
































































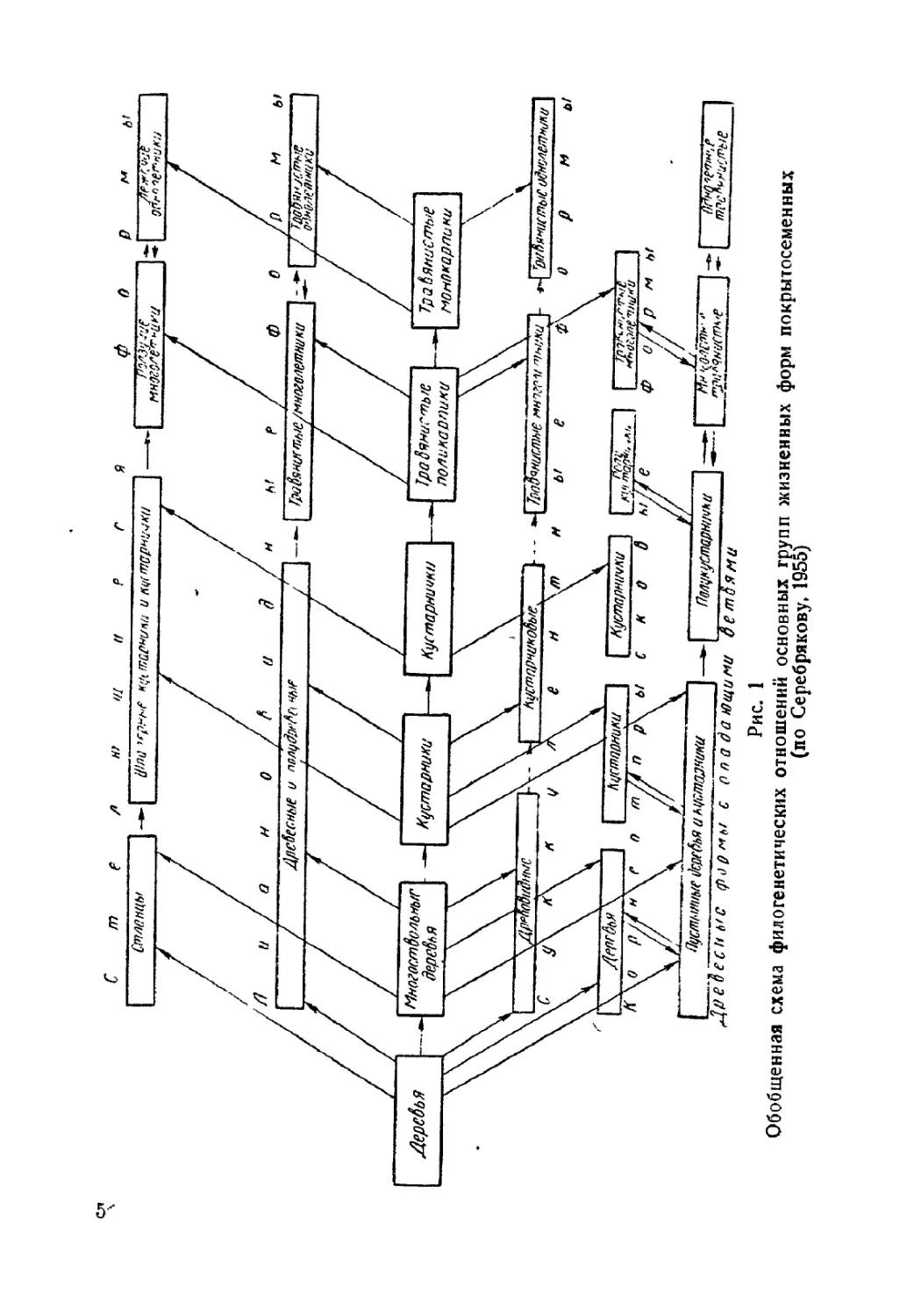









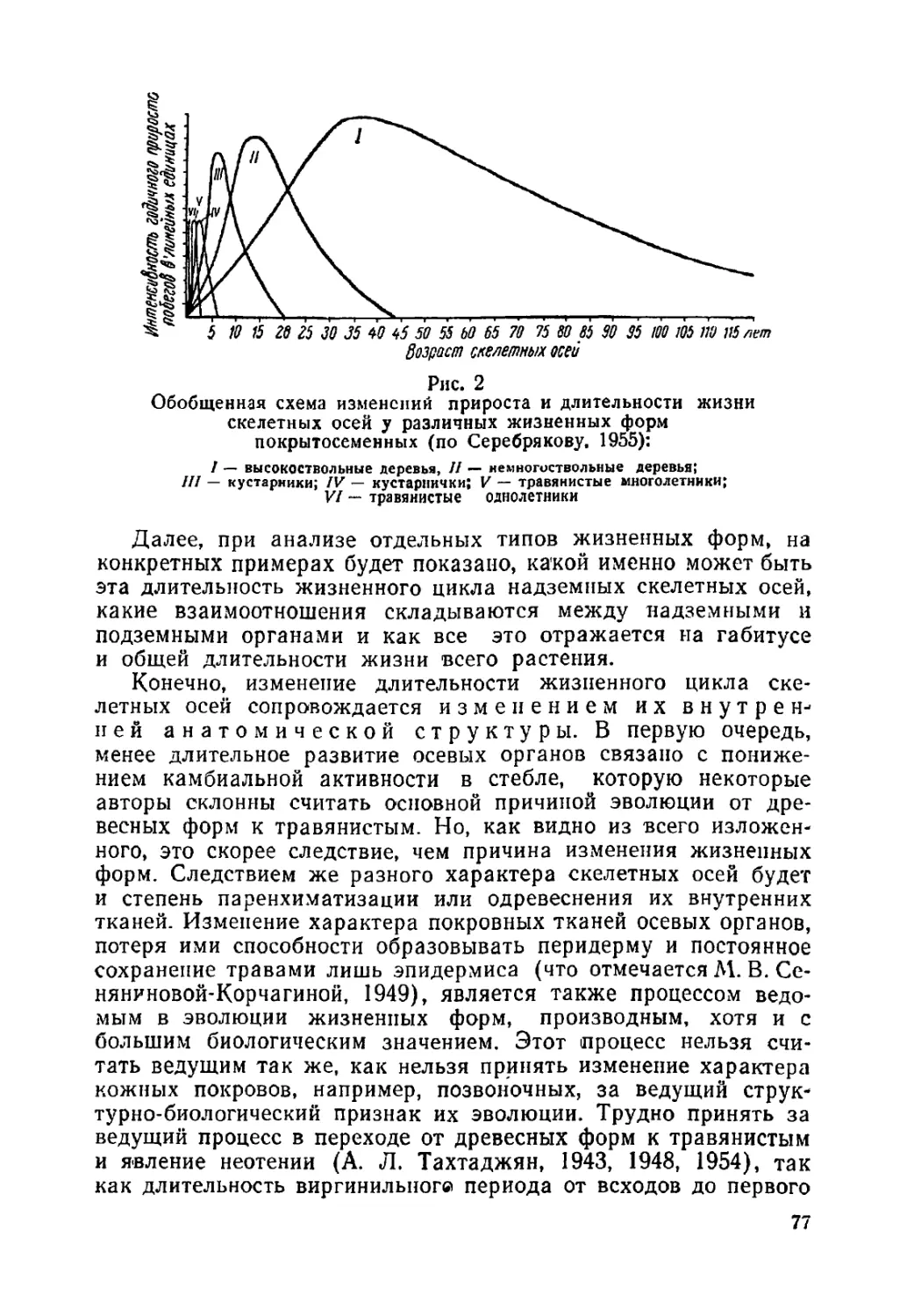





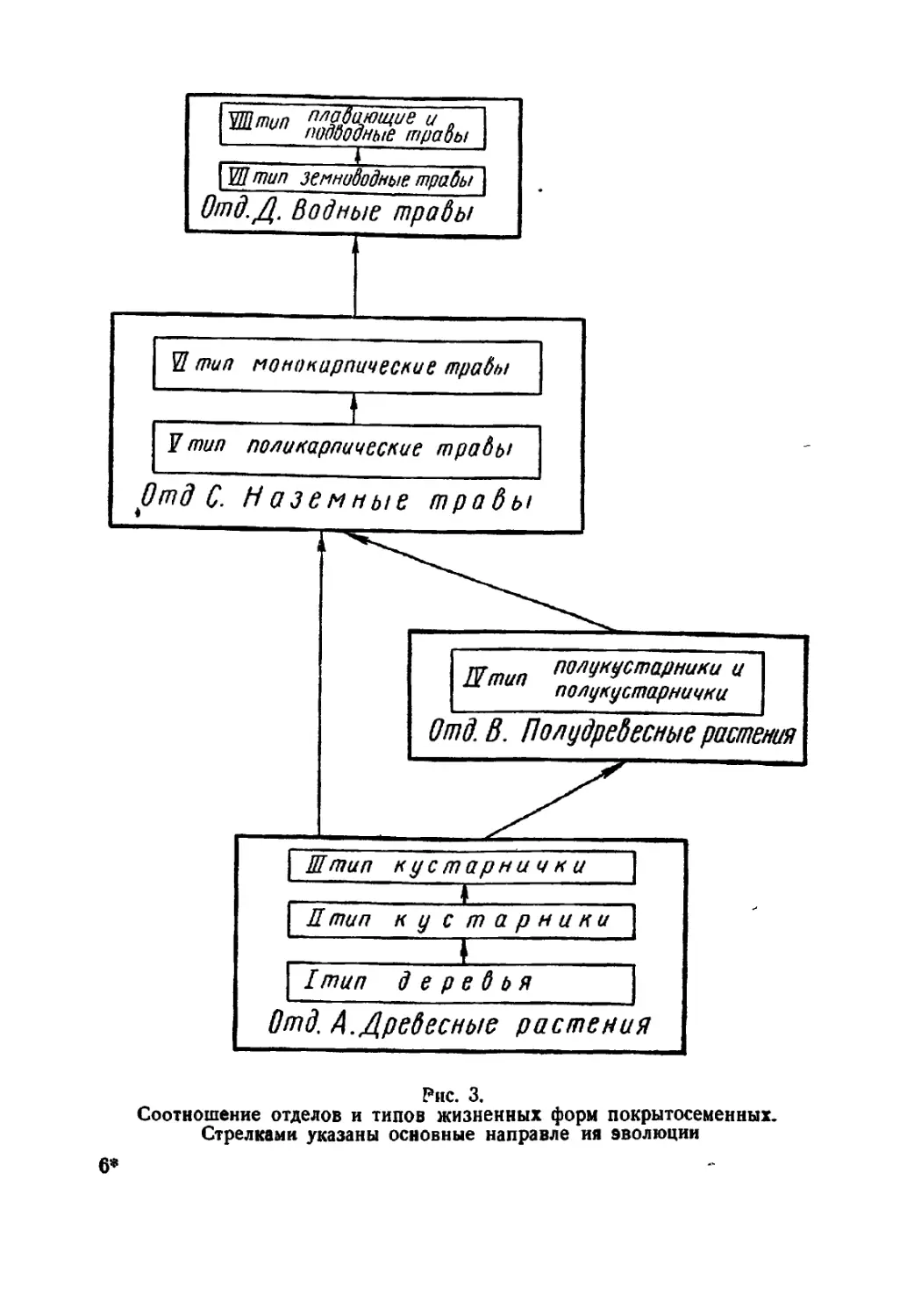
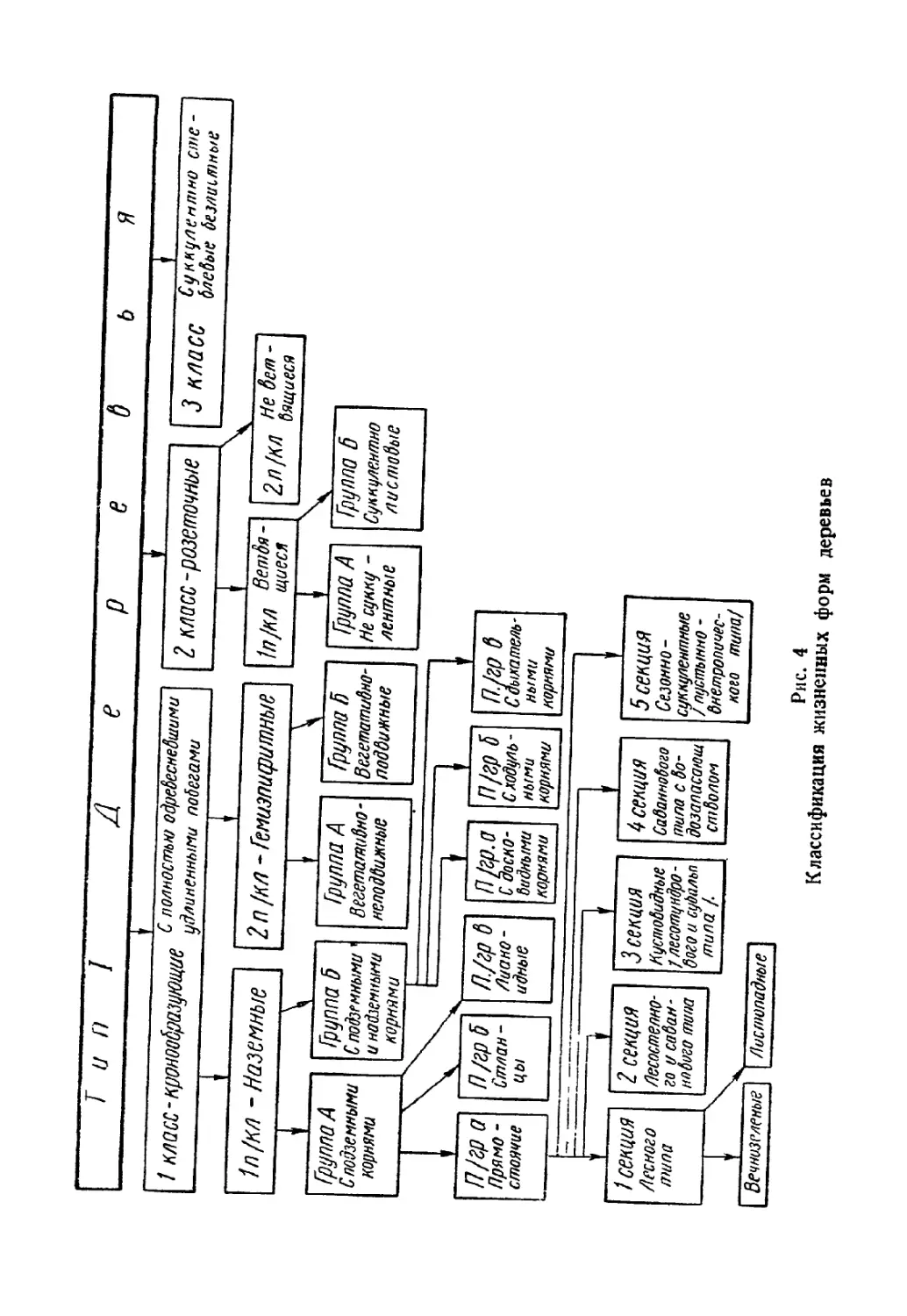


































































