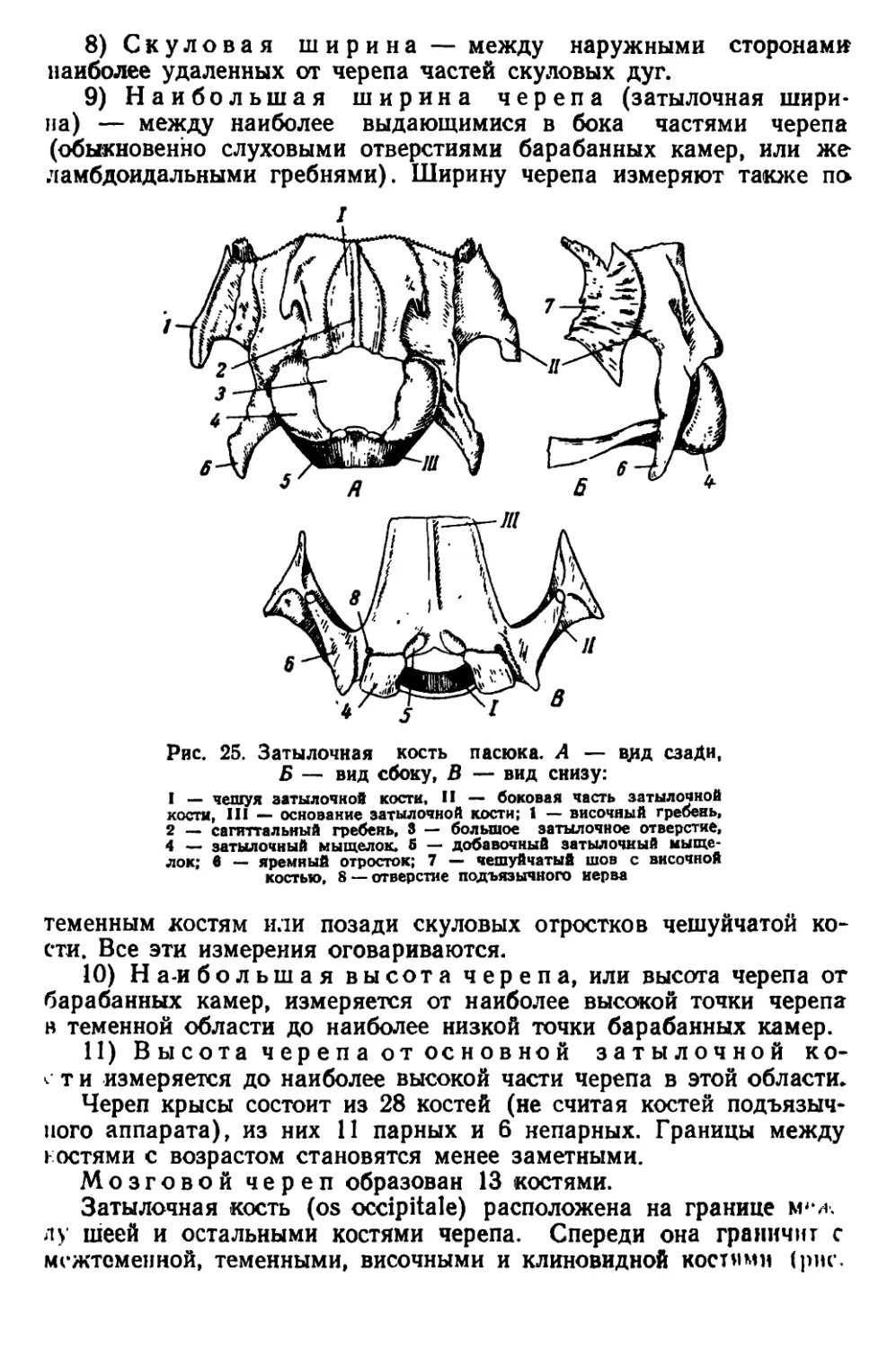
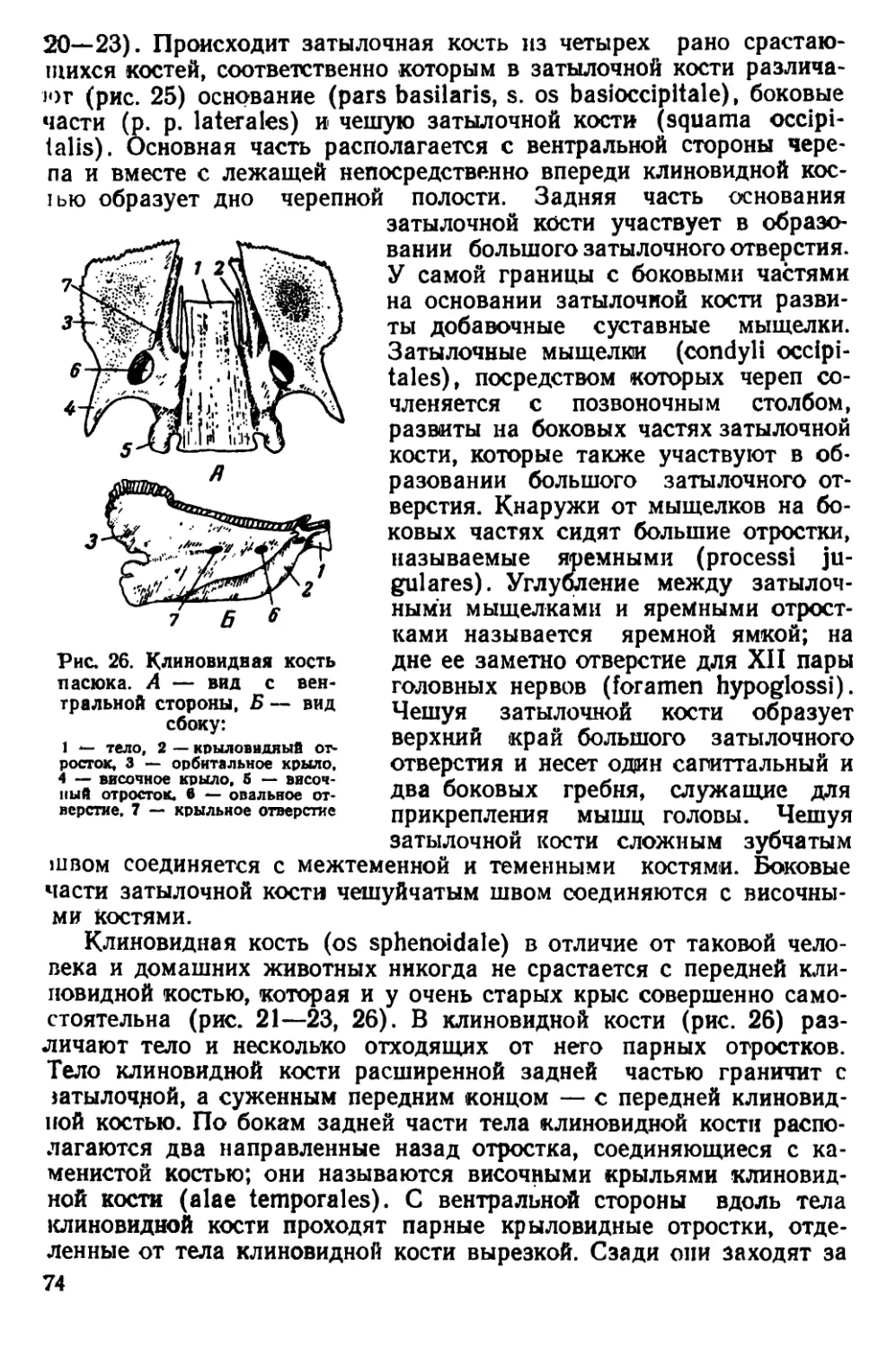
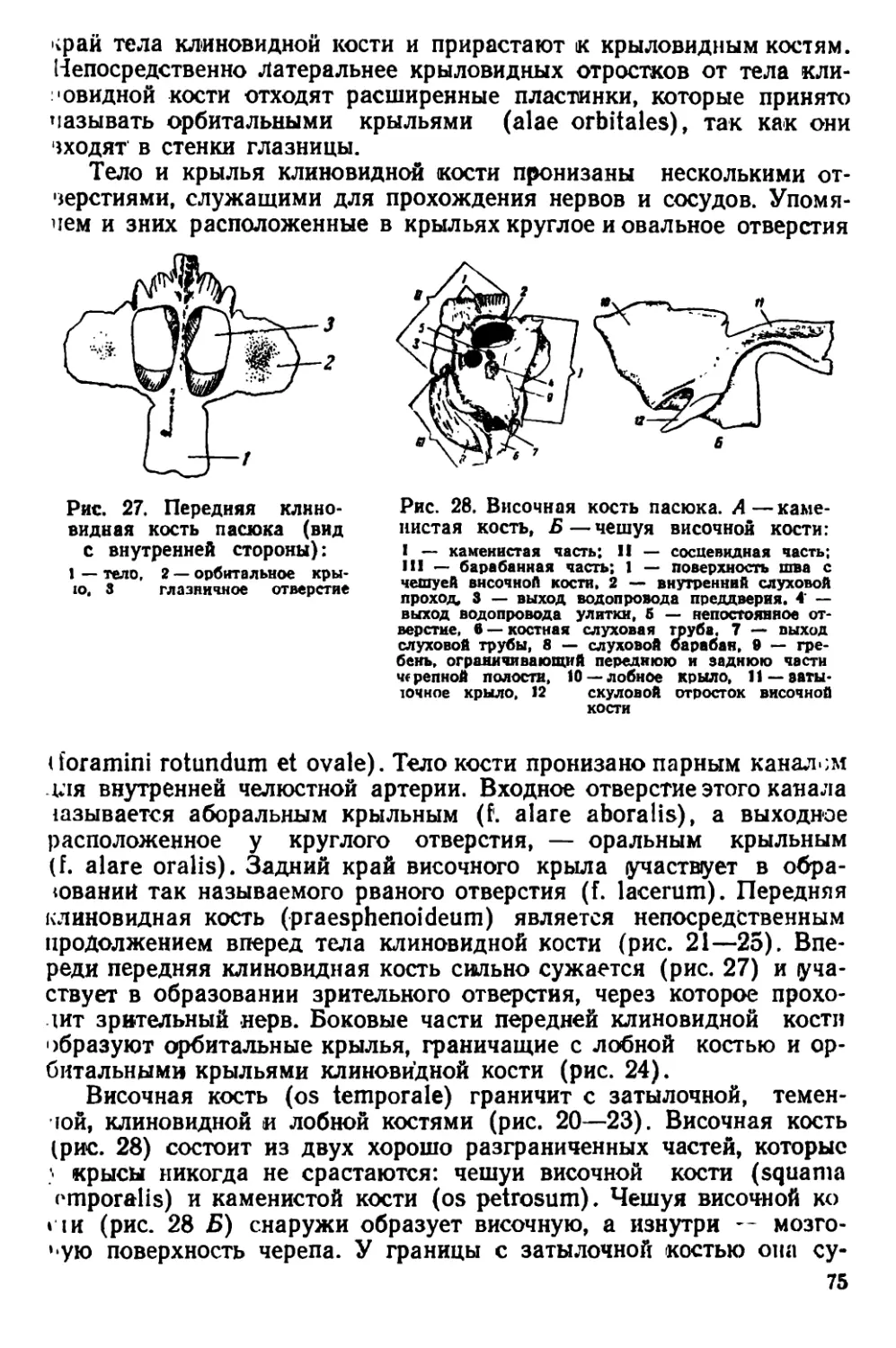
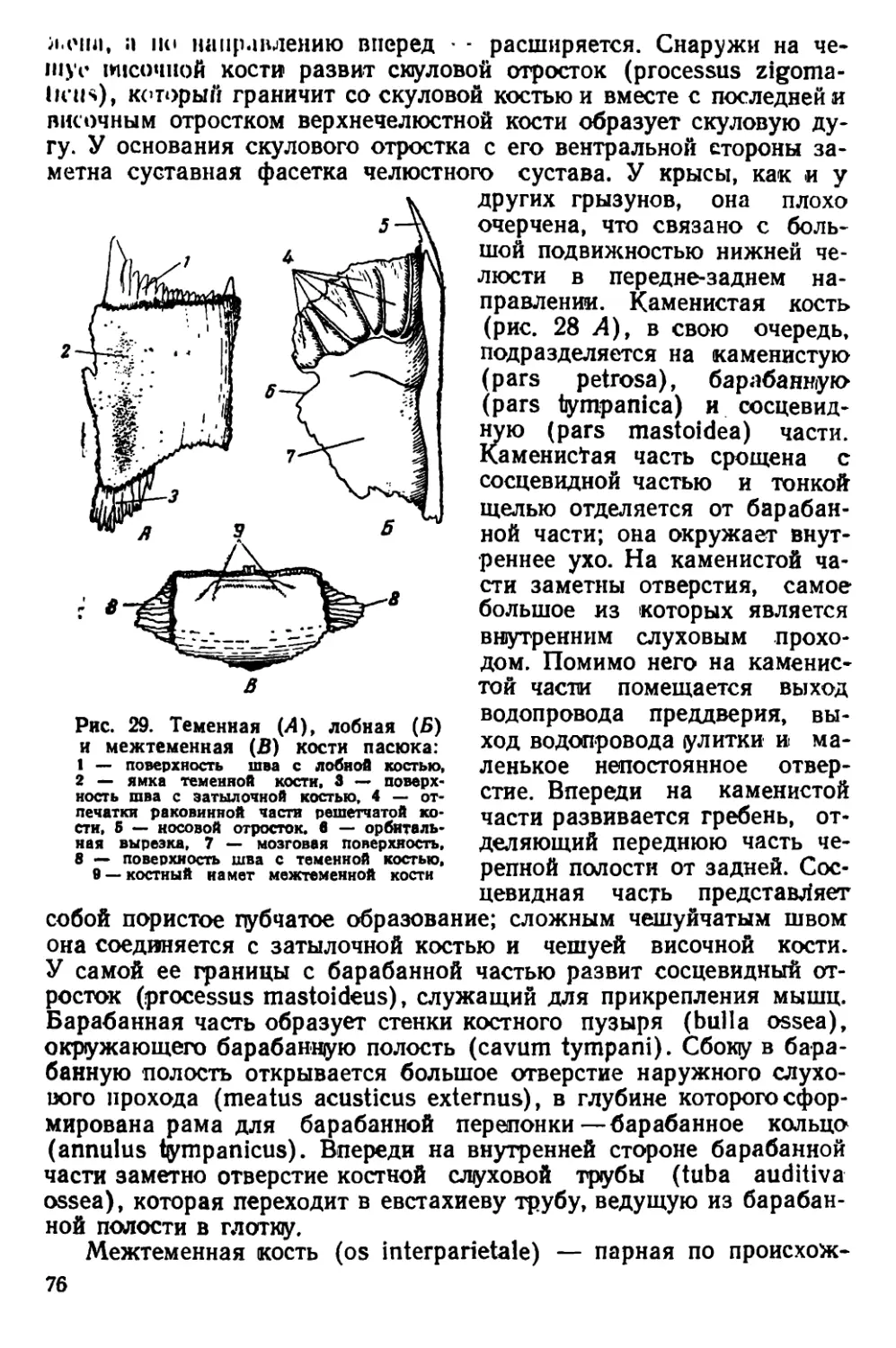
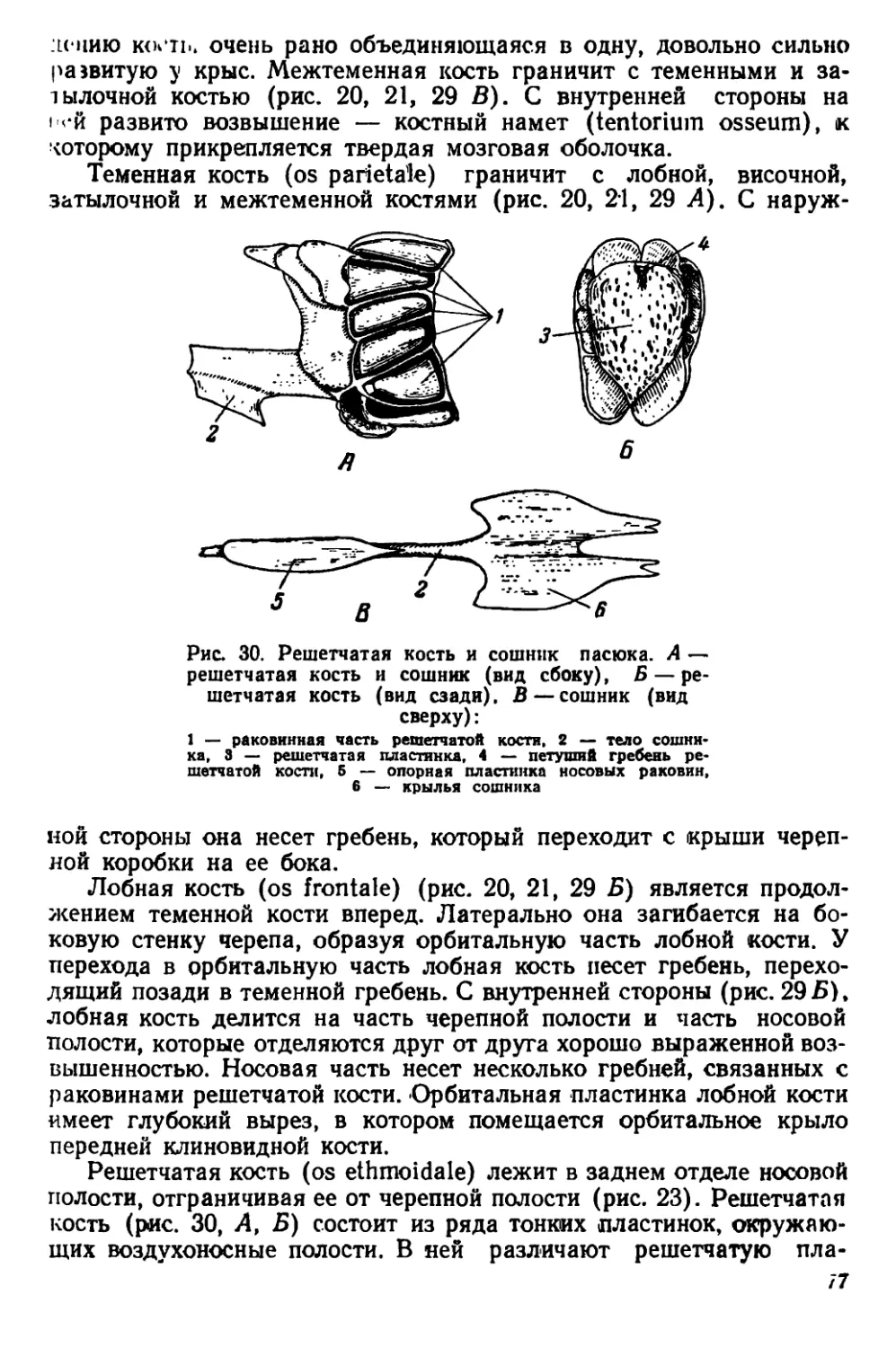
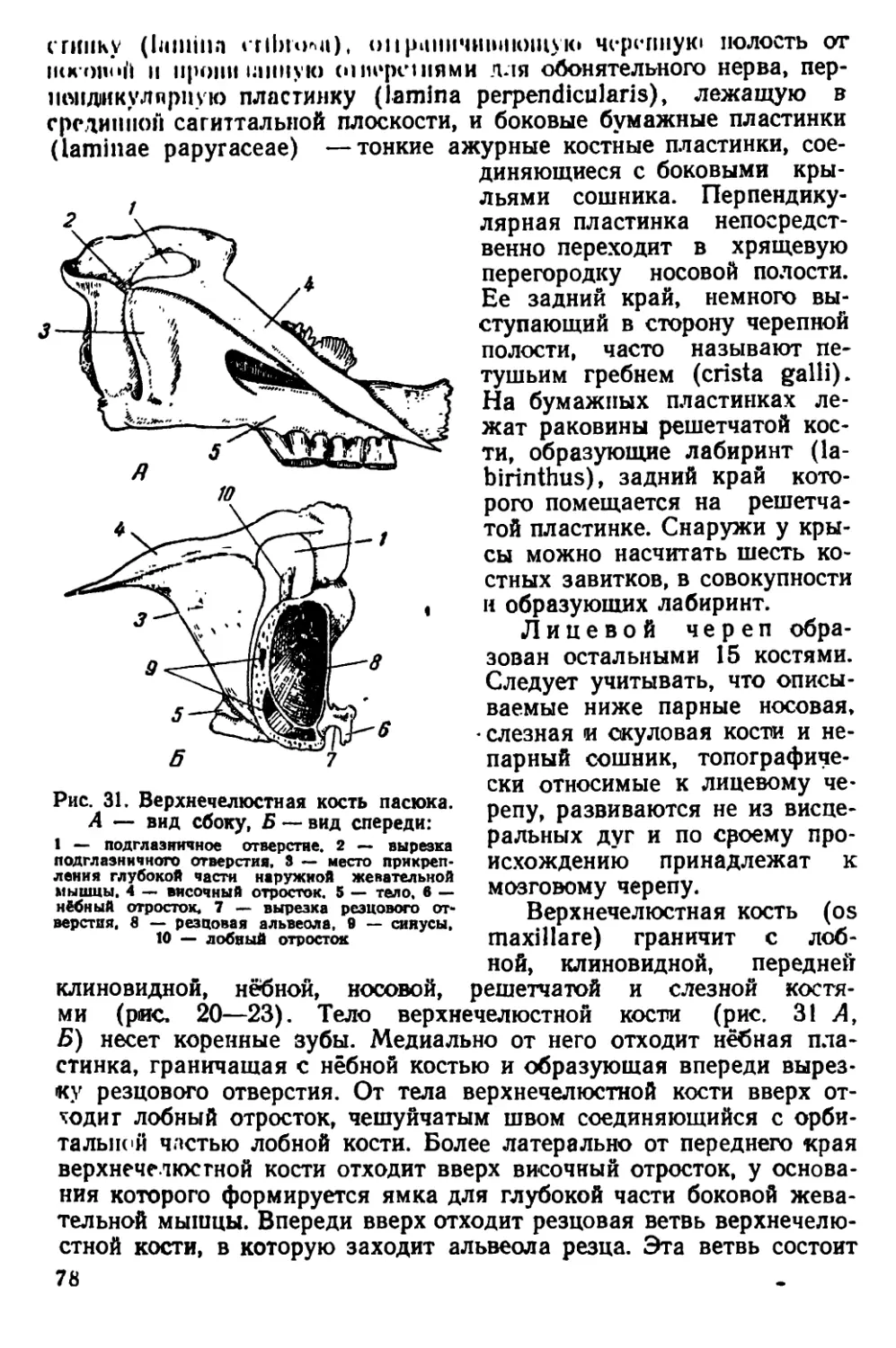













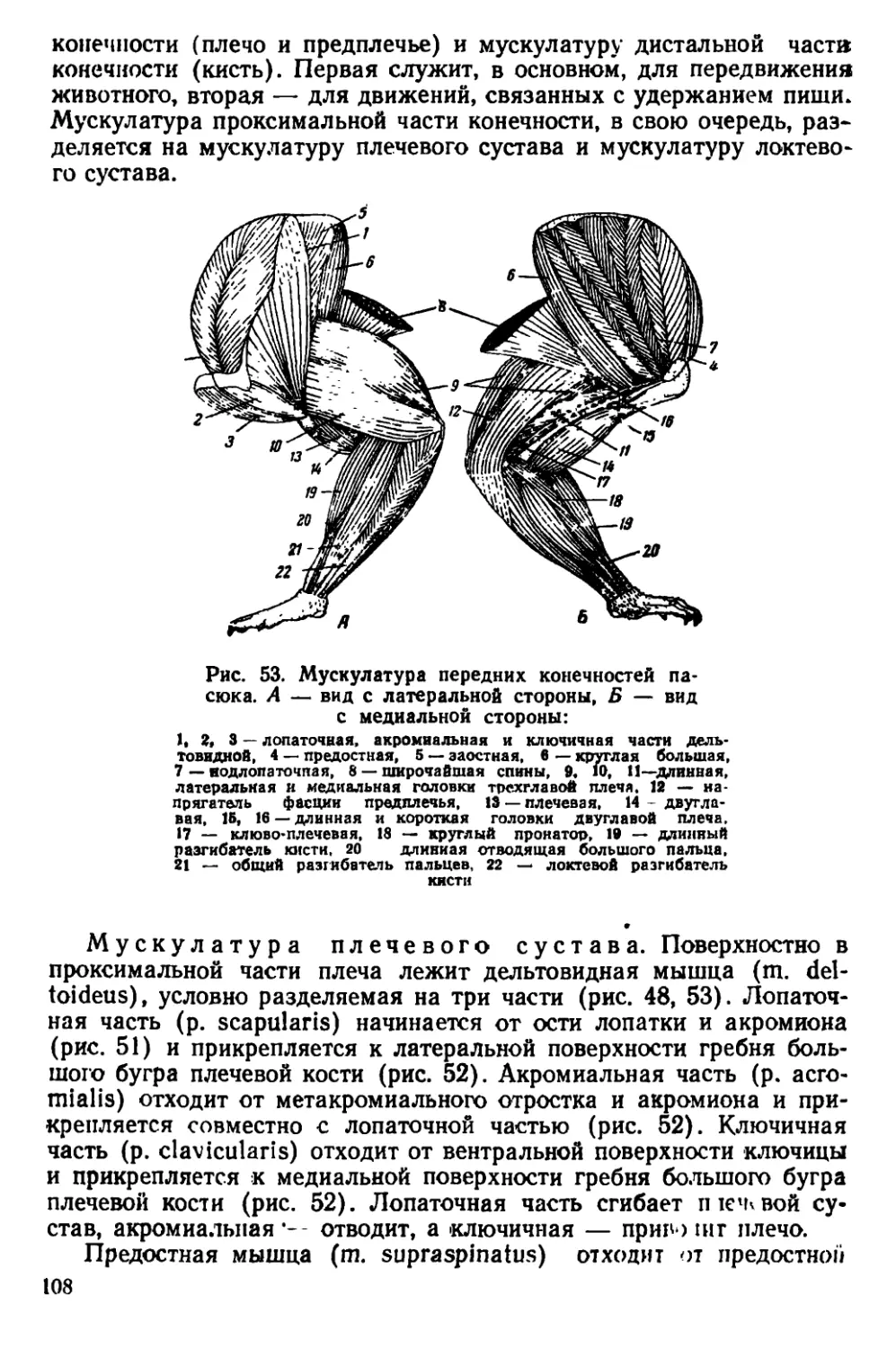

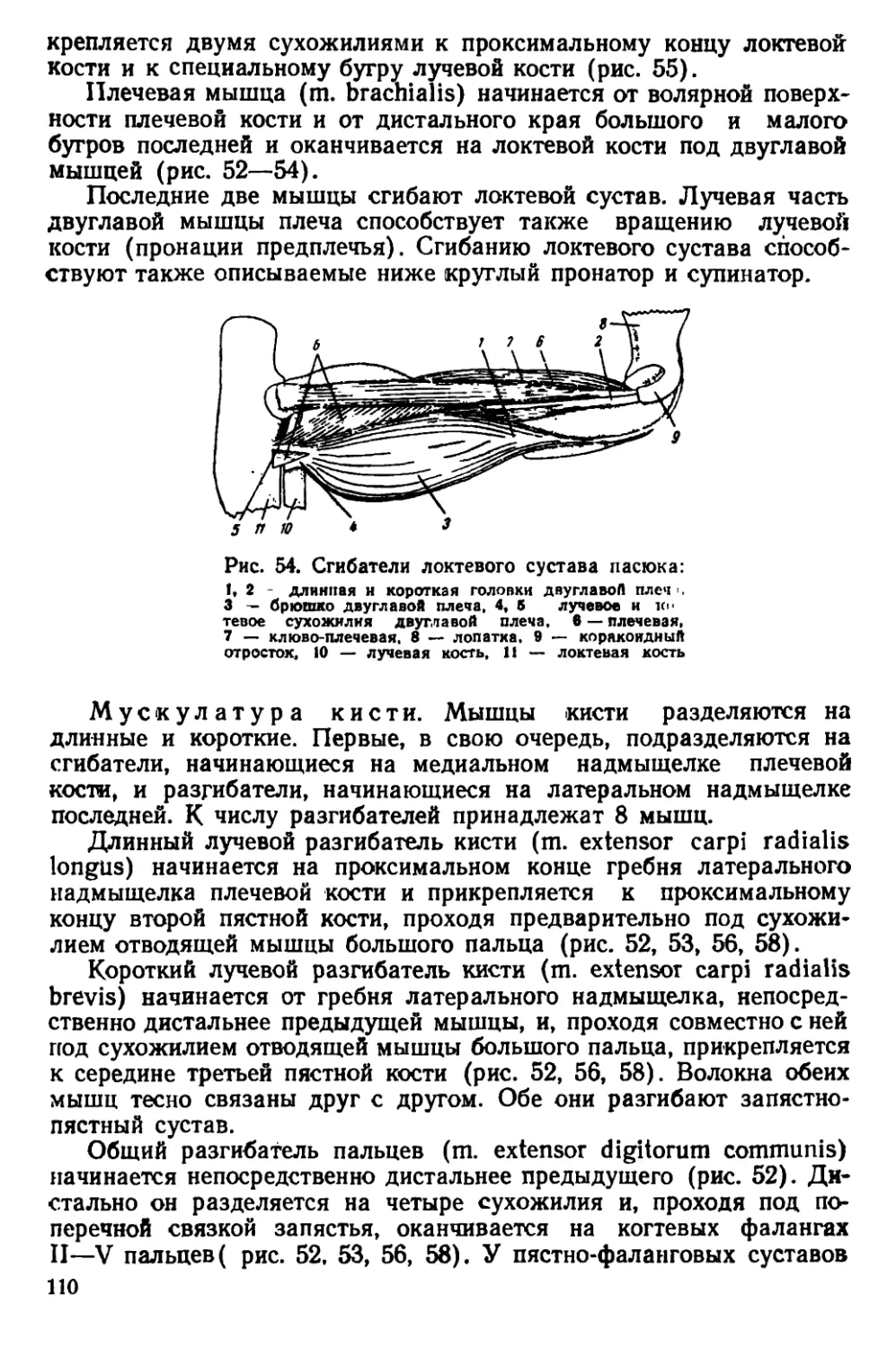
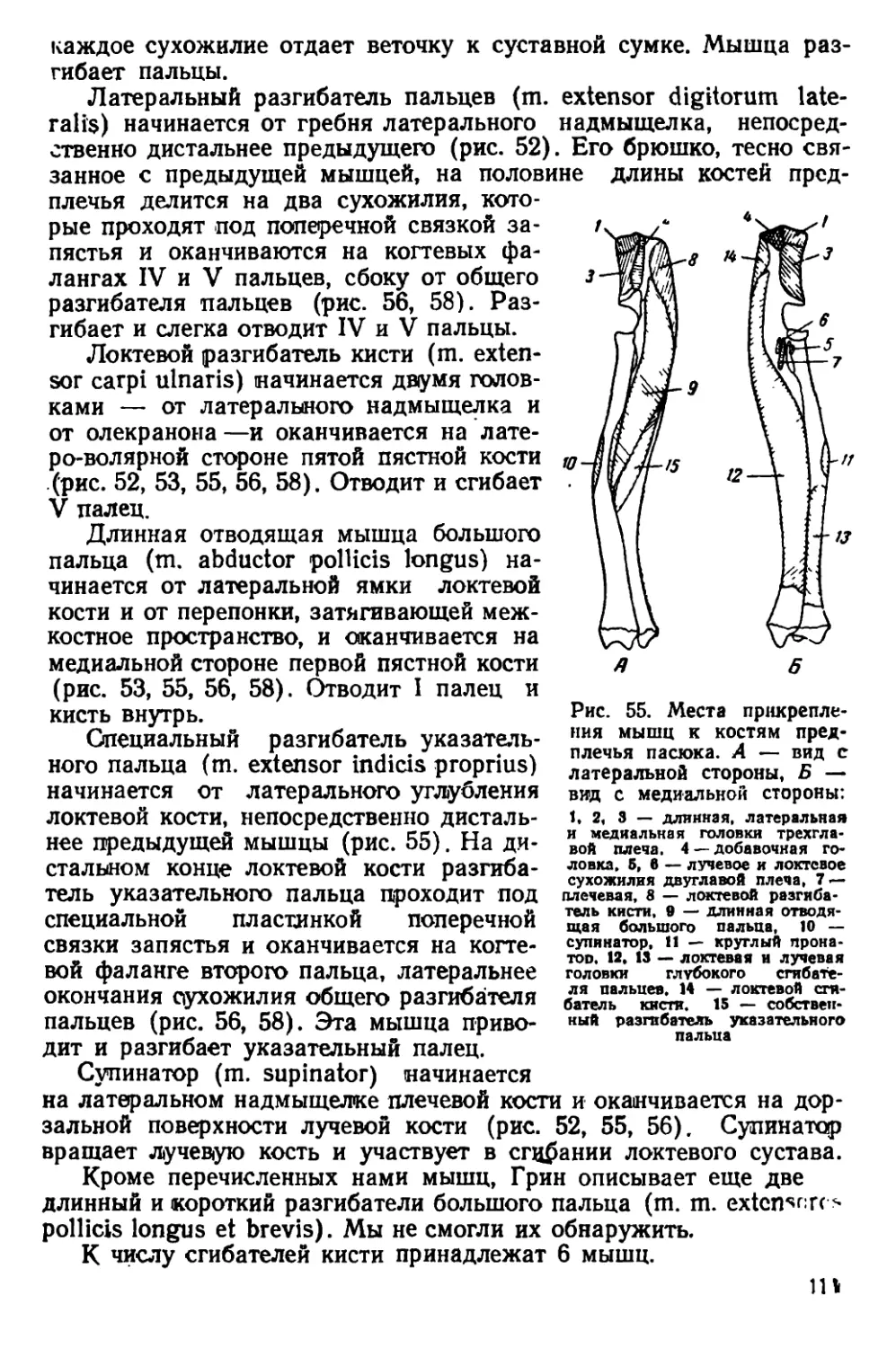
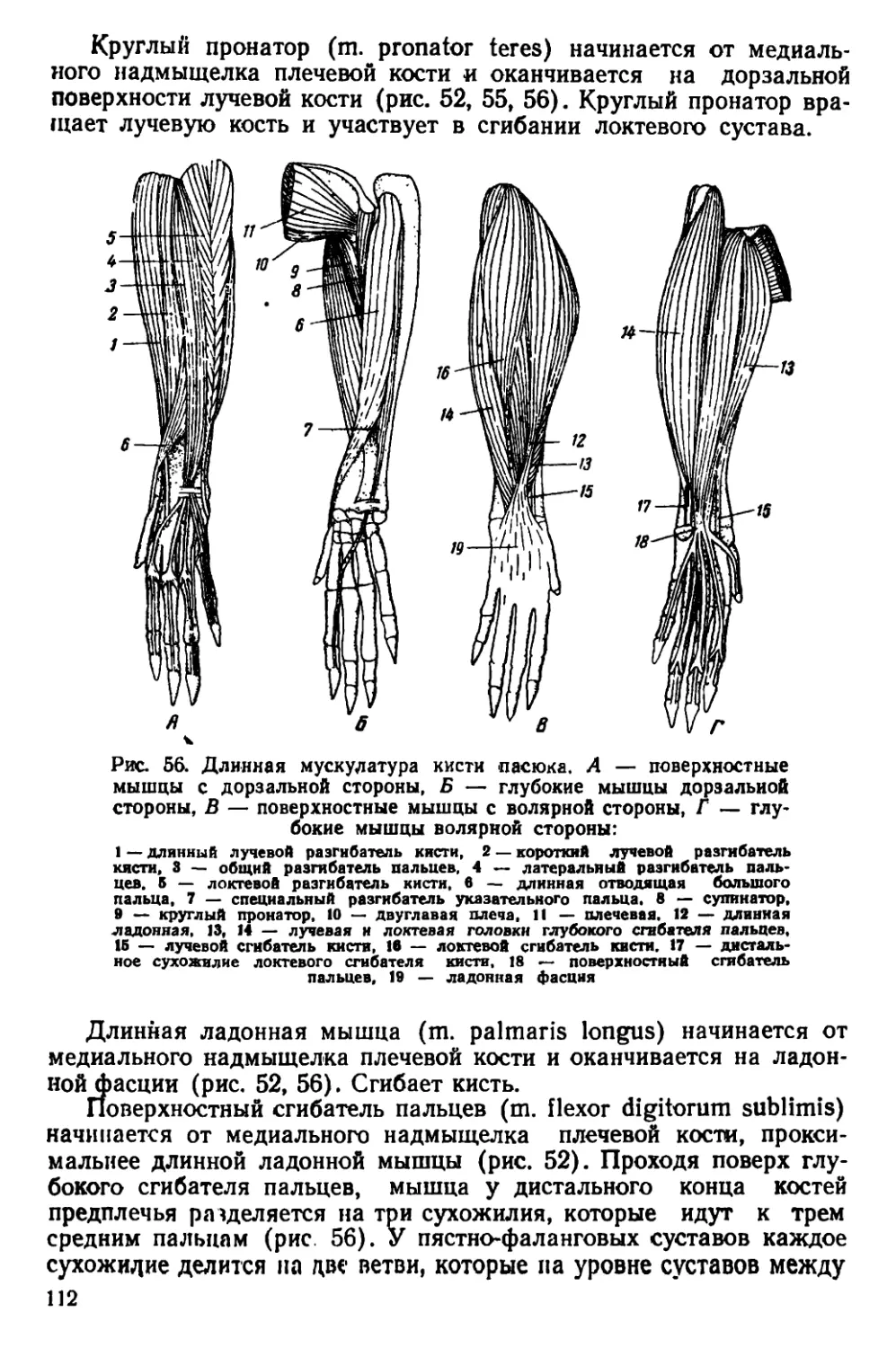
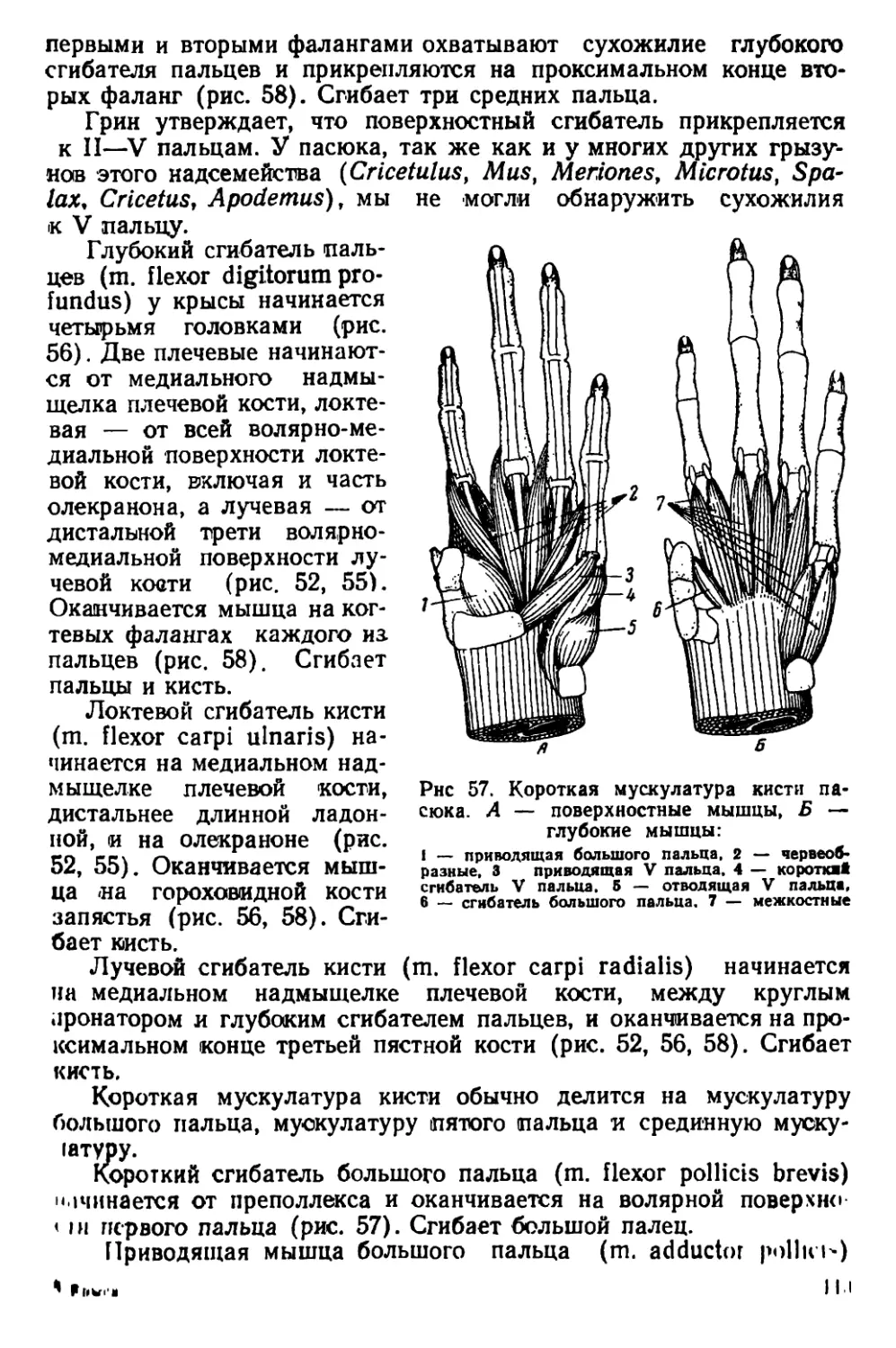
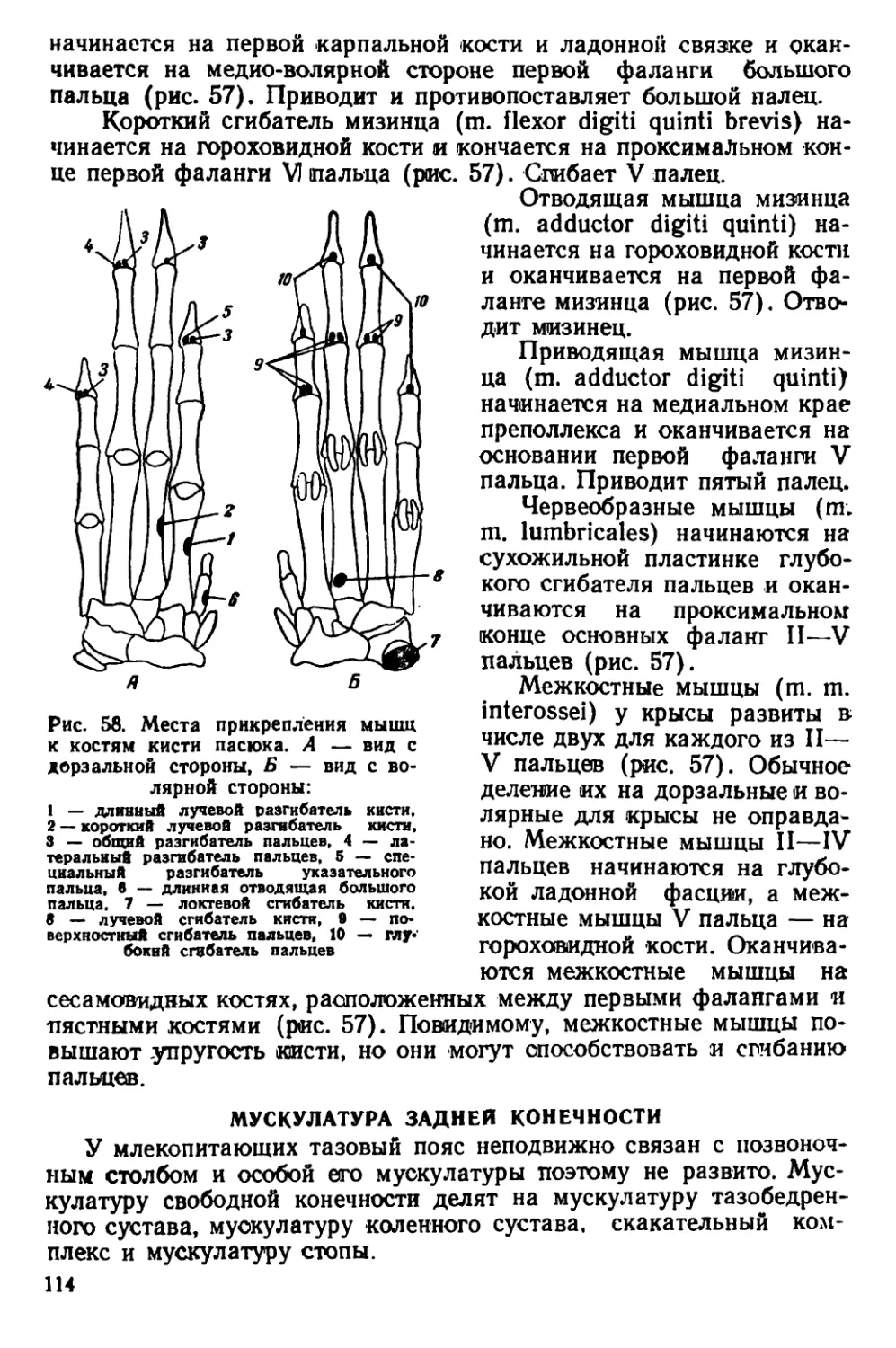

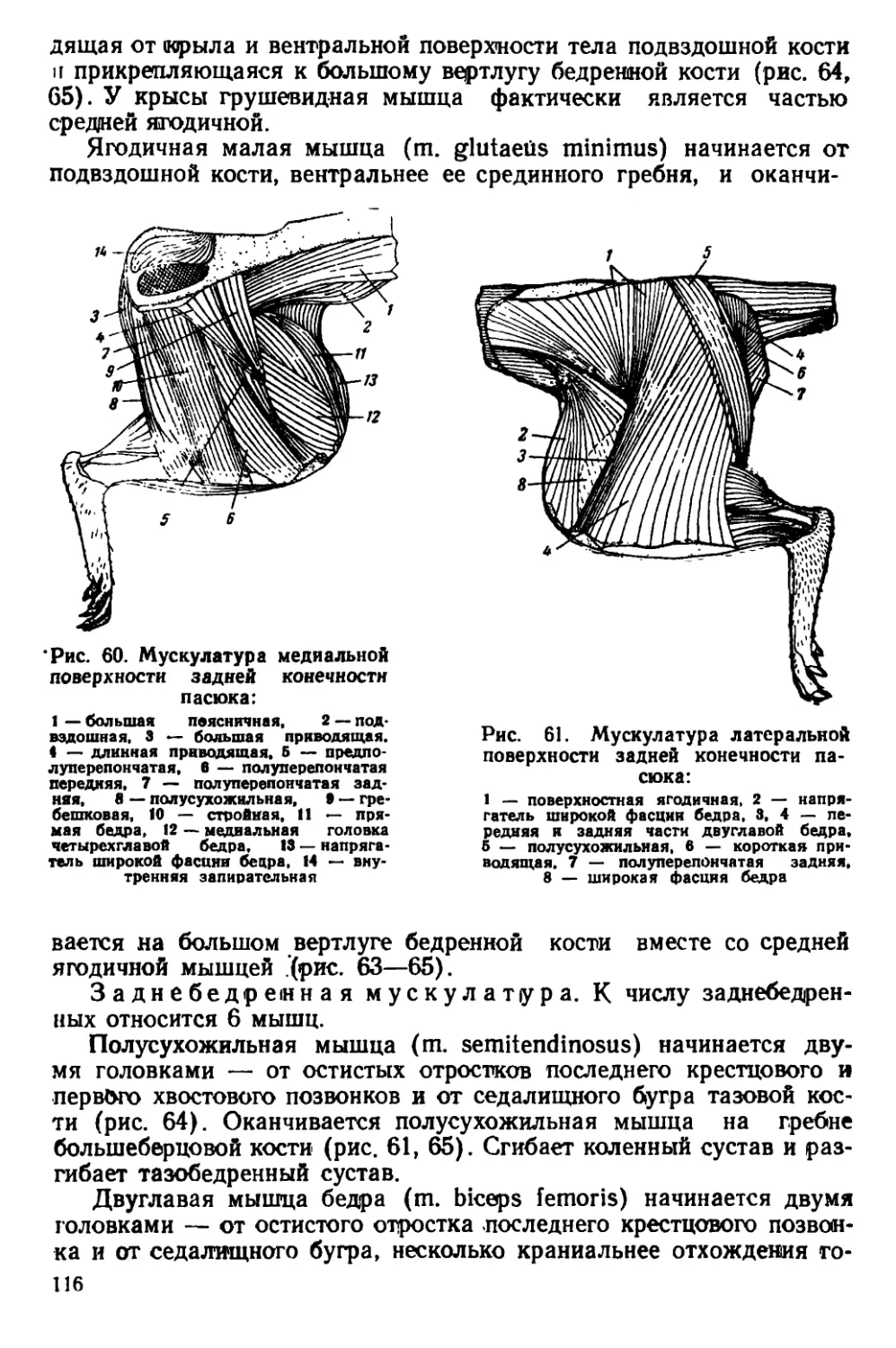
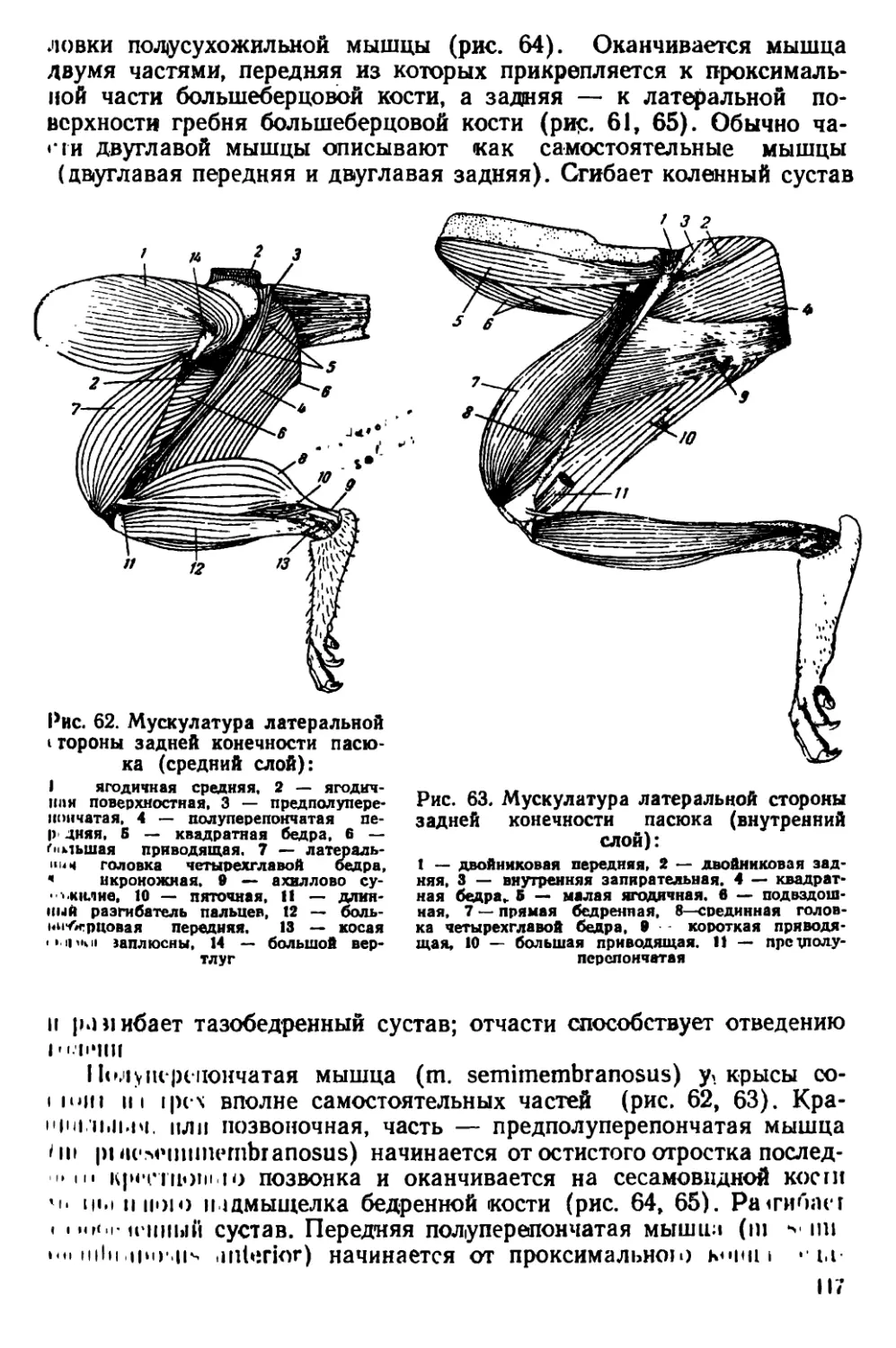




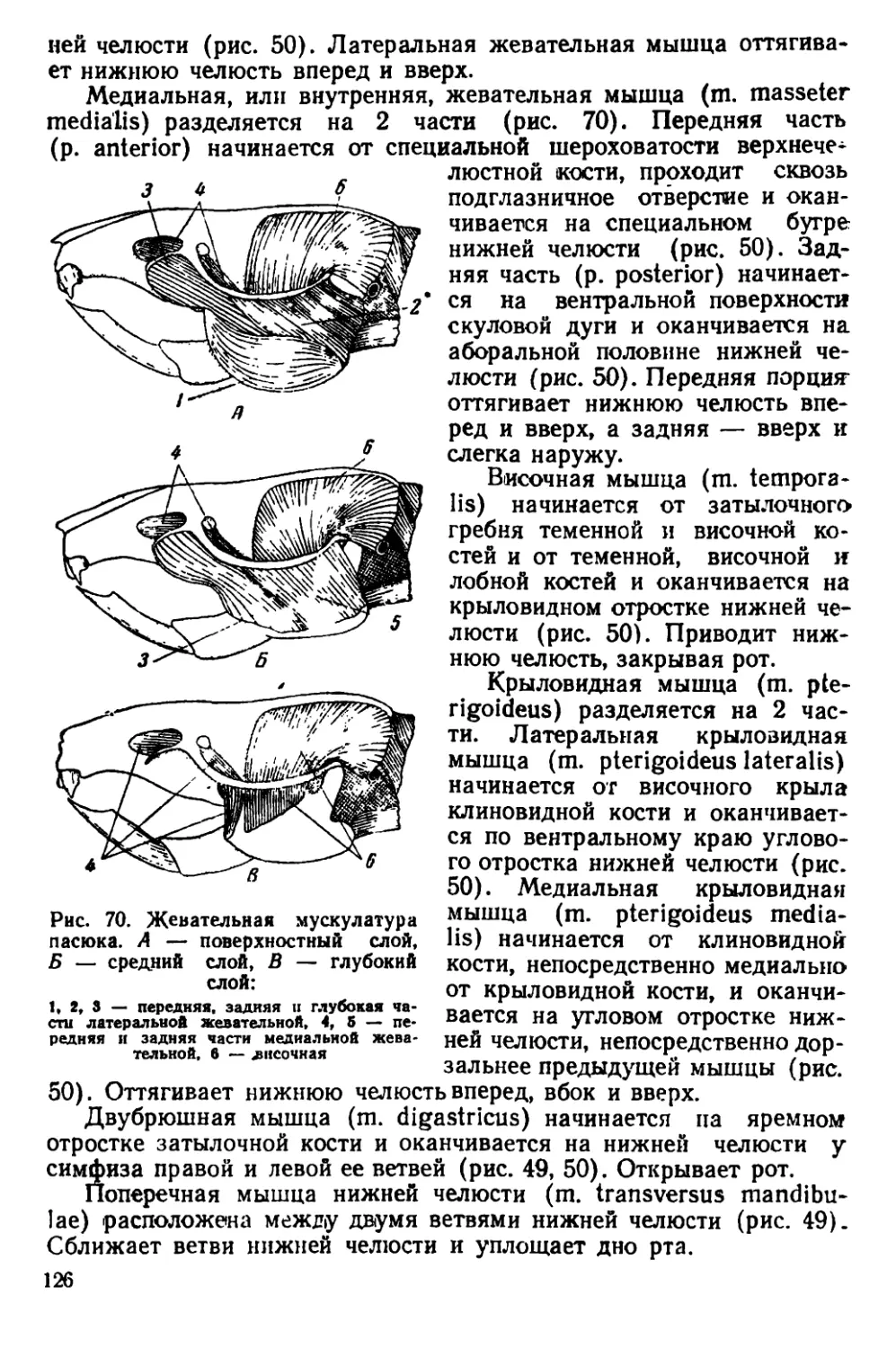
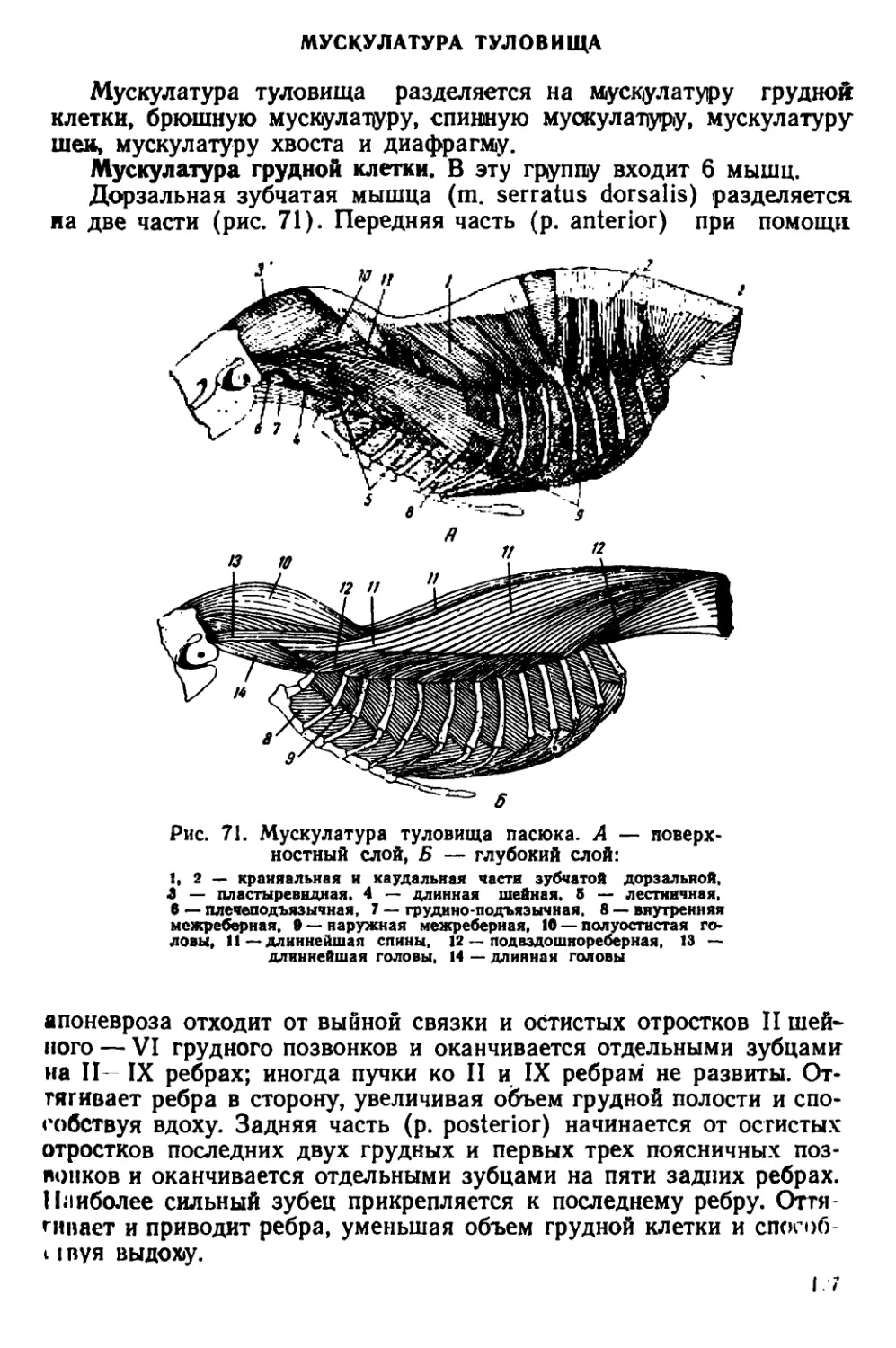








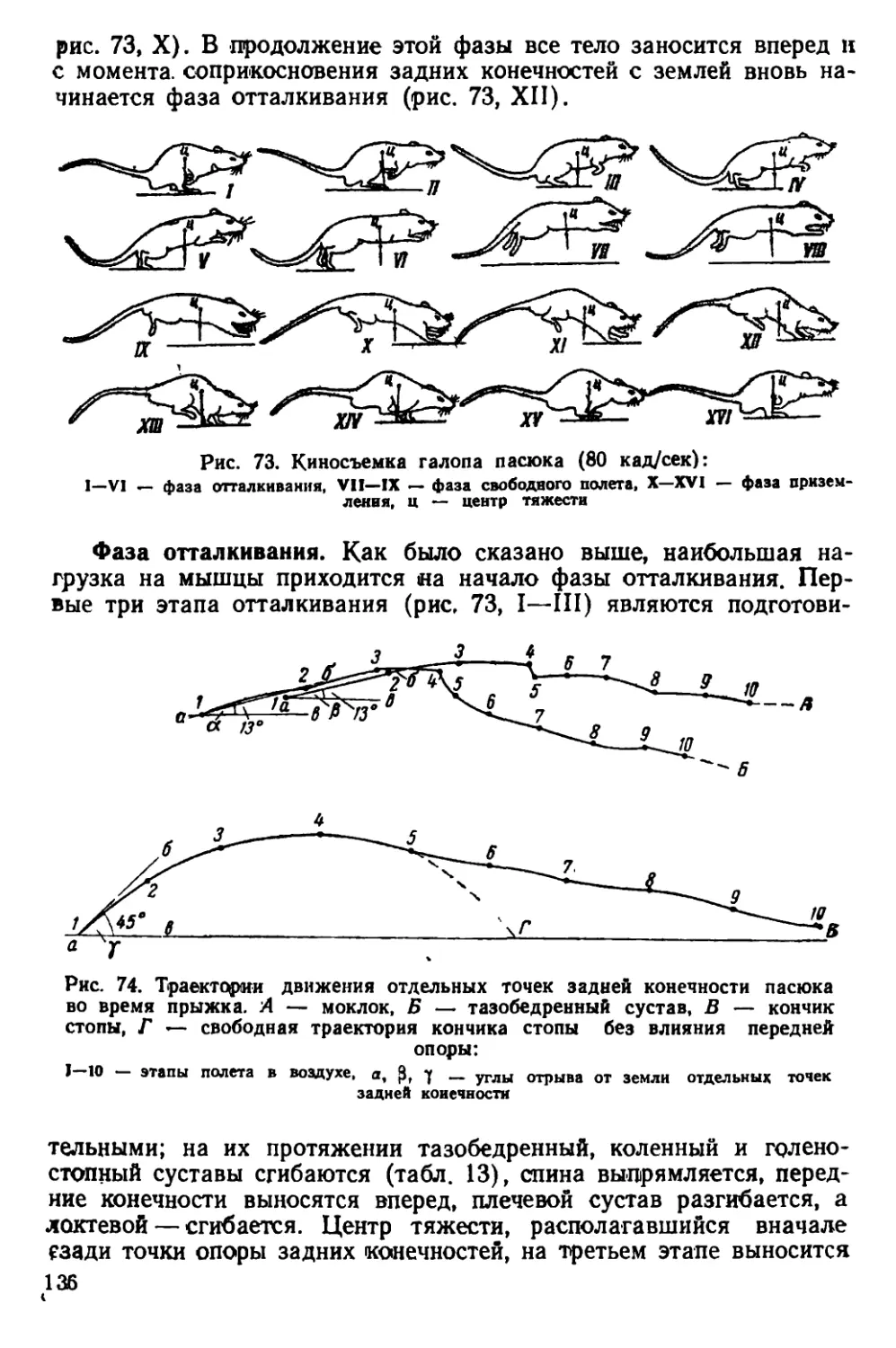




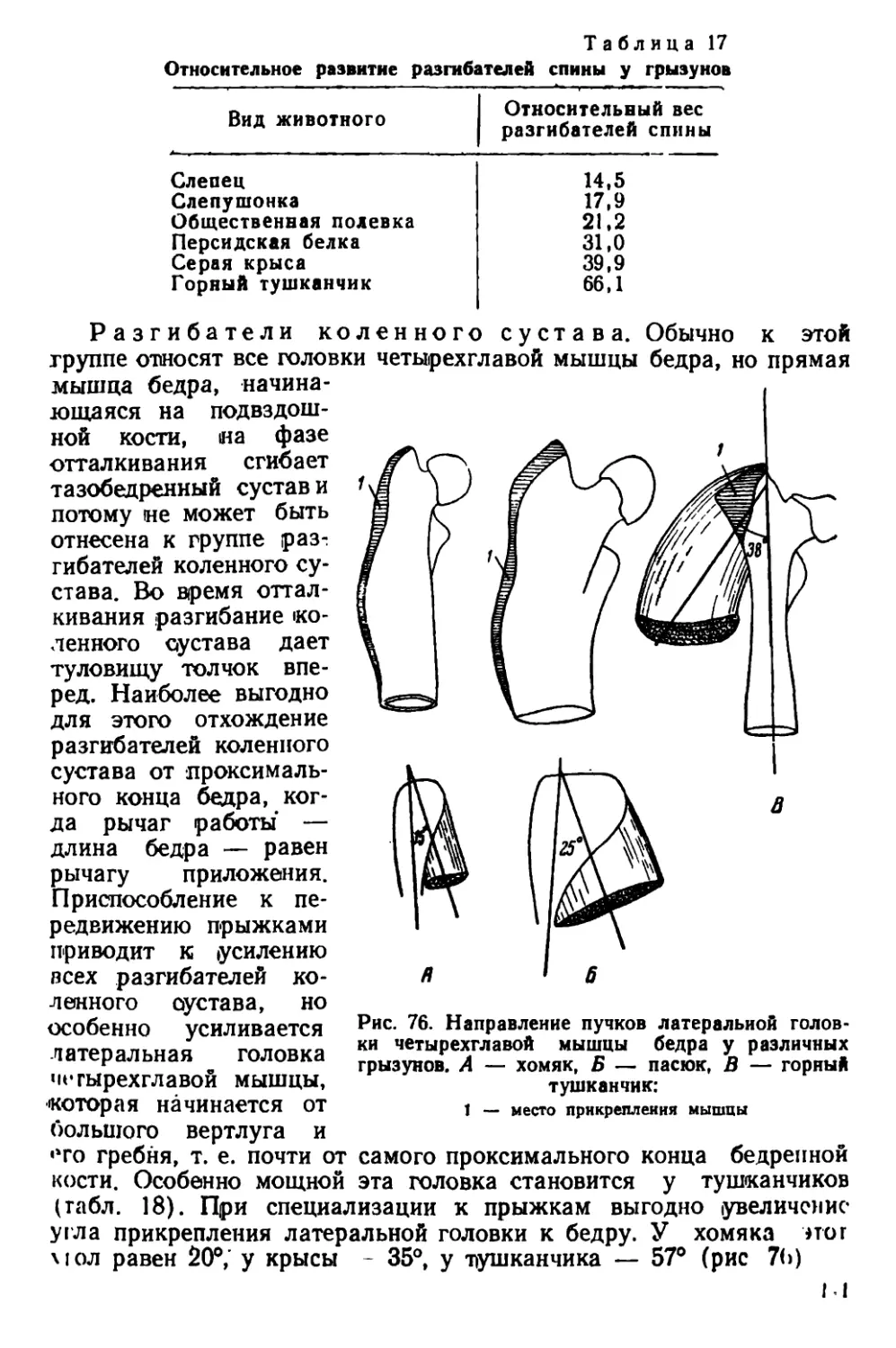



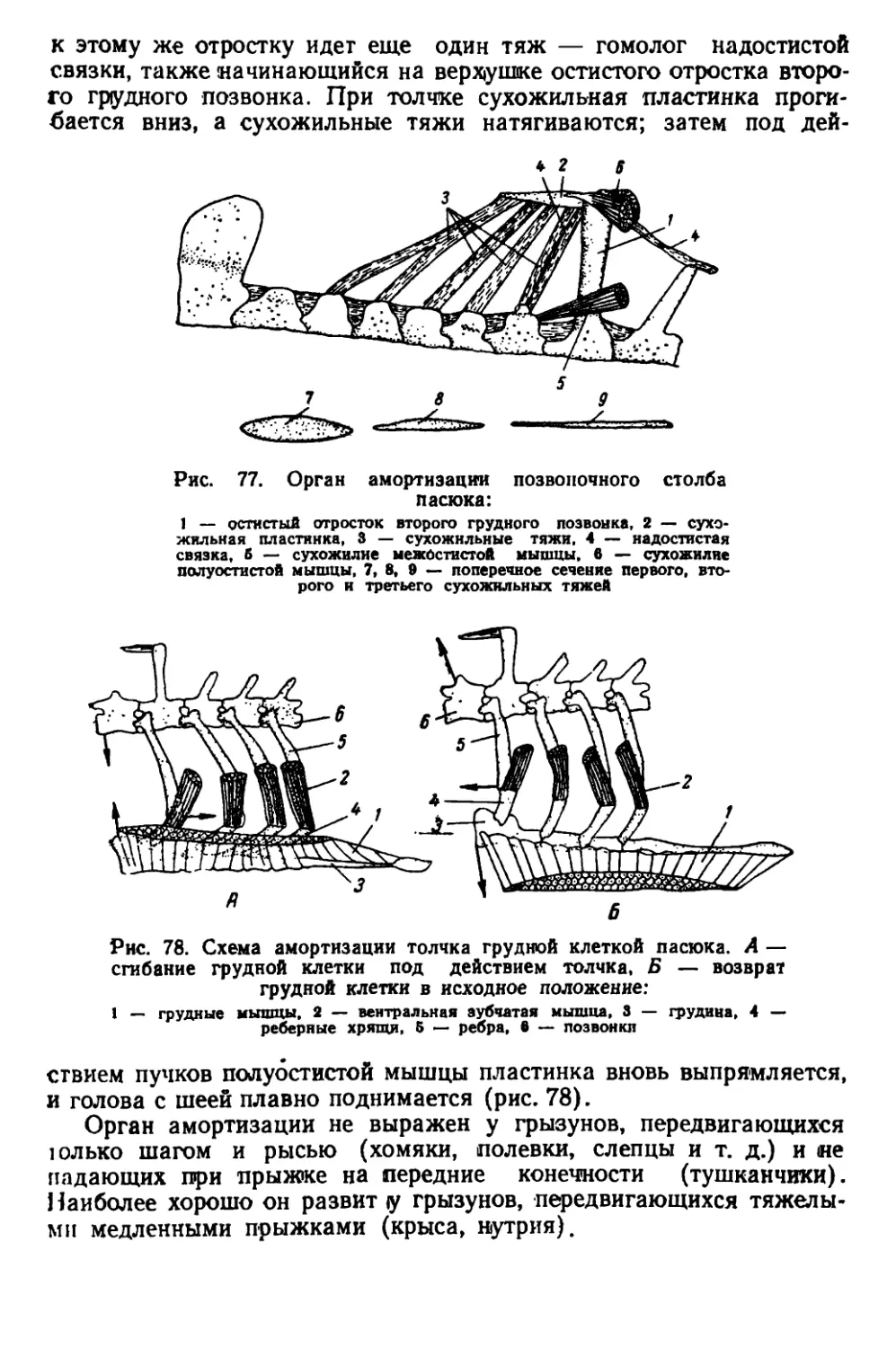


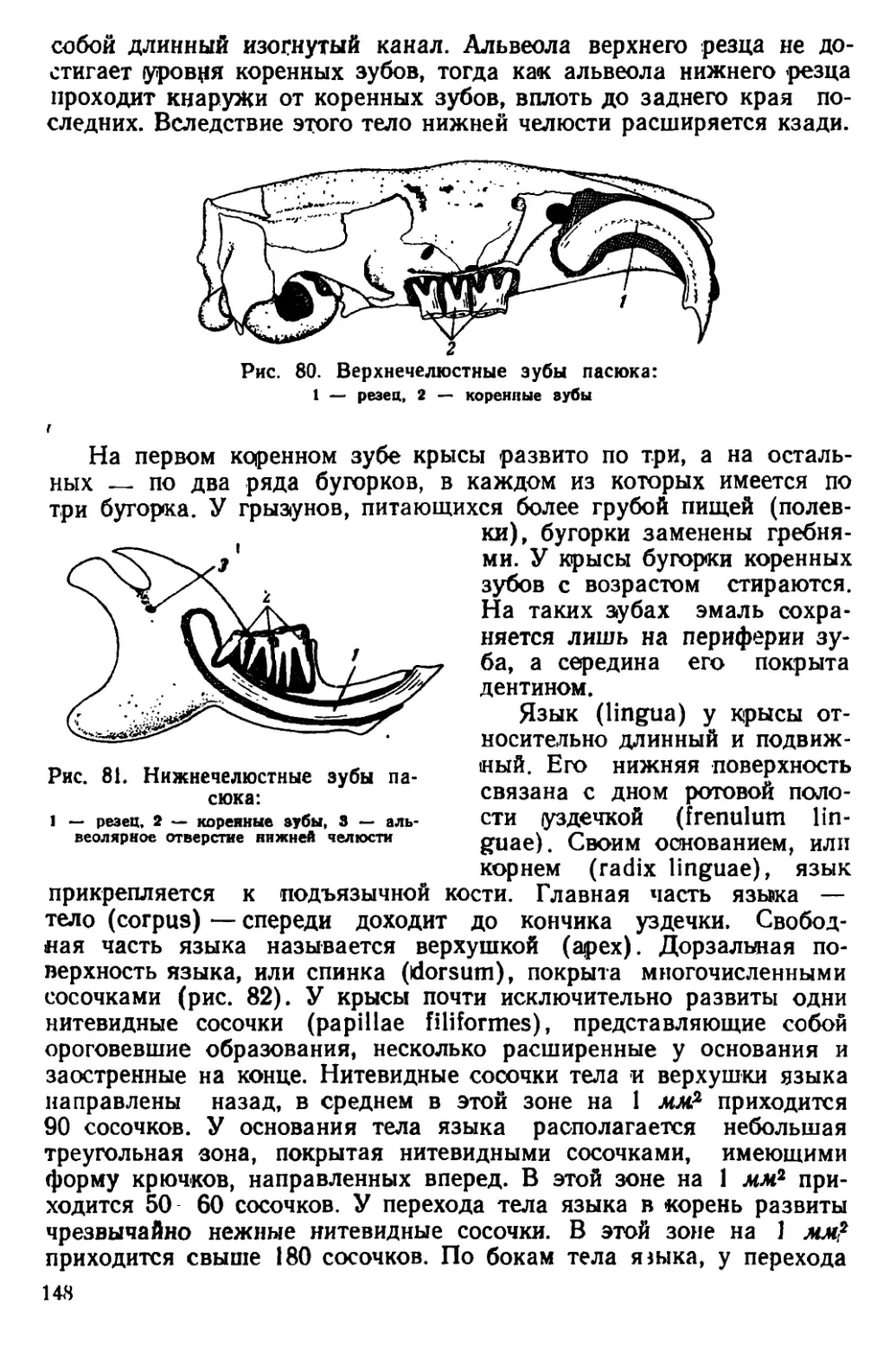
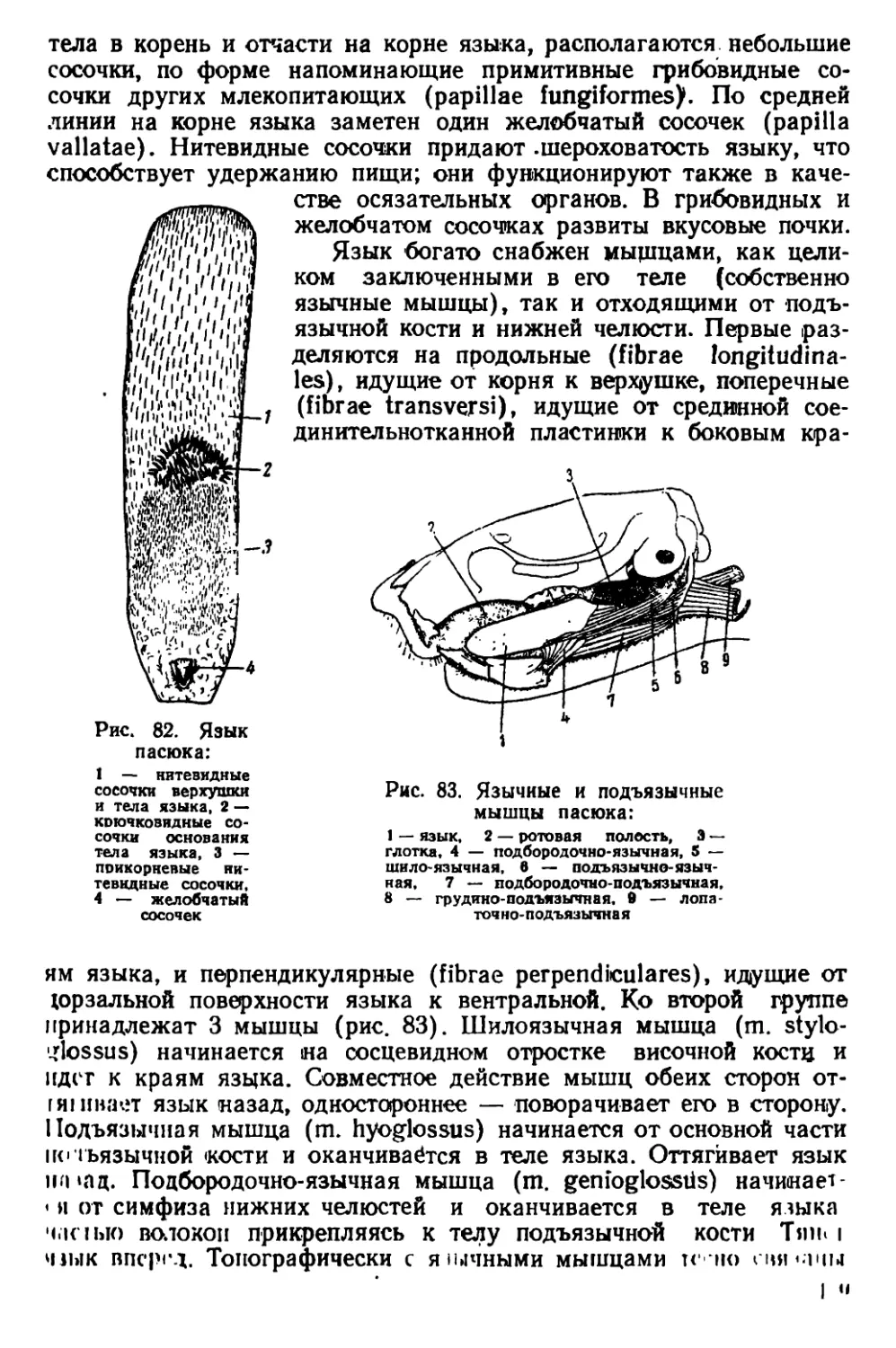
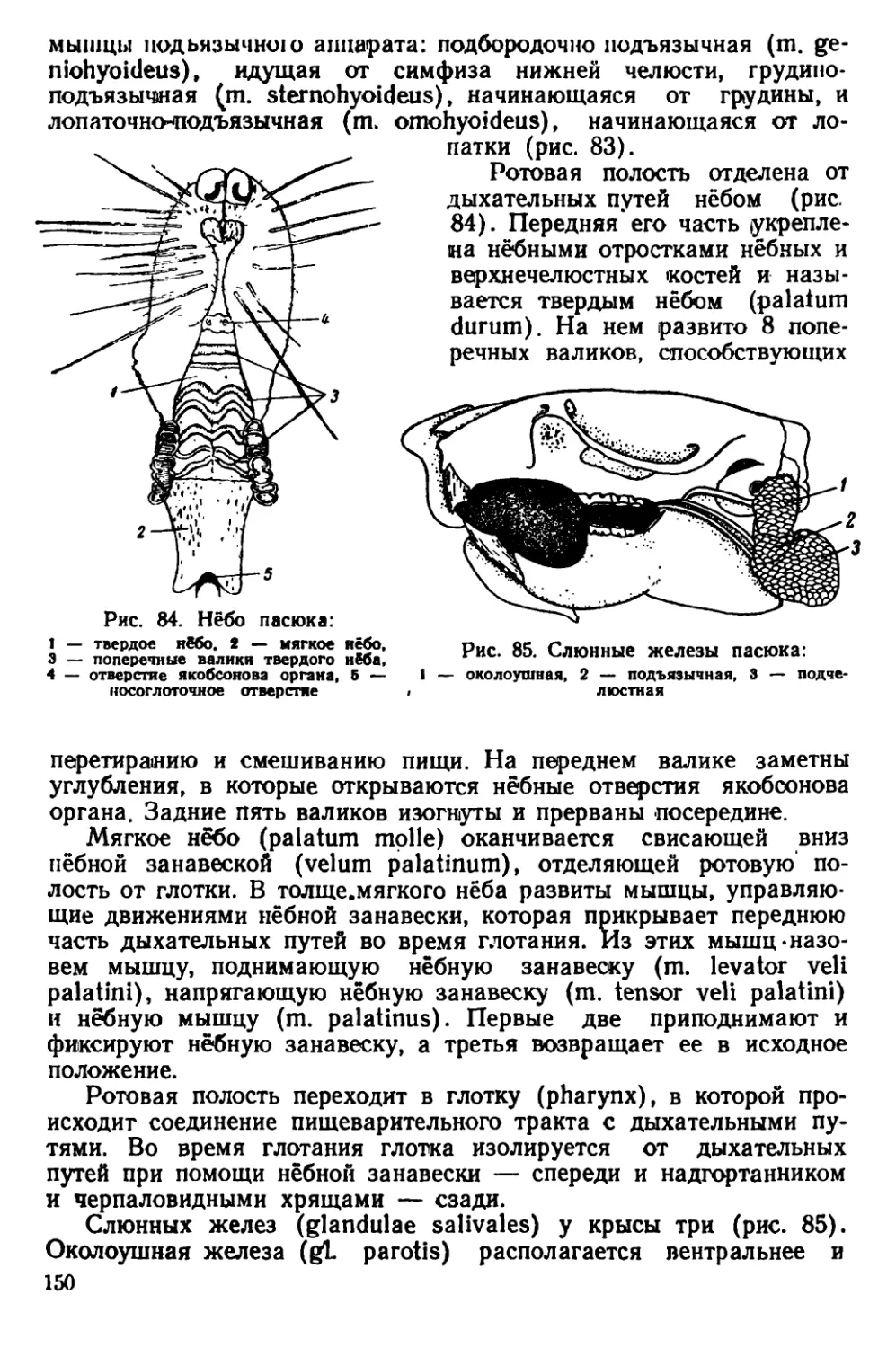

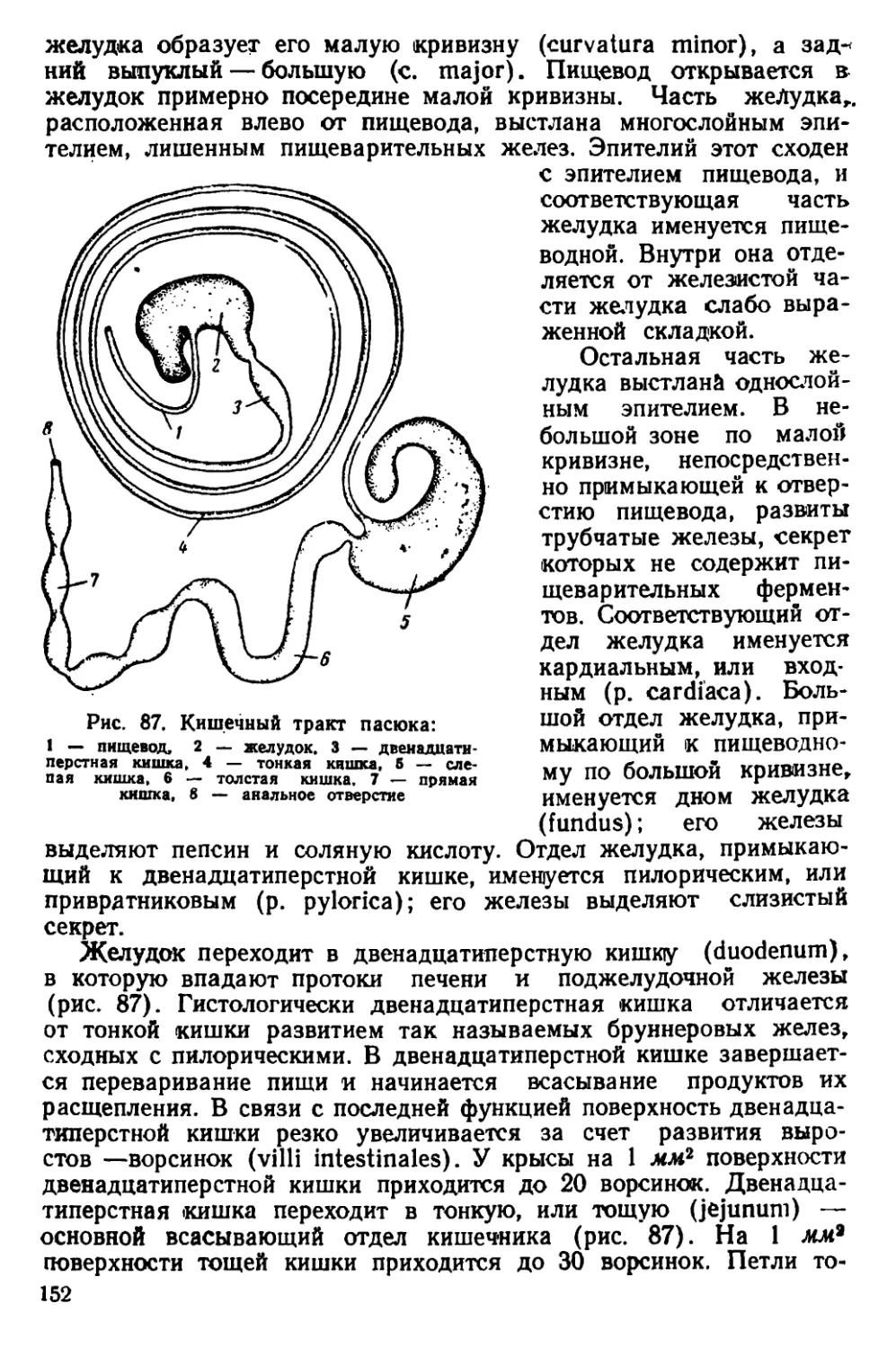
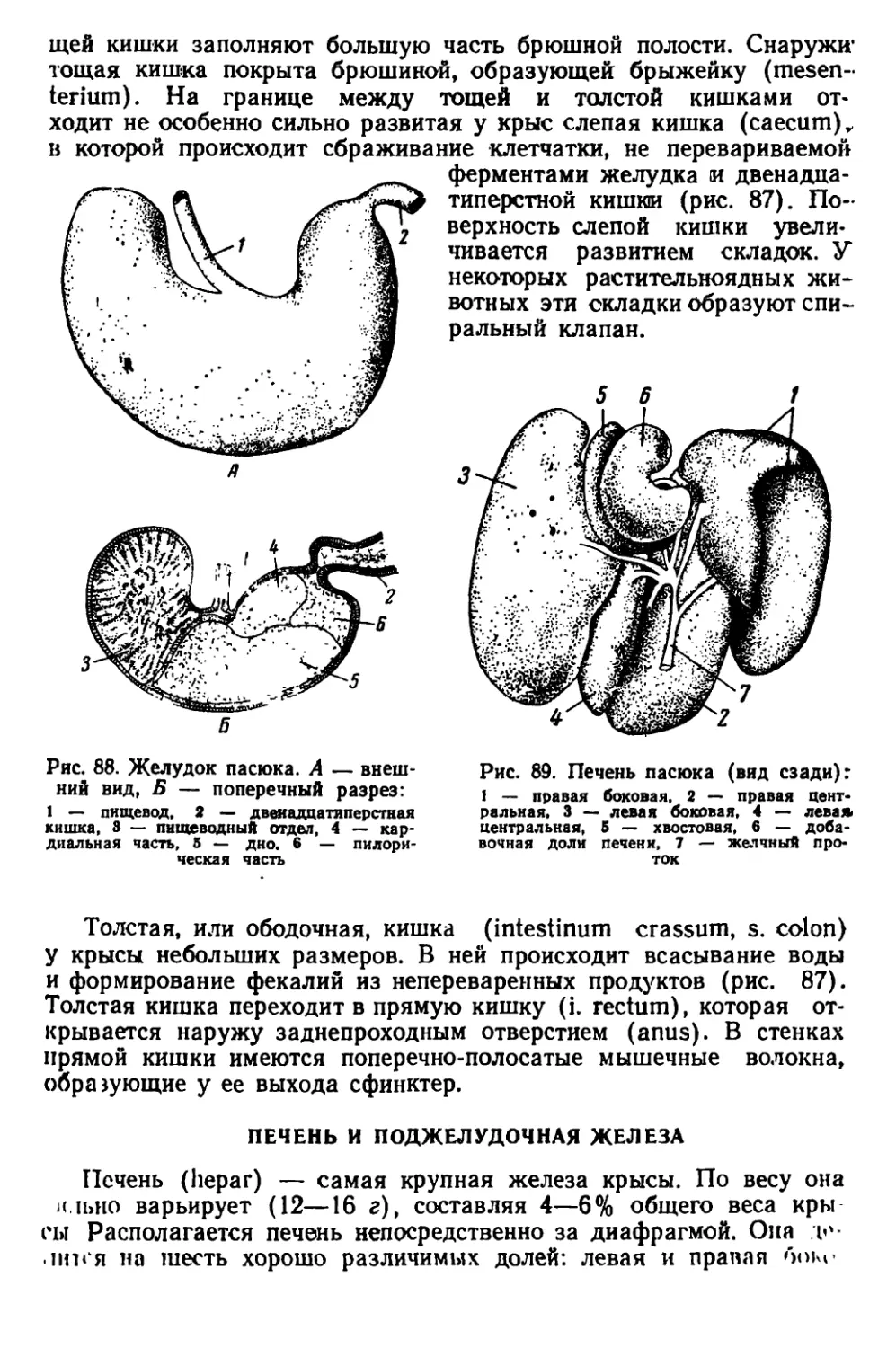


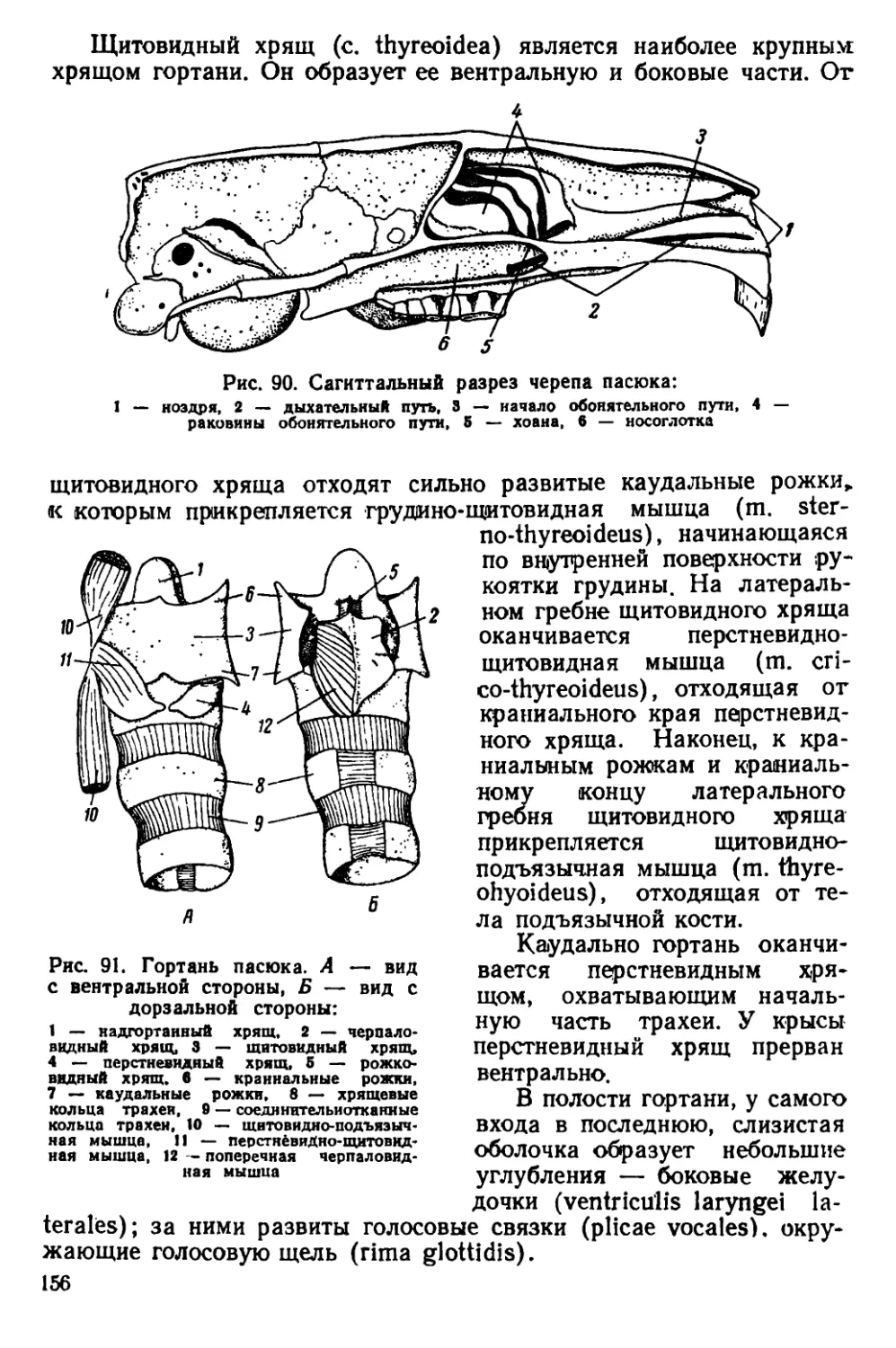
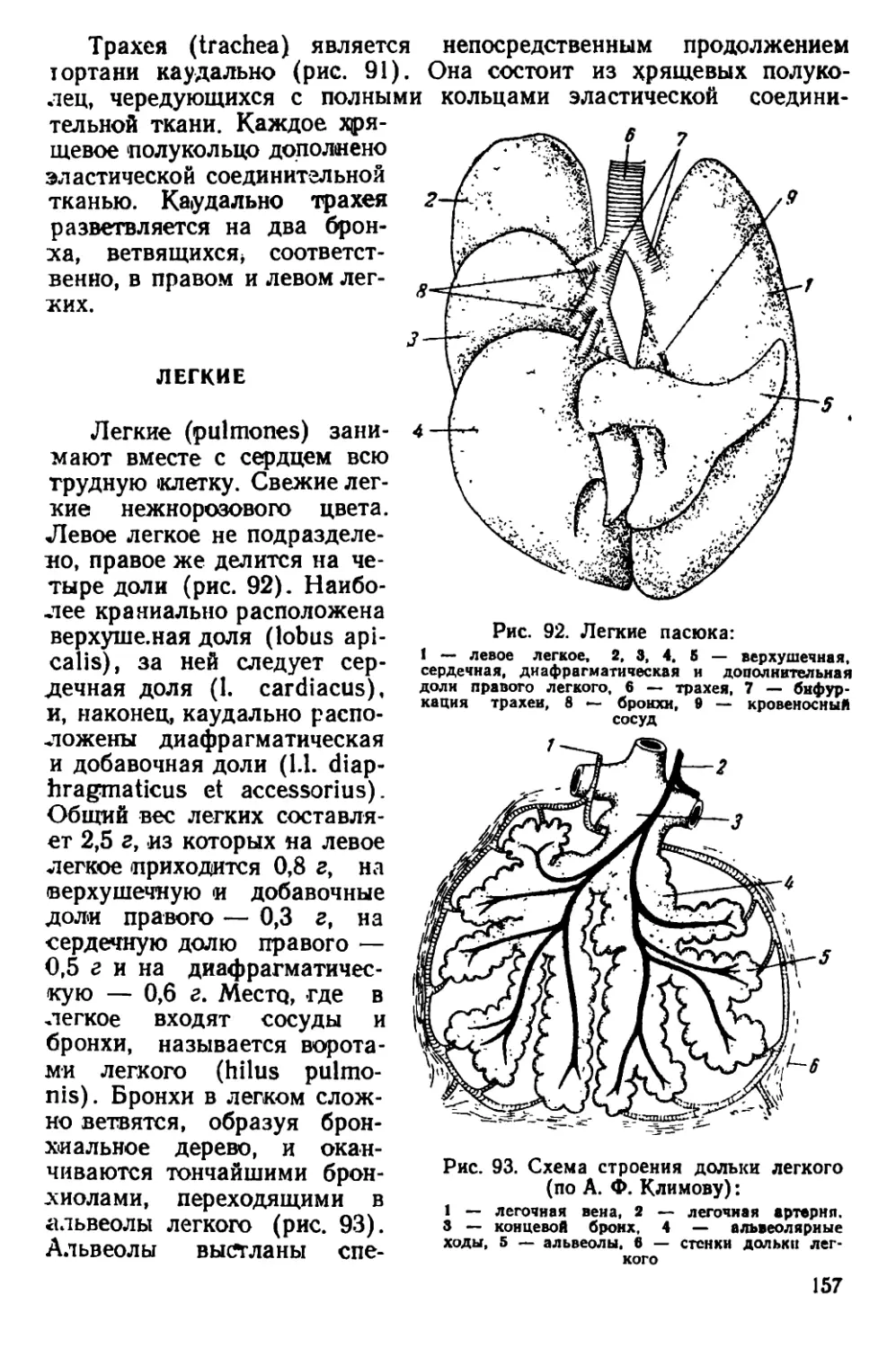

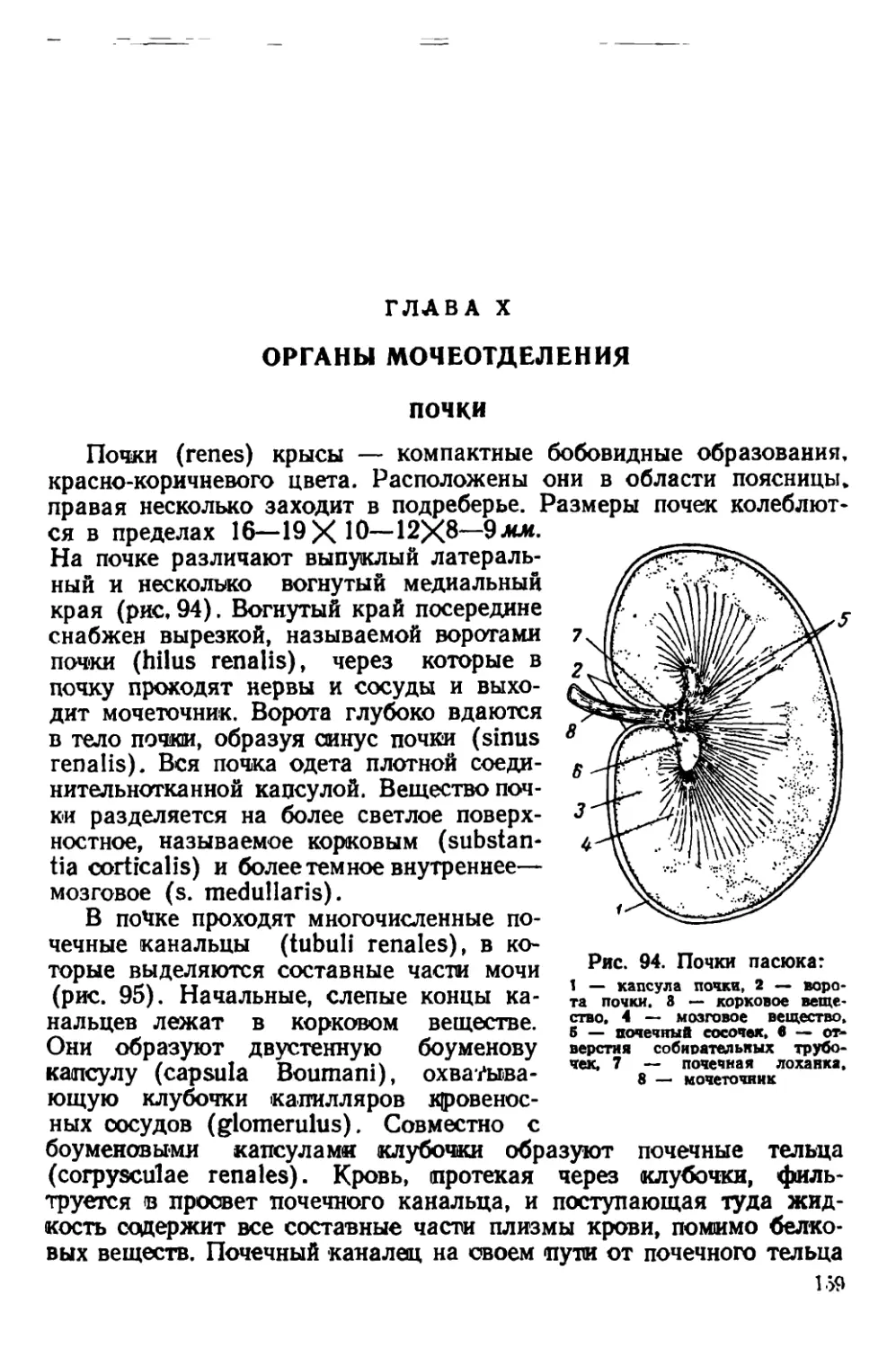
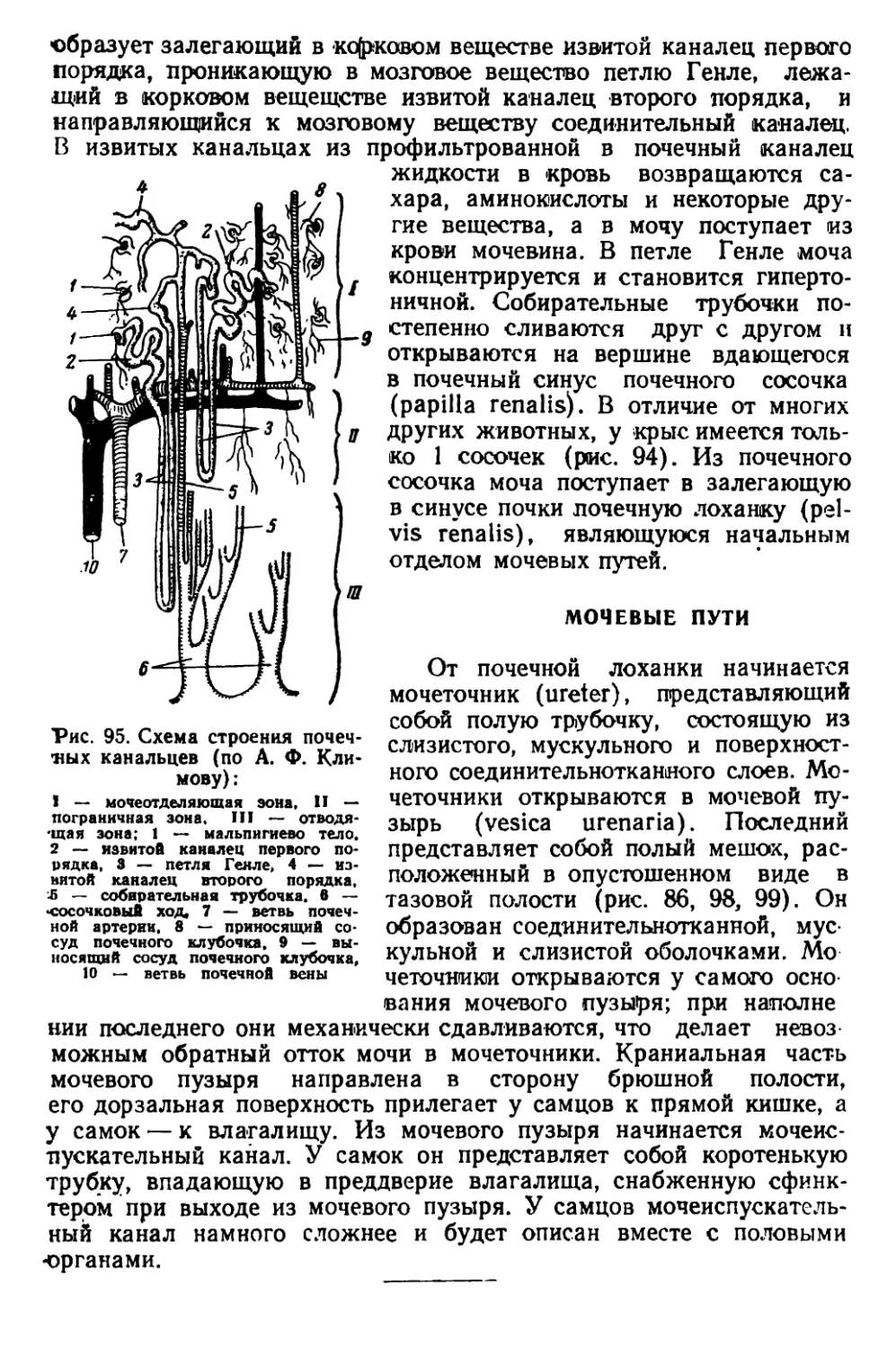

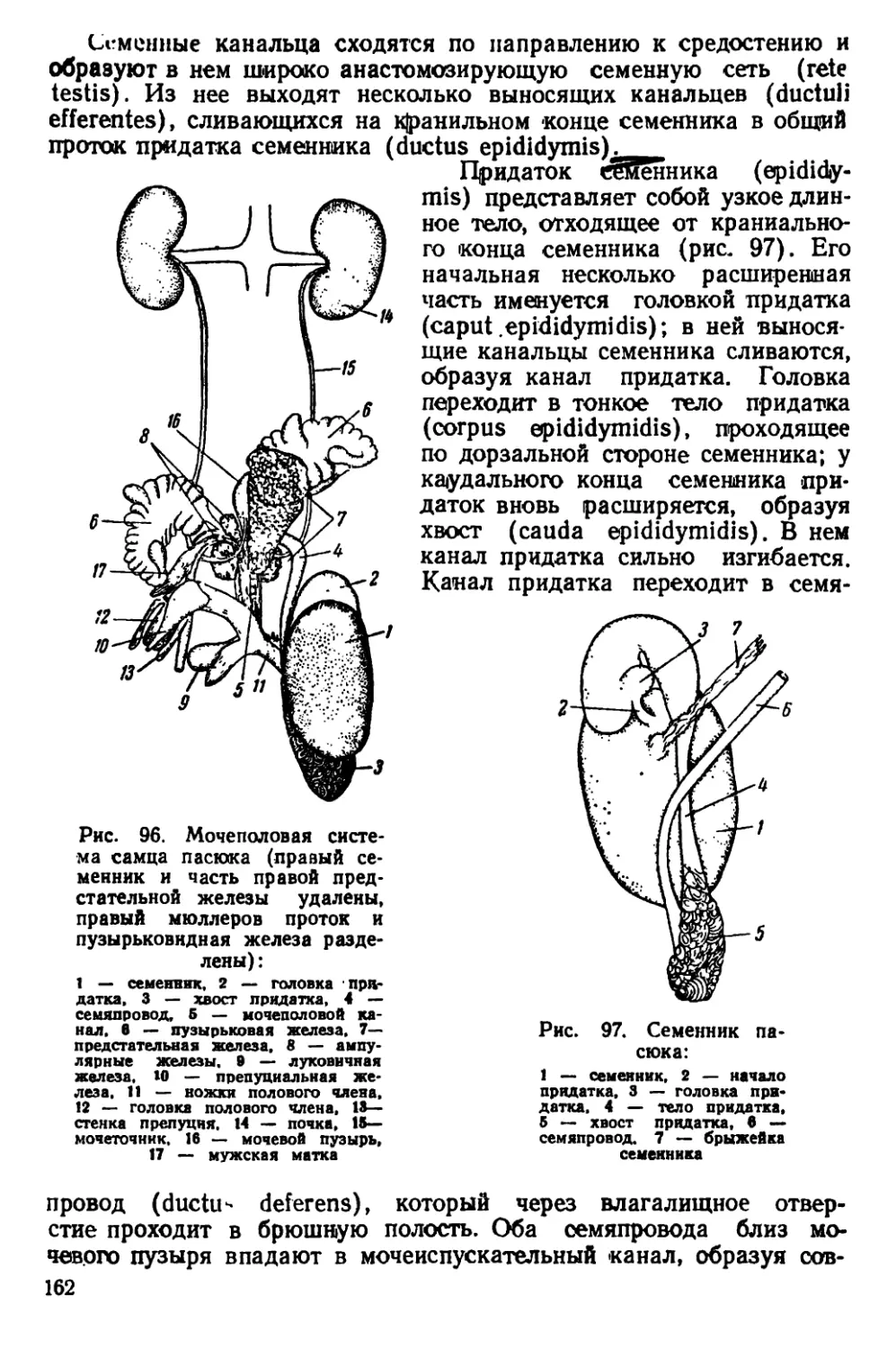
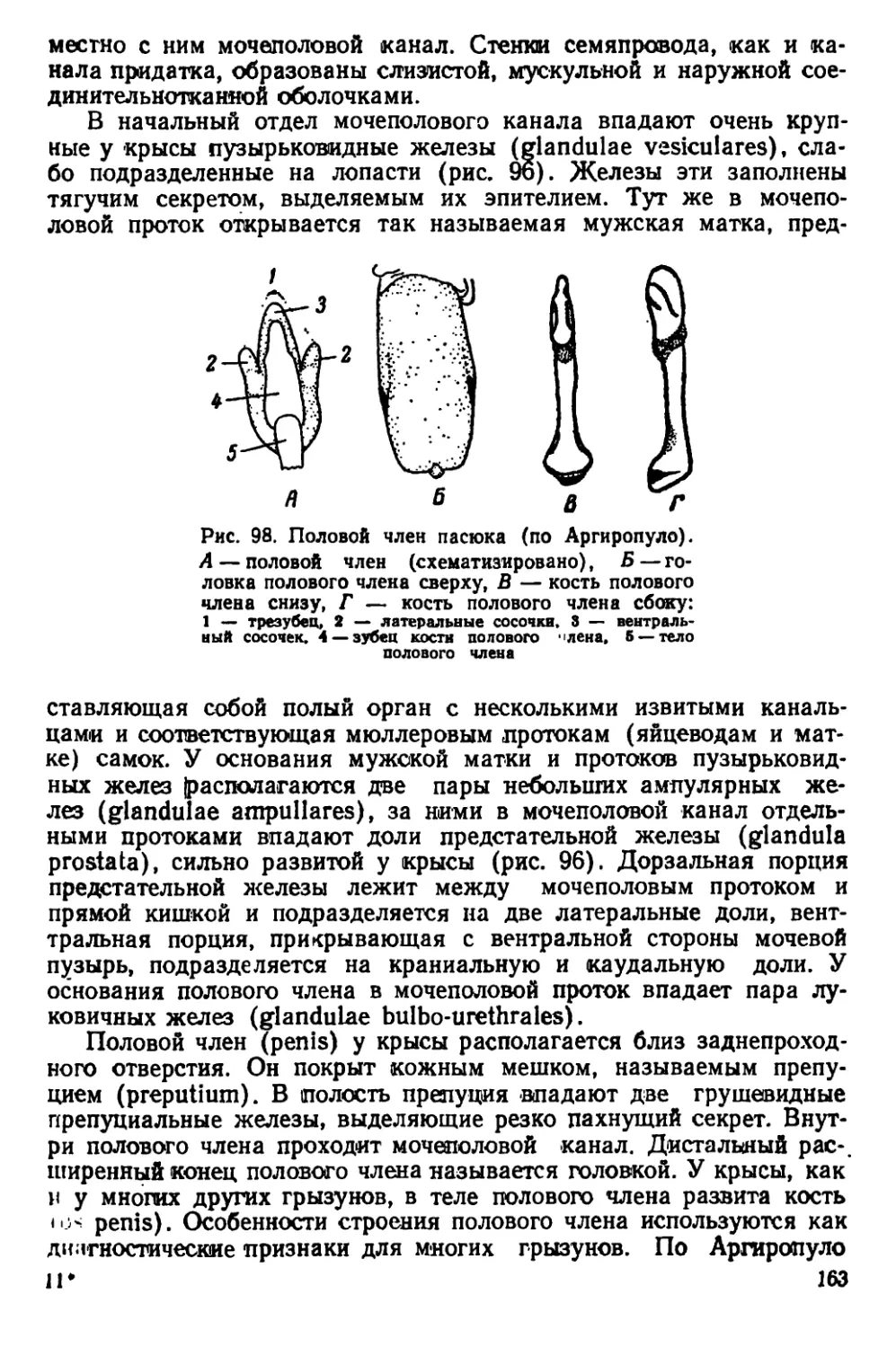








