





















































































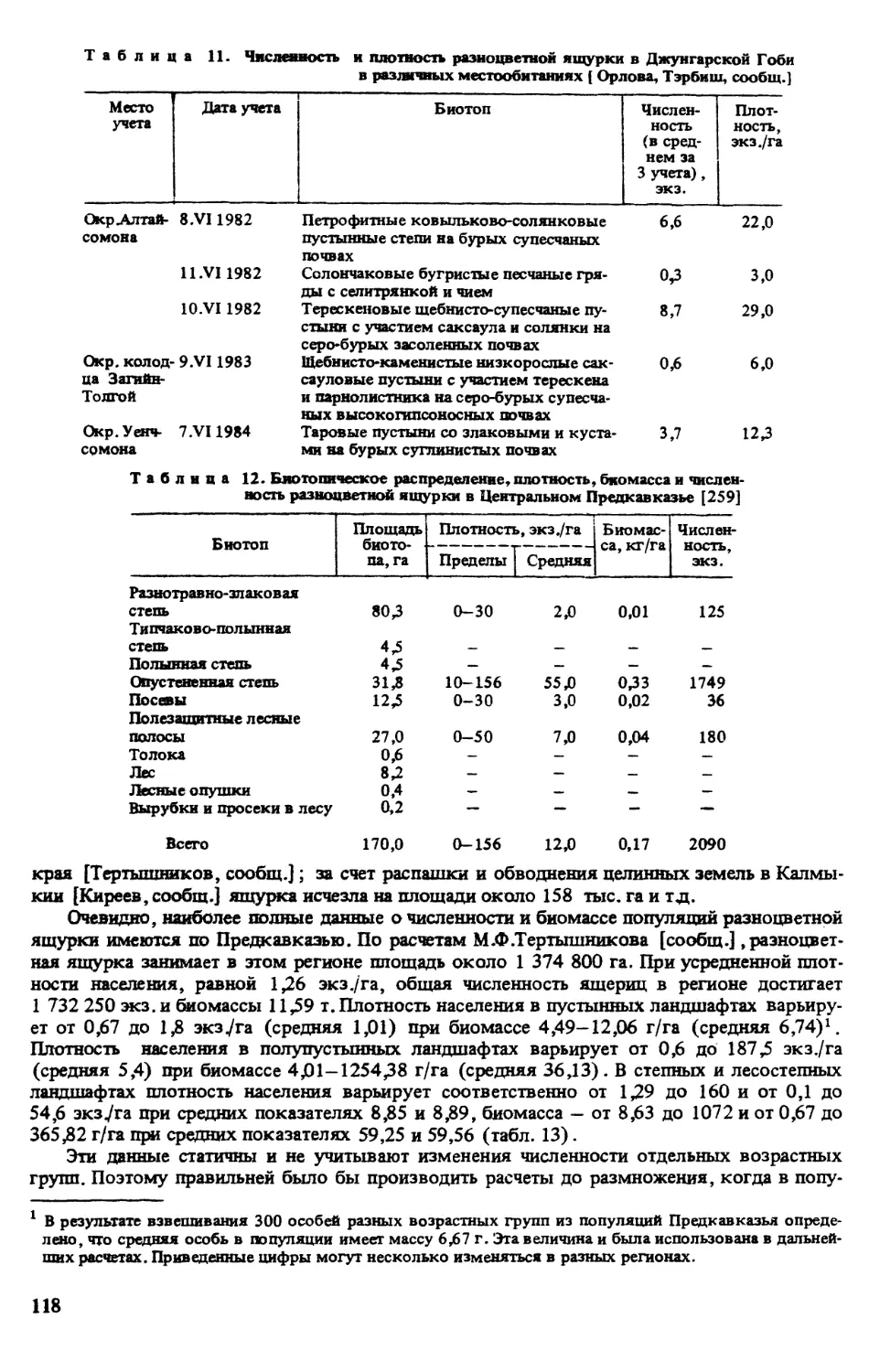




























































































Автор: Щербак Н.Н.
Теги: reptilia рептилии пресмыкающиеся герпетология биология зоология физиология животных издательство наукова думка академия наук украины
ISBN: 5-12-0032494
Год: 1993




Разноцветная
ЯЩУРКА
АКАДЕМИЯ НАУК УКРАИНЫ
Институт зоологии им. И. И. Шмальгаузена
Разноцветная
Я ЩУРКА
Под редакцией
доктора биологических наук Н. н. ЩЕРБАКА
КИЕВ
НАУКОВА ДУМКА
1993
Авторы: НЛЛЦербак, Т.И .Котенко, М.Ф.Тертышников, [ЯС.КЬгок|, Г.И.Василевская, М£ Весе-
ловский. Н.Н.Иорданский, СЛЛьвова, В.В.Неручев, Н.М.Окулова, В.Ф.Орлова, В.И.Горовая, B.nJlIapnu-
ло, CL4 .Шарыгин, ЛВ.Гербильский, В.С.Усенко
УДК 598.113.6
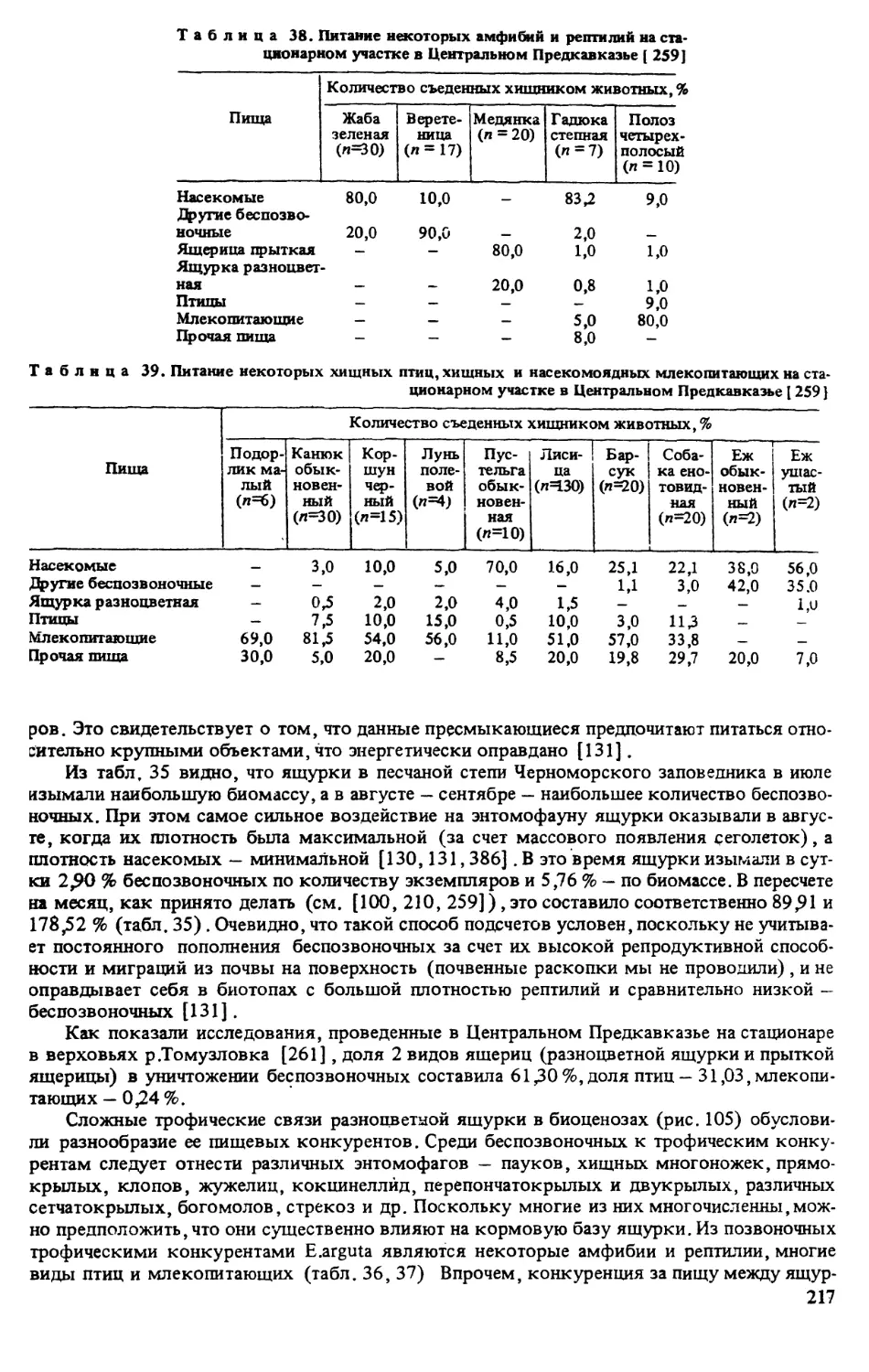
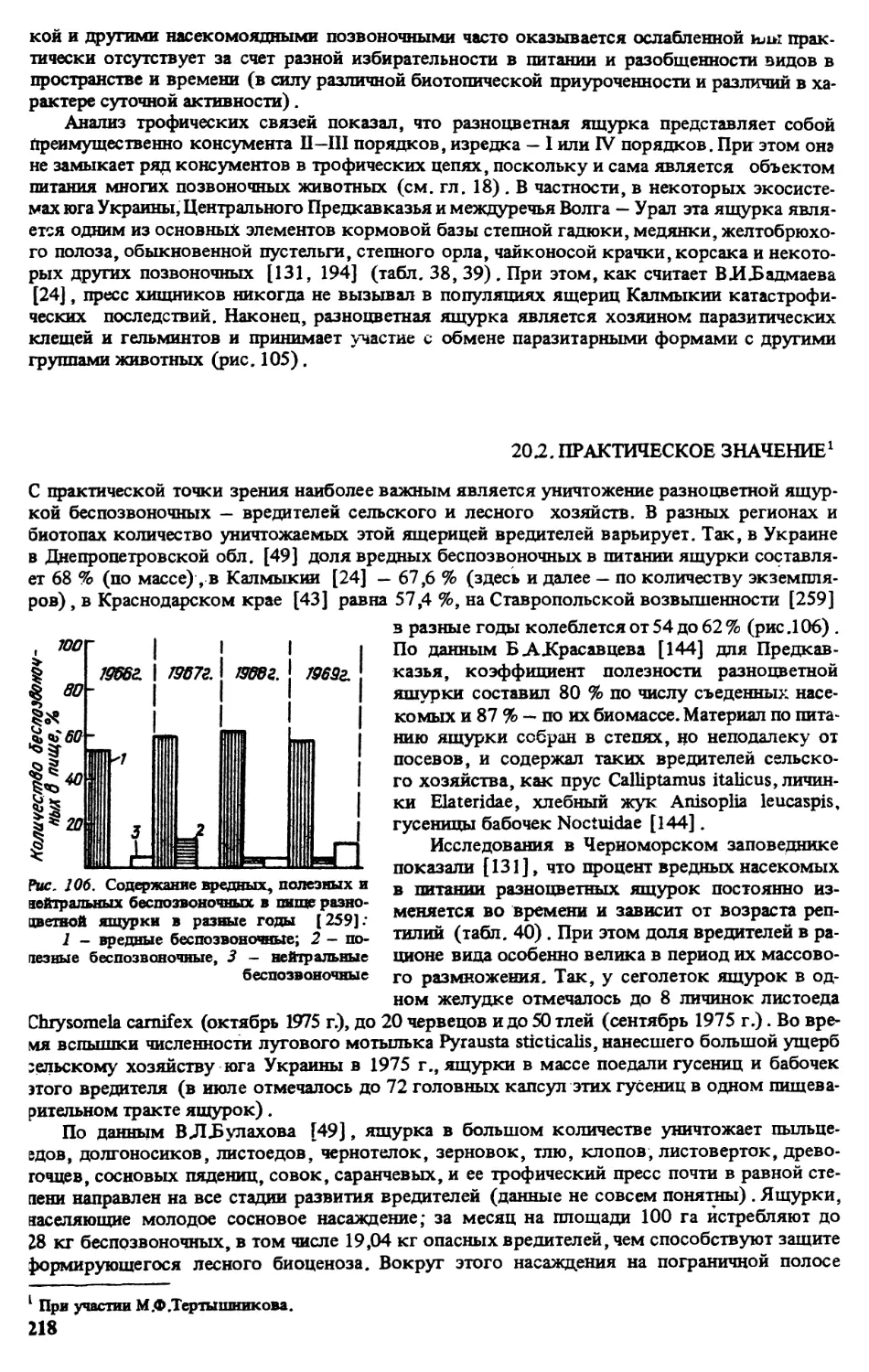
Монография посвящена одному из видов рептилий фауны Палеарктики - разноцветной ящурке.
Рассмотрены особенности ее распространения, систематика, географическая изменчивость, морфоло-
гия, экология (численность, стации, термобиология, питание, размножение, структура популяций и др.),
поведение, практическое и биоценотическое значение.
Для герпетологов, систематиков, морфологов, экологов, биогеоценологов, преподавателей и сту-
дентов вузов.
Монограф1я приев ячена одному з видш рептилий фауни Палеарктики - р1знобарвн!й ящурш. Роз-
глянуто особливосп 1Г поширення, систематику, географ тчну мшливклъ, морфолопю, еколопю (чи-
селыпсть, стат;Г, термобюлопю, живлення, розмноження, структуру популящйттощо), поведШку, прак-
тичне та бюденотичне значения.
Для герпетолопв, систематиков, морфолог!в, еколог!в, бюгеоценолопв, викладач!в та студентов
Ву31В.
Утверждено к 'печати ученым советом
Института зоологии им. ИЛ.Шмальгаузена АН Украины
Редакция биологической литературы
Редакторы С.В.ВЕЧЕРСКАЯ, ТЛ.ГОРБАНБ
р19070000(Ю-107?92ц9?
221-93
ISBN 5-12-0032494
© Н.Н.Щербак, Т.И.Котенко, М.Ф.Тертышников,
[В.С.Кото^ Г.И.Василевская, М.В .Веселовский, Н.Н.Иор-
данский, СЛ.Львова, В.В.Неручев, Н.М.Окулова,
В.Ф.Орлова. В.И.Горовая, В. П. Шар пил о, СА.Шарыгин,
Л.В.Гербильский, В.С.Усенко, 1993
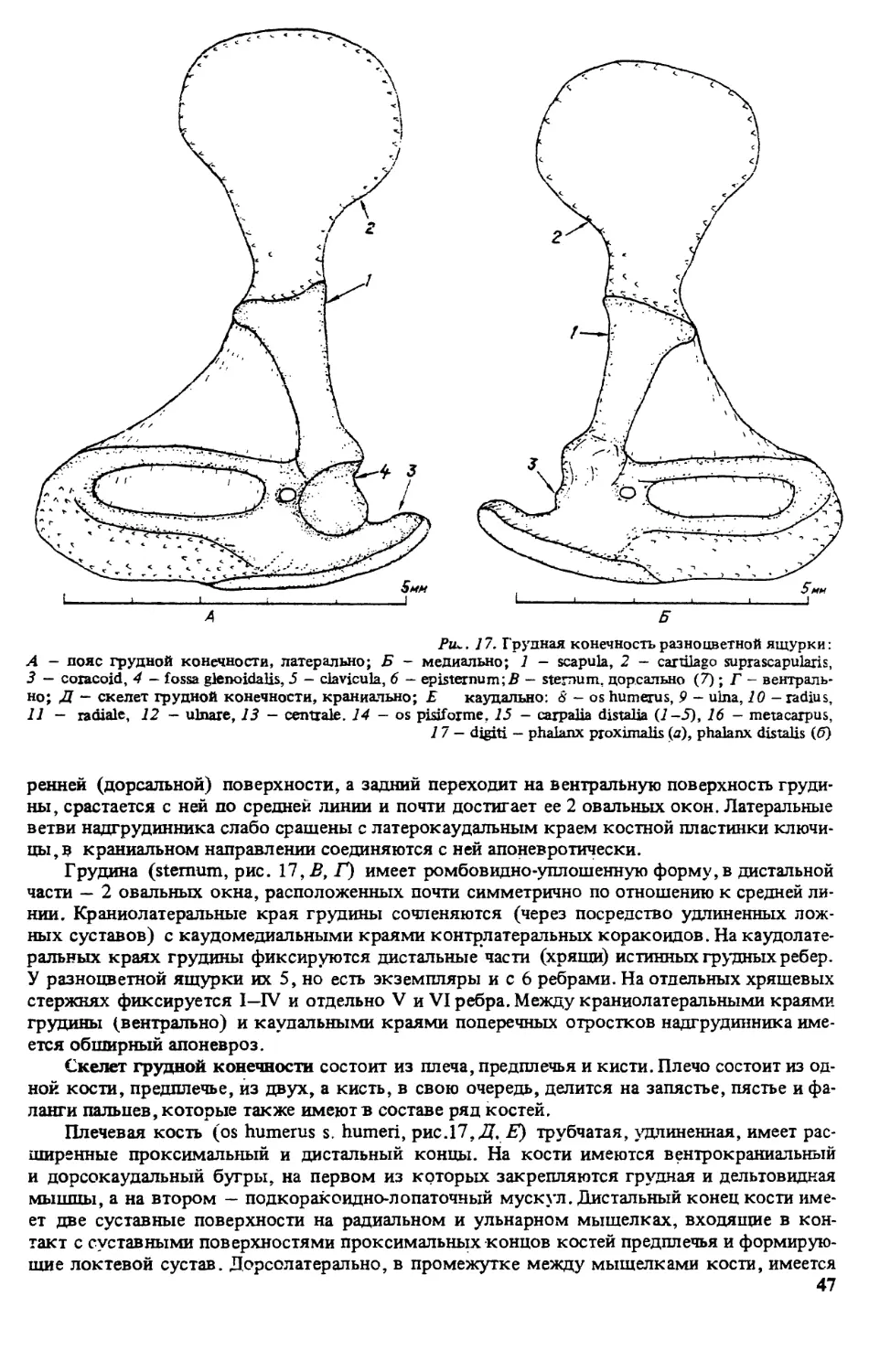
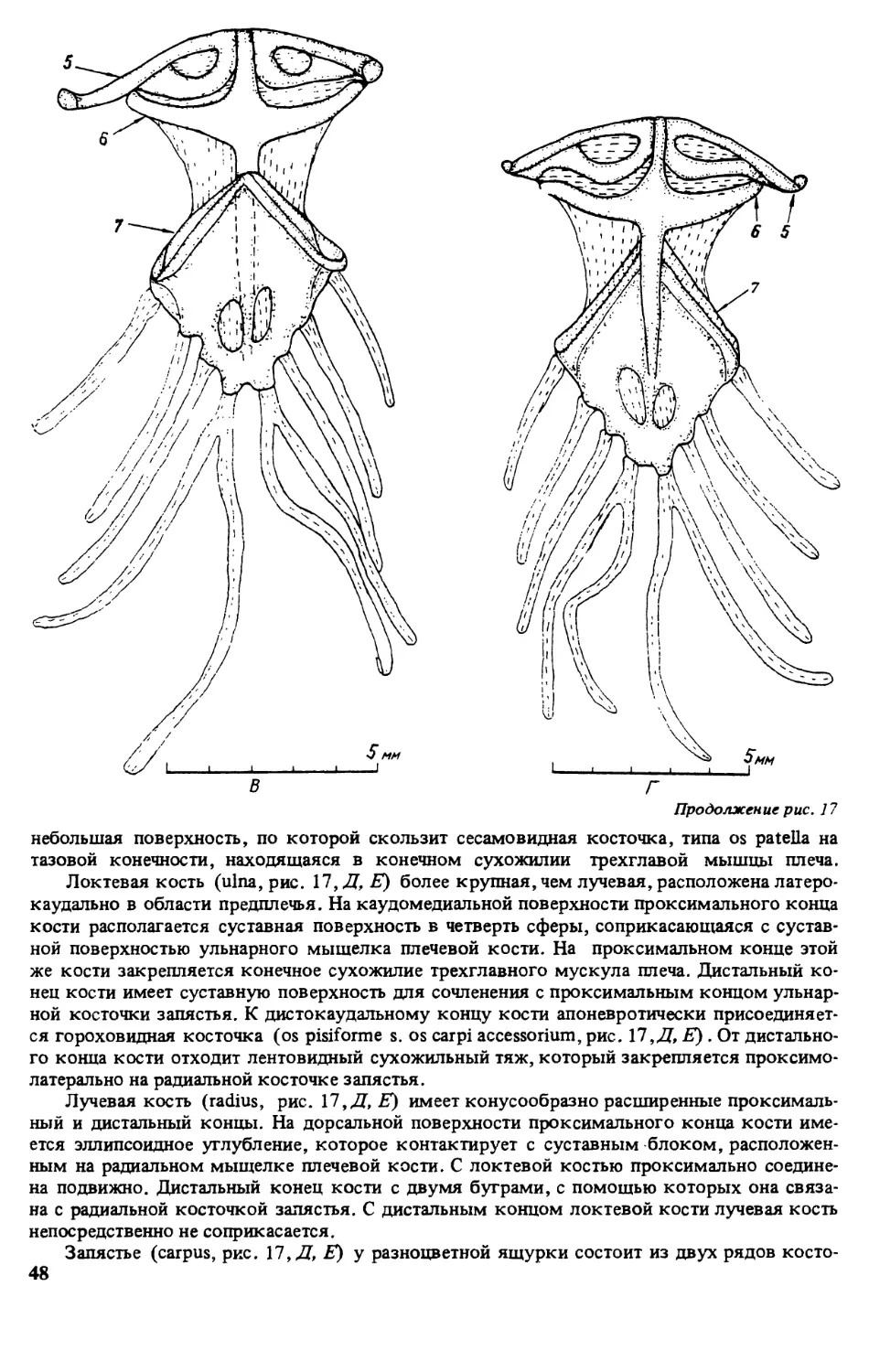
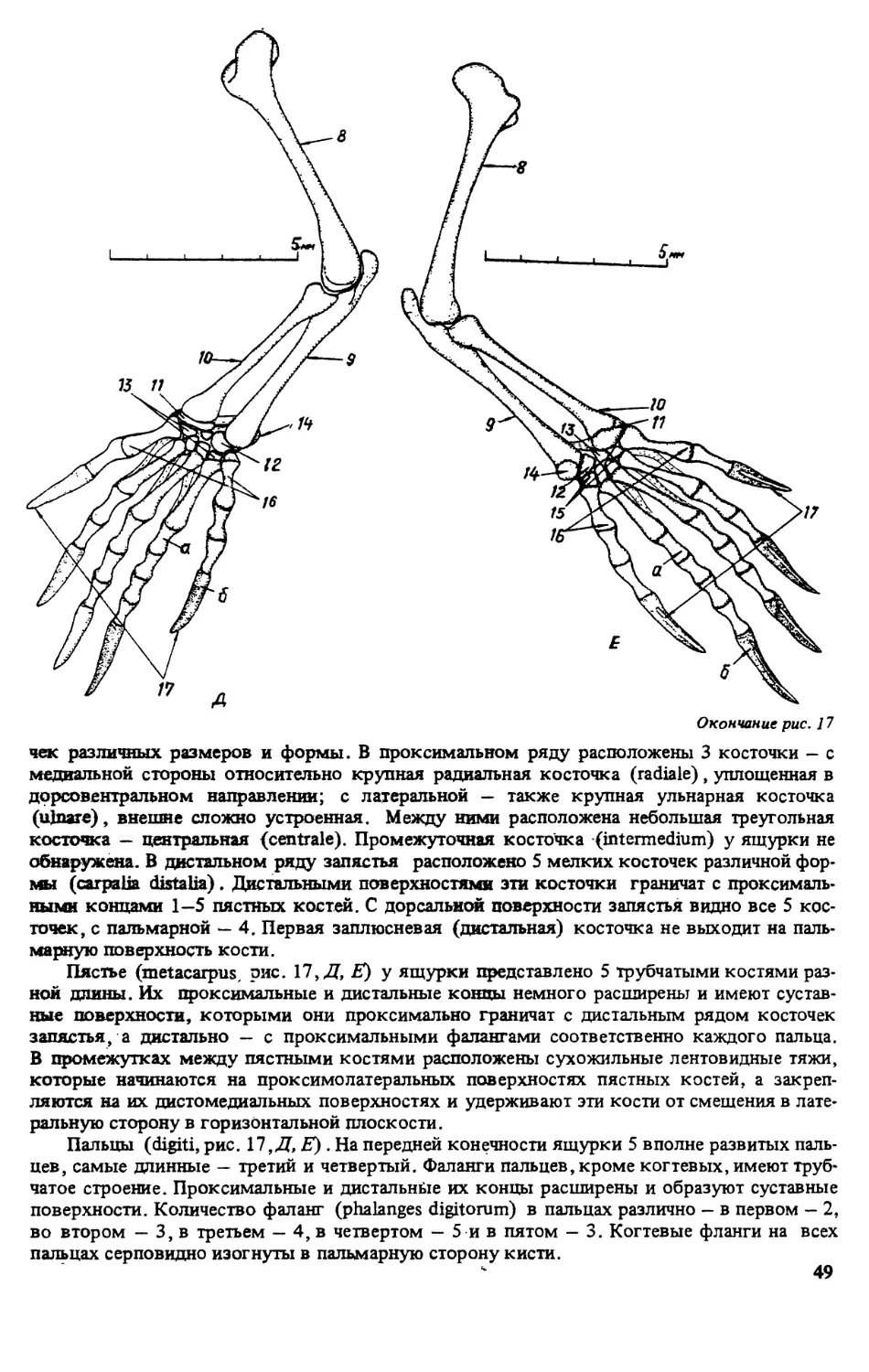
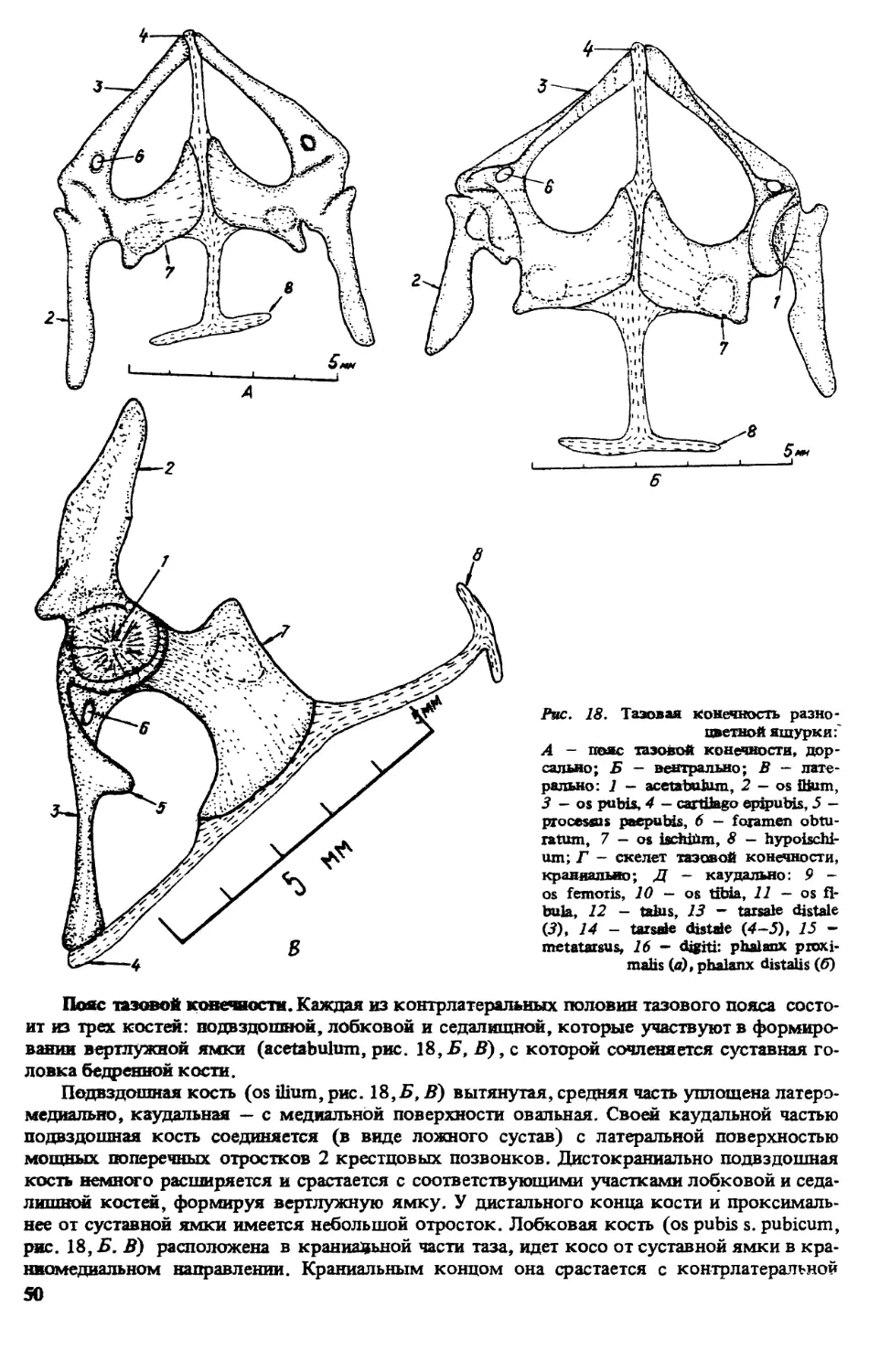
ПРЕДИСЛОВИЕ
Первой из монографий, открывшей серию книг ’’Виды фауны СССР и сопредельных стран”
выполненную по программе ЮНЕСКО ’’Человек и биосфера” (МАБ, проект 86), была
’’Прыткая ящерица” [322]. Второй вид, которому посвящена данная серия — разноцвет-
ная яшурка (Eremias arguta Pall.). Этот вид имеет обширный ареал, но распространен преи-
мущественно в бывшем Советском Союзе, встречается на территории с различными физи-
ко-географическими условиями, вариабелен (известно 6 подвидов, статус некоторых по-
пуляций неясен), в результате хозяйственной деятельности человека быстро сокращает
свой ареал. С 1977 г. проводились плановый сбор материала и наблюдения в 12 пунктах:
в Украине, Предкавказье, Калмыкии, Азербайджане, Грузии, Армении, Казахстане. Узбе-
кистане и др. Нами рассмотрены морфология (фолидбз, пропорции, особенности рисун-
ка) на популяционном уровне и анатомия различных органов и систем, их видовая специ-
фичность, изменчивость в пределах ареала.
При выполнении настоящего исследования использовались большей частью не опубли-
кованные материалы по ящуркам Палеарктики (Н.Н.Щербак), диссертация Э.В.Вашеткс
по ящуркам юга Средней Азии, региональные рукописные сводки: по Степи Украины -
Т.И.Котенко, Предкавказью — М.Ф.Тертышникова, а также были представлены новые ори-
гинальные материалы. В сборе и предоставлении материала приняли участие авторы: Ин-
ститут зоологии АН Азербайджана, г .Баку (Алиев Теймур Рза оглы); Азербайджанский
пединститут, г .Баку (М .Ахмедов); Калмыкский университет, г. Элиста (В И. Бадмаева),
Институт зоологии АН Украины (ИЗ АНУ),г.Киев (НЯЛПербак. Т.Н Котенко. ГВ?С.Коток;,
М .В .Веселовский, Г.ИБасилевская, СБ.Таращук, ВЛ.Шарпило); Ставропольский педин-
ститут, г.Ставрополь (АГБысотин, В .И Г оровая, М .Ф .Т ертышников); Нижегородский
пединститут, гНижний Новгород (НЯИванова, |Е.ЮКудакина], ВВ.Неручев, А.Ф.Ши-
ряев); Кубанский университет, г Краснодар (М.П.Зинякова, П.ПРуденко); Зоологиче-
ский музей Харьковского университета, г Харьков (В.ИВедмедеря, A М Рудик); Даге-
станский госуниверситет, г .Махачкала (С.П Львова, ЗЛХонякина); Институт зоологии
АН Туркменистана, г Ашхабад (С А.Шаммаков); Институт зоологии АН Узбекистана.
г.Ташкент (ЭВВашетко); Институт зоологии АН Казахстана, г.Алма-Ата (ЗКВрушко):
Центральная санэпидстанция Министерства здравоохранения России, г Химки (Д.А.Бонда-
ренко); Зоологический институт АН РАН (ЗИН РАН), г.Санкт-Петербург (И.С.Дарев-
ский); Зоологический музей Московского университета, г Москва (В.Ф.Орлова); Инсти
тут экспериментальной морфологии и экологии животных РАН (ИЭМЭЖ РАН) , гМосква
(ННИорданский); Противочумная станция, г .Элиста (ВАКиреев); Аксу-Джабаглин-
ский заповедник, Чимкентская обл., Казахстан (ВТКолбинцев) ; Монголия (Х.Тэрбиш);
Ивановский университет, г.Иваново (Н.М.Окулова); Никитский ботанический сад, г.Ял-
та (С.А.Шарьп'ин); Адыгейский пединститут, г.Майкоп (Э.А.Шебзухова); Ростовский
университет, гРостов-на-Дону (Г.ПЛукина); Днепропетровский мединститут, г .Днепро-
петровск (Л.В.Гербильский, В.С.Усенко).
Авторский коллектив выражает благодарность за постоянную поддержку и помощь
при проведении исследований президенту Герпетологического общества им.А.М.Николь-
ского, член.-кор. РАН И.С.Даревскому, председателю рабочей группы бывшего националь-
ного комитета Международной биологической программы кандидату биологических наук
Р.С.Вольскису, а также сотрудникам Института зоологии АН Украины Н.П.Бондаревой
и В.И.Вакаренко.
ГЛАВА 1
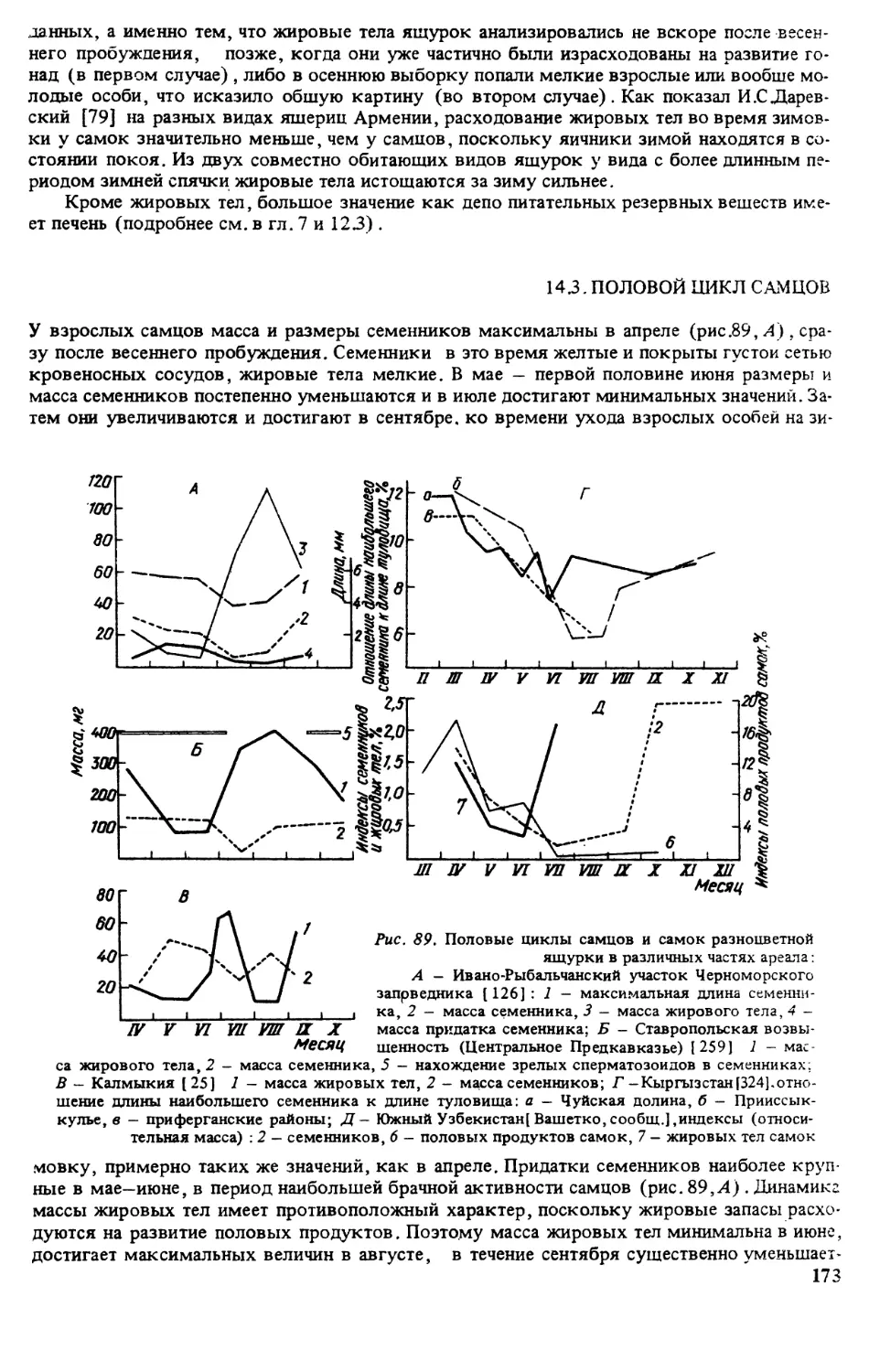
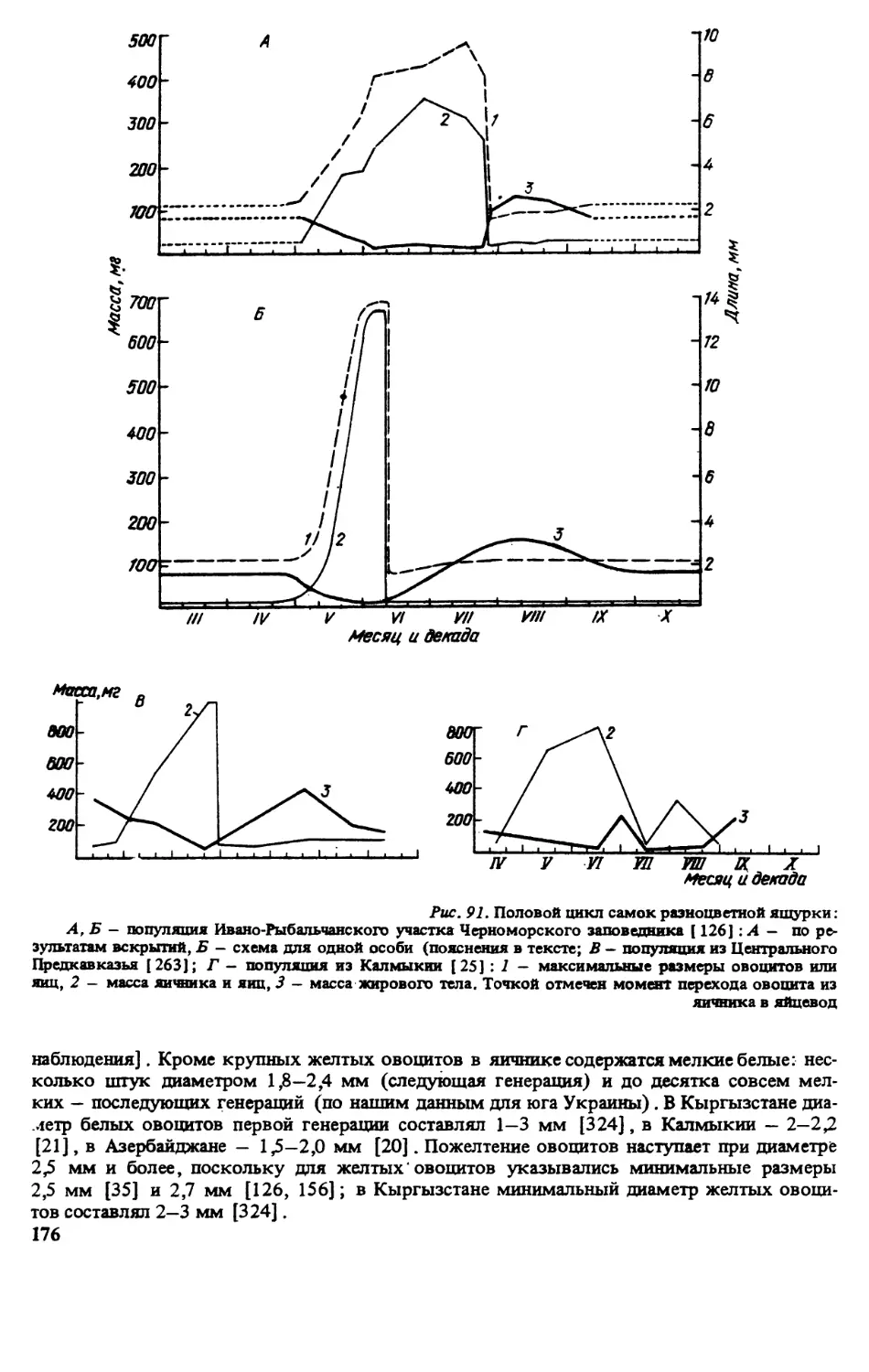
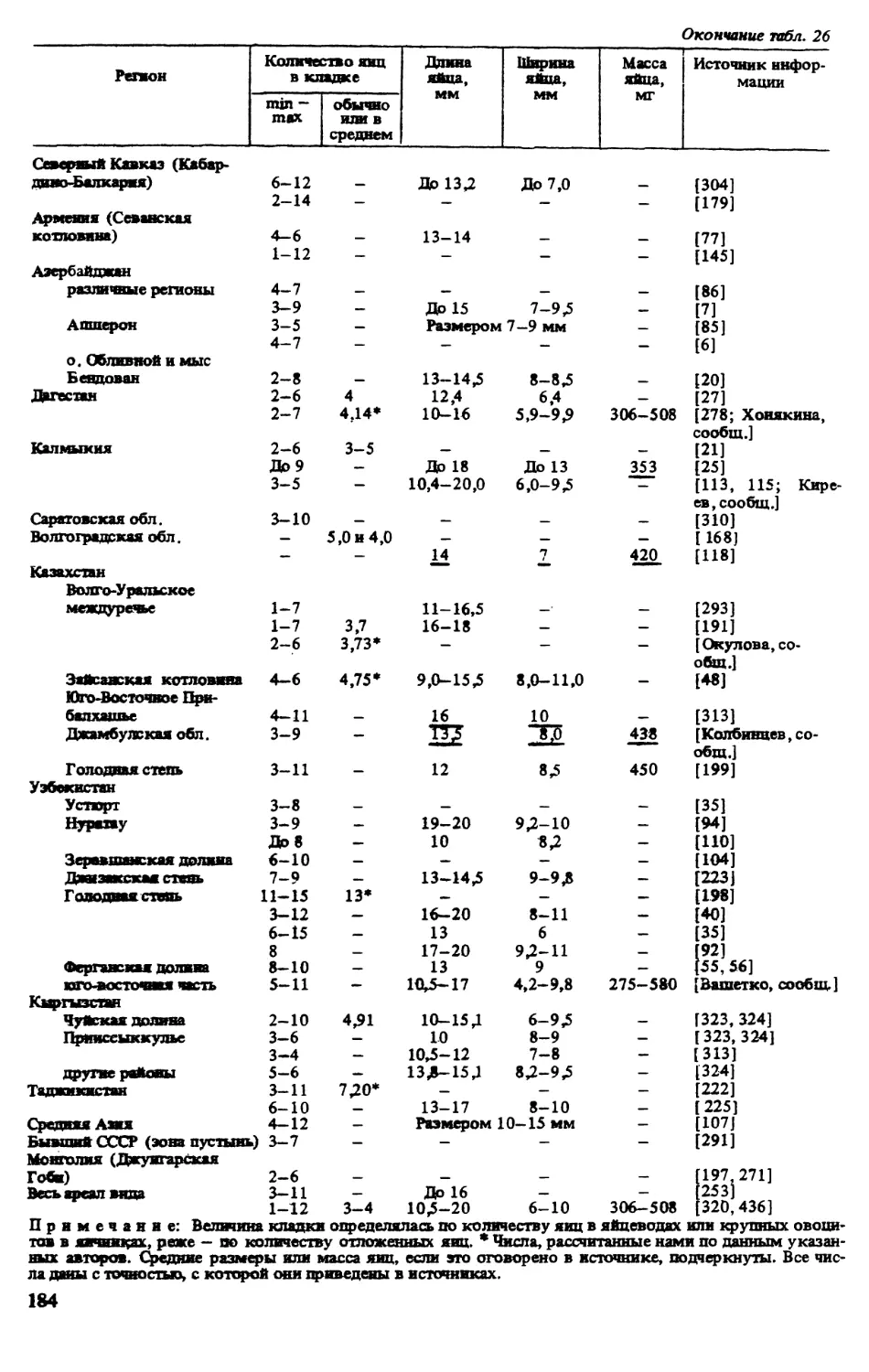
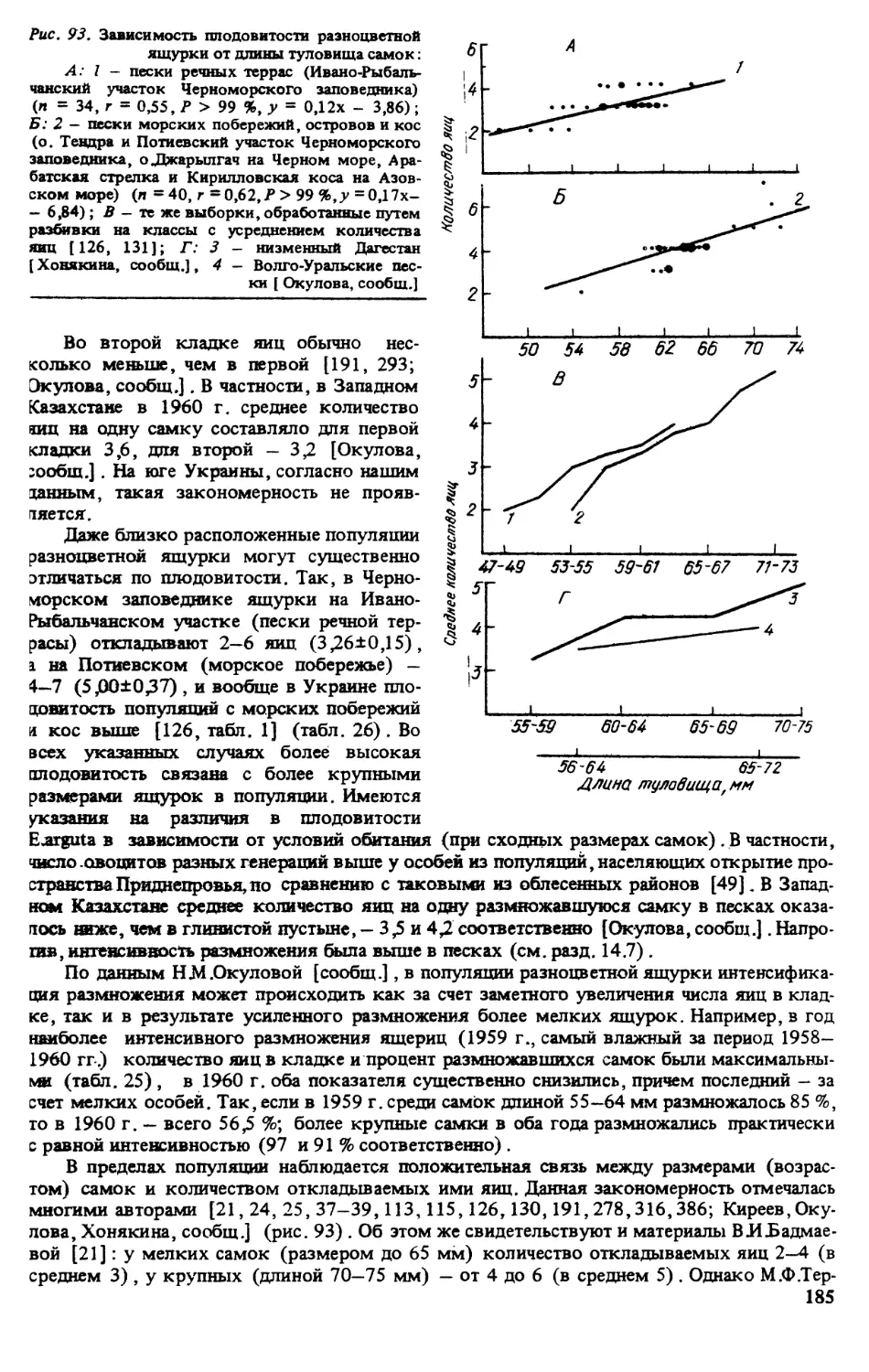
ИСТОРИЯ ИЗУЧЕНИЯ ВИДА
Впервые разноцветная ящурка была описана П.С Лапласом под названием Lacetra arguta
в 1773 г. Позже этот вид приобрел довольно много синонимов, причиной чего являлась
его значительная вариабельность на обширном ареале. Родовая принадлежность данного
вида достаточно полно освещена в работе, посвященной ящуркам Палеарктики [320].
Видовая синонимика представлена ниже:
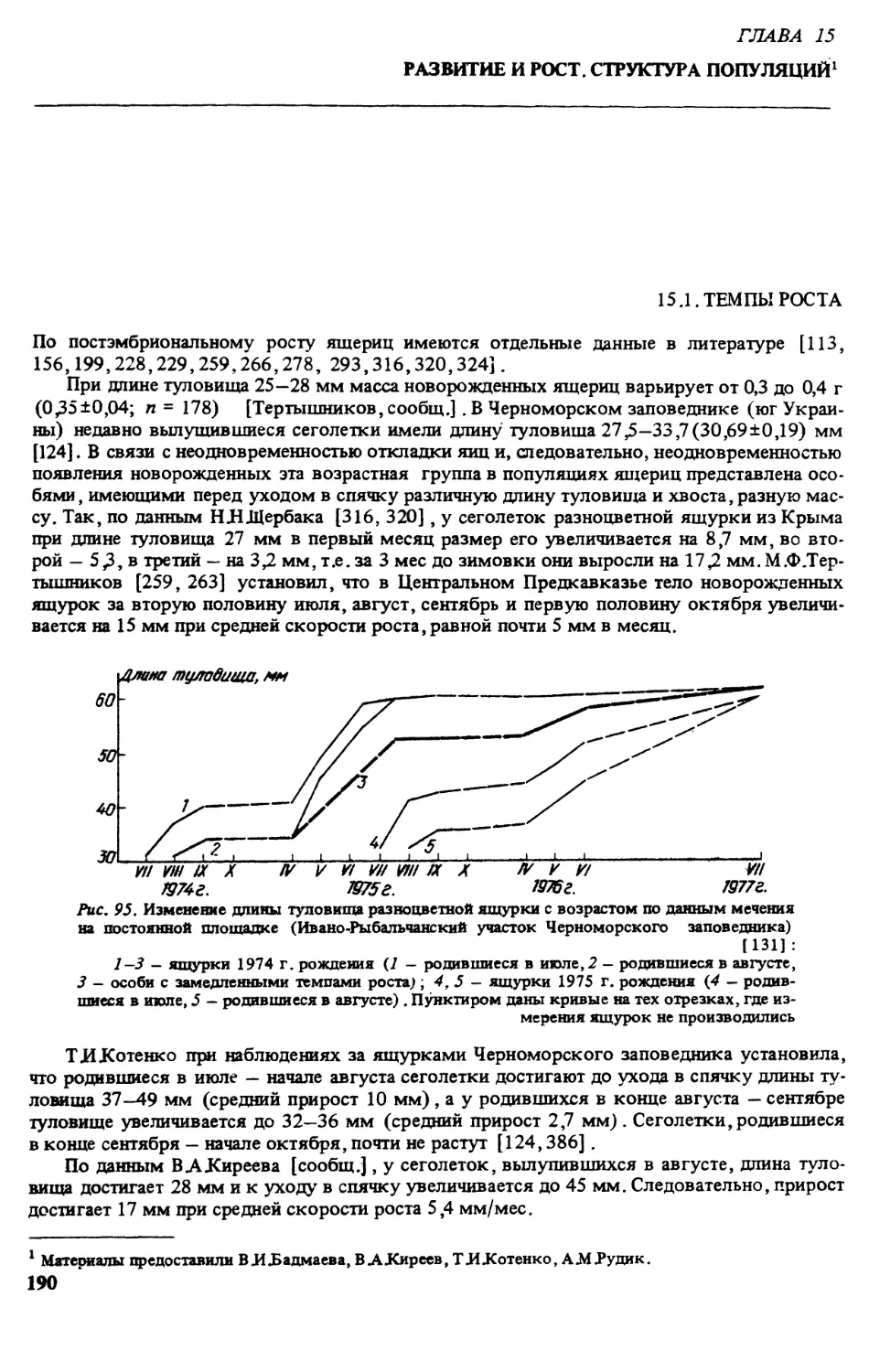
1789 - Lacerta deserti, Gmehn, Linn. Syst. Nat., ed. 13,1: 1076.
1814 - Lacerta variabilis, Pallas, Zoogr. Rosso-Asiat, III: 31.
1823 - Lacerta leucosticta, Lichtenstein, Evcrsmann, Reise Orenb. Buchara: 142.
1830 - Podarcis variabilis Wagler, Nat. Syst. d. Amphib, : 156.
1831 - Lacerta pustulata Eichwald, Zool. Spec., Ill: 187.
1832 - Podarcis irritans, Menetries, Catal. rais. Ob. Zool. Caucas: 62.
1832 - Lacerta boristhenis, Двигубсккй, опыт, естеств. истор., 3: 14.
1834 - Eremias variabilis, Wiegmann, Herpet, mexic, 1:9.
1841 — Podarcis deserti, Eichwald, Fauna Casp.-Cauc.: 96.
1845 — Eremias arguta. Gray, Cat. Lis.: 39. - 1915, Никольский, Фауна Росс. Преем., I: 447. - 1921,
Boulenger, Monogr. Lacert., 2: 342. - 1934, Чернов, Докл. АН СССР, Ш, 8-3: 666. - 1949,
Терентьев и Чернов, Опр. преем, и эемн.: 207. - 1960, Mertens u. Wermuth, D. Amph. und
Rept. Europas: 98.
1876 - Podarcis (Eremias) arguta, Strauch, Voy. Przewalsky, Pept.: 27.
1884 - Podarcis arguta, Boettger, Arch. f. Naturgesch., II: 388.
1928 — Eremias (Ommateremias), Lantz, Bull. Mus. George, IV-V: 95. - 1961, Fuhn si Vancea, Repti-
lia, Fauna RPR, 2: 253. - 1966, Щербак, Земн. и преем. Крыма: 161; - 1974, Щербак, Ящ.
Палеарктики; 147.
В 1912 г. ЯЗЗедряга, изучавший ящериц Центральной Азии, описал вариацию ”ро-
tanini”, которая без должных оснований в 1922 г. была сведена ЛЛантцем в синоним но-
минативной формы. Он же впервые выделил две формы: Еа. arguta и Е.а.deserti, описан-
ные ранее как самостоятельные виды. Неудачная попытка описать форму Earguta occi-
dentalis была предпринята ПЗ.Терентьевым [452]. В работе С.А.Чернова [287] анализиру-
ется форма Earguta и описывается новый подвид Earguta uzbekistanica. Позже И.СДарев-
ским [76] были описаны подвиды E.arguta transcaucasica из Армении и С.И.Царуком
[279] E.a.darevskii из Иссык-Кульской котловины. Упорядочение внутривидовой номен-
клатуры и составление таблиц для определения, диагнозов и описаний форм Ел.arguta,
Ea.deserti, E.a.potanini, Ел.uzbekistanica и Е.a.transcaucasica проведены Н.НЩербаком
[320]. С тех пор во внутривидовой системе разноцветных ящурок, кроме упомянутого
выше описания ОД Дарука [279], ничего не изменилось. Коротко отметим исследования
данного вида на популяционном уровне.
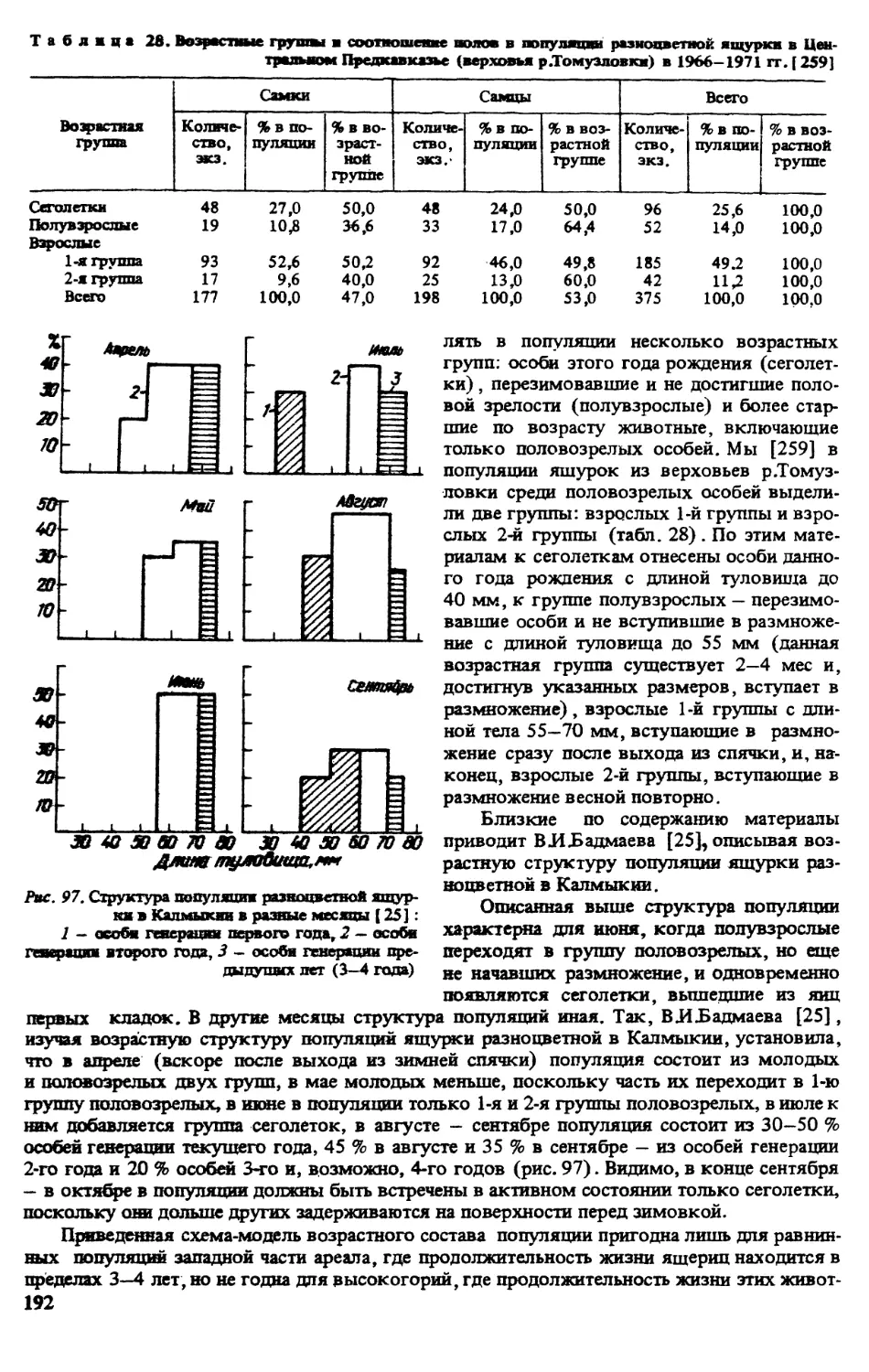
В монографии И.Д.Яковлевой [324], посвященной пресмыкающимся Киргизии, дан
детальный анализ переходных популяций между Е.а. arguta и Е.а. uzbekistanica из Чуйской
долины и прилежащих к Кыргызстану р-нов Казахстана. Несколько ранее в Румынии
И.Фуун и СЗаньча [359] изучали популяции E.arguta из провинций Молдова и Добруджи.
Это дало возможность [316, 320] сравнить различные популяции и выявить клинальную
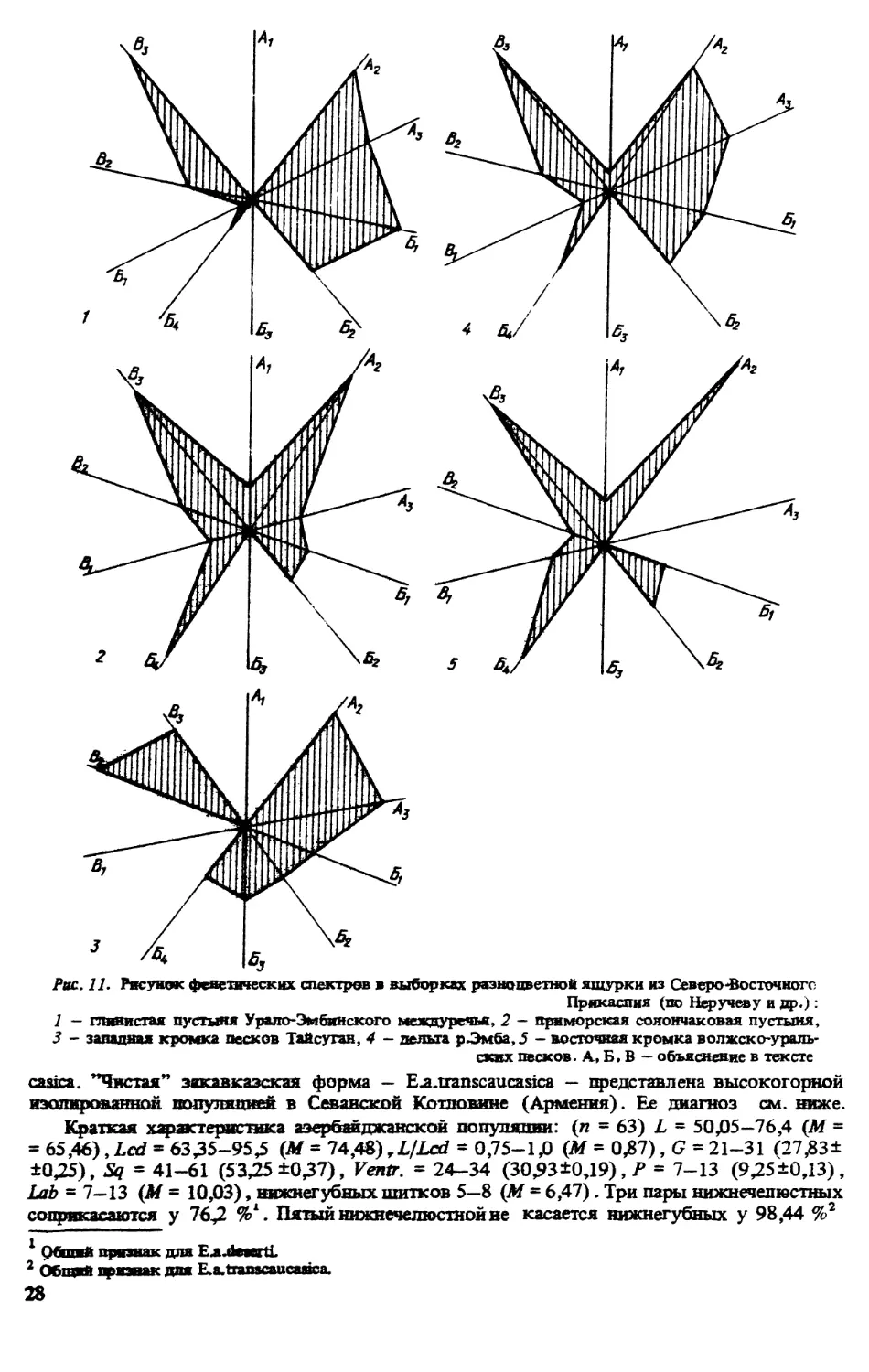
изменчивость некоторых признаков. ВЗ.Неручев и др. [181] дали феногеографическую
характеристику популяций Е.arguta из Северного Прикаспия. В том же году были опубли-
кованы данные В.Ф.Орловой, в которых показаны отличия ящурок Монголии от восточно-
казахстанских и джунгарских (Ea.potanini).