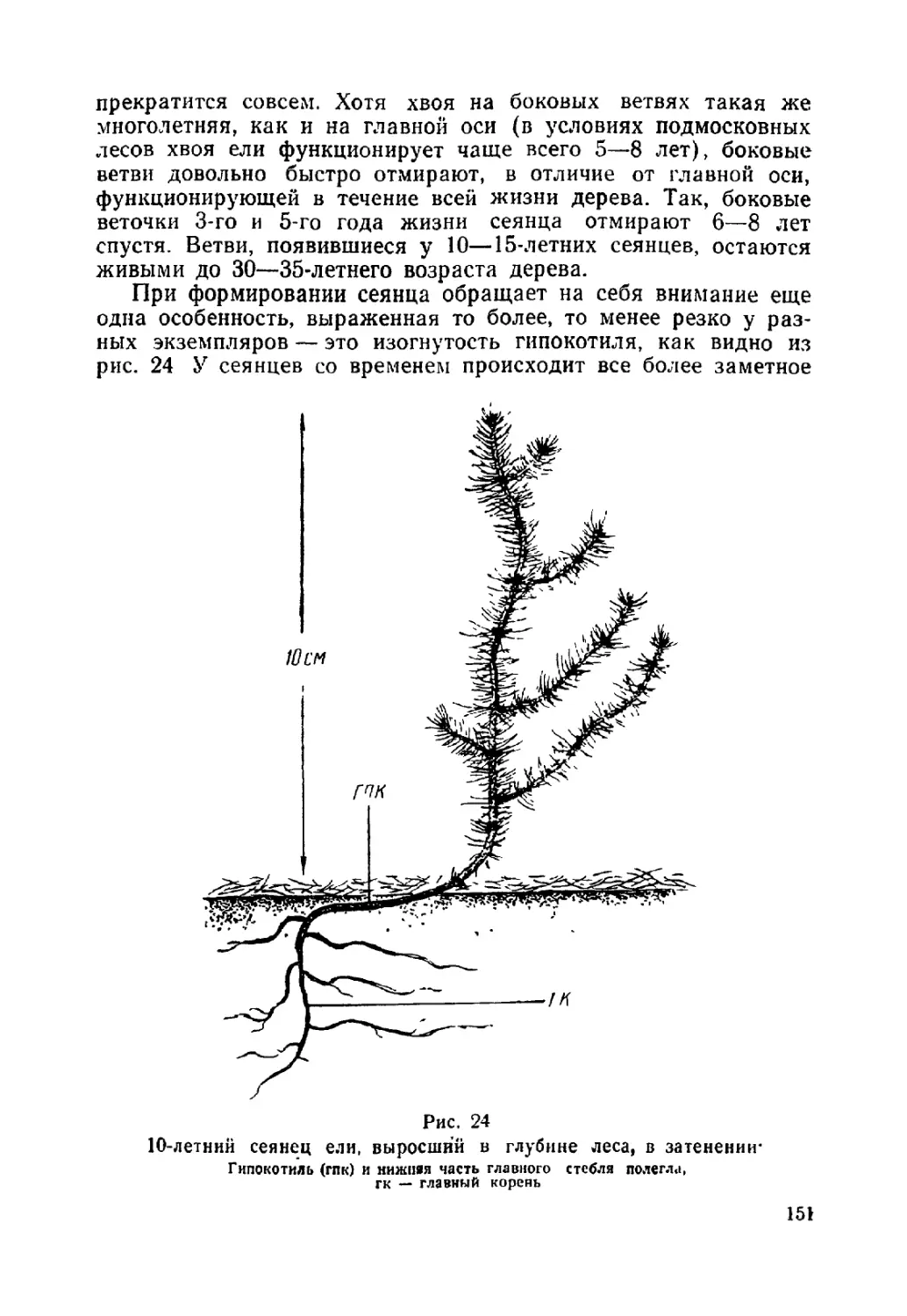



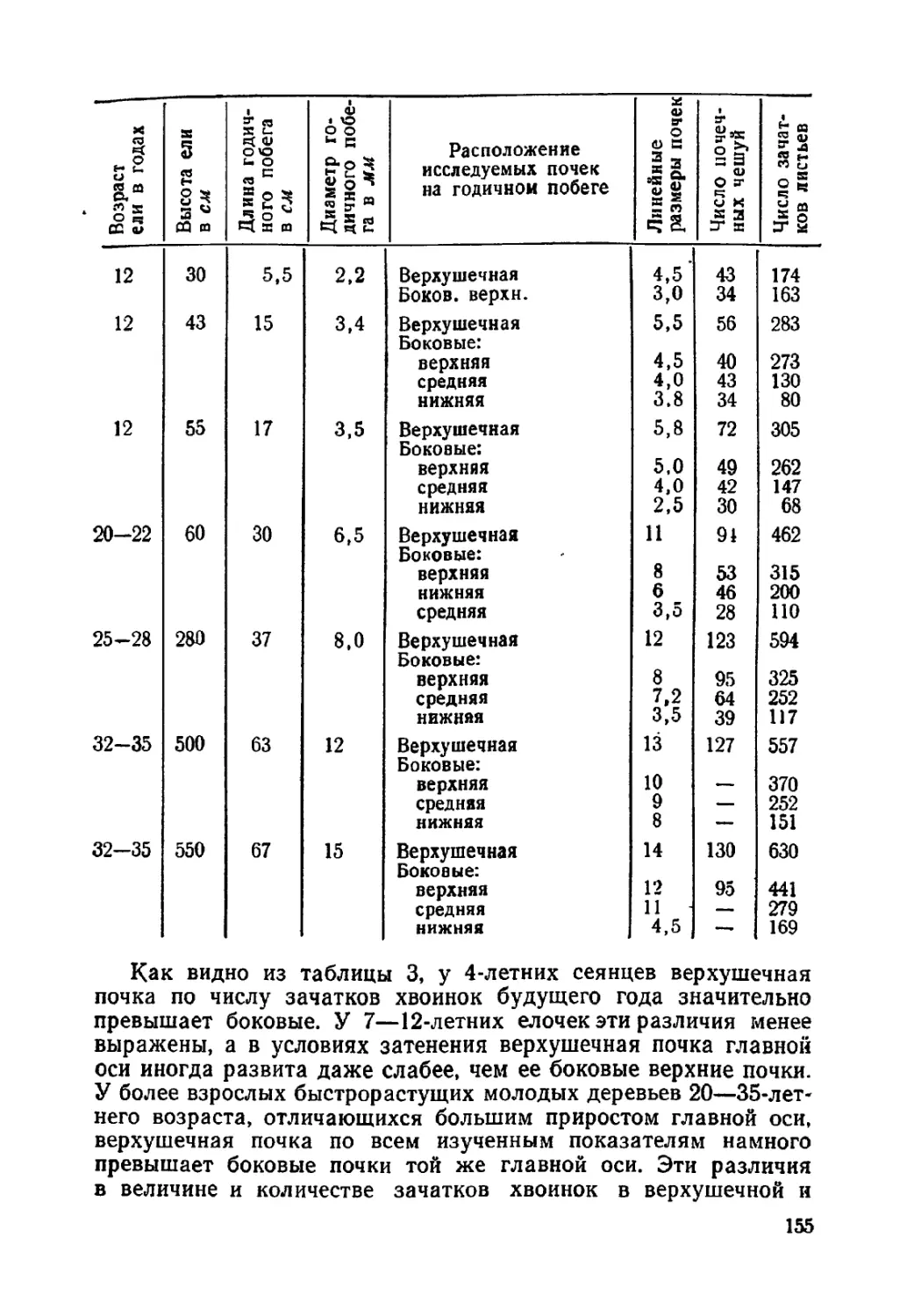

























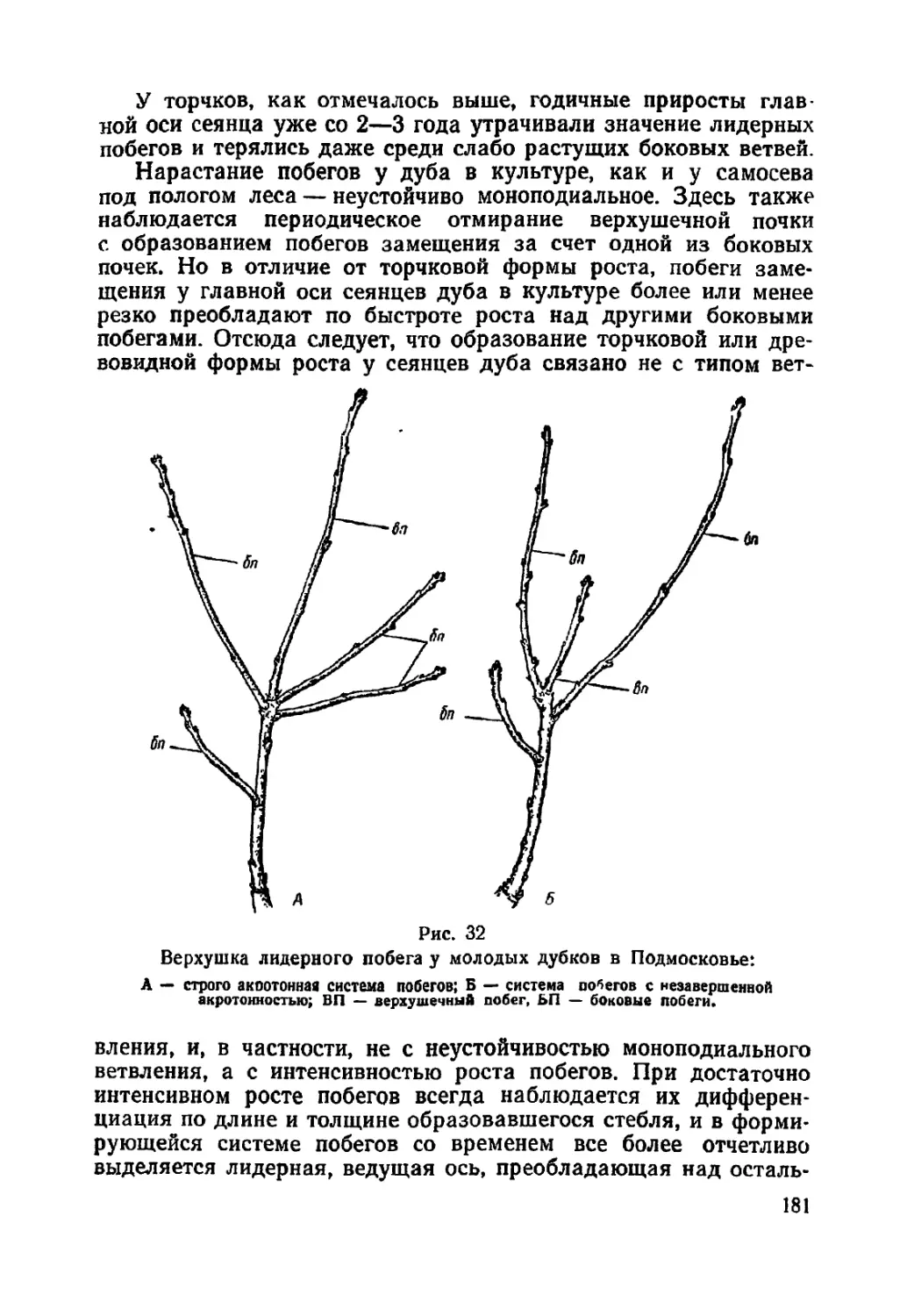
















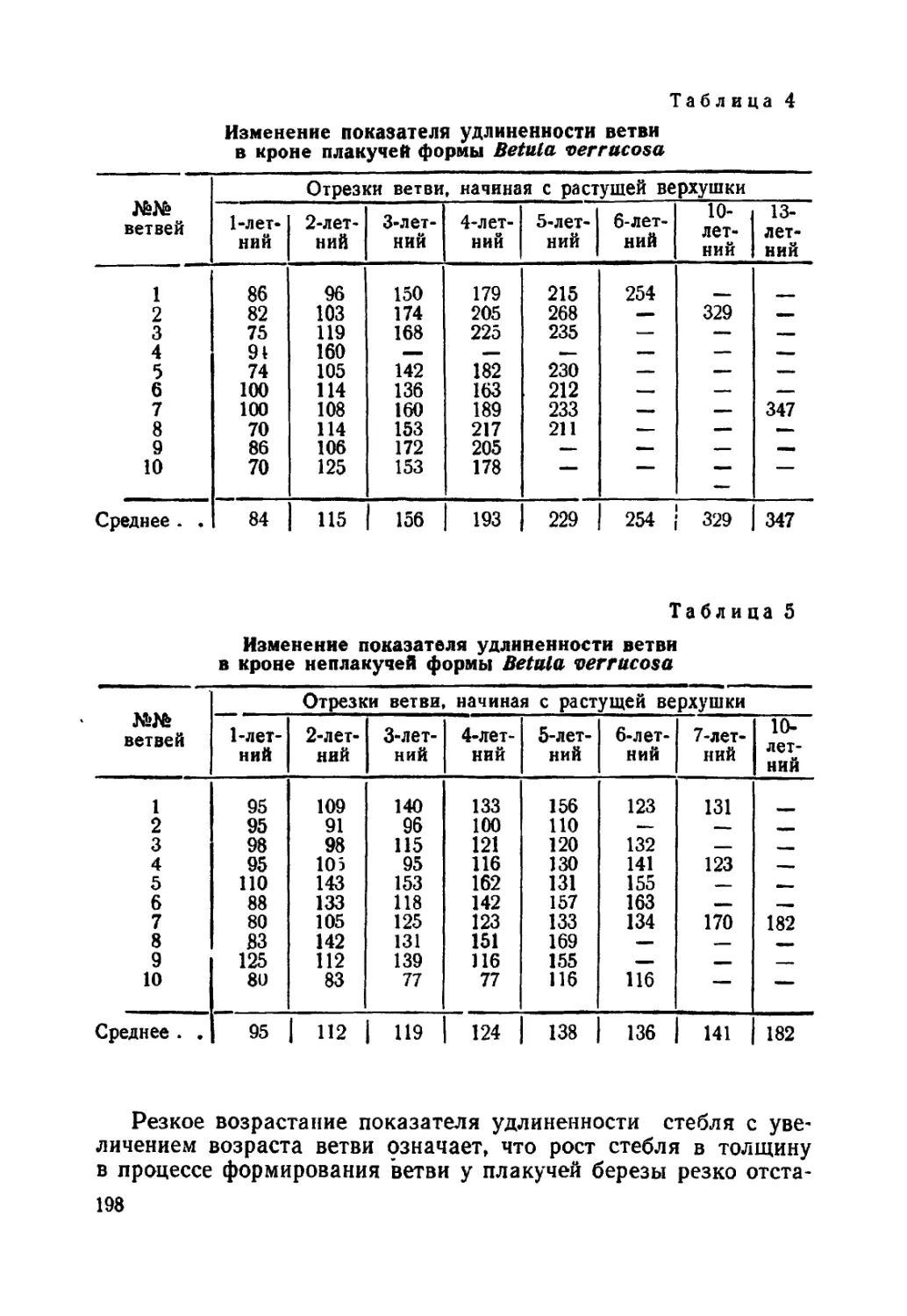
































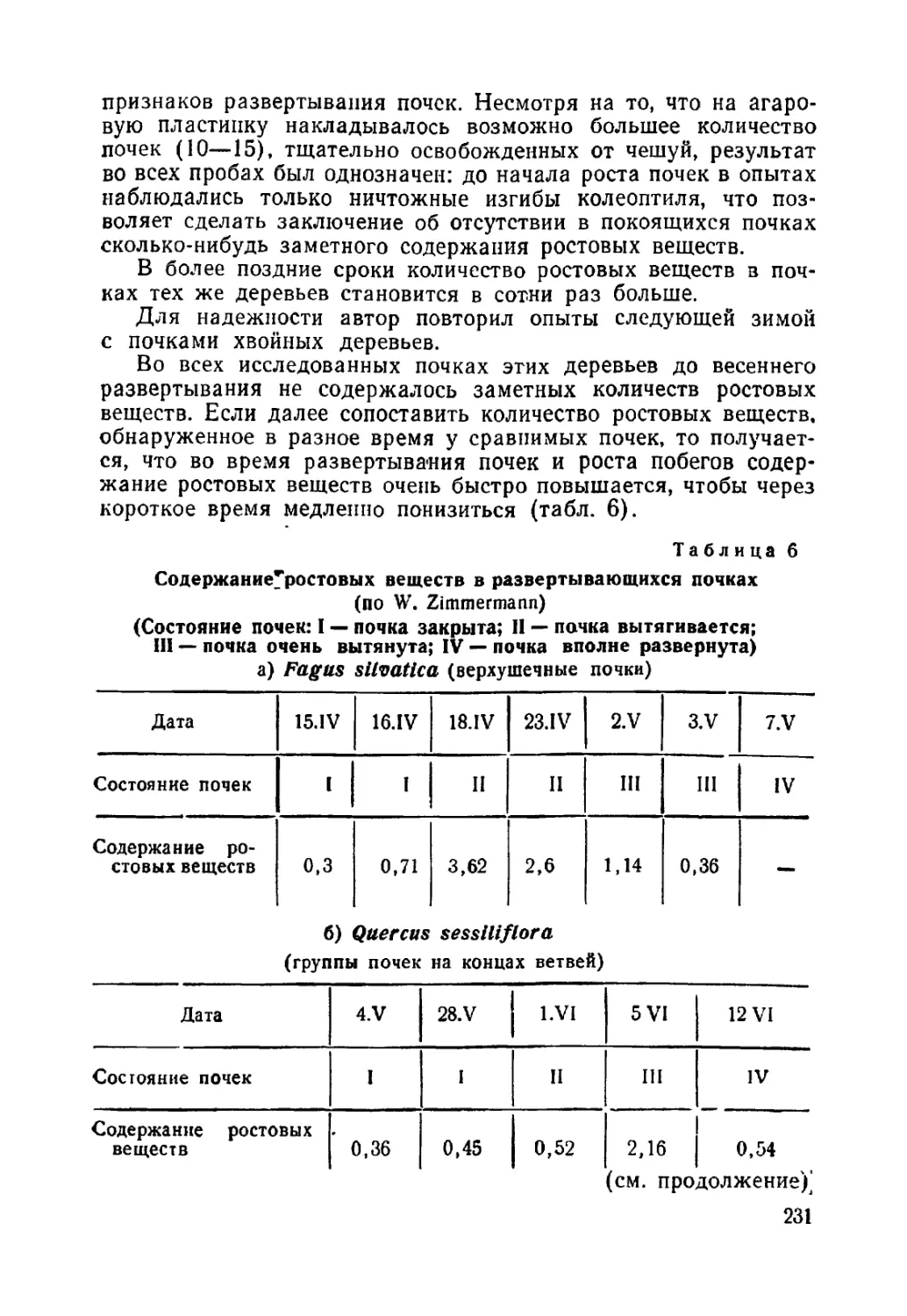







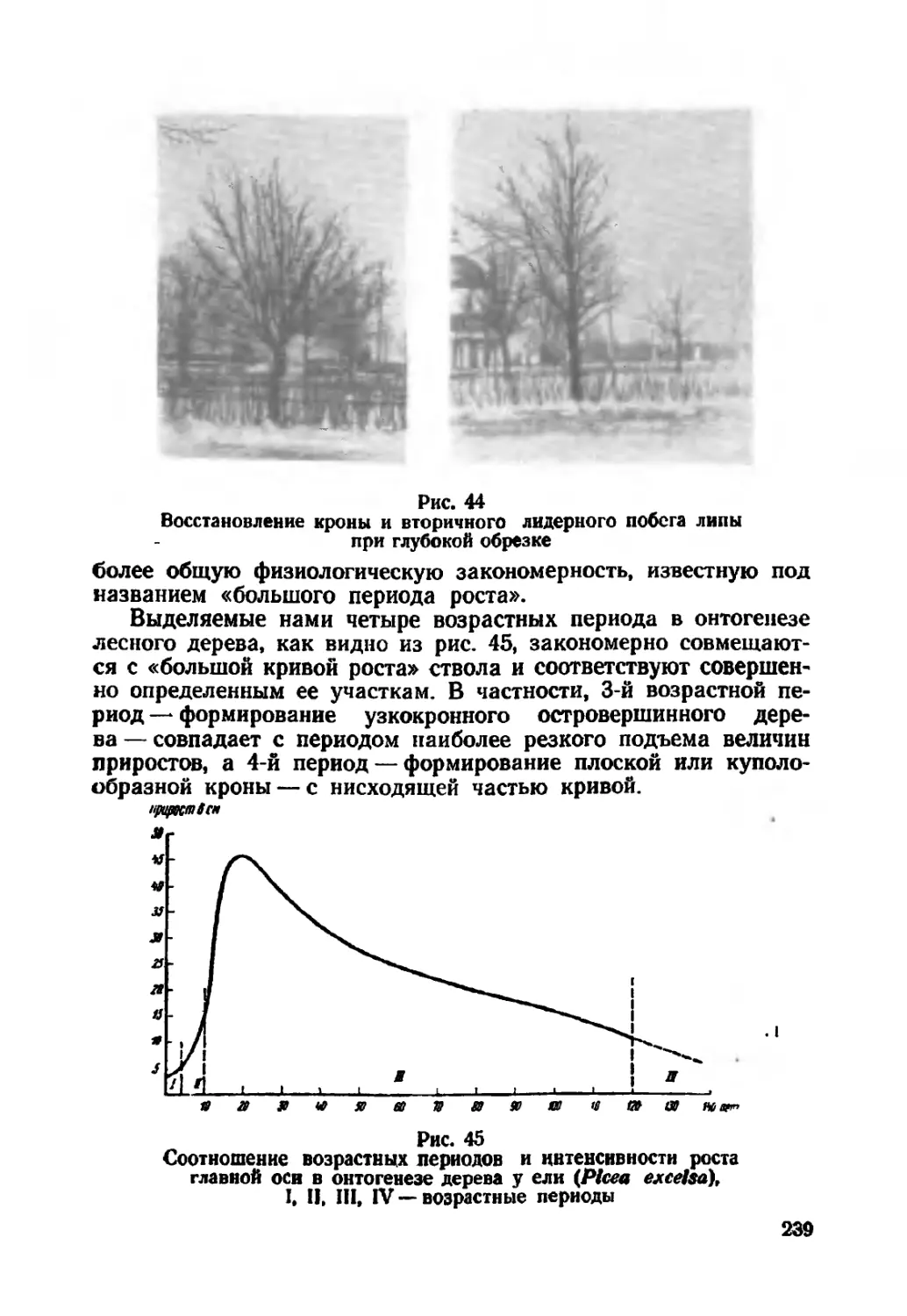















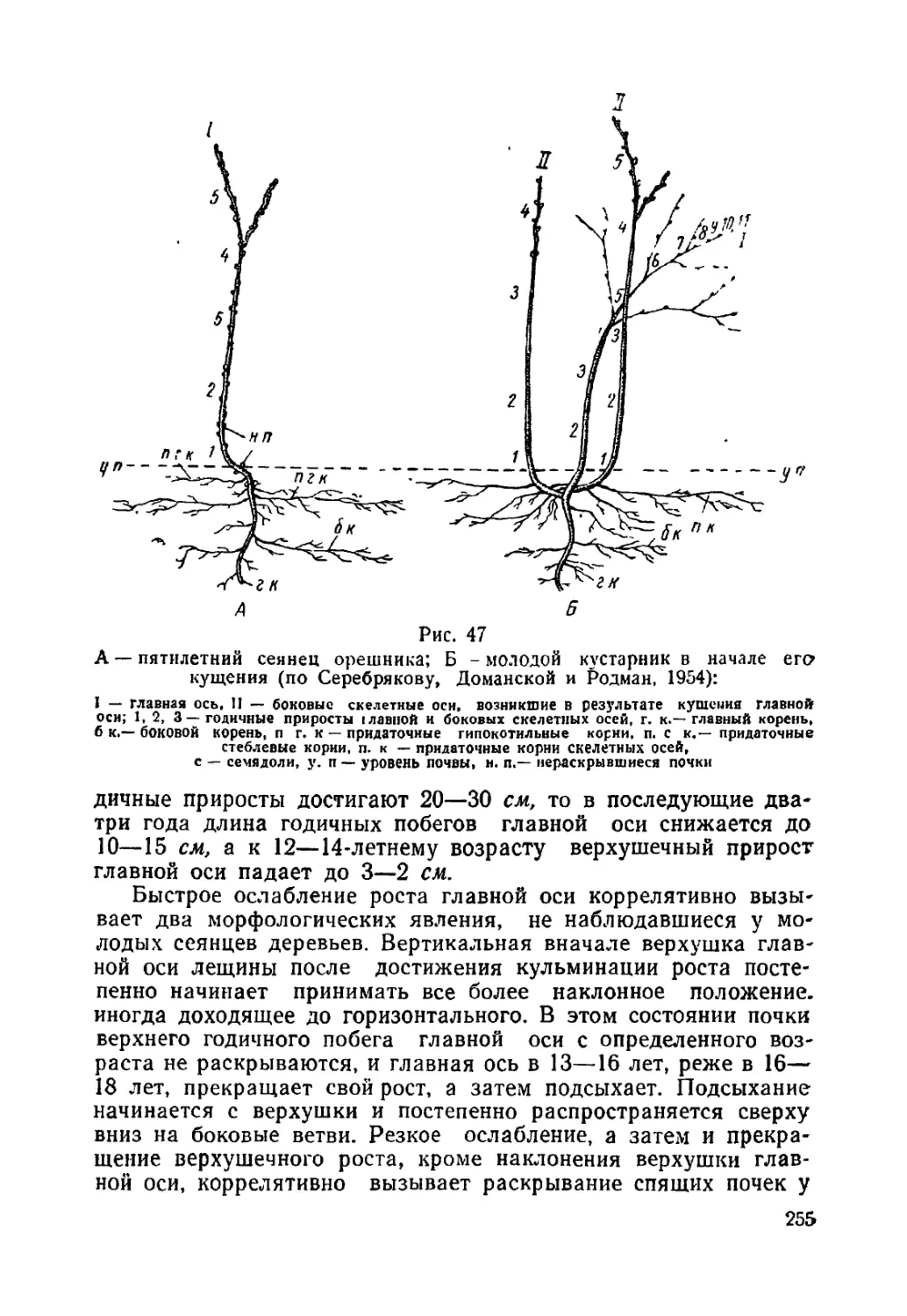
















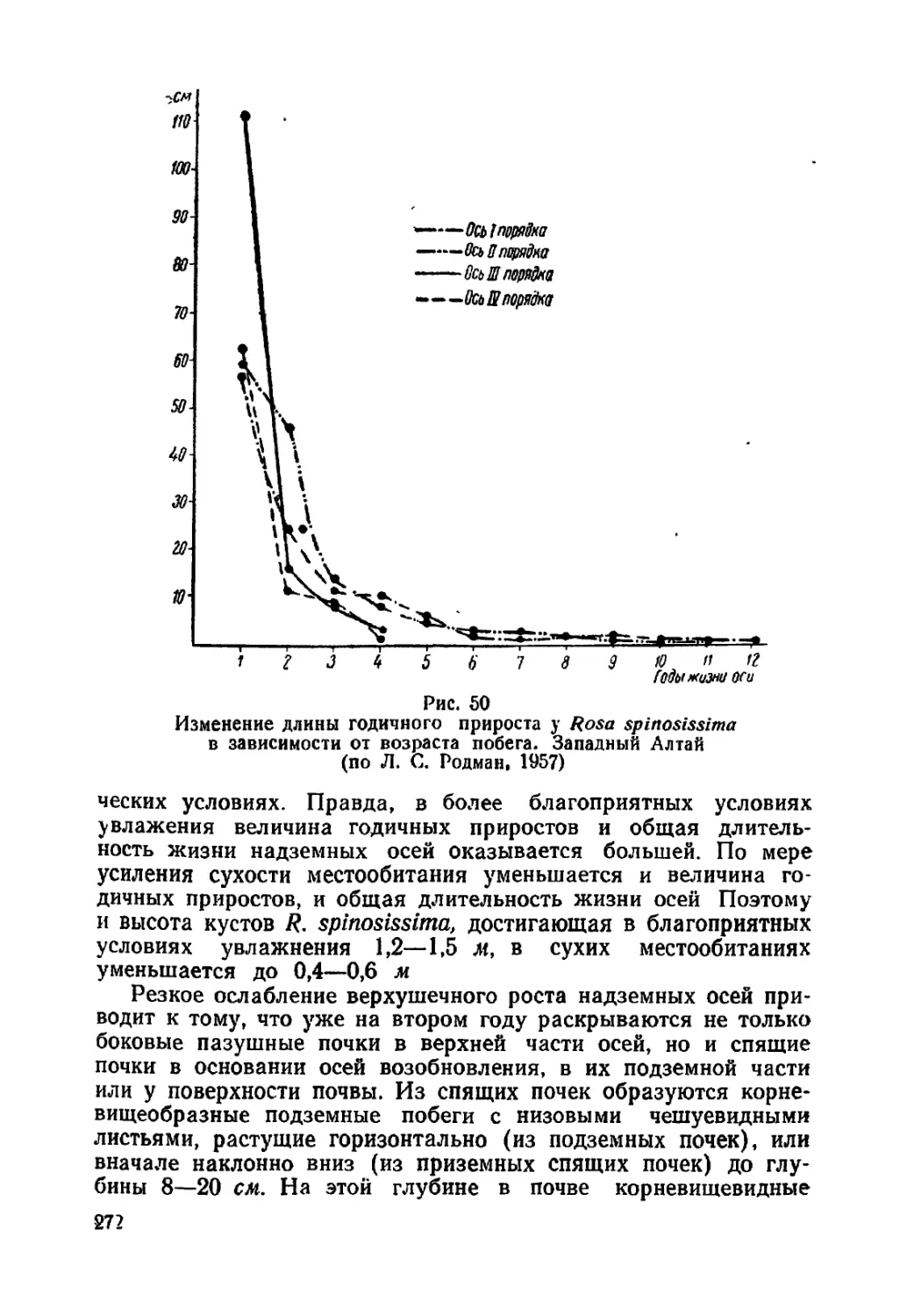

















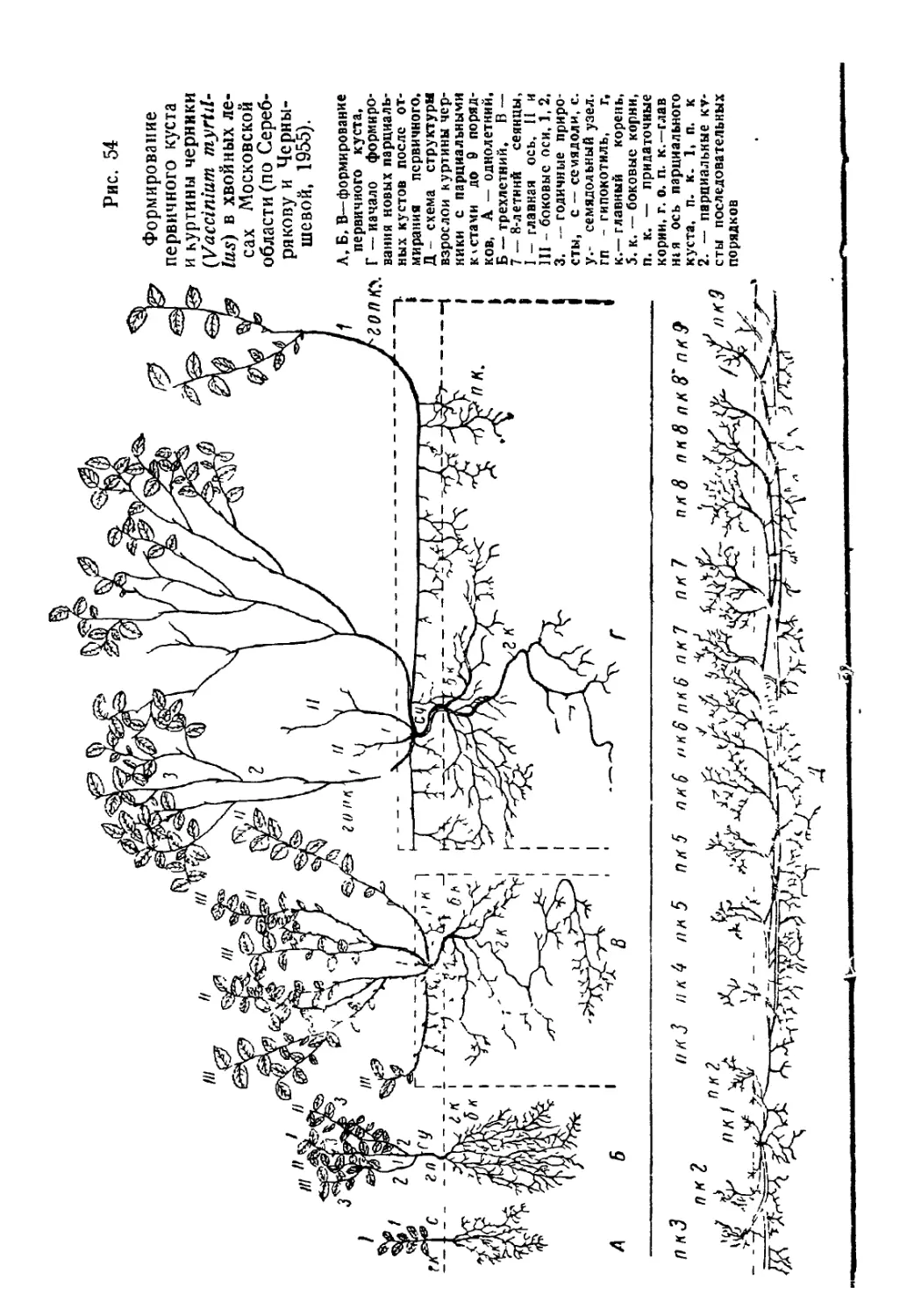













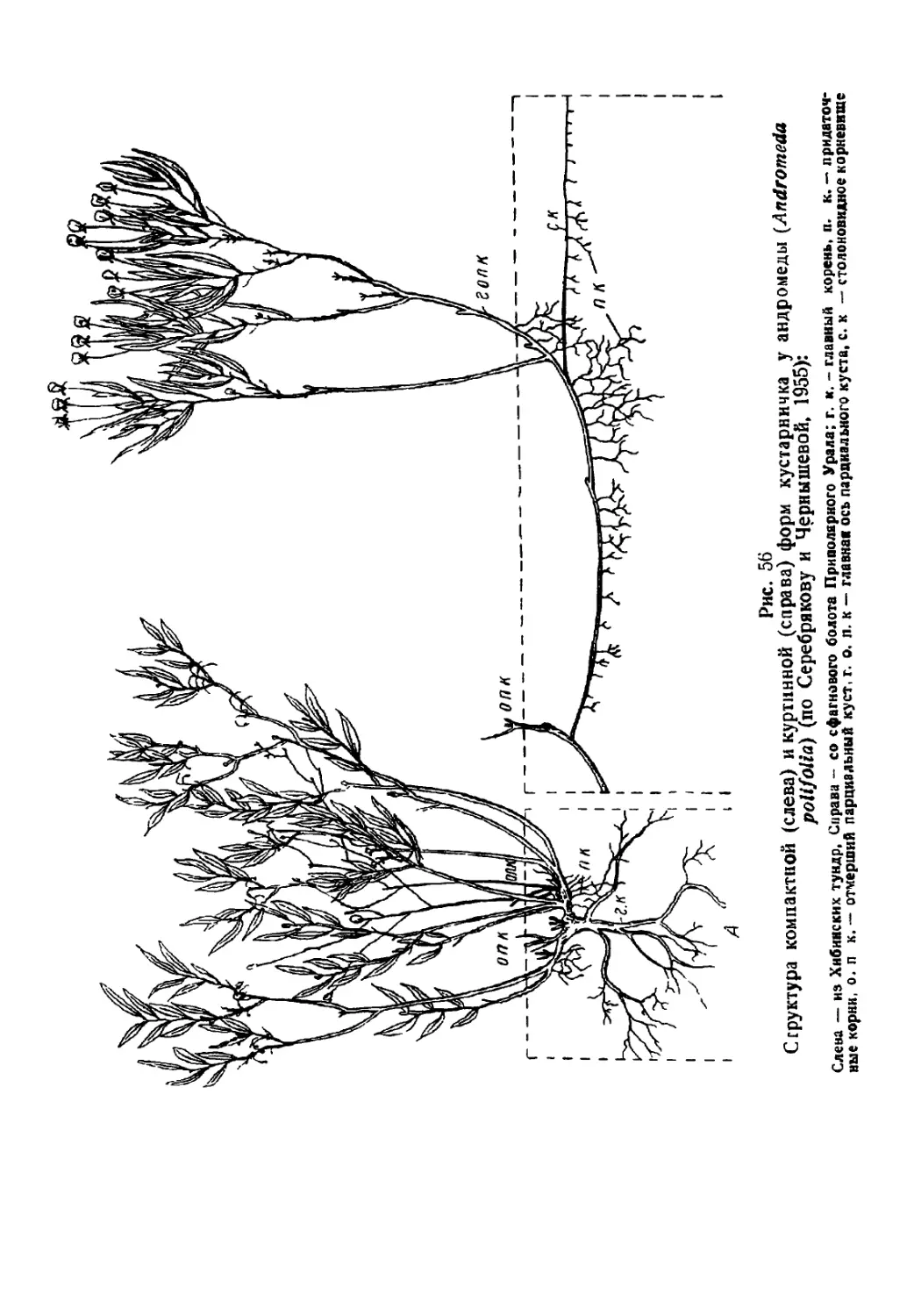



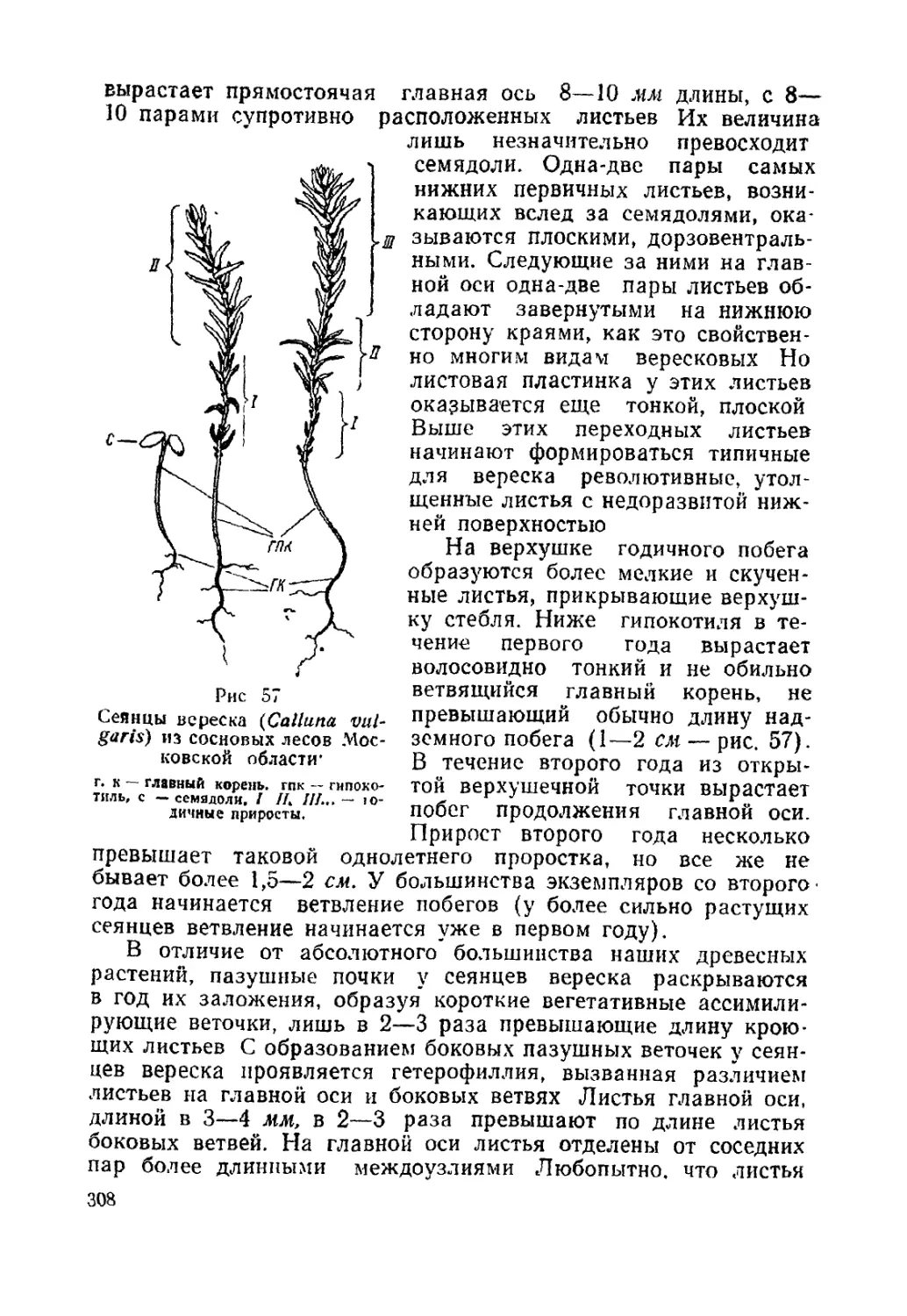









































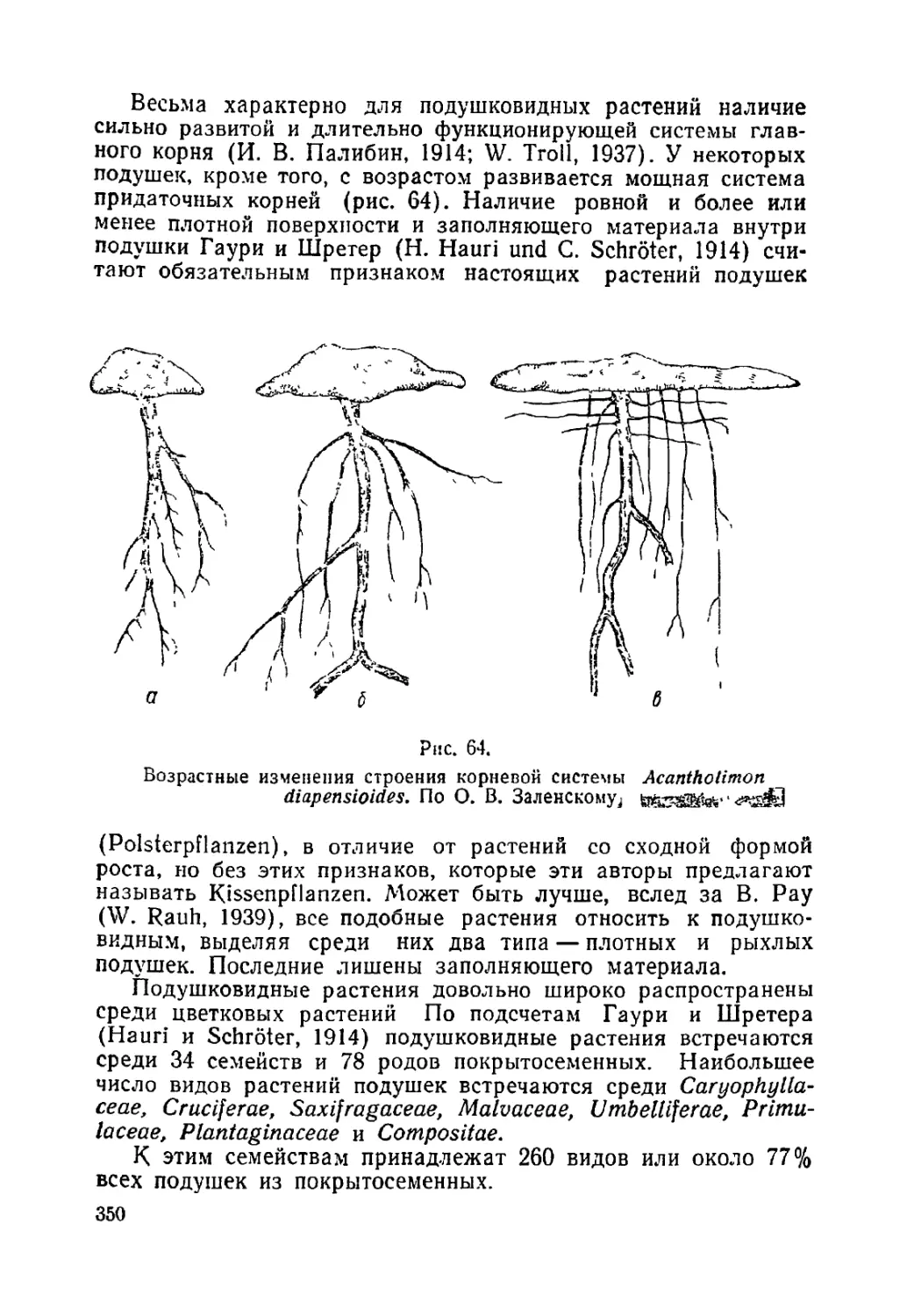


































Текст
Проф. И. Г. СЕРЕБРЯКОВ
ЭКОЛОГИЧЕСКАЯ
МОРФОЛОГИЯ
РАСТЕНИЙ
ЖИЗНЕННЫЕ ФОРМЫ
ПОКРЫТОСЕМЕННЫХ И ХВОЙНЫХ
Допущено
Министерством высшего и среднего
специального образования СССР
в качестве учебного пособия
для государственных университетов,
педагогических и лесотехнических вузов СССР
&
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
«ВЫСШАЯ ШКОЛА»
Москва —1962
ПРЕДИСЛОВИЕ
Настоящее учебное пособие не претендует на полное изло-
жение экологической морфологии растений. Оно посвящено
лишь одному из важных разделов этой науки — учению о жиз-
ненных формах у растений.
Эта книга — итог пятнадцатилетних исследований автора с
сотрудниками и учениками. Необходимость глубокого анализа
проблемы жизненных форм растений не раз подчеркивалась
в советской ботанической литературе и, в частности, была
отмечена в решениях II делегатского съезда Всесоюзного Бо-
танического Общества в 1957 г
Истоки этой книги — в студенческой работе автора на севе-
ро-восточном Тянь-Шане (1939—1940 гг.), в многолетних бота-
нических экскурсиях и наблюдениях «в дебрях» Подмосковья,
обычно вместе со студентами, аспирантами и сотрудниками, и,
в частности, в окрестностях Звенигородской биостанции МГУ,
в экспедициях в Хибины (1946, 1955 г), Западный Таймыр
(1949 г.), на трассы степного лесоразведения Камышин —
Волгоград и Волгоград — Черкесск (1950 г.), на поемные
луга нижнего течения Дона (1950 г), среднего течения Оки
(1947 г.), низовий Северной Двины (1951 г), в полупустыни
Прикаспия (1952 г.), в центральный Тянь-Шань (1953 г.), вос-
точные Кара-Кумы (1954 г.), во Францию, Пиренеи и тропи-
ческую Западную Африку (1954 г), па южный берег Крыма
(1954 г), в дубравы и степи Курского Государственного Запо-
ведника (1957 г).
Незабываемые картины природы этих стран и областей,
разнообразные формы растений, определяющие своеобразие
ландшафта и растительного покрова, детальное изучение исто-
рии развития (онтогенеза) типичных представителей разных
жизненных форм невольно создавали определенный строй
мыслей, и перед мысленным взором все более четко и ясно
3
вырисовывались контуры всего изложенного на страницах этой
книги.
Значительная часть исследований была опубликована в виде
отдельных статей; в этой книге они сведены и дополнены.
Как продолжение этого пособия автор предполагает напи-
сать книгу, посвященную жизненным формам наземных
травянистых растений и теоретическим вопросам, связанным
с ними (происхождение травянистых форм в различных систе-
матических группах покрытосеменных, классификация и эво-
люция жизненных форм наземных травянистых растений,
возникновение монокарпичности у растений и развитие моно-
карпических жизненных форм до образования однолетников
и т. д ). Отдельная работа должна быть посвящена жизненным
формам растений — амфибий и водных растений. Автор на-
деется, что отклики ботаников на предлагаемую работу послу-
жат стимулом для продолжения и завершения труда.
Настоящая книга — результат коллективного труда многих
ботаников, связанных в своей творческой работе с автором.
И я рад предоставившейся возможности выразить искреннюю
признательность и благодарность сотрудникам и аспирантам
(настоящим и бывшим) кафедры ботаники Московского город-
ского педагогического института имени В. П. Потемкина, сту-
дентам (тоже настоящим и бывшим), членам ботанического
кружка этого педагогического института, а также бывшим сту-
дентам кафедры высших растений и геоботаники Московского
Государственного университета имени М. В. Ломоносова, рабо-
тавшим под руководством автора. Имена многих из них упо-
минаются с благодарностью на страницах этой книги
Чувство особой благодарности хочется выразить моей вер-
ной помощнице и другу — Т. И Серебряковой, принимавшей
большое участие в создании этой книги, от сборов материалов
и наблюдений в природе до редактирования рукописи и подго-
товки многих рисунков. Опа была первым читателем, критиком
и редактором этой книги. Без преувеличения можно сказать,
что созданием этой книги автор во многом обязан ей.
Август 1959 г., Москва И. Г. Серебряков.
Глава J
ИСТОРИЯ УЧЕНИЯ О ЖИЗНЕННЫХ ФОРМАХ
У РАСТЕНИИ
1. У истоков УЧЕНИЯ о ЖИЗНЕННЫХ ФОРМАХ
Едва ли не первое, что обращает на себя внимание при
обзоре окружающего растительного мира, — это господство
определенных биологических типов растений, или жизнен-
ных форм, создающих ландшафтные картины растительного
покрова земли от тропиков до полярных стран.
Деревья и кустарники, кустарнички и травы, подушки и
стланцы, полукустарники и суккуленты, лианы и эпифиты опре-
деляют не только физиономию, но и структуру растительных
формаций различных широт — лесов и степей, пустынь и са-
ванн, лугов и болот, альпийских ковров и тундр. Не удивитель-
но, поэтому, что жизненные формы растений стали объектом
внимания исследователей уже на первых этапах развития бо-
таники, на заре ее становления Изучение их вызывалось
потребностями человека, использовавшего в своей жизни раз-
нообразные формы растений, от гигантских деревьев до эфе-
мерных однолетников
История учения о жизненных формах довольно сложна. Ее
истоки уходят в древний мир, к творениям Феофраста, а в
Новое время к трудам Цезальпина, Морисопа, Турпефора Но
самостоятельное бытие это учение приобрело лишь с начала
XIX в., с появления трудов основателя ботанической географии
Александра Гумбольдта. Развитие представлений о жизненных
формах и их системах у растений не раз излагалось в спе-
циальных монографиях (О Drude, 1913, Н. Gams, 1918;
Е. Du Rietz, 1931). Однако в отечественной литературе отсут-
ствует сколько-либо полный очерк этого вопроса. К тому же в
сводках зарубежных авторов остались не учтенными труды
русских и советских исследователей, а после выхода в свет
этих сводок появились новые труды. Поэтому представляется
вполне целесообразным в настоящей книге изложение основ
пых направлений исторического развития учения о жизненных
формах Разумеется, предлагаемый очерк не претендует на
исчерпывающую полноту.
Литература вопроса очень обширна, и ее полный обзор
потребовал бы весьма значительного объема.
Анализ показывает, что с начала XIX в до настоящего вре-
мени исследование жизненных форм у растений проводилось
главным образом в трех основных направлениях* 1) изучение
многообразия жизненных форм и решение вытекающей отсюда
проблемы их классификации, 2) изучение отношений жизнен-
ных форм к окружающей внешней среде и 3) изучение взаим-
ных филогенетических отношений отдельных групп жизненных
форм. Преимущественно с первым направлением совпадает
исследование закономерностей морфогенеза у разных групп
жизненных форм, приобретшее теперь сравнительно-морфоло-
гический и онтогенетический характер. В сущности, наибольшее
развитие до настоящего времени получило первое, системати-
ческое направление Экспериментальная экология, филогенез,
закономерности онтогенетического развития жизненных форм в
последнее время привлекают все большее внимание исследова-
телей. Их расцвета, несомненно, следует ожидать в будущем
Как видим, развитие учения о жизненных формах в значи-
тельной мерс повторяет историю учения о видах Преобладание
систематического направления на первых этапах, развертыва-
ние других и, в частности, экспериментально-аналитических и
филогенетических направлений в последующем наблюдается и
там и здесь Современный период и характеризуется расшире-
нием фронта исследований жизненных форм в направлении
морфолого-онтогенетическом, экологическом и филогенети-
ческом
Вряд ли можно указать дату возникновения понятия жиз-
ненной формы у растений, она, несомненно, теряется в глубине
веков. Об этом свидетельствует наличие уже широкого обзора
жизненных форм в трудах Фсофраста. Более чем за три столе-
тия до нашей эры в «Исследованиях о растениях» Феофраст
приводит подробное описание отдельных видов растений, при-
надлежащих к разным группам жизненных форм, начиная от
деревьев и кончая однолетними травами /Аы не находим у него
термина «жизненная форма растения», он возникнет более, чем
две тысячи лет спустя. Но содержание этого понятия у
Феофраста есть. «Первыми и самыми главными видами, охва-
тывающими почти все растения, или большинство из них, —
пишет Феофраст, — будут деревья, кустарники, полукустарники
и травы Дерево — это то, что дает от корпя один ствол со
6
множеством веток и узлов и не легко погибает; таковы, напри-
мер, маслина, смоковница, виноградная лоза. Кустарник даег
множество веток прямо от корня, например, ежевика и держи-
дерево Полукустарники дают от корня много стеблей и мно-
жество веточек, например, чабер и рута У травы листья идут
от корня, а ствола пет вовсе, таковы, например, хлеба и ово-
щи» (Феофраст, 1950, стр. 18—19). Эти «главные виды», по
Феофрасту, в свою очередь подразделяются на широколистные,
узколистные и игольчатые, с глубоко расположенной или по-
верхностной корневой системой, прямостоячими или лежачими
побегами, обладающие или нс обладающие корнеотпрыско-
востью. Среди травянистых он описывает клубненосные и луко-
вичные, розеточные и полурозеточные, многолетние и однолет-
ние растения. В «Исследованиях о растениях» излагаются
наблюдения над зависимостью формы роста от климата, почвы
и способов возделывания растений «Деревья в горах пизко-
ствольны, сучковаты, при пересадке в долины становятся боль-
ше и красивее на вид» (кн. III). Представляет безусловный
интерес положение Феофраста о том, что, несмотря на различие
отдельных «главных видов» растений, «есть нечто общее в при
роде и у деревьев, и у кустарников, и у кустарничков и трав»
(кн 1, стр 20) «Исследования» Феофраста представляют
энциклопедию ботанических знаний его времени, синтез всего
известного о жизни растений в науке и практике Эллады Уро-
вень знаний этого периода был неожиданно высоким Любо-
пытно, что группы жизненных форм у Феофраста служили
основой сто систематики растений Описание многочисленных
видов (около 500) ведется им с подразделением их на группы
деревьев, кустарников, кустарничков (фриганоидпых форм)
трав многолетних и однолетних.
Использование жизненных форм в таксономических целях
было свойственно и ботаникам Нового времени Так, в системе
Цезальпина (A Caesalpino, 1583), где все растения делятся на
15 классов, два первых класса относятся к деревьям (Arbores),
остальные к травянистым (НсгЬас). Среди последних два клас-
са выделяются по структуре их подземных органов — волокни-
стых (Triplici principia fibrosae — 8-ой класс) и клубненосных
(Triplici principia btilbosae — 9-ый класс) (цит по С Linnaeus,
1751). В системе растений Морисона (R. Morison, 1715) из 18
выделяемых им классов три первые относятся к древесным
1) деревья (Lignosae arbores), 2) кустарники (Lignosae fruti-
ces) и 3) полукустарники (Lignosae suffrutices) Остальные
классы включают лишь травянистые растения Среди послед-
них некоторые классы выделяются также по физиономическому
принципу, например, лиановидные травы (Herbae scandentes —
1-й класс) и злаковидные (Plantae culmiferae, calamiferac —
7
8-ой класс) В системе Турнефора (I. Р Tournelort, 1719) из
22-х классов растений, выделяемых автором, первые 17 обоз-
начаются как травы и полукустарники (Herbae et suffrutices) и
последние 5 относятся к деревьям и кустарникам (Arbores et
fiutices). Интересно, что розоцветные в этой системе оказыва-
ются в двух классах — 6-ом и 21-ом, потому что среди них есть
травянистые и древесные формы То же относится и к бобовым,
разбитым между двумя классами 10-ым и 22-ым.
Использование жизненных форм в систематике растений
удерживалось до Линнея, который, как известно, в основу так-
сономии положил генеративные признаки (С Linnaeus, 1751).
В последующем использование жизненных форм в таксономии
стало рассматриваться как грубая ошибка, а вегетативные при-
знаки утратили интерес и значение для систематиков
Но интерес к жизненным формам вновь возрос в начале
XIX в. На этот раз он пришел не от систематики, а со стороны
ботанической географии В 1806 г выходит небольшая, но за-
мечательная по своим последствиям статья А. Гумбольдта
«Идеи о физиономичпости растений», написанная им по живым
впечатлениям от пятилетнего путешествия по странам тропи-
ческой Америки. «Хотя характер различных местностей земного
шара находится в зависимости одновременно от всех внешних
явлений, если синева неба, строение облаков и прозрачность
воздуха обусловливают совокупность впечатления, то тем не
менее нельзя отрицать, что главное, определяющее это впечат-
ление, принадлежит растительному покрову» (1936, стр. 83).
Отметив решающую роль растительности в определении харак-
тера ландшафта, Гумбольдт пишет: «Если окинуть взором раз-
личные виды явнобрачных растений, имеющиеся в настоящее
время , то в этом удивительном множестве можно найти изве-
стные основные формы, к которым можно свести все остальные.
При выделении этих типов, красотой, распределением и груп-
пировкой которых определяется физиономия растительности
страны, надо обращать внимание нс на мелкие признаки, как
органы размножения, околоцветники и плоды (как это делает-
ся из других побуждений в ботанических системах), а па то,
что массой общего впечатления индивидуализирует местность»
(А. Гумбольдт, 1936, стр. 83—84) Ботаническая география, по
Гумбольдту, и должна начинать с изучения основных физионо-
мических типов растений, сочетания которых создают неповто-
римые ландшафтные картины от тропиков до полярных стран.
Гумбольдт выделяет вначале 16, а затем 19 «основных
форм» (Grundformen), резко отличающихся друг от друга фи-
зиономически.
1. Форма пальм — с высокими, стройными кольчатыми,
иногда колючими стволами, несущими на своей вершине бле-
8
стящие веерные или перистые листья. По мере удаления от
экватора убывают красота и размеры пальм; в Европе сохра-
няется лишь одна карликовая пальма Chamaerops Находимые
в северной Европе стволы пальм, погребенные в недрах земли,
дают возможность судить о грандиозных революциях в истории
нашей планеты, менявших климаты и физиономию природы.
2. Форма пизангов или бананов с низким, сочным, почти
травянистым стволом и облегающими его шелковисто-блестя-
щими листьями.
3. Форма мальвовых и баобабовых с колоссально толстыми
стволами и мягкими крупными листьями; эта форма придает
растительности своеобразный южный характер.
4. Вересковые, внешне сходные с хвойными от древовидных
до небольших кустарников, преимущественно встречающиеся в
Африке
5. Форма кактусовых в виде шаров или членистых стволов,
иногда канделябровидно разветвленных, покрытых иглами, но
с сочной внутренностью; по преимуществу в Америке.
6. Орхидеи, растущие, по преимуществу, на стволах тропи-
ческих деревьев и в расселинах пустынных скал.
7. Казуариновые деревья с ветвями, напоминающими хвощи,
по преимуществу в Австралии и Ост-Индии.
8 Форма ароидных — на стволах тропических деревьев с
мясистыми стеблями и крупными, часто стреловидными толсто-
нервными листьями.
9. Форма лиан — с максимальным развитием в тропиках.
10. Форма алоэ — с сочными мясистыми заостренными
листьями, скученными на верхушке стебля.
11 Форма злаков — с легкими и гибкими стеблями, древо-
видные характерны для тропиков.
12. Папоротники — древовидные в жарких странах, сходные
с пальмами, но с более короткими стволами и нежными
листьями в умеренных горных странах
13. Лилейные — с мечевидными листьями и роскошными
цветками.
14 Ивовые — во всех частях земного шара.
15. Миртовые.
16. Меластомовые.
17. Хвойные.
18. Мимозовые.
19 Потосовые.
Выделенные физиономические группы не идентичны систе-
матическим, как это можно было бы предполагать, судя по их
названиям. «Систематики выделяют множество групп расте-
ний, которые физиономист вынужден объединять одну с дру-
гой» (А. Гумбольдт, 1936, стр. 84). В основе выделения этих
9
групп лежит сходство во внешнем облике растений Как видно
из перечисленных характеристик «основных форм» Гумбольдта,
в них много от эстетического взгляда па природу. Но наиболее
интересно то, что уже здесь высказывается положение о зави-
симости распространения физиономических форм растений от
господствующих климатических условий. Работы Гумбольдта
послужили исторической основой для развития современного
учения о жизненных формах у растений.
2. О ПЕРИОДИЗАЦИИ ИСТОРИИ УЧЕНИЯ
О ЖИЗНЕННЫХ ФОРМАХ У РАСТЕНИЙ
В последующем идеи Гумбольдта были развиты многими
исследователями. В разное время и разными авторами было
предложено большое количество систем жизненных форм у
растений, с использованием различных принципов их выделе-
ния и классификации. При изложении истории учения о жиз-
ненных формах у растений появилась необходимость ее перио-
дизации. Впервые такая периодизация была приведена в мо-
нографии Дю Рие (Du Rietz, 1931), который выделяет 5 перио-
дов изучения и классификации жизненных форм у растений:
1) ранний период чисто физиономических систем жизненных
форм (с 300 лет до н. э. до 1884 г.);
2) ранний период эфармонических систем жизненных форм
(1884—1905 гг.);
3) период первых работ Раункиера по жизненным формам
(1905—1913 гг.);
4) период начавшихся реакций против односторонне эфармони-
ческой точки зрения в классификации жизненных форм
(1913—1920 гг);
5) новый период восстановления чисто физиономических систем
жизненных форм (1921 —1930 гг.).
Несколько иная периодизация была предложена Г. Мёйзе-
лем (Н. Meusel, 1951). Мёйзель выделяет три эпохи в исследо-
ваниях жизненных форм, или форм роста (Wuchsform), отли-
чающиеся друг от друга направлением исследований: 1) фи-
зиономическое направление Гумбольдта, Гризебаха и
др.; 2) экологическое направление в систематике жизнен-
ных форм, связанное прежде всего с трудами Раункиера, рас-
пространенное еще и в настоящее время, иЗ) сравнитель-
но-морфологическое направление в изучении жизнен-
ных форм, куда включаются, в частности, работы самого Мёй-
зеля, Рау, Тролля.
Однако с периодизацией, предложенной этими авторами,
вряд ли можно согласиться Начиная с первых десятилетий
XIX в. и до настоящего времени постоянно и одовременно су-
10
ществовали различные направления в изучении и классифика-
ции жизненных форм — и физиономические, и экологические, а
чаще смешанного эколого-физиономического характера. Что
касается сравнительно-морфологического подхода, то и он
ведет свое начало с замечательных работ Т. Ирмиша
(Th. Irmisch, 1846—1864), отчасти А. Брауна (А. Braun, 1849,
1851), продолженных впоследствии Е. Вармингом (Е. Warming,
1884—1918), Раункиером (С Raunkiaer, 1907—1918) и другими
авторами.
В весьма обстоятельных работах Г. Мёйзеля (Н. Meusel,
1935, 1951, 1952) и В. Pay (W. Rauh, 1937, 1938, 1939, 1940 и др.),
действительно проводится углубленный сравнительно-морфоло-
гический анализ процессов морфогенеза в разных группах
жизненных форм. Однако принципы классификации жизненных
форм, принятые Л1ёйзелем и Рау, не отличаются достаточно
резко от ряда их предшественников. Резко отличались в раз-
личные периоды лишь воззрения на связь жизненных форм со
средой и эволюцию их Решающую роль здесь, как и в других
областях биологии, сыграла победа дарвинизма Лишь в после-
дарвиновский период учение о жизненных формах стало по-
настоящему экологическим и эволюционным.
Что касается проблемы классификации жизненных форм,
то, вероятно, лучше говорить не об исторических периодах раз-
вития этого учения, а о направлениях в систематике
жизненных форм, развивавшихся в течение XIX и XX вв. Изу-
чение истории вопроса склоняет к тому, чтобы выделить два
основных направления в классификации жизненных форм:
1) эколого-физиономическое и 2) морфолого-
биологическое. Эти направления почти идентичны
отмеченным Г. Гамсом (Н. Gams, 1918) физиономическому и
экологическому. Мы предпочитаем все же говорить для первого
об эколого-физиономическом направлении, потому что все авто-
ры этого направления выделенные ими формы связывают с
господствующими климатическими условиями среды. Название
«морфолого-биологическое», по нашему мнению, лучше отра-
жает сущность подхода, чем «экологическое».
3. ЭКОЛОГО-ФИЗИОНОМИЧЕСКОЕ НАПРАВЛЕНИЕ
В ИЗУЧЕНИИ И КЛАССИФИКАЦИИ ЖИЗНЕННЫХ ФОРМ
Его родоначальником не без основания считается А. Гум-
больдт, впервые выделивший группу форм по физиономическим
признакам. Большое значение в развитии этого направления,
как и вообще учения о жизненных формах в целом, имели
труды Кернера и Гризебаха. На них лежит печать нового1,
11
эволюционного учения Дарвина, ставшего основой экологи-
ческого направления в ботанике.
В известной книге «Жизнь растений Дунайских стран'»
(1863) Кернер говорит, что всюду растительность приспособле-
на к местным климатическим условиям И если мы раскроем
большую зеленую книгу растительного царства, то в ней мы
можем прочитать о климате гораздо более полно и верно, чем
в пожелтевших листах метеорологических фолиантов. Расти-
тельный мир всюду — отражение климата. Но из тысяч расте-
ний, составляющих ландшафт, лишь единицы обусловливают
физиономический облик растительного покрова И если мы хо-
тим описать растительные формации, необходимо выделить
основных строителей ее, назвать основные формы («Grund-
fornien») растительного мира. Кернер выделяет 12 «основных
форм», бросающихся в глаза благодаря своей массовости
1) деревья; 2) кустарники; 3) крупные травы (Stauden);
4) дернистые растения (Filzpflanzen), 5) низкие травы, не дер-
нистые (Krauter); 6) листовые растения, типа корневищных
папоротников (Blattpflanzen); 7) лазящие и вьющиеся расте-
ния; 8) нитчатые растения, паразиты (Fadenpflanzen); 9) труб-
чатые растения (Rohres); 10) злаковидные (Halmgewachse);
И) грибы; 12) лишайники
Как видно, в отличие от Гумбольдта, «основные формы»
Кернера отражают специфику растительности не тропических,
а умеренных широт. Второе отличие от Гумбольдта заключает-
ся в том, что Кернер считает ошибочным использование родо-
вых названий для выделенных им «основных форм». Система-
тика и физиогномика растений идут совершенно разными
путями, поэтому, во избежание путаницы, не стоит использо-
вать термины одной дисциплины в другой.
Более сложная и детально разработанная система жизнен-
ных форм (или растительных форм — «Vegetationsforni») была
предложена Гризебахом в его книге «Растительность земного
шара» (1872) ’. Гризебах также исходит из того, что нужно
различать физиономическую и естественную системы растений.
Первая указывает зависимость организации растений от физи-
ческих условий и, в частности, климата, вторая — пути их про-
исхождения Лишь иногда группы «растительных форм» сов-
падают с систематическими группами Гризебах выделяет 7
основных групп, подразделяемых в свою очередь па 54 «расти-
тельные формы». Каждая из выделенных форм, или групп их,
по мнению автора, отражает особенности климата страны, а
также ее историю.
1 Переведена на русский язык и вышла с предисловием А Н Бекетова
в 1874 и 1877 гг
12
1 . Д е р е в я н и ст ы е растения
А — С простым стволом, на верхушке несущим пучок листьев (6 форм—
например, форма пальм, форма древовидных папоротнков, форма
пандаобразных и др.).
В —С простым стволом, но с боковыми пучками листьев (1 форма бамбуко-
образных).
С —С ветвистым стволом и массивной кроной (10 форм, например, формы
хвойных, лавровидных, и вообрази ых и др.).
D — С несколькими стволами, взаимно соединенными кроной (2 формы —
баньянов и манговообразных).
Е— Со стволом, ветвящимся от основания — кустарники (7 форм —вере-
скообразные, миртообразные, олеандры и др.)
F — С недоразвитой листвой (3 формы — казуарины, кипарисовые, спарции).
G — Без видимых стволов и разветвления (1 форма стелющихся пальм).
II . Сочные растения (три формы: лебедовые — кустарники и травы
с сочными листьями, агавы —без заметного ствола и кактусообразные
—без листьев).
III Вьющиеся растения (3 формы — лианы древесные двудоль-
ные, ротанги и лианы травянистые).
IV Эпифиты (2 формы — лорантообразные-чужеядные и орхидеи)
V. Травы.
А — Стебель облиственный (3 формы — полукустарники, гнафалиеобразные,
бессмертницы).
В— Стебель безлистный, листья пучками из почвы (1 формы — лукович-
ные, бромелиевые и др.).
С — Пучки листьев без заметного стебля (1 форма — папоротников).
VI. 3 л а к о о б р а з н ы е (6 форм — луговые, степные, однолетние,
осоки и др.).
VII. Бессосудистые растения (2 формы — мхи и лишайники)
Отдельные «растительные формы» в пределах каждой груп-
пы автор выделяет прежде всего по типу их листьев Например,
среди кронообразующих деревьев им выделяются хвойные с
жесткими вечнозелеными листьями, лавровидные — с широки-
ми глянцевитыми, ивообразпые — с узкими летнезелеными.
Среди кустарников — всрескообразные, с жесткими, часто игло-
видными листьями, олеандры, с крупными листьями больше
2,5 см (1 дюйм) и т. д
Система Гризебаха охватывает почти все наземные расте-
ния различных географических широт. Некоторые из выделен-
ных им групп используются в системах различных авторов и до
настоящего времени (О Drude, 1913, Е Du Rietz, 1931 и др).
Наиболее удачной здесь представляется классификация древес-
ных растений Недостатком системы Гризебаха является не
дробность ее подразделений, как это отмечают Варминг и
Гребнер (Е. Warming и Н Graebner, 1918), а несоответствие
частных признаков, используемых для выделения «раститель-
ных форм», с основной идеей его классификации — зависимо-
стью формы и структуры растений от климатических условий.
Именно поэтому многие «растительные формы» Гризебаха
оказываются «формальными» и экологически неоправданными.
13
Это впечатление усиливается из-за применения автором так-
сономической терминологии, с использованием родовых назва-
ний растений
Но из-за этого не следует систему Гризебаха расценивать
как чисто физиономическую. Он неоднократно подчеркивает
связь господствующих форм растений с климатом местности.
«Узкий лист лилейных может развиваться и в сухом воздухе,
но широкий лист бананов, пронизанный бесчисленными спираль-
ными сосудами, ... требует тропической сырости» (1877). Говоря
об оригинальной форме розеточного дерева Xanthorrhoea из
лилейных, он пишет: «Именно эти-то травянистые пучки на
верхушках стволов лучше всего указывают на климатические
особенности Австралии» (1877, стр. 181). Далее: «Самое большое
богатство форм (подчеркнуто нами — И. С.) представляют как-
тусовые под влиянием равномерной и высокой температуры
в скалистых саваннах Мексики и в Андах Южной Америки»
(там же, стр. 273). Как видим, автор пытается даже самую
форму растений поставить в зависимость от господствующих
климатических условий. Конечно, с высоты биологических дости-
жений за минувшие 85 лет эти попытки нам кажутся иногда
наивными. Но они свидетельствуют о четко выраженном эколо-
гическом подходе Гризебаха к проблеме жизненных форм.
Сходную систему растений, но применительно лишь к Скан-
динавии, разработал Хульт (R. Hult, 1881). Хульт выделяет
43 «растительные формы», или жизненные формы, объединен-
ные в 10 групп «основных форм». Среди них он различает:
А. С древесным стволом:
I Деревья (Вйшпе),
1 — основная форма хвойных деревьев (Pinusform, Abiesform)
2 — основная форма лиственных деревьев (Betulaform).
IJ Кустарники. (StrHucher).
3 —основная форма собственно кустарников — Gebusche (Salixform)
4 — основная форма кустарничков — Reiser (Ericaform, Ledumform, Oxycoccus-
form, Myrtusrorm, Myrtillusform).
В. С травянистыми воздушными побегами:
I. Стебли и листья летнезеленые.
5 — основная форма — узколистные
♦) поликарпичные (Airaform, Festucaform, Miliumform, Eriophorumform
и др.),
**) монокарпичные (Hordeumform, Роа annua-form).
6 — основная форма — широколистные.
*) поликарпичные (Geraniumform, Solidagoform и др.),
♦*) монокарпичные (Stellaria media-form, Galeopsisform и др.)
II. Стебли и листья многолетние.
7—10 основные формы включают мхи и лишайники (Sphagnumform, Poly-
trlchumform, Cladoniaform и т. п.).
14
Конечно, «основные формы» Хульта далеко не исчерпывают
разнообразия жизненных форм Скандинавии, особенно среди
травянистых растений. Расчленение последних у Хульта слиш-
ком схематично. Оставляет чувство неудовлетворенности и
использование родовых названий для отдельных «растительных
форм». Так, в сущности, можно перечислить весь состав флоры
страны по родам и система жизненных форм окажется излиш-
ней Но по сравнению с системой Гризебаха мы видим здесь
и нечто новое:* более полное использование собственно биоло-
гических признаков — длительность жизни растения и его над-
земных органов и др. Система Хульта более биологична, чем
Гризебаха. Но отделять мхи и лишайники от цветковых по дли-
тельности жизни листьев не имеет смысла.
Выше говорилось о глубоком влиянии дарвинизма на разви-
тие учения о жизненных формах. Уже в работах Кернера, а
также Гризебаха отмечается адаптивный характер «основных
форм» у растений Однако не все органы одинаково четко реа-
гируют на изменение внешних условий, вегетативные органы
более лабильны, генеративные более стабильны. На этом осно-
вании К Негели (К- Nageli, 1884) были введены понятия об
«организационных» и «приспособительных» признаках у орга-
низмов. Первые, по Негели, развиваются независимо от внеш-
ней среды, вторые в процессе своего развития могут изменяться
при соответствующих воздействиях среды, приспособляясь
к окружающим условиям Это подразделение признаков орга-
низма оказало существенное влияние на многих исследователей
жизненных форм у растений
Так, О. Друде, начавший с 1887 г. большую серию работ по
«биологической системе» растений, следует Негели, строго раз-
личая признаки «морфологические» и «биологические», т. е.
организационные и приспособительные, по Негели. Таксономи-
ческая система, по Друде, базируется на признаках первого
рода, биологическая — на вторых, и, в частности, на способах
роста. Он критикует системы Гумбольдта и Гризебаха за то, что
в них смешиваются «биологические» и «морфологические»
признаки, что ставит в затруднение при их использовании и фло-
риста и биолога. В биологической системе 1887 г. О Друде вы-
деляет 7 основных групп «растительных форм»:
1. Облиственные древесные р а с т е н и я: А — деревья и кустар-
ники; В —лианы; С —древесные паразиты.
11. Безлистные д р е в е с н ы е ра с т е и и я: А — стеблевые суккуленты
и В — безлистные кустарники
III. Полукустарники.
IV. Травы с сохраняющимися несколько лет подземными органами (ро-
зеточные, листовые суккуленты, эпифиты)
V. Гапаксантные растения: 1 — наземные: а) мхи; Ь) кустистые
15
травы (Stauden); с)дву-и однолетние травы, 2 — пресноводные и
3 — морские.
VI. Автотрофные, безлистные: лишайники.
VII. Па ра з ит и ы е и сапрофитные: а) семенные, Ь) споровые —
грибы.
Довольно близка к этой системе система Паунда и Кле-
ментса (Pound and Clements, 1898)
В 1896 г. Друде несколько дополняет свою систему Но в
наиболее разработанном виде «биологическая классификация»
растений была представлена им в 1913 г. в книге «Экология
растений». Автор отмечает, что он стоит на позиции дарвинизма
и рассматривает жизненные формы у растений как физиологи-
ческие приспособления, становящиеся в течение геологических
периодов морфологическими. Но каждая филогенетическая
группа, по Друде, обладает лишь определенным «модусом при-
способляемости», вытекающим из характера ее наследственной
организации. Поэтому приспособляемость проявляется по-
разному в разных филетических рядах, что и обусловливает
специфику их жизненных форм В связи с этим жизненные
формы растений будут различными у видов, отличающихся
положением в филогенетической системе, хотя и находящихся
в сходных условиях внешней среды. Автор ратует за связь
биологических типов с систематическими группами в целях
придания первым филогенетического значения Было бы поверх-
ностным и необоснованным, говорит Друде, объединять в одну
группу древовидных папоротники, цикадовые, пальмы и драцены
лишь на том основании, что все они несут розетку крупных
листьев на верхушке ствола Они все различны. По крайней
мере, папоротники должны рассматриваться независимо от се-
менных, так как они встречаются там, где есть условия для раз-
вития их обоих поколений Основой для выделения жизненных
форм, по Друде, является их роль и энергия расселения в био-
сфере, в борьбе за пространство, за сохранение вида. Но при
выделении «биологических групп» Друде стоит за использование
физиономических особенностей, понимая под последними мор-
фологические признаки, сходные у разных растений габиту-
ально и по их функциям в данных экологических условиях.
Необходимо подчеркнуть, что в этот период были широко рас-
пространены системы жизненных форм «биологического» или
«эфармонического» характера в связи с работами Раункиера
и Варминга (см ниже). Автор отмечает односторонний харак-
тер этих работ и говорит о том, что физиономические жизнен-
ные формы лучше характеризуют ландшафт и могут быть
полнее использованы в ботанической географии
Друде предложил чрезвычайно подробно разработанную
систему физиономических жизненных форм для наземных и вод-
16
ных растений. По существу она продолжает систему Гризебаха,
детализируя ее, подразделяя сходные формы по систематиче-
ским группам. Автор выделяет свыше 40 жизненных форм на-
земных и водных высших растений, а включая и таллофиты —
55 жизненных форм.
Воззрения Друде и его подход к выделению жизненных
форм без сомнения являются большим шагом вперед Впервые
с полной отчетливостью прозвучала мысль о филогенетическом
подходе к исследованию жизненных форм. К сожалению, в
предложенной системе идея историзма и развития жизненных
форм не нашла отражения Что касается связи жизненных форм
с систематическими группами, то здесь выявился интересный
параллелизм древесных и травянистых форм растений у споро-
вых и семенных, двудольных и однодольных Этот параллелизм
свидетельствует о копвергентности процессов -развития жизнен-
ных форм в разных филогенетических рядах растительного
мира
Оригинальная система жизненных форм была предложена
Линдманном (Lindmann, 1914); к сожалению, она охватывает
лишь незначительный круг древесных форм.
Наименее экологичной системой в рассматриваемом ряду
явилась, пожалуй, система Дю Рие (Du Rietz, 1921) Дю Рие
правильно отмечает субъективность в разделении признаков на
«приспособительные» и «индиферентныс», и призывает не счи-
таться с ним при выделении жизненных форм Но он скепти-
чески относится к адаптивной оценке жизненных форм И по-
этому вместо термина «жизненные формы» предпочитает гово-
рить «основные формы» (Grundformen), выделяемые с чисто
физиономической точки зрения Понятие «жизненная форма»,
по Дю Рие, связано с различными гипотезами приспособления
и вырастает и сменяется вместе с ними Физиономические «ос-
новные формы» выделяются индуктивно и поэтому носят более
устойчивый характер Дю Рие предлагает возвратиться к воз-
зрениям Гумбольдта, Гризебаха, Хульта А вместо старых, раз-
лично толкуемых народных терминов предлагает использо-
вать новые, научные
Десять лет спустя в известной монографии, специально
посвященной анализу жизненных форм у наземных цветковых
растений, Дю Рие (Du Rietz, 1931) продолжает развивать мысль
о необходимости индуктивного изучения жизненных форм,
отвлекаясь от предвзятых теорий адаптаций « Только жизнен-
ные формы, выделяемые независимо от какой-либо теории адап-
тации, могут быть использованы всеми как основа для индук-
тивного изучения проблемы приспособления и действительных
отношений между жизненной формой и средой» (Du Rietz, 1931,
стр 42)
В-113 И Г Серебряков —2
17
Дю Рие отмечает, что у экологов нет согласия и в общей
теории и в практике классификации жизненных форм: одни
выделяют для классификации эфармонические, другие — физио-
номические особенности растений. Даже у авторов близких тео-
ретических воззрений при разработке биологических систем
наблюдаются резкие различия в выборе главных и соподчинен-
ных признаков. Дю Рие считает, что в зависимости от постав-
ленных целей может быть создано несколько параллельных
систем жизненных форм. В частности, для характеристики рас-
тительного покрова и его отношений со средой им рекомен-
дуются следующие системы:
1. Система основных жизненных форм (Main life-forms),
базирующаяся на общей физиономии растений в течение вегетационного
периода, без анализа деталей их морфологии и переживания неблагоприят-
ного времени года; соответствуют «основным формам» Кернера, Хульта,
отчасти Друде.
2. Система форм ро ста (Growth-forms), жизненных форм, выде-
ляемых по структуре побегов в смысле Варминга (1903).
3. Система жизненных форм по периодичности развития
(Periodicity life-forms) и, в частности, по различию в облике растений в
разные сезоны года, преимущественно в вегетативной сфере (в плане работ
J. Massart, 1908; I. Jeswiet, 1914; К. Linkola, 1922).
4. Система жизненных форм по высоте расположения почек
возобновления (Bud-height life-forms) во время неблагоприятного
сезона года, как это использовано Раункиером (1903, 1907 и др.).
5. Система жизненных форм по типам защиты почек, перено-
сящих неблагоприятное время года (Bud-type life-forms).
6. Система жизненных форм по характеру листьев (Leaf life-
forms)—их величине, форме, структуре, длительности жизни и т. д.
Имеются в виду работы Раункиера (1916), A. Seybold (1927) и др.
Желательна была бы, по Дю Рие, также система жизненных
форм, основанная на структуре корневых систем растений,
однако, знания в этой области у нас, к сожалению, еще недо-
статочны
В монографии Дю Рие (1931) приводятся две первые сис-
темы. При изложении системы основных жизненных форм
Дю Рие, в отличие от своей первой монографии (1921), ратует
за возможно более простую терминологию, так как системой
будут пользоваться не только профессионалы ботаники, но и
географы, лесоводы и др. Дю Рие различает:
А. Древесные растения (Holoxyles)
I — Деревья. Очень высокие — больше 30 м; высокие — 8—30 м\ низкие
— 2-8 м; карликовые —0,8—2,0 м и пигмейные — ниже 0,8 м.
И — Кустарники — выше 0,8 м'. а) хтонофиты (живущие на земле), очень
высокие — более 8 м\ высокие —2—8 м и обычные —0,8—2.0 л
Ь) эпифитоиды (живут на деревьях и кустарниках, не паразитируя);
с) паразиты-деревья и кустарники.
III — Кустарнички — ниже 0,8 м, обильно ветвящиеся, но не подушковид-
18
ные: а) хтонофиты — высокие 0,25—0,8 м', низкие — 0,05—0,25 м я
очень низкие — менее 0,06 м; Ь) эпифитоиды; с) паразиты.
IV — Древесные подушковидные растения.
V — Древесные лианы разной высоты и типов прикрепления.
В. Полудревесные растения — полукустарники
(Н е m i х у 1 е s)
I — высокие, более 0,8 м и II — низкие, менее 0,8 м.
С. Травянистые растения.
I — Хтонофиты, не лианоидные. Очень высокие, более 2,0 м; высокие
0,8—2,0 м; средние 0,25—0,8 м и низкие —менее 0,25 м.
11 — Эпифиты.
III — Паразиты.
IV — Травянистые лианы.
Система весьма простая и достаточно полная в своих глав-
ных подразделениях. Но внутри классов границы разделения
растений по высоте, в сущности, условны и остаются неаргу-
ментированными ни биологически, ни фитоценотически.
Более детально разработана у Дю Рие система форм роста,
основанная на структуре побегов Ее изложению автор предпо-
сылает краткий, но весьма содержательный очерк, посвященный
анализу типов стебля у цветковых растений. Он правильно от-
мечает, что, вероятно, нет другой области в новой ботанике, где
царил бы такой же терминологический хаос, как в морфологии
стебля и особенно подземного стебля у цветковых растений
Автор предлагает детальную классификацию побегов, которую
затем и использует в своей системе жизненных форм. В системе
выделяются:
А. Древесные растения (Holoxyles)
I. Деревья
а) Деревья кронообразующие с удлиненными побегами: 1) Эухтонофиты
— неподвижные, надземноподвижные (например, ель при укоренении
ветвей), корнеотпрысковые; 2) Гемиэпифиты — неподвижные и распро-
страняющиеся воздушными корнями (например, баньян).
Ь) Розеточные деревья с укороченными побегами: 1) простые (неразвет-
вленные) и 2) ветвящиеся.
с) Безлистные суккулентно-стеблевые деревья — простые и ветвящиеся.
II. Кустарники.
а) Хтонофиты:
1. С удлиненными побегами:
а) Прямостоячие: 1) аэроксильные — без подземного ветвления (Prunus
padas, Sorbus aucuparia) и 2) геоксильные — с подземным ветвлением
побегов (виды Rosa, Corylus avellana).
0) Полупростратные: 1) аэроксильные (Metrosideras umbellata) и 2) геок-
сильные (например, Lonicera xylosteum).
2. С укороченными побегами, розеточные (тропические)
2*
19
3. Безлистные суккулентно-стеблевые (виды Opuntia, Euphorbia).
bi Эпифиты (виды Plttosporum).
с) Паразиты (из Loranthaceae).
Ill, Кустарнички.
а) Хтонофиты.
1. Неподвижные, без укореняющихся побегов.
а) Прямостоячие: 1) аэроксильные (виды Epacris, Leucopogon);
2) геоксильные (Catectasia суапеа и др.).
?) Полупростратные: 1) аэроксильные (например. Epacris alpina):
2) геоксильные.
2. Подвижные, с укореняющимися побегами,
а) Ползучие посредством надземных побегов:
1) Полуподвиэюные, с сохраняющимся главным корнем.
(Cassiope tetragona, Phyllodoce coerulea и др.),
2) Настоящие ползучие, с отмирающим главным корнем
(Empetrum nigrum, Linnaea borealis и др.). Здесь же имеются некоторые
розеточные и суккулентные,
₽) Ползучие посредством подземных побегов (Vaccinium vitis idaea, V
myrtillus и др.).
b) Эпифиты.
с) Паразиты (Viscum album).
IV. Древесные подушковидные растения с переходными
формами к ползучим кустарничкам и травянистым подушкам.
V. Древесные лианы, близкие к кустарникам и ползучим кустар-
ничкам: а) лазящие посредством шиповатых или гладких побегов;
Ь) корнелазящие; с) вьющиеся; d) лианы, образующие усики,
В. Полудревесные (Hemixyles)
I. Настоящие полукустарники (Suffrutices).
а) Неподвижные* 1) прямостоячие: а) аэроксильные (Euphrasia paludosa)
и р) геоксильные (Genista tinctorial 2) полупростратные (Fumana
vulgaris, Helianthemum canum и др.).
в) Надземно-ползучие, с переходами к травам (Thymus seipylluni, Coma-
rum palustre, виды Helianthemum).
с) Подземно-ползучие (Potentilla tridentata).
II. Прутьевидные полукустарники (Virgulta)—
Pubus idaeus например.
В монографии приводится довольно подробная характери-
стика каждой группы форм с указанием ее географической
распространенности В повой экологической литературе система
Дю Рие, пожалуй, наиболее детально разработана. Но она
осталась незаконченной: в ней отсутствует классификация
травянистых растений В системе сочетается использование раз-
личных признаков физиономического и биологического порядка.
Из последних особое внимание уделяется вегетативному рас-
пространению и размножению. В тексте автор не раз говорит
об экологической и, в частности, климатической приуроченности
различных групп выделяемых жизненных форм Значительным
недостатком системы, как отмечает ряд авторов (R. S Adamson,
1939; J. Braun-Blanquet, 1951 и др), является ее сложность,
20
а также, по нашему мнению, ее статичность Хотя в системе
и указывается ряд переходов между отдельными группами,
автор не пытается делать выводы о развитии жизненных форм,
оставаясь до конца на позициях их описательного, «индуктив-
ного» изучения.
В отличие от Дю Рие, система которого охватывает растения
всего земного шара, рядом авторов были предложены системы
жизненных форм регионального характера или охватывающие
лишь отдельные типы растительности. Например, Коккейн (Coc-
kayne, 1921, 1928) при изучении растительности Новой Зеландии
выделяет 4 класса жизненных форм. 1) деревья, 2) кустарники,
3) травянистые и полудревесные растения и 4) лианы, эпифиты
и паразиты. Каждый из классов им подразделяется далее по
физиономическим признакам, высоте растений (деревья), вели-
чине листьев, листопадности или вечнозелепос-ти и т. д.
С. Scottsberg (1910, 1914), отмечая недостатки системы
жизненных форм Друде и Раункиера, полагает, что может быть
лучше сейчас совсем оставить попытки создания всеобщей
системы жизненных форм, выделяя в силу необходимости спе-
циальные типы для специальных целей.
Ряд систем жизненных форм был предложен для растений
нашей страны.
Для еловых лесов таежной зоны СССР В Н Сукачев (1928)
приводит характерные группы жизненных форм: теневыносли-
вые микотрофные хвойные (ель, пихта); длиннокорневищные
микотрофные травянистые растения теневого типа; длинно-
корневищные травянистые микотрофы с листьями брусничного
типа; злаковидные зимнезеленые растения теневого типа;
теневые травы с клейстогамными цветками, вечнозеленые
иглолистные микотрофные травы; зимнезеленые микотрофные
кустарнички; летнезеленые кустарнички; полусапрофиты; сапро-
фиты, полупаразиты.
Перечисленные В. Н. Сукачевым группы растений отражают
разнообразие форм и путей приспособления растений к усло-
виям произрастания в тенистых еловых лесах. Здесь учтены
кроме признаков физиономического характера, способ питания,
сезонность развития листвы, наличие или отсутствие корневищ
и др
Большое внимание в отечественной литературе уделялось
изучению жизненных форм степных и пустынных растений
Б. А. Келлер (1923—26) приводит для степей, пустынь и полу-
пустынь- 1) деревья, 2) кустарники; 3) полукустарники и полу-
кустарнички, 4) многолетние травы, двудольного и однодоль-
ного типа, 5) летние и осенние однолетники; 6) весенние и
осенне-весенние эфемеры; 7) низшие растения на почве.
21
Перечисленные группы Келлера остаются слишком обобщен-
ными, что особенно относится к многолетним травам, составляю-
щим основу степного травостоя. В 1933 г. Б. А. Келлер не-
сколько видоизменил систему, выделяя в степях, полупустынях
и пустынях СССР: 1) многолетние двудольные травы; 2) много-
летние рыхло-кустовые и корневищные травы; 3) многолетние
дерновинные степные злаки с узкими щетинистыми листьями;
4) карликовые полукустарнички, обыкновенно сильно опушён-
ные; 5) весенние эфемеры; 6) низшие растения на почве.
Здесь, как видим, группа травянистых многолетних растений
дифференцируется значительно глубже.
Более детальная система жизненных форм для степной рас-
тительности разработана В В. Алехиным (1936). Автор выде-
ляет:
А. Кустарники и полукустарники.
I. Степные кустарники.
II. Полукустарники.
В. Травянистые двулетники и многолетники.
1П. Растения с листьями по всей длине стебля. Стебель вертикальный,
прочный.
1 — высокие растения верхнего степного яруса;
2 — растения средних и более низких ярусов.
IV. То же, но стебель слабый, приподнимающийся или цепляющийся.
V. Растения с листьями, сближенными у поверхности земли в розетки.
VI. Растения, образующие перекати-поле.
VII. Растения луковичные и клубневые эфемеры.
VIII. Растения луковичные летнего цикла.
IX. Дерновинные злаковидные растения с узкими листьями—злаки иосоки.
X. Недерновинные злаковидные растения с более широкими листьями.
С, Однолетники.
XI. Весенние эфемеры.
XII. Летне-осенние однолетники.
XIII. Мхи.
XIV. Лишайники.
XV. Водоросли.
Автор считает желательным проведение и дальнейших под-
разделений групп, например по способам защиты от излишнего
испарения, различиям фенологического цикла.
В С. Закржевским и Е. П. Коровиным (1935) дается пере-
чень основных экологических (жизненных) форм растений для
глинистых пустынь Бетпак-Дала. Авторы различают:
I. Группу склероморфных кустарников.
II. Группу ксерофитных полукустарников:
1) форму афильную или с редуцированными листьями; 2) форму с эфе-
22
мерными листьями; 3) с суккулентными листьями; 4) с склероморфными
листьями; 5) с плоскими листьями.
III. Группу травянистых суккулентных форм.
IV. Группу эфемерных трав: 1) эфемеры; 2) эфемероиды.
Коровин и Закржевский отмечают, что каждая из назван-
ных форм обладает своими биологическими особенностями,
отличаясь ритмом развития, динамикой роста и т. д. В то же
время они обнаружили и определенную экологическую приуро-
ченность выделенных групп растений к различным условиям
в пределах Бетпак-Дала. В описаниях отдельных растений в
тексте вскрываются чрезвычайно интересные биологические осо-
бенности, к сожалению, не нашедшие отражения в характери-
стике и названиях групп
Весьма оригинальная и стройная система жизненных форм
для пустынной области Средней Азии была предложена
А В Прозоровским (1936). Среди высших цветковых растений
автор выделяет пять основных типов жизненных форм-
I тип. Древесные растения, все стеблевые побеги которых много-
летние и древеснеющне.
Подтипы: 1) деревья, 2) кустарники, 3) кустарнички.
И тип. Полудревесные растения, часть стеблевых побегов ко-
торых однолетняя и ежегодно опадающая, а другая часть много-
летняя, древеснеющая.
Подтипы: 1) полудеревья, 2) полукустарники и 3) полукустарнички.
Ill тип. Пол у травянистые растения, обладающие однолетними
древеснеющими надземными побегами и многолетними деревяни-
стыми подземными.
IV тип. Травянистые растен'ия (многолетние), обладающие одно-
летними травянистыми надземными побегами и многолетними тра-
вянистыми подземными.
V тип. Эфемерные растения — однолетники.
В 1940 г. автор дополняет систему тремя новыми типами,
выделяя: VI — мхи, VII—слоевищные растения и VIII — одно-
клеточные Среди первых пяти групп автор делит растения на
самостоятельно-поддерживающие свои наземные побеги и тре-
бующие поддержки — лианы.
Более детальное подразделение, по мысли автора, должно
пойти по линии учета длительности жизни ассимилирующих
частей растений и их экологической принадлежности (мезофиты,
ксерофиты, галофиты и т д)
Система Прозоровского подкупает своей последователь-
ностью и логичностью. Расположение типов в системе в значи-
тельной степени соответствует этапам эволюционного развитии
жизненных форм Ее недостатком является слабая разработан-
ность последних двух типов, отсутствие подразделения на более
мелкие группы у полутравянистых и травянистых растений
Для зональной растительности СССР в целом и ее
23
характеристики Б. А. Келлер (1938) рекомендует выделять
следующие группы жизненных форм:
1. Мхи и лишайники.
2. Вечнозеленые карликовые кустарнички брусничного и верескового типов.
3. Летнезеленые карликовые кустарнички и полукустарнички.
4. Многолетние травы — осоки и злаки с чертами ксероморфии,
5. Хвойные деревья.
6. Летнезеленые лиственные деревья, широколиственные и мелколиственные.
7. Весенние эфемеры.
8. Карликовые полукустарнички.
9. Летние однолетники.
Эти группы жизненных форм, по мнению автора, закономер-
но сменяются в растительном покрове СССР при движении от
тундр к пустыням. Кроме перечисленных, Келлер выделяет
и некоторые другие формы, например, лесные многолетние
травы, степные длительно вегетирующие злаковидные и дву-
дольные травы и др Этот список представляет собой лишь
перечисление наиболее распространенных жизненных форм
зональной растительности. Системы жизненных форм растений
здесь еще нет.
Попытка создания такой системы была сделана В. В. Алехи-
ным (1944) В равнинах Европейской и Азиатской частей СССР
В. В Алехин различает следующие жизненные формы:
А. Жизненные формы с деревенеющими стеблями: 1) деревья, 2) кустар-
ники, 3) кустарнички.
Б. Жизненные формы переходного типа между деревянистыми и травяни-
стыми, или имеющие представителей обоих типов: 1) полукустарники,
2) растения-подушки, 3) суккуленты, 4) вечнозеленые травы, 5) зимне-
зеленые травы, 6) лианы.
В. Жизненные формы с травянистыми стеблями: I. Наземные—1) высокие
двудольные травы; 2) травы средней высоты; 3) злаковидные травы;
4) ситниковидные травы; 5) папоротниковидные травы; 6) розеточные
травы; 7) перекати-поле.
II. Эпифитные.
III. Водные—1) плавающие; 2) погруженные
Г. Жизненные формы мхов и низших растений.
Формы: 1) кукушкина льна, 2) гипновых мхов, 3) сфагновых мхов,
4) кустистых лишайников, 5) эпилиты.
В своих главных подразделениях система В В Алехина от-
личается определенной направленностью и в известной степени
сходна с системой А. В. Прозоровского. Но в пределах некото-
рых групп эта классификация не может быть признана удачной.
Как справедливо отмечает М В. Сенянинова-Корчагина (1949),
лианы не могут быть отнесены к переходным формам между
деревянистыми и травянистыми, так как среди них есть те и
другие. То же самое относится к подушкам и суккулентам
(см., например, системы Друде, 1913, и Дю Рие, 1931). Неясно
24
также, почему вечно- и зимнезеленые травы должны считаться
промежуточными по отношению к древесным. Малоэкологично
подразделение травянистых наземных растений, где не чувст-
вуется ведущий принцип классификации.
Как видно из только что изложенного, среди сторонников
эколого-физиономического направления классификации наблю-
даются весьма значительные различия в подходе к выделению
отдельных жизненных форм, их объединению в группы и к ис-
пользованию определенной терминологии. Это вполне естествен-
но и попятно, учитывая громадное разнообразие жизненных
форм растений в окружающем растительном мире. Вместе
с тем нельзя не заметить, что у многих авторов, особенно XX в.,
наблюдается известная повторяемость при выделении крупных
и даже более мелких групп жизненных форм. Наибольший раз-
мах, амплитуда различий в подходах к выделению и группи-
ровке жизненных форм, вероятно, пройдены.’ Начался новый
этап, синтетический и обобщающий, в создании таких систем
жизненных форм, каждая из которых будет приниматься все
большим кругом исследователей.
4. МОРФОЛОГО-БИОЛОГИЧЕСКОЕ НАПРАВЛЕНИЕ
В КЛАССИФИКАЦИИ ЖИЗНЕННЫХ ФОРМ РАСТЕНИИ
Жизненные формы, как выражение приспособленности расте-
ний к господствующим условиям их произрастания, характери-
зуются не только габитуально-физиономическими признаками,
по и рядом биологических свойств, таких как длительность
жизни, ритм развития, способ питания, способы вегетативного
размножения и пр Поэтому естественно стремление ботаников
использовать эти признаки в классификации жизненных форм,
создать «биологические системы растений», отличающиеся не
только от таксономических, ио и от физиономических.
Конечно, все биологические свойства высших растений
проявляются так или иначе в структуре и внешней форме орга-
низмов, т. е снова оказываются связанными с физиономиче-
скими особенностями растений И в настоящее время нам уже
не кажутся столь категоричными различия этих двух типов
признаков Однако в истории науки эти два направления сло-
жились как самостоятельные, особенно на первых этапах их
развития, и отдельное изучение их вполне целесообразно.
Вероятно, первой попыткой биолого-морфологической класси-
фикации растений была система, предложенная А П Де Кан-
доллем (А Р. De Candolle, 1818) Все растения Де Кандоллсм
были подразделены на 8 групп, в зависимости от длительности
их жизни, повторяемости плодоношения и структуры побегов:
25
1. Plantae monocarpicae annuae — однолетние монокарпиче-
ские растения
2. Plantae monocarpicae biennes — двулетние монокарпики.
3 Plantae monocarpicae perennes—многолетние монокар-
пики.
4. Plantae rhizocarpicae — растения с ежегодно плодонося-
щими и отмирающими побегами, но в целом длительно сохра-
няющиеся.
5. Plantae caulocarpicae suffrutescentes полукустарники высо-
той в 30—60 см (1—2 фута).
6. Plantae caulocarpicae frutescentes — кустарники высотой
в 60 см — 3 м (2—10 футов).
7 Plantae caulocarpicae arbusculiformes — деревца 3 м —
7,5 м (10—25 футов) высоты.
8. Plantae caulocarpicae arborescentes — деревья высотой
более 7,5 м (25 футов).
Позже (1832) Де Кандолль группы «rhizocarpicae» и «caulo-
carpicae» в свою очередь объединил в единую группу «polycar-
picae», т. е многократно плодоносящих растений. Эти подразде-
ления Де Кандолля сохранились в ботанике и до настоящего
времени и используются в исследованиях жизненных форм
с большей или меньшей полнотой
Большое влияние на развитие биолого-морфологического
направления в изучении жизненных форм оказали многочислен-
ные исследования Т. Ирмиша (Th. Irmisch, 1849, 1851, 1864) и
отчасти А. Брауна (А. Braun, 1849, 1852). Биологические очерки
Ирмиша по истории развития многих травянистых лесных рас-
тений отличаются исключительной точностью и не утратили
своего значения и до настоящего времени В одной из статей
по истории развития травянистых бобовых Ирмиш (1859) кроме
описания, предложил также биологическую классификацию
клеверов, основанную на морфологии побегов и способах пере-
зимовки растений В сокращенном виде эта классификация
такова:
I. Растения, зимующие с надземными побегами
с зелеными листьями.
А. С укороченной главной осью в форме центральной розетки:
а) в пазухах зеленых листьев главной оси развиваются ассимилирующие
побеги, несущие соцветия (растения 4-осные);
а Цветоносные ассимилирующие побеги лежачие, растет плагиотропно
(например, Trifolium repens).
р Цветоносные ассимилирующие побеги не лежачие, с ограниченным
ортотропным ростом (например, Tri folium pratense).
b) В пазухах зеленых листьев главной оси развиваются побеги с терми-
нальным соцветием (растения 3-осные, например, Trifolium alpestre).
В. С удлиненной главной осью, несущей соцветия:
а) главная ось и ее дочерние ветви зимуют в зеленом состоянии (на-
пример, Trifolium fragiferum)-,
26
Ъ) главная ось не лежачая и не зимующая (например, Trifolium
nybridum).
II. Растения, зимующие с подземными побегами,
несущими чешуевидные листья.
А. Подземные побеги корневищевидные (например, Trifolium medium).
Б. Побеги располагаются плотно на материнской оси (Trifolium rubens,
Т. pannonicumj.
Автор отмечает, что различия в структуре побегов и спосо-
бах перезимовки растений имеют большое значение для ботани-
ческой географии и хозяйства, так как определяют характер
растительного покрова страны В то же время для систематики
эти особенности имеют подчиненное значение, ибо она базирует-
ся на признаках структуры цветка.
Как отмечалось выше, решающее значение в развитии уче-
ния о жизненных формах имела победа эволюционной теории
Ч. Дарвина (1859). В сущности лишь под влиянием дарвинизма
жизненные формы у растений стали рассматриваться как формы
приспособления растения к среде. Да и сам термин «жизненные
формы» («Lebensformcn» и «Life-forms») был предложен одним
из основателей этого учения Е. Вармингом (Е. Warming) лишь
в 1884 г., т е после утверждения эволюционного учения Дар-
вина. Под жизненной формой Варминг понимал «форму, в кото-
рой вегетативное тело растения (индивида) находится в гармо-
нии с внешней средой в течение всей его жизни, от колыбели
до гроба, от семени до отмирания» (1884). В этой же статье
Вармингом приводится детально разработанная система жиз-
ненных форм с подразделением растений по длительности
жизни, циклу развития побегов, способности к вегетативному
размножению, структуре корневой системы и другим биолого-
морфологическим признакам. Особое значение Варминг придает
активности расселения растений. Отсюда становится понятным
большое внимание, которое он уделяет в своей системе подзем-
ным органам Цель Вармипга — не выражение физиономии
растительности, как у Гумбольдта, Гризебаха, Хульта, а отра-
жение эфармонических, т е приспособительных черт этой
растительности Автором выделяются.
I главная группа. Гапаксантные растения (монокарпические по Де
Кандоллю)
А — Моноциклические (однолетние); побеги «моноцикличны» .... Гр 1.
В — Дициклические (двулетние); побеги «дицикличны»........Гр. 2.
С — Плейо-полициклические (многолетние); побеги «три-полицик-
личны»................................................Гр. 3.
II главная группа. Многолетние поликарпические растения
А —Растения без или со слабой способностью к пе-
редвижению.
27
1. В течение всей жизни сохраняется главный корень или стеблевой клу-
бень. Размножение только семенами.
а) Побеги одревесневшие, длительно-живущие. Обычные двудольные
деревья и кустарники.........................................Гр. 4.
Ь) Травянистые растения.
х) с «многоглавым корнем»....................................Гр. 5.
а Побеги ограниченные моноциклические (например, Lotus, Agrimo-
nia и др.).
р Побеги ограниченные дициклические (Taraxacum, Hypochaeris).
7 Ограничены только боковые побеги (Plantago media, Trifolium pratense,
T. montanum и др.).
хх) С многолетними клубнями..................................Гр. 6.
а Клубнеобразен главный корень.
аа Побеги ограничены (Bryonia и др.).
РР Главная ось не ограничена (Rhodiola rosea и др.).
р Клубнеобразен гипокотиль.
аа Побеги ограничены (Eranthis hiemalis и др.).
рр Главная ось не ограничена (Corydalis cava и др.).
7 Клубнеобразен эпикотиль (Tamus и др.).
2. Главный корень быстро (через 1—3 года) отмирает, и за ним отми-
рает главный стебель и последующие побеги. Способны к вегетативному
размножению.
а) Из оснований побегов, живущих более одного астрономического
года, возникают вертикальные или наклонные ризомы системы оснований
побегов) ................-...................................Гр. 7.
аа) Формы побегов обычные
а Криптогамы (например, Aspidium filix mas).
р Фанерогамы с ограниченными моноциклическими побегами (напри-
мер Cynanchum vincetoxicum, Hieracium umbellatum).
7 Фанерогамы с ограниченными дицикличсскими побегами (например,
Leontodon autumnalis, Spiraea, Filipendula, Cardamine).
5 Фанерогамы с неограниченным главным побегом и ограниченными
боковыми цветоносными (например. Succisa pratensis, Geum rivale).
bb) Побеги клубневидные (подразделения, как и в аа).
сс) Побеги луковицеобразующие (подразделения как и в аа).
Ь) Сохраняющихся систем оснований побегов (ризомов) не образуется,
так как моноциклические побеги ежегодно полностью отми-
рают ....................................................Гр. 8.
аа) Побеги омоложения перезимовывают как мелкие укоренившиеся ко-
роткочленные листоносные побеги (например, Samolus Valerandi).
bb) Побеги омоложения перезимовывают как замкнутые почки с запа-
сающими корнями (например, Orchis). Переходная форма аа — bb —
Anthriscus silvestris
В. Виды, обладающие часто значительной способ-
ностью к передвижению.
1. Передвигающиеся наземно.
а) С длительно сохраняющимся главным корнем. Некоторые, например,
Calluna Arctostaphylos, Emvetrum, с одревесневшими побегами. Гр. 9.
b) С быстро отмирающим главным корнем...................Гр. 10.
аа) Криптогамы (например Polypodium vulgare).
bb) Фанерогамы.
а Собственно столонов (Auslaufer) не образует.
х) Побеги моноцикличны (например, Asarum europaeum).
хх) Побеги моно- или слабо дицикличны, но живут 2 и более лет (на-
пример, Comarum palustre, Menyanthes, Iris).
xxx) Побеги дицикличны; живут побеги 2 и более лет (например, Anten-
naria dioica, Hieracium pilosella).
28
р Кроме укорененных побегов или их частей образуются столоны
(Auslaufer).
х) Все побеги ограниченные (например, Saxifraga flagellaris, Ranuncu-
lus repens. Fragaria)
xx) Главная ось вертикальная укорененная, столоны ограничены соцве-
тием (например, виды Potentllla).
ххх) Вертикальные плодущие побеги ограничены. Ползучие побеги не-
ограничены (например, виды Lycopodium, Linnaea borealis, Veronica
officinalis).
2. Передвигающиеся подземно. Главный корень обычно быстро отмирает.
а) Передвижение посредством горизонтально растущих побегов, обра-
зующих систему оснований побегов................................Гр. 11
аа) Оси все подземные, несущие зеленые листья (например, Pieris
aquilina).
bb) Побеги с подземными ползучими частями и отвесными надземными;
все ограничены.
а Надземные части однолетние (моноцикличны),
аа Ветвление подземных частей побегов сильное и не регулярное (на-
пример, Equisetum, Phragmites communis и др. злаки, Urtica dioica).
РР Ветвление подземных частей побегов слабое, ло регулярное посред-
ством мощных пазушных почек на определенных местах симподия.
х) Ежегодно образуется лишь одна генерация подземных побегов (на-
пример, Polygonatum, Anemone nemorosa, Epipactis).
xx) Число ежегодных генераций подземных побегов более одной (с од-
ним междоузлием — Hippuris vulgaris, с двумя — виды Potamogeton,
Juncos, с несколькими — виды Scirpus).
р Подземные части двух-многолетни.
аа Побеги дицикличны и травянисты; подземное ветвление нерегуляр-
ное (например, Tussilago, Achillea, Aegopodium', переходное к рр~ Pirola
rotundifolia).
РЗ Побеги дицикличны и одревесневшие (например, Vactinium myrtillus,
V. vitis idaea).
cc) Побеги двух типов — подземные ползучие неограничены; надземные
ограничены (например, Adoxa, Paris)
b) Передвижение благодаря горизонтальным подземным побегам, быстро
отмирающим и поэтому не образующим систему оснований побегов. Над-
земные части побегов однолетние или более длительно живущие . Гр 12
х) По отмирании материнского растения новые подземные побеги пе-
резимовывают целиком, отмирая в следующем году вместе с надземными
(например, виды Mentha, Lycopus europaeus, Oxalis stricta).
xx) Перезимовывают только специальные верхушечные части подземных
побегов, отмирая при образовании надземных (например, Solanwn tubero-
sum, Trientalis europaea, Stachys palustus, Epilobium palustre).
с) Корнеотпрысковые растения, перезимовывающие, размножающиеся
и передвигающиеся благодаря побегообразующим корням.............Гр. 13.
х) Размножение только корневыми почками, побеги не ветвятся (напри-
мер, Pirola uniflora).
хх) Размножение преимущественно побегами на корнях, ветвление сла-
бое или для передвижения несущественное (например, Linaria vul-
garis, Epilobium angustlfolium, Rumex acetosella, Convolvulus arvensis).
3. Плавающие водные растения.................................Гр. 14.
а) Розеточные формы (например, Siratiotes, Hydrocharis, Lemna).
b) Горизонтально лен^ащие с удлиненными побегами.
аа) Без особых зимующих почек (например, Hottonia, Ceratophyllum).
bb) С таковыми (например, Myriophyllum, Utricularia).
29
Как видно из приведенного, система Варминга отличается
весьма большой детальностью и разработанностью. Она, пожа-
луй, наиболее полно охватывает биологические типы травяни-
стых растений лесной зоны Европы. Недостатком системы
является отсутствие единого ведущего принципа классифика-
ции, что невольно вызывает впечатление ее незаконченности,
несмотря на значительную сложность. Варминг не аргумен-
тирует выделение различных биолого-морфологических призна-
ков как признаков различного ранга. Поэтому соподчинение
групп в системе кажется произвольным В сущности это морфо-
лого-таксономическая система. Детализируя схему Варминга,
мы дойдем до видов, создав ключ к определению растений по
вегетативным признакам Как отмечает сам автор (Warming,
1908), эта система «скорее морфологическая, чем биологиче-
ская».
С иных позиций подошел к биологическому подразделению
растений шведский ботапик Аршуг (F W. Areschoug, 1896).
В зависимости от положения почек возобновления и способов
перезимовки все растения подразделяются им на аэрофиты
(Aerophyten) и геофиты (Geophyten). «Геофильные растения»
характеризуются тем, что почки возобновления их заклады-
ваются под землей, там же протекают и основные этапы форми-
рования воздушных побегов. К ним относятся, например, многие
лесные травянистые многолетники По способу перезимовки
среди травянистых растений им выделяются пять групп.
1. Rasenperennen— с зимующими побегами, образующими дер-
нину; 2 Brutknospenperennen — с зимующими придаточными
почками; 3. Stengelbasisperennen — с зимующими основаниями
побегов, 4. Rosettenperennen — с зимующими розетками и
5. Rhizomperennen — с зимующими корневищами
В начале XX в появляется серия работ К Раункиера
(С Raunkiaer, 1903, 1905, 1907 и др.), определивших на ряд
десятилетий общее развитие учения о жизненных формах Им
была создана система жизненных форм наземных сосудистых
растений, резко отличающаяся от предыдущих своей простотой,
ясностью и законченностью. Он с самого начала отказался от
выделения главных групп жизненных форм с различных эколо-
гических точек зрения. Принцип подразделения по Раункиеру,
должен быть один, но большого приспособительного значения;
выделяемый признак должен быть легко доступен исследова-
телю В основу подразделения жизненных форм Раункиер по-
ложил лишь одно — различия в приспособлении растений к
переживанию неблагоприятного времени года Но и здесь из
всего комплекса адаптивных признаков им выбран также лишь
один — положение почек или верхушек побе-
гов в течение неблагоприятного времени года
30
ПО отношению к поверхности почвы. Все расте-
ния, в соответствии с этим принципом, были подразделены
вначале (1903) на пять типов.
I. фанерофиты —с почками или верхушками побегов,
отрицательно геотропичными и расположенными в течение
неблагоприятного времени года более или менее высоко в воз-
духе (подразделяются на четыре подтипа).
П. Хамефиты — с почками или верхушками побегов,
расположенными в этот же период близ поверхности почвы
(также подразделяются на четыре подтипа).
III. Г е м и к р и п т о ф и т ы — с почками или верхушками
побегов, расположенными непосредственно на поверхности
почвы (подразделяются на три подтипа).
IV Криптофиты — почки или верхушки побегов сохра-
няются под землей на разной глубине у разных видов (три под-
типа).
V. Однолетники или терофиты — растения бла-
гоприятного времени года
Понятно, что степень приспособленности растений к пере-
живанию неблагоприятного времени года возрастает от пер-
вого к четвертому типу.
В 1907 г. эта система была значительно детализирована.
Фанерофиты были подразделены на 15 подтипов — по высоте
растений (мегафанерофиты — выше 30 м, мезофаперофиты 8—
30 м, микрофанерофиты 2—8 м. нанофанерофиты — ниже 2 лг),
по ритму развития листвы (вечнозеленые и листопадные), по
степени защищенности почек (с чешуями и без защитных че-
шуй), по консистенции стебля (кроме древесных, выделены
травянистые и суккулентные фанерофиты). Более детальные
подразделения предложены и для других типов В общем виде
система Раункиера такова.
I. Фанерофиты (Phanerophytes) — почки возобновления выше 30 см
над уровнем почвы:
1. Травянистые фанерофиты.
2. Вечнозеленые мегафанерофиты без почечных чешуи.
3. Вечнозеленые мезофанерофиты без почечных чешуи.
4. Вечнозеленые микрофанерофиты без почечных чешуй
5. Вечнозеленые нанофанерофиты без почечных чешуй.
6. Элифитные фанерофиты,
7. Вечнозеленые мегафанерофиты с почечными чешуями,
8. Вечнозеленые мезофанерофиты с почечными чешуями
9 Вечнозеленые микрофанерофиты с почечными чешуями.
10. Вечнозеленые нанофанерофиты с почечными чешуями.
11. Суккулентно-стеблевые фанерофиты.
12. Листопадные мегафанерофиты с почечными чешуями.
13. Листопадные мезофанерофиты с почечными чешуями.
14. Листопадные микрофанерофиты с почечными чешуями
15. Листопадные нанофанерофиты с почечными чешуями.
31
II. Хамефиты (Chamaephytes) — почки возобновления на поверхности
почвы, или не выше 20—30 см-.
16. Полукустарники (Suffruticose Ch.). Воздушные побеги отрицательно
геотропичны, в конце вегетативного периода отмирают до нижнем части их,
где сохраняются почки возобновления.
17. Пассивные хамефиты (Passive Ch ). Воздушные побеги отрицательно
геотропичны, но полегают от собственной тяжести, верхушка приподнимаю-
щаяся (вечнозеленые и листопадные).
18. Активные хамефиты (Active Ch.). Побеги трансверзально геотропич-
ны и плагиотропны (вечнозеленые и листопадные).
19. Подушковидные растения (Cushion plants).
III. Гемикриптофиты (Hemikryptophytes) — почки возобновления в по-
верхностном слое почвы, покрытые часто подстилкой.
20. Прото-гемикриптофиты (Proto-Hemikryptophytes)— без розеток листьев
в основании надземных побегов: А — без столонов, В — со столонами.
21. Полурозеточные растения (Partial rosette plants)— с розеткой у осно-
вания побега и облиственным стеблем: А — без столонов, В — со столонами
22. Розеточные растения (Rosette plants) — с базальной розеткой листьев
и безлистным или почти безлистным стеблем.
х) Симподнальные: А —без столонов, В — со столонами.
хх) Моноподиальные.
А — Моноподиальная ось несет только зеленые листья, не образуя че-
шуевидных:
а) Воздушные боковые побеги с зелеными листьями.
в) Воздушные боковые побеги без зеленых листьев:
О — без столонов, 00 — со столонами.
В — Моноподиальная ось образует н чешуевидные и зеленые листья:
а) без столонов, в) — со столонами.
С — Моноподиальная ось несет только чешуевидные листья.
IV. Криптофиты (Kryptophytes) — почки возобновления в неблагоприят-
ное время скрыты в почве или под водой.
х) Геофиты (Geophytes) — почки возобновления в почве.
23. Корневищные геофиты (Rhizome geophytes/
24 Стебле-клубневые геофиты (Stem-tuber geophytes).
25. Корне-клубневые геофиты (Root-tuber geophytes).
26. Луковичные геофиты (Bulbose geophytes).
27. Корневые геофнты (Root geophytes).
хх) Гелофиты и гидрофиты (Helophytes a. Hydrophytes).
28. Гелофиты — вегетативные побеги обычно воздушные.
29. Гидрофиты — вегетативные побеги погружены в воду.
V. Терофиты (Therophytes) — возобновление после неблагоприятного
времени года только семенами.
В своих главных подразделениях (на типы) система Рауп-
киера скорее может быть названа эколого-физиономической.
Но классификация в пределах каждого типа, особенно начиная
с хамефитов, базируется на признаках морфолого-биологиче-
ского характера. Автор и сам признает, что его система осно-
вывается в значительной степени на биолого-морфологических
исследованиях Ирмиша и Варминга
Жизненные формы Раункиер рассматривал как итог и ре-
зультат приспособления растений к климатическим условиям
страны, возникшие в процессе исторического развития. В связи
32
с этим жизненные формы, свойственные той или иной стране,
могут служить индикатором климата, а само учение о жизнен-
ные формах, по Раункиеру, является основой ботанической
1еографии Он выделяет как самостоятельную область зна-
ния _ биологическую географию растений, исследующую отно-
шения между растительностью и климатом, в целях получения
фито-биологического показателя климата той или иной страны.
Мысль о связи господствующих- жизненных форм и климата
сама по себе не нова, так как в общей форме она высказыва-
лась еще Гумбольдтом (1806), и особенно Кернером (1869) и
Гризебахом (1872) (см. выше). Но Раункиер впервые в эколо-
гии и ботанической географии применил статистический метод
при изучении закономерностей распределения жизненных форм
в зависимости от климата. С этой целью все виды той или иной
страны он распределял по 10 группам жизненных форм — стеб-
левых суккулентов (S), эпифитов (Е), мега- и мезофанерофитов
(ММ), микрофанерофитов (М), нанофанерофитов (N), хаме-
фиюв (Ch), гемикриптофитов (Н), геофитов (G), гело- и гидро-
фитов (НН) и терофитов (Th). Процентное распределение ви-
дов изучаемой территории по этим десяти группам жизненных
форм автор назвал биологическим спектром, или фито-климати-
ческим спектром страны.
Сравнение биологических спектров различных стран — от
тропиков до приполярий — обнаруживает в них весьма резкие
различия, закономерно изменяющиеся Но как эталон для срав-
нения, автор строит так называемый «нормальный спектр»,
отражающий распределение видов растений всей земли по
указанным выше группам жизненных форм. Такой нормальный
спектр был им построен вначале для 400, а затем 1000 видов
растений, произвольно взятых из «Index kewensis» (1907,
1918).
В течение двадцати с лишним лет Раункиер провел гро-
мадную работу по изучению биологических спектров различных
областей земного шара с различным растительным покровом —
от влажных тропических лесов до тропических и впетропиче-
ских пустынь, арктических тундр и альпийских лугов. Итоги
этих работ широко известны и вряд ли стоит о них детально
говорить здесь Они действительно выявили «климат фанеро-
фитов» (влажные тропические области), «климат гемикрипто-
фитов» (умеренно-холодные области), «климат хамефитов»
(полярные страны) и т д. Раункиер считает, и это весьма с\-
щественно, что сходства и различия биологических спектров
определяются климатом и не зависят от флористических осо-
бенностей той или иной страны.
В отличие от предшествующих система Раункиера нашла
широкое применение в ботанико-географических и экологиче-
В-113. и Г. Серебряков —3 33
ских работах различных авторов. Но в то же время основные
положения классификации Раункиера и ее ботанико-географи-
ческие следствия подверглись весьма резкой критике.
Серьезные возражения против взглядов Раункиера были
выдвинуты Вармингом (Warming, 1908) По Вармингу, соотно-
шения различных жизненных форм в той или иной стране
определяются не только климатом, но и характером почв.
Поэтому он скептически оценивает значение жизненных форм
как показателей климата Зависимость жизненных форм от
почв особенно очевидна, например, в кампосах (саваннах)
Бразилии, где при сравнительно одинаковом климате на лес-
ных и степных территориях наблюдаются совершенно иные на-
боры жизненных форм1 Единственное отличие этих террито-
рий— в характере почв. В отличие от Раункиера, Варминг
считает, что для характеристики естественных условий важно
не соотношение видов, а соотношение индивидов каждого вида.
Виды доминирующие лучше говорят о климате местности, чем
многие, по редко встречающиеся растения Бореалыгые области
Сибири и влажные тропики Южной Америки одинаково отно-
сятся к климату деревьев Однако процентные соотношения
жизненных форм там и здесь весьма различны. Небольшое
число видов хвойных деревьев в Сибири и богатство ими до-
лины Амазонки, по Вармингу, определяется не только клима-
том, по и историческими факторами (ледниковый период и
т д). Словом, флора страны зависит нс только от ее климата,
но и истории, а между тем исторический фактор игнорируется
в сопоставлениях Раункиера.
Наконец, в подсчетах Раункиера совершенно не учитывают-
ся изменения состава жизненных форм, вносимые в раститель-
ный покров человеком. А между тем многие виды, например, во
флоре Дании, являются иммигрантами после нарушения се
естественного растительного покрова.
В ответ на критические замечания Варминга Раункиер
(1934) пишет, что для характеристики климата той или иной
области в целом необходимо брать растения различных почв и
фитоцепозов Лишь обобщенные спектры различных формаций
позволяют характеризовать фитоклимат той или иной области.
Человек действительно может изменить соотношение домини-
рующих видов в растительном покрове и, таким образом, изме-
нить характер растительности. Но флористический состав при
этом остается более или менее постоянным и процентное соот-
ношение жизненных форм практически не изменяется. В то же
время натурализующиеся виды, например, европейские в Се-
верной Америке, по соотношению жизненных форм не отлича-
ются от аборигенных. Что касается исторического фактора, то
исследования автора показали, что различия в геологической
34
истории страны с одинаковым климатом не отражаются на
соотношении жизненных форм.
Серьезные критические замечания в адрес системы Раункие-
ра были сделаны Скоттсбергом (Scottsberg, 1912, 1913, 1914).
Он впервые отмечает гетерогенный характер типа хамефитов,
отдельные подтипы которых несут черты приспособления к раз-
личным климатическим условиям. Различия между хамефитами
и гемикриптофитами на Фолклендских островах часто носяг
морфологический, но не экологический характер, так как
осенью у гемикриптофитов развиваются облиственные побеги,
которые и зимуют без особой защиты Скоттсберг (1914) счи-
тает ошибочным утверждение Раункиера о том, что абориген-
ная и интродуцированная флора относятся к одному и тому же
биологическому спектру. В то время как аборигенная флора
островов Хуан Фернандес относится к фанерофитному спектру,
интродуцированная — скорее к гемикриптофитному и терофит-
ному. Последняя, по Скоттсбергу, более соответствует тепереш-
нему климату, чем аборигенная, обусловленная историческими
факторами
Аллен (Allan, 1927) отмечает, что биологический спектр
растений Новой Зеландии, построенный по системе Раункиера,
не отражает действительных климатических условий. На
Южном и Северном островах, изученных автором, спектры
оказываются сходными, несмотря на значительные различия
климата Это несовпадение биологических спектров и климата
Аллен объясняет тем, что классы жизненных форм и, в част-
ности, хамефиты, слишком обширны и включают растения с
разным отношением к климату. Имеет значение и историче-
ский фактор, т. с. происхождение флоры
На слишком большую обширность класса хамефитов ука-
зывает Хагеруп (О Hagerup, 1930), говоря о том, что этот
класс жизненных форм свойственен не только арктическому
климату, но и полупустынному Гетерогенен он и морфологи-
чески, так как включает растения деревянистые и травянистые.
Дю Рие (G. Е. Du Rietz, 1931) также отмечает неестествен-
ность этой группы и отделяет древесные и травянистые хаме-
фиты друг от друга. Вносит путаницу в систему жизненных
форм Раункиера, по мнению Адамсона (Adamson, 1939), и дру-
гой класс — суккуленты, образующие различные группы, отно-
сящиеся к нанофанерофитам, хамефитам, гемикриптофитам и
терофитам. Адамсон считает, что существенное значение в воз-
никновении специфики биологического спектра той или иной
страны, набора ее жизненных форм, имеет история флоры.
Исследование биологических спектров различных пустынь
Азии, Южной и Северной Африки, Америки и Австралии при-
водят этого автора к выводу о том, что при сходстве климата
35
часто обнаруживаются значительные различия в составе жиз-
ненных форм, что объясняется особенностями флоры и се про-
исхождением Это относится, в частности, к высокому проценту
гемикриптофитов в Закаспийских пустынях, окруженных сте-
пями, к высокому проценту фанерофитов в пустынях Австра-
лии, окруженных формациями древесных растений и т д. Бу-
дучи производным от основного флористического состава стра-
ны, пустыни отражают его влияние в наборе жизненных форм
Разнообразие жизненных форм в пустынных областях будет
определяться не только суровостью климата, но и временем, в
течение которого протекало заселение пустынь. Чем древнее
пустыни, тем более разнообразны жизненные формы па их тер-
ритории. Автор считает, что необходимо создавать новые систе-
мы жизненных форм с учетом не только надземных, ио и под-
земных органов и, в частности, корневых систем. Учет послед-
них особо важен в аридных областях.
Возможность использования системы Раункиера при харак-
теристике растительного покрова страны обсуждается также в
ряде работ отечественных ботаников (А Э. Жадовский, 1925;
Б. Н. Городков, 1936, 1937; М П. Петров, 1936, М. В Сеняни-
нова-Корчагипа, 1949). М. В. Сенянинова-Корчагина (1949) счи-
тает правильным замечание Варминга о необходимости учиты-
вать почвенные условия, так как они могут оказать влияние на
географическое распространение растений (например, деревьев
на севере) и, таким образом, на соотношение жизненных форм.
.Что касается исторического фактора, то его необходимо учиты-
вать, но не переоценивать, как Варминг и Скоттсберг, ибо в
процессе флорогенеза сохраняются только такие формы, кото-
рые соответствуют климату Не следует переоценивать, по Се-
няниновой-Корчагиной, и влияние человека на состав жизнен-
ных форм растений той или иной страны.
Необходимо признать, что критика воззрений Раункиера в
значительно?! мере справедлива. В настоящее время представ-
ляется, что соотношение жизненных форм любой страны зави-
сит не только -от ее современного климата, но также от ком-
плекса почвенно-литологических условий; состав жизненных
форм в то же время отражает историю страны, т. е. изменение
почвенно-климатических условий и флоры ее по крайней мере
с третичного периода; все более ощутимым становится и влия-
ние человеческой культуры
Обсуждение системы Раункиера сопровождалось одновре-
менно ее модификациями, а также разработкой новых систем,
базирующихся на иных принципах, с использованием других
структурно-биологических признаков.
В интересной статье «Биологическая особенность осота
(Cirsium arvense Scop.)» И Пачоский (1916) выдвигает ориги-
35
нальную биологическую систему, созданную под влиянием идей
Раункиера. Растения подразделяются в ней на группы в зави-
симости от степени отмирания и потери частей во время небла-
гоприятного периода года:
1. Вечнозеленые растения (все органы многолетние).
2. Деревья и кустарники с опадающими лиаьями (все органы много-
летние, за исключением листьев, которые являются однолетними).
3. Полукустарники (однолетними являются не только листья, но и вер-
хушки побегов).
4. Многолетние травянистые растения — гемикриптофиты (однолетними
являются все надземные части).
5. Многолетние травянистые растения — криптофиты (однолетними яв-
ляются не только надземные части, но и подземные до известной глубины).
6. Однолетники (включая и «многолетники», от которых к зиме оста-
ются лишь почки, заменяющие семена).
Этот ряд типов рисует переходы от полных многолетников к
однолетникам. Он близок к системе Раункиера, по в основу
его положены размеры потерь в неблагоприятный период года,
но не положение почек возобновления По Пачоскому, нельзя
в одной группе объединять вечнозеленые и листопадные формы.
С другой стороны, он объединяет многолетники, отмирающие
к зиме целиком, за исключением почек, с типичными однолет-
никами В принципе деление Пачоского во многом естественнее
и последовательнее, чем Раункиера, но оно осталось без даль-
нейшей разработки и ботанико-географического применения.
Детально разработанная система жизненных форм у расте-
ний была предложена известным австрийским ботаником
Г. Гамсом (Н Garns 1918) Излагая историю развития учения
о жизненных формах, Гаме отмечает путаницу в использовании
признаков в классификациях различных авторов Он возражает
против использования физиономических и систематических
признаков, говоря, что нужно следовать таксономии или систе-
матической, или экологической, нс путая их. Все признаки,
применяемые в разных системах жизненных форм, им подраз-
деляются на три группы:
А —чисто эфармонические (как различия в местообитании,
способ питания ит. д);
В — смешанные (органографические и фенологические) и
С — чисто конституционные — филогенетические и онтоге-
нетические В системе жизненных форм, по Гамсу, должны
быть использованы эфармонические и, частично, смешанные
признаки1 характер местообитания, способ питания, способ-
ность к передвижению и др Его система в кратком изложении
такова:
57
L Тип прикрепленных организмов, живущих в однородной среде
(Ephaptomenon);
А —Водные (1 класс) Nereiden.
а) Автотрофные (подкласс) Autonereiden; в) Гетеротрофные; а) Сапро-
фитные (подкласс) Sapronereiden; р) Паразитные (подкласс) Рагапе-
reiden; 7) Хищные (подкласс) Phagonereiden.
В — Амфибии (2 класс) Amphinereiden.
а) Автонереиды (подкласс) Autamphinereiden, в) Гетеротрофные (под-
класс) Heteramphinereiden;
С — Воздушные (3 класс) Antephaptomenon.
Дальнейшие подразделения, как в А.
Д — Более или менее погруженные в субстрат:
а) В твердый (5 класс) а) В неорганический (подкласс) Endopetren;
Р) В органический (подкласс) Endoxylen.
b) В организмы: а) Автотрофы (подкласс) Endophyten; (3) Гетеротрофы
(7 класс) Endoxen (подкласс Endosaproben, подкласс Endoparasiten).
IL Тип укореняющихся организмов, т. е. живущих в разнородной* среде
(Rhizumenon):
А — Органы переживания высоко над землей (8 класс) Phanerophyten.
а) Полностью самостоятельны (подкласс) — стволовые (древовидные,
кустарниковидные, суккулентно-стеблевые, травянисто стеблевые);
Ь) Нуждающиеся в опоре (подкласс) древесные лианы —Holzlianen;
с) Паразитирующие (подкласс) древесные паразиты — Parasiten,
В —Органы переживания невысоко над землей (9 класс)
а) Воду воспринимают преимущественно над землей (подкласс) Вгуос-
hamaephyten; в) то же —надземно и подземно (подкласс) Polster-
pflanzen — подушковидные; с) то же, преимущественно подземно (под-
класс) Euchamaephyten.
С —Органы переживания на поверхности почвы (10 класс) Hemikryptophyta.
а) Листья узкие (подкласс) Pooidea; в) Листья широкие;
а) Только розеточные (подкласс) Basiphylla; 0) Только стеблевые
(подкласс) Thyrsophylla; 7) Розеточные и стеблевые (подкласс) Basit-
hyrsophylla
Д — Органы переживания в земле (Kryptophyta):
а) Вегетативные органы постоянно в воде—11 класс HydroKryptophyta;
b) Вегетативные органы земноводные —12 класс Amphikryptophyta;
с) Вегетативные органы лишь как исключение в воде Geophyta;
а') Автотрофные; а) Покоящиеся в земле не из-за недостатка воды —
13 класс Engeophyta; ₽) Покоящиеся в земле из-за недостатка воды —
14 класс Xerogeophyta; в') Гетеротрофные—15 класс Heterogeophyta
(подкласс Saprogeophyta и подкласс Parageophyta).
HL Тип подвижных организмов (Planometion):
А —Движение преимущественно пассивно
а) В открытых водах —16 класс Plankton, я Автотрофные — подкласс
Phytoplankton; р сапробные — подкласс Saproplankton; у паразит-
ные— подкласс Paraplankton; 8 фагонты — подкласс Phagoplankton.
b) На поверхности воды—17 класс Pleuston. я Под или над поверх-
ностной пленкой — подкласс Macropleuston; 0 в поверхностной
пленке — подкласс Micropleuston.
с) Преимущественно в снегу и на снегу и льду — 18 класс Kryoplankton.
d) В почвенной воде—19 класс Edaphon
В —Движение преимущественно активное —20 класс Tacheioru
38
а) Плавающие — подкласс Neicton; b) Ползающие — подкласс Herpon;
с) Бегающие или прьпающие — подкласс Trechon; летающие — под-
класс Petomenon. »
Дальнейшие подразделения подклассов Гаме проводит по степени по-
движности, потребности в кислороде, степени ксероморфности.
Система Гамса охватывает не только растения, но и группы
животных, особенно детально — низшие филетические типы Ее
основное назначение — обзор и систематизация организмов, в
зависимости от их распространения, в целях изучения законо-
мерностей сложения сообществ. Последнее особенно подчерки-
вается автором. Система Гамса—синэкологическая, по преиму-
ществу, и для биоценологии она имеет существенное значение.
Аутэкологически и биологически она явно недостаточна, осо-
бенно для высших растений Нельзя согласиться с автором и в
его подразделении признаков и, в частности, с отрицательным
отношением к использованию в классификации жизненных
форм длительности жизни растений и их фенологических осо-
бенностей Эти признаки столь же эфармоничны, как и спосо-
бы питания. В связи с этим соединение терофитов и криптофи-
тов в одну группу биологически не обосновано. Система Гамса
представляет большой интерес с точки зрения биоценологи-
ческого подхода к классификации жизненных форм.
Оригинальная система жизненных форм была предложена
в 1915 г Г. Н. Высоцким в известном ботанико-географическом
очерке «Ергепя» Автор выделяет ряд групп растений на основе
их способов вегетативного размножения и распространения.
Среди господствующих в Ергенях многолетников он различает.
Первый отдел — многолетники, не обладающие способностью свобод-
ного вегетативного размножения.
1. Осевые растения:
а) Стержнекорневые (например, прутняк, белая полынь),
Ь) Кистекорневые (например, лютиковые).
II. Дерновые растения. Переходные к ползучим. Образуют рас-
ходящиеся круговины (например, ковыли, типчак, ромашник). Сюда же
относятся клубнекорневые (например, Ficaria ranunculoides, Filipendula he-
xapetala и др.).
Второй отдел — многолетники с активным вегетативным размноже-
нием.
111. Ползучие растения:
а) Корневищные, с подгруппами — по глубине прохождения корневищ,
их ветвлению, направлению роста конечной почки — горизонтальному
(моноподиальные) и с выходом на поверхность (симподиальные);
Ь) Корнеотпрысковые, с подгруппами — по глубине почкообразующих
корней.
Кроме этих групп отмечаются луковичные многолетники (Tulipa, Al-
lium) и клубне-луковичные (виды Crocus, Gladiolus и др.).
Третий отдел — многолетники с надземными луковицами и пазушными
клубнями (виды Gagea, Ficaria, Dentaria bulbifera и др.).
39
Четвертый отдел — малолетников, куда входят двулетники, озимые
и яровые однолетники.
Пятый отдел — низших тайнобрачных: мхи, лишайники, грибы.
Шестой отдел (с очень редкими представителями в сухих степях) над-
земных многолетних ксилофоров, т. е. деревьев, кустарников, полукустар-
ников и подземных ксилофитов, у которых деревянеет только корень (Ljtfft-
rum virgatum, Geranium sanguineum). Автор отмечает любопытную гр}ппу
однолетних ксилофоров, например, Salsola, Suae da, Agriophyllum arenarium.
В заключение Высоцкий предлагает схему взаимных отношений
выделенных групп растений.
Система Г. Н. Высоцкого была первой, предложенной для
многолетних растений сухих степей на основе их способности к
вегетативному размножению. Она базируется на глубоком изу-
чении ее автором биологии многих степных растений.
В 1922 г. Л. И Казакевич предложил новую биологическую
классификацию травянистых многолетников, взяв за основу
систему Г. Н. Высоцкого (1915). Он принимает его принцип
подразделения типов и групп многолетних растений, но считает
более удобным группу кистекорневых отнести к дерновым, так
как у них размножение все-таки идет за счет деления коротко-
го вертикального корневища. В результате им выделяется
5 групп.
1. Стержнекорневые — вегетативное размножение отсутствует.
2. Дерновые — вегетативное размножение слабо выражено.
3. Луковичные и клубне-луковичные — вегетативное размножение
слабо выражено
4. Корневищные (включая стелющиеся и укореняющиеся) — с сильно
выраженным вегетативным размножением.
5. Корнеотпрысковые — вегетативное размножение интенсивно
. Как и Высоцкий, Казакевич отмечает наличие переходных
форм в природе; количество их увеличивается под влиянием
внешних условий, воздействующих па общий габитус растения
и способность к вегетативному размножению. Каждая из групп,
по Казакевичу, может подразделяться далее в зависимости от
длительности жизни их побегов, длительности сохранения под-
земных частей последовательно возникающих побегов Быстрое
и полное отмирание побегов после завершения их развития
приводит к появлению «вегетативных малолетников» (термин
Высоцкого).
Большое достоинство работы Казакевича заключается в
том, что автор статистически изучил распределение выделенных
им биологических групп в разных типах растительности быв-
шей Саратовской губернии от сухо-степных и меловых террито-
рий до лесных. В работе Казакевича мы встречаемся с интерес-
ным исследованием экологических закономерностей распреде-
ления жизненных форм. Им выявлен ряд закономерностей —
уменьшение процента стержнекорневых растений по мере пе-
40
рехода от меловых и сухо-степных участков к лесным, увеличе-
ние в том же направлении процента корневищных растений и
т. д 1
В том же 1922 г. оригинальная система жизненных форм лу-
говых растений была предложена В. Р. Вильямсом В своих
построениях Вильямс исходит из теории сложения и развития
луговой формации, как растительного сообщества, в связи с
процессами почвообразования Он отмечает прежде всего, что
всякая растительная формация, и, в частности, луговая, по
необходимости должна включать как автотрофные, так и гете-
ротрофные организмы, обеспечивающие круговорот веществ в
природе Среди тех и других Вильямс выделяет биологические
группы, отличные от систематических и хозяйственных В ряде
автотрофов им выделяются: I группа — Злаки; II группа —
Осоки; III группа — Зеленые мхи и IV группа — Торфяные мхи.
Господство тех или иных групп, по Вильямсу, совпадает с опре-
деленными этапами развития луговой формации — от лугового
до сфагново-болотного периода. Лхговые злаки им подразде-
ляются по длительности жизни — на одно-, дву- и многолетние,
последние, в свою очередь, делятся на группы по длительности
жизненного цикла их побегов Наибольшее распространение в
нашей литературе получило подразделение Вильямсом злаков
по типу их кущения. Йм выделяются три типа. I — Корневищ-
ные злаки с боковыми побегами, отходящими перпендикуляр-
но от материнского и с длинными горизонтальными участками,
распространяющимися под землей, II — Рыхлокустовые злаки
с боковыми побегами, отходящими от материнских под углом
менее 90°; III - Плотпокустовые злаки с боковыми побегами,
растущими параллельно материнским, горизонтальная часть
побегов почти отсутствует Эти группы, выделенные по внешне-
морфологическим признакам, отличаются друг от друга биоло-
гически и тонко сопряжены с особенностями почв, на которых
они произрастают Переход от I к III группе сопровождается
уплотнением почвы и ослаблением ее аэрации.
А. М. Дмитриев (1947) выделяет четвертою группу — кор-
невищеворыхлок} стовых злаков, широко распространенную во
флоре лугов. На корневищах этих растений образуются группы
побегов, как у рыхлокустовых злаков Классификация Вильям-
са-Дмитриева широко используется в работах геоботаников,
экологов и луговедов нашей страны
1 Позже закономерности распределения биологических групп Высоцкого,
Казакевича в разных ботанико-географических зонах—от арктических тундр
до пустынь Средней Азии и в горах Тянь-Шаня были изучены И. Г. Се
ребрякоиым вместе с сотрудниками Е Ф Кошкиной, Т. И Серебряковой,
Г. И Денисовой, М А Борисовой, Г. П. Белостоковым, Р А Ротовым и
И. Н Былинкиной (Серебряков — 1954)
41
С фитоценологической точки зрения классифицирует тра-
вянистые формы растений Л. Г. Раменский (1938). Видоизме-
няя систему Высоцкого — Казакевича, Раменский разделяет
травянистые растения по их способности к вегетативному
возобновлению и разрастанию на три группы: 1) вегетативно-
неподвижные и не разрастающиеся, (стержнекорневые, кисте-
корневые и малолетники); 2) разрастающиеся, вегетативно-
слабоподвижные (плотнодернинные, рыхлодернинпые и корот-
ко-ползучие); 3) вегетативно-подвижные (ползуче-корневищные,
стелющиеся и шагающие и корнеотпрысковые).
В другом плане построена биологическая система травяни-
стых растений К. Линкола (К. Linkola 1922). Изучая способы
перезимовки сорняков Южной Финляндии, автор установил
существенные различия между их отдельными видами В ка-
честве принципа подразделения групп он использовал различия
зимнего и летнего состояний у разных видов растений. Им вы-
деляются пять основных групп.
I. Растения, перезимовывающие в виде семян.
П. Растения, перезимовывающие посредством почек, более или менее
замкнутых, находящихся в почве или на ее поверхности.
III. Растения, перезимовывающие с зелеными розетками листьев.
IV. Растения, перезимовывающие с зелеными ползучими побегами, от-
личающимися от летних
V. Растения, перезимовывающие в летнем состоянии, включая даже
цветки.
А. И. Мальцев (1931) дал другое подразделение сорняков
на биологические типы, основываясь в первую очередь на про-
должительности жизни, характере жизненного цикла и спосо-
бах размножения и связав свою классификацию со способами
борьбы с разными типами сорняков. Он выделяет-
I. Малолетники. Плодоносят один раз в жизни, живут не более
2-х лет, размножаются только семенами.
1. Эфемеры, дающие в лето несколько поколений.
2. Настоящие яровые однолетники.
3. Зимующие однолетники (могут цвести и без перезимовки).
4. Озимые однолетники (цветут только после перезимовки).
5. Двулетники.
II. Многолетники. Плодоносят несколько или много раз в жизни
и могут размножаться, кроме семян, вегетативно.
1. Стержнекорневые.
2. Кистекорневые.
3. Дерновые.
4. Ползучие (с плетями и усами).
5. Луковичные.
6 Корневищные.
7. Корнеотпрысковые
42
Последние два типа, где особенно сильно развито вегетатив-
ное размножение, Мальцев относит к наиболее тяжелым сор-
някам.
Необходимо отметить, что ритм развития ассимилирующих
листьев широко использовался в биологической классификации
и более ранними, чем Линкола, авторами (Grisebach, 1872;
Warming, 1884, Massart, 1908; Jeswiet, 1914). Два последних
автора для побережья Бельгии и Голландии выделяют: 1) зим-
незеленые растения, вегетирующие с осени до весны; 2) ранне-
весенние; 3) летнезеленые и 4) вегетирующие беспрерывно в
течение всего года. Сходные группы в лесах Южной Англии
отмечал и Солсбери (Salisbury А., 1916). Классификация тра-
вянистых растений по способам их перезимовки была предло-
жена также Е. И Лапшиной (1928). Она придерживается
системы Раункиера. Но при этом хамефиты и гемикриптофиты
ею подразделяются на две группы — перезимовывающие поч-
ками и в зеленом состоянии, а зимне-зеленые гемикриптофи-
ты — па факультативно и собственно-розеточные формы.
При экологическом анализе растительности Репетекского
песчано-пустынного заповедника в Кара-Кумах М П. Петров
(1935) использует систему жизненных форм Раункиера-Гамса,
несколько ее видоизменяя. Автор подразделяет гемикрипто-
фиты на две группы — с периодом покоя, зависящим от недо-
статка воды и не зависящим от него. Для Кара-Кумов Петров
принимает следующую систему: А — Фанерофиты; Б — Хаме-
фиты; В — Гемикриптофиты а) покой не зависит от недостатка
воды и б) покой зависит от недостатка воды; Г — Криптофиты*
а) вегетативные органы всегда в воде; б) вегетативные органы
земноводны, в) вегетативные органы в земле, геофиты а ) авто-
трофные* 1) период покоя не зависит от недостатка воды,
2) период покоя зависит от недостатка воды; р)—гетеро-
трофные
Интересная работа по классификации жизненных форм
(форм роста) у лиственных мхов была проведена Г. Мёйзелем
(Н Meusel, 1935). Мёйзель установил замечательный паралле-
лизм для многих форм роста у лиственных европейских мхов
и высших сосудистых растений. Все лиственные мхи он разде-
ляет на два типа: А — Ортотропные и В — Плагиотропные.
Среди ортотропных им выделяются группы: а) протонемных
мхов, Ь) ризоидных мхов, с) ризоидошнуровидных, d) с бази-
тонными инновациями, е) с акротонными инновациями, вклю-
чая дерновые и подушкообразные Основная линия в этом
ряду — изменение способа возобновления побегов от прото-
немы и ризоидов до боковых ветвей подземных побегов —
базальных и субтерминальных Среди плагиотропных мхов
различают: f) нитевидные мхи, g) ползучие, h) гребенчатые,
43
i) вертикально-ветвистые. В этой сисюме Мёйзсля крайне реду-
цированы экологические моменты; в центре внимания — при-
знаки морфологического порядка.
В ряде работ В. Pay (W. Rauh, 1936, 1937, 1938, 1940 и др )
рассматриваются жизненные формы (формы роста, по автору)
отдельных групп растений: кустарников, шпалерных кустарни-
ков, подушкообразных и др Цель Рау — установить организа-
ционно-морфологические типы у растений, относящихся к раз-
ным группам жизненных форм. В. Рау, как и Г. А4ейзель, пыта-
ется выявить морфологический механизм образования разных
«форм роста» в-связи с закономерностями ветвления, симмет-
рии побегов, направления их роста и т д Как указывалось
(см стр. 10), Мейзель работы свои и Рау выделял в особое
сравнительно-морфологическое направление классификации
жизненных форм Однако внимательное изучение их систем
показывает, что основания для выделения их из более общего
биолого-морфологического направления не имеется Напри-
мер, классификация подушковидных растений, предлагаемая
В. Рау (1940), скорее несет черты физиономического, чем био-
лого-морфологического характера. Рау резко расходится с пре-
дыдущими авторами в трактовке жизненных форм, но об этом
будет сказано ниже Система Рау 'интересна тем, что в ней
показано многообразие подушковидных растений — от древо-
видных и кустарниковых до однолетних (Более детальный
обзор и анализ системы В Рау’ см. в гл. VIII).
В В Скрипчинский (1940) делит растения на монокарпи-
ческие и поликарпическис Среди монокарпических растений он
выделяет подгруппу многолетних мопокарпиков (бамбуки,
пальмы), а однолетники и двулетники (моноциклические и
дициклические растения) объединяет в другой подгруппе
монокарпиков. Среди однолетников он выделяет эфемеры как
своеобразный тип Неэфемеры делятся им на три группы по
световой стадии- а) растения длинного дня, б) растения корот-
кого дня и в) нейтральные.
М В. Сенянинова-Корчагина (1949) при анализе фито-
климата Ленинградской и смежных с ней областей, исполь-
зует систему жизненных форм Раункиера, дополняя ее Ею
выделяются 20 жизненных форм
1—Фанерофиты вечнозеленые, 2—Фанерофиты летнезеленые, 3—Хаме-
фиты вечнозеленые, 4— Хамефиты летнезеленые, 5— Гемикриптофиты вечно-
зеленые розеточные с корневищем, б— Гемикриптофиты вечнозеленые ро-
зеточные с корнем, 7— Гемикриптофиты вечнозеленые с побегами, 8— Ге-
микриптофиты вечнозеленые дерниниые, 9— Гемикриптофиты вечнозеленые
рыхлокустовые, 10— Гемикриптофиты летнезеленые розеточные и рыхлоку-
стовые корневищные, 11— Гемикриптофиты легнезеленые розеточные и дер-
нинные с корнями, 12— Гемикриптофиты летнезеленые с побегами, 13— Ге-
микриптофиты летнезеленые, зимующие с почками на корневище, 14— Ге-
44
микриптофиты летнезеленые, зимующие с почками на корне. 15— Геофиты
корневищные, 16— Геофиты л\ковично-корневищные, 17— Геофиты с клуб-
нями, .8—Гелофиты или гигрофиты с корневищем, 19— Гидрофиты' и
20— Терофиты.
Как видно, в этой системе детально расчленены гемикрип-
тофиты. Автор говорит, что среди них имеется ряд хорошо раз-
личимых морфологических и экологических форм, характерных
для разных ассоциаций растительности с особым экологическим
режимом. М. В. Сенянинова-Корчагина приводит интересные
данные о закономерностях распространения перечисленных
жизненных форм по разным растительным формациям. Следует
отметить лишь неудачную терминологию в названиях отдельных
групп гемикриптофитов (например, групп б и 7, 11 и 12)
Детальный статистический анализ жизненных форм (Раун-
киера) в отдельных ассоциациях широколиственных лесов при-
водится также в работе С. А Кейна (S. A. Cain, 1950) по мате-
риалам ряда авторов и оригинальным данным Кейн приходит
к выводу, что спектры отдельных ассоциаций достаточно тонко
реагируют на особенности микроклимата.
Своеобразно изменяет систему жизненных форм Раункиера
применительно к растениям Восточного Памира К. В. Станю-
кович (1949). Станюкович различает:
I. Фанерофиты.
И. Хамефиты.
1. Кустарнички; 2 Полукустарнички; 3. Растения-подушки: а) плотные
подушки, б) рыхлые подушки, в) воздушные подушки. В каждой из трех
видов подушек различаются подтипы — с неукореняющимися побегами и
укореняющимися побегами.
III. Гемикриптофиты
1. Суккулентные (листостебельные); 2. Розеточные; 3. Дернистые;
4. Плогнокустовые злаки, 5. Обитатели осыпей щебня (путешествующие
со щебнем, переползающие по щебню, пронизывающие щебень, запружи-
вающие щебень); 6. Подземные кустарнички.
IV. Криптофиты
1. Луковичного типа; 2 Корневищного типа.
V. Терофиты
В работе Станюковича приводятся данные об экологическом
распространении и биологических особенностях растений Па-
мира, относящихся к различным группам жизненных форм.
Укажем также на систему жизненных форм, приводимую
в «Фитосоциологии» Браун-Бланке (J. Braun-Blanquet, 1951)
Автор присоединяется к воззрениям Раункиера на классифика-
цию жизненных форм и приводит следующую систему (мы ее
излагаем с некоторыми сокращениями).
I Фитопланктон.
а) аэрофиты; Ь) гидрофиты; с) криофиты.
II. Фитоэдафон.
45
а) аэробионты; b) анаэробионты.
III. Эндофиты.
а) эндолитофиты; b) эндоксилофиты; в) эндозоофиты.
IV. Терофиты.
I) таллотерофиты; 2) микотерофиты; 3) бриотерофиты; 4 птеридотеро-
фиты; 5) эутерофиты (а — ползучие терофиты, b — цепляющиеся терофиты,
с — приподнимающиеся, d — пучковые).
V. Гидрофиты.
а) плавающие, Ь) прикрепленные, с) укореняющиеся.
VI. Геофиты.
1. грибные; 2. паразитные; 3. эугеофиты (а — клубневые, Ь— ризомные,
с — корне-почковые).
VII. Гемикриптофиты.
1. талломные, 2. укореняющиеся: а) дерновинные, Ь) розеточные, с) стеб-
лестержневые, d) лазящие).
VIII. Хамефиты.
а) ползучие мхи, Ь) кустистые лишайники, с) ползучие травы, d) сук-
кулентные, е) подушковидные, f) лазящие многолетники, g) сфагновые мхи,
h) дерновинные злаки, i) шпалерные кустарники, j) полукустарники, к) ку-
старнички.
IX. Фанерофиты:
а) кустарники, Ь) деревья, с) стеблевые суккуленты, d) розеточные де-
ревья, е) травяно-стеблевые, f) лианы.
X. Древесные эпифиты.
Очевидно, что система Браун-Бланке охватывает весьма
широкий круг организмов. По своей, направленности и подраз-
делениям классов она в значительной степени сходна с системой
Гамса (1918). Это сходство вполне естественно, так как и та
и другая имеет синэкологическое, фитоценологическое назначе-
ние.
Рядом исследователей разработаны системы значков и сим-
волов для обозначения жизненных форм при описании расти-
тельности. П. Данзеро (Р. Dansereau, 1951) разработал систему
обозначений жизненных форм, предназначенную главным обра-
зом для специалистов неботаников, которой можно пользовать-
ся при полевом описании растительности (ландшафта), не при-
бегая к определению видов растений. Предлагаемая система,
по мнению Dansereau, отражает структуру растительных сооб-
ществ, являясь в то же время индикатором условий среды. Эти
значки могут быть использованы также при изучении варьиро-
вания растительных сообществ
Система обозначений Данзеро является дальнейшим разви-
тием подобной же системы, предложенной Кюхлером (Kuchler,
1949). Она основана на четырех сериях обозначений, комби-
нируя которые можно изображать различные типы раститель-
ности. В этой системе выделяются и получают буквенное обоз-
начение следующие физиономические группы. Большими бук-
вами изображаются:
46
Древесная растительность
В — вечнозеленые крупнолистные;
D — листопадные крупнолистные;
Е — вечнозеленые игольчатолистные;
N — листопадные игольчатолистные;
О — безлистные
Травянистая растительность
G — граминоидные;
U — forbs (прочие травы);
L — лишайники и мхи.
Малыми буквами обозначаются:
I группа. Высота растений
t — высокие (деревья и травы);
m — средние (деревья и травы);
1 — мелкие (деревья и травы).
s — кустарники;
z — кустарнички.
II группа. Густота стояния растений
(Буквы с, i, р, г, b указывают разную степень густоты, умень-
шающуюся от с к Ь);
III группа. Специальные формы.
е — эпифиты;
j — лианы;
к — суккуленты;
q — растения-подушки;
и — пальмы;
v— бамбуки;
w — водные растения;
у — древовидные папоротники.
При описании растительности Dansereau предлагает раз-
личать следующие жизненные формы: деревья, кустарники,
травы, бриоиды, эпифиты, лианы. Первые три категории трак-
туются в обычном смысле. К бриоидам относятся не только мхи
и лишайники, но и водоросли и растения с бриоидным обликохм
вроде Selaginella, Cassiope hypnoides, Raoulia australis. Среди
эпифитов могут быть цветковые растения, мхи, папоротники
и лишайники. Лианы подразделяются на вьющиеся и лазящие.
Жизненные формы изображаются специальными значками.
Кроме этого, имеются особые значки для обозначения формы,
величины и консистенции листьев.
Своей системой обозначений Dansereau широко пользуется
в книге «Biogeography» (1957).
Оригинальная система символов для изображения жизнен-
ных форм (форм роста) разработана Е. Schmid (1956). Жизнен-
47
ные формы, по этому автору, намного упрощают изучение
структуры растительных сообществ, особенно тропических
стран, где набор видов мало известен.
При классификации типов жизненных форм Е. Шмид исхо-
дит из анализа растительности экваториальной зоны, как наи-
более богатой ими. Признаки выделяемых типов связаны
с вегетативными органами и находятся в определенной зави-
симости от среды Комбинации избранных признаков автор
обозначает символами, которые легко воспринимаются; в них
сразу видна вся совокупность признаков. Важнейшим призна-
ком, выявляющим реакцию растительности на внешнюю среду,
согласно автор}, является степень одревеснения. Е. Шмид
различает- акс ильные растения (неодрсвеснсвшие),
олигоксиявные растения (мало одревесневшие), на-
пример, виды Senecio восточно-африканских вулканов, г е-
миксильпые (полуодревесневшие) — например, мягко-
ствольные Carica papaya, Fatsia japonica, гол окси л ь и ы е
(полностью одревесневшие, как большинство древесных розо-
цветных, бобовых и др ), п е р к с и л ь н ы е (жесткодревесные,
например виды Eugenia, Buxus и др.) , мероксильные
(лишь с одревесневшими участками стебля, например полу-
кустарники) .
Вторым основным признаком, легко прослеживающимся
у растений, Шмид считает соотношение между длительно живу-
щими и сезонными осями. Шмид отмечает явно выраженную
тенденцию к увеличению роли сезонных осей у растений холод-
ного и аридного климата вплоть до полного исчезновения дли-
тельно живущих осей. Эта тенденция совпадает с тенденцией
к уменьшению одревеснения осей, т. е к образованию вторично-
аксильных травянистых форм вместо древесных.
Кроме того, Е Шмид использует в своей системе признаки
ветвления, длины междоузлий, величины и сезонности развития
листьев и некоторые другие. Этот автор приводит схематиче-
ское изображение структуры изученных им типов раститель-
ности (в виде вертикальных профилей), на которых хорошо
видны биоморфологические особенности основных видов, вхо-
дящих в их состав.
Детальная и многоплановая система жизненных форм степ-
ных растений разработана М. С. Шалытом (1955) В основу
системы положено несколько признаков: форма роста, продол-
жительность жизни, количество плодоношений, способ вегета-
тивного размножения, характер корневой системы и т д.
Цветковые растения в системе Шалыта разделяются па
многолетники (в большинстве поликарпики) и малолетники.
Среди поликарпиков выделены: 1 Кустарники и кустарнички.
II Полукустарники и полукустарнички III. Полутравы.
48
IV. Травы, длительно вегетирующие и коротко вегетирующие
(весенние, летние и осенние) При дальнейшем подразделении
выделяются более узкие группы осевых растений (стержнекор-
невые, кистекорневые, у трав — клубнскорпсвые), дернистых
(отсутствуют у кустарников и кустарничков) и ползучих (с над-
земными укореняющимися побегами, корневищные и корне-
отпрысковые). К малолетникам отнесены двулетники и однолет-
ники (озимые, яровые и зимующие)
Кроме этой основной системы автор выделяет группы сукку-
лентов, перекати-поле и подушкообразных форм Интересны
данные Шалыта о переходных жизненных формах и их измене-
нии под влиянием внешних условий.
В фитоценологическом плане выдержана система жизнен-
ных форм Г. М. Зозулина (1959) По способу удержания пло-
щади за особью все растения им разбиваются на четыре типа
жизненных форм I Реддитивные растения (от лат. reddere —
уступать)—многолетние виды, уступающие при росте особей
свое место в случае уничтожения по какой-либо причине над-
земной части К этому типу отнесены, например, многие деревья
тропических лесов, у которых отсутствуют спящие почки у осно-
вания стволов. В наших широтах к этому типу относится боль-
шинство хвойных пород 2 Рестативпые растения (от лаг.
restare — оставаться)—многолетние виды, возобновляющиеся
от спящих почек или почек возобновления, в случае уничтоже-
ния надземной части Сюда относится большинство деревьев,
кустарников и трав умеренного пояса 3 Ирруптивные растения
(от лат irrumpcre — вторгаться) — многолетние виды, не только
вегетативно возобновляющиеся при уничтожении надземной
части, но и вторгающиеся посредством новых надземных побе-
гов в места, занятые другими видами В умеренных и холодных
широтах представлены главным образом корневищными и
корнеотпрысковыми формами 4 Вагативпые растения (от лат.
vagare— кочевать)—однолетние и двулетние виды, размно-
жающиеся только семенами, не удерживающие за собой пло-
щади, кочующие путем разноса семян и прорастания на новом
месте Далее эти типы делятся па постоянные вечнозеленые,
постоянные листопадные и периодические. Дальнейшее подраз-
деление жизненных форм Зозулин проводит по тому же прин-
ципу— способу удержания за растительной особью площади
обитания. Единство принципа классификации жизненных форм
придает системе Зозулина черты целенаправленности и строй-
ности Вместе с тем эта система искусственная Деревья в ней,
например, отнесены к трем типам (1 -у, 2-у и 3-у), а во втором
типе оказываются деревья, кустарники и травы
Обзор приведенных систем жизненных форм демонстрирует
их чрезвычайное многообразие Но как и в первом, эколого-
В 413 И Г Серебряков —4 49
физиономическом, направлении, здесь также видна общность
многих групп жизненных форм, выделяемых в системах разных
авторов под разными названиями
Приведенный исторический обзор классификаций жизнен-
ных форм позволяет не только выявить разные пути решения
этой проблемы в прошлом, но и заглянуть в завтрашний день,
увидеть основные черты будущих систем жизненных форм.
В биолого-морфологическом направлении сделан шаг вперед
по сравнению с предыдущим. Появилась система, принимаемая
с теми или иными видоизменениями значительным числом
исследователей (система Раункиера). Сравнивая системы пер-
вого и второго направлений, можно в свою очередь констати-
ровать выделение многих общих групп жизненных форм там
и здесь. Различия остаются, главным образом, в терминологии,
объеме выделенных групп и их субординации Таким образом,
вновь подтверждается мысль о том, что период поисков разно-
образия жизненных форм в основном, вероятно, пройден. Насту-
пает новый период создания систем, приемлемых для все более
широкого круга исследователей.
Вследствие большого разнообразия и сложности отношений
жизненных форм уже теперь целесообразно вначале строить их
системы для ограниченных систематических групп, например,
двудольных или однодольных или даже отдельных порядков
и семейств. Можно согласиться с мнением Дю Рие (1931) о
целесообразности одновременной разработки нескольких систем
жизненных форм различного служебного назначения. Характе-
ристику жизненных форм целесообразно основывать на струк-
турно-биологических особенностях не только надземных и под-
земных побегов, но и корневых систем, еще недостаточно
используемых теперь. Новым в системах будущего, очевидно,
будет ясное представление о закономерностях онтогенеза раз-
личных жизненных форм. В будущих системах найдут отраже-
ние основные пути эволюции жизненных форм, их взаимные
филогенетические отношения. Короче говоря, вместо статичных,
появятся системы эволюционные.
5. ПОНЯТИЕ «ЖИЗНЕННАЯ ФОРМА РАСТЕНИЙ»
И ЕГО ОПРЕДЕЛЕНИЕ У РАЗНЫХ АВТОРОВ
Если все виды растений в процессе своей жизнедеятельности
принимают ту или иную жизненную форму, и все группы жиз-
ненных форм состоят из определенных видов, то, казалось бы,
можно говорить о полном совпадении этих понятий. Однако
у высших растений они далеко не идентичны Один и тот же
вид в различных частях своего ареала или в разных экологиче
ских условиях нередко принимает различные жизненные формы,
50
иногда значительно отличающиеся друг от друга. Так, многие
виды деревьев близ пределов своего распространения образуют
кустарниковые, нередко стелющиеся формы. Широко известно
образование стелющихся форм у многих хвойных, например,
сибирской лиственницы на Таймыре (Л. Н. Тюлина, 1937),
обыкновенной ели на Крайнем Севере (В. Н. Сукачев, 1938),
сибирской ели на Южном Урале (Л. Н. Тюлина, 1929),
в Хибинах (И Г. Серебряков, 1952), тяньшанской ели в Заи-
лийском Алатау (И. Г. Серебряков, 1945) и т. д. Стелющиеся
формы образуются иногда у бука близ верхней границы леса
в Крыму и на Кавказе (А. В. Кожевников, 1935, А. Я. Орлов,
1953, Г. И. Поплавская, 1927, 1929, В Н. Сукачев, 1938). Тур-
кестанский можжевельник — Juniperus turkestanica Кот.—
в Терскей Алатау в лесном поясе (2 600—2 800 м н. у. м.)
растет как дерево 5—6 м высоты, а в субальпийском поясе того
же хребта (3 200—3 300 м н у. м.) образует форму распластан-
ных куртин высотой 30—60 см и диаметром 8—10 м (И. Г. Се-
ребряков, 1954).
Китайский лимонник — Schizandra chinensis — на Дальнем
Востоке в различных экологических условиях образует то
форму лианы, то кустарника (И. С. Михайловская, 1952). Мно-
гие растения Центральной Европы и Средиземья образуют то
форму терофитов, то гемикриптофитов или форму фанерофитов
и хамефитов (J. Braun-Blanquet, 1951).
В еще большей степени изменение жизненных форм наблю-
дается у растений при интродукции их за пределами естествен-
ного ареала. Б. М. Козо-Полянский (1944) сообщает, что неко-