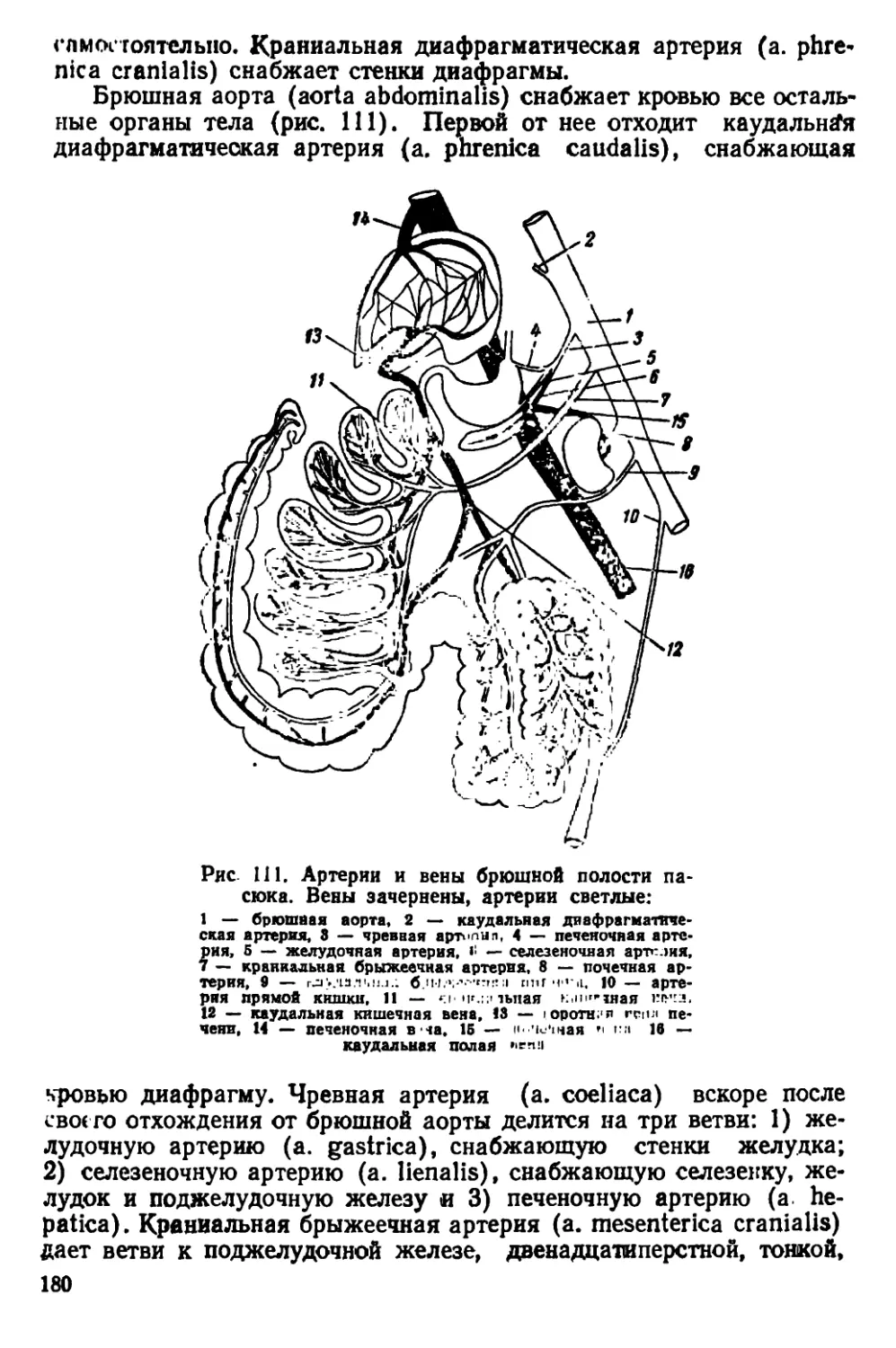
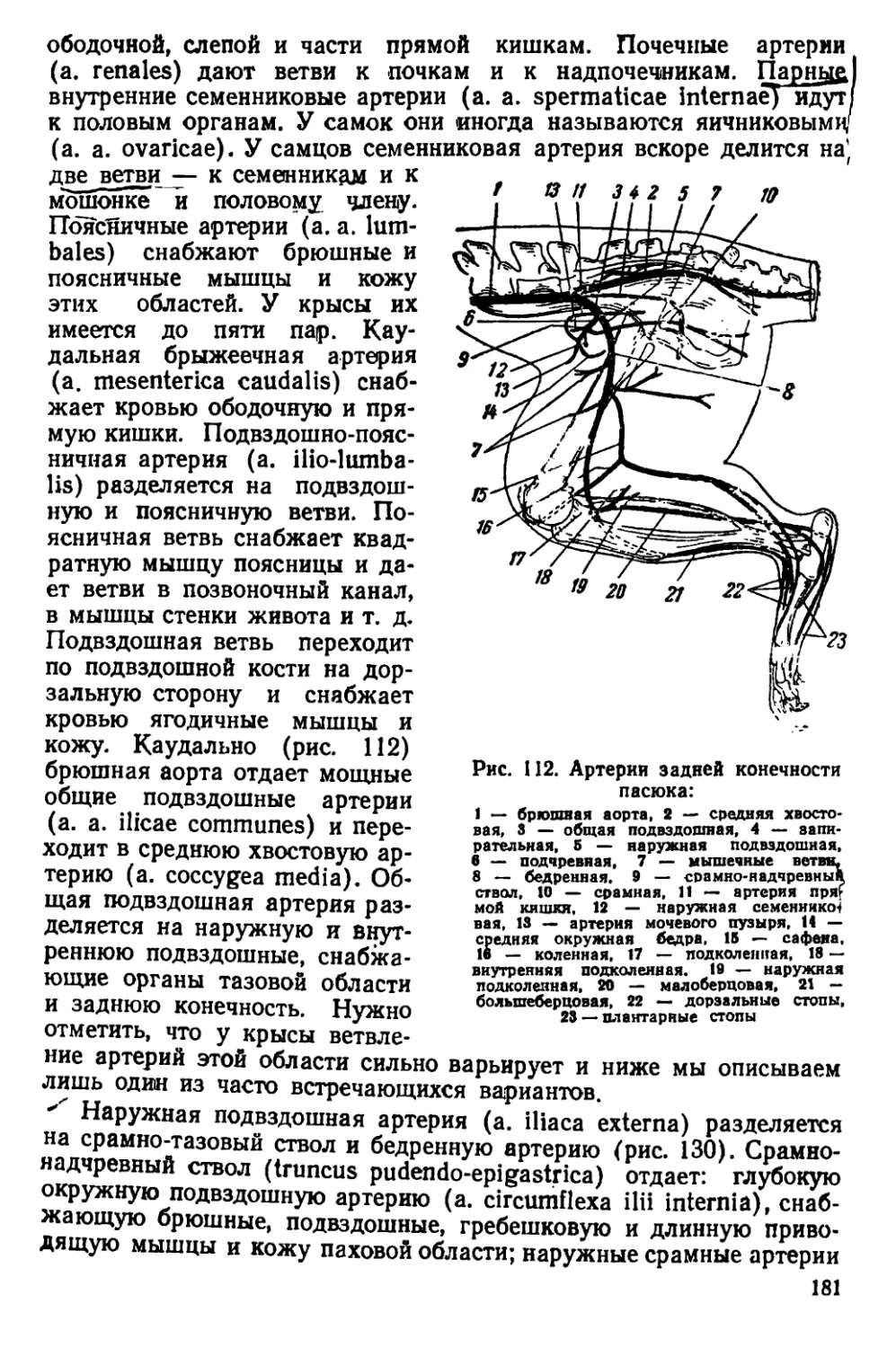


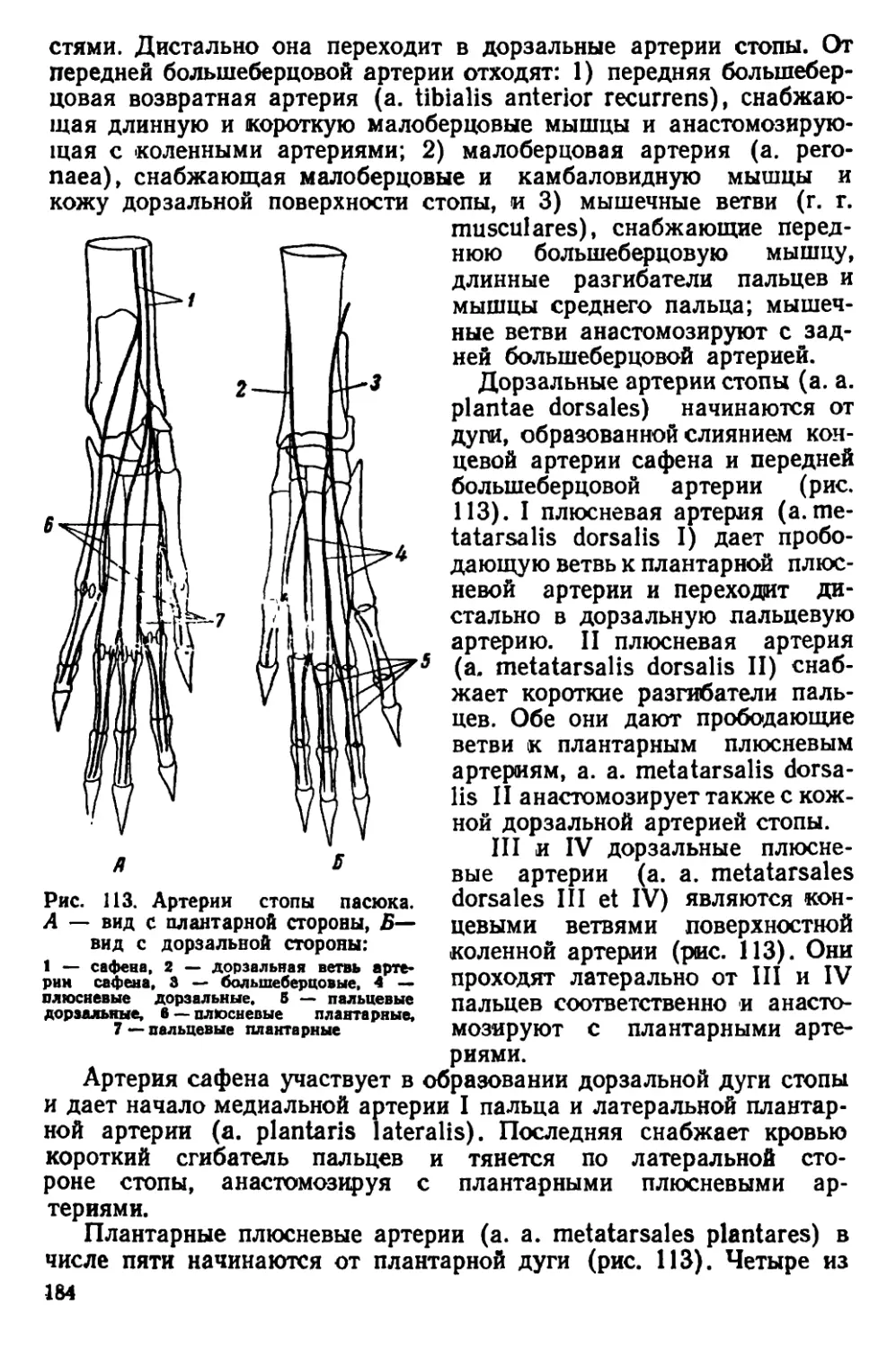

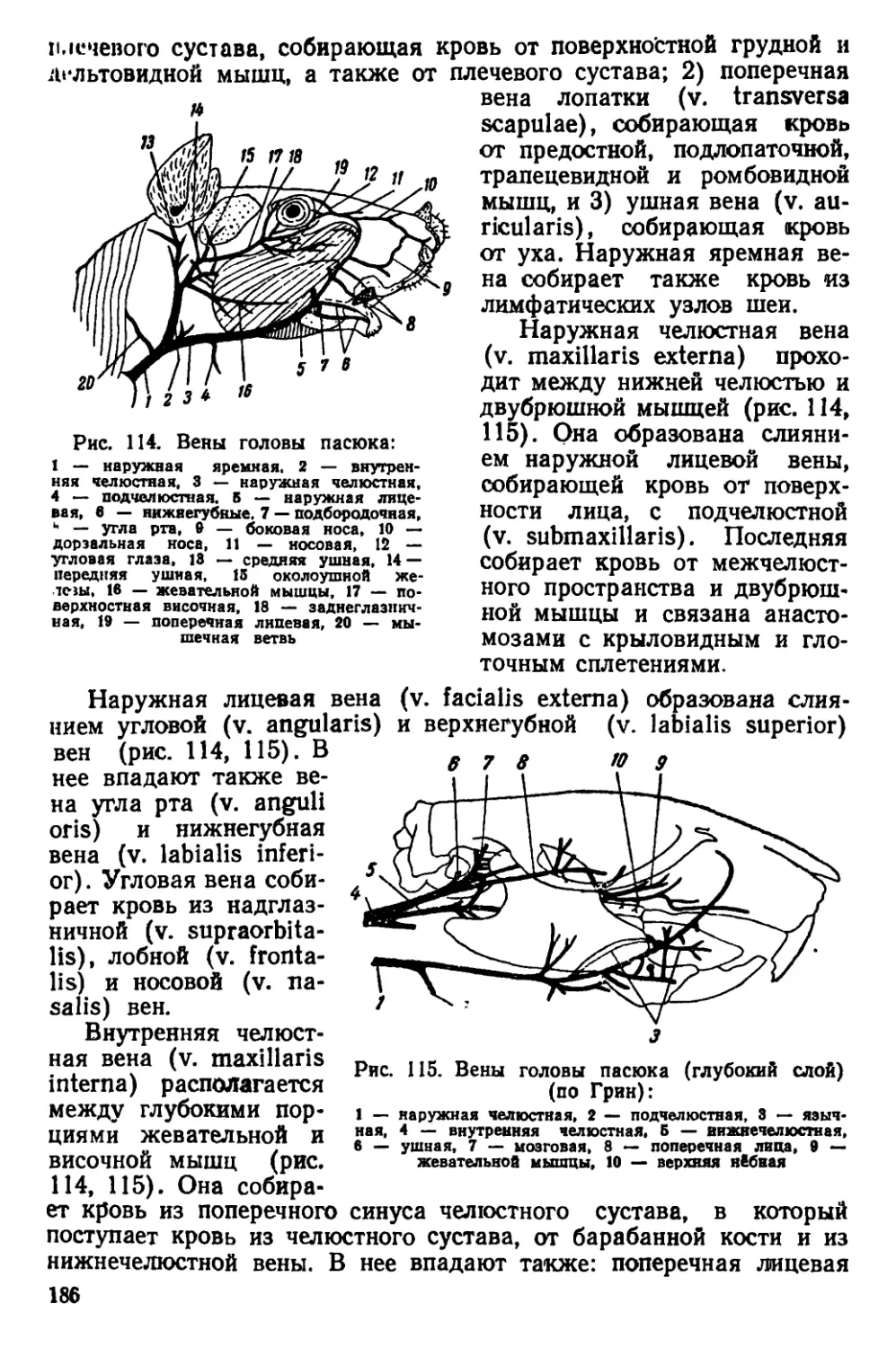




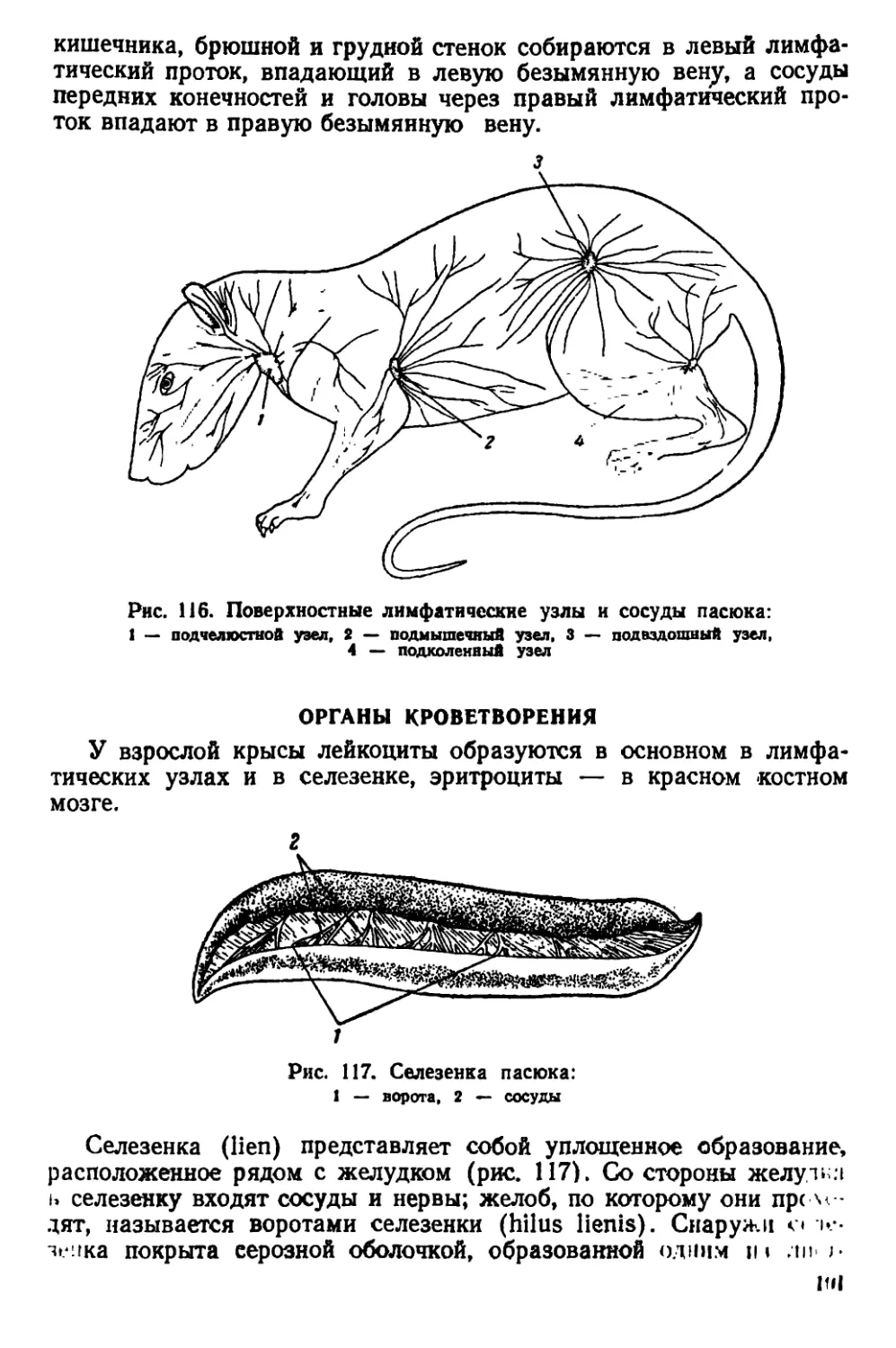



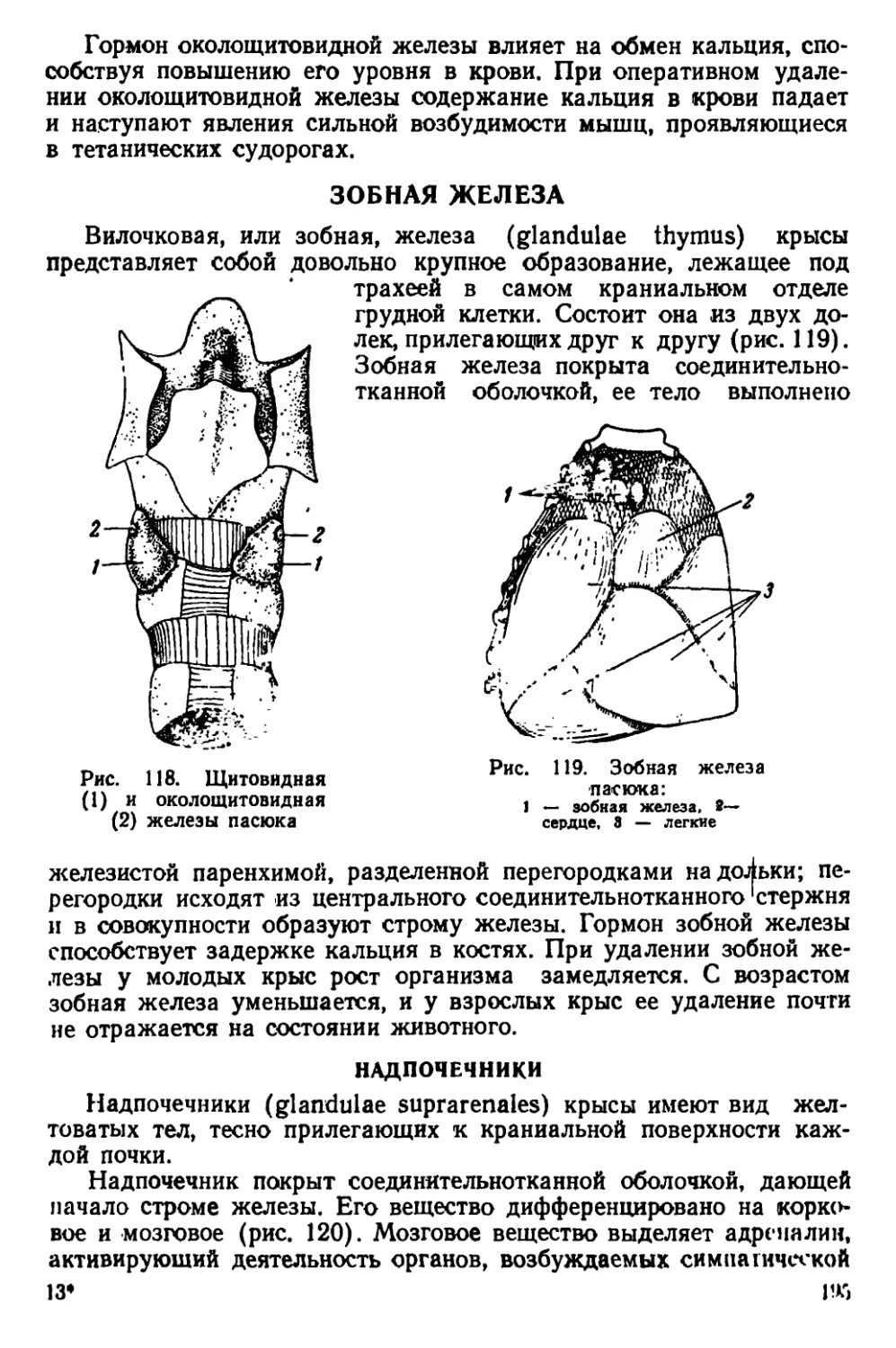





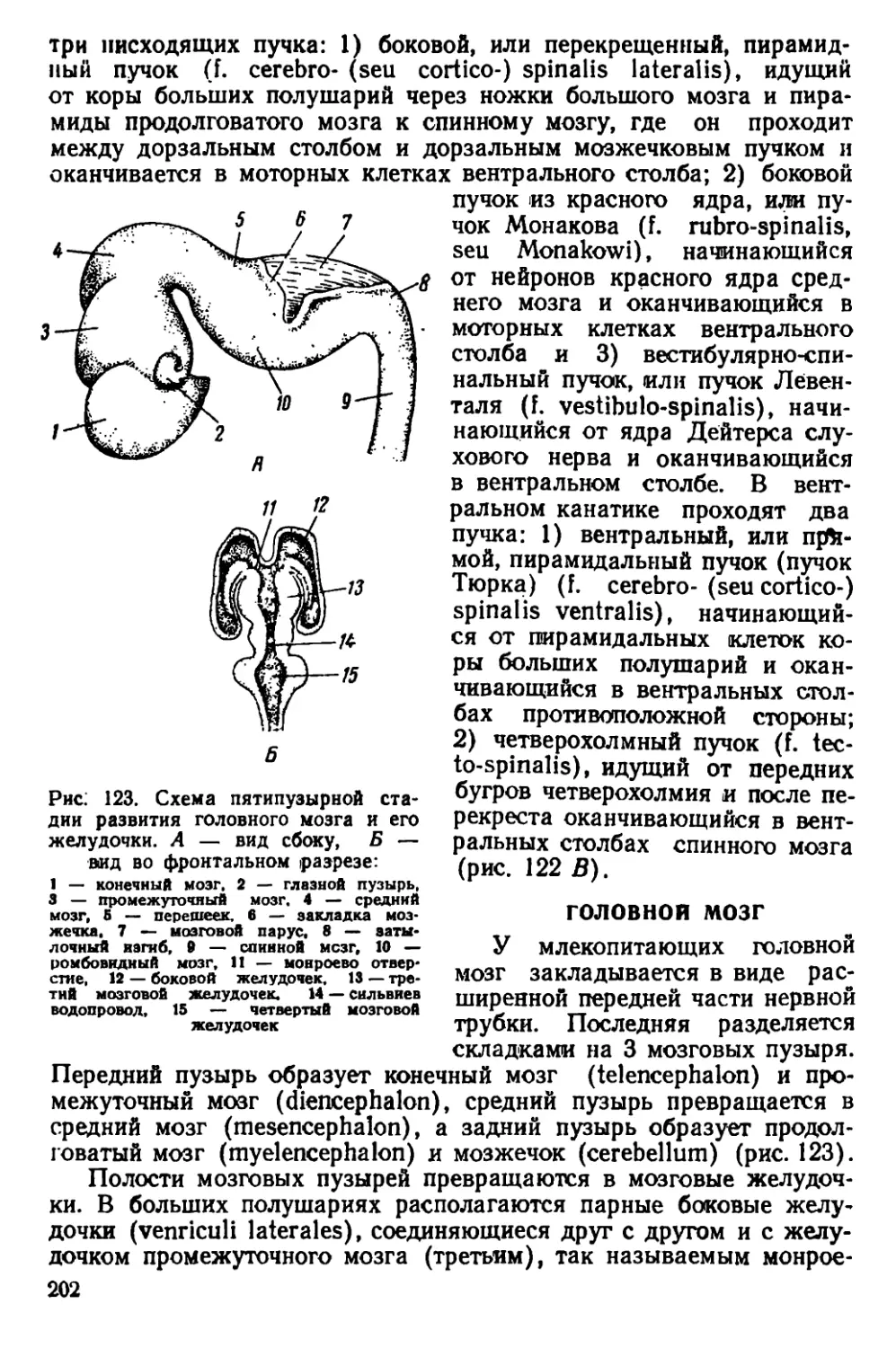







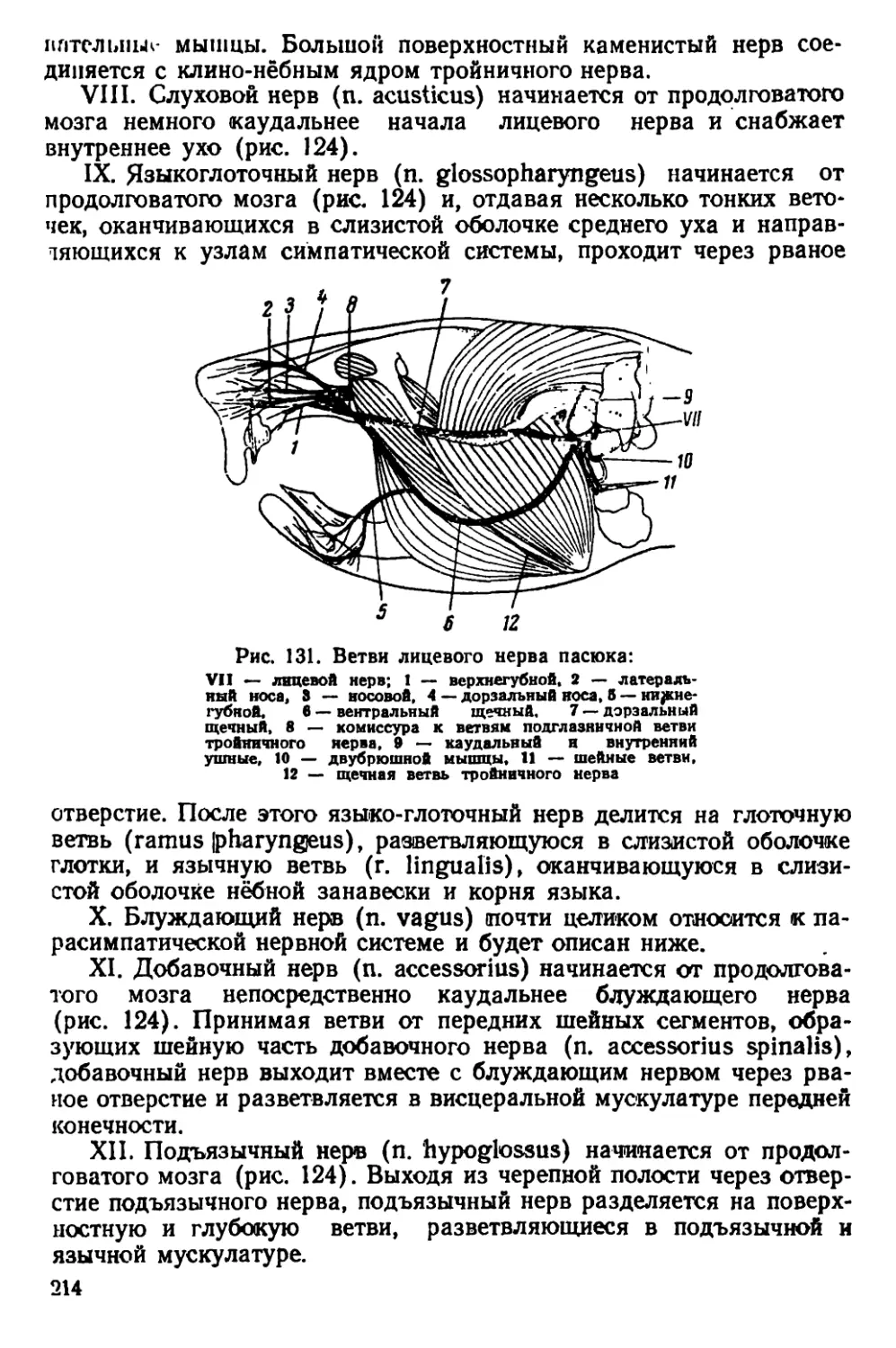
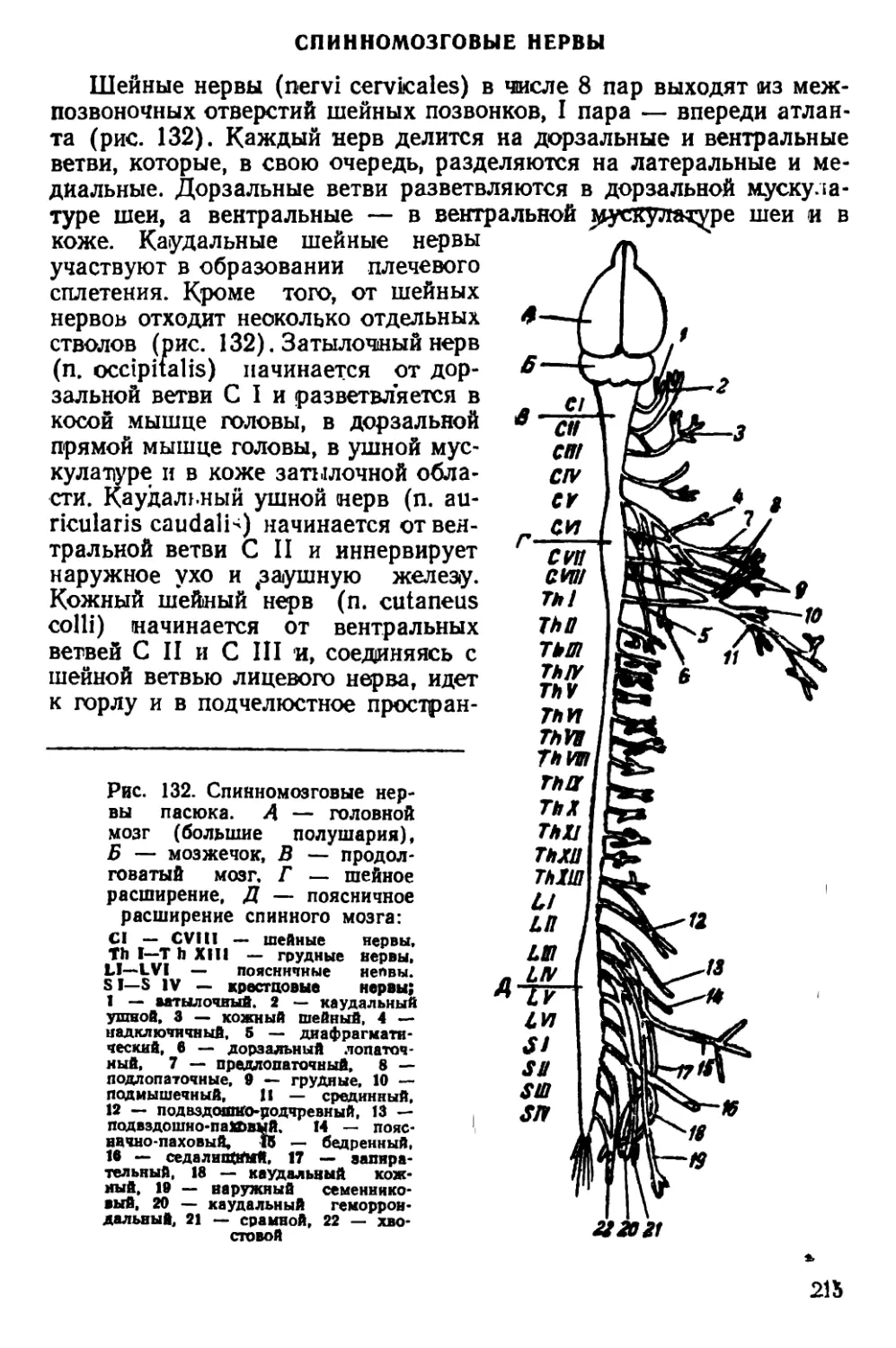

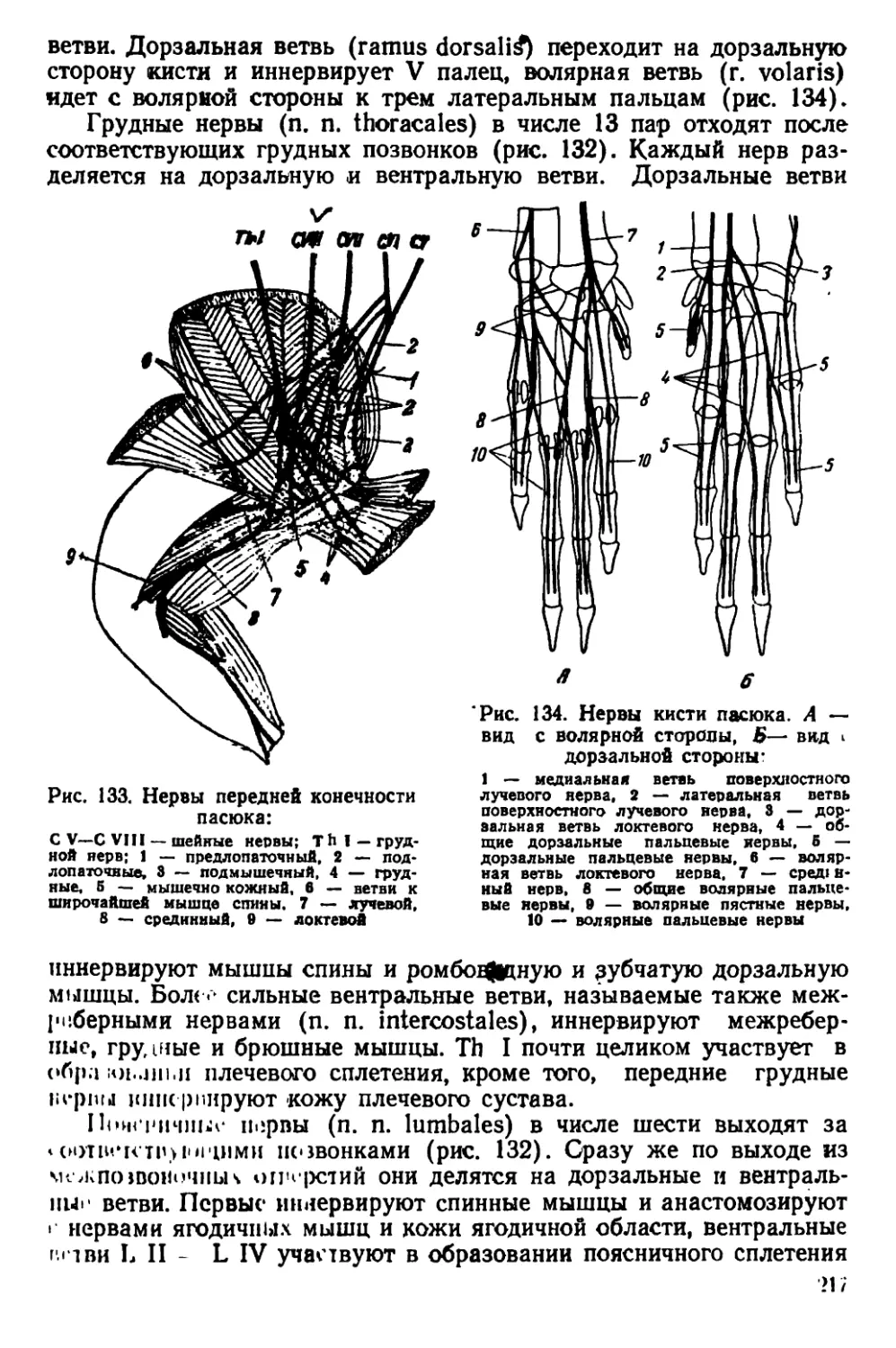
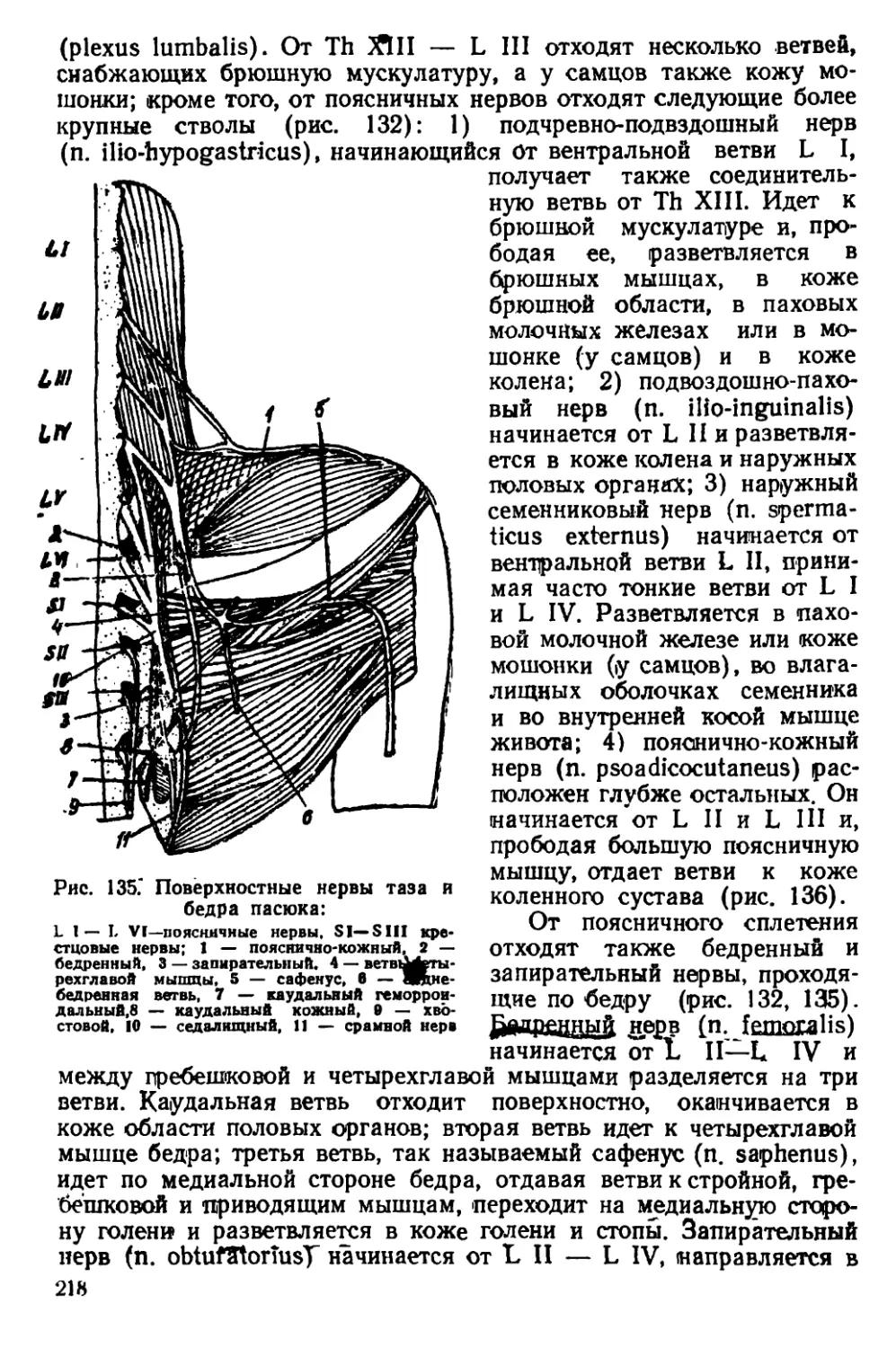
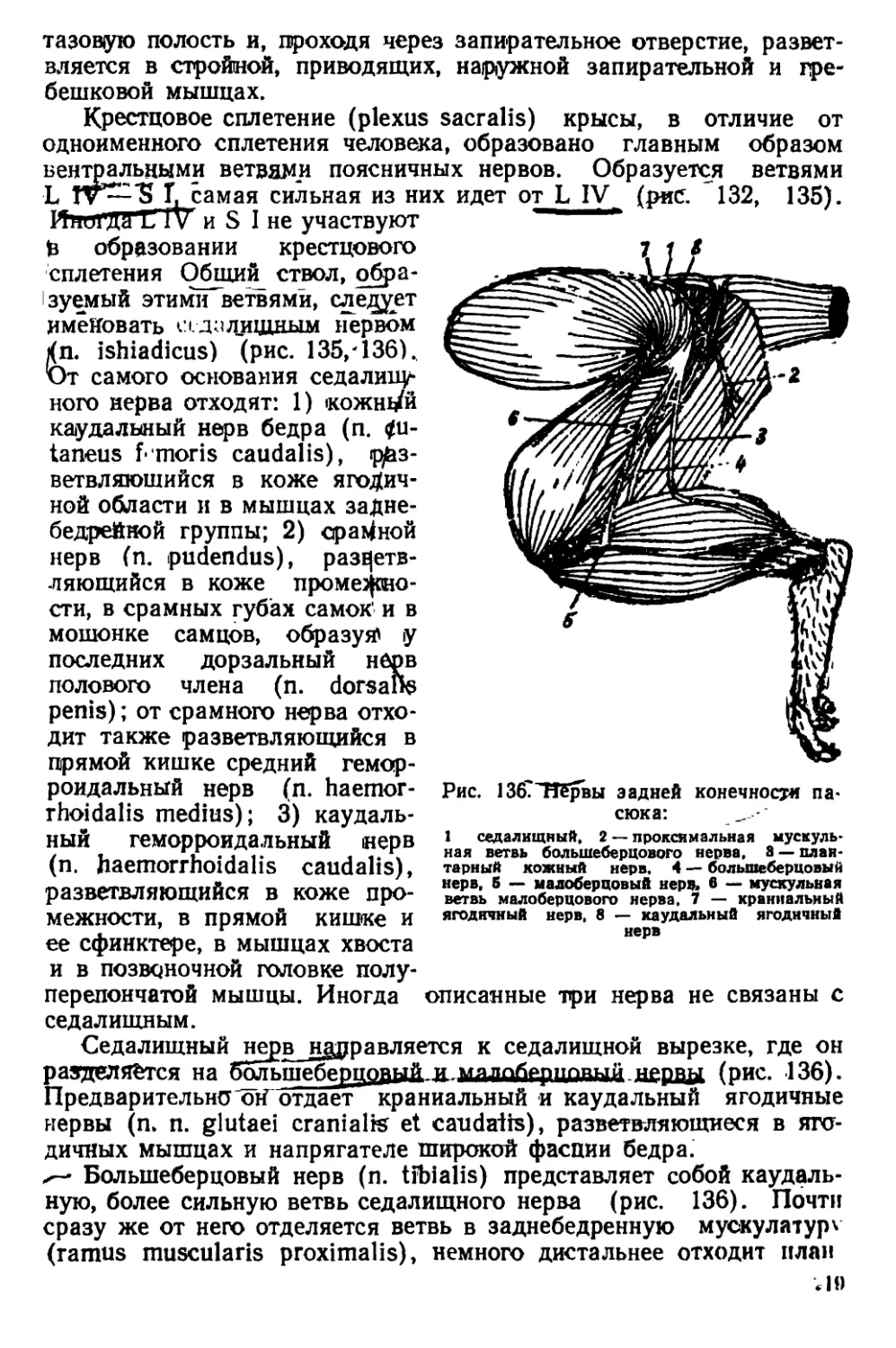
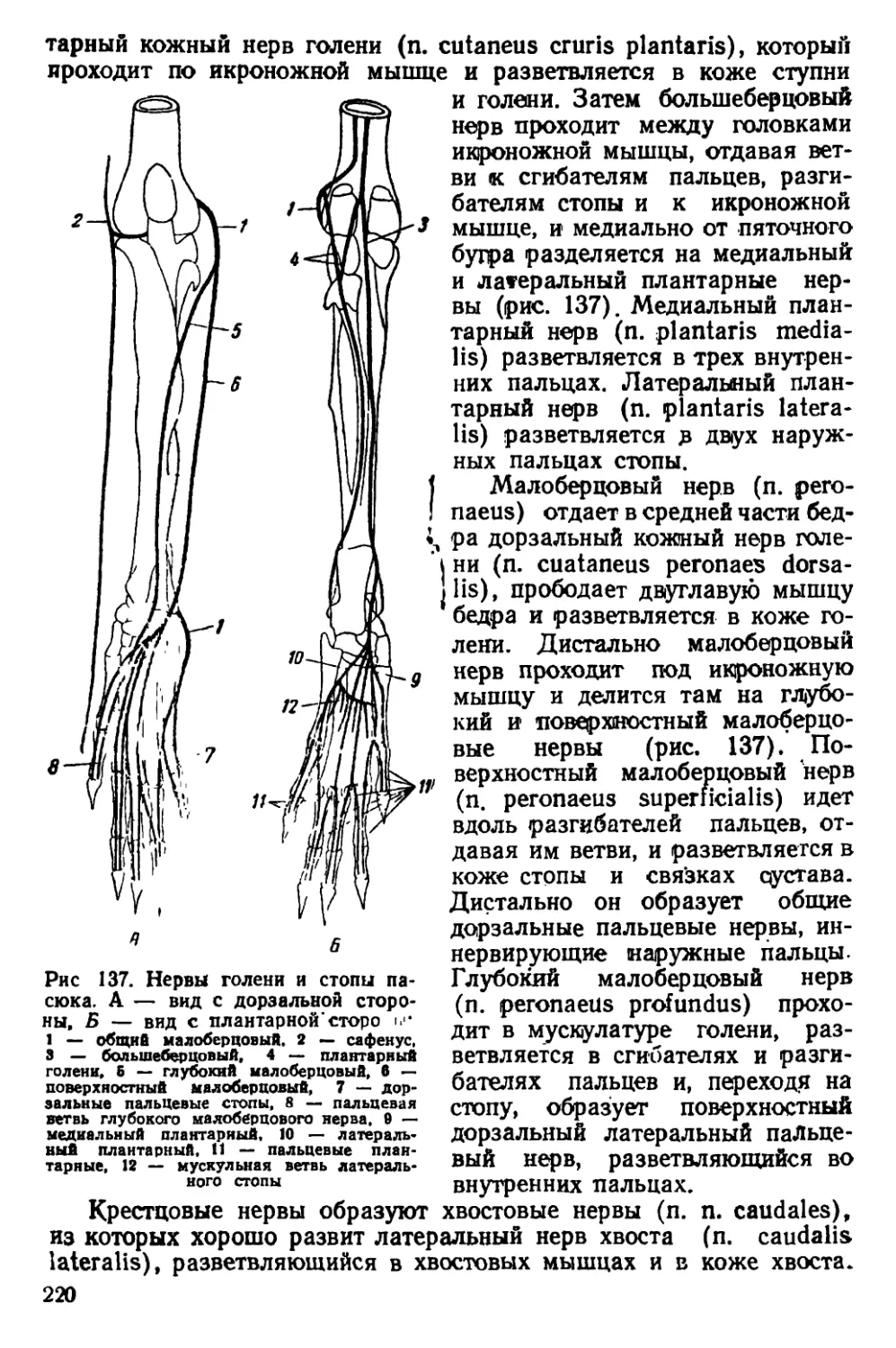
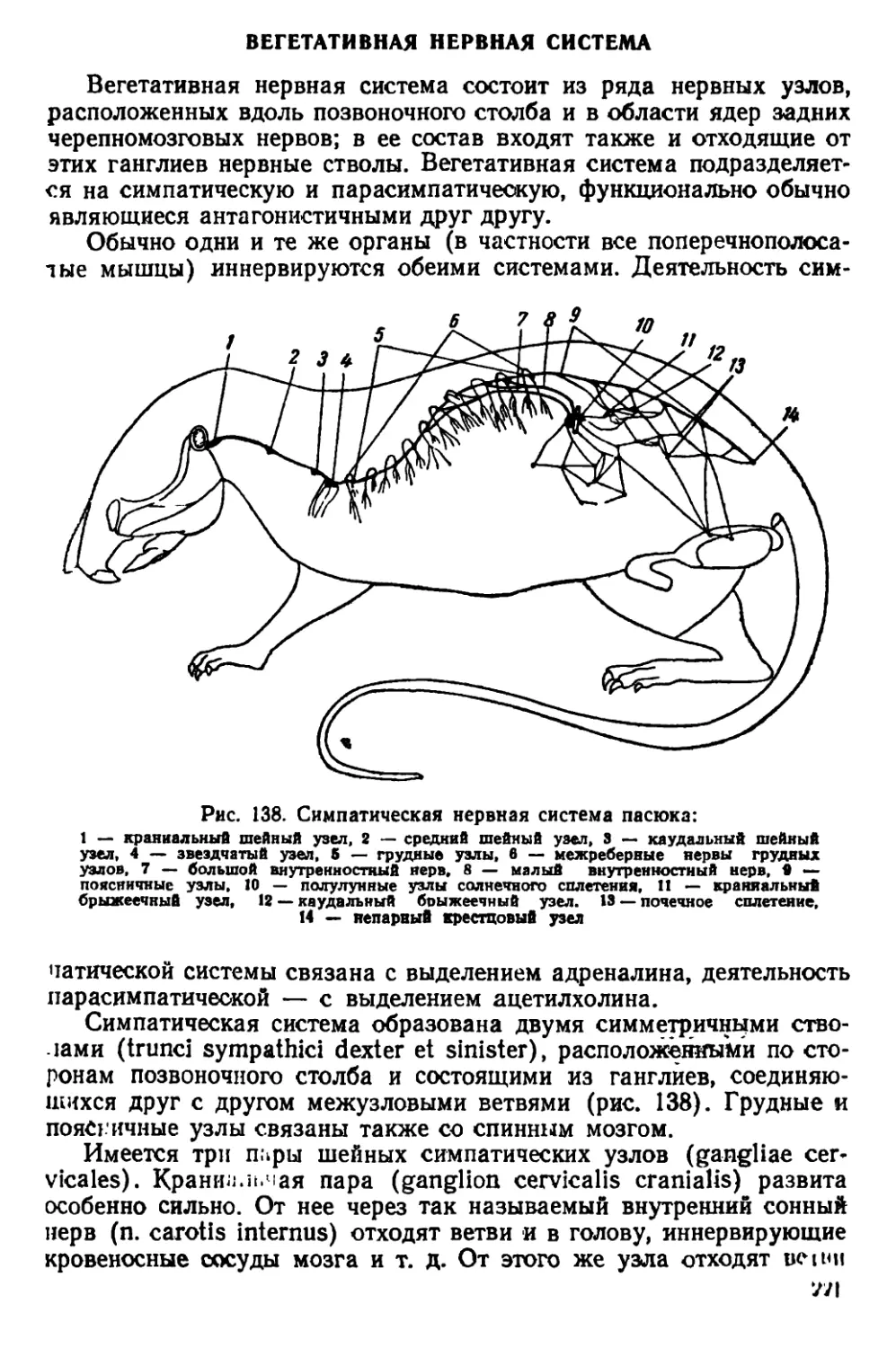



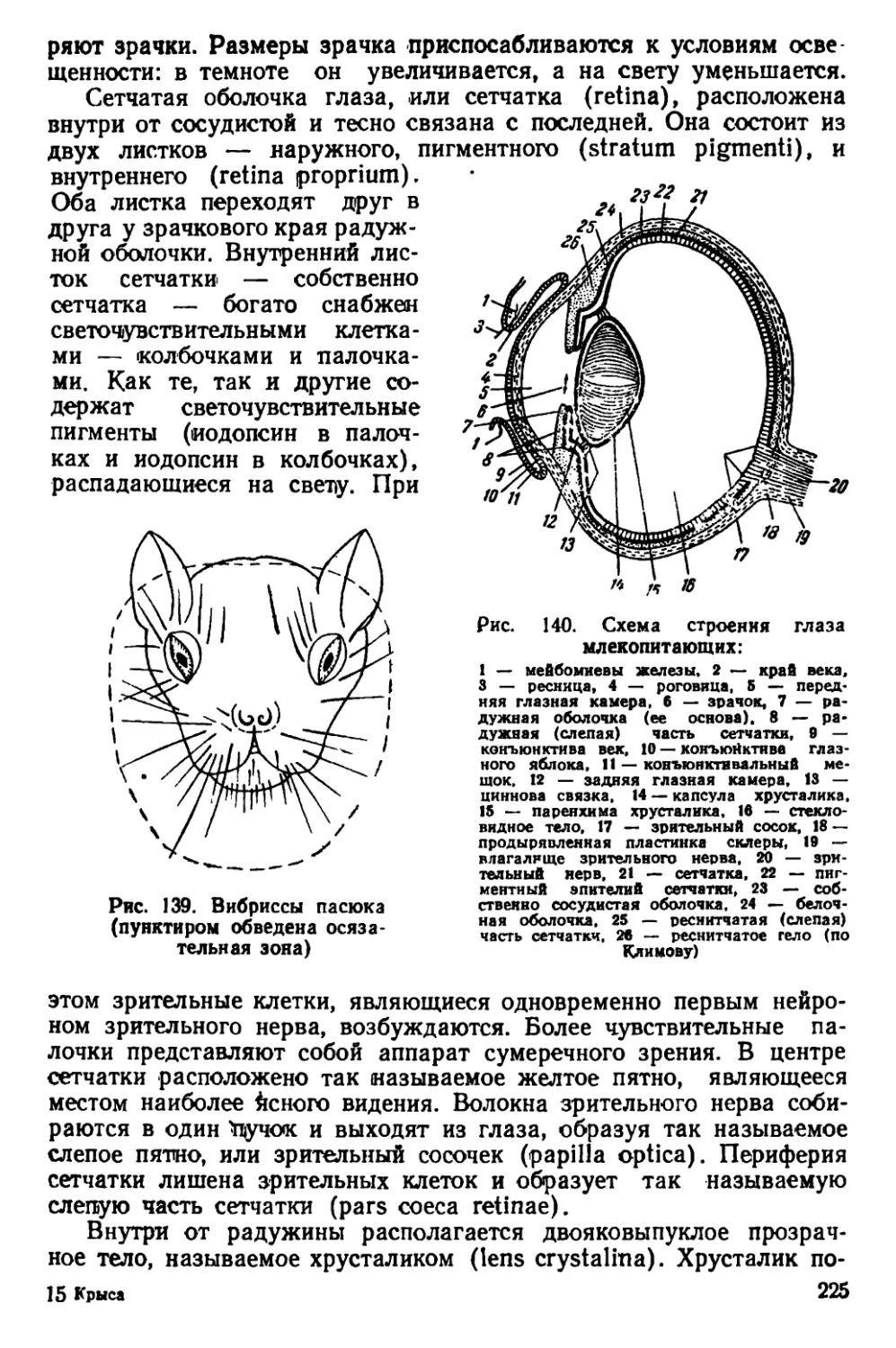



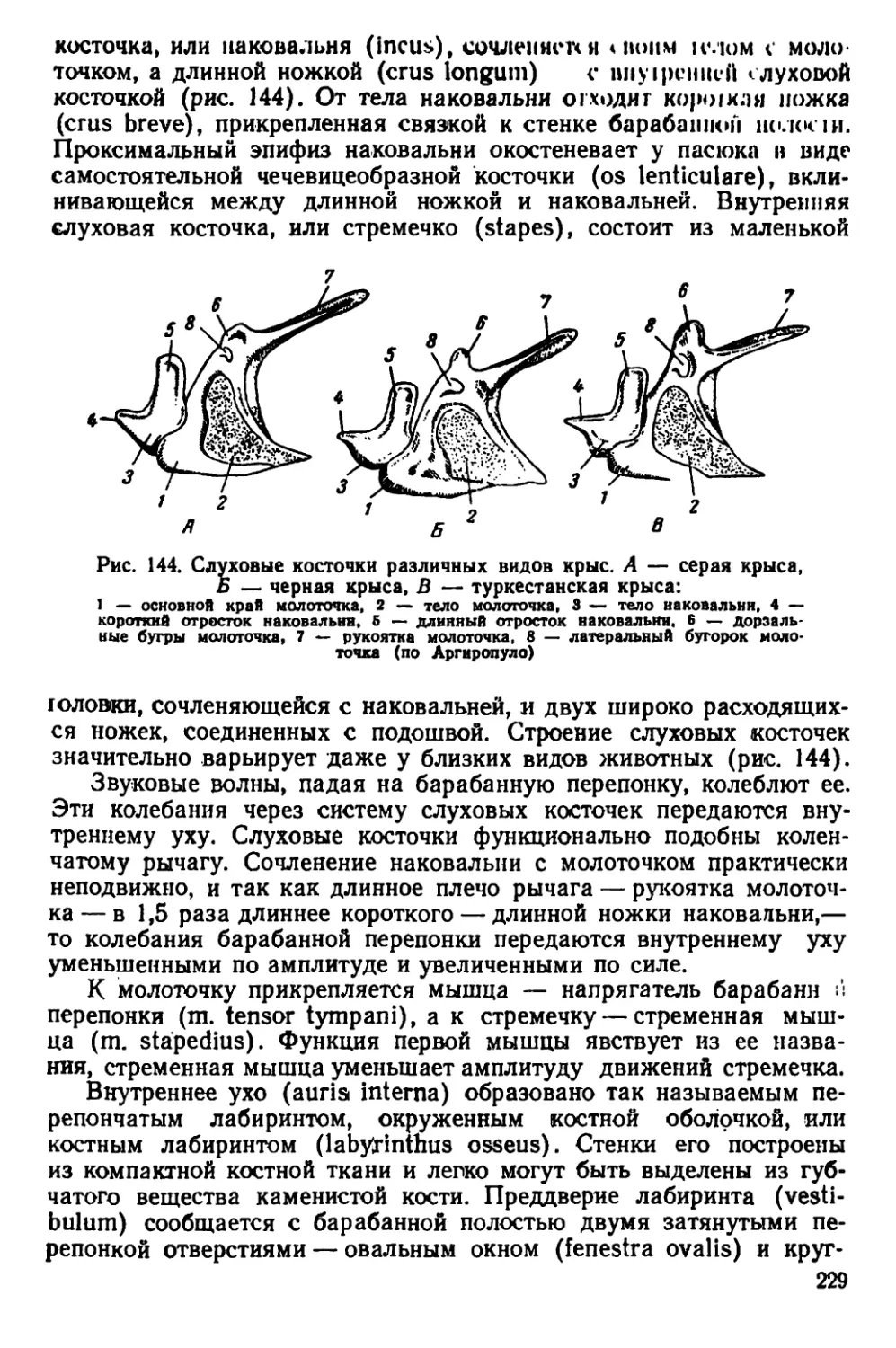
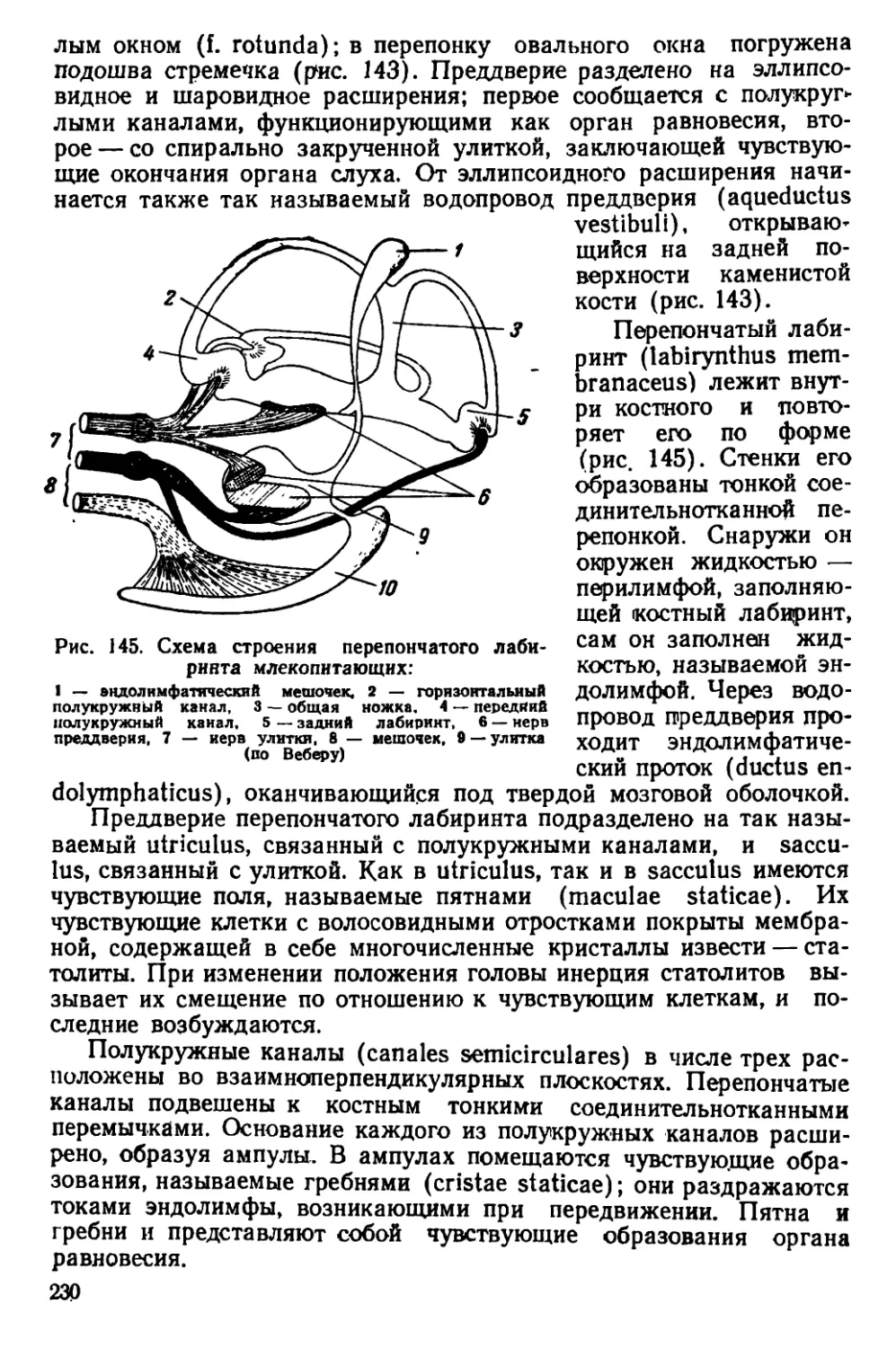





























