












































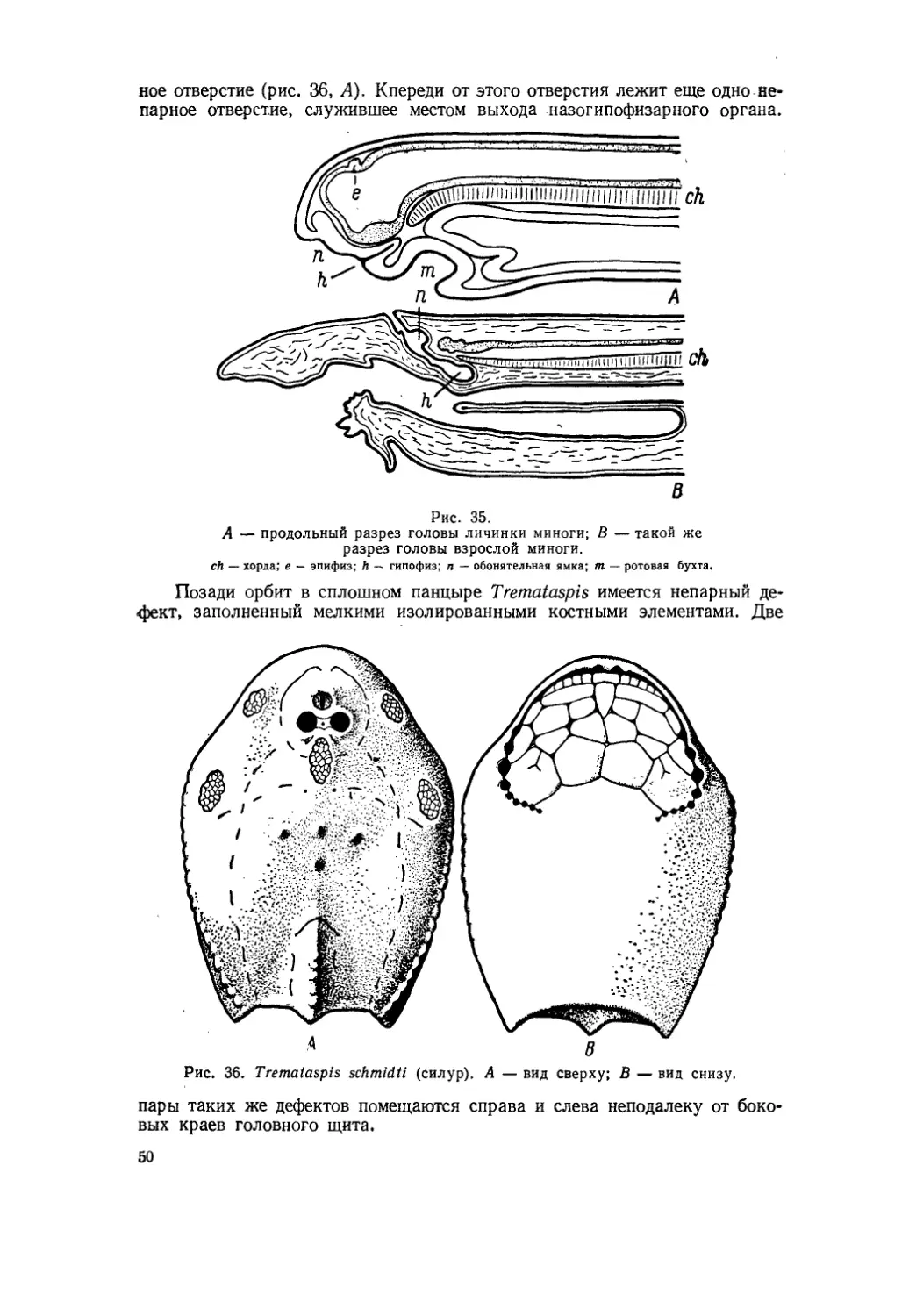






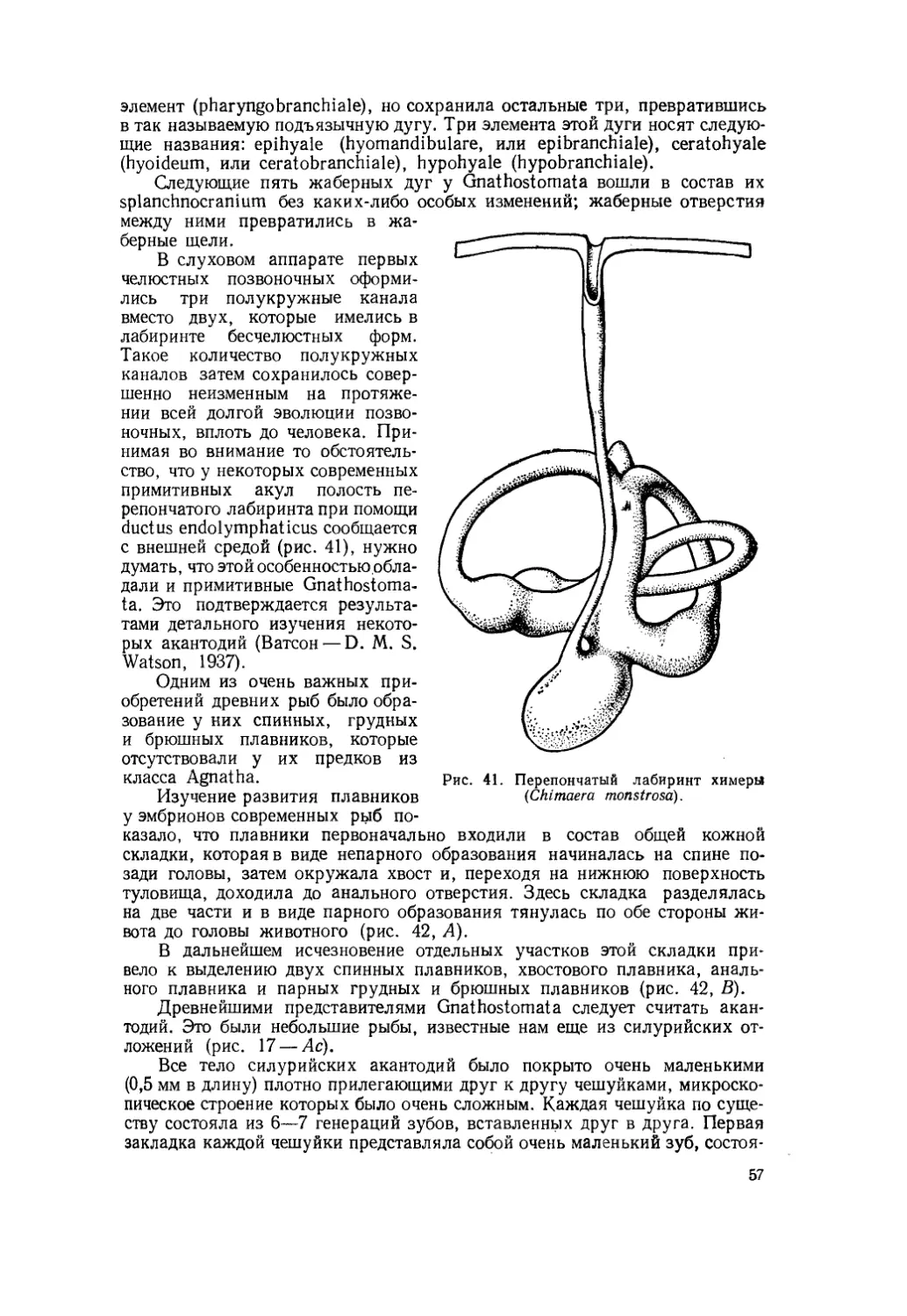




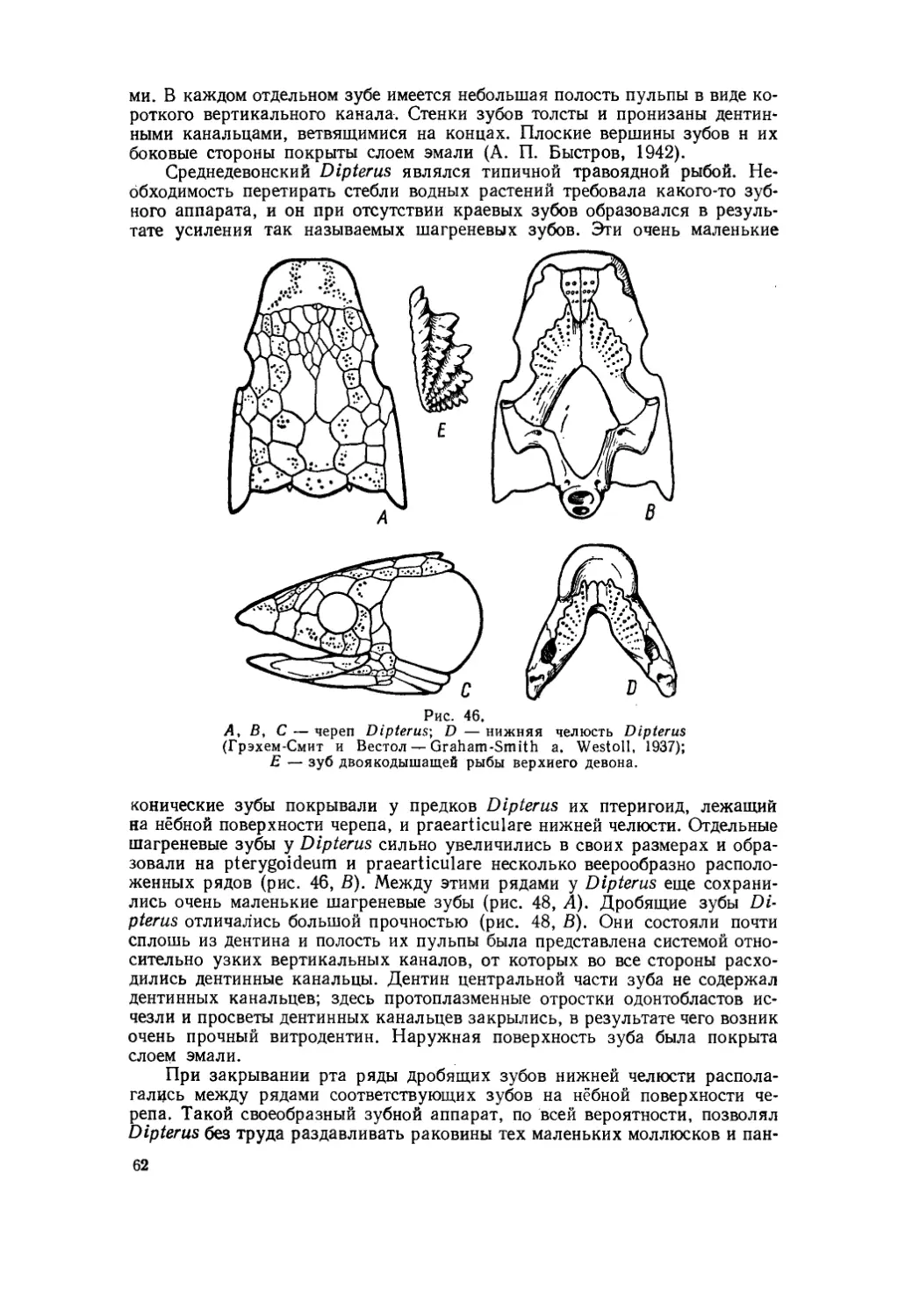







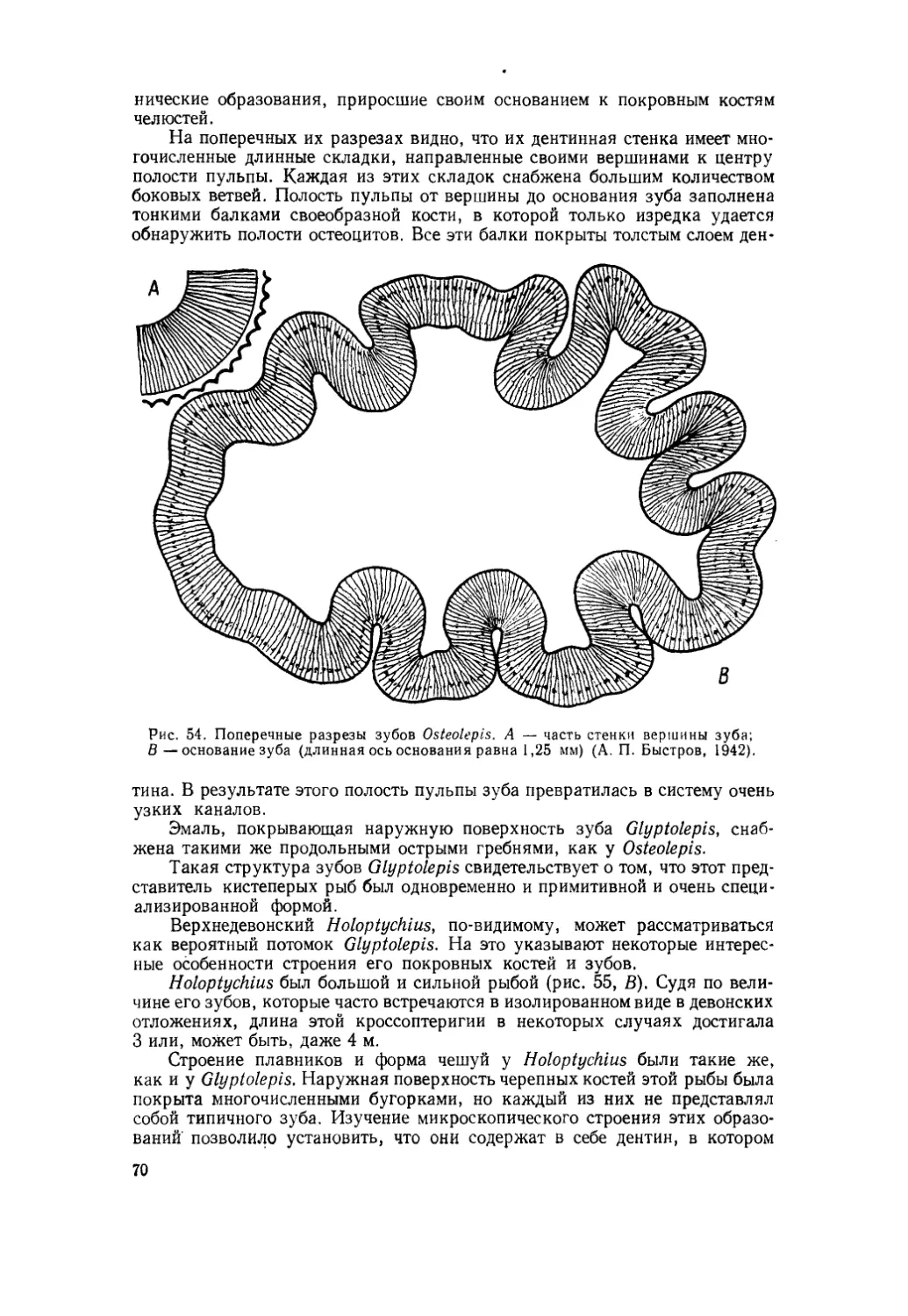

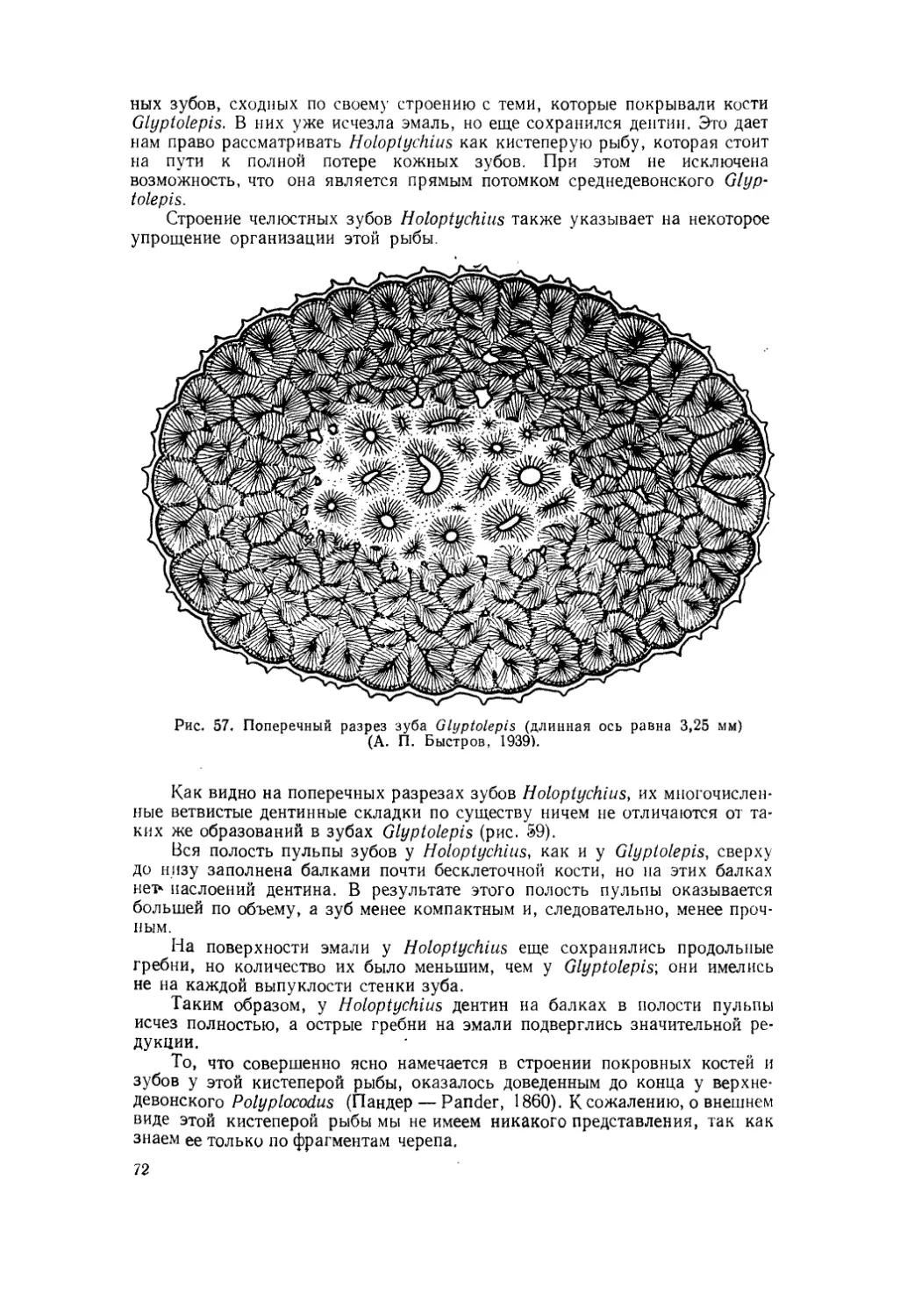




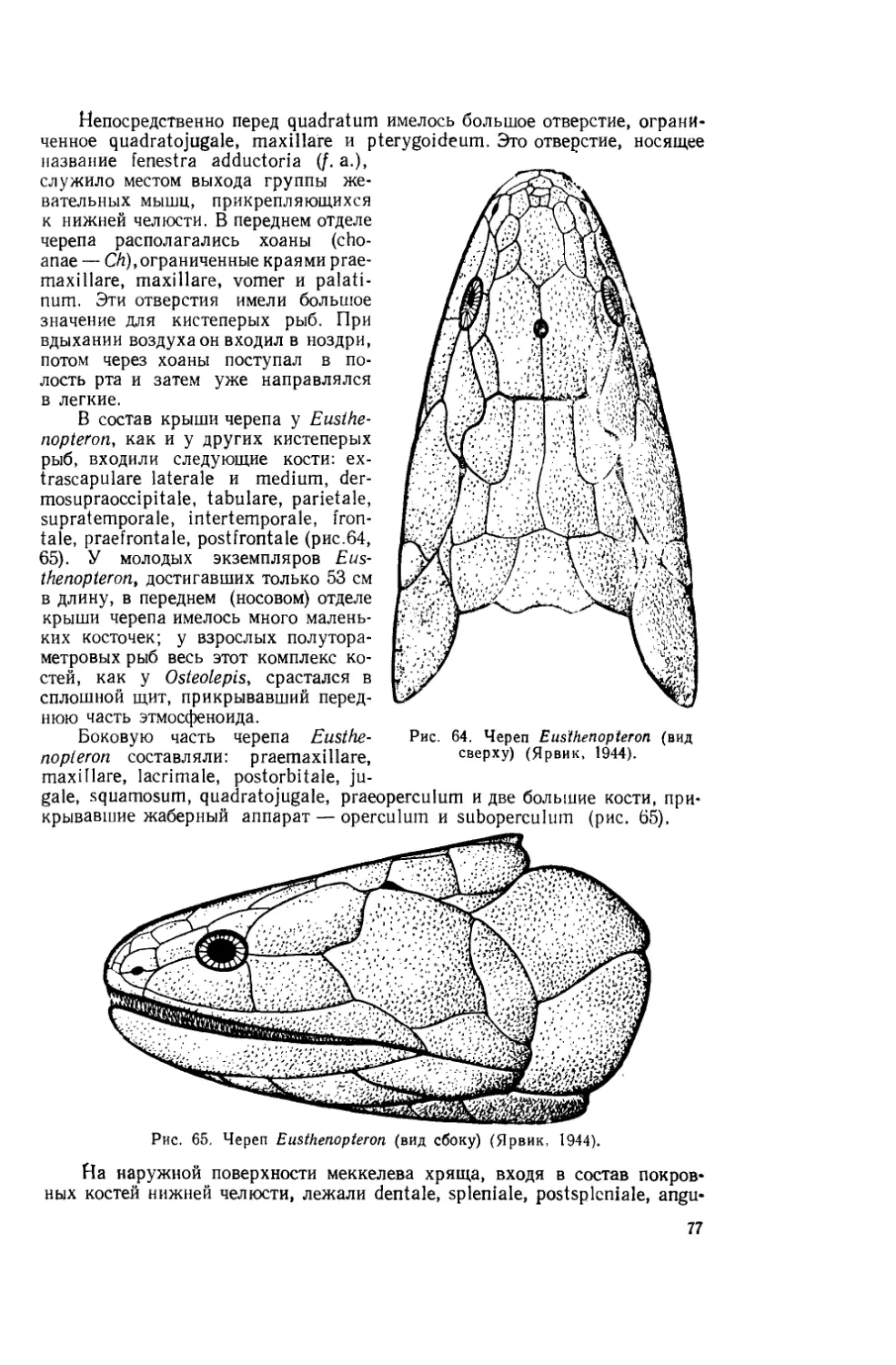
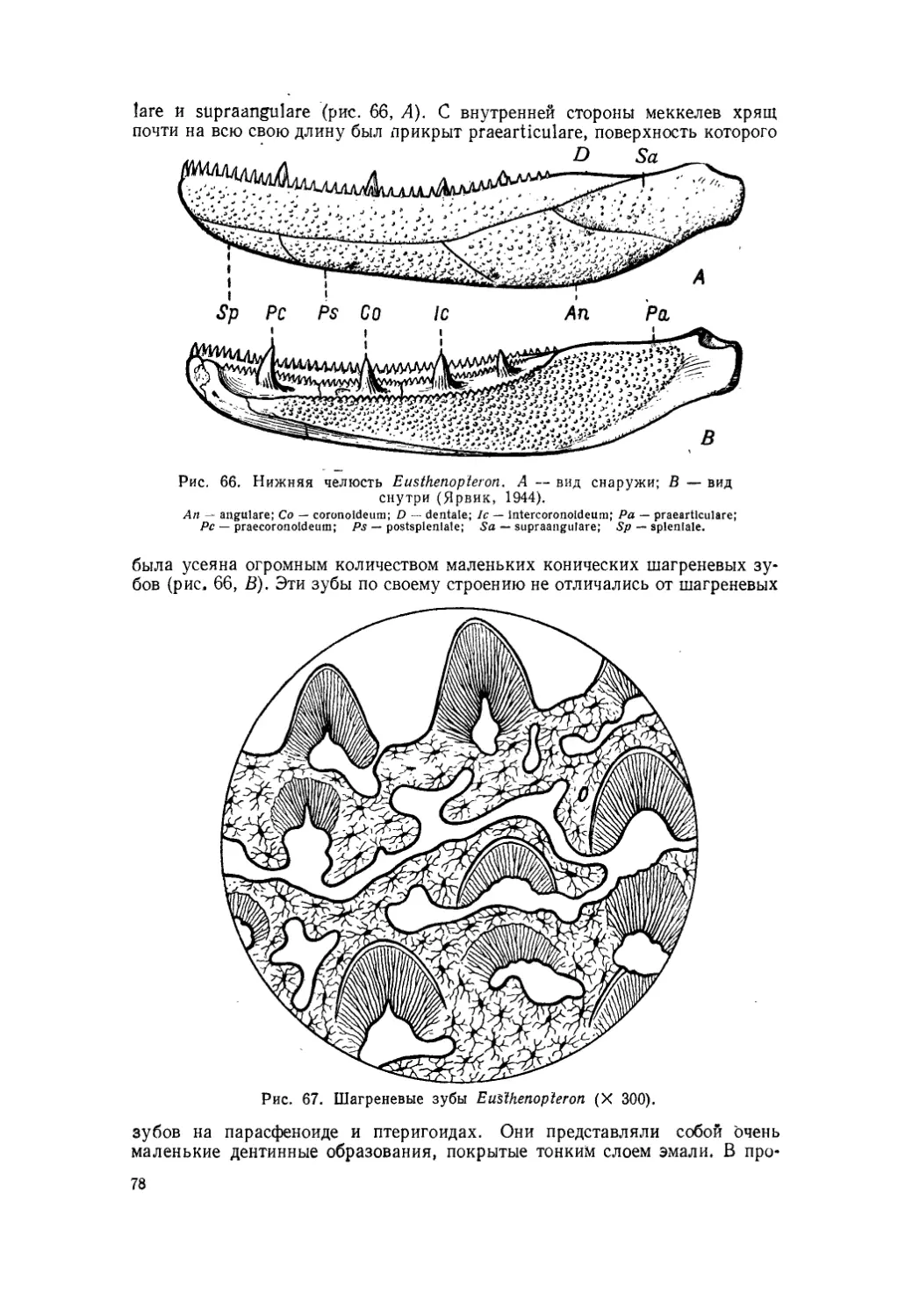

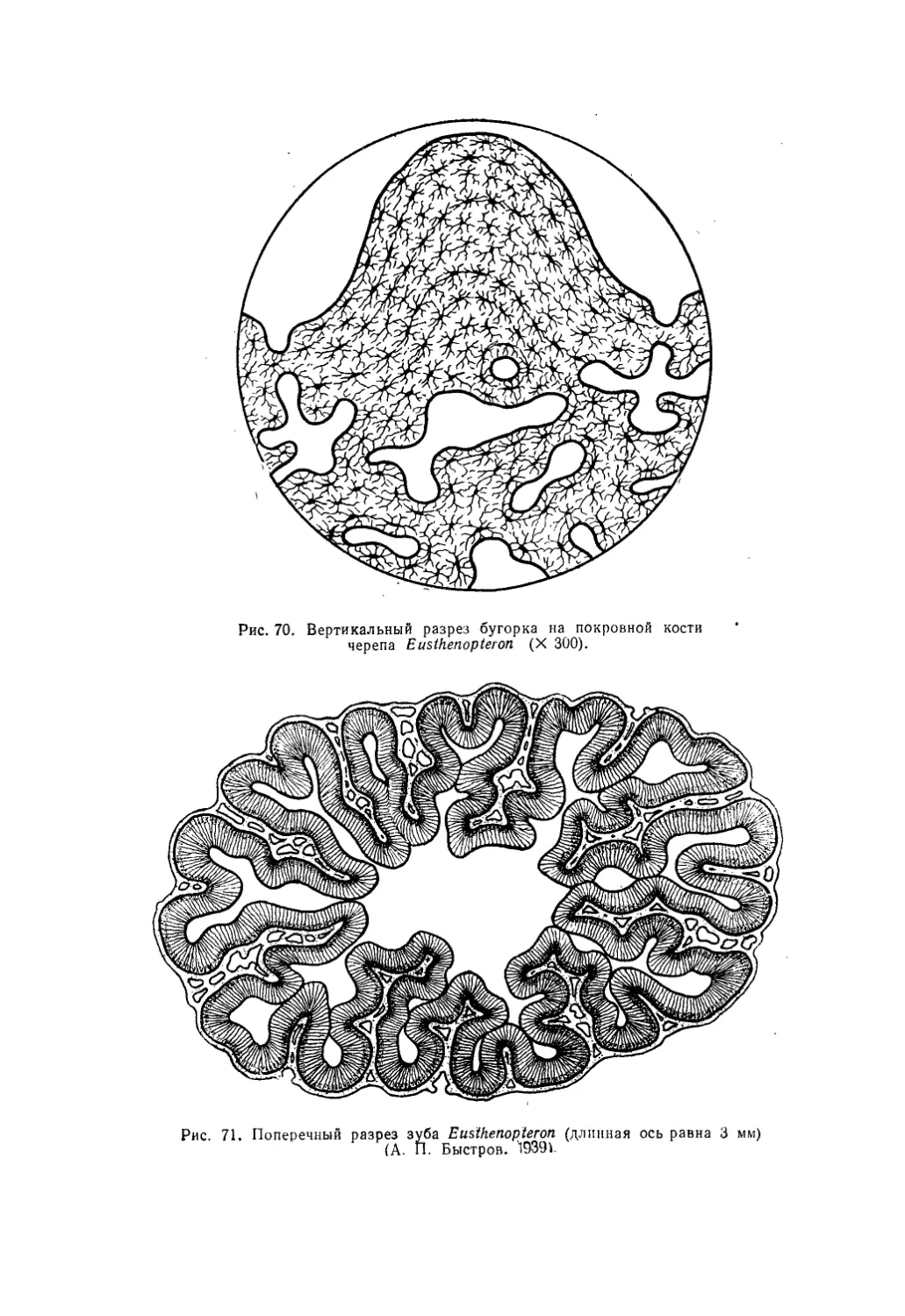


























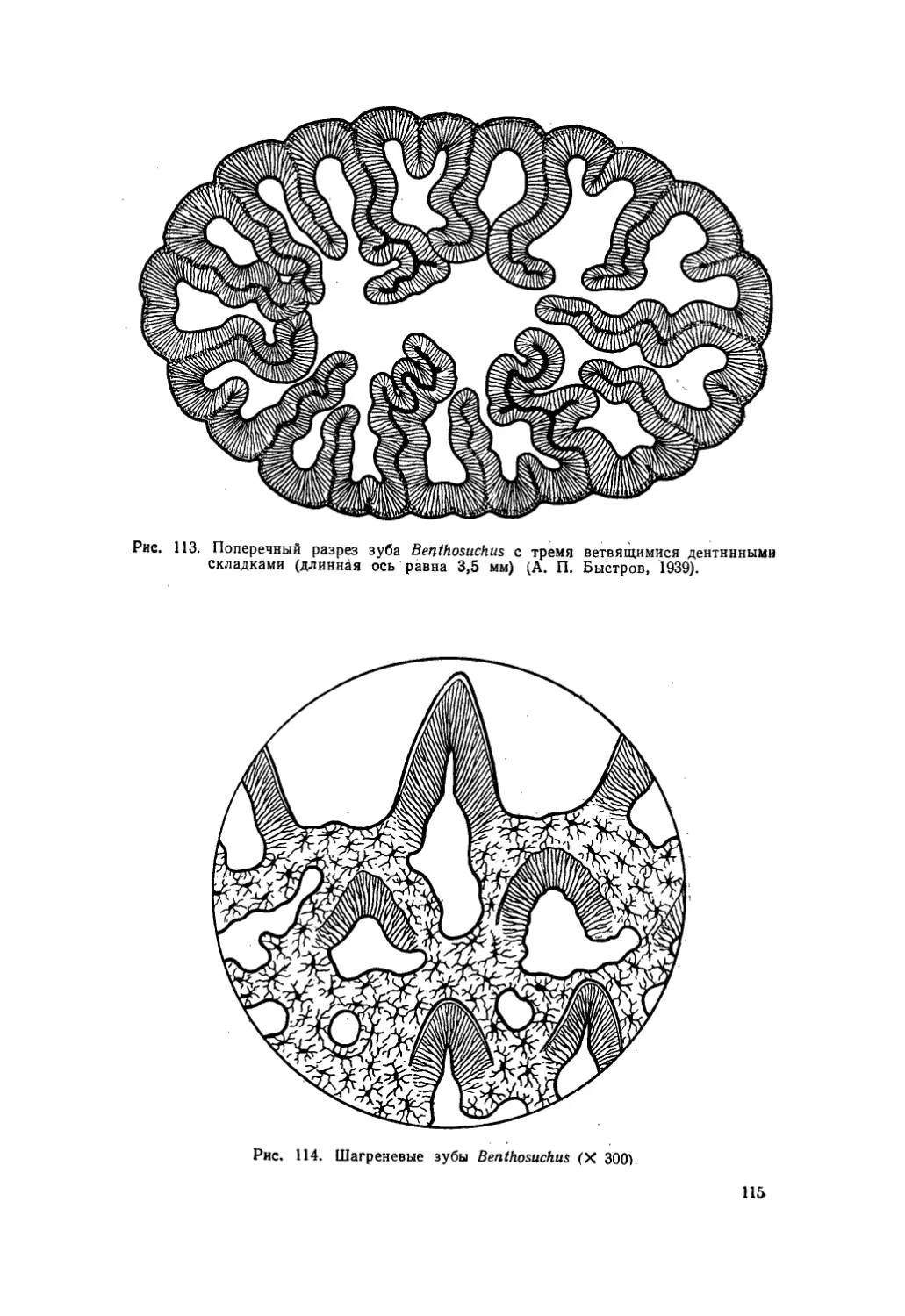
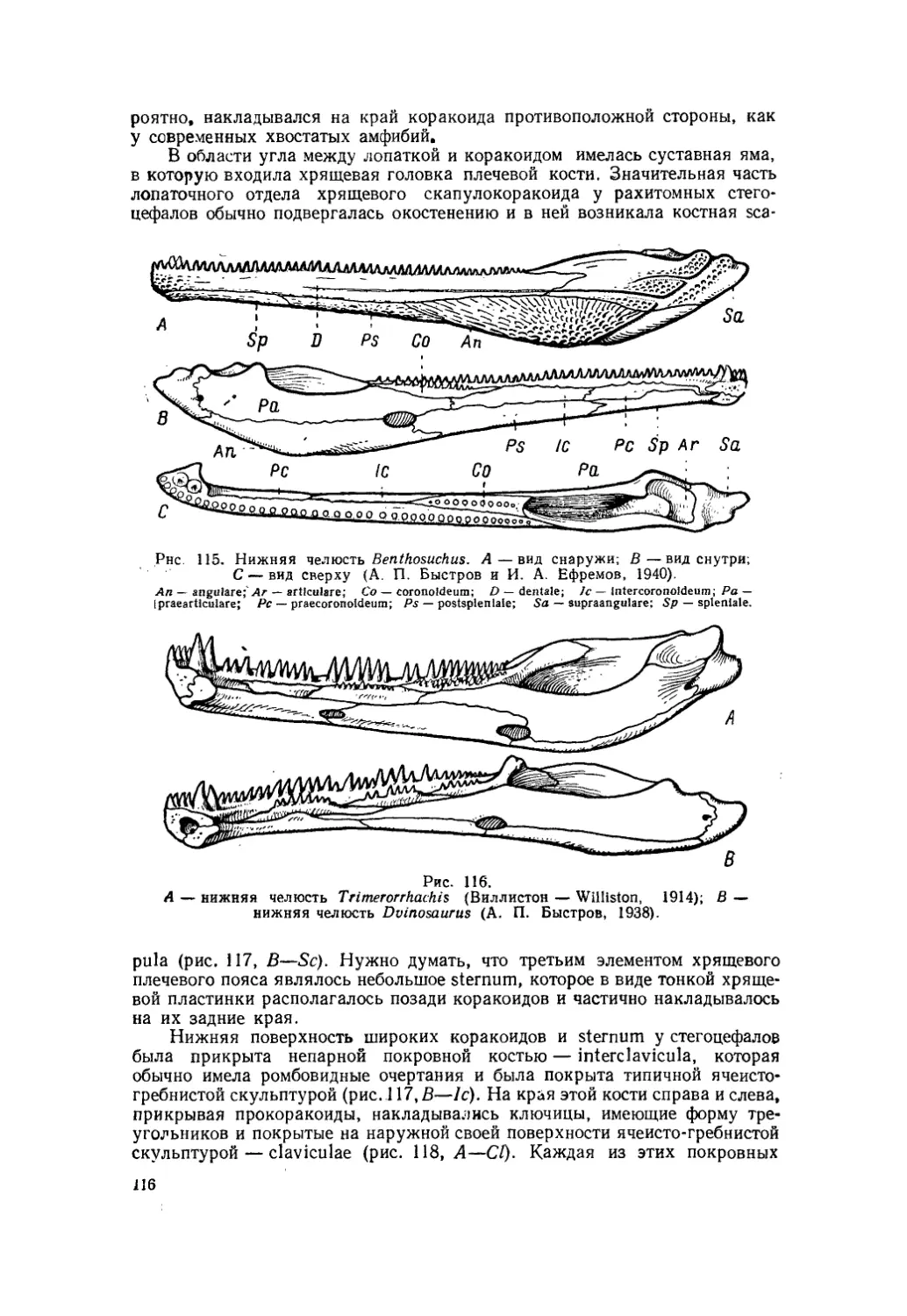
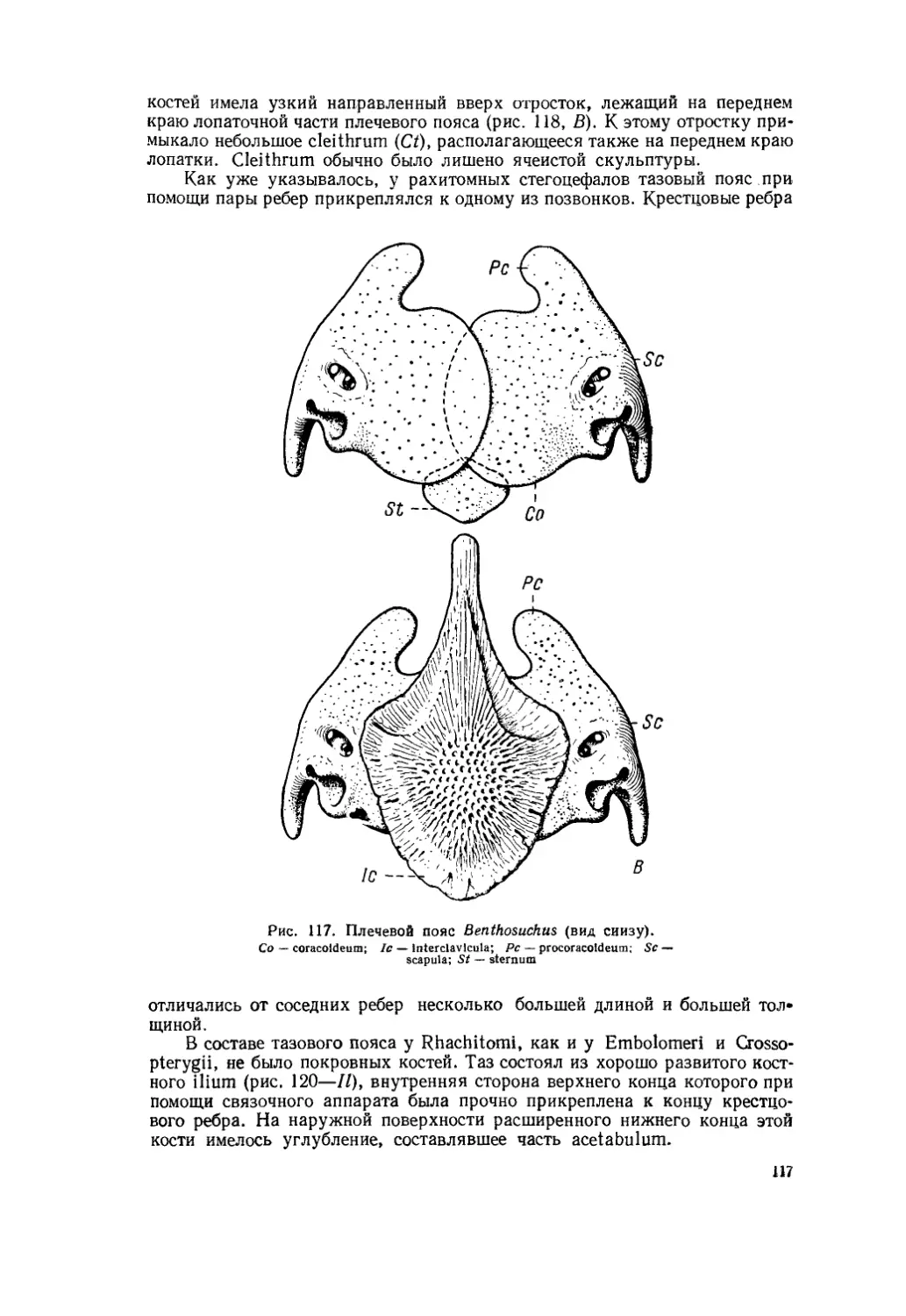


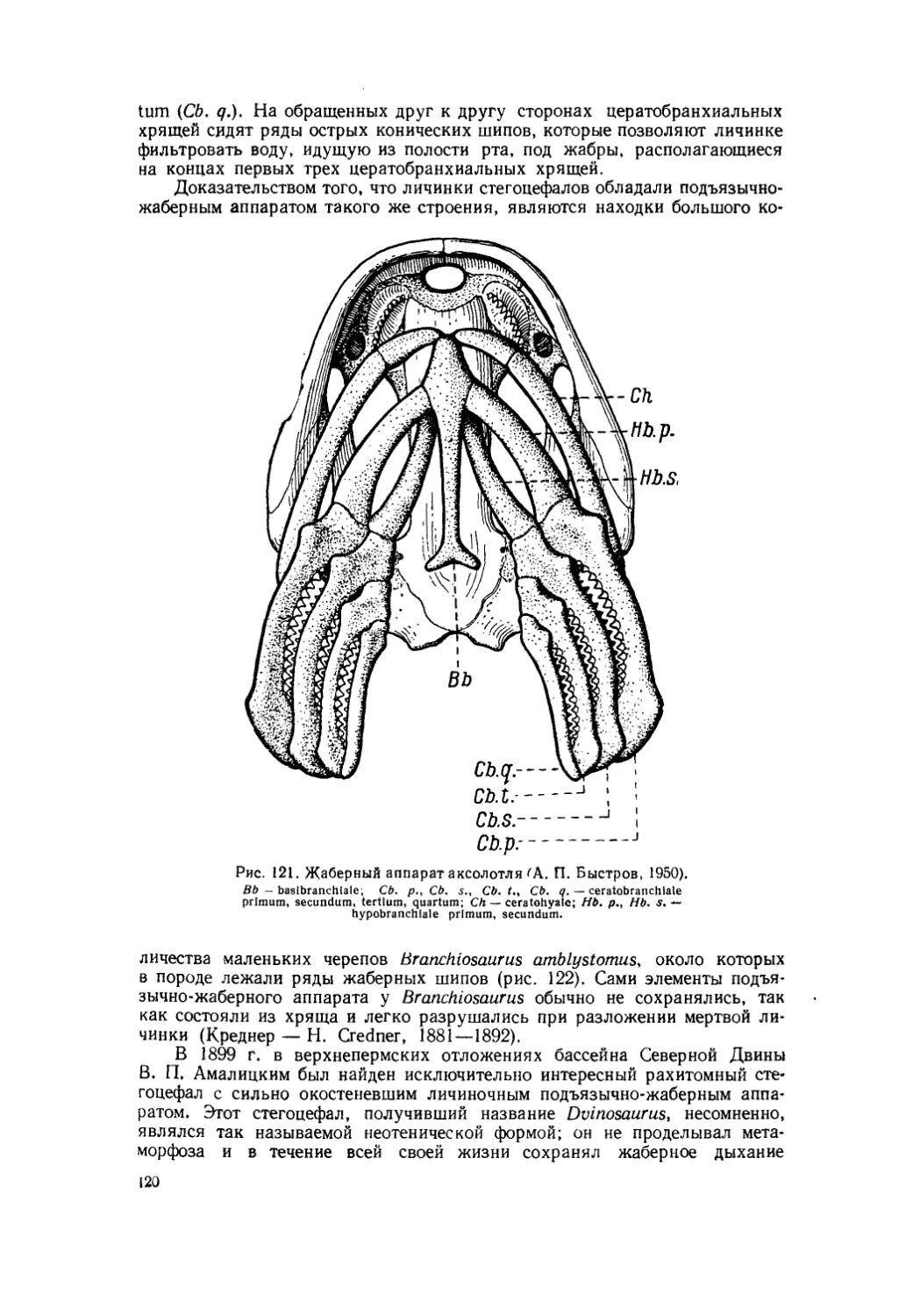












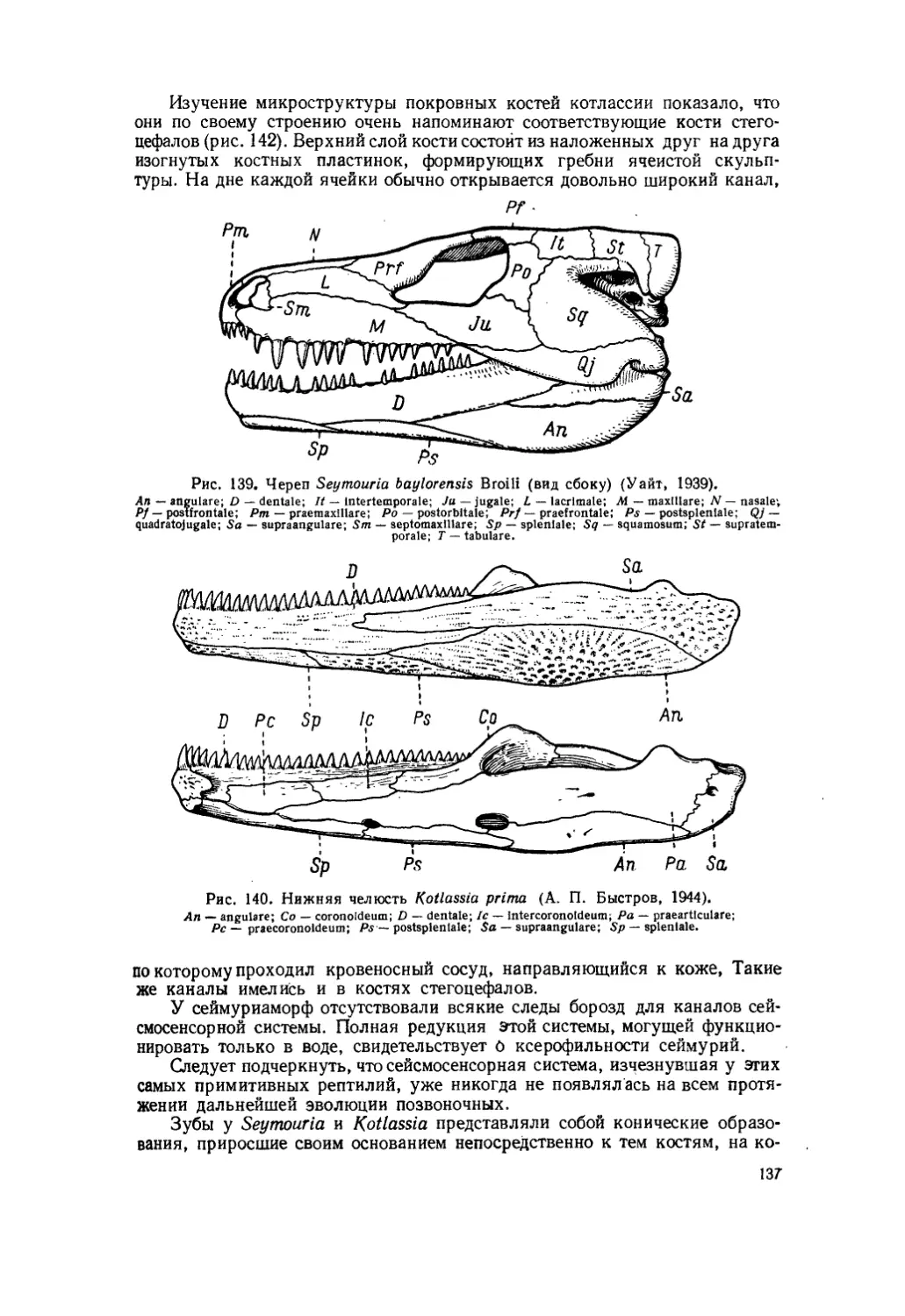




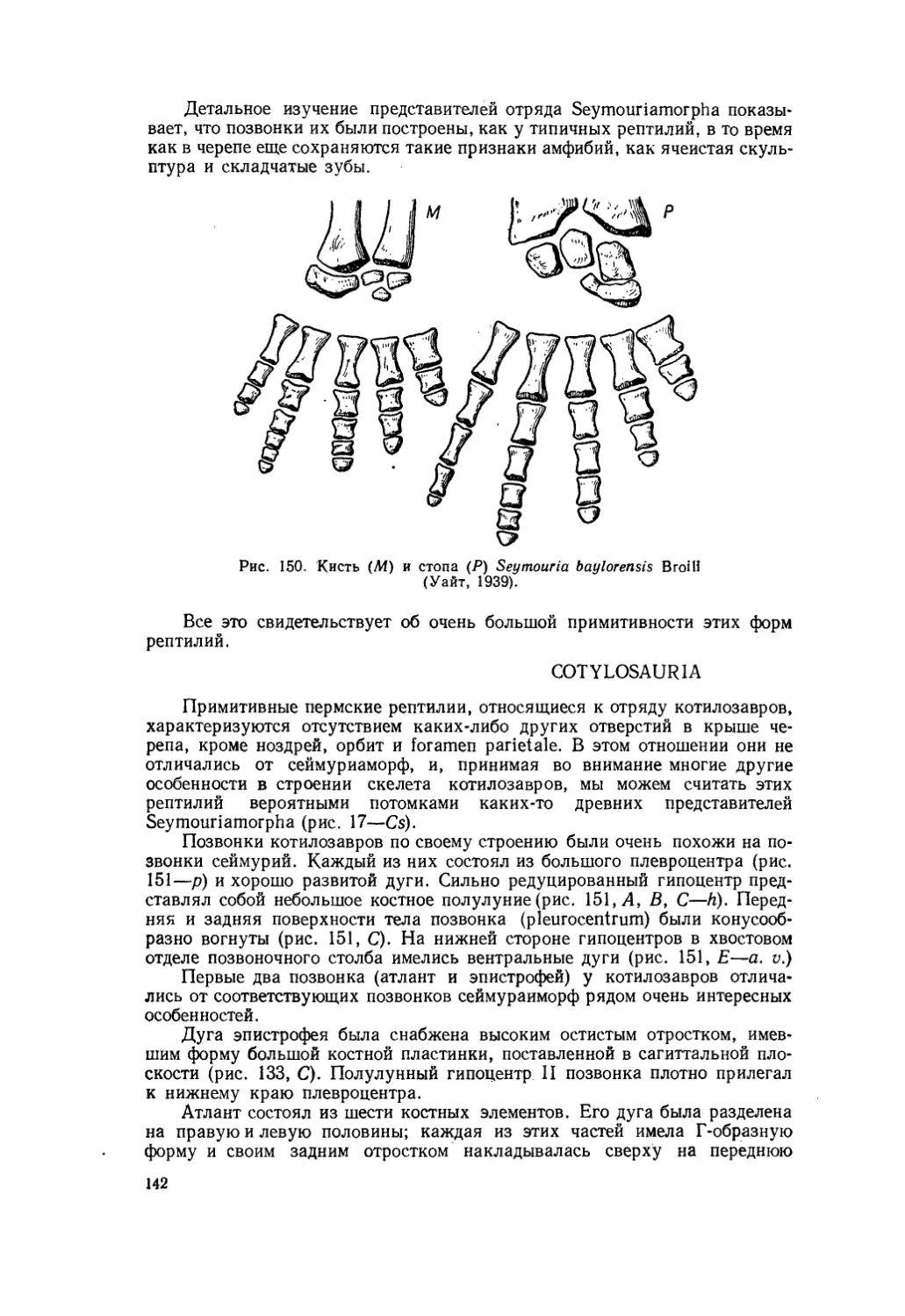
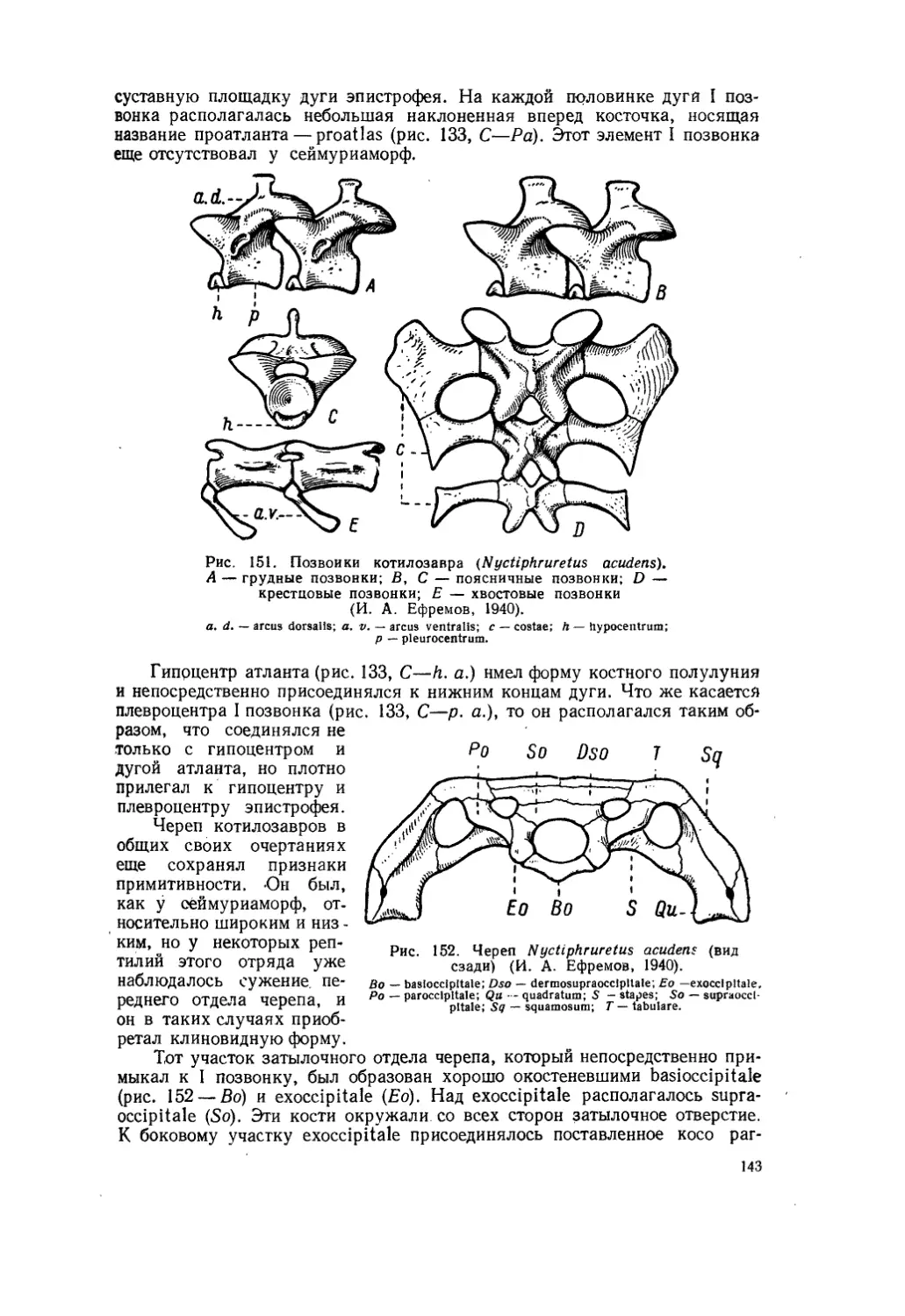



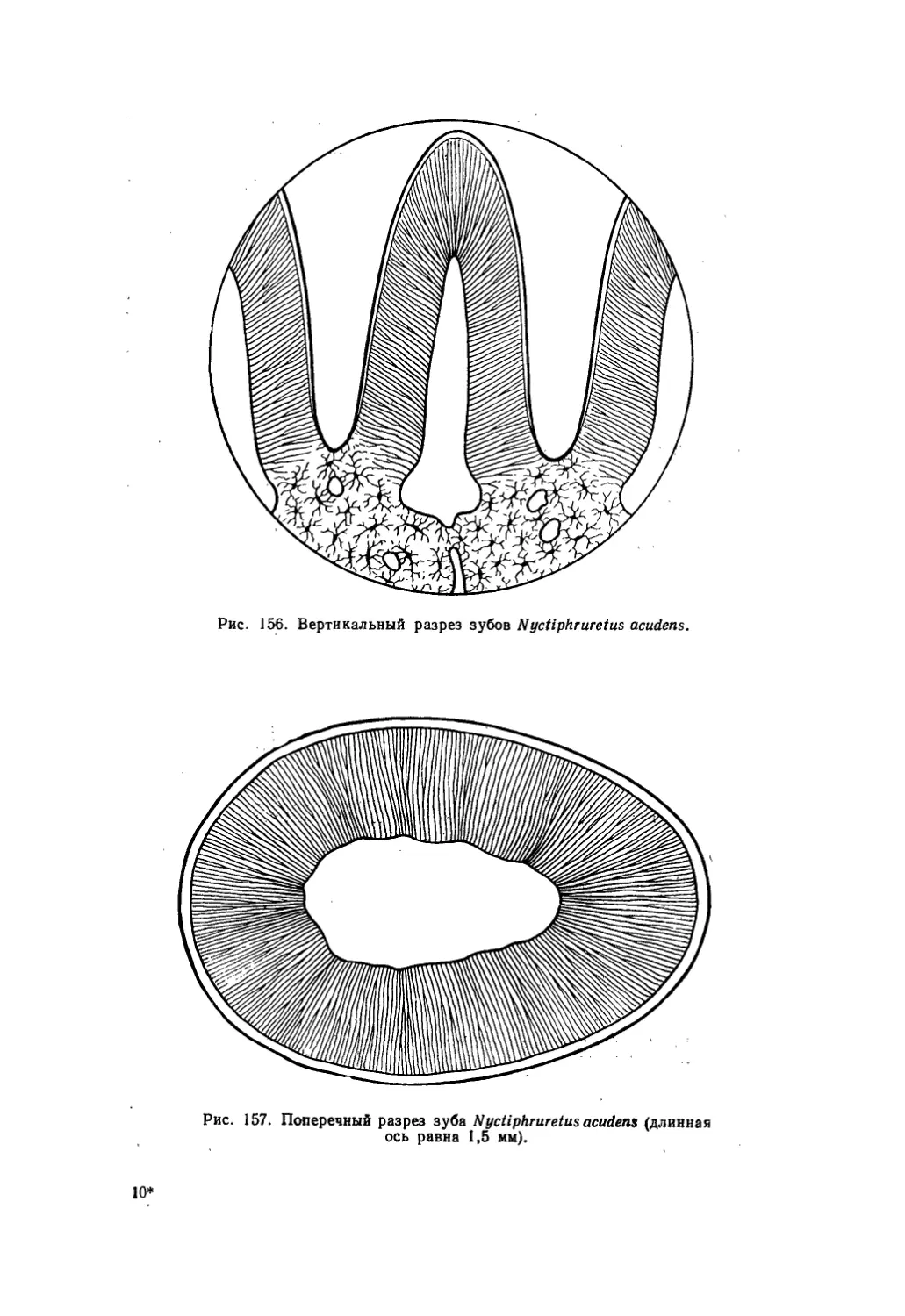












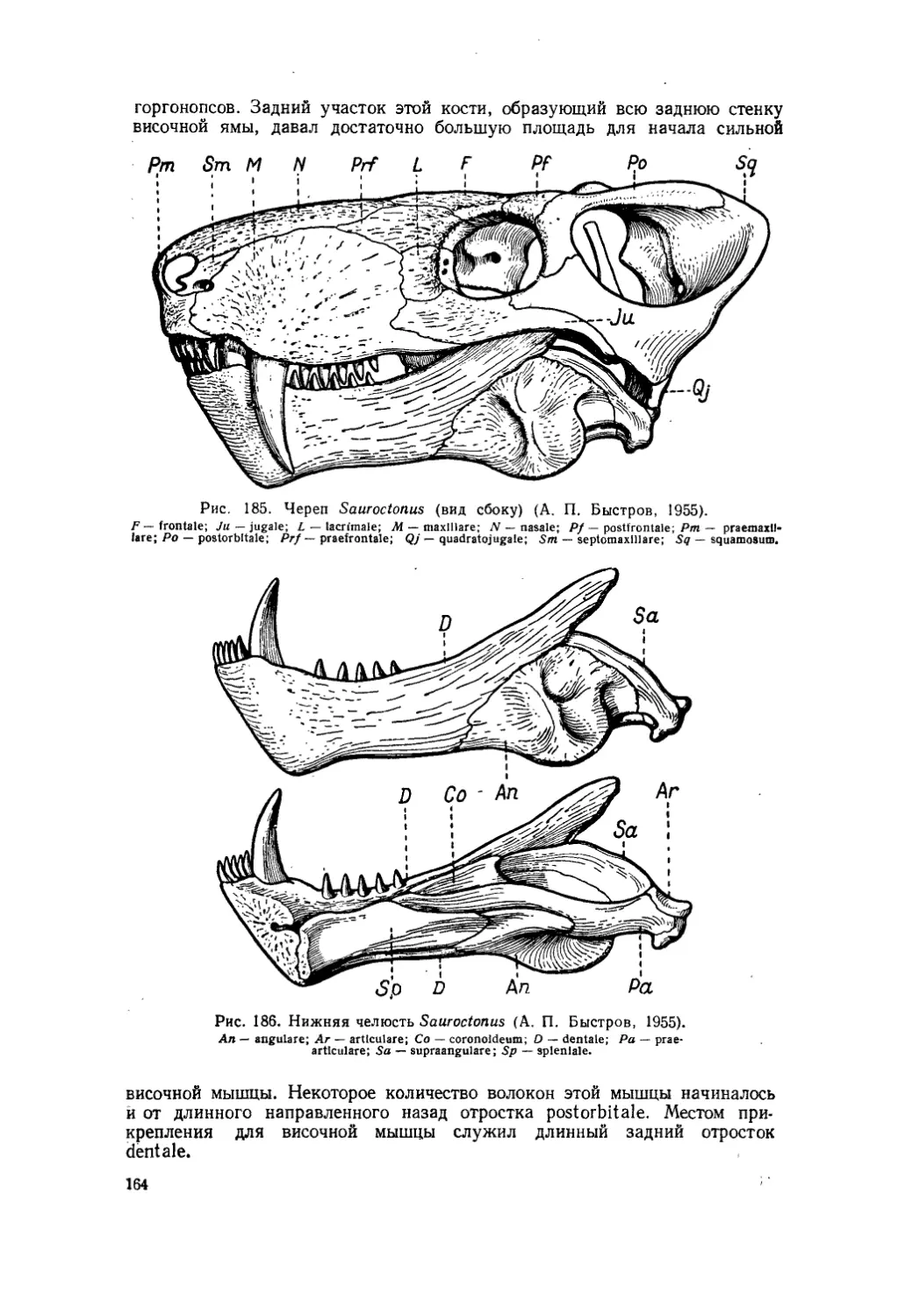
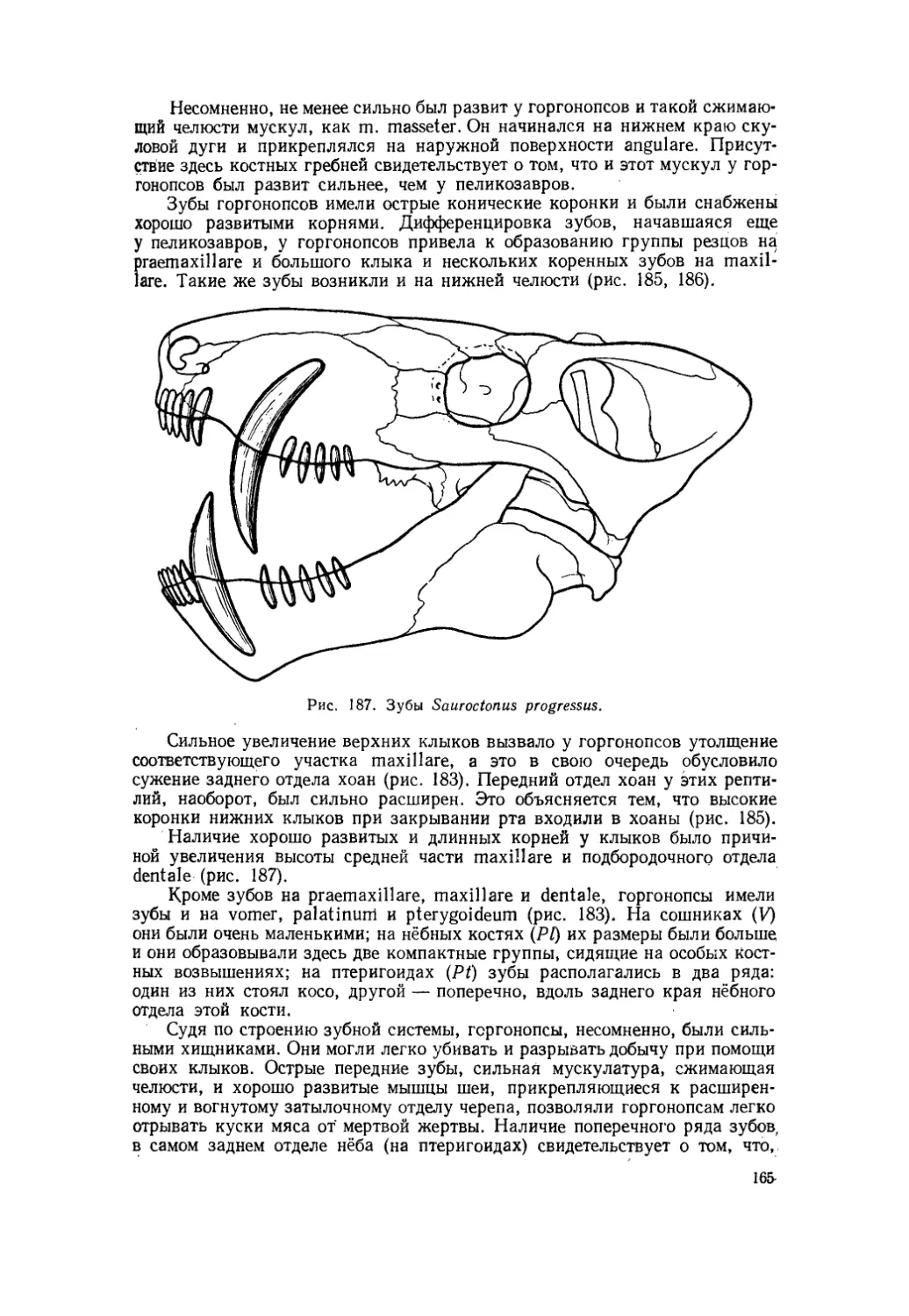

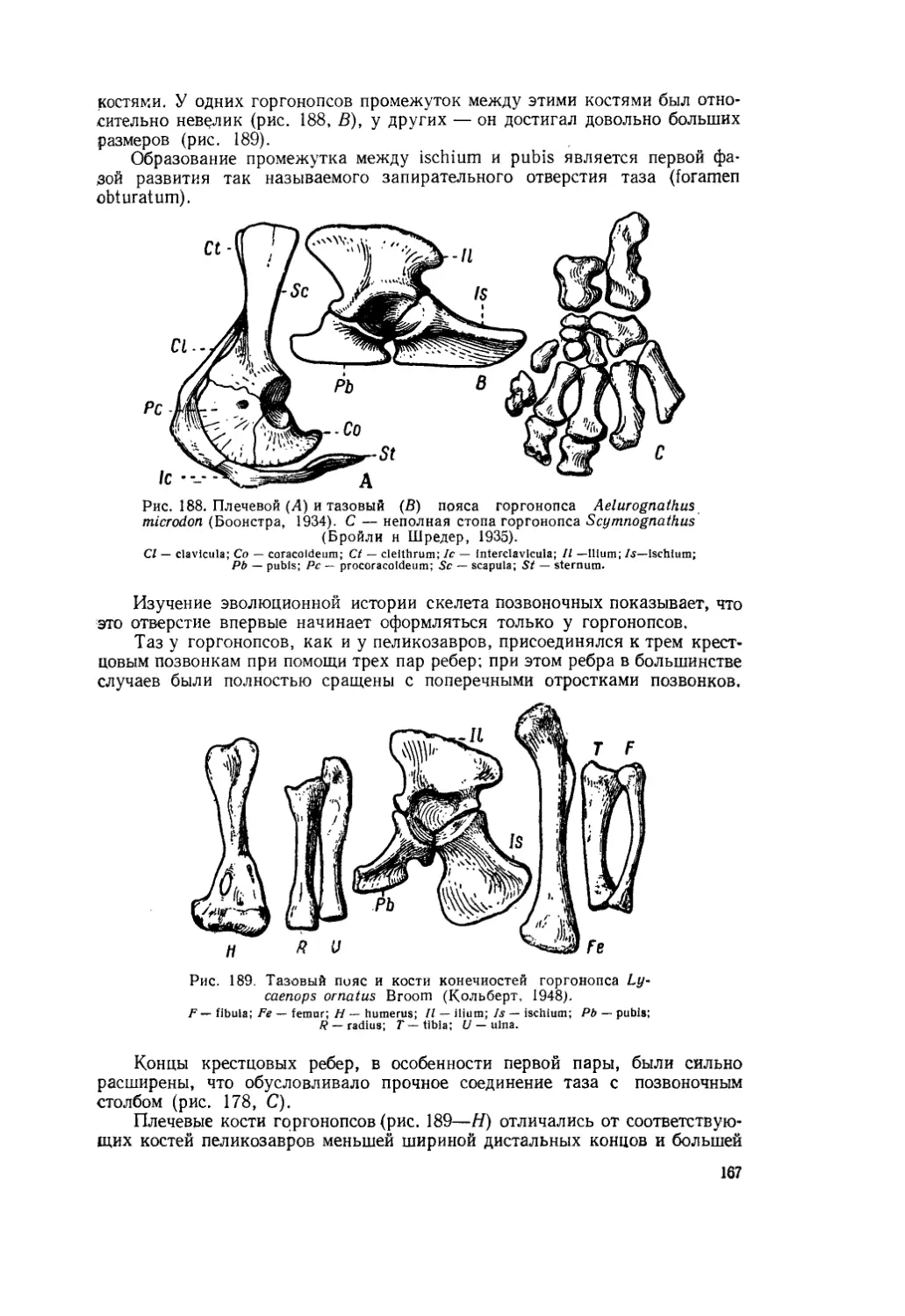



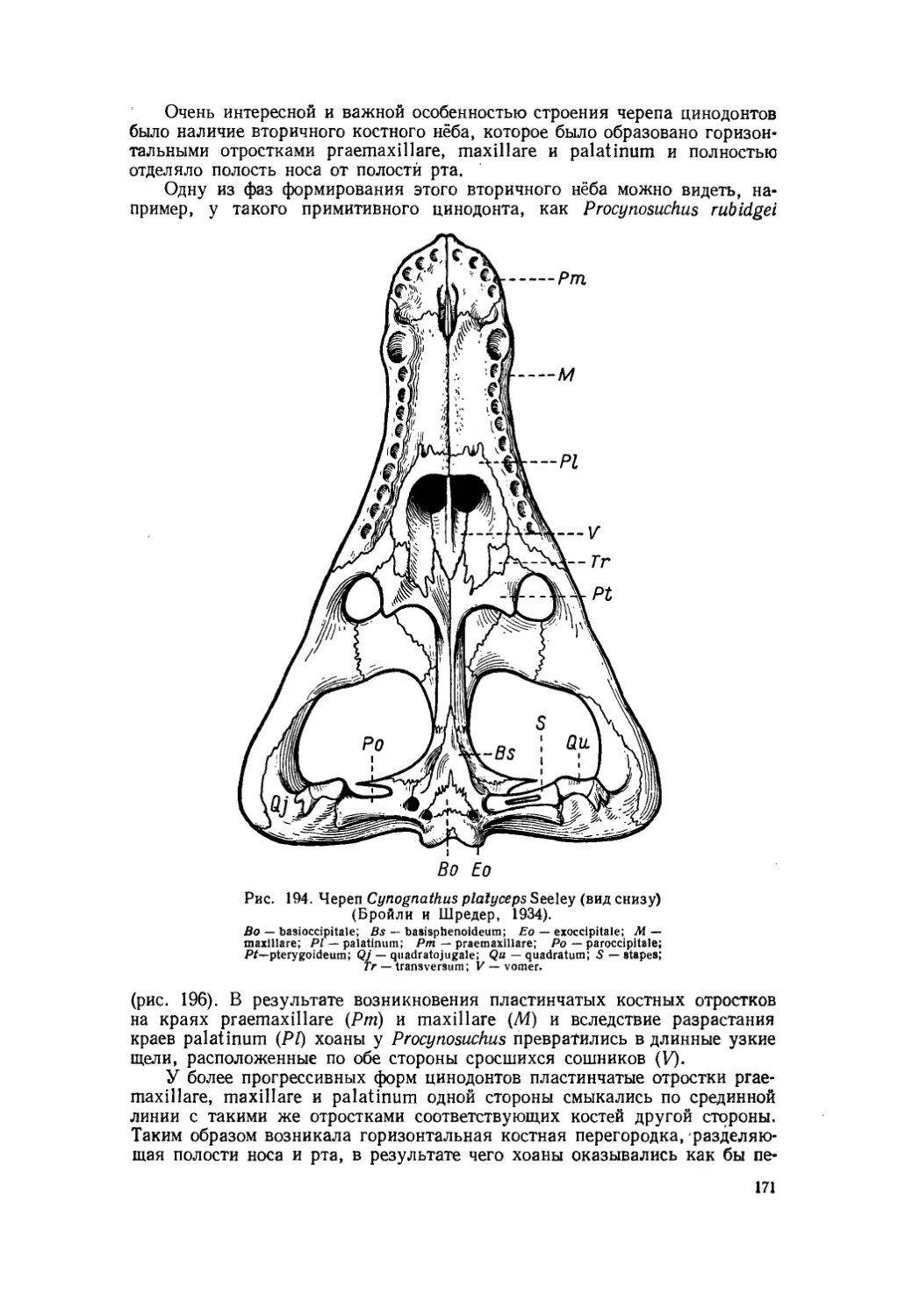




















































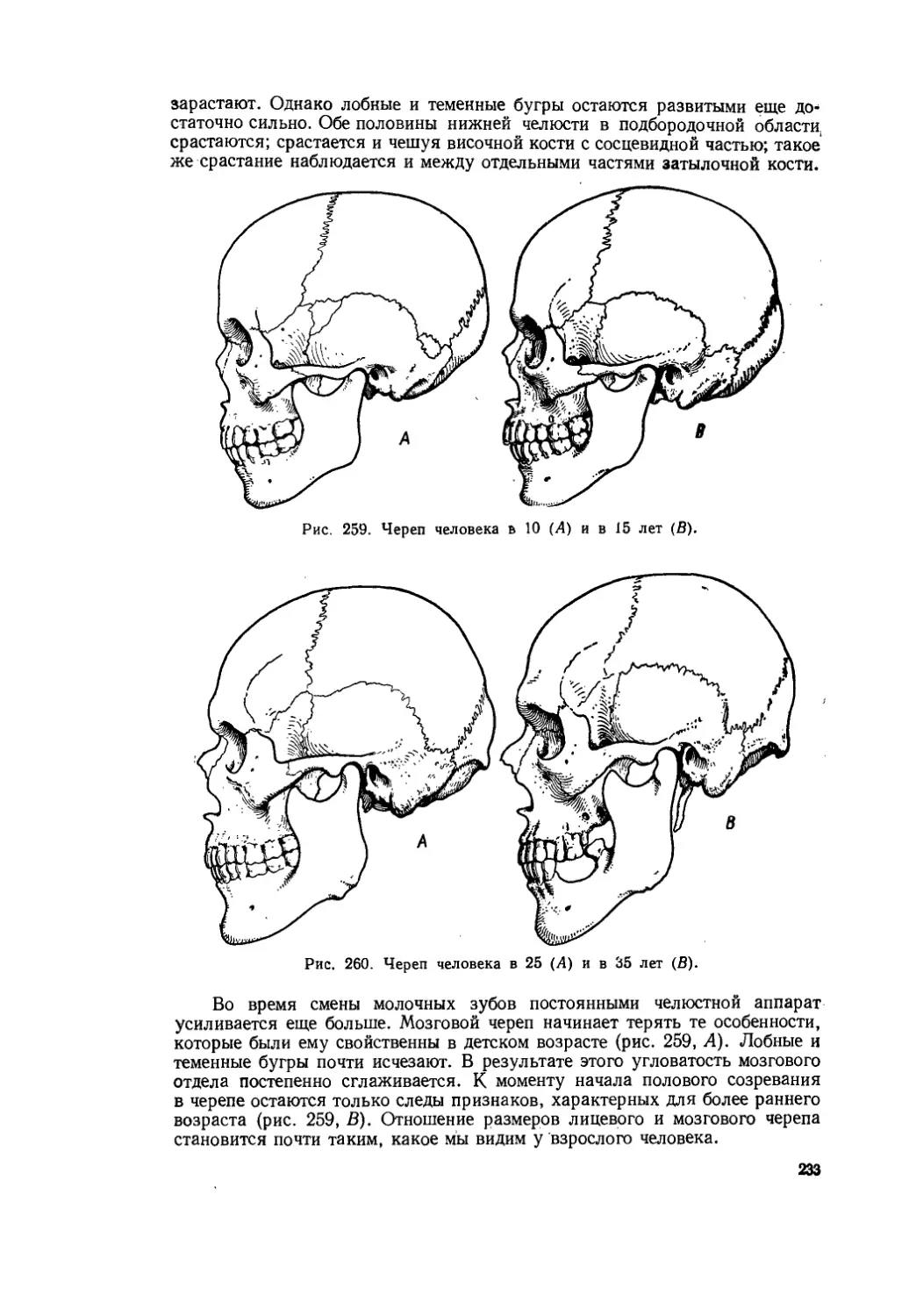
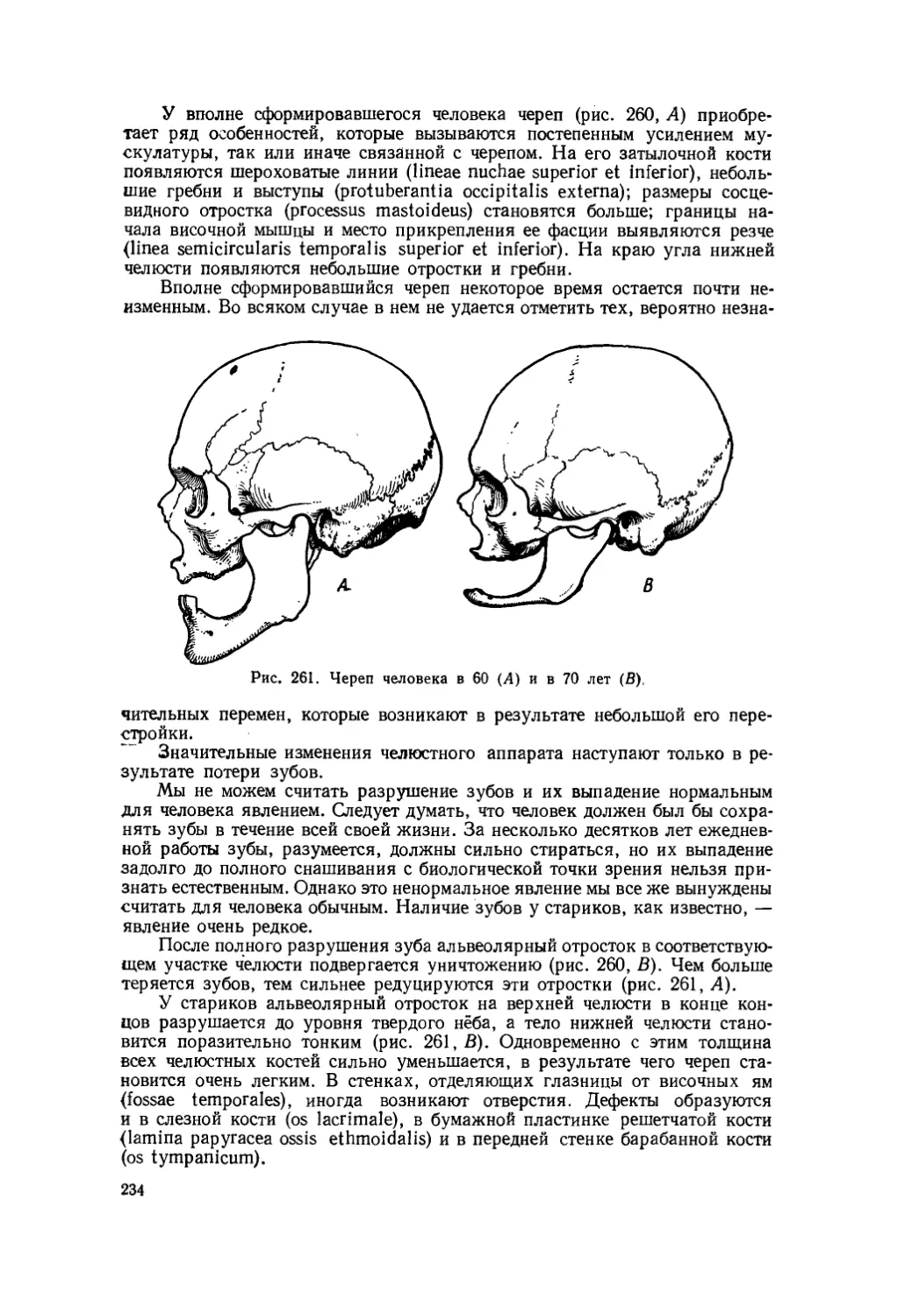

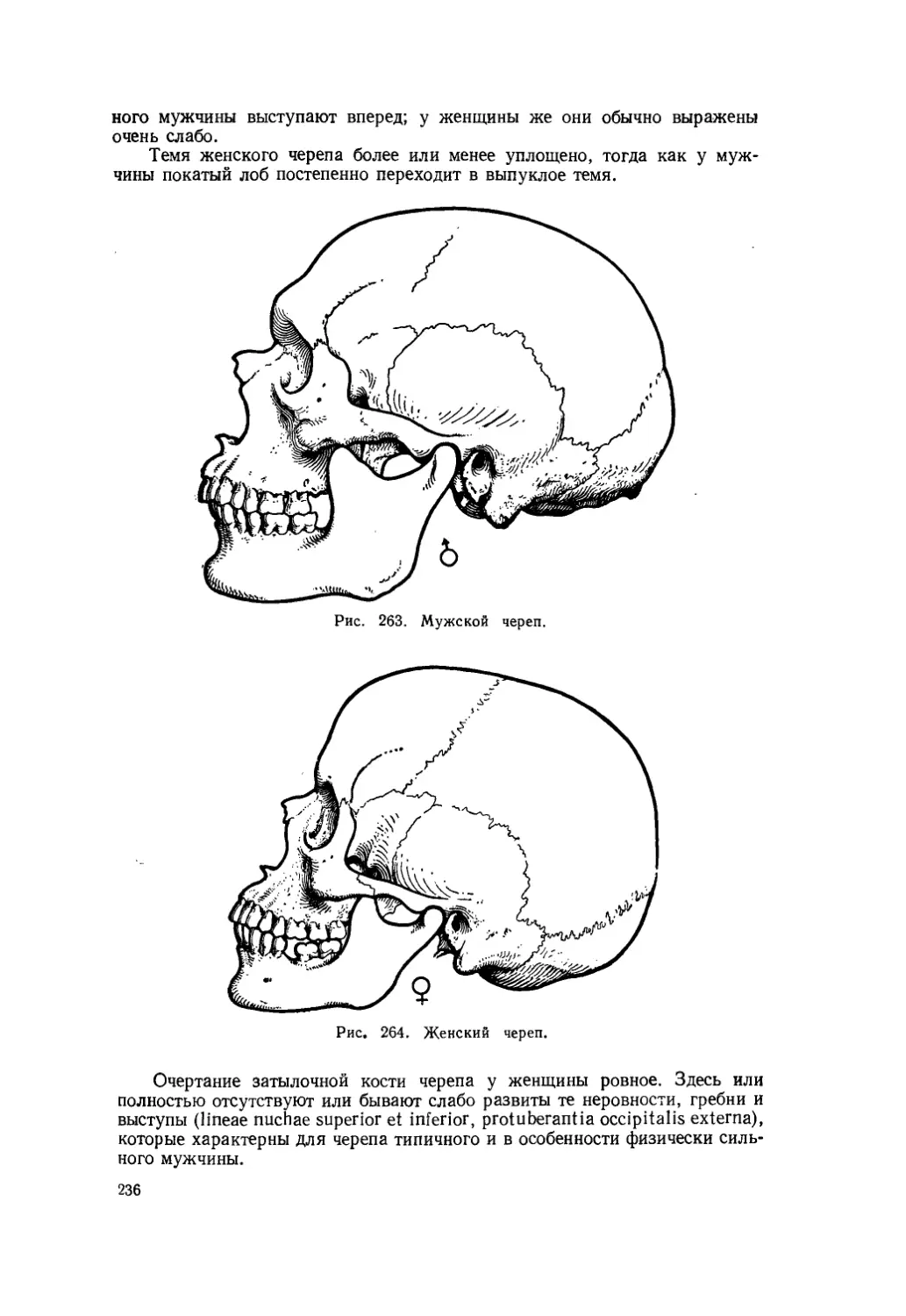
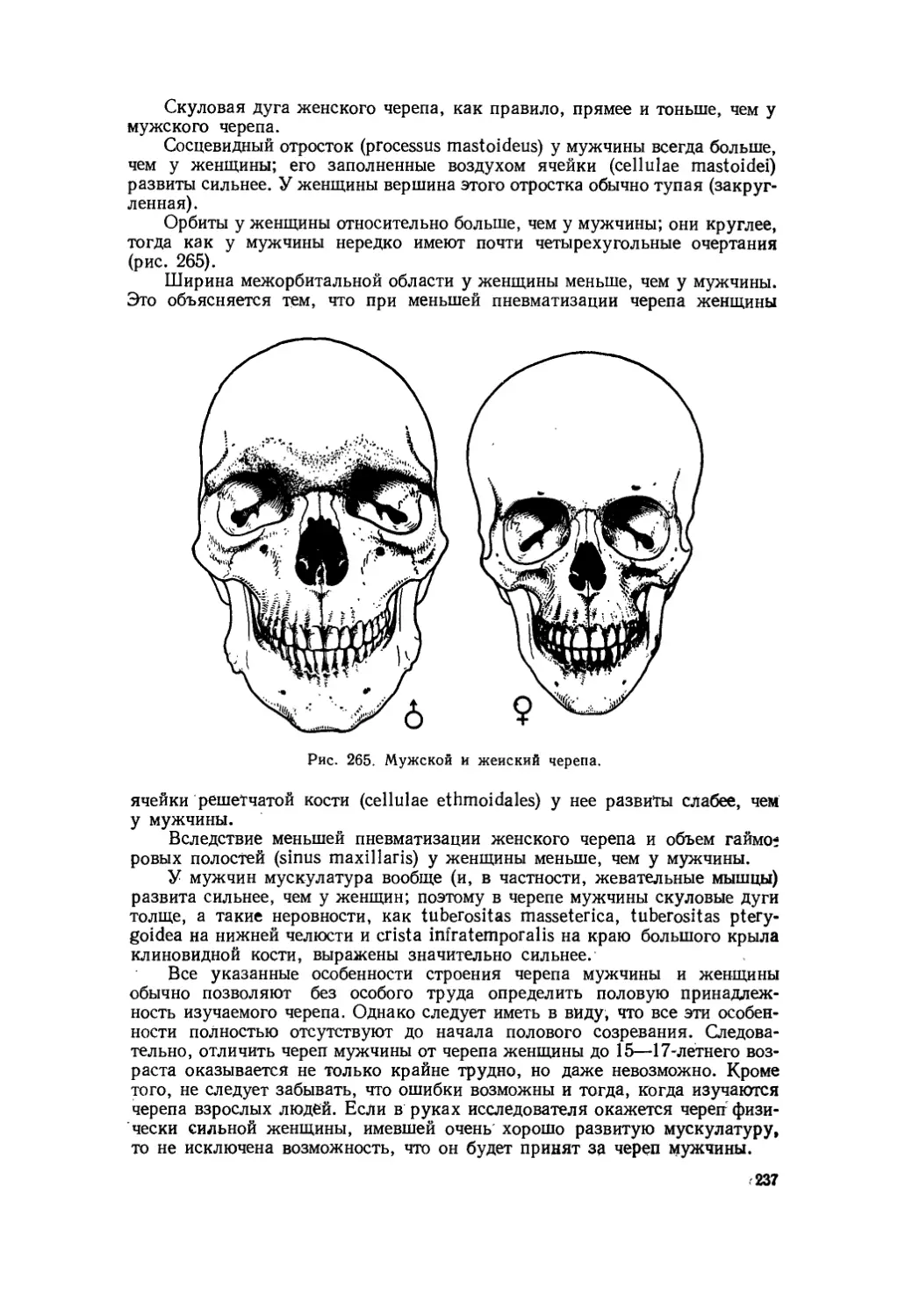