торые древесные растения субтропических и умеренно-теплых
областей в Воронежском и Алма-Атинском ботанических садах
приобретают (из-за ежегодного обмерзания побегов почти до
поверхности почвы и последующего возобновления их за счет
приземных почек) новую «биоморфу», близкую к полукустар-
никам. Подобным же образом ведет себя Amorpha fruticosa
в открытом грунте Ботанического сада МГУ в Москве. Нам
приходилось видеть в Гвинее в Ботаническом саду Шевалье
чайпые кусты, ставшие небольшими деревьями до 5—6 м
высоты с диаметром стволов в 6—7 см под влиянием влажного
тропического климата. На Кольском полуострове в Полярно-
альпийском ботаническом саду, в опытах Н. А. Аврорина, клен
и липа (Acer platanoides и Tilia cordata), приближались по био-
логии и жизненной форме к низким кустарникам и даже полу-
кустарникам В многолетних капитальных работах по интро-
дукции растений в Хибинах Н. А. Аврориным (1956) выявлены
многие примеры изменения морфологических признаков интро-
дуцируемых растений, вплоть до изменения их жизненной
формы
4^5
51
Как видим, под влиянием резких изменений климатических,
а иногда и почвенных и цепотических условий один и тот же
вид может значительно изменять свою жизненную форму.
Вполне понятно, что изменения жизненных форм всегда необ-
ходимо иметь в виду в работах по интродукции растений и при
организации защитного лесоразведения за пределами лесной
зоны.
С другой стороны, сходные или весьма близкие жизненные
формы могут развиваться в разных флористических областях
земного шара у разных, часто систематически далеких видов,
но при условии близкого сходства климата, почвы и характера
растительного покрова страны. Достаточно вспомнить влажно-
тропические леса долины Амазонки, Конго, Западной Явы с их
гигантскими деревьями с досковидными выростами у основания
колонновидных стволов, с их лианами и эпифитами. Историче-
ское развитие каждой группы жизненных форм в этих лесах шло
независимо и копвергенгно в разных областях на базе разлого
флористического материала. И досковидиые выросты корней
у основания стволов тропических деревьев, в сущности, являют-
ся не систематическими, а экологическими признаками. Широко
известно также биолого-морфологическое и габитуальное сход-
ство в вегетативной сфере под} шковидных, розеточпо-сукку-
лентных и сукку лентно-стеблсвых жизненных форм, включаю-
щих виды из различных семейств покрытосеменных. Конечно,
степень копвергентности не следует преувеличивать, имея в виду
специфичность проявления приспособительных наследственных
свойств в каждой из филогенетических групп (О. Drude, 1913,
Б А. Келлер, 1933; J Braun-Blanquet, 1951). Но так или
иначе, понятия о группах жизненных форм и видов не совпа-
дают, и классификация их, как мы видели, построена на раз-
личных основаниях.
Жизненные формы подавляющее большинство авторов, как
мы видели, понимает как выражение приспособленности расте-
ний к господствующим условиям произрастания Но это толко-
вание подходит в значительной степени и к так называемым
экологическим группам, например, мезофитам, ксерофитам,
психрофитам, псаммофитам, литофитам и т д Отношения
между экологическими группами и жизненными формами, нам
думается, правильно оценивают Гл И. Поплавская (1948)
и А П. Шенннков (1950) По мнению этих авторов, первые
отражают приспособленность организмов лишь к отдельным
факторам внешней среды, например, условиям увлажнения
(мезофиты, ксерофиты и т д), почв или грунтов (псаммофиты,
литофиты и др ), освещения и т д Вторые, т. е. жизненные
формы, отражают приспособленность растений ко всему ком-
плексу факторов внешней среды, т. е специфике данного место-
52
обитания в целом Поэтому признаки экологических групп могут
входить лишь как часть общей характеристики жизненных форм,
не заменяя их целиком В таком плане признаки экологических
групп рекомендуют использовать в системах жизненных форм
Г. Гаме (1918) и А В Прозоровский (1938) Выделение же
жизненных форм по одному из экологических или биологиче-
ских признаков невольно ведет к путанице, к замене жизненных
форм экологическими группами, замене общего частным
Развитие учения о жизненных формах сопровождалось вве-
дением определенной терминологии и, в частности, названия
самого объекта Гумбольдт (1806) и Кернер (1863) выделенные
ими группы растений называли «основными формами» («Grund-
formen»); Гризебах (1872), Хульт (1881), Рейтер (1885), Друде
(1887),— «растительными формами» («Vegetationsformen»); Вар-
минг (1884), Раупкиср (1903, 1906), Друде (1913), Гаме (1918),
Дю Рие (1931), Келлер (1933), Алехин (1936, 1944), Коровин
(1934), Кульгиасов (1950), Браун-Бланке (1951),—«жизнен-
ными формами» («Lebensformen», «Life-forms»); Бекетов (1898),
Вильямс (1922) —«биологическими группами», Дю Рие (1921),
Мейзель (1935, 1951, 1952), Рау (1936, 1937, 1940) — «формами
роста» (Wuchsformen)
Как указывалось выше, термин «жизненная форма» был
предложен Вармингом в 1884 г, поэтому названия «основные
формы» Гумбольдта и Кернера, «растительные формы» Гризе-
баха и других исследователей являются как бы его предшест-
венниками Что касается других терминов, применяемых позд-
нейшими авторами вместо «жизненных форм», то они имели
у них определенное обоснование Так, Дю Рце (1931) «жизнен-
ными формами» называет группы, выделяемые на основе об-
щего габитуса растений во время вегетационного периода
Группы растений, выделяемые по структуре их побегов, он
предлагает называть «формами роста» Как справедливо ука-
зывает Адамсон (1938), такое подразделение вряд ли целе-
сообразно’ оно искусственно, эти понятия взаимозаменяемы
Совершенно иначе мотивируют свой отказ от термина «жизнен-
ная форма» Мейзель (1936), Рау (1937, 1940), Тролль (1936)
Рау (1940) квалифицирует экологическое, адаптивное толкова-
ние жизненных форм как поверхностное и необоснованное
В жизненных формах он видит, как Тролль и Мейзель, проявле-
ние не экологической, т е. приспособительной конвергенции,
а конвергенции чисто морфологического характера 1. В связи
1 Следует отметить, что Г Мейзель в позднейших работах отошеч от
этих взглядов, и рассматривает жизненные формы как итог исторического
развития растений в определенных условиях среды (Н Meusel, 1951, 1952).
53
с отказом от экологической трактовки «жизненных форм», Рау
и применяет термин «формы роста». «Формы роста», по Рау,—
это проявления «типа», «организационного плана» растений, в
основе которого лежат симметрия и типы ветвления. Рау (1940),
как и его учитель Тролль (1936), говорит, правда, о тесной
связи организма и среды, и даже о немыслимости существова-
ния первого без второго. Но среда, по Троллю и Рау, лишь спо-
собствует выявлению внутренних потенций организма, его
«плана строения», «типа». Таковы взгляды представителей
современной идеалистической или типологической морфологии
растений. Но критерий действительности — не на стороне идеа-
листической морфологии.
Термин «формы роста» применяет и Р. Шарфеттер (1953)'.
Каждый вид, по Шарфеттеру, для нормальной жизнедеятель-
ности должен находиться в соответствии с окружающей внеш-
ней средой, т. е. обладать определенной «жизненной формой».
Но «жизненная форма» объемлет по Шарфеттеру и внешний
облик растения и его жизненные проявления, т. е. фенологиче-
ские особенности развития Для обозначения лишь внешней
морфологической структуры растения автор предлагает упот-
реблять название «форма роста». Однако это предложение
вряд ли можно назвать удачным Фенологические и морфоло-
гические признаки представляют часто лишь разные стороны
одного и того же процесса жизнедеятельности растений. Изме-
нения в ритме развития неизбежно влекут за собой изменения
в морфологии, в структуре растительного организма (Серебря-
ков, 1948, 1949). Поэтому разные «формы роста» будут неиз-
бежно и разными «жизненными формами».
В истории учения о жизненных формах менялась не только
терминология, применяемая для их наименования, но и содер-
жание, вкладываемое в это понятие разными авторами.
У Гумбольдта и Гризебаха подход к жизненным формам
был скорее географическим, чем биологическим. Никаких адап-
тационных представлений с этим термином авторы не связы-
вали
Впервые жизненные формы как объект специальных эколого-
ботанических исследований стали рассматриваться Вармингом
(1884), который определяет их как форму, в которой вегета-
тивное тело растения (индивида) находится в гармонии с внеш-
ней средой в течение всей его жизни, от семени до отмирания.
Иной оттенок придает этому понятию Рейтер (Н. Reiter,
1885). К той или иной жизненной форме («Vegetationsform»),
Рейтер относит все растения, которые в отношении своей жизне-
деятельности и относящихся сюда признаков структуры суще-
ственно сходны друг с другом, независимо от того, близки они
систематически или нет. В этом определении жизненная форма
54
выступает как группа растений, т. е. единица классификацион-
ного порядка, единица экологической системы растений. Это
дополнение нельзя не признать целесообразным.
Но это уже совершенно другая сторона дела Таким обра-
зом, термин «жизненная форма» приобретает двоякий смысл,
обозначая с одной стороны эфармопию растения, с другой —
классификационную единицу
Далее речь идет, главным образом, о толковании термина
«жизненная форма» именно в первом, более широком смысле.
Что касается единиц классификации, то из обзора приведенных
систем жизненных форм разных авторов видно, что все они
понимали обьем «жизненных форм» по-разному.
Весьма близкое к Вармипгу, но в то же время более узкое
определение жизненным формам дается Раункиером (1906),
Он рассматривает жизненные формы как итог и результат
приспособления растений к внешней среде. Но из всего ком-
плекса внешних условий решающее значение он придает кли-
мату. Отсюда и жизненные формы, по Раункиеру, являются
отражением климата различных областей земного шара; они
совершенно не связаны с систематическим положением расте-
ний Несмотря на простоту и законченность системы жизненных
форм у Раункиера, его определение и толкование жизненных
форм односторонне, что неминуемо влечет к искусственности
всей его концепции. Отсюда понятна справедливость критиче-
ских замечаний многих исследователей в адрес его системы.
Иное содержание вкладывает в понятие жизненных форм
Друде (1913). Он рассматривает жизненные формы как эколо-
гически сходные группы растений главнейших филогенетиче-
ских рядов растительного мира, выделенные с морфолого-
физиономической точки зрения В отличие от Варминга и Раун-
киера он говорит о необходимости выделения параллельных
жизненных форм по систематическим группам, так как приспо-
собительные признаки в каждой из них проявляются специ-
фично, несмотря на одинаковые воздействия внешних условий.
Одновременно Друде подчеркивает целесообразность выделения
жизненных форм по физиономическим, но не чисто биологиче-
ским признакам, ибо лишь физиономические жизненные формы
имеют особое значение для ботанической географии. Замечание
о специфичности жизненных форм в разных систематических
группах представляется безусловно интересным.
В весьма широком экологическом плане дается определение
жизненных форм Гамсо.м (1918), который считает их адекват-
ным выражением условий жизни организмов. Жизненные
формы, по Гамсу, могут служить наиболее надежным средством
экологической оценки местообитания. Он особенно настаивает,
в противоположность Друде, на освобождении учения о
55
жизненных формах от «систематическогобалласта», на учете при
выделении жизненных форм только эфармонических признаков.
В воззрениях Гамса экологическое направление в изучении
жизненных форм находит наиболее резкое и крайнее выраже-
ние. Уже говорилось, например, о том, что классы криптофитов
и терофитов Раункиера Гаме обьединяет в один на том осно-
вании, что у того и другого органы, переживающие неблаго-
приятное время года, располагаются в почве. Что касается раз-
личия этих органов — у одних клубни и луковицы, у других
семена — то, по Гамсу, это экологически несущественно, ибо
различия между семенами и луковицами морфологические, а не
биологические. С этим, конечно, вряд ли можно согласиться.
Несколько сходно с Друде понимание жизненных форм
Б. А. Келлером (1933). Под жизненными формами, по Келлеру,
надо понимать определенную систему экологических приспособ-
лений, тесно связанных с организационным типом растения,
с принадлежностью его к определенному классу, семейству,
а часто и роду. В этом определении, как видим, подчеркивается
как экологизм жизненных форм, так и их историзм, последнее
находит свое выражение в их систематической приуроченности.
Мысль о связи экологических приспособлений с систематиче-
ским положением растений, но в еще более резкой форме, под
черкивается Ю С. Григорьевым (1954), который считает, что
таксономические единицы, например, подсекции, секции, роды
есть «жизненные формы высшего порядка» (стр 141), ибо отно-
сятся к тому же «эколого систематическому типу» Для многих
случаев .это, вероятно, правильно Но в общем виде этот тезис
вряд ли может быть принят
Совпадения систематических групп и жизненных форм да-
леко не обязательны.
Е II Коровин (1934) рассматривает жизненную форму как
приспособление растений к местообшанию, выраженное в фор-
мативных, биологических и физиологических признаках, соче-
тающихся друг с другом. При этом физиологические признаки
могут и не иметь, по Коровину, морфологического проявления
Позже он, совместно с Закржевским, определяет жизненную
форму как форму накопления растением органического веще-
ства (Б С Закржевский и Е П Коровин, 1936). Но последнее
определение имеет скорее биогеохимический характер, в смысле
В. И. Вернадского (1940), чем биологический.
Большой четкостью отличается определение В В Алехина
(1944), который считает, что «жизненная форма — это резуль-
тат длительного приспособления растения к местным условиям
существования, выраженный в его внешнем облике» (стр 55)
В этом определении подчеркивается физиопомичноегь жизнен-
ных форм, рассматриваемая в экологическом и историческом
56
пианах. Здесь отмечается одновременно независимость жизнен-
ных форм от систематического положения составляющих их
растений. Но это простое и емкое определение не учитывает
изменения жизненной формы в пределах вида, возникающие
в онтогенезе растений в разных экологических условиях
По А. П Шенникову (1950), отнесение к одной и той же
жизненной форме означает одинаковый путь и способ приспо-
собления растений к жизненной среде. Судя по этому опреде-
лению, жизненные формы могут выделяться по самым различ-
ным признакам, и не обязательно структурно-морфологическим
В то же время Шеппиков подчеркивает таксономическое значе-
ние жизненных форм, которые являются систематическими еди-
ницами в экологии.
Близкое к этому определение жизненных форм дает
М В. Культиасов (1950). Под жизненной формой автор пони-
мает совокупность, т е. объединение растений, сходных по
исторически сложившимся приспособлениям к условиям суще-
ствования, с помощью которых эти растения утверждаются
к жизни и, размножаясь, прогрессируют
В экологическом плане трактует жизненные формы И Браун-
Бланке (1951) как организмы, в общем строении которых более
или менее отчетливо выражены одинаковые приспособления
к условиям жизни Это определение близко к пониманию жиз-
ненных форм Гамсом Но в отличие от последнего, Браун-
Бланке считает, что приспособление может найти выражение
не только в эфармонических, по и организационных признаках.
Господствующие условия, по Браун-Бланке, отражаются
в жизненных формах лишь в известной степени, в пределах
конституционных границ. В определении Браун-Бланкс, как
видим, отсутствует момент историзма, который подчеркивается
в формулировках советской школы, например, Келлера, Алё-
хина, Шеппикова, Культиасова
Экологическая и историческая трактовка жизненных форм
отражена в определении их у И Г Серебрякова (1954), по
которому жизненная форма «это своеобразная внешняя форма
растений (или их габитус), неразрывно связанная с биологией
развития и внутренней структурой органов растения, возник-
шая в определенных почвенно-климатических и ценотических
условиях и отражающая приспособленность растений к этим
условиям» (стр. 54) Так как общий облик или габитус расте-
ния есть итог и внешнее выражение процессов его жизнедея-
тельности в определенных условиях среды, автор в последую-
щем несколько видоизменил определение (Серебряков, 1955).
«.. под жизненной формой можно понимать своеобразие тех
или иных групп растений, выраженное в специфике их сезонно-
го развития, в способах ежегодного нарастания и возобпов-
57
ления, во внутренней и внешней структуре их органов и, в
конечном итоге, как следствие, во внешнем облике, или габиту-
се растения, исторически возникшем в определенных почвенно-
климатических и ценотических условиях и отражающем при-
способленность растений к этим условиям» (стр. 71). Жизнен-
ные формы в этих определениях рассматриваются с точки
зрения биолого-морфологической, автор пытается осмыслить
многообразие жизненных форм как следствие историче*
ского развития растений в определенных условиях внешней
среды
Учение о жизненных формах, возникшее в ботанике, нахо-
дит все большее распространение и в зоологии. Особое внима-
ние этой проблеме уделяется в отечественной зоологической
литературе в последние два десятилетия. Как и у ботаников,
здесь наблюдаются разнообразные подходы к созданию систем
и трактовке жизненных форм Так, по Д. Н. Кашкарову (1944),
«тип животного, находящегося в соответствии с окружающими
условиями, мы называем «жизненной формой». В «жизненной
форме», как в зеркале, отражаются главнейшие доминирующие
черты местообитания видов и основные черты климата и
... почв, и биотических отношений» (стр. 173). Выделение жиз-
ненных форм, по Кашкарову, необходимо вести не на филоге-
нетических, а на адаптивных признаках, — лишь они харак-
теризуют сред) Автор приводит данные (1937) о жизненной
форме нырца, на примере баклана, у которого вся структура
тела приспособлена к добыче пищи в воде. При построении
системы жизненных форм для животных Кашкаров, подобно
тому как Варминг для растений, использует самые различные
биологические признаки и свойства организмов. Им выделяют-
ся группы по отношению к климату, по способам передвижения,
по роду пищи, по отношению к влаге и т д
По Г. Я. Бей-Биеико (1950), в жизненной форме «реали-
зуются все специфические особенности вида: его биология, по-
ведение, экология, физиология и пр» (стр. 134). В предложен-
ной им классификации прямокрылых насекомых Средней Азии
выделяются 4 класса: фитофилы, геофилы, засадники и сим-
бионты, подразделяющиеся далее па более мелкие группы.
Автор вслед за Б. П. Уваровым (1938), указывает, что среди
ширина
саранчевых закономерно изменяется соотношение ’ высота в
самой широкой части тела в зависимости от условий место-
ширина
обитания У пустынных форм высота обычно более еди-
ницы и покровы тела часто шиповаты. У живущих в густом
ширина
травостое отношение высота " менее единицы, т. е. тело сжато
58
с боков и покровы гладкие; это облегчает им передвижение
Как видим, и у насекомых габитуально-морфологические при-
знаки одновременно являются адаптивными, экологическими.
По Н. И. Калабухову (1955), характеристика жизненных
форм отражает непосредственные связи организма и среды,
которые определяются повседневным и глубоким влиянием
условий существования и факторов воздействия на растения и
животных. Выделение жизненных форм служит весьма полез-
ным и существенным способом оценки сходного влияния внеш-
ней среды на различные организмы Как пример конвергирую-
щего влияния одинаковых условий местообитания автор при-
водит группы степных и полупустынных грызунов различных
стаций
А. К Рустамов (1955) возражает против дробности при вы-
делении жизненных форм, считая, что «каждой ландшафтной
зоне в широком биологическом и эволюционном аспекте соот-
ветствует определенная жизненная форма» (стр. 717), отра-
жающая морфо-физиологический ответ различных организмов
на сходные условия существования С этой точки зрения автор
рассматривает характеристику животных средне-азиатских
пустынь.
А. Г. Банников (1955) считает неверным исходить из ланд-
шафта при выделении жизненных форм и определяет жизнен-
ные формы (биологические группы) как группы животных,
объединенных суммой общих морфо-биологических адаптивных
черт, исторически возникших у видов, сходных по характеру
питания Разбирая биологическую группу копытных, Банников
показывает, что все черты биологии их так или иначе связаны
с питанием Автор приводит довольно стройную систему жиз-
ненных форм копытных, исходя из способа питания, оговари-
вая, правда, что выделенные группы до известной степени
условны из-за недостаточной их изученности
По М Н Акимову (1955), жизненные формы животных
представляют собой тот или иной тип приспособления живот-
ных к определенной среде обитания и к определенному образу
жизни в ней Он выделяет многочисленные группы насекомых
по отношению к местообитанию, по способу питания и т. д., а
среди растений — по способам опыления, расселения. Но выде-
ленные им группы, отражая лишь одну из сторон жизни вида,
скорее являются экологическими группами (см. выше), чем
жизненными формами
Иначе трактует жизненные формы Н. П. Наумов (1955).
Автор согласен с С. А. Северцовым (1937), что в понятии жиз-
ненная форма внимание центрируется на особом специфическом
именно для определенной систематической группы отношении
к окружающей среде; жизненная форма, по Наумову, это
экологическая характеристика вида или любой другой систе-
матической группы. Поэтому можно говорить о жизненной
форме «млекопитающее», «птица», «рыба», «насекомое».
Сходные концепции в экологии растений развивает
Ю. С. Григорьев (1954).
И может быть прав В. Мазинг (1958), говоря, что предло-
жение рассматривать экологические особенности каждого вида
как самостоятельную жизненную форму нельзя считать целе-
сообразным, так как такая узкая трактовка жизненной формы
не способствует выявлению основных направлений эволюции,
свойственных многим видам в определенных условиях суще-
ствования. Такая трактовка жизненных форм у растений недо-
статочно учитывает внутривидовую изменчивость жизненных
форм и конвергснтность развития сходных жизненных форм в
разных систематических группах
Большое разнообразие определений жизненных форм, про-
иллюстрированное в приведенном выше очерке, свидетельствует
о большой сложности и многогранности этого биологического
явления и разнообразии подходов к исследованию жизненных
форм разными авторами В каждом из приведенных определе-
ний нашли отражение те или иные существенные особенности
жизненных форм растений.
Несмотря на калейдоскопическую пестроту определений
жизненных форм у разных авторов, общий обзор их позволяет
обнаружить несомненные закономерности развития этого поня-
тия в течение XIX и XX столетий С течением времени понятие
о жизненных формах становится все более полным, вскрыва-
ются все новые черты, выявляются новые связи и зависимости
жизненных форм от окружающей среды.
Вначале трактовка жизненных форм носит внешний описа-
тельный характер Первое, что было отмечено со времен Гум-
больдта, это решающее значение вегетативных органов расте-
ний в архитектонике жизненных форм. Это положение прини-
мается всеми исследователями от зарождения учения о жиз-
ненных формах до настоящего времени Правда, одни авторы
предпочитают использовать для выделения жизненных форм
признаки физиономического порядка, подчеркивая их значение.
Другие берут морфолого-биологические особенности вегетатив-
ной сферы растений. Но эти различия, в сущности, носят
частный характер, будучи связанными с направлением работ
того или иного исследователя — ботанико-географическим,
морфолого-биологическим или биоценотическим. Страстные
споры, иногда разгоравшиеся между сторонниками того или
другого направления, лишены принципиальной основы, ибо все
признаки вегетативной сферы — и «физиономические» и «био-
логические»— суть выражение определенного характера жизне-
60
деятельности растительных организмов, возникшие в истории
развития различных видов в определенных условиях среды.
Задача заключается в том, чтобы вскрыть сущность отношений
между теми или иными признаками жизненных форм и средой.
А выбор признаков определяется целевым назначением созда-
ваемых систем жизненных форм.
Затем, с победой дарвинизма, возникает и все более распро-
страняется убеждение в приспособительном характере жизнен-
ных форм, крепнет мысль о зависимости их от господствующих
внешних условий. В связи с этим обогащается и определение
жизненных форм, приобретая экологическое содержание. Прав-
да, вопрос о путях приспособления растений — прямом, в
смысле Ламарка, или косвенном, в смысле Дарвина — раз-
лично решается различными исследователями. Но экологи-
ческая концепция жизненных форм стано-
вится в настоящее время общепринятой, за
исключением немногих сторонников «типологической» школы в
ботанике, ратующих за возврат учения о жизненных формах к
концепциям ушедшей в прошлое додарвиповской эпохи в био-
логии
Наконец, как неизбежное следствие экологической концеп-
ции, все более кристаллизуется мысль исследователей об
историзме жизненных форм, о путях возникновения
их в процессе длительного развития различных филетических
групп растений в условиях различного климата, почв и биоти-
ческого окружения. При этом неизбежно поднимается вопрос о
пределах конвергентности, о необходимости учета систематиче-
ского положения растений при выделении жизненных форм и
их классификации. Думается, что решение этого комплекса
вопросов должно быть отнесено к недалекому будущему
Таким образом, в настоящее время достаточно полное опре-
деление жизненных форм должно неминуемо включить описа-
ние предмета исследования, отразить его связи с внешней сре-
дой и его историзм
6. РАЗВИТИЕ ПРЕДСТАВЛЕНИЙ ОБ ЭВОЛЮЦИИ ЖИЗНЕННЫХ
ФОРМ У РАСТЕНИЙ
«Везде базальт образует двойные купола и усеченные кону-
сы, везде трапп-порфир представлен в виде причудливых мае.
сивов скал, а гранит в виде закругленных куполов Точно так
же и сходные растительные формы — ели и дубы венчают
горные склоны как Швеции, так и южных частей Мексики»
(стр 82). В этих словах А. Гумбольдта (1806) с предельной
четкостью выражены его взгляды на жизненные формы у
растений. Физиономические формы деревьев или трав, эпифитов
61
или суккулентов присущи растительным организмам, По-
добно тому, как присуща любому материальному телу опреде-
ленная форма. В воззрениях Гумбольдта отсутствуют даже на-
меки на возможность исторического развития форм. Формы
могли лишь сменяться под влиянием величайших революций,
происходивших время от времени на нашей планете, как об
этом можно судить по сохранившимся остаткам растений в
земных слоях.
Историзм во взглядах на жизненные формы возник срав-
нительно поздно даже после триумфального распространения
теории Дарвина на все области ботаники и зоологии. Поэтому
и в настоящее время вопрос об эволюции жизненных форм
нельзя считать достаточно разработанным.
Идеи об эволюции жизненных форм у цветковых растений
возникли па рубеже XIX и XX вв., более или менее одновре-
менно в морфологии, систематике и географии растений. Вна-
чале лишь в общем виде обсуждался вопрос о соотношении
древесных и травянистых форм и, главным образом, у
покрытосеменных растений.
Многие исследователи, на основе совершенно различных
данных, пришли к выводу о первичности древесных и вторич-
ности травянистых форм у ангиосперм Так, А Н Краснов
(1899) писал: «Однолетние и травянистые типы растительности
суть новейшие типы. Флора древнейших геологических эпох с
ними не была знакома. Они являются со средины третичного
периода, получая все больше и больше преобладания по мере
приближения к современной нам эре» (стр. 56). К этому
заключению Краснов приходит при изучении истории развития
растительности с появления первых ископаемых покрытосемен-
ных в меловом периоде Фактором развития покрытосеменных
от древесных к травянистым, по Краснову, было изменение
внешних условий «Развитие это шло рука об руку с эволюцией
самих условий жизни па пашей планете, с развитием ее суши,
ее гор и плоскогорий, ее климатов Растительные формы и их
изменение были следствием такого развития Они не могли
быть иными» (стр 52—53) Он приводит значительное количе-
ство примеров «вырождения» древесных типов в низкорослые
и наконец травянистые формы в различных семействах покры-
тосеменных, по мере перехода их от тропических областей Явы,
Цейлона, Филиппин к умеренным и холодным зонам Азии и
Европы (А Н. Краснов, 1894, 1897).
Первичность древесных форм в структурном и историческом
плане у покрытосеменных признают Е. Джеффри (Е. Jeffrey,
1889), Г Галлир (И. Hallier, 1905), Р. Веттштейн (R. Wettstein,
1912), а вслед за ними И. Бейли (I. Bailey, 1911, 1912, 1914),
А. Имс (A. Eames 1910, 1911), Комптон (Compton 1914), Вэрн-
62
хем (Wernchem, 1914), Е. Синнот (Е. Sinnott, 1914), Синнот и
Бейли (Sinnott a. Bailey, 1914, 1922), Ирмшер (Irmscher, 1922),
Б. М. Козо-Полянский (1922, 1937); М. И. Голенкин (1927),
В. Г. Александров и О. Г. Александрова (1928), В. Циммер-
ман (W. Zimmermann, 1930, 1949), М Г. Попов (1940),
Е П Коровин (1940), А. А. Федоров (1943), А Л Тахтаджян
(1943, 1948, 1954), В В. Алехин (1944), А. А. Яценко-Хмелев-
ский (1947), А. А. Гроссгейм (1950), М. В. Культиасов (1950),
М. В. Сенянинова-Корчагина (1949), Г. Мейзель (Н. MeuseU
1952), Р. Шарфеттер (R. Scharfetter, 1953) и др.
Признание первичности древесных форм у покрытосеменных
аргументируется различно (см. гл. III). Анализ данных палео-
ботаники, филогенетической систематики, анатомии растений и
ботанической географии склоняет к мысли, что основным
направлением эволюции жизненных форм покрытосеменных
было их развитие от древесных к травянистым формам И
лишь в отдельных группах покрытосеменных ограниченно про-
цесс протекал в обратном направлении.
Но тезис о первичности древесных и вторичности травяни-
стых форм ангиосперм носит слишком общий характер, и
поэтому естественно стремление исследователей проследить
более детально пути эволюционных преобразований и истори-
ческого развития жизненных форм. В связи с этим делались
попытки указать основные этапные формы развития от деревь-
ев к травам Галлир (Н. Hallier, 1905) отмечает непрерывную
цепь постепенной редукции от древнейших древовидных форм
Magnoliaceae к лианам, кустарникам, многолетним травам,
однолетним и водным растениям Подобный ряд для покрыто-
семенных в целом принимает и М. Г. Попов (1940)
Весьма интересные соображения об эволюции жизненных
(рорм высказаны Раункиером (1906). Он говорит для того,
чтобы расположить жизненные формы покрытосеменных в
естественный ряд, соответственно с их эволюцией, необходимо
выяснить, какая из форм может быть принята за самую при-
митивную, первичною, родоначальную для других Если мы
примем, что первичная форма та, которая приспособлена к
жизни в условиях климата наиболее древнего для покрытосе-
менных, то вопрос может быть изменен так: какой из клима-
тов, ныне существующих, наиболее близок к климату, господ-
ствовавшему в эпоху возникновения цветковых растений? Есть
все основания полагать, что климат в этот ранний период исто-
рии был более равномерным, теплым и влажным, чем теперь.
Допустив, что постоянно теплый и влажный климат, который
теперь господствует в некоторых тропических областях, более
древний, чем другие из современных, мы должны призвать, что
и жизненные формы, приспособленные к нему, более древние.
63
От таких жизненных форм должны возникнуть и все другие,
как приспособление к климатам, возникшим позже.
В постоянно влажном и теплом климате рост растений
ничто не тормозит, поэтому развиваются формы с побегами,
уходящими высоко в воздух над поверхностью почвы, как у
деревьев и кустарников. Здесь господствуют формы фанерофи-
тов. Они дифференцируются на вечнозеленые без почечных
чешуи и листопадные с почечными чешуями Различны они и
по высоте, в зависимости от влажности, так что здесь возни-
кает ряд от мега- до нанофанерофитов.
В условиях климата с сухим или холодным периодом года,
фанерофиты уменьшаются в размерах, их рост все более
затрудняется и возникает новый тип жизненных форм — хаме-
фитов, с почками возобновления у поверхности почвы. При
дальнейшем ухудшении климата возникает третий тип жизнен-
ных форм — гемикриптофитов, с еще более защищенными поч-
ками возобновления, находящимися на поверхности почвы, или
прикрытыми подстилкой. В областях с длительным сухим
периодом года возникает четвертый тип жизненных форм —
криптофиты, с почками возобновления, скрытыми более или
менее глубоко в почве В степях и пустынях появляется и пя-
тый тип — терофитов, наиболее приспособленный к пережива-
нию длительного сухого и жаркого периода года в виде семян.
Таким образом, система Раункиера отражает, по мнению ее
автора, определенные исторические отношения жизненных
форм, но не только эколого-географические, как это часто при-
нято отмечать в экологической и ботанико-географической ли-
тературе.
Сходная концепция развивается и Р. Шарфеттером (1953).
В 1956 г. вышел в свет в виде небольшой монографии труд
соотечественника Раункиера Г. М. Ганзена (Н. Hansen, 1956),
посвященный изучению геологического возраста основных
групп жизненных форм, понимаемых в смысле Раункиера.
Исследования современных растений различного возраста
(четвертичного, или известных в ископаемом состоянии в тре-
тичном или меловом периодах), а также ископаемых флор
привело Ганзена к кардинальному выводу — р а з л и ч и ы е
жизненные формы, каки виды, имеют различ-
ный возраст: их время существования и мас-
сового развития различны. Наиболее молодые
жизненные формы — хамефиты, геофиты и терофиты с макси-
мумом распространения в четвертичном периоде. Более ранни-
ми оказываются нанофанерофиты и гемикриптофиты, развитие
которых происходило в неогене. Еще более ранними следует
считать микрофаперофиты и лианы, максимум развития кото-
рых приходился на палеоген Самыми древними были мега- и
64
мезофанерофиты с максимумом развития в меловом периоде.
Следует подчеркнуть, что эти выводы Ганзена основаны на
колоссальном фактическом материале. В его таблицах отмече-
но свыше 38 000 видов сосудистых, преимущественно покрыто-
семенных растений. Интересно, что общие выводы для всех
покрытосеменных оказываются применимыми и к отдельным
родам, виды которых резко отличаются по жизненным формам,
например, к роду Salix.
Схему, близкую к Галлиру и Попову, принимает В. В. Але-
хин (1944) Отношения между главнейшими группами жизнен-
ных форм покрытосеменных он иллюстрирует следующим
образом:
Более схематично отношения основных жизненных форм
ангиосперм отображает А Л. Тахтаджян (1948): деревья—►кус-
тарники—> полукустарники—► многолетние травы—► однолетники
М. В. Сенянинова-Корчагина (1949) приводит классифика-
цию жизненных форм покрытосеменных холодных и умеренно-
холодных областей северного полушария Ею выделяются де-
ревья, кустарники, кустарнички, полукустарники, полукустар-
нички, полутравы и собственно травы. Этот ряд форм генети-
чен Переход от деревьев к травам сопровождается изменением
целого комплекса признаков — уменьшением длительности
жизни, высоты растения, характера ветвления, но ведущим
признаком в этом ряду М В Сеняпиновой-Корчагипой призна-
ется редукция пробковой ткани и накопления древесины в
надземных стеблях.
По мнению Андреанского (1954), общее направление разви-
тия жизненных форм установить трудно. Но для некоторых
групп это возможно, например, о кактусовидных известно, что
они происходят от колючих кустарников и являются или моно-
филетическими или происходят от малого количества исходных
единиц, например, в Америке от семейства Cactaceae, в Африке
от рода Euphorbia и т д Мангровые, по Андреанскому, разви-
лись из вечнозеленых тропических деревьев. Для возникнове-
ния жизненных форм требуются не только специфические
условия среды, но и соответствующие исходные типы.
В-413 И Г. Серебряков —5 65
Более де7альную схему филогенетических отношений неко-
торых групп жизненных форм умеренных и холодных зон при-
водит Серебряков (1954). Здесь также за первичную принята
форма дерева Ведущей силой развития жизненных форм от
деревьев к однолетним травам признается изменение условий
жизни. На схеме показано возникновение сложного комплекса
жизненных форм в процессе географического распространения
покрытосеменных из умеренных зон в полярно-высокогорные и
пустынно-степные области В 1955 г. тот же автор предложил
обобщенную схему филогенетических отношений основных
групп жизненных форм покрытосеменных (рис 1). В этой схе-
ме основным направлением эволюции признан переход от дре-
весных к травянистым. Как видно из схемы, путь эволюции от
древесных к травянистым оказывается свойственным не только
прямостоячим растениям с обычной для современных цветко-
вых структурой стебля (с удлиненными побегами), но и розе-
точным формам, стелющимся, лиановидным, суккулентным, а
также корнеотпрысковым формам. Таким образом, в процессе
расселения по земной поверхности образуются взаимно связан-
ные друг с другом, параллельно развивающиеся ряды жизнен-
ных форм. Каждый из рядов предпочитает определенный ком-
плекс почвенно-климатических и фитоцснотических условий,
обнаруживая определенные закономерности в своем географи-
ческом распространении.
Представления об эволюции жизненных форм развивались
не только в общем плане, но и при монографической обработке
отдельных групп растений Укажем в качестве примеров рабо-
ты М Г Попова (1929, 1940) по родам Cicer и Eremosiuchys\
П А Баранова (1950) по роду Vitis; Н. Meusel (1941, 1952)
по семействам Araceae, Lemnaceae, родам Carlina, Echium,
Sonchus, Campanula, монографию «Биографии групп растений»
Шарфеттера (R Scharfetter, 1953) Это направление работ
имеет замечательные перспективы, так как заключения о ходе
исторического морфогенеза жизненных форм в пределах рода
или семейства отличаются большой аргументированностью п
«документальностью».
Большое значение для суждения об эволюции жизненных
форм и их экологической интерпретации имеют также многие
работы по биологии и экологии отдельных видов растений.
Подобные работы весьма многочисленны Многие из них цити-
руются ниже, в последующих главах, при анализе отдельных
групп жизненных форм. Отметим здесь некоторые интересные
работы отечественных авторов
В серии исследований Т. А. Работнов (1945, 1946, 1950,
1953 и др) установил возрастные этапы развития многих,
преимущественно луговых, травянистых многолетников и дли-
66
Рис. 1
Обобщенная схема филогенетических отношений основных групп жизненных форм покрытосеменных
(по Серебрякову, 1955)
тельность их жизни, разработав методы возрастного анализа
популяций травянистых видов в фитоценозах. Большой интерес
представляют данные Т. А. Работнова о ювенильном состоянии
травянистых многолетников в природных сообществах.
Широтой анализа жизненных форм, с привлечением физио-
логических, эколого-морфологических и ботанико-географиче-
ских методов исследования, отличается работа О. В. Заленско-
го (1948), посвященная подушковидным формам растений вы-
сокогорий Памира. Под руководством О. В. Заленского выпол-
нен ряд весьма интересных исследований по морфогенезу
жизненных форм полукустарничков и подушковидных растений
(М. Г. Зайцева, 1949; А. П. Стешенко, 1956). Интересные дан-
ные по биологии полукустарничков приводятся в работе
Н. А. Козловой (1953).
В. Н. Голубев (1956—1959) в ряде интересных исследований
установил четко выраженные возрастные изменения в морфо-
генезе различных жизненных форм у травянистых многолет-
ников (преимущественно лесной зоны Европейской части
СССР) и их изменчивость в разных условиях произрастания.
Полученные данные В. Н. Голубев рассматривает эволюцион-
но-экологически, основываясь, главным образом, на биогенети-
ческом принципе.
В исследовании А. П. Пошкурлат (1940), выполненном под
руководством А. А. Уранова, детально прослежен большой
жизненный цикл чия (Lasiagrostis splendens). Работой Пош-
курлат начата большая серия исследований возрастной биоло-
гии растений.
Исследования онтогенетического развития растений различ-
ных ботанико-географических зон нашей страны чрезвычайно
важны для дальнейшего развития учения о жизненных формах.
Вероятно, можно не сомневаться в том, что ближайшие
десятилетия дадут многое для понимания эволюции жизненных
форм, ознаменовав новую эпоху в учении о жизненных формах
у растений.
Глава II
ОПЫТ ЭВОЛЮЦИОННО-ЭКОЛОГИЧЕСКОЙ СИСТЕМЫ
ЖИЗНЕННЫХ ФОРМ ПОКРЫТОСЕМЕННЫХ
И ХВОЙНЫХ РАСТЕНИЙ
1. ОПРЕДЕЛЕНИЕ ЖИЗНЕННОЙ ФОРМЫ
У СЕМЕННЫХ РАСТЕНИИ
Обзор определений жизненных форм растений разными
авторами позволяет заключить, что учение о жизненных фор-
мах растений приобрело к настоящему времени по крайней
мере два аспекта — эколого-морфологический и эколого-цено-
тический. Они тесно связаны и в то же время достаточно
отличаются друг от друга. Поэтому могут быть два подхода
и к определению жизненных форм.
Эколого-морфологически жизненную форму у высших расте-
ний мы определяем как своеобразный общий облик (габитус)
определенной группы растений (включая их надземные и под-
земные органы — подземные побеги и корневые системы), воз-
никающий в их онтогенезе в результате роста и развития в опре-
деленных условиях среды. Этот габитус исторически возникает
в данных почвенно-климатических условиях как выражение
приспособленности растений к этим условиям.
Именно это определение приходит на ум, когда мы говорим
«дерево», «кустарник», «полукустарник», «подушковидное расте-
ние», «корневищный многолетник», «древесная лиана» и т. д.
С эколого-ценотической точки зрения жизненные формы
исторически возникли как приспособления к наиболее полному
использованию всего комплекса условий местообитания. Жиз-
ненная форма, с этой точки зрения, есть выражение способ^
ности определенных групп растений к пространственному рассе-
69
лению и закреплению на территории, к их участию в формиро-
вании растительного покрова. Понятно, что эколого-морфологи-
ческий и эколого-ценогичсский подход к жизненным формам
дополняют друг друга, отражая две грани единого сложного
явления.
Каждое растение в онтогенезе меняет свою жизненную
форму Однолетние сеянцы ели и сосны, так же как и «торчки»
дуба под пологом леса, еще не деревья. То же относится и к
травянистым многолетникам: стержнекорневые в первые годы
жизни, они затем могут стать корневищными, кистекорневыми,
столонообразующими, клубневыми и т п (В. Н. Голубев, 1957,
1959)
Но как бы пи изменялось растение в онтогенезе, для завер-
шения своего жизненного цикла оно постоянно должно быть
приспособлено к среде обитания. При этом направление и ин-
тенсивность роста органов, длительность жизни и общий габи-
тус растения будут определяться наследственными свойствами
организма и комплексом внешних условий, непрерывно воз-
действующих на живое растение в течение его онтогенеза.
Различия в ходе роста и развития растений одного и того
же вида в разных условиях могут приводить к более или менее
резко выраженным габитуальным различиям особей — к внутри-
видовой изменчивости жизненных форм.
Многочисленные опытные данные (Bonniet, 1883, Kraszan,
1900—1901; Clements, 1923, П. А Баранов, 1931; О. В Зален-
ский, 1948; Н. А. Аврорин, 1953, 1955 и др ) говорят о том, что
резкое изменение условий роста и развития радикально изме-
няет и размеры, и ритм роста, и габитуальные особенности
растений. Самый значительный вывод из подобных работ,
вероятно, заключается в том, что постоянные воздействия сход-
ных факторов среды, длящиеся в течение ряда поколений, вызы-
вают сходные физиологические и биолого-морфологические
реакции у растений разных видов В конечном итоге быть
может именно таким путем образуются сходные формы у видов
с разной наследственной основой, т е разного систематического
положения
Известно, что приспособление растений к окружающим
условиям проявляется не только в физиологических особенно-
стях (например, в засухо-, холодо- или солеустойчивости), не
только в анатомической структуре их органов (например, ксеро-
морфности или гигроморфности), но и во внешних морфологи-
ческих признаках, определяющих общий габитус или внешнюю
форму растений, следовательно,— в их жизненных формах
1 Понятно, что степень конвергентности будет снижаться то мере увели-
чения систематического гиатуса.
70
Но если это так, то из сказанного следуют два существен-
ных вывода Во-первых, понятие «жизненная форма» резко отли-
чается, как указывалось выше, от понятия «экологическая
группа» у растений, например, мезофиты, ксерофиты, литофиты,
псаммофиты, галофиты и т д Особенности последних, в отли-
чие от жизненной формы отражают приспособленность растений
только к одной группе факторов среды При этом габитуальные
признаки далеко не всегда здесь будут ведущими. Чаще тако-
выми в экологических группах оказываются признаки внутрен-
ние, физиологические. Во-вторых, если жизненные формы суть
морфологические категории в экологии растений, то характери-
зовать их необходимо в первую очередь признаками морфоло-
гическими.
Не следует смущаться при возможных упреках в «форма-
лизме» подобной трактовки жизненных форм Мы глубоко убеж-
дены в том, что у высших растений габитуальные особенности
отражают черты их биологии, ритма развития и роста, функций
отдельных органов Форма организмов у высших растений —
наиболее чуткий индикатор своеобразия их жизненных процес-
сов, особенностей их роста и развития Благодаря постоянному
нарастанию все новых органов и метамеров в течение онтоге-
неза, во внешней форме сосудистых растений и структуре их
органов запечатлена история их жизни, отношения со средой,
пути воздействия среды на организм Этим высшие растения
выгодно отличаются от животных, не обладающих столь выра-
женной хронологической мета мерностью структуры И можег
быть там выделять жизненные формы целесообразно по другим
признакам i высших растений выделение жизненных форм по
габитуальпым признакам оказывается не только простым и
удобным, но и биологически обоснованным
Говоря о том, что жизненные формы несут печать глубокого
воздействия внешних условий на виды растений разного систе-
матического положения, уместно сделать следующие замечания
Во-первых, жизненные формы отражают особен-
ности среды, например, климата или почв,
через специфику роста и развития растений
в господствующих почвенно-климатических
и ценоз и чес к их условиях. Среда действует на
форму через изменение жизнедеятельности
организма и особенно через изменение интен-
сивности и направления роста, а также дли-
тельности жизни его вегетативных органов.
Во-вторых, было бы ошибочно думать, что жизненные формы
возникают только под влиянием неблагоприятных факторов
среды или неблагоприятного времени года, как это подчерки-
вается при анализе жизненных форм в системе Раункиера.
71
Жизненные формы есть результат всей жизнедеятельности
организма как в онтогенетическом плане, так и в плане их
сезонного развития, т. е отражают не только неблагоприятные,
но и благоприятные условия роста и развития растений.
Далее в этой книге жизненные формы растений будут рас-
сматриваться только в одном, эколого-морфологическом плане
В последующих главах рассматриваются в эколого-морфо-
логическом аспекте типы древесных растений — деревья, кус-
тарники и кустарнички, а также древесные лианы и подушко-
видные растения На некоторых конкретных примерах анали-
зируется изменение жизненной формы вида в онтогенезе и ста-
новление той жизненной формы, которая свойственна данному
виду во взрослом состоянии в определенных условиях обитания.
В этой главе приводится общий обзор системы жизненных форм
древесных и наземных травянистых растений
2. ПРИНЦИПЫ ПОСТРОЕНИЯ ЭВОЛЮЦИОННОЙ СИСТЕМЫ
ЖИЗНЕННЫХ ФОРМ РАСТЕНИЙ
Всякая попытка классификации биологических явлений
встречает почти непреодолимые трудности из-за большого мно-
гообразия их и наличия переходных форм и типов в природе.
Это положение в полной мере относится и к классификации
жизненных форм, оно не раз отмечалось создателями их систем.
Трудности подобного рода несомненно возрастают при созда-
нии систем эволюционных, вследствие слабой изученности путей
филогенеза жизненных форм растений, особенно среди травя-
нистых многолетников. И все же, развитие учения о жизненных
формах привело теперь к необходимости создания именно эво-
люционных систем. Первые попытки их построения, вероятно,
будут мало удачными; но они послужат отправным этапом для
создания все более совершенных систем, все более верно отра-
жающих реальный ход истории развития жизненных форм
растений.
Уже отмечалось, что изучение эволюции жизненных форм
у растений велось и ведется двумя путями. Первый путь лежит
через детальные исследования филогенеза жизненных форм в
пределах отдельных небольших систематических групп, напри-
мер, секций или родов; второй — через широкие обобщения
хода исторического развития жизненных форм в крупных систе-
матических группах, например, однодольных и двудольных, или
покрыто-семенных в целом. Каждый JJ3 этих путей имеет свои
преимущества и недостатки И не имеет смысла их противо-
поставлять друг другу. Очевидно, создающиеся филогенетиче-
ские системы должны аккумулировать знания, накопленные
как на первом, так и на втором пути исследования.
72
Создаваемые системы должны быть одинаково и физиономи-
ческими и биологическими, а выделяемые жизненные формы
легко доступными для изучения в полевой обстановке. Специ-
фические габитуальные особенности жизненных форм должны
отражать существенные биологические черты этих растений,
возникшие под влиянием господствующих факторов внешней
среды Не стоит повторять ошибку Гризебаха и выделять жиз-
ненные формы по морфологическим признакам, биологическая
значимость и зависимость от условий жизни которых еще
недостаточно выяснены. Без учета этого обстоятельства системы
неминуемо рискуют стать «формалистическими».
Создание эволюционной системы жизненных форм ставит
вопрос о соотношении ее с эволюционной системой видов, о
путях и факторах возникновения новых жизненных форм, о
соотношении этого процесса с видообразованием.
Уже говорилось выше о том, что эволюция высших растений
шла по пути образования не только новых видов, сменяющихся
в течение геологической истории, но и определенных эколого-
морфологических групп, или жизненных форм растений, несу-
щих в своей биологии и внешнем облике неизгладимую печать
воздействия господствующих почвенно-климатических и цено-
тических условий Но так как видообразование у высших расте-
ний в конечном итоге представляет собой также морфологи-
ческое выражение эволюционного процесса, связанного с освое-
нием новых экологических ниш, то невольно возникает сомне-
ние— закономерна ли вообще постановка проблемы эволюции
жизненных форм у растений, наряду с проблемой эволюции
видовЕще со времени Дарвина остается непоколебимым поло-
жение о том, что видообразование есть процесс приспособи-
тельный, и каждый вид поэтому является своеобразной жизнен-
ной формой Однако, несмотря на очевидцую близость, а иногда
и единство процессов видообразования и развития жизненных
форм, эти процессы отличаются своеобразием Видообразование
всегда протекает как процесс дивергентный в том смысле, что
каждый вид в истории жизни возникает лишь однажды и от
одного или определенного сочетания родительских видов (по-
следнее при процессах гибридизации). Науке неизвестны случаи
независимого появления одного и того же нового вида от раз-
ных родительских видов, или одного и того же вида в разные
геологические эпохи При появлении новых видов, вызываемом
в конечном итоге изменением внешних условий, у них всегда
сохраняются систематические черты большей или меньшей
древности, выражающиеся в родовых, семейственных и т. д
признаках, определяющих неповторимость вида в истории
жизни
В отличие от эволюции видов и таксонов более высокого
73
ранга, в эволюции жизненных форм, кроме дивергенции, ши-
роко распространено явление конвергенции. Дивсргентно новые
жизненные формы возникают в пределах узкой систематической
группы, например, в процессе дифференциации единого вида
на близко родственные самое! оятельпые, при расширении
ареала и заселении новых районов с иными условиями, или
изменении климата, или занятии новых экологических ниш
в пределах старого ареала. Формирование новых видов и разно-
видностей и новых жизненных форм в таких случаях совпадает.
Достаточно вспомнить соотношение древесной формы Pinus
sibirica (Rupr) Моуг. и стланцевой Р. pumila (Pall.) Rgl.,
равнинного древовидного Jumperus communis L, и горного
стелющегося J. clepressa Stev, высокотравной луговостепной
формы Sanguisorba officinalis L., и полурозеточной приполярной
Sanguisorba polygama Nyl , лесной высокотравной Solidago
virga aurea L, и низкотравной горно-тундровой 5 lapponica
With, луговолесной высокотравной формы Polygonum bistorta
L , и тундровой полурозеточной Р. elliptica Willd, и т. д. Изуче-
ние эволюции жизненных форм в подобных группах, как пра-
вильно указывает Г Мейзель (Н. Meusel, 1951, 1952) t открывает
перспективы причинного исследования процессов видообразо-
вания Но сходные жизненные формы, как указывалось выше,
могут возникнуть и независимо друг от друга в далеких систе-
матических группах, например, разных семействах и порядках,
т. е конвергентно, под влиянием одинаковых факторов внешней
среды, действующих из поколения в поколение
Правда, тождественные жизненные формы в разных систе-
матических группах не образуются1 к одной и той же жизнен-
ной форме, понимаемой как основная классификационная еди-
ница (см ниже), относятся особи лишь одного вида, да и то
во взрослом состоянии в определенных условиях произраста-
ния Но конвергентно возникают существенные эколого-морфо-
логические признаки и даже их комплексы, лежащие в основе
выделения таксономических групп жизненных форм более или
менее крупного объема. Поэтому в одних и тех же группах
жизненных форм оказываются виды растений разного система-
тического положения. Достаточно вспомнить примеры много-
численных вечнозеленых деревьев влажных тропиков с колон-
новидными стволами и досковидными выростами или ходуль-
ными корнями в их основании, принадлежащих к разным
семействам двудольных; подушковидные растения высокогор-
ных, арктических и субантарктических областей; суккулентно-
стеблевые и суккулентпо-листовые формы растений аридных
стран, весьма различные по своему' систематическому положе-
нию и т д. Родовые, семейственные и другие систематические
признаки сохраняются преимущественно в генеративной сфере,
74
ле отражаясь существенно в признаках, по которым выделяют-
ся эти группы жизненных форм.
Способность растений одного и того же вида образовывать
разные жизненные формы (в разных условиях произрастания),
а растений разных систематических групп конвергентно образо-
вывать сходные жизненные формы, позволяет рассматривать
эволюцию жизненных форм растений как самостоятельную
проблему. Конечно, изучение эволюции жизненных форм долж-
но основываться на филогенетической системе видов В свою
очередь знание эволюции жизненных форм растений от-
кроет новые возможности перед их филогенетической система-
тикой
Движущей силой исторического развития жизненных форм
растений являются изменения условий существования, включая
факторы как абиотической, так и биотической среды, вызываю-
щие сходные структурно-биологические реакции растений раз-
ных систематических rpjnn, например, видов и родов. В оценке
действия среды наиболее полная и правильная формула, по-
видимому, предложена Б. А Келлером (1948). Согласно его
формулировке среда воздействует па организмы как формирую-
щий и отбирающий фактор.
Но какие же именно изменения условий существования
оказали наиболее серьезное влияние на эволюцию жизненных
форм современных семенных растений и что принять за веду-
щий биологический процесс в бесконечных преобразованиях
жизненных форм"* Как найти здесь «нить Ариадны», которая
могла бы провести через сложный лабиринт эволюционных
путей развития жизненных форм от гигантских деревьев влаж-
ных тропиков до прижимающихся к земле шпалерных кустар-
ничков тундры и эфемерных пигмеев-однолетников азиатских
и африканских пустынь?
История формирования современных флор различных
областей земного шара, начиная с мелового периода, с очевид-
ностью показывает, что основным стимулом эволюции расти-
тельного мира, оказывающим свое влияние на растения с мо-
мента выхода их на сушу, была борьба за влагу (Е. В Вульф,
1944). Изменения режима увлажнения, в сторону ли физиче-
ской сххости или физиологической сухости, чаще всего связан-
ной со снижением температур, всегда неизбежно влекли за
собой изменения состава флоры, появление новых видов расте-
ний, несущих черты приспособления к постоянному или времен-
ному недостатку влаги, изменение внешнего облика растений,
их жизненной формы
А Н Краснов, наблюдавший изменение растительности тро-
пической Азии при подъеме в горы, где сильно сказывается
влияние низких температур, писал (1894), что в верхнем поясе
75
склонов гор Явы невольно видишь элементы тундры, увеличен-
ные до деревьев, или, правильнее, наоборот — элементы тропи-
ческого леса, собранные в формацию тундры, но еще не успев-
шие уменьшиться и выродиться в настоящие тундровые формы.
После подъема на вершины гор Цейлона автор отметил, что
там растительность представляла собой как бы недоконченный
луг, где господствующие семейства наших луговых растений
еще имели вид деревьев и кустарников. Это все подтверждало
мысль Краснова о том, что растительность холодного пояса
нашей планеты образовалась путем отбора из элементов тропи-
ческой растительности видов, могущих расти при условии
низкой температуры. Действуя веками на этих избранников,
она содействовала их вырождению в арктических пигмеев.
Изменения влажности и температуры, как показываю!
многочисленные факты, вызывают у древесных растений прежде
всего изменения характера роста и размеров. При недостатке
тепла формируются все более низкорослые деревья, кустар--
ники, наконец, кустарнички В сухих областях тропиков с резко
выраженным засушливым периодом тропический высокостволь-
ный лес заменяется низкоствольным лесом, зарослями колючих
кустарников и саваннами. В областях с еще большим периодом
засухи, в пустынях и полупустынях, появляются мелкие тра-
вы — эфемеры с чрезвычайно кратким жизненным циклом
Анализ морфогенеза различных типов жизненных форм
покрытосеменных, от крупных деревьев до многолетних и одно-
летних трав, позволяет заключить, что важнейшим структурно-
биологическим признаком, определяющим габитус растения,
является сравнительная длительность жизнен-
ного цикла главной оси и заменяющих ее основных
надземных скелетных осей растения В длительности
жизненного цикла скелетных осей и их струк-
туре концентрируется как в фокусе та самая
специфика роста и развития растений, которая
и Я-влястся видимой реакцией па воздействие
условий внешней среды, главным образом, на
изменение влажности и температуры, о которых
шла речь выше.
лМожно определенно говорить, что главное содержание про-
цесса эволюции жизненных форм у покрытосеменных от де-
ревьев к травам связало с сокращением длительности жизнен-
ного цикла надземных скелетных осей (рис 2).
Длительность жизни (в годах) у надземных скелетных осей
(выше места возникновения инноваций) обычно совпадает
с длительностью их нарастания в длину (мопоподиального или
симподиального). С прекращением нарастания начинается их
подсыхание.
76
возраст скелетных осей
Рис. 2
Обобщенная схема изменений прироста и длительности жизни
скелетных осей у различных жизненных форм
покрытосеменных (по Серебрякову. 1955):
/ — высокоствольные деревья, II — немногиствольные деревья;
HI — кустарники; IV — кустарнички; V — травянистые многолетники;
VI — травянистые однолетники
Далее, при анализе отдельных типов жизненных форм, на
конкретных примерах будет показано, какой именно может быть
эта длительность жизненного цикла надземных скелетных осей,
какие взаимоотношения складываются между надземными и
подземными органами и как все это отражается на габитусе
и общей длительности жизни всего растения.
Конечно, изменение длительности жизненного цикла ске-
летных осей сопровождается изменением их внутрен-
ней анатомической структуры. В первую очередь,
менее длительное развитие осевых органов связано с пониже-
нием камбиальной активности в стебле, которую некоторые
авторы склонны считать основной причиной эволюции от дре-
весных форм к травянистым. Но, как видно из всего изложен-
ного, это скорее следствие, чем причина изменения жизненных
форм. Следствием же разного характера скелетных осей будет
и степень паренхиматизации или одревеснения их внутренних
тканей. Изменение характера покровных тканей осевых органов,
потеря ими способности образовывать перидерму и постоянное
сохранение травами лишь эпидермиса (что отмечается М. В. Се-
няниновой-Корчагиной, 1949), является также процессом ведо-
мым в эволюции жизненных форм, производным, хотя и с
большим биологическим значением. Этот процесс нельзя счи-
тать ведущим так же, как нельзя принять изменение характера
кожных покровов, например, позвоночных, за ведущий струк-
турно-биологический признак их эволюции. Трудно принять за
ведущий процесс в переходе от древесных форм к травянистым
и явление неотении (А. Л. Тахтаджян, 1943, 1948, 1954), так
как длительность виргинилыюг® периода от всходов до первого
77
цветения у травянистых растении нередко не меньше такового
у древесных (Т. А Работпов, 1950).
Наряду с изменением длительности жизни надземных ске-
летных осей для появления и развития многих классов жизнен-
ных форм имело большое значение также изменение на-
правления роста скелетных осей (ортотропное
или плагиотропное), появление суккулентности,
появление специальных органов — цепляющихся,
запасающих и т д Так, стелющиеся и шпалерные формы кус-
тарников и кустарничков смогли приспособиться к жизни в
условиях низких температур воздуха, не сокращая длитель-
ности жизни своих побегов по сравнению с прямостоячими фор-
мами, живущими в более теплых условиях; так, стеблевые сук-
куленты в пустынях смогли сохранить длительность жизни
своих осей, характерную для деревьев или кустарников более
влажных областей Однако эти классы и группы растений обыч-
но носят более частный, специализированный характер, обра-
зуя ряды жизненных форм — стелющихся, лианоидных, сукку-
лентных и других, входящих каждая в определенный тип
жизненных форм деревьев, кустарников, кустарничков, полу--'
кустарников, поликарпических или монокарпических трав
(рис 1). К обзору этих типов и рядов, образующих систему
жизненных форм, мы и переходим.
3. ОСНОВНЫЕ КОНТУРЫ ЭВОЛЮЦИОННО-ЭКОЛОГИЧЕСКОЙ
СИСТЕМЫ ЖИЗНЕННЫХ ФОРМ ПОКРЫТОСЕМЕННЫХ
И ХВОЙНЫХ РАСТЕНИЙ
Всякая система жизненных форм должна иметь определен-
ное назначение Предлагаемая здесь система в известной сте-
пени сходна с системой «главных жизненных форм» Дю Рие
(1931) Она ставит своей целью обзор и ориентировку в много-
образии жизненных форм покрытосеменных и хвойных и иллю-
страцию эволюционных отношений их таксономических групп
крупного ранга Система носит эколого-морфологический харак-
тер Но если она правильно отражает реальные отношения
жизненных форм, она может быть полезной в ботанико-геогра-
фических исследованиях и исследованиях фитоцснологического
характера.
Обзору системы жизненных форм мы предпосылаем анализ
единиц классификации различного ранга, принятых в нашей
системе Как было показано в историческом обзоре классифи-
каций жизненных форм (см. главу 1), в системах ряда авторов
выделенные группы разного объема получили определенные
названия. Так, в системе жизненных форм Гамса (Н. Gams,
78
1918) наиболее крупные группы называются типами Типы жиз-
ненных форм подразделяются па классы, в свою очередь в ряде
случаев подразделяемые на подклассы. Крупные группы в сис-
теме жизненных форм Варминга (Е Wanning, 1923) — классы
подразделяются на более мелкие группы без особых названий
Дю Рие (Du Rietz, 1931) некоторые группы выделяемых им
форм роста называет типами, не придавая, впрочем, этому тер-
мину смысла таксономической единицы определенного объема
В системе А В Прозоровского (1936) наиболее крупные таксо-
номические группы — типы (древесные, полудревесные, полу-
травянистые, травянистые и эфемерные); в первых двух типах
выделены подтипы В системе жизненных форм растений це-
линных степей юго-востока России Г. Н Высоцкого (1915) наи-
более крупные единицы системы — отделы, подразделены на
группы и в ряде случаев на подгруппы У многих авторов
выделяемые группы жизненных форм разного объема и сопод-
чинения не имеют никакого названия В общей форме, насколь-
ко нам известно, вопрос об единицах классификации жизнен-
ных форм, их объеме, принципах выделения и названий в фито-
экологической литературе не обсуждался.
Для общей системы жизненных форм покрытосеменных и
хвойных здесь предлагаются следующие единицы классифика-
ции: отделы, типы, классы, подклассы, группы, подгруппы, сек-
ции жизненных форм и собственно жизненные формы. Послед-
няя единица классификации, или первая, если ряд начать
с наиболее мелких единиц, собственно жизненная форма, может
быть уподоблена виду в систематике и является основной еди
ницей экологической системы растений Под жизненной
формой, как основной таксономической единицей автор
понимает совокупность взрослых особей дан-
ного вида в определенных условиях произ-
растания, обладающих, как уже было сформули-
ровано выше, своеобразным общим обликом
(габитусом), включая надземные и подземные
органы (подземные побеги и корневую сис-
тему). Онтогенетически этот габитус возни-
кает в результате роста и развития в данных
условиях среды, а исторически — в определен-
ных почвенно-климатических и ценотических
условиях, как выражение приспособленности
растений к этим условиям Такое определение позво-
ляет более или менее строго ограничивать объем и границы
каждой жизненной формы, в связи с возрастной и экологической
изменчивостью жизненных форм, обычной среди ангиосперм
Самой крупной классификационной единицей в нашей системе
является отдел, подразделяемый на типы Выделение отделов
79
основано в первую очередь на структуре надземных осей (дре-
весные, полудревесные и травянистые), а выделение типов в
отделе древесных растений — на относительной длительности
жизни надземных осей, в отделе же наземных травянистых
растений — на длительности жизни растений в целом, связан-
ной, как правило, с различной способностью к вегетативному
возобновлению и размножению после плодоношения. Последо-
вательность отделов и типов, принятая в нашей системе, пока-
зывает основное направление эволюции жизненных форм по-
крытосеменных — от деревьев к травам, вплоть до однолетних.
В эволюционном плане отделы и типы обозначают боль-
шие этапы основного пути эволюции жизненных форм анги-
осперм.
Классы в пределах типа выделяются на основании призна-
ков большого экологического и эволюционного значения, на-
шедших свое проявление в структуре побегов (например, расте-
ния с суккулентпыми, ползучими, розеточными и т. д. побе-
гами) В ряде типов выделяются классы на основе специали-
зации питания растений (сапрофиты и паразиты) и образа
жизни (эпифиты). Но выделение этих классов не нарушает
принятого принципа, так как своеобразие питания и образа
жизни этих растений находит свое выражение и в структуре их
побегов. Особенности структуры побегов придают жизненным
формам каждого класса резко выраженное морфологическое
и экологическое своеобразие. Классы жизненных форм в ряде
случаев иллюстрируют направления экологической эволюции
в пределах типа, их приуроченность и приспособление к раз-
личным почвенно-климатическим и ценотическим условиям.
Вероятно можно говорить о разной древности классов в преде-
лах типа, например, о древности розеточных форм деревьев
и кустарников (Corner, 1949, 1955). Но эти вопросы еще мало
исследованы, данных для суждения о них совершенно недоста-
точно
Более мелкие таксономические единицы (подклассы, группы,
подгруппы, секции жизненных форм) определяются по отдель-
ным более частным экологическим признакам, не имеющим
столь крупного эволюционного значения Исключение состав-
ляют лишь поликарпические травы, самый обширный класс
которых — многолетние травы с ассимилирующими побегами
песуккулентного типа — подразделяется на подклассы, харак-
теризующие основное направление их эволюции
Уже говорилось, что система Раункиера отражает, и в зна-
чительной степени правильно, основные этапы эволюционного
развития жизненных форм от деревьев до травянистых одно-
летников. Однако нельзя не согласиться с рядом критических
замечаний в адрес этой системы, служащих основанием для ее
80
существенных изменений и создания новых эволюционных
систем. В частности, прав Г. Гаме (1918), говоря, что подраз-
деление фанерофитов у Раункиера по высоте (от мега- до нано-
фанерофитов) носит произвольный характер и мало аргументи-
ровано. Поэтому в нашей системе все древесные растения,
многие из которых входят в группу фанерофитов, подразделе-
ны на деревья, кустарники и кустарнички, как это принято в
системах многих исследователей (Друде, 1913; Дю Рие, 1931,
Алехин, 1944 и др). Кустарники и кустарнички, как отмеча-
лось, можно рассматривать как самостоятельные типы и этап-
ные формы на эволюционном пути от деревьев к травам. Обра-
зующийся ряд деревья—кустарники — кустарнички характери-
зуется все большим сокращением жизненного цикла их скелет-
ных осей, как это будет показано ниже (главы III, IV и V).
Группа хамефитов Раункиера — сборная и искусственная;
в нее входят древесные и травянистые растения, приуроченные
к разным климатическим областям (например, подушки, полу-
кустарники). Резко различаются разные хамефиты и эколого-
морфологически. Растения, рассматриваемые в системе Раун-
киера как хамефиты, отнесены в нашей системе в более есте-
ственные типы кустарничков, полукустарников и полукустар-
ничков, поликарпических трав и подушковидных растений.
Выделение гемикриптофитов и криптофитов Раункиера, как
этапных групп в широком эволюционном плане, вряд ли
оправдано. Целесообразнее говорить о типе поликарпических
травянистых растений, весьма разнообразном в экологическом
и морфологическом отношениях. Эволюция травянистых поли-
карпиков в наиболее обширном классе — многолетников с асси-
милирующими побегами несуккулептного типа — шла не только
в сторону дальнейшего сокращения длительности жизни ске-
летных осей и образования вегетативных и семенных малолет-
ников и однолетников, но и по пути изменения типов окорене-
ния и способов вегетативного размножения растений В эволю-
ции травянистых многолетников этого класса проявляются две
противоположные тенденции1 1) усиление вегетативного размно-
жения с образованием различных специализированных органов
такого размножения при сокращении жизни побегов материн-
ского растения (образование «вегетативных малолетников» по
Г Н Высоцкому, 1915) и 2) ослабление вегетативного размно-
жения и возобновления после плодоношения с образованием
олигокарпиков, монокарпиков, вплоть до однолетников. Обе
тенденции экологически связаны с ухудшением условий произ-
растания. Но вторая из них несомненно всегда связана с
усилением аридности климата, первая же — только иногда
(например, вивипария у Роа bulbosa, образование воздушных
луковичек у видов Allium и т. д.). Гораздо чаще усиление веге-
В-413 И Г. Серебряков—6 81
тативного размножения и специализация его органов бывают
связаны с холодным климатом и избыточным увлажнением,
с созданием условий типа олиготрофных болот и тундр.
Система жизненных форм завершается типом монокарпи-
ческих травянистых растений, проходящих свой жизненный
цикл в течение нескольких, двух лет или одного года. Форма-
ми, переходными от травянистых поликарпиков к монокарпи-
ческим травам и, в частности, к однолетникам будут олигокар-
пики, многолетние монокарпики и двулетние растения.
Водные и земноводные травы, представляющие большое
разнообразие жизненных форм, в нашей системе детально не
расчленены. Она включает только формы наземные.
Жизненные формы покрытосеменных и хвойных
Эволюционные отношения отделов и типов жизненных
форм покрытосеменных даны на рис. 3
Отдел А. Наземные и эпифитные древесные растения
(Holoxyles, Du Rietz, 1931)
Всегда несут более или менее возвышающиеся над уров-
нем почвы многолетние скелетные оси, в большей или мень-
шей степени одревесневшие и одетые вторичной покровной
тканью (перидермой, коркой) включают три типа: I Деревья,
II Кустарники, III Кустарнички.
1 тип. Деревья
Всегда обладают более или менее развитым многолетним,
в разной степени одревесневшим стволом, разветвленным или
неветвящимся, сохраняющимся в течение всей жизни рас-
тения; длительность жизни измеряется десятками и сотнями
лет; высота от 3—5 до 150 м. Обитатели влажных и частич-
но аридных областей от экваториального до умеренно-хо-
лодных поясов, подразделяются на три класса. Классифика-
цию деревьев иллюстрирует рис. 4.
1 класс. Кронообразующие деревья
с полностью отревесневшими удлиненными побегами
(Long-shoot trees, Du Rietz, 1931)
Ствол сильно ветвящийся, несущий в своей верхней части
крону из ветвей разного порядка (вегетативных и цветоносных).
82
Рис. 3.
Соотношение отделов и типов жизненных форм покрытосеменных.
Стрелками указаны основные направле ия эволюции
6*
Рис. 4
Классификация жизненных форм деревьев
Эволюционно молодые (Comer, 1949, 1955), широко распро-
страненные в настоящее время «обычные» деревья, разделя-
ются на два подкласса.
1 подкласс. Наземные кронообразующие деревья
В течение всей жизни связаны с почвой корневой систе-
мой.
Группа А. Деревья с «обычной», лишь подзем-
ной корневой системой
Подгруппа а—деревья с прямостоячими
стволами.
Рост стволов всегда ортотропен.
Секция 1. —одноствольные деревья — деревья лесного
типа.
Ствол (моноподиальный или симподиальный), единствен-
ный в течение всего онтогенеза дерева, длительное время со-
храняет резкое преобладание в интенсивности роста в длину
и толщину над боковыми ветвями. Крона начинается более или
менее высоко над землей из-за отмирания нижних слабых
ветвей («очищения ствола» по терминологии лесоводов). Да-
же в кроне главная ось заметно выделяется по толщине
среди боковых ветвей. Сюда относятся, например, дуб —
Quercus robur, липа — Tilia cordata и очень многие деревья
влажных умеренно-холодных, умеренно-теплых и тропических
областей. При спиливании дерева из спящих почек могут вы-
расти два или несколько вторичных стволов, например, у ви-
дов березы, дуба и др. По ритму развития листвы подразде-
ляются на листопадные и вечнозеленые с разной длительно-
стью жизни листьев. Деревья следующих трех секций также
могут быть разделены на листопадные и вечнозеленые.
е . 9 Кустовидные или немногоствольные (плей-
Секция 2. окормные) деревья, деревья субарктического
и субальпийского типа.
Во взрослом состоянии имеют несколько стволов, возни-
кающих из спящих почек у основания материнского главного
ствола. В отличие от лесных деревьев боковые стволы воз-
никают здесь в результате естественного процесса старения,
но не в связи с удалением материнского ствола. Предста-
вляют собой переходные формы от типичных деревьев к ку-
старникам. Наиболее характерны для субальпийской и субарк-
тической границы лесной области. Хорошим примером может
служить Betula, tortuosa Ldb,, описанная ниже, в гл. Ш.
с о Деревья лесостепного или caeaHHOBoVo типа,
секция б. или «дерева плодового типа» (A. F. Schimper,
С. Faber, 1935).
85
В онтогенезе дерева ствол рано теряет преобладание в
росте над боковыми ветвями, поэтому крона начинается близ
поверхности почвы, а в самой кроне главная ось не выделя-
ется среди мощных боковых ветвей. В онтогенезе и архи-
тектуре дерева есть черты кустарников (см. главу IV). При-
мер—лесная яблоня (Maias silvestris, см. главу III), многие
деревья саванновых лесов Африки, Южной Америки и Азии
из разных семейств двудольных (Parkia biglobosa, рис. 5).
Рис. 5
Типичное саванновое дерево Parkia biglobosa
в травянистой саванне Африки (по П. А. Баранову,
с . Саванновые деревья с водозапасающим рез-
Секция 4. ко утолщенным (иногда бочковидно) мощным
стволом. Например, баобаб (Adansonia dig it ata), каваниллезия
(Cavanillesia arborea) и др. Деревья этой секции имеют чер-
ты, сближающие их с суккулентно-стеблевыми деревьями (см.
класс 3).
г - Сезонно-суккулентные деревья, деревья вне-
Секция о. тропических аридных областей с сезонно-сук-
кулентными безлистными ветвями. Суккулентны лишь специ-
ализированные однолетние побеги, так называемые «ассими-
лирующие веточки», опадающие в течение сухого и жаркого
лета и осени первого года их жизни. Сохраняющиеся долго-
86
летние побеги, образующие скелет кроны, не суккулентны,
а наоборот, сильно одревесневают. Обитатели пустынь и
полупустынь с жарким летом и морозной зимой, например —
виды саксаула (Haloxylon persicum и Н. aphyllum), см. гл. III.
Подгруппа б —Деревья с лежачим укоре-
няющимся стволом и главными ветвями —
с тла нцы
Главный ствол довольно рано переходит от ортотропного
роста к плагиотропному, полегая на землю и укореняясь.
Сюда относятся стланцы, нередко встречающиеся среди хвой-
ных в субальпийском поясе гор, близ северных границ леса
и иногда на торфяниках и песках в таежной зоне. Типичный
пример таких деревьев — кедровый стланик (Pinus pamlla) и
туркестанский можжевельник (Juniperus turkestanica), опи-
санные ниже, в гл. VI.
Подгруппа в —Д еревья с лианоидн ы.м ство-
лом. Неспособны к прямостоячему росту и используют
соседние древесные растения как опору. Распростране-
ны исключительно во влажно-тропических лесах, напри-
мер, Lonchocbrpus cyanescetis (рис. 6).
Группа Б. Деревья, обладающие, кроме «обыч-
ных» подземных корней, еще и специализиро-
ванными надземными корнями.
Тропические деревья из рщных семейств. Широко описаны
в литературе.
Подгруппа а — Деревья с «ходульными» над-
земными корнями в нижней части ствола.
Преимущественно деревья мангровых зарослей и невы-
сокие деревья влажно-тропических лесов (рис. 7), на-
пример, Uapaca sp. в лесах Гвинеи.
Подгруппа б — Деревья с «дыхательными»
корнями, обладающими отрицательным геотропизмом
и высовывающимися из почвы. Встречаются в мангро-
вых зарослях, например, виды Avicennia.
Подгруппа в —Деревьяс «д о с к о в и д н ым и»
выростами — подпорками корневого происхождения
в основании ствола. Главным образом гигантские де-
ревья влажно-тропических лесов (рис. 8), например, Ceiba
pentandra во влажно-тропических лесах Западной Афри-
ки.
2 подкласс. Гемиэпифитные кронообразующие деревья
Начинают развитие как типичные эпифиты, но с течением
времени воздушные корни их достигают поверхности почвы,
87
Рис. 6
Древовидная лиана Lonchocarpus в
Ботаническом саду Конакри (Гвинея)
Рис. 7
Uар аса guineensls с ходульными
корнями
проникают в нее, сильно утолщаются над землей и образуют
ложные стволы — «корневые стволы» (Cockayne, 1928; Comer,
1951).
Рис. 8
Ceiba pentandra с досковидными придаточными
корнями
Группа А. Вегетативно неподвижные гемиэпи-
фиты. Воздушные корни—ложные стволы растут лишь
вниз по стволу дерева-хозяина, тесно окружая его со всех
сторон; утолщение их приводит к повреждению и отмиранию
ствола дерева-хозяина. Сюда относятся «деревья-душители»
различных типов, например виды Ficus влажно-тропических
лесов юго-восточной Азии (Ан. А. Федоров, 1959) (рис. 9).
89
А, Б, В, Г, Д,
Рис. 9
Фикусы-душ ители:
Развитие фикуса-душителя по Корнеру (Corner. 1958) и Федорову (1959)
А — фикус-эпифит пророс на ветви дерева-хозяина и выпустил вертикальный
канатообразный корень; Б — корень фикуса достиг почвы и укоренилса в ней,
В — образование разветвлений и анастомозов корня фикуса, охватывающих ствол
дерева-хозяина, Г — образовался корневой ствол типа решетчато корзинообразного
чехла, дерево-хозяин отмирает, Д — ф^кус-душитсль в стадии независимого
дерева, дерево-хозяин погибло.
Группа Б. Вегетативно подвижные гемиэпифи-
т ы. Воздушные корни растут не только вниз по главному
стволу, но и вдоль ветвей кроны дерева-хозяина, спускаясь
к земле под всей его кроной и образуя множество колонно-
видных «стволов» корневого происхождения. Сюда относятся
некоторые виды Ficus влажнотропических лесов юго-восточ-
ной Азии, например баниан {Ficus benghalensis, F. religiose)
Это — «баниан-форма» Гризебаха (1882).
2 класс. Розеточные деревья (Rosette trees, Du Rietz, 1931)
Деревья co стволом слабо или совсем неветвящимся, по-
стоянно образующим на своей вершине лишь укороченное
побеги, несущие розетки крупных листьев. В отличие от де-
ревьев предыдущего класса, образующих крону из ветвей,
розеточные деревья несут листовую крону. Недоразвитие
междоузлий у розеточных деревьев связано со слабым одре-
веснением и недостаточной механической прочностью стебля
(Corner, 1949; Schmid, 1956). Иногда, по-видимому, это эволю-
ционно древние формы (в пределах соответствующих систе-
матических групп), сохранившиеся в тропических и субтро-
пических областях.
1 подкласс. Неветвящиеся розеточные деревья
Крайние формы этого класса, имеющие лиш^ простой не-
90
Рис. 10
Неветвящиеся розеточные деревья Klngia australis из
Западной Австралии (по Дю Рие, 1931)
ветвящийся ствол с розеткой листьев на верхушке. Виды
пальм, Ravenala, Strelitzia: некоторые Lobeliaceae, Composi-
tae и т. д., (рис. 10). (Е. Du Rietz, 1931).
2 подкласс. Ветвящиеся розеточные деревья
Образуют ряд форм, от очень слабо ветвящихся (напр.
Xanthorrhoea Preissil (рис. 11), виды Pandanus) до регулярно
ветвящихся. Хорошим примером могут служить различные
древесные лилейные (Dracaena, Yucca, Cordyline, некоторые
Composltae). Можно различать группу ветвящихся розеточ-
ных деревьев с несуккулентными листьями и с суккулент-
ными листьями, например, виды А1оё.
Рис. 11
Ветвящиеся розеточные деревья со слабым монопоциальным
ветвлением Xanthorrhoea Prelssii из Западной Австралии
(по Дю Рие, 1931)
3 класс. Суккулентно-стеблевые безлистные деревья
(Leafless succulent-stammed trees. Du Rietz, 1931). Деревья
co стволом и ветвями (где они есть) мясистой консистенции,
с укороченными междоузлиями и недоразвитыми листьями,
нередко превращенными в колючки. Стебли выполняют водо-
запасающую и ассимилирующую функции, слабо одревесне-
вают, но имеют мощно развитую водоносную паренхиму из
сильно вакуолизированных клеток. Характерно резко ослаб-
ленное ветвление, иногда оно совершенно отсутствует. Оби-
татели тропических и субтропических аридных областей без
холодного (зимнего) времени года. Сюда относятся много-
численные виды Cactaceae, Euphorblaceae (рис. 12) и ряд
других. ______________________
II тип. Кустарники
Главный ствол имеется лишь в начале жизни растения, а
затем он теряется среди равных ему или более сильных над-
земных скелетных осей нескольких порядков, возникающих
последовательно из спящих почек у основания материнских
осей, и отмирает. Во взрослом состоянии кустарник всегда
обладает несколькими или многими надземными скелетными
осями, последовательно сменяющимися в онтогенезе расте-
ния и связанными друг с другом своими базальными участками.
92
Рис 12
Безлистные суккулентно-стеблевые деревья Carnegia gigantea:
Аризона. США (по Дю Рие, 1931)
В отличие от деревьев, длительность жизни надземных скелет-
ных осей у кустарников в большинстве случаев невелика:
10—20 лет (хотя может варьировать от 2—3 до 30—40 и
более лет). Высота кустарников — от 0,8—1,0 м до 5—6 м\
диаметр надземных скелетных осей от 1—2 см до 5—8 см.
Широко распространены от экваториальных областей до хо-
лодных зон. Количество видов и эколого-морфологическое
разнообразие кустарников возрастает близ континентальных,
субальпийских и субарктических границ лесной раститель-
ности, где они нередко становятся эдификаторами раститель-
ного покрова. Кустарники подразделяются на четыре класса.
Три первых класса аналогичны таковым деревьев.
Систему жизненных форм кустарников иллюстрирует
рис. 13.
1 класс. Кустарники с полностью одревесневшими удлинен-
ными побегами
Этот класс аналогичен подобному классу деревьев. Сюда
относится большинство кустарников. Скелетные надземные
оси ортотропные, плагиотропные или лианоидные, более или
менее обильно ветвящиеся, с удлиненными междоузлиями
годичных побегов.
1 подкласс. Прямостоячие кустарники
Надземные оси ортотропны, они образуют более или ме-
нее плотный пучок, при слабом развитии их плагиотропных
93
Рис. 13.
Классификация^жизненных форм кустарников
подземных частей. При наличии более или менее длинных
подземных одревесневших стеблей — ксилоподиев или обшир-
ной корневой системы с корневыми отпрысками — они рас-
полагаются рыхло, образуя куртины в несколько десятков
квадратных метров. Наиболее богатый представителями
подкласс кустарников.
Они распространены в различных климатических областях,
как гумидных, так и аридных.
Группа А. Аэроксильные кустарники (Shrub-trees
Lindman’a, 1914; Aeroxylic erect long shoot shrubs Du Rietz, 1931).
Неспособны к подземному ветвлению. Под землей новые оси
могут возникать лишь как корневые отпрыски. Ветвление осей
надземное, начинается близ поверхности земли образованием
нескольких вертикальных или наклонных осей. Аэроксильные
кустарники —переходные формы между деревьями и кустар-
никами.
Подгруппа а—Рыхлые аэроксильные ку-
старники. Довольно близки к секции немногостволь-
пых или кустовидных деревьев, описанных на примере
Betula tortuosa. Примером аэроксильных рыхлых кустар-
ников могут служить Sorbus aucuparia, Padus racemosa,
Salix caprea в неблагоприятных заболоченных место-
обитаниях таежной зоны Европейской части СССР. В бо-
лее благоприятных условиях эти же виды образуют ку-
стовидные деревья.
Подгруппа б—Подушковидные аэроксиль-
ные кустарники. Характеризуются ничтожными и
одинаковыми для всех побегов годичными приростами
и плотным расположением скелетных осей и ветвей.
Верхушки ветвей образуют более или менее ровную по-
верхность подушки. Примером могут служить Bupleurum
fruticosum,Poterium spinosum (W. Rauh, 1939), виды рода
Astragalus из секции 7ragacantha (М. С. Шалыт, 1955)
и др.
Группа Б. Геоксильные или настоящие кустар-
ники (True shrubs, Geoxyles по Lindman, 1914; Geoxylic erect
long shoot shrubs Du Rietz, 1931). Растения с подземным
ветвлением осей, образующих толстые и долговечные под-
земные одревесневшие оси — ксилоподии, от которых отхо-
дят более тонкие и менее долговечные надземные оси. Сюда
относится большинство кустарников европейской флоры.
Хорошими примерами могут служить виды Rosa, Corylus avel-
lana, Berberis vulgaris и др. К этой же группе принадлежат
многие виды бамбуков тропических и субтропических стран.
Аэроксильные и геоксильные кустарники в свою очередь
95
могут быть подразделены на подгруппы вегетативно непод-
вижных и вегетативно подвижных (посредством корневых
отпрысков и столонов). Примерами корнеотпрысковых аэрок-
сильных кустарников могут служить Prunus spitiosa (см. гл. IV),
Hippo phae rhamnoides\ среди геоксильных кустарников спо-
собны к возобновлению при помощи корневых отпрысков
и столонов, например, Philadelphus coronarius, Syringa 'vul-
garis, Symphoricarpus racemosa (Du Rietz, 1931).
2 подкласс. Полупростратные
и стелющиеся кустарники.
Главная ось и боковые ветви лежачие, укореняю-
щиеся, приподнимающиеся у верхушки. В связи с уко-
ренением лежачих осей система главного корня отмирает
или сохраняется (при слабом окоренении осей). Однако
эти отношения первичной и вторичной корневых систем
остаются мало изученными. Большинство полупрострат-
ных кустарников, вероятно, ветвится только надземно.
В противоположность прямостоячим кустарникам, полу-
простратные редко встречаются в аридных областях, но
оказываются доминирующими в субальпийской и субарк-
тической областях, образуя криволесье (например, в
Альпах — Krummholz из Pinas mughus и Ainas viridis (по
С. Schroeter, 1908); заросли ивняков в горах Сканди-
навии; кустарниковые виды Juniperus в горах Европы
(по Du Rietz, 1931 и т. д).
3 подкласс. Лианоидные кустарники
Обладают скелетными осями, неспособными к самостоя-
тельному ортотропному росту и использующими соседние
древесные растения как опору. Отличаются от лианоидных
деревьев более тонкими стеблями (диаметром не более
5—8 см) Преимущественно обитатели влажных тропических
и субтропических лесов, реже —саванновых и колючих лесов
и кустарников. К этому подклассу относят примитивные
древесные лианы. Классификация в пределах лиан основы-
вается на способах лазания (см. гл. VII).
2 класс. Розеточные кустарники с укороченными побегами
(Rosette shrubs, Du Rietz, 1931)
Аналогичны классу розеточных деревьев. Надземные оси
с укороченными междоузлиями и розетками крупных листьев.
Встречаются сравнительно редко; например, кустарниковая
пальма Badris major (О. Drude, 1913), Dracophyllum fiordense,
Senecio Stewartiae (E. Du Rietz, 1931). Имеются переходные
формы между розеточными кустарниками и кустарниками с
96
удлиненными побегами, например, Olearia operina, О. Lyallii
(Du Rietz).
3 класс. Суккулентно-стеблевые безлистные кустарники
Аналогичны подобному классу деревьев. Скелетные оси
мясистой консистенции, выполняющие водозапасающую
и ассимиляционную функции. Преобладают прямостоячие
формы. Сюда относятся виды Opuntia и другие кактусы,
виды Euphorbia и др.
4 класс. Полупаразитные и паразитные кустарники
С почвой не связаны. Живут на других древесных растениях,
присасываясь к ним специальными органами— гаусториями.
В большинстве случаев имеют зеленые листья и способны
сами к фотосинтезу. Сюда относятся виды Loranthaceae
(главным образом, в тропических странах, в умеренной зоне—
омела — Viscum album) и некоторые Santalaceae (например,
Osyris alba, из Средиземноморской области, по Веттштейну,
1912). ________________________
Ш тип. Кустарнички
Древесные растения, у которых главная ось имеется лишь
в начале онтогенеза, а затем сменяется боковыми надзем-
ными осями из спящих почек базальной части материнской
оси, во взрослом состоянии несут значительное количество
ветвящихся скелетных осей (парциальных кустов), связанных
друг с другом надземно или подземно и последовательно
сменяющихся в течение онтогенеза растения. В отличие от
кустарников, длительность жизненного цикла ортотропных
надземных осей у кустарничков обычно не превышает 5—10
лет, а высота растений колеблется от 5—7 см до 50—60 см.
Преимущественно обитатели высокогорных областей, уме-
ренно-холодных и холодных зон. Система жизненных форм
кустарничков дана на рис. 14.
1 класс. Кустарнички с полностью одревесневшими
удлиненными побегами
Надземные оси более или менее обильно ветвятся, годич-
ные побеги имеют удлиненные междоузлия.
1 подкласс. Вегетативно неподвижные прямостоячие
кустарнички с полностью одревесневшими удлиненными по-
бегами
В-413 И Г. Серебряков — 7 97
Рис. 14
Классификация жизненных форм кустарничков
Образуют формы довольно плотных кустарничков, с тесно
сближенными надземными осями, без длинных подземных
ризомов, неспособны к вегетативному размножению.
Группа А. Аэроксильные вегетативно-непод-
вижные кустарнички. Ветвление лишь в надземной
части, близ поверхности почвы.
Подгруппа а — Рыхлые аэроксильные ку-
старнички. К этой подгруппе мсгут быть отнесены
некоторые экземпляры вереска Calluna vulgaris. Регу-
лярно развивающихся по этому типу кустарничков в Ев-
ропе не имеется. По Дю Рие (Du Rietz, 1931) такие
кустарнички весьма обычны среди зарослей кустарнич-
ков Южной Австралии и, вероятно, других областей
с подобным климатом (например, в Капской области).
Хорошие примеры таких форм кустарничков встречаются,
по Дю Рие, среди Epacridaceae в окрестностях Сиднея,
например, Epacris microphylla, Е. obtusifolia, Leucopo-
gon microphyllus (рис. 15). Только аэроксильные кустар-
нички известны в пределах рода Euphrasia* например,
Е. formosissima с островов Хуан Фернандес.
Подгруппа б — Подушковидные аэрок-
сильные кустарнички. Годичные приросты всех
побегов одинаковы и ничтожны по величине. Скелетные
оси и ветви расположены плотно и образуют подушку
с более менее ровной поверхностью. Распространены
в субальпийских и альпийских, субантарктических
и арктических областях. Примером может служить
Loiseleuria procumbens на ветровых местообитаниях
горных и равнинных тундр.
Группа Б. Геоксильные вегетативно непод-
вижные прямостоячие кустарнички
Аналогичны геоксильным неподвижным кустарникам.
Новые оси возникают от подземного побега типа ксилопо-
дия. В Европе такие кустарнички отсутствуют, но весьма
обычны, по Дю Рие, среди кустарничков Австралии, например,
Calectasia cyanea (Llliaceae, рис. 16), Beckea linifolia (Му г fa-
сеаё) и др.
2 подкласс. Вегетативно подвижные ползучие кустарнички
с полностью одревесневшими удлиненными побегами.
Группа А. Вегетативно подвижные кустар-
нички с ортотропными надземными побегами.
Образуют обширные рыхлые куртины и латки площадью
в десятки кв. м\ парциальные кусты куртин соединены длин-
7Х 99
Рис. 15
Аэроксильный прямостоячий вегетатив-
ночгеподвижный кустарничек Leucopo-
gon mlcroihyllus из окрестностей Сид-
нея (по Дю Рие, 1931)
ными одревесневшими подземными ризомами. Размножаются
вегетативно.
Подгруппа а — По л унеподвижные кустар-
нички. В течение всей жизни сохраняется главный
корень. . Поэтому растение в целом неподвижно, но
каждый ползучий побег подвижный. Хорошим примером
может служить вереск (Calluna vulgaris, см. гл. V),
Cassiope tetragona (Е. Warming, 1908), Erica tetralix
(E. Warming, 1916).
Подгруппа б —Настоящие ползучие кус-
100
Рис. 16
Геоксильный прямостоячий вегетативно-неподвижный
кустарничек Catectasia суапеа из Западной Австралии
(по Дю Рие, 1931)
тарни'чки, Главный корень рано или поздно отмирает,
поэтому в целом образуется настоящее ползучее расте-
ние. Сюда относятся Vactinium vitis idaeu, V. myrtillus,
Empetrutn nigrum (E. Warming, 1916) и др.
Группа Б. — Вегетативно-подвижные кустар-
нички шпалерного типа. В отличие от предыдущей
группы, имеют только плагиотропные надземные побеги,
101
прижатые к земле и укореняющиеся на всем своем протя-
жении. Приурочены, главным образом, к наиболее неблаго-
приятным местообитаниям равнинных и горных тундр,
а также сфагновым болотам. Примерами могут служить
Arcious alpina, Dryas octopetala, Dryas punctata, виды
Oxycoccus.
3. подкласс. Вегетативно-неподвижные лианоидные ку-
старнички. Побеги неспособны к постоянному самостоятель-
ному ортотропному росту и нуждаются в опоре. В случае
отсутствия опоры стелются по земле и укореняются, обра-
зуя обширные куртины. Типичный пример — плющ (Hedera
helix) — см. гл. VII.
2 класс. Розеточные кустарнички
По структуре побегов аналогичны розеточным кустар-
никам и деревьям. Встречаются редко. Пример — Dracophyl-
turn Menziesii из Новой Зеландии (Du Rietz, 1931).
3 класс. Суккулентно-стеблевые безлистные кустарнички
Надземные побеги с водозапасающей и ассимилирующей
функцией и поэтому мясистой консистенции. Встречаются
редко, например, среди Chenopodiaceae Среднеазиатских
пустынь (напр. Halocnemum strobilaceum, М. С. Шалыт,
1955); чаще в этих условиях встречаются формы, переход-
ные к суккулентным полукустарничкам, характеризующиеся
ежегодным отмиранием более или менее значительной части
годичных побегов.
4 класс. Паразитные и полупаразитные кустарнички
Живут на ветвях и сучьях деревьев, используя их пласти-
ческие вещества. Пример — некоторые виды Loranthaceae.
Отдел Б. Полудревесные растения (Hemixyles 1
___________________Е. Du Rietz, 1931) _____________|
В этом отделе имеется лишь один тип — полукустарники
и полукустарнички. Характеристики отдела и типа совпадают.
IV тип. Полукустарники и полукустарнички
Удлиненные ортотропные надземные оси на значительной
части их длины остаются травянистыми и отмирают ежегод-
но
но. Сохраняются и одревесневают лишь базальные части
надземных осей. Многолетние осевые органы с почками во-
зобновления располагаются, как правило, близ поверхности
почвы, частично оказываются даже под землей. Цикл разви-
тия монокарпических побегов — не более 3—5 лет, общая
высота растений обычно до 50—80 см, иногда до 1,5—2 м,
редко более. Преимущественно обитатели аридных областей.
1 класс. Полукустарники и полукустарнички с удлиненными
несуккулентными побегами
1 подкласс. Прямостоячие полукустарники и полукустарнички
Группа А. Прямостоячие полукустарники. Веге-
тирующие побеги высотой до 1,5—2,0 м. РМноголетние одре-
весневшие базальные участки побегов до 20—30 см высоты.
Пример — Artemisia arenaria в песчаных пустынях и полу-
пустынях Средней Азии и некоторые другие виды полыней.
Группа Б. Прямостоячие полукустарнички. Веге-
тирующие побеги высотой обычно в 20—30 см. Многолетние
одревесневшие базальные участки побегов с почками возоб-
новления обычно не выше 3,5—7 см. Обычная в среднеази-
атских пустынях группа полукустарничков (виды Artemisia
подрода seriphidium), Chenopodiaceae (напр., Kochia prostrata)
и др. Возможно выделение подгруппы подушковидных полу-
кустарничков (виды Silene, Astragalus, Шалыт М. С., 1955).
2 подкласс. Стелющиеся полукустарники с удлиненными
несуккулентными побегами. Базальные многолетние участки
с почками возобновления обычно засыпаны почвой (песком);
однолетние побеги — приподнимающиеся. Сюда относятся
некоторые Chenopodiaceae и виды Astragalus среднеазиатских
пустынь, а также Artemisia procera на берегах рек средней
полосы Европейской части СССР.
3 подкласс. Лианоидные полукустарники. Побеги слабые,
опирающиеся на соседние растения, а в случае отсутствия
опоры стелющиеся по земле и укореняющиеся. Типичный
пример — Solanum dulcamara в зарослях кустарников по бе-
регам рек.
2 класс. Полукустарнички с побегами суккулентного типа
Многолетние виды Chenopodiaceae среднеазиатских
пустынь («солянки»).
103
Розеточные полукустарнички, аналогичные розеточным
деревьям, кустарникам и кустарничкам, в литературе не опи-
саны и, вероятно, отсутствуют из-за различия условий мор-
фогенеза полукустарников, с одной стороны, и розеточных
форм — с другой.
Паразитные полукустарнички в литературе тоже не
описаны.
Отдел В. Наземные травянистые растения1
Совершенно лишены надземных сильно одревесневших
многолетних осей, со вторичной древесиной, формирующейся
в течение нескольких или многих лет; все надземные стебли
травянистые; покровные ткани их первичные, без перидермы.
Тип V. Травянистые поликарпики
Многолетние многократно плодоносящие травы. Надзем-
ные удлиненные ортотропные побеги отмирают в конце пер-
вого вегетационного периода; укороченные надземные или
удлиненные, но плагиотропные надземные побеги могут со-
храняться несколько лет. Подземные части побегов функцио-
нируют как органы возобновления или запасающие органы в
течение нескольких или многих лет, или также отмирают
ежегодно, дав начало новым подземным и надземным побе-
гам. Распространены от экваториальных до арктических и
субантарктических стран.
1 класс. Травянистые поликарпики с ассимилирующими
побегами несуккулентного типа (многолетние травы
«обычного» типа)
Побеги с зелеными листьями или зеленым стеблем, орто-
тропные или плагиотропные, удлиненные или укороченные.
Собственно многолетними являются обычно подземные орга-
ны, обеспечивающие вегетативное возобновление и размно-
жение растений этого класса. В связи с этой жизненно важ-
1 Предлагаемая здесь система жизненных форм травянистых растений
рассматривается автором как предварительная. В ней совсем не отражены
весьма своеобразные и мало известные нам травы влажно-тропических
и субтропических лесов. В нее также не включены водные и земноводные
травы. (Примеч. автора)
104
ной ролью подземные органы здесь отличаются большим
разнообразием, дающим основание для дальнейшего подраз-
деления класса.
Многолетние травы этого класса отличаются весьма ши-
роким распространением от тропиков до арктических и суб-
антарктических стран и наибольшей изученностью.
1 подкласс. Стержнекорневые травянистые поликарпики
В течение всей жизни растения сохраняют стержневой
главный корень нередко значительной длины и диаметра с
системой боковых корней; стеблеродные корни (стеблевое
окоренение) отсутствуют или выражены слабо. Вегетативно
неподвижны и неспособны к вегетативному размножению, за
исключением корнеотпрысковых форм.
Группа А. Одноглавые стержнекорневые много-
летники
Выше главного корня развивается неразветвленный, резко
укороченный подземный побег, каудекс, переходящий в над-
земный ортотропный стебель, более или менее разветвленный
в верхней части. В надземной части развивается обычно один,
редко 2 или 3 крупных ветвящихся побега. Одноглавые
стержнекорневые обычно относятся к высоким или средне-
высоким травам. Стержневой главный корень, иногда очень
длинный, идущий на глубину более 1 м. Примерами могут
сложить Palitnbia racemosa, Ferula tatarica, F. caspica, Scor-
zonera strlcta, S. hispanica в степях Северного Казахстана
(И. В. Борисова, i960).
Группа Б. Многоглавые стержпекорне’вые много-
летники
Выше главного корня развивается несколько, иногда
много, наклонных укороченных подземных побегов, (разветв-
ленный каудекс), переходящих в надземные ортотропные
стебли.
Подгруппа й—Длинностержнекорневые мно-
гоглавые многолетники. В надземной части об-
разуется до нескольких десятков побегов. По данным
И. В. Борисовой (1960) в Северном Казахстане подоб-
ные растения, например, Sllene sibirlca, Medicago roma-
nica, Onobrychis sibirlca, Seseli Ledebourii, Echinops ritro,
Limonium sareptanum и др., характерны для всех типов
степей.
Подгруппа б — Короткостержнекорневые
многоглавые многолетники. Короткостержне-
вые, выделенные впервые Е. М. Лавренко (1935) среди
растений Нижнеднепровских песков, отличаются не
только экологической приуроченностью к особым усло-
виям почвенного увлажнения, но и структурно-биологи-
ческими чертами, сближающими эту группу с под-
классом кистекорневых растений. Главный корень корот-
костержнекорневых сравнительно короткий, сильно сбе-
жистый, уходит в почву обычно менее чем на 1 м.
Иногда он отмирает у взрослого растения, заменяясь
системой боковых и стеблеродных (придаточных) корней,
например, у Senecio Jacobaea, S. eruciformis, видов
Pulsatilla (И. В. Борисова, 1960). Отмирание или сохра-
нение главного корня зависит также от условий место-
обитания. Как и среди длинностержнекорневых, здесь
наблюдается образование корнсотпрысковых форм от
примитивных с побегами лишь на главном корне (Silene
multiflora, Viola rupestris), до типичных корнсотпрыско-
вых с особыми «корнями размножения» (виды Linaria,
Achillea gerberi и т. д.). И. В. Борисова (1960) отмечает
среди этой подгруппы ряд растений с различной дли-
тельностью жизни от типичных многолетников (виды
Pulsatilla, Scussurea сапа) до переходных к двулетникам
(Silene parvifolia, S. wolgensis, Senecio Jacobaea, Erige-
ron acer и т. д.). В этой же подгруппе, по данным
И. В. Борисовой, имеются формы, близкие к кистекор-
невым растениям (виды Pulsatilla'), к кустарничкам
(Saussurea сапа, некоторые гвоздики и астрагалы), по
душковидным (Dianthus acicularis).
Группа В — Стержнекорневые травянистые мно-
голетники, образующие форму «перекати-поле»
Специализированная группа стержнекорневых поликарпи-
ков с чертами приспособления к распространению семян на
большие расстояния и удалению массы надземных побегов
с места произрастания растения (В. В. Алехин, 1936). Широ-
ко распространены в открытых степных, полупустынных и
пустынных формациях. Примерами могут служить Gypsophila
paniculata, Statice tatarica, Nepeta ucralnica и т. д. Подоб-
ные специализированные формы перекати-поле образуются и
в других подклассах травянистых поликарпиков, например,
среди кистекорневых и короткокорпевищных (Asparagus
officinalis) и даже луковичных (Hyacinthus ciliatus,
В. В. Алехин, 1936).
106
Группа Г — Подушковидные травянистые поли-
ка рпики
Характеризуются ничтожным и одинаковым у всех веге-
тативных побегов приростом, вследствие чего поверхность
подушки оказывается более или менее ровной и над ней мо-
гут возвышаться лишь цветоносные побеги. Приурочены к
альпийским, арктическим и реже аридным областям. Приме-
ры: Silene acaulis, описанная в гл. VIII, некоторые виды
Saxifraga, Draba, Arabis и др.
2 подкласс. Кистекорневые и короткокорневищные
травянистые поликарпики
В отличие от стержнекорневых система главного корня
рано отмирает в онтогенезе растения и заменяется системой
стеблеродных корней. Вегетативное размножение слабо выра-
жено или совсем отсутствует.
Группа А. Кистекорневые травянистые мно г о-
л етн ики
Система главного корня отмирает более или менее рано
в онтогенезе растения и во взрослом состоянии функциони-
рует лишь система стеблеродных (придаточных) корней. Над-
земные побеги и стеблеродные корни отходят от корневище-
подобного подземного органа, каудекса, с короткими годичны-
ми приростами, живущими каждый не более 2—3 (4) лет.
Таким образом, вся живая часть каудекса состоит не более
чем из 2—3 (4) годичных приростов и всегда остается ко-
роткой и лишена отходящих в стороны укореняющихся от-
ветвлений. Пластические вещества распределены диффузно
не только в стебле, но и в основаниях корней, нередко утол-
щенных и мясистых в связи с этой функцией. Из всех имею-
щихся почек возобновления ежегодно раскрывается одна,
реже 2—3, в связи с чем находится их одпостебельность
или немногостебельность. Зато стебли часто обладают силь-
ным ростом; кистекорневые обычно относятся к высоким
или средневысоким травам. Вновь образующиеся надземные
монокарпические побеги несут весьма короткую плагиотроп-
ную часть. Поэтому типичные кистекорневые многолетники
относятся к «сидячим» растениям и обладают весьма слабо
выраженной способностью к вегетативному распространению
107
и размножению. Широко распространены от тундровой до
степной зоны СССР, наиболее обычны среди луговых много-
летников лесной зоны; например, Caltha palustris, виды
Ranunculus, Veratrum Lobellanum, Succlsa praemorsa, Trollius
europaeus.
Группа Б. Короткокорневищные травянистые
многолетники
Формы, экологически и морфологически близкие к пре-
дыдущей группе. Во взрослом состоянии обычно лишены
системы главного корня и живут за счет вторичной корне-
вой системы из стеблеродных корней. Из почек возобновле-
ния на ризоме ежегодно развертывается одна, реже 2—3,
образуя средне-высокие надземные побеги. В отличие от
кистекорневых короткокорневищные многолетники всегда
имеют четко выраженный подземный мясистый ризом (корне-
вище), состоящий из значительного числа коротких годичных
приростов, сохраняющихся живыми до 10—15 и более лет.
Ризом постоянно нарастает у верхушки и отмирает у осно-
вания. Способность к ветвлению ризома и вегетативному
размножению выражена слабо. Примеры: Polygonatum offi-
cinale, Р. multiflorum, Anemone ranunculoides, Paris quadri-
folia и др.
3 подкласс. Дерновые многолетники
Во взрослом состоянии лишены системы главного корня,
который отмирает раньше, чем у растений предыдущего
подкласса. Обладают лишь вторичной корневой системой,
состоящей из стеблеродных (чаще «узловых» —W. Rauh, 1937)
корней. От кистекорневых и короткокорневищных сущест-
венно отличаются тем, что у них ежегодно раскрывается не
одна или немного, а большое количество почек возобновле-
ния и образуются более или менее крупные «кусты» или
куртины. Кусты и куртинки развиваются по принципу «центро-
бежного роста» (П. А. Смирнов, 1958), т. е. дочерние побеги
всегда направлены к периферии куста или куртины.
Группа А. Плотнокустовые дерновые многолет-
ники
В основании каждого ортотропного монокарпического
надземного побега образуется зона укороченных междоузлий
с расположенными на ней почками возобновления и стебле-
родными корнями. Симподиальная система таких укорочен-
ных оснований монокарпических побегов образует многолет-
108
нюю часть плотнокустового растения/ расположенную на
поверхности почвы (иногда немного погруженную в почву
или, наоборот, чуть приподнятую над ней). Вегетативно эти
растения почти неподвижны, за некоторыми исключениями
неспособны к вегетативному распространению и размножению.
Весьма типичны для этой группы плотнокустовых злаки,
например виды Stipa, Festuca sulcata, F. ovina, Deschampsia
caespitosa, Nardas stricta и т. д. Обычно плотнокустовые
растения, особенно злаки, приурочены к местообитаниям с
плохой аэрацией почвы (В. Р. Вильямс, 1922) — степям,
сильно задерненным лугам, иногда болотам и т. п.
Группа Б. Рыхлокустовые дерновые многолет-
ники
Монокарпические побеги имеют ясно выраженную плагио-
тропную часть, образующую недлинное подземное горизон-
тальное или наклонное корневище. Обычны среди растений
тундровой и лесной зоны СССР, особенно на лугах. В сте-
пях и пустынях становятся более редкими. Типичные при-
меры: Festuca pratetisis, Alopecurus pratensis, Phleum praten.se,
Luzula pilosa, Carex leporlna и т. д.
Группа В. Длиннокорневищные дерновые мно-
голетники
В отличие от предыдущих групп подкласса обладают
длинными подземными ветвящимися корневищами, обусловли-
вающими образование крупных рыхлых кура ин надземных по-
бегов. Резко выражена способность к вегетативному рас-
пространению и размножению. Сильное развитие вегетатив-
ного размножения иногда коррелятивно связано с резко
ослабленным семенным воспроизведением.
Куртины, как правило, представляют собой систему пар-
циальных кустов, соединенных между собой более или менее
длинными подземными корневищами. В зоне кущения каждого
парциального куста возникают побеги с разной длиной пла-
гиотропной подземной части (от нескольких мм до десятков
см), и наиболее длинные из них становятся материнскими
побегами новых парциальных кустов.
Различные виды обладают корневищами разной длины.
Увеличение длины корневища в серии видов оказывается кор-
релятивно связанным с уменьшением их толщины и долго-
летия. Вместе с тем длина корневищ тесно связана с плот-
ностью почвы и ее аэрацией. Длиннокорневищные экологи-
чески приурочены к хорошо аэрируемым, достаточно увлаж-
ненным почвам (В. Р. Вильямс, 1922). Весьма обычны среди
травянистых растений хвойных и широколиственных лесов,
109
лугов, довольно обычны в тундрах и становятся реже в сте-
пях и пустынях (за исключением песчаных пустынь).
Особенно типичны для этой группы злаки—Agropyrott
repens, Bromus inermis, Digraphis arundlnacea\ осоки — Carex
pilosa, Carex physodes и др.
4 подкласс. Столонообразующие и ползучие травянистые мно-
голетники
Многолетники с резко выраженной способностью к веге-
тативному размножению при помощи надземных или подзем-
ных столонов или стелющихся побегов.
Группа А. Столонообразующие травянистые
многолетники
Резко выражена способность к вегетативному размноже-
нию посредством образования столонов (надземных или под-
земных), не связанных, подобно корневищам (ризомам), с
запасающей функцией, и поэтому обычно довольно тонких и
ежегодно отмирающих. В связи с образованием столонов
иногда подавлено семенное воспроизведение, особенно в
крайних условиях произрастания (например, Saxifraga flagel-
laris в арктических тундрах Таймыра, Б. А. Тихомиров,
1951). Ежегодное отмирание надземных и подземных органов
материнских побегов позволяет говорить о таких растениях
как о своеобразных «вегетативных малолетниках» (Г. Н. Вы-
соцкий, 1915). Некоторые столонообразующие растения (на-
пример, Lysitnachia vulgaris, Mentha arvensis) в соответ-
ствующих условиях произрастания вместо столонов образуют
длинные корневища, сохраняющиеся в жизнедеятельном to-
стоянии до 3—4 лет (В. Н. Голубев, 1957). Эти факты гово-
рят о морфологической близости столонообразующих и длин-
нокорневищных форм травянистых * многолетников. Экологи-
чески столонообразующие многолетники приурочены к влаж-
ным и избыточно увлажненным местообитаниям с рыхлой,
часто заторфованной почвой — лесам, болотам.
П-одгруппа а — Подземно-столо иные расте-
ния, например, Adoxa moschatellina, Trientalis euro-
paea,
Подгруппа б— Надземно-столонные расте-
ния, например, Fragaria vesca, Saxifraga flagellaris,
Rubus saxatilis.
Группа Б. П о лзу чи е т р а в я н и с т ы е м н о го л е тн и к и
Сюда относятся многолетние травы, все побеги которых рас-
тут плагиотропно, по поверхности почвы, регулярно образуя
стеблеродные корни. Как и столонообразующие, и дерновые
НО
растения, ползучие многолетники рано теряют систему глав-
ного корня и во взрослом состоянии обладают лишь вторич-
ной корневой системой. В силу этого у ползучих многолет-
ников сильно развито вегетативное распространение и размно-
жение. В отличие от ортотропных удлиненных побегов тра-
вянистых многолетников, отмирающих ежегодно, плагиотроп-
ные удлиненные побеги ползучих травянистых поликарпиков
сохраняются живыми в течение нескольких лет. Подобная
же тенденция к удлинению жизненного цикла ползучих по-
бегов отмечается и для стелющихся древесных растений,
начиная от древовидных стланцев (Pinus pumila и т. п.) и
кончая арктоальпийскими шпалерными кустарничками различ-
ных видов Ericaceae. У многих ползучих растений нарастание
плагиотропных побегов моноподиальное (например, Veronica
officinalis, Lysimachia nummularia). Экологически они при-
урочены к местообитаниям с достаточно влажной почвой,
что необходимо для стеблевого окоренения побегов. Коли-
чественных данных об экологическом и ботанико-гео!рафиче-
ском распределении ползучих трав не имеется.
5 подкласс. Клубнеобразующие травянистые многолетники
Обладают специализированными запасающими органами —
клубнями корневого, стеблевого и листового происхождения,
часто выполняющими и функцию вегетативного размножения.
Свойственны областям с прерывающейся вегетацией вслед-
ствие наступления засушливого времени года. Но обстоятель-
ных исследований экологической и ботанико-географической
приуроченности клубневых многолетников в районах их мак-
симального видового и морфологического разнообразия и
наибольшего участия в растительном покрове не имеется.
В пределах подкласса четко выражен морфогенетический ряд
от растений с запасающими клубнями, неспособных к веге-
тативному размножению и расселению, до растений, у кото-
рых клубни оказываются специализированными органами ве-
гетативного размножения.
Группа А. Клубневые многолетники с клубнями
корневого происхождения (корне клубневые
многолетники)
Подгруппа а —Редьковидные корнеклуб-
невые многолетники с клубневидно утолщенным
главным корнем. Обычно у этих многолетников разраста-
ние в толщину не ограничивается главным корнем, но
переходит на гипокотиль и базальные междоузлия глав-
ного стебля, образуя клубень смешанной корнестеблевой
природы. Хорошим примером могут служить некоторые
ill
Crassulaceae — Sedum Telephium, S. maximum (W. Troll,
1937) и др.
Подгруппа б — Корнеклубневые растения с клубня-
ми,. образующимися из стеблеродных (придаточных) кор-
ней, возникающими на коротких корневищах (например,
Ficaria ranunculoides, Filipendula hexapetala, виды Orchis,
Platanthera bifolia и др.), или на длинных корневищах
(Potentilla alba, В. Н. Голубев, 1959). Экологическая
приуроченность этих растений не изучена. Как и в пре-
дыдущей подгруппе, корневые клубни этих растений
являются запасающими органами.
Группа Б. Клубневые травянистые многолетни-
ки с клубнями стеблевого происхождения (стеб-
леклубневые многолетники)
Подгруппа а —Стеблеклубневые много-
летники с клубневидно утолщенным осно-
ванием главной оси. В образовании клубня прини-
мает участие или только гипокотиль (гипокотильно-
клубневые растения, вроде Carum bulbo casta пит, виды
Cyclamen — W. Troll, 1937), или вместе с гипокотилем
также и эпикотильная часть главной оси с укороченны-
ми междоузлиями (Corydalis cava — W. Troll, 1937).
По участию гипокотиля и базальных междоузлий
главной оси в образовании клубня эти растения примы-
кают к редьковидным многолетникам. Но в отличие от
последних, у стеблеклубневых растений главный корень
рано отмирает, и во взрослом состоянии функционирует
вторичная корневая система из стеблеродных корней.
Этот признак свойствен, вероятно, всем стеблеклуб-
невым многолетникам. Клубни главной оси, по W. Troll
(1937), многолетни и в разной степени способны к вто-
ричному утолщению.
Подгруппа б — Стеблеклубневые расте-
ния с клубнями на боковых побегах —кор-
невищах (корневищно-клубневые растения). В отличие
от клубней главной оси, корневищные клубни функцио-
нируют лишь один год, и ежегодно, после использова-
ния пластических веществ, сменяются новыми. Типич-
ными примерами могут служить Arum maculatum, виды
Crocus, Gladiolus, Colchicum.
Подгруппа в — С т е бл е к л у б н е в ы е расте-
ния с клубнями на верхушках столонов (сто-
лонно-клубневые многолетники). Образование клубня на
верхушке столона влечет за собой отмирание его не-
утолщенного начала, благодаря чему клубень изолирует-
112
ся от материнского растения и дает начало новой особи.
Столонные клубни поэтому оказываются не только за-
пасающими органами, но и специализированными орга-
нами вегетативного размножения. Как и корневищные,
столонные клубни функционируют и сохраняются от
одного вегетационного периода до другого и после по-
требления их питательных веществ на развитие новых
побегов отмирают и заменяются новыми, возникающими
из почек возобновления. Типичными примерами могут
служить картофель (Solatium tuberosum), Hellanthus tu-
berosus, Stachys tubiferns, Sagittaria sagittae folia.
Подгруппа г — Надземно-клубневые мно-
голетники. Клубневидны пазушные почки надземных
побегов, включая область соцветия. Клубни способны
отделяться от материнского растения и давать начало
новым особям, оказываясь таким образом весьма спе-
циализированными органами вегетативного размножения.
Примером может служить Polygonum viviparum, у ко-
торого основная масса клубенька образована первым
междоузлием бокового побега (гипоподием). В области
соцветия клубеньки могут возникать или только сл>чай-
но, или образуются регулярно наряду с образованием
плодов и семян, или образование клубеньков сопровож-
дается недоразвитием плодов из цветков, или в со-
цветии формируются только клубеньки, а образование
цветков подавлено. Все эти степени преобразования со-
цветия в органы вегетативного размножения можно ви-
деть у растений одного и того же вида, например, у
того же Polygonum vivlparum в горно-лесном и горно-
тундровом поясах Приполярного Урала, что вероятно
связано с ухудшением экологических условий, становя-
щихся все более неблагоприятными для развития цвет-
ков и семян.
Группа В. Клубневые многолетники с клубнями
листового происхождения. Встречаются редко. Примером
может служить описанный Д. Е. Янишевским и Б. А. Тихо-
мировым Cardamine tenuifolia (Ldb) Turcz. (1939).
6 подкласс. Луковичные травянистые много-
летники
Обладают специализированными запасающими органами в
форме подземных и надземных луковиц, выполняющих одно-
временно функции органов вегетативного размножения. Эко-
логически наиболее характерны для аридных областей, где
вегетация прерывается наступлением сухого и жаркого вре-
В-413 И Г. Серебряков—8 113
мени года. Возможно выделение луковичных эфемероидов —
с очень короткой длительностью жизни надземных органов.
Из-за слабой эколого-морфологической изученности подраз-
деление луковичных многолетников на группы и подгруппы
не приводится.
7 подкласс. Корнеотпрысковые травянистые
поликарпики (корнеотпрысковые многолетники)
Растения с более или менее подавленным надземным вет-
влением и соответственно недоразвитой системой главного
побега, способные к внепазушному ветвлению за счет обра-
зования почек и побегов на гипокотиле (гипокотильных по-
бегов) и на корнях (корневых побегов). Образуют морфоге-
нетический ряд форм от растений с факультативным появле-