Исследования экологии разноцветной яшурки были начаты в России еще в прошлом
веке. Уже А.М .Никольский [189] собрал сведения об экологии этого пресмыкающегося
4
[Ill, 112, 231, 328, 355]. Разноцветной ящурке в Украине посвящены работы ААБрау-
нера [45, 47], Г.Ф.Сухова [246], ЛБАренса [15], НБ.Шарлеманя [299], ПЛКуцевола
[151], И.А.Цемша [280], ЮЛ Лащенко и В.АМежжерина [202], ВЛЛончаровой [66],
ВИ.Таращука [248], Н.Н.Щербака [314-317, 319, 320, 436], В Л.Булахова [49], ТИКо-
тенко (Ситко) [121—138, 140, 141, 232, 385, 386]. Ящурок в Предкавказье изучали
ДКрасовский [146], А.Г.Банников [27], СИМедведев и ВБЛетров [170], 3 ЛХоня-
кина [278], Г Л Лукина [156], Э.А.Шебз ухова [305], М.Ф.Тертышников [254—270],
в Прикаспии — НКосарева [118], С .А.Чернов [293], СМ.Федоров [273], НМ.Окулова
[192, 193], В И Б адм а ев а [21, 22], В.В.Неручев и др. [180, 182, 183—185], Ю.ЮКудаки-
на [148], Н Л Иванова [99]. Сведения о ящурках Кавказа есть в трудах Р Джафарова
[86], AM Алекперова [5, 7], И.СДаревского [77], ТАМусхелишвили [176]. Обширна
литература о разноцветной ящурке Средней Азии и Казахстана [35, 39, 55, 92, 94, 96,
120, 199, 222, 290, 294, 298, 313, 323, 324]. Материалы по разноцветной ящурке из зару-
бежных стран — Румынии [359, 397], Монголии [196, 197] — фрагментарны и немного-
численны.
Сведения о систематике и экологии ящурок обобщены П.В .Терентьевым и СА.Чер-
новым [251—253], АЛ Банниковым, И.СДаревским и А. К. Рустамовым [28], АЛБан-
никовым и др. [29], НИ Щербаком [320,436].
Морфологию разноцветных ящурок описали Г.И .Василевская и МББеселовский
[52], микроэлементный состав — СА.Шарыгин [302] и В.ЯЛассо [62], энергетический
метаболизм — С Л Львова [161].
ГЛАВА 2.
МАТЕРИАЛ И МЕТОДИКА ИССЛЕДОВАНИЙ
Основные методики, используемые при выполнении настоящей работы, описаны довольно
подробно {219, 320], поэтому мы отсылаем читателя к указанным работам, здесь остано-
вимся на некоторых методиках, ранее не применяемых.
Отметим более глубокие и регулярные (мониторинговые) полевые исследования, про-
веденные в Украине (Т.И.Котенко), Предкавказье (М.Ф.Тертышников, В.И.Горовая,
МЛЗинякова), Прикаспии (В.И.Бадмаева, В.В.Неручев, Н.Н.Иванова, ВА-Киреев,
Н.М.Окулова), а также в Средней Азии и Казахстане (ДА.Бондаренко, В Г Колбинцев,
ЗКЬрушко, С.Шаммаков). Это позволило уточнить ареал, получить данные о морфоло-
гии ряда неисследованных ранее популяций, сравнить их. Всего обработано около 10 тыс.
ящурок.
Впервые исследованы анатомия и морфология большинства систем данного вида
(Г.И.Василевская, М.В.Веселовский, Н.Н.Иорданский, В. С. Кото к). Описания методик содер-
жатся в тексте соответствующих разделов. Значительно пополнился и арсенал экологи-
ческих методик. Так, было изучено воздействие разноцветной ятцурки на видовой состав,
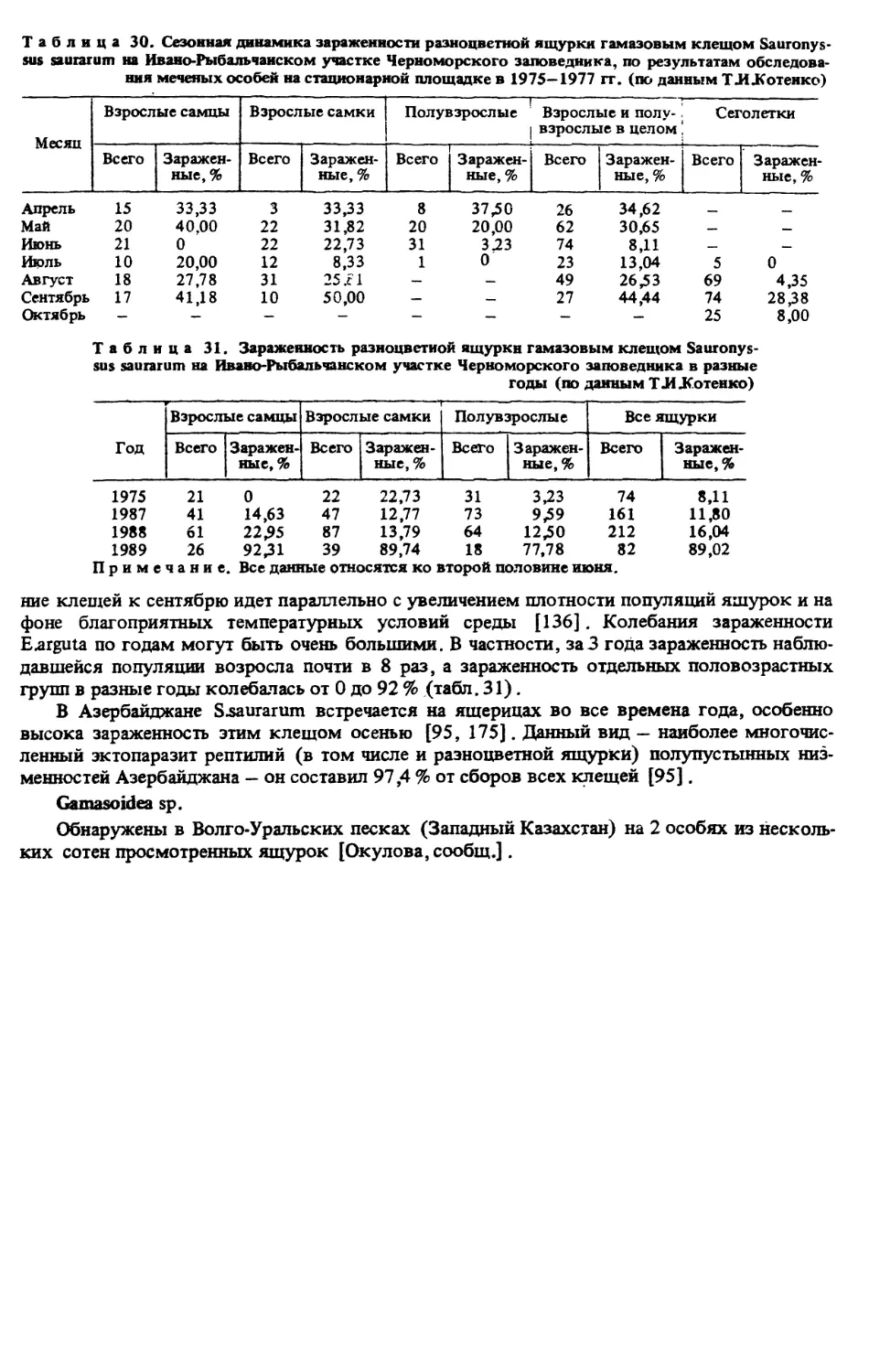
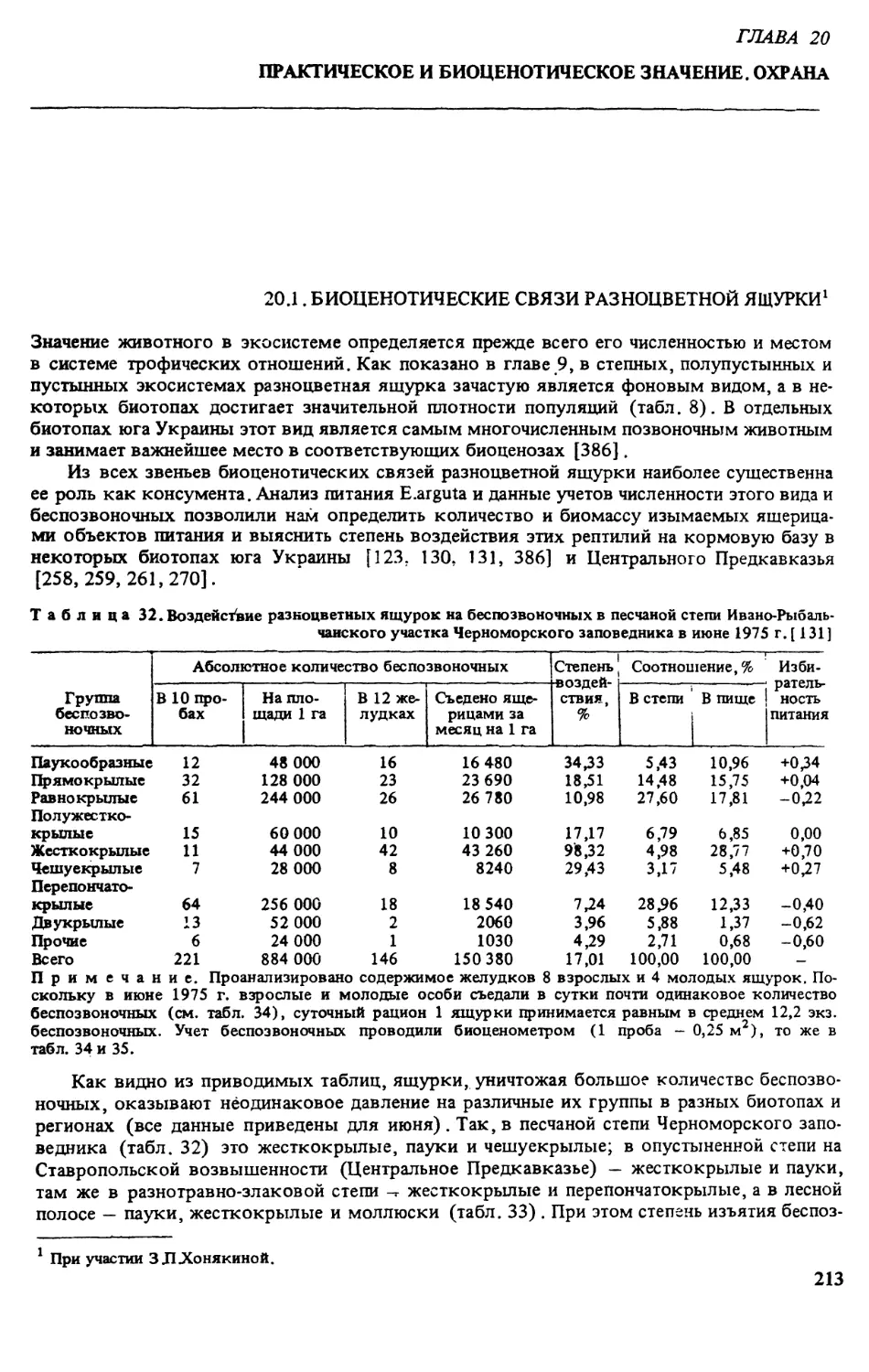
плотность населения и биомассу беспозвоночных. ТЛ.Котенко [139] с помощью биоцено-
метра в песчаной степи Черноморского государственного биосферного заповедника1 —
стации разноцветной ящурки — учитывала животных на стационарной площадке ежеме-
сячно с апреля по октябрь (в июле учет проводился в 19 биотопах). Пробы, преимуще-
ственно насекомых, обрабатывали сразу после их фиксации — во избежание потери массы
вследствие высыхания. Разобранных по видам или группам беспозвоночных взвешивали
на торсионных весах с точностью до 1 мг, а затем хранили на вате и в спирте.
Биоценометр представляет собой сборный дюралюминиевый каркас (параллелепи-
пед) с чехлом из мельничного газа высотой 1 м, основанием 50 х 50 см. Вертикальные
стойки несколько выступают ниже основания (они заострены), что позволяет устойчиво
фиксировать биоценометр на почве. Сбоку биоценометра имеется рукав, позволяющий
исследователю проникать внутрь прибора для сбора насекомых. При их обилии биоцено-
метр можно превратить в фотоэклектор. Для этого с противоположной стороны в верти-
кальной плоскости есть небольшой рукав, куда вставляют (привязывают) стеклянный
сосуд, а остальную поверхность биоценометра затемняют специальным чехлом из черной
ткани. Насекомые собираются в сосуд, после чего чехол снимают, рукав завязывают и
продолжают сбор ручным способом. Затем биоценометр переносят на следующий учас-
ток. При этом двигаются осторожно, против солнца и ветра, набрасывают прибор быстро,
держа на вытянутых руках, и плотно фиксируют на почве, чтобы между его основанием
и поверхностью почвы не было щелей. Описанный биоценометр легок (масса 2,8 кг) и
портативен — в разобранном виде его размер 100 х 10 см. На сборку биоценометра тре-
буется 10—15 мин. Время отбора пробы зависит от плотности беспозвоночных и густоты
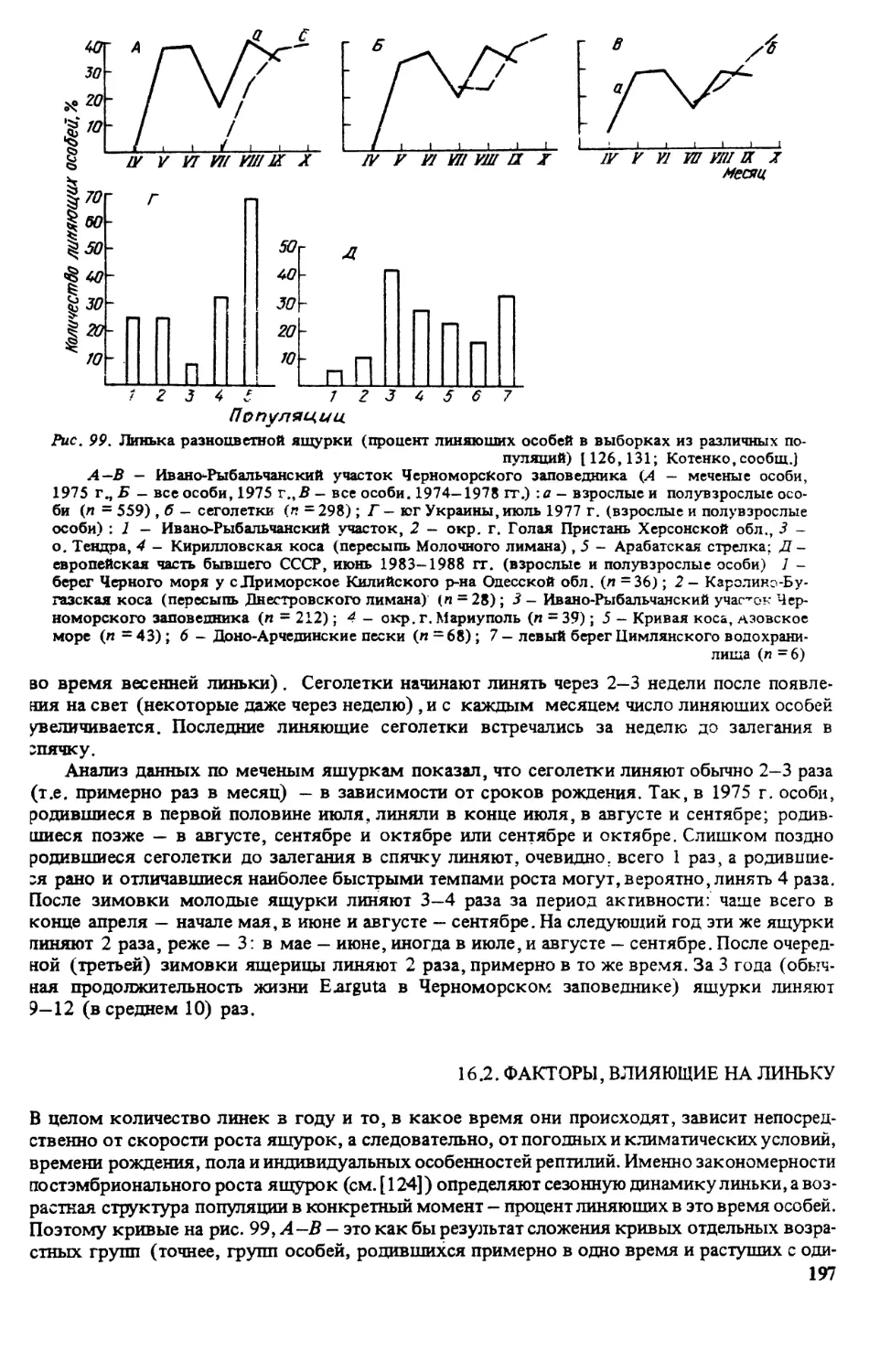
растительности. На взятие одной пробы на площади 0,25 м2‘затрачивалось от 10 мин
(слабозадерненные пески и солончаки) до 2 ч (разнотравно-типчаково-ковыльные и
типчаково-ковыльные степи). При этом учитывали всех беспозвоночных травяного яруса
и подстилки размером более 2 мм — объекты питания пресмыкающихся. Для получения
удовлетворительного уровня точности достаточно брать 10 проб в биотопах с густым
травостоем и высокой плотностью насекомых (настоящие степи) и 20 проб в биотопах
с редким травостоем (песчаные степи, солончаки). В итоговых оценках ошибка колеба-
лась от 6 до 19 %.
* Далее в монографии - Черноморский заповедник.
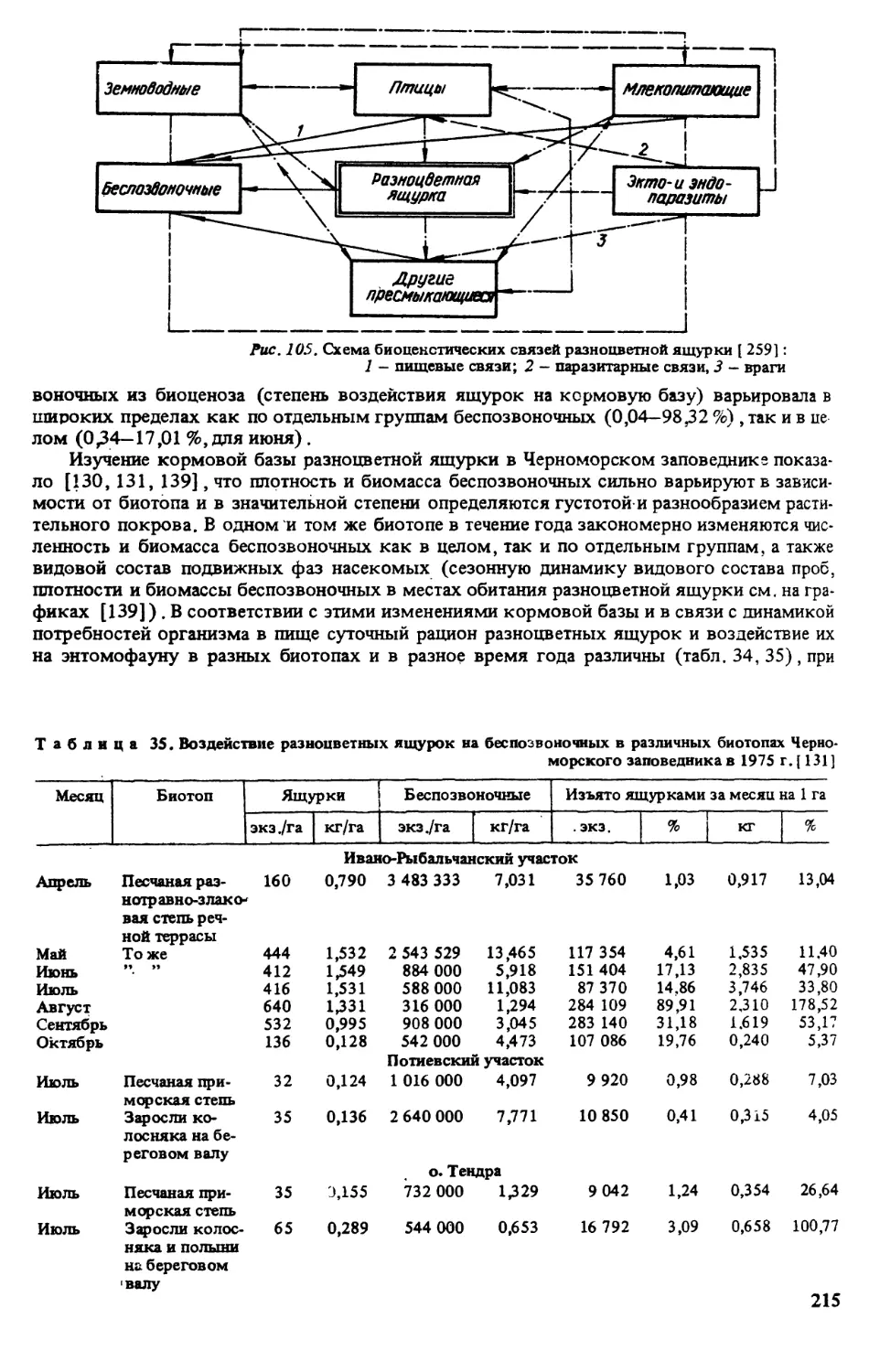
Анализ полученного материала показал [139], что плотность и биомасса беспозвоноч-
ных сильно варьируют в зависимости от биотопа и в значительной степени определяются
густотой и разнообразием растительного покрова, а также зависят от климатических осо-
бенностей года, В одном и том же биотопе в течение года закономерно изменяются числен-
ность и биомасса беспозвоночных в целом и по отдельным группам, а также видовой со-
став подвижных фаз насекомых.
В песчаной степи наибольшее количество видов было в майских пробах, плотность
максимальна — в апреле (за счет цикадок), биомасса — в мае (за счет многих групп) и в
июле (в результате массового появления взрослых прямокрылых). В августе наблюдал-
ся минимум по всем трем показателям, что обусловлено выгоранием степной растительно-
сти. В сентябре — октябре отмечен некоторый подъем численности и биомассы насеко-
мых, связанный с осенним отрастанием растений. В целом плотность беспозвоночных варь-
ировала по месяцам от 300 тыс. до 3,5 млн экз./га, биомасса — от 1,3 до 13,5 кг/га с мини-
мумом в августе.
Исходные данные о составе и количестве беспозвоночных при наличии сведений о чис-
ленности ящурок, массе съеденной ими пищи позволили решить следующие вопросы:
воздействие пресмыкающихся на беспозвоночных и их биоценологическое значение, изби-
рательность кормов (видовая, в зависимости от пола и возраста, климатических особенно-
стей и сезона) , практическое значение ящурки.
Сведения о размножении получены в результате вскрытий животных, при обследова-
нии ежемесячно отлавливаемых меченых особей и во время наблюдений за ящурками в
природе и террариуме. Гонады, яйца и жировые тела взвешивали на торсионных весах с
точностью до 1 мг, размеры семенников, фолликулов и зрелых яиц измеряли штангенцир-
кулем с точностью до 0,1 мм. При определении размеров и массы яиц брали лишь готовые
к откладке или только что отложенные яйца. Выясняя плодовитость, учитывали самок с
яйцами в яйцеводах и особей с крутыми развивающимися фолликулами в яичниках.
Достоверность различий между средними арифметическими определяли по критерию Стью-
дента с учетом объемов выборок (т.е. по дисперсии) [126].
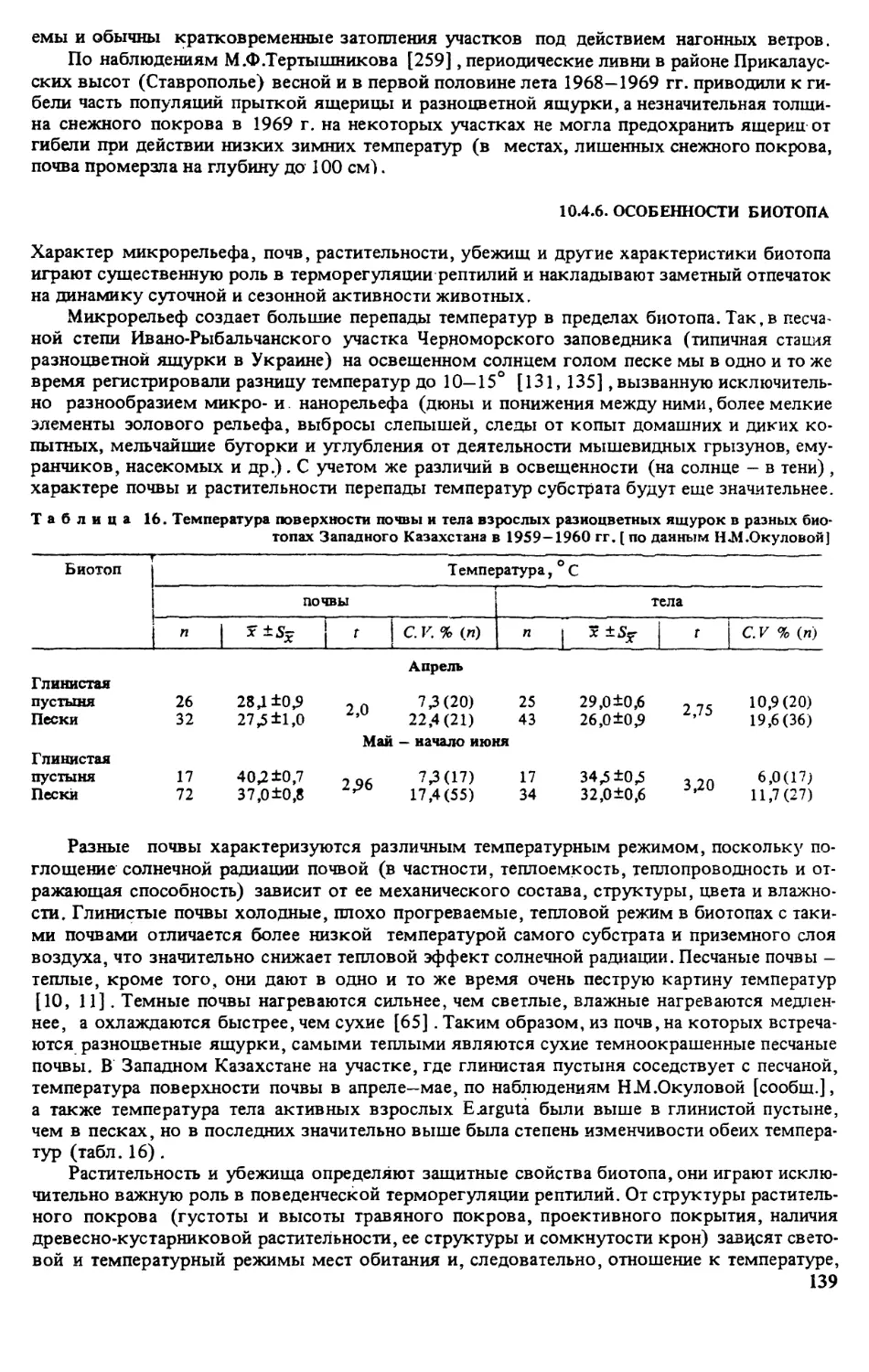
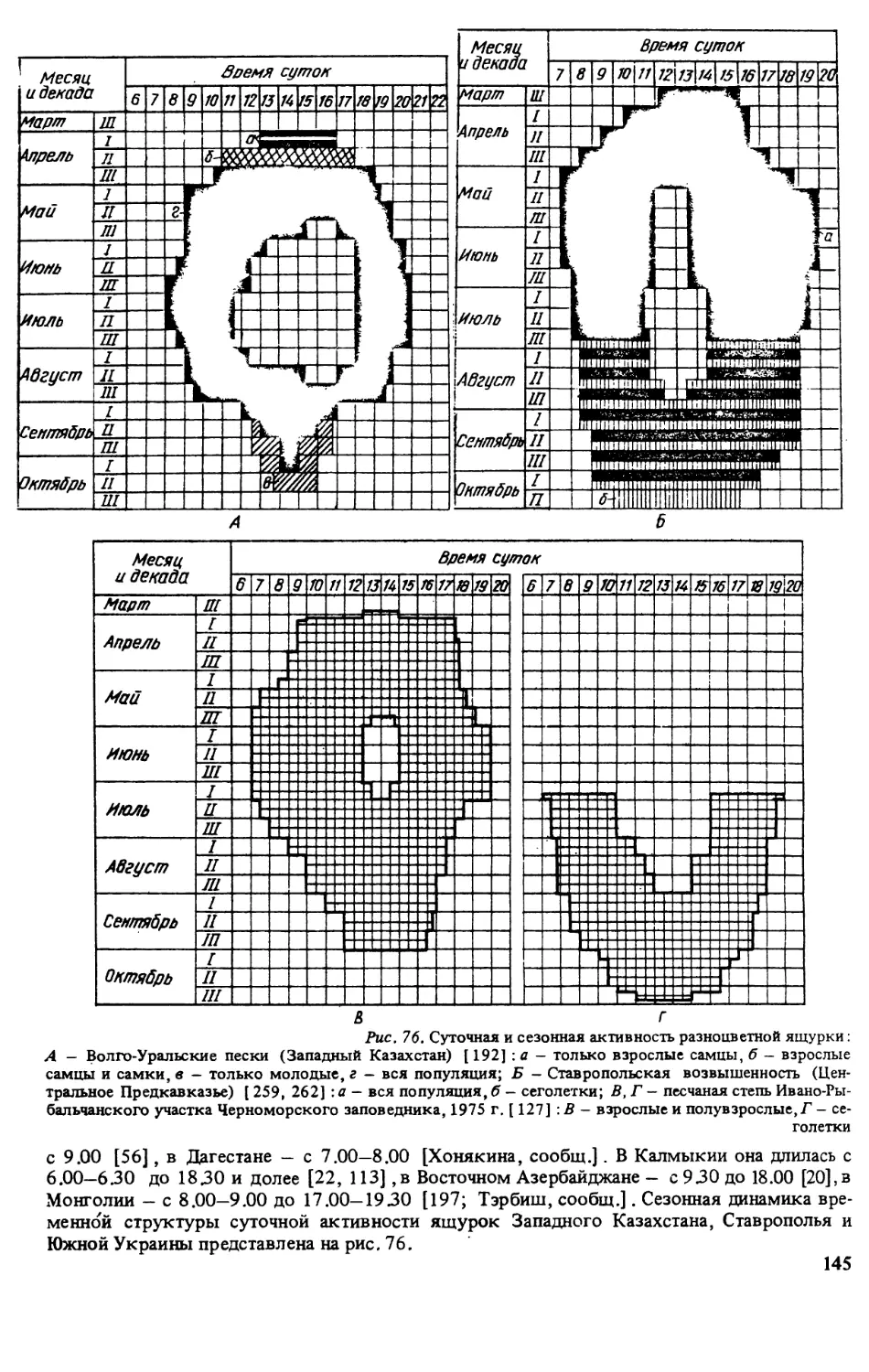
При изучении суточной и сезонной активности и отношения ящурок к погодным фак-
торам фиксировали время поимки каждого животного и учитывали относительную числен-
ность ящурок на постоянных маршрутах или площадках (через каждый час в течение всего
периода суточной активности и в разные сезоны) . При этом отмечали температуру и отно-
сительную влажность припочвенного слоя воздуха, температуру поверхности почвы, атмо-
сферное давление, освещенность, скорость ветра, а также облачность (в баллах) иосадки
(визуально) [127].
Для изучения влияния на ящурок температуры в песчаной степи на участке с высокой
их плотностью была выделена стационарная площадка размером 0,25 га, на которой ежеме-
сячно учитывали относительную плотность ящурок, измеряли температуру и относитель-
ную влажность припочвенного слоя воздуха (аспирационным психрдметром Ассмана на
высоте 3—5 см от поверхности субстрата) и температуру поверхности почвы (ртутным или
электротермометром). Периодически измеряли также атмосферное давление, освещен-
ность и скорость ветра, отмечали облачность и осадки. Затем проводили отлов-выпуск ме-
ченых ящериц, при этом регистрировали время и место отлова, измеряли температуру тела
(перорально, медицинским электротермометром ТПЭМ-1 с точностью до 0,1 ° х) и темпе-
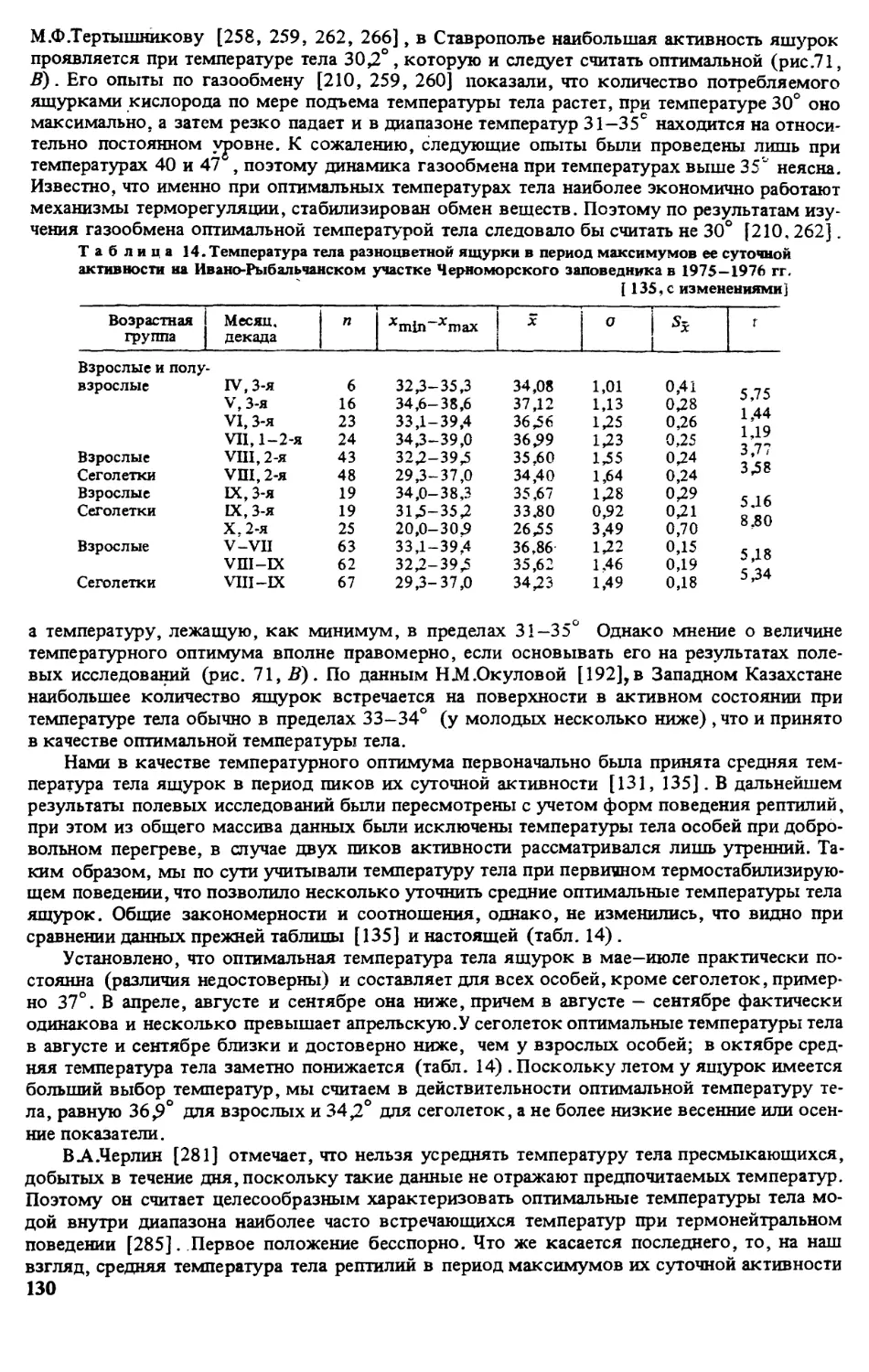
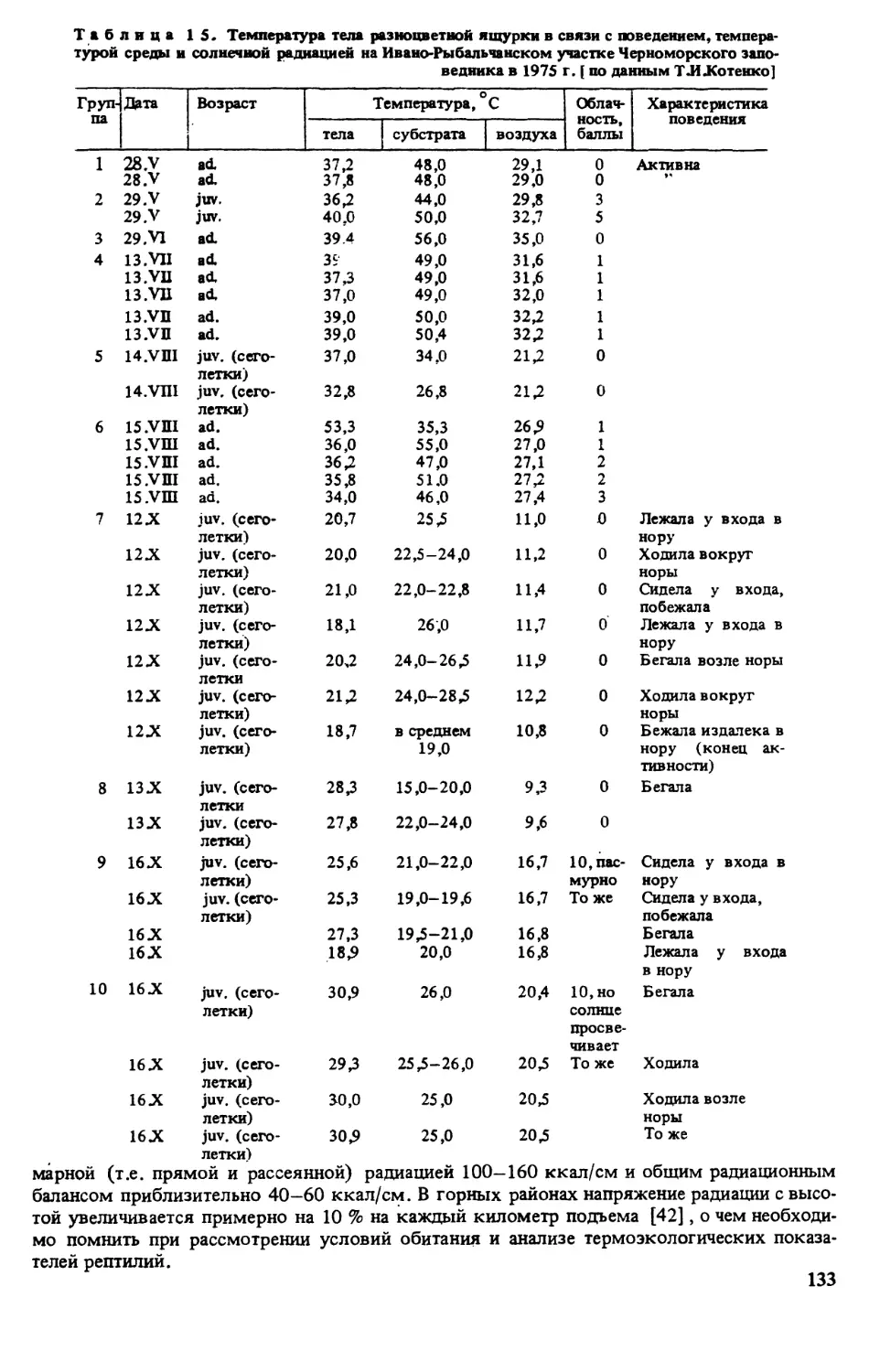
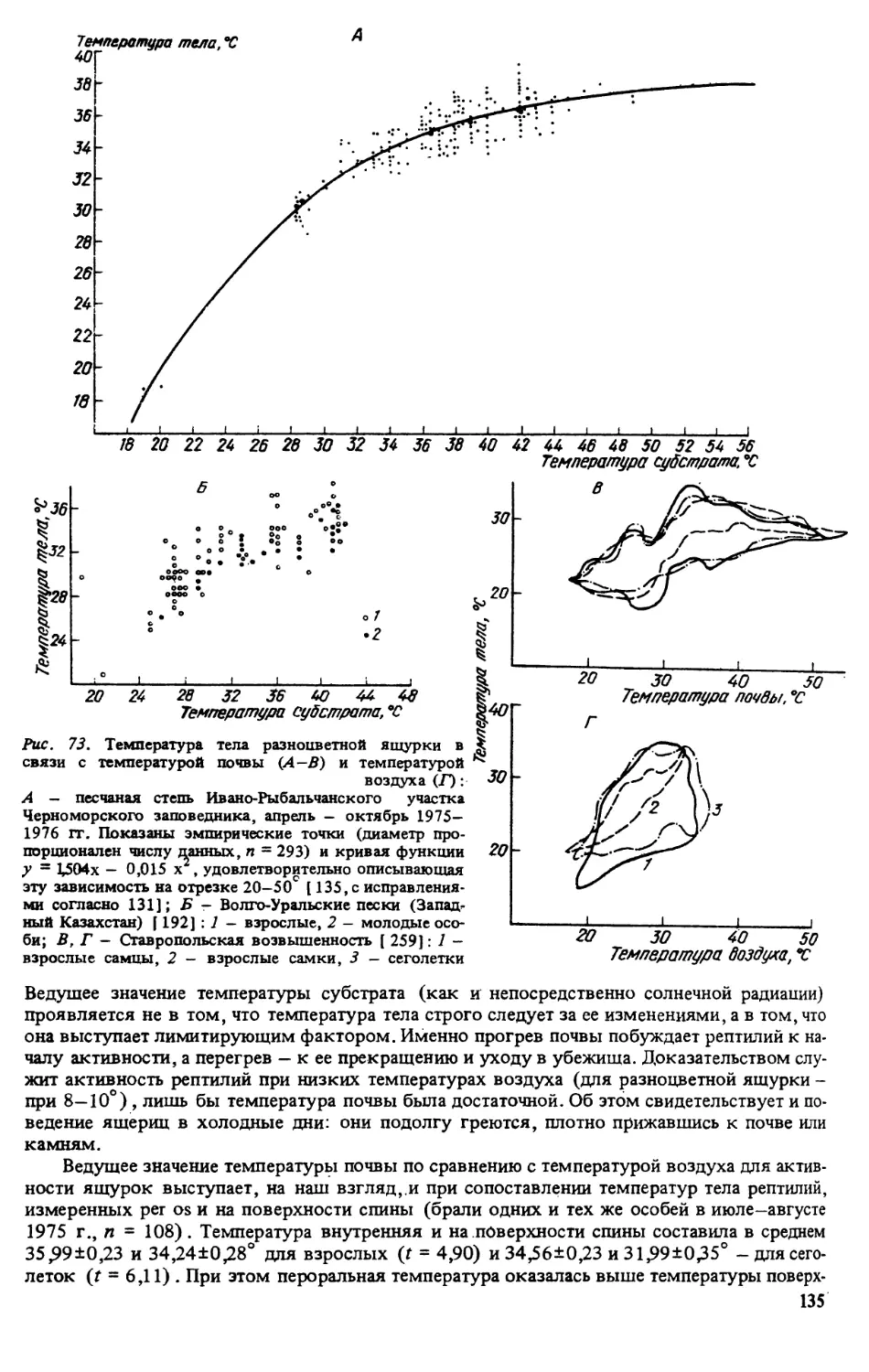
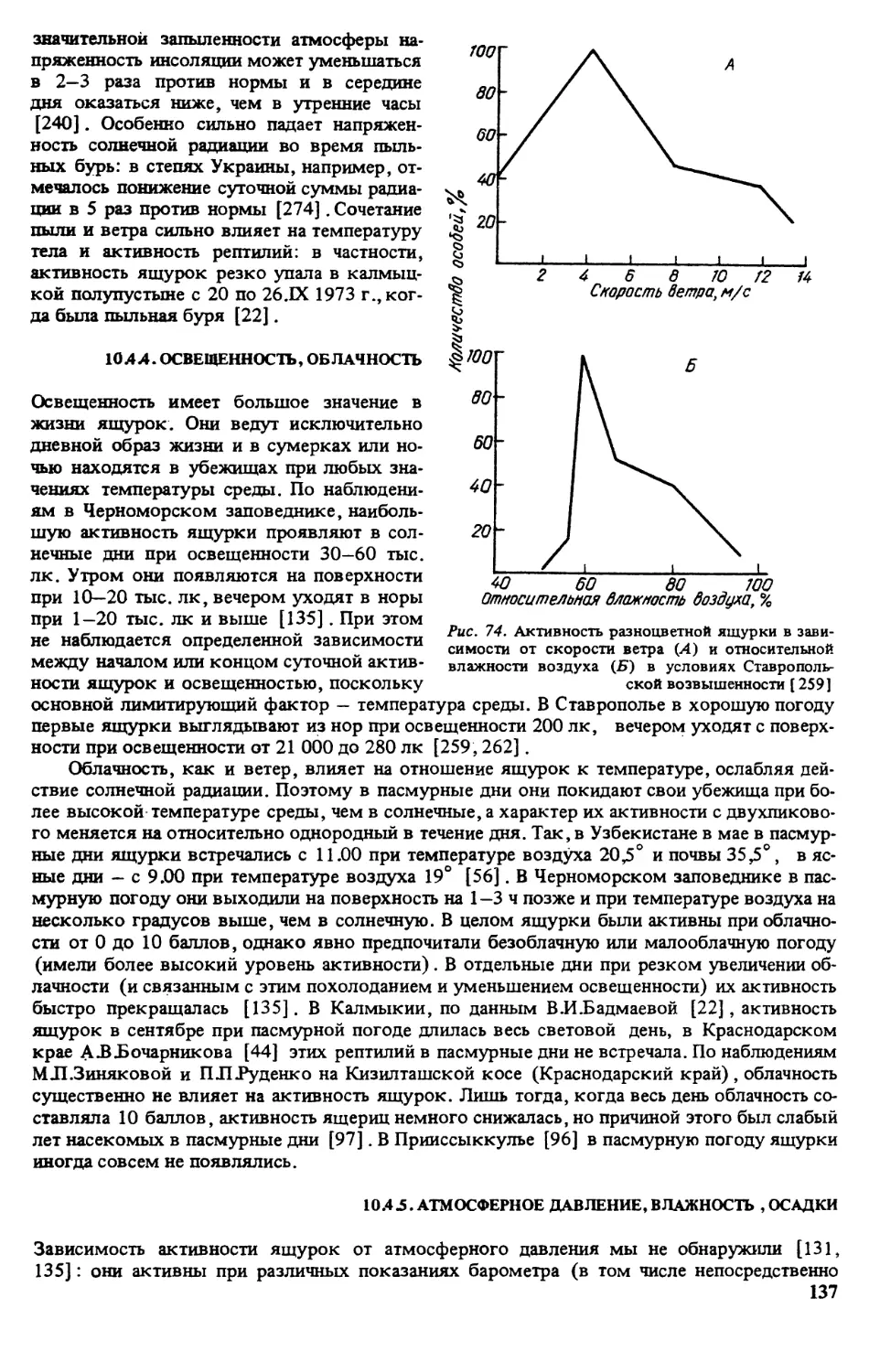
ратуру поверхности субстрата в месте отлова животного [135]
Метили ящурок отсечением пальцев в различных комбинациях (постоянные метки) и
окрашиванием головы или спины ярким лаком (временные метки) . Все норы, служив-
шие убежищем для ящурок, нумеровали и картировали. Это позволило параллельно полу-
чать сведения о поведении отдельных особей в различных условиях, а также миграциях и
индивидуальном участке [135].
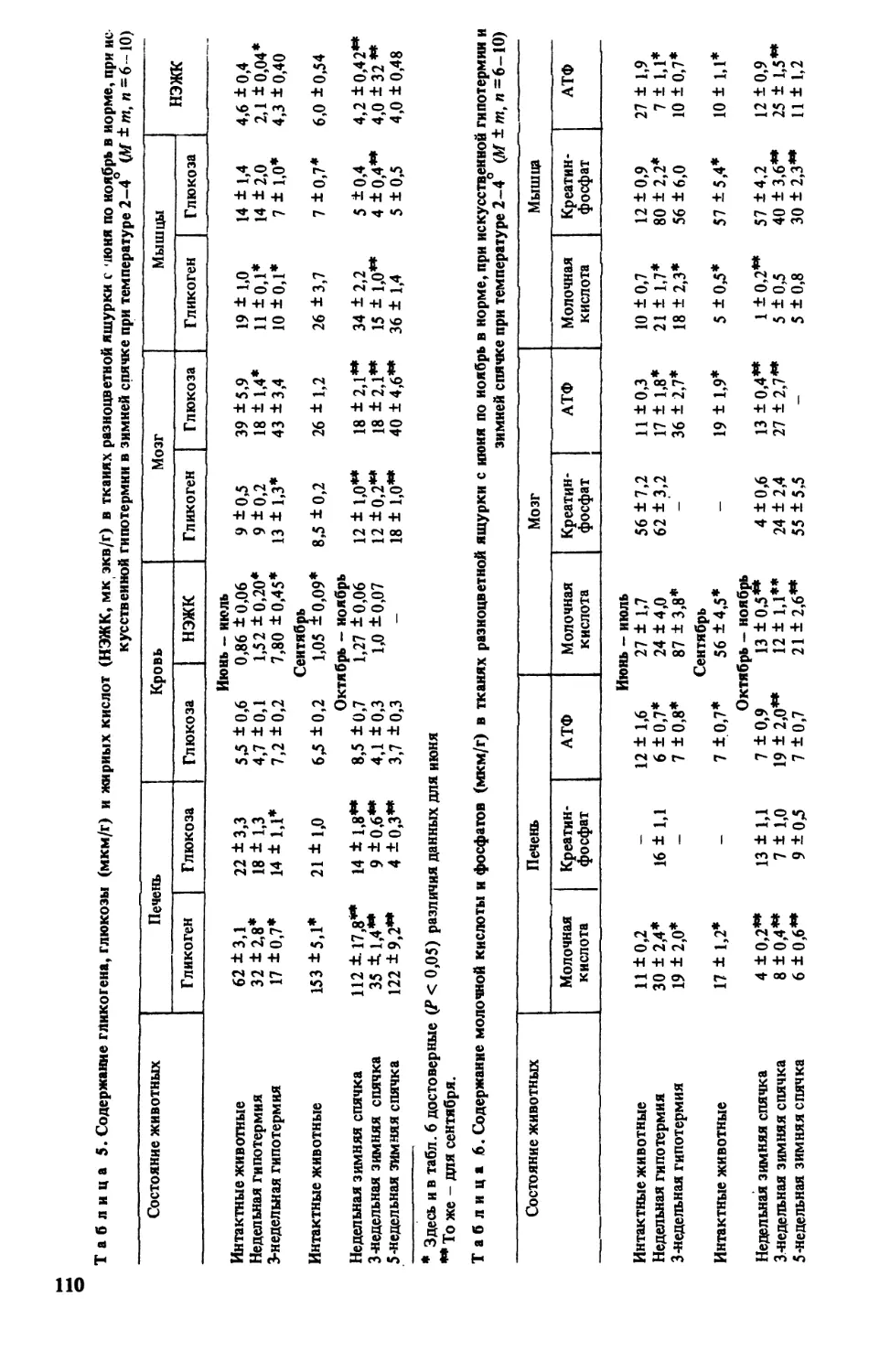
Содержание некоторых энергетических метаболитов в тканях разноцветной ящурки
при зимней спячке определяла СП-Львова [161]. Исследовалось содержание гликогена,
глюкозы, неэтерифицированных жирных кислот (НЭЖК), молочной кислоты, АТФ, креа-
тинфосфата, неорганического фосфора в печени, скелетных мышцах, мозге, а также глюко-
зы и НЭЖК в крови. Опыты проведены на 135 особях взрослых разноцветных ящурок
обоего пола, отловленных в окрестностях г Махачкала. Более детально использованная ме-
1 В монографии температура приведена в градусах Цельсия.
7
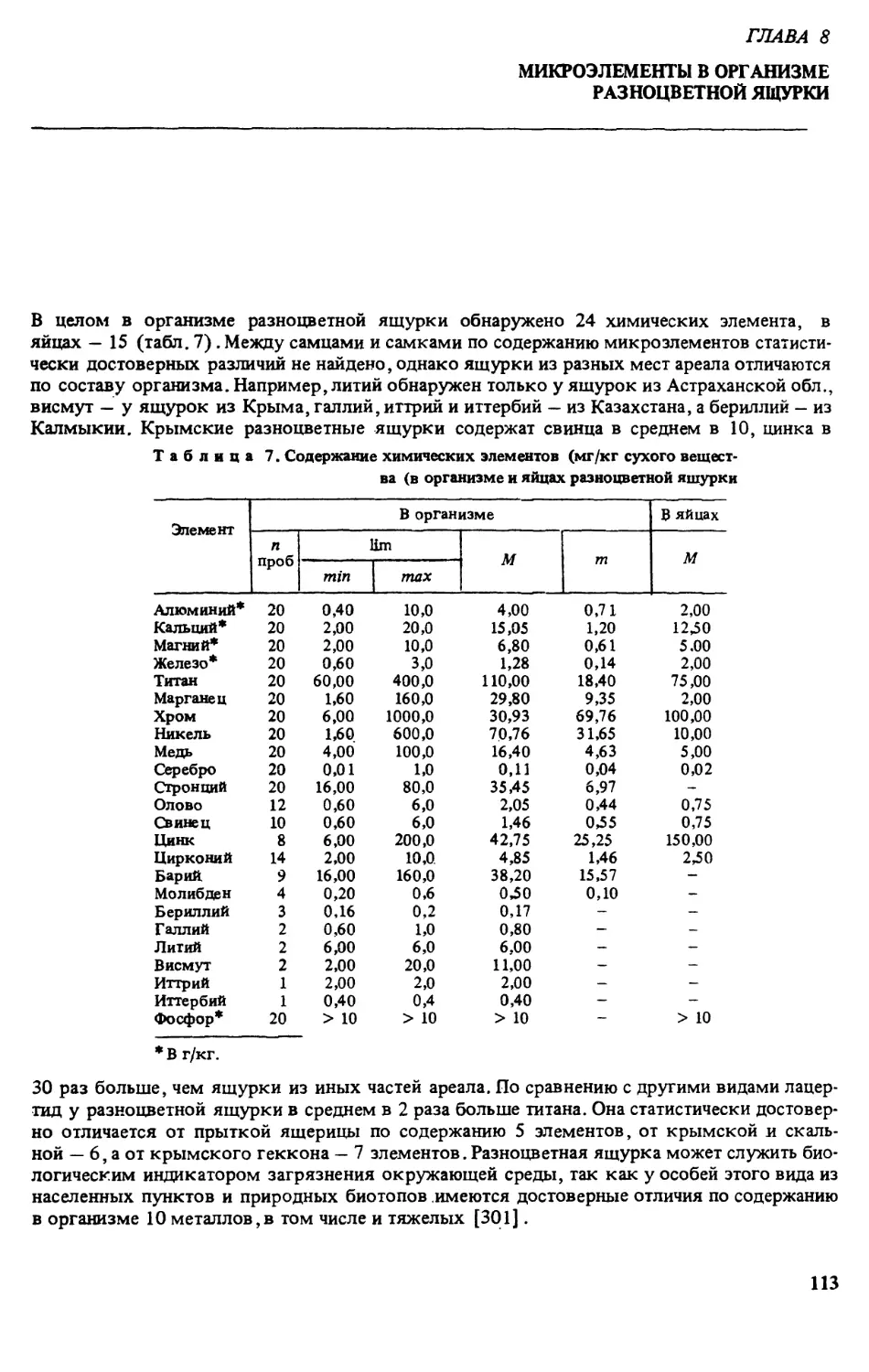
тодика описана в соответствующем разделе. САДИарыгин методом полуколичественного
эмиссионного спектрального анализа на спектрографе ДФС-8 исследовал содержание неко-
торых макро- и микроэлементов в организме разноцветной ящурки (использовано 22 осо-
би, проанализировано 20 проб).
Избирательность питания определялась по формуле В.С. Ивлева (1955) а—в/а+в, где
а — отношение вида, выраженное в процентах, к количеству съеденных животных; в — от-
ношение вида, выраженное в процентах, к количеству учтенных животных.
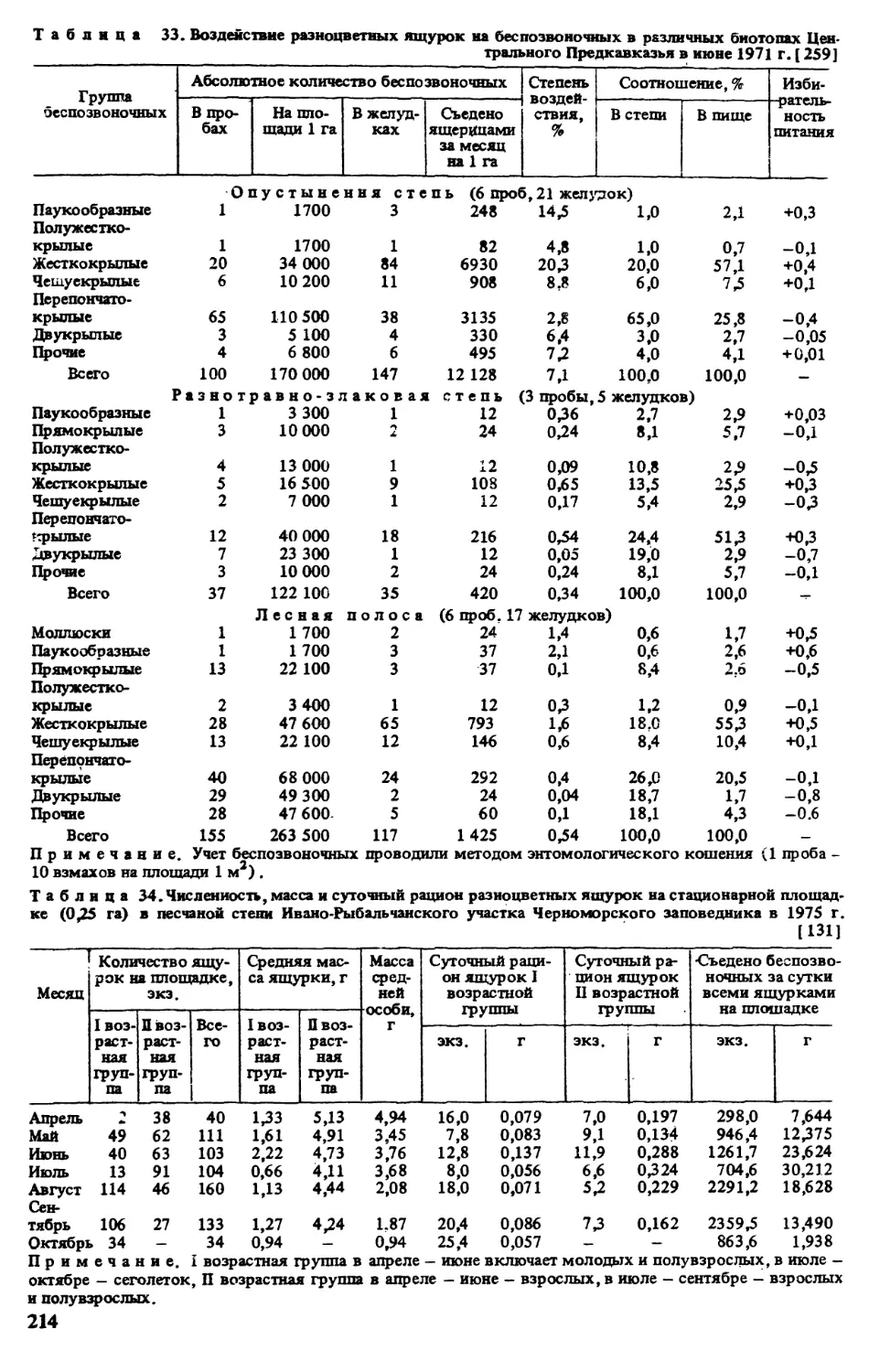
Функциональную роль ящурки в биогеоценозах выясняли определением потока энер-
гии в сутки и в течение года, проходящего через ее популяции. Для этого мы использовали
материалы по сезонной (годовой) и суточной активности ящерицы, структуре популяции
(количестве половозрелых самок — ad 9 , половозрелых самцов — ad д, полувзрослых
особей — зиЬасЦсеголетков — juv), плодовитости, отходу яиц и сеголетков, биомассе одной
условной особи в популяции, яйца и сеголетка в первые сутки жизни, количеству усред-
ненной пищи одной усредненной особи в течение сезона активности. Следуя методическим
обоснованиям оценки роли рептилий в потоке энергии [60, 61,348], единой мере измере-
ния численности ящерицы, массы пищи, эффективности размножения был избран энерге-
тический эквивалент в джоулях (Дж). Ввиду недостаточно репрезентативного материала
по некоторым сторонам биологии разноцветной ящурки нами сделаны следующие допу-
щения. Так, считали, что 1 г сырой зоомассы равен 0,2 г сухого вещества и соответствует
4,2 Дж; зольность сухой зоомассы равна 15 %; степень ассимиляции близка к 90 % [60,
216]; беззольное вещество яйца равно 6,37 кал/мг [456]; за первоначальную биомассу
популяции принимали биомассу половозрелых и неполовозрелых особей весной до наступ-
ления откладки яиц; суточное потребление пищи определяли по результату наблюдений за
животными в вольерных условиях, а также по уравнению зависимости энергообмена от
массы тела, в частности по формуле М = 39,6 W0*75 [90], где М — потребление пищи в 1 мг
сухой массы при 20°, И7 — живая масса животного; наконец, эмбриональную смертность,
гибель сеголетков и ящериц других возрастных групп приравнивали к нулю.
Биомассу ящурки рассчитывали по методу ’’объединенного гектара” [213], т.е. с уче-
том доли площади каждого биотопа на территории региона.
Для изучения индивидуальной территории ящериц и перемещения их использовали
следующую методику [149, 255,258]. В различных биотопах находили обитаемые норы ящу-
рок разного пола и возраста, вокруг них размечали участок размером 20 х 20 м, который
затем разбивали на квадраты площадью 1x1, составляли подробные картосхемы. Перед
началом работ определяли пол и возраст ящериц — хозяев убежищ. Для удобства наблюде-
ния им на спину наносили индивидуальные метки белой или красной нитрокраской. За каж-
дой ящерицей следили с помощью бинокля в течение нескольких дней с 5.00—7.00 до
18.00—21.00 с расстояния 10—15 м. Все дневные перемещения ящериц хронометрировали
и картировали. В конце каждого дня работы в картосхемы вносили уточнения после ос-
мотра следов ящериц, оставшихся на песке (в полупустынных и пустынных биотопах).
Подобные наблюдения проводили в период максимальной активности животных (май) и
во время ее затухания (август, сентябрь). Выясняя направление и дальность передвижения
ящериц, их привязанность к месту обитания, нами помечено большое количество (несколь-
ко тысяч) особей путем сечения фаланг пальцев в разных комбинациях [34]. Животных
выпускали на месте первоначального отлова. В дальнейшем раз в неделю в течение 1—3 лет
проводили повторные отловы. Полученные данные о первоначальном и повторных отло-
вах меченых ящериц также наносили на картосхему местности.
Эндопаразитов ящурок изучали традиционным методом паразитологических вскры-
тий [300].