Текст
П. П. ГАМБАРЯН и Н. М. ДУКЕЛЬСКАЯ
КРЫСА
Допущено
Главным управлением университетов,
экономических и юридических вузов
Министерства высшего образования СССР
в качестве учебного пособия
для государственных университетов
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО «СОВЕТСКАЯ НАУКА»
Москва — 1955
ОТ АВТОРОВ
В лабораторной практике в высших учебных заведениях и
научно-исследовательских институтах в качестве подопытных
животных широко используют белых крыс. Они необходимы
для экспериментальных работ при вирусологических, бактерио-
логических и токсикологических исследованиях и при изуче-
нии физиологии теплокровных животных. Белые крысы служат
объектом для вскрытий на практических занятиях по зоологии
при ознакомлении со строением тела млекопитающих. Кроме
того большой интерес имеет изучение диких крыс, так как
они являются носителями ряда инфекционных заболеваний и
вместе с тем наносят значительный экономический ущерб на-
родному хозяйству.
Несмотря на то, что еще Пастер, Мечников и Кох исполь-
зовали в экспериментальных работах белых крыс, литература
по нормальной анатомии этих животных, знакомство с кото-
рой необходимо при лабораторных исследованиях, совершенно
недостаточна. В отечественной литературе до настоящего вре-
мени отсутствует не только руководство по анатомии крыс, но
пс т и сводки разрозненных материалов по экологии и систе-
матике этой группы грызунов, опубликованных в различных
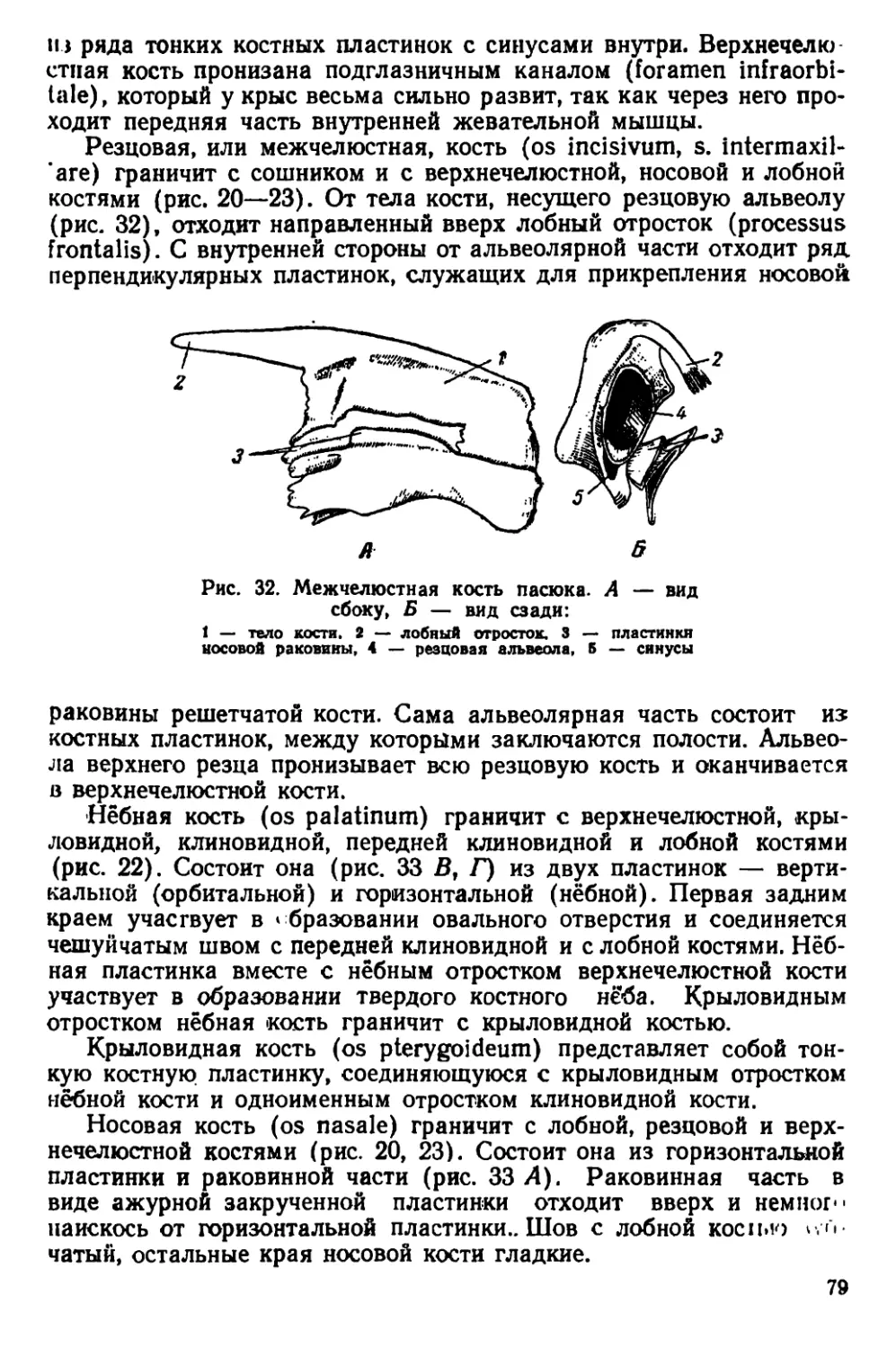
журналах.
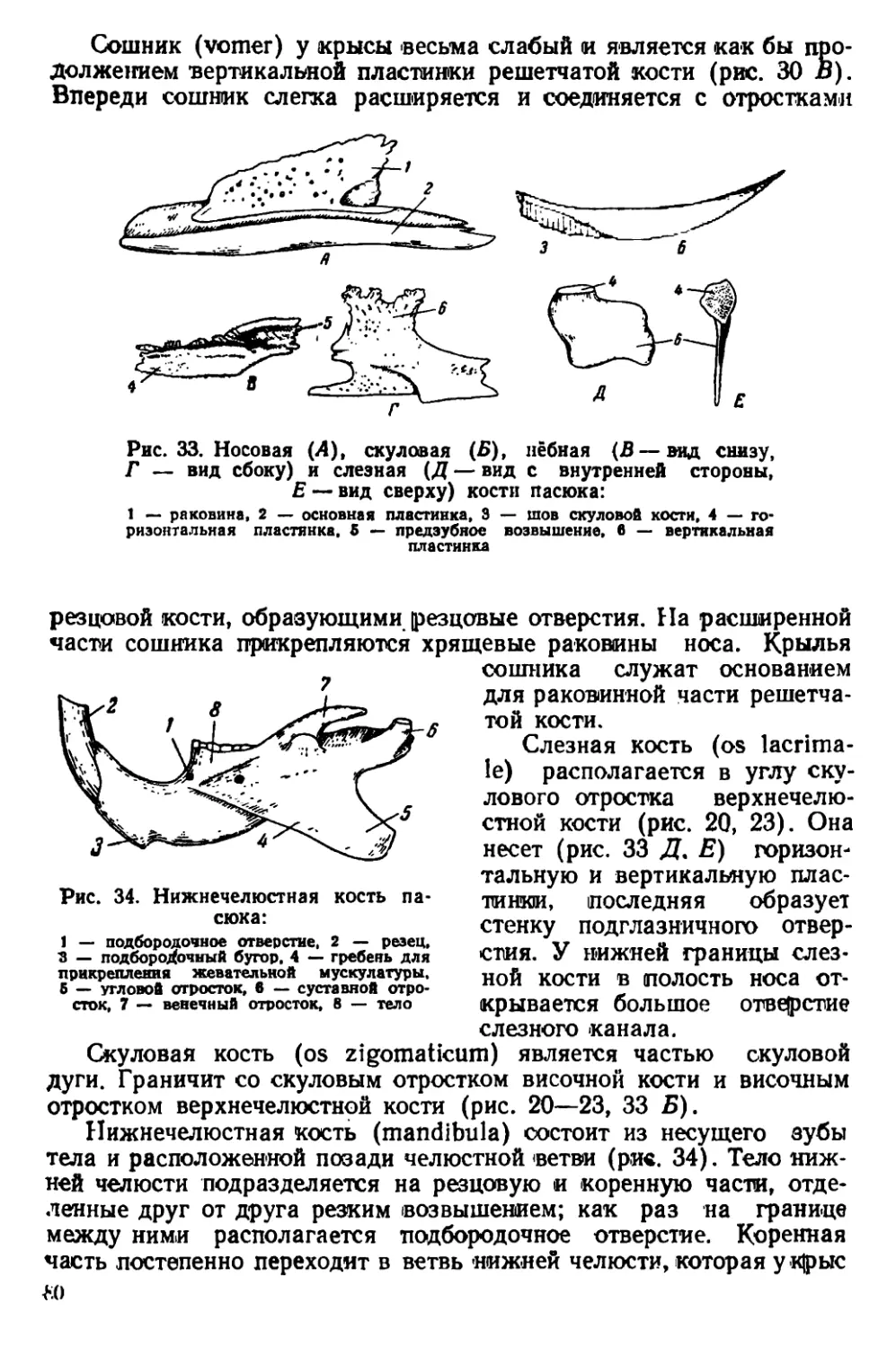
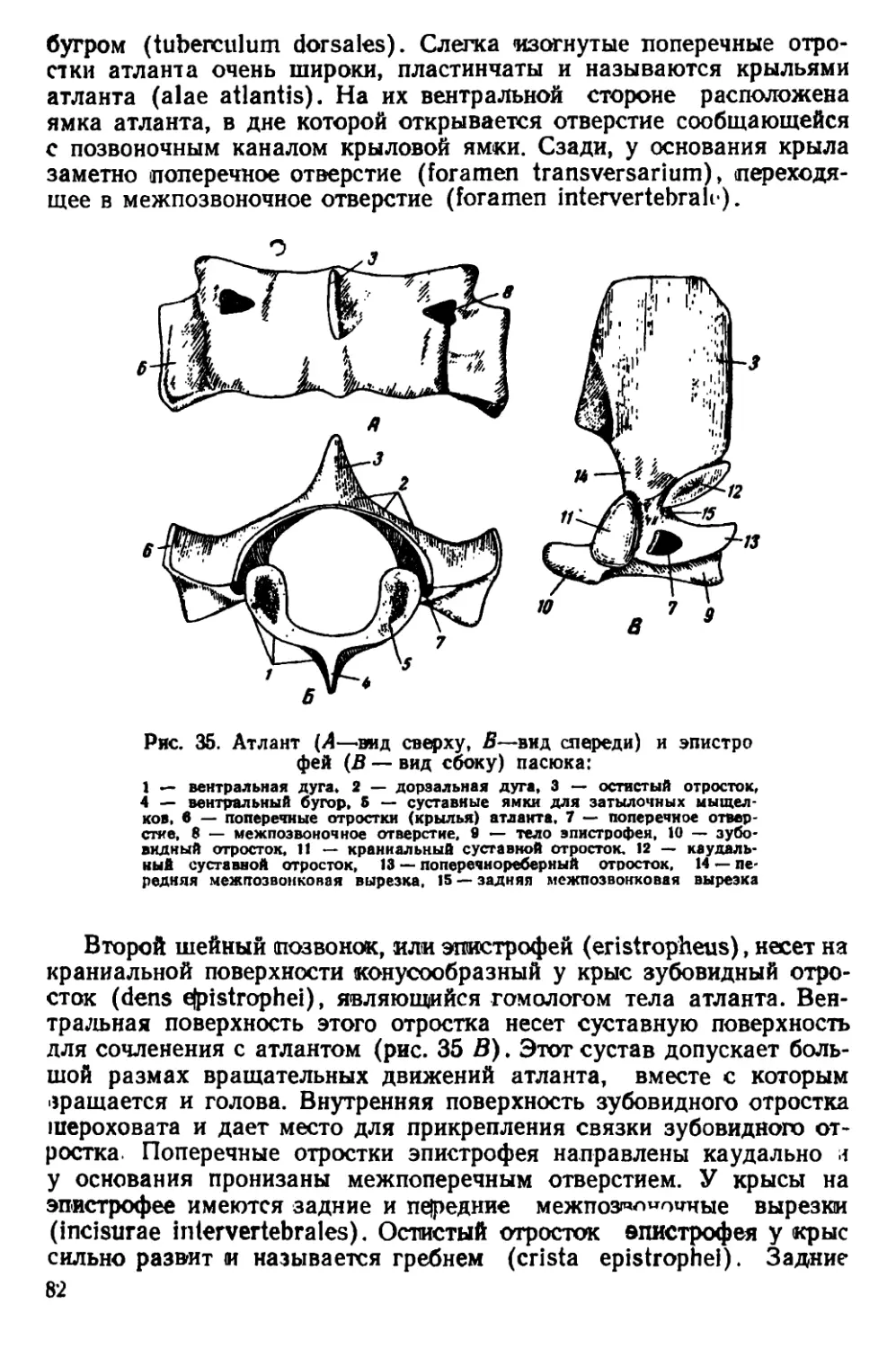
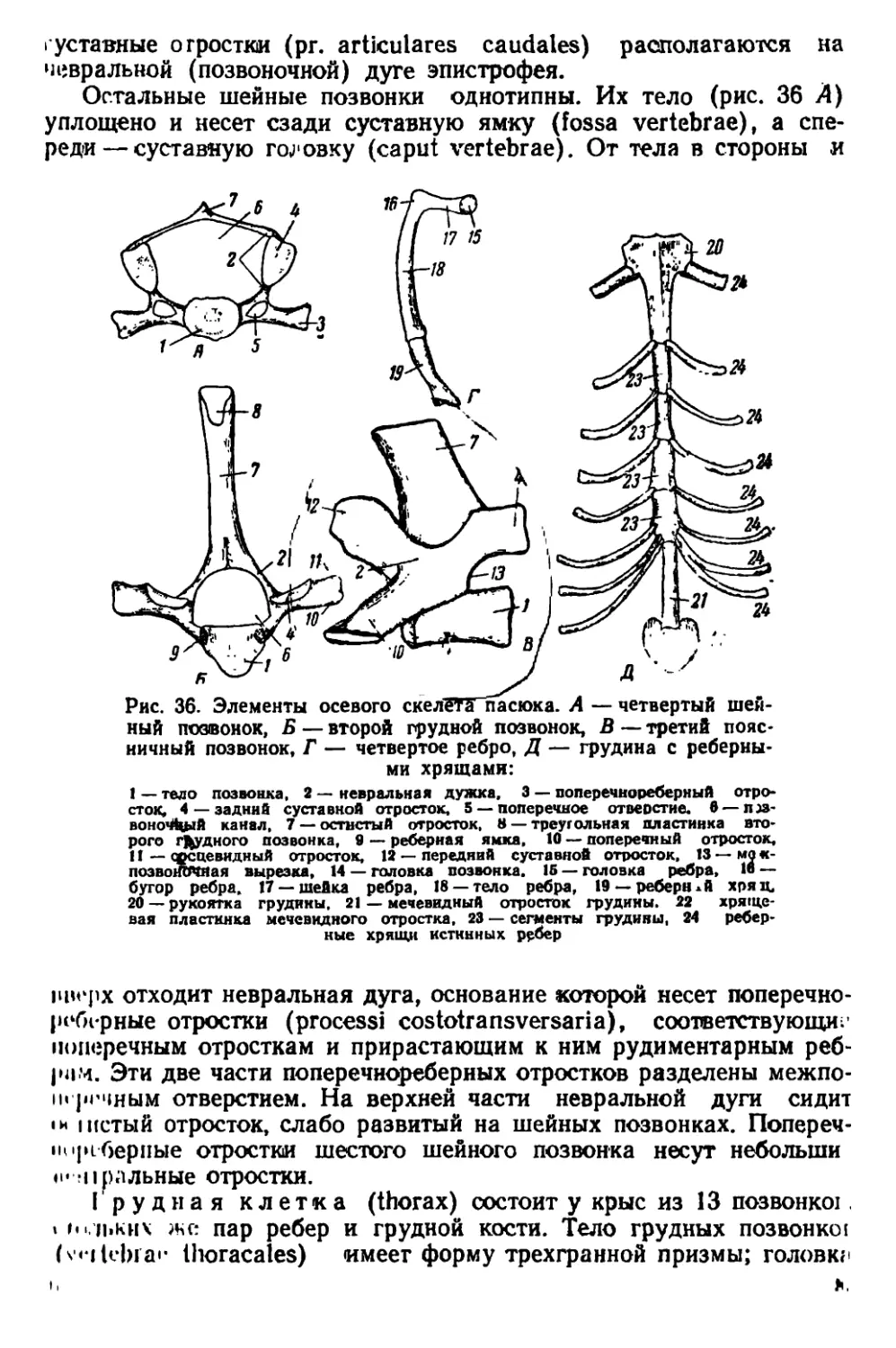
Указанные причины определяют необходимость издания ру-
ынюдства по систематике, экологии и анатомии крыс, которое
। м<> л»ст служить дополнительным учебным пособием для сту-
1Н’Ч1 дипломников и аспирантов. Такое руководство может
г«мн. ip пол».«iiuiiio и iui'K4-(bo справочника также физиолога-
ми, пни।<«мимн и тоологами.
При <1 н гпвлейии раздела, посвященного анатомии крыс,
II. II. I лмбарян пользовался консультацией Б. С. Виноградо-
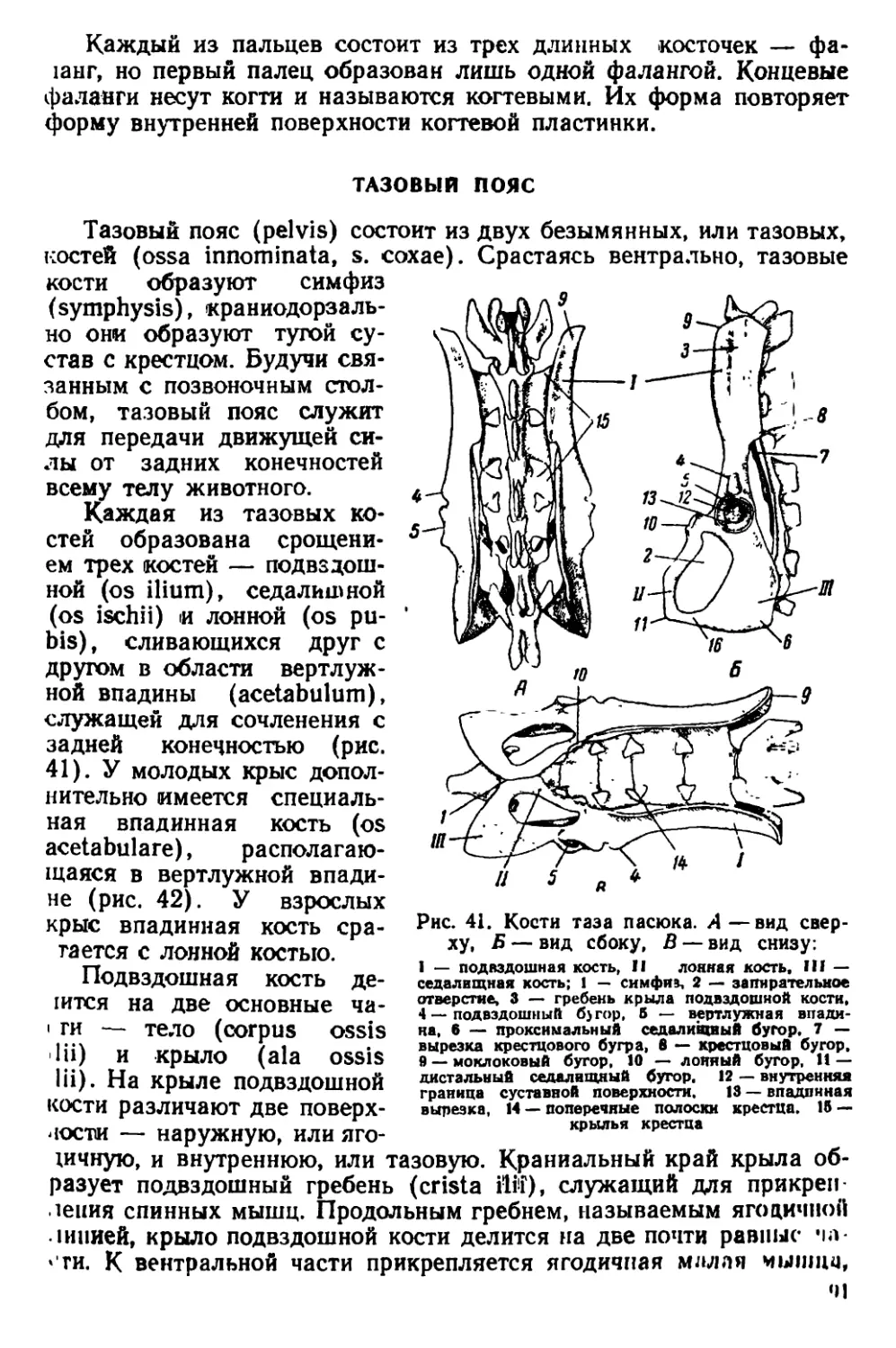
IHI, (•. М Смнренского и Г. А. Новикова; ему была оказана
помощь также со стороны А. С. Аветян, А. Е. и Е. Е. Терте-
рян. Всем этим товарищам он приносит свою глубокую бла-
1одарность.
ЧАСТЬ ПЕРВАЯ
ЭКОЛОГО-СИСТЕМАТИЧЕСКИЙ ОЧЕРК
ГЛАВА I
СИСТЕМАТИКА, ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
И МЕСТА ОБИТАНИЯ
СИСТЕМАТИКА РОДА RATTUS FISCHER
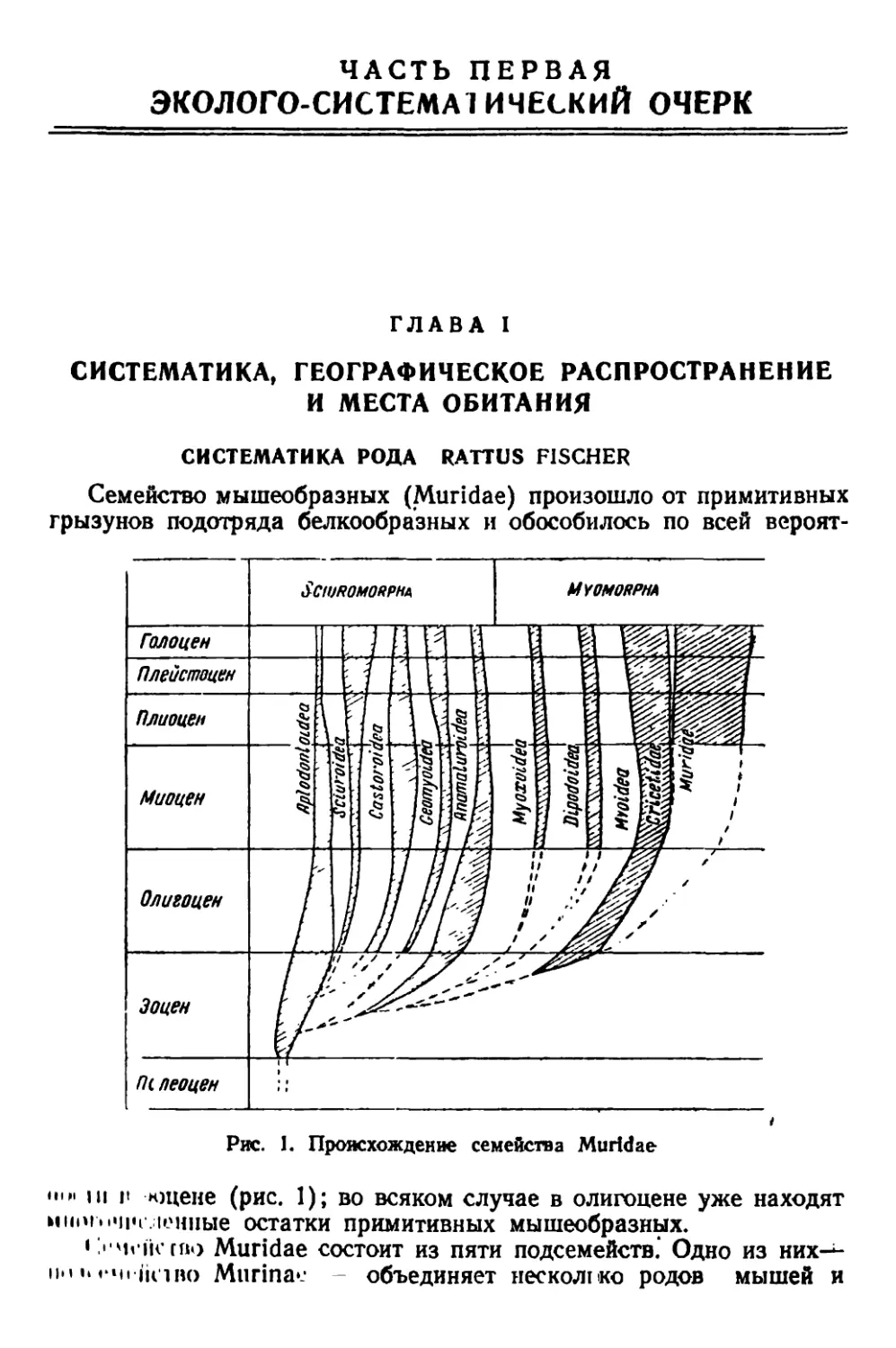
Семейство мышеобразных (Muridae) произошло от примитивных
грызунов подотряда белкообразных и обособилось по всей вероят-
Scujromorpha
My ом or ph а
Плиоцен
Миоцен
Эоцен
Голоцен
Плейстоцен
Олигоцен
ГК леоцен
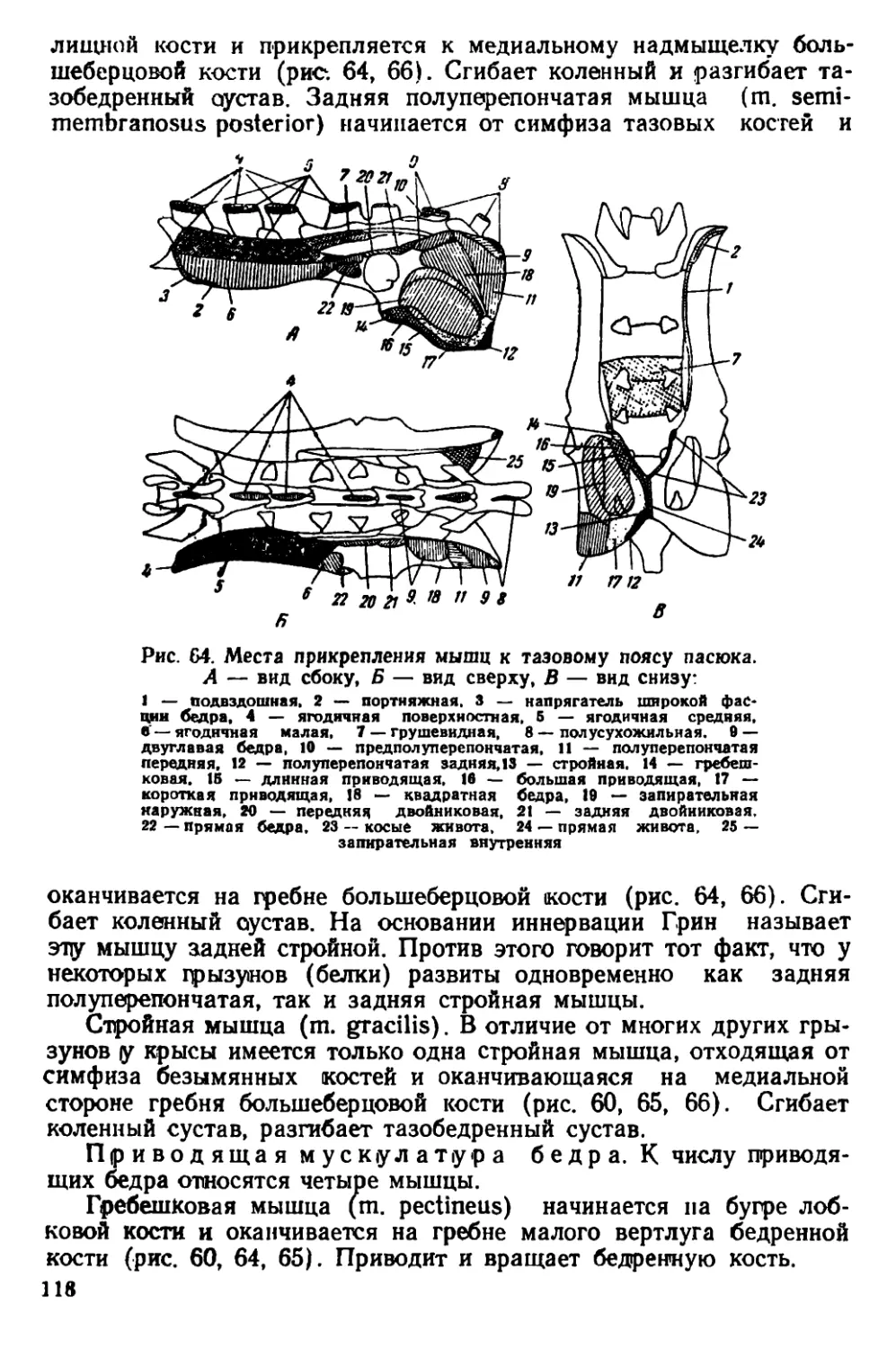
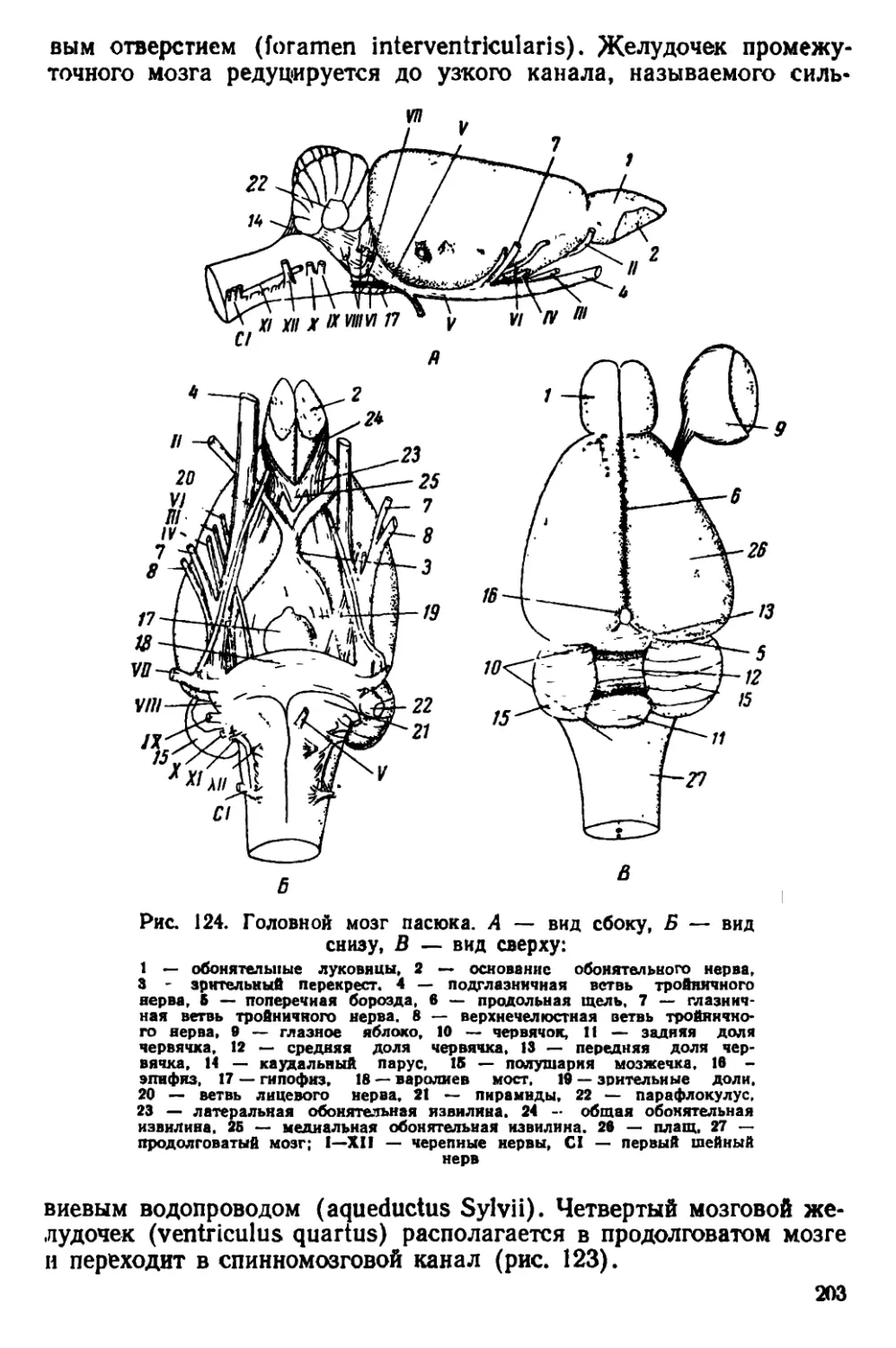
Рис. 1. Происхождение семейства Muridae
..hi г нщене (рис. 1); во всяком случае в олигоцене уже находят
иiii'b "ни-лепные остатки примитивных мышеобразных.
'j-'b iH rno Muridae состоит из пяти подсемейств.' Одно из них—
iiciBo Miirina'.- - объединяет несколико родов мышей и
i.pin, к !«•.» чм. -’ll* род Rallm I isch. (крысы) и род Ncsokia Gray
(ii.*t<K*ntn<i4to<y6uc крысы). Представители этих двух родов встре-
•i.uoi< я па территории СССР.
Водяные крысы только по внешним признакам несколько напо-
минают настоящих крыс. На самом деле они принадлежат к под-
семейству полевок (Microtinae) и их правильнее называть водяны-
ми полевками.
Род Rattus 6oiaT представителями, но в СССР встречаются толь-
ко три вида с подвидами: серая крыса, или пасюк (Rattus norve-
Рис. 2. Серая крыса
gicus Berk.), дальневосточный пасюк, или крыса-карако (Rattus
norvegicus caraco Pall.), черная крыса (Rattus rattus L.), александ-
рийская крыса (Rattus rattus alexandrinus Geofr.), рыжая крыса
(Rattus rattus rufescens Gray) и туркестанская крыса (Rattus
turkestanicus Sat.). В качестве лабораторных животных используют
альбиносов серой и черной крысы, которых разводят в специальных
питомниках или вивариях.
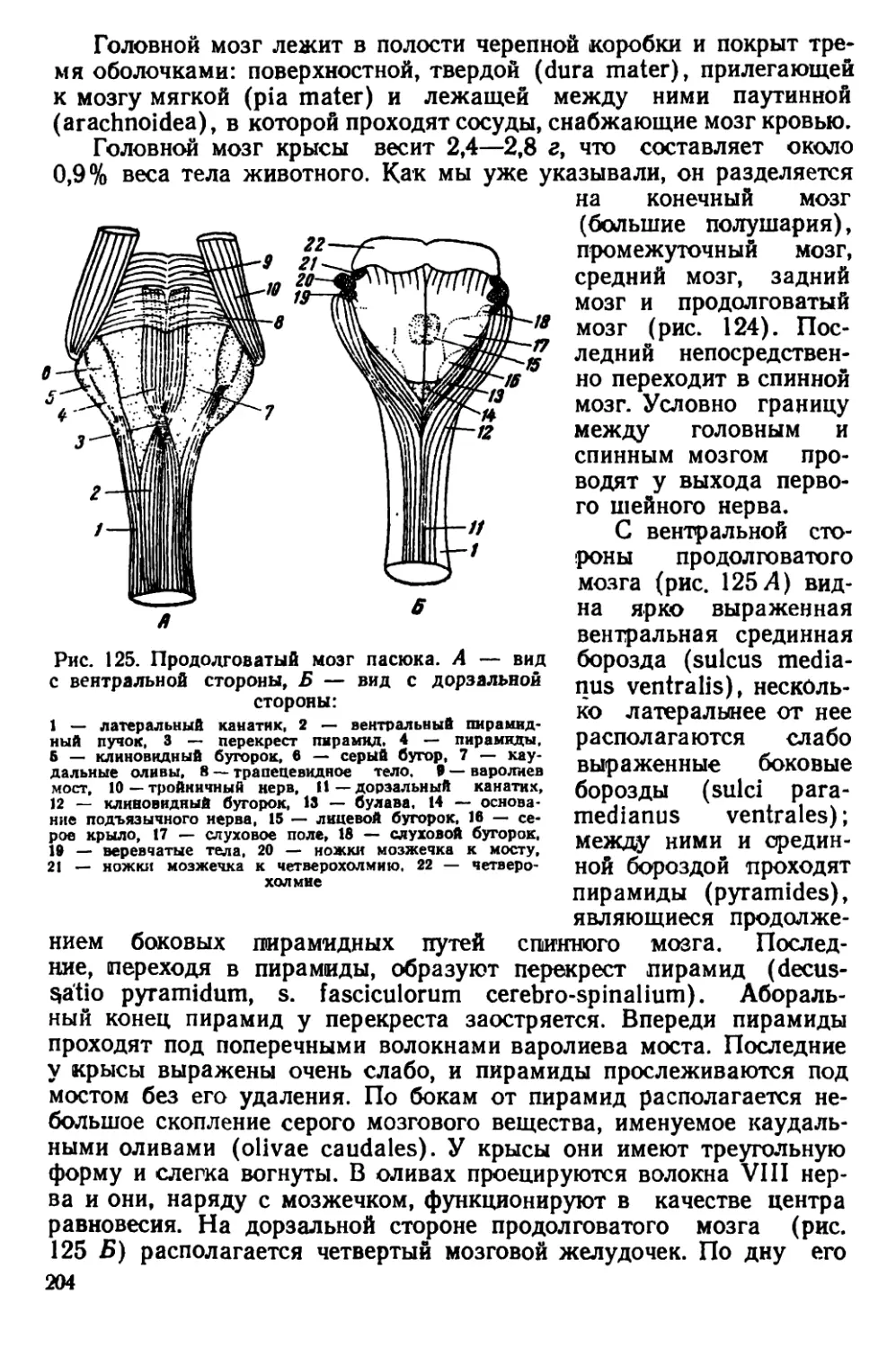
Серая крыса, или пасюк (Rattus norvegicus Berk.). Один из са-
мых крупных представителей подсемейства Murinae. Этот вид крыс
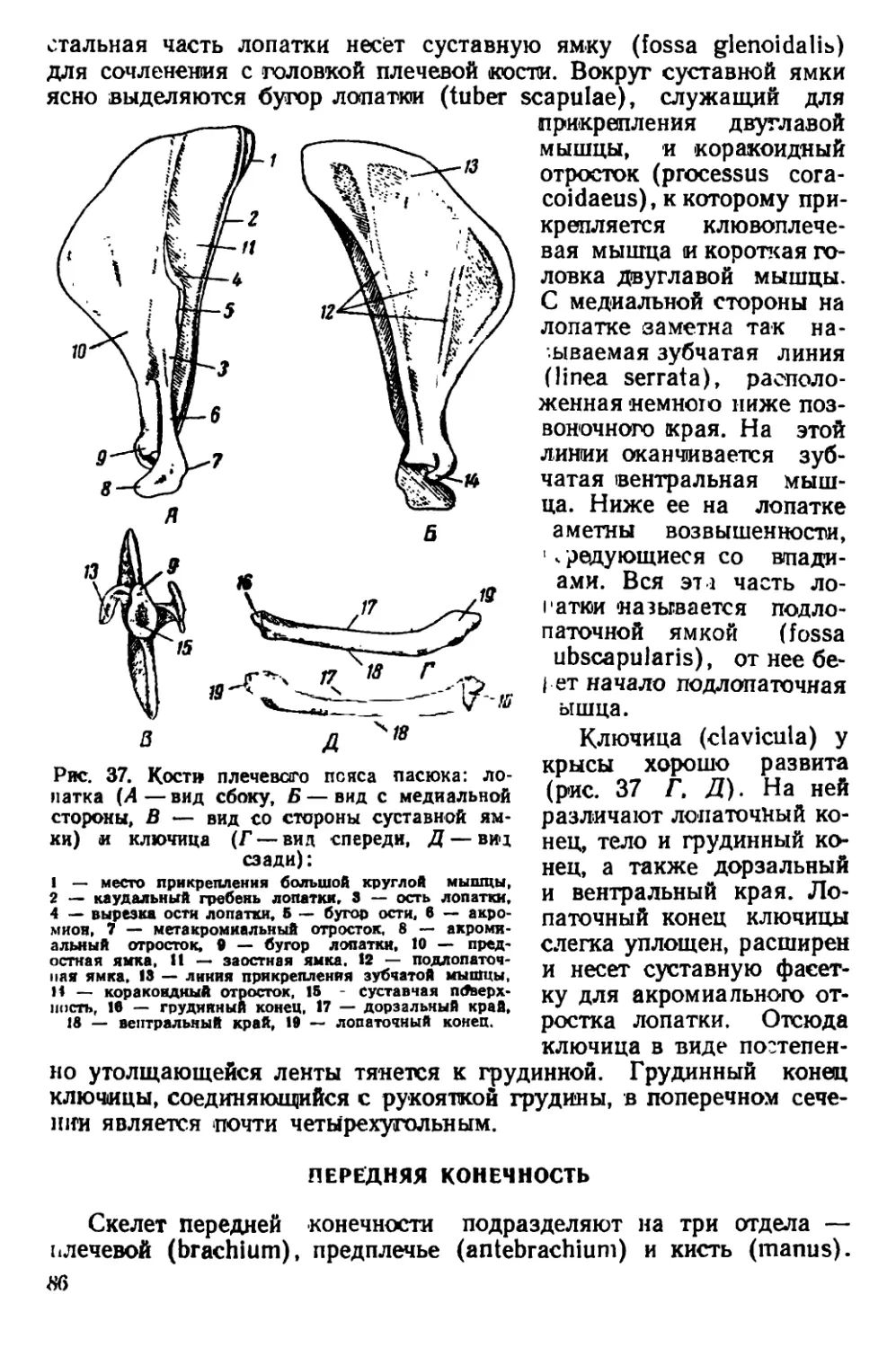
характеризуется плотным телосложением и широкой, тупой головой
(рис. 2, ЗД). Мех жесткий с длинными остевыми волосами. Окра-
«ка довольно изменчива. Верхняя сторона тела окрашена в рыже-
вато-бурый цвет с различно выраженными серыми и охристыми
тонами. Нижняя сторона светлее — серовато-белесая, с бурым от-
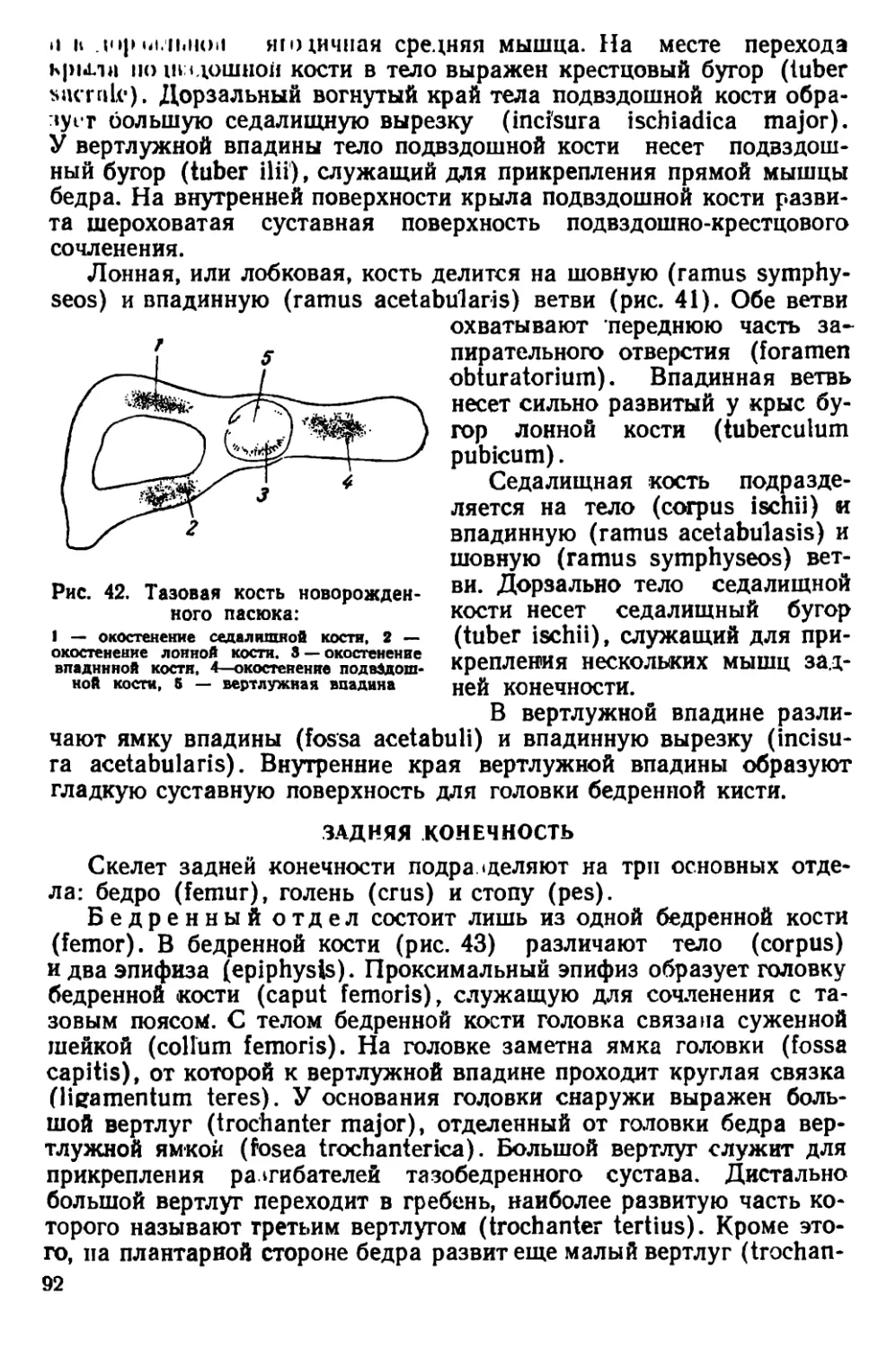
тенком. Хвост голый, покрыт чешуйчатыми кольцами и редкими
волосками между ними. Хвост обычно короче туловища, иногда
равен ему. Сосков 10—12. Ушная раковина небольшая; если ее
отогнуть вперед, то она не достигает края глаза. Между основания-
ми пальцев на задних конечностях имеются небольшие кожистые
перепонки.
Размеры тела: длина тела 150—248 мм, длина хвоста 100 —
120 мм, длина ступни (без когтей) 27—44 мм, высота уха 17 —
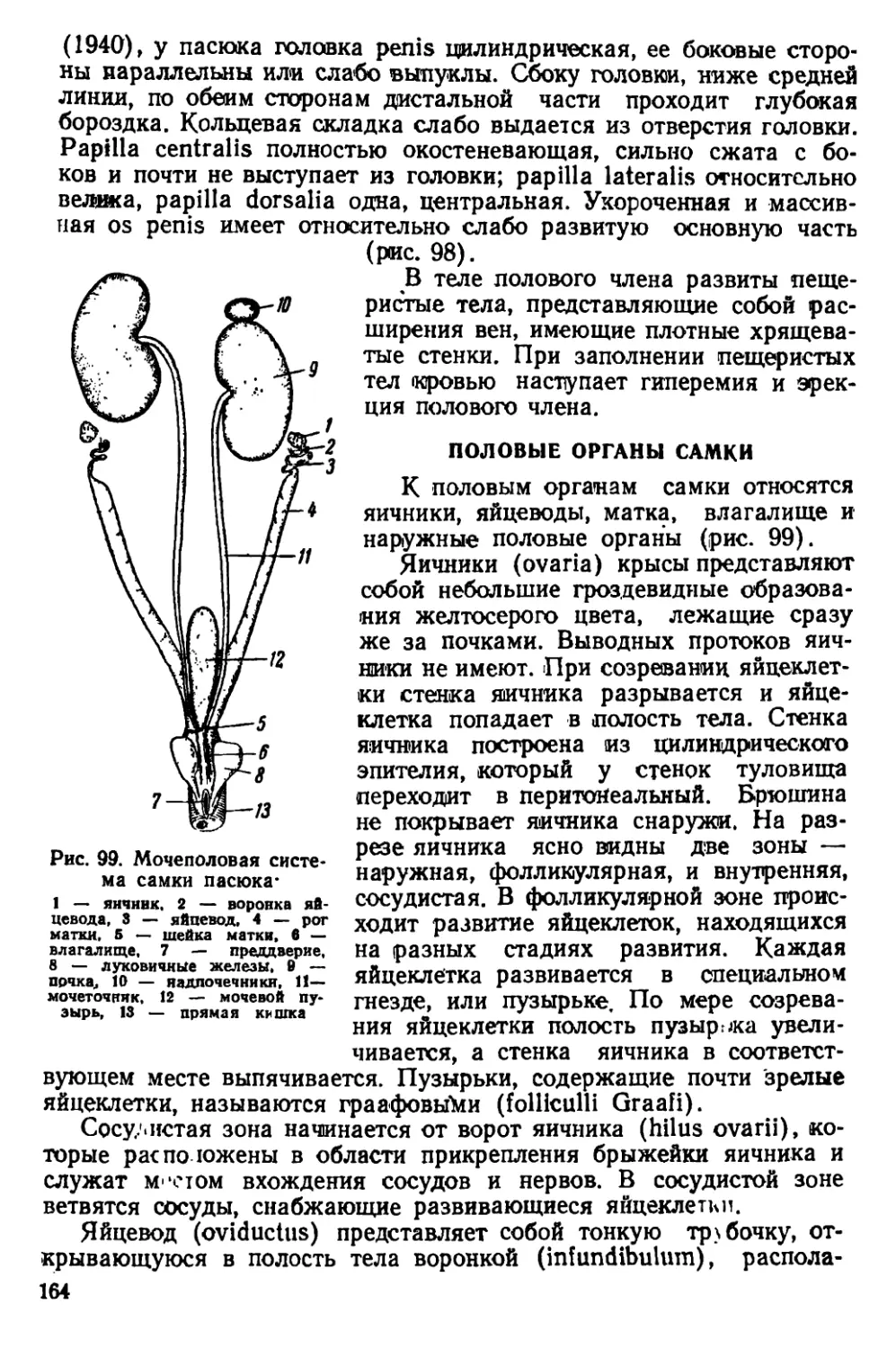
22 мм.
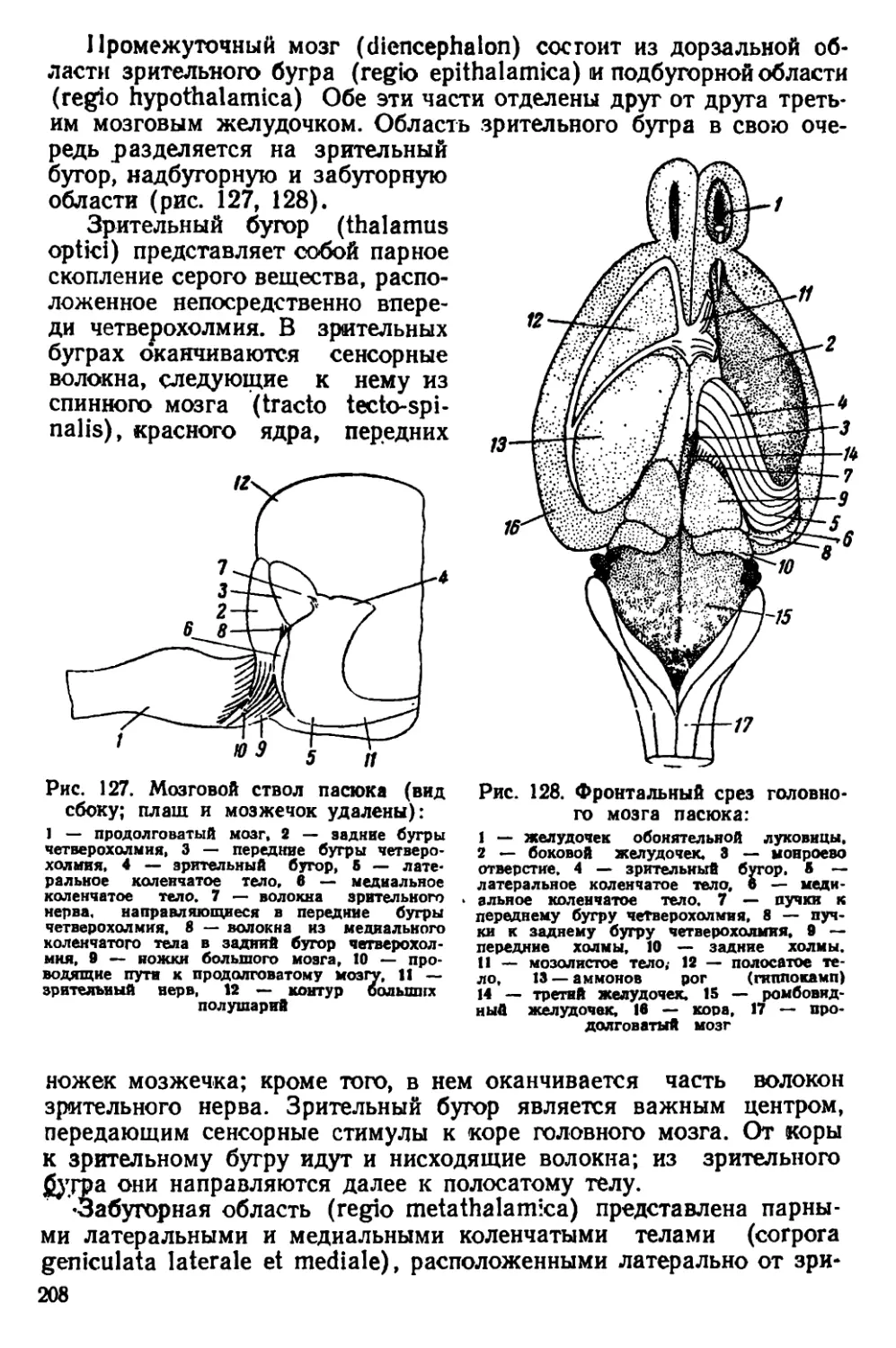
б
Ряс. 3. Головы крыс. А — голова серо*
крысы, Б — голова черной крысы
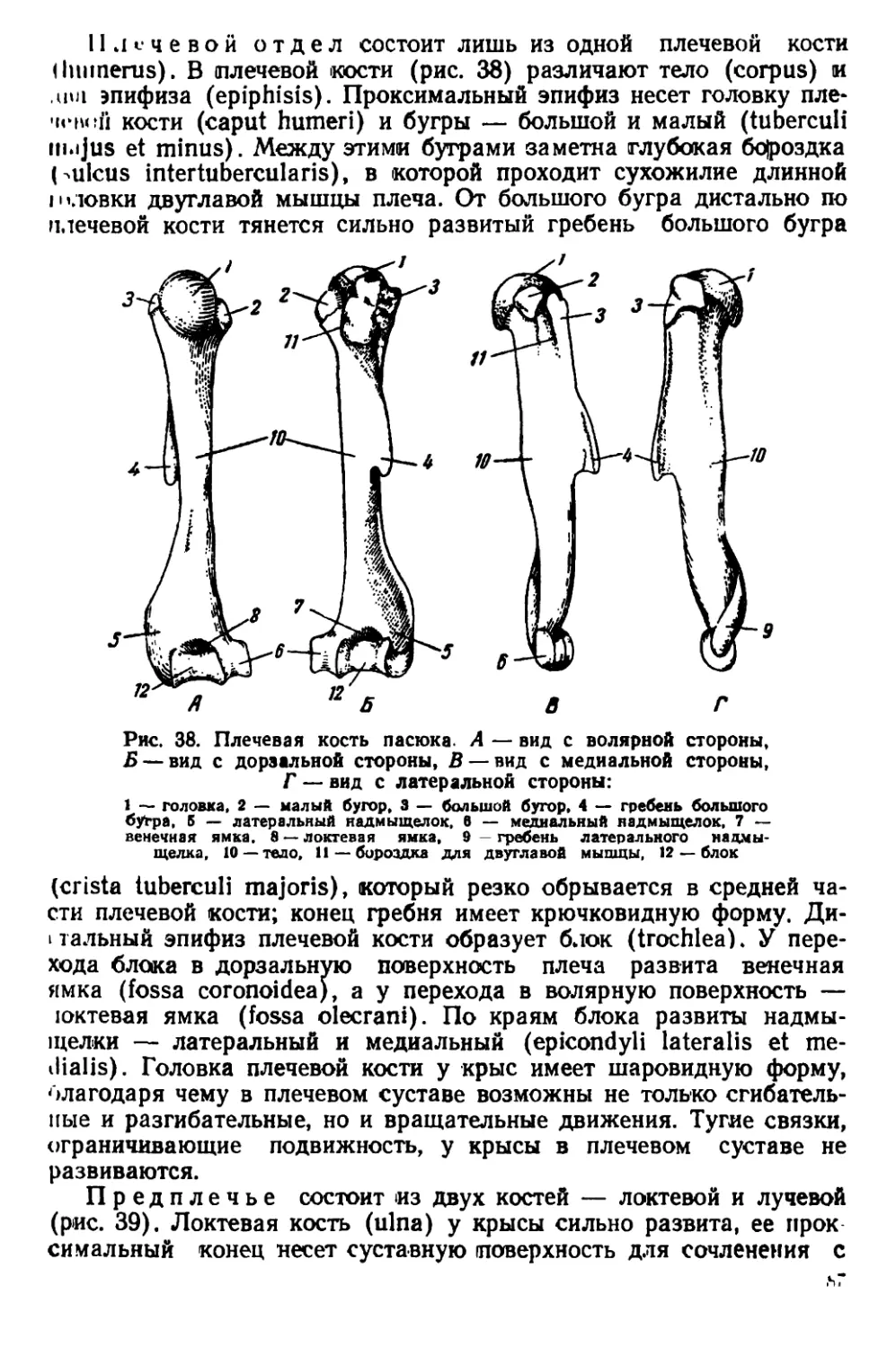
У взрослых особей на черепе развиты костные гребни, придаю-
щие ему угловатый вид (рис. 44). Гребни расположены почти па-
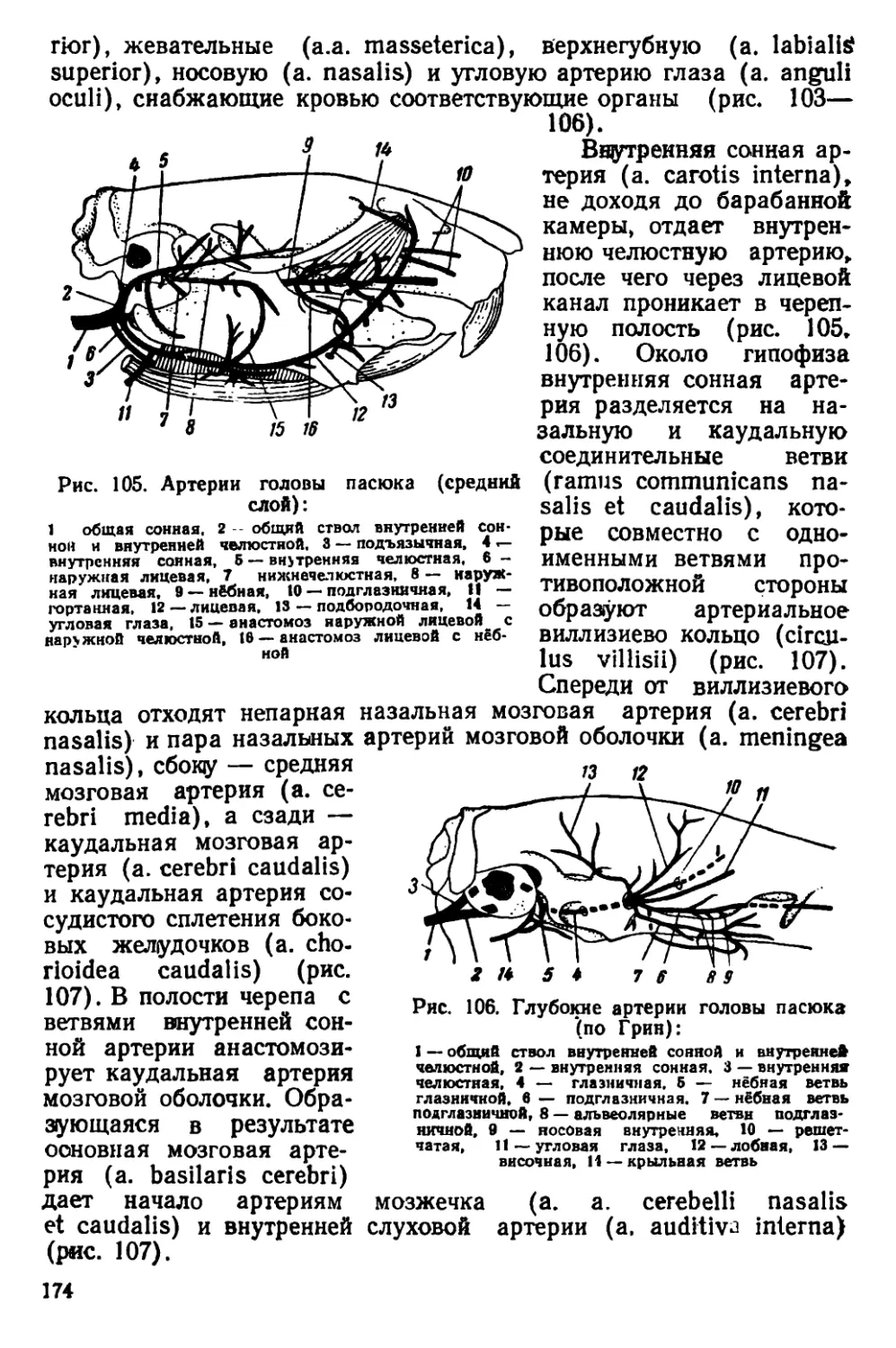
раллельно друг другу. Они идут вдоль теменных костей, изгибаются-
на уровне задней трети лобных костей и сближаются друг с другом
у переднего края скуловых дуг. Теменные кости плоские. Наиболь-
шей высоты череп достигает в области коренных зубов. Задний край
костного нёба расположен позади последних коренных зубов. Ску-
ловые дуги прижаты к
черепу; наибольшей шири-
ны они достигают в своей
задней трети. Слуховые
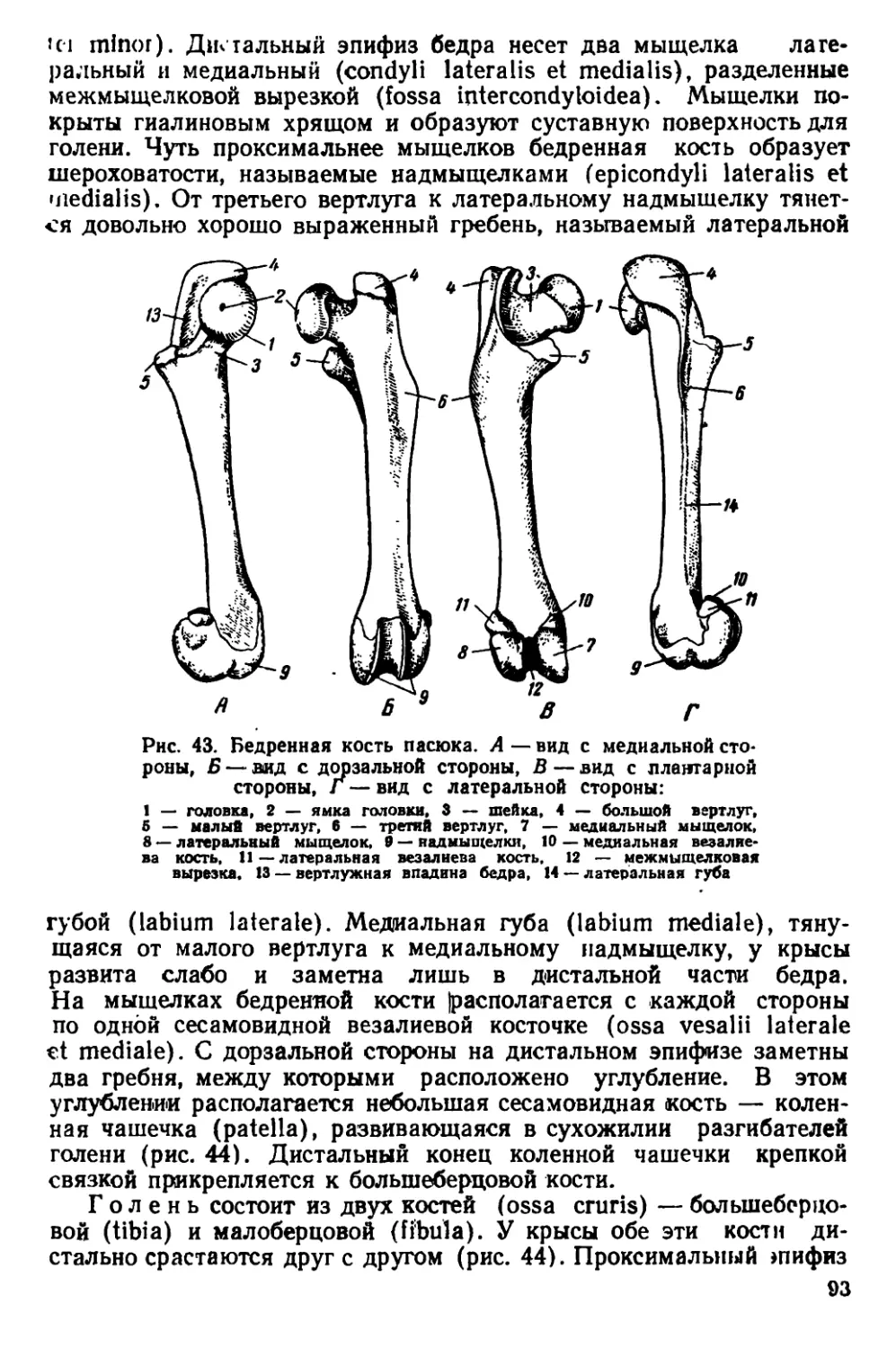
капсулы небольшие. Рез-
цы узкие и длинные, с
стороны эмаль
окрашена в желтовато-
оранжевый цвет.
У молодых особей че-
реп округлый. Гребни на
верхней его стороне слег-
ка намечаются лишь в
возрасте 20 дней. Сначала
гребни появляются по бо-
кам передней части те-
менных костей. Ширина
межглазничного про-
странства по отношению к
общей длине черепа боль-
ше, чем у взрослых. Те-
менные и лобные кости
выпуклые. Скуловые ду-
ги сильно прижаты к че-
репу.
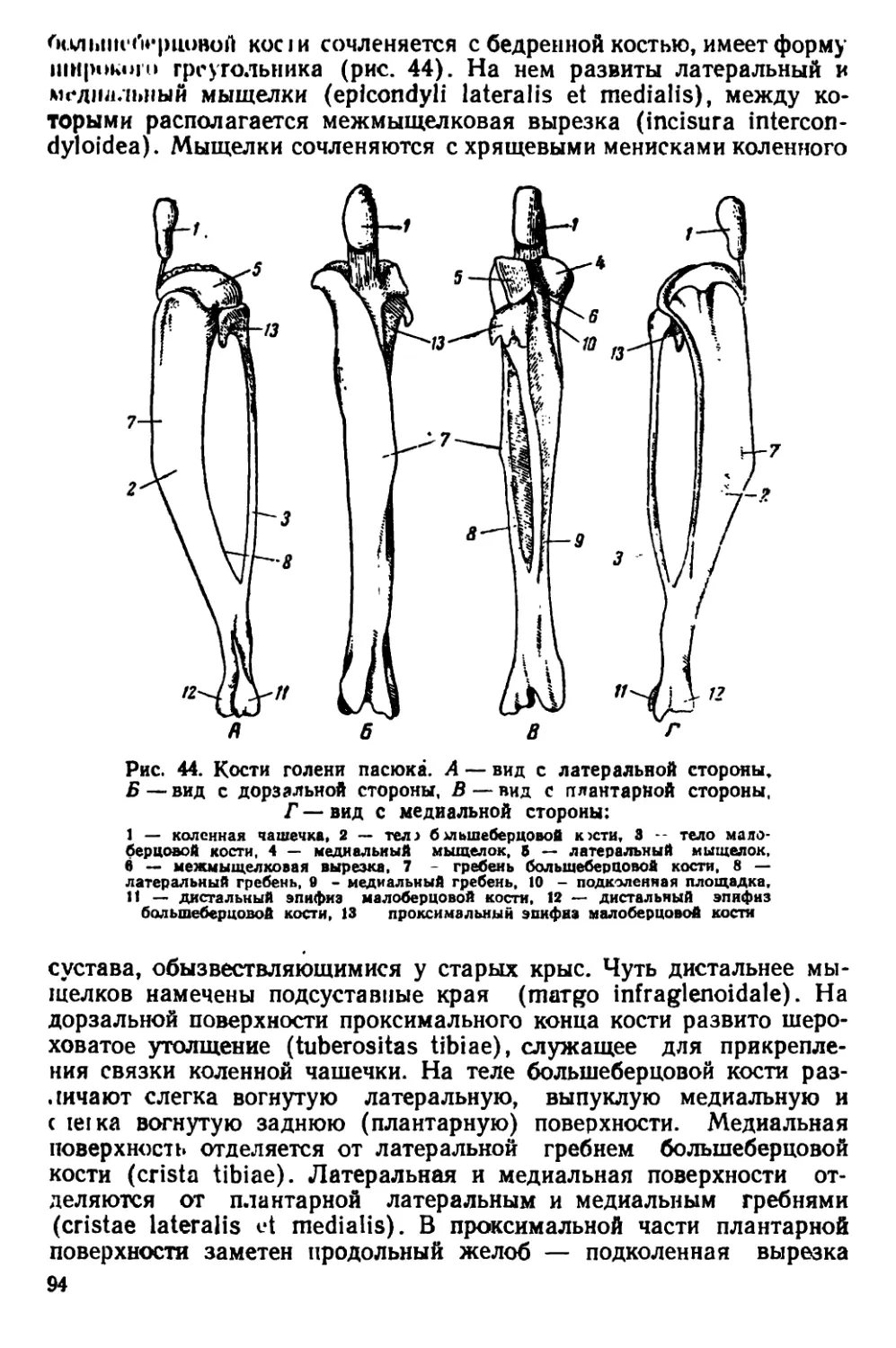
Крыса карако, или дальневосточный пасюк (Rattus norvegicus
caraco Pall.). Отличается от типичной формы серой крысы мень-
шими размерами и более коротким хвостом. Сосков 10. Краниоло-
гически дальневосточный пасюк характеризуется большей шириной
-4' репной коробки и большей шириной скуловых дуг.
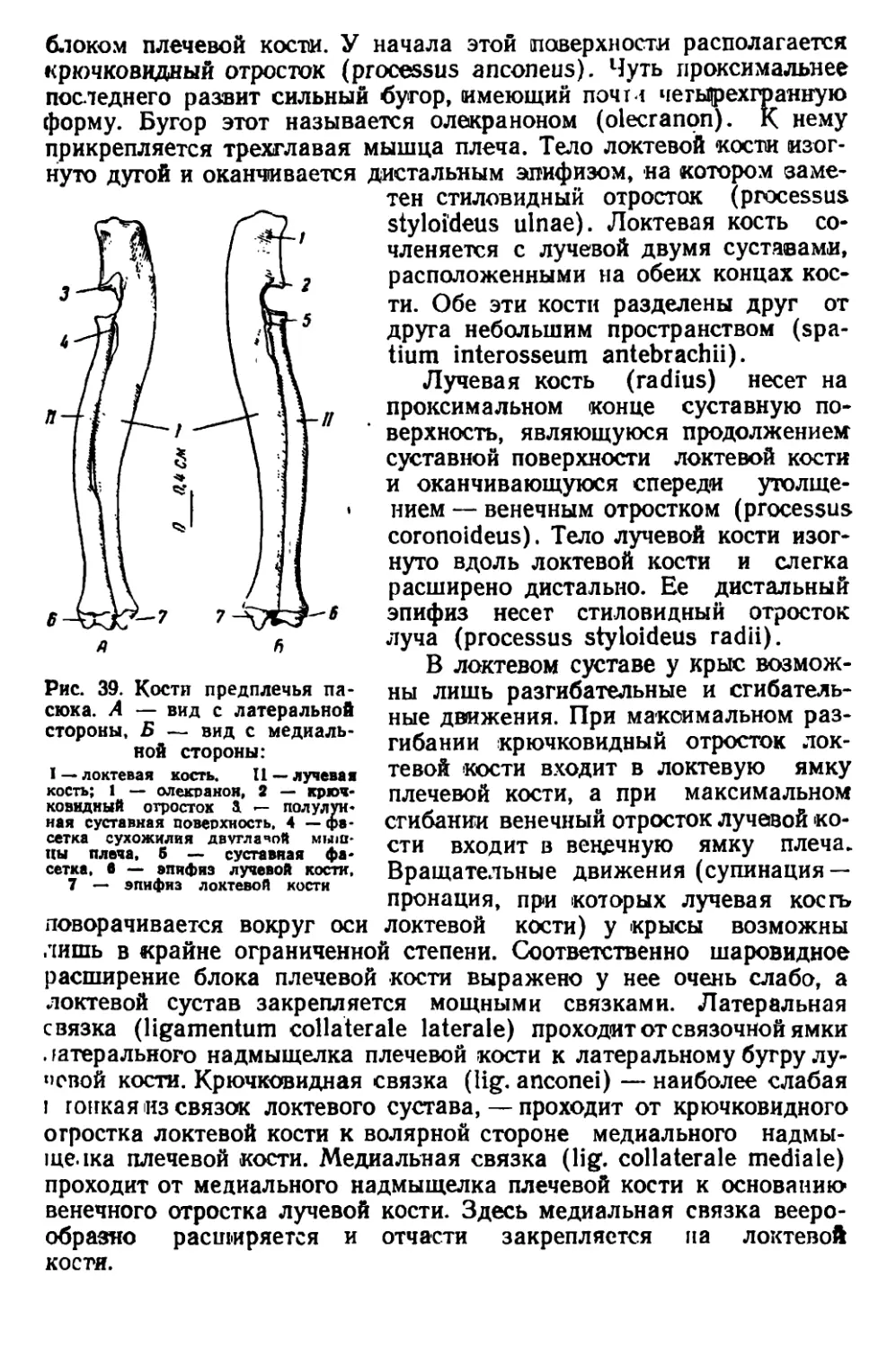
Черная крыса (Rattus rattus L.). Размеры тела меньше, чем у
•рой крысы; морда узкая и более вытянутая (рис. 3, 5). Хвост
обычно длиннее туловища, в редких случаях равен ему. Волоски
между чешуйками, покрывающими хвост, гуще, чем у пасюка.
Ушная раковина крупная, округлой формы. Если отогнуть ухо впе-
!'< г. »<> оно достигает края глаза; этот признак характерен не толь-
••• । иг взрослых, но и для молодых черных крыс. Сосков 10. Окра-
ь.| меча черной крысы сильно варьирует. Преобладают два типа
•нЧ'.ь «и у крыс первого типа тело сверху почти черное, с коричне-
ншым оттенком и металлическим зеленоватым блеском. Бока тела
'•|iifiii' Hiii п'етлее, нижняя сторона дымчато-серого цвета. Черные
7
крысы nгорою inii.i no окраске меха ноши нс ог.ппыклси uj пасю-
ков. Верхняя сторона тела у них серо-бурого цвета, без коричневых
оттенков, но с тем же металлическим блеском, который характерен
для крыс первого типа. Снизу тело свинцово-серого цвета с желто-
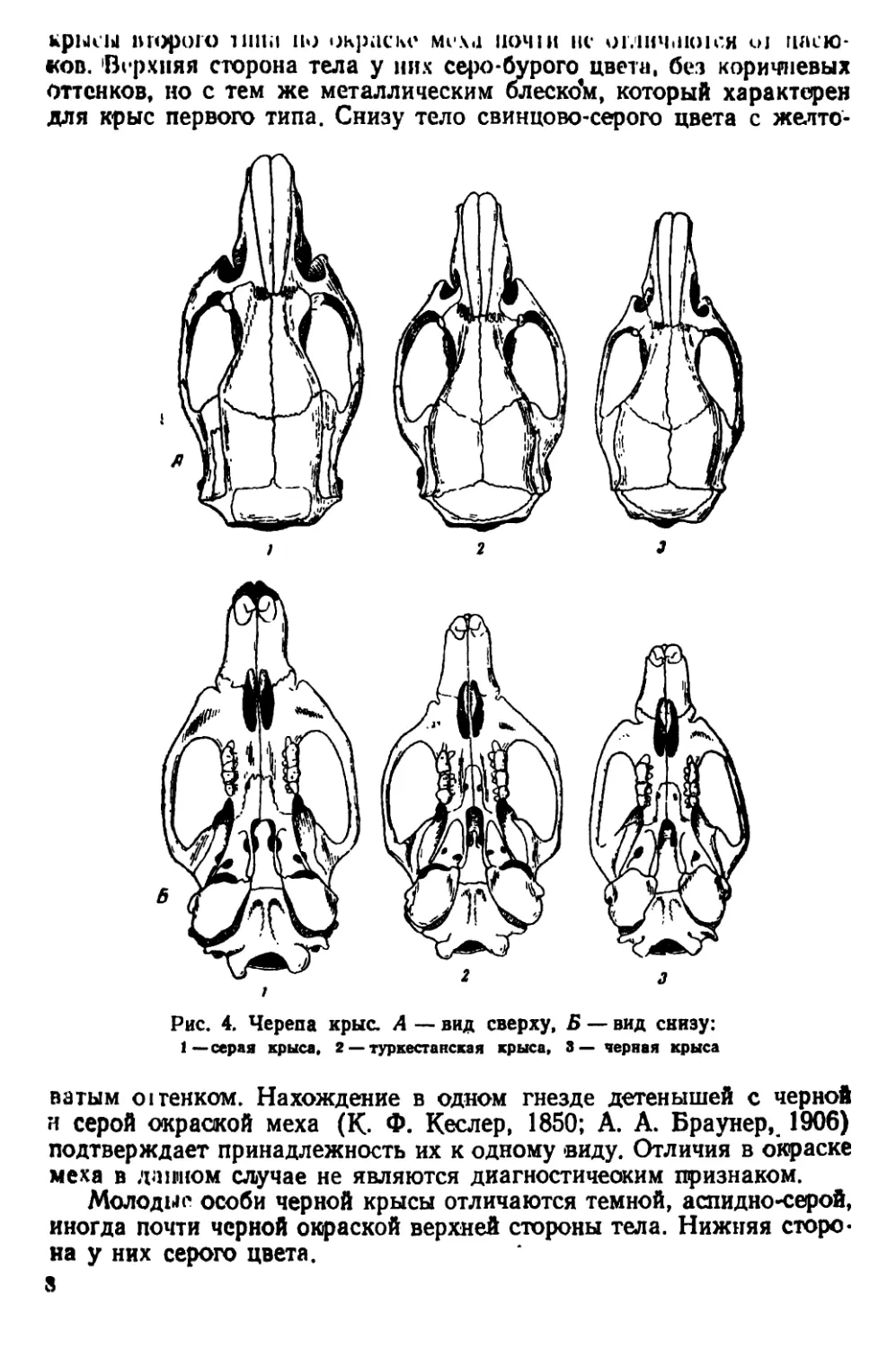
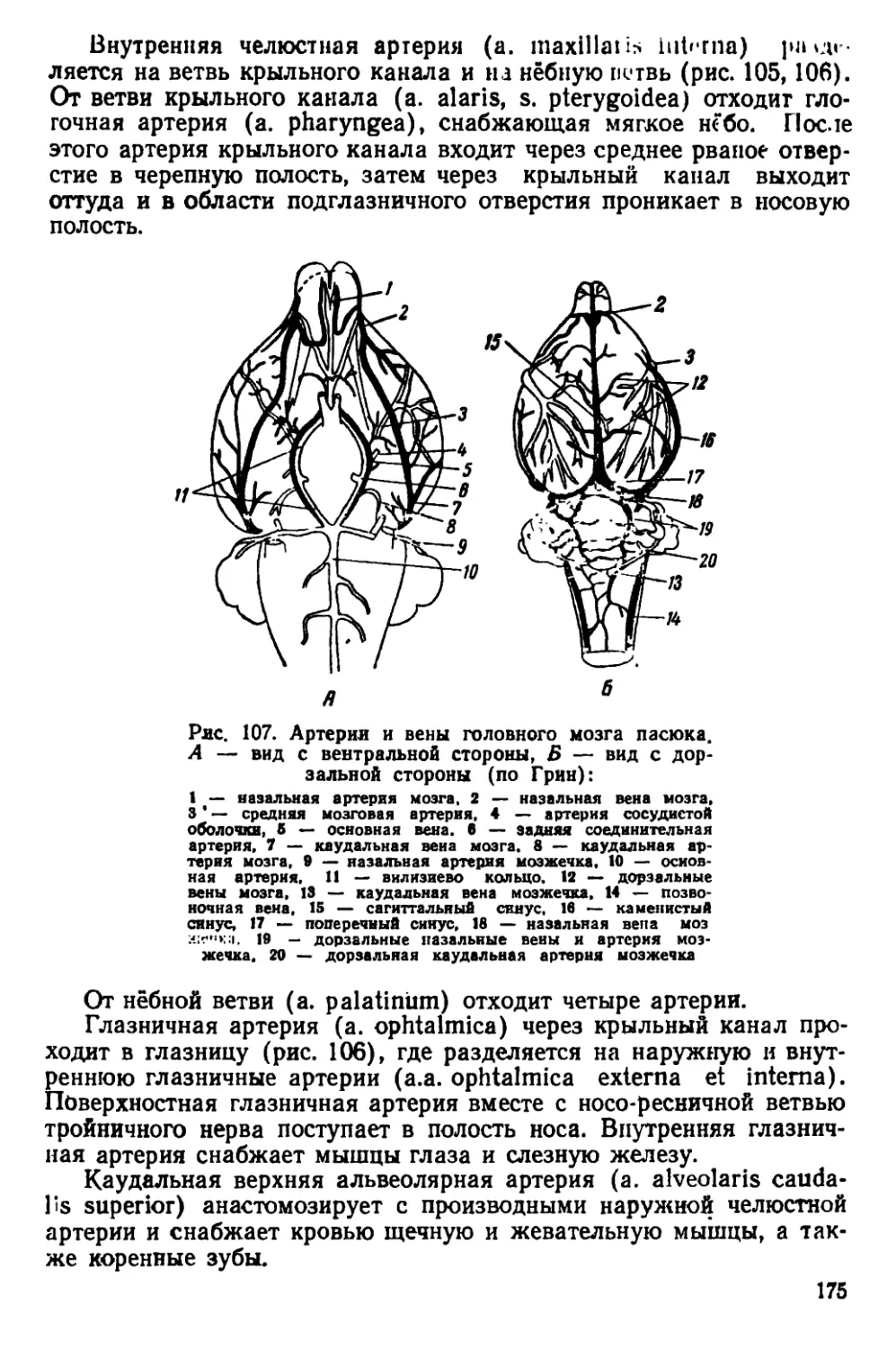
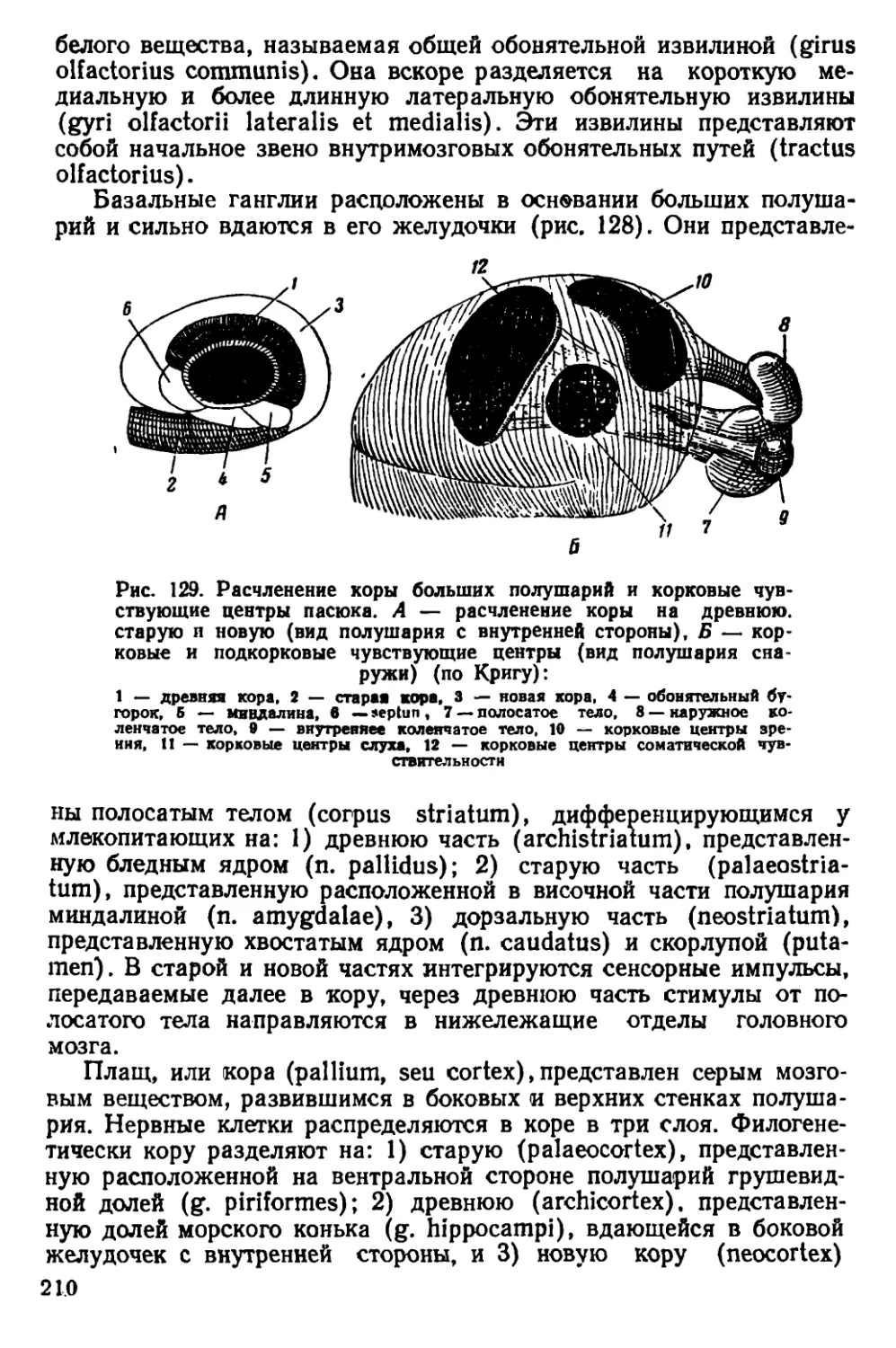
Рис. 4. Черепа крыс. А — вид сверху, Б — вид снизу:
1— серая крыса, 2 — туркестанская крыса, 3— черная крыса
ватым огтенком. Нахождение в одном гнезде детенышей с черной
и серой окраской меха (К. Ф. Кеслер, 1850; А. А. Браунер,. 1906)
подтверждает принадлежность их к одному виду. Отличия в окраске
меха в данном случае не являются диагностическим признаком.
Молодые особи черной крысы отличаются темной, аспидно-серой,
иногда почти черной окраской верхней стороны тела. Нижняя сторо-
на у них серого цвета.
8
I'.inppH ic.ia: лтна ii ia 130 190 мм, длина хвоста 134 —
“*н мм, высота уха 18,0- 25,5 мм, длина сгупни без когтей
?'),1 36,5 мм.
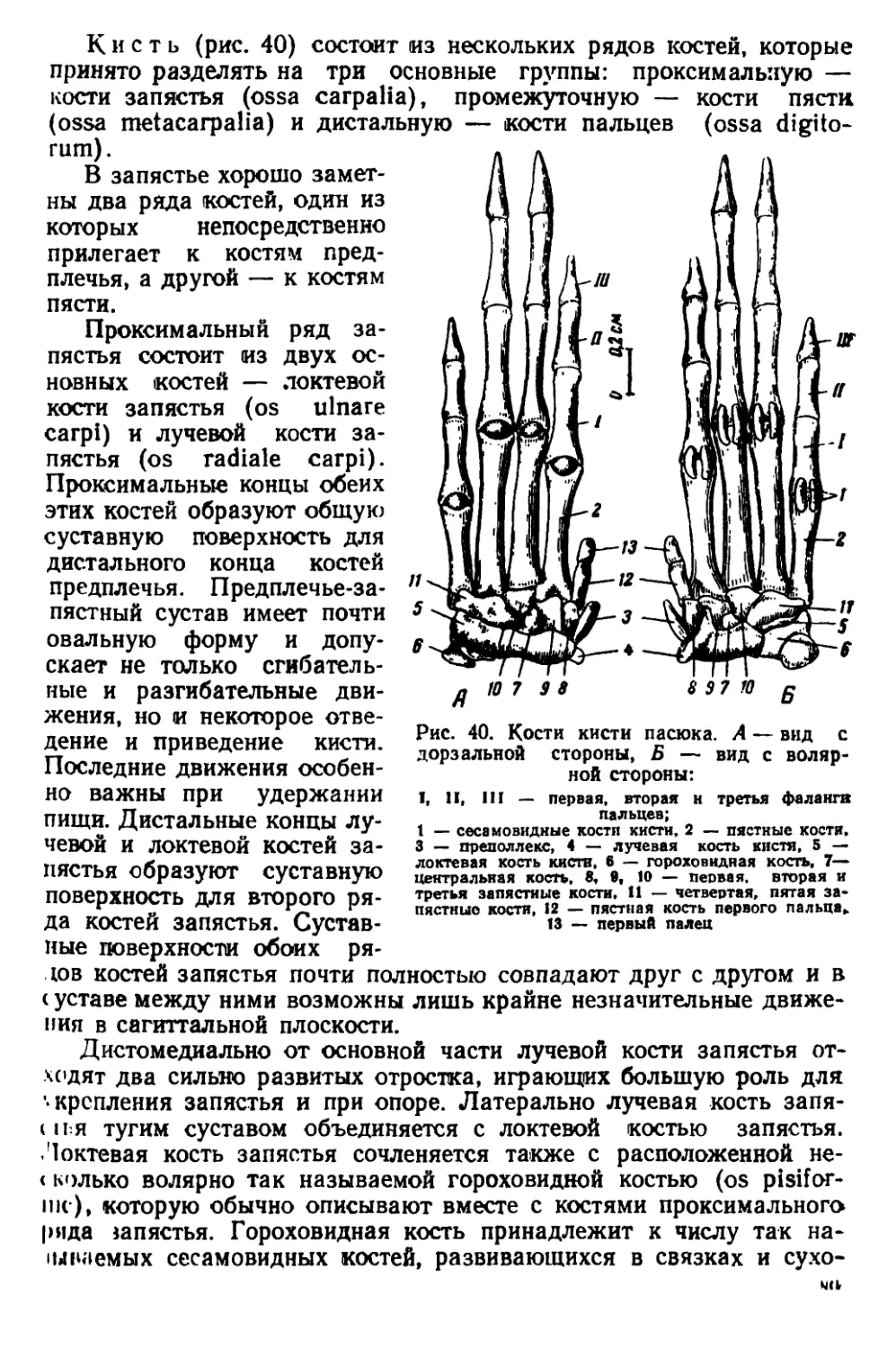
По строению черепа /?. rattus отличаются от R. norvegicus тем,
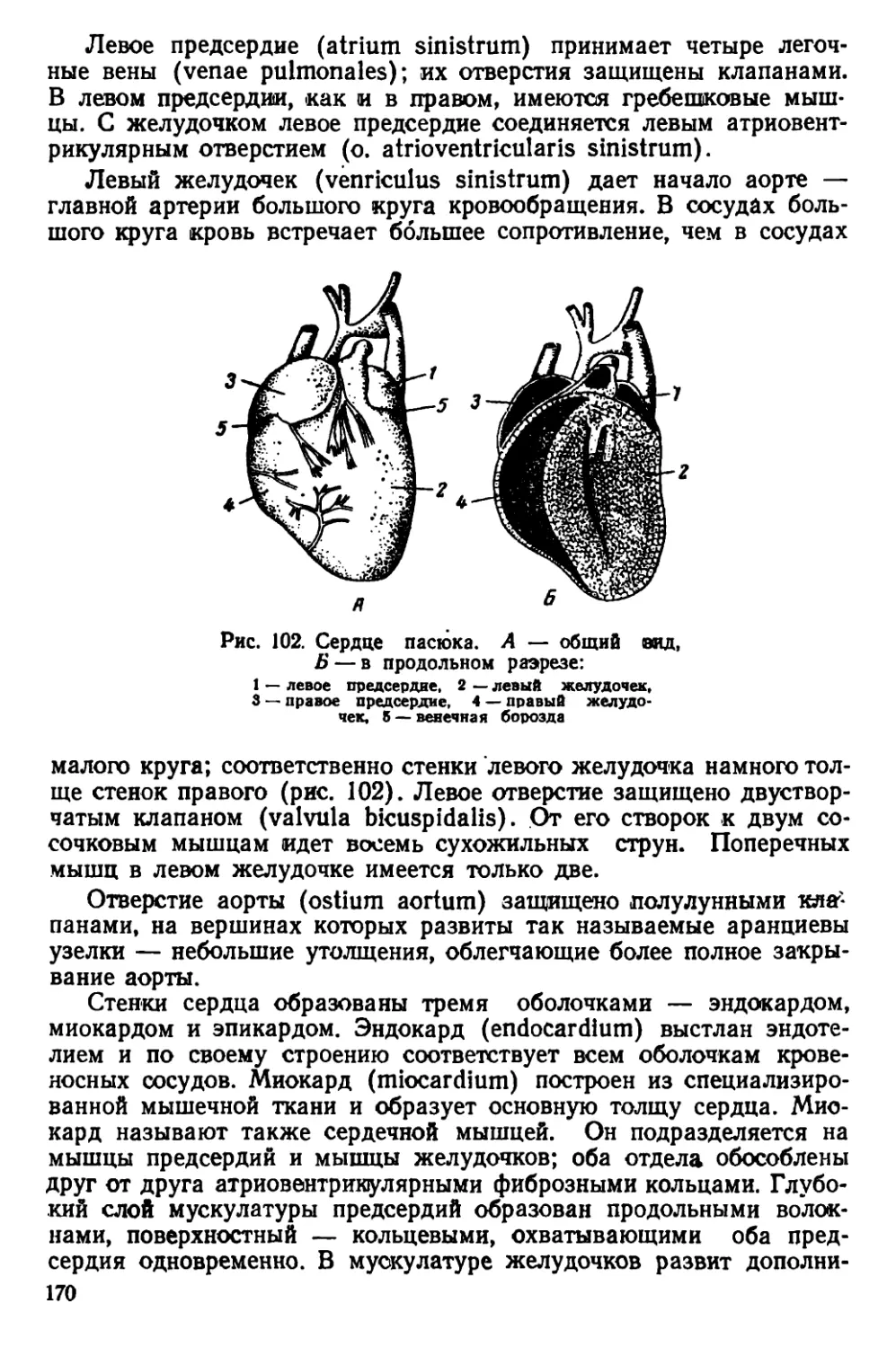
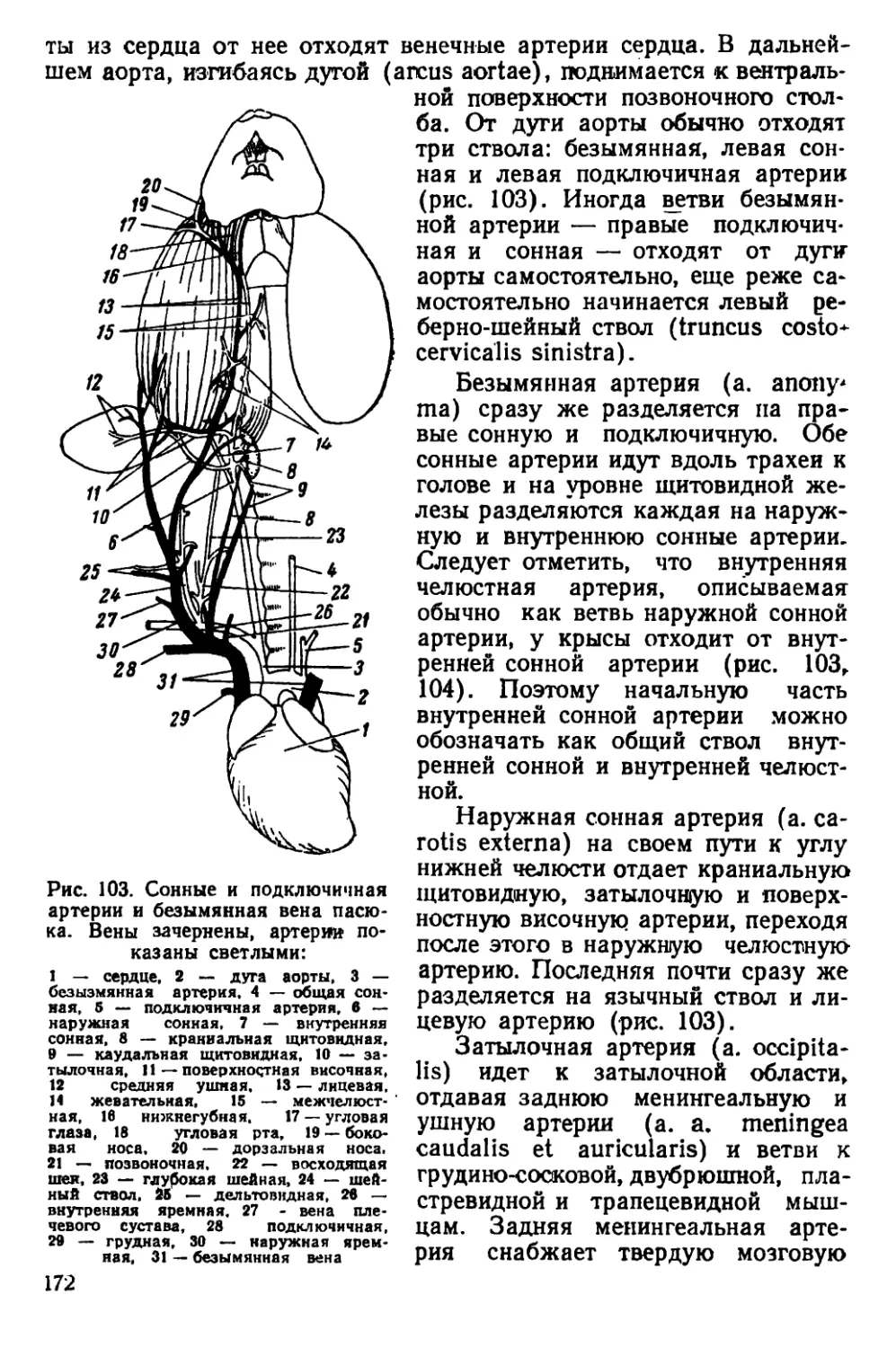
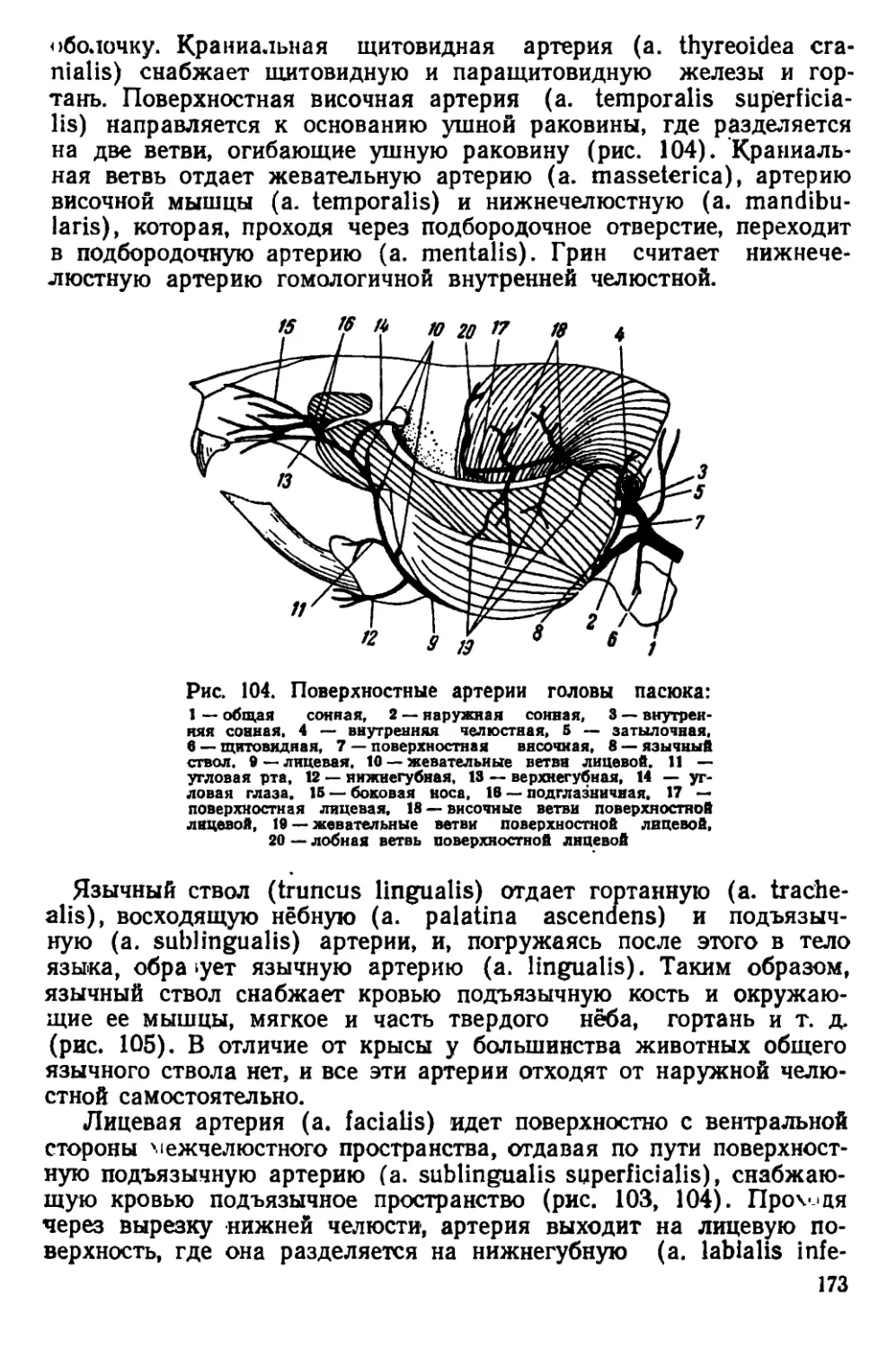
чio черепная коробка у них более округлая, так как лобные, темен-
ные и межтеменная кости несколько вздуты. Скуловые дуги дости-
пнот наибольшей ширины в области своих задних концов. Гребни
пл теменных костях дугообразно изогнуты.
Рис. 5. Черная крыса
Черепа молодых особей серых и черных крыс сходны по своему
строению; некоторые различия имеются только в форме межтемен-
ной кости (рис. 6).
На материале из Смоленской области С. И. Огнев и С. У. Стро-
ганов описали новый подвид Rattus rattus ruthenus Ogn. et Strog.,
распространенный в юго-западных областях Европейской части
СССР. Отличительными признаками этого подвида является доми-
нирующий коричневатый оттенок в окраске меха на верхней сто-
роне тела и несколько более крупные размеры тела по сравнению
< типичной формой, к которой авторы относят черных крыс из
Ленинградской области.
Александрийская крыса {Rattus rattus alexandrinus Geofr.).
Ц|)/1ицд описан из Египта (окрестности Александрии). В СССР эта
форма встречается в портовых городах, расположенных на Черном
норе. По окраске меха алексадрийские крысы, благодаря преобла-
UIHIHO серых тонов близки к серым крысам;'от типичных R. rattus
umi огчнчаются также и большими размерами тела.
9
В pa t.iiriHin пунктах Европы, а также в пгдгш СССР были
имАдены крысы, сходные по своей окраске с александрийскими. Не-
которые авторы ошибочно относили их к подвиду Rattus rattus
alexttntlrinus. В действительности же эти экземпляры представляют
гобой цветовую вариацию типичных черных крыс.
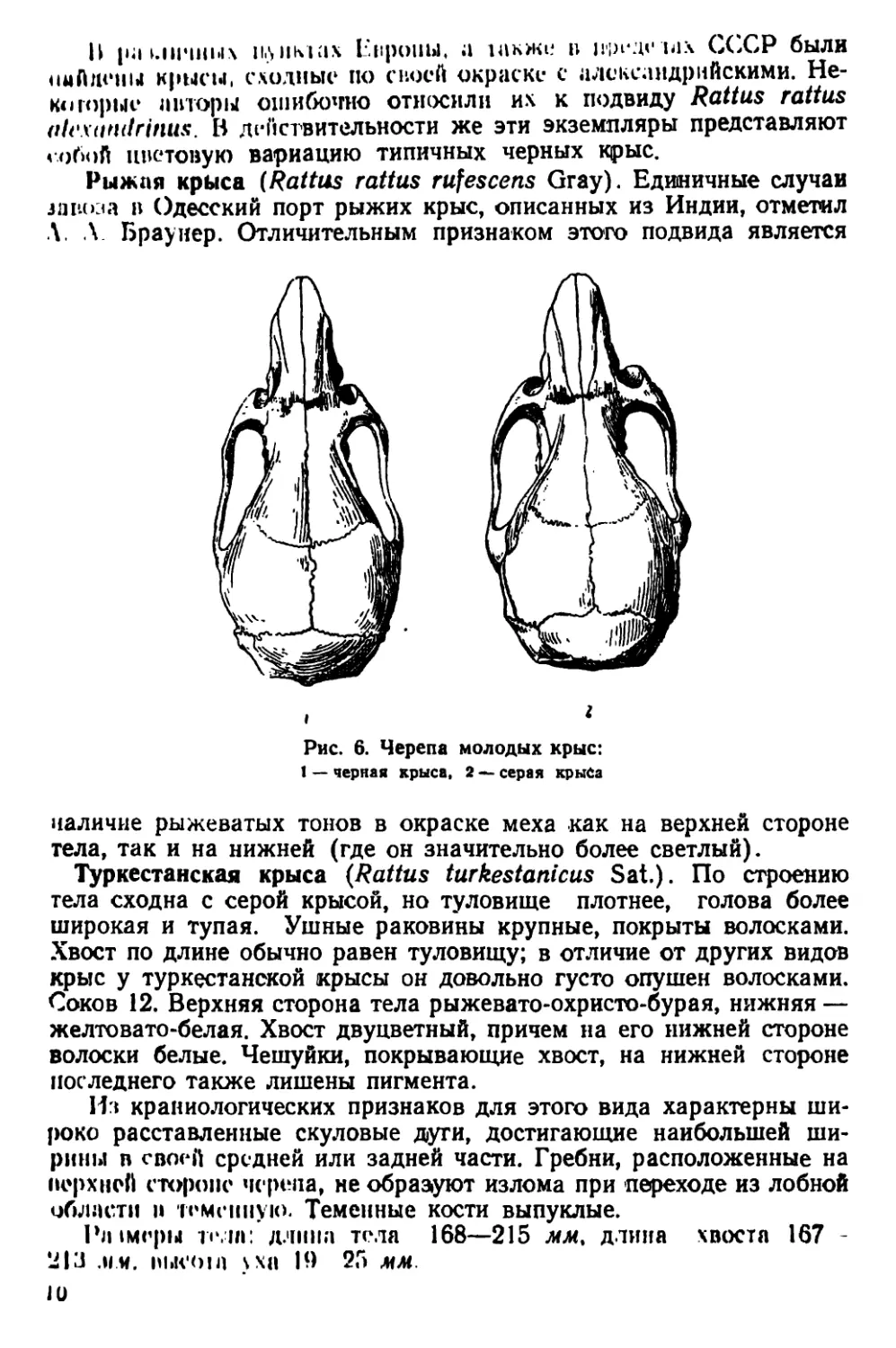
Рыжая крыса (Rattus rattus rufescens Gray). Единичные случаи
завоза в Одесский порт рыжих крыс, описанных из Индии, отметил
Л. Л Браунер. Отличительным признаком этого подвида является
Рис. 6. Черепа молодых крыс:
1 — черная крыса, 2 — серая крыса
наличие рыжеватых тонов в окраске меха как на верхней стороне
тела, так и на нижней (где он значительно более светлый).
Туркестанская крыса (Rattus turkestanicus Sat.). По строению
тела сходна с серой крысой, но туловище плотнее, голова более
широкая и тупая. Ушные раковины крупные, покрыты волосками.
Хвост по длине обычно равен туловищу; в отличие от других видов
крыс у туркестанской крысы он довольно густо опушен волосками.
Соков 12. Верхняя сторона тела рыжевато-охристо-бурая, нижняя —
желтовато-белая. Хвост двуцветный, причем на его нижней стороне
волоски белые. Чешуйки, покрывающие хвост, на нижней стороне
последнего также лишены пигмента.
Из краниологических признаков для этого вида характерны ши-
роко расставленные скуловые дуги, достигающие наибольшей ши-
рины в своей средней или задней части. Гребни, расположенные на
верхней стороне черепа, не образуют излома при переходе из лобной
области и теменную. Теменные кости выпуклые.
Рл1меры толп: длина тола 168—215 мм, длина хвоста 167 -
213 .ии. eiiicoin >хп 19 25 мм.
J0
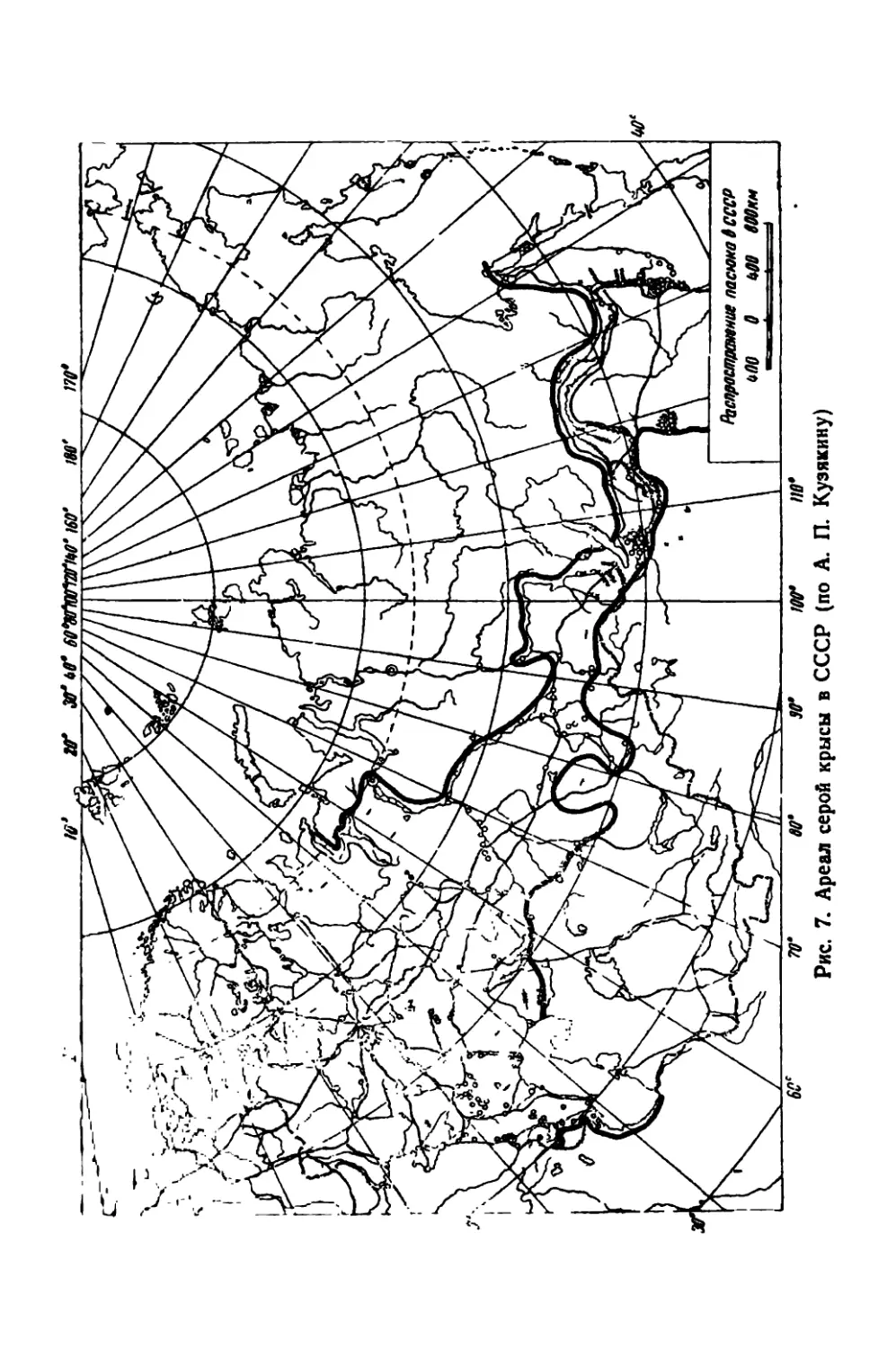
rev* гл* ня г. иное ГДСПРОСТРАМГ.НИЕ СЕРО* КРЫСЫ
Первые факыи нахо/ьленпи серых крыс (Railus norvcguu:i) на
ivppniopitn .Ешадной Европы относятся к XVI в. За три столетия
»ти 1рызуны широко расселились на восток и в настоящее время
нс|рсчаются в населенных пунктах Европейской части СССР,
начиная от крайних северных портовых городов (Архангельск —
Мурманск) до Крыма и Закавказья.
В Сибири ареал пасюка ограничен сравнительно узкой полосой
вдоль линии железной дороги; на Дальнем Востоке он охватывает
южные районы Приморья.
Громадные пространства тундры и таежной полосы Сибири, Се-
верного Урала и северо-востока Европейской часта Союза, так же
как Средняя Азия и почти вся зона полупустынь юго-востока СССР,
свободны от крыс. На карте (рис. 7) вычерчен ареал серой крысы
по крайним пунктам ее нахождения, но следует учитывать, что
численность серой крысы в отдельных городах различна; некоторые
населенные пункты, особенно в сельской местности, вообще свободны
от крыс.
Расселение крыс на восток по территории Сибири началось в
самом конце прошлого века.
Первые данные о нахождении серых крыс в Западной Сибири
мы находим у Н. Я. Словцова (1892), который указывает, что они
изредка встречаются около г. Тюмени. М.*Д. Рузский (1897) отме-
чает пасюков в незначительном количестве в южной части б. То-
больской губернии. По Н. Ф. Кащенко первые экземпляры серых
крыс были зарегистрированы в Омске и в Томске в 1907 году.
М. Д. Зверев считает, что впервые крысы попали в Сибирь в 1900 г.,
в период Русско-Китайской войны.
Массовому расселению серых крыс по Сибири на восток содей-
ствовало проведение Сибирской железной дороги. Особенно благо-
приятная в этом отношении обстановка сложилась во время Русско-
Японской войны в связи с усиленными перевозками на восток про-
довольствия и фуража для армии. Развитие речного судоходства
также ускорило расселение крыс по Сибири. В настоящее время
пасюками заселены главным образом города и населенные пункты
вдоль линии железной дороги и в бассейнах судоходных рек.
Дальнейшее расселение R. norvegicus по Западной Сибири‘на-
блюдалось в период гражданской войны, когда, благодаря разрухе
.'ошйства и увеличению скоплений продовольствия на армейских
складах, для крыс создались благоприятные условия существова-
ния. Так. М. Д. Зверев отмечает, что в Кузнецкой степи крысы
поягилнс. в 1919—1920 гг. — после того, как по этой местности
прош ди ш)й>'ка.
В По । очной Сибири ареал типичной формы R. norvegicus Вегк.
гоприклгиегсн с ареалом другого подвида R. п. caraco Pall. На
Дальнем Востоке встречаются обе формы, причем в больших горо-
дах преобладаю! .трые крысы, а в населенных пунктах, удаленных
12
ОТ H Ul'Hl и»|Н'ГЛ, обычны IIIIIIPUIhH' <К И'М||,1Я|)Ы Крысы lilipjlhl*. П
• iiitiiin 1яч ।<>6111.1101 оба полнила. i . ip. чп'1 я большое количс
|И IIOH'dl МСЖЛ> НИМИ. ЧТО Ш1РУДПЯСТ опредс К1П1Г
) ранними пунктами нахождения серых крыс на гюстоке являют-
II Камчатка и Сахалин. По данным А. П. Державина (1916), се-
рки крысы известны в Большерецке, Петропавловске, Усть-Камчат-
। м-, 11ижне-Камчатске и в Камаках. В Ключи их завозили несколько
ра >, по в этом населенном пункте они не обосновались. На Саха-
лине А. М. Никольский отмечал наличие большого количества крыс
к Александровоке, в Дуэ и на Корсаковском посту.
В связи с освоением Севера граница распространения пасюка в
Сибири продвигается и в этом направлении. К- К. Флеров (1933)
указывает, что R. norvegicus обычны в поселках по р. Оби и изред-
ка попадаются в населенных пунктах по р. Сосве и р. Ляпине.
11. Ф. Егорин (1939) считает, что в Западной Сибири самым север-
ным пунктом постоянного обитания пасюка на Оби является
р. Сургут.
В. Н. Скалой (1939) предполагает, что современный ареал серой
крысы не переходит за пределы северного полярного круга, хотя
известны случаи завоза их речным транспортом и дальше на север.
Так, в бассейне р. Оби они были завезены до Салехарда, но не
закрепились ни там, ни в поселке Сальмер-Седе в устье р. Таз;
в населенных пунктах, расположенных в бассейне р. Таз, крыс нет.
Не смогли обосноваться крысы, завезенные в Яновстан (среднее
течение р. Турухан) и в Дудинку на Енисее (71° сев. ш.) На восток
от Енисея крысы не встречаются до бассейна р. Лены, где они
обнаружены в г. Олекминске.
В условиях сурового климата севера серые крысы нашли благо-
приятную обстановку для зимовки в Якутске и в Игарке, где име-
ются хорошо утепленные благоустроенные жилые помещения;
последнее обстоятельство и позволило пасюку закрепиться в этих
городах.
В Западной Сибири, по М. Д. Звереву (1935), серые крысы на
юге доходят до западных районов (б. Рубцовского, Барнаульского,
Каменского, Новосибирского и Томского районов). С началом
лксплуатации Ачинско-Минусинской железной дороги крысы рас-
। <лились на юг и встречаются теперь в населенных пунктах, рас-
положенных вдоль этой дороги до Минусинска и Абакана включи-
n ibHO. А. П. Разоренова (1939) отмечает, что пасюки довольно
"бычны в предгорьях и в северной части Алтая. В населенных
'нпмах Центрального Алтая серой крысы нет.
Т<’ последнего времени в литературе обычно излагалась гипо-
согласно которой серые крысы заселили Европу с востока —
побережья Каспийского моря. Подобного рода указания даже с
>рмложением карты, на которой изображены пути и центры рассе-
яния крыс, можно найти почти во всех больших руководствах по
11 |i<i 1 и пщии (Я. Л. Окуневский, 1936; X. Д. Архипианц, 1939;
И II Шорохов и С. И. Шорохов, 1938, и др.). Б. М. Житков ука-
13
i. 'i>i< <ih uiiiiiiiv i<> пинаны на неправильном переводе и тол-
. i< к< 1,1 llaii.ic.i (Zoographia rosso-asiatica), где указан
•| и । ni'|H iip uiH крыс черен Волгу из Европы в Азию, а не из При-
|><н ниц, ниц « б.bu rn на Запад.
I in<’ II. Ф Кащенко (1912У в результате анализа современного
р.|| п|"н ।ранения R. norvegicus пришел к выводу, что первоначально
>i' i инд расселялся из Восточной Азии на Запад двумя путями —
«1И.Ц1- медленно по суше и сравнительно быстро по морским путям
и п»я hi с развитием торговли между странами Восточной Азии и
Енропой. Такое предположение о путях расселения серых крыс,
пониднмому, более правильно. Вместе с тем большой интерес пред-
еывляют археологические находки остатков пасюков в голоценовых
пещерных отложениях Западной Европы. Эти данные, так же как и
наличие постоянных поселений R. norvegicus вне связи с человеком,
свидетельствуют о том, что эти грызуны являются древними компо-
нентами европейской фауны (Д. С. Айзенштадт). Возможно, что
широкое расселение серых крыс по Европе с запада на восток сле-
дует рассматривать как вторичное явление.
Приуроченность серых крыс к населенным пунктам, расположен-
ным вдоль линий железных дорог, привело к заключению о пассив-
ном расселении этих грызунов путем завоза их железнодорожным
1 ранспортом. Более подробное исследование этого вопроса показы-
вает, что такой способ расселения не мог бы привести к столь
широкому распространению серых крыс, которое характерно для
этого вида грызунов. В этом отношении заслуживают внимания
наблюдения Д. С. Айзенштадта (1950) на Кишиневской железной
дороге, которые показали, что пасюки обитают не только в по-
стройках, но и вдоль железнодорожного полотна, где они питаются
главным образом зерновыми продуктами, просыпанными во время
перевозок, и отбросами из пассажирских вагонов. Эти наблюдения
представляют большой интерес, так как на основании подобных
фактов можно найти правильное объяснение быстрого расселения
пасюков вдоль линий железной дороги, в частности вдоль Сибир-
ской ж.-д. магистрали, которое нельзя относить за счет пассивного
завоза крыс, поскольку нахождение их в вагонах известно только
в единичных случаях. Расселению пасюков вдоль железной дороги
способствуют не только усиленные перевозки продовольствия и
фуража, но и интенсивное освоение новых территорий в районах,
примыкающих к железной дороге. В станционных поселках и дру-
гих населенных пунктах крысы при продвижении на новые терри-
тории находят для себя благоприятные условия существования, что
способствует их быстрому размножению. Д. С. Айзенштадт пред-
полагает, что значительная часть Сибирской железной дороги была
заселена крысами, следовавшими за поселками строительных рабо-
чих еще до начала движения поездов.
При расселении крыс одним из факторов, регулирующих их про-
движение в новые районы, является влажность климата. Изучая
расселение серых крыс в восточных районах Ростовской области,
U
I II yo.ipeh (1941) пришел к выводу, что неблагоприятные или
Минск > кис условия засушливой зоны, с характерным для нее
in ы точным водным режимом, является одним из основных фак*
i.ipui», определяющих медленное продвижение серых крыс с запада
hi н'и'юк. Даже наличие железной дороги и передвижение вой-
• к.п.1.1 \ и продовольственных обозов в период гражданской войны
«и привело к расселению крыс по этой области. Поэтому «не слу-
к|iiiKt. пишет автор, — что расселение крыс шло более интен-
< IHIHO по населенным пунктам, расположенным вдоль рек и речек,
। при заселении у крыс наблюдается стремление к таким помеще-
ниям, как свинофермы, конюшни, погреба, где имеется большая
|>.1ажность, чем в амбарах, сараях, которые заселяются крысами
только в исключительных случаях».
То обстоятельство, что от крыс свободны Туркестано-Сибирская.
Ташкентская и Ашхабадская железные дороги, расположенные в
безводной местности, Д. С. Айзенштад также объясняет отсут*
чвием воды, исключающим возможность расселения крыс вдоль
полотна этих дорог.
Пассивный завоз по железной дороге также способствует рас-
селению крыс, но в меньшей степени. Например, единичные случаи,
нахождения крыс (/?. norvegicus и /?. rattus) в г. Фрунзе, сообщен-
ные Д. П. Дементьевым (1950), можно объяснить только завозом
их по железной дороге, так как эти виды не встречаются в ближай-
ших к городу областях.
Водный транспорт играет большую роль в расселении крыс, так
как суда различного тоннажа и назначения являются местами их.
постоянного обитания. Поэтому при передвижении судов, заселен-
ных крысами, происходит их пассивный завоз на большие расстоя-
ния. Этим и объясняется наличие крыс не только вдоль линии же-
лезной дороги в Сибири, но и по бассейнам судоходных рек.
Активное расселение серых крыс может происходить достаточно
быстро. В период военных действий в одном из районов северо-
птпадной части РСФСР разрушение населенных пунктов и эвакуа-
ция скота в тыл послужили причиной расселения крыс по всему
району (Айзенштадт, 1945). Крысы передвигались вдоль дорог и
илетилов, где находили пищу в виде просыпанного фуража и про-
дуктов, непереваренных зерен в навозе и т. п. Встречая по дороге
л io гы и блиндАжи, крысы заселяли эти постройки. По данным авто-
ра, крысам потребовалось около года для того, чтобы расселиться
hi разрушенных населенных пунктов в прилежащие леса и обосно-
IHH).<’« в расположенных там складах и постройках, для чего им
и у лпю было пройти по прямой расстояние более 20 км.
МЕСТА ОБИТАНИЯ СЕРОЙ КРЫСЫ
<л*рая крыса в своем распространении связана с человеком. Оби-
1'14 и постройках различного хозяйственного назначения, она обес-
iiiHiriin благоприятными защитными условиями, особенно в зимнее
npi мм. и имеет постоянную кормовую базу.
15
!i |ородц\ крысы обитают в постройках разнообразного типа и
м) miiciвенного использования: в жилых домах, в магазинах, скла-
•irtx, на предприятиях пищевой промышленности, в конторских по-
мещениях, в различных подземных сооружениях (канализация, во-
доснабжение), в зоопарках и т. д. Обычно пасюки живут в
подвальных и первых этажах зданий, но при высокой численности
проникают и в верхние этажи. Как правило, наиболее охотно крысы
заселяют помещения предприятий пищевой промышленности, мага-
зины, продуктовые и фуражные склады, где они постоянно находят
корм.
(.•гры" крысы могут приспосабливаться к различным температур-
ным условиям. В качестве примера можно указать на наблюдения
Д. П. Дементьева (1950), который отметил единичные случаи на-
хождения крыс в тушах мороженого мяса в холодильниках, где
температура воздуха достигает —13°. Устраивая гнезда внутри мяс-
ных туш, крысы используют для их постройки обрывки веревок,
клочки бумаги, а также рассеченные ими сухожилия и пленки.
Вместе с тем крысы могут жить и при весьма высокой темпера-
туре. Так, тот же автор находил гнезда крыс на пивоваренном заво-
де в термоизоляционной прокладке парового котла, обшитого сна-
ружи деревянным каркасом, где температура достигала 40—50°.
В пределах города или населенного пункта отдельные популя-
ции серых крыс разобщены друг от друга, численность их на смеж-
ных объектах бывает различна и много помещений свободно от них.
На железнодорожном транспорте серые крысы заселяют не толь-
ко помещения вокзалов, складов, различных подсобных помещений,
вплоть до путевых будок, но изредка встречаются и в подвижном
составе (в пассажирских, почтовых и багажных вагонах, чаще в
вагонах-ресторанах).
Водный транспорт также не свободен от серых крыс. Они могут
встречаться не только в портах, но и на всех видах судов: грузовых,
пассажирских и других, — куда крысы попадают или пассивно,
вместе с упакованными грузами, или забегают с берегов во время
стоянок у причалов. В некоторых южных портовых городах отмече-
но, что серые крысы на судах держатся главным образом в глубо-
ких нижних трюмах, тогда как черные крысы предпочитают засе-
лять верхние подпалубные помещения. По данным И. В. Степанова
(1949), собранным в одном из южных портовых городов, серые
крысы встречаются как в самом городе, так в порту и на парохо-
дах. Из общего числа исследованных крыс большинство составляли
пасюки (92,7%).
В населенных пунктах сельской местности места обитания серых
крыс также разнообразны. Они заселяют жилые дома и подполья,
скотные дворы, помещения птицеферм и животноводческих хо-
зяйств, мельницы, различные хранилища и склады. В теплое время
года пасюки нередко выселяются на огороды, в заросли сорняков
и т. п. места, где они живут в земляных норах. С наступлением
холодов они возвращаются в постройки. Такие сезонные перекочев-
16
।.11 чмечаюг Б. II. Богачев и Н. М. Дукельская (1936) даже для
.. IH.JI4. и} районов Архангельской области, где зимой крысы засе-
р| ч| помещения молочной фермы, а весной, когда начинался выпас
м>ы, уходили на окрестные поля. К- Плятер-Плохоцкий (1935) ука-
<и|1.ц?т на регулярные сезонные передвижения крыс-карако в усло-
• ihi\ Дальнего Востока.
Н пределах ареала R. norvegicus на территории СССР известны
шьие популяции серых крыс, не связанные с населенными пункта-
ми и приспособившиеся к обитанию в природных стациях. Б. К. Фе-
шок (1941) перечисляет несколько фактов нахождения пасюков вне
нс селений человека в Волго-Ахтубинской пойме. В январе 1938 г.
I" сколько экземпляров R. norvegicus были добыты в стогах и скир-
IX, в норах на лугах близ воды и в пойменном лесу на расстоянии
ос 800 м до 4 км от жилья. Пасюков ловили также у нор водяных
иолевок в пойме р. Волги в 40 км севернее Саратова (в пойменном
лесу и на‘заливном лугу). Вне связи с жильем человека серую
.рысу добыли и в пойме р. Аткары. В. А. Хлебников (1924) писал,
что в Астраханском крае в долине Волги серые крысы встречаются
вдали от жилищ человека вплоть до взморья.
А. Н. Формозов (1945) приводит данные о нахождении диких
популяций серых крыс в дельте Волги, где, по опросным сведениям,
шалаши и зимницы рыбаков заселяются крысами во время зимнего
лова рыбы. В восточной части дельты Волги А. Н. Формозов добыл
беременную самку /?. norvegicus 2 ноября 1941 г. Добывали серых
крыс, в том числе и молодых, и на территории Астраханского запо-
ведника, а также на берегу ерика в дельте Волги. Кроме того,
А. Н. Формозов ссылается на сообщение Н. Шатуновской о нахож-
тении пасюков в пойме р. Волги в Сталинградской области.
Наиболее подробные сведения о поселениях диких крыс В дельте
Волги мы находим у Г. А. Кондрашкина (1949), который указывает,
что эти грызуны встречаются по всей территории дельты, причем
•основная масса их сосредоточена вдоль высоких берегов речек и
ериков с зарослями тальника и ветлы, где они живут в норах между
корнями деревьев. Во время паводка пасюки выселяются во времен-
ные норы водяных полевок, расположенные у воды, а после затоп-
ления всей поверхности земли заселяют дупла деревьев. А. Н. Фор-
мозов совместно с Н. П. Наумовым отмечают, что в 1934 г. в Бер-
(янском районе УССР на р. Обиточной не было водяных полевок,
I встречались обычные пасюки. Известны факты добычи пасюков
и пойме среднего и нижнего течения Дона.
М. К. Серебренников (1929) сообщает, что в Южном Зауралье
I Челябинская область) он находил серых крыс вне поселений чело-
пжа, по берегам озер и болот в зарослях прибрежных кустарников,
г> есть в стациях, характерных для водяных полевок. Н. К» Вере-
щ1цин находил колонии пасюков на берегу р. Шексны. Этот же
•luiop наблюдал их также на побережье Апшеронского полуострова,
> и- па песке были видны следы крыс, подбирающих выбросы моря.
Ь. А. Кузнецов (1928) отмечает, что в г. Орске пасюки довольно
17
Рис. 8. Гнездо серых крыс, устроен-
ное во время наводнения (рис. Д. С.
ины иг юлько в юродских домах, но и в зарослях тала гь-
берегам р. Урала и его стариц. В окрестностях г. Уральска серая
крыса и «редка попадается по берегам речек, например р. Чагана.
II Пог точном Забайкалье это же автор (1929) нашел колонию
днкоживутих пасюков на берегу старицы р. Нерчи. Многочислен-
ные колонии серых крыс в природных условиях наблюдал Д. С. Ай-
зенштадт (1945) в лесах северо-западных районов РСФСР в период
военных действий, имевших место на этой территории. Здесь
Я. norveg icus встречались в наибольших количествах в ельниках на
слабо заболоченной почве и в сос-
новом бору на песчаных возвы-
шенностях; иногда пасюки попа-
дались и на сфагновых болотах.
В летнее время норы серых крыс
были обнаружены под корнями
деревьев.
В природных условиях много-
численные поселения пасюков на-
блюдал Н. К. Верещагин (1949)-
в некоторых районах Закавказья:
в Абхазии — на озере Бабестри,
на р. Гагиди и ее притоках и
вдоль побережья Черного моря от
устья р. Гагиди до устья р. Ин-
гуры; в Западной Грузии — в бас-
сейне р. Пикеры и по другим
речкам до р. Ингуры; на р. Су-
же и озерах Джадана — близ
ст. Самтреди. Наиболее плотно
заселены серыми крысами ни-
зовья р. Гагиди и болота, окружающие озеро Палеостом. Много-
численны пасюки и в тростниково-рогозовых болотах юго-восточной
Мугани и в Приталышской низменности. О численности R. norve-
gicus в Закавказье можно было судить по тому, что в Колхиде в
сезон 1938/39 гг. во время промысла нутрии заготовляли и шкурки
пасюков. Здесь пасюки настолько связаны с водоемами (озерами
и реками), что Б. М. Житков (1944) считал возможным назвать
их «водяными пасюками». По данным Н. К. Верещагина, в Грузии
серые крысы заселяют следующие стации: 1) ложбины, залитые
юдой и заросшие колючим ситником (Juncus maritimus); 2) ма-
рискусовые луга с участками сфагновых болот (Cladium mariscus,
Juncus sp., Carex sp., Sphagnum sp.); 3) тростниково-осоковые за-
росли (Phragmites communis. Carex gracilis); 4) топкие ольховые
кустарники по пересекающим их протокам.
В прирщны'. условиях серые крысы обитают также в низовьях
'Хуная, Прута и Днестра. Во время паводка они устраивают на
деревьях гнезда, располагая их на высоте 0,25—1,5 м от уровня
воды (рис. 8). Материалом для постройки гнезд служат свежие
18
>•< I mi того дерева, на котором они устроены; внутреннюю камеру
ы выстилают листьями (Д. С. Айзенштадт, 1951).
Пл Дальнем Востоке R. norvegicus сагасо встречается не толь-
»<> и населенных пунктах, но обычна также на полях и на огородах;
нобенно многочисленна она на рисовых плантациях, где эти гры-
> . пы заселяют дамбы и валики оросительной системы. Этот подвид
' рых крыс можно встретить и в тайге на расстоянии 20—30 км
"I поселений человека.
Но данным А. С. Фетисова, в южных и западных районах
Восточной Сибири серые крысы круглый год обитают в ивово-топо-
’|-вых и черемуховых зарослях по берегам рек и в осоковых и тро-
шиковых зарослях по берегам рек, стариц и озер. В бесснежный
период они встречаются на полях, залежах и огородах, в камени-
11ых россыпях лесостепной и степной полос, на осоково-моховых и
шшайниковых болотах разнотравных степей.
Многочисленные факты нахождения серых крыс вне построек
че ловека представляет большой интерес, но образ жизни их в при-
родных условиях изучен еще далеко недостаточно, несмотря на то,
что здесь они имеют большое эпидемиологическое значение, так
как легко могут вступать в контакт с другими видами грызунов.
В этом отношении особое внимание должно быть обращено на изу-
чение зимовок пасюков в природных условиях, так как для сохране-
ния тепла они в больших количествах скучиваются в одной норе.
Так, Б. К. Фенюк (1941) в начале января обнаружил в одной норе
j молодых и 12 взрослых крыс, а в другой норе он добыл 13
взрослых крыс.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ И МЕСТА ОБИТАНИЯ
ЧЕРНОЙ крысы
В Европе черная крыса появилась раньше, чем пасюк; возмож-
но, она проникла в Европу из стран Малой Азии еще в XII в. —
и период крестовых походов.
Вместе с тем заслуживают внимания находки в Закавказье ко-
гтей черной крысы во время раскопок в 200 м от правого берега
Куры у Мингечаура. Кости были обнаружены вместе с кувшинными
погребениями, которые относятся к концу I и ко II тысячелетию
до н. э. (Н. К. Верещагин, 1949). Эти археологические находки,
j также довольно широкое распространение диких популяций чер-
ной крысы в Закавказье в наши дни позволяют предполагать, что
<ш грызуны входили в состав древней фауны.
В пределах СССР R. rattus встречается спорадически в различ-
ил районах, не связанных друг с другом, главным образом в Ев-
ропейской части Союза. Б. С. Виноградов и И. М. Громов (1952)
И .ИЫН.1ЮТ следующие места нахождения черной крысы: Архан-
и h i г., Вологда, Молотов, Ленинградская область, Псковская об-
пк и.. Калининская область, Великолукская область, Смоленская
"ii i.u и.. Белоруссия, Московская область, Тула, Курская область,
19
Калужская область. Орловская область, Казань, Горьковская об-
ласть, Ростовская область, юго-западная часть Украины, Черномор-
ское побережье Кавказа, Закавказье. В Сибири черные крысы от-
сутстуют; они встречаются только в нескольких городах Дальнего
Востока: во Владивостоке, Благовещенске, Хабаровске, Комсомоль-
ске, Советской гавани, а также на Командорских островах.
Черные крысы обитают как в городах, так и в сельских местно-
стях. Они отличаются от серых крыс тем, что более сухолюбивы,
в связи с чем предпочитают заселять верхние этажи зданий и чер-
дачные помещения.
В портовых городах преобладают серые крысы, но черных крыс
туда завозят на судах также в достаточно больших количествах.
В одном портовом городе на Дальнем Востоке отмечено следующее
распределение крыс по видам (А. М. Колосов):
Черная крыса Серая крыса
(в %) (я %)
Город 1 99
Порт 6 94
Суда 46 54
На Черноморском побережье Кавказа, в портовых городах с
оживленным движением торгового флота, также отмечается завоз
черных крыс. В Батуми среди нескольких десятков тысяч исследо-
ванных крыс черные крысы составляли только 4,7%, александрий-
ские крысы — 2,6%. Встречаются черные крысы главным образом
на окраинах города, в порту, а также и на пароходах. Александрий-
ские крысы чаще всего вылавливаются на пароходах и в меньших
количествах в порту: в городе эти крысы попадаются редко
(И. В. Степанов, 1949).
В Закавказье черные крысы часто встречаются в садах и лесах.
Они хорошо лазают по деревьям, и их гнезда можно найти в дуп-
лах.
В противоположность серым крысам J?. rattus характеризуется
привязанностью к определенным местам обитания. Черная крыса
активно не расселяется и не расширяет свой ареал. Этим объяс-
няется значительное сокращение их распространения в Европе по
сравнению с первоначальным, когда они представляли собой до-
минирующий вид. В то же время серая крыса, легче приспосабли-
вающаяся к различным условиям существования, за короткий срок
расселилась по огромным территориям.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ И МЕСТА ОБИТАНИЯ
туркестанской крысы
Туркестанская крыса в пределах СССР встречается только в
Средней Азии. Она известна в западной части Памира по Алаю,
Каратегину и Гиссарскому хребту, по Туркестанскому хребту до
Самарканда, в Ферганской долине и окружающим ее горам, на
20
' >лных склонах Таласского Ала-Тау и западном Кара-Тау. Обычна
<-па также в Ташкенте и его окрестностях. Найдена туркестанская
<«рыса и в некоторых пунктах Киргизии. Описан этот вид крысы
о экземпляру, добытому из Арсланбобского лесного массива.
Туркестанские крысы обитают как в постройках человека, так и
п природных условиях, где они заселяют горные лиственные леса,
юлины горных рек, а также и возделанные земли: сады, при-
иадебные участки и т. п.
ГЛАВА II
ЭКОЛОГИЯ
ПОВАДКИ КРЫС
Широкое распространение крыс не только в пределах СССР, но
и по всему земному шару свидетельствует о большой экологической
пластичности этих грызунов, легко приспосабливающихся к самым
разнообразным условиям существования.
В основном крысы — ночные животные, но суточный ритм их
активности зависит от характера деятельности человека в тех по-
мещениях, где они обитают или добывают пищу. Крысы активны
преимущественно в те часы суток, когда их меньше тревожит при-
сутствие людей. Только при высокой численности они мало осто-
рожны и не боятся человека.
Крысы очень подвижны; они хорошо лазают и прыгают, а также
свободно плавают и ныряют. Для проникновения внутрь помещений
они используют имеющиеся дефекты в стенах и полу или активно
прогрызают входные отверстия; в помещения крысы проникают
также через различные люки и отверстия, легко передвигаясь по
вертикально расположенным трубам и по столбам. Они забираются
и на деревья, откуда могут попадать в дома через окна. При за-
возе их на пароходах крысы перебегают на берег по причальным
канатам.
Внутри помещений крысы нередко имеют определенные пути
г зредвижений, которые иногда ясно заметны в виде хорошо про-
топтанных тропинок; здесь же бывают рассеяны и их экскременты.
Обычно крысы бегают вдоль стен или по ходу труб. В производ-
• твенных помещениях пищевых предприятий крысы передвигаются
। чавпым образом по трубам и подвесным путям, расположенным
под потолком.
Пути подхода крыс к месту кормежки были установлены непо-
'•редс! вашими наблюдениями за их жизнедеятельностью в жилой
квартире, где постоянным местом кормежки служило мусорное
ведро, стоявшее из года в год на одном и том же месте. Предва-
рительное опыление пола тальком позволило проследить по сле-
там, что в течение ночи крысы несколько раз проходили вдоль степ
22
• । входного отверстия норы в углу помещения к мусорному ведру,
•I котором находились кухонные отбросы. При недостатке корма в
ipe крысы начинали беспорядочно бегать по помещению в по-
исках пищи. При перемещении ведра с отбросами на новое место
оказалось, что крысы проделывали сначала обычный для них путь,
I затем приступали к поискам кормушки. Такого маршрута крысы
придерживались в течение 5 суток, но некоторые особи посещали
га рое место кормежки более долгое время — до 11 суток.
Значительно затрудняет проведение истребительных мероприя-
|ий большая настороженность крыс по отношению ко всяким новым
предметам — к расставленным капканам или ловушкам, а также
к разложенным отравленным приманкам. Преодолевать эту насторо-
женность удается путем предварительного приучения крыс к неза-
ряженным ловушкам, снабженным приманками, и подкормкой перед
.(травкой неотравленными продуктами.
Присутствие крыс в помещении определяется по различным
следам их жизнедеятельности — по наличию входных отверстий в
полу, у основания стен или по ходу технических труб, по скопле-
ниям экскрементов, по следам на запыленных участках пола и, на-
конец, по наличию повреждений на продуктах питания и на пред-
метах домашнего обихода. Все эти признаки указывают на присут-
ствие крыс в помещении, но по ним нельзя определить расположе-
ние гнезда, где крысы обитают и выводят детенышей. Для своих
убежищ крысы выбирают наиболее укромные места, нередко зна-
чительно удаленные от мест кормежки. Для выстилки и утепления
гнезда крысы используют обрывки бумаги, тряпки, перья и т. п.
Поселяясь в подвалах и подпольях, они роют в земле глубокие
норы.
ПИТАНИЕ КРЫС
Изучение питания серых крыс имеет большое практическое зна-
чение для правильного использования приманочного метода борьбы,
позволяя подбирать при изготовлении отравленных приманок до-
статочно привлекательные для крыс пищевые продукты и рассчи-
тывать расход приманок.
Наблюдениями в естественных условиях обитания серых крыс
.авно установлена всеядность этих грызунов. Они употребляют в
пищу не только все виды продуктов животного и растительного
происхождения, но при недостатке кормов используют также раз-
личные отбросы, нечистоты и поедают трупы животных.
Следует отметить, что организм серой крысы нуждается в бел-
.< г >й пище животного происхождения. Так, в опытах В. К. Шепе-
u it )й (I960) серые крысы, которых содержали в термокамере при
। юянной температуре воздуха 36° и относительной влажности 75 %,
при кормлении их овсом за 15—20 суток теряли в весе 0,4 7,2%.
IliiiaiiHe сырым картофелем дало за те же сроки потерю в весе в
||.|.мере 13 24%. Питание сырой столовой свеклой приводило
>.ч
подопытных животных к гибели от голода через 2- 3 суток, причем
они за >го время теряли в весе 20—22%. Только кормление варе-
ным мясом дало у подопытных крыс прибавление в весе (б—13%
w 10 суток).
Опыты в условиях клеточного содержания показывают, что па-
сюки едят хлеб (ржаной и пшеничный), каши, мясной и рыбный
фарш, овощи, творог; хуже берут зерно и продукты его переработ-
ки. Из зерновых продуктов крысы предпочитают овес и овсяную
крупу, очень плохо поедают пшено и муку. Методика проведения
подобного рода опытов не сложна. Предварительно крыс содержат
н течение 5 суток на каком-нибудь однородном корме, например
па ржаном хлебе. После этого им дают одновременно 4—5 видов
других пищевых продуктов в различных комбинациях. По количе-
ству съеденной за сутки пищи определяют сравнительную поедае-
мость крысами различных видов корма.
Более точные данные были получены при испытании реакции
серых крыс на различные виды корма при помощи специального
Прибора — ольфактометра конструкции П. А. Свириденко. Он пред*
ставляет собой два больших, укрепленных один над другим бараба-
на, сообщающихся друг с другом при помощи трубы. В верхнем
барабане по его окружности расположено 10 кормушек, соединен-
ных с самопишущими регистрирующими приборами, что позволяет
точно учитывать число посещений крысой той или иной кормушки и
длительность времени, затраченного на кормежку. Количество
съеденного корма определялось взвешиванием его остатков. Под-
опытную крысу пускали в нижний барабан, откуда она в поисках
пищи переходила в верхний барабан. Опыты проводились на кры-
сах, добытых на различных объектах города и питавшихся, следо?
вательно, разнообразной пищей. Поэтому перед опытом крыс в те-
чение трех недель выдерживали на одном из видов зернового корма
(овес, пшеница или пшено). Такая подкормка создавала однообраз-
ные условия питания в течение достаточно длительного срока, что
позволяло сравнивать результаты опытов.
Второй этап опыта начинался после пересадки крысы, выдержан-
ной в течение трех недель на однотипном корме, в ольфактометр.
Чтобы подопытная крыса освоилась с новой для нее обстановкой
в ольфактометре, могущей в первые дни привести к сокращению
количества поедаемого ею корма, крыса в течение трех дней полу-
чала тот же -вид корма, который был использован для предвари-
тельной подкормки. Заключительный этап опыта состоял в том, что
в три кормушки из десяти в течение 5 суток помещали новый вид
корма, который крысы не получали во время подкормки при под-
готовке к опыту.
Через ольфактометр удалось пропустить 12 серых крыс, пока-
завших различную реакцию на те виды зерна, которые им давали
в качестве корма (рис. 9). Крыса № 1 И течение первых двух эта-
пов опыта питалась исключительно овсом, поедая его в среднем
около 20 г в сутки. При •введении в рацион пшеницы, крыса в пер-
21
i-un день оказала предпочтение этому новому для нее виду зерна,
hi р последующие дни резкой количественной разницы в поедании
• чп а и пшеницы установить не удалось. Крыса № 2 после питания
И' м в течение 25 дней при свободном выборе между овсом и пше-
iiineft явное предпочтение оказала последнему виду зерна. Инте-
,ц ено отметить, что после интервала в 5 недель, в течение которого
। рыса № 2 получала смешанную пищу, причем пшеница в ее состав
। входила, эта крыса вновь оказывала предпочтение пшенице и
и называлась от овса.
Рис. 9. Количество овса и пшеницы, съеденное крысами во время опытов.
1 — пшеница, 2 — овес
Такое же предпочтение пшенице оказывала и крыса № 8. Для
проверки полученных результатов опыт в ольфактометре был про-
должен, причем пшеницу помещали в различные кормушки. Не-
• мотря на это, крыса продолжала питаться пшеницей и не трогала
овса.
Противоположные результаты были получены в опыте с крысой
•Mb 3. На первых двух этапах опыта эта крыса также питалась
овсом, но при введении в рацион пшеницы она продолжала почти
исключительно поедать овес. При повторном опыте через 40 дней
>та крыса опять не трогала пшеницу и питалась в основном только
овсом.
Пять подопытных крыс не обнаружили сколько-нибудь ясно вы-
раженного предпочтения определенному виду зерна и при свобод-
1ом выборе корма в ольфактометре в равной мере питались овсом,
пшеницей и пшеном.
Из двух крыс, получавших в период подкормки до опыта пшени-
цу, одна оказывала явное предпочтение этому виду корма и при сво-
бодном выборе почти совсем не трогала овса. Вторая крыса в за-
i. почительном этапе опыта в течение 5 суток поедала оба вида
юрна примерно в равном количестве.
По записям регистрирующих приборов видно, что крысы в
•льфактометре активно выбирают корм. Отмечено посещение ими
•тех кормушек, но длительное пребывание около них, связанное
। кормежкой, зарегистрировано только в тех кормушках, где был
помещен более привлекательный для них вид зерна. Количество
поедаемой крысами пищи в течение суток колебалось иногда в зна-
«чительных пределах, но у некоторых животных реакция по отноше-
нию к определенному виду зерна была весьма устойчивой, и коли-
чество потребляемых пшеницы и овса за сутки соответствовало друг
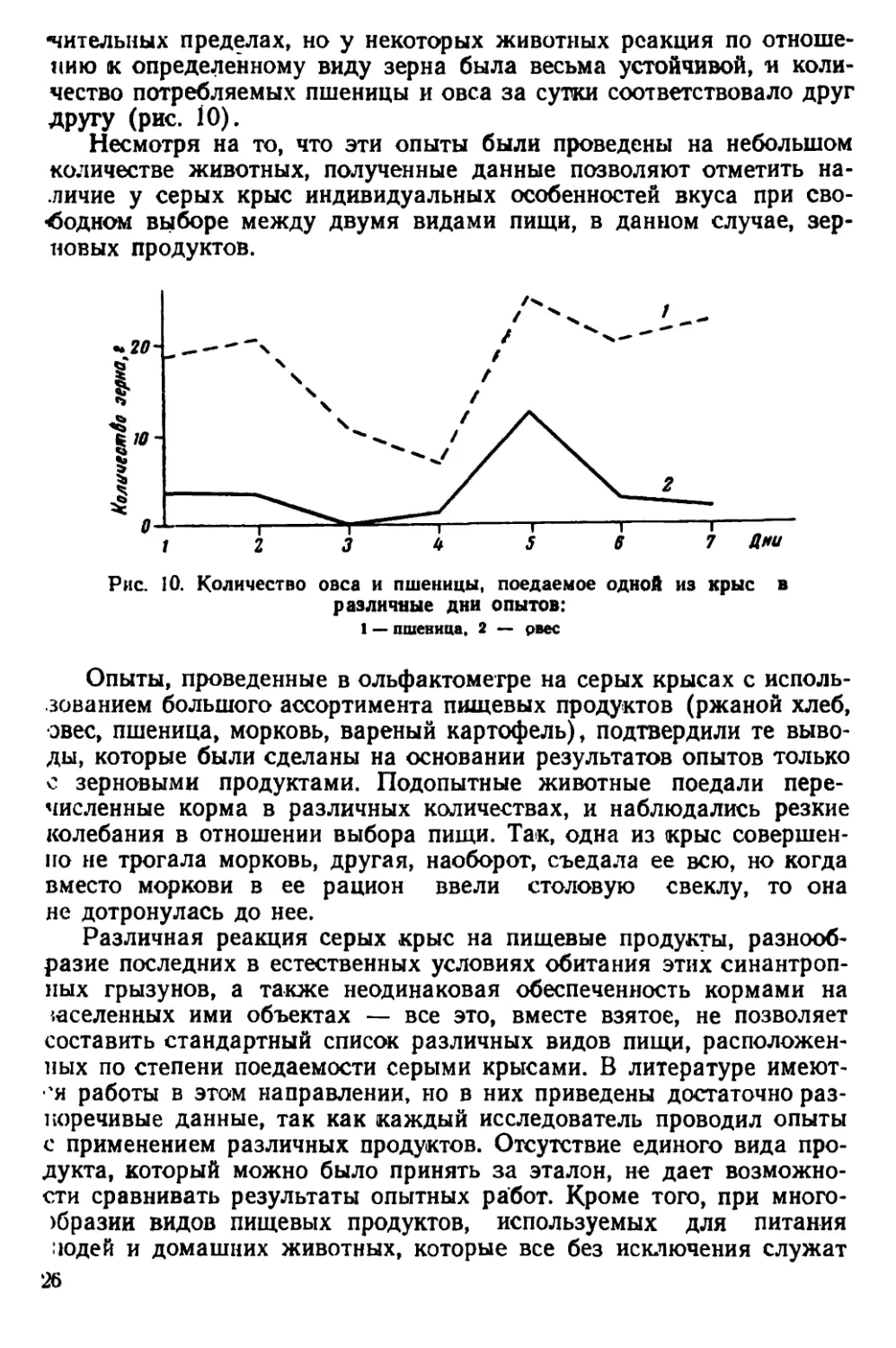
другу (рис. 10).
Несмотря на то, что эти опыты были проведены на небольшом
количестве животных, полученные данные позволяют отметить на-
личие у серых крыс индивидуальных особенностей вкуса при сво-
бодном выборе между двумя видами пищи, в данном случае, зер-
новых продуктов.
Опыты, проведенные в ольфактомегре на серых крысах с исполь-
зованием большого ассортимента пищевых продуктов (ржаной хлеб,
овес, пшеница, морковь, вареный картофель), подтвердили те выво-
ды, которые были сделаны на основании результатов опытов только
с зерновыми продуктами. Подопытные животные поедали пере-
численные корма в различных количествах, и наблюдались резкие
колебания в отношении выбора пищи. Так, одна из крыс совершен-
но не трогала морковь, другая, наоборот, съедала ее всю, но когда
вместо моркови в ее рацион ввели столовую свеклу, то она
не дотронулась до нее.
Различная реакция серых крыс на пищевые продукты, разнооб-
разие последних в естественных условиях обитания этих синантроп-
ных грызунов, а также неодинаковая обеспеченность кормами на
’.вселенных ими объектах — все это, вместе взятое, не позволяет
составить стандартный список различных видов пищи, расположен-
ных по степени поедаемости серыми крысами. В литературе имеют-
ся работы в этом направлении, ио в них приведены достаточно раз-
норечивые данные, так как каждый исследователь проводил опыты
с применением различных продуктов. Отсутствие единого вида про-
дукта, который можно было принять за эталон, не дает возможно-
сти сравнивать результаты опытных работ. Кроме того, при много-
>бразии видов пищевых продуктов, используемых для питания
юдей и домашних животных, которые все без исключения служат
26
।инн и также u серой крысе, очередность, в смысле предпочтения
ti'n> ми иного корма, может значительно изменяться.
Существует предположение, что ддя успеха истребительных ме-
роприятий отравленные приманки должны изготовляться из таких
Мидов продуктов, которых нет на данном объекте. Это утверждение
>н новывается на том, что крысы будто бы лучше берут пищу, от-
шчающуюся от привычных для них видов корма. Противоположная
•очка зрения сводится к тому, что отравленные приманки следует
юговить из тех пищевых продуктов, которые серые крысы обычно
находят в местах своего обитания, так как у крыс вырабатывается
привычка к определенному виду корма, которую необходимо иметь
к виду при выборе приманочного продукта. Опыты в лабораторных
н практических условиях показали, что такая закономерность от-
сутствует и поедаемость приманок зависит не только от вида
пищевого продукта, но и от ряда факторов внешней среды.
Надо также отметить, что положительный результат проведения
перед затравкой предварительной подкормки крыс зависит не только
<>г того, что крысы привыкают к определенному виду корма, кото-
рый в дальнейшем используется для изготовления отравленных
приманок, большое значение имеет также расположение подкормки
па определенном месте, так как крысы проявляют большую насто-
роженность к пище, расположенной там, где они обычно ее не
находили. Поэтому в первые дни подкормки крысы берут ее в
меньших количествах, чем в последующие, и максимум поедания
нищи достигается лишь на 9-й, а иногда даже на 10—12-й день.
Наряду с поисками хорошо поедаемых крысами видов пищи
неоднократно делали попытки найти пахучие вещества, привлекаю-
цие крыс; использование таких веществ позволило бы повысить
>ффективность приманочного метода борьбы. В старой рецептуре
ложно найти указания, что добавление анисового и тминного масел
। отравленным приманкам повышает их поедаемость крысами.
)днако это положение дальнейшими опытами не подтвердилось.
> 1е удалось найти и другие пахучие вещества, привлекающие крыс,
оторые можно было бы использовать на практике. В настоящее
(>емя проводятся исследования по определению роли обонятельного
'щептора у грызунов при выборе ими пищи. Подобные исследо-
вания помогут правильно решить эту задачу.
Изучение сравнительной поедаемости различных видов кормов
показало, что при введении в рацион пищевых отходов или брако-
"1нпых продуктов (плесневелый хлеб, рыба или мясо с гнилостным
шахом, отходы продуктов переработки зерна, мучные сметки
1 т. п.) серые крысы явное предпочтение оказывают доброкаче-
iBi иным, свежим продуктам. Опыт по скармливанию плесневелого
!(<’ а. проведенный в условиях естественного обитания пасюков,
попал. что крысы не брали его не только при наличии доброкаче-
|||> иных кормов, но и при их отсутствии. Этот вывод имеет боль-
't. . практическое значение для правильного подбора продуктов
<1>н и п тов 1ении отравленных приманок.
Характер питания серых крыс оказывает влияние на их физио-
логическое состояние. Так, по данным В. К. Шепелевой (1950),
пасюки, обитающие на элеваторе, загруженном овсом, несмотря на
благоприятные температурные условия (10—15°), отличались мень-
шим весом тела по сравнению с крысами, обитающими в холодиль-
нике при температуре —5° в камерах, загруженных яйцами. Крысы
с холодильника отличались значительным развитием жировых от-
ложений и плотным волосяным покровом; их вес превышал вес
крыс с элеватора в среднем на 21,5%. Питание белковым кормом
компенсирует большую теплоотдачу, благодаря чему крысы в хо-
лодильниках не только приспосабливаются к обитанию при отри-
цательной температуре, но и размножаются в течение круглого
года.
Количество пищи, поедаемое серой крысой, зависит от веса
тела и от калорийности корма. При клеточном содержании крысы
весом от 150 до 400 г съедают в сутки от 25 до 75 г серого пшенич-
ного хлеба; в единичных случаях крупные экземпляры съедают в
сутки до 100 г хлеба.
Пасюки очень чувствительны к голоданию и при отсутствии
пищи погибают в течение 3—4 суток. Хуже переносят они недоста-
ток воды и без питья и корма гибнут в еще более короткие сроки.
В естественных условиях обитания крысы не только пьют воду из
различных источников внутри построек, но и слизывают влагу с во-
допроводных и канализационных труб, с запотевших стекол и т. д.
Количество потребляемой крысами воды зависит от количества
влаги, которая содержится в кормах. Питание сухими кормами
резко повышает потребность в воде (табл. 1). Из таблицы видно,
что добавление сухих кормов к влажным увеличивает потребление
воды. Еще более резкая разница может быть отмечена при сравне-
нии потребности в воде крыс, питающихся влажным кормом, и
крыс, питающихся одним зерном.
Таблица t
Потребление серыми крысами пищи и волы в лабораторных условиях
(по А. А. Пасешнику)
Число под- опытных крыс Средний вес, г Среднее суточное потребле ние пищи, г Суточное потребле- ние воды, мл Вид корма
влажные корма зерновая пища
78 217 53 _ 2,5 Хлеб, каше
11 282 88,4 14,6 9,94 Горох
12 259 60,3 7,0 8.1 К нопляное семя
40 м 30,4 32,6 < вес
24 — м» 28,8 37,6 Пшеница
Интересные результаты дали опыты, проведенные С Л. Янсо-
ном (1937), для установления роли и значения влаги в кормовом
28
«" I uiHce серых крыс. Автор указывает, что кормление подопытных
шинных пищей, содержащей 14% влаги (рожь), без дополнитель-
<i hi дачи воды приводило к острому водному голоданию и гибели
11 среднем через 5—6 суток; при скармливания пищи, с влажностью
и 32% (белая булка), гибель наступала через 17 суток, а при кор-
•1г, содержащем 45% влаги (ржаной хлеб) — через 27,7 суток.
Корм с влажностью около 65% обеспечивал нормальное физиоло-
। ическое состояние животных в течение всего времени опытов
(55 дней).
Эти данные позволяют объяснить описанные в литературе слу-
чаи массовых передвижений пасюков из зерновых складов к местам
s одопоя. Питание одними зерновыми продуктами настолько увели-
чивало потребность в воде, что в поисках ее серые крысы, невзирая
па опасность, двигались по открытым пространствам.
В опытах С. Л. Янсона было также отмечено, что содержание
•-ерых крыс на однообразном корме (ржаной хлеб) не сказывалось
на длительности жизни подопытных животных, но такое авитами-
нозное питание вызвало у крыс ряд функциональных расстройств.
Поэтому в опытах с кормом, содержащем 65% воды, в пищевой
рацион были введены цельное молоко, сырая морковь и сырой кар-
тофель; это положительно сказалось на физическом состоянии жи-
ь иных, которые в течение периода опытов прибавили в весе в
среднем на 40—50 г.
Высокая потребность серых крыс в воде объясняет то обстоя-
тельство, что они при свободном выборе пищи всегда предпочитают
влажные корма. Опыт истребительных работ подтверждает это
положение. Известно, что наиболее охотно поедаются крысами отрав-
ленные приманки, приготовленные из мякиша хлеба, каши, мясного
или рыбного фарша, то есть из продуктов с высоким содержанием
влаги; сухие (зерновые, мучные, сахарные и т. п.) приманки крысц
берут плохо, и применение их мало эффективно.
Серым крысам, обитающим в природных условиях (вне поселе-
ний человека), свойственна привязанность к водоемам. Питание
диких серых крыс изучено недостаточно. Известно, что они пита-
ются растительными кормами, но поедают также пищу животного
происхождения, например, мышевидных грызунов, обитающих в
скирдах и ометах, мелкую рыбу, пресноводных моллюсков, лягушек
н пр. По наблюдениям Н. К. Верещагина (1949), в Западной Гру-
ши серые крысы питаются преимущественно листьями осоки (Carex
<>racilis) и молодыми побегами тростника (Phragmites communis).
В сентябре и октябре основной корм их составляют ядра водяного
ореха (Trapa colchica).
Данные противоположного характера приводит Г. А. Кондраш-
кнн (1949), который отмечает, что крысы, обитающие в дельте
Волги, питаются почти исключительно животной пищей (грызунами,
птенцами, лягушками, двухстворчатыми моллюсками, мелкой рыбой
и насекомыми). Автор достаточно обоснованно предполагает, что
<<|>и совместном обитании пасюков с водяными полевками остатки
•и
пищи на кормовых столиках водяных полевок были ошибочно*
приняты Н. К. Верещагиным за иоеди пасюков; это и привело к
неправильному выводу о характере питания серой крысы в природ-
ных условиях.
Черные крысы относятся к преимущественно растительноядным
животным. В. К. Шепелева (1950) провела сравнительное исследо-
вание питания серых и черных крыс в лабораторных условиях.
Опыты показали, что при пересчете на единицу веса тела оба вида
крыс поедают примерно одинаковые количества одноименного»
корма. При питании овсом, ржаным хлебом и столовой свеклой,
черные крысы чувствуют себя нормально; следовательно, они яв-
ляются типично растительноядными животными. Черные крысы,
содержащиеся на вареном мясе, мало ели и потеряли за 2 суток
4—12% веса, тогда как серые крысы при этом рационе за 10 суток
прибавляли в весе на 5—13%. Следовательно, пасюки, в противо-
положность черным крысам, нуждаются в животной пище.
Различия в питании отражаются на морфологических и физиоло-
гических особенностях пищеварительного аппарата черных и серых
крыс (табл. 2).
Таблица 2
Морфологические и физиологические особенности пищеварительного
аппарата крыс (по В. К. Шепелевой)
Животное Длина слепой кишки п<> от- ношению к длине кише- чника, % ( 'тношенк е веса печени к весу тела, % pH желудоч- ного сока Содержание кислоты в же- лудочном соке, %
Серая крыса Черная крыса 4,5 6,4 6,5 5,0 1 1,58 | 5,3 0,16 0,35
Туркестанские крысы питаются главным образом растительной
нищей. По данным Д. И. Прутенского (1947), содержимое желуд-
ков туркестанской крысы состоит из плодов древесно-кустарниковой
растительности — грецкого ореха, яблони, жимолости, боярышника
и т. п. Крайне редко в желудках попадаются остатки животной
пиши: перья птиц, хитиновые остатки насекомых, кусочки раковин
моллюсков и т. д. Основной корм туркестанской крысы составляют
грецкие орехи. Во время осенних раскопок в подземных камерах
были обнаружены запасы грецких орехов весом до 12 кг. Для хра-
нения таких запасов туркестанские крысы используют также дупла
деревьев. По подсчетам П. М. Сеславиной и П. П. Алексеева (1936),
в старых рощах грецкого ореха грызуны уничтожают не менее 20%
урожая ореха.
РАЗМНОЖЕНИЕ КРЫС
Серые крысы, как и многие другие виды мышевидных грызунов,
отличаются высокой плодовитостью. При благоприятных темпера-
турных и кормовых условиях они могут размножаться в течение
30
hpxr.ioro года. Длительность жизни пасюков равна трем, а в редких
। лучаях даже четырем годам. Максимальный вес самок достигав!
•1К5—529 г, самцов — 560—570 г (А. П. Кузякин, 1952). Крысята
родятся голыми, слепыми. На 10-й день жизни они обрастают
шерстью, на 15-й день у них открываются глаза. Самостоятельную
<м1знь они начинают вести в возрасте 25—30 дней, когда вес их
достигает 30—35 г.
Молодые крысы приступают к размножению в возрасте около<
3 месяцев. Беременность длится 22—26 дней. Плодовитость самок
г возрастом сначала повышается, но по мере старения организма
угасает. Первый помет у крыс относительно невелик, два последую-
щих — самые многочисленные. По данным А. С. Фетисова (1945),
плодовитость крыс-карако находится в прямой зависимости от воз-
раста самок (табл. 3).
Таблица .
Зависимость плодовитости самок крысы-карако от возраста*
(по А. С. Фетисову)
Соотношение полов у крыс колеблется в пределах, близких 1:1.
По данным А. П. Кузякина (1952), самки несколько преобладают,
составляя 55,1—56% от общего количества пасюков, выловленных
п Москве в 1944—1946 гг. И. В. Степанов (1946) приводит для Ба-
гуми близкие цифры (54,4%). Определение пола у 963 пасюков,
выловленных в Ленинграде в 1933 г., показало, что соотношение
полов изменяется по месяцам (табл. 4). В среднем за год оно при-
ближалось к 1:1, причем отмечалось незначительное преоблада-
ш*<’ самок.
Таблица I
Процентное соотношение самцов и самок по месяцам (по С. Л. Янсону)
Месяц Число самцов» % | Число самок, % Месяц Число самцов, % Число самок, %
\| рель 50,0 50,0 W j Сентябрь 45,0 55,0
м и 42,2 ’ 57,8 Октябрь 63,3 36,7
II • 411. 55,4 44,6 Ноябрь 46,0 54,0
Ill 41. 16,6 83,4 Декабрь 48,4 51,6
5 м|vri 61,1 38,9 Январь 54,9 15.1
В течение года самка приносит от 2 до 8 пометов, таким образом
потенциальные возможности размножения чрезвычайно велики. Вы-
числения показывают, что одна пара крыс может дать за год около
800 особей потомства. Практически, численность потомства от одной
пары крыс бывает значительно меньше.
Размножение серых крыс, обитающих в городах и населенных
пунктах сельской местности, происходит в течение всего года, но
интенсивность его сильно колеблется по месяцам. Так, по данным
С. М. Щастного (1912), основанным на вскрытии большого числа
серых крыс, в Одессе размножение достигает наибольшей интенсив-
ности в апреле и в сентябре. Наименьшее число самок приносят
приплод в октябре, ноябре и декабре. Исследование 466 самок, вы-
ловленных в Ленинграде в 1933—1934 гг., также показало наличие
в течение года двух пиков размножения в апреле-мае и в сен-
тябре (С. Л. Янсон, 1934). Интересные цифры, характеризующие
размножение серых крыс, получил А. А. Пасешник (1936). Из
16 000 самок, выловленных в Москве в 1933—1934 гг., почти две
трети оказались беременными, причем наибольшее число беременных
самок было добыто в апреле-мае и в сентябре-ноябре. Наличие двух
пиков размножения подтверждается также и тем, что среди вылов-
ленных крыс наибольшее число неполовозрелых особей отмечалось
в июне, июле и августе, а также в ноябре и декабре то есть в
те месяцы, которые следуют за периодами наибольшей добычи
беременных самок. По данным М. М. Слепцова, в Хабаровском
крае в 1944—1945 гг. в условиях закрытых местообитаний размно-
жение серых крыс достигало наибольшей интенсивности в весенне-
Таблица 5
Суммарные данные о размножении крыс в Батуми за 18 лет
(по И. В. Степанову)
Месяцы Па ючи Черная круга % Алек ачдпчйска* пы -я
всего самок ИЗ них беремен- ных % всего самок из них беремен- ных всего самок ИЗ НпХ беремен- ных %
I 1147 47 4,1 99 11 11,1 44 6 13,6
II 1232 86 7,0 107 13 12,1 38 1 2,7
III 1634 174 10,6 133 14 10,6 79 12 15.2
IV 1955 215 11,0 169 29 17,2 55 10 18,2
V 2129 170 8,0 173 13 7,5 90 13 14,4
VI 2200 134 6,1 232 21 9,0 100 8 8,0
VII 2065 116 5,7 144 20 13,8 51 23 45,0
VIII 1812 135 7,4 165 20 12,1 69 4 5,8
IX 1770 153 8,1 114 12 10,5 52 8 15,4
X 3029 211 6.9 174 15 8,6 43 8 18,6
XI 1585 62 3.9 185 7 3,8 59 8 13,6
XII 1185 41 3.4 179 10 5,6 76 2 2,6
Всего 21756 1544 7,1 1874 185 9,9 856 103 12,9
32
'—।—г—।—। ~г~ г~1—।—гп—।
/ и /// IV V V! W Vga X X! XX
Месяцы
Рис. 11. Изменение среднего ко-
личества эмбрионов у самок па-
сюка в течение года (по А. П.
Кузякину)
летний и осенний периоды, когда беременные самки составляли
<5—40% всех отловленных самок. Зимой среди отловленных самок
беременных было не более 10%. Во Владивостоке максимум интен-
сивности размножения пасюков падает на весенне-летний период.
Такие же сезонные изменения интенсивности размножения подтвер-
ждаются и большим фактическим материалом, собранным в Москве
в 1944—1946 гг. (А. П. Кузякин, 1952).
Сравнительные данные по плодовитости тех видов крыс, кото-
рые встречаются на юге СССР (табл. 5), свидетельствуют о нали-
чии пиков размножения в весенне-
летний и осенний периоды.
Крысы-карако, обитающие в
природных условиях, приносят
обычно два помета: весной и ле-
том; третий помет — в летне-осен-
ний период — удается наблюдать
только в единичных случаях. Г. А.
Кондрашкин (1949) отмечает, что
самки пасюков, добытые в дель-
те Волги за период с октября по
март, были яловые. В конце июня
в дупле была найдена самка с 8
голыми слепыми крысятами.
Число эмбрионов, находящих-
ся на разных стадиях развития, колеблется у одной самки от 4 до
22. В нашей практике при многократных “Вскрытиях беременных са-
мок серых крыс у одной особи ни разу не удалось видеть больше
16—17 эмбрионов — и то только в единичных случаях. В среднем
число зародышей у одной самки колеблется от 6 до 11.
С. Л. Янсон (1934) отмечает, что численность зародышей в од-
ном помете достигает максимума в периоды массовой беременности
(апрель, май и сентябрь), когда, повидимому, условия для разви-
1 ня плода наиболее благоприятны. А. П. Кузякин (1952) также
о I метил, что в среднем наибольшее число эмбрионов у одной самки
приходится на весенние и осенние месяцы, когда увеличивается
процент самок, участвующих в размножении (рис. 11).
Интересные сравнительные данные о числе эмбрионов у бере-
менных самок по отдельным видам крыс мы находим у И. В. Сте-
панова (1946). Данные эти основаны на исследовании громадного
фактического материала, накопленного автором в течение его мно-
। плетней работы в г. Батуми. Среднее число эмбрионов у одной
' амки оказалось приблизительно одинаковым для всех видов крыс,
in 1речающихся в Батуми (табл. 6).
Плодовитость крыс зависит не только от численности помета,
। огорая приближенно может быть установлена путем подсчета
1брионов у беременных самок, но и от числа пометов в год, кото-
рые приносит каждая половозрелая самка. По этому вопросу в
чпературе имеются довольно разноречивые данные. Все они осно-
' I |>Ш» I *
Таблица б>
Числи амбриоиов у 8 888 беременных самок трех видов крыс, выловленных
в Батуми* (по И. В. Степанову)
Вид крысы Число эмбрионом
' 1 1 2 3 ! 4 1 5 6 7 8 | 9 10 11 |12 13114| |_1_5
Серая крыса 4 12 31 46 79 138 195 266228 207155 94 60 ]22 7
Черная крыса — 2 5 11 21 25 31 32 23 10 10 5 2 1 —
Александрийская крыса — — 1 5 12 17 20 27 12 6 2 — —- ——
* Цифры в колонках означают частоту встреч
ваны на материалах, полученных в результате вылова крыс при
дератизации. Как известно, молодые зверьки легче попадаются в
различные ловушки и по сравнению с ними взрослые отличаются
гораздо большей осторожностью и поэтому выловить их труднее.'
Этим и объясняется завышенный процент молодых крыс в уловах.
Так, по материалам, использованным А. П. Кузякиным (1952),.
30,9% от общего числа самок составляли крысы в возрасте от 2
до 6 месяцев. Возможно, что взрослые беременные самки так же
менее осторожны, чем яловые, и отлавливаются в больших количе-
ствах. Поэтому крысы весом от 421 до 529 г, которые имеют воз-
раст более двух лет и составляют, по данным А. П. Кузякина, 1,1 %
От общего числа отловленных самок, дают очень высокий процент
беременных, равный 30,5.*
Эти цифры наглядно показывают, что материалы, основанные на
вылове крыс, даже при условии его массовости, когда количество
добытых зверьков исчисляется свыше 10000, не отражают действи-
тельные возрастные группировки популяции и не дают твердой
почвы для определения плодовитости. Таким образом, если мы бу-
дем принимать за основу процент беременных самок от общего1
числа всех добытых самок, то цифра, характеризующая годовой
процент размножения самок будет занижена.
По данным А. П. Кузякина (1952) молодые крысы в возрасте-
до 2-х месяцев составляют 12,1 % от общего числа выловленных
самок, причем среди них не встречаются беременные. Следующая
возрастная группа, которую составляют самки в возрасте от 2 до
6 месяцев, еще более многочисленна — к ней относятся 30,9% всех
добытых самок, но процент беременных среди них очень низок
(1,1%). А. П. Кузякин в своих вычислениях использовал все воз-
растные группы самок, в том числе и в возрасте до 6 месяцев,
среди которых имеется ничтожный процент беременных. Используя
методику вычисления автора, можно получить совершенно различ-
ные результаты при определении плодовитости крыс, в зависимости
от того, какие возрастные группы самок принимать в расчет. При-
веденные А. К. Кузякиным данные о наличии у серых крыс в сред-
нем 1,8 помета в год нам кажутся преуменьшенными. Аналогичные
34
данные И. В. Степанова (1946) по материалам, собранным в Ба-
туми, получены в результате использования метода, предложенного
П. Кузякиным. Поэтому выводы обоих авторов о плодовитбсТТг
|.рыб совпали друг с другом.
Зависимость результатов вычислений от методики использова-
гия материала можно видеть и по тем данным, которые приводит
V А. Пасешник (1936). По его расчетам беременные самки состав-
ляют в среднем 63,8% (от 36,8 до 72% в различные месяцы) от
>бшего числа добытых взрослых самок (при вычислениях во вни-
мание не была принята большая группа самок весом до 100 г.).
Невидимому, наиболее правильно при определении интенсивности
размножения крыс исключать группу молодых крыс примерно до
6-месячного возраста; тогда в среднем приходится в год около 3 по-
метов на каждую самку. Не следует забывать, что несмотря на
такую плодовитость численность крыс постоянно сокращается в
результате планомерной борьбы с этими грызунами, которая при-
водится в городах и других населенных пунктах. Велика и есте-
твенная смертность крыс (особенно молодняка) от различных ин-
фекционных заболеваний и влияния неблагоприятных условий су-
ществования.
ЭКОНОМИЧЕСКОЕ И ЭПИДЕМИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ КРЫС
Многие виды мышевидных грызунов, в том числе и крысы, яв-
ляются серьезными вредителями. Экономическое значение крыс
определяется стоимостью уничтоженных и приводимых ими в негод-
ность продуктов питания, промышленных товаров и сырья, а также
юрчей построек, в которых они обитают. Убытки, причиняемые
крысами народному хозяйству, очень велики. Исходя из среднего
количества пищи, поедаемой парой крыс, и численности их приплода
ia год, можно подсчитать, что крысы уничтожают и приводят в
негодность тысячи тонн продуктов. Кроме тогорв птицеводческих и
животноводческих хозяйствах, где обилие корма в помещениях для
птиц или свиней создает благоприятные условия существования
। рыс, они наносят большой ущерб, уничтожая молодых птиц и
чйца, а в свинарниках, — нападая на поросят. Нередки случаи
' кусов крысами и взрослых свиней.
Большую роль играют крысы и как носители ряда инфекцион-
ных заболеваний, опасных для человека и домашних животных.
( вязь между эпидемиями чумы у человека и эпизоотиями среди
||>ы»унов известна давно. Передача инфекции от крысы к крысе, а
;n»4v и человеку происходит при контакте с больными животными
। мере j блох. Чумные бактерии попадают в пищеварительный тракт
• < кнп при сосании крови крысы, больной чумой. Переходя на ново-
। • мвяина, блохи заражают его своими испражнениями, содержа
ним чумных бактерий, которые попадают на расчесы кожи иди в
>.|пки от укусов. Возможен и другой путь заражения, осункт i нляю-
<пн11ся, когда чумные бактерии размножаются в пии1сплрщсл|.ч« t
I к ои-'ii it i.ikin ко.шт стах, что происходит закупорка желуд-
I I Г«>| д.1 при поцыже сосать кровь нового хозяина брызги послед-
ши иона laioi обратно в ранку; при этом с брызгами захватывают-
н ч\мпыс бактерии, закупорившие желудок.
Одним из основных перёносчиков чумы служит блоха Xenopsilla
theopis, которая обычно паразитирует на серых и черных крысах,
но может также сосать кровь человека. Кроме эндемичных по
чуме районов, вспышки этой болезни наблюдаются главным образом
в портовых городах, куда иногда случайно завозят зараженных чу-
мой крыс. Такая опасность заставила обратить особое внимание на
дератизацию судов. Соответствующие мероприятия, обязательные
для всех стран, регламентируются специальными постановлениями
Международной конвенции. В пределах СССР, благодаря правиль-’
ной организации профилактических мероприятий, не бывает случаев
заболевания чумой.
Серые крысы передают человеку также спирохету возбудителя
инфекционной желтухи, или болезни Вейля. Загрязняя продукты
питания и предметы домашнего обихода, крысы рассеивают этих
спирохет, которые у больных животных выделяются со слюной и с
мочой. При укусе крыса может заразить человека и другой бо-
1езнью так называемой болезнью укуса крыс, или содоку. Кроме
того, серые крысы являются носителями бешенства и поэтому лю-
дям, укушенным крысами, приходится проходить курс специального
лечения.
Загрязняя продукты питания и водоемы, серые крысы могут
распространять ряд кишечно-тифозных заболеваний человека, в том
числе холеру, дезинтерию, брюшной тиф, паратифы А и Б и др.
Крысы служат также переносчиками бацилл группы паратифов и
палочки Гартнера, а также В. botulinus.
На серых крысах паразитирует крысиный клещ Bdellonyssus
bacoti S. Hirst, который благодаря широкому распространению свое-
го хозяина встречается почти во всех странах света. Его можно
нстретить не только на самих крысах, но и в местах обитания этих
|рызунов. Bd. bacoti паразитирует на R. norvegicus, но напала зт
также и на человека. Эпидемиологическое значение крысиных
клещей заключается в том, что они распространяют среди людей
крысиный тиф. Кроме того, укусы их вызывают раздражение кож-
ных покровов. Крысиные блохи Xenopsylla cheopis и Ceratophyllus
tasciatus также являются переносчиками риккетсий крысиного сып-
шио тифа, который передается от крысы к крысе и от крысы к че-
ловеку. Такое же значение имеют кошачьи и собачьи блохи Cieno-
< ephalus canis и Ct. fells, которых можно найти и на крысах.
В Средней Азии в биотопах, связанных с постройками человека,
туркестанские крысы служат основным источником, откуда клеши
Ornithodorus papillipes Bir. получают спирохет среднеазиатского
клещевого возвратного тифа. Спирохеты передаются со слюной кле-
ща во время сосания крови. Этот вид клещей встречается в норах
R. turkestanicus, в помещениях для скота, а реже и в жилых по-
t (ройках. Благодаря чрезвычайной длительности сохранения ви-
рулентных спирохет клещи О. papillipes служат резервуаром этой
инфекции (Е. Н. Павловский, 1948).
Нападают на человека, а также на некоторых животных и
« греча ющиеся на александрийских крысах мелкие клещики Trombi-
. ч1а akamuschi Вг., служащие переносчиками возбудителя речной
лихорадки.
Большое значение имеют крысы в распространении трихинелеза.
Среди крыс широко распространено заражение трихинами Trichi-
nella spiralis Ow, во взрослой стадии паразитирующими в кишеч-
жке крыс, свиней, собак и человека. Самки трихин в течение корот-
। зго срока пребывания в тонких кишках хозяина отрождают боль-
шое количество мелких личинок, которые, попадая в ток крови и
лимфы, разносятся по всему организму и в конечном итоге внедря-
ются в поперечнополосатые мышцы, где инкапсулируются. В таком
виде они годами сохраняются в мышцах, а при поедании мяса с
живыми личинками происходит заражение трихинами нового хо-
зяина. Таким образом, полный жизненный цикл трихин протекает
в двух хозяевах, которые могут принадлежать к одному и тому же
виду. Среди крыс заражению здоровых животных способствуют яв-
ления каннибализма. Одним из источников заболевания свиней яв-
ляется поедание ими трихинелезных крыс или корма, загрязненного
экскрементами последних.
У крыс паразитируют и некоторые виды ленточных червей и не-
матод. Так, у них в переднем отделе тонкой кишки найден крысиный
цепень Hymenolepis diminuta Rud. Яйца этого червя рассеивают
крысы, затем их проглатывают личинки или взрослые насекомые
(личинки мучного хрущака, черных тараканов, прусаков, крысиных
блох), которые встречаются в местах обитания крыс. В полости
т«*ла промежуточных хозяев личинки превращаются в цистицерко-
пдов. Крысы проглатывают зараженных насекомых, и в их кишеч-
1 ике цистицеркоиды превращаются во взрослых червей.
Из нематод в печени серых и черных крыс паразитирует Hepati-
cola hepatica Вапсг., где она откладывает яйца, из которых, пови-
цимому, здесь же развиваются личинки У крыс, съевших животных,
сраженных Н. hepatica, в кишечнике вылупляются личинки, кото-
) ые вбуравливаются в стенку кишки, а затем с током крови попа-
:ают в печень.
Помимо всего этого, крысы, поедая фекалии человека, прогла-
(ывают яйца анкилостомид, что содействует их рассеиванию во
। пешней среде; при этом анкилостомиды проходят через кишечник
животного без всякого для себя вреда.
Работами многих авторов установлено, что крысы содействуют
। «кже распространению среди домашних животных таких инфек-
ционных заболеваний, как ящур, сибирская язва, инфлюэнца ло-
шадей, рожа свиней, холера птиц, ложный туберкулез, кокцидиоз
ч 1 ц. Через этих грызунов передается также парша, микоз кожи,
•юлос и ногтей.
I lpin»i цинь», it диких популяций R. norvegicus к водоемам также
MbKi'r имен. лщдемноло! ичеокое значение; отметим в связи с этим,
•но и некоторых приречных городах установлен факт носительства
крысами лептоспироза. Загрязняя воду своими выделениями, крысы
итосят в нее инфекцию, которая может распространяться на людей
н домашних животных.
ИСТРЕБЛЕНИЕ КРЫС, ИЛИ ДЕРАТИЗАЦИЯ
В связи с большим экономическим и эпидемиологическим зна-
чением серых крыс во всех странах мира ведется интенсивная борь-
ба с этими вредными грызунами.
В СССР дератизационные мероприятия в городах и населенных
пунктах проводятся силами органов Министерства здравоохранения
и ряда других заинтересованных организаций — Министерства сель-
ского хозяйства, Министерства путей сообщения и др. Опыт истре-
бительных работ показал, что наилучшей организационной формой
борьбы с серыми крысами является проведение в городах сплошной
плановой дератизации, предусматривающей систематическую обра-
ботку всех строений и обязательное наблюдение за площадью, не-
заселенной грызунами или освобожденной от них в результате
истребления крыс.
Дератизация предусматривает проведение комплекса мероприя-
тий, который включает в себя профилактические и истребительные
мероприятия. Многолетняя практика дератизационных работ пока-
зала большое значение профилактических мероприятий, без которых
не представляется возможным ни предотвратить заселение различ-
ных построек крысами, ни правильно организовать истребление
грызунов и закрепить результаты дератизации. Профилактические
мероприятия должны быть направлены в сторону создания для
грызунов неблагоприятных условий существования, что сводится, в
основном, к лишению их кормовой базы и защищенных убежищ.
Достигается это путем содержания всех строений и прилежащих
территорий с надворными постройками в надлежащем санитарном
состоянии, правильной организацией сбора и вывоза мусора, обес-
печения максимальной крысонепроннцаемости зданий и организа-
цией хранения пищевых продуктов в местах, недоступных грызунам.
Истребление грызунов осуществлятся применением нескольких
методов борьбы — химических, механических и бактериальных.
Химические методы борьбы. Для истребления серых крыс в горо-
дах и других населенных пунктах из химических методов борьбы
применяют, главным образом, отравленные ядами пищевые при-
манки. Сущность этого метода заключается в том, что ядовитые
вещества сочетают с пищевым продуктом. При этом приманки
должны остараться достаточно привлекательными для грызунов,
чтобы те съедали их в количествах, содержащих летальную дозу
яда; кроме того, необходимо стремиться, чтобы отравленные при-
манки сохраняли токсические свойства в течение возможно более
33
;.niтельных сроков. Особенно большое значение имеет первое усло-
вие, так как в тех случаях, когда в организм вводятся только суб-
тетальные дозы яда, к некоторым из них вырабатывается повышен-
ная устойчивость, в результате чего при повторных обработках для
отравления необходимо увеличивать дозировку яда. Вместе с тем
у серых крыс, взявших пищевые приманки в недостаточном для
отравления количестве, повышается настороженность по отношению
ко всем видам приманок и тем более к приманкам с повышенными
дозировками яда. Поэтому особое внимание следует обращать на
высокую эффективность первичных истребительных работ.
Поедаемость приманок серыми крысами в значительной мере
зависит от обеспеченности их кормами в местах обитания. В тех
случаях, когда не удается добиться положительных результатов,
увеличение поедаемости приманок может быть достигнуто путем
предварительной подкормки крыс. Для этого их в течение 5—7 су-
ток подкармливают положенной в определенных местах в избытке
неотравленной пищей; после этого крысы утрачивают осторожность
по отношению к тому же виду пищевого продукта, положенному в
ом же месте, и чаще поедают его даже при добавлении к нему яда.
Для истребления серых крыс в дератизации применяют ряд ядо-
витых веществ. Остановимся на наиболее употребительных из них.
Фосфид цинка (2п3Рг) — тонко размолотый порошок черного
цвета с сильным чесночным запахом. Нерастворим в воде. Под
действием слабых кислот, в том числе и соляной кислоты, содержа-
щейся в желудочном соке, разлагается с выделением фосфористого
водорода, который и является действующим началом при отравле-
нии. Токсичен для всех теплокровных животных. Летальная доза
для серых крыс 15—30 мг.
Крысид («- нафтилтиомочевина — C10H7NHCSNH2) — свет-
лосерый тонкий порошок, горький на вкус, нерастворимый в воде.
Этот препарат получил название крысид в связи с тем, что он об-
ладает избирательной токсичностью по отношению к серым крысам,
а также и некоторым другим видам грызунов, и практически безопа-
сен для домашних животных и человека. Летальная доза для серых
крыс равна 25—30 мг на I кг веса, тогда как для собаки она состав-
ляет 60—150, а для цыпленка — 750—2500 жг на 1 кг веса.
Углекислый барий (ВаСОз) — белый порошок без вкуса и ‘за-
паха, нерастворимый в воде. Менее токсичен, чем фосфид цинка и
'рысид; летальная доза для серых крыс — 120—200 мг.
Сернокислый таллий (TISO4) — белый кристаллический поро-
шок без вкуса и запаха, хорошо растворимый в воде. Токсичен для
всех теплокровных животных; летальная доза для серых крыс --
20 л«г.
В зависимости от токсических свойств ядов установлены следую-
щие нормы содержания их в отравленных приманках: фосфида цин-
3—5%; крысида — 1 %-, сернокислого таллия — 1 %, углекислого
'ария — 7—10% (концентрация яда указана в процентах по отно-
шению к весу приманочного продукта).
39
Icxiuih.i mi отопления отравленных приманок зависит от свойств
uiuiii-Bpx приманочных продуктов и применяемых ядов. Однородную
rniinrnvio массу, как например, раскрошенный хлеб, каши и т. п.,
чех.ншчески смешивают с ядовитым веществом, стремясь достигнуть,
равномерного распределения яда в пищевом продукте. Зерновую
приманку с нерастворимыми в воде порошкообразными ядами гото-
вят путем приклеивания их к поверхности зерен при помощи аппли-
каторов, или приклеивателей, в качестве которых используют ра-
стительное масло, крахмальный или мучной клейстер или казеино-
вый клей. Зерновую приманку готовят также путем вымачивания
пли вываривания зерна в ядовитом растворе, для чего используют
растворимые в воде яды. Как указывалось, сухие зерновые приман-
ки серые крысы берут плохо и предпочитают приманки, приготов-
ленные из пищевых продуктов, содержащих достаточное количество
влаги.
Меньшее распространение в практике дератизации имеют различ-
ные пасты с растворимыми и нерастворимыми в воде ядами. Изве-
стны паста с сернокислым таллием, основой которой служит раз-
бухающий в воде коллоид, и пасты с фосфидом цинка, приготовляе-
мые па глицерине или вазелине. Ядовитую пасту намазывают на
кусочки хлеба, после чего приманка готова к употреблению.
Отравленные приманки закладывают в количестве 10—15 г во
все входные отверстия, обнаруженные в обрабатываемом помеще-
нии, после чего производят заделку отверстий. Открытая раскладка
приманок допускается только в тех помещениях, в которых ограни-
чен допуск людей. Остатки несъеденной приманки в обязательном
порядке должны быть собраны и уничтожены. В тех помещениях,
где не безопасна открытая раскладка отравленных приманок, пх
помещают в специальные ящики с отверстиями в боковых стенках,
через которые крысы могут пролезть внутрь ящика, но не имеют
возможности растаскивать из них приманку.
Кроме пищевых приманок в дератизации применяют водные при-
манки. Этот способ борьбы основан на влаголюбивост и серых крыс.
Водные приманки особенно привлекательны для крыс в помещениях,
где отсутствует вода, но опытные работы в практических условиях
показали, что даже на объектах, где крысы имеют свободный до-
ступ к воде, они охотно берут водные приманки. Техника изготов-
ления водных приманок не сложна. На поверхность воды, налитой
в какую-нибудь плоскую, достаточно устойчивую посуду, наливают
чистую воду и высыпают отвешенную дозу крысида. Слегка покачи-
т а я сосуд с водой, достигают равномерного распределения частичек
порошкообразного яда по поверхности воды, который устойчиво
держится на ней. Крысы, выпивая воду, заглатывают одновременно
п ядовитый препарат. Опыты по использованию ядовитых раство-
ров дали отрицательные результаты, так как присутствие в воде рас-
творенных ядов серые крысы легко обнаруживают и не пьют их.
К новым методам борьбы с серыми крысами относится опылива-
ние порошкообразными илами входных отверстий или путей по-
40
< iчинного передвижения крыс. Проходя по опыленной ядом поверх-
ности, зверек пачкает мех и лапки, а при чистке или при принятии
пищи заносит частицы яда в ротовую полость. Летальная доза кры-
. и да настолько незначительна, что такое его применение оказывает-
। ч достаточно эффективным. В городских условиях при использо-
вании метода опыливания можно работать только с крысидом, так
как другие порошкообразные яды, как фосфид цинка или препа-
раты мышьяка, для этих целей непригодны, поскольку они токсичны
ия всех теплокровных животных.
Рис. 12. Давилка типа «Геро»
Распиливание крысида производят при помощи специальных
приспособлений с дозирующими наконечниками, а при отсутствии
hs входные отверстия заделывают тампонами из пакли, мочалы или
ваты, на которые нанесен ядовитый порошок. Яд попадает в рот
крысы при попытке вскрыть отверстие, заделанное тампоном.
Механический метод борьбы. Одним из старейших способов
П' гребления крыс является применение механических орудий лова.
-)тот метод сохранил свое значение и до настоящего времени и
и пользуется как в комплексе с химическими и бактериальными
•модами борьбы, так и самостоятельно в тех объектах, где проти-
опоказано применение ядов или бактериальной культуры. Для вы-
чва серых крыс пользуются ловушками различных систем.
В дератизационной практике в больших масштабах используют
.1 вилки типа «Геро», которые изготовляют двух размеров - боль
пне для отлова крыс и маленькие — для домовых мышей
(рис. 12). Когда зверек потянет приманку, надетую на крючок, на
о гором при зарядке ловушки укрепляется сторожок, проволочная
iv/кка захлопывается. Чувствительность давилки повышается при
iwene крючка подвижной пластинкой, которая приводит ловушку
(ействие, как только крыса до нее дотронется.
Для отлова крыс применяют также дуговые капканы № 0 п № I.
>"юрые ставят без приманки около нор или в тех мепач. где
||)ысы часто бегают. Кроме того, в практической рабоо* испои»
11
.) ни i uMt’.'ioBKii * верш И' и ловушку Тишлеева. Такие ловушки дей-
’ план ши ома 1ичееки и нет надобности перезаряжать их после
поимки крысы.
Верша (рис. 13) представляет собой проволочную клетку с пло-
< ким дном и полукруглой верхней стенкой (длина 50 см, шири-
на - 32 см, высота — 25 см). С торцовой стороны вмонтирован
проволочный конус, узкой частью направленный внутрь клетки. В
горловине этого конуса укреплена металлическая площадка на про-
Рис. 13. Верша
тивовесах, которая закрывает отверстие в нижней части горловины.
Внутри верши подвешивают на крючке или кладут на дно пище-
вую приманку. Пробираясь к ней, крыса попадает «а качающуюся
площадку, которая опускается под тяжестью ее тела, и крыса со-
скальзывает в клетку. Площадка под действием противовесов зани-
мает первоначальное положение и закрывает выход из клетки. С
противоположной от горловины стороны имеется дверца, через ко-
торую пересаживают крыс из ловушки. В одну вершу можно пой-
мать до 10 и более пасюков.
Ловушка Тишлеева (рис. 14) представляет собой проволочную
клетку прямоугольной формы (длина — 50 см, высота — 18 см,
ширина — 35 см). Внутри клетки у входа в ловушку расположена
качающаяся площадка, по которой крыса подходит к приманке,
расположенной у стенки, противоположной входу. При этом под
тяжестью тела зверька площадка опускается и закрывает собой
входное отверстие. Крыса в поисках выхода попадает. в коридор,
расположенный внутри клетки. В нем имеются два металлических
клапана, которые приподнимаются только в одну сторону при пе-
редвижении крысы вперед. Когда поднимается второй клапан,
соединенный при помощи рычагов с падающей площадкой, послед-
няя занимает первоначальное положение, и входное отверстие авто-
12
нически открывается. Дверца для пересадки пойманных пасюков
к.', положена в торцовой стенке ловушки, напротив входного отвер-
тя. В ловушку Тишлеева, как и в вершу, можно одновременно
ь ймать более 10 зверьков. Наличие в ней коридора, снабженного
। планами, один из которых соединен с падающей площадкой,
>'»<‘спечивает надежность действия ловушки и, благодаря этим при-
иособлениям, пойманные крысы не имеют возможности выбраться
нее. Это является большим преимуществом ловушки Тиш-
|> ева, так как падающая площадка, установленная в горловине
iii-рши, не всегда плотно закрывает отверстие, и пойманные крысы
• ' югда уходят из верши.
Рис. 14. Ловушка Тишлеева
Вылов крыс производят также и при помощи различных само-
«• чьных приспособлений, изготовляемых населением. Такие ловуш-
ки обычно представляют собой ловчие бочки с качающимися крыш-
ками, вертушками и т. п.
Эффективность механического метода борьбы с крысами зависит
ч правильной расстановки ловушек в местах, наиболее часто посе-
цаемых грызунами, и использования достаточно привлекательных
|риманок. Для того, чтобы вылов был успешным, рекомендуется в
ачение нескольких (3—5) дней ставить незаряженные ловушки,
пабженные приманкой, в результате чего крысы приучаются к рас-
гагленным орудиям лова. Кроме того, необходимо следить за без-
тазностью работы спусковых механизмов, для чего их следует
'.'.мывать каким-нибудь жиром или маслом. После мытья ловушек
|> надо тщательно просушить, иначе они заржавеют. Пятна крови
• сгатки шерсти пойманных зверьков не отпугивают крыс, и опыт
сказал, что в одну и ту же ловушку подряд можно в течение ко-
ми кого срока поймать несколько крыс.
Бактериальный метод борьбы. Сущность бактериального метода
1 >рьбы заключается в распространении инфекции среди крыс путем
1' кладки приманок, зараженных бактериями крысиного тифа (бак-
• рии Исаченко). Распространению инфекции способствует контакт
» н-ных крыс со здоровыми. В России бактериальный метод борьбы
крысами в большом масштабе впервые был применен в 1902 г. в
’юсе. В настоящее время культура бактерий Исаченко исполь-
!3
♦усчся для дератизации в городских условиях лишь в ограниченных
размерах.
Смертельный исход наступает при введении в организм крысы
?•> мл культуры, выращенной в течение суток в термостате при 37°