нием почек в подсемядолыюй части растения до облигатно
корнеотпрысковых растений, с образованием несколько ме-
таморфизированных «корней размножения» (Л. И. Казакевич,
1921). У растений последнего типа ветвление главной оси в
надсемядольной части нередко редуцировано. Типично кор-
неотпрысковые растения с побегами на боковых корнях спо-
собны к интенсивному вегетативному размножению и рассе-
лению. Экологическая приуроченность корнеотпрысковых
многолетников недостаточно выяснена.
Группа А. Факультативно корнеотпрысковые
растения. Корневые отпрыски могут появляться, напри-
мер, в условиях улучшенного роста {Anemone silvestris, Alli-
aria officinalis} или при поранении главного побега (Сус-
lamen persicum, Linum austriacum).
Группа Б. Облигатно корнеотпрысковые расте-
ния. Многие виды, Linaria, Euphorbia, Cirsium и др.
Подгруппа а — Корнеотпрысковые расте-
ния с побегами лишь на главном корне.
Подгруппа б — Корнеотпрысковые расте-
ния с побегами на боковых корнях, несколько
видоизмененных в связи с функцией вегетативного раз-
множения (например, Mulgedium tataricum, А. М. Бы-
лова, 1959).
2 класс. .Травянистые поликарпики с ассимилирующими
побегами суккулентного типа
Характерные растения аридных областей тропических и
внетропических стран.
1 подкласс. Суккулентно-стеблевые травя-
нистые многолетники. Примерами могут служить Pelar-
gonium tetragonum. Р. echinatum (Geraniaceae), Cissus cacti-
114
for mis {Vitae eae)\ Kleinia articulata (Compositae) {W. Troll,
1937), а также мелкие представители кактусов.
2 подкласс. Суккулентно-листовые травянистые много-
летники. Вероятно, виды Mesembryanthemum (если они
поликарпичны), некоторые Chenbpodiaceae Среднеазиат-
ских пустынь; представители Crassullaceae {Sempervi-
vum, Sedum).
3 класс. Сапрофитные и паразитные травянистые
многолетники
Лишены зеленых ассимилирующих побегов. Резко специа-
лизированная группа.
1 подкласс. Травянистые поликарпики сапрофитного типа
питания, например, Coralliorhiza innata, Neottia nidus
avis, Epipogpn aphyllus, Monotropa hypopitys.
2 подкласс. Травянистые поликарпики паразитного типа
питания. Виды Orobanchaceae (например, Lathraea Squa-
maria, виды Orobanche, Boschniakia rossica)', Balanopho-
raceae-, Rafflesiaceae. Внешний вид этих паразитов чрез-
вычайно разнообразен.
4 класс. Эпифитные травянистые поликарпики
С Почвой не связаны. Побеги (и корни) метаморфизированы
в^связи с эпифитным образом жизни. Чрезвычайно разнооб-
разны по облику. Исключительно виды влажно-тропических
и субтропических лесов. По Шимперу и Фаберу (A. Schimper
и С. Faber, 1935) этот класс можно разделить на:
1 подкласс. Гемиэпифиты, характеризуются незначитель-
ной специализацией побегов. Сюда относятся многие
орхидеи, среди которых интересны ризомоклубневые
формы: у них в основании симподиально нарастающих
воздушных побегов образуются клубни, где кроме ре-
зервных пластических веществ находится и запасная
вода.
2 подкласс. Гнездовые эпифиты, имеющие приспособле-
ния для накопления мелкозема.
3 подкласс. Цистерновые эпифиты с весьма своеобраз-
ными морфологическими приспособлениями для запаса-
ния воды (Bromeliaceae, Dischidia).
5 класс. Лианоидные травянистые поликарпики
(многолетние травянистые лианы)
Надземные стебли слабые, неспособны сохранять само-
стоятельно прямостоячее положение и нуждающиеся в опоре.
8+ 115
Возможно, что подземные органы травянистых лиан тоже
очень многообразны и могут быть представлены как в форме
корневищ, так и в виде клубней и других запасающих ор-
ганов (например, виды Ipomaea, Calystegla, Gloriosa superba,
и т. д.) Но классификация травянистых лиан по подземным
органам не разработана.
VI тип. Монокарпические травы
Жизненный цикл длится в течение одного, двух или не-
скольких лет и завершается цветением и плодоношением,
после чего растения отмирают из-за отсутствия способности
к вегетативному возобновлению. Широко распространены в
аридных областях умеренной зоны северного полушария;
к ним же принадлежат многие культурные и сорные растения.
1 класс. Монокарпические травы с ассимилирующими побега-
ми несуккулентного типа, не нуждающимися в опоре («обыч-
ные» монокарпические травы).
Этот наиболее обширный класс подразделяется на под-
классы по длительности жизни растений:
1 подкласс. Многолетние и двулетние монокарпики, на-
пример, различные виды из Unibelllferae и Cruelferae\
иногда подземные органы у них представлены сильно
утолщенным корнем или стеблем.
2 подкласс. Однолетние монокарпики, куда относится
большинство травянистых монокарпиков. Растения этого
подкласса подразделяются на:
1 группу — длительно вегетирующих однолетников с
подгруппами озимых и яровых и
2 группу — эфемеров.
По структуре побегов растения этого класса могут
быть разделены на безрозеточные, полурозеточные и
розеточные формы. Подобные формы могут быть выде-
лены и среди поликарпиче*1ких трав. Правда, в отличие
от древесных растений, у которых наличие лишь розе-
точного типа побегов, вероятно, нередко является па-
леоморфным признаком (Corner, 1949, 1955; Schmid, 1956)
и может служить основанием для выделения класса,
у травянистых растений розеточный тип побегов (напри-
мер, у видов Draba, А. И. Толмачев, 1958) является
признаком неоморфным. Экологическое значение розе-
точной, полурозеточной и безрозеточной структуры
116
побегов остается недостаточно выясненной, поэтому
автор не использует этот признак при выделении круп-
ных таксономических единиц среди травянистых ра-
стений.
Кроме того, в пределах класса*монокарпических трав,
как и среди поликарпических, может очень сильно варьи-
ровать степень ветвления главного побега, так что
имеются все переходы от «одностебельных» форм, до
форм, образующих «перекати-поле» и подушкообразных.
2 класс. Лианоидные травянистые монокарпики, с побегами,
нуждающимися в опоре, Нам известны лишь однолетние
представители этого класса: Vicia saliva, Polygonum, dume-
torum, Polygonum convolvulus и др.
Могут быть разделены на вьющиеся и лазящие.
3 класс. Монокарпические травы с суккулентными побегами
Растения областей с резко выраженным аридным климатом.
Примером может служить Salicornia herbacea.
4 класс. Полупаразитные и паразитные травянистые монокар-
пики. Подразделяются на подклассы:
1 подкласс — Полупаразитные монокарпические травы,
к которым относятся, например, многие Scrophulariaceae
умеренной зоны северного полушария, имеющие зеленые
листья, но присасывающиеся корнями к корням других
растений: виды Melampyrum, Euphrasia, Rhinanthus,
Odontites и др.
2 подкласс — Паразитные монокарпические травы, к ко-
торым, вероятно, относятся некоторые виды Cuscuta.
Монокарпические эпифиты в литературе не описаны.
Внимательное изучение предлагаемой системы жизненных
форм обнаруживает, что эволюционный ряд жизненных форм
деревья—кустарники—кустарнички—многолетние травы—одно-
летники наблюдается не только среди растений с удлинен-
ными прямостоячими ассимилирующими побегами несукку-
лентного типа, наиболее широко распространенных среди
современных покрытосеменных. Подобные ряды жизненных
форм объединяют и растения, обладающие побегами своеоб-
разной структуры и специфичным направлением роста, напри-
мер, суккулентно-стеблевые, стелющиеся, лиановидные расте-
ния, древесные растения с розеточными побегами, подушко-
видные, эпифитные, сапрофитные и паразитные растения и т. п.
Одни из этих «специализированных гомологов» встречают-
ся во всех типах жизненных форм, от деревьев до травяни-
стых однолетников, например, стелющиеся и лиановидные
формы; другие представлены более короткими рядами, на-
пример, подушковидные формы. В пределах разных типов
одни и те же специализированные формы образуют класс,
подкласс или более мелкие группы в зависимости от мно-
гообразия таких специализированных форм. Каждый класс,
подкласс или группа специализированных гомологов приспо-
соблены к развитию в особых почвенно-климатических
условиях и отличается своеобразном географическим и эко-
логическим распространением.
Если принять деревья — кустарники—кустарнички (или па-
раллельные последним полукустарники и полукустарнички) —
травянистые многолетники—травянистые однолетники за этап-
ные формы основного эволюционного пути жизненных форм
покрытосеменных, то можно говорить об экологическом
многообразии жизненных форм на каждом из этапов эволю-
ции от деревьев к однолетним травам. Так как наблюдается
повторяемость перечисленных гомологов среди деревьев,
кустарников, кустарничков, многолетних и однолетних трав,
то эволюционная система может быть представлена как
система параллельных рядов жизненных форм,
своеобразно и с различной полнотой повторяющих все этап-
ные формы от деревьев до травянистых однолетников (см.
рис. 1). В одном направлении размещаются этапные эволю-
ционные формы, нашедшие выражение в типах жизненных
форм; перпендикулярно к нему размещаются специализиро-
ванные гомологи в пределах каждого типа.
В связи с этим выявляется необходимость выделения осо-
бого понятия — рядов жизненных форм, по объему (рангу)
примерно соизмеримых с типами жизненных форм. Вопрос
об относительном возрасте различных рядов жизненных форм
растений ждет своей разработки.
Наличие параллельных эколого-морфологических рядов
жизненных форм на каждом из эволюционных этапов (от
деревьев до однолетних трав) напоминает принцип парал-
лельной изменчивости близких видов, родов и семейств,
отмеченный Дарвином и сформулированный Н. И. Вавило-
вым (1920) в форме закона гомологических рядов наслед-
ственной изменчивости. В сущности, здесь также можно
говорить о параллельных гомологических рядах наследствен-
ных признаков в сфере вегетативных органов покрытосе-
менных растений. Сходные ряды жизненных форм, например,
суккулентных, лианоидных, стелющихся среди деревьев, ку-
старников, кустарничков и трав обусловлены, главным обра-
зом, закономерной изменчивостью их побегов. Благодаря
этому образуются своеобразные гомологические ряды наслед-
ственно закрепленных форм. Это явление можно трактовать
118
как частный случай закона гомологических рядов наслед-
ственной изменчивости, открытого Дарвином и Вавиловым.
Закон параллельной изменчивости близких видов Дарвина —
Вавилова, видимо, является основой конвергентности жиз-
ненных форм покрытосеменных. В отличие от гомологических
рядов признаков в генеративной сфере растений, изученных
Н. И. Вавиловым, отмеченные здесь признаки вегетативных
органов и жизненных форм обнаруживают более явственную,
зависимость от экологических условий.
Глава III
ЖИЗНЕННЫЕ ФОРМЫ ДЕРЕВЬЕВ
1. РАЗВИТИЕ древовидной формы
у древних папоротникообразных растений
Жизненная форма дерева у высших растений возникла в
процессе длительной эволюции в условиях наземного образа
жизни. Первые достоверно известные наземные растения —
псилофиты, в частности, их примитивные формы из риниевых
(Rhynlaceae) были, как известно, сравнительно мелкими тра-
вянистыми растениями, до полуметра высотой (А. Л. Тахтад-
жян, 1956; Л. М. Кречетович, 1955; К. И. Мейер, 1948, 1957:
А. Н. Криштофович, 1957 и др.). Наличие лишь верхушечных
меристем, при полном отсутствии боковых, неспособность ко
вторичному утолщению стебля, которое обеспечивает необхо-
димую биологическую корреляцию в росте стебля в высоту и
толщину, привело к тому, что псилофиты так и не продвину-
лись сколько-нибудь значительно в направлении образования
древесных форм с силура до среднего девона. Правда, со сред-
него девона среди псилофитов появляются растения с некото-
рыми признаками древовидных форм. Но это были уже расте-
ния переходные к Pteropsida, Lycopsida и Sphenopsida.
Значительный интерес представляют в связи с этим первич-
ные папоротники—Primofitices— среднего и верхнего девона,
содержавшие почти непрерывный ряд промежуточных форм от
примитивных псилофитов до «типичных» папоротников. В этой
группе встречались формы, приближавшиеся к древовидным.
Из последних может быть назван, например, аневрофитои
(Aneurophyton germanicutn Kraus, et. Weyl.) из среднего девона
Германии и среднего и верхнего девона Америки (рис. 17).
120
Aneurophyton germanicutn
имел четко выраженный
тонкий ствол высотой около
полутора метров, на вер-
шине которого располага-
лись крупные, сильно рас-
сеченные «листья», так что
растение в целом походило
на современные древовид-
ные папоротники (W. Got-
han и Н. Weyland, 1954).
Более близкое изучение об-
наруживает, что «листья»
Aneurophyton germanicum
в действительности пред-
ставляли собой осевые ор-
ганы, ветвящиеся и прости-
рающиеся в одной плоско-
сти Первые ветвления
рахиса таких «листьев»
проходили еще объемно, в
нескольких плоскостях про-
странства, и только потом,
дважды или трижды, в од-
ной плоскости (К. Magdef
ran, 1956). Стебель Апеигор-
hyton germanicutn имел
трехлопастную первичную
ксилему и обладал хоро-
шо выраженным вторичным
ростом в толщину.
Переходной формой от
рипиевых к папоротникам
W Zimmermann (1949) счи-
тает Pseudosporochnus Krej-
с ii. Pot. et Berthr., средпеде-
вонское деревцо, высотой
в 2—3 м (рис. 18). Нераз-
в-етвленная часть ствола из
шапп, 1949) утолщалась
Рис. 17
Реконструкция древовидного
Aneurophyton germanicutn (по
Krausel и Weyland, из Magdefrau, 1956)
сросшихся мезомов (W. Zimmer-
В нижней части утол-
к основанию.
щенного ствола отходили корневидные выросты, а верхний ко-
нец ствола завершался кроной более или менее одинаковых
крепких ветвей, возникавших в результате многократно повто-
рявшихся в разных направлениях дихотомических ветвлений.
Наличие в среднем девоне растений типа Aneurophyton
germanicutn свидетельствует о том, что в этот период у назем-
121
Рис. 18
Реконструкция Pseudosporochmis K/ejcii
(no W. Zimmermann, 1949)
ных растений уже на-
чалось образование
настоящего древесного
стебля со вторичным
ростом в толщину. Та-
кой тип стебля в верх-
нем девоне достигнет
почти полного выра-
жения, а в карбоне
станет обычным явле-
нием
Типичной высоты и
формы, характерных
для настоящего дере-
ва, наземные растения
достигли лишь в верх-
нем девоне и получили
широкое распростране-
ние в карбоне. Струк-
турно появление дре-
вовидных форм в де-
воне и карбоне было
связано с появлением
у растений способно-
сти ко вторичному
росту стебля, т. е. с
появлением вторичной
ксилемы, возникающей
в результате более или
менее длительной дея-
тельности камбия Бла-
годаря этой способно-
сти многие ископаемые
формы в карбоне бы-
ли настоящими дре-
весными растениями
(А. Л. Тахтаджян,
1956). Развитие дре-
вовидных форм в кар-
боне проходило независимо друг от друга в разных филетиче-
ских линиях сосудистых растений. Экологически широкое раз-
витие древовидных форм было, вероятно, связано с господством
в карбоне влажно-тропического климата на обширных терри-
ториях земного шара. Возникшие типы древовидных форм
карбона значительно отличались друг от друга по своей внеш-
ней и внутренней структуре.
122
Едва ли не доминирующее положение по разнообразию
форм, размерам и распространению в древесной растительности
карбона занимали древовидные Lycopsida, а среди них выде-
лялись Lepidodendraceae и Sigillariaceae.
Интересным предшественником древовидных Lycopsida кар-
бона было древовидное растение верхнего девона Европы, Вос-
точной Сибири и Северной Америки Cyclostigma kiltorkense.
Cyclostigtna kiltorkense представляла собой дерево до 8 и более
метров высоты. Ствол этого растения диаметром до 30 см в
верхней части дихотомически ветвился, образуя сильно раз-
ветвленную крону.
Свыше ста видов древовидных растений из рода Lepido-
dendron были широко распространены с конца девона до
верхнего карбона в различных районах земного шара, будучи
одними из наиболее характерных растений болотистых тропи-
ческих лесов, образовавших впоследствии толщи каменного
угля. Особенности лепидодендронов широко известны Это
были высокие деревья 40—45 м высоты с прямым колонновид-
ным стволом, несущим на высоте 20 и более метров мощную,
сильно разветвленную крону дихотомирующих ветвей (рис. 19).
Несмотря на наличие ясно выраженного вторичного роста
стебля, у лепидодендронов, как и \ большинства современных
деревьев влажнотропических лесов, отсутствовали годичные
кольца
В толстых стволах лепидодендронов, диаметром часто более
1 м, водопроводящая система по сравнению с современными
деревьями была развита очень слабо, стель не превышала
10 см в поперечнике и составляла от 9 до 14% от общей по-
верхности поперечного среза стебля. От основания ствола отхо-
дили в разные стороны четыре горизонтальных ризофора-стиг-
марии, образующиеся при дихотомическом ветвлении двух вет-
вей ствола. Ризофоры, в свою очередь, дихотомически ветвились.
Подобно стеблю, стигмарии обладали способностью ко вторич-
ному росту На стигмариях придаточно, как на ризофорах
селагинеллы, образовывались корни
По общему строению, характеру роста и ветвлению ствола
и ветвей весьма близок к лепидодендронам другой, широко
распространенный в карбоне род — Lepidophloios Sternb. Габи-
туально виды Lepidophloios отличались от лепидодендронов
более слабым ветвлением кроны и способом расположения
органов спороношения. Стробилы Lepidophloios располагались
вертикальными рядами в верхней части ствола и на старых
частях ветвей кроны, уже лишенных листьев, т е по типу кау-
лифлории современных деревьев влажнотропических лесов
Значительные морфологические и габитуальные отличия
свойственны древовидны.м формам другого распространенного
123
Рис. 19
Lepldodendron. aculeatum Sternb.,
I — реставрация целого растения (по Смиту из А Л Тахтадляна), 2 — часть поверх-
ности ствота с листовыми подушками; 3—5 — детали строения листьев и стробила
семейства Lycopsida — сигилляриевым (Sigillariaceae) и, в част-
ности, их основному роду — сигиллярии. Сигиллярии достигали
30 м высоты и обладали толстым колонновидным стволом,
часто утолщенным в основании, от которого отходили ризофо-
ры типа стигмарий, но менее ветвистые В отличие от лепидо-
дендронов, верхушка ствола сигиллярий ветвилась слабо, или
оставалась совершенно неветвистой
В отличие от гигрофитных лепидодендронов, вымерших в
конце карбона, некоторые виды сигиллярий были более или
менее мясистыми ксерофитами, что и дало им, вероятно, воз-
можность приспособиться к более суровым условиям пермского
периода (А Л. Тахтаджян, 1956).
В другой филетической линии сосудистых растений — Sphe-
nopsida, как и в пределах Lycopsida, в верхнем девоне и кар-
боне также возникли крупные древовидные формы (Hirmer,
1926). Из верхнего девона и нижнего карбона Европы, Сибири
и Малой Азии известны древовидные Asterocalamites с раз-
ветвленным стволом диаметром до 16 см В отличие от совре-
менных представителей Sphenopsida они обладали крупными —
до 10 см длины — и многократно вильчатыми, свободными до
основания мутовчато расположенными листьями.
Своеобразные древовидные формы, внешне напоминавшие
гигантские хвощи, но со вторичным ростом стебля в толщину
представляли собой каламиты Стволы каламитов достигали
20—30 м высоты и до 1 м в диаметре. Несмотря на древовид-
ный облик, каламиты, подобно современным хвощам, имели
подземные, горизонтально идущие и ветвящиеся под землей
корневища. На их узлах, как и па нескольких нижних узлах
воздушных побегов, располагались мутовки придаточных кор-
ней Простые и большей частью мелкие листья, сидевшие му-
товчато на стебле, в отличие от современных хвощей, содержа-
ли хлорофилл и служили фотосинтезирующими органами
Корни каламитов были густо покрыты корневыми волоска-
ми. Кора у старых корней разрушалась, а образовавшиеся
полости служили для проведения воздуха к подземным орга-
нам, что имело большое значение в связи с болотистыми место-
обитаниями каламитов Древесина их обладала вторичным
ростом и была покрыта, как и у хвощей, двуслойной эндодер-
мой Молодые стебли каламитов были сходны со стеблями
хвощей. Но стебли взрослых каламитов отличались сильным
развитием вторичных тканей. Вторичная ксилема каламитов
толщиною 6 см и больше, как и у лепидодендронов, была ли-
шена годичных колец. Сердцевина молодых стеблей была за-
полнена паренхимой, но у взрослых растений становилась
полой в междоузлиях и служила для проведения воздуха к
подземным органам. Сильно развитая кора с мощной перидер-
125
мой в ее внутренней части, выполняла как защитную, так и
механическую функцию.
Кроме древовидных форм, перечисленных выше, в карбоне
и особенно в пермском периоде получили широкое распростра-
нение древовидные папоротники из порядка Marattiales. Сюда
относятся роды: Asierotheca, Acitheca, Crossotheca, Ptychocarpus
и некоторые другие. Ископаемые остатки листьев этих расте-
ний получили сборное название Pecopteris, а окаменелые ство-
лы описаны под названием Psaronius. Пермо-карбоновый вид
Asterotheca arborescens Schleth (рис. 20) имел габитус, типич-
ный для современных древовидных папоротников. Прямостоя-
чий неветвящийся ствол этого растения высотой до 10 и более
метров и диаметром в 10—15 см на своей верхушке нес розет-
ку крупных трижды перистых листьев длиной свыше 2 м и ши-
риной более 1 м.
Ствол снаружи был покрыт чехле м из воздушных корней,
расположенных продольными рядами. Места отхождения кор-
ней образовывали на поверхности ствола округлые рубцы. На
стволах можно видеть также более редкие и крупные подково-
образные листовые рубцы. По анатомической структуре стебли
Psaronius близки к структуре стебля современных древовидных
папоротников (W. Gothan и Н Weyland, 1954).
Из приведенного краткого очерка развития древовидной
формы роста у древних папоротникообразных растений видно,
что первичной формой роста древнейших Pteridophyta была
форма невысоких травянистых растений, а древовидные жиз-
ненные формы были у них вторичными, возникшими в течение
длительной эволюции от силура да верхнего девона и получили
особо широкое распространение в карбоне. Интересно, что
появление и развитие древовидных форм в течение девонского
и каменноугольного периодов проходило независимо друг от
друга в разных филетических линиях сосудистых наземных
растений. Образовавшиеся древовидные формы резко отлича-
лись друг от друга по внешней и внутренней структуре.
2. СООТНОШЕНИЕ ДРЕВЕСНЫХ И ТРАВЯНИСТЫХ ФОРМ
У ПОКРЫТОСЕМЕННЫХ
До конца XIX столетия в ботанической литературе (включая
учебную, см., например, Sachs, Lehrbuch der Botanik, 1874;
A. Gray, Structural Botany, 1881) было широко распространено
мнение, что древнейшим типом покрытосеменных были травя-
нистые формы, а древесные растения возникли от травянистых
благодаря усилению деятельности камбия. Это мнение основы-
валось, главным образом, на сравнении онтогенеза и филоге-
126
Рис. 20
Asterotheca. (Pecopteris) arborescens
(по K. Magdefrau, 1956)
неза и исходило из общего предположения, что сложное должно
происходить от простого Лишь в конце XIX и в первой поло-
вине XX столетия различные исследователи на основе данных
анатомии растений, филогенетической систематики растений,
палеоботаники и ботанической географии пришли к заключе-
нию, что в отличие от папоротникообразных, где древовидные
формы являются вторичными, возникшими от травянистых
предков, у покрытосеменных первичные формы были древес-
ными, а травянистые возникли от них как вторичные. Эта точка
зрения была высказана и аргументирована в трудах многих
авторов (Jeffrey— 1898, 1912, 1921; Hallier — 1905, 1912; Wett-
stein—1912; Eames—1910, 1911, 1921; Bailey—1910, 1912,
1914; Sinnott a. Bailey— 1914, 1922; В Г Александров и
О. Г. Александрова — 1928; Zimmermann — 1930, 1949; М. Г. По-
пов— 1940; А. Л. Тахтад,._ян — 1943, 1948, 1956; А. А. Яценко-
Хмелевский— 1947 и др.). При обсуждении происхождения
покрытосеменных Галлир (Hallier, 1905) отмечал, что древес-
ные Magnoliaceae, без сомнения, являются древнейшим семей-
ством среди существующих покрытосеменных и от них через
Schizandraceae, Lardizabalaceae, Berberldaceae, Ranunculaceae
и Nytnphaeaceae шло развитие к однодольным, образуя непре-
рывную линию редукции от древних древесных форм к лианам,
кустарникам, травам и водным травам. В этом ряду просле-
живаются изменения как морфологической, так и анатомиче-
ской структуры.
Веттштейн (R. Wettstein, 1912) признавал, как известно, за
древнюю и первичную совсем другую группу покрытосемен-
ных— однопокровные (Monochlatnydeae), и одним из самых
важных доказательств их примитивности считал преобладание
в этой группе древесных растений.
Более детально гипотеза редукции древесных форм и воз-
никновения травянистого типа стебля у покрытосеменных была
разработана Джеффри (Jeffrey, 1899, 1917) и его учениками
(Eames, 1910, 1911, 1912, Bailey, 1910, 1911, 1912, 1914; Sinnott
a. Bailey, 1914, 1915, 1922 и др.). Современные научные пред-
ставления об относительной древности и первичности древесных
покрытосеменных и сравнительной молодости и вторичном
характере травянистых ангиосперм, обоснованные в трудах
перечисленных выше и многих других исследователей, бази-
руются па данных палеоботаники, анатомии растений, филоге-
нетической систематики растений и ботанической географии.
а) Доказательства из области палеоботаники
Современные группы сосудистых растений — Lycopodiales,
Isoetales и Equisetales — сплошь травянистые, со следами кам-
128
бия у Isoetales и Equisetales, что можно рассматривать, как
показатель активности этой ткани у их предков. Весьма веро-
ятно, хотя и не вполне достоверно (Bancroft, 1928; АгЬег, 1929),
что в этих группах ныне живущие травянистые формы проис-
ходят от древних древесных типов. Древние представители
плауновых и хвощовых, как отмечалось выше, имели хорошо
развитую вторичную древесину, крупные размеры и часто-
древовидный облик От процветавших в палеозое древесных
форм сохранились лишь мелкие травянистые потомки, лишен-
ные вторичного роста в толщину. Редукция вторичного роста
оказалась биологически очень важной У криптогамов, по Имсу
(Eames, 1912), опа вызвала появление более устойчивых расте-
ний, лучше приспособленных для жизни в новых, менее благо-
приятных условиях и в соседстве с новыми прогрессирующими
группами растений Подобно этим группам папоротникообраз-
ных, среди ангиосперм травы также оказались более адаптив-
ными и более совершенными биологически формами. У них рано
наступает зрелость, меньше тратится пластических веществ на
построение вегетативных органов, достигается большая продук-
тивность семян по отношению к величине особи и занятому ею
пространству Эволюционный процесс у травянистых форм идет
быстрее, чем у древесных Однолетники среди них — высший
тип. Большинство ископаемых покрытосеменных третичного и
особенно мелового периодов относятся к древесным формам,
хотя известны в ископаемом состоянии и некоторые травы
(А. Н Криштофович, 1957). Последнее обстоятельство может,
наряду с нижеприводимыми фактами, служить возражением
против мнения о том, что количество ископаемых трав из более
древних периодов мало только лишь потому, что их остатки
хуже сохраняются, чем древесные. В середине мелового периода
(в Потомаке) встречаются отпечатки травянистых растений,
относящиеся к Plantago, Xyrtdaceae, Сурегасеае\ в эоцене встре-
чаются листья, относимые к Carduus (Sinnott a. Bailey, 1914).
С миоцена по исследованиям Неег’а в отложениях Шпицбергена
и Швейцарии прогрессивно возрастает количество травянистых
форм. В третичной флоре Швейцарии Неег отмечал среди по-
крытосеменных 24'% травянистых видов. В отложениях, им изу-
ченных, количество травянистых форм больше в позднейшее
время, чем в более древних формациях. К подобным выводам
пришли и другие исследователи ископаемых покрытосеменных
(А. Н Криштофович, 1957).
О преобладании фанерофитов, т е. древесных форм, среди
более древних — меловых и палеогеновых покрытосеменных
свидетельствуют и данные М. Hansen (1956).
В-413 И Г Серебряков —9
129
б) Доказательства из области анатомии
растений
Главное различие между древесными растениями и травами
состоит в наличии или отсутствии активного длительно рабо-
тающего камбия. Большинство травянистых растений, по край-
ней мере из внетропических двудольных, имеют камбий, дея-
тельность которого в надземных стеблях ограничена обычно
одним годом, тогда как у древесных она продолжается многие
десятки и даже сотни лет. При этом, если для древесных рас-
тений характерно заложение сплошного непрерывного камби-
ального кольца, откладывающего сплошное же кольцо вторич-
ной ксилемы, то у травянистых часто встречается камбий, под-
разделяемый на пучковый (где он и продуцирует вторичную
ксилему и флоэму) и межпучковый (продуцирующий парен-
химу, а также л\б и древесину, как и пучковый камбий). Пуч-
ковый камбий формируется и начинает * свою деятельность
раньше, чем межпучковый.
Но существуют также древесные растения, у которых хо-
рошо выражены первичные пучки и широкие сердцевинные
лучи, и камбий у них формируется «по травянистому типу».
И, наоборот, у некоторых трав (Hypericum, Caryophyllaceae,
Lythrutn и др ) кольцо первичных проводящих тканей и кам-
биальное кольцо сплошные.
Как отмечалось выше, до конца XIX столетия в ботаниче-
ской литературе было широко распространено мнение, что
структура стебля травянистых растений является первичной и
примитивной, а «древесная» структура — производной. Это
могло быть доказано тем фактом, что в ранних стадиях разви-
тия стебли многих древесных растений имеют «травянистую»,
пучковую структуру Однако, Джеффри (Jeffrey, 1899, 1911,
1917), Hallier (1905, 1912), Eames (1910, 1911), Bailey (1910,
1911, 1912), Sinnott a Bailey (1914, 1915, 1922), Александров
(1928), Яценко-Хмелевский (1947). Тахтаджян (1948) и дру-
гие приводят доказательства обратного- первичности древесного
типа формирования камбия и вторичной ксилемы и вторич-
ности пучкового «травянистого» типа.
Как известно, у древних древовидных папоротникообразных
(лепидодендронов) кольцо камбия и вторичной ксилемы было
непрерывным, сплошным. У тех же ископаемых форм, которые
обладали отчетливыми первичными пучками (Lyginodendron
и Cordaites) вторичная ткань все-таки была тоже совершенно
непрерывной
Доказательством вторичности проводящей системы травя-
нистой структуры можно считать и образование в семействах
с преобладанием травянистых форм такого несомненно специа-
130
лизированного признака как сосуды с простыми перфорациями
(а не лестничными, как у древесных, Яценко-Хмелевский, 1947).
По анатомическому строению травы более специализированы,
чем родственные древесные формы.
Наконец, если молодые веточки древесных и похожи по
анатомической структуре на стебли трав, то у многих трав,
наоборот, в нижних частях их стебля сохраняется «древесная'»
структура со сплошным кольцом проводящей ткани и камбия,
работающего здесь более долго и активно, чем в верхних ча-
стях. Более мощное развитие древесины в основании стебля
многих травянистых растений (например, подсолнечника) объ-
ясняется как сохранение свойств, присущих предкам. (Earns
1911, Sinnott a. Bailey 1914, Александров 1928), в чем про-
является «консерватизм нижних ярусов» (Тахтаджян 1948).
Возникает вопрос, как появился травянистый тип стебля
у покрытосеменных растений? Наиболее четко разработаны две
гипотезы преобразования древесной структуры стебля в травя-
нистую — гипотеза «листовых запасающих лучей» (Jeffrey, 1899,
1912, 1917; Jeffrey a. Torry, 1921; Earns 1910, 1911; Bailey, 1910,
1911, 1912) и гипотеза «снижения камбиальной активности»
(Sinnott a. Bailey, 1914, 1915, 1922; Тахтаджян, 1948).
Уменьшение камбиальной активности, по мнению Синнотта
и Бейли, является главнейшим анатомическим процессом, при-
водящим к возникновению травянистой структуры стебля.
Сравнительное изучение анатомии стебля у большого числа
видов деревьев и трав, относящихся к различным семействам,
привело этих авторов к выводу, что травянистые стебли в раз-
ных систематических группах представляют как бы однолетний
стебель их древесных предков Развивая это положение Синнот-
та и Бейли с позиций биогенетического принципа, А. Л Тах-
таджян (1943, 1948) пришел к заключению, что травы можно
рассматривать как фиксированную ювенильную фазу дерева.
Травы возникли из деревьев, по Тахтаджяну, путем ярусной
неотении, главным образом, путем уменьшения камбиальной
активности. По Scharfetter’y (1953) интенсивность деятельности
камбия тесно связана с климатом. Это подтверждается тем фак-
том, что некоторые травянистые растения в соответствующем
климате могут проявлять гораздо большую, чем обычно, интен-
сивность работы камбия, что приводит к одревеснению их стебля;
примером служит Cheiranthus cheiri в Средиземноморье.
в) Доказательства из области
филогенетической систематики растений
Филогения покрытосеменных говорит о том, что предпола-
гаемые предки их из голосеменных — древние беннеттиты, се-
9” 131
менпые папоротники или цикадовые были сплошь древесными
растениями с хорошо выраженным вторичным ростом стебля,
и среди них отсутствовали травянистые формы. Поэтому первые
покрытосеменные едва ли могли быть травянистыми.
В систематике покрытосеменных нет единства мнений при
выделении их более примитивных групп. Наиболее древними
покрытосеменными считают или сережкоцветные, или рана-
лиевые. Но сережкоцветные — сплошь древесные растения, за
исключением Saururaceae, и отдельных Piperaceae и Chlorant-
haceae. Раналиевые также почти сплошь древесные, за исклю-
чением Ranunculaceae, некоторых Berberidaceae и нескольких
специализированных семейств. Таким образом, при принятии
за наиболее примитивную и той и другой группы подтверждает-
ся взгляд, что наиболее примитивные покрытосеменные были
древесными или обладали по меньшей мере хорошо выражен-
ным вторичным ростом и не были травянистыми.
Наиболее сильным аргументом в пользу относительной
древности древесных и молодости трав у покрытосеменных
является то, что в порядках и семействах, а часто и в родах,
в которых встречаются как древесные, так и травянистые фор-
мы, и где с известной уверенностью можно говорить об относи-
тельной древности форм, судя по строению цветка, примитивные
типы оказываются древесными, а более новые травянистыми
{Leguminosae, Violaceae, Umbelliferae, Borraginaceae, Rosaceae,
роды Viola, Potentilla, Paeonia, Astragalus и др).
Так, в порядке Leguminosae два семейства Mimosaceae
и Сае salp iniaceae с правильными или почти правильными цвет-
ками и многочисленными свободными тычинками без сомнения
более примитивны, чем семейство Papilionaceae, с неправиль-
ным венчиком и сросшимися тычинками. Характерно, что из 121
рода первых двух семейств 113 родов сплошь древесные, а ос-
тальные 8 содержат травянистые и древесные формы В этих
семействах нет сплошь травянистых родов. Из 304 родов се-
мейства Papilionaceae 187 сплошь древесные, 55 имеют древес-
ные и травянистые виды и 62 рода (29%) сплошь травянистые
Следовательно, почти все примитивные Leguminosae древесные,
в то время как более новые группы имеют высокий процент
трав
Такие же отношения древесных и травянистых растений
имеются в семействе Violaceae, Polemoniaceae и Boraginaceae.
Два крупных семейства Araliaceae и Umbelliferae, образую-
щие порядок Umbelliflorae, имеют такое же соотношение дре-
весных и травянистых видов. Первое семейство более примитив-
ное по структуре цветка и вероятно более древнее. За неболь-
шим исключением сюда относятся деревья и кустарники Второе
семейство с резко выраженным постоянством плана строения
132
цветка, исторически более новое, представлено почти сплошь
травами.
Для суждения об относительной древности древесных
и травянистых ангиосперм представляют интерес данные об
общем распределении трав среди семейств двудольных Из 240
семейств, по Энглеру, 121 семейство, или более 50%, сплошь
древесные, 35 семейств, или 14%, сплошь травянистые, и 84 се-
мейства, или 35%, имеют древесные и травянистые виды К тра-
вянистым семействам относятся Saururaceae, Balanophoraceae,
Rafflesiaceae, Hydnoraceae, Cynocrambaceae, Basellaceae, Nymp-
haeaceae, Ceratophyllaceae, Tovariaceae, Resedaceae, Sarracenla-
ceae, Nepenthaceae, Droseraceae, Podostemonaceae, Hydrostachya-'
ceae, Cephalotaceae, Tropaeolaceae, Catlitrichaceae, Limnanthaceae,
Staekonsiaceae, Balsatninaceae, Hippuridaceae, Cynomoriaceae,
Pirolaceae, Lennoaceae, Primulaceae, Pedallaceae, Martyniaceae,
Orobanchaceae, Lentibulariaceae, Globulariaceae, Phrymaceae,
Adoxaceae, Brunnoniaceae и Calyceraceae. Почти все эти се-
мейства представлены или паразитами, или насекомоядными,
или водными растениями, т е. резко специализированы, или же
монотипны и поэтому также не могут считаться примитивными.
Практически все наземные типичные травы относятся к семей-
ствам с древесными видами, но больше половины семейств
с древесными растениями совсем не имеют трав. Этот факт
трудно объяснить, если признать травы более древними.
Интересно также распределение древесных и травянистых
растений среди Archichlamydeae и Metachtamydeae. По Син-
нотту и Бейли в 3551 роде первой группы 2397 родов древесных,
или 68%, и 1154 рода, или 32%, травянистых. Среди 3002 родов
Metachtamydeae 1254 рода, или 42'%, древесных и более поло-
вины (58%) травянистых Деревья составляют 53% от всех
древесных Archichlamydeae и только 23% у древесных Metach-
lamydeae. Таким образом, большинство древесных и, в част-
ности, древовидных растений встречается среди более прими-
тивных из этих двух подразделений двудольных покрыто-
семенных.
г) Доказательства из области ботанической
географии
Современное географическое распространение древесных
и травянистых растений дает также ряд важнейших доказа-
тельств относительной древности древесных и сравнительной
молодости травянистых покрытосеменных. Как видно из приво-
димой здесь таблицы (табл. 1), в северных умеренных областях
преобладают травянистые двудольные, а в тропиках — древес-
ные
133
Таблица 1
Число и процент трав во флоре различных областей
(по Синнотту и Бейли, с дополнением, 1914)
Умеренная зона
Общее число видов Число ВИДОВ трав 0/0»/0 травянист, видов
Великобритания (Hooker) 927 821 89
Франция (Cusin et Ansberqtie) . . . 3924 3492 89
Германия (Engler) Швейцария (Schinz и Keller) . . . СССР (Флора СССР, первые 24 1117 1899 947 1726 85 91
тома) 13896 12119 87,2
Норвегия (Blytt) о. Элсмира (Simmons) 857 741 90
76 71 93
Тропиче ская зона
Бразилия (Muller) 15991 4092 26
Амазонская область Бразилии . . . Тропическая Африка (Oliver a. Thi- . 2209 .265 12
selton-Dyer) . Цейлон (Tri men) 8577 3560 42
1793 670 37
Ява (Koorders) 3188 867 27
Общий вывод о преобладании в умеренных зонах трав,
а в тропических — древесных подтверждается также распре-
делением трав по отдельным семействам (табл. 2).
Таблица 2
Распределение трав по отдельным семействам
в умеренных и тропических странах
(по Синнотту и Бейли)
(в процентах)
Семейства Северная умерен- ная зона Брази- лия Индия Тропи- ческая Африка
1. Amaranthaceae 100 79 44 84
2 Legumlnosae 90 22 38 41
3. Euphorbiaceae 95 14 10 29
4. Violaceae 100 6 52 18
5. Lythraceae 100 30 42 76
6. Gonvolvulaceae 100 42 52 65
100 20 4 7
8. Rubiaceae 100 20 30 19
9. Composltae 100 39 91 80
134
Из общего количества древесных двудольных большинство
древовидных растений также сосредоточено в тропических стра-
нах, где они составляют от 25 до 40%, а в умеренных зонах —
лишь 1—5%.
Но тропические флоры Азии и Южной Америки, в меньшей
степени — Африки, являются наиболее древними из дошедших
до нас флор покрытосеменных растении, имевших уже в тре-
тичном периоде современный состав О древности этих флор
свидетельствует, в числе прочих доказательств, богатство этих
флор родами и видами, указывающее на спокойное развитие
с самого начала их возникновения (Е. В. Вульф, 1944).
Если в наиболее древних флорах преобладают именно дре-
весные формы покрытосеменных, то напрашивается и вывод
о большей древности самих древесных форм по сравнению
с травами.
Данные о современном географическом распространении
древесных и травянистых ангиосперм имеют большое значение
в связи с палеоклиматологическими данными периода возник-
новения покрытосеменных.
Появление многих групп ангиосперм и их широкое распро-
странение в растительном покрове земли начинается с середины
мелового периода (А. Л. Тахтаджян, 1954). Ископаемые остатки
древних покрытосеменных найдены в различных местах, и еди-
ного мнения о месте их возникновения нет Галлир считал
местом их появления гипотетический материк Пацифик, То-
мас— Гопдвану, Арльдт — Ангариду, очень распространенной
в настоящее время точкой зрения является теория полярного
происхождения покрытосеменных (в пределах современной
арктической зоны) (Криштофович, 1957). Но где бы террито-
риально ни находилась «колыбель» первых покрытосеменных,
климат этих стран, судя по составу ископаемых форм, очевидно,
был достаточно влажным и теплым. Одни авторы считают, что
это были страны с теплым субтропическим климатом (Тахтад-
жян, 1954), другие — что с настоящим влажнотропическим
(Sinnott a. Bailey, 1914; Irmscher, 1922; Е. В Вульф, 1944 и др).
Но если в настоящее время равномерный тропический кли-
мат, а также и влажный субтропический, более благоприятен
для развития древесных форм растений, то можно думать, что
и в период становления покрытосеменных такой климат благо-
приятствовал образованию преимущественно древесных расте-
ний и что ангиоспермы в самом начале были главным образом
или сплошь древесными.
Географическое распространение современных семейств и
родов покрытосеменных свидетельствует о том, что центры
происхождения многих из них находятся в тропической зоне,
откуда они распространились на север и юг, дав начало видам
135
умеренных флор. Примером может служить история антаркти-
ческой флоры по Е. В. Вульфу (1944). Этот ботанико-географи-
ческий вывод, будучи связан с данными сравнительной морфо-
логии, также может служить доказательством первичности
древесных форм. В тропических центрах происхождения систе-
матических групп преобладают древесные растения, а в уме-
ренных флорах — роды тех же семейств или виды тех же родов
имеют облик кустарников или трав. Например, семейство
Leguminosae имеет основных представителей в тропиках в виде
деревьев с перистыми листьями, а в умеренной зоне образова-
лись травянистые трибы Trifolieae, Vicieae и Loteae. Порядок
Ranales включает древовидные, распространенные, главным
образом, в тропиках и субтропиках, семейства Magnoliaceae
и Апопасеае, а в умеренных зонах обоих полушарий — произ-
водные травянистые Ranunculaceae. В семействе злаков древ-
няя триба Bambuseae с ее древесными формами распространена
тоже в тропической и субтропической зоне
По мнению Быоса (Bews, 1925), растительные типы (физио-
номические — И С ) могут быть расположены в систему, отве-
чающую их эволюции. Исходным в ней будет тип растений
влажнотропического леса. Внетропические роды и виды, свя-
занные экологически с влажными условиями (болота, побе-
режья рек и морей) сохраняют физиопомически древний тип
растений. Типы же растений саванн, степей, зарослей ксеро-
фитпых кустарников и полупустынных суккулентов будут дери-
ватами, возникшими из первого типа
Признание древесных покрытосеменных за первичные и
более древние, в травянистых — за более новые вызывает есте-
ственный вопрос о причинах трансформации древесной расти-
тельности в травянистую, причинах, вызвавших возникновение
трав. Закономерности современного географического распро-
странения трав в разных областях Земли указывают пути ре-
шения и этой проблемы.
Наивысшего развития травянистые растения достигли лишь
в настоящее время на больших материках северного полушария
Сильное развитие трав в умеренной зоне северного полуша-
рия, очевидно, было вызвано прогрессирующим похолоданием
климата в течение третичного периода. Как показывают пале-
оботанические данные, в меловом и начале третичного периода
в северной умеренной зоне процветали многие древесные роды
и даже семейства, теперь отсутствующие здесь, но обычные
в более теплых областях (например, Ficus, Eucalyptus, Diospy-
ros, Cinnamomutn, Aralia и др.). Вымирание этих растений свя-
зано с похолоданием, которое шло медленнее в более южных
районах и прогрессивно возрастало в более северных.
Травянистые формы выработались, очевидно, как приспособ-
136
ленив к этому прогрессирующему похолоданию, главным обра-
зом, в северной умеренной зоне.
Подавляющее большинство трав, вероятно, развилось на
материках северной умеренной зоны, и в первую очередь, как
‘считают Синнотт и Бейли, Шарфеттер и др.— в горных обла-
стях, где скорее всего сказывается неблагоприятное действие
климатических факторов
Наиболее существенным фактором распространения травя-
нистых форм и отступления древесных явился ледниковый
период. Именно различиями в характере оледенения можно
объяснить некоторые любопытные различия в соотношении
жизненных форм в современной флоре Северной Америки
и Европы.
Флора восточной части Северной Америки сейчас насчиты-
вает 24 процента древесных видов, а флора Европы (севернее
Альп) лишь 13 процентов. Большая бедность европейской
флоры древесными видами связана с тем, что Альпы, Пиренеи,
Балканы и т д послужили препятствием для передвижения
европейской флоры (особенно древесной) на юг, в область
Средиземья, при наступлении ледника. Поэтому, как показы-
вают данные палеоботаники, многие семейства флоры Европы
середины и третичного периода в последующем нацело исчезли.
В Северной же Америке некоторые из этих семейств (например,
Ebenaceae, Menispermaceae, Lauraceae сохранились, так как
там перигляциальная флора беспрепятственно могла мигриро-
вать па юг до Флориды и лучше сохранилась от вымирания
Иллюстрацией того, насколько глубоко повлияло оледенение
на соотношение травянистых и древесных форм во флоре боре-
альных стран, могут служить следующие цифры, приводимые
Sinnott и Bailey. До ледникового периода в этих странах соот-
ношение древесных и травянистых видов было равным при-
мерно 1 : 1 (как показывают палеоботанические данные).
В настоящее время флора Европы насчитывает 12 процентов
древесных и 88 процентов травянистых видов, флора Северной
Америки — 24 процента древесных и 76 травянистых, а Япония,
растительный покров которой меньше всех пострадал от лед-
ника— 46%'древесных и 54% травянистых (соотношение, наи-
более близкое к третичному).
Похолодание климата в третичном периоде в северном полу-
шарии было главнейшим, но не единственным фактором обра-
зования трав Столь же мощным фактором развития травяни-
стых растений явилась аридизация климата
Как пишет Е. В. Вульф (1944), нельзя делать заключение,
что травянистые формы присущи только умеренным зонам.
Тропические флоры имеют также травянистые виды или целые
роды, например, в семействах Cucurbitaceae, Lobeliaceae, Cotnpo-
137
sitae, Solanaceae, Campotiulaceae, Liliaceae и др. Это подтверж-
дает возникновение жизненных форм умеренного климата
в пределах 1ропической зоны в результате приспособления
к ксерофитным условиям обитания в областях с периодической
сменой влажного периода сухим и при подъеме в горы Это в то
же время делает понятным распространение травянистых по-
томков тропических деревянистых исходных форм в пределы
умеренных зон
В отличие от холодных стран, в аридных областях выраба-
тывались массово однолетние травянистые формы, наиболее
приспособленные к условиям засушливого климата
По Шарфеттеру (1953) можно различать
1 . Травы северной умеренной зоны, возникшие вследствие
похолодания климата во второй половине третичного
периода
2 Горные травы, возникшие в холодных высокогорных
поясах: Veronica, Gentiana, Primula.
3 Травы, возникшие из-за сухости климата например, травы
Южной Африки, Западной Австралии.
4 . Тропические муссонные травы, возникшие из-за смены
дождливого и сухого времени года, например Peperotnia.
Как правильно отмечает А Л. Тахтаджян (1948), многие
травы возникли в тропических лесах в результате приспособле-
ния к лазящему, вьющемуся и эпифитному образу жизни,
а также в результате гидрофильной эволюции Таким образом,
фитоценотические факторы тоже сыграли немаловажную роль
в возникновении некоторых жизненных форм травянистых ра-
стений
3. ЭВОЛЮЦИЯ ЖИЗНЕННОЙ ФОРМЫ ДЕРЕВА
У ПОКРЫТОСЕМЕННЫХ
Разносторонняя и убедительная аргументация, приведенная
в предыдущем разделе, делает почти несомненным, что первые
покрытосеменные представляли собой древесные растения и
вероятнее всего — деревья. Судя по палеоботаническим и срав-
нительно—морфологическим данным, первичные древовидные
покрытосеменные значительно отличались от их современных
представителей.
Первичные покрытосеменные, вероятно, представляли не-
большие деревья с толстым стволом, очень слабо разветвлен-
ной кроной и относительно немногочисленными толстыми вет-
вями (Н Hallier, 1912; А Л Тахтаджян, 1948; Е J. Corner,
1949, Е. Schmid, 1956) По мнению последних двух авторов,
особенности роста и внешнего облика этих примитивных древо-
видных ангиосперм были связаны со спецификой внутренней
структуры их стебля Первичные древовидные покрытосеменные
138
обладали, в отличие oi большинства современных деревьев,
слабым одревеснением стеблей; их стебли отличались мягко-
древесностыо и небольшой механической прочностью. Ветви
первых покрытосеменных были «олигоксильными» (Schmid,
1956) или «толстостебельной» (Pachycaul) структуры (Е J Cor-
ner, 1949), сочными и сильно паренхиматизированпыми, как
> современных Carica papaya, древовидных однодольных и т. д
И-5 за слабого одревеснения (механической непрочности) они
и имели значительную толщину и укороченные междоузлия.
Образование типичных современных кронообразующих де-
ревьев с удлиненными побегами, будь то гигантские тропиче-
ские, или величественные и стройные деревья умеренных широт,
было бы невозможным без полного одревеснения их стеблей,
обеспечивающего быстрый рост и образование удлиненных
побегов, интенсивное ветвление и формирование сильно раз-
ветвленной кроны, поднятой на 20—40 и более метров над
уровнем почвы.
Вопрос о путях эволюции жизненных форм деревьев у по-
крытосеменных разработан еще очень слабо и ждет своих иссле-
дователей Интересные данные по эволюции деревьев у по-
крытосеменных приводятся в исследованиях Корнера (Е. J Cor-
ner, 1949, 1953, 1955) Оставляя без обсуждения систематико-
филогенетические выводы автора, следует отметить, что морфо-
логические заключения Корнера представляют большой инте-
рес.
Свою концепцию о развитии современных деревьев у покры-
тосеменных Корнер называет Durian — Theory, используя на-
звание одного из видов деревьев влажнотропических лесов
Малайской области — Durio zibethinus Durio zibethinus — это
высокое дерево из семейства Botnbacaceae, теперь широко куль-
тивируемое от Индии до Новой Гвинеи Оно имеет небольшие
простые листья и тонкие ветви, на которых развиваются круп-
ные (с человеческую голову величиной) мясистые плоды с круп-
ными, светло-коричневыми семенами, покрытыми мясистым и
толстым белым или кремовым присемянником — ариллусом,
отличающимся высокой питательностью и замечательным вку-
сом. В незрелом состоянии плоды надежно защищены креп-
кими многочисленными шипами. Поэтому незрелые плоды
редко поедаются дикими зверями Но ко времени созревания
плоды становятся доступными для животных и птиц При соз-
ревании плоды Durto Zibethinus начинают издавать сильный
и отвратительный запах чеснока и скатола, привлекающий
диких зверей, которые распространяют семена
Анализируя степень распространенности у покрытосеменных
крупных и замкнутых плодов и такого полезного биологически
признака, как мясистый присемянник, автор приходит к выводу,
134
что этот признак сравнительно редкий. Лишь в 50 семействах
всех ангиосперм имеется более или менее развитый ариллус.
Все эти семейства — главным образом тропические; и в боль-
шинстве из них семена с ариллусом свойственны тропическим
деревьям и древесным лианам. Интересно, что у некоторых
родов встречаются виды с различной степенью развития арил-
луса от крупного до зачаточного и полностью отсутствующего
Пытаясь объяснить причины этого, автор приводит ряд
аргументов, из которых следует, что плоды типа массивной
мясистой коробочки или листовки и семена с присемянником
свойственны древним покрытосеменным и являются реликтовым
морфологическим признаком. Признание таких плодов и семян
реликтовыми позволяет автору сделать ряд следующих заклю-
чений:
1. Сочные тяжелые плоды могли развиваться лишь на мас-
сивных прочных ветвях. В частности, травы не могли продуци-
ровать плоды типа Durio, мускатного ореха, или плоды типа
Апопасеае. Поэтому можно заключить, что предковые цветко-
вые растения были деревьями.
2 Крупные семена с присемянником не обладают периодом
покоя и не имеют защиты от высыхания и холода; такие виды
растений не могут быть географически распространены за пре-
делы тропических или субтропических влажных лесов. Поэтому
первичные покрытосеменные должны быть тропическими (или
субтропическими) деревьями.
3. Тропические деревья с массивными ветвями имеют в типе
крупные, сложные, спирально расположенные листья, перистые
или пальчатые; у таких деревьев есть переходы к простым
листьям. Поэтому архаичные формы тропических деревьев
должны были также иметь сложные листья (вначале перистые,
а затем при редукции оси листа — пальчатые) В настоящее
время такие отношения наблюдаются у некоторых деревьев
тропиков с пазушными плодами на боковых ветвях (например,
Meliaceae, Sapindaceae, Legutninosae, Connaraceae, Sterculia,
Aesculus). В связи с этими заключениями автор напоминает
о двух морфологических принципах структуры цветковых ра-
стений:
а) Соподчинение осей разного порядка (Axial
conformity). Чем крепче и массивнее ось у данного вида, тем
крупнее и сложнее ее боковые органы.
б) Уменьшение при ветвлении (Deminution on
ramification). Усиление ветвления всегда вызывает соответствен-
ное уменьшение ветвей и их боковых органов — листьев, соцве-
тий, цветков, плодов и семян.
При учете этих принципов, по Корнеру, становится понят-
ным, почему архаичные формы покрытосеменных деревьев для
140
образования своих громадных плодов должны были обладать
слабым ветвлением главного толстого ствола Становятся по-
нятными также пути развития современных покрытосеменных
деревьев, сильно разветвленных, с тонкими боковыми ветвями
и относительно мелкими боковыми органами
Слабое ветвление древнего перистолистного дерева, иногда
имевшего лишь один главный ствол, сопровождалось превра-
щением верхушечной почки в простой терминальный цветок
В результате, по Корнеру, выявляется форма цикадоподоб-
ного дерева с розеткой стерильных и фертильных листьев
на верхушке ствола, с терминальной розеткой гигантских
плодолистиков, которые морфологически сходны с мегаспоро-
филлами цикадовых От этого цикадоподобного прототипа
покрытосеменных развился, по Корнеру, тип современного дву-
дольного дерева в процессе усиления ветвления всех его орга-
нов и вызванного этим уменьшения размеров и сложности
структуры ветвей, листьев, цветков, плодов и семян и при пере-
мещении цветков в пазушные почки.
В процессе эволюции плоды чаще, чем другие органы, сохра-
няют тенденцию сохранения предковых форм, будучи по своей
природе консервативными органами.
Автор обращает большое внимание на структуру вегетатив-
ных органов. Типичные цикадовые лишены длинных междо-
узлий. Крупные листья их сосредоточены на верхушке стебля,
как у древовидных папоротников В процессе эволюции по-
крытосеменных проявляется тенденция к образованию более
длинных междоузлий и удлиненных побегов с одновременным
ослаблением оси листа и формированием более мелких листьев
Но признаки цикадоподобной фазы эволюции (укороченные
междоузлия на толстых побегах деревьев) еще ясно проявляют-
ся в форме роста некоторых современных деревьев, например,
Carica papaya, пальм, видов Pandanus и у некоторых деревьев со
сложными листьями в молодом возрасте (Araliaceae, Caesalpinia-
ceae, Bignoniaceae и др). Терминальные соцветия Pandanus
и деревьев из Bignoniaceae, вызывающие симподиальный рост
побегов, также являются, по Корнеру, примитивными чертами
структуры, предшествующими появлению пазушных соцветий
Среди деревьев Корнер различает два основных типа:
А. Толстостебельные (Pachycaul). Деревья с массивным
мягкодревесным, неветвистым или слабо ветвистым стволом,
с короткими междоузлиями, крупнолистные и крупносеменные.
Б. Тонкостебельные (Leptocaul) Деревья с относительно
тонким главным стволом и ветвями, более или менее жестко-
древесные, с более или менее развитыми междоузлиями, крупно-
листные или мелколистные, крупносеменные или мелкосемен-
ные
Последний тип (Leptocaul) эволюционно более новый и сов-
ременный. Тонкие ветви с длинными междоузлиями делаю г
возможным более быстрый рост в высоту или в длину и бла-
годаря этому тонкостебельные деревья получают явное преиму-
щество перед древними неуклюжими толстостебельными фор-
мами с массивными и медленно растущими стволом и ветвями.
Появление тонкостсбелыюй структуры обеспечило новым фор-
мам деревьев и более широкое, по сравнению с толстостебель-
ными, географическое распространение за пределы влажных
тропиков и субтропиков, так как мелкие более многочисленные
почки оказываются лучше защищенными, а мелкие ветви легче
замещаются при повреждении. Деревья тонкостебепытой фор-
мы доминируют в настоящее время и количественно, и по вы-
сотному, и географическому распространению над современ-
ными толстостебельными формами типа пальм, Carica, Panda-
nus, а также цикадовыми и древовидными папоротниками,
которые в силу их конституции сохранились в сравнительно
небольших количествах в тропических и частично в субтропи-
ческих влажных областях.
Интересную интерпретацию дает Корнер явлению каули-
флории, нередко встречающейся у тропических деревьев Явле-
ние каулифлории, по Корнеру, получает простое и естественное
объяснение1 это деревья с развитой новой формой ветвей, по
в ряде случаев сохранившие как атавизм древний облик своих
крупных плодов
На основании вышеизложенного Корнер говорит об опре-
деленном направлении и последовательности эволюции де-
ревьев у покрытосеменных, выделяя две стадии и семь фаз в
процессе эволюции цветковых деревьев
А Толстостебельная стадия (Pachycaul Stage) Стволы мас-
сивные, сочные, мягкодревесные, не ветвящиеся, или слабо
ветвящиеся, со слабым развитием междоузлий, крупнолистные,
крупносемеипые
1. Фаза одностебельного или цикадообраз-
и о го дерева (Monocaul or Cycad Phase). Низкие, толсто-
ствольные, неветвящиеся, вероятно монокарпические деревья,
без удлиненных междоузлий. Листья сложноперистые, вероят-
но, с колючим рахисом, цветки примитивно верхушечные,
крупные, одиночные Плоды крупные с несколькими крупными
семенами, покрытые ариллусом Эта фаза i ипотетическая. Ей
близко соответствуют цикадовые, но некоторые черты структу-
ры, и, в первую очередь, отсутствие ветвления, сближает эту
фазу с древовидными папоротниками. Как форма дерева эта
фаза сохранилась не только у цикадовых, но в модифициро-
ванном виде, с более развитыми междоузлиями, также у
пальм и в молодом состоянии у некоторых- двудольных деревьев
142
со сложными листьями (Carica, Cecropia, Schizolobium, Bombax,
Aralia и др.), где наблюдается рекапитуляция предковых при
знаков.
2. Фаза однодольного дерева (Monocotyledonous
Phase). Деревья типа пальм, древовидных злаков — Bambusa.
Scitamineae (Ravenala) и др.
3. Фаза Carica (Carica Phase). Сходна с предыдущей
формой цикадообразных деревьев, но с более длинными меж-
доузлиями и слабым ветвлением менее массивного ствола. Эта
форма типична, кроме Carica для Pandanus, в известной степени
для Hyphaene, древовидных Cactaceae, На этом уровне раз-
вития дерева находятся некоторые тропические деревья, встре-
чающиеся среди Simarubaceae (Euricoma), Campanulaceae, So-
lanaceae, Compositae, Bignoniaceae и т. д. Трудно провести рез-
кую границу между третьей и четвертой фазой. Промежуточное
положение между ними занимают, например, Adansonia и
Brachychiton. Главные вариации в этой фазе касаются поло-
жения соцветий, то терминального (Bignoniaceae, Pandanus), то
латерального (Carica, Cecropia, Macaranga и др )
Б. Тонкостебельная стадия (Leptocaul Stage). Деревья силь-
но ветвистые. Ветви более или менее тонкие, более или менее
жестокодревесные, с более или менее длинными междоузлиями,
крупнолистные или мелколистные, крупносеменные или мелко-
семенные
Крупносеменные (Macrospermous). Даже сильно ветвящиеся
деревья нс могут выйти за пределы влажных тропиков из-за их
крупных, быстро прорастающих семян. Заселению сухих тро-
пиков и умеренных областей способствовало эволюционное
развитие холодоустойчивых и засухоустойчивых плодов (как
костянка и орех) или мелких семян с плотной кожурой и
частично высыхающим зародышем, с сильно выраженным
периодом покоя. Поэтому крупиосеменные деревья с быстро
прорастающими семенами (и, в частности, так называемые жи-
вородящие формы деревьев, столь характерные для мангровых
зарослей) являются, по Корнеру, примитивными формами, с
сохранившимися предковыми свойствами (как видно, явление
живорождения у мангровых и других тропических деревьев с
точки зрения излагаемой теории представляется реликтовым
признаком).
4. Фаза Dysoxylon (Dysoxylon Stage) Dysoxylon
(Meliaceae) типичен для этой важной формы тропических де-
ревьев с крупными сложными листьями, спирально или супро-
тивно расположенными на более или менее массивных, наклон-
но вверх растущих ветвях. Dysoxylon сохраняет характерные
плоды с мясистым околоплодником и семена с присемянником.
Сюда относятся также некоторые Leguminosae, Sterculiaceae,
143
а также Bocconia, Hevea, Artocarpus. Соцветия, как и у пре-
дыдущих форм, боковые или верхушечные. Травянистых де-
риватов нет
5. Фаза Magnolia (Magnolia Phase). Деревья, сход-
ные с предыдущими формами, но с простыми листьями и без
массивных ветвей. Листья крупные или мелкие. Здесь типичны
Magnoliaceae, Dilleniaceae, Sapotaceae, Persea, Quercus и т. д
Имеются переходы к предыдущим и последующим формам
(Terminalia, Achras и др.)
6. Фаза Myristica (Myristica Phase) Формы сход
ные с предыдущими, с простыми листьями, но расположенными
очередно или супротивно в одной плоскости, образуя горизон-
тально-облиственные ветви. Этот тип олиствения является бо-
лее совершенным типом расположения листьев у деревьев
Ветви типично тонкие, более или менее мелколистные. Харак-
терны среди Myristtcaceae, Апопасеае и некоторых современных
родов тропических и деревьев умеренних широт, как Fagus,
Carpinus, Durio, Diospyros, Eugenia и т. д.
7. Фаза каулифлорного дерева (Cauliflorous
Phase). Такие деревья могут быть производными от некоторых
деревьев вышеперечисленных форм с тонкими ветвями, но
крупными цветками или плодами.
Мелкосеменные (Microspermous). Этот более новый тип
деревьев повторяет формы 4, 5 и 6 фазы, типично представ-
ленные, например, у Cassia, Hibiscus и Vernonia, образующие
обычно также травянистые дериваты. (Хотя есть примеры
среди подобных деревьев и только древесных форм, например,
Welnmannia, Populus, Salix).
На рис. 21 приводятся некоторые формы современных тро-
пических деревьев, иллюстрирующие, по Корнеру, различные
предполагаемые фазы эволюции дерева у покрытосеменных.
Воссоздаваемый Корнером облик первичных покрытосемен-
ных совпадает с образом древнейших ангисперм, созданным
ранее другими исследователями (например, Галлиром
(Н. Hallier, 1912), Б. М Козо-Полянским (1922), А. Л. Тахтад-
жяном (1948). Сходный результат попыток разных исследова-
телей, исходивших из разных оснований, реконструировать
облик древнейших покрытосеменных, говорит, по-видимому, о
правдоподобности этих реконструкций.
Нам кажется хорошо аргументированным также выделение
двух морфологических и экологических типов деревьев у покры-
тосеменных — толстостебельных и тонкостебельных (Pachycaui
и Leptocaul — тип), интерпретируемых Корнером в эволюцион-
ном аспекте.
Выделяемые Корнером фазы эволюции дерева у покрытосе-
менных можно рассматривать как подтипы или более мелкие
144
Рис. 21
Главнейшие формы деревьев и две формы их травянистых
дериватов — А и F (по Корнеру. 1949):
А — розеточная агава, редуцированная форма С; В — крупносеменное жест-
кодревеснос дерево со спирально расположенными перистыми листьями; С —
гипотетическая цикадовая форма, одноосная и монокарпическая, с верхушеч-
ным пучком перистых мегаспорофиллов, семена с ариллусом; D — каулифлор-
ное дерево, жесткодревесное, с простыми очередными листьями (Durio); Е —
Carfca-форма, мягкодревесная, с немногими ветвями, сложными листьями и ко-
роткими междоузлиями; семена с ариллусом, F — каулифлорное однодольное
травянистая корневищная форма с простыми листьями, аналогичная каулиф-
лорным однодольным деревьям, дерИватное от Н (fingiberaceae); О— пагодооб-
разное дерево с замкнутыми плодами и семенами с ариллусом (типа Sloanea),
простые листья в розетках; Н — однодольное мягкодревесное монокарпичес-
кое дерево со сложными листьями, имеющее ариллус (типа Ravenala), I — круп-
носемениая предковая жесткодревесная форма дерева Leguminosae со спи-
рально расположенными сложными листьями, семена с ариллусом (типа Parkia
В-413 И Г Серебряков —10
эколого-морфологические 1р>ппы жизненных форм с различным
сочетанием в их структуре палеоморфных и неоморфных при-
знаков
Интерпретация этих гр^пп, как фаз развития деревьев у
покрытосеменных, мало аргументирована и носит произволь-
ный характер. В частности, странное впечатление производит
выделение второй фазы — однодольного дерева (Monocotyledo-
nous phase), поставленной межд> цикадоподобными деревьями
и фазой Carica (Carrca-phase) Здесь совершенно не учтены
своеобразие и высокая специализация анатомической структу-
ры осевых органов однодольных с их атактостелией. Вряд ли
уместно также выделение специальной фазы каулифлорного
дерева (Cauliflorous phase).
Эти критические замечания, однако, не снижают большого
интереса дурьянной теории Корнера. Теория Корнера эволю-
ционно и филогенетически трактует многие признаки, которые
обычно рассматриваются систематиками как чисто морфоло-
гические и экологические (структура вегетативных побегов,
листорасположение, структура околоплодника — сухие и сочные
плоды, наличие или отсутствие периода покоя у семян и т. д.).
Теория Корнера расширяет «ассортимент» признаков, которые
могут быть использованы филогенетической систематикой.
После ознакомления с вопросами эволюции и филогенеза
жизненной формы дерева целесообразно осветить становление
жизненной формы дерева покрытосеменных и хвойных в онто-
генезе. Ниже это и будет сделано на некоторых конкретных
примерах.
4. ОСОБЕННОСТИ МОРФОГЕНЕЗА ЖИЗНЕННОЙ ФОРМЫ
ПРЯМОСТОЯЧЕГО ДЕРЕВА У НЕКОТОРЫХ ХВОЙНЫХ И
ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ
Морфогенез жизненной формы дерева у ели
Picea excels a Link.
Из методических соображений обзор разнообразия структур
и процессов образования жизненной формы дерева в онтогене-
зе целесообразнее начать с хвойных деревьев, у которых эти
процессы особенно иллюстративны
Анализ онтогенеза дерева как жизненной формы мы начи-
наем с описания его у ели — Picea excelsa Link основной ле-
• При составлении данного очерка автор пользовался, кроме литера-
турных источников и собственных наблюдений, еще неопубликованными дан-
ными о развитии корневой системы ели аспирантки Т Й Вещиковой, кото-
рой приносит искреннюю благодарность
146
сообразующей породы смешанных и хвойных лесов Европей-
ской части СССР. Структурные и биологические особенности
жизненной формы дерева у ели выражены с очень большой
отчетливостью.
Прорастание семян ели, как известно (Kirchner, 1908), про-
исходит по надземному типу. Зародышевый корешок, становя-
щийся ьтавным корнем, по выходе из семени очень быстро
обнаруживает резкую положительно-геотропическую реакцию и
начинает расти отвесно вниз Затем вытягивается из семени
гипокотиль и за ним семядоли Последние окрашены в зеленый
цвет, подобно хвоинкам имеют игловидную форму, достигая
15—25 мм длины. От настоящих хвоинок однолетних сеянцев
они отличаются внешне несколько большей длиной и трехгран-
ным поперечньнм сечением (хвоинки в поперечном сечении че-
тырехгранны, чаще всего ромбовидны) Основания семядолей
срастаются, образуя короткое влагалище — орган защиты вер-
хушечной почки. Семядоли ели имеют большое биологическое
значение, выполняя функцию фотосинтеза в течение первых
2 -3 лет жизни сеянца Они отмирают и опадают лишь на 3—
5 году жизни сеянца Под материнским пологом деревьев, в
лесах Подмосковья, семядоли остаются часто едино венными
фотосинтезирующими листьями в течение первого года жизни,
так как на конусе нарастания выше семядолей начинают фор-
мироваться почечные чешуйки новой верхушечной почки Но
при более благоприятных условиях, при меньшем затенении и
на рыхлых богатых почвах, уже в первое лето развивается эпи-
котильная часть главного стебля с зелеными листьями — хвоин
ками, вырастая в высоту на 1—3 см Величина прироста глав-
ной оси тесно связана с освещением и богатством почвы Хвоин-
ки сеянца первого года жизни мелки, их длина 10—15 мм, а
диаметр обычно не превышает 0,4—0,5 мм.
По форме поперечного сечения, небольшому числу устьиц и
структуре мезофилла, часто со значительным развитием меж-
клетников, хвоинки первого года жизни сеянца отличаются
резко выраженной теневой структурой. Как показали исследо-
вания Шрамма (Schramm, 1912), теневая структура листьев сеян-
цев ели сохраняется вне зависимости от освещения местообита-
ния сеянца и представляет собой признак возрастного характера
Главный стебель сеянца первого года заканчивается верхушеч-
ной почкой, покрытой несколькими десятками почечных чешуй.
Кроме верхушечной почки на стебле могут сформироваться
еще одна или две-три боковые пазушные почки Большинство
же листьев лишено пазушных почек
На главном корне через 1.5—2 месяца после прорастания
появляются боковые корни, развивающиеся по типу всасываю-
щих. К концу лета главный корень вырастает до 5—10 см, а
10*
147
боковые корни, числом 10—15, расположенные более или менее
равномерно по длине главного, достигают длины около 1 см.
Весной второго года жизни, в подмосковных лесах обычно
во второй половине мая, раскрываются верхушечные почки
сеянцев ели. Боковые почки у таких перезимовавших сеянцев
под пологом смешанных и еловых лесов Подмосковья обычно
не раскрываются, так что и в течение второго года сеянец ели
несет неветвящуюся главную ось. Побег второго года на глав-
ной оси по величине и морфологии мало отличается от эпико-
тильной части годичного побега первого года. Длина его в
смешанных и еловых лесах Подмосковья III—IV бонитета ко-
леблется от 2 до 3 см. Хвоинки второго года обладают также
резко выраженной теневой структурой. В течение второго веге-
тационного периода на главном стебле опять формируется
верхушечная почка и 2—3 боковые, обычно расположенные в
верхней части годичного побега, ближе к верхушечной почке
(рис 22У. В корневой системе, кроме небольшого прироста
главного корня (до 3—5 см), происходит образование новых.
Рис. 22.
Проростки и сеянцы ели — Picea excelsa'.
1—3 — проростки, 4 — двухлетним сеянец, 5— четырехлетия!»,
6 — шестилетний; г к. — главный корень с боковыми корнями,
гпк — гипокотиль; см - семядоли; эп — эпикотильная часть годичного
побега 1 года
148
боковых корней II и III порядков. На боковых корнях, по
наблюдениям Т. И. Вещиковой, появляются типичные микориз-
ные окончания.
В условиях значительного затенения и на третьем году
жизни сеянца раскрывается только верхушечная почка и на-
блюдается только прирост главной оси, идущий также слабо и
сопровождающийся образованием годичного побега такого же
типа, что и на втором году. В корневой системе, кроме слабого
прироста старых корней, образуются новые боковые, вплоть
до всасывающих корней IV порядка. Главный корень иногда
отмирает, заменяясь одним-тремя боковыми близ отмершей
верхушки В более благоприятных условиях роста (лучшее
освещение, благоприятные почвенные условия), кроме верху-
шечной почки, раскрывается одна или две-три боковые почки
главной оси; начинается ветвление сеянца, образование боко-
вых побегов.
течение ч—о-го года
жизни сеянца растет не
только главная ось, но и бо
ковые ветви; каждая из них
в свою очередь ветвится.
Каждая боковая ветвь по-
добно главной оси имеет,
кроме верхушечной почки,
несколько боковых почек,
образующихся на каждом
годичном побеге в его верху-
шечной части Однако уво
личение ветвистости сеянца
происходит медленно. Ветви
третьего порядка появляют-
ся под пологом леса у 5—
6-летних сеянцев, а ветви
IV порядка лишь в 10—
15-летнем возрасте, да и то
при достаточном освещении,
например, в лесных окнах,
на опушках. Под пологом
материнских деревьев не
только рост, но и ветвление
оказываются сильно подав-
ленными. 10—12-летние
елочки в сомкнутых еловых
лесах Подмосковья едва до-
стигают 15—20 см' и, кро-
ме главной оси, несут
Рис. 23
Ширококронная форма молодого (10—
12-летнего) дерева ели, выросшего в
лесном окне при хорошем освещении
149
лишь несколько боковых ветвей II порядка (рис. 24). Как видно
из рис 23, 10—15-летние сеянцы ели, с освещенных местообита-
ний, в том же возрасте (10—15 лет) мог} г достигать высоты в
100—150 см и более и нести хорошо развитую крону боковых
ветвей, преимущественно II и III порядков.
В течение первых трех лет в корневой системе главный ко-
рень растет быстрее боковых Но с третьего года боковые начи-
нают расти быстрее главного Корневая система становится все
более поверхностной, так как и главный корень, достигнув 10—
15 см длины, приобретает горизонтальное направление
С 3—5 лет начинается образование придаточных корней,
вначале на гипокотиле, а позже и выше по главной оси Обра-
зование придаточных корней облегчается полеганием гипоко-
тиля и нижних частей главной оси и погружением их в мохо-
вой покров и лесную подстилку
Из сказанного следует, что прирост ыавнон оси сеянцев в
этот период (от 2—3 до 10—15 лет) в очень сильной степени
определяется комплексом экологических условий, среди кото-
рых решающим фактором является свет. Как показали изме-
рения, проведенные у mhoihx десятков сеянцев, в сомкнутых
еловых лесах Подмосковья средний прирост главной оси с
2,8 см на 3—5 году жизни сеянца увеличивается к 12 годам до
4,6 см За пределами сомкнутого древостоя на достаточно рых-
лых (иезадернованных) почвах уже на 6—7 году жизни при-
рост главной оси увеличивается до 8—10 см, а у 10 — 12-летних
сеянцев до 18—20 см в год
Таким образом, в течение . первых 15 лет жизни сеянца в
благоприятных условиях роста более или менее отчетливо
проявляется тенденция к увеличению годичною прироста
главной оси В меньшей степени эта тенденция выражена в
условиях значительного затенения
Прирост боковых ветвей в решающей степени зависит от
их положения на главной оси. Самые нижние ветви, возникшие
на сеянцах 3—5-летнего возраста, отличаются очень незначи-
тельным ростом Величина их годичного прироста не превыша-
ет 1—2 см Боковые ветви, возникшие на главной оси в 10—
15-летнем возрасте сеянца, отличаются более интенсивным
приростом; но где бы ни располагались боковые ветки молодых
елочек, их рост всегда несколько уступает приросту главной
оси близ места отхождения от нее. Кроме более слабого роста,
боковые ветви отличаются от главной оси еще одной весьма
существенной особенностью Если главная ось в исследуемый
период жизни ели обнаруживает увеличение годичных при-
ростов с возрастом, то боковые ветви сеянца такой способности
к увеличению прироста не проявляют. Наоборот, с возрастом
величина их прироста становится все меньше, пока прирост не
150
прекратится совсем. Хотя хвоя на боковых ветвях такая же
многолетняя, как и на главной оси (в условиях подмосковных
лесов хвоя ели функционирует чаще всего 5—8 лет), боковые
ветви довольно быстро отмирают, в отличие от главной оси,
функционирующей в течение всей жизни дерева. Так, боковые
веточки 3-го и 5-го года жизни сеянца отмирают 6—8 лет
спустя. Ветви, появившиеся у 10—15-летних сеянцев, остаются
живыми до 30—35-летнего возраста дерева.
При формировании сеянца обращает на себя внимание еще
одна особенность, выраженная то более, то менее резко у раз-
ных экземпляров — это изогнутость гипокотиля, как видно из
рис. 24 У сеянцев со временем происходит все более заметное
Рис. 24
10-летний сеянец ели, выросший в глубине леса, в затенении-
Гипокотиль (гпк) и нижняя часть главного стебля полегла,
гк — главный корень
151
изгибание гипокотиля. Оно начинается в его базальной части
и в зоне корневой шейки, а затем распространяется выше, иног-
да захватывая и облиственную часть главной оси на несколько
сантиметров. Этот изгиб базальной части главной оси и погру-
жение его в почву имеет большое биологическое значение, облег-
чая, как отмечалось, образование придаточных корней.
Образование н рост придаточных корней приводит к сниже-
нию функциональной значимости, ослаблению роста н, наконец,
отмиранию системы главного корня. Отмирание системы глав-
ного корня у 5—8-летних сеянцев начинается с корней высшего
порядка, прогрессивно усиливаясь. У 15—20-летних сеянцев
значение системы главного корня резко снижается, а иногда
она к этому времени почти полностью отмирает. Функция поч-
Рис. 25
Островершинная крона ели на третьем
возрастном этапе ее онтогенеза
152
Рис 26
Акротонное усиление
боковых ветвей у ели
венного питания и водоснабжения переходит к системе прида-
точных стеблевых корней (по наблюдениям Т. И. Вещиковой).
Нарастание и ветвление боковых побегов идет моноподиаль-
но, как и у главной оси. Кроме верхушечной почки, разверты-
• ваются 2—3 боковые почки. Прирост ветвей III порядка обычно
бывает меньше, чем у ветвей II порядка, а ветви IV по-
рядка обладают приростом более слабым, чем побеги III
порядка. Словом, в величине годичных приростов главной оси
и боковых ветвей все более высшего порядка наблюдается
вполне определенная соподчиненность. Чем выше порядок осей,
тем слабее их годичные приросты. В результате этой соподчи-
ненности к 15 годам на освещенных местах у ели формируется
вокруг главного ствола правильная ширококоническая крона
(см. рис. 23).
По мере дальнейшего роста преобладание главной оси над
боковыми ветвями резко возрастает. С 20—30-летнего возраста
у ели резко возрастают годичные приросты главной оси. На
достаточно богатых слабооподзолеппых почвах при полном сол-
нечном освещении в Подмосковье годичные приросты главной
оси достигают 50—70 см. В то же время приросты боковых
ветвей возрастают не столь значительно, сохраняясь на уровне
20—40 см. В начале лета, из-за слабого еще развития механи-
ческих тканей, новые длинные годич-
ные приросты главной оси нередко
поникают или по крайней мере слегка
изгибаются на верхушке.
В результате резкого несоответ-
ствия в интенсивности роста главной
оси и боковых ветвей заметно меняет-
ся форма кроны: из ширококонической
крона становится узкоконической и
островершинной (рис. 25) В этот пе-
риод усиленного прироста главной оси
у ели с особой отчетливостью видна
еще одна структурно-биологическая
особенность, а именно — акротонность
(W. Rauh, 1937) в развитии боковых
ветвей Чем выше на годичном побеге
располагается боковая ветвь, тем
сильнее ее рост Наибольшей энергией
роста в силу акротонности обладает
побег, развивающийся из верхушечной
почки (рис. 26). Следствием акротон-
ности является «этажированность»
расположения боковых ветвей на
главной оси ели. «Этажированность»
153
в расположении боковых ветвей усиливается вследствие того, что
нижние боковые ветви каждого годичного побега главной оси
отличаются не только слабым ростом и незначительной величи-
ной, но и небольшой длительностью жизни. Они довольно
быстро отмирают и опадают с дерева, тогда как верхние боко-
вые ветви каждого годичного побега имеют сильный рост и
долго сохраняются на дереве. Образующаяся мутовчатость
расположения ветвей у ели и других хвойных (с особой отчет-
ливостью она выражена у видов араукарий, кедров, сосны)
дает возможность довольно просто определять возраст деревьев,
так как мутовки ветвей возникают на главной оси ежегодно по
одной.
Акротонность проявляется не только у взрослых ветвей, но
гораздо раньше, еще в состоянии почек. Величина почек на
годичном побеге довольно строго соответствует величине буду-
щих побегов, вырастающих из них. Исследование верхушечной
и боковых почек одного и того же годичного побега главной оси
ели показало, что соотношения в их структуре и величине не
остаются постоянными в разных возрастах дерева и в различных
экологических условиях (табл. 3).
Таблица 3
Величина и число чешуй и зачатков листьев
в верхушечной и боковых почках в пределах одного
и того же годичного побега главной оси у сеянцев, подроста
и молодых деревьев
Возраст ели в годах Высота ели в см Длина годич- ного побега в см Диаметр го- дичного побе- га в мм Расположение исследуемых почек на годичном побеге Линейные размеры почек । Число почеч- । ных чешуй Число зачат- ков листьев
4 7 1.5 1,2 Верхушечная 1,5 16 54
Боков, верхи. 1,2 12 36
4 6,5 1.3 1,5 Верхушечная 2,1 18 91
Боков верхи. 1,6 14 52
7 14 2,6 1,8 Верхушечная 2,8 21 104
Боков, верхи. 2,5 16 104
7 15 3,4 1,5 Верхушечная 3,0 29 104
Боков, верхи 2,8 21 117
7 18 2,5 1.6 Верхушечная 2,8 22 65
Боков, верхи. 2,2 20 91
12 20 2,7 1,8 Верхушечная 2,8 27 85
Боков, верхи. 2,2 20 104
12 18 2,2 1,3 Верхушечная 2,8 27 52
Боков, верхи. 2,2 20 91
154
Возраст ели в годах 1. Высота ели в см Длина годич- ного побега в см Диаметр го- дичного побе- га в мм Расположение исследуемых почек на годичном побеге Линейные размеры почек Число почеч- ных чешуй Число зачат- ков листьев
12 30 5,5 2,2 Верхушечная 4,5' 43 174
Боков, верхн. 3,0 34 163
12 43 15 3,4 Верхушечная 5,5 56 283
Боковые:
верхняя 4,5 40 273
средняя 4,0 43 130
нижняя 3.8 34 80
12 55 17 3,5 Верхушечная 5,8 72 305
Боковые:
верхняя 5,0 49 262
средняя 4,0 42 147
нижняя 2,5 30 68
20-22 60 30 6,5 Верхушечная 11 91 462
Боковые:
верхняя 8 53 315
нижняя 6 46 200
средняя 3,5 28 НО
25-28 280 37 8,0 Верхушечная 12 123 594
Боковые:
верхняя 8 95 325
средняя 7,2 64 252
нижняя 3,5 39 117
32-35 500 63 12 Верхушечная 13 127 557
Боковые:
верхняя 10 — 370
средняя 9 — 252
нижняя 8 — 151
32-35 550 67 15 Верхушечная 14 130 630
Боковые:
верхняя 12 95 441
средняя И — 279
нижняя 4,5 —’ 169
Как видно из таблицы 3, у 4-летних сеянцев верхушечная
почка по числу зачатков хвоинок будущего года значительно
превышает боковые. У 7—12-летних елочек эти различия менее
выражены, а в условиях затенения верхушечная почка главной
оси иногда развита даже слабее, чем ее боковые верхние почки.
У более взрослых быстрорастущих молодых деревьев 20—35-лет-
него возраста, отличающихся большим приростом главной оси,
верхушечная почка по всем изученным показателям намного
превышает боковые почки той же главной оси. Эти различия
в величине и количестве зачатков хвоинок в верхушечной и
155
боковых почках вполне соответствуют различиям в росте побегов,
из них вырастающих. Таким образом, различия в величине
побегов в кроне ели, а, стало быть, и сама форма кроны, в ре-
шающей степени детерминированы уже в состоянии почек.
И здесь в изучении причин различного развития почек нужно
искать раскрытие законов сложения кроны древесных пород, в.
частности, хвойных.
Из таблицы видно также, что и различия боковых побегов,
расположенных на разной высоте по материнскому годичному
побегу, проявляются уже в состоянии почек.
Резкое преобладание в росте главной оси над боковыми
побегами не продолжается постоянно в течение жизни дерева:
к 80—100 годам рост главной оси ели значительно ослабевает,
в то время как боковые ветви сохраняют еще значительную
величину своих годичных приростов В этот период жизни де-
рева и диаметр боковых ветвей лишь немного уступает диаметру
тлавной оси. Так, у просмотренных деревьев 100—120-летнего
возраста в смешанных лесах Подмосковья диаметр сохраняю-
щейся главной оси непосредственно под верхушечной почкой
равен 12—15 мм, а у боковых ветвей, развивающихся на этом
же побеге главной оси, равен 10—15 мм. Величина годичного
прироста главной оси падает с 50—70 см у молодых деревьев
до 10—15 см к 100—120 годам. К 150—180 годам прирост глав-
ного ствола снижается до 5—7 см.
Интересно, что в период максимального роста дерева
(с 20—30 до 70—90 лет) у него имеется и функционирует лишь
система придаточных корней Система главного корня, функ-
ционировавшая в молодом возрасте ели (до 20—30 лет), уже
отмерла. Крупные скелетные стеблевые придаточные корни,
ветвящиеся до IV—V порядка, по наблюдениям Т. И. Вещико-
вой, функционируют 15—20 лет. По мере нарастания мохового
покрова и погружения старых корней в глубь более плотной
почвы с ослабленной аэрацией, рост их замедляется, затем
прекращается полностью, и корни начинают отмирать. На смену
отмершим образуются новые придаточные корни выше по
стволу, располагаясь в поверхностных, богатых и хорошо аэри-
руемых горизонтах почвы Так происходит регулярная смена
стеблевых придаточных корней в течение всего периода макси-
мального роста дерева у ели Примечательно, что утрата спо-
собности к новообразованию придаточных корней, прекращение
их регулярной смены у деревьев старше 90—100 лет совпадает
с ослаблением прироста дерева вплоть до полного его прекра-
щения и появления суховершинности
Снижение интенсивности роста главной оси приводит к тому,
что главная ось становится мало заметной и теряется среди
сильных боковых ветвей Заметно меняется форма кроны: вер-
156
хушка ее становится все более
округлой, дерево теряет остро-
вершинность (рис. 27); ослаб-
ление роста главной оси у ста-
реющих деревьев совпадает
с многократным отмиранием
верхушечной почки ствола,
ель переходит от моноподиаль-
ного нарастания главной оси
к симподиальному. Резкое
ослабление роста верхушки
ствола и переход к симпо-
диальному нарастанию глав-
ной оси с образованием округ-
лой вершины кроны у ели
означает наступление старче-
ского периода в жизни дерева.
Окончательное отмирание
главного ствола на его вер-
хушке и верхних боковых вет-
вей приводит к отмиранию
всего дерева, так как у ели
спящие почки функционируют
на ветвях и стволе не более
7—10 лет, и поэтому нижняя.
Рис. 27
Верхушка кроны старою дерева ели
в начале четвертого возрастного
периода онтогенеза
еще живая часть ствола ко времени отмирания верхушки дерева
бывает уже лишена жизнеспособных резервных точек роста.
Наблюдения за формированием кроны у Picea excelsa от
сеянцев до старческого состояния позволяют выделить четыре
возрастных периода в жизни этого дерева, отличающиеся
и морфологически.
1. Период роста только главной оси В лесах
Подмосковья он длится от прорастания до 4—6-летнего воз-
раста сеянцев. Боковые почки в течение этого периода остаются
в нераскрытом состоянии. Биологически этот период характе-
ризуется чрезвычайно слабым ростом надземных органов, в
результате чего раскрывается на побеге только одна верху-
шечная почка. В подземной сфере в этот период формируется
и функционирует только система главного корня. Рост корней
не превышает роста побегов. Экологически сеянцы ели в этот
период отличаются наибольшей теневыносливостью.
2. Период широко кронного деревца. Растение
имеет небольшую интенсивность роста главной оси, лишь не-
много превышающую рост боковых ветвей. В подмосковных
лесах этот период продолжается от 4—6-летнего до 15—20-лет -
.него возраста сеянцев. Биологически для него характерна
157
несколько большая интенсивность роста побегов по сравнению
с предыдущим периодом. В подземной сфере в этот период
происходит формирование системы придаточных гипокотильных
и стволовых корней и одновременно постепенное отмирание
системы главного корня. Возможно, что ширококоническая
форма кроны в этот период способствует более полному погло-
щению света молодыми елочками под пологом древесных по-
род
3. Период узкокронного островершинного де-
рева Характеризуется большой интенсивностью роста глав-
ного ствола, которая резко превосходит рост боковых ветвей.
Годичные приросты главной оси достигают 50—70 см. Потреб-
ность в освещении в этот период у ели значительно возрастает.
Ассимилирующая часть кроны ели выносится па прямой сол-
нечный свет С началом этого периода совпадает начало семе-
ношения В подземной сфере в этот период у ели полностью
сформирована система придаточных стеблевых корней и наблю-
дается их регулярная смена.
Наряду с отмиранием старых придаточных корней в глубо-
ких плотных и слабоаэрируемых слоях почвы образуются новые
корни выше по стволу, в верхних богатых и рыхлых горизонтах
почвы. В условиях Подмосковья этот период—наиболее дли-
тельный в жизни дерева: он продолжается от 20—30-летнего
до 80—100-летнего возраста, а иногда и несколько долее. К концу
этого периода в корневой системе дерева утрачивается способ-
ность к образованию новых придаточных скелетных корней,
сменяющих старые отмирающие придаточные корни. Вероятно,
в связи с этим начинается ослабление роста главной оси, а за-
тем и боковых ветвей.
4 Период округловершинного стареющего
дерева с последующим появлением суховершинности. Харак-
теризуется замедлением роста главной оси при сохраняющемся
значительном приросте боковых ветвей. Величина годичных
приростов главной оси падает до 7—5 см. Как отмечалось выше
процесс старения дерева в этот период, наряду с резким ослаб-
лением роста, сопровождается переходом от моноподиального
нарастания главной оси к симподиальному, потерей ее преобла-
дания над боковыми ветвями, что вызывает изменение формы
кроны от островершинной к округловершинной. Вследствие
отмирания придаточных корней и утраты способности к образо-
ванию на стволе новых корней, зона поглощения, ранее удален-
ная на 20 и более метров от ствола, в период старения дерева
постепенно приближается к стволу. Площадь корневого пита-
ния сокращается. В конце четвертого периода, в связи с начав-
шейся суховершинностью и отсутствием спящих почек на старых
сучьях и стволе, дерево отмирает.
158
Разумеется, в онтогенезе дерева, в зависимости от скла-
дывающихся условий, могут наблюдаться различные отклонения
от предложенной здесь схемы и возрастной периодизации. Одно
из наиболее часто встречающихся уклонений — выпадение треть-
его периода, если рост дерева протекает беспрерывно в условиях
сильного затенения под пологом древесных пород. Такие экзем-
пляры ели из состояния подроста как бы непосредственно пере-
ходят в старческий период жизни деревца с зонтиковидной фор-
мой кроны Подобные формы роста ели неоднократно отмеча-
лись в лесоводствепной литературе (Г. Ф. Морозов, 1926).
Выделенные четыре биолого-морфологические периода в онто-
генезе ели наиболее четко проявляются в оптимальных условиях
роста У деревьев, выросших в затенении и на бедных почвах,
островершинность кроны бывает выражена слабо.
Морфогенез жизненной формы дерева
у лесной сосны (Pinas Silvestris L.)
Прорастание семян сосны происходит в основных чертах
так же, как и у ели. Зародышевый корешок по выходе из се-
мени начинает расти отвесно вниз и заякоривается в почве.
Благодаря растягиванию гипокотиля семядоли выносятся на
свет, и лишь верхушки их некоторое время остаются в семени
и потребляют питательные вещества эндосперма. Выйдя из
семени, семядоли оказываются над землей и начинают функцио-
нировать как фотосинтезирующие листья. Семядоли сосны,
чаще всего в числе шести, как и у ели, имеют саблевидную
форму, несколько изогнуты вверх, трехгранны в поперечном
сечении. Длина семядолей сосны несколько больше, чем у ели
и достигает, в среднем, 20 мм вместо 15—20 мм у Picea excelsa.
Длина гипокотиля колеблется от 4 до 7 см. Выше мутовки
семядолей у слабых сеянцев, особенно под пологом леса, начи-
нают сразу же формироваться чешуйки верхушечной почки
главной оси, и семядоли оказываются, таким образом, един-
ственными ассимилирующими листьями сеянца первого года,
У более сильных сеянцев уже на первом году выше семядолей
образуется стебель высотой в 4—6 см (в очень хороших усло-
виях до 8—10 см), на котором спирально (по 5/13) распола-
гаются единичные линейные ассимилирующие листья. Эти
первичные листья сеянцев сосны отличаются от листьев взрос-
лого дерева меньшим диаметром (0,4—0,8 мм вместо 1—2 мм),
наличием тонкой зубчатости по краям, а анатомически —
тонкостенпостью эпидермиса, слабым развитием гиподермы,
и склеренхимного влагалища Формированием верхушечной
почки завершается рост главного стебля.
159
В единичных случаях в лесах Подмосковья у однолетних
сеянцев сосны начинается ветвление главной оси за счет появле-
ния боковых побегов в пазухах ее первичных ассимилирующих
листьев. Эти боковые побеги имеют структуру укороченного
побега. На коротком стебле их длиной 3—5 мм вначале разви-
ваются 2 низовых листа, располагающихся латерально. Выше
этих листьев развиваются расположенные тесной спиралью
зеленые листья, все более увеличивающиеся в размерах (от
1—1,5 см до 5—8 см длины) Число зеленых листьев на побеге
достигает 6—8. Ювенильные боковые побеги сосны резко отли-
чаются от пазушных побегов — брахибластов, взрослых де-
ревьев сосны своей более слабой специализацией
В отличие от стебля, главный корень сеянца в течение пер-
вого года отличается большей интенсивностью роста и за лето
вырастает в лесах Подмосковья на 15—20 см и более, при
обильном ветвлении, особенно в ого основании, у корневой
шейки. Уже на первом году7 на корнях сеянцев сосны появляет-
ся микориза (Kirchner, 19*08). Как и у ели, у сосны в конце
первого вегетационного периода обычно формируется лишь
одна верхушечная почка, из которой после перезимовки про-
должается нарастание главной оси
На втором году, при раскрытии верхушечной почки, на
удлиненном побеге главной оси вначале вновь образуются сидя-
щие поодиночке ассимилирующие листья, которые по длине
годичного побега постепенно переходят в короткие треугольные
коричневые чешуйчатые листья В пазухах первых 10—20 асси-
милирующих первичных листьев не образуется пазушных по-
чек. Но выше сидящие переходные и чешуйчатые листья несут
пазушные почки, которые раскрываются почти одновременно
с весенним ростом их кроющих листьев Из этих почек обра-
зуются крайне укороченные побеги весьма своеобразного строе-
ния— брахибласты Появление на удлиненном стебле чешуе-
видных листьев с пазушными почками, раскрывающимися в год
их формирования, приводит к тому, что у сосны весной обра-
зуется не простой удлиненный годичный побег, а целая система
годичного побега, где ассимилирующие листья располагаются
не на главной оси, а на укороченных боковых побегах. Начало
формирования сложных годичных побегов с зелеными листьями
на брахибластах характеризует у Pinus Silvestris завершение
ювенильного периода в ее развитии
Окончание ювенильной фазы в развитии сосны и прекраще-
ние образования зеленых листьев на удлиненных побегах ее
может быть ускорено при изменении условий освещения. Так,
в опытах В. М Лемана (1948) сеянцы Pinus silvestris в питом-
нике, при освещении их ночью лампами в 100—500 вт, более
чем вдвое увеличили интенсивность роста главной оси и уже
160
в течение первого года переходили к образованию брахибла-
стов
В течение второго года побег сосны вырастает от 5—7 см до
1‘5—20 см. Меньшие приросты наблюдаются у естественного
подроста сосны под пологом леса. Резко отрицательное влияние
на рост сеянцев сосны оказывает густой моховой и травяни-
стый покров. Как и в ’первый год, у сеянцев сосны в течение
второго лета рост корневой системы идет сильнее, чем рост
сгсбля. На рыхлых супесчаных почвах главный корень двухлет-
них сеянцев углубляется до 60—80 см.
Правда, на заболоченных торфянистых почвах в лесах Под-
московья главный корень уходит в глубину не более чем на
10—15 см, и затем растет горизонтально, подобно боковым
корням На таких почвах различие в толщине и длине главного
и боковых корней проявляется слабо; диаметр главного корня
быстро уменьшается до размеров диаметра боковых корней
Но и здесь длина главного корня значительно превышает вы-
соту главного стебля В конце лета рост побега завершается
образованием верхушечной почки и группы крупных боковых
пазушных почек, в числе от 2—3 до 5—7, расположенных почти
мутовчато, чуть ниже верхушечной почки.
С третьего года на всех удлиненных побегах сосны обра-
зуются только низовые чешуевидные листья, расположенные
спирально по 13 34. Их нисходящие влагалища образуют на
стебле листовые подушки, выделяющиеся на коре стебля в виде
продольных ребер, а верхняя свободная часть скоро подсыхает
и опадает. В пазухах этих листьев, как отмечалось выше, фор-
мируются брахибласты. Брахибласты Pinus silvestris имеют
недоразвитый стебель длиной в 0,2—0,5 мм и диаметром около
0,5 мм. У основания стебля развивается 8—10 налегающих друг
на друга низовых чешуевидных или пленчатых листьев, из ко-
торых 2 самые нижние располагаются латерально по отношению
к кроющему листу, а 2 последующих — медианно, на задней
и передней стороне укороченного побега. Нижние чешуевидные
листья — наиболее мелкие, но они имеют хорошо развитую
среднюю жилку. Следующие за ними пленчатые листья почти
стеблеобъемлющие и более крупные, но с недоразвитой сред-
ней жилкой Самые верхние листья вновь становятся мельче.
За этими низовыми листьями следуют два крупных зеленых
листа — хвоинки длиной в 60—80 мм. В почкосложении эти
листья плоские и плотно примыкают друг к другу своими
пластинками. На верхушке стебля между зелеными листьями
располагается очень маленький конус нарастания, иногда от-
мерший у живых побегов.
Изредка у молодых деревьев близ верхушки годичного по-
бега верхушечный конус нарастания брахибластов разрастается
В-4В, И. Г Серебряков—И 161
образуя хорошо развитую верхушечную почку до 6—8 мм
высоты и 3—5 мм в диаметре.
Кроме брахибластов, с третьего года у сеянцев сосны выра-
стают боковые удлиненные побеги из мутовки почек, располо-
женных несколько ниже верхушечной почки Начинается вет-
вление побегов и образование кроны Для Picea excelsa отме-
чалась акротопия в развитии побегов, т е акропетальное уси-
ление боковых побегов. У сосны явление акротонии выражено
в еще более резкой форме Годичные побеги главной оси и бо-
ковых ветвей почти на всем протяжении лишены боковых по-
чек, дающих начало удлиненным побегам. Боковые почки все
сосредоточены в виде мутовки у самой верхушки побега Такое
расположение почек вызывает четкую ярусность в расположе-
нии ежегодно образующихся боковых ветвей Как следствие
ярусного расположения боковых ветвей на главной оси и ветвях
кроны и сдвига их в верхушечную область годичных побегов,
наблюдается постоянное отодвигание ветвей высшего порядка
и ассимилирующей листвы дальше от главного ствола, на пери-
ферию кроны и на ее верхнюю поверхность, где имеются более
благоприятные условия освещения. Акротония ветвления таким
образом оказывается морфологическим проявлением приспособ-
ления древесных растений к наилучшему использованию заня-
того воздушного пространства и наилучших условий освещения.
W Rauh (1937) считает, что акротонность ветвления пред-
ставляет самый существенный признак деревьев и определяет
у растений образование жизненной формы дерева.
Рост сеянцев сосны в течение первых 10—15 лет идет более
быстро, чем у ели Но, как и у ели, сеянцы у сосны весьма чув-
ствительны к интенсивности освещения и условиям почвенного
питания Под пологом леса 1 11 интенсивность роста сеянцев сос-
ны возрастает незначительно, увеличиваясь с 5—7 см у 2—
3-летних сеянцев до 10—12 см у 12—15-летних. Любопытно, что
изучение хода роста у каждого из сеянцев обнаруживает зигза-
гообразный характер кривой роста в течение ряда смежных
лет. Судя по характеру кривой, можно было думать, что эти
колебания связаны с колебаниями метеорологических усло-
вий в соответствующие годы, оказывающихся то более, то
менее благоприятными для роста. Однако сравнение величины
1 Л1осковская область, лесной массив Звенигородской биостанции МГУ,
березово-сосновый лес полнотой 0,6 с кустарниковым ярусом из Lonicera
xylosteum, Rhatnnus frangula, Juniberus communis, густым ярусом кустар-
ничков из Vaccinium myrtillus и Vaccinium vitis idaca, и сплошным мохо-
вым покровом из Hylocomium proliferum и Pleurozium Schreberi (почва
слабо оподзоленная, легко суглинистая, дренаж хороший, условия увлажне-
ния средние).
162
годичных приростов у рядом стоящих сеянцев в одни и те же
астрономические годы показывает более сложную картину.
Различный характер роста рядом стоящих сеянцев сосны в
.одни и те же годы означает, что колебания величин годичных
приростов определяются не только колебанием метеоусловий, а
связаны с изменением физиологического состояния самих сеян-
цев Максимальные и минимальные приросты повторяются
через каждые 2—3 года.
С величиной годичного прироста главной оси тесно связана
мощность боковых побегов. В годы с наибольшим приростом
1лавной оси развиваются и наиболее сильные боковые ветви с
большим диаметром, более интенсивным ростом и, как показы-
вает изучение более взрослых деревьев, дольше сохраняющие-
ся в кроне Наоборот, в годы с минимальным приростом глав-
ной оси вырастают слабые ветви, тонкие, медленно растущие и
менее долговечные.
Несмотря па сильное угнетение сеянцев сосны под пологом
леса, они все же развиваются здесь как типичные деревья,
постоянно имеют вертикально растущую главную ось и отходя-
щие от нее почти под прямым углом боковые ветви.
Наиболее быстрый рост сеянцев в высоту и по толщине
ствола наблюдается на открытых местах, особенно в искусст-
венных посадках. Здесь у каждого из сеянцев проявляется
зигзагообразный характер кривой роста, колебания приростов
в течение ряда смежных лет. Как показало изучение, и здесь
эти колебания нельзя свести только к разногодичным колеба-
ниям метеорологических условий Колебания интенсивности
роста проявляются здесь как биологическая особенность самих
сеянцев сосны Правда, здесь эти колебания не затемняют
общего восходящего характера кривой роста сеянцев в тече-
ние первых 15—20 лет их жизни.
Одновременно с увеличением годичных приростов главной
оси в этот период наблюдается другой, не мепее важный про-
цесс — усиление мощности боковых ветвей. Ветвление сосны
начинается с третьего, реже второго года жизни При этом вет-
ви, возникшие на 2—3 году, отличаются ничтожной величиной
прироста; общий период их роста и их жизни обычно не пре-
вышает 5—8 лет После этого верхушечная почка их не раскры-
вается, хвоя подсыхает, и побег опадает с дерева Ветви, воз-
никшие на 4—6-м годах жизни сеянца, отличаются большей
интенсивностью роста; общий ростовой период и длительность
жизни этих ветвей увеличивается до 10—20 лет.
Боковые ветви, появившиеся в более старшем возрасте
жизни сеянца, характеризуются большей быстротой роста, бо-
лее поздним наступлением максимума прироста, большей дли-
тельностью общего периода роста и большей длительностью
П" ]ЬЗ
жизни самой ветви. Ветви, возникшие в 15—20-летием возрасте,
сохраняются как сучья в кроне взрослого и старого дерева. Их
длительность жизни равна многим десяткам лет. Интересно,
что отмирание нижних ветвей, возникших в первые годы жизни
сеянца, не обязательно связано непосредственно с сильным их
затенением. Быстрое отмирание таких ветвей наблюдается и в
разреженных искусственных посадках с регулярно удаляемыми
сорняками, где прямой солнечный свет свободно доходит до
нижних ветвей. Рано возникшие ветви быстро отмирают под
влиянием внутренних биологических причин, в связи с недо-
статочной возрастностыо этих побегов (Н П. Кренке, 1940),
или в связи с сильным коррелятивным торможением роста,
идущим от растущей верхушки главной оси (Зединг, 1955, см
ниже)
Нарастание и ветвление главной оси и боковых ветвей в
описываемый период жизни дерева у сосны происходит по
строго моноподиальному типу, в результате чего образуется
очень правильная система побегов, в которой главная ось
сохраняет преобладающее положение (по величине годичных
приростов) до самой верхушки кроны и заметно выделяется
над кроной дерева. В этой одноствольной кроне обращает на
себя внимание строгая соподчиненность побегов главной оси и
боковых ветвей различного порядка. Чем выше порядок вет-
вей, тем слабее их рост, тем сильнее они наклонены по отно-
шению к материнской оси В этот период сосна характеризует-
ся островершинной формой кроны с главной осью, легко
прослеживаемой до верхушки дерева. Особенно отчетливо в
этот период проявляется роль главной оси как органа, ре-
гулирующего рост остальных боковых побегов, регу-
лирующего морфогенез кроны дерева в целом Годичные при-
росты главной оси в среднем достигают 40—50 см Описывае-
мый период в онтогенезе сосны в лесах Подмосковья длится до
70—90 лет.
В последующие десятилетия из-за резкого ослабления интен-
сивности роста главной оси, ее годичные побеги становятся все
более короткими и тонкими В 120—150 лет они не превышают
в среднем 20 см длины Главная ось на верхушке кроны вслед-
ствие этого теряется среди боковых ветвей, годичные побеги
которых к этому времени не отличаются существенно по интен-
сивности роста от побегов верхушки главной оси. Ослабление
роста главной оси завершается тем, что ее верхушечная почка
в некоторые годы не раскрывается и подсыхает, вследствие
чего нарастание теряет строго моноподиальный характер и
становится симподиальным. Отмершая главная ось замещается
или одним из боковых побегов самой верхней мутовки, при-
обретающим вертикальное направление роста, или побегом,
164
вырастающим из верхушечной почки одного из брахибластов,
расположенных близ верхушки главной оси. В результате
ослабления роста главной оси при сохранении достаточной
интенсивности прироста боковых ветвей лидерное положение
главной оси на верхушке старого дерева постепенно утрачи-
вается. Сильные боковые ветви сохраняют не только большие
годичные приросты, но и моноподиальный способ нарастания
Форма кроны из островершинной конической постепенно прев-
ращается в куполообразную, более или менее плосковершин-
ную Изменение формы кроны в этот период совпадает с отми-
ранием более слабых и топких ветвей в ее внутренней части,
так что в кроне, сосредоточенной теперь близ верхушки дерева,
сохраняются лишь наиболее толстые и длинные сучья, распо-
ложенные на стволе через 2—3 мутовки. Основание этих сучьев
оказывается так же лишенным боковых ветвей, как и главный
ствол
Отмирание слабых ветвей кроны и очищение оснований
сильных сучьев свидетельствует о понижении теневыносливости
хвои и повышении ее светового порога у старых деревьев
Именно поэтому создавшееся ранее при формировании кроны
затенение становится чрезмерным и вызывает отмирание
охвоенных побегов Куполообразная, более или менее плоско-
вершинная форма кроны характеризует последний, старческий
период в онтогенезе сосны Подсыхание верхушечной почки
главной оси, вначале происходящее редко и нерегулярно, ста-
новится все более частым и постоянным, и нарастание главной
оси становится строго еймподиальным. С дальнейшим увели-
чением возраста дерева отмирание верхушечной почки стано-
вится обязательным не только для главной оси, но и для бо-
ковых ветвей, что приводит в конце концов к подсыханию вер-
шины кроны таких перестойных деревьев В результате отми-
рает все дерево, так как сосна, как и ель, не имеет в нижней
части ствола спящих почек, способных образовать новые вет-
ви кроны.
Все, сказанное выше о формировании жизненной формы де-
рева у сосны, дает возможность выделить в непрерывно иду-
щем процессе морфогенеза ее 4 периода, каждый из которых
характеризуется вполне определенными биологическими и
морфологическими чертами
I возрастной период одноосного, нсветв ящегося
сеянца, с одной лишь главной осью и сидящими
на пей ювенильными ассимилирующими ли-
стьями В природных условиях под пологом леса этот период
длится два года В культуре при непрерывном освещении этот
период может быть сокращен до одного года. Годичный прирост
обычно не более 3—5 см Биологически растения в этот период
165
характеризуются очень слабой способностью к росту, вслед-
ствие чего на побеге не только раскрывается, но и заклады-
вается только одна верхушечная почка. Боковые почки недораз-
виваются. Характерна значительная теневыносливость. Этот
период сходен с первым
Рис. 28
Сосна {Pinus silvestris) во
втором возрастном периоде ес
онтогенеза в искусственных
посадках в Московской области; в
основании ствола еще сохранились
подсыхающие слабые нижние ветви
периодом жизни у ели
II возрастной период усиле-
ния роста боковых вет-
вей и главной оси. В этот
период формируется моноподи-
ально нарастающая одностволь-
ная крона широко веретеновид-
ной формы, с острой вершиной
(рис. 28) Быстрое усиление роста
главной оси и верхних боковых
ветвей выносит крону на прямой
солнечный свет Сильное преоб-
ладание главной оси по интенсив-
ности роста над боковыми побе-
гами (особенно нижними) при-
водит к резкому коррелятивному
торможению нижних боковых по-
бегов и постепенному их отмира-
нию и очищению ствола снизу
В условиях Подмосковья этот
период длится от 3—5-летнего до
15—20-летнего возраста сеянцев.
III возрастной период фор-
мирования дерева с мак-
симальным ростом глав-
ной оси и боковых вет-
вей. Характерно строго моно-
подиальное нарастание побегов и
формирование очень правильной
островершинной кроны со стро-
гим соподчинением в росте побегов разного порядка, от главной
оси до ветвей III—IV порядков. Крона коническая, главная ось
в ней легко'прослеживается от основания до верхушки и замет-
но возвышается над кроной (рис. 29). Этот период длится
у сосны в лесах Подмосковья от 15—20 до 70—90-летнего воз-
раста.
IV возрастной период ослабления роста главной
оси и образования куполообразной кроны. Значи-
тельное ослабление роста главной оси (в среднем менее 20 см
в год) при сохранении достаточной интенсивности роста боко-
вых ветвей сопровождается переходом ее от моноподиального
к симподиальному нарастанию. Изменение в соотношении роста
166
Рис. 29
Сосна (Pinus silvestris) в 1ретьем
возрастном периоде ее онтогенеза
в лесах Московской области
главной и боковых осей приводит постепенно к изменению
формы кроны от конической островершинной к куполообразной,
более или менее плосковершинной (рис. 30). В течение этого
периода наблюдается отмирание более слабых ветвей кроны
‘с сохранением наиболее сильных сучьев, расположенных через
2—3 годичных мутовки друг от друга. Основания оставшихся
сучьев очищаются от мелких
ветвей. Отмирание ветвей в
кроне в этот период свиде-
тельствует о снижении тене-
выносливости хвои старых де-
ревьев. С увеличением воз-
раста уменьшение интенсив-
ности роста и переход к
симподиальному нарастанию
наблюдается и на боковых
ветвях, что завершается от-
миранием верхушек побегов.
Подсыхание верхней части и
периферии кроны приводит к
отмиранию дерева в целом
из-за отсутствия длительно
сохраняющихся спящих почек
на стволе. Этот период длит-
ся от 90—100-лстнсго воз-
раста до 200—250-летнего.
Разнообразие формы крон
и особенности их формирова-
ния у некоторых хвойных
деревьев
По отдельным наблюде-
ниям можно заключить, что
установленные возрастные пе-
риоды и закономерности формирования кроны в онтогенезе сосны
лесной (Pinus silvestris) свойственны и другим видам сосны и,
в частности, Pinus Pallasiana в горах южного побережья Кры-
ма и Р. pinea в странах Средиземноморья. У молодых деревьев
Pinus Pallasiana крона более или менее островершинна. Но
в более старом возрасте у нее происходит быстрое уменьшение
интенсивности роста главной оси, сопровождающееся более
или менее частым подсыханием верхушечной почки В отличие
от Pinus silvestris у Pinus Pallasiana ослабление роста и под-
сыхание верхушки главной оси происходит быстрее. В резуль-
тате у старых деревьев Р. Pallasiana, биологически относя-
1G7
шихся к IV возрастному периоду их онтогенеза, формируется
резко выраженная плосковершинная крона.
У Pinus Pinea — пинии — с ее зонтиковидной кроной, столь
характерной для ландшафтов Средиземноморья, по наблюде-
ниям автора, на юге Франции молодые быстрорастущие де-
ревья имеют коническую, более или менее островершинную
крону, не отличающуюся сильно от кроны молодых деревьев
Pinus silvestris. Пре-
вращение кроны в зон-
тиковидную и плоско-
вершинную происхо-
дит лишь с известно-
го возраста и пред-
ставляет биологиче-
ский возрастной при-
знак. Это возрастное
изменение формы кро-
ны у Pinus pinea, как
и у Р. silvestris, свя-
зано с резким ослаб-
лением роста главной
оси дерева в извест-
ном возрасте при
продолжающемся бо-
М лее или менее интен-
сивном приросте бо-
ковых ветвей кроны,
растуших наклонно
или почти горизон-
тально. Отличие от
более северной Pinus
silvestris заключается
в том, что ослабление
роста главной оси
•—--------------------------------после достижения
Рис. 30
Сосна (Pinus silvestris) в четвертом
возрастном периоле ее онтогенеза в лесах
Московской области
кульминации у пинии
происходит более бы-
стро. Это резкое ос-
лабление роста глав-
ной оси у пинии и при-
водит к столь же резкому переходу от островершинной к зонти-
ковидной кроне. Вероятно, сухой и жаркий климат Средиземья
с длительным вегетационным периодом вызывает быстрое ста-
рение главной оси пинии, что проявляется в резком снижении
ее прироста.
168
Прямую противоположность зонтиковидной кроне пинии
представляет сильно вытянутая, узко пирамидальная крона
ели Шренка (Picea Schrenkiana), растущей в горах Тянь-Шаня
(рис. 31).
* Количественно форма кроны дерева может быть охаракте-
ризована отношением диаметра кроны в ее наиболее широкой
части к высоте кроны. У зрелых деревьев Pinus Pinea это отно-
шение доходит до 2—3, у Picea excelsa в лесах Подмосковья
этот коэффициент в среднем колеблется от У2 до У3. В горных
лесах северо-восточного Тянь-
Шаня (Заилийский Алатау) у
Picea Schrenkiana это отноше-
ние равно У5—Уб, а иногда
доходит до У8—’/э. Впрочем,
и здесь наблюдаются резкие
возрастные изменения формы
кроны. У елочек 10—15 лет
это отношение колеблется от
1 до Vs, у деревьев 30—40
лет —от ’/2 до '/з, у 60—80-
летних елей ‘Д—’А, и лишь у
зрелых деревьев 150 лет и
старше — оно может дохо-
дить до У8—Уэ. Столь резко
вытянутая форма кроны яв-
ляется не только видовой осо-
бенностью Picea Schrenkiana,
но в значительной мере опре-
деляется спецификой горных
условий. Так, судя по фото-
графии Neger в известной
книге «Biologie der Pflanzen»
(1913), у Picea excelsa в
Южном Тироле, на высоте
1800 м над уровнем моря,
кроны имеют отношение диа-
метра к высоте, равное в сред-
нем 76 Значит, в горах Южного
тает форму кроны, по степени суженности не уступающую кроне
Picea Schrenkiana. Подобное изменение формы кроны у видов
ели наблюдается и близ полярных пределов их распростране-
ния. Так, на Кольском полуострове, в предгорьях и нижнем
поясе Хибинских гор у Picea obovata v. fennica образуется силь-
но вытянутая крона со средним отношением диаметра кроны к
ее высоте у зрелых деревьев, равным Уз—У?, в то время как у
южного предела распространения ели форма кроны у Picea
169
Рис. 31
Крона тяныпанской ели (Picea
Schrenkiana) в Заили иском Алатау
Тироля Picea excelsa приобре-
obovatu широко коническая со средним отношением диамшра к
высоте V2—'А
Чем определяется возникновение \ хвойных деревьев столь
резко выраженной узко пирамидальном формы кроны? J Wies
пег (1907), Neger (1913) и В Н Любименко (1924) объясняют
ее появление как приспособление к защите от вредного деист
вия прямых солнечных лучей в горах, а также как приспособ-
ление для лучшею использования диффузного света в север-
ных районах Действительно, частая встречаемость хвойных
деревьев с узко пирамидальной кроной в горах и северных
районах подтверждает это объяснение.
Но как возникает подобная форма кроны и каковы непо-
средственные факторы ее образования? Структурно степень су-
женности кроны определяется соотношением быстроты и про
должительности роста главной оси и боковых ветвей Ясно, что
ослабление роста боковых ветвей при достаточно сильном росте
главной оси повлечет за собой формирование узко пирами-
дальной кроны. Поэтому вопрос о причинах возникновения узко
пирамидальной кроны сводится к вопросу о причинах слабого
роста боковых ветвей при интенсивном росте главной оси.
А. Энглер (A. Engler, 1893) связывал слабый рост боковых
ветвей у ели с действием весенних заморозков Акропетальный по-
рядок развертывания почек на побегах ели приводит к тому, что
боковые почки, раскрывшиеся ранее верхушечной, подвергаются
действию заморозков, тогда как верхушечная почка, разверты-
вающаяся позже, такому действию не подвергается Однако наши
наблюдения в Заилийском Алатау не обнаружили подмерзания
боковых ветвей у тяпьшанской ели.
Один из факторов слабого роста боковых ветвей связан с
особенностями их охвоения Известно, что между верхушечной
почкой побега и количеством зеленых листьев на побеге суще-
ствуют обратные коррелятивные отношения (К Goebel, 1908,
Зединг, 1955) Растущие и функционирующие зеленые
листья в известные периоды оказывают тормозящее влияние
на рост верхушечцой почки побега И чем больше зеленых
листьев на побеге, тем сильнее коррелятивное торможение вер-
хушечной почки Но количество зеленых листьев на побегах
зависит, кроме других показателей, от длительности жизни зе
леных листьев (хвои) Отсюда можно ожидать, что форма крон
у хвойных деревьев будет в известной степени связана с дли-
тельностью жизни хвои Действительно, у сосны (Pinus silvest-
ris) хвоя живет всего 2—4 года, верхушечные приросты ветвей
значительные и крона широкая У ели обыкновенной в южной
части зоны хвойных и смешанных лесов Европейской части
СССР хвоя живет в среднем 6—9 лет, и соответственно крона
более узкая, чем у сосны Несколько большая длительность
170
жизни хвои наблюдается у сибирской пихты (Abies sibirica) —
Ч—12 и более лет, и крона ее еще уже. Тяньшапская ель с
узкопирамидальной кроной отличается в то же время чрезвы-
чайно большой длительностью жизни хвои до 28—29 лет
Но приведенное сравнение все же мало убедительно, так
как приходится сравнивать растения, далекие по своим наслед-
ственным свойствам Больший интерес поэтому представляет
сравнение в пределах одного и того же вида. Длительность
жизни листьев в пределах вида изменяется в различных геогра-
фических областях и высотных поясах. Так, у Picea Schrenkiana
в Заилийском Алатау наблюдается резкое увеличение продол-
/кительности жизни хвои по мере увеличения абсолютной вы-
(. О I ы
Абсолютная высота Длительность жизни хвои
1400 м над уровнем моря 8—11 лет
1800 м » » » 12—1ч »
2600 л? » » » 18—22 года
В соответствии с этим форма кроны зрелых деревьев также
меняется от нижнего к верхнему пределу распространения ели
в горах У нижней границы ели форма ее кроны более широ-
кая, чем в верхней полосе лесного пояса.
Формирование узкоконической кроны у Picea obovata в Хи-
бинах также связано со значительным увеличением длитель-
ности жизни хвои. Вместо 6—9 лет у южного предела хвойных
лесов, хвоя ели на Кольском полуострове в районе Хибин
функционирует до 16—18 лет.
Приведенные данные говорят о наличии определенной связи
между длительностью жизни хвои у видов ели с формой их
кроны Чем больше длительность жизни хвои у ели, тем слабее
рост боковых ветвей, тем более узкой и вытянутой оказывается
крона Любопытно, что увеличение длительности жизни хвои у
тяныпаяской ели по мере поднятия в горы наблюдается только
па боковых ветвях, но не на главной оси. Можно по-разному
толковать коррелятивные отношения между листьями и ростом
верхушечной почки побега (влияние отводящих токов
J Wiesner, 1881. трофические отношения между зелеными
листьями и вновь растущим побегом — К. Goebel, 1908; гормо-
нальные отношения — R. Dostal, 1913; Зединг, 1955), но нали-
чие самой корреляции — установленный факт.
Увеличение долголетия хвои означает в то же время увели-
чение ее теневыносливости, а последнее приводит к замедлению
отмирания нижней части кроны. Действительно, у тяньшанской
ели крона часто доходит почти до поверхности земли, и даже в