ГЛАВА 3
АРЕАЛ1
Общий ареал разноцветной ящурки впервые наиболее достоверно описан в начале XX в. в
работах А.М.Никольского [186, 187,189]. В последующие годы уточняли границы распро-
странения вида [15]. Графическое изображение ареала E^rguta на территории бывшего
СССР опубликовано П.В. Терентьевым иСА.Черновым [253], А Г Банниковым и др. [28],
в Евразии — И.Фууном и С.Ваньчем [367], НЛ.Щербаком [316]. Сведения о распростране-
нии данного вида содержатся во многих региональных сводках [7, 35, 36, 176, 199, 248,
324]. Наиболее детальную схему ареала с указанием мест находок и их кадастром в 1974 г,
опубликовал ННЛЦербак.
Позже был уточнен ареал вида, его изменения под влиянием хозяйственной деятель-
ности человека [132, 140]. Все эти данные позволили нам дополнить известные материалы,
высказать суждение о причинах изменения ареала, представить его в перспективе и прогно-
зировать изменения. Ряд новых находок на востоке ареала свидетельствует о еще недоста-
точной изученности некоторых регионов. Поскольку места находок сравнительно недавно
были подробно опубликованы [320], мы намерены здесь критически рассмотреть их по
регионам, дополнить новыми пунктами.
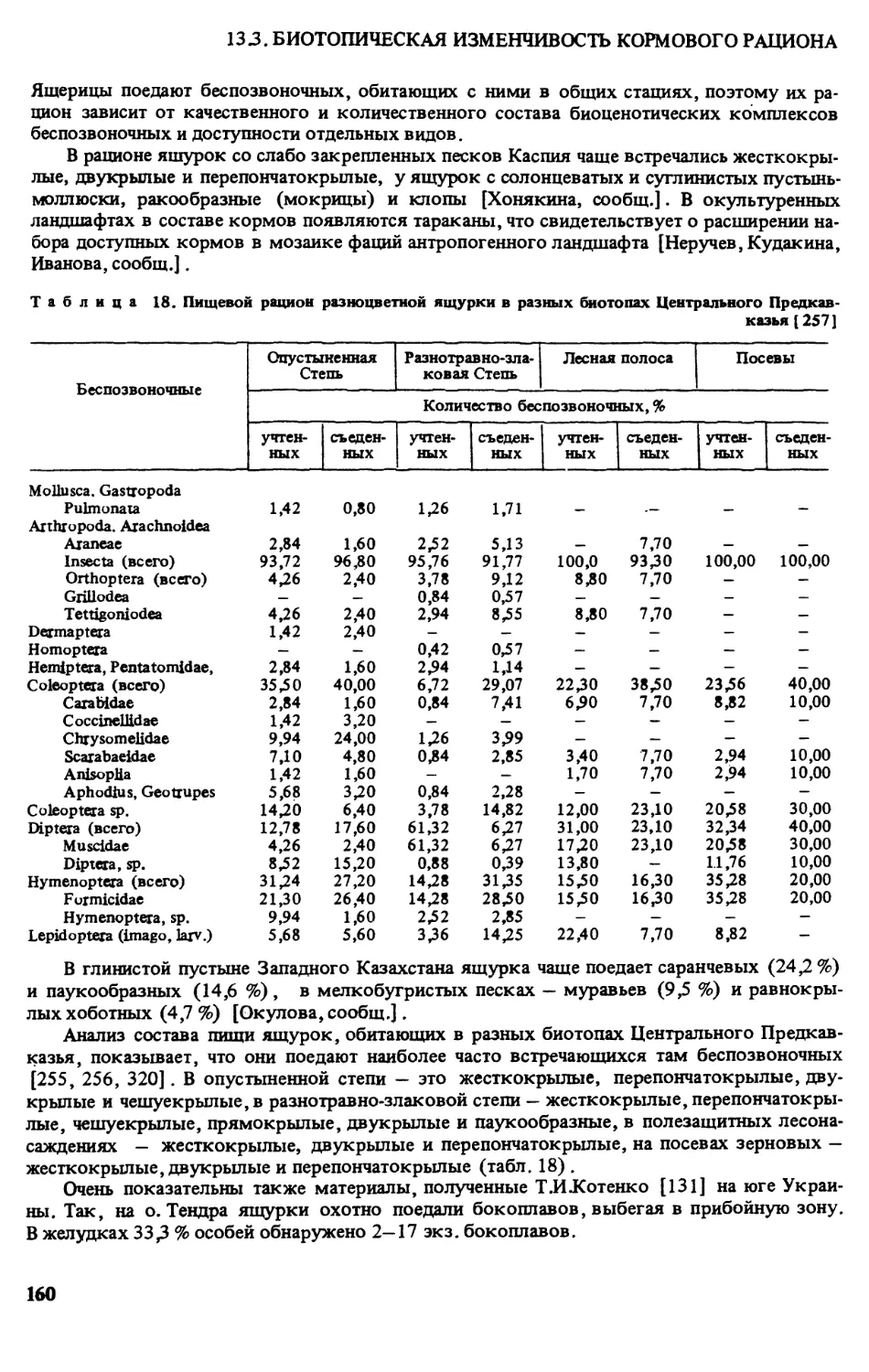
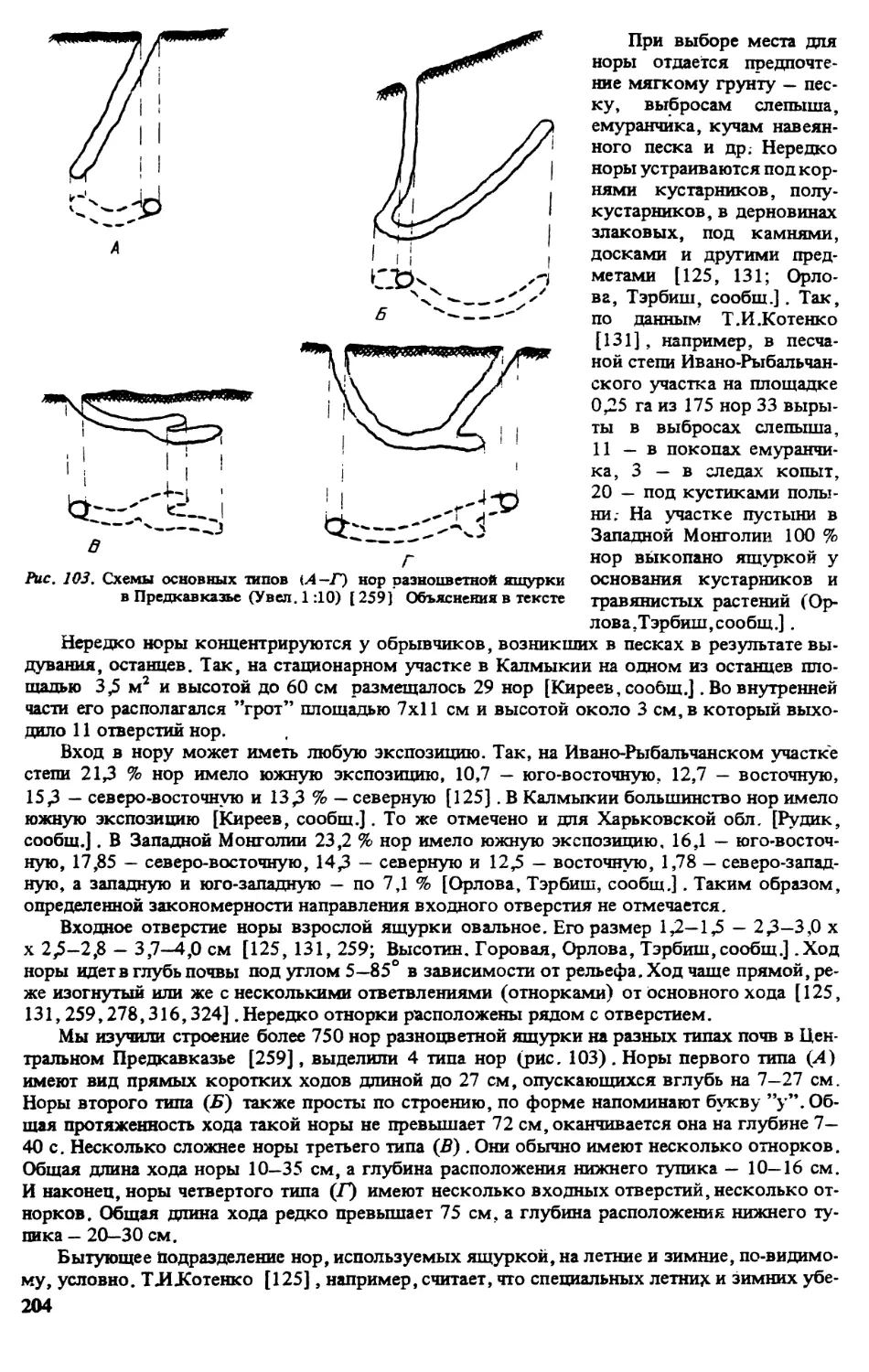
3.1.РАСПРОСТРАНЕНИЕ (МЕСТА НАХОДОК)
Румыния. За последние 15 лет новые сведения о распространении разноцветной ящур-
ки на крайнем западе ее ареала не публиковались. Можно только отметить устное сообще-
ние Т И Котенко о том, что их количество сокращается, особенно в курортных р-нах (гМа-
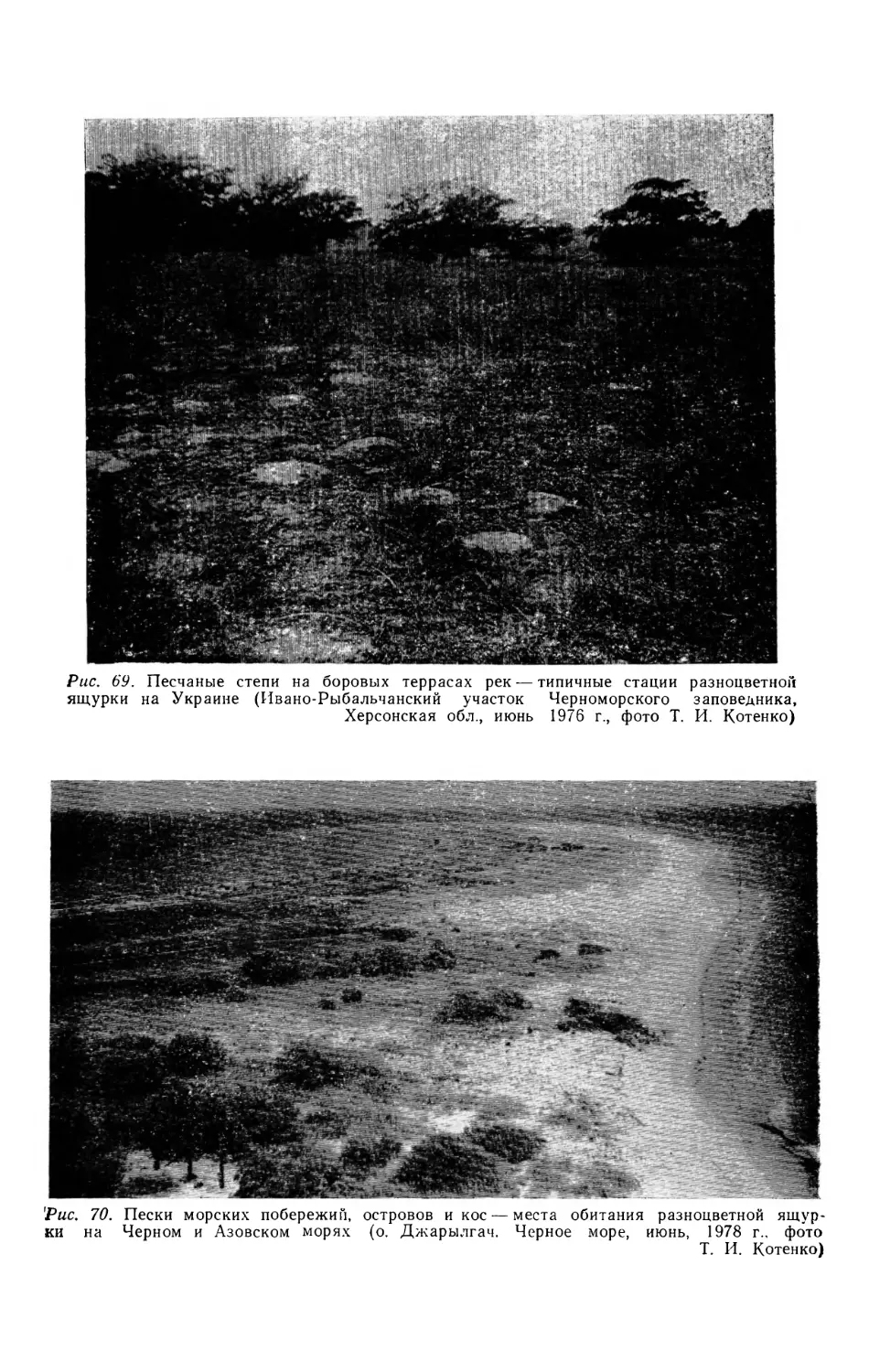
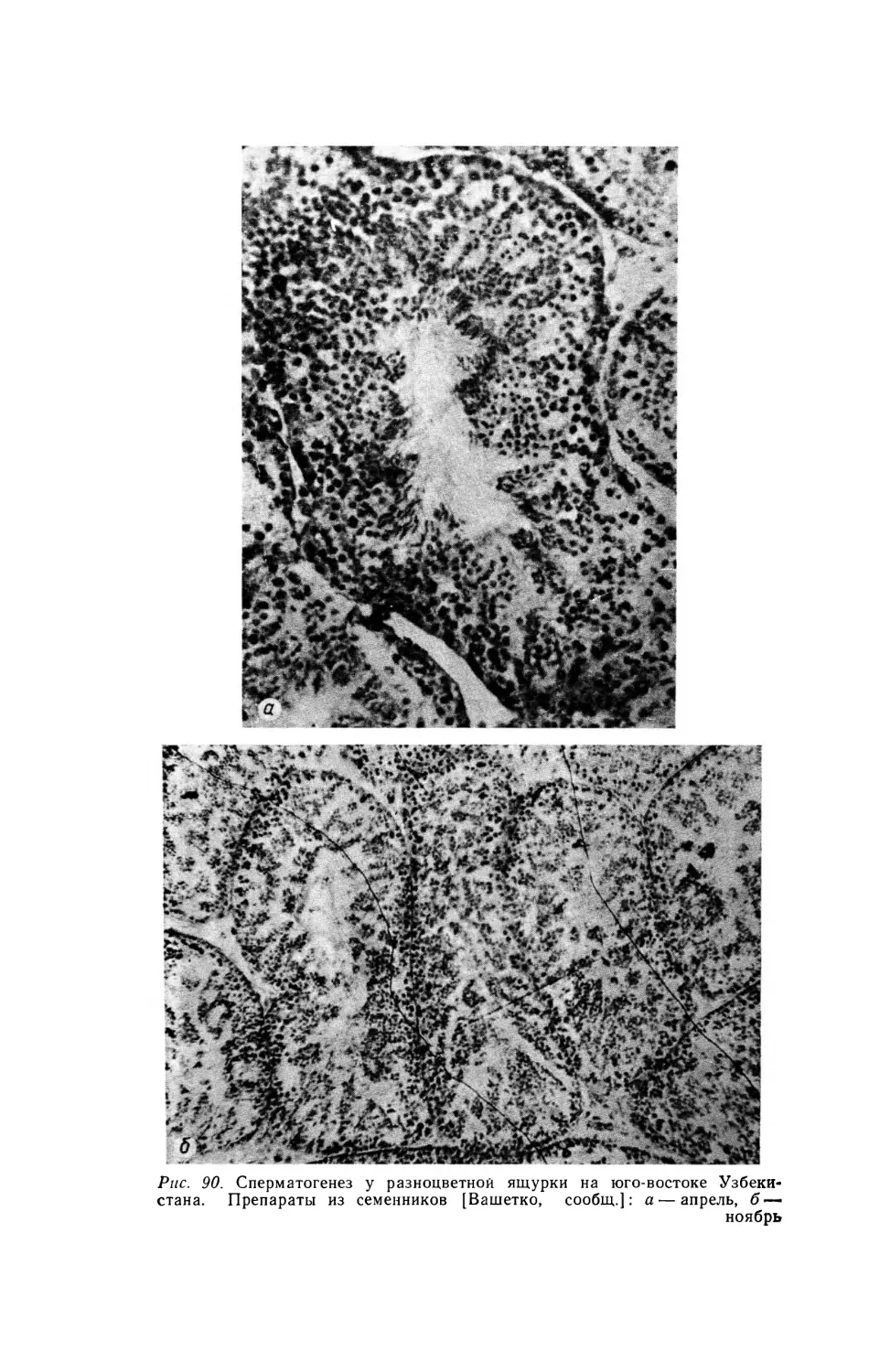
майа) (рис. 1).
2
Кадастр к рис. 1:
Здесь и далее на картах 2-7: а — современные места находок; б - места находок, где этот вид уже
не встречается: 1 - пос. Ганул Кон аки у г.Текуча,р-н Галац, 2 - с. Летя, 3 пос, Караорман, 4 - с.Пери-
тяшка, 5 — оз.Синое, 6 - г.Мамайа, 7 — г.Энисала, 8 - пос. Сфынтул Георге (дельта р.Дунай), 9 -
с. Журилофка, р-н Добруджа [ 359].
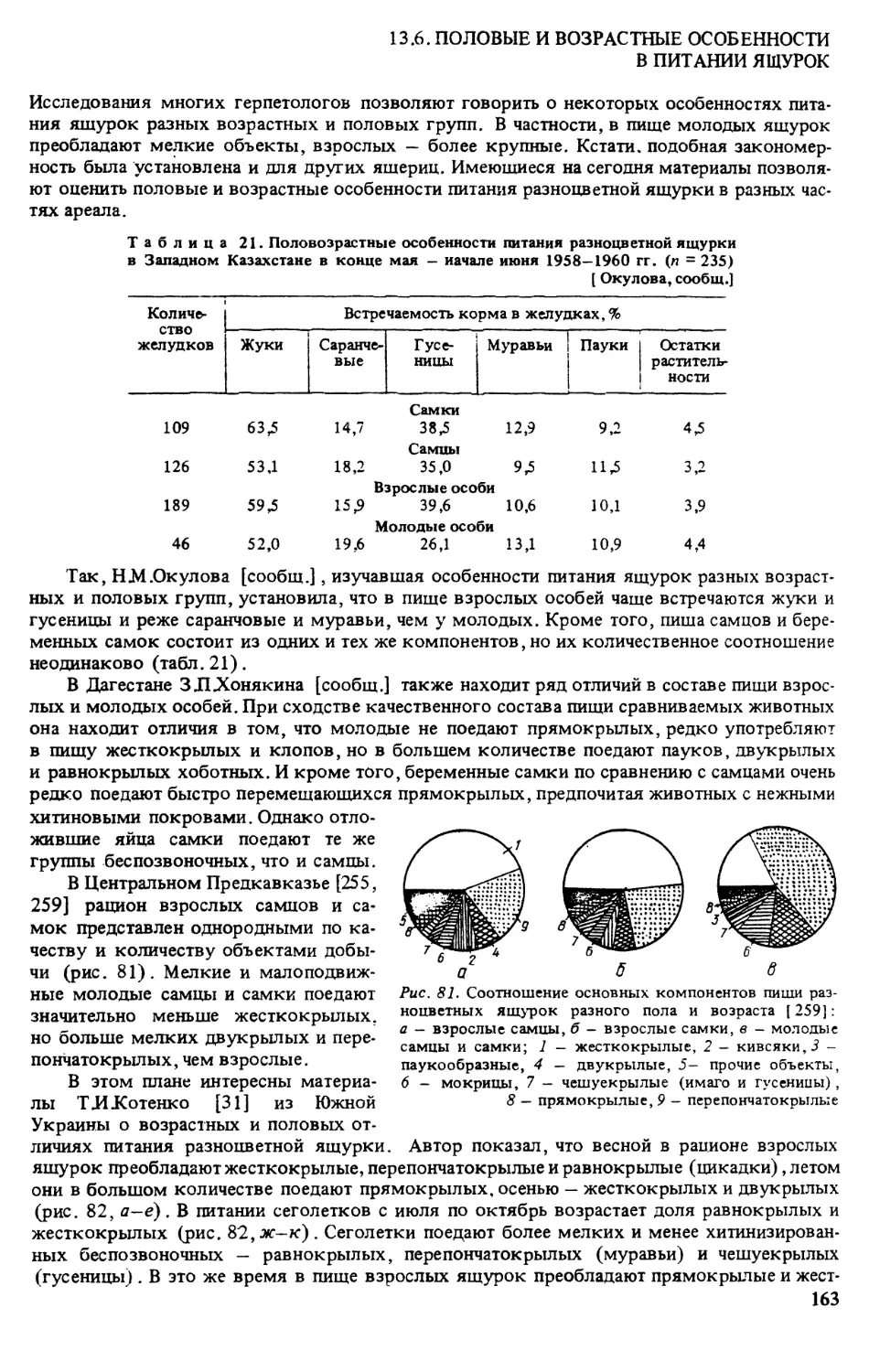
Украина. Распространение разноцветной ящурки в Украине детально исследовала
ТИКотенко [140]. Так, северные пункты границы ареала на западе бывшего СССР -
г Котовск [320, рис. 50, 4} и г Первомайск (там же, 5) — как выяснилось, обозначены
1 Материалы предоставили ТАлиев, Д.А.Бондаренко,З.К.Брушко, Э.В.Вашетко,В.И.Ведмедеря,И.С.Да-
ревский, М.П.Зинякова, Н.Н.Иванова, В .А .Киреев, В .Г .Колбинцев, Т.И.Котенко, Е.Ю.Кудакина,
ВЗ.Неручев, Н.М.Окулова, В.Ф.Орлова,Е.М.Писанец,П.П.Руденко,С.В.Таращук,СШаммаков,ЭА.Шеб-
зухова.
В кадастрах используются следующие сокращения: кол. ЗИН — по коллекциям Зоологического ин-
ститута РАН, г.Санкт-Петербург; кол. ИЗ АНУ- по коллекциям музея Института зоологии АН Украи-
ны, г Киев; кол. ИЗ АН Казахстана — по коллекциям Института зоологии АН Казахстана; кол. ХГУ-
по коллекциям музея Харьковского государственного университета, г.Харьков; кол. МГУ - по кол-
лекциям Московского государственного университета, г .Москва; кол. САГУ — по коллекциям Сред-
неазиатского государственного университета, г.Самарканд; кол. УзГУ - по коллекциям Узбекского
государственного университета, г.Ташкент. Приведен автор находки или источника литуратуры.
Приняты такие сокращения: бывщ. — бывший, окр. - окрестность, вост. - восточный, зап. — запад-
ный, ж.-д. ст. - железнодорожная станция, кол. — колодец, набл. - наблюдение, письм. - письмен-
ный, п. — пункт, прав. — правый, сев. — северный, сев .-вост. — северо-восточный, сев .-зап. — северо-
западный, свх. - совхоз, сообщ. - сообщение, ст. - станция, стан. - станица, ур. — урочище, ущ. -
ущелье, х. — хутор, хр. — хребет, юго-вост. — юго-восточный, юго-зап. — юго-западный, юж.—южный
9
ошибочно. Возможно, г Котовск [248] — это при-
нятое автором с-Котовка Магдалиновского р-на
Днепропетровской обл. [66]. По данным Т.ИКо-
тенко, в окр. городов Котовск и Первомайск
отсутствуют соответствующие биотопы для дан-
ного вида. Достоверные места находки разно-
цветной ящурки (по наблюдениям в 1986 г.):
берега рКучурган в Одесской обл. (окр. с.Но-
вопетровка Великомихайловского р-на, сел Гаев-
ка и Павловка Раздельнянского р-на), низовья
р.Савранка (с.Байбузовка Савранского р-на Одес-
ской обл.) , правый берег р.Южный Буг ниже пос.
Саврани (окр. сТолосково Кривоозерского р-на
Николаевской обл.), правый берег р.Кодыма
Рис. 1. Распространение E<arguta в Румынии (с.Сырово Врадиевского р-на Николаевской
[367] обл.). Указания на находки вида в окр. г.Чер-
кассы [151, 248] также уточнены [140]. В ре-
зультате затопления биотопов Кременчугским водохранилищем здесь на правом берегу
Днепра ящурка исчезла, но сохранилась на левом берегу р.Тясмин от с.Вергуны Черкас-
ского р-на до места впадения ее в Днепр (окр. сБитово Чигиринского р-на) . Окр. пгт Гра-
дижск [320, рис. 50,21} по-прежнему заселены ящуркой (от указанного пункта до сМак-
симовка Кременчугского р-на Полтавской обл.); кроме того, этот вид обнаружен нес-
колько севернее, на левом берегу р.Сула у ее устья (окр. сЛиповое, Глобинского р-на
Полтавской обл.). Самый северный пункт на Левобережье — окр. г.Миргород [320, рис.50,
7] следует после уточнения [140] заменить на окр. с.Великие Сорочинцы Миргородско-
го р-на Полтавской обл. (правый берег рЛсел). Однако и здесь разноцветная ящурка
вследствие облесения песков ныне исчезла и сохранилась несколько южнее, ниже по тече-
нию р.Псел, в Великобагачанском р-не Полтавской обл.: у с. Матяшевка и между пгт Вели-
кая Багачка и с. Яреськи Шишакского р-на. В окр. г. Полтава [320, рис. 50, 75] ящур-
ка была известна еще с дореволюционных времен [189], там же она сохранилась до сих
пор по берегам рЗорскла. Однако в окр. гТрайворон Борисовского р-на и пгт Борисов-
ка Белгородской обл. России [15] этот вид исчез вследствие сплошного залесения со-
сной [140].
В окр. г Харьков [320, рис. 50, 10} ящурка известна издавна [286], и она сохрани-
лась до сих пор у южных окраин города (пески боровой террасы р.Уды). Этот Вид обитает
также на левом берегу р.Оскол в окр. сЗКовтневое Двуреченского р-на Харьковской обл.,
самый северный пункт находки в Украине — на левом берегу р.Северский Донец на гра-
нице с Россией [140].
Относительно находки разноцветной ящурки у г.Киев [320, рис. 50, 75]. Этот пункт
подтверждается коллекционным материалом (ЗИН РАН, кол. Миддендорф, 1838).
и на левом берегу у г Киев имелись соответствующие биотопы, поэтому нельзя данное
место находки не учитывать или подвергать сомнению (см. [140]). Вид здесь мог исчезнуть
не только в результате хозяйственной деятельности человека, но и в силу других факто-
ров, например вымерзания в одну из суровых зим еще в дореволюционные годы.
Можно привести здесь аналогичный пример. К.Ф.Кесслер [111] сообщал о находках
в окр. г Киев зеленой ящерицы (Lacerta viridis). До войны 1941-1945 гг. киевские натура-
листы знали, что севернее гКанев этот вид уже не встречается. После войны поисками
зеленой ящерицы занимался и Н.НЛЦербак. В это время существовало твердое мнение, что
К.ФКесслер ошибся, принял яркого самца прыткой ящерицы за L.viridis. Особенно часто
мы экскурсировали в ур. Конча-Заспа под Киевом. И вот однажды именно там обнаружи-
ли в середине 60-х годов зеленую ящерицу. Спустя несколько лет в этом месте данный вид
стал обычным. Почему же не появилась под Киевом разноцветная ящурка? Причина этого,
видимо, заключается в том, что между Каневом и Киевом биотоп зеленой ящерицы не пре-
рывается (холмы с кустарником, лес) , стации разноцветной ящурки (участки песков)
давно уже прерваны распашкой и посадкой леса, а также водохранилищами; они сохрани-
лись в виде редких островков. Поэтому ящурка, в отличие от зеленой ящерицы, не смогла
воспользоваться представленным ей шансом.
Таким образом, при изображении исторического ареала разноцветной ящурки Киев
10
Рис. 2. Распространение E.arguta в Украине
следует отмечать в качестве его одной из северных точек. Однако северная граница совре-
менного распространения E.arguta в Украине проходит примерно через пгт Великую Ми-
хайловку (рис. 2. 22) — г.Саврань (4) г .Черкассы (2) — устье р.Сула (25) — сТПишаки
(28) — г .Полтава (75) — г.Харьков (7(7) — р.Северский Донец на границе Украины с Белго-
родской обл. России [140].
Кадастр к рис. 2:
7 - с .Великие Сорочинцы Миргородского р-на Полтавской обл. [ 140]; 2 - берег р.Тясмин от
с.Вергуны Черкасского р-на до устья (там же); 3 — правый берег р.Кодыма, с.Сырово Врадиевского
р-на Николаевской обл. (там же); 4 — с.Байбузовка Савранского р-на Одесской обл, (там же); 5 —
окр. г.Вилково (кол. ИЗ АНУ); 6 — Черноморский заповедник, пгт Голая Пристань Херсонской обл.
[наши данные]; 7 - окр. с.Железный Порт Херсонской обл. [наши данные]; 8 — Оле шковские пески
Херсонской обл. [наши данные]; 9 - окр. г.Одесса (кол. ЗИН) ; 10 — окр. г.Харьков [ 140]; 11 - окр.
г.Николаев (кол. ИЗ АНУ); 12 - окр. г.Славянск (кол. ЗИН); 13 - окр. г.Киев (кол. ЗИН); 14 -
окр. г.Херсон (кол. ЗИН) ; 15 — окр. г.Полтава [ 140]; 16 — г. Белгород-Днестровский (кол. ИЗ АНУ);
17 - коса обиточная, Азовское море (кол. ИЗ АНУЗ ; 18 - о.Бирючий, Азовское море (кол. ИЗ АНУ);
19 - окр. г. Павлоград Днепропетровской обл. (кол. ЗИН); 20 - о.Хортица Запорожской обл. (кол.
ЗИН) ; 21 - окр. пгтГрадизк Полтавской обл. [ 140]; 22 - с. Максимовка Киеменчггского р-на (там
же); 23 - устье р.Сула (там же); 24 - с.Жовтневе Двуреченского р-на Харьковской обл. (там же);
25 - окр. г .Днепродзержинск (кол. ХГУ) ; 26 - села Гаевка и Павловка Раздельнянского р-на Одес-
ской обл. [ 140]; 27 - прав, берег р. Южный Буг ниже г.Саврани, окр. с. Голосково Кривоозерского
р-на Николаевской обл. (там же); 28 — между пгт Великая Багачка и с.Яреськи Шишакского р-на
Полтавской обл. (там же); 29 — р.Северский Донец на границе с Россией (там же); 30 — Зачепилов-
ский р-н Харьковской обл. (кол. ХГУ); 31 - Нововодолажский р-н Харьковской обл. (там же); 32 -
г.Балаклея (там же); 33 - пгт Печенеги Чугуевского р-на Харьковской обл. (там же); 34 - окр.
г.Изюм (там же); 35 - сЛески Радьковские Боровского р-на Харьковской обл. (там же). Крым (наши
данные) : 36 - окр. г Керчь; 37 - окр. г.Феодосия; 38 - мыс. Казантип; 39 - Арабатская стрелка; 40 -
Советский р-н, окр. с.Урожайное; 41 - окр. г.Саки; 42 - окр. г.Евлатория; 43 - оз.Донузлав; 44 -
Раздольненский р-н, берег моря; 45 — Красноперекопский р-н, берег моря; 46 - Нижнегорский р-н;
47 - окр. пос. Мирный Мелитопольского р-на [ Котенко, Писанец, сообщ.]; 48 — окр. с.Кирилловка,
Степановская коса [ 126]; 49 - Федотова коса [Писанец, сообщ.]; 50 — Будакская коса [Таращук,
сооб.]; 51 - окр. г.Вознесенск [ Котенко, сообщ.]; 52 - Бердянская коса [ Котенко, сооб.]; 53 - окр.
г.Геническ [ Котенко, сообщ.]; 54 - окр. пгт Октябрьское, с Лиманы Октябрьского р-на Николаевской
обл. [ Таращук, сообщ.]; 55 — сЛриморское (бывш. Жебрияны) Килийского р-на Одесской обл. [наши
данные; 386]; 56 - окр. г.Новоазовск, Кривая коса ( 386].
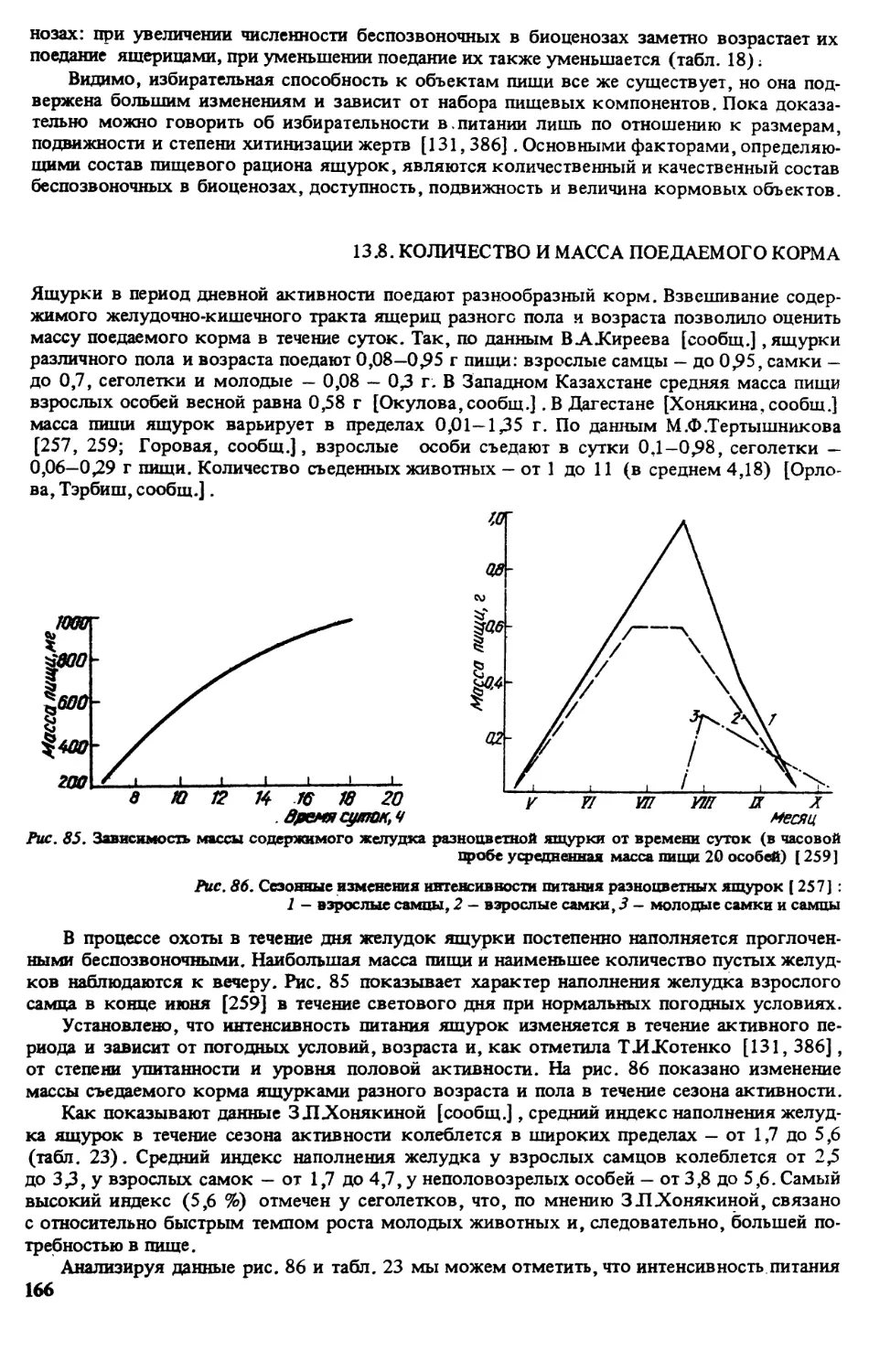
Рас. 3. Распространение E^rguta в России
Россия: Черноземье—Северный Кавказ —Поволжье. К сожале-
нию, новых сведении из западных регионов Российской Федерации в последние годы прак-
тически не публиковалось. Благодаря исследованиям ТИКотенко [140] известно, что у
гХраиворон Белгородской обл. [320, рис. 50, 7] в настоящее время разноцветные ящурки
исчезли, однако они сохранились еще у г .Белгород (сЛЛаслова Пристань Щебекинско-
го р-на); сведения из Воронежском обл. вообще отсутствуют, очень скудные данные о
Саратовской обл., нет таковых из Самарской, мало материала из Оренбургской обл. В.В.Не-
ручев и др. сообщили нам, что Ejirguta не обнаружена в долине р.Урал в окр. р.Оренбург
[320, рис. 50,9] иИлека [то же, 30].
Сведения о находасах в Приазовье, Предкавказье и Дагестане почти на прежнем уров-
не [320].
По Краснодарскому краю подтверждаются находки на Таманском п-ове [320, рис. 50,
14]. Этот вид кчез, по письменному сообщению М ЛЗиняковой и ПЛ Руденко, в окр.
г .Темрюк [320, рис. 50, 75] и хутора Садки после 1970 г. в результате ирригационных
работ. В 1973 и 1977 гг. авторы наблюдали Earguta на юге Таманского п-ова у Кизилташ-
ского лимана и станицы Благовещенской.
В Ставропольском краю (по современным наблюдениям М.Ф.Тертышникова [письм,
сообщ.] отмечено шесть мест находок в Ставропольском р-не (рис. 3, , кроме того, до-
полнительные пункты — на западе края в окр. пгт Новоалександровка (рис. 3,31), окр.
гСветлограда (рис. 3,32), окр. пгт Александровское (рис. 3,33).
ВАЖиреев [письмен, сообщ.] уточняет распространение E^rguta в Калмыкии следу-
ющим образом: вид встречается в республике повсеместно, но неравномерно. Террито-
рию хребта Ергени ящурки заселяют поселениями, расположенными в виде лент, по бал-
кам и оврагам, на склонах и террасах высхоших русел рек, на территории Черных земель
встречаются мозаичные поселешкя.
В Волгоградской обл. (по материалам В.В.Неручева, ЕЛОХудакинойиН Ливановой)
подтверждаются места находок в окр. оз. Баскунчак [320, рис. 50, 73] и окр. пос. До-
санг [то же, рис. 50,27].
12
По сведениям ЭАДИебзуховой [письмен, сообщ.] в Кабардино-Балкарии ящурки
добыты в окр. сел Кахуна, Приближный, Васильевка, Черниговка (долины рек Терека,
Малки и Урвани). Южная граница ареала проходит здесь по линии Урвань — Нальчик —
Баксан (рис. 3).
Кадастр к рис. 3:
1 - рХалка [ Шебзухова, письм. сообщ., 1990 г.); 2 - вост. г.Нальчик и ж-д. ст. Докшукино
(там же) ; 3 - г .Прохладный (там же); 4 - между пгт Залукокоаже и г.Прохладным (там же); 5 -
окр. г.Пятигорск [273] (М.Ф.Тертышников не указывает это место в числе пунктов Ставрополья.
соо(5щ., 1990 г.); 6 - окр. г.Ставрополь [Тертышников, письм. сообщ.]; 7 - гТрайворон Белгород-
ской обл. [ 140]; 8 - г .Астрахань (кол. ЗИН) ; 9 - окр. г. Оренбург, ж.-д. ст. Донгуз [ Неручев и др.,
сообщ.]; 10 — г.Борок Балашсвского р-на Саратовской обл. [189]; 11 — дорога Элиста - Астра-
хань [ Киреев, сообщ.]; 11, 12 - дорога Элиста - Каспийский (тот же); 13-13 - граница Калмыкии с
Дагестаном до Ставропольского края (тот же) ; 14 - Таманский п-ов, стан. Благовещенская [ Зиняко-
ва, сообщ.]; 15 — окр. г.Темрюк (она же) ; 16—11 — дорога Элиста — Цаган-Аман [Киреев, сообщ.);
17 - 1 .Красноармейск (кол. ЗИН); 18 - оз. Баскунчак [Неручев и др.^сообщ.] 19 - г.Новочеркасск
(кол. ЗИН); ±0 - г.Таганрог (кол. ЗИН, [158]); 21 - г Анапа (кол. ЗИН); 22 - окр. г.Моздок
(кол. ЗИН); 23 - р.Маныч, стан. Старогладовская (кол. ЗИН); 24 - оз.Эльтон (кол. ЗИН); 25
окр. г.Волгоград [118]; 26 - с.Замьяны Астраханской обл. (кол. ЗИН); 27 - с.Досанг [Неручев и
др., сообщ.]; 28 — Пугачевский р-н Саратовской обл. [ 253]; 29 — п.Колту банов ский, Бузулукский
бор (Котенко, набл. 1986 г.) (кол. ЗИН) ; 30 - долина р.Илск [ Неручев и др., сообщ.], 31 - Новоалек-
сандровский р-н Ставропольского края [Тертышников, сообщ.]; 32 — г.Светлоград Ставпропольског^
края (он же); 33 — с.Александровское Ставропольского края (он же); 34 - окр. г.Бслгород [ 140]
35 - оз.Дадынское [Тертышников, сообщ.]; 36 - юж. Чограйского водохранилища (он же); 37
окр. с Дивное Ставропольского края (он же), Дагестан [Хонякина, сообщ.]; 38 - с.Тсрекли-Мектеб,
39 — с.Червленная; 40 — г.Кизляр; 41 — сЛасавюрт; 42 - Аграханский п-ов; 43 - г.Главный Судак
44 - г .Махачкала, Кумтор-Кала; 45 - г.Буйнакск; 46 -с.Кутиш Левашинского р-на; 47 - с .Крайнов •
ка; 48 — о.Чечень; 49 - с Леваши в дельте р.Самур; 50 — с.Бажиган; 51 - с.Главный Кут; 52 - г.Ка -
пийск; 53 - пЛИамхал; 54 - сЪерикей; 55 - стан. Вешенская [ 158]; 56 - с.Маргаритовка [46,158]
57 - Калмыкия, окр. оз.Сарпа (Котенко, набл. 1986 г.); 58 - Доно-Арчединские пески, окр. хЛебя
жий Фроловского р-на Волгоградской обл. (Котенко, набл. 1986 г.); 59 - Цимлянские пески, оку
с.Тормосин Волгоградской обл. (Котенко, набл. 1986 г.); 60 - берег р.Чир, окр. стан. Облив ска.
Ростовской обл. (Котенко, набл. 1986 г.); 61 - окр. с. Дьяковка на р.Еруслан Саратовской обл. (Ко
тонко,набл. 1986 г.) ; 62 - с.Садки [ Зинякова, сообщ.).
Дагестан. По дополнитель-
ным материалам 3 Л Лонякиной,
ящурка широко распространена в Но-
гайской степи, Терско-Сулакской и
Приморской низменностях, места-
ми заходит в предгорья до высоты
1600 м H.yjM. Как новые пункты ука-
зываются села Главный Кут, Алмало и
Темиргое, г.Каспийск, села Шамхал и
Берик ей, некоторые из них из-за не-
соответствия масштаба не отмечены
на рис. 3.
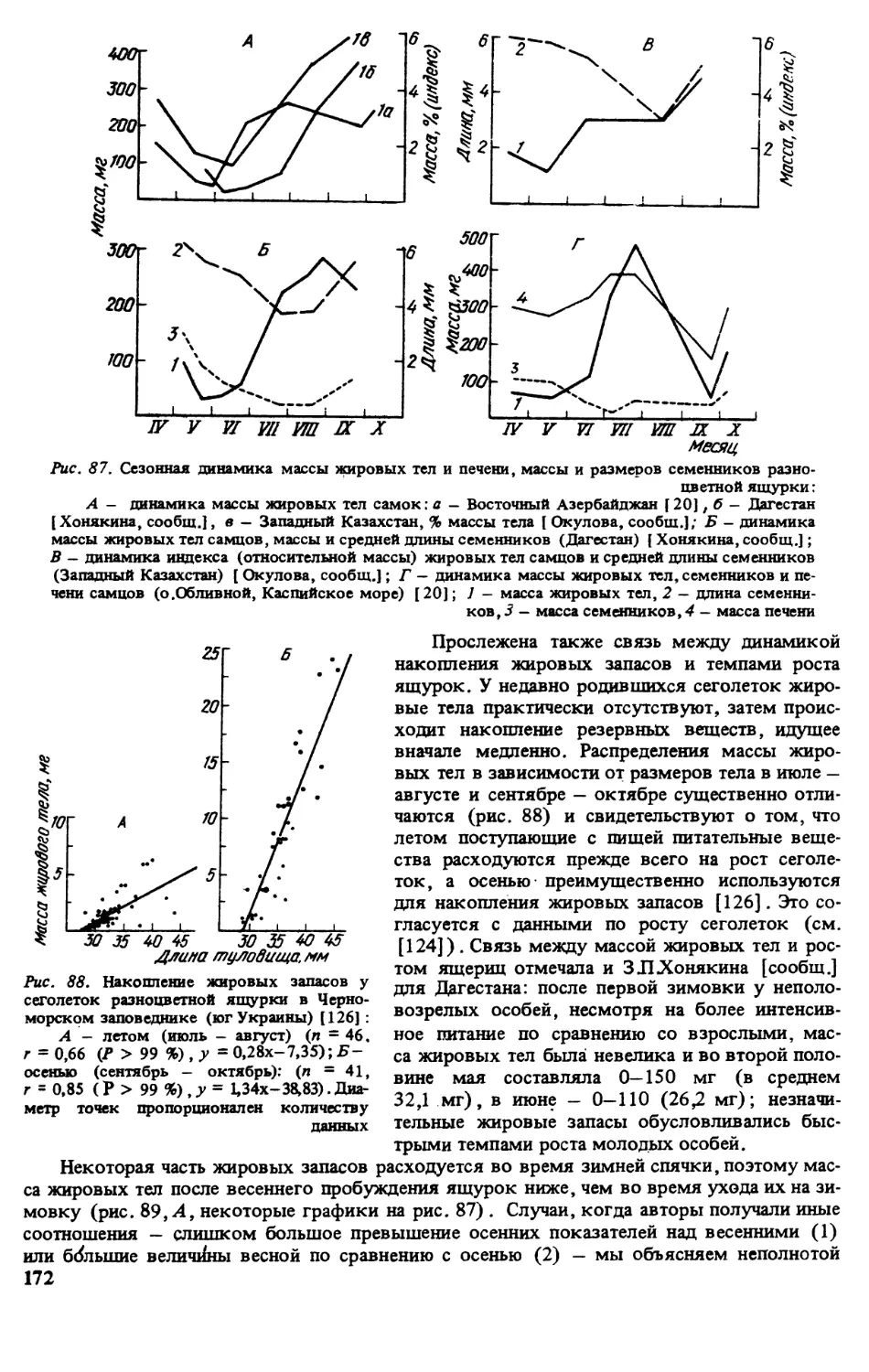
Азербайджан (рис.4). До
сих пор отсутствуют находки между
устьем р.Самур, Дагестан [320, рис .50,
Рис. 4. Распространение E.arguta в Азербайджане. Арме-
нии, Грузии и Турции
12] и Апшеронским п-овом вдоль побережья Каспия. Видимо, они исчезли там в период одной
из трансгрессий моря. Иначе трудно представить, как проник этот вид в Азербайджан. Пс
сведениям Т Алиева, он не нашел ящурку данного вида, известную ранее из Зуванда, Кяль-
ваза и Дуздага. Далее автор утверждает, что она полностью отсутствует в Ленкорань-Та-
лышском р-не, хотя в довоенные годы киевский герпетолог ИЦемш добывал ее у пос. Де-
ман в Ленкоранском р-не и делал попытку описать отсюда форму E.arguta lencoranica, но
война помешала ему это сделать (он погиб на фронте в первые месяцы войны). П-ов Калу-
гина из Ленкоранского р-на (по кол, ЗИН) [320, рис. 50] в действительности пост Кулаги-
на в Сальянском р-не. В посДуванный (ныне Гобустан) и в настоящее время обитают
ящурки. Найдены они также на о,Обливном в Бакинском архипелаге (рис.4, 13}. Кроме
того, данный вид обитает в Бендованском заказнике, близ пос.Шихов а, в Ширванском гос
заказнике, а также в Центральном Азербайджане — Аджыноурской, Бозгольской,
Эльдарской, Джайрангольской (Шамхорский р-н) и Сальянской степях. Ящурка ветре
чается по берегам рек Куры и Аракса, а также Мингечаурского водохранилища
13
в Карабахской степи, Джебраильском и Зангеланском районах, в других низменных райо-
нах республики. По мнению ТАлиева, разноцветная ящурка совсем не встречается в гор-
ных районах, нет ее и в Нахичевани (хотя в Армении она обитает высоко в горах, в Севан-
ской котловине).
Кадастр крис. 4:
Азербайджан: 1 — окр. г.Баку (кол. ИЗ АНУ); 2- Апшеронский п-ов, п.Мардакяны, п.Зых,
п.Бузовна, п.Бинагади [ 7, 86) (кол. ЗИН) ; 3 — ж.-д. ст. Пута [ 84]; 4 — п.Гобустан (бывш. Дуванный)
[ Алиев, сообщ.]; 5 - мыс Бендован (он же); 6 - о.Обливной (20]; 7 - ж.-д. ст. Али-Байрамлы [ 188];
8 - с.Джейранчол [ Алиев, сообщ.]; 9 - г.Евлах, с.Халдан [ 7); 10 - г.Сгепанакерт [ 7]; 11 - г.Джебра-
ил [ 188]; 12 - Мильская степь, оз. Ахчель (7); 13 - с.Дуздаг [ 188]; 14 - г.Сальяны [ 189]; 15 - Бан-
ковски промысел [ 188]; 16 - пост Кулагина Сальянский р-н [ Алиев, сообщ.); 17 - с.Кяльваз, близ
границы с Ираном (кол. МГУ); 18 - г.Зуванд, пос. Деман (7); 19 - г.Гянджа [7]; 20 - Ширванский
заповедник (Алиев, сообщ.]; 21 - Шамхорский р-н (тот же) ; 22 - оз.Аджиноур (тот же); 23 - окр.
ст.Ахмедли G от же); 24 - Зангеланский р-н (тот же) ; 25 - Минчегаурское водохранилище (тот же);
26 - Эвдарская стега» [ 7].
Армения. По письменному сообщению И.СДаревского, область современного
распространения разноцветной ящурки в Армении ограничена высокогорным изолятом,
расположенным на высоте 2000—2100 м н.у.м. в бассейне оз.Севан и отделенным от ос-
новного ареала вида в Закавказье системой горных хребтов на севере и северо-востоке.
В этой высокогорной котловине разноцветная ящурка, как и некоторые другие типично
равнинные виды пресмыкающихся, изолирована, видимо, в раннем плейстоцене в резуль-
тате сводового поднятия Малого Кавказа и связанной с этим перестройки речной системы
Палеокуры [78].
Впервые для Армении разноцветная ящурка была указана ОЪетгером [464] по сбо-
рам экспедиции Валентина и Радде, сделанным якобы в окр. сМец-Мазра в долине р Араке
близ гМегри [464]. Однако, как показал И.С. Даревский [77], скорее всего данная наход-
ка происходит из окрестностей одноименного сМец-Мазра в бассейне оз.Севан где также
проходил маршрут этой экспедиции. Во всяком случае в настоящее время разноцветная
ящурка в долине р Араке не живет. Из окр, с. Мец-Мазра с берегов оз.Севан имеется серия
ящурок также в Зоомузее МГУ. По материалам коллекции Института зоологии АН Арме-
нии СА.Чернов [289] указывает разноцветную ящурку для окр. с. Сатанахач на северо-
восточном берегу оз. Севан, где ее находил позднее и С.К Даль [73]. На берегах Севана в
окр. сАлучалу и в низовьях впадающей в озеро рДахкар ящурку обнаружил И.С.Дарев-
ский [77]. Более поздние местонахождения этого вида были сделаны в окр. сел Норадуза
и Памбак соответственно на юго-западном и северном берегах озера (рис. 4) .
В перечисленных выше местонахождениях разноцветная ящурка обитала в условиях
каменистой разнотравно-злаковой с преобладанием полыни горной степи, местами заходя
также на сильно закрепленные береговые пески. В настоящее время повсеместная распаш-
ка степи под зерновые культуры привела почти к полному исчезновению этого вида в бас-
сейне оз.Севан, чему способствовало также зарастание лесной растительностью берегов
озера, обнажившихся в связи с искусственным понижением его уровня [82]. Например,
береговые песчаные участки с выходами травертинов, где наблюдал ящурок СЛСДаль
[73], в настоящее время находятся в глухом лесу. Для сохранения уникальной высоко-
горной популяции разноцветной ящурки в бассейне оз.Севан в 1959 г. около 30 полово-
зрелых особей, отловленных в низовьях рДахкар на окраине зерновых полей, были пере-
несены на береговые закрепленные пески на юго-западном берегу озера в окр. сЯорадуз
[81]. В дальнейшем яшурки наблюдались здесь в 1967,1970 и последний раз — в 1983 гг.,
после чего, видимо, исчезли в связи с разросшейся лесной растительностью [82]. Сохраня-
ется слабая надежда, что разноцветная ящурка в бассейне оз.Севан уцелела где-либо на це-
линных участках горных степей, оставленных под пастбища для овец.
Кадастр к рис. 4:
Армения;] - с.Мец-Мазра Вардениского р-на (кол. МГУ); 2 - с.Сатанахач на берегу оз.Севан
[77}, окр. с.Мартуни, низовья рДахкар (кол. ЗИН); 3 — г.Мегры, долина р.Аракс [ 288]; 4 - с.Алу-
чалу Мартуниского р-на; 5 - с.Норадуз Камоского р-на; 6 - г.Памбак Вардениского р-на [ Даревский,
сообщ.].
Из Грузии, Турции и Ирана новых сведений о распространении ящурок
не поступало. В монографии МЪажоглу и И .Барана [334] наличие E.arguta в Турции от-
рицается.
14
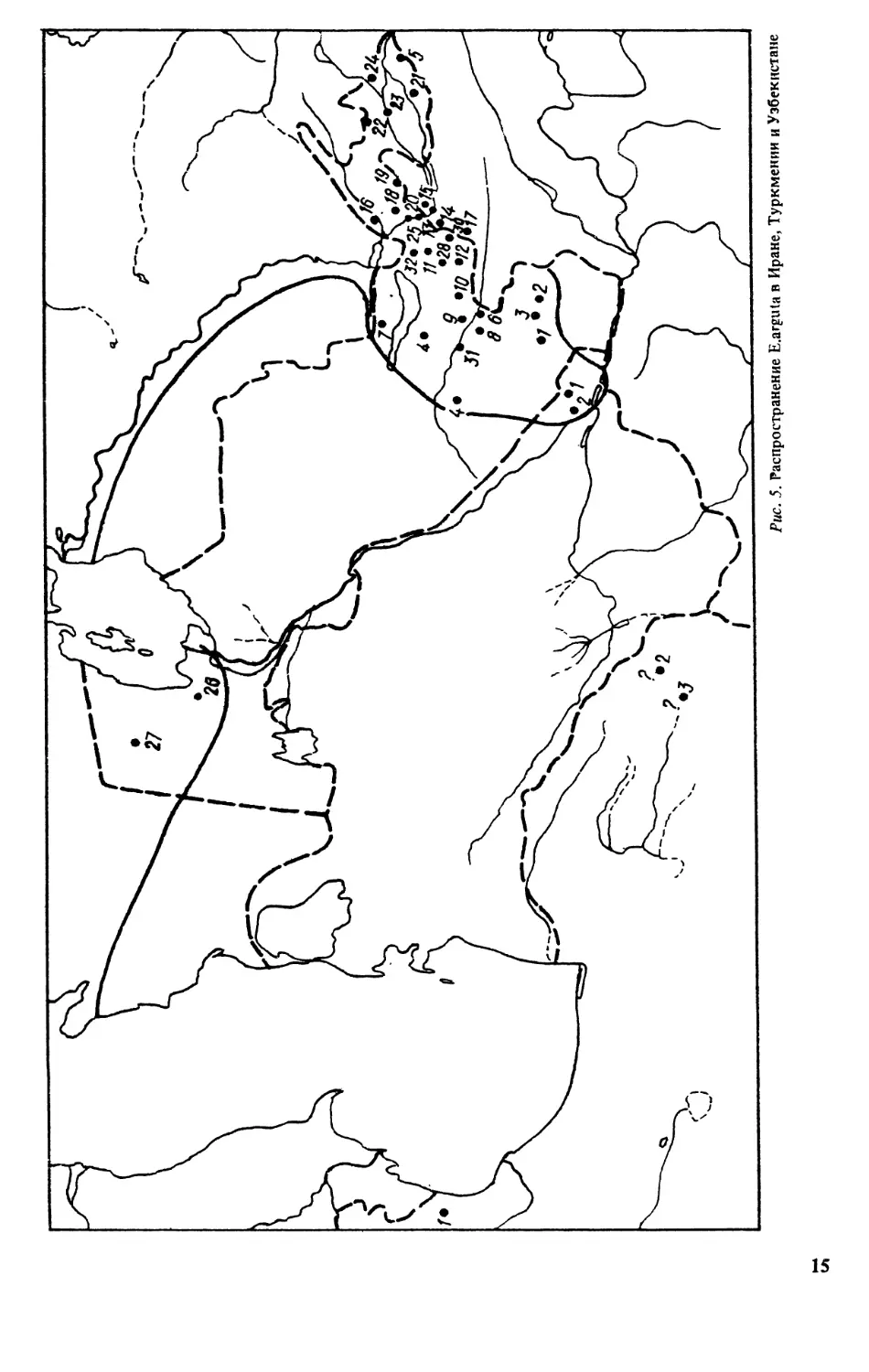
th
Рис. 5. Распространение E.arguta в Иране, Туркмении и Узбекистане
№. 6. Распространение Е Jttguta в Таджикистане, Кыргызстане и Монголии
Кадастр к рис. 4:
Грузия: 1- ьСямухя, Эпшрскля степь [176b Турция: 7 - с.Мнченгардере у оз. Ван (317] -
ошибочное одредпятие.
Туркменистан. Вее известш>1е ранее находки разноцветном ящурки в данном ре-
гионе ошибочны [320]. В новейшей сводах по Туркменистану {298] указаны два место-
обитания вида на крайнем юго-востоке республики (с. Кыркойдии пос Амударья между
сХоджагурлук к ст. Тавммврдявн! Ходжимбасскспго р-на Туркменистана). Предполагаются
находаи на Устюртском плато (на северо-шшде республики) (рис. 5).
В позже опубдаковатаой статье [ 17} сообщается о находке разноцветной ящурки в
Юго-Зяоддаюм Туркменистане (пески, 13 км северовостотаее пос. Мадау Кмзыл-Атрек-
ского р-на Крвснсводской обл.). При ознакомлении с материалом ЧАтаева выяснилось,
что здесь обитает блюкда мд — Е intermedia, ж поэтому этот пункт на карту не наносится.
Кадастр к рис.5:
Иран: 1 - эея. стога яр- Богроидаг (кол. ЙШ); 2 - 66 км от г. Мяяжсда до направлению к
г.Сераксу [373] ;5 - сЛЫМ, ЗЪ км ик. Торбете-Хейдервв {373} (прашяыккзь адредепедан особей из
пужтов 2 и 3 ньттвмт гомкипт).
Т у р к м е нвстан: 1 - сЖыргяйта [298] ;2 - пос. Амударья между с.Ходжагурлук м ст.Тали-
мярджяк Хсцгамв&ссжсзкго p-ea [298].
Узбекистан* В сводке ОШБогданова [40] приведешь данные о распространении
разноцветной ящурки: Голодная ствяь, Цевтрашшй Устюрт, северные предгорья Нуратву,
Зерабулакскме высоты, Каршвиская степь, что в целом соответствует ранее известным
пужтам. По сведениям ДА*Бондаремсо [швсьм. сооб.], севернее Каршннского оазиса
(долша рХаоаодфьк) Е^н^да набшодаткав жр.посешсм Алачбоф иДжаркум.
Кадастр к ряс. 5:
Узбекистан: 1 - г Жяршя (ведя. ЗИН), дамою ДАБоддарсяко - окр. поселков Алтебоф и
дааркум; 2 - г.Ямкабаг [38]; 3 - г.Чярмсти (мол. ЗИН); 4 - г ЗСяттакургян (кол. УзГУ); 5 - с.Свза-
ган (мол.УэГу); б - дорога Самяркяцц - Педгагат; 7-сеж предгорья Нур«тау[ 94]; 8- гХ^маркаяд
(кт. ЗМН, МГУ); 9 - Шт. Яягжурган [169]; /47 - г. ЦраошйардоДОк [35]; 11 - с*Бсшкубе Зеелин-
ского ржа [3SJ; 12 - рОиэар идее с^швдар {72]; 13 - ж.^,. ст. Обручеве (кол. САГУ); 14 - ст.
Уреятьевская (кол. Э1Ш); 15 - г.Чянаэ (код. Ж); М - рХ>шр у г.Ташкект [ 189]; 17 - с.Невич
[35]; 78 -гЛскевг [35]; 19 - оБегсшт (там до); 2t> -сЛвяак (тамже); 21 - г.Ферпва (кол. ЗИН);
22 - г.КасвяеМ [33]; 23 - с.Юкьфг^ Балык^насого р<на [35]; 24 - г.У«сурган [324] ;25 - г.Гули-
спн (бывш. Голодая» Ставь, жал. ЗЙН), Кара-Калпакия; 26 - оз. Судошье [35];27 - крепость Белеулм [35].
Южная граница ареала проходит по чинку Устюрт [35], Джмздаская обл. [Вашетко, сообщ.}; 28 —
окр. поселков Каряташ, Аччн, Бу стан и Галля-арял, городов Дкизмс иПахтакор; 29 - г.Фариш, сзх. ’’Кы-
зылкумский", подножьехр. Нуратау, оз. Тузкан, пос. ДжоЙлянгар; 30 — Заамннский p-в Самаркандской
обл.; 31 - нос. Кутел Сырдарьпнской обл.; 32 - свх. "Социализм”, ЮЗ км от г.Ташюжт.
Таджикистан. Добыта в окр. кишлаков Авзыкент, Ялтак (южньш склон Кура-
ьвшекого хребта), Аккутан (северный склон Моголтау), на Дегмаискои и Иснисорской
возвышенностях, у г-Кайракум,а также в 20 км к северо-западу г Ура-Тюбе. Кроме того, от-
16
Рис, 7. Распространение E.arguta в Казахстане и Китае
мечена в окр. ж.-д. ст. Обручеве [222]. Хотя все находки сделаны в Северном Таджики*
стане, в Ленинабадском р-не, они существенно дополняют наши сведения о распростране-
нии разноцветной яшурки в Таджикистане (рис. 6).
Кадастр к рис.6:
Таджикистан [222] :1-е. Авзыкент (юж. склон Кураминского хр.) ; 2 - с.Аккутан (сев.
склон хр. Моголтау); 3 — г.Кайраккум; 4—20 км свх. Ура-Тюбе; 5 — окр. ж.-д. ст. Обручеве,
Кыргызстан: 1 — окр. г.Бишкек (кол. ЗИН) ; 2 — Чуйская долина [324]; 3 — Таласская долина
(ущ. Кара-Какн. долина р.Чон-Кемин) (там же) ; 4 - с.Чоктал (там же) ; 5 - г. Ош (там же) ; 6 - юж.
берег оз. Иссык-Куль (там же) ; 7 - зап. берег оз.Иссык-Куль (там же) ; 8 - г.Пржевальск (кол, ЗИН).
Монголия [197]: 1 - 12 км юго-вост. Уэнч-Сомона; 2 - 3 км вост. Уэнч-Сомона; 3 - Алтай-Сомон,
берег р.Бодончийн-гол; 4 - такыр Бамбагарын-шал, 10 км у Алтай-Сомона; 5 - котловина Хурэн-
Дэлийн-хотгор; 6 - зап. склон Майхан-хара-ула; 7 - 14-16 см сев.-сев.-вост. хр. Байгаг-Богдо; 8 -
прав, берег р.Уэнч-гол; 9 - 25 км вост, родника Олог-булак; 10 - сомон Jiyj^raH. родник Ушигийнус
(кол. x.Tepf’пи); 11 - 50 км юго-зап. с.Ьиджи-Алтая (кол. МГУ) ; 12 - 45^26 с,ш. 9j 18* вщ.,45°32
сдд.. 93 12 в.д. (кол. МГУ); 13 - Гобийский заповедник, между хребтами Тахийн-Шара-Нуру, Монг,
Алтаем и Хувгийн-Нуру (кол. МГУ); 14 - с.Хонин-У сны-Гоби [437], 15 - 25 км вост, родника Та-
хийн-Ус (Монголия, кол. х.Тербиш).