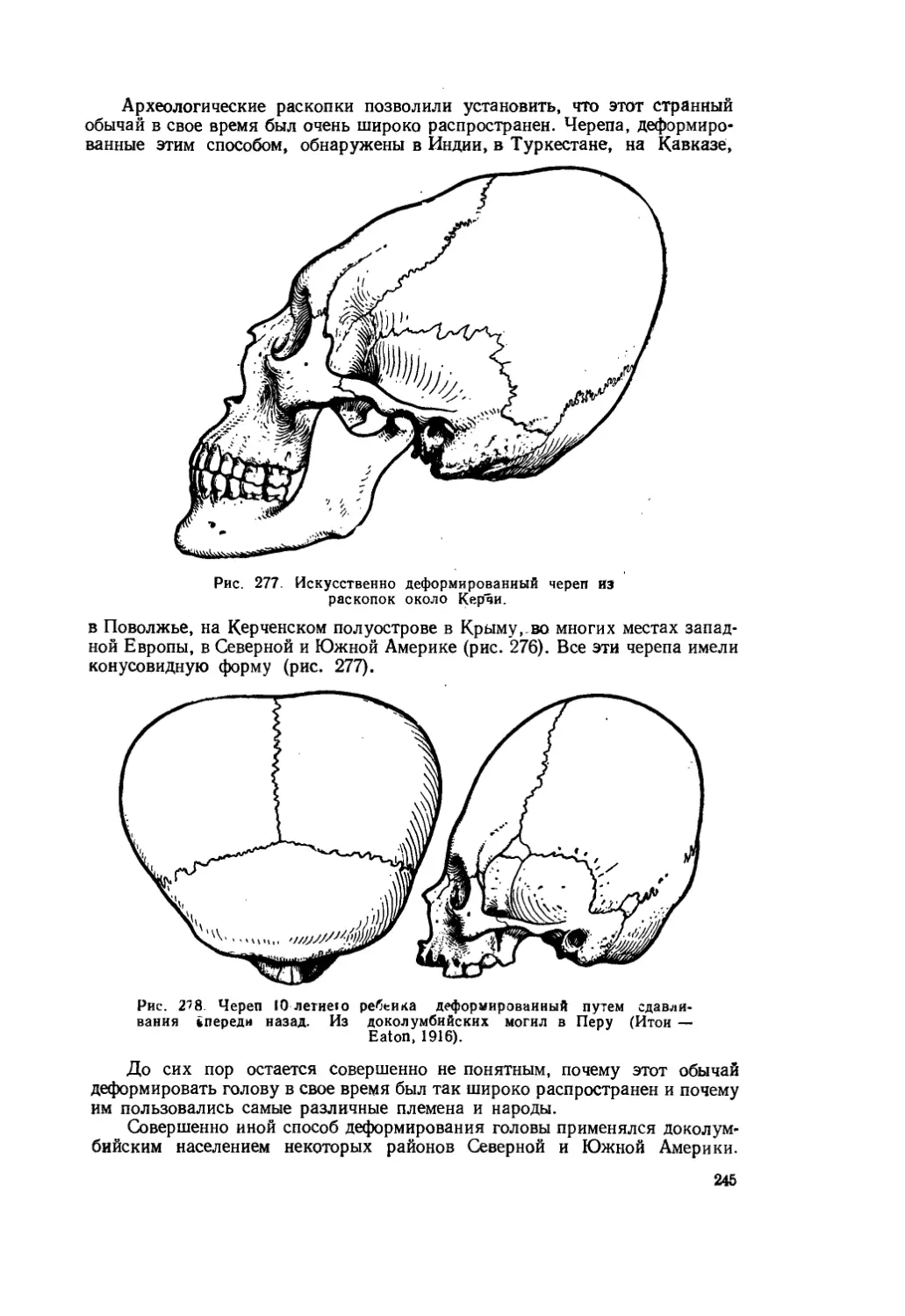
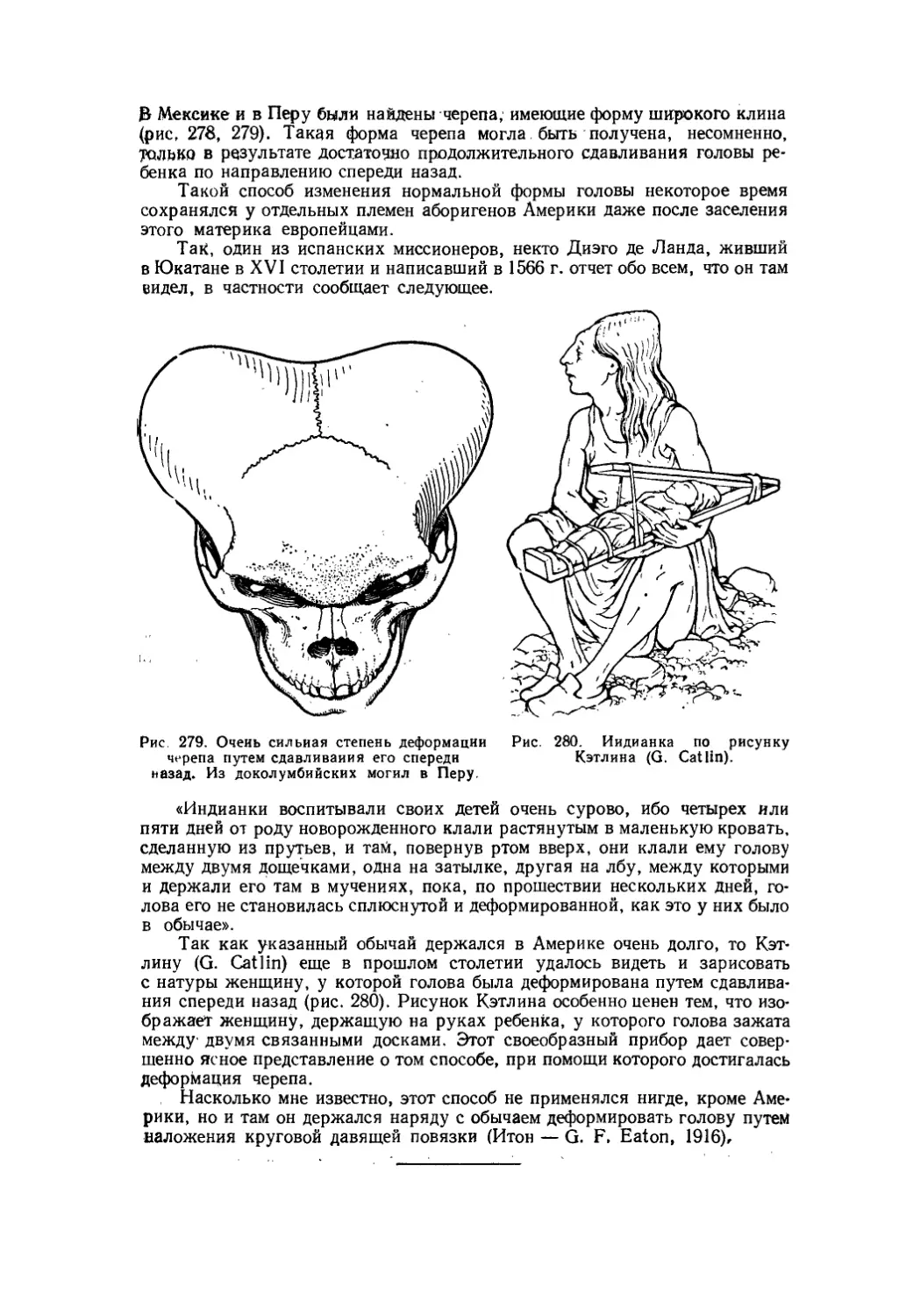
























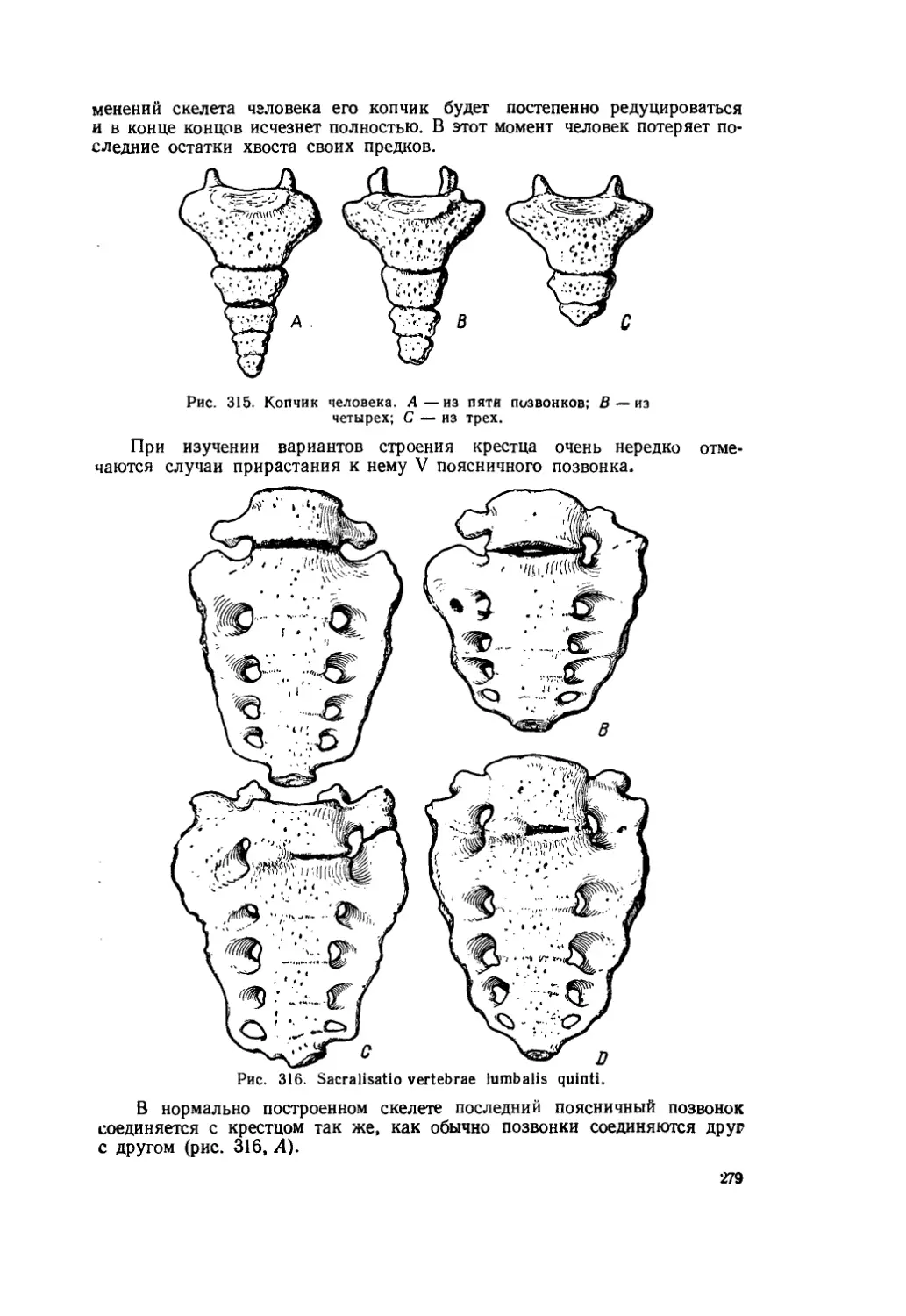
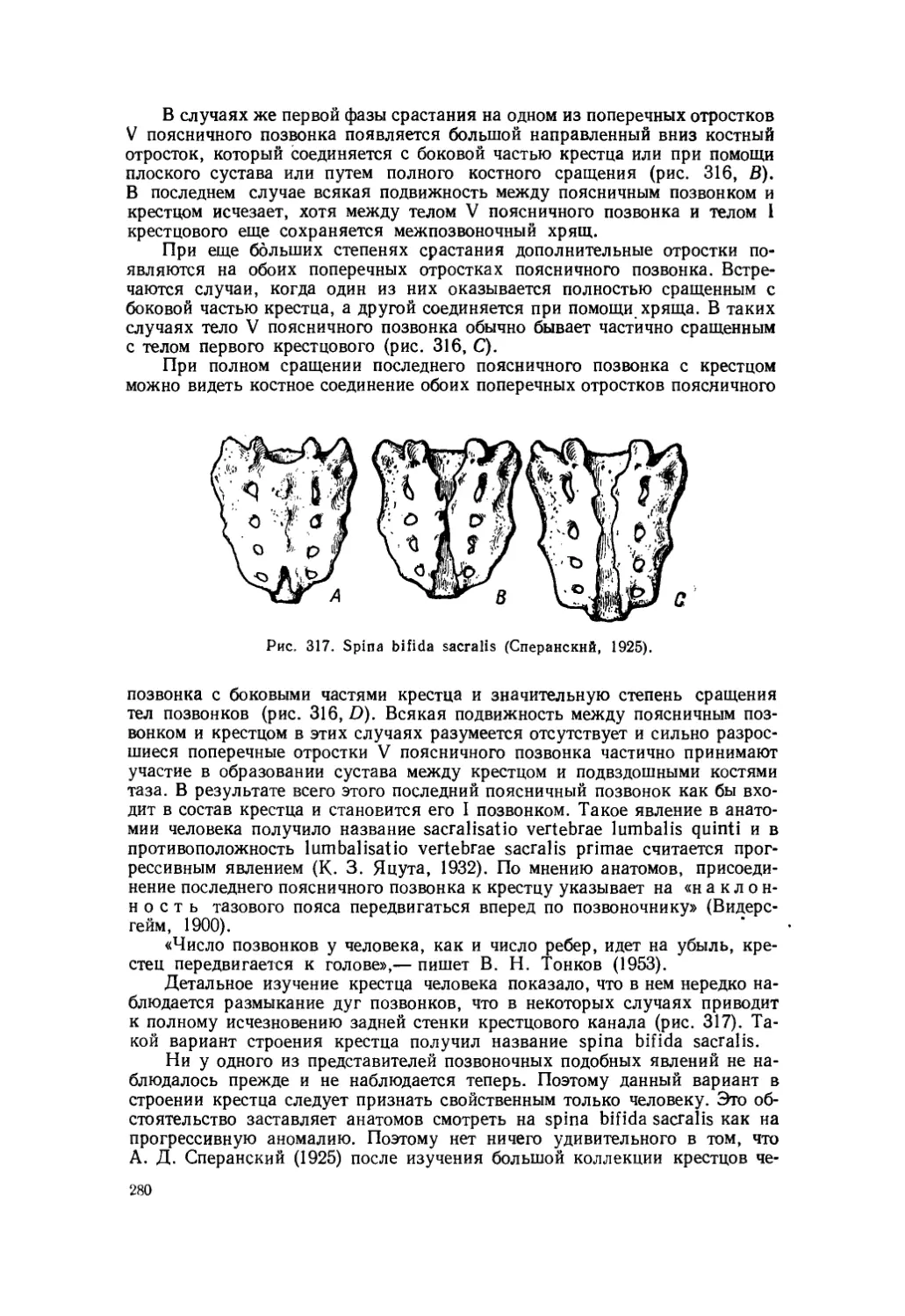
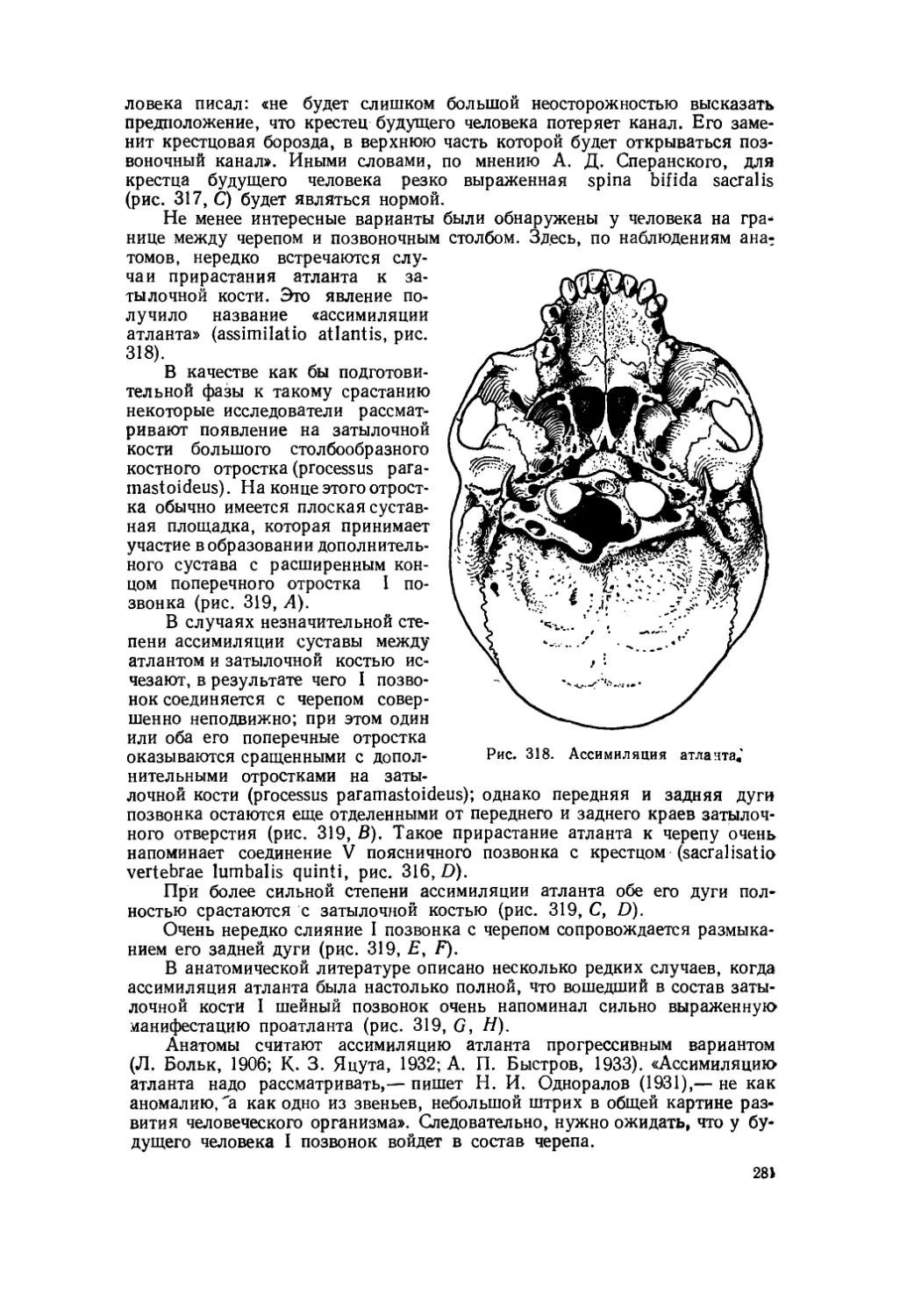
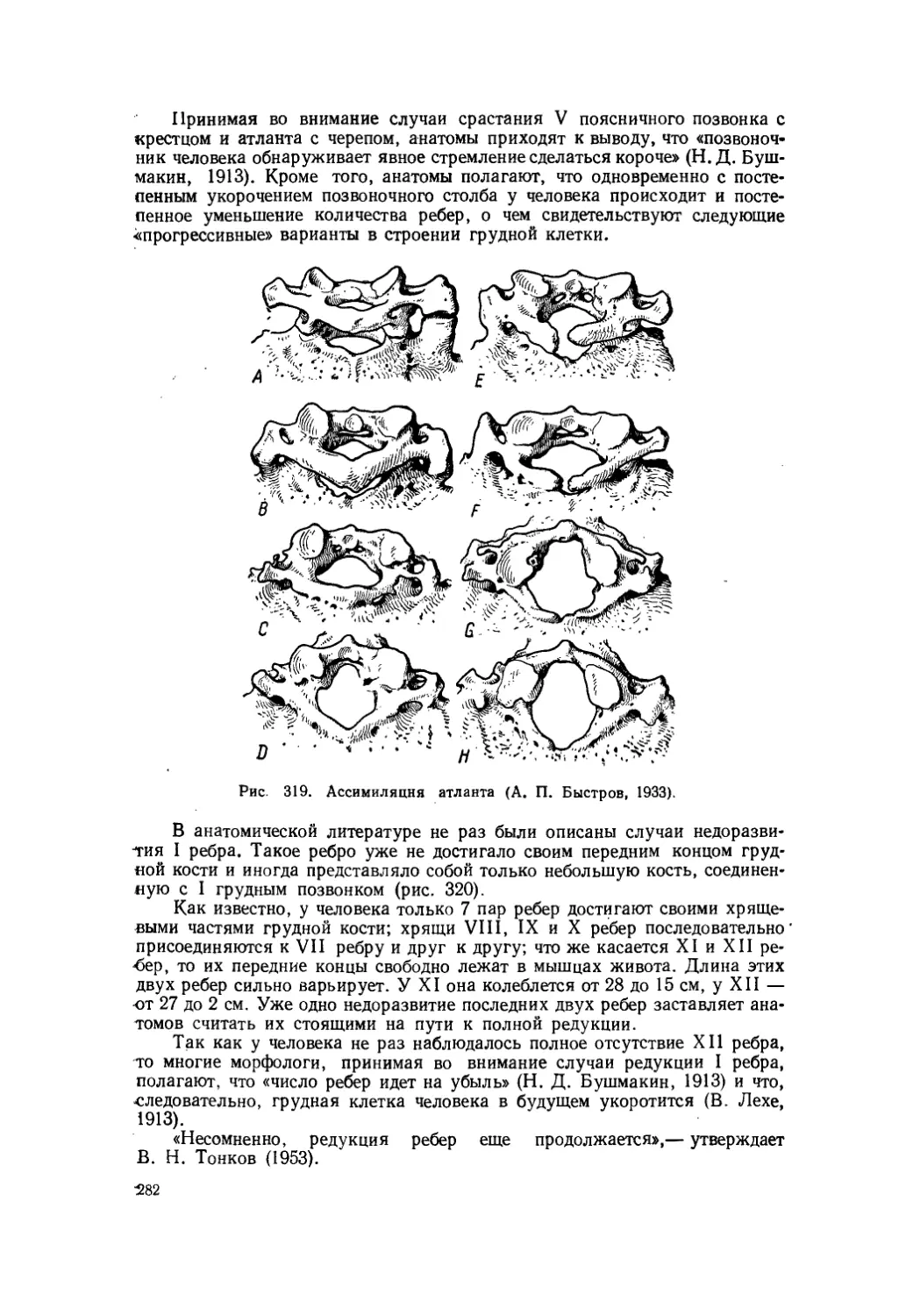