сомкнутых насаждениях крона начинается на высоте 5—7 м.
Это обстоятельство также сильно увеличивает степень сужен-
ности кроны, так как увеличивается ее длина.
Кроме рассмотренной зависимости формы кроны от долго-
летия хвои, едва ли не большее значение в формировании кро-
ны имеет другой биологический фактор — постепенность старе-
ния главной оси дерева после достижения им максимума роста
Показателем быстроты старения главной оси у дерева в этот
период является скорость падения интенсивности ее роста. Чем
быстрее стареет дерево, тем круче нисходящая ветвь кривой ее
роста. В отличие от Pinus Pinea, быстро стареющей и резко
меняющей форму кроны, у Picea Schrenkiana и других видов
ели высокогорных и приполярных стран наблюдается медлен-
ное старение и медленное падение интенсивности роста главной
оси. Интенсивно растущая главная ось не только сама по себе
создаст островершинность кроны, но и вызывает коррелятивное
торможение роста боковых ветвей.
Медленное старение деревьев в приполярных и высокогор-
ных странах, вероятно, связано с особенностями климата —
кратким вегетационным периодом с пониженными температу-
рами. Известно, что комплекс экологических условий высоко-
горных и приполярных районов вызывает у многих видов
растений увеличение длительности жизни и превращение видов
однолетних в двулетние и многолетние, а монокарпических —
в плейо- и даже в поликарпические. Эти изменения в биологии
развития растений также связаны с замедлением процессов
старения в онтогенезе травянистых форм. У хвойных деревьев
в приполярных и высокогорных условиях также наблюдается
замедление процесса старения, но проявляется оно в длитель-
ном сохранении достаточно сильного роста главной оси и в кор-
релятивном торможении роста боковых ветвей; благодаря
этому здесь и формируются узкопирамидальные и островершин-
ные формы кроны и наблюдается длительное сохранение этой
формы в онтогенезе дерева. Правда, уменьшение островершин-
ности кроны у старых деревьев наблюдается и здесь, хотя
в целом крона остается резко вытянутой и узкоконической.
Формирование жизненной формы дерева
у черешчатого дуба (Quercus robur L)
В отличие от сравнительно мелкосемянных ели и сосны,
развитие сеянца дуба начинается с весьма крупного семени
с большим запасом питательных веществ. В среднем вес же-
лудей у дуба в Подмосковье равен 5—7 граммам. Желуди
с двумя и более семенами, из которых развивается соответст-
венное количество проростков, встречаются сравнительно
172
редко ’. Питательные вещества семени дуба сосредоточены
в зародыше, в его массивных семядолях, нацело заполняющих
желудь. Паренхимные клетки семядолей сплошь забиты крах-
мальными зернами. Запасающая ткань испещрена сетью прово-
дящих пучков (Д. Е. Янишевский, 1937) Между семядолями,
у верхушечного полюса желудя, располагается очень короткий
гипокотиль с зародышевым корешком (radicula) и почечкой
(plumula). В почечке зародыша заложены две, редко три пары
зачаточных нерасчлененных листьев Длина гипокотиля с ко-
решком (их трудно разграничить в этом состоянии) не превы
шает 1 —1,5 мм, а длина почечки 0,5—0,7 мм.
При наличии благоприятных условий зрелые желуди про-
растают. Период прорастания длится 30—40 дней В подмосков-
ных лесах проростки дуба появляются с конца апреля и до
июня включительно К середине лета сеянцы дуба в подмосков-
ных лесах имеют вполне сформированный побег с зелеными
листьями и почками На полезащитных полосах от Камышина
до Волгограда в 1950 г. нами наблюдалось прорастание желу-
дей до середины августа. В лабораторных условиях прораста-
ния желудей можно добиться в любое время года. Для уско-
рения прорастания целесообразно снять околоплодник
Прорастание семян дуба происходит по подземному типу
и начинается с вытягивания гипокотиля и зародышевого ко-
решка, дающего начало главному корню дуба При просмотре
анатомического строения осевого органа, вырастающего из
гипокотиля и зародышевого корешка, обнаруживается, что по
всей длине этот осевой корневидный орган имеет вначале ти-
пичную стеблевую структуру, он несет мощно развитую сердце-
вину, сплошное кольцо ксилемы и флоэмы, равномерно рассеян-
ные сосуды и трахеиды Стеблевая структура этого органа
у сеянцев дуба более подробно описана Н. П. Доманской
(1958), подобная структура прослеживается в условиях Под-
московья на протяжении 8—10 см ниже семядолей, а в засуш-
ливых условиях окрестностей Волгограда — более, чем на
15 см. Ниже стеблевая структура постепенно переходит в ти-
пично корневую Сохранение стеблевой структуры много ниже
семядолей означает, что на первых этапах прорастания проис-
ходит резкое вытягивание гипокотиля, обладающего положи-
тельным геотропизмом Благодаря этому корень сеянца дуба
быстрее углубляется в почву, что имеет определенное приспосо-
бительное значение, особенно в засушливых местообитаниях
Корни, отходящие от гипокотиля, обладают типичной корневой
1 По данным А. М Шутяева (1957), в дубравах Курского Государст-
венного Заповедника такие желуди составляют примерно 11% всех желудей.
173
анатомической структурой. Описанные факты позволяют заклю-
чить, что утверждение М. Бюсгена (1913) о наличии короткого
гипокотиля у проростков черешчатого дуба (не более 3—4 мм)
не соответствует действительности. Если же образующийся при
начале прорастания осевой корневидный орган представляет
собой не корень, а имеет на большом протяжении гипоко-
тильную структуру, то боковые корни, отходящие в этой части
сеянца морфологически представляют собой придаточные гипо-
котильные корни.
Когда гипокотиль и корень проростка дуба вместе достигают
уже 10 п более сантиметров в длину, начинает расти почечка
дающая начало главному стеблю сеянца Но иногда рост
почечки зародыша задерживается еще более. Г. Р Эйтинген
(1915) отметил случай, когда почечка начала расти лишь при
достижении корнем длины 30 см
В наших лабораторных опытах 1951 г. выращивались дубки
из желудей, снятых со зрелых деревьев дуба в Лосиноостров-
ском лесном массиве под Москвой в конце августа и начале
сентября. В декабре этого же года началось прорастание, при-
чем наблюдался нормальный рост гипокотиля и корневой
системы, а почечка начала расти лишь 3—4 месяца спустя после
появления корня.— когда последний вместе с гипокотилем
достигал уже 40 см длины и обильно ветвился. Попытки уско-
рить рост почечки подогреванием проростков или их охлаж-
дением (их выносили из помещения на несколько суток, когда
температура воздуха была — 3°—5° С) дотгое время не приво-
дили к желаемым результатам
Рост почечки, а затем и стебля вначале проявляется в рас-
тяжении междоузлий и увеличении размеров зачатков плен-
чатых низовых листьев, заложенных еще в зародыше Но очень
скоро на конусе нарастания верхушечной почки стебля начи-
нается заложение новых зачатков листьев, сначала низовых,
а затем и зеленых ассимилирующих Первые два листа выше
семядолей представлены в виде мелких раздвоенных прилист-
ников около 2 ли: длины, они располагаются почти супротивно
Выше их развиваются еще 2—3 низовых листа, лишенных пла-
стинки или несущих ее недоразвитые зачатки, а затем форми-
руются зеленые ассимилирующие листья.
Эти ювенильные зеленые листья обыкновенно отличаются
от листьев взрослых деревьев тем, что основание их пластинки
клиновидное, как у взрослых листьев Quercus sessiliflora, и они
не имеют ушков при основании пластинки, характерных для
взрослых листьев Quercus robur. Этот факт послужил для
оценки филогенетических отношений Q sessiliflora и Q. robur
(Малеев, 1937) Однако у сеянцев Q. robur довольно часто
174
встречаются также ювенильные листья с ушками у основания
пластинки, п вряд ли этот признак является столь строго
закономерным
Весь комплекс листьев сеянца, образующих первую гене-
рацию, оказывается заложенным в верхушечной почке, когда
•стебель достигает 2—3 см. В дальнейшем происходит разра-
стание зачатков и формирование побега, включая новые боко-
вые и верхушечную почки. Пазушные почки проростка дуба
закладываются очень рано Их можно видеть в пазухах семя-
долей и низовых пленчатых листьев, когда стебель достигает
1—2 см В это время в пазушных почках величиной в 0,2 -
0,3 мм сформировано по 2—3 шчатка листа
Обычно в течение первого года формируется один главный
стебель с группой зеленых листьев, сосредоточенных в верхней
части побега Верхушечная почка в последующем дает продол-
жение плавной оси В отличие от сеянцев ели и сосны с их
небольшими приростами стебля, у дуба в первый период роста
образуется за счет питательных веществ желудя главный сте-
бель высотой в 10—15 см. Еще большая интенсивность роста
в течение первого года паблюдас1ся в корневой системе. Глав-
ный корень сеянца в условиях Подмосковья уходит на глубину
до 60 см, а в засушливых условиях степей и полупустынь (на-
пример, под Волгоградом) до 80—100 см На всем протяжении
главный корень несет большое количество боковых корней,
расположенных более или мепее равномерно от гипокотиля до
верхушки корня, пли сосредоточенных в горизонтах почвы,
наиболее благоприятных для росла корней
Сеянцы дуба обнаруживают большую лабильность внешней
и внутренней структуры побегов в различных почвенно-клима-
тических условиях. Так, сравнение однолетних сеянцев дуба из
окрестностей Москвы, Камышина и Волгограда показало, что
в лесах Подмосковья в среднем развивается в течение первого
периода роста побегов 5 листьев, под Камышином — 7, под
Вол1 оградой — 9 В последнем пункте сеянцы имели большую
толщину стволика, чем под Москвой В листьях сеянцев из
Подмосковья насчитывалось в среднем 622 устьица на 1 мм2
листовой поверхности, под Камышином — 824 В степных усло-
виях (Камышин и Волгоград) у сеянцев дуба развивается
более мощная проводящая система в стебле, более ксероморф-
ная структура листьев, более мощная и глубокоидущая корне-
вая система, чем в условиях Подмосковья (Серебряков И. Г,
1953) Различие структурных признаков сеянцев д^ба в лесах
Подмосковья, степях и полупустынях районов Камышина
и Волгограда говорят о наличии приспособительных реакций,
отчетливо выраженных у сеянцев дуба. Это не подтверждает
мнения Шрамма (Schramm, 1912), согласно которому сеянцы
Рис. 31
Система годичного побега черешчатого
дуба (Quercus robur).
ВП — весенний прирост, ЛП — летние приросты
лесных древесных пород всегда отличаются теневой структурой
листьев и побегов, вне зависимости от внешних условий.
Способность к интенсивному росту побега у сеянцев дуба
в течение первого года проявляется не только в достижении им
сразу значительной высоты (10—15 слг), но и в образовании
нескольких приростов за год Рост побега, начавшийся при
прорастании желудя, длится не более 1,5—2 месяцев и завер-
шается формированием верхушечной почки длиною в 2—4 мм.
покрытой почечными чешуями из метаморфизированных при-
листников Верхушечная почка несет сформированные зачатки
зеленых листьев побега
следующей генерации.
После небольшого пе-
риода покоя (2—4 недс
ли) верхушечная почка
вновь трогается в рост,
давая начало второму
приросту однолетнего
сеянца. Этот второй при-
рост получил название
Иванова побега. По дли-
не стебля и числу листь-
ев Иванов побег неред-
ко превышает побег
первой, весенней генера-
ции (рис. 31). Иногда
на открытых местах и
богатых почвах в тече-
ние первого года наблю-
дается и третий прирост
так что общая высота
сеянца к концу лета до-
стигает 60—70 см. Число
приростов в течение ле-
та в степных районах
Европейской части
СССР больше, чем в
Подмосковье
В лесах Подмосковья
образование Ивановых
побегов часто определяют условия освещения. Под пологом де-
ревьев, при значительном затенении, Ивановы побеги обычно не
образуются, и сеянцы дуба имеют лишь один прирост в тече-
ние года.
Годичный побег сеянцев с несколькими приростами в тече-
ние лета в сущности представляет собой систему годичного
176
побега, так как слагается из элементарных побегов нескольких
генераций, образующихся в несколько периодов роста.
Условия освещения определяют не только образование Ива-
новых побегов, но и некоторые другие существенные процессы
формирования дерева. Так, характер роста дуба под пологом
леса и на открытых местах совершенно различен.
Под пологом леса при значительном затенении годичный
прирост побегов на 2-м году резко уменьшается по сравнению
с первым годом Вместо 10—15 см на первом году, прирост
побега второго года составляет всего 1—3—5 см. На втором
году под пологом леса раскрывается верхушечная почка глав-
ного стебля, а у сильных растений, кроме того, одна или две-
три боковые почки в верхней части материнского побега. При-
рост всех побегов колеблется от 1 до 3 см. По интенсивности
роста верхушечный и боковые побеги мало отличаются друг
от друга. На третьем и четвертом годах также раскрываются
обычно верхушечные почки; годичные приросты остаются столь
же небольшими, как и на втором году. Резкое снижение роста
побегов у сеянцев со второго года связано с полным использо-
ванием пластических веществ семени в течение первого лета
По истечении 2—3, реже 4—5 лет, верхушечная почка главной
оси отмирает, а одна или две-три боковые почки дают начало
соответственно побегам замещения, которые в отличие от глав-
ной оси растут не строго вертикально, а под острым углом
вверх. Побеги замещения не отличаются заметно от других
боковых ветвей не только по величине, но и по направлению
роста. Ивановы побеги в этих условиях не образуются
Любопытно, что через 2—3 года и у побегов замещения
также подсыхает верхушечная почка, и дальнейшее нарастание
идет за счет одной или нескольких боковых почек, дающих
начало побегам замещения третьего порядка. Отмирание верху-
шечных почек происходит не только на главной оси, но и на
боковых ветвях, повторяясь периодически через несколько лет.
Таким образом, под пологом леса сеянцы дуба характеризуются
не только слабым ростом, но и периодическим отмиранием
верхушечных почек, т е неустойчиво-моноподиальным нараста-
нием и ветвлением.
Периодическое отмирание верхушечных почек и образование
побегов замещения приводит к формированию значительного
количества побегов, растущих почти вертикально или несколько
наклонно Величина годичного прироста побегов в течение пер-
вых 10—15 лет остается при затенении очень небольшой
(1—3—5 см) и не обнаруживает обычного для деревьев увели-
чения с возрастом. Поэтому часто 12—15-летние сеянцы не
превышают 30—50 см в высоту. Толщина ветвей таких дубков
измеряется в 3—5 мм.
В-413 И. Г Серебряьов —12
177
Описанные процессы роста и ветвления побегов приводят
к образованию своеобразной формы сеянца дуба с более или
менее значительным количеством ветвей на верхушке, сходных
между собой по интенсивности и косо вверх наклонному направ-
чснию роста, с корозкими и тонкими годичными побегами
Книзу эти ветви сходятся к одной тонкой главной оси, образо-
вавшейся в течение первых лет жизни сеянца Впрочем, и глав-
ная ось, толщиной в 5—8 мм, не отличается резко о г более
верхних боковых ветвей Такая форма роста дуба под пологом
леса получила название торчков (Корнаковский, 1882, Юпиц-
кий В. А, 1924; Пятницкий С. С, 1949; Т. Н. Астапова, 1952)
Иногда (в еловых лесах большой сомкнутости без кустар-
никового яруса, на подзолистых почвах), оказывается подавлен-
ным не только рост, ио и ветвление побегов у сеянцев, и торчки
дуба поэтому слабо разветвлены пли даже на значительном
протяжении одпостебельны, причем их тонкий стебель 2—4 мм
толщины состоит из коротких (1—3 см) годичных побегов.
Однако чаще торчкам дуба свойственна форма со значительным
количеством ветвей Общая высота таких торчков в 12—15-лет-
нем возрасте достигает 40—60 см. Весьма характерно для них
наличие более или менее одинаковых слабо растущих ветвей,
вследствие чего среди них нельзя выделить одну в качестве
ведущей осп (лидера). Именно по этим признакам торчки дуба
сближаются с кустарниковыми и подушковидными формами
растений (см. гл VIII)
Таким образом, при формировании торчковой формы дуба
наблюдается потеря ведущей роли главной оси, неустойчивое
моноподиалыюе ветвление побегов, образование системы ветвей
с более или менее одинаковым слабым ростом и сглаживание
различий между главной осью или замещающими ее побегами
и боковыми ветвями Последний процесс особенно заметно от-
даляет торчковую форму от формы дерева.
В состоянии торчков сеянцы дуба могут пребывать в тече-
ние 15—20 и более лет. В этот период дуб обладает значитель
ной теневыносливостью, и можно думать, что торчки дуба с их
низкой интенсивностью роста есть приспособительная форма,
обеспечивающая выживание дуба в течение первых десятков
лет жизни в условиях значительного затенения под пологом
леса. Биологически торчки дуба могут быть уподоблены так
же слабо растущим ювенильным формам травянистых растений,
способным значительное время сохраняться в густом травостое
естественных лугов, степей и т д (Т. А Работнов, 1950)
Обладая большой теневыносливостью, торчки дуба весьма
чутко реагируют на увеличение интенсивности освещения Если
верхушка одного из побегов торчка оказалась под влиянием
каких-либо причин в лучших условиях освещения, она резко
178
увеличивает интенсивность роста Длина юдичиых побегов
такой ветви в течение 1—2 лет может увеличиться с 3—5 до
30—40 см, и в течение 2—3 лет образуется мощная ветвь дли-
ной’ до метра и более Резкое увеличение интенсивности верху-
шечного роста одного из побегов торчка приводит к весьма
интересному результату Сильный верхушечный прирост побега
вызывает столь же резкое усиление камбиальной деятельности
в стебле ниже растущей верхушки на всем протяжении стебля,
соединяющего верхушку этого быстро растущего побега с глав-
ным корнем растения Наблюдается быстрое утолщение этого
стебля, причем оно начинается наверху, непосредственно в ин-
тенсивно растущем побеге и ниже его, и постепенно переме-
щается вниз.
Через 3—4 года вследствие такого утолщения образуется
толстый стебель, соединяющий верхушку быстро растущей
ветви с главным корнем сеянца. Этот стебель теперь резко
выделяется по толщине среди других Он становится ведущим
побегом, лидером в общей системе побегов торчка. Усиление
роста лидерного побега вызывает в дальнейшем коррелятивное
ослабление и торможение в росте других побегов торчка Веду-
щий побег превращается в главную ось с вертикально ориенти-
рованным сильным ростом, а другие побеги, гомологичные ему
превращаются в боковые ветви Когда лидерный побег стано-
вится биологически главной осью, дуб переходит из формы
торчка в форму деревца с четко обозначившимся стволом и
кроной более слабо развитых ветвей Дальнейшее коррелятив-
ное торможение роста боковых ветвей, идущее от верхушки
появившейся главной оси, приводит к отмиранию этих ветвей
и очищению ствола
Таким образом, у дуба под пологом леса наблюдается свое-
образное явление, которое не наблюдалось в процессе роста
и формирования дерева у ели и сосны — базипетальное усиле-
ние ствола, вызванное усиленным верхушечным ростом одного
из побегов
Как видно из изложенного, сеянцы дуба под пологом леса
имеют два явно выраженных периода в развитии: период
торчкообразной формы и период формирования одноствольного
дерева.
Как показывают наблюдения Н. П. Доманской (1958), в те-
чение периода торчкообразной формы у сеянцев дуба в под-
земной сфере формируется отвесно растущий главный корень
е системой всасывающих и переходных к скелетным корней,
в результате чего корневая система растет главным образом
вглубь, но не в стороны Усиление роста побегов п переход
сеянцев дуба к форме одноствольного дерева сопровождается
12-
179
усиленным ростом боковых скелетных корней, приводящим
к резкому разрастанию корневой системы дуба вширь.
। *
*
Значительные отличия в росте и формировании молодого
дерева наблюдаются у сеянцев дуба в благоприятных условиях
произрастания и, в частности, при культуре дуба в искусствен,-
ных посевах. По размерам побегов первого года резких разли-
чий между дубками в культуре и естественным самосевом дуба
не наблюдается Как и под пологом леса, рост стебля однолет-
них дубков в культуре колеблется обычно от 7—15 до 20—30 см
Вероятно, здесь сказывается значение больших запасов пласти-
ческих и других веществ в семени, используемых в течение
первого лета и обеспечивающих меньшую зависимость сеянцев
от складывающихся условий среды. В связи с этим большое
влияние на рост дуба в первое лето оказывает вес желудя Так,
по данным А М. Шутяева (1957) из крупных желудей сеянцы
достигали 17—18 см высоты, из мелких желудей в тех же усло-
виях культуры — 9—13 см.
На втором году раскрывается верхушечная почка и нередко
2—3 боковые близ верхушки. Следовательно, уже со второго
года наблюдается не только нарастание главной оси, но и ее
ветвление Характерной особенностью роста сеянцев в культуре
(в хороших условиях освещения и почвенного питания) по
сравнению с самосевом дуба под пологом леса является отсут-
ствие резкого снижения прироста главной оси па втором году
сравнительно с первым годом. При хорошем уходе в лесных
питомниках Подмосковья размеры побега второго года в два
и более раз превышают размеры однолетних сеянцев В росте
годичных побегов — боковых и верхушечного — заметно про-
является акротония. По длине, толщине стебля и величине
почек побег главной оси превосходит боковые. На третьем и
четвертом году развертываются верхушечные почки главного
и боковых- побегов и несколько боковых почек на главной оси.
Рост главной оси с возрастом сеянца усиливается, и к 5—7 го-
дам годичные побеги главного стебля достигают 30—40 см
длины Акротонность ветвления, заметное преобладание годич-
ных побегов главной оси по толщине и длине над боковыми
вертикальное направление роста главного стебля при наклон
ном росте боковых ветвей уже с первых лет приводит к фор-
мированию лидерной главной оси и кроны боковых ветвей, т. е
к образованию настоящей кустовидной формы у сеянцев дуба,
в культуре в отличие от формы торчка под пологом леса.
180
У торчков, как отмечалось выше, годичные приросты глав-
ной оси сеянца уже со 2—3 года утрачивали значение лидерных
побегов и терялись даже среди слабо растущих боковых ветвей.
Нарастание побегов у дуба в культуре, как и у самосева
под пологом леса — неустойчиво моноподиальное. Здесь также
наблюдается периодическое отмирание верхушечной почки
с образованием побегов замещения за счет одной из боковых
почек. Но в отличие от торчковой формы роста, побеги заме-
щения у главной оси сеянцев дуба в культуре более или менее
резко преобладают по быстроте роста над другими боковыми
побегами. Отсюда следует, что образование торчковой или дре-
вовидной формы роста у сеянцев дуба связано не с типом вет-
Рис. 32
Верхушка лидерного побега у молодых дубков в Подмосковье:
А — строго акоотонная система побегов; Б — система побегов с незавершенной
акротонностью; ВП — верхушечный побег, ЬП — боковые побеги.
вления, и, в частности, не с неустойчивостью моноподиального
ветвления, а с интенсивностью роста побегов. При достаточно
интенсивном росте побегов всегда наблюдается их дифферен-
циация по длине и толщине образовавшегося стебля, и в форми-
рующейся системе побегов со временем все более отчетливо
выделяется лидерная, ведущая ось, преобладающая над осталь-
181
ними (с более длинными юдичными побеыми и более толстым
стеблем). Чем интенсивнее рост побегов, тем быстрее и более
резко проявляется их дифференциация, тем быстрее и отчет-
ливее становятся отличия самой сильной, биологически главной
оси от боковых, более слабых ее ветвей. При симподиальном
ветвлении такая главная ось у сеянцев лишь в своей базальной
части является собственно главной осью в строго морфологи-
ческом смысле, а выше она состоит из побегов замещения т о
боковых побегов все более высокого порядка Подобная биоло-
гически главная ось и образуется у сеянцев дуба в культуре.
Характерной особенностью древовидной формы у молодых
дубков в культурах является относительно сильное развитие
боковых ветвей. В результате образуется ширококронная древо-
видная форма, более или менее обильно разветвленная, но
с различимой главной осью Иногда у таких кустовидных моло-
дых дубков наблюдается смена лидерных осей, если верхушка
функционирующей «главной оси» ослабляет свой рост, а одна
из ветвей начинает более сильно расти в высоту и толщину.
Подобная смена лидерных побегов может быть вызвана по
вреждением верхушки главной оси (особенно неоднократным)
Но часто такая смена представляет результат естественных
коррелятивных отношений в росте побегов молодого дуба.
Как видно из рисунков, у сеянцев дуба в искусственных
посадках в Подмосковье, наряду с образованием строго акро-
юнной системы побегов (с самым сильным верхушечным, рис.
32 А), можно наблюдать системы побегов с незавершенной
акротонностью, когда самым сильным в системе вновь образо-
вавшихся побегов оказывается не верхушечный, а один из боко-
вых (рис 32 Б) Кустовидная форма дуба в молодом возрасте
является следствием недостаточно интенсивного роста побегов
и связанной с этим нерезко выраженной дифференциации побс-
юв на лидерный и боковые. Длительность периода кустовидного
роста дуба в решающей степени определяют условия культуры
(богактво почвенного питания, наличие или отсутствие боко-
вого затенения). В хороших условиях культуры в Подмосковье
уже к 7—10 годам дуб образует четко выраженную древовид-
ную форму с главным стволом и кроной боковых ветвей
В росте и формировании боковых ветвей у дуба наблюдает-
ся известное сходство с явлениями, описанными у ели и сосны.
Ветви перъых двух-трех годичных побегов, расположенные близ
поверхности почвы, с самого начала отличаются более слабым
ростом сравнительно с главной осью. У них не наблюдается
увеличения годичных приростов в последующие годы их жизни
т е типичной для главной оси восходящей ветви кривой роста.
В противоположность главной оси, на нижних боковых ветвях
через несколько лет начинается ослабление роста, а через
182
7— 10 лет рост их прекращается совсем Эги ветви постоянно
остаются в наклонном, почти горизонтальном положении и не
поднимаются над поверхностью почвы Опп отмирают, достигая
в среднем 0,5—1 ж длины и 1 см в диаметре
Ветви, возникающие позже, в течение последующих лет, и
потому расположенные выше по главной оси сеянца, отличают
ся более интенсивным ростом Как отмечалось выше, ветви,
возникшие в 5—10-летием возрасте, иногда могут превосходить
по интенсивности роста в длину и толщину биологически глав-
ную ось (лидерный побег) и сами становятся лидерными
осями Однако с усилением роста биологически тлавнои оси
и образованием древовидной формы с четко выраженным глав
ным стволом, рост этих боковых ветвей все более замедляется
и они подобно самым нижним ветвям, через некоторое время
также отмирают Так, в молодых смешанных лесах Подмо-
сковья с полнотой насаждения 0,6—0,7 у молодых деревьев
дуба высотой в 5—6 м и диаметром ствола у основания в
8—10 см боковые ветви отмирают и большей частью опадают
со ствола на протяжении полутора метров от поверхности
почвы. Годичные побеги биологически главной оси у таких
деревьев на верхушке кроны имеют длину от 20—30 до 40—
50 см Ветвление деревьев в этом возрасте и у зрелых деревьев
можно характеризовать как неустойчиво моиоподиальное
с отмиранием верхушечной почки побегов через 2—3 года или
несколько реже
По данным Т. Н Астаповой (1954) отмирание почек дуба
вызвано весенними заморозками. Несмотря на отмирание
верхушечной почки, продолжение ствола в кроне в виде лидер-
ной оси прослеживается до ее верхушки Устойчивое сохране-
ние лидерного положения биологически ктавной оси в кроне
связано с акротонным усилением побегов при их ветвлении,
с преобладанием в росте в длину и толщину над боковыми вет-
вями не только самой главной оси в молодом возрасте сеянцев,
по и ее побегов замещения в последующие годы
Смена моноподиального гг симподиального ветвчения про-
является в непостоянстве вертикального роста главной оси
Каждый раз при отмирании верхушечной почки побш замеще-
ния растет под некоторым углом к вертикали. В соответствии
с этим главный стебель довольно длительное время остается
коленчато изогнутым и выпрямляется медленно, лишь при до-
стижении стеблем достаточной толщины.
Отмирание нижних ветвей на стволе дуба зависит от густо-
ты насаждения и связано со значительным затенением В мо-
лодых густых насаждениях стволы дуба несут боковые ветви
с зелеными листьями почти до поверхности земли, но благо-
даря значительному затенению эти ветви постепенно начинают
18
-отмирать, так что в более взрослых и уже разреженных насаж-
дениях стволы деревьев оказываются очищенными от ветвей
или несут отмершие и отмирающие ветви на большом протяже-
нии от поверхности почвы. Продолжающееся отмирание ниж-
них боковых ветвей у дуба в более благоприятных условиях
освещения, но в более позднем возрасте деревьев, когда рост
главной оси становится более сильным, свидетельствует о том,
что очищение ствола у этого дерева определяется не столько
непосредственным затенением нижних ветвей, сколько корреля-
тивными отношениями побегов, складывающимися к этому
времени в корне дерева. Эти отношения проявляются в корре-
лятивном торможении роста боковых побегов, идущем от
растущей верхушки биоло-
гически главной оси и при-
водящим в конечном итоге
к полному прекращению
роста этих побегов и их
отмиранию.
Как известно (М. Е. Тка-
ченко, 1952), черешчатый
дуб отличается, в общем,
медленным ростом, хотя
в некоторые годы при-
рост побегов может дости-
гать у него 120—140 см.
Так, в 1957 г. в лееах Кур-
ского Государственного за-
поведника у 8-летних сеян-
цев дуба нами были отме-
чены годичные побеги те-
кущего года в 106 и 129 см
с тремя приростами за ле-
то. Эти годичные побеги нес-
ли более 10 боковых ветвей,
выросших из почек, зало-
жившихся в текущем году.
С медленным ростом
главной оси связано силь-
ное развитие боковых вет-
вей кроны, и поэтому дуб характеризуется мощной кроной, осо-
бенно на открытых местах. У молодых деревьев дуба до 60—80
лет, а иногда и в большем возрасте, в кроне хорошо прослежи-
вается до ее верхушки лидерная ось, и поэтому общая форма кро-
ны в этом возрасте может быть более или менее островершинной
(рис. 33). Но у деревьев с более слабым ростом главная ось
теряется на верхушке кроны среди боковых ветвей. В связи с
184
4
Рис. 33
Молодое дерево черешчатого дуба
(Quercus robur) с островершинной
кроной в смешанных лесах Подмосковья
этим и крона подобных деревьев отличается большей округло-
вершинностью. Как у ели и сосны, такое изменение формы
кроны тесно связано с возрастом дерева (рис. 34). Но в связи
с медленным ростом в высоту у молодых деревьев и медлен-
ным ослаблением роста у старых деревьев (рост дуба в высо-
ту длится до 150—200 лет) возрастные изменения кроны, пере-
ход от островершинной формы кроны к куполообразной и
плосковершинной с изменением возраста выражен слабо и
более четко проявляется лишь в благоприятных почвенных
и климатических условиях произрастания В гораздо большей
степени подобные различия в структуре кроны в лесах Москов-
ской области обусловлены особенностями различных место-
Рис. 34
Старое дерено мережчатого дуба (Quercus robur)
с куполообразной кроной в Эстонии (по Мазингу, 1959)
обитаний и представляют, таким образом, не столько возраст-
ной, сколько экологический признак.
Естественный ход ослабления верхушечного роста, как и у
ранее рассмотренных деревьев — ели и сосны, — приводит к
прекращению роста в высоту и подсыханию почек и побегов на
вершине кроны, к суховершинности.
В широколиственных и смешанных лесах Подмосковья пре-
кращение роста и явление суховершинности у старых деревьев
дуба наступает не ранее 150—200 лет (если суховершинность
не вызвана в более раннем возрасте внешними факторами). В
более южных и особенно юго-восточных районах Европейской
части СССР в условиях степей и полупустынь, за пределами
лесной зоны, в искусственных посадках суховершинность дуба
наступает в более раннем возрасте и наносит большой ущерб
лесному хозяйству
185
П. Д. Никитин (1950) отмечает в Куйбышевской области
усыхание посадок дуба 45—55-летнего возраста, связывая его
с сочетанием засух и сильных морозов 1938—1942 гг. По
С. Н Андрианову (1953), па водоразделах в Сальских степях дуб
начинает суховершинить в 17—18 лет, в то время как по днищам
балок хорошо сохраняется до зрелого возраста, образуя хоро-
шие насаждения. Дьяченко А Е., Землянский Л. Т. и Ханбе-
ков И И (1949), а также И. С Матюк (1950) приходят к вы-
воду, что в условиях полупустыни ускорение суховершинности
дуба и его дальнейшее усыхание определяется почвенными
условиями, в частности, засолением. В районе Волгограда в
посадках дуба на водоразделах суховершинность также насту-
пает часто в 15—20 лет, в то время как по днищам и склонам
балок можно видеть мощные деревья высотой до 30 м с диа-
метром ствола до 1 м без следов суховершинности.
По данным О. Н. Чистяковой (1959), под Волгоградом и
Камышином у дуба отмечается более быстрый темп образова-
ния кольцесосудистости, что, по мнению автора, может быть
одним из симптомов более раннего созревания дуба в степных ус-
ловиях по сравнению с более северными широтами. Действитель-
но, под Волгоградом и Камышином встречаются многочислен-
ные дубки, плодоносящие в 4—6 лет, тогда как в Тульских за-
секах плодоношение у дуба начинается только в 30 лет и позже
В отличие от ели и сосны, подсыхание вершины кроны и
верхушек боковых ветвей у дуба может не повлечь за собой
отмирания дерева в целом На ветвях и стволе дуба длитель-
ное время сохраняются (в благоприятных условиях 60—80 лет,
в неблагоприятных — дольше) спящие почки. Спящие почки
дуба возникают как обычные пазушные почки на однолетних
побегах (начиная от однолетнего сеянца и далее в течение всей
жизни дерева) в пазухах почечных чешуй (чешуепазушные и
венечные чешуепазушные — М. И Падеревская, 1951) Они
сохраняются на коре ветвей и ствола в живом состоянии мно-
гие десятки лет. На их верхушке ежегодно формируются очень
мелкие, морфологически слабо дифференцированные чешуйча-
тые листья, опадающие на второй — третий год. Ежегодное
верхушечное нарастание приводит к образованию хорошо вы-
раженного стебля, и поэтому спящие почки д>ба с увеличени-
ем возраста поднимаются над поверхностью коры ветвей и
ствола в виде крайне укороченных бурых побегов, высотой в
5—8 мм и диаметром около 1 мм. От основания до верхушки
они покрыты горизонтальными валиками — листовыми рубца-
ми опавших чешуйчатых листьев. Иногда верхушки этих по-
чек— побегов отмирают, что вызывает их ветвление, происхо-
дящее за счет пазушных почек этих крайне укороченных побе-
гов. Благодаря ветвлению спящие почки расположены обычно
186
Рис. 35
Старое суховершипяшее дерево
черешчатого дуба с водяными побегами
из спящих почек, формирующих
вторичную крову
на коре ствола и толстых сучьев небольшими группами — в 5—
10 почек или несколько больше.
Прекращение роста и подсыхание побегов на вершине кроны
и по ее периферии вызывает коррелятивно усиление деятельности
верхушечной меристемы спящих почек; вместо мелких чешуй-
чатых листьев в них начинают формироваться зеленые ассими-
лирующие, отделенные друг от друга удлиненными междоуз-
лиями. В результате интенсивного роста спящих почек обра-
зуются новые побеги. За счет их роста и жизнедеятельности
поддерживается дальнейшая жизнь дерева в целом.
Побеги из спящих почек (водяные побеги, «волчки») отли-
чаются обычно быстрым
ростом, на них в боль-
шом числе образуются
крупные зеленые листья,
за их счет отмершая на
периферии кроны асси-
милирующая поверхность
быстро восстанавливает-
ся (рис. 35).
Следует, однако, под-
черкнуть, что образова-
ние побегов из спящих
почек дуба не гаранти-
рует их быстрого роста
во всех случаях. Неред-
ко из спящих почек на
сучьях и стволе выра-
стают мелкие недораз-
витые побеги с мелкими
листьями, с приростом
за лето в 3—5 см. Такие
побеги обычно через не-
сколько лет отмирают.
Скорость роста, мощ-
ность и длительность
жизни побегов, возни-
кающих из спящих почек
непосредственно связаны
со степенью отмирания
или повреждения кроны
дерева. Чем больше по-
вреждена или подсохла
крона, тем быстрее и тем более крупные побеги развиваются из
спящих почек Лишь к этим побегам и применимо народное наз-
187
вание «водяных побегов». Скорость роста водяных побегов быстро
падает, и через 5—8 лет на их верхушках формируются годич-
ные побеги, мало отличающиеся от побегов первичной кроны
Спящие почки характерны для многих деревьев умеренных
широт (В Н Любименко, 1900; И Г. Серебряков, 1952) К со-
жалению, в ботанической литературе очень мало данных о спя-
щих почках деревьев влажно-тропических лесов (Sandt, 1923;
Schimper — Faber, 1935; Richards, 1957).
Судя по небольшим наблюдениям группы советских ботани-
ков-участников VIII Международного ботанического кон-
гресса, в тропических лесах Гвинеи в Африке у крупных де-
ревьев влажно-тропических лесов и деревьев мангровых
зарослей пет спящих почек или они довольно быстро отмирают
(Серебряков, 1955, П. А Баранов, 1957). По Ричардсу же
(1961), деревья дождевого леса обыкновенно очень легко дают
порослевые побеги, если главный ствол был повален или сгнил
Но происхождение этих побегов не выяснено. Неясно также,
насколько распространено это явление. Можно полагать, что
спящие почки встречаются у деревьев тропических лесов не-
сравненно реже, чем у деревьев наших широт.
Появление в процессе эволюции спящих почек у деревьев
послужило, очевидно, таким морфологическим признаком, ко-
торый явился биологической предпосылкой дальнейшего хода
эволюции жизненных форм покрытосеменных от деревьев к
кустарникам, кустарничкам и травянистым многолетникам Без
возникновения спящих почек этот эволюционный ряд жизнен-
ных форм был бы невозможен.
Биологическое значение спящих почек деревьев заклю-
чается в том, что они служат для замещения поврежденных
или подсохших побегов и для восстановления кроны в целом
Как видно из сказанного выше, в онтогенезе дуба также
представляется возможным выделить несколько возрастных
этапов, характеризующихся определенными биолого-морфоло-
гическими признаками
Первый возрастной период. Рост сеянца за счет
запасных веществ семени В связи с этим находится
интенсивный рост побега, часто с несколькими ростовыми
периодами, с образованием системы годичного побега до 60—
80 см высоты Интенсивному росту побегов здесь соответствует
столь же интенсивный рост корневой системы, с образованием
главного корня до 60—80 см длины и системой боковых и при-
даточных всасывающих и переходных корней.
Второй возрастной период. Формирование торчко-
вой, или к у с т о в и д н о й, формы Под пологом леса этот
период характеризуется резко ослабленным ростом побегов после
исчерпания запасных веществ семени (со второго года жизни).
188
Под пологом материнских деревьев образуется форма торчков,
с короткими годичными побегами (не более 1—3—5 см) и тон-
кими стеблями (2—3 льи), без заметной дифференциации побе-
гов на лидерную ось и боковые ветви, со слабым ростом бо-
ковых и придаточных скелетных корней По слабости роста и
значительной теневыносливости форма торчков дуба является
приспособительной для сохранения под пологом материнского
покрова Этот период длится до 15—20 лет и иногда, вероятно,
дольше.
При хорошем освещении, за пределами лесного полога, в
течение этого периода формируется кустовидная форма дуба,
с заметной лидерной осью и мощными боковыми ветвями,
иногда перерастающими в лидерную ось На богатых рыхлых
почвах при боковом затенении (например, в культурах) этот
период сокращается до 5—8 и даже 3—5 лет
Третий возрастной период. Формирование дерева с
быстрым ростом главной оси и более или менее
островершинной кроной. Характеризуется усилением
роста побегов, что сопровождается дифференциацией побегов
с выделением лидерной оси и боковых ветвей и завершается
образованием древовидной формы. В развитии побегов наблю-
дается четко выраженная акротонность ветвления. В сформи-
ровавшемся и быстро растущем дереве вплоть до вершины
кроны легко прослеживается лидерная ось, возвышающаяся
над кроной и делающая ее островершинной. Период длится
от 15—30-летнего возраста до 60—90-летпего
Четвертый возрастной период. Ослабление роста
главной оси и формирование куполообразной
кроны. Характеризуется ослаблением роста побегов и лидер-
ной оси, потерей биологически главной осью лидерного положе-
ния в кроне. Главная ось теряется в верхней части кроны среди
мощных боковых ветвей, форма кроны становится все более
куполообразной и плосковершинной. Ослабление роста побегов
завершается полным прекращением роста и появлением сухо-
вершинности кроны В корневой системе преобладают процес-
сы отмирания корней, начиная с питающих корней высшего
порядка, расположенных на периферии корневой системы.
Степень ветвления скелетных придаточных корней снижается,
поглощающая поверхность корневой системы уменьшается и
приближается к стволу, а уменьшение площади корневого пи-
тания влечет за собой ослабление роста побегов вплоть до
появления суховершинности. Период длится от 80—100 — до
150—200-летнего возраста и иногда дольше. При наличии
сохранившихся жизнеспособных спящих почек на сучьях и
стволе дерева возможно выделение пятого возрастного пе-
риода.
189
Пятый возрастной период. Формирование вторич-
ной кроны. Характеризуется формированием вторичной
кроны за счет водяных побегов из спящих почек, пробуждаю-
щихся при появлении суховершинности и подсыхании первич-
ной кроны.
Морфогенез жизненной формы дерева у бородавчатой
и пушистой березы (Betula verrucosa Ehrh.
и В. pubescens Ehrh )
Рис 37.
Однолетние сеянцы березы
(Betula verrucosa):
гк — главный корень, гпк — гипокотиль,
см — семядоли
Из не вполне двугнездной завязи с двумя семяпочками у
березы при созревании образуется односемянный плод — ши-
рококрылатая семянка, или-орешек. Хорошо дифференцирован-
ный зародыш в созревшем семени бородавчатой березы имеет
две овальные, плосковыпуклые, мясистые семядоли около 1 мм
длины и 0,5 мм ширины, явственно обособленный зародышевый
корешок 0,1—0,2 мм длины и очень маленькую, почти недиффе-
ренцированную почечку. Прорастание семян, чаще наблюдае-
мое в лесах Подмосковья после перезимовки их, в апреле и
мае, происходит по надзем-
ному типу. Гипокотиль, вытя-
гивающийся до 10—15 мм,
выносит семядоли над уров-
нем почвы По выходе из се-
мян семядоли вытягиваются до
3—4 мм длины, несколько
расширяются, зеленеют и
функционируют как ассими-
лирующие листья почти до
конца вегетационного перио-
да. Выше овальных цельно-
крайних семядолей вскоре по-
являются мелкие трехлопаст-
ные первичные листья с тре-
мя-пятью жилками, за которы-
ми следуют уже листья, форма
и величина которых становит-
ся все более близкой к листь-
ям взрослых деревьев (рис 37).
В первый и второй год
листья Betula verrucosa Ehrh.,
как и В. pubescens Ehrh,
сильно опушены. Но у В. verrucosa уже в первый и второй
юд на стебле появляются клейкие железки, количество кото-
190
рых в следующие годы возрастает, а опушение полностью
исчезает на третий-пятый год Несмотря на небольшие раз-
меры семени и отсутствие в нем значительных запасов, необ-
ходимых для роста веществ, сеянцы березы в благоприятных
условиях могут в первый же год достигнуть высоты 30 см
и образовывать не менее 6 зеленых ассимилирующих листьев
Но в природных условиях, под пологом леса или в густом
травостое за пределами лесных насаждений, по наблюдениям
в Московской области, сеянцы березы к концу года обычно
вырастают на 2—3 см, а иногда и меньше, образуя 2—5
мелких зеленых листьев. Главный корень за это время выра-
стает также лишь на 2—3—4 см, образуя боковые корни
на всем своем протяжении (Т И. Воробьева, 1959). Рост побе-
га завершается формированием верхушечной почки, а также
боковых в пазухах зеленых листьев, включая семядоли.
На первом же году боковые корни заражает микориза, они
ветвятся (Н В. Лобанов, 1949)
Весной второго года раскрывается верхушечная почка, а
у сильных растений в хороших условиях (на рыхлой почве без
травостоя) — и 2—3 пазушные боковые почки в средней части
побега Среди новых побегов наибольшим оказывается верху-
шечный Его длина резко колеблется у разных растений — от
3—5 см в густом травостое до 30 и более см на почве, лишен-
ной наземного покрова. Но обычно в природных условиях под-
московных лесов прирост березы и на втором году остается
очень небольшим, а боковые почки совсем не раскрываются. В
последующие 2—3 года рост сеянцев в подобных условиях
также незначителен и мало отличается от первых двух лет.
Боковые пазушные почки у сеянцев, находящихся в глубине
травостоя или под пологом леса, остаются пераскрывшимися и
переходят в состояние спящих почек.
На 3—4-м году у бородавчатой и пушистой березы появля-
ются придаточные корни, вначале на гипокотиле, а позже и на
эпикотильной части стебля (Т И. Воробьева, 1959).
Скорость роста побегов березы быстро возрастает после
выхода из-под затеняющего влияния травостоя. Нарастание и
ветвление побегов в этот период переходит от моноподиального
к неустойчиво моноподиальному типу. Главная ось березы,
образующаяся в этот период, как и в течение первых лет жизни,
не отличается резко по высоте и особенно по толщине от боко-
вых ветвей Последние по выходе из травостоя характери-
зует гонкий рост, по интенсивности нс уступающий главной
оси
В результате усиления роста побегов березы в возрасте 6—
8 лет в условиях Подмосковья образуется кустовидная
форма, несколько напоминающая сеянцы дуба Высота
191
кустов в этот период достигает 1 —1,5 м, а в течение нескольких
лет увеличивается до 2—2,5 м.
У кустовидных сеянцев березы, которые нередко встречают-
ся на опушках лесов, в молодых возобновляющихся лесах и
зарослях кустарников в Подмосковье, наблюдается иногда
смена лидерных осей в пределах одного и того же растения,
что связано в этот период с нерезкой дифференциацией побегов
на главную ось и боковые более слабые ветви Часто такая
смена лидерных побегов вызвана повреждением верхушки ли-
дерной оси
В корневой системе к этому времени сформированы и функ-
ционируют две части — система главного корня, состоящая из
собственно главного корня 60—70 см длиной и боковых, расту-
щих вниз, до 60—80 см длиной — и система придаточных стеб-
левых корней, приповерхностных и горизонтальных, 40—60 см
длиной Характерна двуярусность в расположении корней: в
глубоких горизонтах — главный и боковые, ближе к по-
верхности— придаточные стеблевые (Т. И. Воробьева, 1959).
Период к)стовидной формы в онтогенезе березы длится не-
долго (от 6—8 до 10—15-летнего возраста). Она менее постоян-
на, чем у сеянцев дуба, и, вероятно, часто вызывается повреж-
дением верхушек побегов. Быстрое усиление роста побегов (у
10—15-летних берез годичные побеги главной оси достигают
50—70 см длины) приводит к четкой дифференциации биологи-
чески главной оси и образованию древовидной формы
с главным стволом и кроной боковых ветвей С образованием
древовидной формы в корневой системе резко снижается роль
системы главного корня из-за ослабления ее роста, а функция
корневого питания все более и более переходит к системе при-
даточных стеблевых корней
Обычно в этот период в лесах Подмосковья нарастание по-
бегов у березы становится строго симподиальным, вследствие
того, что верхушечная почка удлиненных побегов недоразвива-
ется и подсыхает уже в середине лета Вместе с верхушечной
почкой подсыхает обычно и стебель на протяжении одного-
двух верхних междоузлий, так что к осени одна из боковых па-
зушных почек оказывается на верхушке побега. Вследствие сдви-
гания в сторону подсохшей верхушки побега эта пазушная поч-
ка занимает положение верхушечной и после перезимовки
образует ложно-верхушечный удлиненный побег — продолже-
ние материнского.
Удлиненные побеги березы начинаются одним-двумя
очень короткими междоузлиями в 1—3 мм длины; пазушные
почки в этой части побегов остаются обычно недоразвитыми
или совсем не образуются. Затем на побеге следуют более
длинные междоузлия в 2—3 см, а ближе к верхушке вновь
192
образуются более короткие. Пазушные почки в этой части
побега развиты хорошо и дают начало удлиненным или укоро-
ченным побегам. Иногда на удлиненных побегах вместо недо-
развитой верхушечной почки формируется мужская сережка
(рис. 38). Нередко на верхушке побега образуется не одна, а
две-три мужские сережки, из которых одна верхушечная и
одна-две боковые, образовавшиеся в пазухах зеленых листьев.
Пазушные почки в средней и верхней части побега развиты
хорошо и образуют укоро-
ченные или удлиненные
боковые побеги По отцве-
тании мужские сережки
подсыхают, а вместе с
ними подсыхает верхушка
побега до верхней боковой
вегетативной почки. Из
этой почки следующей вес-
ной вырастает новый лож-
новерхушечный побег про-
должения материнской оси
Формирование терминаль-
ных мужских сережек и
образование псевдотерми-
нальных побегов продол-
жения на удлиненных по-
бегах березы может повто-
ряться в течение ряда
смежных лет, в результате
чего образуются симподи-
альные ветви, слагающиеся
д
Рис 38
Удлиненные вегетативные побеги
бородавчатой березы (Betula verrucosa):
А — С верхушечной почкой, подсыхающей к кон-
цу вегетационного периода, Б — с верхушечным
мужским соцветием — сережкой, 1 — верхушка
подсохшего побега прошлого года, 2 — подсы-
хающая верхушечная почка текущего года,
3 — мужская сережка.
из удлиненных годичных
побегов Симподии в кроне
березы, таким образом, -
возникают двумя способа-
ми,— при подсыхании ве-
гетативной верхушечной
почки побега и при обра-
зовании терминального муж-
ского соцветия — сережки.
Иногда вместо удлиненных ложно-верхушечных побегов
продолжения из верхней боковой почки удлиненного побега
вырастает укороченный побег. Укороченные побеги березы не-
сут, как правило, по два зеленых листа, разделенных междо-
узлием в 1 мм На верхушке укороченного побега образуется
зимующая верхушечная вегетативная почка или терминальное
женское соцветие — сережка, также покрытая почечными че-
В-413 И. Г. Серебряков —13
193
шуйками (рис. 39). В первом случае следующей весной разви-
вается новый терминальный укороченный побег, и подобное
моноподиальное нарастание укороченного побега может про
должаться от 2—3 до 6—10 лет Ветвление укороченных побе-
гов наблюдается очень редко.
После указанного периода моноподиального нарастания из
верхушечной почки вместо укороченного вновь может образо-
ваться удлиненный побег В север-
ных районах лесной зоны период
моноподиалыюго нарастания побе-
гов березы (обычно В. pubescens)
может значительно удлиняться.
Так, в Хибинских горах и на При-
полярном Урале мы наблюдали
укороченные побеги В. pubescens,
нараставшие моноподиально 15—
20 и более лет и лишь затем пере-
ходившие к образованию удлинен-
ных побегов Чередование удли-
ненных и укороченных годичных
побегов на одной и той же ветви в
этих районах наблюдается доволь-
но часто и представляет характер-
ную особенность ветвей березы.
При образовании на верхушке
укороченных побегов женской се-
режки нарастание происходит
симподиально, за счет пазушной
почки верхнего листа. Если из нее
вторично образуется женская се-
режка, в побег развивается почка
в пазухе нижнего листа (обычно
недоразвитая).
В кроне березы, таким обра-
зом, можно видеть 4 типа побе-
гов с различным типом нараста-
ния и ветвления* 1) удлиненные симподиально ветвящиеся
побеги с подсыхающей терминальной почкой. 2) удлиненные
симподиально ветвящиеся побеги с мужским терминальным
соцветием-ссрежкой; 3) укороченные симподиально ветвящие-
ся побеги с терминальным женским соцветием-сережкой и
4) укороченные моноподиально нарастающие побеги
В период интенсивного роста у березы образуются ветвя-
щиеся годичные побеги за счет распускания пазушных почек
средней части побега в год их заложения. Образование ветвя-
щихся годичных побегов у деревьев умеренной зоны представ-
194
Рис. 39
Укороченные побеги бородав-
чатой березы (Betiila
verrucosa)’
А — с верхушечной почкой; Б — с
верхушечной женской сережкой,
ПП — пазушная почка
ляет редкое явление; оно чаще встречается у деревьев тропи-
ческих лесов (R. Scharfetter, 1953).
Удлиненные побеги березы превосходят побеги других де-
ревьев своей относительной длиной и тонкостью стебля. Wigand
(1854) для удлиненных побегов деревьев ввел особый показа-
тель — коэффициент стройности (Schlankheitkoefficient) — отно-
шение длины побега к толщине его стебля. По данным Виган-
да это отношение у березы достигает 396, у дуба — 82, у
ольхи — 48—114. Как видно, по этому показателю побеги бе-
резы намного отличаются от побегов дуба и ольхи.
Увеличение относительной
длины побегов улучшает усло-
вия освещения зеленых листь-
ев Кроме того, образующиеся
удлиненные прутьевидные по-
беги березы обладают боль-
шой подвижностью Наличие
длинных и тонких побегов в
кроне березы не только улуч-
шает условия освещенности
листьев, но и способствует
рассеиванию семян и, наконец,
создает возможность схлесты-
вания ветвями березы ветвей
и вершин рядом растущих де-
ревьев, увеличивая тем самым
в известной степени ее конку-
рентную способность в отно-
шениях с другими порода-
ми (В. Н. Сукачев, 1938;
М. Е Ткаченко, 1952).
Как отмечалось выше, пе-
реход от кустовидной к дре-
вовидной форме в онтогенезе
березы неразрывно связан с В
усилением роста главной оси.
Характерной особенностью
березы, отличающей ее от
ранее рассмотренных деревь-
ев — ели, сосны и дуба, яв- рис> 49
ляется быстрое достижение Молодое дерево бородавчатой березы
ею кульминации роста, кото- с островершинной кроной, с главной
рая на хороших почвах насту- 0Cbl°- заметной до верхушки кроны
пает во втором, а на более бедных — на третьем десятилетии ее
жизни (О. Kirchner, 1914; Г. Р. Эйтинген, 1940); прирост побе-
гов достигает в среднем в год 60—70 см.
13*
195
Рис. 41
Старое дерево бородавчатой березы с ку-
полообразной кроной, в которой главная
ось теряется на вершине кроны среди
сильных ветвей
Как видно на рис. 40, у молодых деревьев березы, как и у
ранее рассмотренных ели и сосны, формируется довольно узкая
островершинная крона. Правда, у главной оси березы, как и у
боковых, медленно увеличивается диаметр стебля, и главная
ось у вершины кроны мало отличается от боковых ветвей.
Вследствие этого у березы образуется ажурная изящная крона,
без особенно выраженных закономерностей в росте и располо-
жении ветвей разных порядков. Быстрое достижение максимума
роста вызывает в последующем и быстрое замедление роста в
высоту, который практически прекращается у березы к 60—70 го-
дам; позже заметно продолжается лишь прирост в толщину. Зна-
чительное ослабление роста главной оси в высоту вызывает
постепенное изменение формы кроны от островершинной к
широко раскидистой и
более или менее куполо-
образной. В Подмосковье
такая форма кроны обра-
зуется у деревьев с 50—
60-летнего возраста (рис.
41), у Betula verrucosa в
этом возрасте достигает
полного выражения пла-
кучесть кроны. У более
молодых деревьев пла-
кучесть кроны выражена
гораздо слабее, сосредо-
точиваясь, главным обра-
зом, в нижней части
кроны.
Дютроше объяснял све-
шивание побегов плаку-
чего ясеня отрицательным
геотропизмом (см. Бара-
нецкий, 1901). W. Hof-
meister (1868) усматри-
вал причину этого явле-
ния в механическом
влиянии собственной тя-
жести побегов с их бо-
лее длинными и тонкими
междоузлиями по срав-
нению с обыкновенной
разновидностью того же дерева. К этому объяснению позже
присоединились А. В. Frank и Н. Vochting. Исследования
О. В. Баранецкого (1901) плакучих форм Caragana arborescens,
UImus montana и Fraxinus excelsior подтвердили такой вывод.
196
У Caragana arborescens var pendula молодые растущие побе-
ги обладают отчетливо выраженным отрицательно геотропи-
ческим изгибом вверх. Позднейшее свешивание побегов обу-
словлено их собственной тяжестью. Об этом говорит, по Бара-
нецкому, внешний вид побегов обыкновенной и плакучей формы
желтой акации У последней побеги более длинные и тонкие. Кро-
ме этого, по данным О. В. Баранецкого, волокна вторичной древе-
сины у обыкновенной формы более толстые, чем у плакучей
формы Древесина плакучей формы рыхлая. Таким образом,
заключает Баранецкий, в самом строении древесины дано усло-
вие, благодаря которому побеги плакучей формы не в состоя-
нии удержать тяжесть листьев, и по мере удлинения должны
пассивно свешиваться. Подобная особенность отмечена у
Ultnus montana \аг pendula, а также Fraxinus excelsior var.
pendula. Любопытно, что у плакучей формы ясеня толщина по-
бегов больше, чем у обычной, но в плакучих стеблях преобла-
дает паренхима, в них относительно сильнее развита сердцеви-
на, а образование механических тканей начинается позже
Уже внешний вид плакучей формы кроны Betula verrucosa
наводит на мысль, что пассивное свешивание побегов вниз
обусловлено их собственной тяжестью. Свисающие вниз ветви
в кроне В. verrucosa морфологически представляют собой оси
высоких порядков (обычно 4-го, 5-го и более высоких), в ко-
торых, как известно, отрицательно геотропическая реакция по
сравнению с главной осью и осями второго порядка значи-
тельно ослаблена. Сравнение структуры ветвей плакучей и
неплакучей формы березы обнаруживает в них существенные
различия. Ветви плакучей формы, как и у деревьев, изученных
О В. Баранецким (1901), более тонкие и длинные Количе-
ственной мерой удлиненности ветвей и тонкости их стебля
может служить отношение длины ветви к диаметру ее стебля
(коэффициент стройности побега по Wigand’y, 1854). Только, мо-
жет быть, его лучше называть показателем удлиненности стебля.
Ниже приведены показатели удлиненности 10 типичных по-
бегов плакучей и неплакучей формы В. verrucosa, взятых из
крон старых деревьев в Лосиноостровском лесном массиве под
Москвой (табл. 4 и 5).
Как видно из данных, приведенных в таблицах 4 и 5, у по-
бегов плакучей и пеплакучей формы В. verrucosa различия в
отношении их длины к толщине стебля становятся все более
четко выраженными и резкими по мере увеличения возраста
ветви. Если у неплакучей формы березы показатель удлинен-
ности стебля несущественно увеличивается на всем протяжении
многолетней ветви в течение 10 и более лет, то у плакучей фор-
мы березы с увеличением возраста ветви показатель удлинен-
ности стебля увеличивается резко. С 80—100 у однолетних по-
бегов он возрастает до 320—350 у 10—13-летних ветвей.
Таблица 4
Изменение показателя удлиненности ветви
в кроне плакучей формы Betula 'verrucosa
№№ ветвей Отрезки ветви , начиная с растущей верхушки
1-лет- ний 2-лет- ний 3-лет- ний 4-лет- ний 5-лет- ний 6-лет- ний 10- лет- ний 13- лет- ний
1 86 96 150 179 215 254
2 82 103 174 205 268 — 329 ___
3 75 119 168 225 235 — —
4 91 160 — — —
5 74 105 142 182 230 — — —
6 100 114 136 163 212 — — —
7 100 108 160 189 233 — — 347
8 70 114 153 217 211 —- —
9 86 106 172 205 — — мм
10 70 125 153 178 — — — —
Среднее . . 84 115 156 193 229 254 i 329 347
Таблица 5
Изменение показателя удлиненности ветви
в кроне неплакучей формы Betala verrucosa
№№ ветвей Отрезки ветви, начиная с растущей верхушки
1-лет- ний 2-лет- ний 3-лет- ний 4-лет- ний 5-лет- ний 6-лет- ний 7-лет- ний 10- лет- ний
1 95 109 140 133 156 123 131
2 95 91 96 100 ПО —
3 98 98 115 121 120 132 —
4 95 105 95 116 130 141 123 —
5 НО 143 153 162 131 155
6 88 133 118 142 157 163 —В
7 80 105 125 123 133 134 170 182
8 83 142 131 151 169 —
9 125 112 139 116 155 — —
10 80 83 77 77 116 116 — —
Среднее . . 95 112 | 119 124 138 136 141 182
Резкое возрастание показателя удлиненности стебля с уве-
личением возраста ветви означает, что рост стебля в толщину
в процессе формирования ветви у плакучей березы резко отста-
198
ет от роста в длину. Из таблиц следует, что утолщение стебля
у ветвей плакучей березы идет слабее, чем у нсплакучей
формы
Анатомический анализ показывает, что у однолетних побе-
гов ширина годичного кольца ксилемы у плакучих и неплаку-
чих форм более или менее одинакова В последующие годы
толщина годичного кольца ксилемы у ветвей неплакучей фор-
мы остается такой же, как и у одногодичного стебля, т. е
утолщение идет примерно равномерно из года в год На ветвях
плакучей формы уже со второго года и в течение последующих
лет до 10—15-лстнего возраста наблюдается резкое уменьше-
ние ширины годичного кольца ксилемы. Обычно ширина го-
дичного кольца, образующегося на 2-м, 3-м, 4-м и т д годах
жизни ветвях плакучей формы березы в 3—4 раза меньше го-
дичного кольца однолетнего побега Биологически это означает,
что в стеблях плакучей формы березы образование длинных и
тонких ветвей связано с резким ослаблением в них камбиаль-
ной деятельности
Таким образом, формирование свисающих ветвей у плаку-
чей формы березы вызывается недоразвитием в них тканей вто-
ричной ксилемы стебля на 2, 3, 4 и последующих годах жизни
ветви и свисание ветвей происходит под влиянием их собствен-
ной тяжести так же, как это было показано О. В. Баранецким
для плакучих форм желтой акации, вяза и ясеня
Известно, что камбиальную деятельность вызывают и сти-
мулируют ростовые вещества, притекающие от растущей вер-
хушки стебля Чем сильнее верхушечный рост стебля, тем
интенсивнее деятельность камбия и тем больше прирост в тол-
щину стебля. В связи с этим представляет интерес тот факт,
что рост в длину у свисающих вниз ветвей плакучей формы бе-
резы идет слабее, чем у ветвей, растущих вверх Исходя из
этого свешивание ветвей плакучей березы можно рассматри-
вать, как следствие общего ослабления роста побегов у старею-
щих деревьев Одним из косвенных доказательств этого явля-
ется полное развитие плакучести кроны лишь у старых деревь-
ев березы Любопытно, что и у более молодых деревьев сви-
сающие ветви представляют собой обычно оси высоких поряд-
ков (5-го, 6-го, 7-го) с ослабленным ростом и признаками ста-
рения
Ослабление и прекращение роста у стареющих деревьев бе-
резы (обычно не позже 100—120-летнего возраста) приводит к
появлению суховершинности и все более увеличивающемуся
подсыханию кроны Но, как и у черешчатого дуба, подсыхание
кроны у старых деревьев березы может и не вызвать отмира-
ния всего дерева, так как береза обладает длительно сохра-
няющимися спящими почками
Спящие почки березы располагаются по всем ветвям и
сучьям, а также по стволу вплоть до его основания. Они воз-
никают из нераскрывшихся пазушных почек удлиненных побе-
гов, будучи приурочены обычно к их нижней половине. Выше
спящих почек на материнском годичном побеге находятся па-
зушные почки, образующие укороченные побеги описанной
выше структуры. Как и у дуба, спящие почки березы отличают-
ся довольно интенсивным верхушечным ростом (даже более
интенсивным, чем у дуба, так как на их конусе нарастания мо-
гут формироваться даже зачатки зеленых листьев, Серебря-
ков И. Г, 1952); они способны к обычному аксиллярному
ветвлению за счет своих пазушных почек. Верхушечное нара-
стание приводит к тому, что спящие почки березы постоянно
возвышаются над поверхностью коры, постепенно увеличиваясь
в размерах (от 0,5—1 до 8—10 мм) По своей структуре они
представляют как бы миниатюрные укороченные побеги, ли-
шенные развернувшихся зеленых листьев Впрочем, между ти-
пичными укороченными побегами и спящими почками у березы
имеются переходные образования и часто между ними нелегко
установить границы (после опадения зеленых листьев с укоро-
ченных побегов) Располагающиеся вначале поодиночке, спящие
почки вследствие ветвления на 10—15-летних ветвях уже встре-
чаются по 2—3, а на более старых сучьях и стволе — все более
крупными группами В основании стволов старых деревьев спя-
щие почки образуют крупные гру ппы, занимающие площадь в 5—
8 см диаметром, и состоящие из нескольких десятков почек.
Как видно, на поперечных спилах ствола, такие группы по-
чек возникают в результате ветвления одной единственной
почки, возникшей в течение первых лет жизни сеянца березы.
На хороших почвах спящие почки сохраняются у березы до
60—80 лет, а на бедных — до 100—120 лет (Г. Ф Морозов,
1922, Н. А Асосков, 1931; М. Е. Ткаченко, 1952)
Все изложенное выше позволяет выделить несколько воз-
растных периодов в онтогенезе березы (Betula verrucosa и
В. pubescens) в природных условиях подмосковных лесов
Первый возрастной период (до 5—8 лет). Одностебель-
ный с е я н е ц с очень слабым ростом стебля (годичные побеги
в 1—3 см) и подавленным ветвлением (образовавшиеся пазуш-
ные почки нс раскрываются, переходя в состояние спящих).
В подземной сфере формируется система главного корня, со
слабым ростом корней, не превышающим резко прирост стебля.
Отличается значительной теневыносливостью, что дает возмож-
ность сеянцам сохраниться в течение ряда лет в травостое или
под пологом кустарников и деревьев.
Второй возрастной период. Образование кустовид-
нон формы (от 5—8 до 10—15-летнего возраста). Характе-
200
ризуется более сильным ростом побегов по сравнению с первым
периодом, особенно после выхода из-под затенения травостоем.
Из-за слабой дифференциации в росте побегов лидерная ось не
формируется или выделяется не надолго (сменяясь другими
лидерными осями), что приводит к образованию кустовидпой
формы высотой в 1—2 м В корневой системе двуярусной струк-
туры примерно одинаково развиты система главного корпя,
располагающаяся более глубоко в почве (от 40—60 см до 1 —
1,5 к), и система придаточных стеблевых корней, более поверх-
ностных (от 5—10 до 30—40 см глубины и длиной до 1,5—2 it)
Третий возрастной период. Узкокронное и островер-
шинное дерево с лидерной осью, прослеживаю-
щейся до вершины кроны (от 10—15 до 40—50-летнего
возраста). Период максимального роста побегов (особенно глав-
ной оси) В корневой системе преобладающее значение приоб-
ретает система придаточных стеблевых корней, обильно ветвя-
щихся и интенсивно растущих, с регулярной сменой всасываю-
щих корней 5—7 и более высоких порядков.
Четвертый возрастной период. Формирование раски-
дистой куполообразной кроны (с 50—70 до 100—120-
летнего возраста). Характеризуется ослаблением роста побегов,
начиная с главной оси, вследствие чего лидерная ось теряется
среди ветвей в верхней части кроны, а сама крона постепенно
становится все /юл ее раскидистой с куполообразной вершиной
В корневой системе среди корней высших порядков начинают
преобладать процессы отмирания над новообразованием. При
наличии сохранившихся жизнеспособных спящих почек на
сучьях и стволе дерева наступает пятый возрастной период.
Пятый возрастной период. Формирование вторич-
ной кроны. Характеризуется прекращением роста дерева
в высоту, появлением суховершинности, пробуждением спящих
почек на сучьях и стволе дерева и образованием вторичной
кроны.
Особенности морфогенеза жизненной формы дерева
у березы извилистой (Betula tortuosa Ldb )
Betula tortuosa Ldb. распространена на Алтае, Приполяр-
ном Урале, Кольском полуострове, произрастая у северной и
верхней границы леса в горах (Флора СССР, т. VI). От Betula
pubescens отличается низкорослым, сильно корявым стволом
(Сукачев В. Н. 1938). Ниже приводятся данные об особен-
ностях морфогенеза жизненной формы дерева у Betula tortuosa
на основе наблюдений автора у верхнего предела лесного пояса
в Хибинских горах, где эта береза образует неширокий, но
четко выраженный пояс березового криволесья (Флора Мур-
манской области, т. III, 1956; Мишкин, Флора Хибинских гор,
Как и у выше рассмотренных видов березы — Betula verru-
cosa и В pubescens, прорастание семян В. tortuosa происходит
по надземному типу. В конце июля в Хибинах, в поясе березо-
вого криволесья, на осыпях, по щебнистым участкам, лишен-
ным сомкнутого растительного покрова, можно видеть молодые
всходы извилистой березы еще с остатками семени на красно-
ватых или уже зеленых семядолях. Их гипокотиль, 5—8 мм
длины, у корневой шейки имеет резкий перехват и переходит
в более тонкий главный корень, начинающий ветвиться вскоре
после прорастания, еще при наличии семядолей В течение
первого года всходы вырастают не более чем на 1 см и обра-
зуют 4—5 первичных листьев 2—3 мм длины, с одной-двумя
боковыми жилками. Формированием зимующей верхушечной
почки завершается рост побега первого года. Главный корень
вырастает за первое лето не больше чем на 2—3 см.
После перезимовки верхушечная почка раскрывается, обра-
зуя побег продолжения главной оси. Листья второго года, по
крайней мере в нижней части побега, сохраняют ювенильную
форму, с широко-яйцевидной пластинкой, ширина которой пре-
вышает длину, с одной-двумя боковыми жилками и зубцами.
По величине (1—3 мм) и форме ювенильные листья Betula
tortuosa очень близки к листьям сеянцев карликовой березки
Betula папа. После образования 4—5 мелких зеленых листьев
на миниатюрном побеге второго года дпиной в 5—8 мм форми-
руется вновь зимующая верхушечная почка.
В 3—4-летнем возрасте у В. tortuosa начинается образова-
ние придаточных корней вначале на гипокотиле, затем в над-
семядольной части стебля
Дальнейший рост сеянцев В. tortuosa происходит по-разно-
му, в зависимости от условий произрастания. В тени камней и
под кронами материнских деревьев рост сеянцев первые три-
шесть лет сильно замедлен. Главная ось сеянцев или образует
удлиненные годичные побеги до 8—10 мм длины, или на тре-
тий-четвертый год переходит к росту по типу укороченных по-
бегов, так что 6—8-летние сеянцы едва достигают высоты 3
реже 4 см. Все эти годы на побеге раскрывается лишь одна
верхушечная почка, боковые почки не раскрываются и ветвле-
ние отсутствует. Нераскрывшиеся почки становятся спящими
Спящие почки хорошо видны уже у молодых сеянцев на
приростах прошлых лет главной оси, особенно в ее основании
на побегах первых двух-трех лет В 6—8 летнем возрасте сеян-
цев спящие почки бывают удлиненной формы, с заостренной
верхушкой зеленоватого цвета и имеют длину 1—2 мм Они
направлены в разные стороны под прямым углом от стебля и
напоминают торчащие гвозди. На хорошо освещенных местах,
лишенных растительности, сеянцы Betula tortuosa растут
202
быстрее. С 3—5-го года их годичные побеги увеличиваются до
3—5 см, к 8—10 годам — до 10 см, так что 10-летние экземпля-
ры на открытых местах достигают до 40—50 см высоты. Боль-
шая длительность жизни и роста в высоту у В. tortuosa в Хи-
бинах наблюдается в лесном поясе па незаболоченных почвах,
более раннее прекращение роста и отмирание главной оси
характерно для верхнего предела лесного пояса в березовом
криволесье
Рост боковых ветвей главной оси В. tortuosa происходит
также по-разному у экземпляров, произрастающих в разных
экологических условиях В лесном поясе боковые ветви
В. tortuosa растут относительно слабо (по сравнению с главным
стволом), причем, нижние скорее отмирают, а верхние усили-
ваются. Такое акротопное усиление ветвей главной оси уже
отмечалось выше у ели, сосны, дуба и березы бородавчатой и
пушистой. Этот процесс, как известно, приводит к очищению
ствола в его нижней части и формированию кроны па некото-
рой высоте над уровнем почвы. В таких условиях у В. tortuosa
формируется типично одноствольная форма дерева.
Но близ верхнего предела лесов, и особенно в поясе березо-
вого криволесья, боковые ветви отличаются относительно более
сильным ростом. Они мало уступают главной оси. Вследствие
этого возникают ветвящиеся от основания деревца с двумя-
тремя мощными ветвями Основание ствола со временем засы-
пается щебнем, лесной подстилкой или зарастает мхами, обра-
зующими здесь слой толщиной 10—20 см. В результате обра-
зуется кустовидная форма дерева, и общее происхождение
ствола и отходящих ветвей может быть установлено только при
раскопке Такие кустовидные деревья составляют подавляющее
большинство экземпляров в поясе криволесья (рис 42). Хотя
интенсивность роста нижних ветвей у кустовидных форм отно-
сительно велика по сравнению с интенсивностью роста главной
оси, абсолютные приросты в этом случае значительно меньше,
чем у одноствольной формы В. tortuosa
Образующаяся у верхнего предела лесного пояса Хибин
кустовидная форма В. tortuosa представляет собой дерево со
слабо или почти не выраженным главным стволом, и кроной,
начинающейся у поверхности почвы Нижние вегви кроны
вскоре начинают укореняться, особенно если они покрыты мо-
ховым покровом или лесной подстилкой. Образование самостоя-
тельной системы придаточных корней усиливает независимость
нижних боковых ветвей от главной оси и ее корневой системы
Укоренение ветвей сопровождается усилением их роста в высо-
ту и толщину. Вскоре по высоте и диаметру стебля они не
только сравниваются с главной осью, но и превосходят ее Под-
счеты показывают, что и по длительности жизни они превосхо-
дят главную ось 0
Отмирание главной оси сопровождается постепенным отми->
раннем системы главного корня. Поэтому через некоторое время
у дерева остаются лишь укоренившиеся ветви, становящиеся
самостоятельными стволами вегетативного происхождения.
Рис. 42
Взрослое дерево извилистой березы (Betula tortuosa)
из пояса березового криволесья Хибин
каждый со своей корневой системой (из стеблеродных при-
даточных корней). Количество таких стволов, возникших от
одного дерева, в криволесье Хибин чаще всего колеблется от
2 до 4, иногда увеличиваясь до 6—8. Стволы достигают 45—SO-
204
летнего возраста (редко более), 3,5—4,5 м высоты и 10—12 см
в диаметре у основания.
Ослабление роста у стареющих стволов вегетативного про-
исхождения вызывает пробуждение спящих почек. Своеобраз-
ной особенностью В. tortuosa, отличающей ее от всех ранее рас-
смотренных деревьев, является то, что побеги, возникшие из
спящих почек, способны к длительному росту при наличии жи-
вой материнской оси с неповрежденной кроной Рост побегов из
спящих почек, расположенных у основания материнского
ствола, со временем не только не ослабевает, но даже усили-
вается, вследствие образования в основании этих побегов при-
даточных корней. Укоренение этих побегов усиливает их неза-
висимость от материнской оси В результате у В. tortuosa можно
наблюдать одновременное развитие многолетних надземных
осей двух порядков, отличающихся друг от друга по возрасту,
как материнская и дочерняя оси; такие соотношения осей ти-
пичны для кустарников.
Интересно, что у одноствольных деревьев В. tortuosa в горно-
лесном поясе Хибин также наблюдается раскрытие спящих по-
чек ствола в его основании и длительный рост побегов, из них
возникающих, при наличии растущей и неповрежденной кроны
материнской оси Правда, рост побегов из спящих почек начи-
нается лишь у старых деревьев с резко ослабленным ростом
главной оси Как и у деревьев В. tortuosa из пояса березового
криволесья, побеги, возникшие из спящих почек, укореняются,
и два-три из них, обладающие наиболее быстрым ростом, со
временем образуют новые дочерние стволы второго порядка
Подобные оси второго и более высших порядков в виде стволов
вегетативного происхождения, как известно, образ} ются и в
более южных районах лесной зоны у В. verrucosa и В pubescens.
Но если у пушистой и бородавчатой березы образование новых
стволов из спящих почек может происходить только после уда-
ления материнской оси или очень сильного ее повреждения,
у В. tortuosa рост и формирование боковых стволов со своими
кронами может происходить и в присутствии материнской оси
с живой кроной Исключения из этого правила наблюдаются
единично у В. pubescens на моховых болотах, где она может
образовывать кустовидную форму. В результате образуется
дерево с 2—3—4 (редко больше) стволами, каждый из которых
имеет собственную систему придаточных корней. Морфологи-
чески эти оси могут быть или одного порядка, например, вто-
рого, возникшими из спящих почек главной оси, или несколь-
ких порядков, например двух — первого и второго, или второго
и третьего, как это наблюдается в типе у кустарников.
Формирующаяся в зрелом возрасте у В. tortuosa жизненная
форма значительно отличается от формы типичного дерева
рассмотренной выше на ряде примеров По наличию нескольких
одновременно растущих стволов, обычно разных порядков (ма-
теринских и дочерних), по способу возникновения новых стволов
из спящих почек базальной части материнской оси и способ-
ности к вегетативному возобновлению надземных осей по мере
старения и ослабления материнской оси, форма дерева
у В. tortuosa сходна с кустарниками (см гл. IV)
В то же время здесь ярко выражены и черты отличия от
кустарников — большая длительность жизни стволов (40—60
лет, иногда более), отсутствие регулярной смены осей низших
порядков осями все более высшего порядка в онтогенезе расте-
ния, наличие лишь небольшого количества осей, относящихся не
более, чем к двум порядкам, большое тормозящее коррелятив-
ное воздействие материнской оси на рост дочерних осей из
спящих почек. Подобная жизненная форма может быть названа
немногоствольным деревом (плейокормпое дерево).
Это название подчеркивает постоянное наличие у этой формы
небольшого количества стволов (2—3—4), но не десятков над-
земных осей, характерных для кустарников, которые по этому
признаку могли бы быть названы полнкормными древесными
растениями.
Это сравнение выявляет ряд жизненных форм древесных
растений- монокормные древесные растения — типичные де-
ревья, плейокормные древесные растения — немногоствольные
деревья типа В. iortuosa и поликормные древесные растения —
типичные кустарники. Филогенез в этом ряду шел от монокорм-
ных к поликормным древесным растениям
В. tortuosa представляет высокогорную и северную расу
В. pubescens (Сукачев, 1938), и образование у В. tortuosa жиз-
ненной формы немпогостволыюго дерева, являющейся переход-
ной от дерева к кустарникам, полно эколого-филогенетическою
смысла. Образование формы пемногостволыюго дерева у В. tor-
tuosa показывает исторический путь и факторы эволюции жиз-
ненных форм в пределах этих близких видов березы от деревьев
к кустарникам
Как видно из данных И Л. Крыловой (1958), немногостволь-
ная форма дерева образуется также у бука (Fagus taurica)
у верхней границы леса в Крымских горах, а по данным
В И Запрягаевой (1953)—у некоторых плодовых деревьев
в горах Киргизии. Форма немногоствольного дерева возникает
в неблагоприятных экологических условиях у границы распро-
странения древесной растительности.
В Хибинах нижняя часть стволов у В tortuosa коленчато
изогнута, что обусловлено рядом причин. Здесь имеет значение
и пригибание стволов тяжестью снега, сползающего со склонов,
и удары осыпающихся камней Кроме того, коленчатость ствола
206
В. tortuosa обусловливается биологическими особенностями
роста этого дерева: отмирание главной оси в молодом возрасте
близ ее верхушки сопровождается образованием осей замеще-
ния, которые в первое время растут наклонно и даже горизон-
тально
В кроне у В tortuosa, как и у В verrucosa и В. pubescens,
образуются удлиненные и укороченные побеги. Последние могут
возникать в любом возрасте дерева. Но они становятся особенно
частыми у стареющих деревьев с ослабленным ростом побегов
скелетной оси Тогда укороченные побеги появляются обильно
по всей кроне. В укороченном состоянии побеги В tortuosa, как
отмечалось, могут расти до 15—20 лет. У В. tortuosa в отличие
от В. verrucosa и В. pubescens, на укороченных побегах можег
развиваться не 2, а 3—4 зеленых листа Замечательна их спо-
собность снова возвращаться к образованию удлиненных побе-
гов, которые в свою очередь опять могут перейти в укороченные
побеги и т. д. Закономерная смена па одной и той же ветви
укороченных и удлиненных годичных побегов позволяет гово-
рить не о типах побегов, а о состоянии одних и тех же
побегов, развивающихся у В. tortuosa по типу укороченных или
удлиненных.
В процессе морфогенеза дерева в индивидуальном развитии
извилистой березы, в условиях пояса березового криволесья
Хибин можно выделить несколько возрастных периодов, как и у
двух предыдущих видов березы.
Первый возрастной период. О д постебел ьн ый сеянец
с очень слабым ростом стебля и корней. Период
длится от появления проростков до 6—8-летнего возраста их.
В этот период раскрывается лишь верхушечная почка, образуя
ежегодно побеги в 1—2 см высоты. Боковые пазушные почки не
раскрываются и переходят в состояние спящих. В корневой
системе формируется система главного корня, а в конце пе-
риода появляются первые придаточные корни Рост корней идет
так же слабо, как и рост стебля.
Второй возрастной период. Низкорослое дерево,
ветвящееся от основания ствола. В этот период у
невысокого (до 2,5—4 дт) кустовидного дерева нет заметных
отличий главной оси и нижних ветвей, отходящих близ поверх-
ности почвы и укореняющихся. В поясе березового криволесья
Хибин этот период длится от 10—15- до 30—40-летнего воз-
раста В лесном поясе Хибин боковые нижние ветви не отлича-
ются сильным и длительным ростом, и поэтому в этот период
здесь формируется одноствольное дерево.
Третий возрастной период. Формирование дочерних
вегетативных стволов. Характеризуется ослаблением
роста и отмиранием главной оси с образованием нескольких
207
новых стволов, вырастающих из спящих почек в основании
ствола (в лесном поясе) или нижних укореняющихся ветвей
с интенсивным и длительным ростом (в поясе березового
криволесья) Начинается с 25—40 лет. Система главного корня
сменяется системой придаточных (стеблеродных) корней в ос-
новании каждого ствола вегетативного происхождения.
Морфогенез жизненной формы дерева
у дикой яблони (Mallis silvestris Mill)
Maias silvestris произрастает в широколиственных и смешан-
ных лесах Европейской части СССР, доходя на севере, при-
мерно, до Ленинграда, Новгорода, Осташкова, Ярославля,
Костромы, Казани, Уфы, встречаясь чаще в южных дубравах
подзоны лесостепи (В Н. Сукачев, 1938). В лесостепи дикая
яблоня растет по только в лесных массивах как дерево второй
величины, но выходит одиночно или небольшими группами на
открытые степные пространства Поэтому обзор морфогенеза
дикой яблони интересен тем, что здесь мы встречаемся с дре-
весной формой лесостепного типа.
Ниже приводятся краткие данные о морфогенезе жизненной
формы дерева у дикой яблони на основе наблюдений автора и
Т И Серебряковой на территории Курского Государственного
Центрально-Черноземного заповедника имени В. В. Алехина
Прорастание семян М. silvestris происходит по надземному
типу Интенсивно растущий гипокотиль, достигающий длины
3—5 см, выносит мясистые и толстые округло-яйцевидные семя-
доли на свет, и они вскоре зеленеют и начинают фотосинтези-
ровать Выше семядолей на стебле вскоре появляются первич-
ные клиновидно заостренные зеленые листья, покрытые с обеих
сторон волосками У взрослых деревьев листья сверху голые,
при основании пластинки округлые, с сильно выступающими
снизу боковыми жилками Количество зеленых листьев, появ-
ляющихся в течение первого лета, может достигать 15—20. В те-
чение первого года сеянцы дикой яблони в питомнике Курского
Государственного заповедника вырастают в среднем на 20—30 см.
По интенсивности роста сеянцы яблони проявляют большую
изменчивость: высота их в конце первого лета колеблется от
10—15 до 60—70 см. Как и у сеянцев дуба, у сильных экзем-
пляров яблони уже в первый год наблюдается два, а иногда
даже три прироста, сильно затрудняющие в последующем
определение возраста сеянцев. Рост побега завершается форми-
рованием зимующей верхушечной почки. Но нередко уже с пер-
вого года моноподиальность нарастания нарушается подсыха-
нием терминальной почки и верхней части побега.
208
На второй год рост сеянцев яблони становится еще более
интенсивным, прирост достигает в среднем 30—40 см, а у от-
дельных экземпляров более 60 см. Нередко со второго года
начинается ветвление за счет верхних боковых почек побега
первого года. Но столь же часто первые боковые ветви по-
являются из почек побегов второго года жизни сеянца лишь на
третьем году. В последующие годы интенсивность роста замет-
но увеличивается
На величине годичного прироста у молодых яблонь сказы-
вается также и сочетание метеорологических условий, склады-
вающихся в течение смежных лет Так, в 1957 г, благоприятном
в метеорологическом отношении, годичные побеги у семилетних
яблонь в посадках Курского Государственного заповедника
в среднем имели длину 70—80 см, а у отдельных экземпляров
достигали 130—140 см (2—3 прироста в лето). В результате
интенсивного роста у М. silvestris уже в первые 5—7 лет фор-
мируется древовидный габитус с четко выраженным вертикаль-
ным главным стволом и наклонными, слабее растущими и более
тонкими ветвями.
В росте боковых ветвей у молодых деревьев М. silvestris
проявляется свойство, уже отмеченное у сеянцев ели, лесной
сосны, дуба и березы — возрастное усиление боковых ветвей.
Ветви, возникшие на побегах первого и второго года жизни
сеянца, растут слабо и недолговечны. Вследствие этого
у 10—12-петних деревьев высотой около 3 м и диаметром ствола
у основания 5—6 см, растущих на открытых полянах, эти ветви,
расположенные па высоте 15—25 см от земли, едва достигают
длины 40 см. Они тонкие, подсыхающие, коряво ветвятся Ветви,
расположенные выше, на приростах следующих лет, отличаются
большей интенсивностью роста в длину и толщину и большим
долголетием Сильные боковые ветви, составляющие скелет
кроны, появляются на 5—6-м году, или у некоторых экземпля-
ров на 8-м году. Скелетные ветви кроны у 10—12-летних де-
ревьев достигают 1,5 м длины, при диаметре 2,5—3 см и почти
не отличаются от главной оси Г равная ось в этом возрасте
дерева еще может быть прослежена до верхушки кроны Годич-
ные приросты ее резко колеблются в смежные годы, вероятно
в связи с колебанием метеоусловий, от 15—25 до 60—70 см
Нижние слабые ветви первых лет жизни сеянца в последую-
щем отмирают, и у 20—30-летних деревьев нижняя часть ствола
до высоты 50—80 см оказывается очищенной от отмерших вет-
вей На высоте 70—100 см начинаются скелетные ветви кроны,
отходящие от главной оси в количестве 15—20 Они распола-
гаются на расстоянии 10—12 см друг от друга по главному
стволу, иногда группами, почти мутовчато Их диаметр дости-
гает 2,5—3 см, а длина — 2,5—3 м. Эти ветви вначале растут
В-413 И Г Серебряков—14 209
почти горизонтально, затем приобретают почти вертикальное
положение.
От 10—12-летних деревьев, где главная ось может быть
прослежена до верхушки кроны, а крона имеет суженную форму
с более или менее острой вершиной, отличаются 20—30-летние
зрелые деревья М. silvestris, распщие на больших лесных
полянах и открытых степных участках Курского заповедника.
Их главная ось теряется среди сильных ветвей на высоте 2 м.
Ее диаметр от 10—12 см у основания снижается до 2—2,5 см
(на высоте 2 м)\ здесь главная ось не отличается по толщине
от боковых ветвей. Одновременно с уменьшением толщины
главной оси ослабляется ее рост в высоту.
Ослабление роста главной оси проявляется не только отно-
сительно, при сравнении с боковыми ветвями, но и абсолютно,
по сравнению с ростом ее от 5—8 до 15—20 лет. Позже 20 лет
наблюдается резкое ослабление роста дерева в высоту. Как
и у рассмотренных ранее деревьев — дуба и березы, ослабление
и прекращение роста кроны и начинающееся подсыхание кроны
у дикой яблони не влечет за собой отмирания дерева в целом
благодаря способности к образованию новых осей вместо ста-
реющих и прекративших рост материнских. Правда, образова-
ние новых осей происходит за счет придаточных почек на кор-
нях, дающих корневую поросль (П. Г. Шитт, 1953), но не спя-
щих почек ствола, как это характерно для дуба и рассмотрен-
ных видов березы.
Как видно из краткого описания роста и формирования
дерева у дикой яблони, здесь наблюдается много общего с по-
добными процессами в морфогенезе дерева у дуба и трех видов
березы: изменение интенсивности роста главной оси в онтоге-
незе дерева по типу одновершинной кривой, усиление боковых
побегов с возрастом дерева у молодых формирующихся де-
ревьев, изменение формы кроны от более или менее вытянутой
с заостренной верхушкой к раскидистой кроне с округлой вер-
шиной, способность к замене стареющих и прекративших рост
материнских осей новыми, развивающимися из спящих или
придаточных почек На фоне общих с рассмотренными выше
лесными деревьями черт морфогенеза у дикой яблони четко
проявляются черты своеобразия.
Опа отличается от ранее рассмотренных деревьев более
коротким жизненным циклом главной оси Быстрое усиление
интенсивности роста главной оси приводит к раннему наступле-
нию кульминации роста (в 5—10-летнем или 10—15-летнем воз-
расте), а затем к столь же раннему ослаблению и прекращению
роста Ранней кульминации роста главной оси соответствует
раннее достижение максимума роста и долголетия боковыми
ветвями главной оси.
210
Как отмечалось выше, максимальным ростом и долголетием
обладают боковые ветви главной оси, возникшие на 5—6 году
жизни дерева. Именно эти ветви составляют скелет кроны
взрослого дерева. Напомним, что у дуба максимальным ростом
и долголетием обладают боковые ветки главной оси, возникшие
во 2-м, 3-м и даже 4-м десятилетии их жизни. В этой связи
интересно отметить, что у домашней яблони максимум в росте
боковых ветвей наступает еще раньше.
Как известно (Курындин, 1948), крона домашней яблони
формируется из боковых побегов 3-го и 2-го годов жизни
дерева В этом, вероятно, проявляется влияние условий куль-
туры, ускоряющих развитие растений в онтогенезе и вместе
с тем ведущих к сокращению общей длительности жизни расте-
ния Вообще в условиях культуры у растений далеко не еди-
ничны примеры сокращения онтогенеза, потери поликарпич-
пости и перехода к монокарпичности, перехода многолетних
растений к развитию по типу двулетников, а последних —
к однолетникам (Работнов Т. А , 1950, Купцов А. И , 1953 и др ).
Видимо, в этом же направлении изменялось развитие и форми-
рование дерева у дикой яблони в процессе ее одомашнивания
В отличие от ранее рассмотренных деревьев, у дикой яблони
очень рано теряется главная ось среди мощных ветвей форми-
рующейся кроны В результате образуется своеобразная форма
дерева с коротким стволом, низко расположенной кроной, ли-
шенной четко выраженной лидерной оси. У домашней яблони
потеря лидерной оси при формировании кроны наступает даже
раньше, чем у дикой лесной яблони. Удаление главной оси
в 3—4-летнем возрасте при формировании у нее кроны уже не
приводит к образованию вторичной лидерной оси за счет побе-
гов замещения
Ранняя потеря лидерной оси и потеря способности к ее вос-
становлению за счет побегов замещения значительно отличает
формирование дерева у дикой яблони от ранее рассмотренных
лесных деревьев, у которых главная ось (моно- и симподиаль-
ная) не только сохраняет свое лидерное положение в кроне
многие десятки лет, но и восстанавливается за счет боковых
побегов замещения при глубокой обрезке даже у старых де-
ревьев. Этот же признак—ранняя потеря лидерной оси — при
ближает форму дерева, наблюдаемую у яблони, к кустарникам,
у которых, как известно, главная ось рано теряет лидерное
положение в онтогенезе растения
Отмеченные своеобразные черты жизненной формы дерева
у дикой яблони (низко расположенная раскидистая крона,
отсутствие лидерной оси в кроне, округлая или чаще уплощен-
ная вершина кроны) свойственны не только видам яблони, но
и множеству других видов деревьев различного систематиче-
14' 211
ского положения Подобная форма дерева весьма характерна
для сухих саванновых лесов и саванн (Schimper и Faber, 1935).
Широкое распространение деревьев подобной формы у гра-
ниц, лесной растительности, обусловленных аридностью, и ее
наличие в разных систематических группах (Rosaceae, Legumi-
nosae, Proteaceae и др.) позволяет думать, что жизненная форма
«плодового дерева» экологически возникла в аридных условиях
как следствие ускорения развития и сокращения жизненного
цикла главной оси. Дальнейшее усиление этого процесса сокра-
щения длительности жизни главной оси привело к появлению
новой группы жизненных форм — кустарников (см. гл. IV).
Форма «плодового дерева» является как бы переходной между
деревьями и кустарниками, наряду с отмеченной выше формой
немногоствольного дерева типа Betula tortuosa. Но форма «пло-
дового дерева» чаще встречается у континентальной границы
леса и связана с аридностью климата, а форма немногостволь-
ного дерева приурочена к субарктической и субальпийской гра-
нице леса.
Морфогенез жизненной формы дерева
у черного саксаула — Haloxylon aphyllum Bge
и белого саксаула Haloxylon persicum (Minkw) Iljin
Haloxylon aphyllum и H. persicum имеют довольно близкие
ареалы и на территории СССР распространены на востоке
и юге Арало-Каспийской низменности, в Прибалхашье, Кзыл-
Кумах, Кара-Кумах и по долинам Аму-Дарьи и Сыр-Дарьи
(М. ЛА Ильин, 1936). При общем сходстве области распростра-
нения эти виды обнаруживают значительные различия в эколо-
гии Haloxylon persicum приурочен к бугристым и грядовым
пескам, Н. aphyllum — к солончаковым окраинам песков, лес-
сово-солончаковым почвам, такырам, солончаковым и солонце-
ватым сероземам (М М. Ильин, 1936; М. П. Петров, 1936).
Ниже приводятся краткие данные по морфогенезу жизненной
формы дерева у черного и белого саксаула на основе наблюде-
ний, проведенных на территории Репетекского государственного
песчано-пустынного заповедника В сборе полевых материалов
принимал участие Р. А. Ротов, котором} автор приносит искрен-
нюю благодарность.
Черный саксаул (Haloxylon aphyllum) во взрослом состоянии
на территории Репетекского госзаповедника образует типичную
форму дерева высотой 5—6 я, с толстым стволом диаметром
в 40—60 см у его основания У зрелых деревьев черного сак-
саула ствол обычно расщеплен почти до основания на 2—3
части, так что каждая из этих частей несет самостоятельно
свою часть кроны.
212
Белый саксаул (Haloxylon persicum) в Репетекском госзапо-
веднике во взрослом состоянии достигает 3—4 м высоты с диа-
метром ствола у основания 15—20 см. Наряду с одноствольной
древовидной формой у белого саксаула и во взрослом состоя-
нии иногда сохраняется 2—3 ствола, каждый со своей кроной,
так что получается своеобразная форма кустовидного дерева.
Своеобразие формы дерева у саксаула проявляется не
только в расщеплении его ствола у зрелых деревьев, но и в осо-
бенностях его побегов. Годичные побеги Я. persicum и Н. ophyl-
turn, как известно, несут крайне редуцированные листья. Они
лишены ассимилирующей зеленой пластинки и имеют лишь
слабо развитое влагалище в виде узенькой каймы, охватываю-
щей основание междоузлий. Отсутствие зеленых ассимилирую-
щих листьев влечет за собой усиленное развитие ассимиляцион-
ной ткани стебля (Радкевич О. Н и Василевская В К., 1933).
Зеленые прутьевидные стебли у саксаула оказываются един-
ственными органами фотосинтеза Хлорофиллоносная ткань
побега саксаула сосредоточена во внутренних слоях первичной
коры Как и у всех Chenopodiaceae, у саксаулов имеется два
слоя ассимиляционной ткани, наружный — палисадного типа и
внутренний — из более округлых клеток. Последний соответ-
ствует эндодерме
Безлистность побегов саксаула сочетается с членистой струк-
турой стебля и с наличием интеркалярной зоны роста в осно-
вании каждого членика — междоузлия (как у злаков) При
делении паренхимных клеток каждого узла образуется отделяю-
щий слой, благодаря чему отдельные членики стебля легко опа-
дают по мере их усыхания.
В кроне черного и белого саксаула наблюдается четко выра-
женный диморфизм годичных побегов, но иного типа, чем у
ранее рассмотренных деревьев. Например, резкие различия
удлиненных и укороченных побегов наблюдаются у сосны и ви-
дов березы. Менее резко эти типы побегов выражены у дуба
и ели У всех рассмотренных деревьев четко отличимы друг от
друга побеги вегетативные и генеративные. У черного и белого
саксаула диморфизм побегов проявляется в ином плане и, как
показано в исследовании Радкевич О. Н. и Василевской В. К.
(1933), соответствует климатическим особенностям пустынь
Средней Азии.
В кроне саксаула (Н. aphyllum и И. persicum) можно видеть
побеги двух типов. Одни — тонкие, мягкие, зеленые, несколько
сочные, однолетние, опадающие с середины лета до осени; дру-
гие — более толстые и жесткие, серые или коричневые, с более
энергичным ростом, обычно ортотропные, с большей продолжи-
тельностью жизни. Первые получили название «ассимиляцион-
ных веточек» (см. Радкевич О. Н. и Василевская В. К., 1933).
213
Посредством побегов второго типа осуществляется нараста-
ние кроны дерева; они образуют скелет кроны, поэтому их
целесообразно назвать побегами нарастания или ростовыми
побегами.
Различия между ассимиляционными и ростовыми побегами
заметны уже в мае, так как первые в основном заканчивают
рост в конце мая, а вторые продолжают его до августа (Арци-
ховский В М, 1928). В июле диаметр ассимилирующих веточек
не превышает 3 мм, у ростовых он нередко достигает 7—9 мм.
Наряду с более слабым и кратковременным ростом в длину
ассимиляционные веточки характеризуются задержкой вторич-
ного роста в толщину. У них до самого опадения сохраняется
фаза первичного пучкового строения стели, что сильно сбли-
жает их структуру с побегами травянистого типа. Наличие тон-
костенной хлоренхимы, крупноклетной водоносной ткани пер-
вичной коры, пучкового типа стели и общая слабая склерифика-
ция тканей не только приближают структуру ассимилирующих
однолетних побегов к травянистой, по и придают им некоторые
черты суккулептности
В отличие от ассимилирующих, побеги нарастания разви-
ваются как типично древесные- они рано переходят к образова-
нию сплошного широкого кольца ксилемы, в их лубе образуют-
ся лубяные волокна, а в паренхимных частях перицикла непо-
средственно под склеренхимными пучками образуется пробко-
вая ткань (Арциховский, 1928)
Раннее образование перидермы в побегах нарастания спо-
собствует переживанию ими неблагоприятных сезонов года
(летней сухости и низких зимних температур), чем обеспечи-
вается их жизнедеятельность в кроне саксаула в течение
нескольких или даже многих лет Но вместе с тем образование
пробки на этих побегах влечет за собой отмирание ассимили-
рующей ткани, утрату зеленого цвета побегами и потерю спо-
собности к фотосинтезу. Вместе с ассимилирующей тканью при
образовании перидермы отмирает и водоносная ткань, имев-
шаяся в первичной коре
Таким образом, в кроне саксаула наблюдается своеобразный
диморфизм годичных побегов, не встречавшийся у ранее рас-
смотренных деревьев В кроне саксаула развиваются однолет-
ние ассимилирующие побеги с чертами травянистой структуры
и признаками суккулептности, и многолетние побеги с типичной
древесной структурой, с ранним опробковением и потерей пер-
вичных неодревесневших тонкостенных и крупноклетных тка-
ней, свойственных суккулентным формам. Потеря саксаулом
в течение лета ассимилирующих веточек и сбрасывание неодре-
весневших мягких первичных тканей приводит к тому, что на
зиму деревья становятся-типичными склерофитами и по зимо-
214
стойкости сходными с лесными деревьями более холодных ши-
рот. Саксаул, как и другие древесные псаммофиты среднеазиат-
ских пустынь (древовидные солянки, виды Calligonum), отно-
сится к своеобразной группе сезонно суккулентных растений.
Последние резко отличаются от постоянных суккулентов жар-
ких пустынь, где отсутствует зимний период (Радкевич и Ва-
силевская, 1933).
Это своеобразие саксаула обусловлено экологическими осо-
бенностями среднеазиатских пустынь — низким температурным
минимумом зимой и резко пониженным, но постоянным запа-
сом влаги в песчаном грунте в течение летнего периода Лет-
ний минимум влаги обеспечивает лишь небольшой процент по-
бегов, возникших весной в кроне саксаула, это обстоятельство
и обусловливает длительный и постепенный летний стеблепад,
свойственный саксаулу и другим древесным псаммофитам с
безлистными членистыми стеблями (И. М. Васильев, 1931).
Прорастание семян солончакового и песчаного саксаула,
как и других растений из Chenopodiaceae, происходит по над-
земному типу (И. Т. Васильченко, 1938). Зрелые семена после
перезимовки в природных условиях Репетекского госзаповед-
ника прорастают в марте и апреле Как показали наблюдения
Н Т Нечаевой и С Я. Приходько (1952), семядоли еще до вы-
хода на поверхность песка имеют зеленую окраску. По выходе
на свет очи оказываются первыми ассимилирующими органа-
ми проростка В противоположность семядолям, первичные
листья, развивающиеся выше семядолей на главной оси, начи-
ная с эпикотипыюго узла, имеют характер бесцветных пленча-
тых чешуек. Вследствие этого уже с самого начала главная
ось проростка приобретает черты афилльного членистого побега
Наилучшее прорастание семян- саксаула, по данным
М И Сусловой (1936), происходит на глубине 2—3 см. В этих
условиях гипокотиль проростка, достигающий 4—5 см длины,
легко выносит семядоли и ассимилирующий побег на свет,
обеспечивая дальнейшее нормальное развитие. Главная ось
проростка отличается довольно быстрым ростом, и к концу
мая, когда рост побега в основном заканчивается, достигает
высоты 10—15 см В отличие от побега, корень проростка сак-
саула отличается большей интенсивностью роста и в течение
первого года достигает глубины 80—90 см, т е. горизонта с
постоянно увлажненным песком (М П Петров, 1936).
В течение первого же лета начинается ветвление побегов
саксаула (у сеянцев Haloxylon aphyllum оно происходит силь-
нее, чем } Н. persicum), причем в росте боковых ветвей сеянцев
саксаула хорошо выражено базитонное усиление — наиболее
сильными оказываются самые нижние ветви. В течение летнего
периода не только останавливается верхушечный рост главной
215
оси сеянца, но и наблюдается подсыхание ее верхней части,
иногда до половины длины побега Из-за отмирания верхушки
тлавиой оси дальнейшее нарастание ее в высоту происходит по
симподиальному типу, за счет одной или двух-трех боковых
ветвей, приобретающих ортотропный рост. Симподиальность
ветвления, отмеченная Арциховским В. М (1928, 1933) для
взрослых растений Н. aphyllum и Н. persicum, начинается, как
видим, с однолетнего возраста.
Представляет безусловный интерес тот факт, что побеги мо-
лодых сеянцев саксаула (от однолетнего до 5—7-летнего воз-
раста) развиваются только по типу побегов нарастания Побе-
ги типа ассимиляционных веточек появляются в онтогенезе со-
лончакового и песчаного саксаула значительно позже (с 5—
7-летнего возраста). Напрашивается биогенетическое толкова-
ние такой последовательности появления побегов древесного
и травянистого типа. Однако из-за сложности филогенетических
отношений среди Chenopodiaceae подобный вывод был бы
преждевременным
С первых же лет жизни сеянцев саксаула обращает на себя
внимание еще одна особенность — отмирание значительной
части побега в его верхней половине (часто до половины го-
дичного прироста и даже более). Как известно, подобное раз-
витие побегов (с отмиранием верхней половины) свойственно
не только саксаулу, но и другим пустынным древесным расте-
ниям Кара-Кумов и, в частности, кустарникам (например,
Astragalus paucljugus, A. unifoliatus, Smirnovia turkestana и др )
Оно проявляется еще сильнее у полукустарников, где побеги
отмирают почти до поверхности земли
Вследствие базитонного характера ветвления двух-, трех-,
четырехлетние сеянцы саксаула приобретают кустовидный га-
битус У них развивается одновременно несколько сильных
ветвей, обычно 5—7 у черного саксаула и до 8—12 у белого.
Ежегодный прирост сильных ветвей, несмотря на отмирание
верхних частей побегов, уже в 3—4-летнем возрасте увеличи-
вается до 30—35 см, и общая высота сеянцев саксаула к 3—
4 годам достигает 80—120 см. Быстрое усиление роста саксаула
приводит к тому, что уже с 5—7-летнего возраста начинается
период максимального роста. Достигнутая к этому времени
большая интенсивность роста (в среднем 50—80 см в год, без
к чета подсыхающей части побега) сохраняется до 15—20-лет-
него возраста, а затем начинает постепенно снижаться. К 7—8
годам высота саксаула достигает 1,5—2 м. В этом возрасте
начинает резко проявляться различие в интенсивности роста
отдельных скелетных осей куста саксаула, приводящее к пол-
ному прекращению роста некоторых из них и последующему их
отмиранию. Происходит интенсивный процесс дифференциации
216
«выбраковки» (изреживания) скелетных осей В результате
изреживания остается сначала 2—4 наиболее крупные оси и,
наконец, только лишь одна, которая и становится стволом фор-
мирующегося дерева. Усиление интенсивности роста в высоту
сопровождается быстрым утолщением скелетных осей
К 10—15-летпему возрасту Н. aphyllum достигает 2,5 м вы-
соты и 12—15 см в диаметре у основания; в 20—25 лет черный
саксаул достигает 3—3,5 м в высоту при диаметре 20—25 см
Нередко уже у 20—25-лстних деревьев черного саксаула по-
являются признаки суховершинности, особенно в центральной
части кроны
Примерно одновременно с этим па стволе черного саксаула
появляются продольные трещины, которые в последующие годы
быстро расширяются и углубляются, доходя до центральной
части ствола. Под влиянием собственной тяжести ствол черно-
го саксаула в конечном счете разваливается на 2—3 отдельные
части, так что общая часть ствола оказывается лишь ниже по-
верхности песка. При продольном расщеплении ствола и неко-
тором полегании образовавшихся частей на них неч наблюда-
ется образования новых придаточных корней, хотя часто при
этом распавшиеся части ствола оказываются несколько засы-
панными песком.
Расщепление ствола у черного саксаула совпадает с рез-
ким ослаблением и практическим прекращением роста в высо-
ту и в свою очередь приводит к дальнейшему ускорению про-
цессов старения дерева. Через некоторое время распавшиеся
части ствола и принадлежащие им кроны полностью отмирают.
Распадение ствола представляет своеобразную особенность
стареющих деревьев саксаула и очевидно является одним из
проявлений процесса старения дерева По внешнему проявле-
нию и биологическим последствиям этот процесс напоминает
широко распространенное среди полукустарников Средней
Азии явление партикуляции
Раннее усиление роста главной оси в онтогенезе сеянцев чер-
ного саксаула вызывает не только быстрое наступление куль-
минации роста, но и сопровождается ускоренным развитием и
переходом в генеративное состояние Черный саксаул начинает
плодоносить с 5—7-летнего возраста, а иногда и раньше
В течение периода, предшествующего цветению, наблюдает-
ся, как отмечалось, усиление быстроты роста годичных побегов
главной оси, изменение общего габитуса растения от кустовид-
ного, с несколькими скелетными осями, к древовидному с
одним толстым стволом и низко начинающейся сильно ветви-
стой кроной, появление наряду с древесными побегами нара-
стания однолетних травянистых ассимилирующих веточек.
Интересно, что развитие древовидной формы саксаула обуслов-
217
лсно не только резкой дифференциацией в быстроте роста
между отдельными ветвями, но и изменением характера ветвле-
ния Наблюдающееся на первом и втором году жизни сеянца
базитониое усиление боковых ветвей в последующие годы ста-
новится нс столь четко выраженным, а когда среди образую-
щихся годичных побегов наряду с ростовыми многолетними
появляются ассимилирующие — однолетние веточки, начинает
проявляться характерное для всех рассмотренных деревьев
акротонное усиление ветвей. Слабо растущие ассимилирующие
однолетние ветви развиваются из нижних почек материнского
годичного побега, а более сильные многолетние и быстро расту-
щие ростовые побеги возникают из верхних почек годичного
побега, располагающихся близ его отмирающей верхушки.
Быстрое достижение саксаулом периода максимальной ин-
тенсивности роста сопровождается соответственно и ранним
началом замедления его для главной оси К 20—25 годам рост
саксаула в высоту практически прекращается, и деревья начи-
нают суховершинить. Первые отмирающие верхние ветви появ-
ляются в центральной части кроны Как и при партикуляции
полукустарников, отмирание центральных веток влечет за собой
отмирание более или менее широкого сектора тканей главного
стебля (ниже отхождения отмерших веток) и последующее рас-
щепление стебля. Как отмечалось выше, расщепление и распа-
дение ствола ускоряет дальнейшее усыхание кроны саксаула
В результате этого уже к 25—30-летнему возрасту заросли
саксаула относят к возрастной категории перестойных
(В. Л. Леонтьев, 1954), а к 40—60 годам деревья обычно пол-
ностью усыхают Вероятно, лишь в отдельных случаях деревья
саксаула могут достигать большей длительности жизни (до
70—90 лет)
Как и ранее рассмотренные лесные деревья (дуб, береза),
саксаул имеет спящие почки Но они не оказывают существен-
ного влияния на формирование кроны дерева в его онтогенезе
Спящие почки обычно не «просыпаются» и при усыхании кро-
ны и не дают начала новым стволам и кронам при старении
материнской главной оси, как это наблюдалось, например, у
Betula tortuosa Лишь при сильном повреждении кроны саксау-
ла или ее полном удалении из спящих почек появляется пневая
поросль, обеспечивающая вегетативное возобновление дерева.
5. ФИЗИОЛОГИЧЕСКИЙ АНАЛИЗ ФОРМИРОВАНИЯ КРОНЫ ДЕРЕВА
(Факторы, определяющие направление и интенсивность роста
побегов. Корреляции в росте побегов в кроне дерева)
Различие в форме кроны у разных деревьев, равно как и
само образование кроны у каждого дерева, становится возмож-
218
ним лишь в результате различий в интенсивности и направле-
нии роста отдельных побегов, образующих крону Поэтому
анализ морфогенеза дерева был бы незавершенным без рас-
смотрения причин, обусловливающих это разнообразие побегов,
без физиологического андлиза формирования кроны.
Классические опыты Найта в начале XIX в впервые пока-
зали влияние силы земного притяжения на вертикальное по-
ложение главных осей растения, доказав экспериментально,
что естественное вертикальное направление главного стебля и
корня определяется воздействием силы тяжести.
Достаточно отклонить молодой растущий стебель на
несколько градусов от вертикали, чтобы вызвать возвращение
его верхушки вновь в вертикальное положение и образовать
искривление, получившее название гсотропического, из-за выз-
вавшей его причины.
Способность главного побега постоянно расти вертикально
вверх в поле действия силы земного притяжения получила наз-
вание отрицательного геотропизма, а главные оси растения,
растущие вертикально, были названы ортотропными.
По поводу положения боковых ветвей, обычно наклонного, а
не вертикального, Найт высказал предположение, что эти вет-
ви обладают одинаковым с главной осью свойством расти вер-
тикально вверх под влиянием силы тяжести; но они не дости-
гают этого положения вследствие более слабого роста и меха-
нического воздействия их собственной тяжести
Этот взгляд Найта впоследствии поддержали А Декан-
долль, Дютрошс, Мейен, Сакс и Гофмейстер, не проводя,
однако, новых опытов для более детального изучения этого
вопроса
А Б Франк (А. В. Frank, 1870) провел специальное
исследование физиологических свойств боковых побегов и при-
чин, обусловливающих их наклонное положение. Франк прида-
вал боковым растущим побегам хвойных и лиственных деревь-
ев, а также стелющимся стеблям вертикальное положение, а
часть побегов оставлял в горизонтальном положении, но пере-
вертывая их морфологически нижней стороной вверх. Верти-
кально ориентированные растущие побеги при этом изгиба-
лись, пока не возвращались в горизонтальное положение, а
перевернутые горизонтально расположенные побеги перекручи-
вались на 180°, снова поворачиваясь вверх морфологически
верхней стороной. Из этих опытов Франк заключил, что поло-
жение боковых побегов деревьев также определяется земным
притяжением, но нормальное положение для этих побегов
(положение равновесия) — горизонтальное, и определяется оно
особым физиологическим свойством этих побегов, их трансвер-
зальным или поперечным геотропизмом Кроме того, боковые
219
побеги по Франку обладают физиологической двусторопностью,
так как всегда обращаются вверх определенной стороной.
Исследования Франка послужили исходным пуктом для ра-
боты Дс Фриза с листьями и ветвями деревьев. Де Фриз
подтвердил предположение Сакса (1874) о неравномерности
роста верхней и нижней стороны черешка и главной жилки
листьев. По выходе листа из почки на черешке и главной жилке
листа сильнее растет верхняя сторона и листья отгибаются от
стебля (эпинастия), а в более раннем возрасте листа, в почке
сильнее растет морфологически нижняя сторона листа (гипо-
настия), вследствие чего лист оказывается прижатым к стеблю,
прикрывая конус нарастания.
Де Фриз обрезал на растущих листьях пластинку парал-
лельно главной жилке и помещал эти листья во влажный воз-
дух в горизонтальном положении, ориентируя к зениту морфо-
логически верхней, нижней или боковой стороной Растущие
листья образовывали изгибы черешка и главной жилки. Опыты
Де Фриза показали, что листья, как и побеги, обладают отри-
цательным геотропизмом, но с известного возраста ему проти-
водействует эпинастия, и листья получают наклонное положе-
ние. Такое же заключение сделано Де Фризом и в отношении
ветвей деревьев и кустарников
Таким образом, если по Франку наклонное положение вет-
вей и листьев определяется их своеобразным отношением к
силе тяжести — поперечным геотропизмом, то по Де Фризу
наклонное положение этих органов есть следствие взаимодей-
ствия противоположно направленных отрицательного геотро-
пизма и эпинастии. Эпинастия, по Де Фризу, представляет
внутреннее свойство боковых органов растений, не зависящее
ни от каких внешних условий. У растений с гипонастией
наклонное положение ветвей обусловлено собственной тя-
жестью облиственного побега, которое противостоит сов-
местному действию отрицательного геотропизма и гипо-
настии.
Обширные исследования физиологических свойств и причин
наклонного направления ветвей деревьев и кустарников были
проведены О. В Баранецким (1901). Этим автором было откры-
то у побегов хвойных и лиственных деревьев и кустарников
особое физиологическое свойство «противоизгиба».
Фехтинг (1882) впервые заметил, что стебли и корни с геот-
ропическим изгибом в процессе роста на клиностате выпрям-
ляют изгиб и продолжают расти прямо. Это свойство Фехтинг
назвал ректипетальностью, или внутренним свойством осевых
органов выпрямлять изгиб и расти в прямолинейном направле-
нии Это свойство основано на усиленном росте вогнутой сто-
роны осевых органов.
В опытах с крупными ветвями клена, каштана, черемухи,
липы, чубушника, вяза, с почти нормальным ростом опытных
побегов, О. В Баранецкий обнаружил, что на клипостате дей-
ствительно происходит выпрямление геотропического изгиба.
Но при этом ветви не только разгибались, но и сгибались в
противоположную сторону в той же плоскости, затем снова сги-
бались в первоначальном направлении и т. д несколько раз.
Эти опыты подтвердили, что выпрямление побегов на клино-
стате зависит от усиленного роста на вогнутой стороне. Но
если Фехтинг, а затем Чапек думали, что этот рост происходит
до выпрямления, опыты О В. Баранецкого показали, что этот
рост происходит и после выпрямления, до изгиба в обратную
сторону. Затем усиленный рост переходит на внутреннюю сто-
рону нового изгиба, пока не уменьшится амплитуда изгибов и
стебель не выпрямится окончательно. Следовательно, дело не в
свойстве выпрямлять изгиб, а в свойстве, на всякий изгиб, выз-
ванный внешними или внутренними причинами, реагировать
стремлением к изгибу в противоположную сторону, усилением
роста на противоположной стороне стебля. Это стремление к
противоизгибу столь резко, что проявляется даже при действии
фактора, вызвавшего первоначальный изгиб Стремление к про-
тивоизгибу вызывает ряд попеременных колебаний роста на
противоположных сторонах стебля в плоскости первоначаль-
ного изгиба.
Таким образом, всякий односторонний рост стебля вызыва-
ет стремление к усилению роста на противоположной стороне,
вследствие чего всякий изгиб служит исходной точкой для по-
переменных колебаний роста, пока не исчезнет или не умень-
шится изгиб
Наблюдения и опыты О В. Барапецкого привели его к вы-
воду, что по характеру побегов среди лиственных деревьев и
кустарников можно выделить два физиологических типа:
1) «тип черемухи» и 2) «тип липы» «К «типу черемухи» отно-
сятся виды черемухи, клена, ясеня, а также конский каштан,
европейский бересклет и др Боковые побеги этих растений не
отличаются от главной оси и физиологическая двусторопность
у них отсутствует Все побеги этих растений обладают нор-
мальным отрицательным геотропизмом и при всяком не верти-
кальном положении стремятся образовать изгиб вверх Но с
образованием геотропического изгиба в побеге возникает про-
тиводействие этому изгибу, которое, наконец, уменьшает гео-
тропический изгиб. Образующийся наклон у многолетних ветвей
представляет положение равновесия между отрицательным
геотропизмом и противодействующим ему' стремлением к про-
тивоизгибу К этим двум силам у многолетних боковых ветвей
присоединяется также влияние собственной тяжести ветви,
221
из-за чего у опустившихся тяжелых старых ветвей верхушки
получают дугообразный изгиб вверх.
К деревьям и кустарникам физиологического «типа липы»
относятся виды липы, вяза, а также граб и лещина. Их моло-
дые побеги обладают свешивающимися верхушками и дуго-
образно изогнуты у тургесцептных побегов. Эти побеги эпина-
стичны в смысле Де Фриза, тогда как побеги «типа черемухи»
по Баранецкому — гипонастичны, и их наклонное положение
определяется свойством противоизгиба.
Из опытов О В Баранецкого видно, что побеги липы физио-
логически двусторонни, эпинастичны, но эпинастия их связана
не с морфологически верхней стороной побега, а присуща
каждый раз физически верхней стороне побега Свойство про-
тивоизгиба есть и у липы (в ответ на отрицательный геотро-
пизм), но эта реакция противоизгиба здесь получает характер
органического свойства, сохраняясь долгое время (но все же
затем ослабевая и исчезая совсем) Поэтому различия между
побегами «типа черемухи» и «типа липы» по Баранецкому —
количественные. Побегам липы и вяза свойственна эпинастия,
особенно в их молодых частях, но затем она пересиливается
геотропизмом, под влиянием которого и происходит выпрямле-
ние эпинастического изгиба Как видно, и у побегов «типа
липы» наклонное положение является равновесным от воздей-
ствия двух противоположных сил — отрицательного геотропиз-
ма и физиологического свойства побегов к противоизгибу (как
и у побегов «типа черемухи»), но в отличие от побегов «типа
черемухи» свойство противоизгиба у побегов «типа липы»
сохраняется длительное время и проявляется как эпинастия
У сосны и ели, по данным О В Баранецкого, как и у побе-
гов «типа черемухи», нельзя обнаружить признаков физиоло-
гической двустороннее™ боковых побегов Но у сосны расту-
щие побеги обладают сильным отрицательным геотропизмом,
и поэтому лишь позже, по окончании роста, становится замет-
ным воздействие собственной тяжести побега Побеги ели,
напротив, в первое время роста отличаются слабым отрицатель-
ным геотропизмом, и поэтому подчиняются главным образом
действию собственной тяжести, и только в конце периода роста
обнаруживается влияние геотропизма, которое затем проявля-
ется и на многолетних ветвях, вызывая искривление вверх их
верхушек.
*
* t
Корреляция роста побегов в кроне дерева. Последующие
исследования физиологов и морфологов растений обнаружили,
что меньшая интенсивность роста боковых побегов в кроне де-
222
рева (по сравнению с главным стеблем), их наклонное положе-
ние и морфологическая дорзовентральность (в отличие от ра-
диальной структуры главного побега) обусловлены наличием в
кроне дерева главной оси, которая оказывает физиологическое
и формативное воздействие на боковые побеги, вызывая спе-
цифические особенности их роста и формирования Эти данные
вновь подтвердили потенциальную физиологическую равноцен-
ность главной оси и боковых ветвей. Лишь в процессе взаимо-
влияния и взаимодействия (корреляции) главной и боковых
осей возникают физиологические и морфологические различия
между ними
Как отмечалось выше, при формировании кроны ели
выявляются резкие различия в интенсивности и направлении
роста, морфологии и долголетии главного ствола и боковых
ветвей. Чем определяется это своеобразие в росте и морфоге-