Кыргызстан. Новые сведения о находках отсутствуют.
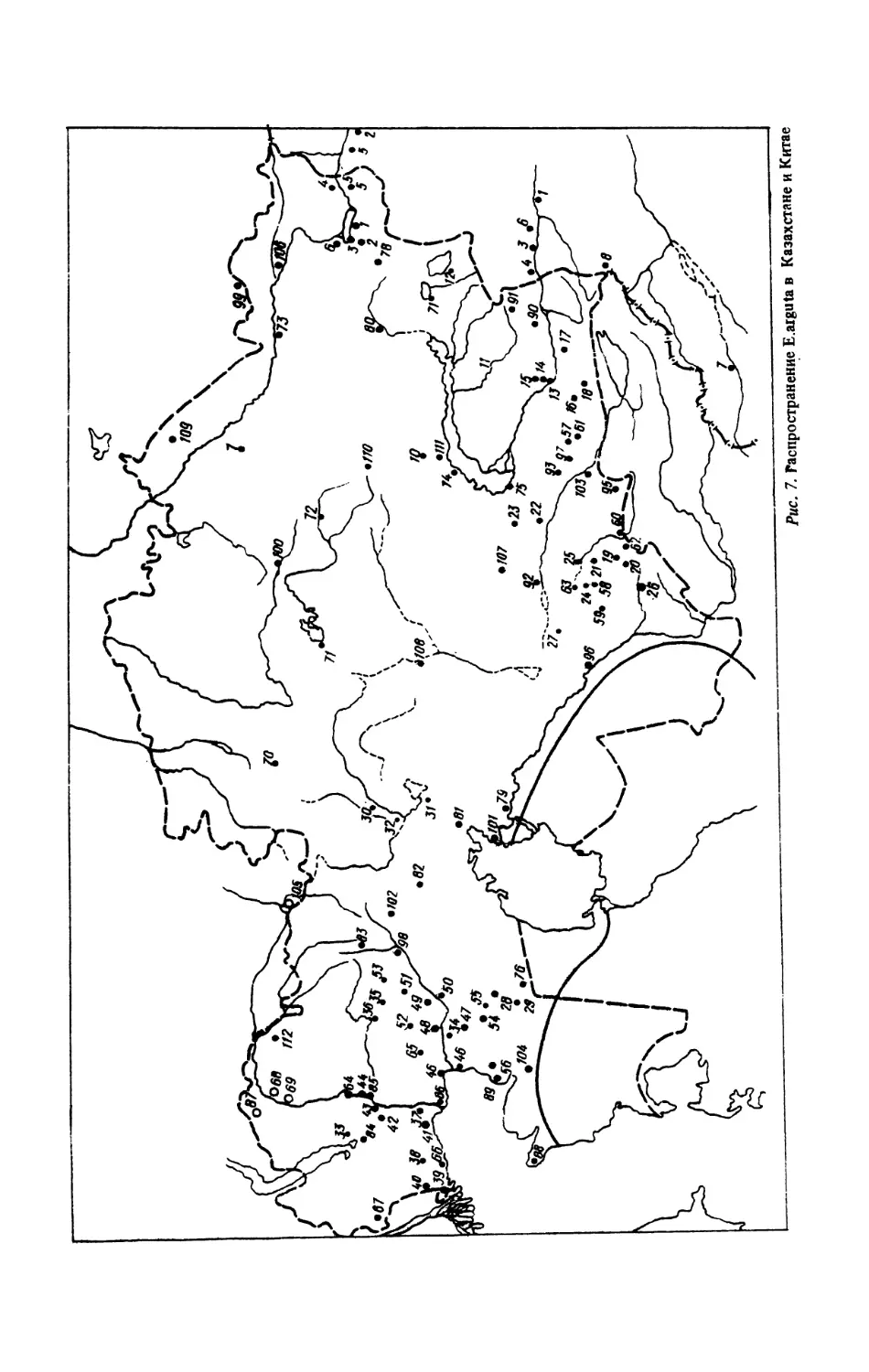
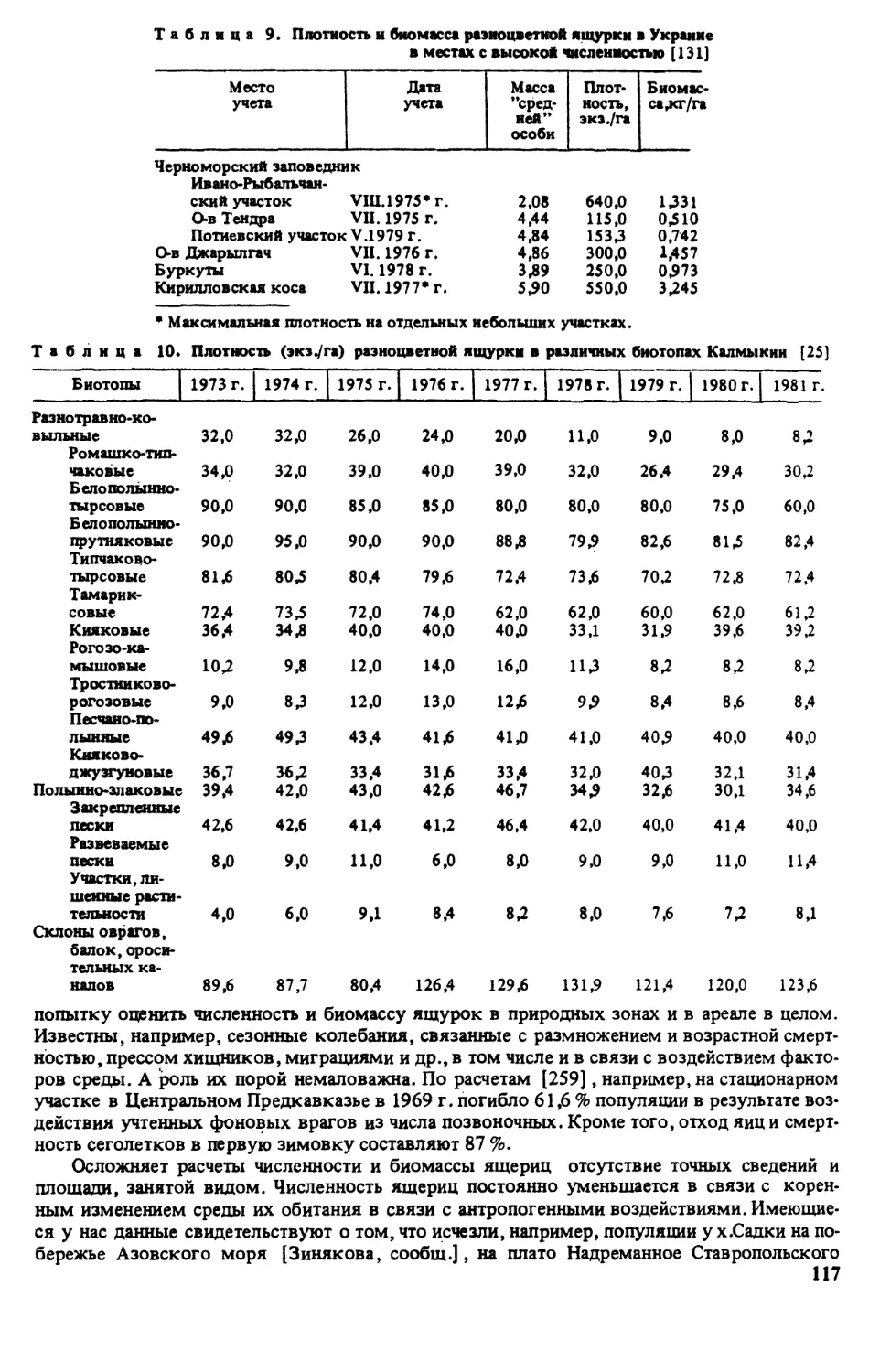
Казахстан (рис. 7) . В 1972—1985 гг. детальные исследования в Северо-Восточном
Прикаспии провели В.В.Неручев, Е.ЮКудакина, НЛИванова [письм. сообщ.]. Как оказа-
лось, разноцветная ящурка уже исчезла в окр. г.Уральска [320, рис. 50,1] и даже южнее
его у сКоловертное в долине р.Урал. Здесь отмечено отступление на юг северной границы
ареала примерно на 120 км. Подтверждены новейшими находками следующие пункты
Прикаспия (приведены без нумерации, так как их нельзя поместить вследствие мал ©мас-
штабности картосхемы): берег оз.Индер и горы; пос. Гребенщиков в долине р.Урал; на
восточной кромке Волго-Уральских песков у пос.Кулагино; на восточной кромке Волго-
Уральских песков урКамбакты; у пос. Зеленый, долина р.Урал; с Танюшкино и ур.Кара-
Узек на южной кромке Волго-Уральских песков; бывш. пос. Ракуша, вост. г .Гурьев ;
гКулкары, дельта р.Эмба; ур. Терень-Узек, зап. г.Кульсары; ур.Прорва, вост, побережье
Каспийского моря; г Лман-Кара, склоны горы сев г.Кульсары; пойма р.Сагиз; ypJCapa-
бау, кромка песков Таисуган; междуречье Уил-Сагиз; юго-западная кромка песков Ак-
кум-Сагиз; плато Ак-Толагай; ур. Ушкан, сев. кромка Прикаспийских Каракумов; сЖан-
бике, долина р.Эмба; села Шолькара и Жанаси, 100 км юго-вост. г.Кульсары; с. Кара-Ар-
на, западная кромка Прикаспийских Каракумов.
Кадастр к рис. 7:
Восточно-Казахстанская обл.: Тарбагатайский р-н. Зайсанская котловина (кол.
ИЗ АН Казахстана, данные З.К.Брушко): 1 - 14-15 км на зап. п. Приозерный; 2 - 12-15 км яа
сев .-вост. с^Белая Школа; 3-20 км на север от с.Крупское; 4 - Маркакольский р-н, 45 км на запад
от с. Буран; 5 - Зайсанский р-н, 20 км сев.-зап. с.Майкапчагай; 6 — Самарский p-и, Буконьские пески;
Павлодарская обл.; 7 - Ермаковский р-н, Малый Калкаман, юго-зап, г.Ермак; Целиноградская обл.;
8 - Тенгизский р-н. Кургальджино, озЛСыпшак, Тургайская обл.; 9 - Джангилвдинский р-н. метеостан-
ция Брали; Джезказганская обл.; 10 - Актогайский р-н, окр, г.Бектауата Талды-Курганской обл.,
h - Каратальский р-н, пл. Ушколь; 12 - Алакульский р-н, оз.Алакуль; Алма-Атинская обл.; 13 -
Талгарский р-н, окр. с.Чингильды; 14 - окр. с.Кускудук, 15 - перевал Архарды; 16 — Илийскийр-н,
оз.Сорбулак; 17 — Чиликский р-н, долина р.Чилик; 18 — Алма-Ата; Джамбульская обл., Джувадин-
ский р-н, 19 — Терс-Апшбулакское водохранилище; 20 — истоки р.Терс, Джамбульский р-н; 21 — Ма-
лый Каратау, ущ.Алмалы, Мойынкумский р-н; 22 - Бетпакдала, Андасайский заказник; 23 — горы
Курманчиты; Таласский р-н; 24 - оз.Аккуль; 25 - 12 км юго-вост, с.Бостандык; Курдайский р-н;
26 - окр. с.Черная речка; Чимкентская обл., Суэакский р-н; 27 - с. Шу курой Бейнеуского р-на Ман-
гышлакской обл.; 28 - с. Акжигит; 29 - пески Сам Актюбинская обл., Иргизский р-н; 30 - 15 км.
сев.-зап. с.Мамыр, ур. Улькаяк; 31 — 25 км юж. с. Жайсамбай; 32 — вост, часть оз.Кумколь. Ураль-
ская обл.; 33 - с.Борки Джангалинский р-н; Гурьевская обл., 34 - с.Кульсары Эмбенского р-на;
35 - Кзылкогинский р-н, пески Тайсойган, окр. с.Сармайчагыл; 36 - Кзылкопшский р-н, окр. с.Ка-
рабау; 37 - окр. с. Махамбет Махамбетского р-на; 38 - Новобогатинский р-н, Волго-Уральские пески,
По В.В.Неручеву, Е.Ю.Кудакиной и Н.Н.Ивановой [письм. сообщ., набл. 1972-1985 гг.]: 39 - окр, пос.
Танюшкино; 40 - ур.Кара-Узек; 41 - ур.Камбакты; 42 - окр. пос. Кулагине; 43 — окр. пос. Гребен-
щиков; 44 - оз.Индер, Индерские горы; 45 - бывш. пос. Ракуша, вост. г.Гурьева; 46 - ур.Терень-
Узек; 47 - ур. Ушкан; 48 - ур.Иман-Кара; 49 - пл. Ак-Толагай; 50 - ур.Жанбике в долине р.Эмба;
51-пески Аккум-Сагиз; 52 - долина р.Сагиз; 53 — междуречье Уил-Сагыз; 54 - ур.Шолькара; 55 -
урЛСапасу; 56 — урЛрорва, приморская солончаковая пустыня. По сведениям В.Г.Колбинцева из
Джамбульской обр. (1981—1986 гг.): 57 — Чуйский р-н, 15-20 км к востоку сев.^оср?, oi сЛенирское,
74°00 вщ„ 43°36 сдц.; 58 - Таласский р-н, 5 км к юго-юго-зрп, от ст.Коктал 70 14 в.д.,43 17 ст.;
59 - Сарысусский р-н, 15 км зап? с.Бугунь, 6?°5б' вщ., 43°30 с ди., зап. ст.Байкадан; 60 - Джамбуль-
ский р-н, окр. г.Джамбул, 71°43 вд, 42°55 с.ш.; 61 - Красногорский р-н, окр, ст.Кенеп (Умтыл);
62 - Джамбульский р-н, 20 км зап. с.Головачевка, 15 км к север-сев.-зап. от с.Головачевка, южное под-
ножье Улькен-Бурултау; 63 - Таласский р-н, окр. оз. Ащикуль. По данным Н.М.Окуловой [письм.
сообщ.) ~4 - пос.Калмыково; 65 - ст.Макат; 66 - с.Сазде (Сазды); 67- Суюндик (Суйиндик). По
материалам Н.Н.Щербака [320]: 68 - окр. г.Уральск (с поправкой В.В.Неручева и др.) ; 69 - Коловерт-
ное, долина р.Урал; 70 - НаурзумскиЙ заповедник; 71 - оз.Тенгиз, 72 - окр. Караганды; 73 - окр.Се-
мипалатинска; 74 - Таргыл; 75 - Мын-Арал; 76 - колодец Кюще; 77 - оз. Сасыккуль; 78 - с.Бакты,
г.Тарбагатай; 79 - окр. Казалинска; 80 - окр. Сергиополя; 81 - сев. ст. Аральск; 82 - окр. г.Челкар;
83 — пески Кок-Джада, р.Темир; 84 — Камыш-Самарские озера; 85 — пос. Индеборский; 86 - г.Гурь-
ев; 87 - с.Семиглавый Мар; 88 - г.Форт-Шевченко; 89 - ур.Кара-Арпа; 90 - хр. Алтынэмель; 91 -
с.Капал; 92 - низовья р.Чу; 93 - с.Хантау; 94 - г. Чимкент; 95 - с.Мерке; 96 - с.Яны-Курган; 97-
с.Эспе; 98 - между р.Эмбой и р.Темиром; 99 - окр. г.Змеиногорска, граница Казахстана и Алтайского
края; 100 — г.Целиноград (бывш. Акмолинск); 101 — устье р.Сырдарья; 102 — горы Мугоджары;
103 - ст. Чу; 104 - Сор. Кайдак; 105 г.Орск; 106 - г.Усть-Каменогорск; 107 - Голодная Степь
(Бет-Пак-Дала) ; 108 - г.Джезказган; 109 - Кулундинская степь; ПО - Шетскийр-н; 111 - Коунрад-
ский p-Hj 112 - окр. с. Миргородка, Аксайского р-на Уральской обл. (Котенко, набл, 1987 г.).
Китай (кол. ЗИН) : 1 - р.Кунгес. 2 — Джунгария, с. Булун-Тохой. 3 - тьКульджа. 4 - рЛалка, 5 -
горы Сал бур ты, 6 - г.Мазар, 7 - г.Кашгар, 8 - оз. Эби-Нур.
Ценные дополнительные сведения о распространении разноцветной ящурки б Запад-
ном Казахстане — междуречье Волги и Урала, предоставила нам Н.М.Окулова [письмен,
сообщ.]. Среди указанных автором пунктов следующие урочища: Абл-Кзыл, Айгыр,
Ак-Мола, Алтапан, Арен, Аще-Кудук, Байгазы, Байтерек, Б ал тай-Пике, Басчикан, Бекет,
Беспай, Бисагач, Большой Иман, Бустандык (Казталовского р-на), Городище, Девлятк-
стау, Джантемир, Дынкзыл I, Егоров-Куль, Жира, Жума, Калмыков© в долине р.Урал, Ка-
рабас, Караканат, Кара-Кудук, Кармыза, Кати-Чагл, Кенджали-Мола, Кирале, Колбай,
Колтума, Конор-Кудук, Косак, Косчеген, Кунак, Май-чагл, Макат, Мантыково, М штатов о.
Огыз-Кура, Орл-Кудук, Полевой Стан, Сазде,Салык-Мола,Саргужа, Сары-Иван.Суюндык,
Сыиргумак, Текей. Текей-Кзыл, Терен-Кудук, Тсун-Аурган. Тумар-Кудук, Чагр-Чигр,
Шошай-Кубук, Шошакты. Эти места находок, как правило, нельзя указать на картосхеме
из-за ее масштаба, однако мы приводим их здесь, надеясь, что они будут полезны для бу-
дущих исследователей региона.
Нами получена карта с кадастром Казахстана от ЗКБрушко, на которой отмечены
пункты коллекционных сборов E.arguta за 1975—1985 гг. Приводим их, кроме перечислен-
ных выше: Уральская обл.: сБорки Джангалинского р-на Гурьевской обл.: пески Тайсо-
ган, окр. с.Сармайчагыл [320, рис. 50,18] и окр. сКарабау; окр. с. Махамбет Махамбет-
ского р-на; Волго-Уральские пески, Новобагатинский р-н Актюбинской обл.: Иргиз-
ский р-н, окр. с.Мамыр, ур.Улькаяк, 25 км южнее сЖайсамбай, восточная часть оз. Кум-
коль. Мангышлакская обл.: Бейнеускйй р-н, сАкжигит; пески Сам. Чимкентская обл.:
с.Шукурой Сузакского р-на. Джамбульской обл.: Джувалинский р-н, Терс-Ашибулакское
водохранилище; верховье р.Терс, Джамбульский р-н, Малый Каратау, уш.Алмалы. Из это-
го района имеются также сведения от ВТ.Колбинцева, который добывал здесь ящурок в
окр. г .Джамбул — Мойынкумский р-н, Бетпакдала (Голодная степь), Апдасайский заказ-
ник; горы Курманчиты; Таласский р-н, оз. Аккуль; 12 км юго-вост. сБостандык; а также
данные В.Г.Колбинцева — окр. ст. Коктал Курдайского р-на, окр. с.Черная речка. Сведения
(дополнительные) по Джамбульской обл.: Чуйский р-н, западный склон Чу-Илийских гор,
20 км к сев .вост, от с.Ленинское Сарысусского р-на, 15 км западнее стБугунь. Алма-Атин-
ская обл.: Талгарский р-н, окр. сел. Чингильды и Кускудук, перевал Архарлы; Илийский
р-н, оз. Сорьулак; Чиликский р-н, долина р.Чилик; Алмаатинский р-н, г.Алма-Ата. Талды-
Курганская обл.: Каратальскийр-н,платоУшколь, Алакульскийр-н, оз.Алакуль [320,рис.50,
70]. Джезказганская обл.: Актогайский р-н, окр. г.Бектауат. Тургайская обл.: Джангиль-
динский р-н, метеостанция Брали. Целиноградская обл.: Тенгизский р-н, Кургальджино,
оз. Кыпшак. Павлодарская обл.: Ермаковский р-н, Малый Калкаман, юго-зап. г.Ермак.
Восточно-Казахстанская обл.: Тарбагатайский р-н, Зайсанская котловина, 14—15 км на за-
пад от сЛрйозерный; 12—15 км на северо-восток от сБелая Школа; 20 км севернее
с-Крупское [320, рис. 50, 6], Маркакольский р-н. 45 км на запад от с.Буран Зайсанского
р-на; 20 км северо-западнее сМайкапчагай Самарского р-на Буконьские пески.
Приведенный список показывает, что большинство данных — новые. К сожалению,
отсутствуют многие известные северные находки, на границе ареала. Если в тех местах
уже не встречаются ящурки, то можно говорить о заметном сдвиге северной границы аре-
ала на юг.
Новые данные о распространении Е. argutaB Китае отсутствуют (рис. 7). Сведения о
19
находках разноцветных ящурок в Монголии дополнены (рис .6) благодаря исследовани-
ям В.Ф.Орловой. Как уже указывалось [320], находка в Хиргиснурской котловине
(кол. ЗИН) вызывает сомнение, а первая достоверная находка из Монголии принадлежит
ГПетерсу (Джунгария, Уэнг-Сомон). В.Ф.Орлова и Х.Тербиш [197] описали серию нахо-
док (16) в данном районе, что дает основание предполагать о сплошном ареале вида от
г.Зайсан в Казахстане, китайской и монгольской частях ареала. Сведения о Earguta допол-
нены также работами монгольских коллег [174,271]. Ниже приведены карты с кадастра-
№ мест находок разноцветной ящурки.
3 2. ОБЩАЯ ХАРАКТЕРИСТИКА АРЕАЛА
Разноцветная ящурка распространена от Северо-Восточной Румынии на западе до Юго-За-
падной Монголии на востоке. Северная граница ареала часто совпадает с границей степной
зоны, доходит до 52—53° сдп. на востоке европейской части бывшего СССР, южная ограни-
чена Черным морем, далее идет по степным районам и предгорьям Северного Кавказа, про-
никает в Западном Прикаспии на Кавказ, заходит в Северо-Западный Иран и в Закаспии
обходит пустыни и высокогорья Средней Азии (однако на востоке проникает в ряд долин
Тянь-Шаня), далее ареал включает степи Казахстана до Кашгарии и Джунгарии. Как видно,
это (кроме некоторых краевых форм) типичный степной вид, и если он встречается в гор-
ных районах Кавказа и Тянь-Шаня, то занимает там остепненные биотопы. Горные популя-
ции этого вида часто изолированы от основного ареала, хотя и в равнинных районах ареал
чаще состоит из разобщенных поселений.
Большое значение (в европейской части ареала — определяющее) для распространения
вида имеет характер почв. Так, в европейской части бывшего СССР ареал E.arguta опреде-
ляется наличием песчаных почв. Поэтому поселения этого вида связаны с приморскими
пляжами, он Проникает на север только по пойменным пескам речных долин, которые в
прошлом былг распространены значительно более широко. Видимо, в ксеротермическое
время в голоцене, когда этот вид проник из Азии в Европу, в результате смены стаций
(из глжшсто-щебнистых местообитаний разноцветная ящурка проникла в песчаные био-
топы как более открытые и прогреваемые), он расселялся по западному побережью Кас-
пийского моря, пескам Кумо-Манычской впадины и Предкавказья и далее вдоль Азово-
Черноморского побережья. Одновременно с этим разноцветная ящурка продвигалась на
север по пойменным пескам речных долин. В это время ее ареал представлял собой поло-
су, протянувшуюся с востока на запад (вдоль моря) с ветвистыми ’’отростками'’ на север
соответствующим речным бассейнам. Необходимо помнить и о вековых колебаниях гра-
ниц зон, связанных с периодичностью солнечной акпшности.
Не исключено, что по этой причине ящурка исчезла из-под Киева и Оренбурга, в доли-
не р-Урал. Расчленение поселений ящурки происходило по разным причинам. Прежде все-
го это залесение песков, образование сосновых боров. Подобные явления происходят и
на наших глазах (например, ящурки уходят из облесенных участков Олешковских песков
в Херсонской обл., окр. сЭеликие Сорочинцы Полтавской обл., окр. гТрзйворон Белго-
родской обл. России) [140].
Способствовала этому явлению и другая хозяйственная деятельность человека. Многие
местообитания Eargut а в Украине были затоплены при зарегулировании Днепра. По со-
общению МЛ.Зиняковой и ПЛ .Руденко, ящурка в результате ирригационных работ ис-
чезла в окр. г.Темрюк. Многие местообитания этого вида были распаханы, особенно губи-
тельно для него орошаемое земледелие. Мы наблюдали влияние последнего на пресмы-
кающихся в долине р.Терек под г. Кизляр. Все побережье Черного моря от Одесской обл.
до г Херсона застроено санаториями и пансионатами, где, по нашим наблюдениям, разно-
цветная ящурка уже почти исчезла. Аналогичная картина отмечена и на Азовском побе-
режье. Поэтому в Украине встает вопрос о включении данного вида в Красную книгу
Украины. Не случайно в последние годы отсутствуют находки мест обитания и в ряде об-
ластей России.
Азиатская часть ареала вида находится в более благоприятном состоянии. Здесь Елг-
guta населяет обширные пространства целинных степей Казахстана, где она обитает на раз-
режешнях глинистых и лессовых, а в горных районах — на каменисто-щебнистых почвах,
20
менее подверженных хозяйственной деятельности человека. И хотя ящурка местами про-
никает в полупустыню (Казахстан, Узбекистан, Таджикистан) , в Европе ее ареал ограни-
чен изотермами июля 18—24 °, в Азии — 24—26, изотермами января соответственно — 3...—
8 и —14° и годового количества осадков 300—500 и 100—300 мм. Суммарная солнечная
радиация территории ареала между 100 и 140 ккал/см2 [9].
Связь дагестанских и азербайджанских популяций, видимо, нарушилась в результате
трансгрессий Каспийского моря. Об этом свидетельствуют и находки вида на о.Обливной в
Бакинском архипелаге на Каспии [20] В Азербайджане Earguta распространена узкой
лентой от Апшеронского п-ова вдоль моря до границ Ленкоранского р-на, а от нее вдоль
рек Кура (проникает здесь и в Грузию) и Аракса, по низменным районам республики,
где ее ареал также разорван в результате хозяйственной деятельности человека. Совершен-
но изолирована высокогорная севанская популяция. На востоке горные популяции (Талас-
ская долина, Иссык-Кульская котловина) сейчас тоже изолированы. Вероятно, это связа-
но с общим поднятием Тянь-Шаня в четвертичное время. Таким образом, на современное
формирование ареала E^rguta в разное время влияли исторические, климатические (темпе-
ратура, влажность) , почвенные и антропогенные факторы.
Современный ареал E^rguta можно очертить следующим образом. Его северная грани-
ца начинается в Добрудже (Румыния), проходит через дельту Дуная и далее по Украине
через Раздельнянский и Савранский районы Одесской обл., Черкасский р-н, устье р.Сулы —
Шищакский р-н Полтавской обл., окр. г Ларьков; в России — окр. гБелгород, стан. Вешен-
скую на рДон, г .Борок и Пугачевский р-н Саратовской обл., Бузулукский бор (самая се-
верная точка ареала) и затем южнее г.Уральск продолжается на территории Казахстана,
через пос. Калмыков о на рУрал, южнее р.Уил, пески Кок-Джида, Иргизский р-н Актюбин-
ской обл., Наурзумский заповедник, оз. Тенгиз, окр. г. Целиноград, окр. г.Ермак Павло-
дарской обл., Кулундинскую степь, затем граница ареала резко спускается к югу через
окр. г. Змеиногорск (Россия) , г. Усть-Каменогорск, через Зайсанскую котловину уходит
в Джунгарию (Китай), горы Салбурты и Б улун-Тох ой и заходит на востоке в монголь-
скую часть Джунгарии до окр. с. Уэнчсомона. Южная граница ареала: на западе распростра-
нение Earguta ограничено Черным морем, в Крыму она занимает только степные - при-
морские райошя на западе и востоке полуострова, встречается на юге Таманского п-ова,
затем проходит по Ставрополью (Ново-Александровский и Ставропольский районы) и
вдоль предгорий обходит Кавказский хребет (долина рМалки,окр.гМоздок,г. Терекли-
Мекбет, ст. Червленая, г Хасавюрт и далее до дельты р.Самур) .
Изолированная часть популяции находится в Азербайджане от Апшеронского п-ова до
устья рКура и далее на запад по равнинам доходит до Грузии. Современное распростране-
ние в Зуванде и Иране вызывает сомнения. Еще одна изолированная популяция Earguta
в Севанской котловине и (возможно) у оз. Ван в Турции. Ареал вида далее на восток об-
ходит Каспий и от г.Форт Шевченко, с.Кайдак (Казахстан), пгт Белеули и оз.Судочье
(в Каракалпакии) доходит почти до устья р Амударья.
Далее она продолжается вдоль р.Сырдарья, доходит до предгорий Нуратау, Чиракчи в
Узбекистане и на юге проникает на территорию Туркменистана (пос. Амударья Ходжам-
басского р-на), затем возвращается в Узбекистан (города Карши и Яккабаг), обходит
горные системы Таджикистана и проникает в Ферганскую долину (окр. г.Ходжент, Таджи-
кистан; г.Фергана, Узбекистан), с севера распространение разноцветной ящурки ограни-
чено Кураминским хребтом. Обходя систему гор Тянь-Шаня, южная граница ареала прохо-
дит через ее северные предгорья (однако заходит в Таласскую долину и Иссык-Кульскую
котловину), Чуйскую долину, г Алма-Ату, долину рЛли и по р.Кунгес проникает в Ки-
тай, где обнаружена в Кашгарии. На востоке южная граница обходит Алакульскую и Зай-
санскую котловины и уходит в Джунгарию (Китай), где смыкается с северной границей
(рис. 6,7) .
ГЛАВА 4
СИСТЕМАТИКА, ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
И ВНУТРИВИДОВАЯ СТРУКТУРА1
Разноцветная ящурка имеет широкий видовой ареал, населяет различные климатические и
ландшафтные зоны. Известен ряд изолированных популяций, обитающих в горных райо-
нах, и обширные равнинные территории, занятые четко выраженными формами, в ряде
случаев они постепенно переходят в другие, при наличии зон интерградации. Этот вид ха-
рактеризуется как политипический. В настоящее время еще не утратило своей актуально-
сти описание вида, данное Н.Н-Щербаком [320]. Выборки взяты со всей территории ареала
(Добруджа, Румыния; Крым и Южная Украина; междуречье Волга — Урал; Западный Ка-
захстан, левобережье р.Урал; Восточный Казахстан, Семиречье; Джунгария; Кыргызстан,
Чуйская долина, Прииссыккулье; Узбекистан; Калмыкия; Дагестан; Азербайджан; Арме-
ния) . Описание составлено но экземплярам из коллекций ЗИН РАН, ИЗ АНУ и Бер-
линского музея.
4.1. ОПИСАНИЕ ВИДА
Ящурка разноцветная — Eremias aiguta (Pallas, 1773)
Кариотип: 2л = 38 35А + Зт, отмечен гетероморфизм половых хромосом 18-й пары
[150].
Неотип: № 13 205 (кол. ЗИН РАН) , добыт в Западном Казахстане, в песчаном масси-
ве Кок-Джида по р.Темир, сентябрь 1932 г., Гожев, в, L = 34,1 мм,Led = 45 Д мм, G = 30,
Sq =45, Ventre 30,Р/. = 7/8.
Общее ошеание: (п = 270) ; L = 28,3—96,6 мм (М = 61Д мм) ; Led = 38,5—122Д мм
(М = 70,4 мм). L/Lcd = 0,7-1,09 (3f=0,84); G = 20-33(М±т = 27,03*0,19); Sq = 37-
64(51,18*0,34, половой диморфизм отсутствует); Ventr. = 27—35(30,5*0,1); P.f = 5—
15 (9,68*0Д9), зачаточных бедренных пор 1—3; выражен половой диморфизм (г = 3,36) •
Pf. 9 = 9^4 * 0Д5; d= 9,98*0,12. Верхнегубных щитков (Lofr)7—13 (М = 10,15). Под-
глазничный щиток не касается рта у 99,53 %, касается (аномальный) у 0,47 %. Нижне-
губных 5—9 (М = 6,83) .
Пятый нижнечелюстной щиток доходит до нижнегубных щитков (11,28 %), не дохо-
дит (88,72 %). Лобноносовых цдетков один (99,53 %), два (1,44 %). Длина лобноносового
меньше его ширины (98,56 %), равна ей (1,44 %) . Надглазничные не отделены рядом зер-
нышек от лобного и лобнотеменных щитков (90,4 %), не полностью отделены (9,52 %) .
Между предлобшям нет дополнительных щитков (68 ,93 %) , имеется один дополнитель-
ный щиток (18,18 %), два (9,56 %), три (2,39 %), четыре и пять (по 0,47 %). Длина нижне-
го края межчелюстного щитка больше длины площадки перед первым надглазничным,
занятой зернышками (31,03 %) , равна (31,46 %), меньше (37,51 %) . Промежуток между
рядами бедренных пор укладывается в длине одного ряда 0,9—2,5, в среднем 1,5 раза. Ряд
бедренных пор не доходит до сгиба колен (99,06 %) , доходит (0,94 %) . Количество чешуй
вокруг 9—10-го кольца хвоста 20—31 (М ±т = 24,27*0,12) . Верхнехвостовые чешуи глад-
кие (40£ %), слаборебристые (41,43 %), ребристые (18,57 %). В преанальной области 5—
15 чешуй, увеличенных 0-3, у самцов (в основном) один (72,75 %) , у самок отсутствуют
(52,53 %) или один (42,42 %) .
1 Матервазы предоставили Э.В.Вашетко, М.П.Зинякова, Н.Н.Иванова, В.А.Киреев, В.Г.Колбинцев,
Е.ЮЛСудакина, П.П.Руденко.
22
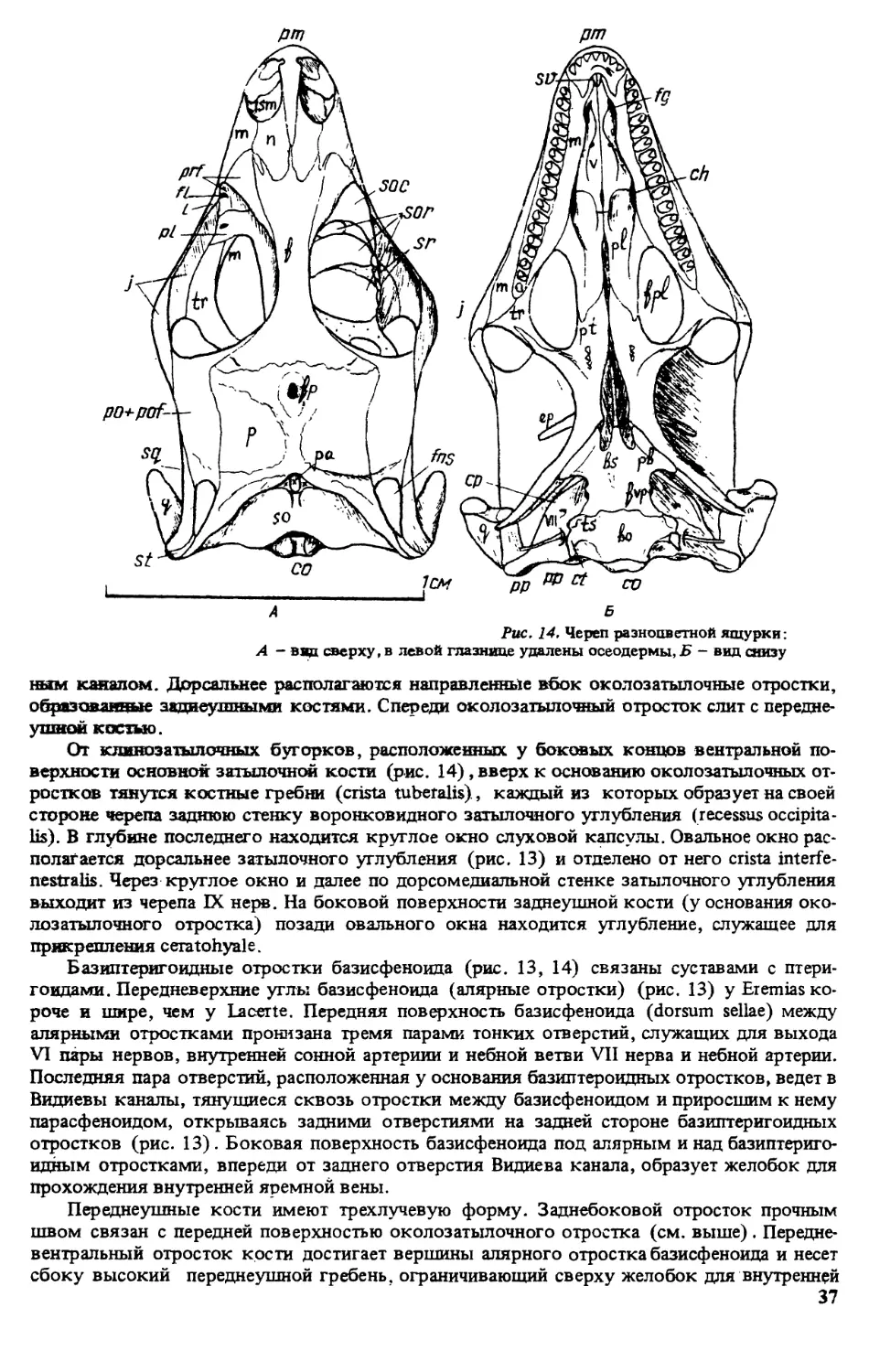
Рис. 9. Щиткование головы разноцветной ящурки [320]. форма E.a.deseni. имеются дополнитель-
ные шитки между предлобными (сверху, сбоку и снизу)
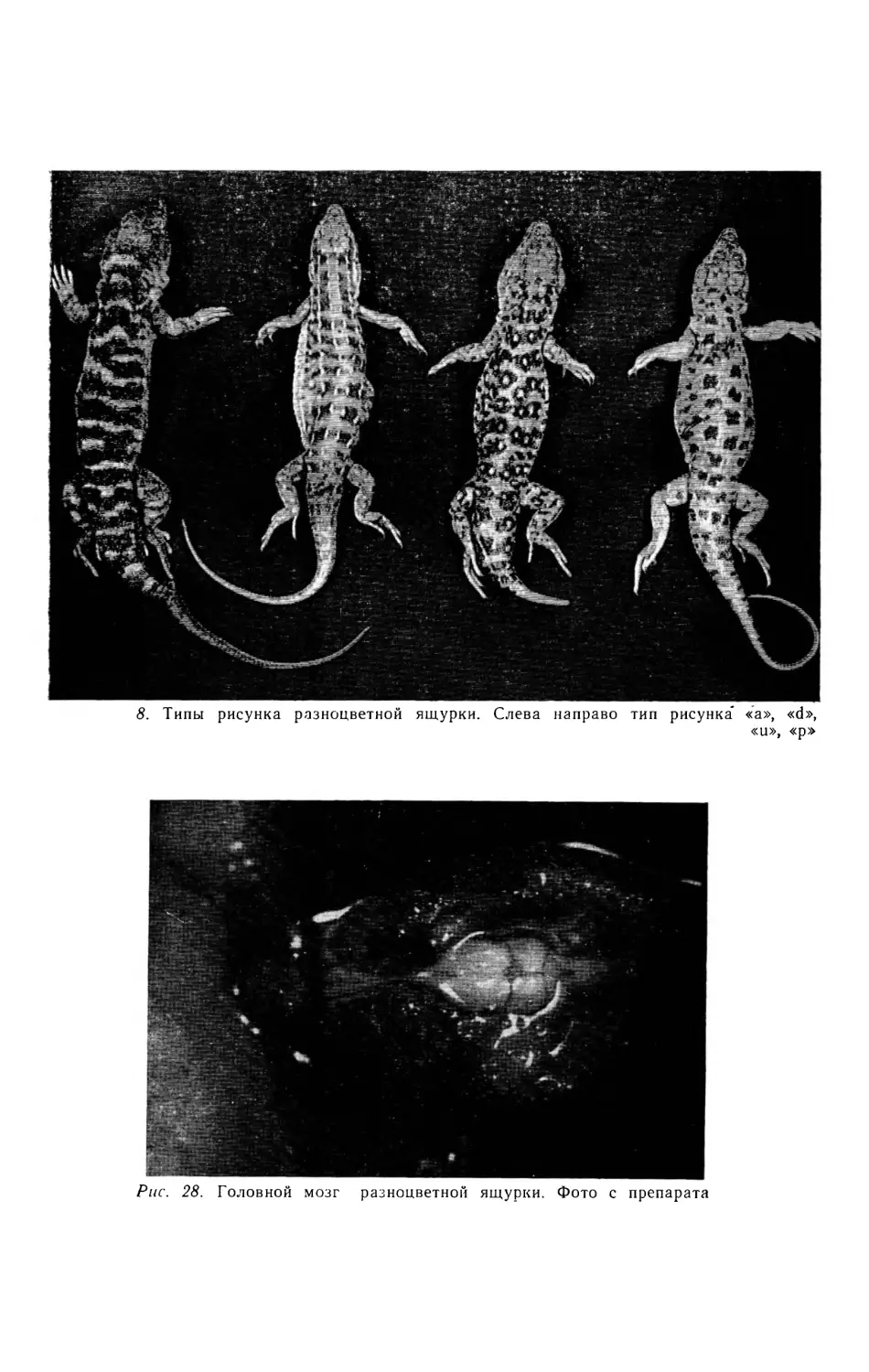
Рисунок молодых ящурок в общем не отличается от такового взрослых, но более чет-
кий. Основной фон туловища сверху серый с оливковым или буроватым оттенком. четко
соответствует цвету субстрата, на котором обитают ящурки. Наблюдается четыре типа ри-
сунка, хотя встречаются особи с рисунком, имеющим промежуточный характер: ”а” — на
спине неправильные черноватые или черные поперечные пятна и полосы (характерен для
многих представителей формы E.a.arguta); ”d” — на спине продольные ряды светлых то-
чек и черточек, между которыми расположены темные пятна (характерен для большинства
представителей формы E^.deserti); ”u” — на спине округлые светлые, отороченные черным,
пятна, расположенные более или менее правильными поперечными рядами (часто встреча-
ется у особей из Средней Азии (E^a.uzbekistanica), Вост. Казахстана и Закавказья) ; ”р” -
на спине редкие беспорядочно разбросанные светлые и довольно крупные темные пятна,
вытянутые чаще в поперечном направлении. Самый редкий тип рисунка, впервые описан у
формы Eapotanini (рис. 8) . Внешний вид фолидоза головы разноцветной ящурки пред-
ставлен на рис. 9.
42. ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
На неоднородность вида E.arguta впервые обратил внимание ЛЛантц1 [391.392], который
выделил на основании морфологического и географического критериев две формы Еалг-
guta (Pall.) и E^.desertii (Gm.). Последняя форма наиболее четко очерчена, хорошо выделя-
ется наличием ряда признаков фолидоза, в частности характеризуется наличием дополни-
тельного щитка между предлобными, и специфического рисунка (тип ”d”). Нами [320]
был показан ряд общих закономерностей географической изменчивости. Так, обнаружена
кливальная изменчивость размеров туловища половозрелых особей в направлении с запа-
да (Lm =59,7, Молдова) на восток (Lm = 68,3, Семиречье, Восточный Казахстан). а также
увеличение количества чешуи вокруг середины туловища половозрелых особей в направ-
лении с востока (Sqm = 45,9, Джунгария) на запад (Sqm = 53,3, Молдова) , а также от ме-
ждуречья Волга—Урал (Sqm - 50,0) до Дагестана (Sqm =54,3) . Еще более наглядны приме-
ры дискретной изменчивости некоторых признаков E.arguta. Так, по признаку ’’наличие
дополнительных щитков между предлобными или их отсутствие” отмечается четкая гра-
ница, которая на юге европейской части ареала проходит по предгорьям Кавказа (импе-
дентная граница) , на востоке — в междуречье Волга — Урал.
Таким образом, эта группа популяций четко обособлена от других (позднее будут по-
казаны признаки, характерные для западных популяций) . По признаку соприкасания тре-
тьей пары нижнечелюстных щитков (% 50 %) четко выделяются узбекистанские попу-
ляции и заметно отличается от остальных изолированная высокогорная армянская попу-
ляция [320]. В результате дополнительных исследований, проведенных в последующие го-
ды, мы получили описания некоторых популяций, ранее нами не изученных. Так, по дан-
1 Рис. 8, 28, 33-40,45-70, 90 см. на вклейке.
23
шм МЛЗиняковой и ПЛ .Руденко [письм. сообщ.] приводим описания выборки (л =
= 63 зкз.), добытой на Таманском п-ове — Кызылташской косе у стан. Благовещенской:
L = 31—75 мм (Л/ = 59,48), Led = 41-100 мм (М = 7421). L/Lcd = 0,68-0,94 (М = 0,81),
G - 23-33 (28,05 ±0,45), Sq = 44-61 (52,84±0,85) , Ventr. = 27-35 (31,59±0,51) , P.f. =
= 8-12(10,08±0,16), Lab = 7-11 (M = 9,4) , Sublad. = 6-10 (M = 7,46) ,Sq. c. cd. =9-10 -
23—24 (29 /56±0,48). Пятый нижнечелюстной щиток касается нижнегубных у 20,64% осо-
бей. нс касается у 7936. Три пары нижнечелюстных щитков не разделены у 93,65 % осо-
бей, разделены у 635. Промежуток между рядами бедренных пор укладывается в длине
одного ряда 0^—137, в среднем — 1,17. Ряд бедренных пор не доходит до коленного сги-
ба на 2 чешуйки у 7,94 %; на 3 — 47,63; на 4 — 33,09; на 5 — 634 %. Между предлобными
щитками имеется дополнительный щиток: 1 — у 61,91 %; 2 — у 31,75; 3 — у 3,16; 4 —
у 139 %, нет щитков у 139 %. Рисунок: тип ”4” — у 54,55 %, тип ”и” — у 25,45, переходной
tjhi ~d/u” — у 20,0 %. Все сказанное выше свидетельствует о том, что данная популяция
принадлежит к подвиду Е.a .deserti.
В А Кире ев исследовал выборки половозрелых ящурок из Калмыкии (п d= 45 зкз.,
п 9 = 34 зкз.). Как показывают приведенные материалы, ящурки из данной местности так-
же относятся к указанной форме: L d= 63,5-71,3 мм (66,8 ± 13); L 9 = 58,5—68,2 мм
(643*0,7), Led d= 75,9-97,5 мм (903 *2,2); Led 9 = 56,4-78,1 (65,1 ±2,2) , L/Lcd =0,7,
G = 24—33 (283*0Д), Ventr. = 28-33 (30,1 ±0,1) , P.f. = 8-12 (93*03) ♦ Дополнитель-
ный щиток между предлобными имеется: Lab = 7 (26 %), 8(56 %) и 9 (18 %). Нижнече-
люстных щитков 4 (28 %) , 5 (72 %) . Ряд бедренных пор не доходит до коленного сгиба,
встречается 1—3 недоразвитые поры. Вокруг 9—10-го кольца хвоста — 22—30 (253*0,1) .
Масса 43,8—9,1 г (6,9* 1,8), 9 - 4,04-9,9 г (63*0,7).
Ящурки из Калмыкии серые, соломенно-желтые или охристые, рисунок спины состо-
ит из продольно расположенных рядов светлых пятен, окруженных полосками темного
или светло-коричневого цвета. У молодых особей рисунок выражен значительно ярче.
Брюшная сторона светлая. Окраска ящурок соответствует их месту обитания. В одно-
родных условиях у ящурок почти отсутствуют изменения в окраске и образуется моно-
морфная группа. Так, у ящурок, населяющих кромки песчаных массивов — пески Маш-
так, Тавкаца и солончюсовые пески (побережье Каспия), окраска, как правило, более
светлая, чем у ящурок из других районов Калмыкии. Потемнение окраски отмечено у
ящурок, обитакндах во влажных биотопах (урочище Андра-Ата). Более тусклые по окрас-
ке ящурки, населяющие северные районы республики.
Наиболее интересный район взаимоотношения различных популяций яшурок находит-
ся в Западном Казахстане, особенно междуречье Волги и Урала. Этот вопрос детально ис-
следовали Н.М .Окулова, а также ВЗЗЯеручев, ЕЯЗКудакина и НЛ-Иванова [письм. со-
обцЦ * НМ .Окулова обследовала территорию общей площадью около 5 тыс. км2 на севе-
ро-востошой окраине Волго-Уральских песков и сравнила две микропопуляцми разноцвет-
ной ящурки в ландшафтах глшжстой и песчаной пустынь (отловлено соответственно
261 и 430 зкз.). Западный подвид Ел .deserti характеризуется обитанием на песчаных грун-
тах, йоммнативнда форма придерживается плота глинистых почв. На исследуемой тер-
риторми отмечено наличие и тех м других биотопов. Характеристика половозрелых ящу-
рок Западного Казахстана: L d= 65,73±O,27, L 9 = 6431*^38; d= 73 Д9±035,£с^ 9 =
= 6433±0,49, L/Lcd d = 1,16±0,01, L/Lcd 9 = 1 Д>3±0ДЭ1. Среднее тасло дополшггельных
щитков на одну особь: 3— 0,69—0,03; 9 — 0Д9±0,05. Масса тела 3— 7,11 ±0,23,9 — 6,15±
± 0,75 г., особи с глазчатой окраской: d— 39,1 ±3,1 %, 9 — 103*23; то же с поперечно-
павосчатой окраской: в— 353*3,0%, 9 — 40Д±3,6.
До рассмотренным признакам западаэказахстанская выборка занимает промежуточ-
ное положение между Ел arguta и Ел. deserti. Автор также установила, что обитатели пес-
чагых грунтов отличаются от особей, обитающих на твердых грунтах, следующими черта-
ми: более длинным хвостом, светлее окраской с преобладанием ’’глазчатого” типа и боль-
шйм количеством бедренных пор. Эти черты характеризуют, таким образом, более строй-
ных подвижных и светлых особей, что соответствует специфике жизни на песках по сравне-
нию с глинистыми участками, позволяет избегать перегрева, быстрее скрываться и быть
менее заметными.
Для данного вида, кроме того, характерна возрастная изменчивость. Рассмот-
рим кривые распределения длины тела E.arguta. Вся популяция нами разделена на че-
24
тыре размерные группы, из которых две первые объединяют неполовозрелых молодых
особей, а две последние — половозрелых взрослых особей: 1) L <48 ммг2) L = 49-55;
3) L = 56—64;4) L - 65—76 мм.
У ящурок было выделено четыре типа окраски. Молодая — все новорожденные ящур-
ки не старше 1 мес (длина до 45 мм) имеют окраску с преобладанием продольных рядов
светлых и белых пятнышек и со светлой полосой вдоль хребта. С их ростом возрастает
пигментация покровов, они приобретают окраску глазчатого (светлые пятна, отороченные
черным, разбросаны по всей спине) или поперечнополосатого типа (темные пятна сливают-
ся в поперечные черные полосы, иногда разделенные более светлой полосой вдоль хребта).
Промежуточные варианты окраски относятся к неопределенному типу. В данной работе
рассматриваются главным образом морфологические особенности самцов; близкие зако-
номерности характерны и для самок.
Рассмотрение окраски у 259 самцов позволило обнаружить, что окраска поперечнопо-
лосатого типа свойственна главным образом более крупным, особенно самым крупным
особям. У ящурок третьей размерной группы уже почти не встречается окраска молодого
типа, тогда как в первой не найдено ни одной особи с окраской поперечнополосатого ти-
па. Иными словами, с возрастом окраска ящурок изменяется в сторону преобладания
типа окраски, характерного для Ед .arguta. Глазчатая окраска встречается лишь незначи-
тельно чаще у самых крупных (наиболее старых) особей. Относительная длина хвоста
уменьшается с ростом ящурок. Следовательно, с возрастом у самцов разноцветной ящур-
ки начинают преобладать свойства, характерные для номинативного подвида: поперечно-
полосатая окраска и меньшая длина хвоста. Это подтверждает положение о том, что номи-
нативный подвид Еа arguta по сравнению с Ea.deserti более примитивный, так как с воз-
растом именно такие черты начинают проявляться более четко.
Как показывают ландшафтные особенности морфологических признаков, в глинистой
пустыне рисунок поперечнополосатого типа встречается почти вдвое чаще, чем в песках,
среди молодых и взрослых особей. По относительной длине хвоста молодые ящурки не
различаются: у старых в глинистой пустыне гораздо чаще, нежели в песках, встречаются
особи с хвостом короче тела, редко — с хвостом длиннее тела.
В старшей возрастной группе, среди половозрелых особей, в песках больше длинно-
хвостых. чем на глинистых грунтах, молодые по этому признаку не различаются. Следова-
тельно, в песках хвост растет сильнее или особи с длинным хвостом выживают лучше,
чем в глинистой пустыне. По числу дополнительных предлобных щитков и по длине тела
ящурки здесь четко не различаются.
Проверка достоверности различия эдафических вариантов популяций по критерию
Стьюдента показала, что особи с размерами тела менее 65 мм достоверно различаются толь-
ко по одному признаку — процент особей с поперечнополосатой окраской — в степи этот
показатель достоверно выше, чем в песках (t = 2,15). Для крупных ящурок t значительно
выше (4,9). В группе крупных особей (65 мм и более) достоверно реже встречается глаз-
чатая окраска в стели, чем в песках (/ = 4,6); в степи достоверно ниже процент особей с
хвостом длиннее тела (г = 2,15), что также прослежено только у более крупных особей.
Таким образом, ландшафтные особенности изученных микропопуляций аналогичны
особенностям подвидов; для песков характерны более длиннохвостые и светлые ящурки,
с большей долей глазчатого типа C’d”) рисунка, а для глинистой пустыни — короткохво-
стые, более неуклюжие ящурки, с более темной окраской, чаще поперечнополосатого
типа (”а”).
Из полученного материала можно сделать вывод, что приспособление к характеру
грунта — ведущий фактор в формообразовании в пределах вида Е arguta. Это можно про-
следить в пределах одной популяции (Западный Казахстан) в различных ландшафтах, а
также на разных подвидах, для которых характерна приуроченность к разным грунтам.
Эдафические варианты в популяции западноказахстанских ящурок можно рассматри-
вать как фенокопии [163] географических подвидов (вариант глинистой пустыни —
Е.а. arguta, песков — Ea.deserti). Очевидно, подвид Ea.deserti образовался вследствие обед-
нения генофонда, возможно, из-за временной изоляции западной части популяции во
время трансгрессии Каспия. В европейской части ареала Earguta не отмечено сколько-
нибудь значительной примеси типа ’’arguta” в популяции Ea.deserti. Экологически несом-
ненно, что ящурки типа’’arguta” могли отыскать пригодные для обитания твердые грунты
25
в европейской части ареала и эдафические фенотипы были бы найдена и там. ягп,< бы не
отсутствие соответствующих генов. Видимо, обеднение генотипа привело к тому, что на за-
паде этот вид стал обитателем исключительно песков. Во всяком случае все сказанное вы-
ше свидетельствует о наличии в данном районе границы двух форм с наличием небольших
зон интерградации.
Подвидовому статусу и изучению популяционных различий ящурок в Северном При-
каспии посвящена работа также В-В.Неручева, Е.Ю.Кудакиной и И.Н.Ивановой [письм.
сообщ.]. Материалы первых экспедиций авторов подтвердили то обстоятельство, что
условная граница между ареалами Ел .desert! и E^^rguta расположена в восточной части
Волго-Уральского междуре^я. в долине р.Урал. В то же время Е.Ю.Кудакина [148] уточ-
няет, что ящурки с западной и восточной кромок Волго-Уральских песков морфологиче-
ски и экологически сходны; население вида между песками и поймой Урала крайне разре-
жено, поэтому условной границей можно считать долину его. Поскольку на левобережье
Урала преобладают глинистые пустыни (E-a.arguta экологически связана преимущественно
с ними), что переходная зона (она представляет собой зону вторичной интерградации, свя-
заикую с восстановлением целостности ареала после трансгрессии Каспия) представля-
лась узкой, заключенной между песками и Уралом. В этой зоне пр ед полагал ось относитель-
ное пространственное разобщение вторично контактирующих подвидов, обладающих раз-
ной экологией.
Таблица 1- Встречаемость фенов рисунка свины у рамоцветной явдгрвд жз Северо-Восточного Прикаслыя Материалы, полученные в 1980— 1985 гг., позволили оценить картину несколько по-иному. Были взяты ящурки из пяти точек: глинистой пу- стыни урало-эмбинского между- речья - 40 экз.. приморской солонча-
Фен Глини- стая пу- стыня Примор- ские со- лон- чаки Пески Тайсуган Дельта р.Эмба Волго- Ураль- ские пески
A j 15,0 2,0 16 ковой пустыни — 26, западной кромки
^2 57,0 66,0 50,0 53,0 84 песков Тайсуган — 38, дельты р .Эм-
Аз 43,0 194) 50,0 45,0 — бы — 49, восточной кромки Волго-
Б, б2 524 324 214 24,6 294 24,0 33,2 33,2 22 28 Уральских песков — 38 экз. Изучал-
Б3 264) ся тип рисунка спинной стороны те-
БЛ 154) 534 21,0 344 50 ла с выделением его элементарных
Б1 24 114 — 104) 18 составляющих, приблизительно в том
В2 274 23,1 55,3 264 10 плане, как это сделано А.С .Барано-
Бз 704) 66Л 44,7 644 71 вым [31] для прыткой ящерицы. Просмотр серий показал, что суще-
ствует, по крайней мере, три группы фенов, свободно комбинирующихся между собой.
В каждой из жх есть два альтернативных варианта я один-два промежуточных. Группа
А — продольная светлая полосатость спины и шеи (Аг — ,4 сплошных полосы вдоль все-
го тела, А2 - полосы только на шее и передней части спины, А3 - полос нет). Группа
Б — светлые пятна на спинной стороне тела (Б^ — округлые светлые ’тлазки” по всему
телу, Бъ — ’’глазки” преимущественно в задней части спины, Б3 — светлые "глазки”
сливаются в пятна неправильной формы, Б* — светлых "глазков” нет). Группа В — тем-
ные пятна на спине (Вi — округлые пятна по всей спине, В2 ~ округлые пятна, сливающие-
ся в поперечные цепочки, В3 — бесформенные темные пятна, нередко выстраивающиеся в
поперечные ряды) - Схемы их даны на рис. 10, соотношение отдельных фенов — в табл. 1.
На этой основе построены графические изображения фенетических спектров (рис. 11).
Каждая выборка имеет свой фенетический спектр, характеризующийся как представлен-
ностью отдельных фенов, так и их частотами, что свидетельствует об относительной попу-
ляционной специфичности изучаемых группировок. Заметна также разная степень разли-
чий между выборками, свидетельствующая о возможных родственных связях между по-
пуляциями. Из табл. 1 видно, что выборки из глинистых пустынь и дельты Эмбы близки,
приморская популяция тоже имеет ряд общих с ними черт, но отличается высокой концен-
трацией фена Ь4, что сближает ее с серией из Волго-Уральских песков. Серия из песков
Тайсуган очень специфична (отсутствие и появление фена 53). Различия обнаружены
также по некоторым деталям фолидоза (табл. 2), а также в отношении экологии.
Приведенные факты свидетельствуют о сложности процесса вторичного контактиро-
вания подвиде®, между которыми возникли довольно четкие экологические различия. Аб-
26
солютной изоляции между ними нет, гибридный характер популяций в довольно широкой
зоне Северного Прикаспия не вызывает сомнений. Однако в этой зоне нет постепенного,
плавного перехода от одной формы к другой. Внутри нее выделяются гибридные популя-
ции с различной степенью сходства с тем или иным подвидом. Основой для этой дифферен-
цировки может служить ландшафтная неоднородность территории, различные части кото-
рой соответствуют экологическим требованиям той или другой формы. Отсюда - разно-
направленность отбора, формирующая различный (сходный с той или иной формой) фено-
тип популяций. Завершая обзор европейских и пограничных популяций разноцветной
ящурки, следует остановиться на особенностях кавказских E.arguta. Нами {320] доказано,
что форма ’’deserti” распространена до Южного Дагестана включительно (впоследствии это
подтвердил ОЛЦарук [279]), а также, что территория Азербайджана населена изолиро-
ванными популяциями, отнесенными к промежуточной форме E.a.deserti apt E.a.transcau-
27
Рас. 11. Рисунок фснетических спектров в выборках разноцветной ящурки из Северо-Восточного
Прмкаспия (по Неручеву и др,) :
1 — глинистая пустыня Урале-Эмбинского междуречья, 2 - приморская солончаковая пустыня,
3 - западная кромка песков Тайсуган, 4 - дельта рЭмба,5 - восточная кромка волжско-ураль-
ских песков. А, Б, В — объяснение в тексте
casica. „Чистая” закавказская форма — Ел-transcaucasica — представлена высокогорной
изолированной популяцией в Севанской Котловине (Армения). Ее диагноз см. шоке.
Краткая характеристика азербайджанской популяции: (п = 63) L = 50,05—76,4 (М =
= 65,46), Led = 63,35—95,5 (М = 74,48) ,£/£«/ = 0,75-1,0 (№ 0,87), G = 21—31 f27,83 ±
±0,25), Sq = 41-61 (53,25 ±0,37), Ventr. = 24-34 (30,93±0,19), Р = 7-13 (9,25±0,13),
Lab = 7—13 (М = 10,03), нижнегубных щитков 5—8 (М = 6,47). Три пары нижнечелюстшдх
соприкасаются у 76^2 %1. Пятый нижнечелюстной не касается нижнегубных у 98,44 %1 2
1 артик, для ЕллЗезегИ.
2 Обдай признак для E.a.transcaucaeica.
28
Таблица 2. Сравнение некоторых морфологических признаков разноцветной ящурки из разных
популяций Северного Прикаспия
Но- мер при- знака E.a.de- serti (по Н.Н.Щер- баку) западная кромка Волго- Уральских | песков (п=35) Восточная кромка Волго- | Уральских песков (л=^1) 1 Уральск о- Эмбинское между- речье, глинистые пустыни (м=40) Уральско- Эмбин- ское ме- ждуречье, примор- ский со- лончак 1 (л=26) Уральск о- Эмбинское между- речье, пески Тайсуган (и=25) Дельта р.Эмба (л =49) Е.а.ат- guta (по Н.Н.Щер- баку)
Дополнительный щиток между предлобными имеют
75,3 % 9,22 ±0,11 57 % 9,5 ±0,15 53% 18% 46% 41% Количество бедренных пор 9Д7±0Д4 74±025 9,39±0,19 8Д2±0,13 •21 % 9,17 ±0,23 22,7 % 8,35*0.23
3 4 40% ’Ъ”=0?. Пятый нижнечелюстной щиток касается нижнегубкых 3,3% 10% 5% 3.8 % 0% 6% Тип рисунка* спинной стороны тела "а" - ** "а" - 2,4 % "а" -75 % "а" - 20% "а" - 82 % "а" - 35 % 4,3% - 28,6%
”<Г=70% ”d”-He ме- "d"- 87 Л % "d" -12,5% ”</”-65% ”</” - 6% ”</” - 35 % "d” - 14,3%
”U’* = 30% нее 66 % ”и” - 1 "и" - 9,8 % ”М” -12,5% ”и” - 15% "и" -12 % "и" - 30 % ’V’ - 31,3%
* Типы рисунка по Н.Н.Щербаку [320].
** Плохая сохранность серии не позволила подробно анализировать рисунок.
Между предлобными нет дополнительного щитка у 87,31 %, имеется оданщитоку И, 11,2—2
у 1,58 %г. Вокруг 9— 10-го кольца хвоста 22—31 (25,85 ±0,26) чешуй. Промежуток между
рядами бедренных пор укладывается в длине одного ряда 0,9—2 (Л/ = 1,45) раза2. Ряд бед-
ренных пор не доходит до сгиба колен на 1 чешуйку у 1,16 %, на 2 — 17,44, на 3 - 51,16.