па нормальном мясо-пептонном бульоне. Обычно крысы гибнут
через 5—12 суток после заражения, но некоторые особи обладают
природным иммунитетом по отношению к бактерии Исаченко и не
восприимчивы к инфекции. Кроме того, часть крыс, переболев кры-
синым тифом, приобретает иммунитет. Поэтому повторное исполь-
зование для дератизации культуры бактерий Исаченко менее эф-
фективно.
При практическом использовании бактериальных культур решаю-
щее значение имеет их вирулентность, которая зависит от многих
причин и довольно непостоянна. Она снижается при хранении куль-
туры свыше 45 дней, при пребывании ее на свету или на холоде,
при засорении посторонними микроорганизмами и пр.
Данные А. Д. Касицкой, В. Г. Полежаева и П. С. Гречищева
(1950) показали, что эффективность бактериального метода борьбы
повышается при введении в пищевые приманки вместе с бактери-
альной культурой и сублетальных доз крысида (0,3 % по отношению
к весу приманки). Введение в организм небольших доз яда сни-
жает устойчивость крысы к бактерии Исаченко, чем и объясняется
большая эффективность такого комбинированного метода истребле-
ния крыс.
СОДЕРЖАНИЕ И РАЗВЕДЕНИЕ КРЫС
Правила содержания крыс в неволе
Экспериментальная работа с лабораторными животными может
быть правильно организована только при условии использования
здоровых, полноценных подопытных животных. В связи с этим,
при содержании и разведении крыс в питомниках, или в вивариях,
должны соблюдаться определенные требования.
Обычно научно-исследовательские институгы приобретают белых
крыс через кооперативные организации, ведающие поставкой раз-
личных лабораторных животных и другого биологического материа-
ла. Крупные институты иногда имеют собственные питомники. При
незначительных масштабах экспериментальных работ белых крыс
можно разводить и в условиях сравнительно небольшого вивария.
Виварий для крыс должен быть помещен в сухом, отапливае-
мом помещении, обеспеченном естественным освещением и венти-
ляцией. В сыром, темном или холодном помещении животные не
могут нормально развиваться и подвержены различным губитель
вым заболеваниям. Воспитание при высокой температуре также
снижает устойчивость белых крыс к различным инфекциям.
Даже небольшой виварий, рассчитанный на несколько сотен жи-
вотных, должен помимо основного помещения, иметь изолятор, или
бокс, для отсадки больных животных и для карантишвацип вновь
поступающих в виварии, отдельную комнату тля хранении продук-
41
Рис. 15. Клетка для содержания
белых крыс
i ов и приготовления кормов, и, наконец, помещение для мытья и
дезинфекции клеток, кормушек и поилок.
Крыс содержат в клетках, размещаемых на стеллажах, но не
больше, чем в 2—3 яруса. Емкость вивария исчисляется из расчета
’.5 л3 на 15 взрослых крыс. Клетки должны быть достаточно вме-
тптельными — животные должны в них свободно двигаться — и
удобными при обслуживании, т. е., при чистке клеток и пересадке
животных.
Для содержания белых крыс используют клетки из деревянных
jpycKOB или углового железа, стенки которых затянуты металличе-
«кой сеткой (размер ячеек
1 см2). Клетка должна иметь
1ва отделения с отдельными
терцами, в одно из которых
помещают корм, а в другое —
шездо. Ту часть клетки, где
расположено гнездо, следует
сатемнять, так как крысы, осо-
бенно в период размножения,
i пльно реагируют на различ-
ные внешние раздражители.
(по клетки должно быть двой-
ным, причем верхнее дно долж-
но выдвигаться. Над выдвиж-
ным дном делают решетчатый
настил. Дверцы клетки снаб-
жают металлическими наклад-
ками. Деревянных вертушек для запора следует избегать, так как
<>ни быстро ослабевают и могут быть случайно повернуты при
м5орке вивария.
В одну большую клетку можно помещать не больше 10 самок
I 1 самца. Такая клетка должна иметь в длину — 1 ж, в ширину —
к) см и в высоту — 35 см (рис. 15).
Серых крыс рекомендуется содержать только в металлических
и )етках, так как деревянный каркас они легко перегрызают и ухо-
1ят из клеток. Для изготовления клеток мягкие сорта металла, на-
пример дюралюминий, непригодны, ибо пасюки легко их перегры-
1ЮТ.
Серые крысы не приручаются даже при длительном содержании
неволе и остаются столь же дикими и злобными животными, как
। в условиях естественного обитания. Поэтому клетки для их со-
। ржания имеют некоторые конструктивные особенности. К ним ot-
ic сится, в первую очередь, выдвижная поперечная перегородка,
г своляющая изолировать зверьков в одной половине клетки, без
'с ю нельзя производить чистку клетки и дачу корма и питья. Двер-
цы удобнее располагать с торцовых сторон клетки, что облегчает
• ресадку крыс, техника которой будет описана ниже. Практика
> । Ы1НЫХ работ с серыми крысами показала, что наиболее рацио
1 >
h i 1Ы1Ы h. iv I mi. имеющие следующие размеры: длина — 50 см, ши-
рина ;<5 гн, высота — 32 см. высота с ножками — 35 см
(рис. 16). Пасюков рассаживают в клетки поодиночке, так как в
ц-\ случаях, когда они пойманы в разных местах, между ними воз-
ни кп юг драки, причем более сильные нередко загрызают слабых.
Чистку клеток производят ежедневно, удаляя при помощи метал-
лического скребка или лопатки экскременты животных и остатки
корма, а также меняя подстилку. После окончания опытов клетки
тщательно моют горячей водой с содой. При длительных опытах
такую обработку клеток производят не реже, чем 2 раза в месяц.
Один раз в 2—3 месяца все клетки дезинфицируют, для чего их по-
гружают в ванну с дезинфици-
рующим раствором. В случае по-
явления в виварии инфекционных
заболеваний клетки, в которых со-
держались больные животные, не-
обходимо дезинфицировать. Убор-
ку помещения вивария производят
ежедневно, протирая для этого
пол мокрой тряпкой или вымывая
его.
Работа с крысами требует не-
которых навыков со стороны об-
служивающего персонала, так
Рис. 16. Клетка для содержания как при неосторожном обращении
серых крыс со зверьками легко получить уку-
сы, и, кроме того, крысу, неумело
взятую в руки, легко травмировать.
Белых крыс рекомендуется брать пальцами (большим и указа-
1ельным), удерживая зверька со спинной стороны в подмышечной
области так, чтобы пальцы на брюшке крысы почти сходились друг
с другом. Так взятый зверек не может укусить и, вместе с тем, его
нельзя травмировать, даже сильно сжимая пальцы. Этот способ
применяют главным образом при пересадке животных. В тех слу-
чаях, когда нужно крысам ввести какое-либо вещество per os или
' делать влагалищный мазок, зверька схватывают сначала за хвост,
опускают на какую-нибудь горизонтальную поверхность, а затем
другой рукой берут со стороны спины так, чтобы голова крысы рас-
полагалась между указательным и средним пальцами, а передние
1апы — между большим и безымянным. Белые крысы, если их
брать; не причиняя им боли, довольно быстро привыкают к рукам
и перестают кусаться.
Пасюков, вследствие их злобности, брать голыми руками не
представляется возможным. Для пересадки их удобнее всего поль-
зоваться мешком из плотной прорезиненной ткани. Такой мешок
надевают на ловушку или клетку с той стороны, где имеется двер-
ца, и перегоняют в него зверьков. Затем открытую сторону мешка
собирают складками, зажимают у горловины рукой и, приоткрыв
46
дверцу новой клетки, просовывают в нее зажатую часть мешка и
перегоняют туда крысу. Размеры мешка должны соответствовать
величине клеток, чтобы можно было надеть его на торцовую сто-
рону клетки.
В лабораторной практике серых крыс используют главным об-
разом для испытания различных средств и методов их истребления;
при проведении опытов такого рода нет необходимости брать подо-
пытных животных в руки. То же самое относится к эксперименталь-
ному изучению поведения и отдельных вопросов биологии серых
крыс. В зависимости от задач, которые разрешаются в ходе опытов,
в конструкцию клеток вносятся специальные изменения, что позво
ляет регулировать доступ к корму, освещение и т. п.
В исследованиях экологического характера правильная методи
ка работ может быть соблюдена только в том случае, если их про-
изводить в условиях, максимально близких к обстановке естествен-
ного обитания серых крыс; это достигается проведением опыто: в
специально оборудованных больших вольерах.
Если белых крыс сравнительно легко разводить в неволе, то
необходимых для вивария серых крыс приходится отлавливать в
местах их естественного обитания. Для этого пользуются живолов-
ками — вершами или ловушками Тишлеева. Поскольку пасюки с
большой настороженностью относятся ко всем новым предметам,
рекомендуется маскировать ловушки бумагой, мешками, рогожей
п т. п., а также приучать брать приманку из незаряженных лову-
шек. Это увеличивает эффективность вылова и сокращает его
роки.
Кормление крыс
Лишение крыс возможности свободного передвижения снижает
их обмен веществ и делает зверьков более восприимчивыми к раз-
личным инфекциям. Поэтому одним из главных условий успешного
содержания и разведения крыс в неволе является правильная орга-
низация кормления.
Кормовые продукты должны содержать белки, жиры, углеводы,
минеральные вещества, витамины и воду в определенных соотноше-
ниях. Отсутствие в рационе какой-нибудь составной части и недо-
статок или преобладание одной из них приводят к различным нару-
шениям нормальной жизнедеятельности организма. Для питания
крыс используют концентрированные корма — различного вида
зерно, продукты его переработки в виде отрубей, жмыхов, круп, а
также печеный на дрожжах хлеб. В качестве источника витаминов
даются корнеплоды, среди которых наиболее ценным пишевь|м
продуктом является морковь. В пищевой рацион входят и корм$
животного происхождения — молоко и молочные продукты, мясо
к субпродукты — печень, почки, селезенка. Вода для питья во из-
бежание какой-либо инфекции должна даваться кипяченая. Мине-
ральные вещества добавляются в виде поваренной соли, мела, др-
веского угля и костяной муки. Корма, используемые для питания
(Г
крыс, обязательно должны быть доброкачественными. Данные о со-
ставе суточного рациона крыс различного возраста и физиологиче-
ского состояния приведены в табл. 7.
Таблица 7
Суточные кормовые нормы для крыс (по К. Л. Ковалевскому)
Животное Концентри- рованный корм, г Белый хлеб, г Мясо, е Моло- ко, г
Самец и самка в со- 17 23 10 —
стоянии покоя -Самка во время случки 19 25 12 —
Самка беременная до 10 дней 22 27 24
Самка беременная до 22 дней 25 30 15
Молодняк в возрасте 1 недели 5 6 5
Молодняк в возрасте 2 недель 7 10 7
Молодняк в возрасте 3 недель 12 14 10
Молодняк в возрасте , 4 недель 15 18 15
Молодняк в возрасте 6 недель 17 23 10 —
При содержании крыс в неволе большое значение имеет обес-
печенность пищи витаминами. При витаминном голодании разви-
ваются различные нарушения жизнедеятельности организма, делаю-
щие крыс непригодными для использования в экспериментах. Дача
витаминных кормов особенно необходима самкам в период размно-
жения, что обеспечивает нормальный рост и развитие молодняка.
Данные о составе полноценного рациона для кормящей самки кры-
сы приведены в табл. 8.
Таблица 8
Суточный рацион для самки, кормящей 8 детенышей, на первые 7 дней
(по К. Л. Ковалевскому)
Наименование корма Количество, а Витамины
А В С 1 f д ! 1 В
Овес 27 + ++ — — —
Кукуруза желтая ^цельное зерно) 15 ++ + — 0-+
Просо 7 +++ — —
Пшеница 8 ; ++ — — +
Рыбий жир Белый хлеб 0,2 71 ++++ -г ^-4-* — 1 Н 1 —
Молоко 40 + + + — —
48
Зоогигиенические правила кормления лабораторных животных
предусматривают дачу кормов в установленные часы и в определен-
ной последовательности, а также соблюдение чистоты при изготов-
лении и раздаче кормов. Несмотря на то, что серые крысы в усло-
виях естественного обитания бывают достаточно неразборчивы в
пище, при содержании их в неволе следует соблюдать те же пра-
вила кормления, какие установлены для кормления белых крыс.
Это позволяет сохранять животных здоровыми и пригодными для
проведения экспериментальных работ.
Размножение крыс в неволе
Белые крысы хорошо размножаются в неволе. Подбор племен-
ного материала определяет продуктивность стада, поэтому в каче-
стве производителей используют вполне здоровых, сильных живот-
ных хорошего телосложения, устойчивых к инфекционным забо-
леваниям. Правильная организация случки и тщательный уход за
беременными самками и молодняком позволяет получить здоровое
потомство. Учет размножения проводят путем записей в племенной
книге, в которой по определенной форме регистрируют дату случ-
ки, число беременных самок и дату отсадки их в отдельные клетки,
время рождения и численность молодняка, характеристика детены-
шей, данные о лактации самок и их материнских качествах и т. п.
По данным племенных записей производят выбор производите-
лей. К. Л. Ковалевский (1948) рекомендует ежемесячно произво-
дить замену старых крыс молодыми на 10—13% из расчета, чтобы
крысы находились в размножении 8—10 месяцев. Случка крыс из
)дного гнезда не допускается, так как длительное внутрисемейное
спаривание может привести к вырождению, выявляющемуся в
уменьшении плодовитости, роста и т. п.
К первой случке не следует допускать слишком молодых крыс;
обычно для этого используют зверьков в возрасте от 14—-16 недель
до 8—10 месяцев, которые дают наиболее сильное и многочислен-
ное потомство. При одновременном содержании в клетке самок и
амца (5 самок и 1 самец) происходит произвольная случка. Со
второй половины беременности (которая продолжается 16—22 дня)
беременных самок рассаживают в отдельные клетки. Следует учи-
1ывать, что при недостатке влаги в корме и питьевой воды самка
зачастую поедает своих детенышей. В обычных условиях явления
каннибализма у белых крыс наблюдаются редко.
Плодовитость белых крыс очень высока — самка за год может
принести 4—7 пометов по 5—12 детенышей в каждом. Рождаются
крысята голыми, слепыми. На 8—10-й день у них прорезаются рез-
цы, и они начинают обрастать шестью; глаза открываются на 14—
17-й день. Крысята до месячного возраста не имеют устойчивой
1емпературы тела и их обогревает своим телом находящаяся в гнез-
де мать. Начиная с 6-недельного возраста, при перемещении кры-
< ят в более холодное помещение, температура их тела повышается.
I Крыса 'Ч
। । и. с .... noipiH ia крысята постепенно переходят на само-
. и.»««и нн।«тис н перестают питаться молоком матери, так что
к и- Mi-» мца их можно отсаживать от матери. Обычно их no-
ч. hi пн и п специальные клетки до 25 штук в каждую. Через две не-
IH nt после того, как отнимают молодых, самка может быть исполь-
зована для случки.
П. П. Сахаров и др. (1952) приводят следующие биологические
особенности, характеризующие размножение белых крыс: продол-
жительность жизни — 2—2,5 года продолжительность беременно-
сти — 20—30 дней; количество пометов в год — 4—7; число дете-
нышей в помете — 5—9; предельная длительность питания моло-
ком матери — 30 дней; срок отделения детенышей от матери —
4 недели; срок отделения самцов от самок — 1—1,5 месяца; срок
допущения к первой случке — 3,5—4 месяца; предел производи-
тельности самцов — I—1,5 года; предел производительности са-
мок — 1,5—2 года; продолжительность половой охоты — 5 дней;
отношение числа самцов к числу самок в производстве 1 : 2—4;
вес взрослых животных — 150 а и выше.
Половой диморфизм у белых крыс выражается главным образом
в размерах: самцы крупнее и тяжелее самок. Наиболее полные дан-
ные об изменении веса белых крыс с возрастом приводит Г. Д. Кинг
(1923) (табл. 9).
Таблица 9
Вес белых крыс по возрастным группам (по Г. Д Кинг)
Возраст, дни Самцы Самки
число крыс нес, ? число крыс вес, г
макси- мальный средний мини- мальный макси- мальный средний мини- мальный
13 50 21 17,2 13 67 20 15,7 13
30 50 КО 48,5 41 50 57 45,7 39
60 50 170 122,9 64 50 153 107,0 66
90 50 238 184,8 103 39 178 148,0 95
120 50 284 223,9 146 42 212 173,4 1 125
151 50 307 244,8 169 45 225 186,3 147
182 50 343 258,4 172 42 245 116,5 147
212 48 366 268,0 176 42 245 197,3 196
243 44 355 279,7 194 43 256 209,6 141
273 41 349 280,9 202 38 254 210,8 158
304 36 379 216,1 198 38 262 219,1 165
334 33 385 300,8 238 35 276 222,4 170
365 28 377 307,1 245 31 276 223,1 176
395 24 381 314,1 246 31 284 220,5 175
Г25 23 397 312,2 243 30 271 215,8 169
155 Г. 414 323.9 249 18 264 220,2 196
ж> 1? 137 326,0 255 13 324 234,7 197
Орг.тннюшит. рптслепие серых крыс в неволе очень трудно.
Рождение мн.п'Лыч muzmio наблюдать чаше всего тогда, когда в ви-
варий доставляю! бг|ц-М1’||ц|<|м симок В болмпипстгн* случаев мать
после родов сейчас же • ы nni'r • пишмспю и лини, в редкие сту-
50
чаях удается его сохранить. Для этого до родов в клетке должно
иметься достаточное количество подстилки, из которой самка может
сделать гнездо. Родившей самке необходимо обеспечить максималь-
ный покой, чтобы ее не раздражал излишний свет и шум. В обыч-
ный рацион добавляют корма с большим содержанием влаги и да-
ют в достаточном количестве питьевую воду. Чистку клетки лучше
производить не раньше, чем через 7—8 дней после появления па
свет новорожденных.
Болезни белых крыс
Белые крысы подвержены ряду заболеваний, которые иногда
приводят к гибели значительную часть поголовья. В деле предот-
вращения эпизоотий большую роль играет комплекс профилактиче-
ских мероприятий, который включает соблюдение на должном уров-
не санитарного состояния помещения и инвентаря вивария, систе-
матический осмотр животных, организацию карантина для посту-
пающих в виварий белых крыс, изоляцию больных животных и т. п.
Высокая восприимчивость белых крыс к инфекционным заболе-
ьлпиям, а также распространение среди них бацилло- и вирусоноси-
тсльства обычно бывают причинами вспышек различных заболева-
нии Поэтому для организации противоэпидемических мероприятий
в вивариях должен быть организован ветеринарный надзор, что
iic'iih наблюдение за состоянием здоровья животных, контроль
лн « aiiiiiарным состоянием вивария, а также за качеством кормов
и приин.пин <П opt дни »ацией питания.
u.'ihiim hi iiaiifioiu распространенных заболеваний белых крыс
нк ни г ч 1Ь1рл1Н(||, по iO\ ди।ель которого относится к группе Salmo-
I, и,). |»рнг|.1, Он в.пне паратифом, делаются вялыми, пере*
• .... in, шгр< п> у них г рче| блеск, развивается конъюнктивит,
xri.ii । in । < к.ц inwiioh ii Ocipaii форма заболевания дает быструю
ин» и. ах pi ioin, при \р..ЦНЧГ1 мм к.чепии болезнь затягивается на
. чп и б и.их и |пп>1 ы прппи.ни к выздоровлению животных, ко-
H'piJi к in п.1|<-111>1< м Mi.ni 01О1ЫИ4 Н бациллоносителями.
Ixi'n 1110. р.п up... ipaii. ши < р< ди бг.тыч крыс можег иметь пасте-
pr.iivi, в. мГц in । г н’м in 11. .pul ।> ini.'iнс г vii luiciepe.i.ia Pasteurella
nttuisiptuu lap.izh< line iipiiin ii jiiii при вдыхании воздуха, зара-
ж« iiiioi (I паc i<‘p.'.'i. iaми. Kxiiipui’ попадают в него при чихании и
мчите больных ЖПВ01НЫХ. У белых крыс пастереллез чаще всего
inn '-г форму инфекционного ренита.
1‘< । о заболевают белые крысы инфекционной эктромелией, вы-
1ын.1см‘1Й фильтрующимся вирусом. Болезнь эта выражается в по-
ражении кожи на передних конечностях и на хвосте, что вызывает
1ангренозные воспаления. Заражение происходит через пищевари-
тельный канал.
Псевдотуберкулез, вызываемый Bad. pseudotuberculosis, также
не имеет большого распространения среди белых крыс, ра >витке
этого заболевания наблюдается спорадически. Заражение ирписхо-
дит через пищеварительный аппарат. Признаками заболевания яв-
ляется общее истощение, которое в конце концов приводит живот-
ных к гибели.
Из грибковых заболеваний у белых крыс наблюдается парша,
। оторая поражает кожу (главным образом на голове и конечно-
стях). Реже встречается стригущий лишай, характеризующийся
появлением на голове и на других частях тела небольших очажков
выпадения волос. Заражение грибковыми заболеваниями происхо-
дит при контакте с больными животными, через подстилку или через
инвентарь вивария (клетки, кормушки и пр.).
Наконец, у белых крыс можно наблюдать поражение ушей и
хвоста чесоточным зуднем. Наличие этого заболевания препят-
(твует использованию крыс в лабораторных условиях. Кроме того,
у белых крыс известны заболевания, возбудителем которых являют-
ся паразитические простейшие.
Кроме инфекционных и инвазионных заболеваний в условиях
вивария белые и серые крысы болеют воспалением легких, которое
возникает главным образом вследствие простуды, и расстройствами
пищеварения, обязанными неправильному кормлению животных.
С едует учитывать также, что при скученном содержании крысы
чогут ранить друг друга и пораженные места иногда нагнаиваются.
Л ИТЕРАТУРА
Авакян А.. Наблюдения над простейшим» паразитами московских крыс
Mus decumanus, Тр. Ill Закавказского съезда по борьбе с малярией, 1939.
Авакян А.. Наблюдения над простейшими паразитами московских крыс
(Mus decumanus), Тр. Москв. обл. ин-та инфекционных болезней нм. И. И.
Мечникова, 1939.
Авакян А., Наблюдения над кишечными цростейшилси у крыс. Сообщение
ДП, Тр. Моск. обл. ин-та инфекционных болезней им. И. И. Мечникова, 1939.
Айзенштадт Д. С.. Некоторые данные по распространению и образу
жизни серой крысы (Rattus norvegicus Berk.) лесного района северо-западной
части РСФСР. «Зоол. жури.». т. XXIV. вып. 3. 1945.
Айзенштадт Д. С., Методика обследования и учет эффективности де-
ратизации, «Военно-мед. журю.», № 11, 1949.
Айзенштадт Д. С. Расселение серых крыс вдоль железнодорожного
полотна, <Природа», 4. 1950.
Айзенштадт Д. С., Гнезда серых крыс, устраиваемые во время поло-
водья, «Природа», 6, 1950.
АйзенштадтД. С. и Арсеньев А. А., Истребление серых крыс в ка-
нализации, «Гигиена и санитария», № 8, 1954.
А й с е л и А. А. Экспериментальная туляремия у серой крысы. Иэв. Ир-
кутского гос. Н.-И. противочумного ин-та Сибири и Дальнего Востока, т. IX,
1951.
А р г и р о п у л о А. И., К вопросу о систематическом положении туркестан-
ской крысы (Rattus turkestanicus Sat.), «Докл. АН СССР», 1928.
Аргнропуло А. И. (Argyropulo A.)» Kurze Bestimmungstabelle
der R&fen Russlands, Zeits. fur Saugetierk., Bd. II. Heft °, 35, 1928.
Аргпр о пу ло А. И.,Материалы к познанию рода RaWnsFitzinger (1867).
Изв. курсов прикл. зоол. и фитопат., вып. 4. 1929.
АргиропулоА И., Определение крыс рода Rattus фауны СССР. Н •»
Северо-западной г,бл. станции зашиты растений. 1929.
52
Аргиропуло А. И., Крысы Средней Азии и прилежащих частей Ка-
захстана, Тр. зоол. ин-та АН СССР, в. IV. 1935.
Аргиропуло А. И., Мыши (Фауна СССР, Млекопитающие, т. III,
выл. 5). Изд. АН СССР. 1940.
Архипианц X. Д.» Рациональная постановка дератизационного дела,
Тр. VIII Всероссийского съезда бактер.. эпидем. и сан. врачей в Ленинграде
20—26 мая 1924 г., 1925.
Архипианп X. Д., Год практики на дератизационных работах, Тр. VIII
Всероссийского съезда бактер., эпидем. и сан. врачей в Ленинграде 20—26 мая
1924 г.. 1925.
А р х и п и а н ц X. Д., Практическое руководство по дератизации изд. 2-е,
Л., 1939.
Бабалова Е.. Крысиный риккетсиоз. наблюдавшийся в Б.. «Журн.
микроб., эпидем. и иммунол.>, № 10—11, 1945.
, Бажанов В. С.» Проникновение крыс на северные морские побережья,
Изв. Иркутского гос. н.-и. противочумного ин-та Сибири и Дальнего Востока,
т. VI. 1946.
Белугина К. и Крей пер С.» К вопросу об эпидемическом вирусе
сыпного тифа у серых крыс, 'Изв. Азово-Черноморского н.-и. ин-та микроб, и
эпвдем., вып. 16. 1937.
Берман В., С л ав с к а я Е. и Попов Н., О носительстве дикими кры-
сами микробов из группы Salmonella, Вопросы возрастной иммунол., вып. I,
1947.
Берта Л., Резистентность крыс, иммунизированных вирусом острого эн-
цефаломиелита человека. «Bonn мед. вирус.», вып. 3. 1950.
Б и рул я А., Предварительное сообщение о грызунах (Rodent? а) из чет-
вертичных отложений Крыма. «Докл. АН СССР», 1930.
Близниченко А.» Характеристика лептоспир, выделенных от крыс,
«Жуон. микроб., эпидем. и иммунол.», № 8. 1949.
Бобринский Н. А., Кузнецов Б. А. и Кузякин А. П.. Опреде-
литель млекопитающих СССР М. 1944.
Богачев Б. П. иДукельская Н. М., Мышевидные грызуны южной
части Северного края, Защита растений, сб. II. 1936.
Боголюбов О. М., К вопросу о плодовитости и способности к вскарм-
ливанию детенышей у самок крыс при некоторых видах гиповитаминоза, «Арх.
биюл. наук», т. XXXI. вып. 5. 1931.
Бондарь Е., Грызуны населенных пунктов Туркменистана, Изв. Турк-
мен. филиала АН СССР, № 1 1946.
Борю С. И. и Фердинанд Я. М. Крысы как носители возбудителей
желудочно-кишечных заболеваний человека. Тр. Куйбышевск. военно-мед. ака-
демии Красной Армии, т IV. 1940.
Браунер А. А.. Млекопитающие Новороссии н Крыма. Крысы, водящие-
ся в г. Одессе, «Записки Новорос, об-ва естествомспыт.». т. XXX. 1906.
В а н д о в а С.. Кокцидии серых крыс (Rattus norvegicus Berk.) из Баку и
окрестных поселений, Изв. АН АзербаЙдж. ССР, № 9, 1949.
В а ш к о в В. И., Избирательное токсическое действие препарата «крысид-
ЦНИДИ» (альфа^нафталтномочевииа) на некоторых животных, Тр. Центр, н.-и.
дезинф. ин-та. вып. 2, 1946.
В а ш к о в В. И., Устойчивость сеоых крыс к крысиду при повторном его
применении, Тр. Центр, н.-и. дезинф. ин-та. вып. 4, 1948.
Вашков В. И., Руководство по дезинфекции, дезинсекции и дератизации,
М- 1952
Вашков В. И., Поликарпов В. Н. и Пасешник А. А.. Токсич
worn фгораиетата бария и натрия для серых крыс, белых мышей и кроликов,
Тр Центр, н.-и. дезинф. ин-та, вып. 5, 1949.
Вашков В. И. и Полежаев В. Гм Возможность применение в дера-
пгзационной практике заделочяых материалов, обработанных ядовитыми пре-
паратами, Тр. Центр, н. и. дезинф. ин-та, вып. 6, I960.
В е рш а г и н Н. К.. О происхождении крыс рода Rattus в Закавказье,
* Природа» И, 1949.
Виноградов Б.. С.. Грызуны. Л.. 1933.
Виноградов Б. С. и Аргиропуло А. И., Определитель грызунов.
Изд. АН СССР, Л., 1936.
В и н о г п а д о в Б. С. иАргиропулоА. И.. Определитель грызунов
(Фауна СССР. Млекопитающие.). Изд. АН СССР. М.-Л.. 1941.
R и» огр ап о в Б. С. » Громов И. М., Грызуны фауны СССР. Изд. АН
СССР, М -Л., 1952.
Виноградов Б. С. и Оболенский С. И., Вредные и полезные в
сельском хозяйстве млекопитающие. М.-Л.. 1932.
Воронин В. В Кпысоистребление; организация и результаты, Сб. <Чума
в Одессе в 1910 г.», СПБ, 1912.
Гамалея Н. Ф. Чумз и крысы. Одесса, 1902.
Гвелесианп И.. Крысы как резервуар Balantidium coli «Мед. парази-
юл. и паразит, заболев», № 6. 1949.
Гептя.еп В. Г.. М о о о з о в а-Т у р о в а Л. Г. и Цалкнн В. И., Вред-
ные и полезные звери районов полезащитных насаждений. М, 1950.
Г е р ш к о в и я Н., Матипалы по фауне блох серой крысы Хабаровского
края. Сообщение I, Изв. Иркутского тос. н.-и. противочумного ин-та Сибири и
Дальнего Востока, т. VII. 1949.
Гиренко Л. Л., К вопросу о распространении черной крысы в УССР,
«Научи зап. Киевск. гос. ун-та», т. IX. вып. VI. 1950.
Голещянин А., Опыт сплошной дератизации «Врач. дело». № И. 1950.
Грязнов Н, О восприимчивости диких крыс (пасюков) к вирусу гриппа,
«Дохл. АН СССР», т. IX, № 3. 1948.
Губарев Л. Д.. Распространение серой крысы (Rattus norvegicus) в
восточных районах Ростовской области, Тр. Ростовского-на-Дону гос. н.-и. про-
тивочумного ин-та. т. II. 1941.
Дементьев Д. П.» Расселение крыс по железным дорогам. «Природа»,
4. 1950.
Демидова А. А. и Конькова К. В., К вопросу об избирательном от-
ношении домовых грызунов к различным видам кормов, Изв. Ипкутского гос.
н.-и. противочумного ин-та Сибири и Дальнего Востока, т. ХШ 1954.
Д и н н и к Ю. и Каменев В., Паразитические черви кпыс и мышей
г. Краснодара, Тр. Краснодарск. гос. пед. ин-та, т. VI, вып. 1, 1937.
Диомидова Н., Закономерности сперматогенеза крыс вне организма,
«Докл. АН СССР», т. XXVIII. № 6, 1940.
Долманова А.. К вопросу об особенностях бактериологической диагно-
стики псевдотуберкулеза у серых крыс, Тр. Ростовского-на-Дону гос. н.ни. про-
тивочумного ин-та, т. II. 1941.
Доппельмаир Г. Г., Заметки о диагностических признаках Mus rattus
и Mus decumanus Pali.. Изв. Лесного ин-та т. XI. 1914.
Дукельская Н. М., Крысы и мыши я борьба с ними. «Фельдш. и
акуш.», № 11, 1946.
Дукельская И. М., Распространение и биология серой крысы. Тр.
Центр, н.-и. дезинф. ин-та. вып. 3. 1947.
» Дукельская Н. М., Подбор пищевых продуктов для изготовления сгг-
равлеиых приманок для серых крыс, Тр. Центр, н.-и. дезинф. ин-та. вып. 4,
1948.
Дукельская Н. М.. Эффективность применения в дератизация жидких
приманок, Тр. Центр, н.-и. дезинф. ин-та, вып. 4. 1948.
Дукельская Н. М.» Применение в дератизации метода опыливания ки-
шечными ядами, Тр. Центр, н.-и. дезинф. ин-та, вып. 5, 1949.
Житков Б. М.» Замечание о крысах н некоторых условиях их исследо-
вания, «Зоол. жури.», т. XXIII, вып. 2—3. 1944.
Жуховицкий И., Случай нифекцио1Н1юй болезни от укуса крыс (бо-
лезнь сод оку), «Врач. дело». № 10, 1940.
54
Зверев М. Д.. К изучению серых крыс в Сибири и методика борьбы с
ян мн» Изв. Сибирск. краевой станции защиты растений, № 4—7, 1930.
Зверев М. Д., Распространение серых крыс в Сибири, Изв. Гос. н.»ж
противочумного ин-та Сибири и Дальнего Востока, т. II, 1935.
Зверев М. Д., Распространение серых крыс в Сибиои. Изв. Гос. н.-и.
противочумного ин-та Сибири и Дальнего Востока, т. II, 1935.
Зейтленок Н. А. и Колосов Н. И., Наблюдения над инвазией чело»
века крысиным цепнем, «Мед. паразитол, и паразит, болезни*, т. V, вып. 5. 1936,
Зотова А. А. О задачах борьбы с грызунами в животноводческих хозяй-
ствах, «Ветеринарная дезинфекция», 1954.
Иванов А. В. и Джараева Е. К.. О результатах дератизации веще-
ствами, безвредными для человека., «Гигиена и санитария». № 9—10, 1937.
К а л а б у х о в Н. Щ Применение фосфористого цинка (ZnaPj) в борьбе
с грызунами, «Фармакол. и токсикол.», вып. 2.. 1946.
К а Л а б у х о в Н. И., Новейшие достижения в области дератизации,
«Военно-мед. жури.», № 12. 1946.
Калита См Блохи серых крыс (Rattus norvegicus Berk.) Прикаспийской
полосы Северного Ирана, «Зоол. журн.», т. XXVII, вып. 6, 1948.
Кащенко Н. Ф., Крысы и заместители их в Западной Сибири и Турке-
стане, Ежегодник Зоол. музея Акад, наук СССР, т. XVII, 1912.
Клиентова А., Крысы как возможный резервуар вируса сыпного тифа,
«Арх. биол. наук», т. XXXV, серия Б, вып. 2, 1934.
Клодницкий Н. Н. и Кац Л. Я., Роль крыс в эпидемиологии кишеч-
ных инфекций, «Жури, микроб.» эпидем. и иммунол.», т. XVII. вып. 4. 1936.
Ковалевский К. Л.. Лабораторные крысы и мыши, М., 1948.
Ковалевский К. Л., Содержание мелких лабораторных животных в
вивариях, М.. 1949.
Ковалевский К- Л.» Лабораторное животноводство. М., 1951.
* КондрашкинГ. А., О серых крысах (Rattus norvegicus Berk.) дельты
Волги, «Бюлл. Моск, общ-ва испыт. прир.», отд. биол., т. IV, вып. I, 1949.
Кузнецов Б. А., Млекопитающие степной полосы Южного Урала,
сБюлл. Моск, общ-ва испыт. прир.», 1928.
Кузнецов Б. А., Черная крыса в Мало-Ярославском уезде Калужской
губернии, Тр. по лесному опытному делу, Центр, лесная опытная станция,
вып. VII, 1930.
Кузякин А. П., Проблемы городской и транспортной дератизации, Тр.
научи, конференции к 25-летию института «Микроб», Саратов, 1944.
Кузякин А. П., К методике учета и вылова серых крыс в городских
объектах, Сб. «Грызуны и борьба с ними», вып. 3, Саратов, 1950.
Кузякин А. П., История расселения, современное распространение и ме-
ста обитания пасюка в СССР, Фауна и экология грызунов, вып. 4, 1950.
Кузякин А. П., О размножении пасюков в городах, «Бюлл. Моск,
общ-ва испыт. прир.», отд. биол., т. VII, вып. 3, 1952.
Липатов Л., Об экспериментальном бруцеллезе у крыс, «Арх. биол.
наук», т. VI, вып. 1 (10), 1939.
Литвинов Н. И. и Карнаухова Н. Г., О видовом составе крыс,
населяющих морские суда порта Владивосток, Изв. Иркутского Гос. н.-и. про-
(ивочумного ин-та Сибири и Дальнего востока, т. XIII, 1954.
Львов Г. И., Колхидский пасюк* Уч. зап. Горьковск. гос. ун-та» вып. 14,
1949.
Майзелис М. Г. и Руттенберг С. О., Суточный ритм активности
н температуры* тела у крыс, «Опыт изучения периодических изменений фено-
логических функций в организме», М.. 1949.
Малевич И. И., Резистентность мышей и крыс к повторному заражению
личинками трихинеллы, «Бюлл. эксп. биол. и мед.», № 3, 1941.
Михайлов А. И., Опыты по подбору наиболее эффективных приманок для
борьбы с домовыми грызунами, Изв. Иркутского Гос. н.-и. противочумного
ин-та Сибири и Дальнего востока, т. XIII» 1954.
Ч •• p I '• । I11* i ь J я II I , Ц i i .1.1 . a 3. К. и М»н?сяи A. А., Гры-
ц>|14 1ч|н.дн 1гЧм’чн, Tp Зоол. им та Акад, наук Груз. ССР. т. VII, 1947.
II» д II Н.» Избирательное отношение некоторых мышевидных гры-
ънов ь различным кормам. Изв. Иркутского Гос. н.-я. Противочумного ин-та
Сибири и Дальнего -востока, т. XIII. 1954.
Никитин В. П., О распростарнени» чеоной крысы на Дальнем Востоке.
Вопросы географии Дальнего Востока. № 1. 1949.
Никитин В. П., К биологии домовой крысы. «Природа*. 3, 1950.
Никитин В. П. Борьба с домовыми грызунами с помощью отравленной
клеевой массы. Сб. «Грызуны и борьба с ними*, вып. VI. Саратов. 1955.
Никитин П. И.. Борьба с грызунами на железнодорожном транспорте,
М., 1939. | ' 1 ,П,Я
Николаев И., Лептоспироз среди крыс крупного города, «Журн. микроб.,
эпидем. и иммунол.», № 12, 1945.
Огнев С. И., и Строганов С. У.. Новый подвид черной крысы
R rattus ruthenus subsp. nov. «Сб. н.-и. ин-та зоол. Москов. гос. ун-та». № 3,
1936
Огнев С. И. (О g n е f f S. I.), A synopsis of Russian Rats, «Journal of
Mammology», 8, 1927.
Огнев С. И., Новые данные о крысах (рода Rattus} Дальнего Востоку
«Бюлл. Моск, общ-ва испит. пряр.», отд. биол., вып. 2, 1950.
Окуневский Я. Л., Практическое руководство по дезинфекции, т. IV
Дезинфекция, дератизация. М.—Л.. 1936.
Организационно-методические материалы по дезинфекции, дезинсекции и ле-
ратяэации. М.. 1950.
Панкратов А., О бруцеллезе у крыс. «Арх. биол. наук», т. Ш. вып 1
1939.
П а с е ш н и к А. А., К вопросу о размножаемости крыс в Москве, Тр
Моск, дез ив ф. ин-та вып. I, 1936.
П а ее ш ник А. А.. Применение препарата «коысид-ЦНИДИ» для отрав*
ления серых крыс, Тр. Центр, н.-и. дезинф. ин-та, вып. 2. 1946.
Пикуль Н., Материалы к изучению рол» кэыс в распространении бешен-
ства, «Журя, микроб., эпмдем. и иммунол.». т. ХХП вып. 1, 1939.
Плятер-Плохоцкий К., Вредные грызуны южных районов ДВК в-
1935 году. «Вести. Дальневосточного филиала Акад, наук», 1936.
г Подхватильнев П. Ф. и Девятова А. П.. Новая крысоловка-
давилка, «Грызуны и борьба с ними», вып. IV, 1955.
Полежаев В. Г., Организация дератизационных мероприятий органами
здравоохранения.. Тр. центр, н.-и. дезинф. ин-та, вып. 4. 1948.
Полежаев В. Г., Способность кпыс перегрызать заделочные материалы,
Тр. Центр, н.-и. дезинф. ин-та, вып. 5. 1949.
Полежаев В. Г., Изучение поведения серой крысы в зависимости» от
факторов внешней среды, Тр, Центр, н.-и. дезинф. ин-та, вып. 6, 1950.
Полежаев В. Г. и Греч и щев П. G, Практическое значение примене-
ния комбинированного метода (яда совместно с культуоой Данича) в борьбе
с грызунами. Тр. Центо. н.-и. дезинф. ин-та. вып. 6. 1950.
Прохоров М. И.. Использование бактериальных культур в борьбе с
мышевидными грызунами в животноводческих хозяйствах, «Ветеринарная дезин-
фекция», 1954.
Прутенский Д.» Туркестанская крыса (Rattus turkestanicus Sat), ее
образ жизни и экономическое значение в Аослаибобском лесном массиве,
Тр. Ьиол. ин-та Киргизского филиала АН СССР, вып. 2. 1948.
Решетник Е.. О возможности применения препарата ДДТ в борьбе с
серой крысой (Rattus norvegicus Berk.), «Врач, дело», № И—12, 1946.
Р у ц е в и ч А., Крысиный клещ Liponyssus — переносчик эпидемического
сыпного тифа. «Природа», 12. 1947.
Сахаров П. П., Важнейшие заболевания лабораторных животных,
/кроликов, морских свинок, мышей и крыс) и изучение мер борьбы с ними*
«Журн. микроб., эпидем. и иммунол.». № 6. 1939.
56
. \ а г о в П Пм Метелкин А. И., I у д к о в а I И., Лабораторные
• in иные. М.. 1952.
Серебренников М. К.. Материалы по . истематике и экологии гры-
нов Южного Зауралья, Ежегодник Зоол. музея Акад, наук СССР, 1929
Сеславина П. М. иАлексеев П. П., Грецкий орех в Средней Азии,
I »
Сильвере И. Л., Некоторые биологические особенности черных крыс
N rattus L. по материалам Тульской экспедиции с 25/II по 1/V 1935. <Вестн.
•|икроб., эпидем. и паразитол.», т. ,Х1Х» вып. I. 1940.
Скалой В. Н., Северная граница домовой крысы в Сибири, «Природа»,
6 1939.
Солитерман П.. Заражение лабораторных грызунов риккетсиями кры-
* иного тифа через слизистые и кожные покровы. «Журн. микроб., эпидем. и
и'мунол.», № 8. 1950.
Степанов И. В., Крысы и мыши Батуми, «Вести, микроб и эпидем.»,
X, вып. I. 1931.
Степанов И. В.. Материалы по биологии размножения крыс и мышей
•• Батуми. «Бюлл. Моск, общ-ва испыт. пртф.», отд. биол., вып. 6, 1946.
Степанов И. В.. Блохи на крысах г. Батуми. «Бюлл. Моск, общ-ва
пспыт. прир.», отд. биол., вып. 4. 1939.
Степин Е. К., О крысах порта Николаевск-на-Амуре, Изв. Иркутского
юс. н.-и. противочумного ин-та Сибири и Дальнего Востока, т. VI, 1946.
Строганов С. У., О таксономическом положении Rattus rattus и о рас-
пространении ее в СССР. «Зоол. журн.», т. ХШ, № 4. 1935.
Траут И. И.. Численность и размещение мышевидных грызунов в насе-
ленных пунктах городского типа, Сб. «Грызуны и борьба с ними», вып. 3,
Саратов, 1950.
Турин М. Л., Рациональные методы дератизации. «Фельдш. и акуш.»,
№ 5. 1949.
Турин М. Л. и Соловьев В. В.. Дезинфекция, дезинсекция и дерати-
зация. М., 1951.
Фенюк Б. В., Массовое размножение мышевидных грызунов на Юго-
Востоке РСФСР в 193? г.. Сб. «Грызуны и борьба с ними». № 1, Алма-Ата,
1941.
Фетисов А. С„ Видовой состав грызунов населенных пунктов Западного
Забайкалья, «Вести, микроб., эпидем. и паразитол.», т. XIX, вып. 3—4, 1940.
Фетисов А. С., Крыса-карако в Восточной Сибири. Иркутск, 1945.
Формозов А. Н., Несколько дополнений к статье Б. М. Житкова «За-
мечания о крысах и некоторых условиях их исследования», «Зоол. жуон»
т. XXIV, вып. Z 1945. F ’
Хлебников Б. А., Позвоночные враги промысловых птиц и зверей Астра-
ханского края. 1924.
Цибулевская Ф. С., К вопросу о кровепаразитах серой крысы, Изв.
Иркутского гос. н.-и. противочумного ин-та Сибири и Дальнего Востока, т. IX,
1951.
Шепелева В. К, Некоторые особенности питания пасюка (Л. norve-
fficus Berk.) и черной крысы (R. rattus L.), Тезисы докл. второй эколог, кон-
ференции, ч. 2. Киев. 1950.
Шорохов П. И. и Шорохов С. И., Вредители запасов зерна и зер-
иопродуктов. М.. 1938.
Щастный С. М., Отчет по исследованию крыс на Одесской бактериоло-
гической станции с сентября 1910 г. по декабрь 1911 г., Сб. «Чума в Одессе
в 1910 г.». СПБ. 1912.
Яковлев М., Некоторые экологические особенности серой крысы в ни-
зовьях р. Дона, Рефераты иаучно-исслед. работ Ростовского-на-Дону гос. н.-и
противочумного ин-та. т. XIII, 1949.
Янсон С. Л.. Заметки о биологии серой крысы (R. norvegicus Erxi),
Ьюлл. нвучно-эксперим. лаб. Осоавиахима, № 1, 1934.
Янсон С. Л., Потребность серой крысы в воде и рентабельность приме
57
нения для целей дератизации жидких отравленных приманок, Тр. научно-экспе-
рим. лаб. Осоавиахима, т. И, 1937.
Boelter W. R., The Rat problem, London, 1909.
Chitty D.t A relative census method for brown rats (/?. norvegicus).
Nature, vol. 150. 1942.
Chitty D. and Shorten M., Techniques for the study of the norway
rat (R. norvegicus), Journ. of Mammol., vol. 27, № 1, 1946.
Control of rats and mice. Vol. I, II. Rats, edited by D. Chitty, Oxford. 1954.
Dawson A., Rat destruction.. Agricult., 51, 5, 1944.
Donaldson H., The Rat. 2. ed., Philadelphia, 1924.
Ell erm an J. R., The families and genera of the living rodents, vol. 1—2,
Brit. Mus. Nat. Hist. London, 1940—1941.
Ell erm an J. R. and T. G. S. M о r r i s о n-S co tt. Checklist of palaear-
ctic and Indian mammals. Brit. Mus. Nat. Hist. London. 1951.
Kemper H.. Die Haus- und Gesundhelfsschadlinge und ihre Bekampfung,
Berlin, 1943.
К i n g H. D., The growth arid variability in the bodi weigth of the Norway
Rat (Mus norwegicus), Anat., Record, v. 25, H. 79, 1923.
King H. D., Life processes in gray Norway rats during fourteen Jears in*
captivity, Amer. Anat. Memoirs, 17, Philadelfia, 1939.
Schaub S.. TertiSre und quartfire Murine, Abk. Schweiz. palSont. Ges.,
1928. «I
Silver J., The house rat. 1941.
Simpson G., The principles of classification and a classification of Mam-
mals, Bull. Amer. Museum Nat Hist., vol. 85. 1945.
Steiniger F., Rattenbiologie und Rattenbekfimpfung einschliesslich der
Toxikologie gebrauchlicher Rattengifte, Stuttgart, 1952.
Ulmer F., Unusual behavior of the Norway rat, Journ. of Mammal., 32(1),
1951.
Venables L. S. and Leslie P. H., The rat and mouse populations of
corn ricks, Journ anim. ecol, VI. 11, № 1, 1942.
Weber M.. Die Saugetiere, Bd. II, 2. Aufl., Jena, 1928.
ЧАСТЬ ВТОРАЯ
АНАТОМИЯ
ГЛАВА III
ВНЕШНЯЯ МОРФОЛОГИЯ И ТЕРМИНОЛОГИЯ,
УПОТРЕБЛЯЕМАЯ ПРИ ОПИСАНИИ
Как и у всякого другого млекопитающего, тело (corpus) кры-
ы принято подразделять на осевую часть и конечности. Осевую
часть в свою очередь делят на голову (caput), шею (cervix, s. col-
lum), туловище (truncus) и хвост (cauda) (рис. 17 А).
Голову подразделяют на мозговую (pars cranialis) и лицевую
•| iCth (pars facialis). Мозговую часть подразделяют на затылочную
(regio occipitalis), теменную (г. parietalis), лобную (г. frontalis) и
височную (г. temporalis) области. Затылочная область расположена
на границе с шеей, теменная и лобная — по верхней поверхности
। лловы, а височная занимает боковую сторону головы. Лицевую
•асть головы подразделяют на носовую (г. nasalis), ротовую, или
«убную (г. oralis, s. labialis) и щечную (г. buccalis) области.
Шею обычно делят на верхнюю — выйную область (г. nuchalis)
и нижнюю — собственно шею (г. colli ventralis).
Туловище подразделяют на спинно-грудной, пояснично-брюшной
и крестцово-ягодичный отделы. В верхней части спинно-грудного от-
.«ла различают переднюю— межлопаточную (г. interscapularis) и
। |днюю — спинную (г. dorsalis) области. Боковые части этого от-
1ела называют реберными областями (г.г. costales), а нижнюю
часть — грудной областью (г. stemalis). Пояснично-брюшной от-
г*л подразделяют на поясницу (f. lumbalis) и на брюхо (abdo-
men), которое в свою очередь подразделяют на области мечевид-
юго хряща (г. xiphoidea), пупочную (г. umbilicalis), паховую
(г inguinalis) и подвздошную (г. iliaca). Крестцово-ягодичный от-
ц'л состоит из крестцовой области (г. sacral is) и ягодичной, или
ивовой, области (г. glutaea).
Передняя конечность состоит из лопаточной области (г. scapula-
Г1.н), плеча (brachium), предплечья (antebrachium) и лапы, или ки-
। in (manus). Задняя конечность ягодичной областью граничит с
шальной (г. analis). Последняя впереди гранична с промежно-
• п»ю (perineum). Сама конечность делится на бедро (femur), го-
лень (cru>) и стопу (planta). Положение всех этих отделов можно
видеть на схеме, приведенной на рис. 17 А.
Для облегчения описания формы органов в анатомии принято
давать определенные наименования различным плоскостям, поверх-
Рис. 17. Терминология, употребляемая при описании стро-
ения крысы. А — схема деления тела крысы на области,
Б — направления в теле крысы:
1 — ротовая, 2 — носовая, 3 — щечная, 4 — лобная, б — глаз. 6 —-
теменная, 7 — ухо, 8 — затылочная, 9 — височная. 10 — жевательной
мышцы, 11 — собственно шеи. 12 — выйная, 18 — межлопаточная,
14 — спинная, 15 — грудная, 18 — реберная, 17 — мечевидного
хряща, 18 — пупочная. 19 — подвздошная, 20 — поясничная. 21 —
крестцовая, 22 — хвост, 23 — тазовая. 24 — анальная. 25 —лопаточ-
ная. 26 — плечевая. 27 — предплечье, 28 — кисть. 29 — бедро, 30 —
голень, 31 — стопа; а — краниальное, б — каудальное, в — дорзаль-
ное, г — вентральное, Д — оральное, е —* аборальное, ж - прокси-
мальное, з - - дистальное, и — вол яркое, к — плантарное
ноет ям и направлениям тела животного (рис. 17 Б). Продольные
плоскости, разделяющие тело на правые и левые части, называют
сагиттальными. Плоскости, рассекающие тело на верхние и нижние
части, называют фронтальными, а поперечные плоскости называют
сегментальными, или трансверзальными. Всякий орган имеет ось,
направленную к срединной сагиттальной плоскости. Это направле-
ние называю) метни.иным, а противоположное направление — ли-
бо
тральным. Всякое направление вверх называют дорзальным, или
пикным, а всякое направление вниз — вентральным, или брюш-
ным. Всякое направление вперед, в сторону головы, называют кра-
ниальным, или передним, а назад, в сторону хвоста — каудальным,
или задним. Кроме того, для головы и конечностей приняты спе-
циальные термины. Для головы направление ко рту называют
оральным, а противоположное — аборальным. В конечностях же
направление в сторону пальцев называют дистальным, а противо-
положное — проксимальным. Ладонную поверхность передней ко-
нечности называют также волярной, а подошвенную поверхность
> лдней конечности—плантарной. Противоположные поверхности
конечностей называют дорзальными, спилковыми или тыльными.
ГЛАВА IV
КОЖНЫЕ ПОКРОВЫ И ИХ ПРОИЗВОДНЫЕ
КОЖА
Кожа, (derma), покрывающая все тело крысы, не только защи-
щает организм от внешних воздействий, но и выполняет много дру-
гих функций. Она служит выделительным органом, воспринимает
различные внешние стимулы и т. д.
Кожа состоит из трех слоев (рис. 18 Л): наружного, или
эпидермиса (epidermis) ; собственно кожи (cutis, s. derma prop-
rium) и подкожной жировой клетчатки (hypoderma, s. tela subcu-
tanea).
Эпидермис тонок, прозрачен; на местах постоянного механиче-
ского раздражения (подошва, ладонь) он сильно утолщается. Эпи-
дермис состоит из нескольких слоев. Самый поверхностный роговой
слой представляет из себя ороговевшие клетки, постепенно слущи-
вающиеся под влиянием механических воздействий. Сброшенные
клетки заменяются новыми, возникающими путем деления в глубо-
ком мальпигиевом слое. В мальпигиевом слое клетки имеют цилин-
дрическую форму, но по мере перехода в поверхностный слой они
все более и более уплощаются, превращаясь в тонкие роговые
пластинки.
Собственно кожа состоит из плотной опорной соединительной
ткани. Подразделяется она на богатый нервами и сосудами сосоч-
ковый слой, лежащий непосредственно под эпидермисом и питаю-
щий его, и на более глубокий сетчатый слой, состоящий из сложно
переплетенной сети коллагеновых и эластических волокон. Эласти-
ческие волокна развиты и в сосочковом слое. Они и придают эла-
стичность всей коже. Толщина собственно кожи сильно варьирует и
наиболее велика на подошвах.
Подкожная клетчатка построена из рыхлой соединительной тка-
ни. В неподвижных участках кожи подкожная клетчатка более
плотна, в подвижных же она очень рыхла и слабо тормозит сме-
щение участков кожи. Этим в известной мере предупреждаются ме-
ханические повреждения кожи.
62
ПРОИЗВОДНЫЕ КОЖИ
К производным кожи относятся волосы, кожные железы (пото-
вые и сальные), когти, мозоли и некоторые другие образования.
Волос (pilus) представляет собой тонкую, эластичную роговую
Рис. 18. Кожа и производные кожных покровов (схематизи-
ровано). А — вертикальный срез кожи, Б — вертикальный
срез волосяной сумки, В — потовая железа, Г — коготь, Д—
млечная железа:
1 — эпидермис. 2 — дерма. 3 — жировая клетчатка. 4 — сосочки дер-
мы. б — жировые клетки. 6 — стержень волоса, 7 •— луковица волоса,
в — волосяной сосочек. 9 — волосяная сумка, 10 — сальная железа.
И — потовая железа. 12 — проток потовой железы. 13 — мальпи-
гиевый слой эпидермиса, 14 — секреторные клетки. 15 — гладкие мы-
шечные клетки, 10 — когтевая фаланга, 17 — когтевая пластинка,
13 — подошвенная пластинка, 19 — ногтевой валик. 20 — ногтевой
желобок. .21 — пальцевая подушечка. 22 — выводной проток млечной
железы
пить (рис. 18 Б). Свободный, выступающий над поверхностью ко-
жи участок волоса называется стержнем волоса (scapus pili), часть,
погруженная в кожу — корнем волоса (radix pili), а ее утолщен-
ное основание — луковицей волоса (bulbus pili). В луковицу про-
никает сосочек собственно кожи. У крысы все тело, кроме кистей и
/тупней, густо покрыто волосами. Мало'волос на хвосте, где они
имеют вид щетинок, расположенных между чешуйками, и на ушах.
63
Волосы iy крыс подразделяются на грубые и длинные, называе-
мые ocibKi. короткие нежные и тонкие, называемые подшерстком, и
расположенные между ними промежуточные по строению волосы
шерсти. Кроме того, различают так называемые осязательные ще-
тинки, или вибриссы — длинные грубые волосы, к корням которых
подходят многочисленные нервные окончания.
У крыс периодически происходит линька волос — выпадение
старых и замена их новыми. В течение года у крыс бывает две
линьки — весенняя и осенняя. Во время первой выпадает значи-
тельная часть подшерстка, а во время второй происходит сильное
отрастание последнего. При осенней линьке кожа сильно набухает
за счет прилива крови в сосуды собственно кожи. У крыс в это вре-
мя кожа бывает более теплая, чем обычно, а при снятии шкурки
линяющие места выступают в виде темносиних пятен, которые на
ощупь грубее участков, еще не начавших линять. По темным пятнам
на мездре судят о ходе линьки у млекопитающих. Обычно линька
начинается с головы и спины, позднее вылинивают брюхо и ко-
нечности. Линька эпидермиса у млекопитающих происходит непре-
рывно — в виде шелушения его поверхностных ороговевших слоев.
На кончиках пальцев у крыс формируются когти (unguicola),
тоже являющиеся производными кожи. Когти представляют собой
твердые крючкообразные роговые образования, сидящие на тыль-
ной поверхности концевых фаланг пальцев. В когте (рис. 18 Г)
различают: ногтевую пластинку (выпуклую верхнюю стенку когтя),
образованную твердым ротовым веществом и являющуюся наиболее
прочной рабочей частью когтя; подошвенную пластинку, занимаю-
щую вогнутую подошвенную часть когтя, представленную у крысы
слабо развитой полоской плотной кожи, и корень когтя, сидящий в
глубокой складке кожи — когтевом желобке. Благодаря глубине
желобка, коготь прочно связан с фалангой, что очень важно при
рытье, удержании добычи и т. д. Основание когтя охвачено полу-
круглой складкой кожи когтевым валиком. С подошвенной сто-
роны под когтем развита так называемая подошвенная подушечка.
К производным кожи относят также кожные железы. Для млеко-
питающих характерно наличие трех основных типов кожных желез:
потовых, сальных и млечных. Потовые железы (рис. 18 В) — труб-
чатого типа; в толще кожи они обычно закручены клубком. Пото-
вая железа образована кубовидным железистым эпителием, выст-
ланным снаружи тонким слоем гладких мышечных волокон.
Сальные железы (рис. 18 Б) — альвеолярного типа. Они обра-
юваны многослойным железистым эпителием и не выстланы снару-
жи слоем мускульных волокон. Сальные железы обычно открыва-
ются в волосяные сумки и смачивают своим секретом стержень во-
лоса, который, благодаря этому, становится более эластичным и
менее гигроскопичным.
Млечные железы крыс (рис. 18 Д) имеют сложное альвеоляр-
ное строение. lit г< вые ж( лезы являются трубчатыми ж-лезами, во
(Ч
тем не менее гомологию млечных и потовых желез можно считать
твердо установленной. У однопроходных (утконос и ехидна) млеч-
ные железы еще сохраняют трубчатое строение.
У крысы выводные протоки млечной железы открываются непо-
средственно на поверхности соска и общего выводного протока не
образуется. Таким образом, у крысы, как и у человека, сосок истин-
ный. Число сосков у крыс варьирует. Обычно их бывает от 8 до 12,
реже — больше. Среди них обычно насчитывается четыре грудных
(иногда их два), четыре брюшных и два паховых. Считается уста-
новленным, что прямой связи между числом сосков и числом дете-
нышей в помете нет.
ГЛАВА V
СКЕЛЕТ
ОБЩИЕ ЗАМЕЧАНИЯ
У крысы скелет состоит из 265—285 костей. Число их варьирует
за счет изменчивости в числе позвонков (в основном хвостовых),
ребер и сесамовидных костей. Кости объединяются мышцами и
связками в единую систему, выполняющую опорную, защитную и
локомоторную функции. Вес свежего скелета составляет 10—12%
общего веса туловища взрослой крысы. Свежие кости содержат
50% воды, 28—30% органических веществ и 20—22% неорганиче-
ских веществ. У молодых животных неорганических веществ в ко-
стях содержится значительно меньше, чем у старых.
Кость (рис. 19) богато снабжена сосудами и нервами. Снаружи
она покрыта плотной соединительной тканью — надкостницей.
Сама кость состоит из компактного (substantia compacta) и губча-
того (substantia spongiosa) вещества. Рост костного вещества идет
под влиянием механических сил, действующих на кость. Пластинки
губчатого вещества всегда ориентированы по линиям сжатия и рас-
тяжения (рис. 19 Л). При изменении направления этих сил костное
вещество перестраивается соответственно новым механическим ус-
ловиям. Внутри кости находится костный мозг, являющийся крове-
творным органом. К старости в большинстве случаев происходит
его жировое перерождение.
По своей форме кости делятся на 4 вида: 1) длинные кости
(ossa longa). Длина этих костей значительно превышает как ши-
рину, так и толщину, обычно они цилиндрические. В длинных ко-
стях различают сидящие по концам головки — эпифизы (epiphysis)
и тело — диафиз (diaphysis). Каждая из этих частей окостеневает
самостоятельно; 2) короткие кости (ossa brevia). Эти кости имеют
приблизительно одинаковые размеры во всех направлениях. Корот-
кие кости выполняют обычно опорную функцию. Чаще всего они
имеют многогранную форму; 3) плоские кости (ossa plana) имеют
вид тонких широких пластинок. Основная их функция защитная;
4) смешанные кости (ossa mixta). Эти кости имеют признаки не-
скольких типов костей, но не могут быть отнесены ни к одному иг
66
пих. К смешанным костям относят позвонки и некоторые другие
кости.
В онтогенезе животного кости возникают по-разному. Одни из
них проходят три стадии развития — соединительнотканную, хря-
щевую и костную, другие, так называемые
тельнотканной стадии переходят прямо к
костной.
Вес костей скелета различных отделов
туловища позволяет до некоторой степени
судить о значении каждого из них. Так,
например, у крысы скелет задних конеч-
ностей тяжелее скелета передних; это ста-
новится понятным, если учесть способ-
ность крысы передвигаться прыжками.
Данные о весе костей различных отделов
тела крысы приведены в табл. 10 (кости
взвешивались в свежем состоянии).
ЧЕРЕП
При взгляде на череп крысы заметно,
что он сильно вытянут, скуловые дуги
расставлены не широко. Более широкая
задняя часть черепа называется мозговой.
Она сужена в области глазниц и вновь
расширяется впереди последних. Впереди
предглазничного расширения начинается
лицевая часть черепа, к которой отно-
сится также нижняя челюсть.
На верхней стороне черепа (рис. 20)
спереди лежат парные межчелюстная и
носовая кости, окружающие наружное
носовое отверстие. Сзади "носовые кости
граничат с лобными, а межчелюстные —
«• верхнечелюстными костями. У перед-
него края верхнечелюстных костей обра-
>уется большое подглазничное отверстие
(foramen infraorbitale), через которое
покровные, из соедини-
Рис. 19. Строение ко-
сти (схематизировано).
А — продольный разрез
бедренной кости, Б —
кусок трубчатой кости со
вскрытой надкостницей:
1 — губчатое вещество эпи-
физов бедренной костя. 2 —
губчатое вещество дисталь-
ной части тела бедренной к<
сти, 3 — надкостница. 4 —
компактное вещество кости
проходят нервы, сосуды и часть наруж-
н it жевательной мышцы. В углу между
• looHofl и верхнечелюстной костями имеется маленькая слез-
ная кость, пронизанная открывающимся в носовую полость
слгшым каналом (canalis lacrimalis). Сбоку верхнечелюстная
кп-п, почт сильно (развитый скуловой отросток, образующий
переднюю часть скуловой дуги и граничащий со скуловой
|«ч Т1,к>. Последняя составляет среднюю часть скуловой д)Тн,
I 1И1НЯЯ сс часть образована скуловым отростком височной к"сщ.
1 нпи лобные кости граничат с теменными, а последние — с мс
м>*иной костью. Межтеменная кость граничит позади с kw i< ы
тылочной кости. Сбоку (рис. 21) позади межчелюстной и челюстной
костей заметна передняя клш
челюстной располагается реш
1е), окаймляемое сверху лобн
передней клиновидной кости,
Рис. 20. Череп пасюка (вид
сверху).
А — носовая кость, Б — межче-
люстная кость, В — верхнечелю-
стная кость, Г — слезная кость,
Д — лобная кость. Е — скуло-
вая кость, Ж — височная кость,
3 — теменная кость. И — межте-
менная кость, К. — затылочная
ювидная кость. Между нею и верхне-
етчатое отверстие (foramen ethmoida-
эй костью. На орбитальном крыле
граничащем с пластинкой лобной
кости, располагается большое зри-
тельное отверстие (foramen opticum)
для зрительного нерва. Орбитальные
крылья передней клиновидной кости
граничат позади с одноименными
крыльями основной клиновидной
кости; на границе между ними и
верхнечелюстной костью располага-
ется круглое отверстие (foramen го-
tundum), через которое в глазницу
проходят III, IV и VI черепномозго-
вые нервы, иннервирующие мышцы
глаза, и ветвь тройничного нерва
(V). Основная клиновиная кость
граничит сзади с височной костью,
которая вместе с клиновидными,
лобной и задней частью верхнечелю-
стной кости составляет боковую
стенку черепа. На границе между
чешуей височной кости и ее каменис-
той частью располагается видимая
сбоку часть рваного отверстия (fora-
men lacerum), на самой же камени-
стой части заметно большое отвер-
стие наружного слухового прохода
(meatus acusticus externus). Снизу
(рис. 22), на переднем конце черепа
располагаются межчелюстные кости,
граничащие сзади с верхнечелюст-
ными костями. Между межчелюст-
ными и челюстными костями замет-
кость:
1 — подглазничное отверстие, 2 — ску-
ловой отросток верхнечелюстной кости,
8 — гребень лобной кости, 4 — скуло-
вой отросток височной кости, 5 — гре-
бень теменной кости. 6 — затылочные
мыщелки
ны сильно развитые у крысы резцо-
вые, или передненёбные, отверстия
(f. incisivum, s. palatinum anterior).
Сзади верхнечелюстные кости гра-
ничат с нёбными, которые про-
низаны одной-двумя парами нёбных
отверстий (f. palatinum). Нёбные кости совместно с верхнечелюст-
ными составляют твердое нёбо, образующее дно дыхательного трак-
та, соединяющего носовую полость с задней частью ротовой. Крышу
дыхательного тракта сзади образует тело передней клиновидной
кости, по бокам от которого располагается глубокая вырезка. Нёб-
ные кости граничат сзади с крыловидными отростками основной
68
Таблица 10
Вес костей различных отделов тела пасюка
Вес кости
Наименование кости Вес костинг ' в % от об- । щего веса скелета
Скелет конечностей 10,48- tl,43 32,8±0,5
Передняя конечность 1,384 НО,32 4,56±0,18
Лопатка 0,314 НО,07 0,93±0,8
Ключица 0,044 НО,02 0,12±0,05
Плечевая 0,524 Н0,07 0,61 -ЬО,05
Локтевая 0,224 10,04 0,69±0.03
Лучевая 0,144 г0,03 0,45 ±0,05
Кисть 0,244 НО,04 0,73 ±0,03
Задняя конечность 3,794 40,63 11,96±0,48
Тазовая . 0,964 Ё0.23 3,08±0,42
Бедро 1,144 НО, 14 3,634 0,15
Коленная чашечка 0,054 -0,01 0,15±0,02
Голень 0,874 НО,13 2,71±0,09
Стопа 0,77+0,13 2,41±0,14
Осевой скелет 21Д1 ±2,44 67,2+0,5
Череп 4^8±0,72 14,99±0,89
Нижняя челюсть 0,66+0,03 2,09±0,17
Шейные позвонки 1,06±0,17 3,28±0,16
Грудные позвонки 2,13+0,16 6.70±0.30
Поясничные позвонки 2,83+0,43 8,85±0,27
Крестец 1,69±0,28 6,00±0,30
Хвостовые позвонки 3,17+1,06 13,10±1,10
Ребра 1,41+0,14 4,48±0,22
Грудина 0,62±0,09 1,95±0,02
Итого 31,89±3,88 100
Рис. 21. Череп пасюка (вид сбоку).
А — носовая кость, Б — межчелюстная кость, В — верхнече-
люстная кость, Г — слезная кость, Д — лобная кость, Е —
височная кость, Ж — межтеменная кость, 3 — теменная
кость, И — затылочная кость, К — передняя клиновидная
кость (орбитальное крыло), Л —скуловая кость, М — основ-
ная клиновидная кость:
1 — подглазничное отверстие, 2 — решетчатое отверстие, 3 — глазиич
ное отверстие, 4 — круглое отверстие, 5 — крыловидный отрос! ни
основной клиновидной кости, 6 — рваное отверстие. ’ 7 — наружный
слуховой проход, 8 — яремный отросток, 9 — затылочный мыщс кл
клиновидной кости. Сбоку от крыловидных костей на орбитальном
крыле основной клиновидной кости располагается крыльное отвер-
стие (f. alare), а на височном крыле — овальное отверстие
(f. ovale). На границе между основной клиновидной и височной
костями видно рваное отверстие. Сзади основная клиновидная кость
граничит с затылочной; между последней и барабанной частью ос-
новной клиновидной кости располагается яремное, или заднее рва-
ное, отверстие (f. jugulare, s, lacerum posterior). Затылочная кость
окружает большое затылочное отверстие (f. occipitale magnum), а
несколько спереди несет небольшое отверстие для подъязычного
нерва (f. hypoglossi).
При вскрытии крыши черепа можно видеть, что на границе меж-
ду черепной и носовой полостями располагается решетчатая кость
(рис. 23). Она пронизана многочисленными отверстиями, через ко-
торые проходят волокна обонятельного нерва.
Череп служит не только для защиты головного мозга и органов
чувств (обоняния, зрения, слуха); лицевая часть черепа служит
также для захватывания пищи. Последняя функция приводит к
развитию мощной мускулатуры, прикрепляющейся на черепе. Части
черепа, выполняющие различные функции, развиваются независи-
мо друг от друга. Мозговая часть черепа развивается из передних
склеротомов, образующих соединительнотканную оболочку вокруг
головного мозга и органов чувств. Лицевая (точнее, висцеральная *)
часть черепа происходит из висцеральных дуг, появляющихся в
стенках передней части эмбрионального кишечника. В своем разви-
тии череп проходит три стадии: перепончатую (соединительноткан-
ную), хрящевую и костную.
При изучении черепа грызунов, в частности крыс, обычно про-
изводят некоторые измерения. Укажем наиболее распространенные
из них (рис. 24). Приводимые определения заимствованы из рабо-
ты Б. С. Виноградова и И. М. Громова, 1952.
1) Общая длина черепа — от наиболее выдающейся впе-
ред точки черепа (резцы, верхнечелюстная кость, носовые кости)
до наиболее выступающей назад части (сочленовные мыщелки, за-
тылочная кость и пр.).
2) Кондилобазальная длина — от наиболее выдаю-
щейся вперед части межчелюстной1 2 кости (между резцами) до
задней стороны затылочных мыщелков.
3) Длина лицевой части — от выдающейся вперед меж-
ду резцами точки межчелюстных костей до задней стороны альвео-
лы последнего коренного.
1 Топографически череп подразделяется на лицевой и мозговой, а по про-
исхождению — на висцеральный и мозговой. Этя деления не вполне совпадают
друг с другом, так как в состав лицевого черепа, помимо висцерального, входит
и передняя часть мозговою черепа (см. стр. 67). При описании черепа мы
придерживаемся деления ого по топографическому признаку.
2У Б. С. Виноградова и И М. Громова «верхнечелюстной-
70
4) Длина мозговойчасти — от задней стороны альвеолы
последних коренных до затылочных мыщелков.
5) Ширина межглазничного промежутка — наи*
более узкое расстояние между глазницами.
Рис. 22. Череп пасюка (вид снизу).
1 — носовая кость, Б — межчелю-
стная кость, В — верхнечелюстная
кость, Г — скуловая кость. Д — ви-
* очная кость, Е — основная клино-
видная кость, Ж — затылочная
кость, 3 — нёбная кость, И — лоб-
ная кость, К — передняя клиновид-
ная кость:
I — носовое отверстие. 2 — резцовое, или
। редненёбное, отверстие, 3 — вырезка
надглазничного отверстия. 4 — ямка глу-
бокой части наружной жевательной мыш-
• - 5 — задненёбное отверстие, 6 — ску-
овой отросток височной кости. 7 — ка-
зистая часть височной кости, 8 —
< сильное отверстие. 9 — вырезка клнно-
'*дной кости, 10 — овальное отверстие,
II непостоянное отверстие, 12 наруж-
‘ПйЛ слуховой проход, 13 — яремное от-
f'P( 1ие, 14 — яремный отросток, 16 —
। "лвочиый затылочный мыщелок, 16 —
• |гмлочный мыщелок, 17 — большое за-
-|)чиое отверстие, 18 — сагиттальный
ннАгнь основания затылочной кости. 19 —
nil»..г отверстие, 20 — отверстие подъя-
зычного нерва
Рис. 23. Череп пасюка (вид сверху;
черепная полость вскрыта):
А — решетчатая кость, Б — перед-
няя клиновидная кость, В — основ-
ная клиновидная кость, Г — височ-
ная кость (каменистая часть), Д~
затылочная кость, Е — крыловид-
ная кость:
1 — раковины решетчатой кости, 2 — ре-
шетчатая пластинка решетчатой кости,
3 — отверстие зрительного нерва, 4 —
круглое отверстие, 5 — орбитальное крыло
клиновидной кости, 6 — височное крыло
клиновидной кости, 7 — непостоянное от-
верстие, 8 — крыльное отверстие, 9 —
овальное отверстие, 10 — височный отро
сток клиновидной кости. II - синуии за-
тылочной и височной костей, 12 отвер-
стие ПОДЪЯЗЫЧНОГО ‘KjUi.i
71
6) Длина носовой части (диастема) — от задней сто-
роны основания верхних резцов до основания первого ложнокорен-
ного или коренного (если ложнокоренные отсутствуют) зуба.
Рис. 24. Измерения черепа крысы:
1 — общая длина черепа, 2 — конднлобазальная длина. 3 — длина лицевой части.
4 — длина мозговой части, б — ширина межглазничного промежутка, 6 — длина
носовой части (диастема). 7 — длина ряда коренных зубов (а — альвеолярная,
б — по краям коронок крайних коренных зубов), 8 — скуловая ширина, 9 — наи-
большая ширина черепа, 10 — наибольшая высота черепа, 11 — высота черепа от
основной затылочной кости, 12 — длина носовых костей
7) Длина ряда коренных зубов: а) альвеолярная (от
основания крайних зубов) и б) по краям коронок крайних корен-
ных зубов. Наиболее употребительно первое измерение, так как при
стирании зуба площадь коронки изменяется*. Способ измерений
коренных всегда следует оговаривать.
1 У Б. С. Виноградова и И, М. Громова «уменьшается*.
72
8) Скуловая ширина — между наружными сторонам»
наиболее удаленных от черепа частей скуловых дуг.
9) Наибольшая ширина черепа (затылочная шири-
на) — между наиболее выдающимися в бока частями черепа
(обыкновенно слуховыми отверстиями барабанных камер, или же
ламбдоидальными гребнями). Ширину черепа измеряют также по
Рис. 25. Затылочная кость пасюка. А — вид сзади,
Б — вид сбоку, В — вид снизу:
I — чешуя затылочной кости, II — боковая часть затылочной
кости, III — основание затылочной кости; 1 — височный гребень,
2 — сагиттальный гребень, 8 — большое затылочное отверстие,
4 — затылочный мыщелок» 6 — добавочный затылочный мыще-
лок; в — яремный отросток; 7 — чешуйчатый шов с височной
костью, 8 —отверстие подъязычного нерва
теменным костям или позади скуловых отростков чешуйчатой ко-
сти. Все эти измерения оговариваются.
10) На-и большая высота черепа, или высота черепа от
барабанных камер, измеряется от наиболее высокой точки черепа
в теменной области до наиболее низкой точки барабанных камер.
11) Высота черепа от основной затылочной ко-
сти измеряется до наиболее высокой части черепа в этой области.
Череп крысы состоит из 28 костей (не считая костей подъязыч-
ного аппарата), из них 11 парных и 6 непарных. Границы между
костями с возрастом становятся менее заметными.
Мозговой череп образован 13 костями.
Затылочная кость (os occipitale) расположена на границе М'-я.
лу шеей и остальными костями черепа. Спереди она граничит с
межтеменной, теменными, височными и клиновидной костями (рис.
20—23). Происходит затылочная кость из четырех рано срастаю-
щихся костей, соответственно которым в затылочной кости различа-
ют (рис. 25) основание (pars basilaris, s. os basioccipitale), боковые
части (p. p. laterales) и чешую затылочной кости (squama occipi-
talis). Основная часть располагается с вентральной стороны чере-
па и вместе с лежащей непосредственно впереди клиновидной кос-
ого образует дно черепной полости. Задняя часть основания
затылочной кости участвует в образо-
Рис. 26. Клиновидная кость
пасюка. А — вид с вен-
тральной стороны, Б — вид
сбоку:
1 — тело, 2 — крыловидный от-
росток, 3 — орбитальное крыло.
4 — височное крыло, 5 — височ-
ный отросток. 6 — овальное от-
верстие. 7 — крыльное отверстие
вании большого затылочного отверстия.
У самой границы с боковыми частями
на основании затылочной кости разви-
ты добавочные суставные мыщелки.
Затылочные мыщелки (condyli occlpi-
tales), посредством которых череп со-
членяется с позвоночным столбом,
развиты на боковых частях затылочной
кости, которые также участвуют в об-
разовании большого затылочного от-
верстия. Кнаружи от мыщелков на бо-
ковых частях сидят большие отростки,
называемые яремными (process! ju-
gulares). Углубление между затылоч-
ными мыщелками и яремными отрост-
ками называется яремной ямкой; на
дне ее заметно отверстие для XII пары
головных нервов (foramen hypoglossi).
Чешуя затылочной кости образует
верхний край большого затылочного
отверстия и несет один сагиттальный и
два боковых гребня, служащие для
прикрепления мышц головы. Чешуя
затылочной кости сложным зубчатым
швом соединяется с межтеменной и теменными костями. Боковые
части затылочной кости чешуйчатым швом соединяются с височны-
ми костями.
Клиновидная кость (os sphenoidale) в отличие от таковой чело-
века и домашних животных никогда не срастается с передней кли-
новидной костью, которая и у очень старых крыс совершенно само-
стоятельна (рис. 21—23, 26). В клиновидной кости (рис. 26) раз-
личают тело и несколько отходящих от него парных отростков.
Тело клиновидной кости расширенной задней частью граничит с
затылочной, а суженным передним концом — с передней клиновид-
ной костью. По бокам задней части тела клиновидной кости распо-
лагаются два направленные назад отростка, соединяющиеся с ка-
менистой костью; они называются височными крыльями клиновид-
ной кости (alae temporales). С вентральной стороны вдоль тела
клиновидной кости проходят парные крыловидные отростки, отде-
ленные от тела клиновидной кости вырезкой. Сзади они заходят за
74
край тела клиновидной кости и прирастают к крыловидным костям.
Непосредственно Латеральнее крыловидных отростков от тела кли-
новидной кости отходят расширенные пластинки, которые принято
называть орбитальными крыльями (alae orbitales), так как они
чходят' в стенки глазницы.
Тело и крылья клиновидной кости пронизаны несколькими от-
верстиями, служащими для прохождения нервов и сосудов. Упомя-
нем и зних расположенные в крыльях круглое и овальное отверстия
Рис. 28. Височная кость пасюка. А — каме-
нистая кость, Б—чешуя височной кости:
I — каменистая часть: II — сосцевидная часть;
III — барабанная часть; 1 — поверхность шва с
чешуей височной костя. 2 — внутренний слуховой
проход, 3 — выход водопровода преддверия. 4 —
выход водопровода улитки, 5 — непостоянное от-
верстие, 6 — костная слуховая труба. 7 — выход
слуховой трубы, 8 — слуховой барабан, 9 — гре-
бень, ограничивающий переднюю и заднюю части
черепной полости, 10—- лобное крыло, 11 —заты-
ючное крыло, 12 скуловой отросток височной
кости
Рис. 27. Передняя клино-
видная кость пасюка (вид
с внутренней стороны):
1 — тело, 2 — орбитальное кры-
ю. 3 глазничное отверстие
(toramini rotimdum et ovale). Тело кости пронизано парным каналам
для внутренней челюстной артерии. Входное отверстие этого канала
шзывается аборальным крыльным (f. alare aboralis), а выходное
расположенное у круглого отверстия, — оральным крыльным
(f. alare oralis). Задний край височного крыла участвует в обра-
зовании так называемого рваного отверстия (f. lacerum). Передняя
клиновидная кость (praesphenoideum) является непосредственным
продолжением вперед тела клиновидной кости (рис. 21—25). Впе-
реди передняя клиновидная кость сильно сужается (рис. 27) и уча-
ствует в образовании зрительного отверстия, через которое прохо-
дит зрительный нерв. Боковые части передней клиновидной кости
образуют орбитальные крылья, граничащие с лобной костью и ор-
битальными крыльями клиновидной кости (рис. 24).
Височная кость (os temporale) граничит с затылочной, темен-
юй, клиновидной и лобной костями (рис. 20—23). Височная кость
(рис. 28) состоит из двух хорошо разграниченных частей, которые
' крысы никогда не срастаются: чешуи височной кости (squama
cmporalis) и каменистой кости (os petrosum). Чешуя височной ко
«ди (рис. 28 Б) снаружи образует височную, а изнутри — мозго-
вую поверхность черепа. У границы с затылочной костью она су-
75
ji.oiui, а пи направлению вперед расширяется. Снаружи на че-
шуе |*11сочпой кости развит скуловой отросток (processus zigoma-
Ih iis), который граничит со скуловой костью и вместе с последней и
височным отростком верхнечелюстной кости образует скуловую ду-
гу. У основания скулового отростка с его вентральной стороны за-
метна суставная фасетка челюстного сустава. У крысы, как и у
других грызунов, она плохо
очерчена, что связано с боль-
шой подвижностью нижней че-
люсти в передне-заднем на-
правлении. Каменистая кость
(рис. 28 Л), в свою очередь,
подразделяется на каменистую
(pars petrosa), барабанную
(pars tympanica) и сосцевид-
ную (pars mastoidea) части.
Каменистая часть срощена с
сосцевидной частью и тонкой
щелью отделяется от барабан-
ной части; она окружает внут-
реннее ухо. На каменистой ча-
сти заметны отверстия, самое
большое из которых является
внутренним слуховым прохо-
дом. Помимо него на каменис-
той части помещается выход
водопровода преддверия, вы-
ход водопровода (улитки и ма-
ленькое непостоянное отвер-
стие. Впереди на каменистой
части развивается гребень, от-
деляющий переднюю часть че-
репной полости от задней. Сос-
цевидная часть представляет
собой пористое губчатое образование; сложным чешуйчатым швом
она соединяется с затылочной костью и чешуей височной кости.
У самой ее границы с барабанной частью развит сосцевидный от-
росток (processus mastoideus), служащий для прикрепления мышц.
Барабанная часть образует стенки костного пузыря (bulla ossea),
окружающего барабанную полость (cavum tympani). Сбоку в бара-
банную полость открывается большое отверстие наружного слухо-
вого прохода (meatus acusticus externus), в глубине которого сфор-
мирована рама для барабанной перепонки—барабанное кольцо
(annulus tympanicus). Впереди на внутренней стороне барабанной
частя заметно отверстие костной слуховой трубы (tuba auditiva
ossea), которая переходит в евстахиеву трубу, ведущую из барабан-
ной полости в глотку.
Межтеменная кость (os interparietale) — парная по происхож-
в
Рис. 29. Теменная (Л), лобная (Б)
и межтеменная (В) кости пасюка:
1 — поверхность шва с лобной костью.
2 — ямка теменной кости. 3 — поверх-
ность шва с затылочной костью. 4 — от-
печатки раковинной части решетчатой ко-
сти, 5 — носовой отросток, в — орбиталь-
ная вырезка, 7 — мозговая поверхность.
8 — поверхность шва с теменной костью.
9 — костный намет межтеменной кости
76
яению кость, очень рано объединяющаяся в одну, довольно сильно
развитую у крыс. Межтеменная кость граничит с теменными и за-
тылочной костью (рис. 20, 21, 29 В). С внутренней стороны на
(ей развито возвышение — костный намет (tentorium osseum), к
которому прикрепляется твердая мозговая оболочка.
Теменная кость (os parieta’le) граничит с лобной, височной,
затылочной и межтеменной костями (рис. 20, 21, 29 А). С наруж-
Рис. 30. Решетчатая кость и сошник пасюка. А —
решетчатая кость и сошник (вид сбоку), Б — ре-
шетчатая кость (вид сзади), В — сошник (вид
сверху):
1 — раковинная часть решетчатой кости. 2 — тело сошни-
ка, 3 — решетчатая пластинка. 4 — петуший гребень ре-
шетчатой кости. 5 — опорная пластинка носовых раковин.
6 — крылья сошника
ной стороны она несет гребень, который переходит с крыши череп-
ной коробки на ее бока.
Лобная кость (os frontale) (рис. 20, 21, 29 Б) является продол-
жением теменной кости вперед. Латерально она загибается на бо-
ковую стенку черепа, образуя орбитальную часть лобной кости. У
перехода в орбитальную часть лобная кость песет гребень, перехо-
дящий позади в теменной гребень. С внутренней стороны (рис. 29 Б),
лобная кость делится на часть черепной полости и часть носовой
полости, которые отделяются друг от друга хорошо выраженной воз-
вышенностью. Носовая часть несет несколько гребней, связанных с
раковинами решетчатой кости. Орбитальная пластинка лобной кости
имеет глубокий вырез, в котором помещается орбитальное крыло
передней клиновидной кости.
Решетчатая кость (os ethmoidale) лежит в заднем отделе носовой
полости, отграничивая ее от черепной полости (рис. 23). Решетчатая
кость (рис. 30, А, Б) состоит из ряда тонких пластинок, окружаю-
щих воздухоносные полости. В ней различают решетчатую пла-
77
сгкнку (lamina i ribtoMi), 011paiiii’iHiiiiioniyio черепную полость от
поеопой и пропн uiiniyio (niiepeiiiHMH для обонятельного нерва, пер-
иоиднкулярпую пластинку (lamina perpendicularis), лежащую в
срединной сагиттальной плоскости, и боковые бумажные пластинки
(laminae раругасеае) —тонкие ажурные костные пластинки, сое-
диняющиеся с боковыми кры-
Рис. 31. Верхнечелюстная кость пасюка.
А — вид сбоку, Б — вид спереди:
1 — подглазничное отверстие. 2 — вырезка
подглазничного отверстия, 3 — место прикреп-
ления глубокой части наружной жевательной
мышцы, 4 — височный отросток, 5 — тело, в —
нёбный отросток, 7 — вырезка резцового от-
верстия, 8 — резцовая альвеола, 9 — синусы,
10 — лобный отросток
льями сошника. Перпендику-
лярная пластинка непосредст-
венно переходит в хрящевую
перегородку носовой полости.
Ее задний край, немного вы-
ступающий в сторону черепной
полости, часто называют пе-
тушьим гребнем (crista galli).
На бумажных пластинках ле-
жат раковины решетчатой кос-
ти, образующие лабиринт (1а-
birinthus), задний край кото-
рого помещается на решетча-
той пластинке. Снаружи у кры-
сы можно насчитать шесть ко-
стных завитков, в совокупности
и образующих лабиринт.
Лицевой череп обра-
зован остальными 15 костями.
Следует учитывать, что описы-
ваемые ниже парные носовая,
• слезная и скуловая кости и не-
парный сошник, топографиче-
ски относимые к лицевому че-
репу, развиваются не из висце-
ральных дуг и по сроему про-
исхождению принадлежат к
мозговому черепу.
Верхнечелюстная кость (os
maxillare) граничит с лоб-
ной, клиновидной, передней
клиновидной, нёбной, носовой, решетчатой и слезной костя-
ми (ряс. 20—23). Тело верхнечелюстной кости (рис. 31 А,
Б) несет коренные зубы. Медиально от него отходит нёбная пла-
стинка, граничащая с нёбной костью и образующая впереди вырез-
ку резцового отверстия. От тела верхнечелюстной кости вверх от-
водит лобный отросток, чешуйчатым швом соединяющийся с орби-
тальной частью лобной кости. Более латерально от переднего края
верхнечелюстной кости отходит вверх височный отросток, у основа-
ния которого формируется ямка для глубокой части боковой жева-
тельной мышцы. Впереди вверх отходит резцовая ветвь верхнечелю-
стной кости, в которую заходит альвеола резца. Эта ветвь состоит
78
из ряда тонких костных пластинок с синусами внутри. Верхнечелю
стпая кость пронизана подглазничным каналом (foramen infraorbi-
tale) , который у крыс весьма сильно развит, так как через него про-
ходит передняя часть внутренней жевательной мышцы.
Резцовая, или межчелюстная, кость (os incisivum, s. intermaxil-
are) граничит с сошником и с верхнечелюстной, носовой и лобной
костями (рис. 20—23). От тела кости, несущего резцовую альвеолу
(рис. 32), отходит направленный вверх лобный отросток (processus
frontalis). С внутренней стороны от альвеолярной части отходит ряд
перпендикулярных пластинок, служащих для прикрепления носовой
Рис. 32. Межчелюстная кость пасюка. А — вид
сбоку, Б — вид сзади:
1 — тело кости. 2 — лобный отросток. 3 — пластинки
носовой раковины, 4 — резцовая альвеола, Б — синусы
раковины решетчатой кости. Сама альвеолярная часть состоит из
костных пластинок, между которыми заключаются полости. Альвео-
ла верхнего резца пронизывает всю резцовую кость и оканчивается
в верхнечелюстной кости.
Нёбная кость (os palatinum) граничит с верхнечелюстной, кры-
ловидной, клиновидной, передней клиновидной и лобной костями
(рис. 22). Состоит она (рис. 33 В, Г) из двух пластинок — верти-
кальной (орбитальной) и горизонтальной (нёбной). Первая задним
краем участвует в < бразовании овального отверстия и соединяется
чешуйчатым швом с передней клиновидной и с лобной костями. Нёб-
ная пластинка вместе с нёбным отростком верхнечелюстной кости
участвует в образовании твердого костного нёба. Крыловидным
отростком нёбная кость граничит с крыловидной костью.
Крыловидная кость (os pterygoideum) представляет собой тон-
кую костную пластинку, соединяющуюся с крыловидным отростком
нёбной кости и одноименным отростком клиновидной кости.
Носовая кость (os nasale) граничит с лобной, резцовой и верх-
нечелюстной костями (рис. 20, 23). Состоит она из горизонтальной
пластинки и раковинной части (рис. 33 А). Раковинная часть в
виде ажурной закрученной пластинки отходит вверх и немног"
наискось от горизонтальной пластинки.. Шов с лобной хоснио '
чатый, остальные края носовой кости гладкие.
79
Сошник (vomer) у крысы весьма слабый и является как бы про-
должением вертикальной пластинки решетчатой кости (рис. 30 В).
Впереди сошник слегка расширяется и соединяется с отростками
Рис. 33. Носовая (Л), скуловая (Б), нёбная (В — вид снизу,
Г — вид сбоку) и слезная (Д — вид с внутренней стороны,
Е — вид сверху) кости пасюка:
1 — раковина, 2 — основная пластинка, 3 — шов скуловой кости, 4 — го-
ризонтальная пластинка, 5 — предзубное возвышение, 6 — вертикальная
пластинка
Рис. 34. Нижнечелюстная кость па-
сюка:
1 — подбородочное отверстие, 2 — резец,
3 — подбородочный бугор, 4 — гребень для
прикрепления жевательной мускулатуры,
Б — угловой отросток, в — суставной отро-
сток, 7 — венечный отросток, 8 — тело
резцовой кости, образующими (резцовые отверстия. На расширенной
части сошника прикрепляются хрящевые раковины носа. Крылья
сошника служат основанием
для раковинной части решетча-
той кости.
Слезная кость (os lacrima-
1е) располагается в углу ску-
лового отростка верхнечелю-
стной кости (рис. 20, 23). Она
несет (рис. 33 Д. Е) горизон-
тальную и вертикальную плас-
тинки, последняя образует
стенку подглазничного отвер-
стия. У нижней границы слез-
ной кости в полость носа от-
крывается большое отверстие
слезного канала.
Скуловая кость (os zigomaticum) является частью скуловой
дуги. Граничит со скуловым отростком височной кости и височным
отростком верхнечелюстной кости (рис. 20—23, 33 Б).
Нижнечелюстная кость (mandibula) состоит из несущего зубы
тела и расположенной позади челюстной 'ветви (ри«. 34). Тело ниж-
ней челюсти подразделяется на резцовую и коренную частя, отде-
ленные друг от друга резким возвышением; как раз на границе
между ними располагается подбородочное отверстие. Коренная
часть постепенно переходит в ветвь нижней челюсти, которая у крыс
М)
сильно наклонена назад. Ветвь нижней челюсти несет два отрост-
ка — венечный (processus coronoideus) и суставной (рг. condyloi-
deus). К первому прикрепляется височная мышца, а вторым нижняя
челюсть сочленяется с черепом. От тела нижней челюсти вниз и на-
зад отходит хорошо развитый у крыс угловой отросток, по которому
«идет вертикальная, так называемая жевательная линия.
Подъязычная кость (os hyoideum) располагается между ниж-
ними челюстями и служит основой для языка, глотки и гортани.
Она разделена на непарное тело (corpus) и парные ветви (rami).
I ело служит для прикрепления корня языка, глотки и прилежащей
к ней гортани. Парные ветви представлены малыми (cornua mino-
ra) и большими (cornua majora) рогами. К большим рогам при-
крепляется гортань. Подъязычная кость прикрепляется к черепу на
основании клиновидной кости при помощи так называемого
tympanohyale, который у них никогда не окостеневает. От tympa-
nohyale к малым рогам тянется костный stylohyale, соединяющийся
с малыми рогами связкой.
ОСЕВОЙ СКЕЛЕТ
Осевой скелет образует опорную ось туловища. Позвоночный
столб — основа осевого скелета — состоит из отдельных позвонков.
От каждого тела позвонка отходят парные •невральные дужки
tarcus neurales), которые окружают спинной мозг. Совокупность
невральных дужек всех позвонков образует спинномозговой канал.
Невральные дужки каждого позвонка сливаются над спинным моз-
гом, образуя так называемый остистый отросток (рг. spinosus). В
грудном отделении от каждого позвонка отходит пара ребер, сово-
купность которых составляет грудную клетку, защищающую вну-
тренние органы животного. Весь позвоночный столб принято подраз-
делять на 5 отделов: шейный, грудной, поясничный, крестцовый и
хвостовой.
Шейный отдел, как и у большинства других млекопитаю-
щих, состоит из семи позвонков (vertebrae cervicales). Передние два
позвонка обеспечивают подвижность головы и сильно изменены в
связи с этим. Первый шейный позвонок, или атлант (atlas), имеет
вид кольца (рис. 35 А, Б) и дает большие возможности сгиба-
сельных и разгибательных движений в атланто-головном суставе.
У эмбриона тело атланта прирастает к телу второго шейного позвон-
i.a, образуя зубовидный отросток последнего (рис. 35 В). Атлан
(рис. 35 А, Б) состоит из двух дуг — дорзальной и вентральной
с arcus dorsalis et ventralis). Последняя несет сильно выступающий
вентральный бугор (tuberculum ventrale), а на внутренней поверх-
ности — суставную поверхность для сочленения с зубовидным от-
рчстком эпистрофея. На краниальной поверхности атланта располо
л»сны суставные ямки для сочленения с головой (foveae articulare--
I iniales). Дорзальная дуга сильно выпукла и на верхней поп«*рхно
11и несет остистый отросток, который принято называть дор >>ими-1ч
(ИДСА
бугром (tuberculum dorsales). Слегка изогнутые поперечные отро-
стки атланта очень широки, пластинчаты и называются крыльями
атланта (alae atlantis). На их вентральной стороне расположена
ямка атланта, в дне которой открывается отверстие сообщающейся
с позвоночным каналом крыловой ямки. Сзади, у основания крыла
заметно поперечное отверстие (foramen transversarium), переходя-
щее в межпозвоночное отверстие (foramen intervertebralc).
Рис. 35. Атлант (А—вид сверху, В—вид спереди) и эпистро
фей (В — вид сбоку) пасюка:
1 — вентральная дуга, 2 — дорзальная дуга, 3 — остистый отросток,
4 — вентральный бугор, 6 — суставные ямки для затылочных мыщел-
ков, в — поперечные отростки (крылья) атланта, 7 — поперечное отвер-
стие, 8 — межпозвоночное отверстие, 9 — тело эпистрофея, 10 — зубо-
видный отросток, 11 — краниальный суставной отросток. 12 — каудаль-
ный суставной отросток, 13 — поперечнореберный отоосток, 14 — пе-
редняя межпозвонковая вырезка, 15 — задняя межпозвонковая вырезка
Второй шейный позвонок, или эпистрофей (eristropheus), несет на
краниальной поверхности конусообразный у крыс зубовидный отро-
сток (dens efpistrophei), являющийся гомологом тела атланта. Вен-
тральная поверхность этого отростка несет суставную поверхность
для сочленения с атлантом (рнс. 35 В). Этот сустав допускает боль-
шой размах вращательных движений атланта, вместе с которым
вращается и голова. Внутренняя поверхность зубовидного отростка
шероховата и дает место для прикрепления связки зубовидного от-
ростка. Поперечные отростки эпистрофея направлены каудально и
у основания пронизаны межпоперечным отверстием. У крысы на
эпистрофее имеются задние и пе|редние межпозто^оичые вырезки
(incisurae intervertebrales). Остистый отросток эпистрофея у крыс
сильно развит и называется гребнем (crista epistrophei). Задние
82
суставные отростки (pr. articulates caudales) располагаются на
невральной (позвоночной) дуге эпистрофея.
Остальные шейные позвонки однотипны. Их тело (рис. 36 Л)
уплощено и несет сзади суставную ямку (fossa vertebrae), а спе-
реди— суставную готовку (caput vertebrae). От тела в стороны и
ный позвонок, Б — второй орудной позвонок, В — третий пояс-
ничный позвонок, Г — четвертое ребро, Д — грудина с реберны-
ми хрящами:
1 _ тело позвонка. 2 — невральная дужка. 3 — поперечнореберный отро-
сток, 4 — задний суставной отросток, 5 — поперечное отверстие. 6 — п»-
воно'Йцлй канал, 7 —остистый отросток, 8 — треугольная пластинка вто-
рого грудного позвонка, 9 —- реберная ямка, 10 — поперечный отросток,
11 — сюсневидный отросток, 12 — передний суставной отросток, 13 — мо «-
позвоночная вырезка, 14 — головка позвонка. 15 — головка ребра, w —
бугор ребра. 17 — шейка ребра, 18 —тело ребра, 19 —реберной хряц,
20 —рукоятка грудины, 21 — мечевидный отросток грудины. 22 хряще-
вая пластинка мечевидного отростка, 23 — сегменты грудины, 24 ребер-
ные хрящи истинных ребер
пвсрх отходит невральная дуга, основание которой несет поперечно-
рсГн-рные отростки (process! costotransversaria), соответствующие
поперечным отросткам и прирастающим к ним рудиментарным реб-
ром. Эти две части поперечиореберных отростков разделены межпо-
п> речным отверстием. На верхней части невральной дуги сидит
<» (истый отросток, слабо развитый на шейных позвонках. Попереч-
иирсберпые отростки шестого шейного позвонка несут неболыпи
<!• :((рольные отростки.
Грудная клетка (thorax) состоит у крыс из 13 позвонко!,
। (ч.ц.кнх же: пар ребер и грудной кости. Тело грудных позвонко(
(vci|cbra«’ thoracales) имеет форму трехгранной призмы; головке
1| й.
(caput vertebrae) и ямка тела (fossa vertebrae) уплощены еще
больше, чем на шейных позвонках (рис. 36 Б). На краях тела за-
метны передняя и задняя реберные ямки (foveae costales cranialis
et caudaiis). Задние ямки совместно с передними следующих поз-
вонков образуют полную суставную ямку для головки ребер. От
нервных дужек отходят поперечные отростки, у основания которых
имеются передний и задний суставные отростки. Грудные позвонки,
в особенности второй, несут сильно развитые остистые отростки.
На остистом отростке второго грудного позвонка имеется сустав,
соединяющий его с направленной вперед треугольной костной плас-
тинкой, которая служит для прикрепления нескольких связок. Кон-
цы поперечных отростков развилены. Нижняя часть развилки обра-
зует суставную фасетку для реберного бугра, верхняя часть — так
называемый сосцевидный отросток (processus mammillaris). На
грудных позвонках хорошо развиты межпозвоночные вырезки, кото-
рые в сумме образуют межпозвоночные отверстия для выхода нервов
и сосудов из позвоночного канала. Ребра (costae) представляют со-
бой костные пластинки, имеющие хрящевые концы (рис. 36 Г).
Костная часть, называемая костным ребром (os costale), имеет вид
изогнутой внутрь ленты. У проксимального конца ребра на месте
его изгиба заметно возвышение — бугорок ребра (tuberculum cos-
tae). От основания бугорка внутрь тянется суженная шейка ребра
(collum costae), которая оканчивается головкой ребра (capitulum
costae). Шейка лучше выражена на первых ребрах, а на последних
она исчезает вместе с реберным бугорком. Вниз от реберного бу-
горка тянется тело ребра (corpus costae), которое па конце остается
хрящевым и слегка расширено. Конец ребра связан с реберным
хрящем (cartilage costalis). Реберные хрящи передних ребер, на-
зываемых истинными, оканчиваются на рукоятке или же на теле
грудины, а реберные хрящи задних ребер, называемых ложными, не
достигают грудины. У крысы семь истинных и шесть ложных
ребер.
Грудина (sternum) образует вентральную основу грудной клетки
(рис. 36 Д). Она состоит из рукоятки (manubrium sterni), тела
(corpus sterni), состоящего из ряда сегментов, и мечевидного хряща
(cartilago xiphoidea). Передний конец рукоятки грудины сильно