незе главной и боковых осей^ Заложено ли оно в самой орга-
низации побегов, образующих ствол и ветви кроны, и поэтому
постоянно и стабильно, или в
определяется какими-то причи-
нами и может быть видоизме-
нено? В молодых насаждениях
ели, особенно в условиях, бла
гоприятных для роста, можно
видеть, как у некоторых дерев-
цев вместо одной вертикально
растущей верхушки главной
осп и отходящих от нее под
углом в 20—45° мутовок боко-
вых ветвей поднимаются более
или менее вертикально две или
более боковых ветви, как бы
образуя дополнительные глав
ные оси (рис 43) Вниматель-
ный осмотр показывает, что у
всех подобных деревьев с рез-
свою очередь это своеобразие
Рис. 43
Верхушка молодого деревца
ели — Picea excelsa
с поврежденной верхушечной
почкой главной оси’(слева)
ко приподнимающимися боко-
выми ветвями наблюдается
повреждение верхушки глав-
ной оси, ее надлом или подсы-
хание верхушечной почки.
В этих случаях проявляется известная зависимость роста боко-
вых ветвей от наличия главной оси.
Эта зависимость выявляется еще более четко в опытах с
искусственным удалением или повреждением верхушки глав-
ной оси ели Боковые побеги в таких случаях приподнимаются,
приобретая вертикальное положение, рост их усиливается,
223
достигая интенсивности, свойственной главной оси, меняется их
симметрия с дорзовентральной (как у боковых ветвей) на
радиальную (свойственную главной оси). Интересно,что подня-
тие боковых ветвей при повреждении верхушки главной оси
наблюдается только в период роста побегов ели. Если верхуш-
ка ели повреждается в конце лета или осенью, поднятие боко-
вых ветвей происходит лишь следующей весной (К. Гебель,
1908).
Боковые ветви при повреждении верхушки деревца приобре-
тают свойства главной оси, и обычно одна из них, наиболее
интенсивно растущая, вскоре полностью замещает удаленную
или поврежденную главную ось. Через несколько лет ветвь,
заместившую главную ось, оказывается трудно отличить от
настоящей оси первого порядка. Если удаляется не однолет-
ний, а многолетний участок верхушки главной оси ели, то
наблюдается поднятие соответственно и многолетних боковых
ветвей до 8—10-летнего возраста Подобные случаи можно
нередко наблюдать в подмосковных лесах в результате среза-
ния верхушек молодых деревьев на новогодние елки.
Нижние участки многолетних приподнимающихся ветвей
сохраняют дорзовентральную структуру, верхняя же, вновь на-
растающая часть становится ортотропной и радиальной
Изгиб боковых ветвей, особенно многолетних, сопровождаю-
щийся преодолением значительного механического сопротив-
ления, осуществляется благодаря явлению гипонастии, т. е
более быстрого разрастания тканей на нижней поверхности
ветви
Обычно лишь одна из боковых ветвей, наиболее быстро
растущая, образует продолжение главного ствола, замещая его
Но изредка образуются две или более вертикально растущие
ветви, замещающие главную ось. Тогда возникают двух- г ли
трехствольные ели, единично встречающиеся в лесных масси-
вах. Из сказанного вытекает, что боковые побеги, по крайней
мере в молодом возрасте, способны приобретать особенности
тлавной оси и образовывать новую верхушку дерева Но это
произойдет только в том случае, если главный побег удален
или сильно поврежден. Мы видим, таким образом, что главный
побег и боковые побеги находятся во взаимной зависимости
или корреляции, которая проявляется в том, что главный побег
так воздействует на боковые побеги, или «индуцирует» их, что
они по направлению своего роста и морфологии отличаются ог
главного побега.
Главный побег тормозит рост боковых и как бы мешает им
формироваться по типу, свойственному самому главному стволу.
Но каким путем протекает это торможение? В чем заключается
тормозящее воздействие главного побега^
224
Еще в начале двадцатого столетия Еггега (1904) высказал
предположение, что от органов растения, в данном случае от
верхушки главного побега, исходит «внутренняя секреция»,
которая и действует тормозяще на боковые побеги так, что
мешает им формироваться по типу главной оси Однако это
предположение в то время нс имело никакого научного обосно-
вания
К. Goebel (1908) выдвинул другое объяснение явлению кор-
реляции в росте главного и боковых побегов у деревьев, и в
частности у ели, которое казалось более обоснованным и вер-
ным. Гебель отмечает, что если весной верхушечный побег в те-
чение немногих недель из маленькой почки вырастает до 1 м
длины, то понятно, что для этого необходимо много строитель-
ного материала, который не может быть обеспечен в столь ко-
роткий срок ассимиляционной деятельностью самого растущего
побега Строительные материалы, по Гебелю, идут в главный
ствол и корни из боковых ветвей, которые функционируют как
ассимилирующие органы. Там они потребляются для вторич-
ного роста в толщину и для образования эмбриональных тканей
на верхушке Эти эмбриональные ткани живут за счет ассими-
лятов дерева и могугт быть уподоблены семенам, которые также
целиком или частично потребляют резервные вещества Можно
образно сказать, что эмбриональные точки роста дерева, подобно
созревающим семенам, являются в организме как бы местами
притяжения строительных веществ Гебель подчеркивает, что
точка роста главной оси отличается от точек роста боковых
ветвей большей величиной и большей физиологической актив-
ностью Отсюда и ее более интенсивный рост
Отношения, свойственные главной оси и верхним боковым
ветвям, свойственны также верхним и нижним боковым ветвям,
развивающимся в кроне одного и того же дерева Верхние
ветви, как более сильные, ускоряют отмирание нижних ветвей,
что особенно сильно проявляется в густых насаждениях В ниж-
них затененных ветвях образование ассимилятов идет более
медленно, чем отток пластических веществ, поэтому рост их
прекращается и они отмирают. С другой стороны, изобилие
питательных веществ приводит к образованию на верхушке
сильных вертикально растущих радиальных побегов Об этом
свидетельствует, по Гебелю, образование радиальных побегов
вместо боковых дорзовентральных у пихты при поселении гриба
Melatnpsorella caryophyllacearum, вызывающего приток пласти-
ческих веществ.
Одним из важнейших доказательств трофической природы
явления корреляции в росте главной оси и боковых побегов
у ели Гебель считает поведение укореняющихся побегов ели.
Если нижние ветви кроны ели, например, на болотах или в
В-413 И Г Серебряков—15 225
замоховелых лесах, укореняются, то верхушки этих ветвей при-
поднимаются и начинают интенсивный рост с образованием
самостоятельных стволов, несмотря на наличие связи с главной
осью материнского дерева и продолжающийся приток «внут-
ренней секреции» от главной оси Укоренение боковых побегов
делает их менее зависимыми от главной оси
Поведение укоренившихся боковых побегов, по Гебелю,,
является убедительным аргументом против принятия гипотезы
Эррера, так как связь этих побегов с главной осью не поры-
вается и передача секреторного торможения продолжается
Гебель высказывает мысль, что корреляции в росте главной оси
и боковых ветвей определяются, быть может, в большей мере
не просто количественным распределением питательных ве-
ществ в тех и других побегах, а различием в содержании ве-
ществ ферментативного порядка в главной и боковых осях,
веществ, которые способствуют более быстрому усвоению пита-
тельных материалов, имеющихся в этих побегах и идущих на
построение новых эмбриональных тканей. Однако эта мысль
Гебеля о роли и распределении ферментативных веществ в по-
бегах древесных растений в дальнейшем не была эксперимен-
тально подтверждена и разработана.
Воздействие («индукция») главной оси па рост и формооб-
разование боковых ветвей сохраняется различное по длитель-
ности время у разных видов деревьев. Это обстоятельство
послужило основанием для выделения двух типов «индукции»,
исходящих от главной оси растения (К. Гебель, 1908). Первый
тип Гебель назвал «лабильной индукцией», проявляющейся,
например, у ели (Picea excelsa). Боковые ветви кроны у расте-
ний этого типа испытывают воздействие главной оси только
при наличии и жизнедеятельности ее верхушки в кроне. При
повреждении верхушки или удалении главной оси ее воздей-
ствие прекращается, верхушки боковых ветвей приподнимают-
ся, и боковые ветви начинают приобретать физиологические и
морфологические особенности, свойственные главной оси.
У других деревьев (например, Araucaria excelsa) воздействие
главной оси на рост и формообразование боковых ветвей сохра-
няется и после удаления главной оси, и даже при росте боко-
вых ветвей при их укоренении Этот тип воздействия главной
оси на боковые К. Гебель назвал «стабильной индукцией».
Вопросы корреляции роста и морфогенеза главного и боко-
вых побегов у растений привлекали внимание многих исследо-
вателей и породили значительную специальную литературу.
Обширное исследование корреляции роста между почками
и побегами главной оси и боковых ветвей проведено было
W. Mogk (1914) В. Мок проводил свою работу как продолже-
ние исследования своего учителя Пфеффера (Pfeffer, 1893,
226
1904). Как показал Pfeffer (1893) на проростках Vicia faba,
механическое торможение роста главного побега коррелятивно
действует на зависящие от него боковые побеги таким же обра-
зом, как и его удаление: нормально покоящиеся пазушные
почки при этом развертываются и растут, как если бы главный
побег был обрезан. Опыты проводились на ветвях хвойных де-
ревьев (виды Picea, Torreya), лиственных деревьев (Tilia, Salix,
fragilis), а также па проростках бобовых, отличающихся моно-
подиальным ростом и четко выраженными корреляциями в
росте главного побега и боковых пазушных побегов. Для тор-
можения роста Мок применял гипсовую повязку, которая, по
Моку, имеет ряд преимуществ перед другими методами механи-
ческого торможения роста. У деревьев Мок загипсовывал почки
в кроне ранней весной до начала их набухания. Для изучения
корреляций в росте побегов в пределах одной ветви эта ветвь
отрезалась от кроны материнского дерева.
Мок выявил различные типы корреляции в росте материн-
ских и боковых побегов, вегетативных и генеративных почек
у разных видов растений Чтобы выяснить влияние ассимиля-
ционной деятельности материнского и боковых побегов на
взаимные коррелятивные отношения в их росте, Мок видоизме-
нял условия проведения опытов Часть опытов он проводил
в темноте, в другой части опытов растения вначале были в тем-
ноте, затем выставлялись па свет; один из вариантов опыта
полностью проводился на свету; в двух вариантах растения
вначале были на свету, а затем переносились в темноту.
В результате проведенных опытов автор приходит к выводу, что
различия в ассимиляционной деятельности не меняют сущест-
венно характера коррелятивных отношений в росте главного
и боковых побегов
В своих опытах Мок видоизменял также условия транспи-
рации подопытных побегов Все эти опыты привели Мока
к принципиально важному выводу, что коррелятивные отноше-
ния в росте побегов в пределах одного растения не зависят пи
от ассимилирующей деятельности побега, ни от их транспира-
ции. Эти выводы Мока находятся в резком противоречии с об-
щей концепцией К. Гебеля (1908) о трофических отношениях
как основе коррелятивных отношений роста побегов.
Еще более резкой критике трофическая концепция корреля-
тивного торможения роста подвергалась в работах Me Callum,
Child and Bellamy, R Snow (1925). R. Snow (1925) говорит
о торможении аксилляриев верхушкой. Относительно природы
этого торможения возможны три теоретических предположения:
1) возможно, что растущая верхушка каким-то образом
оттягивает запасы питательных веществ, или веществ, необхо-
TZ 227
димых для роста, и таким образом, оставляет аксиллярии уми-
рающими от голода и неспособными к росту,
2) торможение может быть вызвано каким-то тормозящим
веществом, возникающим в точке роста и спускающимся по
стеблю,
3) это может быть какой-то физиологический процесс неиз-
вестной природы, передающийся вниз по побегу
Первая из этих трех теорий — «перехват питания» — кажется
на первый взгляд наиболее простой и приемлемой, но с физико-
химической точки зрения она очень неясна. Крайне трудно
представить себе механизм, одна часть которого тянет себе
питательные вещества от других частей, истощая их. Согласно
Me Callum, нужно ожидать, что питательные вещества диффун-
дируют от мест большей концентрации к местам их меньшей
концентрации и этим автоматически выравнивают отдельные
местные недостатки в таких веществах Кроме того, экспери-
ментальные данные резко противоречат трофической теории
корреляций.
Из трех гипотез, высказанных выше, эти авторы считают
наиболее вероятной и отвечающей имеющимся эксперименталь-
ным данным гипотезу тормозящих веществ
Дальнейшая разработка проблемы коррелятивного тормо-
жения роста отдельных органов у растений привела к призна-
нию решающего значения в этих процессах ростовых ве-
ществ.
Участие и посредничество ростовых веществ в коррелятив-
ном торможении роста побегов в пределах одного растения было
открыто исследованиями Тимана и Скуга (Thiniann a. Skoog,
1934). Эти авторы вначале проводили обычные опыты с про-
ростками Vicia Faba\ удаляя верхушку главного побега, они
вызывали развертывание аксиллярных побегов, не разверты-
вающихся при наличии верхушки. Однако если вместо удален-
ной верхушки на место среза насадить агаровый кубик с росто-
выми веществами и сменять этот кубик свежим через каждые
6 часов, то у таких опытных растений пазушные почки не про-
растают, так же как у нормальных контрольных растений
с сохраняющейся верхушкой главного побега. Среди ростовых
веществ, используемых этими авторами, были гетероауксин
и ауксин В опытах эти авторы установили, что для торможения
пазушных побегов в верхушечной почке должно быть больше
ростовых веществ, чем их необходимо для роста побега в длину,
так как декапитированные стебли бобов содержат еще доста-
точное количество ростовых веществ, чтобы обеспечить на неко
торое время рост побега, но уже недостаточно для того, чтобы
подавлять пазушные почки Эти результаты были затем под-
тверждены разными исследователями.
228
Так, Мюллер обнаружил тормозящее влияние ростовых ве-
ществ на прорастание пазушных почек у растений нескольких
семейств Весьма обширные и тонкие опыты по изучению влия-
ния ростовых вещсс!в при корреляциях в росте пазушных почек
и кроющих листьев, в росте материнского и пазушных побегов
были проведены чешским ботаником Досталем (Dostal, 1915,
1936, 1937). Посредничество ростовых веществ в коррелятивном
торможении роста побегов делает понятным, почему сильное
тормозящее действие исходит от молодых сильно растущих по-
бегов Чем сильнее растет побег, тем больше он содержит росто
вых веществ, тем сильнее он тормозит рост других побегов,
и тем в меньшей мере тормозится его собственный рост Наобо-
рот, чем слабее ветвь, тем меньше она содержит ростовых
веществ, тем большее торможение испытывает она сама и тем
слабее исходящее от нее тормозящее влияние На эти соотно-
шения взаимозависимого роста и торможения оказывает влия-
ние свет, сильно освещенные почки оказываются наиболее мощ-
ными и тормозят или подавляют рост почек, находящихся
в тени Однако, как правильно подчеркивает Зедипг (1955),
недостаток света сам по себе не может быть причиной того, что
затененные почки больше не прорастают На отрезанных ветвях
почки развертываются и побеги растут, будучи полностью за-
крыты от света Это было показано в опытах Носта (lost)
(1894) па буке и в опытах Молиша (Н. Molisch, 1930) Но если
на дереве затемняются лишь отдельные ветви, то эти bcibh уже
не дают побегов Обьяснение влияния света по Зедингу (1955)
заключается в том, что сильнее всего освещенные почки стано-
вятся самыми сильными и, благодаря идущему от них корреля-
тивному торможению, все более и более подавляют прораста-
ние затененных почек Такая закономерность, вероятно, харак-
терна для всех древесных видов, хотя более подробные иссле-
дования по этому вопросу еще отсутствуют. Зединг полагает
(1955, стр. 233), что эту регуляцию роста в кроне деревьев
можно рассматривать как приспособление, чтобы новый прирост
всегда снова попадал па свет, а внутренняя часть крон не ста-
новилась слишком густой. Без этого управляемого светом тор-
можения не могла бы возникнуть гармоничная и биологически
приспособленная форма дерева Вследствие того, что потреб-
ность в свете у разных видов древесных растений различна, у
одних (теневыносливых) почки прорастают и при умеренном
освещении, и образуется густая крона (ель, бук и др.); у дру-
гих (светолюбивых) тормозящее действие периферических
сильно растущих побегов приводит скорее к подавлению почек
внутренней части кроны, и крона формируется рыхлой и скво-
зистой.
Понятно, конечно, что тормозящее влияние оказывает не
229
только главный верхушечный побег на боковые. Эти последние,
в свою очередь, тормозят развитие главного побега Это делает
понятным, почему у деревьев, выросших на открытых местах,
главный ствол в кроне скорее теряет свое ведущее положение
при мощном развитии боковых побегов, что приводит к образо-
ванию более широких и плосковершинных форхМ кроны. По мне-
нию Зединга (1955), изменение формы кроны у ели, в процессе
ее старения, переход от узкоконической и островершинной
кроны к плосковершинной кроне, в какой-то мере происходит
под влиянием длительно нарастающих постоянно суммирую-
щихся коррелятивных торможений со стороны боковых ветвей.
Согласно этим гипотетическим представлениям, коррелятивное
торможение принимает участие также и в процессах старения
деревьев Следует, однако, отметить, что сложные биологиче-
ские вопросы старения растений остаются еще недостаточно
разработанными.
Значение ростовых веществ для роста и морфогенеза от-
дельных побегов или их целых систем в кроне дерева вызвали
необходимость изучения распределения и динамики ростовых
веществ в различных частях кроны дерева, изучения их дина-
мики во времени и распределения в пространстве. Из исследова-
ний подобного рода обширностью материала и методическим
совершенством отличается работа W. Zimmermann (1936).
Этот автор исследовал содержание ростовых веществ в поч-
ках и побегах различных хвойных и лиственных деревьев, в
разных частях их кроны и в разные сроки вегетационного
периода. Среди исследованных видов деревьев были Picea
excelsa, Abies europaea, Pseudotsuga Douglasii; Carpinus betulus,
Fagus silvatica, Fraxinus excelsior, Acer pseudoplatanus, Tilia
tomentosa, Quercus sessiliflora, Aesculus Hippocastanum. Росто-
вые вещества извлекались из почек и побегов в агаровые
пластинки толщиной в 1 мм и площадью в 1 кв. см (или в
0,25 кв. см, если почки были слишком мелки). Агаровые пластин-
ки прикладывались к поверхности среза, сделанного у основания
почки, или к нижнему срезу взрослых побегов. Ростовые веще-
ства диффундировали при этом из почек и стеблей в агаровые
кубики. Большое внимание уделялось чистоте препарирования
почки и среза стебля, очищению от тканей, содержащих дубиль-
ные вещества и смолы, так как эти вещества инактивируют рос-
товые вещества и тормозят их выход из почек и побегов в агар.
Количество ростовых веществ, выделившихся в агаровые ку-
бики, определялось по общепринятой методике, так называемых
овсяных единиц.
Исследование покоящихся почек па содержание ростовых
веществ автор начал в середине апреля, когда в окрестностях
Дрездена (где проводились исследования) еще не замечалось
230
признаков развертывания почек. Несмотря на то, что на агаро-
вую пластинку накладывалось возможно большее количество
почек (10—15), тщательно освобожденных от чешуй, результат
во всех пробах был однозначен: до начала роста почек в опытах
наблюдались только ничтожные изгибы колеоптиля, что поз-
воляет сделать заключение об отсутствии в покоящихся почках
сколько-нибудь заметного содержания ростовых веществ.
В более поздние сроки количество ростовых веществ в поч-
ках тех же деревьев становится в сотни раз больше.
Для надежности автор повторил опыты следующей зимой
с почками хвойных деревьев.
Во всех исследованных почках этих деревьев до весеннего
развертывания не содержалось заметных количеств ростовых
веществ. Если далее сопоставить количество ростовых веществ,
обнаруженное в разное время у сравнимых почек, то получает-
ся, что во время развертывания почек и роста побегов содер-
жание ростовых веществ очень быстро повышается, чтобы через
короткое время медленно понизиться (табл. 6).
Таблица б
Содержание^ростовых веществ в развертывающихся почках
(по W. Zimmermann)
(Состояние почек: I — почка закрыта; II — почка вытягивается;
III — почка очень вытянута; IV — почка вполне развернута)
a) Fag us sllvatica (верхушечные почки)
Дата 15.IV 16.IV 18.IV 23.IV 2.V 3.V 7.V
Состояние почек I I II II III III IV
Содержание ро- стовых веществ 0,3 0,71 3,62 2,6 1,14 0,36 —
б) Quercus sessiliflora
(группы почек на концах ветвей)
Дата 4.V 28.V 1.VI 5 VI 12 VI
Состояние почек I I II Ш IV
Содержание ростовых веществ 0,36 0,45 0,52 2,16 0,54
(см. продолжение)’
231
в) Picea excelsa
(верхушечная почка)
Дата I 1 29.IV 1 21 .V 29.V(>-14 С.)
Состояние почек | I II 11
Содержание ростовых веществ . . 1 ’ 6,9 | 1 3,36
Из приведенной таблицы видно, что максимальное количе-
ство ростовых веществ в верхушках побегов наблюдается в мо-
мент весеннего развертывания почек.
Как показали определения Циммермана, в верхушечной
почке вновь образующегося годичного побега никогда не содер-
жится ростовых веществ в количестве, которое можно устано-
вить в материнской почке при се развертывании.
Наблюдениями Губера (В. Huber, 1925) установлено, что
побеги, срезанные с разной высоты дерева и поставленные
в воду, развертывают свои почки с разной быстротой, причем
верхние побеги развертывают листья раньше, чем нижние. Воз-
никло предположение, что эти различия обусловлены разным
содержанием ростовых веществ. Проведенные Циммерманом
определения показали, что побеги из верхней части кроны
всегда отличаются большим содержанием ростовых веществ по
сравнению с нижними побегами того же дерева
Большой интерес представляют исследования Циммермана
о содержании ростовых веществ в почках, развивающихся на
разной высоте одного и того же годичного побега. Исследования
были проведены на побегах различных деревьев, но особенно
удобными для исследований подобного рода оказались побеги,
обладающие большой концентрацией ростовых веществ Все
проведенные этим автором определения показали, что содержа-
ние ростовых веществ больше всего в верхушечной почке побега.
В боковых почках всех испытанных объектов содержание росто-
вых веществ от верхних боковых почек до нижних неуклонно
снижается, причем содержание ростовых веществ находится
в полном соответствии с величиной почки и величиной побега,
который из нее разовьется следующей весной Это соответствие
доходит до того, что архитектура побега до мелочей точно, по
Циммерману, отражается в содержаний ростовых веществ.
Особенно четкие закономерности в распределении ростовых
веществ в почках и побегах были установлены Циммерманом
у хвойных У ели, как и у пихты, боковые почки развертываются
раньше верхушечных; верхушечная почка развертывается
232
несколько позже, когда боковые побеги достигают уже замет-
ной величины. Это явление имеет большое биологическое значе-
ние как защита важного верхушечного побега от весенних позд-
них заморозков. В соответствии с этим содержание ростовых
веществ в верхушечной почке главной оси начинает увеличи-
ваться несколько позже, чем в почках боковых побегов. Но при
сравнимом состоянии побегов количество ростовых веществ
в верхушечных почках главной оси и боковых ветвей закономер-
но снижается от верхушки кроны к ее основанию.
Столь же закономерное снижение количества ростовых ве-
ществ наблюдается и в почках каждого годичного побега. В
связи с этим находится и мощность развития побегов в раз-
ных частях кроны и в пределах годичного побега, так называе-
мая ярусность в расположении ветвей, особенно отчетливо вы-
раженная у хвойных деревьев
Любопытно, что у кустарников и многоствольных деревьев,
представляющих переходную форму от деревьев к кустарни-
кам, эта закономерность не выражена.
Большой интерес представляет обширное эксперименталь-
ное исследование явлений корреляции роста в кроне хвойных
деревьев, проведенное Е. Мюнхом (Е. Munch, 1938).
Мюнх отмечает, что все организмы отличаются определен-
ной гармоничностью структуры У растений строгая соподчи-
ненность отдельных частей и, в частности побегов, наиболее
ясно выражена у деревьев. Гармоничность и строгая соподчи-
пенность у них проявляется в соотношении роста побегов в
длину и толщину, в соотношении роста главной оси и отдельных
боковых ветвей между собой и в направлении роста. Опыты
этого автора направлены к тому, чтобы выяснить процессы и
причины, которые приводят к таким корреляциям и гармонич-
ности строения дерева.
При изучении соотношений между ростом надземных орга-
нов в длину и толщину Мюнх исходит из современных пред-
ставлений, согласно которым в растущих почках и молодых
побегах, и особенно в молодых растущих листьях, образуются
ростовые вещества, которые делают возможным рост этих орга-
нов, а также идут к камбию ствола и вызывают там вторичный
прирост в толщину. Передвижение ростовых веществ идет по
лубу и почти исключительно в базипетальном направлении.
Это не представляет препятствий для передвижения ассимиля-
тов в любом направлении — вверх и вниз, смотря по тому, в
каком месте ростовые вещества вызвали рост. Исходя из этого
представления, автор и ставил свои опыты.
Значительный интерес представляют опыты этого автора по
изучению соотношения роста надземных органов дерева в дли-
ну и толщину.
233
Первый опыт. У веймутовой сосны (Pimis strobus) удаля-
лись все почки на опытных побегах При удалении почек рост
ветвей и ствола в толщину, по сравнению с контролем, не
уменьшился, ио наблюдалось смещение роста в толщину в ниж-
нюю часть ветвей и ствола
Второй опыт. Обрезались все почки, кроме одной в верхней
мутовке Из этой единственной почки развивался побег, заме-
щавший верхушку Прирост в толщину па верхнем отрезке
стебля резко уменьшался, а ниже побега, замещавшего вер-
хушку, становился значительно сильнее нормального. Расту-
щая верхушка главной оси или замещающая ее ось стимули-
рует рост в толщину верхних частей ствола и препятствует па-
дению прироста вниз по стволу, которое наступает при удале-
нии верхушки.
Третий опыт — главный и комплексный, очень сложный В
нем изучались ход роста в толщину во времени, роль запасных
веществ, влияние корней па распределение роста, влияние ветв-
ления на рост в толщину и т. д Особое внимание уделялось
изучению распределения прироста по длине ствола Опыт про-
водился на десятилетних деревьях веймутовой сосны В опыте
были следующие варианты, контрольные растения, растения с
удаленной хвоей и почками на главной оси, растения с удален-
ными почками, но оставленной хвоей, растения с окольцован-
ным у основания стволом в целях выявления роли корней,
растения с удаленными почками и нижними ветвями, растения
с удаленными нижними ветвями, растения с затененной верху-
шечной почкой, растения с затененной верхушечной почкой при
освещении боковых, растения с окольцованной и затененной
верхушкой; растения, окольцованные на охвоенной и неохвоен-
иой части одного и того же ствола
У всех опытных деревьев, а также контрольных, прирост
ствола в толщину измерялся в линейных единицах и в квадрат-
ных (площадь) в разных частях ствола.
У контрольных деревьев линейный прирост ствола шел бо-
лее или менее одинаково и вверху, и в середине, и внизу ство-
ла Лишь в средней части ствола (по длине) прирост был
чуть-чуть больше Колебания между отдельными деревьями
невелики.
Опыт с удалением хвои и почек у деревьев был сделан для
того, чтобы испытать, как будет идти рост в толщину только
за счет запасных веществ Опытные деревья жили все лето, ио
прирост был крайне ничтожен. В конце опыта наблюдалось
отмирание В следующем опыте хвоя была удалена, но почки
оставлены Почки без хвои развивались очень медленно, давая
очень короткие щеткообразные побеги Хвоинки на этих побе-
гах росли долго, но медленно Прирост в толщину был не
234
намного больше, чем в предыдущем Новых хвоинок и запас»
пых веществ оказалось недостаточно, чтобы обеспечить нор-
мальную деятельность камбиальных клеток При удалении по-
чек, но с сохранением старой хвои прирост в толщину почти не
уменьшался по сравнению с контролем Но этот прирост шел
совсем иначе, чем \ неповрежденных контрольных деревьев.
Наиболее сильный и резко отчетливый прирост был в нижней
части ствола. Особенно резко при этом возрастал объем ствола
в его нижней части. Таким образом, удаление почек на вер-
хушке кроны у главной оси перемещает прирост в нижние части
ствола и приводит здесь к увеличению объема выше нормаль-
ного Два дерева в опыте с удалением почек в верхней части
кроны были окольцованы в основании ствола, чтобы узпать,
не от корня ли идет усиленное утолщение ствола в его основа-
нии при отсутствии почек наверху Однако кольцевание ствола
у его основания не повлияло отрицательно на его утолщение в
нижней части Значит, это усиленное утолщение ствола идет не
от корня, и во всяком случае идет не по коре
Чтобы испытать, не от большого ли количества боковых вет-
вей внизу зависит сильный прирост ствола при основании, были
удалены кроме почек на верхушке кроны и нижние боковые
ветви Оказалось, что и в этом случае наибольший прирост в
толщину был сдвинут к основанию ствола, хотя и не столь
сильно, как при наличии нижних ветвей Если же при удалении
нижних ветвей почки на верхушке кроны были оставлены, то
при этом наибольший прирост в толщину смещался по стволу
кверху Следовательно, распределение роста в толщину по
длине ствола определяется, главным образом, наличием или
отсутствием растущих побегов и почек в верхней части кроны,
и особенно на верхушке растущего ствола Чем интенсив-
нее верхушечный рост и чем дольше он про-
должается в жизни дерева, тем равномернее
распределяется прирост в толщину по всей
длине ствола от его основания до верхушки.
Этот вывод, вытекающий из опытов Мюнха, представляет
большой экологический интерес, так как объясняет колонновид-
ные формы стволов у деревьев влажно-тропических лесов
Наиболее ярко выраженной цилиндрической формой ствола
отличаются, как известно, тропические и субтропические розе-
точные деревья, для которых характерно наличие моноподиаль-
ного верхушечного нарастания.
Автор наблюдал также распределение прироста, или ход
камбиальной деятельности во времени, в течение вегетацион-
ного периода. У контрольных деревьев прирост в толщину идет
главным образом в мае, июне и июле, в августе он ничтожен,
а в сентябре прекращается одинаково во всех частях ствола.
233
У деревьев с удаленными почками в верхней части кроны при-
рост ствола на верхушке и в средней части его кончается много
раньше, и лишь в нижней части протекает так же, как у кон-
трольных деревьев.
Опыты Мюнха показывают, что развертывающиеся почки и
растущие побеги стимулируют деятельность камбия в стеб-
левых органах, расположенных ниже этих почек и побегов.
Однако стимулирующая деятельность растущих побегов рас-
пространяется лишь на стебли, соединяющие эти побеги с
главным корнем растения Все боковые по отношению к дан-
ному побегу стебли и почки тормозятся. Стимулирование кам-
биальной деятельности в системе стеблей, соединяющих расту-
щий побег с главным корнем, находит яркое подтверждение у
молодых деревьев дуба черешчатого и березы при формирова-
нии у них ведущих лидерных побегов. Торможение боковых
побегов сопровождается одновременно усилением их «эпипа-
стизма» (Мюнх), который приводит к их отклонению в сторону,
плагиотропному росту и дорзовентральности. Как показывают
опыты с кольцеванием главного и боковых стеблей, стимули-
рующее и тормозящее влияние растущих побегов распростра-
няется по коре в базипетальном направлении Наиболее сильно
эти влияния проявляются непосредственно под развертываю-
щимися почками и побегами и ослабевают по мере удаления ог
них
Если участие ростовых веществ в системе коррелятивных
явлений роста и торможения в кроне деревьев не вызываем
сомнения, так как базируется на многочисленных точно постав-
ленных экспериментах, то механизм действия ростовых ве-
ществ и, в частности, механизм коррелятивного торможения,
остаются недостаточно выясненными. Зедипг (1955) указывает,
что нельзя считать установленным, будто при коррелятивных
торможениях дело идет всегда об одних и тех же физиологи-
ческих процессах Форма проявления торможения меняется от
едва заметной задержки роста до отмирания органов и, следо-
вательно, в конечном счете, речь идет здесь о разных явлениях.
Нарушение роста может происходить, очевидно, вследствие
самых разнообразных вмешательств в цепь ростовых процес-
сов. Поэтому весьма возможно, что совершенно разные гипоте-
зы правомерны как для объяснения различных случаев, так и
для объяснения общего конечного эффекта у одного и того же
растения. Из числа гипотез, приводимых Зедингом, наиболее
широкий круг явлений, изученных экспериментально, объясня-
ет гипотеза тормозящих веществ, разработанная рядом иссле-
дователей (R. Snow, 1937, 1940; Laibach, 1933; Muller, 1935 и
др.). По данным этих исследователей в растениях, под влия-
нием ростовых веществ, возникают тормозящие рост вещества,
236
распространяющиеся по всему растению R. Snow (1940) ука-
зывает, что в растениях неоднократно обнаруживались тормо-
зящие рост вещества Некоторые из этих веществ, например,
антиауксин, обладают способностью к передвижению по расте-
нию в обоих морфологических направлениях, в отличие от сти-
мулирующих рост веществ, передвигающихся только в базипе-
тальном направлении. По этой гипотезе в растущей верхушеч-
ной почке образуются не только стимулирующие, но и тормо-
зящие рост вещества, которые по мере образования передви-
гаются из почки вниз по стеблю, вызывая разрастание стебля
в толщину. Но вследствие того, что ростовые вещества способ-
ны передвигаться только в базипетальном направлении, они не
могут подняться акропетально из главного побега в верхушки
боковых побегов, тогда как тормозящие вещества свободно
проникают к верхушкам боковых побегов, подавляя их рост
Если боковые побеги мощные и обладают достаточным коли
чеством ростовых веществ, то действие тормозящих веществ,
пришедших из верхушки главного побега, нейтрализуется, или
проявляется слабо. Если же боковой побег слабый и содержи';
мало ростовых веществ, их оказывается недостаточно для ней-
трализации подавляющего действия тормозящих веществ, при-
ходящих из верхушки главного побега, и боковой побег обна-
руживает резкое торможение роста вплоть до полного не-
раскрытия почки.
Эта гипотеза хорошо объясняет, почему передвижение
ростовых веществ из верхушечной почки в боковые тормозит
их рост, когда, казалось бы, эти вещества должны усиливать
рост боковых побегов, так как эти побеги содержат ростовых
веществ меньше, чем верхушки главной оси (см. выше, данные
I (иммермана).
Таким образом, ростовые вещества, включая стимуляторы и
ингибиторы роста, вследствие их чрезвычайно большой физио-
логической активности и своеобразию передвижения по живым
тканям листовых и осевых органов, оказываются не только
посредниками между отдельными побегами, но и регуляторами
их роста, определяющими соотношение роста стеблей в длину
и толщину, различную интенсивность роста отдельных побегов,
вызывая этим самым бросающуюся в глаза соподчинепиость
побегов и гармоничное строение кроны дерева Деятельность
этих веществ, образующихся в процессе жизнедеятельности
растения, обусловливает физиологическое единство всех побе-
юв в кроне дерева, биологическую целостность дерева как
единого организма. Старение и отмирание одних побегов,
сопровождающееся возникновением и развитием других, их
смена во времени выражают собой единый процесс развития
организма древесного растения в целом.
237
6. БИОЛОГО-МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ФОРМИРОВАНИЯ
НАЗЕМНЫХ КРОНООБРАЗУЮЩИХ ПРЯМОСТОЯЧИХ ДЕРЕВЬЕВ
Приведенный выше обзор морфогенеза дерева у разных
видов и физиологический анализ формирования кроны позво-
ляют выделить основные биолого-морфологические особенности,
лежащие в основе формирования типичного кронообразующего
прямостоячего дерева из секции лесных деревьев.
В. Рау (1937) считает, что главным свойством деревьев яв-
ляется акротонность ветвления побегов, делающая возможным
само формирование дерева. Акротонность ветвления побегов
действительно является одним из важнейших свойств деревьев
Но этот тип ветвления побегов свойствен не только деревьям,
но также кустарникам, кустарничкам и даже стелющимся и
подушковидным формам (см главы IV, V, VI, VIII) н, следо-
вательно, одной акротонности ветвления недостаточно для фор-
мирования растений по типу дерева
Более правильным будет считать, что основой формирова-
ния дерева является процесс, тесно связанный с акротонностью
ветвления побегов, а именно дифференциация расту-
щих побегов с обязательным выделением ли-
дерного побега, наиболее интенсивно растущего и при-
нимающего вертикальное направление. Так формируется ствол
дерева. Более слабые побеги с менее интенсивным ростом и
наклонные становятся боковыми ветвями, формирующими
крону.
Весьма характерно для лесного дерева то, что дифферен-
циация растущих побегов с выделением лидерной оси сохраня-
ется в течение большей части его онтогенеза, утрачиваясь лишь
в старческом возрасте, в связи с резким ослаблением роста У
лесных деревьев биологически главная ось восстанавливается
даже у старых деревьев после глубокой обрезки (рис 44)
Этот процесс характеризует секцию лесных деревьев, у которой
черты дерева выражены наиболее сильно
Возрастная . периодизация в онтогенезе дерева, предложен-
ная выше для ели, сосны, березы, дуба, пожалуй, может счи-
таться применимой ко всей секции лесных деревьев. Возраст-
ные периоды, выделенные у описанных деревьев, неразрывно
связаны с особенностями роста главного ствола и формирова-
ния кроны.
Основная закономерность роста главного ствола деревьев,
укладывающаяся в одновершинную кривую приростов, была
выявлена многочисленными исследованиями лесоведов и ден-
дрологов (см. Г Ф. Морозов, 1923; В. Н. Сукачев, 1948;
М Е. Ткаченко, 1952 и др.), а также физиологов растений (см.
Л А Иванов, 1931). Эта кривая в свою очередь выражает
238
Рис. 44
Восстановление кроны и вторичного лидерного побега липы
при глубокой обрезке
более общую физиологическую закономерность, известную под
названием «большого периода роста».
Выделяемые нами четыре возрастных периода в онтогенезе
лесного дерева, как видно из рис. 45, закономерно совмещают-
ся с «большой кривой роста» ствола и соответствуют совершен-
но определенным ее участкам. В частности, 3-й возрастной пе-
риод-формирование узкокронного островершинного дере-
ва— совпадает с периодом наиболее резкого подъема величин
приростов, а 4-й период — формирование плоской или куполо-
образной кроны — с нисходящей частью кривой.
Соотношение возрастных периодов и интенсивности роста
главной оси в онтогенезе дерева у ели (Picea excelsa),
I. II, III, IV — возрастные периоды
239
Зависимость формы кроны у разных видов деревьев от воз-
раста и условий произрастания, в том числе и в первую оче-
редь — ценотических, была давно известна в лесоведении и
лесоводстве (Г Ф Морозов, 1923; В Г. Нестеров, 1949). Как
явствует из всего вышесказанного, условия произрастания
влияют па общий облик дерева именно через основную законо-
мерность роста дерева — дифференциацию растущих побегов с
выделением из них лидерного Эта дифференциация наблюда
ется у деревьев, например, у дуба и видов березы, только при
достаточно интенсивном росте их побегов, что зависит от воз-
раста и условий среды В начале онтогенеза, при слабом при-
росте побегов, у дуба и березы образуется не форма дерева, а
или кустовидная форма, или — у дуба — еще более далекая от
дерева форма торчка, несущая некоторые черты структуры
подушковидных растений
Таким образом, дифференциация растущих побегов с выде-
лением лидерной оси непосредственно связана с большой
интенсивностью роста побегов, и в то же время сама приводит
к максимально быстрому росту в высоту и в стороны, обеспе-
чивает захват максимального воздушного пространства.
Главная ось дерева служит не только механической основой
всех его надземных органов и не только органом, обеспечиваю-
щим >наилучшее расположение ассимилирующих органов, но
регулирующим рост боковых ветвей, она определяет соподчи-
непность в росте побегов, образующих крону, и вследствие
этого обусловливает формирование кроны дерева как единого
гармонического целого
Дерево в биологическом отношении оказывается жизненной
формой растений, более всех других приспособленной к тому,
чтобы занимать максимальное жизненное пространство и
верхние ярусы фитоценозов. Исторически эта форма сложилась
в климатических условиях, благоприятствующих длительному
росту растений, и в ценотических условиях, вызывающих необ-
ходимость роста органов ассимиляции вверх Как писал
Ирмшер (Irmscher, 1922), «дерево —это идеальнейшее вопло-
щение беспрепятственного хода жизненных процессов», на ко-
торое, впрочем, резкая смена влажного и сухого, теплого и хо-
лодного времени года, безусловно наложила свой отпечаток
Другие жизненные формы, от кустарников до однолетних трав
по Ирмшеру, суть выражение реакции на те или иные неблаго-
приятные для роста условия.
Формой, противоположной в морфологическом и экологи-
ческом плане жизненной форме дерева, является подушковид
ная форма растений (см. гл. VIII), с ничтожным приростом
побегов, с отсутствием дифференциации побегов по интенсив-
ности их роста, прижатая к субстрату и имеющая ровную
240
поверхность В отличие от дерева, эта форма выработалась в
условиях, крайне неблагоприятных для роста из-за низких
температур, сухости воздуха и иссушающего ветра, и пол-
ностью этим условиям соответствует
7. МНОГООБРАЗИЕ И РАСПРОСТРАНЕНИЕ ДРЕВОВИДНЫХ
ЖИЗНЕННЫХ ФОРМ
Описанные в этой главе формы деревьев иллюстрируют
лишь небольшую часть древовидных жизненных форм покрыто
семенных и хвойных.
Разнообразие форм деревьев чрезвычайно велико. Попытка
классификации их была дана в главе II. Эта классификация в>
крупных единицах близка, как упоминалось, к классификации
деревьев Дю Рие (Du Rietz, 1931), так как нам кажется пра-
вильным основной принцип классификации деревьев по харак
теру побегов
В пределах типа деревьев выделено 3 класса по характеру
роста побегов: 1) кронообразующие деревья с полностью одре-
весневшими удлиненными несуккулентными побегами, 2) розе
точные деревья и 3) суккулентностеблевые безлистные деревья
Характер побегов у разных форм деревьев не только хорошо
отражает экологические и морфологические особенности раз
ных классов древовидных растений, но, вероятно, может слу-
жить в ряде случаев показателем уровня их филогенетического
развития Как уже указывалось, по Корнеру (Corner, 1949).
розеточный характер побегов деревьев в сочетании с толстосте-
белыюстью и слабым одревеснением стеблей можно считать
более древним признаком по сравнению с «нормальной струк-
турой» вполне одревесневшего удлиненного побега
Наиболее обширный класс — несуккулептных кронообра-
зующих «обычных» деревьев.
В пределах этого класса наблюдается крайнее разнообразие
групп, подгрупп и секций жизненных форм, что безусловно
связано с большой экологической амплитудой и их широким
географическим распространением
Если подкласс гемиэпифитпых деревьев, группа деревьев с
корнями-подпорками разных типов, подгруппа деревьев с лиа-
новидным стволом иллюстрируют разнообразие приспособи-
тельных черт, главным образом, деревьев влажно-тропических
лесов, где находится максимум видового разнообразия деревь
ев вообще и, возможно, центр возникновения всего типа жиз-
ненных форм деревьев, то секции в пределах подгруппы прямо-
стоячих «обычных» деревьев показывают приспособительные
особенности, пути изменения роста и ветвления деревьев при
В-Ш И 1 Серебряков-16 241
освоении ими областей земного шара, расположенных за пре-
делами влажных тропиков и субтропиков.
По существу только жизненная форма лесного дерева, осо-
бенности формирования которой были подчеркнуты в предыду-
щем разделе, развивающаяся в различнейших лесных областях,
ют экваториальной до таежной, сохраняет во всех своих ва-
риантах характерные черты дерева как жизненной формы,
«воплощающей в себе условия беспрепятственного роста»
(Irmscher). Во всех этих лесных областях, так или иначе,
несмотря на разницу климатов, условия увлажнения оказыва-
ются достаточно благоприятными Листопадные формы здесь
представляют реакцию на периодический физический или фи-
зиологический недостаток влаги. Все остальные секции — де-
ревьев лесостепных и саванновых типа «плодовых» деревьев,
саванновых с водозапасающими стволами, деревьев полу
пустынных и пустынных с сезонно-суккулентными веточками,
наконец, многоствольных или кустовидных деревьев северных
и верхних границ леса, — представляют собой дальнейшее
своеобразное выражение реакции на неблагоприятные условия,
впрочем еще оставляющего возможность для древовидного
роста растений
Как видно из перечисления секций, ухудшение условий про-
израстания вызывает весьма различные приспособления, отра-
жающиеся в габитусе растений.
Один путь приспособления — запасание влаги. Здесь в ка-
честве приспособлений выступают или утолщение стволов с раз-
витием в них водозапасающей ткани, или полная безлистность
и суккулентность специализированных, опадающих па зиму
ассимилирующих побегов. Крайним выражением перехода к
суккулентности служат уже деревья особого класса — сукку-
лентно-стеблевые, теряющие все характерные черты «обычного»
дерева, кроме основных: наличия долговечного главного ствола
и крупных размеров
С другой стороны, потеря именно этих черт — крупных раз-
меров и постоянного наличия долговечного главного ствола —
приводит от жизненной формы дерева к жизненной форме кус-
тарника. Это другой путь приспособления к неблагоприятным
условиям роста — сокращение длительности роста главного
ствола и скелетных ветвей, коррелятивно связанное с более
обильным ветвлением побегов, вплоть до пх подземной части
(геоксильные или настоящие кустарники).
Глава IV
ЖИЗНЕННЫЕ ФОРМЫ КУСТАРНИКОВ
1. ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ КУСТАРНИКОВ
Кустарники обладают несколько большей, чем деревья, эко-
логической амплитудой и отличаются более широким, сравни-
тельно с деревьями, географическим распространением. Ареал
кустарников перекрывает полностью ареал деревьев, выходя за
пределы последнего у верхней границы распространения лесов
в горах тропической и умеренных зон, у полярных пределов
распространения деревьев, и отчасти у континентальной гра-
ницы лесов, в тропиках и субтропиках, где ксерофитные колю-
чие леса при усилении сухости климата переходят в заросли
колючих кустарников. Области наибольшего видового и мор-
фологического разнообразия форм деревьев и кустарников не
«совпадают. Для деревьев наиболее благоприятные условия во
влажно-тропическом и субтропическом климате, где они дости-
гают максимального разнообразия форм (особенно во влажно-
тропических лесах). Господство кустарников в растительном
покрове и их наибольшее многообразие в пределах тропической
и умеренных зон наблюдается в областях с аридным климатом,
а в холодно-умеренных зонах — у полярных пределов лесов и
также близ континентальной границы лесной растительности,
т е. в климатических условиях, неблагоприятных для развития
высокоствольных деревьев.
В пределах тропической зоны кустарники встречаются в
различных типах растительности, от влажно-тропических лесов
до пустынь. В дождевых тропических лесах кустарники отмече-
ны во всех областях распространения этого типа раститель-
ности. Так, во влажно-тропических лесах Западной Явы в
16? 243
форме кустарников или кустовидных деревьев (Baum-Straucher
по S. Н Koorders, 1914) произрастают некоторые виды Сот-
melina, Mussaenda, Sauravla (Dillemaceae) виды Boehme-
iia, Laportea (Urticaceae), некоторые виды Piper, виды Phyllant-
hus, Ostodes, Homalanthus и др. Кустарникам влажно-тропиче-
ских лесов свойственна значительная высота (до 6—8 м и бо-
лее), слабое ветвление побегов и развитие крупных листьев,
обычно гигрофильной структуры. Многие из них, вероятно,
относятся к группе аэроксильных кустарников, без ветвящихся
долголетних стеблей под землей
Во влажно-тропических лесах Бирмы к прямостоячим на
земным кустарникам относятся некоторые виды Alsodeja
(Violaceae), виды Mussaenda, Morinda, Ixora (Rubiaceae), виды
Boehmeria из Urticaceae, Clerodendron из Verbenaceae, Unona
из Апопасеае, Capparis из Capparidaceae, виды Maesa и Ardisia
из Myrsinaceae, виды Diospyros, Cannarus и др (Scliinipei u.
Faber, 1935).
Во влажно-тропических лесах провинции Юньнань Ан. А. Фе-
доров (1958) среди кустарников отмечает Antidesma, Briedelia,
Croton (Euphorbiaceae), Ardisia, Embetia, Maesa (Myrtaceae)
Psychotria, Uncaria (Rubiaceae), Rauwolfia (Apocynaceae).
В дождевых лесах Африки кустарники и кустовидные де-
ревья образуют иногда густой подлесок, состоящий из предста-
вителей Acanthaceae, Mimosaceae, Commelinaceae, Maranthaceae,
Labiatae, Convolvulaceae, Melastomataceae.
Во влажно-тропических лесах Меланезии подлесок состоит
из многих видов Pittosporum, Psyochordia, Dracaena, Наппе-
mannia и т. д.
Однако фитоценотически кустарники не играют сколько-
нибудь значительной роли в сложении влажно-тропических ле-
сов в ненарушенном состоянии При нарушении лесов вырубка-
ми или пожарами кустарники могут разрастаться па значи-
тельных территориях, образуя вторичные труднопроходимые за-
росли. Многие • виды кустарников влажно-тропических лесов
относятся к формам, переходным от деревьев к кустарникам
(Baum-Straucher — S Н Koorders, 1914, A. Schimper и F Faber,
1935).
Изучение распространения кустарников выявляет относи
тельную флористическую бедность кустарников влажно-тропи-
ческих лесов. Так, в детальном флористическом списке древес-
ных растений влажно-тропических горных лесов Западной Явы
(S. Н. Koorders, 1914) отмечено 35—40 видов кустарников из
15 семейств и 140 видов деревьев из 45 семейств. Две трети
всех семейств и родов древесных цветковых растений этого ти-
пичного влажно-тропического леса представлены только разно
244
образными формами деревьев и совершенно лишены кустар-
ников.
Кустарники произрастают и в различных растительных фор-
мациях тропических областей с сухим периодом года. Так, в
муссонных тиковых лесах Восточной Явы, между высокостволь-
ными деревьями обильны многочисленные виды низких деревьев
и кустарников
Роль кустарников в сложении ценозов значительно возра-
стает при усилении аридности климата тропических областей,
удлинении засушливого времени года, в саванновых и колючих
лесах Колючие леса в тропической Африке Энглер (A. Engler,
1895) называет формацией «колючих кустарников», или
«кустарниковыми лесами» (Buschwalder), из-за господства в
этих формациях ксерофитных кустарников. Энглер отмечает
систематическое сходство «кустарниковых лесов» тропической
Африки с подобными формациями Индии и физиономи-
ческое сходство «кустарниковых лесов» тропической Афри-
ки, Центральной и Южной Америки. В систематическом
отношении для формации «колючих кустарников» характерно
постоянное наличие разнообразных видов Acacia, а также дру-
гих родов Leguminosae с дважды перистыми листьями, напри-
мер, рода Albizzia и др. Изредка доминируют древесные с
просто перистыми листьями, например Adina из Anacardiaceae,
Harrisonia из Sitnarubaceae, некоторые Rutaceae, Burseraceae,
Bignoniaceae и др
Большинство кустарников и деревьев этих формаций воору-
жены колючками, что послужило поводом для названия их
«колючими лесами», или «колючими кустарниками»
Кустарники колючих и саванновых лесов, как и произра-
стающие здесь деревья, обладают резко выраженными чертами
ксероморфизма В колючих и саванновых лесах появляется
форма кустарников с подземным ветвлением побегов, так на-
зываемые геоксильные кустарники. Их подземные побеги в
сравнении с надземными развиты сильнее, имеют большую тол-
щину и более долговечны. Некоторые из таких кустарников,
например, Andira laurifolia и Anacardiutn humile из кампосов
Бразилии получили название «подземных деревьев» (A. Schim-
рег, F Faber, 1935) У Andira, например, ветвящаяся система
подземных осей толщиной в руку занимает площадь до 10 м в
диаметре, в то время как надземные листоносные побеги тонкие
и не превышают одного метра в высоту. Толстые подземные
оси служат водозапасающими органами, подобно клубневид-
ным одревесневающим ризомам других мелких кустарников и
полукустарников, произрастающих в кампосах.
Господство кустарников в таких ксерофитных древесных
формациях проявляется не только фитоценотически, но и
245
флористически Так, в кампосах Бразилии с резко разграниченной
лесной и безлесной травянистой ксерофитной растительностью
Е. Варминг (Е. Warming, 1892) отметил 170—180 видов кус-
тарников и всего лишь 70—80 видов деревьев Среди кустарни-
ков особенно обильно представлены Myrtaceae и Malpighiaceae,
за которыми следуют Melastomataceae и Composiiae. Резкое
преобладание кустарников по сравнению с деревьями во флоре
кампосов Бразилии особенно наглядно, если эти данные срав-
нить с соотношением числа видов деревьев и кустарников во
флоре влажно-тропических лесов Западной Явы, приводимым
Кордерсом (S. Н. Koorders, 1914).
Обзор растительных формаций тропической зоны обнаружи-
вает, что роль кустарников в растительном покрове возрастает
по мере усиления аридности климата, удлинения засушливого
времени года Наряду с усилением фитоценотической роли
кустарников в аридных областях тропиков наблюдается уве-
личение их видового многообразия, появляются и оказываются
наиболее распространенными «настоящие кустарники», с под-
земным ветвлением побегов, т. е. кустарники геоксильного типа
В областях тепло-умеренной зоны северного и южного по
лхшарий кустарники также встречаются в различных расти-
тельных формациях от дождевых субтропических и умеренных
лесов до формации жестколистной древесной растительности
Так, в тепло-умеренных влажных лесах Южной Японии, со-
стоящих из многих видов высокоствольных вечнозеленых де-
ревьев, по данным Миошп (Mioschi, 1906—1926), густой подле
сок образован Thea japonica, Eurya japonica, Pitiosporutn Tobira
i. некоторых других
Таким образом, во влажных субтропических лесах Японии
соотношение между количеством видов деревьев и кустарников
остается примерно таким же, как и во влажно-тропических ле-
сах (деревья резко преобладают).
Влажные леса южной части Чили на скалистых склонах по
бережья переходят в густые заросли кустарников из видов
Drymis, родов Myrtaceae, Lomatia ferruginea и др. В умеренных
и влажных лесах южных областей Чили, по данным К. Reiche
(1907), развит густой подлесок, иногда в виде труднопроходи-
мых зарослей кустарников. На вырубках разрастаются кустар-
ники Однако и в областях тепло-умеренных зон, подобно тро-
пической зоне, наибольшее значение в растительном покрове,
наибольшее флористическое и эколого-морфологическое разно-
образие кустарники получают в областях с сухим временем
2'16
года, и особенно с зимними осадками и сухим жарким летом.
Таким областям умеренно-теплых зон свойственны, как извест-
но, формации жестколистной древесной растительности, с пре-
обладанием склерофильных вечнозеленых кустарников Широко-
распространены формации жестколистной древесной раститель-
ности, например, в прибрежных странах и на островах Среди-
земного моря, в формациях маквиса, гариги, фриганы Изда-
лека маквис и гарига производят впечатление серо-зеленых
однообразных зарослей, монотонных на фоне сухих склонов
холмов и невысоких гор Болес близкое изучение обнаруживает
большее флористическое и морфологическое разнообразие
входящих в их состав растений (A Schiniper u. F Faber, 1935)
Маквис состоит из кустарников более высоких, чем гарига
Высота кустарниковых зарослей, состоящих из вечнозеленых
растений с листьями склероморфного эрикоидного типа колеб
лется от полутора до четырех метров Иногда здесь встреча-
ются низкорослые вечнозеленые деревья с уплощенно-округлы-
ми кронами, например, Quercus ilex, Ceratonia siliqua, Pinus.
halepensis В различных районах Средиземья, на разной высоте
над уровнем моря и на почвах разного состава преобладают
различные виды кустарников В маквисах Испании доминируют
виды ладанника (Cistiis), ьлавным образом, Cistus inonspelien-
sis, красивый вечнозеленый кустарник с белыми, красными или
золотистыми цветками, на острове Сардиния — Erica arboreu —
высокий кустарник 6 м высоты, иногда древовидный В более
восточных областях, например, на побережье Адриатического
моря, довольно обычен Juniperus oxycedrus, который здесь, как
правило, растет в форме кустарника, по иногда приобретает
форму дерева с пирамидальной кроной (L Adamovicz, 1929)
Довольно часто в состав маквиса входит дикая маслина —
Olea oleaster, которая в сухих маквисах образует форму кус-
тарника с колючими побегами. В культуре маслина (культур-
ный вид—Olea еигораеа) всегда образует форму дерева.
Из большого числа обычных для маквиса кустарников мо-
гут быть названы1 Myrtus italica, теплолюбивый кустарник, при-
уроченный к теплым прибрежным местообитаниям, Arbutus
unedo (земляничное дерево) — кустарник, внешне сходный с
камелией, Daphne gnidium, Phyllirea media с узкими серо-зеле-
ными листьями, Cneorum tncoccum, Erica verticillata — высотой
в 1,5—2 я, образующий иногда густые заросли Кроме перечис-
ленных видов, выделяются своими перистыми листьями
фисташки вечнозеленая Pistacia Lentiscus и листопадная Pista-
cia Terebinthus Характерен и своеобразен в маквисах испанский
дрок (Spartium junceum) — кустарник с гибкими прутьевидны-
ми зелеными стеблями и ярко-желтыми крупными цветками
Довольно обычна также вечнозеленая роза — Rosa sempervirens
247
Rhamnus Alaternus, Colutea arborescens, Smilax aspera, Lonicera
implexa, Lonicera etrusca и др.
Растительность, экологически и физиономически сходная с
только что описанными зарослями склерофильных вечнозеле-
ных кустарников Средиземья, им'еет широкое распространение
в юго-западной Капландии (R. Marloth, 1908, A. Schimpcr и.
F Faber, 1935) Климат этой части Капской области весьма
близок к средиземноморскому. Виды, слагающие заросли кус-
тарников капского маквиса весьма обычны. Rhus tomentosa с
желтовато-серыми с нижней стороны листьями, Rhus rosmarini-
folia, Rhus villosa, Philicia paniculata, Ph obtusifolia, Berrelia
abrotanoides с мелкими игольчатыми листьями, В intermedia,
Colleonema album с ветвями типа игольчатых можжевельников,
Diosma succulentum с мелкими мясистыми листьями, Agathosma
capitatum с гусго опушенными ветвями и ряд других своеобраз-
ных видов Весьма характерны в кустарниковых зарослях юго-
западной Капской области разнообразные виды Erica, например
Erica matntnosa, Е. concina, Е. baccans, Е cor if oil a, E mucosa,
E. pulviformis, E. rumentacea и др.
В отличие от других областей с жестколистной древесной
растительностью, в Австралии, в юго-западных и юго-восточных
областях более важную роль играют деревья, образуя нередко
сомкнутые жестколистные леса с господством различных видов
эвкалипта, например Eucalyptus marginata, Eucalyptus diversico-
lor и др, и участием некоторых древовидных Proteaceae
(A Schitnpet и F Faber, 1935). Но и в Австралии жестколист-
ные леса обладают сильно развитым, часто трудно проходимым
и разнообразным по видовому составу кустарниковым под-
леском В более сухих районах при изреживании древесного
поло! а образуются на обширных территориях густые заросли
кустарников, известные австралийские скрэбы Кустарники этой
формации экологически близки к кустарникам маквиса Среди-
земья и Капской области Они редко достигают высоты 2 м,
обычно не превышая одного метра
Среди них.наиболее часто встречаются красиво цветущие
Proteaceae, затем Myrtaceae и Sterculiaceae.
Как известно, вечнозеленые низкорослые склерофильные
кустарники, экологически сходные с только что рассмотренными
кустарниками Средиземья, Капской области и Австралии, свой-
ственны также калифорнийскому побережью Северной Америки.
Здесь на больших пространствах кустарники образуют трудно
проходимые чащи, состоящие из кустарников одного-трех метров
высоты и известные под названием чаппараль. Область распро-
странения чаппараля совпадает с климатической областью, где
летом (с апреля до конца сентября) выпадает не более 20%
юдового количества осадков. В центре распространения этой
248
формации летние осадки составляют лишь 10% от их общей
годовой суммы
Интересно, что одиночные склерофильные деревья, произ-
растающие в зарослях чаппараля, на более сухих местах обра-
зуют форму кустарника Это наблюдается, например, у Quercus
agrifolia, Q. chrysolepis, Q. Wislinezii, Q. dumosa, Q oblongifolia,
Q emoryi, обладающих мелкими жесткими цельнокрайними или
колючезубчатыми листьями Заросли кустарников чаппараля
образованы представителями различных семейств и отличаются
большим флористическим разнообразием Очень характерны
для чаппараля, например, розоцветное, Adenostoma fasciculatum,
вечнозеленый кустарник с эрикоидными листьями и цветками,
сходными со Spiraea Часто встречается Ceanothus cuneatus
(Rhamnaceae) до 2 м высоты, образующий густые гр\ппы,
Ericameria arborescens из сложноцветных, внешне весьма сход-
ная со средиземноморскими видами Cistus.
Обзор жестколистной древесной растительности показывает,
что в различных областях тепло-умеренных зон северного и
южного полушарий с зимними дождями и сухим и жарким ле-
том, на основе различных флор сформировались экологически
сходные, сравнительно низкорослые (от 0,5—0,8 до 2—3 м вы-
соты), вечнозеленые, жестколистные, сильно ветвящиеся геок-
сильные (см ниже) кустарники Эти области, наряду с арид-
ными тропическими областями, явились одним из центров фор-
мирования кустарников во многих семействах покрытосемен-
ных
Интересно, что подобные же формации склерофильных
кустарников характерны для высокогорий тропической и тепло-
умеренных зон, где такие заросли кустарников распространены
у верхней границы древесного пояса, например, в горных масси-
вах Малайского архипелага, Цейлона, Килиманджаро, Кор-
дильер, и в умеренной зоне южного полушария — например,
в горах Чили (A Schimper u F. Faber, 1935)
Географическое распространение кустарников позволяет
заключить, что кустарники в тропической зоне и тепло-умерен-
ных зонах северного и южного полушарий приобретают наи-
большее ценотическое значение (доходящее до господства в рас-
тительном покрове) и наибольшее многообразие в областях
с периодически сухим и жарким временем года (в саванновых
и колючих лесах тропической зоны и в областях с жестколист-
ной древесной растительностью в умеренно-теплых зонах),
а также у верхнего предела лесной растительности в горных
массивах тех же широт. В перечисленных областях кустарники
становятся обычно эдификаторами растительных ценозов, дости-
гают максимального флористического и эколого-морфологиче-
ского разнообразия. Эти области, вероятно, явились родиной
249
кустарников во многих семействах покрытосеменных. В част-
ности, вероятно, здесь возникли в результате приспособления
к неблагоприятным условиям увлажнения наиболее типичные
для кустарников геоксильные формы с сильно развитыми и наи-
более долговечными подземными ветвящимися стеблями.
t *
В холодно-умеренной зоне северного полушария кустарники
также находят широкое распространение. Так, в хвойных лесах
Центральной Канады хорошо развит подлесок и ярус кустар-
ников. В западных борах США в хорошо развитом подлеске
преобладают виды Opulaster, Rosa, Viburnum, Ribes, Jamesia,
Acer glabrum, Prunus pensileanica, Cornus amotnum и др
(А П Ильинский, 1937).
В темнохвойных лесах Европейской части СССР в кустар-
никовом ярусе, хорошо развитом в менее сомкнутых насажде-
ниях довольно обычны виды Salix, Rosa acicularis, Alnus incana,
Sorbus aucuparia. В северной подзоне тайги появляется Betula
папа в форме кустарника (В. Б. Сочава, 1956).
Хорошо развит ярус кустарников также в широколиствен-
ных лесах Европейской части СССР Его состав варьирует
в различных областях. В юго-западных областях Европейской
части СССР представлены относительно термофильные и засу-
хоустойчивые кустарники — кизил — Cornus mas, клекачка —
Staphylea pinnata, скумпия — Cotinus coggygria, гордовина —
Viburnum lantana, виды Crataegus. В средней и восточной части
Русской равнины доминируют более холодостойкие виды.
Corylus avellana, Frangula alnus, Lonicera xylosteum, а также
Evonymus verrucosa, Daphne mezereum. На Дальнем Востоке
эти виды кустарников сменяет Corylus heterophylla и Lespedeza
bicolor, нередко доминирующие в подлеске. В дубравах Кавказа
на осветленных участках густые заросли образует Rhododendron
luteum (В. Б. Сочава и А. М. Семенова-Тянь-Шанская, 1956)
Интересно, что и в холодно-умеренной зоне северного полу-
шария, как и в рассмотренных ранее тепло-умеренных зонах
и тропической зоне, роль кустарников в растительном покрове
усиливается у пределов распространения лесной раститель-
ности. Так, в степных районах Европейской части СССР нередко
развиваются заросли ксерофитных степных кустарников, изве-
стные под названием дерезняков или кустарниковых степей.
Здесь довольно часто доминируют Prunus spinosa, Amygdalus
папа, Cerasus stepposa, Spiraea cremfolia и Spiraea hypericifolia.
Доминируют кустарники иногда и в более сухих областях, на-
пример, в кустарниковых пустынях пустынной зоны СССР, где
250
отмечается значительное разнообразие кустарников, своеобраз-
ных в экологическом и флористическом отношении. Наибольшее
флористическое разнообразие и широкое распространение за-
росли ксерофитпых кустарников получают в сухих горных стра-
нах, например, в Передней и Средней Азии В пределах СССР,
в низкогорных и среднегорных поясах Закавказья и гор Средней
Азии нередко образуют густые заросли в арчевых и фисташни-
ковых редколесьях (Е. М. Лавренко, 1956). Так, в кустарни-
ковых арчевниках Закавказья преобладают Paliurus Spina
Christi, Lonicera iberica, Cotoneaster racemiflora, Spiraea hyperi-
ctfolia, Jasminum fruticans, виды Rosa (H И Рубцов, 1956)
В ксерофильных зарослях кустарников Тянь-Шаня доминируют
различные виды Rosa, Lonicera, Cotoneaster, Crataegus, Spiraea,
Berberis. В горах Средней Азии и Кавказа в родах Rosa, Loni-
cera, Cotoneaster отмечается значительно большее число видов,
по сравнению с обширными территориями равнинных лесных
районов Европейской и Азиатской части СССР.
Доминируют кустарники в растительном покрове и севернее
полярного предела лесной растительности, где выделяется осо-
бая подзона тундровых кустарников (Б. Н. Городков, 1956),
образованная, главным образом, видами Salix, а также Betula
и Alnus Подобный пояс тундровых кустарников, то более, то
менее ясно выраженный, имеется и в горных тундрах Кольского
полуострова, Урала, Средне- и Восточно-Сибирских гор, Алтая
и Саяна.
Изучение географического распространения кустарников
выявляет вполне определенные ботанико-географические и эко-
логические закономерности. Кустарники приобретают господ-
ствующее положение в растительном покрове в условиях, не-
благоприятных для развития высокоствольных древесных расте-
ний — в саванновых и колючих лесах тропической зоны, в об-
ластях с жестколистной древесной растительностью тепло-уме-
ренных зон северного и южного полушарий, в ксерофиль-
ных редколесьях и формациях горно-степных кустарников
Передней и Средней Азии, и севернее полярного предела лесной
растительности холодно-умеренной зоны в подзоне тундровых
кустарников. Усиление фитоценотической роли кустарников в
этих областях сопровождается увеличением видового разно-
образия кустарников и эколого-морфологического их многооб-
разия. В этих областях наблюдается увеличение числа видов
кустарников в отдельных систематических группах (например,
родах, семействах). В перечисленных областях наблюдается
наибольшее разнообразие в характере роста кустарников’ по
высоте, интенсивности ветвления надземных осей, структуре
листьев и т д В этих областях наиболее часто наблюдаются
случаи варьирования жизненных форм в пределах одного и
251
того же вида, особи которого образуют то форму дерева, то
кустарника Эти явления в свою очередь могут указывать на
пути и факторы исторического метаморфоза древовидных жиз-
ненных форм в кустарники
2. МОРФОГЕНЕЗ ЖИЗНЕННОЙ ФОРМЫ КУСТАРНИКА
У НЕКОТОРЫХ ТИПИЧНЫХ КУСТАРНИКОВЫХ ВИДОВ РАСТЕНИИ
Морфогенез жизненной формы кустарника
у лещины — Corylus avellana L
Род Corylus — типичный представитель флоры умеренных
областей северного полушария, с 8 видами во флоре СССР
(Е. Г. Бобров, 1936), из которых 5 видов — типичные кустар-
ники (Corylus avellana L., С heterophylla Fisch., C. pontica
C. Koch., C, colchica Allon., C. manschurica Maxim) и лишь
3 вида могут образовывать настоящие деревья (С. colurna L.,
С. maxima Mill., С. breviluba Кот).
С. avellana широко распространен в широколиственных
лесах Европейской части СССР и других европейских странах,
будучи типичным спутником дуба и обладая сходным с ним
ареалом. По структуре надземных и подземных органов, по
экологическим свойствам С. avellana может служить типичным
представителем лесных кустарников, часто образующих хорошо
выраженный ярус в листопадных летпезеленых лесах холодно-
умеренной зоны северного полушария.
Биология и морфология лещины изучалась рядом исследова-
телей, выявивших основные особенности структуры куста
(Blomquist, 1911, С Lindman, 1914, Е Du Rietz, 1931) и его ста-
новление в онтогенезе (М Biisgen, 1913; W. Rauh, 1939; Сереб-
ряков И. Г., Доманская Н. П. и Родман Л. С, 1954). Семена
лещины прорастают подземно, в лесах Подмосковья обычно
в начале мая Под давлением набухшего семени каменистая
стенка плода раскалывается на две половинки Вследствие
удлинения черешков семядолей зародышевый корешок и по-
чечка выходят за пределы плода. Сквозь треснувший околоплод-
ник вначале выходит зародышевый корень, а затем гипокотиль,
также обладающий положительно-геотропической реакцией
и подобно корню врастающий в почву. Гипокотиль у проростков
лещины достигает от 3,5 до 8,5 см длины и постепенно пере-
ходит в главный корень. Семядоли остаются внутри плода не
только при прорастании, но и позже, сохраняясь у проростка
в течение всего первого лета
Наличие значительных запасов пластических веществ в се-
мени обусловливает интенсивный рост проростка в течение пер-
вого года К концу лета побег у сеянца лещины достигает вы-
252
соты 12—18 см. Выше мясистых семядолей, сохраняющихся
вместе с плодом у проростка лещины до осени в лесной под-
стилке или под подстилкой, на стебле образуется 3—5 недораз-
витых чешуевидных, затем 4—6 зеленых ассимилирующих
листьев (рис. 46) (Серебряков, Доманская, Родман, 1954).
Побег завершается верхушечной почкой, которая в лесах Под-
московья иногда подсыхает осенью и поэтому нарастание побега
со второго года происходит симподиально.
В пазухах всех листьев, начиная от семядолей и кончая
верхним ассимилирующим, формируются почки. На второй год
Рис. 46
Сеянцы орешника (Corylus avellana) (по Серебрякову,
Доманской и Родман, 1954):
А — схема проростка, Б— двухлетний сеянец, гк — главный корень, бк — бо-
ковые корни, гпк — гипокотиль, пгк — придаточные гипокотильные корни,
уп — уровень почвы, с — семядоля, СП — пазушная почка семядоли, ил — ни-
зовой лист (опавший), пп — пазушная почка, нп— иераскрывшиеся почки, вп —
верхушечная почка; 1 и 2 — годичные приросты первого и второго года
253
раскрывается только верхушечная почка или, при ее подсыха-
нии, верхняя боковая почка, заменяющая верхушечную; обра-
зуется побег продолжения главной оси. Остальные пазушные
почки остаются в нераскрывшемся состоянии, переходя в кате-
горию спящих почек. Длина побега второго года резко колеб-
лется, в зависимости от условий освещения и богатства почвы.
Но обычно он меньше побега первого года и не превышает
10—15 см. При значительном затенении прирост второго года
падает до 3—5 см. У слабо растущих побегов верхушечная
почка обычно отмирает, и ее заменяет пазушная почка верхнего
зеленого листа.
Под густым пологом кустарников при сомкнутом древостое
в лесах Подмосковья в течение третьего и четвертого года
у сеянцев лещины развертывается лишь верхняя пазушная
почка, образуя побег продолжения главной оси 5—8 см длины.
Но в более благоприятных условиях освещения и почвенного
питания годичные побеги третьего и четвертого года достигают
15—20 см длины. На них нередко сохраняется верхушечная
почка, и рост главной оси происходит моноподиально Интен-
сивно растущие сеянцы лещины с 3-го—4-го года начинают
ветвиться В последующие два-три года прирост главной оси
лещины резко возрастает, достигая 30—40 см в год Главная
ось четко выделяется в системе побегов, отличаясь от слабее
растущих наклоненных в стороны боковых ветвей своей боль-
шей толщиной и вертикальным положением.
В результате описанных процессов роста надземных органов
у сеянцев лещины в течение первых 5—7 лет жизни образуется
форма (рис 47), близкая к форме молодых сеянцев дерева,
например, пушистой и бородавчатой березы, а также сеянцев
дуба в хороших условиях роста (см. гл III). Вначале обра-
зуется вертикально растущая неветвящаяся главная ось, затем
она сменяется системой побегов с заметно выделяющейся по
силе роста главной осью и более слабыми, наклоненными в сто-
роны боковыми ветвями, как это имеет место у деревьев
В последующие годы рост и формирование надземных орга-
нов у лещины начинают все более и более отличаться от тако-
вых у деревьев Если у лесных деревьев (см гл. III) усиление
роста главной оси продолжается в течение первого и второго
десятилетий жизни, а у некоторых видов (например, у ели —
Picea excelsa) продолжает усиливаться в третьем и четвертом
десятилетиях, то у лещины кульминация роста главной оси
наступает в благоприятных экологических условиях в 4—6 лет
(в неблагоприятных условиях роста, например, при значитель-
ном затенении, максимум роста наступает на несколько лег
позже), а затем интенсивность роста главной оси начинает
снижаться. Если в период кульминации роста (рис. 47 Б) го-
254
2
Рис. 47
А — пятилетний сеянец орешника; Б - молодой кустарник в начале его
кущения (по Серебрякову, Доманской и Родман, 1954):
I — главная ось, 11 — боковые скелетные осн, возникшие в результате кушеиня главной
оси; 1, 2, 3—годичные приросты (лавной и боковых скелетных осей, г. к.—главный корень,
б к,— боковой корень, п г. к — придаточные гипокотильные корни, п. с к,— придаточные
стеблевые корни, п. к — придаточные корни скелетных осей,
с — семядоли, у. п — уровень почвы, и. п.— нераскрывшиеся почки
дичные приросты достигают 20—30 см, то в последующие два-
три года длина годичных побегов главной оси снижается до
10—15 см, а к 12—14-летнему возрасту верхушечный прирост
главной оси падает до 3—2 см.
Быстрое ослабление роста главной оси коррелятивно вызы-
вает два морфологических явления, не наблюдавшиеся у мо-
лодых сеянцев деревьев. Вертикальная вначале верхушка глав-
ной оси лещины после достижения кульминации роста посте-
пенно начинает принимать все более наклонное положение,
иногда доходящее до горизонтального. В этом состоянии почки
верхнего годичного побега главной оси с определенного воз-
раста не раскрываются, и главная ось в 13—16 лет, реже в 16—
18 лет, прекращает свой рост, а затем подсыхает. Подсыхание
начинается с верхушки и постепенно распространяется сверху
вниз на боковые ветви. Резкое ослабление, а затем и прекра-
щение верхушечного роста, кроме наклонения верхушки глав-
ной оси, коррелятивно вызывает раскрывание спящих почек у
255
ее основания, обычно оказывающегося к этому времени
покрытым лесной подстилкой. Из спящих почек возникают
быстро растущие побеги, биологически подобные водяным
побегам деревьев. Обычно в основании главной оси рас-
крываются 1—2, реже 3 спящие почки в течение одного лета
или 2—3 лет, следуя друг за другом За первый год побеги из
спящих почек вырастают до 20—30 см; они несут 8—10 зеле
ных листьев Их рост завершается образованием крупной вер-
хушечной почки, обычно сохраняющейся живой до следующего
вегетационного периода
Нарастание этих побегов в течение первых 3—4 лет идет по
моноподиалыюму типу Интересно, что побеги лещины с моно-
подиальным нарастанием имеют радиальное листорасположе-
ние, близкое к 2/г„ С переходом побега к симподиальному на-
растанию, в связи с ослаблением роста и плагиотропным на-
правлением его, радиальное листорасположение сменяется ла-
теральным, близким к ’/а
В 2—4-летнем возрасте у надземных осей, образовавшихся
из спящих почек, наблюдается резко выраженная кульминация
роста Годичные побеги их в этот период достигают длины 60—
70 см и в течение лета на них формируется до 15—20 зеленых
листьев, расположенных радиально. Междоузлия на этих го-
дичных побегах изменяются по типичной одновершинной кри-
вой, а формирование побега завершается образованием круп-
ной верхушечной почки
С 4—5 года происходит резкое снижение интенсивности
роста этих осей Годичные приросты их уменьшаются до 20—
30 см, на побеге от основания до верхушки можно видеть лишь
увеличивающиеся по длине междоузлия, верхушечная почка
отмирает, нарастание становится симподиальным Годичные
побеги их становятся подобными нижней части годичных побе-
гов тех же осей в период кульминации их роста. Вначале невет-
вящиеся, с 3—4-летнего возраста стебли начинают ветвиться
в верхней части Как и у главной оси, ослабление роста боко-
вых осей, возникающих из спящих почек, сопровождается изме-
нением направления их верхушек с вертикального ко все более
наклонному.
Быстрый рост надземных боковых осей из спящих почек
приводит к тому, что через 6—8 лет они достигают высоты и
толщины главной оси кустарника, а затем и превосходят ее
Эти надземные оси, возникающие из спящих почек главной оси
кустарника лещины и в будущем дающие начало подобным же
осям более высокого порядка, целесообразно назвать осями
возобновления кустарника.
Ветвление, начавшееся на 3—4-м году жизни, приводит к
тому, что с 7—8-го года верхушка оси возобновления теряется
256
среди ее быстро растущих боковых ветвей. Как и у главной
оси, переход осей возобновления от моноподиального к симпо-
диальному нарастанию сопровождается изменением листорас-
положения от радиального, близкого к 2/5, к латеральному, по
’/а К 13—16-летнему возрасту осей возобновления их верху-
шечный прирост снижается до 3—2 см и, наконец, прекраща-
ется совсем Годичные побеги таких стареющих осей возобнов-
ления представляют как бы базальную часть годичного побега
тех же осей в период кульминации их роста, отмершие в их
верхней части, не завершившие свой цикл развития.
Как и у главной оси сеянца, ослабление верхушечного роста
осей возобновления вызывает коррелятивно пробуждение спя-
щих почек в их основании, обычно прикрытых лесной подстил-
кой Из спящих почек вырастают побеги, формирующиеся по
типу материнских осей возобновления, т е осн возобновления
следующего порядка
Из подземных спящих почек вырастают вначале подземные
горизонтально растущие белые мясистые побеги корневище-
образного характера, несущие мелкие чешуйчатые низовые
листья На поперечных срезах подземных стеблей видна мощ-
ная паренхима первичной коры и сердцевины В подземных
стеблях слабее, чем в надземных, развита вторичная древеси-
на, а в конце первого вегетационного периода они остаются
без вторичной покровной ткани. Подземный моноподиальный
рост таких корневищеобразных побегов продолжается до 2—3-х
лет. Затем их верхушка приобретает вертикальное положение
'и они выходят на поверхность почвы. Вместо чешуйчатых на
них начинают формироваться зеленые ассимилирующие листья
Весьма характерна для таких белых подземных побегов лещи-
ны крючковидно загнутая верхушка. В дальнейшем из таких
подземных побегов вырастают надземные оси, формирующиеся
по типу материнских осей возобновления. Раскрытие в основа-
нии каждой оси двух-трех спящих почек приводит соответ-
ственно к двух-трехкратному увеличению числа осей возобнов-
ления в общей системе куста.
Рост и формирование осей возобновления происходит в
основном так же, как и у главной оси сеянцев Жизненный
цикл осей возобновления от раскрытия спящих почек до прек-
ращения их верхушечного прироста и начала подсыхания, как
и у главной оси, длится 13—16 лет, изредка достигая 17—
20 лет. Оси возобновления отличаются от главной оси лишь
более быстрым ростом в течение первых 3—4 лет, более ран-
ним наступлением кульминации роста (в 2—4-летнем возрасте),
более резкой выраженностью максимума на кривой роста, бо-
лее быстрым падением интенсивности верхушечного роста
после достижения кульминации. Следует отметить также, что
В-413 И Г Серебряков-17 257
оси возобновления II, III и IV порядков отличаются от главной
оси несколько большей высотой и диаметром стебля. Главная
ось лещины, вырастающая из семени, обычно достигает высоты
2 м, с диаметром стебля у поверхности почвы в 2—3 см Оси
возобновления II, III и IV порядков вырастают до 3—4 в вы-
соту, с диаметром стебля у поверхности почвы в 3—4 см.
Подземная часть осей возобновления располагается в лесах
Подмосковья на глубине 5—15 см в рыхлых верхних горизон-
тах почвы в виде дуговидно вниз изогнутых подземных стеблей
Подземные участки осей возобновления лещины отличаются ог
надземных стеблей большей долговечностью, они продолжаю1’
рост в толщину и после подсыхания и обламывания надземной
части осей возобновления Такие подземные стебли соединяют
в почве пеньки, остающиеся от подсохших надземных стеблей,
располагаясь обычно под землей в радиальном направлении от
центра куста к его периферии. Диаметр подземных стеблей ле-
щины в подмосковных лесах нередко достигает 8—10 см Под-
земные одревесневшие стебли у кустарников получили на-
звание ксилоподиев (Е. Warming, 1923, Е. Du R>etz,
1931).
В ксилоподиях лещины обнаруживается большее количество
юдичных колец, чем в надземных стеблях Так, в одном из
старых кустарников лещины из подмосковных смешанных ле-
сов с надземными стеблями диаметром у поверхности почвы в
2,5 см, с 12 годичными кольцами, в ксилоподиях диаметром
6—7 см имелось 26—27 годичных колец, а па отрезке того же
ксилоподия, ближе к центру куста, было отмечено 32 годичных
кольца Иногда у лещины наблюдается срастание соседних ра-
диальных подземных стеблей с образованием горизонтально
уплощенных досковидных одревесневших стеблевых органов,
расположенных неглубоко под поверхностью почвы.
Образование у кустарников подземных одревесневших стеб-
лей — ксилоподиев, более толстых и долговечных по сравне-
нию с надземными стеблями тех же кустарников, представляет
один из наиболее характерных признаков геоксильных
или настоящих кустарников
Общая длина каждого из подземных стеблей равна радиусу
куста и увеличивается с возрастом куста в целом Структурно
каждый подземный стебель представляет симподий, состоя-
щий из 01 дельных дуговидно изогнутых участков, каждый из
которых представляет собой подземную базальную часть осей
возобновления Каждый изогнутый участок подземных стеблей
лещины, длиной до 15—20 см, несет от 3—5 до 10—12 толстых
придаточных корней диаметром в 3—4 см, растущих горизон-
тально и ветвящихся, образующих придаточную корневую сис-
тему каждой оси возобновления
258
Как отмечалось выше, в основании главной оси, а затем бо-
ковых осей возобновления раскрываются по две и даже по три
спящих почки, что приводит к увеличению общего количества
надземных многолетних осей по мере увеличения возраста кус-
та В лесах Подмосковья у старых кустов лещины иногда
насчитывается до 60—80 и даже 120—150 многолетних надзем-
ных осей (осей возобновления). Если принять, что в основании
каждой материнской оси раскрывается по две спящих почки и
образуются две дочерние оси возобновления, то число осей в
ку сте возрастает с появлением каждого нового поколения осей
следующим образом (теоретически): главная ось—1; с появ-
лением осей возобновления II порядка — 3 оси; с появлением
осей III порядка — 6 осей; с появлением осей IV порядка —
12 осей; с появлением осей V порядка — 24 оси; с появлением
осей VI порядка — 48 осей; с появлением осей VII порядка —
96 осей.
В действительности, увеличение числа живых осей возоб-
новления в общей системе куста лещины происходит несколько
медленнее, чем в этом теоретическом ряд), из-за прекращения
верхушечного роста и подсыхания верхушек осей с их 16—
18-летнего возраста. Но подсохшие оси еще долго сохраняются
в общей системе надземных осей кустарника и обламываются
спустя длительное время после подсыхания.
Образующиеся оси возобновления располагаются около ма-
теринской оси более или менее плотной группой, обычно от 5
до 15—20 см друг от друга у поверхности почвы, образуя округ-
лую или овальную картину с самыми старыми осями в ее
центре. Вследствие того, что вновь вырастающие оси возобнов-
ления чаще располагаются по периферии куртины, с увеличе-
нием возраста куста происходит увеличение диаметра такой
куртины У молодых кустов лещины вся куртина состоит из
живых надземных многолетних осей, включая располагающую-
ся в центре куртины главную ось. У более старых кустов про-
исходит подсыхание и обламывание надземных осей в центре
куртины, с сохранением живых надземных осей по периферии
куртины. В результате образуется своеобразная форма курти-
ны, типа лысого куста.
В дальнейшем, наряду с подсыханием и обламыванием
старых осей в центре куртины, происходит формирование но-
вых осей по периферии куртины, что приводит к кольцеобраз-
ному расположению надземных осей. У наиболее старых кус-
тов лещины наблюдается распад кольца из надземных много
летних осей на отдельные небольшие группы, представляющие
последний старческий этап в отногенезе куста лещины. Над-
земные оси в таких старческих группах отличаются от осей
более молодых кустов меньшей высотой и диаметром стебля, а
И* 259
также ослабленным плодоношением Надземные оси в этот
период онтогенеза куста лещины характеризуются свойствами,
типичными для сенильного этапа в онтогенезе многолетних
растений (Т А Работнов, 1951).
Как видно из сказанного, в онтогенезе лещины наблюдает-
ся смена главной оси осями возобновления все более высокого
порядка При этом вначале имеет место некоторое усиление
роста осей возобновления, хорошо заметное при сравнении
главной оси с осями возобновления II, III и IV порядков. За-
тем, начиная с осей возобновления VI—VIII и с образованием
осей все более высокого порядка происходит ослабление их
роста, что проявляется в меньшей высоте и диаметре стебля у
взрослых осей В дубравах и липовых лесах Подмосковья
надземные оси лещины в старческом состоянии, даже в бла-
гоприятных условиях, достигают высоты не более 2—2,5 м, при
диаметре стебля у основания в 2—2,5 см, т с примерно в
2 раза менее высоты и диаметра надземных осей молодых
кустов лещины, произрастающих в этих же условиях Внешне
надземные оси старческих кустов лещины напоминают еще не
выросшие молодые кусты, и лишь наличие толстых, отмираю-
щих с дистальной части ксилоподиев, говорит о том, что здесь
мы имели дело со слабо растущими осями старческих кустов
лещины.
Все сказанное о росте и формировании куста лещины поз-
воляет выделить два крупных возрастных этапа в онтогенезе
Corylus avellana:
1. Этап древовидного роста — от прорастания
семени до 5—8-летнего возраста, т е до раскрытия спящих
почек в основании главной оси В течение первого этапа проис-
ходит образование вертикально растущей главной оси, сначала
неветвящейся, затем образующей систему боковых ветвей, по-
добно боковым ветвям в кроне дерева В связи с ослаблением
верхушечного роста главной оси в конце первого этапа наблю-
дается раскрытие спящих почек в основании главной оси с
образованием осей возобновления, т е осей II порядка, знаме-
нующих переход ко второму возрастному этапу в формирова-
нии кустарника у лещины.
П. Этап кустовидного роста —от раскрытия
спящих почек в основании главной оси до сенильного периода
в формировании куста и его последующего полного подсыха-
ния Второй возрастной этап в онтогенезе лещины характери-
зуется последовательной сменой осей возобновления все более
высокого порядка за счет раскрытия спящих почек в основа-
нии материнских осей при ослаблении их верхушечного роста.
В первой половине этого этапа происходит увеличение числа
осей возобновления в общей системе куста, так как новообра-
260
зование осей происходит быстрее их подсыхания и выпадения
из системы куста. Во второй половине этого этапа увеличение
числа осей возобновления прекращается и наступает уменьше-
ние числа надземных осей в общей системе куста, так как
процесс образования живых осей уже не компенсирует отмира-
ния и выпадения старых В пределах второго возрастного этапа
выделяются более частные периоды, отличающиеся друг о г
друга расположением надземных осей в общей системе
куста.
1) Период, характеризующийся равномерным и сплошным
расположением осей возобновления в форме округлой кур-
тины от ее периферии до центра.
2) Период, характеризующийся кольцеобразным
расположением надземных осей в общей системе
куста, с отмиранием старых осей на внутренней стороне кольца
и появлением новых осей на внешней стороне кольца
3) Период группового расположения слабо
растущих и плодоносящих осей возобновления, возникающих в
результате распада общего кольца надземных осей на отдель-
ные небольшие группы и образования клона Этот период ха-
рактеризуется как наиболее старческий из наблюдавшихся
автором в широколиственных и смешанных лесах Европейской
части СССР. В указанный период у лещины, вероятно, резко
снижается способность к образованию новых надземных осей
за счет раскрытия спящих почек материнских осей возобновле-
ния. В конце этого сенильного периода наступает полное под-
сыхание куста.
Изучение структуры куста лещины и его формирования в
отногенезе позволяет предложить некоторые методы определе-
ния возраста куста
Вследствие того, что в онтогенезе куста лещины происходит
многократная смена осей возобновления все более высокого
порядка, о возрасте куста в целом нельзя судить по возрасту
его отдельных надземных осей, так как их возраст всегда
будет меньше общего возраста куста
Как отмечалось выше, возраст надземных осей у лещины
достигает 15—18 лет, тогда как кустарник в целом живет много
десятков лет Более близкие данные о возрасте куста можно
получить при определении возраста (по числу годичных колец)
подземных одревесневших стеблей — ксилоподиев, более долго-
летних, чем надземные оси. В подмосковных лесах, как указы-
валось выше, в ксилоподиях лещины насчитывается до 32 го-
дичных колец Однако и возраст ксилоподиев не отражает пол-
ностью общего возраста к>ста, вследствие того, что в старых
базальных частях ксилоподия годичные кольца откладываются
нерегулярно, или даже совсем не откладываются, при возник-
261
новении на ксилоподиях мощных функционирующих придаточ-
ных корней.
Возраст кустарника у лещины можно определить путем
анализа общей структуры куста (Серебряков, Доманская.
Родман, 1954). Для определения возраста куста лещины необ-
ходимо подсчитать число порядков осей' возобновления у изу-
чаемого куста (лучше подобный подсчет произвести по несколь-
ким радиусам от центра куста до его периферии, взяв наиболь-
шее число порядков из ряда полученных чисел). Затем необ-
ходимо определить среднюю для данного куста разность в воз-
расте материнских и дочерних надземных осей. Разность в воз*
расте осей смежного порядка легко устанавливается у живых
растущих осей возобновления в периферийной части куста
Наконец, необходимо определить возраст осей самого высшего
порядка для изучаемого куста. Имея эти данные, общий воз-
раст куста лещины можно определить по формуле:
А = а -|- в X с,
где А — общий возраст куста;
а — возраст оси высшего порядка;
в — средняя разность в возрасте осей смежного порядка;
с— число порядков осей в данном кусте.
Указанным методом удается определить возраст кустов
лещины в подмосковных лесах до 60—80 лет Так, у одного из
изученных старых кустов было отмечено 12 порядков осей и
средняя разность в возрасте осей смежного порядка в 6,5 лет
при возрасте осей высшего порядка 4 года. При подставлении
числовых значений в формулу А = а + в X с, общий возраст
данного куста оказался равным 82 годам (Д = 4 12 X 6,5 =
= 82).
По данным М. Бюсгена (М. Biisgen, 1911) длительность
жизни лещины может достигать 200 лет.
В заключение общего обзора морфогенеза кустарника у ле-
щины целесообразно кратко охарактеризовать особенности
роста и формирования корневой системы этого кустарника.
При прорастании семени лещины в условиях смешанных
лесов Подмосковья на оподзоленных супесчаных почвах фор-
мируется хорошо выраженный стержневой корень, растущий
отвесно вниз на 20—25 см. Уже на первом году главный ко-
рень ветвится, особенно обильно в зоне, переходной к стеблю,
в рыхлых горизонтах почвы. На втором, а чаще третьем-
четвертом году жизни сеянца появляются придаточные корни в
надсемядольной части стебля, засыпанной к этому времени
лесной подстилкой. Отмирание верхушки главного корня вызы-
вает более сильный рост боковых и придаточных корней. В ре-
262
зультате роста и ветвления главного корня формируется систе-
ма главного корня, функционирующая до 6—8 лет, т. е. до
раскрытия спящих почек в основании главной оси, и до по-
явления на образовавшихся побегах придаточных корней. С
появлением боковых осей возобновления корневая система ле-
щины обогащается за счет их придаточных корней, возникаю-
щих в зоне перехода их из горизонтального к вертикальному
положению Придаточные корни осей возобновления отличают-
ся быстрым ростом и обычно направлены горизонтально. У 12—
14-летних боковых осей возобновления II, III и более высоких
порядков придаточные корни, обильно ветвящиеся, достигают
3,5 м длины и диаметра 2—3 см в основании.
Придаточные корни осей возобновления простираются ра-
диально от центра куста, на глубине 10—20 см, намного вы-
ходя за пределы кроны кустарника Мощное развитие прида-
точных корней боковых осей возобновления уменьшает функцио-
нальное значение главного корня, который постепенно отмира-
ет, начиная с верхушки. Но и сами придаточные корни осей
возобновления относительно недолговечны. У 13—15-летних
осей возобновления корни оказываются отмершими на значи-
тельном протяжении в их верхней части. На смену отмершим
корням данного поколения осей возобновления приходят новые
придаточные корни осей возобновления более высокого по-
рядка.
Хотя корни функционируют гораздо дольше побегов, их
породивших, все же в онтогенезе куста лещины происходит
регулярная смена придаточных корней по мере смены надзем-
ных осей возобновления.
Исходя из вышеописанного, можно сказать, что корневая
система лещины представлена системой главного корня и сис-
темой придаточных корней осей возобновления все более высо-
кого порядка, последовательно сменяющих друг друга в отно-
генезе кустарника. Остатки отмершего главного корня можно
видеть у лещины до 50—80-летнего возраста. Формирование
корневой системы лещины таким образом в известной степени
оказывается подобным формированию системы надземных
осей.
Сохранение в онтогенезе лещины фазы древовидного роста,
вероятно, отражает черты предков С. avellana. По Е. Г. Боб-
рову (1936), С. avellana обнаруживает непосредственную связь
через вымершее звено с С. column, которая может рассматри-
ваться как отдаленный предок С. avellana. С. column отличает-
ся древовидной формой и достигает высоты 20 м. С. column и
С. avellana образуют особую серию Lucerne в секции Diphyllon.
В этой серии древовидная С. colurna — древняя форма, распро-
страненная в настоящее время в лесах Кавказа и Балкан, а
263
C. avellana по происхождению наиболее молодая кустовидная
форма с широким современным распространением и крайне по-
лиморфная (установлено 50 разновидностей и форм).
Восточный отрезок ареала С. colurna занимает область
Центральных Гималаев и здесь этот вид связывается с другой
серией видов (Incisae) той же секции Diphyllon, ближайшим из
которых является С. Potanini, довольно широко распространен-
ный в Китае С. Potanini — дерево, достигающее 20 м высоты,
представляет собой довольно древний тип, непосредственно с
С. colurna не связанный, а параллельный ему и имеющий, оче-
видно, общего с ним предка
К С Potanini близок типичный кустарник С. heterophylla,
распространенный от провинций Северного Китая и Кореи до
Восточного Забайкалья, Амурской и Уссурийской области,
и Японии. Е. Г. Бобров (1936) считает С. heterophylla видом,'
возникшим непосредственно от С. Potanini Анализ эволюции
видов в секции Diphyllon рода Corylus показывает, что в двух
сериях (Lacerae и Incisae) образование новых видов сопро-
вождалось метаморфозом жизненных форм от деревьев
к кустарникам Этот исторический метаморфоз проходил в пре-
делах лесных областей умеренного пояса. Вероятно, фактором
подобного метаморфоза явилось похолодание климата этих
областей в позднетретичное время.
Особенности морфогенеза жизненной формы кустарника
у бересклета бородавчатого (Evonymus verrucosa Scop )
Семейство Celastraceae, к которому относится род Evonymus,
обладает широким географическим распространением, преиму-
щественно в субтропическом и тропическом поясах Семейство
включает разнообразные жизненные формы, деревья, кустар-
ники, древесные лианы, стелющиеся кустарники и др Род
Evonymus представлен во флоре СССР 10-ыо видами, преиму-
щественно кустарникового типа, среди которых нс только листо-
падные, но и вечнозеленые формы, например, Е sempervirens
(Медведев, 1883), Е. japonica (Я И Проханов, 1949; Л А Гросс-
гейм, 1949). Характер географического распространения и пре-
обладание в семействе Celastraceae древесных, часто древовид-
ных форм, а также наличие вечнозеленых видов в роде Evony-
mus делают понятными некоторые морфологические и биологи-
ческие особенности в морфогенезе кустарника у Е verrucosa
Scop. Бородавчатый бересклет — типичный спутник дуба на
территории Европейской части СССР, входит в состав кустар-
никового яруса дубрав. Ареал бородавчатого бересклета лишь
немного отличается от ареала черешчатого дуба, не доходя
несколько до северо-восточных пределов последнего.
Изучение структуры кустов Е. verrucosa в широколиствен-
964
пых и смешанных лесах Подмосковья обнаружило некоторые
специфические особенности, отличающие его от структуры кус-
тарника у лещины. Большинство надземных осей бородавчатого
бересклета более или менее рассеяно отходит от лежачих и за-
сыпанных лесной подстилкой материнских осей; происхождение
последних удалось выяснить лишь, при изучении онтогенеза
кустарника бересклета от проростков до взрослого состояния
В работе Н. Н Иорданской и И Г Серебрякова (1954) просле-
жены основные особенности формирования кустарника у бере-
склета бородавчатого.
Семена бородавчатого бересклета прорастают надземно
Очень часто проростки и молодые сеянцы встречаются под по-
логом леса небольшими тесными группами, в числе 3—7 экзем-
пляров. Это обусловлено, вероятно, особенностями плодоноше-
ния бересклета бородавчатого, семена которого не высыпаются
при опадении плодов
В течение первого года у сеянцев бородавчатого бересклета
в лесах Подмосковья, кроме двух ассимилирующих овальных
семядолей, успевают сформироваться 2—3 пары небольших
зеленых листьев и верхушечная почка. Высота сеянца, обуслов-
ленная, главным образом, вытянутым гипокотилем, в конце лета
не превышает 3—4 см. В течение 2-го, 3-го и последующих
нескольких лет под пологом леса, при значительном затенении,
раскрывается только верхушечная почка, с образованием тон-
кой неветвящейся оси. Боковые пазушные почки остаются в со-
стоянии спящих Прирост на втором, третьем и четвертом годах
незначительный, он даже меньше прироста первого года (рис.
48). В культуре, на черноземных почвах при обильном удобре
нии, сеянцы бересклета растут значительно быстрее (И. А Ива-
нов, 1939)
Обращает на себя внимание характерная особенность сеян-
цев бересклета — их вечнозеленость. Семядоли и зеленые листья
сеянцев живут до 3-х лет. Под пологом леса вечнозеленость
сеянцев бересклета сохраняется до 7—8-летнего возраста Эта
биологическая особенность сеянцев бересклета бородавчатого
является отголоском свойств его древних предков, произрастав-
ших в условиях более теплого климата третичного периода Как
отмечалось выше, некоторые виды бересклета в более южных
широтах и в настоящее время являются вечнозелеными Ч
1 Интересно, что у бересклета бородавчатого вечнозелены не только
сеянцы, но и корнеотпрысковые побеги и побеги, возникшие из спящих почек
полегших материнских осей, в тс генме первых лет их жизни
В начале зимы в лесах Подмосковья нередко можно видеть торчащие
из-под снега небольшие веточки (до 20—25 см) с зелеными листьями. Эти
листья краснеют и опадают лишь весной следующего года. Таким образом,
эти побеги обнаруживают свойства ювенильных побегов сеянцев
265
Рис. 48
Сеянцы бересклета бородавчатого {Evonymus verrucosa)
в широколиственных лесах Московской области (по Иорданской
и Серебрякову, 1954):
а — однолетний сеянец; б — двухлетний; в — трехлетний, г — четырехлетний,
д — пятилетний; прерывистой линией обозначены границы годичных приростов
С 4—5-го года интенсивность роста главной оси у сеян-
цев возрастает, достигая к 10—15 годам у некоторых экземпля-
ров 15—18 см в год; но в общем интенсивность роста под поло-
гом леса в условиях затенения остается низкой. Рост главной
оси бородавчатого бересклета идет по типу одновершинной
кривой, со слабо выраженным максимумом в 10—15 лет. По
сравнению с лещиной главная ось бородавчатого бересклета
отличается более поздним наступлением кульминации роста и
в соответствии с этим значительно большим долголетием Глав-
ная ось бересклета бородавчатого живет до 30 лет, вместо
13—16 у лещины. Эти особенности роста главной оси бересклета
выражают тенденцию к образованию у него древовидной фор-
мы. Иногда действительно отмечается способность бересклета
бородавчатого расти в виде небольшого деревца (Я- С. Медве-
дев, 1883) Таким образом, жизненная форма кустарника у бо-
266
родавчатого бересклета структурно и биологически стоит ближе
к форме дерева, чем у рассмотренного выше кустарника —
Corylus avellana.
Своеобразная особенность бересклета проявляется в поле-
гании его главной оси. К 15—20 годам, когда главная ось
сеянца достигает 70—100 см, она полегает на значительном
своем протяжении. При этом вертикальное положение иногда
сохраняет лишь ее верхушка. Лесная подстилка покрывает сте-
бель; на засыпанном стебле вскоре появляются придаточные
корни (рис. 49). Окоренение главной оси и ослабление роста ее
верхушки приводит вскоре к раскрытию спящих почек, распо-
ложенных по длине погребенного стебля. Полегание стеблей
Рис 49
Формирование боковых скелетных осей бересклета бородавчатого
при полегании материнской главной оси:
1 — главная ось, 2 — боковые скелетные оси, возникающие от спящих почек
материнской оси; 3 — скелетная ось корнеотпрыскового происхождения
(по Иорданской и Серебрякову, 1954)
у бересклета связано со слабым ростом стеблей в толщину,
с резким ослаблением камбиальной деятельности при затене-
нии под пологом леса. Здесь образуются тонкие годичные
кольца, недоразвивается ксилема. Стебель бересклета полегает
в сомкнутых лесах, а на открытых местах, например на опуш-
ках, лесных полянах и т. д., формируются прямостоячие кусты
с более толстыми осями
От основания и до верхней части полегшей укоренившейся
главной оси из раскрывшихся спящих почек развиваются новые
267
надземные оси в числе 2—4. Биологически эти оси равнозначны
осям возобновления в кустах Corylus avellana. Разрастание этих
осей из спящих почек способствует дальнейшему ослаблению
роста и, наконец, отмиранию верхушки главной оси Боковые
многолетние оси, возникающие из спящих почек, отличаются
от главной оси сеянцев интенсивным ростом, особенно в тече-
ние первых 5—7 лет. Вероятно, их быстрый рост обеспечивается
сформировавшейся к этому времени корневой системой и запа-
сом пластических веществ Прирост боковых осей из спящих
почек идет, как и у главной оси, по типу одновершинной кривой
Только максимум роста их приходится на второй, реже третий
год жизни оси после выхода ее на поверхность почвы. Период
максимального роста’ у боковых осей, в отличие от главной оси,
выражен чрезвычайно резко. Но интенсивный рост боковых
осей продолжается всего лишь несколько лет, после чего они нс
отличаются от слабо растущей главной оси по величине своих
годичных приростов Длительность жизни боковых осей, как
и главной оси сеянцев, достигает 30—35 лет А И. Страто-
нович (1938) отмечает длительность жизни осей у кустов бере-
склета до 40 лет По длительности жизни осей — важнейшему
биологическому признаку — кустарник бородавчатого береск-
лета стоит ближе к форме дерева, чем кустарник С. avellana.
Боковые оси возобновления у бересклета, как и у лещины,
в течение первых лет остаются неветвящимися Лишь к 8—10
годам их жизни начинают разрастаться верхние боковые ветви
Эти ветвящиеся оси возобновления бересклета в сомкнутых
лесах, в свою очередь, обычно полегают, подобно главной оси
При полегании они покрываются лесной подстилкой и укоре-
няются Из спящих почек полегших стеблей возникают оси
возобновления нового поколения, развивающиеся по типу мате-
ринских Подобный процесс развития осей возобновления и
последующего их полегания повторяется неоднократно в онто-
генезе бересклета бородавчатого. В результате образуются
заросли бересклета на значительной площади со сравнительно
редким расположением многолетних надземных осей (осей
возобновления). По этому признаку бересклет надо отнести
к полупростратным геоксильным кустарникам (см. гл. II)
Таким образом, кусты бересклета бородавчатого представ-
ляют собой систему сменяющих друг друга осей возобновления
с длительностью жизни до 30 и более лет, образующих каждый
раз свою систему придаточных корней Полегание осей бере-
склета под пологом леса, вызываемое значительным затенением,
влечет за собой усиление его вегетативного размножения
Последнее имеет большое биологическое значение в связи с тем
что семенное размножение бересклета под пологом леса значи-
тельно подавлено Кроме того, полегание надземных осей
бересклета бородавчатого имеет значительный хозяйственны/'
интерес, потому что в коре полегших стеблей, покрытых лесной
подстилкой, значительно возрастает содержание г>тты. На этом
основан метод обогащения стеблевой коры бересклета гуттой
(А. Л. Кащеев, 1953; Н В. Мацкевич и А. Л. Кащеев, 1953).
Бересклет бородавчатый способен к вегетативному возоб-
новлению и размножению не только за счет спящих почек
побегов, но и за счет придаточных почек корней, т. е корневых
отпрысков (рис 49). Корнеотпрысковое возобновление береск-
лета довольно широко распространено, например, в лесах Под-
московья Оно имеет существенное биологическое значение,
потому что у бересклета бородавчатого под пологом леса обра-
зование новых надземных осей из спящих почек побегов проис-
ходит не столь регулярно и часто, как например у. лещины.
У бересклета образование новых осей происходит не всегда за
счет почек базальной части материнских осей, т е. возрастно-
молодых эмбриональных тканей. И возобновление за счет поле-
гающих стеблей неминуемо должно приводить к довольно бы-
строму старению. Корнеотпрысковое возобновление бересклета
бородавчатого поэтому можно рассматривать как путь биологи
ческого омоложения растений, обычно возобновляющихся за
счет стеблевых почек
Морфогенез корневой системы бересклета бородавчатого
протекает весьма различно, в зависимости от характера почвы,
типа развития куста и, в частности, степени полегания его глав-
ной и боковых осей. В смешанных и широколиственных лесах
Подмосковья у сеянцев бересклета бородавчатого формируется
главный корень, ветвящийся уже на первом году жизни сеянца
С приближением к горизонту вымывания на оподзоленных
почвах в лесах Московской области верхушка главного корня
принимает горизонтальное положение и в дальнейшем продол-
жает рост параллельно поверхности почвы. Так же растут и
боковые корни В результате формируется поверхностная
система главного корня, располагающаяся в лесах Подмосковья
на глубине до 10 см По мере полегания главной оси и засыпа-
ния ее почвой на ней образуются стеблевые придаточные корни,
ветвящиеся и растущие горизонтально Если главная ось и бо-
ковые оси возобновления полегают незначительно, образуя
компактный куст, система главного корня функционирует до
40 и более лет При сильном полегании главной оси и боковых
осей возобновления наблюдается формирование сильно разви-
той системы придаточных корней, вследствие чего функциональ-
ное значение системы главного корпя снижается, и она отми
рает быстрее Ее сменяет система стеблевых придаточных кор
ней. Смена поколений надземных осей возобновления в таких
случаях сопровождается последующей сменой их корневой
269
системы. Это биологическое свойство бересклета бородавчатого
позволяет в течение онтогенеза куста проводить искусственно
неоднократную смену его корневой системы. Удаление части
ростовых корней для промышленного использования (получение
гутты) сопровождается формированием новых ростовых корней
(М. Д Данилов и В. И Пчелкин, 1953)
Особенности морфогенеза жизненной формы кустарника 1
у шиповников — Rosa spinosissima L. и Rosa Alberti Rgl.
Политипный род Rosa насчитывает в настоящее время нс
менее 380—400 видов, широко распространенных от тропиков
до областей холодно-умеренных зон, где некоторые виды этого
рода, например, Rosa acicularis, в Евразии доходят до северного
предела лесов (В. Г Хржановский, 1952). По форме роста
большинство видов Rosa представляет многостебельные прямо-
стоячие кустарники, высотой в 1—2 м Однако в пределах рода
имеются деревья, древесные лианы, а также карликовые кус-
тарники 15—30 см высоты, со слабо одревесневающими и не-
долговечными стеблями, переходные к травам.
Древовидные формы роз и древесные розы-лианы широко
распространены в субтропической флоре; они преобладают
в крупнейшей секции рода — Synstylae Стволы роз-деревьев
обычно несколько изогнуты и свилеваты, что, по мнению
В. Г. Хржановского (1952), объясняется их происхождением от
лианоидных предков. Среди лианоидных древесных видов Rosa
выделяются более древние вечнозеленые формы, первичные по
В Г. Хржановскому (секция Synstylae) и листопадные розы-
лианы, вторичные по В Г. Хржановскому (секции Cinnamomeae,
Pimpinellifoliae, в которых преобладают прямостоячие кустар-
никовые формы) Высота кустов у видов Rosa может быть
более или менее постоянной в пределах целой секции (напри-
мер, виды секции Pimpinellifoliae имеют в среднем высоту 80—
120 см, в секции Caninae — около 2 м), или резко различной
в пределах небольшой систематической группы, например,
серии. Так, Rosa Fedtschenkoana достигает 3—4 м высоты, а
близкий вид — R. nanothamnus — карликовый кустарник, высо-
той 15—30 см Высота куста, кроме видовой обусловленности
и возраста растения, зависит от условий произрастания Так,
R. cinnamomea, имеющая высоту обычно около 1,5 м, по данным
В. Г. Хржановского (1952), в пойме р Иртыш, от Усть-Камено-
горска до Павлодара, на выходах известняков образует карли-
ковые кусты в 20—40 см, а на галечных конусах выноса рек
Большой и Малой Алма-Атинок (Заилийский Ала-тау) — в 30—
50 см.
Rosa spinosissima L из секции Pimpinellifoliae — невысо-
кий кустарник каменистых горных склонов, с общим распро-
270
странением в Скандинавии, Средней Европе, Средиземноморье,
Западной Сибири (Алтай) и в горах Средней Азии (С. В. Юзеп-
чук, 1949)
В общей структуре куста R. spinosissima четко выделяются
парциальные кусты, т е. группы тесно расположенных
надземных осей, в числе от 3—5 до 10—15 (вместе с подсох-
шими) В общей системе куста или куртины насчитывается до
<50 парциальных кустов. В пределах каждого парциального
куста оказываются побеги 3—4 порядков Как отмечает
Л. С. Родман (1957), в кустарниковой степи западных предгорий
Алтая парциальные кусты /?. spinosissima могут быть удалены
друг от друга на значительное расстояние, диффузно распро-
страняясь среди остальной массы кустарников В условиях недо-
статочного увлажнения в блюдцевидных понижениях парциаль-
ные кусты располагаются близко друг к другу и образуют
плотные куртины 1,5—5 jh в диаметре. Форма куртины соответ-
ствует форме микропонижения, которое она занимает.
В формировании осей возобновления /?. spinosissima имеется
ряд своеобразных черт, отличающих этот кустарник от Corylus
avellana и Evonymus verrucosa Основное своеобразие надзем-
ных осей R. spinosissima заключается в том, что максимум
роста их достигается уже в первое лето после их выхода из
почвы па дневную поверхность.
Рост надземных осей возобновления R. spinosissima проис-
ходит по нисходящей кривой (рис. 50), тогда как рост осей
возобновления Corylus avellana проходит по кривой одновер-
шинного типа, хотя и у С. avellana восходящая ветвь кривой
роста надземных осей более короткая, чем ее нисходящая
часть.
Резкое уменьшение интенсивности верхушечного роста над-
земных осей R. spinosissima приводит к быстрому (в 5—7 лет)
отмиранию оси Из-за отмирания верхушки побега нарастание
надземных осей уже со второго года происходит симподиально,
за счет пазушных почек. Лишь изредка моноподиальное нара-
стание сохраняется до третьего года. Со второго года обычно
начинается ветвление осей, боковые ветви образуются на вы-
соте 20—40 см от поверхности почвы Боковые ветви, подобно
материнской оси, наиболее интенсивно растут в первый год
жизни. Максимум роста в течение первого года свойствен
боковым ветвям и более высокого порядка. Эта особенность
роста приводит к тому, что уже со второго года жизни боковые
ветви осей возобновления превышают по высоте материнскую
ось.
Резкое ослабление роста надземных осей уже со второго
года и общий характер роста надземных осей по типу нисходя-
щей кривой наблюдается у R. spinosissima в разных экологи-
271
Рис. 50
Изменение длины годичного прироста у Rosa spinosissima
в зависимости от возраста побега. Западный Алтай
(по Л. С. Родман, 1957)
ческих условиях. Правда, в более благоприятных условиях
увлажения величина годичных приростов и общая длитель-
ность жизни надземных осей оказывается большей. По мере
усиления сухости местообитания уменьшается и величина го-
дичных приростов, и общая длительность жизни осей Поэтому
и высота кустов R. spinosissima, достигающая в благоприятных
условиях увлажнения 1,2—1,5 м, в сухих местообитаниях
уменьшается до 0,4—0,6 м
Резкое ослабление верхушечного роста надземных осей при-
водит к тому, что уже на втором году раскрываются не только
боковые пазушные почки в верхней части осей, но и спящие
почки в основании осей возобновления, в их подземной части
или у поверхности почвы. Из спящих почек образуются корне-
вищеобразные подземные побеги с низовыми чешуевидными
листьями, растущие горизонтально (из подземных почек), или
вначале наклонно вниз (из приземных спящих почек) до глу-
бины 8—20 см. На этой глубине в почве корневищевидные
272
побеги растут горизонтально В течение второго года, или через
несколько лет, верхушка подземных побегов приобретает орто-
тропное направление роста и выходит на поверхность почвы,
давая начало новой надземной оси. В зависимости от длины
подземного корневищеобразного побега, новые надземные оси
располагаются на большем или меньшем удалении от материн-
ской надземной оси. Если подземные побеги короткие, из них
вырастают надземные оси, входящие в состав того же парциаль-
ною куста, к которому относится их материнская ось. Если же
подземные корневищеподобные побеги длинные, они выходят
за пределы материнского парциального куста и, выйдя на по-
верхность почвы, дают начало материнской оси нового пар-
циального куста Вследствие этого, материнская ось каждого
парциального куста обладает длинной подземной частью, и в
связи с этим — сильным окоренением Изучение показало, что
в пределах каждого парциального куста оси более высокого
порядка, входящие в его состав, отличаются более слабым
окоренением, они несут меньше жизнеспособных спящих почек,
из которых вырастают новые надземные оси Вследствие такого
понижения жизнеспособности осей все более высокого порядка
в пределах парциального куста, последний всегда имеет лишь
ограниченное количество надземных осей Вслед за отмиранием
осей I и II порядков наступает отмирание всего парциального
куста в целом. Но из спящих почек осей I и II порядков старею-
щих парциальных кустов возникают длинные подземные корне-
вищеобразные побеги, дающие начало новым материнским
побегам новых парциальных кустов
Таким образом, у R spinosissima, подобно Corylus avellana,
во взрослом состоянии наблюдается последовательная смена
не только надземных многолетних осей (осей возобновления)
все более высокого порядка, но и целых групп их, или пар-
циальных кустов Как долго в онтогенезе куста в целом у
R. spinosissima происходит подобная последовательная смена
парциальных кустов и какова общая длительность жизни
куста — остается невыясненным
Rosa Alberti Rgl., из секции Cinnamomeae,— сильно ветви-
стый кустарник, высотой более 1 м, с длинными дуговидно изог-
нутыми ветвями, распространенный в горных лесах, по опушкам
и кустарникам гор Западной Сибири (Алтай) и Средней
Азии — Джунгарский Алатау, Тарбагатай, Тянь-Шянь (С В
Юзепчук, 1941)
Ниже приводятся некоторые данные о структуре кустов и
морфогенезе жизненной формы кустарника у R. Alberti по на-
блюдениям автора летом 1953 г в Центральном Тянь-Шане
(хребет Терскей Алатау, долина р Чон-Кызыл-су) В долине
р Чон-Кызыл-су R Alberti довольно обычна для лугово-лесного
В-413 И Г. Серебряков —18 273
и субальпийского пояса от 1700—1800 до 2800 м над уровнем
моря. Высота кустов R. Alberti в лугово-лесном поясе обычно
достигает 2—3 м В росте и формировании надземных осей
R- Alberti имеется много общего с R. spinosissima.
Максимум роста надземных осей приходится па первый год
после выхода их из почвы. В лесном поясе Терскей Алатау на
высоте 2300—2600 м над уровнем моря прирост первого года
надземных осей достигает в среднем 145 см (среднее из изме-
рений 50 надземных осей, колебания прироста от 120 до 160 см)
Диаметр молодых стеблей равен в среднем 10—12 мм. На вто-
ром году интенсивность верхушечного роста надземных осей
резко снижается и в среднем составляет (по данным 50 изме-
рений) 65 см. На третьем году, в результате дальнейшего ослаб-
ления верхушечного роста, длина годичных побегов снижается
в среднем до 25 см, на четвертом — до 15,6 см, пятом — до
8,6 см, шестом — до 3,7 см, седьмом — до 3 см
Как видно из приведенных цифр, рост надземных осей
у R. Alberti, как и у R. spinosissima, идет по типу нисходящей
кривой, с максимумом роста на первом году их жизни. На пя-
том году жизни оси начинается плодоношение, которое продол-
жается в течение 4—5 лет, прекращаясь в более старом воз-
расте В отличие от R. spinosissima, рост надземных осей
у R. Alberti продолжается не 5—7, а 8—10 лет. и подсыхание
•осей начинается после 11—12 лет Большая длительность жиз-
ненного цикла надземных осей R. Alberti и, в частности, их
периода роста определяет большую высоту кустов R Alberti по
сравнению с R. spinosissima.
Ветвление надземных осей у R. Alberti, как и у R. spino-
sissima, начинается со второго года их жизни Вследствие
отмирания верхушки побега оно происходит симподиально.
В течение всего весенне-летнего периода роста на конусе нара-
стания годичных побегов надземных осей формируются только
зачатки зеленых ассимилирующих листьев.
Если в этот период на конусе нарастания стебля форми-