на 4 — 23,25 и на 5 — 6,99 % особей2. Недоразвитые поры имеются у 76,08 % особей, по 1 -
38,04; по 2 — 34,78, по 3 — 3.26 %2. Рисунок: тип — 19,36, — 16,13, промежуточ-
ный вариант ”u/d” — 64,51 %2 Как видно, у азербайджанских популяций имеются общие
признаки и с западной, и с армянской популяциями, однако последние преобладают. От-
сутствие контактов в настоящее время азербайджанских и дагестанских популяций позво-
ляет нам отнести первых условно к форме Е. transcaucasica. Сложные взаимоотношения
популяций наблюдаются в юго-восточных районах ареала, особенно краевых.
ЭББашетко [письм. сообщ.] предоставила неполное описание интересной ферганской
популяции: = 96,5 мм, L/Lcd = 0,7-1,0 (М = 0,8) , G = 20-28 (24,16*0,5), Sq = 40-
51 (44,8*0,25), Ventr. = 27-32(29,06*0,35) , Lab = 8-11 (9,5 ±0,2). Количество чешуй на
9-10-м кольце хвоста 19-27 (22,6*0,39),?./. = 10-14(11,8*0,16) .Между рядами бедрен-
ных пор .6—8 чешуй. Окраска спины зеленовато-серая. Рисунок спины из поперечных по-
лос черного цвета с 2 округлыми серовато-фиолетовыми пятнами. Черные полосы между
светлыми в форме ”Х”. По краям полос бежеватые тонкие полоски. На голове светлые
пятна сливаются в линию. Молодые особи не отличаются по рисунку от взрослых, только
он у iBix обычно ярче. Насколько можно судить по данному описанию, ферганская попу-
ляция близка к форме Е.а. uzbekistanica.
В.Г.Колбинцев [письм. сообщ.] проанализировал две выборки из популяций Джам-
бульской обл. (Южный Казахстан). Одна из них (п = 33) собрана в предгорьях Каратау (юго-
запад области, западнее г .Джамбул), другая (и = 45), обозначенная исследователями как
”курдайская”, происходит из юго-востока области (на северо-восток от г Бишкек). Он
указал, что между ними по ряду признаков отмечены существенные различия. Приводим
здесь сокращенные описания указанных выборок.
Каратауская предгорная популяция: L = 59—81,3 (68,68*2,25) ,Lcd = 69-109 (85.56*
±2,07), L/Lcd = 0,72-1,07 (0,81*0,02), Sq = 40-52 (45,09*0,57),?./. = 10-14 (11,67*
*0,19); между рядами бедренных пор 5—10 (7,28* 17) чешуй. Длина ряда бедренных пор
не доходит до сгиба колен на 1—3 чешуйки. Lab =8—11 (М = 9,52) ; нижнегубных щитков
5—9 (6,58*0,17). Пятый нижнечелюстной не касается нижнегубных или незначительно ка-
сается внешним передним углом у 100 %. Дополнительный маленький щиток между пред-
* Общий признак для Е.а. transcaucasica.
Промежуточный признак между E.a.desertiH Е.а.transcaucasica,
29
лобными у 3,03 %. G = 20-26 (22,62±0,74), Ventr. =28-34 (32,0±0^7), количество че-
шуи вокруг 9-104*0 кольца хвоста 21-25 (23,25±0,45). Рисунок типа ”и” - у 24,24 %,
промежуточный тип ”u/d” — у 72,72. Основной фон спины от серого до песочного с розо-
вато-винным оттенком. Как видно из приведенных данных, каратауская популяция, оби-
тающая на границе гор (предгорий) и казахской степи, по большинству признаков (L,
Sq, G, отношению ряда P.f. к сгибу колена, отсутствию недоразвитых PJ.) относится к
форме E.a.uzbeckistanica.
Курдайская популяция: L = 60-72 (68,25±2,78) , Led = 66-81 (73,0±3,39), L/Lcd =
= 0,81-1,09 (М = 0,94±0Д5),А? =44-54 (48.8±2,25),PJ. = 10-12 (11,25 ±0,25). Между
рядами бедренных пор 7-9 (8,2±0,37) чешуи, Lab = 9-11 (10,6±0,25). Нижнегубных
щитков 7—8 (7,4±025), пятый нижнечелюстной не касается нижнегубных у 100 %. Допол-
нительный маленький щиток между предлобными у 40 %. G = 24—26 (25,4±0,4) . Ventr. =
= 31-32 (За,2±0,2). Рисунок промежуточного типа ”u/d” у 100 %, основной фон спины
серо-зеленоватый (d) и песочный ( 9) .
К сожалению, автор не указывает важных признаков, насколько далеко до коленного
сгиба доходит ряд бедренных пор, наличие недоразвитых бедренных пор. Заметно увели-
чение количества дополнительных щитков между предлобными до 40 %, что резко отли-
чает их от каратауских. Как показал О Л Царук [279], данный признак характерен д ля
ящурок Прииссыккулья (до 62,82 %) и ранее отмечен [324] для особей из Чуйской доли-
ны (33,3 %) что свидетельствует об их генетической близости.
Популяция из Прииссыккулья, которая благодаря изоляции обладала рядом особен-
ностей фолидоза [320], впоследствии [279] была возведена в ранг подвида Ел. darev-
skii. Ее описание будет приведено ниже.
Наименее изученными до сих пор остаются восточные популяции (Джунгария. Кашга-
рия). Однако и этот пробел в наших знаниях в последние годы исчезает благодаря работам
В.Ф.Орловой и ее мошольских коллег. В.Ф.Орлова отмечает следующую изменчивость у
особей из Кобдосского и Гоби-Алтайского аймаков Монголии (Джунгарская Гоби).
43. ПОЛОВОЙ ДИМОРФИЗМ
Взрослые особи отличаются от ювенильных и неполовозрелых абсолютными и относитель-
ными размерами тела и его частей, неровной поверхностью головы и, как правило, отсут-
ствием глазчатого рисунка дорсальной поверхности; максимальная длина туловища отме-
чена дня самцов, хотя по средним значениям этого признака взрослые обоих полов почти
не отличаются (у 9 - 51,0-72,2; 62,09±0,96, п = 27; у d= 52,7-75,0; 64,95±1,31,и = 26).
Мощное телосложение взрослых самцов, исключительно грубо пятнистый черный рисунок,
сильно развитая мускулатура в височной области хорошо отличают их от самок. Самцы и
самки в выборке из окр. Алтай-Сомона и Боданчийн-Гол отличаются также достоверно
знгшмыми разлмтаями количества чешуи вокруг 9—10-го кольца хвоста (yd— 27—34;
29<90±Q,24, п = 41; у 9 — 25—32; 27,77 ±0,29, п = 31; Р < 0ДО1) и числа подпальцевых
пластинок (yd— 17-24; 20,46±0,26; у9- 17-22; 19^5±0^5; Р<0Д5).
Часть взрослых самок сохраняет в полном или редуцированном виде рисунок дор-
сальной поверхности, характерный для неполовозрелых особей. Незначительные разлитая
отмечены в относительной длине передних в задних конечностей, в отношении ширины го-
ловы к ее высоте. Географическая изменчивость на собранном материале с изолированной
территории Джунгарской Гоби не прослеживается. Возможно, это объясняется не только
небольшой площадью исследованного района, но и тем небольшим количественным мате-
риалом, который был собран на юге, в предгорьях Байтак-Богло-Нуру. Очень заметна
сизо-голубая окраска боков головы, шеи, туловища и хвоста крупных самцов. В централь-
ной части Джунгарской Гоби, из окр. с. Уенча и с надпойменных террас Боданчийн-гола ни
одного экземпляра с подобной окраской не было и все самцы были мельче.
4.4. ВНУТРИВИДОВАЯ СТРУКТУРА
Элементы продольного рисунка наиболее выражены на западе ареала, глазча-
тый рисунок особешю часто встречается на юге его (Закавказье, Средняя Азия),
30
Рис. 12. Общий ареал разноцветной ящурки и распространение ее подвидов .
1 — Е.а.deserti, 2 - Е.а. transcaucasica, 3 — E.a.arguta. 4 - Е.а.uzbekistanica, 5 - E.a.darewskii, 6 —
E.a.potanini
I
в Казахстане выражена в той или иной степени поперечная полосатость, которая на край-
нем востоке сохраняется в виде отдельных темных пятен. Чаще подвергаются географи-
ческой изменчивости признаки GnP.f. [320], затем Ventr. nSq.
Наряду с клинальной существует дискретная изменчивость признаков разноцветной ящур-
ки. Ряд близких популяций можно сгруппировать в категории высшего порядка — подви-
ды. Реально существуют подвиды Ea.arguta, E.a.deserti, E.a.potanini, Е.a.uzbekistanica.
Е.а. darevskii и Ел .transcaucasica (рис. 12) . Отмечены зоны интерградации подвидов: Север-
ный Прикаспий, Азербайджан, Голодная Степь в Казахстане. Обнаружены популядии-изо-
ляты в Армении и Иссыккульской котловине. Центром становления вида предполагают-
ся степи Центрального Казахстана, где отмечается высокая степень полиморфизма особей,
сходятся границы ареалов по крайней мере трех форм.
Таблица для определения подвидов Eremias arguta
1 (2) . Три пары нижнечелюстных щитков всегда (около 100 % особей) соприкасаются по средней ли-
нии горла.................................................................. 3
2 (1) . Три пары нижнечелюстных щитков чаще не соприкасаются (более 50 % особей).7
3 (4). Дополнительный щиток между ппедлобными, как правило, имеется (у 75 % особей) .
. . E.a.deserti (также: &? =42 (52)62. G =23 (29) 36,ЛС. =6(10)13; тело стройное,£тах =77,5,
чаше - 60—66 мм) .Рисунок: из светлых черточек, расположенных продольными рядами.
Распространение: от Румынии до р.Урал и на юг до Азербайджана.
4(3). Дополнительный щиток между предлобными, как правило, отсутствует (может быть у 20-
22 % особей)............................................................... 5
5(6). Ряд бедренных пор не доходит до сгиба колен на 1-2 чешуи, дополнительные недоразвитые
поры часто отсутствуют......................................... E.a.potanini
(также: Sq = 37 (46)55, G = 22(25)32, P.f. = 8(11)14 ; тело массивнее и коренастее, чем у
Е.а. deserti, = 84,чаще - 61-68 мм) . Рисунок: чаще светлые отороченные черным пят-
на, расположенные поперечными рядами. Распространение: от Прибалхашья до Юго-
Западной Монголии, Джунгария. Кашгария.
6(5). Ряд бедренных пор не доходит до сгиба колен на 3-5 чешуй, дополнительные недоразвитые
бедренные поры имеются.......................... ................... E.ajirguta
(также: Sq = 42 (50)56, G = 23 (28) 32,P.f. = 5 (8) 12. Пропорции тела и рисунок спины промежу-
точные между E.a.deserti и E.a.rotanini. хотя часто (до 28 %) встречаются особи с неправильны-
ми черноватыми поперечными пятнами без светлых глазков (с Zmax = 85 мм, чаще - 67 мм).
Распространение: от р.Урал до Прибалхашья, на юг до пустынь Каракум, Кызылкум и
зоны гор Средней Азии.
См. также описание подвида
31
7 (8). Дополнительный щиток между предлобными часто имеется (до 70 % особей).......9
8 (7). Дополнительный щиток между предлобными, как правило, отсутствует ...........10
9 (10). Ряд бедренных пор не доходит до сгиба колен на 2-3 чешуи, дополнительные недоразвитые по-
ры, как правило, отсутствуют ..............................................ЕлДатеуакн
(также: &? = 34(43)51; <7 = 24(29)34; Р/ = 8(10), 12; тело стройное,Lw = 76 мм)\Р и с у-
н о к: светлые, отороченные черным пятна, расположенные поперечными рядами. Распро-
странение: Иссыккульская котловина.
10 (11) .Рад бедренных пор не доходит до сгиба колен на 1 —2 чешуи, недоразвитых, как правило, нет . .
- Ел. ияМсЫжса (также: Sq « 41 (47)62; G 3 20 (22) 26,P.f = 9 (11) 151 тело массивное и коре-
настое. = 96 мм, чаще — 67—68 мм) . Рисунок: светлые, отороченные черным пятна,
расположенные поперечным! рядами. Расп ро странение: юго-восточная часть Средней
Азии, зона гор.
11 (10) .Ряд бедретых пор не доходит до сгиба колеи на 3-5 чешуй, недоразвитые поры имеются ....
ЕлЛпмсавсаака (также: Sq 3 52 (54)56; G = 24(26)29; P.f. = 5 (6)8. По пропорциям тела и ри-
с>дку похожи на E^jutekbtsaica, = 85 мм, чаще - 70-72 мм) .Распространение:
Закавказье, Севанская котловжа.
Ящурка разноцветная обыкновенная -
Eremias arguta arguta (Pallas, 1773)
Тепа typiea - Междуречье Урала и Эмбы.
1773 - Lacerta arguta, Pallas, Rebe versch. Prov.russ. Reich, 2: 718.
1922 - Eremias argutaarguta, Lantz, Bull See. Zool France, 47.
1974 - Eremias arguta arguta, Щербак, Ящурки Палеарктики : 167.
Описание: Тип видовой, (п = 22), 34Д5 (5035) 85,1; Led =423(55,4) 15^,L/Lcd=
= 0,75(0,82)035, G = 23-32(28,68*0,45), Sq = 42-56(50,23*0,84), Р/. = 5-12 (835*
±0ДЗ), Lab - 9 (1037) 12, Ventr. = 29—35 (3137*30). Три пары нижнечелюстных не разде-
вши у 100 %. Между предлобными нет дополнительного щитка у 77,28 %, имеется — у
22,72. Количество чешуи вокруг 9—10-го кольца хвоста 21-29 (М ±т = 2437*0,45) . Про-
межуток между радами бедренных пор укладывается в диете одного рада 0,9—13, в сред-
нем — 1,16 раза. Рад бедренных пор не доходат до сгиба колен на 2 чешуйки у 6,66 %, на
3 — 50 Д на 4 — 26,66. на 5 — 16,68 %. Недоразвитые поры имеются у 85,29 % особей. Ри-
сунок: тип ”а” - 45,47, ”и” - 9Д9, "<Г - 18,18, "a/u” - 9,09, ”u/d” - 13,63, ”a/d” -
434%.
Распространение: Западшлй Казахстан (рис. 12).
Ящурка разноцветная завидная -
Eremias azgttta deserti (Gmetin, 1789)
Тя» typiea - ”Яипкие степи” (межцуре’ье Волги и Урала).
1789 - Lacerta deserti Gmettn, Linn. Sys. Nat. Ed. 13, L 1076.
1922 - Eremias arguta deserti, Lantz, Bulk Soc. Zool France, 47,193.-1974, Щербак, Ящурки Палеарк-
тжи : 168.
Описание: Тип утерян, (и = 93),£ =31 (5EJS)TI £,Lcd L/Lcd =
= 0,68(031) 1Д G = 23-36(29,59*0,27), Sq = 42-64(52^1 *0,41), Ventr. = 27-33(29,63*
+0Д4), Pf. = 6—13(9,92*0,11), Lab. = 7(9)12. Три пары шокнечешостных не разделены
у 100 %. Между предлобными дополнительный цщток имеется у 75,27 %, отсутствует у
24,73 %. Количество чешуи вокруг 9— 1&го кольца хвоста 22—31(26,47*0Д9), Промежу-
ток между радами бедоеншх пор укладывается в длине одного рада 0,9—23,® среднем -
1,64 рвза. Рад бедрсятах шэр не доходит до сгиба колета на 1 чешуйку у 3,06 %, на 2 -
30,79, на 3 — 46,42, та 4 — 15,57, на 5 — 4,16 % особей. 1—3 недоразвитые поры у 53,41 %.
Рисунок: тип "сГ - 66,81, - 16,53, ”d/u” - 16,66%.
Распространение: Видовой ареал на западе от р.Урал, на юг до Дагестана вклю-
чительно.
Ящурка разиоцвеппя армянская —
Eremias aiguta transcaucasica Darevskii, 1953
1953 - &emias argute transcaucasica
Двревский, Докл. АН АрмССР, 164: 117.
1974 - Eremiasarguta transcaucasica Щербак,Ящурки Палеарктики: 169.
О п и с а ни е. Тип: Зоомузей МГУ, № 2811, Армения,Басаргечарскии р-н, окр. сЛ4ец-
Мазра, 2100 м нули., 8JX 1951 г. кол. И.СДаревский. (п = 13).£ = 39,6(54,9)85 мм,
Led = 48,6(59,7)88 мм, L/Lcd = 0,76(0,88) 1,09, G = 24-29(26,0*0,39), Sq = 52-56 (54,4*
*036), Ventr. = 29-33(30^*035) ,Р/. = 5-8(637*0,22) .Lab = 8(10) 12. Три пары нижне-
челюстных не разделены у 46,15 %. По И.С.Даревскому [76], горловые чешуйки разделя-
32
8. Типы рисунка разноцветной ящурки. Слева направо тип рисунка* «а», «d»,
«и», «р»
Рис. 28. Головной мозг разноцветной ящурки. Фото с препарата
Рис. 33. Шейный отдел truncus sympathicus: 1 — первый грудной узел, 2 — нервы, форми-
рующие truncus sympathicus, 3—узел nervus vagus, 4 — nervus larvngeus, 5 — ramus caro-
ticus, 6 — ramus cardiacus, 7 — nervus hypoglossus, 8 — hyoideus, 9 — trachea, 10 — ramus
pulmonalis, 11 — основной ствол n. vagi
Puc. 34. Варианты строения начального
отрезка truncus sympathicus thoracalis,
ganglion stellatus и два последующих
грудных узла: а Е. a. uzbekistanica, б,
в — Е. a. deserti
Рис. 35. Варианты строения truncus sympa-
thicus в области желудка: а — Е. a. uzbeki-
stanica; б, в — Е. a. deserti
Рис. 36. Варианты строения проксимальной части plexus brachialis: а —
Е. a. deserti, б — Е. a. uzbekistanica
Рис. 37. Строение plexus lumbo-sacralis. Е. a. uzbekistani-
ca, XXIII спинальный нерв
Рис. 38. Строение plexus brachialis. Е. a. deserti
Рис. 39. Варианты строения
plexus lumbo-sacralis Е. a. de-
serti, XXIII спинальный нерв
Рис. 40. Варианты строения plexus lumbo-sacralis.
Е. a. deserti, $, XXIII спинальный нерв
Рис. 45. Гортань разноцветной ящурки: 1 — перстневидный хрящ,
2— черпаловидные хрящи (3,5Х^)
Рис. 46. Центральный отдел гортани разноцветной
ящурки: 1 — слизистая оболочка гортани, 2— му-
скулатура гортани, 3—перстневидный хрящ, 4 —
цилиндрический мерцательный эпителий.
Фото с препарата (3,5X10)
Рис. 47. Трахея разноцветной ящурки: 1 — хрящи
трахеи (10X10)
Рис. 48. Легкие взрослой разноцветной ящурки
Рис. 49. Искусственно раздутые легкие взрослой разноцветной
ящурки. Видны соединительные септы, образующие каркас лег-
кого
Рис. 50. Схема септ легкого разноцветной ящурки. Септы I—III порядков
Рис. 51. Микрофотография легочных септ: / — соединительная
септа первого порядка (3,5X8)
Рис. 52. Микрофотография центральной части языка раз-
ноцветной ящурки (поперечный срез). Гематоксилин-эозин
(3,5X10)
Рис. 53. Подъязычный аппарат разноцветной
ящурки: 1—entoglossum, 2 — жаберные дужки,
3 — гиоидные рожки
Рис. 54. Желудок разноцветной ящурки. Фото с препарата (3,5X10)-
Рис. 55. Двенадцатиперстная кишка ящурки: 1 — мышечный слой, 2 — склад-
ки слизистой оболочки, 3—ворсинки слизистой оболочки (3,5X10)
Рис. 56. Тонкий кишечник разноцветной ящурки: 1 — адвентиция, 2 —
мышечный слой, 3 — подслизистый слой, 4 — слизистый слой
(3,5X10)
Рис. 57. Участок печени разноцветной ящурки: 1 — си-
стема кровеносных капилляров, заполненная эритроци-
тами. Микрофото (10X10)
'Рис. 58. Выделительная система разноцветной ящурки: 1 — Capsula glomeruli, 2 —
ячейки соединительнотканного остова почки. Гематокснлин-эозин (Х2ОО)
Рис. 59. Nephron. Гематоксилин-эозин (Х200)
Рис. 60. Иннервация капсулы железы, микроганглий. Импрегнация солями
серебра по Бильшовскому-Грос
Рис. 61. Мочеполовая система самца: 1 — orchis, 2 —
epididymus, 3— ductus deferens, 4 — hemipenis, 5 —
ostium uro-genitalis, 6 — ren
Рас. 62. E. arguta deserti. Паренхима epididymus. 1 ематоксилин-эозин
(X200)
Рис. 63. Е. arguta uzbekistanica. Паренхима epididymus. Гема-
токсилин-эозин (Х200)
Рис. 64. Мочеполовая система самки: 1 — infundibulum
oviducti, 2— oviductus, 3— ostium genitalis, 4 — ostium
uretris, 5 — ovarium, 6 — cloaca, 7 — vesica urinaris, 8 —
ran
Рис. 65. Щитовидная железа неполовозрелой Е. arguta. Фолликулы среднего
размера. Окраска по Слинченко (Х120)
Рис. 66. Щитовидная железа половозрелой Eremias arguta. Крупные фоллику-
лы. Окраска по Слинченко (X 120)
Рис. 67. Участок опустыненной степи. Место обитания ящурки разноцветной (Прика-
лаусские высоты, Центральное Предкавказье, май 1984 г., фото М. Ф. Тертышникова)
Рис. 68. Полоса выветривания в разнотравно-злаковой степи.Место обитания ящур-
ки разноцветной (Прикалаусские высоты, Центральное Предкавказье, май 1984 г.,,
фото М. Ф. Тертышникова)
Рис. 69. Песчаные степи на боровых террасах рек — типичные стации разноцветной
ящурки на Украине (Ивано-Рыбальчанский участок Черноморского заповедника,
Херсонская обл., июнь 1976 г., фото Т. И. Котенко)
Рис. 70. Пески морских побережий, островов и кос — места обитания разноцветной ящур-
ки на Черном и Азовском морях (о. Джарылгач. Черное море, июнь, 1978 г., фото
Т. И. Котенко)
Рис. 90. Сперматогенез у разноцветной ящурки на юго-востоке Узбеки-
стана. Препараты из семенников [Вашетко, сообщ.]: а — апрель, б*—
ноябрь
ют третью, а иногда и вторую пару нижнечелюстных у 76 % особей. Между предлобными
отсутствует дополнительный щиток у 83,34 %, имеется один у 16,66 %. Количество чешуй
вокруг 9—10-го кольца хвоста 21—29 (26,2±0,61) . Промежуток между рядами бедренных
пор укладывается в длине одного ряда 0,8—(1,0) 13 раза. Ряд бедренных пор не доходит
до сгиба колена у 100 %: на 2 чешуйки у 5,88 %, на 3 — 70,6, на 4 — 23,52 % особей. Имеют-
ся недоразвитые поры: 1 — 4,09, 2 — 36,37, 3 - 22,73 %. Рисунок: тип ”и” — 30,76 ”d” -
38,48 и ”d/u”— 30,76 %.
Распространение: Закавказье.
Ящурка разноцветная узбекская -
Eremias arguta uzbekistanica Cernov, 1934
Terra typica - Узбекистан, Гулистан (бывш. ст. Голодная Степь)
1934 - Eremias arguta uzbekistanica, Чернов, Докл. АН СССР,!!!, 8-9: 666. - 1974, Щербак, Ящурки
Палеарктики: 171.
Описание: Голотип: № 10093, кол. ЗИН РАН, Узбекистан, ст. Голодная Степь.
Среднеаз. жд., Г Якобсон, V, 1903, (и = 14). L = 34(60,8)96,6; Led = 38,5(78,0) 122:
L/Lcd = 0,74(0,78)0,88; G = 20-26(22,7±0,45) ; Sq = 41-62 (47,22±1,22) ; Ventr. = 29-
34(313±039), P.f = 9-3 (1136±0,2). По С.А.Чернову (294), Р/. = 7-16; Gmax = 33;
Lab = 8(9,8) 12. Три пары нижнечелюстных не соприкасаются (92,86 %) . Между предлоб-
ными нет дополнительного щитка у 78,58 %. Количество чешуй вокруг 9—10-го кольца
20—16(21,93±0,48) . Промежуток между рядами бедренных пор укладывается в длине од-
ного ряда 1,4—23 (М = 1 »73) раза. Ряд бедренных пор доходит до сгиба колен у 6,25 %, не
доходит на 1 чешуйку у 81,25 %, на 2 — 12,5. Недоразвитых бедренных пор (по одной) у
9,09 %. Рисунок: тип ”и” - 85,72, ”u/d” - 14,28 %.
Распространение: Узбекистан, пограничные районы Южного Казахстана, Кыр-
гызстана и Таджикистана.
Ящурка разноцветная восточная -
Eremias arguta potanini (Bedriaga, 1912)comb. nova Szczerbak, 1974.
Тепа typica — Джунгария, у гор Салбурты на юго-запад от Булун-Тохоя.
1912 — Eremias arguta var. potanini, Bedriaga, Rept. Przewalsk., Ill, 624.
1974 — Eremias arguta potanini, Щербак, Ящурки Палеарктики: 172; - 1986 Орлова. Тэрбиш, Матер,
по герпетофауне Джунгарской Гоби 103.
Кариотип разноцветной ящурки из Монголии внешне не отличается от описанных из дру-
гих частей ареала (см. выше) . Череп относительно высокий, массивный, отношение шири-
ны черепа к его длине (п = 12) составляет 0,42—0,49 (0,45 ±0,06) мм, отношение ширины
черепа к его высоте равно 1,29—1,65 (1,43±0.03) . Количество зубов на предчелюстной кос-
ти — 6—7, в нижней челюсти — 14—18/14—18. Имеются птеригоидные зубы.
Описание. Лектотип № 5165, кол. ЗИН РАН, Джунгария, пустыня у гор Салбурты,
1878 г., кол. Г.Н.Потанин. 9 L = 63,7 мм, Led =R; G = 28, Sq = 51, Ventr. 30,P.f =8/7 (и=36 экз.
из Семиречья, Джунгария и Кашгарии) . L = 283 (53,81)83,7 мм [по данным В.Ф.Орло-
вой, у особей из Монголии (п = 72) Lwax = 88 мм], Led = 42,0(64,5)77,7 мм (поданным
В.Ф.Орловой, у особей из Монголии Zctfmax = 83 мм) ,L/Lcd = 0,75 (0,85/1,04) (по данным
В.Ф.Орловой, 0,86—1,26), G = 22—32(25,31 ±0,35) [по данным В.Ф.Орлов ой, 26—34
(30,22±0,49)], Sq = 37-55(45,89±0,19) [по данным В.Ф.Орловой, 44-57(52,29±0,30)],
Ventr, = 28—32(29,64±6,15) [по данным В.Ф.Орловой, 28—33(31,07±0,12)], Pf = 8-
14(11,07±0,15) [по данным В.Ф.Орловой, 7-12 (9,75±0,13)], Lab = 8(10,5)12 [по дан-
ным В.Ф.Орловой, 9—15(11,53 ±0,21)]. Три пары нижнечелюстных соприкасаются у
91,66 %. Между предлобными отсутствует дополнительный щиток у 80,56 %. Вокруг 9—
10-го кольца хвоста 21—28 (243±032) чешуй [по данным В.Ф.Орловой, 25—34(28,99±
±0,22)]. Промежуток между рядами бедренных пор укладывается в длине одного ряда
1,3(1,81)2,2 раза [по данным В.Ф.Орловой, 0,9—2,25 раза].Слабо выраженные бедренные
поры имеются у 14,28—81,81 %. Ряд бедренных пор не доходит до коленного сгиба на 1 че-
шуйку у 56,43 %, на 2—35,89, на 3 — 2,56, на 4 — 2,56 %, доходит до сгиба колена 2,56 %
[по данным В.Ф.Орловой, у особей из Монголии бедренные поры не доходят до коленного
сгиба на 3—6 чешуй (?) ]. Рисунок: тип ”и” — 77,15, ”d” — 5/71, ”d/u” — 17,14 %.
По данным В.Ф.Орловой, у особей из Монголии окраска верхней стороны туловища
серая, нижняя белая, цветных глазков по бокам туловища нет; у некоторых самцов бока
головы и шеи (включая верхне- и нижнегубные щитки, височную область), бока туловища
и хвоста окрашены в сизо-голубой цвет. На шее, спине и в верхней части хвоста по основно-
33
му фону расположены крупные черные пятна неправильной формы, часто сливающиеся в по-
лосы, вытянутые в поперечном направлении. Отдельные темные пятнышки могут встречать-
ся сверху и по бокам головы. Ювенильные и неполовозрелые особи имеют рисунок, состоя-
щий из двух рядов белых глазков, отороченных черным, середина спины без рисунка. Та-
кой же рисунок сохраняется у 53,06 % самок; у других взрослых особей на крупных
черных пятнах различимы 1—2 маленьких белых глазка, иногда редуцированных (18,37%),
а доминируют особи с крупными черными пятнами. Такой тип рисунка (по нашей схеме
”р”) был характерен для серии, собранной Г Л.Потаниным в Джунгарии. Видимо, количе-
ство особей с таким рисунком увеличивается при движении от Семиречья на восток. Не-
которые расхождения данных по фолидозу (например, признак на сколько чешуи не до-
ходят бедренные поры до коленного сгиба и некоторые другие) могут свидетельствовать
о том, что на территории Кобдоского и Гоби-Алтайского аймаков Монголии обитает свое-
образная популяция, достойная пристального внимания и дальнейшего изучения.
Распространение: от Прибалхашья до Юго-Западной Монголии, Джунгарии и
Кашгарии (Северо-Западный Китай).
Ящурка разноцветная иссыккульская —
Eremias arguta darevskii Tsaruk, 1986
Тепа typiea - Пржевальск, Иссык-Кульская котловина, Кыргызстан.
1986 - Eremias arguta darevskii, Царук, Тр. ЗИН РАН, 157: 209.
Описание. Голотип — ЗИН РАН № 20018, Ad ЛТржевальск, Иссык-Кульская кот-
ловина Кыргызстана, 1906, кол. ДЛедашенко. L = 67 мм, Lcd(R) = 79 мм, Sq = 47, G =
= 30; Ventr, = 29, P.f. = 10/9. Между предлобными 2 дополнительных мелких щитка. Па-
ратипы (л = 51) ЗИН РАН № 10584, добыты там же, кол. тот же. L = 53—68 мм£/Led =
= 0,72—1,26 (0,91 ±0,04), Sq = 43-59 (51,0±0,5 8) , G = 24-35 (30,22±0,25) , P.f. = 8-12
(9,78±0,15). Три пары нижнечелюстных разделены, касаются у 5,56 %. Между предлоб-
ными дополнительные щитки имеются у 70,83 %, 1 — 56,25, 2 — 12,5, 3 — 2,0 %. Ряд бед-
ренных пор не доходит до коленного сгиба на 1 чешуйку у 7,41 %, на 2 — 44,44. на 3 —
44,44, на 4 - 3,71 %. Недоразвитые бедренные поры у 6,9 %. Рисунок: тип ”и”-
47,22 % особей, ”и/а” — 2,78 %, у остальных рисунок близок к типу ”и”, но темные
поперечные полосы с глазками не сливаются посередине спины — вдоль хребта остается
светлая продольная полоса из 3—6 рядов чешуи (такой тип рисунка встречается уже у
ювенильных особей).
Распространение: Иссык-Кульская котловина (побережье вокруг озера), на
западе проникает в долину р.Чу, доходя до Боомского ущелья и долины р. Чонг-Кемин
[324].
ГЛАВА 5
МОРФОЛОГИЯ
Представители рода Eremias. объекты нашего исследования, принадлежат к наиболее рас-
пространенной, процветающей группе современных рептилий, демонстрирующей значи-
тельные адаптационные возможности. Это и предопределило акценты в исследованиях
особенностей морфологии указанных рептилий: основное внимание было уделено опорно-
двигательному аппарату и нервной системе. Транспортные, пищеварительная и выдели-
тельная системы рассмотрены с меньшей степенью детализации.
5.1. СКЕЛЕТ
5ЛЛ. ЧЕРЕП
Вследствие специфики издания мы не можем привести здесь полное описание строения
черепа разноцветной ящурки. Поэтому последующий текст составлен так, чтобы служить
дополнением к достаточно детальным иллюстрациям (рис. 13—15): в нем приводятся
в первую очередь те сведения, которые трудно передать графически.
Детальное описание черепа Lacertidae, не потерявшее значения до настоящего време-
ни, было сделано Ф.Зибенроком [445], исследовавшим 18 видов этого семейства, в том
числе Eremias arguta и E.velox. В литературе можно найти общую характеристику черепа
лацертид [428], данные об онтогенезе [335. 336, 369. 422] и строении различных отделов
черепа и подъязычного аппарата [370, 420, 421, 435]. Наиболее полное описание различ-
ны! структур головы ящериц (на примере игуаниды Ctenosaura) было сделано Т.Элричем
[404] . Особенности черепного кинетизма у некоторых видов лацертид рассмотрены Т.фра-
зеттой [358] и НЛИорданским [101,102].
Невральный череп. Строение его у разноцветной яшурки в целом типично для лацер-
тид. Общая форма черепа Eremias отличается от таковой Lacerta относительно более узкой
мордой (в наибольшей степени у Е.gram mica) с несколько вздутыми участками позади
отверстий наружных ноздрей и перед глазницами. В результате морда оказываемая слегка
вогнутой дорсально. Межглазничное пространство очень узкое, а глазницы относительно
больше, чем у Lacerta (особенно у E.arguta) , и глазничная область черепа в целом расшире-
на (рис. 13, 14). Череп E.arguta в целом более низкий и плоский, чем у E^rammica и La-
certa.
Остеодермы на черепе ящурок хорошо развиты, особенно крупные и грубые у E.gram-
mica. У E.arguta они более тонкие и, прирастая к костям, не закрывают швов между ними.
Имеются крючковидная supraorbitale, 4 supraocularia и ряд мелких supraciliaria (рис. 13).
Остальные остеодермы, обычно удаляемые вместе с кожей, располагаются на восходящем
отростке скуловой и у дорсального конца барабанного гребня квадратной костей. Остео-
дермы нераздельно сливаются с наружной поверхностью носовых, лобной, теменной, за-
глазничных и предлобных костей (у E.grammica также на скуловых и верхнечелюстных
костях) .
Небо лацертид псевдоновохоанового типа [390]. Отверстия хоан и Якобсонова органа
кажутся разделенными костью (рис. 14), но над небными отростками верхнечелюстных
костей связаны щелевидным пространством. Верхние височные окна у всех лацертид ре-
дуцированы и оттеснены назад разрастанием заглазничных и заднелобных костей (рис. 13,
14). Межптеригоидное окно у всех лацертид очень узкое, тянется вперед как небный синус
(рис. 14) , разделяющий узкой щелью задние концы сошников.
35
f
a - над сбоку; б — осевой череп, вид сбоку (здесь и на рис. 14,15: ап — угловая кость,
ар - переднеуиаюе крыло, аг - сочленовная кость, bh - тело гиоида. Ьо - основная
затьиюадвя кость, bs - баэиофеноид, ch - хоакь, chc - cartilago hypochiasmatica, cl -
слуховой столбик (стремечко), со — затылочный мыщелок, ср — переднеушной гребень.
crb (2, 2 — жаберные рожки гиоида), crh - гиоидные рожки, ст - Меккелев хрящ,
ст - веиешяк кость, ct — crista ttrtseralis, d — зубная кость, ер - эпипгеригов^ ехс -
эксзракалумезхлзь/— лобиад кость, fad - аддукторная яма нюкней челюсти, jet — отвер-’
езж дай chorda tympaoi, fee — fenestra epioptica, fl - отверстие Якобсонова органа.
fl - слезное отверстие, fin — fenestra metoptica, fap - заднее височное окно, fns - верх-
нее височное окно, fb - fenestra optica, fov — овальное окно, fp — теменное отверстие.
fpl - небное окно, fpo - fenestra piootica, fsi - fenestra septalis mferior, fss — fenestra sep-
ta}» superior, /sz - fissura supratrabecularis, fvp - заднее отверстие Видиева канала, hph -
hypofeyale, in - вставочный хрящ» / - скуловая кость, I - слезная кость, т ~ верхнече-
люстная кость, л-носовая кость, от - орбигосфеновд, р - теменная кость, ра - восхо-
дящий отросток, poo - piia airtotica, pb — базиптеригоидный отросток, pl — небная
кость, рт - предчелюстная кость, ро — загдазнячная кость, pol — заднелобная кость,
pot - передне ушная кость, рр — околозатылочный отросток, рга - предсочленошвя
кость, prf - предлобная кость, рза - алярный отросток базисфеноида, psr - парасфено-
адный ростр ум; pt - гггеригоад, q - квадратная кость, го — tbcossjls oticus, san - надуг-
ловая кость, sb — subiculum infundibulum, яс — supraciliaria, si - межглазничная септа,
sm - септомакс илла, яо — верхняя затылочная кость, soc - supraocularia, sor — supraor-
bitalia, sp - крышечная кость, sq - чешуйчатая кость, ss — solum supraseptale, st — надви-
сочная кость, jv — spina vomcrina, tb — trabecula cranii, tm — taenia marginal», tpm — taenia
parietalis media, tr — зктоптериговд, ts — клинозатылочный бугорок, v - сошник; I-XII -
отверстия выхода соответствующих черепномозговых нервов
Парные предчелюстные, лобные и теменные кости срастаются в соответствующие не-
парные элементы. Кроме того, слиты друг с другом, образуя комплексные кости, базисфе-
нвоид с парасфеноидом; основная, боковые и верхняя затылочные и заднеушные; заглаэ-
ничная и заднелобная (у молодых ящурок между ними еще заметны швы); в нижней
челюсти — сочленовная и предсочленовная кости.
Задняя поверхность каждой боковой затылочной кости пронизана 3—4 отверстиями
(их число может варьировать на двух сторонах одного черепа). Дорсальное (более круп-
ное) из них служит для выхода из черепа X нерва, 2—3 вентральных отверстия — для вы-
хода корней XII нерва. Сбоку от отверстий поверхность кости вздута задним полукруж-
36
pm pm
Рис, 14, Череп разноцветной ящурки:
А - вид сверху, в левой глазнице удалены осеодермы, Б - вид снизу
шям каналом. Дорсальное располагаются направленные вбок околозатылочные отростки,
образование эадаеушными костями. Спереди околозатылотаый отросток слит с передне-
ушной костью.
От клинозатылочных бугорков, расположенных у боковых концов вентральной по-
верхности основной затылочной кости (рис. 14), вверх к основанию околозатылочных от-
ростков тянутся костные гребни (crista tuberalis), каждый из которых образует на своей
стороне черепа заднюю стенку воронковидного затылочного углубления (recessus occipita-
lis). В глубине последнего находится круглое окно слуховой капсулы. Овальное окно рас-
полагается дорсальнее затылочного углубления (рис. 13) и отделено от него crista interfe-
nestralis. Через круглое окно и далее по дорсомедиальной стенке затылочного углубления
выходит из черепа IX нерв. На боковой поверхности заднеушной кости (у основания око-
лозатылочного отростка) позади овального окна находится углубление, служащее для
прикрепления ceratohyale.
Базиптеригоидные отростки базисфеноида (рис. 13, 14) связаны суставами с птери-
гоидами. Передневерхние углы базисфеноида (алярные отростки) (рис. 13) у Eremias ко-
роче и шире, чем у Laceite. Передняя поверхность базисфеноида (dorsum sellae) между
алярными отростками пронизана тремя парами тонких отверстий, служащих для выхода
VI пары нервов, внутренней сонной артериии и небной ветви VII нерва и небной артерии.
Последняя пара отверстий, расположенная у основания базиптероидных отростков, ведет в
Видиевы каналы, тянущиеся сквозь отростки между базисфеноидом и приросшим к нему
парасфеноидом, открываясь задними отверстиями на задней стороне базиптеригоидных
отростков (рис. 13) . Боковая поверхность базисфеноида под алярным и над базиптериго-
идным отростками, впереди от заднего отверстия Видиева канала, образует желобок для
прохождения внутренней яремной вены.
Переднеушные кости имеют трехлучевую форму. Заднебоковой отросток прочным
швом связан с передней поверхностью околозатылочного отростка (см. выше). Передне-
вентральный отросток кости достигает вершины алярного отростка базисфеноида и несет
сбоку высокий переднеушной гребень, ограничивающий сверху желобок для внутренней
37
Рис. 15. Череп разноцветной ящурки:
а - над черева сзадм, б — «пеня челюсть (7 — сбоку, 2 - медиально), е - подъязычный аппарат
^имкм вша. В задней части этого желобка находится отверстие для выхода VII нерва.
Бежевая поверхность переднеушной кости воздута валиком над парасагиттальным верти-
кальным полукружным каналом. Впереди него кость несет широкий передний отросток —
переднеушное крыло, располагающееся сбоку от перепончатой стенки мозговой коробки.
В узком щепевидаом пространстве между ними начинаются мышцы — подниматель птери-
гонда и подниматель глазного яблока. Между переднеушным крылом и передневентраль-
ным отростком кости располагается широкая вырезка, разделенная выступающим вперед
гонким супратригемшгальным отростком на вентральную часть для выхода V нерва и дор-
сальную — для выхода медиальной мозговой вены.
Глзэиичио-иисочная часть стенок мозговой коробки у ящериц образована ажурной
хрящевой решеткой с болыпнади окнами, затянутыми перепонками (рис. 13). Ее основа-
ние поддерживается тонким костным парасфеновдным рострумом, дорсальнее которого
тянется пара трабекулярных хрящей, впереди сливающихся в интертрабекулу. Последняя
образует вентральный край хрящевой межглазничной септы, впереди пронизанной неболь-
шим нижним септальным окном. Верхнее септальное окно дорсально отделяет септу от
хрящевого solum supraseptale, расположенного под нисходящими флангами лобной кости
и образующего вместе с последними стенки переднего продолжения черепно-мозговой по-
лости (которое вмещает обонятельные тракты переднего мозга) .
Позади черепно-мозговая полость резко распшряется, здесь от solum supraseptale отхо-
дят два хрящевых тяжа. Дорсальный тяж (taenia marginalis) тянется под нисходящими
флангами лобной и теменной костей и по дорсальному краю переднеушной кости до верх -
Э8
ней затылочной кости. Вентральный тяж (taenia parietalis media) отделяет супрасептальное
окно от надглазничного и связан вентрально с заднедорсальным отростком межглазничной
септы и с переднедорсальным конном pila met optica. Последняя идет вниз — назад и у
взрослых ящурок почти целиком окостеневает как орбитосфеноид, имеющий форму изог-
нутого стержня с расширенным дорсальным концом. Pila metoptica и задний край межглаз-
ничной септы сзади и спереди ограничивают глазничное окно, большая часть которого за-
нята отверстием для II нерва. Вентрально это окно ограничено cartilago hypochiasmatica,
задний конец которого отогнут вниз как короткий тупой отросток — subiculum infundi-
bulum. От последнего начинаются прямые мышцы глаза.
Между cartilago hypochiasmatica и трабекулярным хрящом сзади вдается узкая супра-
трабекулярная вырезка. Pila metoptica отделяет глазничное окно от большого заднеглаз-
ничного окна, в переднедорсальном углу которого выходит IV. а примерно в середине —
Ш нерв. Дорсально это окно ограничено хрящевой pila antotica. сзади — передним краем
базифеноида, а вентрально — трабекулярным хрящом. Над pila antonica находится передне-
ушное окно, которое спереди отделено от надглазничного окна pila accessoria, а дорсально
ограничено taenia marginalis. Некоторые хрящи глазнично-височной стенки мозговой ко-
робки обызвествляются (у взрослых ящурок — дорсальная часть межглазничной септы
и taenia parietalis media).
Переднечелюстная кость (рис. 13, 14) у ящурок имеет узкий межносовой отросток,
у Еarguta — очеш> длинный. Он отделяет передние половины носовых костей друг от дру-
га. достигая у отдельный особей переднего медиального отростка лобной кости. Вентраль-
но позади зубного гребня имеется пара резцовых отростков, вклинивающихся спереди в
верхнечелюстные кости. Между этими отростками на середине заднего края предчелюстной
кости находится небольшой полумесяцеобразный гребень, выступающий вентрально
(spina praemaxillaris). Сзади в него упираются узкие передние концы сошников.
Боковая поверхность верхнечелюстной кости пронизана шестью тонкими отверстиями
для выхода кожных веточек верхнего альвеолярного нерва и верхнечелюстной артерии.
Отверстие основных стволов кровеносного сосуда и нерва находится медиальнее гребня,
ограничивающего задний край ноздри (т.е. в ее полости). Шов верхнечелюстной кости со
скуловой у Eremias не имеет зубца, характерного для Lacerta. Сошники (рис. 13) имеют
характерные для лацертид особенности: сзади разделены узким небным синусом, вдоль
которого посылают назад узкие отростки; передние концы вытянуты и сужены, заканчива-
ясь у сочленения с предчелюстной костью небольшим вентральным отростком (сошни-
ковой остью). Медиально и позади от наружных отверстий Якобсонова органа вентраль-
ная поверхность сошника приподнята в виде острого гребня, ограничивающего глубокий
желобок для небной ветви VII нерва, которая выходит через отверстие на медиальной сто-
роне этого гребня. Сбоку от него в узкой щели над небным отростком верхнечелюстной
кости тянется слезный желобок,
На дорсальной поверхности максиллярного отростка небной кости вблизи шва со ску-
ловой костью (рис. 14) расположено отверстие для верхнего альвеолярного нерва (перед-
нее продолжение верхнечелюстной ветви тройничного нервд) и верхнечелюстной артерш.
Небный отросток птеригоида вытянут вдоль медиального края небной кости, с кото-
рым связан прочным швом. Поперечный (боковой) отросток птеригоида вклинен в край
эк то птеригоида. Задний конец птеригоида связан с медиальной поверхностью вентрального
конца квадратной кости очень рыхлым синдесмозом, допускающим значительную взаим-
ную подвижность этих костей. У основания заднего (квадратного) отростка птеригоида
на его дорсальной поверхности находится маленькое углубление, вмещающее вентральный
конец эпиптеригоида. Эпиптеригоиды — тонкие костные столбики, связывающие птери-
гоиды с переднеушными костями (рис. 13), являясь подвижными распорками черепного
механизма.
Эктоптеригоиды имеют широкий контакт с медиальной поверхностью верхнечелюст-
ных костей, впереди достигая контакта с небными костями (рис. 14). Квадратные кости
(рис. 13—15) у Eremias отличаются от таковых Lacerta прямоугольным (а не закруглен-
ным) , резко выступающим дорсальным концом барабанного гребня, образующего лате-
ральный край кости. Дорсальный конец квадратной кости образует выпуклую вверх пло-
щадку, медиальная часть которой оттянута назад и закруглена, образуя так называемый
головчатый мыщелок. Последний служит для связи квадратной кости с чешуйчатой и над-
височной, а также (через посредство вставочного хряща) с околозатылочным отростком.
39
Все связи квадратной кости с прилежащими костями - рыхлые синдесмозы, допускающие
возможность движения.
Септомаксиллы образуют крышу полости для Якобсонова органа и расположены в
носовой капсуле у межносовой септы. Снаружи кости видны в глубине наружных носовых
отверстий (рис. 14). Лобная кость глубоко заходит под носовые, у некоторых особей
может полностью разделять последние по средней линии, достигая предчелюстной. Нисхо-
дящие фланги лобной кости впереди вытянуты в нисходящие отростки, тянупдаеся вдоль
заднемедиальных краев предлобных костей (рис. 13). Между нисходящим отростком лоб-
ной кости и межглазжчной септой располагается глазнично-носовое отверстие, затя-
нутое хрящом, за исключением небольшого канала для глазничной ветви тройнич-
ного нерва.
Предлобные кости состоят из поверхностной и нисходящей (глазнюшой) частей. У ос-
нования последней (иногда между предлобной и лобной костями) имеется отверстие, че-
рез которое веточка глазничной ветви тройничного нерва выходит из носовой полости в
глаэшщу. Сбоку исходящая часть предлобной кости образует медиальный край слезного
отверстия, латералы&хм краем которого является прилежащий участок небольшой слез-
ной кости (рис. 14),
Скуловые кости Eremias отличаются от таковых Lacerta отсутствием заднего отростка
и формой шва с верхнечелюстными костями (см. выше). Узкий восходящий отросток об-
разует основную часть заглазничной пряжки (рис. 13),
Заднелобные кости у взрослых ящурок срастаются с заглазничными, хотя у молодых
особей (с длиной черепа до 15 мм) следы шва еще хорошо заметны. Заднелобные кости у
всех лацертид разрастаются назад в виде прямоугольных пластин, перекрывающих боль-
шую часть верхних височных окон. Заглаэничная кость имеет форму узкого треугольника
с длинным задним отростком, сзади прилежащим к медиальной стороне переднего угла
чешуйчатой кости. Последняя у всех лацертид представлена маленьким плоским элемен-
том, образующим латеральный край верхнего височного окна.
Надвисочшае кости у всех лацертид — маленькие костные клинья между теменной и
чешуйчатыми костяади, околозатылочными отростками и вставочными хрящами
(рис.13—15).
Задняя часть теменной кости у Eremias, отделенная от основной ее пластины парой
невысоких поперечных гребней, скошена к затылку. В середине заднего края теменной
кости расположено небольшое отверстие, ведущее в канал, который тянется в теле кости
кпереди по средней линшз к заканчивается слепо. В этот канал входит хрящевой восходя-
щий отросток, связанный с дорсальным краем верхней затылочной кости. Вблизи передне-
го края теменной кости находится относительно большое теменное отверстие. По бокам
нейтральной поверхности теменной кости тянутся невысокие костные валики, ограниад-
ваюоде дорсолатеральио задавав часть черепно-мозговой полости. Веятралшо ат перепон-
ками связаны с taeniae marginate.
Нвкш wshdctk Ветви нижней челюсти впереда связаны гибким симфизом, образо-
наадым свиховдрозом Меккелевых хрящей и рыхлой соещетгтелъной тканью. Задние кон-
цы ветвей нижней челюсти, образованные компнексюймм костями (сочленовше 4 предсо-
чдеиовные), несут короткие засочленовиые отростки (рис. 15) с сильно вогнутой дорсаль-
ной поверхностью, пронизанной тонкжи отверстием для веточки падевого нерва chorda
tympani. Последняя проходит сквозь комплексную кость в аддукторную яму нижней че-
люсти, выходя туда через отверстие вентральнее челюстного сустава. Сочленовная ямка
челюстного сустава обращена назад и дорсально. Передняя часть комплексной кости обра-
зует медиальную стенку аддукторной ямы и примордиального канала нижней челюсти,
по дау которых стержневидньш Меккелев хряш тянется от сочленовной кости до нижне-
челюстного симфиза.
Надугловые кости образуют латеральную стенку аддукторной ямы нижней челюсти.
У переднего и заднего концов они пронизаны отверстиями для кожных веточек нижнече-
люстной ветви тройничного нерва. Угловые кости образуют дао задней части приморди-
ального канала нижней челюсти. На медиальной поверхности кости имеется отверстие
(рис. 15) для ramus myiohyoideus posterior нижнечелюстной ветви тройничного нерва.
Ежовая поверхность зубной кости несет 6—7 отверстий для кожных веточек того
же нервного ствола, тянущегося в примордиальном канале внутри зубной кости, который
разделен на дорсальную часть — для нерва, и вентральную — для Меккелева хряща. Меди-
40
альную стенку примордиального канала образует крышечная кость (рис. 15, б), прони-
занная двумя отверстиями: большим дорсальным — для лингвальной веточки нижнечелю-
стной ветви тройничного нерва, и меньшим вентральным — для ramus mylohyoideus anterior
последней. У E.grammica вентральное отверстие смещено значительно вперед относительно
дорсального, у E^rguta они располагаются одно над другим.
Венечные кости (рис. 15) имеют дорсальный (венечный) и 3 вентральных отростка,
латеральный, передне- и заднемедиальные. От вершины венечного отростка вниз тянутся
3 гребня: латеральный, задний и медиальный (последний продолжается по заднемедиаль-
ному отростку кости, ограничивая на заднемедиальной поверхности ее желобообразное
углубление).
Зубы. По внутреннему краю зубных (альвеолярных) гребней предчелюстной, верхне-
челюстных и зубных костей укреплены плевродонтные зубы (рис. 13,15) , На предчелюст-
ной кости 7 зубов (срединный зуб непарный) . В области шва между пред- и верхнечелюст-
ной костями зубные ряды этих костей разделены небольшим промежутком. Передние зу-
бы заостренные, задние несколько притупленные, некоторые с неясно обозначенными
двумя вершинами (задняя из них — большая) . На вентральной поверхности тела птеригои-
да находится 3—6 неправильно расположенных маленьких и тупых небных зубов. Зубная
Форму» аа*г£Ят.т/>(о)*.<(о) ♦ <з-о х 2.
о (1о—19)
Подъязьгавдй аппарат (рис. 15). У Eremias хрящевой. Небольшое непарное тело гиоипа
(basihyale) продолжается кпереди длинным тонким клиновидным hypohyale. Заднелате-
рально от тела гиоида отходят 3 пары длинных и тонких отростков, или рожек гиоида.
Дистальные концы передних гиоидных рожек (ceratohyalia) срастаются с заднеушными
костями, входя в небольшие углубления у основания околозатылочных отростков поза-
ди овального окна. Две задние пары отростков — жаберные рожки (ceratobranchialia 1 и
2) — не связаны с черепом и располагаются в толще подъязычных мышц.
Кииетнзм черепа у Eremias сходен с таковым Lacerta [101]. Особенностями черепного
кинетизма лацертид являются: относительно малая подвижность основной части мак сил-
лярного сегмента, в которой метакинетическая гибкость преобладает над слабо выражен-
ной мезокинетической; череп фирмипалатинарный — небные отделы представляют собой
жесткие стержни, не изгибающиеся поперечно; связь квадратных костей с птеригоидами
очень рыхлая, в связи с этим квадратные кости обладают высокой подвижностью (гипер-
стрептостилия), поворачиваясь на своих дорсальных связях (с околозатылочными отро-
сткаьш, надвисочными и чешуйчатыми костями; см. выше). Вместе с квад ратными костя-
ке! смещается нижняя челюсть. При поворотах квадратных костей вперед (протракция)
нижняя челюсть двигается вперед относительно верхней, при поворотах назад (ретрак-
ция) — возвращается в исходное положение. Такое продольное движение нижней челюсти
было отмечено на кинокадрах поедания пищи быстрой ящуркой E.velox. ЭЮЗоробьева
и TJ0 .Чугунова полагают [59], что это движение используется ящурками для первичной
переработки (’’перекатывания”) пищи между зубами верхней и нижней челюстей. Сход-
ство строежя и кинетизма черепа у разных видов ящурок позволяет предполагать, что и
разноцветная ящурка аналогично использует указанную подвижность квадратно-мандибу-
лярной системы.
5.12. ОСЕВОЙ СКЕЛЕТ
Осевой скелет у Eremias arguta разделяется на отделы: шейный, первый позвонок которо-
го подвижно сочленен с черепом; туловищный, или пояснично-грудной, передние позвон-
ки которого соединены с ребрами, образующими грудную клетку; крестцовый, позвон-
ки которого сочленяются с костями таза; хвостовой, состоящий из ряда позвонков,
упрощающихся по форме в каудальном направлении хвоста (рис. 16, А —Е).
Шейный отдел (рис. 16, Б) содержит 8 позвонков, на контрлатеральных поверхно-
стях которых имеются в разной степени развитые поперечные отростки (processus trans-
versus s.diapophysis) (рис. 16,Ж, 3), к которым у каудальных 5 позвонков подсоединяют-
ся шейные ребра. Последние имеют разную форму. Поперечные отростки контрлатераль-
ных поверхностей IV—VIII позвонков меняют свою ориентацию, т.е. расположение самой
их латеральной части по отношению к телу позвонка. Так, если у IV позвонка овально-
продолговатый латеральный край отростка идет от дорсальной к вентральной поверхности
41
42
ж з
Рис. 16. Осевой скелет разнодоетной ящурки:
А — cofainma vertebraiis, Б — vertebrae cervicates, В — vertebrae thoracicae, Г - vertebrae lumbalis, Д - ver-
tebrae aerates, E — vertebrae crudaies, Ж — atlas, краниально, 3 - каудально, И - epistropheus, кранналь-
но, К - каудально, Л — латерально; 1 - processus tzansversus, 2 - arcus neuralis. 3 - processus spinosus,
4 — processus articularis (zygapophysis) anteriors, cranialis, 5 — processus articularis (zygapophysis) posterior
s. caudalis, 6 — hypapophysis, 7 - processus odontoideus, 8 - costae,9 -arcus haemaiis
и в каудокраниальном направлении, а у VII позвонка он расположен вертикально, то у
УШ уже наблюдается его поворот в краниокаудальное направление.
Все гпетные позвожи (рис. 16, Ж, 3) имеют хорошо развитые невральные дуги (ar-
ena neuiatis). Проксимально невральные дуги переходят в остистые отростки (processus
spinosus). На крашо и каудолатеральных краях невральных дуг расположены сочленов*
иые отростки (zygapophysis). Краниально расположены передние сочленовные отростки
(processus articularis anterior), сочленовные поверхности которых расположены прокси-
мально. Каудально расположены задние сочленовные отростки (processus articularis poste-
rior), сочленовные поверхности которых расположены вентрально. У каждого из шейных
позвонков с вентральной поверхности имеется нижний остистый отросток (hypapophysis).
Форма этих отростков варьирует. Проксимальные остистые отростки у 6 каудальных шей-
ных позвонков имеют сходное строение, а их угол наклона к продольной оси позво-
ночника слегка увеличивается. Первые, краниально расположенные, шейные позвонки,
именуемые атлантом и эпистрофеем (atlas et epistropheus), отличаются от каудально распо-
ложенных .
Атлант (atlas, рис. 16,Ж) представляет собой подобие кольца неправильной формы,
с несколькими отростками по периметру. Проксимо-каудально на теле невральных дуг
позвонка расположены 2 задних сочленовных отростка, которые соединяются с соответ-
ствующими передними сочленовными отростками эпистрофея. Краниолатеральнее от них
расположены дополнительные отростки. Дорсальная часть невральной дуги позвонка рас-
ширена и уплощена и по средней линии почти не срастается. На контрлатеральных поверх-
ностях, около середины тела позвонка, имеются поперечные отростки. На вентральной
поверхности тела позвонка имеется мощный нижний отросток, который дистальным кон-
цом направлен чуть каудовентрально,
Дистальная половина тела позвонка имеет утолщенные стенки, а сам он, по внутрен-
нему периметру, разделен поперечным плоским сухожильным тяжем на две неравноцен-
ные части. Проксимальная — большая, овальная и служит для прохождения ствола спин-
ного мозга, а нижняя — меньшая, имеет форму полукольца и служит для соединения за-
тылочной кости (os occipitale) с затылочным мыщелком (condylus occipitalis). Поперечная
43
и
Окончание рис. 16
сухожильная перемычка ат-
ланта каудаиьио срастается
с дорсокзудальным краем
краниального ”зубя” эпи-
строфея, что, однако, не ме-
шает его полувращательным
движениям вокруг продоль-
ной оси ”зуба”. Кравиально
атлант по всему периметру
и через посредство обшир-
ного плоского апоневроза
соединяется с каудальной
поверхностью затылочных
костей, вокруг большого
затылочного отверстия (for-
amen occipitale magnus) и за-
тылочного мыщелка.
Эпистрофей (epistrophe-
us, рте. 16, 5) имеет также
сложное внешнее строе-
ние. Тело позвонка вытя-
нуто в дррсовентральном
направлении и чуть расши-
рено в контрлатеральные
стороны. Невральные дуги
44
проксимально срастаются между собой, образуя расширенный в краниокаудальном
направлении остистый отросток* На латерокраниальной поверхности невральных
дут расположены передние сочленовные отростки. Чуть проксимальнее от них рас-
полагаются задние сочленовные отростки. Дистальная половина позвонка в дорсальной
части (средняя линия позвонка) вытянута в краниокаудальном направлении. Краниально
на позвонке расположен сочленовный отросток (”зуб” — processus odontoideus), к которо-
му дорсокаудально прирастает поперечная сухожильная перегородка первого шейного
позвонка.
Краниальный сочленовный отросток позвонка дорсально почти плоский, а вентраль-
но имеет поверхность конуса, которая конгруирует с каудальной поверхностью дистально-
го отдела атланта и с желобом на дорсокаудальной поверхности затылочного мыщелка.
Каудально на теле позвонка имеется сочленовная головка в виде почти полусферы, частич-
но обособленной от тела позвонка. Дистальнее передних сочленовных отростков на лате-
ральных поверхностях тела позвонка располагаются поперечные отростки.
На самой вентральной точке поверхности тела позвонка расположен нижний остистый
отросток, уплошенный в контрлатеральных направлениях с овальными краями, направлен
вейтрокаудально по отношению к телу позвонка. На дистокраниальной половине тела
позвонка видны швы, по которым между собой срастаются краниальный сочленовный от-
росток и средняя часть тела позвонка. Остальные 6 шейных позвонков имеют сходную внеш-
нюю структуру (рис. 16, Б), На дистокраниальных поверхностях тел III—VIII шейных
позвонков заметны полушаровидные углубления, которые служат для конгруэнтного
сочленения с полусферными головками, расположенными на каудальных поверхно-
стях II—VIII шейных позвонков. Лишь у III и IV шейных позвонков имеются довольно
выраженные лопатовидные остистые отростки, уплощенные в латеральном направлении
и расположенные в вертикальной плоскости.
Туловищный отдел у разноцветной ящурки состоит из 17 позвонков (рис. 16, В, Г),
практически все несут на своих контрлатеральных поверхностях ребра. Этот отдел можно
разделить на грудной и поясничный. Непосредственно на грудине закрепляются 4-5 груд-
ных ребер (всего их 9) . Остальные 8 позвонков являются поясничными. Ребра на пояснич-
ных позвонках более чем в 2 раза короче грудных и расположены почти все в горизон-
тальной плоскости; они постепенно уменьшаются линейно от краниального до каудаль-
ного позвонка. На дорсальной поверхности всех туловищных позвонков имеются хорошо
выраженные остистые отростки, направленные проксимокаудально под острым углом.
Нижних остистых отростков нет. На контрлатеральных поверхностях всех туловищных
позвонков имеются хорошо развитые передние и задние сочленовные отростки. Дистокра-
ниальнее от передних сочленовных отростков на контрлатеральных поверхностях всех ту-
ловищных позвонков расположены поперечные отростки, сориентированные в проксимо-
каудальном направлении и имеющие эллипсоидную форму. К этим поперечным отросткам
и крепятся все ребра. Проксимальные расширенные концы ребер фиксируются на туло-
вищных позвонках 2 короткими сухожильными тяжами — нейтральнее и дорсальнее от
поперечных отростков. Дистальнее места соединения краниальных и каудальных сочленован-
ных отростков позвонков имеются контрлатеральные овальные окна, через которые выхо-
дят дорсальные и вентральные корешки спинного мозга. Невральные дуги у всех позвон-
ков хорошо развиты и формируют сквозной канал для спинного мозга.
Крестцовый отдел у разноцветной ящурки состоит из 2, аналогичных туловищным,
позвонков (рис. 16, Д) , которые, однако, имеют хорошо развитые поперечные отростки,
За счет ложных суставов последние соединяются с подвздошными костями (по их меди-
альным поверхностям). Латеральные концы поперечных отростков сближены. Краниаль-
ная пара поперечных отростков слабо уплощена в краниокаудальном направлении и рас-
положена почти в горизонтальной плоскости, а каудальная — в дорсовентральном и на-
правлена латеровентрально. Проксимальнее от поперечных отростков на невральных ду-
гах имеются передние и задние сочленовные отростки. На вентральных поверхностях
обоих позвонков отсутствуют нижние остистые отростки.