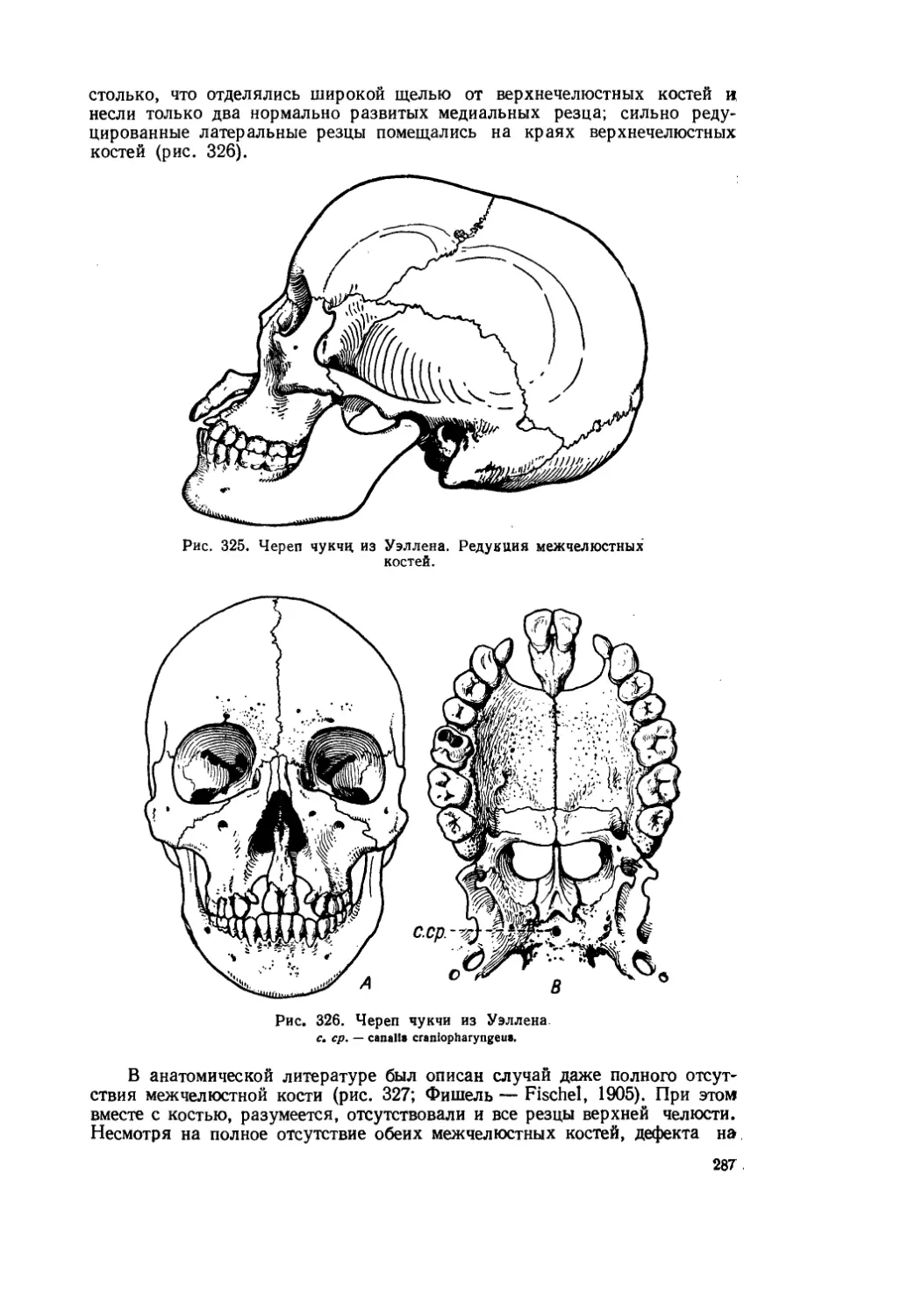



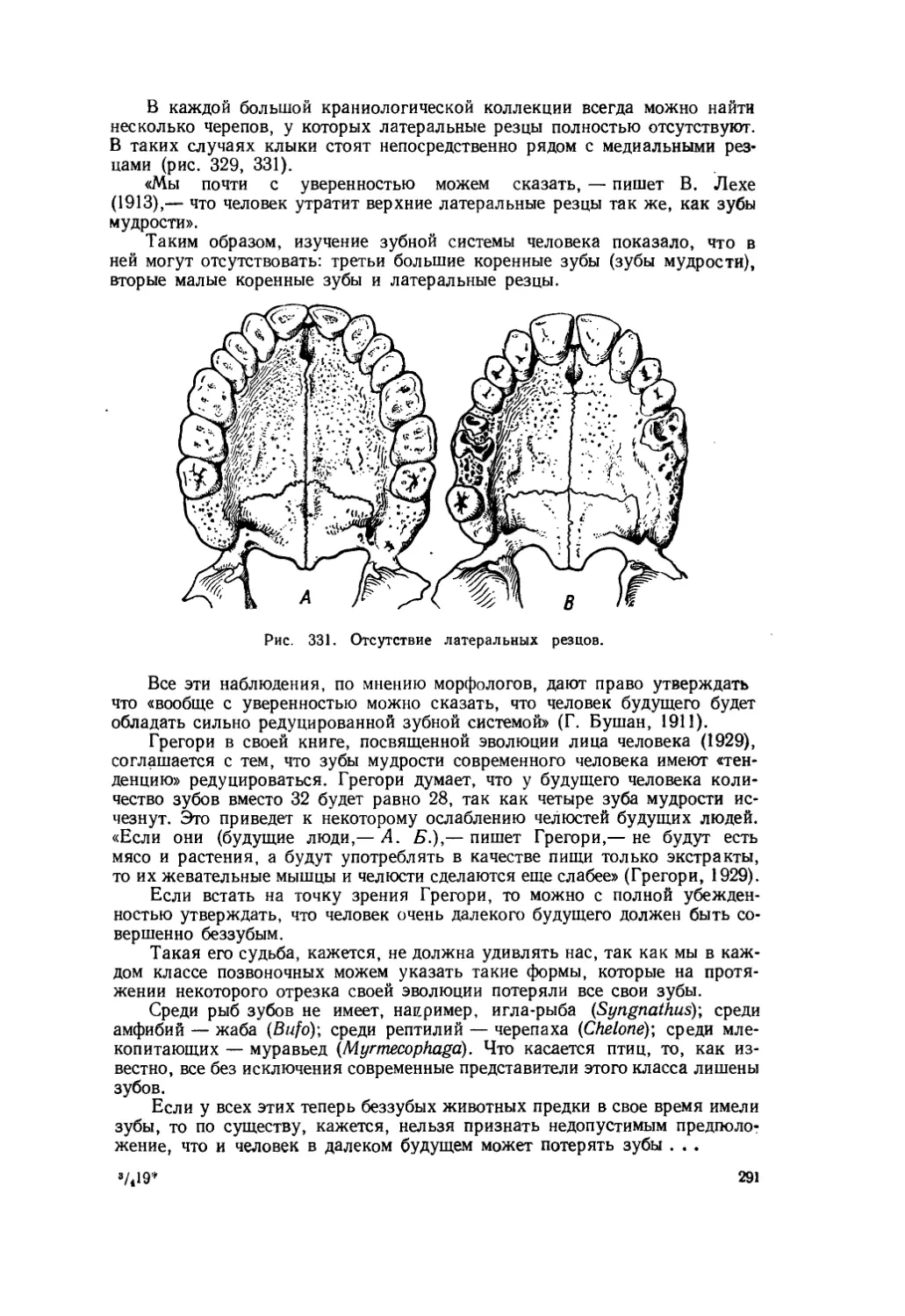




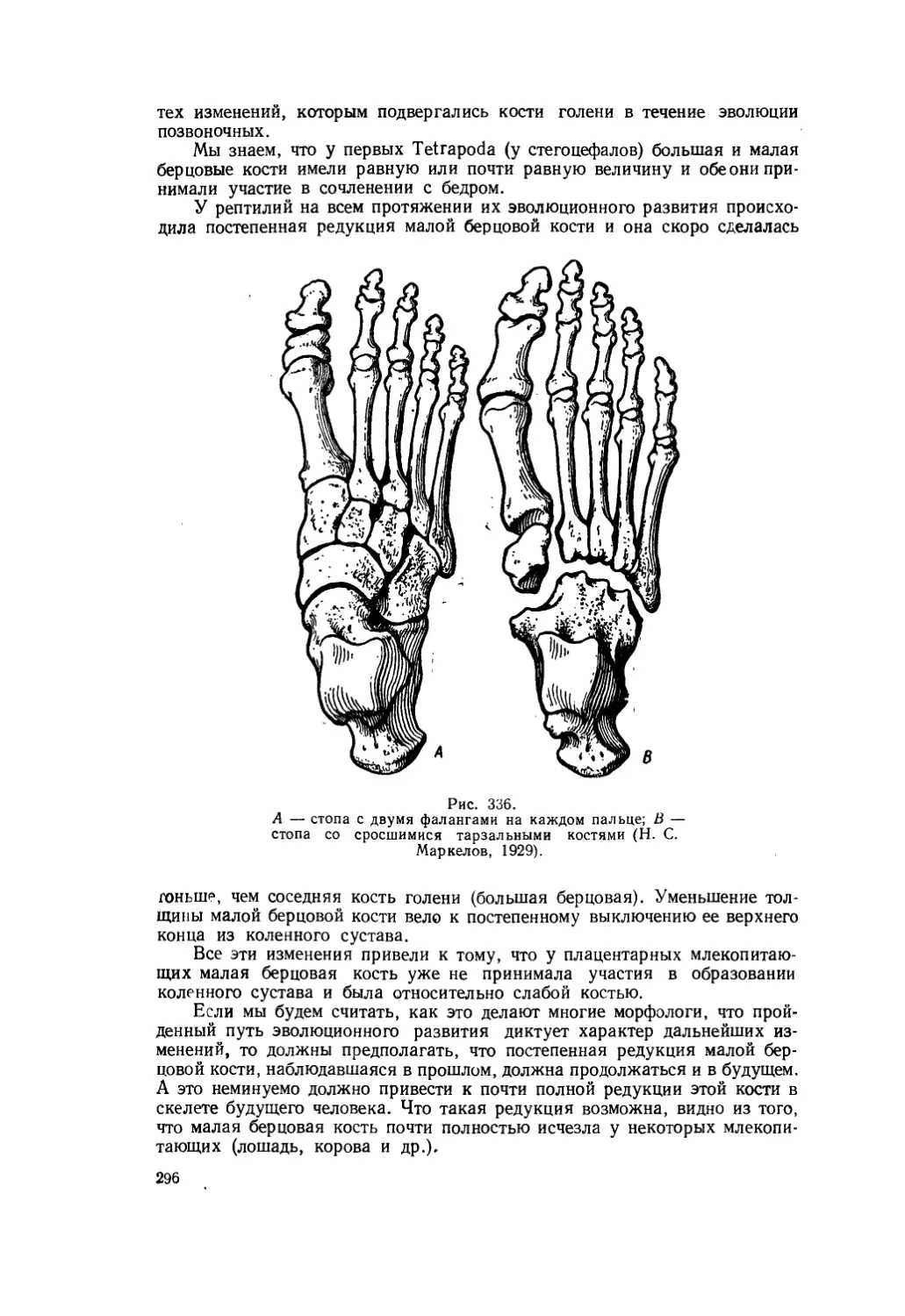







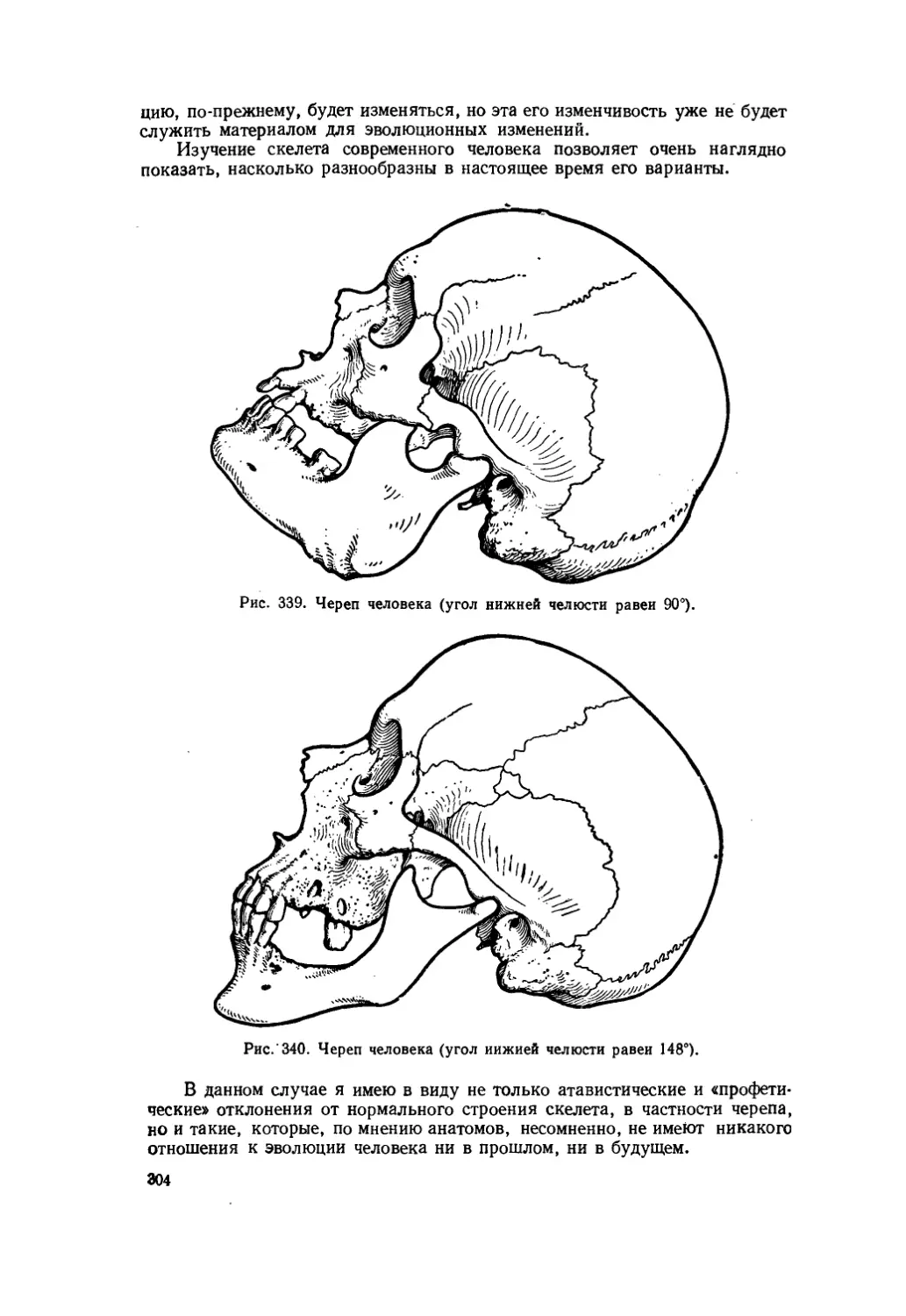










Текст
А. П. БЫСТРОВ
ПРОШЛОЕ, НАСТОЯЩЕЕ, БУДУЩЕЕ ЧЕЛОВЕКА
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО МЕДИЦИНСКОЙ ЛИТЕРАТУРЫ МЕДГИЗ
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ . 195?
ПРЕДИСЛОВИЕ
Я решил написать эту книгу после того, как мне несколько раз пришлось принять участие в горячих спорах по вопросам, касающимся эволюции человека.
Во время обсуждения прошлого человека у многих моих оппонентов иногда возникали некоторые сомнения; не все они соглашались с тем или иным объяснением некоторых явлений в эволюционной истории человека. Но их возражения обычно касались частных, а не общих вопросов. Большинство участников наших споров совершенно не сомневалось в происхождении человека от животных и признавало его близкое родство с обезьянами.
Настоящее человека обычно не вызывало каких-либо серьезных споров, хотя не все участники наших дружеских диспутов одинаково оценивали значение некоторых морфологических деталей в строении современного человека.
Что касается будущего человека, то обсуждение его всегда приводило к принципиальным разногласиям.
Никто из участников наших бесед не сомневался в том, что биологическая эволюция человека происходит и в настоящее время. При этом одни сторонники такого взгляда утверждали, что факторами этой эволюции являются те же причины, которые обусловливали филогенетические изменения человека в прошлом. Другие находили, что эти факторы в настоящее время должны быть в какой-то мере иными или даже совершенно иными.
Защитники такого взгляда делились на две группы.
Одни из них признавались, что не могут конкретно указать факторов» обусловливающих современную эволюцию человека, и считают их пока еще невыясненными.
Другие — смело утверждали, что человек в настоящее время эволюционирует под влиянием «био-социальных факторов», как они выражались. Несмотря на полную убежденность в своей правоте, сторонники такого взгляда не могли разъяснить нам, что они разумеют под этими новыми факторами эволюционного развития.
Все это приводило к тому, что будущее человека казалось нам гораздо1 более темным, чем его далекое прошлое.
В своей книге я попытался использовать имеющийся в настоящее время> морфологический материал для того, чтобы осветить прошлое, на -1* а
стоящее и будущее организма человека на основании изучения истории его скелета.
Я старался сделать свою книгу доступной по возможности широкому кругу читателей. Однако я не мог отказаться от довольно большого количества латинских анатомических терминов, так как они не всегда могли быть заменены русскими названиями. Кроме того, употребление специальных терминов оказалось совершенно неизбежным во всех тех случаях, когда мне приходилось говорить об ископаемых животных, так как ни одно из них никаких названий, кроме греческих и латинских, не носит. Наряду с этим, пытаясь проследить эволюционную историю различных элементов скелета, я во многих случаях был вынужден останавливаться на ряде таких анатомических деталей, которые неизвестны неподготовленному читателю.
Все это привело к тому, что моя книга оказалась требующей от чита-теля некоторой морфологической эрудиции.
Считаю необходимым подчеркнуть, что в этой книге идет речь только о формировании организма человека (точнее, только его скелета) в прошлом, о современном состоянии человека (собственно его скелета) и о предполагаемом строении человеческого скелета в будущем. В этой книге нет ничего о прошлом, настоящем и будущем человечества. Поэтому читатель в ней не найдет никаких сведений о первых этапах развития человеческого общества, о современном состоянии населения земного шара, о будущих судьбах человечества и его культуры. Все эти интересные и важные вопросы, разумеется, не могут решаться на основе морфологических данных-
ВВЕДЕНИЕ
1
Как известно, слово «эволюция» происходит от латинского глагола evolvo (evolvi, e vol ut um, evolvere) — раскатываю, развертываю. Но для понимания этого термина одного его перевода с латинского языка недостаточно, так как в понятие «эволюция» вкладывается значительно больше содержания, чем имеется в указанном латинском глаголе.
Дать определение понятия «эволюция» в немногих словах нелегко.1
Эволюция — это не простое «развертывание» чего-то уже имеющегося; она сопровождается появлением того, чего прежде не было.
Эволюция — это длинный ряд последовательных изменений, которые приводят к так называемому филогенетическому развитию живых организмов, или к филогенезу (®oXov — phylon — род, племя; -feveai? — genesis — происхождение).
Эволюция — это процесс изменений, сам по себе не свойственный живым организмам, но связанный с определенными их свойствами: способностью к изменчивости и способностью передавать свои морфологические и физиологические особенности по наследству. Эти свойства приводят к эволюции только при наличии определенных условий. При отсутствии этих условий они сами по себе не могут вызвать эволюционных изменений. '
Эволюция — это процесс не направленный, так как эволюционные изменения живых организмов могут совершаться в любых направлениях. Поэтому все и всякие эволюционные изменения никогда и ничем не бывают предрешены и никогда и никем не могут быть предсказаны. Однако все они и всегда оказываются обусловленными рядом причин и никогданебывают самопроизвольными (спонтанными). Evolutiospontanea abes t— самопроизвольной эволюции не существует.
Эволюция — это процесс необратимый, так как всякий организм, достигнув той или иной ступени своего развития, уже не может вновь вернуться к своему прежнему состоянию. Evolutio irreversa е s t — эволюция необратима. Это приводит к тому, что никакой организм не может быть предком такой новой формы, которая была бы более примитивной, чем он сам.
Во всех этих определениях мы не один раз называли эволюцию живых организмов развитием и изменением. Однако эволюция, развитие и изменение— не одно и то же.
1 Я считаю, что «эволюция» — понятие биологическое; это понятие выработалось при изучении живой природы. Использование термина «эволюция» в других дисциплинах я считаю незаконным. Если астрономы по недоразумению говорят об эволюции звезд, то это не значит, что она действительно происходит.
а
Эволюция есть изменение, но не всякое изменение является эволюцией, так как мы не имеем оснований называть эволюцией, например, старческие изменения живых организмов.
Эволюция — это развитие, но не всякое развитие — эволюция, так как мы не можем считать эволюцией, например, развитие зародыша или развитие болезненного процесса.
Развитие сопровождается изменениями, но изменения не обязательно являются развитием. Изменения культурных растений и домашних животных под влиянием искусственного отбора, применяемого человеком, — не развитие и не эволюция. Поэтому все те изменения, которые получает человек у находящихся в его распоряжении животных и растений, нельзя считать результатом «направленного овладения эволюцией», как думают некоторые.
Таким образом, не всякие изменения и не каждое развитие есть эволюция. Однако эволюция представляет собой изменения живых организмов и является процессом их филогенетического развития.
Факторами, обусловливающими эволюцию, как известно, являются: наследственность, изменчивость, борьба за существование и естественный отбор.
Изучение роли каждого из этих факторов представляет собой очень нелегкую задачу, так как в процессе эволюции все эти факторы действуют не последовательно один за другим, а одновременно. Поэтому приходится намеренно и искусственно расчленять единый эволюционный процесс.
До появления первого живого вещества на нашей планете эволюционного процесса на ней не происходило, так как никакие изменения тел мертвой природы не могут быть названы эволюцией.
Живое вещество, возникшее в мертвой природе, конечно, могло состоять только из неживых элементов. Но живые организмы не являются простым скоплением неживых частиц. Если бы они представляли собой только скопление мертвых элементов, то нам не пришлось бы говорить о возникновении живого. Мы говорим о живом веществе только потому, что оно, состоя из неживых элементов, отличается от тех мертвых вещей, в состав которых входят те же химические элементы. Таким образом, живое вещество, очевидно, может быть названо живой комбинацией неживых частиц.
После своего появления живые организмы прошли очень длинный путь эволюционного развития, прежде чем на Земле появились первые представители позвоночных.
Так как человек относится именно к этой группе животных, то мы при разборе основных движущих сил эволюционного процесса будем иметь в виду главным образом позвоночных.
Уже давно установлено, что всякое позвоночное развивается из половых клеток его родителей. Ни эпителиальные, ни мышечные, ни нервные, ни соединительнотканные клетки позвоночных никогда не служат исходным материалом для развития нового поколения. Это обстоятельство дает нам основание не сомневаться в том, что все особенности родителей в тех случаях, когда они в той или иной степени обнаруживаются у их детей, передаются по наследству только через половые клетки, так как только эти клетки преемственно связывают следующие друг за другом поколения.
Несмотря на то, что дети в силу наследственности бывают сходны со своими родителями, никакие особенности тела родителей, приобретенные ими в течение индивидуальной жизни, не могут передаваться по наследству.
Это верно, по крайней мере, по отношению к животным, о которых идет .речь в настоящей книге.
6
Совершенно невозможно представить себе, каким образом вызванные непосредственными воздействиями внешней среды особенности тела родителей могли бы отразиться на их половых клетках и вызвать в них такие изменения, которые обусловили бы появление у потомков подобных же особенностей.
Несмотря на всю невероятность подобных явлений, признание наследственной передачи приобретенных признаков было основным положением теории Ламарка.
Ламарк (1744—1829) учил (1809), что изменения особенностей окружающей природы всегда вызывают у животных появление новых привычек; новые привычки и, следовательно, новое поведение животных в свою очередь обусловливают упражнение или, наоборот, неупотребление некоторых органов; продолжительное употребление любого органа приводит к его усилению, неупотребление, наоборот, — к постепенной редукции.
По мнению Ламарка, результаты упражнения или неупражнения органов родителей всегда передаются их потомству. Без этого утверждения теория Ламарка ничего не смогла бы «объяснить».
Несостоятельность теоретических взглядов Ламарка становится совершенно очевидной при изучении эволюционной истории любого органа любого животного. Например, в процессе филогенетического формирования птиц происходило постепенное развитие грудных мышц. Это развитие выражалось в увеличении количества мышечных волокон, в результате чего мышцы, связанные с летательным аппаратом, становились все более и более мощными.
Все это не могло быть вызвано упражнением мышц и передачей по наследству результатов этого упражнения, так как никакое упражнение не способно вызвать увеличения количества поперечнополосатых мышечных волокон: эти волокна у взрослых животных уже не могут размножаться путем деления.
Однако известно, что упражнение мышц всегда приводит к их увеличению: каждая из них в таком случае становится толще и сильнее. Но это увеличение размеров и силы мышцы объясняется только утолщением входящих в ее состав мышечных волокон, что обусловливается увеличением в них количества саркоплазмы и количества миофибриллей.
Таким образом, если усиленная функция не способна вызвать в мышце увеличение количества мышечных волокон, то, очевидно, упражнение не могло играть никакой роли в филогенетическом развитии летательного аппарата птиц.
Ламарк пытался объяснить отсутствие зубов у муравьеда привычкой не жевать пищу. Но неупотребление зубов не может вызвать в них каких-либо изменений.
Известно, что эмаль и дентин не способны ни восстанавливаться после разрушений, ни срастаться после переломов зубов. Поэтому всякое усиленное употребление (упражнение) зубов всегда приводит только к одному неизбежному результату — к непоправимому стиранию их коронок. Если бы такие результаты упражнения передавались по наследству, то это давно бы привело к тому, что все позвоночные были бы беззубыми. . .
Таким образом, если теоретические положения Ламарка считать правильными, то следует признать, что не привычка не жевать пищу, а, наоборот, усиленное употребление зубов привело к тому, что муравьеды стали беззубыми. Но в таком случае становится совершенно непонятным, почему то же самое не случилось со многими другими млекопитающими.
Так как зубы не могут изменять свою форму под действием упражнения или неупражнения, то мы не можем объяснить с позиций Ламарка их эволю
7
ционные изменения и поразительное соответствие их формы всем особен? ностям пищи, которой питаются различные позвоночные.
Ламаркизм немыслим без признания возможности передачи по наследству приобретенных особенностей. Что же касается современной эволюционной теории, то, по моему мнению, она вообще не нуждается в учении о наследовании приобретенных признаков, так как для нее достаточно одного признания возможности возникновения у животных в силу их постоянной изменчивости разнообразных наследующихся особенностей. Таким образом, вместо наследования приобретенного признака необходимо приобретение наследуемого признака.
Интересно, что при развитии зародышей животных их половые клетки обособляются от других клеток тела очень рано. У некоторых животных первую половую клетку можно видеть уже у зародыша, состоящего только из 32 клеток. Это явление было обнаружено, например, у Sagitta.
При наличии непрерывного метаболизма структура всяких клеток, в том числе и половых, очевидно, не может оставаться неизмененной. Иными словами, обмен веществ должен вызывать непрерывные изменения в структуре клеток. Поэтому половые клетки не могут быть неизменными. Но всякие изменения в половых клетках неминуемо должны отразиться на строении того организма, который разовьется из них.
Таким образом, следует признать, что метаболизм является не только основной, но и единственной причиной изменений в структуре половых клеток. И именно эти изменения приводят к возникновению у взрослых животных тех или иных наследуемых особенностей.
Все живое не может даже короткое время оставаться абсолютно неизменным. Это обстоятельство делает понятным, почему дети одних и тех же родителей при всем их семейном сходстве никогда не бывают совершенно одинаковыми. Семейное сходство детей является следствием того, что они происходят от одних и тех же родителей, а индивидуальные различия этих детей представляют собой результат метаболизма в половых клетках их родителей. В данном случае я не имею в виду тех не передающихся по наследству особенностей, которые могут возникнуть у живых существ в течение их жизни в результате воздействия тех или иных внешних причин.
Всякий новый признак, вызванный метаболизмом в половых клетках родителей, разумеется, является наследуемым, но он в то же время подвергается изменению у представителей нового поколения в результате обмена веществ в их собственных половых клетках.
Таким образом, метаболизм обусловливает и появление нового признака, и его изменение. В результате этого всякий наследуемый признак никогда не бывает неизменным. При этом следует иметь в виду, что только эти признаки имеют значение для эволюционного процесса. Все и всякие ненаследуемые изменения живых существ в их эволюционном развитии никакой роли не играют.
Все это приводит нас к заключению, что постоянные изменения в структуре половых клеток, обусловливающие появление новых наследуемых особенностей, являются необходимым условием эволюционного процесса. Однако все изменения в половых клетках, обусловленные протекающим в них обменом веществ, все же не приводят за короткое время к коренной перестройке всей структуры половых клеток.
Если бы не было такой относительной стабильности структуры половых клеток, то существование каких бы то ни было таксономических единиц было бы совершенно невозможно, так как это привело бы к исчезновению даже семейного сходства.
Отсутствие некоторой стабильности в структуре половых клеток сделало бы эволюционный процесс невозможным, так как приводило бы к по-1,
8
явлению во всяком новом поколении таких животных, которые были бы совершенно непохожи на своих непосредственных предков. Это, разумеется, сразу уничтожало бы все ценные особенности, приобретенные в течение предыдущего развития.
Таким образом, некоторая стабильность структуры половых клеток животных является совершенно необходимым условием эволюции. Без этой стабильности никакое эволюционное развитие немыслимо. Но эволюционное развитие невозможно и без постоянных изменений в структуре половых клеток.
Вся эволюционная история позвоночных была историей изменений стабильности структуры их половых клеток. Эти клетки оставались неизменными постольку, поскольку позвоночные оставались позвоночными, но они одновременно с этим и изменялись постольку, поскольку в процессе эволюции позвоночных появляются новые их представители.
Допустим, что под влиянием окружающей природы у какого-нибудь животного возникли некоторые новые морфологические особенности. Чтобы эти изменения приобрели значение для эволюции, необходимо, чтобы они были переданы потомству. Как известно, эта передача может быть осуществлена только через половые клетки путем соответствующих их изменений. Можно предположить, что изменения в теле животного вызовут изменения в его общем обмене веществ, а это неминуемо отразится и на метаболизме в половых клетках и приведет к структурным изменениям в них. Эти структурные изменения, несомненно, обусловят появление новых признаков у потомства, но эти признаки не могут быть тождественными новым признакам у родителей. Следовательно, воздействие факторов внешней среды на животных не может вызвать и никогда не вызывает у них такие изменения, которые являлись бы прямым эволюционным приспособлением к вызвавшим их особенностям мертвой природы.
II
В течение почти столетия после опубликования книги Ч. Дарвина о происхождении видов (1859) в биологической литературе накопилось очень много разнообразных и интересных данных, указывающих на то, что и животные, и растения производят потомства несравненно больше, чем его выживает.
Так, подсчитано, что один экземпляр инфузории Paramaecium, делящейся в среднем один раз в сутки, при беспрепятственном размножении и достаточном питании в течение 4 месяцев мог бы дать потомство в 4- 10за экземпляров. Общий объем такой массы инфузорий был бы равен объему земного шара.
Подсчитано, что в плодовом теле гриба-дождевика (Lycoperdon gigan-teum), иногда достигающем размеров арбуза, образуется 7,5 триллионов спор. Если бы каждая спора такого гриба могла прорасти и дать подобное же плодовое тело, а затем каждый из этих 7,5 триллионов грибов вновь образовал бы соответствующее количество спор, то грибы, развившиеся из спор только второго поколения, по своему общему объему оказались бы больше земного шара в 800 раз.
Подсчитано, что одуванчик при беспрепятственном размножении через 10 лет мог бы дать такое количество экземпляров, которое смогло бы сплошным ковром покрыть площадь, в 15 раз превышающую поверхность всей суши земного шара.
Потомство одной мухи в течение 4 месяцев могло бы достигнуть 4- 10*t особей. Такое количество мух покрыло бы площадь, равную 2000 км8.
»
Потомство одной пары полевых мышей при условии беспрепятственного размножения и достаточного количества пищи могло бы в течение 2 лет достигнуть 14 000 особей.
Однако в обычных условиях ни одно живоё существо не имеет возможности размножаться с такой быстротой, на которую оно способно, так как огромное количество живых существ обычно погибает. Поэтому все цифры, показывающие большую интенсивность размножения, являются доказательством наличия в природе беспощадной борьбы за существование, которая охватывает все живое. В такой борьбе за жизнь уничтожается огромное количество растений и животных. При этом легче погибают те формы, которые по тем или иным особенностям своего строения оказываются менее «удачными», чем их конкуренты. А это приводит к так называемому естественному отбору.
Быть менее «удачно» организованным — это значит иметь такие морфологические и физиологические особенности, которые в определенных условиях не дают преимуществ в борьбе за жизнь. Однако следует подчеркнуть, что ценность всех и всяких морфологических и физиологических особенностей любого организма всегда относительна. Если в одних условиях некоторые морфо-физиологические особенности какого-нибудь животного оказываются в процессе борьбы за жизнь, скажем, полезными, то в других условиях эти же особенности могут оказаться в равной мере вредными.
Наряду с разнообразными относительно полезными и относительно вредными особенностями все живые существа обладают одним общим, безусловно полезным для них свойством — свойством производить потомство в количестве, всегда превышающем число родительских форм. Это полезное свойство является одной из непосредственных причин борьбы за существование и в то же время представляет собой основное средство, при помощи которого ведется борьба за сохранение вида. Таким образом, быстрое размножение одновременно и вызывает борьбу за жизнь и является одним из способов борьбы за существование.
Некоторые биологи различают несколько видов борьбы за существование и говорят о межвидовой, внутривидовой и конституциональной борьбе за жизнь (о «борьбе» с мертвой природой). Я думаю, что в такой детализации нет большой нужды, так как в природе все виды борьбы настолько слиты между собой, что по существу становятся неразделимыми. В частности, межвидовая и конституциональная борьба проявляется главным образом через внутривидовую.
В жизни природы, несомненно, бывают такие моменты, когда все эти виды борьбы за существование действуют одновременно, но бывают и такие условия, при которых тот или иной вид борьбы становится особенно напряженным, в то время как другой значительно ослабевает.
Когда, например, млекопитающие впервые начали заселять Северный полярный океан, то они, разумеется, не боролись ни с водой, ни со льдами, ни с низкой температурой в буквальном смысле этого слова. Но вся сумма особенностей полярного океана производила жесточайший отбор среди проникающих в его воды мигрантов. Конечно, полярные условия были способны уничтожить неограниченное количество живых существ, в то время как сам полярный океан и его льды и снега не могли быть уничтожены никакими мигрантами. Однако не только полярные условия производили беспощадный отбор среди мигрантов, но и сами мигранты вели жестокую борьбу за существование друг с другом в условиях полярного океана. При этом, несомненно, многие животные вымерли, но некоторые в результате длительного естественного отбора приобрели все те особенности, которые позволяли им жить в своеобразных условиях Крайнего Севера.
10
Миграция тех же животных в иной ареал, например в жаркую пустыню, несомненно, привела бы к другим результатам. Возможно, что в этом случае победителями в борьбе за существование оказались бы не те виды, которые приспособились к жизни на Крайнем Севере. Но в обоих этих случаях длительный естественный отбор привел бы к возникновению таких животных, которые были бы удивительно приспособлены к жизни в заселенном ими ареале.
Таким образом, борьба за жизнь между многочисленными и разнообразными представителями флоры и фауны происходит на фоне различных условий, созданных мертвой природой и при непосредственном воздействии со стороны сил самой мертвой природы. При этом жестокая борьба за существование и естественный отбор в конце концов приводят к тому, что вся масса животных и растений оказывается в какой-то степени приспособленной и к особенностям мертвой природы данного ареала, и к совместному существованию в нем. Это приводит к возникновению так называемого биоценоза (pioc — bios — жизнь; xotvoc — coenos — община).
Все члены любого биоценоза всегда оказываются в достаточной степени приспособленными к совместному существованию и к особенностям мертвой природы. Следует подчеркнуть, что эта приспособленность никогда не бывает и не может быть абсолютной, так как непрерывная изменчивость животных и растений постоянно приводит к появлению новых форм. А это вызывает нарушение установившейся гармонии отношений. Впрочем, установившейся «гармонии отношений» в биоценозах по существу никогда не бывает. Все они находятся как бы в неустойчивом равновесии Непрерывное появление новых особенностей у различных членов биоценозов постоянно меняет их отношения и характер борьбы за существование. При этом естественный отбор, уничтожая менее приспособленных к данной конкретной обстановке и в данное время, приводит к эволюционным изменениям членов биоценоза.
Поэтому можно сказать, что творцом всех живых существ на нашей планете является смерть — Mors creator vitae est (творцом жизни является смерть).
Для того чтобы понять ход эволюционного процесса в каком-нибудь ареале, необходимо изучить все детали взаимоотношений между всеми без исключения живыми существами, его населяющими.
Учесть и изучить влияние одних животных на других, учесть и изучить влияние друг на друга различных растений, учесть и изучить влияние разных представителей флоры на представителей фауны и наоборот — задача, разумеется, необычайно трудная. Следует иметь в виду, что в каждом ареале находится очень большое количество разнообразных видов живых существ, если принять во внимание и те из них, размеры которых микроскопически малы. Не учитывать этих членов биоценозов нельзя потому, что и они играют в жизни и в эволюции органического мира совсем не малую роль.
Если мы предположим, что эта сложная задача будет нами как-то разрешена, то все же вопрос о взаимоотношениях между членами изучаемого биоценоза останется по-прежнему невыясненным, так как одних этих знаний недостаточно. Ясное представление о видовом составе населения данного ареала позволит нам выяснить только характер взаимоотношений, обусловливающих различные формы борьбы за существование между членами биоценоза, но не больше. А для понимания хода эволюционного процесса совершенно необходимо ясное представление и о степени напряженности борьбы за жизнь.
Острота или напряженность борьбы за существование не обусловливается только одним тем фактом, что те или иные живые существа так или иначе вытесняются другими представителями биоценоза. Напряженность
II
борьбы зависит главным образом от количества индивидуумов, находящихся в состоянии борьбы друг с другом. При этом не следует забывать, что количественное отношение между многочисленными членами любого биоценоза никогда не остается стабильным. Оно постоянно меняется вследствие непрерывных изменений числа представителей тех или иных живых форм, входящих в состав биоценоза.
Все это должно быть учтено с достаточной точностью и полнотой. Только в этом случае мы получим возможность выяснить те условия, в которых протекает эволюция данной группы живых организмов.
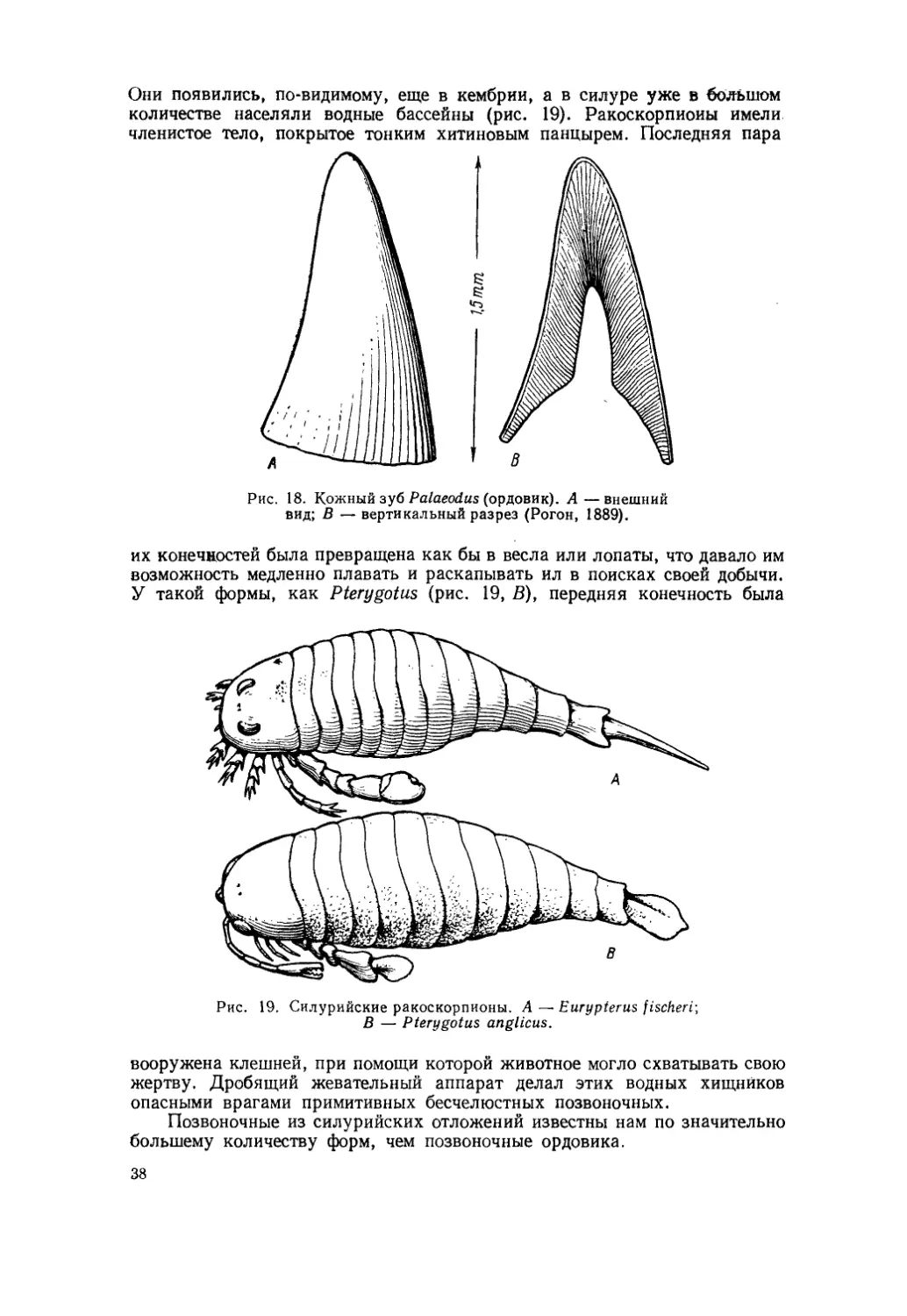
Рис. 1. Alligator mississipplensis и Trionyx ferox.
Следует признать, что такая задача необычайно трудна и, пожалуй, практически неразрешима. Поэтому в настоящее время мы можем составить себе некоторое представление о всей сложности взаимоотношений между членами биоценоза только на основании чисто теоретических соображений, основанных на отрывочных наблюдениях.
Крайне сложные и запутанные взаимоотношения в биоценозах и их роль в эволюционном процессе легче всего понять при помощи упрощенной схемы биоценоза, в котором количество видов животных намеренно и искусственно уменьшено до минимума.
Известно, например, что североамериканский крокодил, Alligator mississipplensis, преследует и уничтожает черепаху Trionyx ferox, но также установлено, что Trionyx истребляет молодых аллигаторов. Таким образом, взаимоотношения между этими двумя рептилиями образуют как бы замкнутый круг, так как они преследуют и истребляют друг друга (рис. 1).
Trionyx, уничтожая молодых аллигаторов, наносит вред этим рептилиям и приносит себе пользу, так как спасает самого себя от будущих врагов. При этом уменьшение числа крокодилов, естественно, приводит к увеличению количества черепах. Но размножение Trionyx не является для него абсолютно полезным, так как среди представителей этого вида 12
черепах тотчас же возникают некоторые затруднения в добывании пищи. Таким образом, истребляя потомство своих врагов, черепахи одновременно с пользой приносят себе и вред. Следовательно, взрослые крокодилы, истребляя Trionyx, с известной точки зрения становятся не врагами, а друзьями черепах, точнее, будучи врагами, приносят им пользу. Но истребление Trionyx полезно и самим аллигаторам, так как, уничтожая черепах, они не только питаются, но и спасают от гибели свое собственное потомство. Но. . . при этом увеличивается количество крокодилов и уменьшается запас пищи,- что неминуемо влечет за собой обострение борьбы за жизнь и гибель многих аллигаторов. В результате этого польза становится вредом.
При таком непрерывном взаимном истреблении всякая польза для крокодилов одновременно является вредом для них и пользой для черепах и наоборот. Иными словами, польза и вред в этом случае неразличимы, или: польза вредна, а вред полезен. Следовательно, польза и вред в биоценозе — это одно и то же. . . Во всяком случае они являются понятиями совершенно относительными.
Такая борьба за существование, при которой и польза и вред становятся понятиями относительными, обусловливает среди аллигаторов и черепах жестокий естественный отбор. А так как у всех живых организмов в силу изменчивости постоянно появляются новые наследующиеся особенности, то этот отбор неминуемо приведет к появлению новых форм и среди крокодилов и среди черепах. Эти новые формы будут отличаться от своих предшественников самыми разнообразными особенностями, которые могут в некоторых случаях давать им преимущества перед их конкурентами. А это всякий раз будет вызывать новое обострение борьбы за жизнь.
Понятно, что возникновение в таких условиях всяких новых форм представляет собой последовательные этапы эволюционного процесса. Очевидно, что, непрерывно осуществляя взаимный отбор, эта пара форм рептилий не может не эволюционировать. Появление новых приспособлений то у крокодилов, то у черепах неминуемо и постоянно будет нарушать неустойчивое равновесие во взаимоотношениях между этими животными, а это повлечет за собой отбор, который будет закреплять новые приспособления, являющиеся новыми этапами эволюционного процесса.
Совершенно ясно, что до тех пор, пока эта пара животных форм будет существовать в одном ареале и их взаимоотношения останутся неизмененными, их эволюция не может остановиться.
Следует признать, что в природе таких изолированных эволюционирующих пар не существует. Дело в том, что Alligator mississippiensis питается не только Trionyx ferox и пищей для этих черепах служит не только беззащитное потомство крокодилов. И те и другие питаются, кроме того, и многочисленными видами рыб. Но всякое введение в сферу борьбы за жизнь новых живых организмов сразу сильно усложняет картину. Эго можно показать хотя бы на таком менее упрощенном примере (рис. 2).
Известно, что гадюки уничтожают мышей; ежи — гадюк и мышей; филин — ежей, гадюк и мышей. Но указанные животные, конечно, не исчерпывают биоценозов наших лесов. Филин, например, нападает, кроме того, на зайцев, истребляет многих птиц и нередко уничтожает летучих мышей; зайцы в свою очередь истребляют многие формы растений, птицы питаются их семенами и плодами, одновременно уничтожая большое количество насекомых, которыми питаются летучие мыши, и т. д. По существу мы не можем установить объема биоценоза до тех пор, пока не введем в него всех без исключения животных и все растительные формы данного ареала.
В любом биоценозе все его многочисленные и очень разнообразные члены всегда оказываются так или иначе связанными друг с другом. В ре
13
зультате этого каждый биоценоз представляет собой такую систему живых организмов, в которой имеет значение и состав флоры и фауны, и количество экземпляров каждого вида растений и животных.
Изменение количества экземпляров какого-либо вида, входящего в состав биоценоза, всегда в какой-то мере нарушает неустойчивое равновесие в сложных взаимоотношениях огромной массы организмов, образующих биоценоз.
Основной причиной таких нарушений равновесия является непрерывная изменчивость живых существ, в конце концов приводящая к тому,
Рис. 2. Схема связей между различными животными в биоценозе.
что в процессе естественного отбора среди представителей какого-либо вида появляются новые формы, которые оказываются несколько более приспособленными к окружающей обстановке, чем их непосредственные предшественники. А это вызывает немедленное нарушение неустойчивого равновесия в биоценозе. Борьба за существование и естественный отбор через некоторое время приводят к оформлению новых приспособлений и к появлению новых видов. А это в конце концов обусловливает и изменение всего состава биоценоза.
Все это создает такие условия, при которых члены биоценоза не эволюционировать не могут, так как эволюция является прямым следствием их совместного существования. При этом эволюция, представляющая собой непрерывную и последовательную смену живых форм, сопровождается уничтожением одних видов и заменой их другими. Иными словами, всякая эволюция неминуемо должна сопровождаться вымиранием. Если эволюция по ряду причин оказывается для живых организмов неизбежностью, то такой же неизбежностью следует признать и вымирание. Таким образом, вымирание и эволюция неотделимы друг от друга. Эволюция не может протекать без вымирания, и вымирание является непосредственным результатом эволюции.
14
Поэтому всякий вымерший вид следует рассматривать как вид, в свое время вытесненный из состава какого-то древнего биоценоза.
Многочисленные попытки биологов объяснить вымирание живых существ непосредственно теми или иными изменениями в мертвой природе нужно признать излишними.
В мертвой природе, конечно, наблюдаются такие явления, которые способны уничтожить не только огромное количество живых существ, но и целые биоценозы. Но если эти явления наступают быстро, то они приводят не к вымиранию, а к простому уничтожению живых форм. Если же они протекают медленно, то они могут и не ускорять вымирания, обусловленного самим эволюционным процессом.
Если эволюционные изменения постепенно меняют состав биоценоза, то ни один вид, являвшийся членом прежнего биоценоза, не может суще-
Рис. 3. Череп Mesenosaurus romeri Efr.
ствовать в условиях нового биоценоза. Такой вид не смог бы успешно бороться за жизнь с более совершенными членами нового биоценоза. Это обстоятельство делает понятным, почему эволюционный процесс является необратимым.
Эволюция оказалась бы обратимой только в том случае, если бы все без исключения члены биоценоза в своем эволюционном развитии одновременно и в одинаковой степени пошли бы назад, что привело бы к восстановлению прежнего биоценоза в целом. Так как такие явления для всех членов биоценоза невозможны, то и эволюция оказалась необратимой для каждого в отдельности члена биоценоза и, следовательно, для всех членов всяких биоценозов вообще.
Однако следует иметь в виду, что необратимость эволюции не является каким-то свойством живых организмов. Возвращение утраченного признака вообще возможно. И если старый признак в новой обстановке окажется более полезным, чем его отсутствие, то он может сохраняться достаточно долгое время.
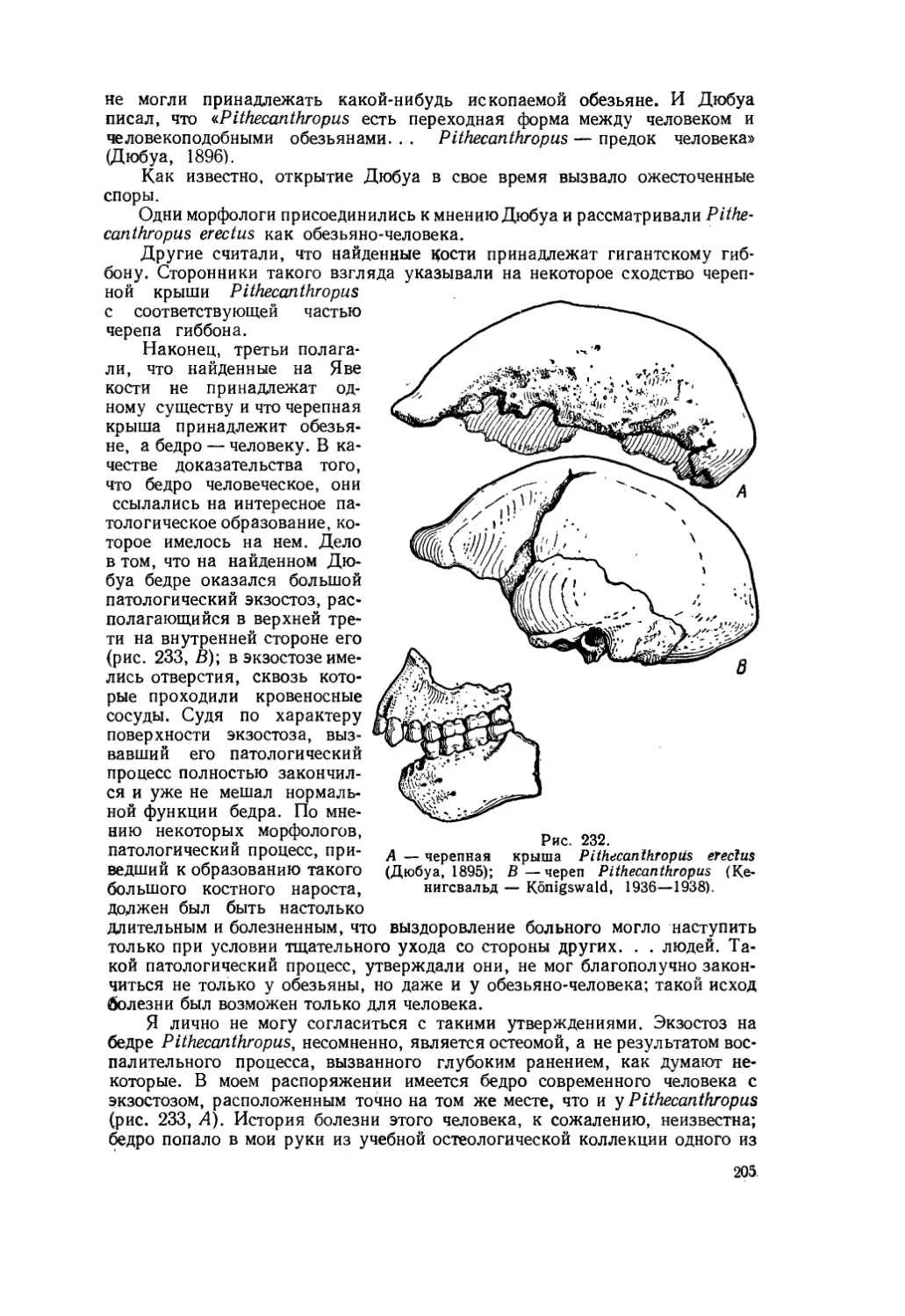
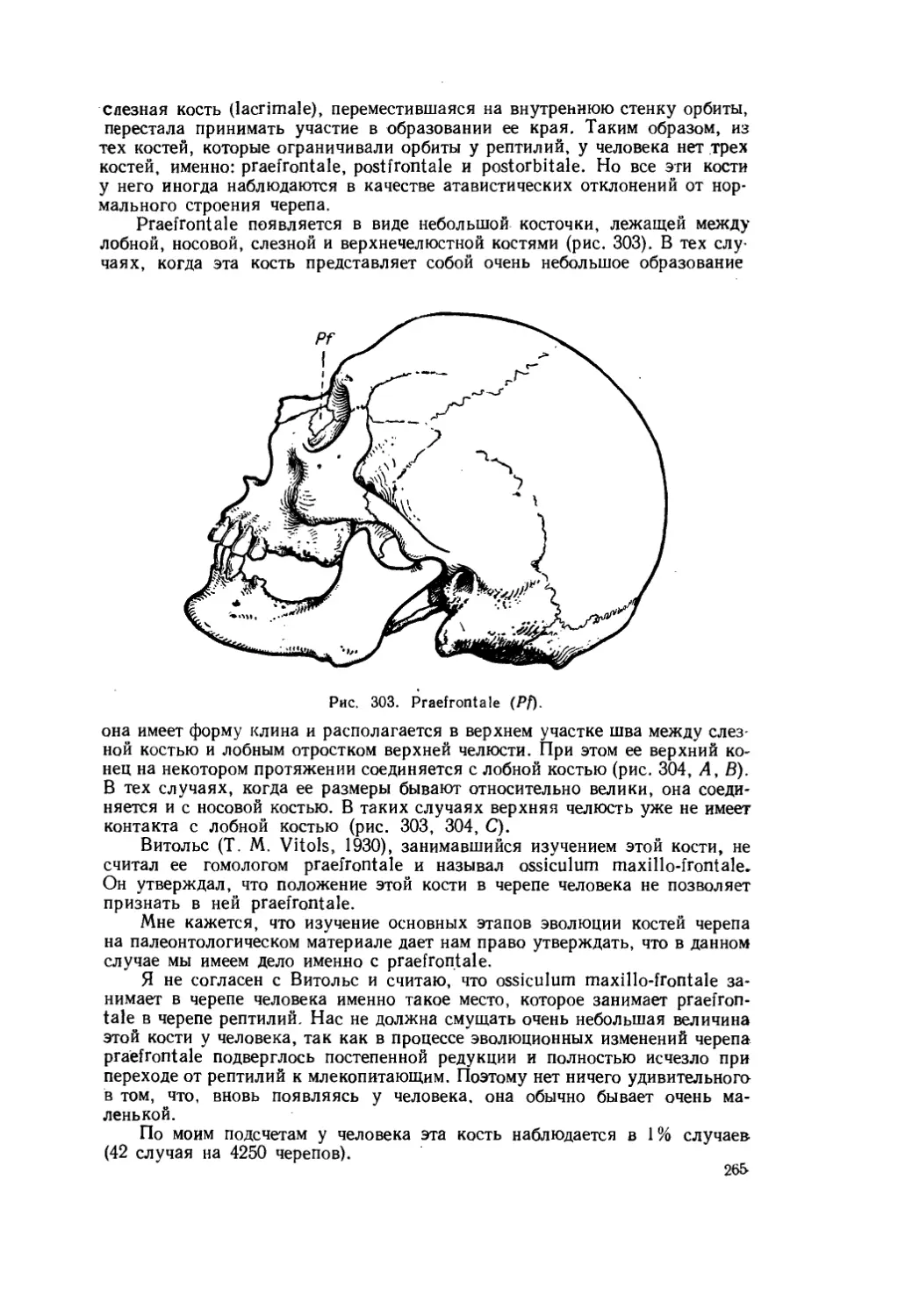
В качестве примеров можно указать на следующие явления. Известно, что у всех амфибий и рептилий передние концы носовых костей (nasale) соединены с межчелюстными костями (praemaxillare = intermaxillare); в результате этого носовые отверстия оказываются разделенными костной перемычкой (рис. 3). У млекопитающих такой контакт отсутствует, что привело к образованию в их черепе одного, непарного, входа в носовую полость. Разумеется, все случаи, когда у какого-либо млекопитающего вновь возникает соединение передних концов носовых и межчелюстных костей, следует считать возвращением предковой особенности, в частности повторением того, что было в черепах рептилий.
Такие явления не один раз наблюдались у различных ископаемых млекопитающих. Например, у южноамериканского ископаемого наземного
15
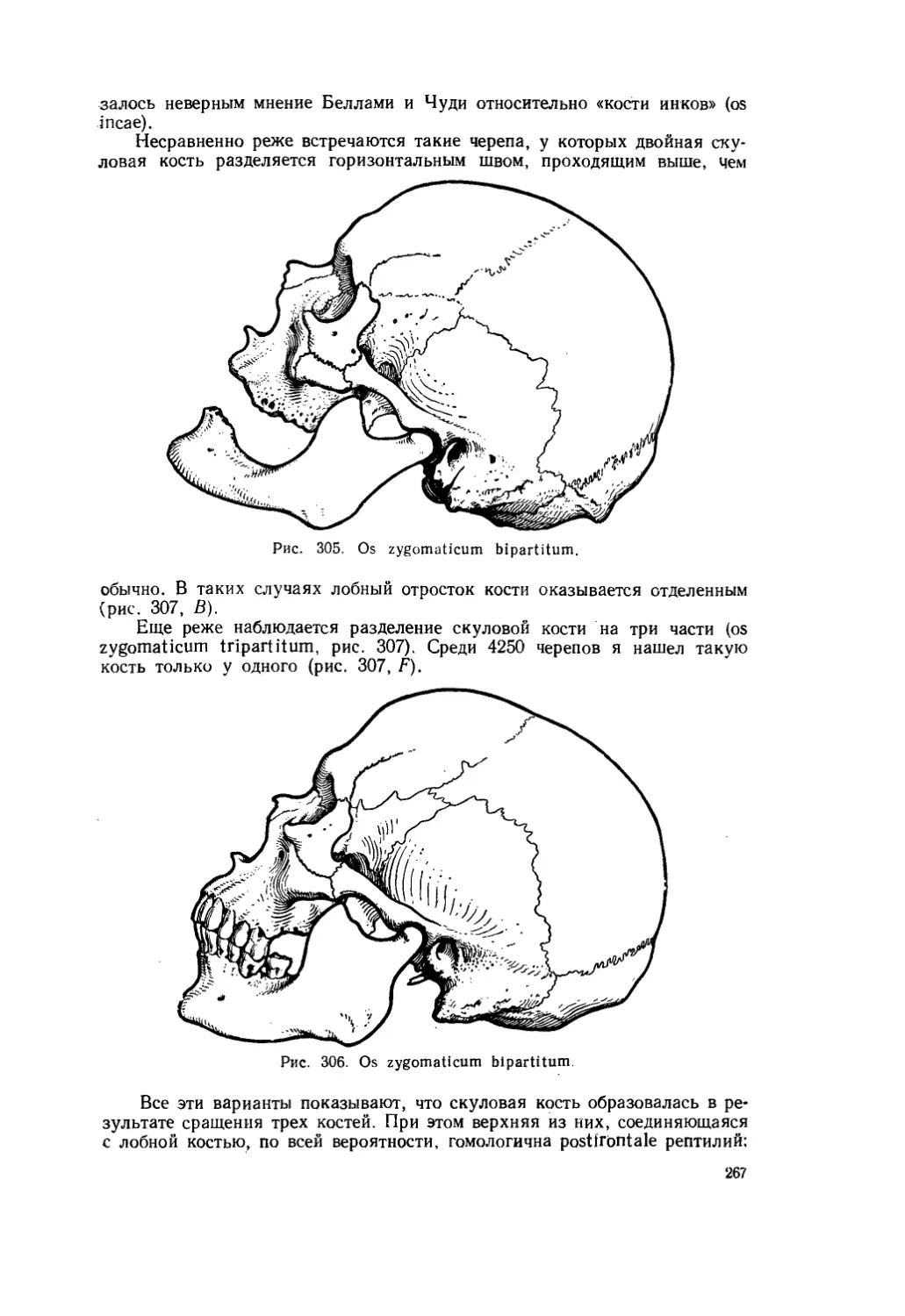
ленивца Mylodon robustus (рис. 4), как у обычного млекопитающего, контакт между передними концами носовых и межчелюстных костей отсутствует, а у другого наземного ленивца Grypotherium darwini, жившего одновременно с милодоном, эти кости соединены так же, как они соединены в черепе всех рептилий (рис. 5).
Такое же соединение носовых и межчелюстных костей имелось и в черепе своеобразного эоценового млекопитающего Gobiatherium mirificum, у олигоценового Arsinoitherium zitteli и у некоторых других ископаемых млекопитающих.
Восстановление одного или даже нескольких уже утраченных признаков, разумеется, еще не является полным возвратом к строению предков.
Рис. 4. Череп Mylodon robustus Owen
Возникновение вновь всех без исключения предковых особенностей у всякого организма и невозможно (невероятно) и неминуемо привело бы к немедленному его уничтожению, так как он не смог бы отстоять свою жизнь в борьбе с более прогрессивными членами нового биоценоза.
Все это, несмотря на возможность возвращения отдельных уже исчезнувших особенностей, делает эволюцию всех живых существ необратимой.
Всякий живой организм всегда поражает нас удивительной согласованностью формы и функции всех входящих в его состав органов. В результате такой согласованности организм в целом представляет собой нечто единое. Нарушение работы какого-нибудь одного из его органов в некоторых случаях может привести к гибели всего организма.
Эволюция такого организма сопровождается многочисленными и разнообразными изменениями во всех его участках, при этом все его ткани, органы и системы органов на каждом новом этапе эволюционного развития остаются согласованными друг с другом не в меньшей степени, чем прежде, что дает организму возможность сохранять жизнеспособность и в новых условиях. Таким образом, филогенез любого сложно построенного организма сопровождается изменениями как макроскопических, так и микроскопических его структур. Организм всегда эволюционирует как нечто целое, как система неразрывно связанных между собой частей. Именно как единая система он вступает в борьбу за жизнь и подвергается естественному отбору. При этом неудовлетворительная работа даже одного органа может повести к гибели всего организма.
Это положение может быть проиллюстрировано следующим примером.
Представим себе какого-нибудь хищника, например тигра, выслеживающим ночью свою добычу.
Физиология установила, что в автономной нервной системе, состоящей из sympathicus и parasympathicus, в данном случае sympathicus будет на
16
ходиться в состоянии возбуждения. Это приведет к тому, что зрачки животного расширятся, что даст возможность большему количеству лучей света проникать в глаз; работа слюнных желез затормозится, так как выделение слюны до овладения добычей является преждевременным; слюна сделается более вязкой; сердцебиение ускорится, так как усиление работы сердца в данном случае, естественно, будет иметь большое значение для всего организма; кровяное давление повысится вследствие воздействия симпатической нервной системы на надпочечники, из которых будет выделяться большее количество адреналина; бронхи в легких расширятся, что даст возможность усилить газообмен и улучшить снабжение организма кислородом; сосуды кожи сузятся и это понизит теплоотдачу тела; секреция
Рис. 5. Череп Grypotherium darwini Owen.
желез желудка прекратится; движение (перистальтика) кишок замедлится до полной остановки.
Таким образом, организм хищника будет подготовлен к нападению и все его силы будут мобилизованы.
После того как животное овладеет своей добычей, возбуждение симпатической части его автономной нервной системы прекратится и наступит возбуждение парасимпатической, которая выполняет совершенно противоположные функции.
В результате этого зрачки хищника сузятся, слюнные железы будут выделять обильную и жидкую слюну, сердцебиение замедлится, кровяное давление понизится, бронхи в легких сузятся, дыхание сделается спокойным, секреция желез пищеварительной системы возобновится, перистальтика кишечника восстановится.
Таким образом, весь организм хищника будет подготовлен к усвоению той пищи, которую ему удалось добыть.
Добывание пищи для каждого живого существа является одной из самых важных сторон его деятельности. Поэтому во время выслеживания добычи и при нападении на нее все органы хищника должны работать в высшей степени согласованно и точно. Всякое нарушение этой согласованности может привести к гибели всего организма.
На протяжении долгой эволюции живых существ всегда в конце концов погибали все те особи, у которых оказывались те или иные дефекты в функции отдельных органов или недостатки в согласованности их работы с работой других органов. Все это приводило к тому, что на каждом отдельном этапе своего развития все организмы представляли собой относительно совершенные живые системы, в которых форма и функция составляющих их частей были достаточно точно согласованы. Однако, существуя в составе биоценозов, живые организмы непрерывно вовлекались в борьбу за жизнь,
2 А. П. Быстров 17
так как непрерывная изменчивость членов биоценозов постоянно меняла обстановку и предъявляла новые требования. Но при непрерывной борьбе за существование естественному отбору подвергался каждый организм как нечто единое, как нечто целое. И это неминуемо приводило на основе непрерывной изменчивости, охватывающей не только системы органов и отдельные органы, но и образующие их ткани, к тому, что эволюционные изменения охватывали весь организм вплоть до формы и функции каждой его клетки.
В результате этого естественный отбор у сложно построенных живых существ в конце концов приводил к отбору и всех разнообразных тканевых структур организма и обусловливал их «гистологическую» эволюцию.
Мало того, отбору подвергались не только взрослые, вполне оформившиеся организмы, но и все молодые формы на всех фазах их развития. Поэтому всякое выгодное своеобразие развития эмбриона или личинки также закреплялось отбором, как и выгодная структура взрослого организма. При этом по понятным причинам всякие гибельные пороки развития эмбрионов или личинок делали невозможным формирование взрослого организма, а гибельные недостатки в строении и функции взрослого организма делали невозможным появление зародышей.
Все это приводило к тому, что естественный отбор на протяжении многих миллионов лет контролировал строение и функцию живых существ на всех фазах их развития и во всех формах проявления. Всякое несоответствие строения и функции любого живого существа той обстановке, в которой оно жило, всегда приводило только к одному результату — к его уничтожению.
Таким образом, в процессе эволюции все живые организмы в конце концов «платили» жизнью за неудовлетворительную функцию любой части своего тела. Здесь следует подчеркнуть, что всякая структура всегда «оценивалась» только по той роли, которую она играла в жизни организма. Следовательно, значение любого органа «определялось» по его функции. Это обстоятельство послужило причиной того, что некоторые биологи стали считать, что функция в процессе эволюции является «ведущей» и изменяется в первую очередь, а ее изменения затем вызывают соответствующие изменения формы. Такой взгляд на форму и функцию встречается у тех, кто в той или иной мере признает ламаркизм.
Однако несомненно, что функция любого органа не может изменяться без изменений его формы, и, наоборот, форма органа не может изменяться, не вызывая изменений в его функции.
Таким образом, ни форма, ни функция не могут в своих изменениях опережать друг друга; их изменения всегда бывают совершенно одновременными (абсолютно синхронными).
В биологической литературе можно найти немало примеров, показывающих, что всякий орган, почему-либо потерявший значение для организма, постепенно редуцируется и, наконец , исчезает. Все подобные явления обычно считаются доказательством, что функция в эволюционном процессе играет ведущую роль и что неупотребление (отсутствие функции) является достаточной причиной для исчезновения органа. Такой взгляд следует признать явной данью ламаркизму.
Причиной исчезновения всякого нефункционирующего органа, конечно, не является только то обстоятельство, что он бездействует. Если бездействующий орган не приносит вреда организму, то, по существу, нет причин, которые могли бы вызвать его исчезновение. Но всякий бездействующий и, следовательно, функционально бесполезный орган всегда оказывается в какой-то мере вредным, так как он использует некоторое количество питательных веществ, вводимых в организм. Поэтому всякий живой организм, 18
у которого в силу вариабильности нефункционирующий орган будет уменьшен в своих размерах или будет совсем отсутствовать, получит некоторое преимущество в борьбе за жизнь по сравнению с теми организмами того же вида, у которых эти органы еще имеются. При таких условиях естественный отбор, действуя в течение очень долгого времени, неминуемо приведет к тому, что сохранятся именно те животные, у которых нефункционирующие органы отсутствуют.
Постепенная редукция бездействующих органов, приводящая в конце концов к их исчезновению, еще раз показывает нам, что всякий организм является очень сложной системой. Форма и функция всех органов этой системы строго согласованы; и эта согласованность является необходимым условием существования всякого организма и непрерывно поддерживается естественным отбором.
III
Для того, чтобы добиться совершенно ясного представления о значении тех факторов, которые обусловливают эволюционный процесс, я считаю очень полезным поставить перед собой и попытаться разрешить несколько теоретических задач.
Допустим невероятное и предположим, что в нашем распоряжении имеется два совершенно равных по своей площади ареала. Предположим, что они расположены на одной параллели, но в диаметрально противоположных пунктах земного шара, и, таким образом, совершенно изолированы друг от друга. Предположим, что геологическое строение этих ареалов и их климат совершенно одинаковые. Предположим, что каждый из этих ареалов мы заселили равным количеством животных и растений. Предположим, что для этого мы имели возможность взять совершенно одинаковое количество видов и одинаковое количество экземпляров каждого вида. Таким образом, в данном случае мы допускаем, что исходный материал для дальнейшей эволюции в обоих ареалах оказался совершенно одинаковым как в количественном, так и в качественном отношении; одинаковы и все особенности мертвой природы.
В этой теоретической задаче нас интересует следующий вопрос: что случится с нашей флорой и фауной в обоих этих ареалах, скажем, через миллион лет, если в течение этого времени мертвая природа останется совершенно неизменной?
Казалось бы, что эволюционное развитие животных и растений в одинаковых условиях должно будет привести к появлению в обоих ареалах вполне тождественных форм, так как в обоих случаях организмы будут вести борьбу за жизнь с одними и теми же членами биоценоза и приспособляться к одинаковым и неизменным особенностям мертвой природы.
Однако можно уверенно сказать, что одинаковых растений и животных в этих условиях, несомненно, не разовьется. Через некоторое время после начала нашего «опыта» биоценозы обоих наших ареалов окажутся совершенно различными по своему составу.
Причиной этого в данном случае будет только один из четырех факторов эволюции, именно: изменчивость. Дело в том, что беспорядочная изменчивость животных и растений, разумеется, не может постоянно давать одинаковые формы живых существ. Поэтому рано или поздно в обоих изолированных друг от друга биоценозах среди представителей одного и того же вида появятся несходные формы. И это послужит первым толчком к возникновению различий в составах обоих биоценозов. Различные наследующиеся особенности, появившиеся в силу изменчивости, немедленно подвергнутся естественному отбору. А это через некоторое время приведет к появлению в одном из ареалов таких форм, которых нет в другом.
2*
19
На следующих этапах эволюции различия в обоих наших биоценозах будут постепенно увеличиваться, так как появление первых различий неминуемо вызовет и появление ряда новых различий. В результате этого различия в биоценозах будут постепенно накопляться. В конце концов оба ареала окажутся занятыми двумя биоценозами, вполне приспособленными к условиям существования, но состоящими из совершенно несходных растений и животных.
Все это показывает, что если мертвая природа в течение даже очень долгого времени останется совершенно неизменной, то это не значит, что живые организмы, обитающие в соответствующих ареалах, за это время не будут эволюционировать. Они вследствие непрерывно действующего естественного отбора будут улучшать свои приспособления даже к неменяю-щимся особенностям мертвой природы. Эти приспособления могут быть очень разнообразными, так как одна и та же задача может быть разрешена различными способами.
Для всякого эволюционирующего биоценоза (а неэволюционирующих биоценозов не существует), конечно, не безразлично, где он находится, т. е. какой ареал занимает. Изменения мертвой природы в любом ареале, разумеется, не могут не отразиться на строении членов биоценозов.
Возникновение гор там, где прежде была равнина, конечно, всегда приводило к появлению животных и растений, приспособленных к жизни в горах; равным образом возникновение равнины на месте разрушившихся гор, разумеется, всегда приводило к образованию равнинных биоценозов. Однако никакие изменения в мертвой природе сами по себе никогда не были основными причинами прогрессивной эволюции живых существ.
Возникновение горных массивов на различных участках суши в течение палеозойской эры всегда приводило к появлению горных биоценозов. То же самое происходило и на протяжении мезозойской и кайнозойской эр. Но горные биоценозы палеозоя, мезозоя и кайнозоя были составлены, конечно, совершенно различными формами живых существ, которые, однако, были в достаточной мере приспособленными к жизни в горах.
Все это показывает, что все члены эволюционирующих биоценозов всегда оказываются вынужденными приспосабливаться ко всем особенностям мертвой природы ареала своего обитания. Но изменения в мертвой природе не являются ведущей причиной эволюции. Все изменения, происходящие в мертвой природе, не протекают и не протекали в такой последовательности, которая сама по себе могла бы обеспечить прогрессивную эволюцию живых существ на нашей планете. Из этого можно сделать только один вывод: эволюционное развитие всего живого на земле не вызывалось изменениями в мертвой природе.
Изучение окружающей нас живой природы показывает, что животные и растения всюду образуют разнообразные биоценозы, в которых наблюдаются очень сложные связи и тесные взаимоотношения, обусловленные борьбой за существование и естественным отбором.
Однако допустим невероятное и скажем, что по каким-то причинам борьба за жизнь среди членов биоценоза прекратилась и исчез естественный отбор.
При таких условиях среди живых существ в силу их непрерывной изменчивости стали бы по-прежнему возникать формы, обладающие новыми, многочисленными и разнообразными наследующимися признаками. Однако отсутствие естественного отбора исключило бы возможность закрепления любой из этих новых особенностей. В результате этого, несмотря на непрерывные колебания в строении организмов, они прекратили бы свое эволюционное развитие.
.20
Таким образом, решение этой теоретической задачи приводит нас к заключению, что прекращение действия таких факторов эволюции, как борьба за существование и естественный отбор, немедленно вызвало бы остановку эволюционного процесса.
Допустим, что мы не будем нарушать сложных взаимоотношений, вызванных борьбой за существование и естественным отбором между многочисленными членами биоценозов, но предположим невероятное, что у всех живых существ, составляющих эти биоценозы, исчезла изменчивость. Если бы у живых организмов перестали непрерывно появляться многочисленные, разнообразные, новые, передающиеся по наследству особенности, то естественный отбор был бы лишен материала для своей «работы». А это неминуемо привело бы к тому, что эволюция организмов немедленно остановилась бы. Биоценозы различных ареалов навсегда остались бы такими, какими их застало допущенное нами исчезновение изменчивости, и мы не смогли бы обнаружить в них каких бы то ни было изменений даже на протяжении многих миллионов лет.
Все это показывает, что изменчивость живых существ является необходимым условием эволюции и отсутствие изменчивости делает эволюционный процесс невозможным.
Сделаем еще одно недопустимое допущение. Предположим, что все живые существа потеряли способность передавать по наследству потомству свои морфологические и физиологические особенности.
Если бы исчезла наследственность и те особенности, которыми обладает данное поколение живых организмов, перестали бы передаваться потомкам, то и при таких условиях эволюция немедленно прекратилась бы. Это случилось бы потому, что отбор по ненаследуемым признакам, разумеется, не смог бы привести к каким-либо стойким изменениям организации живых существ. Всякие полезные особенности животных были бы безразличны для их потомства, так как они не могли бы у них сохраниться.
При таких условиях в процессе борьбы за жизнь выживало бы всякое удачно организованное существо, но это не давало бы решительно никаких преимуществ его непосредственным потомкам, так как они не имели бы возможности сохранить выгодные особенности своих родителей. .
Таким образом, прекращение действия любого из четырех факторов эволюции должно приводить к одному и тому же результату — к остановке эволюционного развития. В то же время ни один из этих факторов в отдельности не в состоянии обусловить эволюционное развитие живых существ. Для такого развития совершенно необходимо одновременное действие всех четырех факторов эволюции.
Некоторые биологи считают, что эволюционное развитие возможно только при условии наследования приобретенных признаков, возникающих в результате воздействия окружающей среды.
Это положение нуждается в некоторых уточнениях.
Многообразная изменчивость живых существ, чем бы она ни вызывалась, всегда приводит к возникновению двух видов признаков: ненасле-дуемых и наследуемых. Ненаследуемые особенности никакого значения для эволюции не имеют, так как естественный отбор не может закрепить и сохранить их. Что же касается наследуемых признаков, то эволюционную ценность приобретают только те из них, которые сохраняются естественным отбором. Таким образом, даже наследуемые признаки в организации живых существ сами по себе не имеют эволюционного значения. Они приобретают это значение только в результате воздействия естественного отбора, так как в процессе борьбы за жизнь сохраняются те организмы, которые обладают полезными при данных условиях признаками.
21
Вследствие этого в конце концов возникают живые формы, поразительно приспособленные ко всем особенностям окружающей среды.
Следует иметь в виду, что беспорядочная, ненаправленная изменчивость живых существ приводит только к возникновению огромного количества разнообразных вариантов строения, из которых только немногие случайно оказываются соответствующими «требованиям» окружающей обстановки.
Такая изменчивость, разумеется, не может привести к эволюционному развитию живых форм.
Только естественный отбор, сохраняя на протяжении многих миллионов лет удачные организации живых существ, мог обусловить прогрессивную эволюцию.
Все это позволяет нам прийти к заключению, что никакая изменчивость сама по себе, без естественного отбора, не в состоянии обусловить эволюционный процесс. Но и естественный отбор при отсутствии изменчивости, разумеется, не смог бы вызвать эволюционное развитие.
IV
Как известно, Ч. Дарвин придавал большое значение данным палеонтологии, которые позволяют проследить изменения организации живых существ на протяжении многих миллионов лет. Но в 1859 г. он имел все основания жаловаться на «неполноту геологических летописей».
После выхода в свет книги Ч. Дарвина о происхождении видов в течение почти столетия в результате усиленных поисков палеонтологи накопили очень большой материал по истории позвоночных. Огромная теоретическая ценность этого материала, разумеется, не подлежит никакому сомнению. Однако, непрерывно пополняя соответствующие музеи все новыми и новыми находками, палеонтологи получают возможность изучать только фактическую сторону эволюционного процесса. В некоторых случаях ряд ценных находок позволяет им выяснить основные этапы филогенетического развития той или иной группы животных. Но при этом исследователи обычно не находят никаких указаний на те конкретные причины, которые заставили данное животное пройти именно такой, а не иной путь эволюционного развития.
В отличие от палеонтологов зоологи, изучающие современных животных, имеют возможность выяснить не только все их особенности как живых организмов, но и исследовать те сложные взаимоотношения, которые обусловливают нх эволюционное развитие. Однако зоологи не в состоянии непосредственно наблюдать ход эволюционного процесса.
Таким образом, изучение прошлого привело к констатации эволюции без выяснения ее факторов, а изучение настоящего привело к выяснению факторов эволюции без достаточно очевидной ее констатации.
Такое положение потребовало допущения, что при изменениях древних животных и растений действовали те же факторы эволюции, которые действуют и в современном органическом мире, и что эволюционный процесс, констатированный для прошлого, продолжается и в настоящее время. Иными словами, в прошлом мы констатируем факты эволюции, а в настоящем мы констатируем факторы эволюции; в прошлом мы предполагаем факторы эволюции, в настоящем мы предполагаем факты эволюции.
Все это приводит к тому, что одни палеонтологи верят, другие допускают, третьи отрицают, что причинами эволюционных изменений являются факторы, установленные Ч. Дарвином. Так как эти факторы пока еще не могут быть доказаны ископаемым материалом, то именно среди палеонтологов чаще всего встречаются противники теории Дарвина. Не имея возмож-22
ности проследить влияние дарвиновских факторов на филогенетические изменения вымерших форм, антидарвинисты обычно утверждают, что теория Дарвина неверна. Для ее замены они нередко создают, хотя и совершенно неубедительные, но новые гипотезы.
Впрочем, не все антидарвинисты полностью отвергают теорию Дарвина. Некоторые из них пытаются внести в нее ряд дополнений и исправлений.
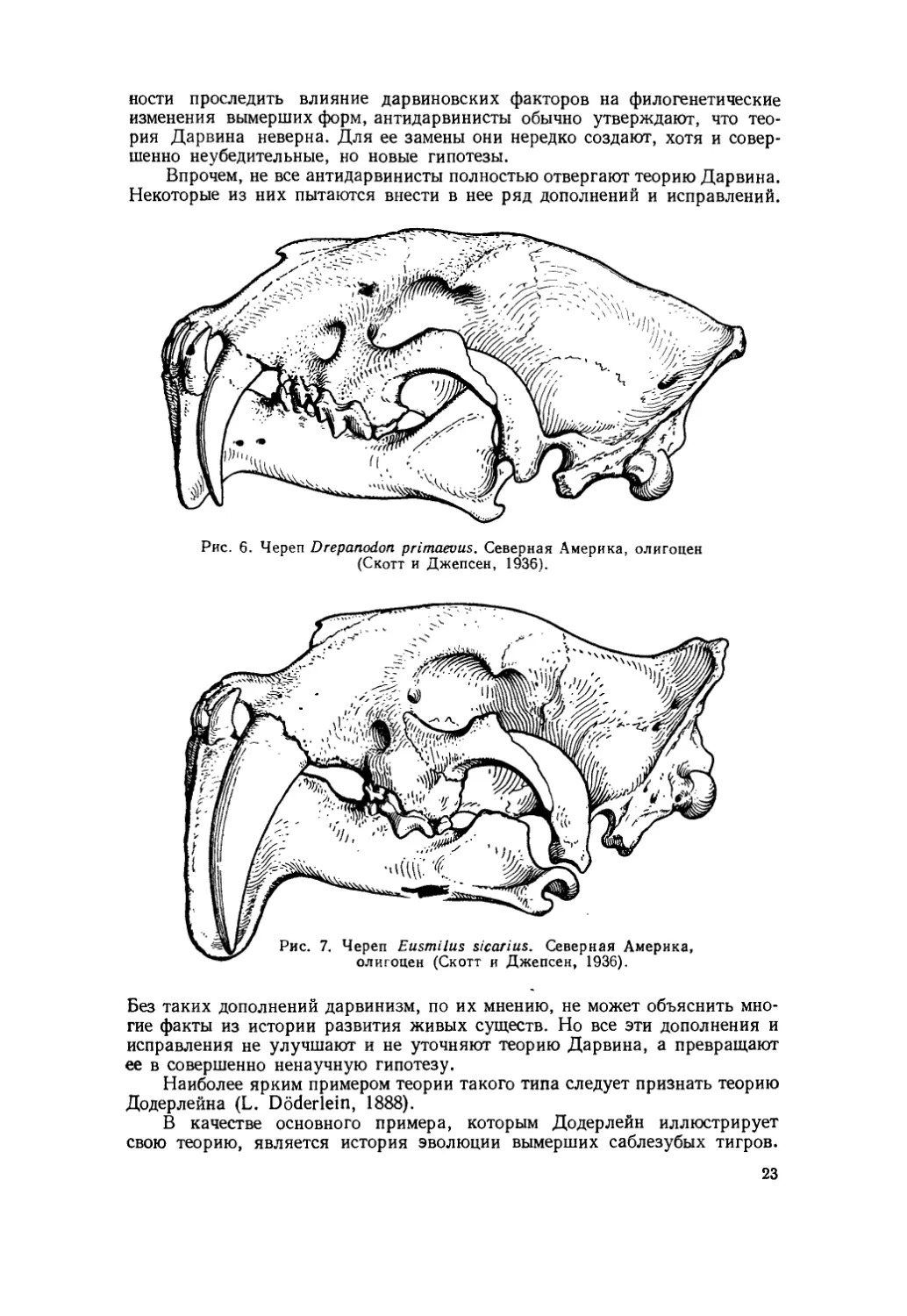
Рис. 6. Череп Drepanodon primaevus. Северная Америка, олигоцен (Скотт и Джепсен, 1936).
Без таких дополнений дарвинизм, по их мнению, не может объяснить многие факты из истории развития живых существ. Но все эти дополнения и исправления не улучшают и не уточняют теорию Дарвина, а превращают ее в совершенно ненаучную гипотезу.
Наиболее ярким примером теории такого типа следует признать теорию Додерлейна (L. Doderlein, 1888).
В качестве основного примера, которым Додерлейн иллюстрирует свою теорию, является история эволюции вымерших саблезубых тигров.
23
Судя по довольно богатому ископаемому материалу, эти тигры прошли очень своеобразный путь эволюционного развития, на протяжении которого у них постепенно увеличивались размеры верхних клыков и шла редукция коренных зубов. Одновременно с этим произошли некоторые изменения и в челюстных суставах (рис. 6 и 7; Скотт и Джепсен — Scott and Jepsen, 1936).
Л. Додерлейн согласен с тем, что образование больших клыков на верхних челюстях у саблезубых тигров было вызвано естественным отбором,
Рис. 8. Череп Eusmilus sicarius (рот открыт).
закрепляющим полезные варианты строения. Но при этом постепенное увеличение клыков под влиянием отбора, по его мнению, продолжалось до тех пор, пока эти мощные зубы не дали тиграм возможности вытеснить и полностью истребить всех своих конкурентов.
«Можно и даже должно считать, — говорит Додерлейн, — что если у ряда животных важные органы, служащие, например, оружием, достигают высшей целесообразности, то это настолько вредит их прежним конкурентам, что они уступают им место».
Таким образом, сильное развитие клыков у саблезубых тигров, по мнению Додерлейна, привело к тому, что борьба за существование сначала значительно ослабла для них, а затем совсем исчезла. Одновременно с этим, разумеется, должен был прекратить свое действие и естественный отбор. Это, казалось бы, должно было повлечь за собой и немедленную остановку дальнейшей эволюции зубов. Однако Додерлейн полагает, что дальнейшее
24
развитие клыков и после этого продолжалось, но теперь уже только под влиянием одной «силы инерции».
«Здесь имеется своего рода vis inertiae» — говорит он. Таким образом, по Додерлейну, естественный отбор обусловливал эволюционный процесс только до момента возникновения у животного максимально целесообразного строения органа. Все же последующее развитие совершалось только по инерции, причем направление развития в этом случае диктовалось уже пройденным эволюционным путем.
«Я считаю возможным думать, — пишет Додерлейн, — что после того, как у длинного ряда предков упорно сохранялось определенное направление развития, пока, например, не достигалось состояние наибольшей целе-
Рис. 9. Череп Thylacosmilus atrox. Сумчатый хищник из плиоцена Аргентины (Риггс, 1933).
сообразности, потомки наследуют не это определенное состояние, а только тенденцию развиваться именно в этом определенном направлении».
Следовательно, по мнению Додерлейна, развитие под влиянием «силы инерции» являлось не только непосредственным продолжением развития, протекавшего под действием факторов Ч. Дарвина, но и совершалось оно в том же направлении, в котором шло прежде. В результате этого клыки животного продолжали увеличиваться. Это увеличение с некоторого момента было уже ненужным и нецелесообразным. В первое время при отсутствии уже уничтоженных конкурентов, а следовательно, и при отсутствии борьбы за существование и естественного отбора, большие, но бесполезные клыки не мешали хищнику жить. Но потом дальнейшее увеличение этих зубов привело к такой их длине, что животное лишилось возможности есть (рис. 8).
«Эти зубы, — говорит Додерлейн, — сами по себе представлявшие ужасное оружие, одновременно с этим вследствие своей необыкновенной длины чрезвычайно мешали при еде, действуя как намордник». Это обстоятельство, по мнению Додерлейна, и было причиной вымирания саблезубых тигров.
Таким образом, эволюционная сила инерции или унаследованная тенденция развития в конце концов привела этих своеобразных хищников к гибели.
Такие мощные клыки, как у наиболее специализированных саблезубых тигров (Eustnilus, рис. 7), были обнаружены и у найденного в 1926 г.
25
в плиоценовых отложениях Аргентины сумчатого хищника Thylacostnilus atrox (рис. 9; Риггс — Riggs, 1933). Если бы это своеобразное животное было найдено лет на 40 раньше, то Л. Додерлейн, несомненно, воспользовался бы им в качестве хорошей иллюстрации своей гипотезы.
Мы не знаем, какими животными питался Thylacosmllus atrox и какую пользу приносили ему его огромные клыки. Однако можно уверенно сказать, что они для него, несомненно, представляли большую ценность. Доказательством этого является то, что подбородочная^часть нижней челюсти живот-
Рнс. 10. Скелет мамонта.
ного сильно разрослась вниз и, помещаясь при закрытом рте между клыками, предохраняла их от случайных переломов. Такое же приспособление, как известно, имелось и у многих саблезубых тигров, например у Drepan-odon (рис. 6), и было сильно развито у Eusmilus (рис. 7).
Кроме ссылки на эволюцию саблезубых тигров, Л. Додерлейн в подтверждение своей гипотезы указывает на огромные бивни мамонта (рис. 10) и на необычайно сильное развитие рогов у плейстоценового оленя Mega-ceros (рис. 11).
По мнению Л. Додерлейна, бивни мамонта и рога оленя достигали таких больших размеров в силу тенденции к увеличению. Эта тенденция была выработана предками мамонта и предками оленя. Л. Додерлейн считает, что в течение длительного периода эволюционного развития увеличение размеров бивней было полезно предкам мамонта, полезно было и увеличение рогов предкам оленя. Но после достижения состояния наибольшей целесообразности дальнейший рост и бивней и рогов был уже не только бесполезным, но даже вредным. Однако он продолжался до тех пор, пока не оказался причиной вымирания и мамонта и гигантского оленя.
Наконец, для доказательства правоты своих взглядов Л. Додерлейн указывает на сильное развитие верхних и нижних клыков у современной
26
дикой свиньи бабируссы (Babirussa babirussa), живущей на острове Целебес (рис. 12). Он считает, что после того, как бабирусса была изолирована от крупных хищников на острове и перестала подвергаться нападениям, ее клыки продолжали развиваться в прежнем направлении только под влиянием vis inertiae и скоро достигли настолько нецелесообразных размеров, что й некоторых случаях стали своими концами врастать в череп (рис. 13).
Рис. 11. Скелет Megaceros hibernicus Owen из торфяных болот Ирландии.
Изучение гипотезы Додерлейна показывает, что его vis inertiae представляет собой некое дополнение к теории Дарвина, такое дополнение, без которого будто бы совершенно невозможно объяснить некоторые факты из истории эволюционного развития животных. По существу, все новое в гипотезе Л. Додерлейна сводится к утверждению, что эволюционные изменения являются в значительной степени внутренним свойством живых организмов, с чем нельзя согласиться.
В 1897 г. Т. Эймер (Th. Eimer) выдвинул типично антидарвиновскую теорию прямого развития, или «ортогенеза» (6р&6<: — orthos — прямой; •yeveatc — genesis — происхождение). «Именно в силу того, — говорит он, —
27
что органическое образование формы основывается на физико-химических процессах, оно является, как и форма неорганических кристаллов, определенным и при всяком новообразовании может идти только по определенным направлениям».
Из этого можно сделать вывод, что orthogenesis как эволюционный процесс является одним из свойств живых организмов. Однако, по мнению Эймера, «внешние воздействия, т. е. климат, свет, тепло, влажность, и различия в питании изменяют организмы без какого-либо участия отбора, и так как возникающие этим путем изменения наследующиеся, то они обусловливают возникновение новых видов».
«Отбор, — говорит Эймер, — не может создать решительно ничего нового».
Рнс. 12. Череп бабируссы (Babirussa babirussa).
Таким образом, эволюционный процесс, по Эймеру, оказывается обусловленным в значительной мере тем, что «плазма имеет свойство, будучи благодаря воздействию внешних сил измененной в физиологическом или в морфологическом отношении, принимать иной вид».
Следовательно, основной причиной появления изменений в плазме Т. Эймер считает внешние воздействия. Именно они (и только они) способны так или иначе изменять плазму живых организмов. Но эволюционные изменения вызываются не одним тем фактом, что плазма под влиянием внешних воздействий изменяется, а тем, что последовательные фазы этих изменений обусловливают соответствующую последовательность в этапах эволюционного развития организмов.
«Согласно моим исследованиям,— говорит Т. Эймер, — главной причиной эволюции является обусловленный внешними влияниями, климатом и пищей органический рост в плазме, выражением которого. . . является определенно направленное развитие (ортогенез)».
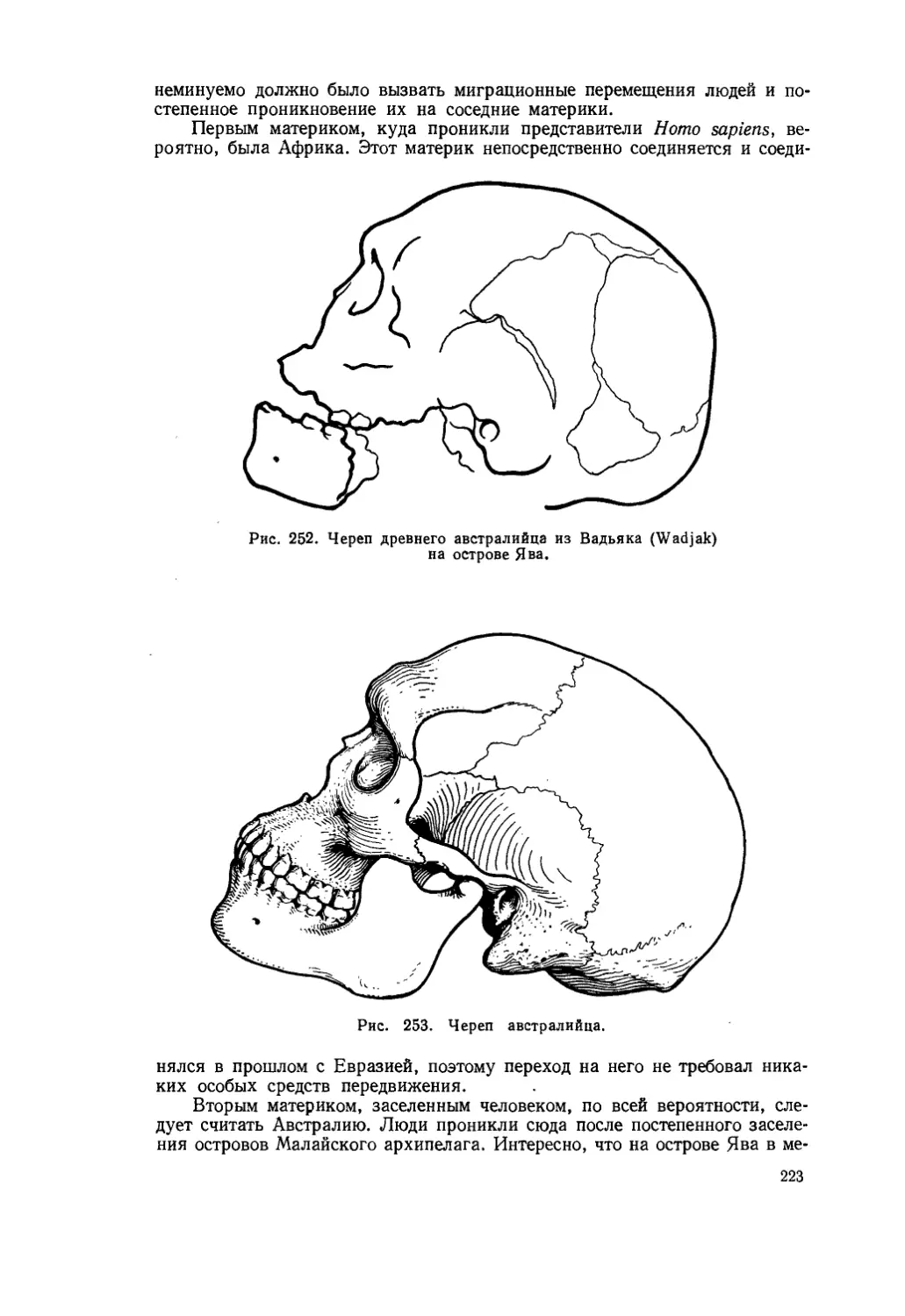
Таким образом, основным фактором эволюционных изменений Эймер считает прямое воздействие внешней среды, вызывающее в плазме такие изменения, которые приводят к направленной эволюции.
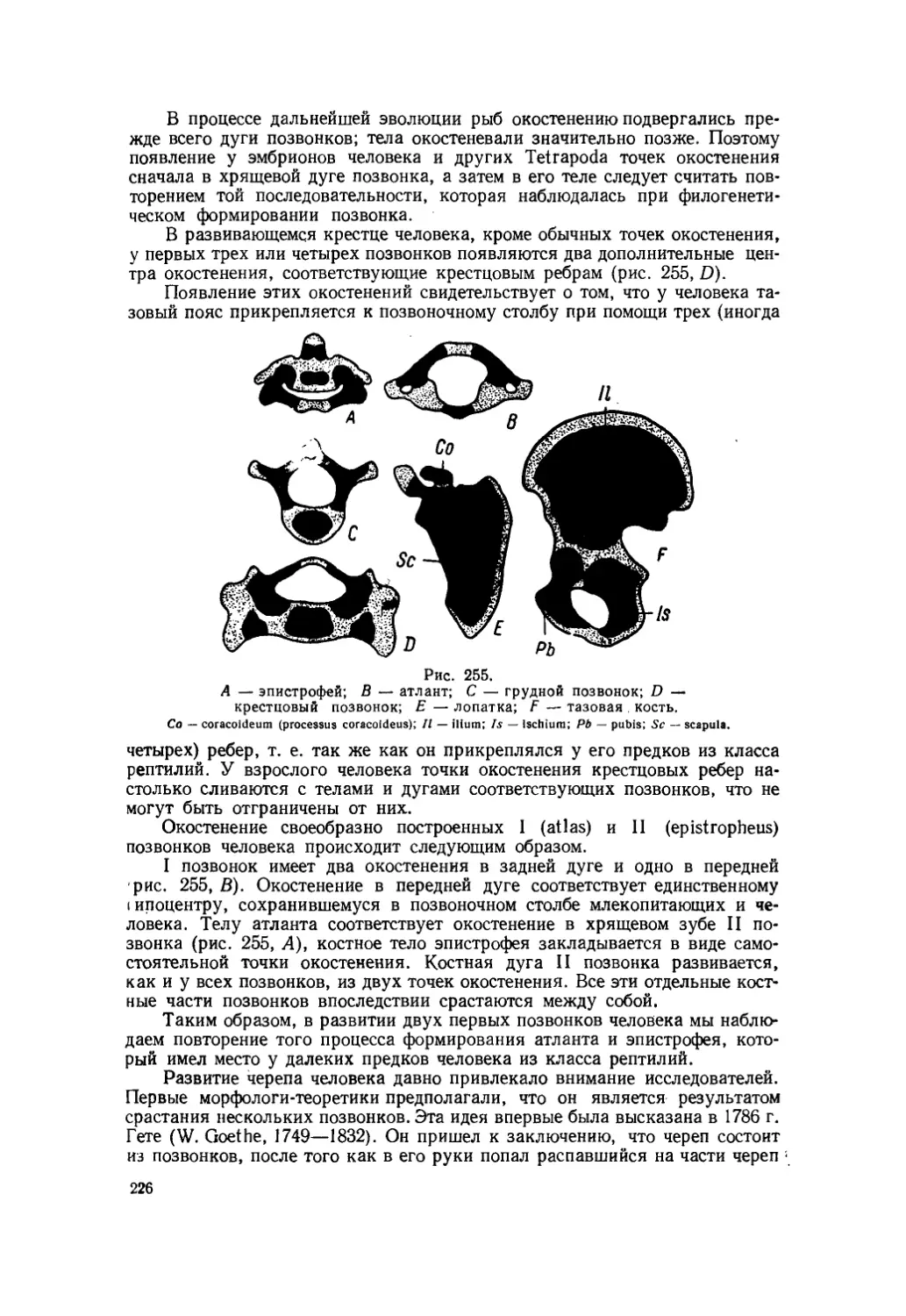
В 1922 г. Л. С. Берг опубликовал свою теорию «номогенеза» (v6p.oc — nomos — закон; yeveaic — genesis — происхождение). Он утверждал, что «эволюция организмов есть результат некоторых закономерных процессов, 28
протекающих них. Она есть номогенез, развитие по твердым '«законам».
По мнению Л. С. Берга, основными причинами эволюции являются изменения стереохимических свойств белков протоплазмы живых существ. Эти изменения приводят к развитию в определенном направлении.
Л. С. Берг писал, «что процесс эволюции идет закономерно, что он покоится на основе номогенеза. Как проявляются эти закономерности, мы видим, но почему они таковы, — это пока скрыто от нас. Равным образом, почему организмы в общем прогрессируют в своей организации, мы не знаем». Однако Л. С. Берг утверждал, что при этом «природа работает исключительно при помощи законов физики и химии».
Рис. 13. Череп бабнруссы (Babirussa babirussa).
Свою теорию Л. С. Берг иллюстрирует многочисленными примерами» взятыми из самых различных областей морфологии, и эти примеры, ко' нечно, во всех случаях оказываются подтверждением его номогенеза.
В частности, ссылаясь на описанный им в 1916 г. интересный экземпляр .аральской плотвы (Ratlins rutilus aralensis monstrositas exocoetoides, рис. 14, В) с необычайно длинными грудными плавниками, напоминающими соответствующие плавники летающей рыбы Exocoetus volitans (рис. 14, Л), он приходит к заключению, что планирующий полет у летающих рыб развился «на основе закономерностей, вне всякого участия как естественного отбора, так и употребления и неупотребления органа». «Удлиненные плавники, — говорит он, — образуются потому, что они должны образоваться».
Таким образом, эволюционный процесс, по мнению Л. С. Берга, представляет собой непосредственный результат особых закономерных изменений в самих живых организмах. Эти изменения, независимо от окружающей среды, всегда ведут эволюцию по такому пути, который приводит к тому, что должно было быть.
Если Додерлейн в 1888 г. рассматривал vis inertiae только как необходимое дополнение к дарвиновским факторам эволюции, то Абель (О. Abel) в 1929 г. считал, что «сила инерции» является во всех случаях единственной причиной эволюционного процесса. По его мнению, причиной филогенетических изменений живых существ является так называемый «биологический закон инерции» (Biologisches Tragheitsgesetz). Этот закон некоторые биологи называют «законом Абеля».
29
Хотя Абель, как и Додерлейн, говорил об инерции, однако он давал совершенно иное обоснование своим взглядам.
Абель считал, что в настоящее время у нас нет никаких оснований сомневаться, что все процессы в живых организмах подчинены законам физики и химии, а все физические и химические процессы могут рассматриваться, как движение. Поэтому все без исключения функции живых организмов представляют собой явления не только физико-химические, но и механические. Но так как одним из основных законов механики следует считать закон инерции, то, по мнению О. Абеля, всё функции животных и растений
Рис. 14.
А — Exocoetus volitans; В — R uti.1 us rutilus aralensis monstrositas exocoetoides (по Л. С. Бергу).
и их эволюционное развитие полностью подчинены этому закону. Поэтому однажды взятое направление эволюции затем на всех своих этапах в силу этого закона остается неизменным. Таким образом, по теории О. Абеля эволюционные изменения^ являются внутренним свойством t живых организмов.
«В настоящее время мы знаем, — говорит Абель, — что всякий функционирующий организм, так же как его отдельные части и даже сама протоплазма, находится в состоянии движения, потому что функция есть деятельность и, следовательно, — движение. Но если это так, то все части организма должны подчиняться законам физики и, следовательно, механики, к которым в первую очередь относится закон инерции. Этот закон и для органического мира следует понимать в механическом, а не в переносном смысле».
Разумеется, мы не можем отрицать наличия физических и химических процессов в живых организмах. Но такие факторы эволюции как борьба за существование и естественный отбор не могут быть признаны ни физическими, ни химическими явлениями и они не встречаются в мертвой природе. А без этих факторов никакие физические и химические процессы, протекающие в живых организмах, не смогут вызвать их эволюционных изменений.
30
Таким образом, можно уверенно сказать, что физико-химические процессы в живых организмах также не приводят и не приводили к эволюцион
ным изменениям, как они не превращали в эволюцию никакие изменения
в неорганическом мире.
В 1933—1934 гг. Осборн (Н. F. Osborn) опубликовал свою теорию «аристогенеза» (арюто<: — aristos — наилучший; тЫак; — genesis — происхо
ждение).
Изучая в течение многих лет титанотериев и ископаемых хоботных,
Осборн на большом материале проследил основные этапы эволюции этих
млекопитающих. При этом у наиболее древних форм титанотериев он наблюдал появление небольших костных выступов на переднем отделе черепа, которые у более поздних форм превратились в хорошо выраженные рога (рис. 15).
У хоботных Осборн особенно детально изучал постепенное увеличение количества бугорков и гребней на жевательной поверхности зубов.
Постепенное увеличение размеров рогов у титанотериев и постепенное усложнение строения зубов у хоботных Осборн считал результатом особых изменений в плазме половых клеток этих животных. Эти изменения, по его мнению, выражались в появлении так называемых «ари-стогенов», которые затем медленно развивались и были причиной возникновения у животных различных полезных и притом постепенно улучшающихся особенностей.
«Мы знаем, — говорит Осборн, — как появляются эти аристогены, но мы не знаем, почему они появляются, т. е. причины их возникновения». «То, что вызывает аристоген в гене-плазме, — пишет он, — для нас также таин
ственно, как и то, что вызывает зачаток рога на
черепе титанотерия».
Однако, несмотря на всю эту таинственность, Осборн признавал, что «аристогены, конечно, не появляются самопроизвольно или
Рис. 15. Эволюционные изменения титанотериев.
без какой-либо прежде
существующей или побуждающей причины». Они, по его мнению, возни-
кают в результате способности животного реагировать приспособительно на различные особенности внешней среды и на борьбу с другими животными. В результате этого изменение пищи вызвало у хоботных появление ари-стогенов, обусловливающих постепенное усложнение зубов, а борьба
с другими животными привела у титанотериев к возникновению аристоге-нов, которые были причиной появления и постепенного увеличения рогов.
Несмотря на то, что появление аристогенов в генеплазме животных, по мнению Осборна, как-то связано с внешней средой, сам эволюционный процесс (aristogenesis) все же, по его убеждению, является только результатом развития аристогенов.
«Аристогенез,— говорит он, — есть творческий процесс, вызванный совершенно новым биохимизмом зародышевой плазмы, процесс направленный, постепенный, прямой, идущий по пути к будущему приспособлению».
Таким образом, по теории Осборна, всякие эволюционные изменения предрешены биохимическими реакциями, происходящими в плазме половых клеток живых существ.
31
Все авторы, предлагающие ту или иную теорию эволюции для замены неудовлетворяющего их дарвинизма, в подтверждение своих взглядов, разумеется, всегда приводят некоторое количество фактических данных.
По моему мнению, нет ничего удивительного в том, что это оказывается возможным. Строение как современных, так и ископаемых животных настолько разнообразно, что если не пытаться выяснить в каждом отдельном случае ту конкретную обстановку, в которой формировались все морфологические особенности данного организма, то при желании можно без особого труда найти неограниченное количество доказательств в подтверждение правильности любой антидарвиновской гипотезы. Мало того, наши знания
Рис. 16.
А — Dactylopterus orientalis; В — Comephorus dybowskii.
конкретной истории формирования многих анатомических особенностей животных настолько недостаточны, что одну и ту же морфологическую структуру можно легко трактовать с различных точек зрения.
Это можно показать хотя бы на следующем примере.
В свое время в озере Байкал была обнаружена своеобразная рыба Comephorus dybowskii (рис. 16, В), у которой имелись очень большие грудные плавники, напоминающие соответствующие плавники летающей рыбы Dactylopterus orientalis (рис. 16, Л). Причина образования и назначение, т. е. функция, этих плавников пока еще совершенно не выяснены. Поэтому ничто не мешает нам утверждать, что под влиянием внешней среды в плазме этой рыбы в свое время произошли такие изменения, которые привели к увеличению грудных плавников, которые, однако, аппаратом для планирующего полета пока еще не служат. Но дальнейшее их развитие, обусловленное дальнейшим органическим ростом плазмы, надо думать, будет идти в том же направлении, а это в конце концов неминуемо должно будет привести к появлению нового вида летающих рыб. Таким образом, очевидно, что Comephorus подтверждает ортогенез Эймера.
Однако в данном случае мы можем, соглашаясь с мнением Эймера, что развитие грудных плавников у Comephorus обусловлено физико-химическими изменениями плазмы рыбы, в то же время утверждать, что эти изменения наступили совершенно независимо от каких бы то ни было особен-
32
ностей внешней среды. Мы можем предполагать, что они были обусловлены закономерными процессами, постоянно протекающими в плазме живых существ. Поэтому длинные плавники образовались только потому, что они должны были образоваться. Следовательно, байкальский Cotnephorus dybowskii является хорошей иллюстрацией не ортогенеза Т. Эймера, а номогенеза Л. С. Берга.
Впрочем, если принять во внимание, что все физико-химические процессы сопровождаются движением, а основным законом движения является закон инерции, то никто не может лишить нас права считать, что эволюционный процесс направляется именно этим законом и что развитие плавников у Cotnephorus шло по существу под влиянием только силы инерции. Эта сила и обусловливала и направляла эволюцию рыбы по строго определенному пути, что, в частности, привело к образованию у нее больших грудных плавников, а в будущем даст ей возможность превратиться в летающую рыбу. . .
Если это действительно так, то Cotnephorus dybowskii представляет собой прекрасный пример, подтверждающий не номогенез Л. С. Берга, а биологический закон инерции О. Абеля.
Наконец, у нас имеются все основания ссылаться на Cotnephorus как на очевидное доказательство того, что в генеплазме живых существ возникают аристогены, обусловливающие эволюцию. В данном случае мы имеем дело с аристогеном, вызвавшим появление больших грудных плавников. Дальнейшая эволюция этого аристогена неминуемо должна будет привести к возникновению байкальской летающей рыбы, вполне приспособленной ко всем особенностям жизни в этом озере. Поэтому мы можем считать, что Cotnephorus dybowskii является подтверждением не закона Абеля, а аристо-генеза Осборна.
Все это можно утверждать, но ничего этого нельзя доказать. При этом следует иметь в виду, что интересные морфологические особенности Соте-phorus dybowskii, с одинаковой легкостью подтверждающие различные анти-дарвиновские гипотезы, в то же время не могут служить доказательством правильности только одной теории, именно: теории Ч. Дарвина, так как для этого совершенно необходимо всестороннее и детальное знание той конкретной и, несомненно, очень сложной обстановки, в которой на протяжении не одного миллиона лет совершалась эволюция этой рыбы.
ЛИТЕРАТУРА
Берг Л. С. Номогенез или эволюция на основе закономерностей. Труды Географического института, т. 1, 1922.
Ламарк Ж. Б. Философия зоологии. 1809. М.—Л., 1933.
Abel О. Palaeoblologie und Stammesgeschichte. Jena, 1929.
Doderlein L. Phylogenetische Betrachtungen. Biologisches Centralblatt, Bd. 7, 1888.
E i m er Th. Orthogenesis der Schmetterlinge. Ein Beweis bestimmt gerichteter Entwicklung und Ohnmacht der natiirlichen Zuchtwahl bei der Artbildung. Leipzig, 1897.
Osborn H. F. Aristogenesis, the creative principle in the origin of species. The American Naturalist, vol. 68, N 716, 1934.
Riggs E. S. Preliminary description of a new marsupial sabertooth from the pliocene of Argentina. Geological Series of Field Museum of Natural History, vol. 6, 1933. Scott W. B. and J e p s e n G. L. The mammalian fauna of the White River Oli-gocene. Part I. Insectivora and Carnivora. Transactions of the American Philosophical Society, vol. 28, 1936.
3
А. П. Быстров
ЧАСТЬ ПЕРВАЯ
ПРОШЛОЕ
Прошлое с точки зрения историка охватывает такой период жизни человечества, в течение которого люди уже оставляли после себя те или иные документы. Это были или напечатанные книги, или рукописные летописи, или надписи, высеченные на каменных стенах дворцов и храмов. По этим документам историки пытаются восстановить политические события прошлого и изучить социальные отношения у различных народов на протяжении их так называемой исторической жизни.
Такое понимание прошлого не может быть принято нами, так как за указанный промежуток времени в анатомическом строении человека по существу никаких изменений не произошло. При изучении скелетов представителей, например, такого древнего народа, как египтяне, мы не обнаруживаем решительно никаких отличий от скелетов людей XX века. Таким образом, историческое прошлое человека для морфолога, очевидно, слишком коротко.
Прошлое с точки зрения антрополога является несравненно более продолжительным. Антрополог, изучающий современного человека, интересуется и доисторическими людьми, выясняя и вопрос о их происхождении; но при этом он обычно ограничивается поисками их предков только среди высших форм млекопитающих — обезьян.
Следует признать, что и такое понимание прошлого для морфолога-эволюциониста неприемлемо, так как оно в данном случае совершенно произвольно и искусственно ограничивается моментом появления примитивного человека среди древних обезьян.
Так как у нас нет никаких оснований сомневаться в том, что обезьянообразные предки человека в свою очередь произошли от каких-то иных форм млекопитающих, то мы можем уверенно считать, что среди этих млекопитающих были и «предки» человека.
«Предки» человека должны были быть и среди представителей класса рептилий, которые дали начало млекопитающим. Мало того, мы вынуждены согласиться, что наши «предки» были и среди тех примитивных амфибий, которые послужили исходным пунктом для развития рептилий. Наконец, мы не можем отрицать существования очень далеких «предков» человека и среди древнейших рыб того времени, когда на земле позвоночные были представлены только этими формами животных.
Таким образом, история формирования человека, очевидно, не может быть отделена от эволюционной истории позвоночных вообще. Поэтому с точки зрения морфолога прошлое человека является необычайно продолжительным. Оно по своей длительности равно по крайней мере прошлому позвоночных, т. е. приблизительно 500 000 000 лет.
34
«Предки» человека, несомненно, прошли длинный путь развития, последовательно находясь в составе класса бесчелюстных позвоночных, класса рыб, класса амфибий, класса рептилий и, наконец, класса млекопитающих. При этом многие органы человека складывались не на последних этапах его долгой эволюции; не малое их количество оформилось в различные и нередко в очень древние периоды; некоторые из них возникли в самом начале эволюционного развития позвоночных и затем в том или ином виде сохранились до настоящего времени. Поэтому в строении человека мы обня-
Рис. 17. Филогенез позвоночных.
А—Anaspida; Ас — Acanthodii; С — Coelolepida; Ср—Crossopterygii; Cs — Cotylosauria; D — Dipnoi; E — Embolomeri; H — Heterostraci; L — Lepo-spondyli; M—Mammalia; О — Osteostraci; P — Pelycosauria; R— Rhachitomi;
S — Stereospondyli; T — Therapsida.
О — ордовик; S — силур; D — девой; С — карбои; Р — пермь; Т — триас; J — юра; Сг — мел; Сг — кайнозой; 55—480 — миллионы лет.
руживаем немало несомненных доказательств его родственных связей не только с относительно поздними, но и с очень древними животными.
Изучение такого прошлого человека вынуждает нас широко пользоваться палеонтологическим материалом, так как именно он дает необходимое количество ценных данных, позволяющих проследить основные этапы биологической истории человека.
Накопленный в настоящее время ископаемый материал свидетельствует о том, что родословное дерево позвоночных было необычайно сложным. Оно имело большое количество ветвей, направленных в разные стороны. Огромное большинство из них не имело никакого отношения к истории формирования человека, так как представляло собой такие эволюционные пути, которые вели далеко в сторону и нередко заканчивались полным вымиранием тех животных, развитие которых шло в данном направлении. Такие боковые ветви при изучении биологической истории человека иас, разумеется, интересовать не могут. Наше внимание должен привлекать
3*
35
только тот непрерывный путь, который, в конце концов привел к появлению человека^ При этом мы должны детально изучить все особенности строения тех вымерших животных, которые занимали , в сложном родословном дереве позвоночных такие места, через которые прошел эволюционный путь человека (см. прерывистую линию на рис, 17).
•: -Несмотря, на то, что палеонтология дает в наше распоряжение очень большое: количество разнообразных ископаемых форм, история развития человека все же не может быть прослежена с исчерпывающей полнотой. При изучении этой истории мы, к сожалению, не сможем указать конкретных предков человека среди представителей отдельных вымерших групп позвоночных. Эту задачу в настоящее время следует признать невыполнимой. Она, вероятно, останется для нас непосильной и в течение очень долгого времени. Больше того, возможно, что найти конкретных древнейших предков человека среди многочисленных давно вымерших животных нам никогда не удастся, так как мы не имеем никаких оснований надеяться, что даже в очень отдаленном будущем в распоряжении палеонтологии окажется такой материал, который позволит проследить с достаточной детальностью историю эволюционного развития всех без исключения групп позвоночных. Изучение условий, при которых было возможно окаменение остатков древних животных, показало, что многие из них вообще не могли сохраниться и должны считаться потерянными для нас навсегда. Таким образом, следует признать, что нам известно значительно меньшее количество вымерших форм, чем их существовало в действительности; огромное большинство из них исчезло бесследно и искать их в земле — это все равно, что искать скелет Савонаролы на кладбищах Флоренции.
Детальное изучение тех ископаемых позвоночных, которые в настоящее время найдены и которые позволяют выяснить отдельные фазы формирования человека, всегда сводится к изучению только скелетов, так как никакие мягкие ткани животных в ископаемом состоянии, как правило, не сохранились.
Поэтому, естественно, что при всякой попытке восстановить фактическую историю развития человека морфолог всегда оказывается вынужденным ограничиваться только историей скелета. Следует, конечно, признать, что самая детальная история скелета не является историей всего крайне сложного организма человека, но она все же дает нам достаточно ясное общее представление о ходе его долгой эволюции.
ГЛАВА I
БЕСЧЕЛЮСТНЫЕ ПОЗВОНОЧНЫЕ (AGNATHA)
В отложениях кембрийского периода было найдено немало остатков самых разнообразных беспозвоночных (губки, иглокожие, моллюски, трилобиты), но следов позвоночных обнаружено не было. Это указывает на то, что в это время позвоночные или еще не появились, или их тело еще не имело таких твердых образований, которые могли бы сохраниться в ископаемом состоянии.
Несомненные остатки древнейших позвоночных в настоящее время известны из ордовикских отложений.
В 1889 г. Рогон (Rohon) описал найденные им в глауконитовом песчанике Прибалтики конические зубы, высота которых была равна только 1,5 мм (рис. 18, А). Древность их была около 480 000 000 лет (начало ордовика).
Изучение микроскопического строения этих зубов показало, что их стенка построена из типичного дентина, пронизанного тонкими дентинными канальцами, а наружная поверхность покрыта слоем эмали; внутри зубов имелась довольно большая полость пульпы (рис. 18, В).
Животное, которому принадлежали эти зубы, получило название Palaeodus и должно быть признано самым древним из известных нам в настоящее время ископаемых позвоночных.
Можно уверенно сказать, что Palaeodus был бесчелюстной формой, т. е. являлся представителем класса Agnatha. Найденные зубы не помещались во рту этого животного, а без сомнения сидели в его коже и играли роль чешуи.
Мы не имеем никакого представления о внешнем виде Palaeodus, так как его тело, кроме кожных зубов, несомненно, не содержало никаких твердых образований, которые могли бы сохраниться в ископаемом состоянии. Что же касается чешуек-зубов, то они оказались разбросанными среди зерен глауконитового песчаника и не давали возможности судить о форме тела животного. Нужно думать, что предки Palaeodus не только во внутреннем скелете, но и на своей коже не имели никаких твердых образований, поэтому у нас по существу нет надежды на то, что мы когда-нибудь найдем их остатки. Таким образом, первые этапы эволюции позвоночных, по всей вероятности, никогда не будут выяснены путем изучения ископаемых форм.
Многочисленные кожные зубы у Palaeodus, несомненно, следует рассматривать как органы защитного значения. Покрывая все тело животного,1 они в какой-то мере защищали его от врагов. Так как сами бесчелюстные позвоночные не были хищниками и не могли нападать друг на друга, то их преследователей нужно искать среди древних беспозвоночных. Предполагается, что такими врагами могли быть ракоскорпионы (Gigantostraca).’
37
Они появились, по-видимому, еще в кембрии, а в силуре уже в большом количестве населяли водные бассейны (рис. 19). Ракоскорпионы имели членистое тело, покрытое тонким хитиновым панцирем. Последняя пара
Рис. 18. Кожный зуб Palaeodus (ордовик). А—внешний вид; В — вертикальный разрез (Ротон, 1889).
их конечностей была превращена как бы в весла или лопаты, что давало им возможность медленно плавать и раскапывать ил в поисках своей добычи. У такой формы, как Pterygotus (рис. 19, В), передняя конечность была
Рис. 19. Силурийские ракоскорпионы. А — Eurypterus fischeri-, В — Pterygotus anglicus.
вооружена клешней, при помощи которой животное могло схватывать свою жертву. Дробящий жевательный аппарат делал этих водных хищников опасными врагами примитивных бесчелюстных позвоночных.
Позвоночные из силурийских отложений известны нам по значительно большему количеству форм, чем позвоночные ордовика.
38
Подобно современным миногам, все древнейшие формы Agnatha дышали при помощи жабр, которые представляли собой мешки со складчатыми
Рис. 20. Схема строения головы Agnatha.
b - basibranchiale; с — ceratobranchlale; ch — chorda; е — eplbranchlale; h — hypobranchiale; p — pharyngobranchlale.
стенками, расположенные по обе стороны глотки. Каждый из этих мешков
с одной стороны сообщался с глоткой, с другой — открывался на поверхность кожи. Число жаберных мешков у различных
древних форм колебалось от 6 до 15.
Все эти примитивные позвоночные имели не-сохранявшийся в ископаемом состоянии хрящевой скелет (рис. 20). Их осевой скелет был представлен хорошо развитой хордой (chorda dorsalis). Возможно, что на верхней стороне хорды располагались хрящевые дуги позвонков, каждая из которых состояла из правой и левой половин. Эти половины не были сращены друг с другом. При этом одному позвонку соответствовали, как у современных миног, две такие дуги. Таким образом, каждый позвонок был по существу двойным (дип-лоспондильным) и состоял из четырех самостоятельных хрящевых элементов. Тела позвонков, по всей вероятности, еще отсутствовали, как они отсутствуют у современных круглоротых (миноги и миксины).
Лишенный челюстного аппарата череп (chon-drocranium) древнейших позвоночных состоял из хрящевой мозговой коробки (neurocranium) и ряда жаберных дуг, между которыми располагались отверстия жаберных мешков (splanchnocranium). Каждая из жаберных дуг, вероятно, состояла из четырех элементов: pharyngobranchiale, epibranchiale, ceratobranchiale и hypobranchiale. Послед-
Рнс. 21. Thelodus
ний из этих элементов своим нижним концом
присоединялся к непарному хрящу, носящему на- (силур),
звание basibranchiale.
Приспособление бесчелюстных позвоночных к условиям жизни в пресноводных бассейнах уже в силуре привело к тому, что многие из них превратились в придонные формы и по своему внешнему виду стали напоминать скатов. Они рылись в илистом дне водоемов, питаясь продуктами распада растений и жившими в илу мелкими беспозвоночными.
39
Такой йридонной формой был, например, Thelodus (рис. 21). Он имел широкую уплощенную голову, относительно узкое туловище, заканчивающееся гетероцеркальным хвостом. Размеры этого животного были невелики: он имел около 8 см в длину. Его тело было покрыто многочисленными
Рис. 22. Чешуя Thelodus. А — вид сверху; В — вид снизу; С — вид сбоку; D — вертикальный разрез (силур) (А. П. Быстров, 1955).
и хорошо развитыми кожными зубами, которые по своей форме сильно отличались от кожных зубов Palaeodus.
Каждый кожный зуб Thelodus имел овальные очертания с заостренным концом, который был направлен назад (рис. 22, Л); длинная ось овала
Рнс. 23. Чешуя одного из представителей целолепид (Bystrowia aspidinella Berg.). А — вид сбоку; В — вертикальный разрез (силур) (А. П. Быстров, 1955).
равнялась только 0,75 мм. Верхняя поверхность зуба была относительно плоской. Основание зуба, входившее в кожу животного, было сужено (рис. 22, В, С). Как показало изучение шлифов под микроскопом, стенка такого зуба состояла из типичного дентина, содержащего многочисленные дентинные канальцы. Сверху и с боков зуб был покрыт слоем эмали (рис. 22, D).
Такие чешуйки-зубы, несомненно, в какой-то мере защищали Thelodus от современных ему хищных форм беспозвоночных. Однако такая защита, 40
по-видимому, не была достаточно надежной, поэтому у некоторых представителей силурийских Agnatha вместо многочисленных кожных зубов возник сплошной прочный панцырь, покрывающий переднюю часть их тела.
Рис. 24. Щит Tolypelepis undulata (силур).
Этот панцырь, по всей вероятности, образовался следующим путем.
Изучение строения кожных зубов у различных силурийских Agnatha, относящихся, как и Thelodus, к отряду Coelolepida (рис. 17—С), показало, что у некоторых из них каждый кожный зуб имел на переднем краю своего основания более или менее сильно развитый отросток (рис. 23, А). Этот отросток не состоял из кости (в нем никогда не удавалось обнаружить по
41
лостей остеоцитов); он не был построен и из дентина (в нем не было дентинных канальцев); но вся его масса была пронизана беспорядочно разбросанными тонкими канальцами, иногда собранными в пучки (рис. 23, В). Судя по расположению этих канальцев и их ширине, можно уверенно сказать, что у живого животного в них помещались коллагеновые волокна соединительной ткани. Эти каналы по существу ничем не отличаются от шар-пеевских каналов кости.
Такое твердое вещество — не дентин и не кость, — уже не встречающееся в скелетных образованиях у современных позвоночных, получило название аспидина.
Рис. 25. Вертикальный разрез панцыря Tolypelepis undulata (Х300) (А. П. Быстров, 1955).
Возможно, что появление аспидиновых наростов на основаниях кожных зубов у некоторых представителей целолепид было первым шагом по пути к образованию сплошного панцыря. Если мы допустим, что затем последовало срастание аспидиновых отростков отдельных зубов друг с другом, то это естественно должно было привести к возникновению на теле животного сплошного панцыря.
Подтверждением этого, мне кажется, может служить головной щит представителя отряда Heterostraci (рис. 17—Н), Tolypelepis undulata Pander.
Tolypelepis был небольшим животным; его головной щит имел немного больше 2 см в ширину. Толщина щита была незначительной, она равнялась только 0,7 мм. Вся наружная поверхность панцыря Tolypelepis была покрыта многочисленными гладкими валиками, собранными в отдельные группы (рис. 24).
Изучение микроскопического строения этого панцыря позволило установить, что он состоит из сильно видоизмененных и сросшихся друг с другом кожных зубов (рис. 25). Оказалось, что каждый валик на панцыре
42
представляет собой изогнутую пластинку дентина, пронизанную дентинными канальцами и покрытую тонким слоем эмали. Места сращений каждой пары таких зубов соединены с тонкими вертикальными перегородками, которые в свою очередь сращены с такой же тонкой базальной пластинкой. И перегородки и базальная пластинка в панцыре Tolypelepis построены из аспидина.
Несмотря на тонкость аспидиновых перегородок и базальной пластинки, которые ограничивали заполненные рыхлой соединительной тканью камеры, головной щит Tolypelepis все же мог лучше защищать животное от нападений хищников, чем изолированные кожные зубы, покрывавшие тело целолепид.
Сходное строение имели панцырные щиты и у таких Heterostraci силура, как Poraspis polaris и Pteraspis rostrata. Обе эти формы, несомненно, на-
Рис. 26,
А — Poraspis polaris; В — Pteraspis rostrata (силур).
ходились в родственной связи с Tolypelepis, но были значительно более прогрессивными.
Poraspis (рис. 26, Л) достигал 8 см в длину, его голова сверху была прикрыта сплошным щитом, на переднем конце которого справа и слева имелись небольшие вырезки для глаз. На каждой стороне позади глаза располагалась длинная относительно узкая панцырная пластинка, прикрывавшая жаберный аппарат. Жаберным отверстием у Poraspis служила щель между задним концом этой пластинки и головным щитом. Снизу голова животного была защищена большим вентральным щитом. Все тело Poraspis было покрыто большими чешуями; хвост был гипоцеркальным.
Длина Pteraspis (рис. 26, В) колебалась от 13 до 15 см. Это животное обладало большим количеством панцырных элементов на голове, чем Poraspis, и. было снабжено острым шипом, сидящим на заднем краю головного щита. Туловище Pteraspis было покрыто мелкими ромбическими чешуями.
На наружной поверхности панцырных элементов обеих этих форм можно видеть узкие длинные блестящие валики, ширина которых равняется только 0,2 мм (рис. 27). Изучение вертикальных разрезов панцыря под микроскопом показало, что эти валики построены, как зубы. Они состоят из дентина, пронизанного ветвящимися на концах дентинными канальцами, и покрыты эмалью (рис. 28). Под слоем этих зубов в щитах Poraspis и Pterasp's имеются, как и у Tolypelepis, вертикальные перегородки, присоединяющиеся к базальной пластинке. И перегородки, и базальная пластинка у них несравненно толще, чем у Tolypelepis, поэтому под микроскопом хорошо видно, что эти образования построены из типичного
43
аспидина, в котором легко обнаруживаются многочисленные шарпеевские каналы.
Более прочный панцырь Poraspis и Pteraspis, несомненно, являлся более надежной защитой от нападений хищников, чем панцырь Tolypelepis.
Рис. 27. Щит Poraspis polaris (Киэр и Гейнц, 1935).