расширен и соединен с ключицами. Тело грудной кости состоит из
четырех сегментов, между которыми имеются углубления для при-
крепления реберных хрящей истинных ребер. Первое истинное ребро
прикрепляется на рукоятке грудины, а последние два — к послед-
нему сегменту тела грудины. От тела грудины назад отходит мече-
видный хрящ, который у крыс представляет собой костную пла-
стинку с лопатообразным хрящевым расширением.
Передний конец грудной клетки образует грудной вход (apertura
thoracis cranialis), а задний конец — грудной выход (apertura
thoracis caudaiis).
Поясничный отдел состоит из 7 или 6 поясничных по-
звонков (vertebrae Jumbales). У всех поясничных позвонков сильно
84
развиты поперечнореберные отростки и передний и задний
суставные отростки (рис. 36 В). Тело поясничного позвонка на кон-
цах имеет хорошо выраженные головку и ямку. Межпозвоночная
вырезка особенно хорошо развита в задней части позвонка, и
межпозвоночные отверстия почти целиком образованы задними меж-
позвоночными вырезками. Поперечнореберные отростки шести пе-
редних поясничных позвонков несут задний отросток, особенно силь-
ный на четвертом поясничном позвонке.
Крестцовый отдел (os sacrum) состоит из четырех слив-
шихся позвонков (рис. 41). Иногда к нему присоединяется первый
хвостовой позвонок, и тогда в состав крестцовой кости входит пять
позвонков. Разделение на отдельные позвонки намечено попереч-
ными полосками (lineae transversales). Слившиеся поперечноребер-
иые отростки крестцовых позвонков образуют крылья крестцовой
кости (alae sacra les), служащие для сочленения с тазовыми костями.
Наиболее расширенная часть крыльев крестцовой кости образована
поперечнореберным отростком первого крестцового позвонка. Пос-
ледние 2 позвонка непосредственно не связаны с подвздошной костью.
Остистые отростки крестцовых позвонков у крысы хорошо развиты.
Хвостовой отдел состоит из 25—30 хвостовых позвонков
(vertebrae caudales). Из этих позвонков только первые два или три
имеют невральную дужку и хорошо развитые поперечные и остистые
отростки. Последующие позвонки теряют сначала невральную дуж-
ку, а затем и все отростки, и уже с шестого позвонка получают
вид шестигранников с легкими возвышениями на концах. Послед-
ние хвостовые позвонки имеют вид столбиков.
ПЛЕЧЕВОЙ ПОЯС
У крыс плечевой пояс состоит из двух костей: лопатки и клю-
чицы.
Лопатка (scapula) имеет форму слегка вытянутой треугольной
пластинки (рис. 37, А, Б. В). Основанием треугольника служит
позвоночный край, а вершиной — суставной угол лопатки. По на-
ружной поверхности лопатки тянется гребень — ость лопатки (spina
rapulae), которая делит лопатку на две неравные части. Часть,
лежащая перед остью, называется предостной ямкой (fossa supraspi-
•ata), а часть, лежащая за остью — заостной ямкой (fossa
htfiaspinata). Каудальный край ’заостной ямки поворачивает в ла-
’•ральную сторону и образует каудальный Гребень лопатки (crista
। audalis). У позвоночного края, за каудальным гребнем, имеется
"-большая площадка для прикрепления большой круглой мышцы.
< ’«гь лопатки у позвоночного края очень слабо развита. Дистально
"и । становится осе выше и наконец образует бугор ости. За бугром
'» и- постепенно переходит в акромион (acromion), переходя при
чим и-« вертикального положения в горизонтальное. Акромион
шчивается расширением, краниальная часть которого назыпастсч
тпенно акромиальным отростком (processus acromialls). u kv.)
• in кая часть — мет акромионом (processus metacromialh) .'In
i r.
стальная часть лопатки несёт суставную ямку (fossa glenoidalis)
для сочленения с головкой плечевой кости. Вокруг суставной ямки
ясно выделяются бугор лопатки (tuber scapulae), служащий для
Рис. 37. Кости плечевого пояса пасюка: ло-
патка (4 — вид сбоку, Б — вид с медиальной
стороны, В — вид со стороны суставной ям-
ки) и ключица (Г — вид спереди, Д — вид
сзади):
1 — место прикрепления большой круглой мышцы,
2 — каудальный гребень лопатки. S — ость лопатки,
4 — вырезка ости лопатки, Б — бугор ости, 6 — акро-
мион, 7 — мета премиальный отросток, 8 — акроми-
альный отросток, 9 — бугор лопатки, 10 — пред-
остная ямка, И — за ости а я ямка. 12 — подлопаточ-
ная ямка, 13 — линия прикрепления зубчатой мышцы,
К — коракоидный отросток, 15 - суставная поверх-
ность, 10 — грудинный конец, 17 — дорзальный край,
18 — вентральный край, 19 — лопаточный конец.
прикрепления двуглавой
мышцы, и коракоидный
отросток (processus сога-
coidaeus), к которому при-
крепляется клювоплече-
вая мышца и короткая го-
ловка двуглавой мышцы.
С медиальной стороны на
лопатке заметна так на-
зываемая зубчатая линия
(1 i nea serrata), располо-
женная немного ниже поз-
воночного края. На этой
линии оканчивается зуб-
чатая вентральная мыш-
ца. Ниже ее на лопатке
аметны возвышенности,
1.. редующиеся со виади-
ами. Вся эта часть ло-
патки называется подло-
паточной ямкой (fossa
ubscapularis), от нее бе-
I ет начало подлопаточная
ышца.
Ключица (clavicula) у
крысы хорошо развита
(рис. 37 Г. Д). На ней
различают лопаточный ко-
нец, тело и грудинный ко-
нец, а также дорзальный
и вентральный края. Ло-
паточный конец ключицы
слегка уплощен, расширен
и несет суставную фасет-
ку для акромиального от-
ростка лопатки. Отсюда
ключица в виде постепен-
но утолщающейся ленты тянется к грудинной. Грудинный конец
ключицы, соединяющийся с рукояткой грудины, в поперечном сече-
нии является почти четырехугольным.
ПЕРЕДНЯЯ КОНЕЧНОСТЬ
Скелет передней конечности подразделяют на три отдела —
плечевой (brachium), предплечье (antebrachium) и кисть (manus).
Л6
Плечевой отдел состоит лишь из одной плечевой кости
Hniinerus). В плечевой кости (рис. 38) различают тело (corpus) м
дна эпифиза (epiphisis). Проксимальный эпифиз несет головку пле-
•hMMtfl кости (caput humeri) и бугры — большой и малый (tuberculi
ni.ijus et minus). Между этими буграми заметна глубокая бо|роздка
(-ulcus intertubercularis), в которой проходит сухожилие длинной
и «ловки двуглавой мышцы плеча. От большого бугра дистально по
плечевой кости тянется сильно развитый гребень большого бугра
Рис. 36. Плечевая кость пасюка. А — вид с велярной стороны,
Б — вид с дорзальной стороны, В — вид с медиальной стороны,
Г — вид с латеральной стороны:
1 головка. 2 — малый бугор. 3 — большой бугор. 4 — гребень большого
бугра, Б — латеральный надмыщелок, в — медиальный надмыщелок, 7 —
венечная ямка, 8 — локтевая ямка, 9 — гребень латерального надмы-
щелка. 10 —тело» И —бороздка для двуглавой мышцы, 12 — блок
(crista tuberculi majoris), который резко обрывается в средней ча-
сти плечевой кости; конец гребня имеет крючковидную форму. Ди-
। тальный эпифиз плечевой кости образует блок (trochlea). У пере-
хода блока в дорзальную поверхность плеча развита венечная
ямка (fossa coronoidea), а у перехода в волярную поверхность —
юктевая ямка (fossa olecrani). По краям блока развиты надмы-
щелки — латеральный и медиальный (epicondyli lateralis et me-
dians). Головка плечевой кости у крыс имеет шаровидную форму,
благодаря чему в плечевом суставе возможны не только сгибатель-
ные и разгибательные, но и вращательные движения. Тугие связки,
ограничивающие подвижность, у крысы в плечевом суставе не
развиваются.
Предплечье состоит из двух костей — локтевой и лучевой
(рис. 39). Локтевая кость (ulna) у крысы сильно развита, ее прок
симальный конец несет суставную поверхность для сочленения с
нуто дугой и оканчивается
п
6
д
39. Кости предплечья па-
блоком плечевой кости. У начала этой поверхности располагается
крючковидный отросток (processus anconeus). Чуть проксимальнее
последнего развит сильный бугор, имеющий почта четырехгранную
форму. Бугор этот называется олекраноном (olecranon). К нему
прикрепляется трехглавая мышца плеча. Тело локтевой кости изог-
дистальным эпифизом, на котором заме-
тен стиловидный отросток (processus
styloi'deus ulnae). Локтевая кость со-
членяется с лучевой двумя суставами,
расположенными на обеих концах кос-
ти. Обе эти кости разделены друг от
друга небольшим пространством (spa-
tium interosseum antebrachii).
Лучевая кость (radius) несет на
проксимальном конце суставную по-
верхность, являющуюся продолжением
суставной поверхности локтевой кости
и оканчивающуюся спереди утолще-
нием — венечным отростком (processus
coronoideus). Тело лучевой кости изог-
нуто вдоль локтевой кости и слегка
расширено дистально. Ее дистальный
эпифиз несет стиловидный отросток
луча (processus styloideus radii).
В локтевом суставе у крыс возмож-
ны лишь разгибательные и сгибатель-
ные движения. При максимальном раз-
гибании крючковидный отросток лок-
тевой кости входит в локтевую ямку
плечевой кости, а при максимальном
сгибании венечный отросток лучевой ко-
сти входит в венечную ямку плеча.
Вращательные движения (супинация —
пронация, при которых лучевая косгь
[ локтевой кости) у крысы возможны
лишь в крайне ограниченной степени. Соответственно шаровидное
расширение блока плечевой кости выражено у нее очень слабо, а
локтевой сустав закрепляется мощными связками. Латеральная
связка (ligamenturn collaterale laterale) проходит от связочной ямки
латерального надмыщелка плечевой кости к латеральному бугру лу-
чевой кости. Крючковидная связка (lig. anconei) — наиболее слабая
I гонкая из связок локтевого сустава, — проходит от крючковидного
отростка локтевой кости к волярной стороне медиального надмы-
щелка плечевой кости. Медиальная связка (lig. collaterale mediate)
проходит от медиального надмыщелка плечевой кости к основанию
венечного отростка лучевой кости. Здесь медиальная связка вееро-
образно расширяется и отчасти закрепляется па локтевой
кости.
Рис.
сюка. А — вид с латеральной
стороны, Б — вид с медиаль-
ной стороны:
I — локтевая кость. 11 — лучевая
кость; 1 — олекранон, 2 — крюч-
ковидный отросток а — полулун-
ная суставная поверхность, 4 — фа-
сетка сухожилия двуглазой мыш-
цы плеча, 6 — суставная фа-
сетка. • — апяфяз лучевой кости,
7 — эпифиз локтевой кости
Кисть (рис. 40) состоит из нескольких рядов костей, которые
три основные группы: проксимальную —
carpalia), промежуточную — кости пясти
дистальную — кости
замег-
к костям
ряд за-
двух ос-
локтевой
пальцев (ossa digito-
ш
и
I
и
п
5
иг
з
3 37 Ю
с
13
12
принято разделять на
кости запястья (ossa
(ossa metacarpalia) и
rum).
В запястье хорошо
ны два ряда костей, один из
которых непосредственно
прилегает к костям пред-
плечья, а другой —
пясти.
Проксимальный
пястья состоит из
новных костей —
кости запястья (os ulnare
carpi) и лучевой кости за-
пястья (os radiale carpi).
Проксимальные концы обеих
этих костей образуют общую
суставную поверхность для
дистального конца костей
предплечья. Предплечье-за-
пястный сустав имеет почти
овальную форму и допу-
скает не только сгибатель-
ные и разгибательные дви-
жения, но и некоторое отве-
дение и приведение кисти.
Последние движения особен-
но важны при удержании
пищи. Дистальные концы лу-
чевой и локтевой костей за-
пястья образуют суставную
поверхность для второго ря-
да костей запястья. Сустав-
ные поверхности обоих ря-
дов костей запястья почти полностью совпадают друг с другом и в
< уставе между ними возможны лишь крайне незначительные движе-
ния в сагиттальной плоскости.
Б
кисти пасюка. А — вид
Д 10 7 9 8
Рис. 40. Кости
дорзальной стороны, Б — вид с воляр-
ной стороны:
1f II, III — первая, вторая н третья фаланга
пальцев;
1 — сесамовидные кости кисти, 2 — пястные кости,
3 — преполлекс, 4 — лучевая кость кистя, 5 —
локтевая кость кисти, 6 — гороховидная кость, 7—
центральная кость, 8, 9, 10 — первая, вторая и
третья запястные кости, 11 — четвертая, пятая за-
пястные кости, 12 — пястная кость первого пальца,.
13 — первый палец
Дистомедиально от основной части лучевой кости запястья от-
ходят два сильно развитых отростка, играющих большую роль для
’•крепления запястья и при опоре. Латерально лучевая кость запя-
(it я тугим суставом объединяется с локтевой костью запястья.
Локтевая кость запястья сочленяется также с расположенной не-
< колько волярно так называемой гороховидной костью (os pisifor-
пк), которую обычно описывают вместе с костями проксимального
ряда запястья. Гороховидная кость принадлежит к числу так на-
1ываемых сесамовидных костей, развивающихся в связках и сухо-
жилиях. У крысы, как и у других стопоходящих животных, горохо-
видная кость хорошо развита и служит твердой основой для боль-
шой ладонной мозоли. Концы гороховидной кости, один из которых
сочленяется с локтевой костью запястья, а другой служит твердой
i новой ладонной мозоли, несколько расширены.
Кроме этих трех костей в состав проксимального ряда заня-
тья входит и кость так называемого предпервого пальца — пре-
поллекс (praepollex). У крыс преполлекс имеет вид тонкой пластин-
ки, прилегающей к запястью с внутренней стороны, и служит твер-
юй о новой ладонной мозоли. Предполагают, что преполлекс яв-
ляется рудиментом самого внутреннего пальца кисти, лучше раз-
витого у некоторых примитивных наземных позвоночных. У эмбриог
чов крысы имеется также так называемая промежуточная кость (os
intermedium), срастающаяся позднее с лучевой костью кисти.
В состав дистального ряда запястья входят четыре запястные
кости и одна центральная кость. Из запястных костей самой боль-
шой является наиболее латеральная кость, представляющая собой
росшиеся в единое целое четвертую и пятую запястные кости (os
। arpale IV-j-V). Она имеет неправильную форму и несет несколько
' уставных поверхностей, из которых дистальная связана с пятой и
четвертой пястными костями, а проксимальная — с локтевой и лу-
чевой костями запястья. Третья запястная кость (os carpale III),
имеет форму почти правильной треугольной призмы, на гранях ко-
торой развиты суставные поверхности, соединяющие третью запяст-
ную кость с четвертой и второй костями запястья, центральной ко-
стью (os centrale) и с третьей костью пясти. Центральная кость —•
амая маленькая кость запястья крысы — сдвинута несколько про-
ксимально и внутрь по отношению к третьей кости запястья. Вто-
>ая и третья кости запястья имеют неправильную форму и сочле-
няются дистально с соответствующими пястными костями. Дисталь-
но кости запястья тугим суставом соединяются с костями пясти.
Кости запястья несут многочисленные, направленные в разные
тороны суставные поверхности. Передаваясь по ним, сила толчка
i ри падении на передние конечности разлагается по разным на-
правлениям. Тем самым ее действие на животное несколько смяг-
чается.
В состав пясти у крысы входит пять костей, из которых первая
ильно редуцирована. Кости пясти принадлежат к числу длинных
костей и имеют тело и эпифизы. Проксимально пяетные кости со-
членяются с костями дистального ряда запястья, дистально они не-
сут шаровидную головку для сочленения с основными фалангами
соответствующих пальцев. Помимо сгибательных и разгибательных
движений в сочленениях пястных костей и основных фаланг воз-
можны и боковые движения в пределах до 30°. У основания четы-
рех латеральных пальцев, на границе между ними и пястными ко-
(тями, возникают парные сесамовидные кости, образующие совме-
• гно с синовиальной сумкой канал, облегчающий скольжение сухо-
жилий сгибателей палыкш.
<:о
Каждый из пальцев состоит из трех длинных косточек — фа-
1анг, но первый палец образован лишь одной фалангой. Концевые
фаланги несут когти и называются ногтевыми. Их форма повторяет
форму внутренней поверхности когтевой пластинки.
ТАЗОВЫЯ ПОЯС
Рис. 41. Кости таза пасюка. А —вид свер-
ху, Б — вид сбоку, В — вид снизу:
1 — подвздошная кость, II лонная кость, III —
седалищная кость; 1 — симфиз, 2 — запирательное
отверстие, 3 — гребень крыла подвздошной кости,
4 — подвздошный б) гор, 5 — вертлужная впади-
на, 6 — проксимальный седалищный бугор, 7 —
вырезка крестцового бугра, в — крестцовый бугор.
9 — моклоковый бугор, 10 — лонный бугор, И —
дистальный седалищный бугор. 12 — внутренняя
граница суставной поверхности, 13 — впадпнная
вырезка, 14 — поперечные полоски крестца. 13 —
крылья крестца
Тазовый пояс (pelvis) состоит из двух безымянных, или тазовых,
костей (ossa innominata, s. coxae). Срастаясь вентрально, тазовые
кости образуют симфиз
(symphysis), краниодорзаль-
но они образуют тугой су-
став с крестцом. Будучи свя-
занным с позвоночным стол-
бом, тазовый пояс служит
для передачи движущей си-
лы от задних конечностей
всему телу животного.
Каждая из тазовых ко-
стей образована срощени- '
ем трех костей — подвздош-
ной (os ilium), седалищной
(os ischii) и лонной (os pu- ’
bis), сливающихся друг с
другом в области вертлуж-
ной впадины (acetabulum),
служащей для сочленения с
задней конечностью (рис.
41). У молодых крыс допол-
нительно имеется специаль-
ная впадинная кость (os
acetabulare), располагаю-
щаяся в вертлужной впади-
не (рис. 42). У взрослых
крыс впадинная кость сра-
тается с лонной костью.
Подвздошная кость де-
ются на две основные ча-
। ги — тело (corpus ossis
1 Hi) и крыло (ala ossis
lii). На крыле подвздошной
кости различают две поверх-
ности — наружную, или яго-
гичную, и внутреннюю, или тазовую. Краниальный край крыла об-
разует подвздошный гребень (crista ilif), служащий для прикреп-
ления спинных мышц. Продольным гребнем, называемым ягодичной
линией, крыло подвздошной кости делится на две почти равные 'i.v
'Ти. К вентральной части прикрепляется ягодичная малая мышца,
<И
ii h jnpui.ii.iioii ягщичная средняя мышца. На месте перехода
hpiaLni по 1нздоишоН кости в тело выражен крестцовый бугор (tuber
sacrnk*). Дорзальный вогнутый край тела подвздошной кости обра-
зует большую седалищную вырезку (incisura ischiadica major).
У вертлужной впадины тело подвздошной кости несет подвздош-
ный бугор (tuber ilii), служащий для прикрепления прямой мышцы
бедра. На внутренней поверхности крыла подвздошной кости разви-
та шероховатая суставная поверхность подвздошно-крестцового
сочленения.
Лонная, или лобковая, кость делится на шовную (ramus symphy-
seos) и впадинную (ramus acetabularis) ветви (рис. 41). Обе ветви
Рис. 42. Тазовая кость новорожден-
ного пасюка:
1 — окостенение седалищной кости, 2 —
окостенение лонной кости. 3 — окостенение
впадннной кости, 4—окостенение подвздош-
ной кости, 5 — вертлужная впадина
охватывают переднюю часть за*
пирательного отверстия (foramen
obturatorium). Впадинная ветвь
несет сильно развитый у крыс бу-
гор лонной кости (tuberculum
pubicum).
Седалищная кость подразде-
ляется на тело (corpus ischii) и
впадинную (ramus acetabulasis) и
шовную (ramus symphyseos) вет-
ви. Дорзально тело седалищной
кости несет седалищный бугор
(tuber ischii), служащий для при-
крепления нескольких мышц зад-
ней конечности.
В вертлужной впадине разли-
чают ямку впадины (fossa acetabuli) и впадинную вырезку (incisu-
ra acetabularis). Внутренние края вертлужной впадины образуют
гладкую суставную поверхность для головки бедренной кисти.
ЗАДНЯЯ КОНЕЧНОСТЬ
Скелет задней конечности подразделяют на три основных отде-
ла: бедро (femur), голень (crus) и стопу (pes).
Бедренный отдел состоит лишь из одной бедренной кости
(femor). В бедренной кости (рис. 43) различают тело (corpus)
и два эпифиза (epiphysis). Проксимальный эпифиз образует головку
бедренной кости (caput femoris), служащую для сочленения с та-
зовым поясом. С телом бедренной кости головка связана суженной
шейкой (collum femoris). На головке заметна ямка головки (fossa
capitis), от которой к вертлужной впадине проходит круглая связка
(ligamentum teres). У основания головки снаружи выражен боль-
шой вертлуг (trochanter major), отделенный от головки бедра вер-
тлужной ямкой (fosea trochanterica). Большой вертлуг служит для
прикрепления разгибателей тазобедренного сустава. Дистально
большой вертлуг переходит в гребень, наиболее развитую часть ко-
торого называют третьим вертлугом (trochanter tertius). Кроме это-
го, па плантарной стороне бедра развит еще малый вертлуг (trochan-
92
<ci minor). Дистальный эпифиз бедра несет два мыщелка лате-
ральный и медиальный (condyli lateralis et medialis), разделенные
межмыщелковой вырезкой (fossa intercondyloidea). Мыщелки по-
крыты гиалиновым хрящом и образуют суставную поверхность для
голени. Чуть проксимальнее мыщелков бедренная кость образует
шероховатости, называемые надмыщелками (epicondyli lateralis et
uiedialis). От третьего вертлуга к латеральному надмыщелку тянет-
ся довольно хорошо выраженный гребень, называемый латеральной
Рис. 43. Бедренная кость пасюка. А — вид с медиальной сто-
роны, Б — вид с дорзальной стороны, В — вид с плантарной
стороны, Г — вид с латеральной стороны:
1 — головка, 2 — ямка головки, 3 — шейка, 4 — большой вертлуг,
5 — малый вертлуг, 6 — третий вертлуг, 7 — медиальный мыщелок,
8 — латеральный мыщелок, 9 — надмыщелки, 10 — медиальная везалие-
ва кость, 11—латеральная везалиева кость, 12 — межмыщелковая
вырезка, 13 — вертлужная впадина бедра, 14 — латеральная губа
губой (labium laterale). Медиальная губа (labium mediale), тяну-
щаяся от малого вертлуга к медиальному надмыщелку, у крысы
развита слабо и заметна лишь в дистальной части бедра.
На мыщелках бедренной кости (располагается с каждой стороны
по одной сесамовидной везалиевой косточке (ossa vesalii laterale
et mediale). С дорзальной стороны на дистальном эпифизе заметны
два гребня, между которыми расположено углубление. В этом
углублении располагается небольшая сесамовидная кость — колен-
ная чашечка (patella), развивающаяся в сухожилии разгибателей
голени (рис. 44). Дистальный конец коленной чашечки крепкой
связкой прикрепляется к большеберцовой кости.
Голень состоит из двух костей (ossa cruris) — большеберцо-
вой (tibia) и малоберцовой (fibula). У крысы обе эти кости ди-
стально срастаются друг с другом (рис. 44). Проксимальный эпифиз
93
OcMiiiiiti'i'ifpuoHoft кос J и сочленяется с бедренной костью, имеет форму
тиром>|ч» треугольника (рис. 44). На нем развиты латеральный и
медиальный мыщелки (epicondyli lateralis et medial is), между ко-
торыми располагается межмыщелковая вырезка (incisura intercon-
dyloidea). Мыщелки сочленяются с хрящевыми менисками коленного
Рис. 44. Кости голени пасюка. А — вид с латеральной стороны.
Б—вид с дорзальной стороны, В—вид с плантарной стороны,
Г — вид с медиальной стороны:
1 — коленная чашечка, 2 — тел> большеберцовой юсти, 3 -- тело мало-
берцовой кости, 4 — медиальный мыщелок, 6 — латеральный мыщелок,
6 — межмыщелковая вырезка. 7 - гребень большеберцовой кости. 8 —
латеральный гребень, 9 - медиальный гребень, 10 - подколенная площадка,
11 — дистальный эпифиз малоберцовой кости. 12 — дистальный эпифиз
большеберцовой кости. 13 проксимальный эпифиз малоберцовой кости
сустава, обызвествляющимися у старых крыс. Чуть дистальнее мы-
щелков намечены подсуставные края (margo infraglenoidale). На
дорзальной поверхности проксимального конца кости развито шеро-
ховатое утолщение (tuberositas tibiae), служащее для прикрепле-
ния связки коленной чашечки. На теле большеберцовой кости раз-
личают слегка вогнутую латеральную, выпуклую медиальную и
с ieiKa вогнутую заднюю (плантарную) поверхности. Медиальная
поверхность отделяется от латеральной гребнем большеберцовой
кости (crista tibiae). Латеральная и медиальная поверхности от-
деляются от плантарной латеральным и медиальным гребнями
(cristae lateralis et medialis). В проксимальной части плантарной
поверхности заметен продольный желоб — подколенная вырезка
94
(incisura poplilea). Дистально большеберцовая кость образует су
ставную поверхность для сочленения со стопой, с медиальной сто-
роны которой образуется бугорок — медиальная лодыжка (malleo-
lus medialis).
Малоберцовая кость (рис. 44) несет на проксимальном конце
головку (caput fibuli), сочленяющуюся с большеберцовой костью.
Дистальный эпифиз малоберцовой кости целиком срастается с боль-
шеберцовой костью, образуя
латеральную лодыжку (malleo-
lus lateralis), выступающую
латерально от голеностопного
сочленения.
Стопа (рис. 45) состоит из
нескольких рядов костей, кото-
рые принято разделять на три
основные группы: проксималь-
ную — кости предплюсны (ossa
tarsalia), промежуточную —
кости плюсны (ossa metatarsa-
lia) и дистальную кости паль-
цев (ossa digitorum). У крысы
в стопу входит 36 костей.
В состав предплюсны входит
восемь костей. С голенью соч-
леняется лишь одна из них —
так называемая таранная, или
большеберцовая кость пред-
плюсны (os astragalus, s. tibiale
tarsi). Она несет на своем про-
ксимальном конце сильно раз-
витую суставную поверхность,
имеющую форму блока (рис.
46). Медиальная грань этого
блока короче латеральной и
поэтому он несколько скошен.
Дистально тело таранной кости
переходит в головку, сочленяю-
щуюся с центральной костью
предплюсны (os centra’e). Соч-
ленение это мало подвижно.
Головка таранной кости отде-
ляется от тела сужением, называемым шейкой. Изучение эмбрио
нального развития показывает, что таранная кость происходит сра-
станием двух костей — большеберцовой и промежуточной пред
плюсны (ossa tibialeintermedium).
Несколько латерально от таранной кости располагается слм;| •
большая кость предплюсны — пяточная, или малоберцовая прел
плюсна (os calcaneus, s. fibulare tarsi). Совместно с таранной ко
А 6
Рис. 45. Кости стопы пасюка. А —
вид с дорзальной стороны, Б — вин
с плантарной стороны:
I, II, III — первая, вторая и третья
фаланги пальцев;
! таранка •« кость. 2 - пяточная кость
3 ъбавочная кость. 4 центральная
кость. Б первая предплюсневая кость,
в — вторая предплюсневая юсть, 7 —
третья предплюсневая кость. 8 четвертая
пятая предплюсневая кэсть, 9 плюс
невые кости. 10 — сесамовнднъ с кости
стью она образует проксимальный ряд предплюсны (рис. 45). От
тела пяточной кости отходит направленный назад пяточный бугор
(tuber calcanei), служащий для прикрепления икроножной мышцы.
11а теле пяточной кости видны две суставные фасетки для таранной
кости, образующие совместно держатель последней (sustentaculum
astragali). Соответственно на таранной кости развиты две сустав-
ные фасетки (facies articulares medialis et lateralis). Двойное сочле-
нение этих костей делает их почти неподвижными по отношению
друг к другу. Движение между ними возможно лишь в плоскости
обеих суставных поверхностей, причем по дуге не более 5—8°.
Рис. 46. Таранная (А—вид сверху) и пяточная (Б —
вид сверху, В — вид снизу) кости пасюка:
1 — пяточный бугор, 2, 3 — держатели тарзияой кости. 4 —
тело пятой кости, 5 — шейка таранной кости, б — головка та-
ранной кости, 7 — медиальный гребень блока таранной кости,
в — латеральный гребень блока таранной кости, 9, 10 — сухо-
жильные бугры, 11, 12 — суставные поверхности пяточкой кости
С внутренней стороны с обоими только что описанными костями
сочленяется характерная для грызунов так называемая дополни-
тельная кость предплюсны (os accessorius). Ее происхождение не-
ясно.
Между проксимальным и дистальным рядами костей предплюс-
ны вклинивается центральная кость (см. рис. 45). Дистально с ней
сочленяются первая, вторая и третья кости предплюсны (ossa tarsa-
lia I, II et III). Четвертая и пятая кости предплюсны (os tarsale
IV4-V) сращены друг с другом и сочленяются с пяточной
костью.
Плюсневые кости (см. |рис. 45) у крысы довольно длинные и
хорошо развиты. На дистальном эпифизе каждой из пяти плюсне-
вых костей располагается по паре сесамовидных костей, которые
совместно с сухожильным влагалищем образуют канал для су-
хожилий сгибателей пальцев. Первый палец состоит из двух, а ос-
тальные из трех фаланг (см. рис. 45). Концевые фаланги несут
когти и называются когтевыми.
96
ТЕХНИЧЕСКИЕ СОВЕТЫ
Для изучения скелета пользуются как изолированными костями,
так и скелетом, скрепленным связками. Чтобы получить скелет в
разобранном виде, можно применять метод выварки костей в воде.
Для этого свежий труп крысы очищают от внутренностей и по воз-
можности срезают с костей мышцы, а затем вываривают скелет до
тех пор, пока остатки мышц не будут легко отделяться от костей.
Вываренные кости окончательно очищают от мягких частей грубой
зубной щеткой, скальпелем и пинцетом. Изолированные кости мож-
но отбелить, помещая их на одни сутки в бензин, причем удаляют-
ся остатки жира, а затем на 2—3 часа в 3—5%-ный пергидроль. Но
наиболее хорошие результаты дает мацерация, при которой можно
достичь любой степени очистки скелета. Для мацерации свежий
труп крысы по возможности тщательно очищают от внутренностей
и от мышц и помещают в воду на 3—15 дней в зависимости от
температуры. При комнатной температуре (20°) мацерация длится
около недели. Ежедневно воду нужно менять, лучше же всего
держать скелет в проточной воде. При этом сроки мацерации не-
сколько удлиняются, но скелеты получаются особенно чистые. Ма-
церацию продолжают до тех пор, пока не начнут отделяться остат-
ки мышц. После этого щеточкой счищают остатки мышц, тонкой
проволокой удаляют спинной мозг и ваткой под струей воды уда-
ляют из черепной коробки головной мозг. Затем маленькими нож-
ницами обрезают торчащие концы сухожилий и скелет кладут су-
шить. Таким образом получают скелет на связках. Если же маце-
рацию немного продолжить, то сухожилия разлагаются и скелет
получается разобранным. Отмацерированный скелет обычно бывает
настолько чистым, что специальной отбелки не требуется.
При приготовлении нескольких скелетов выварку и мацерацию
можно проводить в одном сосуде, помещая каждый неочищенный
скелет в особый марлевый мешочек; если мацерацию желательно
довести до полного разделения костей, то пользуются мешочками
из бязи. В мешочек вместе со скелетом кладется этикетка цз плот-
ной бумаги, надписанная простым карандашом; еще лучше выши-
вать на кусочке бязи номерки. При обработке скелетов молодых
крыс от костей легко отделяются эпифизы. После того как скелет
высохнет, их можно подклеивать.
Наиболее удобен для работы скелет, одна половина которого со-
храняется на связках, а вторая полностью разобрана. Для хранения
скелетов обычно пользуются картонными коробками. При хранении
скелеты обязательно нужно пересыпать нафталином.
ГЛАВА VI
МУСКУЛАТУРА
ОБЩИЕ ЗАМЕЧАНИЯ
Основной функциональной частью мышцы является мышечное
волокно. Мышечные волкна одеты тонкой собственной оболочкой —
сарколеммой (sarcolemma) и образуют пучки, покрытые более гру-
бой соединительнотканной оболочкой — внутренним перимизием
(perimysium internum). Мышца образована несколькими пучками
волокон и покрыта общей соединительнотканной оболочкой — на-
ружным перимизием (perimysium externum). По внутреннему пери-
мизию в мышцу проникают кровеносные сосуды. По концам мышца
переходит в плотные соединительнотканные тяжи — сухожилия, об-
разованные параллельными пучками колагеновых волокон. Каждое
волокно покрыто собственной оболочкой, а пучки их одеты peritendy-
neum internum. Все сухожилие покрыто наружной оболочкой (peri-
tendyneum externum). Концы сухожилий проникают в костное ве-
щество и прочно связывают с ним мышцу. Сухожилие менее эла-
стично, чем мышца, но обладает огромной сопротивляемостью на
растяжение.
Во многих мышцах сухожилие не является непосредственным
продолжением мышечных пучков, и последние прикрепляются к не-
му под углом (рис. 47 В). Если сухожилие проходит вдоль края
мышцы, то она получает вид одноперистой (musculus unipennatus).
При внедрении сухожилия в глубь мышцы получается двуперистая
мышца (m. bipennatus). Если же сухожилие разделяется в мышце
на несколько полосок, то получается многоперистая мышца
(m. multipennatus).
По форме мышцы принято делить на три группы: 1) широкие,
пластинчатые; обычно они располагаются на туловище и оканчива-
ются на конечностях; 2) кольцеобразные, располагаются около от-
верстий (ротового, анального и пр.); 3) округлые, имеющие вид
веретена, цилиндра, конуса; образуют главную массу мускулатуры
конечностей.
Работу мышц облегчают некоторые вспомогательные образова-
ния: фасции, синовиальные сумки и синовиальные влагалища. Фас-
98
ции представляют собой соединительнотканные оболочки, одеваю-
щие мышцы. Поверхностная, или подкожная, фасция (fascia subcu-
tanea, seu superficialis) прикрывает под кожей все тело. Глубокая
фасция (f. profunda) лежит под поверхностной и непосредственно
покрывает все мышцы с наружной стороны. От внутренней фасшн
к каждой мышце отходят специальные фасции (f. propria), которые
Рис. 47. Строение мышцы и вспомогательных органов
(по А. Ф. Климову). А — поперечный срез мышцы, Б —
синовиальное влагалище и сухожилие в продольном (I) и
поперечном (II) разрезе, В—типы мышечных брюшек:
1 — пучка мышечных волокон. 2 — внутренние перимвзяй, 3 —
Жжный перимизий. 4 — кровеносные сосуды, 5 — сухожилие, в —
.юзный пласт. 7 — листки синовиального пласта. 8 — синовиаль-
ная полость, 9 — кость; а — брюшко с параллельными волокнами,
б — одноперистая мышца, в — двупергстая мышца, г — много-
перистая мышца
как бы прибинтовывают мышцы к костям. Специальные фасции за-
крепляются и на костях.
На местах трения мышц о скелет или о соседние мышечные пуч-
ки образуются синовиальные сумки. Они представляют собой соеди-
нительнотканные мешочки, заполненные жидким секретом (сино-
вией) , облегчающим скольжение стенок сумки, а тем самым и рас-
положенным по соседству мышцам и сухожилиям. Синовиальные
влагалища имеют сходное строение, но охватывают сухожилие со
всех сторон (рис. 47 Б). Поверхностный фиброзный слой влага-
лища (stratum fibrosum) образован из плотной соединительной
ткани; глубокий синовиальный слой (stratum sinoviale). носгроен-
7f
иый из рыхлой соединительной ткани, имеет полость, заполненную
синовией.
Эффективность мышцы зависит от угла, под которым ее волок-
на прикрепляются к кости. Если этот угол не равен 90°, то сила,
развиваемая мышцей, разлагается пэ нескольким составляющим и
потому используется неполно. В таких случаях зачастую разви-
ваются костные образования — сесамовидные кости и блоки. Сухо-
жилия проходят через них и прикрепляются к кости под более вы-
годным углом.
Мышц в теле крысы насчитывают более трехсот. Естественно,
что их желательно объединять в группы по какому-либо принципу.
Многие авторы за основу для выделения' групп мышц принимают
их топографическое положение. Мы в основном придерживаемся
здесь этого же принципа, но все же считаем необходимым провести
деление мускулатуры на две основные естественные группы — осе-
вую мускулатуру и мускулатуру парных конечностей.
Принятое деление мышц не совсем отвечает истории их проис-
хождения. Так, в число мышц передней конечности входят две рав-
ные по происхождению группы мышц, производные так называемой
висцеральной мускулатуры (трапецевидные и грудино-ключично-
сосковая мышцы) и производные соматической мускулатуры, в
число которых входят все остальные мышцы передней конечности.
Соматическая мускулатура передней конечности в свою очередь
разделяется по происхождению на первичную и вторичную и т. д.
Вес мышцы почти прямо пропорционален ее силе и поэтому
для более полной характеристики мускулатуры крысы было прове-
дено взвешивание почти всех ее мышц в отдельности (табл. 11).
ПОДКОЖНАЯ МУСКУЛАТУРА
Подкожная мускулатура туловища представляет собой производ-
ное поверхностных пучков грудной мышцы (m. pectoralis) и широ-
чайшей мышцы спины (m. latissimus dorsi). Совместно они обра-
зуют большую подкожную мышцу (m. panniculus carnosus). У кры-
сы хорошо развит лишь тонкий мышечный листок, являющийся про-
изводным широчайшей мышцы спины. Волокна этого листка отхо-
дят от средней линии спины, начиная от четвертого грудного поз-
вонка и до крестца позади. Сближаясь друг с другом, они прикреп-
ляются к коракоидному отростку лопатки и к плечевой кости — у
малого бугра последней.
Подкожная мускулатура шеи и головы является производным
так называемого сжимателя шеи (m. sphincter colli). Эти мышцы
принадлежат к так называемой висцеральной мускулатуре и иннер-
вируются не спинномозговыми, а лицевым нервом. Вентральная
часть сжимателя кожи у крысы развита плохо и представляет собой
тонкую пленку, включающую мышечные волокна.
У млекопитающих от сжимателя шеи обосабливается поверхно-
стный слой (platysma myoides) и оба слоя разрастаются вперед,
100
Таблица 11
Вес мышц пасюка (в % к общему весу скелета)
Наименование мышцы Относи- тельный вес Наименование мышцы Относи- тельный вес
Мускулатура передней 75,26±3,50 Разгибатель пальцев ла-
конечности
1,46±0,2 теральный 0,08+0,01
Трапецевидная шеи Разгибатель пясти лок-
Трапецевидная головы 0,66±0,1 тевой 0,17±0,02
Трапецевидная спины 1,12=1=0,1 Отводящая большого 0,13±0,03
Ромбовидная шейная и пальца
грудная 1,18±0,2 Длинная ладонная 0,13±0,01
Ромбовидная головы 0,8±0,1 Сгибатель кисти луче-
Широчайшая спины 3,89±0,4 вой 0,12±0,02
Ключично-сосковая Г рудино-сосковая Грудная поверхностная 0,36±0,1 0,8940,1 3,83±0,3 Сгибатель кисти локте- вой Сгибатель пальцев глу- бокий 0,34+0,04 1,08±0,03
Грудная глубокая 1,75=1=0,1 Сгибатель пальцев по-
Грудная живота 0,85±0,2 верхностный 0,37±0,03
Подключичная 0,18±0,02 Мускулатура задней ко-
Зубчатая вентральная 2,95±0,3 нечности 113,93±10,0
Атланто-лопаточная 0,23d 0,02 Поясничио-подвздош-
Дельтовидная 1,49 0,1 пая 6,60±1,3
Предостная Злостная Подлопаточная 1.58-10,05 1,55+0,06 1.71+0,05 Ягодичная поверхност- ная Ягодичная средняя 3,12+0,13 5,60±0,23
Круглая большая 1,35±0,15 Ягодичная малая Г рушевидная 0,01+0,05 0,60±0,05
Круглая малая 0,07±0,02 Двойниковая 0,29 И),07
Клюво-плечевая 0,11 ±0,02 Запирательная наружная 0,73+0,05
Трехглавая, длинная голэв- ка 3.144=0,12 Запирательная внутрен- няя 1 0,24+0,06
Трехглавая, латеральная го- ловка 0,84 ' 0,05 I Г ребешковая । Четырехглавая бедра 0,81±0,1 7,35 fcO,8
Трехглавая, медиальная го- ловка 0,85 1 0,06 1 Двуглавая передняя 1,34+0,09
Напрягатель фасции пред- плечья 0,27±0,05 Двуглавая задняя Полусухожильная 6,04± 1,0 2,68 ! 0,42
Добавочная головка 0,06±0,01 Полуперепончатая перед-
Двуглавая плеча 0,53 ±0,03 няя I 3,73+0,4
Плечевая 0,70±0,06 Полуперепончатая задняя' 0,94±0,1
Круглый пронатор 0,20±0,03 Предполуперепончатая 1 0,62±0,1
Супинатор 0,09±0,02 Приводящая длинная j 0,41 ±0,0?
Разгибатель пясти длин- Приводящая короткая 1 1,83±0,21
ный лучевой 0,27±0,04 Приводящая большая 2,32 i 0,1
Разгибатель пясти ко- Квадратная бедра । 0,69 0,04
роткий лучевой 0,54+0,11 Стройная 0,57 ! 0,04
Разгибатель пальцев об- Подколенная 0 22 f 0,02 0,91 _L0>01 101
щий 0,20±0,02 Большеберцовая задняя
Продолжение табл. II
Наименование мышцы Относи- 1 ельный вес Наименование мышцы Относи- тельный вес
Большеберцовая перед- Двубрюшная 1 0,48±0,02
няя 1,52±0,13 Крыловидная 0,76 fcO, 1
Икроножная латераль-
ная 1,89±0,5 Спинная мускулатура 39,90
Икроножная медиальная 1.69+0,2 Дорзальная шеи 2.7
Пяточная 0,88±0,09 Полуостистая голозы 1,80
Камбаловидная 0,43±0,04 Длиннейшая головы 0,65
Оибатель пальцев длин- ' 1 Зубчатая дорзальная 2,26
ный 1,35±0,03 Лестничная 1,64
Разгибатель пальцев Диафрагма 5,32
длинный 0,39+0,06
Осевая мускулатура Брюшная мускулатура 27,8
Жевательная 5,67+0,5 Хвостовая дорзальная 1,64
Височная 2,68±0,6 । Хвостовая вентральная 9,02
образуя подкожную мускулатуру головы. Глубокий сжиматель шеи
дает начало мышцам рта, а поверхностный листок мышцам на-
ружного уха и век. Подробнее подкожная мускулатура головы бу-
дет описана в разделах, посвященных соответствующим органам.
МУСКУЛАТУРА ПЛЕЧЕВОГО ПОЯСА И ПЕРЕДНИХ
КОНЕЧНОСТЕЙ
Мускулатура плечевого пояса. Мышцы плечевого пояса соеди-
няют плечевой пояс с туловищем. У животных, утративших ключи-
цу, передние конечности связаны с туловищем только посредством
этих мышц. У крысы к числу мышц плечевого пояса принадлежит
7 основных мышц.
Трапецевидная мышца (m. trapezius) у крысы развита весьма
сильно (рис. 48, 49) в подразделяется на три части. Головная часть
(р. capitis, s. m. clavo-trapezius) отходит от затылочного гребня
височной кости и идет вдоль краниального края шеи к ключице,
прикрепляясь к дорзальному краю последней. Мышца тянет клю-
чицу вперед и вверх и участвует в разгибании головы.
Шейная часть (р. cervicalis, s. m. acromio-trapezius) отходит от
средней линии шеи, достигая позади треугольной пластинки второго
грудного позвонка. Прикрепляется шейная часть трапецевидной
мышцы к ости лопатки, не доходя 4—5 мм до позвоночного края
последней, и к акромиальному отростку (рис. 51). Краниальные
пучки мышцы двигают лопатку суставным углом вперед, каудаль-
ные пучки являются антагонистами первых и двигают лопатку
позвоночным краем вперед.
102
Спинная часть (р. spinalis, s. пк ppino-trapezius) отходит от
остистых отростков IV грудного — III поясничного позвонков; II и
III поясничного позвонка мышца достигает при помощи апоневроза.
Прикрепляется на ости лопатки, вплоть до ее бугра, не доходя
3—4 мм до позвоночного края лопатки (рис. 51). Мышца тянет
лопатку позвоночным краем назад.
Грудино-ключично-сосковидная мышца (m. sterno-cleido-mas-
toideus) отходит от сосцевидного отростка височной кости (рис. 50).
Рис. 48. Мышцы пасюка (вид сбоку):
1, 2, 3, — головная, шейная п спинная порции трапецевидной мышцы, 4 —
предостная, 5 — ромбовидная, в — дельтовидная, 7 — разгибатель спины,
8 — широчайшая спины. 9 — поверхностная ягодичная. 10 — напрягатель
широкой бедренной фасции, 11 — широкая бедренная фасция, 12 — полусу-
хожильная, 13 — латеральная хвоста, 14 — вентральная хвоста. 15 — наруж-
ная косая живота, 16 — зубчатая вентральная, 17 — поверхностная грудная,
18 — брюшная порция грудной, 19 — прямая живота, 20 — большая круглая,
21 — лестничная, 22. 23 — латеральная и длинная головка трехглавой плеча,
24, 25, 26 — передняя, задняя и глубокая порции наружной жевательной.
27 — передняя порция внутренней жевательной, 28 — височная, 29 — атланто-
плечевая, 30 — двуглавая бедра
и сразу же разделяется на два пучка (рис. 49). Более поверхно-
стная, грудино-соковидная часть (р. sterno-mastoideus) прикреп-
ляется к рукоятке грудины. Глубокая, ключично-сосковидная часть
(р cledo-mastoideus) прикрепляется к средней трети дорзального
грач ключицы. Мышца разгибает голову и поднимает ключицу.
Об'- описанные выше мышцы относятся к висцеральной мускула-
туре. У многих млекопитающих (копытные и некоторые другие)
грудипоключично-сосковидная мышца сливается с головной частью
трапецевидной мышцы и с акромиальной частью дельтовидной
мышцы. Образующаяся комплексная мышца называется плече-го-
1овной мышцей (m. brachiocephalicus).
Широчайшая мышца спины (m. latissimus dorsi) oixouii от
ОСТИСТЫХ ОТРОСТКОВ V XIII ГруДНЫХ ПОЗВОНКОВ И, ПОСрС 11'11"'М по-
им
Рис. 49. Мышцы пасюка (вид снизу):
1 — поперечная нижней челюсти. 2 — жевательная. 3. 4 — передняя и задняя части дву-
брюшной мышцы. S — плече-подъязычная, 6 — головная часть трапецевидной, 7 — под-
бородочно-подъязычяая, 8 — атлано-лопаточная, 9 — грудная глубокая, 10 — подключич-
ная. И — клюво-плечевая, 12 — двуглавая плеча, 13 — грудная живота, 14 — широчайшая
спины, 15 — прямая живота, 18 — зубчатая вентральная, 17 — лестничная. 18 —межребер-
ная наружная, 19 — межреберная внутренняя, 20 — грудная поверхностная, 21 — коса»
внутренняя живота, 22 — белая линия, 23 — подвздошная. 24 — поясничная большая. 25 —
прямая бедра, 26 — четырехглавая бедра (медиальная головка), 27 — полуперепончатая пе-
редняя, 28 — полуперепончатая задняя, 29 — грудино-подъязычная, 30 — вентральная хвоста,
31 — приводящая короткая, 32 — гребешковая, 33 — приводящая длинная, 34 — стройная, 35 -
полусухоЖИльная, 36 — приводящая большая, 37 — трехглавая голени, 38 — портняжная,
39 — мечевидный хрящ, 40 — косая наружная живота, 41 — передняя большеберцовая, 42 —
трехглавая плеча, 43 — напрягатеть фасции предплечья, 44, 45 — ключичная и акромиальная
порция дельтовидной, 46 -- нлючнца, *7— ключично-сосковая, 48 груднно сосковая,
49 — височная
ясничной фасции, от остистых отростков I—III поясничных позвон-
ков (рис. 48, 49). Прикрепляется широчайшая мышца спины на во-
лярной стороне плечевой кости, у головки последней (рис. 52).
Мышца оттягивает переднюю конечность назад и сгибает плечевой
сустав.
Ромбовидная мышца (m. rhomboideus* отходит от сосцевидного
отростка и затылочного гребня височно'" кости (рис. 50), гребня
Рис. 50. Места прикрепления мышц к черепу пасюка
(А — вид сбоку, Б — вид снизу):
I трапецевидная (головная часть), 2 — грудино-ключично-соско-
вая, 3 — ромбовидная (головная часть), 4, 6, 6 — передняя, глубокая
и задняя части наружной жевательной, 7, 8 — передняя и задняя
части внутренней жевательной, 9 — височная, 10. 11 — латеральная
н медиальная части крыловидной, 12 — двубрюшная, 13 — пластыре-
видная, 14 — полуостисгая головы, 15 — длиннейшая головы, 16 —
косая i-оловы (краниальная часть), 17. 18, 19, 20 — дорзальная боль-
шая, дор сальная малая, латеральная и вентральная части прямой
головы, 21 — длинная головы.
эпистрофея и остистых отростков шейных и передних (I—II) груд-
ных позвонков. Снаружи она прикрыта трапецевидной мышцей
(рис. 48). Мы разделяем ромбовидную мышцу на три части: голов-
ную (р. capitis), шейную (р. cervicalis) и грудную (р. thora-
*<tlis). Обычно головную часть описывают как самостоя-
тельную мышцу (Грин, 1935; Хилл, 1930; Хоуэлл, 1936), однако у
многих беличьих она не обособлена от остальных частей ромбовид-
ной мышцы. У крысы головная часть ромбовидной мышцы отходит
от черепа и прикрепляется по верхней трети ости и по кранипль
пому концу позвоночного края лопатки (рис. 51). Мышца unci
лопатку вперед и способствует подниманию головы. Шейная чю и
отходит от дорзального бугра атланта, гребня эпистроф’ ч и •" til
иг.
in ।и -кг и четвертого) шейного позвонка.
.........к и ни мгшильной стороне краниальной трети позвоноч-
ник* крци лопатки (рис. 51). Тянет лопатку позвоночным краем
ппср' Д. I рудная часть отходит от остистых отростков четырех
последних шейных и первых двух; грудных позвонков и прикрепляет-
ся по медиальной стороне позвоночного края лопатки (рис. 51). Тя-
п
№
6
А
Рис. 51. Места прикрепления мышц к ло-
патке пасюка. А — вид с латеральной сто-
роны, Б — вид с медиальной стороны:
нет лопатку позвоночным краем вперед.
Грудная мышца (m. pectoralis) представлена у крысы тремя
самостоятельными мышцами, отходящими от грудины и от белой
линии живота (рис. 49).
Грудная поверхностная
мышца (ш. pectoralis su-
perficialis) отходит от ру-
коятки тела и мечевидно-
го отростка грудины,
вплоть до основания рас-
ширенной части мечевид-
ного отростка, и оканчи-
вается на шероховатости
большого бугра плечевой
кости (рис. 52). Фикси-
рует туловище между ко-
нечностями, поворачивает
и сгибает плечевой сустав.
Грудная глубокая
мышца (m. pectoralis pro-
fundus) отходит от первых
четырех сегментов тела
грудины и оканчивается
1—2 — шейная н спинная части трапецевидной
мышцы, 8 — атланто-лопаточная. 4—5 — лопагоч-
мая и акромиальная части дельтовидной, 6, 7, 8—
головная, шейная и грудная части ромбовидной.
• — грудная глубокая, 10 — грудная живота,
1!) 12 — шейная и грудная части вентральной
зубчатой, 13 — предостная, 14 — заостная, 15 —
подлопаточная, 16 — круглая большая, 17 —
круглая малая, 18 — трехглавая плеча (длинная
головка), 19 — двуглавая плеча (длинная голов-
ка), 20 — клюво-плечевая
па большом и малом буг-
рах плечевой кости и на
коракондном отростке ло-
патки (рис. 51, 52). Фик-
сирует туловище между
конечностями; оттягивает
гшечность назад и участ-
вует в повороте плечевого
сустава. Мышца снаружи прикрыта поверхностной грудной мышцей
(рис. 49). Грудная мышца живота (m. pectoralis abdominalis) отхо-
дит от белой линии живота и тянется вперед, прикрывая каудаль-
ные пучки поверхностной грудной мышцы (рис. 49). Постепенно
она переходит в почти округлое брюшко, проходящее по груди под
обеими описанными выше частями грудной мышцы и оканчиваю-
щееся на коракоидном отростке лопатки (рис. 51). Оттягивает
переднюю конечность назад.
Подключичная мышца (m. subclavius) отходит от первого ребра
я прикрепляется в средней части каудо-вентральной поверхности
ключицы. Тянет ключицу вниз и назад.
106
Зубчатая вентральная мышца (m. serratus ventralis) по про-
исхождению является, по всей вероятности, двойной. Следуя Кли-
мову, мы здесь разделяем ее на шейную и грудную части (рис. 48,
19), однако по новейшим данным такое деление не отвечает исто-
рии происхождения мышцы (Максименко). Шейная часть (р. сег-
v icalis) отходит от поперечнореберных отростков последних четырех
шейных позвонков и оканчивается на медиальной поверхности ло-
Рис. 52. Места прикрепления мышц к плечевой кости пасюка.
А — вид с волярной стороны, Б — вид с дорзальной стороны,
В — вид с медиальной стороны, Г — вид с латеральной
стороны:
1 — широчайшая спины, 2 — грудная поверхностная. 3 — грудная глу-
бокая, 4—5 — лопаточная и ключичная части дельтовидной, 6 — пред-
метная, 7 — заостиая, 8 — подлопаточная, 9 — круглая малая, 10 —
круглая большая, 11, 12 — латеральная и медиальная головки трех-
главой плеча. 13 — добавочная головка, -14 — плечевая. 15 — длинный
лучевой разгибатель кисти, лучевой разгибатель кисти, 10 — короткий,
17 — общий разгибатель пальцев, 18 — латеральный разгибатель паль-
цев, 19 — локтевой разгибатель запястья, 20 — супинатор, 21 — круг-
лый пронатор, 22 — длинная ладонная, 23 — локтевой сгибатель за-
пястья, 24 — поверхностный сгибатель пальцев. 25 — лучевой сгиба-
тель кисти, 26, 27 — наружная п внутренняя головки глубокого сги-
бателя пальцев
натки (рис. 51). Грудная часть (р. thoracal is) отходит от первых
семи ребер и оканчивается на медиальной стороне лопатки (рис.
51). Зубчатая мышца подвешивает тело между лопатками и являет-
ся основным фиксатором туловища.
Атланто платочная мышца (m. atlanto-scapularis) отходит от
вентральной дуги, вентрального бугра и крыла атланта и оканчи-
вается на метакромиальном отростке лопатки (рис. 48, 49, 51).
Тянет лопатку суставным углом вперед.
Мускулатура свободной конечности. Мускулатура свободной
передней конечности делится на мускулатуру проксимальной чл-тп
|о>
конечности (плечо и предплечье) и мускулатуру дистальной част»
конечности (кисть). Первая служит, в основном, для передвижения
животного, вторая — для движений, связанных с удержанием пиши.
Мускулатура проксимальной части конечности, в свою очередь, раз-
деляется на мускулатуру плечевого сустава и мускулатуру локтево-
го сустава.
Рис. 53. Мускулатура передних конечностей па-
сюка. А — вид с латеральной стороны, Б — вид
с медиальной стороны:
1, 2, 3 — лопаточная, акромиальная и ключичная части дель-
товидной, 4 — предостная, 5 — заостная, в — круглая большая,
7 — нодлопаточпая, 8 — широчайшая спины, 9, 10, 11—длинная,
латеральная н медиальная головки трехглавой плеча. 12 — на-
прягатель фасции предплечья, 13 —плечевая, 14 - двугла-
вая, 16, 16 —длинная и короткая головки двуглавой плеча,
17 — клюво-плечевая, 18 — круглый пронатор, 19 — длинный
разгибатель кисти, 20 длинная отводящая большого пальца,
21 — общий разгибатель пальцев, 22 — локтевой разгибатель
кисти
Мускулатура плечевого сустава. Поверхностно в
проксимальной части плеча лежит дельтовидная мышца (m. del-
toideus), условно разделяемая на три части (рис. 48, 53). Лопаточ-
ная часть (р. scapularis) начинается от ости лопатки и акромиона
(рис. 51) и прикрепляется к латеральной поверхности гребня боль-
шого бугра плечевой кости (рис. 52). Акромиальная часть (р. асго-
mialis) отходит от метакромиального отростка и акромиона и при-
крепляется совместно с лопаточной частью (рис. 52). Ключичная
часть (р. clavicularis) отходит от вентральной поверхности ключицы
и прикрепляется к медиальной поверхности гребня большого бугра
плечевой кости (рис. 52). Лопаточная часть сгибает птеч^вой су-
став, акромиальная'-- отводит, а 'ключичная — притч шг плечо.
Предостная мышца (m. supraspinatus) отходит от предостно»
108
ямки лопатки и прикрепляется к проксимальному концу большого
бугра плечевой кости (рис. 51—53). Разгибает плечевой сустав.
Заостиая мышца (m. infraspinatus) отходит от всей заостной
ямки лопатки и оканчивается на большом бугре плечевой кости
(рис. 51—53). Вращает и отводит плечевую кость.
Подлопаточная мышца (m. subscapularis) начинается от подло-
паточной ямки на медиальной поверхности лопатки и оканчивается
на малом бугре плечевой кости (рис. 51—53). Вращает и отводит
плечевую кость.
Круглая большая мышца (m. teres major) начинается от спе-
циальной площадки каудального угла лопатки и оканчивается на
медиоволярной стороне плечевой кости, около окончания широчай-
шей мышцы спины (рис. 48, 51—53). Сгибает плечевой сустав и
способствует приведению плеча.
Круглая малая мышца (тп. teres minor) начинается от каудаль-
ного гребня лопатки и оканчивается на большом бугре плечевой
кости (рис. 51—53). Отводит плечевую кость.
Клюво-плечевая мышца (m. coraco-brachlalis) отходит от кора-
коидного отростка лопатки и оканчивается на медиальном надмы-
щелке плечевой кости и по медиальному краю последней (рис. 53,
54). Приводит плечевую кость.
Мускулатура локтевого сустава. К этой группе мы
относим 5 мышц.
Трехглавая мышца плеча (m. triceps brachii) разделяется на
три головки (рис. 53). Длинная головка (с. longum) начинается от
дистальной трети каудального края лопатки и оканчивается на вер-
хушке олекранона. Латеральная головка (с. laterale) начинается от
дистального края большого бугра плечевой кости и оканчивается
по латеральному гребню олекранона. Медиальная головка начинает-
ся от волярной поверхности плечевой кости и оканчивается по дор-
зальной поверхности олекранона и по медиальному краю последнего
(рис. 51, 52, 55).
Добавочная головка (m. epijtrochleo-anconeus) начинается от ме-
диального надмыщелка плечевой кости и оканчивается на олекра-
ноне (рис. 52, 55).
Напрягатель фасции предплечья (m. dorsoepitrochlearis) начи-
нается на широчайшей мышце спины и оканчивается на медиальной
поверхности олекранона и на фасции предплечья (рис. 53).
Все эти три мышцы разгибают локтевой сустав. Напрягатель
фасции предплечья способствует также натяжению фасции предпле-
чья. Наиболее мощной из описанных мышц является трехглавая
мышца плеча, особенно ее длинная головка.
Двуглавая мышца плеча (m. biceps brachii) длинной головкой
(с. longum) начинается от бугра лопатки, а короткой (с. breve) -
от коракоидного отростка лопатки. Отсюда короткая головка идет
совместно с клюво-плечевой мышцей до середины длины плсчевои
кости, где соединяется с длинной головкой (рис. 54). Munniii при
ion
крепляется двумя сухожилиями к проксимальному концу локтевой
кости и к специальному бугру лучевой кости (рис. 55).
Плечевая мышца (m. brachialis) начинается от волярной поверх-
ности плечевой кости и от дистального края большого и малого
бугров последней и оканчивается на локтевой кости под двуглавой
мышцей (рис. 52—54).
Последние две мышцы сгибают локтевой сустав. Лучевая часть
двуглавой мышцы плеча способствует также вращению лучевой
кости (пронации предплечья). Сгибанию локтевого сустава способ-
ствуют также описываемые ниже круглый пронатор и супинатор.
Рис. 54. Сгибатели локтевого сустава пасюка:
1, 2 - длинная и короткая головки двуглавой плеч.
3 — брюшко двуглавой плеча, 4, б лучевое и
тевое сухожилия двуглавой плеча, 6 — плечевая,
7 — клюво-плечевая, 8 — лопатка, 9 — коракоидный
отросток, 10 — лучевая кость, 11 — локтевая кость
Мускулатура кисти. Мышцы кисти разделяются на
длинные и короткие. Первые, в свою очередь, подразделяются на
сгибатели, начинающиеся на медиальном надмыщелке плечевой
кости, и разгибатели, начинающиеся на латеральном надмыщелке
последней. К числу разгибателей принадлежат 8 мышц.
Длинный лучевой разгибатель кисти (m. extensor carpi radialis
longus) начинается на проксимальном конце гребня латерального
надмыщелка плечевой кости и прикрепляется к проксимальному
концу второй пястной кости, проходя предварительно под сухожи-
лием отводящей мышцы большого пальца (рис. 52, 53, 56, 58).
Короткий лучевой разгибатель кисти (m. extensor carpi radialis
brevis) начинается от гребня латерального надмыщелка, непосред-
ственно дистальнее предыдущей мышцы, и, проходя совместно с ней
под сухожилием отводящей мышцы большого пальца, прикрепляется
к середине третьей пястной кости (рис. 52, 56, 58). Волокна обеих
мышц тесно связаны друг с другом. Обе они разгибают запястно-
пястный сустав.
Общий разгибатель пальцев (m. extensor digitorum communis)
начинается непосредственно дистальнее предыдущего (рис. 52). Ди-
стально он разделяется на четыре сухожилия и, проходя под по-
перечной связкой запястья, оканчивается на когтевых фалангах
II—V пальцев ( рис. 52, 53, 56, 58). У пястно-фаланговых суставов
ПО
каждое сухожилие отдает веточку к суставной сумке. Мышца раз-
гибает пальцы.
Латеральный разгибатель пальцев (m. extensor digitorum late-
ralis) начинается от гребня латерального надмыщелка, непосред-
ственно дистальнее предыдущего (рис. 52). Его брюшко, тесно свя-
занное с предыдущей мышцей, на половине длины костей пред-
плечья делится на два сухожилия, кото-
рые проходят под поперечной связкой за-
пястья и оканчиваются на когтевых фа-
лангах IV и V пальцев, сбоку от общего
разгибателя пальцев (рис. 56, 58). Раз-
гибает и слегка отводит IV и V пальцы.
Локтевой разгибатель кисти (m. exten-
sor carpi ulnaris) начинается двумя голов-
ками — от латерального надмыщелка и
от олекранона —и оканчивается на лате-
ро-волярной стороне пятой пястной кости
(рис. 52, 53, 55, 56, 58). Отводит и сгибает
V палец.
Длинная отводящая мышца большого
пальца (m. abductor pollicis longus) на-
чинается от латеральной ямки локтевой
кости и от перепонки, затягивающей меж-
костное пространство, и оканчивается на
медиальной стороне первой пястной кости
(рис. 53, 55, 56, 58). Отводит I палец и
кисть внутрь.
Специальный разгибатель указатель-
ного пальца (m. extensor indicis proprius)
начинается от латерального углубления
Рис. 55. Места прикрепле-
ния мышц к костям пред-
плечья пасюка. А — вид с
латеральной стороны, Б —
вид с медиальной стороны:
локтевой кости, непосредственно дисталь-
нее предыдущей мышцы (рис. 55). На ди-
стальном конце локтевой кости разгиба-
тель указательного пальца проходит под
специальной пластинкой поперечной
связки запястья и оканчивается на когте-
вой фаланге второго пальца, латеральнее
окончания сухожилия общего разгибателя
пальцев (рис. 56, 58). Эта мышца приво-
дит и разгибает указательный палец.
Супинатор (m. supinator) начинается
1, 2, 3 — длинная, латеральная
и медиальная головки трехгла-
вой плеча. 4 —добавочная го-
ловка. Б, в — лучевое и локтевое
сухожилия двуглавой плеча. 7 —
плечевая. 8 — локтевой разгиба-
тель кисти. 9 — длинная отводя-
щая большого пальца, 10 —
супинатор, 11 — круглый прона-
тор. 12, 13 — локтевая н лучевая
головки глубокого сгибате-
ля пальцев. 14 — локтевой сги-
батель кисти. 15 — собствен-
ный разгибатель указательного
пальца
на латеральном надмыщелке плечевой кости и оканчивается на дор-
зальной поверхности лучевой кости (рис. 52, 55, 56). Супинатор
вращает лучевую кость и участвует в сгибании локтевого сустава.
Кроме перечисленных нами мышц, Грин описывает еще две
длинный и короткий разгибатели большого пальца (m. т. extcnsnn"
pollicis longus et brevis). Мы не смогли их обнаружить.
К числу сгибателей кисти принадлежат 6 мышц.
11»
Круглый пронатор (m. pronator teres) начинается от медиаль-
ного надмыщелка плечевой кости и оканчивается на дорзальной
поверхности лучевой кости (рис. 52, 55, 56). Круглый пронатор вра-
щает лучевую кость и участвует в сгибании локтевого сустава.
Рис. 56. Длинная мускулатура кисти пасюка. А — поверхностные
мышцы с дорзальной стороны, Б — глубокие мышцы дорзальной
стороны, В — поверхностные мышцы с волярной стороны, Г — глу-
бокие мышцы волярной стороны:
1 — длинный лучевой разгибатель кисти, 2 — короткий лучевой разгибатель
кисти, 3 — общий разгибатель пальцев, 4 — латеральный разгибатель паль-
цев. 5 — локтевой разгибатель кисти, в — длинная отводящая большого
пальца, 7 — специальный разгибатель указательного пальца, в — супинатор,
9 — круглый пронатор, 10 — двуглавая плеча, II — плечевая. 12 — длинная
ладонная, 13, 14 — лучевая и локтевая головки глубокого сгибателя пальцев,
15 — лучевой сгибатель кисти, 16 — локтевой сгибатель кисти. 17 — дисталь-
ное сухожилие локтевого сгибателя кисти, 18 — поверхностный сгибатель
пальцев, 19 — ладонная фасция
Длинная ладонная мышца (m. palmaris longus) начинается от
медиального надмыщелка плечевой кости и оканчивается на ладон-
ной фасции (рис. 52, 56). Сгибает кисть.
Поверхностный сгибатель пальцев (m. flexor digitorum sublimis)
начинается от медиального надмыщелка плечевой кости, прокси-
мальнее длинной ладонной мышцы (рис. 52). Проходя поверх глу-
бокого сгибателя пальцев, мышца у дистального конца костей
предплечья разделяется на три сухожилия, которые идут к трем
средним пальцам (рис. 56). У пястно-фаланговых суставов каждое
сухожилие делится на две ветви, которые па уровне суставов между
112
первыми и вторыми фалангами охватывают сухожилие глубокого
сгибателя пальцев и прикрепляются на проксимальном конце вто-
рых фаланг (рис. 58). Сгибает три средних пальца.
Грин утверждает, что поверхностный сгибатель прикрепляется
к П—V пальцам. У пасюка, так же как и у многих других грызу-
нов этого надсемейства (Cricetulus, Mus, Meriones, Microtus, Spa-
lax, Cricetus, Apodetnus), мы не могли обнаружить сухожилия
к V пальцу.
Глубокий сгибатель паль-
цев (ш. flexor digitorum pro-
fundus) у крысы начинается
четырьмя головками (рис.
56). Две плечевые начинают-
ся от медиального надмы-
щелка плечевой кости, локте-
вая — от всей волярно-ме-
диальной поверхности локте-
вой кости, включая и часть
олекранона, а лучевая — от
дистальной трети волярно-
медиальной поверхности лу-
чевой кости (рис. 52, 55).
Оканчивается мышца на ког-
тевых фалангах каждого из
пальцев (рис. 58). Сгибает
пальцы и кисть.
Локтевой сгибатель кисти
(m. flexor carpi ulnaris) на-
чинается на медиальном над-
мыщелке плечевой кости,
дистальнее длинной ладон-
ной, и на олекраноне (рис.
52, 55). Оканчивается мыш-
ца на гороховидной кости
запястья (рис. 56, 58). Сги-
бает кисть.
Рнс 57. Короткая мускулатура кисти па-
сюка. А — поверхностные мышцы, Б —
глубокие мышцы:
I — приводящая большого пальца, 2 — червеоб-
разные, 3 приводящая V пальца, 4 — коротки!
сгибатель V пальца, Б — отводящая V пальца,
6 — сгибатель большого пальца. 7 — межкостные
Лучевой сгибатель кисти (m. flexor carpi radialis) начинается
на медиальном надмыщелке плечевой кости, между круглым
пронатором и глубоким сгибателем пальцев, и оканчивается на про-
ксимальном конце третьей пястной кости (рис. 52, 56, 58). Сгибает
кисть.
Короткая мускулатура кисти обычно делится на мускулатуру
большого пальца, мускулатуру пятого пальца и срединную муску-
(атуру.
Короткий сгибатель большого пальца (m. flexor pollicis brevis)
н.1чинается от преполлекса и оканчивается на волярной поверхно
< in первого пальца (рис. 57). Сгибает большой палец.
Приводящая мышца большого пальца (гл. adductor pollici-)
11.1
я Б
Рис. 58. Места прикрепления мышц
к костям кисти пасюка. А — вид с
дорзальной стороны, Б — вид с во-
лярной стороны:
1 — длинный лучевой разгибатель кисти.
2 — короткий лучевой разгибатель кисти,
3 — общий разгибатель пальцев, 4 — ла-
теральный разгибатель пальцев, 5 — спе-
циальный разгибатель указательного
пальца, в — длинная отводящая большого
пальца, 7 — локтевой сгибатель кисти,
в — лучевой сгибатель кястя, 9 — по-
верхностный сгибатель пальцев, 10 — глу.
бокий сгибатель пальцев
начинается на первой карпальной кости и ладонной связке и окан-
чивается на медио-волярной стороне первой фаланги большого
пальца (рис. 57). Приводит и противопоставляет большой палец.
Короткий сгибатель мизинца (m. flexor digiti quinti brevis) на-
чинается на гороховидной кости и кончается на проксимальном кон-
це первой фаланги V пальца (рис. 57). Сгибает V палец.
Отводящая мышца мизинца
(m. adductor digiti quinti) на-
чинается на гороховидной кости
и оканчивается на первой фа-
ланге мизинца (рис. 57). Отво-
дит мизинец.
Приводящая мышца мизин-
ца (ш. adductor digiti quinti)
начинается на медиальном крае
преполлекса и оканчивается на
основании первой фаланги V
пальца. Приводит пятый палец.
Червеобразные мышцы (пт.
т. lumbricales) начинаются на
сухожильной пластинке глубо-
кого сгибателя пальцев и окан-
чиваются на проксимальном
конце основных фаланг II—V
пальцев (рис. 57).
Межкостные мышцы (т. т.
interossei) у крысы развиты в
числе двух для каждого из II—
V пальцев (рис. 57). Обычное
деление их на дорзальные и ве-
лярные для крысы не оправда-
но. Межкостные мышцы II—IV
пальцев начинаются на глубо-
кой ладонной фасции, а меж-
костные мышцы V пальца — на
гороховидной кости. Оканчива-
ются межкостные мышцы на
сесамовидных костях, расположенных между первыми фалангами и
пястными костями (рис. 57). Повидимому, межкостные мышцы по-
вышают .упругость кисти, но они могут способствовать и сгибанию
пальцев.
МУСКУЛАТУРА ЗАДНЕЙ КОНЕЧНОСТИ
У млекопитающих тазовый пояс неподвижно связан с позвоноч-
ным столбом и особой его мускулатуры поэтому не развито. Мус-
кулатуру свободной конечности делят на мускулатуру тазобедрен-
ного сустава, мускулатуру коленного сустава, скакательный ком-
плекс и мускулатуру стопы.
114
Мускулатура тазобедренного сустава. Включает основную мае*
< у мышц задней конечности. Мы разделяем ее на поясничную, яго-
i и иную, заднебедренную, приводящую и глубокую тазобедренную
।руппы.
Поясничная мускулатура. К числу поясничных при-
надлежат 2 мышцы. Их можно видеть лишь после удаления всех
нпутренностей.
Поясничная большая мышца (m. psoas major) начинается от
«антральной поверхности тел последних двух грудных и всех пояс-
ничных позвонков и оканчивается на малом вертлуге бедренной ко-
< ги (рис. 59, 60, 65). Сгибает тазобедренный сустав.
Рис. 59. Поясничная мускулатура пасюка:
1 большая поясничная, 2 — подвздошная. 3 — поясничная
квадратная, 4 — тазовая кость, 6 — бедренная кость
Подвздошная мышца (m. iliacus) начинается только от под-
ь тошной кости (рис. 64). Она сливается с большой поясничной
мышцей и оканчивается вместе с ней на малом вертлуге бедренной
•)сти (рис. 59, 65). Сгибает тазобедренный сустав.
Поясничная малая мышца (m. psoas minor), описываемая Грин,
• крыс отсутствует.
Ягодичная мускулатура. К числу ягодичных принадле-
.кат 6 мышц. Ягодичная поверхностная, напрягатель широкой фас-
ции бедра и портняжная мышцы (m. m. glutaeus superficialis, ten-
or fasciae latae et sartorius) у крысы тесно связаны друг с другом
(рис. 60, 61). Они берут начало от вентрального, краниального и
юрзального краев подвздошной кости, а ягодичная поверхност-
«я также и от остистых отростков последнего поясничного и
| " р''дних крестцовых позвонков (рис. 64). Поверхностная ягодич-
III идет к гребню большого вертлуга бедренной кости, напряга-
• п. широкой бедренной фасции — к широкой бедренной фасции, а
портняжная мышца — к коленной чашечке (рис. 65, 66). Все эти
•плицы сгибают бедренный сустав, а портняжная влияет на разги-
'.iiiiir коленного сустава.
Яютчиая средняя мышца (m. glutaeus mediu?) начинается от
ip 1.1ЛЫЮЙ ямки подвздошной кости и остистых и сосцевидных от-
1»х'1Ко|1 кр. отцовых позвонков (рис. 64). Прикрепляется эта мыш-
п । к проксимальному концу большого вертлуга и, отчасти, к кау-
। ц.Ф‘ч краю последнего (рис. 62, 65). Является одним из ’1мы
р пых разгибателей бедренного сустава. С ягодичной ц» .!<• и
. uiim ii х 1рысы тесно связана грушевидная (m. piriforini' ) ipi
II
дящая от оврыла и вентральной поверхности тела подвздошной кости
и прикрепляющаяся к большому вертлугу бедренной кости (рис. 64,
65). У крысы грушевидная мышца фактически является частью
средней ягодичной.
Ягодичная малая мышца (m. glutaeiis minimus) начинается от
подвздошной кости, нейтральнее ее срединного гребня, и оканчи-
Рис. 61. Мускулатура латеральной
поверхности задней конечности па-
сюка:
’Рис. 60. Мускулатура медиальной
поверхности задней конечности
пасюка:
1 — большая поясничная. 2 — под-
вздошная, 3 — большая приводящая.
4 — длинная приводящая. 6 — предпо-
луперепончатая. 6 — полуперепончатая
передняя. 7 — полуперепончатая зад*
няя. 3 — полусухожильная, 9 — гре-
бешковая. 10 — стройная. 11 — пря-
мая бедра. 12 — медиальная головка
четырехглавой бедра, 13 — напряга-
тель широкой фасции бецра. 14 — вну-
тренняя запирательная
1 — поверхностная ягодичная, 2 — напря-
гатель широкой фасции бедра. 3. 4 — пе-
редняя и задняя части двуглавой бедра»
б — полусухожильная, в — короткая при-
водящая. 7 — полуперепончатая задняя,
8 — широкая фасция бедра
вается на большом вертлуге бедренной кости вместе со средней
ягодичной мышцей .’(рис. 63—65).
3 а д и е бедр енн а я мускулатура. К числу заднебедрен-
ных относится 6 мышц.
Полусухожильная мышца (m. semitendinosus) начинается дву-
мя головками — от остистых отростков последнего крестцового и
первого хвостового позвонков и от седалищного бугра тазовой кос-
ти (рис. 64). Оканчивается полусухожильная мышца на гребне
большеберцовой кости (рис. 61, 65). Сгибает коленный сустав и раз-
гибает тазобедренный сустав.
Двуглавая мышца бедра (m. biceps femoris) начинается двумя
головками — от остистого отростка последнего крестцового позвон-
ка и от седалищного бугра, несколько краниальнее отхождения го-
116
ловки полусухожильной мышцы (рис. 64). Оканчивается мышца
двумя частями, передняя из которых прикрепляется к проксималь-
ной части большеберцовой кости, а задняя — к латеральной по-
верхности гребня большеберцовой кости (рис. 61, 65). Обычно ча-
сти двуглавой мышцы описывают «сак самостоятельные мышцы
(двуглавая передняя и двуглавая задняя). Сгибает коленный сустав
Рис. 62. Мускулатура латеральной
i тороны задней конечности пасю-
ка (средний слой):
I ягодичная средняя, 2 — ягодич-
ная поверхностная, 3 — предполупере-
шшчатая. 4 — полуперепокчатая пе-
Р дняя, 6 — квадратная бедра, 6 —
Гпныиая приводящая. 7 — латераль-
144*4 головка четырехглавой бедра,
" икроножная. 9 — ахиллово су-
• к|ыне, 10 — пяточная, 11 — длин-
ный разгибатель пальцев, 12 — боль-
нич>рцовая передняя. 13 — косая
1 • 'ii'Mi заплюсны. 14 — большой вер-
тлуг
Рис. 63. Мускулатура латеральной стороны
задней конечности пасюка (внутренний
слой):
1 — двойниковая передняя, 2 — двойниковая зад-
няя, 3 — внутренняя запирательная. 4 — квадрат-
ная бедраF 5 — малая ягодичная, в — подвздош-
ная, 7 — прямая бедренная, 8—срединная голов-
ка четырехглавой бедра, 9 короткая приводя-
щая, 10 — большая приводящая. 11 — прецюлу-
перспончатая
и р.В1ибает тазобедренный сустав; отчасти способствует отведению
Полуперепончатая мышца (m. semimembranosus) у, крысы со-
। 1>‘И1 hi ipcx вполне самостоятельных частей (рис. 62, 63). Кра-
I'lui ii.ii.iq, или позвоночная, часть — предполуперепончатая мышца
Ни pi iicM'iminernbranosus) начинается от остистого отростка послед-
.... К|»'сти<)||| ю позвонка и оканчивается на сесамовидной косш
'ь цы и ною |цдмыщелка бедренной кости (рис. 64, 65). Ра«гибас1
< ><».г к нный сустав. Передняя полуперепончатая мыший (ш пн
• и, hiIii ii'ii 'ii'' .interior) начинается от проксимальной) b<>niu • bi-
ll;
лищной кости и прикрепляется к медиальному надмыщелку боль-
шеберцовой кости (рис. 64, 66). Сгибает коленный и разгибает та-
зобедренный сустав. Задняя полуперепончатая мышца (m. semi-
membranosus posterior) начинается от симфиза тазовых костей и
Рис. 64. Места прикрепления мышц к тазовому поясу пасюка.
А — вид сбоку, Б — вид сверху, В — вид снизу:
1 — подвздошная, 2 — портняжная, 3 — напрягатель широкой фас-
ции бедра, 4 — ягодичная поверхностная, 5 — ягодичная средняя,
6’—ягодичная малая, 7 — грушевидная, 8 — полусухожильная. 9 —
двуглавая бедра, 10 — предполуперепончатая, 11 — полуперепончатая
передняя, 12 — полуперепончатая задняя, 13 — стройная. 14 — гребеш-
ковая. 16 — длинная приводящая, 16 — большая приводящая, 17 —
короткая приводящая, 18 —- квадратная бедра, 19 — запирательная
наружная, 20 — передняя двойниковая, 21 — задняя двойниковая,
22—прямая бедра, 23 — косые живота, 24 — прямая живота, 25 —
запирательная внутренняя
оканчивается на гребне большеберцовой кости (рис. 64, 66). Сги-
бает коленный сустав. На основании иннервации Грин называет
эцу мышцу задней стройной. Против этого говорит тот факт, что у
некоторых грызунов (белки) развиты одновременно как задняя
полуперепончатая, так и задняя стройная мышцы.
Стройная мышца (m. gracilis). В отличие от многих других гры-
зунов у крысы имеется только одна стройная мышца, отходящая от
симфиза безымянных костей и оканчивающаяся на медиальной
стороне гребня большеберцовой кости (рис. 60, 65, 66). Сгибает
коленный сустав, разгибает тазобедренный сустав.
Приводящая мускул ат ура бедра. К числу приводя-
щих бедра относятся четыре мышцы.
Гребешковая мышца (m. pectineus) начинается па бугре лоб-
ковой кости и оканчивается на гребне малого вертлуга бедренной
кости (рис. 60, 64, 65). Приводит и вращает бедренную кость.
118
Длинная приводящая мышца (m. adductor longus) начинается
от лонной кости и от симфиза безымянных костей, непосредственно
каудальнее гребешковой мышцы, и оканчивается на гребне малого
вертлуга бедренной кости (рис. 60, 64, 65). Приводит бедро и спо-
собствует разгибанию тазобедренного сустава. Большая приводя-
maximus) начинается от симфиза тазо-
щая мышца (in. aaauci
вых костей и от лон-
ной кости и оканчи-
вается на плантарной
стороне бедренной ко-
сти, непосредственно
дистальнее гребня боль-
шого вертлуга (рис. 60,
65). Дистально волокна
мышцы достигают обо-
их надмыщелков бед-
ренной кости, а с ме-
диальной стороны один
из пучков мышцы до-
стигает большеберцо-
вой кости (рис. 66).
Разгибает тазобедрен-
ный сустав, приводит
бедренную кость, сги-
бает коленный сустав.
Короткая приводя-
щая мышца (m. adduc-
tor brevis) начинается
от седалищной кости и
оканчивается на план-
1лрной поверхности
||'1'бня большого верт-
iyia бедренной кости
Рнс. 65. Места прикрепления мышц к бедренво*
кости пасюка. А — вид с медиальной стороны,
Б — вид с дорзальной стороны, В — вид с план-
тарной стороны, Г —вид с латеральной стороны:
1 — поясничная большая и подвздошная. 2 — ягодичная
поверхностная. 3 — ягодичная средняя и малая. 4 —
предполуперепончатая, Б — гребешковая. 6 — длинная
приводящая. 7 — большая приводящая. 8 — короткая
приводящая. 9 — квадратная бедра, 10 — наружная за*
пирательная. 11 — внутренняя запирательная. 12 — двой-
никовая передняя. 13 — двойниковая задняя. 14, 15. 16—
медиальная, срединная и латеральная головки четырех*
главой бедра. 17. 18 — медиальная и латеральная го*
ловки трехглавой голени, 19 — подошвенная, 20 — длин*
ный разгибатель пальцев, 21 — подколенная
(рис. 63—65). Разгибает тазобедренный сустав, вращает и, отчасти,
пр иводит бедренную кость.
Глубокие тазобедренные мышцы. К числу глубоких
। । обе.тренных относится 5 мышц.
Квадратная мышца бедра (m. quadratus femoris) начинается от
1.1.1НШНОЙ кости, непосредственно краниальнее передней полупе-
||' ||()11Ч.п<'й мышцы, и оканчивается на малом вертлуге бедренной
tn и по плантарному соединению малого вертлуга с большим
|«*|>1лу|ом бедренной кости (рис. 63—65). Разгибает тазобедренный
' ’• hiii
I Ги|п жнчя запирательная мышца (m. obturator externus) начи*
.... м си окружности запирательного отверстия и на самой мем-
|||>лн< чширшсльного отверстия и оканчивается в ямке большого
г» ши бедренной кости (рис. 64, 65). Отводит бедро и пиблет
'•| •(" >шый сустав.
II'»
Внутренняя запирательная мышца (m. obturator internus) начи-
нается от медиальной поверхности седалищной кости и оканчивает-
ся в ямке большого вертлуга бедренной кости (рис. 60, 63—65).
Отводит бедренную кость и сгибает тазобедренный сустав.
Двойниковые мышцы (m. m. gemelli) состоят из двух частей
(рис. 63). Краниальная начинается от седалищной и подвздошной
я баг
Ряс. 66. Места прикрепления мышц к костям голени
пасюка. А — вид с латеральной стороны, Б — вид с дор-
зальной стороны, В — вид с плантарной стороны.
Г —вид с медиальной стороны:
1 — полусухожильная, 2, 3 — передняя н задняя двуглавая бед*
ра, 4 — передняя полуперепончатая, S — задняя полуперепонча-
тая, в — стройная, 7 — камбаловидная, 8 — подколенная, 9 —
передняя большеберцовая, 10 — разгибатель большого пальца,
И — длинная малоберцовая, 12 — короткая малоберцовая, 13 —
малоберцовая IV пальца, 14 — малоберцовая V пальца, 1Б —
задняя большеберцовая, 18 — длинный сгибатель пальцев, 17 —
длинный сгибатель большого пальца
костей, непосредственно, каудальнее малой ягодичной мышцы, и
оканчивается в ямке большого вертлуга бедренной кости (рис. 64,
65). Она отводит бедренную кость и способствует сгибанию тазо-
бедренного сустава. Каудальная часть начинается от седалищной
кости, непосредственно каудальнее краниальной части и сухожилия
внутренней запирательной мышцы, и оканчивается в ямке большого
вертлуга бедренной кости (рис. 64, 65). Она сгибает тазобедрен-
ный сустав.
Мускулатура коленного сустава. К мускулатуре коленного су-
става относится 5 мышц.
120
Четйрехглавая мышца бедра (m. quadriceps femoris) имеет че-
тыре головки (рис. 60, 62). Одна из них, начинающаяся на бугре
тазовой кости (рис. 64), называется прямой мышцей бедра (m. rec-
tus femoris). Остальные головки отходят от бедренной кости и соот-
ветственно своему отхождению именуются латеральная, ме-
диальная п срединная головки (с. с. laterale, mediale et interme-
dium). Латеральная головка начинается от края и гребня большого
вертлуга и дистальнее — от латеральной губы бедренной кости, ме-
диальная головка — от дорзальной поверхности малого вертлуга и
от медиальной губы бедренной кости, срединная головка — от
дорзальной поверхности тела бедренной кости, начиная от ее шей-
ки и почти до дистального эпифиза (рис. 65). Все четыре части
четырехглавой мышцы оканчиваются на коленной чашечке и явля-
ются мощным разгибателем коленного оустава.
Подколенная мышца (m. popliteus) начинается от латерального
надмыщелка бедренной кости и оканчивается на медиальном крае
проксимального конца большеберцовой кости (рис. 65—67). Сги-
бает коленный сустав и, отчасти, вращает голень.
На движения коленного сустава влияют также четырехглавая
мышца голени и длинный разгибатель пальцев.
Скакательный комплекс. В эту группу входят трехглавая голени
и подошвенная мышца.
Трехглавая мышца голени (m. triceps surae) состоит из трех го-
товок (рис. 67). Латеральная головка (m. gastrocnemius lateralis)
начинается от сесамовидной кости латерального надмыщелка бед-
ренной кости, медиальная головка (m. gastrocnemius medialis) —
от сесамовидной кости медиального надмыщелка бедренной кости,
а третья головка, называемая пяточной мышцей (m. soleus) — от
головки малоберцовой кости (рис. 65, 66). Все части трехглавой
мышцы переходят в общее сухожилие, называемое ахилловым, кото-
рое прикрепляется к бугру пяточной кости (рис. 69). Трехглавая
мышца голени является самым мощным разгибателем голеностоп-
ного сустава.
Подошвенная мышца (m. plantaris) берет начало от сесамовид-
ной косточки латерального надмыщелка бедренной кости, несколь-
ко медиальнее латеральной головки трехглавой бедра, и оканчивает-
l я на пяточном бугре у места отхождения короткого сгибателя
пальцев (рис. 65, 67, 69). Сгибает голеностопный сустав.
Мускулатура стопы. Разделяется на длинную и короткую.
Члннная мускулатура стопы. В эту группу вхо-
1м । 10 мышц, которые в свою очередь разделяются на разгибате-
in и < шпатели пальцев. К числу разгибателей принадлежит
’11411111-
II'pt щяя большеберцовая мышца (m. tibialis anterior) начи-
'>’• I-ч tn лаифо-проксимального края гребня большеберцовой кости
о • ‘к ин'.кия на латеральном крае дистального конца первой плюс-
...... • in (рис 66, 67, 69). Сгибает и отчасти приводит стону.
Г?1
Длинный разгибатель пальцев (m. extensor digitorum longue)
начинается на латеральном надмыщелке бедренной кости и, прохо-
дя под косой связкой голени, разделяется на три сухожилия
(рис. 65, 67); латеральное сухожилие вскоре в свою очередь раз-
деляется на два. Сухожилия длинного разгибателя пальцев окан-
Рис. 67. Длинная мускулатура стопы. А — поверхностная му-
скулатура дорзальной стороны, Б — глубокая мускулатура дор-
зальной стороны, В — поверхностная мускулатура плантарной
стороны, Г — глубокая мускулатура плантарной стороны:
1—длинный разгибатель пальцев, 2 — малоберцовые, 3 — подколенная,
4 — задняя большеберцовая, Б — длинный сгибатель пальцев, 6 — длин-
ный разгибатель большого пальца, 7 — малоберцовая четвертого паль-
ца, 8 — малоберцовая пятого пальца. 9 — малоберцовая длинная, 10 —
подошвенная. 11 — латеральная головка икроножной. 12 — медиальная
головка икроножной, 13 — передняя большеберцовая, 14 — камбаловид-
ная, 15 — поперечная связка заплюсны, 16 — короткая малоберцовая
чиваются на суставных сумках плюсно-пальцевых суставов II—V
пальцев, отдавая веточки к ногтевым фалангам II—IV пальцев.
Сухожилия II и III пальцев отдают также веточки к суставным
сушам плюсно-пальцевых суставов III и IV пальцев соответствен-
но (рис. 69). Мышца разгибает пальцы задней конечности и сгибает
голеностопный сустав.
Разгибатель большого пальца (m. extensor pollicis longus) на-
чинается от проксимального конца латеральной поверхности мало-
берцовой кости и оканчивается на дорзальной стороне когтевой фа-
ланги I пальца (рис. 66, 67, 69). Разгибает большой палец.
122
Длинная малоберцовая мышца (m. peronaeus longus) начинает-
ся от дистального края головки малоберцовой кости и оканчивает-
ся на проксимальном конце четвертой плюсневой кости, с ее план-
тарной стороны (рис. 66, 67, 69). Мышца сгибает голеностопный су-
став и отводит стопу. " ----- ~
Короткая малоберцовая мышца (гл. peronaeus brevis) начинает-
ся от внутреннего края малоберцовой кости и оканчивается на пя-
той плюсневой кости (рис. 66, 67, 69). Сгибает голеностопный су-
став.
Малоберцовая мышца V пальца (m. peronaeus digiti quinti) на-
чинается на латеральной стороне головки малоберцовой кости и
оканчивается на латеро-дорзальном крае ногтевой фаланги V паль-
ца (рис. 66, 67, 69). Разгибает V палец.
Малоберцовая мышца IV пальца (m. peronaeus digiti quarti) на-
чинается на каудальном крае малоберцовой кости и оканчивается
па латеро-дорзальной стороне когтевой фаланги IV пальца (рис. 66,
67, 69). Разгибает IV палец.
К числу сгибателей пальцев принадлежит 3 мышцы.
Длинный сгибатель пальцев (m. flexor digitorum longus) начи-
нается от плантарной поверхности большеберцовой кости и от ла-
теральной поверхн!ости малоберцовой кости и оканчивается на ког-
тевых фалангах I—V пальцев (рис. 66, 67, 69). Сгибает пальцы и
влияет на разгибание голеностопного сустава.___
Задняя большеберцовая мышца (m. tibialis posterior) начинает-
ся от плантарной стороны большеберцовой кости, охватывая с обе-
их сторон длинный сгибатель пальцев и подколенную мышцу (рис.
66, 67). Оканчивается задняя большеберцовая мышца у прокси-
мального конца первой предплюсневой кости (рис. 69). Разгибает
голеностопный сустав, являясь антагонистом передней большебер-
цовой мышцы.
Дополнительная задняя большеберцовая мышца (m. tibialis pos-
terior accessorius) у крысы развита слабо (рис. 67). Она начинает-
ся непосредственно под подколенной мышцей и оканчивается на
плантарной стороне первой плюсневой кости (рис. 66, 69). Сгибает
большой палец и влияет на разгибание голеностопного сустава.
У человека длинный сгибатель пальцев расположен медиально
от задней большеберцовой мышцы, а латерально от последней про-
ходит длинный сгибатель большого пальца. У крысы на месте по-
следнего развит общий сгибатель пальцев (часто смешиваемый из-
'.I <то положения с длинным сгибателем большого пальца), а на
м"гт<’ длинного сгибателя пальцев — мышца, прикрепляющаяся к
••ормсэй плюсневой кости. Мы называем ее дополнительной больше-
'нрцовой.
Короткая мускулатура стопы. Разделяется на дор-
< 11Ы1ую (разгибатели пальцев) и плантарную (сгибатели пальцев).
I top члыюй относится лишь одна мышца.
Короткий разгибатель пальцев (m. extensor digitorum) отходит
' литеральных костей предплюсны (III и IV + V). Имеет два
123
небольших брюшка, сухожилия которых проходят по латеральной:
стороне II и III пальцев (рис. 68). Оба сухожилия кончаются на
суставных сумках плюсно-пальцевых суставов, но отдают веточки к
суставным оумкам первых межфаланговых суставов (рис. 69). Раз-
гибает второй и третий пальцы.
К вентральной короткой мускулатуре стопы принадлежат сле-
дующие мышцы.
Рис. 68. Короткая мускулатура стопы. А — мускулатура дорзальной
стороны, Б — поверхностная мускулатура плантарной стороны, В —
средний слой мускулатуры плантарной стороны, Г — глубокая муску-
латура плантарной стороны:
1 — сухожилие длинного сгибателя пальцев, 2 — короткий сгибатель* пальцев.
3 — квадратная стопы. 4 — червеобразные, 5 — дорзальные сухожилия межко-
стных, 6 — межкостные, 7 — короткий разгибатель пальцев
Короткий сгибатель пальцев (m. flexor digitorum brevis) начи-
нается на пяточном бугре пяточной кости и вскоре разделяется на
четыре брюшка, каждое из которых оканчивается сухожилием, иду-
щим к четырем латеральным пальцам (рис. 68, 69). Сухожилие к
V пальцу обычно очень слабо и нередко вовсе отсутствует. На уров-
не плюсно-фаланговых суставов сухожилия раздваиваются и тянут-
ся до первых межфаланговых суставов, 'Прикрепляясь к основанию
вторых фаланг II—V пальцев. Сгибает четыре латеральных пальца.
Квадратная мышца стопы (m. quadratus plantae) начинается от
пяточного бугра пяточной кости и оканчивается на основании пя-
той плюсневой кости (рис. 68, 69).
124
Червеобразные мышцы (m.m. lumbricales) начинаются на сухо*
жильной пластинке длинного сгибателя пальцев и имеют четыре
брюшка, из которых три начинаются с плантарной стороны сухо-
жильной пластинки, а четвертое — с его дорзальной стороны
(рис. 68). Первые 3 брюшка оканчиваются на медиальной стороне
II—IV пальцев, а последнее -
на медиальной стороне V паль-
ца. Сухожилия червеобразных
мышц до некоторой степени пе-
реходят на дорзальную сторону
пальцев (рис. 68). Червеобраз-
ные мышцы сгибают пальцы. 6
Межкостные мышцы (т. т.
interossei) развиты в числе че- g
тырех для каждого из пальцев.
Два пучка оканчиваются на се-
самовидных костях плюсно-фа-
ланговых суставов, а два —
переходят с плантарной сторо-
ны на дорзальную и прикреп-
ляются по сторонам дорзаль-
ной поверхности когтевых фа-
ланг. Отходят межкостные
мышцы от костей плюсны. (рис.
68). Сгибают и разгибают
пальцы.
Описываемые Грин специ-
альные мышцы большого паль-
ца и мизинца ничем не отлича-
ются от соседних межкостных
мышц и нами не выделяются.
МУСКУЛАТУРА ГОЛОВЫ
В этом разделе мы описыва-
ем .тишь челюстную мускулату-
ру Подкожная лицевая муску-
тагура рассматривается в раз-
има посвященных органам
Рис. 69. Места прикрепления мышц на
стопе. А — дорзальная сторона, Б —
плантарная сторона:
1 — четырехглавая мышца голени, 2 — пе-
редняя большеберцовая, 3 —• разгибатель
большого пальца, 4 — длинная малоберцовая,
5 — короткая малоберцовая. 6 ~ малоберцо-
вая пятого пальца, 7 — малоберцовая четвер-
того пальца, 8 — длинный разгибатель паль-
цев, 9 — задняя большеберцовая, 10 — корот-
кий сгибатель пальцев, 11 — длинный сгиба-
тель пальцев, 12 —длинный сгибатель боль-
шого пальца, 13 — квадратная стопы
пищеварения, слуха и зрения.
.Латеральная, или наружная, жевательная мышца (m. masseter
l.ih i-ill ) разделяется на 3 части (рис. 70). Передняя часть (р. ап-
hiloi) начинается от специальной шероховатости на вентральной
• i ороне верхнечелюстной кости и оканчивается на конце углового
"ipo’ iba нижней челюсти (рис. 50). Задняя часть (р. posterior)
н ruin-к гея от вентральной поверхности скуловой дуги и оканчивает-
' по ./>' |'-1 тьному краю кижней челюсти (рис. 50). Глубокая
। " и. ср profundus) качинается от предокуловой площадки вср\не-
....... . кости и оканчивается по шероховатой линии на г< мс пиж-
Рис. 70. Жевательная мускулатура
пасюка. А — поверхностный слой,
Б — средний слой, В — глубокий
слой:
1, 2, 3 — передняя, задняя п глубокая ча-
сти латеральной жевательной, 4, 5 — пе-
редняя и задняя части медиальной жева-
тельной, 6 — височная
ней челюсти (рис. 50). Латеральная жевательная мышца оттягива-
ет нижнюю челюсть вперед и вверх.
Медиальная, или внутренняя, жевательная мышца (m. masseter
media'lis) разделяется на 2 части (рис. 70). Передняя часть
(р. anterior) начинается от специальной шероховатости верхнече-
люстной кости, проходит сквозь
подглазничное отверстие и окан-
чивается на специальном бугре
нижней челюсти (рис. 50). Зад-
няя часть (р. posterior) начинает-
ся на вентральной поверхности
скуловой дуги и оканчивается на
аборальной половине нижней че-
люсти (рис. 50). Передняя порция-
оттягивает нижнюю челюсть впе-
ред и вверх, а задняя — вверх и
слегка наружу.
Височная мышца (m. tempora-
lis) начинается от затылочного
гребня теменной и височной ко-
стей и от теменной, височной и
лобной костей и оканчивается на
крыловидном отростке нижней че-
люсти (рис. 50). Приводит ниж-
нюю челюсть, закрывая рот.
Крыловидная мышца (m. pte-
rigoideus) разделяется на 2 час-
ти. Латеральная крыловидная
мышца (m. pterigoideus lateralis)
начинается or височного крыла
клиновидной кости и оканчивает-
ся по вентральному краю углово-
го отростка нижней челюсти (рис.
50). Медиальная крыловидная
мышца (m. pterigoideus media-
lis) начинается от клиновидной
кости, непосредственно медиально
от крыловидной кости, и оканчи-
вается на угловом отростке ниж-
ней челюсти, непосредственно дор-
зальнее предыдущей мышцы (рис.
50). Оттягивает нижнюю челюсть вперед, вбок и вверх.
Двубрюшная мышца (m. digastricus) начинается па яремном
отростке затылочной кости и оканчивается на нижней челюсти у
симфиза правой и левой ее ветвей (рис. 49, 50). Открывает рот.
Поперечная мышца нижней челюсти (m. transversus mandibu-
lae) расположена между двумя ветвями нижней челюсти (рис. 49).
Сближает ветви нижней челюсти и уплощает дно рта.
126
МУСКУЛАТУРА ТУЛОВИЩА
Мускулатура туловища разделяется на мускулатуру грудной
клетки, брюшную мускулатуру, спинную мускулатуру, мускулатуру
шеи, мускулатуру хвоста и диафрагму.
Мускулатура грудной клетки. В эту группу входит 6 мышц.
Дорзальная зубчатая мышца (m. serratus dorsalis) разделяется
на две части (рис. 71). Передняя часть (р. anterior) при помощи
Рис. 71. Мускулатура туловища пасюка. А — поверх-
ностный слой, Б — глубокий слой:
1, 2 — краниальная и каудальная части зубчатой дорзальной,
3 — пластыревидная, 4 — длинная шейная, 5 — лестничная,
6 — плечеподъязычная, 7 — грудино-подъязычная. 8 — внутренняя
межреберная, 9 — наружная межреберная, 10 — полуостистая го-
ловы, 11 — длиннейшая спины, 12 — подвздошнореберная, 13 —
длиннейшая головы, 14 — длинная головы
апоневроза отходит от выйной связки и остистых отростков II шей-
ного — VI грудного позвонков и оканчивается отдельными зубцами