руются зачатки почечных чешуй, ветвление будет идти моно-
подиально Если же в течение весенне-летнего периода роста
побега на конусе нарастания продолжают образовываться но-
вые зачатки зеленых листьев, верхушечная почка не образует-
ся и ветвление приобретает симподиальный характер (Серебря-
ков И. Г, 1952).
Ослабление верхушечного роста надземных осей вызывает
коррелятивно раскрытие спящих почек в приземной и подзем-
ной части материнских осей и образование на них новых над-
земных осей. Как и у R. spinosissima, из базальных спящих
почек вначале формируются подземные корневищеобразные
побеги, белые, несущие низовые чешуевидные листья, с сильным
274
развитием паренхимных тканей в стебле. На них уже на второй
или третий год их жизни вырастают стеблевые придаточные
корни, отличающиеся быстрым ростом и обильным ветвлением
По выходе на поверхность почвы эти корневищеобразные по-
беги дают начало новым надземным осям, растущим и форми-
рующимся по вышеописанному типу. Как показали массовые
подсчеты, разность в возрасте материнских и дочерних надзем-
ных осей в пределах одного куста равна обычно 5—6 годам
Как и у двух рассмотренных выше кустарников — Corylus
avellana и R. spinosissima, из подземных ризомообразных побе-
гов R. Alberti, являющихся базальной частью каждой надземной
оси, по мере роста последней образуются толстые одревеснев-
шие подземные стебли — ксилоподии, соединяющие под землей
в единое целое все надземные оси куста У R. Alberti* как видно
из рис. 51, диаметр ксилоподиев в 3—4 раза и даже более
превосходит диаметр старых надземных осей. Ксилоподии
R. Alberti отличаются от надземых осей большим долголетием.
Рис. 51
Нижние части стеблей Rosa Alberti, соединяющие их
подземные стебли-ксилоподии
(в Центральном Тянь-Шане)
Таким образом, у R. Alberti, как и у других кустарников,
рассмотренных выше, морфогенез куста сопровождается обра-
зованием многочисленных надземных одревесневших осей и
еще более долговечных и толстых подземных одревесневших
стеблей — ксилоподиев. Дифференциация стеблей па более
тонкие и менее долговечные надземные и более толстые и дол-
18*
275
говечные подземные (ксилоподии), которые оказываются орга-
нами возобновления надземных осей, является одним из наи-
более характерных признаков подобных, геоксильных кустарни-
ков
Особенности морфогенеза жизненной формы кустарника
у терна — Prunus spinosa L.
Prunus spinosa L — сильно разветвленный, очень колючий
кустарник, реже небольшое деревце до 4—8 м высоты. Растет
преимущественно в зоне лесостепи, на полянах, лесных опуш-
ках. по оврагам, в лес заходит редко. В северной части ареала
встречается на лугах. Высотные пределы распространения —
800—1 200 м над уровнем моря в Закавказье (С В Юзепчук,
1941). По Lindman (1914) и Du Rietz (1931) Р. spinosa относит-
ся к аэрокс ильным кустарникам, вегетативно рас-
пространяющимся посредством корневых отпрысков. Ниже при-
водятся некоторые данные о росте и формировании кустарника
у Р. spinosa по наблюдениям автора на территории Курского
Центральночерноземного государственного заповедника летом
1957 г.1.
В лесах Курского заповедника Р. spinosa встречается очень
часто в качестве одного из- главнейших видов подлеска Терн
приурочен к опушкам и лесным полянам, где он образует плот-
ные заросли или куртины, до нескольких десятков метров в
диаметре. По периферии куртины терна располагаются обычно
более молодые ветвящиеся надземные оси, а вокруг куртины
довольно часто можно встретить надземные оси различного
возраста, включая однолетние и двулетние стебли, обычно
наиболее удаленные от куртины (на 10—20 ж).
При раскапывании стеблей обнаруживается, что все молодые
стебли отходят от горизонтально растущих корней толщиной
около 1 см на глубине 10—15 см от поверхности почвы. В тече-
ние первого года по выходе стебля на поверхность почвы побеги
вырастают от 50—60 до 90—100 см От поверхности почвы и до
верхушки стебли покрыты зелеными листьями. Изредка уже в
конце первого лета у верхушки побега появляются несколько
боковых наклонно растущих ветвей в 20—30 см длины. В тече-
ние второго года интенсивность роста надземной оси несколько
снижается, достигая обычно 40—60 см При этом прирост глав
пой оси и боковых ветвей оказывается примерно одинаковым и
различия главной оси и боковых ветвей по длине годичных побе-
гов и толщине их стебля становятся несущественными. На треть
1 В сборах и анализе полевых материалов принимала участие Т И Се
ребрякова; автор приносит ей искреннюю благодарность
276
ем году рост надземных осей не только не ослабевает, но срав-
нительно с предыдущим годом часто даже несколько возрастает,
достигая 60—70 см. Но уже с четвертого года рост побегов
заметно уменьшается. С этим ослаблением роста совпадает и
изменение направления растущей верхушки главной оси: из вер-
тикальной она становится наклонной, оказываясь незаметной
среди боковых ветвей
Рост надземных осей Р. spinosa проходит в общем по типу
нисходящей кривой с максимумом роста в течение первых 2—3
лет. В эти годы приросты достигают в среднем 60—70 см В те-
чение четвертого года прирост побегов снижается до 30—40 см,
на пятом году жизни оси годичные побеги имеют среднюю длину
32,5 см, в течение двух следующих лет годичный прирост падает
до 24 см В дальнейшем быстрота роста побегов снижается
медленнее, и у 12—14-летних надземных осей годичные приросты
нередко остаются равными 15—25 см. Верхушечный рост над-
земных осей сохраняется и в 15—18-летнем возрасте их. Наши
попытки найти прекратившие верхушечный рост и подсыхающие
надземные оси терна в его куртинах в лесах Курского государ-
ственного заповедника не увенчались успехом. Жизненный цикл
надземных осей Р spinosa здесь, вероятно, длится не менее
20—30 лет
Уменьшение интенсивности роста надземных осей по мере
увеличения их возраста сопровождается резким изменением мор-
фологической структуры и дифференциацией их годичных побе-
гов В течение первых 4—5 лет на надземных осях терна фор-
мируются лишь удлиненные вегетативные побеги, несущие до
Ю—20 и более зеленых листьев. С 5-го года жизни надземных
корнеотпрысковых осей на них начинают формироваться, наря-
ду с удлиненными вегетативными, укороченные специализиро-
ванные цветоносные побеги Последние возникают как боковые
побеги на 2, 3 и 4-летних годичных удлиненных побегах
материнской оси Характерное для терна сочетание вегетатив-
ных и генеративных побегов и расположение цветоносных побе-
гов показано на рис. 52.
Укороченные вегетативные побет связаны переходными фор-
мами с удлиненными вегетативными. В последовательной смене
боковых побегов от основания до верхушки материнского у
Р. spinosa находит своеобразное проявление акротонность ветв-
ления, отмеченная ранее у деревьев (см гл III) Верхушечная
почка на удлиненных побегах формируется редко. Поэтому
нарастание побегов у Р. spinosa в основном симподиальное
Укороченные вегетативные побеги Р. spinosa несут от 1 до
5—6 зеленых листьев, на верхушке они заканчиваются (как и
Удлиненные вегетативные) более или менее длинной и острой
колючкой (рис 52).
277
В подземной части стебли
Р, spinosa отличаются от ранее
рассмотренных кустарников
Corylus avellana и видов Rosa
неспособностью к ветвлению
Это типичный аэроксильный
кустарник. Как видно из рис
53, побег, возникший из прида-
точной почки корня, растет от-
весно вверх, не образуя боко-
вых ветвей,выходит на поверх-
ность почвы и продолжает рост
как надземный ассимилирую-
щий, а затем и цветоносный
побег с обильным ветвлением
в своей верхней части, с акро-
тонным развитием боковых
ветвей. Новые надземные оси
образуются не за счет ветвле-
ния материнских осей в их
подземной части, как у Corylus
avellana, Rosa spinosissima и
R. Alberti (см. выше), а за счет
придаточных почек на гори-
зонтально или наклонно расту-
щих корнях. Наиболее часто
такие почки образуются на
верхней выпуклой стороне
корней, при переходе корней
от горизонтального к верти-
кальному росту
Рост надземных побегов из
Рис. 52
Цветоносные и укороченные
вегетативные побеги терна
(Prunus spinosa),
развивающиеся на двухлетних
юдичных удчиненных побегах
придаточных корневых почек вызывает разрастание в толщину
материнского корня. Как и у других корнеотпрысковых древес-
ных растений, корень утолщается только на участке, идущем от
основания придаточного побега в сторону растущей верхушки
корня. В противоположном направлении, т. е на участке от
основания придаточного побега до основания материнского кор-
ня, прироста в толщину не происходит. Поэтому диаметр мате-
ринского корня по обе стороны от места прикрепления корно-
отпрыскового стебля оказывается резко различным (рис. 53)
Как видно из приведенного краткого очерка, основное свое-
образие морфогенеза кустарника у Р. spinosa, отличающее его-
от рассмотренных выше кустарников, заключается в отсутствии
у пего ветвящихся подземных долголетних стеблей — ксилопо-
диев, дающих начало новым надземным осям. Возобновление
278
надземных осей в онтогене-
зе Я. spinosa происходит за
счет придаточных почек на
корнях. Морфогенез кус-
тарника у Р. spinosa во
взрослом состоянии пред-
ставляет в сущности после-
довательную смену надзем-
ных многолетних осей
(с жизненным циклом в
20—30, а, может быть, и
более лет) корнеотпрыско-
вого происхождения, приво-
дящую к образованию более
или менее густых зарослей
или куртин в несколько де-
сятков метров в диаметре.
Быстрота разрастания кур-
тин и образование новых
куртин в первую очередь
связаны с быстротой роста
горизонтальных корней, на
которых формируются при-
даточные почки, дающие
начало новым надзем-
ным осям В лесах Курско-
в дубравах Центрально-черноземного
государственного заповедника
Курской области
у. п. - уровень почвы, к — корень
о п. — подземная часть побега
го государственного заповедника вегетативное корнеотпрыско-
вое размножение происходит у терна так интенсивно, что его
заросли, как отмечалось выше, оказываются одним из главней-
ших компонентов подлеска. В то же время столь интенсивное
вегетативное размножение Р spinosa повлекло, видимо, к резко-
му ослаблению семенного воспроизведения. Наши попытки най-
ти проростки и молодые кусты семенного происхождения окончи-
лись неудачно. Поэтому цикл развития главной оси сеянцев
Р. spinosa и переход их к корнеотпрысковому возобновлению
остался неизученным.
3. ВАЖНЕЙШИЕ МОРФОЛОГО-БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
НАЗЕМНЫХ ПРЯМОСТОЯЧИХ КУСТАРНИКОВ С УДЛИНЕННЫМИ
ПОБЕГАМИ
Габитуально кустарники отличаются от деревьев значительно
меньшей высотой и одновременным наличием нескольких или
даже многих крупных «скелетных» осей — надземных ветвящихся
стеблей, среди которых первая ось — главная, не выделяется по
размерам и недолговечна
Приведенные выше очерки морфогенеза кустарниковой формы
27<*
у Corylus avellana, Evonymus verrucosa, Rosa spinosissima и
R Alberti, равно как и наблюдения за десятками видов тундро-
вых, лесных, степных и пустынных кустарников флоры нашей
страны, позволяют заключить, что меньшая высота кустарников
по сравнению с деревьями определяется вовсе не более слабым
ростом их стеблей. Выше было показано, что величина годичных
приростов в период кульминации роста осей у ряда кустарников,
например, С. avellana и видов Rosa превышает 60—70 см, иногда
достигая 140—160 см Рост такой интенсивности и такие длин-
ные годичные побеги встречаются редко даже у высокостволь-
ных деревьев. Меньшая высота кустарников, по сравнению с
деревьями, связана не с более слабым ростом кустарников, а с
меныиим долголетием и резким сокращением жизненного цикла
их надземных осей по сравнению с осью дерева.
Если у деревьев жизненный цикл главной оси (ствола) и
период ее роста (моноподиального или симподиального) длится
до 80—100 и более лет, то у кустарников главная ось и заме-
няющие ее оси возобновления (тоже часто симподиальные)
завершают свой жизненный цикл (от появления над поверх-
ностью почвы до прекращения верхушечного роста) в течение
10—15, иногда 20—30, редко более, лет. Интенсивность роста
надземных осей у кустарников, как и \ деревьев, изменяется в
онтогенезе оси по типу одновершинной кривой, и лишь у неко-
торых кустарников, например видов Rosa, от одновершинной
кривой в надземной области сохраняется лишь ее нисходящая
часть Длительность общего периода роста надземных осей
у кустарников в среднем в 10—15 раз меньше таковой у де-
ревьев. В связи с этим кульминация роста, наступающая
у наших лесных деревьев в возрасте от 10—20 до 40—60 лет
(см. гл III), у кустарников наблюдается на 3—5 году жизни
оси. Быстрое достижение кульминации роста надземных осей
с необходимостью вызывает соответственно раннее ослабление
и прекращение верхушечного роста осей и, как следствие этого,
раннее коррелятивное раскрытие сохранившихся жизнеспособ-
ных спящих почек в базальной части осей, ослабивших или уже
прекративших свой верхушечный рост.
Раскрытие спящих почек в основании надземных стеблей и
образование из них новых осей возобновления у кустарников
(процесс кущения) является, таким образом, следствием быст-
рого ослабления и прекращения роста материнских осей.
У деревьев спящие почки на разных частях ствола тоже мо-
1ут пробуждаться при прекращении роста ствола по естествен-
ной или искусственной причине. У кустарников это происходит
естественно и закономерно, обычно только у основания побегов
и весьма рано по сравнению с деревьями (образование вторич-
ных крон происходит лишь у старых деревьев!).
280
Вероятно, следствием резкого сокращения жизненного цикла
главной оси и замещающих ее осей возобновления оказывается
и вторая габитуальная особенность кустарников—наличие у
них одновременно нескольких или многих надземных осей
Говоря об образовании куста, не нужно путать явлений за-
мещения главной оси новыми осями возобновления у кустарни-
ка (или срубленного дерева) и симподиально!о формирования
самой главной оси (ствола) или скелетной оси любого порядка,
где также происходит замещение одних осей другими, более
высокою порядка
При формировании симподия верхушечная почка или вер-
хушка побега отмирает, но главная или скелетная ось продол-
жает рост (иногда весьма интенсивный) в том же направлении,
за счет одной ближайшей к верхушке боковой почки, разверты-
вающейся сразу и не впадающей в состояние спящей
При замещении главной оси осями возобновления раскры-
ваются одна или несколько почек, наиболее удаленных от
верхушки, находящихся очень близко к корневой системе и пре-
бывавших до этого некоторое время в состоянии спящих Про-
исходит кущение При этом одной из главнейших особенностей
кустарников служит одновременное функционирование надзем-
ных осей нескольких поколений (обычно двух-трех, например, у
Corylus avellana, видов Rosa и др ) в общей системе кустарника
Если принять за первичную для покрытосеменных жизнен-
ную форму дерева (хотя бы и розеточного, своеобразного, о чем
говорилось в гл III), то жизненную форму кустарника надо
считать исторически вторичной, эволюционно возникшей от более
древней формы дерева
Формирование у кустарников менее долговечных, чем у де-
ревьев, надземных стеблей сопровождается дифференциацией их
стеблей на надземные, выполняющие функции ассимиляции и
плодоношения, и более долговечные и толстые, также одревес-
невшие подземные стебли — ксилоподии, выполняющие функции
вегетативного возобновления, а также функцию запасающих
органов У кустарников аридных областей подземные стебли
иногда выполняют водозапасающую функцию Развитие у кус-
тарников наиболее долговечных побегов, служащих органами
возобновления, под землей, позволяет отнести большинство
кустарников к геофильным древесным растениям с подземным
способом возобновления Это так называемые геоксильные, или
«настоящие» кустарники
Историческое преобразование древовидных жизненных форм
у покрытосеменных в кустарниковые, связанное с отмеченными
выше сокращением жизненного цикла главной оси и подземным
способом возобновления, могло вызываться в разных группах
покрытосеменных различными климатическими факторами —
281
похолоданием климата, а чаще, вероятно, аридизацией клима-
та), которые в общем можно охарактеризовать как изменение
климатических условий в сторону, неблагоприятную для разви-
тия высокоствольных деревьев. Иллюстрацией такого воздей-
ствия условий обитания может служить географическое рас-
пространение кустарников.
Причины возникновения кустарниковых форм, вероятно, мог-
ли быть и ценотическими.
Судя по тому, что и в начальный период возникновения по-
крытосеменных условия жизни этих растений могли быть очень
неоднородными (обязательное существование горных и аридных
областей даже в тропическом климате, Е. В. Вульф, 1944),
жизненная форма кустарника возникла, вероятно, филогенети-
чески очень рано и эволюционировала одновременно с жизнен-
ной формой дерева, завоевав впоследствии более широкий ареал
благодаря своей большей экологической лабильности.
Приведенный в этой главе обзор морфогенеза некоторых
кустарников позволил выявить среди кустарников определен-
ный биологический ряд форм, отличающихся по длительности
жизненного цикла надземных скелетных осей На одном флан-
ге этого ряда находятся формы, надземные оси которых отли-
чаются большой продолжительностью жизненного цикла, для-
щегося 30 и более лет, как у терна — Prunus spinosa, бородав-
чатого бересклета — Evonymus verrucosa и др. Подобные фор-
мы кустарников непосредственно примыкают к немногостволь-
ным деревьям типа Beiula tortuosa. Ближе к противоположно-
му флангу этого ряда стоят кустарники вроде Rosa spinosissima,
жизненный цикл надземных осей у которых длится 5—7 лет.
Крайняя форма этого ряда — малина — Rubus idaeus, надзем-
ные оси которой завершают жизненный цикл в 2 года. Эта
форма непосредственно граничит с травами, у которых удли-
ненные надземные ортотропные побеги живут один год 1
Интересно, что различные формы этого ряда кустарников
имеются в пределах одной и той же систематической группы,
например, семейства розоцветных, мотыльковых и др., и даже
в пределах рода, например, Rubus, Potentilla, Geum, Astragalus
и др.
Наличие подобных форм в пределах рода или близких ро-
дов позволяет проследить эволюцию жизненных форм в кон-
кретных систематических группах.
1 Чрезвычайно интересные данные о росте и возобновлении у многих
видов кустарников в условиях горного Алтая приводятся в недавно вышед-
шей книге 3 И Лучник «Обрезка кустарников» (I960). Автор ее тоже при-
ходит к выводу о существовании закономерного ряда форм кустарников
с разной длительностью жизни скелетных осей и разными способами возоб-
новления, и дает на этом основании свою классификацию кустарников, свя-
зывая ее с практическими рекомендациями по их обрезке
Глава V
ЖИЗНЕННЫЕ ФОРМЫ КУСТАРНИЧКОВ
1. ЭКОЛОГИЧЕСКИЕ, ОСОБЕННОСТИ И РАСПРОСТРАНЕНИЕ
КУСТАРНИЧКОВ
Кустарнички образуют довольно разнообразную группу
жизненных форм древесных растений, чаще с прямостоячими
или приподнимающимися надземными побегами. Отличитель-
ной габитуальной особенностью кустарничков является их
низкорослость. Кустарнички, как правило, не превышают высоты
20—30 см и лишь в редких случаях поднимаются над землей
на 60—80 см. Побеги кустарничков располагаются в приземном
слое воздуха, обычно не превышая высоты яруса травянистых
растений. Наибольшее экологическое значение низкорослости
кустарничков проявляется в неблагоприятное время года, зи-
мой, когда побеги кустарничков оказываются прикрытыми сне-
гом. Под защитой снегового покрова кустарнички в тундрах и
высокогорных альпийских поясах выходят за пределы распро-
странения деревьев и кустарников. До таежных лесов и тундр
сохраняют некоторые биологические черты тропических и суб-
тропических древесных растений, например, вечнозеленость.
Как правильно отмечает М. В. Сенянинова-Корчагина
(1949), высота этих растений (которые этот автор называет
полукустарничками) соответствует мощности снегового покро-
ва, более или менее постоянной для той или иной географиче-
ской области. В тундрах, на ровных местах, глубина плотного
снегового покрова (по Б. Н. Городкову, 1935) в среднем равна
20—30 см. В связи с этим и высота тундровых кустарничков не
превышает 20—30 см, Таким образом, низкорослость кустар-
ничков является не внешним формальным признаком этой
283
группы растений, а признаком, связанным с экологией кустар-
ничков: она позволяет им использовать в умеренных и холод-
ных зонах и высокогорных поясах лучше прогреваемые и за-
щищенные от иссушающего действия холодных ветров призем-
ные слои воздуха.
Отличительная экологическая и габитуальная особенность
кустарничков, отмеченная выше, тесно связана с их географи-
ческим распространением. Наибольшее значение в раститель-
ном покрове, доходящее до степени эдификаторов, кустарнички
получают за пределами полярной границы лесной растите пь-
ности, в северном полушарии, где они образуют так называе-
мые кустарничковые тундры, широко распространенные в зоне
тундр Евразии от Кольского полуострова до Чукотки (В. Б Со-
чава и Б. Н Городков, 1956) ’Здесь широко распространены:
Empetrum nigrum, Vaccinium myrtillus, V uliginosum, V. vitis
idaea, Cassiope tetragona, Cassiope hypnoides, Andromeda polifo-
lia, Ledum decumbens, Dryas octopetala, D. punctata, Arctous
alpina, Loiseleuria procumbens и др. В различных областях
тундровой зоны, на разных почвах, в различных элементах
рельефа, преобладают те или иные виды Подобные формации
кустарничковых тундр,.с преобладанием тех же кустарничков
из семейства вересковых, характерны для тундровой зоны Се-
верной Америки и Гренландии (A. Schimper и F. Faber, 1935;
А. П. Ильинский, 1937). Так, в растительности Южной Грен-
ландии преобладают формации вечнозеленых мелколистных
кустарничков, в которых доминируют Empetrum nigrum, Cassio-
pe tetragona, C. hypnoides, Loiseleuria procumbens, Vaccinium
uliginosum var. microphyllum, Ledum palustre var decumbens,
L. groenlandicum, Phyllodoce coerulea, Rhododendron lapponicutn
и др. Вечнозеленые кустарнички из вересковых (Colluna vulga-
tis, а иногда Erica vagans, Е. ciliaris, Е ietralix. Empetrum nig-
rum, Arctostajphylos uva ursi, Vaccinium vitis idaea, V, myrtillus,
V. uliginosum и др ) образуют своеобразный тип раститель-
ности верещатников или пустошей. В странах с океаническим
климатом, на бедных почвах, например, в прибрежных облас-
тях и северо-западной Европы, верещатники часто занимают
обширные территории
Четко выраженный ярус кустарничков из вересковых
характерен для таежных лесов Евразии и Канады
В таежных лесах Европейской части СССР по доминан-
там кустарничкового яруса — Vaccinium vitis idaea nV myrtil-
lus выделяются ельники-брусничники и ельники-черничники и
параллельные им типы боров В состав кустарничкового яруса
европейских таежных лесов входят также V uliginosum,
линнея — Linnaea borealis, Ledum palustre, Empetrum nigrum,
Arctostaphylos uva ursi.
284
Для влажных лесов умеренных, умеренно-теплых зон и тро-
пической зоны кустарнички не характерны. В умеренных влаж-
ных лесах южных провинций Чили отмечается развитие низ-
ких зарослей кустарников из Vgni molinae (Myrtaceae) и раз-
личных видов Pernettya и Gaultheria (A Schimper и F. Faber,
1935). Во влажных тропических лесах южного Китая Ан А Фе*
доров (1958) отмечает развитие среди подроста деревьев мелких
кустарников из Melastomataceae, Acanthaceae, Rubiaceae и видов
Piper Но вряд ли эти виды мелких кустарников умеренных
влажных и тропических лесов могут быть отнесены к группе
типичных кустарничков, хотя следует подчеркнуть, что резкую
границу между кустарничками и кустарниками установить
трудно. Иногда один и тот же вид, в зависимости от условий
произрастания, образует то форму кустарничка, то кустарника,
(например, болотный багульник — Ledum palustre).
Довольно широкое распространение кустарнички получают
также в высокогорных поясах различных горных стран, от тро-
пиков до холодно-умеренных зон Так, на вершинах Западно-
Яванских вулканов, выше пояса лесной растительности и криво-
лесья, весьма обычны кустарнички с мелкими, плотными
листьями—Rhododendron reiusum, Gaultheria fragrantissima,
Anaphalis javanica, Gaultheria nummularioides и др. В высоко-
горьях восточной Явы на каменистых почвах находят широкое
распространение сильно ветвящиеся кустарнички с кожистыми
несколько ксероморфными листьями — Photinia notoniana, Сор-
rosma sundana, Vaccinium variingifolium, эндемичный кустарни-
чек восточной Явы — Styphelia pungens и др В высокогорьях
Борнео, на высотах 3660—3960 м над уровнем моря, отмечены
кустарнички — Symplocos buxtfolia (Styracaceae), Drapetes eri-
coides (Thymelaeaceae) и Coprostna Hookeri (Schimper и Faber,
1935).
В высокогорьях Южной Америки, от северного Перу до
северо-западной Аргентины, в пустынеподобных формациях
пуны, па высоте 3700—4200 м над уровнем моря, среди ксеро-
фитных злаков, особенно на каменистых почвах, встречаются
многочисленные, мелколистные ксероморфные кустарнички эри-
коидного типа, преимущественно из сложноцветных, например,
Lepidophyllum quadrangulare, Baccharis Tola, Micromeria quad'
ra&gulare и др
В высокогорных формациях пуны, на засоленных почвах
у разных видов растений проявляется тенденция к возможно
меньшему выносу побегов на поверхность почвы. Подобная тен-
денция проявляется также у некоторых видов кустарничков
в высокогорных пустынных формациях восточного Памира
(К В. Станюкович, 1949). К- В Станюкович выделяет на вос-
точном Памире особую группу жизненных форм, названную им
285
подземными кустарничками (например, Dasiphora dryadanthol-
des, Lonicera Semenovii, Ephedra Fedtschenkoana). Тенденция
к подземному росту побегов проявляется также у некоторых
арктических кустарничков, например у полярной ивы — Salix
polaris.
В высокогорных поясах умеренно-теплых и умеренных зон
также значительно развиты кустарнички. Так, большие терри-
тории Армянского нагорья заняты кустарничковой полупусты-
ней Здесь часты заросли подушковидных кустарничковых форм
астрагалов {Astragalus) и Acantholimon, а также виды Zygop-
hullum, Calligonum и др.
В альпийском поясе Новой Зеландии на восточных склонах
Новозеландских гор господствуют высокогорно-пустынные фор-
мации, сходные с высокогорной растительностью Анд Южной
Америки.
По данным К. В Станюковича (1949), в субальпийском
поясе и нижней части альпийского пояса Восточного Памира
кустарнички обычны. Здесь они встречаются по самым узким
каньонообразным долинам Но даже в этих более или менее
защищенных местообитаниях кустарнички Восточного Памира
представлены шпалерными формами. Один из наиболее обыч-
ных памирских кустарничков — Salix Fedtschenkoi встречается
на лугах между кочек, образуя лежащий на земле стволик
с боковыми ветвями, не превышающими высоты трав, полегаю-
щих в зимнее время.
В альпийском поясе северо-восточного Тянь-Шаня (напри-
мер, в Заилийском Алатау и Кунгей Алатау) изредка ветре*
чается шпалерный кустарничек Arctous alpina. В более северных
горных системах Азии — на Алтае, Саянах, в Яблоновом хребте
и др., выше лесного пояса располагается пояс гольцов или гор-
ных тундр, в котором кустарнички становятся одним из доми-
нирующих типов жизненных форм. У полярного предела рас-
пространения лесов горные тундры непосредственно смыкаются
с зональными типами тундровой растительности и, в частности,
с кустарничковыми тундрами.
Из приведенного краткого обзора географического распро-
странения кустарничков видно, что кустарнички встречаются
в областях умеренных и холодных зон, а также альпийских
поясов, т. е. на территориях с холодным зимним временем года
и снеговым покровом.
Наибольшее значение в сложении растительного покрова
кустарнички приобретают за пределами распространения лесной
растительности, где они образуют ряд формаций так называе-
мых кустарничковых тундр На бедных почвах в пределах
лесной зоны в странах с океаническим климатом кустарнички
286
образуют на обширных территориях так называемые верещат-
никовые пустоши.
В областях с преобладанием кустарничков в растительном
покрове наблюдается в то же время наибольшее видовое и эко-
лого-морфологическое разнообразие кустарничков. В кустар-
ничковом ярусе таежных лесов и болот северного полушария
получает наибольшее распространение группа ползучих кустар-
ничков с ранним отмиранием главного корня, вегетативно под-
вижных, обладающих резко выраженным вегетативным размно-
жением посредством укореняющихся ползучих побегов.
2. МОРФОГЕНЕЗ ЖИЗНЕННОЙ ФОРМЫ КУСТАРНИЧКА
У НЕКОТОРЫХ ТИПИЧНЫХ ПРЕДСТАВИТЕЛЕЙ
Морфогенез жизненной формы кустарничка
у черники (V actinium myrtillus L.)
Семена черники прорастают надземно (S. Grevillius и
О. Kirchner, 1923; А Авдошенко, 1949). В лесах Московской
области всходы черники появляются в мае. На гипокотиле
10—15 мм высоты развиваются две овальные поЬти сидячие
семядоли 2,5—3 мм длины и около 1 мм ширины. Они сохра-
няются на стебле в зеленом состоянии в течение 2—3 лет Выше
семядолей на главной оси проростка за лето образуется от
5—7 до 12—15 мелких, слабо зубчатых зеленых ассимилирую-
щих листьев, отмирающих, как и семядоли, через 2—3 года.
Зеленые листья расположены довольно тесной спиралью на
коротком стебле, вырастающем за первое лето примерно на
1 см. В конце лета верхушка стебля отмирает с образованием
небольшого шипика Поэтому уже со второго года нарастание
побега происходит симподиально. Главный корень за первое
лето вырастает лишь на 1,5—2 см, но довольно обильно ветвит-
ся до корней 3-го порядка.
Из всех пазушных почек главной оси (от семядолей до верх-
него зеленого листа) весной следующего года раскрывается
лишь самая верхняя боковая почка, дающая начало побегу
продолжения главной оси. Остальные пазушные почки остаются
в состоянии нераскрывшихся спящих почек. Годичный побег
второго года, как и в первый год, не превышает 1,5—2 см. В его
основании формируются 3—4 чешуевидных, а выше 5—7 асси-
милирующих зеленых листьев, сохраняющихся на стебле жи-
выми (как и семядоли) в течение 2—3 лет. Свойство вечно-
зелености ювенильных растений черники сохраняется до 10—
12-летпего возраста, и лишь позже, с образованием более круп-
ных кустов из укореняющихся подземных ползучих побегов,
черника становится летне-зеленым растением с листвой, еже-
годно отмирающей в конце вегетационного периода. Переход от
287
вечнозелености к летнезелености в онтогенезе черники, вероятно,
следует объяснять биогенетически, как следствие первичности
вечнозеленых древесных форм среди Ericaceae (Н Sleumer,
1941, F. Niedenzu, 1889).
Верхушка побега, как и в первый год, в конце лета отмирает
с образованием характерного для годичных побегов черники
шиповидного бурого выроста. Длительное сохранение этого
шипика на ветвях позволяет легко определить возраст сеянцев
черники, а у взрослых растений — возраст ветвей до 8—10 лет,
а иногда и более. Из пазушной почки верхнего зеленого листа
весной 3-го года образуется побег продолжения главной оси, по
величине, структуре и свойству вечнозелености не отличающийся
заметно от годичного побега предыдущего лета Наряду с побе-
гом продолжения в течение третьего года раскрываются одна
или две спящие почки присемядольной зоны главной оси, из
которых образуются боковые наклонно вверх растущие ветви
Годичные побеги боковых ветвей не превышают 1,5—2 см
длины и не отличаются существенно от годичных побегов сим-
подиальной главной оси.
С появлением боковых ветвей сеянцы черники начинают
приобретать облик миниатюрных вечнозеленых кустарничков.
Веспой четвертого года у некоторых экземпляров раскрываются
верхние боковые почки главной оси и боковых ветвей, обеспечи-
вая симподиальное нарастание этих осей. У других экземпляров
пазушная почка верхнего годичного побега главной оси не
раскрывается и главная ось, достигнув 5—8 см высоты, на чет-
вертом, реже пятом или шестом году жизни сеянца, теряет
способность к дальнейшему нарастанию в высоту Дальнейший
рост кустовидных сеянцев черники, по прекращении роста глав-
ной оси, идет за счет нарастания их боковых ветвей. Послед-
ние, как и главная ось, нарастают симподиально, с ежегодным
отмиранием па верхушке и образованием характерного буро-
ватого шиповидного выроста Как и у главной оси, побеги нара-
стания их формируются из пазушной почки верхнего зеленого
листа
На втором или третьем году после их появления, на некото-
рых ветвях раскрывается не только верхняя пазушная почка
с образованием побега продолжения, но и одна или две спящие
почки в основании этих ветвей. С раскрытием спящих почек
формируются ветви третьего порядка в общей системе миниа-
тюрного кустика сеянца черники. По интенсивности роста ветви
третьего порядка не отличаются существенно от главной оси
и ее боковых ветвей (осей второго порядка). Они оказываются
лишь еще более наклоненными, нередко растущими горизон-
тально, и обладают еще более кратким периодом роста в вы-
соту, который у них продолжается обычно 2—3 года
288
Рост за счет формирования побегов продолжения главной оси
и боковых ветвей и раскрытия спящих почек в основании этих
осей приводит к тому, что к 8—10 годам жизни сеянца у чер-
ники образуется миниатюрный, карликовый вечнозеленый кус-
тарничек, высотой в 6—10 см и такого же диаметра. Он несет
около десятка боковых наклонных или почти вертикальных
боковых ветвей второго и третьего порядка, сходящихся у сво-
его основания к главной оси. Верхняя часть главной оси к этому
времени оказывается уже отмершей на большем или меньшем
протяжении. Корневое питание этого миниатюрного кустарничка
осуществляется за счет сильно разветвленного главного корня
достигающего 20—30 см длины, и образующего систему боковых
корней до V—VII порядков. Карликовый кустарничек, форми-
рующийся в онтогенезе черники при семенном воспроизведении
целесообразно назвать ее первичным кустом (Серебряков И. Г
и Чернышева М. Б., 1955). Структурно первичный куст черники
действительно представляет собою ветвящуюся от основания до
III порядка главную ось проростка, снабженную первичной кор-
невой системой. Наличие главной оси и главного корня —
весьма характерный признак кустарничка у черники на этом
возрастном этапе (рис. 54).
Дальнейший рост первичного куста черники сопровождается
не только образованием новых ассимилирующих побегов Из
спящих почек, расположенных в базальной части главной оси.
и побегов второго порядка первичного куста, прикрытых к этому
времени лесной подстилкой или моховым покровом, начинают
формироваться длинные подземные ползучие побеги Они вна-
чале имеют розовато-белую окраску и несут спирально располо-
женные мелкие пленчатые низовые листья, отделенные междо-
узлиями в 2—3 см В пазухах пленчатых листьев формируются
почки, образующие боковые подземные ползучие побеги, по
типу материнских. Чуть выше пазушной почки (на 0,5—1 льч),
на ползучих подземных побегах черники, формируется по типу
сериальных образований по одному стеблевому надузловому
корню. Формирование надузловых стеблеродных корней чуть
выше пазушной почки характерно и для других Ericaceae
(Е Warming, 1908; W. Rauh, 1937)
Ползучие подземные побеги некоторое время растут гори-
зонтально, удаляясь в разные стороны по радиусам от первич-
ного куста. Их длина в рыхлой лесной подстилке или моховом
покрове достигает в лесах Московской области от 10—15 до
30—40 см После периода горизонтального роста верхушка пол-
зучих подземных побегов приобретает ортотропное направление,
выходит на поверхность почвы, продолжая рост как ассимили-
рующий воздушный побег. Ассимилирующие побеги, формирую-
щиеся на ползучих подземных побегах при выходе их верхушки
В-413. И Г. Серебряков -19 289
Рис. 54
Формирование
первичного куста
и куртины черники
(Vaccinium myrtil-
lus) в хвойных ле-
сах Московской
области (по Сереб-
рякову и Черны-
шевой, 1955).
Л, Б, В—формирование
первичного куста,
Г — начало формиро-
вания новых парциаль-
ных кустов после от-
мирания первичного,
Д - схема структуры
взрослой куртины чер-
ники с парциальными
юстами до 9 поряд-
ков, А — однолетний,
Б — трехлетний, В —
7 — 8-летннй сеянцы,
I - главная ось, II и
III - боковые оси, 1, 2,
3. — годичные приро-
сты, с — семядоли, с.
у.- семядольный узел,
гп - гипокотиль, г,
к.— главный корень,
5. к. — боковые корни,
п. к. — придаточные
корни, г. о. п. к.—глав
hi я ось парциального
куста, п. к. 1, п. к
2. — парциальные ку-
сты последовательных
порядков
на поверхность почвы, отличаются несколько большей интен-
сивностью роста, в сравнении с побегами первичного куста
У взрослых экземпляров черники они вырастают за первое лето
на 5—8 см и несут более десятка зеленых листьев. Но, как и в
первичном кусте, верхушка этих годичных побегов в конце ве-
сенне-летнего роста отмирает, с образованием характерного для
черники буроватого острого шипика. Зеленые листья осенью на
некоторых побегах отмирают, и под снег уходят лишь стебли
с закрытыми почками. Но у других побегов еще сохраняются
на зиму зеленые листья, которые продолжают ассимилировать
и в течение следующего вегетационного периода После пере
зимовки на этих воздушных побегах раскрывается обычно две,
реже три верхние пазушные почки, давая начало побегу про-
должения и боковым ветвям новых кустов, сходных по струк-
туре с первичным материнским кустом черники, но несколько
больших размеров (10—15с.и высоты притойже ширине куста)
Кусты, развивающиеся терминально на пол-
зучих подземных побегах, по выходе верхушек
этих побегов на поверхность почвы, целесо-
образно назвать парциальными кустами чер-
ники, так как каждый из них действительно
оказывается частью целого материнского рас-
тения, представляющего к этому времени не-
большую куртину или латку (Серебряков и Черны-
шева, 1955).
Молодая куртина черники состоит из системы отдельных
надземных кустов с материнским первичным кустом в 1&нтре
и несколькими парциальными кустами, окружающими материн-
ский первичный куст, причем все эти кусты соединены с первич-
ным кустом подземными ползучими побегами типа удлиненных
горизонтальных корневищ. Парциальные кусты черники, внешне
сходные с материнским первичным кустом, отличаются от по-
следнего рядом существенных признаков. Биологически главная
ось парциальных кустов, в отличие от первичного куста, пред-
ставляет в морфологическом отношении не главную ось расте-
ния, а боковой побег, являющийся продолжением подземных
ползучих побегов, возникших из спящих почек главной оси или
боковых осей первичного куста В отличие от первичного куста,
окоренение парциальных кустов происходит не за счет главного
корня, а посредством придаточных стеблевых корней подземных
ползучих побегов; их корневая система — вторичная
В формировании и росте парциальных кустов черники есть
некоторые общие черты с се первичным кустом Как отмечалось
выше, по выходе на поверхность почвы, ползучий подземный
побег приобретает характерную для стеблей черники зеленую
окраску, а вместо низовых на стебле формируются зеленые
19* 291
ассимилирующие листья. Из-за отмирания верхушки стебля по
окончании весеннего роста нарастание биологически главной
оси парциального куста уже со второго года происходит сим-
подиально. Максимум роста биологически главной оси парци-
ального куста достигается на втором году его жизни, а общее
нарастание главной оси парциальных кустов, как и в первичном
кусте черники, продолжается не более 4—5 лет, после чего начи-
нается ее отмирание с верхнего годичного побега После отми-
рания верхушки главной оси нарастание парциальных кустов
продолжается за счет боковых ветвей Ветвление главной оси
парциальных кустов начинается со второго года ее жизни и
кончается на четвертом году Каждая из боковых ветвей вто-
рого порядка также нарастает 4—5 лет, после чего отмирает,
начиная с верхушки. В отличие от главной оси парциального
куста, интенсивность нарастания боковых ветвей снижается уже
после первого года Вследствие того, что отмирание боковых
ветвей второго порядка начинается позже, чем у главной оси,
длительность жизни парциальных кустов возрастает до 6—7 лет.
Боковые ветви второго порядка в свою очередь ветвятся, обра-
зуя ветви третьего порядка, а последние — четвертого и пятого
порядков Но ветви высших порядков прекращают нарастание
быстрее, чем главная ось парциального куста, уже на 2-м или
3-м году их жизни Поэтому за счет образования ветвей IV
и V порядков общая длительность жизни парциального куста
возрастает незначительно — до 8, редко 9 лет. Парциальные
кусты стареют и отмирают, едва превышая по высоте их мате-
ринскую ось (20—30 см)
Но отмирание куста происходит лишь в его верхней и сред-
ней части, так как в основании парциального куста, из спящих
почек первого годичного побега его главной оси вырастает
один, реже два новых побега, обладающие относительно более
мощным ростом Они были названы побегами замещения глав-
ной оси парциальных кустов (Серебряков И Г. и Черны-
шева М Б, 1955). Общий период их нарастания, как и у глав-
ной оси парциальных кустов, длится не более 5 лет. Ветвление
побегов замещения начинается со 2 и прекращается на 4-м году
их жизни В результате нарастания и ветвления побегов заме-
щения в основании парциального куста формируется новый
парциальный куст замещения, структурно подобный материн-
скому Длительность жизни его также достигает 8—9 лет.
В общем жизненном цикле парциальных кустов взрослых
экземпляров черники в лесах Московской области плодоношение
начинается с 4—5 лет, а заканчивается в 12—15 лет каждого
парциального куста.
Но с отмиранием первых генераций парциальных кустов чер-
ники общее отмирание растения в целом не наступает Еще
292
задолго до отмирания парциального куста, на 4—6-м году его
жизни, в подземной части главной оси парциального куста из
спящих почек возникают новые подземные ползучие побеги,
растущие горизонтально в верхних слоях почвы. Моноподиаль-
ное нарастание этих подземных побегов может продолжаться
в почве до 6—7 лет, а общая длина их достигает 120—140 см,
пока верхушка этих побегов не примет ортотропного положения
и не выйдет на поверхность почвы, чтобы дать начало новому
парциальному кусту.
Интенсивный рост подземных ползучих побегов обеспечивает
довольно быстрое разрастание куртин черники в радиальных
направлениях, захват парциальными кустами черники все новой
территории Отмирание системы главного корня, а затем и всего
первичного куста, делает чернику с известного возраста веге
тативно подвижным растением, распространяющимся при по-
мощи подземных ползучих побегов, дающих начало новым
поколениям парциальных кустов, по мере старения и отмирания
старых. «Расползание» куртин черники усиливается благодаря
тому, что большинство подземных ползучих побегов в пределах
куртины растет радиально от центра к периферии куртины При
этом максимальной длины подземные ползучие побеги дости-
гают у парциальных кустов, расположенных на периферии кар-
тины О быстроте разрастания куртин черники можно судить,
например, по тому, что в ельниках-зеленомошниках 60—70-лет-
него возраста на территории Звенигородской биостанции МГУ
(Московская область) иногда можно видеть куртины черники
диаметром 6—8 м из нескольких сотен парциальных кустов,
возникших в результате разрастания одного семенного экзем-
пляра
Изучение подземных структур таких куртин показывает, что
каждая из них представляет единое структурное целое (комп
леке парциальных кустов, связанных сетью подземных ползучих
побегов) В конечном итоге все подземные побеги ведут к од-
ному биологическому центру — первичному материнскому
кусту, отмирающему на определенном возрастном этапе форми-
рования куртины
Как видно из описанного выше, с начала образования на
материнском первичном кусте подземных ползучих побегов и
формирования на них, после выхода из верхушек на поверх-
ность почвы, парциальных кустов, замещающих первичный куст,
дальнейший онтогенез черники в морфолого-биологическом
плане выражается в последовательной смене парциальных
кустов все более высокого порядка. Сменяющиеся одно за дру-
гим поколения парциальных кустов в общей куртине черники
сходны между собою структурно и биологически Однако по-
следующие поколения парциальных кустов не являются точной
293
копией предыдущих Первые парциальные кусты, возникшие
непосредственно из первичного материнского куста, а иногда
и второе поколение парциального куста, сохраняют еще свой-
ства вечнозелености Парциальные кусты, возникающие позже,
свойством вечнозелености уже не обладают Смена первых
поколений парциальных кустов (не менее 10—15 поколений)
сопровождается усилением интенсивности роста побегов и, как
следствие этого, увеличением общей высоты парциальных
кустов. Если первое и второе поколение парциальных кустов
в ельниках-зеленомошниках Московской области (Звенигород-
ская биостанция МГУ) достигают высоты 10—15 см, то пар-
циальные кусты последующих поколений (порядка 8—10 и
выше) у более старых куртин, диаметром в 6—8 м, достигают
высоты 30—40 см.
Наряду с увеличением высоты надземных побеюв, смена
парциальных кустов в этот период формирования куртины
сопровождается также увеличением длины подземных ползучих
побегов. Так, при формировании двух-трех первых поколений
парциальных кустов длина подземных ползучих побегов в ель-
никах-зеленомошниках Московской области достигает от 10—
15 до 30—40 см, а у более старых куртин их длина может
достигать 100—150 см Наконец, в описываемый период форми-
рования куртины наблюдается повышение интенсивности плодо-
ношения у парциальных кустов более высокого порядка.
Наряду с изменением высоты и интенсивности роста и плодо-
ношения у парциальных кустов в течение описываемого воз-
растного периода у черники наблюдается также изменение
общей структуры куртины. В начале куртина черники пред-
ставляет систему парциальных кустов, связанных при помощи
подземных ползучих побегов с центральным первичным кустом,
их породившим В таких молодых куртинах черники, по иссле-
дованию Ю. А. Злобина (1960) все парциальные кусты обла-
дают одинаковой способностью к вегетативному возобновлению
Затем, по мере разрастания куртины, по данным Злобина,
наблюдается дифференциация парциальных кустов, в резуль-
тате которой центральные более старые парциальные кусты
обладают меныпей способностью к вегетативному размножению
по сравнению с парциальными кустами на периферии куртины.
Старение парциальных кустов в центре куртины приводит в ко-
нечном итоге к их отмиранию в центре куртины и формирова-
нию старых куртин В дальнейшем, по данным Ю. А Злобина,
наблюдается ослабление вегетативного возобновления парци-
альных кустов и на периферии куртины.
Естествен вопрос — как долго может повторяться после
довательная смена парциальных кустов в онтогенезе черники?
Образование своей придаточной корневой системы у каж-
294
лого из парциальных кустов обеспечивает значительную само
стоятсльность и затушевывает признаки старения в ряду после-
довательных поколений парциальных кустов в разрастающейся
куртине черники. Большой жизненный цикл черники, вероятно,
длится много более сотни лет, и по возрасту куртины черники
приземного яруса наших хвойных лесов может быть ничуть не
уступают старым деревьям, господствующим в верхнем ярусе,
а иногда даже и превосходят их. О значительной длительности
жизни черники свидетельствует, кроме сказанного, также
и позднее начало плодоношения ее. В общем онтогенезе чер-
ники, в лесах Московской области, ее плодоношение начинается
не ранее 15—20-летнего возраста
Анализ морфогенеза черники от прорастания и до форми-
рования зрелых и старых куртин позволяет выделить в ее онто-
генезе в лесах Московской области два основных морфолого-
биологических периода.
I возрастной этап — формирование первичного куста — от
проростков до вполне сформированного первичного куста, высо-
той и диаметром в 6—8 см Структурно первичный куст состоит
из симподиальной ортотропной главной оси, являющейся про-
должением главной оси проростка, и более или менее наклонных
боковых осей II и III порядков, возникающих из спящих почек
базального годичного побега материнской оси Жизненный цикл
главной и боковых осей первичного куста (от начала и до конца
нарастания в длину) длится 4—5 лет. Корневая система пер-
вичного куста представлена системой главного корня, ветвяще-
гося до корней V—VII порядков, с расположением большой
части поглощающих корней в поверхностных горизонтах почвы,
не глубже 10—15 см. Завершение первого возрастного периода
и переход ко второму основному периоду в онтогенезе черники
связан с появлением в основании первичного куста, из спящих
печек подземной главной или боковых осей длинных подземных
ползучих побегов, верхушки которых, по выходе их на поверх-
ность почвы, дают начало новым парциальным кустам, заме-
няющим первичный куст, и образующим в общей совокупности
форму латки, а затем и куртины, характерной для второго воз-
растного периода в онтогенезе черники Этап первичного куста
в онтогенезе черники относится к виргинильному периоду.
II возрастной этап — формирование все более разрастаю-
щейся куртины, состоящей из системы парциаль-
ных кустов, связанных между собой под землей ползучими
укореняющимися побегами Морфогенез черники в течение
И возрастного этапа представляет по существу последователь-
ную смену парциальных кустов все более высокого порядка, с
образованием наиболее интенсивно растущих парциальных
кустов по периферии куртины Второй возрастной этап начи-
295
нается с момента выхода на поверхность почвы верхушек пол-
зучих подземных побегов, возникших на первичном к\сте
и дающих начало парциальным кустам II порядка (считая пер-
вичный материнский куст за куст I порядка). Второй возраст-
ной период заканчивается отмиранием всего растения в целом.
Большая длительность II возрастного периода и изменение в
структуре куртины позволяют разделить его на два периода.
1 период — характеризуется более или менее равно-
мерным распределением парциальных кустов на всей
площади разрастающейся куртины от периферии до центра.
В начале этого подпериода центр куртины бывает занят сохра-
няющимся некоторое время материнским первичным кустом.
В течение этого подпериода наблюдается усиление интенсив-
ности роста побегов и соответственно увеличение высоты пар-
циальных кустов все более высокого порядка.
2 п е р и о д — характеризуется тем, что в центральной части
куртины первичный куст отмирает, а вместе с ним и самые
старые парциальные кусты. Живые парциальные кусты распо-
лагаются в периферийной части куртины, образуя подобие
«лысого куст а». В течение второго подпериода наблю-
дается ослабление роста побегов парциальных кустов и пони-
жение способности их к вегетативному возобновлению.
Материалы по биолого-морфологическим изменениям струк-
туры куста и куртины черники в целом в онтогенезе этого рас-
тения позволяют высказать некоторые соображения относи-
тельно определения возраста черники в природных условиях.
Следует отметить, что определение возраста черники, сравни-
тельно легкое и простое в начале ее онтогенеза (в течение пер-
вого возрастного периода — формирования первичного куста),
у более старых экземпляров, уже перешедших к формированию
разрастающейся куртины, становится все более сложным, осо-
бенно с момента отмирания первичного куста в центре куртины.
Возраст таких старых куртин может быть определен лишь
приблизительно.
Возраст первичного куста может быть легко определен по
годичным побегам главной оси, если она еще нарастает, или
по годичным побегам одной из боковых нарастающих осей с
прибавлением возраста главной оси, с которого началось
ветвление. Вследствие симподиальности нарастания, границы
юдичных побегов у черники остаются заметными до 5—8 лет.
Для контроля можно параллельно использовать метод подсче-
та годичных колец в основании главной оси — в присемядоль-
ной зоне или па гипокотиле
С переходом черники во второй возрастной период—фор-
мирования куртины, состоящей из системы парциальных кус-
тов, связанных подземными и укореняющимися побегами, ста-
296
ловится >же недостаточным определение возраста отдельных
парциальных кустов для суждения о возрасте всей куртины в
целом Последняя слагается из сменяющих друг друга во
времени парциальных кустов все более высоких порядков
Можно было думать, что более близкие данные о возрасте
куртины можно получить при изучении подземных побегов,
связывающих отдельные парциальные кусты. Вследствие того,
что границы годичных побегов у подземных осей черники не-
заметны, оставалось обратиться к подсчету годичных колец
Однако подсчеты годичных колец и общего количества слоев
ксилемы на поперечных срезах стеблей показали, что ежегод-
ное нарастание вторичной ксилемы в результате камбиальной
деятельности наблюдается только в надземной части побегов,
и наибольшее количество годичных колец насчитктвается в
приземной части каждого парциального куста.
По мере удаления от основания парциального куста по
подземному стеблю в базипетальном направлении диаметры
стебля и количество слоев вторичной ксилемы в нем часто за-
метно уменьшаются. Особенно заметно уменьшается толщина
стебля и мощность вторичной ксилемы позади многолетних,
хорошо развитых придаточных корней Нарастание ксилемы и
соответственно числа годичных колец в стеблях парциальных
кустов происходит до зоны окоренения стеблей Дальше этой
зоны в базипетальном направлении нарастание вторичной кси-
лемы не распространяется.
На этом основании можно говорить об определенном бло
кирующем действии придаточных корней, которые задержи-
вают идущее сверху нарастание ксилемы и «поглощают» его
(нарастание ксилемы продолжается в этих корнях). Корни нс
пропускают физиологические импульсы, необходимые для кам-
биальной деятельности, далее по длине подземных стеблей в
базипетальном направлении. Вероятно, воздействие мощных
придаточных корней, расположенных близ выхода подземных
стеблей на поверхность почвы, препятствует нарастанию кси-
лемы в подземных стеблях черники и они остаются тонкими,
несмотря на их значительный возраст. Исключением из всех
подземных органов в куртине черники оказываются лишь гипо-
котиль и главный корень первичного куста, в которых долго
сохраняется способность к увеличению числа годичных колец,
длительно сохраняется камбиальная активность. Иначе гово-
ря, эти органы долго сохраняют предковые черты подобных же
органов настоящих деревьев.
Отмеченная способность гипокотиля и основания главного
корня первичного куста черники может быть использована при
определении возраста сравнительно молодых куртин черники с
еще сохранившимся первичным кустом. В лесах Московской
297
области таким путем удается определить возраст куртин чер-
ники 25—30-летнего возраста, достигающих к этому времени
диаметра в 2,5—3 м. При применении этого метода надо обя-
зательно найти первичный куст в изучаемой куртине, взять его
главную ось и главный корень, и на поперечных срезах гипо-
котиля или основания главного корня подсчитать число годич-
ных колец
Примерный возраст куртины черники может быть опреде-
лен и иным методом, основанным на подсчете числа поколений,
или порядков парциальных кустов, входящих в состав кур-
тины.
Как отмечалось выше, спящие почки в основании главной
оси парциального куста пробуждаются и дают начало главной
оси нового парциального куста только на 5—6-м году жизни
главной оси материнского парциального куста после выхода ее
на поверхность почвы в связи с прекращением ее роста в
длину. Поэтому каждый последующий порядок парциальных
кустов возникает позже предыдущего на 5—6 лет. Другими
словами, разность в возрасте парциальных кустов смежных
поколений в лесах Московской области равна 5—6 годам.
Отсюда, очевидно, возраст куртины будет равен числу генера-
ций парциальных кустов куртины, умноженному на разность в
возрасте парциальных кустов смежных порядков. Для опре-
деления возраста куртины черники необходимо подсчитать
число порядков (или поколений) парциальных кустов в кур-
тине (обнажая подземные ползучие побеги) по разным ра-
диусам ее и взять наибольшее число из полученных. Одновре-
менно следует проверить начало раскрытия спящих почек в
основании главных осей парциальных кустов для уточнения
разности в возрасте парциальных кустов смежных порядков в
данных экологических условиях Если середина куртины уже
лишена парциальных кустов, необходимо к полученному числу
(произведению от числа порядков парциальных кустов на раз-
ность в возрасте смежных порядков) прибавить примерный
возраст срединной части изучаемой куртины с отмершими
парциальными кустами. Последнее число должно быть полу-
чено в тех же экологических условиях при определении воз-
раста более молодых куртин.
Особенности морфогенеза кустарничка
у брусники (Vaccinium vitis idaea L.)
Данные о морфогенезе первичного куста брусники изложены
по наблюдениям автора на территории Полярно-Альпийского
ботанического сада в Хибинских тундрах, а по структуре и
298
росту взрослых куртин брусники — по наблюдениям в еловых
и сосновых лесах Московской области. Прорастание семян
брусники, как и у черники, происходит по надземному типу. На
гипокотиле 10—15 мм длины формируются ланцетно-овальные,
почти сидячие семядоли 2—3 мм длины и около 1 мм ширины.
На эпикотильной части опушенного стебля, вырастающего за
первое лето примерно на 1 см, формируется 5—8 зеленых
листьев. Рост годичного побега проростка завершается форми-
рованием зимующей верхушечной почки, следующей весной
дающей начало побегу продолжения главной оси. Семядоли и
зеленые листья проростка функционируют, как и листья взрос-
лых растений, по типу вечнозеленых, сохраняясь на растении в
течение 3—4 лет. Главный корень вырастает за первое лето
на 2—3 см.
Весной 2-го года из верхушечной почки развивается побег
продолжения главной оси высотой в 1—2 см, несущий в осно
вании несколько недоразвитых низовых, а выше их 5—10 зе-
леных ассимилирующих листьев. В конце лета формируется
верхушечная почка, дающая весной третьего года новый побег
продолжения главной оси Подобное моноподиальное нара-
стание главной оси продолжается 3—4 года. Затем верхушеч-
ный рост главной оси прекращается и верхушка ее отмирает.
Но в течение 3-го года раскрывается одна или две спящие поч-
ки главной оси, образуя побеги замещения главной оси, с не-
большой плагиотропной частью в своем основании. Плагиотроп-
ная часть боковых побегов оказывается тем большей, чем ниже
располагается боковой побег на материнской оси. Нарастание
боковых побегов у сеянцев брусники происходит также моно-
подиально, а верхушечное нарастание боковых побегов также
длится не более 3—4 лет, после чего их верхушка отмирает,
когда боковые побеги достигают примерно высоты главной оси.
Подобным же образом из спящих почек боковых побегов
II порядка возникают побеги III порядка, сходные по разме-
рам, структуре, способу и длительности нарастания с материн-
скими побегами.
В результате ветвления образуется небольшой первичный
куст брусники, по величине и структуре сходный с таковым у
черники (рис. 55 А). Как и у последней, корневая система пер-
вичного куста брусники представлена системой главного корня,
обильно разветвленного до корней 5—7 порядков. Отличие от
первичного куста черники проявляется в моноподиальном на-
растании побегов, которое длится у брусники до цветения и
плодоношения, после чего цветоносные побеги уже нарастают
симподиально.
На 4—5-м годах жизни у сеянцев брусники, кроме образо-
вания боковых ассимилирующих побегов одна, реже две спящие
299
Рис 55
Структура парциального куста и куртины у брусники (Vaccinium vitis idaea) в лесах Московской
области и Хибинских тундрах (по Серебрякову и Чернышевой. 1955):
А, Б — формирование первичного куста брусники и подземного столоновидного корневища (Хибины); В — отрезок взрослой
куртины (Подмосковье); А — трехлетии», Б — 8 - 10-летний сеянцы; 1 и II — главная и боковые оси; г. п. - гипокотиль;
г. п. — гипокотиль, г. к. — главный корень, п. к.— придаточные корни, с. к. — столоновидное корневище, з. а. п. — зачатки
ассимилирующих побегов, пр. к. — парциальные кусты
почки главной оси дают начало подземным, горизонтально
растущим ползучим побегам, диаметром в 1—2 мм (см.
рис. 55 Б), из которых позже возникают парциальные кусты
ассимилирующих и цветоносных побегов, образуя куртину
брусники, несколько сходную с таковой у черники. Вначале
подземные побеги имеют белый или слабо-желтоватый, или
розовый цвет, а в более старом возрасте приобретают светло-
коричневую окраску. Ползучие подземные побеги у брусники,
как и у черники, несут мелкие пленчатые, прижатые к стеблю
низовые листья, расположенные более или менее правильной
спиралью и удаленные друг от друга междоузлиями длиной от
1 до 2—3 см
В пазухах низовых листьев формируются довольно быстро
раскрывающиеся почки, дающие начало новым подземным
ползучим побегам, растущим горизонтально, или анизотроп-
ным побегам, вскоре выходящим на поверхность почвы и
образующим зеленые листья и соцветия. Чуть выше пазушной
почки (около 1 мм), по типу сериального образования, форми-
руется зачаток надузлового корня, который позже вырастает в
более или менее тонкий всасывающий корень. Длинные подзем-
ные ползучие побеги брусники отличаются от черничных тем.
что они никогда не выходят на поверхность почвы Их верхуш-
ка постоянно сохраняет горизонтальное положение, обеспечи-
вая длительное моноподиалыюе нарастание этих побегов
После некоторого периода роста верхушка таких подземных
побегов все же может отмереть В таком случае ее заменяют
одна или две верхние боковые почки, ближайшие к отмершей
верхушке, которые дают начало новым подземным постоянно
горизонтально растущим побегам
Парциальные кусты надземных побегов брусники, в отли-
чие от черники, всегда возникают из боковых пазушных почек,
значительно удаленных от растущей верхушки горизонтальных
подземных побегов Надземные побеги парциальных кустов
нарастают в течение 4—5 лет. Затем, чаще после плодоноше-
ния, они заменяются боковыми побегами, с жизненным циклом
в 3—4—5 лет Смена побегов происходит не более 4—5 раз,
так что общая длительность жизни парциального куста не пре-
вышает 10—15 лет (см рис 55 В). Подземные побеги у брус-
ники нс только дают начало новым парциальным кустам, но
выполняют и запасающую функцию
В стеблях брусники, надземных и подземных, более или
менее четко выражены годичные кольца Особый интерес в
этом отношении представляют длинные многолетние подземные
побеги, которые позволяют путем подсчета годичных колец
определить возраст куртины брусники По длине подземного
укореняющегося побега брусники на поперечных срезах,
301
сделанных близ его верхушки и далее, в базипетальном направ-
лении, удается констатировать последовательное увеличение
числа годичных колец.
Возрастание числа годичных колец по длине подземного
ползучего побега позволяет установить, что средний прирост в
длину подземных стеблей брусники в изученных экологических
условиях равен около 20 см в год. В одной из куртин брусники
(в сосновом лесу с господством брусники в наземном ярусе на
территории Звенигородской биостанции МГУ) на отпрепариро-
ванном подземном побеге длиной в 3,5 м (с сохранившейся
верхушкой), в основании побега было насчитано 27 годичных
колец. У этой куртины средний прирост подземного ползучего
стебля в длину равен 13 см в год. Увеличение числа годичных
колец по длине подземного ползучего побега, т. е. длительность
нарастания вторичной ксилемы, свидетельствует о» том, что
подземный ползучий стебель у брусники выполняет активную
проводящую функцию на значительном протяжении и значи-
тельное время. Длительное нарастание ксилемы и продолжаю-
щаяся активность камбия в таких ползучих стеблях, вероятно,
•связаны с тем, что на этих побегах брусники не образуется
мощных блокирующих придаточных корней, которые развива-
ются, например, в подземной части каждого парциального кус-
та у черники.
В сосновых лесах на территории Звенигородской биостанции
МГУ (Московская область) автору вместе со студентами МГУ
удавалось отпрепарировать подземные ползучие побеги брус-
ники до 18 м длины. К сожалению, тогда не было подсчитано
количество годичных колец у этого стебля, в целях определения
возраста куртины, из которой был взят этот побег; однако,
приняв средний прирост подземного стебля брусники в данных
экологических условиях за 15—20 см (на основе определения
прироста у растений, произрастающих рядом с данной курти-
ной), находим, что возраст данной куртины достигает 90—
120 лет.
Приведенные данные о возрастании числа годичных колец
по длине подземного ползучего побега у брусники позволяют
этот признак рекомендовать для определения возраста ее кур-
тин
Как модификацию этого метода можно рекомендовать
следующее: путем подсчета числа годичных колец по длине
подземного ползучего стебля вначале определить среднюю ве-
личину годичного прироста в длину в данных экологических
условиях. Затем, по нахождении величины годичного прироста
в длину, необходимо общую длину подземного стебля разде-
лить на величину ею ежегодного среднего прироста.
302
Особенности структуры кустарничка
у андромеды (Andromeda polifolia L.)
Приводимые сведения основаны на наблюдениях и сборах
андромеды на сфагновых болотах Московской области и в гор-
ных тундрах Приполярного Урала, в верховьях р. Ляпин.
На сфагновых болотах Московской области семенных про-
ростков андромеды обнаружить не удается; размножение и
воспроизведение андромеды в этих условиях происходит веге-
тативным путем. Во взрослом состоянии на сфагновых болотах
Московской области кустарнички андромеды представлены раз-
бросанными по поверхности сфагновых болот, главным обра-
зом на микроповышениях, отдельными слаборазветвленными
вечнозелеными парциальными кустами или их небольшими
группами, высотой в 20—40 см Из-за сильной разбросанности
парциальных кустов вначале даже не создается впечатления о
развитии у этого кустарничка куртин. А между тем.парциаль
ные кусты андромеды связаны между собой подземными пол-
зучими побегами, подобно тому как это имеет место у черники
Во взрослом состоянии на сфагновых болотах Московской
области кустарнички андромеды образуют рыхлую куртищ.
структурно представляющую собой систему парциальных кус-
тов все более высоких порядков, сменяющихся последовательно
во времени. Как и у черники, новые парциальные кусты андро
меды развиваются из специализированных подземных укоре-
няющихся побегов, возникающих из спящих почек подземной
части главной оси материнского парциального куста (рис. 56).
Подземные побеги андромеды диаметром около 1 мм, бело-
розовые, с низовыми пленчатыми прижатыми к стеблю листь-
ями, вначале растут горизонтально, а затем, по ослаблении
роста материнского парциального куста, выходят на поверх-
ность сфагнового покрова. Длина подземных ползучих побегов
андромеды колеблется от 40—70 см до 3—5 м. Как и у черни-
ки, выше каждой пазушной почки на подземном стебле фор-
мируется зачаток придаточного корня, из которого вырастает
тонкий разветвленный питающий корень. По выходе на по-
верхность сфагнового покрова надземные побеги начинают
расти по типу ассимилирующих, становясь в ходе последую-
щего роста главной осью нового парциального куста. Нараста-
ние главной оси парциального куста происходит симподиально.
завершаясь терминальным соцветием на 3—4 году жизни. За
счет ветвления длительность жизни парциального куста увели-
чивается до 6—8 лет, после чего он отмирает, начиная с вер
хушки.
Нарастание ксилемы наблюдается, как и у черники, только
в надземной части куста, прекращаясь в зоне укоренения.
303
Рис. 56
Структура компактной (слева) и куртинной (справа) форм кустарничка у андромеды (Andromeda
polifolia) (по Серебрякову и Чернышевой, 1955):
Слева — из Хибинских тундр, Справа - со сфагнового болота Приполярного Урала; г. к. - главный корень, п. к. — придаточ-
ные корни, о. л к. — отмерший парциальный куст, г. о. л. к — главная ось парциального куста, с. к — столоновидное корневище
Структура и морфогенез куртины андромеды на сфагновых
болотах Московской области в основных чертах сходны с та-
ковыми у черники.
Однако подобный тип морфогенеза кустарничка у андроме-
ды — не единственный. На минеральных почвах горных тундр
Хибин и Приполярного Урала (верховья р. Ляпин) у андроме-
ды во взрослом состоянии формируются компактные кустар-
нички, лишенные длинных горизонтально растущих побегов,
удаляющих парциальные кусты на значительное расстояние
друг от друга В подобных условиях у андромеды развивается
мощный главный корень, функционирующий в течение всей
жизни растения, наряду с придаточными стеблевыми корнями
Однако значение стеблевых придаточных корней здесь срав-
нительно невелико.
Сильное развитие главного корня и его длительное функцио-
нирование приводит к длительному формированию годичных
колец в основании главного побега и главного корня. У ком-
пактного кустарничка из горных тундр Хибин, на гипокотиле
диаметром в 6,5 мм было обнаружено 21 годичное кольцо
Очевидно у таких форм кустарничков андромеды, с постоянно
функционирующим главным корнем, подсчет годичных колец
в основании главного стебля и главного корня является падеж-
ным способом определения возраста
Особенности структуры кустарничка
у голубики (Vaccinium uliginosum L.),
болотного багульника (Ledum palustre L.)
и Кассандры (Cassandra calyculata D Don)
На сфагновых болотах Московской области весьма свое-
образно проходит морфогенез кустарничка у голубики, болот-
ного багульника и Кассандры В отличие от черники, андроме-
ды и брусники, у этих кустарничков на сфагновых болотах со-
вершенно отсутствуют специализированные подземные побеги,
возникающие из спящих почек. Их побеги ежегодно нарастают
верхушкой, поднимаясь все выше над уровнем сфагнового
покрова. Но одновременно нижние части ветвей ежегодно по-
гружаются в беспрерывно нарастающий сфагновый покров,
что облегчается и ускоряется вследствие придавливания этих
стеблей снегом. Погребенные в сфагновом покрове ветви вскоре
укореняются, причем образование придаточных корней ежегод-
но продвигается все выше по стеблям, по мере их погружения
в моховой покров Такое верхушечное нарастание ветвей, с
одновременным погружением их старых частей в сфагновую
толщу и последующим укоренением, может продолжаться
в-41 i и г Серебряков -20 305
многие десятки лет. Погруженные части ветвей довольно долго
-сохраняются живыми, затем отмирают.
На Луцинском сфагновом болоте, близ Звенигородской био-
станции МГУ (Московская область) удавалось извлекать из
сфагнового покрова 40-летние подземные стебли багульника и
56-летние стебли Кассандры. Возраст погребенных ветвей (от
верхушки) легко определяется по хорошо выраженным грани-
цам годичных побегов, с ежегодно отмирающим соцветием на
верхушке годичного побега. Извлеченные побеги почти до осно-
вания были живыми, в сердцевине и сердцевинных лучах их
сохранялось (в июне) значительное количество крахмала Но
ни в одном случае не удавалось обнаружить у этих растений
на болотах главного корня и первичного куста. Не было обна-
ружено здесь и семенных проростков
Нарастание ксилемы в стеблях багульника, Кассандры и го-
лубики происходит только в надземной части У болотного ба-
гульника на сфагновых болотах не обнаруживается более 6—
7 годичных колец; у Кассандры — более 8—9 годичных колец
у голубики — более 8—10 годичных колец, независимо от их
возраста.
Однако описанный здесь способ нарастания и структура
кустарничка у голубики и багульника — не единственный. В
горных тундрах Хибин, в горных лесах и тундрах Приполяр-
ного Урала, в тундрах западных отрогов хребта Бырранга на
Таймыре автору приходилось наблюдать голубику и багульник
в форме компактных кустарничков, типа описанных выше для
андромеды
В горных тундрах Приполярного Урала (долина р Старик
Шор) на каменистых почвах кустарнички голубики достигают
высоты 5—10 см. Сближенные парциальные кусты их функцио-
нируют 8—10 лет, заменяясь со временем другими парциаль-
ными кустами за счет спящих почек в основании побегов ма-
теринских кустов. В мощном моховом покрове из этих спящих
почек образуются иногда подземные ползучие побеги, типа по-
бегов черники, но более короткие (20—30 см). По выходе па
дневную поверхность подземные побеги дают начало новым
парциальным кустам. Система главного корня у голубики в
горных тундрах Приполярного Урала функционирует многие
десятки лет. Поэтому в основании главного побега и главного
корня у голубики в горных тундрах длительное время наблю-
дается нарастание вторичной ксилемы. На собранных в При-
полярном Урале экземплярах голубики в основании главного
побега и главного корня мы насчитывали до 78 годичных ко-
лец У багульника на минеральных почвах в горных лесах
Приполярного Урала система главного корня функционируег
также длительное время. На каменистых осыпях в Хибинах, у
306
верхнего предела лесного пояса, голубика образует форму
шпалерных кустарничков, с мощным развитием главного кор-
ня. Лежачие побеги на камнях при этом почти не укореняются.
Но длительность нарастания побегов в длину у таких шпалер-
ных форм голубики значительно возрастает по сравнению с
прямостоячими кустами (с 4—5 лет до 12—13 лет). В воронич-
но-черничных ельниках Хибин, у верхнего предела лесного
пояса, с мощным развитием мохового покрова, у голубики на-
блюдается длительное нарастание побегов в длину, при полега-
нии их базальной части и укоренении, по мере погребения в
моховом покрове. При этом резко проявляется различие в тол-
щине подземных и надземных стеблей. Погребенные длительно
сохраняющиеся побеги достигают толщины 1,5—2 см, тогда
как воздушные побеги остаются тонкими, обычно не превышая
3—5 мм в диаметре. В результате в таких условиях образуется
форма своеобразного подземного кустарника с толстыми и
длинными подземными стеблями (до 3—3,5 м длины и 1,5—
2 см толщины) и тонкими короткими надземными ветвями, еще
возвышающимися над моховым покровом Возраст подземных
побегов достигает 25—30 лет
Особенности морфогенеза жизненной формы кустарничка
у вереска (Calluna vulgaris (L.) Hill.)
Географическое распространение вереска ограничено Евро
пой, где он встречается в различных экологических условиях,
от 71,5° сев широты до Португалии, Корсики, Средней Италии,
Далмации и западных берегов Черного моря на юге Европы,
заходя далее в северную часть Малой Азии (Флора СССР,
т. 20, 1955) Центром массового распространения вереска явля-
ются северо-западные Приатлантические области, где на зна-
чительных территориях вереск оказывается важнейшим эдифи-
катором так называемых верещатниковых пустошей.
Ниже приводятся некоторые данные о морфогенезе кустар-
ничка у вереска, по наблюдениям и сборам в сосновых лесах
Московской области (главным образом близ ст. Ромашково
Кунцевского района). Проростки и ювенильные растения ве-
реска встречаются нередко на открытых участках песчаной
почвы, вблизи от взрослых плодоносящих зарослей вереска.
Прорастание семян вереска, как и других растений и семейства
вересковых, рассмотренных выше, происходит по надземному
1ипу. Тонкий гипокотиль в 5—7 мм высоты выносит на свет
зеленые сидячие овальные семядоли длиной в 1—1,5 мм и ши-
риной около 1 мм Выше семядолей в течение первого года
20*
307
вырастает прямостоячая главная ось 8—10 мм длины, с 8—
10 парами супротивно расположенных листьев Их величина
Рис 57
Сеянцы вереска (Calluna vul-
garis) из сосновых лесов Мос-
ковской области*
г. к — главный корень, гпк — гипоко-
тиль, с — семядоли, I Ih Ш... — ю-
дичные приросты.
лишь незначительно превосходит
семядоли. Одна-две пары самых
нижних первичных листьев, возни-
кающих вслед за семядолями, ока-
зываются плоскими, дорзовентраль-
ными. Следующие за ними на глав-
ной оси одна-две пары листьев об-
ладают завернутыми на нижнюю
сторону краями, как это свойствен-
но многим видам вересковых Но
листовая пластинка у этих листьев
оказывается еще тонкой, плоской
Выше этих переходных листьев
начинают формироваться типичные
для вереска резолютивные, утол-
щенные листья с недоразвитой ниж-
ней поверхностью
На верхушке годичного побега
образуются более мелкие и скучен-
ные листья, прикрывающие верхуш-
ку стебля. Ниже гипокотиля в те-
чение первого года вырастает
волосовидно тонкий и не обильно
ветвящийся главный корень, не
превышающий обычно длину над-
земного побега (1—2 см — рис. 57).
В течение второго года из откры-
той верхушечной точки вырастает
побег продолжения главной оси.
Прирост второго года несколько
превышает таковой однолетнего проростка, но все же не
бывает более 1,5—2 см. У большинства экземпляров со второго
года начинается ветвление побегов (у более сильно растущих
сеянцев ветвление начинается уже в первом году).
В отличие от абсолютного большинства наших древесных
растений, пазушные почки у сеянцев вереска раскрываются
в год их заложения, образуя короткие вегетативные ассимили-
рующие веточки, лишь в 2—3 раза превышающие длину крою-
щих листьев С образованием боковых пазушных веточек у сеян-
цев вереска проявляется гетерофиллия, вызванная различием
листьев на главной оси и боковых ветвях Листья главной оси,
длиной в 3—4 мм, в 2—3 раза превышают по длине листья
боковых ветвей. На главной оси листья отделены от соседних
пар более длинными междоузлиями Любопытно, что листья
308
главной оси отмирают осенью или
ветвей они сохраняются зелеными
В течение 3—4-го года жиз-
ни сеянца продолжается моно-
подиальное нарастание главной
оси и боковых ветвей.
В росте боковых ветвей наблю-
дается определенная диффе-
ренциация по длине материнско-
го годичного побега. Нижние
боковые веточки каждого годич-
ного побега растут медленнее
и через 3—4 года обычно прекра-
щают рост и подсыхают. Верх-
ние боковые ветви годичного по-
бега растут сильнее и более
долголетни, мало уступая по ин-
тенсивности роста и долголетию
главной оси (рис 58). Границы
между годичными побегами на
надземных осях вереска морфо-
логически выражены довольно
слабо, и лишь при внимательном
осмотре могут быть разли-
чимы благодаря развитию здесь
более мелких листьев, отде-
ленных от соседних пар междо-
узлиями более короткими, чем
в середине годичного побега.
Наличие более мощных боковых
ветвей в верхней части годич-
ных побегов (акротония ветвле-
ния) также может быть исполь-
зовано для подсчета числа го-
дичных побегов на боковых вет-
вях или главной оси.
В 5—7-летнем возрасте у ве-
реска в изученных экологических
условиях начинается цветение
и плодоношение. Первые цветки
появляются вначале на главной
оси, в средней или верхней
части годичного побега текуще-
го года. На 1—2 года позже
цветки появляются также и на
сильно растущих верхних боко-
зимой, тогда как у боковых
в течение 2—3 лет.
Рис. 58
Структура ветвящегося годичного
побега вереска из сосновых лесов
Московской области:
/ — неветвяшаяся базальная часть годич-
ного прироста, //—зона годичного побега
с вегетативными боковыми веточками.
/// — зона годичного побега с цветками в
пазухах листьев. IV — верхушечная невет-
вящаяся часть годичного прироста.
309
вых ветвях, развиваясь, как это свойственно вереску, на верх-
них годичных побегах Ко времени зацветания сеянцы вереска
в лесах Московской области достигают высоты 8—12 см. В об-
щей системе побегов к этому времени наиболее мощной ока-
зывается вертикально растущая главная ось, на которой распо-
лагаются супротивные боковые ветви, по преимуществу в ниж-
ней части главной оси Такое соотношение интенсивности и на-
правления роста главной оси и боковых ветвей создает в этом
возрасте вереска облик миниатюрного деревца Развитие си-
стемы побегов у вереска в сухих местообитаниях происходит по
аэроксильному типу Под землей происходит лишь рост глав-
ного корня и его боковых ветвей. Интенсивность роста корневой
системы не превышает таковой у побегов. Главный корень
к началу цветения достигает длины 10—12 см Из боковых кор-
ней наиболее интенсивно растут поверхностные, отходящие от
основания главного корня, на границе его с гипокотилем.
С началом цветения у вереска более четкой и сложной ста-
новится дифференциация боковых пазушных побегов, развиваю-
щихся на разной высоте материнского годичного побега. В ниж-
ней части годичного побега продолжают формироваться корот-
кие ассимилирующие веточки, с плотным расположением более
мелких (чем на удлиненных побегах) и вечнозеленых листьев,
функционирующих 2—3 года Эти веточки прекращают рост на
3—4-й год и подсыхают Выше их, в средней части и частично
верхней половине ветвящихся удлиненных годичных побегов
располагаются цветоносные пазушные побеги. На каждом та-
ком побеге в 2—3 мм длиной развивается терминальный цве-
ток, под чашечкой которого формируется одна пара зеленых
листьев — прицветничков или предлистьев В верхней части
удлиненного годичного побега вновь формируются вегетативные
пазушные побеги, отличающиеся от пижних ассимилирующих
коротких веточек более интенсивным ростом и большим долго-
летием. Они в дальнейшем дают начало сильным боковым вет-
вям, по росту и долголетию не резко отличающихся от мате-
ринских удлиненных побегов Как видим, у вереска наблюдает-
ся своеобразное сочетание разделения функций (дифференциа-
ции) и акротонности в росте боковых ветвей.
Переход к цветению и плодоношению в онтогенезе вереска
сопровождается значительным усилением интенсивности роста
побегов
Во взрослом состоянии вереск в сосновых лесах Московской
области обычно достигает высоты 30—40 см. У экземпляров
с прямостоячим главным стеблем и слабее растущими наклон-
ными боковыми ветвями, формирующимися с небольшой
высоты над уровнем почвы, в течение всей жизни сохраняется
и функционирует только система главного корня.
Но более обычной и чаще встречающейся формой кустар-
ничка у вереска в сосновых лесах Московской области оказы-
вается кустарничек с сильно развитыми и полулежачими боко-
выми ветвями, среди которых трудно определить главную ось.
В основании полегшей главной оси и боковых ветвей обра-
зуется значительное количество придаточных стеблевых корней,
которые функционируют одновременно с системой главного
корня. Укореняющиеся полупростратные боковые оси могут
достигать 1—1,5 м в длину, а кустарничек в целом приобретает
характер небольшой куртины, диаметром в 2—3 м, с сохраняю-
щейся главной осью и функционирующей системой главного
корня. С образованием придаточных корней боковые оси кус-
тарничка становятся хрупкими и ломкими. При обламывании
и отделении от материнского растения боковые побеги продол-
жают дальнейший рост как самостоятельные растения, ис-
пользуя систему придаточных стеблевых корней. Однако подоб-
ное вегетативное размножение и расселение у вереска в лесах
Московской области не является регулярным и обязательным.
Подобную же структуру имеют взрослые кустарнички вереска,
собранные на сфагновых болотах Талдомского района Москов-
ской области
Как видно из приведенного очерка, морфогенез и структура
кустарничка у вереска значительно отличаются от таковых
у рассмотренных выше черники и брусники. Основное своеобра-
зие морфогенеза и структуры кустарничка вереска, по сравне-
нию с только что названными видами растений проявляется
в тол, что-
1) у вереска в течение всей жизни сохраняется система
главного корня, а система придаточных корней или не обра
зуется совсем, или развита сравнительно слабо и имеет второ-
степенное значение;
2) у вереска все побеги, от заложения и до конца роста,
формируются как надземные древесные побеги, т е. аэрок-
с ильн о;
3) у вереска отсутствуют специализированные подземные
побеги, лишенные зеленых листьев и выполняющие функции
вегетативного размножения или запасающих органов. По дан-
ным Гревиллиуса и Кирхнера (S. Grevillius и О. Kirchner,
1923) у вереска па влажных местообитаниях с мощным разви-
тием мохового покрова наблюдается геоксильный тип развития
побегов, начинающих свой рост под землей и завершающих его
по выходе на поверхность почвы или мохового покрова В лесах
Московской области подобный тип морфогенеза у вереска не
встречен
311
з. главнейшие биолого-морфологические
ОСОБЕННОСТИ КУСТАРНИЧКОВ
Отмеченная в начале настоящей главы Основная габитуаль-
но-экологическая особенность Кустарничков — их крайняя
низкорослость — в типе довольно четко отделяет их от кустар-
ников.
Обзор кустарников, приведенный в IV главе, позволил заклю-
чить, что их относительная низкорослость по сравнению с де-
ревьями связана с резким сокращением длительности жизнен-
ного цикла главной оси и сменяющих ее осей возобновления
(в среднем до 15—20 лет). В то же время отмечалось, что ин-
тенсивность роста надземных осей кустарников и величина их
годичных приростов не меньше, чем у деревьев, а в ряде слу-
чаев даже превосходит их Изучение различных групп кустар-
ничков выявило, что для них характерно дальнейшее
сокращение длительности жизненного цикла
их надземных осей, по сравнению с кустарниками
(в среднем до 4—5 лет). Эта биологическая особенность оказа-
лась одной из основных причин их низкорослости.
Но наряду с резким сокращением длительности жизни их
надземных осей, низкорослость кустарничков определяется
также резким снижением интенсивности роста их
побегов. Даже в период кульминации скорости роста над-
земных осей годичные побеги их обычно не превышают 8—
10 см. За пределами периода кульминации ежегодный прирост
побегов у кустарничков, как правило, не превышает 2—3 см
Таким образом, крайняя низкорослость кустарничков обуслов-
лена как резким сокращением длительности жизненного цикла
надземных осей, так и столь же резким уменьшением интен-
сивности роста побегов. Но при всем этом кустарнички остаются
типичными древесными растениями, удлиненные ортотропные
побеги которых способны к верхушечному нарастанию (моно-
или симподиальному) в течение нескольких лет. В ветвлении
надземных побегов кустарничков четко проявляется акротон-
ность роста вновь возникающих ветвей, столь характерная для
рассмотренных выше деревьев и кустарников
В типе кустарнички хорошо отличаются и от полукустар-
ничков (группы жизненных форм, характерной для многих
аридных областей), годичные побеги которых ежегодно отми-
рают на значительную длину (обычно до базальной части по-
бега с укороченными междоузлиями). Отмечая характерные
биолого-морфологические особенности группы кустарничков,
отличающие эту группу от других, следует в то же время под-
черкнуть наличие переходных форм как от кустарников к кус-
тарничкам, так и от кустарничков к полукустарничкам.
312
Отмеченные выше признаки, небольшая длительность жиз-
ненного цикла надземных осей (в среднем 4—5 лет) и низкая
интенсивность роста побегов (годичные побеги обычно не более
2—3 см, и даже в период кульминации роста осей не превышаю-
щие 8—10 см) являются, по-видимому, наиболее общими чер-
тами кустарничков; с ними связана крайняя низкорослость этой
группы древесных растений. Что касается других эколого-мор-
фологических признаков, как развитие подземных ползучих
побегов, способность к вегетативному расселению, образование
шпалерных форм и т. д, то они оказываются более частными
признаками, свойственными лишь некоторым группам кустар-
ничков. В частности, отмирание системы главного корня, обра-
зование вторичной корневой системы, формирование подземных
ползучих побегов типа удлиненных горизонтальных корневищ
или столонов свойственны кустарничкам сфагновых болот и
торфяников, а также лесных и тундровых формаций с мощным
развитием мохового покрова. Шпалерные формы кустарничков
свойственны преимущественно различным тундровым форма-
циям и обычно приурочены к суровым климатическим условиям.
Обзор кустарничков, приведенный в настоящей главе и главе
II показал, что эта группа древесных жизненных форм встре-
чается в различных семействах, главным образом, двудольных
покрытосеменных (Ericaceae, Empetraceae, Salicaceae, Rosaceae,
Compositae, Myrtaceae, Epacridaceae, Dilleniaceae и др). Среди
однодольных кустарнички встречаются редко (например, розе-
точный ползучий кустарничек Dracophyllum Menziesii (Liliaceae)
из субальпийского пояса Новой Зеландии
Географическое распространение кустарничков показывает,
как уже говорилось выше, что наибольшее значение в сложении
растительного покрова (доходящее до степени эдификаторов),
наибольшее флористическое и эколого-морфологическое разно
образие кустарнички получают за пределами распространения
Деревьев и кустарников, и в частности, в тундрах северного
полушария, где выделяется особая подзона кустарничковых
тундр, а такж-е в альпийском поясе различных горных систем
Старого и Нового света. Концентрация кустарничков в этих
областях, наибольшее флористическое и эколого-морфологиче-
ское разнообразие кустарничков здесь дает основание полагать,
что эти области были главными центрами формирования кустар-
ничков в различных семействах покрытосеменных, а сама жиз-
ненная форма кустарничков возникла как приспособление
растений к развитию в неблагоприятных климатических усло-
виях этих стран и использованию особой экологической ниши,
создающейся в приземном слое воздуха, наиболее прогреваемом
и защищенном от иссушающего действия ветра. Появление
кустарничков связано с господством условий, которые в общем
313
можно охарактеризовать, как неблагоприятные для развития
деревьев и кустарников.
Из всех перечисленных семейств двудольных наиболее много-
численны и разнообразны кустарнички среди Ericaceae. Это
семейство интересно также наличием ряда форм, переходных
от настоящих кустарничков к типичным корневищным и столо-
нообразующим травянистым лесным растениям, характерным
для близкого к этому семейству грушанковых. В семействе
брусничных и грушанковых, естественно, прослеживается путь
метаморфоза одной из групп древесных жизненных форм в одну
из групп травянистых многолетних растений, свойственную
лесам бореальной области.
Глава VI
ЖИЗНЕННЫЕ ФОРМЫ СТЕЛЮЩИХСЯ РАСТЕНИЙ
1. ЭКОЛОГО-МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ СТЕЛЮЩИХСЯ
РАСТЕНИИ, ИХ ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
И ПОЛОЖЕНИЕ В СИСТЕМЕ ЖИЗНЕННЫХ ФОРМ
Стелющиеся растения образуют разнообразный, но довольно
четко очерченный ряд жизненных форм, специфическим призна-
ком которых является развитие горизонтальных или приподни-
мающихся на верхушке скелетных надземных осей, занимающих
приземное положение, или даже прижатых к почве.
По особенностям роста побегов стелющиеся растения суще-
ственно отличаются от прямостоячих деревьев, кустарников и
кустарничков. Как показано в главах III, IV и V, у прямостоя-
чих деревьев, кустарников и кустарничков интенсивность роста
побегов неразрывно связана с их вертикальным положением.
Чем ближе растущие побеги или ветки к вертикальному поло-
жению, тем интенсивнее их рост. Наоборот, чем больше отклоне-
ны от вертикали побеги или ветви в кроне деревьев, кустарни-
ков и кустарничков, тем слабее их рост, тем менее долговечны
такие ветви.
У стелющихся древесных растений наблюдаются совершенно
иные коррелятивные связи между направлением и интенсив-
ностью роста побегов. Наибольшей интенсивностью роста
в длину, толщину и наибольшей долговечностью обладают над-
земные горизонтальные побеги В общей системе побегов
горизонтальные оси оказываются скелетными, наиболее долго-
летними осями. Вертикально растущие побеги у стелющихся
древесных растений отличаются слабым ростом в длину и тол-
315
щину и обладают небольшим долголетием. На скелетных гори-
зонтальных осях вертикальные побеги развиваются как корот-
кие тонкие ветви, ассимилирующие или цветоносные.
Экологически стелющиеся древесные растения близки к кус-
тарничкам тем, что их надземные органы также располагаются
в приземном слое воздуха, получая здесь защиту от иссушаю-
щего действия ветра. Но в отличие от кустарничков, приземное
положение побегов у стелющихся древесных растений дости-
гается не за счет резкого ослабления роста и сокращения дли-
тельности жизненного цикла надземных осей, а за счет изме-
нения направления интенсивно растущих побе-
гов с ортотропного на плагиотропное.
Стелющиеся растения занимают своеобразное положение
в системе жизненных форм покрытосеменных и хвойных, входя
в состав типов деревьев, кустарников и кустарничков, встре-
чаясь и у травянистых форм, включая однолетние. Они пред-
ставляют собой обычно подкласс или более мелкую группу
в пределах типа, отличающуюся особой специализацией строе-
ния и роста побегов Как подчеркивалось в главе II, при изло-
жении системы жизненных форм покрытосеменных и хвойных,
стелющиеся растения составляют один из рядов специализиро-
ванных гомологов от деревьев до травянистых однолетников.
В главах III, IV и V при характеристике деревьев, кустарников
и кустарничков имелись в виду исключительно или преиму-
щественно прямостоячие формы. В настоящей главе стелю-
щиеся древесные растения будут рассмотрены более подробно.
Как бы ни были разнообразны древесные стелющиеся растения,
их всех объединяют особенности роста их побегов, приводящие
к занятию этими растениями приземной экологической ниши,
но способом существенно иным, по сравнению с прямостоячими
кустарничками.
Рассматривая стелющиеся древесные растения как дериваты
прямостоячих деревьев, кустарников и кустарничков, теорети-
чески естественно и не трудно выделение стелющихся деревьев,
стелющихся кустарников и стелющихся кустарничков Практи-
чески выделение таких групп стелющихся древесных растений
встречает большие затруднения. Метаморфоз прямостоячих
деревьев в стелющиеся и полупростратные сопровождается,
кроме значительного полегания надземных осей, некоторым
ослаблением роста главной оси при сохранении интенсивного
роста боковых ветвей. В результате этого вместо древовидных
простратных форм образуются кустовидные приподнимающиеся
формы, которые трудно отграничить от настоящих ползучих
или приподымающихся кустарников. Стелющиеся кустарники,
в свою очередь, приобретают черты, сближающие их с про-
стратными кустарничками.
316
Экологическое своеобразие стелющихся древесных растений
•оказывается тесно связанным с их географическим распростра-
нением
Стелющиеся или приподымающиеся деревья широко рас-
пространены у верхнего предела лесного пояса различных
горных стран, от тропиков до холодно-умеренных зон, а также
у полярных пределов лесной растительности, образуя здесь
разнообразные формации криволесья (Krummholz), занимаю-
щие то более, то менее узкий пояс в горах, то более или менее
обширные территории на равнинах.
По широте географического распространения и значению
в растительном покрове среди стелющихся древесных растений
нашей страны следует прежде всего назвать кедровый стла-
ник — Pinus pumila Rgl. Формации кедрового стланика в СССР
занимают громадные территории от бассейна Лены до Охот-
ского моря, от границ Монголии и до тундр Восточной Сибири
и Чукотки (Б. А Тихомиров, 1949)
У верхней границы лесной растительности Карпат, Альп,
Балканских гор широкое распространение находят стелющиеся
формы горной сосны — Pinus montana var pumilio, var. uncinata
и var. tnughus В этом же поясе и частично выше, в альпийском
поясе этих гор весьма обычны также стелющиеся формы Alnus
viridis, Rhododendron hirsutum, R. ferrugineum, а в пределах
альпийского пояса в более неблагоприятных условиях произра-
стания — шпалерные кустарники и кустарнички:! Juniperus
папа, Salix retusa, S. reticulata, S herbacea, Empetrum nigrum
и др Шпалерные кустарники и кустарнички, как известно,
составляют одну из характерных и широко распространенных
жизненных форм в различных формациях тундровой раститель-
ности. Шпалерные кустарнички в тундрах занимают обычно
наиболее неблагоприятные для древесных растений местообита-
ния
Формы, экологически и габитуально близкие к стелющимся
формам горной сосны, наблюдаются у разных видов деревьев
близ верхнего предела лесной растительности тропических гор-
ных систем Так, в горах западной Явы, на высоте около
3 000 м над уровнем моря развит че^ко выраженный пояс криво-
лесья (A Schimper и F. Faber, 1935). Стволы карликовых де-
ревьев этого пояса короткие и толстые, косые или даже горизон-
тальные, несут толстые, мало разветвленные ветви винтообраз-
но изогнутые и переплетающиеся между собой. Только на кон-
цах ветвей располагаются кожистые листья, образуя негустую,
резко уплощенную крону. В поясе криволесья западно-яванских
гор доминируют: V actinium varingiifolium, Eurya acuminata,
Rhododendron retusum, Symplocos sessilifolia, Rapanea avenis,
Schefflera rugosa, Viburnum coriaceum и др.
317
В горах восточной Явы у верхнего предела лесного пояса
(2800 м над уровнем моря) габитус, типичный для деревьев
криволесья, приобретают Quercus pruinosa и Vaccimum varingii-
folium. Их ствол становится коротким, ветви изогнутыми и длин-
ными, а крона становится зонтикообразной, плоской В горах
Западной Африки (Камерун), выше 2700 м над уровнем моря,
у верхнего предела лесного пояса образуют чащи криволесья
виды Adenocarpus Одиночные карликовые узловатые деревца
Polylepis lanuginosa, габитуально близкие к формам криволесья,
встречаются в Кордильерах Северного Перу, в формациях
парамо. В высокогорьях Юньнани в субальпийском поясе в
составе криволесья доминируют виды Rhododendron
Широкое распространение стелющиеся древесные растения
имеют в различных хребтах Тянь-Шаня. В отличие от многих
других горных стран, на Тянь-Шане стелющиеся древесные
формы встречаются не только выше предела лесной раститель-
ности, в субальпийском поясе, но и ниже лесного пояса, среди
горно-степных формаций, заходя также в пределы лесного
пояса. Распространение стелющихся древесных форм ниже
лесного пояса на Тянь-Шане связано, вероятно, со значитель-
ной аридностью климата. Наибольшее значение среди стелют
щихся древесных растений Тянь-Шаня имеют различные виды
можжевельника или арчи, например, Juniperus turkestanica,
J. sabina и др Кроме видов Juniperus, здесь встречаются и дру-
гие стелющиеся древесные растения, например Caragana pleiop-
hylla.
Как видно из приведенных данных и материалов, излагаемых
ниже в этой главе, стелющиеся древовидные формы (стланцы)
и шпалерные кустарники и кустарнички приурочены к высоко-
горным и полярным областям, получая распространение в край-
них для древесных растений условиях произрастания При этом
большая степень простратиости форм у разных видов древесных
растений связана с господством более неблагоприятных усло-
вий. Анализ распространения стелющихся древесных растений
выявляет особенно близкую зависимость такой формы роста
растений от особенностей местообитания. В неблагоприятных
условиях высокогорных областей простратные формы образуют
часто те же самые виды, которые в более благоприятных усло-
виях растут как прямостоячие.
В. Pay" (W. Rauh, 1937) среди аркто-альпийских шпалерных
кустарников различает две группы — активные и пассивные
шпалерные кустарники.
К первым (активным шпалерным кустарникам) относятся
растения, шпалерные формы которых—наследственные, и
внешние условия могут лишь усиливать или ослаблять их
особенности, но не устранять полностью; высокогорные
318
шпалерные кустарники этой группы при переносе в равнины
сохраняют особенности своего роста, за исключением неко-
торого удлинения междоузлий К этой группе относятся, на-
пример, Dryas octopetala, Arctostaphylos alpina и др Вторая
группа (пассивные шпалерные кустарники), по Рау, возникают
в результате воздействия неблагоприятных климатических фак-
торов, как низкая температура воздуха, ветер и сухость При
устранении этих тормозящих рост факторов шпалерный тип
роста в короткое время теряется; у растений развиваются
прямостоячие формы.
Примером второй группы, по Рау, может служить Betula
pubescens var tortuosa, образующая, по описанию Кильмана
(Kihlnian, 1890), формы шпалерных кустарников за пределами
лесной растительности в открытых тундрах Подобные же
формы (по Рау) образуют и другие деревья у полярного пре-
дела их распространения, например, сосна.
Образование стелющихся форм у деревьев близ полярной
границы лесов и верхнего предела распространения деревьев
в горах неоднократно отмечалось в отечественной ботанической
литературе Стелющиеся формы у сибирской лиственницы —
Larix siblrica — на Таймыре описала Л Н. Тюлина (1937)
В. Н Сукачев (1934, 1938) отмечает стелющиеся формы 5
обыкновенной ели на крайнем Севере, Л Н Тюлина (1929) —
у сибирской ели на Южном Урале, а И Г. Серебряков (1945) —
у 1янь-шанской ели в Заилийском Алатау и Кунгей Алатау
Подобные, формы образуются иногда и у лиственных де-
ревьев, например, у бука близ верхней границы леса в Крыму
и на Кавказе (Поплавская Г. И , 1927, 1929; Кожевников А. В..
1935; Сукачев В Н, 1938, Орлов А Я., 1953). Судя по резуль-
татам географических посевов сосны, лиственницы и дуба (Де-
рюжкин Р. Р, 1952; Самофал С , 1929) и культуры древесных
пород из лесотундры в условиях, благоприятных для роста де-
ревьев в лесной зоне (В Н Андреев, 1948), стелющиеся формы
у этих деревьев вряд ли можно считать наследственно закреп-
ленными.
В процессе исторического освоения древесными растениями
новых территорий с суровыми климатическими условиями и
недоразвитыми почвами выработались своеобразные формы
стланцев, наследственно закрепленные, сохраняющие свою
форму при переносе в более благоприятные почвенно-климати-
ческие условия.
Среди хвойных в качестве примера может быть назван кед-
ровый стланик — Pinus pumila Rgl При интродукции в Евро-
пейской части СССР кедровый стланик обычно сохраняет
чашевидную форму куста (Б. А. Тихомиров, 1947; В Н Вехов,
1957, 1958)
319
2. МОРФОГЕНЕЗ ЖИЗНЕННОЙ ФОРМЫ СТЛАНЦА
У НЕКОТОРЫХ РАСТЕНИИ
Некоторые биологические особенности
кедрового стланика — Pinus putnila Rgl.
Несмотря на широкое распространение кедрового стланика
и большое значение в сложении растительного покрова,” его
биолого-морфологические особенности изучены недостаточно.
В частности, представляло бы большой интерес сравнительное
изучение морфогенеза у сибирского кедра и кедрового стла-
ника
Проростки кедрового стланика по форме и величине мало
отличаются от проростков кедровой сосны — Pinus sibirica Мауг
Их гипокотиль в 3—5 см длины выносит зеленые семядоли на
свет и они оказываются первыми ассимилирующими листьями
в течение первых лет жизни сеянца, как и у других видов сос-
ны. Выше семядольного узла главный стебель вырастает в те-
чение первого года на 1—2 см В природных условиях произ-
растания стланика, по D А. Тихомирову (1949), сеянцы кедро-
вого стланика обычно встречаются пучками, наиболее часто в
числе 7—15 сеянцев в пучке. Расположение сеянцев пучками
приводит к тому, что в процессе дальнейшего роста происходит
тесное контактирование отдельных экземпляров и образуется
единый куст стланика.
Пучковое расположение сеянцев, как показали наблюдения
многих исследователей, связано с заготовками кедровых орехов
впрок кедровкой (Nucifraga caryocatactes).
В течение первых лет жизни сеянца кедрового стланика
главная ось его остается ортотропной и отличается небольшой
интенсивностью роста. По С. А. Пивник (1958) в 6—10 лет
сеянцы на гольцах достигают 5—7,5 см, а/ в лиственничных ле-
сах 14—17 см высоты. По данным Б. А. Тихомирова (1949),
12—15-летние экземпляры стланика имели высоту 30—45 см.
После некоторого периода ортотропного роста, по С А Пив-
ник (1958), уже с 5—7 лет (на гольцах) главная ось растения
становится плагиотропной. К 15—20 годам сеянцы приобретают
типично стелющуюся форму
В подгольцовом и лесном поясах главная ось кедрового
стланика сохраняется прямостоячей до 10—12 лет, в после-
дующем происходит ослабление роста в высоту и постепенное
полегание главной оси, с погружением ее в подстилку.
В более благоприятных условиях сомкнутых лесных со-
обществ полегают лишь базальные части стволиков. Обра-
зующиеся боковые ветви растут почти вертикально. К 25—
30 годам происходит дальнейшее ослабление роста главной
320
оси, которая вскоре теряется среди быстрее растущих боковых.
Сеянцы теряют форму дерева. Образуется чашевидный куст.
Пивник отмечает, что чашевидная и стелющаяся форма роста
зависит от интенсивности и направления роста главной и боко-
вых осей и не связана с первоначальным групповым располо-
жением всходов.
Прижатость к почве ствола усиливается благодаря форми-
рованию на нем придаточных корней, притягивающих его к
поверхности почвы Развитие сильных, обильно ветвящихся
придаточных корней на стволе приводит к ослаблению функ-
ционального значения системы главного корня, который через
некоторое время отмирает В последующем отмирание распро-
страняется и на базальную часть полегшей главной оси Но
от.мирание в базальной части главной оси сопровождается
непрерывным нарастанием ветвей и побегов продолжения в
приподымающейся верхушке кроны стланца, а также образо-
ванием новых придаточных корней ближе к живой кроне.
Отмирание главного корня и формирование на стелющихся
осях придаточных корней приводит к тому, что стланец Pinus
pumila становится вегетативно подвижным, наподобие стелю-
щихся кустарничков (см ы IV) Под влиянием факторов сре-
ды (например, господствующих ветров) в перемещении стлан-
ца иногда наблюдается определенная направленность В резуль-
тате этого вновь образующиеся укоренившиеся ветви кедрового
стланца могут занимать более благоприятные местообитания,
защищенные от ветра (D А. Тихомиров, 1949)
Развитие системы придаточных корней на стелющихся осях
приводит к значительному увеличению длительности жизни и
общего периода нарастания кроны. Придаточные корни как бы
снимают признаки старения и пи резко замедляют старение в
растущей верхушке кроны В литературе но отмечены явпения
суховершинности и естественного отмирания кедрового стлани-
ка (Б. Л Тихомиров, 1949). Все случаи отмирания отдельных
кустов были связаны с нарушением нормальных условий роста
каким-либо посторонним фактором (чаще всего пожаром)
На форму роста кедрового сту^нйка"и, в частности, чг ст^
ег° иТтли простратности, влияют условия его
произрастания. ~В условиях лесных ценозов где кедровый
стланик растет в подлеске, у него образуется форма с прямо-
стоячими, слабо изогнутыми стволами. На безлесных равнинах,
в условиях субальпийского пояса образуются ползучие формы
(Б А Тихомиров, 1949)
Г. Э Гроссет (1959), как и другие исследователи, отмечает
зависимость формы роста кедрового стланика от условии сре-
ды В очень плохих условиях роста, на тощих, сухих и холод-
ных почвах образуются наиболее простратные формы шпалер-
91 321
В-413 И Г Серебряков—Л
ного типа, со стволиком, лежащим на всем его протяжении В
лучших условиях роста верхняя часть стволика в большей или
меньшей степени приподымается. Изгиб стволика и поднятие
его верхушки связано, по Гроссету, с образованием крепевой
древесины, которая формируется на нижней стороне стволика в
верхушечной его части.
В литературе давно отмечена связь между образованием
креневой древесины, интенсивностью роста и распределением
ростовых веществ
Известно, например, что в стволах хвойных, имеющих
наклонное положение, ростовых веществ содержится больше
на нижней стороне ствола, т. е. там, где образуется креневая
древесина. Связь крепевой древесины и ростовых веществ уда-
ется доказать также экспериментально, путем обработки росто-
выми веществами. Если обрабатывать ростовыми веществами
лишь одну сторону стволика или ветви, то креневая древесина
образуется только здесь, и стволик изгибается в противополож-
ную сторону. Креневая древесина образуется при достаточно
высокой концентрации ростовых веществ (Е. W. Sinnot, 1952;
Necesanij 1955). При образовании креневой древесины проис-
ходит односторонний усиленный рост не только в толщину, но
и в длину, отчего происходит изгиб стволика или ветвей
Одной из наиболее интересных экологических особенностей
кедрового стланика является способность его ветвей к актив-
ному полеганию при наступлении морозов. Пригнувшиеся или
даже прижавшиеся к поверхности почвы кусты засыпаются
выпадающим снегом. Благодаря этой способности даже мало-
мощный снежный покров совершенно скрывает стланик там,
где летом поднимаются трудно проходимые заросли, высотой
в 4—5 м
Механизм полегания стланика при наступлении морозов
разные исследователи объясняли различно По мнению
Л Н Тюлиной (1936), полегание происходит вследствие неоди-
накового промерзания разных сторон веток, связанного с
неодинаковым охлаждением соков верхней и нижней стороны
ветнсй. Пр В If Васильеву полегание происходит,
вероятно, вследствие уменьшения ''тургор^^.^^’ З.?
влаги в начале зимнего периода, когда испарение еще продоч-
и оеЛС\Я’ 3 ДОСТУП о ВОДЫ от к°Рпей прекратился В. И. Андреев
(1У49), изучивший движение вегвей в кроне ряда деревьев в
течение зимнего времени, полагает, что движение боковых вет-
вей при колебаниях температуры обусловлено неодинаковым
расширением воды и древесной массы, находящихся в различ-
ных количествах в древесине верхней и нижней стороны ветви.
сЭтот вывод В. Н. Андреева, по Б. А. Тихомирову (1949) при-
меним и для объяснения пригибания ветвей и стволиков’стла-
322
ника. Близкое объяснение этому явлению дают также Стари-
ков Г. Ф. и Дьяконов П. Н. (1954).
Г. Э. Гроссет (1959) установил, что ветви стланика не реа-
гируют на колебания температур, если они положительные
(выше 0°). Движение ветвей начинается только с частичного
замерзания воды, содержащейся в древесине. При этом
наиболее значительное движение ветвей имеет место при отри-
цательных температурах, близких к 0°. Ветви пригибаются не
только при падении температуры, но и при потере влаги. Не-
пременным условием движения ветвей при изменении темпера-
туры, по Г. Э Гроссету, является дорзовентральность строения
ветвей и, в частности, развитие в них креневой древесины.
Образование креневой древесины, по этому автору, имеет очень
большое значение в жизни стланика, определяя как его способ-
ность к полеганию, так и форму роста, о чем уже говорилось
выше
Какое бы объяснение механизма пригибания ветвей стла-
ника ни было принято, несомненным остается большое биоло-
гическое значение этой способности. Стланцевая форма роста,
в соединении со способностью к активному полеганию, позво-
ляет кедровому стланику обитать в крайне суровых условиях.
Интересно также, что движение ветвей в кроне деревьев
при колебании температуры в зимнее время, отмеченное для
ряда видов, у кедрового стланика приобретает ярко выражен-
ное экологическое значение. Экологическое значение этого
явления у прямостоячих лесных деревьев вряд ли может счи-
таться столь существенным и вообще остается невыясненным
Морфогенез жизненной формы стланца
у туркестанского можжевельника — Juniperus turkestanica Korn.
Флора СССР, по В Л. Комарову (1934), насчитывает 21
вид из рода Juniperus, широко распространенные в лесной зоне
и лесотундре равнинной части СССР и особо разнообразные в
горных районах Крыма, Кавказа, Средней Азии и Дальнего
Востока 12 видов можжевельника — низкорослые, по типич-
ные деревья, высотой от 4 до 18 м {Juniperus communis L.,
J. oblonga M. B., J. rigida Siel., J. foetidissima Wil id , J. excelsa
M. В , /. isophyllus C. Koch., J. polycarpus C. Koch., J. turkomani-
ca В Fedtsch., J. seravschanica Kom , J. semiglobosa Rgl., J. ta-
lassica Lipsky, J. schugnanica Kom Семь видов представлены
типичными «стелющимися кустарниками» (В. Л. Комаров) —
/. depressa Stev., J. sibirica Burqsd., /. littoralis Maxim., J. pseu-
dosabina F. et M., J. sargenii T., J. dahurica Pall., J sabina L.).
Эта группа видов обычно распространена за пределами лесного
пояса. Наконец, два вида можжевельника — J. o^ycedrus L. и
21* 323
J. turkestanica Кош мог)Т образовывать и древовидные и кус-
тарниковые формы Особый интерес представляет / turkestani-
ca, широко распространенный в различных хребтах Тянь-Шаня;
он может образовывать деревья до 18 м высоты и стланцевые
формы, возвышающиеся лишь на 30—50 см над уровнем почвы.
Как очевидно, туркестанский можжевельник, или туркестан-
ская арча, способен образовывать переходные формы от де-
ревьев к стланцам
Приводимые ниже данные о морфогенезе туркестанского
можжевельника основаны на наблюдениях и описаниях автора
в одном из хребтов Центрального Тянь-Шаня — Тсрскей Ала-
тау, в долине р. Чон-Кызыл-су (Серебряков И. Г, 1954).
Juniperus turkestanica в пределах Терскей Алатау распро-
странен преимущественно в верхней части луговолесного пояса,
выходя за его пределы до альпийских лугов Высотные грани-
цы распространения I. turkestanica в долине р. Чон-Кызыл-су —
от 2300 до 3300 м над уровнем моря В отличие от казацкого
можжевельника (J. sabina), произрастающего в долине Чон-
Кызыл-су только в виде прижатых к почве куртин, /. turkesta-
nica в пределах одной и той же речной долины, на разных абсо-
лютных высотах имеет различную форму роста — от типичных
стланцев, в виде прижатых к почве куртин, до настоящих дре-
вовидных форм высотой в 4—6 м.
Семена /. iurkestanica прорастают надземно У проростка
в течение первого года формируется довольно обильно развет-
вленный главный корень. Гипокотиль имеет длин) около 4 см.
На семядольном узле образуются две длинные лентовидные
семядоли длиной 20—25 мм, резко отличающиеся от последую-
щих листьев Выше семядолей в течение первого года разви-
вается еще 3—5 мутовок зеленых ассимилирующих листьев
длиной до 7—8 мм На второй год раскрывается лишь верху-
шечная почка, образуя побег продолжения главной оси Моно-
подиалыюе нарастание продолжается и в последующие годы
Годичные побеги первых 10 лет — небольшие, и средний прирост
главной оси не превышает 7—10 мм Границы годичных побегов
не очень отчетливы Они узнаются по чуть укороченным междо-
узлиям и более мелким зеленым листьям Ветвление главной
оси начинается с 2—4 лет, боковые ветви образуются из пазуш-
ных почек верхних листьев каждого годичного побега. Пазуш-
ные почки, расположенные в нижней части годичных побегов,
остаются в нераскрывше.мся состоянии Таким образом, в вет-
влении туркестанского можжевельника проявляется акротонное
усиление побегов, свойственное, как отмечалось выше, деревьям
и кустарникам. Боковые побеги начинаются с двух латерально
расположенных предлистьев, несколько меньших по размеру,
чем последующие листья.
324
В онтогенезе /. turkestanica наблюдается четко выраженная
смена ювенильных игловидных листьев чешуевидными, свой-
ственными взрослым растениям. Игловидные листья 7—8 мм
длины сохраняются на главной оси до 8—12 и более лет.
Позже они сменяются более короткими, но еще игловидными
листьями, до 2,5—3 мм длины, а затем, несколько лет спустя
после начала формирования коротких игловидных листьев, на
побега к начинают формироваться чешуевидные листья. Смена
ювенильных игловидных листьев чешуевидным и начинается
раньше у сильно ветвящихся экземпляров. Одновременно с изме-
нением формы листьев с возрастом у /. turkestanica происходит
также изменение листорасположения. Вначале четырехлистное
мутовчатое, на 3—8-м году жизни сеянца оно сменяется трех-
и затем двухлистным, т. е. супротивным.
Рост надсемядольной части стебля сопровождается, как и
у деревьев и кустарников, полеганием гипокотиля, а нередко
и более верхних участков 1лавной оси. На полегшем стебле,
который скоро покрывается подстилкой или моховым покровом,
образуются придаточные корни
В течение первых 12—15 лет у туркестанского можжевель-
ника сохраняется облик сеянцев типичных деревьев, более или
менее вертикальная главная ось несет отклоненные в сторону
слабые боковые веточки Но уже в конце второго десятилетия
у сеянцев /. turkestanica в условиях лесного пояса долины Чон-
Кызыл-су наблюдается некоторая дифференциация.
У одних экземпляров остается доминирующей в системе
побегов главная ось, сохраняющая вертикальное положение
Интересно, что даже отмирание верхушки главной оси у этих
экземпляров не вызывает ее ослабления,— отмершая верхушка
заменяется одной из боковых ветвей, как это наблюдается у
симподиально нарастающих лиственных деревьев (липы, вяза,
древовидных ив и т. д) К 20—25 годам сеянцы древовидного
типа достигают высоты-50—60 см. В этом возрасте у J. turkes-
tanica в пределах лугово-лесного пояса долины р. Чоп-Кызыл-
су, на высоте 2600—2800 м выше уровня моря начинается
семеношение.
У других экземпляров, встречающихся чаще, наблюдается
довольно резкое ослабление роста главной оси, хотя она и оста-
ется моноподиальной Ослабление главной оси у этих экземп-
ляров обычно сопровождается более или менее сильным ее по-
леганием и разрастанием боковых осей (рис 59) Развитие мощ-
ных боковых ветвей вызывает усиление формирования прида-
точных корней в основании главной оси.
Пока трудно сказать, чем вызывается подобная дифферен-
циация сеянцев — индивидуальными особенностями их, или
воздействием внешних условий. Нередко удается отмечать, что
32.5
СОсмг
Рис. 59.
Начало дифференциации у подроста туркестанского
можжевельника (Juniperus turkestanica) на древовидную (%)
и стланцевую (б) форму в возрасте 20—22 лет
(по Серебрякову, 1954)
слабое развитие боковых ветвей у прямостоячих древовидных
экземпляров является результатом механического повреждения
ветвей падающими со склона камнями. Но этот фактор имеет
лишь частное значение. Замечается также некоторая приурочен-
ность древовидных прямостоячих сеянцев к участкам склонов
с более развитым почвенным покровом, т. е. к более благо-
приятным микроусловиям. В связи с этим вопросом представ-
ляется интересным провести специальные опыты по изучению
влияния обработки почвы и удаления нижних ветвей на форму
роста этого важного высокогорного растения, а также по отбору
сеянцев в целях получения устойчивых древовидных форм.
С возрастом дифференциация между отдельными экземпля-
рами J. turkestanica усиливается (рис. 60). К 35—40 годам неко-
торые экземпляры сохраняют древовидный облик, с ясно выра-
женным главным вертикальным стволом и отходящими от него
более или менее тонкими, несколько изогнутыми вверх ветвями.
От настоящих деревьев габитуально их отличает лишь развитие
наиболее мощных ветвей у самой поверхности почвы, в основа-
нии главной оси. Но у других экземпляров того же возраста
главная ось становится плагиотропной, теряясь среди мощных
326
150см
Рис. 60.
Дифференциация у туркестанского можжевепьника на древовидную (а)
и стланцевую форму (б) в возрасте 35—40 лет
(по Серебрякову, 1954)
боковых приподымающихся ветвей Длина боковых ветвей у
таких экземпляров нередко превышает длину главной оси.
Иногда уже в этом возрасте главная ось отмирает с верхушки
на значительном протяжении. Куст приобретает типичную та-
релковидную форму.
Из-за прекращения роста главного и боковых корней наи-
большее функциональное значение приобретают придаточные
корни в основании главного ствола. Древовидные прямостоячие
и простратные кустовидные экземпляры данного возраста в
лесном поясе долины р. Чон-Кызыл-су нередко встречаются
рядом. Лишь в известной степени их распространение опреде-
ляется характером почвенного покрова — более щебнистым
почвам, в отличие от богатых мелкоземом, соответствуют более
простратные формы.
К 50—60 годам различия между древовидными и стланце-
выми формами J. turkestanica становятся более очевидными.
У типичных стланцевых форм к этому времени не только теряет-
ся среди ветвей или отмирает почти до основания главная ось,
но и наблюдается укоренение боковых лежачих ветвей. Послед-
нее обстоятельство коренным образом меняет дальнейший ход
развития стланца. Наибольшее разрастание получают укоренив-
шиеся ветви. Их верхушки выше зоны укоренения приобретают
вертикальное положение. Но довольно скорое отмирание вер-
327
хушек неминуемо влечет за собой усиление ветвления укоренив-
шихся побегов. Высота стланцев в этом возрасте в лесном поясе
достигает 0,5—0,8 м, при диаметре куста 2,5—3 м. Древовидные
формы того же возраста в тех же условиях достигают высоты
2,5—3 м У древовидных экземпляров в основании главного
ствола иногда наблюдается также разрастание одной-двух или
трех нижних ветвей, позже укореняющихся в местах их сопри-
косновения с почвой.
Различие между стланцевыми и древовидными формами
у J. turkestanica становится максимальным в его зрелом и ста-
ром возрасте (рис. 61)