У Poraspis и Pteraspis, как, вероятно, и у всех представителей силурийских Agnatha, была хорошо развита так называемая сейсмосенсорная система. Она была образована каналами, проходящими в самом веществе пан-
44
цыря и открывающимися при помощи ряда коротких трубок на наружную поверхность (рис. 29, Л). После удаления верхнего слоя щита сейсмосен-сорная система Poraspis и Pteraspis обнаруживается в виде сети узких каналов (рис. 29, В), а на поперечных разрезах панцыря ее отдельные участки представляются отверстиями, располагающимися в аспидиновых перегородках под слоем кожных зубов (рис. 29, С).
В этих каналах помещались эпителиальные трубочки, в некоторых участках которых имелись группы особых чувствительных клеток, снабженных волосками.
О структуре этих своеобразных органов мы можем судить, разумеется, только на основании изучения строения таких же образований у современных рыб.
Сейсмосенсорная система рыб образована эпителиальными каналами, заложенными в покровных костях черепа и под чешуей (рис. 30). Эти каналы при помощи коротких ветвей открываются на поверхность кожи. В некоторых участках стенки каналов имеются скопления эпителиальных клеток, снабженных волосками и образующих своеобразные рецепторы.
К каждому такому рецептору направляется нерв и охватывает своими ветвями его чувствительные клетки (рис. 31, Л).
На голове рецепторы сейсмосенсорной системы иннервируются ветвями nervus facialis (VII) и на очень небольшом протяжении — n. glossor pharyngeus (IX); на туловище — ветвями nervus vagus (X) (рис. 31, В),
Функция сейсмосенсорной системы до сих пор является для нас физиологической загадкой. Мы не можем ясно представить себе ее действие;, так как у человека от нее не сохранилось никаких следов. Однако нужно
45
думать, что рецепторные органы этой своеобразной системы реагируют на вибрационные раздражения.
Так как вся сейсмосенсорная система при помощи многочисленных отверстий сообщалась с внешней средой (с водой), то легко себе представить, что всякие колебательные движения в ней в той или иной мере приводили в движение волоски клеток сейсмосенсорных рецепторов и возбуждали их,
Рис. 29.
А — Poraspis polaris; В — каналы сейсмосенсорной системы в его дорзальном панцырном щите; С — вертикальный разрез щита; SSS — сей-смосенсорный канал.
что тотчас же по соответствующему нерву передавалось в головной мозг животного. Если сейсмосенсорная система функционировала именно так, то она должна быть признана очень важной для всех водных форм позвоночных. Она позволяла им воспринимать разнообразные колебательные движе-
ния, например волны, вызванные движением других животных, и, может быть, даже слабые отраженные волны, возникающие в результате собственных перемещений позвоночных. Такая функция сейсмосенсорной системы позволяла водным позвоночным даже при слабом освещении легко ориентироваться в окружающей обстановке.
Кроме Agnatha, покрытых изолированными кожными зубами (Coelo-lepida) или снабженных панцырем, состоящим из сильно видоизмененных и сросшихся друг с другом зубов (Heterostraci), в силуре, появились и такие формы бесчелюстных позвоночных, у которых все тело было покрыто несросшимися элементами наружного скелета, нигде не образующими сплошного щита.
46
Эти представители древних бесчелюстных позвоночных, относящиеся к отряду Anaspida (рис. 17 — А), были небольшими рыбообразными формами, их длина колебалась от 2 до 20 см (рис. 32, А, В, С). Голова их была покрыта маленькими чешуйками, а туловище — несколькими рядами
Рис. 31.
А — поперечный разрез сейсмосенсорного канала акулы (Mustelus canis); В — сейсмосенсорная система и ее иннервация у костистых рыб; С — перепончатый лабиринт акулы Scyllorhynchus-, D — чувствительные клетки в перепончатом лабиринте мыши.
длинных узких пластинок. Глаза у Anaspida были хорошо развиты и располагались на боковых сторонах головы. Края беззубого рта были покрыты панцырными пластинками. На боковой стороне переднего'отдела туловища располагались в виде косого ряда круглые жаберные отверстия; их число колебалось от 6 до 15. На спине у Anaspida вместо дорзальных плавников помещался ряд своеобразных шипов. Грудные и брюшные плавники у них
47
отсутствовали; анальный плавник был более или менее развит. Хвост у всех Anaspida был гипоцеркальным.
Как показало изучение под микроскопом, маленькие панцирные элементы Anaspida не содержат ни дентинных канальцев, ни полостей кост-
Рис. 32. Anaspida: Birkenia elegans (A), Pterolepis nitida (В), Pharyngolepis oblonga (С) (силур).
ных клеток и, следовательно, состоят из аспидина; в нем удается обнаружить большое количество шарпеевских каналов (рис. 33).
Anaspida, по всей вероятности, произошли от каких-то примитивных и древних представителей целолепид Эти целолепиды не были придонными
Рис. 33. Поперечный разрез чешуи одного из представителей аиаспид (Х300) (А. П. Быстров, 1955).
формами; их тело было покрыто многочисленными кожными зубами, снабженными аспидиновыми отростками. Срастание этих отростков у первых анаспид привело к образованию на их теле многочисленных коротких и длинных панцырных пластинок, состоящих из аспидина. Кожные зубы на поверхности этих панцырных элементов затем подверглись полной редук-
48
ции. Все Anaspida, несомненно, были очень подвижными животными. Будучи способными быстро плавать, они, по всей вероятности, относительно легко избегали нападений преследующих их врагов. Этим, видимо, и объяс-
няется отсутствие у них такого хорошо развитого панцыря, какой имелся
у менее подвижных Heterostraci.
На верхней поверхности головы у Anaspida между их большими орбитами располагались два непарных отверстия (рис. 34). В заднем меньшем
по своим размерам отверстии помещался пинеальный орган (эпифиз мозга—
epiphysis cerebri). Что же касается большего переднего отверстия, то назначение его, вероятно, навсегда осталось бы для нас непонятным, если бы мы не имели возможности изучить строение и развитие так называемого назогипофизарного органа у современных миног.
У личинки миноги между ротовой бухтой (рис. 35, А — т) и обонятельными ямками образуется так называемый гипофизарный мешок (/г). Его вершина направлена косо вверх и назад к переднему концу хорды (с/г). По мере развития миноги происходит сильное разрастание того участка эктодермы, который располагается между гипофизарным мешком и ротовой бухтой. Это в конце концов приводит к тому, что гипофизарный мешок (/г) и обонятельные ямки (п) смещаются с вентральной стороны головы на дорзальную и у взрослого животного оказываются расположенными следующим образом. Конец гипофизарного мешка (/г) лежит непосредственно под передним концом хорды (с/г); его выводной проток идет косо вверх и вперед, а сближенные обонятельные ямки (/г) открываются в него сзади. Так формируется и так построен на-зогипофизарный орган у современных миног.
У Anaspida строение назогипофизарного органа, несомненно, было таким же, и он
Рис. 34. Голова Pharyngolepis oblonga (вид сверху).
е — отверстие для эпифиза; nh — отверстие для назогипофизарного органа; о — орбита.
открывался у них на верхней стороне головы через отверстие, расположен^ ное кпереди от foramen pineale (рис. 34 — н/г).
В отличие от Heterostraci и Anaspida силурийские Agnatha, относящиеся к отряду Osteostraci, были снабжены панцырем, состоящим из настоящей кости. Эти бесчелюстные позвоночные по всей вероятности являлись потомками анаспид (рис. 17 — О). Представители Osteostraci были сходны с Anaspida тем, что также имели хорошо развитый назогипофизарный орган. Но все они отличались от Anaspida тем, что были типичными придонными формами и по своему виду напоминали скатов.
Костный панцырь Osteostraci, по-видимому, образовался из аспиди-
новых панцырных элементов анаспид вследствие того, что соединительнотканные клетки стали замуровываться в образованный ими аспидин и тем самым превратили его в кость.
Одна из форм Osteostraci — Tremataspis schmidti (рис. 36), имела сильно уплощенную голову, сверху и снизу покрытую панцырными щитами, сросшимися по краям. Длина панцыря равнялась 3,5 см, ширина — 2,5 см. На верхней стороне головного щита Tremataspis можно видеть сближенные друг с другом орбиты, между которыми располагается небольшое пинеаль-
4 А. П. Быстров
49
ное отверстие (рис. 36, Л). Кпереди от этого отверстия лежит еще одно непарное отверстие, служившее местом выхода назогипофизарного органа.
Рис. 35.
А — продольный разрез головы личинки миноги; В — такой же разрез головы взрослой миноги.
ch — хорда; е — эпифиз; h — гипофиз; п — обонятельная ямка; т — ротовая бухта.
Позади орбит в сплошном панцыре Tremataspis имеется непарный дефект, заполненный мелкими изолированными костными элементами. Две
Рис. 36. Tremataspis schmidti (силур). А — вид сверху; В — вид снизу.
пары таких же дефектов помещаются справа и слева неподалеку от боковых краев головного щита.
50
Детальное изучение некоторых Osteostraci дало возможность Стеншио (Stensio, 1927) высказать предположение, что под этими полями, состоящими из мелких костных элементов, у остеостраков располагались электрические органы.
Позади центрального электрического поля у Tremataspis имеются два маленьких отверстия, которые служили местом выхода ductus endolympha-ticus на поверхность кожи животного.
Изучение развития перепончатого лабиринта у современных позвоночных показало, что он образуется в результате впячивания эктодермы в виде кармана в область слуховой капсулы хрящевого черепа (рис. 37, А, В, С, D).
Из этого эктодермального мешка затем формируются перепончатые полукружные каналы и другие отделы внутреннего уха. На начальных
Рис. 37. Последовательные фазы развития перепончатого лабиринта (схема).
стадиях своего развития лабиринт еще сохраняет сообщение с наружной средой при помощи так называемого ductus endolymphaticus (рис. 37, Е, F), но впоследствии этот проток у большинства позвоночных закрывается и лабиринт внутреннего уха совершенно изолируется (рис. 37, G).
Присутствие пары маленьких отверстий на щите Tremataspis свидетельствует о том, что полости его перепончатых лабиринтов еще сообщались с внешней средой. Интересно, что Стеншио (1927) удалось показать, что лабиринт у представителя Osteostraci — Kiaeraspis, имел только два полукружных канала —передний и задний (рис. 38, А); латеральный канал отсутствовал. Такую же особенность мы наблюдаем и у миноги (рис. 38, В).
В нашем распоряжении нет прямых морфологических доказательств того, что все без исключения силурийские (и девонские) Agnatha имели-в своем перепончатом лабиринте два полукружных канала, но у нас нет оснований и отрицать это.
Интересно, что на перепончатый лабиринт позвоночных мы можем смотреть как на образование, по своему происхождению связанное с их сейсмосенсорной системой. Он возник, по всей вероятности, в результате погружения части сейсмосенсорной системы в полость слуховой капсулы хрящевого черепа. Доказательством этого являются следующие факты.
Лабиринт развивается, как и сейсмосенсорная система, из эктодермы,» на первых фазах формирования его полость сообщается с внешней средой. Таким образом, ductus endolymphaticus можно считать гомологичным тем трубочкам, при помощи которых сейсмосенсорные каналы открываются-на поверхности кожи. (У некоторых акул это сообщение лабиринта сохраняется в течение всей жизни, рис. 31, С.)
4*
51
Некоторые участки эпителиальной стенки лабиринта имеют такие же клетки с волосками, какие находятся и в рецепторных аппаратах сейсмосенсорной системы (рис. 31, D).
Построенные таким образом чувствительные поля в перепончатом лабиринте у всех позвоночных иннервируются ветвями n. statoacusticus (VIII). Этот нерв выходит из мозга между n. facialis (VII) и n. glossopha-ryngeus (IX).
Все это дает нам право предполагать, что в процессе развития позвоночных некоторый участок сейсмосенсорного канала отделился от всей остальной системы, погрузился в хрящевой череп и здесь превратился в лабиринт. Это привело к тому, что нерв, выходящий из мозга между n. facialis (VII) и п. glossopharyngeus (IX) и иннервирующий этот участок, превратился в п. statoacusticus (VIII). Таким образом, сохраняя по-прежнему
Рис. 38.
А — левый лабиринт одного из представителей Osteost-raci (Kiaeraspis, Стеншио, 1927); В — левый лабиринт миноги (вид сверху).
способность реагировать на колебательные раздражения, часть сейсмосенсорного канала после очень больших анатомических изменений превратилась в перепончатый лабиринт — орган слуха и сохранения равновесия.
На нижней стороне панцыря Tremataspis можно видеть два ряда жаберных отверстий (рис. 36, В). Передний отдел вентральной части щита у этого животного состоит из отдельных кусков панцыря. Отсутствие сращений между ними позволяло животному опускать и поднимать дно рта во время дыхательных движений и при захватывании пищи. Рот у Tremataspis представлял собой узкую поперечную щель, ограниченную спереди краем головного щита, а сзади — рядом мелких костных пластинок, которые приводились в движение при захватывании пищи.
Толщина панцыря Tremataspis была равна 0,3 мм. При изучении строения панцыря под микроскопом в его веществе легко удается обнаружить полости остеоцитов, соединяющиеся друг с другом тонкими канальцами (рис. 39). Следовательно, панцырь Tremataspis состоял из костной ткани.
В среднем слое панцыря имеются довольно обширные полости, от которых отходят вертикальные каналы, открывающиеся на поверхности щита отверстиями или порами. Эти поры хорошо видны при рассматривании поверхности панцыря под лупой.
Костный панцырь силурийских Osteostraci, несомненно, не был вполне надежной защитой от врагов, так как его толщина колебалась от 0,1 до 0,3 мм. Нужно думать, что именно это обстоятельство было причиной того, что у всех представителей этой группы Agnatha возникли электрические органы, способные своим разрядом оглушить нападающего врага. Таким
образом, наличие электрических органов следует рассматривать как компенсацию недостаточной прочности панцыря.
Тот факт, что сближенные глаза у Osteostraci располагались на дорзальной стороне головного щита, является несомненным доказательством того, что эти своеобразные представители Agnatha, прячась от своих врагов, обычно зарывались в ил на дне водного бассейна и там занимались поиском своей пищи. При этом те хищники, которые случайно наталкивались на них, получали такой электрический удар, который, вероятно, был способен заставить их прекратить преследование.
Рис. 39. Вертикальный разрез панцыря Tremataspis schmidti (Х300) (А. П. Быстров, 1955).
Детальное изучение всех известных нам силурийских Agnatha показывает, что ни один из представителей этого класса не может рассматриваться в качестве возможного предка позвоночных, снабженных челюстным аппаратом (Gnathostomata). Все эти формы были в той или иной степени специализированными животными.
Большинство силурийских целолепид было придонными формами, и их жизнь на дне сильно отразилась на их строении.
Heterostraci, снабженные хорошо развитым панцырем, состоящим из сросшихся и сильно видоизмененных кожных зубов, уже не могли дать лишенного панцыря предка Gnathostomata.
Наличие назогипофизарного органа у Anaspida не дает нам права искать среди них предков рыб, у которых, как известно, перемещения гипофизарного кармана (карман Ратке) на дорзальную сторону головы никогда не происходит.
Появление у Osteostraci костной ткани нельзя рассматривать как указание на их близость к рыбам. Эти Agnatha, как и Anaspida, имели назогипо-
53
физарный орган и, следовательно, предками примитивных рыб .быть не могли. .•
Кроме того, следует иметь в виду, что остатки примитивных Gnatho-stomata найдены в силурийских отложениях. Следовательно, они были современниками многих древних форм Agnatha.
Все это свидетельствует о том, что непосредственных предков Gnatho-stomata следует искать где-то среди ордовикских неспециализированных бесчелюстных позвоночных. Но, к сожалению, о них мы знаем очень мало и, возможно, никогда не будем знать достаточно. Если в скелете этих форм твердыми образованиями, как у Palaeodus, были только кожные зубы, игравшие роль чешуи, то в таком случае мы не имеем оснований надеяться когда-нибудь найти в нижнесилурийских отложениях что-нибудь более ценное, чем рассыпанные в породе кожные зубы.
ЛИТЕРАТУРА
Быстров А. П. Микроструктура панцыря бесчелюстных позвоночных силура и девона. Сборник памяти акад. Л. С. Берга, изд. АН СССР, 1955.
К i а е г I. and Heintz A. The Downtonian and Devonian vertebrates of Spitsbergen. Skrifter om Svalbard og Ishavet, № 40, 1935.
Pander Ch. Monographie der fossilen Fische des Silurischen Systems. St-Petersburg, 1856.
R ohon I. W. Uber untersilurischen Fische. Melanges Geologiques et paleonto-logiques Theses du Bulletin de i’Academie Imperial des Sciences. T. I, St. Peters-bourg, 1889.
Stensio E. The Downtonian and Devonian vertebrates of Spitsbergen. Scrifter om Svalbard og Nordishavet, N 12, 1927.
ГЛАВА II
ЧЕЛЮСТНЫЕ ПОЗВОНОЧНЫЕ (GNATHOSTOMATA). РЫБЫ
Отсутствие в ордовикских отложениях хорошо сохранившихся остатков древнейших представителей Agnatha не дает нам возможности выяснить на палеонтологическом материале все те изменения, которые имели место при превращении бесчелюстных позвоночных в позвоночных, снабженных челюстями.
Однако это обстоятельство не лишает нас права считать, что позвоночные, имеющие челюсти, произошли от каких-то бесчелюстных форм, и пытаться теоретически представить себе все те изменения, которые совершились в скелете позвоночных при переходе от Agnatha к Gnathostomata.
Внутренний скелет первых Gnathostomata, несомненно, был, как и у их бесчелюстных предков, хрящевым, но в нем произошли следующие очень важные изменения.
В позвоночном столбе, по всей вероятности, уже появились хрящевые элементы тел позвонков. Они располагались на нижней стороне хорды в количестве двух пар на каждый позвонок. Таким образом, в состав одного позвонка у Gnathostomata входили две пары дуг и две пары тел. Так построены позвонки у эмбрионов современных акул и это дает нам право предполагать, что именно такое строение имели хрящевые позвонки и у примитивных Gnathostomata (рис. 40).
Одновременно с этими усложнениями в позвоночном столбе происходила и очень сильная перестройка хрящевого скелета жаберного аппарата. По существу именно эта перестройка и превратила Agnatha в Gnathostomata.
В процессе формирования хрящевого черепа Gnathostomata первая и вторая жаберные дуги Agnatha подверглись сильной редукции; они потеряли свои верхние и нижние элементы (pharyngobranchiale и hypobranchiale), а две сохранившиеся части (epibranchiale и ceratobranchiale) превратились в тонкие хрящи, заложенные в кожной складке угла рта. Таким образом возникли так называемые губные хрящи, еще сохраняющиеся у многих современных акул.
После редукции двух первых висцеральных дуг жаберные отверстия, располагающиеся позади каждой из них, закрылись.
Значительно более сильным изменениям подверглась третья жаберная дуга. Ее pharyngobranchiale и hypobranchiale редуцировались. Что же касается epi- и ceratobranchiale, то сни, наоборот, сильно увеличились в своих размерах и превратились в челюстные хрящи; при этом epibran-chiale дало так называемое palatoquadratum, a ceratobranchiale — mandi-bulare или cartilago Meckelii (меккелев хрящ). Место соединения этих элементов превратилось в челюстной сустав.
55
Так как слизистая оболочка рта у зародыша образуется в результате вворачивания эктодермы, то, естественно, она оказывается способной образовывать во рту такие же зубы, какие создавала кожа на теле животного. В результате этого покровы, выстилающие полость рта, у примитивных челюстных позвоночных оказались снабженными зубами. Но здесь зубы уже не могли играть прежнюю роль и с момента образования челюстных хрящей (palatoquadratum и mandibulare) приобрели новую функцию. Прикрепившись своими основаниями к хрящам челюстей, зубы стали теперь уже не орудиями пассивной защиты, а органами, помогающими животному
Рис. 40. Схема строения головы бесчелюстного (Л) и челюстного (В) позвоночного.
b — baslbranchlale; с — ceratobranchiale; ch — chorda; е — eplbranchiale; h — hyp obran chlale; p — pharyngobranchiale.
добывать пищу. Это обстоятельство имело огромное значение для всей дальнейшей эволюции Gnathostomata. Эти животные впервые получили возможность не только использовать недоступные их предкам виды пищи, но и более активно вести борьбу за свое существование. Однако следует иметь в виду, что зубы только что возникшего челюстного аппарата на первых этапах своего развития еще не давали рыбам возможности во всех случаях легко отражать нападения современных им хищников. Поэтому все тело древнейших рыб было покрыто, как у их бесчелюстных предков, многочисленными кожными зубами, игравшими роль чешуи и выполнявшими защитную функцию.
Только в процессе дальнейшей эволюции рыб их челюстные зубы постепенно приобретали все большее и большее значение, в то время как кожные зубы, наоборот, теряли свою ценность и, как мы увидим, подвергались постепенной редукции.
После превращения третьей висцеральной дуги в челюстной аппарат располагающееся позади нее жаберное отверстие превратилось в так называемое «брызгальце» (spiraculum), при помощи которого глотка сохраняла сообщение с внешней средой.
Четвертая дуга жаберного аппарата Agnatha у Gnathostomata подверглась относительно небольшой перестройке. Она потеряла свой верхний
56
элемент (pharyngobranchiale), но сохранила остальные три, превратившись в так называемую подъязычную дугу. Три элемента этой дуги носят следующие названия: epihyale (hyomandibulare, или epibranchiale), ceratohyale (hyoideum, или ceratobranchiale), hypohyale (hypobranchiale).
Следующие пять жаберных дуг у Gnathostomata вошли в состав их splanchnocranium без каких-либо особых изменений; жаберные отверстия
между ними превратились в жаберные щели.
В слуховом аппарате первых челюстных позвоночных оформились три полукружные канала вместо двух, которые имелись в лабиринте бесчелюстных форм. Такое количество полукружных каналов затем сохранилось совершенно неизменным на протяжении всей долгой эволюции позвоночных, вплоть до человека. Принимая во внимание то обстоятельство, что у некоторых современных примитивных акул полость перепончатого лабиринта при помощи ductus endolymphaticus сообщается с внешней средой (рис. 41), нужно думать, что этой особенностью обладали и примитивные Gnathostomata. Это подтверждается результатами детального изучения некоторых акантодий (Ватсон — D. М. S. Watson, 1937).
Одним из очень важных приобретений древних рыб было образование у них спинных, грудных и брюшных плавников, которые
отсутствовали у их предков из
класса Agnatha. Рис. 41. Перепончатый лабиринт химеры
Изучение развития плавников (Chimaera monstrosa).
у эмбрионов современных рыб по-
казало, что плавники первоначально входили в состав общей кожной
складки, которая в виде непарного образования начиналась на спине по-
зади головы, затем окружала хвост и, переходя на нижнюю поверхность туловища, доходила до анального отверстия. Здесь складка разделялась на две части и в виде парного образования тянулась по обе стороны живота до головы животного (рис. 42, А).
В дальнейшем исчезновение отдельных участков этой складки привело к выделению двух спинных плавников, хвостового плавника, анального плавника и парных грудных и брюшных плавников (рис. 42, В).
Древнейшими представителями Gnathostomata следует считать акантодий. Это были небольшие рыбы, известные нам еще из силурийских отложений (рис. 17 — Ас).
Все тело силурийских акантодий было покрыто очень маленькими (0,5 мм в длину) плотно прилегающими друг к другу чешуйками, микроскопическое строение которых было очень сложным. Каждая чешуйка по существу состояла из 6—7 генераций зубов, вставленных друг в друга. Первая закладка каждой чешуйки представляла собой очень маленький зуб, состоя
57
щий из дентинной основы и покрытый сверху и с боков тонким слоем эмали (рис. 43). Основание этого первого зуба было сращено с подстилающим его слоем аспидина. На следующем, этапе развития чешуи вокруг первого.
Рис. 42. Образование плавников у рыб (схема).
центрального, зуба возникал новый слой дентина. Он в виде кольца охватывал зуб со всех сторон и наслаивался непосредственно на эмаль, покры-
Рис. 43. Вертикальный разрез чешуи Acanthodes.
вающую его боковые стороны. При этом дентин заходил и на верхнюю поверхность зуба; он откладывался на эмаль и здесь, оставляя свободной только центральную часть ее. Тонкие, снабженные большим количеством ветвей дентинныр канальцы. во вновь образованном участке дентина обычно переги
бе
бались через острый край эмали центрального зуба. Вскоре после образования этого кольцевидного, или, точнее, цилиндрического, слоя дентина на нем возникал тонкий эмалевый колпак. При этом в центральной своей части эмаль ложилась непосредственно на эмаль первого зуба. Одновременно с этим происходило разрастание аспидинового основания чешуи.
Последовательный ряд таких наслоений дентина, эмали и аспидина приводил к тому, что чешуя акантодий превращалась в очень сложное образование, состоящее из 6—7 вставленных друг в друга зубов и сращенных со слоистым аспидиновым основанием (рис. 43). Сложность строения чешуи является, разумеется, результатом специализации акантодий, но тот факт, что каждая чешуйка все же представляет собой только видоизмененный зуб (или зубы), одновременно указывает на примитивность этих рыб.
Рис. 44. Euthacanthus macnicoli Powrie. Нижний девон (Ватсон, 1937).
Все плавники у этих древнейших рыб были снабжены острыми шипами. На спине, например, у нижнедевонского Euthacanthus (рис. 44) помещались два шипа, позади которых имелись кожные складки. Такими же складками у этой рыбы были снабжены и анальный и брюшные шипы. Грудные шипы Euthacanthus были лишены кожных складок; не имели кожных складок и пять парных шипов, располагающихся на животе рыбы между ее грудными и брюшными шипами.
Наличие на брюхе акантодий двух рядов шипов, несомненно, указывает на то, что эти шипы развились в парной кожной складке.
Все акантодий были хорошо плавающими рыбами. При этом очень мелкие чешуйки, сидевшие в их коже, не мешали их телу изгибаться при движении. Что же касается острых плавниковых шипов, то они, по-видимому, служили наряду с чешуей неплохими орудиями пассивной защиты от нападений хищников.
Палеонтология в настоящее время не располагает таким материалом, который позволил бы проследить тот эволюционный путь, который прошли Gnathostomata от примитивных форм, сходных с акантодиями, до настоящих костных рыб (Osteichthyes). В силурийских отложениях мы не находим ничего, что давало бы нам возможность выяснить историю развития рыб, снабженных костями.
Для объяснения этого странного явления высказывалось предположение (Ромер — A. S. Romer, 1933), что основные этапы своей эволюции Osteichthyes проделали в верховьях рек, куда в начале девона мигрировали из дельт и озер.
59
Такую миграцию могли проделать, разумеется, только хорошо плавающие формы, так как жизнь в верховьях рек требовала от рыб способности бороться с быстрым течением. Но это переселение в небольшие притоки рек оказалось очень выгодным для рыб, так как избавляло их от преследований тех врагов, которые, будучи плохими пловцами, не могли проделать такой же миграции (Gigantostraca).
Жизнь в быстро текущих водах верховий рек, по-видимому, была главной причиной все большего усовершенствования плавательных движений, что привело к появлению среди рыб очень сильных пловцов. Кроме того, скоро возникло разделение этих рыб на две группы по способу питания. Одни из них стали питаться в основном растительной пищей, другие превратились в типичных хищников. К сожалению, все этапы этого процесса не могут быть прослежены нами на ископаемом материале, так как никаких остатков силурийских представителей костных рыб мы не находим. Это объясняется тем, что ручьи и небольшие реки, размывающие свои русла, не оставляют осадочных отложений. Поэтому приходится признать, что история развития костных рыб потеряна для нас навсегда. Мы, по-видимому, никогда не сможем найти такие ископаемые остатки, которые позволили бы проследить все интересующие нас фазы развития Osteichthyes. Их кости были перетерты до полного уничтожения в быстро текущих водах верховий рек.
В девоне костные рыбы, уже проделавшие длинную эволюцию в притоках больших рек, неожиданно появились в дельтах и озерах.
Причиной такого возвращения Osteichthyes в места обитания их предков могло быть перенаселение верховий рек. Однако возможно и даже наиболее вероятно, что это было вызвано климатическими особенностями девонского периода. Ряд геологических данных свидетельствует о том, что климат девона был сухим и жарким, по крайней мере в районе нынешних Канады, южной Гренландии, Шпицбергена, Англии, Прибалтики, Воронежской области, Южного Китая, Северной Австралии. На это указывают и некоторые особенности в строении девонских растений (Криштофович, 1941).
Сухой и жаркий климат, естественно, привел к уменьшению сети водных потоков, что вынудило рыб мигрировать в сохранившиеся водные бассейны, т. е. в большие озера и низовья рек.
О сухом климате девона говорят и некоторые особенности самих костных рыб.
Появившиеся в девоне Osteichthyes были частично представлены двоякодышащими (Dipnoi) и кистеперыми (Crossopterygii) рыбами (рис. 17 — D, Ср). И Dipnoi и Crossopterygii, кроме жабр, имели легкие и были снабжены хорошо развитыми грудными и брюшными плавниками, в которых находились наборы хрящевых (или костных) элементов. Эти элементы, к которым прикреплялись мышцы, делали плавники достаточно сильными органами передвижения (рис. 45, А, В).
Легкие двоякодышащих и кистеперых рыб, несомненно, образовались в качестве приспособлений, дающих возможность использовать для газообмена кислород воздуха. Совершенно очевидно, что древнейшие костные рыбы некоторое время находились в таких условиях, в которых одного жаберного дыхания для снабжения организма кислородом было совершенно недостаточно. Такие условия могли создаться только в периодически пересыхающих бассейнах, в воде которых нередко оказывался недостаток кислорода.
Периодическое пересыхание водоемов, по всей вероятности, было причиной появления у двоякодышащих и кистеперых рыб и сильных грудных и брюшных плавников, которые позволяли им в некоторых случаях переползать из пересыхающих водоемов туда, где вода еще сохранялась.
60
Двоякодышащие и кистеперые рыбы для истории эволюции позвоночных имеют исключительно большое значение, так как изучение их дает возможность выяснить многие вопросы, связанные с проблемой происхождения амфибий.
Dipterus — среднедевонский представитель Dipnoi — был небольшой рыбой; его длина колебалась от 30 до 35 см. В строении этой интересной фор* мы были смешаны как признаки примитивности, так и признаки высокой специализации.
Все тело Dipterus было покрыто циклоидной чешуей, что указывает на некоторую прогрессивность этой рыбы по сравнению с древнейшими формами кроссоптеригий.
Dipterus имел два спинных плавника и гетероцеркальный хвост. Его трудный и брюшные плавники были очень длинными и в средней своей части
Рис. 45.
А — двоякодышащая рыба Dipterus (Форстер-Купер — Forster-Cooper, 1937);
В — кистеперая рыба Osteolepis. Средний девон (Ярвик, 1948).
были покрыты несколькими рядами чешуй (рис. 45, А). В хрящевом черепе подвергалась окостенению только задняя часть neurocranium (рис. 46, В). Покровные кости, лежащие на chondrocranium, были очень многочисленными, а в переднем отделе черепа срастались в один сплошной щит (рис. 46, Л). На нёбе имелось только пять костей: два сошника (vomer), Два птеригоида (pterygoideum) и непарный парасфеноид (parasphenoideum) (рис. 46, В); жаберный аппарат был прикрыт большим operculum. Ргае-maxillare и maxillare у Dipterus подверглись полной редукции, поэтому у него отсутствовали краевые зубы. В составе покровных костей нижней челюсти не было dentale; это привело к тому, что и нижняя челюсть была лишена краевых зубов (рис. 46, D). Каналы сейсмосенсорной системы у Dipterus проходили в самом веществе покровных костей и открывались на их наружную поверхность многочисленными отверстиями (рис. 46, А, С).
Изучение микроскопического строения чешуи Dipterus и покровных элементов его черепа показало, что эти образования состоят из типичной костной ткани. В ней можно видеть многочисленные полости остеоцитов С тонкими канальцами, отходящими от них в разные стороны (рис. 47). На наружной поверхности как чешуй, так и всех покровных костей Dipterus располагаются очень своеобразные по своей форме кожные зубы. Эти зубы сращены друг с другом и образуют слой, в котором проходят каналы, открывающиеся на поверхность кости круглыми отверстия
61
ми. В каждом отдельном зубе имеется небольшая полость пульпы в виде короткого вертикального канала. Стенки зубов толсты и пронизаны дентинными канальцами, ветвящимися на концах. Плоские вершины зубов н их боковые стороны покрыты слоем эмали (А. П. Быстров, 1942).
Среднедевонский Dipterus являлся типичной травоядной рыбой. Необходимость перетирать стебли водных растений требовала какого-то зубного аппарата, и он при отсутствии краевых зубов образовался в результате усиления так называемых шагреневых зубов. Эти очень маленькие
Рис. 46.
А, В, С — череп Dipterus-, D — нижняя челюсть Dipterus (Грэхем-Смит и Вестол — Graham-Smith a. Westoil, 1937);
Е — зуб двоякодышащей рыбы верхнего девона.
конические зубы покрывали у предков Dipterus их птеригоид, лежащий на нёбной поверхности черепа, и praearticulare нижней челюсти. Отдельные шагреневые зубы у Dipterus сильно увеличились в своих размерах и образовали на pterygoideum и praearticulare несколько веерообразно расположенных рядов (рис. 46, В). Между этими рядами у Dipterus еще сохранились очень маленькие шагреневые зубы (рис. 48, А). Дробящие зубы Dipterus отличались большой прочностью (рис. 48, В). Они состояли почти сплошь из дентина и полость их пульпы была представлена системой относительно узких вертикальных каналов, от которых во все стороны расходились дентинные канальцы. Дентин центральной части зуба не содержал дентинных канальцев; здесь протоплазменные отростки одонтобластов исчезли и просветы дентинных канальцев закрылись, в результате чего возник очень прочный витродентин. Наружная поверхность зуба была покрыта слоем эмали.
При закрывании рта ряды дробящих зубов нижней челюсти располагались между рядами соответствующих зубов на нёбной поверхности черепа. Такой своеобразный зубной аппарат, по всей вероятности, позволял Dipterus без труда раздавливать раковины тех маленьких моллюсков и пан-
62
цыри тех ракообразных, которые вместе с растительностью могли попадать в его рот.
У верхнедевонских представителей двоякодышащих рыб радиально расположенные ряды изолированных конических зубов на нёбе и на нижней челюсти срослись таким образом, что образовали гребнистые зубы, которые являлись еще более мощным аппаратом раздавливания (рис. 46, Е).
Dipnoi, часто встречавшиеся в средне- и верхнедевонских отложениях, продолжали существовать, но в меньшем количестве и в карбоне, и в перми.
Рис. 47. Вертикальный разрез покровной кости черепа Dipterus (Х300) (А. П. Быстров, 1942).
Они жили в пресноводных бассейнах и в течение всего мезозоя. Эти рыбы сохранились и до настоящего времени в Южной Америке (Lepidosiren ра-radoxa), в Африке (Protopterus annectens) и в Австралии (Neoceratodus forsteri).
Некоторые морфологи конца прошлого столетия (Коп — Соре; Боас— Boas) предполагали, что предков наземных позвоночных (Tetrapoda) следует искать среди двоякодышащих рыб. В настоящее время можно считать установленным, что Dipnoi, уже на первых этапах своей эволюции потерявшие praemaxillare, maxillare и dentale, и приобретшие высокоспециализированный зубной аппарат, предками первых амфибий быть не могли. Этих предков, несомненно, нужно искать только среди кистеперых рыб. Поэтому детальное изучение различных представителей Crossopterygii имеет очень большое значение для решения вопроса о выходе позвоночных из воды на сушу.
Древний и примитивный представитель кистеперых рыб Osteolepis-был современником Dipterus. Обе эти рыбы обитали в одних и тех же пресноводных бассейнах среднего девона, на это указывает тот факт, что в соответствующих отложениях их кости нередко бывают перемешаны. .
63-
Osteolepis представлял собой рыбу, размеры которой у разных видов колебались от 7 до 45 см (Ярвик — Е. Jarvik, 1948). Тело Osteolepis было покрыто ромбической чешуей (рис. 45, В); его хвост, как у Dipterus, был гетероцеркальным; на спине располагались два дорзальных плавника; грудные и брюшные плавники на некотором протяжении были покрыты чешуей.
Позвоночный столб Osteolepis не окостеневал и поэтому не сохранялся в ископаемом состоянии. Мозговой череп (neurocranium) состоял из двух
Рис. 48.
А — поперечный разрез шагреневого зуба Dipterus (диаметр зуба равен 0,1 мм);
В — поперечный разрез основания нёбного зуба Dipterus (длинная ось зуба равна 0,6 мм) (А. П. Быстров, 1942).
окостеневших частей — передней и задней; передняя, ethmosphenoideum (рис. 49), располагалась на уровне глаз и заключала в себе обонятельные капсулы и передний отдел мозга; задняя, oticooccipitale (рис. 49), нижним своим участком охватывала передний конец хорды и содержала в себе задний отдел головного мозга и перепончатые лабиринты. Обе эти части мозгового черепа были соединены друг с другом подвижно.
Покровных костей, лежавших на окостеневших и хрящевых частях черепа, у Osteolepis было меньше, чем у Dipterus.
Затылочный край черепной крыши был образован тремя костями: парным extrascapulare laterale (рис. 50, В — Е. /.) и непарным extrascapu-lare medium (Е. т.). Кпереди от этих костей располагались dermosupra-occipitale (Dso), tabulare(T)H supratemporale (St). Все эти кости, соединенны e друг с другом швами, лежали на окостеневшем заднем отделе neurocranium и отделялись от остальных покровных костей черепа щелью. Наличие этой щели позволяло переднему и боковым отделам черепа двигаться вверх и вниз по отношению к oticooccipitale и лежащему на нем комплексу покровных костей.
64
В переднем отделе крыши черепа Osteolepis можно различить два ра-rietale (Р), в шве между которыми располагается отверстие для пинеального органа (foramen parietale). По обе стороны от теменных костей помещаются intertemporalia. Остальные покровные кости здесь обычно срастаются в один общий щит, лежащий на ethmosphenoideum. На боковых сторонах этого щита имеются два отверстия, ведущие в полости обонятельных капсул. Под орбитами и позади них располагаются lacrimale (рис. 50, А — L), jugale (Ju) и postorbitale (Ро).
Челюстной край черепа был образован praemaxillare, сросшимся с комплексом костей, вошедших в состав носового щита, и длинным maxil-lare (Л4). На нижних краях обеих этих костей сидели зубы.
Рис. 49. Мозговой череп Osteolepis.
Позади postorbitale, jugale и maxillare помещалось большое squamosum (S7) и quadratojugale (Qj), а за ними praeoperculum (Pop). Между squamosum и tabulare у Osteolepis имелось небольшое отверстие — spira-culum (рис. 50, А, В — S). Задний отдел боковой части черепа был образован двумя большими подвижными костями, прикрывавшими жаберный аппарат operculum (Op) и suboperculum (So).
Нёбная часть черепа Osteolepis еще недостаточно изучена, по она, вероятно, была построена так же, как у других представителей кроссопте-ригий (см. Eusthenopterori).
В состав нижней челюсти Osteolepis входит ряд покровных костей, развившихся вокруг меккелева хряща (mandibulare), задний участок которого окостеневал и в виде кости, носящей название articulare, соединялся с quadratum черепа.
На наружной поверхности меккелева хряща у Osteolepis лежало длинное dentale, на верхнем краю которого имелся ряд острых зубов (рис. 50, А — D). Позади этой кости располагались сросшиеся у Osteolepis в одно образование postspleniale (Ps), angulare (Ап) и supraangulare (Sa). У других кистеперых рыб эти три кости отделены друг от друга.
У Osteolepis, как и у всех кистеперых рыб, на нижней стороне головы располагалась пара больших костей, носящих название gulare (рис. 50, С — G); спереди между ними вклинивалось небольшое непарно^ gulare medium (G. т.). Между gulare и каждой ветвью нижней челюсти лежал ряд покровных костей — branchiostegalia (Вг).
Во многих покровных костях черепа Osteolepis проходили каналы сейсмосенсорной системы. От этих каналов отходили короткие боковые веточки, которые открывались на поверхность кости небольшими отверстиями.
5 А. П. Быстров
65
Каналы сейсмосенсорной системы, переходя из одной кости в Другую, Всегда пересекали их центры окостенений; иными словами, они шли от центра окостенения одной покровной кости к центру окостенения другой.
В системе сейсмосенсорных каналов различают несколько отделов* носящих специальные названия. Надглазничный канал (canalis supraor-bitalis, рис. 51 — с. s.) у Osteolepis на значительном своем протяжении про-
Рис. 50. Череп Osteolepis. А — вид сбоку; В — вид сверху; С — вид снизу (Ярвик, 1948).
Ас — anocleithrum; Ап — angulare; Br —branchlostegale; Cl — clavlcula; Ct — cleithrum; D — dentale; Dso— dermosupraocclpltale; E. I. — extrascapulare laterale; E. m. — extrascapulare medium; О — gulare; O. m. — gulare medium; Ic — Interclavlcula; Ju — jugale; L — 1асг!та1е;Л1— maxlllare; Op — operculum; P — parletale; Po — postorbitale; Pop — praeoperculum; Ps — postsplenlale; Pt — posttemporale; QJ — quadratojugale; 5— splraculum; Sa — supraangulare; Sc — supraclelthrum; So — suboperculum; Sp — spleniale; Sq — squamosum; St — supratemporale; T — tabulare.
ходил в костном щите передней части черепа и только в заднем своем отделе пересекал центры окостенения parletale и intertemporale, где соединялся с подглазничным каналом (canalis infraorbitalis — с. i.), проходившим в ргае-maxillare, lacrimale и jugale.
Как бы непосредственным продолжением надглазничного канала являлся canalis temporalis (с. t.), последовательно пересекавший центры окостенения supratemporale, tabulare и extrascapulare laterale. Правый и левый canales temporales в заднем отделе крыши черепа соединялись друг с другом при помощи поперечного анастомоза, а затем шли в покров-
66
ные кости плечевого пояса, а из них на туловище, где располагались в чешуе.
В области центра окостенения jugale от подглазничного канала начинался canalis jugalis (с. j.), который, пройдя в squamosum и praeopercu-lum, располагался позади челюстного сустава, а затем под названием canalis mandibularis (с. tn.) шел в supraangulare, angulare, postspleniale и spleniale.
Рис, 51. Каналы сейсмосенсорной системы в костях черепа Osteolepis (Ярвик, 1948).
с. I. — canalis Infraorbitalls; с. j. — canalis jugalis; c.m. — canalis mandibula-rls; c. s. — canalis supraorbitalis; c. t. — canalis temporalis.
Изучение микроскопического строения покровных костей и чешуй Osteolepis показало поразительное сходство их структуры со структурой костей и чешуй Dipterus. Основная их масса также состоит из костной ткани, в которой хорошо видны многочисленные полости остеоцитов; наружная поверхность костей покрыта слоем таких же сросшихся друг с другом кожных зубов. По своей форме эти зубы очень похожи на соответствующие зубы у Dipterus. Единственным отличием их является значительно большая густота дентинных канальцев (рис. 52) (А. П. Быстров, 1942).
Это удивительное сходство, несомненно, свидетельствует об общем происхождении обеих этих рыб. Можно уверенно сказать, что Osteolepis и Dipterus ведут свое начало от какого-то общего предка. Расхождение их эволюционных путей по двум направлениям, по всей вероятности, явилось результатом приспособления к питанию различными видами пищи. При этом Osteolepis превратился в типичного хищника, в то время как Dipterus стал в основном травоядной рыбой.
67 5*
Ha pterygoideum и praearticulare у хищного Osteolepis имелись Шагреневые зубы, но они не подверглись таким изменениям, как у Dipterus. Эти зубы остались очень небольшими коническими образованиями, приросшими своими основаниями непосредственно к кости, и помогали Osteolepis удерживать живую добычу в челюстях. Интересно, что шагреневые зубы при утолщении тех костей, на которых они помещались, постепенно замуровывались в костное вещество. После погружения одной генерации шатре.
Рис. 52. Вертикальный разрез покровной кости черепа Osteolepis (X 300) (А. П. Быстров, 1942).
невых зубов в кость на ее поверхности появлялись зубы новой генерации, которые через некоторое время так же замуровывались. Полностью погруженные в костное вещество зубы обычно подвергались разрушению в результате перестройки кости. В шлифах из pterygoideum и praearticulare Osteolepis нередко можно видеть полуразрушенные шагреневые зубы (рис. 53).
Зубы, расположенные на краях praemaxillare, maxillare и dentale, имели форму немного изогнутых острых конусов. Покрывавшая их эмаль была снабжена большим количеством очень острых продольных гребней (рис. 54, Л). Большие зубы, или так называемые «клыки»,1 сидящие на vomer, palatinum, transversum черепа и на короноидах нижней челюсти, у Osteolepis имели складчатую стенку (рис. 54, В).
Примыкая непосредственно к голове, у Osteolepis располагались шесть покровных костей плечевого пояса: posttemporale (рис. 50—Pt), supracleith
1 «Клыки» кистеперых рыб совершенно не соответствуют клыкам рептилий и млекопитающих.
68
rum (Sc), anocleithrum (Ac), cleithrum (Cl), clavicula (С/) и непарная маленькая interclavicula (Ic). Все эти кости лежали на несохранившихся хрящевых элементах плечевого пояса, к которому присоединялись также хрящевые элементы грудного плавника.
Хрящевой тазовый пояс и набор хрящей в брюшном плавнике у Osteolepis не подвергались окостенению, а потому и не были найдены в ископаемом состоянии. О строении этого отдела скелета Osteolepis мы не имеем никакого представления.
Рис. 53. Шагреневые зубы Osteolepis (X 300) (А. П. Быстров, 1942).
Кроме Osteolepis, в среднедевонских отложениях была найдена другая кистеперая рыба, Glyptolepis, размеры которой были по крайней мере в Два раза больше размеров Osteolepis.
Glyptolepis (рис. 55, Л) как типичная кистеперая рыба имел длинные грудные и брюшные плавники, в средней своей части покрытые чешуей. Чешуя Glyptolepis была не ромбической, а циклоидной. Это являлось признаком прогрессивности Glyptolepis по сравнению с Osteolepis.
Череп Glyptolepis был построен в общем так же, как у всех кистеперых
рыб.
На наружной поверхности покровных костей и на чешуе у этой рыбы имелись многочисленные бугорки. Изучение их строения под микроскопом показало, что каждый из них представляет собой зуб, состоящий из большого куска дентина, покрытого очень толстым слоем эмали (рис. 56). Основание такого зуба было непосредственно сращено с костью. Таким образом, чешуя и кости черепа Glyptolepis не были покрыты сплошным слоем сросшихся друг с другом кожных зубов, как у Osteolepis (А. П. Быстров, 1939).
Челюстные зубы Glyptolepis отличались необычайной сложностью рвоего строения (рис. 57). Они, как и у Osteolepis, представляли собой ко
69
нические образования, приросшие своим основанием к покровным костям челюстей.
На поперечных их разрезах видно, что их дентинная стенка имеет многочисленные длинные складки, направленные своими вершинами к центру полости пульпы. Каждая из этих складок снабжена большим количеством боковых ветвей. Полость пульпы от вершины до основания зуба заполнена тонкими балками своеобразной кости, в которой только изредка удается обнаружить полости остеоцитов. Все эти балки покрыты толстым слоем ден-
Рис. 54. Поперечные разрезы зубов Osteolepis. А — часть стенки вершины зуба; В — основание зуба (длинная ось основания равна 1,25 мм) (А. П. Быстров, 1942).
тина. В результате этого полость пульпы зуба превратилась в систему очень узких каналов.
Эмаль, покрывающая наружную поверхность зуба Glyptolepis, снабжена такими же продольными острыми гребнями, как у Osteolepis.
Такая структура зубов Glyptolepis свидетельствует о том, что этот представитель кистеперых рыб был одновременно и примитивной и очень специализированной формой.
Верхнедевонский Holoptychius, по-видимому, может рассматриваться как вероятный потомок Glyptolepis. На это указывают некоторые интересные особенности строения его покровных костей и зубов.
Holoptychius был большой и сильной рыбой (рис. 55, В). Судя по величине его зубов, которые часто встречаются в изолированном виде в девонских отложениях, длина этой кроссоптеригии в некоторых случаях достигала 3 или, может быть, даже 4 м.
Строение плавников и форма чешуй у Holoptychius были такие же, как и у Glyptolepis. Наружная поверхность черепных костей этой рыбы была покрыта многочисленными бугорками, но каждый из них не представлял собой типичного зуба. Изучение микроскопического строения этих образований' позволило установить, что они содержат в себе дентин, в котором
70
хорошо видны дентинные канальцы, но слой эмали в них полностью отсутствует (рис. 58). Вместо него каждый бугорок покрыт слоем типичной ко-
Рис. 55.
А — Glyptolepis (Иекель— Jaekel, 1911); В — Holoptychius. Девон.
сти. В этом слое хорошо видны полости остеоцитов, отростки которых соединяются с концами дентинных канальцев (А. П. Быстров, 1939).
Рис. 56. Вертикальный разрез кожного зуба Glyptolepis (X 300).
Такая структура бугорков на костях Holoptychius, по-видимому, свидетельствует о том, что они являются результатом неполной редукции кож-
71
ных зубов, сходных по своему строению с теми, которые покрывали кости Glyptolepis. В них уже исчезла эмаль, но еще сохранился дентин. Это дает нам право рассматривать Holoptychius как кистеперую рыбу, которая стоит на пути к полной потере кожных зубов. При этом не исключена возможность, что она является прямым потомком среднедевонского Glyptolepis.
Строение челюстных зубов Holoptychius также указывает на некоторое упрощение организации этой рыбы.
Рис. 57. Поперечный разрез зуба Glyptolepis (длинная ось равна 3,25 мм) (А. П. Быстров, 1939).
Как видно на поперечных разрезах зубов Holoptychius, их многочисленные ветвистые дентинные складки по существу ничем не отличаются от таких же образований в зубах Glyptolepis (рис. 59).
Вся полость пульпы зубов у Holoptychius, как и у Glyptolepis, сверху до низу заполнена балками почти бесклеточной кости, но на этих балках нет* наслоений дентина. В результате этого полость пульпы оказывается большей по объему, а зуб менее компактным и, следовательно, менее прочным.
На поверхности эмали у Holoptychius еще сохранялись продольные гребни, но количество их было меньшим, чем у Glyptolepis', они имелись не на каждой выпуклости стенки зуба.
Таким образом, у Holoptychius дентин на балках в полости пульпы исчез полностью, а острые гребни на эмали подверглись значительной редукции.
То, что совершенно ясно намечается в строении покровных костей и зубов у этой кистеперой рыбы, оказалось доведенным до конца у верхнедевонского Polyplocodus (Пандер — Pander, 1860). К сожалению, о внешнем виде этой кистеперой рыбы мы не имеем никакого представления, так как знаем ее только но фрагментам черепа.
72
На покровных костях черепа Polyplocodus также имелись многочисленные бугорки, но все они состояли только из кости и никаких следов дентина уже не содержали (рис. 60) (А. П. Быстров, 1939).
В полости пульпы челюстных зубов Polyplocodus полностью отсутствовали те балки бесклеточной кости, которые еще имелись в зубах Holoptychius (рис. 61).
Дентинная стенка зубов Polyplocodus еще образовывала ветвистые складки, но они не были так многочисленны, как у Glyptolepis и Holoptychius, и имели меньшее количество боковых ответвлений.
Рис. 58. Вертикальный разрез бугорка на покровной кости черепа Holoptychius (X 300).
Эмаль на зубах Polyplocodus была совершенно лишена гребней; они у этого представителя верхнедевонских кистеперых рыб уже подверглись полной редукции.
Очень интересной и совершенно новой особенностью зубов Polyplocodus являлось расщепление периферических участков дентинных складок на протяжении от 1/3 до 2/3 их длины; полного разделения двух слоев дентина на всей длине складки никогда не наблюдалось; также не было разделения этих слоев в боковых ветвях складок. В глубокие щели, образовавшиеся таким образом, с периферии входили две пластинки кости. Эти пластинки во многих местах соединялись друг с другом при помощи коротких перекладин.
Хорошо изученная верхнедевонская кистеперая рыба, Eusthenopte-гоп, достигала по меньшей мере 1,5 м в длину (рис. 62).
Eusthenopteron, как все кроссоптеригии, имел два спинных плавника, но его хвостовой плавник был не гетероцеркальным, а дифицеркальным; тело рыбы было покрыто циклоидной чешуей.
73
Большинство позвонков у Eusthenopteron окостеневало.
Мозговой череп (neurocranium) состоял, как у Osteolepis, из двух окостеневших кусков — ethmosphenoideum и oticooccipitale, соединённых друг с другом подвижно.
В нижней части oticooccipitale имелся канал, в котором проходил передний конец хорды (рис. 63). На боковых поверхностях этой части мозгового черепа у Eusthenopteron. были обнаружены суставные площадки, к которым присоединялось hyomandibulare подъязычной дуги (рис. 63 — f. hm ).
Рис. 59. Поперечный разрез зуба Holoptychius (длинная ось равна 3,25 мм) (А. П. Быстров, 1939).
На нижней стороне oticooccipitale располагались две овальные покровные кости, покрытые шагреневыми зубами. Таких костей до сих пор не было описано ни у одной кроссоптеригии.
На нижней стороне этмосфеноида у Eusthenopteron лежал парасфеноид (Ps), в котором имелось небольшое отверстие — canalis hypophyseos. Кпереди от этого канала поверхность парасфеноида была покрыта шагреневыми зубами.
Передний острый конец парасфеноида входил между парой сошников (vomer — V), снабженных зубами. Самой большой костью неба являлось pterygoideum (Pt), на поверхности которого находилось большое поле шагреневых зубов. Вдоль бокового края птеригоида лежали palatinum (Pl) и transversum (Тг). Обе эти кости несли большие «клыки» и ряд маленьких зубов. Передний и боковые края нёбной части черепа были образованы нижними краями praemaxillare (Рт) и maxillare (Л4), на которых сидели зубы. Непосредственно позади maxillare лежало quadratojugale (Qj). Между этой костью и задним концом птеригоида (Pt) помещалось quadratum (Qu), представлявшее собой результат окостенения заднего участка хрящевого palatoquadratum. Блок quadratum принимал участие в образовании челюстного сустава.
74
Рис. 60. Вертикальный разрез бугорка на покровной кости черепа Polyplocodus (X 300).
Рис. 61. Поперечный разрез зуба Polyplocodus (длинная ось равна 3,5 мм) (А. П. Быстров, 1939).
Рис. 62. Eusthenopteron. Верхний девон.
Рис. 63. Череп Eusthenopteron (вид снизу) (Ярвик, 1944).
ch — choana; f.a. — fossa adductorla;/. hm. — facies hyomandibularls; Af — maxilJare; Pl— pa-latlnum’, Pm — prae-n]axUlare;P$—parasphe-noldeum; Pt — pterygoideum; Qj—quadraioju-gale; Qu — quadratum; Tr — transversum; V — vomer*
Непосредственно перед quadratum имелось большое отверстие, ограниченное quadratojugale, maxillare и pterygoideum. Это отверстие, носящее название fenestra adductoria (f. а.), служило местом выхода группы жевательных мышц, прикрепляющихся к нижней челюсти. В переднем отделе черепа располагались хоаны (choanae — С/г),ограниченные краями ргае-maxillare, maxillare, vomer и palati-num. Эти отверстия имели большое значение для кистеперых рыб. При вдыхании воздуха он входил в ноздри, потом через хоаны поступал в полость рта и затем уже направлялся в легкие.
В состав крыши черепа у Eusihe-nopteron, как и у других кистеперых рыб, входили следующие кости: ех-trascapulare laterale и medium, der-mosupraoccipitale, tabulare, parietale, supratemporale, intertemporale, frontale, praefrontale, postfrontale (рис.64, 65). У молодых экземпляров Eus-thenopteron, достигавших только 53 см в длину, в переднем (носовом) отделе крыши черепа имелось много маленьких косточек; у взрослых полутораметровых рыб весь этот комплекс костей, как у Osteolepis, срастался в сплошной щит, прикрывавший перед-
нюю часть этмосфеноида.
Боковую часть черепа Eusthe- Рис. 64. Череп Eusthenopteron (вид nopteron составляли: praemaxillare, сверху) (Ярвик, 1944).
maxillare, lacrimale, postorbitale, jugale, squamosum, quadratojugale, praeoperculum и две большие кости, прикрывавшие жаберный аппарат — operculum и suboperculum (рис. 65).
Рис. 65. Череп Eusthenopteron (вид сбоку) (Ярвик, 1944).
На наружной поверхности меккелева хряща, входя в состав покровных костей нижней челюсти, лежали dentale, spleniale, postsplcniale, angu-
77
fare и stipraangulare (рис. 66, А). С внутренней стороны меккелев хрящ почти на всю свою длину был прикрыт praearticulare, поверхность которого
Рис. 66. Нижняя челюсть Eusthenopteron. А — вид снаружи; В — вид снутри (Ярвик, 1944).
Ап — angulare; Со — coronoideum; D — dentale; Ic — intercoronoideum; Pa — praearticulare; Pc — praecoronoldeum; Ps — postsplenlale; Sa — supraangulare; Sp —• splenlale.
была усеяна огромным количеством маленьких конических шагреневых зубов (рис, 66, В). Эти зубы по своему строению не отличались от шагреневых
Рис. 67. Шагреневые зубы Eusthenopteron (X 300).
зубов на парасфеноиде и птеригоидах. Они представляли собой очень маленькие дентинные образования, покрытые тонким слоем эмали. В про-
78
Рис. 68. Поперечные разрезы нижних челюстей Eusthenopteron (Д) и Benthosuchus (В). D — dentale; Ic — Intercoronoldeum; Ра — praearticulare;
Ps — postsplenlale
рейсе роста кости при ее постепенном утолщении эти зубы не спадали, а замуровывались вновь образованными слоями костного вещества и затем, находясь в кости, подвергались полному разрушению при ее перестройке. Следы различных фаз разрушения шагреневых зубов обычно удается наблюдать в каждом препарате, представляющем собой вертикальный разрез парасфеноида, птеригоида или praearticulare (рис. 67) (А. П. Быстров, 1939). Такую своеобразную судьбу имели шагреневые зубы у всех без исключения кистеперых рыб.
На верхней поверхности cartilage Meckelii лежали три коро-ноида (praecoronoideum, inter-coronoideum и coronoideum), снабженные рядами маленьких зубов и так называемыми «клыками».
На поперечных разрезах нижней челюсти Eusthenopteron видно, что меккелев хрящ помещался в очень широком канале, проходившем вдоль всей
Между ветвями нижней челюсти на вентральной поверхности головы Eusthenopteron лежали такие же кости, как у Osteolepis и у всех других кистеперых рыб.
Проходящие в костях каналы сейсмосенсорной системы у Eusthenopteron снабжены очень большим количеством боковых ветвей (рис. 69).
нижней челюсти (рис. 68, А).
Рис. 69. Каналы сейсмосенсорной системы в костях черепа Eusthenopteron (Ярвик, 1944).
Структура покровных костей Eusthenopteron и строение его зубов позволяют думать, что эта кистеперая рыба могла быть непосредственным потомком Polyplocodus.
Черепные кости Eusthenopteron были покрыты бугорками, которые состояли только из костной ткани (рис. 70). Следовательно, в этом отношении Eusthenopteron не отличался от Polyplocodus.
Что же касается челюстных зубов, то они являлись как бы результатом дальнейшей эволюции зубов Polyplocodus (рис. 71). Их полость пульпы была совершенно свободна от каких-либо костных балок; эмаль на своей поверх-
79
Рис. 70. Вертикальный разрез бугорка на покровной кости черепа Eusthenopteron (X 300).
Рис. 71. Поперечный разрез зуба Eusthenopteron (длинная (А. П. Быстров. 19391-
ось равна 3 мм)
Рис. 72. Череп Eusthenopteron. А — вид сверху: В — вид снизу (Ярвик, 1944).
Ac — anocleithrum; Cl — clavicula; Ct — cleithrum; Ic — interciavicula; Pt — posttem-porale; Sc — supraclelthrum .
Рис. 73. Череп Eusthenopteron (В — operculum и suboperculum удалены, чтобы показать связь костей плечевого пояса с черепом).
Ас — anoclelthrum; Cl — clavicula; Ct — cleithrum; Pt — posttemporale; Sc — supraclelthrum.
6 А. П. быстро»
ности не имела продольных гребней. Следовательно, в этих деталях зубы Eusthenopteron были сходны с зубами Polyplocodus. Отличием их являлась следующая очень интересная особенность. Все их дентинные складки были расщеплены до конца. При этом два слоя дентина в каждой складке были
раздвинуты и во всех ее боковых ветвях. Такого полного расщепления зуб-
ных складок у Polyplocodus еще не было.
В промежутки между двумя дентинными слоями зубных складок у Eusthenopteron, как и у Polyplocodus, с периферии входили две пластинки кости, соединенные короткими перемычками, но у Eusthenopteron они проникали во все расщепленные боковые ветви дентинных складок.
Как у всех кистеперых рыб, у Eusthenopteron непосредственно позади черепа
располагался ряд покров-
Рис. 74. Cleithrum (Ct) и scapula (S) ных костей плечевого пояса
Eusthenopteron (вид изнутри). (рис. 72, А, В). Самая верх-
няя из этих костей, posttem-porale (Pt), присоединялась к задним краям extrascapulare laterale и medium. Далее следовали supracleithrum (Sc), anocleithrum (Ле), большое cleithrum (Ct), clavicula (Cl) и непарная interclavicula (Ic). Таким
Рис. 75. Плечевой пояс и грудной плавник Eusthenopteron (Грегори и Равен— Gregory and Riven, 1941).
Ac — anoclelthrum; Ct — cleithrum; H — humerus; Pt — posttemporale; R — radius; Sc — supraclelth-rum; U — ulna.
образом, получалось незамкнутое кольцо покровных костей, охватывающее переднюю часть туловища рыбы и соединяющееся своими верхними концами с костями крыши черепа. Сбоку между собственно черепом и плечевым поясом находились две большие подвижные кости, прикрывающие жаберный аппарат — operculum и suboperculum (рис. 73, А). Если удалить эти кости, то перед нами откроется большое пространство, в котором лежали жабры, и анатомическая связь покровных костей плечевого пояса с такими же костями крыши черепа будет совершенно очевидной (рис. 73, В).
82
Cleithrum у Eusthenopteron прикрывало развившуюся в результате окостенения хряща лопатку, scapula. Эта кость прикреплялась к трем шероховатым площадкам на внутренней поверхности cleithrum (рис. 74 — S).
К лопатке присоединялись кости передней конечности. В наборе этих костей можно различить плечевую кость, humerus (рис. 75—Н), лучевую, radius (R), и локтевую, ulna ((7). Остальные кости трудно сравнивать с костями передних конечностей наземных позвоночных. Это, вероятно, объясняется тем, что грудные плавники верхнедевонского Eusthenopteron были в значительной степени измененными в результате специализации и уже не могли дать конечности примитивных Tetrapoda.
Если у кистеперых рыб в состав плечевого пояса, кроме скапулокора-коида, входил еще ряд покровных костей, непосредственно связанных с черепом, то тазовый пояс этих рыб был совершенно лишен покровных костей. Это различие, по всей вероятности, объясняется только непосредственным соседством таких костных и хрящевых элементов плечевого пояса, как scapula и coracoideum, с черепом. Очевидно, плечевой пояс был покрыт костями, развивающимися в соединительной ткани, только потому, что весь передний отдел тела рыбы был защищен такими костями. Поэтому можно считать, что posttemporale, supracleithrum, anocleithrum, cleithrum, clavicula и interclavicula являются по существу покровными костями черепа, распространившимися на плечевой пояс.
Тазовый пояс рыб, находящийся на большом расстоянии от черепа, естественно, не мог иметь никаких покровных костей. Его хрящевые элементы были заключены в мышцах подвижного отдела туловища и прикрыты только чешуей.
В состав тазового пояса Eusthenopteron входила только одна парная кость (os coxae), представлявшая собой результат окостенения хрящевого образования (рис. 76). Она состояла из сросшихся ilium, pubis и ischium. Эта комплексная кость, лежащая в мышцах, не соединялась с позвоночным столбом рыбы.
Скелет задней конечности у Eusthenopteron состоял из бедренной кости — femur (рис. 76—Fe), большой и малой берцовых костей — tibia и fibula (Т, F), и еще четырех костей, соответствие которых костям стопы наземных позвоночных не ясно.
Fe T
Рис. 76. Брюшной плавник. Eusthenopteron (Грегори и Ривен, 1941). Ге — femur; F — fibula; Г— tibia.
ЛИТЕРАТУРА
Быстров А. П. (Bystro w). Zahnstruktur der Crossopterygier. Acta Zoologies. Bd. 20, 1939.
Быстров А. П. Deckknochen und Zahne der Osteolepis und Dipterus. Ibidem, Bd. 23, 1942.
Forster - Cooper C. The Middle Devonian Fish fauna of Achanarras. Transactions of Royal Society Edinburgh, vol. 59, 1937.
Graham-Smith B. A. a. Westoil T. S. Ona new longheaded fish from the Upper Devonian of Scaumenac Bay, P. Q. Canada, Transactions of the Roy. Soc. of Edinburgh, vol. 59, 1937.
Gregory W. K. and R i v e n . Structure of the vertebral column in Eusthenopteron ioordi Whiteaves. Journal of Paleontology, vol. 13, N 1, 1939.
Gregory W. K. and Riven. Studies on the origin and early evolution of paired, fins and limbs. Annal of the New-York Academy of Sciences, vol. 42, N 15, 1941.' J a г v i k E. On the species of Eusthenopteron found in Russia and the Baltic States,.
Bull, of Geol. Instit. of Uppsala, vol. 27, 1937.
J a r v i k E. On the structure of the snout of Crossopterygians and lower Gnathostomes in general. Zoologiska Bidrag fran Uppsala, Bd. 21, 1942.
6* 83.
J a r v i к E. On the dermal bones, sensory canals and pitlines of the skull in Eusthenopteron foordi Whiteaves, with some remarks on Eusthenopteron Save-Soderbergh Jarvik. Kungl. Svenska Vetenskapsakademins Handlinger, Bd. 21, N 3, 1944.
Jarvik E. On the exosceletal shouldergirdle of Teleostomian fishes, with special reference to Eusthenopteron foordi Whiteaves. Ibidem, Bd. 21, N 7, 1944.
J arvik E. On the Morphology and Taxonomy of the Middle Devonian Osteolepid fishes of Scotland. Ibidem, Bd. 25, N 1, 1948.
J a e к e 1 O. Die Wirbeltiere. Eine Obersicht uber die fossilen und lebenden Formen. Berlin, 191.1.
Pander Ch. Ober die Saurodipterinen, Dendrodonten, Glyptolepiden und Cheirole-piden des Devonischen Systems. St. Petersburg, 1860.
Romer A. S. Eurypterid influence on vertebrate history. Science, vol. 78, 1933.
Save-Soderbergh G. The dermal bones of the head and the lateral line system in Osteolepis macrolepidotus Ag. R. Soc. Sci. Ups. N. Acta, Ser. IV, vol. 2, N 2, 1933.
S a v e-Soderb ergh G. Notes on the dermal bones of the head in Osteolepis macrolepidotus Ag. and the interpretation of the lateral line system in certain primi-; five vertebrates. Zoologiska Bidrag fran Uppsala, Bd. 20, 1941.
Watson D. M. S. The Acanthodian fishes, Philosophical Transactions of the Royal Society of London, vol. 228, 1937.
ГЛАВА III
НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ (TETRAPODA). АМФИБИИ
Ископаемые остатки амфибий из карбоновых отложений были известны уже давно (с 1841 г.). И эти находки давали все основания считать, что первые наземные позвоночные (Tetrapoda) появились в начале каменноугольного периода. Это мнение было общепринятым на протяжении ряда десятилетий. Но в 1896 г. американский палеонтолог Марш (Ch. Marsh, 1831 — 1899) описал след так называемого Thinopus antiquus из девонских отложений Северной Америки (рис. 77). Марш предполагал, что это был отпечаток левой стопы какой-то древней амфибии. На плитах из тех же отложений были найдены и круглые ямки, являвшиеся следами капель дождя, и трещины, вызванные высыханием почвы. Все это позволяло думать, что по мягкой сырой почве на берегу девонского водного бассейна- прошла какая-то примитивная амфибия; затем почва высохла и следы животного вместе со следами капель дождя сохранились на поверхности затвердевшей позже породы.
Так как отпечатки ног различных ископаемых животных в более поздних отложениях удавалось находить не один раз, то не было ничего удивительного в том, что подобные следы могли сохраниться и в отложениях девонского периода. Однако отпечаток, описанный Маршем, все же не позволял уверенно сказать, что в данном случае мы действительно имеем дело со следом позвоночного; такое углубление на поверхности породы могло возникнуть и каким-нибудь иным путем. Надо признать, что трактовка подобных образований — вообще дело очень трудное. Поэтому «след» так называемого Thinopus antiquus еще не доказывал, что Tetrapoda существовали уже в девонское время.
Вопрос о существовании девонских наземных позвоночных был разрешен только спустя 35 лет, когда в 1931 г. на восточном берегу Гренландии в верхнедевонских отложениях было найдено семь черепов очень примитивных амфибий. Это обстоятельство заставило палеонтологов уверенно передвинуть момент появления первых Tetrapoda из нижнего карбона по крайней мере в средний девон. Таким образом, древность амфибий оказалась равной приблизительно 325 000 000 лет.
Изучение всех известных в настоящее время кистеперых рыб показывает, что среди них трудно указать непосредственных предков амфибий.
Такие верхнедевонские формы, как Eusthenopteron, Polyplocodus и Holoptychius не могли быть предками первых наземных позвоночных хотя бы потому, что примитивные Tetrapoda уже были их современниками. Что же касается Glyptolepis и Osteolepis, то у нас нет прямых доказательств того, что именно от них произошли амфибии. Glyptolepis, судя по строению его зубов, был высокоспециализированной формой и мы не можем его считать
85
Рис. 77. Thinopus antiquus Marsh (длина следа равна 10 см; Марш— Marsh, 1896).
предком Tetrapoda. Он, по всей вероятности, был непосредственно связан с Holoptychius, который, по-видимому, был его потомком. На это указывает микроскопическая структура покровных костей и зубов обеих этих рыб.
Не менее специализированной формой следует считать и Osteolepis с его слоем своеобразных кожных зубов на наружной поверхности черепных костей и на чешуе.
Казалось бы, что такая специализация могла быть непреодолимым препятствием для происхождения от этой кистеперой рыбы первых амфибий, черепные кости которых лишены всяких следов кожных зубов. Однако нам известен следующий интересный и очень важный факт. Черепные кости и чешуя древнейшей двоякодышащей рыбы Dipterus были покрыты слоем кожных зубов, которые по своему строению почти не отличались от соответствующих образований у Osteolepis. Но это обстоятельство не помешало непосредственным потомкам Dipterus— двоякодышащим рыбам верхнего девона и карбона — быстро потерять всякие следы кожных зубов. Можно допустить, что это же явление могло произойти и при переходе от кистеперых рыб к первым Tetrapoda.
Челюстные зубы Osteolepis не были настолько специализированными, чтобы из них не могли образоваться зубы древних амфибий. Но у Osteolepis покровные кости, входившие в состав переднего отдела черепа, обычно срастались между собой, и это указывает на то, что эта рыба предком Tetrapoda, у которых подобных явлений не наблюдалось, по всей вероятности, все же не была.
Принимая все это во внимание, можно сказать, что ни одну из известных
нам кистеперых рыб непосредственным предком амфибий мы уверенно считать не можем. Но предками амфибий, несомненно, были какие-то неизвестные нам кистеперые рыбы. Эти рыбы покинули пресноводные бассейны среднего девона и, приспособившись к жизни на суше, превратились в первых примитивных амфибий.
Выход кистеперых рыб из воды на сушу не был вызван недостатком пищи в реках и озерах. На это указывает тот факт, что и для первых амфибий пресноводные бассейны по-прежнему оставались единственным местом, откуда они добывали свою пищу (рыб).
Выход рыб из воды на сушу не вызывался и большим обилием пищи на суше. На пустынных берегах девонских водоемов хищные кроссоптери-гии, несомненно, не могли найти для себя подходящего питания.
Выход рыб из воды на сушу не вызывался и наличием в воде каких-либо опасных для них врагов, так как более сильных, чем Crossopterygii, позвоночных в пресноводных бассейнах девона по существу не было.
Выход кистеперых рыб на сушу без сомнения был обусловлен только сухим и жарким климатом девонского периода. Эта особенность климата приводила к периодическим пересыханиям пресноводных бассейнов и делала их во многих случаях совершенно непригодными для жизни рыб.
86
Уже один тот факт, что появившиеся в девоне представители Osteichthyes были двоякодышащими и кистеперыми (по существу также двоякодышащими) рыбами, говорит о том, что и их миграция из верховий рек и образование у них легких было обусловлено постепенным пересыханием водоемов, которое сопровождалось сильным уменьшением количества кислорода в воде. Кислород быстро поглощался гниющими растениями и делал воду совершенно непригодной для жизни рыб. Нужно думать, что периодическое возникновение таких условий сначала вызвало появление у рыб легких, а затем стало вынуждать их мигрировать из тех бассейнов, в которых дальнейшее существование становилось для них уже невозможным.
Можно предполагать, что переселение рыб в низовья рек и в относительно большие пресноводные бассейны в первое время улучшало для них условия существования. Но скоро жаркий и сухой климат стал неблагоприятно отражаться и на этих водоемах; и здесь начались полные или почти полные периодические пересыхания, сопровождающиеся быстрым нарастанием дефицита растворенного в воде кислорода.
В такой тяжелой обстановке у рыб были только три способа сохранить свое существование, именно:
1) мигрировать через дельты рек в море, в воде которого количество кислорода во все сезоны года оставалось неизменным;
2) остаться в высохшем бассейне и как-то приспособиться к переживанию периода полного безводия;
3) переползти из высохшего бассейна в такой, в котором вода еще сохранилась в достаточном количестве.
Все эти способы были использованы рыбами девона.
Те кистеперые рыбы, которые обитали в реках, смогли воспользо* ваться первым способом и уйти через дельты в море. Мигрировавшие в море формы дали группу так называемых Coelacanthini. Эти рыбы появились еще в пресноводных бассейнах верхнего девона и представляли собой боковую ветвь, отделившуюся от Crossopterygii. Их череп по сравнению с черепом кистеперых рыб сильно изменился; многие его покровные кости исчезли (Nesides, Rhabdodermd).
Приспособившись к жизни в соленой воде, Coelacanthini дожили до настоящего времени. Непосредственным потомком морских юрских (Dn-dina — рис. 78, Л) и меловых (Масгорота — рис. 78, В) форм является современная Latimeria chaltitntiae, впервые пойманная 22 декабря 1938 г. около Южной Африки. Эта рыба имела металлически-синий цвет, достигала 150 см в длину и весила больше 57 кг. Ее тело было покрыто циклоидной чешуей; на спине имелись два дорзальных плавника; хвостовой плавник был построен, как у Undincr, грудные и брюшные плавники — как у типичной крос-соптеригии (рис. 78, С).
Легкие, которые были унаследованы Coelacanthini от их предков крос-соптеригий, изменили свою функцию и превратились в плавательный пузырь, служащий гидростатическим аппаратом.
Я присоединяюсь к мнению тех морфологов, которые считают, что плавательный пузырь является видоизменением легких, а не наоборот, и думаю, что легкие возникли раньше, чем плавательный пузырь. Наличие плавательного пузыря у большинства современных рыб указывает на то, что все они являются потомками тех девонских форм, которые были вынуждены жить в периодически пересыхающих пресноводных бассейнах, в которых часто возникал дефицит кислорода. Интересно отметить, что все современные акулы лишены плавательного пузыря. Это объясняется тем, что их предки из группы силурийских акантодий не имели легких.
Двоякодышащие рыбы воспользовались вторым способом сохранения своего существования. Возможно, что причиной того, что Dipnoi не мигри,
87
ровали в море, было то обстоятельство, что они обитали главным образом в таких богатых растительностью замкнутых пресноводных бассейнах, кац небольшие озера и болота. Будучи почти всеядными формами, для которых в пересыхающих бассейнах еще долго сохранялось достаточное количество пищи, они во время засух оставались на месте своего прежнего обитания.
Рис. 78. Coelacanthini. А — Undina (юра); В — Масгорота (мел); С — Latimeria chalumnae (рецент)
По мере того, как уменьшалось количество воды и исчезал в ней кислород, поглощаемый распадающимися растениями, эти рыбы все чаще и чаще пользовались для дыхания легкими, захватывая в них воздух, в котором количество кислорода, несмотря на сухость климата, разумеется, оставалось неизменным. В поисках воды и пищи эти рыбы вынуждены были ползать по илистому дну почти совершенно пересохшего водоема. Это в процессе естественного отбора в конце концов привело к значительному усилению у них тех своеобразных грудных и брюшных плавников, которые как основные органы передвижения были выработаны еще их предками в пересыхающих верховьях рек.
88
В момент полного высыхания водоема эти рыбы стали закапываться в ил и впадать в спячку на весь период засухи. При этом их очень сильно замедленный газообмен происходил через легкие и воздух поступал к неподвижной рыбе через канал, оставшийся в илу. Канал был закрыт слоем
пористого грунта.
Так, несомненно, вели себя девонские двоякодышащие рыбы. Так ведут себя и современные двоякодышащие рыбы экваториальной Африки (Protopterus annectens, рис. 79) и Южной Америки (Lepidosiren para-doxa), впадающие в спячку на 7 — 8
месяцев.
Когда сезон засухи сменялся сезоном обильных дождей, спячка Dipnoi кончалась и эти рыбы снова становились деятельными. С этим временем года был связан и период их размножения, и из икры, отложенной в воду, быстро развивались личинки. По всей вероятности, личинки девонских Dipnoi были снабжены такими же наружными жабрами, как и личинки современных двоякодышащих рыб Африки и Южной Америки (рис. 80, В, С).
Несомненно, только это удивительное приспособление к очень неблагоприятным условиям существования дало возможность двоякодышащим рыбам сохраниться, начиная от девона до наших дней (рис. 81).
Так как постепенное пересыхание замкнутых бассейнов и нарастание дефицита кислорода в их воде, по всей вероятности, сопровождалось массовой гибелью рыб,
Рис. 79. Protopterus annectens — африкан. ская двоякодышащая рыба во время лет ней спячки.
то кроссоптеригии как типичные хищники, по-видимому, начинали чувствовать недостаток пищи значительно раньше, чем Dipnoi, пи
тавшиеся растительностью.
Это обстоятельство вынуждало их делать попытки покинуть те водоемы, в которых еще могли оставаться Dipnoi, и переползать в ближайшие более многоводные бассейны. Наличие сильных брюшных и грудных плавников и наличие легких, которые были выработаны еще во время пребывания кистеперых рыб в пересыхающих верховьях рек, позволило им осуществить эти вначале недалекие миграции.
Таким образом, кистеперые рыбы воспользовались третьим из названных выше возможных путей сохранения своего существования в очень тяжелых условиях.
Если мы допустим, что климат девонского периода постепенно становился все более сухим и жарким, то это должно было в каждый сезон вызывать полное пересыхание все большего и большего количества замкнутых пресноводных бассейнов. Это явление в свою очередь обусловливало непре-
89
рывное увеличение расстояний между высохшими водоемами и теми, которые еще сохраняли воду. Иными словами, увеличение засушливости климата неминуемо вело к удлинению миграционных путей кистеперых рыб.
Рис. 80. Личинки современных двоякодышащих рыб. А — Neoceratodus forsteri (Австралия); В — Protopterus annectens (Африка); С — Lepidosiren paradoxa (Южная Америка).
Совершенно естественно, что подобные миграции были нелегким делом для рыб, и они, несомненно, во многих случаях погибали, не доползая до
Рис. 81. Современные двоякодышащие рыбы. А — Neoceratodus forsteri (Австралия); В — Protopterus annectens (Африка); С — Lepidosiren paradoxa (Южная Америка).
воды. Но в тех ареалах, в которых непересыхающих многоводных бассейнов было относительно много, кроссоптеригии сравнительно легко осуществляли свои сезонные переселения туда, где вода еще сохранилась, или не мигрировали совсем, если оказывались обитателями никогда полностью
«о
не высыхающего водоема. Это обстоятельство, по-видимому, было главной причиной того, что они просуществовали в течение всего среднего и верхнего девона, а одна форма (Megalichthys) даже дожила до перми.
В тех же ареалах, в которых водоемы были небольшими и, следовательно, быстро пересыхающими, поиски воды для кистеперых рыб были крайне затруднительными. Их миграции в таких условиях далеко не всегда приводили к благоприятным результатам и на путях переселений развертывалась жестокая борьба за жизнь.
Большим преимуществом в этой борьбе являлось наличие сильных грудных и брюшных плавников, так как они ускоряли движение рыб по суше и увеличивали их шансы на спасение. Не менее важным было и хорошее развитие легких, так как это удлиняло срок пребывания рыб на суше и помогало им доползти до воды. Поэтому всякая кистеперая рыба, имеющая относительно слабые плавники и недостаточно развитые легкие, легко по-
Рис. 82. Скелет Ichthyostega. Верхний девон Гренландии (Ярвик, 1955).
гибала в пути, и, наоборот, всякая рыба, способная преодолеть длинный путь и долго оставаться вне воды, имела больше шансов на сохранение и, следовательно, на размножение. Кистеперые рыбы, как и Dipnoi, откладывали икру только в период дождей, когда и все пересохшие бассейны вновь наполнялись водой. Личинки кроссоптеригий, как и личинки двоякодышащих рыб, были снабжены, кроме внутренних, и наружными жабрами. В этом можно быть уверенным потому, что такие жабры образуются и у личинок их потомков — амфибий.
У нас нет оснований сомневаться в том, что жестокий естественный отбор на путях вынужденных миграций в конце концов должен был привести к превращению грудных и брюшных плавников кистеперых рыб в передние и задние конечности наземных позвоночных. Одновременно с этим тот же отбор неминуемо должен был сделать легкие единственным органом газообмена у взрослых форм первых амфибий.
Жабры кистеперых рыб, разумеется, не сохранились у их наземных потомков. Они подверглись полной редукции; при этом исчезли и такие прикрывающие жаберный аппарат кости, как operculum и suboperculum.
Ряд покровных костей, которые у кистеперых рыб лежали на нижней стороне головы между ветвями нижней челюсти (gulare и branchiostegale), у амфибий также не сохранились. Полная редукция этих костей сделала более подвижным дно полости рта, и лежащие здесь мышцы стали выполнять роль дыхательных мышц. Опуская дно полости рта, амфибии набирали воздух в рот, втягивая его через ноздри и хоаны; затем, закрывая ноздри и сокращая мышцы дна рта, они вгоняли его в легкие.
После редукции ряда покровных костей черепа оставшиеся образовали сплошной щит на голове древних амфибий, что послужило поводом назвать их стегоцефалами (Stegocephalia), т. е. щитоголовыми.
Древнейшие из известных нам стегоцефалов, верхнедевонские Ichthyo-stegidae (восточная Гренландия), обладали рядом очень интересных осо-
91
Рис. 83. Череп Tchthyostegopsis. Вид сверху (Сэве-Зедерберг — Save-Soderbergh, 1932).
Рис. 84. Череп Ichthyostega (вид сбоку) (Ярвик, 1952).
Ап — angulare; D —dentale; F — frontale; Ju — jugale; L — lacrimale; M — maxillare; iV — nasale; P — arletale; Pf—praefrontale; Pm — praemaxlllare; Po — postorbttale; Pop — praeoperculum; Prf — praefrontale; p spostsplenlale; Qj — quadratojugale; Sa — supraangulare; Sm — septomaxillare; Sop —suboperculum;
Sp — spleniale; Sq — squamosum; St — supratemporale; T — tabulare.
бенностей, которые указывают на несомненную их связь с кистеперыми рыбами (рис. 82).
Каналы сейсмосенсорной системы у ихтиостегид проходили так же в костях, как у всех кистеперых рыб, и открывались на поверхность черепа при помощи отверстий (рис. 83). У этих древнейших амфибий еще сохранялось praeoperculum и даже небольшое suboperculum (рис. 84).
Рис. 85. Череп ichthyostega (вид снизу) (Ярвик, 1952).
М — maxlllare; Pl — palatinum; Pm — praemaxillare; Ps — parasphenoldeum;
Pt — pterygoideum; Qj — quadratojugale; Qu — quadratum; Tr — transver sum; V—vomer.
Нёбная поверхность черепа ихтиостегид во многих отношениях напоминала соответствующую часть черепа кистеперых рыб (рис. 85). Соединение птеригоидов (Pt) с мозговой коробкой располагалось почти на средине длины черепа; небольшой парасфеноид (Ps) имел сквозное отверстие для гипофиза; в вентральной части oticooccipitale имелся широкий канал для хорда.
Позвонки ихтиостегид, впервые изученные Ярвиком (Е. Jarvik, 1952, 1955), были очень похожими на позвонки кистеперой рыбы Eusthenopteron (рис. 86). Тело каждого позвонка состояло из кольцевидного, незамкнутого сверху гипоцентра (hypocentrum), охватывавшего толстую хорду. Позади дуги позвонка располагался небольшой плевроцентр (pleurocentruml. имевший овальную форму. Ребра присоединялись к дуге позвонка и к его гипоцентру.
93
Судя по строению скелета (рис. 82), верхнедевонские ихтиосте-гиды имели наряду с признаками примитивности такие признаки специализации, что Ярвик (1955) не считает возможным рассматривать этих животных в качестве предков более поздних, карбоновых, стегоцефалов и думает, что ихтиостегиды были боковой ветвью в эволюционном развитии амфибий.
Стегоцефалы, появившись в верхнем девоне, просуществовали до конца триаса. В конце триаса они вымерли полностью, и уже ни одного из них мы не находим в юрских отложениях.
В настоящее время нам известно достаточно большое количество самых разнообразных форм этих ископаемых амфибий. Все они обладали способ-
Рис. 86. Позвонки Eusthenopteron (А, В) и Ichthyostega (С, D) (Ярвик, 1952).
а — arcus; с — costa; Л — hypocentrum; р — pleurocentrum.
ностью передвигаться по суше при помощи своих еще не очень сильных четырех конечностей. Их размеры колебались от величины современного тритона почти до размеров нильского крокодила.
Стегоцефалов по строению их позвонков принято разделять на четыре группы: Embolomeri, Lepospondyli, Rhachitomi и Stereospondyli.
Первая группа наиболее примитивных форм появилась в начале карбона или еще в девоне и просуществовала до начала перми (рис. 17—Е).
Одновременно с Embolomeri жили представители группы Lepospondyli, появившиеся в карбоне и вымершие также в начале пермского периода (рис. 17 — L).
Rhachitomi, по-видимому, представляли собой боковую ветвь Embolomeri. Появившись в конце карбона, они жили в течение всего пермского периода и вымерли в начале триаса (рис. 17 — R).
Наконец, группа Stereospondyli, отделившаяся от Rhachitomi, в конце перми полностью вымерла на границе между триасом и юрой. Представители этой группы были последними стегоцефалами (рис. 17 — S).
Эмболомерные стегоцефалы карбона имели хорошо окостеневшие позвонки.
Каждый их позвонок состоял из дуги (arcus dorsalis) (рис. 87—a. d.) и двух дисковидных тел; оба они были двояковогнутыми и в центре имели
94
небольшие отверстия, сквозь которые проходила хорда. Первый костный диск, hypocentrum (Л), был немного тоньше второго, pleurocentrum (р), и на своей боковой поверхности имел небольшую ямку для сочленения с ребром. Дуга позвонка соединялась с обоими телами и имела хорошо развитые остистый, поперечные и суставные отростки.
Двухголовчатые ребра у эмболомерных стегоцефалов присоединялись к позвонку в двух местах: к суставной площадке на конце поперечного отростка дуги и к суставной площадке на боковой поверхности hypocentrum (рис. 87, А, В).
Рис. 87. Эмболомерные позвонки Eogyrinus attheyi (Ватсон).
А. В.—грудные позвонки; D, С — хвостовые позвонки.
а. a.—ыат dorsalis; а. г.—arcus ventralis; с—costa; Л—hypocentrum; р—pleurocentrum.
Весь позвоночный столб этих амфибий может быть разделен только на два отдела: на туловищный и хвостовой, но точную границу между ними у наиболее примитивных форм установить трудно.
Почти все хвостовые позвонки у Embolomeri были снабжены так называемой вентральной дугой, которая представляла собой большой костный отросток, отходящий от нижней поверхности hypocentrum. В основании этого отростка имелось отверстие, сквозь которое проходил хвостовой участок аорты (рис. 87, С, D).
Передняя поверхность первого позвонка примыкала непосредственно к костям мозгового черепа.
Мозговой череп (neurocranium) у эмболомерных стегоцефалов окостеневал только в заднем своем отделе (рис. 88, В); в области обонятельных капсул он оставался в течение всей жизни животного хрящевым. Таким образом, по сравнению с кистеперыми рыбами у древнейших амфибий наблюдалась меньшая степень окостенения хрящевого черепа. Затылочный отдел мозгового черепа у них был образован непарным basioccipitale (рис. 89 — Во) и парным exoccipitale (£о). В месте контакта с позвоночным столбом
95
на этих костях имелось коническое углубление. Таким образом, этот уча сток мозгового черепа напоминает позвонок и дает право сравнивать basi-occipitale с телом, a exoccipitale — с двумя несращенными друг с другом половинками дуги позвонка.
Кпереди от exoccipitale и над ним располагалось костное непарное supraoccipitale (So), а перед ним лежало prooticum (рис. 88—Р). К боковой поверхности exoccipitale присоединялось paroccipitale (рис. 88—Ро), также развивающееся в результате окостенения хряща. Передняя часть окосте-
Рис. 88. Череп Palaeogyrinus decorus (карбон) (Ватсон, 1926).
Во — basiocclpltale; Bs — baslsphenoldeum; Ео — exoccipitale; Ер — eplpterygoldeum; Ju — jugale; L — lacrimale; M—maxillare; P — prooticum; p. bp. — processus basipterygoldeus; Pr — praemaxlllare; Po — paroccipitale; psf — pseudofenestra ovalls; Pt — pterygoldeum; Qj — quadratojugale; Qu — quadratum; Se — sphenethmoldeum; Sq — squamosum.
невшего neurocranium была образована большим костным сфенэтмоидом (sphenethmoideum,рис. 88, В, С — Se),в стенках которого имелись отверстия для выхода некоторых черепномозговых нервов. В основании мозгового черепа лежало костное basisphenoideum (рис. 88, В — Bs), снабженное так называемыми базиптеригоидными отростками (processus basipterygoideus, рис. 88, В — р. вр.).
Весь этот комплекс костей охватывал полость, в которой помещался головной мозг стегоцефала, а в боковых участках заднего отдела мозгового черепа лежали лабиринты внутреннего уха.
Слабо окостеневающий splanchnocranium стегоцефалов, по всей вероятности, был построен так же, как построена соответствующая часть черепа у современных амфибий (рис. 90).
Несколько редуцированное palatoquadratum у них соединялось с мозговым черепом, вероятно, в двух пунктах. Первое из этих соединений, сохранившееся у некоторых современных Urodela (рис. 90), представляло со-
9J
бой эпиптеригоид (epipterygoideum, рис. 88, 90—Ер), направленный от ра-latoquadratum к верхнему краю мозгового черепа; при помощи второго соединения palatoquadratum было связано со слуховым отделом neurocranium. Задний конец palatoquadratum принимал участие в образовании сустава с задним концом mandibulare или cartilago Meckelii (рис. 90—Md).
Некоторые участки этого отдела черепа у стегоцефалов окостеневали. Так, подверглось окостенению epipterygoideum (рис. 88, С—Ер) и задний участок palatoquadratum, превращающийся в сустав- DsO SO 1
ную кость черепа — quad-ratum (рис. 88, 89—Qu). Такому же окостенению подвергался и задний конец хрящевого mandibulare, образуя костное arti-culare (рис. 95—Аг).
У стегоцефалов имелся и еще один очень важный элемент черепа, являющийся результатом окостенения hyomandibulare (epi-
hyale). Рис. 89. Череп Palaeogyrinus (вид сзади) (Ватсон, 1926).
Во—basiocclpltale; Dso — dermosupraocclpitale; Ео — exocclpltale? Pt—pterygoideum; Qy —quadratojugaie; Qu—quadratum; Sq —squamosum; So — supraoccipitale; T— tabulate.
Hyomandibulare у ки-степерых рыб верхним
своим концом упиралось
в наружную стенку слуховой области окостеневшего мозгового черепа, но слуховой косточкой еще не являлось. Следы контакта hyomandibulare с черепом можно видеть, например, у Eusthenopteron (рис. 63).
У примитивных карбоновых стегоцефалов hyomandibulare также еще не играло роли слуховой косточки и, как у кистеперых рыб, одним своим
Рис. 90. Хрящевой череп аксолотля (Siredon pisciformis). Все покровные кости сняты (вид сбоку) (А. П. Быстров, 1939).
Md—mandibulare (cartilago Meckelii); Ep—epipterygoideum; Pq — palatoquadratum.
концом было соединено co стенкой слуховой капсулы. Очень интересным и важным является тот факт, что на боковой стенке окостеневшего мозгового черепа на границе между prooticum и paroccipitale у некоторых карбоновых стегоцефалов было доказано присутствие небольшого углубления, которое являлось местом контакта hyomandibulare со стенкой слухового отдела черепа. Это углубление получило название pseudofenestra ovalis (Ватсон, 1926; рис. 88, В — psf). Другой конец hyomandibulare у этих примитивных эмболомерных стегоцефалов соединялся с quadratum. Поэтому
7 А. П. Быстров 97
в области верхнего угла этой кости у некоторых из них имелось углубление овальной формы.
Отсутствие отверстия в костной стенке слуховой капсулы в месте контакта hyomandibulare с черепом (pseudofenestra ovalis) свидетельствует о том, что hyomandibulare у этих стегоцефалов еще не являлось настоящей слуховой косточкой и еще не может быть названо стремечком (stapes).
У более прогрессивных форм стегоцефалов pseudofenestra ovalis превратилась в отверстие, ведущее в полость внутреннего уха. Таким образом воз-
Рис. 91. Вертикальный разрез покровной кости черепа карбонового стегоцефала Baphetes (X 300).
никла fenestra ovalis, и окостеневшее hyomandibulare стало играть роль слуховой косточки — stapes.
Окостеневшие и неокостеневшие отделы хрящевого черепа стегоцефалов снаружи были покрыты, как и у кистеперых рыб, многочисленными костями, развившимися в соединительной ткани кожи. На наружной поверхности этих костей имелись многочисленные круглые ячейки и иногда радиально расположенные гребни. При микроскопическом изучении структуры всех этих костей видно, что они состоят только из типичной костной ткани. Никаких следов кожных зубов, которые когда-то, несомненно, располагались на черепных костях у предков стегоцефалов, на этих костях уже нет (рис. 91).
Задний край черепной крыши стегоцефалов был образован двумя парными покровными костями — dermosupraoccipitale (рис. 92—Dso) и tabulate (Т). Кпереди от этих костей лежала пара больших parietale (Р), в шве между которыми помещалось пинеальное отверстие (foramen parietale). По обе стороны от parietale располагались supratemporale (St) и in-tertemporale (It), а кпереди на уровне орбит лежало парное frontale (F). Верхние края орбит были образованы двумя костями — praefrontale (Pf)
98
и postfrontale (Pof). В состав переднего отдела крыши черепа входило большое nasale (N) и расположенное между орбитами и носовыми отверстиями длинное lacrimale (L). Передний край черепа был образован praemaxillare (Рт). Боковая сторона черепа состояла из maxillare (М), jugale (Ju), postorbitale (Po), squamosum (Sq) и quadratojugale (Qj).
Позади squamosum и quadratojugale располагалось небольшое quadratum (Qu), представляющее собой результат окостенения заднего конца palatoquadratum. У некоторых стегоцефалов в состав черепной крыши вхо-
дила еще одна очень интересная небольшая покровная косточка, носящая название se-ptomaxillare. Она располагалась между maxillare и nasale непосредственно позади носового отверстия.
Строение черепной крыши эмболомерных стегоцефалов карбона показывает, что в ее состав входит значительно меньшее количество костей, чем у кистеперых рыб. Следовательно, у амфибий многие покровные кости подверглись полной редукции. Так, исчез ряд костей, образующий задний отдел черепа: extrascapulare medium, extrascapulare laterale, operculum, suboperculum k praeoperculum (последние две еще имелись у девонских ихтиостегид); исчезли и все маленькие косточки, лежавшие в переднем отделе черепа. В этом участке черепной крыши сохранились только praemaxillare, nasale и lacrimale.
Покровные кости нёбной части черепа эмболомерных
Рис. 92. Череп Palaeogyrinus decorum (вид сверху) (Ватсон, 1926).
Dso — dermosupraocclpltale; F — frontale; It — Intertemporalei Ju—jugale; L — lacrimale; M —maxillare; TV—nasale; P—parletale; Pf—praefrontale; Pm—praemaxillare; Po —postorbitalel Pof~ postfrontale; Qj —quadratojugale; Qu — quadratum; Sq — squamosum; St — supratemporale; T — tabulare.
стегоцефалов по своей форме и положению были похожи на нёбные кости
у кистеперых рыб.
Непосредственно на мозговом черепе у них располагалось непарное parasphenoideum (рис. 93—Ps). Эта кость имела длинный острый отросток, Направленный вперед — processus cultriformis (р. с.). По обе стороны от парасфеноида лежали птеригоиды (pterygoideum — Pt), которые были соединены с отростками на basisphenoideum (processus basisphenoideus). Птеригоиды имели очень сложную форму. Передняя часть каждого птеригоида представляла собой костную пластинку, лежащую в полости нёба (рис. 93); эадняя часть, направляясь к quadratum, стояла в вертикальной плоскости и была сильно вдавлена с латеральной стороны; при этом ее верхний край
почти достигал крыши черепа.
Вдоль наружного края каждого птеригоида располагались три кости — transversum (Tr), palatinum (Pl) и vomer (V). Наружные края нёбной поверхности черепа были образованы praemaxillare (Pm), maxillare (М) и quadratojugale (Qj). Непосредственно перед quadratum имелось большое от-
99-
верстие (fenestra adductoria), служившее местом выхода группы жевательных мышц. В переднем отделе нёба помещались хоаны, обычно ограниченные краями четырех костей: praemaxillare, maxillare, vomer и pala-tinum.
Нижние края praemaxillare и maxillare у стегоцефалов, как и у кистеперых рыб, были усажены небольшими коническими зубами. Кроме этих зубов, на vomer, palatinum и transversum располагался еще один ряд зубов, среди которых имелись сильно развитые «клыки». Такую же зубную
Рис. 93. Череп Baphetes (вид снизу) (Ватсон, 1926).
/. а. — fenestra adductoria; М — maxillare; р. с. — processus cultrlformfs; Pm — praemaxillare; Pl — palatinum; Ps — pa-rasphenoldeum; Pt — pterygoldeum; Qj — quadratojugale; Qu — quadratum; Tr — transversum; V — vomer.
систему мы видели и у кроссо-птеригий.
Конические зубы стегоцефалов, как и у кистеперых рыб, своими основаниями прирастали непосредственно к тем костям, на которых они располагались.
Изучение поперечных разрезов зубов показало, что все они были снабжены дентинными складками, не имеющими боковых ветвей, как зубы Glyptolepis, Holoptychius, Polyplocodus и Eusthenopteron. Периферические участки дентинных складок не были расщеплены, как у Polyplocodus и Eusthenopteron (рис. 94).
Кроме этих зубов, на парасфеноиде и птеригоидах у стегоцефалов находились поля многочисленных мелких конических шагреневых зубов.
В состав нижней челюсти стегоцефалов входили десять костей (рис. 95). В заднем конце ее помещалось articulare, являющееся результатом окосте
нения хрящевого mandibulare. Эта кость имела на своей верхней поверхности углубление, в которое при образовании челюстного сустава входила выпуклая часть quadratum. К переднему участку articulare присоединялся неокостеневающий отдел меккелева хряща, который проходил вдоль всей нижней челюсти и достигал ее подбородочного конца. Кроме articulare, остальные девять костей нижней челюсти являлись покровными костями и накладывались со всех сторон непосредственно на хрящевое mandibulare (cartilage Meckelii). Поэтому на поперечных разрезах нижней челюсти стегоцефалов на любом ее уровне можно видеть более или менее широкое от-