на П IX ребрах; иногда пучки ко II и IX ребрам не развиты. От-
тягивает ребра в сторону, увеличивая объем грудной полости и спо-
собствуя вдоху. Задняя часть (р. posterior) начинается от остистых
отростков последних двух грудных и первых трех поясничных поз-
вонков и оканчивается отдельными зубцами на пяти задних ребрах.
Наиболее сильный зубец прикрепляется к последнему ребру. Оття-
гивает и приводит ребра, уменьшая объем грудной клетки и способ-
11 вуя выдоху.
1.7
Рис. 72. Дорзальная
мускулатура шеи па-
сюка:
1 — полуостистая шеи,
2 — каудальная косая го-
ловы, 8 — краниальная
косая головы, 4 — боль-
шая дорзальная прямая
головы. 5 — малая дор-
зальная прямая головы,
6 — латеральная прямая
головы
Лестничная мышца (m. scalenus) отходит зубцами от 6 перед-
них ребер и идет к поперечнореберным отросткам шейных позвон-
ков (рис. 71). Поворачивает шею и расширяет трудную клетку.
Короткие подниматели ребер (m.m. levatores costaruni) идут от
сосцевидных отростков грудных позвонков к дорзальной части кра-
ниального края соответствующих ребер. Поднимают ребра.
Межреберные наружные мышцы (m. т. in-
tercostales externi) являются непосредственным
вентральным продолжением коротких подни-
мателей ребер. Пучки их, проходящие спере-
ди — назад и вниз, протягиваются между
смежными ребрами (рис. 71).
Межреберные внутренние мышцы (т. т.
intercostales interni) проходят между смежны-
ми ребрами в направлении спереди — назад и
вверх (рис. 71).
Поперечная грудная мышца (т. transversus
thoracis) представляет собой очень слабую пла-
стинку, проходящую между внутренней поверх-
ностью грудной кости и хрящами истинных ре-
бер.
Брюшная мускулатура. В эту группу входят
4 мышцы.
Косая наружная мышца живота (m. obli-
quus abdominis externus) начинается от восьми
последних ребер и при помощи апоневроза —
от поясничной фасции. Оканчивается двумя
порциями (рис. 49) — на средней линии жи-
вота и на паховой пластинке (ligamentum in-
guinale s. Pouparti), которая тянется от мокло-
ка до лонного бугра.
Косая внутренняя мышца живота (m. obli-
quus abdominis internus) начинается от пахо-
вой пластинки и поясничной фасции и оканчи-
вается на внутренней поверхности фасции пря-
мой брюшной мышцы и на внутренней поверх-
ности последних трех-четырех ребер (рис. 49).
Поперечная мышца живота (m. transversus abdominis) проходит
под косыми мышцами от внутренней фасции поясницы к фасции
прямой мышцы живота.
Прямая мышца живота (m. rectus abdominis) начинается от сим-
физа безымянных костей и от лонного бугра и оканчивается на
первом реберном хряще (рис. 49).
Брюшные мышцы сжимают брюшную полость и, оттягивая
вниз ребра и уплощая диафрагму, способствуют выдоху.
Спинная мускулатура. К спинной мускулатуре относится 7 мышц.
Все они, за исключением подвздошно-реберной и квадратной пояс-
128
ничной, разгибают спину. При одностороннем действии они изгибают
спину в соответствующую сторону.
Подвздошно-реберная мышца (m. iliocostalis) отходит от под-
вздошной кости и остистого отростка первого крестцового позвонка
и прикрепляется к трем задним ребрам; наиболее мощный пучок
прикрепляется к последнему ребру. Начиная от третьего ребра, под-
вздошно-реберная мышца снова постепенно усиливается, перебра-
сывается зубцами через одно-два ребра и прикрепляется к впереди
лежащим ребрам и поперечнореберным отросткам последних трех-
четырех шейных позвонков (рис. 71). Поясничная часть мышцы спо-
собствует выдоху, а спинная — вдоху.
Длиннейшая мышца спины (m. longissimus dorsi) отходит от
подвздошной кости и остистых отростков двух передних крестцо-
вых, поясничных и двух задних грудных позвонков. Мышца разде-
ляется на медиальные пучки, оканчивающиеся на сосцевидных и
поперечных отростках грудных позвонков, и на латеральные пучки,
оканчивающиеся на реберных бугорках и на поперечнореберных
отростках четырех-пяти задних шейных позвонков (рис. 71). Явля-
ется наиболее мощным разгибателем спины. Краниально мышца не-
сколько ослабевает.
Остистая и полуостистая мышцы спины (m. m. spinalis et semi-
spinalis dorsi) начинаются от остистых отростков грудных и первых
поясничных позвонков и оканчиваются на остистых отростках впе-
реди лежащих позвонков (кончая вторым грудным). Каудально
эти мышцы почти полностью вытеснены длиннейшей мышцей спины.
Многораздельная мышца (m. multifidus) начинается на сосце-
видных или на поперечных отростках поясничных, грудных и шей-
ных позвонков и оканчивается на остистых отростках впереди ле-
жащих позвонков, зачастую перебрасываясь через один-два сег-
мента.
Межпоперечные мышцы (m. intertransversarii) располагаются
между поперечнореберными отростками поясничных и шейных по-
звонков. В спинных позвонках они почти отсутствуют.
Межостистые мышцы (m. interspinales) располагаются между
остистыми отростками смежных позвонков, начиная от второго
I рудного впереди и кончая поясничными позади.
Квадратная мышца поясницы (m. quadratus lumborum) распола-
гается с вентральной стороны поясничных позвонков (рис. 59). На-
чинается от поперечнореберных отростков поясничных позвонков и
вентральной поверхности крыла крестцовой кости и оканчивается
на телах поясничных и трех задних грудных позвонков. Сгибает
позвоночный столб.
Мускулатура шеи. В эту группу входит 12 мышц.
Пластыревидная мышца (т. splenius) начинается вдоль лате-
ральной стороны треугольной пластинки второго грудного позвонка
ч краниально — по срединной линии вплоть до эпистрофея. Окан
•ишпется на затылочном гребне, от теменных костей вплоть до конца
1 "I невидного отростка (рис. 50, 71). Поднимает голову.
' ... 129
Длиннейшая мышца головы (m. longissimus capitis) начинается
от поперечных отростков первых шести-семи грудных позвонков и
последних четырех, реже пяти шейных позвонков. Оканчивается на
сосцевидном отростке, непосредственно глубже прикрепления пла-
стыревидной мышцы (рис. 50, 71). Поднимает голову. При одно-
стороннем действии поворачивает голову в сторону.
Длинная мышца головы (m. longus capitis) начинается от попе-
речнореберных отростков шейных позвонков и оканчивается на
основании клиновидной кости (рис. 50, 71). Опускает голову.
Прямая вентральная мышца головы (m. rectus capitis ventralis)
начинается от вентральной дуги атланта и оканчивается на заты-
лочной кости, непосредственно каудальнее длинной мышцы головы
(рис. 50). Опускает голову.
Полуостистая мышца шеи (m. semispinalis cervicis) начинается
от поперечных отростков пяти передних грудных и трех задних шей-
ных позвонков и оканчивается на остистом отростке эпистрофея
(рис. 72). Разгибает шею.
Полуостистая мышца головы (m. semispinalis capitis) начинает-
ся на поперечных отростках шейных и передних грудных позвонков,
и оканчивается на затылочном гребне, глубже и каудальнее пла-
стыревидной мышцы (рис. 50, 71). Поднимает голову.
Длинная шейная мышца (m. longus cervicis) начинается от вен-
тральной поверхности поперечных отростков последних грудных и
нередких шейных позвонков и оканчивается на вентральных гребнях
шейных позвонков (рис. 71). Сгибает шею.
Каудальная косая мышца головы (m. obliquus capitis caudalis)
начинается от гребня эпистрофея и оканчивается на крыле атланта
(рис. 72).
Краниальная косая мышца головы (m. obliquus capitis cranialis)
начинается от крыла атланта и оканчивается на затылочном гребне
(рис. 50, 72). Эти две мышцы обслуживают повороты головы в сто-
рону.
Большая дорзальная прямая мышца головы (m. rectus capitis
dorsalis major) начинается на гребне эпистрофея и оканчивается на
затылочной кости (рис. 50, 72). Поднимает голову.
Малая дорзальная прямая мышца головы (m. rectus capitis
dorsalis minor) начинается от дужки атланта и оканчивается на дор-
зальном крае большого затылочного отверстия (рис. 50, 72). Под-
нимает голову.
Латеральная прямая мышца головы (m. rectus capitis latera-
lis) начинается от крыла атланта и оканчивается на яремном от-
ростке затылочной кости (рис. 50, 72). Поворачивает голову в.
сторону.
Мускулатура хвоста. К мускулатуре хвоста относятся 6 мышц.
Хвостовая мышца (m. coccygeus) начинается от внутренней по-
верхности седалищной кости и оканчивается на поперечных отро-
стках пяти передних хвостовых позвонков. Отводит хвост в сторону
и опускает его.
130
Боковой опускатель хвоста (m. sacrococcygeus ventralis latera-
lis) отходит от вентральной поверхности крыла крестца, начиная
со второго крестцового позвонка, и от поперечнореберных отростков
последних двух-трех поясничных позвонков. Крепкие сухожильные
пучки мышцы оканчиваются на поперечнореберных отростках хво-
стовых позвонков. Опускает хвост.
Внутренний опускатель хвоста (m. sacrococcygeus ventralis те-
dialis) отходит от вентральной поверхности крыла крестца, несколь-
ко краниальнее предыдущей мышцы, и оканчивается на вентральной
поверхности хвостовых позвонков. Опускает хвост.
Боковой подниматель хвоста (m. sacrococcygeus dorsalis latera-
lis) отходит от гребня подвздошной кости и оканчивается на сос-
цевидных и остистых отростках хвостовых позвонков. Поднимает
хвост.
Внутренний подниматель хвоста (m. sacrococcygeus dorsalis
inedialis) начинается от дорзальной поверхности крыла крестцовой
кости и оканчивается на остистых отростках хвостовых позвонков.
Поднимает хвост. •
Межпоперечные мышцы хвоста (m. т. intertransversarii caudi)
представляют собой короткие пучки, располагающиеся между по-
перечными отростками хвостовых позвонков. Обслуживают боковые
движения хвоста
Диафрагма. Выделяется в особую группу, так как не может
быть причислена ни к одной из описанных выше.
Диафрагма (diaphragma) отделяет грудную полость от брюш-
ной. У крыс диафрагма почти целиком мышечная и прикрепляется
к хрящам 5—6-го задних ребер и к мечевидному отростку грудной
кости. Посередине диафрагму прободают отверстия для пищевода
и задней полой вены. Эти отверстия окружены сухожильной частью,
слабо развитой у крысы. От сухожильной части диафрагмы берет
начало столб диафрагмы, представляющий собой мышечный пу-
чок, идущий к вентральному гребню второго и третьего поясничного
юзвонков. У основания столба с его левой стороны проходит спин-
ная аорта.
В заключение приводим данные об иннервации и кровоснаб-
жении описанных мышц (табл. 12).
Таблица 12
Иннервация и кровоснабжение мышц пасюка
Название мышцы Иннервация Кровоснабжение
1рппецевидная головы Трапецевидная шеи h i пецевидная спины 1 1ч дино*ключично-сос- ‘ц)ная Добавочный, С И, III Добавочный, С II, III Добавочный, С II, III Добавочный, С III, IV Поперечная шеи Поверхностная шеи, затылочная Поверхностная подло* паточная Затылочная (шейный ствол)
I 1
Продолжение табл. 12
Название мышцы Иннервация Кровоснабжение
Широчайшая спины Ромбовидная Грудные Подключичная Зубчатая вентральная Атланто-лопаточная Дельтовидная Предостная Заостная Подлопаточная Круглая большая Круглая малая Клюво-плечевая Трехглавая плеча Напрягатель фасции предплечья Двуглавая Длинный подлопаточный, С VI, VII Дорзальный лопаточный, С V Краниальный грудной С VI (специальная ветвь) С III, дорзальный лопа- точный, грудные С III Подключичный Подлопаточный Подлопаточный Подлопаточный Подключичный Подмышечный Кожномускульный, луче- вой Лучевой Локтевой Мускульный (С VI— Подлопаточная Возвратная и поверхно- стная шейные Латеральная грудная, подключичная Подключичная Поперечная, поверхност- ная шейная, подлопа- точная Поверхностная шейная Подключичная, задняя окружная плеча, глу- бокая плечевая Поверхностная шейная, поперечно-лопаточная Поперечно-лопаточная Грудо-спинная, подлопа- точная, поперечно-ло- паточная, Подлопаточная Задняя окружная плеча Лучевая Окружные лопатки и плеча, плечевая Глубокая плечевая Лучевая
Плечевая Разгибатели кисти Кожномускульный (С V—VII), лучевой Лучевой Глубокая плечевая Поперечная локтя, меж- костные, срединно-лу- чевая
Сгибатели кисти Срединный, локтевой Локтевая, срединно-лу- чевая, локтевая, окольная
Поясничная большая и подвздошная L II, III Поясничная, подвздош- но-поясничная, окруж- ная бедра подвздошная, окружная
132
Продолжение табл. 12
Название мышцы Иннервация Кровоснабжение
Ягодичная группа Полусухожильная Двуглавая бедра Полусухожильная Полуперепончат^я пе- редняя > • ,z‘;i Полуперепончатая зад- няя Стройная Портняжная •-< ' <-? ‘ *-$• Отводящие Квадратная бедра Запирательная внутрен- . Двойниковые * ^тыр^сгулавая б^дра 11одколенн^я. Поверхностный, нижний и наружный ягодич- ные Большеберцовый L VII, S I—III Большеберцовый Большеберцовый Запирательный Запирательный Бедренный Запирательный Запирательный Запирательный Ветвь от крестцового сплетения Ветвь от крестцового сплетения Бедренный Большеберцовый Большеберцовый Подвздошно-пояснич- ная, поверхностная, глубокая и нижняя ягодичные, латераль- ные и медиальная окружные бедра Наружная срамная, под- коленная, медиальная окружная бедра Глубокая и поверхност- ная половые, подколен- ная, задняя семенни- ковая, глубокая яго- дичная, окружная бед- ра Глубокая ягодичная, бедренная, задняя се- менниковая, половые, медиальная окружная бедра Бедренная, медиальная окружная бедра Бедренная, срамная Бедренная Медиальная окружная бедра, окружная под- вздошная, лонная Бедренная, медиальная окружная бедра, поло- вая и семенниковая Медиальная окружная бедра, запирательная Медиальная окружная бедра, запирательная Глубокая ягодичная, запирательная Запирательная, поверх- ностная и глубокая ягодичные, латераль- ная окружная бедра Поверхностная половая, окружная подвздош- ная, окружная бедра, подколенная Коленная Задняр большеберцовая и коленная
Продолжение табл. 12
— - , Иннервация Кровоснабжение
Название мышцы
Разгибатели стопы Глубокий и поверхно- Передние большеберцо-
стный малоберцовые вая и малоберцовая
Сгибатели стопы Большеберцовый Задняя большеберцовая
Жевательная мускулату- ра Тройничный и лицевой Внутренняя и наруж- ная челюстные, на- ружная сонная
Мускулатура спины Дорзальные ветви спин- Дорзальные ветви пояс-
номозговых 1 ничной и грудной ар- । теряй
Мускулатура хвоста Ветви от крестцового сплетения I Запирательная. срам- ная, поверхностная 1 ягодрчная
Мускулатура шеи С II, С III и затылочный Реберно-шейная, сон- ная, позвоночная
Мускулатура живота Нижние межреберные, подвздошно-надчрев- ный, подвздошно-па- ховый Надчревные, окружная подвздошная, лате- ральная окружная । бедра, запирательная
Мускулатура грудной клетки Межреберные Межреберные
ТЕХНИЧЕСКИЕ СОВЕТЫ
Препаровку мышц на свежезабитых крысах производить затруд-
нительно, поэтому рекомендуется использовать тушки, фиксирован-
ные в 3—5 %-ном формалине или в 70%-ном спирте. Предпочти-
тельно пользоваться выдержанным в формалине материалом. За
сутки до начала работы формалинный материал помещают в воду.
Для определения мест прикрепления мышц их следует перерезать
поперек волокон и натягивать концы пинцетом. Полученные данные
следует сопоставлять с результатами изучения очищенного скелета,
который необходимо иметь под руками.
При фиксации следует надрезать кожу в области шеи, живота
и ягодиц и, поместив в таком виде крысу в фиксирующую
жидкость, выдерживать ее в ней около месяца. При необходимо-
сти можно использовать материал, выдержанный в фиксаторе в
течение 10—12 дней. Эту процедуру можно ускорить, если фикси-
ровать тушки со снятой кожей.
ГЛАВА VH
ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ОРГАНОВ
ДВИЖЕНИЯ
По функциям отдельных мышц очень трудно представить их
действительную роль при любом конкретном движении. В связи с
этим мы посвящаем функциональному анализу органов движения
крысы особую главу.
Все многообразие движений крысы почти не поддается учету.
Поэтому мы ограничиваемся разбором функции органов движения
лишь при поступательном передвижении прыжками, или галопе.
Выбор наш объясняется тем, что при галопе — наиболее быстром
способе передвижения крысы — функциональные особенности ее
скелета и мускулатуры выявляются особенно ярко. (В своем анализе
мы широко пользовались сравнением крысы с другими грызунами,
.что облегчает понимание функций отдельных органов.
ГАЛОП
Галоп, или передвижение прыжками, слагается из периодически
повторяющихся фаз отталкивания, свободного полета и приземления
(рис. 73). Наибольшая нагрузка на мышцы падает на начало оттал-
кивания (I—III); на следующих ‘этапах отталкивания (IV—VI),
которые можно называть стадией разгона, тело животного получает
скорость, необходимую для отрыва от земли. Длина прыжка зави-
сит от скорости, достигнутой в момент отрыва (VI), и угла вылета.
Чем более угол вылета приближается к 45°, тем дальше летит
животное, однако скорость движения с увеличением угла вылета
замедляется. Этим и объясняется то, что все быстроаллюрные жи-
вотные как бы «стелятся по земле» и передвигаются по очень по-
логой траектории. Если вычертить траектории полета для отдель-
ных точек тела животного, то можно увидеть, что все они в на-
чальной стадии полета имеют очень пологую траекторию (около
13°) и лишь кончики пальцев отрываются от земли под углом 45е
(рис. 74). Фаза свободного полета заканчивается падением крысы
на передние конечности, когда начинается фаза приземления (см.
рис. 73, X). В продолжение этой фазы все тело заносится вперед и
с момента, соприкосновения задних конечностей с землей вновь на-
чинается фаза отталкивания (рис. 73, XII).
Рис. 73. Киносъемка галопа пасюка (80 кад/сек):
1—VI «— фаза отталкивания, VII—IX — фаза свободного полета, X—XVI — фаза призем-
ления, ц — центр тяжести
Фаза отталкивания. Как было сказано выше, наибольшая на-
грузка на мышцы приходится «а начало фазы отталкивания. Пер-
вые три этапа отталкивания (рис, 73, I—III) являются подготови-
Рис. 74. Траектории движения отдельных точек задней конечности пасюка
во время прыжка. А — моклок, Б — тазобедренный сустав, В — кончик
стопы, Г — свободная траектория кончика стопы без влияния передней
опоры:
1“10 “ этапы полета в воздухе, а, Зг 7 — углы отрыва от земли отдельных точек
задней конечности
тельными; на их протяжении тазобедренный, коленный и голено-
стопный суставы сгибаются (табл. 13), спина выпрямляется, перед-
ние конечности выносятся вперед, плечевой сустав разгибается, а
локтевой — сгибается. Центр тяжести, располагавшийся вначале
сзади точки опоры задних конечностей, на третьем этапе выносится
136
вперед, и равновесие не нарушается только благодаря тому, что
тело крысы получает ускорение, направленное вперед и вверх и
противодействующее силе тяжести.
Таблица 13
Изменения величины углов в суставах задней
конечности при прыжке (в град.)
Этапы прыжка Суставы
голеностопный коленный тазобедренный
I 100 118 72
II 90 78 63
III 70 73 69
IV 72 76 83
V 80 94 109
VI 117 108 109
VII 107 107 108
VIII 107 32 92
IX 77 76 87
X 50 60 74
XI 53 46 60
XII 63 50 60
XIII 64 52 53
XIV 65 77 67
XV 83 100 68
XVI 98 120 72
Тело крысы получает ускорение за счет работы мышц задних:
конечностей. Принято считать, что при нижней опоре подвижными
точками для всех мышц являются их верхние прикрепления, однако
Рис. 75. Схема положения задних конечностей пасюка во
прыжка:
I—XVI — этапы прыжка; а—б — таз, в—г — бедро, г—д —
е—ж — стопа
время
голень,
это положение не всегда оправдывается. Прыжок совершается при
одновременной работе всёх мышц задней конечности. Для облег-
чения понимания их роли мы разделим мышцы задней конечности
на группы по сходству функций. Нужно отметить, что функции
мышц, объединяемых в отдельные группы, являются сходными лишь
4<а фазе отталкивания и могут быть различными на дру1их фазах
137
Таблица 14
Величина слагающих мыши задней конечности на фазе отталкивания
Название мышцы Величина угла прик- репления (в град.) Относи- тельный вес Рычаг Слагающая
прило- жения дей- ствия верти- кальная гори- зонталь- ная
Заднебедренная группа
Квадратная бедра 25 0,69 18 10 0,52 0,63
Приводящая длинная 56 0,41 5 10 0,17 0,23
Приводящая короткая 29 1,83 18 10 1,54 1,60
Приводящая большая 50 2,22 12 10 1,89 1,46
Полуперепончатая передняя 58 3,74 14 10 4,31 2,10
Полуперепончатая задняя 58 0,93 17 : 10 1,22 0/2
Предполуперепончатая 62 0,62 10 10 0,54 0,29
Полусухожильная 71 2,68 16 10 4,05 0,87
Стройная 86 0,57 1 10 0,05 0,04
Двуглавая передняя 65 1,34 • 9 : ю 1.08 0,58
Двуглавая задняя 76 6,04 12 10 6,96 1,64
Поверхностная ягодичная 60 1,58 4 10 0,54 0,80
Ягодичная группа
Ягодичная средняя 7 5,60 17 10 0,85 4,90
Ягодичная малая 5 1,01 10 10 0,41 1,00
Грушевидная 5 0,60’ 5 10 0,04 0,59
Фиксаторы тазобедрен-
ного сустава
Подвздошная ' 25 3,30
Поясничная большая 22 3,30
Сгибатели тазобедрен-
ного сустава
Прямая бедра 75 2,39 —3,1 10 -0,62 0,59
Гребешковая 78 0,81 —0,1 I 10 -0,07 0,08
Разгибатели спины 25 39.90 1
Разгибатели коленного
сустава
Четырехглавая латеральная 25 2,21 20 30 —2,00 0,62
Четырехглавая медиальная 35 1,94 30 30 -1,59 1,10
Четырехглавая промежуточная 15 0,91 15 30
Скакательный комплекс 1
Икроножная 86 5,06 6 30 0.2 1,00
Длинные стопы
Сгибатель пальцев длинный 1,39 0.2 — 1,34
Разгибатель пальцев длинный .г 0*49 -0,03 —0,36
Большеберцовая передняя 1,50 -0,49 —1,40
Малоберцовые 0,50 - 0,04 Г°’48
ч шжения. Так, прямая мышца бедра на фазе отталкивания функ-
ционально отличается от остальных частей четырехглавой мышцы,
1 во время свободного полета функционирует так же, как и осталь-
<ые головки. Чтобы сделать описание более наглядным, мы сопро-
вождаем его схемой, на которой нанесены равнодействующие основ-
ных мышц задней конечности (рис. 75).
Заднебедренная группа. В ее состав входят мышцы,
начинающиеся от седалищной и, отчасти, лонной костей и крестца и
оканчивающиеся на бедренной кости или на костях голени; все они
способствуют разгибанию тазобедренного сустава. Действие каж-
юй из этих мышц разлагается на две слагающие — вертикальную,
влияющую на разгибание тазобедренного сустава, и горизонталь-
ную, обслуживающую продвижение туловища вперед. Величина
обеих слагающих зависит от величины угла прикрепления, от точки
приложения и силы каждой из мышц (табл. 14).
Рычагом приложения для заднебедренной группы является рас-
стояние от центра тазобедренного сустава до начала мышцы, а ры-
чагом работы — расстояние от центра тазобедренного сустава до
центра соединения таза с крестцом. Все эти мышцы не только
разгибают тазобедренный сустав, но и имеют вредную для прыжка
сторону действия. Заднебедренные мышцы, оканчивающиеся на
бедре, способствуют оттягиванию дистальной части бедра назад, а
мышцы, оканчивающиеся на голени, — сгибанию коленного суста-
ва; обе эти функции вредны для прыжка. Величина «вредного дей-
ствия» прямо пропорциональна расстоянию от головки бедра до ме-
ста окончания мышцы на бедре и расстоянию от коленного сустава
до места окончания мышцы на голени; поэтому специализация к
прыжку должна сопровождаться усилением тех заднебедренных
мышц, которые прикрепляются на бедре и на голени более прокси-
мально (квадратная бедра и передние двуглавая и полуперепонча-
'ая). Данные, приведенные в табл. 15, в которой различные виды
грызунов расположены по степени их специализации к прыжкам,
подтверждают это положение.
Таблица 15
Изменения в относительной силе заднебедренных мышц в связи
с |фиспособлением к прыжкам
Вид животного Двуглавая .передняя Полупере- пончатая передняя Квадратная бедра Остальные мышцы заднебед- ренной группы
Слепец 0,39 1,18 0,43 7,25
Слепушонка 0,97 2,19 0,71 6,97
Хомяк малоазиатский 2,32 0,51 0,50 26,10
Крыса серая 1,33 3,74 0,69 15,18
Персидская белка 4,57 3,40 0,49 24,95
Горный тушканчик 22.76 12,29 1,50 12,75
139
Интересно, что iy хорошо прыгающих форм происходит также
перемещение проксимально окончания этих мышц. Если у крысы
отношение расстояния от коленного сустава до места прикрепления
передней двуглавой к длине голени равно 0,1, а такая же величина
для передней пол|уперепончатой — 0,26, то у тушканчика вторая
величина равна 0,1, а передняя двуглавая мышца оканчивается на
прямой связке колена и из сгибателя коленного сустава превра-
щается в его разгибатель.
Ягодичная группа. Из трех ягодичных мышц — поверх-
ностной, средней и глубокой — «а фазе отталкивания общей функ-
цией обладают только две последние, и поэтому поверхностную яго-
дичную мы относим здесь к другой группе. Мышцы ягодичной груп-
пы, так же как и заднебедренные, разгибают тазобедренный сустав,
ко их горизонтальная составляющая является отрицательной (на-
правлена назад).
Фиксаторы тазобедренного сустава. В эту группу
мы включаем две мышцы — подвздошную и большую поясничную.
Обе они прижимают головку бедренной кости к краниальной стенке
вертлужной ямки и немного тормозят выпрямление спины и тазо-
бедренного сустава. При прыжке растягиваются.
Сгибатели тазобедренного сустава. К этой группе
относятся прямая бедра и гребешковая мышца. Они препятствуют
чрезмерному разгибанию тазобедренного сустава, будучи в этом
отношении антагонистами заднебедренных мышц; такая их роль
возрастает с увеличением расстояния от места отхождения мышцы
до центра движения тазобедренного сустава. У животных, специа-
лизированных к прыжкам, происходит некоторое перемещение нача-
ла этих мышц в краниальном направлении (табл. 16).
Таблица 16
Перемещение прикрепления сгибателей тазобедренного сустава
в связи со специализацией к прыжкам
Вид животного Отношение длины рычага при- ложения к рычагу действия
прямая бедра | гребешковая .
Хомяк малоазиатский 0,157 0,05
Полчок 0,240 0,01
Крыса серая 0,314 0,01
Горный тушканчик 0,617 0,26
Разгибатели спины. Во время прыжка эти мышцы спо-
собствуют подниманию туловища. Трудность поднимания туловища
прямо пропорциональна скорости движения. При прыжках без па-
дения на передние конечности роль разгибателя спины чрезвычайно
усиливается, так как передние конечности совсем перестают играть
роль тормоза при падении туловища. Этим объясняется чрезвычай-
ное развитие разгибателей спины у тушканчиков (табл. 17).
140
Таблица 17
Относительное развитие разгибателей спины у грызунов
Вид животного Относительный вес разгибателей спины
Слепец Слепушонка Общественная полевка Персидская белка Серая крыса Горный тушканчик 14,5 17,9 21,2 31,0 39,9 66,1
Разгибатели коленного сустава. Обычно к этой
группе относят все головки четырехглавой мышцы бедра, но прямая
мышца бедра, начина-
ющаяся на подвздош-
ной кости, «а фазе
отталкивания сгибает
тазобедренный сустав и
потому не может быть
отнесена к группе раз-:
гибателей коленного су-
става. Во время оттал-
кивания разгибание «о-
ленного сустава дает
туловищу толчок впе-
ред. Наиболее выгодно
для этого отхождение
разгибателей коленного
сустава от проксималь-
ного конца бедра, ког-
да рычаг работы —
длина бедра — равен
рычагу приложения.
Приспособление к пе-
редвижению прыжками
приводит к усилению
всех разгибателей ко-
Рис. 76. Направление пучков латеральной голов-
ки четырехглавой мышцы бедра у различных
грызунов. А — хомяк, Б — пасюк, В — горный
тушканчик:
1 — место прикрепления мышцы
ленного сустава, но
особенно усиливается
латеральная головка
чггырехглавой мышцы,
^которая начинается от
большого вертлуга и
’•го гребня, т. е. почти от самого проксимального конца бедренной
кости. Особенно мощной эта головка становится у тушканчиков
(табл. 18). При специализации к прыжкам выгодно увеличение
угла прикрепления латеральной головки к бедру. У хомяка ног
мол равен 20°, у крысы - 35°, у тушканчика — 57° (рис 7<>)
II
Скакательный комплекс. Сюда мы относим икронож-
ную мышцу, которая наряду с заднебедренными мышцами и разги-
бателями голени является основной мышцей, обусловливающей1
отталкивание. Икроножная мышца является основной мышцей, сооб-
щающей ускорение телу животного. Казалось бы, что при нижней
опоре икроножная мышца должна сгибать коленный сустав, но в
действительности это ее действие нейтрализуется разгибателями ко-
ленного сустава и икроножная мышца разгибает голеностопный су-
Табляца 1ь
Изменения веса головок четырехглавой мышцы в связи
с приспособлением к прыжкам
Вид животного Относительный вес
латераль- ная медиаль- ная промежу- точная прямая
Малоазиатский хомяк 2,44 1, 81 2,75
Суслик 3,82 1,02 0,37 3,00
Полчок 3.35 1,11 0,48 2,30
Серая крыса 2,98 0,93 0,77 2,49
Персидская белка 6,40 1,80 0,72 4,65
Горный тушканчик 10,28 3,07 0,74 3,04
став. Действие икроножной мышцы на коленный сустав ослабляете:
благодаря тому, что она отходит от самого дистального конца бед-
ренной кости; кроме того, у всех прыгающих грызунов развиваются
сесамовидные косточки, связанные с надмыщелками бедра связка-
ми, почти параллельными телу последнего, и начало икроножной
мышцы переносится на эти косточки. В результате основная часть
силы икроножной мышцы, приложенной к бедру, направляется
вдоль тела бедренной кости и не влияет на сгибание колена.
Дистальный конец икроножной мышцы прикрепляется к пяточ
пому бугру. Так как основная роль икроножной мышцы — созда-
ние ускорения, то для прыжка выгодно относительное укорочение
пяточного бугра при усилении самой икроножной мышцы. Это i
наблюдается у прыгающих форм (табл. 19).
' Таким образом, фаза отталкивания разделяется на две стадии:
подготовительную и разгона. Подготовительная стадия (рис. 73, I.
II, III) сопровождается сгибанием тазобедренного, коленного и го-
леностопного суставов с растягиванием соответствующих мышц и
является наиболее тяжелым рабочим моментом. Стадия разгона
(рис. 73, IV, V, VI) сопровождается разгибанием тазобедренного,
коленного и голеностопного суставов. При этом туловище получает
ускорение в основном за счет быстрого разгибания голеностопного,
а отчасти и коленного суставов. Разгибатели коленного сустава дей-
ствуют в основном как силовые мышцы, толкающие туловище впе-
ред. К ним присоединяется также действие горизонтальной слагаю-
щей заднебедренных мышц, которая через разгибатели тазобедре.ч-
U2
ного сустава действует и на подъем туловища. Основным поднима-
телем туловища являются разгибатели спины.
Таблица 19
Изменение пяточного бугра и икроножной мышцы в связи с
приспособлением к прыжкам
Вид животного Отношение а X б
длины пя- точного бу- гра к длине пятки (л) | веса икро- ножной к весу скеле- та (б)
Слепец 0,26 1,64 0,4*3
Слепушонка Малоазиатский 0,21 2,21 0,46
хомяк 0,21 2,35 0,49
Серая крыса 0,15 3,63 0,53
Полчок 0,14 4,10 0,59
Горный тушканчик 0,093 13,03 1,21
Фаза свободного полета. Оттолкнувшись от земли, крыса проле-
тает определенное расстояние в воздухе (рис. 73, VII—IX), после:
чего падает на передние конечности. Во время полета передние ко-
нечности крысы вытягиваются сильно вперед, а задние конечности-
начинают перемещаться в переднее положение. У специализирован-
ных прыгающих животных это перемещение заходит настолько да-
леко, что животное падает на задние конечности; это ведет к пол-
ному отрыву от земли передних конечностей при передвижении.
Перемещение задних конечностей вперед совершается за счет сги-
бания голеностопного, коленного и тазобедренного суставов. При
этом функции ряда мышц оказываются не такими, какими они были
во время отталкивания при задней опоре. Основными сгибателями
коленного сустава являются мышцы заднебедренной группы, окан-
чивающиеся на голени; для этой их функции наиболее выгодны по
положению мышцы, заканчивающиеся на голени более дистально
(полусухожильная, задняя двуглавая и полуперепончатая), но так
как сгибание голени — процесс не особенно трудный, то названные
мышцы развиты у крысы довольно слабо.
Фаза приземления. Крысы после прыжка падают на передние
конечности. В результате толчка уровень тазового пояса резко сни-
жается, а задние конечности перегоняют тазовый пояс. Первое вре-
мя они продолжают сгибаться во всех трех суставах (табл. 13, этап
X), но уже начиная с XI этапа голеностопный сустав начинает
вновь разгибаться, перенос же стопы в более горизонтальное ^поло-
жение вплоть до XIV этапа осуществляется за счет крайнего сгиба-
ния позвоночного столба. На XIV этапе процесс заноса задних ко-
нечностей достигает своего апогея; с этого момента начинают дей-
ствовать разгибатели коленного сустава (все головки четырехглавой
мышцы), сгибатели же голени начинают растягиваться, фиксируя
тазобедренный сустав и облегчая тем самым действие прямой
11»
мышцы 6cipa Передние конечности при приземлении ипрают роль
пассивных амортизаторов толчка. При этом вначале происходит
сгибание локтевого, кистевого и плечевого суставов, а в дальней-
шем — разгибание локтевого сустава, связанное с перемещением
вперед туловища (рис. 73, X—XIV). В первый момент толчка про-
исходит переразгибание позвоночного столба в области соединения
передних конечностей с позвоночным столбом. Амортизация толчка
совершается в основном за счет уступающих движений в локтевом
и плечевом суставах, соединении плечевого пояса с туловищем,
грудной клетке и в позвоночном столбе. Некоторое значение для
амортизации толчка имеет и кистевой сустав.
Кистевой сустав. Образует первое звено амортизации.
При падении первыми с землей соприкасаются пальцы и дистальные
части кисти. Опусканию проксимальной части кисти противодейст-
вуют сгибатели пальцев, что и ведет к смягчению толчка.
Локтевой сустав. Образует второе звено амортизационно-
го аппарата. Сразу же после падения на передние конечности про-
ксимальный конец локтевой кисти поднимается, так как он переме-
щается краниально. Казалось бы, что при этом должно происходить
разгибание локтевого сустава, однако плечевая кость вначале опу-
скается быстрее, чем поднимается олекранон.
Плечевой сустав. Образует третье звено амортизацион-
ного аппарата. Сгибание плечевого сустава продолжается дольше,
чем сгибание локтевого, но начиная с IV. этапа фазы передней
опоры и в нем происходит разгибание с поднятием туловища. У
копытных и хищных разгибание плечевого сустава используется
чля отталкивания, у крысы движение в плечевом суставе служит,
вероятно, лишь для смягчения толчка.
Соединение плечевого пояса с туловищем. Об-
разует четвертое звено амортизации. В качестве амортизаторов
используются мышцы плечевого пояса, связывающие его с тулови-
щем (вентральная зубчатая, подключичная, грудные).
Грудная клетка. Образует пятое звено амортизационного
аппарата. Через грудную и вентральную зубчатую мышцы толчок
передается на ребра. Ребра сгибаются и разгибаются в местах их
соединения с реберными хрящами. Сгибание в ребрах значительно
смягчает толчок (рис. 77).
Орган амортизации позвоночного столба. Обра-
зует шестое звено амортизационного аппарата. Этот орган пред-
ставляет собой разросшийся остистый отросток второго (у крысы)
грудного позвонка, к верхушке которого прикрепляется короткая,
направленная вперед связка или обызвествляющаяся треугольная
сухожильная пластинка. С вентральной стороны к пластинке при-
крепляются несколько сухожильных тяжей, оканчивающихся на
остистых отростках грудных и шейных позвонков (рис. 78). У крысы
и у нутрии таких тяжей пять, у песчанки хорошо заметны лишь три.
У крысы особенно хорошо развит первый тяж, идущий от начала
костной пластинки к остистому отростку первого грудного позвонка;
144
к этому же отростку идет еще один тяж — гомолог надостистой
связки, также начинающийся на верхушке остистого отростка второ-
го грудного позвонка. При толчке сухожильная пластинка проги-
бается вниз, а сухожильные тяжи натягиваются; затем под дей-
Рис. 77. Орган амортизации позвоночного столба
пасюка:
1 — остистый отросток второго грудного позвонка, 2 — сухо-
жильная пластинка, 3 — сухожильные тяжи, 4 — надостистая
связка, б — суъожлянъ межостистой мышцы, 6 — сухожилие
полуостистой мышцы, 7, 8, 9 — поперечное сечение первого, вто-
рого и третьего сухожильных тяжей
Рис. 78. Схема амортизации толчка грудной клеткой пасюка. А —
сгибание грудной клетки под действием толчка, Б — возврат
грудной клетки в исходное положение:
1 — грудные мышцы, 2 — вентральная зубчатая мышца, 3 — грудина, 4 —
реберные хрящи, б — ребра, 6 — позвонки
ствием пучков полуостистой мышцы пластинка вновь выпрямляется,
и голова с шеей плавно поднимается (рис. 78).
Орган амортизации не выражен у грызунов, передвигающихся
юлько шагом и рысью (хомяки, полевки, слепцы и т. д.) и не
падающих при прыжке на передние конечности (тушканчики).
Наиболее хорошо он развит у грызунов, передвигающихся тяжелы-
ми медленными прыжками (крыса, нутрия).
ГЛАВА VIII
ОРГАНЫ ПИЩЕВАРЕНИЯ
ОБЩИЕ ЗАМЕЧАНИЯ
Органы пищеварения служат для механической и химической об-
работки пищи. В органах пищеварения пищевые продукты перево-
дятся в растворимую форму и, проникая в кровь, разносятся по
всему организму. Пищеварительный тракт начинается ротовой по-
лостью, где пища пережевывается и обволакивается слюной. Через
второй отдел — пищевод — пища попадает в желудок. В желудке и
в следующей за ним двенадцатиперстной кишке пища подвергается
основательной химической переработке; переваренные пищевые про-
дукты всасываются в тонких кишках. Непереваренные остатки по-
ступают в слепую кишку, где они подвергаются сбраживанию. (В
толстых кишках происходит всасывание перебродивших продуктов
и формирование фекалий, которые через прямую кишку выводятся
наружу.
Для анатомического исследования кишечника производят вскры-
тие брюшной полости. Для этого делают разрез по средней линии
живота от мечевидного хряща до лонного сращения. Затем, прощу-
пав симфиз тазовых костей, перерезают его. Если исследователя
интересует топография отдельных частей кишечника, тогда произ-
водят еще два разреза: 1-й — у самого основания мечевидного
хряща — по направлению к позвоночному столбу, перерезая попут-
но и ребра, и 2-й — на уровне моклока от середины брюха — к
позвоночному столбу. Чтобы обнаружить пищевод, приходится
вскрыть грудную полость и удалить сердце. Кроме того, желательно
отделить грудино-подъязычную мышцу и отодвинуть трахею. Для
исследования ротовой полости необходимо отделить нижнюю че-
люсть. Для проведения измерений необходимо всю пищеваритель-
ную трубку вывести наружу, для чего пищевод отделяют от глотки,
затем отделяют пищевод от диафрагмы, разрезают диафрагму, от-
деляют желудок от сальника, а кишечник—от брыжейки; в заклю-
чение от стенок тазовой полости отделяют прямую кишку. Измере-
ние пищеварительного тракта желательно производить на более или
менее свежем материале, поскольку при использовании подопре-
вшего материала кишечник легко рвется.
146
Общая длина кишечника крысы достигает 1,3—1,5 м, превышая
длину ее тела в 5—9 раз. Пищевод составляет 4,8%, желудок —
1,4%, двенадцатиперстная и тонкая кишки — 77,5'%(, слепая киш-
ка— 3,5%, толстая кишка — 8,8% и прямая кишка—4,0% общей
длины кишечника.
Рис. 79. Лицевая мускулатура пасюка:
1 — оттягивающая угол рта, 2 — поднимаю*
щая верхнюю губу, 3 — боковая носа, 4 —
опускающая нижнюю губу
РОТОВАЯ ПОЛОСТЬ И ГЛОТКА
Ротовая полость (cavum oris) начинается преддверием рта
(vestibulum oris), расположенным между губами (labii superior et
inferior) и альвеолярными отростками челюстей. Верхняя и нижняя
губы переходят друг в друга по
углам ротового отверстия (rima
oris). С губами связано не-
сколько мышц, являющихся
производными висцеральной
platysma myoideus (рис. 79). К
комиссуре губ с внутренней сто-
роны прикрепляется мышца, от-
тягивающая угол рта (m. ret-
ractor anguli oris). К верхней
губе прикрепляется поднима-
ль верхней губы (m. levator
li'bil superioris) и латеральная
мышца носа (m. lateralis nasi),
соединяющая лубу с крылом
носа; последняя мышца приподнимает верхнюю губу и расширяет
। осовое отверстие. К нижней губе прикрепляется мышца, опускаю-
। (,ая нижнюю губу (m. depressor labii inferioris). Все ротовое отвер-
стие окружено круговой мышцей рта (m. orbicularis oris), сжимаю-
щей губы. У крысы эта мышца развита весьма слабо. Губы пере-
ходят в щеки, в глубине которых развита щечная мышца (m. buc-
cinator).
С внутренней стороны ротовая полость выстлана слизистым
сштелием. На альвеолярных отростках в специальных ямках
(альвеолах) сидят зубы (dentes). Ткань, покрывающая альвеоляр-
ные отростки, называется десной (gingiva). У крысы на каждой
ч;люсти развито лишь по одному резцу и по той коренных зуба
(рис. 80, 81), так что ее зубная формула имеет следующий вид:
, 1 „0 _ О и 3
I ! . С q . Pm 0 . М3.
Резцы служат только для кусания и грызения. Растут они по
с пирали, и если один из резцов ломается, то противостоящий ему
разрастается в такой степени, что его кончик прободает нёбо н
животное гибнет. Эмаль развита лишь на передней поверхности
I <-щов, сзади же они покрыты более мягкой тканью — дентином
|,ептия1 изнашивается быстрее эмали, и вследствие этого |ччцы
<.рысы постоянно затачиваются. Резцовая альвеола прсдмын.(кеч
КГ н.
собой длинный изогнутый канал. Альвеола верхнего резца не до-
стигает уровня коренных зубов, тогда как альвеола нижнего резца
проходит кнаружи от коренных зубов, вплоть до заднего края по-
следних. Вследствие этого тело нижней челюсти расширяется кзади.
Рис. 80. Верхнечелюстные зубы пасюка:
1 — резец, 2 — коренные 8убы
На первом коренном зубе крысы развито по три, а на осталь-
ных — по два ряда бугорков, в каждом из которых имеется по
три бугорка. У грызунов, питающихся более грубой пищей (полев-
ки), бугорки заменены гребня-
Рис. 81. Нижнечелюстные зубы па-
сюка:
1 — резец, 2 — коренные зубы, S — аль-
веолярное отверстие нижней челюсти
прикрепляется к подъязычной
ми. У крысы бугорки коренных
зубов с возрастом стираются.
На таких зубах эмаль сохра-
няется лишь на периферии зу-
ба, а середина его покрыта
дентином.
Язык (lingua) у крысы от-
носительно длинный и подвиж-
ный. Его нижняя поверхность
связана с дном ротовой поло-
сти уздечкой (frenulum lin-
guae). Своим основанием, или
корнем (radix linguae), язык
:ти. Главная часть языка —
тело (corpus) — спереди доходит до кончика уздечки. Свобод-
ная часть языка называется верхушкой (apex). Дорзальная по-
верхность языка, или спинка (dorsum), покрыта многочисленными
сосочками (рис. 82). У крысы почти исключительно развиты одни
нитевидные сосочки (papillae filiformes), представляющие собой
ороговевшие образования, несколько расширенные у основания и
заостренные на конце. Нитевидные сосочки тела и верхушки языка
направлены назад, в среднем в этой зоне на 1 мм2 приходится
90 сосочков. У основания тела языка располагается небольшая
треугольная зона, покрытая нитевидными сосочками, имеющими
форму крючков, направленных вперед. В этой зоне на 1 лслс2 при-
ходится 50 60 сосочков. У перехода тела языка в корень развиты
чрезвычайно нежные нитевидные сосочки. В этой зоне на 1 мм?
приходится свыше 180 сосочков. По бокам тела языка, у перехода
148
тела в корень и отчасти на корне языка, располагаются небольшие
сосочки, по форме напоминающие примитивные грибовидные со-
сочки других млекопитающих (papillae fungiformes). По средней
линии на корне языка заметен один желобчатый сосочек (papilla
vallatae). Нитевидные сосочки придают .шероховатость языку, что
способствует удержанию пищи; они функционируют также в каче-
стве осязательных органов. В грибовидных и
желобчатом сосочках развиты вкусовые почки.
Язык богато снабжен мыщцами, как цели-
ком заключенными в его теле (собственно
язычные мышцы), так и отходящими от подъ-
язычной кости и нижней челюсти. Первые раз-
деляются на продольные (fibrae longitudina-
les), идущие от корня к верхушке, поперечные
(fibrae transversi), идущие от срединной сое-
динительнотканной пластинки к боковым кра-
3
Рис. 83.
ай»
Рис. 82. Язык
пасюка:
1 — нитевидные
сосочки верхушки
и тела языка, 2 —
крючковидные со-
сочки основания
тела языка, 3 —
прикорневые ни-
тевидные сосочки,
4 — желобчатый
сосочек
Язычные и подъязычные
мышцы пасюка:
1 — язык, 2 — ротовая полость, Э —
глотка, 4 — подбородочно-язычная, 5 —
шило-язычная, в — подъязычно-языч-
ная, 7 — подбородочно-подъязычная,
8 — грудино-подъязычна я, 9 — лопа-
точно-подъязычная
ям языка, и перпендикулярные (fibrae perpendiculares), идущие от
щрзальной поверхности языка к вентральной. Ко второй группе
принадлежат 3 мышцы (рис. 83). Шилоязычная мышца (гл. stylo-
glossus) начинается на сосцевидном отростке височной кости и
идет к краям языка. Совместное действие мышц обеих сторон от-
гя1 нвает язык назад, одностороннее — поворачивает его в сторону.
Подъязычная мышца (гл. hyoglossus) начинается от основной части
по тьязычной кости и оканчиваётся в теле языка. Оттягивает язык
шиац. Подбородочно-язычная мышца (m. genioglosstis) начинает
< я от симфиза нижних челюстей и оканчивается в теле языка
налью волокон прикрепляясь к телу подъязычной кости Тян< i
чшк вперед. Топографически с я ночными мышцами тс но гня’.тнй
I "
мышцы подъязычною аппарата: подбородочке подъязычная (m. ge-
niohyoideus), идущая от симфиза нижней челюсти, грудино-
подъязычная (m. sternohyoideus), начинающаяся от грудины, и
лопаточно-подъязычная (m. omohyoideus), начинающаяся от ло-
1 — твердое нёбо. 2 — мягкое нёбо.
3 — поперечные валики твердого нёба.
4 — отверстие якобсонова органа, 5 —
носоглоточное отверстие
патки (рис. 83).
Ротовая полость отделена от
дыхательных путей нёбом (рис.
84). Передняя его часть укрепле-
на нёбными отростками нёбных и
верхнечелюстных костей и назы-
вается твердым нёбом (palatum
durum). На нем развито 8 попе-
речных валиков, способствующих
Рис. 85. Слюнные железы пасюка:
1 — околоушная, 2 — подъязычная, Э — подче-
люстная
перетиранию и смешиванию пищи. На переднем валике заметны
углубления, в которые открываются нёбные отверстия якобсонова
органа. Задние пять валиков изогнуты и прерваны посередине.
Мягкое нёбо (palatum molle) оканчивается свисающей вниз
нёбной занавеской (velum palatinum), отделяющей ротовую по-
лость от глотки. В толще.мягкого нёба развиты мышцы, управляю-
щие движениями нёбной занавески, которая прикрывает переднюю
часть дыхательных путей во время глотания. Из этих мышц-назо-
вем мышцу, поднимающую нёбную занавеску (m. levator veil
palatini), напрягающую нёбную занавеску (т. tensor veil palatini)
и нёбную мышцу (т. palatinus). Первые две приподнимают и
фиксируют нёбную занавеску, а третья возвращает ее в исходное
положение.
Ротовая полость переходит в глотку (pharynx), в которой про-
исходит соединение пищеварительного тракта с дыхательными пу-
тями. Во время глотания глотка изолируется от дыхательных
путей при помощи нёбной занавески — спереди и надгортанником
и черпаловидными хрящами — сзади.
Слюнных желез (glandulae salivales) у крысы три (рис. 85).
Околоушная железа (gL parotis) располагается вентральнее и
150
каудальнее наружного уха. Подъязычная железа (gl. sublingualis)
лежит непосредственно нейтральнее околоушной, подчелюстная же-
леза (gl. mandibularis) располагается вентральнее и каудальнее
подъязычной, в ямке, образованной лопаточно-подъязычной и
грудино-подъязычной мышцами и подъязычной костью. Секрет
Рис. 86. Внутренности пасюка:
1 — пищевод, 2 — трахея, 3 — зобная железа, 4 — легкое, Б — сердце,
6 — ребра, 7 — диафрагма, 8 — печень, 9 — желудок. 10 — селезенка,
11 — почка, 12 — поджелудочная железа, 13 — тонкая кишка, 14 — тол-
стая кишка, 15 — слепая кишка, 16 — прямая кишка, 17 — пузырьковид-
ная железа, 18 — придаток семенника, 19 — семенник. 20 — половой
член, 21 — мочевой пузырь
слюнных желез служит для смачивания ротовой полости и ее со-
держимого; кроме того, слюна содержит фермент птиалин, рас-
щепляющий крахмал до дисахаридов.
КИШЕЧНЫЙ ТРАКТ
Пищевод (oesophagus) представляет собой сравнительно длин-
ную трубку, служащую для проведения пищи из глотки в желудок
(рис. 86, 87). Слегка изгибаясь по левой стороне гортани, пищевод
проходит в грудную полость и, прободая диафрагму, открывается
в желудок. Стенки пищевода образованы слизистой, мышечной и
соединительнотканной оболочками; в переднем отделе пищевода в
мышечной оболочке развиты поперечнополосатые волокна.
Желудок крысы (ventriculus) представляет собой изогнутый
мешок, неразделенный внешне на отделы (рис. 88). Он, как
к кишечник, образован слизистой, мускульной и серозной оболоч-
1.1 ми. У места перехода пищевода в желудок и желудка в две-
Н4 щатиперстную кишку мышечные волокна желудка образуют
б" ivr или менее выраженные сфинктеры. Передний вогнутый край
151
желудка образует его малую кривизну (curvatura minor), а зад-<
ний выпуклый — большую (с. major). Пищевод открывается в
желудок примерно посередине малой кривизны. Часть желудка,,
расположенная влево от пищевода, выстлана многослойным эпи-
телием, лишенным пищеварительных желез. Эпителий этот сходен
87. Кишечный тракт пасюка:
Рис.
пищевод, 2 — желудок. 3 — двенадцати-
перстная кишка, 4 — тонкая кишка, 6 — сле-
пая кишка, 6 — толстая кишка, 7 — прямая
кишка, 8 — анальное отверстие
выделяют пепсин и
с эпителием пищевода, и
соответствующая часть
желудка именуется пище-
водной. Внутри она отде-
ляется от железистой ча-
сти желудка слабо выра-
женной складкой.
Остальная часть же-
лудка выстлана однослой-
ным эпителием. В не-
большой зоне по малой
кривизне, непосредствен-
но примыкающей к отвер-
стию пищевода, развиты
трубчатые железы, секрет
которых не содержит пи-
щеварительных фермен-
тов. Соответствующий от-
дел желудка именуется
кардиальным, или вход-
ным (р. cardi'aca). Боль-
шой отдел желудка, при-
мыкающий к пищеводно-
му по большой кривизне,
именуется дном желудка
(fundus); его железы
'. Отдел желудка, примыкаю-
щий к двенадцатиперстной кишке, именуется пилорическим, или
привратниковым (р. pylorica); его железы выделяют слизистый
секрет.
Желудок переходит в двенадцатиперстную кишку (duodenum),
в которую впадают протоки печени и поджелудочной железы
(рис. 87). Гистологически двенадцатиперстная кишка отличается
от тонкой кишки развитием так называемых бруннеровых желез,
сходных с пилорическими. В двенадцатиперстной кишке завершает-
ся переваривание пищи и начинается всасывание продуктов их
расщепления. В связи с последней функцией поверхность двенадца-
типерстной кишки резко увеличивается за счет развития выро-
стов —ворсинок (villi intestinales). У крысы на 1 леи2 поверхности
двенадцатиперстной кишки приходится до 20 ворсинок. Двенадца-
типерстная кишка переходит в тонкую, или тощую (jejunum) —
основной всасывающий отдел кишечника (рис. 87). На 1 мм*
поверхности тощей кишки приходится до 30 ворсинок. Петли то-
152
щей кишки заполняют большую часть брюшной полости. Снаружи1
тощая кишка покрыта брюшиной, образующей брыжейку (mesen-
terium). На границе между тощей и толстой кишками от-
ходит не особенно сильно развитая у крыс слепая кишка (caecum),
в которой
происходит сбраживание клетчатки, не перевариваемой
ферментами желудка и двенадца-
типерстной кишки (рис. 87). По-
верхность слепой кишки увели-
чивается развитием складок. У
некоторых растительноядных жи-
вотных эти складки образуют спи-
ральный клапан.
Ряс. 88. Желудок пасюка. А — внеш-
ний вид, Б — поперечный разрез:
1 — пищевод, 2 — двенадцатиперстная
кишка, 8 — пищеводный отдел, 4 — кар-
диальная часть, В — дно. 6 — пилори-
ческая часть
Рис. 89. Печень пасюка (вид сзади):
1 — правая боковая, 2 — правая цент-
ральная, 3 — левая боковая, 4 — лева»
центральная, 5 — хвостовая, 6 — доба-
вочная доли печени, 7 — желчный про-
ток
Толстая, или ободочная, кишка (intestinum crassum, s. colon)
у крысы небольших размеров. В ней происходит всасывание воды
и формирование фекалий из непереваренных продуктов (рис. 87).
Толстая кишка переходит в прямую кишку (i. rectum), которая от-
крывается наружу заднепроходным отверстием (anus). В стенках
прямой кишки имеются поперечно-полосатые мышечные волокна,
образующие у ее выхода сфинктер.
ПЕЧЕНЬ И ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
Печень (liepar) — самая крупная железа крысы. По весу она
ic.ibHO варьирует (12—16 г), составляя 4—6% общего веса кры
сы Располагается печень непосредственно за диафрагмой. Опа до-
лин я на шесть хорошо различимых долей: левая и правая бокс
тзые, левая и правая центральные и охватывающие последнюю хво-
статая и добавочная (рис. 89). Правая боковая доля почти полно-
стью подразделена на две. Каждая доля на внутренней стороне
имеет небольшое углубление — ворота (hilus), через которые в пе-
чень входят кровеносные сосуды и выходят печеночные протоки.
Печеночные протоки, сливаясь, образуют общий желчный проток,
открывающийся в двенадцатиперстную кишку на расстоянии 20—
30 мм <уг привратника. Желчного пузыря у крысы нет.
Поджелудочная железа (pancreas) — удлиненное гроздевидное
тело, залегающее в брыжейке кишечника (рис. 86). Часть железы,
обращенная ,к желудку, именуется головкой; она подразделяется
на правую и левую доли. Противоположный конец железы име-
нуется хвостом, а средняя часть — телом. Поджелудочная железа
достигает в длину 30 мм, в ширину — 3 мм, а в толщину — 1 мм.
У крысы имеется два очень тонких протока поджелудочной желе-
зы, впадающих или в желчный проток, или непосредственно рядом
с ним прямо в двенадцатиперстную кишку.
ГЛАВА IX
ОРГАНЫ ДЫХАНИЯ
ОБЩИЕ ЗАМЕЧАНИЯ
Органы дыхания наземных позвоночных представляют собой
разного вида легкие, построенные из эластической ткани. Воздух
проходит в легкие по дыхательным путям, начальным звеном ко-
торых служит носовая полость.
При изучении строения легких необходимо вскрыть грудную
полость, для чего после удаления шкурки перерезают реберные
хрящи и отделяют грудину. Строение носовой полости рекомен-
дуется изучать на продольном и поперечном распилах черепа.
Предварительно черепа необходимо декальцинировать, помещая их
на несколько суток в 5—10%-ный раствор соляной кислоты.
ДЫХАТЕЛЬНЫЕ ПУТИ
У млекопитающих вдыхаемый воздух поступает в основном че-’
рез ноздри и носовую полость (рис. 90). Последняя у них подраз-
деляется на два хода — дыхательный и обонятельный, сообщаю-
щиеся друг с другом по концам. Обонятельный путь выстлан обо-
нятельным эпителием. При нормальных дыхательных движениях
воздух проходит главным образом через дыхательный ход. Далее,
через хоаны поступает в носоглотку, а через глотку — в гортань.
Гортань (larynx) состоит из ряда хрящей, объединенных систе-
мой связок и мышц в единое целое (рис. 91). Гортань служит не
только для проведения воздуха: в ней локализованы также голо-
совые связки, играющие важнейшую роль при производстве звуков.
С вентральной стороны у входа в гортань развит надгортан-
ный хрящ (cartilago epyglottica), выполняющий роль клапана гор-
тани. Дорзально в передней части гортани развит черпаловидный
хрящ (с. arytaenoidea).. Краниально от черпаловидного хряща от-
ходят парные рожковые хрящи (с. с. corniculatae), совместно с
надгортанным хрящом выполняющие роль клапанов гортани.
Сверху на черпаловидном хряще развита черпаловидная попереч-
ная мышца (гп. arytaenoideus transversus), срастающаяся по сред-
ней линии.
1У»
Щитовидный хрящ (с. thyreoidea) является наиболее крупным
хрящом гортани. Он образует ее вентральную и боковые части. Ог
4
Рис. 90. Сагиттальный разрез черепа пасюка:
1 — ноздря, 2 — дыхательный путь, 3 — начало обонятельного пути, 4 —
раковины обонятельного пути, 5 — хоана, 6 — носоглотка
10
11
12
б
Л
Гортань пасюка. А — вид
щитовидного хряща отходят сильно развитые каудальные рожки,,
к которым прикрепляется грудино-щитовидная мышца (m. ster-
no-thyreoideus), начинающаяся
по внутренней поверхности ру-
коятки грудины. На латераль-
ном гребне щитовидного хряща
оканчивается перстневидно-
щитовидная мышца (m. cri-
co-thyreoideus), отходящая от
краниального края перстневид-
ного хряща. Наконец, к кра-
ниальным рожкам и краниаль-
ному концу латерального
гребня щитовидного хряща
прикрепляется щитовидно-
подъязычная мышца (m. thyre-
ohyoideus), отходящая от те-
ла подъязычной кости.
Каудально гортань оканчи-
вается перстневидным хря-
щом, охватывающим началь-
ную часть трахеи. У крысы
перстневидный хрящ прерван
вентрально.
В полости гортани, у самого
входа в последнюю, слизистая
оболочка образует небольшие
углубления — боковые желу-
дочки (ventriculi's laryngei la-
terales); за ними развиты голосовые связки (plicae vocales). окру-
жающие голосовую щель (rima glottidis).
156
Рис. 91.
с вентральной стороны, Б — вид с
дорзальной стороны:
1 — надгортанный хрящ, 2 — черпало*
видный хрящ, 3 — щитовидный хрящ,
4 — перстневидный хрящ, б — рожко-
видный хрящ, в — краниальные рожки,
7 — каудальные рожки, 8 — хрящевые
кольца трахеи, 9 — соединительнотканные
кольца трахеи, 10 — щитовидно-подъязыч-
ная мышца, !1 — перстнёвиДно-щитоввд-
ная мышца, 12 — поперечная черпаловид-
ная мышна
Трахея (trachea) является непосредственным продолжением
гортани каудально (рис. 91). Она состоит из хрящевых полуко-
лец, чередующихся с полными кольцами эластической соедини-
тельной ткани. Каждое хря-
щевое полукольцо дополнено
эластической соединительной
тканью. Каудально трахея
разветвляется на два брон-
ха, ветвящихся; соответст-
венно, в правом и левом лег-
ких.
ЛЕГКИЕ
Легкие (pulmones) зани-
мают вместе с сердцем всю
трудную клетку. Свежие лег-
кие нежнорозового цвета.
Левое легкое не подразделе-
но, правое же делится на че-
тыре доли (рис. 92). Наибо-
лее краниально расположена
верхуше.ная доля (lobus api-
cal is), за ней следует сер-
дечная доля (1. cardiacus),
и, наконец, каудально распо-
ложены диафрагматическая
и добавочная доли (1.1. diap-
hragmaticus et accessorius).
Общий вес легких составля-
ет 2,5 г, из которых на левое
легкое приходится 0,8 г, на
верхушечную и добавочные
доли правого — 0,3 г, на
сердечную долю правого —
0,5 г и на диафрагматичес-
кую — 0,6 г. Место, где в
легкое входят сосуды и
бронхи, называется ворота-
ми легкого (hilus pulmo-
nis). Бронхи в легком слож-
но ветвятся, образуя брон-
Рис. 92. Легкие пасюка:
1 — левое легкое, 2, 3, 4. б — верхушечная,
сердечная, диафрагматическая и дополнительная
доли правого легкого, 6 — трахея, 7 — бифур-
кация трахеи, 8 «- бронхи, 9 — кровеносный
сосуд
Рис. 93. Схема строения дольки легкого
(по А. Ф. Климову):
1 — легочная вена, 2 — легочная артерия.
3 — концевой бронх, 4 — альвеолярные
ходы, 5 — альвеолы, в — стенки дольки лег-
кого
хиальное дерево, и окан-
чиваются тончайшими брон-
хиолами, переходящими в
альвеолы легкого (рис. 93).
Альвеолы выстланы спе-
157
циальным однослойным дыхательным эпителием и богато снабжа-
ются капиллярами от легочной артерии. В стенках альвеол и про-
исходит газообмен.
ДЫХАТЕЛЬНЫЕ ДВИЖЕНИЯ
Вдох (inspiratio) может происходить под действием мышц
грудной клетки, оттягивающих ребра в сторону и вперед (лестнич-
ные, краниальная часть дорзальной зубчатой, межреберные на-
ружные, прямая грудная и передняя часть прямой брюшной), и
при уплощении диафрагмы. Первый тип дыхания называется груд-
ным, второй — брюшным.
Выдох (expiratio) при грудном дыхании осуществляется под
действием мышц, оттягивающих ребра назад (каудальная часть
зубчатой дорзальной мышцы, пояснично-реберная, межреберные
внутренние и поперечная грудная мышцы).
При брюшном дыхании выдох осуществляется под действием
диафрагмы, впячивающейся в грудную полость. Это достигается
при расслаблении диафрагмы и действии всех мышц брюшного
пресса (прямая, поперечная и косая мышцы живота). Нормально
грудной и брюшной типы дыхания в чистом виде не встречаются
и можно говорить лишь о преобладании одного из них.
ГЛАВА X
ОРГАНЫ МОЧЕОТДЕЛЕНИЯ
почки
2
8
в
3
1 ______
та почки. 8
ство, 4 — 1 _
5 _ почечный сосочек, в — от*
верстия
чек, 7
Рис. 94. Почки пасюка:
— капсула почки, 2 — воро-
П — корковое веще-
мозговое вещество,
Почки (renes) крысы — компактные бобовидные образования,
красно-коричневого цвета. Расположены они в области поясницы,
правая несколько заходит в подреберье. Размеры почек колеблют-
ся в пределах 16—19X10—12X8—9 мм.
На почке различают выпуклый латераль-
ный и несколько вогнутый медиальный
края (рис, 94). Вогнутый край посередине
снабжен вырезкой, называемой воротами
почки (hilus renalis), через которые в
почку прокодят нервы и сосуды и выхо-
дит мочеточник. Ворота глубоко вдаются
в тело почки, образуя синус почки (sinus
renalis). Вся почка одета плотной соеди-
нительнотканной капсулой. Вещество поч-
ки разделяется на более светлое поверх-
ностное, называемое корковым (substan-
tia corticalis) и более темное внутреннее—
мозговое (s. medullaris).
В поЧке проходят многочисленные по-
чечные канальцы (tubuli renales), в ко-
торые выделяются составные части мочи
(рис. 95). Начальные, слепые концы ка-
нальцев лежат в корковом
Они образуют двустенную
веществе,
боуменову
капсулу (capsula Boumani), охва?ыва-
ющую клубочки капилляров кровенос-
ных сосудов (glomerulus). Совместно с
боуменовыми капсулами клубочки образуют
(corpysculae renales). Кровь, протекая через
труется в просвет почечного канальца, и поступающая туда жид-
кость содержит все составные части плизмы крови, помимо белко-
вых веществ. Почечный каналец на своем пути от почечного тельца
1 S9
собирательных трубе*
— почечная лоханка,
8 — мочеточник
почечные тельца
клубочки, филь-
•образует залегающий в корковом веществе извитой каналец первого
порядка, проникающую в мозговое вещество петлю Генле, лежа-
щий в корковом вещещстве извитой каналец второго порядка, и
направляющийся к мозговому веществу соединительный каналец.
В извитых канальцах из профильтрованной в почечный каналец
Рис. 95. Схема строения почеч-
ных канальцев (по А. Ф. Кли-
мову) :
I “ мочеотделяющая эона, II —
пограничная зона, III —- отводя*
•щая зона; 1 — мальпигиево тело.
2 — извитой каналец первого по*
рядка, 3 — петля Генле, 4 — из-
витой каналец второго порядка,
-5 — собирательная трубочка. 6 —
«сосочковый ход, 7 — ветвь почеч-
ной артерии. 8 — приносящий со-
суд почечного клубочка. 9 — вы-
носящий сосуд почечного клубочка,
10 — ветвь почечной вены
жидкости в кровь возвращаются са-
хара, аминокислоты и некоторые дру-
гие вещества, а в мочу поступает из
крови мочевина. В петле Генле моча
концентрируется и становится гиперто-
ничной. Собирательные трубочки по-
степенно сливаются друг с другом и
открываются на вершине вдающегося
в почечный синус почечного сосочка
(papilla renalis). В отличие от многих
других животных, у крыс имеется толь-
ко 1 сосочек (рис. 94). Из почечного
сосочка моча поступает в залегающую
в синусе почки почечную лоханку (pel-
vis renalis), являющуюся начальным
отделом мочевых путей.
МОЧЕВЫЕ ПУТИ
От почечной лоханки начинается
мочеточник (ureter), представляющий
собой полую трубочку, состоящую из
слизистого, мускульного и поверхност-
ного соединительнотканного слоев. Мо-
четочники открываются в мочевой пу-
зырь (vesica urenaria). Последний
представляет собой полый мешок, рас-
положенный в опустошенном виде в
тазовой полости (рис. 86, 98, 99). Он
образован соединительнотканной, мус-
кульной и слизистой оболочками. Мо
четочники открываются у самого осно
вания мочевого пузыря; при наполне
нии последнего они механически сдавливаются, что делает невоз
можным обратный отток мочи в мочеточники. Краниальная часть
мочевого пузыря направлена в сторону брюшной полости,
его дорзальная поверхность прилегает у самцов к прямой кишке, а
у самок — к влагалищу. Из мочевого пузыря начинается мочеис-
пускательный канал. У самок он представляет собой коротенькую
трубку, впадающую в преддверие влагалища, снабженную сфинк-
тером при выходе из мочевого пузыря. У самцов мочеиспускатель-
ный канал намного сложнее и будет описан вместе с половыми
-органами.
ГЛАВА XI
ПОЛОВЫЕ ОРГАНЫ
ПОЛОВЫЕ ОРГАНЫ САМЦА
К половым органам самца относятся семенники с их придат-
ком, семявыносящие протоки, половой член и придаточные желе-
зы (рис. 96).
Семенники (testes) закладываются в области поясницы рядом
с почками. С возрастом они перемещаются вентрально и выходят
в мошонку (scrotum) —специальное выпячивание брюшной поло-
сти. Внутренняя оболочка мошонки (tunica serosum) представляет
собой выпячивание брюшины. Ее внутренний, висцеральный ли-
сток тесно связан с оболочкой семенника, наружный, париеталь-
ный листок выстилает внутреннюю поверхность полости мошонки.
Последняя у крысы в течение всей жизни соединяется с брюшной
полостью влагалищным отверстием (foramen vaginale). Кнаружи
от серозной оболочки располагается мышца, втягивающая семен-
ник (m. cremaster), тесно связанная с фасцией (f. crema s ter ica).
Эта фасция, в свою очередь, прикрыта мясистой оболочкой (tunica
dartos), формирующейся из соединительнотканной оболочки
брюшной стенки. Она образует перегородку между полостями мо-
шонки (septum scroti). Снаружи мошонка покрыта кожей (cutis
scroti). Типичного кожного мешка у крысы нет и мошонка зна-
чительно выступает лишь при увеличении размеров семенников в
период усиленного сперматогенеза.
Семенник крысы имеет форму эллипса (рис. 97). Он окружен
плотной соединительнотканной белочной оболочкой (tunica albu-
ginea), которая у выхода семявыносящих канальцев значительно
вдается в тело семенника, образуя так называемое средостение,
или гайморово тело (mediastinum testis s. corpus Highmori). От
' н-достения к белочной оболочке идут перегородки (septulae testis),
разделяющие семенник на отдельные камеры. Камеры заполнены
нежной соединительной тканью с особыми интерстициальным в
клетками, в которой располагаются извитые семенные каналыы; в
этих канальцах формируются сперматозоиды.
11 Криса 161
Семенные канальца сходятся по направлению к средостению и
образуют в нем широко анастомозирующую семенную сеть (rete
testis). Из нее выходят несколько выносящих канальцев (ductuli
efferentes), сливающихся на к|ранильном конце семенника в общий
проток придатка семенника (ductus epididymis)^_^__
Придаток сменника (epididy-
mis) представляет собой узкое длин-
ное тело, отходящее от краниально-
го конца семенника (рис. 97). Его
начальная несколько расширенная
часть именуется головкой придатка
(caput .epididymidis); в ней вынося-
щие канальцы семенника сливаются,
образуя канал придатка. Головка
переходит в тонкое тело придатка
(corpus epididymidis), проходящее
по дорзальной стороне семенника; у
каудального конца семенника при-
даток вновь расширяется, образуя
хвост (cauda epididymidis). В нем
канал придатка сильно изгибается.
Канал придатка переходит в семя-
6
п
Мочеполовая систе-
X?
Ю
Рис. 96.
ма самца пасюка (правый се-
менник и часть правой пред-
стательной железы удалены,
правый мюллеров проток и
пузырьковидная железа разде-
лены) :
1 — семенник, 2 — головка при-
датка, 3 — хвост придатка, 4 —
семяпровод, 5 — мочеполовой ка-
нал, 6 — пузырьковая железа, 7—
предстательная железа, 8 — ампу-
лярные железы, 9 — луковичная
железа, Ю — препуциальная же-
леза, 11 — ножки полового члена,
12 — головка полового члена, 13—
стенка препуция, 14 — почка, 15-
мочеточник, 16 — мочевой пузырь,
17 — мужская матка
3 ?
г
Рис. 97. Семенник па-
сюка:
1 —• семенник, 2 — начало
придатка, S — головка при-
датка, 4 — тело придатка,
5 — хвост придатка, 6 —
семяпровод. 7 — брыжейка
семенника
провод (ductu- deferens), который через влагалищное отвер-
стие проходит в брюшную полость. Оба семяпровода близ мо-
чевого пузыря впадают в мочеиспускательный канал, образуя сов-
162
местно с ним мочеполовой канал. Стенки семяпровода, как и ка-
нала придатка, образованы слизистой, мускульной и наружной сое-
динительнотканной оболочками.
В начальный отдел мочеполового канала впадают очень круп-
ные у крысы пузырьковидные железы (glandulae vesiculares), сла-
бо подразделенные на лопасти (рис. 96). Железы эти заполнены
тягучим секретом, выделяемым их эпителием. Тут же в мочепо-
ловой проток открывается так называемая мужская матка, пред-
Рис. 98. Половой член пасюка (по Аргиропуло).
А — половой член (схематизировано), Б—го-
ловка полового члена сверху, В — кость полового
члена снизу, Г — кость полового члена сбоку:
1 — трезубец, 2 — латеральные сосочки, S — вентраль-
ный сосочек. 4 — зубец кости полового ' лена, S — тело
полового члена
ставляющая собой полый орган с несколькими извитыми каналь-
цами и соответствующая мюллеровым протокам (яйцеводам и мат-
ке) самок. У основания мужской матки и протокой пузырьковид-
ных желез (располагаются две пары небольших ампулярных же-
лез (glandulae ampul lares), за ними в мочеполовой канал отдель-
ными протоками впадают доли предстательной железы (glandula
prostata), сильно развитой у крысы (рис. 96). Дорзальная порция
предстательной железы лежит между мочеполовым протоком и
прямой кишкой и подразделяется на две латеральные доли, вент-
тральная порция, прикрывающая с вентральной стороны мочевой
пузырь, подразделяется на краниальную и каудальную доли. У
основания полового члена в мочеполовой проток впадает пара лу-
ковичных желез (glandulae bulbo-urethrales).
Половой член (penis) у крысы располагается близ заднепроход-
ного отверстия. Он покрыт кожным мешком, называемым препу-
цием (preputium). В полость препуция впадают две грушевидные
препуциальные железы, выделяющие резко пахнущий секрет. Внут-
ри полового члена проходит мочеполовой канал. Дистальный рас-,
ширенный конец полового члена называется головкой. У крысы, как
и у многих других грызунов, в теле полового члена развита кость
• os penis). Особенности строения полового члена используются как
диагностические признаки для многих грызунов. По Аргиропуло
11» 163
Рис. 99. Мочеполовая систе-
ма самки пасюка*
1 — яичник. 2 — воронка яй-
цевода, 8 — яйцевод, 4 — рог
матки, 5 — шейка матки, 6 —
влагалище, 7 — преддверие,
8 — луковичные железы, 0 —
почка., 10 — надпочечники, 11—
мочеточник, 12 — мочевой пу-
зырь, 13 — прямая кишка
(1940), у пасюка головка penis цилиндрическая, ее боковые сторо-
ны параллельны или слабо выпуклы. Сбоку головки, ниже средней
линии, по обеим сторонам дистальной части проходит глубокая
бороздка. Кольцевая складка слабо выдается из отверстия головки.
Papilla centralis полностью окостеневающая, сильно сжата с бо-
ков и почти не выступает из головки; papilla lateralis относительно
велика, papilla dorsalia одна, центральная. Укороченная и массив-
ная os penis имеет относительно слабо развитую основную часть
(рис. 98).
В теле полового члена развиты пеще-
ристые тела, представляющие собой рас-
ширения вен, имеющие плотные хрящева-
тые стенки. При заполнении пещеристых
тел кровью наступает гиперемия и эрек-
ция полового члена.
ПОЛОВЫЕ ОРГАНЫ САМКИ
К половым органам самки относятся
яичники, яйцеводы, матка, влагалище и
наружные половые органы (рис. 99).
Яичники (ovaria) крысы представляют
собой небольшие гроздевидные образова-
ния желтосерого цвета, лежащие сразу
же за почками. Выводных протоков яич-
ники не имеют. При созревании яйцеклет-
ки стенка яичника разрывается и яйце-
клетка попадает в полость тела. Стенка
яичника построена из цилиндрического
эпителия, который у стенок туловища
переходит в перитонеальный. Брюшина
не покрывает яичника снаружи. На раз-
резе яичника ясно видны две зоны —•
наружная, фолликулярная, и внутренняя,
сосудистая. В фолликулярной эоне проис-
ходит развитие яйцеклеток, находящихся
на разных стадиях развития. Каждая
яйцеклетка развивается в специальном
гнезде, или пузырьке. По мере созрева-
ния яйцеклетки полость пузырика увели-
чивается, а стенка яичника в соответст-
вующем месте выпячивается. Пузырьки, содержащие почти 'зрелые
яйцеклетки, называются граафовыми (folliculli Graafi).
Сосудистая зона начинается от ворот яичника (hilus ovarii), ко-
торые расположены в области прикрепления брыжейки яичника и
служат местом вхождения сосудов и нервов. В сосудистой зоне
ветвятся сосуды, снабжающие развивающиеся яйцеклетки.
Яйцевод (oviductus) представляет собой тонкую трубочку, от-
крывающуюся в полость тела воронкой (infundibulum), распола-
164
Рис. 100. Разрез влага-
лища и основания матки
пасюка:
1 — рог матки. 2 — непар-
ная часть матка, 3 — губы
шейки матки. 4 — вход в
матку, Б — влагалище
гающейся близ яичника. На другом конце яйцевод открывается в
матку. Стенки яйцевода выстланы мерцательным эпителием и об-
разованы слизистым, мышечным и соединительнотканным слоями.
Зрелое яйцо, попав в яйцевод, быстро прогоняется по нему дейст-
вием мерцательного эпителия. Оплодотворяется яйцо обычно в на-
чале яйцевода.
Матка (uterus) — орган, в котором происходит развитие заро-
дышей (рис. 99, 100). Матка образована слизистой (endometrium),
мускульной (myometrium) и серозной (pe-
rimetrium) оболочками. Оплодотворенное
яйцо вростает в слизистую оболочку матки
и снабжается сосудами последней. Приро-
дах зародыши действием мышц мускуль-
ной оболочки выталкиваются из матки.
У крысы матка разделена почти до са-
мого влагалища на две части, называемые
рогами. Такая матка называется двураз-
дельной (uterus bipartitus). Непарная
часть матки у входа во влагалище сужена
и снабжена сильно развитым сфинктером;
эта часть матки называется шейкой (col-
lum uteri). Сфинктер полностью закрывает
отверстие матки, которая раскрывается
лишь при родах.
Влагалище (vagina) соединяет матку с
наружными половыми органами (рис. 99,
100). Внутри влагалище выстлано. много-
слойным слизистым эпителием, обнаружи-
вающим сильно развитые складки. Соеди-
нительнотканная оболочка выражена толь-
ко в маточном конце влагалища. Вход во
влагалище прикрыт поперечной складкой
кожи — девственной плевой (hymen), ко-
торая у совокуплявшихся животных исче-
зает.
У входа во влагалище расположено
преддверие (vestibulum urogenitale) (рис. 99). С вентральной сто-
роны в преддверие открывается мочеиспускательный проток; та-
ким образом в преддверии мочевые и половые пути самки соеди-
няются. По происхождению преддверие является впячиванием
задней стенки промежности. Его слизистая оболочка выстлана
многослойным эпителием и богата железами. В стенках преддве-
рия развит сильный сжиматель преддверия (гл. constrictor vesti-
buli), который каудально переходит в сжиматель половой щели
(m. constrictor vulvae). В преддверие открываются две лукович-
ные железы (gl. bulbo-urethrales), соответствующие одноименным
железам самца.
Преддверие открывается наружу половой щелью (rim.i piub'ii-
1(>й
dis), окаймленной кожной складкой — половыми губами (labia pu-
dendis). С внутренней стороны от губ располагется клитор — ми-
ниатюрное подобие полового члена самца. В его теле, отходящем
двумя ножками от седалищных бугров, развито пещеристое тело.
Снаружи клитор окружен кожным препуциальным мешком.
Половые органы самки со времени наступления половозрелости
подвержены циклическим изменениям. Ко времени созревания яйце-
клетки эпителий матки слущивается, ее слизистая оболочка набу-
хает и подготавливается к восприятию яйца. Во время беременности
слизистая матки резко утолщается, млечные железы увеличиваются,
а дальнейшее созревание половых клеток приостанавливается. Пос-
ле родов наступает период восстановления, по окончании которого
вновь начинаются процессы созревания яйцеклеток.
ГЛАВА ХИ
СОСУДИСТАЯ СИСТЕМА
ОБЩИЕ ЗАМЕЧАНИЯ
Сосудистая система представляет собой систему трубок — сосу-
дов, по которым циркулируют жидкости, доставляющие тканям пи-
тательные вещества и уносящие из них продукты жизнедеятельно-
сти. У позвоночных сосудистая система раз-
деляется на кровеносную и лимфатическую;
по лимфатической системе продукты жизне-
деятельности тканей поступают в кровенос-
ную систему. Жидкость, циркулирующая по
кровеносной системе, называется кровью, а
жидкость лимфатической системы — лим-
фой.
Центральным органом сосудистой систе-
мы является сердце, выполняющее роль на-
соса, обеспечивающего циркуляцию жидко-
стей. Сосуды, выходящие из сердца, называ-
ются артериями, а сосуды, приносящие
кровь в сердце, называются венами. Арте-
рии сообщаются с венами' посредством тон-
чайших сосудов — капилляров.
У млекопитающих имеется два круга
кровообращения — большой и малый. По
артериям большого круга кровь разносится
по всему телу; возвращаясь по венам в
сердце, по артериям малого круга (легоч-
ным) она поступает в легкие. Благодаря
этому кровь, оттекающая от тканей, сначала
обогащается кислородом в легких, а затем
уже вновь поступает к тканям.
Кровеносные сосуды изнутри выстланы
эндотелием — соединительнотканными клет-
ками, организованными по типу плоского
эпителия (рис. 101). Далее идет тонкая сое-
Рис. 101. Схема строе-
ния сосудов (по А. Ф.
Климову). А — артерия,
Б — вена:
1 — эндотелий, 2 — яити-
ма. 8 — внутренняя власти-
ческая оболочка, 4 — сред-
няя оболочка, S - наруж-
ная оболочка, fl utana-
HIJ |1Г<|||4
167
динятельнотканная оболочка (intima) и внутренняя эластичная обо-
лочка (membrana elastica interna). Эти три слоя совместно обра-
зуют внутреннюю оболочку (tunica interna). Средняя оболочка
(tunica media) образована гладкими мышечными волокнами, а на-
ружная, или адвентиция (tunica externa), — волокнистой соедини-
тельной тканью. В артериях, ближайших к сердцу, внутренняя обо-
лочка развита особенно сильно, что позволяет им выдерживать осо-
бенно высокое здесь кровеносное давление. В венах средняя оболоч-
ка слабо развита, и кроме того, в них имеются многочисленные
кармановидные клапаны, препятствующие обратному току крови.
Капилляры образованы лишь эндотелием. Более крупные капилля-
ры охватываются сократимыми кольцевыми клетками Руже. При
сокращении последних, капилляр сужается и сопротивление току
крови в нем повышается.
КРОВЬ
Кровь состоит из жидкости — плазмы, в которой свободно пла-
вают форменные клеточные элементы. Последние разделяются на
окрашенные, как правило безъядерные, эритроциты, придающие
крови красный цвет, и бесцветные лейкоциты. Эритроциты имеют
форму двояковыпуклых дисков с диаметром около 6,3 р . Они со-
держат дыхательный пигмент — гемоглобин, и служат для тран-
спорта кислорода. У новорожденных крыс в 1 мм3 крови находится
2,5—3,5 млн. эритроцитов, у взрослых крыс число их повышается
до 9,27 (у самцов) — 9,42 (у самок) млн. в 1 мм3. Число лейкоци-
тов у крыс в норме колеблется от 6 до 18 тыс. в 1 мм3 крови, со-
ставляя в среднем 9 000. Данные о содержании в крови крысы раз-
личных видов лейкоцитов приведены в табл. 20.
Таблица 20
Содержание различных видов лейкоцитов
в крови крысы
Название форменного элемента Среднее ко- личество в % Коле- бания В %
Нейтрофилы 20 8-39
Лимфоциты 78 55-96
Моноциты меньше 1 0-3
Эозинофилы 2 0-4
Базофилы меньше 1 0—3
У новорожденных крыс формула лейкоцитов совершенно другая
(табл. 21). Число лейкоцитов в крови у новорожденных крыс со-
ставляет в среднем 3200 на 1 лл3 крови.
Форменные элементы очень быстро разрушаются и на их место
в кроветворных органах (костный мозг, селезенка, лимфатические
узлы) возникают новые.
168
Таблица 21
Содержание различных видов лейкоцитов
в крови новорожденных крыс
Название форменного элемента Среднее количество В %
Миэлоциты
Метамиелоциты
Гранулоциты (нейтрофилы-}*
базофилы-|-9Озинофилы)
Лимфоциты
25
25
СЕРДЦЕ
Сердце (сот) представляет собой толстостенный мышечный ме-
шок, подразделенный перегородками на четыре камеры правое
и левое предсердия и желудочки (рис. 102). Внешне предсердия
отделены от желудочков поперечной венечной бороздой (sulcus со-
ronarius). Сердце покрыто специальной околосердечной сумкой и
располагается в области II—V ребер. Весит оно у крысы около
1,3 г.
Правое предсердие (atrium dextrum) принимает краниальную и
каудальную полые вены (рис. 102). Отверстия обеих полых вен
разделены небольшим утолщением мышечных стенок предсердия и
защищены клапанами, препятствующими обратному оттоку крови.
Рядом с отверстием каудальной полой вены в предсердие Откры-
вается так называемый венечный синус сердца (sinus coronarius
cordis), собирающий кровь из сердечных вен. На перегородке меж-
ду предсердиями заметно небольшое овальное углубление (fossa
ovalis); у эмбрионов перегородка пронизана в этом месте отвер-
стием. Спереди на внутренней поверхности предсердия развиты мы-
шечные валики — гребешковые мышцы (m. т. pectinati). Предсер-
дие сообщается с правым желудочком правым атриовентрикуляр-
ным отверстием (ostium atrioventricularis dextrum).
Правый желудочек (ventriculus dextrum) в поперечном сечении
имеет форму полумесяца, так как межжелудочная перегородка вдает-
ся в его сторону. Между предсердием и желудочком располагается
правый атриовентрикулярный, или трехстворчатый, клапан (val-
vula tricuspidalis). К каждой из его створок прикрепляются сухо-
жильные струны (chordae tendineae), которые начинаются от трех
сосочковых мышц (m.m. papillares).
От стенок межжелудочковой перегородки к боковой стенке идут
поперечные^мыШЦы се(рдца (m.m. transversi cordis).
Из краниальной части желудочка выходит легочная артерия. В
ее отверстие вдается три полулунных клапана (valvulae semiluna-
res), которые препятствуют обратному поступлению крови п жч-1\
Дочек при диастоле (расширении) последнего.
169
Левое предсердие (atrium sinistrum) принимает четыре легоч-
ные вены (venae pulmonales); их отверстия защищены клапанами.
В левом предсердии, как и в правом, имеются гребешковые мыш-
цы. С желудочком левое предсердие соединяется левым атриовент-
рикулярным отверстием (о. atrioventricularis sinistrum).
Левый желудочек (venriculus sinistrum) дает начало аорте —
главной артерии большого круга кровообращения. В сосудах боль-
шого круга кровь встречает большее сопротивление, чем в сосудах
Рис. 102. Сердце пасюка. А — общий вид,
Б — в продольном разрезе:
1 — левое предсердие, 2 — левый желудочек»
3 — правое предсердие, 4 — правый желудо-
чек, 5 — венечная борозда
малого круга; соответственно стенки левого желудочка намного тол-
ще стенок правого (рис. 102). Левое отверстие защищено двуствор-
чатым клапаном (valvula bicuspidalis). От его створок к двум со-
сочковым мышцам идет восемь сухожильных струн. Поперечных
мышц в левом желудочке имеется только две.
Отверстие аорты (ostium aortum) защищено полулунными кла-
панами, на вершинах которых развиты так называемые аранциевы
узелки — небольшие утолщения, облегчающие более полное закры-
вание аорты.
Стенки сердца образованы тремя оболочками — эндокардом,
миокардом и эпикардом. Эндокард (endocardium) выстлан эндоте-
лием и по своему строению соответствует всем оболочкам крове-
носных сосудов. Миокард (miocardium) построен из специализиро-
ванной мышечной ткани и образует основную толщу сердца. Мио-
кард называют также сердечной мышцей. Он подразделяется на
мышцы предсердий и мышцы желудочков; оба отдела обособлены
друг от друга атриовентрикулярными фиброзными кольцами. Глубо-
кий слой мускулатуры предсердий образован продольными волок-
нами, поверхностный — кольцевыми, охватывающими оба пред-
сердия одновременно. В мускулатуре желудочков развит дополни-
170
тельно поверхностный СЛОЙ продольных IViJloKull, pImIUIII И I...Il'
желудочков, средний, кольцевой слой слмосг«1Н1«'л<*н дли .».<!.* чт«•
желудочка.
Координация сокращений частей миокарда осущспвлчгпи при
помощи специальной системы, проводящей раздражение. Опа "('цм
зована специальными мышечными пучками, богатыми нервными
элементами, которые соединяют различные отделы миокарда. На-
чальное звено проводящей системы представлено синоаурикуляр-
ным узлом (nodulus sinoauricularis), расположенным в правом
предсердии у впадения краниальной полой вены. В конце правого
предсердия расположен узел Тавары (nodulus Tawara), от которого
по перегородке между желудочками спускается атриовентрикуляр-
ный пучок (fasciculus atrioventricularis), объединяющий мышцы
предсердий с мышцами желудочков. Атриовентрикулярный пучок
разделяется на правый и левый, оканчивающиеся в стенках соответ-
ствующих желудочков. Проводящую систему сердца крысы лишь с
трудом удается обнаружить при помощи бинокуляра.
Эпикард (epicardium) представляет собой внутренний (висце-
ральный) листок околосердечной сумки (pericardium), тесно свя-
занный с миокардом. Эпикард продолжается на полые и легочные
вены, легочную артерию и аорту и переходит там в наружный
(париетальный) листок перикарда. Полость между обоими листка-,
ми (cavum pericardii) заполнена серозной жидкостью (liquor peri-
cardii), уменьшающей трение при соприкосновении. Париетальный
листок состоит из внутреннего (серозного) и наружного (фиброз-
ного) слоев. Последний непосредственно переходит в адвентицию
сосудов и посредством связки (iigamenta siernopericardiaca) при-
крепляется к внутренней поверхности грудины.
Сердечная мышца снабжается двумя венечными артериями (а.а.
coronales dextra et sinistra). Левая венечная артерия дает начало
большой сердечной вене (v. cordis magna), а правая венечная ар-
терия — средней сердечной (v. cordis media) и четырем-шести ма-
лым сердечным венам (v.v. cordis parvae). Средняя сердечная вена
впадает в большую сердечную вену.
АРТЕРИИ
Все сосуды разделяются на сосуды малого и большого кругов
кровообращения.
Начальное звено малого круга кровообращения образует легоч-
ная артерия (a. pulmonalis), начинающаяся от правого желудочка
сердца. Непосредственно по выходе из сердца легочная артерия де-
лится на ряд ветвей, которые, идя к легким, образуют в их альвео-
лах сложную сеть капилляров. Обогащаясь здесь кислородом, кровь
собирается в легочные вены (v.v. pulmonales), ^оторые в числе
2—4 впадают в левое предсердие.
Начальное звено большого круга кровообращения образует
аорту (aorta), выходящую из левого желудочка. При выходе яор-
171
ты из сердца от нее отходят венечные артерии сердца. В дальней-
шем аорта, изгибаясь дугой (arcus aortae), поднимается к вентраль-
Рис. 103. Сонные и подключичная
артерии и безымянная вена пасю-
ка. Вены зачернены, артерии по-
казаны светлыми:
1 — сердце, 2 — дуга аорты, 3 —
безызмянная артерия, 4 — общая сон-
ная, 5 — подключичная артерия, в —
наружная сонная, 7 — внутренняя
сонная, 8 — краниальная щитовидная,
9 — каудальная щитовидная, 10 — за-
тылочная, 11 — поверхностная височная,
12 средняя ушная, 13 — лицевая,
И жевательная, 15 — межчелюст-
ная, 16 нижнегубная, 17 — угловая
глаза, 18 угловая рта, 19 — боко-
вая носа, 20 — дорзальная носа.
21 — позвоночная, 22 — восходящая
шеи, 23 — глубокая шейная, 24 — шей-
ный ствол, 25 — дельтовидная, 26 —
внутренняя яремная. 27 - вена пле-
чевого сустава, 28 подключичная,
29 — грудная, 30 — наружная ярем-
ная, 31 — безымянная вена
ной поверхности позвоночного стол-
ба. От дуги аорты обычно отходят
три ствола: безымянная, левая сон-
ная и левая подключичная артерии
(рис. 103). Иногда ветви безымян-
ной артерии — правые подключич-
ная и сонная — отходят от дуги
аорты самостоятельно, еще реже са-
мостоятельно начинается левый ре-
берно-шейный ствол (truncus costo-
cervical is sinistra).
Безымянная артерия (a. anonym
ma) сразу же разделяется па пра-
вые сонную и подключичную. Обе
сонные артерии идут вдоль трахеи к
голове и на уровне щитовидной же-
лезы разделяются каждая на наруж-
ную и внутреннюю сонные артерии.
Следует отметить, что внутренняя
челюстная артерия, описываемая
обычно как ветвь наружной сонной
артерии, у крысы отходит от внут-
ренней сонной артерии (рис. 103,
104). Поэтому начальную часть
внутренней сонной артерии можно
обозначать как общий ствол внут-
ренней сонной и внутренней челюст-
ной.
Наружная сонная артерия (а. са-
rotis externa) на своем пути к углу
нижней челюсти отдает краниальную
щитовидную, затылочную и поверх-
ностную височную артерии, переходя
после этого в наружную челюстную
артерию. Последняя почти сразу же
разделяется на язычный ствол и ли-
цевую артерию (рис. 103).
Затылочная артерия (a. occipita-
lis) идет к затылочной области,
отдавая заднюю менингеальную и
ушную артерии (a. a. meningea
caudalis et auricularis) и ветви к
грудино-сосковой, двубрюшной, пла-
стревидной и трапецевидной мыш-
цам. Задняя менингеальная арте-
рия снабжает твердую мозговую
172
оболочку. Краниальная щитовидная артерия (a. thyreoidea cra-
nialis) снабжает щитовидную и паращитовидную железы и гор-
тань. Поверхностная височная артерия (a. temporalis superficia-
lis) направляется к основанию ушной раковины, где разделяется
на две ветви, огибающие ушную раковину (рис. 104). Краниаль-
ная ветвь отдает жевательную артерию (a. masseterica), артерию
височной мышцы (a. temporalis) и нижнечелюстную (a. mandibu-
laris), которая, проходя через подбородочное отверстие, переходит
в подбородочную артерию (a. mentalis). Грин считает нижнече-
люстную артерию гомологичной внутренней челюстной.
Рис. 104. Поверхностные артерии головы пасюка:
1 — общая сонная, 2 — наружная сонная, 3 — внутрен-
няя сонная, 4 — внутренняя челюстная, 5 — затылочная,
6 — щитовидная, 7 — поверхностная височная, 8 — язычный
ствол, 9 — лицевая, 10 — жевательные ветви лицевой. 11 —
угловая рта, 12 — нижнегубная, 13 — верхнегубная, 14 — уг-
ловая глаза. 15 — боковая носа, 16 — подглазничная, 17 —
поверхностная лицевая, 18 — височные ветви поверхностной
лицевой, 19 — жевательные ветви поверхностной лицевой,
20 — лобная ветвь поверхностной лицевой
Язычный ствол (truncus lingualis) отдает гортанную (a. trache-
alis), восходящую нёбную (a. palatina ascendens) и подъязыч-
ную (a. sublingualis) артерии, и, погружаясь после этого в тело
языка, обра 'ует язычную артерию (a. lingualis). Таким образом,
язычный ствол снабжает кровью подъязычную кость и окружаю-
щие ее мышцы, мягкое и часть твердого нёба, гортань и т. д.
(рис. 105). В отличие от крысы у большинства животных общего
язычного ствола нет, и все эти артерии отходят от наружной челю-
стной самостоятельно.
Лицевая артерия (a. facialis) идет поверхностно с вентральной
стороны межчелюстного пространства, отдавая по пути поверхност-
ную подъязычную артерию (a. sublingualis superficialis), снабжаю-
щую кровью подъязычное пространство (рис. 103, 104). Проходя
через вырезку нижней челюсти, артерия выходит на лицевую по-
верхность, где она разделяется на нижнегубную (a. labialis infe-
173
rfor), жевательные (a.a. masseterica), верхнегубную (a. labials
superior), носовую (a. nasalis) и угловую артерию глаза (a. anguli
oculi), снабжающие кровью соответствующие органы (рис. 103—
106).
Внутренняя сонная ар-
терия (a. carotis interna),
не доходя до барабанной
камеры, отдает внутрен-
нюю челюстную артерию,
после чего через лицевой
канал проникает в череп-
ную полость (рис. 105,
106). Около гипофиза
внутренняя сонная арте-
рия разделяется на на-
8 >5 16 ’ зальную и каудальную
соединительные ветви
Рис. 105. Артерии головы пасюка (средний
слой):
1 общая сонная, 2 - общий ствол внутренней сон-
ною и внутренней челюстной, 3 — подъязычная. 4 «—
внутренняя сонная, Б — внутренняя челюстная, 6 -
наружная лицевая, 7 нижнечелюстная, 8 — наруж-
ная лицевая, 9 — нёбная, 10 — подглазничная, И —
гортанная, 12—лицевая, 13 — подбородочная, 14 —
угловая глаза, 15 — анастомоз наружной лицевой с
наружной челюстной, 16 —анастомоз лицевой с нёб-
ной
(ramus communicans na-
salis et caudalis), кото-
рые совместно с одно-
именными ветвями про-
тивоположной стороны
образуют артериальное
виллизиево кольцо (circii-
lus villisii) (рис. 107).
Спереди от виллизиевого
кольца отходят непарная назальная мозговая артерия (a. cerebri
nasalis) и пара назальных артерий мозговой оболочки (a. meningea
nasalis), сбоку — средняя
мозговая артерия (a. ce-
rebri media), а сзади —