Хвостовой отдел позвоночника состоит из 45—48 внешне различно устроенных позвон-
ков (рис. 16, £) . В каудальном направлении позвонки уменьшаются в размерах и меняют
форму. У всех хвостовых позвонков имеются проксимальные остистые отростки, распо-
ложенные под острым углом к продольной оси позвоночника, в проксимокаудальном
45
направлении, степень выраженности которых уменьшается к каудальному концу хвоста.
Наиболее типично устроены первые 3 хвостовых позвонка, на вентральной поверхности
которых отсутствуют нижние остистые отростки. Поперечные отростки направлены в
контрлатеральные стороны, у I — чуть каудально, VIII — чуть краниально. Начиная с IV
хвостового позвонка на протяжении следующих 20 на вентральных поверхностях имеют-
ся хорошо выраженные гемальные дуги (arcus haemalis, рис. 16,2?) , образующие в прокси-
мальной части гемальный канал. На остальных каудальных позвонках эти дуги переходят
в короткие хрящевые отростки и у самых последних исчезают совсем.
Первые 9 хвостовых позвонков имеют хорошо выраженные, латерально направленные
и уменьшающиеся линейно, в каудальном направлении хвоста, поперечные отростки, ко-
торые представляют собой сплошные треугольные пластинки, расположенные по средней
линии позвонка и в промежутке между его сочленовными отростками. Тела этих позвон-
ков ттельные и не имеют следов срединного поперечного разделения. Следующие за ними
позвонки уже имеют видимое срединно-поперечное разделение. Их поперечные отростки
уменьшаются линейно в каудальном направлении, вплоть до полного исчезновения. По
срединно-поперечной линии разделения позвонка происходит отламывание хвоста ящурки
при аутотомии.
На контрлатеральных поверхностях хвоста также имеются межпозвоночные окна,
через которые выходят корешки хвостовой части нервного ствола. Краниальная часть по-
звонков имеет явно выраженный процельный тип строения. У каудальной части позвон-
ков процельность постепенно сглаживается. т.е. исчезают краниальная ямка и каудальная
головка, а сочленение их происходит по вертикальной плоскости.
5 Л 3. СКЕЛЕТ КОНЕЧНОСТЕЙ
Пояс грудной конс<шости. Пояс каждой из грудных конечностей состоит из двух основных
частей — лопаточной и коракоидной. Кроме этого, в него входят также грудина, надгрудин-
ник и ключица.
Лопатка (scapula, рис. 17, А, Б) состоит из двух элементов — костного и хрящевого.
Костная часть лопатки занимает ее дистальную половину и располагается почти вертикаль-
но, имеет расширенный проксимальный и суженный дистальный концы. Дистально кост-
ный элемент сочленяется с коракоидом, образуя латеровентрально суставную поверхность.
Котор» и входит в состав гленоидной ямки. К последней, с латеральной стороны, причле-
няется плечевая кость. К верхнему расширенному концу костной части лопатки, по види-
мой границе раздела, прирастает уплощенно-веерообразный надлопаточный хрящ (carti-
lage suprascapuhris). Дистокраниальный край лопатки и латеральный край коракоида
расположены под острым углом и соединены между собой обширным плоским апоневро-
зом. От латерокаудалыюго края лопатки отходит лентовидное сухожилие, которое закреп-
ляется на дорсокраниальной поверхности головки плечевой кости.
Коракоид (coracoid, рис. 17,Л, Б) составляет вентральную часть плечевого пояса, име-
ет форму неправильного овала, частично вытянутого в каудальном направлении. На теле
коракоида, краниолатерально от суставной ямки, расположено обширное овальное окно,
Латерокаудальный край коракоида утолщается, срастается с дистальным концом костного
элемента лопатки и способствует формированию нижней половины гленоидной ямки
(fossa glenoidales). Краниомедиально коракоид представлен хрящевой пластинкой. От кау-
дального края коракоида отходит лентовидное сухожилие, которое закрепляется на вен-
тральной поверхности границы между суставной головкой и дельтовидным бугром плече-
вой кости, напротив дорсального закрепления лентовидного сухожилия от костного эле-
мента лопатки.
Ключица (clavicula, рис.17, В, Г) расположена краниолатерально от надгрудинника и
коракоида. В ней выделяются более мощная краниолатеральная часть в виде овального
стержня и более слабая — каудальная в виде костной серповидной пластинки. Они сраще-
ны, а промежуток между ними заполняет овальное окно. Медиальным краем ключица сра-
стается с краниальным краем латеральной ветви надгрудинника и контрлатеральной клю-
чицей. Латеральная часть ключицы срастается с проксимокраниальным краем костного эле-
мента лопатки, т.е. в том месте, где он срастается с надлопаточным хрящем.
Надгрудинник (epistemum, рис. 17, В, Г) имеет крестообразную форму. Его передний
отросток простирается краниально до места сращения контрлатеральных ключиц, с их внут-
46
Piu. 17. Грудная конечность разноцветной ящурки:
А — пояс грудной конечности, латерально; Б - медиально; 1 — scapula, 2 — carulago suprascapularis,
3 — coracoid, 4 — fossa glenoidalis, 5 - clavicula, 6 - episternum;В — sternum, дорсально (7); Г - вентраль-
но; Д — скелет грудной конечности, краниально; Е каудально: 8 — os humerus, 9 — ulna, 10 — radius,
11 — radiale, 12 — ulnare, 13 — centrale. 14 - os pisiforme. 15 - carpalia distalia (2-5), 16 - metacarpus,
17 — digiti — phalanx proximalis (a), phalanx distalis (5)
ренней (дорсальной) поверхности, а задний переходит на вентральную поверхность груди-
ны, срастается с ней по средней линии и почти достигает ее 2 овальных окон. Латеральные
ветви надгрудинника слабо сращены с латерокаудальным краем костной пластинки ключи-
цы, в краниальном направлении соединяются с ней апоневротически.
Грудина (sternum, рис. 17, В, Г) имеет ромбовидно-уплошенную форму, в дистальной
части — 2 овальных окна, расположенных почти симметрично по отношению к средней ли-
нии. Краниолатеральные края грудины сочленяются (через посредство удлиненных лож-
ных суставов) с каудомедиальными краями контрлатеральных коракоидов. На каудолате-
ральных краях грудины фиксируются дистальные части (хрящи) истинных грудных ребер.
У разноцветной ящурки их 5, но есть экземпляры и с 6 ребрами. На отдельных хрящевых
стержнях фиксируется I—IV и отдельно V и VI ребра. Между краниолатеральными краями
грудины (вентрально) и каудальными краями поперечных отростков надгрудинника име-
ется обширный апоневроз.
Скелет грудной конечности состоит из плеча, предплечья и кисти. Плечо состоит из од-
ной кости, предплечье, из двух, а кисть, в свою очередь, делится на запястье, пястье и фа-
ланги пальпев, которые также имеют в составе ряд костей.
Плечевая кость (os humerus s, humeri, рис. 17, Д. Е) трубчатая, удлиненная, имеет рас-
ширенные проксимальный и дистальный концы. На кости имеются вентрокраниальный
и дорсокаудальный бугры, на первом из которых закрепляются грудная и дельтовидная
мышцы, а на втором — подкоракоидно-лопаточный мускул. Дистальный конец кости име-
ет две суставные поверхности на радиальном и ульнарном мышелках, входящие в кон-
такт с суставными поверхностями проксимальных концов костей предплечья и формирую-
щие локтевой сустав. Дорсолатерально, в промежутке между мыщелками кости, имеется
47
небольшая поверхность, по которой скользит сесамовидная косточка, типа os patella на
тазовой конечности, находящаяся в конечном сухожилии трехглавой мышцы плеча.
Локтевая кость (ulna, рис. 17, Д, Е) более крупная, чем лучевая, расположена латеро-
каудально в области предплечья. На каудомедиальной поверхности проксимального конца
кости располагается суставная поверхность в четверть сферы, соприкасающаяся с сустав-
ной поверхностью ульнарного мыщелка плечевой кости. На проксимальном конце этой
же кости закрепляется конечное сухожилие трехглавного мускула плеча. Дистальный ко-
нец кости имеет суставную поверхность для сочленения с проксимальным концом ульнар-
ной косточки запястья. К дистокаудальному концу кости апоневротически присоединяет-
ся гороховидная косточка (os pisiforme s. os carpi accessorium, рис. 17, Д, E) . От дистально-
го конца кости отходит лентовидный сухожильный тяж, который закрепляется прок симо-
латерально на радиальной косточке запястья.
Лучевая кость (radius, рис. 17, Д, Е) имеет конусообразно расширенные проксималь-
ный и дистальный концы. На дорсальной поверхности проксимального конца кости име-
ется эллипсоидное углубление, которое контактирует с суставным блоком, расположен-
ным на радиальном мыщелке плечевой кости. С локтевой костью проксимально соедине-
на подвижно. Дистальный конец кости с двумя буграми, с помощью которых она связа-
на с радиальной косточкой запястья. С дистальным концом локтевой кости лучевая кость
непосредственно не соприкасается.
Запястье (carpus, рис. 17, Д, Е) у разноцветной ящурки состоит из двух рядов косто-
48
8
Л9
IZ
16
Окончание рис, 17
10
11
чек различных размеров и формы. В проксимальном ряду расположены 3 косточки — с
медиальной стороны относительно крупная радиальная косточка (radiale), уплощенная в
дорсовентральном направлении; с латеральной — также крупная ульнарная косточка
(ulnare), внешне сложно устроенная. Между ними расположена небольшая треугольная
косточка — центральная (centrale). Промежуточная косточка (intermedium) у ящурки не
обнаружена. В дистальном ряду запястья расположено 5 мелких косточек различной фор-
№i (carpalia distalia). Дистальными поверхностями эти косточки граничат с проксималь-
ными концами 1—5 пястных костей. С дорсальной поверхности запястья видно все 5 кос*
точек, с пальмарной — 4. Первая заплюсневая (дистальная) косточка не выходит на паль-
марную поверхность кости.
Пястье (metacarpus, рис. 17, Д, Е) у ящурки представлено 5 трубчатыми костями раз-
ной длины. Их проксимальные и дистальные концы немного расширены и имеют сустав-
ные поверхности, которыми они проксимально граничат с дистальным рядом косточек
запястья, а дистально — с проксимальными фалангами соответственно каждого пальца.
В промежутках между пястными костями расположены сухожильные лентовидные тяжи,
которые начинаются на проксимолатеральных поверхностях пястных костей, а закреп-
ляются на их дистомедиальных поверхностях и удерживают эти кости от смещения в лате-
ральную сторону в горизонтальной плоскости.
Пальцы (digiti, рис. 17, Д, Е) . На передней конечности ящурки 5 вполне развитых паль-
цев, самые длинные — третий и четвертый. Фаланги пальцев, кроме когтевых, имеют труб-
чатое строение. Проксимальные и дистальные их концы расширены и образуют суставные
поверхности. Количество фаланг (phalanges digitorum) в пальцах различно - в первом - 2,
во втором — 3, в третьем — 4, в четвертом - 5 ив пятом — 3. Когтевые фланги на всех
пальцах серповидно изогнуты в пальмарную сторону кисти.
49
Пояс тазовой конечности. Каждая из контрлатеральных половин тазового пояса состо-
ит из трех костей: подвздошной, лобковой и седалищной, которые участвуют в формиро-
вании вертлужной ямки (acetabulum, рис. 18, Б, В), с которой сочленяется суставная го-
ловка бедренной кости.
Подвздошная кость (os ilium, рис. 18,Б, В) вытянутая, средняя часть уплощена латеро-
медиально, каудальная — с медиальной поверхности овальная. Своей каудальной частью
подвздошная кость соединяется (в виде ложного сустав) с латеральной поверхностью
мощных поперечных отростков 2 крестцовых позвонков. Дистокраниально подвздошная
кость немного расширяется и срастается с соответствующими участками лобковой и седа-
лищной костей, формируя вертлужную ямку. У дистального конца кости и проксималь-
нее от суставной ямки имеется небольшой отросток. Лобковая кость (os pubis s. pubicum,
рис. 18, Б. В) расположена в краниальной части таза, идет косо от суставной ямки в кра-
ниомедиальном направлении. Краниальным концом она срастается с контрлатеральной
50
костью с помощью надлобкового хряща (cartilago epipubis, рис. 18, А -В) , а каудальным —
с днстокраниальным концом подвздошной, проксимолатеральным — с седалищной, фор-
мируя при этом суставную ямку.
Вентрокаудальная часть кости имеет вид пластинки, на которой сформирован предлоб-
ковый отросток (processus praepubis, рис. 18, А— Е) . На дорсолатеральной поверхности кау-
дального конца кости имеется небольшое запирательное отверстие (foramen obturatum,
рис. 18, А—Е) . От каудального края надлобкового хряща отходит хрящевидный шнур, ко-
51
торый срастается с краниовентральными выступами контрлатеральных седалищных костей
(каудальный край шнура имеет вид пики) и достигает их симфиза. Седалищная кость
(os ischium, рис. 18, А—Е) расположена вентро медиально по отношению к лобковой и под-
вздошной костям, имеет вид костной пластинки, внутри которой просвечивается вееро-
образное жилкование. Вентрокраниальный край вытянут краниально и срастается, совме-
стно с контрлатеральной костью, с каудальным концом хрящевого шнура, идущего от кау-
дального конца надлобкового хряща.
Дистовентрально между контрлатеральными костями имеется хрящевидная прослой-
ка, простирающаяся на месте седалищного симфиза. Каудально хрящевая прослойка пере-
ходит в Т-образный вырост (hypoischium, рте. 18, А-Е), от которого берет свое начало
подседалищный мускул (m. hypoischiadicus) . Проксимолатерально кость срастается с дру-
гими костями таза и формирует вертлужную ямку. Парными лобковыми и седалищными
костями сфоршровано, в пределах таза, сердцевидной формы лобковоседалищное окно
(foramen puboischiadicum).
Скелет тазовой конечности разноцветной ящурки состоит из трех отделов: бедра,
голени и стопы (рте. 18, Г, Д). В состав голени входят два костных элемента, а стопа со-
стоит из заплюеш, плюсны и фаланг пальцев. Бедренная кость (os femoris) трубчатая, с
расширенными проксималышми и дистальными концами, из которых первый формирует
овальную суставную головку для сочленения с суставной ямкой таза, а второй — 2 мы-
щелка с суставныьш поверхностями для сочленения с таранной костью. На вентральной
поверхности проксимального конца кости расположен гребень (большой вертлуг — tro-
chanter major) , на который крепятся мышцы таза. На дорсальной поверхности дистального
конца бедренной кости располагается коленная чашечка (patella)которая срослась с ко-
неадым сухожилием дорсальной группы мышц бедра. Между дистальными суставными
поверхностями бедренной кости и проксимальными костей голени расположен хряшевой
мениск (meniscus).
Большая берцовая кость (tibia) расположена на голени краниомедиально, проксимо-
латеральио гранитах с проксимальным концом малой берцовой кости, но не срастается
с ней. Дистально она с малоберцовой костью не соприкасается, между их дистальными
концат проходагт лентовидный сухожильный тяж. Малая берцовая кость (fibula) имеет
слабо расширенный проксимальный и силыю — дистальный концы, которые формируют
суставные поверхности: прокошально — для бедренной кости, дистально — для таранной.
Заплюсна (tarsus) разноцветной ящурки состоит из трех различной формы косточек: та-
ранной, заплюсневой (днеталыюй) третьей и дистальных заплюсневых четвертой и пятой
(сросшиеся).
Таранная кость (talus, рис. 18, Г, Д) ътшютлыею крупная, занимает полностью про-
ксимальный ряд эаплюевд, проксимально граничит через суставные поверхности с дисталь-
ными концами костей гопеж, дистально — с дорсальными суставными поверхностями 1-й
и 2-й плюсневых костей, 3-й (tarsale distale 3, рис. 18,13, Г, Д), 4—5-й (tarsale distale 4—5,
рис. 18,15, Г, Д) зашиосневых костей с дорсомедиальной поверхностью проксимального
конца 5-й плюсневой кости. На ее краниальной поверхности заметая в разной степени вы-
раженные швы. Это свидетельствует о том, что тело кости состоит из сросшихся централь-
ной, промежуточной, тибиальной и фнбулярной косточек заплюсны. Плюсна (metatarsus,
рис. 18, Г, Д) cQjzppxzn 5 трубчатых костей, разной степени удлиненности; 5-я плюсневая
кость силыю видоизменена (за счет срастания с 5-й заплюсневой костью) .
Проксимальные и дистальные концы плюсневых костей расширены и образуют соот-
ветствующие суставные поверхности для сочленения с заплюсной и проксимальными кон-
цами фаланг. Самая длинная из них 4-я, самая короткая и сложно устроенная — 5-я. На
плюсне, в промежутках между ее костями, имеются лентовидные сухожильные тяжи. Они
не позволяют плюсневым костям расходиться в стороны и укрепляют стопу.
Пяльцы (digiti, рис. 18,7”, Д). На тазовой конечности ящурки, как и на грудной, имеет-
ся 5 пальцев разной длины и ориентации, содержащих разное количество фаланг. Самый
длинный 4-й палеп, самый короткий — 5-и. Все фаланги, кроме когтевых, имеют трубчатое
строение и в разной степени удлинены. Проксимально и дистально фаланги расширены и
образуют суставные поверхности. Количество фаланг в пальцах распределяется следующим
образом: в 1-м — 2, во 2-м — 3,в 3-м — 4,в 4-м — 5, в 5-м — 4, тх. в 5-м пальце на одну фа-
лангу больше, чем на передней конечности. Когтевые фаланги на всех пальцах также сер-
повидно изогнуты в плантарную сторону стопы.
52
5 2. МУСКУЛАТУРА
Ящерицы. ввиду многообразия группы и наличия у них большого числа двигательных
адаптации, являются по существу единственными пресмыкающимися, по сравнительно-
анатомическим данным локомоторного аппарата которых можно с большой достоверно-
стью судить об уровне развития и специализации локомоции у рептилий в целом. В свя-
зи с этим справедливо утверждение ВЪ.Суханова [245. — С. 92], что ”ни черепахи, ни
крокодилы с их узкой защитной или амфибиотической специализацией, ни тем более
змеи не могут дать правильной картины локомоции этой, некогда столь обширной, груп-
пы животных”. Сравнительная анатомия костно-мышечной системы ящериц давно при-
влекала внимание исследователей: систематиков, обосновывающих филогенетические свя-
зи, и функциональных морфологов, анализирующих форму и функцию мышц и мышечных
комплексов, обусловленных спецификой локомоций.
Ранние работы по сравнительной анатомии костно-мышечной системы некоторых ви-
дов рептилий выполнены за рубежом. Были предприняты попытки выяснения гомологии
мышечной системы в ряду позвоночных, изучения эмбрионального развития мускулату-
ры, исследования этапов эволюции опорно-двигательного аппарата [345, 346, 360-362.
365, 366, 379, 400, 405, 410-412, 419. 423-429, 430, 433, 434, 446,447]. Своего расцвета
эта отрасль эволюционной морфологии достигла в 20—30-х годах нашего века, дав ряд
более или менее успешных реконструкции локомоторного аппарата вымерших живот-
ных, включая и его мускулатуру [245, 372, 423, 459]. В настоящее время усилия морфо-
логов направлены на создание общей картины структурно-функциональной эволюции
локомоторного аппарата как в целом, так и в отдельных деталях [152,243-245.325,341,
371,373, 374, 417, 418]. Лишь в нескольких работах [152,243,244] приведены фрагмен-
тарные данные по сравнительной анатомии костно-мышечной системы разноцветной ящур-
ки. Вот почему мы и предприняли попытку хоть частично восполнить этот пробел. Задачей
настоящей работы и было сравнительно-анатомическое и функциональное изучение кост-
но-мышечной системы у разноцветной яшурки (Eremias arguta deserti) с точки зрения при-
способления ее к наземной локомоции.
Материалом для наших исследований послужили морфологические сборы ящурки из
ее естественного местообитания (Херсонская обл.) . Материал фиксирован в 5 %-ном фор-
малине (животные предварительно усыплялись эфиром). Основной методикой в исследо-
вании была макропрепаровка под увеличением (бинокуляр МБС-9). Изучали общую архи-
тектонику посткраниальных мышечных групп и каждой мышцы в отдельности; их прокси-
мальные и дистальные места фиксации; соотношение с близлежащими мышцами; функ-
цию каждой мышцы.
При наименовании костей и мускулов мы придерживались (в основном) номенклату-
ры, применяемой в ’Практикуме зоотомии позвоночных. Земноводные и пресмыкающие-
ся” [69] и отчасти в работе Н£ергер-Дель’моура [342].
5 2.Ь МЫШЦЫ ШЕИ
Подкожный мускул шеи (m.subcutaneus colli, рис. 19, Л) прикрывает m.constrictor colli и
m.cervicomandibularis, которые мышечно срастаются. Мускул начинается расширенным
пластинчатым апоневрозом на дорсолатеральной поверхности мышц шеи. сразу же кау-
дальнее от костей головы и на протяжении трех сухожильных перехватов на дорсальной
поверхности этих мышц. До остистых отростков первых шейных позвонков апоневроз не
достигает, так как они прикрыты дорсальными мышцами шеи. но срастается с проксималь-
ным апоневрозом контрлатеральной мышцы. Мышечное брюшко огибает шею латераль-
но и переходит на ее вентральную поверхность, где заканчивается также обширным апо-
неврозом. При функционировании сжимает шею в вертикальном направлении.
Сжиматель шеи (m.constrictor colli, рис. 19, Л) лентовидный, расположен на латераль-
ной поверхности шеи, прикрыт почти полностью предыдущим мускулом, частично прикры-
вает проксимокраниальную часть m. epistemocleidomastoideus и краниально мышечно сра-
стается с каудальным краем m. cervicomandibularis. Начинается мышпа обширным апонев-
розом на дорсолатеральной поверхности мышц шеи, проксимокраниальнее от краниально-
го угла надлопаточного хряща. Проксимальный апоневроз мышцы достигает уровня дор-
53
A - латеральной — дорсально,В - вентрально; - m. subcutaneus соЩ, 2 — т,constrictor colli, 5-m.cer
nalis, 7 - m. longissimus dorsi, 8 — m. iliocostalis, 9 - m. obliquus abdominis extemus superfidalis, 10 - m. tran
laris, 14 - m. triceps brachii, 15 - m. brachialis, 16 - m. extensor antebrachii radialis, 7 7 - m. extensor digito
22 — m. flexor tibialis extemus, 23 - m. extensor digitorum longus, 24 — m. peroneus longus, 25 - m^astrocne
nis lateralis, 29 - m.biceps brachii, 30 - m.flexor carpi ulnaris,57 - m.sternohyoideus, 32 - m.pecroralis, 33 - m.
chiocaudalis, 31 — m. flexor carpi radia
54
Рис» 19, Поверхностные мышиы тела разноцветной ящурки:
vicomandibuiaris sjn. depressor mandibularis, 4 - m. trapezius, 5 - m. latissimus dorsi. 6 - m. transversospi-
sversospinalis caudae, 11 - m. iliocauaalis, 12 - m. intertransversarius caudae, 13 - m. deltoideus pars scapu-
rum communis, 18 - m. extensor carpi ulnaris, 19 — m. ilio-tibialis, 20 - m. ambiens, 21 - m. iliofibularis,
mius extemus, 26 — m. epistemo-cleidomastoideus, 27 — m. deltoideus pars clavicularis, 28 - m. rectus abdomi-
rectus abdomini ventralis s. medialis, 34 — m. hypoischiadicus, 35 — m. transversus cloacae cranialis, 36 - m. is-
lis, 38 - m. puboischio-tibiaiis. 39 - m. gastrocnemias internus, 40 - m. omohyoideus, 41 - m. tibialis anterior
55
сальной поверхности остистых отро-
стков шейных позвонков, где сраста-
ется с апоневрозом коллатеральной
мышцы. При функционировании спо-
собствует подкожному мускулу шеи и
прижимает глотку снизу.
Шейно-челюстной мускул (m. cer-
vicomandibularis, рис. 19,А) треуголь-
ный, расположен на дорсолатеральной
поверхности краниальной части шеи,
непосредственно за ушным отверсти-
ем, частично прикрыт m. subcutaneus
colli и каудально мышечно срастается
с m. constrictor colli. Сам мускул при-
крывает проксимальную часть m.epi-
stemocleidomastoideus. Начинается мы-
шечно-сухожильно от дорсолатераль-
ной поверхности чешуйчатой и темен-
ной костей, а также от затылочной
фасции. Мышечные пучки брюшка ве-
ером сходятся к каудальному отрост-
ку нижней челюсти (mandibula), где
предварительно закрепляются на удли-
ненном сухожильном тяже. При син-
хронной работе контрлатеральных
мышц нижняя челюсть опускается ди-
стально.
Лопаточно-подъязычный мускул
(m. omohyoideus, рис. 19, В) располо-
жен на латеровентральной поверхно-
сти шеи, прикрывает расположенный
от него дорсомедиально грудинно-подъ-
язычный мускул (m. sternohyoideus),
а сам латерально частично прикрыт
надгрудинно -ключично -сосцевидным
мускулом (m. epistemocieidomastoide-
us) и подкожным мускулом шеи. На-
чинается от латеральной части ключи-
цы и частично от поверхности лате-
рального отростка надгрудинника. За-
крепляется на кауд©вентральной по-
верхности жаберного рожка, на его
2/3 медиальной части, вплоть до сред-
ней линии тела. При функционирова-
нии ретрагирует подъязычный аппарат,
одновременно приподнимая его.
Грудинно-подъязычный мускул
(m. sternohyoideus, рис. 19, В) распо-
ложен на вентральной поверхности
шеи, дорсальнее от лопаточно-подъ-
язычного. Медиальным краем эта
мышца проходит по средней линии
тела, прикрывая частично с вентральной поверхности трахею. Начинается мышечно-сухо-
жильно на вентрокраниальной поверхности латерального отростка надгрудинника и час-
тично от сухожильного апоневроза, соединяющего краниальный край этого отростка и
каудальный край ключицы. Закрепляется на каудальной поверхности медиальных
2/3 длины жаберного рожка. При функционировании ретрагирует подъязычный ап-
парат.
Остистый мускул шеи (m. spinalis cervicis, рис. 21, Л) слабо выражен, состоит из 3-4
56
Рис. 20. Глубокий слой мышц тела разноизетной ящурки:
А - дорсально, Б - латерально, В - вентрально; 1 - m. minaus
capitis, 2 — m. longissimus capitis, 3 — m. obiiquus capitis rnagnug,
4 — m. levator scapulae, 5 — m. serratus ventralis, 6 — m. serratus
dorsalis, 7 - m. obiiquus abdominis extemus profundus, 8 - m. inter-
costales externi brevis, 9 - m. intercostales externi longus, 10 - m. in-
tercostales intemi, 11 - m. scaiares intemi, 12 - m. obiiquus abdomi-
nis intemus, 13 — m. costocoracoideus, 14 — m pyramidalis profundus
sjn. rectus abdominis intemus, 15 — m. protractor penis, 16 - m. re-
tractor cloacae, 1 7 — m. retractor penis
5мм
*—i—L—I——J 8 9 6 3
Б
сросшихся между собой веретеновидных мышечных пучков, расположен на дорсальной
поверхности шеи и дорсально прикрыт остистым мускулом головы. Начинается мышеч-
но-сухожильно на дорсолатеральной поверхности остистых отростков II—III шейных
позвонков. Мышечные пучки направляются каудально, закрепляются на краниодорсальной
поверхности остистых отростков V-VIII шейных позвонков. Мышца является как бы са-
мым краниальным продолжением поперечно-остистого мускула туловища. Мускул осу-
ществляет дорсальное сгибание шеи и поднятие головы вверх.
Остистый мускул головы (m. spinalis capitis, рис.20, А) расположен на дорсальной
поверхности шеи, латеральнее от средней линии трла животного, лентовидно вытянут в
краниокаудальном направлении, в средней области брюшка имеет сухожильный перехват.
Латерально мышечное брюшко срастается с длиннейшим мускулом головы (m. longissimus
capitis), с контрлатеральной мышцей — только апоневротически. Начинается мышечно-су-
хожильно на небольшой каудальной поверхности теменной и верхней затылочной костей.
Вентрально мускул прикрывает задний прямой мускул головы (m. rectus capitis posterior)
и остистый мускул шеи. Кроме этого имеются тонкие сухожильные анастомозы к дорсаль-
ной поверхности апоневроза, покрывающего поперечно-остистый мускул (m. transversospi-
nalis) и длиннейший мускул спины (т. longissimus dorsi). Конечные сухожилия мышцы
срастаются также с вентрально расположенными конечными сухожилиями остистого
мускула шеи. При функционировании мускул поднимает голову вверх.
Длиннейший мускул головы (m. longissimus capitis, рис. 20, А) относительно мощный,
расположен на дорсовентральной поверхности шеи, латеральнее от остистого мускула голо-
вы, с которым срастается по всей длине медиальной поверхности. Вентрально мышечное
брюшко граничит с большим косым мускулом головы (m. obliguus capitis magnus) и нераз-
57
Окончание рис, 20
с ним срастается, а также с задним прямым мускулом головы (m. rectus capitis pos-
terior), но с последним не срастается. Начинается обширно мышечно-сухожильно от кау-
дального края теменной кости (or parietale), Закрепляется на латеральной поверхности
краниального конца длиннейшего мускула спины. При функционировании осуществляет
боковые повороты головы и в большей степени поднятие ее вверх.
Большой косой мускул головы (m. obliguus capitis magnus, рис. 20, Ь, дорсально гра-
ничит и полностью срастается с длиннейшим мускулом головы. Вентрально частично грани-
58
Рис, 21, Глубокий слой мышц области шеи. туловища и хвоста разноцветной ящурки:
А - дорсально, Б - латерально, В - вентрально; 1 - m. spinalis cervicis, 2 - т> rectus capitis posterior
3 — m, obligatus atlantis, 4 — m. obiiquus capitis inferior, 5 * m. longus colli lateralis, 6 ~ m, costocervicalis,
7 - m, rectus capitis anterior, 8 - m. longus colli medialis, 9 — m. transversus dorsalis, 10 - m. transversus ab-
dominis, 11 — m. quadratus lumborum, 12 — m. obiiquus abdominis intemus, 13 - m. caudifemoralis longus,
14 - m. caudi-femoralis brevis
59
чит с нижним косым мускулом головы (m. obliguus capitis inferior) и латеральным длин*
ным мускулом шеи (m. longus colli lateralis) . Начинается мышечно-сухожильно на каудаль-
ной поверхности боковой затылочной кости и частично на самом дистокаудальном конце
отростка теменной кости. Закрепляется брюшко на латеральной поверхности косого сухо-
жильного перехвата на краниальном конце длиннейшего мускула спины. При функциони-
ровании мускул поворачивает голову латерально, а при совместном действии с контрла-
теральной мышцей — фиксирует голову в горизонтальной плоскости.
Задний прямой мускул головы (m. rectus capitis posterior, рис. 21, А) уплощенно-тре-
угольный, расположен кпереди от краниального края остистого мускула шеи. Ни с одним
из окружающих его мускулов не срастается. Дорсально прикрыт длиннейшим мускулом
головы, вентрально граничит с большим косым мускулом головы и частично прикрывает
косой мускул атланта (m. obliguus atlantis). Начинается мышечно-сухожильно от дорсокау-
дальной поверхности боковой и верхнезатылочных костей (exoccipitale et supraoccipitale)
и направляется в каудомедиальном направлении. Закрепляется на латеральных поверхно-
стях остистых отростков П—Ш шейных позвонков. При функционировании поворачивает
голову латерально, а при действии с коллатеральной мышцей поднимает ее вверх.
Косой мускул атланта (m. obliguus atlantis, рис. 21, А) слабый, уплощенный в дорсовен-
тральном направлении, расположен на дорсальной поверхности невральных дуг II—III шей-
ных позвонков, дорсально прикрыт задним прямым мускулом головы, с которым не сра-
стается. Мышечные пучки закрепляются на невральных дугах и на дистальных частях ости-
стых отростков II—Ш шейных позвонков. При функционировании поворачивают атлант
вокруг продольной оси в дорсальную сторону туловиша и частично в горизонтальной плос-
кости.
Нижний косой мускул головы (m. obliguus capitis inferior, рис. 21, Б) относительно
мощный, слабо веерообразный, расположен нейтральнее от большого косого мускула го-
ловы, с которым частично срастается дорсокаудальным краем. Вентрально брюшко грани-
чат! с латеральным длинным мускулом шеи (in. longus colli lateralis) и вентрокаудальным
краем также частично с ним срастается. Краниовентрально брюшко частично граничит с ла-
теральной головкой поднимателя лопатки (m. levator scapulae) и с самым краниальным
концом переднего прямого мускула головы (т. rectus capitis anterior). Начинается на кау-
дальной поверхности основного затылочного отростка черепа, каудальнее от закрепления
на нем переднего прямого мускула головы. Каудальным конном брюшко небольшим
количеством мышечных пучков выходит на латеродорсальную поверхность краниального
конца длиннейшего мускула спины (m. longissimus dorsi) и с ним срастается. Мускул пово-
рачивает голову около продольной оси туловища.
йебершнвейвые мускулы (m. costocervicales, рис. 21, Б) расположены между двумя
последними ребрами и между последним шейным и I грудным. Мышечные пушси идут ко-
со в дистокаудальном направлении. При функционировании способствуют подтягиванию
I грудного ребра в краниальном направлении.
Лкгерелышй длинный мускул шеи (m. longus соШ lateralis, рис. 21,5) расположен на
латеровеятральной поверхности шеи, проксимокраниальнее от дорсального зубчатого мус-
кула (m. serratus dorsalis) и дистокаудально от нижнего косого мускуле головы. Начинает-
ся коротким сухожилием на латеральной поверхности поперечного отростка атланта. За-
крепляется мышца на краниолатеральной поверхности поперечных сухожильных миосепт
шеи. Дорсокаудалыю брюшко снова переходит в тонкое лентовидное сухохопше, кото-
рым дополнительно фиксируется на апоневрозе, покрывающем краниальный край длин-
нейшего мускула спины. При функционировании мускул изгибает латерально шею.
Передний прямой мускул головы (m. rectus capitis anterior, рис. 21,5) мощный, верете-
новидньхй, расположен на вентральной поверхности шейных позвонков, латеральнее от их
вентральных остистых отростков. На протяжении краниальных 2/3 длины мышечного
брюшка имеется внутренний сухожильный тяж, к которому прикрепляются мышечные пуч-
ки. Начинается как от вентральной поверхности тел всех шейных позвонков, так и от
латеровентральной поверхности их вентральных остистых (хрящевых) отростков. Внут-
ренний сухожильный тяж выходит на вентрокраниальную поверхность брюшка, которым
оно и закрепляется на вентрокаудальной поверхности вентрального (контрлатерального)
отростка черепа, расположенного дистокраниальнее от затылочного мыщелка на основной
затылочной кости. Мускул опускает голову и сгибает шею в вертикальной плоскости.
Медиальнълй дтанш^й мускул шеи (m. longus colli medialis, рис. 21, В) слабо выражен-
ньж. С вентральной поверхности прикрыт передним прямым мускулом головы. Начинает-
60
ся на поперечном отростке II шейного позвонка, закрепляется этот мускул на вентролате-
ральной поверхности 6 каудальных шейных позвонков, на вентральной поверхности их
поперечных отростков и оснований соответствующих ребер. Мускул сгибает шею вентро-
латерально.
522. МЫШЦЫ ТУЛОВИЩА
Поперечно-остистый мускул (m.transversospinalis, рис. 19, А, Б) расположен на дорсальной
поверхности позвоночника, латеральнее от остистых отростков позвонков и на дорсальной
поверхности их сочленовных отростков. Мускул на всем протяжении покрыт апоневро-
зом, благодаря чему видна граница раздела между ним и длиннейшим мускулом спины.
Краниально мускул простирается вплоть до 3-го шейного позвонка, каудально — до по-
следнего хвостового. Мышечное брюшко состоит из отдельных лепестков, из которых бо-
лее мощные расположены дорсолатерально, а более слабые — медиально на брюшке. Пер-
вые мышечные лепестки многоперистые, начинаются удлиненными лентовидными сухо-
жильными тяжами на проксимокаудальной поверхности остистых отростков позвонков,
с Ш шейного до предпоследних хвостовых. Через полтора межреберных промежутка на-
чальное сухожилие лепестка переходит в мышечный пласт, прикрывает дорсально кау-
дальную часть краниально расположенного мышечного лепестка и с ним срастается. Началь-
ное сухожилие заходит внутрь лепестка, поэтому мышечные пучки образуют двуперистую
структуру. Вентральная часть удлиненных мышечных пучков как бы частично огибает с ла-
теральной поверхности краниально расположенный мышечный лепесток и закрепляется на
дорсокраниальной поверхности сочленовных отростков позвонков (в краниальной части
позвоночника на II, в каудальной — на III) . Каждый мышечный лепесток простирается на
протяжении V (краниально) или VI (каудально) туловищных позвонков. Дорсолатераль-
ная часть мышечных лепестков переходит на границе с длиннейшим мускулом спины в ко-
роткое поверхностное сухожилие, которое срастается с проксимомедиальным краем соот-
ветствующих сухожильных миосепт. От этих сухожилий берут начало дорсомедиальные
лентовидные сухожильные тяжи мышечных лепестков длиннейшего мускула спины. Меж-
ду каудальным краем каждого остистого отростка и краниолатеральным краем невраль-
ной дуги следующего позвонка располагается уплощенный в коллатеральных направлени-
ях мышечный пласт, который также частично закрепляется и на медиальной поверхности
краниального конца короткого сухожилия мышцы. Этот мышечный пласт, по-видимому,
в какой-то степени соответствует межостистой мышце млекопитающих (m. interspinales) .
При функционировании мускул сгибает позвоночник в вентральном направлении и подни-
мает шею и хвост.
Длиннейший мускул спины (m. longissimus dorsi, рис. 19, А, Б) расположен на дор-
сальной поверхности спины, латеральнее от поперечно-остистого мускула, но грани-
чит с ним через обширный апоневроз, которым он покрыт дорсолатеромедиально. Лате-
рально он граничит с подвздошно-реберным мускулом. Мышца простирается от каудаль-
ной области подвздошной кости до первых 2 шейных позвонков, где краниальным краем
срастается с каудальными концами дорсолатеральных мышц шеи. Каждый мышечный ле-
песток краниально имеет лентовидное сухожильное начало: от дорсального апоневроза
поперечно-остистого мускула., от конечных сухожилий его дорсолатеральных мышечных
лепестков, а также от дорсомедиального конца каждой сухожильной миосепты подвздош-
но-реберного мускула. Самый каудальный конец данной мышцы срастается с краниаль-
ным концом подвздошно-хвостового мускула, который служит как бы его каудальным
продолжением. Данный мускул вызывает латеральное и дорсальное сгибание позвоноч-
ника.
Подвздошнореберный мускул (m. iliocostalis,pHC. 19, Л, Б) расположен на дорсальной
поверхности проксимальных концов шейных и грудино-поясничных ребер, медиально
граничит с латеральным краем длиннейшего мускула спины, латерально — с наружной ко-
сой мышцей, а вентрально прикрывает дорсальное продолжение наружных межреберных
мышц. Четкой границы раздела между мышечными пучками подвздошно-реберного мус-
кула и наружных межреберных мышц нет, и их продольное направление плавно переходит
от дистокраниального у первых к каудолатеральному у вторых. Мышца простирается от
подвздошной кости, где начинается почти по всей длине вплоть до I шейного позвонка;
здесь закрепляется на каудальной поверхности поперечного отростка. Этот мускул разде-
лен поперечными сухожильными миосептами на 24—25 частей. Все поперечные сухожиль
ные миосепты его срастаются с дорсальной поверхностью ребер и расположены косо, соот-
ветственно каудолатеральному ходу последних. При функционировании мускул оттяги-
вает ребра в дорсокаудальном направлении.
Прямая мыпща живота (m. rectus abdominis) в краниальной половине отчетливо делит-
ся на две части: медиальную (m. rectus abdominis ventralis s. medialis) и парные латеральные
(m. rectus abdominis lateralis) H.OЛанге [152] выделяет и третью часть — глубокую (т.
rectus adbominis intemus). НЛТуртовой и др. [69] рассматривают эту третью часть мыш-
цы как глубокий пирамидальный мускул (m. pyramidalis profundus), а МРомер [429]
и Н.Бергер-Дель’моур [341] — как прямой треугольный мускул (m. rectus triangularis).
М, rectus abdominis ventralis s. medialis (рис. 19, В) обширная, удлиненная, простирает-
ся от вентрокаудальной поверхности дистальных концов последних грудных ребер и от
каудального конца самой грудины до вентральной поверхности таза, где переходит в
короткое расширенное сухожилие, которым и закрепляется на ligamentum pubo-ischiadi-
cum. Самым каудальным концом мышца закрепляется и на вентральной поверхности се-
далищной кости, латерокаудальнее от начала хряшевидного отростка (hypoischium). Ла-
теральными краями мышечное брюшко граничит с прямыми латеральными мышцами жи-
вота и срастается с ними (а также срастается с дистальным краем наружной глубокой
косой мышцы живота). При функционировании мышца поднимает живот и оттягивает
конечные грудные ребра в каудальном направлении против действия грудной мыпщы.
М. rectus abdominis lateralis (рис. 19,Л,В) лентовидная,уплощена,расположенанавен-
тролатеральной поверхности брюшка ящурки, медиальным краем (каудальная половина
брюшка) срастается полностью с латеральным краем прямой медиальной мышцы живота.
Начинается удлиненным апоневрозом на вентрокаудальной поверхности латерального от-
ростка надгрудинника, где сращена с конечным сухожилием надгрудинно-ключично-сос-
цевидного мускула. Латеромедиально апоневроз имеет дополнительное начало и на вен-
тральной поверхности грудной мышцы (m. pectoralis). Почти по всему латеральному краю
брюшко срастается с дистальным концом наружной поверхностной косой мыпщы тулови-
ща (m. obiiquus abdominis extemus superficialis). Вентральной поверхностью брюшко при-
крывает дистальный конец наружной глубокой косой мышцы туловища (m.obiiquus ab-
dominis extemus profundus). Закрепляется мышца совместно с медиальной прямой мышцей
живота на вентромедиальной поверхности лобковоседалищной связки. Мышца способ-
ствует средней прямой мышце живота в поддержании внутренностей тела с вентролате-
ральной поверхности.
Глубокий дарамидальный мускул (m. pyramidalis profundus, или, по Н.О Ланге [152],
т. rectus abdominis intemus, рис. 20, В) уплощенно-треугольный, расположен вентрокрани-
ально от таза, вентрально прикрыт каудальной частью прямой медиальной мышцы живота.
Начинается прямолинейными мышечными пупсами (латеральные более длинные, чем ме-
диальные) на вентральном конце предлобкового отростка (processus praepubis) и на вен-
трокрантальной поверхности всей длины лобковоседалищной связки (ligamentum pubo-
iachiadicum). Мышечные пучки брюпжа направляются краниомедиально к средней линии
туловища, где закрепляются на медиальном ашшевротяческом тяже, который продолжа-
ется краниально, и срастаются с контрлятеральными пучкаэет. При фушашонировашш мус-
кул, совместно с другими, укрепляет стенку живота.
Наружная косая мышца (m. obiiquus abdominis extemus) расположена латеровентрально
на туловище и свободно разделяется на 2 самостоятельных мышечных слоя, соединяющих-
ся нежной соединительной тканью, — поверхностный (superficialis) и глубокий (profundus).
М. obiiquus abdominis extemus superficialis (рис. 19, А —В) состоит из 10 лентовидных
N&imewx полосок, соединенных апоневротически. Мускул своими лепестками наедна-
ется на латеральной поверхности последнего шейного и 9 последующих туловищных ре-
бер, а также на дистальных концах соответствующих inscriptiones tendineae подвздошно-
реберного мускула, т.е. у самой дистальной границы. По направлению к хвосту, тх. на про-
тяжении 7 сухожильных перегородок подвздошно-реберного мускула, мышечные полоски
исчезают, переходя в сплошной обширный апоневроз, который латерально покрывает кау-
дальную часть мышечных полосок глубокой порции наружного косого мускула.
М.obiiquus abdominis extemus profundus (рис. 20, Б, В) состоит из 10 мышечных удли-
ненных лепестков, расположенных на латеральной поверхности туловища, прикрытых ла-
терально поверхностной наружной косой мышцей, направленных косо, как и мышечные
лепестки предыдущей мышцы. Первые 2 мышечных лепестка начинаются на латеральной
поверхности последних 2 шейных ребер, проксимальнее и дистальнее от начала на них
62 !
вентрального зубчатого мускула (m. serratus ventralis). Остальные 8 мышечных лепестков
начинаются от латеральной поверхности 5 стернальных и следующих 3 туловищных ребер,
непосредственно у дистального конца подвздошно-реберного мускула. Все лепестки за-
крепляются по латеровентральному краю медиальной прямой мышцы живота, вплоть до
ее каудального, конца, а первые 2 лепестка закрепляются на вентрокраниальной поверхно-
сти дистальных (хрящевых) концов IV—V стернальных ребер. Обе части наружной косой
мышцы поддерживают брюшную стенку тела.
Наружные межреберные мышцы (mm. intercostales externi. рис. 20,2>) расположены на
латеральной поверхности туловища от шейных ребер на протяжении 10 туловищных ребер.
Однако если в пределах последних 4 туловищных ребер расположены четко выраженные
мышечные пучки коротких наружных межреберных мышц (mm. intercostales externibre-
vis), то в области стернальных ребер (и с заходом на шейные) расположены мышечные пуч-
ки длинных наружных межреберных мышц (mm. intercostales externi longus), под котота
ми расположен тонкий слой коротких межкостных мышц. Короткие мышцы располжс
ны в пределах 2 соседних ребер. Длинные мышечные пучки соединяют 3—4 стернальных
ребра. У дистальных концов каудальных 4 туловищных ребер пучки коротких межребер-
ных мышц располагаются полого, т.е. ближе к горизонтальной плоскости. От каудальной
поверхности IV туловищного ребра отходит относительно мощный лентовидный мышеч-
ный пласт (ramus abdominalis mm. intercostales externi), который направляется дистокау*,
дально и закрепляется совместно с брюшной частью наружного косого мускула на пред-
лобковом отростке.
Внутренние межреберные мышцы (mm. intercostales intemi, рис. 20,Б, 2?) расположены
на латеральной поверхности туловища, в промежутках между ребрами, соединяя их внут-
ренние поверхности в косом направлении (противоположно направлению мышечных пуч-
ков наружных межреберных мышц) . У дистальных концов ребер мышечные пучки распо-
лагаются более полого, т.е. ближе к горизонтальной плоскости. Мышечные пучки ниже
дистальных концов 4 туловищных ребер как бы сливаются с внутренней косой мышцей
(m. obliquus abdominis internus). В предкрестцовой области имеется плоский мышечный
треугольный пласт, который тянется от дистальных концов первых 3 поясничных ребер
и от соответствующих сухожильных перемычек подвздошно-реберного мускула и закреп-
ляется на каудальной поверхности последнего туловищного ребра. Его закрепление при-
крыто началом брюшной ветви наружного межреберного мускула. Обе межреберные мыш-
цы играют значительную роль в дыхательном акте ящурки.
Внутренних лестничных мышц (mm. scalares intemi, рис. 20,В) 3, тонкие, лентовидные,
их проксимокаудальные концы расширены, а дистокраниальные — сужены, расположены
на латеральной поверхности брюшка, дистальнее от4 торакальных и дистокаудальнее от
V стернального ребер. Латерально мышечные лепестки прикрыты глубокой наружной ко-
сой мышцей, сами располагаются на латеральной поверхности внутренней косой мышцы.
Начинаются от краниальной поверхности дистальных концов 3 каудальных торакальных
ребер. У латерального края медиальной прямой мышцы живота они переходят в тонкие
лентовидные сухожилия, которыми и закрепляются на ней.
Поперечная мышца (m. transversus) у разноцветной ящурки состоит из двух отдельных
мышц: поперечной дорсальной (m. transversus dorsalis) и поперечной брюшной (т. trans-
versus abdominis).
Поперечная дорсальная мышца (т. transversus dorsalis, рис. 21,2?) плоская, обширная,
состоит из 13 сросшихся лентовидных мышечных пучков, расположена на вентральной
поверхности туловища, латеральнее от позвоночного столба. Начинается на вентролате-
ральной поверхности тел 13 туловищных позвонков (включая 9 грудных и 4 поясничных),
закрепляется на каудомедиальной поверхности ребер (9 грудных и 2 последних шейных),
проксимокаудальнее начала мышечных пучков поперечной брюшной мышцы. Каудальный
край мышцы достигает уровня межлобкового симфиза и частично прикрывает квадратный
мускул поясницы (m. quadratus lumborum), а краниальный — граничит с каудальным кон-
цом переднего прямого мускула головы.
Поперечная брюшная мышца fm. transversus abdominis, рис. 21,В) занимает самую ме-
диальную поверхность брюшной стенки и простирается от последних 2 шейных ребер до
лобково-седалищной связки. Брюшко состоит из отдельных удлинённых мышечных ле-
пестков, сросшихся и расположенных в дорсовентральном направлении. Латеральной по-
верхностью мышца граничит с внутренней косой мышцей живота (m. obliquus abdominis
internus) и апоневротически с ней срастается. Мышечные лепестки брюшка начинаются каж-
63
дый отдельно на краниомедиальной незакрытой поверхности 2 последних шейных и всех
туловищных ребер, медиальнее от начала аналогичных лепестков внутренней косой мыш-
цы живота на них; в поясничной области начинаются от дистальных концов всех пояс-
ничных ребер и соответствующих поперечно-сухожильных миосепт. Дистальным концом
мышца срастается плоским обширным апоневрозом с контрлатеральной мышцей и с меди-
альной прямой мышцей живота. Обе поперечные мышцы способствуют поддержанию внут-
ренностей брюшка.
Внутренняя косая мышца (m. obliquus abdominis intemus, рис. 20,Б, В\ 21,2?) обшир-
ная, расположена на внутренней поверхности туловищных ребер и с медиальной поверхно-
сти прикрыта поперечной брюшной мышцей, состоит из сросшихся апоневротически ряда
мышечных пучков. Каудальная часть мышцы имеет вид сплошного тонкого мышечного
пласта, закрывающего пространство между последним туловищным ребром и областью
лобково-седалищной связки. В поясничной области мышечные полоски начинаются от ди-
стальных концов всех поясничных ребер и соответствующих сухожильных миосепт под-
вздошно-реберного мускула. В грудной области мышечные полоски начинаются на крани-
альной поверхности последнего шейного и 9 грудных ребер, у самой дистолатеральной
границы подвздошно-реберного мускула и проксимолатеральнее от начала мышечных
пучков поперечной брюшной мышцы, с которыми апоневротически срастаются. Дисталь-
ным концом мышца апоневротически срастается с латеральным краем медиальной
прямой мышцы живота. Таким образом, мышца простирается по всей внутренней
поверхности туловищных ребер, начиная от 2 последних шейных вплоть до лобко-
во-седалищной связки. Мышца способствует выдоху, а также укреплению вентральной
части туловища.
Квадраттшй мускул поясницы (m. quadratus lumborum, рис. 21, В) расположен на вен-
тральной поверхности позвоночника, уплошен в вентродорсальном направлении, слабо
веерообразный, простирается почти от каудальных шейных ребер до подвздошной кости.
Дорсально граничит и срастается с наружными межреберными мышцами, которые дохо-
дят до тел позвонков, вентрально прикрыт дорсальной поперечной мышцей. Своим каудо-
латералъным краем мускул граничит с проксимальными концами поперечной брюшной и
внутренней косой мышцами. Каудально брюшко закрепляется мышечно-сухожильно на
краниальном конце подвздошной кости с частичным заходом на краниовентральную по-
верхность поперечного отростка первого крестцового позвонка. В краниальном направле-
на мышечные пучки расходятся веером, достигая последнего грудного ребра. При функ-
ционировании мускул предотвращает провисание позвоночника, а при одностороннем дей-
ствии изгибает позвоночник в латеровентральном направлении.
Реберио-кораковдный мускул (m. costocoracoideus, рис. 20,Б) веерообразный. Начи-
нается брюлвсо на мощном сухожилии, расположенном на медиальной поверхности плече-
вого пряса. Вертикальная ветвь этого сухожилия берет начало от проксимального конца
лопатки (ближе к краниальному ее краю), дистальная — от медиальной поверхности кора-
коида (ближе к его каудальному краю), каудальная — от внутренней сухожильной ветки
трехглавого мускула плеча. Мылдошое брюшко ориентировано в вертикальной плоско-
сти. Закрепляется брюшко мышечно-сухожильно на краниомедиальной поверхности пер-
вого грудного ребра, в области соединения вертикальной (проксимальной) части ребра
с горизонтальной. При функционировании мускул ретрагирует коракоид в его сочленении
с грудиной и отводит первое грудное ребро в латеральную сторону и вперед, тем самым
способствуя вдоху.
Грудино-коракоидный внутренний поверхностный мускул (m. stemocoracoideus in-
terims superficialis, рис. 22, Б) слабо развит, плоский, имеет вид тупоугольного треуголь-
ника. Начинается мышечно-сухожильно от дорсальной (внутренней) поверхности кора-
коида, краниомедиальнее от начала на нем медиальной ветви m. subcoraco-scapularis pars
coracoideus и медиальнее от овального коракоидного окна. Закрепляется на кранио дор-
сальной поверхности дорсального валика (ложного сустава) краниомедиальной части гру-
зины. Мышца удерживает коракоид и грудину в статодинамическом режиме.
Грудашо-коракоидный внутренний глубокий мускул (m. stemocoracoideus intemus
profundus, рис. 22,2?) веретеновидно-уплощенный, удлинен в краниокаудальном направ-
лении, расположен на внутренней (дорсальной) поверхности грудины и частично на кау-
цомедиальном крае коракоида. Начинается от дорсальной поверхности каудомедиального
края грудины, в местах срастания ее с самыми дистальными хрящевыми концами II—V
И
Рис 22. Поверхностный слой мышц грудной конеч-
ности разноцветной ящурки:
А - латерокраниально, Б — дорсомедиально; 1 —
m. deitoideus pars scapularis, 2 - т. deltoideus pars
ciavicularis, 5 — m. scapulohumeralis anterior, 4 — m. sup-
racoracoideus, 5 - m. biceps brachii, 6 - m. brachialis,
7 — m.coracobrachialis longus, 8 — m. coracobrachialis
brevis, 9 - m. triceps brachii, 10 - in. extensor ante-
brachii radialis, 11 - m. extensor digitorum communis.
12 - m. extensor carpi ulnaris, 13 - m. subcoracoscapu-
laris pars scapularis, 14 - m. subcoracoscapularis pars co-
racoidea, 15 — m. serratus dorsalis, — m. serratusven-
tralis, 17 — m. stemo-coracoideus. internus superficialis,
18 - m.stemo-coracoideus internus profundus, 19 —
m. flexor carpi ulnaris + m. flexor palmaris superficialis.
20 — m. flexor carpi radialis, 21 — mm. flexores digito-
rum breves superficiaies
65
стернальных ребер и на самых дистальных концах этих ребер. По всему медиальгэ,. у краю
мышечное брюшко только апоневротически срастается с коллатеральной мышцей. Самый
краниолатеральный край брюшка постепенно переходит в короткое обширное и плоское
сухожилие, которым и закрепляется на дорсальной поверхности коракоида, по узкой по-
лоске (в краниокаудальном направлении) и медиальнее от крупного овального его окна и
начала m. subcoracoscapularis pars coracoideus. Краниальная треть брюшка прикрывает собой
грудино-коракоидный внутренний поверхностный мускул. Мускул содержит в динамиче-
ском режиме грудину и коракоид.
523. МЫШЦЫ ХВОСТА
Поперечно-остистый мускул хвоста (m. transversospinalis caudae, рис. 19, А, Б) — непосред-
стгэнное продолжение поперечно-остистого мускула спины, не обнаруживает структурных
изменений по сравнению с ним и способен вызывать иг гибы хвоста в вертикальной
плоскости.
Подвздошно-хвостовой мускул (m. iliocaudalis, рис. 19,Л, Б) относительно мощный,
веретеновидный, двуперистьш, расположен на дорсальной поверхности краниальной час-
ти хвоста, латеральнее от каудальной части поперечно-остистого мускула. Вентральной по-
верхностью брюшко расположено на дорсальной поверхности поперечных отростков кра-
ниальных хвостовых позвонков. Краниально брюшко срастается своей дорсомедиальной
частью с каудальным концом длиннейшего мускула спины, дистолатеральной частью — с
подвздошно-реберным мускулом. На дорсальной поверхности краниальной четверти брю-
шка просматривается мощный сухожильный тяж, которым оно и начинается на каудоме-
диальном конце подвздошной кости. Часть мышечных пучков брюшка, идущих в каудоме-
диальном направлении, закрепляется на латеральном апоневрозе поперечно-остистого мус-
кула. Часть мышечных пучков, идущих каудовентрально и латерайьно, закрепляется на
дорсокраниальной поверхности поперечных отростков 7—8 краниальных хвостовых по-
звонков. При функционировании мускул изгибает хвост дорсолатерально.
Седалищно-хвостовой мускул (m. ischiocaudalis, рис. 19, В) веретеновидный, располо-
жен на вентральной поверхности хвоста, у его медиальной (средней) линии и кзади от кло-
аки. Начинается крепким шнуровидным сухожилием от каудальной части седалищной кос-
ть, на ее внутренней (дорсальной) поверхности. Это сухожилие идет до уровня подседа-
лищного хрящевого выроста (hypoischium), а затем переходит в мышечную головку, мы-
шечные пучки которой закрепляются на латеральной и вентральной поверхностях гемаль-
ных дуг хвостовых позвонков (на расстоянии 20 мм) При функционировании мускул
прижимает хвост к земле и участвует в пропульсивном движении тела от конечностей.
Межпоперешый мускул хвоста (m. intertransversarius caudae, рис. 19, Л, Б) мощный,
длинный, расположен на латеровентральной поверхности хвоста и простирается от первого
до последнего хвостовых позвонков. Мышечное брюшко расширено в дорсовентральном
направлении. Краниальная часть брюшка медиально граничит с подвздошно-хвостовым
мускулом и частично охватывает его каудальную половину; вентромедиально граничит с
хвостово-бедренными мускулами и протрактором копулятивного органа (у самцов). По
всей латеральной поверхности брюшка имеется борозда, которая проходит на уровне ла-
теральных концов поперечных отростков хвостовых позвонков. Медиальные сухожильные
тяжи дорсальной мышечной полосы срастаются своими каудальными концами с сухожиль-
ными тяжами от мышечных лепестков поперечно-остистого мускула, а дистолатеральные —
срастаются с дорсолатеральными сухожильными тяжами вентральной мышечной полосы.
На вентральной поверхности хвоста коллатеральные мышечные брюшка срастаются по
средней линвд позвонков с помощью дополнительных V-образной формы сухожилий, к
которым прикрепляются мышечные пучки с контрлатеральных сторон, а сами сухожилия
срастаются с одной стороны с каудальными концами медиальных сухожильных тяжей кол-
латеральных мышечных брюшек, с другой — через посредство мышечных пучков с вентро-
каудальным краем гемальных дуг каждого хвостового позвонка. При функционировании
мышца сгибает хвост в латеральную сторону.
Протрактор копулятивного органа (m. protractor penis рис, 20, В) имеет вид полой
трубы с поперечным расположением мышечных пучков, в которой располагается копуля-
тивный орган. Медиальный край брюшка переходит в тонкий обширный апоневроз, кото-
рым оно крепится к дистальным концам 5—6 гемальных дуг. Его дорсолатеральный край
66
также переходит в обширный тонкий апоневроз, которым оно и срастается с дорсолате-
ральной поверхностью ретрактора клоаки (m. retractor cloacae) и закрепляется на вен-
тральной поверхности латеральных концов 4 хвостовых позвонков. Краниально-мышечное
брюшко через посредство апоневроза соединяется с подвздошно-седалищной связкой.
Краниомедиально от мышечного брюшка имеется небольшой мышечный пласт, который
лежит кауцальнее клоаки и срастается медиально с таким же коллатеральным мышечным
пластом. Он усиливает работу самого протрактора копулятивного органа, который выдви-
гает его краниально.
Ретрактор клоаки (m. retractor cloacae, рис. 20,2?) веерообразный, расположен дррсола-
терально от копулятивного органа, но находится как бы внутри трубки, сформированной
его протрактором. Дорсомедиально брюшко граничит с длинным хвостово-бедренным
мускулом, но с ним не срастается. Лишь своей латеральной поверхностью брюшко сраста-
ется с тонким дорсолатеральным апоневрозом протрактора копулятивного органа и та-
ким образом закрепляется на поперечных отростках хвостовых позвонков. В краниальном
направлении брюшко немного утолщается и закрепляется на каудолатеральном крае ниж-
ней губы клоаки и на каудолатеральном крае подседалищного мускула (большей своей
частью) .При функционировании мускул ретрагирует клоаку.
Ретрактор копулятивного органа (m. retractor penis, рис. 20, В) удлиненный, располо-
жен в глубине хвоста, каудальнее от копулятивного органа, и является как бы его кау-
дальным продолжением. Сам копулятивный орган сложно устроен: самую его краниаль-
ную часть занимает объемное дольчатое тело, от которого через посредство обширного тон-
кого, но короткого апоневроза отходят относительно длинные мышечные пучки, сгруппи-
ровавшиеся в веретеновидную форму. Снаружи эти пучки покрыты тонким апоневрозом.
По всей латеральной поверхности мышечной части органа расположена уплотненная ткань
в виде сухожильной пластинки, к которой частично присоединяются мышечные пучки (с
вентральной и дорсальной поверхностей). Медиальная часть мышечных пучков простирает-
ся от дольчатого тела непосредственно к каудальному концу сухожильной пластинки. На
уровне 7-й гемальной дуги хвостового позвонка от каудального конца пластинки и мышеч-
ных пучков начинается посредством поперечной сухожильной перемычки мышечное брюш-
ко ретрактора копулятивного органа. Он закрепляется на вентролатеральной поверхности
8—9-й гемальной дут и тел соответствующих хвостовых позвонков. При функционирова-
нии мускул втягивает копулятивный орган в боковую полость основания хвоста. В месте
расположения данного мускула имеется значительное количество жировой ткани желтого
цвета.
5.2.4.МЫШЦЫ ГРУДНОЙ КОНЕЧНОСТИ
Трапециевидный мускул (m. trapezius, рис. 19, Л, Б) в виде тупоугольного треуголь-
ника, расположен на латеральной поверхности туловища, частично над лопаткой и прокси-
мокаудальнее от нее, прикрывает проксимокраниальный угол широчайшего мускула спи-
ны. Мышечные пучки брюшка имеют слабую веерообразность в дорсокаудальном направ-
лении. Начинается брюшко коротким обширным апоневрозом на остистых отростках по-
следних шейных и первых грудных позвонков. Закрепляется же оно на проксимокраниаль-
ном крае лопатки, в месте срастания последней с латеральным концом ключицы. При
функционировании мускул фиксирует и частично смещает лопатку в дорсомедиальном
направлении.
Надгрудинно-ключично-сосцевидный мускул (m. episterno-cleidomastoideus, рис. 19,А)
расположен на дорсолатеровентральной поверхности шеи, в косом направлении, прикрыт
с латеральной поверхности мускулами — подкожным шеи и сжимателем шеи. По отноше-
нию к лопатке и трапециевидному мускулу брюшко расположено краниально, с ним мы-
шечно срастается. Начинается от латерокаудальной поверхности теменной и частично заты-
лочной костей, проксимокаудальнее ушного отверстия. Проксимокраниальная часть мыш-
цы имеет начало также и от латерального отростка теменной кости относительно мощным
сухожильным тяжем. Проксимальная часть брюшка имеет апоневротическое начало oi
дорсальной поверхности апоневроза мышц шеи — на протяжении от каудолатерального
края головы и вплоть до начала трапецевидного мускула. Закрепляется мышца коротким
апоневрозом на краниовентральной поверхности ключицы, латеральной ветви надгрудин-
ника и частично доходит до грудины, где граничит с краниальным краем грудного муску-
67
ла. При функционировании отводит шею в латеральную сторону и поворачивает голову во-
круг продольной оси.