верстие, в котором когда-то находилась соответствующая часть меккелева хряща (рис. 68, В). На наружной поверхности articulare лежало sup-raangulare (рис. 95, А — Sa), а на внутренней — длинное praearticulare (рис. 95, В — Ра). Обе эти кости образовывали стенки большого углубления, в которое входила группа жевательных мышц. Снизу, накладываясь на cartilago Meckelii, помещалось angulare (Лп). Самой длинной костью нижней челюсти являлось dentale (D). Верхний край этой кости был снабжен рядом конических зубов. На внутренней стороне меккелева хряща параллельно dentale лежали три короноида: coronoideum (Со), intercoronoideum (1с) и praecoronoideum (Рс).
400
Перед angulare, образуя нижний край челюсти, располагались post-spleniale (Ps) и spleniale (Sp).
Покровные кости плечевого пояса кистеперых рыб, располагающиеся непосредственно позади костей, прикрывавших жаберный аппарат, как известно, соединялись с задним отделом крыши черепа.
Так как при переходе от кистеперых рыб к примитивным Tetrapoda одновременно с редукцией жабр полной редукции подверглись и такие ко-
Рис. 94. Поперечный разрез зуба карбонового стегоцефала Megalocephalus (Эмблтон и Этзи — Emblton a. Etthey, 1874).
сти, как operculum и suboperculum, то в результате этого между черепом и плечевым поясом возник промежуток. Редукция operculum и subopercu-lum, конечно, не наступила сразу: обе эти кости, несомненно, постепенно уменьшали свою величину, что сопровождалось постепенным приближением плечевого пояса к черепу. В результате этого промежуток между черепом и плечевым поясом после полной редукции operculum и suboperculum оказался не очень большим; однако его наличие все же сделало и голову и плечевой пояс у амфибий значительно более подвижными, чем у их предков — кроссоптеригий.
Очень интересно, что плечевой пояс примитивных эмболомерных стегоцефалов карбона еще сохранял непосредственную связь с черепом (рис. 96).
Процесс уменьшения количества покровных костей, которым сопровождались первые этапы развития амфибий, отразился и на строении их плечевого пояса: в нем исчезла такая кость, как anocleithrum. В результате этого в состав плечевого пояса стало входить только пять покровных костей. Самой верхней из них являлось небольшое posttemporale (Pt).
101
Эта кость одним своим концом соединялась с tabulare, другим — с supracleithrum (Set), вследствие чего плечевой пояс стегоцефала, как у кистеперых рыб, еще сохранял непосредственный контакт с затылочным отделом крыши черепа. Далее следовали большие cleithrum (Ct) и clavicula (Cl). Между правой и левой ключицами располагалась interclavicula (Ic).
Рис. 95. Нижняя челюсть Megalocephalus (Ватсон, 1926).
Ап — angulare; Аг — articulare; Со — coronoldeum; D — dentale; Ic — Intercoronoldeum;
Pa — praearticulare; Pc — praecoronoldeum; Ps — postsplenlale; Sa — supraangulare;
Sp — spleniale.
Cleithrum и clavicula прикрывали хорошо развитую лопатку (scapula), являющуюся результатом окостенения некоторых хрящевых элементов плечевого пояса. Coracoideum и procoracoideum у эмболомерных стегоцефалов оставались в течение всей жизни хрящевыми.
Рис. 96. Череп и плечевой пояс Eogyrinus attheyi (Ватсон, 1926).
Cl — clavicula; Ct — cleithrum; Ic — Interclavicula; 5c — scapula; Set — supraclelthrum; Pt — post-temporale.
Как указывалось выше, кости тазового пояса у кистеперых рыб лежали в мышцах туловища и не имели непосредственного контакта с какими-либо костями осевого скелета. У эмболомерных стегоцефалов каждая половина тазового пояса состояла из трех костей — ilium, ischium, pubis (рис. 97 — II, Is, Pb).
Эти три кости представляли собой результат окостенения хряща и каждая из них соответствовала определенной части хрящевого отдела плечевого пояса, именно: ilium было гомологично scapula; pubis — procoracoideum и ischium — coracoideum.
102
У наиболее примитивных карбоновых стегоцефалов ilium имело два длинных отростка, которые лежали непосредственно на ребрах, но не соединялись с ними при помощи суставов. Об этом свидетельствует то, что на их внутренней стороне нет никаких следов скрепления с ребрами (рис. 97).
Два длинных отростка на ilium давали возможность достаточно хорошо фиксировать эту кость на ребрах только при помощи мышц.
В скелете эмболомерного стегоцефала карбона Eogyrinus attheyi (рис. 98)
типичные крестцовые ребра, при помощи которых тазовый пояс мог бы присоединяться к одному из позвонков, отсутствовали. Ребра того отдела его скелета, где располагался таз, построены, как обычные грудные ребра.
Таким образом, тазовый пояс у некоторых древних стегоцефалов был скреплен с осевым скелетом почти так же непрочно, как и у кистеперых рыб.
В то время как относительная подвижность плечевого поя-
са для всякого наземного поз- Рис. 97. Таз карбонового стегоцефала (вид воночного была, несомненно, изнутри) (Стин —Steen, 1934).
полезной особенностью, недо- 11 ~ lllum; Is - ischium; Pb — pubis.
статочно прочное прикрепление
тазового пояса к осевому скелету, напротив, было невыгодным, так как задние конечности для Tetrapoda являлись основным аппаратом, толкающим тело животного вперед.
Передние и задние конечности примитивных наземных позвоночных (стегоцефалов), развившиеся из набора костей или хрящей своеобразных плавников кистеперых рыб, были относительно длинными образованиями.
Рис. 98. Неполный скелет Eogyrinus attheyi. Карбон (Ватсон, 1926).
Первой костью передней конечности являлась плечевая кость (humerus), соединяющаяся с суставной ямкой на плечевом поясе. К этой кости присоединялись лучевая (radius) и локтевая (ulna) кости, входящие в состав предплечья. Скелет кисти распадался на три отдела: carpus, metacarpus и phalanges.
В состав carpus входили небольшие хрящевые или костные элементы, образующие два ряда. В первом ряду помещались: radiale, intermedium и ulnare; во втором — carpale primum, secundum, tertium, quartum и quin-tum. Между этими рядами располагались два centrale. К каждому из элементов дистального ряда carpus присоединялось по одной метакарпальной кости, которые на своих концах несли фаланги пальцев.
103
Задняя конечность стегоцефалов была построена по тому же плану, как и передняя. Первой ее костью являлся femur, сочленяющийся с особой суставной ямкой, помещающейся на границе между тремя элементами тазового пояса. Далее следовали tibia и fibula — кости голени.
Стопа, как и кисть, разделялась на три отдела: tarsus, metatarsus и phalanges.
В первом ряду tarsus располагались tibiale, intermedium и fibulare. Далее следовали два центральных элемента и ряд tarsalia, к которым присоединялись пять metatarsalia, несущие фаланги пальцев.
Для того, чтобы подчеркнуть сходство в строении передней и задней конечностей у наземных позвоночных, мы приводим таблицу, показывающую соответствие составляющих их элементов.
Плечевой пояс и передняя конечность
Тазовый пояс и задняя конечность
Posttemporale Supracleithrum Cleithrum Clavicula Interclavicula Scapula Coracoideum Procoracoideum Humerus Radius Ulna Radiale Intermedium Ulnare Centrale
I. II. III. IV. V Metacarpalia Phalanges
покровные кости
carpale
Покровных костей нет
Ilium
Ischium
Pubis
Femur
Tibia
Fibi la
Tibiale
Intermedium
Fibulare
Centrale
I. II. III. IV. V tarsale
Metatarsalia
Phalanges
Одновременно с эмболомерными стегоцефалами в течение каменноугольного периода существовала еще одна группа древних амфибий. Эти амфибии, имевшие очень своеобразные позвонки, известны под названием лепоспондильных (Lepospondyli, рис. 17). Они, по всей вероятности, представляли собой боковую ветвь, отделившуюся в начале карбона от эмболомерных стегоцефалов.
Каждый позвонок лепоспондильных амфибий имел хорошо развитое тело с коническим углублением спереди и сзади (амфицельные позвонки); дуга позвонка была полностью сращена с телом (рис. 99, А). Такие позвонки по-видимому, образовались в результате полного слияния гипоцентра (Л) и плевроцентра (р) позвонка эмболомерного типа. Ребро на туловищном позвонке в типичном случае присоединялось своим бугорком к концу поперечного отростка дуги, а головкой — к площадке на переднем краю тела (рис. 99, В) — и это дает нам право рассматривать передний отдел тела позвонка как гипоцентр. У последнего, туловищного, у крестцового (S) и у первых хвостовых позвонков суставная площадка для головки ребра была смещена на средину тела позвонка (рис. 99, С). Хвостовые позвонки, начиная с III или IV, были снабжены хорошо развитыми вентральными дугами, каждая из которых являлась частью гипоцентра (рис. 99, А, С).
В черепе лепоспондильных стегоцефалов нередко наблюдалось срастание друг с другом некоторых покровных костей (рис. 100). Сейсмосенсор-ная система у тех форм, у которых она имелась, была представлена эпите
104
лиальными каналами, лежащими на наружной поверхности покровных костей; они вызывали на них образование углублений в виде борозд или ряда ямок. Нёбо у наиболее примитивных Lepospondyli было построено, как у эмболомерных стегоцефалов.
Поздние, пермские, представители лепоспондильных стегоцефалов нередко имели очень своеобразную форму черепа и, несомненно, являлись высокоспециализированными амфибиями (рис. 101, А, В).
Маленькие лепоспондильные амфибии, населявшие пресноводные бассейны каменноугольного периода (рис. 102), по всей вероятности, были предками современных хвостатых амфибий (Urodela).
Рис. 99. Лепоспондильные позвонки Urocordylus (Стин, 1938).
a. d. — arcus dorsalis; a. v. — arcus ventralls; h — hypocentrum; p — pleurocentrum; S — крестцовый позвонок.
Изучение многочисленных ископаемых амфибий показало, что еще в конце карбона появились так называемые рахитомные стегоцефалы. В начале пермского периода они полностью вытеснили своих предков — эмболомерных стегоцефалов — и просуществовали до начала триаса (рис. 17, Rhachitomi)..
Позвонки рахитомных стегоцефалов отличались следующими интересными особенностями.
Гипоцентр каждого позвонка имел форму костного полулуния, которое охватывало хорошо развитую хорду снизу (рис. 103, Л, В). На боковой поверхности этой части тела позвонка имелась небольшая площадка для соединения с ребром. Таким образом, гипоцентр рахитомного позвонка по сравнению с соответствующим образованием в позвонке эмболомерного типа был в значительной степени редуцированным: он не представлял собой полного двояковогнутого диска — верхняя его часть отсутствовала.
Плевроцентр в рахитомном позвонке подвергся еще более сильной редукции. Он распался на два отдельных костных элемента ромбовидной формы, которые располагались по обе стороны хорды непосредственно позади полулунного гипоцентра (рис. 103, А, С — р). Дуга позвонка была хорошо развита и имела высокий остистый отросток; она обычно не срасталась с телом позвонка.
Ребра присоединялись к поперечному отростку дуги и к небольшой суставной площадке на заднем краю полулунного гипоцентра.
Так как у всех рахитомных стегоцефалов тазовый пояс присоединялся к концам пары ребер одного из позвонков, то в позвоночном столбе выделялся единственный крестцовый позвонок (рис. 104).
105
Позвонки шейные, грудные и поясничные не отличались друг от друга по своему строению и обычно все были снабжены ребрами той или иной длины (рис. 105).
Хвостовые позвонки рахитомных стегоцефалов имели на своих гипоцентрах хорошо развитые вентральные дуги (рис. 103 ,С, D—а. о.). Несколько
Рис. 100. Череп Keraterpeton galvani Huxley (вид сверху). Карбон (по фотографиям Стин, 1938).
хвостовых позвонков, располагающихся непосредственно позади крестцового позвонка, обычно были снабжены короткими ребрами (рис. 105).
Таким образом, у рахитомных стегоцефалов можно было различить три отдела позвоночного столба: предкрестцовый, состоящий у разных видов из различного количества позвонков, крестцовый, всегда представленный только одним позвонком, и хвостовой, содержащий различное число позвонков.
Хрящевой neurocranium у рахитомных стегоцефалов окостеневал значительно слабее, чем у Embolomeri.
106
Затылочный его отдел был образован костным exoccipitale, на котором имелся отросток с суставной площадкой для соединения с I позвонком (рис. 106, А, В — Ео). Basioccipitale (Во) и supraoccipitale (So) представляли собой обычно очень небольшие кости; при этом basioccipitale не принимало
Рис. 101. Черепа Diplocaulus limbatus (Д) и Diplocaulus magnicornis (В). Пермь (Кейз — Case).
Рис. 102. Скелеты лепоспондильных стегоцефалов Urocordylus scalaris Fritsch (А) и Keraterpeton galvani Huxley (В). Карбон (Стин, 1938).
участия в образовании суставной поверхности для сочленения с позвоночным столбом. У многих рахитомных стегоцефалов basioccipitale и supraoccipitale в течение всей жизни животного оставались хрящевыми (рис. 106, В).
Окостенение переднего отдела мозгового черепа нередко приводило у рахитомных стегоцефалов к возникновению костного сфенэтмоида (sphen-
107
ethmoideum, рис. 107, 108), обычно имеющего форму трехгранной призмы, в задней части которой имелась полость для переднего отдела головного мозга; от этой полости вперед направлялись два канала, в которых проходили обонятельные нервы, а в боковых стенках располагались относительно большие отверстия для зрительных нервов (foramen opticum, рис. 107).
Хрящевой splanchnocranium рахитомных стегоцефалов подвергался незначительному окостенению.
Palatoquadratum окостеневало только в двух местах. Одно из них после окостенения давало epipterygoideum, обычно прочно прирастающее к пте-ригоиду, другое — превращалось в костное quadratum (рис. 108, 109).
Рис. 103. Рахитомные позвонкиDvinosaurus. А, В—грудные позвонки; С, D — хвостовые позвонки (А. П. Быстров, 1938). a. d. — arcus dorsalis; a. v. — arcus ventralls; c — costa; h — hypocentrum;
p — pleurocentrum.
Mandibulare у рахитомных стегоцефалов, как и у эмболомерных, окостеневало только в своем заднем конце, образуя костное articulare.
В отверстие в хрящевой стенке слухового отдела мозгового черепа у рахитомных стегоцефалов было вставлено основание слуховой косточки — stapes (рис. 106 — S). Stapes, имеющий форму немного изогнутой костной колонки, обычно располагался косо, под углом 30—40° к плоскости основания черепа. Его верхний конец был направлен в центр слуховой вырезки (incisura oticalis, рис. 108), образованной краями tabulare и squamosum, и прикреплялся к натянутой здесь барабанной перепонке (membrana tympani).
Таким образом, у рахитомных стегоцефалов в отличие от эмболомерных stapes уже играл роль настоящей слуховой косточки и передавал звуковые колебания от барабанной перепонки во внутреннее ухо через fenestra ovalis, возникшее в боковой стенке хрящевой слуховой капсулы. Таким образом, pseudofenestra ovalis примитивных эмболомерных стегоцефалов у рахитомных превратилась в fenestra ovalis, и это, несомненно, было большим шагом вперед.
108
Черепная крыша у рахитомных стегоцефалов (рис. ПО) была образована таким же набором покровных костей, как и у эмболомерных стегоцефалов карбона, но intertemporale обычно отсутствовало. Впрочем, эта кость иногда отсутствовала и у Embolomeri.
Очень интересным следует считать строение septomaxillare, которое имелось не у всех представителей Rhachitomi. В тех случаях, когда эта
маленькая кость входила в состав крыши черепа, она располагалась, как у Embolomeri, позади ноздри между nasale и lacrimale (рис. 111— Srn). У некоторых форм от нижней .поверхности septomaxillare отходил небольшой пластинчатый отросток, который соединялся с краем vomer на нёбной поверхности черепа. Такая интересная особенность была
Рис. 105. Скелет Micropholis stowi Huxley (вид сверху). Триас (Бройли и Шредер, 1937).
Рис. 104. Позвонки Micropholis stowi (вид сверху) (Бройли и Шредер—Broil! und Schroder, 1937). j — крестцовый позвонок; с — крестцовое ребро; I — подвздошная кость.
обнаружена, например, на черепе Edops (рис. 111, В; Ромер).
Нёбная сторона черепа рахитомных стегоцефалов была образована такими же костями, как и у Embolomeri, именно: parasphenoideum, pterygo-ideum, vomer, palatinum, transversum (рис. 112). Но птеригоид соединялся при помощи зубчатого шва непосредственно с парасфеноидом. Соединение птеригоида с базисфеноидом у Rhachitomi не имело такого механического значения, как у примитивных эмболомерных стегоцефалов. Однако хрящевой базисфеноид все же сохранял контакт с птеригоидом и в виде конического отростка входил в особое углубление в птеригоиде. Это углубление — recessus conoideus — располагалось над шовным соединением птеригоида с парасфеноидом и непосредственно под тем местом, к которому прирастал эпиптеригоид (рис. 109, А).
Не менее важной особенностью нёба рахитомных стегоцефалов было наличие больших пространств между передними крыльями птеригоидов
109
и prosessus cultriformis parasphenoidei. У Embolomeri передние крылья птеригоидов обычно были почти вплотную прижаты к processus cultriformis. У Rhachitomi крылья птеригоидов настолько далеко были отодвинуты от парасфеноида, что на нёбе образовались два больших окна, носящие на-
Рис. 106. Черепа Dvinosaurus (Л) (А. П. Быстров, 1938) и Benthosuchus (В) (вид сзади).
Во — basioccipitale; Dso — dermosupraocclpitale; Ео — exocclpitale; L. qs. — ligamentum quadra-tostapediale; Ps — paraspbenoideum; Pt — pterygoldeum; Qu — quadratum; 5 — stapes; So — supraoccipitale; T — tabulare.
звание интерптеригоидных отверстий (fenestrae interpterygoideae). У живых стегоцефалов эти отверстия были закрыты слизистой оболочкой, лежавшей на крыше полости рта. На нижних краях praemaxillare и гпахП'аге у рахи-
Рис. 107. Sphenethmoideum стегоцефала Benthosuchus. А — вид сверху; В — вид снизу; С — вид сбоку; D — внд сзади; Е — вид спереди; F — схема строения (А. П. Быстров и И. А. Ефремов, 1940).
с, о. — canalis olfactorlus; f. о, — foramen optlcum.
томных стегоцефалов, как и у всех Embolomeri, помещался ряд небольших конических зубов. Изучение их строения на поперечных разрезах позволило установить у них наличие дентинных складок, обычно не имеющих таких боковых ответвлений, которые мы видели в зубах большинства кистеперых рыб. Однако при исследовании большого количества зубов рахитомных стегоцефалов мне удалось в 2% случаев найти зубы, в которых одна, две или три складки обладали боковыми ветвями (рис. 113). Этот ва
110
риант указывает на то, что в родословной стегоцефалов были предки, снабженные зубами с ветвящимися дентинными складками.
Кроме praemaxillare и maxi Hare, у рахитомных стегоцефалов зубы имелись и на таких костях нёба, как vomer, palatinum и transversum. Здесь,
Рис. 108. Череп Benthosuchus, Часть покровных костей удалена (вид сверху).
Ер — epipterygoideum; th с. — processus cultriformts; Pl — palatinum; Se — sphenethmo-laeum; Tr — transversum; V — vomer.
кроме больших так называемых «клыков» (рис. 112), у некоторых форм располагались ряды мелких зубов. В результате этого возникал второй ряд зубов, параллельный на большей части своего протяжения краевым зубам.
У многих стегоцефалов нёбная поверхность парасфеноида и передних крыльев птеригоидов, как и у кистеперых рыб, была покрыта так называемыми шагреневыми зубами (рис. 112). Эти зубы представляли собой очень
Ш
Рис. 109. Птеригоид и эпиптеригоид Benthosuchus. А — вид снутри; В — вид снаружи.
Ер — eplpterygoldeum; Pt — pterygoldeum: Qu — quadratum; r. c. — recessus condldeus.
Рис. ПО. Череп Benthosuchus (триас, СССР) (А. П. Быстров и И. А. Ефремов, 1940).
маленькие конические образования, состоящие из дентина и покрытые тонким слоем эмали (А. П. Быстров, 1938).
Как у всех без исключения кистеперых рыб, шагреневые зубы стегоцефалов в процессе роста соответствующих костей замуровывались в костное вещество и при его перестройке постепенно подвергались полному разрушению (рис. 114). Совершенно очевидно, что стегоцефалы унаследовали эту удивительную особенность от своих предков — кистеперых рыб.
Нижняя челюсть рахитомных стегоцефалов состояла из таких же костей, как и у Embolomeri (рис. 115). Зубы располагались вдоль верхнего
Рис. 111.
А — передний участок черепа Trematosaurus, В — хоана Edops (Ромер, 1942).
М — maxlllare; Pl — palatinum; Pm — praemaxillare; Sm—septomaxlllare; И — vomer.
края длинного dentale; строение их не отличалось от строения таких же зубов на краях praemaxillare и maxillare. У некоторых форм зубами были снабжены и короноиды. Так, у Dvinosaurus зубы имелись на всех трех ко-роноидах; у Trimerorhachis — на короноиде и интеркороноиде; у Bentho-suchus — только на короноиде (рис. 115, 116).
На переднем конце dentale обычно помещались два хорошо развитых конических «клыка». В заднем отделе нижней челюсти перед articulare всегда имелась глубокая яма, куда входила группа жевательных мышц, сжимающих челюсти. Эта яма продолжалась вперед в виде канала, проходившего под покровными костями вдоль всей нижней челюсти (рис. 68, В). В этом канале располагался не сохраняющийся в ископаемом состоянии меккелев хрящ, окостенение которого в заднем отделе приводило к образованию articulare — единственной кости нижней челюсти, развивающейся на месте хрящевой ткани. Все покровные кости нижней челюсти, которые лежали на ее наружной стороне, были покрыты такой же ячеистой скульптурой, как и наружная поверхность костей крыши черепа (рис. 115, А); поверхность костей, располагающихся на внутренней стороне нижней челюсти, была лишена скульптуры.
Эпителиальные каналы сейсмосенсорной системы у пермских стегоцефалов обычно лежали не в покровных костях, как у кистеперых рыб и у девонских ихтиостегид, а в особых желобках на наружной поверхности Этих костей.
В А. П. Быстров
113
Плечевой пояс рахитомных стегоцефалов уже не имел непосредственного соединения с черепом, так как в нем подвергались полной редукции posttemporale и supracleithrum. Одновременно с этим размеры cleithrum и clavicula значительно уменьшились.
Рис. 112. Череп Benihosuchus (вид снизу) (А. П. Быстров и И. А. Ефремов, 1940).
Ео -г- exocclpitale; Ju — jugale; Л1—maxillare; p. c.—processus cultrlformls; Р/— palatinum; Pm — praemaxillare; Ps — parasphenoldeum; Pt — pterygoldeum; QJ — quadratoju-gale; Qu — quadratum; Se — sphenethmoldeum; T — tabulare; Tr — transversum; V — vomer.
Плечевой пояс рахитомных стегоцефалов был образован двумя хрящевыми скапулокоракоидами (scapulocoracoideum) (рис. 117, А). Лопаточные части этих образований были поставлены под прямым углом к тонкой пластинчатой коракоидной части, у которой можно было различить два неравных по своей величине отростка:.procoracoideum (рис. 117, А, В—Рс) и со-racoideum (рис. 117, А—Со). Один из коракоидов своим тонким краем, ве-
114
Рис. 113. Поперечный разрез зуба Benthosuchus с тремя ветвящимися дентинными складками (длинная ось равна 3,5 мм) (А. П. Быстров, 1939).
Рис. 114. Шагреневые зубы Benthosuchus (X 300).
115
роятно, накладывался на край коракоида противоположной стороны, как у современных хвостатых амфибий.
В области угла между лопаткой и коракоидом имелась суставная яма, в которую входила хрящевая головка плечевой кости. Значительная часть лопаточного отдела хрящевого скапулокоракоида у рахитомных стегоцефалов обычно подвергалась окостенению и в ней возникала костная sca-
Рнс 115. Нижняя челюсть Benthosuchus. А—вид снаружи; В—вид снутри; С — вид сверху (А. П. Быстров и И. А. Ефремов, 1940).
Ап — angulare;\Ar — artlculare; Со — coronoldeum; D — dentale; Jc — Intercoronoldeum; Pa — [praearticulare; Pc — praecoronoldeum; Ps — postsplenlale; Sa — supraangulare; Sp — spleniale.
Рис. 116.
A — нижняя челюсть Trimerorrhachis (Виллистон — Williston, 1914); В — нижняя челюсть Dvinosaurus (А. П. Быстров, 1938).
pula (рис. 117, В—Sc). Нужно думать, что третьим элементом хрящевого плечевого пояса являлось небольшое sternum, которое в виде тонкой хрящевой пластинки располагалось позади коракоидов и частично накладывалось на их задние края.
Нижняя поверхность широких коракоидов и sternum у стегоцефалов была прикрыта непарной покровной костью — interclavicula, которая обычно имела ромбовидные очертания и была покрыта типичной ячеистогребнистой скульптурой (рис. 117, В—1с). На края этой кости справа и слева, прикрывая прокоракоиды, накладывались ключицы, имеющие форму треугольников и покрытые на наружной своей поверхности ячеисто-гребнистой скульптурой — claviculae (рис. 118, А—С1). Каждая из этих покровных 116
костей имела узкий направленный вверх отросток, лежащий на переднем краю лопаточной части плечевого пояса (рис. 118, В). К этому отростку примыкало небольшое cleithrum (Ct), располагающееся также на переднем краю лопатки. Cleithrum обычно было лишено ячеистой скульптуры.
Как уже указывалось, у рахитомных стегоцефалов тазовый пояс при помощи пары ребер прикреплялся к одному из позвонков. Крестцовые ребра
Рис. 117. Плечевой пояс Benthosuchus (вид снизу).
Со — coracoideum; Ic — Interclavicula; Pc — procoracoideum; Sc —• scapula; St — sternum
отличались от соседних ребер несколько большей длиной и большей тол» щиной.
В составе тазового пояса у Rhachitomi, как и у Embolomeri и Crosso-pterygii, не было покровных костей. Таз состоял из хорошо развитого костного ilium (рис. 120—II), внутренняя сторона верхнего конца которого при помощи связочного аппарата была прочно прикреплена к концу крестцового ребра. На наружной поверхности расширенного нижнего конца этой кости имелось углубление, составлявшее часть acetabulum.
117
Второй костью таза являлось ischium (рис. 120—/s); что касается pubis, то оно у рахитомных стегоцефалов обычно в течение всей жизни оставалось хрящевым.
Передние и задние конечности Rhachitomi были построены по обычному плану.
Рис. 118. Плечевой пояс Benthosuchus. А — вид снизу; В — вид сбоку (А. П. Быстров и И. А. Ефремов, 1940).
Cl — clavlcula; Ct — cleithrum; Ic — interclavlcula; Sc — scapula.
Передняя конечность состояла из humerus, radius, ulna, карпальных элементов — хрящевых или частично окостеневших, метакарпальных костей и фаланг пальцев (рис. 119).
В состав задней конечности входили: femur, tibia, fibula, хрящевые или костные tarsalia, metatarsalia и phalanges (рис. 120).
Все длинные, так называемые трубчатые, кости конечностей стегоцефалов (humerus — femur, radius — tibia, ulna—fibula) имели хрящевые эпифизы, т. е. их верхние и нижние концы (головки), принимавшие участие в образовании суставов, в течение всей жизни животного не окостеневали.
Изучение стегоцефалов как примитивных представителей Tetrapoda показывает, что с выходом позвоночных из воды на сушу в их организме произошли многочисленные и очень важные изменения. Эти изменения уже
118
давали возможность древнейшим амфибиям во взрослом состоянии обитать на суше, однако еще не позволяли им полностью оторваться от водных бассейнов: амфибии еще нуждались в воде как среде для размножения; они еще продолжали, как их предки — кистеперые рыбы,— откладывать икру в пресноводные бассейны, где их личинки проходили все фазы своего развития. Только после метаморфоза, во время которого у личинок редуциро-
Рис. 119. Плечевой пояс и передняя конечность Dvinosaurus (А. П. Быстров, 1938)
Cl — clavicula; Ct — cleithrum; Н — humerus; Ic — Interclavicula; M — manus; £ — radius; Sc — scapula; U — ulna.
вались жабры и заменялись легкими, молодые животные получали возможность покинуть воду.
В распоряжении палеонтологии имеются совершенно несомненные доказательства того, что стегоцефалы в личиночной фазе своего развития дышали при помощи жабр и по своему внешнему виду, вероятно, были очень
Рис. 120. Тазовый пояс и задняя конечность Dvinosaurus (А. П. Быстров, 1938).
F — fibula; Ре — femur; II — Ilium; Is — Ischium; P — pes; T — tibia.
похожи на личинок современных амфибий. Во всяком случае мы можем уверенно сказать, что они обладали таким же сложным подъязычно-жаберным аппаратом, каким обладают личинки современных Urodela.
У личинок современных хвостатых амфибий имеется длинное непарное basibranchiale (рис. 121—ВЬ); к переднему концу этого элемента на каждой стороне присоединено короткое hypohyale, которое в свою очередь соединено с длинным ceratohyale (Ch). К наиболее широкой части basibranchiale справа и слева примыкает по два немного изогнутых хряща: hypobranchiale primum (Hb. р.) и hypobranchiale secundum (Hb. s.). Задние концы этих хрящей соединяются с ceratobranchiale primum и secundum (Cb. р., Cb. s.), к которым с внутренней стороны прилегают еще два таких же элемента — ceratobranchiale tertium (Cb. t.) и ceratobranchiale quar
119
turn (Cb. q.). На обращенных друг к другу сторонах цератобранхиальных хрящей сидят ряды острых конических шипов, которые позволяют личинке фильтровать воду, идущую из полости рта, под жабры, располагающиеся на концах первых трех цератобранхиальных хрящей.
Доказательством того, что личинки стегоцефалов обладали подъязычножаберным аппаратом такого же строения, являются находки большого ко-
Рис. 121. Жаберный аппарат аксолотля ГА. П. Быстров, 1950).
Bb — basibranchialei Cb. р., Cb. s., Cb. Cb. q. — ceratobranchiale prlmum, secundum, tertium, quartum; Ch — ceratohyaie; Hb. p., Hb. s. — hypobranchiale prlmum, secundum.
личества маленьких черепов Branchiosaurus amblystomus. около которых в породе лежали ряды жаберных шипов (рис. 122). Сами элементы подъязычно-жаберного аппарата у Branchiosaurus обычно не сохранялись, так как состояли из хряща и легко разрушались при разложении мертвой личинки (Креднер — Н. Credner, 1881—1892).
В 1899 г. в верхнепермских отложениях бассейна Северной Двины В. П. Амалицким был найден исключительно интересный рахитомный стегоцефал с сильно окостеневшим личиночным подъязычно-жаберным аппаратом, Этот стегоцефал, получивший название Dvinosaurus, несомненно, являлся так называемой неотенической формой; он не проделывал метаморфоза и в течение всей своей жизни сохранял жаберное дыхание
120
(П. П. Сушкин, 1936). Двинозавр достигал почти 2 м в длину и напоми-
нал гигантского аксолотля.
В подъязычно-жаберном аппарате у относительно молодых экземпляров двинозавра окостеневали следующие элементы: ceratohyale (рис. 123 — Ch), оба hypobranchialia (Hb. р., Hb. s.) и все четыре ceratobranchialia (Cb. р., СЬ. s., Cb. t., Cb. q.). У вполне взрослого животного той или иной степени окостенения подвергались все без исключения части подъязычно-жаберного аппарата (рис. 124).
Поразительное сходство в строении подъязычно-жабепного аппарата
двинозавра с таким же аппаратом у личинок всех современных хвостатых амфибий позволяет уверенно сказать, что стегоцефалы в личиночном состоянии дышали при помощи жабр. В нашем распоряжении имеется доказательство и того, что они, подобно личинкам современных амфибий, обладали тремя наружными жабрами.
В 1938 г. в Сибири на берегу реки Нижней Тунгуски была сделана очень редкая находка остатков личинки стегоцефала (Тип-gussogyrinus bergi, рис. 125) с обуглившимися жабрами. Жабр было по три на каждой стороне, как у личинок современных амфибий (И. А. Ефремов, 1939).
Неотеническое состояние двинозавра, естественно, не могло не отразиться на многих деталях его строения, в частности на характере скульптуры его покровных костей. Наружная поверхность этих костей у
>———--.---- 5 тпт
Рис. 122. Череп личинки Branchiosaurus amblystomus (вид снизу). Пермь (Креднер, 1881 — 1892).
него покрыта вместо ячеек ра-
диально расположенными высокими гребнями (рис. 126). Такой скульптуры не было обнаружено ни у одного из известных в настоящее время стегоцефалов. Эта особенность, по-видимому, могла возникнуть только в результате очень сильной гиперплазии личиночной скульптуры.
Покровные кости личинок стегоцефалов, еще дышавших жабрами, были лишены ячеистой скульптуры. Первые ячейки, по-видимому, появлялись только после того, как животное проходило метаморфоз.
Кроме необычной скульптуры и наличия окостеневшего подъязычножаберного аппарата, двинозавр обладал еще одной интересной особенностью. В отличие от обычных рахитомных стегоцефалов он не имел слуховой вырезки и был лишен барабанной перепонки. Его stapes имел три отростка; один из них закрывал fenestra ovalis в хрящевой стенке слухового отдела мозгового черепа, второй был направлен вверх и соединялся с хрящевым paroccipitale, а третий был связан при помощи длинной связки с quadratum (рис. 106, А). Отсутствие барабанной перепонки и наличие непосредственной связи stapes с quadratum делали слуховой аппарат двинозавра похожим на такой же аппарат примитивных эмболомерных стегоцефалов карбона.
121
После того как личинки стегоцефалов проделывали все фазы метаморфоза и вместо жабр приобретали легкие, они становились в значительной мере наземными животными. Однако полностью оторваться от водных бассейнов стегоцефалы все же не могли, так как именно здесь они добывали свою пищу — рыб.
Cb, р., Cb. s., Cb. I., Cb. q. — ceratobranchiale primum, secundum, tertium, quartum; Ch — ceratohyale; fib. p.t fib. s. — hypobranchiale primum, secundum.
Очень разнообразные рахитомные стегоцефалы вымерли в начале триасового периода. На их смену пришли стереоспондильные лабиринтодонты, являющиеся потомками рахитомных (рис. 17 — S).
Первые представители Stereospondyli появились еще в конце перми. Нужно думать, что их позвонки развились из позвонков рахитомного типа
122
путем постепенного усиления гипоцентра. При этом полулунные гипоцентры в позвонках стереоспондильных стегоцефалов превратились в массивные диски, на верхних краях которых сохранились только небольшие вырезки для хорды (рис. 127, А, В).
У некоторых стереоспондильных стегоцефалов (Buettneria bakeri Case) верхние концы гипоцентра замкнулись над хордой полностью и только ко-
роткая вертикальная борозда, идущая от верхнего края дисковидного тела позвонка, указывает на его развитие из гипоцентра полулунной формы (рис. 127, С).
Передняя и задняя поверхности тел стереоспондильных позвонков стали почти плоскими или очень слабо вогнутыми.
Плевроцентры у Stereospondyli отсутствовали и массивные гипоцентры соседних позвонков примыкали непосредственно друг к другу. Дорзаль
123
ные дуги позвонков были развиты хорошо. Среди стереоспондильных стегоцефалов встречались формы, у которых эти дуги соединялись одновременно с двумя соседними телами позвонков, а двухголовчатые ребра имели контакт с поперечным отростком дуги и с суставной площадкой, образованной двумя соседними гипоцентрами (рис. 127).
Рис. 125. Tungussogyrt-nus bergi. Видны три пары обуглившихся жабер (b.p.d.,b. s.d., b. t. d., b. p. s., b. s. s., b. t. s.) (А. П. Быстров. 1939).
Как и рахитомные стегоцефалы, стегоцефалы со стереоспондильными позвонками имели только три отдела позвоночного столба, именно: пред-крестцовый, крестцовый, представленный только одним позвонком, и хвостовой (рис. 128). Их тазовый пояс соединялся с позвоночным столбом по-прежнему только при помощи пары хорошо развитых крестцовых ребер.
124
Череп стереоспондильных стегоцефалов триасового периода по своему строению принципиально ничем не отличался от черепа рахитомных стегоцефалов перми (А. П. Быстров, 1935).
Некоторые формы триасовых стереоспондильных стегоцефалов имели совершенно своеобразную форму головы. Их черепа были необычайно ко-
Рис. 126. Череп Dvinosaurus (вид сверху) (А. П. Быстров, 1947).
роткими, очень широкими и плоскими, причем орбиты достигали очень большой величины (рис. 129). Эти особенности, несомненно, следует рассматривать как результат приспособления к каким-то своеобразным условиям существования. Скульптура у такого широкоголового лабиринтодонта, как Gerrothorax (рис. 129, А), резко отличалась от скульптуры, характерной для других стегоцефалов; она была образована не ячейками, а высокими бугорками, которые на многих костях располагались в виде радиально идущих рядов.
125
Рис. 127. Стереоспоидильные позвонки. А, В — позвонки Plagio-Saurus (Хюне — Ниеле, 1922); С — Buettneria (Кайз —Case, 1932). h — hypocentrum.
Рис. 128. Скелет Metoposaurus diagnosticus. Триас (Фраас — Fraas, 1890).
126
Плечевой пояс и передние конечности у стереоспондильных стегоцефалов были построены так же, как у Rhachitomi.
В тазовом поясе некоторых Stereospondyli pubis окостеневало. Таз прикреплялся при помощи пары больших крестцовых ребер к одному сакральному позвонку, как и у рахитомных лабиринтодонтов.
30 СП-------*-
Рис. 129. Черепа Gerrothorax pulcherrimus Fraas (Л) й'Plagiostec-num granulosum (В). Триас (Фраас, 1913).
В строении задних конечностей не было никаких новых особенностей.
ЛИТЕРАТУРА
Амалицкий В. П. Dvinosauridae. Изд. АН СССР, 1921.
Быстров А. П. Двинозавр из верхнепермских отложений на р. Волге. Вопросы палеонтологии, т. 1, 1950.
Быстров А. П. (В у s t г о w). Morphologische Untersuchungen des Schadels der Wirbeltiere. I Mitt. Schade! der Stegocephalen. Acta zoologica, Bd. 16, 1935. Быстров А. П. Dvinosaurus als neotenische Form der Stegocephalen. Ibidem, Bd. 19, 1938.
Быстров А. П. Zahnstruktur der Labyrinthodonten. Ibidem, Bd. 19, 1938.
Быстров А. П. Blutgefassystem der Labyrinthodonten. Ibidem, Bd. 20, 1939.
БыстровА. П. Hydrophilous and xerophilous Labyrinthodonts Ibidem, Bd. 28, 1947. Быстров А. П. и Ефремов И. A. Benthosuchus sushkini — лабиринтодонт из эотриаса реки Шарженги. Труды Палеонтологического института АН СССР, т. 10, 1940.
Ефремов И. А. (Е f г е m о V). Benthosuchus sushkini, ein neuer Labyrinthodont aus den permo-triassischen Ablagerungen des Scharschenga Flusses. Известия АН СССР, N 8, 1929.
Ефремов И. А. Первый представитель наземных позвоночных из Сибири. Доклады АН СССР, т. 23, 1939.
127
С у ш к и н П. П. (S u s h к i n). Notes on the permo-jurassic Tetrapoda from Russia. Ill, Dvinosaurus Amalytzki a perennibranchiate stegocephalian from the Upper Permian of North Dvina. Труды Палеонтологического института АН СССР, т. 4. 1936.
В г о i 1 i F. und S.c h r 6 d e r J. Beobachtungen an Wirbeltieren der Karrooformation, XXV. Uber Micropholis huxlei. Sitzungsberichte der Bayerischen Akade-mie der Wissenscheften. Miinchen, 1937.
Case E. A Collection of Stegocephalians from Scurry County Texas. Contribution from the Museum of Paleontology, Univ, of Michigan, vol. 4, 1932.
Credner H. Die Stegocephalen und Saurier aus dem Rotliegenden des Plaunschen Grund bei Dresden. I—X. Zeitschrift der Deutsch. Geol. Geselschaft, Bd. 33—45, 1881—1892.
E m b 1 t о n and A t t h e y. On the skull and other bones of Loxomma. Ann. and Mag. Nat. History, ser. 4, vol. 14, 1874.
Fraas E. Labyrinthodonten aus der Schwabischen Trias. Palaeontographica, Bd. 36, 1890.
Fraas E. Neue Labyrinthodonten aus der Schwabischen Trias. Ibidem. Bd. 60, 1913.
H u e n e F. Beitrage zur Kenntnis der Organisation einer Stegocephalen der Schwabischen Trias. Acta Zoologica, Bd. 3, 1922.
J a ek e 1 O. Ober Ceraterpeton, Diceratosaurus und Diplocaulus. Neues Jahrbuch fur Min., Geol. und Palaeont., Bd. 1, 1903.
J arv i k E. On the fish-like tail in the Ichthyostegid Stegocephalians. Meddelelser om Gronland, Bd. 114, 1952.
J a г v i k E. The oldest Tetrapoda and their forerunners. The Scientific Monthly, vol. 80, 1955.
Marsh C. Amphibian footprints from the Devonian. American Journal of Science, 1896. Nilsson T. Uber einige postcraniale Skeletreste der triassischen Stegocephalen Spitz-bergens. Bull, of the Geol. Inst, of Uppsala, vol. 30, 1943.
Romer A. Edops, a primitive rhachitomous Amphibian from the Texas Red Beds. The Journal of Geology, vol. 50, N 1942.
Save-Soderbergh G. Preliminare Notes on Devonian Stegocephalian from East Greenland. Meddelelser om Gronland, Bd. 97, 1932.
Steen M. Amphibian Fauna from the South Jogging Nova Scotia. Proceedings of the Zool. Society of London, 1934.
Steen M. On the fossil Amphibia from Gas Coal of Nyrany and other deposits in Chechoslovakia. Ibidem, vol. 108, 1938.
Watson D. M. S. The structure, evolution and origin of the Amphibia. Philosophical Transactions, vol. 209, 1919.
Watson D. M. S. The evolution and origin of the Amphibia. Philosonhical Transactions, vol. 216. 1926.
Williston S. The osteology of some American Permian Vertebrates. The Journal of Geology, vol. 22, 1914.
ГЛАВА (V
РЕПТИЛИИ
У нас нет никаких оснований сомневаться в том, что рептилии произошли от амфибий.
Принимая во внимание тот факт, что ископаемые рептилии известны уже из карбоновых отложений, мы можем уверенно считать, что их предками были какие-то стегоцефалы этого периода. Таким образом, древность рептилий оказывается очень большой; она равна приблизительно 300 000 000 лет (рис. 17).
Наиболее примитивные из древних рептилий, по всей вероятности, были очень похожи на стегоцефалов.
Но нужно думать, что даже самые примитивные из древних представителей этого класса были’в еще большей степени приспособлены к жизни на суше, чем стегоцефалы, и это так или иначе отражалось на многих деталях строения их скелетов. Однако основным отличием первых рептилий от их непосредственных предков — амфибий были не особенности строения тела взрослых форм, а различия в способах размножения и в развитии зародышей.
Эти различия заключались в том, что в то время как амфибии, подобно своим предкам — рыбам, размножались путем откладывания икры в пресноводные бассейны, в которых затем и развивались их личинки, рептилии стали откладывать свои яйца непосредственно на сушу и перешли к развитию без личиночной стадии, без метаморфоза.
Это оказалось возможным только потому, что яйца рептилий приобрели сначала плотную кожистую оболочку, способную предохранить их от высыхания, а затем и скорлупу, состоящую из слоя своеобразных известковых сферокристаллов (рис. 130, А). Кроме того, каждое яйцо было снабжено большим количеством желтка, которого в качестве питательного материала для эмбриона хватало на весь период его развития.
Доказательством того, что вымершие рептилии размножались путем откладывания именно таких яиц, является находка в пермокарбоновых отложениях Северной Америки небольшого окаменелого яйца какой-то рептилии (рис. 130, В). Это яйцо имело овальную форму и было покрыто типичной для рептилий скорлупой; длина.его равнялась 59 мм. Древность яйца определяется приблизительно в 225 000 000 лет.
Изучение онтогенеза современных рептилий показывает, что их зародыш имеет ряд своеобразных приспособлений, позволяющих ему развиваться в яйце, находящемся на суше.
На некоторой стадии развития зародыша у него возникает своеобразная оболочка, образующая наполненный жидкостью пузырь, носящий название амниона (рис. 131—Ат). В результате этого эмбрион оказывается заключенным как бы в маленький бассейн. Таким образом, рептилии, отор-
9 А П Быстров
12?
ванные от тех водоемов, в которых развивались зародыши их предков -амфибий, оказались вынужденными создать в яйце свой собственный водоем.
По понятным причинам амнион не образуется ни у одного представителя бесчелюстных позвоночных, рыб и амфибий. Их зародыши, развиваясь
А
, 59 тт
Рис. 130.
А — вертикальный разрез скорлупы яйца черепахи;
S — ископаемое яйцо рептилии (Ромер и Прайс— Romer and Price, 1939).
в воде, не нуждаются в этом образовании. Поэтому эти три класса позвоночных принято объединять в одну группу Anamnia (лишенных амниона), в то время как все классы позво
В
ночных, начиная с рептилий, образуют группу Amniota (снабженных амнионом).
Хотя амниотическая жидкость у рептилий и являлась подходящей средой для развивающегося эмбриона, однако она не могла служить средой для газообмена при помощи жабр. По-видимому, именно это обстоятельство оказалось главной причиной того, что жабры у зароды-и вместо них вне амниона
шей рептилий подверглись полной редукции образовался снабженный большим количеством кровеносных сосудов аллантоис (рис. 131—А/), который и взял на себя функцию органа газообмена.
Рис. 131. Зародыш рептилии.
Al — allantois; Ат — amnion; V — желточный мешок.
Появление всех этих своеобразных органов у зародыша, несомненно, было результатом вынужденного приспособления рептилий к неблагоприятным условиям жизни. При этом необходимость откладывать яйца
130
на сушу могла возникнуть только вследствие пересыхания тех бассейнов, в которых прежде откладывалась икра, а образование на поверхности яйца сначала кожистой оболочки, а затем и скорлупы являлось защитным приспособлением, предохраняющим яйцо от высыхания.
Поэтому можно уверенно сказать, что причиной всех этих изменений в строении яиц и в развитии зародыша, а следовательно, и причиной появления рептилий мог быть только сухой и жаркий климат. В момент появления рептилий он, очевидно, был настолько сухим, что нередко вызывал полное пересыхание тех пресноводных бассейнов, в которых обычно развивалась икра древних позвоночных, и настолько жарким, что делал возможным развитие зародыша в яйце, отложенном на сушу.
Климат-карбона обычно принято считать влажным и теплым. Накопление мощных слоев каменного угля в'течение этого периода свидетельствует об очень благоприятных условиях для бурного расцвета самых разнообразных растений. Однако в каменноугольное время, несомненно, существовали и такие ареалы, климат которых был сухим. На это указывают как палеоботанические, так и геологические данные. Изучение ископаемой флоры показало, что в конце карбона в некоторых достаточно обширных ареалах появились растения с такими особенностями строения, которые являлись приспособлением к сухому климату. На это же указывает и накопление гипса и каменной соли в соответствующих отложениях.
Доказательством того, что в карбоне было немало таких областей, в которых пресноводные бассейны подвергались периодическим пересыханиям, является^ тот факт, что в это время жили и типичные двоякодышащие рыбы, как Uronemus, Ctenodus, Gnathorhiza и Sagenodus, способные зарываться в ил и впадать в спячку на весь период засухи.
По-видимому, именно такие области и были теми местами, в которых происходило формирование первых рептилий.
Таким образом, если сухой и жаркий климат девона был основной причиной первого этапа приспособления позвоночных к жизни на суше и привел к превращению некоторых кистеперых рыб в амфибий, которые во взрослом состоянии уже могли жить на суше, но еще были вынуждены откладывать свои яйца в воду, то такой же климат в некоторых ареалах в каменноугольное время обусловил и второй этап приспособления позвоночных к жизни на суше и привел к таким изменениям в строении их яиц и в развитии зародышевых оболочек, которые позволили Tetrapoda полностью оторваться от водоемов и превратиться в рептилий.
SEYMOUR I AMOR РИА
Представители отряда Seymouriamorpha являются самыми примитивными из известных в настоящее время ископаемых рептилийВ их скелете еще сохранились некоторые признаки стегоцефалов. Это обстоятельство' было причиной того, что одни палеонтологи считали сеймуриаморф рептилиями (Бройли — F. Broili, 1904; Ватсон, 1918; Уайт — Т. Е. White, 1939), другие — амфибиями (Брум — R. Broom, 1913; П. П. Сушкин, 1925; А. П. Быстров, 1944).1
В настоящее время известны только две формы сеймуриаморф, именно: Seymouria baylorensis Broili из нижнепермских отложений Северной Америки и Kotlassia prima Amalitzky, найденная в верхнепермских отложениях, на берегах Северной Двины {Lanthanosuchus watsoni, описанный И. А. Ефремовым, 1946, по черепу из верхнепермских отложений Поволжья и отне-
1 Сейчас я отказываюсь от прежнего своего взгляда и считаю сеймуриаморф реп--тилиями
9*
13L'
Рис 132. Позвонки Kotlassia prima. А, В — грудные позвонки; С — крестцовый позвонок; D — хвостовой позвонок
(А. П. Быстров, 1944).
a. d.—arcus dorsalis; a. ч>. —arcus ventralls; c—costa; Л—hypocentrum; p—pleurocent-rum.
Рис. 133.
A — три первых позвонка Seymouria; В — первый позвонок Seymouria (Уайт 1939); С — три первых позвонка котилозавра (Procolophon trigoniceps.
а. а. — arcus atlantis; h. а. — hypocentrum atlantls; h. e. — hypocentrum epistrophei; p. a. — pleurocentrum atlantis; p. e, — pleurocentrum epistrophei; pa — proatlas.
сенный им к Seymouriamorpha, по моему мнению, не является представителем этого отряда).
Позвонки Seymouria и Kotlassia были построены, как позвонки рептилий. Тело каждого позвонка представляло собой сильно увеличенный
2
Ра
Рис. 134. Черепа Seymouria baylorensis Broil! (Л) и Kotlassia pritna Amalitzky (вид сзади).
Во — baslocclpltale, Dso — dermosupraocclpltale; Во — exocclpltale; Po — parocclpl-tale; Pt — pterygoideum; Qj — quadratojugale; Qu — quadratum; 5 — stapes; Sy— squamosum; T — tabulate.
плевроцентр (рис. 132, A—p). Хорошо развитая дуга позвонка была сращена с его телом. На передней и задней сторонах плевроцентра имелись конусообразные углубления, соединенные небольшим отверстием, сквозь которое
Рис. 135. Мозговой череп Kotlassia ptima (вид сбоку)
;А. П. Быстров, 1944).
Bs — baslsphenoldeum; /. о. — fenestra ovalis; Po — parocclpitale; Pro — prootlcum: Ps — parasphenoldeum; Se — sphenethmoideum.
проходила хорда (рис. 132, В). Гипоцентр, хорошо развитый у стегоцефалов, у сеймуриаморф подвергся сильной редукции; он имел форму небольшого костного полулуния и располагался непосредственно перед телом позвонка (рис. 132, А, В). На нижней стороне гипоцентров’ хвостовых позвонков имелись вентральные дуги (рис. 132, D—a. v.).
133
Ко всем* туловищным позвонкам и к первым 5—7 хвостовым позвонкам у сеймуриаморф присоединялись двухголовчатые ребра. При этом одна из головок сочленялась с поперечном отростком дуги, другая — с суставной площадкой на боковой поверхности маленького гипоцентра (рис. 132). Таким образом, гипоцентр, несмотря на свою сильную редукцию, у этих примитивных рептилий не потерял контакта с ребром.
Совершенно очевидно, что такие позвонки могли возникнуть только из позвонков эмболомерных стегоцефалов. Это произошло путем постепен-
ного увеличения плевроцентров при одновременном сильном .уменьшении гипоцентров. Позвонки сеймуриаморф не могли развиться из позвонков рахитомного или сте-реоспондильного типа, так как плевроцентры в рахитомных позвонках уже подверглись значительной редукции, а в стереоспондильных — полностью отсутствовали. Что же касается гипоцентров, то они, наоборот, были очень сильно развиты у Rhachitomi, а у Stereospondyli составляли все тело позвонка.
Два первые позвонка, атлант и эпистрофей, у сеймуриаморф были : построены очень своеобразно. Дуга ат-I ланта состояла . из двух не-сросшихся друг с другом половин и представляла собой изолированное образование ! (рис'. 133, Л, В). Плевроцентр
Рт
Я/-
i.o.
Prf
Ро
t
Dso
Рис. 136. Череп
Seymouria baylorensis (вид сверху) (Уайт, 1939).
Dso — dermosupraocclpl.tale; d. nl. — ductus nasolacrimalls; /’•—frontale; It — Intertemporale; i. o. — Incisura otlcalls; Ju — jugale; L— lacrimale; M — maxillare; N — nasale; P— parietale; Pf — postfrontale; Pm — praemaxillare; Po — postorbitale; /V/—praefrontale; Qj — quadratojugale; Qu — quadratum; Sm~ septomaxlllare; Sq— squamosum; St — supratem-porale; T — tabulare.
Broili
этого'позвонка располагался несколько позади дуги и имел отверстие для хорды [(рис.
’ 133,В—р. а.). Гипоцентр был относительно большой костью и помещался непосредственно под дугой. Двухголовчатое ребро атланта присоединялось к поперечному отростку дуги и к. суставной площадке на гипо
центре.
Эпистрофей отличался от обычных туловищных позвонков клиновидной формой своего плевроцентра (рис. 133, А—р.е.). Его дуга была сращена с телом позвонка; гипоцентр имел обычную форму костного полулуния. Эпистрофей, как и атлант, был снабжен парой ребер, головки которых присоединялись к его дуге и гипоцентру.
Череп сеймурии по своей общей форме был очень похож на череп карбонового стегоцефала.
В состав мозгового черепа (neurocranium) у сеймуриаморф входило несколько костей.
Basioccipitale (рис. 134, Д, В—Во), примыкавшее к плевроцентру атланта, имело на своей задней поверхности небольшое углубление для пе
134
реднего конца хорды. Рядом с этой костью находилось exoccipitale (Ео), нижний край которого также принимал участие в сочленении с I позвонком. Supraoccipitale у сеймурии было хрящевым; у котлассии оно представляет собой небольшое костное образование, прикрытое сзади dermosupra-occipitale. Paroccipitale (Ро) представляло собой большую кость, лежащую латерально от exoccipitale.
Перед paroccipitale располагалось хорошо развитое prooticum (рис. 135—Pro). На границе между этими костями имелось большое отверстие,
ведущее во внутреннее ухо (fenestra ovalis, рис. 135). .
Непосредственно к переднему краю prooticum примыкало небольшое костное basisphenoideum (Bs), на некотором расстоянии от которого помещалось хорошо развитое sphenethmoideum (Se). Эта кость по своей форме была очень похожа на соответствующую кость в черепе стегоцефалов (рис. 107).
В хрящевом splanchno-cranium у сеймуриаморф развивались три костные элемента, именно: quadratum (рис. 134, А, В—Qu), articulare (в нижней челюсти) и stapes (рис. 134, А, В—S). Stapes своим основанием закрывал fenestra ovalis, а вершиной был направлен косо вверх и соединялся с барабанной перепонкой, помещавшейся в слуховой вырезке на заднем краю черепной крыши.
В состав крыши черепа у сеймуриаморф входили те же покровные кости, что и у карбоновых стегоцефалов
Рис. 137; Череп Seymouria baylorensts Broili (вид снизу) (Уайт, 1939).
/. a.— fenestra adducioria, М - maxillare, р. с. — processus cultrlformls; Pl — palatlnum; Pm — praemaxillare; Ps — pa* rasphenoideum: Pt — pterygoideum; Qj — quadratojugale; Qu — quadratum; Tr —transversum; V — vomer.
(рис. 136). Затылочный край
был образован tabulare (Т) и dermosupraoccipitale (Dso). Перед этими костями лежали parietale (Р), supratemporale (St) и intertemporale (It).
В шве между парой parietalia имелось небольшое пинеальное отверстие. Далее кпереди располагались frontale (F), nasale (N) и praemaxillare (Pm). Орбиты были окружены praefrontale (Prf), postlrontale (Pf), postorbitale (Po), jugale (Ju) и lacrimale (L).
Lacrimale у сеймуриаморф, как у примитивных стегоцефалов, доходило своим передним концом до носового отверстия. У Seymouria baylorensis в этой кости имелся длинный слезно-носовой канал (ductus nasolacrimalis—
d. nl.), который, пройдя сквозь небольшое septomaxillare (Sm), открывался
в полость носа.
Боковой край черепа был образован относительно узким maxillare (М), к которому сзади примыкало quadratojugale (Qj), прикрывавшее вме*: сте со squamosum (Sq) квадратную кость.
Слуховая вырезка (incisura oticalis — i. о.), в которой натягивалась
135
барабанная перейонка, была ограничена краями трех костей: tabulare, supratemporale и squamosum.
Нёбная часть черепа у сеймуриаморф была построена так же, как соответствующая часть черепа у карбоновых стегоцефалов (рис. 137, 138). Сильно развитые птеригоиды (Pt) были сближены друг с другом и соединялись с особым отростком базисфеноида (processus basipterygoideus); между ними входил processus cultriformis parasphenoidei (р. с.). Vomer, ра-
Рис. 138. Череп Kotlassia prima (вид снизу) (А. П. Быстров, 1944), /. а. — fenestra adductorla; М — maxlllare; Pl —palatinum; Pm — praemaxillare; Ps — parasphenoldeum; Pt — pterygoideum; QJ — quadratojugale; Qu — quadratum; Tr — transversum; V — vomer.
latinum и transversum располагались перед птеригоидом и сбоку от него. Хоаны были далеко отодвинуты друг от друга.
Новой особенностью нёба сеймуриаморф по сравнению с нёбом стегоцефалов было появление большого бокового отростка у птеригоида, что вызвало сильное сужение переднего отдела fenestra adductoria (f. а.).
В состав нижней челюсти у сеймурии и котлассии входило такое же количество костей, как и у стегоцефалов (рис. 139, 140). На ее наружной стороне лежали supraangulare (Sa) и dentale (D), на верхнем краю которого сидел один ряд острых конических зубов. Вдоль нижнего края челюсти помещались spleniale (Sp), postspleniale (Ps) и angulare (Лп). На внутренней стороне нижней челюсти располагались большое praearticulare (Ра), сого-noideum (Со), intercoronoideum (Ic) и praecoronoideum (Рс). Все три коро-ноида у сеймуриаморф были лишены зубов.
Те кости черепа и нижней челюсти, которые были непосредственно покрыты кожей, на своей наружной поверхности имели многочисленные ячейки, по своей форме не отличавшиеся от таких же образований на черепах стегоцефалов (рис. 140, 141),
136
Изучение микроструктуры покровных костей котлассии показало, что они по своему строению очень напоминают соответствующие кости стегоцефалов (рис. 142). Верхний слой кости состоит из наложенных друг на друга изогнутых костных пластинок, формирующих гребни ячеистой скульптуры. На дне каждой ячейки обычно открывается довольно широкий канал,
Pf-
Рис. 139. Череп Seymouria baylorensis Broili (вид сбоку) (Уайт, 1939).
Ап — angulare; D — dentale; It — Intertemporale; Ju — jugale; L — lacrimale; M — maxillare; W — nasale; P/ —postfrontale; Pm — praemaxillare; Po — postorbitale; Prf — praefrontale; Ps — postsplentale; Qj — quadratojugale; Sa — supraangulare; Sm — septomaxillare; Sp — splenlale; Sq — squamosum; St — supratemporale; T — tabulare.
Рис. 140. Нижняя челюсть Kotlassia prima (А. П. Быстров, 1944).
An — angulare; Co — coronoldeum; D — dentale; Ic — intercoronoldeum; Pa — praearticulare; Pc — praecoronoideum; Ps— postsplenlale; Sa — supraangulare; Sp — splenlale.
по которому проходил кровеносный сосуд, направляющийся к коже, Такие же каналы имелись и в костях стегоцефалов.
У сеймуриаморф отсутствовали всякие следы борозд для каналов сейсмосенсорной системы. Полная редукция этой системы, могущей функционировать только в воде, свидетельствует 0 ксерофильности сеймурий.
Следует подчеркнуть, что сейсмосенсорная система, изчезнувшая у этих самых примитивных рептилий, уже никогда не появлял ась на всем протяжении дальнейшей эволюции позвоночных.
Зубы у Seymouria и Kotlassia представляли собой конические образования, приросшие своим основанием непосредственно к тем костям, на ко-
13Z
Рис. 141. Череп Kotlassia prima (вид сверху) (А. П. Быстров, 1944).
Рис. 142. Вертикальный разрез покровной кости черепа Kotlassia prima (X 300).
торых они располагались. Изучение поперечных разрезов зубов показало, что их дентинная стенка, как у типичных стегоцефалов, была снабжена складками (рис. 143) (Ф. Бройли, 1927; А. Быстров, 1944).
В состав плечевого пояса у сеймуриаморф входили те же элементы, что и у стегоцефалов, но лопатки и коракоиды окостеневали полностью и, срастаясь друг с другом, образовывали большое костное scapulocoraco-ideum (рис. 144, 145).
Тазовый пояс был образован хорошо окостеневшими и сросшимися между собой ilium, ischium и pubis. Крылья ilium были отклонены назад и, как у стегоцефалов, соединялись при помощи пары сильно развитых ребер с одним крестцовым позвонком (рис. 146). Тот конец крестцового
Рис. 143. Поперечный разрез зуба Kotlassia prima (длинная ось равна 3 мм) (А. П. Быстров, 1944).
ребра, который соединялся с крылом позвоночной кости (ilium), был сильно расширен, что обеспечивало прочную связь между этими костями.
Интересно, что у одного из экземпляров котлассии на левой стороне имелось одно крестцовое ребро, а на правой — два (рис. 147), при этом оба они своими расширенными концами были соединены с правым ilium. Такой вариант в строении, несомненно, свидетельствует о том, что сейму-риаморфы уже стояли на пути к включению двух позвонков в состав крестца.
Седалищная и лонная кости у сеймуриаморф лежали, как у стегоцефалов, в горизонтальной плоскости.
Передние и задние конечности сеймурий были развиты сильнее, чем у стегоцефалов, и были в достаточной степени приспособлены к передвижению животного по земле.
На плечевой кости Seymouria baylorensis имелось небольшое отверстие (foramen entepicondyloideum, рис. 148—Н), которое, однако, отсутствовало у Kotlassia.
В коленном суставе сеймурий бедро опиралось, как у стегоцефалов, одновременно на большую и малую берцовые кости (рис. 149).
Некоторые элементы carpus и tarsus у сеймурий окостеневали; кисть и стопа у Seymouria baylorensis Broili имели по пяти пальцев (рис. 150). Фаланговые формулы были: для кисти — 23453, для стопы — 23454.
139
Рис. 144. Плечевой пояс Kotlassia prima (вид свизу) (А. П. Быстров, 1944).
С —coracoldeum; Cl — clavlcula; Ic — Interclavl-cula; Sc — scapula.
Рис. 145. Плечевой пояс Kotlassia prima (вид сбоку) (А. П. Быстров, 1944).
Ci — clavlcula; Ct — cleithrum; !c — Interclavlcula; Sc-scapula.
Рис. 146. Таз Kotlassia prima (вид слева) (А. П. Быстров, 1944). Л — Ilium; Is — Ischium; Pb — pubis,
Рис. 147. Крестцовые позвонки Kotlassia prima (А. П. Быстров. 1944).
Рис. 148. Кости конечностей Seymouria baylorensis (Уайт, 1939).
Г — fibula; Fe — femur; H—humerus; R — radius; U —ulna; T — tibia.
Рис. 149. Скелет Kotlassia prima. Пермь <А. П. Быстрое, 1944).
Детальное изучение представителей отряда Seymouriamorpha показывает, что позвонки их были построены, как у типичных рептилий, в то время как в черепе еще сохраняются такие признаки амфибий, как ячеистая скульптура и складчатые зубы.
Все это свидетельствует об очень большой примитивности этих форм рептилий.
COTYLOSAURIA
Примитивные пермские рептилии, относящиеся к отряду котилозавров, характеризуются отсутствием каких-либо других отверстий в крыше черепа, кроме ноздрей, орбит и foramen parietale. В этом отношении они не отличались от сеймуриаморф, и, принимая во внимание многие другие особенности в строении скелета котилозавров, мы можем считать этих рептилий вероятными потомками каких-то древних представителей Seymouriamorpha (рис. 17—Cs).
Позвонки котилозавров по своему строению были очень похожи на позвонки сеймурий. Каждый из них состоял из большого плевроцентра (рис. 151—р) и хорошо развитой дуги. Сильно редуцированный гипоцентр представлял собой небольшое костное полулуние (рис. 151, А, В, С—h). Передняя и задняя поверхности тела позвонка (pleurocentrum) были конусообразно вогнуты (рис. 151, С). На нижней стороне гипоцентров в хвостовом отделе позвоночного столба имелись вентральные дуги (рис. 151, Е—a. v.)
Первые два позвонка (атлант и эпистрофей) у котилозавров отличались от соответствующих позвонков сеймураиморф рядом очень интересных особенностей.
Дуга эпистрофея была снабжена высоким остистым отростком, имевшим форму большой костной пластинки, поставленной в сагиттальной плоскости (рис. 133, С). Полулунный гипоцентр II позвонка плотно прилегал к нижнему краю плевроцентра.
Атлант состоял из шести костных элементов. Его дуга была разделена на правую и левую половины; каждая из этих частей имела Г-образную форму и своим задним отростком накладывалась сверху на переднюю 142
суставную площадку дуги эпистрофея. На каждой половинке дуги I позвонка располагалась небольшая наклоненная вперед косточка, носящая название проатланта — proatlas (рис. 133, С—Ра). Этот элемент I позвонка еще отсутствовал у сеймуриаморф.
Рис. 151. Позвонки котилозавра (Nyctiphruretus acudens). А — грудные позвонки; В, С — поясничные позвонки; D — крестцовые позвонки; Е — хвостовые позвонки (И. А. Ефремов, 1940).
a. d. — arcus dorsalis; a. v. — arcus ventralls; c — costae; h — hypocentrum; p — pleurocentrum.
Гипрцентр атланта (рис. 133, С—h. а.) имел форму костного полулуния и непосредственно присоединялся к нижним концам дуги. Что же касается плевроцентра I позвонка (рис. 133, С—р. а.), то он располагался таким об
разом, что соединялся не только с гипоцентром и дугой атланта, но плотно прилегал к гипоцентру и плевроцентру эпистрофея.
Череп котилозавров в общих своих очертаниях еще сохранял признаки примитивности. 'Он был, как у сеймуриаморф, относительно широким и низ -ким, но у некоторых рептилий этого отряда уже наблюдалось сужение, переднего отдела черепа, и он в таких случаях приоб-
Рис. 152. Череп Nyctiphruretus acudens (вид сзади) (И. А. Ефремов, 1940).
Во — basiocclpltale; Dso — dermosupraocclpltale; Ео —exoccipitale. Po — parocclpltale; Qu — quadratum; S — stapes; So — supraocct-pltale; Sq — squamosum; T — tabulate.
ретал клиновидную форму.
Тот участок затылочного отдела черепа, который непосредственно примыкал к I позвонку, был образован хорошо окостеневшими basioccipitale (рис. 152 — Во) и exoccipitale (Ео). Над exoccipitale располагалось supra-occipitale (So). Эти кости окружали со всех сторон затылочное отверстие. К боковому участку exoccipitale присоединялось поставленное косо раг-
143
occipitale (Po). Все эти элементы черепа представляли собой окостеневшие' части заднего отдела хрящевого neurocranium.
У некоторых представителей Cotylosauria окостеневали и такие кости мозгового черепа, как basisphenoideum и sphenethmoideum.
Окостенение splanchnocranium у . котилозавров, как и у сеймуриаморф, вызывало образование таких важных когтей, как quadratum (рис. 152—Qu), articulare (в нижней челюсти) и stapes (рис. 152—S).
Очень важным являет-
’ис. 153. Череп Nyctiphruretus acudens (вид сверху) (И. А. Ефремов, 1940).
>so — dermosupraoccipitale; F — irontale; Ju — jugale; L — lacrlma-;; M — maxillare; N — nasale; P — parietale; Pf — postfrontale; Fit — praemaxillare; Po — postorbitale; Prf — praefrontale; Qj — quadratojugale; Sq — squamosum; St —supratemporale; T — tabulare.
ся тот факт, что stapes у этих рептилий одним своим концом входил в fenestra ovalis, ведущее во внутреннее ухо, а другим — присоединялся непосредственно к quadratum (рис. 152). Такое положение stapes ясно свидетельствует о том, что барабанная перепонка у котилозавров отсутствовала. Это подтверждается и тем, что на заднем краю черепной крыши для нее не было соответствующей вырезки (рис. 153).
В состав черепной крыши котилозавров входило меньшее количество костей, чем у сеймуриаморф, но некоторые из этих костей имели интересные и важные особенности.
Такая парная у всех стегоцефалов и сеймурий покровная кость, как der-mosupraoccipitale (Dso), у котилозавров в результате срастания превратилась й непарное образование. Эта впервые возникшая особен
ность затем удержалась на всем протяжении эволюции позвоночных. Tabulare (Т), располагавшееся по обе стороны непарного dermosup-raoccipitale, у котилозавров не образовывало вместе со squamosum (Sq) слуховой вырезки.
Parietale (Р), frontale (Г) и nasale (N) помещались на обычных своих местах.
Supratemporale у примитивных котилозавров имелось. Полное сращение этой кости с parietale у N yctiphruretus acudens (рис. 153—St) не является особенностью, свойственной всем котилозаврам.
Intertemporale, входившее в состав крыши черепа у примитивных стегоцефалов и у сеймуриаморф, у котилозавров подверглось полной редукции. Эта кость в течение всей дальнейшей эволюции рептилий уже никогда не появляется вновь.
Вокруг орбит у котилозавров располагались praefrontale (Prf), postfrontale (Pf), postorbitale (Po), jugale (Ju) и lacrimale (L). При этом lac-
144
rimale у большинства форм уже не достигало носового отверстия. Края черепа были образованы, как обычно: praemaxillare (P/n), maxillare (М) и quadratojugale (Qj).
Передние отделы носовых отверстий у котилозавров были сближены друг с другом настолько, что nasale и praemaxillare между ними оказывались сильно суженными. Эта особенность в строении черепа рептилий была Новой.
Рис. 154. Череп Nyctiphruretus acudens (вид снизу) (И. А. Ефремов, 1940).
Во — basloccipltale; /. а. — fenestra adductorla; М — maxillare; Pl — palatlnum; Pm — praemaxillare; Po — paroccipitale; Ps — paraspheno-Ideum; Pt — pterygoldeum; QJ — quadratojugale; Qu — quadratum; S— stapes; Tr — transversum; V — vomer.
Нёбная часть черепа у котилозавров имела такое же строение, как у сеймуриаморф.
Хорошо развитые птеригоиды (рис. 154 — Pt) своими внутренними краями вплотную прилегали к парасфеноиду (Ps), поэтому межптеригоидные отверстия отсутствовали. Вследствие развития боковой части птеригоида передний отдел fenestrae adductoriae (f. а.) был сужен. Сошники (vomer— V) были широкими, поэтому хоаны оказывались так же отодвинутыми друг от друга, как у сеймуриаморф. Palatinum (РГ) и transversum (Tr) были почти одинаковой величины.
На наружной стороне нижней челюсти располагались dentale (рис. 155— D), снабженное одним рядом зубов, supraangulare (Sa), angulare (Ап) и spleniale (Sp). Postspleniale отсутствовало. Эта кость, еще хорошо развитая у сеймуриаморф, у котилозавров подверглась полной и окончательной
10 А. П. Быстров
145
редукции и на протяжении всей дальнейшей эволюции рептилий уже никогда не появлялась.
На внутренней стороне нижней челюсти у котилозавров имелось довольно хорошо развитое praearticulare; что же касается короноидов, то они уже начали подвергаться редукции.
Зубы наиболее примитивных котилозавров представляли собой Дентинные конусы, покрытые тонким слоем эмали. Их основания были, как у стегоцефалов и сеймурий, непосредственно сращены с теми покровными костями, на которых они располагались (рис. 156). Поперечные разрезы таких зубов показывают, что они уже не имели никаких следов дентинных
Рис. 155. Череп Nyctiphruretus acudens (вид сбоку) (И. А. Ефремов, 1940).
Ап — angulare; D — dentale; Л —frontale; Jw —jugale; L — lacrlmale; M — maxil-lare; TV — nasale; Р/— postfrontale; Pm — praemaxillare; Po — postorbitale; Prf — praefrontale; Qj — quadratojugale; Sa — supraangulare; Sp — spleniale; Sq — squamosum; T — tabulare.
складок (рис. 157). Следовательно, можно считать, что при переходе от сеймуриаморф к котилозаврам произошла полная редукция дентинных складок; и это, несомненно, было важным шагом вперед. Интересно, что у некоторых котилозавров мы видим следующую фазу эволюции зубов. Эта фаза сопровождалась образованием корня зуба (рис. 158). Такие зубы уже не прирастали непосредственно к костям, а были вставлены своими длинными корнями в глубокие ячейки (альвеолы).
Как указывалось выше, у стегоцефалов на parasphenoideum, pterygoideum, а иногда на vomer и palatinum располагались поля очень маленьких так называемых шагреневых зубов.
У котилозавров эти зубы на парасфеноиде исчезли, но сохранились на птеригоидах, сошниках и небных костях.
У некоторых представителей этого отряда рептилий нёбные зубы располагались не беспорядочно, а рядами (рис. 154). Наиболее постоянным из этих рядов был поперечный ряд относительно больших зубов, сидящих на заднем краю бокового отростка птеригоида.
Плечевой пояс у котилозавров был образован такими же элементами, как и у сеймурий. В состав его входило полностью окостеневшее scapulo-coracoideum (рис. 159, А, В — Sc, Со, Рс) с хорошо оформленной суставной ямкой для головки плечевой кости. На края коракоидов с вентральной стороны накладывалась несколько редуцированная interclavicula,
146
Рис. 156. Вертикальный разрез зубов Nyctiphruretus acudens.
10*
снабженная длинным отростком (рис. 159, А — 1с). К переднему краю этой кости и к переднему к{ чипа (clavicula, рис. 159, А,
Рис. 158. Зубы парейазавра.
лопатки присоединялась тонкая клю-В). Cleithrum представляло собой еще более тонкую кость, лежащую вдоль переднего края лопатки (рис. 159, В — Ct).
Если сравнить плечевой пояс котилозавров с плечевым поясом сеймуриаморф и в особенности с плечевым поясом стегоцефалов, то можно видеть, что у типичных рептилий такие его покровные кости, как interclavicula, clavicula и cleithrum, подверглись сильной редукции и в значительной степени утратили свое прежнее значение, в то время как полностью окостеневший скапулокоракоид стал играть роль основного элемента этого отдела скелета.
Тазовый пояс котилозавров, как и у сеймурий, был представлен тремя полностью окостеневшими элементами: ilium (рис. 160 — II), ischium (Is) и pubis (Pb). При этом ischium и pubis сохраняли то же
положение, которое они занимали в тазу сеймурий, т. е. располагались горизонтально (рис. 160, В, С). В месте соединения этих трех костей помещалась хорошо выраженная суставная ямка для головки бедра.
Рис. 159. Плечевой пояс Nyctiphruretus acudens. А — вид снизу; В — вид сбоку (слева) (И А. Ефремов, 1940). Cl — clavicula; Со — coracoldeum; Ct — cleithrum; Ic — Intercla-vlcula; Pc — procoracoldeum; Sc — scapula.
Рис. 160. Тазовый пояс Nyctiphruretus acudens. A — вид сверху;
В — вид снизу; С — вид сбоку (слева) (И А. Ефремов, 1940).
II — Ilium; Is — ischium; Pb — pubis.
Верхний конец ilium у котилозавров был сильно расширен. Эта особенность была связана с тем, что тазовый пояс у этих рептилий прикреплялся к трем позвонкам при помощи трех пар ребер (рис. 160, Я).
148
Концы этих ребер были расширены и соединены с крылом ilium, а головки сочленялись с соответствующими позвонками (рис. 151, D).
Прочное соединение таза с тремя крестцовыми позвонками, несо
мненно, является очень важным шагом вперед в эволюции скелета наземных позвоночных. Три пары крестцовых ребер хорошо фиксировали таз на позвоночном столбе и это делало воз-
можным дальнейшее усиление задних конечностей как основного аппарата, толкающего тело животного вперед.
Передние и задние конечности котилозавров, построенные по тому же глзну, что и у сеймурий, отличались только большим развитием и, несомненно, большей подвижностью (рис. 161).
Таким образом, оформление скелета типичных рептилий привело к сильным изменениям в строении первых двух позвонков, к слиянию парного der-mosupraoccipitale в одну кость, к полной редукции intertemporale и post-spleniale, к исчезновению дентинных складок в зубах, к некоторой редукции покровных костей плечевого пояса и к соединению таза при помощи трех пар ребер с тремя крестцовыми позвонками.
PELYCOSAURIA
Рептилии, относящиеся к отряду Pelycosauria, по-видимому, являются непосредственными потомками котилозавров (рис. 17 — Р).
Главной отличительной их особенностью было наличие в черепной крыше дополнительного отверстия, расположенного непосредственно позади орбиты (fenestra postorbitalis). В результате этого у пеликозавров впервые появилась скуловая дуга, в образовании которой у примитивных форм этого отряда принимали участие jugale, squamosum и quadratojugale.
По своей общей форме черепа пеликозавров обычно сильно отличались
Рис. 161. Скелет N yctiphruretus acudens Efr. Пермь (И. А. Ефремов, 1940).
от черепов котилозавров. Они были не низкими и широкими, а относительно
высокими и узкими.
Позвонки пеликозавров отличались от позвонков котилозавров большей высотой и более сильным развитием остистых отростков (рис. 162,В, С). Дуги позвонков были полностью сращены с телами, которые представляли собой плевроцентры с вогнутыми передней и задней поверхностями; отверстие для хорды в них было очень мало, а иногда совсем отсутствовало. Гипоцентры имели форму небольших костных полулуний, вставленных в виде жлиньев между плевроцентрами соседних позвонков (рис. 162).
Все шейные, грудные, поясничные, крестцовые и первые хвостовые позвонки у пеликозавров были снабжены ребрами. Ребра, имеющие в раз
149
личных отделах разную длину, соединялись с поперечными отростками и с гипоцентрами позвонков.
Атлант и эпистрофей у пеликозавров были построены так же, как у котилозавров, но гипоцентр II позвонка (рис. 162, А—Л. е.) у некоторых
Рис. 162. Позвонки пеликозавра Ophiacodon retroversum. А — атлант и эпистрофей; В —шейные позвонки, вид сбоку; С — шейный позвонок, вид спереди.
а. а. — arcus atlantls; Л — hypocentrum; Л. а. — hypocentrum atlantls; Л. е. — hypocentrum epistrophei; р — pleurocentrum; pa — proatlas; p. a.pleuro-l> centrum atlantls; p. e. — pleurocentrum epistrophei.
форм почти полностью срастался с плевроцентром атланта (р. а.). В результате этого возникала большая комплексная кость, которая своей задней
Рис. 163. Ophiacodon retroversum. А —I и II позвонки, вид спереди; В — атлант, вид спереди (Ромер и Прайс, 1940).
а. а. — arcus atlantls; h. а. — hypocentrum atlantls; Л. e. — hypocent-rum epistrophei; p. a. — pleurocentrum atlantls; p. e, — pleurocentrum epistrophei.
поверхностью плотно прилегала к плевроцентру эпистрофея. Дуга атланта имела Г-образную форму (рис. 162, А—а. а.); к ней присоединялся проатлант (ра).
150
Вогнутые суставные площадки на обеих половинках дуги атланта и такая же площадка на его гипоцентре образовывали полушаровую поверхность, которая принимала участие в образовании сустава с черепом (рис.
DS0
Рис. 164. Череп Dimetrodon (вид сзади) (Ромер и Прайс, 1940).
Во -f- Ео — basioccipitale -j- exoccipitale; Dso — dermosupraoccipitale; Po — paroccipitale; Qj — quadratojugale; Qw — quadratum; S — stapes; So — supraoccipitale; Sq — squamosum; St — supratemporale; T — tabulare.
163, В). В углубление, образованное этими тремя элементами I позвонка, входил шаровидный суставной отросток черепа (рис. 164). Он был образован сросшимися друг с другом basioccipitale и двумя exoccipitale. На
Рис. 165. Продольный разрез черепа Dimetrodon (Ромер и Прайс, 1940).
Ер — eplpterygoideum; Pro — prootlcum; Ps — parasphenoideum; Pt — pterygoideum; Qu — quadratum; Se — sphenethmoldeum.
верхнем участке каждого exoccipitale имелась небольшая суставная площадка, к которой присоединялся передний конец проатланта. Таким образом, череп сочленялся не только с гипоцентром и дугой атланта, но и с проатлантом.
151
Рис. 166. Череп Dimetrodon (вид сверху) (Ромер и Прайс, 1940).
F — frontale; Ju — jugale; L — lacrimale; M — maxillare; A'— nasale; P — parletale; Pf — postfrontale; Pm — praemaxillare; Po — postorbitale; Prf — praefrontale; Sq — squamosum.
Над exoccipitale в черепе специализированных пеликозавров располагалась большая кость, являвшаяся результатом слияния supraoccipitale (So) и парного paroccipitale (Ро), к которому спереди прирастало prooticum (рис. 165 — Pro). Basisphenoideum у пеликозавров было сращено с prooticum и с parasphenoideum (Ps). Небольшое sphenethmoideum (Se) располагалось между processus cultriformis parasphenoidei (Ps) и крышей черепа.
Частичное окостенение palatoquadratum привело к образованию большого quadratum (рис. 164, 165—Qu) и высокого узкого epipterygoideum "или columella cranii (рис. 165 — Ер); окостенение заднего конца mandibulare, как обычно, дало articulare в нижней челюсти.
Окостенение hyomandibulare вызвало у пеликозавров образование очень большого stapes, снабженного тремя отростками (рис. 164 — S). Один из этих отростков закрывал fenestra ovalis, другой соединялся с нижней поверхностью paroccipitale и третий был направлен в сторону сустава. Этот отросток, несомненно, имел хрящевое продолжение, которое упиралось в небольшую ямку на задней поверхности quadratum. Функция такого stapes, как слуховой косточки, несомненно, была очень ограничена.
Количество покровных костей, входящих в состав черепной крыши, у пеликозавров было такое же, как у котилозавров, но у многих форм эти кости имели иные размеры, иные очертания,, а в некоторых случаях и иное положение.
Например, непарное dermosupra-occipitale (Dso), располагавшееся у котилозавров на заднем краю черепной крыши, у пеликозавров сместилось в область затылка и приняло вертикальное
положение (рис. 164). То же случилось с tabulare (Т) и supratemporale (S/)-Наружная поверхность всех этих костей у пеликозавров стала служить местом прикрепления сильных мышц шеи. Parietale (Р), сильно сдвинутое кзади, сделалось коротким и широким; postorbitale (Ро) приобрело длинный отросток, ограничивающий верхний край позадиглазничного отверстия (fenestra postorbitalis). Frontale (F) и postfrontale-(Pf) подверглись относительно небольшим изменениям (рис. 166).
В переднем отделе черепа покровные кости praefrontale (Prf), lacrimale (L) и nasale (N) у многих пеликозавров были сильно вытянуты в длину, а размеры maxillare (М) увеличены.
Носовые отверстия у пеликозавров были так же сближены друг с другом, как у котилозавров. Поэтому разделявший их отросток praemaxillare (Pm) был относительно узким. В заднем отделе носового отверстия помещалась небольшая, но сложная по своей форме кость — septomaxillare (Sm). Она соединялась швами с nasale, maxillare и praemaxillare и имела отвер
152
стие, сквозь которое проходил конец слезно-носового канала (ductus nasolacrimalis).
Jugale (рис. 167 — Ju) и squamosum (Sq) не изменили своих положений в черепе, но стали принимать участие в образовании впервые появившейся скуловой дуги; quadratojugale (рис. 164—Qj) значительно уменьшилось в своих размерах.
Очень интересным и важным изменениям подверглись кости нёбной части черепа пеликозавров — vomer и transversum. Сошники (рис 168 — V) у них превратились в пару узких длинных костей, отделяющих
Рнс. 167. Череп Dimetrodon (вид сбоку) (Ромер и Прайс, 1940).
Р — frontale; Ju — jugale; L — lacrimale; M — maxlllare; ЛГ — nasale; Pf — postfrontale; Pm — praemaxillare; Po — postorbitale; Prf — praefrontale; Sm — septomaxillare; Sq — squamosum; St — supratemporale.
друг от друга сближенные и сильно вытянутые в длину хоаны. Такая форма сошников была новой особенностью в строении черепа рептилий.
Размеры palatinum и transversum у стегоцефалов, сеймурий и котилозавров были почти одинаковыми; у пеликозавров transversum (рис. 168 — Тг) стало отставать в росте и обычно оказывалось меньшим образованием, чем palatinum. Птеригоиды у пеликозавров были снабжены поперечным рядом конических зубов (рис. 168). Передний отдел fenestrae adductoriae был так же сужен, как у котилозавров.
Нижняя челюсть примитивных пеликозавров (рис. 169) в своем строении почти ничем не отличалась от нижней челюсти котилозавров, но у прогрессивных форм в ней появились очень важные изменения. Они привели к тому, что размеры angulare (Лп) сильно увеличились и на нижнем краю этой кости образовался большой отросток в виде пластинки. Ргаесогопо-ideum в челюсти отсутствовало; intercoronoideum (Ic) превратилось в длинную узкую кость; таким же длинным и узким стало и coronoideum (Со); большое spleniale (Sp) почти полностью лежало на внутренней стороне нижней челюсти; dentale (О) изменилось в связи с новым этапом развития зубов; praearticulare (Ра) подверглось значительной редукции и приобрело форму узкой изогнутой костной пластинки; supraangulare (Sa) сохранило свое обычное положение, но его задний конец вместе с задним концом praearticulare и angulare был отогнут вниз в результате смещения в том же направлении articulare (Аг).
153
Многие особенности в строении черепа пеликозавров, несомненно, были связаны с теми очень важными изменениями, которым подверглась их зубная система.
Зубы у всех пеликозавров были снабжены хорошо развитыми корнями.
Образование у зубов корней, несомненно, было очень полезным для позвоночных приобретением. Конические зубы, непосредственно сращенные с костью (у рыб, стегоцефалов и у неспециализированных котилозавров), были значительно менее надежными органами, чем зубы, снабженные длинными корнями и сидевшие в альвеолах (у некоторых специализированных
котилозавров, у всех пеликозавров и У Других прогрессивных форм рептилий). В первом случае зуб мог сравнительно легко отламываться от кости, так как при этом нарушалась только целость костной ткани, во втором — животное теряло зуб только
Рис. 168. Череп Dimetrodon (вид снизу) (Ромер и Прайс, 1940).
М — maxillare; Pl — palatinum; Pm — praemaxillare; Pt — pterygoideum; Tr — transversum; V — vomer.
Рис. 169. Нижняя челюсть Dimetrodon (Ромер и Прайс, 1940).
Ап — angulare; Ar — articulare; Со — coronoideum; D — dentale; Ic — intercoronoideum; Pa — praearticulare; Sa — sup-raangulare; Sp—spienlale.
тогда, когда переламывалась стенка самого зуба, состоящая из более прочного, чем кость, дентина.
У наиболее примитивных пеликозавров зубные ряды на челюстях, как у котилозавров, состояли из конических зубов, размеры которых постепенно уменьшались по направлению к челюстному суставу (рис. 170, А). Но у многих прогрессивных представителей этого отряда уже отмечалось начало дифференцировки зубов. На первых своих этапах она выражалась в том, что размеры верхних зубов становились неодинаковыми. Начиная с первого зуба на praemaxillare высота конических коронок в зубном ряду постепенно уменьшалась, затем на maxillare увеличивалась и, наконец, снова уменьшалась. Таково, например, строение верхних зубов у примитивного нижнепермского пеликозавра Mycterosaurus (рис. 170, В). Несмотря на наличие у этой рептилии больших конических зубов, мы, однако, еще не можем говорить о появлении у нее клыков, так как на это название могли бы претендовать по меньшей мере четыре больших зуба. Но у пеликозавра Dimetrodon. (рис. 167) особенно большими размерами отличались только
154
два зуба — один на maxillare и один на dentale; эти зубы являлись настоящими клыками. Эти особенности дают нам возможность различить у Dimetr-odon три группы зубов: резцы, клыки и «коренные» зубы.
Увеличение размеров некоторых зубов у пеликозавров сопровождалось
не только увеличением высоты их конических коронок, но и соответствующим удлинением корней. Это обстоятельство неминуемо вызывало и увеличение размеров тех костей, в которые входили корни зубов.
Поэтому по мере развития больших зубов на maxillare размеры этой кости постепенно увеличивались; при этом наибольшая высота этой кости всегда соответствовала положению корней самых больших зубов (рис. 170, Л,В). Интересно, что dentale, несущее зубы нижней челюсти, не увеличивалось в своих размерах в такой степени, как maxillare. Это объяснялось тем, что нижние клыки располагались в переднем участке этой кости, а «коренные» зубы не отличались большой величиной. Однако длинный корень хорошо раз
витого клыка все же вызывал разрастание переднего конца dentale (рис. 167).
В состав плечевого пояса пеликозавров входили те же кости, что и у котилозавров (рис. 171, А, В). Scapula (Sc), procoracoideum (Рс) и cora-
В
Рис. 170. Черепа пеликозавров Edaphosaurus (А) и Mycterosaurus (В).
Рис. 171. Плечевой пояс Dimetrodon. А — вид слева; В — вид снизу (Ромер и Прайс, 1940).
Cl — clavicula; Со — coracoideum; Ct — cleithrum; Ic — interclavicula; Pc — procoracoideum; Sc — scapula.
coideum (Co) у них окостеневали полностью. Cleithrum (Ct) было сильно редуцировано, но clavicula (Cl) и interclavicula (Ic) были развиты еще достаточно хорошо.
155
при помощи трех ребер,
Хорошо окостеневающий тазовый пояс пеликозавров отличался от тазового пояса котилозавров тем, что ischium и pubis одной стороны соединялись с такими же костями противоположной стороны под некоторым углом. Иными словами, эти кости у пеликозавров уже не лежали в горизонтальной плоскости, как это было у их предков из отряда котилозавров. Эти изменения в положении указанных двух костей таза были новым шагом вперед в эволюционном развитии скелета позвоночных (рис. 172 — Is, Pb).
Верхний расширенный конец ilium у пеликозавров присоединялся к позвоночному столбу, как и у котилозавров, имеющих сильно расширенные концы. У некоторых пеликозавров можно наблюдать первые фазы оформления крестца путем полного срастания его позвонков друг с другом и путем неподвижного присоединения к ним соответствующих ребер. Например, у Dimetrodon (рис. 173) подвижное соединение III крестцового позвонка со II уже исчезло. Тело
--IL
Рис. 173. Три крестцовых позвонка Dimetrodon (вид слева) (Ромер и Прайс, 1940).
Рис. 172. Таз Dimetrodon (вид слева) (Ромер и Прайс, 1940).
II — ilium; Is — Ischium; Pb — pubis.
и суставные отростки этих позвонков были соединены при помощи зубчатых швов. Такими же швами, исключающими всякую подвижность, были соединены и головки трех ребер с поперечными отростками соответствующих позвонков.
Передние и задние конечности пеликозавров были еще в большой степени приспособлены к передвижению животного по суше, чем у котилозавров (рис. 174).
На дистальных концах таких костей, как плечевая (рис. 175—Н) и бедренная (Fe), уже оформились соответствующие суставные поверхности в результате полного окостенения хрящевых эпифизов. То же произошло и с проксимальными концами костей предплечья (radius и ulna) и костей голени (tibia и fibula). На нижней стороне дистального конца бедра у пеликозавров впервые возникли два суставных отростка, которые при образовании коленного сустава опирались только на верхний расширенный конец большой берцовой кости. Головка малой берцовой кости соединялась с особой дополнительной суставной площадкой на латеральной поверхности дистального конца бедра. Таким образом, в коленном суставе пеликозавров мы видим как бы первую фазу вытеснения малой берцовой кости из состава этого сочленения.
156
'о
0“ ^/^£Г ^^VPftMjX|W^)il^l,tT*iril|iL||i|iru I I I ii~>i|~i,1'Y>rt<*'*'*rtr"‘F"™
Рис. 174. Скелет Ophiacodon rostratum (длина скелета равна 227 см). Пермь (Ромер и Прайс, 1940).
Рис. 176. Левая кисть Ophiacodon (Ромер и Прайс, 1940). I — Intermedium; R — radlale; U — ulnare.
Рис. 175. Кости конечностей Dimetrodon (Ромер и Прайс, 1940).
F — fibula; Fe — femur; Н — humerus; R — radius; T — tibia; V — ulna.
Рис. 177. Левая стопа Ophiacodon (Ромер и Прайс, 1940).
Г + I — сросшиеся tibiale и Intermedium; F — fibulare.
Все элементы кистей и стоп у пеликозавров были вполне окостеневшими. В кисти в состав проксимального ряда carpus входили radiale, intermedium и ulnare; дистальный ряд был образован пятью carpalia; между этими рядами располагались два центральных элемента — centrale mediate и laterale (рис. 176). Ни одна из названных костей не отличалась какой-либо особой формой или необычными размерами. Следовательно, кисть у пеликозавров сохраняла все оссбенности примитивного строения.
Что же касается стопы, то она, несомненно, уже довольно сильно уклонялась от примитивного типа. Проксимальный ряд tarsus в стопе пеликозавров был образован двумя большими костями. Одна из них представляла собой сильно увеличенное iibulare, другая являлась результатом слияния intermedium и tibiale. Две центральные кости и пять тарзальных были развиты нормально (рис. 177).
Сильное увеличение размеров fibulare у пеликозавров без сомнения было началом развития у позвоночных пяточной кости (calcaneus), а слияние intermedium и tibiale — первым этапом оформления таранной кости (talus).
И кисть, и стопа у пеликозавров имели по пяти пальцев. Количество фаланг пальцев осталось таким же, как у котилозавров; фаланговая^фор-мула для кисти была 23453, для стопы — 23454.
THERAPSIDA
Рептилии, относящиеся к отряду Therapsida, были непосредственными потомками пеликозавров. Они появились в начале пермского периода и вымерли в конце триаса (рис. 17 — Т).
Рис. 178. Позвонки горгонопса Scymnognathus. А, В — грудные позвонки; С — крестцовые позвонки (Бройли и Шредер, 1935). h — hypocentrum; р — pleurocentrum.
Therapsida имеют для нас исключительно большое значение, так как изучение строения скелета у различных представителей этого отряда дает возможность проследить постепенное оформление и накопление у них разнообразных признаков млекопитающих.
Позвонки у таких относительно примитивных представителей Therapsida, как горгонопсы, уже напоминали позвонки млекопитающих.
158
стали присоединяться к попе-
Рис. 179. Три первых''позвонка гор-гонопса Sauroctonus progressus (А. П. Быстрой, 1955).
а. а. — arcus atlantls; h. а. — hypocentium atlantis; Л. e. — hypocentrum epistrophei', pa— proatlas; p. a. —pleurocentrum atlantia; p. e.— pleurocentrum epistrophei.
Хорошо развитые дуги позвонков были снабжены высокими^ остистыми отростками и длинными поперечными. Телами позвонков являлись большие плевроцентры, полностью сращенные с дугами. Передняя и задняя поверхности тела каждого позвонка были еще конусообразно вогнутыми, но отверстие для хорды обычно отсутствовало (рис. 178, В). Гипоцентры имели форму маленьких костных полулуний, вставленных снизу между телами соседних позвонков. Их размеры обычно были настолько малы, что ребра уже не могли соединяться с ними. Мало того, в некоторых участках позвоночного столба гипоцентры у многих горгонопсид совсем отсутствовали (рис. 178, А). Сильная редукция и в иных случаях полное исчезновение гипоцентров привели к тому, что ре' речному отростку дуги и к суставной площадке на переднем краю плевроцентра, играющего роль тела позвонка (рис. 178, А, В). Ребрами у горгонопсов были снабжены все позвонки, кроме хвостовых.
Атлант и эпистрофей у горгонопсов были построены так же, как у пеликозавров. Дуга атланта (рис. 179—а. а.) состояла из двух несросшихся половин; на каждой из них располагался небольшой парный элемент проатланта (ра). Гипоцентр I позвонка (/г. а.) представлял собой небольшое костное полулуние, присоединенное к нижним концам половинок дуги. Плевроцентр (р. а.) примыкал к дуге и гипоцентру сзади. Двухголовчатое ребро атланта присоединялось к суставным площадкам на его дуге и гипоцентре.
Эпистрофей был хорошо развитым позвонком. Его дуга, сращенная с плев
роцентром (р. е.), была снабжена высоким пластинчатым остистым отростком, который имел, как у пеликозавров, треугольные очертания. Гипоцентр эпистрофея (Л. е.) в виде небольшого костного клина был выдвинут снизу между плевроцентрами I и II позвонков. Ребро эпистрофея в одних случаях присоединялось, как у атланта, к дуге и гипоцентру (рис. 179), в других — его головка соединялась с суставной площадкой на переднем краю плевроцентра или тела II позвонка.
Строение двух первых позвонков еще у пеликозавров показывает, что у так называемых тероморфных рептилий начала постепенно вырабатываться большая подвижность дуги и гипоцентра атланта по отношению к его плевроцентру и одновременно с этим стала возникать меньшая подвижность плевроцентра I позвонка по отношению к телу (к плевроцентру) II. Все эти изменения представляли собой первые этапы в формировании своеобразных по своему строению атланта и эпистрофея млекопитающих.
Среди Therapsida имелись и такие рептилии, у которых оформление атланта и эпистрофея сделало новый шаг вперед. Например, у Jonkeria. оба эти позвонка подверглись следующим изменениям. Проатлант полностью редуцировался; правая и левая половины дуги атланта срослись друг с другом, но еще сохранили соединение с передними суставными отростками дуги эпистрофея (рис. 180, А, В); контакт гипоцентра атланта (Л. а.) с его дугой усилился, сделавшись по существу неподвижным; плевроцентр атланта (р. а.) приобрел форму большого костного цилиндра с конусооб-
159
разным передним концом и почти полностью прирос к плевроцентру эпистрофея (р. е.); оба эти элемента отделялись друг от друга узкой щелью только снизу; в эту щель между ними, как клин, входил небольшой гипоцентр II позвонка (h. е.).
Рис. 180. Два первых позвонка Jonkeria (Брум, 1934). А — вид сбоку; В — атлант, вид спереди (обозначения те же, что на рис. 179).
Dso
I
Рис. 181. Череп Sauroctonus (вид сзади) (А. П. Быстров, 1955).
Во + Ео — basioccipitale 4* exoccipitale; Dso — dermosupraoccipitale; Po — paroccipitale; Qj — quadratojugale; Qu —-quadratum; S — stapes; So — supraoccipitale; Sq — squamosum; T— tabulare.
Черепа горгонопсов по общей своей форме напоминают черепа хищных млекопитающих; однако, несмотря на это, они по своему строению все же являются черепами типичных рептилий.
Суставной отросток, при помощи которого череп у горгонопсов соединялся с I позвонком, был образован сросшимися basioccipitale (рис. 181 — Во) и exoccipitale (Ео). На верхнем отделе каждого exoccipitale имелась небольшая площадка, к которой присоединялся передний конец проатланта. Над exoccipitale располагалось сильно вогнутое большое supraoccipitale (So), а по сторонам в поперечном направлении лежали paroccipitale (Ро). Все эти кости представляли собой результат окостенения заднего отдела мозгового черепа (neurocranium). Остальные кости затылочного отдела являлись покровными. Непосредственно над supraoccipitale помещалось вогнутое непарное dermosupraoccipitale (Dso), представлявшее собой, как одноименных костей. Справа
и слева к dermosupraoccipitale примыкали большие tabularia (Т), соединявшиеся швами с supraoccipitale и paroccipitale. В шве между tabulare и paroccipitale у горгонопсов, как и у пеликозавров, имелось отверстие для vena capitis lateralis.
Непосредственно перед paroccipitale лежало prooticum (рис. 182 — Pro), которое, продолжаясь вверх, достигало крыши черепа. Между этими ко-160
пеликозавров, результат слияния двух
стями на нижней стороне черепа располагалось fenestra ovalis (рис. 183, f. о.), в которое входило основание stapes.
Базисфеноид, лежавший перед basioccipitale, у горгонопсов был прикрыт снизу парасфеноидом (Ps). Передняя часть парасфеноида (processus cultriformis), имевшая форму узкой пластинки, с нижней стороны в свою очередь была закрыта сближенными ветвями птеригоидов (Pt)', концы этих ветвей кзади расходились в стороны, и достигали quadratum (Qu).
На верхней поверхности каждого птеригоида, на уровне basispheno-ideum, располагались узкие и высокие эпиптеригоиды (рис. 182—Ер). Они были направлены вверх и почти достигали крыши черепа. По своей форме
Рис. 182. Череп Sauroctonus. Часть костей удалена (А. П. Быстров, 1955).
Ер — eplpterygoideum; F — frontale; М — maxillare; N — nasale; Pf — posttrontale; Pm — praemaxillare; Pro — prooticum; Pt — pterygoldeum; Qu — quadratum; Se — sphenethmoldeum; Sm — septomaxlllare; Sq — squamosum; v. c. I. — борозда venae capitis lateralis.
эти кости у горгонопсов были поразительно похожи на соответствующие образования у пеликозавров.
Принимая участие в образовании заднего отдела нёба, птеригоиды у горгонопсов сильно расширялись, но здесь они были значительно короче, чем у пеликозавров.
Над этими участками птеригоидов, примыкая к крыше черепа, располагался сфенэтмоид (рис. 182—Se), в боковых стенках которого имелись довольно широкие отверстия для зрительных нервов.
Transversum (рис. 183 — Тг) у горгонопсов имело такие же размеры, как у пеликозавров, но palatinum (Pl) сильно уменьшилось. Что касается сошников (vomer, V), то они были узки и в отличие от сошников пеликозавров обычно срастались друг с другом. Длина узких хоан у горгонопсов достигала половины длины первичного нёба.
Вследствие присутствия очень больших позадиглазничных отверстий (fenestrae postorbitales) черепная крыша у этих рептилий в заднем своем отделе была относительно узкой (рис. 184). Главными костями этого участка являлись два parietale (Р), в шве между которыми помещалось пинеальное отверстие (foramen parietale). Задний край теменных костей был сйльно вогнут; по обе стороны от parietalia располагался длинный узкий отросток postorbitale (Ро).
Supratemporale, которое в виде небольшой кости еще Сохранялось в составе черепа пеликозавров, у горгонопсов уже отсутствует. '
Между орбитами лежали. три парные покровные кости: postfrontale (Pf), frontale (F) и praefrontale (Prf).
11 А. П. Быстров
161
Длинное nasale (V) своим передним концом соединялось с узким отростком praemaxillare (Рт).
Lacrimale (L), образующее передний край орбиты, у горгонопсов было относительно небольшой костью; в нем проходил слезно-носовой канал (ductus nasolacrimalis), начинающийся двумя отверстиями (рис. 184, 185). Передний конец этого канала проходил сквозь septomaxillare и открывался в полость носа (рис. 184 и 185 — Sm).
Рис. 183. Череп Sauroctonus (вид снизу) (А. П. Быстров, 1955).
Во — basioccipitale; /. о. — fenestra ovalis; Ju — jugale; M — maxillare; Pl — palatinum; Pm — praemaxillare; Ps — parasphenoldeum; Pt — pterygoideum; Qj — quadratojugale; Qu — quadratum; Sq — squamosum; Tr — transversum; V — vomer.
Maxillare (рис. 185 — M) было самой большой костью переднего отдела черепа горгонопсов. К его заднему краю примыкало jugale (Ju); дальше следовало сильно развитое squamosum (Sq). Задний отдел этой кости представлял собой большую изогнутую пластинку, которая образовывала заднюю стенку височной ямы и достигала postorbitale (Ро) и prooticum (Pro). Внизу к squamosum присоединялось quadratum, на наружной стороне которого лежало сильно редуцированное quadratojugale (Qj).
Нижняя челюсть горгонопсов имела ряд интересных особенностей (рис. 186).
162
Самой большой из входящих в ее состав костей являлось dentale, (D). Размеры этой кости были настолько велики, что она полностью,закрывала spleniale (Sp), которое оказалось лежащим только на внутренней стороне нижней челюсти. Задний участок dentale был сильно вытянут назад и вверх и надел форму длинного отростка.
Supraangulare (Sa) и praearticulare (Ра) у горгонопсов почти не отличались от соответствующих костей у пеликозавров. Что же касается angu-
Рис. 184. Череп Sauroctonus (вид сверху) (А. П. Быстров, 1955).