каудальная мозговая ар-
терия (a. cerebri caudalis)
и каудальная артерия со-
судистого сплетения боко-
вых желудочков (a. cho-
rioidea caudalis) (рис.
107). В полости черепа с
ветвями внутренней сон-
ной артерии анастомози-
рует каудальная артерия
мозговой оболочки. Обра-
зующаяся в результате
основная мозговая арте-
рия (a. basilar!s cerebri)
1 — общий ствол внутренней сонной и внутренней
челюстной, 2 — внутренняя сонная, 3 — внутренняя
челюстная, 4 — глазничная, Б — нёбная ветвь
глазничной, 6 — подглазничная. 7 — нёбная ветвь
подглазничной, 8 — альвеолярные ветви подглаз-
ничной, 9 — носовая внутренняя, 10 — решет-
чатая, 11— угловая глаза, 12 —лобная, 13 —
височная, 11 —крыльная ветвь
дает начало артериям мозжечка (a. a. cerebelli nasalis
et caudalis) и внутренней слуховой артерии (a. auditiva interna)
(ряс. 107).
174
Внутренняя челюстная артерия (a. maxlllaii> lut< rna) pnu<-
ляется на ветвь крыльного канала и на нёбную ветвь (рис. 105,106).
От ветви крыльного канала (a. alaris, s. pterygoidea) отходит гло-
точная артерия (a. pharyngea), снабжающая мягкое небо. После
этого артерия крыльного канала входит через среднее рваное отвер-
стие в черепную полость, затем через крыльный канал выходит
оттуда и в области подглазничного отверстия проникает в носовую
полость.
Рлс. 107. Артерия и вены головного мозга пасюка.
А — вид с вентральноб стороны, Б — вид с дор-
зальной стороны (по Грин):
1 — назальная артерия мозга. 2 — назальная вена мозга,
3 * — средняя мозговая артерия, 4 — артерия сосудистой
оболочки, б — основная вена. 6 — Задняя соединительная
артерия, 7 — каудальная вена мозга. 8 — каудальная ар-
терия мозга, 9 — назальная артерия мозжечка, 10 — основ-
ная артерия, И — вилизиево кольцо. 12 — дорзальные
вены мозга, 13 — каудальная вена мозжечка, 14 — позво-
ночная вена, 15 — сагиттальный синус, 16 — каменистый
синус, 17 — поперечный синус, 18 — назальная вела моз
19 — дорзальные назальные вены и артерия моз-
жечка. 20 — дорзальная каудальная артерия мозжечка
От нёбной ветви (a. palatinum) отходит четыре артерии.
Глазничная артерия (a. ophtalmica) через крыльный канал про-
ходит в глазницу (рис. 106), где разделяется на наружную и внут-
реннюю глазничные артерии (а.а. ophtalmica externa et interna).
Поверхностная глазничная артерия вместе с носо-ресничной ветвью
тройничного нерва поступает в полость носа. Внутренняя глазнич-
ная артерия снабжает мышцы глаза и слезную железу.
Каудальная верхняя альвеолярная артерия (a. alveolaris cauda-
lis superior) анастомозирует с производными наружной челюстной
артерии и снабжает кровью щечную и жевательную мышцы, а так-
же коренные зубы.
175
Клино-нёбная артерия (a. sphenopalatinum) входит в носовую
полость, а оттуда проникает в альвеолу резца. Снабжает кровью
слизистую носа и раковины.
Подглазничная артерия (a. infraorbitalis) отдает небольшие вет-
ви к мышцам глаза и, проходя через подглазничное отверстие, раз-
деляется на снабжающую зубы краниальную верхнюю альвеоляр-
ную артерию (a. alveolaris cranialis superior) и на носовую ветвь
(ramus nasalis) (рис. 106). Послед-
няя анастомозирует с ветвями лице-
вой артерии и дает ветви к вибрис-
сам.
Отдав все эти ветви, нёбная ар-
терия проходит по твердому нёбу в
альвеолу резца, где она анастомо-
зирует с ветвями клино-нёбной ар-
терии.
Подключичные артерии (а. а.
subclavia), отделившись от дуги
аорты и безыменной артерии, идут
симметрично. Их ветвление сильно
варьирует. Ниже мы описываем ос-
новные ветви подключичной арте-
рии.
Плече-шейный ствол (truncus bra-
chio-cervicalis) обычно отходит сов-
Рис. 108. Артерии и вены грудной
полости пасюка (по Грин):
1 — общая сонная артерия, 2 — шей*
ный ствол, 3 — подмышечная арте-
рия, 4 — наружная грудная артерия,
5 — реберно-шейный ствол, в — вен-
тральные межреберные артерии и ве-
ны, 7 — мышечно-диафрагматические
артерия и вена, 8 — дорзальные меж-
реберяые артерия и вены, 0 — непар-
ная вена. 10 — внутренняя грудная
вена
местно с подмышечной артерией
(a. axillaris) (рис. 103, 108). Плече-
шейный ствол идет в области клю-
чицы и выше на шею, отдавая ветви
к дельтовидной, трапецевидной, гру-
дино-ключично-сосковой и глубоким
мышцам шеи.
Реберно-шейный ствол (truncus
costo-cervicalis) обычно отходит от
подключичной артерии совместно с
внутренней грудной и позвоночной артериями (рис. 108). Дает
первую межреберную артерию (a. intercostalis cranialis), снабжаю-
щую кровью первые три межреберные пространства. Кроме того, от
реберно-шейного ствола отходят передняя перикардиальная арте-
рия (a. pericardialis), артерия средостения (a. mediostenalis) и
бронхиальные ветви.
Позвоночная артерия (a. vertebralis), проникая в поперечно-ре-
берное отверстие VI шейного позвонка, проходит вперед и, прони-
кая в черепную полость, образует вместе с затылочной и внутрен-
ней сонной артериями основную мозговую артерию.
Глубокая шейная артерия (a. cervical is profunda) чаще начи-
нается от подключичной артерии самостоятельно. Она проходит по
шее, снабжая кровью глубокие мышцы шеи, пищевод, гортань,- щи-
176
товидную железу и другие органы шеи (рис. 103). Иногда отходит
совместно с реберно-шейным стволом.
Внутренняя грудная артерия (a. thoracalis interna) нередко
отходит от подключичной артерии самостоятельно. Дает несколько
ветвей (рис. 108). Вентральные межреберные артерии (a. a. inter-
costales ventrales) отходят в числе восьми-девяти и снабжают
межреберные пространства, начиная со второго-третьего. Грудинная
артерия (a. sternalis) фактически является общим стволом вен-
тральных реберных артерий. Диафрагмо-перикардиальная артерия
(a. pericardiaco-phrenica) вместе с артерией средостения участвует
в кровоснабжении перикарда, а кроме того, дает ветвь к диафраг-
ме. Средост'-но-перикардаальная артерия (a. pericardiaco-meaios-
tenalis) участвует в кровоснабжении перикарда и средостения. Про-
бодающая артерия (a. perforans) прободает стенки межреберных
пространств и снабжает кровью грудные мышцы. Краниальная
надчревная артерия (a. epigastrica cranialis) является продолжени-
ем внутренней грудной артерии на брюшную стенку, где она, про-
ходя между косой и прямой мышцами живота, анастомозирует с ка-
удальной надчр-. аноп a;i и ;?ией. Мышечно-диафрагматическая ар-
терия (a. musculo-phrenica) снабжает реберную часть диафрагмы.
Кроме того, от подключичной артерии отделяется шейный ствол
(truncus cervicalis), который дает начало: 1) восходящей шейной
(a .cervicalis ascendens), снабжающей мышцы шеи, пищевод и тпл-
хею, дорзальную зубчатую и лестничную мышцы; 2) повсрхш ст-
ной шейной артерии (a. cervicalis superficialis),снабжающей груди-
но-ключично-сосковую, грудино-щитовидную, грудино-подъязыч-
ную, плече-поцъязычную мышцы и шейные лимфатические узлы;
3) поперечной артерии лопатки (a. transverse scapularis), снаб-
жающей предостную, заостную, подлопаточную и лопаточно-подъ-
язычную мышцы и 4) поперечной шейной артерии (a. transversa
cervicalis), снабжающей трапецевидную, ромбовидную, пластыре-
видную и предостную мышцы (рис. 103).
По отхождении всех этих ветвей подключичная артерия перехо-
дит в подмыш-чную (a. axillaris), которая дает три ветви
(рис. 109). Грудо-йг; ^мчильная артерия (a. thoraclco-acromuUis)
t набжает кровью прямую мышцу живота, грудные и дельтовидное
мышцы. Латеральная грудная л]перия (a. thoracalis laterr-lis)
снабжает кровью широчайшую сп нпл, большую подкожную, груд-
ную, Прямую брюшную И межреберную МЫШЦЫ И ПОЛМЫ1ПечГ|'.1;|
.hrt’Niiи«-ескчн узел. Окру.'КРая под.юплтпчпая .ipti’piiH (я. с r-.’i'г>-
П v.i subseaрularis) разделяется на: 1) краинлльнур» <кру.нвю
артерию шеча (a. circumflexa brachialis cranialis), снаб-
жающую плечевой сустав и длинную головку дау гл а в-vs
мышцы плеча: 2) каудальную окружную артерию плечм
(a. circumflexa brachielis caudalis), снабжающую двуглавую,
трехглавую, дельтовидную и круглые мышцы и плечевой
сустав и 3) подлопаточную артерию (a. subscapularis). lloi-
лопаточная артерия в свою очередь делится на окружную лмпа-
12 Крыса 177
Гоч и ую артерию (a. circumflexa scapularis), идущую к подлопаточ-
ной, заостной и круглым мышцам и к длинной головке трехглавой
мышцы, и грудо-спинную артерию (a. thoracico-dorsalis), идущую
к подмышечному лимфатическом)' узлу, подлопаточной, круглой
большой, широчайшей спины, зубчатой вентральной, трапецевид-
ной и большой подкожной мышцам и к грудным позвонкам.
По отхождении всех этих ветвей подмышечная артерия переходит
в плечевую (a. brachialis), проходящую по плечевой кости (рис. 109).
У локтевого сустава плечевая артерия разделяется на лучевую,
срединную и локтевую артерии. До локтевого сустава от плечевой
артерии отходят несколько ветвей, в том числе: 1) глубокая плече-
вая артерия (a. brachialis profunda), снабжающая кровью трехгла-
вую мышцу плеча, m. epitrochleo-anconeus и плечевую мышцу; кроме
того, эта артерия дает анастомозы к локтевой артерии; 2) локте-
вая коллатеральная артерия (a. collateralis ulnaris), снабжающая
локтевой сустав, длинные сгибатели кисти, трехглавую и пле-
чевую мышцы; 3) поперечная локтевая артерия (a. ulnaris trans-
verse), снабжающая двуглавую мышцу и оба лучевых разгибателя
запястья; 4) локтевая возвратная артерия (a. ulnaris recurrens),
дающая ветви к локтевой мышце, локтевому сгибателю запястья и
локтевой головке глубокого сгибателя пальцев.
Лучевая артерия (a. radialis) снабжает кровью длинные сгиба-
тели кисти. Дистально она соединяется с межкостными пястными
артериями.
Срединная артерия (a. mediana) располагается между лучевым
сгибателем кисти и поверхностной частью глубокого сгибателя паль-
цев. Проходя под поперечной связкой запястья, срединная артерия
разделяется на две ветви. Латеральная анастомозирует с локтевой
артерией, медиальная — с срединно-лучевой. Вместе они образуют
волярную дугу, которая дает ряд пястных волярных артерий, пере-
ходящих в пальцевые артерии. От каждой пальцевой артерии к дор-
зальным пальцевым артериям отходят прободающие ветви.
Проксимально от срединной артерии отходит срединно-лучевая
артерия (a. mediano-radialis). Она участвует в образовании воляр-
ных пальцевых артерий и дает ветвь на дорзальную поверхность
кисти, образующую анастомоз с дорзальными межкостными арте-
риями. Срединно-лучевая артерия анастомозирует также с локтевой
артерией и дает ряд ветвей к мышцам предплечья.
Локтевая артерия (a. ulnaris) участвует в образовании волярнрй
дуги вместе со срединной и срединно-лучевой артериями. Прокси-
мальнолоктевая артерия отдает мышечную ветвь (ramus muscu-
laris), снабжающую длинные сгибатели кисти, сгибатели пальцев
и пястный сустав, и общую межкостную артерию (a. interossea
communis), разделяющуюся на два ствола, из которых один анасто-
мозирует с срединно-лучевой, а другой — с лучевой артериями. От
локтевой артерии отходят также четыре дорзальные артерии пясти и
артерия пятого пальца (рис. ПО).
178
Дуга аорты, отдав подключичные артерии, подходит к вентраль-
ной поверхности позвоночного столба и тянется по ней в виде груд-
ной, а затем брюшной аорты.
Грудная аорта (a. thoracica) дает несколько ветвей, снабжаю-
щих кровью органы грудной полости и ряд пристеночных ответвле-
ний. Межреберные артерии (a. a. intercostales) отходят от грудной
Рис. 109. Артерии передней
конечности пасюка:
1 — подмышечная. 2 — грудо-
акромиальная, 8 — подлопаточ-
ная. 4 — окружная лопаточная,
5 — краниальная окружная плеча,
в — грудо-спинная. 7 — плечевая,
8 — каудальная окружная плеча,
9 — двуглавой мышцы, 10 —мышеч-
ные ветви плечевой. 11 — лучевая.
12 — общая межкостная, 13 —сре-
динная. 14 — локтевая, 16 — сре-
динно-лучевая. 16 — срединно-лок-
тевая, 17 — дорзальная сеть кисти,
18 — волярная сеть кисти
Рис. НО. Артерии кисти пасю-
ка. А — вид с волярной сто-
роны, Б — вид с дорзальной
стороны:
1 — срединная, 2 — локтевая, 3 —
межкостные воляоные, 4 — паль-
цевые волярные, Б — срединно-лу-
чевая, 6 — лучевая. 7 — дорзаль-
ная ветвь волярной межкостной,
8 — межкостные дорзальные, 9 —
пальцевые дорзальные
аорты и направляются в IV—XII межреберные пространства. Каж-
дая межреберная артерия отдает вентральную, дорзальную и спин-
номозговую ветви. Дорзальная ветвь идет к мышцам спины, спин-
номозговая ветвь попадает через межпозвоночные отверстия в
спинномозговой канал, а вентральная ветвь идет у каудального
края каждого ребра, снабжая межреберные мышцы, кожу и под-
кожные мышцы. Пищеводобронхиальный ствол (truncus ofesophago-
bronchialis) делится на бронхиальную (a. bronchialis) и пищевод-
ную (a. oesophagea) артерии; иногда они отходят от грудной аорты
I,- 179
глмоггоятелыю. Краниальная диафрагматическая артерия (a. phre-
nica cranialis) снабжает стенки диафрагмы.
Брюшная аорта (aorta abdominalis) снабжает кровью все осталь-
ные органы тела (рис. Ш). Первой от нее отходит каудальная
диафрагматическая артерия (a. phrenica caudalis), снабжающая
Рис. 111. Артерии и вены брюшной полости па-
сюка. Вены зачернены, артерии светлые:
1 — брюшная аорта, 2 — каудальная ди а ф par магиче-
ская артерия, 8 — чревная арп'пып, 4 — печеночная арте-
рия, Б — желудочная артерия, — селезеночная артклия,
7 — краниальная брыжеечная артерия, 8 — почечная ар-
терия, 9 — '.ни/. и гшгч-»'^ Ю — арте-
рия прямой кишки, 11 — нг.:;>1ьпая мучная
12 — каудальная кишечная вена, 18 — юротнпл тепл пе-
чени, 14 — печеночная в ча. 15 — и- чи'тая * пи 16 —
каудальная полая »>епн
кровью диафрагму. Чревная артерия (a. coeliaca) вскоре после
свое го отхождения от брюшной аорты делится на три ветви: 1) же-
лудочную артерию (a. gastrica), снабжающую стенки желудка;
2) селезеночную артерию (a. lienalis), снабжающую селезенку, же-
лудок и поджелудочную железу и 3) печеночную артерию (a. he-
patica). Краниальная брыжеечная артерия (a. mesenterica cranialis)
дает ветви к поджелудочной железе, двенадцатиперстной, тонкой,
180
ободочной, слепой и части прямой кишкам. Почечные артерии
(a. renales) дают ветви к почкам и к надпочечникам. Парный
внутренние семенниковые артерии (a. a. spermaticae internae) идут
к половым органам. У самок они иногда называются яичниковыми,'
(a. a. ovaricae). У самцов семенниковая артерия вскоре делится на'(
две ветви — к семенникам и к
мошонке и половому члену.
Поясничные артерии (а. а. 1шп-
bales) снабжают брюшные и
поясничные мышцы и кожу
этих областей. У крысы их
имеется до пяти пар. Кау-
дальная брыжеечная артерия
(a. mesenterica caudaiis) снаб-
жает кровью ободочную и пря-
мую кишки. Подвздошно-пояс-
ничная артерия (a. ilio-lumba-
lis) разделяется на подвздош-
ную и поясничную ветви. По-
ясничная ветвь снабжает квад-
ратную мышцу поясницы и да-
ет ветви в позвоночный канал,
в мышцы стенки живота и т. д.
Подвздошная ветвь переходит
по подвздошной кости на дор-
зальную сторону и снабжает
кровью ягодичные мышцы и
кожу. Каудально (рис. 112)
брюшная аорта отдает мощные
общие подвздошные артерии
(a. a. ilicae communes) и пере-
ходит в среднюю хвостовую ар-
терию (a. coccygea media). Об-
щая подвздошная артерия раз-
деляется на наружную и внут-
реннюю подвздошные, снабжа-
ющие органы тазовой области
и заднюю конечность. Нужно
отметить, что у крысы ветвле-
ние артерий этой области сильно варьирует и ниже мы описываем
лишь один из часто встречающихся вариантов.
Наружная подвздошная артерия (a. iliaca externa) разделяется
на срамно-тазовый ствол и бедренную артерию (рис. 130). Срамно-
надчревный ствол (truncus pudendo-epigastrica) отдает: глубокую
окружную подвздошную артерию (a. circumflexa ilii internia), снаб-
жающую брюшные, подвздошные, гребешковую и длинную приво-
дящую мышцы и кожу паховой области; наружные срамные артерии
Рис. 112. Артерии задней конечности
пасюка:
1 — брюшная аорта, 2 — средняя хвосто-
вая, 3 — общая подвздошная, 4 — запи-
рательная, Б — наружная подвздошная,
6 — подчревная, 7 — мышечные ветви,
8 — бедренная. 9 — срамно-надчревный
ствол, 10 — срамная, 11 — артерия npnf
мой кишки. 12 — наружная семенников
вая, 13 — артерия мочевого пузыря, 14 —
средняя окружная бедра, 1Б — сафеиа,
16 — коленная, 17 — подколенная, 18 —
внутренняя подколенная. 19 — наружная
подколенная, 20 — малоберцовая, 21 —
большеберцовая, 22 — дорзальные стопы,
23 — плантарные стопы
181
(a. pudendae externae), снабжающие наружные половые органы,
полусухожильную и стройную мышцы, у самок — млечные железы;
и наружную семенниковую артерию (a. spermatica externa), снаб-
жающую подниматель пениса, наружную окружную артерию бедра
(a. circumflexa femoris lateralis), снабжающую четырехглавую мыш-
цу бедра, и каудальную надчревную артерию (a. epigastrica cauda-
lis), идущую в прямую мышцу живота; у самцов каудальная над-
чревная артерия снабжает также m. cremaster.
Внутренняя подвздошная, или подчревная, артерия (a. iliaca in-
terna, s. hipogastrica) снабжает остальные органы тазовой области
(рис. 112). Краниальная артерия мочевого пузыря (a. vesical is
cranialis) у самок дает артерию матки, которая анастомозирует с
яичниковой артерией и снабжает кровью стенки матки и влагали-
ща. У самцов эта артерия анастомозирует с запирательной арте-
рией. Средняя артерия прямой кишки (a. haemorrhoidalis media)
снабжает кровью прямую кишку. Запирательная артерия (a. obtura-
toria) проходит через запирательное отверстие и дает слабые ветви
к матке, влагалищу (у самок) или к предстательной железе (у сам-
цов). Соединена анастомозами с окружной бедра и со средней арте-
рией прямой кишки. У крыс часто отходит от общей подвздошной
артерии. Срамная артерия (a. pudenda) проходит вместе с запира-
тельной артерией, снабжая кровью тазобедренный сустав, наруж-
ную и внутреннюю запирательные, квадратную бедра, близнецовые,
грушевидную и хвостовые мышцы, а также длинную приводящую
мышцу. Анастомозирует с каудальной надчревной артерией. Сред-
няя окружная артерия бедра (a. circumflexa femoris media) выхо-
дит из полости таза между гребешковой и большой (поясничной)
мышцами. Снабжает гребешковую, приводящие, двуглавую, полу-
перепончатую, полусухожильную и квадратную бедра. Латеральная
окружная артерия бедра (a. circumflexa femoris lateralis) проходит
по большой поясничной мышце и дает нисходящую и восходящую
ветви. Первая снабжает кровью ягодичные, четырехглавую и под-
вздошную мышцы и тазобедренный сустав, вторая идет в прокси-
мальные части ягодичных мышц и в краниальную близнецовую
мышцу. Внутренняя срамная артерия (a. pudenda interna) разде-
ляется на каудальную ягодичную артерию и артерию пениса (кли-
тора). Каудальная ягодичная артерия выходит из тазовой полости
каудальнее грушевидной мышцы и снабжает кровью таз и ягодич-
ные мышцы. Краниальная ягодичная артерия (a. glutaea cranialis)
снабжает кровью близнецовые, грушевидную, среднюю ягодичную
и хвостовую мышцы, а также кожу каудальной части спины и яго-
дичной области.
Бедренная артерия (a. femoralis) является непосредственным
продолжением наружной подвздошной артерии. Проходит по меди-
альной поверхности бедра, переходя дистально в подколенную арте-
рию (рис. 112). По дороге от нее ответвляются: 1) поверхностная
окружная подвздошная артерия (a. circumflexa ilii superficialis),
снабжающая кровью большую подвздошную, гребешковую и четы-
182
рехглавую мышцы; 2) мышечные ветви (rami musculares), снаб*
жающие приводящие, стройную и полуперепончатую мышцы;
3) поверхностная надчревная артерия (a. epigastrica superficialis),
снабжающая кровью прямую и косую мышцы живота и паховую
млечную железу, и 4) коленная проксимальная артерия (a. genu
suprema), снабжающая кровью коленный сустав и медиальную и
промежуточную головку четырехглавой, большую и короткую приво-
дящие мышцы.
Кроме того, от бедренной артерии отходит довольно крупная
артерия сафена (a. saphena). Эта артерия снабжает кровью боль-
шую приводящую, полуперепончатую, полусухожильную и стройную
мышцы и образует коленное сплетение. Дистально от нее отходит
коленная артерия. Около полусухожильной мышцы от артерии са-
фена отходят две дистальные ветви (рис. 112). Одна из них прохо-
дит по дорзальной поверхности голени, а другая — между задней
большеберцовой мышцей и большеберцовой костью, переходя на
стопу. Эта ветвь подробнее описана ниже.
Подколенная артерия отдает несколько ветвей, снабжающих
кровью дистальную часть бедра, голень и стопу, в том числе: 1) по-
верхностную мышечную артерию (a. muscularis superficialis), снаб-
жающую двуглавую и полусухожильную. мышцы и-коленный сустав
и отдающую поверхностную коленную артерию (a. genu superfi-
cialis). Последняя, проходя по дорзальной поверхности голени,
образует четыре межкостные артерии, расположенные в межпаль-
цевых пространствах (рис. 113); 2) наружную и внутреннюю колен-
ные артерии (a. a. genu intemas et externas), снабжающие
кровью икроножную мышцу; 3) латеральную и медиальную про-
ксимальные колейные артерии (a. a. genu proximales lateralis et
medialis), участвующие в образовании сосудистой сети колена. Ла-
теральная снабжает также че^ырехглавую, двуглавую и полупере-
пончатую мышцы, а медиальная — медиальную головку четырех-
главой мышцы; 4) среднюю коленную артерию (a. genu media),
снабжающую кровью короткую приводящую и икроножную мышцы,
а также капсулу коленного сустава, и 5) латеральную и медиальную
дистальные коленные артерии (a. a. genu distales medialis et late-
ralis). Из них латеральная участвует в образовании сосудистой сети
коленной чашечки и снабжает кровью латеральные головки икро-
ножной и двуглавой мышц, а медиальная снабжает кровью голов-
ки берцовых костей.
Дистально подколенная артерия разделяется на переднюю и зад-
нюю большеберцовые артерии (рис. 112).
Задняя большеберцовая артерия (a. tibialis posterior) распола-
гается м< жду длинным сгибателем пальцев и длинным сгибателем
большого пальца. Она снабжает кровью длинные сгибатели паль-
цев, заднеберцовую мышцу и медиальную головку икроножной
мышцы.
Передняя большеберцовая артерия (a. tibialis anterior) идет по
межкостной мембране между большеберцовой и малоберцовой кл-
стами. Дистально она переходит в дорзальные артерии стопы. От
передней большеберцовой артерии отходят: 1) передняя большебер-
цовая возвратная артерия (a. tibialis anterior recurrens), снабжаю-
щая длинную и короткую малоберцовые мышцы и анастомозирую-
щая с коленными артериями; 2) малоберцовая артерия (а. рего-
паеа), снабжающая малоберцовые и камбаловидную мышцы и
кожу дорзальной поверхности стопы, и 3) мышечные ветви (г. г.
musculares), снабжающие перед-
Рис. 113. Артерии стопы пасюка.
А — вид с плантарной стороны, Б—
вид с дорзальной стороны:
1 — сафена, 2 — дорзальная ветвь арте-
рии сафена. 3 — большеберцовые. 4 —
плюсневые дорзальные. 5 — пальцевые
дорзальные, в — плюсневые плантарные*
7 — пальцевые плантарные
нюю большеберцовую мышцу,
длинные разгибатели пальцев и
мышцы среднего пальца; мышеч-
ные ветви анастомозируют с зад-
ней большеберцовой артерией.
Дорзальные артерии стопы (а. а.
plantae dorsales) начинаются от
дуги, образованной слиянием кон-
цевой артерии сафена и передней
большеберцовой артерии (рис.
113). I плюсневая артерия (a. me-
tatarsalis dorsalis I) дает пробо-
дающую ветвь к плантарной плюс-
невой артерии и переходит ди-
стально в дорзальную пальцевую
артерию. II плюсневая артерия
(a. metatarsalis dorsalis II) снаб-
жает короткие разгибатели паль-
цев. Обе они дают прободающие
ветви к плантарным плюсневым
артериям, a. a. metatarsalis dorsa-
lis II анастомозирует также с кож-
ной дорзальной артерией стопы.
III и IV дорзальные плюсне-
вые артерии (a. a. metatarsales
dorsales III et IV) являются кон-
цевыми ветвями поверхностной
коленной артерии (рис. 113). Они
проходят латерально от III и IV
пальцев соответственно и анасто-
мозируют с плантарными арте-
риями.
Артерия сафена участвует в образовании дорзальной дуги стопы
и дает начало медиальной артерии I пальца и латеральной плантар-
ной артерии (a. plantaris lateralis). Последняя снабжает кровью
короткий сгибатель пальцев и тянется по латеральной сто-
роне стопы, анастомозируя с плантарными плюсневыми ар-
териями.
Плантарные плюсневые артерии (a. a. metatarsales plantares) в
числе пяти начинаются от плантарной дуги (рис. 113). Четыре из
184
них разветвляются в межпальцевых пространствах и образуют план-
тарные пальцевые артерии, артерия V пальца сливается с дорзаль-
ной плюсневой артерией.
ВЕНЫ
Вены большого круга кровообращения собирают кровь со всего
организма в правое предсердие. У крысы в него впадают нижняя
полая вена, парные безымянные и сердечные вены. Краниальной по-
лой вены, образующейся при слиянии безымянных, у крысы нет.
Сердечные вены уже описывались выше и здесь мы останавливаем-
ся лишь на системе безымянных и нижней полой вен.
Безымянная вена (v. innominata) образуется слиянием подклю-
чичной и внутренней яремной вен. По пути в нее впадает несколько
вен (см. рис. 103, 108), в том числе: 1) позвоночная вена (v. ver-
tebralis), проходящая через отверстия поперечнореберных отростков
шейных позвонков; 2) внутренняя грудная вена (v. thoracicus), со-
бирающая кровь из краниальной надчревной, диафрагматической,
прободающих, грудной и передней межреберной артерий; 3) средо-
стено-перикардиальная вена (v. pericardiaco-interstenalis), собираю-
щая кровь от перикарда и от средостения; часто впадает непосред-
ственно в безымянную вену; 4) бронхиальные вены (v. v. bronchia-
les), собирающие кровь от трахеи, бронхов и пищевода, и от лим-
фатических бронхиальных узлов; 5) краниальная межреберная вена
(v. intercostalis cranialis), собирающая кровь от первых трех-че-
тырех межреберных пространств; 6) краниальная диафрагматиче-
ская вена (v. frenica cranialis), добирающая кровь от диафрагмы, и
7) непарная вена (v. azygos), проходящая вдоль позвоночного стол-
ба слева от грудной аорты и собирающая кровь от последних семи
межреберных пространств, от квадратной мышцы поясницы, а, по
всей вероятности, и от позвонков. В непарную вену впадает также
полунепарная (v. hemiazygos), собирающая кровь от краниальных
сегментов грудной клетки.
Внутренняя яремная вена (v. jugularis interna) выходит из
черепной полости через заднее рваное отверстие и проходит по шее
вдоль общей сонной артерии (рис. 103). Она собирает кровь от
затылочного и глубокого каменистого синусов и тем самым — от го-
ловного мозга. Во внутреннюю яремную вену впадают затылочная
вена (v. occipitalis), собирающая кровь от области затылочной кос-
ти, щитовидная вена (v. thyroidea), собирающая кровь от области
щитовидной и паращитовидной желез, и глоточная вена (v. pharyn-
gea), собирающая кровь от подъязычных мышц, глотки, гортани
и двубрюшной мышцы. Внутренняя яремная вена собирает также
кровь от ромбовидной, атланто-плечевой, трапецевидной, пластыре-
видной, длинной полуостистой и глубоких шейных мышц (рис. 103).
В подключичную вену впадает наружная яремная вена (v. jugu-
laris externa), собирающая кровь от остальных частей головы. Она
образуется слиянием наружной и-внутренней челюстных веп и про-
ходит по наружной стороне шеи (рис. 103). В нее впадают: 1) йена
1%
№
15 1718
2D
IS
Рис. 114. Вены головы пасюка:
1 — наружная яремная, 2 — внутрен-
няя челюстная, 3 — наружная челюстная,
4 — подчелюстная. 5 — наружная лице-
вая, 6 — нижнегубные, 7 — подбородочная,
u — угла рта, 9 — боковая носа, 10 —
дорзальная носа, 11 — носовая, 12 —
угловая глаза, 13 — средняя ушная, 14 —
передняя ушная, 15 околоушной же-
лезы, 16 — жевательной мышцы, 17 — по-
верхностная височная, 18 — заднеглазпнч-
ная, 19 — поперечная лицевая, 20 — мы-
шечная ветвь
плечевого сустава, собирающая кровь от поверхностной грудной и
дельтовидной мышц, а также от плечевого сустава; 2) поперечная
вена лопатки (v. transversa
scapulae), собирающая кровь
от предостной, подлопаточной,
трапецевидной и ромбовидной
мышц, и 3) ушная вена (v. au-
ricularis), собирающая кровь
от уха. Наружная яремная ве-
на собирает также кровь из
лимфатических узлов шеи.
Наружная челюстная вена
(v. maxillaris externa) прохо-
дит между нижней челюстью и
двубрюшной мышцей (рис. 114,
115). Она образована слияни-
ем наружной лицевой вены,
собирающей кровь от поверх-
ности лица, с подчелюстной
(v. submaxillaris). Последняя
собирает кровь от межчелюст-
ного пространства и двубрюш-
ной мышцы и связана анасто-
мозами с крыловидным и гло-
точным сплетениями.
(v. facialis externa) образована слия-
и верхиегубной (v. labialis superior)
В 7 8
4
Рис.
115. Вены головы пасюка (глубокий слой)
(по Грин):
1 — наружная челюстная, 2 — подчелюстная, 3 — языч-
ная, 4 — внутренняя челюстная, Б — нижнечелюстная,
в — ушная, 7 — мозговая, 8 — поперечная лица, 9 —
жевательной мышцы, 10 — верхняя нЗбная
Наружная лицевая вена
нием угловой (v. angularis)
вен (рис. 114, 115). В
нее впадают также ве-
на утла рта (v. anguli
oris) и нижнегубная
вена (v. labialis inferi-
or) . Угловая вена соби-
рает кровь из надглаз-
ничной (v. supraorbita-
lis), лобной (v. fronta-
lis) и носовой (v. na-
salis) вен.
Внутренняя челюст-
ная вена (v. maxillaris
interna) располагается
между глубокими пор-
циями жевательной и
височной мышц (рис.
114, 115). Она собира-
ет кровь из поперечного синуса челюстного сустава, в который
поступает кровь из челюстного сустава, от барабанной кости и из
нижнечелюстной вены. В нее впадают также: поперечная лицевая
186
пена (v. facialis trans versa), собирающая кровь от височной и же-
вательной мышц, ушные вены (v.v. auricuiares), вены жевательной
н височной мышц (v.v. masseterica et temporalis) и некоторые дру-
। не второстепенные вены. Она собирает также кровь из глоточного
и крыловидного сплетений.
Нижнечелюстная вена (v. mandibularis) проходит по нижнече-
люстному каналу и собирает кровь из альвеолярной вены (v. alveo-
laris) и подбородочной вены (v. mentalis) (рис. 114). Последняя
собирает кровь из нижней губы и связана анастомозами с нижне-
губной веной.
Глоточное сплетение (plexus venosus pharhyngeu) образовано
венами, окружающими глотку и гортань; оно собирает также кровь
от крыловидной мышцы. Крыловидное сплетение (venosus pterigoi-
deu) располагается медиально от нижней челюсти. Оно образова-
но задней верхней альвеолярной, задней менингеальной и крыло-
видной венами, иногда в его образовании участвует глубокая bi -
сочная и нижняя альвеолярная вены. Задняя верхняя альвеолярная
вена (v. alveolaris superior posterior) собирает кровь от альвеол
верхних коренных зубов и из подглазничной вены. Задняя менян
геальная вена (v. meningaea posterior) собирает кровь из оболочек
головного мозга и выходит из черепной полости через вырезку ка-
менистой кости.
Посредством мозговой вены (v. cerebraiis) внутренняя челюст-
ная вена собирает кровь также от головного мозга. Вены головного
мозга связаны с подглазничной веной посредством поперечного и
каменистых синусов (рис. 107). Вентральный и дорзальный камени-
стые синусы (s.s. petrosus ventralis et dorsalis) собирают кровь
от боковых сторон больших полушарий. Поперечный синус (sinus
transversus) располагается на границе между большими полуша-
риями и мозжечком. Каудально от поперечного синуса отходят вен-
тральные и дорзальные вены мозжечка (v.v. cerebelli ventralis et
dorsalis). Дорзальные мозжечковые вены анастомозируют друг с
другом, вентральные мозжечковые вены собирают кровь из кау-
дальной мозжечковой вены (v. cerebelli caudalis) и дают анасто-
моз к позвоночной вене. Краниально от поперечного синуса отхо-
дит сагиттальный синус (sinus sagittalis), располагающийся в бо-
розде между большими полушариями. В сагиттальный синус прино-
сят кровь дорзальные вены большого мозга (v.v. cerebri dorsa-
les). Впереди сагиттальный синус образуется слиянием двух перед-
них (назальных) вен большого мозга (v. cerebri nasalls), прохо-
дящих между обонятельными долями и большими полушариями. В
сагиттальный синус впадают также вентральные вены большого
мозга (v. cerebri ventralis), собирающие кровь с вентральной
стороны больших полушарий. Вентральные вены большого мозга
анастомозируют с дорзальными венами и в области выхода глаз-
ного нерва образуют вентральный синус (s. ventralis).
От передней конечности и плечевого пояса кровь собирается в
подмышечную вену (v. axillaris), переходящую далее в подключнч-
187
ную (v. subceavia). В подмышечную вену впадают: 1) окружная
лопаточная вена (v. circumflexa scapularis), собирающая кровь от
круглой большой мышцы и от широчайшей спины; 2) акромиально-
грудная вена (v. thoracico-acromialis), собирающая кровь от лате-
ральных сторон грудной клетки и от подлопаточной мышцы, и
3) латеральная грудная вена (v. thoracica lateralis), собирающая
кровь от широчайшей мышцы спины и от большой подкожной
мышцы.
Сама подмышечная вена является непосредственным продолже-
нием плечевой (v. brachialis), проходящей по плечевому отделу
передней конечности. Из вен, впадающих в плечевую вену, назовем
дорзальную и волярную окружные вены плеча (v.v. circumflexae
brachii volaris et dorsalis), собирающие кровь от мышц, располо-
женных вокруг плечевой кости. Плечевая вена образуется в резуль-
тате слияния нескольких крупных стволов предплечья, наиболее по-
стоянными из которых можно считать срединную (v. media), об-
щую межкостную (v. interossei communis) и локтевую (v. ulnaris)
вены. Локтевая вена выносит кровь из волярной венозной сети ки-
сти (rete manus volaris), а общая межкостная и срединная — из
дорзальной венозной сети кисти (tere manus dorsalis).
С каудальной стороны в сердце впадает каудальная полая века
(v. cava caudalis), образующаяся путем слияния общих подвздош-
ных вен. Она поступает в грудную полость, пробочая диафрагму.
У самой диафрагмы в каудальную полную вену впалает печеночная
вена (v. hepatica). Последняя собирает кровь, приносимую в печень
воротной веной печени (v. porta hepatis).
Воротная вена печени образуется слиянием селезеночной, кра-
ниальной брыжеечной и каудальной брыжеечной вен (рис. 111). Се-
лезеночная вена (v. lienalis) собирает кровь от селезенки, желудка
и двенадцатиперстной кишки. Краниальная брыжеечная вена
(v. mesenterica cranialis) начинается от тонкой, ободочной и сле-
пой кишок. Каудальная брыжеечная вена (v. mesenterica caudalis)
собирает кровь от толстой и от прямой кишок.
Воротная вена образует в печени капиллярную сеть, от которой
и берет начало печеночная вена.
Каудальная полая вена образуется слиянием общих подвздошных
вен. На своем пути к сердцу каудальная полая вена принимает не-
сколько парных вен, в том числе: 1) каудальную диафрагматиче-
скую вену (v. phrenica caudalis), собирающую кровь от диафраг-
мы и надпочечника; 2) почечную вену (v. renalis); 3) семеннико-
вую (яичниковую) вену (v. spermatica s. ovarii); 4) поясничные
вены (v.v. lumbales), собирающие кровь от поясничных позвонков,
спинных мышц и от квадратной мышцы поясницы, и 5) подвздош-
но-поясничную вену (v. ilio-psoas), собирающую кровь от брюшных
мышц, от большой подкожной мышцы, и от большой поясничной и
квадратной мышц поясницы.
У крысы каудальные диафрагматические и почечные вены обыч-
но не симметричны. Левая каудальная диафрагматическая вена у
188
нее собирает кровь, главным образом от надпочечника, и впадает в
левую почечную вену; в последнюю нередко впадает и левая се-
менниковая вена. Правая почечная вена собирает кровь только от
почки, а иногда еще и от надпочечника.
Общая подвздошная вена (v. iliaca communis) собирает кровь
от органов тазовой области и задних конечностей. Образуется она
слиянием наружной подвздошной и подчревной (внутренней под-
вздошной) вен.
Подчревная вена (v. hipogastrica) принимает: 1) краниаль-
ную вену мочевого пузыря (v. vesicalis cranialis), собирающую
кровь от мочеиспускательного канала и от мочевого пузыря. У са-
мок она принимает маточную вену (v. uretrae); 2) запирательную
вену (v. obturaioria), собирающую кровь от глубокой мускулатуры
таза. В нее впадает средняя вена прямой кишки (v. haemorrhoi i.i-
iis media), которая собирает кровь от хвостовых мышц и от прямой
кишки; 3) медиальную окружную вену бедра (v. circumflexa L пю-
ris medialis), собирающую кровь от мышц области малого вертлу-
га, т. е. приводящих, гребешковой, стройной, поясничной и запира-
тельной; 4) лат. ри. п яую окружную вену бедра (v. circumflexa fe-
monis lateralis), собирающую кровь от четырехглавой, ягодичных
поверхностной и средней, близнецовых и большой поясничной
мышц; 5) внутреннюю срамную вену (v. pudenda interna), соби-
рающую кровь от половых органов. В эту вену впадают дорзальная
вена полового члена (клитора у самок) и вена пещеристых тел по-
лового члена. Внутренняя срамная вена анастомозирует с геморрои-
дальным сп.’н ъ Ши м, в которое собирается кровь от хвостов: :х
мышц и от прямой кишки; 6) внутреннюю ягодичную вену (v. izlu-
taea interna), собирающую кровь от мошонки самцов и от про-
межности у самок. Нередко впадает в подчревную вену само-
стоятельно; и 7) наружную ягодичную вену (v. glutaea externa),
собирающую кровь от ягодичных, грушевидной, хвостовой и близ-
нецовых мышц.
Наружная подвздошная вена (v. iliaca externa) образуется
• сиянием надчрспчо-срамной и бедренной вен. Надчревно-срН’ ’ " я я
вена (v. pudendo-epigastrica) принимает следующие вены: 1) глубо-
кую окружную подвздошную вечу (v. circumflexa ilii profunda), со-
бирающую кровь от мускулатуры живота и подвздошио-пояс*'!р-чой
мышцы; 2) каудальную надчревную вену (v. epigastrica each'-'-),
собирающую кровь от прямой мышцы живота; 3) наружную се-п'-ч-
иую tv ну (v. spermatica externa), собирающую кровь от пошпча-
теля полового члена, и 4) краниальную и каудальную наружные
срами: и- вены (v. pudenuae externae cranialis et caudalis). ~Kay*
лмычая собирает кровь от геморроидального венозного сплетения
и половых <>:• .'i.t vi, расположенных в полости таза, краниалы яя ::<е
у . .(мцов собирает кровь от препуция, мошонки, полового члена и
семенников, а у самок — от преддверия влагалища и паховых мо-
лочных желез.
Бедренная вена (v. femoraiis) проходит с медиальной стороны
мышц бедра. Она образуется близ коленного сустава слиянием под-
коленной вены и вены сафена. Бедренная вена принимает мышеч-
ные ветви (rami musculares), собирающие кровь из заднебедренных
мышц и из разгибателей голени, и поверхностную надчревную вену
(v. epigastrica superficialis), собирающую кровь от промежности,
подвздошной и паховой областей.
Вена сафена (у. saphena) тянется вдоль медиальной поверхно-
сти голени до стопы и собирает кровь из дорзального и латераль-
ного венозных сплетений стопы (plexus lateralis et plantaris pedis).
Вена сафена принимает также кровь из латеральной вены стопы
(v. lateralis pedis) и глубоко# плантарной дуги (arcus venosus
plantaris pedis pipfundus), в которую несут кровь четыре плантар-
ные плюсневые вены (v.v. metatarsales plantaresi). Кроме того, в
вену сафена впадает медиальная вена стопы (v. pedis medialis),
собирающая кровь от медиальной стороны стопы и пальцев и ана-
стомозирующая с ветвями дорзальной венозной дуги.
Подколенная вена (v. poplitea) образуется в результате слия-
ния передней и задней большеберцовых вен. Передняя большебер-
цовая вена (v. tibialis anterior) собирает кровь из дорзальной ве-
нозной дуги стопы (arcus venosus dorsalis pedis)). В последнюю
впадают дорзальные плюсневые вены I и II (v.v. metatarsales dor-
sales I et II). Задняя большеберцовая вена (v. tibialis posterior)
собирает кровь от икроножной и других мышц плантарной стороны
голени и от дорзальных плюсневых вен III и IV.
ЛИМФАТИЧЕСКАЯ СИСТЕМА
Лимфатическая система образует в тканях систему слепых ка-
пиллярных сосудов, объединяющихся в более крупные сосуды. По-
следние открываются в безымянные вены. Лимфа свободно фильт-
руется сквозь стенки капилляров и непосредственно связана с
межтканевой жидкостью. По своему строению лимфатические сосу-
ды близки к венам, но на своем пути они образуют так называемые
лимфатические узлы — соединительнотканные образования, в кото-
рых лимфатические сосуды распадаются на капилляры.
Лимфатические сосуды делятся на поверхностные, собирающие
лимфу от кожи и подкожных пространств, и глубокие, собирающие
лимфу от остальных органов. Лимфатические сосуды поступают в
регионарные лимфатические узлы, располагающиеся в брыжейке,
в ямках межмышечных пространств и в сгибах (рис 116). Лимфа-
тические сосуды передних конечностей проходят через подмышеч-
ный узел, сосуды задних конечностей — через подколенный, пахо-
вый и подвздошный, сосуды головы — через подчелюстной и
подъязычный лимфатические узлы. В лимфатические узлы бры-
жейки собирается лимфа со стенок кишок. Соответствующие лим-
фатические сосуды именуются млечными, так как лимфа в них ок-
рашена в белый цвет капельками жира, поступающими из содер-
жимого кишечника. Лимфатические сосуды задп'-й половины тела,
190
кишечника, брюшной и грудной стенок собираются в левый лимфа-
тический проток, впадающий в левую безымянную вену, а сосуды
передних конечностей и головы через правый лимфатический про-
ток впадают в правую безымянную вену.
Ряс. 116. Поверхностные лимфатические узлы и сосуды пасюка:
1 — подчелюстной узел, 2 — подмышечный узел, 3 — подвздошный узел,
4 — подколенный узел
ОРГАНЫ КРОВЕТВОРЕНИЯ
У взрослой крысы лейкоциты образуются в основном в лимфа-
тических узлах и в селезенке, эритроциты — в красном костном
мозге.
г
1
Рис. 117. Селезенка пасюка:
1 — ворота, 2 — сосуды
Селезенка (lien) представляет собой уплощенное образование,
расположенное рядом с желудком (рис. 117). Со стороны желулнз
i> селезенку входят сосуды и нервы; желоб, по которому они пре v -
дят, называется воротами селезенки (hilus lienis). Снаружи <• к-
^ика покрыта серозной оболочкой, образованной одним in ли >
ИИ
ков большого сальника — частью брыжейки желудка, богатой жи-
ровыми клетками.
Селезенка имеет и собственную соединительнотканную оболочку,
продолжающуюся в толщу органа в виде сети перекладин.
Последние образуют строму селезенки. Перекладины широко ана-
стомозируют друг с другом, промежутки между ними заполнены
нежной лимфоидной тканью — пульпой. Пульпа и является тканью,
продуцирующей лейкоциты. Кровеносные сосуды селезенки сообща-
ются с межклеточными пространствами, и по ним образующиеся
лейкоциты легко поступают в кровяное русло.
В селезенке осуществляется также распад отживших эритроци-
тов. В ней, как и в лимфатических уздах, кровь очищается от по-
павших в нее бактерий. Последние захватываются и уничтожаются
лейкоцитами и другими клетками, способными к внутриклеточному
пищеварению. Во время внутриутробного развитии в селезенке об-
разуются эритроциты. Кроме того, селезенка может переполняться
кровью и служит одним из кровяных депо организма.
Лимфатические узлы (noduli lymphatica по строению близки к
селезенке. Они одеты соединительнотканной оболочкой, от которой в
i«>.'iiiiy узла отходят перегородки, образующие строму лимфатиче-
ского узла. Промежутки между перекладинами заполнены лимфоид-
ной тканью, продуцирующей лейкоциты. Лимфатические сосуды рас-
падаются в лимфатических узлах на капилляры. Функции лимфати-'
веских узлов близки к функциям селезенки, но в них кровеносные
сосуды не сообщаются с межклеточными пространствами и распа-
да эритроцитов не происходит.
Органом образования эритроцитов у взрослой крысы служит
красный костный мозг, располагающийся в плоских костях, напри-
мер, в грудине. Он образован нежной ретикулярной тканью, богато
снабженной капиллярами. Ретикулярная ткань и образует эритро-
циты; кроме того, в ней образуются и зернистые лейкоциты (гра-
нулоциты) .
ТЕХНИЧЕСКИЕ ЗАМЕЧАНИЯ
Изучение кровеносной системы немыслимо без инъекции сосу-
дов. Наиболее хорошие результаты получают при инъецировании
сосудов окрашенным раствором целлулоидина. При инъецировании
вен в качестве красителя принято брать синьку, а при инъецирова-
пси артерий — кармин.
Для растворения целлулоидина берут смесь спирта с эфиром
или ацетоном в соотношении 1:1 или 7:3. Целлулоидин раство-
ри) -т до консистенции жидкого масла. Для инъекции артерий ис-
пользуют шприц емкостью 10 мл. Шприц меньшей емкости недо-
статочен, а ббльшей — неудобен для работы.
Артерии ин ) еитруют через брюшную аорту. На аорту наклады-
вают петлю из хирургического шелка или простой нитки так, что-
бы ее стенки плотно сидели на игле; нужно следить, чтобы нитка
не прорезала стенки сосуда. Наполнение сосудов следует произво-
192
дить легким равномерным нажатием на поршень шприца. Для рав-
номерности давления рекомендуется прикрепить к стенкам цилинд-
ра и к поршню шприца два резиновых тяжа. Когда дальнейшее
наполнение сосудов начинает встречать сильное сопротивление, что
обычно происходит после инъекции по крайней мере 3 мл, нужно
сделать перерыв на полчаса, после чего вновь повторить наполне-
ние сосудов и так в течение 3—5 часов. При изучении артерий все-
го тела лучше проводить инъекцию на двух животных, так как
выполнить двустороннюю инъекцию очень трудно.
Вены перед инъекцией приходится промывать, чтобы удалить
кровь, скапливающуюся в них посмертно. Клапаны вен создают
большое сопротивление инъецирующей жидкости, и потому вены
приходится инъецировать в нескольких пунктах. При этом не сле-
дует брать шприц емкостью более 2 мл, так как венозную систе-
му все равно наполнить за один раз технически невозможно.
Иногда инъекцию сосудов производят так называемым горячим
способом. Для этого одну часть желатины растворяют в течение
3—4 часов в двух частях воды, а затем нагревают и окрашивают в
нужный цвет. Перед инъекцией крысу помещают в воду, подогре-
тую до 50—60°.
Для изучения лимфатической системы в толщу тканей живой
крысы в нескольких местах вводят обыкновенную тушь или бер-
линскую лазурь; инъецируемая жидкость распространяется в ор-
ганизме по лимфатическим сосудам. Таким образом удается выя-
вить области, от которых лимфа собирается в тот или иной регио-
нарный лимфатический узел, и проследить направление лимфатиче-
ских сосудов.
ГЛАВА XIII
ЖЕЛЕЗЫ ВНУТРЕННЕЙ СЕКРЕЦИИ
ОБЩИЕ ЗАМЕЧАНИЯ
Все ткани и органы живого организма выделяют продукты об-
мена, которые, поступая в кровь и димфу, разносятся по всему ор-
ганизму. Такие продукты могут влиять на жизнедеятельность дру-
гих органов. У позвоночных имеются специальные органы, выде-
ляющие в кровь вещества, регулирующие жизнедеятельность орга-
низма. Такие органы называются железами внутренней секреции, а
вещества, выделяемые ими, гормонами. Ниже мы описываем важ-
нейшие железы внутренней секреции крысы, кроме так называемых
лангергансовых островков поджелудочной железы, упоминавшихся
выше.
ЩИТОВИДНАЯ И ОКОЛОЩИТОВИДНАЯ ЖЕЛЕЗЫ
Щитовидная железа (glandula thyreoidea) — небольшое парное
образование треугольной формы. Располагается она симметрично с
вентральной стороны трахеи, у самого основания последней
(рис. 118). Щитовидная железа построена из железистого эпителия,
образующего отдельные мешочки — фолликулы. Железа покрыта
соединительнотканной оболочкой, от которой отходят нежные пере-
городки между отдельными фолликулами. Фолликулы заполнены
секретом — коллоидом, который всасывается в кровь через стенки
сосудов, проходящих в перегородке.
Гормон щитовидной железы повышает обмен веществ и имеет
большое значение для процессов роста и дифференцировки, в част-
ности волос. При удалении щитовидной железы у крыс наблюдает-
ся общее недоразвитие половых желез, замедленный рост костей.
Усиление функции железы также ведет к патологическим явлениям
(базедова болезнь).
Околощитовидная железа, или эпителиальные тельца (glandulae
parathyreoideae), представляет собой небольшие образования, рас-
положенные на теле щитовидной железы в дорзальной части по-
следней (рис. 118).
194
Гормон околощитовидной железы влияет на обмен кальция, спо-
собствуя повышению его уровня в крови. При оперативном удале-
нии околощитовидной железы содержание кальция в крови падает
и наступают явления сильной возбудимости мышц, проявляющиеся
в тетанических судорогах.
ЗОБНАЯ ЖЕЛЕЗА
Вилочковая, или зобная, железа (glandulae thymus) крысы
представляет собой довольно крупное образование, лежащее под
Рис. 116. Щитовидная
(1) и околощитовидная
(2) железы пасюка
трахеей в самом краниальном отделе
грудной клетки. Состоит она из двух до-
лек, прилегающих друг к другу (рис. 119).
Зобная железа покрыта соединительно-
тканной оболочкой, ее тело выполнено
Рис. 119. Зобная железа
пасюка:
1 — зобная железа, 8—
сердце, 8 — легкие
железистой паренхимой, разделенной перегородками на дольки; пе-
регородки исходят из центрального соединительнотканного 'стержня
и в совокупности образуют строму железы. Гормон зобной железы
способствует задержке кальция в костях. При удалении зобной же-
лезы у молодых крыс рост организма замедляется. С возрастом
зобная железа уменьшается, и у взрослых крыс ее удаление почти
не отражается на состоянии животного.
НАДПОЧЕЧНИКИ
Надпочечники (glandulae suprarenales) крысы имеют вид жел-
товатых тел, тесно прилегающих к краниальной поверхности каж-
дой почки.
Надпочечник покрыт соединительнотканной оболочкой, дающей
начало строме железы. Его вещество дифференцировано на корко-
вое и мозговое (рис. 120). Мозговое вещество выделяет адреналин,
активирующий деятельность органов, возбуждаемых симпатической
13* 1%
нервной системой. При его введении в кровь сердцебиения учаща-
ются, кровеносные сосуды сужаются, а кровяное давление повы-
шается и снимается мышечное утомление. Корковое вещество вы-
деляет различные стероидные гормоны, влияющие на обмен ве-
ществ, в частности, калиевый и водный. При удалении коры ткани
обезвоживаются, уровень калия в крови повышается, наступают яв-
Рис. 120. Поперечный срез ле-
вого надпочечника пасюка:
1 — оболочка. 2 — корковый слой,
3 — мозговой слой
Рис. 121. Гипофиз
пасюка:
1 — задняя доля,
2 — передняя доля,
3 — ножка
ления мышечной слабости. При полном удалении коры неизбежна
тибель животного. При гиперфункции коры часто ускоряется поло-
вое созревание.
ПОЛОВЫЕ ЖЕЛЕЗЫ
Половые железы относятся к числу желез внутренней секреции.
Их гормоны стимулируют развитие вторичных половых признаков.
Желтое тело яичников выделяет гормоны, стимулирующие рост
эндометрия и приостанавливающие дальнейшее созревание яйце-
клеток.
ЭПИФИЗ И ГИПОФИЗ
Эпифиз (epiphysis) представляет собой небольшой пузырек, рас-
полагающийся в сагиттальной плоскости между большими полуша-
риями и мозжечком. Он является гомологом так называемого те-
менного глаза, функционирующего у некоторых низших позвоноч-
ных. Строма эпифиза, связанная с его оболочкой, разделяет тело
железы на дольки. Функции эпифиза еще недостаточно выяснены,
повидимому его гормон несколько задерживает половое созревание.
С возрастом эпифиз постепенно перерождается.
Гипофиз мозга (hypophysis) представляет собой небольшое ок-
руглое образование, располагающееся на вентральной поверхности
головного мозга и связанное ножкой с дном промежуточного мозга.
Гипофиз состоит из двух легко различимых долей — задней и перед-
196
ней (рис. 121). Первая по происхождению представляет собой вы-
рост мозга, вторая — производное глоточного эпителия. Обе доли
разделены слабо развитой промежуточной, которую можно обнару-
жить только на срезах.
Гормоны передней доли стимулируют рост организма и дея-
тельность других желез внутренней секреции. Гормон промежуточ-
ной доли у низших позвоночных способствует контракции пигмент-
ных клеток — меланофоров. Его функции у высших позвоночных
неясны.
Гормоны задней доли повышают тонус гладких мышц, кровяное
давление и замедляют мочеобразование. У самок они играют боль-
шую роль при родах, стимулируя сокращение матки.
ГЛАВА XIV
НЕРВНАЯ СИСТЕМА
ОБЩИЕ ЗАМЕЧАНИЯ
По нервным клеткам — невронам — раздражения передаются от
органов, воспринимающих их, к органам, отвечающим на эти раз-
дражения. Совместно с органами чувств нервная система опосре-
дствует связи организма со средой и координирует деятельность
его частей. Эмбрионально нервная система происходит из эктодермы.
Нервная система крысы, как и других позвоночных, разделяется
на центральную (головной и спинной мозг со спинномозговыми
ганглиями) и периферическую (нервные волокна, идущие от голов-
ного и спинного мозга). Кроме того, в особую группу выделяют
так называемую вегетативную нервную систему, включающую пери-
ферические нервы, сплетения и ганглии, связанные с внутренними
органами и более или менее обособленные от центральной нервной
системы. Последняя совместно с периферическими нервами, связан-
ными с органами чувств и поперечнополосатой мускулатурой, обра-
зует соматическую нервную систему.
Нервная клетка (рис. 122 Г) состоит из тела и отростков. Ко-
роткие, сильно ветвящиеся отростки, называемые дендритами, про-
водят раздражение к телу клетки, более длинные отростки — ней-
риты — передают раздражение на периферию. Нейриты одеты бе-
лой миелиновой оболочкой и образуют в совокупности белое веще-
ство головного и спинного мозга. Серое вещество мозга образовано
самими нервными клетками; следует отметить, что нейриты вегета-
тивной системы лишены миелиновой оболочки и отличаются серым
цветом.
Нейрон, воспринимающий раздражение, совместно с нейроном,
передающим его к эффекторному органу, образует рефлекторную
лугу (рис. 122 А). Между рецепторным и эффекторным нейронами
располагается один или несколько вставочных. Нейроны, воспри-
нимающие раздражение от кожи, передают его в дорзальные выро-
сты (рога) спинного мозга; оттуда через вставочный нейрон раз-
198
дражение передается дендритам эффекторной клетки, расположен-
ной в вентральном роге спинного мозга. По нейрону последней
раздражение достигает эффекторного органа.
СПИННОЙ мозг
Центральная нервная система развивается из полоски утолщен-
ной эктодермы, сворачивающейся в трубку (рис. 122 Б). Передняя
часть трубки значительно расширяется и образует головной мозг, а
остальная ее часть превращается в спинной мозг. Спинной мозг
располагается в спинномозговом канале, образованном дугами поз-
вонков. Спинной мозг растет медленнее позвоночного ствола и
оканчивается на уровне 3—4, а у старых — на уровне 1—2 пояс-
ничных позвонков. Так как спинномозговые нервы выходят из
спинномозгового канала посегментно, они на большом протяжении
тянутся внутри спинномозгового канала позади конца спинного
мозга, образуя так называемый конский хвост (cauda equina).
Спинной мозг крысы образует два веретенообразных расшире-
ния— шейное (intumescentia cervicalis), располагающееся в обла-
сти IV шейного — П грудного позвонков, « поясничное (i. lumba-
lis), располагающееся в области X грудного — II поясничного поз-
вонков (рис. 132). От шейного расширения отходят спинномоз-
говые нервы, обравующис плечевое сплетение, от поясничного рас-
ширения отходят нервы, образующие пояснично-крестцовое спле-
тение.
Спинной мозг одет поверхностной твердой (dura mater) и более
глубокой мягкой (pia mater) оболочками. Эти оболочки продолжа-
ются и после окончания спинного мозга, образуя так называемую
концевую нить (filum terminale) (рис. 122 В).
На вентральной поверхности спинного мозга развита вентраль-
ная щель (fissura medi'ana ventralis), а на дорзальной стороне —
дорзальная срединная борозда (sulcus medianus dorsalis). По сто-
ронам спинного мозга проходят дорзальная и вентральная лате-
ральные борозды (sulci laterales dorsalis et ventralis).
Серое вещество (substantia grisea) расположено вокруг спинно-
мозгового канала (canalis centralis). На поперечном разрезе
(рис. 122 В) оно напоминает бабочку, крылья которой именуются ро-
гами (cornua dorsalis et ventralis). От рогов серого вещества отхо-
дят спинные (чувствующие) и брюшные (двигательные) корешки
спинномозговых нервов; соединяясь друг с другом, корешки обра-
зуют спинномозговые нервы. В спинных корешках располагаются
спинномозговые ганглии (ganglia spinales). Рога образуют прохо-
дящие по всему спинному мозгу столбы серого вещества (columnae
grisea). По сторонам спинномозгового канала столбы серого веще-
ства соединяются друг с другом спайками (comissurae grisea)
(рис. 122 В). Серое вещество сильнее развито в области шейного и
поясничного утолщений, особенно толсты здесь брюшные рога, в ко-
торых располагаются тела двигательных нейронов, иннервирующих
конечности.
199
Белое вещество (substantia alba) занимает периферию спинного
мозга. Столбами серого вещества и бороздками на поверхности
спинного мозга белое вещество разделяется на дорзальный, вент-
ральный и боковые канатики (^b^iculi dorsalis, ventralis et late-
rals). В белом веществе проходят нервные волокна, соединяющие
различные отделы спинного мозга, й образующие его проводящие
пути (рис. 122 В), ведущие к головному мозгу и обратно.
Центростремительные (чувствующие) проводящие пучки, иду-
щие к головному мозгу, расположены в дорзальном и отчасти в
боковых канатиках. Чувствующие пучки от задней части тела идут
вдоль дорзомедиальной борозды, образуя пучок Голля (fasciculus
gracilis, seu Golli). Рядом с ним, начиная от уровня передних ко-
нечностей, проходит клиновидный пучок, или пучок Бурдаха (f. cu-
neatus seu Burdachi), граничащий латерально с дорзальным стол-
бом серого мозгового вещества. Первый пучок кончается в продол-
говатом мозге в специальном ядре пучка Голля (nucleus fasciculi
gracilis), ядро пучка Бурдаха (n. fasciculi cuneati) лежит у основа-
ния задних ножек мозжечка. Некоторая часть восходящих волокон
обоих пучков оканчивается у медиальной части основания дорзаль-
ного рога спинного мозга. В боковом канатике проходит три восхо-
дящих пучка: 1) дорзальный мозжечковый jfcL spino-cerebellaris
dorsalis), проходящий через задние ножки моЛпка* в переднюю
долю червячка; 2) центральный мозжечковый "(Г. sj^ho-cerebellaris
ventralis), направляющийся в червячок через передние ножки моз-
жечка; 3) пучок зрительного бугра и четверохолмия (tractus spino-
halamicus), оканчивающийся в сером веществе зрительного бугра и
в переднем холме четверохолмия.
Нисходящие (двигательные) проводящие пути располагаются в
боковом и вентральном канатиках. В боковом канатике различают
Рис. 122. Схема строения и развития нервной^астемы. Л — ррфлек-
торная дуга,'® — развитие спинного мозга/^Х" проводящие
спинного мозга, Г — нейрон:
I—III — последовательные стадии развития мозга (поперечные разрез), IV—
нервная трубка в продольном разрезе; ,1 — дорзальный канатик, 2 — вста-
вочный нейрон, 3—4 — дорзальный рбг, Б — дорзальный корешок, в — бо-
ковой канатик, 7 — центральный спинномозговой канал, 8 — вентральный
рог, 9 — тело рецепторного иейрона, 10 —ТкожаД 11 — рецепторное оконча-
ние, 12 — вентральный канатик, 13 — вентральная ^срединная щ,ель, 14 — те-
ло эффекторного нейрона, 15 — нейрит эффекторного нейрона. 10 — вент-
ральный корешок, 17 — смешанный нерв, 18 — мускульное волокно с эффек-
торными нервными окончаниями,} 19 — боковые нервиые валики, 20 — ган-
глионарная пластинка, 21 — нервная борозда, 22 — нервная пластинка, 23 —
вторичный мозговой пузырь, 24 — пластинка покрышки, 25 — нервная труб-
ка, 26 — боковая пластинка, 27 — центральный канал, 28 — основная пла-
стинка, 29 — пластинка дна, 30 — первичный мозговой пузырь, 31 — невро-
пор, 32 — глазной пузырь, 33 — вентральная складка, 34 — хорда, 35 — заднее
отверстие, 36—четверохолмный пучок, 37 — вентральный пирамидный пучок,
38 — пучок зрительного бугра, 89 — вестибулоспинальный пучок, 40 —• вентраль-
ный мозжечковый пучок бокового канатика, 41 — боковой основной пучок,
42 — пучок Монакова, 43 — пирамидный боковой пучок, 44 — дорзальный моз-
жечковый пучок бокового канатика, 45 — дорзальный основной пучок, 46 — пу-
чок Бурдаха, 47 — пучок Голля, 48 — коллатераль нейрита. 49 — дендриты,
50 —тело нейрона, 51 — нейрит, 52 — сегменты миелиновой оболочки, 53 — пе-
рехваты Ранвье, 54 — телодендрнй (эффекторное нервное окончание), 55 —му-
скульное волокно (по Климову)
200
Рис. 122
три нисходящих пучка: 1) боковой, или перекрещенный, пирамид-
ный пучок (f. cerebro- (seu cortico-) spinalis lateralis), идущий
от коры больших полушарий через ножки большого мозга и пира-
миды продолговатого мозга к спинному мозгу, где он проходит
между дорзальным столбом и дорзальным мозжечковым пучком и
оканчивается в моторных клетках вентрального столба; 2) боковой
Б
Рис: 123. Схема пятипузырной ста-
дии развития головного мозга и его
желудочки. А — вид сбоку, Б —
вид во фронтальном разрезе:
1 — конечный мозг. 2 — глазной пузырь,
3 — промежуточный мозг. 4 — средний
мозг, 6 — перешеек, 6 — закладка моз-
жечка. 7 — мозговой парус. 8 — заты-
лочный изгиб, 9 — спинной мозг, 10 —
ромбовидный мозг, 11 — монроево отвер-
стие, 12 — боковой желудочек, 13 — тре-
тий мозговой желудочек. 14 — сильвиев
водопровод, 15 — четвертый мозговой
желудочек
пучок из красного ядра, или пу-
чок Монакова (f. rubro-spinalis,
seu Monakowi), начинающийся
от нейронов красного ядра сред-
него мозга и оканчивающийся в
моторных клетках вентрального
столба и 3) вестибулярно-спи-
нальный пучок, млн пучок Левен-
таля (f. vestibulo-spinalis), начи-
нающийся от ядра Дейтерса слу-
хового нерва и оканчивающийся
в вентральном столбе. В вент-
ральном канатике проходят два
пучка: 1) вентральный, или пря-
мой, пирамидальный пучок (пучок
Тюрка) (f. cerebro- (seu cortico-)
spinalis ventralis), начинающий-
ся от пирамидальных клеток ко-
ры больших полушарий и окан-
чивающийся в вентральных стол-
бах противоположной стороны;
2) четверохолмный пучок (f. tec-
to-spinalis), идущий от передних
бугров четверохолмия и после пе-
рекреста оканчивающийся в вент-
ральных столбах спинного мозга
(рис. 122 В).
головной мозг
У млекопитающих головной
мозг закладывается в виде рас-
ширенной передней части нервной
трубки. Последняя разделяется
складками на 3 мозговых пузыря.
Передний пузырь образует конечный мозг (telencephalon) и про-
межуточный мозг (diencephalon), средний пузырь превращается в
средний мозг (mesencephalon), а задний пузырь образует продол-
говатый мозг (myelencephalon) и мозжечок (cerebellum) (рис. 123).
Полости мозговых пузырей превращаются в мозговые желудоч-
ки. В больших полушариях располагаются парные боковые желу-
дочки (venriculi laterales), соединяющиеся друг с другом и с желу-
дочком промежуточного мозга (третьим), так называемым монрое-
202
вым отверстием (foramen interventricular is). Желудочек промежу-
точного мозга редуцируется до узкого канала, называемого силь-
Рис. 124. Головной мозг пасюка. А — вид сбоку, Б — вид
снизу, В — вид сверху:
1 — обонятельные луковицы, 2 — основание обонятельного нерва,
3 - зрительный перекрест. 4 — подглазничная ветвь тройничного
нерва, Б — поперечная борозда, в — продольная щель, 7 — глазнич-
ная ветвь тройничного нерва. В — верхнечелюстная ветвь тройнично-
го нерва, 9 — глазное яблоко, 10 — червячок, 11 — задняя доля
червячка, 12 — средняя доля червячка, 13 — передняя доля чер-
вячка, 14 — каудальный парус, 15 — полушария мозжечка. 16 -
эпифиз, 17 — гипофиз, 18 — варолиев мост, 19 — зрительные доли,
20 — ветвь лицевого нерва, 21 — пирамиды, 22 — парафлокулус,
23 — латеральная обонятельная извилина. 24 - общая обонятельная
извилина, 26 — медиальная обонятельная извилина. 26 — плащ, 27 —
продолговатый мозг; 1—XII — черепные нервы, CI — первый шейный
нерв
виевым водопроводом (aqueductus Sylvii). Четвертый мозговой же-
лудочек (ventriculus quartus) располагается в продолговатом мозге
и переходит в спинномозговой канал (рис. 123).
203
Головной мозг лежит в полости черепной коробки и покрыт тре-
мя оболочками: поверхностной, твердой (dura mater), прилегающей
к мозгу мягкой (pia mater) и лежащей между ними паутинной
(arachnoidea), в которой проходят сосуды, снабжающие мозг кровью.
Головной мозг крысы весит 2,4—2,8 г, что составляет около
0,9% веса тела животного. Как мы уже указывали, он разделяется
Рис. 125. Продолговатый мозг пасюка. А — вид
с вентральной стороны, Б — вид с дорзальной
стороны:
1 — латеральный канатик, 2 — вентральный пирамид-
ный пучок, 3 — перекрест пирамид, 4 — пирамиды,
Б — клиновидный бугорок, 6 — серый бугор, 7 — кау-
дальные оливы, 8 — трапецевидное тело. 8 — варолиев
мост, 10 — тройничный нерв, 11 — дорзальный канатик,
12 — клиновидный бугорок, 13 — булава, 14 ~ основа-
ние подъязычного нерва, 15 — лицевой бугорок, 16 — се-
рое крыло, 17 — слуховое поле, 18 — слуховой бугорок,
19 — веревчатые тела, 20 — ножки мозжечка к мосту,
21 — ножки мозжечка к четверохолмию. 22 — четверо-
холмие
на конечный мозг
(большие полушария),
промежуточный мозг,
средний мозг, задний
мозг и продолговатый
мозг (рис. 124). Пос-
ледний непосредствен-
но переходит в спинной
мозг. Условно границу
между головным и
спинным мозгом про-
водят у выхода перво-
го шейного нерва.
С вентральной сто-
роны продолговатого
мозга (рис. 125Л) вид-
на ярко выраженная
вентральная срединная
борозда (sulcus media-
nus ventralis), несколь-
ко латералынее от нее
располагаются слабо
выраженные боковые
борозды (sulci рага-
medianus ventrales);
между ними и средин-
ной бороздой проходят
пирамиды (pyramides),
являющиеся продолже-
нием боковых пирамидных путей спинного мозга. Послед-
ние, переходя в пирамиды, образуют перекрест пирамид (decus-
$a'tio pyramidum, s. fasciculorum cerebro-spinalium). Абораль-
ный конец пирамид у перекреста заостряется. Впереди пирамиды
проходят под поперечными волокнами варолиева моста. Последние
у крысы выражены очень слабо, и пирамиды прослеживаются под
мостом без его удаления. По бокам от пирамид располагается не-
большое скопление серого мозгового вещества, именуемое каудаль-
ными оливами (olivae caudales). У крысы они имеют треугольную
форму и слегка вогнуты. В оливах проецируются волокна VIII нер-
ва и они, наряду с мозжечком, функционируют в качестве центра
равновесия. На дорзальной стороне продолговатого мозга (рис.
125 Б) располагается четвертый мозговой желудочек. По дну его
204
проходит дорзальная срединная борозда (sulcus medianus dorsalis),
проходящая по поверхности продолговатого мозга и за пределами
четвертого желудочка. Эта борозда разделяет дорзальные канатики
белого вещества, являющиеся непосредственным продолжением од-
ноименных канатиков спинного мозга. Слабо выраженной борозд-
кой каждый дорзальный канатик разделяется на две части. Меди-
альный, или стройный, канатик (fasciculus gracilis, seu Golli)
оканчивается небольшим утолщением — булавой (clava). Лате-
ральный. или клиновидный канатик (f. cuneatus) оканчивается
клиновидным бугорком (tuberculum cuneatum). У крысы оба эти
утолщения тесно связаны друг с другом. Впереди они суживаются
и переходят в так называемые веревчатые тела (corpora restifor-
mia), ограничивающие заднюю часть четвертого желудочка. К ла-
теральной стороне продолговатого мозга проходят боковые кана-
тики (fasciculi laterales medullae oblongatae), являющиеся непо-
средственным продолжением одноименных канатиков спинного
мозга. От дорзальных канатиков они отделены едва заметной бо-
роздкой. Боковые канатики огибают сбоку каудальные оливы и об-
разуют впереди утолщение — серый бугорок (tuberculuru cinereum);
у крысы последний покрыт белым веществом. Впереди серого бу-
горка боковые канатики переходят в веревчатые тела. Последние
принимают также перекрещивающиеся волокна из каудальных олив
и образуют задние ножки мозжечка. Четвертый мозговой желудо-
чек (ventriculus rhombocephali) образует расположенную на по-
верхности продолговатого мозга ромбовидную ямку. Позади он пе-
реходит в спинномозговой канал, а впереди суживается и переходит
в сильвиев водопровод среднего мозга. Впереди крыша четвертого
желудочка образована мозговыми парусами, позади она остается
эпителиальной и совместно с сосудистой оболочкой мозга образует
вдающееся в полость желудочка сосудистое сплетение. На дне ром-
бовидной ямки, по краям срединной борозды располагается неболь-
шое возвышение, оканчивающееся впереди так называемым лице-
вым бугорком (colliculus facialis), образованными волокнами лице-
вого нерва. Позади лицевых бугорков располагается поле подъязыч-
ного нерва (area hypoglossi), под которым лежит ядро этого нерва.
Латеральнее поля подъязычного нерва находится серое крыло (ala
cinerea), образованное одним из ганглиев блуждающего нерва.
У бокового угла ромбовидной ямки расположено слуховое поле
(area acustica), граничащее латерально со слуховым бугорком
(tuberculum acustlcum).y заднего угла ромбовидной ямки имеется
слабо выраженное у крысы возвышение, являющееся дыхательным
центром — так называемое писчее перо (calamus scriptorius). В об-
ласти ромбовидной ямки расположены также ядра всех черепно-
мозговых нервов, начиная с V (тройничного).
Задний мозг (metencephalon) состоит из дорзального выроста
(мозжечка) и вентрального пучка поперечных волокон (варолиево
го моста).
Варолиев мост (pons Varolii) у крысы развит очень слабо (ри<
205
124, 125). По сторонам пучки волокон моста переходят в средние
ножки мозжечка (brachia pontis, seu crura cerebelli ad pontum). У
основания ножек располагается ганглий тройничного нерва, назы-
ваемый гассеровым узлом (ganglion Gasseri). У границы продолго-
ватого мозга располагается трапецевидное тело (corpus trapezioi-
des), разделенное продолжением пирамид на две части. Между
волокнами моста располагаются отдельные скопления серого веще-
ства, в том числе краниальная олива (oliva cranialis). Кроме то-
го, через мост проходят проводящие пути головного мозга.
Мозжечок (cerebellum) состоит из двух полушарий (hemisphe-
ria) и непарной срединной части — червячка (vermis) (рис. 124,
126). Мозжечок соединяется с другими частями мозга тремя нож-
ками, по которым проходят нервные волокна. Задние ножки (верев-
чатые тела) соединяют мозжечок с продолговатым мозгом, перехо-
дят в волокна варолиева моста, а передние ножки (brachium conjuc-
tivum, seu crura cerebelli ad corpora quadrigemina) идут к четверо-
холмию. Между передними ножками натянут так называемый пе»
.редний, а между задними ножками — задний мозговые паруса
(veli cerebri anterior et posterior).
Поверхность мозжечка покрыта многочисленными параллельно
расположенными бороздками, разделяющими его на длинные узкие
нзвилины. Группы извилин часто объединяют в дольки мозжечка,
из которых слагаются разграниченные более глубокими бороздами
три крупные доли — передняя, средняя и задняя.
Деление червячка и полушарий на дольки, принятое в анатомии
человека, выражено нечетко (рис. 126 Л), однако и у крысы бороз-
ды частично переходят с полушарий на червячок, не прерываясь.
Сбоку на полушариях хорошо выраженной бороздой отделяется так
называемый клочок (frocculus). Червячок и клочки составляют фи-
логенетически более древнюю часть мозжечка и под названием древ-
ний мозжечок (palaeocerebellum) противопоставляются остальной
части полушарий — новому мозжечку (neocerebellum). На сагит-
тальном разрезе мозжечка (рис. 126 Л) видно, что серое вещество
образует его поверхностную часть, белое же вещество древовидно
ветвится внутри, образуя так называемое древо жизни (arborvitae).
Как указывалось, проводящие пути, связывающие мозжечок с
остальными частями мозга, проходят по его ножкам. По каудаль-
ным ножкам (веревчатым телам) в мозжечок идут волокна от ядер
обоих дорзальных канатиков (Голля и Бурдаха), от каудальных
олив (так называемый пучок Флексига, проходящий по боковым
канатикам спинного мозга), и от ганглия вестибулярной части VIII
нерва (ядра Дейтерса). Из мозжечка по веревчатым телам идут
также нисходящие волокна к спинному мозгу. По средним ножкам
проходят волокна, исходящие из больших полушарий. После пере-
креста эти волокна направляются к коре полушарий мозжечка. Че-
рез средние ножки проходит также и часть нисходящих волокон моз-
жечка. По краниальным ножкам проходят волокна, исходящие из-
206
мозжечка и оканчивающиеся в красном ядре среднего мозга и в
промежуточном мозге. Под четверохолмием эти волокна образуют
перекрест.
Средний мозг (mestencephalon) состоит из ножек большого моз-
га и бугров четверохолмия (рис. 127, 128). Полость среднего мозга
представлена сильвиевым водопроводом (aqueductus Sylvii).
Ножки большого мозга (pedunculi cerebri) составляют вентраль-
ную часть среднего мозга. Они в виде двух сильно развитых стол-
бов проходят от больших полушарий головного мозга к продолгова-
Рис. 126. Мозжечок пасюка. А — сагиттальный раз-
рез через червячок, Б — общий вид сзади:
1 — червячок. 2 — передняя доля червячка (язык, централь-
ные долька и горка), 3 —средняя доля червячка (листок я
бугорок), 4 — задняя доля мозжечка (пирамида, язычок и яд-
рышко). 5 — полушарие мозжечка. 6 — клочок. 7 — каудаль-
ный парус. 8 — древо жизни, 9 — продолговатый мозг
тому мозгу. Ножки разделяются на вентральную часть — основание
(basis) и дорзальную — покрышку (tegmentum). В основании моз-
га проходят нисходящие волокна из больших полушарий в спинной
мозг (пирамидные пути), в ядра варолиева моста и к ядрам дви-
гательных черепно-мозговых нервов. В покрышке расположено
крупное красное ядро (nucleus rubera) серого мозгового вещества,
принимающее волокна из мозжечка и базальных ганглиев больших
полушарий, восходящие и нисходящие волокна к четверохолмию и
исходящий из красного ядра пучок Монакова, направляющийся в
спинной мозг. Кроме того, в покрышке расположены ядра III, IV,
а частично и V черепно-мозговых нервов. Между ножками боль-
шого мозга располагается так называемое заднее продырявленное
пространство (substantia perforata posterior).
Четверохолмие (corpora quadrigemina) образует дорзальную
часть среднего мозга. Оно представлено четырьмя буграми, разде-
ленными продольной и поперечной бороздками. Каждый бугор со-
стоит из серого мозгового вещества, покрытого снаружи белым.
В передних буграх четверохолмия оканчиваются зрительные, а в
задних — слуховые пути среднего мозга. Кроме того, в задних буг-
рах оканчивается часть восходящих волокон, проходящих в боковых
канатиках спинного мозга (tractus spino-tectalis).
207
Промежуточный мозг (diencephalon) состоит из дорзальной об-
ласти зрительного бугра (regio epithalamica) и подбугорной области
(regio hypothalamica) Обе эти части отделены друг от друга треть-
им мозговым желудочком. Область зрительного бугра в свою оче-
редь разделяется на зрительный
бугор, надбугорную и забугорную
области (рис. 127, 128).
Зрительный бугор (thalamus
optici) представляет собой парное
скопление серого вещества, распо-
ложенное непосредственно впере-
ди четверохолмия. В зрительных
буграх оканчиваются сенсорные
волокна, следующие к нему из
спинного мозга (tracto tecto-spi-
nalis), красного ядра, передних
Рис. 126. Фронтальный срез головно-
го мозга пасюка:
1 — желудочек обонятельной луковицы.
2 — боковой желудочек. 3 — монроево
отверстие, 4 — зрительный бугор, Л —
латеральное коленчатое тело, о — меди-
альное коленчатое тело. 7 — пучки к
переднему бугру четверохолмия, 8 — пуч-
ки к заднему бугру четверохолмия. 9 —
передние холмы, 10 — задние холмы.
11 — мозолистое тело,- 12 — полосатое те-
ло, 13 — аммонов рог (гиппокамп)
14 — третий желудочек. 1S — ромбовид-
ный желудочек, 19 — кора, 17 — про-
долговатый мозг
Рис. 127. Мозговой ствол пасюка (вид
сбоку; плащ и мозжечок удалены):
1 — продолговатый мозг, 2 — задние бугры
четверохолмия, 3 — передние бугры четверо-
холмия, 4 — зрительный бугор, 5 — лате-
ральное коленчатое тело, в — медиальное
коленчатое тело. 7 — волокна зрительного
нерва, направляющиеся в передние бугры
четверохолмия, 8 — волокна кэ медиального
коленчатого тела в задний бугор четверохол-
мия, 9 — ножки большого мозга, 10 — про-
водящие пути к продолговатому мозгу, 11 —
зрительный нерв, 12 — контур больших
полушарий
ножек мозжечка; кроме того, в нем оканчивается часть волокон
зрительного нерва. Зрительный бугор является важным центром,
передающим сенсорные стимулы к коре головного мозга. От коры
к зрительному бугру идут и нисходящие волокна; из зрительного
gjTpa они направляются далее к полосатому телу.
•Забугорная область (regio metathalamica) представлена парны-
ми латеральными и медиальными коленчатыми телами (corpora
geniculata laterale et mediale), расположенными латерально от зри*
208
тельных бугров. В латеральном коленчатом теле оканчивается боль-
шая часть волокон зрительного нерва, а в медиальном — волокна
слуховых путей. Соответственно, латеральное коленчатое тело вы-
полняет функции подкоркового центра зрения, а медиальное —
центра слуха.
Надбугорная область (regio epithalamica) представлена эпифи-
зом, или шишковидной железой.
Подбугорная область разделяется на заднюю, собственно под-
бугорную область, и переднюю, зрительную часть, развивающуюся
за счет конечного мозга. Подбугорная часть является непосред-
ственным продолжением вперед покрышки ножек мозга и тесно
связана с основанием зрительных бугров. Из ножек мозга в нее
вдается красное ядро, целиком в подбугорной области расположено
люисово тело (corpus Luyisi), связанное нисходящими волокнами с
полосатым телом. В подбугорной области проходят также восходя-
щие и нисходящие волокна зрительного бугра. Зрительная часть
подбугорной области образована парными сосцевидными телами
(corpora mammilaria), серым бугром (tuberculum cinereum) и во-
ронкой с гипофизом. Впереди она отделяется от конечного мозга
перекрестом зрительных нервов. Сосцевидные тела связаны с обо-
нятельными центрами конечного мозга нисходящими волокнами.
Серый бугор представляет собой непарное выпячивание третьего
желудочка, тесно связанное у крысы с сосцевидными телами. Вер-
шина серого бугра переходит в воронку, к которой прикрепляется
гипофиз.
Третий желудочек (ventriculus tertius) располагается между
зрительными буграми- и подбугорной областью. Позади он перехо-
дит в сильвиев водопровод, а впереди посредством монроевых от-
верстий сообщается с желудочками больших полушарий головного
мозга.
Конечный мозг (telencephalon) представлен большими полуша-
риями, разделяющимися на обонятельный мозг, плащ, или кору, и
базальные ганглии (рис. 124, 128). Центры обоих полушарий сое-
диняются друг с другом комиссуральными волокнами, проходящими
в концевой пластинке, соединяющей оба полушария. Внутри полу-
шарий располагаются боковые желудочки (ventriculi laterales),
сообщающиеся посредством монроева отверстия друг с другом и с
третьим желудочком.
Обонятельный мозг (rhinencephalon) представляет собой перед-
нюю часть больших полушарий, расположенную на вентральной
стороне последних. Там образуются парные выросты — обонятель-
ные луковицы (bulbi olfactorii), соединяющиеся с большими полу-
шариями широким основанием. Спереди в обонятельные луковицы
входят короткие нервные волокна (fila olfactoria), в совокупности
образующие обонятельный нерв (п. olfactorius). Внутри луковиц
располагаются желудочки (ventriculi bulbi olfactorii), сообщающие’-
ся с желудочками больших полушарий. От обонятельных луковиц
по вентральной поверхности большого мозга назад тянется полоска
14 Крыса
белого вещества, называемая общей обонятельной извилиной (girus
olfactorius communis). Она вскоре разделяется на короткую ме-
диальную и более длинную латеральную обонятельную извилины
(gyri olfactorii lateralis et medialis). Эти извилины представляют
собой начальное звено внутримозговых обонятельных путей (tractus
olfactorius).
Базальные ганглии расположены в основании больших полуша-
рий и сильно вдаются в его желудочки (рис. 128). Они представле-
б
Рис. 129. Расчленение коры больших полушарий и корковые чув-
ствующие центры пасюка. А — расчленение коры на древнюю,
старую и новую (вид полушария с внутренней стороны), Б — кор-
ковые и подкорковые чувствующие центры (вид полушария сна-
ружи) (по Кригу):
1 — древняя кора, 2 — старая кора, 3 — новая кора, 4 — обонятельный бу-
горок, Б — мивдалина, в — septun, 7 — полосатое тело, 8 — наружное ко-
ленчатое тело, 9 — внутреннее коленчатое тело, 10 — корковые центры зре-
ния, 11 — корковые центры слуха, 12 — корковые центры соматической чув-
ствительности
ны полосатым телом (corpus striatum), дифференцирующимся у
млекопитающих на: 1) древнюю часть (archistriatum), представлен-
ную бледным ядром (n. pallidus); 2) старую часть (palaeostria-
tum), представленную расположенной в височной части полушария
миндалиной (n. amygdalae), 3) дорзальную часть (neostriatum),
представленную хвостатым ядром (n. caudatus) и скорлупой (puta-
men). В старой и новой частях интегрируются сенсорные импульсы,
передаваемые далее в кору, через древнюю часть стимулы от по-
лосатого тела направляются в нижележащие отделы головного
мозга.
Плащ, или кора (pallium, seu cortex), представлен серым мозго-
вым веществом, развившимся в боковых и верхних стенках полуша-
рия. Нервные клетки распределяются в коре в три слоя. Филогене-
тически кору разделяют на: 1) старую (palaeocortex), представлен-
ную расположенной на вентральной стороне полушарий грушевид-
ной долей (g. piriformes); 2) древнюю (archicortex), представлен-
ную долей морского конька (g. hippocampi), вдающейся в боковой
желудочек с внутренней стороны, и 3) новую кору (neocortex)
210
(рис. 129 Л). Старая и древняя кора представляют собой обоня-
тельные центры коры. Вся остальная часть плаща образует новую
кору. У крысы поверхность ее гладкая, и лишь на основании осо-
бенностей микроструктуры удается выделить различные ее зоны.
На прилагаемой схеме (рис. 129 Б) мы показываем только корко-
вые центры слуха, зрения и соматических ощущений и пути, связы-
вающие эти центры с подкорковыми ганглиями.
Новая кора обоих полушарий соединяется друг с другом силь-
ным пучком волокон, образующим мозолистое тело (corpus callo-
,sum) (рис. 128). Кроме того, древняя кора связана специальной
комиссурой гиппокампа (с. hippocampi), а старая — передней ко-
миссурой (с. pallii anterior). Последние две комиссуры развиты
незначительно и при макроскопическом исследовании мозга их об-
наружить очень трудно.
ГОЛОВНЫЕ НЕРВЫ
У крыс, как и у других млекопитающих, имеется двенадцать
пар головных, или черепномозговых, нервов (nervi cerebrales).
I. Обонятельный нерв (п. olfactorius) представляет собой ряд
коротких нервных волокон, начинающихся от обонятельного эпите-
лия носовой полости. Проходя через отверстия решетчатой кости,
они оканчиваются в обонятельных луковицах (рис. 124).
II. Зрительный нерв (n. opticus) начинается от сетчатки глаза и
проходит в черепную полость через зрительное отверстие. После
перекреста волокна нерва следуют к наружному коленчатому телу,
а частью — к передним буграм четверохолмия и в зрительный бугор
(рис. 124, 127, 130).
III. Глазодвигательный нерв (n. oculomotorius) начинается от
ножек большого мозга. Проходя через верхнюю глазничную щель
вместе с глазничной ветвью тройничного нерва и отводящим нервом,
глазодвигательный нерв оканчивается в глазодвигательных мышцах
(m. т. recti superior, inferior et intemus, m. obliquiis inferior) а в
мышце верхнего века (рис. 124, 130).
IV. Блоковой нерв (n. trochlearis) начинается в области соеди-
нения краниального мозгового паруса с четверохолмием, проходит
через верхнюю глазничную щель и иннервирует m. obliquus supe-
rior (рис. 124, 130).
V. Тройничный нерв (n. trigeminus) — самый крупный из голов-
ных нервов. В отличие от чисто чувствующих I и II и чисто двига-
тельных III и IV включает как двигательные, так и чувствующие
волокна. Чувствующий ганглий отделен от двигательного и назы-
вается полулунным, или гассеровым, узлом (ganglion semilunare,
seu Gasseri). От гассерова узла начинаются три главных ствола
тройничного нерва — глазничный, верхнечелюстной и нижнечелю-
стной; двигательные волокна идут совместно с последним из них
(рис. 124, 130).
1. Глазничный нерв (n. ophthalmicus) является самов слабой
ветвью тройничного нерва. Проходя верхнюю глазничную пв н>. он
14* -Ч ।
сразу же делится на три нерва: лобный, слезный и носо-ресничный
(рис. 130). Лобный нерв (п. frontalis) разветвляется в коже лобной
области и верхнем веке. Слезный нерв (n. lacrimalis) иннервирует
слезную железу, кожу области лба и верхнее веко. Носо-ресничный
нерв (n. naso-ciliaris) иннервирует конъюнктиву глаза и глазное
яблоко; продолжение нерва под названием решетчатого нерва
(п. ethmoidalis) через решетчатую кость входит в носовую полость
и иннервирует ее слизистую оболочку.
Рис. 130. Передние черепномозговые нервы пасюка:
II — зрительный вера, III — глазодвигательный нерв. IV — бло-
ковой нерв. V —тройничный нерв; V, — глазничная ветвь,
V • — верхнечелюстная ветвь. Vа—нижнечелюстная ветвь, VI —
отводящий нерв; 1 — слезный 2 — носоресничный, 8 — лобный,
4 — нижний альвеолярный, 5 — верхний альвеолярный, 6 — под-
глазничный, 7 — скуловой, 8 — клинонёбный, 9 — аборальный но-
совой, 10 — малый нёбный, И — большой нёбный, 12 глубокие
височные, 13 — щёчный, 14 — нерв мышцы, поднимающей нёб-
ную занавеску, 15 — поверхностный височный. 10 — язычный,
17 — жевательный, 18 — межчелюстной
2. Верхнечелюстной нерв (п. maxilaris) выходит из черепа через
круглое отверстие и почти сразу разделяется на 3 ветви: подглаз-
ничную, скуловую и клинонёбную. Подглазничный нерв (n. infra-
orbitalis) — наиболее сильная из них. Она проходит через подглаз-
ничное отверстие, отдавая предварительно альвеолярную и резцовую
ветви (rami alveolaris et incisivum), иннервирующие зубы верхней
челюсти, твердое нёбо и десну. Выходя на лицевую поверхность,
разделяется на верхнегубной нерв (n. labialis superior), иннерви-
рующий вместе с одноименной частью лицевого нерва вибриссы и
верхнюю губу, наружные носовые нервы (n. п. nasales extern!),
иннервирующие кожу носа, и оральный носовой нерв (n. nasa'lis
oralis), разветвляющийся в верхней губе и наружной стенке носа.
Скуловой нерв (п. zygomaticus) идет в нижнее веко, анастомозируя
со слезным нервом. Клинонёбный нерв (n. spheno-palatinus) почти
сразу же разделяется на три ветви. Из них аборальный носовой
нерв (n. aboralis nasalis) через клинонёбное отверстие входит в но-
212
совую полость и разветвляется в слизистой оболочке носовой пере-
городки, большой нёбный нерв (n. palatinus major) разветвляется в
твердом нёбе, а малый нёбный нерв (n. palatinus minor) раз-
ветвляется в мягком нёбе.
3. Нижнечелюстной нерв (n. mandibularis) выходит из черепа
через овальное отверстие и сразу же разделяется на несколько вет-
вей. Из них жевательный нерв (n. massetericus) идет в наружную
жевательную мышцу, отдавая глубокие височные нервы (n. n. tem-
porales profundi), снабжающие височную мышцу. Щечный нерв
(п. buccinatorius) проходит через крыловидную мышцу в слизистую
оболочку щеки и нижней губы, где анастомозирует с вентральным
щечным нервом, идущим от лицевого нерва. Поверхностный височ-
ный нерв (п. temporalis superficialis) делится на дорзальную и вен-
тральную ветви. Вентральная ветвь анастомозирует с вентральным
щечным нервом лицевого нерва и разветвляется в коже лица и
нижней челюсти, дорзальная ветвь разветвляется в коже, в области
скуловой дуги. Нерв мышцы, поднимающей нёбную занавеску (п.
levator veli palatini), иннервирует одноименную мышцу. Язычный
нерв (n. lingualis) идет в язык и делится в нем на вентральную
ветвь, снабжающую грибовидные сосочки кончика языка, и дорзаль-
ную ветвь, разветвляющуюся в слизистой оболочке зева, передних
частей языка и дна ротовой полости. Альвеолярный нерв (n. alveo-
laris mandibulae) иннервирует нижнечелюстные зубы и, выходя че-
рез подбородочное отверстие, образует подбородочный нерв, развет-
вляющийся в коже нижней губы п подбородка. Межчелюстной нерв
(п. mylo-hyofdeus) оканчивается в двубрюшной и межчелюстной
мышцах.
VI. Отводящий нерв (n. abducens) отходит от продолговатого
мозга сбоку от пирамид и, проходя через овальное отверстие, окан-
чивается в m. rectus externus (рис. 124, 130).
VII. Лицевой нерв (n« facialis) начинается от продолговатого
мозга впереди места отхождения слухового нерва, связанного с ним
соединительной ветвью (рис. 124). Отдавая несколько ветвей, и ।
которых мы назовем большой поверхностный каменистый нерв
(п. petrosus superficialis maior) и так называемую барабанную
струну (chorda tympani), лицевой нерв проходит через лицевой ка-
нал. Барабанная струна проходит через полость среднего уха и
снабжает вкусовые почки языка, лицевой же нерв отдает несколько
ветвей (рис. 131), в том числе шейные ветви (rami colli), идущие к
околоушной железе и к подкожным мышцам шеи, каудальный и
внутренний ушные нервы (п. п. auriculares caudalis et internus),
разветвляющиеся в мышцах и коже наружного уха совместно с вет-
вями I и II шейных нервов, веко-ушной нерв (auriculo-palpebralis),
иннервирующий ушные мышцы и круглую мышцу век, и нерв дву-
брюшной мышцы (п. digastricus), снабжающий переднюю часть
двубрюшной мышцы и шилоподъязычную мышцу. В дальнейшем
лицевой нерв разделяется на дорзальный и вентральный щечные
нервы (n. buccales dorsalis et ventralis), которые иннервируют же-
213
пптслышг мышцы. Большой поверхностный каменистый нерв сое-
диняется с клино-нёбным ядром тройничного нерва.
VIII. Слуховой нерв (п. acusticus) начинается от продолговатого
мозга немного каудальнее начала лицевого нерва и снабжает
внутреннее ухо (рис. 124).
IX. Языкоглоточный нерв (n. glossopharyngeus) начинается от
продолговатого мозга (рис. 124) и, отдавая несколько тонких вето-
чек, оканчивающихся в слизистой оболочке среднего уха и направ-
ляющихся к узлам симпатической системы, проходит через рваное
Рис. 131. Ветви лицевого нерва пасюка:
VII — лицевой нерв; I — верхнегубной, 2 — латераль-
ный носа, 3 — носовой, 4 — дорзальный носа, S — низкие*
губной. 6 — вентральный щечный, 7 — дорзальный
щечный, 8 — комиссура к ветвям подглазничной ветви
тройничного нерва, 9 — каудальный и внутренний
ушные, 10 — двубрюшной мышцы, 11 — шейные ветви,
12 — щечная ветвь тройничного нерва
отверстие. После этого языко-глоточный нерв делится на глоточную
ветвь (ramus (pharyngitis), разветвляющуюся в слизистой оболочке
глотки, и язычную ветвь (г. lingualis), оканчивающуюся в слизи-
стой оболочке нёбной занавески и корня языка.
X. Блуждающий нерв (и. vagus) почти целиком относится к па-
расимпатической нервной системе и будет описан ниже.
XI. Добавочный нерв (п. accessorius) начинается от продолгова-
того мозга непосредственно каудальнее блуждающего нерва
(рис. 124). Принимая ветви от передних шейных сегментов, обра-
зующих шейную часть добавочного нерва (n. accessorius spinalis),
добавочный нерв выходит вместе с блуждающим нервом через рва-
ное отверстие и разветвляется в висцеральной мускулатуре передней
конечности.
XII. Подъязычный нерв (n. hypoglossus) начинается от продол-
говатого мозга (рис. 124). Выходя из черепной полости через отвер-
стие подъязычного нерва, подъязычный нерв разделяется на поверх-
ностную и глубокую ветви, разветвляющиеся в подъязычной и
язычной мускулатуре.
214
СПИННОМОЗГОВЫЕ НЕРВЫ