Рис. 61.
Стланцевая и древовидная форма туркестанского
можжевельника (Juniperus turkestanica) с высотой 3200 и 2900 м
н у м с хребта Терскей Алатау (возраст не .менее 250 лет)
(по Серебрякову, 1954)
Выше отмечалось, что форма роста J. turkestanica в преде-
лах Терскей Алатау зависит от абсолютной высоты. Маршрут
по долине р. Чон-Кызыл-су от нижней границы леса до ледни-
ков Кара-Баткак и Кельдеке дает возможность наблюдать
смену формы роста у этого растения. Первые экземпляры тур-
кестанского можжевельника в долине этой реки появляются
с высоты 2300 м над уровнем моря и приурочены к пеболь-
328
шим полянам среди насаждений тяньшанской ели Они пред-
ставлены типичными стланцами Стланцевые формы преобла-
дают и на высоте 2600—2650 м над уровнем моря, особенно на
открытых полянах по долине реки или склонам Лишь изредка
встречаются здесь на небольших полянках древовидные экземп-
ляры /. tarkesianica, обычно с несколькими приподымающимися
стволами, до 3,5—4 м высоты Однако на высоте 2650—2800 м
над уровнем моря, близ верхней границы ельников, туркестан-
ский можжевельник почти сплошь приобретает древовидную
форму Верхушки кроны его поднимаются на 4—6 м над уров-
нем почвы Именно здесь можно видеть мощные деревья с диа-
метром ствола 30—35 см.
Иногда главный ствол дерева на высоте 2—3 м причудливо
изгибается или скручивается; иногда сохраняется и главный
ствол, и разрастаются нижние мощные приподымающиеся
ветви, образуя своеобразную чашу со свечой главной оси в сере-
дине Но по мере дальнейшего подъема в горы, за пределами
еловых • насаждений высота туркестанского можжевельника
вновь начинает падатр. Он снова приобретает все более типич-
ный облик стланца. У верхнего предела своего распространения,
в долине Кара-Баткак и Кельдекс, на высоте 3 300 м над уров-
нем моря, стланцы туркестанского можжевельника приобретают
подчеркнуто простратную форму Его прижатые к почве лепеш-
ковидные куртины не превышают 40—50 см в высоту, достигая
диаметра 5—8 м, а иногда образуя сплошные, трудно проходи-
мые заросли на многие десятки метров
Как видно из предыдущего, у туркестанского можжевель-
ника, как и у других древесных пород, например, бука
(Г И Поплавская, 1929), тяньшанской ели (И Г Серебряков,
1945), наблюдается проявление общебиологической закономер-
ности — наилучший рост древесных растений в горах наблю-
дается в средней части пояса их распространения, по мере
приближения к его пределам высота деревьев падает, иногда
вплоть до перехода к кустовидным стелющимся формам По-
пятно, конечно, что эта закономерность свойственна видам де-
ревьев, действительно имеющим нижние и верхние пределы
своего распространения в данной горной системе У деревьев
равнинных, по мере поднятия в горы, будет наблюдаться лишь
нисходящая ветвь этой одновершинной биологической кривой,
и стелющиеся формы приурочены лишь к верхнему пределу их
распространения в горах
Изучение структуры старых стелющихся куртин туркестан-
ского можжевельника на высоте 3300 м над уровнем моря дает
возможность установить у таких стланцев наличие главной оси
или ее остатков Верхушка главного ствола здесь обычно отми-
рает на высоте 30—40 см. Дальнейший рост стланца продол-
329
жается за счет роста боковых ветвей, расположенных в сохра-
нившейся части ствола. Верхние ветви главного ствола верти-
кальны и поэтому вскоре испытывают судьбу материнского
побега: их верхушки отмирают. Но отмирание верхушки,
в свою очередь, вызывает ветвление этих боковых ветвей с обра-
зованием побегов все более высокого порядка. Вскоре здесь
образуется плотная шапка коротких, направленных вверх вет-
вей. Ветви, расположенные ниже на главной оси, в отличие от
более верхних, растут плагиотропно, и лишь их верхушка па
некотором удалении от главной оси направлена вверх. На боль-
шом своем протяжении нижние ветви оказываются погружен-
ными в моховой покров, что в последующем облегчает их уко-
ренение.
Нижние укореняющиеся ветви в свою очередь ветвятся по
типу материнской главной оси. На верхушке их образуются
плотные группы коротких, более или мен^е вертикально расту-
щих ассимилирующих ветвей, а в более нижней части они так
же, как и главная ось, дают начало системе боковых плагиот-
ропных ветвей, уже III порядка, укореняющихся на некотором
расстоянии от материнских. В ветвлении побегов J. turkestanica
выше отмечалось акротоппое усиление боковых побегов. Акро-
тония ветвления отражает особенности продольной симметрии
побега (W. Troll, 1937; W. Rauh, 1937). Вместе с тем, в про-
цессе ветвления побегов проявляется и тип их латеральной сим-
метрии У J. turkestanica при ветвлении на боковых плагиотроп-
ных побегах наблюдается более интенсивный рост почек и боко-
вых побегов, расположенных па нижней стороне материнского
побега Из них развиваются длинные плагиотропные укореняю-
щиеся ветви Из почек, расположенных на верхней стороне
материнского плагиотроппого побега, образуются более слабые,
вертикально ориентированные боковые ветви
Таким образом, в ветвлении побегов /. turkestanica, наряду
с акротонным усилением побегов, наблюдается явление гипото-
нии Понятно, конечно, что более сильный рост ветвей с нижней
(и наружной) стороны материнских латеральных побегов, т. е.
гипотония ветвления, приводит к усилению краевого роста
образующихся куртин стланца.
У старых экземпляров стланца туркестанского можжевель-
ника главная ось и ее боковые ветви в верхушечной части отми-
рают. В функционирующем состоянии остаются лишь укореняю-
щиеся боковые ветви более высшего порядка. Укоренение вет-
вей, как отмечалось, облегчается тем, что на большем своем
протяжении, за исключением приподымающейся верхушки, они
оказываются погруженными в постоянно влажный’ моховой
покров Горизонтальная погруженная часть подобных осей
может иметь различную длину — от 20—30 до 90—ПО см. У ко-
330
ренение ветвей весьма мощное. На стеблях развивается сильно
разветвленная система питающих корней и единичных корней,
принимающих форму стержневых. Замечательно при этом, что
диаметр стебля, в отличие от главной оси, резко увеличивается
у боковых скелетных осей стланца выше зоны их укоренения.
У тонких ветвей диаметр увеличивается, например, с 0,8 до
2,8 см, а у более толстых — с 2,2 до 4,8 см. Это означает, что
почвенное питание вновь укоренившихся ветвей происходит
преимущественно за счет своих придаточных корней. Зависи-
мость таких укоренившихся ветвей от главной оси и системы
главного корня, равно как и от материнских боковых осей,
постепенно ослабевает Вероятно, вследствие этого древесина
плагиотропных ветвей между материнским побегом и зоной их
хкоренения становится хрупкой, легко ломается в процессе пре-
парирования стланца, а затем, со временем, перегнивает, и до-
черние группы побегов (парциальные кусты стланца) приобре-
тают характер самостоятельных образований.
Как видно из сказанного, стланцевые формы туркестанского
можжевельника в зрелом состоянии приобретают черты, сбли-
жающие их с типичными кустарниками, для которых характер-
на последовательная смена укореняющихся скелетных осей,
а затем и парциальных кустов. Конечно, форма роста туркестан-
ского можжевельника не повторяет форму типичных кустарни-
ков, например, орешника, бересклета или розы (см. гл IV).
У туркестанского можжевельника дольше сохраняется главная
ось, хотя бы в се основании В этом признаке стланцы турке-
станского можжевельника ближе к дереву, чем кустарники.
Гораздо дольше, чем у кустарников, живут у туркестанского
можжевельника укореняющиеся скелетные оси Подсчеты, про-
веденные па экземплярах стланцев с высоты 3300 м над уров-
нем моря, обнаружили длительность жизни скелетных осей
в 80—90 лет, вместо 15—20 у кустарников. У стланцев турке-
станского можжевельника, в отличие от геоксильных кустарни-
ков, отсутствуют специализированные подземные корневищеоб-
разные побеги, дающие начало надземным осям возобновления.
В отличие от кустарников у стланцев туркестанского можже-
вельника укореняющиеся скелетные оси возникают не из спя-
щих почек материнских осей (что так отличает кустарники от
деревьев — см. гл IV), а представляют собой обычные боко-
вые ветви с резко выраженной плагиотропной частью, которая
оказывается погруженной в моховой покров, вследствие их
приземного положения.
Изучение структуры и развития стланцевой формы турке-
станского можжевельника позволяет считать, что здесь мы
встречаемся со своеобразной формой, с чертами деревьев (дли-
тельно сохраняющаяся, хотя бы в основании, главная ось и
331
боковые сильные ветви кроны, отсутствие раскрывающихся при
жизни главной оси спящих почек) и некоторыми чертами кус-
тарников (укоренение боковых ветвей, с превращением их в оси
возобновления, последовательная смена укореняющихся осей
возобновления), сочетающимися с приземным положение^ уко-
реняющихся скелетных осей, с резко выраженной плагиотроп-
ностью их роста. Стланец туркестанского можжевельника пред-
ставляет собой как бы дерево с кроной, начинающейся у самой
поверхности почвы и потому с укореняющимися ветвями
О близости стланца туркестанского можжевельника к де-
реву свидетельствуют и переходные формы к настоящим де-
ревьям, встречающиеся у туркестанского можжевельника в бо-
лее благоприятных условиях роста Стройные высокоствольные
деревья туркестанского можжевельника в районе исследования
нс были обнаружены Такие формы приурочены к более запад-
ным хребтам Тянь-Шаня (например, Киргизский Алатау) По
мере продвижения на восток, точнее на северо-восток, турке-
станский можжевельник постепенно теряет древовидную форму
роста. В условиях Заилийского Алатау автор встречал турке-
станский можжевельник на всех абсолютных высотах только
в стланцевой форме
Особенности морфогенеза стланцевой формы
у казацкого можжевельника
(Juniperus sabina L.)
Как отмечалось выше, казацкий можжевельник (Junipprus
sabina L) представляет, по В. Л. Комарову (1934) типичную
форму стелющегося кустарника Лишь единично в пределах
этого вида встречаются древовидные формы с узко пирами-
дальной кроной высотой до 8 м (f jastigiata Host ) или прямо-
стоячего кустарника (f. erecta Host), приуроченные к европей-
ской части ареала (В П. Малеев, 1949).
Изучение показывает, что форма стланца у казацкого мож-
жевельника ушла значительно дальше от формы дерева, по
сравнению с туркестанским можжевельником В пределах рай-
она нашего исследования казацкий можжевельник представлен
только стланцевой формой Изучение морфогенеза казацкого
можжевельника проводилось автором в нижней части долины
р Чон-Кызыл су, в предгорной части Иссыккульской котло-
вины, в урочище «Красная глина».
Прорастание семян и формирование сеянцев у казацкого
можжевельника в течение первых 10—15 лет проходит в основ-
ном так же, как и у туркестанского можжевельника Вначале
главная ось растет ортотропно, достигая высоты 20—25 см
Затем, к 12—15 годам, в основании главной оси от годичных
332
побегов первых 3—5 лет начинают разрастаться боковые ветви.
Число их обычно небольшое: 3—5. Рост ветвей по интенсивности
не уступает главной оси, а часто и превосходит ее. Главная ось
и мощные боковые ветви с возрастом полегают Уже у 25—30-
летних растений, достигающих в диаметре куста 1 —1,5 м, ске-
летные оси приобретают характер прижатых к почве плагио-
тропных осей со слабо приподымающейся верхушкой Покры-
ваясь подстилкой, полегшие части главной оси и боковых вет-
вей у кореняются
У разных экземпляров казацкого можжевельника судьба
главной оси различна Иногда главная ось довольно быстро
отмирает, и дальнейшее нарастание происходит за счет стелю-
щихся укореняющихся боковых ветвей, растущих в радиальных
направлениях В других случаях главная ось сохраняется
долго в полегшем состоянии, и от других укоренившихся осей
(боковых) отличается ослабленным ростом Иногда главная
ось сохраняется и у старых куртин, достигающих 8 м и более
в диаметре
Количество мощных * боковых осей II порядка находится
в известном коррелятивном отношении с интенсивностью роста
главной оси — при отмирании ее в молодом состоянии количе-
ство боковых мощных ветвей увеличивается
Иногда в молодом состоянии в основании главной оси обра-
зуются лишь одна-две мощные боковые ветви, располагающиеся
под острым углом к материнской оси Тогда формируется одно-
сторонняя куртина казацкого можжевельника В таких случаях
главная ось несет мощные ветви не только в ее присемядоль-
ной части, но и выше, на расстоянии 30—35 см Тяжелые боко-
вые ветви пригибают к земле молодую главную ось и укоре-
няются
В отличие от туркестанского можжевельника, мощные боко-
вые ветви казацкого можжевельника остаются постоянно плаги-
отропными Они или активно растут горизонтально, прижи-
маясь к поверхности почвы своей верхушкой, или имеют харак-
тер приподымающихся побегов, которые укореняются при
последующем полегании. От осей II порядка отходят ветви
III порядка, приподымающиеся, более или менее ортотропные.
В отличие от материнских плагиотропных, эти ветви отличаются
меньшей интенсивностью роста и оказываются плодущими,
тогда как ползучие быстро растущие побеги никогда не плодо-
носят, оставаясь вегетативными Вследствие этого, в отличие
от туркестанского можжевельника, здесь можно говорить о
дифференциации побегов на ползучие быстро растущие вегета-
тивные, образующие приземный скелет стланца, и растущие
вверх, более короткие и генеративные, представляющие собой
боковые ветви III—IV—V и более высоких порядков. Послед-
333
ние составляют основную массу приподымающихся побегов по
всей куртине стланца. Их высота не превышает 50—70 см. Неко-
торые побеги III, а, возможно, и более высоких порядков,
могут также отличаться большой интенсивностью роста, распо-
лагаясь при этом горизонтально. Разделение функций между
побегами и, в частности, развитие ползучих, быстро растущих
скелетных укореняющихся побегов обусловливает формирование
крупных куртин казацкого можжевельника, диаметром до 15—
20 м, занятых одним и тем же экземпляром.
Некоторые данные о формировании
и структуре стланцевой формы у сибирского можжевельника
(Juniperus sibirlca Burgsd.)
Формирование проростка и развитие сеянца сибирского мож-
жевельника по наблюдениям близ верхней границы леса в Хи-
бинах в течение первых 8—10 лет его жизни происходит очень
сходно с тем, что было описано у туркестанского и казацкого
можжевельника. Рост главной оси, вначале ортотропной, про-
исходит по моноподиальному типу В течение первых 5—8 лет
годичные приросты не превышают 1,5—2,5 см и лишь в после-
дующие гоДы длина годичных побегов несколько возрастает,
достигая 3—4 см. К 10—12-летнему возрасту высота сеянцев
достигает 20—25 (30) см. Ветвление главной оси начинается
со 2—3-го года. В 10—15-лстнем возрасте становится все более
резко выраженным полегание базальной части главного побега
с последующим его окоренением за счет стеблевых придаточных
корней. Примерно с 20-летнего возраста начинается плодоно-
шение молодых растений, достигающих к этому возрасту
высоты 50—70 см.
В дальнейшем, вследствие более сильного полегания главной
оси (на протяжении 15—20 см), основания боковых ветвей ока-
зываются также полегшими и погруженными в моховой покров
Вскоре они также укореняются. Сильные укоренившиеся боко-
вые ветви оказываются морфолого-биологически подобными
главной оси Из всех боковых ветвей главной оси, как и у турке-
станского можжевельника, самыми мощными оказываются ниж-
ние ветви. Эти ветви обладают в то же время и наиболее
длинной плагиотропной частью, скрытой в моховом покрове
и обильно укоренившейся Ортотропная верхняя часть этих
ветвей имеет ограниченный рост. По мере поднятия над уров-
нем почвы верхушечный рост побегов ослабевает и, наконец, по
достижении известной высоты, прекращается совсем. В верхней
части лесного пояса Хибин побеги сибирского можжевельника
достигают высоты 100—120 см, а в поясе горных тундр всего
лишь 15—20 см.
334
Укоренившиеся приподымающиеся ветви в свою очередь
ветвятся с образованием самых мощных ветвей в базальной
части материнских. Эти мощные нижние ветви, подобно мате-
ринским осям, полегают на значительном протяжении и \коре-
няются, выполняя функции осей возобновления. С появлением
укоренившихся приподымающихся нижних ветвей на главной
оси сибирского можжевельника дальнейший рост его происхо-
дит в форме последовательной смены полупростратных укоре-
няющихся нижних ветвей. Краевое разрастание куртины при-
водит к тому, что у зрелых экземпляров сибирского можжевель-
ника в Хибинах куртины достигают 5,5—6 м в диаметре.
Формирование скелетных осей у стланца сибирского мож-
жевельника по типу обычных боковых ветвей, но лишь укоре-
няющихся вследствие их приземного положения, позволяет
использовать эту структурную особенность для определения
возраста к.уртины.
При определении возраста куртины необходимо отпрепари-
ровать скелетные оси стланца от центра до периферии (жела-
тельно по нескольким фади}сам), измерить общую длину ске-
летных осей по радиусам, вместе с приподымающейся частью
оси у края куртины, и разделить полеченное число (в см) на
величину среднего годичного прироста скелетных осей в данных
экологических условиях. Последняя величина определяется
путем измерения длины годичных побегов на плагиотропных
скелетных осях у разновозрастных экземпляров данного место-
обитания Измерения длительности жизни куртин сибирского
можжевельника в Хибинах дали следующие результаты. Длина
радиальных скелетных осей старых куртин диаметром 6 м была
определена в 450 см, а средний годичный прирост плагиотроп-
ных осей — в 3—4 см. Возраст подобных картин, таким обра-
зом, достигает 120—150 лет
Особенности морфогенеза стелющихся форм
у некоторых аркто-альпийских шпалерных кустарников
и кустарничков (по W. Rauh, 1937)
Весьма точный обзор и строгий морфологический анализ
морфогенеза шпалерных кустарников и кустарничков на при-
мере широко распространенных аркто-альпийских видов приво-
дится в исследовании В Pay (W. Rauh, 1937)
а) Особенности морфогенеза шпалерного кустарника у Salix
reticulata L. Сетчатая ива — Salix reticulata — широко распро-
странена во всей арктической области земли, а также часто
встречается в альпийских областях бореальных горных систем
(М И. Назаров, 1936). В ювенильном состоянии рост главной
оси сетчатой ивы идет ортотропно Ветвление с первого года
335
происходит симподиально и акротонно Симподиально растущая
главная ось или несколько лет растет ортотропно, или уже со
второго года переходит к плагиотропному росту, вследствие уси-
ленного роста стебля с его верхней стороны. Стебель ложится
на землю и закрепляется в таком положении придаточными
корнями (рис. 62). Только верхушка побега слегка приподы-
мается. Ветвление побегов проходит монохазиально Раскры-
Salix reticulata L. Схема шпалерною роста.
в. с. — верхушки симподиальных побегов, с — соцветия, п п — побеги
продолжения, с — семядоли, гп - гипокотиль, г к.— главный корень,
п к,— стеблеродные корни. По Рау, 1939
Образование боковых ветвей (кроме побега продолжения)
начинается со второго или третьего года Однако в первые годы
боковые ветви остаются короткими и тонкими и вскоре отми-
рают
Лишь с четвертого года или позже появляются более силь-
ные боковые ветви, способные к дальнейшему росту. Но они
растут не ортотропно, а сохраняют плагиотропное направление
роста главной оси. Некоторые из них растут в направлении,
противоположном росту главной оси
Ветвление проходит со схематической правильностью. На
каждом годичном побеге закладывается три почки Базальная
почка остается нераскрывшейся, вторая образует короткую
боковую веточку, верхняя боковая почка дает начало побегу
продолжения Верхушка оси регулярно отмирает. Положение
средней почки на побеге имеет большое значение для развития
боковой ветви. Если эта почка оказывается на верхней стороне
материнской оси, то из нее вырастает слабая веточка, растущая
косо вверх и довольно быстро завершающая свой рост терми-
нальным соцветием. Если эта почка закладывается на нижней
?35
стороне материнского стебля, то ее рост оказывается гипотонно
усиленным, и побег может превосходить по силе самый верхний
побег Так может возникнуть второй побег продолжения. Такие
побеги особенно способствуют усилению краевого роста шпалер.
Если на побеге появляется случайно четвертый лист, то четвер-
тая почка не раскрывается, так что сохраняется только что опи-
санный способ ветвления
б) Особенности морфогенеза шпалерного кустарника у Dryas
octopetala, L. Дриада восьмилепестная (Dryas octopetala L.)
представляет собой циркумполярный вид, встречающийся, кроме
того, также в альпах Западной Европы и Урала. Этот вид обра-
зует резко выраженную шпалерную форму.
Dryas octopetala относится к группе мопоподиальных шпа-
лерных кустарников, нарастающих до начала цветения монопо-
диально, а с появлением цветоносных побегов переходящих к
симподи-альному ветвлению. К этой группе относятся также
L oiseleurla procumbens, Arctostaphylos ииа ursi, A alpina, Globula-
ria cordifolia, Cotonepster humifusa и Juniperus prostrata.
Два последние вида сохраняют в течение всей жизни
моноподиальный способ нарастания побегов D. octopelala. по
данным С Schrdter (1926) представляет весьма ярко выражен-
ный тип шпалерного кустарника В его структуре и морфоге-
незе имеется много существенных признаков, общих с ранее
рассмотренными шпалерными кустарниками и стланцами,
наряду с некоторыми специфическими чертами, отличающими
его от них В отличие от выше рассмотренных видов, имевших
главную ось с длительным более или менее интенсивным
ростом, у D octopetala главная ось остается короткой и недо-
развитой; она нарастает лишь в течение нескольких лет.
Главная ось D. octopelala не испытывает характерного
изгиба и не переходит к плагиотропному росту.
Как видно из рис 63, в течение первого года развивается
довольно сильно разветвленная система главного корпя и уко-
роченный ортотропный главный побег Уже на втором году из
пазушных почек базальной и средней части главного побега
вырастают боковые побеги, которые вскоре превосходят по
длине материнский главный побег, полегают на почву и лишь
их верхушка несколько приподымается Их годичный прирост
вначале небольшой, а междоузлия укорочены Вследствие
этого в первые годы возникает в семядольной области неболь-
шая шпалера типа плоской подушки.
Базальная область главной оси является зоной возобнов-
ления всего растения, отдельные побеги ветвятся более или
менее акротонно, или, скорее, мезотонно Боковые ветви, направ-
ленные вверх, заполняют площадь шпалеры, подобно верти-
кальным побегам в куртине описанного выше Juniperus sabina^
15-413 И I Серебряков —22 337
Рис. 63.
Dryas octopetala L. Развитие шпалерной формы:
I — сеянец в начале второго года жизни, 11 — трехлетнее растение. III — трехлетнее растение с гипотонным усилением боковых
ветвей, с — семядоли, мгпк — гипокотиль, г. к. — главный корень, г. о. — главная ось, б. в. — боковые ветви, с. к. — сгеблерод-
ные корни По Рау, 1939
а нижние побеги продолжения обеспечивают краевой рост
шпалеры
Главная ось D. octopetala, растущая ортотропно, обладает
спиральным листорасположением и поэтому ее боковые ветви
растут более или менее равномерно по всем радиусам. Вслед-
ствие этого образуется более или менее округлая шпалера,
нередко достигающая 50—100 см в диаметре, остающаяся бо-
лее или менее зеленой и в течение зимы, так как листья опа-
дают постепенно
Растение в целом производит впечатление скорее много-
летнего дерновинного травянистого, чем древесного, особенно
потому, что новые побеги длительное время сохраняют травя-
нистый характер. По Drude (1896) дриада — не настоящее
древесное растение, а скорее должна считаться переходным
типом к травянистым. Однако стель Dryas со временем приоб-
ретает сильное одревеснение и отличается длительным ростом
вторичной древесины.
D. octopetala является одним из важнейших пионеров расти-
тельности при заселении голых скал и каменистых субстратов.
Растение обладает сильно развитой системой главного корня и
образует систему мощных придаточных корней, в связи с чем
очень велика его роль в закреплении осыпей на склонах.
в) Особенности морфогенеза шпалерной формы у Arctostap-
hylos uva ursi (L.) Spreng; и A. alpina L. Толокнянка обыкновен-
ная, Arctostaphylos uva ursi (L ) Spreng, вечнозеленый распро-
стертый кустарничек; по Е. А Буш (1952), она широко
распространена в сухих сосновых борах, лиственничных лесах
на песчаных почвах, в березово-лиственничных редколесьях,
среди зарослей кедрового стланика и на сухой песчаной тундре;
типична для лесной зоны северного полушария, немного захо-
дит и в тундру. Толокнянка альпийская (A alpina) по Е А. Буш
(1952) —стелющийся кустарничек с опадающими листьями,
широко распространен в различных тундровых формациях, на
гольцах, по скалам, каменистым склонам, россыпям, на сфаг-
новых торфяниках в тундровой и лесной зонах северного полу-
шария. A. uva ursi и A. alpina могут быть поставлены в ряду
наиболее типичных шпалерных форм
По данным Рау, семепа этих видов должны быть освобож-
дены от семенной кожуры и находиться два года во влажном
состоянии, чтобы началось прорастание и образование про-
ростка. У проростка вначале развивается ортотропный и не-
ветвящийся главный побег. На втором или третьем году в па-
зухах семядолей и базальных зеленых листьев развиваются
боковые ветви, которые интенсивно растут и скоро превосходят
главную ось. К этому времени у побегов проявляется положи-
тельно-гсотропический изгиб, и вскоре главный побег и боко-
22* 339
выс ветви полегают на поверхность почвы Годичные приросты
побегов относительно большие, междоузлия сильно удлинены,
и A. uva ursi в сравнительно небольшое время становится за-
метным компонентом ценоза Питание растения происходит
почти исключительно за счет сильного развития системы глав-
ного корня Придаточные корни у A uva ursi появляются редко
в виде тонких питающих корешков, имея только их, без глав-
ного корпя, растение не может произрастать.
Иначе обстоит дело у A alpina, ветви которой зарываются
в почву или рыхлый гумус На этих ветвях формируются силь-
ные придаточные корни, которые находятся в связи с пазуш-
ными почками, располагаясь выше их Можно видеть часто
отрезки побегов, изолированные от системы главного корня и
продолжающие самостоятельный рост. О длительности такого
роста сказать трудно.
Ветвление всего растения у обоих видов толокнянки идет
базитонно, но отдельные побеги обладают строго гипотонным
ветвлением, нарастая моноподиально в вегетативном состоянии
Из рассматриваемых здесь двух систематически близких
видов A alpina имеет более резко выраженную шпалерную
форму. Все ветви у A. alpina оказываются прижатыми к почве,
и лишь редко их верхушки подымаются на несколько санти-
метров Благодаря ежегодному листопаду между ветвями на-
капливается гумус, покрывающий побеги, так что через ряд
лет ветви оказываются скрытыми, за исключением торчащих
верхушек. Это явление способствует более сильному развитию
придаточных стеблевых корней у A alpina.
У A. uva ursi боковые ветви, за исключением побегов про-
должения, растут косо вверх Они долго сохраняют это направ-
ление роста и через ряд лет или целиком отмирают, или пере-
ходят полностью к плагиотропному росту Интересно, что раз-
ная степень выраженности шпалерного характера роста у этих
родственных видов связана с их зональным распространением
A uva ursi с более слабо выраженным шпалерным ростом
приурочена преимущественно к лесной зоне, в то время как
A. alpina с ее резко выраженной шпалерной формой роста —
типичное аркто-альпийское растение.
В стеблях A. uva ursi долго сохраняется способность к фор-
мированию годичных колец. Kanngisser (1912) насчитывал у
этою растения до 80 годичных колец.
3. ГЛАВНЕЙШИЕ ОСОБЕННОСТИ РОСТА И МОРФОГЕНЕЗА
У СТЕЛЮЩИХСЯ ДРЕВЕСНЫХ РАСТЕНИЙ
Приведенный выше обзор морфогенеза стелющихся форм
на ряде типичных примеров позволяет видеть общие и специ-
340
фичсскис особенности этого процесса у разных видов древес-
ных растений. Приведенные данные подтверждают, что стелю-
щиеся древесные растения являются специализированной фор-
мой роста деревьев, кустарников и кустарничков. Примером
стелющихся деревьев (стланцев) могут служить рассмотрен-
ные выше Pinus pumila и Juniperus turkestanica. (К сожалению,
морфогенез стелющихся кустарников остается слабо изучен-
ным ) Примером стелющихся или шпалерных кустарничков
могут служить рассмотренные выше Dryas octopetala, Arctostap-
hylos alpina, A uva ursi и др.
W. Rauh (1937) различает среди шпалерных форм «шпалер-
ные деревца» («Spalierbaumchen») и «шпалерные кустарники»
(«Spalierstraucher»). Критерием для такого подразделения по
Рау служит морфологический признак — наличие или отсут-
ствие раскрывающихся почек возобновления в базальной части
главной оси У шпалерных деревцев спящие почки в основании
главной оси при ее жизни не раскрываются (как и у прямо
стоячих деревьев), и ветвление растения в целом, как и отдель-
ных побегов, проходит по акротонному типу У шпалерных
кустарников из раскрывающихся спящих почек у основания
главной оси образуются быстро растущие побеги (как и у
прямостоячих кустарников) Поэтому при акротонном вет-
влении отдельных побегов, растение в целом ветвится бази-
тонно
Несмотря па заманчивость подразделения стелющихся дре-
весных растений на базе строго морфологического принципа,
использование лишь его одного приводит к искусственному
выделению групп Думается, что экологически более верным
было бы для подразделения стелющихся древесных растений
на стелющиеся деревья (стланцы), стелющиеся кустарники и
стелющиеся кустарнички использовать комплекс признаков,
таких, как длительность жизни скелетных осей, их длина (вы-
сота) и толщина, общие размеры куртин или шпалер, а также
наличие или отсутствие главного ствола, полегающего, как у
Pinus pumila, или прямостоячего и короткого, но толстого и
сохраняющегося сотни лет, как у Juniperus turkestanica.
Наиболее общим и характерным признаком для всех стелю-
щихся древесных растений оказывается специфика рос-
та их побегов Наиболее интенсивно расту-
щими в длину и толщину и наиболее долго-
летними оказываются горизонтальные пла-
гиотропные побеги Побеги ортотропные или близкие
по направлению роста к вертикали отличаются слабым ростом
в длину и толщину и небольшим долголетием
Эти особенности роста побегов оказываются в свою очередь
связанными с ветвлением и латеральной симметрией побегов
341
Ветвление побегов у стелющихся древесных растений, как
подчеркнуто Рау, проходит с акротонным или мезотонным уси-
лением боковых ветвей. Акротонность и мезотонность ветвле-
ния — это проявление продольной симметрии побега. Акротон-
ность ветвления у стелющихся древесных растений сочетается
с гипотонностью его, т. е. усиленным ростом ветвей, располо-
женных на нижней стороне материнской оси. Гипотонно-акро-
тонное усиление ветвей обусловливает краевое разрастание
шпалер и куртин у стелющихся древесных растений, подобно
тому, как эти же свойства в росте и ветвлении побегов у пря-
мостоячих деревьев обусловливают разрастание кроны в сто-
роны и вверх.
Главная ось у стелющихся древесных растений после непро-
должительного периода ортотропного роста или испытывает
положительно-геотропический изгиб и продолжает рост пла-
гиотропно, горизонтально, как у Juniperus sabina, J. sibirica,
Salix retusa, S. reticulata и др., или остается постоянно орто-
тропной, с укоророченными междоузлиями и довольно скоро
прекращает нарастание вверх в отногенезе растения, как у
Dryas octopetala, Juniperus turkestanica и др.
Наряду с сильным развитием системы главного корня у
многих стелющихся древесных растений развивается более или
менее мощная система придаточных стеблеродных корней, уве-
личивая длительность жизненного цикла и интенсивность роста
простратных осей и всего растения в целом. Полегание скелет-
ных осей у стелющихся древесных растений вызывает рез-
кое увеличение длительности жизненного цикла этих осей.
Так, у стелющегося кустарничка A. uva ursi скелетные про-
стратные оси у верхней границы леса и в горных тундрах Хи-
бин живут до 10—12 лет, тогда как у прямостоячих кустар-
ничков в тех же условиях длительность жизненного цикла
скелетных осей не превышает 4—5 лет.
Подобное же увеличение длительности жизни и верхушеч-
ного нарастания отмечалось выше у кедрового стланика.
Глава VII
ЖИЗНЕННЫЕ ФОРМЫ ЛИАН
Лианы, или лазящие растения, как их называл Дарвин
(1878), могут быть охарактеризованы как укореняющиеся в
почве, обычно автотрофные растения со стеблями, неспособны-
ми постоянно сохранять вертикальное положение и использую-
щие другие растения как опору, чтобы поднять свою ассими-
лирующую листву и цветки над почвой в условия благоприят-
ного освещения.
Стебли лиан отличаются резко выраженным отрицательным
геотропизмом, быстрым ростом в длину и резко вытянутыми
междоузлиями. По данным П. А. Баранова (1949) побеги ви-
нограда (Vitis vinifera) в Средней Азии за один вегетационный
период могут вырастать до 25—30 м в длину. Так как* интенсив-
ному росту стебля в длину не соответствует рост его в толщи-
ну, лианы не в состоянии сохранять вертикальное положение
осей. Пользуясь другими растениями как опорой, они в корот-
кое время достигают значительной высоты, без больших затрат
на построение прочных осей.
Лианы представляют разнообразный и широко распростра-
ненный ряд жизненных форм. По положению в системе жиз-
ненных форм (см. гл. II) лианы сходны со стелющимися расте-
ниями: они образуют своеобразный ряд, начинающийся дре-
вовидными растениями и кончающийся травянистыми
однолетниками. Среди лиан, как и среди стелющихся растений,
имеются древесные и травянистые формы, В пределах древес-
ных лиан имеются лиапоидпые дериваты деревьев, кустарников
и кустарничков. Но наибольшее развитие среди древесных
343
лиан получают различные лазящие кустарники, филогенети-
чески связанные с их прямостоячими формами Некоторые
формы лиан (корнелазящие лианы с придаточными корнями —
прицепками) тесно связаны со стелющимися формами расте-
ний, образующими на стеблях многочисленные придаточные
корни
Лианы характерны не только для современного раститель-
ного покрова. Многочисленные лазящие растения из папорот-
ников и семенных папоротников встречались в лесах каменно-
угольного периода; они произрастали в условиях, близких к
современным во влажно-тропических лесах. Из числа лазящих
папоротников карбона может быть назван Ankyropteris scandens.
Некоторые семенные папоротники из Medulloseae верхнего кар-
бона, например Medullosa anglica, была вьющимся растением;
ее разделенная на три части стель весьма сходна со стслыо
некоторых современных лиан (К. Magdefrau, 1956).
Среди покрытосеменных растений древесные лианы оказы-
ваются также одной из самых древних групп жизненных форм,
уступающих по древности лишь прямостоячим деревьям и кус-
тарникам. По исследованиям Ганзена (Н М Hansen, 1956)
максимальное развитие их происходило в палеогене
В настоящее время лианы широко распространены среди со-
судистых и особенно семенных растений По подсчетам Г. Шен-
ка (Н. Schenk, 1892) в половине всех семейств семенных расте
ний имеются представители лиан Склонность отдельных се-
мейств к лазящему образу жизни различна: то она свойствен-
на большинству родов, то выражена слабо и проявляется лишь
в образовании отдельных лазящих видов и родов Эти разли-
чия связаны с внутренними свойствами групп, а также усло-
виями их развития в настоящем и прошлом
В настоящее время лианы широко распространены в расти-
тельном покрове земли. Они отсутствуют лишь в полярных и
альпийских областях, а также в пустынных и степных, где кли-
матические факторы не благоприятствуют образованию длин-
ных осей Лианы имеют очень широкий ареал, но распределение
их крайне неравномерное В большинстве своем — лианы обита-
тели тропиков. По подсчетам Г Шенка (Н Schenk, 1892), под-
твержденным позднейшими исследователями (A. Schimper
и F Faber, 1935), более 90% всех лиан находится в тропиках
По Palm’y (см Н. Schenk, 1892) в тропиках число лиан превы-
шает 2000 видов, в то время как в умеренных зонах едва дости-
гает 200. Но и в тропиках распространение лиан неоднородно
Большинство древесных лиан оказывается во влажных (дож-
девых) и муссонных лесах, в то время как сухие древесные фор-
мации тропиков и саванны имеют лишь тонкостебельные, глав-
ным образом, травянистые формы лиан
344
Во влажных тропических лесах условия жизни оптимальны
для лиан Наряд\ с эпифитами, лианы во влажно-тропических
лесах являются главнейшими жизненными формами Разнооб-
разие и массовость развития лиан во влажно-тропических лесах
связаны не только с благоприятным климатом, необходимым
для роста длинных побегов лиан (высокая средняя годовая
температура, постоянная высокая влажность воздуха и обилие
осадков), но определяется и борьбой за свет между растениями,
которая в густых влажно-тропических вечнозеленых лесах до-
стигает своего апогея. Последний фактор в развитии лиан ока-
зывается решающим
Начиная с классических трудов Дарвина (1878) и Г. Шенка
(1892, 1893) классификация лиан основывается на способах
лазания. Этот принцип подразделения лиан не только отражает
многообразие лазящих растений и их эколшические особен-
ности, но и позволяет проследить эволюцию приспособлений
к лазящему образу жизни в этой обширной группе растений.
По способу лазания лианы подразделяются на 4 биологиче-
ские группы
I Опирающиеся лианы (Spreizklimmer, Н Schenk, 4892;
Scrambling woody lianes, E Ou Rietz, 1931)
2 . Корнелазяшие лианы (Wurzelkletterer, H. Schenk, 1892;
Root-climbing woody lianes, E Du Rietz, 1931).
3 Вьющиеся лианы (Windenpflanzen, H Schenk, 1892, Win-
dmg woody lianes, E. Du Rietz, 1931).
4 Усиконосные лианы (Rankenpflanzen H Schenk, 1892;
Tendril-climbing woody lianes, E. Du Rietz, 1931).
I\ первой биологической группе опирающихся лиан относят
лазящие растения, не имеющие специальных органов лазания —
вьющихся стеблей, чувствительных усиков, прикрепляющихся
корней Опирающиеся лианы, большей частью кустарники,
а также иногда травы, имеют длинные, отходящие в стороны
ветви, которые из-за слабости их стебля полегают на ветви и
сучья соседних растений, становящихся их опорой. Закрепление
ветвей на опорных растениях часто усиливается с помощью
шипов или колючек Но образование этих последних обычно не
связано с приспособлением к лазящему образу жизни (напри-
мер, у лазящих видов Rosa — см. Сумневич, 1947). Биологически
эта группа лиан представляет низшую ступень развития лазя-
щих растений и не всеми ботаниками признается за лианы.
Действительно, некоторые из относящихся сюда видов вообще
трудно отнести к лианам так мало они отличаются по способу
роста и габитуально от прямостоячих кустарников и трав. Но
эта группа позволяет понять этапы формирования лиан от пря-
мостоячих близко родственных видов.
Корне лазящие лианы взбираются вверх по стволам
315
деревьев, отвесным скалам или стенам с помощью прицепляю-
щихся и прирастающих к опоре придаточных корней. Число
корнелазящих невелико, сравнительно с вьющимися растениями
и усиконосными лианами, хотя они и встречаются среди раз-
личных семейств, и обладают четко выраженными приспособ-
лениями к лазящему образу жизни Развитие корнелазящих
лиан связано с наличием отвесных скал или толстых стволов
деревьев и поэтому сильно локализировано. Исходные формы
для большинства корнелазящих лиан находятся среди растений
с ползучими укореняющимися побегами
Типичным представителем корнелазящих древесных лиан
нашей флоры может служить плющ — Hedera helix.
Вьющиеся растения. Обвивающий рост стебля пред-
ставляет наиболее широко распространенный способ лазания
лиан; по числу вьющиеся растения превышает число лиан дру-
гих категорий
Первые междоузлия проростков у типичных вьющихся расте-
ний, например, Phaseolus, Humulus lupulus, Convolvulus не
вьются и растут отвесно вверх. Их нутация слишком незначи-
тельна, чтобы вызвать обвивающее движение; и только по до-
стижении определенной длины стебля, обычно после образова-
ния 3—4 междоузлий, наступает сильная круговая нутация, и
стебель приобретает характер вьющегося. У некоторых слабо
вьющихся видов отвесное состояние стебля сохраняется дольше.
Подобная смена в онтогенезе растения и отдельного стебля
наблюдается и у усиконосных лиан.
Любопытно, что завивание стебля идет у каждого вида в
строго определенном направлении, или только вправо, или
только влево Этот признак оказывается наследственно закреп-
ленным.
Некоторые виды образуют вьющиеся формы только в благо-
приятных для этого условиях — в густых тенистых лесах, а на
открытых освещенных местах растут как прямостоячие кустар-
ники. По данным И С Михайловской (1953) так ведет себя
лимонник (Schizandra chinensis} на Дальнем Востоке
Чаще всего у растений вьются лишь длинные побеги, и лишь
у сильно вьющихся видов, например Humulus lupulus, Convol-
vulus, Phaseolus вьющимися оказываются все стебли
Кроме резко выраженной круговой нутации, в росте и фор-
мировании побегов вьющихся растений выделяются еще две
черты, отличающие их от обычных прямостоячих побегов —
большая длина растущей зоны стебля и заторможенность роста
листьев в верхней части побега. По данным Тролля (W. Troll.
1937), у вьющихся растений длина растущей зоны побега может
превышать таковую у прямостоячих растений в 15—20 раз и
достигать 40—50 см.
316
Листообразование на вьющихся побегах значительно замед-
ленно. Но это касается не заложения, а роста листьев Вслед-
ствие этого рост междоузлий особенно резко обгоняет рост
листьев Подобный рост вьющихся растений аналогичен росту
затененных этиолированных растений. Быстрое удлинение
междоузлий при замедленном росте листьев приводит к умень-
шению веса растущих верхушек побегов у7 лазящих растений.
У многих тропических вьющихся древесных лиан, например у
видов Dioscorea, листья достигают нормальной величины лишь
в 2—3 метрах ниже верхушки, в нескольких десятках междо-
узлий от нее Такой тип роста приводит к образованию габи-
туально своеобразного бичевидного типа побегов. Развитие
листьев у вьющихся растений часто зависит от наличия опоры
Выше опоры растущие побеги могут засохнуть.
У травянистых вьющихся, как Hamulus lupulus Convolvulus,
виды Phaseolus и др, обычно все побеги развиваются как вью-
щиеся удлиненные с ассимилирующими листьями. Иначе у дре-
весных лиан, которые дают множество примеров диморфизма
побегов Весьма обычна у вьющихся древесных дифференциа-
ция побегов на удлиненные и укороченные, с образованием
фотосинтезирующих листьев лишь на укороченных, тогда как
удлиненные побеги несут недоразвитые чешуевидные листья.
У них произошло разделение функций: удлиненные побеги —
исключительно ищущие и лазящие, а ассимиляция ложится на
невьющиеся укороченные побеги с зеленой листвой. Такой
диморфизм побегов резко выражен, например, у некоторых
древесных лиан из рода Gnetum
Усиконосные лианы. По приспособленности к лазя-
щему образу жизни усиконосные лианы являются наиболее
совершенной группой среди всех лазящих растений Они обла-
дают способностью к очень быстрому и надежному укреплению
тонких и быстро растущих побегов на подпорках. При всем
многообразии усиков у разных видов растений, общим призна-
ком их является контактная чувствительность или резко выра-
женный гаптотропизм, т. е. чувствительность усиков к дли-
тельному соприкосновению с твердой опорой, приводящая
к более или менее быстрому обхватыванию этой подпорки
Морфологически усики у разных видов лазящих растений
могут быть листового или стеблевого происхождения Среди тех
и других выделяются более примитивные и более совершенные,
специализированные формы.
Так, среди листоусиковых лиан выделяется группа
листолазящих растений. Листья этих растений еще обла-
дают листовой пластинкой и способны к фотосинтезу. Но в то
же время им свойственна контактная чувствительность, и они
обвиваются вокруг подпорки при помощи листового черешка
347
или удлиненной верхушки пластинки (например у Gloriosa
superba) От этой группы растений с двойной функцией листьев
в процессе естественного отбора развились собственно листо-
усиковые лианы, с листьями специализированного типа, лишен-
ными пластинки, нитевидными с высокой контактной чувстви-
тельностью, выполняющими лишь функцию лазящих органов.
Листоусиковые известны в различных семействах двудольных.
Во второй группе — стеблеусиковых лиан — выде-
ляется биологически примитивная группа ветвелазящих
растений с боковыми облиственными ветвями, обладающими
контактной чувствительностью, и обвивающими опору при сопри-
косновении с ней. К этой группе относятся многие лазящие
кустарники тропиков От подобных форм возникли более спе-
циализированные лазящие кустарники, на чувствительных вет-
вях которых листья редуцированы, а сами ветви метаморфизи-
рованы в усики. Эти ветви функционируют лишь как лазящие
органы.
Интересные данные о филогенезе стеблеусиковой лианы
у виноградной лозы (Vitis vinifera) приводятся П. А. Барано-
вым (1949). Сравнительно-морфологический и ботанико-геогра-
фический анализ Vitaceae приводит П. А. Баранова к заключе-
нию, что предками винограда были обитатели ярко освещенных
открытых пространств без признаков лианоидной формы роста.
По мнению П. А. Баранова, предки Vitis vinifera и других родов
Vitaceae имели древовидную или кустарниковую форму с моно-
подиальным ростом побегов, но верхушечными соцветиями.
Одним из первых морфогенетических процессов на пути от
прямостоячего кустарника к лиане было преобразование вер-
хушечного соцветия в боковое. Таким путем растения получили
гораздо большую возможность для роста побегов в длину.
Важнейшим этапом на пути исторического морфогенеза лианы
у V. vinifera было преобразование соцветия в специализирован-
ный орган для лазания—усик Такое преобразование облегча-
лось у винограда тем, что ветви соцветия у некоторых видов
винограда обладают гаптотропизмом, и при соприкосновении
с твердым предметом они производят гаптотропическое обхва-
тывающее движение. Вследствие недоразвития цветков в от-
дельных ветвях соцветия происходила их стерилизация и эти
ветви специализировались как органы лазания — усики. Подхва-
ченный естественным отбором процесс стерилизации распро-
странялся на большую и большую часть соцветия, вплоть до
полного преобразования соцветия в усик, свободный от функции
размножения.
Интересно, что в онтогенезе соцветия у винограда не всегда
наблюдается полная дифференциация цветков. При недоразви-
тии цветков в уже заложенном соцветии последнее нередко
вырастает в типичный усик
Глава VIII
ЖИЗНЕННЫЕ ФОРМЫ ПОДУШКОВИДНЫХ РАСТЕНИИ
Как и стелющиеся и лианоидные растения, подушковидные
растения составляют ряд жизненных форм, входящих в типы
жизненных форм от кустарников до травянистых многолет-
ников
1. ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И ГЕОГРАФИЧЕСКОЕ
РАСПРОСТРАНЕНИЕ ПОДУШКОВИДНЫХ РАСТЕНИЙ
Подушковидные растения представляют собой многолетние,
травянистые, реже древесные растения, в большинстве вечно-
зеленые, с едва заметной главной осью (у основания подушки)
и многочисленными, радиально и этажами расположенными
боковыми ветвями, верхушки которых, вследствие одинакового
и всегда ничтожного прироста в длину (несколько мм в год)
образуют общую для всей подушки более или менее плотную
ровную поверхность полушаровидной или плоскокровлеобраз-
ной формы, выше которой могут подниматься лишь отдельные
ветви соцветия и цветки. Стебли подушковидных растений с
крайне короткими междоузлиями плотно покрыты мелкими,
чаще простыми, сидячими ассимилирующими листьями, более
или менее длительно сохраняющимися по подсыхании. Живые
растущие стебли и ассимилирующие листья располагаются
лишь на поверхности подушки, в ее «коровой» части, а внутрен-
ний объем подушки между ее ветвями обычно заполнен торфя-
нистой массой из выветривающихся листьев, пронизанной пи-
тающими придаточными корнями.
349
Весьма характерно для подушковидных растений наличие
сильно развитой и длительно функционирующей системы глав-
ного корня (И. В. Палибин, 1914; W. Troll, 1937). У некоторых
подушек, кроме того, с возрастом развивается мощная система
придаточных корней (рис. 64). Наличие ровной и более или
менее плотной поверхности и заполняющего материала внутри
подушки Гаури и Шретер (Н. Hauri und С. Schroter, 1914) счи-
тают обязательным признаком настоящих растений подушек
Рис. 64.
Возрастные изменения строения корневой системы Acantholimon
diapensioides. По О. В. Заленскому;
(Polsterpflanzen), в отличие от растений со сходной формой
роста, но без этих признаков, которые эти авторы предлагают
называть Kissenpflanzen. Может быть лучше, вслед за В. Рау
(W. Rauh, 1939), все подобные растения относить к подушко-
видным, выделяя среди них два типа — плотных и рыхлых
подушек. Последние лишены заполняющего материала.
Подушковидные растения довольно широко распространены
среди цветковых растений По подсчетам Гаури и Шретера
(Hauri и Schroter, 1914) подушковидные растения встречаются
среди 34 семейств и 78 родов покрытосеменных. Наибольшее
число видов растений подушек встречаются среди Caryophylla-
ceae, Cruciferae, Saxifragaceae, Malvaceae, Umbelliferae, Primu-
laceae, Plantaginaceae и Compositae.
К этим семействам принадлежат 260 видов или около 77%
всех подушек из покрытосеменных.
350
Растения-подушки отличаются широким ареалом. Они
встречаются на всех континентах и во всех географических ши-
ротах, от Арктики до субантарктических островов. Правда, в
тропиках подушковидные формы растений приурочены лишь
к высокогорному альпийскому поясу. В равнинных областях
тропиков подушки отсутствуют, за исключением кампосов Бра-
зилии, где имеются растения, близкие к подушковидным.
Наибольшее число видов подушковидных растений известно
в Южной Америке и особенно в Андах, Патагонии и на
Фальклендских островах. Здесь зарегистрировано наиболь-
шее флористическое многообразие подушковидных растений
{170 видов или более 50% от общего количества подушковид-
ных покрытосеменных, Н. Hauri и. С. Schroter, 1914). Здесь
растения-подушки — важнейшие эдификаторы растительного
покрова и ландшафтные формы (рис. 65).
Рис. 65.
Гигантские подушки азореллы (Azorella, Umbelliferae)
с Малого Кергуелена. По Шенку из И. В. Палибина
Довольно широка экологическая приуроченность подушко-
видных растений. Они распространены на болотах Южной Аме-
рики, Новой Зеландии и субантарктических островов. По Hauri
и Schroter (1914), не менее половины всех подушек относится
к психрофитам.
351
С другой стороны, растения подушки — обитатели крайне
сухих и жарких областей Сахары, крайне иссушенных почв
нагорий Центральной Азии Растения-подушки селятся также
на прибрежных скалах, каменистых осыпях пояса полупустынь
и горных степей Средней Азии, Ирана и Малой Азии, в холод-
ных сухих высокогорных пустынях Памира.
Экологически разнообразные местообитания растений-поду-
шек связывает общий фактор — свободный доступ света
(О В Заленский, 1948) По мнению О. В. Заленского, многие
особенности морфогенеза растений-подушек, вероятно, связаны
с действием света. Все местообитания подушек (особенно
в местах их массового развития) характеризует крайняя не-
благоприятность условий для роста надземных органов. В раз-
личных типах местообитаний подушек ограничивающими рост
могут быть различные факторы то это низкая температура воз-
духа и почвы (высокогорные области, Арктика и субантаркти-
ческие острова), то это крайняя сухость почвы и низкая влаж-
ность воздуха, то это тормозящее рост действие холодных штор-
мовых ветров, то чаще всего сочетание перечисленных неблаго-
приятных факторов, подавляющих рост надземных побегов.
Можно с полным основанием, вслед за многими исследова-
телями, рассматривать подушковидные растения как конечное
звено в приспособительном ряду растений, направленном по
пути редукции ростовых процессов (О. В Заленский, 1948),
а сами подушковидные формы растений — как ярко выражен-
ную приспособительную форму к жизни в крайних условиях
«существования
В экологическом плане жизненные формы подушковидных
растений диаметрально противоположны жизненным формам
деревьев Деревья — это растения, у которых наиболее полно
выражена способность к интенсивному и длительному ростуг их
побегов, вследствие чего они занимают максимальное воздуш-
ное пространство Дерево — жизненная форма, посредством
которой растения наиболее полно используют благоприятные
для роста условия и достигают максимальных для высших
растений размеров Жизненные формы подушковидных расте-
ний — это форма приспособления растений к условиям, в макси-
мальной степени неблагоприятным для роста надземных орга-
нов. а сами подушковидныо растения являются формами много-
летних растений, достигающими при большой длительности
жизни наименьших размеров.
* *
*
Сохранение крайне незначительных размеров при очень
большой длительности жизни подушек не раз отмечалось бота-
352
никами. В отличие от многовековых гигантов растительного
мира — деревьев, подушковидные растения, по образному срав-
нению К. И, Максимовича, могут быть охарактеризованы как
многовековые карлики. «Эти дерновины — подушки,— говорит
И В Палибин (1914),— стойкие ко всем невзгодам климата,
древнее старых толстых деревьев наших лесов, и какой-нибудь
могучий дуб в степном лесу гораздо моложе компактной дерно-
вины (подушки), обитающей па высоких горах Азии» (стр
1442). Известный знаток флоры Тибета К И. Максимович, го-
воря об Androsace ZapeZe Maxim, и других подушковидных расте-
ниях Тибета, заключает, что безлесное Тибетское нагорье имеет
своих, многовековых, но не исполинов, а карликов (из речи,
произнесенной 9 ноября 1888 г. в память ботанических исследо-
ваний Н М Пржевальского — см. И. В. Палибин, 1914).
2. МОРФОГЕНЕЗ ПОДУШКОВИДНОЙ ЖИЗНЕННОЙ ФОРМЫ
У НЕКОТОРЫХ ВИДОВ РАСТЕНИЙ (ПО RAUH 1939).
Биолого-морфологические особенности подушковидных рас-
тений наиболее полно выявляются при изучении их формирова-
ния в онтогенезе
Образование подушковидной формы
у Silene acaulis L.
Сеянцы Silene acaulis уже с первых лет жизни отличаются
резко преобладающим ростом главного корня над ростом глав-
ного стебля Как видно из рис 66, двухлетний сеянец Sdene
acaulis имеет мощный, маловетвящийся главный корень и сла-
бый ортотропный главный побег. Последний вырастает в тече-
ние первого года до 0,5 см. В конце вегетационного периода
рост побега прекращается, и верхушечная почка главной оси,
как и всех позже возникающих боковых ветвей, перезимовы-
вает без особых защитных органов или почечных чешуй по типу
открытых почек. Наличие открытых почек, лишенных почечных
чешуй — характерный признак всех подушковидных растений.
Прирост главного побега остается ничтожным на втором
году и в последующие годы, так что главная ось вскоре оказы-
вается едва заметной. Через несколько лет ее рост прекращает-
ся из-за образования верхушечного цветка. Но часто еще
раньше этого главную ось обгоняют в росте боковые побеги.
Ветвление главной оси начинается на втором, реже третьем
году жизни растения, и всегда происходит определенным обра-
зом, свойственным большинству растений-подушек. Раскры-
ваются лишь самые верхние почки, в то время как почки ба-
зальных листьев и семядолей остаются в покоящемся состоянии,
В-413. И Г Серебряков —23 353
Рис. 66.
Silene acaulis. Формирование плоской подушки:
1 - сеянец в начале второго года, II и III — трехлетние растения,
г о,— главная ось, б. в — боковые ветви, с — семядоли, гпк — гипокотиль,
г. к,— главный корень По Рау, 1939
и в дальнейшем отмирают. Таким образом, в образовании боко-
вых побегов уже с первых лет проявляется резко выраженная
акротонность.
Первичные боковые ветви главной оси трансверзально гео-
тропичны: они располагаются почти под прямым углом к глав-
ной оси и отходят радиально во все стороны, прилегая к суб-
страту и прикрепляясь к почве придаточными корнями по мере
их возникновения. Лишь верхушки этих побегов слегка приподы-
маются. Эти побеги Рау называет периферическими или
краевыми. Боковые ветви второго и следующих лет распо-
лагаются на главной оси под острым углом к ней и вследствие
этого растут более или менее ортотропно. Их Рау называет
354
центральными ветвями подушки, образование их приводит
к заполнению объема и уплотнению молодых подушек.
Все боковые побеги в течение первых лет нарастают моно-
подпально. При достаточном усилении начинается ветвление,
причем у боковых ветвей, как и на главной оси, наблюдается
акротонное усиление образующихся ветвей. Но в то время как
на главной оси, в связи с ее ортотропным ростом и радиальной
структурой, все боковые ветви растут одинаково, на боковых
побегах наблюдается иное, на этих побегах, как трансверзаль-
ио геотропических органах, развиваются различные боковые
ветви: более слабые на верхней и сильнее растущие — на ниж-
ней стороне (гипотония ветвления)
Эта гипотония выражена тем сильнее, чем больше угол
отхождения боковой материнской ветви от главной оси; наи-
более сильно она выражена на периферических плагиотропных
ветвях и менее резко — па растущих вверх центральных ветвях
подушки Центральные побеги подушки образуют примерно
одинаковые боковые ветви, периферические образуют более
сильные ветви на нижней стороне Гипотония проявляется
также в различной величине листьев и цветков на нижней и
верхней стороне ветвей.
В результате ежегодного повторения акротонно-гипотонного
усиления ветвей образуется плоская слабо-куполообразная
подушка, края которой, вследствие усиленного роста нижних
периферических побегов, растут быстрее, чем центральные
части. У плоских подушек, как и у стелющихся растений (см.
гл. V), можно говорить о краевом росте. Краевой рост обусло-
вливает разрыхление края подушки, как это хорошо видно
у плоских подушек Alsine sedoides. A Bosani, A. stellata, Azoretla
trifurcata, A. apola и др.
По достижении известной величины и возраста растений-
подушек начинается их цветение. Каждый побег завершается
образованием верхушечного цветка, только периферические
побеги часто остаются вегетативными Цветки развиваются так
обильно, что почти целиком скрывают зеленую окраску по
душки. С началом цветения прекращается моноподиальное на-
растание и дальнейшее ветвление происходит симподиально
Почки, заложенные непосредственно ниже цветка, дают начало
побегам замещения Как и в вегетативной фазе развития по-
душки, эти побеги в центральной части подушки развиваются
почти одинаково сильно во всех направлениях. На перифериче-
ских ветвях обычно развиваются лишь побеги, закладываю-
щиеся на нижней стороне материнских ветвей.
Так как все побеги растут в длину одинаково, их верхушки
располагаются на одном уровне, образуя общую поверхность
С каждым периодом цветения симпоциальное ветвление повто-
23* 355
ряется и вследствие этого формируется округлая подушка
с радиальным этажеобразным строением; диаметр подушки
достигает 60 см при высоте в 15 см. Ее поверхность образована
верхушками побегов с ассимилирующими листьями, которые
располагаются так тесно, что свет не проходит внутрь подушки.
Схема формирования подушки дана на рис. 67.
Рис 67.
Схема формирования радиально-шаровидной подушки
(по Рау, 1939):
г к.— главный корень, с к — стеблеродные корни
Листья подсыхают довольно быстро, уже в конце первого
вегетационного периода. Но они не опадают с оси, а сохраняют-
ся долго на стеблях внутри подушки; они постепенно выветри-
ваются, образуя торфянистую заполняющую массу, которая не
только придает подушке известную плотность, но играет также
некоторую роль в водном балансе растения Заполняющая
масса сравнима с губкой, которая легко поглощает воду, но
с трудом ее отдает. По Рау, эта масса представляет как бы
временный резервуар для дождевой воды. Запасы этой воды
поглощаются тонкими всасывающими придаточными корнями
Правда, по данным О. В. Заленского (1948), у пустынных
ксерофитных подушек придаточные корни на побегах не обра-
зуются, поэтому говорить здесь о непосредственном использо-
вании воды, накопленной внутри подушки, не приходится
Когда плоская подушка Silene acaulls достигает известной
старости, ее краевой рост начинает ослабевать при сохранении
поверхностного роста Вследствие этого подушка становится
все более выпуклой, вплоть до полушаровидной, высотой до
50 см. Когда краевой рост начинает ослабевать, цветки уже
образуются только на центральных ветвях, в то время как пери-
ферийные остаются вегетативными и неветвящимися
356
3. КЛАССИФИКАЦИЯ ПОДУШКОВИДНЫХ ЖИЗНЕННЫХ ФОРМ
И ИХ ЭКОЛОГО-МОРФОЛОГИЧЕСКАЯ ИНТЕРПРЕТАЦИЯ
Развитие подушковидных растений в различных системати-
ческих группах и разных экологических условиях вызвало
многообразие типов и форм подушек и необходимость их класси-
фикации.
К. Reiche (1893) выделял среди подушковидных рас-
тений кровлеобразные (Deckenpolster, плоские подушки) и
дерновинные растения (Horstpflanzen) из Gramineae, Juncaceae.
Diels (1897) подразделяет подушковидные растения, исходя
не из их общего типа роста, а из структуры листьев, выделяя
Areiia-тип (подушковидные растения с опушенными листьями)
и Лгоге(/а-тип (подушковидные растения с голыми кожистыми
листьями).
В классификации С. Schroter (1908), охватывающей лишь
наиболее типичные подушковидные растения, т. е. растения-
подушки в узком смысле этого слова, выделяются шарообраз-
ные и плоские подушки (Vollkugelpolster и Vollflachpolster).
Более детальна классификация подушковидных растений
Гаури (Н. Hauri, 1912). Он различает подушки-сообщества
и подушки-индивиды (Gesellschaftspolstern и Individualpolstern).
По величине, компактности и наличию заполняющего материала
последние подразделяются на шаровидные кустарники (Kugel-
straucher) и воздушно-шаровидные подушки (Luftkugelpolstern),
с одной стороны, и подушковидные растения в узком смысле —
с другой В свою очередь среди этих растений выделяются
12 групп
Рау (1939) предлагает следующую систему подушковидных
растений (изложено сокращенно).
1. Настоящие п о д у ш к о в и д н ы е растения
Под^шковидный рост настоящих растений-подушек закреп-
лен наследственно, в отличие от ложно-подушковидных расте-
ний, и не меняется существенно при изменении внешних усло-
вий, лишь усиливаясь или ослабляясь. Основные особенности
подушковидных растений связаны с симметрией их побегов,
с ежегодным повторением акротонно-гипотонного усиления вет-
вей. Боковые ветви плотно облиствены и обладают одинаковым
ничтожным ростом. Они располагаются более или менее плотно
в общей поверхности, выше которой поднимаются лишь соцве-
тия. Плотность подушки зависит от накопления заполняющего
материала. Подушки с рыхлым расположением ветвей не имеют
заполняющего материала; он свойствен лишь компактным
подушкам. Вода и минеральные соли поглощаются мощным
357
главным корнем, деятельность которого усиливается придаточ-
ными всасывающими корнями в заполняющей массе подушки
I Радиально-шаровидные подушки. Поверх-
ность подушки сильно выпуклая и имеет форму полушария,
заполненного внутри радиально и поэтажно-расположенными
боковыми ветвями Заполняющий материал имеется или отсут-
ствует
а) Подушковидпые деревья (Polsterbaume). Подушка рас-
полагается на более или менее высоком, лишенном боковых
ветвей стволе. Ветви расположены рыхло. Заполняющая масса
отсутствует. Dracaena Draco, Euphorbia dendroides;
b) Шаровидные кустарники. Достигают 1 м высоты, имеют
едва заметную главную ось и радиально расположенные ветви.
Характерен ежегодный листопад. Листья не превращаются
в заполняющую массу. Примеры:
Bupleurum fruticosum, Daphne glandulosa, Thytnelaea argen-
iea, Daphne sericea, Euphorbia acanthothamnus, Ruta Corsica,
Poterium spinosum и др.
с) Незаполненные шаровидные подушки Древесные или
травянистые растения, сходные с предыдущей группой: виды
Astragalus, Acantholimon и др.
d) Заполненные шаровидные подушки. Подушковидные
растения в узком смысле слова Верхушки радиально располо-
женных ветвей смыкаются вместе в общую поверхность. Отми-
рающие листья остаются на стеблях и выветриваются в торфя-
нистую массу, пронизанную придаточными питающими корнями.
Симподиальные подушки
*) с одревесневшими побегами. Raoulia rubra,
кх) с травянистыми побегами:
Виды Azorella, Saxifraga и др.
?) Моноподиальные подушки, например Oxalis compacta,
виды Androsace
II. Радиально-плоские подушки Поверхность
подушки слабо выпуклая и поднимается лишь на несколько
сантиметров над уровнем почвы Нерезко отграничены от шаро-
видных подушек и связаны с ними переходными формами.
а) незаполненные плоские подушки: Lucllia, Bryomorpha
и др ;
Ь) заполненные плоские подушки. Silene acaulis (в молодом
возрасте), Alsine sedoides, Raoulia Buchanani, Plantago subulata
и др.
Ill Дерновинные подушки
Дерновинным подушкам свойственна организационная струк-
тура подушковидных растений, но они отличаются отсутствием
многолетнего главного корня Он отмирает уже в молодом
358
состоянии растения и заменяется сильными придаточными. По
этому признаку эти подушки становятся сходными с дерновин-
ными растениями. Сюда Рау относит многие осоки и злаки.
IV. Розеточные подушки
Подушка не сохраняется как целое, так как отдельные побе-
ги отмирают по отцветании Они заменяются укореняющимися
побегами возобновления, типа самостоятельных особей (подуш-
ки-сообщества по Hauri). Инновации образуются в таком коли-
честве, что, примыкая друг к другу, формируют компактную
подушку Примером могут служить некоторые виды Saxifraga.
V. Суккулентныс подушковидные растения:
Opuntia floccosa, О. lagopus, Euphorbia hystrix, виды Mamil-
laria
VI. Подушковидные мхи
Grimmia, Sphagnum и др.
VII. Гапаксантные п о д у ш к о в и д н ы с растения —
ложпоподушковидные
К этой группе относят растения без организационных
признаков подушковидных растений. Под воздействием факто-
ров среды, особенно механически повреждающего действия
ветра, они принимают подушковидный рост, что связано боль-
шей частью с отмиранием (верхушек Примеры. Helianthemutn
oelandicum, Hutchinsia andina, Cerastiutn latifolium, Calycoiome
spinosa и др. Подушковидный рост их вызван торможением
роста внешними факторами. Как только тормозящее действие
факторов устраняется, растения начинают развиваться по свой-
ственному им плану строения.
+ *
«
Классификация подушковидных растений В. Рау — наиболее
разработанная и обоснованная из всех известных в литературе.
Безусловный интерес представляют ее наиболее крупные под-
разделения, выделяемые римскими цифрами. Обращает вни-
мание широкая трактовка подушковидных растений, как боль-
шого ряда жизненных форм, подобного, например, ряду стеля-
щихся растений и лиан, начинающихся с дериватов дерева и
заканчивающихся травянистыми монокарпиками (последние у
подушек обладают, по Рау, лишь внешними признаками поду-
359
шек, возникающими под воздействием внешних факторов
среды, но не обусловлены конституционными особенностями са-
мих растений).
Однако вряд ли можно согласиться с такой широкой трак-
товкой растений-подушек и, в частности, с включением в число
настоящих подушек так называемых подушковидных деревьев —
Polsterbdtune. Несмотря на известное своеобразие в структуре
и физиономии крон этих растений, эти жизненные формы все
же с большим основанием (экологическим, физиономическим
и морфологическим) можно рассматривать как своеобразные
формы деревьев, но не как подушки.
Экологически подушки диаметрально противоположны де-
ревьям, хотя морфологически у подушек имеются черты, сбли-
жающие их скорее с деревьями, чем с кустарниками и кустар-
ничками (отсутствие смены скелетных осей). Историческое раз-
витие деревьев и подушковидных форм у покрытосеменных
проходило в диаметрально противоположных условиях: если
деревья возникли и развились во влажно-тропических, субтро-
пических и умеренных областях, наиболее благоприятных для
длительного, интенсивного роста надземных осевых органов, то
подушковидные формы, наоборот, возникли и приурочены к
областям, где почвенно-климатические условия в максимальной
степени неблагоприятны для роста надземных органов рас-
тений
Подушковидные растения в своем массовом распростране-
нии приурочены к областям или (высотным поясам, где деревья
произрастать не могут, поэтому экологически немыслимо суще-
ствование столь противоестественных жизненных форм, как
подушки-деревья
Эколого-морфологически подушковидные растения занимают
своеобразное место в системе жизненных форм покрытосемен-
ных. Среди подушковидных растений имеются и древесные, и
травянистые формы, причем последние преобладают. Древесные
подушки имеют резко ослабленный рост надземных органов,
что и приводит к образованию форм, приспособленных к жизни
в крайних экологических условиях Как было сказано выше,
подушки представляют собой заключительное звено в приспо-
собительном ряду растений, направленном по пути редукции
ростовых процессов.
Поэтому нельзя согласиться также и с общей морфологиче-
ской интерпретацией Рау подушковидных форм растений. По
мнению Рау, подушковидные формы возникают в разных систе-
матических группах покрытосеменных не в результате экологи-
ческой конвергенции, а представляют собой формы проявления
единства внутренней организации этих растений, проявляющей-
ся, прежде всего, в особенностях симметрии их побегов, в един-
360
стве их плана строения в акротонно-гипотонном ветвлении.
Возникновение подушковидных форм, по Рау, вызвано не эко-
логической, а морфологической конвергенцией.
Образование подушковидных растений действительно пред-
полагает у них свойство акротонно-гипотонного ветвления. Но
это свойство может быть квалифицировано как необходимое, но
недостаточное для характеристики подушковидных растений.
Как было показано в предыдущих главах (см. главы III, IV,
V, VI), акротонно-гипотонное ветвление (как следствие соответ-
ствующей симметрии побегов) свойственно не только подушко-
видным растениям, но и деревьям, а также надземным осевым
органам кустарников, кустарничков и стелющихся древесных
растений.
Таким образом, этот важнейший, по Рау, признак не специ-
фичен для подушек, он свойствен и другим группам древес-
ных жизненных форм, и поэтому он один не может обусловли-
вать возникновение особой группы жизненных форм подушко-
видных растений. Главную роль в формировании подушек игра-
ет редукция роста надземных органов.
При морфологическом анализе различных жизненных форм
растений целесообразно руководствоваться принципом, выдви-
нутым одним из основателей современной морфологии растений
В Гофмейстером (W Hofmeister, 1868), согласно которому
форма органов растений и их организма в целом возникает
в результате роста, а своеобразие их форм ярляется
результатом своеобразия их ростовых процессов. С этой точки
зрения и подушковидные формы растений являются резуль-
татом особенностей их роста.
По мнению Райхе (Reiche, 1893), поддержанному в дальней-
шем работами Хелберна (Helborn, 1925) и Вебербауэра (Weber-
bauer, 1931), основной причиной возникновения подушек служит
угнетение верхушечного роста в результате дей-
ствия низкой температуры, иссушающего действия ветра и дру-
гих факторов, угнетающих рост. Угнетение верхушечного роста
коррелятивно вызывает ряд изменений в строении растения
(в первую очередь в ветвлении и компактности расположения
побегов и др.), которые приводят к образованию подушек.
Важнейшим следствием сильного угнетения роста побегов у
подушковидных растений является очень резкое ослабление
дифференциации побегов по интенсивности их роста, приводя-
щее практически к образованию совершенно одинаковых и нич-
тожных по длине годичных побегов. Отсюда и важнейший
признак подушек — образование ровной и более или
менее плотной поверхности подушковидных растений,
образованных облиственными верхушками побегов.
361
Компактное расположение побегов в подушке также обяза-
но не усиленному их ветвлению, а ничтожному приросту и рас-
положению побегов в небольшом пространстве. Как правильно
отмечает М. Г. Зайцева (1949), ветвление у подушек не отлича-
ется высокой интенсивностью У покрытосеменных растений с их
аксиллярным типом ветвления побегов, мерой интенсивности
ветвления может служить отношение числа ежегодно образую-
щихся побегов к числу пазушных точек роста или, практически,
к числу листьев на материнских побегах. По данным
М. Б. Штернберг (1953), у лесных деревьев среднерусской фло-
ры это отношение колеблется от 44% до 88%. По подсчетам
М. Г. Зайцевой, у плотных подушек Памира это отношение не
превышает 3%. Таким образом, у подушек резко подавлен не
только линейный рост побегов, но и его особая форма — ветвле-
ние.
*
* *
Эколого-морфогенетический обзор различных типов и рядов
жизненных форм древесных растений, проведенный в предыду-
щих главах, позволяет заключить, что среди древесных расте-
ний четко выделяются два эволюционно-морфогенетических
ряда жизненных форм.
1 ряд. Деревья — кустарники — кустарнички. Этот эволюци-
онный ряд связан с резким сокращением длительности жизни
надземных осей, а при переходе от кустарников к кустарнич-
кам— и резкой редукцией их роста. Весьма характерно для
этого ряда появление геофильного способа возобновления над-
земных осей. По существу этот ряд является рядом геофильной
эволюции древесных растений.
2 ряд. Деревья — стланцы — подушки. Стланцы в этом ряду
далеко не всегда являются формой, переходной от деревьев к
их антиподам — подушкам Основным содержанием этого ряда
является крайне резкая редукция роста побегов в последнем
звене ряда В отличие от первого ряда, переход от деревьев к
стланцам и подушкам не сопровождается появлением геофиль-
ного возобновления побегов. Все растения этого ряда остаются
аэроксильными У подушек из-за резкой редукции роста полно-
стью исчезает дифференциация побегов по интенсивности роста
Ничтожные и одинаковые по длине годичные побеги их образу-
ют ровную и общую для всей подушки плотную поверхность.
Экологически эволюционно-морфогенетическое преобразова-
ние древесных растений в первом и втором ряду было связано
с ухудшением почвенно-климатических условий, становившихся
все более и более неблагоприятными для развития высоко-
ствольных древовидных форм.
ЛИТЕРАТУРА
АвдошенкоА К Биология северных брусничных «Учен зап Ленингр.
гос пед ин-та им. А. И Герцена», т 82, 1949
Аврорин Н А. Акклиматизация и фенология «Бюлл Главн Бот.
сада», № 16, 1953
Аврорин Н. А. Переселение растений на полярный север М—Л, 1956
Александров В. Г. и Александрова О. Г О сосудисто-волокнис-
тых пучках стебля подсолнечника как объекта экспериментальной мор-
фологии, ч I, «Журн Русск Бот. Об-ва», т. 13, № 3—4, 1928.
Алехин В В География растений, М, 1950
Алехин В В. Растительность СССР в основных зонах В книге Г. Вал fa-
тера и В Алехина. Основы ботанической географии. М., 1936.
Андреев В Н. О движении боковых ветвей древесных пород. Сб :
«Лесоведение и лесоводство», вып 5, изд 1928 ,
Андреев В Н Экологические исследования стелющихся форм древесных
растений «Научн. зап, Молдавск научно исслед. базы АН СССР»,
№ 11, 1949
Андрианов С. И Дуб в Сальской степи «Лес и степь», № 12, 1953
АрциховскийВ М Рост саксаула и анатомическое строение его ствола
«Тр по прикл бот, генет и селекц », т XIX, вып 4, 1928
Асосков Н. А. Порослеобразовательная способность наших древесных
пород Сборник исследований по лесоводству под ред М Е. Ткаченко,
1931
Астапова Т Н Рост и формирование побегов дуба в лесах Подмосковья
«Учен зап. Моск гор. пед ин та им. В. П. Потемкина», т. 37, каф. бота-
пики, вып 2, 1952
Банников А Г О биологических группах копытных «5чей зап Моск,
гор пед ин-та им. В П Потемкина», т 38, 1955
Баранов П А Акклиматизация на Памире Сб «Вопросы экологии и
биоценологии», 3, 1936
Баранов П А. Биологическая наука на службе освоения высокогорного
Памира «Природа», № 2, 1936
Баранов П А. Строение виноградной лозы Ампелография СССР, 1947
Баранов П А Приспособительная эволюция виноградной лозы. «Тр Гл.
бот сада АН СССР», т 1, 1949
Баранов П. А В тропической Африке М, 1957
БаранецкийО В О причинах направления ветвей деревьев и кустар-
ников «Зап Киевск об-ва естествоиспыт» т 17, 1901.
Бекетов А Н. География растений СПб, 1896.
363
Бобров Е Г. История и систематика рода Corylus. «Сов. ботаника»,
№ 1, 1936.
Бобров Е. Г. Corylus L. Флора СССР, т V, 1936
Борисова И. В Основные жизненные формы двудольных млоюлетних
растений степных фитоценозов северного Казахстана. «Бот. журн,», т. 45,
№ 1, 1960
Бы л о в а А М О корневой системе Mulgediutn tataricum DC. «Бот. журн.»,
т 43, № 4, 1958
Васильев В II. Растительность Анадырского края М.—Л., 1956
Веттштейн Р. Руководство по систематике растений, т И, ч. 2.
Покрытосеменные. М, 1912.
Вернадский В И, Биогеохимические очерки, М, 1940
Бехов В. Н. Поведение кедровою стланика (Pinus /umila (Pall.) Rgl.)
в условиях лесостепи. «Научн. докл. высш, школы». «Билд, науки», № 4,
1958
Вильямс В Р. Естественнонаучные основы луговодства, или луговедение.
Общее земледелие, ч 11, 1922
Воробьева! И Некоторые данные по формированию корневой системы
березы пушистой, березы бородавчатой, березы извилистой и березы кар-
ликовой в процессе индивидуального развития «Бюлл. МОИП», отд.
биол., т 64, вып 3, 1959
Вульф Е В. Историческая география растений М—Л, 1944
Высоцкий Г. Н. Ергеня. Культурио-фитологический очерк. «Тр по прикл.
бот.», т. V, М, 1915
Г а б а й В С Бересклет бородавчатый Кишинев, 1952.
Го лен кин М И Победители в борьбе за существование, М, 1927
Голубев В Н. О закономерностях морфогенеза корневищ травянистых
растений и некоторые вопросы их происхождения. «ДАН СССР», т. 106,
№ 2, 1956
Голубев В Н О морфогенезе и эволюции жизненных форм травянистых
растений лесо-луговой зоны. «Бюлл. МОИП», отд. биол, т. 62 (6), 1957.
Голубев В Н. Материалы к эколого-морфологической и генетической
характеристике жизненных форм травянистых растений. «Бот. жури»,
т 42, № 7, 1957
Городков Б. II Растительность тхндровой зоны СССР, растений Запад-
ной Сибири. «ДАН СССР», т 120, № 1, 1936
Городков Б Н Растительность Арктики и горных тундр СССР. Расти-
тельность СССР, т. 1, 1938.
Григорьев Ю С Сравнительно-экологическое исследование ксерофили-
зации высших растений. М.—Л, 1955.
Гризебах А Растительность земного шара, ч 1 и 2. М, 1874—77.
Г р о с с г е й м А. А. Определитель растений Кавказа, М , 1949
Гроссгейм А А. Теория ксероморфогенеза и некоторые вопросы истории
флоры Сб «Проблемы ботаники», т 1, 1950.
ГроссетГ Э. К изучению экологии кедрового стланика. «Бюлл. МОИП»,
отд биол, it 64, вып 2, 1959
ГроссетГ Э Кедровый стланик М, 1959
Гумбольдт Л. Идеи о физиономичности растений М, 1936.
ДамбергЭ И Дуб на крайней северной границе его распространения.
«Лесной журн», вып 1, 1914
Данилов М, Д Классификация деревьев в древостоях на основе ста-
дийного развития «Лесное хоз-во», № 3, 1949.
Данилов М Д и Пч ели и В. И Организация хозяйства с прижизнен-
ным использованием бересклета бородавчатого. «Лесное хоз во».
№ 10, 1953
Дарвин Ч О лазающих растениях, 1878 4
Денисова Г. М Побегообразование и сезонное развитие некоторых злаков
364
на поемных пугах низовий Северной Двины Вопросы биологии расте-
ний. «Уч. зап. Моск гор. пед. ин-та им В П. Потемкина», т. 57, каф.
ботаники, вып 4, 1960.
Де лисов а Л В К морфогенезу извилистой березы. «Бюлл. МОИП», отд.
биол, т 63, вып 5, 1958
ДерюжкинВ В Географические посевы сосны в Воронежской области.
«Лесное коз во», № 12, 1952.
Дмитриев А М Луговодство с основами луговедения. М, 1948.
До майская Н П Формирование корневой системы черешчатого дуба.
«Докл АН СССР», т. 119, № 6, 1958
Досталь Р. Наш вклад в исследование причин роста и развития растений
Сб. Высшей школы земледелия и лесного хоз ва в Брно, 1956
Досталь Р Некоторые тормозящие и стимулирующие явления в морфо-
-генезе растений. София, 1958
Ж ад о в ск и й А Э. Экологический спектр Костромской флоры. «Тр Ко-
стромск. научи об-ва изуч. местного края», XVI, 1920
Зайцева М. Г. О развитии формы у растений-подушек. «Сообщ. Тадж„
фил АН СССР», вып 17, 1949.
Заленский О. В О температурном и водном режиме растений-подушек.
«Бот журн», т. 33, № 6, 1948
Запрягаева В И. Корневая система туркестанского клена. «Сообщ.
Тадж. фил. АН СССР», вып 31, 1951.
Запрягаева В И Биологические особенности фисташки в связи с ее
культурой в Таджикистане. «Бот. журн.», т. 39, № 3, 1954
Зёдинг Г Ростовые вещества растений М, 1955
Злобин Ю. А. Строение и развитие клонов черники «Уч зап. Моск гос.
пед ин-та им. В И. Ленина», каф. ботаники, вып. 4, 1961
Зозулин Г. М. Подземные части основных видов травянистых растений
и ассоциаций плакоров Средне-русской лесостепи в связи с вопросам»
формирования растительного покрова «Труды Центр -черноз гос запо-
веди им В. В. Алехина», вып. 5, 1959
Иванов Л А Физиология растении М, 1931
Ильин М. М Chenopodiaceae Less Флора СССР, т VI, 1936 ’
Ильин М. М. Поликамбиальность и эволюция Сб «Проблемы ботаники»,
вып. 1, 1950.
Ильинский А. П. Растительность земного шара. Л., 1937
Иорданская Н Н и Серебряков И Г О морфогенезе жизнен-
ной формы 'Кустарника у бересклета бородавчатого «Бот. журн.», т 39,
№ 5, 1954
К а з а к е в и ч Л И Материалы ik биологии растений юго востока России.
«Изв Сарат обл с, х опытн ст», т Ill, 1921, вып 2—3, 1922.
Казарян В О Физиологические основы онтогенеза растений Ереван. 1959,
Ка лабух о в Н И Эколого-физиологические особенности «жизненных
форм» грызунов лесостепи и степей левобережья Украины и Европейской
части РСФСР. Третья экологии конференция Тезисы докладов Киев,
1954
К'ашкаровД Н Экология животных М, 1944.
Келлер Б А Растительный мир русских степей, полупустынь и пустынь.
Вып. 1, 2 Воронеж, 1923, 1926
Келлер Б. А Степи Центрально-Черноземной области М.—Л., 1931.
Келлер Б. А Об изучении жизненных форм при геоботанических иссле-
дованиях «Сов бот », № 2, 1933
К е л л е р Б А. Главные типы растительности СССР. «Растительность
СССР», т. 1. М—Л, 1938
Келлер Б. А. Основы эволюции растений. М— Л., 1948.
Кожевников А В Материалы по экологии буковых лесов, «Сов. бот.»г
№ 5, 1935
365
Козлова Н А. Анатомо экологическая характеристика полукустарников
Крыма. «Бот жури », т. 38, № 4, 1953
К о з о • П о л янский Б М. Введение в филогенетическую систематику
выофих растений, 1922
К озо - П о л ян ск ий Б М Биогенетический закон с ботанической точки
зрения. М, 1937
К оз о-Пол янс кий Б М Случай превращения биоморф культурных
растений и их значение «Тр. Воронежск. Гос. ун-та», 1945.
Комаров В Л. Juniperus L. Флора СССР, т. 1, 1934
Коровин Е. П. Растительность Средней Азии. М., 1934
КоровинЕ П иЗакржевскийБ С Экологические особенности
главнейших растений Бетпакдала «Тр Ср. Азиат. Гос ин-та, сер 8,
бот., вып 23, 1935
Кошкина Е. Ф. Ритм развития луговой растительности низовий Дона.
«Уч зап. Моск гос. пед. ин-та им В П Потемкина», т 37, каф бот ,
вып 2, 1954
Кощеев А Л Обогащение стеблевой коры бересклета гуттой «Тр
Ин-та леса АН СССР», XI, 1953.
Кравченко Р. В Рост и производительность бересклета бородавча-
того и европейского в условиях открытых плантаций «Лесное хоз во»,
12, 1952
Красно в, А Н Из поездки на Дальний Восток Азии. «Землеведение»,
№ 2-3, 1894
Краснов А Н География растений, 1899
КренкеН П Теория циклического старения и омоложения растений
М, 1940
Кречетович Л М Вопросы эволюции растительного мира. М, 1952
Криштофович А. Н Палеоботаника. 1957.
Крылова И. Л. Развитие замещающих побегов у сосны и бука «Бюлл.
МОИП», отд биол, т 63, вып 3, 1958
КультиасовМ В Проблема становления жизненных форм у расте-
ний. Сб. «Проблемы ботаники», 1, 1950
Купцов А. И Превращение диких многолетних растений в культурные
малолетники Докл АН СССР, т. 86, № 5, 1952
Курындин И И, М ал и н ко в с к ий В В, Беньяминов А Н,
Белохонов И В, Плодоводство М, 1947.
Лавренко Е. М. О Сахаро-Гобийской пустынной ботанико-географи-
ческой области и ее разделении Докл АН СССР, т 134, № I, 1960
Лавренко Е М Некоторые наблюдения над корневой системой, эколо
гией и хозяйственным значением псаммофитов нижнеднепровских песков
Сб «Пробл растениеводч. освоения пустыни», 1935.
Лавренко Е М Основные закономерности растительных сообществ и
пути их изучения. Полевая геоботаника, т 1, изд АН СССР, 1959.
Лапшина Е И О перезимовывании высших растений по наблюдениям
в окрестностях Петергофа «Тр Петергофск. ест -научного ин-та», вып 5,
1928.
Леонтьев В Л. Саксауловые леса пустыни Кара-Кум М—Л, 1954
Лобанов Н. В Микотрофный тип питания лесных деревьев. «Лесное
хоз-во», № 1, 1949.
Лобановы В Микотрофность древесных растений М, 1953.
Лучник 3. И Обрезка кустарников М, 1960.
ЛюбименкоВ Н. О спящих почках «Труды СПБ об-ва естеств», т 30,
отд бот., вып 3, 1900.
ЛюбименкоВ Н. Курс общей ботаники. 1923
МазингВ О понятии «жизненная форма» в экологин высших растений,
«Учен зап Тартуск гос ун та», вып 64 Тарту, 1958
МалашкинаН С Анатомо-морфологическое исследование суховсршин-
366
ности дуба летнего (Quercus ro^ur L.) в условиях Камышина «Бюлл
МОИП», отд биол, т. 63, вып 3, 1958
Малеев В. П Можжевельник. Деревья и кустарники СССР, т. 1, 1949
Мальцев А И. Сорно-полевая растительность и меры борьбы с нею.
М—Л., 1931.
М а т ю к И И. Рост лесных насаждений в зависимости от условий среды
Полезащитное лесоразведение. М, 1950
Мацкевич Н. В, Кощеев А Л. Укоренение и гуттон а копление зазем-
ленных стеблей бересклетов «АН СССР», т. 92, № 5, 1953.
Медведев Я. С. Деревья и кустарники Кавказа, 1883
Мейер К И Происхождение наземной растительности М, 1946
Мейер К- И. Морфогения высших растений М, 1958.
Мильков Ф Н. Стелющаяся форма дуба на меловых холмах юго восто-
ка Русской равнины «Природа», № 6, 1951
Михайловская И С Особенности приспособительной эволюции лимон-
ника китайского «Учен зап Моск гос. пед ин-та им В И Ленина»,
т. 73, каф бот, вып. 2, 1953
Молиш Г. Физиология растений как теория садоводства М, 1933.
Морозов Г. Ф Учение о лесе, 1926
Назаров М И. Salix L Флора СССР, т. V, 1936.
Наумов Н. П Экология животных М, 1955
Нестеров В. Г. Общее лесоводство М, 1949
Нечаева Н. Т. и Приходько С. Я Всхожесть семян и характеристика
всходов некоторых пустынных растений, пригодных для улучшения
пастбищ. «Изв АН Туркменск ССР», № 1, 1952
Никитин И Н Возрастная стадийность и методы ускорения роста дуба
в первые годы его жизни. «Лес и степь», № 2, 1949.
Орлов А. Я. Буковые леса Севере западного Кавказа. Сб. «Широколист-
венные леса Северо Западного Кавказа». М, 1953
П адеревскаяМ И Биологические особенности почек дуба Уч зап.
Моск гос пед ин-та им В И Ленина», т 73, каф ботаники, вып 2, 1953
ПалибинИ В Растения-подушки «Природа», № 12, 1914 t
Па ч ос кий И К. Биологическая особенность осота (Cirsium arvensc).
«Труды бюро по прикл бот.», т. IX, № 1.
Пачоский И. К Основы фитосоциологии Херсон, 1921
Петров М П. Экологический очерк растительности Репетекского песчано-
пустынного заповедника в гою восточных Кара-кумах Проблемы расте
ниеводческого освоения пустынь, вып. 4, 1935
ПетровМП Об экологии солончакового и песчаного саксаулов. Вопро-
сы экологии и биоценологии, вып 3, 1936
Пив ник С А Эколого биологические особенности кедрового стланика
(Pinus pumfta (Pall.) Rgl) в основных растительных ассоциациях
северо-западной часть его ареала Автореф канд дисс Ленинград, 1958.
Пономарева И Н (Былинкина). Ритм сезонного развития растений
Иссык-Кульской котловины Вопросы биологии растений «Уч зап Моск
гос пед ин та им. В. П Потемкина», т 100, 1959.
Поплавская Г. И Материалы по изучению изменчивости крымскою
бука «Журн русск. бот общ ва», т. 1’2, № 1—2, 1927
Поплавская Г. И О некоторых взаимозаменяющих буковых ассоциа-
циях в Крыму Очерки по фитосоциологии и фитогеографии, 1929
Поплавская Г. И Экология растений М, 1948.
П о п о в М Г Род Cicer и его виды «Тр. по прикл бот, генет и селекц »,
т. 21, вып. 1, 1928-1929
П о п о в М Г Опыт монографии рода Eremostachys Bge. Нов мем
МОИП, т 19, 1940
Попов М Г. О применении ботанико-географического метода в система-
тике растений Сб «Проблемы ботаники», т. 1, 1950
367
ПошкурлатА. П Строение и развитие дерновины чия. «Уч зап Моск,
гос пед ин-та им. В. И. Ленина», т. 30, каф ботаники, вып 1, 1941
Прозоровский А. В О биологических типах растений пустыни «Бот.
журн», т. 21, вып. 5, 1936
Прозоровский А. В Полупустыни и пустыни СССР. Растительность.
СССР, т II, М, 1940,
Проханов Я И. Celastraceae Lindl Флора СССР, т XIV, 1949.
Пятницкий С. С. Спящие почки в пазухах семядолей дубовых всходов,
«Природа», № 7, 1947.
Работнов Т. А Биологические наблюдения на субальпийских лугах
Северного Кавказа «Бот. журн », т. 30, № 4, 1945
Работнов Т. А. Опыт определения возраста у травянистых растений.
«Бот журн», т. 31, № 5, 1946
Работнов Т. А. Длительность виргинильного периода жизни травянистых
растений в естественных ценозах. «Бюлл МОИП», отд. биол, т 51, 2,
1946.
Работнов Т. А. Определение возраста и длительность жизни у много-
летних травянистых растений. «Усп. совр. биол», XXIV, 1 (4), 1947
Работнов Т. А Жизненный цикл многолетних травянистых растений.
«Тр. БИН АН СССР, Геоботаника», вып 6, 1950
Работнов Т. А О жизненном цикле борщевика сибирского Бюлл МОИП,
отд. биол, т. LXI, 2, 1956
Радкевич О. Н и В ас и леве кая В К Анатомическое строение побе-
гов первого года жизни у древесных форм песчаной пустыни Кара-Кум.
«Тр. по прикл бот. генетике и селекц», сер. 1, № 1, 1933.
Раменский Л. Г Введение в комплексное почвенно геоботаническое
изучение земель, М., 1938.
Ричардс П Тропический дождевой лес. М, 1961.
Родман Л. С Образование кустарниковой формы у Rosa spinosissima L.
в условиях кустарниковых степей западных предгорий Алтая. «Бюлл.
МОИП», отд. биол, т. 62, вып 1, 1957.
Ротов Р. А Биолого-морфологические особенности древесно-кустарниковых
жизненных форм растений песчаной пустыни Кара-Кум «Учен зап
Моск гос пед ин-та им В П Потемкина», т. 100, каф бот, вып. 5,
1959
Рубцов Н И Горные плодовые леса и горные кустарники Растительный
покров СССР. Пояснительный текст к геоботанической карте СССР, т. II,
1956
Рустамов А К- К вопросу о понятии «жизненная форма» в экологии
животных. «Зоол. жури », т. 34, вып 4, 1955
СамофалС К. К изучению климатических рас сибирской лиственницы.
«Тр. по лесн. опытн. делу», вып. 1 (75), 1929.
Сенянинова-Корчагина М В К вопросу о классификации жиз-
ненных форм «Учен зап ЛГУ», сер геогр наук, вып 5, 1949
Серебряков И Г. Биология тяньшанской ели и типы ее насаждений в
пределах Заилийского и Кунгей Алатау. «Учен зап МГУ», т 82, «Тр.
Бот сада ЛАГУ», вып 5, 1945
СеребряковИ Г О ритме сезонного развития растений Подмосковных
лесов, «Вестник Моск, ун-та», № 6, 1947
СеребряковИ Г. Структура и ритм в жизни цветковых растений.
1 Запасающие органы, положение цветков на побеге и время зацветания.
«Бюлл МОИП», отд. биол, т. 53(2), 1948
СеребряковИ Г Структура и ритм в жизни цветковых растений.
II «Бюлл. МОИП», отд биол, т. 54(1), 1949.
СеребряковИ Г. Морфология вегетативных органов высших растений.
М, 1952.
СеребряковИ Г. О морфогенезе жизненной формы дерева у лесных
368
пород средней полосы Европ. части СССР «Бюлл МОИП», отд. биол,
т 59(1), 1954
Серебряков И Г. О морфогенезе жизненной формы стланца у турке-
станского и казацкого можжевельников, «Бюлл МОИП», отд биол,
т. 59, вып. 5, 1954.
Серебряков И. Г. Биолого-морфологический и филогенетический ана-
лиз жизненных форм покрытосеменных «Учен зап Моск Гор. пед ин-та
им В. П Потемкина», т 37, каф бот, вып. 2, 1954
Серебряков И. Г. VIII Международный Ботанический конгресс
Экскурсии по Франции и Африке. «Бюлл. МОИП», отд. биол., т. 60,
вып 1, 1955
Серебряков И. Г. Основные направления эволюции жизненных форм
у покрытосеменных растений «Бюлл. МОИП», отд. биол., т. 60, вып 3,
1955
Серебряков И. Г, Дома вс к а я Н П, Родман Л. С. О морфо-
генезе жизненной формы кустарника на примере орешника «Бюлл
МОИП», отд биол, т. 59, вып. 2, 1954
Серебряков И. Г. и Чернышева М. Б. О морфогенезе жизненной
формы кустарничка у черники, брусники и некоторых болотных
Ericaceal «Бюлл. МОИП», отд биол, т 60, вып 2, 1955.
Серебрякова Т. И. Побегообразование и ритм сезонного развития
растений заливных лугов Средней Оки. «Учен зап Моск. гос. пед ин-та
им. В. И. Ленина», т. 97, каф. бот, вып. 3, 1956
Смирнов П. А О HieradilQa^dorata авторов среднерусской флоры.
«Бюлл. МОИП», отд.биол, т. 63, в. 5, 1958
Сочава В. Б и Городков Б Н. Арктические пустыни и тундры.
Растительный покров СССР, пояснительный текст к геоботанической
карте СССР, т 1, М.—Л., 1956
Станюкович К В Растительный покров Восточного Памира М, 1949.
Стариков Г Ф. и Дьяконов П. Н Леса полуострова Камчатки.
Хабаровск, 1954.
Ст еш ен ко А. П. Формирование структуры полукустарничков в условиях
высокогорий Памира. Автореф. дисс. на соиск. степ. капд. биол наук,
1953
Стешенко А. П. Формирование структуры полукустарничков в условиях
высокогорий Памира «Тр Памирск биол ст т. 50, 1956
Сукачев В. Н. Растительные сообщества, изд. 4, 1928
Сукачев В. Н Наши древесные породы. Хвойные М, 1928
Сукачев В Н Дендрология с основами лесной геоботаники М, 1938
СумневичГ П. Розы — лианы ореховых лесов Ферганы «Бюлл МОИП»,
отд. биол, т. 52, в. 3, 1947.
Суслова М И Рост побегов и опадение ассимилирующих органов у
деревьев и кустарников песчаной пустыни Кара-Кум Проблемы растение-
водч. освоения пустынь, вып 4, 1935.
Тахтаджян А. Л. Морфологическая эволюция покрытосеменных. М ,
1948
ТахтаджянА Л Происхождение покрытосеменных растений М., 1954
Тихомиров Б. А. К характеристике флоры западного побережья Тай-
мыра Петрозаводск, 1948.
Тихомиров Н А. Кедровый стланик, его биология и использование.
М., 1949
Тихомиров Б А. и ЯнишевскийД Е. Новое в морфологии листа
подземного побега у многолетнею двудольного растения «Сов бот»,
№ 4, 1939.
ТкаченкоМ Е. Общее лесоводство М, 1952.
Толмачев А И. К истории развития и географического распространения
рода Drala «Бот. журн », т. 42, № 9, 1957
JB-413. И Г Серебряков —24
369
Т ю л и н а Л И Из высокогорной области Южного Урала. Очерки по фи-
тосоциологии и фитогеографии, 1929.
Тюлина Л И О лесной растительности Анадырского края и ее взаимо-
отношениях с тундрой «Тр. Арктич ин та», т. 15, геоботаника, 1936
Тюлина Л И. Лесная растительность Хатангского р-на у ее северного
предела. Труды Аркт. ин-та, т 63, 1937
Федоров Ан А Влажнотропические леса Китая. «Ботанич журнал»,
т 43, № 10, 1958
Федоров Ан. А Древесные эпифиты и фикусы удушители в тропических
лесах Китая «Бот журн », т 44, № 10, 1959
Феофраст, Исследования о растениях М, 1950
Флора СССР, тт I—XXIX
ХржановскийВ Г. Розы. Филогения и систематика М, 1958
Чистякова О. Н О возникновении кольцесосудистости у дуба черешча-
того (Quercus robur L.) «Научи, докл высш, школы». Биол. науки,
№ 1, 1959.
Чистякова О. Н. О формировании трахей древесины дуба Quercus ro-
bur L.) в онтогенезе в различных экологических условиях. «Научн.
докл. высш, школы» Биол. науки, № 2, 1960.
Шалит М. С Система жизненных форм степных растений «Уч зап Тадж.
гос универе», т. VI, вып 1, 1955
Шенников А. П Экология растений М, 1950
Шитт П Г. Биологические основы агротехники плодоводства, 1953
Штернберг М Б К вопросу о строении кроны моноподиальных и сим-
подиальных древесных пород «Бюлл МОИП», отд биол, т 58, вып. 1,
1953
Шут я ев А М Влияние величины желудей на рост сеянцев дуба в пер-
вый год жизни «Тр Центр-Черноземн гос заповеди им проф.
В. В Алехина», вып IV, 1957
Эйти и ген Г. Р. Рост дуба в зависимости от веса желудей «Советская
экономия», № 8—9, 1946
Э йт и н ген Г Р. Лесоводство М , 1949
Юзепчук С. В. Rosaceae. Matus Less. Флора СССР, т. IX, 1939.
Ю ниц к и й А А. О возобновлении дуба торчками «Лесоведение и лесо-
водство», вып 4, 1927
Яценко-Хмелевский А А Строение древесины кавказских предста-
вителей сем. Ericaceae и его систематическое и фило!енетическое
значение «Изв АН Арм ССР», № 9, 1946
Яценко-Хмелевский А А Принципы систематики древесины «Тру-
ды Бот ин-та АН Арм ССР», вып 5, 1948
AdamsonR S. The classification of the lifeforms plants «Bot Rev», 5, 10,
1939
Allan H New Zealand trees and Shrubs and How to identify them.
Wellington, 1928
Anoreanszky G. Az eletforma fejlodesi kozpontokrol «Bot kozl», 45,
№ 1—2. Реф «Реферат, журн , Биология», т. 18, 1956, реф. 76169 (Об
очагах возникновения жизненных форм)
А г be г А., 1920. Water-plants. Cambridge.
Ar be г A, 1928. The tree habit in angiosperms: its. Origin and meaning.
«New Phytologist», v. 27, № 1.
A r b e r A., 1934. The Gramineae.
Areschoug F. W., 1896. Beitrage zur Biologie der geophilen Pflanzen. «Acta
. Reg. Soc. Phys. Lund», t. VI.
Bailey 1, 1911. The relation of the leaftrace to the formation of compound
rays in the lower dicotyledons «Ann. Bot.», 25.
Bailey I., 1912 The evolutionary history of the foliar ray in the mood of
the dicotyledons and its phylogenetic significance. «Ann. Bot », 26.
370
Ba пег о f t, 1930. The Arborescent Habit in Angiosperms- a rexiew. «New
Phytologist», v. 29.
Bews I. W., 1925. Plant forms and their evolution in South Africa London.
Bews 1 W„ 1927. Studies in the ecological evolution of angiosperms. «New
Phytologist », 26.
Bonnie t G, 1888. Etude expcrimentale de 1’influence du climat alpin sur la
vegetation et les fonctions des plantcs. «Bull, de la Soc. hot. de France»,
35. Paris.
Braun A., 1851. Betrachtungen fiber die Erscheinung der Verjungung in der
Natur. Leipzig.
Braun A„ 1853. Das Individuum der Pflanzen.
Braun-Blanquet I., 1951. Pflanzensoziologie Vien
Busgen M., 1913. Cupuliferae, В кн. Lebensgeschichte der Blutenpflan-
zen Mitteleuropas O. Kirchner, E. Loew, C. Schroeter.
Busgen M., 1917. Ban und Leben unserer Waldbaume 2 Aufl, Jena.
Cain S. A., 1950 Life-forms and phytoclimate Bot review, v. XVI, № 1.
Clements F P, 1920. Plant Indicators. The Relations of plant communities
to process and practice. CarneWash
Cockayne L, 1921, 1928. The Vegetation of New Zealand. В книге: Die
Vegetation der Erde, 14. 1. u 2 Ed Leipzig.
Cockayne L, 1928. Monograph of the new Zealand beech forests. Part 11.
The forests fron the practical and economical standpoints. «.New Zeal.
State Forest Ser\ ice Bull.», 4. Wellington.
Corner E. S., II, 1949. lhe Durian-Theorie of the origin of the modern
tree. «Ann. Bot.», N. S., v. XIII, № 52.
Corner E. S. H., 1954. The Durian-theory. «Phytomorphology», 3.
Dansereau P., 1951 Description and recording of vegetation "upon a structural
basis «Ecology», v. 32, № 2
Dansereau P, 1957 Biogeography, Alonreal
De Candolle A. P., 1818. Regni vcgctabilis systema naturalis. Vol J Pari-
shs.
Dos tai R, 1914. Uber innere, das Wachsturn der Pilanze regtilierende Fak-
toren. Ref «Bot Zentralbl », CXXV. ’
Dos tai R., 1926 Uber die wachstumsregulierende Wirkung der Laulblattes,
«Acta Soc. Scient. nat. Morav.», T. 3.
Dos tai R, 1959 Versuche zur apikalen Dominanz in der Pfianzenmorpho-
genese «Prace Brnenske Zakladny Ceskosfovenske Akademie Ved »•
R. 31
Drude O, 1887. Die systematischc und geographische Anordnung der Phane-
rogamen Schenk Handbuch der Botanik. Bd 111, 2 Breslau ’
Drude O., 1896. Handbiicher zur Deutschen Landes-und Volkskunde. Bd.
4, T 1.
Drude O„ 1913. Okologie der Pflanzen. Die Wissenschaft, Bd. 50 Braunsch-
weig.
D r u d e O., 1928. Pflanzengeographische Okologie. Abderhalden’s Handbuch
der biologischen Arbeitsmethoden. Abt. XI, V. IV.
Du Rietz G. E., 1921. Zur metodologischen Grundlage der modernen Pflan-
zensoziologie Uppsala
Du Rietz G. E., 1931 Life-forms of terrestrial flowering plants. Uppsala.
Eames A., 1910. On the origin of the ray in Quercus. «Bot Gaz », 49.
Eames A, 1911 On the origin of the herbaceous type in the angiosprems.
«Ann Bot.», 25
Engler A., 1895 Die Pflanzenwelt Ostafrikas und Nachbargebiete. Grundzuge
der Pflanzenverbreitung in Deutsch-Afrika. Berlin
Engler A., 1908—1925. Pflanzenwelt Afrikas, 1-5. Engler u. Drude. Die Ve-
getation der Erde, 9. Leipzig.
24-
371
Er re r a L., 1904. Comflits de presence et excitation inhibitors chez les vege-
taux «Bull. Soc. Roy. Bot Belgique», 42, 27
Frank A., 1870. Die natiirliche wagerechte Richtung von Pflanzenteilen und
ihre Abhangigkeit vom Lichte und der Gravitation. Leipzig.
Gams H., 1918 Prinzipienfragen der Vegetationsforschung. Ein Beitrag zur
Begriffsklarung und Methodik der Biocoenologie. «Vierteljahrsschrift der
Naturforsch. Ges. in Zurich», 63.
Gams H„ 1950. Importance of Growth Forms for Taxonomical and Eco-
logical Systematics. Proceed, of the 7-th Intern. Bot. Congress Stock-
holm.
Gluck H , 1904, 1906, 1910, 1924. Biologische und morphologische Untersu-
chungen uber Wasser — und Sumpfgewachse Jena
Goebel K., 1908. Einleitung in die experimentelle Morphologie der Pfbanzen
Berlin und Leipzig.
Goebel К.» 1928 Organographie der Pflanzen, 3. Aufl Jena.
Gothan W. u. Weyland H., 1954 Lehrbuch der Palaobotanik Berlin.
Gray A., 1884. Structural Botany.
Grevillius S. u. Kirchner 0, 1923. Ericaceae in Lebensgeschichte der
Blutenpflanzen Mitteleuropas von O. Kirchner, E. Loew u. C. Schroeter,
Lief, 23—24. Stutgart.
Ha ger up O., 1930 Etudes des types biologiques de Raunkiaer dans la
fiore autour Timbouctou. «Kgl. Dansk. Vidensk. Selsk Bid Medd.»; 9,4.
Hallier H, 1905. Ein zweiter Entwurf des natiirlichen (phylogenetischen)
Systems der Blutenpflanzen. «Вег. d. Deutsch». Bot. Ges., 23
Hallier H., 1912. L’origine et le systeme phyletique des Angiusperms
exposes a 1’aide le leur arbre genealogique. «Archives Neerland.» Ser
11 В (Sci. Nat., 1)
Hansen H. M, 1956. Life-forms as age indicators. Riuckjbing.
Hauri H„ 1912 Anabasis aretioides Mog. et Coss.—eine Polsterpflanze der
algerischen Sahara. «Beih. zum Bot Ctbl », 28, 1.
H el born O, 1925 Contribution to the ecology of the Ecuadorian paramos
with special reference to cushion plants and osmotic pressure. «Svensk
Botanisk Tidskrift», Bd. 19.
Hirmer M., 1927. Handbuch der Palaobotanik. Miinchen und Berlin.
Hofmeister W., 1868. Allgemeine Morphologie der Gewachse Leipzig.
Huber B., 1926 Okologische Probleme der Baumkrone. «Planta», № 2.
Hull R., 1881. ForsOk till analitisk behandling af vaxtformationerna. «Meddel
af Soc. pro Fauna et Flora fennica», 8.
Hutchinson I., 1926. The families ot flowering plants. London.
Jeswiet L, 1914. Eine Einteilung der Pflanzen der niederlandischen Kusten-
dunen in okologische Gruppen. «Beih zur Bot. Centralblatt», 31, 2 Dres-
den
Jost L., 1936. Baum und Wald. Berlin
Irmisch Th., 1850. Zur Morphologie der monokotyli;chen Knotlen-und
Zwiebelgewachse Berlin.
I rm is ch Th., 1859. Bemerkungen uber einige Wassergewach'e. «Bot. Ztg», 17.
Irmisch Th., 1860. Ober einigen Ranunculaceen. «Bot Ztg», 18.
Irmisch Th., 1860. Ober einigen Cra«sulaceen, Bot. Ztg. 18.
Irmisch Th, 1861. Ober Polygonum amphibium. Lysimachia vulgaris, Co-
marum palustre und Menyanthes trifoliata. «Bot. Ztg.», 19.
IrmscherE., 1922. Pflanzenverbreitung und Entwicklung der Kontinente.
«Mitt. Inst. f. allg. Bot. im Hamburg», V.
Jeffrey E., 1899. The morphology of the central cylinder in the angiosperms.
«Trans Canadian Institute», 6.
Jeffrey E., 1917. The anatomy of woody plants. Chicago.
Jeffrey E. a. Torrey R., 1921. Physiological and morphological correlation
in herbaceous angiosperms. Bot. Gaz, 71.
372
Jeffrey E. a. Torrey R., 1921. Transitional herbaceous dicotyledons. «Ann.
Bot.», 35.
Kanngisser F.. 1906, Ober Alter und Dickenwachstum von Calluna vulga-
ris. «Naturw. Zeitschr. Land — und Forstwirtsch.»
Kanngisser F, 1907. Ober Lebensdauer der Straucher. «Flora», Bd. 97
und 98.
Kanngisser F., 1910. Ober Lebensdauer einigen Zwergstiauchern. «Garten-
flora».
Kanngisser F., 1911. Lebensdauer von Ericaceen. «Naturwissensch. Wochen-
schrift», N. F., Bd. 9, № 40.
Kanngisser F., 1912. Ober Lebensdauer Polarstrauchern. «Cartenflora».
Kanngisser F., 1917. Ein Beltrag zur Kenntnis der Lebensdauervon Zwerg-
strauchern aus hOhen Hohen der Schweiz. «Mitteil. Deut. dendrol. Ges »,
№ 26.
Kanngisser F., 1923. Ein Beltrag zur Kenntnis der Lebensdauer arktischer
und alpiner Holzgewachse. «Mitt. Deut. dendrol. Ges », № 23.
Kerner A., 1863. Das Pflanzenleben der Donaulander. Innsbruck.
К 1 h 1 m a n A. 0., 1890. Pflanzenbiologische Studien aus Russisch-Lappland.
«Acta Soc. pro Fauna et Flora Fennica», 6.
Kirchner 0., LoewE., SchroeterC. Lebensgeschlchte der Bliitenpflanzen
Mitteleuropas. Bd. 1. Abt. I, 1908.
Koorders 8. H., 1914. Florlstischer Uberblick fiber die Bliitenpflanzen des
Urwaldes von Tjibodas im Westjawa. «En6lers Bot. Jahrb.», B. 50.
Kraszan F. 1900—1901. Ergebnisse m'einer neuesten Untersuchungen fiber
die Polymorphic der Pflanzen «Englers bot. Jahrb», Bd. XXVIII.
Krazan F„ 1900—1901. Weitere Beobachtungen an breiwachsenden und an
versetzen Pflanzen. «Englers Bot. Jahrb». Bd. 28.
Krause E., 1891. Die Einteilung der Pflanzen nach ihrer Dauer. Ber. d. Deut.
Bot. Ges, 9.
L i n d m a n С. A. M., 1914. Magra bidrag till fragan: buske eller trad? K. Ve-
tenskapsakademiens Arsbok, 12.
Lin kola K., 1922. Zur Kenntnis der Oberwinterung der Unkrauter und Ru-
deralpflanzen in der Gegend von Helsingfors. «Soc. Zoolog.—Bot^n. Fenn.
Vanamo». Tom 1, № 7.
Linnaeus C„ 1753. Species plantarum. Holmiae.
Massart I„ 1908. Essal de geographic botanlque des districts littoraux et
alluviaux de la Belgique. «Rec. de 1’lnstit. Bot. Leo Errere, 7. Bruxelles.
Magdefrau K., 1953. Palaobiologie der Pflanzen. Jena.
MeuselH., 1935 Wuchsformen und Wuchstypen der europaischen Laub-
moose, «Nova acta Leopoldina», N. F., Bd. 3, № 12.
Me us el H., 1951. Die Bedeutung der Wuchsform ffir die Entwicklung der
natiirlichen Systems der Pflanzen. «Feddes Repertor», Bd. 54, H. 2/3.
xMeusel H, 1952. Ober Wuchsformen, Verbreitung und Phylorenie einlger
mediterran — mitteleuropaischen Angiospermen-Gattungen/ «Flora», Bd.
139, H. 3.
Mi о sc hi, 1906. Atlas of Japanese vegetation. Tokio.
Mioschi, 1926. Naturdenkmalpflege in Japan. «Nachrlchtebl. f. Naturdenk*
malpflege», 3.
Mog к W., 1914. Untersuchungen der Korrelationer. von Knospen und Spros-
sen. «Archiv d. Entwicklungsmechanik d. Organismen», v. 38
Morison R., 1669 Preludia botanica. London!.
Morison R„ 1715. Plantarum histories universalis Oxanlensis... Oxania.
Miinch E., 1938. Untersuchungen liber die Harmonie der Baumgestalt. «Jahrb.
f. wiss. Bot.». Bd. 86, 4.
Nageli C., 1884. Mechanisch-physiologische Theorie der AbstammungSlehre.
Mfinchen und Leipzig.
37.1
Nefcesanij V., 1955. Vztah tnezi reakcnim drevem listnatych a JehliSnatych
drevin. Biologia, X, № 6.
Neg er, 1913. Biologie der Pflanzen auf experimenteller Grundlage. Stuttgart.
Nt edenzu F., 1889. Ober den anatomischen Bau der Laubblatter der Arbuto-
ideae und Vaccinioideae in Beziehung zu ihrer systematischen Gruppierung
und geographischen Verbreitung, «Bot. Jahrb. f. Syst» (A. Engler), Bd.
Pfeffer W„ i904. Pflanzenphysiologie. Leipzig.
Pound R. and Clements F. E., 1898. The Phytogeography of Nebraska. 1.
Lincoln, Nebr.
Rauh W„ 1937. Die Blldung von Hypocotyl-und Wurzelsprossen und ihre
Bedeutung fUr die Wuchsformen der Pflanzen. «Nova Acta Leopoldina»,
N. F„ 4.
R a u h W., 1937—39. Ober die Verzweigung auslauferbildender Straucher mil
besonderer BerQckslchtigung ihrer Beziehungen zu den Stauden. «Hercinia,
Abhandl. d. bot. Verein Mitteldeutsch.», 1.
Rauh W., 1938. Beitrage zum Morphologic und Biologie der Holzgewachse. 1
Entwicklungsgeschichte und Verzweigungsverhaltnisse arktlsch-alpiner
Spalierstraucher «Nova Acta Leopoldina», N. F„ 5.
Rauh W., 1939. Ober Qesetzmafiigkeit .der Verzweigung und deren Bedeutung
fiir die Wuchsformen der hoheren Pflanzen. «Mitteil. d Deut. dendrol. Ges.»,
№ 52.
Rauh W., 1939. Ober polsterfdrmigen Wuchs. Ein Beitrag zur Kenntnis
der Wuchsformen der hdheren Pflanzen. «Nova Acta Leopoldina.»,
N. F., 7.
Raunkiaer C., 1904. Om biologiske Typer, med Hensyn til Planternes Til-
pasning til at overleve ugunstige Aarstider. «Bot. Tidsskrift», 26.
Raunkiaer C., 1905. Types biologiques pour la geographic botanique Over-
sigt over det Kgl. «Danske Videnskabernes Selsk. Forhandl.» № 5.
Raunkiaer C., 1907. Planterigets Livsformer og deres Betydning for Geog-
rafien. Koebenhavn og Kristiania.
Raunkiaer C„ 1916. Om Bladstcerelsens Anvendelse I den biologiske Planten-
geographie. «Botanisk Tidsskrift»,34, Koebenhavn.
Raunkiaer C, 1934. The life form of plants and statistical Plant geography.
Oxford.
R e i c h e К , 1893. Ober Polster — und Deckenformigwachsende Pflanzen. «Verh.
wiss. Verz Santjago», Bd. II, H. 5—6.
Reiter H., 1885. Die Consolidation der Physiognomik als Versuch einer
Okologie der Gewachse. Graz.
Richards P. W., 1952. The tropical rajn forest. Cambridge.
Sachs J, 1873—77 Ober orthotrope und plagiotrope Pflanzenteilen. «Arb.
Bot. Inst. Wiirzburg», I.
Sachs J., 1874. Lehrbuch der Botanik. 4. Aufl. Leipzig.
Salisbury E., 1916. The Oak-Horn beam woods of Hertfordshire. «Jotirn.
of Ecology», vol. IV.
S a n d t W„ 1923. Zur Kenntnis. der Beiknospen. «Bot. Abhandl », hgb v
K. Goebel, H. 7.
Scharfetter R., 1953. Biographien von Pflanzensippen. Wien.
Schenk H., 1892, 1893. Beitrage zur Biologie und Anatomie der Lianen.
«Bot. Mitteil. aus den Tropen», hgb. von A. F. W. Schimper, H 4.
SchimperA. F. W. and Faber C , 1935. Pflanzengeographie auf physiolo-
gischer Grundlage, 3. Aufl, Jena.
Schmid E, 1956. Die Wuchsformen der Dicotyledonen. Bericht fiber der geo *
botanische Forschungsinstitut Riibel in Zurich, f. d. Jahr 1955
Schmid E., 1957. Ein Vergleich der Wuchsformen im illyrischen Buchen-
und Laubmlschwald. Bericht Uber das geobot. Forschungsinst. Rflbel in
Zfirich.
374
Schramm, 1912. Ober die anatomischen Jugendformen der Blatter einhei-
mischer Holzpflanzen. 4Flora», Bd. 104.
Sc h roe ter C., 1908. Das Pflanzenleben der Alpen.
S c h г о e t e r C., 1926. Das Pflanzenleben der Alpen.. 2. Aufl. Ztirich.
S c h r 0 e t e r C. u. H a u r i H., 1914. Versuch einer Ubersicht der Siphonogamen
Polsterflanzen. «Englers Bot Jahrb», 50, Suppl. 618.
SeyboldA., 1927. Untersuchungen Uber die Formgestaltung der Blatter
der Angiospermen. Bibl. Gen., Bd. XII, Leipzig.
S 1 n n 0 11 E W., 1914. Investigations of the phylogeny of the angiosperms.
1 The anatomy of the node as an aid in the classification of angiosperms
«Amer. Journ of Bot.», v 1.
Sinnott E. W 1952. Reaction wood and the regulation of tree form. «Amer
Jour, of Botany», v. 39, № 1.
Sinnott E. W. a. В a i 1 e у I. W., 1914 The origin and dispersal of herbaceous
angiosperms. «Ann. Bot.», 28.
Sinnott E W. a Bailey I. W., 1922. The significance of the «foliar ray» in
the evolution of herbaceous angiosperms. «Ann. Bot» 36.
SkottsbergC., 1910 Vegetationsbilder von den Juan — Fernandez Inseln
Karsten u Schenk Vegetationsbilder. 8. Reihe, H. 2. Jena.
S kottsberg C., 1912, The vegetation of South Georgia. Wiss. Ergebn. d.
Sc hued Siidpolar—Expedition 1901 — 1903. Bd. IV, Lief. 12.
Skottsberg C, 1913, A botanical Surxey of the Falkland Island. Ill
К Svenska Vetensk. Akad. Hande, Bd 50, № 3
Skottsberg C, 1914 Studien tiber die Vegetation der Juan-Fernandez
Inseln «Bot. Erhebn.», IV, Bd. 51, № 9.
Sleumer H, 1941. Vaccinioideen-Studien Bot. Jahrb. f. Syst., Bd. 71.
Snow R, 1925. Correlative ingibition of the growth of axillary buds. «Ann
Bot », v. 39.
Troll W, 1937. Vergleichende Morphologie der hOheren Pflanzen. Bd. I
Berlin
ThimannK. a. Skoog T., 1933. The inh'bition action of the growth sub-
stance on bud development. «Proc. Nat. Akad. Sci», v. 19.
T 0 u r n e f о r t I. P., 1719. Institutiones rei herbariae. Ed. 3. Paris.
V d c h t i n g H. 1884 Ober Organbildung im Pflanzenreich. Bonn. ♦
Warming E, 1884. Ober perenne Gewachse. «Bot. Centralbl». Bd. 18, № 19.
Warming E, 1892 Lagoa Santa. «К. Danske Vid. Selk. Skriftber». 6.
W a r m i n g E , 1908. Ericineae {Ericaceae, Pirolaceae) Morphology and Bio-
logy. The Structure and Biology of Arctic Flowering Plants. I. «Medd. om
Grdnland», 36.
Warming E., 1909. Oecology of plants. Oxford.
Warming E , 1916. Dansk Plantevaekst. 3 Skovene. 1 och 2 Hefte. «Botanisk
Tidsskrift» 35, 1-2.
Warming E, 1916. Bemacrkningen om Livsform og Standplads. «Forhandl.
ved det Skandinav-ke Naturf.», 16 (Ref.: Englers Bot. Jahrb, 56, 1920).
W arming E., 1923. 0kologiens Grundformer Udkast til en systematisk Ord-
ning «D. Kgl Danske Vidensk. Selsk. Skrifter», Naturvidensk. og Mathem
Afd , 8, Raekke, IV, 2,
U arming E. u. G r a e b n e r P., 1918. Lehibuch der okologische Pflanzengeog-
raphie 3. Aufl. Berlin.
Weberbauer A, 1931. Ober die Polsterpflan’e Picnophyllum aristatum und
Polsterpflanzen im allgememen. «Вег. d. Deut. Bot. Ges.», 49.
Werdermann E„ 1930- 31. Die Pflanzenwelt Nord — und Mittelchiles. Ve-
getationsbilder v. Karsten u. Schenk., 21 Reihe, H. 6/7.
WlgandA., 1854. Der Baum. Betrachtungen tiber Gestalt und Lebensge-
schichte der Holzgewachse
Zimmermann W., 1930. Phylogenie der Pflanzen. Jena.
Zimmermann W., 1949. Geschichte der Pflanzen. Stuttgart.
Zimmermann W, 1936 Untersuchungen tiber die raumliche und zeitliche
Verteilung der Wuchsstoffer. Zeitschr. Bot., Bd. 30, H 5—6.
ОГЛАВЛЕНИЕ
Стр.
Предисловие ...............• .7.............................. 3
Глава I. История учения о жизненных формах у растений ... Ь
1. У истоков учения о жизненных формах..................... 5
2. О периодизации истории учения о жизненных формах у ра-
стений .................................................. 10
3. Эколого-физиономическое направление в изучении и классифи-
кации жизненных форм...................................... И
4. Морфолого-биологическое направление в классификации жиз-
ненных форм растений..................................... 25
5. Понятие «жизненная форма растений» и его определение у
разных авторов........................................... 50
6. Развитие представлений об эволюции жизненных форм у ра-
стений ................................................ 61
Глава II. Опыт эволюционно-экологической системы жизненных
форм покрытосеменных и хвойных растений.................. 69
1. Определение жизненной формы у семенных растений....... 69
2. Принципы построения эволюционной системы жизненных форм
растений................................................. 72
3. Основные контуры эволюционно-экологической системы жиз-
ненных форм покрытосеменных и хвойных растений........... 78
Глава IIL Жизненные формы деревьев...........................120
1. Развитие древовидной формы у древних папоротникообразных
растений ................................................120
2. Соотношение древесных и травянистых форм у покрытосе-
менных ..................................................126
3. Эволюция жизненной формы дерева у покрытосеменных ... 138
4. Особенности морфогенеза жизненной формы прямостоячего де-
рева у некоторых хвойных и покрытосеменных растений . . 146
Морфогенез жизненной формы дерева у ели {Picea excelsa,
Link) .................................................146
Морфогенез жизненной формы дерева у лесной сосны {Pinus
silvestris L.).........................................159
Разнообразие формы крон и особенности их формирования у
некоторых хвойных деревьев.............................167
Формирование жизненной формы дерева у черешчатого дуба
{Quercus robur L.).....................................172
376
стр.
Морфогенез жизненной формы дерева у бородавчатой и пуши-
стой березы (Betula verru osa Ehrh и Betala pubescens Ehrh.) . 190
Особенности морфогенеза жизненной формы дерева у березы
извилистой (Betula tortuosa Ldb.)..... ...............201
Морфогенез жизненной формы дерева у дикой яблони (Malus
Silvestris Mill)............................. ... 208
Морфогенез жизненной формы дерева у черного саксаула (Ha-
lo iylon aphyllum Bge) и белого саксаула (Haloxylon persicum
5. Физиологический анализ формирования кроны дерева..........218
6 Биолого-морфологические особенности формирования назем-
ных кронообразующих прямостия шх дег евьев..................238
7. Многообразие и распространение древовидных жизненных
форм...................................................... 241
Глав» IV Жизненные формы кустарников...........................243
1. Географическое распространив кустарников.................243
2. А1орфо1енез жи «ценной формы кустарника у некоторых типич-
ных кустарниковых видов растений....................... ... 252
Морфогенез жизненной формы кустарника у лещины — Corv-
ins avellana L. . .... .... .........252
Особенности морфоге, еза жизненной формы кустарника у
бересклета бородавчатого (Evonymus verrucosa S«op.) . . . 264
Особенности морфогенеза жизненной формы кус।арника у ши-
Н 'вников Rosa spinosissima L. и Rosa Alberti R.l. . . . 270
Особенности морф генеза жизненной формы кустарника у
терна — Prunus spinosa L...................................276
3. Важнейшие морфолого-биологические особенности наземных
прямостоячих кустарников с удлиненными побегами.............279
Глава V Жизненные формы кустарничков . ... ... 283
]. Экологические особенности и распространение кустарничков . 283
2. Морф )генез жизненной формы кустарничка у некоторых ти-
пичных представителей . . ... ...... 287
Морфогенез жизненной формы кустарничка у черники (Vaccinium
mvrtillus L) . ........... . . . ... 287
Особенности морфогенеза кустарничка у брусники (Vaccinium
Ос Ценности структуры кустарничка у андромеды (Andromeda
Особенности структуры кустарничка у голубики (Vaccinium
uliginosum L.), болотного балльника (Ledum } alustre L.) и
Кассандры (Cassandra calycuhta D. D п.) . . *. . . . 305
Особенности морф >генеза жизненной формы кустарничка у ве-
реска (Calluna vulgaris (L) Hill)........................ 307
3. Главнейшие морфолого-биологич ские особенности кустар-
ничков ................................................... 312
Глава VI. Жизненные формы стелющихся растений . . . . 315
1. Экологические особенности мелющихся растений, их геогра-
фическое распространение и положение в системе жизненных
форм........... ........................... . . 315
2. Морфогенез жизненной формы стланца у некоторых рас-
тений ......................................................320
Некоторые биологические особенности кедрового стланика (Pinus
pumila RhI).................................................320
. Морфогенез жизненной формы стланцт у туркестанского мож-
жевельника (Juniperus turkestanica Kom.)...................323
377
S § ЛгаЗ §
Особенности морфогенеза стланцевой формы у казацкого мож-
жевельника (Juniperus sabina L.). . . . ................332
Некоторые данные о формировании и структуре стланцевой
формы у сибирского можжевельника (Juniperus sibirica
Burgsd.).......................................* . . . .
Особенности морфогенеза стелющихся форм у некоторых
аркто-альпийских шпалерных кустарников (по Rauh, 1937). . .
3. Главнейшие особенности роста и морфогенеза у стелющихся
древесных растений.......................................
Глава VII. Жизненные формы лиан.............................
Глава VIII. Жизненные формы подушковидных растений ....
1. Экологические особенности и географическое распространение
подушковидных растений...................................
2. Морфогенез подушковидных жизненных форм у некоторых
видов растений (по Rauh).................................
Образование подушковидной формы у Silene acaulis L.....
3. Классификация подушковидных жизненных форм и их морфо-
лого-биологическая интерпретация.........................
Литература...............................................
Иван Григорьевич Серебряков
Экологическая
морфология растений
Редактор Г. М. Проскурякова
Художественный редактор Н. Л. Кузнецова
Технический редактор Л, Л. Ежова
Корректор Г. П. Меркулова
Сдано в набор 13/Х-61 г. Подписано к печати 14/IV-62 г. Бумага 60Х90*/ю.
.23,75 печ. л. 24,17 уч.-нзд. л. Тираж 3500. Т —03642. Изд. № Е/13. Зак. В-413.
Цена 83 коп,
Государственное издательство «Высшая школа»,
Москва, К-62, Подсосенский пер., 20.
Казань, «Татполиграф», ул. Миславского, 9.
Замеченные опечатки
Страница Строка Напечатано Следует читать
180 2 снизу кустовидной древовидной
308 17 снизу точки почки
338 2 снизу мгпк гп