F — frontale; Ju — jugale; L — lacrimale; M — maxillare; W — nasale; P — parletale; Pf — postfrohtale; Pm — praemaxillare; Po —postorbitale; Prf — praefrontale; Sm — septomaxillare; squamosum.
lare (An), то oho было несколько меньше по своим размерам, чем у пеликозавров, и имело на наружной поверхности костные гребни.
Articulare (Аг) у горгонопсов было небольшой костью; intercorono-ideum подвергалось полной редукции, но coronoideum (Со) еще сохранялось и имело форму костного клина, лежащего между dentale и praearticulare.
Присутствие в черепе горгонопсов большого позадиглазничного отверстия (fenestra postorbitalis), превратившегося в обширную височную яму. свидетельствует о том, что помещавшаяся здесь височная мышца (m. temporalis) была развита у этих рептилий гораздо сильнее, чем соответствующий мускул у пеликозавров. На это указывает и строение squamosum
11* 16»
горгонопсов. Задний участок этой кости, образующий всю заднюю стенку височной ямы, давал достаточно большую площадь для начала сильной
Рис. 185. Череп Sauroctonus (вид сбоку) (А. П. Быстров, 1955).
F— frontale; Ju — jugale; L — lacnmale; M — maxillare; N — nasale; Pf — postfrontale; Pm — praemaxillare; Po — postorbitale; Prf — praefrontale; QJ — quadratojugale; Sm — septomaxlllare; Sq — squamosum.
Рис. 186. Нижняя челюсть Sauroctonus (А. П. Быстров, 1955). An — angulare; Ar — articulare; Co — coronoideum; D — dentale; Pa — praearticulare; Sa — supraangulare; Sp — spleniale.
височной мышцы. Некоторое количество волокон этой мышцы начиналось й от длинного направленного назад отростка postorbitale. Местом прикрепления для височной мышцы служил длинный задний отросток dent ale.
164
Несомненно, не менее сильно был развит у горгонопсов и такой сжимающий челюсти мускул, как m. masseter. Он начинался на нижнем краю скуловой дуги и прикреплялся на наружной поверхности angulare. Присутствие здесь костных гребней свидетельствует о том, что и этот мускул у горгонопсов был развит сильнее, чем у пеликозавров.
Зубы горгонопсов имели острые конические коронки и были снабжены хорошо развитыми корнями. Дифференцировка зубов, начавшаяся еще у пеликозавров, у горгонопсов привела к образованию группы резцов на praemaxillare и большого клыка и нескольких коренных зубов на maxillare. Такие же зубы возникли и на нижней челюсти (рис. 185, 186).
Сильное увеличение верхних клыков вызвало у горгонопсов утолщение соответствующего участка maxillare, а это в свою очередь обусловило сужение заднего отдела хоан (рис. 183). Передний отдел хоан у этих рептилий, наоборот, был сильно расширен. Это объясняется тем, что высокие коронки нижних клыков при закрывании рта входили в хоаны (рис. 185).
Наличие хорошо развитых и длинных корней у клыков было причиной увеличения высоты средней части maxillare и подбородочного отдела dentale (рис. 187).
Кроме зубов на praemaxillare, maxillare и dentale, горгонопсы имели зубы и на vomer, palatinurri и pterygoideum (рис. 183). На сошниках (V) они были очень маленькими; на нёбных костях (Р[) их размеры были больше и они образовывали здесь две компактные группы, сидящие на особых Костных возвышениях; на птеригоидах (Pt) зубы располагались в два ряда: один из них стоял косо, другой — поперечно, вдоль заднего края нёбного отдела этой кости.
Судя по строению зубной системы, горгонопсы, несомненно, были сильными хищниками. Они могли легко убивать и разрывать добычу при помощи своих клыков. Острые передние зубы, сильная мускулатура, сжимающая челюсти, и хорошо развитые мышцы шеи, прикрепляющиеся к расширенному и вогнутому затылочному отделу черепа, позволяли горгонопсам легко отрывать куски мяса от мертвой жертвы. Наличие поперечного ряда зубов, в самом заднем отделе нёба (на птеригоидах) свидетельствует о том, что,
165-
разрывая свою добычу, горгонопсы в некоторый момент захватывали куски мяса так, что они заполняли всю их пасть.
Слабо развитые конические коренные зубы у горгонопсов не были приспособлены к измельчению или перетиранию пищи. Группы небольших зубов на сошниках, нёбных костях и птеригоидах, не имевшие антагонистов, также не давали горгонопсам возможности в какой-нибудь мере раздроблять пищу. Поэтому можно уверенно сказать, что эти рептилии, отрывая от убитого ими животного куски, глотали их без предварительного пережевывания. Это они делали не только потому, что при свойственных им особенностях строения зубной системы они не могли ничего пережевывать, но и потому, что вообще не были в состоянии долго держать пищу во рту. Это объяснялось тем, что всякий достаточно большой кусок пищи закрывал у них не только рот, но и расположенные в переднем отделе нёба хоаны, а это делало дыхание совершенно невозможным.
Указанное положение хоан было причиной того, что наиболее сильно развитыми зубами у горгонопсов были конические резцы и клыки, т. е. зубы, расположенные перед хоанами и на уровне их переднего отдела.
Когда горгонопсы, разрывая добычу, действовали этими зубами, задний участок хоан оставался открытым, что еще позволяло животному дышать; кроме того, воздух мог проходить и через не вполне закрытый рот. Но хоаны и рот закрывались полностью, как только куски добычи захватывались и птеригоидными зубами. С этого момента дыхание становилось уже невозможным. Это обстоятельство вынуждало хищника поспешно оторвать кусок мяса и быстро проглотить его.
Работа коренных зубов по раздроблению пищи у горгонопсов по существу была совершенно невозможной, так как такую функцию эти зубы могли бы выполнять только в том случае, если бы животное могло достаточно продолжительное время не дышать.
Невозможность одновременного пережевывания пищи и дыхания являлась основной причиной того, что коренные зубы у всех горгонопсов были слабо развитыми, а у такой специализированной формы, как иностран-цевия (/nostrancevia alexandri), коренные зубы на нижней челюсти совершенно отсутствовали. Это указывает на то, что этот страшный хищник нападал на свою жертву, действуя только своими мощными острыми резцами и клыками, и, разрывая убитое животное, поспешно глотал окровавленные куски мяса, не пережевывая их. . .
Плечевой пояс горгонопсов состоял из хорошо окостеневших лопатки (рис. 188, А—Sc), коракоида (Со) и прокоракоида (Рс). Суставная ямка для головки плечевой кости, образованная этими костями, была направлена латерально и назад, что указывает на то, что в отличие от пеликозавров горгонопсы могли ставить переднюю конечность под углом около 30—40° по отношению к продольной оси тела.
Межключица (1с) и ключицы (С/) у горгонопсов были развиты слабее, чем у пеликозавров. Обе эти кости прикрывали только края прокоракоидов.
Cleithrum (Ct) у многих горгонопсов подверглось почти полной редукции и представляло собой очень маленькое образование, приросшее к переднему краю лопатки.
У некоторых горгонопсов (Aelurognathus, Hipposaurus, Lycaenops) было обнаружено окостеневшее sternum (St), которое представляло собой вогнутую сверху костную пластинку, на краях которой имелись следы контакта с ребрами (Боонстра — L. Boonstra, 1934; Кольберт — Е. Colbert, 1948).
Тазовый пояс горгонопсов, состоящий из хорошо окостеневших ilium (II), ischium (Is) и pubis (Pb), отличался от тазового пояса пеликозавров тем, что в нем отсутствовало полное соединение между седалищной и лонной 166
костями. У одних горгонопсов промежуток между этими костями был относительно невелик (рис. 188, В), у других — он достигал довольно больших размеров (рис. 189).
Образование промежутка между ischium и pubis является первой фазой развития так называемого запирательного отверстия таза (foramen obturatum).
Рис. 188. Плечевой (Д) и тазовый (В) пояса горгонопса Aelurognathus microdon (Боонстра, 1934). С — неполная стопа горгонопса Scymnognathus (Бройли н Шредер, 1935).
Cl — clavicula; Со — coracoideum; Ct —- cleithrum; Ic — interclavlcula; Il —Ilium; Is—Ischium; Pb — pubis; Pc — procoracoideum; Sc — scapula; St — sternum.
Изучение эволюционной истории скелета позвоночных показывает, что это отверстие впервые начинает оформляться только у горгонопсов.
Таз у горгонопсов, как и у пеликозавров, присоединялся к трем крестцовым позвонкам при помощи трех пар ребер; при этом ребра в большинстве случаев были полностью сращены с поперечными отростками позвонков.
Рис. 189. Тазовый пояс и кости конечностей горгонопса Ly-caenops ornatus Broom (Кольберт, 1948).
F — fibula; Fe — femur; H — humerus; Il — ilium; Is — ischium; Pb — pubis; R — radius; T— tibia; U — ulna.
Концы крестцовых ребер, в особенности первой пары, были сильно расширены, что обусловливало прочное соединение таза с позвоночным столбом (рис. 178, С).
Плечевые кости горгонопсов (рис. 189—Н) отличались от соответствующих костей пеликозавров меньшей шириной дистальных концов и большей
167
уплощенностью проксимальных. Foramen entepicondyloideum имелось. Кости предплечья горгонопсов очень напоминали такие же кости пеликозавров (рис. 189 — 2?, U).
Интересными особенностями строения кисти горгонопсов было появление дополнительной кости pisiforme в проксимальном ряду carpus (рис. 190)
Рис. 190. Кисть (Л) и стопа (В) горгонопса Lycaenops ornptus Broom (Кольберт, 1948).
С — calcaneus; I — intermedium; W — naviculare; P — pisiforme; /? — radiale; T — talus; U ~ ulnare.
и слияние костей дистального ряда — carpale quarturn и carpale quintum. Это срастание привело к образованию одной кости, соответствующей hama-tum высших форм позвоночных. Кисть горгонопсов была снабжена пятьк? пальцами с фаланговой формулой 23343.
Рис. 191. Скелет горгонопса Lycaenops ornatus Broom (длина скелета равна 120 см). Пермь (Кольберт, 1948).
Задняя конечность горгонопсов во многих отношениях отличалась от задней койечности пеликозавров. Прежде всего следует обратить внимание на то, что бедренная кость горгонопсов уже обладала достаточно хорошо выраженной шейкой, поставленной под углом 160° к продольной оси кости, и хорошо оформленной закругленной головкой (рис. 189). Эти особенности позволяли горгонопсам поворачивать бедро кпереди и держать его под углом 30—40° по отношению к продольной оси тела (рис. 191).
На нижней стороне дистального конца бедра у горгонопсов имелись два круглых суставных отростка, которыми эта кость опиралась на верхний конец большой берцовой кости. Эти образования не являлись новопри-16®
обретением для горгонопсов: как указывалось выше, они возникли еще у пеликозавров.
Большая и малая берцовые кости не отличались какими-нибудь новыми особенностями.
В стопе горгонопсов проксимальный ряд, как и у пеликозавров, состоял только ‘Из двух костей, так как tibiale и intermedium полностью срослись друг с другом и образовали talus, в то время как сильно увеличенное fibu-lare превратилось в calcaneus (рис. 190, В).
Два центральных элемента, которые были еще изолированными в кисти горгонопсов, в их стопе срослись и образовали naviculare. Такого явления в стопе пеликозавров еще не наблюдалось.
ро. р.е.
Рис. 192. Позвонки цинодонта Galesaurus. А —три первых позвонка; В — грудные позвонки (Паррингтон — Parrington, 1934).
а. а. — arcus atlantis; Л. а. — hypocentrum atlantis; Л. е. — hypocentrum epistrophei; pa — proatlas; p. a. — pleurocentrum atlantis; p. e. — pleurocentrum epistrophei; p — pleurocentrum.
Полному слиянию подверглись и tarsale quartum и quintum, в результате чего возникло cuboideum, к которому присоединялись метатарзальные кости четвертого и пятого пальцев. При таких изменениях в строении tarsus первая, вторая и третья тарзальные кости уже могут быть названы клиновидными (cuneiforme primum, secundum, tertium).
Фаланговая формула стопы горгонопсов, как и их кисти, выражалась следующими цифрами: 23343.
Слияние некоторых элементов в составе carpus н tarsus и уменьшение количества фаланг в кисти и стопе горгонопсов свидетельствуют о большей прогрессивности этих рептилий по сравнению с пеликозаврами.
Непосредственными потомками горгонопсов были цинодонты (Супо-dontia) — еще более прогрессивные рептилии конца перми и начала триаса. Многими особенностями строения своего скелета эти рептилии очень напоминали млекопитающих.
Каждый туловищный позвонок у цинодонтов состоял из хорошо развитого плевроцентра и дуги; гипоцентр отсутствовал, поэтому ребро присоединялось к поперечному отростку дуги и к «уставной площадке на переднем краю плевроцентра (рис. 192, В). Тела (плевроцентры) позвонков еще сохраняли такой признак рептилий, как амфицельность, т. е. имели спереди и сзади конические углубления.
Между затылочным отделом черепа и I шейным позвонком у цинодонтов еще имелся парный костный элемент проатланта (рис. 192, А—Ра). Атлант состоял из дуги, разделенной на правую и левую половины (а. а.), и небольшого непарного гипоцентра (Л. а.). Плевроцентр (р. а.) I позвонка
169
имел закругленный передний конец; его задний конец был почти полностью сращен с плевроцентром II позвонка (эпистрофея). В щель между этими элементами снизу был вдвинут гипоцентр эпистрофея (h. е.). Почти полное срастание плевроцентра атланта с плевроцентром II позвонка привело по существу к образованию так называемого «зуба» эпистрофея (dens epistrophei).
Ill и IV позвонки еще сохраняли свои гипоцентры, остальные же позвонки цинодонтов были уже лишены этих элементов.
Череп цинодонтов отличался от черепа горгонопсов многими очень важными и интересными особенностями, которые свидетельствуют о том, что именно среди этих рептилий мы должны искать предков млекопитающих.
Рис. 193. Череп цинодонта Cynognathus platyceps Seeley (вид сзади) (Бройли и Шредер, 1934).
Во — basioccipitale; Dso — dermosupraoccipitale; Ео — exoccipitale; Po — paroccipitale; QJ — quadratojugale; Qu — quadratum; So — sup-raoccipitale; Sq — squamosum; T — tabulare.
Затылочный отдел черепа у цинодонтов не был так сильно вогнут, как у горгонопсов, и у некоторых форм был почти плоским (рис. 193).
Вокруг затылочного отверстия располагались basioccipitale (Во), парное exoccipitale (Ео) и supraoccipitale (So). С I позвонком соединялись своими суставными отростками только exoccipitalia. Этой особенностью цино-донты уже напоминали млекопитающих.
Непосредственно перед basioccipitale (рис. 194 — Во) располагалось basisphenoideum (Bs), на котором у примитивных цинодонтов еще лежал парасфеноид (parasphenoideum) с сильно редуцированным processus cult-riformis. У более прогрессивных форм этих рептилий парасфеноид полностью отсутствовал.
С basisphenoideum (Bs) соединялись птеригоиды (Pt).
Эти кости у прогрессивных форм цинодонтов были развиты слабее, чем у горгонопсов, и на своей нёбной поверхности никогда не имели зубов.
Развившееся в результате окостенения хряща paroccipitale (Ро) у цинодонтов, как и у горгонопсов, имело форму костной балки, направленной от exoccipitale к суставному углу черепа (рис. 193, 194).
Над задней ветвью птеригоида располагались две кости мозгового черепа: prooticum (Pro) и epipterygoideum (Ер) (рис. 195).
Эпиптеригоид у цинодонтов имел форму довольно широкой костной пластинки. Расширение эпиптеригоида привело к тому, что у прогрессивных форм цинодонтов он стал принимать большое участие в образовании боковой стенки мозгового черепа, соединяясь с теменной костью посредством шва.
170
Очень интересной и важной особенностью строения черепа цинодонтов было наличие вторичного костного нёба, которое было образовано горизонтальными отростками praemaxillare, maxillare и palatinum и полностью отделяло полость носа от полости рта.
Одну из фаз формирования этого вторичного нёба можно видеть, например, у такого примитивного цинодонта, как Procynosuchus rubidgei
Во Ео
Рис. 194. Череп Cynognathus plalyceps Seeley (вид снизу) (Бройли и Шредер, 1934).
Во — basioccipitale; Bs — basisphenoideum; Ео — exoccipitale; М — maxillare; PZ — palatinum; Pm — praemaxillare; Po — paroccipitale; Pt—pterygoideum; Qj — quadratojugale; Qu — quadratum; S — stapes; Tr — transversum; V — vomer.
(рис. 196). В результате возникновения пластинчатых костных отростков на краях praemaxillare (Р/n) и maxillare (М) и вследствие разрастания краев palatinum (Pl) хоаны у Procynosuchus превратились в длинные узкие щели, расположенные по обе стороны сросшихся сошников (V).
У более прогрессивных форм цинодонтов пластинчатые отростки praemaxillare, maxillare и palatinum одной стороны смыкались по срединной линии с такими же отростками соответствующих костей другой стороны. Таким образом возникала горизонтальная костная перегородка, разделяющая полости носа и рта, в результате чего хоаны оказывались как бы пе
171
ремещенными далеко назад (рис. 194). При этом правый и левый сошники (V) полностью срастались друг с другом и, вытянувшись в сагиттальной
Рис. 195. Череп Cynognathus platyceps Seeley (вид сбоку) (Бройли и Шредер, 1934). Скуловая дуга удалена.
£р-е pipterygoideum; Л — lacrimale; М — maxillare; N — nasale; Р — parletale; Pm — praemaxillare; Po — postorbitale; Prf — praefrontale; Pro — prooticum; Pt — pterygoideum; Sm — septomaxillare; Sq‘— squamosum.
плоскости, образовывали - непарную носовую перегородку. Сошники такого строения по существу ничем не отличаются от сошника в черепе млекопитающего.
Рис. 196. Череп Procynosuchus rubidgei Broom (Брум, 1938).
М — maxillare; Pl — palatinum; Pm — praemaxillare; V — vomer.
Все эти изменения в строении черепа сопровождались полным исчезновением нёбных зубов на pterygoideum, palatinum и vomer.
172
Эти зубы, появившиеся в виде многочисленных очень мелких конических зубов еще у кистеперых рыб, оставались такими же у стегоцефалов, затем увеличились в размерах, но уменьшились в количестве у котилозавров, пеликозавров и горгонопсов и, наконец, полностью исчезли в момент образования вторичного нёба у примитивных цинодонтов, после чего они уже больше никогда не возникали вновь на всем протяжении дальнейшей эволюции позвоночных.
Transversum (Тг) у цинодонтов сильно уменьшилась в размерах и потеряла то значение, которое она еще имела в черепе горгонопсов.
В поперечном шве между горизонтальным отростком palatinum и таким же отростком maxillare у цинодонтов возникло небольшое отверстие, сквозь которое проходили сосуды и нервы для слизистой оболочки нёба. Эти отверстия сохранились также в черепе млекопитающих.
В переднем отделе твердого нёба между praemaxillare и maxillare также образовалось отверстие для сосудов и нервов; и это отверстие можно видеть в черепе всех млекопитающих, не исключая и человека (foramen incisivum).
Возникновение твердого нёба у цинодонтов, несомненно, явилось очень большим шагом вперед в эволюционном развитии рептилий. .Наличие горизонтальной костной перегородки между полостью рта и полостью носа впервые позволило животным привлечь к раздроблению пищи’ и коренные зубы, не затрудняя при этом акта дыхания. Это привело к тому, что у цинодонтов впервые появились сложно построенные коронки коренных зубов. В то время как резцы и клыки цинодонтов еще сохраняли примитивно коническую форму, их моляры приобрели коронки, снабженные несколькими острыми вершинами. Однако, несмотря на такое усложнение коренных зубов, их корни оставались еще одиночными.
Перемещение хоан назад, обусловленное образованием твердого нёба,' и появление у коренных зубов коронок с несколькими острыми бугорками давало цинодонтам возможность хорошо пережевывать пищу. Эта особенность без сомнения была их огромным преимуществом по сравнению с их предками из группы горгонопсов.
Покровные кости крыши черепа у цинодонтов по своему положению, а во многих случаях и по форме очень напоминали соответствующие кости млекопитающих.
Непарное dermosupraoccipitale (рис. 193 — Dso) стояло вертикально во фронтальной плоскости; справа и слева от него в той же плоскости помещались два tabulare (Т).
Парное parietale (рис. 197—Р), прикрывавшее мозговую полость сверху, еще сохраняло в сагиттальном шве теменное отверстие (foramen parietale), которое, однако, у более прогрессивных цинодонтов было очень небольшим.
Перед parietale располагалось хорошо развитое frontale (F). Сзади к нему примыкало postorbitale (Ро), еще сохранившее узкий задний отросток, но он у цинодонтов уже не достигал squamosum, что наблюдалось у всех горгонопсов.
Postfrontale (Pf) исчезло. Эта кость, по всей вероятности, слилась с postorbitale и уже не появляется в качестве самостоятельного образования при дальнейшей эволюции черепа позвоночных.
Praefrontale (Prf) в составе черепной крыши цинодонтов еще имелось.
Lacrimale (L) представляло собой относительно небольшую кость и в своем заднем краю имело два отверстия для слезно-носового канала (рис. 195, 197).
Nasale (N) было длинным и несколько суженным в переднем своем отделе. Его передний конец соединялся с узким отростком praemaxillare, в результате чего у цинодонтов еще сохранялось костное разделение обоих
,173
носовых отверстий, характерное для всех рептилий. В углу между передними концами nasale (N) и maxillare (М) помещалось небольшое septoma-xillare (Sm). Maxillare у цинодонтов было хорошо развито, так как в него входили длинные корни зубов (рис. 198).
Jugale (Ju) вместе со squamosum (Sq) образовывало хорошо развитую скуловую дугу, служившую местом начала такой жевательной мышцы.
Рис. 197. Череп Cynognathus platyceps Seeley (вид сверху) (Бройли и Шредер, 1934).
F — frontale; Ju — jugale; L — lacrimale; M — maxillare; W — nasale; P — parietale; Pm — praemaxillare; Po — postorbitale; Prf — praefrontale; Pro —prooticum; Sq — squamosum.
как m. masseter. Задний отдел squamosum был сильно расширен и, как у горгонопсов, образовывал заднюю стенку очень большой височной ямы. Находившаяся здесь височная мышца (m. temporalis) в отличие от соответствующего мускула у горгонопсов начиналась не только от заднего участка squamosum, но и от расширенного эпиптеригоида и от сильно разросшегося книзу края теменной кости. Иными словами, эта мышца начиналась от образовавшейся у цинодонтов костной боковой стенки мозговой коробки.
Quadratum (рис. 193, 194—Qu) у цинодонтов было редуцировано. Еще большей редукции подверглось quadratojugale (Qj).
Stapes (рис. 194 — S) представлял собой небольшую кость с большим отверстием и, располагаясь горизонтально, одним своим концом закрывал fenestra ovalis, другим соединялся с quadratum.
174
Нижняя челюсть цинодонтов отличалась от нижней челюсти горгонопсов сильной редукцией всех входящих в ее состав костей, кроме dentale, которое, напротив, у цинодонтов было развито значительно сильнее, чем у горгонопсов.
У такой прогрессивной формы, как Cynognathus platyceps Seeley (рис. 199) огромное dentale (D) прикрывало снаружи все остальные кости, кроме маленького articulare (Лг). На внутренней стороне dentale у Cynognathus располагались supraangulare (Sa), praearticulare (Ра) и angulare (Ап), превратившиеся в длинные и узкие кости. Несколько более сильно
Рис. 198. Череп Cynognathus platyceps Seeley (вид сбоку) (Бройли и Шредер, 1934). D — dentale; Ju — jugale; L — lacrimale; M — maxillare; N — nasale; Pm — praemaxillare; Po — postorbitale? Prf — praefrontale; Sm — septomaxillare; Sq — squamosum.
было развито spleniale (Sp), а короноид (С) представлял собой небольшую короткую костную пластинку.
Необычайно сильное развитие dentale в нижней челюсти цинодонтов, несомненно, было связано с образованием у них вторичного твердого нёба и с появлением возможности раздроблять пищу при помощи коренных зубов. Усложнение строения коронок коренных зубов в свою очередь было связано с той новой функцией, которую они могли теперь выполнять.
У сеймуриаморф, котилозавров, пеликозавров и горгонопсов высота последних зубов всегда постепенно уменьшалась по направлению к челюстному суставу. То же наблюдалось и у всех стегоцефалов и кроссоптеригий. Что же касается цинодонтов, то у этих рептилий впервые мы видим обратное явление — увеличение размеров зубов по мере приближения к челюстному суставу (рис. 198, 200). Эта особенность объясняется, конечно, только тем, что цинодонты имели возможность пользоваться своими коренными зубами для измельчения пищи, причем самая большая работа выполнялась именно самыми задними из них, так как они располагались ближе к челюстному суставу и, следовательно, могли быть сильнее сжаты жевательными мышцами.
Принимая во внимание сложное строение коронок коренных зубов, размеры dentale, ширину скуловой дуги (jugale и squamosum) и большие площади начала и прикрепления жевательных мышц, мы можем считать, что челюстной аппарат прогрессивных форм цинодонтов обладал немалой
175.
силой и отличался некоторым,совершенством. Однако одновременно с этими положительными качествами он имел один большой недостаток. Этот недо-
Рис. 199. Нижняя челюсть Cynognathus platyceps Seeley (Бройли и Шредер, 1934).
Ап — angulare; Ar — articulare; Со — coronoideum; D — dentale; Ра — praearticulare; Sa — supraangulare; Sp — spleniale.
статок заключался в том, что мощное dentale было сочленено с черепом при помощи очень небольшого articulare, образующего сустав с quadratum.
Рис. 200. Зубы Cynognathus platyceps Seeley.
И нужно удивляться, как могло это очень слабое подвижное соединение без повреждений выдерживать ту большую нагрузку, которая, несомненно, возлагалась на челюстной сустав цинодонтов.
176
В состав плечевого пояса цинодонтов входили scapulocoracoideum, clavicula, interclavicula и sternum. Лопатка представляла собой высокую
Рис. 201. Кости плечевого пояса цинодонта {Cynognathus craterono-tus Seeley) (Грегори и Камп — Gregory and Kamp, 1918).
Cos— coracoideum; Pc — procoracoideum; Sc — scapula.
костную пластинку, на наружной поверхности которой имелся продольный гребень (spina scapulae, рис. 201). Этот гребень еще отсутствовал на лопатке горгонопсов. Коракоид и прокоракоид у цинодонтов были развиты значительно слабее, чем у горгонопсов. Cleithrum отсутствовало. Clavicula и interclavicula были относительно небольшими костями.
Рис. 202. Грудной позвонок (Л), таз (В) и крестцовые позвонки (С) Permocynodon sushkini (Е. Д. Коижукова, 1949).
II — ilium; Is — ischium; Pb — pubis.
Тазовый пояс цинодонтов по своему строению был очень близок к тазу млекопитающих. Ilium было гораздо более широким, чем у горгонопсов. Оно прочно скреплялось с четырьмя крестцовыми ребрами и давало большую площадь для начала сильных мышц задней конечности (рис. 202).
Рнс. 203. Таз н кости конечностей Galesaurus. Бедро Permocynodon (Л).
Fe — femur; Н — humerus; Il — ilium; Is — ischium; Pb — pubis; R — radius; Г-tibia; U - ulna.
В результате разрастания нижних концов седалищной и лонной костей навстречу друг другу между этими костями образовалось довольно большое отверстие (foramen obturatum). У примитивных цинодонтов нижние концы ischium и pubis еще не были сращены друг с другом (рис. 203), но у более прогрессивных форм это сращение уже имелось и запирательное отверстие оказывалось совершенно замкнутым (рис. 202).
12 А. П. Быстров
177
Плечевая кость цинодонтов мало чем отличалась от такой же кости горгонопсов. Но бедро у более прогрессивных представителей имело головку, гораздо яснее выраженную, чем у горгонопсов (рис. 203, А). В костях предплечья, голени, кисти и стопы каких-либо новых и важных особенностей у цинодонтов не наблюдалось (рис. 204).
Рис. 204. Скелет Cynognathus crateronotus Seeley. Триас (Грегори, 1918).
Детальное изучение скелетов известных в настоящее время цинодонтов показало, что именно среди этих рептилий следует искать предков млекопитающих.
ЛИТЕРАТУРА
Быстров А. П. (В у s t г о w). Kotlassia prima Amalitzky. Bull, of the Geol. Society of America, vol. 55, 1944.
Быстров А. П. Горгоиопс из верхнепермских отложений иа Волге. Вопросы палеонтологии, т. 2, 1955.
Ефремов И. А. (Е f г е m о v). Die Mesen-Fauna der permischen Reptilien. Neues Jahrbuch fur Mineralogie etc., Bd. 84, 1940.
Ефремов И. А. О подклассе Batrachosauria — группе форм, промежуточных между земноводными и пресмыкающимися. Известия АН СССР, № 6, 1946.
Конжукова Е. Д. К морфологии Permocynodon и эволюции зубного аппарата Cynodontia. Труды Палеонтологического института АН СССР, т. 20, 1949.
Сушкин П. П. (S u s h k i n). On the representatives of the Seymouriamorpha, supposed primitive reptile from the Upper Permian of Russia, and on their phylogenetic relations. Boston soc. Nat. Hist. occ. papers, vol. 5, 1925.
Boonstra L. A contribution to the morphology of the Gorgonopsia. Annals of the South African Museum, vol. 31, 1934.
Broili F. Permische Stegocephalen und Reptilien aus Texas. Palaeontographica, Bd. 51, .1904.
Broili F. Uber Zahnbau von Seymouria. Anatomischer Anzeiger, Bd. 49, 1927.
Broili F. und Schroer J. Zur Osteologie des Kopfes von Cynognathus. Sitzungs-berichte der Bayerischen Akademie der Wissenshaften, 1934.
Broili F. und Schroder J. Beobachtungen an Wirbeltiere der Karrooformation. Ibidem, 1935.
Broom R. On the structure of affinities of Bolosaurus. Bulletin of American Museum of Natural History, vol. 32, 1913.
Broom R. The mammal-like reptiles of South Africa. 1934.
Broom R. The origin of the Cynodonts. Annals of the Transvaal Museum, vol. 19, 1938.
Colbert E. The mammal-like reptile Lycaenops. Bulletin of the American Museum of Natural History, vol. 89, 1948.
Gregory W. K. and Camp C. L. Studies in comparative myology and osteology, N 3. Ibidem, vol. 38, 1918.
Parrington F. On the Cynognath genus Galesaurus, with a note on the functional significance of the changes in the evolution of the theriodont skull. Annal and Magazine of Natural History, vol. 13, 1934.
Parrington F. On the cranial anatomy of Cynodonts. Proceedings of the Zoological Society, vol. 116, 1946.
Romer A. and Price L. The oldest vertebrate egg. American Journal of Science, vol. 237, 1939.
Romer A. and Price L. Review of Pelycosauria. Geol. Soc. of America, 1940.
Watson D. M. S. On Seymouria, the most primirive reptile. Proceedings of Zoological Society, London, 1918.
White T. Osteology of Seymouria. Bull, of the Museum of Comparative Zoology, vol. 85, 1939.
ГЛАВА V
МЛЕКОПИТАЮЩИЕ
Судя по очень немногочисленным находкам отдельных частей скелета, первые млекопитающие появились в юрском периоде.
Существует достаточное количество фактов, здесь не рассматриваемых, которые заставляют думать, что одновременно с млекопитающими в юре возникли и птицы,
Главнейшей особенностью млекопитающих и птиц, отличающей их от рептилий, является способность сохранять постоянную температуру тела, не зависящую от колебаний температуры окружающей среды.
Эта очень важная особенность обусловливается наличием в головном мозгу представителей обоих этих классов особого терморегуляторного цен* тра, который дает возможность животным так регулировать теплоотдачу, что при данной теплопродукции температура тела остается всегда неизменной.
То или иное количество тепла, вырабатываемого в теле всякого животного, зависит исключительно от окисления в его тканях тех пищевых веществ, которые поступают в организм.
Этих веществ обычно вводится в каждый организм столько, сколько необходимо для его нормальной жизнедеятельности. Таким образом, теплопродукция оказывается величиной более или менее постоянной или во всяком случае подвергающейся относительно небольшим колебаниям. Что же касается теплоотдачи, то она непрерывно изменяется в зависимости от температуры окружающей среды. Регулирование этой теплоотдачи и осуществляется терморегуляторным центром головного мозга.
У млекопитающих и птиц потеря тепла уменьшается благодаря наличию шерсти и перьев, которые состоят из плохо проводящего тепло кератина. Кроме того, при понижении температуры окружающей среды и птицы и млекопитающие имеют возможность путем сокращения особых мышц поднимать перья и волосы, в результате чего толщина слоя неподвижного воздуха, непосредственно окружающего их тело, увеличивается.
У млекопитающих между мускулом, поднимающим волос (musculus erector pili), и самим волосом располагается сальная железа. При сокращении мускула в момент поднимания волоса эта железа оказывается сдавленной. В результате этого из нее выжимается некоторое количество сала, которое покрывает тонким слоем кожу и уменьшает испарение воды с ее поверхности, а это значительно уменьшает теплоотдачу, так как испарение воды всегда сопровождается потерей тепла.
Появление на коже у млекопитающих волос, а у птиц — перьев и возникновение терморегуляторного центра в головном мозгу, несомненно, является результатом приспособления этих животных к жизни в ареале
12*
179
с относительно низкой температурой. А это дает нам право уверенно говорить, что только холодный климат мог быть причиной превращения некоторых рептилий в млекопитающих и птиц. У нас имеются все основания предполагать, что в юрский период в каком-то достаточно обширном ареале на земной поверхности установился относительно холодный климат и этот ареал оказался именно тем местом, где впервые и одновременно возникли примитивные млекопитающие и примитивные птицы.
Холодный климат родины птиц и млекопитающих не мог не отразиться и на их размножении.
Птицы, подобно их предкам — рептилиям, продолжали откладывать яйца, но так как яйца, положенные непосредственно на землю, легко погибали в результате воздействия низкой температуры, то птицы оказались вынужденными согревать их теплом своего тела; только это могло обусловить нормальное развитие их зародышей. Однако высиживание яиц стало возможным только потому, что птицы обладали постоянной и довольно высокой температурой тела.
Первые примитивные млекопитающие, по всей вероятности, также были яйцекладущими. Но свои яйца они, как современная ехидна, помещали в кожную.сумку на животе, что позволяло зародышу развиваться вне вредного влияния низкой температуры окружающей среды и под непосредственным воздействием высокой температуры тела самки.
Одновременно или почти одновременно с этим у примитивных млекопитающих появилась и возможность вскармливать свое слабое потомство выделением кожных желез.
На следующем этапе своего эволюционного развития млекопитающие етали рождать живых, но еще недоразвитых детенышей, которых они помещали в ту сумку, в которую их предки клали яйца.
В дальнейшем более прогрессивные формы древнейших млекопитающих перешли к рождению уже вполне развитых детенышей; после этого их кожная сумка исчезла.
Холодный климат того ареала, в котором оформлялись птицы и млекопитающие, создал совершенно неподходящие условия для жизни стегоцефалов. Эти амфибии могли сохраняться только в том случае, если бы они были в состоянии прятаться под камнями или зарываться в землю и там впадать в спячку на очень продолжительный срок. Однако их большие размеры мешали прибегнуть к такому приему защиты от часто наступающих холодов.
Неспособность к быстрому и длительному передвижению по суше мешала стегоцефалам покинуть холодный ареал, несмотря на то, что существование в нем было для них крайне тяжелым. Очень возможно, что миграции стегоцефалов в более теплые области во многих случаях мешали и моря, так как соленая вода являлась непреодолимым препятствием для амфибий.
Следует думать, что именно эти причины обусловили полное вымирание стегоцефалов в течение триасового периода.
Небольшие размеры таких амфибий, как лягушки, жабы, саламандры, тритоны, дали им возможность находить себе в холодные сезоны года такие места, где они могли, впадая в спячку, пережить неблагоприятное время. Именно это обстоятельство и было причиной того, что только небольшие амфибии смогли сохраниться до наших дней.
Тот холодный ареал, в котором впервые появились млекопитающие и птицы, по-видимому, был местом вымирания не только стегоцефалов, но и крупных мезозойских рептилий. Их большие размеры также не позволяли им находить места для сезонной спячки. Развитие зародыша в яйце, отложенном непосредственно на землю, по-видимому, было также очень затруд
180
нительно. Холодный климат, заставивший птиц высиживать яйца, а млекопитающих помещать их в кожную сумку, по всей вероятности, сделал размножение крупных рептилий совершенно невозможным.
Интересно, что многие современные рептилии холодных и высокогорных районов обычно размножаются путем живорождения. В северных районах Евразии живут только живородящие виды ящериц; только живородящие формы обитают и в горных районах от высоты в 3800 м до верхней границы распространения рептилий — 5400 м.
Но живорождение у них по существу сводится к задержке яйца в теле самки до того момента, когда из него выведется детеныш, который и выбрасывается как уже вполне жизнеспособное существо.
Эта особенность размножения, по-видимому, вызывается следующими причинами.
Температура тела рептилий, как известно, полностью зависит от температуры окружающей среды и колеблется вместе с ней. Для того, чтобы сохранить температуру своего тела более или менее высокой и неизменной, рептилии вынуждены при понижении температуры окружающей среды искать более теплые места, забираясь под камни, под корни деревьев и в норы. В холодных ареалах яйца рептилий, отложенные непосредственно на землю, разумеется, легко гибнут от резких и сильных понижений температуры. В таких ареалах рептилии могут сохранить возможность размножаться только при том условии, если самка, не откладывая яиц, будет постоянно менять свое местопребывание и таким образом сохранять температуру своего тела относительно высокой и неизменной. Это даст возможность эмбрионам нормально развиваться в соответствующих температурных условиях.
Подобные изменения в способе размножения были бесполезны для крупных древних рептилий, так как они не могли при своих больших размерах находить такие места, относительно высокая температура которых позволяла бы им избегать чрезмерного охлаждения тела. Иными словами, крупные и гигантские рептилии мезозоя не'могли прятаться под корни деревьев и под камни. Поэтому температура их тела непрерывно колебалась, находясь в полной зависимости от температуры окружающей среды. Все это делало крупных представителей класса рептилий неспособными приспособиться к жизни в ареалах с холодным климатом.
Таким образом, общее понижение температуры на достаточно большой площади суши неминуемо должно было привести и привело к полному вымиранию крупных амфибий (стегоцефалов) и крупных рептилий, к сохранению маленьких амфибий и рептилий и к появлению первых представителей птиц и млекопитающих.
В настоящее время палеонтология накопила очень много данных, которые говорят и о том, что триасовый, юрский и меловой периоды были не столько временем вымирания рептилий, сколько временем их расцвета. Рептилии юры и мела были многочисленны, очень разнообразны и во многих случаях достигали необычайно больших размеров.
Это обстоятельство свидетельствует о том, что холодный климат сначала в триасе, а затем в юре и мелу охватывал не всю сушу земного шара; на многих континентах имелись достаточно обширные области, где продолжали жить и развиваться самые разнообразные рептилии в достаточно благоприятной обстановке. Климат этих областей оставался настолько теплым, что эмбрионы в яйцах, отложенных рептилиями непосредственно на землю, развивались нормально; сами же рептилии не имели нужды зарываться в норы и впадать в спячку на тот или иной срок.
Теплокровность и живородность млекопитающих, позволяющие им обитать в холодных областях, разумеется, не мешали им жить и в районах
181
с теплым климатом. Поэтому млекопитающие вместе с птицами скоро стали появляться и там, где до тех пор жили только рептилии.
Постепенное расширение или смещение ареала холодного климата неминуемо приводило к сужению областей обитания крупных мезозойских рептилий, но не мешало миграциям млекопитающих и захвату ими новых территорий, независимо от климата, который в них наблюдался.
Все те особенности млекопитающих, которыми они отличались от своих предков, давали им настолько большие преимущества в борьбе за жизнь при всяких условиях, что они не могли не выйти победителями из конку-
Рис. 205. Позвонки млекопитающего (Meles taxus). А — шейный; В, С — грудные; D — поясничный; Е — крестцовый; F — хвостовой.
ренции с сильными и многочисленными представителями класса рептилий. Это привело к тому, что все гигантские рептилии мезозоя вымерли в конце мелового периода и начало кайнозоя оказалось началом господства и расцвета млекопитающих.
В скелете млекопитающих наряду с признаками, свидетельствующими об их генетической связи с рептилиями, имеются и такие особенности строения, которые являются совершенно новыми.
Тела позвонков млекопитающих представляют собой сильно увеличенные плевроцентры (рис. 205); их передняя и задняя поверхности плоские; характерная для рептилий амфицельность в позвонках млекопитающих не сохранилась. Хорошо развитые дуги срослись с телами позвонков. Гипоцентры у всех позвонков, кроме I, полностью исчезли. Результатом этого явилось то, что головка ребра, соединявшаяся у предков млекопитающих с гипоцентром, теперь стала соединяться с двумя суставными площадками на краях тел соседних позвонков (рис. 205, С). Таким образом, головка каждого ребра у млекопитающих направлена в промежуток между плевроцентрами, т. е. туда, где когда-то находился гипоцентр, с которым ребро соединялось у всех низших Tetrapoda. Бугорок ребра у млекопитающих, как и у их предков из класса рептилий, образует сустав с концом поперечного отростка дуги позвонка (рис. 205, В).
Полное срастание ребер с позвонками в переднем участке позвоночного столба привело к появлению у млекопитающих типичных шейных позвон-182
ков, количество которых обычно равняется семи (рис. 205, Л). Редукция ребер у ряда позвонков предкрестцового отдела обусловила оформление поясничных позвонков (рис. 205, D).
В состав крестца (sacrum) млекопитающих входит обычно не меньше трех позвонков. Но в некоторых случаях число крестцовых позвонков увеличивается за счет присоединения к ним нескольких хвостовых. Крестцовые ребра у взрослых млекопитающих полностью срастаются с соответствующими позвонками (рис. 205, Е)\ у молодых животных они бывают еще свободными (рис. 206, В—с). Эти крестцовые ребра у млекопитающих, как и у их предков из класса рептилий, служат для соединения тазового пояса с позвоночным столбом. Таз млекопитающих обычно присоединяется толь-
Рис. 206. Позвонки млекопитающих. А — III шейный позвонок молодой ехидны (Echidna aculeata); В — I крестцовый позвонок медведя (Ursus arctos); С — хвостовые позвонки молодого муравьеда (Myrmecophaga jubata).
a. d. — arcus dorsalis; a. v. — arcus ventralls; c — costa.
ко к трем первым позвонкам крестца; другие позвонки не имеют непосредственного контакта с подвздошной костью.
Среди млекопитающих встречаются формы, у которых, несмотря на полное отсутствие гипоцентров, в хвостовом отделе позвоночного столба сохраняются вентральные дуги. Это можно видеть, например, у муравьеда (Myrmecophaga jubata) (рис. 206, С — a. v.).
Своеобразное строение первых двух позвонков рептилий сохранилось и у млекопитающих. Однако проатлант (proatlas) исчез полностью. Атлант млекопитающих представляет собой костное кольцо, состоящее из дуги, сросшейся с гипоцентром. Гипоцентр атланта является единственным гипоцентром, сохранившимся в позвоночном столбе млекопитающих.
По обе стороны к дуге и гипоцентру I позвонка приросла пара рудиментарных ребер. У молодых животных все эти части бывают еще изолированными образованиями (рис. 206, А).
Тот сустав между дугами атланта и эпистрофея, который имелся у рептилий, у млекопитающих не сохранился. Вместо него между I и II позвонками возникло новое соединение, расположенное ниже прежнего (рис. 207, А).
Тело атланта (плевроцентр) у млекопитающих полностью приросло к телу эпистрофея, превратившись в так называемый «зуб» II позвонка —
183
dens epistrophei (рис. 207, В). Гипоцентр между двумя плевроцентрами (двумя телами) II шейного позвонка подвергся полной редукции.
Очень своеобразной и совершенно новой особенностью позвонков млекопитающих является образование у молодых животных так называемых
Рис. 207. Позвонки млекопитающих. А — три первых шейных позвонка барсука (Meles taxus); В—II шейный позвонок барсука; С—три первых шейных позвонка молодого медведя (Ursus arctos).
а — atlas; ad. а. — arcus dorsalis atlantis; e — epistropheus; h. a. — hypocentrum atlantis; p. a. — pleurocentrum atlantis; p. e. — pleurocentrum epistrophei.
эпифизарных окостенений. Эти дополнительные окостенения возникают в виде небольших косточек на вершинах остистого, поперечного и сустав-
Рис. 208.
А, В—поясничные позвонки молодого человека; С — поясничный позвонок взрослого человека; D — большая берцовая кость молодого человека.
ных отростков и в виде костных пластинок на передней и задней поверхностях тела позвонка (рис. 208, А, В). Эпифизарные окостенения отделяются от соответствующих элементов позвонка слоем непрерывно растущего и
184
непрерывно окостеневающего хряща. Наличие этих хрящевых прослоек дает позвонку возможность увеличивать длину своих отростков и размеры тела несколько более интенсивно, чем это можно было бы осуществить только путем одного периостального окостенения, т. е. равномерного наслоения костного вещества на всю поверхность позвонка,
У взрослых млекопитающих эпифизарные хрящи исчезают, что приводит к прирастанию всех дополнительных окостенений к соответствующим участкам позвонка (рис. 208, С).
Очень интересно «поведение» эпифизарных пластинок на телах позвонков в тех случаях, когда соседние позвонки срастаются друг с другом, на*
Рис. 209. Крестцы медведей (Ursus arctos) различного возраста.
пример при образовании крестца. При этом прежде всего исчезают межпозвоночные; хрящи (f i brocart i Iago intervertebral is), что приводит к срастанию задней эпифизарной пластинки одного позвонка с передней эпифизарной пластинкой следующего позвонка (рис. 209, А). В результате этого между телами позвонков у молодого животного появляются толстые костные диски, отделенные прослойками эпифизарных хрящей от тел позвонков (рис. 209, В).
Через некоторое время подвергаются окостенению и эпифизарные хрящи. В этот момент тела позвонков полностью срастаются друг с другом (рис. 209, С). Таким образом, при срастании двух позвонков прежде всего исчезают межпозвоночные хрящи и только после этого исчезают эпифизарные хрящи. Все это свидетельствует о том, что во время срастания позвонки млекопитающих еще сохраняют все характерные признаки своей молодости.
Так как у атланта нет тела и его дуга не имеет хорошо выраженных отростков, то он лишен эпифизарных пластинок. Но такие пластинки имеются в эпистрофее, обладающем двумя телами. Они располагаются следующим образом. Одна помещается на вершине «зуба», две — между зубом и телом и одна — на задней поверхности тела позвонка (рис. 207, С).
Пара эпифизарных пластинок, находящаяся между зубом и телом эпистрофея, очень рано срастается между собой и у молодых животных обычно образует относительно толстый диск, на дорзальной и вентральной сторонах которого хорошо видны борозды, указывающие на образование его в результате сращения двух пластинок.
При переходе от рептилий к млекопитающим в черепе" позвоночных произошли еще более важные изменения, чем в строении позвоночного столба.
185
Рис. 210.
A —затылочная часть черепа дикого кабана (Sas scrofa); В—затылочная часть черепа молодого муравьеда (Myrmecophaga jubata).
р. b. — pars basilaris; р. I. — pars lateralis; р. s.— pars squamosa ossis occipitalis.
Такие кости, как непарные basioccipitale и supraoccipitale и парные exoccipitale, dermosupraoccipitale и tabulare, срослись между собой и образовали одну затылочную кость (os occipitale). У молодых млекопитающих эта сложная комплексная кость бывает еще разделена на четыре части, именно: pars basilaris (basioccipitale), pars lateralis (exoccipitale) и pars squamosa (supraoccipitale, dermosupraoccipitale и tabulare) (рис. 210, A). Некоторое уменьшение размеров basioccipitale еще у цинодонтов привело к тому, что эта кость почти перестала принимать участие в образовании сустава между черепом и I позвонком. Эта особенность сохранилась и у млекопитающих; суставные отростки на их черепе образованы в основном выступами на pars lateralis затылочной кости, т. е. на exoccipitale.
Интересно, что у молодых млекопитающих нередко наблюдаются эпифизарные пластинки на суставных отростках затылочной кости (рис. 210, В).
Теменные кости (os parietale) у млекопитающих увеличились в своих размерах и foramen parietale между ними исчезло (рис. 211).
«Лобная кость (os frontale) вместе с теменной стала принимать участие в образовании мозговой коробки.
Носовая кость (os nasale) несколько укоротилась и потеряла контакт с praemaxillare (os interma-xillare), в результате чего возник один непарный вход в полость носа.
Слезная кость (os lacrimale) сильно уменьшилась в своих размерах и оказалась лежащей наполовину на внутренней стенке орбиты. В этой кости сохранились два отверстия для слезно-носового канала.
Septomaxillare полностью переместилось в полость носа и превратилось в нижнюю носовую раковину — concha nasalis inferior (рис. 212 — с. п. i.).
Таким образом, эта покровная кость перестала быть наружной костью лицевого черепа, но еще сохранила прежнюю свою связь с передним концом слезно-носового канала (canalis nasolacrimal is). Этот канал у млекопитающих входит в отверстие в слезной кости, затем идет в полость носа и открывается под нижнюю носовую раковину.
Межчелюстная кость (os intermaxillare = praemaxillare), потерявшая контакт с передним концом носовой кости, у млекопитающих, как и у всех Therapsida, несет резцы (рис. 213). Верхнечелюстная кость (maxilla, maxillare) сохранила свое положение и размеры. В ее нижний край входят корни клыка и коренных зубов.
Jugale рептилий, получившее у млекопитающих название os zygo-maticum, также не изменило своего положения (рис. 213).
Squamosum сильно увеличилось в той своей части, которая у Therapsida принимает участие в образовании задней стенки височной ямы, и вошло в состав боковой стенки мозговой коробки. Эта разросшаяся часть squamosum, получившая у млекопитающих название squama temporalis, по-прежнему служит местом начала для одной из жевательных мышц (mu-sculus temporalis) и сохраняет контакт с jugale (zygomaticum) при помощи так называемого processus zygomaticus ossis temporalis.
186
Prooticum подверглось полному окостенению и сильно изменило свою форму и положение. Эта кость включила в себя перепончатый лабиринт
внутреннего уха и, расположившись в основании черепа под сильно уве-
личившимся головным мозгом, приросла к squamosum (squama temporalis) в виде ее пирамиды (pyramis s. pars petrosa ossis temporalis).
Paroccipitale у млекопитающих стало принимать участие в образовании задней стенки среднего уха и превратилось в так называемый сосцевидный отросток височной кости (pars mas-toidea ossis temporalis).
Basisphenoideum, примыкающее непосредственно к basioccipitale, образовало задний отдел тела клиновидной кости (corpus ossis sphenoidalis).
Epipterygoideum приросло к basisphenoideum и превратилось в большое крыло клиновидной кости—ala magna ossis sphenoidalis (рис. 214 — a. tn.).
Sphenethmoideum дало передний отдел тела и малые крылья клиновидной кости (alae parvae ossis sphenoidalis).
Твердое нёбо (palatum durum) в черепе млекопитающих образовано такими же отростками maxillare и palatinum, как и в черепе цинодонтов. Но transversum у млекопитающих исчезло.
Сошник представляет собой непарную костную пластинку, которая разделяет полость носа на правую и левую половины.
Парасфеноид (paraspheno-ideum) у млекопитающих под-
Рис. 211. Череп опоссума (Didelphys virgini-апа) (вид сверху).
F — frontale; Im — intermaxillare (praemaxillare); L — lacrimale; M — maxillare; N — nasale; P — parietale; Sq — squama temporalis (squamosum); Z — zygomatlcum (jugale).
вергся полной редукции; от этой кости не сохранилось никаких следов (рис. 215).
Птеригоид (pterygoideum)
потерял то значение, которое он имел в черепе рептилий, и превратился в небольшую костную пластинку — lamina medialis processus pterygoidei ossis sphenoidalis (рис. 215). Несмотря на очень сильную редукцию, птеригоид у млекопитающих по-прежнему служит местом начала такой жевательной мышцы, как musculus pterygoideus (internus).
Нижняя челюсть у млекопитающих (mandibula) образована только одной костью, именно: очень сильно увеличенным dentale. Эта кость соеди-
няется посредством сустава с височной костью (squamosum, squama ossis temporalis). Для черепа млекопитающих этот сустав (articulatio mandibu-laris) является совершенно новым анатомическим образованием. Его не было ни у кого из представителей класса рептилий. Он отсутствует и в черепе птиц, которые в этом отношении не отличаются от рептилий.
187
Образование у первых млекопитающих подвижного контакта между dentale и squamosum сопровождалось очень сильными изменениями в строении среднего уха позвоночных.
Стремечко (stapes), которое у предков млекопитающих было единственной слуховой костью и одним своим концом закрывало fenestra ovalis,
а другим — соединялось c qua-dratum, у млекопитающих сильно уменьшилось в размерах, но не изменило своего отношения к соседним костям и сохранило прежнюю функцию.
Quadratum, принимавшее у рептилий участие в образовании челюстного сустава, у млекопитающих сильно изменило свою форму, размеры и функцию. Эта кость, сохраняя контакт со stapes, переместилась в среднее ухо и превратилась в слуховую косточку — в наковальню (incus).
Подвижное соединение между quadratum и articulare, которое играло роль челюстного сустава у всех позвоночных, начиная с рыб и кончая рептилия-
Рис. 212. Полость носа в черепе человека. с. п. I. — concha nasalis inferior (septomaxillare); Z. m. — lamina medians processus pterygoidei ossls sphenoidalis (pterygoideum).
ми (и птицами), у млекопитающих сохранилось, но articulare выключилось из состава сложной нижней челюсти и переместилось в среднее ухо. Здесь оно, так же как quadratum,
Рис. 213. Череп опоссума (Didelphys virginiana) (вид сбоку).
Л — frontale; Im — intermaxillare (praemaxillare); L — lacrimale; M — maxillare; N — nasale; О — occlpitale; P — parletale; T — temporale; Z — zygomaticum (jugale).
превратилось в маленькую слуховую кость. У млекопитающих эта кость получила название молоточка (malleus).
Таким образом, тот сустав, который у рептилий являлся челюстным, у млекопитающих превратился в сустав между двумя новыми слуховыми косточками (incus и malleus).
188
Такие коренные изменения в устройстве уха могли наступить, разумеется, только после возникновения сустава между dentale и squamosum.
Рис, 214. Череп опоссума (Didelphys olrginiana) (вид сбоку); скуловая дуга удалена. л. т. — ala magna ossis sphenoidalis (eplpterygoldeum); F — frontale; Im — Intermaxlllare; L — lacrlmale; M — maxillare; N — nasale; О — occipitale; P — parietale; T — temporale (squama temporalis, squamosum).
И образование этого сустава было одним из очень важных изменений при оформлении черепа млекопитающих.
Одновременно с превращением quadratum в incus, a articulare в malleus сильно редуцированное praearticulare нижней челюсти, сохраняя свое непосредственное соединение с articulare, приросло к malleus и образовало его processus anterior Folii.
Angulare нижней челюсти рептилий в процессе развития черепа млекопитающих потеряло контакт с dentale и, сильно изменив свою форму, приросло к височной кости в виде os tym-panicum.
В результате ряда перестроек височная кость млекопитающих оказалась одной из самых сложных костей черепа; в ее состав вошли следующие восемь костей черепа рептилий: squamosum, prooticum, paroccipitale, stapes, quadratum, articulare, praearticulare и angulare.
В период оформления среднего уха у млекопитающих у них заново возникла барабанная перепонка (membrana tym-pani), которая и стала воспринимать звуковые волны и про-
Рис. 215. Череп опоссума (Didelphys virginiana} (вид снизу).
Im — Intermaxlllare (praemaxillare); М — maxillare; Pl — palatinum; Pt — pterygoideum; T — temporale (squamosum); Z — zygomatlcum (jugale).
водить их в виде колебаний через цепь костей (malleus-*incus-* st apes) в лабиринт внут-
реннего уха.
189
Кости нижней челюсти рептилий — supraangulare и coronoideum —
у млекопитающих исчезли полностью.
Рис. 216> Нижние челюсти зародышей Didelphys aurata (А) и Erinaceus europaeus (В).
Ап — angulare; с. М. — cartilago Meckelii; D — dentale; i — incus; m — malleus; Md — mandlbula; Pa — praearticulare; p. F. — processus anterior Foil!; s — stapes; t — tympanicum.
Изучение эмбрионального развития черепа современных млекопитающих с совершенной очевидностью доказывает соответствие молоточка и наковальни млекопитающих articulare и quadratum рептилий.
Рис. 217. Нижние челюсти зародыша человека (А) и ребенка (В).
с. М. — cartilago Meckelii; I — incus; Afd — mandlbula; m — malleus; p. F. — processus anterior Folii; 5“ — stapes; t — os tympanicum.
У зародышей около хорошо развитого меккелева хряща (cartilago Meckelii) в соединительной ткани образуется большое dentale (mandibula) и закладываются сильно редуцированные praearticulare и angulare (рис. 216, Л).
190
При дальнейшем развитии зародыша задний отдел меккелева хряща окостеневает, превращаясь в молоточек (malleus), к которому прирастает praearticulare и образует его processus anterior Folii. Так как окостенение заднего участка меккелева хряща у рептилий дает articulare, то мы можем быть совершенно уверены в том, что эта кость гомологична молоточку млекопитающих.
Angulare у зародышей млекопитающих приобретает кольцевидную форму и у взрослых животных прирастает к squama temporalis (squamosum) в виде os tympanicum, служа как бы рамкой, в которой натягивается барабанная перепонка.
Изолированный кусок палатоквадратного хряща, соединенный при помощи сустава с задним концом cartilage Meckelii, у млекопитающихжпри
Рис. 218. Зубы опоссума (Didelphys virginiana).
своем окостенении образует наковальню (incus). Так как у рептилий окостенение этого хряща дает quadratum, то у нас нет никаких оснований сомневаться, что quadratum рептилий соответствует incus млекопитающих.
Таким образом, изучение развития среднего уха у современных млекопитающих показывает, что костные элементы челюстного сустава рептилий (quadratum и articulare) у млекопитающих переместились в среднее ухо и превратились в слуховые косточки (incus и malleus) (рис. 217).
Зубы примитивных млекопитающих во многих отношениях очень напоминают зубы цинодонтов; их резцы и клыки еще сохраняют конические коронки и имеют по одному корню.
Однако малые коренные зубы уже приобретают по два корня, а большие коренные верхние челюсти — даже по три. Коронки коренных зубов, в особенности больших, снабжены на своей вершине несколькими бугорками (рис. 218).
После своего оформления череп млекопитающих на всем протяжении дальнейшей эволюции изменился только в своей общей форме. При этом отдельные кости меняли свои размеры и очертания, но количество их оставалось неизменным.
Для подведения общих итогов рассмотренной нами истории формирования черепа Tetrapoda ниже приводится таблица, которая дает представление о составе черепа на различных этапах его эволюции.
191
Названия костей Обозначения Stegocephalia Seymouria-morpha Cotylosauria Pelycosauria Qorgonopsia Cynodontla Mammalia Homo
Basioccipitale . . . Bo + + + 4- 4- 4- 4- Pars basilaris ossis occipitalis
Exoccipitale .... Eo + + 4- 4- 4- 4- 4- Pars lateralis ossis occipitalis
Supraoccipitale . . . So + + 4- 4- 4- 4- 4- Squama ossis occipitalis (часть)
Paroccipitale .... Po + + 4- 4- 4- 4- 4- Pars mastoidea ossis temporalis
Prooticum Pro + + 4- 4- 4- 4- 4- Pyramis ossis temporalis
Basisphenoideum . . Bs + + 4- 4- 4- 4- 4- Corpus ossis sphenoidal is (задний отдел)
Sphenethmoideum . Se + + 4- 4- 4- 4- 4- Corpus et alae par-vae ossis sphenoidalis
Epipterygoideum . . Ep + + 4- 4- 4- 4- 4- Ala magna ossis sphenoidalis
Parasphenoideum . . Ps + 4- 4- 4- 4- ± —— Нет
Pterygoideum . . . Pt 4- + 4- 4- 4- 4- 4- Lamina medialis processus pterygoi deus ossis sphenoidalis
Transversum .... Tr + + 4- 4- 4- 4- —• Нет
Palatinum Pl + + 4- 4- 4- 4- 4- Os palatinum
Vomer Dermosupraoccipi- V + + 4- 4- 4- 4- 4- Vomer
tale Dso + 4- 4- 4- 4- 4- 4- Верхняя часть squamae occipitalis
Tabulare ...... T + + + 4- 4- 4- 4- Верхняя часть squamae occipitalis
Supratemporale . . . St 4- 4- 4- 4- — — Нет
Intertemporale . . . It -t- + — — — — — »
Parletale P + 4- 4- 4- 4- 4- 4- Os parietale
Front ale F + + 4- 4- 4- 4- Os frontale
Nasale N + + 4- 4- 4- 4- Os nasale
Postorbitale .... Po + + 4- 4- 4- 4- —. Нет
Postfrontale .... Pf + + 4- 4- 4- —— »
Praefrontale .... Prf • + + 4- 4- 4- 4- »
Lacrimale L + + 4- 4- 4- 4- 4- Os lacrimale
Septomaxillare . . . Sm + + 4- 4- 4- 4- 4- Concha nasalis inferior
Praemaxillare . . . Pm + 4- 4- 4- 4- 4- 4- Os intermaxillare = os incisivum
Maxillare M + + 4- 4- 4- 4- 4- Maxilla
Jugale Ju + + 4- 4- 4- 4- 4- Os zygomaticum
Squamosum .... Sq + 4- 4- 4- 4“ 4- 4- Squama temporalis
Quadratojugale . . . QJ + 4- 4- 4- 4- 4- — Нет
Stapes S + 4- 4- 4- 4- 4- 4- Stapes
Quadratum Qu + 4- 4- 4- 4- 4- 4- Incus
Articulare Ar + 4- + 4- 4- 4- Malleus
Praearticulare . . . Pa + 4- 4- 4- 4- 4- 4- Processus anterior mallei Folii
Supraangulare . . . Sa + 4- -+- 4- 4- 4- — Нет
Angulare An + 4- 4- 4- 4- 4- 4- Pars tympanica ossis temporalis
Dent ale D + 4- 4- 4- 4- + 4- Mandibula
Postspleniale .... Ps + 4- — — Нет
Splenlale Sq + 4- + 4- 4- + — »
Coronoideum .... Co + 4- 4- 4- -j- 4- — »
Intercoronoideum . . Ic + 4- 4- — — »
Praecoronoideum . . Pc + 4- — — — — »
Примечание. Плюсом (+) отмечено наличие кости, минусом (—) ее отсутствие.
192
В состав плечевого пояса наиболее примитивных несомненно, входило такое же количество костей, как
млекопитающих, и у цинодонтов.
Рис. 219. Плечевой пояс ехидны. А — вид снизу. В — вид сверху.
Cl — clavicula; Со — coracoldeum; Ic — interclavicula; Pc — procoracoldeum; Sc — scapula; St — sternum.
Рис. 220.
A — плечевой пояс ехидны, вид сбоку; В —левая плечевая кость ехидны, вид снизу; С — левое бедро ехидны, вид сверху.
Cl ~ clavicula; Со — coracoldeum; Pc — procoracoldeum; Sc — scapula: St — sternum.
Доказательством этого является строение плечевого пояса у современных яйцекладущих млекопитающих — утконоса и ехидны. Их плечевой пояс
13 А. П. Быстрое 193
Рис. 221.
А — левая лопатка вомбата; В — левая лопатка и ключица человека.
CZ — clavicula; р. г. — processus coracoideus scapulae (coracoideum); 5г — scapula.
образован следующими костями: scapula, coracoideum, procoracoideum, sternum, clavicula и interclavicula (рис. 219, 220). У взрослых животных обе ключицы и межключица обычно срастаются.
При дальнейшем своем развитии млекопитающие скоро потеряли прокоракоид и межключицу, но ключица у них сохранилась и оказалась единственной покровной костью плечевого пояса; коракоид прирос к лопатке и превратился в так называемый processus coracoideus scapulae (рис. 221).
Передний край лопатки цинодонтов и примитивных форм млекопитающих (рис. 220, Л) у более прогрессивных представителей Mammalia ' превратился в длинный лопаточный гребень, расположенный на наружной поверхности кости (spina scapulae, рис. 221). Такое разрастанием лопатки кпереди,
К нижнему участку переднего края лопатки у цинодонтов и у примитив-
изменение в строении было обусловлено дальше ее прежнего переднего края.,
Рис. 222.
А — бедро и кости голени ехидны; В — таз ехидны.
Г — fibula; Ге — femur; 11 — Ilium; Is — ischium; M — os marsuplale; Pb — pubis; P — patella; T — tibia.
Рис. 223.
A — плечевая кость саблезубого тнгра. В — плечевая кость барсука.
(Масштабы различные.)
ных форм млекопитающих и привел к образованию сустава между ключицей и так называемым acromion (рис. 221, В),
194
Тазовый пояс млекопитающих по своему составу не отличается от та- зового пояса низших позвоночных; он образован теми же тремя костями: ilium, ischium и pubis. У сумчатых форм к pubis присоединялась особая парная кость, поддерживающая кожную сумку на животе, — os marsu-piale (рис. 222). После редукции сумки эта кость исчезла.
Длинные кости передней и задней конечностей млекопитающих отличаются от соответствующих костей рептилий тем, что их концы (эпифизы) окостеневают независимо от средней, диафизарной, части. И это приводит к появлению у молодых животных так называемых эпифизарных хрящей, которые позволяют костям расти в длину даже тогда, когда их концы уже
Рис. 224. Нижние челюсти юрских млекопитающих (Осборн, 1907). А — Phascolotherium (нижняя юра);
В — Amblotherium (верхняя юра).
полностью окостенели (рис. 208, D). Таким образом, кости молодых животных могут функционировать с полной нагрузкой и в то же время не прекращать своего роста в длину. Такие же дополнительные окостенения, отделенные от основной кости прослойками хряща, мы отмечали и у позвонков. Следовательно, эпифизарные окостенения появились почти у всех элементов посткраниального скелета млекопитающих.
На плечевой кости у многих млекопитающих сохранилось foramen entepicondyloideum (рис. 223). На бедре развились хорошо выраженные trochanter major и trochanter minor, служащие местом прикрепления сильных тазовых мышц (рис. 220, С). У древнейших и примитивных форм шейка бедра, вероятно, была также слабо выражена, как у современной ехидны (рис. 220), и эта часть кости напоминала соответствующий участок бедра цинодонта.
Совершенно новой особенностью скелета задней конечности млекопитающих было появление костной коленной чашки — patella (рис. 222, Л) — и полное выключение верхнего конца малой берцовой кости из коленного сустава (этого еще не произошло в скелете ехидны, рис. 222, Л).
В кисти и стопе млекопитающих установилась следующая формул» фаланг: 23333.
Судя по неполным и разрозненным остаткам скелетов, млекопитающие юрского периода были очень маленькими и слабыми животными. По своим размерам они не превосходили величины крысы. Такие животные, разумеется, не могли претендовать на господствующее положение в природе. Однако эти примитивные млекопитающие обладали хорошо развитым
13*
195
зубным аппаратом; их коренные зубы были снабжены несколькими острыми вершинами и давали им возможность хорошо размельчать пищу (рис. 224); количество зубов у них было обычно довольно велико. Судя по нижним челюстям, которые сохранялись лучше, чем какие-либо другие части скелета, зубная формула у юрских млекопитающих выражалась следующими цифрами: 4. 1. 3. 4 (Phascolotherium, нижняя юра); 4. 1. 4. 7 (Amblotherium, верхняя юра).
Юрские млекопитающие были очень подвижными маленькими хищниками, не боящимися низких температур и быстро расселяющимися повсюду.
В течение всего мелового периода млекопитающие оставались такими же небольшими и незаметными обитателями степей и лесов.
Только в кайнозое они заняли господствующее положение в природе и, постепенно заселяя всю сушу, стали приобретать самое различное строение. Некоторые из них уже в начале кайнозоя приспособились к жизни в воде и к полету по воздуху. Об этом свидетельствуют находки скелетов зубатых китов (Zeuglodon) и летучих мышей (Palaeochiropteryx) в эоценовых отложениях.
ГЛАВА VI
ОБЕЗЬЯНЫ И ЧЕЛОВЕК
Филогенетическое развитие большинства разнообразных млекопитающих кайнозоя, разумеется, не вело к человеку. Но эволюционный путь древнейших предков человека прошел через те примитивные формы, которые, будучи обитателями лесов, приспособились к жизни на деревьях. Такие формы появились уже в самом начале кайнозоя.
Древнейшим из таких животных был среднеэоценовый Notharctus, найденный в Северной Америке (штат Вайоминг). Это маленькое древесное млекопитающее очень напоминало современных полуобезьян — лемуров. Череп Notharctus был длинным и узким; орбиты располагались сбоку; зубная формула выражалась следующими цифрами:
2, 1, 4. 3
2. 1. 4. 3
(рис. 225).
Notharctus обладал длинным хвостом; тонкими костями конечностей, его кисти и стопы были приспособлены к передвижению по ветвям деревьев (рис. 226).
Сходное животное, Adapts parisiensis, было найдено в верхнеэоценовых отложениях Франции. Череп Adapts (рис. 227) был более коротким и более высоким, чем у Notharctus, но зубная формула была такой же.
Мы имеем все основания полагать, что именно такие формы, как Notharctus и Adapts, могли быть предками настоящих обезьян.
Древнейшие обезьяны на всем протяжении своей эволюции оставались жителями лесов.
К сожалению, в распоряжении палеонтологов нет достаточного количества ископаемого материала, который позволил бы проследить все этапы развития этой очень важной для нас группы животных. Отсутствие хорошо сохранившихся скелетов ископаемых обезьян объясняется тем, что остатки всех обитателей лесов, как правило, полностью разрушаются и очень редко попадают в такие условия, которые позволяют им окаменеть. Поэтому у нас по существу не может быть больших надежд, что и в будущем мы сумеем изучить все те морфологические изменения в скелете обезьян, которые сопровождали их эволюцию на протяжении всего кайнозоя. По-видимому нам придется примириться с тем, что очень ценные для нас скелеты этих форм погибли безвозвратно.
Это обстоятельство лишает нас возможности проследить шаг за шагом тот эволюционный путь, который непосредственно вел к человеку.
Что же касается хорошо изученных современных высших обезьян, то, разумеется, ни одна из них не может считаться предком человека. Но, принимая во внимание их строение, мы можем составить себе некоторое
197
представление и о строении тех форм, которые были нашими непосредственными предками.
Из четырех форм современных человекообразных обезьян гиббоны, живущие на островах Малайского Архипелага и в Индокитае, несомненно, представляют собой наиболее далеких родственников человека.
Рис. 225. Череп Notharctus osborni (средний эоцеи).
Череп гиббона (рис. 228) по своей общей форме еще очень похож на череп более примитивных обезьян, хотя имеет достаточно большую мозговую
Рис 226 Скелет Notharctus osborni (Грегори, 1935)
часть. Надглазничные дуги (arcus supraorbitalis) у этой обезьяны развиты слабо; гребни на черепе отсутствуют; клыки большие; подбородочная часть нижней челюсти тупая.
198
Гиббоны появились, по всей вероятности, еще в олигоцене и за время своей долгой эволюции очень хорошо приспособились к жизни на деревьях, превратившись в высокоспециализированных обезьян. Об этом свидетельствует необычайная длина их рук, которые при вертикальном положении
Рис. 227. Череп Adapts parisiensis (верхний эоцеи).
тела животного достигают земли. Такие руки позволяют гиббонам передвигаться по лесу с ловкостью изумительных акробатов.
Древние менее специализированные формы этих обезьян, вероятно, находились в непосредственной родственной связи с миоценовыми предками оранг-утана,
Рис. 228. Череп гиббоиа.
Современные оранг-утаны, живущие на Суматре и Борнео, по своему строению стоят ближе к человеку, чем гиббоны.
Череп оранг-утана (рис. 229) имеет необычайно сильно развитую лицевую часть, по сравнению с которой мозговой отдел кажется очень маленьким; лицевая поверхность черепа уплощена и вогнута; орбиты обычно высокие и узкие; надглазничные дуги невелики.
199
Оранг-утаны, как и гиббоны, ведут древесный образ жизни, но их руки относительно не так длинны, как у гиббонов, и при вертикальном положении тела достигают только голеностопных суставов.
Африканские формы антропоморфных обезьян — горилла и шимпанзе — являются без сомнения самыми близкими родственниками человека (рис. 230, 231).
Размеры мозговой части черепа у этих обезьян больше, чем у орангутана. На крыше черепа шимпанзе гребни отсутствуют; у гориллы, в особенности у старых самцов, они достигают большой величины. Надглазничные дуги у обеих этих обезьян сильно развиты.
Рис. 229. Череп оранг-утана.
Руки у шимпанзе короче, чем у оранг-утана, и спускаются только немного ниже колен; у гориллы они немного длиннее, чем у шимпанзе.
Обе эти обезьяны появились, по всей вероятности, в конце миоцена или в начале плиоцена и развивались своими собственными путями.
Шимпанзе и горилла, конечно, не являются предками человека, но какие-то далекие обезьянообразные предки человека, вероятно, были одновременно и предками этих обезьян. Это обстоятельство дает нам право считать, что непосредственные предки человека были очень похожи на такую обезьяну, как шимпанзе. Они обладали относительно большим мозгом; на их черепе отсутствовал такой сагиттальный гребень, который имеется у самцов горилл; надглазничные дуги были развиты не слабее, чем у шимпанзе; челюсти были сильными, вследствие чего лицевая часть черепа была выдвинута вперед, подбородочная часть нижней челюсти была тупой; зубная формула была такой же, как у всех современных человекообразных обезьян, 2 12 3
именно: ~ руки были длинными, ноги же, напротив,— относительно
короткими; кисти и стопы были приспособлены к передвижению по ветвям деревьев. Эти наши предки были обитателями лесов, подобно современным древесным формам обезьян.
Превращение таких обезьян более миллиона лет тому назад в человекоподобные существа могло быть обусловлено только тем, что какие-то причины вынудили их покинуть деревья и приспособиться к жизни на земле. Только приспособление к этой новой среде обитания могло превратить обезьяну в человекообразное существо, 200
Обезьяны, оказавшиеся предками человека, покинули деревья, несомненно, не потому, что на деревьях стало меньше пищи. На земле для них пищи было еще меньше.
Обезьяны покинули деревья и не потому, что на деревьях было много врагов. На земле для них врагов было несравненно больше и защита от них там была гораздо труднее.
Рис. 230. Череп гориллы.
Рис. 231. Череп шимпанзе.
Обезьяны покинули деревья и не потому, что были лучше приспособлены к жизни на земле. На земле они были очень беспомощными существами.
Обезьяны покинули деревья, конечно, и не потому, что превратились в человекоподобные существа. Наоборот, они стали человекоподобными потому, что спустились на землю и только после того, как сделали это. Все и всякие обезьяны, сохранившие древесный образ жизни, по-прежнему остались обезьянами.
Обезьяны покинули деревья, несомненно, только потому, что леса исчезли. Всюду, где этого не случилось, они, как правило, остались жите-
20)
лями деревьев, но не всюду, где исчезли леса, обезьяны, жившие в них, превратились в предков людей. Предки павианов, живущих в горных рай* онах Абиссинии, несомненно, в свое время обитали на деревьях, но, приспособившись к жизни на земле, они все же остались сравнительно низко -организованными формами обезьян, не имеющими никакого права претендовать на почетное звание предка человека. Следовательно, не всякая обезьяна, покинувшая лес как среду обитания, способна дать начало человеку. Его предками были обезьяны, несравненно более высоко организованные, чем предки павианов.
Человек, несомненно, не мог бы появиться, если бы его предки — обезьяны — к определенному моменту не оказались подготовленными к тому, чтобы встать на задние ноги и выработать новый способ передвижения. Но человек и при этих условиях не появился бы, если бы его предки в то же время не оказались вынужденными покинуть деревья и спуститься на землю. Таким образом, для того, чтобы понять, как мог возникнуть человек, мы должны предположить, что в каком-то достаточно обширном аре-~але лес стал исчезать и постепенно заменяться степью, покрытой редкими деревьями и кустарником.
И именно это обстоятельство вынудило обезьян спуститься на землю. Для наших предков жизнь в этих совершенно новых условиях, несомненно, была очень тяжелой, что сильно обострило для них борьбу за существование.
Обезьяны, впоследствии оказавшиеся нашими предками, не могли спасаться от преследования путем быстрого убегания: их движения на земле были неловкими и относительно медленными. При таких условиях всякая подобная обезьяна, способная подняться на задние ноги, могла своевременно обнаружить приближение хищника и так или иначе укрыться от него. Жизнь на деревьях уже выработала у наших предков способность держаться более или менее вертикально, поэтому в попытке встать на задние ноги не было ничего принципиально нового. Вертикальное положение тела освободило руки и они могли быть использованы для борьбы с нападающим врагом наряду с зубами. Таким образом, в новой обстановке вертикальное положение тела и использование для передвижения, только ног давали обезьянам некоторое преимущество в борьбе за жизнь.
Жестокий и длительный естественный отбор в новых условиях неминуемо должен был привести к появлению таких существ, которые оказались способными не только легко подниматься на задние ноги, но и уверенно и относительно быстро передвигаться на них.
Однако этот новый способ передвижения наших предков в дальнейшем не был развит ими до высокой степени совершенства. Эволюция их ног не шла по пути развития способности к быстрому бегу, как это случилось у многих жителей равнин, например у лошади, страуса и т. д. Это свидетельствует о том, что не стремительное бегство от хищников давало возможность ндшим предкам сохранить свое существование.
При относительно слабо развитой зубной системе наших далеких предков их руки, освобожденные от функции передвижения, оказались по существу единственными и наиболее надежными органами защиты от врагов. Но и они сами по себе не были приспособлены к такой роли: на пальцах наших предков отсутствовали крепкие острые когти, которые могли бы •помочь им в борьбе с нападающими. Однако их руки в то же время обладали и некоторыми очень ценными особенностями: они были сильными, подвижными и были способны крепко схватывать. Все это было результатом приспособления к жизни на деревьях в прошлом.
Поэтому нет ничего удивительного в том, что обломок сука в руках наших предков мог превратиться в орудие защиты. Взять в минуту опасности 202
палку для нашего предка было так же естественно, как схватиться за ветвь дерева, на котором он прежде искал спасения. Вначале это могло быть чисто инстинктивным движением, но потом к нему присоединился и разум.
Защита от нападения при помощи такого примитивного орудия, как тяжелая дубина, для наших предков оказалась, если не единственным, то во всяком случае достаточно надежным способом обороны. Однако этим способом могли воспользоваться только такие обезьяны, которые были В состоянии понять и оценить все его значение. При этом та обезьяна, интеллект которой позволял ей быстрее и лучше использовать новый способ защиты, получала в борьбе за индивидуальную жизнь, вероятно, не малые преимущества по сравнению с менее одаренными представителями того же вида.
Все это неминуемо должно было привести к появлению существа, передвигающегося на задних конечностях и защищающегося при помощи тяжелой дубины. Это существо, разумеется, еще не было человеком, но оно уже не было и человекообразной обезьяной. Суровые условия, в которых оно жило, обусловливали то, что в процессе жестокого и непрерывного естественного отбора сохранялись только те, которые отличались наиболее развитым мозгом. И это обстоятельство было основным фактором развития интеллекта.
Прогрессивное развитие мозга давало нашим предкам возможность все более и более целесообразно применять даже и такое примитивное орудие защиты, как тяжелый обломок сука. В то же время борьба за жизнь настойчиво требовала от наших предков улучшения искусственных способов защиты, а естественный отбор, приводящий к общему повышению интеллекта, делал возможным усовершенствование орудий и техники их применения.
С того момента как наши предки впервые воспользовались обломком сука в качестве орудия защиты, этот сук превратился для них и в орудие нападения. Палка дала им возможность убивать относительно небольших животных, чаще всего, вероятно, грызунов и насекомоядных, которых они подстерегали у их нор.
Нелегкое добывание пищи, по всей вероятности, в свою очередь явилось мощным фактором отбора. Не погибал от голода только тот, кто имел достаточный интеллект, чтобы суметь выследить и убить небольшое животное. Таким образом, не только необходимость защиты от врагов, но и трудности добывания пищи должны были обусловливать прогрессивную эволюцию головного мозга наших предков.
Нет ничего удивительного в том, что при таких условиях они в конце концов заменили легкую палку тяжелым камнем, которым и стали убивать небольших животных около их нор.
Искусственное соединение палки с камнем, несомненно, было огромным шагом вперед в развитии примитивной техники наших предков. Такое соединение дало в их руки сильное орудие, при помощи которого можно было наносить очень серьезные повреждения всякому врагу и смелее нападать на более крупную добычу.
Мне кажется, что только тогда, когда наши предки оказались способными изготовить такое орудие, как примитивный каменный топор, они получили право называться примитивными людьми.
Применение искусственных орудий для защиты и нападения было по существу обязательным условием, при котором наши предки могли сохранить свое существование. Без этого их судьба была бы такой же, как и судьба всех тех обезьян, которые прежде жили в том же ареале, именно — полное вымирание.
203
Наши обезьянообразные предки, бывшие стадными животными во время своей жизни в лесах, остались стадными и в безлесной равнине. По всей вероятности, они в течение долгого времени были кочующими и постояннб меняли места своего обитания в поисках более обильной пищи.
К моменту изготовления примитивных искусственных орудий они прошли в своем эволюционном развитии ряд настолько важных этапов, что уже не смогли бы вернуться к жизни на деревьях, если бы вновь оказались в местности, сплошь покрытой лесом.
Полное отсутствие ископаемых форм человека в Южной и Северной Америке указывает на то, что оба эти континента не были родиной человека.
Родиной человека не была и Австралия. На этом материке на протяжении всего кайнозоя до наших дней жили и развивались только однопроходные и сумчатые млекопитающие; обезьяны там не появлялись.
Несмотря на то, что в Африке найдено несколько форм ископаемых обезьян, а в настоящее время там обитает такая высокоорганизованная обезьяна, как шимпанзе, мы все же не можем считать, что первые люди возникли на этом континенте. Климатические особенности экваториальной Африки не могли создать такие условия, которые привели бы к превращению обезьян в людей. Для этого требовалась несравненно более суровая окружающая обстановка.
Родиной человека, несомненно, был Евразиатский материк. Здесь* в Центральной Азии, по всей вероятности там, где теперь находится пустыня Гоби, возникла та обстановка, которая обусловила появление первых обе-зьяно-людей.
В конце мезозоя, в меловом периоде, в Центральной Азии обитали многочисленные травоядные и хищные динозавры и маленькие примитивные млекопитающие. Климат здесь в это время был, несомненно, теплым и вся обширная область была покрыта богатой растительностью. Но в кайнозое* начиная с олигоцена, обстановка изменилась и здесь постепенно стали образовываться травянистые равнины в результате отступания тропической флоры на юг. Эти данные палеоботаники дают нам право думать, что именно Центральная Азия была родиной человека.
Здесь впервые появились примитивные обезьяно-люди и отсюда они стали путем медленной миграции расселяться на юго-восток, юг, юго-запад и запад.
Ископаемые остатки примитивных форм этой первой миграционной волны в настоящее время обнаружены на Малайском архипелаге и в Европе, это — Pithecanthropus erectus Dubois и так называемый гейдельбергский человек.
Первые остатки Pithecanthropus были найдены на острове Ява. Здесь на берегу реки Бенгаван (Bengavan, или Solo) при раскопках в 1891—1894 гг. голландским врачом Дюбуа (Dubois) были обнаружены на глубине 15 м черепная крыша, два коренных зуба и левое бедро.
Черепная крыша (рис. 232, А) отличалась сильно покатым лбом, хорошо развитыми надглазничными дугами и заостренным затылком; на ней отсутствовал сагиттальный гребень, характерный для самцов таких человекообразных обезьян, как оранг и горилла.
Коренные зубы по своему строению были похожи на соответствующие зубы человека.
Бедро было очень длинным (45,5 см) и прямым. На основании детального изучения этой кости Дюбуа пришел к заключению, что «бедро Pithecanthropus выполняло ту же механическую работу, что и бедро человека». Следовательно, это ископаемое существо имело, подобно человеку, прямое положение тела и такую же походку на двух ногах.
Таким образом, изучение черепной крыши и бедра показало, что они
204
Наши обезьянообразные предки, бывшие стадными животными во время своей жизни в лесах, остались стадными и в безлесной равнине. По всей вероятности, они в течение долгого времени были кочующими и постояннд меняли места своего обитания в поисках более обильной пищи.
К моменту изготовления примитивных искусственных орудий они прошли в своем эволюционном развитии ряд настолько важных этапов, что уже не смогли бы вернуться к жизни на деревьях, если бы вновь оказались в местности, сплошь покрытой лесом.
Полное отсутствие ископаемых форм человека в Южной и Северной Америке указывает на то, что оба эти континента не были родиной человека.
Родиной человека не была и Австралия. На этом материке на протяжении всего кайнозоя до наших дней жили и развивались только однопроходные и сумчатые млекопитающие; обезьяны там не появлялись.
Несмотря на то, что в Африке найдено несколько форм ископаемых обезьян, а в настоящее время там обитает такая высокоорганизованная обезьяна, как шимпанзе, мы все же не можем считать, что первые люди возникли на этом континенте. Климатические особенности экваториальной Африки не могли создать такие условия, которые привели бы к превращению обезьян в людей. Для этого требовалась несравненно более суровая окружающая обстановка.
Родиной человека, несомненно, был Евразиатский материк. Здесь, в Центральной Азии, по всей вероятности там, где теперь находится пустыня Гоби, возникла та обстановка, которая обусловила появление первых обе-зьяно-людей.
В конце мезозоя, в меловом периоде, в Центральной Азии обитали многочисленные травоядные и хищные динозавры и маленькие примитивные млекопитающие. Климат здесь в это время был, несомненно, теплым и вся обширная область была покрыта богатой растительностью. Но в кайнозое, начиная с олигоцена, обстановка изменилась и здесь постепенно стали образовываться травянистые равнины в результате отступания тропической флоры на юг. Эти данные палеоботаники дают нам право думать, что именно Центральная Азия была родиной человека.
Здесь впервые появились примитивные обезьяно-люди и отсюда они стали путем медленной миграции расселяться на юго-восток, юг, юго-запад и запад.
Ископаемые остатки примитивных форм этой первой миграционной волны в настоящее время обнаружены на Малайском архипелаге и в Европе, это — Pithecanthropus erectus Dubois и так называемый гейдельбергский человек.
Первые остатки Pithecanthropus были найдены на острове Ява. Здесь на берегу реки Бенгаван (Bengavan, или Solo) при раскопках в 1891—1894 гг. голландским врачом Дюбуа (Dubois) были обнаружены на глубине 15 м черепная крыша, два коренных зуба и левое бедро.
Черепная крыша (рис. 232, А) отличалась сильно покатым лбом, хорошо развитыми надглазничными дугами и заостренным затылком; на ней отсутствовал сагиттальный гребень, характерный для самцов таких человекообразных обезьян, как оранг и горилла.
Коренные зубы по своему строению были похожи на соответствующие зубы человека.
Бедро было очень длинным (45,5 см) и прямым. На основании детального изучения этой кости Дюбуа пришел к заключению, что «бедро Pithecanthropus выполняло ту же механическую работу, что и бедро человека». Следовательно, это ископаемое существо имело, подобно человеку, прямое положение тела и такую же походку на двух ногах.
Таким образом, изучение черепной крыши и бедра показало, что они
204
не могли принадлежать какой-нибудь ископаемой обезьяне. И Дюбуа писал, что ^Pithecanthropus есть переходная форма между человеком и человекоподобными обезьянами. . . Pithecanthropus — предок человека» (Дюбуа, 1896).
Как известно, открытие Дюбуа в свое время вызвало ожесточенные
споры.
Одни морфологи присоединились к мнению Дюбуа и рассматривали Pithecanthropus erectus как обезьяно-человека.
указывали на некоторое сходство череп-
А
Рис. 232.
крыша Pithecanthropus erectus В—череп Pithecanthropus (Ке-
— черепная (Дюбуа, 1895);
нигсвальд — Konigswald, 1936—1938).
ли,
Другие считали, что найденные Кости принадлежат гигантскому гиббону. Сторонники такого взгляда ной крыши Pithecanthropus с соответствующей частью черепа гиббона.
Наконец, третьи полага-что найденные на Яве
кости не принадлежат одному существу и что черепная крыша принадлежит обезьяне, а бедро — человеку. В качестве доказательства того, что бедро человеческое, они ссылались на интересное патологическое образование, которое имелось на нем. Дело в том, что на найденном Дюбуа бедре оказался большой патологический экзостоз, располагающийся в верхней трети на внутренней стороне его (рис. 233, В); в экзостозе имелись отверстия, сквозь которые проходили кровеносные сосуды. Судя по характеру поверхности экзостоза, вызвавший его патологический процесс полностью закончился и уже не мешал нормальной функции бедра. По мнению некоторых морфологов, патологический процесс, приведший к образованию такого большого костного нароста, должен был быть настолько
длительным и болезненным, что выздоровление больного могло наступить только при условии тщательного ухода со стороны других. . . людей. Такой патологический процесс, утверждали они, не мог благополучно закончиться не только у обезьяны, но даже и у обезьяно-человека; такой исход болезни был возможен только для человека.
Я лично не могу согласиться с такими утверждениями. Экзостоз на бедре Pithecanthropus, несомненно, является остеомой, а не результатом воспалительного процесса, вызванного глубоким ранением, как думают некоторые. В моем распоряжении имеется бедро современного человека с экзостозом, расположенным точно на том же месте, что и у Pithecanthropus (рис. 233, Л). История болезни этого человека, к сожалению, неизвестна; бедро попало в мои руки из учебной остеологической коллекции одного из
205
анатомических институтов. Но мне кажется, что написанной врачом история болезни, связанной с экзостозом на бедре, у этого человека вообще не было. Я Думаю так потому, что мне приходилось обнаруживать подобные экзостозы на бедрах при рентгеноскопических обследованиях больных. Несмотря на очень большие размеры костных разращений, пациенты не жаловались на них и даже не подозревали об их присутствии. Все это дает мне право утверждать, что медленно увеличивающаяся остеома на бедре Pithecanthropus никогда не вызывала у него болезненных ощущений и не мешала нормаль-
ной функции его левой ноги.
В 1907—1908 гг. вдова немецкого зоолога Зелёнки (Selenka) организовала новую научную экспедицию на остров Ява и провела обширные раскопки в том месте, где Дюбуа нашел кости Pithecanthropus. Эта экспедиция за 18 месяцев работы просмотрела 10 000 м3 земли, но никаких новых остатков обезьяно-че-ловека не обнаружила. Несмотря на такую неудачу, раскопки Зеленки дали много очень ценного материала. В тех слоях, в которых Дюбуа нашел Pithecanthropus, были обнаружены многочисленные кости современных ему животных. Изучение их показало, что они не отличаются от костей ископаемых животных, найденных у подножий Гималаев. Это, несомненно, свидетельствует не только о том, что острова1 Малайского архипелага когда-то соединялись с Азиатским материком, но и о том, что фауна Азии в свое время мигрировала в юго-восточном направлении.
Все споры о значении находки Е. Дюбуа потеряли всякий смысл после того, как в 1936—1939 гг. Кенигсваль-ду (G. Konigswald) удалось найти на острове Ява четыре неполных мозговых черепа Pithecanthropus и фрагменты его верхней и нижней челюсти (рис. 232, В).
Судя по этим остаткам, можно не
Рис. 233.
А — бедро человека; В — бедро Pithecanthropus (Дюбуа, 1895).
сомневаться, что яванский обезьяно-человек существовал и что он имел относительно небольшой мозговой отдел черепа, низкий черепной свод, покатый лоб, хорошо развитые надглазничные дуги, заостренный затылок; его челюстной аппарат был слабее, чем у обезьян, но значительно более сильным, чем у человека; коронки клыков не имели такой конической формы, как соответствующие зубы всех обезьян; подбородочная часть нижней челюсти была тупой.
В тех же слоях, в которых были найдены новые остатки Pithecanthropus, Кенигсвальд обнаружил кости следующих животных: оранг-утана (Simia satyrus), гиббона (Hylobates leuciscus и Symphalangussyndactylus), ма-’ кака (Macacus irus), носорога (Rhinocerus sondaicus), тапира (Tapirus indicus), свиньи (Sus brachygnathus), бегемота (Hippopotamus antiquus), оленя (Cercus zwaani), антилопы (Antilope saatensis), быка (Bos banteng), слона (Ele-phas и Stegodon trigonicephalus), выдры (Lutra), медведя (Ursus malaynaus),
206
гиены {Hyaena bathygnatha), обычного тигра {Fells tigris) и саблезубого тигра (Epimachairodus zwierzyckii).
Обычный и саблезубый тигры, входившие в состав фауны, окружавшей обезьяно-человека, несомненно, являлись наиболее опасными врагами его. Однако эти хищники, как правило, преследовали многочисленных копытных, животных, и, вероятно, не часто нападали на обезьяно-человека. Яванский Pithecanthropus мог относительно благополучно и долго существовать В условиях теплого климата и обилия легко добываемой пищи. Не исключена возможность, что он еще сохранялся на островах Малайского архипелага даже тогда, когда туда пришла новая волна более совершенных мигрантов. На это указывает, в частности, интересная находка, сделанная в 1908 г. экспедицией Зеленки: при раскопках в 3,5 км к западу от Триниля (место раскопок Дюбуа) был обнаружен коренной зуб ниЖней челюсти человека.
•Таким образом, морфологическая примитивность яванского обезьяночеловека, по-видимому, не соответствует его небольшой геологической древности (начало плейстоцена).
Pithecanthropus erectus Dubois, мигрировавший из Центральной Азии на острова Малайского архипелага, вероятно, является близким родственником того существа, нижняя Челюсть которого была найдена около Гейдельберга в Германии.
Эта находка была сделана 21 октября 1907 г. Шэтенсаком (Schoetensack). В песках на глубине 24 м была обнаружена хорошо сохранившаяся очень мощно развитая нижняя челюсть с тупым обезьяньим подбородком, но с человеческими зубами (рис. 234, 235).
Такая челюсть могла принадлежать только существу, очень похожему на Pithecanthropus. Возможно, что в данном случае мы имеем дело-с европейской формой обезьяно-человека. Он пришел в Европу также из Центральной Азии, являясь представителем тех форм обезьяно-людей, которые мигрировали на запад.
Древность Гейдельбергской находки равна приблизительно 500 000 лет.
В состав фауны, среди которой жил гейдельбергский обезьяно-чело-век, входили: древний слон {Elephas antiquus), этрусский носорог {Rhino-cerus ethruscus), лошадь {Equus masbaehensis), дикий кабан {Sus scrofa), лось {A Ices), олень {Cervus elaphus), первобытный бык {Bos pritnigenius), зубр {Bison priscus), медведи {Ursus deningeri и Ursus arvernensis), лев {Lea leo), волк {Canis lupus).
Саблезубый тигр в тех слоях, в которых была найдена гейдельбергская нижняя челюсть, не был обнаружен, но он входил в состав синхронной фауны, обнаруженной в древних пещерах Пиренеев.
Дальнейшее эволюционное развитие человекообразных форм в Центральной Азии через некоторое время привело к появлению того существа, остатки которого были обнаружены в 1927—1938 гг. около Пекина. Эта ископаемая форма получила название Sinanthropus pekinensis.
Были найдены различные фрагменты больше чем десяти черепов Sinanthropus. Это дало возможность Вайденрайху (F. Weidenrejch) реконструировать весь череп этого представителя древних людей (рис. 236).. Мозговой череп у синантропа был развит сильнее, а лицевой — слабее, чем у питекантропа. Синантроп еще обладал очень низким лбом, невысоким черепным сводом, хорошо развитыми надглазничными дугами, выступающими вперед челюстями, тупым подбородком и заостренным затылком. Однако его processus mastoideus был развит сильнее, чем у Pithecanthropus, что указывает на более вертикальное положение тела.
Вместе с фрагментами скелетов Sinanthropus удалось обнаружить очень
207
примитивные каменные орудия, остатки костров, разбитые и обожженные
кости различных млекопитающих, чашки
Рис. 234. Гейдельбергская нижняя челюсть (1907).
Рис. 235.
А — нижняя челюсть современного человека;
В—гейдельбергская нижняя челюсть; С — нижняя челюсть шимпанзе.
нз черепов животных.
Таким образом, можно уверенно сказать, что синантроп уже не только изготовлял примитивные каменные орудия, при помощи которых охотился на различных животных, но и умел добывать и поддерживать огонь.
Не один раз высказывалось предположение, что древние люди впервые познакомились с огнем, наблюдая его в тех случаях, когда молния зажигала лес. Это возможно, но мне кажется более вероятным, что открытие огня было связано с обработкой примитивных каменных орудий. При отбивании краев от куска кремня при изготовлении из него грубого топора наши предки видели, что при каждом ударе камня о камень отскакивают искры. Но от этого постоянного и привычного наблюдения до использования огня для бытовых нужд было еще очень далеко. Примитивные люди увидели настоящий огонь только в тот момент, когда при обработке кремня от случайно попавшей искры загоралась сухая трава. Возможно, что наблюдение над огнем в таких случаях было только забавой для любопытного и уже достаточно интеллектуального древнего человека. Наши предки, вероятно, не раз делали попытки взять огонь, как всякий другой незнакомый предмет, в руки, но всякий раз с удивлением и испугом убеждались в очень странных и непонятных его свойствах. Нет ничего удивительного в том, что очень скоро в этих «экспериментах» рука была заменена суком дерева. Это было не менее любопытно, но более безопасно. Подобные
занятия, естественно, не могли не привести к следующим очень важным открытиям. Оказалось, что сук, всунутый в пламя, сам покрывается огнем и что огонь при помощи горящего сука может быть перенесен в другое место — на другой сук или даже на кучу сухих ветвей, и таким образом размножен в неограниченном ко
208
личестве. Возможно, что человек довольно скоро по своему личному печальному опыту понял, что допускать «размножение» огня в очень больших количествах — дело опасное, но что огонь, «размноженный» в умеренном количестве, дает приятное тепло и не причиняет болезненных ожогов, если с ним обращаться осторожно. Словом, примитивные люди в конце концов должны были понять, что огонь при умелом с ним обращении может быть очень полезным.
Дальнейшее постепенное и невольное изучение свойств огня неминуемо должно было привести к ряду других открытий. Например, к такому, что не всякий предмет, положенный в огонь, легко загорается. Свежая кость
Рис. 236. Череп Sinanthropus pekinensis.
животного с остатками мяса на ней не горит, но мясо нагревается и изменяет свой запах.
Вероятно, несколько позже наши предки заметили, что подгоревшее мясо приобретает иной вкус, легче отделяется от кости и с меньшим трудом пережевывается.
В результате всех этих ценнейших наблюдений перед пещерами примитивных людей в конце концов запылали костры, на которых люди стали поджаривать мясо убитых ими животных. . .
Следующей после синантропа фазой развития человека было появление неандертальца. Человек неандертальского типа возник, по всей вероятности, также в Центральной Азии и оттуда медленно мигрировал в Европу, Африку и на острова Малайского архипелага. Остатки его скелетов в настоящее время найдены в Туркестане, Крыму, Германии, Бельгии, Франции, Италии, на Гибралтарской скале, в Палестине, в районе озера Танганайка, в Южной Африке и на острове Ява. Это, по-видимому, была очень мощная миграционная волна людей, способных жить в довольно разнообразных условиях.