Широчайший мускул спины (m. latissimus dorsi, рис. 19, А, Б) в виде тупоугольного
треугольника, расположен на дорсолатеральной поверхности туловиша, проксимальнее от
плечевой кости и на протяжении 12 сухожильных миосепт подвздоггнс.?бсрного мускула,
начиная от каудального края лопатки. Начинается от остистых отростков последних шей-
ных и от IX—X туловищных позвонков, из которых первые 5 - грудные. У проксималь-
ного конца плечевой кости мышечные пучки веерообразно сходятся, переходят в короткое
мощное сухожилие, которым и закрепляк/гся на переднедорсальной поверхности самой
плечевой кости. Мышца ретрагирует конечности и пронирует плечо.
Вентральный зубчатый мускул (m. serratus ventralis, рис. 20/>; 22, Б) веерообразный.
Начинается от дистальных концов 2 последних шейных ребер, прикрывая частично межре-
оерные мышцы, а сам латерально прикрыт широчайшим мускулом спины. Мускул идет
к каудальной поверхности надлопаточного хряща, где и закрепляется, частично на каудаль-
ной поверхности выемки этого хряща, а частично на его медиальной поверхности — про-
ксимокаудально. При функционировании совместно с поднимателем лопатки фиксируй'
лопатку относительно туловища, нагружая ее частично за счет массы тела животного.
Подниматель лопатки fm. levator scapulae, рис. 20,Б) состоит из 2 краниально сросших-
ся головок разной длинхм и мощности и со слабо выраженной веерообразностью. Обе го-
ловки начинаются совместно с латеральным длинным мускулом шеи на поперечном отро-
стке и на самом теле атланта: дорсальная головка — на коротком сухожилии латерального
длинного мускула шеи, а вентральная — на теле позвонка. Дорсальная головка закрепля-
ется мышечно-сухожильно на латеральной поверхности прсксимокраниальной части надло-
паточного хряща, а вентральная — на краниомедиальной поверхности надлопаточного хря
ща. Латерально обе головки прикрыты лопаточно-подъязычным мускулом. При функцио-
нировании мышца протрагирует лопатку, а при одностороннем действии сгибает шею в ла-
теральном направлении.
Дорсальный зубчатый мускул (m. serratus dorsalis, рис. 20, Б; 22, Б) состоит из двух
порций — латеральной и медиальной, расположенных под углом 55—60°. Латеральная часть
мыпшы начинается от одноименной поверхности дистальных концов 2 шейных ребер, кпе-
реди от начала вентрального зубчатого мускула, а закрепляется к его проксимомедиальной
поверхности надлопаточного хряща, ближе к его проксимальному краю. Медиальная часть
мышцы имеет вид прямоугольной трапеции, ее мышечные пучки располагаются вертикаль-
но. Начинается от дистальных концов 3 шейных ребер, расположенных кпереди от 2 по-
следних ребер. Мышечные пучки закрепляются на медиальной поверхности надлопаточного
хряща. Мышца подвешивает лопатку к осевому скелету, передавая вертикальные и гори-
зонтальные усилия.
Дельтовидный мускул (m. deltoideus) состоит из двух порций; лопаточной и клю-
чичной.
Лопаточная порция дельтовидного мускула <m. deltoideus pars scapularis, рис. 19, Л;
22. А) частично прикрыта трапецевидным и широчайшим мускулами спины. Начинается
на латеральной поверхности надлопаточного хряща, а также от проксимальной половины
лопатки, закрепляется на проксимокраниальной поверхности дельтовидного бугра, под
прикрытием ключичной порции данного мускула. При функционировании отводит и супи-
нирует плечо.
Ключичная порция дельтовидного мускула (m. deltoideus pars clavicularis, рис. 19, Л;
22, А) расположена дистомедиальнее от расположения его лопаточной порции. Начинается
на вентральной и медиальной поверхностях основной части ключицы, закрепляется, при-
крывая закрепление лопаточной порции данного мускула. При функционировании протра-
гирует плечевую кость и всю конечность в целом
Подкоракоидно-лопаточный мускул (m. subcoracoscapularis, рис. 22) — мощный, вееро-
образный. состоит из 2 сросшихся мышечных брюшек, которые можно условно считать
как лопаточная и коракоидная части.
Поцкоракоидно-лопаточный мускул, часть лопаточного (гп. subcoracoscapularis parssca-
pularis, рис. 22,Б) также состоит из двух сросшихся брюшек и краниально, в области кора-
коида и лопатки, граничит с коракоидной частью данного мускула. Кауд©латеральное
бсюцрсо начинается от одноименного края лопатки и частично на дистокаудальном крае
68
надлопаточного хряща. Медиальное брюшко начинается от медиальной поверхности надло-
паточного хряща и частично от проксимомедиальной части лопатки, в месте соединения ее
с надлопаточным хрящом. Мышечные пучки сливаются с таковыми каудолатерального
брюшка, переходят в удлиненное ухожилие, которым и закрепляются на дорсокаудаль-
ном бугорке плечевой кости. При функционировании поднимает и ретрагирует плечо.
Подкоракоидно-лопаточный мускул, часть коракоидная (m. subcorascapularis pars cora-
coidea, рис. 22, Б) сложноперистый, проксимокаудально граничит с лопаточной частью дан-
ной мышцы. Начинается от дорсальной поверхности коракоида, дистального конца лопат-
ки, от апоневроза, закрывающего коракоидное окно, и от апоневроза, прикрывающего
пространство между краниальным краем прокоракоида, медиальным — лопатки и каудаль-
ным — ключицы. Мышечные пучки закрепляются на каудальной поверхности специально-
го бугра головки кости. Поднимает, пронирует и сильно ретрагирует плечо.
Грудной мускул (m. pectoralis, рис. 19,В) мощный, веерообразный, расположен на вен-
тральной поверхности грудины, прикрыт с вентральной поверхности краниальным апонев-
розом латеральной ветви прямой мышцы живота. Начинается от латеровентральной по-
верхности грудины и от дистальных концов всех 5 грудных ребер. Мышечные пучки закре-
пляются на вентральной поверхности дельтовидного бугра плечевой кости. Краниальная
часть мышцы покрыта обширным апоневрозом, простирающимся от поперечного отростка
надгрудинника к каудальномедиальному краю грудной кости. При функционировании
мышца приводит плечевую кость, а также пронирует и ретрагирует ее.
Надкоракоидный мускул (m. supracoracoideus, рис. 22, А) веерообразный, состоит из
2 сросшихся брюшек, расположен на вентральной поверхности коракоида. Начинается от
вентральной поверхности коракоидной пластинки, прикрывая коракоидное отверстие,
и частично от дистального конца лопатки. Закрепляется на проксимальном конце дельто-
видного бугра плечевой кости. С медиальной стороны граничит с двуглавым мускулом
плеча, а каудально — с передним лопаточно-плечевым мускулом. При функционировании
протрагирует и приводит плечо.
Передний лопаточно-плечевой мускул (m. scapulohumeralis anterior, рис. 22, А) 2-пери-
стый, начинается на латеральной ветви коракоида, на краниолатеральной поверхности ло-
патки, а также на каудальной поверхности проксимальной части ключицы. Конечное су-
хожилие брюшка закрепляется между дельтовидным и задним буграми плечевой кости,
проксимальнее от закрепления широчайшего мускула спины. Протрагирует, отводит и
пронирует плечо.
Трехглавый мускул плеча (m. triceps brachii, рис. 19,В\ 22, А) занимает дорсальную
поверхность названного отдела и состоит из трех головок: латеральной, медиальной и
длинной.
Латеральная головка трехглавого мускула плеча (m. triceps brachii. caput laterale) мощ-
ная, веретеновидная, расположена краниодорсально на плечевой кости, латерально частич-
но срастается с длинной головкой данной мышцы. Начинается от каудальной поверхности
дельтовидного бугра и вплоть до ее дистального конца. Закрепляется мощным сухожиль-
ным тяжем, имеющим сесамовидную косточку, вместе с длинной головкой, на прокси-
мальном конце локтевой кости.
Длинная головка трехглавого мускула плеча (m. triceps brachii caput longum) мощная,
веретеновидная,расположена на дорсальной поверхности плечевой кости, между латераль-
ной и медиальной головками мускула. Начинается коротким уплощенным сухожильным
тяжем медиальнее от дельтовидного бугра. Дистально головка мышечно срастается с лате-
ральной и медиальной головками и вместе с ними закрепляется коротким мощным сухо-
жилием на проксимальном конце локтевой кости.
Медиальная головка трехглавого мускула плеча (m. triceps brachii caput mediale) разви-
та слабо, расположена на каудодорсальной поверхности плечевой кости, частично прикрыта
длинной головкой и по латеральной поверхности с ней срастается. Начинается от каудаль-
ного бугра плечевой кости вплоть до ее дистального конца. Закрепляется головка вместе
с остальными головками мышцы на проксимальном конце локтевой кости. При функцио-
нировании разгибает локтевой сустав, а с помощью длинной головки поднимает конеч-
ность в целом.
Двуглавый мускул плеча (m. biceps brachii, рис. 19, В\ 22, А) расположен вентральнее
и каудальнее от дельтовидного бугра плечевой кости, состоит из двух неравноценных го-
69
ловок, соединенных лентовидным сухожилием, которое прикрывает вентрально плечевой
сустав. Медиальная (проксимальная) головка более слабая, уплощена; начинается от вен-
тральной поверхности коракоида, проксимальнее от начала короткого и длинного корако-
идно-плечевых мускулов, частично прикрывая их с вентральной поверхности. На уровне
дельтовидного бугра мышечная головка переходит в лентовидное сухожилие (2 i мм)
Сухожилие тут же переходит в латеральную (дистальную) мышечную головку, которая
идет по вентрокаудальной поверхности плечевой кости, краниальнее от плечевого мускула,
с которым срастается мышечно. Закрепляется коротким пластинчатым сухожилием вме-
сте с плечевым мускулом на внутренней поверхности проксимальных концов лучевой и
локтевой костей, у самого локтевого сустава и на суставной сумке. Сгибает конечность
в локтевом суставе и вентрально приводит ее.
Плечевой мускул (ш. brachialis, рис. 19, Б; 22, А) веретеновидный, расположен про-
ксимальнее от двуглавого мускула плеча. Начинается от латеральной поверхности дельто-
видного бугра и далее по узкой полоске от всей плечевой кости. У дистального конца
брюшко срастается с двуглавой мышпей плеча и закрепляется на внутренней поверхности
проксимальных концов костей предплечья. При функционировании сгибает конечность
в локтевом суставе.
Длинный коракоидно-плечевой мускул (m. coracobrachialis longus,pnc. 22, А) располо-
жен ближе к каудальному краю плеча, с вентральной поверхности частично прикрыт дву-
главым мускулом плеча. Краниовентрально граничит и срастается с коротким коракоид-
но-плечевым мускулом. Начинается от вентральной поверхности коракоида и закрепляется
на ульнарном мышелке плечевой кости, чуть проксимальнее начала мышц предплечья. Ре-
трагирует и слегка пронирует плечевую кость.
Короткий коракоидно-плечевой мускул (m. coracobrachialis brevis, рис. 22, А) располо-
жен на вентральной поверхности каудальнее конца коракоида, откуда и начинается мышеч-
но-сухожильно; вентрально частично прикрыт краниальной головкой двуглавой мышцы и
надкоракоидной мышцей. Закрепляется мышечно-сухожильно на вентрокраниальной по-
верхности головки плечевой кости, краниальнее ее дельтовидного бугра, и на вентральной
пср.пхности самой кости. При функционировании ретрагирует и пронирует плечо.
Лучевой разгибатель предплечья (m. extensor antebrachii radialis, рис. 19,£, 22, А)
мощный, состоит как бы из 2 сросшихся брюшек. Начинается на краниальной поверхности
радиального надмыщелка плечевой кости, проходит по латеромедиальной поверхности лу-
чевой кости к ее дистальному концу, где и закрепляется. При функционировании отводит
предплечье дорсально и супинирует его.
Лучевой разгибатель кисти (m. extensor carpi radialis) частично прикрывает лучевой раз-
гибатель предплечья и отводящий мускул первого пальца. Начинается на краниальной по-
верхности радиального надмыщелка плечевой кости, совместно с лучевым разгибателем
предплечья, но латеральнее последнего. Закрепляется на поверхности проксимальных кон-
цов 2, 3 и 4-й пястных костей, в промежутке между началами дорсальных межкостных
мышц. Разгибает кисть в интеркарпальном суставе.
Общий разгибатель пальцев (m.extensor digitorum communis, рис. 19, Л— В; 22, А) час-
тично прикрывает лучевой разгибатель предплечья и отводящий мускул первого пальца.
Начинается на краниальной поверхности радиального надмыщелка плечевой кости вместе
с лучевым разгибателем предплечья, но латеральнее него, закрепляется на поверхностях
проксимальных концов 2—4-й пястных костей, в промежутке между началами дорсальных
межкостных мышц. Разгибает кисть в интеркарпальном суставе.
Отводящий мускул первого пальца (m. abductor pollicis, рис. 23, Л) треугольный, на-
чинается от латерокраниальной поверхности дистальной половины локтевой кости, закреп-
ляется на краниомедиальной поверхности проксимального конца пястной кости первого
пальца. Яри функционировании отводит первый палец и супинирует кисть.
Локтевой разгибатель кости (m. extensor carpi ulnaris, рис. 19, А, Б; 22, А) медиально
граничит с общим разгибателем пальцев, а с латерокаудальной стороны — с локтевым сги-
бателем кисти. Начинается от латеральной поверхности радиального надмыщелка плечевой
кезти и от латерокраниальной поверхности проксимального конца локтевой кости, закреп-
ляется на дорсолатеральной поверхности гороховидной косточки (вместе с локтевым сги-
бателем кисти) . При функционировании отводит и разгибает кисть.
Локтевой сгибатель кисти (m. flexor carpi ulnaris, рис. 19, Л: 22, Б) с медиальной по-
верхности срастается с поверхностным ладонным сгибателем. Начинается на каудолате-
70
Рис. 23. Глубокий слой мышц грудкой конечности разноцветной ящурки:
А — краниально, Б - каудально; 1 — m. abductor policis, 2 — mm. extenso-
res digitorum breves, 3 - mm. interossei dorsales, 4 - m. flexor palmaris profun-
dus, 5 — m.pronator teres, 6 - m. pronator profundus
ральной поверхности ульнарного надмыщелка плечевой кости, совместно с поверхностным
ладонным сгибателем, но латеральнее от него. Закрепляется мышечно-сухожильно на про-
ксимокаудальной поверхности гороховидной косточки запястья, но дает сухожильный ана-
стомоз на ладонный апоневроз и особенно к 5-й пястной кости. Сгибает кисть.
Поверхностный ладонный сгибатель (m. flexor palmaris superficialis, рис. 22, Б) лате-
ральной поверхностью срастается с локтевым сгибателем кисти, с глубоким ладонным сги-
бателем срастается только апоневротически. Начинается на ульнарном надмыщелке плече-
вой кости, проксимальнее от начала глубокого ладонного сгибателя. У проксимального
конца запястья мышечное брюшко переходит в ладонный апоневроз, который прикрывает
ладонь с пальмарной поверхности и закрепляется на пальцах. При функционировании сги-
бает кисть.
Глубокий ладонный сгибатель (m. flexor palmaris profundus, рис. 23, Б) с латеральной
поверхности граничит с поверхностным ладонным сгибателем, медиально брюшко грани-
чит с лучевым сгибателем кисти. Начинается двумя головками: от ульнарного надмыщел-
ка плечевой кости, рядом, но латеральнее от начала лучевого сгибателя кисти; от каудаль-
ной поверхности локтевой кости, почти на всем ее протяжении. В области запястья оба
брюшка, до этого сросшихся, переходят в мощный сухожильный апоневроз, армирован-
ный двумя мощными уплощенными сесамовидными окостенениями. Далее он распадается
на 5 мощных сухожильных ветвей. Конечные сухожилия закрепляются на пальмарной по-
верхности проксимальных концов конечных фаланг 5 пальцев. При функционировании
сгибает пальцы и кисть.
Помимо этого на пальмарной поверхности ладони расположен мышечный пласт, начи-
нающийся от пальмарной поверхности ульнарной косточки запястья и 5-й запястной кос-
точки, фиксирующейся на дорсальной поверхности проксимомедиальной части конечного
сухожилия глубокого ладонного сгибателя. При функционировании он подтягивает конеч-
ное сухожилие сгибателя в проксимолатеральном направлении.
71
5мм
Рис. 24, Мышцы пальмарной поверхности
кисти разноцветной ящурки:
1 - mm. lumbricales, 2 - m. abductor digiti
minimi, 5 - m. adductor digiti minimi, 4 —
m. carpo-digitalis ventralis communis, 5 -
mm. interossei volares, 6 — m. carpo-digita-
Hsvolaris I
-Л Лучевой сгибатель киста (m. flexor carpi
radialis, рис. 19, В; 22, Б) граничит с глубоким
ладонным сгибателем, однако не срастается с
ним. Начинается от медиального надмыщелка
плечевой кости, медиальнее от начала глубокого
ладонного сгибателя, закрепляется на медиаль-
ной поверхности лучевой кости, достигает каудо-
медиальной поверхности радиальной косточки
запястья и частично доходит своим небольшим
конечным сухожилием до каудомедиальной по-
верхности проксимального конца 1-й пястной
кости. При функционировании приводит и прони-
рует предплечье.
Круглый пронатор (m, pronator teres , рис.23,
Б) прикрыт лучевым сгибателем кисти. Начина-
ется коротким овальным сухожилием на каудо-
медиальной поверхности ульнарного надмыщел-
ка плечевой кости, под прикрытием начала луче-
вого сгибателя кисти; закрепляется на медиаль-
ной поверхности лучевой кости. При функциони-
ровании пронирует предплечье.
Глубокий пронатор (m. pronator profundus,
рис. 23, Б) расположен между внутренними по-
верхностями костей предплечья, по всей их дли-
не. Мышечные пучки идут от внутренней поверх-
ности локтевой кости косо вниз к внутренней
поверхности лучевой кости. Часть мышечных
пучков начинается от вентральной поверхности
ульнарного надмыщелка плечевой кости и за-
крепляется на внутренней поверхности лучевой
кЪсти. При функционировании пронирует луче-
вую кость.
Поверхностные короткие сгибатели пальцев (mm. flexores breves superficiales, рис. 24)
в количестве пяти начинаются мышечно-сухожильно на пальмарной поверхности ладонного
апоневроза который соединяет гороховидную косточку и радиальную кость запястья.
В области пястно-фаланговых суставов переходят в удлиненные и уплощенные сухожилия,
закрепляются на проксимальных концах почти всех фаланг пальцев, за исключением кон-
цевых (когтевых). Сгибают пальцы.
Червеобразные мышцы (пип. lumbricales, рис. 24) в количестве трех начинаются на дор-
сальной поверхности конечного апоневроза глубокого ладонного сгибателя и направляют-
ся к основаниям 1-х фаланг и частично к следующим за ними второго-четвертогопальцев,
где и закрепляются. При функционировали брюшка способствуют передаче усилий глубо-
кого ладонного сгибателя на проксимальные фаланги.
Абдуктор пятого пальца (m. abductor digit! minimi, рис. 24) начинается мышечно-сухо-
жильно от пальмарнолатеральной поверхности гороховидной косточки и направляется к
пястно-фаланговому суставу пятого пальца, закрепляется на латеральной поверхности ди-,
стального конца 1-й фаланги и проксимального — 2-й фаланги пятого пальца. Отводит лате-
рально пятый палец.
Приводящая мышца пятого пальца (m. adductor digiti minimi, рис. 24) начинается ко-
ротким лентовидным сухожилием на пальмарнолатеральной поверхности радиальной кос-
точки запястья и частично на такой же поверхности проксимального конца 1-й пястной кос-
ти, закрепляется на медиопальмарной поверхности дистального конца 1-й фаланги пятого
пальца и частично на проксимальном конце 2-й фаланги. Приводит пятый палец.
Общая запястно-пальцевая мышца (m. carpo-digitalis ventralis communis, рис. 24) состоит
из 5 мышечных брюшек, которые начинаются на пальмарной поверхности костей запястья
и закрепляются на пальмарнобоковых поверхностях дистальных концов 1—5-й пястных
костей. При функционировании сгибает кисть в пястно-запястных суставах.
Подопшеншле межкостные мышцы (mm. interossei volares, рис. 24) наиболее глубокая
часть мышечных брюшек, расположенных на пальмарной поверхности пястных костей. На-
72
чикаются на пальмарномедиальной поверхности 2—5-й пястных костей; закрепляются они
на латеропальмарной поверхности 1—4-й пястных костей и частично — на проксимальных
концах 1-х фаланг соответствующих пальцев. При функционировании стягивают и отво-
дят латерально пальцы.
Подошвенная запястно-пальцевая мышца 1 (m. сагро-digitalis volaris 1,рис. 24) располо-
жена на пальмарной поверхности кисти косо, ближе к ее медиальному краю. Начинается на
пальмарной поверхности 1—2-й запястных костей и частично от радиальной косточки за-
пястья. Закрепляется на пальмарнолатеральной поверхности дистального конца 1-й пяст-
ной кости и частично на проксимальном конце 1-й фаланги. Приводит первый палец.
Короткие разгибатели пальцев (mm. extensores digitorum breves, рис. 23) в количестве
пяти начинаются от дорсальной поверхности локтевой кости запястья, закрепляются на
проксимодорсальной поверхности последних фаланг каждого пальца. Непосредственно на
дорсальной поверхности пястных костей расположены мышечные брюшка дорсальных
межкостных мышц (mm. intefossei dorsales) (рис. 23) . Начинаются они мышечно-сухожиль*
но на дорсолатеральной поверхности 1—5-й пястных костей, а закрепляются также на дор
сальной и боковых поверхностях соответствующих 1—5-й пястных костей.
523. МЫШЦЫ ТАЗОВОЙ КОНЕЧНОСТИ
Подвздошно-большеберцовый мускул (mjlio-tibialis, рис. 19, Л, Б; 24, Л) каудально
граничит с подвздошно-малоберцовым мускулом, а краниально — с охватывающим. С вен-
тральной поверхности прикрывает бедренно-большеберцовый мускул. Начинается упло-
щенным апоневрозом от дорсолатеральной поверхности подвздошной кости. Нижняя часть
мышечных пучков направляется к краниальной части подвздошной кости, а верхняя —
к началу подвздошно-седалищной связки. Заканчивается брюшко плоским сухожильным
тяжем, которым и закрепляется на дорсальной поверхности проксимального конца боль-
шеберцовой кости, совместно с охватывающим и бедренно-большеберцовым мускулами.
На вентральной поверхности конечного сухожилия имеется небольших размеров колен-
ная чашечка. Отводит конечность в тазобедренном и разгибает в коленном суставах.
Охватывающий мускул (m. ambiens, рис. 19, Б; 24, А) начинается крепким уплощен-
ным сухожильным тяжем на латерокраниальной поверхности капсулы тазобедренного су-
става. У коленного сустава брюшко переходит в короткое уплощенное сухожилие, которое
срастается с конечными сухожилиями подвздошно-большеберцовою и бедренно-больше-
берцового мускулов и закрепляется на проксимолатеральной поверхности большеберцо-
вой кости. Выносит конечность вперед и разгибает в коленном суставе.
Бедренно-большеберцовый мускул (m. femorotibialis, рис. 26, А) обширный, мощный,
начинается мышечносухожильно от большей части бедренной кости, начиная от самой ее
головки вплоть до дистального конца (область коленного сустава), закрепляется совме-
стно с предыдущими мышцами на дорсальной (латеральной) поверхности проксимально-
го конца большеберцовой кости. Совместно с предыдущими мышцами разгибает конеч-
ность в коленном суставе.
Подвздошно-малоберцовый мускул (m. iliofibularis, рис. 19, А> Б\ 25, Л) начинается
от латеральной поверхности подвздошной кости, кпереди от начала подвздошно-седалищ-
ной связки. Начало мышечного брюшка прикрыто каудальным лентовидным сухожиль-
ным тяжем, идущим от проксимального конца подвздошно-большеберцового мускула.
Каудальнее от мышечного брюшка расположен наружный большеберцовый сгибатель.
У проксимального конца голени брюшко переходит в короткое уплощенное сухожилие,
которым и закрепляется на латеральной поверхности проксимального конца малоберцо-
вой кости, под прикрытием начала длинного малоберцового мускула. Супинирует голень,
отводит конечность вверх и сгибает ее в коленном суставе.
Лобково-седалищно-большеберцовый мускул (m. pubo-ischio tibialis, рис. 19, В; 25
Б) треугольноуплошекный, мышечные п\*чки конвергируют в короткий сухожильный
тяж, который расположен большей частью с подкожной стороны дистальной половины мы-
шечного брюшка. Мышечное брюшко прикрывает область бедра с вентрокаудальной по-
вехности и граничит с охватывающим мускулом и наружным большеберцовым сгибате-
лем. Начинается от латерального лобкового бугорка, лобково-седалищной связки, дисталь-
ного конца подвздошно-седалишной связки и частично на вентральной поверхности седа-
лищной кости. Дистальным сухожильным тяжем брюшко крепится к медиальной поверх-
ности проксимального конца большеберцовой кости, у дистального конца медиальной
73
Рис. 25. Поверхностный слой мышц тазовой конечности разноцветной ящурки;
А — дорсолатерально, Б — вентрально; I — m. ilio-tibialis, 2 — m. ambiens, 3 — m. ilio-
fibularis, 4 — m. flexor tibialis ex tenuis, 5 — m. pubo-ischjo-femoralis internus, 6 — m. tibia-
lis anterior, 7 - m. extensor digitorum longus, 8 - m. peroneus longus, 9 - m. pubo-ischio-
femoralis externus, 8 - m. peroneus longus, 9 - m. pubo-ischio-femoralis extemus, 10 -
m. pubo-tibialis, 11 — m. pubo-ischio-tibialis, 12 — m. flexor tibialis internus, 13 - m. gastro-
cnemius internus, 14 — m. gastrocnemius extemus, 15 — m. hypoischiadicus, 16 — m. trans-
versus cloacae cranialis, 17 — m. caudifemoralis longus, 18 — m. caudufemoralis brevis, 19 —
mm. extensores digitorum breves
контрлатеральной связки колена. Приводит конечность в целом и пронирует голень.
Внутренний большеберцовый сгибатеш» (m. flexor tibialis internus, рис. 25, Б) располо-
жен венТрокаудально в области бедра. Обычно выделяют в мышце три головки: заднюю,
медиальную и латеральную; у разноцветной ящурки их две, одна более массивная, а вто-
рая — поменьше, но имеет конечное лентовидное сухожилие. Они срастаются слабо апонев-
ротически. Более мощная головка закрепляется на медиальной поверхности проксималь-
ного конца большеберцовой кости и очень коротким сухожильным тяжем на ее медиаль-
ном мыщелке, что рядом с медиальной контрлатеральной связкой колена' Эта часть мыш-
цы приводит конечность, разгибает ее в тазобедренном и сгибает в коленном суставах и
пронирует голень и бедро. Меньшая головка этой мышцы начинается, как и предыдущая
часть, от латеральной поверхности седалищно-подаздощной связки. Мышечные пучки за-
крепляются на слабом внутреннем сухожильном зеркале, которое переходит в конечное
сухожилие в виде шнура. Им головка и закрепляется на латеральной поверхности прокси-
мального конца большеберцовой кости вместе с мышечным сухожилием наружного боль-
шеберцового сгибателя. Функция сходна с таковой у предыдущей части мышцы.
Подседалищный мускул (гл. hypoischiadicus, рис. 19,В; 25,Б) расположен вентрокра-
ниально от клоаки и вентролатерально от hypoischium. Мышечные пучки начинаются на
вентролатеральной поверхности hypoischium и от каудальной поверхности седалищной кос-
ти, закрепляются на вентромедиальной поверхности седалищно-подвздошной связки. При
функционировании прижимает с вентральной поверхности hypoischium, а через него и клоа-
ку, смещая одновременно медиально подвздошно-седалищную связку.
Передний поперечный мускул клоаки (m. transversus cloacae cranialis,рис. 19,В; 25,В)
расположен поверхностно на передней губе клоаки, позади предыдущего мускула, и про-
стирается между контрлатеральными подвздошно-седалищными связками, но слабо выра-
жен. Прижимает клоаку и сближает связки.
Лобково-большеберцовый мускул (m. pubotibialis, рис. 25,В) веретеновидно уплощен-
ный, начинается от латеровентральной поверхности лобково-седалищной связки (каудаль-
ная часть брюшка) и на сухожильном перехвате внутреннего лобково-седалищно-бедрен-
ного мускула. Закрепляется мышечно-сухожильно на латеральной поверхности ее прокси-
74
16 &
мального конца, совместно с конечным сухожилием внутреннего большеберцового а лба-
теля. При функционировании протрагирует и супинирует бедро и частично сгибает конеч-
ность в коленном суставе.
Наружный большеберцовый сгибатель (m. flexor tioialis externus, рис. 19, Л, Б, 25,А)
расположен каудальнее от подвздошно-малоберцового мускула. Начинается мышечно-су-
хожильно на латеральной поверхности подвздошно-седалишнои связки, дорсальнее начала
внутреннего большеберцового сгибателя, закрепляется на латеральной поверхности про-
ксимального конца большеберцовой кости. Перед закреплением брошка от его дистально-
го сухожилия отходит дополнительное шнуровидное сухожилие, которое направляется к
голени и сливается с внутренним икроножным мускулом. Пои функционировании ретра-
гирует бедро, супинирует голень и сгибает конечность в коленном суставе
Подвздошно-бедренный мускул (mjliofemoralis, рис. 26, А) расположен каудальнее
проксимального конца бедренной кости и бедрено-большеберцового мускула, дорсально
прикрыт подвздошно-большеберцовым мускулом. Начинается мышечно-сухожильно от ла-
терокраниальной поверхности подвздошной кости, непосредственно за началом охватыва-
вшего мускула, закрепляется на протяжении ее проксимальных 2/3. При функционирова
нии отводит и пронирует бедро.
Внутренний лобково-седалишно-бедренный мускул (m. рио о ischiofemoral is internus,
рис. 26, Л) довольно мощный, расположен на дорсальной (внутренней) поверхности таза
(на лобковых и седалищных костях контрлатеральных сторон). Мышечное брюшко гори-
зонтально разделено на дорсальную и вентральную части. Начинается (дорсальная часть
мышцы) на дорсолатеральной поверхности предлобкового хряща, самой лобковой коси:
75
Рис. 26. Глубокий слой мышц тазовой конечности разноцветной ящурки:
А — дорсолатерально. Б — вентрально; 1 — тп. femorotibialis, 2 — m. iliofemoralis, 3 — m. adductor femoris,
4 - m. ischiotrochantericus, 5 - m. peroneus brevis, 6 - m. flexor digitorum longus, 7 - m. pronator profun-
dus. 8 - m. fibulo-tibialis superior s.m. popliteus, 9 - m. fibulo-tibialis inferior, 10 - m. abductor et extensor
hallucis et indicis
и на сухожильном тяже, который простирается от предлобкового хряща и к началу дорсо-
каудальной части мышцы, связывая их таким образом в одно целое. Закрепляется на дор-
сокраниальной поверхности бедренной кости, у ее проксимального конца. Вентральная
часть мышцы состоит из нескольких уплощенных мышечных слоев, которые на 2/3 началь-
ной длины свободно отделяются друг от друга; перед закреплением они срастаются. Осо-
бенностью этой части мышцы является то, что наблюдается своеобразный перекрест мы-
шечных слоев контрлатеральных сторон поочередно. Левая сторона мышцы представлена
четырьмя нормально развитыми мышечными лепестками, правая — пятью.
Каудальная порция вентральной части мышцы имеет вид треугольника. Начинается от
дорсолатеральной поверхности каудального края сухожильной перемычки между симфи-
зами, от симфиза и тела седалищной кости, кроме каудальной поверхности, где начинается
седалищно-вертлужный мускул. Далее мышечные волокна идут по внутренней (дорсальной)
поверхности седалищной кости, до места сращения ее в суставной ямке, огибают сустав
дорсально и переходят в лентовидный сухожильный тяж, которым и закрепляются на дор-
сокаудальной (латеральной) поверхности шейки бедренной кости. При функционировании
протрагирует и пронирует бедренную кость.
Наружный лобково-седалищно-бедренный мускул (m. puboisciofemoralis extemus,
рис. 25, Б) начинается от вентро-медиальной поверхности лобковой кости, сухожильной
перемычки между симфизами и вентральной поверхности седалищной кости, закрепляет-
ся в области шейки головки и на проксимальном конце большого трохантера. Окончание
мышцы прикрыто конечным сухожилием длинного хвостово-бедренного мускула. Приво-
дит и супинирует бедро.
Аддуктор бедра (m. adductor femoris, рис. 26, Б) начинается коротким плоским сухо-
жилием от латеральной поверхности каудальной части седалищной кости и лобково-седа-
лищной связки, а также от дистальной части подвздошно-седалищной связки. До лобково-
го бугорка брюшко не доходит. Закрепляется мышечно-сухожильно на вентральной по-
76
верхности бедренной кости, в ее проксимальной половине, заходя частично на ее каудаль-
ную поверхность. Вентрально полностью прикрыт лобково-Сидалищно-большеберцо-
вым мускулом. Разгибает конечность в тазобедренном суставе, привод^ и прони-
рует бедро.
Длинный хвостово-бедренный мускул (m. caudifemoralis longus. рис. 21, В'. 25. Ь)
треугольный, расположен на вентральной поверхности основания хвоста, медиальнее от
начала короткого хвостово-бедренного мускула, но латеральнее от седалишно-хвостового.
Начинается от тел и поперечных отростков первых 7—8 хвостовых позвонков.Мышечные
пучки переходят в пластинчатый сухожильный тяж, которым и закрепляются на кранио-
вентральной поверхности большого вертела бедренной кости. Сильный ретрактор и прона-
тор бедра.
Короткий хвостово-бедренный мускул (m. caudifemoralis brevis, рис. 2*. В; 25,
обширный,сложно устроенный. Начинается мышечйо-сухожильно от тел и боковых отрост-
ков первых 5 хвостовых позвонков, а также через посредство седалищно-подвздошной
связки и от подвздошной кости, близ ее каудального конца. Часть мышечных пучков начи-
нается на 1-И хвостовых позвонках и затем закрепляется мышечно-сухожильно на лате-
ральном крае трохантера, рядом и под прикрытием конечного сухожилия от длинного хво-
стово-бедренного мускула. Каудальная часть пучков начинается от остальных 3—4 хвосто-
вых позвонков и закрепляется на медиальной поверхности подвздошно-седалищной связ-
ки, а также на каудальной поверхности седалищной кости, вблизи ct-далищного бугра и
под прикрытием этой связки. Краниальная порция мышцы ретраглрует и пронирует бед-
ренную кость, а каудальная — натягивает подвздошно-седалищную связку в каудомеди-
альном направлении, противодействуя закрепленным на ней большеберцовым сгибателям,
а через посредство седалищной кости прижимает хвост книзу.
Седалищно-вертлужный мускул (m. ischiotrochantericus, рис. 26. Б) расположен кау-
дальнее от внутреннего лобково-седалишко-бедренного мускула. Начинается от каудаль-
ной части симфиза седалишных костей и дорсокаудального края седтпищной кости. Далее
брюшко направляется проксимолатерально, переходит в лентовидное ?vxoжилиfc. которое
направляется через суставную сумку к дорсальной поверхности суставной головки бедрен-
ной кости, где и закрепляется. Супинирует бедренную кость укрепляет тазобедренный
сустав, прижимая головку к суставной ямке.
Передний большеберцовый мускул (m. tibialis anterior, пне. 19,В\ 4) мощный, пе-
ристый. латерально граничит с общим разгибателем пальцев, с каудомедиальной поверхно-
сти - с внутренним икроножным и подколенным мускулами. Начинается от краниомеди-
77
альной поверхности проксимального конца большеберцовой кости, в промежутке между
прямой и медиальной (коллатеральной) связками колена. Дистальный конец брюшка
полностью перемещается на медиальную поверхность голени и превращается в лентовид-
ный сухожильный тяж, который следует на медиальную поверхность проксимального кон-
ца первой плюсневой кости, где и закрепляется. Кроме этого, дистальный конец брюшка
отдает сухожильный анастомоз и к каудомедиальной поверхности проксимального конца
1-й фаланги первого пальца. Отводит медиально и частично сгибает в плюсне-фаланговом
суставе первый палец.
Длинный малоберцовый мускул (гл. peroneus longus, рис. 19, Л,Б; 25,А) расположен
латеральнее от общего разгибателя пальцев, частично прикрывает короткий малоберцо-
вый мускул и срастается с ним. Начинается коротким уплощенным сухожильным тяжем
от латерального мышелка бедренной кости, кпереди от латеральной коллатеральной связ-
ки коленного сустава, а также мышечно-сухожильно от латеропроксимальной части мало-
берцовой кости. Закрепляется на латеральной поверхности 5-й плюсневой кости. На уровне
таранной кости от конечного сухожилия мышцы отходит плоский сухожильный анастомоз,
который срастается с конечным сухожилием длинного сгибателя пальцев и отдает сухо-
жильную ветвь к медиальному краю интертарсального сустава. Отводит латерально и про-
нирует стопу.
Короткий малоберцовый мускул (m. peroneus brevis, рис. 26. Л) латерокаудально час-
тично прикрыт длинным малоберцовым мускулом. Начинается почти от всей латеральной
поверхности малоберцовой кости, кроме ее проксимальной части. Дистально брюшко пе-
реходит на латерокаудальную поверхность малоберцовой кости и в области интертарсаль-
ного сустава переходит в шнуровидное сухожилие, которым и закрепляется на латерокау-
дальной поверхности 5-й плюсневой кости, чуть проксимальнее от закрепления длинного
малоберцового мускула. При функционировании пронирует и разгибает стопу.
Длинный разгибатель пальцев (m. extensor digitorum longus. рис. 19, А, Б\ 25.Л) распо-
ложен на краниальной поверхности голени, в промежутке между передним большеберцо-
вым мускулом и малоберцовыми мускулами. Начинается лентовидным сухожильным
тяжем на латеральном мышелке бедренной кости. Закрепляется на дорсолатеральных по-
верхностях проксимальных концов 2-й и 3-й плюсневых костей и частично на брюшке аб-
дуктора и разгибателя первого и второго пальцев, которое она своим дистальным концом
прикрывает. Сгибает конечность в интертарсальном,суставе и частично супинирует стопу.
Внутренний икроножный мускул (m. gastrocnemius internus, рис. 19,2?; 25,Б) апонев-
ротически срастается с наружным икроножным мускулом и частично прикрывает его. На-
чинается от подколенной области медиального мыщелка бедренной кости и частично от
мениска, а также от каудомедиальной поверхности проксимальной половины большой бер-
цовой кости; мышечные пучки закрепляются на ветви конечного сухожилия наружного
большеберцового сгибателя, которая и выполняет роль конечного сухожилия данного мус-
кула. От конечного сухожилия брюшка отходит дополнительный лентовидный апоневроз
к медиальной поверхности дистального конца наружного икроножного мускула, капсуле
интертарсального сустава и Проксимальному концу первой плюсневой кости. Разгибает ко-
нечность в интертарсальном суставе; частично сгибает и отводит в латеральную сторону
пятый палец.
Наружный икроножный мускул (m. gastrocnemius externus. рис. 19,2>; 25, Б) частично
прикрыт внутренним икроножным мускулом, расположен на каудальной поверхности го-
лени (ближе к латеральному краю). Начинается мощным шнуровидным сухожилием на
каудальной (вентральной) поверхности бедренной кости, чуть проксимальнее латерально-
го мышелка. К этому сухожилию косвенно подсоединяется длинный сухожильный тяж.
идущий от длинного хвостово-бедренного мускула, именно в том месте, где в сухожилии
имеется сесамовидная косточка. Закрепляется на плантарной поверхности проксимального
_сонца 5-й плюсневой кости и дальше следует к проксимальным концам первых фаланг
третьего и четвертого пальцев, а также на 5-й плюсневой кости вентрально (плантарно), у
ее проксимального конца, и соединяется расширенным поперечным апоневрозом с меди-
альной стороной интертарсального сустава и с заплюсневой и плюсневой областями. При
функционировании сгибает конечность в интертарсальном суставе, способствует сгибанию
пальцев и эффективно отводит стопу в латеральную сторону.
Длинный сгибатель пальцев (m. flexor digitorum longus, рис. 26, Б) состоит как бы из
двух веретеновидно уплощенных сросшихся головок. Более поверхностная часть брюшка
78
начинается коротким шнур обидным сухожилием на латеральном мыщелке бедренной
кости, ниже начала наружного икроножного мускула, но чуть выше и каудальнее от начала
длинного малоберцового. Более глубокая часть начинается мышечно-сухожильно от каудо-
медиальной поверхности проксимального конца малоберцовой кости, а также от апоневро-
за подколенного мускула. На каудальной поверхности брюшка расположена дополнитель-
ная мышечная головка, которая начинается на конечном сухожилии наружного большебер-
цового сгибателя, а затем сливается с медиальной поверхностью дистального конца. Основ-
ная часть мышцы в области плюсны делится на 4 ветви — к концевым фалангам первого —
четвертого пальцев. При функционировании мышца сгибает пальцы и конечность в колен-
ном и интертарсальном суставах.
Глубокий пронатор (m. pronator profundus, рис. 26, Б) расположен на каудальной по-
верхности голени, в промежутке между берцовыми костями и краниальнее ci длинного
сгибателя пальцев, прикрывает верхний и нижний межберцовые мускулы. Начинается на
каудомедиальной поверхности малоберцовой кости, почти на всем ее прстяжении. За-
крепляется на плантарной поверхности проксимальных концов 1 -й и 2-й плюсневых костей
и частично на заплюсне. При функционировании пронирует стопу и сгибает интертарсаль-
ный сустав.
Верхний межберцовый, или подколенный, мускул (m. fibulotibialis superior sjn. popli-
teus, рис. 26, Б) начинается мышечно-сухожильно от медиокаудальной поверхности про-
ксимального конца малоберцовой кости. Мышечные пучки направляются под острым уг-
лом к каудальной поверхности большеберцовой кости, где и закрепляются узкой полос-
кой почти по всей длине, вплоть до медиального мыщелка. Пронирует большеберцовую
кость.
Нижний межберцовый мускул (m. fibulotibialis inferior, рис. 26,Б) направляется чуть
наклонно сверху вниз от малоберцовой к большеберцовой кости. Дистальная часть
мышечных пучков идет почти горизонтально от кости к кости. Каудально мускул
частично прикрыт подколенным мускулом и глубоким пронатором. Пронирует кости
голени.
Короткие разгибатели пальцев (mm. extensores digitorum bicves, рис. 25, Л; перистые,
составляют два апоневротически сросшихся ряда, расположенных друг над другсм. Дор-
сальная часть начинается узкими уплощенными сухожильными тяжами на дорсальной по-
верхности заплюсны, на границе между дистальными и проксимальными рядами костей,
ближе к дистальному концу большеберцовой кости, Более глубоко лежащие брюшка начи-
наются на дорсальной поверхности дистального конца заплюсны и нз дорсолатеральных
поверхностях плюсневых костей первого-четвертого пальцев, а закрепляются все они, пере-
ходя в уплощенные сухожильные тяжи, на капсулах плюсне-ф ал антовых суставов и далее
на основаниях всех фаланг пальцев, вплоть до последней. При функционировании разги-
бают, отводят вверх и сводят к центру стопы первый—четвертый пальцы.
Абдуктор и разгибатель первого и второго пальцев (m. abductor et extensor hallicus
etindicis, рис. 26, А) состоит из 2 сросшихся брюшек. Брюшко к первому пальцу начина-
ется на дистолатеральном конце малоберцовой кости и частично на проксимальном конце
таранной кости, закрепляется на медиальной поверхности дистальных 2/3 1-й плюсневой
кости и отдает слабые сухожильные апоневрозы к дорсальной поверхности 2-й плюсневой
кости и к конечному сухожилию переднего большеберцового мускула. Брюшко ко вто-
рому пальцу начинается на дорсолатеральной поверхности таранной кости и направляется
на дорсальную поверхность 2-й плюсневой кости, где и закрепляется дистальнее от закреп-
ления медиальной головки общего разгибателя пальцев. При функционировании супиниру-
ет стопу, отводит первый и второй пальцы дорсально и способствует тыльному сгибанию
конечности в интертарсальном состав
Поверхностные короткие сгибатели пальцев (mm. flexores digitorum breves superficiales,
рис. 27) веретеновидно уплощенные, расположены на плантарной поверхности стопы и
служат как бы продолжением наружного икроножного мускула. Мышечные брюшка име-
ются к первому—третьему и пятому палылам. которые перед плюснефаланговыми сустава-
ми переходят в тонкие парные сухожильные тяжи, дающие отводы ко всем фалангам паль-
цев, кроме концевых, и идущие по бокам от конечных сухожилий длинного сгибателя
пальцев. К четвертому пальцу имеется самостоятельное брюшко, которое начинается шну-
рсвидным сухожилием на дистоплантарной поверхности таранной кости, которое затем
пооходит в промежутке между проксимальными концами 4—5-й плюсневых костей. Это
79
Рис. 27. Мышш стопы разноцветной ящурки (Д, 2>) - плантарно:
1 - mm. flcxores djgitorun breves superficiales, 2 - mm. ftexores digitorum breves superficiaies profundus, 3 -
mm. lumbricales, 4 — mm. contrachentes digitorum, 3 — mm. interossei piantares
брюшко отводит четвертым палец латерально и частденго сгибает его; остальные брюшка
сгибают пальцы плантарно.
Глубоюю иширттви гии короткие сгибатели вш&оеа (пкп. flexores digitorum breves
superficial^ profundus, рве. 27) — 3 мышечных брюшка веретеновидно уплощенные, соеди-
няют дорсальную поверхность конечного сухожшшя длинного сгибателя пальцев и основа-
ния первых фаланг второго—четвертого пальцев. При функционировании сгибают пальцы
в плюснефалетговых суставах и сводят их к центру стопы.
Чфвеобраяиые мыешы (шт. lumbricales, рис. 27) в количестве 2 брюшек начинаются
между конездыми сухожилиями длинного сгибателя пальцев ко второму—третьему и
третьему—четвертому пальцам. Закрепляются короткими лентовидными сухожилиями на
основаниях предпоследних фаланг соответственно третьего и четвертого пальцев. Способ-
ствует сгибанию последних.
Мускулы, сютшагарю лшьвд (mm. contrachentes digitorum, рис. 27) , в количестве 4
вгашаются на плантарной поверхности 5-й плюсневой кости, со стороны пальцев, и направ-
ляются на латероплантарную поверхность проксимальных концов 1-х фаланг первого—чет-
вертого пальцев, где и закрепляются короткими сухожилиями. При функционировании
стягивают пальцы.
Межкостные мышцы (mm. interossei piantares, рис. 27) идут только к первому—чет-
вертому пальцам. Начинаются на плантарномедиальных поверхностях 2—4-й плюсневых
костей, почти по всей длине, а закрепляются на латероплантарных поверхностях дисталь-
ных концов 1— 3-й плюсневых костей и частично переходят на капсулы плюснефаланговых
Ю
суставов и проксимальные концы 1-х фаланг названные пальцев. Мышечное брюшка . че-
твертому пальцу начинается уплощенным сухожильным тяжем на медис плантарной повер-
хности 5-й плюсневой кости и от заплюсневых 4—5-й. Закрепляется на латеральной поверх-
ности проксимального конца первой фаланги четвертого пальца и дает сухожильную ветвь
к дальнейшим фалангам пальца. При функционировании стягивают и отводят латерально
плюсневые кости; частично способствуют сгибанию стопы в плюснефаланговых суставах
первого — четвертого пальцев.
Особенностью межкостной мышцы первого пальца является то, что брюшко ее более
отчетливо дифференцировано на три части, из которых средняя — истинно межкостная
мышца, медиальная отводит палец в медиальную сторону, а латеральная приводит его к се-
редине стопы.
5 3. НЕРВНАЯ СИСТЕМА
53.1. ГОЛОВНОЙ МОЗГ
Головной мозг рептилий интенсивно исследовался в историческом и сравнительном аспек-
тах [32, 226, 275, 329,330, 382,394]. В структурной организации вентральной нервной си-
стемы (ЦНС) пресмыкающихся наблюдаются различия, свойственные представителям
определенного подкласса или отряда, но наряду с ними сохранились и особенности, общие
для всего класса рептилий. Это закономерно, если учесть особенности филогенеза и обпзг
жизни составляющих этот класс специфических групп.
Мы использовали литературные данные и результаты изучения 3 подвидов разноцвет-
ной ящурки (E.a.deserti. Ед. arguta, Е.а. uzbekistanica). Большинство образований, о ко-
торых будет идти речь, даже если им присуши видовые отличия, имеют микроскопические
размеры и скрыты в глубине мозга.
Передний мозг — самый крупный отдел головного мозга разноцветной ящурки пред-
ставлен хорошо выраженными полушариями и центральными узлами полушарий — к'члп-
Рис. 29. Головной мозг разноцветной ящурки:
1 - обонятельные луковицы. 2 - обонятельный тракт. 3 - передний мозг. 5 - мозжечок.
6 - гипоталамус (воронка), 7 - продолговатый мозг. 8 - гипофиз; II-XII - черепномоз-
говые нервы
лексами нервных ядер и проводящих афферентных и эфферентных путей. Вместе с пла-
щом, или мантией (pallium), они составляют конечный мозг (telencephalon) Передний
мозг (рис. 28, 29) имеет грушевидную форму, под ним скрыты промежуточный мозг, под-
бугорная область, таламус и зрительные тракты. Поверхность полушарий лишена борозд
и.извилин. Поперечные срезы теленцефалона, сделанные на разных уровнях, четко показы-
вают преобладание по объему базального отдела над паллиальным (рис. 30) Важнейшая
особенность паллиума всех пресмыкающихся в том, что строение поверхностного слоя пла-
ща полностью гомологично коре (серому веществу) полушарий мозга млекопитающих.
Кора (cortex cerebri) состоит в свою очередь из трех слоев — плексиформного (молеку-
лярного, зонального) , клеточного и слоя белого вещества, в целом являясь еще примитив-
ной. Образующие кору нейроны и их отростки охватывают в виде пластинки дорсальную к
боковые поверхности полушария; у чешуйчатых рептилий (ящерицы хамелеоны) кора со-
стоит из нескольких обособленных, но перекрывающих одна другую пластинок [32] .Ме-
диальный отдел дорсальной коры содержит клетки Гольджи I типа. Видовые особенней ,
строения латерального отдела у ящурки не выяснены.
81
1
2
Рис. 30. Поперечные срезы переднего мозга :
1—4 - через 0,5 мм, начиная от краниальной границы переднего мозга
В настоящее время общепризнано, что с точки зрения филогенеза медиодорсальная зо-
на коры рептилий относится к архикортексу, латеральная — к палеокортексу, а один из
участков коры — паллиальное утолщение — часть исследователей рассматривают как зача-
точный неокортекс. Найдены цитоархитектонические и гистологические различия между
этими областями коры, но такое подразделение следует учитывать также в связи со специа-
лизацией переработки информации мозговой корой, ято является одним из показателей
степени (высоты) организации нервной системы. В новой коре рептилий отчетливо прояв-
ляется тенденция к специализированному восприятию сигналов обонятельных афферентов,
с одной стороны, и зрительных и соматических — с другой [108]. Этим свойством обладает
и мозг разноцветной ящурки. Возможно, что в передних 2/3 общей коры паллиума подоб-
но Emys orbicularis [32] локализуется представительство зрительной, слуховой и сомати-
ческой сенсорных систем.
В толще каждого полушария большого мозга расположен боковой мозговой желудо-
чек. Оба желудочка сообщаются между собой, а также с третьим мозговым желудочком,
который принадлежит промежуточному мозгу, но проникает и в передний мозг (рис. 31).
К щелевидной полости третьего желудочка прилегают полосатые тела (corpora striata);
они же выступают в виде выпуклостей на вентролатеральной поверхности мозга. В эти
структуры поступает информация от обонятельных и необонятельных экстерорецепторов.
В вентромедиальной части полушария расположена перегородка (septum), в которой за-
ключены ядра, где, как полагают, у рептилий переключаются импульсы, идущие от эксте-
рорецепторов в сетевидную формацию мозга [32]. Дорсомедиальная часть полушария —
место расположения обширной структуры, называемой гиппокампом, или аммоновым
комплексом.
Для большинства дневных ящериц главное значение имеют цветные зрительные сигна-
лы, для ночных — звуки [344], у них, наряду с этим, как у всех рептилий, хорошо развита
обонятельная система. Ее первичные центры — обонятельные луковицы (ОЛ) , к ним из пе-
редних областей полушарий тянутся тонкие обонятельные тракты (рис. 28,29) . ОЛ полые
и хорошо развиты у всех разноцветных ящурок независимо от места их обитания. Крани-
82
Рис. 31. Парасагиттальные срезы головного мозга:
1-4 через 0,15 мм от латеральной поверхности мозга
альная поверхностная зона ОЛ примыкает к наружной поверхности обонятельной мем-
браны. Средняя мозговая оболочка, которой покрыты луковицы, как и мозговые полуша-
рия, значительно пигментирована, но распределение хроматофоров неравномерно.
Строение ОЛ рептилий, а также связи их нейронов с рецепторами и структурами юлов-
ного мозга освещены в ряде работ [32, 58, 226, 247]. Здесь отметим лишь, что ольфактор-
ные раздражители вызывают в нервном аппарате ОЛ яшериц сложные процессы, как мест-
ные межнейронные взаимодействия, так и реакции, включающие контроль со стороны цен-
тральных образований в мозге. Большое число главных дендритов митральных клеток, ин-
нервирующих клубочки, и малый диаметр обонятельных трактов свидетельствуют в поль-
зу суммации афферентных импульсов в ОЛ. Сигнализация луковиц распространяется в
структурах обоих мозговых полушарий, вызывая обратные потоки импульсов из вторич-
ных обонятельных центров переднего мозга. В генерацию обратных потоков импульсов во-
влечены такие структуры, как габенулярный комплекс, пириформная (грушевидная) ко-
ра, амигдалярный комплекс; последний получает информацию и от ОЛ, и от Якобсонова
органа — по вомероназальному нерву. Кроме того, от ОЛ в кору обоих полушарий следу-
ют прямые волокна. В передаче ольфакторной информации у ящериц и черепах обязатель-
но участвует габенулярный комплекс. Главный вторичный обонятельный центр - пири-
формная кора. Гиппокампальная кора активируется помимо обонятельных висцеральны-
ми и экстерорецепторными импульсами [32]; следовательно, ее интегративная деятель-
ность непосредственно связана с поведенческими реакциями.
В толще мозговых полушарий расположена значительная группа центральных нервных
узлов, среди которых можно отметить, например, бледный шар, скорлупу, хвостатое ядро.
Эти узлы выполняют функцию высших двигательных центров, поскольку у рептилий нет
пирамидной системы (корковые центры и проводники), осуществляющей у высших по-
звоночных произвольную координацию моторики.
К полушариям переднего мозга прилегает средний мозг (mesencephalon, рис. 32).
В дорсальной части его лежит важная структура — крыша (tectum) - центр переработки
зрительной и двигательной информации. Глубже расположены развивающиеся задние буг-
ры четверохолмия. У разноцветной ящурки они не выступают за пределы гектума, крыши
среднего мозга. В задней части зрительных бугров имеется октаволатеральное ядро (ком-
понент зрительной системы).
В краниовентральной части среднего мозга отчетливо выделяется хиазма, образован-
ная обоими короткими мощными зрительными нервами, точнее — каждый из нервов раз-
деляется на 5—6 пучков волокон, которые перекрещиваются подобно пальцам рук, следу-
ют латеродорсально и в виде оптических трактов ясно различимы на боковых поверхно-
стях мозга.
Вентральный отдел среднего мозга — ствол — является продолжением заднего мозга
(metencephalon). В центре ствола проходит сильвиев водопровод, с помошью которого
сообщаются между собой мозговые желудочки, хорошо развитые у разноцветной яшурки
на уровне заднего и межуточного, слабее — переднего мозга. В субэпендиме бокового же-
лудочка низших позвоночных расположена зона матрикса, где образуются нервные клетки
мозговых полушарий [217].
83
72J4S6
Рис. 32. Средний мозг:
1—6 — поперечные срезы, 7 — уровень срезов
Краниальный отдел разросшегося переднего мозга прикрывает межуточный мозг
(diencephalon). К его важнейшим структурам относится парный зрительный бугор (thala-
mus opticus) — система ядерных образований и связующих волокон вблизи бокового же-
лудочка мозга. В специализированные ядра таламуса поступают афферентные импульсы
от зрительного, слухового и соматических рецепторов. В аборальной части крыши межу-
точного мозга расположены несимметричные ганглии, или габенулярный комплекс (gan-
glia habenulae). В его ядрах импульсы с обонятельной системы переключаются на двига-
тельные ядра ствола мозга; ядра комплекса соединены габенулярной комиссурой.
84
В гипоталамусе (вентральной части межуточного мозга) имеется несколько ядер свя-
занных с корой полушарий, таламусом и другими подкорковыми центральными узлами
переднего и заднего мозга. Структуры оказывают существенное нервное и гормональное
влияние на протекание вегетативных функций (терморегуляция, обмен веществ и др.), с по-
мощью которых достигается эффективная адаптация рептилий к условиям среды обитания.
В центре гипоталамической области расположены воронка гипофиза и сам гипофиз -
важнейшая железа. Воронка у Eremias — компактное образование, выступающее
на вентральной поверхности мозга и тонким тяжем соединяющееся с гипофизом.
Глубоко в воронку проникает сильвиев водопровод. У молодых особей E.arguta (гНико-
лаев) воронка и гипофиз относительно ббльших размеров, чем у крупных половозрелых.
Для мозга ящериц характерно наличие своеобразных структур, развивающихся как
полые выросты крыши промежуточного мозга, которые постепенно отшнуровываются
и превращаются в два пузырька, соединенных ножкой с мозгом [311]. Передний пузырек
называют теменным, или париетальным, органом; у высших рыб, особенно у Amia, у двоя-
кодышащих рыб и у многих ящериц он приобретает строение более или менее сформиро-
ванного теменного, или третьего, галаза или теменного органа. Задний пузырек превраща-
ется в железу внутренней секреции эпифиз (epiphisis cerebri s. gl. pinealis).
Теменной глаз наиболее развит у Ед. uzbekistanica; к овальному отверстию в теменной
кости подходит теменной нерв, по диаметру превышающий многие черепномозговые нер-
вы, он следует под сильно пигментированной мозговой оболочкой. В отверстии теменной
кости располагается линзообразная, прозрачная в центре структура, окруженная пигменти-
рованным кольцом. Часть пигментных клеток мигрирует от края к центру линзы. У Е.а.
deserti теменной нерв развит слабее, после выхода на поверхность мозга он тонким тяжем
направляется к париетальному органу. Теменной орган городской популяции Е.a.arguta
рудиментарен, так как отсутствует сформированный концевой отдел (линза), теменной
нерв не выходит за пределы мозга, а заканчивается расширением на дорсальной границе
среднего и переднего мозга далеко позади foramen parietale черепа, что исключает попада-
ние в такой третий глаз прямых лучей света.
Третий глаз построен у разноцветной ящурки по принципу прибора прямого изобра-
жения. В отличие от инвертированных глаз позвоночных в нем отсутствует сложная систе-
ма нейронов сетчатки. Информация рецепторов теменного органа по париетальному нер-
ву поступает к nucleus habenulae, расположенному перед задней комиссурой мозга. Учиты-
вая то, что в габенулярных узлах заканчиваются аксоны рецепторов теменного органа,
можно предположить, что этим же путем к двигательным центрам поступает информация
как непосредственно от теменного органа, так и после ее интеграции с ольфакторно-лим-
бической .
Теменной орган развит не у всех лацертид. Обычно он сохраняется у роющих ящериц,
у древесных форм часто редуцируется, и в области пинеального комплекса откладывает-
ся хрящ [376]. Третий глаз не был обнаружен у бразильских семейств Gekkonidae и Tei-
idae. тогда как у игуановых, особенно у Topinambis torquatus, в этом органе имеются хру-
сталик, сетчатка, развитый нерв и хороидная оболочка. Наряду с этим в нем отсутствуют
мышечные волокна, т.е. аккомодация таким глазом не достигается [409]. С помощью те-
менного органа Sceloporus olivaceus (игуаны) продолжал различать изменения интенсивно-
сти постоянного освещения после хирургического удаления глаз, но Lacerta sicula (настоя-
щие ящерицы) такую способность утратила [457]. На основании сравнений выдвинуто
представление, дающее более широкое толкование распространения третьего глаза среди
семейств ящериц [377]. Установлено, что практически все геккониды и тейиды лишены
третьего глаза; они обитают в экваториальных широтах. Агами ды, игу аниды, л ацсртиды и
сцинковые ящерицы, у которых третий глаз имеется, распространены в более высоких ши-
ротах, но те немногие из них, у которых нет теменного органа, — животные экваториаль-
ных областей. Согласно предложенной теории, деятельность фоторецепторов третьего гла-
за, принимающих участие в терморегуляции и синхронизации репродуктивных циклов (см.
ниже) , является одним из факторов, позволивших адаптироваться ящерицам к относитель-
но суровому климату.
Фоторецепторная деятельность (активность) теменного органа влияет на организм реп-
тилий различно. Эпифиз найден у всех изучавшихся ящериц, также у видов, не имеющих
теменного глаза [457]. Чередование освещенности и темноты в процессе жизненного цикла
(и у разноцветной ящурки), воспринимаемое третьим глазом, отражается на продукции
85
гормона эпифиза — мелатонина, что в конечном итоге ведет к изменениям размеров тес-
тикулов и интенсивности сперматогенеза. В последнее время подтверждено участие те-
менного органа в терморегуляции. Так, ящерицы Anolis caroliensis с удаленным теменным
органом выбирали в термоградиенте (15—25 °, фотопериод постоянный) более высокую
температуру во все часы суток [383].
Длина продолговатого мозга (myelencephalon) у крупных взрослых особей разноцвет-
ной ящурки 5 мм, средняя ширина — 2—2,5. Каудальная граница продолговатого мозга —
место выхода первой пары спинномозговых нервов, краниальная — перешеек (isthmus).
Снизу проходит только вентральная серединная борозда, на дорсальной поверхности име-
ется глубокая ромбовидная ямка (четвертый мозговой желудочек) . Центральная дорсаль-
ная борозда продолговатого мозга выражена у Eremias слабо. Продолговатый мозг имеет
дяа изгиба: передний — на границе со средним мозгом, задний — на уровне VIII пары череп-
номозговых нервов. В пределах продолговатого мозга расположены ядра V—XII пар череп-
номозговых нервов.
Серединная часть продолговатого мозга ящурки занята сетевидной, или ретикулярной,
формацией. Сетевидной формации заднего мозга, особенно у наземных позвоночных, при-
надлежит важная роль в координации движений; так, она обеспечивает быстрые и резкие
движения (схватывание добычи, например).
В определенных участках нейроны сетевидной формации сгруппированы в ядра с раз-
ветвленной системой контактов между их телами, аксонами и дендритами соседних кле-
ток и более отдаленных нервных клеток. У рептилий нервные волокна, принадлежащие се-
тевидной формации, собраны как бы в 3 пучка, которые тянутся вдоль мозгового ствола
в сторону продолговатого мозга. Ядра сетевидной формации продолговатого мозга, в
том числе черепномозговых нервов, являются не только посредниками между системами
рецепторов, прежде всего органов равновесия, слуха, внутренних органов, мышц и кож-
ных покровов в зонах, где ветвятся эти нервы, и вышележащих отделов ЦНС, но и центра-
ми, которые регулируют (модифицируют) протекание процессов на периферии. В качестве
примеров таких функций ядер можно назвать формирование реакции на звуковые раздра-
жители; хотя ящерицы, по-видимому, как и аллигаторы, способны воспринимать комп-
лекс звуков, у них импульсы слуховых рецепторов поступают в дорсальное крупнокле-
точное и латеральное мелкоклеточное ядра (охватывает подобно чаше наружный край ром-
бовидной ямки), тогда как у амфибий имеется только одно ядро [226].