Шейные нервы (nervi cervicales) в числе 8 пар выходят из меж-
позвоночных отверстий шейных позвонков, I пара — впереди атлан-
та (рис. 132). Каждый нерв делится на дорзальные и вентральные
ветви, которые, в свою очередь, разделяются на латеральные и ме-
диальные. Дорзальные ветви разветвляются в дорзальной мускула-
туре шеи, а вентральные — в вентральной ^ускулмуре шеи и в
коже. Каудальные шейные нервы
участвуют в образовании плечевого
сплетения. Кроме того, от шейных
нервов отходит несколько отдельных
стволов (рис. 132). Затылочный нерв
(л. occipitalis) начинается от дор-
зальной ветви С I и разветвляется в
косой мышце головы, в дорзальной
прямой мышце головы, в ушной мус-
кулатуре и в коже затылочной обла-
сти. Каудальный ушной нерв (n. аи-
ricularis caudalh) начинается от вен-
тральной ветви С II и иннервирует
наружное ухо и раушную железу.
Кожный шейный нерв (п. cutaneus
colli) начинается от вентральных
ветвей С II и С III и, соединяясь с
шейной ветвью лицевого нерва, иде
к горлу и в подчелюстное простран
Рис. 132. Спинномозговые нер-
вы пасюка. А — головной
мозг (большие полушария),
Б — мозжечок, В — продол-
говатый мозг. Г — шейное
расширение, Д — поясничное
расширение спинного мозга:
Cl — CVIII — шейные нервы.
Th I—Т h XIII — грудные нервы,
LI-LVI - поясничные непвы.
SI—S IV — крестцовые нервы;
1 — затылочный. 2 — каудальный
ушной, 3 — кожный шейный, 4 —
надключичный, 5 — диафрагматн-
ческий, 6 — дорзальный лопаточ-
ный, 7 — предлопаточный, 8 —
подлопаточные, 9 — грудные, 10 —
подмышечный, II — срединный,
12 — подвздошко-родчревный, 13 —
подвздошно-па»вый, 14 — пояс-
ннчно-паховый, 15 — бедренный,
16 — седалиЩНый, 17 — запира-
тельный, 18 — каудальный кож-
ный, 19 — наружный семеннико-
вый, 20 — каудальный геморрои-
дальный, 21 — срамной, 22 — хво-
стовой
&2021
215
ство. Диафрагматический нерв (n. phrenicus) начинается от
вентральных ветвей С V, С VI, С VII и иннервирует мышцы
диафрагмы. Дорзальный лопаточный нерв (п. scapularis dorsalis)
начинается от вентральных ветвей С V и С VI и разветвляется в
ромбовидной и вентральной зубчатой мышцах. Надключичный нерв
(n. supraclavicularis) начинается от вентральной ветки С VI и идет
в кожу плечевого сустава.
Плечевое сплетение (plexus brachialis) образуется из вентраль-
ных ветвей V—VIII шейных и I грудного нервов (рис. 132, 133).
Часто ветвь от V шейного нерва отсутствует. Наиболее сильно раз-
виты ветви С VII — Th I. От плечевого сплетения в области лопатки
отделяется ряд нервов, разветвляющихся в мышцах плечевого пояса.
Предлопаточный нерв (и. suprasca'pularis), образованный ветвями
от С VI и С VII, входит в щель между предостной и подлопаточной
мышцами и разветвляется в предостной и заостной мышцах. Подло-
паточный нерв (п. subscapularis) начинается от С VII и С VIII и
оканчивается в подлопаточной мышце. Подмышечный нерв (и. axi-
laris) начинается от С VII и С VIII и, проходя между круглой боль-
шой и подлопаточной мышцами, разветвляется в них, отдавая также
ветви к малой грудной и дельтовидной мышцам и к коже. Грудные
нервы (п. п. pectorales) начинаются от Th I и оканчиваются в по-
верхностной и глубокой грудных мышцах.
Передняя конечность снабжается четырьмя нервами, также на-
чинающимися от плечевого сплетения (рис. 134). Мышечно-кожный
нерв (n. musculocutaneus) начинается от С VIII, получая также
веточку от срединного нерва. Этот нерв проходит между коракоидо-
плечевой и круглой большой мышцами и разветвляется в двуглавой
и плечевой мышцах и в коже плеча. Лучевой нерв (n. radialis)
начинается от С III, проходит между длинной и медиальной голов-
ками трехглавой мышцы плеча, отдавая им ветви, и, выходя на
латеральную поверхность предплечья, разделяется на глубокий лу-
чевой нерв (и. radialis profundus),’снабжающий разгибатели паль-
цев, и на поверхностный лучевой нерв (п. radialis superficialis),
иннервирующий кожу предплечья и пальцев. Переходя на кисть,
поверхностный лучевой нерв разделятся на медиальную и латераль-
ную ветви, образующие дорзальные пальцевые нервы (n. n. di pita-
les dorsales) (рис. 134) — по два для каждого пальца. Латераль-
ный пальцевой нерв V пальца является производным локтевого
нерва.
Срединный нерв (n. medianus) начинается от Th I и отчасти от
С VIII и проходит по медиальной поверхности плечевой кости ме-
диальнее двуглавой и коракоидоплечевой мышц. Переходя на пред-
плечье, он дает межкостный нервен, interosseus) и ветви к сгиба-
телям пальцев, а сам переходит на кисть, где образует волярные
пальцевые нервы (n. n. digitales volares) (рис. 134).
Локтевой нерв (n. ulnaris) отделяется от общего ствола со сре-
динным нервом на середине плеча, проходит под малой локтевой
мышцей на волярную поверхность предплечья, где разделяется на
216
ветви. Дорзальная ветвь (ramus dorsali^) переходит на дорзальную
сторону кисти и иннервирует V палец, волярная ветвь (г. volaris)
идет с волярной стороны к трем латеральным пальцам (рис. 134).
Грудные нервы (n. n. thoracales) в числе 13 пар отходят после
соответствующих грудных позвонков (рис. 132). Каждый нерв раз-
деляется на дорзальную и вентральную ветви. Дорзальные ветви
Рис. 133. Нервы передней конечности
пасюка:
С V—С VIII — шейные нервы; Т h I — груд-
ной нерв; 1 — предлопаточный, 2 — под-
лопаточные, 3 — подмышечный, 4 — груд-
ные, Б — мышечно кожный, 6 — ветви к
широчайшей мышце спины. 7 — лучевой,
8 — срединный, 9 — локтевой
л 6
Рис. 134. Нервы кисти пасюка. А —
ВИД С волярной стороны, 5— ВИД L
дорзальной стороны*
1 — медиальная ветвь поверхностного
лучевого нерва, 2 — латеральная ветвь
поверхностного лучевого нерва, 3 — дор-
зальная ветвь локтевого нерва, 4 — об-
щие дорзальные пальцевые нервы, б —
дорзальные пальцевые нервы, 6 — воляр-
ная ветвь локтевого нерва, 7 — сред> н-
ный нерв, 8 — общие велярные пальце-
вые нервы, 9 — вол яря ые пястные нервы,
10 — волярные пальцевые нервы
иннервируют мышпы спины и ромбоидную и зубчатую дорзальную
мышцы. Бол< > сильные вентральные ветви, называемые также меж-
рсберными нервами (п. n. intercostales), иннервируют межребер-
пыс, грудные и брюшные мышцы. Th I почти целиком участвует в
обра юьшц.и плечевого сплетения, кроме того, передние грудные
нерпы иннервируют кожу плечевого сустава.
I!<>т'|Ч1ч)1К\- нервы (п. п. lumbales) в числе шести выходят за
«ooinef<Ti')i<«i'UiMH позвонками (рис. 132). Сразу же по выходе из
мокпо юоночньн опнрстий они делятся на дорзальные и вентраль-
ный ветви. Первые иннервируют спинные мышцы и анастомозируют
। нервами ягодичных мышц и кожи ягодичной области, вентральные
г.и ви L II - L IV участвуют в образовании поясничного сплетения
>1;
(plexus lumbalis). От Th — L III отходят несколько ветвей,
снабжающих брюшную мускулатуру, а у самцов также кожу мо-
шонки; кроме того, от поясничных нервов отходят следующие более
крупные стволы (рис. 132): 1) подчревно-подвздошный нерв
(п. ilio-hypogastricus), начинающийся От вентральной ветви L I,
Рис. 135.' Поверхностные нервы таза и
бедра пасюка:
Ll-L VI—поясничные нервы, SI— SIII кре-
стцовые нервы; 1 — пояснично-кожный, 2 —
бедренный, 3 — запирательный. 4 — ветв1Мвгы-
рехглавой мышцы. 5 — сафенус, 6 — Япне-
бедренная ветвь, 7 — каудальный геморрои-
дальный, 8 — каудальный кожный, 9 — хво-
стовой. 10 — седалищный, 11 — срамной нерв
получает также соединитель-
ную ветвь от Th XIII. Идет к
брюшной мускулатуре и, про-
бодая ее, разветвляется в
брюшных мышцах, в коже
брюшной области, в паховых
молочных железах или в мо-
шонке (у самцов) и в коже
колена; 2) подвоздошно-пахо-
вый нерв (п. ilio-inguinalis)
начинается от L II и разветвля-
ется в коже колена и наружных
половых органах; 3) наружный
семенниковый нерв (и. sperma-
ticus externus) начинается от
вентральной ветви L II, прини-
мая часто тонкие ветви от L I
и L IV. Разветвляется в пахо-
вой молочной железе или коже
мошонки (у самцов), во влага-
лищных оболочках семенника
и во внутренней косой мышце
живота; 4) пояснично-кожный
нерв (n. psoadicocutaneus) рас-
положен глубже остальных. Он
начинается от L II и L III и,
прободая большую поясничную
мышцу, отдает ветви к коже
коленного сустава (рис. 136).
От поясничного сплетения
отходят также бедренный и
запирательный нервы, проходя-
щие по бедру (рис. 132, 135).
^мпенный дерв (и. femotalis)
начинается от L II—I* IV и
между гребешковой и четырехглавой мышцами разделяется на три
ветви. Каудальная ветвь отходит поверхностно, оканчивается в
коже области половых органов; вторая ветвь идет к четырехглавой
мышце бедра; третья ветвь, так называемый сафенус (п. saphenus),
идет по медиальной стороне бедра, отдавая ветви к стройной, гре-
бешковой и приводящим мышцам, переходит на медиальную сторо-
ну голени и разветвляется в коже голени и стопы. Запирательный
нерв (n. obtufStonusT начинается от L II — L IV, направляется в
218
тазовую полость и, проходя через запирательное отверстие, развет-
вляется в стройной, приводящих, наружной запирательной и гре-
бешковой мышцах.
Крестцовое сплетение (plexus sacralis) крысы, в отличие от
одноименного сплетения человека, образовано главным образом
вентральными ветвями поясничных нервов. Образуется ветвями
L Tv^^> Т, самая сильная из них идет от L IV (рис. 132, 135).
Иногда L IV и S 1 не участвуют ~~
р образовании крестцового
сплетения Общий ствол, обра-
1зуемый этими ветвями, следует
именовать <ч д;1л_ищным нервом
i(n. ishiadicus) (рис. 135,-136).
От самого основания седалищ-
ного нерва отходят: 1) кожнж/й
каудальный нерв бедра (n. ф1-
taneus f-moris caudaiis), раз-
ветвляющийся в коже ягодич-
ной области и в мышцах задне-
бедреймой группы; 2) срайной
нерв (n. pudendus), разветв-
ляющийся в коже промежно-
сти, в срамных губах самок’ и в
мошонке самцов, образуя) у
последних дорзальный нерв
полового члена (п. dorsalis
penis); от срамного нерва отхо-
дит также разветвляющийся в
прямой кишке средний гемор-
роидальный нерв (n. haemor-
rhoidalis medius); 3) каудаль-
ный геморроидальный нерв
(n. haemorrhoidalis caudaiis),
разветвляющийся в коже про-
Рис. 13б7~Нёрвы задней конечности па-
сюка:
1 седалищный. 2 — проксимальная мускуль-
ная ветвь большеберцового нерва, 8 — план*
тарный кожный нерв, 4 — большеберцовый
нерв, б — малоберцовый нерж в - мускульная
ветвь малоберцового нерва, 7 — краниальный
межности, в прямой кишке и
ее сфинктере, в мышцах хвоста
ягодичный нерв, 8 — каудальный ягодичный
нерв
и в позвоночной головке полу-
перепончатой мышцы. Иногда описанные три нерва не связаны с
седалищным.
Седалищный нерв направляется к седалищной вырезке, где он
разделяется на бблыпебердояый и мялпбррпппый нервы (рис. 136).
Предварительно oH отдает краниальный и каудальный ягодичные
нервы (и. п. glutaei cranialis et caudaiis), разветвляющиеся в яго-
дичных мышцах и напрягателе широкой фасции бедра;
z— Большеберцовый нерв (n. tibialis) представляет собой каудаль-
ную, более сильную ветвь седалищного нерва (рис. 136). Почти
сразу же от него отделяется ветвь в заднебедренную мускулатура
(ramus muscularis proximalis), немного дистальнее отходит план
тарный кожный нерв голени (n. cutaneus cruris plantaris), который
проходит по икроножной
8
/7
па-
Рис
сюка. А — вид с дорзальной сторо-
ны, Б — вид с плантарной'сторо ।
1 — общие малоберцовые. 2 — сафенус,
3 — большеберцовый, 4 — плантарный
голени, 6 — глубокий малоберцовый, в —
поверхностный малоберцовый, 7 — дор-
зальные пальцевые стопы, 8 — пальцевая
ветвь глубокого малоберцового нерва. О —
медиальный плантарный, 10 — латераль-
ный плантарный. 11 — пальцевые план-
тарные, 12 — мускульная ветвь латераль-
ного стопы
6
137. Нервы голени и стопы
мышце и разветвляется в коже ступни
и голени. Затем большеберцовый
нерв проходит между головками
икроножной мышцы, отдавая вет-
ви к сгибателям пальцев, разги-
бателям стопы и к икроножной
3 мышце, и медиально от пяточного
бугра разделяется на медиальный
и латеральный плантарные нер-
вы (рис. 137). Медиальный план-
тарный нерв (и. plantaris media-
ns) разветвляется в трех внутрен-
них пальцах. Латеральный план-
тарный нерв (n. plantaris latera-
lis) разветвляется в двух наруж-
ных пальцах стопы.
| Малоберцовый нерв (в. рего-
I naeus) отдает в средней части бед-
4, ра дорзальный кожный нерв голе-
’ i ни (п. cuataneus peronaes dorsa-
jlis), прободает двуглавую мышцу
бедра и разветвляется в коже го-
лени. Дистально малоберцовый
нерв проходит под икроножную
мышцу и делится там на глубо-
кий и поверхностный малоберцо-
вые нервы (рис. 137). По-
верхностный малоберцовый нерв
(и. peronaeus superficialis) идет
вдоль разгибателей пальцев, от-
давая им ветви, и разветвляется в
коже стопы и связках сустава.
Дистально он образует общие
дорзальные пальцевые нервы, ин-
нервирующие наружные пальцы.
Глубокий малоберцовый нерв
(п. peronaeus profundus) прохо-
дит в мускулатуре голени, раз-
ветвляется в сгибателях и разги-
бателях пальцев и, переходя на
стопу, образует поверхностный
дорзальный латеральный пальце-
вый нерв, разветвляющийся во
внутренних пальцах.
Крестцовые нервы образуют хвостовые нервы (n. n. caudales),
из которых хорошо развит латеральный нерв хвоста (n. caudalis
lateralis), разветвляющийся в хвостовых мышцах и в коже хвоста.
220
W
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Вегетативная нервная система состоит из ряда нервных узлов,
расположенных вдоль позвоночного столба и в области ядер задних
черепномозговых нервов; в ее состав входят также и отходящие от
этих ганглиев нервные стволы. Вегетативная система подразделяет-
ся на симпатическую и парасимпатическую, функционально обычно
являющиеся антагонистичными друг другу.
Обычно одни и те же органы (в частности все поперечнополоса-
тые мышцы) иннервируются обеими системами. Деятельность сим-
Рис. 138. Симпатическая нервная система пасюка:
1 — краниальный шейный узел, 2 — средний шейный узел, 3 — каудальный шейный
узел, 4 — звездчатый узел, 5 — грудные узлы, в — межреберные нервы грудных
узлов, 7 — большой внутренностный нерв, 8 — малый внутренностный нерв, й —
поясничные узлы, 10 — полулунные узлы солнечного сплетения, 11 — краниальный
брыжеечный узел, 12 — каудальный боыжеечный узел. 13 — почечное сплетение,
14 — непарный крестцовый узел
патической системы связана с выделением адреналина, деятельность
парасимпатической — с выделением ацетилхолина.
Симпатическая система образована двумя симметричными ство-
лами (trunci sympathies dexter et sinister), расположёнными по сто-
ронам позвоночного столба и состоящими из ганглйев, соединяю-
щихся друг с другом межузловыми ветвями (рис. 138). Грудные и
поясничные узлы связаны также со спинным мозгом.
Имеется три пары шейных симпатических узлов (gangliae сег-
vicales). Крайни.п ^ая пара (ganglion cervicalis cranialis) развита
особенно сильно. От нее через так называемый внутренний сонный
нерв (n. carotis internus) отходят ветви и в голову, иннервирующие
кровеносные сосуды мозга и т. д. От этого же узла отходят dpi ни
771
к органам шеи (трахее, пищеводу и др.), к сердцу и передним
артериальным стволам. Средняя * пара шейных узлов (ganglion
cervicalis medius) дает ветви к сердцу и щитовидной железе. Кау-
дальная пара шейных узлов (ganglion cervicalis caudalis) часто
объединяется с краниальным грудным узлом, образуя так называе-
мый звездчатый узел (ganglion stellatum). От этого узла идут вет-
ви к сердцу и другим органам грудной клетки.
Грудные узлы (gangliae thoracales) дают начало двум внутрен-
ностным нервам — краниальному и каудальному (п. n. splanchnic!
cranialis et caudalis) (рис. 138). Оба эти нерва прободают диа-
фрагму и входят в брюшную полость, иннервируют кишечник. Кро-
ме того, грудные симпатические узлы иннервируют органы грудной
полости и заходят в конечности.
Поясничные узлы (gangliae lumbales) дают начало нервам, раз-
ветвляющимся в пищеварительных органах. В краниальной части
брюшной полости из нервов, выходящих от поясничных симпатиче-
ских нервных узлов, образуется солнечное сплетение (plexus Sola-
ris); в нем расположено два крупных нервных узла. Поясничные
симпатические нервы образуют также краниальный и каудальный
брыжеечные нервные узлы (gangliae mesentericae cranialis et cau-
dalis) .
Парасимпатическая нервная система разделяется на головной и
крестцовый отделы. Головной отдел, в свою очередь, делится на
среднемозговой и продолговатомозговой. Среднемозговой отдел
представлен вегетативной частью глазодвигательного нерва, иннер-
вирующей мышцу, суживающую зрачок. Интересно, что мышцу,
расширяющую зрачок, иннервирует ветвь симпатического внутрен-
него сонного нерва. Продолговатомозговая часть парасимпатической
нервной системы представлена вегетативными волокнами VII, IX и
X нервов. Парасимапатические волокна лицевого нерва разветвля-
ются в слезной железе и образуют так называемую барабанную
струну, разветвляющуюся в подчелюстной слюнной железе. Языко-
глоточный нерв содержит парасимпатические волокна, идущие к
околоушной слюнной железе. Основным нервом парасимпатической
системы является блуждающий нерв. Он дает ветви к сердцу и,
проходя по шее, образует ряд ганглиев на трахее, от которых отхо-
дят волокна к органам шеи и грудной полости. Волокна блуждаю-
щего нерва участвуют также в иннервации кишечника и других
органов брюшной полости. Раздражение парасимпатической системы
ведет к ускорению перистальтики кишечника и замедлению сердце-
биений.
Ганглии крестцового отдела парасимпатической системы тесно
связаны с боковыми рогами крестцового отдела спинного мозга.
Они отдают тазовый нерв, который иннервирует половые органы,
мочевой пузырь и прямую кишку. Раздражение этих волокон ведет
к опорожнению мочевого пузыря и кишечника и к гиперемии поло-
вых органов.
ГЛАВА XV
ОРГАНЫ ЧУВСТВ
ОРГАНЫ ОСЯЗАНИЯ
Среди различных чувствующих образований, расположенных в
коже и воспринимающих температурные, болевые и другие раздра-
жения, имеются тельца Мейсснера (corpusculi Meissneri), являю-
щиеся специализированными органами осязания. Они имеют оваль-
ную форму и состоят из поперечно ориентированных клеток, между
которыми ветвятся нервные окончания. Снаружи они окружены
кожей. Тельца Мейсснера особенно обильны на кончиках пальцев.
Большую роль в осязании играют также волосы, корни которых
окружены нервными окончаниями, зачастую связанными со спе-
циальными осязательными тельцами Меркеля (с. Merceli). Волосы
морды, специализированные в качестве органов осязания, именуют-
ся вибриссами (vibrissae). Это весьма длинные и жесткие волоски,
расположенные в основном на губах (рис. 139). Их длина у крысы
колеблется от 6 до 56 мм, общее число вибрисс достигает 150, из
них над глазом с каждой стороны располагается 2—3, между ухом
и глазом — 3—4, на верхней губе — 23—25 и на нижней — 6—9.
Вибриссы раздражаются как при непосредственном соприкосно-
вении с твердыми телами, так и колебаниями воздуха; поэтому жи-
вотное может ориентироваться, воспринимая воздушные токи, отра-
жающиеся от твердых тел. Даже слепые крысы передвигаются,
почти не наталкиваясь на препятствия.
ОРГАНЫ ЗРЕНИЯ
Органом зрения служит глаз (oculus). Он составляется из глаз*
иого яблока (bulbus oculi) и вспомогательных образований. Глаз-
ное яблоко представляет собой чуть сдавленный шар, с несколько
выпуклой наружной стенкой. Его передне-задний диаметр у крысы
достигает 6,2—6,5 мм, вертикальный — 6,2—6,4 мм-, глубина глаза
(глазная ось) крысы равна 5,7—6,0 мм. Помещается глаз в полости
орбиты, лежащей в средней части головы. Снаружи глазное яблоко
окружено шаровой подушкой, выполняющей роль, буфера.
223
Глазное яблоко образовано тремя оболочками: фиброзной, со-
судистой и сетчатой (рис. 140).
Фиброзная оболочка (tunica fibrosa) окружает глазное яблоко
снаружи. Внутренние 2/з глаза окружает белочная часть фиброзной
оболочки, или склера (sclera), снаружи склера переходит в проз-
рачную выпуклую роговицу (cornea), проходя через которую лучи
света попадают в глаз. Снаружи роговица покрыта плоским много-
слойным эпителием, изнутри — эндотелием. Роговица почти лишена
сосудов, но богата нервными окончаниями. Латеро-вентрально от
заднего полюса глазного яблока располагается продырявленная
пластинка склеры (lamina cribrosa slclerae), через отверстия которой
из глазного яблока выходят волокна зрительного нерва. У перехода
в роговицу склера несколько утончается.
Сосудистая оболочка (tunica vasculosa) расположена под фиб-
розной оболочкой. В ней различают собственно сосудистую оболочку
(chorioidea), реснитчатое тело (corpus ciliare) и радужную оболочку
(iris). Собственно сосудистая оболочка лежит во внутренней части
глаза непосредственно под склерой. Она образована рыхлой сое-
динительной тканью, в которой обильно ветвятся сосуды. Содержа-
щимся в ней пигментом сосудистая оболочка окрашена в коричне-
вый цвет. На небольшом участке сосудистой оболочки откладывает-
ся гуанин — пигмент, обладающий металлическим блеском. Соответ-
ствующий участок называется зеркальцем (tapetum fibrosum). Лучи
света, отражающиеся от зеркальца, дважды проходят через свето-
чувствительную сетчатку, благодаря чему чувствительность в усло-
виях сумеречного зрения повышается. Сосудистая оболочка соеди-
нена со склерой очень неплотно.
Реснитчатое тело представляет собой утолщенный край сосуди-
стой оболочки, образующий кольцевой валик, расположенный у
места перехода склеры в роговицу. Реснитчатое тело несет около
100 радиально расположенных складок, образующих реснитчатую
корону (corona ciliaris). В сторону роговицы эти складки сильно
возвышаются и образуют так называемые реснитчатые отростки
(process! ciliares). К ним прикрепляются тонкие волоконца (fibrae
zonula res), образующие совместно реснитчатый пояс, или циннову
связку (zonula ciliaris, seu Zinnii), посредством которой хрусталик
прикрепляется к реснитчатому телу. В реснитчатом теле расположе-
на гладкая кольцевая мышца (m. ciliaris).
Радужная оболочка представляет собой непосредственное про-
должение сосудистой, подстилающее роговицу. Окраска радужной
оболочки обусловливает цвет глаза. Радужная оболочка по краям
тесно связана с реснитчатым телом. Поверхность радужной оболоч-
ки несет нежные складки (plicae iridis). Посередине радужная
оболочка пронизана круглым отверстием — зрачком (pupilia).
Зрачковый край радужной оболочки снабжен радиальными и коль-
цевыми мышечными волокнами. Кольцевые волокна (m. sphincter
pupillae) сужают, а радиальные (m. dilatator pupillae) — расши-
224
ряют зрачки. Размеры зрачка приспосабливаются к условиям осве
щенности: в темноте он увеличивается, а на свету уменьшается.
Сетчатая оболочка глаза, или сетчатка (retina), расположена
внутри от сосудистой и тесно связана с последней. Она состоит из
двух листков — наружного, пигментного (stratum pigmenti), и
внутреннего (retina proprium).
Оба листка переходят друг в
друга у зрачкового края радуж-
ной оболочки. Внутренний лис-
ток сетчатки — собственно
сетчатка — богато снабжен
светочувствительными клетка-
ми — колбочками и палочка-
ми. Как те, так и другие со-
держат светочувствительные
пигменты (иодопсин в палоч-
ках и иодопсин в колбочках),
распадающиеся на свепу. При
гзг? и
Рис.
140.
глаз-
ме-
Ряс. 139. Вибриссы пасюка
(пунктиром обведена осяза-
тельная зона)
Схема строения глаза
млекопитающих:
1 — мейбомиевы железы, 2 — край века,
3 — ресница, 4 — роговица, Б — перед-
няя глазная камера, 6 — зрачок, 7 — ра-
дужная оболочка (ее основа). 8 — ра-
дужная (слепая) часть сетчатки, 9 —
конъюнктива век, 10 — конъюИктивв
ного яблока, 11 — конъюнктивальный
щок, 12 — задняя глазная камера, 13 —
циннова связка, 14 — капсула хрусталика,
15 — паренхима хрусталика, 16 — стекло-
видное тело, 17 — зрительный сосок, 18 —
продырявленная пластинка склеры, 19 —
влагалище зрительного нерва, 20 — зри-
тельный нерв, 21 — сетчатка, 22 — пиг-
ментный эпителий сетчатки, 23 — соб-
ственно сосудистая оболочка, 24 — белоч-
ная оболочка, 25 — реснитчатая (слепая)
часть сетчатки, 26 — реснитчатое тело (по
Климову)
этом зрительные клетки, являющиеся одновременно первым нейро-
ном зрительного нерва, возбуждаются. Более чувствительные па-
лочки представляют собой аппарат сумеречного зрения. В центре
сетчатки расположено так называемое желтое пятно, являющееся
местом наиболее йеною видения. Волокна зрительного нерва соби-
раются в один йучок и выходят из глаза, образуя так называемое
слепое пятно, или зрительный сосочек (papilla optica). Периферия
сетчатки лишена зрительных клеток и образует так называемую
слепую часть сетчатки (pars соеса retinae).
Внутри от радужины располагается двояковыпуклое прозрач-
ное тело, называемое хрусталиком (lens crystalina). Хрусталик по-
15 Крыса 225
крыт тонкой оболочкой, к которой прикрепляется реснитчатая связ-
ка. Промежутки между волокнами этой связки выполнены жидко-
стью и образуют так называемый петитов канал (canalis Petiti).
Пространство между радужиной, хрусталиком и роговицей вы-
полнено жидкостью и называется передней камерой глаза (camera
oculi anterior). Пространство между радужиной и реснитчатой
связкой также выполнено жидкостью и называется задней камерой
глаза (camera oculi posterior). Вся остальная часть глаза внутри
от хрусталика заполнена желеобразным стекловидным те-
лом (рис. 140).
Роговица и хрусталик представ-
ляют собой важнейшие светопре-
ломляющие среды глаза. Лучи све-
та, преломляясь в них, фокусируются
на сетчатку. У близоруких живот-
2 5
Рис. 142. Вспомогательные ор-
ганы глаза пасюка:
1 — слезная железа. 2 — глазное
яблоко, 3 — роговица, 4 — веко, 5—
косая каудальная мышца глаза.
/ 2
Рис. 141. Разрез гЛаза пасюка:
1 — хрусталик, 2 — стекловидное
тело, 3 — передняя камера
6 — слезная кость. 7 — косая кра-
ниальная мышца глаза, 8 — прямая
дорзальная мышца. 9 — прямая
вентральная мышца. 10 — прямая
латеральная мышца, И — прямая
медиальная мышца
ных, в том числе и у крысы, глаз становится более короткофокус-
ным. Соответственно, размеры и выпуклость хрусталика у них уве-
личиваются. У крысы хрусталик заполняет почти все глазное ябло-
ко и имеет почти шарообразную форму (рис. 141). Его эквато-
риальный диаметр 4,0 мм, осевой — 3,6 мм; отношение обоих
диаметров составляет 1,1 : 1, в то время как у человека оно равно
почти 3:1.
Выпуклость хрусталика может увеличиваться, благодаря чему
глаз может приспосабливаться к видению более близко располо-
женных предметов. Этот процесс называется аккомодацией. При
аккомодации реснитчатая мышца сокращается и реснитчатое тело
приближается к хрусталику. При этом реснитчатая связка, растя-
226
гивающая хрусталик, расслабляется, и последний становится болег
выпуклым.
Вспомогательный аппарат глаза представлен слезной железой,
глазными мышцами и веками.
Глаз движется при помощи шести мышц, из которых четыре на-
зываются прямыми, а две — косыми (рис. 142). Прямые мышцы
глаза прикрепляются к четырем полюсам глазного яблока и назы-
ваются, соответственно, вентральной, дорзальной, латеральной и ме-
диальной прямыми мышцами (m. т. recti ventralis, dorsalis, latera-
lis et medialis). Одна из косых мышц глаза называется вентральной,
а вторая — дорзальной (m. т. obliqui ventralis et dorsalis). Все
эти мышцы, кроме вентральной косой, начинаются от сухожиль-
ного кольца, окружающего глазной нерв. Вентральная косая начи-
нается от костных стенок глазницы. При рассматривании более
близких предметов глаза поворачиваются передним краем внутрь,
так что глазные оси сближаются (конвергируют).
Веки (palpebrae) представляют собой кожные складки, защи-
щающие глазное яблоко от повреждений. Различают верхнее и
нижнее веко, разделенные друг от друга глазной щелью (рис. 142).
На внутренней поверхности век открываются так называемые мей-
бомиевы железы (glandulae tarsales), увлажняющие края век. На
веках имеются также сальные и апокриновые железы. По краям
век растут небольшие щетинкообразные волосы — ресницы. Они
служат органами осязания, предупреждающими глаза об опасности.
При прикосновении к ресницам веки рефлекторно закрывают глаз.
Это происходит под действием развитой в обоих веках кольцевой
мышцы глаза (m. orbicularis oculi). Верхнее веко снабжено спе-
циальной мышцей, поднимающей его (m. levator palpebrae supe-
rior). Внутренняя поверхность век покрыта соединительнотканной
оболочкой — конъюнктивой, которая с внутренних стенок век пере-
ходит на поверхность глазного яблока; в месте перехода образуют-
ся глубокие складки, называемые сводом. Эти складки допускают
значительную подвижность глаз. Конъюнктива построена из много-
слойного эпителия и содержит дополнительные слезные железы.
Она непосредственно переходит в эпителий век.
Слезная железа (gl. lacrimalis) представляет собой альвеоляр-
ную железу, секрет которой — слезы — служит для смачивания
поверхности глазного яблока (рис. 142). Из слезной железы слезы
поступают во внутренний угол между верхним и нижним веками.
Через носослезный проток слезы поступают в носовую полость и
выводятся наружу. При мигании веки увлекают накапливающиеся
в их углу слезы и смачивают ими глазное яблоко.
ОРГАНЫ СЛУХА И РАВНОВЕСИЯ
У позвоночных орган равновесия тесно связан с органом слуха
и составляет ча< гь внутреннего уха. Поэтому оба органа будут
описаны совместно.
1S* 227
Топографически орган слуха и равновесия разделяется на на-
ружное, среднее и внутреннее ухо. Наружное ухо (auris externa)
состоит из ушной раковины (auricula) и наружного слухового про-
хода (meatus acusticus externus). Ушная раковина представляет
собой кожную складку, поддерживаемую эластическим ушным хря-
щом. У перехода ушной раковины в наружный слуховой проход
правая и левая ее половины разделены вырезкой. У серой крысы
она узкая, всегда в виде острого угла, а у туркестанской и черной
Рис. 143. Схема строения органа слуха и
равновесия млекопитающих:
1 — лицевой нерв, 2 — эндолимфатический ме-
шок, 3 — слуховой нерв, 4 — лицевой нерв, 6 —
перепончатый лабиринт, 6 — костный лабиринт,
7 — мешочек, 8 — улиточный ход, 9 — костная
улитка, 10 — водопровод улитки. II — евстахиева
труба, 12 — напрягатель барабанной перепонки,
13 — среднее ухо, 14 — круглое окно, 13 — бара-
банная перепонка, 16 — наружное ухо, 17 — стре-
мечко, 18 — мышца стремечка, 19 — барабанная
струна, 20 — молоточек. 21 — наковальня (по
Веберу)
крысы — широкая и округ-
лая. На оральном крае уш-
ной раковины имеется не-
большой вырост — завиток
(helix). У пасюка он слабо
загнут внутрь, достигает
трех четвертей высоты уха и
не вырезан по наружному
краю, у туркестанской и у
черной крыс завиток всег-
да вырезан (Аргиропуло,
1940). Наружный слуховой
проход крысы представляет
собой небольшую хрящевую
трубку, заканчивающуюся
барабанной перепонкой (рис.
143).
Среднее ухо . (auris me-
dia) состоит из барабанной
полости (cavum tympani),
сообщающейся с глоткой
евстахиевой трубой; снаружи
она закрыта барабанной пе-
репонкой. Цепью из трех
слуховых косточек барабан-
ная перепонка соединяется с
так называемым овальным окном, ведущим во внутреннее ухо
(рис. 143).
Барабанная перепонка (membrana tympani) построена из трех
слоев. Толща ее состоит из фиброзной соединительной ткани, сна-
ружи она докрыта утонченным продолжением кожного покрова, а'
изнутри — слизистым эпителием. С барабанной перепонкой сра-
стается отросток наружной слуховой косточки, проходящий от края
перепонки до ее середины.
Наружная слуховая косточка, или молоточек (malleus), являет-
ся самой крупной из всех слуховых косточек (рис. 144). Его ру-
коятка (manubrium mallei) срощена с барабанной перепонкой, а
головка (caput) сочленяется со средней слуховой косточкой. У пере-
хода рукоятки молоточка в тело располагаются дорзальные отро-
стки молоточка (protuberantia mallei dorsales). Средняя слуховая
228
косточка, или наковальня (incus), сочленнепя «нонм н'.юм с моло-
точком, а длинной ножкой (crus longum) с nnyipciutvil слуховой
косточкой (рис. 144). От тела наковальни отходит коршкам ножка
(crus breve), прикрепленная связкой к стенке барабанной полосiи.
Проксимальный эпифиз наковальни окостеневает у пасюка в виде
самостоятельной чечевицеобразной косточки (os lenticulare), вкли-
нивающейся между длинной ножкой и наковальней. Внутренняя
слуховая косточка, или стремечко (stapes), состоит из маленькой
Рис. 144. Слуховые косточки различных видов крыс. А — серая крыса,
о — черная крыса, В — туркестанская крыса:
1 — основной край молоточка, 2 — тело молоточка, 3 — тело наковальни, 4 —
короткий отросток наковальня, 5 — длинный отросток наковальня, 6 — дорзаль-
ные бугры молоточка, 7 — рукоятка молоточка, 8 — латеральный бугорок моло-
точка (по Аргиропуло)
головки, сочленяющейся с наковальней, и двух широко расходящих-
ся ножек, соединенных с подошвой. Строение слуховых косточек
значительно варьирует даже у близких видов животных (рис. 144).
Звуковые волны, падая на барабанную перепонку, колеблют ее.
Эти колебания через систему слуховых косточек передаются вну-
треннему уху. Слуховые косточки функционально подобны колен-
чатому рычагу. Сочленение наковальни с молоточком практически
неподвижно, и так как длинное плечо рычага — рукоятка молоточ-
ка — в 1,5 раза длиннее короткого — длинной ножки наковальни,—
то колебания барабанной перепонки передаются внутреннему уху
уменьшенными по амплитуде и увеличенными по силе.
К молоточку прикрепляется мышца — напрягатель барабаня st
перепонки (m. tensor tympani), а к стремечку — стременная мыш-
ца (m. stapedius). Функция первой мышцы явствует из ее назва-
ния, стременная мышца уменьшает амплитуду движений стремечка.
Внутреннее ухо (aurisi interna) образовано так называемым пе-
репончатым лабиринтом, окруженным костной оболочкой, или
костным лабиринтом (labyTinthus osseus). Стенки его построены
из компактной костной ткани и легко могут быть выделены из губ-
чатого вещества каменистой кости. Преддверие лабиринта (vesti-
bulum) сообщается с барабанной полостью двумя затянутыми пе-
репонкой отверстиями — овальным окном (fenestra oval is) и круг-
229
лым окном (f. rotunda); в перепонку овального окна погружена
подошва стремечка (рис. 143). Преддверие разделено на эллипсо-
видное и шаровидное расширения; первое сообщается с полукруг*
лыми каналами, функционирующими как орган равновесия, вто-
рое— со спирально закрученной улиткой, заключающей чувствую-
щие окончания органа слуха. От эллипсоидного расширения начи-
нается также так называемый водопровод I. . '
открываю’
задней по-
10
Рис.
145. Схема строения перепончатого лаби-
ринта млекопитающих:
1 — эндолимфатический мешочек. 2 — горизонтальный
полукружный канал, 3 — общая ножка. 4 — передний
полукружный канал, 5 — задний лабиринт, 6 — нерв
преддверия, 7 — нерв улитки, 8 — мешочек, 9 — улитка
(по Веберу)
преддверия (aqueductus
vestibuli),
щийся на
верхности каменистой
кости (рис. 143).
Перепончатый лаби-
ринт (labirynthus mem-
branaceus) лежит внут-
ри костного и повто-
ряет его по форме
(рис. 145). Стенки его
образованы тонкой сое-
динительнотканной пе-
репонкой. Снаружи он
окружен жидкостью —
перилимфой, заполняю-
щей костный лабиринт,
сам он заполнен жид-
костью, называемой эн-
долимфой. Через водо-
провод преддверия про-
ходит эндолимфатиче-
ский проток (ductus еп-
dolymphaticus), оканчивающийся под твердой мозговой оболочкой.
Преддверие перепончатого лабиринта подразделено на так назы-
ваемый utriculus, связанный с полукружными каналами, и saccu-
lus, связанный с улиткой. Как в utriculus, так и в sacculus имеются
чувствующие поля, называемые пятнами (maculae staticae). Их
чувствующие клетки с волосовидными отростками покрыты мембра-
ной, содержащей в себе многочисленные кристаллы извести — ста-
толиты. При изменении положения головы инерция статолитов вы-
зывает их смещение по отношению к чувствующим клеткам, и по-
следние возбуждаются.
Полукружные каналы (canales semicirculares) в числе трех рас-
положены во взаимноперпендикулярных плоскостях. Перепончатые
каналы подвешены к костным тонкими соединительнотканными
перемычками. Основание каждого из полукружных каналов расши-
рено, образуя ампулы. В ампулах помещаются чувствующие обра-
зования, называемые гребнями (cristae staticae); они раздражаются
токами эндолимфы, возникающими при передвижении. Пятна и
гребни и представляют собой чувствующие образования органа
равновесия.
230
Чувствующее образование, воспринимающее звуковые раздра-
жения, называется кортиевым органом (organon Corti). Расположен
он в перепончатой улитке (cochlea membranous), представляющей
собой спиральный канал. У крысы этот канал имеет три завитка.
Одна из стенок перепончатой улитки срастается со стенкой костной
улитки, другая прикрепляется к спиральному выросту костной улит-
ки, вдающемуся в ее просвет. Перилимфатический ход улитки под-
разделяется перепончатой улиткой на дорзальную лестницу пред-
дверия, сообщающуюся с овальным окном, и вентральную барабан-
ную лестницу, сообщающуюся с круглым окном; обе лестницы на
вершине костной улитки сообщаются друг с другом. Кортнев орган
располагается по всей длине перепончатой улитки. Он состоит из
поддерживающих и чувствующих клеток с волосовидными отрост-
ками, расположенных в несколько рядов. Дорзально над ними нави-
сает отходящая от стенок перепончатой улитки кроющая мембрана
(membrana tectorii), снизу кортиев орган подстилает так называе-
мая основная мембрана (membrana basilaris), состоящая из попе-
речно ориентированных волокон. Длина последних уменьшается в
соответствии с уменьшением диаметра хода улитки. По теории Гель-
мгольца звуковая волна, распространяющаяся по перилимфе улит-
ки, вызывает резонансные колебания в волокнах основной мембра-
ны, имеющих соответствующую длину. Последние вызывают коле-
бания сидящих на них чувствующих клеток, раздражаемых при со-
прикосновении с кроющей мембраной. Согласно Гельмгольцу, часть
кортиева органа, расположенная ближе к вершине улитки, в кото-
рой волокна основной мембраны короче, связана с восприятием
более высоких звуков.
ОРГАНЫ ОБОНЯНИЯ
Обонятельные клетки располагаются в слизистом эпителии но-
совой полости, покрывающем верхнюю (обонятельную) раковину и
противоположную ей стенку носовой полости. Соответствующую
часть носовой полости называют обонятельной, а остальную — ды-
хательной.
Обонятельные клетки, представляющие собой первые нейроны
обонятельного нерва, принадлежат к типу ганглионарных клеток и
имеют на наружном конце волосовидный отросток. В слизистом
эпителии дыхательной части носовой полости обонятельных клеток
не бывает. В отличие от светлорозового эпителия дыхательной
области, обонятельный эпителий крысы окрашен в светложелтый
цвет.
По степени развития обонятельного эпителия можно до изве-
стной степени судить о развитии обоняния у животного. У крысы
на сагиттальном разрезе носовой полости обонятельный эпителий
занимает примерно такую же часть, как и дыхательный; однако,
учитывая сложность поверхности обонятельной раковины, мы долж
пы признать, что общая поверхность обонятельного эпителия h.i
.'31
много превышает общую поверхность дыхательной части носовой
полости.
Эмбрионально от носовой полости отделяется так называемый
якобсонов орган (organon Jacobsonii), от чувствующего эпителия
которого начинаются волокна, присоединяющиеся к обонятельному
нерву. У крысы якобсонов орган представляет собой узкую трубку
длиной 12—15 мм, расположенную у основания сагиттальной пере-
городки носа и открывающуюся в ротовую полость на переднем
валике твердого нёба. Кроме того, якобсонов орган обычно соеди-
няется и с носовой полостью, в которую открывается непосред-
ственно рядом с ноздрями.
ОРГАНЫ ВКУСА
Органы вкуса крысы представлены так называемыми вкусовы-
ми почками, расположенными в сосочках языка и отчасти в сли-
зистой мягкого нёба. Вкусовые почки представляют собой оральные
тельца, состоящие из покровных клеток и чувствующих клеток с
волосовидным отростком. Вкусовые почки погружены в слизистую
оболочку, но сообщаются с полостью рта так называемыми вкусо-
выми порами, представляющими собой небольшие отверстия в по-
верхностном слое эпителия.
ЛИТЕРАТУРА
Общие работы
Алезе Г.. Морская свинка. 1911.
Беляев М. М., Зоотомия. М., 1947.
Кюкенталь В., Руководство к практической зоологии. М., 1919-
М о ж а й к о Л. Н.» Крыса. М.» 1936.
Паркер Т., Курс зоотомии. М., 1904.
Сахаров П. П., Лабораторные животные, М.» 1937.
Терентьев П. В., Дубинин В. Б. и Новиков Г. А-, Кр^™^- М.»
1952.
Терентьев П. В., Малый практикум зоологии позвоночных. М» 1948.
Donaldson Н., The rat 2-nd ed*, Philadelphia, 1924.
F a r i s s E., The rat in laboratory investigations, 2-nd ed, London, 1942.
Green E., Anatomy of the rat, «Trans. Amer. Philos. Soc.>, № i27, 1935.
Кожа и ее производные
Арутюнов А. И. и Семенов Н. И., Кожная температура при раз-
личных воздействиях на симпатическую нервную систему, «Вести, хирургии»,
т. 56. вып 2. 1938.
Бабенышев В. П., Значение мехового покрова и размера тела некото-
рых видов грызунов для теплоотдачи и их стойкость к действию внешних фак-
торов, «Зоол, журн.>, т. XVII, вып. 3. 1938.
Синельников Я. С., Результаты исследования методами макроскопии
вегетативной нервной системы и желез слизистых оболочек и кожи. «Врач.
дело>, № 1—2. 1946.
Скелет
А н ич кя и А., Челюстное сочленение человека и животных, «Из». С -П.
биол. лаб.>, т. I, вып. L СПБ, 1896,
Гамбарян П. П.» Роль остистого отростка второго грудного позвонка
некоторых грызунов, «Здол. журн.>, т. LXXX, вып. 2. 1951.
232
Герцен И. Г.. Материалы по сравнительной анатомии голеностопного су*
става, «Арх. анат. гист. и эмбриол.», т. XX, вып. 2. 1939.
Домбровский Б. В., Антнклиния у млекопитающих и ее функциональ-
ное освещение, «Зоол. журн.», т. XIII, вып. 1. 1934.
Долг о-С а б у р о в Б. А., Роль мышц в морфогеюии скелета в местах на*
чала и прикрепления мышц, «Изв. Ленингр. научн. ин-та им. Лесгафта», т. XVI,
вып. 1 и 2. 1930.
Дружинин А. Н., К вопросу о строении и эволюции локтевого сустава
•млекопитающих, «Докл, АН СССР», нов. сер., т. Ill (XII), Хе 5 (100). 1936.
Дубинин В. Б., Изменения шейного отдела позвоночника животных в
филогенезе и онтогенезе, «Журн. общ. биол.», т. X, вып. 2. 1949.
М а из ий С. Praepollex млекопитающих в свете эволюции их конеч-
ностей, Тр. ihct. Зоол. АН УРСР, т. V. 1950.
КасьяненкоВ. Г., Про особливости спещализацп колшого суглобли
у деяких ссявщв, Тр. 1нст. Зоол. АН УРСР, т. I. 1948.
Касьяненко В. Г., Анал1з скелета заплесна деяких ссавщв, Тр. 1нст.
Зоол. АН УРСР, т. I. 1948.
Касьяненко В. Г., Об особенностях строения и функции стопы, Тр.
1нст. Зоол., т. II. 1949.
Касьяненко В. Г., К сравнительной анатомии и функции стопы млеко-
питающих, Тр. 1нст. Зоол. АН УРСР, т. III, 1950.
Касьяненко В. Г., Функциональный анализ суставов тазовой конечно-
сти некоторых млекопитающих, Тр. нн-та Зоол. АН УССР, т. V, 1950.
Ковешникова А. К., Изменения лопатки животных в связи с перехо-
дом от горизонтального положения на четырех к вертикальному положению на
двух конечностях, «Изв. Ленингр. научн. ин-та им. Лесгафта», т. XIV, вып. 1
и 2. 1928.
Кравец Л. П., История развития грудины и эпистернального аппарата
млекопитающих, Тр. сравнительноанатомического ин-та МГУ, вып. 5. 1905.
Лесгафт П. Ф., О различных типах конечностей млекопитающих, «Изв.
С.-П. биол. лаб.», т. 1 и 2. 1896—97.
Малеев Е. А., Морфо-функциональный анализ.затылочной области чере-
па и шейных позвонков млекопитающих (двое.), 1950/
Страхов Н. Н., О костях запястья млекопитающих. СПБ, 1857.
Якимов В. П., Развитие скелета передних конечностей человека и неко-
торых млекопитающих, «Зоол. журн.», т. XXVI, вып. 4. 1947.
Вгопп К., Klassen und Ordnungen des Tier-Rrfchs, Bd. VI. Abt. V. 1874—
1900.
Howell B., Anatomy of the wood rat, 1927.
Мускулатура
Александровский В. Н.» Глубокая мускулатура и подвздошноребер-
ный мускул шеи с их иннервацией у ряда млекопитающих домашних животных,
Тр. Моск, зоовет. ин-та, т. 4. 1940.
Гамбарян П. П., Мускулатура переднего пояса некоторых грызунов,
Изв. АН Арм. ССР», № 7, 1946.
Гамбарян П. П., Мускулатура проксимального отдела передней конеч-
ности некоторых грызунов, «Иэв. АН Арм. ССР», т. I, № 3. 1948.
Гамбарян П. II., Адаптивные особенности передних конечностей горного
слепца Spalax leucodon nehrlngi Sat., «Зоосб. АН Арм. ССР», т. 8. 1952.
ИвановС. В., Сравнительная анатомия и физиология m. т. biceps brachii
и brachialis internus млекопитающих, «Русск. арх. анат., гист. и эмбриол.»,
т IX, вып. 2. 1930.
Ковешникова А. К-, Сравнительноанатомические исследования m. bi
1 • ds brachii млекопитающих и анатомия нижнего прикрепления его у человека,
«Изв. Ленингр. научн. нн-та им. Лесгафта», т. IX, вып. 2. 1924.
233
Лебедева М. В., Соотношение между окраской* и строением попереч-
нополосатых мышц в связи с их деятельностью и возрастом животного, «Изв.
Ленннгр, яаучн. ин-та ям. Лесгафта», т. XVI, вып. 1 и 2. 1930.
Лерхе А. В., Наблюдения над добавочными головками двуглавой мышцы
плеча у животных и человека, «Виол. н.-и. «н-т Ростовского-на-Дону универ-
ситета», вып. 2. 1938.
М а к р о в И. К., Сравнительноанатомическое исследование мускулов, дей-
ствующих на тазобедренный сустав у млекопитающих, Тр. Моск, зоовет. ин-та
т. 3. 1938.
Преображенский П. П., Сравнительноанатомнческие данные по дор-
зальной мускулатуре поясничного отдела домашних животных (внутренняя
структура и статодинамика). Тр. Вологодск. с.-х. ин-та, вып. 3. 1941.
Прозоров Н. Н., Номенклатура скелетных мышц домашних млекопитаю-
щих и их группировка по функции, Казань. 1914.
С п и р и х о в И. К.» Сравнительная анатомия мускулатуры предплечья, дей-
ствующей на карпальный сустав, с изучением внутренней структуры мускулов
Тр, Бурят-Монгол. зоовет. ин-та, вып. 2. Улан-Уде. 1941.
Ю с т о в К. Р., Подкожная мускулатура шеи у млекопитающих, Уч. зап.
Ростовского-на-Дону гос. ун-та, Бюлл. н.-и. ин-та, вып. 2. 1934.
В i d i n о s t L. E., Rate of formation of myosin in the muscle of the rat,
«Journ. Biol. Chem.», vol. 190. 1951.
Gregory W., Notes on the principles of quadrupedal locomotion «Ann.
New. JOrk Ak, Set», vol. 22. 1912.
Hill J. E.t Morphology of the pocket-gopher mammalian genus Thomomys,
«Univ, of Californ. Public. Zool.,» vol. 42. № 2. 1937.
Hol liger C. D., Anatomical adaptation in the thoracic limb of the Cali-
fornia Docket goDher and other rodents, «Univ, of Calif. Public Zool», vol.
13. № 12. 1926. ’
Howell B. A., Phylogeny of the distal musculature of the pectoral apen-
dage, «J. Morph.», vol. 60. 1930.
Parsons F. G., Some point in the myology of rodents, «Proc. Zool. Soc.
Lond.», 1892.
Parsons F., On the myology of the Sciuromorphinae and Hystricomorphi-
nae rodents, «Proc. Zool. Soc. bond». 1894.
Parsons F. G. Myology of Rodents, II, «Proc. Zool. Soc. Lond.». 1896.
Функциональная характеристика аппарата движения
Бернштейн Н. А., О построении движений. Медгиз, 1947.
Бернштейн Н. А. (ред.), Исследования по биодинамике локомоций, кн. I.
1935.
Гамбарян П. П., Роль остистого отростка второго грудного позвонка
некоторых грызунов, «Зоол. журн.», т. XXX, вып. 2. 1951.
Попова Т. С., М о г и л я п с к а я 3. В., Техника изучения движений,
1935.
Н i г s с h W., Zur physiologischen mechanik des Froschsprunges, «Zeitsch. f.
vergleich. Physiol.», Bd. 15. 1931.
Howell A. B., Speed in animals. 1942.
Howell A. B., The saltatorial rodents Dipodomys, the functional and com-
parative anatomy of its muscular and osseous systems, «Proc. Amer. Acad, of
Arts and Science», vol. 67. № 10. 1932.
Органы питания
В e л и ч к о M. А., О некоторых особенностях строения пищевода и желуд-
ка некоторых грызунов, «Арх. анат., систол, и эмбриол.», т. XX, вып. 2, сер.
анатом., кн. 2. 1939.
234
Величко М. А., Кишечник грызунов (Rodentia). Уч. зап. фак-та естеств.
Ленингр. пед. ин-та, вып. 2. 1939,
Величко М. А.» Кишечник грызунов (Rodentia): II желудок, Уч. зап.
фак-та естеств. Ленингр. пед. ин-та, т. 5, вып. 1. 1949.
Величко М. А. и Макеева Т. М, О некоторых характерных w/i н-
ностях строения и функции кишечника грызунов, Тр. Всес. им-Tai защиты расте-
ний, вып. 2. 1949,
Окунь М. М. и Дмитренко М. И.. Влияние острого отравления бен-
золом на превращение гликогена в печени крыс, «Фармакол и токсикол.», № 3.
1950,
Gilson G. and Wiseman G., Selective absorption of stereoisomers
of a min a acid from loops of the small intestive of the rat. «Bioch. Journ.», vol.
48, 1951.
G 1 a s t о n e S.. A comparative study of the development in vivo and in
vitro of Rat and Rabbit molars, «Proc. Royal Soc. London». 1938.
Lagerstedt S. and S t er n am N., Selective acetilholine phosphatase
activity within the epithelia of the small intestine and Kindey in rat, «Acta Anat.
Bozel», Bd. 10. 1950.
M i 1 s t e i n В.» Regeneration in the submacsillary gland of the rat, «Brit.
Journ. Exp. Path.», vol. 31, 1950.
Органы дыхания
Арандаренко М., Обонятельный орган и его функция у животных и
человека, «Вести, болезн. уха, носа, горла». 1913.
Ж е д е н о в В. Н.» О классификации типе» дольчатости легких у млекопи-
тающих, «Докл. АН СССР», т. LXXIV, № 2. 1950.
Жеденов В. Н., Сравнительноанатомические типы дольчатости легких v
млекопитающих в эволюционном освещении, «Докл. АН СССР», т. LXXXI,
№ 4. 195L
Жеденов В. Н., Образование вторично-недольчатых легких у млекопи-
тающих в процессе истории развития», «Докл. АН СССР», т. LXXXV1I № 5 1951.
Мухамедгалиев Ф. С., Биоморфология дыхательной системы млеко-
питающих (докторск. диес.). Алма-Ата, 1948.
Сахарова Т. М., Адаптивные особенности морфологии носовой полости
грызунов (канднд. дисс.). М., МГУ, 1948.
Сергиевский М. Н., Дыхательный центр млекопитающих животных и
регуляция его деятельности. М., 1951.
Яхтмн Г. А., Сравнительная характеристика носа у белой мыши и белой
крысы. Материалы по анатомии и гистологии носа у лабораторных животных,
«Вести, ото-рино-ларннгол.», № 3. 1939.
Мочеполовая система
Завадовский М. М. и> Юдинцев С. Д., Взаимодействие между се-
менниками и простатой с семенными пузырьками у крыс, «Сб. труд. НИИ
Зоол., МГУ», № 3. 1936.
Ильина А. А. и Смелов Н. К., Патолого-ан атомические изменения во
внутренних органах крыс и кроликов при применении различных доз осарсола,
«Веста, венерол. и дерматол.», № 12. 1934.
Кукушкина* Е. И., Возрастные изменения в половой системе самцов
крыс и влияние на них половых стимуляторов, «Док. Всес. акад. с.-х. наук
имени В. И. Ленина», вып. 18. 1939.
Кудряшов Б. А и Беляева Е. А., Действие продуктов распада жи-
ров на беременную матку крысы, «Сб. труд. НИИ Зоол МГУ», № 3. 1936.
Кудряшов Б. А. и Поликарпова Е. А.. К вопросу о временной
стерилизации самок крыс и кроликов продуктами распада жиров, «Сб. труд
НИИ Зоол. МГУ», № 3, 1936.
735
Рихтер И. Д., Об особенностях смены клеток вагинального эпителия
грызунов, «Бюлл. эксп. биол. и мед.», т. 8, вып. 2 (8). 1939.
Тереза С. Н., О механизме стерилизующего действия избытка тиреоиди-
на на беременность белых крыс «Сб. труд. НИИ Зоол. МГУ», № 3. 1936.
Э с к н и И. А., Вызывание ложной беременности у крыс фолликулином,
«Бюлл. эксп. бнол. и мед.», т. 5, вып. 3. 1938.
Органы кровообращения и внутренней секреции
Крылова П. С., Сравнительноанатомические данные об артериях кожи,
Тр, V-ro Всес. съезда анат., гистол и эмбриол. 1951.
Луков Б. Н., К вопросу о кровяном давленини при носовом, ротовом и
трахеальным дыхании, Тр. ин-та физиол. Саратовск. мед. мн-та. т. I, № 1. 1934.
Никитин В. Б., Атлас клеток крови сельскохозяйственных и лаборатор-
ных животных. Мч 1949.
Пегельман С. Г., Влияние температуры и литания на внутренние ор-
ганы и реакцию крови у некоторых грызунов, Тр. Всес. 1ин-та защиты растений,
вып. 2, 1949.
Evans G. Adrenal corten and endogenous carbohydrate formation, «Amer.
Journ. Physiol.», vol. 114, 1936.
Jugle D., Higgins G. and Nilson H., Homoplastic transplantation
of adrenal glands in rats of inbred strains. «Amer. Journ. Physiol.», vol. 121.
1938.
M a г k e e J.. Rhythmic vascular tissue changes, «Amer. Journ. Physiol.»,
vol. 100, 1932.
Shapiro R. and Pincus G., Pancreatic diabetes and hypophysectoray in
the rat, «Proc. soc. exper. Biol, and Med.», vol. 34. 1936.
Stendell W. Zur vergleichenden Anatomie und Histologie des Hypophy-
sis cerebri, «Arch, fur mikr. An at.», Bd. 82, 1913.
Нервная система
Боровский M. Н., Методика исследования нервного аппарата щитовид-
ной железы у крыс, морских свинок и кроликов, «Пробл. эндокринол.», Кв 4,
1940.
By л И. М., Исследование на крысах (о функциональных особенностях
нервномышечной системы в онтогенезе. Сообщ. I), «Физиол. журн.», т. XVI, № 1.
1937.
Гамаков С. Г., Влияние отсутствия носового дыхания на сосуды мозга,
Тр. Сарат. ин-та физиол., т. I. 1934.
Кононова Е. А., Атлас мозгового ствола человека и животных, М., 1947
Матвеев Б. Си Биоморфология головного мозга позвоночных. Тр.
V Всес. съезда анатом., гистол. и эмбриол. 1951.
Полежаев В. Г., Изучение поведения серой крысы в зависимости от
факторов внешней среды, Тр. ЦНИДИ, вып. 6. 1950.
С а р к и с я н А. А. и Фанарджян В. В., К вопросу о значении нерв-
ной системы в возникновении злокачественных опухолей в эксперименте, «Изв.
АН Арм. ССР», т. IV, № 9, 1951.
С е п п Е., История развития нервной системы позвоночных от бесчерепных
до человека. М„ 1949.
Фаворский Б. В., Внутриствольное строение головного мозга у чело-
века, собаки, кошки, кролика, морской свинки и белой мыши, «Арх. биол.
наук», т. XLII. вып. 1/2. 1936.
Gurevitsch М. und andere, Zur vergleichenden Zytoarchitektonik der
Grosshirurinde der Saugetiere; 1—Nager, «Zeitschr. Anat. Entw.», Bd. 90. 1929.
Herrick G. The brain of rats and man, 1926.
Kappers С.» Huber G. and G г о s b у E.. The comparative anatomy of
the wervous system of vertebrates. New Jork. vol. 1—2. 1936.
236
Органы чувств
Арандаренко Е. Н., Обонятельный орган и его функция *
и человека, «Вести, болезн. уха, носа, горла». 1913.
Аргиропуло А. И., Мыши и крысы. Л., 1940 (Фауна СССР, Miexonii
тающие, т. III, вып. 5).
Турке ви ч Б. П., Новая методика изготовления коррозионных препа чм •
костного лабиринта млекопитающих и птиц, Тр. Узб. гос. ун-та, т. 5. 1936.
В е г 1 е г i с h and Nussbaum, Nase Anatomie und Pathologic der Spon-
tanerkranknngen der kleinen Laboratoriumstiere, Berlin. 1931.
Broman, Das Organon vomeronosale Jacobsonen’s — ein Wassergeruch-
sorgan, «Anat. Hefte». Bd, 58. 1920.
D e *t w i 11 e r, The vertebrate photoreceptors. 1943.
Walls G., The vertebrate aye and its adaptive radiation. 1942.
Meyer M., Die normale Anatomie der Labyrinthkapsel, Untersuchungen an
Menschen und Laboratoriumtieren, «Zeitsr. Hals. u. Heilkunde». Bd. 34. 1933.
РУССКИЙ УКАЗАТЕЛЬ
А
Анальное отверстие 152 Ф1, 153
Аорта 170, 171, 172 *
— брюшная 179, 180 *, 181 ♦
— грудная 179, 185
Артерия (ии) альвеолярная 175. 176
— бедерная 181 *, 182
— безымянная 172 *
— большеберцовая 181 *, 183, 184
— бронхиальная 179
— брыжеечная 180 *, 181
— венечная 171
— верхнегубная 173 *, 174
— височная 173, 174 *
— глаза угловая 172 * — 174 •
— глазничная 174*, 175
— глоточная 174 *
— гортанная 173, 175
— грудинная 177
— грудная 176*, 177
— грудо-акромиальная 177, 179 *
— грудо-спинная 178, 179 *
— дельтовидная 172 *
— диафрагматическая 180 *
— диафрагмо-перикардиальная 177
— жевательная 172 ♦, 173. 174
— желудочная 180*
— запирательная 181 *, 182
— затылочная 172 *, 173 *, 176
Артерия (ин) клино-нёбная 176
— коленная 181 *, 183
— крыльного канала 175
— легочная 157*. 158, 169, 171
— лицевая 173*, 174, 176
— лобная 174 *
— локтевая 178, 179 *
Артерия (ни) лопатки 177
— лучевая 178, 179 *
— малоберцовая 181 *, 184
— матки 182
— межкостные 178, 179 *, 183
— межреберные 176 *, 177, 179
— межчелюстная 172 *
— менингеальная 172, 173
— мозговая 174, 175 *, 176
— мозговой оболочки 174
— мозжечка 174, 175*
— мочевого пузыря 181 *, 182
— мышечно-диафрагматическая
176*, 177
— надчревная 177, 182, 183
— нёбная 173, 174 *
— ниокнегубная 172 *, 173 *
— нижнечелюстная 173, 174 *
— носовая 172*—174*
— пальцевые 178
— перикардиальная 176
— печеночная 180 *
— пищеводная 179
— плечевая 177, 178, 179*
— плюсневые 184
— подбородочная 173, 174 *
— подвздошная 181 *, 182
— подвздошно-поясничная 181
— подглазничная 173 *, 174 *
— подколенная 181 *, 183
— подключичная 172 *, 176, 177, 179
— подлопаточная 177, 179*
— подмышечная 176 *, 177, 179 *
— подчревная 181 *, 182
— подъязычная 173, 174*
— позвоночная 172 *, 176
1 Звездочкой * отмечены страницы, на которых помещены рисунки.
238
Артерия (ни) почечная 180 *, 181 *
— поясничные 181
— прямой кишки 180*, 181 *, 182
— пястные 178
— рта угловая 172*, 173*
— сафена 181 ♦, 183, 184
— селезеночная 180 •
— семенниковая 181 *
— сердца венечные 171, 172
— слуховая 174
— сонная 172 *—174 *, 176 *
— сосудистой оболочки мозга 175 *
— срамная 181 *, 182
— срединная 178, 179 *
— срединно-локтевая 179 *
— срединно-лучевая 178, 179 ♦
— средостения 176, 177
— стопы 181 *, 183
— хвостовая 181 *
— челюстная 172, 173 *, 174, 175
— чревная 180*
— шейная 172 ♦, 176, 177
— щитовидная 172 *
— ягодичная 182
— язычная 173
- яичниковая 181, 182
— яремная 172*
Артериальное виллизиево кольцо 174,
Атлант 81, 82*, 105, 107, 130
Б
Барабанная перепонка 228, 229
- полость 228, 229
Бронхи 157* 185
Брыжейка 146, 153
Брюшина 153
В
Варолиев мост 204—207
Веки 227
Вена (ы) альвеолярная 186, 187
— бедренная 189
— безымянная 172 *, 185, 190
— большеберцовая 190
— бронхиальные 185
— брыжеечная 188
— верхнегубная 186
височная 186 ♦, 187
— глоточная 185
— головного мозга 187
— грудная 172 *, 176*, 185. 188
— диафрагматическая 185, 188, 189
— жевательной мышцы 186*, 187
— заднеглазничная 186*
— запирательная 189
затылочная 185
Вена (ы) кисти 188
— кишечная 180*
— краниально-грудная 188
— крыловидная 187
— легочные 157*, 169, 171
— лицевая 186 *, 187
— лобная 186
— локтевая 188
— лопаточная 186, 188
— маточная 189
— межкостная 188
— межреберные 176*. 185
— менингеальная 187
— мозговая 175 *, 186 *, 187
*— мозжечковая 175 *, 187
— мочевого пузыря 189
— мышечно-диафрагматическая 176
— надглазничная 186
— надчревная 190
— надчревно-срамная 189
— непарная 176 *, 185
— нижнегубная 186*
— нижнечелюстная 186 *
— носовая 186 *
— околоушной железы 186 *
—- пальцев 190
— печени воротная 180 *, 188
— печеночная 180 ♦, 188
— плечевая 187, 188
— плюсневые 190
— подбородочная 186 *
— подвздошная 188, 189
— подвздошно-поясничная 188
— подглазничная 187
— подключичная 172*, 185
— подколенная 189, 190
— подмышечная 187, 188
— подчелюстная 186*
— позвоночная 175 *, 185, 187
— полового члена 189
— полунепарная 185
— полые 131, 171, 180 ♦, 185, 188
— почечная 180*, 188, 189
— поясничные 188
— прямой кишки 189
— сафена 189, 190
— селезеночная 188
— семенниковая 188
— сердечные 171, 185
— срамная 189
— срединная 188
— средостено-перикардиальная 185
— стопы 190
— угла рта 186 ♦
— угловая глаза 186
— ушная 186 *, 187
— челюстная 185. 186*, 187
— щитовидная 185
— ягодичная 189
— язычная 186 *
239
Вена (ы) яичниковая 188
— яремная 172, 185, 186 е
Венозное сплетение геморроидальное
189
— — глоточное 186, 187
---крыловидное 186, 187
---стопы 190
Вибриссы 64, 223, 224, 225 *
Влагалище 164*, 165*, 182, 189
Волос 63 *, 64, 223
Г
Гипофиз 196 *, 197, 209
Глаз 223, 224, 225*, 226*
Глаза задняя камера 226*
— передняя камера 226 *
Глазного яблока оболочка радужная
224
-------сетчатая 224—226
-------сосудистая 224
--- фиброзная 224
---оболочки 224, 225
Глазное яблоко 223, 224, 227
Глотка 81, 146, 147, 150, 155, 185, 187,
228
Головной мозг 185, 187, 199
Гортань 81, 155, 156*, 157, 173, 185,
187
Грудина 83, 84, 103, 106, 150, 155, 156
Губы 147
д
Дерма 63*
Дермы сосочки 63 *
Диафрагма 102, 131, 146, 151 *, 153,
158, 177, 180, 185, 188
Е
Евстахиева труба 228
Ж
Железа (ы) ампуллярные 163
— апокриновые 227
— бруннеровы 152
— вилочковая, или зобная (тимуо)
195 *
— заушная 215
- - зобная 151 *
— кожные 64
— луковичные 163, 165
— мейболиевы 227
— молочные 63*, 64, 65, 183, 189
— надпочечная 181, 188
— околощитовидная 194, 195 ♦
— паращитовидная 173, 185
240
Железа (ы) пилорические 152
— поджелудочная 151 *, 152, 154,
180
— половые 196
— потовые 63 *, 64, 66
— предстательная 163, 182
— препуциальные 163
— придаточные 161
— пузырьковидная, 151*,
162*, 163
— сальные 63 *, 64, 227
— слезная 212, 227
— слюнные 150 *, 151
— щитовидная 172, 173, 177, 185,
104 10Ч 999
Желудок i46, 147, 151 *, 152 *, 153 *,
180, 188, 191
Желчный пузырь 154
3
Зрительный бугор 200, 208, 209, 212
Зубная формула 147
Зубы 78, 147, 148. 175, 187, 212, 213
— верхнечелюстные 148*
— нижнечелюстные 148 *
К
Канал слезный 67
Капилляры 167
Кисть 89, 90
Клитор 166
Ключица 69, 86 *
Кишка двенадцатиперстная 146, 147,
151, 152 *, 153 *, 154, 180, 188
— ободочная 181, 188
— подвздошная 153
— прямая 146, 147, 151 ♦, 152 ♦, 153,
163, 164*, 188, 189
— слепая 146, 147, 151.*, 152*, 153,
181 188
— толстая 152 ♦, 153, 181, 188
— тонкая 146, 147, 151 *, 152 *, 153,
180, 188
Когтевая пластинка 63*, 64
— фаланга 63 *
Когтевой валик 63 *, 64
— желобок 63 *, 64
Когти 64, 91, 96
Когтя корень 64
— подошвенная часть 64
Кожа 62, 63*
— собственно 62
Кожи производные 63 *, 64
Коленчатое тело 208, 209, 211
Конечности передней плечевой отдел
86, 87
Конечность задняя 91, 92
Конечность передняя 86
Ковыоктмва 227
Крови форменные элементы 168, 169
Кровь 168, 169
Кортнев орган 231
Костный мозг 66. 192
Кость (и) барабанная 187
— бедренная 92, 93 *, 94, 115 е, 116,
118—122, 139, 140, 142
— безымянные 118, 119, 128
— большеберцовая 93, 94 *, 95, 116—
119, 121, 123, 183
— верхнечелюстная 67, 68, 69 *, 71 *,
78*, 80, 125, 126
— височная 68, 69 *, 71 *, 75 *, 76,
77, 102, 103, 105, 125, 149
— впадинная 91
— голени 93, 94 *, 139
— гороховидная 89, 90, 114
— грудная (грудина) 128, 131
— длинные 66
— запястья 89, 90, 113
-----локтевая 89, 90
----- лучевая 89
— затылочная 67, 69 *, 70, 71 *, 73 *,
74, 75, 77, 126, 130
— каменистая 75, 187, 229, 230
— клиновидная 74 *, 75, 78, 79, 81,
126, 130
-----основная 68, 69 *, 71 * 70
---передняя 67—69 *, 71 *, 74,
75 *, 78, 79
— короткие 66
— крестцовая 85, 129, 131
— крыловидные 70, 71 *, 79
— лобковая 118
— лобная 67—69 *, 71 *, 75, 76 *, 77,
79, 126
— локтевая 69, 88 *, ПО *, 111 ♦, 144
— лонная 91 *, 92, 119, 139
— лучевая 69, 88 *, ПО*, 111 ♦, 112,
113
— малоберцовая 93, 94 *, 95, 121—
123, 183
— межтеменная 68, 69 *, 76 *, 77, 79,
80
— межчелюстная 67, 68, 69 *, 71 *
— нёбная 68, 69, 71 ♦, 78, 79, 80 •
— нижнечелюстная 80 е, 81
— носовая 67,’ 69 *, 71 *, 78—80
— пальца предпервого 90
— пальцев 89, 91, 95
— плечевая 69, 87*, 88, 100, 105, 106,
107*, 108—113, 144, 178, 216
— плоские 66
— плюсны 95*, 96*, 121, 123, 124,
'125
— подвздошная 85, 91 *, 92, 115, 116,
120, 129, 131, 141
16 Крысз
Кость (и) подъязычная 81, 148, 149*,
173
— полового члена 163
— предплюсны 95, 96, 123
дополнительная 96
центральная 96
— пястья 89, 90, ПО, 111, ИЗ, 114
— пяточная 95, 96 *, 121, 124
— решетчатая 70, 71 ♦, 77 ♦, 78, 79,
80, 211
— седалищная 91 *, 92, 117—120, 130,
139
— сесамовидные 66, 89, 90, 96, 114,
121, 125, 142
— скуловая 67, 71 *. 80 *
— слезная 67, 69 *, 78, 80 *
— смешанные 66, 67
— стопы 95 *
— тазовые 91 *, 92 *, 115*, 116, 118,
121
— таранная 95, 96 •
— теменная 68, 69 *, 75, 76*, 77, 126.
129
— челюстная 67
— щитовидно-подъязычная 156
Л
Лабиринт 78
— костный 229, 230
— перепончатый 229, 230 *
Легкие 151 *, 155, 157 *, 158, 171
Лейкоциты 191, 192
Лимфа 190
Лимфатические протоки 190
— сосуды 190
— узлы 190—192
----бронхиальные 185
----паховые 190
----подвздошные 190, 191 *
----подколенные 190, 191 *
----подмышечные 177, 178, 190,
191 *
----подчелюстные 190, 191 *
----подъязычные 190
----шейные 177, 186
Лопатка 69, 85, 86*, 100, 103, 105,
107, 109, 150
Люисово тело 209
М
Матка 182
— мужская 163
Мозг головной 202, 203 *, 204, 208 *,
215*
— задний 204, 205
— конечный 202, 204, 209
— обонятельный 209
241
Мозг продолговатый 202, 203, 204 *,
206. 208 •, 214, 215
— промежуточный 202. 204, 207, 208
— средний 202, 207
Мозга белое вещество 207
— головного большие полушария
206, 207, 209
----кора 208. 209. 210 ♦, 211
— серое вещество 198, 207, 208, 210
Мозговые желудочки 202—205, 208—
210
Мозжечек 200, 202, 205, 206, 207 *
Мозолистое тело 211
Мочевой пузырь 151 *, 160, 162, 163,
164*, 189
Мочеиспускательный канал 160, 162,
189
Мочеполовой канал 163
Мочеточник 160, 162 ♦, 164 *
Мошонка 161, 189
Мышца (ы) атланто-лопаточная 101,
104*, 106*, 107, 132
— атланто-плечевая 103, 185
— бедра двуглавая 103*, 116*,
118 *, 120*. 133
----квадоатная 101, 117*, 118,
119*, 133, 138, 139, 182
----прямая 92, 104, 116*, 117*,
118*, 121, 137, 140, 141
----четырехглавая 101, 104*. 116*,
117*, 119*, 121, 134, 139, 141 *,
143, 182
— близнецовая 182, 189
— большеберцовая 101, 104 *, 117*,
120*, 121, 122*, 123, 125*, 138.183
— век 213
— височная 102, 103 *, 104 *, 105 *,
126 *, 187, 213
— втягивающая семенник 161
— глазные 227
— голени трехглавая 104 *, 119 *,
121, 133
----четырехглавая 121, 125 *
— головы длинная 105 *, 127, 130
----длиннейшая 102, 105 *, 127 *,
130
----косая 105 *, 128, 130, 215
----полуостистая 102, 105*, 127*
130
----прямая 105 * 128 • 130, 215
----ромбовидная 101
----трапецевидная 101, 103 *, 104*,
132
— гребешковая 101, 104*, 118*,
119* 138, 140, 170, 181. 182, 189,
218, 219
— грудино-ключично-сосковая 105 *,
132, 176, 177
—^грудвдоподъязычяая 104 *, 150 *,
242
Мышца (ы) грудино-сосковая 101,
104*. 172
— грудино-щитовидная 156, 177
— грудная 100, 101, 103 *, 104 \
107 *, 109 *, 144, 177, 186, 216
—. — живота 101, 104 *, 106 *
-----поперечная 128, 158
-----прямая 158
— грушевидная 101, 115, 116, 118*,.
138, 182, 189
— двойниковая 101, 117*, 118*,
119* 120, 133
— двубрюшная 102, 104 •, 105, 126^
172. 185, 186, 213
— двуглавая 86, 101, 139, 140, 143„
216
— дельтовидная 101, 103 •, 104*,
106*, 107*, 108» 176, 186, 216
— жевательная 67, 102, 103 *, 104 *.
105 *, 125, 126 *, 175, 187, 213, 214
— живота косая 103 *, 104 *. 118%
128, 177, 183
----- поперечная 128
-----прямая 118*, 128, 158, 177, 182г
183, 189
— зяднеберцовая 183
— заостная 101, 106 *, 107*, 108 *г
109, 132, 177, 178, 216
— запирательная 101, 116*, 117 *г
118, 119*, 120, 133, 182, 189
— зубчатая 86, 101, 102, 103 *, 104 *г
106*, 107, 127, 132, 144, 145*, 158,
177, 178, 216
V— икроножная 101, 117, 122*. 142,
. 143, 183. 220
V— камбаловидная 101, 120 *, 122,
184
— квадратная 115*, 128, 129, 185,
188
— клюво-плечевая 86, 101, 104*,
106*, 108*, 109, ПО*
— ключично-сосковая 101, 104 *
— ко ракоидо-плечевая 216
— круглая большая 101, 106* —
108*, 109, 132, 178
— круглая малая 101, 106 *, 107 *,
109, 132
— крыловидная 102, 105*, 126. 187,
213
— ладанная 101, 107*, 112*
— лестничная 102, 103 *, 104 *, 127,
128, 158, 177
— локтевая 216. 217
• — лопат'очно-подьязычная 149 ♦, 150,
177
— малоберцовая 120 *, 122 *, 123,
125*, 183
-----IV пальца 120*. 122*, 123,125*
-----V пальца 120*. 122*. 123,125*
— межкостные 113*, 114, 125, 124*
Мышца (ы) межостистые 129
— межпоперечные 129
— межреберные 104 *, 127 *, 128, 158,
177
— межчелюстная 213
— многораздельная 129
— напрягатель барабанной перепой-
к» 229
---- нёбной занавески 150
----фасции предплечья 101, 104 ♦,
108», 132
----широкой фасции бедра 115,
116*, 118
— нёбная 150
— опускатель нижней губы 147 *
---- хвоста 131
— отводящая большого пальца 101,
ПО, 111 *, 112 ♦, 114 *
----мизинца 114
— оттягивающая угол рта 147 •
— перстневидно-щитовидная 156
~ пластчпеви^дняя 105 ♦, 127 •, 129,
130. 172, 177, 185
— плеча двуглавая 87, 101, 104 *,
106е, 108*, 109, ПО*—112*, 177
----трехглавая 88, 103 *, 104 *,
106* 107*, 108*, 109, 111 * 132
178, 216
----плечевая 101, ПО, 111*, 112,
132, 178, 216
—- плече-головная 103
— пттече-подъязычная 104 • 127 *
177
— подбородочно-подъязычная 104 ♦,
149 *, 150
— подвздошная 104 ♦, 115*, 116 *
117 118 *, 119*, 132, 133, 140,’
182
— подвздошно-реберная 127 •, 128,
129
— подключичная 101, 106. 132. 144
— подкожная 100, 177, 178, 188
— подколенная 101, 119*, 120*, 121,
122*, 123. 134
— подлопаточная 86, 101, 106 *—
108 *, 109. 132, 177, 178, 186, 216
— подниматель верхней губы 147*
----нёбной занавески 150
----пениса 182
---- ребер 128
---- хвоста 131
— подошвенная 119*, 121, 122*
— подъязычная 149
— подъязычно-язычная 149 *
— полуперепончатая 101, 104 *, 116*,
117, 118*, 119, 120*, 138, 133, 139,
140, 143, 182, 183
- полусухожильная 101, 103 *, 104 *,
116*, 117, 118*, 133, 138, 143, 182,
183
16*
Мышца (ы) портняжная 104*, 115,
118*, 133
— поясничная 104*, 115*, 116*,
119* 133, 138, 140, 182, 188, 189
— пояснично-подвздошная 102, 189
— пояснично-реберная 168
— предостная 101, 103*, 106*, 107*,
108, 132, 177, 186, 216
— предполуперепончатая 101, 116*,
117*, 118*, 119*, 138
— приводящая большая 116*—119*
-----большого пальца 113*
-----длинная 101, 116*, 118*, 119,
138, 181, 182
-----короткая 101, 116* — 119*,
138
-----V пальца, 113*, 114
— пронатор 107*, 108*, 111*, 112,
113
— пяточная 101, 117*, 121
— разгибатель кисти 107, 108*, ПО,
111, 112 • 114*
-----пальцев 101, 107*, 108*, ПО,
111 *, 112*, 114*, 117*, 119*, 120,
121, 122* 123, 124 *, 125*, 138
-----пясти 101
— ромбовидная 105*, 106*, 132, 177,
185, 186
— сгибатель кисти лучевой 101, 102,
107*, 111*—114*
— пальпев 101, 102, 107*, 112*—
114*, 120*, 121, 122 *, 123, 124 *,
125*, 138
— сердечная 169—171
— сосочковые 169
— спины длиннейшая 127*, 129
----- остистая 129
-----полуостистая 129
-----трапецевидная 101, 103 *
-----широчайшая 100, 101, 103 ♦,
104*. 105, 107*, 108*, 109, 132,
177, 178, 188
— стопы 124 *, 125 *
— стременная 229
— стройная 101, 104*, 116*, 118*,
120*, 133, 138, 182, 183» 218, 219
— супинатор 111 *, 112*, 107*
— тпапепевидная 101, 102, 103 *,
105 *, 106. 132, 172, 176, 178, 186
— ушные 213
— хвостовые 102, 103 *, 104 *, 130,
131, 182
— червеобразные 133* 114, 124 *г
125
— чеппаловыдная поперечная 155,
156*
— шейная длинная 127*, 130
-----полуостистая 128 *, 130
-----ромбовидная 101. 103 •
— трапецевидная 101, 103 *
.’1 <
Мышца (ы) шилоподеьязычная 213
— щечная 175
— ягодичные 91, 92, 102, 103 *, 115—
118*, 119*. 120, 138, 140, 149, 189
Н
Надгортанник 150
Надпочечники) 164 *, 195, 196 *
Нёбо 150*, 212
— твердое 68
Нерв (ы) альвеолярный 213
— • бедра 218, 219
— блоковый 212
— блуждающий 205, 214, 222
— большеберцовый 219. 220
— веко-ушной 213
— верхнегубной 212
— верхнечелюстной 212
— височные 213 г
— внутренностные 222
— геморроидальный 219
— глазничный 212
— глазодвигательный 212
— голени кожный 220
— грудные 216, 217, 218
— двубрюшной мышцы 213
— диафрагматический 216
— добавочный 214
— жевательный 212
— запирательный 218, 219
— затылочный 215
— зрительный 75, 208, 209, 211, 225
— клвнонёбный 212
— кожный шейной 215
— крестцовые 220
— лицевой 100, 205, 213, 214 ♦, 215
— лобный 212
— локтевой 216
— лопаточный 216
— лучевой 216
— малоберцовый 219, 220
— межкостные 216
г- межреберные 217
— межчелюстной 213
— мышечно-кожный 216
— надключичный 216
— нижнечелюстной 212
— носовой 212
— носо-ресничный 212
— обонятельный 209, 211, 231, 232
— отводящий 213
— пальцевые 216, 217, 220
— плантарные 220
— подбородочный 213
— подвздошно-паховый 218
— подглазничный 212
— подлопаточный 216
— подмышечный 216
241
Нерв (ы) подчревно-подвздошный
218
— подъязычный 70, 205, 214
— поясничные 217, 219
— предлопаточный 216
— решетчатый 212
— сафенус 218
— седалищный 219
— семенниковый наружный 218
— скуловой 212
— слезный 212
— слуховой 213, 214
— сонный 221
— спинно-мозговые 199, 215*
— срамной 219
— срединный 216
— тройничный 68, 175, 206, 212, 214
— ушные 213, 215
— хвостовые 220
— черепномозговые 68, 205, 207, 211*
— шейные 213, 215, 216
— щечный 213
— ягодичные 219
— языко-глоточный 214
— язычный 213
Нервная клетка 198
— система вегетативная 198, 221
----парасимпатическая 214, 221,
222
----периферическая 198
----сипатическая 214, 221 *
----соматическая 198
----центральная 198, 199
Нервное сплетение крестцовое 219
----плечевое 199, 215—217
---- поясничное 218
----пояснично-крестцовое 199
---- солнечное 222
Ноздри 155, 156 *
Носовая полость 155
Носовые кости см. Кости носовые
Носоглотка 155, 156*
О
Оливы краниальные 204—206
Органы вкуса 232
— обоняния 231, 232
— осязания 223
— равновесия 227, 228, 230
— слуха 227, 228
— чувств 223
П
Пищевод 146, 147, 151 *, 152*, 153*,
176, 177, 185
Печень 151 *, 152, 153 *, 154
Подкожная клетчатка 62
Позвонки 66, 67, 81, 83, 145 *, 185,
199
Позвонки грудные 69, 83 *, 84, 100,
103, 105, 106, 115, 127, 128, 129, 130,
144, 145, 178, 217
— крестцовые 85, 115—117, 129, 131
— поясничные 69, 84, 85, 103, 105,
115, 127, 129, 131, 188, 217, 218
— хвостовые 66, 69, 85, 116, 130, 131
— шейные 69, 81, 82, 83 •, 84, 105—
107, 127—130, 144, 185, 215
Позвоночного столба крестцовый от-
дел 85
---поясничный отдел 84
--хвостовый отдел 85
---шейный отдел 81
Позвоночный столб 81, 91, 114, 179,
185
Половой член 151, 161, 163 *, 164,
189
Полосатое тело 208, 210
Полукружные каналы 230
Почки 151, 159*, 160, 161, 162*,
164 *» 181
Пояс плечевой 85, 86, 91, 114
Предплечье 88, 89
Преполлекс 90
Препуций 163, 189
Пучок Бурдаха 200
— Голля 200
— зпительного бугра 200
— Левенталя 202
— мозжечковый 200
— Монакова 202, 207
— перекрещенный пирамидальный
202
— прямой пирамидальный 202
— флексита 206
— четверохолмия 200
Р
Ребра 66, 69, 83, 84, 106, 131, 144,
145 *, 151 ♦, 127—129
— ложные 84
Роговица 224, 226
Ромбовидная ямка 205
Ротовая полость 146, 147, 150
С
Сальник 146
Селезенка 151 *, 180.. 188, 191 *, 192
Семенники 151, 161, 162 *, 163*, 189
Семявыносящие протоки 161, 162*
Семяпровод 162, 163
Сердца желудочки 169, 170 *, 171
— камеры 169
— клапаны 169, 170
— предсердия 169, 170 *, 171
Сердце 151 ♦, 157, 167, 169, 170*,
172*, 222
Сильвиев водопровод 205, 207, 209
Синовиальные влагалища 99 *
Синовиальные сумки 99
Слуховой проход 228
Слуховые косточки 228, 229*.
Соски брюшные 65
— грудные 65
— паховые 65
Сошник 77 *, 79, 80
Спинного мозга белое вещество 200
-----проводящие пути 200
-----серое вещество 199, 200
Спинной мозг 199. 202. 204, 205, 207
Спинномозговой канал 199, 203, 205
Спинномозговые ганглии 199
— нервы см. Нервы спинномозговые
Стекловидное тело 226
Сустав атланто-головной 81
— голеностопный 121—123; 136, 137,
142, 143
— запястно-пястный ПО
— запястный 89
__ кистевой 144
— коленный 94, 114—116, 118, 119,
121, 136, 137, 140—143, 183, 189
— локтевой 88, 110—113, 136, 144
— плечевой 87, 105, 106, 108, 109,
136, 144, 178, 186, 216
— пястно-фаланговый ПО, 112
Сустав тазобедренный 92, 114—116,
118, 119, 120, 136, 137, 139, 140—
144, 182
— челюстной 187
Суставы межфаланговые 124
— плюсно-пальцевые 122, 124
— плюсно-фалаиговые 125
Сухожилие ахиллово 121
Сухожилия пальцев 122
Т
Тазовый пояс 143
Тельца Мейсснера 223
— Меркеля 223
Тркат дыхательный 68
Трахеи 151 *, 157*, 172, 177, 185
У
Узел звездчатый 222
Узлы нервные 221
— симпатические грудные 222
-----поясничные 222
-----шейные 221, 222
Улитка 230, 231
Ухо 186
— внутреннее 214, 227, 228
— наружное 213, 215, 228
— среднее 214. 228
Ушная раковина 228
ЛП
ф
Фаланги пальцев 96
Фасции специальные 99
Фасция бедренная широкая 115
Фасция внутренняя 99
— глубокая 99
— ладонная 114
— подкожная 99
— поясничная 105
— предплечья 109
X
Хоаны 155. <56 *
Хрусталик 225, 226 •» 227
Ч
Челюсть нижняя 67, 69, 125. 126, 149,
150, 186
Череп 67
Черепа боковая стенка 68
Черепа крышка 70
лицевая часть 67. 70
— мозговая часть 67
Четверохолмие 200, 206—208, 212
Щ
Щеки 147
3
Эпидермис 62, 63 •
Эпидермиса мальлигиевый слои 63 *
Эпистрофей 81. 82 *, 105, 129, 130
Эпифиз 196, 209
Эритроциты 191, 192
Я
Язык 81, 148, 213
Яичники 164*
Яйцеводы 164 ♦, 165
ЛАТИНСКИЙ
А
Anus 153
Aorta 171
- abdominalis 180
— thoracia 179
Arteria (ae) alveolaris caudalis superior
175
--cranialis 176
— anguli oculi 174
— anonyma 172
— auditive 174
— axillaris 176, 177
— basilaris cerebri 174
— bingualis 173
— brachialis 178, 179
— carotis externa 172
--interna 174
— cerebelli 174
--cerebri caudalis 174
--media 174
-----nasalis 174
— cervicalis ascendens 177
-----profunda 176, 178
•----superficialis 177
— chorioidae 174
— circumflexa brachialis caudalis 177
---------cranialis 177
-----femoris 182
-----iliacus 182
— circumflexa ilii 181
-----scapularis 178
246
УКАЗАТЕЛЬ
Arteria circumflexa subscapularis 177
— coccigea media 181
— coeliaca 180
— collateralis ulnaris 178
— coronales 171
— epigastrica cranialis 177
----superficialis 183
— facialis 173
— femoralis 182
— gastrica 180
— genu distalis 183
----media 183
----proximales 183
----suprema 183
— glutaea 182
— haemorrhoidalis 182
~ hepatica 180
— hypogastrica 182
----communis 181
— — externa 181
----interna s. hipogastrica 182
— ilio-iumbalis 181
— infraorbitalis 176
— intercostalis cranialis 176
----ventralis 177 I
— interossea 178
— labialis inferior 173
----superior 174
7- lienalis 180
— lumbales 181 5
— mandibularis 173
Arteria (ae) rnasseterica 173, 174
— maxillaris 174
- mediana 178
— mediano-radialis 178
— mediostenalis 176
~ meningea 174
• mentaUs 173
— mesenterica caudaiis 181
----cranialis 180
— metatarsales 184
— muscubphrenica 177
— muscularis 183
— nasalis 174
— obturatorius 182
— occipitalis 172
— oesophagea 179
— ophtalmica 175
— ovarica 181
— palatina 175
----ascendens 173
— perforans 177
— pericardlaco- mediostenalis 174
— pericardiaco-phrenlca 177
— pericardialls 176
— peronaeus 184
— pharingealis 175
— phrenica caudaiis 180
----cranialis 180
— plantaris dorsalis 184
— — lateralis 184
— pterygoidea 175
— pudenda 182
— pulmonafis 171
— radialis 178
— renalis 181
— saphena 181
— spermatica 181
— sphenopalatinum 176
— sternalis 177
— subclavia 176
— sublingualis 173
— subscapularis 177
— temporalis 173
— thoracal is intent 177
----lateralis 177
— thoracico-acromialis 177
— thoracico-dorsalis 178
— thyreoidea cranialis 173
— tibialis anterior 183
----posterior 183
— trachealis 173
— transversa cervicalis 177
----scapularis 177
— ulnaris 178
----recurrens 178
----transversa 178
— vertebralis 176
— vesicalis cranialis 182
Atlas 81
Atrium 169, 170
Auris externa 228
— interna 229
— media 228
В
Bulbi olfactorii 209
Bulbus oculi 223
C
Caecum 153
Cerebellum 202, 206
Circulus villisii 174
Clavicula 86
Cor 169
D
Dentes 147
Diaphragma 131
Diencephalon 202, 208
Duodenum 152
E
Epididymis 162
Epiphysis 196
Epistropheus 82
О
Gangliae cervicales 221
— lumbales 222
— mesentericae 222
— spinales 199
— thoracales 222
Ganglion cervicalis caudaiis 222
-----cranialis 221
-----medius 222
— stellatum 222
Gingiva 147
Glandule (ae) ampullares 163
— bulbo-urethrales 163, 165
— lacrimalis 227
— mandibularis 150
— parathyreoideae 194
— parotis 150
— prostata 163
— sali vales 150
— sublingualis 150
— suprarenales 195
— tarsales 227
— thymus 195
— thyreoidea 194
— vesiculares 163
H
Hepar 153
Hilus 154
Hymen 165
Hypophysis 196
247
I
Intestinum crassum s. colon 153
— rectum 153
Intumescentia cervicalis 199
— lumbalis 199
J
Jejunum 152
L
Larynx 155
Lien 191
Lingua 148
M
Mandibula 80
Mesencephalon 202, 207
Mesenterium 153
Musculus (li) adductor brevis 119
------digiti quinti 114
------longus 119
------maximus 119
— adductor pollicis 113
------longus 111
— arytaenoideus transversus 156
— atlanto^scapularis 107
— biceps brachii 109
------femoris 116
— bipennatus 98
— brachialis 110
— brachiocephalicus 103
— buccinator 147
— coccygeus 130
— constrictor vestibuli 165
------vulvae 165
— coraco-brachialis 109
— cremaster 161
— cricothyreoideus 156
— deltoideus 108
— depressor labii inferioris 147
— digastricus 126
— dorsoepitrochlearis 109
— epitrochleo-anconeus 109, 178
— extensor carpi radialis brevis 110
------------longus 110
---------ulnaris 111
------digitorum 123
---------communis 110
---------lateralis 111
---------longus 122
------pollicis longus 122
— — indicis proprius 111
------pollicis brevis 111
---------longus 111
— flexor carpi radialis 113
248
Musculus flexor carpi ulnaris 113
----digiti quinti brevis 111
— — digitorum brevis 124
---------longus 123
---------profundus 113
---------sublimis 112
----pollicis brevis 113
— gastrocnemius lateralis 121
• medialis 121
— gemelli 120
— genioglossus 149
— geniohyoideus 150
— glutaeus medius 115
----minimus 116
----superficialis 115
— gracilis 118
— hyoglossus 149
— iliacus 115
— iliocostalis 129
— infraspinatus 109
— intercostales extern! 128
----interni 128
— interossei 114, 125
— inter spinales 129
— intertransversarii 129
— — caudi 131
— lateralis nasi 147
— latissimus dorsi 100, 103
— leva tores costar ium 128
— levator labii superioris 147
----palpebrae superior 227
----veli palatini 150
----capitis 130
— longissimus dorsi 129
— longus capitis 130
----cervicalis 130
— lumbricales 114, 125
— masseter lateralis 125
----medialis 126
— multifidus 129
— multipennatus 98
— obliqui 227
— obllquus abdominis externus 128
---------internus 128
----capitis caudalis 130
---------cranialis 130
----inferior 212
— obturator externus 119
----internus 120
— omohyoideus 150
— orbicularis oculi 227
----oris 147
— palatinus 150
— palmaris 112
— panniculus carnosus 100
— papillaris 169
— pectinati 169
— pectineus 118
— pectoralis 100, 106
— — abdominalis 106
Musculus pectoralie profundus 106
------superficialis 106
— peronaeus brevis 123
------digiti quart! 123
---------quinti 123
------longus 123
— piriformis 115
— plantaris 121
— popliteus 121
— praesemimembranosus 117
— pronator teres 11*2
— psoas major 115
------ minor 115
— pterigoideus lateralis 126
------medialis 126
— quadratus femoris 119
------limborum 129
------planatae 124
— quadriceps femoris 121
— recti 212, 227
— rectus abdominis 128
------capitis dorsalis major 130
------------minor 130
---------lateralis 130
---------ventralis 130
------femoris 121
— retractor anguli oris 147
— rhomboideus 105
— sacrococcygeus dorsalis lateralis
131
---------medialis 131
------ventralis lateralis 131
---------medialis 131
— salenus 128
— semimembranosus 117
— semispinalis capitis 130
------cervicis 130
— semitendinosus 116
— serratus dorsalis 127
------ventralis 107
— soleus 121
— sphincter colii 100
------pupillae 224
— spinalis 129
— splenius 129
— stapedius 229
— sterno- cleido-mastoideus 103
— sternohyoideus 150
— sternothyreoideus 156
— styloglossus 149
— subclavius 106
— subscapularis 109
— supinator 111
— suprasplnatus 108
— temporalis 126
— tensor fasciae latae et sartorius 115
------tympanl 229
------veli palatini 150
— teres major 109
------minor 109
Musculus thyreo-hyoideus 156
- tibialis anterior 121
------posterior 123
— transversi cordis 169
— trans versus abdominis 128
------mandibulae 126
------thoracis 128
— trapezius 102
— triceps brachii 109
------surae 121
— unipennatus 98
Myelencephalon 202
N
Nervus (vi) abducens 213
— accessorius 214
— acustlcus 214
— alveolaris mandibulae 213
— auriculares 213, 215
— axilaris 216
— buccales 213
— buccinatorius 213
— carotis 221
— caudales 220
— cutaneus colli 215
------cruris plantaris 220
------femoris caudalis 219
------peronaeus dorsalis 220
— supraclavicularis 216
— digastricus 213
— digitales 216
------volares 216
— ethmoldalis 212
— facialis 213
— femoralis 218
— frontalis 212
— glossopharyngeus 214
— glutaei 219
— haemorrhoidalis caudalis 219
------medius 219
— hypoglossus 214
— ilio-hypogastricus 218
— ilio-inguinalis 218
— infraorbitalis 212
— intercostales 217
— interosseus 216
— isehiadicus 219
— labialls 212
— lacrimalis 212
— levator veli palatini 213
— lingualis 213
— lumbales 213
— mandibular is 213
— massetericus 213
— maxilaris 212
— medlanus 216
— musculocutaneus 216
— mylo-hyoideus 213
— nasalis aboral is 212
24fr
Hervi nasales extern! 212
—- nasalis oralis 212
— obturatorius 218
— occipitalis 215
— oculomotorius 212
— olfactorius 209, 211
— ophthalmicus 212
— opticus 211
— palatinus major 213
— palatinus minor 213
— pec tor ales 216
— penis 219
— peronaeus 220
----profundus 220
----superficialis 220
— petrosus superficialis 213
— phrenicus 21₽
~ plantaris lateralis 220
----medialis 220
— psoadicocutaneus 218
— pudendus 219
— radialis 216
----profundus 216
----superficialis 216
— saphenus 218
— scapularis 216
— spermaticus 218
— spheno-palatinus 212
— splanchnic! 222
— suprascapularis 216
— temporales profundi 213
— temporalis superficialis 213
— thoracales 217
— tibialis 219
— trigeminus 212
— trochlearis 212
— ulnaris 216
— vagus 214
— zygomaticus 212
Nodull lymphatici 192
О
Ос ulus 223
Oesophagus 151
Oris cavum 147
— rima 147
— vestibulum 147
Os accessorius 96
— acetabulare 91
— astragalus s. tibiale tarsi 95
— basioccipitale 74
— calcaneus s. fibulare tarsi 95
— carpale 90
— ethmoid ale 77
— femoris 92
— frontale 77
— humerus 87
— hyoideum 81
— ilium 91
— incisivum s. intermaxillare 79
— intermedium 90
250
Os interparietale 76
.Os ischii 91
— lacrimale 80
— maxillare 78
— nasale 79
— occipitale 73
— palatinum 79
— parietale 77
— penis 163
— petrosum 75
— pisiforme 89
— pterygoideum 79
— pubis 91
— radiale carpi 89
— sacrum 85
— sphenoidale 74
— tarsalia 96
— temporale 75
— ulnare carpi 89
— zigomaticum 80
Ossa carpalia 89
— cruris 93
— digitorum 89, 95
— innominate s, coxae 91
— metacarpalia 89
— metatarsalis 95
— tarsalia 95, 96
— vesalis 93
Ovaria 164
Oviductus 164
P
Palatum durum 150
— molle 150
Palpebrae 227
Pancreas 154
Pelvis 91
Penis 163
Pharynx 150
Pilus 63
Praesphenoideum 75
Praepoliex 90
Preputium 163
Pudendis labia 165
— rima 165
Pulmones 157
R
Radius 88
Regio epitalamica 209
— metathaiamica 208
Renes 159
Retina 225
Rhinencephalon 209
S
Scapula 85
Scrotum 161
Sternum 84
Substantia alba 200
— grisea 199
т
Telencephalon 202, 209
Testes 161
Thalamus optid 208
Trachea 157
Truncus lingualis 173
Tibia 93
U
Ulna 87
Unguicola 64
Ureter 160
Uterus 165
V
Vagina 165
Valvulae semilunares 169
Velum palatinum 150
Vena (ae) alveoiarls 186
— angulares 156
— auricularis 186, 187
— axillaris 187
— azygos 185
— brachialis 188
— bronchiales 185
— cave 188
— cerebelles 187
— cerebri 187
— circumflexa brachiales 188
----femoris 189
- — iliaca 189
----scapularis 188
— cordis magna 171
----media 171
— epigastrica caudalis 189
----superficialis 190
— facialis externa 186
----transversa 187
— femoraiis 189
— frenica cranialis 185
— frontalis 186
— glutaea 189
— haemorrhoidalis 189
— hepatica 188
— hemiazygos 185
— hipogastrica 189
— iliaca 188, 189
— ilio-psoas 188
— innominata 185
— intercostaiis 185
Vena (ae) interossei 188
— jugularis 185
— labialis inferior 186
----superior 186
— lateralis pedis 190
— lienalis 188
— mandibularis 186
— masseterica 187
— temporalis 187
— maxillaris 186
— medialis 188
— meningaea 187
— mentaHs 186
— mesenterica caudalis 188
----cranialis 188
— metatarsales plantares 190
— nasalis 186
— obturatoria 189
— occipitalis 185
— pedis medialis 190
— pharyngea 185
— phrenica 188
— poplitea 190
— pericardlaco-interstenalis 185
— porta hepatis 188
— pudenda externa 189
----interna 189
— pudendo-epigastrica 189
— pulmonales 170, 171
— renalis 188
— saphena 190
— spermatica 188
— spermatica externa 189
— subclavia 188
— submaxillaris 186
— - superior bitalis 186
— thoracica lateralis 188
— thoracico-acromialis 188
— thoracicus 185
— thyroidea 185
— tibialis 190
— transversa scapulae 186
— ulnaris 188
— uretrae 189
— vertebralis 185
— vesicalis cranialis 189
Ventriculus 151, 169, 170
Vertebrae caudales 85
— cervicales 81
— lumbales 84
— thoracales 83
Vesica urenaria 160
Vestibulum urogenitale 165
Vomer 80
ОГЛАВЛЕНИЕ
От авторов 3
ЧАСТЬ ПЕРВАЯ
ЭКОЛОГО-СИСТЕМАТИЧЕСКИЙ ОЧЕРК
Глава 1. Систематика, географическое распространение и места обитания 5
Систематика рода Rattus Fischer.............................5
Географическое распространение серой крысы . . 12
Места обитания серой крысы.................................15
Географическое распространение и места обитания черной
крысы................................................19
Географическое распространение в места обитания турке-
станской крысы . .... . . 20
Глава П. Экология................................................. 22
Повадки крыс.............................................. 22
Питание крыс.............................................. 23
Размножение крыс.......................................... 30
Экономическое и эпидемиологическое значение крыс 35
Истребление крыс, или дератизация 38
Содержание и разведение крыс ... 44
Правила содержания крыс в неволе . 44
Кормление крыс .... 47
Размножение крыс в неволе . 49
Болезни белых крыс ... 51
Литература .......................................................... 52
ЧАСТЬ ВТОРАЯ
АНАТОМИЯ
Глава III. Внешняя морфология н терминология, употребляемая при
описании .... ... . . 59
Глава IV. Кожные покровы и их производные . 62
Кожа..................................................... 62
Производные кожи ... 63
Глава V. Скелет ... .66
Общие замечания . 66
Череп ... .67
Осевой скелет . .81
Плечевой пояс * 85
252
1К ре шяя К'ЖечШ’ИЬ
Тазовый пояс .
Задняя конечность
Технические советы
Глава VI. Мускулатура
Общие замечания ......................................
Подкожная мускулатура......................
Мускулатура плечевого пояса и передних конечностей
Мускулатура задней конечности ....
Мускулатура головы
Мускулатура туловища
Технические советы
Глава VII. Функциональная характеристика органов движения
Галоп ................................
Глава VIII. Органы пищеварения
Общие замечания ....
Ротовая полость и глотка
Кишечный тракт ....
Печень и поджелудочная железа
Глава IX. Органы дыхания
Общие замечания ’
Дыхательные пути
Легкие ....
Дыхательные движения
Глава X. Органы мочеотделения
Почки
Мочевые пути
Глава XI. Половые органы
Половые органы самца
Половые органы самки
Глава XII. Сосудистая система
Общие замечания .
</Кровь ,
Сердце ....
Артерии
Вены........................
Лимфатическая система
Органы кроветворения
Технические замечания
Глава XIII. Железы внутренней секреции .
Общие замечания ...
Щитовидная и околощитовидная железы
Зобная железа ....
Надпочечники ....
Половые железы ....
Эпифиз и гипофиз ....
Глава XIV. Нервная система .
Общие замечания .
Спинной мозг
Головной мозг
Головные нервы
Спинномозговые нервы .
Вегетативная нервная система
м-
’И
92
97
9£
9€
10С
102
114
12£
127
1&
13’
13Е
141
141
14;
151
15S
15’
lUi
15;
151
15<
15<
16(
16
16
1ft
16;
16
161
16!
17:
181
19(
191
192
19'
19'
19'
191
191
191
19(
191
191
191
205
211
211
22!
У5.
Глава XV. Органы чувств . 223
Органы осязания . . 223
Органы зрения 223
Органы слуха и равновесия . 227
Орган обоняния 231
Органы вкуса . 232
Литература .... 232
Русский указатель . 238
Латинский указатель . 246
П. П. Гамбарян
и Н. М. Дукельская
КРЫСА
Редактор Л. П. Татаринов
Техн, редактор А. Губер
Сдано в набор 10/VI—55 г. Подо, к печати 7/IX—55 г.
Бумага 60 х 92*,, 16 печ. л.
18,51 уч.-изд. л. Тираж 3000. Т—05275.
Издательство «Советская наука». Заказ ИЗ.
Ц е н а б руб. 55 коп.
Тип. иэд*ва «Советская наука», Неглинная, 29/14. Зак. 795.