Первая находка черепа неандертальца была сделана в 1848 г. лейтенантом Флинтом (Flint) на северной стороне Гибралтарской скалы. Здесь в пещере был найден хорошо сохранившийся череп без нижней челюсти. Он отличался небольшими размерами мозгового отдела, низким лбом и за
14 А. П. Быстров 209
остренным затылком; судя по относительно слабому развитию надглазничных дуг, череп принадлежал женщине (рис. 237, В).
Гибралтарский череп, попавший в частную коллекцию, в течение 20 лет оставался не изученным и не описанным; только в 1868 г. он был передан Баском (Busk) в один из лондонских музеев.
За 12 лет до этого, в 1856 г., в Германии, в долине Неандер (Neanderthal), была сделана вторая находка костей первобытного человека. Здесь в древней пещере рабочие нашли, по всей вероятности, полный скелет, но
разрушили его; сохранилась только черепная крыша (рис. 237, Л) и несколько обломков костей посткраниального скелета.
Все эти кости попали в руки д-ра Фульрота (Fuhlrott), который в 1857 г. сделал о них доклад на съезде натуралистов в Бонне. Фульрот считал, что найденные в долине Неандера кости принадлежат первобытному человеку, когда-то обитавшему в Европе. Мнение Фульрота не было поддержано участниками съезда. «Слушатели,— говорит Фульрот,— конечно, удивлялись и делали большие глаза при виде того, что им показывали, но в то же время пожимали плечами, слушая то, что им говорили, и во всем собрании не нашлось человека, который встретил бы мои сооб-
Рис. 237. ражения относительно древности
А — черепная крыша из Неандерталь (1856); моей находки одобряющим сло-В — череп с Гибралтарской скалы (1848). вом».
Несмотря на то, что неандертальская черепная крыша обладала ясными признаками черепа примитивного человека: низким сводом, покатым лбом и хорошо развитыми надглазничными дугами, — о ее принадлежности были высказаны самые разнообразные мнения.
Р. Вагнер (R. Wagner) считал, что она является частью черепа древнего голландца; Прунер-Бей (Pruner-Вау) находил в ней признаки кельта; Хами (Нашу) утверждал, что он встречал на улицах Брюсселя людей, черепа которых не отличались от неандертальского; Мейер (Meyer), указывая на то, что над правым надглазничным краем черепной крыши имеется косой шрам, а на правой теменной кости — небольшая яма, высказал предположение, что найденный обломок черепа принадлежит русскому солдату из монголов, который при преследовании войск Наполеона в 1814 г. был ранен в голову, затем заполз в пещеру и там умер . . . Р. Вирхов (R. Virchow) нашел в неандертальском черепе признаки рахита и утверждал, что все его особенности вызваны длительным болезненным процессом. Мнение такого авторитета, как Р. Вирхов, было решающим, поэтому неандертальская находка надолго потеряла всякий интерес для морфологов. Ее значение вновь было оценено только после того, как в 1887 г. в Бельгии в пещере около Спи (Spy) были найдены два черепа и неполные скелеты
210
первобытного человека того же типа. Черепа из Спи имели такие же покатые лбы, низкие черепные своды и сильно развитые надглазничные дуги; подбородочная часть нижней челюсти была тупой (рис. 238).
Рис. 238. Череп неандертальца нз Спи (Spy, 1887).
После находок в Бельгии первобытный человек получил название Homo neanderthalensis — «Неандертальский человек» — по месту находки 1856 г.
Рис. 239. Череп неандертальца
из La Chapelle-aux-Saints (1908).
Если бы череп, найденный в 1848 г. на Гибралтарской скале, был в свое время изучен и описан, то мы теперь, вероятно, говорили бы не о неандертальском, а о гибралтарском человеке.
Череп неандертальца наилучшей сохранности был найден 3 августа 1908 г. в Ла Шапель-о-Сэн (La Chapelle-aux-Saints) во Франции (рис. 239).
14*
211.
Мозговая часть черепа у всех неандертальцев была значительно больше, чем у питекантропа и синантропа; лоб был развит сильнее, но оставался еще покатым; большие надглазничные дуги были хорошо выражены; затылочный отдел черепа был заостренным. Вся лицевая часть была немного выдвинута вперед; на подбородочной части нижней челюсти выступ отсутствовал, вследствие чего подбородок был таким же тупым, как у синантропа.
Вейнерт (Weinert, 1930,1932) на основании изучения известных в настоящее время находок сделал удачную попытку восстановить полный скелет неандертальца (рис. 240). Это был человек небольшого роста (155—165 см). Его позвоночный столб еще не имел хорошо выраженных изгибов, которые так характерны для современных людей. Ребра неандертальцев были треугольными в поперечном сечении; лопатки — короткими и широкими; кости рук — массивными; кисти — грубо построенными; ноги — относительно короткими.
Неандертальцы появились в Европе около 100—120 тысяч лет тому назад. Они жили в естественных пещерах, владели огнем и умели изготовлять примитивные каменные орудия. Одновременно с ними жили бизон, дикая лошадь, олень, лось, шерстистый носорог, мамонт, волк, пещерный медведь. Такие животные, как лошадь, олень, лось и бизон, были для неандертальцев предметом охоты. Что же касается пещерного медведя, то с этим опасным зверем неандертальцам приходилось вести постоянную борьбу, отвоевывая у него пещеры.
Пещерные медведи (Ursus spelaeus) были очень большими и сильными животными (рис. 241); старые самцы, поднявшись на задние ноги, достигали 2,5 м в высоту. Несмотря на это, неандерталь-
Рис. 240, Скелет неандертальца цы все же осмеливались нападать на них. (Вейнерт — Weinert, 1930). Изучение многих пещер в Европе
показало, что неандертальцы не только старались изгнать из них медведей, но даже систематически охотились на них.
При раскопках некоторых пещер были обнаружены скопления костей больше чем 1000 убитых медведей. В одной небольшой пещере (Kummet-loch, Франконская Юра) было найдено огромное количество костей медведей. При этом все трубчатые кости оказались расколотыми для извлечения мозга; они были перемешаны с большим количеством разбитых черепов, нижних челюстей и зубов, среди которых, однако, не было обнаружено ни позвонков, ни ребер, ни тазовых костей.
Совершенно очевидно, что такой отбор частей добычи мог сделать только человек. Он приносил в пещеру только отделенные от убитых медведей конечности и головы вместе со шкурой, остальное оставляя на месте охоты.
212
Охота на такого сильного зверя, как пещерный медведь, несомненно, являлась очень опасной, и плохо вооруженные неандертальцы, вероятно, не во всех случаях оказывались победителями. То, что они нападали на медведя не в одиночку, а небольшими группами, разумеется, облёгчало задачу. Однако и при этих условиях печальные для неандертальцев исходы охоты, по всей вероятности, были нередкими.
Постепенное понижение температуры в Евразии, вызванное началом ледникового периода, сделало борьбу за жизнь для неандертальцев очень тяжелой.
Рис. 241.
А —череп неандертальца; В —череп бурого медведя (Ursus arctos); С — череп пещерного медведя (Ursus spelaeus).
Ютясь в сырых пещерах, одеваясь в шкуры убитых на охоте животных и согреваясь у костров, они с трудом поддерживали свое существование.
Возможно, что именно наступившее похолодание климата было причиной вынужденной миграции неандертальцев на юг.
Постепенно расширяя область своего обитания, неандертальцы- через Палестину проникли в Африку и дошли до южной ее части.
В 1931 г. в Палестине было найдено несколько скелетов неандертальцев (рис. 242), которые, однако, отличались от неандертальцев Европы тем, что имели более высокий черепной свод и менее тупой подбородок; их ноги были длиннее и рост выше. Эти люди в морфологическом отношении были развиты выше, чем типичные неандертальцы Европы.
Плохо сохранившиеся черепа неандертальцев были обнаружены и в районе озера Танганайка (1935).
В 1921 г. в Южной Африке (Родезия) на дне пещеры, уходящей под скалы Брокен-Хилл (Broken-Hill) на глубину 27 м, был найден хорошо сохранившийся череп человека (рис. 243). Родезианский череп поражает необычайно слабым развитием лба и очень большими надглазничными дугами. Человек, которому принадлежал этот череп, в морфологическом отношении был развит, несомненно, ниже, чем европейские неандертальцы. Это становится совершенно очевидным, если сравнить череп, найденный
213
в Родезии, с черепом неандертальца, например, из Ла Шапель-о-Сэн (рис. 239, 243, 244). Однако тот факт, что в той пещере, где был найден
Рис. 242. Обломок черепа неандертальца из Галилеи.
Рис. 243. Череп неандертальца из Родезии (Rhodesia Broken-Hill, 1921).
родезианский череп, было обнаружено скопление большого количества костей современных африканских животных (слон, бегемот, носорог, зебра,
Рис. 244.-Черепа неандертальцев. А —из La Chapelle-aux-Saints; В — из Rhodesia (Broken-Hill).
антилопа, лев, леопард), свидетельствует о небольшой древности южноафриканского человека. Этот первобытный человек жил в Родезии, по всей
214
вероятности, относительно недавно. Он, по-видимому, являлся представителем той первой миграционной волны неандертальцев, которая из Евразии докатилась до Южной Африки и здесь сохранялась в течение довольно продолжительного времени. Родезианский человек, сохраняя многие признаки примитивности, по-видимому, еще продолжал существовать на юге Африки в то время, как на севере этого материка уже появились более совершенные представители неандертальцев. Как указывалось выше, неандертальцы ближайшего к Африке участка Евразии — Палестины^— были^ развиты
Рис. 245. Черепа неандертальцев с острова Ява (Кеиигсвальд, 1931).
даже выше, чем типичные неандертальцы Европы. Они, по всей вероятности, были представителями последней миграционной волны неандертальцев, идущей из Евразии в Африку.
Кроме Африки, азиатские неандертальцы мигрировали и по направлению к Австралии, однако не достигли этого материка. Они дошли только до островов Малайского архипелага. Здесь на острове Ява в 1931 г. было найдено одиннадцать их черепов без лицевых частей (рис. 245). На затылочной кости у яванских неандертальцев имелись сильно выраженные поперечные гребни, служившие местом прикрепления шейных мышц. Такой же гребень можно видеть и на затылке черепа из Родезии. Общее сходство мозгового черепа родезианского неандертальца с соответствующей частью черепа неандертальца с острова Ява свидетельствует о том, что эти люди принадлежат к одной фазе развития человека. По-видимому, миграционная волна примитивных неандертальцев Евразии пошла по крайней мере по двум направлениям: по направлению к Африке и по направлению к Австралии. Проникнув в Африку, она дошла до южной ее части. Что же касается Австра
215
лии, то этого материка примитивным неандертальцам достигнуть не удалось; они задержались на островах Малайского архипелага. Возможно, что здесь они сохранялись так же долго, как родезианский человек на юге Африки.
Неандертальцы Центральной Азии не смогли достигнуть Северной Америки, так как путь из Сибири на Аляску проходил слишком далеко на севере. Очень суровый климат этих областей сделал Америку для неандертальцев недоступной. Поэтому совершенно естественно, что все попытки ученых — «патриотов» Северной и Южной Америки — найти на своей родине какие-нибудь остатки первобытного человека не привели ни к чему. На американском континенте неандертальцев никогда не было.
Рнс. 246. Череп ископаемого человека из Combe-Capelle (1909).
В процессе своей дальнейшей эволюции неандертальцы Евразии оказались предками более высоко организованных людей. Около 50 000 лет тому назад здесь появился новый человек. Он был выше ростом, чем неандертальцы; его скелет был менее грубым. Череп этого человека сильно отличался от черепа типичного неандертальца: лоб был хорошо развит, надглазничные дуги были выражены слабо, на подбородочной части нижней челюсти уже намечался выступ. Череп человека этого типа был найден во Франции (Combe-Capelle) в 1909 г. (рис. 246).
Вслед за этим человеком в Евразии появился так называемый кроманьонец, получивший свое название по месту первой находки его черепа (1868 г. — Cro-Magnon во Франции).
Кро-маньонцы по существу ничем не отличались от современных людей. Они были высокого роста, сильные и стройные. Их мозговой череп был развит так же, как у современного человека; надглазничные дуги были не больше, чем у людей нашего времени; лицевой череп был развит слабо; подбородочный выступ был выражен очень хорошо (рис. 247).
Таким образом, можно сказать, что с момента появления кро-маньонца в Евразии впервые возник человек современного типа — Homo sapiens fossilis (рис. 248).
216
Рис. 247. Череп из Cro-Magnon (1868).
Рис. 248. Черепа кро-маньоннев из Obercassel (1914).
Д1О ОИШ>И пл>л>л>г»оо'"_ ои2 110 ZTer’®'
ЛИТЕРАТУРА
и мен к оПП. П. Первобытное общество. 1938.
el О. Die Stellung des Menschen in Rhamen der Wirbeltiere. Jena, 1931.
ick D. Fossil man in China. Geological Memoirs. Ser. A. N 11, Peiping, 1933.
ul M. Les hommes fossiles, Paris, 1923.
b о i s E. Pithecanthropus erectus, eine Stammform des Menschen. Anat. Anz. Bd. 12, 1896.
egory W. K. Our face from fish to man. N.-York, 1929.
user O. Urgeschichte. Jena, 1925.
enigswald G. Neue Pithecanthropus — Funde 1936—1938. Ein Beitrag zur Kenntnis der Praehominiden. Wetenschappelijka Mededeelingen, Bd. 28, Batavia, 1940.
born H. F. Men of the old stone age. OSA, 1925.
mer A. S. Vertebrate paleontology. USA, 1945.
mer A. S. Man and the vertebrates. USA, 1947.
i d en rei ch F. Der Schadelfund von Weimar—Eringsdorf. Jena, 1928.
idenreich F. Apes, giants and man. USA, 1946.
inert H. Menschen der Vorzeit. Stuttgart, 1930.
inert H. Ursprung der Menschenheit. Stuttgart, 1932.
rth E. Der fossile Mensch. Berlin, 1921.
ЧАСТЬ ВТОРАЯ
НАСТОЯЩЕЕ
С абстрактно-философской точки зрения настоящее — это мгновение, не имеющее длительности во времени и отделяющее прошлое от будущего; настоящее — это передвигающаяся граница между прошлым и будущим; настоящее — это никогда не останавливающийся процесс превращения будущего в прошлое. Таким образом, настоящее — не период, а только крайне короткая фаза, длительность которой по существу равна нулю; словом, настоящее как период не существует.
Такое представление о настоящем не может быть принято нами, так как в таком настоящем изучать человека мы не можем.
С точки зрения историка настоящее является периодом, продолжительность которого не может быть точно определена. Но историк не считает настоящее мгновением и некоторую группу явлений рассматривает как события своей современности.
Относительно длительное, хотя и совершенно неопределенное настоящее историка также неприемлемо для нас, так как люди исторического настоящего в морфологическом отношении ничем не отличаются от людей исторического и даже доисторического прошлого.
С точки зрения морфолога «настоящим» человека следует считать несравненно более продолжительный период, именно такой период, в течение которого в организме человека не появилось ни одного нового, важного и общего для всех людей признака, такого признака, который позволил бы разделить человеческий род на предков и потомков, отличающихся друг от друга теми или иными анатомическими особенностями. Такое «настоящее» человека началось с того момента, когда на Земле оформился Homo sapiens, а это, как известно, случилось при возникновении кро-маньонца, т. е. около 50 000 лет тому назад, и . . . будет продолжаться до тех пор, пока не появится такой потомок Homo sapiens, который будет признан систематиками новым видом человека и будет назван, скажем, Homo sapientissimus. Только в этот момент Homo sapiens уйдет в прошлое.
Таким образом, с морфологической точки зрения мы не можем «настоящее» человека ограничить только настоящим моментом, но должны прибавить к нему часть прошлого и часть будущего. Словом, биологическое «настоящее» человека — это такой период, в течение которого существует Ното sapiens как потомок Homo primigenius и как возможный предок гипотетического Homo sapientissimus.
Такое понимание «настоящего» противоречит не только взглядам иных философов, но и представлениям историков, однако для морфологов оно оказывается единственно приемлемым.
ГЛАВА I
РАСЫ
Анатомическое строение живущих сейчас людей показывает, что все человечество образовано представителями одного рода (Ното), в состав которого входит единственный вид (sapiens).
Детальное изучение физических особенностей современного населения Земли позволило установить наличие в человечестве нескольких рас, которые, однако, не соответствуют ни расам, ни подвидам в мире животных.
Наиболее резко отличаются друг от друга представители так называемых больших рас. По цвету кожи они называются белыми, желтыми и черными, а в зависимости от ряда других признаков — европеоидами, монголоидами и негроидами. В посткраниальном скелете представителей этих рас нет ясных и бесспорных отличительных признаков, но в черепе мы находим такие особенности строения, которые позволяют обычно без труда определить его расовую принадлежность.
Череп представителя белой расы (европеоида) характеризуется прямым положением альвеолярных отростков и таким же положением передних зубов (ортогнатизм), высоко поставленными носовыми костями и относительно небольшой шириной скул (рис. 249).
Особенностями строения черепа представителя желтой расы (монголоида) являются уплощение носовых костей, уплощение лица, широкие скулы, ортогнатизм (рис. 250).
У представителей черной расы (у негроидов) мы находим в качестве отличительных признаков косое положение альвеолярных отростков и передних зубов (прогнатизм), немного уплощенные носовые кости, обычно удлиненную форму мозгового черепа — долихоцефалию (рис. 251).
В настоящее время все антропологи разделяют большие расы на ряд более мелких и говорят о существовании микрорас. Можно сказать, что чем детальнее изучается человечество и чем большее значение придается каждой отдельной морфологической особенности человека, тем больше выделяется малых рас. Так, некоторые антропологи насчитывают до 25 рас, а В. В. Бунак (1941) полагает, что «рас столько, сколько существует различных сочетаний признаков на больших территориях». При таком взгляде количество рас оказывается совершенно неопределенным и, вероятно, бесконечно большим.
При установлении расовой принадлежности какого-нибудь человека антропологи учитывают, кроме цвета кожи и строения черепа, и такие признаки, как особенности волос, лба, носа, губ, подбородка, верхнего века (эпикантус) и многое другое.
Несомненно, ни одна из этих особенностей никогда не могла дать и не давала человеку никаких преимуществ в борьбе за жизнь. Поэтому можно 220
уверенно сказать, что расовые признаки не являются результатом естественного отбора.
Расовые признаки впервые стали появляться только у кро-маньонцев и, по-видимому, не на первых этапах их развития.
Рис. 250. Череп эскимоса.
Причины, которые могли бы вызвать появление расовых особенностей, до сих пор еще не выяснены.
В настоящее время предполагается, что в процессе расообразования немалую роль играла миграция небольших групп людей в новые места обитания (Монтагю — М. F. Montagu, 1945). Эти миграционные передвижения р поисках лучших районов охоты могли привести при немногочисленности
221
древнего человечества к изоляции отдельных племен. Такая изоляция небольших групп людей могла обусловить превращение некоторых семейных особенностей в особенности, свойственные всем представителям данного племени. Это могло случиться тогда, когда племя развивалось цз одной семьи.
Таким образом, миграция и следующая за ней изоляция отдельных групп людей могли оказаться причинами закрепления тех или иных признаков. Но ни миграционные перемещения, ни достаточно длительная и полная изоляция отдельных групп людей не могли быть причинами появления даже слабо выраженных расовых особенностей.
Существование представителей различных рас в одинаковых климатических условиях (негроиды в экваториальной Африке и монголоиды в эква-
ториальной Америке) доказывает, что расовые признаки выработались независимо от особенностей среды обитания. Об этом же свидетельствует и тот факт, что одна и та же раса может занимать ареалы, резко отличающиеся по своим климатическим особенностям. Это мы видим, например, в Северной и Южной Америке, где одна и та же большая раса занимает территорию почти от северного и почти до южного полюса.
Великий творец эволюционного учения Дарвин, по-видимому, не считал естественный отбор фактором, способным создать расовые особенности человека. В 1867 г. в своем письме к А. Уоллесу Дарвин писал следующее: «Я продолжаю верить (хотя мне не удалось убедить Вас, а это для меня самый тяжелый удар), что половой отбор был главной действующей силой в образовании человеческих рас».
Однако трудно допустить, что только «мода» на темную кожу могла путем длительного полового отбора постепенно привести к возникновению негров. Так же трудно думать, что такой же половой отбор был способен обусловить образование плоского лица монголов.
Следует признать, что в настоящее время мы не знаем причин, вызвавших появление первых слабо выраженных расовых особенностей, и не понимаем, что заставило эти расовые особенности затем постепенно усилиться.
Если Евразия действительно была родиной человека, то этот материк должен был быть и той территорией, на которой впервые появился Ното sapiens. Образование достаточно многочисленного населения в Евразии
222
неминуемо должно было вызвать миграционные перемещения людей и постепенное проникновение их на соседние материки.
Первым материком, куда проникли представители Homo sapiens, вероятно, была Африка. Этот материк непосредственно соединяется и соеди-
Рис. 252. Череп древнего австралийца из Вадьяка (Wadjak) на острове Ява.
нялся в прошлом с Евразией, поэтому переход на него не требовал никаких особых средств передвижения.
Вторым материком, заселенным человеком, по всей вероятности, следует считать Австралию. Люди проникли сюда после постепенного заселения островов Малайского архипелага. Интересно, что на острове Ява в ме-
223
стечке Вадьяк (Wadjak) был найден древний череп (рис. 252) с характерными признаками черепа австралийца (рис. 253).
Значительно позднее того момента, когда Австралия была заселена людьми, человек впервые проник в Америку.
Начиная с 1492 г. и до настоящего времени на территории Северной и Южной Америки ни разу не было найдено никаких следов неандерталь
Рис. 254. Калаверасский череп (1866).
цев. Это дает нам право уверенно говорить, что неандертальцы не смогли проникнуть на эти материки.
Все остатки древнего населения Америки свидетельствуют о том, что это были люди уже не первобытного типа. Наиболее интересной и важной находкой древнего человека в Америке является так называемый калаверасский череп (рис. 254), найденный в 1866 г. в Калифорнии в одном из рудников (Calaveras), где добывалось золото. В этом черепе отсутствуют какие-либо следы примитивности; «калаверасский человек» был настоящим Homo sapiens.
В самое последнее время, применяя особый метод изучения изменений радиоактивного углерода (Libby’s radiocarbon method), американские
ученые пришли к выводу, что люди заселили западное полушарие не раньше 10 000 лет тому назад.
ГЛАВА II
ЭМБРИОГЕНЕЗ СКЕЛЕТА И ПОЗВОНОЧНАЯ ТЕОРИЯ ЧЕРЕПА
Состояние человека в настоящее время — это, несомненно, результат его прошлого. Поэтому в настоящем человек должен сохранять в организме большое количество следов своего прошлого. Мало того, он на основании гак называемого «биогенетического закона» должен в процессе своего индивидуального развития, как и все животные, повторять в сокращенном и измененном виде основные фазы своей эволюции в прошлом. И действительно, изучение эмбрионального развития человека дает большой материал, подтверждающий это положение. Этот материал очень интересен и очень важен, так как с совершенной очевидностью показывает, что человек прошел длинный эволюционный путь. Это можно видеть при изучении развития любого его органа. Но ввиду того, что, прослеживая формирование человека в Прошлом, мы говорили только о скелете, то и теперь должны ограничиться только историей формирования костей, оставляя в стороне все другие системы органов.
Окостенение хрящевых позвонков'зародыша человека начинается в начале третьего месяца и идет по направлению от головного конца позвоночного столба к копчику.
В каждом хрящевом позвонке появляются три точки окостенения: две в дуге (правая и левая) и одна в теле. При этом точка окостенения в теле появляется. несколько позже, чем точки, окостенения в дуге (рис. 255, G). У всех предков человека из класса млекопитающих, класса рептилий и класса амфибий костная дуга позвонка развивалась также из двух точек окостенения, а затем обе эти костные закладки сливались друг с другом, образуя непарную дугу.
Самостоятельное окостенение правой и левой половины дуги у всех наземных животных, не исключая и человека, по-видимому, следует рассматривать как повторение строения дуги позвонка каких-то древних рыб.
Мы не знаем строения позвонков у древних представителей бесчелюстных позвоночных (Agnatha), так как позвонки у них не окостеневали и поэтому не сохранялись в ископаемом состоянии. Однако, судя по строению позвонков у современных круглоротых (Cyclostomata — миноги и миксины), позвонки вымерших Agnatha, по всей вероятности, состояли только из хрящевых дуг, разделенных на правую и левую половины; тел позвонков у них еще не было.
Хрящевые тела позвонков впервые появились, по-видимому, только у примитивных представителей челюстных позвоночных — у древнейших Gnathostomata. Об этом мы можем судить на основании строения позвоночника современных акул. Хрящевые позвонки таких древних челюстных форм, как силурийские Acanthodii, в ископаемом состоянии, разумеется, не сохранились.
15 А. П. Быстров
225
В процессе дальнейшей эволюции рыб окостенению подвергались прежде всего дуги позвонков; тела окостеневали значительно позже. Поэтому появление у эмбрионов человека и других Tetrapoda точек окостенения сначала в хрящевой дуге позвонка, а затем в его теле следует считать повторением той последовательности, которая наблюдалась при филогенетическом формировании позвонка.
В развивающемся крестце человека, кроме обычных точек окостенения, у первых трех или четырех позвонков появляются два дополнительные центра окостенения, соответствующие крестцовым ребрам (рис. 255, D).
Появление этих окостенений свидетельствует о том, что у человека тазовый пояс прикрепляется к позвоночному столбу при помощи трех (иногда
Рис. 255.
А — эпистрофей; В — атлант; С — грудной позвонок; D — крестцовый позвонок; Е — лопатка; F — тазовая кость.
Со — coracoideum (processus coracoideus); Il — ilium; Is — Ischium; Pb — pubis; Sc — scapula.
четырех) ребер, т. e. так же как он прикреплялся у его предков из класса рептилий. У взрослого человека точки окостенения крестцовых ребер настолько сливаются с телами и дугами соответствующих позвонков, что не могут быть отграничены от них.
Окостенение своеобразно построенных I (atlas) и II (epistropheus) позвонков человека происходит следующим образом.
I позвонок имеет два окостенения в задней дуге и одно в передней рис. 255, В). Окостенение в передней дуге соответствует единственному । ипоцентру, сохранившемуся в позвоночном столбе млекопитающих и человека. Телу атланта соответствует окостенение в хрящевом зубе II позвонка (рис. 255, Л), костное тело эпистрофея закладывается в виде самостоятельной точки окостенения. Костная дуга II позвонка развивается, как и у всех позвонков, из двух точек окостенения. Все эти отдельные костные части позвонков впоследствии срастаются между собой.
Таким образом, в развитии двух первых позвонков человека мы наблюдаем повторение того процесса формирования атланта и эпистрофея, который имел место у далеких предков человека из класса рептилий.
Развитие черепа человека давно привлекало внимание исследователей. Первые морфологи-теоретики предполагали, что он является результатом срастания нескольких позвонков. Эта идея впервые была высказана в 1786 г. Гете (W. Goethe, 1749—1832). Он пришел к заключению, что череп состоит из позвонков, после того как в его руки попал распавшийся на части череп
226
барана. Сначала Гете полагал, что в состав черепа вошли три позвонка. Первый из них, по его мнению, образовал затылочную кость, второй — заднюю часть клиновидной кости и третий — переднюю часть той же кости.
Позднее Гете решил, что к этим трем позвонкам можно прибавить еще гри. Он был склонен рассматривать как видоизмененные позвонки нёбную (palatinum), верхнечелюстную (maxilla) и межчелюстную (intermaxlllare s. praemaxillare) кости.
Хотя Гете может считаться творцом позвоночной теории черепа, однако наиболее полно она была разработана в 1807 г. Океном (L. Oken, 1779— 1851).
Окен строил свою теорию, рассуждая как типичный натурфилософ. В его теории наряду с совершенно правильными положениями, основанными на анатомических наблюдениях, встречается немало выводов, достаточно логичных, но совсем не соответствующих действительности. Это объясняется тем, что длинная цепь следующих друг за другом теоретических положений и выводов нередко приводила Окена к совершенно ложным взглядам.
Исходным пунктом для теории Окена служило следующее положение: число позвонков равно числу пар спинномозговых нервов; поэтому каждому позвонку соответствует одна пара нервов и наоборот.
Так как эти нервы связаны с чувствительными окончаниями в коже, то все позвонки позвоночного столба (Rumpfwirbel) Окен называл «кожными» (Hautwirbel) или «чувствительными» (Gefiihlwirbel). По его мнению, позвоночный столб человека должен быть разделен на три больших отдела по числу систем органов, которые иннервируются соответствующими группами нервов.
Таких систем Окен различал три: дыхательная, пищеварительная и система органов размножения; поэтому весь посткраниальный скелет он делил на грудь, живот и таз.
«К груди относится шея, руки и все ребра», — писал Окен. Во всем этом отделе, по его мнению, имеется 15 позвонков (Brustwirbel), которые разделяются на три группы по пяти позвонков в каждой: пять шейных (Halswirbel), пять ручных (Armwirbel) и пять легочных позвонков (Lungenwirbel).
«Шейными» позвонками Окен называл атлант, зуб эпистрофея, который он считал самостоятельным, но недоразвитым позвонком, эпистрофей (без зуба), III и IV позвонки шеи.
Спинномозговые нервы, соответствующие этим позвонкам, по мнению Окена, иннервируют гортань, состоящую из пяти хрящей и развившуюся, из пяти жаберных дуг. Поэтому указанные пять «шейных» позвонков (Halswirbel) он называл, кроме того, «жаберными» (Kiemenwirbel).
«На руки должно приходиться пять позвонков, — писал Окен, —, потому, что они имеют пять пальцев и пять нервов».
«Ручными» позвонками (Armwirbel) Окен считал три нижних шейных и два верхних грудных позвонка.
Пять грудных позвонков (от III до VII), ребра которых достигают грудной кости, по Окену, являются «легочными» позвонками (Lungenwirbel)., Непосредственно за грудным отделом позвоночного столба следуют, пять «брюшных» позвонков (Bauchwirbel). Каждый из этих позвонков снабжен парой ребер, но эти ребра своими концами не доходят до грудной, кости. Таким образом, грудные позвонки от VIII до XII, по Окену, являются брюшными и оказываются связанными с кишечником. Это — «кишечные» позвонки (Darmwirbel).
Последний отдел позвоночного столба состоит, как и грудной отдел, из пятнадцати позвонков, нервы которых связаны с органами размноже-. ния. Это — так называемые «половые» позвонки (Geschlechtswirbel).
15* 227
Этот отдел Окен разделял Также на три участка по пяти позвонков в каждом.
Пять поясничных позвонков (Lendenwirbel) он называл «ножными» позвонками (Fusswirbel), так как соответствующие им нервы иннервируют ноги.
Пять крестцовых позвонков (Kreuzwirbel) он считал истинными «половыми» позвонками (Geschlechtswirbel).
Наконец, пять копчиковых позвонков (Steisswirbel, или Schwanzwir-bel) Окен рассматривал как позвонки, соответствующие шейным, но помещенные на противоположном конце тела.
Так как согласно теории Окена шейные позвонки при помощи нервов связаны с гортанью, образовавшейся из пяти жаберных дуг, что дало ему Повод назвать их «жаберными» (Kiemenwirbel), то и сильно редуцированные копчиковые позвонки, по его мнению, были когда-то связаны с исчезнувшими в настоящее время «половыми жабрами» (GeSchlechtskiemen).
Такой неожиданный и ни на чем не основанный вывод является очень характерным для теоретических рассуждений Окена.
Продолжая так же смело строить свою теорию, ОкеН пришел к заключению, что слуховые, вкусовые, зрительные и обонятельные ощущения представляют собой только видоизменение чувств туловища. Поэтому нервы, связанные с органами чувств головы, по его мнению, должны соответствовать каким-то позвонкам. Эти позвонки изменились в такой же степени, в какой изменились и сами органы чувств и передний отдел центральной нервной системы, превратившейся в головной мозг. Все это заставило Окена прийти к убеждению, что череп должен состоять из нескольких сильно расширенных позвонков. «Головные позвонки есть только расширенные шейные позвонки», — писал он.
Количество позвонков, вошедших в состав черепа, по теории Окена, должно быть равно количеству органов чувств, т. е. четырем (слух, вкус, зрение и обоняние).
Окен полагал, что I позвонком черепа является затылочный позвонок (Hinterhauptswirbel). Он образован затылочной костью — os occipitale. При этом телом позвонка является pars basilaris ossis occipitalis (рис. 256, A— c. v.)t дугой — partes laterales (a. v.) и остистым отростком — squama occipitalis (p. s.).
Мозжечок, помещающийся в этом «позвонке», по мнению Окена, связан при помощи нервов с органом слуха. Поэтому указанный позвонок может быть назван «ушным» (Ohrwirbel).
Далее II позвонок, «теменной» (Scheitelwirbel), располагается непосредственно перед затылочным. Его телом является задняя часть клиновидной костй — corpus ossis sphenoidalis (рис. 256, В—с. v.), дугой — большие крылья (alae magnae) той же кости (a. v.) и остистым отростком — теменные кости (os parietale—р. s.). «Теменной» позвонок связан с органом вкуса, так как сквозь его дугу (ala magna ossis sphenoidalis) проходит тройничный нерв (n. trigeminus), принимающий участие в иннервации языка. Поэтому Окен назвал этот позвонок «язычным» (Zungenwirbel), или «вкусовым» (Geschmakswirbel).
Следующий III позвонок Окен называет «лобным» (Stirnwirbel). Тело его образовано передней частью клиновидной кости (рис. 256, С — с. v.), дуга — малыми крыльями (alae parvae) этой кости (a. v.) и остистый отросток — лобной костью (os frontale—р. s.). Так как сквозь отверстие в основании малых крыльев (foramen opticum) проходит зрительный нерв, то Окен считал этот пбЗбоПок связанным с органом зрения и назвал его «глазным» (Augen-wirbel), или «зрительным» (Gesichtswirbel).
Последний IV позвонок Окен назвал «носовым» (Nasenwirbel). В его
228
состав, по его мнению, входят следующие кости: сошник (vomer), представляющий собой тело позвонка (рис. 256, D—с. v.), правая и левая половины решетчатой кости (os ethmoidale), соответствующие дуге позвонка (a. v.), и носовые кости (os nasale), являющиеся остистым отростком (р. s.). Так как через отверстия в lamina cribrosa решетчатой кости проходят обонятельные нервы, то Окен считал этот позвонок связанным с органом обоняния и назвал его «обонятельным» (Geruchswirbel).
Рис. 256.
А — «затылочный» позвонок; В — «теменной» позвонок; С — «лобный» позвонок; D — «носовой» позвонок.
а. V. — arcus vertebrae; с. v . — corpus vertebrae; p. s. — processus splnosus.
Позвоночная теория черепа Гете—Окена была опровергнута в 1864 г. работами Т. Г. Гексли (Т. Huxley).
Гексли доказал, что такие кости черепа, как носовая, лобная, теменная и верхняя часть затылочной, не закладываются в виде хрящевых образований, а развиваются непосредственно в рыхлой соединительной ткани. Поэтому указанные кости не могут быть частями позвонков, которые на первом этапе своего развития всегда состоят из хряща. Что же касается хрящевой закладки основания черепа, то она не разделяется на сегменты и в ней нельзя обнаружить отдельные части, соответствующие хрящевым позвонкам (рис. 257).
«Череп, — писал Гексли, — уже не представляется нам видоизмененным позвоночником, равно как и позвоночник не есть видоизмененный череп». В хрящевом основании черепа зародыша развивается несколько костей. Их образование начинается с появления точки окостенения в хрящевой массе. Затем кость постепенно заменяет хрящ, который некоторое время сохраняется только в промежутках между костными образованиями.
229
Затылочная кость человека, как и соответствующая кость черепа млекопитающих, развивается из четырех точек окостенения, появляющихся в соответствующем участке хрящевого основания черепа зародыша. Основная часть затылочной кости (pars basilaris ossis occipitalis) возникает как непарное костное образование в хряще непосредственно перед затылочным отверстием (рис. 257—р. Ь.).
Боковые части затылочной кости (partes laterales ossis occipitalis) развиваются в виде изолированных парных костных образований по обе стороны затылочного отверстия (рис. 257—р. I.). Нижняя часть чешуи затылочной кости образуется в результате окостенения вогнутой хрящевой пла
стинки позади затылочного отверстия (рис. 257—sq). Здесь возникает широкая костная пластинка.
Окостенение боковых участков хрящевого основания черепа приводит к образованию костной пирамиды — pyramis (рис. 257—ру) и сосцевидной части височной кости — pars mastoidea ossis temporalis (рис. 257—p. m.).
Непосредственно перед основной частью затылочной кости (pars basilaris ossis occipitalis) в хрящевом черепе зародыша образуется задняя часть тела клиновидной кости — pars posterior corporis ossis sphenoidalis (рис. 257 — p. p.). Окостенение примыкающих к этому участку хрящевых пластинок приводит к образованию больших крыльев клиновидной кости — alae magnae ossis sphenoidalis (рис. 257 — a. m.).
В результате окостенения хря
щевых образований, расположенных кпереди от турецкого седла (sella turcica), в развивающемся черепе возникают передняя часть тела и малые крылья клиновидной кости — pars anterior et alae parvae ossis sphenoidalis (рис. 257—a. p.). Окостенение самого переднего участка хрящевого основания черепа приводит к образованию решетчатой кости — os ethmoidale (рис. 257—eth).
Все другие кости черепа человека, как и у всех млекопитающих, развиваются непосредственно в эмбриональной соединительной ткани и предварительной хрящевой стадии не проходят. Началом развития каждой из них служит появление так называемой точки окостенения (punctum ossi-ficationis). Эта первая закладка покровной кости затем постепенно увеличивается в своих размерах.
Элементы плечевого, тазового поясов и свободного отдела конечностей у человека, как и у всех позвоночных, закладываются в виде хрящевых образований. Впоследствии они подвергаются окостенению. В лопатке человека появляются две точки окостенения; одна из них соответствует scapulae, другая—коракоиду (coracoideum) низших позвоночных (рис. 255, Е— Sc, Со). Дополнительная (третья) точка окостенения, возникающая у человека и млекопитающих в хрящевом acromion, является новообразованием.
Тазовая кость (os coxae) человека закладывается как хрящевое образование, в котором затем появляются одна за другой три точки окостенения 230
(рис. 255, F). Сначала начинает окостеневать подвздошная кость (ilium), затем седалищная (ischium) и, наконец, лонная (pubis). У взрослого человека все эти три части срастаются друг с другом и образуется одна тазовая кость (os coxae).
Не только образование тазовых костей из трех отдельных частей, но и та последовательность, в которой они возникают в общей хрящевой закладке у человека, является повторением того, что наблюдалось у предков млекопитающих из классов рептилий и амфибий. У стегоцефалов сначала окостеневала подвздошная кость, затем седалищная; лонная кость у многих из них в течение всей жизни оставалась хрящевой. У рептилий тазовые кости окостеневали в такой последовательности: ilium, ishhium, pubis.
Все кости конечностей закладываются у зародыша как хрящевые образования; позже они подвергаются окостенению. В костях плеча и предплечья, бедра и голени возникает по три самостоятельных точки окостенения: одна в средней части (в диафизе) и две в обоих концах (в эпифизах).
ГЛАВА III
ОБЩАЯ ФОРМА ЧЕРЕПА
Как известно, в течение индивидуальной жизни человека его череп подвергается довольно сильным изменениям.
Череп новорожденного отличается очень большой мозговой частью по сравнению с лицевой (рис. 258, Д). Лобные и теменные бугры черепа сильно
Рис. 258. Череп новорожденного (А) н череп 5-летнего ребенка (В).
выступают; около каждого из четырех углов теменных костей имеются дефекты. закрытые только соединительнотканной перепонкой, — так называемые «роднички» (fonticulus frontalis, f. occipitalis, f. sphenoidalis и f. mastoideus); лобная кость разделена швом на правую и левую половины; лобные пазухи (sinus frontalis) еще отсутствуют; гайморовы пазухи (sinus maxillaris) развиты слабо; барабанная кость (pars tympanica ossis temporalis) имеет форму незамкнутого кольца; сосцевидная часть (pars masto-idea) височной кости отделена от чешуи (squama temporalis); правая и левая половины нижней челюсти не сращены в подбородке; затылочная кость состоит из четырех отдельных частей.
По мере увеличения размеров черепа отношения мозгового и лицевого его отделов изменяются. Рост мозгового черепа отстает от роста лицевого. После прорезывания молочных зубов челюстной аппарат становится сильнее, чем был прежде (рис. 258, В). К этому времени все роднички уже
232
зарастают. Однако лобные и теменные бугры остаются развитыми еще до* статочно сильно. Обе половины нижней челюсти в подбородочной области, срастаются; срастается и чешуя височной кости с сосцевидной частью; такое же срастание наблюдается и между отдельными частями затылочной кости.
Во время смены молочных зубов постоянными челюстной аппарат усиливается еще больше. Мозговой череп начинает терять те особенности, которые были ему свойственны в детском возрасте (рис. 259, Л). Лобные и теменные бугры почти исчезают. В результате этого угловатость мозгового отдела постепенно сглаживается. К моменту начала полового созревания в черепе остаются только следы признаков, характерных для более раннего возраста (рис. 259, В). Отношение размеров лицевого и мозгового черепа становится почти таким, какое мы видим у взрослого человека.
233
У вполне сформировавшегося человека череп (рис. 260, Л) приобретает ряд особенностей, которые вызываются постепенным усилением мускулатуры, так или иначе связанной с черепом. На его затылочной кости появляются шероховатые линии (lineae nuchae superior et inferior), небольшие гребни и выступы (protuberantia occipitalis externa); размеры сосцевидного отростка (processus mastoideus) становятся больше; границы начала височной мышцы и место прикрепления ее фасции выявляются резче (linea semicircularis temporalis superior et inferior). На краю угла нижней челюсти появляются небольшие отростки и гребни.
Вполне сформировавшийся череп некоторое время остается почти неизменным. Во всяком случае в нем не удается отметить тех, вероятно незна-
Рис. 261. Череп человека в 60 (Д) и в 70 лет (В),
чительных перемен, которые возникают в результате небольшой его перестройки.
Значительные изменения челюстного аппарата наступают только в результате потери зубов.
Мы не можем считать разрушение зубов и их выпадение нормальным для человека явлением. Следует думать, что человек должен был бы сохранять зубы в течение всей своей жизни. За несколько десятков лет ежедневной работы зубы, разумеется, должны сильно стираться, но их выпадение задолго до полного снашивания с биологической точки зрения нельзя признать естественным. Однако это ненормальное явление мы все же вынуждены считать для человека обычным. Наличие зубов у стариков, как известно, — явление очень редкое.
После полного разрушения зуба альвеолярный отросток в соответствующем участке челюсти подвергается уничтожению (рис. 260, В). Чем больше теряется зубов, тем сильнее редуцируются эти отростки (рис. 261, Л).
У стариков альвеолярный отросток на верхней челюсти в конце концов разрушается до уровня твердого нёба, а тело нижней челюсти становится поразительно тонким (рис. 261, В). Одновременно с этим толщина всех челюстных костей сильно уменьшается, в результате чего череп становится очень легким. В стенках, отделяющих глазницы от височных ям (fossae temporales), иногда возникают отверстия. Дефекты образуются и в слезной кости (os lacrimale), в бумажной пластинке решетчатой кости (lamina papyracea ossis ethmoidalis) и в передней стенке барабанной кости (os tympanicum).
234
В моем распоряжении был череп старухи 99 лет (1844—1943). Зубы у нее, разумеется, полностью отсутствовали; альвеолярные отростки верхней челюсти были атрофированы настолько, что твердое нёбо сделалось плоским. Такой же сильной атрофии подвергся и альвеолярный отросток нижней челюсти. Череп был очень легким; все его швы были заросшими. Скуловая дуга была очень тонкой. В костной стенке, отделяющей глазницу от височной ямы, имелись многочисленные отверстия. Кости мозгового черепа были настолько истонченными, что во многих местах их толщина равнялась только 1,5 мм. Это привело к тому, что борозды артерии мозговой оболочки (sulci arteriae meningeae mediae) стали заметными на наружной поверхности черепа в виде валиков (рис. 262). В некоторых местах костная
Рис. 262. Череп старухи 99 лет.
стенка этих валиков имела даже дефекты; следовательно, при жизни arte-ria meningea media здесь была прикрыта только надкостницей. Каналы venae diploicae превратились в борозды, видимые с внутренней стороны черепа.
Несколько большая толщина кости (около 3 мм) сохранилась только по ходу полукружной линии (linea semicircularis temporalis). В результате по этой линии возник широкий костный вал, который образовал выступающую дугу. Эта дуга служила местом прикрепления fasciae temporalis, внутренняя сторона которой, как известно, служит местом начала для височной мышцы.
В период полового созревания человека в его черепе возникают признаки полового диморфизма. Это приводит к тому, что в строении черепа мужчины и женщины появляется ряд отличительных особенностей (рис. 263, 264, 265).
Возникновение этих особенностей объясняется главным образом двумя причинами.
Это — во-первых, большее у мужчины развитие мускулатуры и, во-вторых, — большая степень пневматизации черепа мужчины по сравнению с черепом женщины.
Лоб у мужчины больше наклонен назад, чем у женщины; у женщины он почти прямой. Эта особенность является результатом значительно более сильного развития лобных воздушных полостей (sinus frontalis) у мужчины, чем у женщины. Поэтому надглазничные дуги (arcus supraorbitalis) у силь
235
ного мужчины выступают вперед; у женщины же они обычно выражены очень слабо.
Темя женского черепа более или менее уплощено, тогда как у мужчины покатый лоб постепенно переходит в выпуклое темя.
Рис. 263. Мужской череп.
Рис. 264. Женский череп.
Очертание затылочной кости черепа у женщины ровное. Здесь или полностью отсутствуют или бывают слабо развиты те неровности, гребни и выступы (lineae nuchae superior et inferior, protuberantia occipitalis externa), которые характерны для черепа типичного и в особенности физически сильного мужчины.
236
Скуловая дуга женского черепа, как правило, прямее и тоньше, чем у мужского черепа.
Сосцевидный отросток (processus mastoideus) у мужчины всегда больше, чем у женщины; его заполненные воздухом ячейки (cellulae mastoidei) развиты сильнее. У женщины вершина этого отростка обычно тупая (закругленная).
Орбиты у женщины относительно больше, чем у мужчины; они круглее, тогда как у мужчины нередко имеют почти четырехугольные очертания (рис. 265).
Ширина межорбитальной области у женщины меньше, чем у мужчины. Это объясняется тем, что при меньшей пневматизации черепа женщины
Рис. 265. Мужской и женский черепа.
ячейки решетчатой кости (cellulae ethmoidales) у нее развиты слабее, чем у мужчины.
Вследствие меньшей пневматизации женского черепа и объем гаймо; ровых полостей (sinus maxillaris) у женщины меньше, чем у мужчины.
У мужчин мускулатура вообще (и, в частности, жевательные мышцы) развита сильнее, чем у женщин; поэтому в черепе мужчины скуловые дуги толще, а такие неровности, как tuberositas masseterica, tuberositas ptery-goidea на нижней челюсти и crista infratemporalis на краю большого крыла клиновидной кости, выражены значительно сильнее.
Все указанные особенности строения черепа мужчины и женщины обычно позволяют без особого труда определить половую принадлежность изучаемого черепа. Однако следует иметь в виду, что все эти особенности полностью отсутствуют до начала полового созревания. Следовательно, отличить череп мужчины от черепа женщины до 15—17-летнего возраста оказывается не только крайне трудно, но даже невозможно. Кроме того, не следует забывать, что ошибки возможны и тогда, когда изучаются черепа взрослых людей. Если в руках исследователя окажется череп физически сильной женщины, имевшей очень хорошо развитую мускулатуру, то не исключена возможность, что он будет принят за череп мужчины.
237
Таким образом, определение половой принадлежности черепа не представляет каких-либо трудностей по существу только в тех случаях, когда все особенности черепа мужчины или женщины достаточно хорошо выражены. В тех же случаях, когда признаки мужчины в изучаемом черепе по тем или иным причинам ослаблены или когда сильное развитие мускулатуры оказало соответствующее влияние на череп женщины, возможны ошибочные решения.
Если череп не дает возможности определить пол скелета с абсолютной достоверностью, если позвонки и кости конечностей далеко не всегда имеют
Рис. 266. Мужской и женский тазы.
необходимые для этого признаки, то изучение строения таза позволяет сделать определение пола совершенно безошибочно.
Таз женщины по понятным причинам сильно отличается от таза мужчины (рис. 266). Он ниже и шире мужского; лонные кости у него сходятся ниже лонного соединения под углом 95—100°, тогда как в мужском тазу этот угол обычно равен только 75°.
Точное измерение костей, образующих правую и левую половины черепа, показывает, что абсолютной симметрии в строении черепа человека нет. Обычно (в 97% случаев) правая его половина развита немного сильнее, чем левая. На этой стороне размеры костей несколько больше, чем на противоположной. Кроме того, различные гребни, шероховатости и отростки справа выражены сильнее, чем слева. Однако эта асимметрия обычно не очень велика и может быть обнаружена только при детальном изучении черепа. Тем не менее, можно уверенно сказать, что абсолютно симметрично построенного черепа не существует; такой череп следует признать парадоксальным (рис. 267, В).
Более сильное развитие правой половины черепа, как и всех костей правой половины скелета, очевидно, является результатом того, что у человека мускулатура справа обычно сильнее, чем слева.
238
В больших краниологических коллекциях иногда встречаются черепа с резко выраженной асимметрией. При этом более развитой чаще оказывается правая сторона, чем левая (рис. 267, А).
Рис. 267.
А — резкая асимметрия черепа; В — парадоксальный череп.
Такое очень сильное развитие правой половины черепа, по всей вероятности, следует рассматривать как крайнюю степень обычной асимметрии.
Рис. 268. Асимметричные черепа человека.
В краниологических коллекциях анатомических институтов очень нередко встречаются случаи асимметричного строения черепа только в его мозговом отделе. Наиболее распространенной формой являются различные степени перекосов (рис. 268).
239
' Причины, вызывающие Такие неправильности строения, не выяснены. Высказывалось предположение, что такие формы черепа являются результатом деформации головы вследствие постоянного лежания ребенка на
Рис. 269. Два случая деформации черепа, вызванной преждевременным зарастанием швов.
одной стороне. Однако далеко не все подобные случаи могут быть объяснены таким образом. Мне даже кажется, что ни одна из этих форм асимметрии не может быть вызвана указанной причиной, так как трудно согласиться
Рис. 270. Скафоцефал (scaphocephalia). А — вид сверху; В — вид сбоку.
с тем, что лежание на одной стороне даже в раннем возрасте может так сильно'отразиться на форме черепа.
Очень интересной группой деформаций черепа человека являются деформации, вызываемые преждевременным зарастанием тех или иных швов.
В тех случаях, когда один из швов зарастает и рост краев костей здесь прекращается, в то время как другие швы продолжают функционировать, 240
возникает нарушение гармоничного роста черепа как целого. Это, естественно, приводит к его общей деформации.
На рис. 269 изображены два случая преждевременного зарастания некоторых швов.
Рис. 271. Череп «штеттинского ткача» (Бониет. 1904).
В первом из них (рис. 269, А) левая половина венечного шва (sutura coronalis) полностью заросла, а правая сохранилась. Поэтому левая темен-
Рис. 272. Гипсицефал (hypsicephalia).
ная кость не могла расти так же, как росла правая. В результате этого об* щие размеры левой половины черепа оказались меньше правой.
16 А. П. Быстров
241
Еще, более интересен второй случай (рис. 269, В). Здесь подверглись полному и, по всей вероятности, раннему зарастанию сагиттальный шов (sutura sagittalis) и левая половина венечного шва (sutura coronalis). Сохранились и продолжали функционировать только правая половина венечного шва и незаросший лобный шов (sutura frontalis). Нормальный рост краев только правой половины лобной кости и краев прилегающих к ней костей привел к резкой деформации переднего отдела мозгового черепа.
Преждевременное зарастание только одного сагиттального шва обычно приводит к тому, что увеличение черепа в1 ширину оказывается значительно
Рис. 273. Естественная деформация черепа. А — череп 9-летнего ребенка: В — череп взрослого человека.
ограниченным. При этом деятельность венечного и ламбдовидного швов продолжается и череп растет в продольном направлении. В результате этого возникает деформация, известная под названием скафоцефалии (scaphocepha-Па, рис. 270). Мозговой череп в этом случае бывает длинным и узким и напоминает лодку, обращенную килем вверх.
Та или иная степень скафоцефалии является относительно часто встречающейся деформацией черепа.
Необычайно сильная степень скафоцефалии имелась у так называемого «штеттинского ткача» (Боннет—Bonnet, 1904). Это был рабочий одной из ткацких фабрик в Штеттине. Его череп был настолько длинным и узким, что затылок упирался в спину, когда несчастный пытался поднять голову (рис. 271). О «штеттинском ткаче» говорили, что он никогда не видел неба и всегда мрачно смотрел в землю. Преследуемый за свое уродство насмешками, он скоро превратился в пьяницу и умер от плеврита на 38-м году своей печальной жизни. . .
Иногда раннему зарастанию подвергается не один, а несколько швов. Это также приводит к деформации черепа.
В тех случаях, когда преждевременно закрываются такие швы, как венечный и сагиттальный, небольшие размеры верхнего отдела черепа ребенка как бы фиксируются и их увеличение прекращается. Так как рост костей в сохранившихся швах черепа продолжается, то у взрослого возникает так 242
называемая «башенная» голова. Мозговой чёрен становится вытянутым вверх и немного назад (рис. 272).
Я имел в своем распоряжении череп 9-летнего мальчика, у которого венечный шов зарос почти на всем своем протяжении; при этом сагиттальный, шов был полностью закрыт (рис. 273, Л). Это фиксировало размеры переднего отдела мозгового черепа. Но так как рост краев костей по ходу ламбдовидного шва продолжается, то это вызвало выступание затылка назад.
Череп этого мальчика в будущем, несомненно приобрел бы ту весьма своеобразную форму, которую мы наблюдаем у взрослых в случаях зарастания тех же швов (рис. 273, В).
Рис. 274. Череп акромегалика Я. Лолли (Lolli).
Что вызывает преждевременное срастание некоторых костей черепа неизвестно. Возможно, что это явление связано с каким-то Нарушением нормальной функции одной из желез внутренней секреции. Однако следует признать, что пока еще не наблюдалось ни одного факта, который бы указывал на существование связи между зарастанием швов и деятельностью какой-нибудь из желез внутренней секреции.
В настоящее время можно считать твердо установленным влияние- на общую форму черепа только гормона гипофиза (hypophysis cerebri). Но гормон этой же железы не вызывает преждевременного зарастания черепных швов и, следовательно, не обусловливает деформации черепа.
Влияние гипофиза на рост костей выражается в том, что в случайх гиперсекреции этой железы возникает так называемая акромегалия, при которой высота человека значительно превосходит норму. Описаны великаны, рост которых равнялся 240 см (Махнов), 250 см (Купер), 275 сН* (Ганс Крав) и даже 283 см (финн Каянус).
При акромегалии руки и ноги человека становятся непропорционально длинными, а кисть и стопы — ненормально большими. Одновременно с этим появляются значительные изменения и в черепе.
Общие размеры череца у акромегаликов больше нормальных. Места, начала и прикрепления мышц на черепе превращаются в сильно развитые4, гребни и отростки. Кости черепа становятся массивными, и череп делается
243
16*
очень тяжелым. Но наибольшие изменения наблюдаются в форме нижней
челюсти. Она становится непропорционально большой.
Рис. 275. Искусственная деформация головы путем кругового бинтования (с фотографического снимка, сделанного в 1939 г. в Центральной Америке).
Все особенности черепа акромегалика были хорошо выражены у такого великана, как Яков Лолли (Jacob Lolli, рис. 274). Его рост равнялся 222 см. Лолли выступал в качестве борца в различных цирках Европы и скончался от разрыва аневризмы аорты в Петербурге в 1814 г. Его скелет в настоящее время находится в анатомическом институте Военно-медицинской академии (в Ленинграде).
Кроме естественных деформаций черепа, которые вызываются не выясненными до сих пор причинами, необходимо отметить деформации искусственные, которые создаются по воле самого человека.
Многие племена и народы в сравнительно недалеком прошлом имели странный обычай искусственно деформировать головы. Этот обычай сохраняется у некоторых народов еще и теперь.
На ряде островов Малайского архипелага и в Центральной Африке и в настоящие дни «башенная» голова считается очень красивой. Такая «модная» форма Черепа получается путем кругового бинтования, головы в детском возрасте. Бинтование производится до тех пор, пока мозговой череп не измейит своей обычной формы. Черепные швы при такой деформации, разумеется, не зарастают; рост изуродованного черепа в дальнейшем происходит нормально; но голова сохраняет искусственно приданную ей форму. Соответствующую форму, разу-
меется, приобретает и мозг, но это никак не отражается на интеллекте человека (рис. 275).
Рис. 276. Карта распространения обычая деформировать голову Черными кружками отмечены места, где были найдены черепа, деформированные путем кругового бинтования.
244
Археологические раскопки позволили установить, что этот странный обычай в свое время был очень широко распространен. Черепа, деформированные этим способом, обнаружены в Индии, в Туркестане, на Кавказе,
Рис. 277. Искусственно деформированный череп из раскопок около Керчи.
в Поволжье, на Керченском полуострове в Крыму, во многих местах западной Европы, в Северной и Южной Америке (рис. 276). Все эти черепа имели конусовидную форму (рис. 277).
Рис. 2'/8. Череп 10 летнею ребенка деформированный путем сдавливания впереди назад. Из доколумбийских могил в Перу (Итон — Eaton, 1916).
До сих пор остается Совершенно не понятным, почему этот обычай деформировать голову в свое время был так широко распространен и почему им пользовались самые различные племена и народы.
Совершенно иной способ деформирования головы применялся доколум-бийским населением некоторых районов Северной и Южной Америки.
245
В Мексике и в Перу были найдены черепа, имеющие форму широкого клина (рис, 278, 279). Такая форма черепа могла , быть получена, несомненно, только в результате достаточно продолжительного сдавливания головы ребенка по направлению спереди назад.
Такой способ изменения нормальной формы головы некоторое время сохранялся у отдельных племен аборигенов Америки даже после заселения этого материка европейцами.
Так, один из испанских миссионеров, некто Диэго де Ланда, живший в Юкатане в XVI столетии и написавший в 1566 г. отчет обо всем, что он там видел, в частности сообщает следующее.
Рис 279. Очень сильная степень деформации черепа путем сдавливания его спереди назад. Из доколумбийских могил в Перу.
Рис. 280. Индианка по рисунку Кэтлина (G. Catlin).
«Индианки воспитывали своих детей очень сурово, ибо четырех или пяти дней от роду новорожденного клали растянутым в маленькую кровать, сделанную из прутьев, и там, повернув ртом вверх, они клали ему голову между двумя дощечками, одна на затылке, другая на лбу, между которыми и держали его там в мучениях, пока, по прошествии нескольких дней, голова его не становилась сплюснутой и деформированной, как это у них было в обычае».
Так как указанный обычай держался в Америке очень долго, то Кэт-лину (G. Catlin) еще в прошлом столетии удалось видеть и зарисовать с натуры женщину, у которой голова была деформирована путем сдавливания спереди назад (рис. 280). Рисунок Кэтлина особенно ценен тем, что изображает женщину, держащую на руках ребенка, у которого голова зажата между двумя связанными досками. Этот своеобразный прибор дает совершенно ясное представление о том способе, при помощи которого достигалась деформация черепа.
Насколько мне известно, этот способ не применялся нигде, кроме Америки, но и там он держался наряду с обычаем деформировать голову путем наложения круговой давящей повязки (Итон — G. F. Eaton, 1916),
ГЛАВА IV
АТАВИСТИЧЕСКИЕ ВАРИАНТЫ
Все особенности формы черепа человека, возникшие по естественным причинам или вызванные искусственно, представляют для морфолога немалый интерес, но не имеют прямого отношения к эволюционной истории человека.
Несравненно большее значение для нас имеют так называемые атавистические 1 варианты в строении человеческого скелета, т. е. такие отклонения от обычной нормы, которые являются повторением строения некоторых предков человека. Эти варианты дают возможность иа основании фактического материала доказать происхождение человека от животных и, в частности, показать его генетическую связь не только с ближайшими, но и с очень отдаленными предковыми формами.
Если не отрывать эволюционную историю человека от истории позвоночных вообще, то мы должны признать, что в его родословной были предки из всех классов позвоночных. Поэтому нет ничего невероятного в том, что в некоторых случаях у человека могут быть обнаружены такие атавистические варианты, которые представляют собой очевидное доказательство его генетической связи с теми или иными формами его предков. При этом чем древнее «предок», тем реже его особенности должны повторяться в виде атавистических вариантов у современного человека, и, наоборот, особенности относительно поздних «предков» должны встречаться у человека более часто.
Это общее теоретическое положение вполне подтверждается изучением вариантов строения различных частей скелета.
Таким образом, мы ставим перед собой задачу на основании изучения разнообразных вариантов в строении скелета современного человека выяснить, как он в своем настоящем «вспоминает» свое прошлое.
Во всякой даже небольшой остеологической коллекции можно найти такие первые позвонки человека, в которых задняя дуга оказывается разомкнутой. Этот вариант в анатомической литературе носит название spina biiida atlantis (рис. 281, С, D).
Известно, что дуга каждого позвонка у зародыша развивается из двух (правой и левой) точек окостенения, возникающих в общей хрящевой закладке. Обычно эти окостенения затем полностью срастаются. Следовательно, наличие дефекта в задней дуге атланта является результатом его недоразвития.
1 Atavus — предок.
247
У примитивных наземных позвоночных — стегоцефалов—I позвонок был снабжен дугой, состоящей из правой и левой половин, в то время как все другие их позвонки имели целые дуги.
Эта особенность строения I позвонка сохранялась и у рептилий из отрядов Seymouriamorpha, Cotylosauria, Pelycosauria и Therapsida.
Полное сращение половин дуги этого позвонка наблюдалось у всех без исключения древних и современных млекопитающих.
Таким образом, в процессе филогенетического развития атланта срастание обеих половин его дуги у взрослых позвоночных наступило только при переходе от прогрессивных рептилий к примитивным млекопитающим.
Рис. 281. Варианты строения 1 позвонка человека.
Такое позднее слияние правой и левой половин дуги I позвонка является причиной того, что spina bifida atlantis у человека встречается относительно часто.
Так как близкие к млекопитающим формы рептилий — Therapsida— в большинстве случаев имели атлант с дугой, состоящей из двух половин, то spina bifida atlantis у человека мы можем рассматривать как вариант, повторяющий особенности строения I позвонка именно этих рептилий.
Задняя дуга I позвонка человека, как известно, лишена хорошо развитого остистого отростка. Вместо него на дуге имеется только небольшой бугорок (рис. 281, В). Однако, хотя и очень редко, встречаются атланты с относительно большим остистым отростком (рис. 281, Л).
Остистый отросток на дуге атланта отсутствует у всех представителей класса млекопитающих; его не было и у рептилий из отрядов Therapsida, Pelycosauria и Cotylosauria. Этот отросток имелся только на разделенной на две части дуге I позвонка сеймурий, т. е. у самых примитивных представителей класса рептилий.
Не исключена возможность, что в тех очень редких случаях, когда \ человека на задней дуге его I позвонка возникает остистый отросток, мы имеем дело с повторением того этапа филогенеза, который был пройден нашими далекими предками на начальной фазе развития рептилий.
Несравненно реже, чем spina bifida atlantis, подобное расщепление дуги встречается и в позвонках шейного, грудного и поясничного отделов.
Если мы будем изучать строение позвонков у ископаемых рептилий и амфибий, то у всех взрослых форм найдем целые дуги на всем протяжении позвоночного столба (исключением будет являться только дуга I позвонка).
248
Только у каких-то древних, вероятно, очень примитивных рыб дуги всех позвонков состояли из двух несросшихся половин.
Мне кажется, что это обстоятельство дает право думать, что очень редкие случаи spina bifida в грудных и поясничных позвонках являются повторением строения позвонков именно этих примитивных рыб.
Крестец человека, состоящий из пяти сросшихся друг с другом позвонков, как известно, соединяется с тазовыми костями таким образом, что-только три первые позвонка принимают участие в образовании крестцовотазового сустава (рис. 282, Л). Но встречаются случаи, когда I крестцовый позвонок отделяется от крестца и приобретает все особенности поясничного (рис. 282, В). Такой переход I крестцового позвонка в поясничный отдел (lumbalisatio vertebrae sacralis primae) является атавистическим вариантом.
Рис. 282.
А — нормальный крестец человека; В — lumbalisatio vertebrae sacralis primae.
В процессе формирования крестца у наземных позвоночных происходило постепенное включение в его состав все большего и большего количества позвонков, что приводило к более прочному соединению таза с позвоночным столбом. У амфибий крестец был образован только одним позвонком; у многих рептилий он уже состоял из двух-трех и даже четырех позвонков; у млекопитающих — из трех-четырех. Поэтому выключение одного позвонка из крестца человека следует считать возвращением к особенностям предков.
Изучение филогенетических изменений в строении позвонков показало, что их гипоцентры (hypocentrum) в процессе эволюции- рептилий подверглись постепенной редукции. Дольше всего они сохранялись в хвостовом отделе позвоночного столба. Здесь каждый из них был снабжен хорошо развитой вентральной дугой.
У млекопитающих гипоцентры исчезли во всех позвонках, кроме I. Однако, несмотря на это, у многих представителей этого класса вентральные дуги редуцированных гипоцентров еще долго сохранялись на несколь-ких|ближайших к крестцу хвостовых позвонках.
У человека в его очень слабо развитом хвостовом отделе позвоночника (копчик) обычно не встречается никаких следов не только гипоцентров, но и их дуг.
Однако в анатомической литературе были описаны два случая, когда наг I копчиковом позвонке была обнаружена приросшая к его нижнему краю-половина дуги гипоцентра (рис. 283). В первом случае (И. Э. Шавловский, 1901) это была правая половина дуги, во втором (Л. Крастин, 1928) — левая.
24»
В этих атавистических вариантах человек-повторил особенности, свойственные хвостовым позвонкам каких-то его примитивных предков из класса млекопитающих.
У многих ископаемых рептилий ребрами были снабжены не только грудные позвонки, но и все шейные, поясничные, крестцовые и даже первые
хвостовые.
Рис. 283. Дуга гипоцентра на I копчиковом позвонке человека (Д — по Шавловскому, 1901; В, С — по Крастин, 1928).
У млекопитающих хорошо развитые ребра сохранились только в грудном отделе; в шейном, поясничном и крестцовом отделах небольшие остатки редуцированных ребер полностью приросли к поперечным отросткам соответствующих позвонков.
Поэтому все случаи появления свободных ребер на шейных и поясничных позвонках у человека следует рассматривать как повторение особенностей
строения каких-то его предков из класса рептилий.
Так как редукция шейных ребер у рептилий шла спереди
назад и у последних шейных позвонков их рудименты прирастали к попе-
речным отросткам и телам позвонков позже, чем у первых, то естественно, что у человека хорошо развитые ребра чаще всего наблюдаются на VII шейном позвонке (рис. 284, А, В). В тех редких случаях, когда они по
Рис. 284.
А — ребро на VII шейном позвонке; В — то же; С — ребро иа I поясничном позвонке.
являются и на других шейных позвонках, их размеры бывают обычно очень невелики; такие ребра имеют форму небольших косточек, мало похожих на ребра.
Ребра в поясничном отделе позвоночника человека наблюдаются относительно редко и обычно на I поясничном позвонке (рис. 284, С). Впрочем, был описан случай, когда отверстие, в котором проходила вена, отделяло небольшое ребро, приросшее к телу и поперечному отростку V поясничного позвонка (рис. 285, Л). У того же человека подобное отверстие располага-250
лось между поперечным отростком и ребром и у 1 крестцового позвонка (рис. 285, В).
В анатомической литературе можно найти немало работ, посвященных так называемому «затылочному позвонку» (Свечников—Swj'etschnikow, 1906. 1908; Кольман — J. Kollmann, 1907; Больк — L. Bolk, 1922; А. А. Дешин. 1926; А. П. Быстров — Bystrow, 1931 и др.).
Изучение затылочной кости человека показало, что вокруг большого затылочного отверстия иногда появляются костные образования, по своей форме очень напоминающие отдельные элементы позвонка, похожего на атлант.
На основной части затылочной кости в таких случаях можно видеть разомкнутую или даже целую переднюю дугу позвонка (рис. 286). Иногда на ней имеется даже суставная площадка, принимающая участие в образовании дополнительного сустава с вершиной зубовидного отростка эпистрофея (рис. 286, С). Бывают случаи, когда один или оба суставных отростка затылочной кости оказываются не выпуклыми, а плоскими, и по своим очертаниям напоминают нижние Суставные площадки атланта (рис. 286, В — левый суставной отросток). Гораздо реже встречаются поперечные отростки «затылочного позвонка» (рис. 286, В, D)-, при этом на конце такого отростка иногда имеется суставная площадка, которая принимает участие в образовании дополнительного сустава с концом поперечного отростка атланта (рис. 286, D—на правой стороне). На заднем краю затылочного отверстия можно наблюдать утолщения, которые очень
лочной кости заднюю дугу позвонка. Эта дуга обычно оказывается сомкнутой (рис. 286, С).
Все эти отклонения от нормального строения затылочной кости трактуются анатомами как выявление «затылочного позвонка» (manifestatio vertebrae occipitalis). Для подтверждения такого взгляда они обычно ссылаются на данные эмбриологии, указывающие, что в состав основной части затылочной кости млекопитающих входит по крайней мере три сегмента, и считают, что в случаях «манифестации» мы имеем дело с выявлением последнего из них.
Палеонтология не располагает такими фактическими данными, которые могли бы подтвердить, что задний отдел хрящевого черепа у позвоночных возник в результате слияния нескольких позвонков. Однако у ископаемых рептилий и амфибий basioccipitale очень похоже на тело позвонка, а парное exoccipitale напоминает его разомкнутую дугу.
Затылочная кость человека является сложным образованием. Судя по ее развитию и строению в детском черепе, в ее состав входя! несколько
Рис. 285
А—поясничный позвонок; с — ребро, В — крестец человека (вид сверху) (Шавловский, 1901).
напоминают приросшую к заты-
251
отдельных частей (рис. 287, Л). Основная часть затылочной кости, pars basilaris, развивающаяся из хрящевой закладки, представляет собой basioccipitale низших форм позвоночных. Боковые части, partes laterales,
Рис. 286. Манифестация так называемого «затылочного позвонка».
развивающиеся также путем замены хряща костью, соответствуют exoccipitale рептилий и амфибий. Тот участок чешуи затылочной кости, который закладывается как хрящевое образование, является гомологом sup-
Рис. 287.
А — затылочная кость новорожденного; В — затылочная кость взрослого.
Во — basioccipitale; Dso — dermosupraoccipitale; Ео — exoccipitale; р. b. — pars basilaris; p. I. — pars lateralis; So — supraoccipitale; Sq — squama; T — tabulare.
raoccipitale рептилий и амфибий. Верхний участок чешуи, как известно, развивается непосредственно в соединительной ткани и не проходит предварительной стадии хрящевой закладки. У новорожденного он еще отделяется двумя щелями (рис. 287, Л); у взрослого человека от этих щелей нередко остаются короткие швы — sutura mendosa (рис. 288, Л). Несрав
252
ненно реже оба эти шва соединяются друг с другом, и в таких случаях верхний участок чешуи оказывается отделенным от остальной части затылочной кости (рис. 288, В).
В 1842 г. английский врач Беллами (Р. F. Bellamy) описал два черепа, взятые от перуанских мумий. При этом у одного из них он обнаружил поперечный шов, отделяющий верхнюю часть чешуи затылочной кости; у другого сохранились небольшие участки такого же шва (sutura mendosa). Беллами решил, что этот вариант является типичной особенностью для до;
Рис. 288.
А — череп с нормальной затылочной костью; В — os incae; С — os incae bipartitum; D — os incae tripar-titum.
колумбийского населения Перу — инков. Немного позже Чуди (Tschudi, 1844); согласившись с мнением Беллами, признал эту особенность расовым признаком для инков и назвал изолированную верхнюю часть чешуи затылочной кости os incae. Это название сохранилось в анатомической литературе до сих пор, несмотря на то, что скоро было доказано, что os incae встречается у инков не только не постоянно, но совсем не чаще, чем у представителей любой другой расы или народа.
Изучение os incae показало, что эта кость очень сильно варьирует.
В некоторых случаях она оказывается разделенной швом на правую и левую половины; при этом возникает так называемая os incae bipartitum (рис. 288, С).
Иногда она разделяется при помощи двух швов на три равные по своим размерам участка. Этот вариант получил название os incae tripartitum (рис. 288, D).
Несравненно реже встречаются случаи, когда os incae состоит из четырех частей и средняя ее часть оказывается разделенной швом, являю-
253
щимся.’как бы.. прямым продолжением: сагиттального ;шва,: проходящего между, теменными'костями (рис. 289, А, В). Такие случаи свидетельствуют о том, что средняя часть os incae tripartitum представляет'собой резуль*. тат слияния двух самостоятельных костей.. / ,
Во.всякой большой краниологической коллекции можно найти черепа с самыми: разнообразными комбинациями сращений четырех костей, входя* щих в состав os incae, друг с другом и с нижним участком чешуи; затылоч-ной кости (рис. 289, 290)............................ .
Таким образом, изучение различных вариантов «костей инков» показывает, что в состав верхней части чешуи затылочной кости, развивающейся
Рис. 289. Варианты строения чешуи затылочной кости.
как соединительнотканная кость, входят четыре отдельные элемента (рис. 287, В). ' .
Эти кости, несомненно, соответствуют четырем покровным костям, образующим задний отдел черепной крыши у низших позвоночных, именно: двум dermosupraoccipitale и двум tabulare.
У стегоцефалов эти кости располагаются непосредственно позади parietale и supratemporale и соединяются с supraoccipitale и. exoccipitale; между основаниями exoccipitalia помещается basioccipitale.
У ^рептилий dermosupraoccipitale и tabulare занимают такое же Положение. У наиболее примитивных из них dermosupraoccipitale обычно является еще парной костью, а у более прогрессивных оно уже представляет собой непарное образование.
У млекопитающих basioccipitale, exoccipitale, supraoccipitale, dermosupraoccipitale и tabulare сливаются в одну кость и образуют os occipitale.
Все это дает нам право считать, что появление у человека таких вариантов, каков incae bipartitum и tripartitum, представляет собой отделение der-
254
mosupraoccipitale и tabulare от остальных частей затылочной кости. А по-явление у человека изолированных dermosupraoccipitale и tabulare. мы можем рассматривать как атавистический вариант, при котором наблюдается! возвращение к особенностям строения черепа рептилий.
В тех случаях, когда имеется os incae tripartitum (рис. 288, D), строение затылочной кости человека оказывается сходным со строением затылочной части черепа тех рептилий, у которых оба dermosupraoccipitale уже слились друг с другом, a tabulare представляют собой еще достаточно большие и изолированные кости (Therapsida). Такие случаи у человека встречаются несравненно чаще, чем случаи разделения средней кости на две
Рис. 290. Варианты строения чешуи затылочной кости.
части, т. е. случаи присутствия двух dermosupraoccipitale (рис. 289, А, В). Это, по всей вероятности, объясняется тем, что наличие двух dermosupraoccipitale является возвратом к более древним предкам, именно: к примитивным рептилиям из отряда Seymouriamorpha.
Клиновидная, или основная, кость (os Sphenoidale) человека представляет собой не менее сложное образование, чем затылочная кость. У взрослого человека она является результатом сращения нескольких костных элементов, соответствующих различным костям черепа низших наземных позвоночных (рис. 291).
Задний отдел тела клиновидной кости, развивающейся в результате окостенения соответствующего участка хрящевого основания черепа, гомологичен базисфеноиду (basisphenoideum) рептилий.
Передний отдел тела и малые крылья (alae parvae), также развивающиеся в хряще, являются видоизменением сфенэтмоида (sphenethmoideum) древних Tetrapoda.
Большие крылья (alae magnae) клиновидной кости, формирующиеся из самостоятельных точек окостенение в хрящевом основании черепа, со:
255.
ответствуют эпиптеригбидам (epipterygoideum) древних рептилий и амфибий.
Единственной частью клиновидной кости, которая развивается в соединительной ткани, является медиальная пластинка крыловидного отростка (lamina medialis processus pterygoidei). Эта пластинка представляет собой сильно редуцированный птеригоид (pterygoideum) низших Tetrapoda — амфибий и рептилий.
От парасфеноида (parasphenoideum), который лежал на нижней поверхности основания черепа рептилий, амфибий и рыб, у млекопитающих
Рис. 291. Клиновидная кость человека.
Bs — basisphenoideum; Ер — epipterygoideum; Pt — pterygoideum; Se — sphenethmoideum.
и человека, как известно, не осталось никаких следов; он подвергся полной редукции.
Очень редким вариантом в строении клиновидной кости человека является наличие узкого канала, начинающегося на дне турецкого седла и открывающегося на наружной поверхности основания черепа. Этот канал получил название canal is craniopharyngeus (рис. 326, В). Подобный канал, проходящий сквозь парасфеноид, имелся у девонских кистеперых рыб и у ихтиостегид.
Височная кость (os temporale) человека, как и соответствующая кость у всех млекопитающих, является результатом срастания таких костей древних рептилий, как paroccipitale (pars mastoidea), squamosum (squama temporalis), prooticum (pyramis) и angulare (pars tympanica). Кроме того, к ней прирастает окостеневший верхний участок hyoideum (из второй висцеральной дуги) и приобретает форму шиловидного отростка (processus styloideus, рис. 292, В).
Наиболее интересным атавистическим вариантом в строении височной кости является присутствие на ее чешуе большого отростка, располагающегося непосредственно позади суставной ямки для нижней челюсти (processus postglenoidalis, Кабиббе — G. Cabibbe, 1901).
256
Обычно этот отросток у человека очень мал (рис. 292, А). В тех же случаях, когда он достигает больших размеров (рис. 292, В), он становится очень похожим на соответствующий отросток у антропоморфных обезьян (рис. 293). В таких случаях человек, по-видимому, повторяет особенности строения височной кости своих недавних обезьянообразных предков.
Рис. 292.
А — нормальная височная кость; В — височная кость с сильно развитым processus postglenoidalis Cabibbe (р р.).
Под влиянием Чезаре Ломброзо (С. Lombroso), который, как известно, почти все отклонения от нормального строения в человеческом организме рассматривал как признаки вырождения, Кабиббе (1901) утверждал, что сильно развитый processus postglenoidalis чаще встречается у душевнобольных и преступников, чем у нормальных людей.
При изучении анатомического строения стато-акустического аппарата, помещающегося в пирамиде височной кости, были обнаружены очень редкие и интересные варианты.
Как известно, кроме улитки — органа слуха,— в состав перепончатого лабиринта у человека входят три полукружных канала, несущих функцию органов сохранения равновесия. Эти каналы располагаются в трех взаимно-перпендикулярных плоскостях. Перепончатый лабиринт заключен
257
17 А П. Быстров
в так называемый костный лабиринт, который'в общих чертах повторяет форму перепончатого (рис. 294, Л). Три полукружные канала, проходящие в кости и содержащие н себе соответствующие перепончатые каналы,
Рис. 293. Processus postglenoidalis (р.р.) на височной кости шимпанзе (Л) и гориллы (В).
открываются отверстиями в полость преддверия (vestibulum). Эти отверстия хорошо видны, если распилить пирамиду височной кости в соответствующем месте поперек (рис. 295, А).
Рис. 294. Костные лабиринты человека.
с» L — canalis semlcircularis lateralis; с. р. — canalis semlcircularis posterior; c. s. — canalla semlcircularis superior.
В анатомической литературе описаны только три случая полного отсутствия у человека одного из трех полукружных каналов (3. И. Ибрагимова, 1940; А. П. Быстров, 1944). Во всех этих случаях отсутствовал латеральный (горизонтальный) канал (рис. 294, В) и на задней стенке преддверия не было соответствующей пары отверстий (рис. 295, В). Отверстия двух других каналов располагались в обычных для них местах.
253
Сравнительная анатомия давно установила, что все представители таких классов позвоночных, как млекопитающие, птицы, рептилии, амфибии и рыбы, имеют три полукружных канала. Поэтому можно уверенно сказать, что отсутствие одного из этих каналов у человека не может рассматриваться как атавистический вариант, повторяющий особенности строения уха у древних его предков, относящихся к одному из этих пяти классов. Три полукружные канала оформились уже у самых древних и примитивных Gnathostomata, и, начиная с этих форм, общий план строения органов сохранения равновесия на протяжении всей эволюции позвоночных вплоть до человека оставался неизменным.
Рис. 295. Поперечные распилы пирамид височных костей.
с. f. — canalis facialis; с. s. 1. — canalis semiclrcularls lateralis; c. s. p. — canalis semiclrcularls posterior, c, s. s. — canalis semiclrcularls superior; /. r. — fenestra rotunda; in — Incus; m — malleus; s — stapes.
Но у Agnatha было не три, а два полукружных канала. На это указывает строение внутреннего уха у современных круглоротых (Cyclostomata). У миноги имеются только передний и задний полукружные каналы. Передний соответствует верхнему (фронтальному) каналу человека, задний —г заднему (сагиттальному). >
Как известно, детальное изучение некоторых ископаемых представителей класса Agnatha, блестяще проведенное шведским палеонтологом Стеншио (1927), показало, что перепончатый лабиринт у них имел, как у миног, только два полукружных канала, именно: передний и задний. Это дает нам право уверенно считать, что все многочисленные и разнообразные представители класса Agnatha силура и девона, подобно современным миногам, имели только два полукружных канала. Третий канал — именно горизонтальный (латеральный) — в процессе эволюции позвоночных впервые возник только у примитивных Gnathostomata. Следовательно, он является, хотя и очень древним, но все же более поздним образованием, чем! два другие.
Изучение развития перепончатого лабиринта у эмбрионов высших позвоночных показало, что передний и задний полукружные каналы сначала закладываются как один общий уплощенный выворот стенки слухового пузырька. Затем в этом вывороте путем прорыва его стенок возникают два 17* 259
отверстия. В результате этого образуются одновременно два полукружных канала — передний и задний. Латеральный канал закладывается как самостоятельное образование, имеющее вначале также форму уплощенного выворота (рис. 37).
Такой ход развития указывает на то, что передний и задний полукружные каналы оформились в процессе эволюции совершенно одновременно; латеральный же возник как добавочное образование.
В указанных нами случаях отсутствия одного из полукружных каналов у человека не было именно этого, латерального (горизонтального), канала.
Рассматривая эти случаи как атавистические варианты, мы должны будем признать, что они указывают на генетическую связь человека с его
древнейшими предками из класса бесчелюстных позвоночных (Agnatha). Иного объяснения отсутствия именно латерального канала дать невозможно.
Очень интересной ненормальностью теменной кости человека являются случаи разделения ее горизонтальным швом на две части. Двойная теменная кость (os parietale bipartitum) встречается очень редко; я нашел ее только у одного черепа из 4250 (рис. 296).
Эмбриологические исследования показали, что каждая из теменных костей, развиваясь в соединительной ткани, закладывается в виде двух точек окостенения, которые, однако, очень скоро сливаются друг с другом. Это обстоятельство, несомненно, свидетельствует о том, что в состав теменной кости вошли две ранее самостоятельные кости. Одна из них (верхняя) представляет-собой собственно теменную кость, в то время как вторая (нижняя) является какой-то другой костью, приросшей к теменной.
Швальбе (G. Schwalbe, 1903) описал череп новорожденного с двойной теценной костью. На этом черепе было хорошо видно, что каждая из двух частей теменной кости имела свою собственную точку окостенения (рис. 297). Это наблюдение подтверждает предположение, что разделение теменной
260
кости наступает в тех случаях, когда обе ее точки окостенения не сливаются друг с другом.
Изучение черепов ископаемых амфибий и рептилий показало, что у них между parietale и squamosum (squama temporalis млекопитающих и человека) помещались еще две кости, именно: intertemporale и supratemporale.
Intertemporale у рептилий скоро исчезло, в то время как supratemporale еще долго сохранялось и подверглось полной редукции только у горгонопсов. Однако очень возможно, что в данном случае мы имеем дело не с редукцией supratemporale, а с ее прирастанием к краю parietale. Такое явление наблюдалось в черепах у некоторых реПтилий, например у Nyctiphruretus.
Рис. 297. Череп ребенка с os parietale bipartitum (Швальбе — Schwalbe, 1903).
Все это дает право думать, что в случаях разделения теменной кости на две части мы имеем дело с появлением вновь самостоятельного supratemporale. А если это так, то человек, имеющий os parietale bipartitum, повторяет особенности строения черепа какого-то своего далекого предка из класса рептилий.
Лобная кость (os frontale) на всем протяжении эволюции рыб, амфибий и рептилий была парной костью. Парной костью она оставалась и у всех млекопитающих, за исключением обезьян. У обезьян обе половины лобной кости разделяются швом только в раннем возрасте; у взрослых животных этот шов зарастает и лобная кость становится непарной.
У человека, как и у обезьян, лобный шов (sutura frontalis s. metopica) в норме существует только в раннем детстве (рис. 298, А); на втором году он обычно уже зарастает. Однако наблюдаются случаи, когда этот шов сохраняется в течение всей жизни человека (рис. 298, В). Изучая краниологическую коллекцию в 4250 черепов, я обнаружил его у взрослых людей в 8% случаев.
Лобный шов у взрослого человека, несомненно, является атавистическим вариантом, но в данном случае мы имеем дело с повторением анатоми-
261
веских особенностей относительно недавних предков человека из класса млекопитающих; эти предки были предшественниками обезьян. Этим, по-видимому, и объясняется тот факт, что лобный шов наблюдается так часто: у 8 человек из 100. Нет ничего удивительного и в том, что и у обезьян иногда
Рис. 298.
А — череп новорожденного; В — череп старика с незаросшим лобным швом.
наблюдаются случаи сохранения лобного шва в течение всей жизни (рис. 299). Такие варианты были описаны, например, Швальбе (G. Schwalbe, 1904) и Больком (L, Bolk, 1913).
Рис 299. Лобный шов у взрослых обезьян. А —Colobus guerezza (Швальбе, 1904); В — Macacus cynomolgus (Больк, 1913)
Рассматривая последовательные фазы развития черепа рептилий, мы видели, что появление особых горизонтальных отростков на maxillare и pajatinum у цинодонтов привело к образованию у них вторичного костного нёба. Таким образом, полость носа отделилась от полости рта при 262.
помощи костной перегородки еще у рептилий. Эта особенность затем перешла к млекопитающим.
Изучение эмбрионального развития млекопитающих показывает, что твердое нёбо (palatum durum) у каждого зародыша возникает в результате образования горизонтальных пластинчатых отростков на верхних челюстях и на нёбных костях. Постепенно разрастаясь, эти отростки затем сходятся по срединной линии и полностью отделяют полость носа от полости рта.
Рис.'300. Небольшое
расщепление твердого иёба.
Рис. 301. Дефекты твердого нёба.
У взрослого человека, хотя и редко, но все же встречаются отклонения от нормального строения твердого нёба.
В коллекции, содержащей 4250 черепов, я обнаружил 9 подобных случаев. При этом наиболее часто наблюдалось недоразвитие горизонтальных пластинок нёбных костей и частичное недоразвитие нёбных отростков верхней челюсти. В результате этого возникает треугольный дефект в заднем отделе твердого нёба (рис. 300, А, В). Иногда этот дефект бывает настолько велик, что достигает середины нёба (рис. 301, Л). Очень редко встречаются случаи полного расщепления твердого нёба на всем его протяжении (рис. 301, В). Такая ненормальность, сопровождающаяся дефектом и в слизистой обо
263
лочке крыши рта, почему-то получила название «волчьей пасти», хотя у нормального волка ничего подобного не наблюдается. Гораздо правильнее латинское и греческое названия этого варианта: palatum fissum, urano-schisma, что значит расщепленное нёбо.
Если мы будем смотреть на недоразвитие твердого нёба у человека как на атавистический вариант, то должны будем признать, что в таких случаях человек повторяет особенности строения черепа таких своих предков из класса рептилий, у которых твердого нёба еще не было. Как известно, вторичное нёбо уже имелось у цинодонтов, но оно еще отсутствовало у горгонопсов.
Одна из покровных костей черепа различных ископаемых позвоночных — transversum — в процессе эволюции подвергается постепенной
Рнс. 302. Твердое нёбо. Pl — palatinum; Tr — transversum.
редукции. В черепе кистеперых рыб transversum представляет собой большую и длинную кость, несущую ряд зубов. У стегоцефалов она также велика и также снабжена зубами. У котилозавров и пеликозавров размеры этой кости становятся уже несколько меньшими и у большинства из этих рептилий она не имеет зубов. У горгонопсов и цинодонтов transversum всегда лишено зубов и отличается небольшой величиной. Особенно мала эта кость у цинодонтов.
При переходе от рептилий к млекопитающим transversum полностью исчезло и даже у самых примитивных млекопитающих оно отсутствует. В нормально построенном черепе человека никаких следов этой кости, разумеется, нет. Однако детальное изучение большого количества черепов человека позволило у немногих из них обнаружить небольшую самостоятельную косточку, представляющую собой отделенный швом processus ру-ramidalis нёбной кости (рис. 302, А, В). Эта кость по своему положению точно соответствует небольшому transversum в черепе цинодонтов и, по всей вероятности, является им.
У древних рептилий в образовании краев орбит принимают участие следующие кости: lacrimale, praefrontale, frontale (не у всех), postfrontale, postorbitale и jugale. У наиболее прогрессивных из них (у цинодонтов) postfrontale отсутствует.
У человека орбита ограничена только тремя костями, именно: лобной (frontale), скуловой (zygomaticum=jugale) и верхнечелюстной (maxillare);
264
слезная кость (lacrimale), переместившаяся на внутреннюю стенку орбиты, перестала принимать участие в образовании ее края. Таким образом, из тех костей, которые ограничивали орбиты у рептилий, у человека нет трех костей, именно: praefrontale, postfrontale и postorbitale. Но все эти кости у него иногда наблюдаются в качестве атавистических отклонений от нормального строения черепа.
Praefrontale появляется в виде небольшой косточки, лежащей между лобной, носовой, слезной и верхнечелюстной костями (рис. 303). В тех случаях, когда эта кость представляет собой очень небольшое образование
Рис. 303. Praefrontale (Pf).
она имеет форму клина и располагается в верхнем участке шва между слезной костью и лобным отростком верхней челюсти. При этом ее верхний конец на некотором протяжении соединяется с лобной костью (рис. 304, А, В). В тех случаях, когда ее размеры бывают относительно велики, она соединяется и с носовой костью. В таких случаях верхняя челюсть уже не имеет контакта с лобной костью (рис. 303, 304, С).
Витольс (Т. М. Vitols, 1930), занимавшийся изучением этой кости, не считал ее гомологом praefrontale и называл ossiculum maxillo-frontale. Он утверждал, что положение этой кости в черепе человека не позволяет признать в ней praefrontale.
Мне кажется, что изучение основных этапов эволюции костей черепа на палеонтологическом материале дает нам право утверждать, что в данном случае мы имеем дело именно с praefrontale.
Я не согласен с Витольс и считаю, что ossiculum maxillo-frontale занимает в черепе человека именно такое место, которое занимает praefrontale в черепе рептилий. Нас не должна смущать очень небольшая величина этой кости у человека, так как в процессе эволюционных изменений черепа praefrontale подверглось постепенной редукции и полностью исчезло при переходе от рептилий к млекопитающим. Поэтому нет ничего удивительного в том, что, вновь появляясь у человека, она обычно бывает очень маленькой.
По моим подсчетам у человека эта кость наблюдается в 1% случаев (42 случая на 4250 черепов).
265
У нас нет, разумеется, никаких оснований отрицать возможность появления такого же варианта и у любого представителя современных млеко-питающих, так как рептилии были и их предками. И такие случаи наблюдались. Например, Ле-Дубль (A. F. Le-Double, 1906) в своей сводке, посвященной вариантам черепа, дал рисунок черепа быка (Bos taurus), у которого имелось хорошо развитое praefrontale, а Брэдли (Bradley, 1925) описал череп льва с такой же костью.
Отсутствие большого количества подобных наблюдений объясняется, несомненно, только тем, что варианты строения черепа млекопитающих очень мало изучаются, так как в остеологических музеях нет таких коллекций, в которых каждый вид млекопитающих был бы представлен, если не тысячами, то по крайней мере несколькими сотнями черепов.
Рис. 304. Praefrontale (Prf) различной формы и величины.
F — frontale; L — lacrimale; N — nasale; M — maxillare; Z — zygomaticum.
Уже одно то обстоятельство, что скуловая кость (os zygomaticum) -человека, по данным эмбриологии, закладывается в соединительной ткани в виде двух или трех отдельных центров окостенения, заставляет предполагать, что в ее состав вошли две или три покровные кости предковых форм. Эти центры окостенения рано сливаются друг с другом и в дальнейшем скуловая кость эмбриона развивается как единое образование. Но совершенно естественно, что при таком ходе развития изоляция одного из центров окостенения или неслияние всех трех может привести к разделению скуловой кости на две или на три части.
Наиболее часто наблюдается разделение скуловой кости на две части. Этот вариант получил название os zygomaticum bipartitum (Матигка — Н. Matiegka, 1899; Тольд — К. Toldt, 1903).
В краниологической коллекции, содержащей 4250 черепов, я обнаружил двойную скуловую кость в 20 случаях (0,47%). Кость разделялась горизонтальным швом на две неравные части; верхняя принимала участие в образовании орбиты, нижняя служила местом начала жевательной мышцы (m. masseter) и немного выступала вниз (рис. 270, 305).
Иногда встречаются случаи, когда нижний участок такой двойной скуловой кости оказывается приросшим к верхней челюсти и представляет собой как бы ее длинный отросток, принимающий участие в образовании скуловой дуги (рис. 306).
Некоторые анатомы называют отделившуюся нижнюю часть скуловой кости «японской костью, (os japonicum) или «костью айнов» (os ainonicum), утверждая, что она у японцев и айнов встречается чаще, чем у представителей других народов. Возможно, что это мнение так же неверно, как ока-266
залось неверным мнение Беллами и Чуди относительно «кости инков» (os incae).
Несравненно реже встречаются такие черепа, у которых двойная скуловая кость разделяется горизонтальным швом, проходящим выше, чем
Рис. 305. Os zygomaticum bipartitum.
обычно. В таких случаях лобный отросток кости оказывается отделенным (рис. 307, В).
Еще реже наблюдается разделение скуловой кости на три части (os zygomaticum tripartitum, рис. 307). Среди 4250 черепов я нашел такую кость только у одного (рис. 307, F).
Рис. 306. Os zygomaticum bipartitum.
Все эти варианты показывают, что скуловая кость образовалась в результате сращения трех костей. При этом верхняя из них, соединяющаяся с лобной костью, по всей вероятности, гомологична postfrontale рептилий;
267
средняя, принимающая участие в образовании края орбиты, соответствует postorbitale и, наконец, нижняя представляет собой jugale. (Прежде, 1947, я считал нижнюю часть os zygomaticum bipartitum гомологом quadratojugale рептилий. Теперь я отказываюсь от такой трактовки и думаю, что
Рис. 307. Os zygomaticum bipartitum и tripartitum (Матигка, 1899)
quadratojugale редуцировалось полностью и в черепе млекопитающих никогда не появляется.)
Как известно, в образовании сложной нижней челюсти примитивных рептилий принимало участие девять покровных костей. В процессе эволюционного развития представителей этого
Рис. 308. Ossiculum mentale (Адаши, 1904; Тольд, 1905).
класса позвоночных происходила постепенная редукция всех костей нижней челюсти, за исключением dentale. Эта кость, наоборот, постепенно увеличивалась и при переходе от рептилий к млекопитающим превратилась в единственную кость их нижней челюсти (man-dibula).
Очень интересным является тот факт, что из редуцирующихся покровных костей нижней челюсти рептилий spleniale отставала в своей редукции и была еще достаточно большой костью у таких тероморфных рептилий, как горгонопсы и циноДонты; она располагалась у них на внутренней стороне нижней челюсти и принимала участие в образовании соединения обеих ее ветвей.
У млекопитающих spleniale не сохранилось, но очень возможно, что
именно его остатки мы наблюдаем у человека в виде подбородочных косточек (ossicula mentalia). Эти маленькие
косточки иногда встречаются в качестве добавочных окостенений в нижнем отделе подбородочного шва нижней челюсти у детей (Адаши — В. Adachi, 1904; Тольд, 1905).
268
Они бывают парными (рис. 308, А), частично сращенными друг с другом (рис. 308, В) или имеют форму небольшого костного клина, вставленного снизу в подбородочное соединение (рис. 308, С).
После срастания обеих половин нижней челюсти подбородочные косточки исчезают бесследно и у взрослого человека никогда не наблюдаются.
Сильное развитие dentale и превращение его у млекопитающих в тап-dibula сопровождалось соответствующим изменением зубного аппарата и
дифференцировкой жевательной мускулатуры. Mm. temporales у обезьян обычно прикрывают всю или почти всю мозговую часть черепа. У человека они значительно слабее, и расстояние между верхними краями правого и левого мускула у него оказывается равным 12—15 см.
Однако иногда удается находить такие черепа человека, у которых на темени расстояние между полями, занятыми началом широких веерообразных височных мышц, очень мало. Например, на черепе, изображенном на рис. 309, оно было равно только 4 см.
Такое сильное развитие височных цышц, очевидно, является повторением особенностей обезьянообразных предков человека, которые, несомненно, имели очень мощную жевательную мускулатуру.
Подводя итог сделанному нами обзору атавистических вариантов костей черепа человека, мы должны признать, что изучение разнообразных и иногда редких отклонений от нормы позволяет обнаружить немало давно исчезнувших костных элементов. В виде иллюстрации этого на рис. 310 дано схематическое изображение черепа
человека, на котором нанесены наряду с обычными костями и все те, которые встречаются только в качестве атавистических вариантов.
Такие кости, как parasphenoldeum, intertemporale, quadratojugale, supraangulare, postspleniale, coronoideum, intercoronoideum и praecorono-ideum, в черепе человека никогда не появляются.
Это, по всей вероятности, объясняется различной судьбой редуцирующихся в процессе эволюции покровных костей.
В одних случаях сильно уменьшившиеся в своих размерах покровные кости совсем перестали закладываться даже в виде самостоятельных точек окостенения. Такие кости исчезали окончательно и уже не могли возникать вновь даже в качестве временных вариантов.
В других случаях сильно редуцированные кости сливались с нормально развитыми соседними костями. Такие кости закладываются у эмбриона в виде самостоятельной точки окостенения и обычно присоединяются к точке
269
окостенения своего соседа. Естественно, что такая кость может возникнуть вновь в виде самостоятельного образования, если ее закладка не срастается с закладкой соседней кости. По-видимому, только такие кости мы и обнаруживаем в черепе как атавистические варианты его строения.
В анатомической литературе принято исключать зубы из описания скелета и рассматривать их в главах, посвященных спланхнологии. Все анатомы считают их тесно связанными с системой органов пищеварения и поэтому описывают наряду со слюнными железами и языком.
Рис. 310.
Ап — angulare (pars tympanica ossis temporalis); D — dentale (mandlbula); Dso — dermosupraoccipitale; Eo — exoccipitale: Ep — epipterygoideum (ala magna ossis sphenoidalis); F — frontale; Pm — intermaxillary (praemaxillare); Ju — jugale; L — lacrlmale; M — maxillare; N — nasale; P — parietale; Pf — postfrontale Po — postorbitale; Poe — paroccipitale; Prf — praefrontale; So — supraoccipitale; Sp — spleniale; Sq — squamosum (squama ossis temporalis); St — supratemporale; T — tabulare.
Такое выделение зубов из остеологии следует признать совершенно неправильным.
Как показывает палеонтология, на первых этапах эволюции позвоночных их зубы были разбросаны по всей поверхности тела и не имели никакого отношения к акту захватывания пищи. После появления покровных костей они оказались тесно связанными с их наружной поверхностью, и только тогда, когда возникли челюсти, некоторые зубы стали прикрепляться к ним и переменили свою функцию, превратившись из органов пассивной защиты в органы нападения и обработки пищи.
Если эту .вторичную функцию зубов считать достаточной причиной, чтобы относить их к органам пищеварения, то следовало бы и челюсти исключить из остеологии, тем более, что они никогда и никакой иной функции, кроме функции захватывания пищи, не выполняли. . .
Постепенная дифференцировка зубов на протяжении эволюции рептилий и млекопитающих привела к оформлению резцов, клыков, малых коренных и больших коренных зубов.
270
Зубные формулы человека и узконосых обезьян, как известно, не отли чаются друг от друга и выражаются следующими цифрами:
2. 1. 2, 3
2. 1. 2. 3 ’
Но у более примитивных млекопитающих зубов было больше. Например, представитель сумчатых — опоссум (Didelphys') — имеет:
5. 1. 3, 4
5. 1. 3. 4 '
Рис. 311. Присутствие четвертого большого коренного зуба.
Рис. 312. Присутствие трех резцов (4) и четырех больших коренных зубов (В).
Поэтому всякое появление лишнего зуба у человека мы должны рассматривать как «воспоминание» о каком-то его предке из группы очень примитивных млекопитающих.
Наиболее частым вариантом количества зубов у Человека следует считать появление четвертого большого коренного зуба.
Обычно этот лишний зуб бывает очень небольшим (рис. 311, Л), но в некоторых случаях его размеры оказываются почти равными так называемому «зубу мудрости», т. е. третьему коренному зубу (рис. ЗГ1, В; 312, В).
271
Значительно реже у человека наблюдаются три резца вместо двух (рис. 312, А; 313, В).
Еще реже встречаются такие случаи, когда появляются три нормально развитых малых коренных зуба (рис. 313)
Насколько мне известно, случаев увеличения количества клыков (два вместо одного) у человека никогда не наблюдалось. Это, несомненно, объ-
ясняется тем, что ни у одного из его предков двух клыков не было.
Рис. 313.
А — нижняя челюсть с тремя малыми коренными зубами на каждой стороне; В — нижняя челюсть с шестью резцами.
В костях плечевого и тазового пояса, так же как и в костях верхней и нижней конечности человека, атавистические варианты почти не встречаются. Это, по всей вероятности, объясняется тем, что строение этих частей скелета у человека сохранило очень примитивный характер.
На протяжении всей эволюционной истории человека его кисть не подвергалась сильной специализации и сохранила свое примитивное строение. В ней не уменьшилось количество пальцев до одного, как у непарнокопытных, в ней не увеличилась длина фаланг, как у летучих мышей, и не увеличилось количество фаланг, как у некоторых китов, — кисть сохранила основы того строения, которое она получила в момент выхода позвоночных из воды на
сушу. Она оставалась у различных предков человека неизменной на протяжении всего палеозоя и
мезозоя. В начале кайнозоя она дала возможность нашим предкам перейти от жизни на земле к жизни на деревьях.
Отсутствие односторонней специализации кисти позволило ей выполнять не одну, а несколько различных работ. Именно эта особенность кисти, обусловленная общей примитивностью ее строения, затем дала возможность первым людям с успехом пользоваться ею при самых различных видах труда. А способность трудиться для своей и общей пользы была, пожалуй, одним из важнейших отличий человека от обезьяны. Как известно, Ф. Энгельс был первым, кто понял то значение, которое имел труд на первых этапах формирования человека.
Что касается стопы, то она у человека, несомненно, специализирована и может выполнять только одну функцию. По своему общему строению она очень похожа на стопу тех примитивных древних млекопитающих, которые при ходьбе еще опирались не только на концы метатарзальных костей, но и на пяточную кость. Эта особенность сохранилась и у человека.
272
Древние стопоходящие млекопитающие имели плоскую стопу, т. е. все ее кости при опоре располагались горизонтально по отношению к грунту.
У человека в отличие от этих примитивных форм стопа имеет высокий свод. Поэтому человек опирается не на все кости своей стопы, а только на головки метатарзальных костей и на пяточный бугор. Свод стопы мы унаследовали от своих обезьянообразных предков. Стопа с приподнятым внутренним краем была выработана ими в качестве приспособления к жизни на деревьях. Такая стопа позволяла крепче схватывать вертикально или почти вертикально стоящие стволы деревьев и их сучья.
Рис. 314.
А — нормальная левая плечевая кость человека;
В — плечевая кость с processus supracondyloideus;
С — плечевая кость с foramen entepicondyloideum.
Пожалуй, единственным интересным атавистическим вариантом в строении скелета конечностей человека является так называемый processus supracondyloideus humeri. Этот отросток иногда наблюдается на внутренней стороне плечевой кости в нижнем ее участке и всегда бывает направлен своим концом вниз (рис. 314, В). В очень редких случаях конец отростка прирастает к кости (рис. 314, С). В результате этого на внутренней стороне плечевой кости образуется отверстие, которое по своему положению вполне соответствует такому же отверстию (foramen entepicondyloideum) на плечевой кости многих млекопитающих и рептилий.
ЛИТЕРАТУРА
Бунак В. В., Нестурх М. Ф., Рогннскнй Я. Я- Антропология. 1941.
Быстров А. П. (В у s t г о w). Assimilation des Atlas und Manifestation des Proatlas. Zeitschrift fur Anatomie und Entwiklungsgeschichte, Bd. 95, 1931.
Быстров А. П. О признаках низших позвоночных в черепе человека. Сообщение I. Лабиринт. Труды Военно-морской медицинской академии, т. Ill, 1944.
Быстров А. П. Idem. Сообщение II. Os zygomaticum bipartitum. Труды Воеино медицинской академии, т. 38, 1947.
Быстров А. П. Idem. Сообщение III. Os fronto-naso-maxillo-laerimale. Ibidem т. 38, 1947.
Дешин А. А. К вопросу о выявлении передней дуги последнего затылочного позвонка (склеромера) у человека. Бюллетень Моск, об-ва испытателей природы, 1926.
Ибрагимова 3. И. Редкая аномалия костного внутреннего уха у человека. Труды Узбекского гос. ун-та, т. 5, 1940.
Энгельс Ф. Роль труда в процессе очеловечения обезьяны, 1932.
18 А П. Быстров
273
A d a c h i B. Ober die Knochelchen in der Symphyse der Unterkiefer. Zeitschrift fur Morphol. und Anthropol., Bd. 7, 1904.
Bellamy P. F. A brief account of two Peruvian mummis. The Annals and Magazine of Natural History, vol. 10, London, 1842.
В о 1 к L. Ober die Obliteration der Nahte am Affenschadel. Zeitschrift fur Morph, und Anthropol. Bd. 15, 1913.
В о 1 к L. Ober unvollstandige Assimilation des letzten Occipitalwirbels des Menschen. Anatomischer Anzeiger, Bd. 55, 1922.
Bonnet R. Der Skaphocephalus synostoticus des Stettiner Webers. Eine Studie. Wiesbaden, 1904.
Bradley. Notes on the skull of a lion. Anatomischer Anzeiger, Bd, 27, 1905.
C a b i b b e G. Il processo postglenoideo nei crani di normali, alienati, criminal! in rapporto a quello dei varii ordini di mammiferi. Anatomischer Anzeiger, Bd. 20, 1901.
Eaton G. The collection of osteological material from Machu Picchu. Memoirs of the Connecticut Academy of Arts and Sciences, vol. 5, 1916.
G о e t h e W. Morphologic. Goethes samtliche Werke. Bd. 32, Stuttgart.
Ko 1 1 m a n n J. Variation am Os occipitale. Anatomischer Anzeiger, Bd. 30, 1907.
Krasti n L. Hamapophysis beim Menschen. Ibidem, Bd. 65, 1928.
Де-Л а н д а Сообщение о делах в Юкатане (1566). Изд. АН СССР, 1955.
Le-D о u b 1 е A. F. Traite des variations des os de la face de 1’homme, Paris, 1906.
Matiegka H. Ober das «Os malare bipartitum». Anatomischer Anzeiger, Bd. 16, 1899.
Montagu M. An introduction to physical anthropology. USA, 1945.
Ok en L. Lehrbuch der Naturphilosophie. 3. Auflage. Zurich, 1843.
Schawlowski J. Ober einige seltane Variation an der Wirbelsaule beim Menschen. Anatomischer Anzeiger, Bd. 20, 1901.
Schwalbe G. Ober geteilte Scheitelbeine. Zeitschrift fur Morphol. und Anthropol., Bd. 6, 1903.
Schwalbe G. Ober die Stirnnaht bei den Primaten. Ibidem, Bd. 7, 1904.
Swjetschnikow. Ober Assimilation des Atlas und Manifestation des Occipital-wirbel beim Menschen. Archiv fflr Anatoinie, 1906.
Swjetschnikow. Ober die Variation des Occipitalwirbels. Anatomischer Anzeiger, Bd. 32, 1908.
T о 1 d t K. Die Querteilung des Jochbeines und andere Varietaten des$elben. Sitzungs-berichte der K. Akademie der Wissenschaften, Bd. 112, Abt. Ill, 1903.
T о 1 d t K. Die Ossicula mentalia und ihre Bedeutung fflr die Bildung des menschlichen Kinnes. Ibidem, Bd. 114, 1905.
T s c h и d i V. Ober die Ureinwohner von Peru. Muller’s Archiv, 1844.
V i t о 1 s T. M. Ober das «Nebentranenbein» des Menschen. Anatomischer Anzeigsr, Bd. 69, 1930.
ЧАСТЬ ТРЕТЬЯ
БУДУЩЕЕ
Если понятия прошлого и настоящего в применении к эволюционной истории человека потребовали больших оговорок, то и понятие будущего нуждается в некотором уточнении.
С точки зрения философа будущим нужно считать не только все, что наступит непосредственно вслед за мгновением настоящего, но и все, что случится на протяжении такого времени, длительность которого равна бесконечности.
Мы можем согласиться с таким общим представлением о будущем. Однако следует оговориться, что ближайшее будущее человека не может интересовать морфолога, так как у него нет никаких оснований ожидать на протяжении нескольких предстоящих веков каких-либо изменений в анатомическом строении человека. Наоборот, у него имеются все основания не ожидать таких изменений.
Эволюционный процесс является процессом очень медленным. Судя по поразительно точным изображениям диких животных на стенах египетских и ассирийских дворцов и храмов, эти животные за два или три тысячелетия не подверглись никаким видимым изменениям; они ничем не отличаются от тех, которые существуют и в настоящее время. Это дает нам право думать, что и человек в течение ближайших тысячелетий останется таким же, каким мы знаем его теперь.
Однако человека ожидает не только ближайшее, но и несомненно очень далекое будущее; у нас нет никаких оснований сомневаться в том, что его существование будет достаточно продолжительным. В настоящее время наука не располагает никакими данными, указывающими на то, что жизнь человечества ограничена каким-то неизбежным концом.
Если оставить в стороне все разнообразные и обычно достаточно фантастичные гипотезы о гибели нашей планеты, то мы не можем указать никаких причин, которые могли бы помешать человеку просуществовать на Земле еще не один миллион лет. В сущности нет ничего невероятного и в том, что он будет существовать не один десяток, а может быть и не одну сотню миллионов лет. . .
Постепенно развивая свою технику, он, по всей вероятности, в ближайшие тысячелетия займет все без исключения участки суши и овладеет природой настолько, что станет единственным и полновластным хозяином повсюду. Вытеснив всех мешающих ему животных из мест их обитания, он сохранит их в специальных заповедниках только для того, чтобы иметь-представление о животном мире прошлого.
Имея в виду очень продолжительное существование человека в будущем, мы, естественно, должны поставить вопрос, что будет с ним через-несколько миллионов лет; иными словами, как отразятся предстоящие миллионы лет на анатомическом строении наших далеких потомков.
18* 275-
Если эволюционные изменения позвоночных на протяжении ряда миллионов лет привели к появлению человека, то кажется совершенно естественным, что прожитый им новый ряд миллионов лет не может пройти для его организма бесследно. Поэтому многие находят совершенно логичным Допущение, что Homo futurus (человек будущий) будет отличаться от современного человека так же сильно, как современный человек отличается от своих далеких предков, и что, следовательно, наши далекие потомки не могут быть похожими на нас. Поэтому, если будущие люди будут производить раскопки на наших кладбищах так же, как мы в пещерах, где жили неандертальцы, то они должны будут невольно поражаться грубостью и примитивностью строения наших скелетов.
Все эти соображения не являются чем-нибудь новым; они уже высказывались многими и не один раз. Например, известный французский астроном Фламмарион (С. Flammarion) в своей «Популярной астрономии» писал: «Новая раса, умственно более развитая, займет наше место на Земле, и кто знает, не встретимся ли мы когда-нибудь с вами, серьезный читатель, или с вами, мечтательная читательница, в кабинете какого-нибудь ученого 276-го века в виде белых величественных скелетов с этикетками на лбу. . . На нас будут смотреть как на любопытные экземпляры вымершей расы, довольно грубой и жестокой, но уже обладавшей зачатками культуры и цивилизации и отличавшейся некоторой склонностью к занятию науками. . .»
ГЛАВА I
ЧЕЛОВЕК ДАЛЕКОГО БУДУЩЕГО (HOMO FUTURUS)
Большинство морфологов не сомневается в том, что эволюционные изменения человека происходят в настоящее время и будут продолжаться в будущем.
Так, А. Н. Северцов, всю жизнь занимавшийся изучением эволюционного процесса, по этому вопросу говорил (1910) следующее:
«Эволюционное учение, раскрыв прошлое филогенеза человека, приступило к изучению направления его современного развития и ставит вопрос о том, нет ли возможности ускорить и направить прогрессивную эволюцию человека в будущем» (А. Н. Северцов, 1945).
В. М. Шимкевич (1923) писал: «Если возможен был в прошлом человечества физический прогресс, то ничто не противоречит предположению возможности дальнейшего прогресса в этом направлении». Оценивая результаты этого прогресса, Шимкевич находил, что «нельзя отрицать, что человек будущего может настолько же отличаться от нас физически и психически, насколько мы отличаемся от дикарей».
Наконец, Б. С. Жуков в своей популярной книжке по вопросу о происхождении человека (1925, 1928) уверенно заявлял, что «современная ступень человеческой эволюции еще далеко не последняя. Эта эволюция еще не закончена и человечеству предстоят еще многие тысячелетия жизни и прогресса».
Такие взгляды разделяются и анатомами, которые уже не первое столетие с величайшим усердием занимаются точным описанием все новых и новых деталей строения человека. Работая в этом направлении со времен основоположника анатомии Андрея Везалия (1514—1564), они изучили человеческий организм настолько, что открытие в нем чего-нибудь прежде неизвестного сделалось уже невозможным. Поэтому современные анатомы, цитируя десятки и сотни работ своих предшественников, все снова и снова, все детальнее и детальнее переописывают строение различных органов человека, создавая огромную литературу, в которой можно найти очень много, но уже трудно найти то, что нужно.
Многолетнее и всестороннее изучение трупов привело к тому, что в руках анатомов накопился огромный материал, показывающий очень большую вариабильность строения человека. У него не оказалось ни одного органа, в котором при детальном его исследовании не были бы обнаружены те или иные отклонения от так называемого нормального строения.
«При изучении нормальной анатомии человека,— писал Н. Д. Буш-макин (1913),— встречаются на каждом шагу те или другие уклонения от нормального строения».
277
Это обстоятельство привело к тому, что многие анатомы с особенным увлечением стали заниматься описанием различных ненормальностей.
Сначала эти ненормальности регистрировались под общим названием «аномалий» и никак не трактовались. Но затем под влиянием учения Ч. Дарвина о происхождении человека (1871) исследователи стали подходить к ним как к явлениям, имеющим значение для доказательства генетической связи человека с животными. Однако при этом оказалось, что не все «аномалии» свидетельствуют о происхождении человека от животных; некоторые из них, несомненно, не имеют никакого отношения к эволюционной истории человека в прошлом. Это заставило морфологов разделить все разнообразные отклонения от нормы на две группы.
1. Регрессивные аномалии. К этой группе относятся такие отклонения от нормального строения человека, которые представляют собой «более или менее точное воспроизведение особенностей строения нормально существующих в различных отрядах животных и тесно связанные с филогенезом человека» (В. В. Бунак, М. Ф. Нестурх, Я. Я- Рогинский, 1941). Эти аномалии, как известно, обычно называются атавистическими, так как они являются повторением некоторых особенностей строения тех или иных предков человека. Мы уже рассмотрели их в одной из предыдущих глав (стр. 247—273),
2. Прогрессивные аномалии. Сюда относятся варианты строения человека, «нормально не встречающиеся у каких-либо животных». По мнению анатомов, такие прогрессивные варианты «также теснейшим образом связаны с филогенезом человека» (В. В. Бунак, М. Ф. Нестурх, Я. Я. Рогинский, 1941) и указывают на направление дальнейшей эволюции человека.
«Уже давно было отмечено,— писал В. М. Шимкевич (1923),— что в организации скелета наряду с колебаниями в сторону более или менее отдаленных предков человека — колебаниями атавистическими — существуют колебания в сторону будущего . . . колебания профетические» . . ?
По убеждению анатомов « . . .человеческий организм не представляет собой нечто неизменное, законченное, а является лишь одной из последовательных стадий своего развития; изменения, в нем совершающиеся, протекают так медленно, что не доступны нашему прямому наблюдению, и лишь различные аномалии и уродства, как опознавательные вехи, указывают нам пути его эволюции. Пути эти в настоящий момент лишь только намечаются и поэтому наша обязанность подробно описывать все наблюдаемые нами аномалии и уродства, чтобы на основании этого сырого материала последующие исследователи могли не только предугадывать, но и точно знать то направление, по которому совершаются филогенетические изменения в нашем организме» (Н. Д. Бушмакин, 1913).
Исходя из такого теоретического положения и занимаясь детальным описанием разнообразных «прогрессивных аномалий» отдельных частей скелета человека, анатомы обычно пытаются наметить ту форму, которую, по их мнению, должна будет принять изучаемая часть у человека далекого будущего.
Как известно, от длинного хвоста предков у человека сохранилось только четыре-пять полуредуцированных позвонков копчика (рис. 315, А, В). При этом наблюдения анатомов показывают, что иногда количество копчиковых позвонков у человека уменьшается до трех (рис. 315, С). Этот вариант обычно рассматривается как прогрессивный, т. е. указывающий на то, что у человека далекого будущего три позвонка копчика будут нормальным явлением. Теоретически предполагается, что по мере дальнейших из-
1 Propheta пророк.
278
менений скелета человека его копчик будет постепенно редуцироваться и в конце концов исчезнет полностью. В этот момент человек потеряет последние остатки хвоста своих предков.
Рис. 315. Копчик человека. А—из пяти позвонков; В—из четырех; С — из трех.
При изучении вариантов строения крестца очень нередко отмечаются случаи прирастания к нему V поясничного позвонка.
Рис. 316. Sacralisatio vertebrae lumbalis quinti.
В нормально построенном скелете последний поясничный позвонок соединяется с крестцом так же, как обычно позвонки соединяются друг с другом (рис. 316, А).
279
В случаях же первой фазы срастания на одном из поперечных отростков V поясничного позвонка появляется большой направленный вниз костный отросток, который соединяется с боковой частью крестца или при помощи плоского сустава или путем полного костного сращения (рис. 316, В). В последнем случае всякая подвижность между поясничным позвонком и крестцом исчезает, хотя между телом V поясничного позвонка и телом 1 крестцового еще сохраняется межпозвоночный хрящ.
При еще больших степенях срастания дополнительные отростки появляются на обоих поперечных отростках поясничного позвонка. Встречаются случаи, когда один из них оказывается полностью сращенным с боковой частью крестца, а другой соединяется при помощи хряща. В таких случаях тело V поясничного позвонка обычно бывает частично сращенным с телом первого крестцового (рис. 316, С).
При полном сращении последнего поясничного позвонка с крестцом можно видеть костное соединение обоих поперечных отростков поясничного
Рис. 317. Spina bifida sacralis (Сперанский, 1925).
позвонка с боковыми частями крестца и значительную степень сращения тел позвонков (рис. 316, D). Всякая подвижность между поясничным позвонком и крестцом в этих случаях разумеется отсутствует и сильно разросшиеся поперечные отростки V поясничного позвонка частично принимают участие в образовании сустава между крестцом и подвздошными костями таза. В результате всего этого последний поясничный позвонок как бы входит в состав крестца и становится его I позвонком. Такое явление в анатомии человека получило название sacralisatio vertebrae lumbalis quinti и в противоположность lumbalisatio vertebrae sacralis primae считается прогрессивным явлением (К. 3. Яцута, 1932). По мнению анатомов, присоединение последнего поясничного позвонка к крестцу указывает на «н а к л о н-н о с т ь тазового пояса передвигаться вперед по позвоночнику» (Видерс-гейм, 1900).
«Число позвонков у человека, как и число ребер, идет на убыль, крестец передвигается к голове»,— пишет В. Н. Тонков (1953).
Детальное изучение крестца человека показало, что в нем нередко наблюдается размыкание дуг позвонков, что в некоторых случаях приводит к полному исчезновению задней стенки крестцового канала (рис. 317). Такой вариант строения крестца получил название spina bifida sacralis.
Ни у одного из представителей позвоночных подобных явлений не наблюдалось прежде и не наблюдается теперь. Поэтому данный вариант в строении крестца следует признать свойственным только человеку. Это обстоятельство заставляет анатомов смотреть на spina bifida sacralis как на прогрессивную аномалию. Поэтому нет ничего удивительного в том, что А. Д. Сперанский (1925) после изучения большой коллекции крестцов че
280
ловека писал: «не будет слишком большой неосторожностью высказать предположение, что крестец будущего человека потеряет канал. Его заменит крестцовая борозда, в верхнюю часть которой будет открываться позвоночный канал». Иными словами, по мнению А. Д. Сперанского, для крестца будущего человека резко выраженная spina bifida sacralis (рис. 317, С) будет являться нормой.
Не менее интересные варианты были обнаружены у человека на границе между черепом и позвоночным столбом. Здесь, по наблюдениям ана
томов, нередко встречаются случаи прирастания атланта к затылочной кости. Это явление получило название «ассимиляции атланта» (assimilatio atlantis, рис. 318).
В качестве как бы подготовительной фазы к такому срастанию некоторые исследователи рассматривают появление на затылочной кости большого столбообразного костного отростка (processus рага-mastoideus). На конце этого отростка обычно имеется плоская суставная площадка, которая принимает участие в образовании дополнительного сустава с расширенным концом поперечного отростка I позвонка (рис. 319, А).
В случаях незначительной степени ассимиляции суставы между атлантом и затылочной костью исчезают, в результате чего I позвонок соединяется с черепом совершенно неподвижно; при этом один или оба его поперечные отростка оказываются сращенными с допол-
Рис. 318. Ассимиляция атланта.'
нительными отростками на заты-
лочной кости (processus paramastoideus); однако передняя и задняя дуги
позвонка остаются еще отделенными от переднего и заднего краев затылочного отверстия (рис. 319, В). Такое прирастание атланта к черепу очень напоминает соединение V поясничного позвонка с крестцом (sacralisatio vertebrae lumbalis quinti, рис. 316, D).
При более сильной степени ассимиляции атланта обе его дуги полностью срастаются с затылочной костью (рис. 319, С, D).
Очень нередко слияние I позвонка с черепом сопровождается размыканием его задней дуги (рцс. 319, Е, F).
В анатомической литературе описано несколько редких случаев, когда ассимиляция атланта была настолько полной, что вошедший в состав затылочной кости I шейный позвонок очень напоминал сильно выраженную манифестацию проатланта (рис. 319, G, И).
Анатомы считают ассимиляцию атланта прогрессивным вариантом (Л. Больк, 1906; К. 3. Яцута, 1932; А. П. Быстров, 1933). «Ассимиляцию атланта надо рассматривать,— пишет Н. И. Одноралов (1931),— не как аномалию, "а как одно из звеньев, небольшой штрих в общей картине развития человеческого организма». Следовательно, нужно ожидать, что у будущего человека I позвонок войдет в состав черепа.
28*
Принимая во внимание случаи срастания V поясничного позвонка с крестцом и атланта с черепом, анатомы приходят к выводу, что «позвоночник человека обнаруживает явное стремление сделаться короче» (Н. Д. Буш-макин, 1913). Кроме того, анатомы полагают, что одновременно с постепенным укорочением позвоночного столба у человека происходит и постепенное уменьшение количества ребер, о чем свидетельствуют следующие «прогрессивные» варианты в строении грудной клетки.
Рис. 319. Ассимиляция атланта (А. П. Быстров, 1933).
В анатомической литературе не раз были описаны случаи недоразви--тия I ребра. Такое ребро уже не достигало своим передним концом грудной кости и иногда представляло собой только небольшую кость, соединенную с I грудным позвонком (рис. 320).
Как известно, у человека только 7 пар ребер достигают своими хрящевыми частями грудной кости; хрящи VIII, IX и X ребер последовательно присоединяются к VII ребру и друг к другу; что же касается XI и XII ре-<5ер, то их передние концы свободно лежат в мышцах живота. Длина этих двух ребер сильно варьирует. У XI она колеблется от 28 до 15 см, у XII — -от 27 до 2 см. Уже одно недоразвитие последних двух ребер заставляет анатомов считать их стоящими на пути к полной редукции.
Так как у человека не раз наблюдалось полное отсутствие XII ребра, то многие морфологи, принимая во внимание случаи редукции I ребра, полагают, что «число ребер идет на убыль» (Н. Д. Бушмакин, 1913) и что, следовательно, грудная клетка человека в будущем укоротится (В. Лехе, 1913).
«Несомненно, редукция ребер еще продолжается»,— утверждает В. Н. Тонков (1953).
282
К. 3. Яцута, много занимавшийся изучением всяких вариантов человеческого скелета, совершенно уверенно писал (1932): «Несомненно, что туловище человека проявляет наклонность к укорочению следующим образом: слияние атланта с черепом, . . . выпадение XII (а реже и XI) ребра, ассимиляция крестцом последнего поясничного позвонка. . . Мы лично склонны
считать это явление прогрессивным, связанным с выгодой сохранения равновесия при вертикальном положении человека».
В черепе человека также можно указать некоторое количество '.прогрессивных или профетических (пророческих) вариантов.
Если обратить внимание на носовые кости у ряда ископаемых рептилий — Sey-mouriamorpha -> Cotylosauria -> Pelyco-sauria -> Therapsida, то легко можно убедиться, что в процессе эволюции относительные размеры этих костей постепенно уменьшались. При этом у всех рептилий
‘Рис. 320. Редукция I ребра.
носовые кости еще имели контакте praema-
xillare. Этот контакт исчез у млекопитающих, и носовые кости у представителей этого класса, стоящих на пути, ведущем к человеку, продолжали постепенно уменьшаться. У высших обезьян носовые кости представляют собой очень небольшие элементы черепа.
Рис. 321. Варианты строения носовых костей.
Носовые кости человека являются одними из очень сильно редуцированных покровных костей его черепа. Очень редко они бывают настолько большими и широкими, что лобные отростки верхней челюсти теряют контакт с лобной костью (рис. 321, Л). Такой вариант, по-видимому, следует рассматривать как атавистический, так как отсутствие контакта между frontale и maxillare представляет собой в филогенетическом смысле довольно древнее явление: эти кости не соединяются друг с другом в черепе рептилий и всегда соединены в черепё млекопитающих.
283
Несравненно чаще наблюдаются случаи, когда носовые кости у человека оказываются по своим размерам значительно меньше нормы. При этом они обычно бывают очень узки в своем верхнем отделе (рис. 321, В). Это сужение в некоторых случаях приводит к тому, что одна из носовых костей совсем не достигает лобной кости и представляет собой узкий костный клин, вставленный между лобным отростком верхней челюсти и носовой костью противоположной стороны (рис. 322, А).
В очень редких случаях носовая кость отсутствует совершенно и заменяется разросшимися краями лобных отростков верхнечелюстных костей (рис. 322, В).
Рис. 322.
А — редукция носовых костей; В — полное отсутствие носовых костей.
Я нашел подобные варианты только у двух черепов из 4250.
Уменьшение размеров и полное отсутствие носовых костей у человека, разумеется, не являются атавистическими вариантами, так как у всех предков человека носовые кости не только имелись, но были всегда развиты сильнее, чем у человека. Поэтому кажется совершенно логичным считать эти варианты профетическими и предполагать, что носовые кости у будущего человека должны будут постепенно подвергнуться полной редукции.
Если такие варианты, как отсутствие I и XII ребер, анатомы рассматривают как доказательство того, что грудная клетка человека стремится укоротиться, то отсутствие носовых костей также нужно считать показателем их стремления к исчезновению.
Все, что было сказано о носовых костях, в одинаковой мере относится и к слезной кости. И эта черепная кость на всем протяжении долгой эволюции позвоночных постепенно уменьшалась до тех пор, пока не превратилась у человека в очень небольшую и тонкую костную пластинку, принимающую участие в образовании внутренней стенки орбиты.
Изучение этой кости у человека показало, что она нередко еще более уменьшается в своих размерах и может даже совсем отсутствовать. В таких случаях она заменяется расширенным лобным отростком верхнечелюстной 284
кости, который контактирует непосредственно с передним краем бумажной пластинки решетчатой кости, lamina papyracea ossis ethmoidalis (рис. 323).
Уменьшенные размеры слезной кости по сравнению с ее нормальной величиной и тем более ее полное отсутствие, по-видимому, следует считать профетическими вариациями. «И полное отсутствие и большое разнообразие в степени развития слезной кости у человека свидетельствует о несомненном стремлении ее к редукции»,— пишет Цабель (Е. Zabel, 1900).
Для всех этих пророчеств анатомы могли бы найти теоретические основания в работах В. К. Грегори (W. К. Gregory, 1934, 1935), посвященных
Рис. 323. Отсутствие слезных костей.
так называемому «закону Виллистона» (Williston's Law) и учению о полиизомерии и анизомерии.
В своих общих построениях Грегори прежде всего опирается на наблюдения палеонтолога Виллистона (1852—1918), который отметил, что в процессе эволюции позвоночных количество костей черепа постепенно уменьшается. Подсчеты, произведенные сотрудниками Грегори, дали следующие цифры. Кистеперые рыбы имели в своем черепе 143 кости, стегоцефалы — 90, рептилии (Cotylosauria) — 84, примитивные млекопитающие — 42 и, наконец, человек — 27 костей.
Наличие большого количества отдельных элементов в черепе Грегори называет полиизомерией (poly isomer ism — множество равных частей). Всякое уменьшенео количества костей, вызванное их полной редукцией или срастанием с исседними, Грегори считает проявлением анизомерии (aniso-merism — немного частей — результат нарушения полиизомерии). Таким образом, в процессе эволюции позвоночных примитивная полиизомерия непрерывно заменялась прогрессивной анизомерией, в результате чего количество костей в черепе постепенно уменьшалось («закон Виллистона»).
Грегори убежден, что его полиизомерия и анизомерия проявляются «во всех степенях организации, начиная от внутриатомных структур вплоть до супрагалактик», и что « истематика, палеонтология, сравнительная анатомия, генетика, эксперименстальная биология, физиология и другие науки
285
могут найти для себя новые общие основы в изучении проявления полиизомерии, анизомерии и гиперполиизомерии» (В. К. Грегори, 1934). ,
Если вместе с Грегори верить в силу закона Виллистона и в стремление всех структур развиваться, идя от полиизомерии к анизомерии, то можно не сомневаться в том, что череп будущего человека будет иметь не 27 костей, а значительно меньше.
Крайне редкой аномалией человеческого черепа является отсутствие контакта между скуловой костью и скуловым отростком чешуи височной кости, что приводит к появлению дефекта в скуловой дуге.
Рис. 324. Редкий случай двустороннего дефекта скуловой дуги.
За время своих занятий остеологией я просмотрел несколько тысяч человеческих черепов и встретил такой вариант только однажды (рис. 324). Череп принадлежал взрослому мужчине; скуловые кости (os zygomaticum) у него были недоразвиты и скуловые дуги на обеих сторонах были широко разомкнуты. Во всех других своих частях череп был построен совершенно нормально.
Так как скуловая дуга у всех древних предков человека была построена нормально, то недоразвитие ее мы должны считать профетическим вариантом и предполагать, что для будущего человека разомкнутая скуловая дуга будет нормальным явлением.
Это не будет исключительной особенностью человека, так как эволюционное развитие млекопитающих уже привело к редукции скуловой дуги у некоторых из них и в составе современной фауны мы знаем таких животных, у которых нет скуловой дуги.
Скуловая дуга отсутствует у землероек (Sorex), у муравьедов (Мугте-cophaga) и у некоторых китов. Ее не было и на черепе ископаемого насекомоядного Apternodus из олигоцена Северной Америки.
Почти так же редко, как дефект в скуловой дуге, встречается и недоразвитие межчелюстных костей (os intermaxillare=praemaxillare).
Я видел только один такой случай. Это был череп чукчи из Уэллена (рис. 325). Межчелюстные кости в этом случае были недоразвиты на-286
столько, что отделялись широкой щелью от верхнечелюстных костей и несли только два нормально развитых медиальных резца; сильно редуцированные латеральные резцы помещались на краях верхнечелюстных костей (рис. 326).
Рис. 325. Череп чукчц из Уэллена. Редукция межчелюстных костей.
Рис. 326. Череп чукчи из Уэллена. с. ср. — canalis craniopharyngeus.
В анатомической литературе был описан случай даже полного отсутствия межчелюстной кости (рис. 327; Фишель — Fischel, 1905). При этом вместе с костью, разумеется, отсутствовали и все резцы верхней челюсти. Несмотря на полное отсутствие обеих межчелюстных костей, дефекта на
287
их месте не было, так как края верхних челюстей были сильно сближены и
правый и левый клыки стояли недалеко друг от друга.
Так как в длинной родословной человека никогда не было предков без межчелюстных костей (без praemaxillare), то мы не можем считать отсутствие этих костей атавистическим вариантом. Сильную редукцию и полное отсутствие межчелюстных костей, по-видимому, следует рассматривать как редкий профетический вариант в строении черепа человека, свидетельст-
Рис. 327. Отсутствие межчелюстных костей (Фишель, 1905).
вующии о том, что в очень отдаленном будущем эта кость, подобно слезной и носовой, исчезнет.
Случаи сохранения лобного шва (sutura frontalis s. metopica) у взрослого человека (метопизм, рис. 326, Л) морфологи обычно считают атавистическим вариантом, повторяющим особенности строения лобной кости предков человека из класса млекопитающих. Однако есть мнение, что лобный шов представляет собой не только атавистический, но одновременно с этим и профетический вариант, так как он не только был у предков человека, но и, как предполагают некоторые морфологи, будет у будущих людей.
Впервые эта мысль была высказана П. П. Сушкиным (1915). Он считал, что нередкие случаи сохранения лобного шва у взрослых людей связаны с постепенным увеличением размеров лобных долей головного мозга. Поэтому
у будущего человека, который будет обладать очень большим мозгом, лобный шов будет нормальным явлением.
Некоторые антропологи (Д. Н. Анучин, 1880; Г. Бушан, 1911) утверждают, что у европейцев черепа с сохранившимся лобным швом в настоящее время встречаются чаще, чем у представителей менее культурных на-
родов.
Находясь под влиянием этих антропологов и пользуясь их данными, П. П. Сушкин полагает, что «частота метопизма в культурном человечестве будет возрастать далее» . . . «Таким образом,— пишет он,— из вида Homo sapiens на наших глазах выделяется раса, отличающаяся известным морфологическим признаком».
Взгляды В. М. Шимкевича (1923) на значение лобного шва по существу ничем не отличаются от взглядов П. П. Сушкина. И В. М. Шимкевич считает лобный шов у взрослого человека прогрессивным вариантом.
Таким образом, по мнению этих морфологов, лобный шов у будущего человека не будет зарастать в течение всей его долгой индивидуальной жизни.
У древних людей лобные пазухи (sinus frontalis), как известно, были очень большими. Это обусловило сильное развитие надглазничных дуг (arcus supraorbital is), присутствие которых было характерной особенностью неандертальцев. У кро-маньонцев размеры sinus frontalis были не больше, чем у современного человека.
288
В настоящее время лобные пазухи у человека сильно варьируют. В одних случаях они оказываются очень большими, в других — незначительными. В редких случаях наблюдается полное отсутствие их.
Отсутствие лобных пазух, по-видимому, нужно считать прогрессивным (профетическим) вариантом, и если мы признаем, что в процессе эволюции человека в прошлом объем его лобных пазух постепенно уменьшался, то следует думать, что у будущего человека эти полости в лобной кости совсем исчезнут.
Всем хорошо известен тот факт, что третий большой коренной зуб, так называемый«зуб мудрости», прорезывается с большим запаздыванием.Обычно он начинает функционировать только в 17—18 лет. Однако нередко наблю
Рис. 328.
А — отсутствие зубов мудрости; В — недоразвитие зубов мудрости.
даются случаи, когда его прорезывание задерживается до 30 и 40 лет В некоторых случаях он не развивается совсем (рис. 328, Д).
По своей форме и размерам «зуб мудрости» очень сильно варьирует. В одних случаях он по своей величине бывает почти равен размерам второго большого коренного зуба, в других — приобретает форму небольшого столбика (рис. 328, В; 329, Л).
Все эти наблюдения привели морфологов к выводу, что «третий коренной зуб у нынешнего культурного человечества медленно регрессирует» (Г. Бушан, 1911). Этот взгляд настолько распространен, что почти каждый автор, говоря о зубах, считает себя обязанным заявить, что «зуб мудрости» , человека в будущем обречен на бесследное исчезновение.
«Самый задний из наших коренных зубов,— пишет В. Лехе (1913),— так называемый зуб мудрости, представляет собой пример органа, который, по крайней мере у цивилизованных народов, находится на пути к вырождению». . . «Совершающийся на наших глазах процесс исчезновения зуба мудрости идет. . . с большой постепенностью»,— утверждает В. М. Шимкевич (1923).
«Нет и тени сомнения в том, что мы собираемся потерять наши зубы мудрости, которые в настоящее время исчезают», — говорит Хауэлс (W. Howells, 1946).
Все это в настоящее время считается настолько очевидным и доказанным, что обычно ни у кого не возбуждает никаких сомнений; все давно примирились с предстоящей потерей этих зубов.
3/<19 А. П. Быстров
289
Детальное изучение зубной системы человека показывает, что при отсутствии зуба мудрости второй большой коренной зуб в некоторых случаях оказывается недоразвитым: его размеры бывают меньше обычных (рис. 329, В) и иногда он имеет на своей коронке три бугорка вместо четырех (рис. 330, В). Подобные варианты строения второго моляра анатомы
Рис. 329.
А — недоразвитие зубов мудрости, недоразвитие правого латерального резца и отсутствие левого; В — отсутствие зубов мудрости, недоразвитие вторых больших коренных зубов, отсутствие малого коренного зуба слева, отсутствие латерального резца справа и недоразвитие такого же зуба слева.
считают прогрессивными (Мартин — R. Martin, 1928) и полагают, что после исчезновения зуба мудрости та же судьба постигнет и этот зуб, и он сначала некоторое время будет прорезываться с запозданием, затем сильно уменьшится в своих размерах и, наконец, подвергнется полной редукции.
Рис. 330.
А — отсутствие зубов мудрости, отсутствие вторых малых коренных зубов; В — отсутствие зубов мудрости и недоразвитие вторых коренных зубов.
Прогрессивными же вариантами, по мнению анатомов, являются и случаи полного отсутствия второго малого коренного зуба (рис. 329, В; 330, Д).
Очень нередко у человека наблюдается недоразвитие боковых резцов; при этом они сильно уменьшаются в размерах и их коронки приобретают коническую форму (рис. 329)
290
В каждой большой краниологической коллекции всегда можно найти несколько черепов, у которых латеральные резцы полностью отсутствуют. В таких случаях клыки стоят непосредственно рядом с медиальными резцами (рис. 329, 331).
«Мы почти с уверенностью можем сказать, — пишет В. Лехе (1913),— что человек утратит верхние латеральные резцы так же, как зубы мудрости».
Таким образом, изучение зубной системы человека показало, что в ней могут отсутствовать: третьи большие коренные зубы (зубы мудрости), вторые малые коренные зубы и латеральные резцы.
Рис. 331. Отсутствие латеральных резцов.
Все эти наблюдения, по мнению морфологов, дают право утверждать что «вообще с уверенностью можно сказать, что человек будущего будет обладать сильно редуцированной зубной системой» (Г. Бушан, 1911).
Грегори в своей книге, посвященной эволюции лица человека (1929), соглашается с тем, что зубы мудрости современного человека имеют «тенденцию» редуцироваться. Грегори думает, что у будущего человека количество зубов вместо 32 будет равно 28, так как четыре зуба мудрости исчезнут. Это приведет к некоторому ослаблению челюстей будущих людей. «Если они (будущие люди,— А. Б.),— пишет Грегори,— не будут есть мясо и растения, а будут употреблять в качестве пищи только экстракты, то их жевательные мышцы и челюсти сделаются еще слабее» (Грегори, 1929).
Если встать на точку зрения Грегори, то можно с полной убежденностью утверждать, что человек очень далекого будущего должен быть совершенно беззубым.
Такая его судьба, кажется, не должна удивлять нас, так как мы в каждом классе позвоночных можем указать такие формы, которые на протяжении некоторого отрезка своей эволюции потеряли все свои зубы.
Среди рыб зубов не имеет, например, игла-рыба (Syngnathus'y, среди амфибий — жаба (Bufo)\ среди рептилий — черепаха (С7ге/оле); среди млекопитающих — муравьед (Myrtnecophaga). Что касается птиц, то, как известно, все без исключения современные представители этого класса лишены зубов.
Если у всех этих теперь беззубых животных предки в свое время имели зубы, то по существу, кажется, нельзя признать недопустимым предположение, что и человек в далеком будущем может потерять зубы . . .
’/*19*
291
Многие антропологи путем изучения могильных черепов и измерения голов живых людей пытаются доказать, что древнее население Европы имело больший процент длинноголовых (долихоцефалов), чем современное.
На основании накопленного материала антропологи сделали следующий очень важный вывод: в современном человечестве протекает процесс «брахицефализации», т. е. количество короткоголовых (брахицефалов) непрерывно нарастает, а количество длинноголовых (долихоцефалов) уменьшается. . .
Г. Ф. Дебец (1948) смело заявляет, что «изменение формы черепа в направлении брахикрании является биологическим процессом».
Причина этой очень подозрительной «брахицефализации» антропологами до сих пор не выяснена.
Вайденрайх (F. Weidenreich, 1946) по вопросу о брахицефализации пишет следующее:
«Можно думать, что она объясняется зависимостью формы головы от вертикального положения тела человека. Шаровидная форма мозговой коробки, с ее тенденцией уровнять свою длину, ширину и высоту, является самой подходящей формой для черепа, балансирующего на вершине позвоночного столба. Растущая брахицефализации является показателем того, что эволюция человека еще продолжается».
Если согласиться с антропологами и считать, что брахицефализации, по крайней мере у европейцев, действительно происходит, то можно утверждать, что будущий человек будет обладать почти шаровидной головой.
Я не знаю ни одного анатома или антрополога, который бы осмелился написать в какой-нибудь своей работе, что мозг человека уже не эволюционирует. Поэтому я уверен, что все они согласятся с мнением Вайденрайха (1946), что «достигнутая величина мозга человека не есть последняя ступень его эволюции».
«У культурных наций объем мозга и в будущем не перестанет увеличиваться»,— уверенно пророчествует Лехе (1913).
Если размеры головного мозга человека будут действительно непрерывно увеличиваться, то человек далекого будущего должен иметь гигантский мозг. В результате этого его психика будет сильно отличаться от нашей.
Проф. Холдэн (J. В. S. Haldane — University College, London) в своем доклада на тему об эволюции человека в прошлом и будущем (Man's evolution: past and future), сделанном 3 января 1946 г. на научной конференции в США (Princeton University, New Jersey), дал следующую характеристику будушего человека:
«Он будет иметь большую голову и меньше зубов, чем мы; его движения будут ловкими, но не сильными. Он будет развиваться медленно, продолжая учиться до зрелого возраста, который будет наступать только в 40 лет; жить он будет несколько столетий. Он будет более разумен и менее подчинен инстинктам и сексуальным эмоциям, чем мы. Его побуждения значительно больше, чем наши, будут зависеть от воспитания. Он будет иметь более высокий, по сравнению с нашим, общий уровень интеллекта и многие будущие люди будут обладать в некоторых отраслях знания такими способностями, которые мы называем гениальностью».
В этой характеристике Холдэн мало уделил внимания анатомическим особенностям будущего человека.
«Он будет иметь большую голову и меньше зубов, чем мы» . . . Это все, что было сказано им по этому вопросу.
Немного больше можно прочитать в работе Шапиро (Н. L. Shapiro, 1933), посвященной вопросу о будущем человеке.
292
Рис. 332. Стопа будущего человека (Шапиро, 1933).
«Суммируя наши пророчества о будущем человеке,— писал Шапиро,— мы можем картинно описать его как более высокого, чем мы,с головой большего объема и более круглой. Его лоб будет более вертикальным, надглазничные дуги — гладкими. Его лицо будет меньшим по размерам и менее выступающим, чем у нас. Зубов на его челюстях будет меньше и они будут слабее. Третий моляр, а в некоторых случаях также латеральные резцы будут отсутствовать. Кариес или порча зубов останется серьезной проблемой и зубы будут требовать постоянного наблюдения и контроля. Некоторые представители будущей расы будут ходить на четырехпалых ногах и многие будут рано становиться лысыми» (рис. 332).
Если доводить до логического конца все рассуждения анатомов по поводу различных профетических вариантов в строении современного человека, то можно без труда составить себе более ясное и более детальное представление о многих особенностях скелета будущего человека.
Мозговая часть его черепа по своим размерам будет, вероятно, значительно превосходить соответствующую часть нашего черепа (рис. 333) —это предполагают многие морфологи.
В лицевой части черепа будут отсутствовать такие кости, как носовые, слезные и межчелюстные — это следует из работ некоторых авторов.
Так как анатомы убеждены, что «. . . зубы человека*
вырождению» (В. П. Воробьев и Г. В. Ясвоин, 1936), то можно на основании
. . подвергаются
Рис. 333. Череп современного человека (Homo sapiens) и череп будущего человека (Homo sapientissimus).
этого предполагать, что челюсти будущего человека будут совершенно лишены зубов, как . . . челюсти жабы. Скуловые дуги у него будут прерваны.
Полное исчезновение зубов, разумеется, неминуемо повлечет за собой редукцию альвеолярных отростков обеих челюстей, а это вызовет сильное уменьшение высоты лицевой части черепа (рис. 333).
293
Рис. 334. Скелет будущего человека (Homo sapientialmus).
Все анатомы, изучавшие строение позвоночного столба человека, согласны с тем, что череп, ассимилируя атлант, стремится переместиться вниз, в то время как таз в результате включения V поясничного позвонка в состав крестца имеет тенденцию передвинуться навстречу черепу, вверх. Одновременно с этим, по мнению анатомов, происходит укорочение грудной клетки путем редукции ребер сверху и снизу.
ь Если оба эти процесса действительно происходят, то они в будущем должны привести к очень сильному укорочению позвоночного столба и грудной клетки человека. Если череп будет неудержимо стремиться навстречу тазу, а таз — навстречу черепу, то не исключена возможность, что через несколько миллионов лет у человека останется один шейный позвонок, один грудной, один поясничный и два-три крестцовых. Этот короткий крестец будет иметь резко выраженную spina bifida, т. е. его задняя стенка будет полностью отсутствовать. В грудной клетке сохранится только одна пара ребер. Эти ребра будут присоединяться к единственному грудному позвонку и к небольшому куску сильно редуцированной грудной кости.
Так как ключицы являются последними сохранившимися покровными костями плечевого пояса, то можно предполагать, что у будущего человека они исчезнут, так же как исчезли у многих млекопитающих. Интересно, что в настоящее время у человека уже наблюдаются случай полного отсутствия этих костей (dysostosis cleido-cranialis).
Постепенное укорочение позвоночного столба и грудной клетки приведет к тому, что таз у будущего человека будет находиться настолько близко к огромному черепу, что лопатки лягут непосредственно на края подвздошных костей (рис. 334). В результате всего этого рост человека далекого будущего окажется значительно меньше нашего. Однако Шапиро (1933) приходит к совершенно противоположному выводу и ду
мает, что люди через 500000 лет будут выше ростом, чем мы. В доказательство этого он ссылается на то, что эволюция многих животных сопровождалась увеличением их размеров. Это было, например, у динозавров, f у лошадей, у верблюдов. Это явление Шапиро почему-то считает общим для всех позвоночных, а потому обязательным и для человека.
У современного человека иногда наблюдается уменьшение количества пальцев на кисти (hypodactylia, рис. 335); описаны также случаи уменьшения количества фаланг до двух у каждого пальца (hypopha-langia).
Кисти и стопы у всех предков человека были пятипалыми; при этом II, III, IV и V пальцы имели не меньше трех фаланг.
Это обстоятельство дает нам право думать, что гиподактилия и гипо-фалангия не могут считаться атавистическими вариантами.
Ж
Если придерживаться взглядов и терминологии анатомов, то следует признать, что гиподактилия и гипофалангия, по всей вероятности, являются профетическими отклонениями от нормального строения человека. Но если это так, то можно предполагать, что кисть будущего человека будет иметь только три двухфаланговых пальца. Нужно думать, что такого числа пальцев для будущего человека будет вполне достаточно, чтобы он имел возможность выполнять свою не требующую большого физического напряжения работу.
Кости рук и ног человека далекого будущего, по всей вероятности, будут изящнее и тоньше, чем у нас. К такому выводу можно прийти на основании следующих данных.
Г. Ф. Дебец (1948) в своих исследованиях по палеоантропологии уверенно говорит об «основных направлениях эпохальных изменений черепа». К таким изменениям он относит «процесс брахицефализации» и «процесс грациали-зации» черепа. Г. Ф. Дебец совершенно не сомневается в том, что такие процессы происходят, , и говорит, что «не только возможно, но даже бесспорно, что брахицефализация происходила ... в более ранние эпохи, а грациализа-ция — в более поздние». Такое заявление антрополога дает право уверенно полагать, что в результате действия «направленной эпохальной» брахицефализации и такой же грациали-зации череп будущего человека будет не только коротким и широким, но и более легким и лишенным различных гребней, шероховатостей и не-ровностей.
Если такая судьба ожидает череп, то нужно
думать, что грациализации подвергнутся И все Рис. 335. Трехпалая кисть другие отделы скелета будущего человека, человека.
в частности кости его конечностей. Поэтому мы
можем предполагать, что кости рук и ног будущего человека будут слабее, тоньше и изящнее, чем наши.
Очень возможно, что руки и ноги наших очень далеких потомков будут не только слабее, но и короче, чем наши, так как и в недоразвитом состоянии они легко смогут выполнять ту минимальную работу, которая еще будет выпадать на их долю.
Как указывалось выше, Шапиро (1933) предполагает, что будущий человек будет выше ростом, чем современный. Однако Шапиро считает необходимым оговориться. «Насколько мне известно,— пишет он,— нет причины считать более высокого человека в механическом отношении построенным лучше. В самом деле, низкорослый человек, вероятно, более экономно расходует ту энергию, которая вырабатывается в его теле, чем более высокий человек. Но в эстетическом отношении более высокий человек предпочитается среди европейцев и американцев, и, может быть, у цивилизо-
ванных народов эстетика получит перевес».
В скелете нижних конечностей будущего человека малая берцовая кость (fibula), по всей вероятности, подвергнется почти полной редукции. Возможно, что от этой кости останутся только верхний и нижний концы, которые прирастут к соответствующим концам большой берцовой кости (рис. 334).
Такое предположение может быть сделано на основании изучения
295
тех изменений, которым подвергались кости голени в течение эволюции позвоночных.
Мы знаем, что у первых Tetrapoda (у стегоцефалов) большая и малая берцовые кости имели равную или почти равную величину и обе они принимали участие в сочленении с бедром.
У рептилий на всем протяжении их эволюционного развития происходила постепенная редукция малой берцовой кости и она скоро сделалась
Рис. 336.
А — стопа с двумя фалангами на каждом пальце; В — стопа со сросшимися тарзальными костями (Н. С.
Маркелов, 1929).
тоньше, чем соседняя кость голени (большая берцовая). Уменьшение толщины малой берцовой кости вело к постепенному выключению ее верхнего конца из коленного сустава.
Все эти изменения привели к тому, что у плацентарных млекопитающих малая берцовая кость уже не принимала участия в образовании коленного сустава и была относительно слабой костью.
Если мы будем считать, как это делают многие морфологи, что пройденный путь эволюционного развития диктует характер дальнейших изменений, то должны предполагать, что постепенная редукция малой берцовой кости, наблюдавшаяся в прошлом, должна продолжаться и в будущем. А это неминуемо должно привести к почти полной редукции этой кости в скелете будущего человека. Что такая редукция возможна, видно из того, что малая берцовая кость почти полностью исчезла у некоторых млекопитающих (лошадь, корова и др.).
296
Принимая во внимание то обстоятельство, что средняя и концевая фаланги мизинца ноги у современного человека очень часто (41%) срастаются, Лехе (1913) считает, что мизинец «находится на пути к редукции» . . . Если это так, то этот палец у будущего человека должен быть рудиментарным.
Шапиро (1933) не сомневается в том, что через 500 000 лет на стопе человека сохранится только четыре пальца, и пишет: «Такое уменьшение числа пальцев не должно удивлять нас, так как подобное явление хорошо известно в эволюции многих млекопитающих, например лошади, коровы верблюда, свиньи и т. д.»
Гораздо реже, но все же встречаются случаи, когда все пальцы стопы человека имеют по две фаланги (рис. 336, Д). Такой вариант, разумеется, не является атавистическим, так как ни у кого из предков человека подобной особенности стопы не наблюдалось. Если же этот вариант считать прогрессивным, то можно предполагать, что у будущего человека все пальцы стопы будут иметь по две фаланги.
В морфологической литературе было описано несколько случаев своеобразных врожденных и наследующихся сращений тарзальных костей стопы (Н. С. Маркелов, 1929).
В одном из таких случаев в обеих стопах были сращены все тарзальные кости, кроме первой клиновидной, которая оставалась свободной (рис. 336, В).
Так как сращение некоторых тарзальных костей уже наступило у ряда современных рептилий, то. высказывались предположения, что подобные явления и у человека имеют филогенетическое значение (Н. С. Маркелов). Поэтому можно думать, что у будущего человека все суставы между тарзальными костями стопы исчезнут вследствие полной их ненужности, а это превратит тарзальный отдел стопы в одну кость . ..
Итак, изучение морфологической литературы, посвященной «аномалиям» костей современного человека, позволило нам познакомиться со взглядами анатомов на особенности строения скелета будущего человека.
При этом оказалось, что анатомы совершенно убеждены в том; что эволюционное развитие человека протекает в настоящее время так же, как оно протекало в далеком прошлом. Больше того, они, по-видимому, уверены, что путь дальнейшей эволюции человека давно намечен. Поэтому почти все они смело говорят о тенденции развития в определенном направлении. Получается такое впечатление, что если человек просуществует на Земле еще несколько десятков миллионов лет, то он неминуемо должен будет превратиться в очень своеобразное существо, скелет которого будет сильно отличаться от нашего . . . Если это физически слабое и беззубое существо с коротким туловищем и с огромной головой действительно когда-нибудь будет жить вместо нас на нашей планете, то оно вполне заслужит название Homo sapientissimus . . .
ГЛАВА II
ЗАКЛЮЧЕНИЕ
Начиная эту главу, я считаю себя обязанным прежде всего признаться, что я совершенно не согласен с тем, что написано в предыдущей главе. Я не разделяю взглядов анатомов на предстоящую судьбу человека и не думаю, что его скелет даже в очень далеком будущем может принять такие уродливые формы, какие пророчествуют они (рис. 334). Я вполне убежден, что все то, что анатомы с такой уверенностью предсказывают человеку в будущем, с ним никогда не случится............................
. . . Эволюционное развитие человека, начавшееся стадией питекантропа, протекало, разумеется, в непосредственной связи с окружающей средой. Pithecanthropus входил в состав соответствующего биоценоза и эволюционировал в зависимости от него и вместе с ним.
Существование каждого питекантропа полностью зависело от того, в какой мере он мог добывать себе достаточное количество пищи и защищать свою жизнь от врагов. Это делало его обычным членом того биоценоза, в состав которого он входил, заставляло вести тяжелую борьбу за существование и подвергаться наряду со всеми другими животными беспощадному естественному отбору. Но в отличие от всех окружающих животных питекантроп вел борьбу за жизнь не при помощи зубов и когтей, а при помощи искусственных, хотя и очень примитивных, орудий защиты и нападения. При этом сила его вооружения зависела не столько от технических качеств орудий, сколько от умелого их применения. Иными словами, могущество искусственных орудий в значительной степени зависело от интеллекта того, кто ими владел. Поэтому естественный отбор на первых этапах эволюции человека неминуемо и всегда приводил к сохранению более одаренных индивидуумов. А это обусловливало прогрессивную эволюцию мозга.
В результате эволюционного развития питекантропа появился непосредственный его потомок — синантроп, а затем и неандерталец.
Борьба за существование, которую вел неандерталец, была, по всей вероятности, менее напряженной, чем у его предков. Однако, принимая во внимание плохое вооружение неандертальцев, состав окружающей фауны и их отношение к ней, мы должны признать большое значение этой борьбы и для неандертальцев. Можно уверенно сказать, что и неандерталец подвергался естественному отбору.
Возможно, что этот отбор был менее жесток по сравнению с отбором у питекантропа, но он во всяком случае еще не потерял для неандертальца значения фактора биологической эволюции. При этом естественный отбор касался не столько физической силы неандертальца, сколько его интеллектуальных особенностей. Физическая сила первобытного человека по 298
сравнению с физической силой, например, огромного пещерного медведя была ничтожной. Поэтому не физическое преимущество давало неандертальцам возможность осмеливаться на преследование этого зверя. Несомненно, только интеллектуальное превосходство и совместные действия неандертальцев позволяли им вступать в борьбу с медведем с достаточной уверенностью в успехе. Можно сказать, что неандертальский человек, будучи физически слабее многих его противников, мог сохранить свое существование на всем протяжении своей эволюции только благодаря деятельности своего головного мозга. Поэтому нет ничего удивительного в том, что в течение многих тысячелетий в процессе естественного отбора сохранялись преимущественно те существа, которые обладали более совершенным мозгом.
Высокое развитие головного мозга давало возможность примитивному человеку создавать все более совершенные искусственные орудия защиты и нападения. Так как на всех этапах эволюции человека его существование зависело исключительно от этих орудий, то их постепенное улучшение имело для него огромное значение. Но развитие индустрии не могло осуществляться без развития мозга.
Так как человек в отличие от всех других животных боролся за свою жизнь исключительно при помощи своего разума, то естественно, что отбор вел к постепенному совершенствованию строения и функции его головного мозга. Но чем совершеннее становился мозг человека, тем совершеннее делалось и его оружие и тем легче для человека становилась борьба за жизнь.
Это постепенное ослабление борьбы за существование неминуемо вело к выходу человека из состава окружающего его биоценоза. Этот медленно протекающий процесс в конце концов привел к тому, что естественный отбор для человека сначала сильно ослабел, а затем совсем прекратился. В этот момент человек вышел из состава биоценоза.
Живущий небольшими обществами в пещерах, одевающийся в шкуры животных, владеющий огнем, охотящийся не в одиночку, а небольшими группами, хорошо вооруженный грубо обработанными каменными орудиями человек оказался способным преследовать и убивать всех, кого встречал на своем пути.
Теперь в составе окружающей фауны уже не было такого зверя, который мог бы решиться на постоянное преследование человека, не было такого хищника, который мог бы успешно осуществлять истребление людей.
Таким образом, эволюционное развитие человека привело к тому, что он из слабого и беззащитного обезьянообразного существа превратился для всех окружающих его животных в страшного и по существу непобедимого врага.
Случайные гибели людей во время охоты на то или иное животное или от каких-либо иных причин, разумеется, не имели значения отбора. Естественный отбор как таковой теперь для человека уже не существовал. Но отсутствие естественного отбора было равносильно прекращению действия одного из факторов эволюции. Так как такие факторы, как наследственность и изменчивость сами по себе без естественного отбора не могут обусловить эволюционного развития, то мы должны признать, что с момента прекращения действия естественного отбора должна была остановиться и биологическая эволюция человека.
Это, несомненно, и произошло около 50 000 лет тому назад, когда оформился кро-маньонец.
Изучение найденных в настоящее время скелетов этого человека дает возможность легко убедиться, что он ничем существенным не отличался от современных людей. Это был настоящий Homo sapiens, хотя и fossilis. Следовательно, в течение 50 000 лет в скелете человека не возникло ни од
299
ного нового признака, такого признака, который давал бы нам право говорить о новом этапе эволюционного развития человека.
Выйдя из состава биоценоза и прекратив свою биологическую эволюцию, человек продолжал постепенно развивать свою индустрию и стал все увереннее и властнее вмешиваться в окружающую его природу. Научившись шлифовать свои каменные орудия, он сделал удачную попытку путем приручения волка получить первое домашнее животное — собаку. Позже были приручены и другие животные.
Рис. 337.
А — золотая рыбка (Carassius auratus); В — амурский карась (Carassius carassius).
Все животные, которых человек делал домашними и таким образом выводил из состава биоценоза, прекращали свою биологическую эволюцию. Вскоре они, варьируя, как прежде, в первую очередь теряли защитную окраску и становились в этом отношении мало похожими на своих предков. В дальнейшем под влиянием длительного, хотя и бессознательного искусственного отбора, осуществляемого человеком, они изменялись все более и более. Но все эти изменения уже не были эволюционными, так как возникали в результате вмешательства человека и приводили к тому, что одомашненные животные уже не могли вернуться в тот биоценоз, из которого вышли их дикие предки. Иными словами, сильно изменившиеся в домашних условиях прирученные животные оказывались уже неспособными жить там, где жили их предки, так как не были в состоянии выдержать конкуренцию с дикими формами. Только тесная связь с человеком и постоянная забота с его стороны обеспечивали их существование.
Применение искусственного отбора позволяло человеку быстро добиваться очень сильных изменений строения любого прирученного животного. Примером может служить амурский карась (Carassius carassius, рис. 337, В), измененный китайцами при помощи искусственного отбора до неузнавае
мо
мости и превращенный в так называемую золотую рыбку (Carassius aura-tus, рис. 337, А).
Насколько мне известно, многие сторонники эволюционной теории считали и считают, что современный человек продолжает подвергаться такому же естественному отбору, который прежде действовал на его предков.
Сам творец эволюционного учения Дарвин не сомневался в этом.
«По-видимому много правды лежит в мнении,—писал он (1871),— что изумительное процветание Соединенных Штатов, равно как и характер тамошнего населения, представляет следствие естественного отбора. Наиболее энергичные, предприимчивые и смелые люди всех частей Европы выселялись в продолжение последних десяти или двенадцати поколений в эту обширную страну и преуспевали здесь соразмерно своим дарованиям».
Такой взгляд разделяется некоторыми биологами и анатомами.
Так, В. М. Шимкевич (1923) утверждал, что «естественный отбор является одним из факторов социальной жизни» и что «идея естественного отбора, понимаемая в ее широком смысле, с удобством применяется к объяснению целого ряда социальных явлений».
Признание того, что естественный отбор еще продолжает действовать на людей, совершенно законно приводит к убеждению, что биологическая эволюция человека в настоящее время продолжает протекать так же, как она протекала в прошлом. Однако все, кто думает так, обычно не приводят каких-либо доказательств правильности своих взглядов и не пытаются показать, как естественный отбор закрепляет новые особенности человека. В данном случае нас должны особенно интересовать новые морфологические признаки, а не те сомнительные психологические преимущества которые, по мнению Дарвина, отличают мигрантов в Северную Америку от тех европейцев, которые не решились переплыть Атлантический океан.
Совершенно несомненно, что для того, чтобы биологическая эволюция человека продолжалась и теперь, нужно поставить людей в такие условия, в которых они расплачивались бы жизнью за те или иные свои морфологические особенности. Только в этом случае эти особенности приобретут эволюционное значение.
Некоторые авторы рассматривают войну как биологическое явление и считают, что война действует как фактор отбора и улучшает человечество.
Нельзя отрицать того, что на воине убивают, но всем известно, что там уничтожение людей не производится ни по физическим, ни по психическим признакам. Поэтому война никогда не играла роли естественного отбора и никогда не могла быть фактором биологической эволюции человечества.
Хирургам-ортопедам, занимающимся изучением различных поражений и дефектов костной системы человека, иногда приходится встречаться с одной очень своеобразной ненормальностью в строении V поясничного позвонка, носящей название spondylolysis. Эта неприятная ненормальность заключается в том, что дуга позвонка оказывается несращенной с телом; при этом щель между дугой и телом проходит так, что нижние суставные отростки находятся на отделенной дуге, а верхние — на теле позвонка (рис. 338). Кроме того, во многих подобных случаях дуга позвонка бывает разделенной на две части. >
Эта врожденная ненормальность строения V поясничного позвонка в Детском и юношеском возрасте не дает себя чувствовать, но у взрослого человека она в конце концов приводит к очень тяжелым явлениям (spondylolisthesis).
Так как тело V поясничного позвонка, имеющее нормальное соединение с IV поясничным позвонком, лежит на наклоненной верхней поверхности крестца, то под влиянием давления всего расположенного выше отдела туловища оно начинает постепенно соскальзывать вперед. При этом
20 А. п. Быстров
301
заработать себе на пропитание и
дуга позвонка, не имеющая костного сращения с телом, но соединенная с суставными отростками крестца, остается на месте.
Смещение при спондилолистезе, тела, последнего поясничного позвонка вперед, естественно, сопровождается сильным растяжением связочного аппарата и. травмированием спинномозговых нервов. Все это вызывает очень болезненные ощущения.
В тяжелых случаях тело V поясничного- позвонка «соскальзывает» с угла крестца (promontorium) и «падает» в.таз. Такой результат спондилолистеза делает человека полным инвалидом.
Современная медицина старается всеми доступными ей средствами помочь таким, людям. Ношение легкого, но прочного корсета, опирающегося на кости таза внизу и поддерживающего туловище вверху, дает возможность предупредить, или задержать сползание последнего поясничного позвонка с крестца.
Если в. наших условиях человек, страдающий спондилолизом (spondylolysis), может так или иначе
прожить достаточно долго, то такая ..же...аномалия в строении позвоночного, столба, для..неандертальца, несомненно, была. очень тяжелым и даже гибельным дефектом,
Тот факт, что spondylolysis для современного человека уже. не является тяжелой аномалией, a spondylolisthesis—гибельным следствием.этой аномалии, свидетельствует о том, что никакого естественного отбора для человека уже не существует.
Можно уверенно сказать, что никакое отклонение от средней нормы строения, если оно не вызвало смерти в первые дни жизни, уже не
Рис. 338. Spondylolysis. . мешает человеку благополучно существовать в течение многих лет. При этом всякий человек, имеющий тот или иной дефект. ..строения, обычно. получает необходимую помощь со стороны окружающих его людей. Это обстоятельство вызывало немалую тревогу у некоторых, биологов, несмотря на то, что они признавали наличие у современного. человека борьбы за существование и естественного отбора и верили в благотворное влияние этих .факторов на человечество.
«Есть основание думать,— писал Дарвин (1871),— что оспопрививание сохранило тысячи людей, которые, при своем слабом сложении в.преж.-нее время погибли бы от оспы. Таким образом, слабые члены цивилизованного общества распространяют свой род. Ни один человек, знакомый, с .законами разведения домашних животных, не будет иметь, ни малейшего сомнения в том, что это обстоятельство — крайне неблагоприятно для человеческой расы».
«Естественно,— говорит. Лехе (1913),— что, поддерживая слабых и больных, человечество подвергается опасности вырождения». .
«Неоднократно указывалось,— пишет В. М. Шимкевич (1923),— что поддержание слабых и помощь болезненным грозит человечеству вырождением. Но, конечно, человечество, вступив на путь искусственного отбора, найдет в нем же самом орудие против этого бедствия». «Внося идею искусственного отбора в свои отношения к животным, человечество не могло не внести ее и в отношения между собой».«Можно с уверенностью предсказать.—
302
утверждает В. М. Шимкевич,— что искусственный отбор постепенно вытеснит естественный ...» ,
Таким, образом, В. М. Шимкевич без всякого смущения готов применить к человеку те же методы искусственного отбора, которыми пользуются скотоводы при выведении новых пород домашних животных.
Мы не можем отрицать большого могущества этого фактора и должны согласиться, что если человек в будущем действительно применит по отношению к самому себе искусственный отбор, то, добиваясь своего собственного улучшения, он, конечно, сможет изменить себя не в меньшей степени, чем изменил карася, превратив его в золотую рыбку.
Как известно, забота генетиков о будущем человечества привела к созданию такой псевдо-науки, как евгеника.1 Ю. А. Филипченко (1921) определял евгенику как «науку, изучающую все влияния, которые способны улучшить врожденные качества будущих поколений».
При таком определении евгеника кажется совершенно безвредной «наукой», занимающейся только мирным изучением наследственности у человека.
Однако следует иметь в виду, что мирное накопление наблюдений считается евгенистами только первым этапом развития их «науки». На втором этапе евгеника, по их мнению, перейдет от теории к практике и превратится в весьма воинственную дисциплину, причем каждый правоверный евгенист при первой же попытке приложить свои научные достижения к человеку потребует для себя полицейских прав, так как его взгляды на взаимоотношения между людьми без жестоких насильственных мер не могут быть приняты никаким обществом. Присяжные евгенисты полагают, что после всеобщего признания их учения каждый из них будет считаться таким же непогрешимым судьёй в делах любви, как папа римский — в делах веры, и что все они будут наделены исключительными правами разрешать и запрещать браки между людьми. Евгенисты утверждают, что человечество может быть «облагорожено» (sofevr|i;) только при условии, если каждый человек будет подыскивать себе пару для брака, пользуясь авторитетными советами евгенистов, и если по указанию евгенистов многие люди будут хирургическим путем лишены возможности оставить после себя нежелательное для общества потомство. Таковы претензии евгенистов. Однако их первые попытки (в США) воспользоваться государственной властью для проведения в жизнь своих взглядов, как и следовало ожидать, встретили общее возмущение и были ими же самими признаны «преждевременными и потому ненужными, даже более того — вредными с точки зрения широкого распространения евгенической идеи» (Ю. А. Филипченко, 1921).
Можно совершенно уверенно сказать, что человечество в будущем не будет прибегать к рекомендуемому евгеникой полицейскому вмешательству в частную жизнь человека и не сможет осуществить ничего похожего на искусственный отбор. Оно, несомненно, пойдет по пути широких и смелых социальных реформ, улучшающих общие условия жизни для всех людей.
Это неминуемо поведет к укреплению и духа и тела человека, и лозунг евгеники: Mens sana in corpore sano (здоровый дух в здоровом теле) — будет осуществлен без каких-либо издевательств над человеческой личностью . . .
Не ведя жестокой борьбы за существование и не подвергаясь ни естественному, ни искусственному отбору, человек, однако, не будет чем-то абсолютно неизменным, так как такой фактор его прошлой эволюции, как. изменчивость, и в будущем не утратит своей прежней силы, хотя и потеряет свое прежнее значение. Иными словами, человек, прекратив свою эволю
1 — благородный.
20*
303,
цию, по-прежнему, будет изменяться, но эта его изменчивость уже не будет служить материалом для эволюционных изменений.
Изучение скелета современного человека позволяет очень наглядно показать, насколько разнообразны в настоящее время его варианты.
Рис. 339. Череп человека (угол нижней челюсти равен 90°).
Рис.'340. Череп человека (угол иижией челюсти равен 148°).
В данном случае я имею в виду не только атавистические и «профети-ческие» отклонения от нормального строения скелета, в частности черепа, но и такие, которые, по мнению анатомов, несомненно, не имеют никакого отношения к эволюции человека ни в прошлом, ни в будущем.
304
Как известно, детальное изучение черепа человека дало возможность обнаружить в нем настолько большое количество разнообразных отклонений от нормального строения, что при обследовании краниологических коллекций мы обычно не находим ни одного нормально построенного черепа,— каждый из них имеет то или иное отклонение от нормы. Поэтому надо признать правильным следующее, казалось бы парадоксальное, положение: отклонение от нормы является нормой. Таким образом, «норма» ставится совершенно отвлеченным понятием. «Нормой» мы можем назвать только
Рис. 341.
А — череп с очень большими .орбитами; В — череп с очень маленькими орбитами.
некую схему строения, в которой отсутствуют какие-либо отклонения от некоторого идеального плана строения.
Как известно, пока еще ни один анатом не обнаружил двух совершенно одинаковых черепов и не нашел ни одного черепа, не имеющего какой-нибудь ненормальности, однако каждый анатом с полным основанием может утверждать, что он имеет совершенно ясное представление о том, что такое череп вообще. Именно такой лишенный каких-либо аномалий воображаемый череп и представляет собой идеальную норму, от которой череп любого человека обязательно будет отличаться той или иной особенностью своего строения.
Изменчивость черепа наблюдается во всех без исключения его частях.
Угол, который образован телом и ветвью нижней челюсти, обычно сильно варьирует. Средняя его величина равна 127°. Но иногда он уменьшается до 90° (рис. 339) или увеличивается до 148° (рис. 340).
Очень интересными являются колебания в размерах орбит. В большинстве случаев величина орбит колеблется около некоторой средней нормы. Но в редких случаях можно видеть черепа с огромными • орбитами (рис. 341, А) или, наоборот, с очень маленькими (рис. 341, В).
Конечно, следует считать фантазией утверждение Ломброзо, что очень большие орбиты обычно наблюдаются у воров. Ломброзо объясняет этот
305
мнимый факт тем, что воры, работая ночью в темноте, напрягают свое зрение, а это будто бы вызывает у них увеличение глазных яблок и орбит.
’ Наряду с колебаниями размеров глазниц нередко изменяется и их форма. Вместо обычных, более или менее круглых, они иногда оказываются очень широкими и низкими (рис. 342, Л) или узкими и высокими (рис. 342, В).
Низкие и широкие орбиты человека по своей форме очень напоминают орбиты гориллы, а узкие и высокие — орбиты оранг-утана. Однако это сходство не имеет никакого значения, так как ни та, ни другая обезьяна не является предком человека.
Носовые кости очень сильно варьируют как по своей форме, так и по размерам. В моем распоряжении был череп с носовыми костями необычай-
Рис. 342.
А—череп с низкими и широкими орбитами; В—череп с высокими и узкими орбитами.
ной длины (рис. 343). Я видел также и череп с очень короткими носовыми костями (рис. 344). Эти два черепа представляют собой крайние формы среди коллекции, содержащей 4250 черепов.
Очень нередкими вариантами в строении черепа Человека следует считать появление в нем добавочных костей.
Наиболее часто наблюдаются дополнительные окостенения в соединительнотканной перепонке, закрывающей в черепе ребенка височный родничок (fonticulus sphenoidalis). Это приводит к тому, что у взрослого человека появляется небольшая самостоятельная косточка, расположенная между чешуей височной кости, теменной костью, лобной костью и большим крылом клиновидной кости (рис. 345, Л).
Несколько реже встречаются случаи такого же окостенения в соединительной ткани затылочного родничка (fonticulus occipitalis), что приводит к появлению у взрослого человека дополнительной кости в углу между теменными костями и чешуей затылочной кости (рис. 345, В).
Еще более редко у взрослого человека можно видеть дополнительную кость, возникающую в результате окостенения в лобном родничке (fonticulus frontalis) (рис. 345, С). Такая кость обычно в виде тупого клина входит в край лобной кости там, где у ребенка проходил шов между правой и
306
левой половинами его лобной кости. Интересно, что эта родничковая кость в прежнее время носила оригинальное название: ossiculum antiepilepti-cum. Она была известна знаменитому врачу средневековья Парацельзу (1493—1541), и порошок, изготовленный из нее, рекомендовался им в качестве надежного средства против эпилептических припадков. . .
Рис. 343. Череп европейца с очень длинными носовыми костями.
Рис. 344. Череп европейца с очень короткими носовыми костями.
Дополнительные окостенения в fonticulus mastoideus возникают очень редко (рис. 345, £>).
Так как в раннем детском возрасте края костей мозгового черепа еще не соприкасаются друг с другом и между ними находится соединительнотканная перепонка, то в случае возникновения самостоятельных окостене
307
ний в этой перепонке у взрослого человека могут появиться так называемые шовные косточки (ossa suturarum s. ossa Wormiana).
Наиболее часто подобные окостенения наблюдаются в затылочном шве (sutura lambdoidea). У ребенка они имеют вид ряда очень маленьких костных островков (рис. 346); у взрослого человека эти кости достигают уже довольно больших размеров (иногда 2—3 см в длину). Встречаются случаи, когда дополнительные косточки в очень большом количестве заполняют весь затылочный шов, в результате чего чешуя затылочной кости нигде не соприкасается с теменными костями (рис. 347, В).
Рис. 345. Родничковые
кости в черепе человека.
Несравненно реже можно наблюдать такие же кости в венечном шве (sutura coronalis). В очень редких случаях они располагаются почти на всем протяжении этого шва (рис. 348).
Почти также редко образуются дополнительные кости в чешуйчатом шве (sutura squamosa) (рис. 349). Интересно, что края тех шовных костей, которые лежат в пределах височного поля (planum temporale), всегда бывают так же косо срезаны, как верхний край височной кости и нижний край теменной кости. Иными словами, в пределах начала височной мышцы (mus-culus temporalis) все швы построены по типу sutura squamosa, а не sutura dentata.
Дополнительные окостенения в сагиттальном шве встречаются крайне редко (рис. 347, Д). Среди 4250 черепов я ни разу не встретил случая, чтобы шовные косточки располагались на всем протяжении этого шва.
В анатомической литературе описано несколько очень редких случаев, когда в черепе человека теменные кости были разделены беспорядочно расположенными швами на.большое количество отдельных косточек (рис. 350). Впрочем, возможно, что в этих случаях мы имеем дело с вытеснением теменной кости большим количеством дополнительных костей.
308
Подобные аномалии черепа обычно сопровождаются дефектами в области лобного родничка и полным отсутствием ключиц (Хульткранц — Hultkrantz, 1908). Это обстоятельство свидетельствует о том, что в данном случае недоразвитию подвергаются не только покровные кости че-
Рис. 346. Череп новорожденного с закладками шовных костей.
Рис. 347. Шовные кости (ossa suturarum).
репа, но и покровные кости плечевого пояса. Поэтому подобные пороки развития получили название dysostosis cleidocranialis.
Все эти многочисленные варианты формы лицевого черепа, размеров и формы орбит, носовых костей, разнообразные дополнительные кости мозговой коробки являются результатом обычной изменчивости человека. Они не могут быть признаны ни атавистическими, ни профетическими, поэтому ни один даже самый смелый на пророчества анатом до сих пор не решился сказать, какую форму и какие размеры будут иметь, например, глазницы будущего человека и сколько шовных костей будет в его мозговом черепе.
309
Эти индивидуальные особенности'не вовлекаются ни в; естественный, ' ни в искусственный отбор, так как ^современный человек не'подвергается ни тому, ни другому виду отбора. Это обстоятельство делает все его варианты по существу совершенно безразличными для него. Во всяком случае они
Рис. 348. Шовные кости в веиечиом шве.
Рис. 349. Шовные кости в чешуйчатом шве.
«е приносят ему никакого вреда и никакой пользы. Поэтому подобные отклонения от средней нормы строения не дают нам права предсказать морфологические особенности будущего человека.
Все те варианты, которые анатомы рассматривают как прогрессивные или профетические, также не вовлечены в какой-либо огбор. Поэтому Можно уверенно сказать, что случаи превращения V поясничного позвонка в I крестцовый и случай так называемой ассимиляции затылочной костью I шей
310
ного позвонка совершенно не свидетельствуют о будущем укорочении позвоночного столба.
Случаи отсутствия задней стенки крестца также не могут считаться указанием на то, что крестец будущего человека не будет иметь задней стенки.
Редукция первого и последнего ребер не является доказательством того, что у. будущего человека ребер будет меньше, чем у нас.
, Такие варианты строения, как отсутствие носовых, слезных и межчелюстных костей и размыкание скуловых дуг, не имеют никакого значения для современного человека. Эти недостатки индивидуального развития обычно обнаруживаются только на секционных столах анатомических ин-
Рис. 350. Случай dysostosis cleidocranialis. Череп 59-летиего мужчины (Хульткраиц, 1908).
статутов или только тогда, когда человек в виде скелета попадает в остеологическую коллекцию. При жизни даже и сами владельцы подобных аномалий, как правило, не подозревают их присутствия у себя.
Отсутствие зубов мудрости, вторых премоляров или латеральных резцов, разумеется, не дает человеку никакого преимущества в жизни. Можно уверенно сказать, что никакие варианты в строении зубного аппарата человека не вовлекаются в какой-либо отбор, так как, повторяю, никакому отбору человек уже давно не подвергается. Поэтому у нас нет никаких оснований утверждать, что у будущего человека многие зубы исчезнут.
Тот факт, что зубы мудрости прорезываются по сравнению с другими коренными зубами с запозданием на несколько лет, не является доказательством их «стремления» исчезнуть. Изучение зубной системы неандертальцев показывает, что и у них зубы мудрости нередко были развиты слабее, чем вторые моляры. Мало того, они, несомненно, прорезывались значительно позднее других зубов. На челюстях молодых неандертальцев можно вйдеть зубы мудрости, находящиеся еще глубоко в кости.
Все это свидетельствует о том, что последний коренной зуб начал подвергаться редукции еще у неандертальцев, но не исчез у них полностью. В несколько редуцированном состоянии он перешел от неандертальцев сначала к кро-маньонцам, а затем и к современному человеку. Нужно думать, что полурудиментарное состояние этого зуба и более поздний срок его про
311
резывания сохранятся неограниченно долгое время, так как при отсутствии естественного отбора по существу уже нет причин, которые могли бы обусловить его исчезновение.
Понимая, что ни один из так называемых «профетических» вариантов в скелете человека не дает ему никакого преимущества в жизни по сравнению с людьми, лишенными подобных особенностей, и, может быть, не видя в современном человечестве никаких проявлений естественного отбора, присяжные анатомы в качестве единственной причины дальнейшей эволюции человека выдвигают тенденцию развития в определенном направлении.
Эта эволюционная тенденция или тенденциозная эволюция правоверных анатомов свидетельствует о том, что они вольно или невольно трактуют биологические процессы с идеалистических позиций. Предполагаемая ими тенденция развития не объясняет решительно ничего, но заставляет считать, что все живые существа, в том числе и человек, обладают неким таинственным свойством, которое обусловливает их эволюцию и направляет ее по строго определенному пути. При таких взглядах на эволюционный процесс анатомы находят, что их задача заключается только в том, чтобы путем тщательного изучения современного человека выяснить тот путь, по которому в настоящее время идет его развитие, и на основании этого предсказать анатомические особенности будущего человека.
Так как при отсутствии борьбы за существование и естественного отбора никакая эволюция невозможна, то все попытки анатомов выяснить направление эволюционного развития современного человека и предсказать особенности строения будущих людей совершенно ненаучны. Нельзя изучать те процессы, которые не происходят, и предсказывать то, чего не будет.
Насколько мне известно, очень немногие морфологи готовы признать, что эволюционное развитие человека прекратилось.
Так, А. Семенов-Тян-Шанский в своей работе о границах вида (1910) между прочим упоминает «род Ното, остановленный в своем дальнейшем видообразовательном движении только интеллектуальным развитием человека».
В. В. Бунак, М. Ф. Нестурх и Я. Я. Рогинский (1941) пишут, что «приспособление человека к природе и влияние последней на человека осуществляется, начиная с верхнего палеолита, уже в столь сильной степени через посредство социальной среды, что трудно ожидать сколько-нибудь и в дальнейшем особых перемен в строении человеческого тела, кроме, например, вероятного некоторого усложнения строения головного мозга и т. п.»
Какие органы следует разуметь под словами «и т. п.», решить трудно. «Во всяком случае,—продолжают авторы,—нельзя полагать, чтобы в эволюции физического типа человека в будущем наступила какая-нибудь новая стадия развития». Из этого заявления видно, что В. В. Бунак, М. Ф. Нестурх и Я. Я. Рогинский согласны с тем, что в настоящее время «особых перемен в строении человеческого тела» не происходит. Однако, говоря об этом, они спешат сделать исключение для мозга и очевидно допускают его дальнейшее усложнение. Я думаю, что эволюционное развитие только одного этого органа совершенно невероятно.
Я. Я. Рогинский и М. Г. Левин (1955) не делают никаких оговорок и пишут, что «в лице современного человека процесс биологической эволюции создал обладателя таких видовых свойств, которые привели к затуханию дальнейшей их эволюции».
Мне кажется, что в настоящее время можно совершенно не сомневаться в том, что эволюционное развитие человека давно остановилось. Поэтому, если человечество не применит по совету некоторых евгенистов
312
по отношению к самому себе такого же искусственного отбора, при помощи которого оно получает новые породы домашнего скота, то люди далекого будущего ничем не будут отличаться от нас — людей современности.
ЛИТЕРАТУРА
Анучин Д. Н. О некоторых аномалиях человеческого черепа и преимущественно об их распределении по расам. Известия Общества любителей естествозиаиия, антропологии и этнографии, т. 38, 1880.
Бунак В. В., Нестурх М. Ф., Рогинский Я. Я- Антропология. 1941.
Бушаи Г. Наука о человеке. 1911.
Бушмакии Н. Д. Аномалии позвоночника с точки зрения его эволюции. Казанский медицинский журнал, т. 13, 1913.
Быстров А. П. (Bystrow). Morphologische Untersuchungen fiber die Occipital-region etc. Zeitschr. f. Anatomie u. Entwickl., Bd. 100 и 102, 1933.
Воробьев В.П. и Я с в о и н Г. В. Анатомия, гистология и эмбриология полости рта и зубов. 1936.
Видерсгейм Р. Строение человека. 1900.
Дарвин Ч. Происхождение человека. 1871.
Д е б е ц Г. Ф. Палеоантропология СССР. 1948.
Жуков Б. С. Происхождение человека. 1928.
Лехе В. Человек, его происхождение и эволюционное развитие. 1913.
Маркелов Н. С. О врожденной гипоплязии суставов. Вестник хирургии, 1929.
Одноралов Н. И. К вопросу о сращении атланта с затылочной костью. Архив анатомии, гистологии и эмбриологии, т. 10, 1931.
Рогинский Я. Я., Левин М. Г. Основы антропологии. М., 1955.
Северцов А. Н. Эволюция и эмбриология. Собрание сочинений, т. 3, 1945.
Семенов-Тян - Шанский А. Таксономические границы вида. Записки Имп. АН, т. 25, 1910.
Сперанский А. Д. Происхождение spina bifida occulta в крестцовом отделе позвоночного столба человека. Вестник хирургии, 1925.
Сушкин П. П. Обратим ли процесс эволюции? Новые идеи в биологии, № 8, 1915.
Тонков В. Н. Учебник анатомии человека. Л., 1953.
Филипченко Ю. А. Что такое евгеника. 1921.
Шимкевич В. М. Будущее человечества с точки зрения натуралиста. Берлин, 1923.
Я цута К-3.0 вариациях человеческого тела (В. П. Воробьев. Анатомия человека, т. 1, 1932).
В о 1 k L. Zur Frage der Assimilation des Atlas am Schadel beim Menschen. Anatomi-scher Anzeiger, Bd. 28, 1906.
F i s c h e 1. Ober einen menschlichen Schadel ohne Zwischenkiefer. Anatomischer Anzeiger, Bd. 27, 1905.
Gregory W. K. Our face from fish to man. New-York, 1929.
Gregory W. K. Polyisomerism and anisomerism in cranial and dental evolution among vertebrates. Proceedings of the National Academy of Sciences, vol. 20, N 1, 1934.
Gregory W. K. «Williston’s Law» relating to the evolution of skull bones in the vertebrates. American Journal of Physical Anthropology, vol. 20, N 2, 1935.
Haldane J.B. Man’s evolution: past and future. Princeton University Bicentennial Conference. Genetics, Paleontology and Evolution. Princeton, New Jersey, 1946.
Howells W. Mankind „so far. New-York, 1946.
Hultkrantz Z. W. Uber Dysostosis cleidocranialis. Zeitschrift fur Morphol. und Anthropol., Bd. 11, 1908.
Martin R. Lehrbuch der Anthropologie. Jena, 1928.
Shapiro H. L. Man 500 000 years from now. Natural History. The Journal of the American Museum of Natural History, vol. 33, N 6, 1933.
Weidenreich F. Apes, giants and man. Chicago, 1946.
Zabel E. Verietat und vollstandiges Fehlen des Thranenbeins beim Menschen. Anato-mische Hefte, Bd. 15, 1900.
ОГЛАВЛЕНИЕ
Стр.
Предисловие........................................................... 3
Введение ............................................................. 5
ЧАСТЬ ПЕРВАЯ ПРОШЛОЕ 34
Глава I. Бесчелюстные позвоночные (Agnatha).....................37
Глава II. Челюстные позвоночные (Gnathostomata). Рыбы ...........55
Глава III. Наземные позвоночные (Tetrapoda). Амфибии..............85
Глава IV. Рептилии .............................................129
Глава V. Млекопитающие.........................................179
Глава VI. Обезьяны и человек ...................................197
ЧАСТЬ ВТОРАЯ НАСТОЯЩЕЕ 219
Глава I. Расы .................................................220
Глава II. Эмбриогенез скелета и позвоночная теория черепа.......225
Глава III. Общая форма черепа ...................................232
Глава IV. Атавистические варианты ..............................247
ЧАСТЬ ТРЕТЬЯ БУДУЩЕЕ 275
Глава I. Человек далекого будущего (Homo futurus)..............277
Глава II. Заключение ...........................................298
Быстров Алексей Петрович
Прошлое, настоящее, будущее человека
Редактор А. Г. КНОРРЕ
Техн, редактор М. С. Рулева Корректоры И. М. Брицына и А. Г. Букреева Обложка работы художника М. Н. Свиньиной
Сдано в набор 12/IV 1957 г. Подписано к печати 2/Х 1957 г. Тираж 4 000 экз. Формат бум. jOXlOS’/w Печ. лист. 27,05 Бум. лист. 9,87. Учетно-издат. лист. 25. Заказ № 935. М 33809. Цеиа 12 руб. 50 коп.-|-2 руб. переплет
Типография № 4 УПП Ленсовнархоза
Ленинград, Социалистическая, 14