Импульсы сетевидной формации поддерживают мозг животного в более или менее
бодрствующем, настороженном состоянии, что имеет решающее значение для выживания
слабо защищенных и охотящихся подобно ящурке животных.
В сетевидной формации заднего мозга содержатся два жизненно важных центра, где
замыкаются афферентные и эфферентные нервные пути, при участии которых непрерывно
поддерживается постоянство внутренней среды организма — центр дыхательных движений
и ядра X пары нервов. Ритм и частота дыхательных движении задаются специализирован-
ной частью нейронов сзтевидной формации близ аборального края четвертого желудочка,
активность которых автохтонна; эти же клетки анатомически и функционально связаны с
нейронами ядер блуждающего нерва. Ядра этого нерва обеспечивают также регуляцию
моторной и секреторной функций пищеварительного тракта и сердечных сокращений.
Основной объем продолговатого мозга ящурки занят центральными проводящими
спинномозговыми путями, связывающими головной мозг со спинным и статоакустическим
аппаратом. По ходу путей имеются ядра, среди них — двигательное ядро чепца (начиная с
рыб). Оно связано специальными нервными волокнами со структурами продолговатого
мозга, средним мозгом и мозжечком. За счет этого ядра, начиная с амфибий, обособля-
ются так называемые оливы — пучки волокон, идущих от вестибулярного (VIII пара) нер-
ва к таламусу.
Мозжечок (cerebellum) у разноцветной ящурки, совершающей точные и быстрые,
но тем не менее относительно простые движения, развит слабо по сравнению с другими от-
делами мозга. Он невелик и внешне его строение просто, деление на доли отсутствует. Моз-
жечок имеет вид лепестка с утолщением в центральной части (на сагиттальном срезе),
частично прикрывает ромбовидную ямку, краниально прилегает к задней поверхности
среднего мозга, даже надвигается на него.
У подвидов Ejirguta, обитающих в разных регионах, размеры и форма мозжечка не
86
являются постоянными. Наибольшего развития он достигает у Е л.uzbekistanica, укоторой
заходит на дорсальную поверхность среднего мозга, а его каудальная часть почти полно-
стью закрывает четвертый желудочек. Средняя часть утолщена, выделяется зачаток чер-
вячка. По нашему мнению, размеры и форма мозжечка могут служить таксономическим
признаком. Мозжечек ящериц изучен слишком слабо, чтобы обрисовать его функциональ-
ную роль, хотя известно, что он более развит у рептилий (Crocodilia, Chelonia) с развитыми
конечностями; у них уже присутствуют внутримозжечковые ядра, участвующие в перера-
ботке информации, связанной с осуществлением двигательных актов. Х.С.Коштоянц
[142] вкратце описал наблюдения Гаккер, из которых следует, что мозжечок влияет на
координацию движений и тонус мышц у ящериц, однако слабее, чем у филогенетически
более развитых позвоночных.
532. ЧЕРЕПНОМОЗГОВЫЕ НЕРВЫ
При макроскопическом осмотре головного мозга разноцветной ящурки различимы ко-
решки черепномозговых (головных) нервов, которые, как известно,выполняют исключи-
тельно важную функцию поддержания непрерывной двусторонней связи между разнооб-
разными периферическими рецепторами и головным мозгом. Говоря о рептилиях, следует
учитывать замечание ИИДИмальгаузена [311] о том, что у низших позвоночных постоянно
имеется вместо 12 лишь 10 пар вполне типичных черепномозговых нервов. Поэтому пер-
вым из черепномозговых нервов И.И.П1мальгаузен отмечает концевой нерв (п. termina-
lis) — смешанный (по функциональному значению нервных волокон) очень тонкий нерв,
идущий от орального края переднего мозга в слизистую оболочку Якобсонова органа.
Если этот нерв и имеется у разноцветной ящурки, то его выделить очень трудно.
I. Обонятельные нервы (n. olfactorius) еще не сформированы, так как обонятельные
мешки с их рецепторами у ящурки сильно сближены с обонятельными луковицами. Поэто-
му аксоны обонятельных рецепторов отдельными тонкими пучками подходят к обонятель-
ным луковицам, где вместе с дендритами митральных клеток образуют обонятельные клу-
бочки. От крупных обонятельных луковиц ольфакторные тракты следуют к обонятельным
отделам переднего мозга.
И. Зрительные нервы (n. opticus) у ящурки самые крупные по диаметру из черепно-
мозговых нервов, но относительно короткие из-за латерального положения глаз животно-
го. В хиазме происходит практически полный перекрест волокон. Степень развития этих
нервов отражает роль зрительной информации в жизни животного.
Ш. Глазодвигательные нервы (n. oculomotorius) отходят почти в области перехода
продолговатого мозга в стволовую часть среднего, сразу же за воронкой. Наиболее круп-
ные среди моторных нервов глазной мускулатуры. Диаметр Ш нерва у разноцветной
ящурки почти в 2 раза превышает таковой блокового нерва (TV пара) . Глазодвигательные
нервы на коротком расстоянии следуют рострально под средним мозгом, затем выходят
за пределы черепа через большое заднеглазничное отверстие. Направляясь к глазной муску-
латуре, нерв распадается на две ветви — соответственно к нижцей косой и внутренней пря-
мой мышцам — первая, к верхней прямой и ретрактору глаза — вторая.
IV. Блоковые нервы (n. trochlearis). По литературным данным [226], ядро блокового
нерва располагается за ядром глазодвигательного, однако блоковый нерв выходит не на
вентральной, а на каудальной поверхности среднего мозга (двухолмия) , сразу за зачатком
паруса. Место его выхода прикрыто мозжечком. IV нерв сначала следует латерально, до бо-
ковой поверхности среднего мозга, затем вдоль нее к глазному яблоку через заднеглазнич-
иое окно (см. разд. ’’Череп”) . Внутри орбиты поворачивает медиодорсально, проходя под
сухожилием мигательной перепонки к верхней косой мышце.
V. Тройничные нервы (n. trigeminus) развиты хорошо. Отходят несколькими корешка-
ми от латеральной поверхности передней части продолговатого мозга, за пределы черепа
выходят двумя стволами, развит Гассеров узел. Делится тройничный нерв на три ветви: глаз-
ничную, максиллярную и мандибулярную. Глазничная ветвь (в одном случае выходила
как самостоятельный нерв, имеющий в основании ресничный ганглий) направляется кра-
ниодорсально к орбите, вступает в нее и делится на нерв для вспомогательных структур
глаза и ветвь, следующую вместе с блоковым, нервом под сухожилием медиально, а затем
через отверстие в передней части глазницы, входящую в носовую полость. Вторая ветвь,
87
менее развитая, следует рострально под глазным яблоком к тканям, иннервируемым ею.
Третья, мандибулярная, после отделения от общего ствола направляется вентрально к
нижней челюсти (ее мускулатуре, коже, зубам).
VI. Отводящие нервы (n. abducens) менее развиты, чем блоковые, отходят от вен-
тральной поверхности продолговатого мозга почти на том же уровне, что и тройничный
(на несколько миллиметров каудальнее), следуют под продолговатым и каудальной ча-
стью среднего мозга, проходят через отверстие в базисфеноиде, направляясь к наружной
прямой мышце глаза и мышцам, смещающим третье веко.
УП, Vffl. Статоакустический (n. statoacusticus) (VII пара) и лицевой (n. facialis) (VIII па-
ра) нервы выходят из продолговатого мозга на уровне заднего изгиба, сразу за тройнич-
ным нервом, но дорсальнее. Основная ветвь направляется к области уха. Лицевая ветвь
следует рострально и ветвится в верхней и нижней челюстях.
К. Языкоглоточный нерв (n. glossopharyngeus) отходит от боковой поверхности про-
долговатого мозга, рядом с корешками, образующими блуждающий нерв. Слабо разви-
тый нерв, вместе с блуждающим выходит через отверстие в затылочной кости и направля-
ется к языку и гортани.
X. Блуждающий нерв (п. vagus) достаточно развит, посылает свои ветви к внутренним
органам (легким,сердцу, желудку, пищеводу и др.) .
ХЖ. Подъязычный нерв (п. hypoglossus) отходит от вентральной поверхности продол-
говатого мозга наиболее каудально. Этот тонкий нерв иннервирует преимущественно мус-
кулатуру языка.
533. ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
Ранние исследования периферической нервной системы Lacertidae были посвящены изуче-
нию устройства шейно-головного отдела симпатического ствола. Описаны ’’глубокая” и
’’поверхностная” части этого нерва в области его выхода из полости черепа [368, 379,
453]. По мнению Т.Терни [453], "глубокая” часть симпатического ствола представляет
собой гомолог позвоночного сплетения, свойственного млекопитающим, хотя у ящериц
в вех кровеносного сосуда, сравнимого с позвоночной артерией млекопитающих.
Более подробные и полные описания симпатической системы и структуры иннерва-
ции области аортальных и легочных дуг, а также каротидных сосудов у ящериц появляют-
ся в 30-40-х годах XX в. Однако по сравнению с имеющимися данными по другим клас-
сам позвоночных (особенно млекопитающим) полученных сведений недостаточно [325—
327,343,384,453].
Симпатический ствол в грудном отделе ящериц представлен билатеральной цепочкой
нервных узле», простирающейся в хвостовой отдел. Самый крупный узел ствола — первый
(звездчатый) узел грудной цепочки. В шейном отделе симпатического ствола Lacerta stri-
gata имеется 2—3 нервных узла. Приводя описание устройства грудного отдела симпатиче-
ской системы этой ящерицы, В .С Абашидзе [1-3] в составе ствола выделил выраженный
внутренностный нерв, берущий начало от последних VI—VII грудных позвонков. В.Адамс
[327] этого нерва у L. vfridis не обнаруживает, однако отмечает* хорошо развитые висце-
ральные ветви последних узлов,калибр которых больше, чем у межузловых ветвей симпа-
тического ствола. По мнению ВА.Кальберга [105, 106], исследовавшего некоторых яще-
риц и черепах, названный нерв у рептилий отсутствует.
Из работ, посвященных вопросам строения соматической нервной системы, с точки
зрения функционального и сравнительного анализа, наиболее информативными представ-
ляются исследования, проведенные В.САбашидзе [1—3] и ДЛ.Асатиани [16]. Авторы вы-
деляют ряд доминирующих тенденций в формировании плечевого и пояснично-крестцово-
го сплетений на примере ряда представителей рептилий: проксимальное формирование
нервов обусловлено более дифференцированной функцией конечностей; в филогенезе
плечевого м пояснично-крестцового сплетений происходит каудальное смещение этих
структур; первоначальной формой проявления концентрации ганглиозных элементов яв-
ляется увеличение объема нервных узлов.
Приведенное положение хорошо согласуется с принципом концентрации функции,
предложенным НЗЗСокшайским [41], отражающим процесс топографической локализа-
ции функции, и эмбриологическими данными, указывающими на то, что большая величина
88
нервных стволов и узлов непосредственно связана с периферической нагрузкой на нервы.
Сведений, касающихся строения симпатической и соматической систем у представите-
лей рода Eremias, в доступной литературе нами не обнаружено. Материалом исследования
послужили представители Е. arguta uzbecistanica и E.a.deserti.
Вегетативная нервная система E.a.uzbekistanica. В области выхода симпатического
ствола из полости черепа регистрируются два стволика примерно одинакового калибра
(рис. 33). Медиально расположенный нерв связан тонкими ветвями с шейными нервами
этой области. Этот ствол прилегает к латеральной полуокружности внутренней сонной
артерии. Латеральный ствол, посылая соединительную веточку к подъязычному нерву,
срастается с медиальным на уровне передней 1/3 II шейного позвонка. Видимых контак-
тов между симпатическим стволом и блуждающим нервом в этой области не зарегистриро-
вано. Шейные узлы ствола не обнаружены.
Блуждающий нерв формирует массивный нервный узел в области IV шейного позвон-
ка (рис. 33). Этот узел принимает участие в иннервации краниального отрезка внутренней
сонной артерии, краниальной каротидной дуги и каротидного синуса. Звездчатый узш
(рис. 33) располагается на уровне межреберного промежутка II и III грудных позвонков.
Дорсальная поверхность узла уплощена, вентральная выпуклая. Узел лежит на поверхно-
сти второго ствола плечевого сплетения. Краниальная межузловая ветвь узла утолщена
и укорочена по сравнению с каудальной. Второй грудной узел значительно меньшего раз-
мера, обладает сходной формой, лежит между III и IV грудными позвонками. К каудально-
му полюсу узла присоединяется ветвь V спинального нерва (рис. 34) . На некоторых пре-
паратах этот узел отсутствовал. Форма третьего узла, расположенного между IV и V по-
звонками, вариабельна. Каудальная ветвь узла уплощена, к одноименному полюсу его при-
соединяется длинная ветвь от VII спинального нерва (у одного из исследованных объектов
указанная ветвь превосходила по размерам и диаметру межузловую ветвь симпатического
ствола, по-видимому, являясь местным коллектором).
Следующий нервный узел располагается на уровне тела VI позвонка или смещается
каудальнее. Это мелкий узел, его каудальная межузловая ветвь приближается по размерам
к его диаметру. Пятый узел смещается к основанию позвоночного столба, его каудальная
ветвь круто поднимается дорсовентралъно (до этого симпатический ствол располагался
в основании позвоночного столба) , идет параллельно позвоночнику, вблизи VIII позвонка
ветвь возвращается к основанию позвоночника. У двух из исследованных ящурок, начи-
ная с V узла, симпатический ствол смещался в плоскость ребер, межузловые ветви упло-
щались. У тела VH грудного позвонка расположен крупный округлый узел. Каудальная
ветвь его укорочена, расширяясь, переходит в крупный звездчатый узел у каудального
полюса VIII позвонка. Этот узел служит корешком крупной висцеральной ветви. От кра-
ниального полюса узла отходит длинная ветвь к краниально расположенному спиналь-
ному нерву. Каудальная ветвь дугой, обращенной к основанию позвоночника, подходит
к IX позвонку, у краниального полюса которого расположен седьмой узел. Он отдает две
извитые висцеральные ветви, направленные краниально, к дорсальной поверхности пече-
ни, попутно иннервируя стенку дорсальной аорты.
В каудальном направлении от названного узла идет хорошо выраженный внутренно-
стный нерв (рис. 35). У одного из объектов он появляется на уровне VIII грудного по-
звонка. Следующий узел симпатического ствола может отсутствовать. От межузловой
ветви отходит висцеральная ветвь. У X позвонка имеется узел овальной формы, от него
отходят три висцеральные ветви, пересекая аорту, направляются к краниальному полю-
су желудка. Начиная с VIII (иногда IX—X) грудного узла, ножки узлов ствола удлиняют-
ся, узлы приближаются к иннервируемым органам (печени, желудку, мочеполовому ком-
плексу) . Между XI и XII позвонками узлы симпатического ствола снижаются до уровня
продольных позвоночных связок, имеющих вид сплошной ’’перепонки”. Одновременно
удлиняющиеся ножки узлов симпатического ствола смещаются в латеральном направле-
нии, вдоль соответствующих спинальных нервов. Это связано, по-видимому, с перемеще-
нием узлов симпатического ствола в соединительнотканную перегородку, развивающую-
ся в связи с задней стенкой перикарда (вентральная область), косо подразделяющую
туловище ящурки в направлении от V ребра (дистального края) к телам X -XI грудных
позвонков. Начиная с XII—XIII позвонков узлы симпатического ствола уменьшаются в
размерах, приобретая овальную форму. В пояснично-крестцовом отделе обнаружены тон-
89
кие нервные ветви между краниальными стволами пояснично-крестцового сплетения и
узлами симпатического ствола.
Наблюдалась определенная асимметрия в строении симпатического ствола. В двух слу-
чаях отмечено дорсовентральное смещение ствола одной из сторон по сравнению с контр-
латеральной, начиная со второго грудного узла. Висцеральные ветви узлов этого нерва так-
же смещались значительно краниальнее — они появлялись на уровне второго—третьего
грудных узлов. В то же время внутренностный нерв был смещен в область XIV—XV по-
звонков. Этот нерв имел два корешка и один плоский ствол.
Соматическая нервная система E^.uzbekistanica. Плечевое сплетение (рис. 36) располо-
жено в области I—IV грудных позвонков, в его составе имеются три—четыре первичных
ствола. Основными источниками сплетения являются первые 4 грудных спинальных нер-
ва. а также тонкие веточки от каудальных шейных нервов. Первые два ствола срастаются
в единый ствол (сращение происходит на незначительном удалении от области выхода из
позвоночного столба). Вблизи плечевого сустава ствол расщепляется на два ствола, кау-
дальный присоединяется к стволу, образованному за счет сращения Ш и IV грудных спи-
нальных нервов. Спинальные нервы срастаются значительно дистальнее ранее упомянутых.
Сразу же после присоединения П—IV нервов начинается дифференциация общего ствола на
ряд ветвей, дающих начало нервам конечности. Слева количество нервов, участвующих в
образовании сплетения, может уменьшаться до трех за счет ^выпадения” каудального
ствола.
Пояснично-крестцовое сплетение. Краниальная граница его (рис. 37) проходит в обла-
сти ХХШ (чаще — начиная с XXIV) спинального нерва. Наиболее развитыми источниками
сплетения являются XXIV и XXV нервы. Вблизи капсулы тазобедренного сустава перечис-
ленные нервы формируют дугообразное сращение; в некоторых случаях, наблюдалось бо-
лее сложно устроенное образование. Пояснично-крестцовое сплетение начинает дифферен-
цироваться на больше- и малоберцовые нервы вблизи капсулы сустава.
Вегетативная нервная система Ед.deserti. Краниальный участок симпатического ствола
в шейном отделе практически не отличается по устройству от такового у Ед. uzbecistanica.
В области выхода из черепа подъязычного и языкоглоточного нервов между симпатиче-
ским стволом и названными нервами имеются многочисленные связи, чего не обнаружено
у ранее исследованной ящурки. В этой же области имеется краниальный шейный узел сим-
патического ствола в виде овального утолщения на нерве. Блуждающий нерв залегает ней-
тральнее симпатического ствола. Краниальный участок нерва проходит в миоэпителиаль-
ном утолщешш, богато васкуляризованном. Узел этого нерва располагается вблизи крани-
альной полуокружности каротидной дуги. На некоторых препаратах была зарегистрирова-
на хорошо развитая гортанная ветвь, отходящая от упомянутого нервного узла блуждаю-
щего нерва, распространяющаяся но боковой поверхности трахеи, краниально. От этого
узла также отделяется каротидная ветвь, идущая к краниальной полуокружности дуги
аорты.
Основной ствол блуждающего нерва направляется к вентральной поверхности сердца.
В этой области от нерва отделяется желудочная ветвь, распространяющаяся по боковой по-
верхности аорты. В области звездчатого узла наблюдаются тонкие веточки, соединяющие
желудочную ветвь блуждающего нерва с указанным узлом. Звездчатый узел (рис. 34) мо-
жет быть представлен одиночным очень крупным овальным узлом (впрочем, форма узла
достаточно вариабельна). В 1/3 случаев регистрировался звездчатый комплекс, состоящий
из двух сращенных между собой узлов, при этом степень сращения различна. Звездчатый
узел располагается на мышцах шеи, иннервируя их одной-двумя мелкими веточками;
кроме этого наблюдаются связи между стволами плечевого сплетения и дорсальной по-
верхностью узла. Каудальная ветвь узла удлинена, за счет чего симпатический ствол в
грудной полости смещается на боковую поверхность тел позвонков. На уровне П грудного
позвонка наблюдаются обмен волокнами между стволами контрлатеральных сторон. Уз-
лы сиьшатического ствола имеют форму овалов с заостренными полюсами.
Начиная с VB—VIII позвонков ножки узлов постепенно удлиняются (рис. 35). В этой
области располагается тонкое сплетение, образованное ветвями блуждающего нерва. На
некоторых препаратах узлы симпатического ствола не выражены. На уровне XII—ХШ по-
звонков у некоторых объектов обнаруживается внутренностный нерв (в составе его име-
ется 2—3 коронка, иногда нерв сопровождается несколькими висцеральными ветвями).
90
В пояснично-крестцовом отделе наблюдаются отдельные нервные ветви, идущие от узлов
симпатического ствола к нервам одноименного сплетения. У одного из исследованных эк-
земпляров обнаружен крупный узел, располагающийся на вентральной поверхности позво-
ночного столба; этот узел связывает стволы противоположных сторон, а также иннервиру-
ет продольные связки позвоночника.
Соматическая нервная система E.a.deserti. По сравнению с Е a. uzbecistanica наблюдает-
ся увеличение компонентов шейного сплетения за счет присоединения каудальных сегмен-
тов спинальных нервов и веточек блуждающего нерва. В составе плечевого сплетения име-
ются четыре основных ствола и три—пять тонких ветвей (рис. 36, 38). Передний ствол
сплетения наиболее мощно развит, на уровне плечевого сустава срастается на значительном
протяжении со вторым стволом. Общий ствол распространяется по медиальной поверхно-
сти плечевой кости. Остальные два ствола сращены на коротком участке, уровень сраще-
ния — несколько проксимальнее предыдущих стволов, затем ствол расщепляется на 3 нер-
ва. Краниальный отдает длинную ветвь к общему стволу, образованному передними нер-
вами сплетения. Остальные нервы, древовидно ветвясь, распространяются среди задних
пучков мускулатуры.
Пояснично-крестцовое сплетение выглядит менее дифференцированным по сравнению
с одноименной структурой у Е.а. uzbecistanica (рис. 39,40) . В формировании его принима-
ют участие XXIII спинальный нерв (в ряде случаев это не наблюдалось) , XXIV, XXV и вет-
ви XXVI нерва. Последние из перечисленных нервов могут образовывать сращение на ко-
ротком отрезке перед капсулой тазобедренного сустава, после чего в проксимальных уча-
стках мускулатуры бедра общий ствол дифференцируется на 2 самостоятельных нерва.
В некоторых случаях XXIV нерв связан с XXV за счет тонкой ветви (уровень капсулы су-
става) .
Следовательно, наблюдается смещение плечевого сплетения в каудальном направле-
нии, характерное для всех исследованных форм ящурок. При этом нервы плеча формиру-
ются значительно проксимальнее у Е.а. uzbecistanica, что указывает на возможность осуще-
ствления более дифференцированных движений передних конечностей подвида. Это же каса-
ется и степени развития и дифференцировки пояснично-крестцового сплетения у назван-
ных подвидов разноцветной ящурки. Отмечаются существенные различия в строении и
топографии симпатического ствола и его взаимоотношениях с системой блуждающего
нерва. Хорошо развиты ганглионарные элементы указанных нервов. У исследованных реп-
тилий в той или иной степени развитый внутренностный нерв.
5.4. ОРГАН ЗРЕНИЯ
5.4Л. ГЛАЗНОЕ ЯБЛОКО
Орган зрения — важнейший дистантный рецептор разноцветных ящурок. Это отражено
в строении их глаза, которое обеспечивает эффективное функционирование его при днев-
ном образе жизни, активной охоте, частом рытье убежищ преимущественно в сухих песча-
ных почвах (рис. 41).
Глазные яблоки у ящурки расположены латерально. Такое их расположение свидетель-
ствует о сильно развитом панорамическом зрении и небольшом бинокулярном. Глазное
яблоко относительно крупное (горизонтальный диаметр около 8 % длины туловища) , сжа-
то по передне-задней оси, вертикальный диаметр несколько меньше горизонтального.
Наружная оболочка глаза очень тонкая. Передняя часть — роговица — круглая, слабо
выпуклая, диаметр ее у взрослых особей может достигать 1,6—1,8 мм, Роговица сформи-
рована такими же слоями, как и у других позвоночных. Переход роговицы в склеру обо-
значен корнеосклеральной бороздой (впадиной). За ней располагаются 14 склеральных
косточек, необычайно тонких, гибких и прозрачных, но лишь во влажной среде. Их коли-
чество вряд ли может служить надежным признаком для таксономической диагностики,
так как характерно для ящериц вообще [458]. Косточки своим роговичным краем на-
кладываются друг на друга. Именно за счёт этого и создается впечатление непрерывного
костного кольца. Оно у ящурки занимает практически всю фронтальную поверхность
склеры, укрепляя ее и поддерживая выпуклую форму глазного яблока. Склера, особен-
91
Рис. 41. Схема представительства зрительной си-
стемы в среднем, промежуточном и переднем
мозгу:
1 - тектум оптикум, 2 - круглое ядро, 3 - ко-
ленчатые тела, 4 — общая кора
но в задней полусфере, состоит главным об-
разом из прозрачной хрящевой ткани, не-
сколько уплотненной в области выхода зри-
тельного нерва.
Средняя оболочка незначительной тол-
щины. Как собственно сосудистая оболоч-
ка, так и радужина темные. Окраску раду-
жины полезно учитывать в целях система-
тики, поскольку, хотя ’’мезодермальная
часть ириса сильно окрашена, как если бы
делалась попытка обратить внимание на
глаз” \353. — С. 437], разные представители
ящериц отличаются цветом этой оболочки;
он может быть красным, желтым, бриллиантовым с металлическим блеском (у хамелео-
нов) или почти черным, как у ящурки (благодаря меланину). Радужина утолщается к
зрачковому краю. В ней хорошо развиты волокна кольцевого сфинктера, волокна дилята-
тора залегают тонким слоем под эпителием. Цилиарная мышца хорошо развита. Само ци-
лиарное тело крупное, зрачок округлый.
Хрусталик у ящурки сплющен по оси глаза. У исследованных нами взрослых особей его
максимальный диаметр (горизонтальный) достигал 2—2,5 мм, вертикальный не превышал
2.2, а медиальный — 2 мм. Хрусталик мягкий, его передняя поверхность уплошена, даже
имеется слабая вогнутость в центре, задняя выпукла и окружена развитой экваториальной
подушечкой, которая образована необычайно разросшимися клетками субкапсулярного
эпителия и достигает мощного цилиарного тела, опирающегося на нее. У большинства днев-
ных видов хрусталик более плоский, чем у других рептилий [458]. Наличие подушечки
свойственно и другим рептилиям.
Макро- и микроскопическими методами исследования на внутренней оболочке мож-
но выявить область соска зрительного нерва; у разноцветной ящурки он круглый и сдви-
нут латерально. В центральной зоне расположен длинный тонкий пигментированный конус
(conus papillaris). Это специальная, характерная для ящериц, структура, образованная тон-
кими пигментированными кровеносными сосудами, которые удерживаются вместе ней-
роглией. Конус тянется от зрительного нерва через стекловидное тело в направлении
центра хрусталика (рис. 42). Это образование, питающее сетчатку, отсутствует л инн» у
семейства червеобразных, роющих ящериц [458]. Сама сетчатка толстая, лишена сосудов.
Ее строение типично для всех дневных ящериц: четкая дифференциация клеточных слоев,
зрительные клетки двух типов — палочки и колбочки, содержащие желтый каротиноид-
ный пигмент.
5 А2. ВСПОМОГАТЕЛЬНЫЕ СТРУКТУРЫ ГЛАЗА
Глаз Ел. arguta защищен хорошо развитыми веками. Верхнее веко неподвижно, покрыто
мелкими чешуйками. В центральной части века этого вида располагается вытянутое черное
пятно. Нижнее веко подвижное, с крупным прозрачным окошком у молодых ящурок и
полупрозрачным у крупных взрослых особей. Таким образом, сомкнутые веки, защищая
глаз от механических раздражений и яркого света, не препятствуют функционированию
глаза. Края век пигментированы (состоят из темноокрашенных ресничных чешуек) . Пиг-
мент заходит на смыкателъную поверхность и конъюнктиву век.
Во внутреннем углу глаза располагается прозрачная мигательная перепонка — третье
веко. От нижнего края мигательной перепонки отходит тонкий сухожильный тяж, перехо-
дящий на заднюю поверхность глазного яблока. Здесь он формирует длинное тонкое ци-
линдрическое сухожилие. В области зрительного нерва оно внедряется в специальную
мышцу, сокращение которой вызывает натяжение мигательной перепонки. Часть волокон
сухожильного тяжа подходит к нижней прямой мышце. Сухожильные волокна связаны и
с верхним краем мигательной перепонки.
Под третьим веком в переднем углу глаза залегает одна из двух желез — Гардерова.
У разноцветной ящурки обращают на себя внимание ее большие размеры (крупная у ла-
цертид вообще); железа переходит на назомедиальную поверхность, а затем на заднюю по-
верхность глаза, почти достигая зрительного нерва. Она практически полностью прикры-
92
вает медиальную прямую мышцу гла-
за, наименее развитую из всех прямых
глазных мышц. Размеры, форма и по-
ложение Гардеровой железы отличают-
ся у разных видов Lacertidae [337].
Железа выделяет как слизистый, так и
серозный секреты. Микроскопические
исследования у Lacerta agilis и L. mura-
lis [440] показали относительную обо-
собленность участков, выделяющих
слизистый или серозный секрет, а так-
же их выводных протоков. Слизистые
протоки открываются снаружи от ми-
гательной перепонки, а серозные выде-
ляют секрет на ее внутреннюю поверх-
ность. Большое количество секрета
распределяется на верхнем веке. Ме-
нее развита слезная железа. Это не-
большое образование, расположенное
в области наружного угла глаза.
По нашему мнению, секрет желез
необходим и для постоянного смачи-
вания нижнего века, чем поддержива-
ется прозрачность окошка, так как с
прекращением выделения секрета, не
позднее чем через 30 мин после смер-
ти животного, прозрачность утрачива-
Рис. 42. Схема строения глазного яблока разноцветно?
ящурки:
1 ~ роговина, 2 - склера, 3 - сосудистая оболочка,
4 - радужина, 5 - цилиарное тело {5а - цилиарные от-
ростки, 56 - цилиарная мышца, 5в — основная пластин-
ка) , 6 - сетчатка, 7 - стекловидное тело, 8 - YnycTc-
лик (8а - подушечка (корзинка) хрусталика), 9 - ко-
нус, 10 - зрительный нерв, 11 - склеральная косточга.
12 - Е1леммов канал
ется.
Наружные мышцы глаза у всех подвидов разноцветной ящурки развиты достаточно
хорошо. Прямые мышцы глаза начинаются от заднего конца cartilago hypochiasmatica.
Верхняя и наружная прямые мышцы глаза прикрепляются к глазному яблоку за склераль-
ными косточками соответственно в верхней и темпоральной области задней полусферы
глазного яблока.Внутренняя прямая развита слабее, почти полностью покрыта Гардеровой
железой, начинается тонким сухожилием. Расположение нижней прямой мышцы обычное.
Внутренняя косая мышца начинается коротким сухожилием из внутреннего угла глаза
(от кости орбиты) , затем, огибая глаз медиально, выходит на заднюю поверхность глазно-
го яблока, проходит над Гардеровой железой и крепится к вентрал: ?й части глаза. Верх-
няя косая мышца начинается от передней назальной орбитальной стенки, переходит нс дор-
сальную верхнюю часть глазного яблока, где и крепится, немного не доходя до верхней
прямой мышцы.
Кроме 6 описанных, мышц у ящурки хорошо развит ретрактор глаза и имеются мыш-
цы, через которые проходит сухожилие мигательной перепонки. Сокращение их вызыва-
ет натяжение сухожилия и задергивает мигательную перепонку.
Иннервация мышц описана в разделе ’Черепномозговые нервы”. Д.Третьяков [455],
подробно изучавший периорбитальные образования Lacerta agilis exigua Eihu и Locellata.
подтвердил наличие открытых Вебером (1877) венозных и лимфатических синусов, за-
легающих между стенками глазницы, поверхностью глазного яблока и его наружными
мышцами. По его мнению, такие структуры облегчают трофику мышц, их сокращения;
по-видимому, они имеются и у разноцветной ящурки, у которой, учитывая особенности ее
природной среды, могут также способствовать оптимизации температуры глаза.
В общем в строении глаза Eremias выявлены признаки, типичные для всего класса пре-
смыкающихся и лацертид в частности, причем активных днем. Исходя из того что многие
из живущих форм и подгрупп рептилий вновь стали ночными, в строении глаза разноцвет-
ной ящурки унаследованы особенности, достигнутые предками этого вида, так как ранним
рептилиям был свойствен строго дневной образ жизни [458].Поскольку глаза являются глав-
ными органами световосприятия, импульсные сигналы фоторецепторов сетчатки - через
головной мозг и железы внутренней секреции — воздействуют и на функцию воспроизведе-
ния; в этом воздействии принимает участие и третий, или теменной, глаз. Его строение рас-
смотрено в разделе ’Толовной мозг”
93
53. КРОВЕНОСНАЯ СИСТЕМА
Еще в ранних исследованиях кровеносной системы рептилий [331,460] выдвигались пред-
положения о том, что, несмотря на несовершенство разделения сердца, следует ожидать
приспособлений, обеспечивающих нормальное функционирование двойного кровообраще-
ния. Дальнейшие изыскания в этой области подтвердили высказанные догадки. Сердце реп-
тилий не только обеспечивает функционально разделение артериальной и венозной крови,
выполняя главное требование для позвоночных — перенос газов, но и позволяет создавать
более высокое давление в артериальной системе по сравнению с давлением в капиллярах
жабр низших позвоночных [116, 206, 312, 357]. Некоторое смешивание потоков крови
(наблюдаемое даже в четырехкамерном сердце крокодилов) и неизбежная при этом гипо-
ксии компенсируется за счет эффективного легочного кровообращения (кроме некоторых
групп черепах, иногда использующих клоакальную респирацию) [206,312].
Артериальная система рептилий по сравнению с амфибиями претерпела довольно суще-
ственные преобразования. За исключением гаттерии и некоторых черепах, у большинства
рептилий sinus venosus сильно редуцирован — большая его часть вошла в состав стенки
правого предсердия. Это же произошло и с артериальным конусом [308]. Обе системные
дуги начинаются самостоятельно: левая системная дуга аорты берет начало от правой сто-
роны желудочка и отклоняется на левую сторону, правая — от левой стороны желудочка,
отклоняется вправо. Эти сосуды соединяются за счет небольшого foramen Panizzae. Веноз-
ная система рептилий изменилась в меньшей степени. Исчезновение кожного дыхания при-
вело к повышению роли легочной циркуляции: легочные вены, несущие оксигенированную
кровь, вариабельны по размерам и форме (одна из них может отсутствовать из-за редук-
ции левого легкого у некоторых змей) . Большая часть крови от задних отделов тела отте-
кает через крупную брюшную вену, включающую в себя воротную вену печени. Значение
воротной системы почек ослаблено: кровь от конечностей и хвостового отдела может по-
падать либо через воротную систему почек и далее в заднюю полую вену, либо через та-
зовую вену и далее — через брюшную вену в печень. Рептилийная vena cava posterior, как и
у амфибий, происходит от кардинальных и (частично) желточных вен [308,310].
53.1. СЕРДЦЕ
Сердце (сог) расположено в медиастинальном пространстве, между обоими листками pleu-
ra mediastinalis. Передняя поверхность и верхушка сердца прилегают к антеродорсальной
поверхности грудной клетки, дорсальная сторона перикардиального мешка сердца сопри-
касается с органами заднего средостения. В перикарде можно выделить два отдела: peri-
cardium viscerale, покрывающий сердечную мышцу, а также расположенные здесь сосуды
сердца; pericardium parietale, связанный с окружающими образованиями посредством рых-
лой соединительной ткани — спереди с задней поверхностью corpus stemi, по бокам с pleura
mediastinalis, дорсально с aorta thoracica и каудально с зачаточной fascia diaphragma tica.
Желудочек сердца разделен неполной косо направленной перегородкой на две полости:
более крупную — дорсолатеральную и меньшую — вентролатеральную.
Каждое из предсердий открывается в соответствующую ему часть дорсолатеральной
полости. Ветвящиеся мышечные пучки названной полости имеют вид трабекул, регуля-
ция ритма наполнения сердца предотвращает полное смешивание артериальной и венозной
крови. Тонкостенный венозный синус встроен в стенку правого предсердия. Сюда впадают
3 крупные вены: пара передних w. cavae и непарная v.cava posrerior. Перепончатые клапаны,
имеющиеся в местах впадения перечисленных вен, являются остатками sinus venosus. Ди-
стальная часть практически редуцированного артериального конуса расщепляется на
3 главных ствола, каждый из которых обладает одним рядом клапанов, лежащих в его
основании.
532. АРТЕРИАЛЬНАЯ СИСТЕМА
Наиболее крупным и заметным из 3 упомянутых кровеносных сосудов является правая
системная дуга (arcus aortae dexter), которая подразделяется на 2 ствола еще в пределах
перикарда (рис. 43). Левая системная дуга (arcus aortae sinister) появляется в области про-
екции свободного края горизонтальной перегородки. Легочная артерия (arteriae pulmona-
94
Рис, 43, Артериальная система
1 - наружная сонная артерия. 2 -
внутрення сонная артерия. 3 — лева:,
и правая общие сонные артерии.
4 - правый сонный проток, 5 - ле-
вая и правая системные дуги аорты,
6 — легочная аорта, 7 - грудная ар-
терия, 8 — плечевая артерия, 9 — шей-
ная артерия, 10 - вентральная и
задняя ветви легочной артерии, 11 -
передние желудочные артерии, 12 -
поджелудочная железа, 13 - желу-
док, 14 - задняя желудочная арте-
рия. 15 - селезеночная артерия,
16 - селезенка, 17 - дорсальная аор-
та. 18 - тонкий кишечник, 19 - пе-
ченочно-кишечная артерия, 10 - гени-
тальная артерии. 21 - почечные арте-
рии, 22 — ректальная артерия, 23 -
наружная подвздошная артерия, 24 -
внутренняя подвздошная артерия
les) — третий главный ствол
(порядок появления сосудов:
легочный ствол, правая и левая
системные дуги) . Легочная арте-
рия начинается от вентральной
поверхности желудочка. Ср^эу
после выхода из желудочка ее
ствол распадается на левую и
правую легочные ветви, направ-
ляющиеся к соответствующим
легким. Каждая из перечислен-
ных артерий, в свою очередь,
расщепляется на 2 ветви: вен-
тральная ветвь распространяется
по одноименной поверхности лег-
кого, вблизи его латерального
края, дорсальная располагается
на спинной части лишь частично
(каудальный отрезок этого сосу-
да проникает на внутреннюю по-
верхность легкого). Правая си-
стемная дуга аорты делится на
безымянную артерию (а.апопу-
ша) и левую общую сонную ар-
терию (a. carotis communis si-
nister) . Безымянная артерия идет
в краниальном направлении по
правой стороне и разделяется на
правую общую сонную артерию (a carotis communis dexter) и правую системную дугу. Правая
общая сонная артерия разделяется на 4 основные ветви: правая наружная сонная артерия
(a. carotis externa dexter), идущая к основанию языка; по ходу распространения этот сосуд
снабжает стенки пищевода и глотки за счет аа. pharyngea et buccinator; правая внутренняя
сонная артерия (a. carotis interna dexter) идет параллельно пищеводу, в области глаза пово-
рачивает в дорсальном направлении (этот сосуд снабжает основание головы и мозга) ; пра-
вая мышечная артерия (a.musciilaris) снабжает мышцы шейного отдела; правый сонный про-
ток (ductus caroticus dexter) идет каудально, затем открывается в одноименную дугу аор-
ты. Левая общая сонная артерия делится на ветви, идентичные названным правой общей
сонной артерии.
Правая системная дуга идет в краниальном направлении параллельно правой общей
сонной артерии. В начале ветвления последней она изгибается дорсально в направлении к
пищеводу. На вершине изгиба в этот сосуд впадает правый сонный проток. В каудальном
направлении дуга пронизывает плевральную полость на уровне перехода pleura mediasti-
nalis в pleura costalis, огибает легкое и идет по направлению к правой полуокружности пи-
щевода. Соединение с левой системной дугой и образование дорсальной аорты (aorta dor-
salis) происходят несколько каудальнее передних конечностей. Краниальнее соединения
этих сосудов от правой системной дуги отходят левая и правая подключичные артерии
(аа. subclaviae sinister et dexter), идущие в толще гипаксиальной мускулатуры. От каждой
из названных артерий, в свою очередь, отделяется ряд более мелких сосудов: a. brachialis,
a. axillaris, a. sub scapularis.
Левая системная дуга после впадения в нее одноименного сонного протока отдает не-
сколько пищеводных артерий, затем дорсально изгибается и идет в точку соединения с
правой системной дугой.
533. ДОРСАЛЬНАЯ АОРТА И ЕЕ ВЕТВИ
В качестве основных ветвей этого крупного кровеносного сосуда представлены передние
желудочные артерии (аа. gastricae anterior), снабжающие кардиальный отдел желудка
.имеется 3—4 пары этих сосудов); желудочно-селезеночная артерия (a. gastrolienalis) —
гонкий сосуд, разделяющийся на ветви к селезенке (a. lienalis) , пилорическому отделу же-
лудка (a. gastrica posterior), и сосуд, направляющийся к поджелудочной железе (а. рапсгеа-
йса); печеночно-кишечная артерия (a. hepatomesenterica) делится на заднюю кишечно-бры-
жеечную артерию (a. mesenterica posterior), направляющуюся к толстому кишечнику, и пе-
ченочно-двенадцатиперстную артерию, снабжающую печень и двенадцатиперстную кишку
(a. hepatoduodenalis); передняя кишечно-брыжеечная артерия (a. mesenterica anterior) снаб-
жает петли тонкого кишечника и брыжейку; генитальные артерии (аа. genitales) — пара
тонких сосудов, обеспечивающих кровоснабжение гонад; ректальная артерия (a. recta-
lis) начинается от дорсальной аорты, этот сосуд располагается между почками, снабжает
прямую кишку; тазовая артерия (a. pelvica) обеспечивает питание мышц таза, клоаку и
соответствующий гемипенис самца; подвздошная артерия (a. iliaca) продолжается в зад-
ние конечности в виде бедренной артерии (a. femoralis); почечные артерии (аа. renales)
в количестве нескольких пар впадают в паренхиму почек; хвостовая артерия (a. cauda-
lis) является продолжением дорсальной аорты в хвостовом отделе. Кроме этого отмечают-
ся мелкие париетальные артерии (аа. parietales), обеспечивающие кровоснабжение позво-
ночного столба и мускулатуры.
534. ВЕНОЗНАЯ СИСТЕМА
Этот круг кровообращения может быть условно разделен на четыре функциональные еди-
ницы: передняя венозная система, легочные вены, портальная система печены и задняя ве-
нозная система (рис. 44) . Передняя венозная система содержит левую и правую передние
полые вены (venae cava anterior dexter et sinister), впадающие в венозный синус совместно
с задней полой веной (vena cava posterior). Передняя vena cava несет кровь от 3 главных со-
судов: наружной и внутренней яремных вен, а также подключичных вен. Наружная ярем-
ная вена (v. jugularis externa) представлена тонким сосудом, обеспечивает дренаж языка,
мыши подъязычной области и щитовидной железы; внутренняя яремная вена (v.jugula-
ris interna) несет кровь от остальной части головы за счет челюстной вены (v. mandibula-
ris), идущей из области нижней челюсти, и головной вены (v. cephalica), несущей кровь от
тканей мозга; подключичная вена (v. subclavia) принимает кровь от грудных позвонков
через позвоночные вены и из передних конечностей, включая область лопатки через v.sub-
scapularis. Подкрыльцовая вена (v. axillaris) образована за счет 3 главных ветвей: v. sub-
scapularis — из одноименного участка, v. pectoralis — от грудных мышц и v. brachialis — из
передних конечностей. Легочные вены (w. pulmonales), транспортирующие оксигенирован-
ную кровь из легких, перед впадением в левое предсердие объединяются в общий ствол.
Портальная система печени обеспечивает транспорт крови, содержащей абсорбирован-
ный пищевой материал из органов желудочно-кишечного тракта. Портальная вена направ-
ляется к печени в близком соседстве с желчным протоком. В месте ее впадения в печень
портальная вена соединяется с брюшной веной (v. abdominalis). Вне печени этот сосуд от-
96
Рис. 44. Венозная система :
1 наружная яремная вена, 2 -
правая и левая челюстные вены, 3 —
внутренняя сонная вена, 4 - правая
и левая легочные вены, 5 - под-
крыльцовая вена, 6 — подлопаточ-
ная вена, 7 - плечевая вена, 8 - ор-
ганные ветви легочной вены, 9 ~ зад-
няя полая вена, 10 - пищеводная ве-
на, 11, 15 — воротная вена печени,
12 - передняя и задняя вены желуд-
ка, 13 - брюшная вена, 14 - перед-
няя кишечная вена, 16 - вены семен-
ника, 17 - тазовая вена, 18 - бедрен-
ная вена, 19 - почечные вены и отхо-
дящие от них подчревные вены,
20 — ректальная вена, 21 - хвосто-
вая вена
дает 2 главные ветви: желудоч-
ную (v. gastrica) и кишечную
(v. mesenterica) вены. Желудоч-
ная вена принимает кровь от пе-
редней и задней ветвей, распола-
гающихся на соответственных
поверхностях желудка. К пече-
ни описываемый сосуд идет по
поверхности поджелудочной же-
лезы, к которой посылает нес-
колько одноименных вен. Ки-
шечная вена образована за счет
слияния желудочно -селезеноч-
ных вен, куда оттекает кровь от
желудка и селезенки, а также от
передней части тонкого кишеч-
ника (от двенадцатиперстной
кишки и подвздошной области).
Венозная система задней час-
ти туловища состоит из брюш-
ной вены, почечной портальной
системы и задней полой вены.
Брюшная вена (v. abdominalis)
в передней своей части представ-
лена 2 ветвями, одна из которых берет начало из паренхимы печени, другая же, более круп-
ная по диаметру, — от портальной вены печени. Задний участок этого кровеносного сосуда
сформирован за счет пары тазовых вен (w. pelvicae), в эту область впадают с вентральной
поверхности подчревные вены (w. hypogastrica) и лобковая вена (v. pudenda).
Почечная портальная система собирает кровь от задней части туловища и задних конеч-
ностей к почкам. Пара коротких почечных вен (vv.renales) расположена на задневентраль-
ной поверхности этих органов. В паренхиме почек они представлены в виде венул и капил-
лярных сплетений. Кровь от каждой конечности транспортируется через бедренную вену,
в полости тела представленную наружной подвздошной веной (v. iliaca externa). Кровь из
области клоаки собирается во внутреннюю подвздошную вену (v. il. interna), которая со-
единяется с наружной, образуя короткую общую подвздошную вену. Этот сосуд впадает в
почечную вену. Тазовая вена берет начало в области сращения описанных сосудов.
Задняя полая вена образована за счет слияния правой и левой почечных вен (правая
более короткая и крупная по диаметру) . Несколько краниальнее в эту вену впадают мно-
гочисленные генитальные вены (w. genitales) . Полая вена проникает в паренхиму печени,
проходит ее транзитом, получая печеночные вены (w. hepaticae), пищеводную вену (v. eso-
phageales), а также позвоночную вену (v. vertebrales).
97
5.6. СИСТЕМА ОРГАНОВ ДЫХАНИЯ
Строение дыхательной системы Eremias arguta типично для настоящих ящериц (Lacerti-
dae). Вдыхаемый воздух через наружные носовые отверстия (ноздри) поступает в прегуте-
рие носа, затем — в проходы между завитками максиллярной носовой раковины (con-
cha), причем в легкие направляется часть воздушного потока, следующая через нижние за-
витки раковины, далее — по носоглоточному ходу и сквозь хоаны в глотку, куда откры-
вается вход в гортань. Эта часть системы — верхние дыхательные пути — выполняет две
функции: проведение воздуха при вентиляции легких и обонятельную. Соответственно в
носовой полости различают два отдела, которые разнятся, в частности строением слизи-
стой оболочки: расположенный впереди и каудовентрально дыхательный (pars respirato-
ria) и задний — собственно обонятельную область (pars olfactoria). Под раковиной, в ниж-
ней части обонятельной области заканчивается тянущийся из конъюктивального мешка
слезно-носовой канал. Для более углубленных морфологических сравнений рекоменду-
ется работа М.Габе и Г.Сен Жерона [364], в которой приведены результаты подробного
анатомического и гистологического исследований носовой полости 28 семейств подклас-
са Lepidosauria.
Дыхательная часть носовой полости разноцветной ящурки выстлана эпителием без
развитых желез и обонятельных клеток. Эпителий pars olfactoria образован чувствитель-
ными, опорными секреторными и базальными клетками, которые укреплены на основной
пластинке; в ней же имеются трубчато-альвеолярные Боуменовы железы. С помощью
электронной микроскопии на поверхности всех видов клеток у австралийской Tilique
scincoides обнаружены микроворсинки, а в чувствительных клетках, кроме того, — палоч-
ки и реснички [387].
Почти несомненно, что подобные образования, способствующие усилению контакта
обонятельного эпителия с молекулами вдыхаемых пахучих веществ, частично растворен-
ных в секретах желез, имеются и у разноцветной ящурки, обладающей развитым чув-
ством обоняния. Обонятельный эпителий лацертид, как и гекконов, амфисбен и ксанту-
зий, особенно обильно снабжен сенсорными клетками; у названных животных они состав-
ляют 77—83 % всех клеток эпителия, тогда как у варана обонятельный эпителий слабо раз-
вит и заметно редуцирован у специализированных водных змей (Acrochordus, некоторые
Hydrophidae) [432].
Чувствительные клетки входят также в состав Якобсонова органа, лежащего в рас-
тральном отделе носовой полости. У ящурки это парное образование на фронтальном сре-
зе имеет полулунную форму со сближающимися краями и небольшую щель (полость)
в середине, из которой узкий канал ведет в ротовую полость. Снаружи орган покрыт хря-
щевой выпуклой капсулой, под ней залегает цилиндрический обонятельный эпителий, об-
разованный клетками тех же видов, что и в pars olfactoria носовой полости. В полость ор-
гана с вентральной стороны вдается выступ, покрытый слоем эпителия. С носовой поло-
стью Якобсонов орган не сообщается. Как в других участках pars olfactoria, так и в Якоб-
соновом органе чувствительные клетки являются биполярными нейронами, их аксоны ок-
ружены телами и отростками базальных клеток. На дендритах сенсорных клеток тоже вы-
явлены микроворсинки, содержащие нитевидные структуры [387]. Вероятно, у ящурки,
подобно остальным ящерицам и змеям, Якобсонов орган сохраняется в течение всей
жизни, хотя его обонятельная чувствительность, если судить по числу сенсорных клеток,
несколько ниже, чем у тейид, ангвиноидей, амфисбен и большинства змей [432].
Первым собственно дыхательным органом является гортань (larynx), расположенная
в задней части дна ротовой полости; частично она прикрывается корневой частью языка.
Вход в гортань имеет продольную форму. Общая форма гортани обусловлена перстне-
видным хрящом (cartilago cricoidea) (рис. 45), который частично разомкнут лишь в перед-
нем участке с вентральной стороны; в этом промежутке есть 2 черпаловидных хряща
(cartilago arytaenoidea). Похоже, у Е. arguta они являются если не отростками, то, по
крайней мере, формируются (обособляются) из вентральной части последнего (рис. 45).
Снаружи гортань покрыта тонким слоем соединительной ткани, в целом орган связан
с близлежащей мускулатурой (рис. 46). Слизистая оболочка, покрывающая мускулату-
ру гортани, является продолжением слизистой нижней поверхности корня языка. Полость
гортани выстлана цилиндрическим мерцательным эпителием. Под слизистой оболочкой
98
лежат хорошо развитые сфинктер и мускулы-расширители гортани; они функционируют
в связи с актом дыхания, а не фонации, поскольку у Eremias, как и у прыткой ящерицы,
не обнаруживаются складки слизистой — голосовые связки, хорошо развитые у гекко-
нов, хамелеонов, крокодилов [375], и голосовые способности ящурки не описаны.
Гортань сращена с относительно длинной трахеей, ее остов образован приблизительно
50 хряшевыми кольцами. Несмотря на относительно легкую доступность использования
этого показателя и в полевых условиях, он вряд ли подходит для систематической диагно-
стики, потому что такое же, тоже заметно варьирующее, количество колец свойственно и
прыткой ящерице [30].
Каждое из колец трахеи разноцветной ящурки не является сплошным замкнутым или
незамкнутым, но состоит из нескольких хрящей (рис. 47). Как и в гортани, хорошо за-
метен выстилающий трахею цилиндрический мерцательный эпителий. Наличие реснитча-
тых клеток свидетельствует о защитной функции эпителия трахеи; судя по некоторым не-
давним работам, эта функция у рептилий, в том числе, по-видимому, и у ящурок, развита
достаточно сильно и подкрепляется также деятельностью желез. Так, у большинства пре-
смыкающихся среди клеток трахеи, несущих реснички, разбросаны секреторные, у кроко-
дилов, змей, варанов они образуют скопления [454]. Установлено, что у Testudo graeca
и Pseudemys scripta elegans слизистые клетки продуцируют сиало- и сульфомуцины, но осо-
бый интерес вызывает тот факт, что электронная микроскопия позволила, во-первых, об-
наружить видовые различия в ультраструктуре реснитчатых клеток у этих черепах, во-вто-
рых, выявить наличие эндокринных клеток среди секреторных клеток [408].
В аборальной части трахея ящурки делится на 2 коротких, относительно крупных по
диаметру бронха. В их стенке также имеются кольца из гиалинового хряща. Изнутри брон-
хи покрыты однослойным цилиндрическим эпителием. Внелегочный бронх, внедрившись
в краниальную треть легкого через его медиальную поверхность, не образует дальнейших
ветвлений, т.е. у ящурки нет обычного для млекопитающих бронхиального дерева.
Легкие ящурки мешковидные, вытянутые образования, суживающиеся в каудальной
части (рис. 48). Их краниальные края располагаются прямо под сердцем, каудальные
достигают разного уровня — правое легкое доходит до желудка, левое вытянуто несколь-
ко больше, вследствие чего легкие в общем несколько асимметричны.
Сквозь поверхность легкого видны переплетающиеся белесые тяжи, что в совокупно-
сти создает рисунок крупноячеистой сети. На разрезе его подобие мешку усиливается бла-
годаря незначительной толщине, мягкости стенок и полости в центральной части, тянущей-
ся от устья бронха до каудальной оконечности органа. После сильного искусственного раз-
дувания легкого переплетение толстых и тонких тяжей становится вполне отчетливьш
(рис. 49).
Как показывает увеличение, соединительнотканные перегородки-септы образуют кар-
кас легкого и служат в то же время стенками ячеек (камер) . Дном ячейки является стен-
ка легкого, которая сформирована очень тонким слоем респираторного эпителия (рис.50,
51). В этом слое и осуществляется газообмен, так как в нем ветвится сеть кровеносных
капилляров — производных крупных сосудов, идущих от бифуркации трахеи по поверх-
ности легкого. Легкие разноцветной ящурки определенно не покрыты сплошным листком
висцеральной плевры, как, например, у крокодилов и варанов [375]. По данным сравни-
тельного исследования [415], форма и строение межкамерных септ в легких разных пре-
смыкающихся сходны; основу септ составляют волокна и клетки соединительной ткани,
свободные концы утолщены и кроме немногочисленных эластиновых волокон содержат
гладкомышечные клетки. Развитие мускулатуры в легких рептилий подчеркивал и А.Жюй-
ес-Пелисье [375].
Однако если строение септ ящурки в общем соответствует приведенному выше, то
вопрос о легочной мускулатуре требует дальнейшего подробного выяснения, поскольку у
других ящериц ей придается большое функциональное значение. Так, у Calotes versicolor
(Agamidae) описана сеть гладких мышц, которые иннервируются блуждающим нервом и
обеспечивают сокращение самих легких [389]. В перегородках легкого австралийской
черепахи Chelonia longicollis вдоль ветвей легочной артерии, вне легкого и в его ткани тоже
установлено наличие пучков гладких мышц, иннервированных адренэргическими волок-
нами [448].
Существенный компонент легких — сеть их кровеносных сосудов—у разноцветной ящурки
99
представлен правой и левой легочными артериями, несущими из правой части желудочка
сердца венозную кровь. Делясь, артерии 3-го порядка образуют капиллярную сеть у наруж-
ной поверхности ячеек и здесь же соединяются с венами 3-го порядка. Правая и левая
легочные вены объединяются в общую легочную вену, которая доставляет в левое пред-
сердие артериальную кровь. Это описание несколько схематизировано, так как для Е. аг-
guta желательно уточнить взаимоотношения легочных сосудов и плевры, вопрос о возмож-
ном сопровождении сосудов мышечными пучками, ход артерий 1-го порядка. Основыва-
ясь на особенностях этих образований, например, сосудистая система легкого гаттерии
оценена как очень простая и стоящая ближе к Anura, чем к другим рептилиям [461].
От функционирования дыхательной системы зависят, исходя из строения верхних ды-
хательных путей, не только способность ориентации ящурки по запахам, но и удовлетворе-
ние энергетических нужд, связанных с общей повседневной активностью животного, и
осуществление терморегуляторных реакций1. Выживание ящурок — пойкилотермных живот-
ных — требует предотвращения тканевой гипоксии и глубоких сдвигов кислотно-щелочно-
го равновесия (последние могут наступить вследствие перегрева животного). Компенса-
ция этих состояний первоначально достигается с помощью вентиляции легких и при учас-
тии кислородтранспортной функции крови. Как видно из опытов М.Ф.Тертышникова
[260], потребление кислорода прыткой ящерицей и разноцветной ящуркой отчетливо из-
меняется в зависимости от температуры окружающей среды и у ящурки потребление выше.
Такая реакция может быть ответом на изменение температуры, как у рептилии Thamnophis
из провинции Манитобы (но не из Флориды) , у которой потребление кислорода уменьша-
ется по мере снижения температуры от 30 до 20 ° и резко падает при температуре ниже
15° [208]. ЛЛроссер сообщает [208], что жабовидная ящерица Phrynosoma, предпочитав-
шая в террариуме условия, при которых ее температура тела удерживалась на уровне
35°, тяжело дышит при 43,2°. Многие рептилии, например сцинки, при приближении тем-
пературы тела к верхней критической границе начинают тяжело дышать, а другие просто
держат рот открытым [208]. Очевидно, в природе пресмыкающиеся изыскивают возмож-
ности избегать критических ситуаций, при этом соответствующим образом изменяют дыха-
ние. По наблюдениям за ящуркой в природе на Украине эти животные появляются весной
при прогревании не ниже чем 17°, в Приуралье — до 21°, а при температуре почвы 45—
48° активность прекращается [320]. НЛ.Щербак установил также, что диапазон переноси-
мых разноцветной ящуркой температур лежит в пределах от —7,8 до 40—42° при средней
предпочитаемой температуре 35,9°
Переносчиком кислорода у рептилий служит гемоглобин, но детали его молекулярно-
го строения и физико-химические особенности, которые определяют его функциональные
свойства, у ящурки неизвестны. Этот вопрос возникает в связи с тем, что у черепахи Pseu-
demys обнаружены два типа гемоглобина, многие рептилии имеют гемоглобины разных ти-
пов, что проявляется в их разной электрофоретической подвижности [208]. Кроме того,
среди 54 исследованных видов черепах были найдены различия в размерах молекул гемо-
глобина, иногда значительные [451]. Не установлены также особенности насыщения кис-
лородом гемоглобина ящурок, тя. эффективность функционирования гемоглобина как
переносчика кислорода.
В качестве меры сродства гемоглобина к кислороду принята величина Р$о — то парци-
альное давление кислорода Pq2 , при котором кислородом насыщено 50 % крови. Обычно
гемоглобин полностью насыщается кислородом при более низком Ро2, чем то, какое су-
ществует в воздухе на уровне моря (155 мм рт. ст.), однако адап^йвные возможности
определяются прежде всего тем, при каких величинах (в каком диапазоне) Pq2 гемогло-
бин данного животного может отдавать и присоединять кислород. В 100 мл крови репти-
лий содержится 6—10 г гемоглобина, но адекватно он функционирует лишь в тех случаях,
когда способен присоединять кислород в количествах, достаточных для удовлетворения
потребностей организма. Показателем этой способности служит кислородная емкость кро-
ви — наибольшее возможное содержание кислорода в крови, полностью им насыщенной.
ЛЛроссером [208] приведены обширные данные для дыхательной характеристики крови
разных классов животных — рыб, амфибий, рептилий, птиц и млекопитающих. В пределах
1 Терморегуляторное поведение ящерицы, по-видимому, складывается с учетом температуры головно-
го мозга и всего тела [401].
100
каждого класса животных Р50 гемоглобинов варьируют не только под влиянием Рсо2 кро-
ви (благодаря эффекту Рута) и уровня термоадаптации, особенно у рыб и земноводных,
но и вследствие генетически закрепленных видовых особенностей.
В сводной таблице [208] приведены характеристики для систематических групп реп-
тилий (Pseudemys, Crocodylus, Alligator, Heloderma, Eumeces, Iguana, Agama, Squaromales).
Из этого следует, что гемоглобины черепахи Chrysemys способны обеспечить 50 %-ное на-
сыщение крови кислородом при Ро 2 15 мм рт. ст., но у ящерицы Iguana Р50 наступало лишь
при Pq2 51 мм рт. ст. По кислородной емкости крови перечисленные пресмыкающиеся
ближе всего, пожалуй, к исследованным амфибиям — 6,6—12,5 у первых и 8,1—10,4 мл
О2/Ю0 мл у вторых. Кровь наземных млекопитающих может полностью насыщаться
в несколько больших количествах — 15,6—17,4 О2/100 мл, тогда как у тюленя кислород-
ная емкость равна 36 мл О2/100 мл.
Ориентируясь на рассмотренные дыхательные характеристики крови, можно ожидать,
что у Е. arguta кислородная емкость близка к 10 мл О2/100 мл, однако нужны специаль-
ные исследования, поскольку на способность гемоглобина насыщаться кислородом на-
кладывает отпечаток и образ жизни вида. У сухопутной коробчатой черепахи Рзд состав-
ляет 28,5 мм рт. ст., у ложной каретты, живущей в воде, — 12,5, у малоподвижной Тег-
гарепе — И, у активно плавающей Chelydra — 32 мм рт. ст. [208]. В среднем черепахой ис-
пользуется 44 % кислорода, поступившего в артериальную кровь, человеком — 27, ска-
том — 66 % [333]. Очевидно, величина Р50 разноцветной ящурки отражает ее большую
подвижность.
В эритроцитах позвоночных содержатся соединения, которые регулируют сродство ге-
моглобина к кислороду. У млекопитающих таким регулятором является дифосфоглице-
рат [ДФГ], у рыб — аденозинтрифосфат, в эритроцитах птиц и черепах — инозитгексафос-
4>ат. У людей, адаптированных к большим высотам, эритроциты богаче ДФГ, чем у жи-
вущих на уровне моря [208]. Подобно млекопитающим у рептилий синтез гемоглобина регу-
лируется соответственно содержанию кислорода в атмосферном воздухе. Так, с увеличением
высоты мест обитания одной из североамериканских ящериц содержание гемоглобина в ее
крови возрастало с 6,62 до 12,18. Наблюдения над прыткой ящерицей и растительнояд-
ной игуаной [402] подтверждают зависимость интенсивности газообмена и метаболизма в
целом от сезона года. Например, поздней весной, в период активного питания, игуана по-
требляла 0,22 мл О2 на 1 г массы в 1 ч, летом — 0,06, зимой — 0,036.
В настоящее время дыхание разноцветной яшурки нельзя охарактеризовать с подоб-
ной конкретностью, дыхательные процессы у лацертид отечественной фауны вообще изу-
чены недостаточно. Для последующих сравнений может послужить исследование влияния
изменений температуры на кинетику артериального pH, углекислого газа и содержание
бикарбонатов у Topinambus nigropunctatus.
Вероятно, вентиляция легких у E^rguta, как и у прыткой ящерицы, осуществляется
благодаря грудной, брюшной и, возможно, шейной мускулатуре и, кроме того, сокраще-
ниям и расслаблениям гладких мышц в легких. Частота, сила и ритм активности этой ске-
летной и висцеральной мускулатуры, безусловно, требуют координации при участии дыха-
тельного центра. Активность образующих этот центр нервных клеток должна отражать вос-
ходящую сигнализацию от хеморецепторов, рецепторов легких и, возможно, других тка-
ней. На существенное значение хеморецепторной регуляции указывает и торможение ак-
тивности центра в периоды спячек. Следовательно, у ящурки, с момента вылупления ды-
шащей атмосферным воздухом, на протяжении всей жизни в продолговатом мозге дей-
ствует центр, анатомически связанный с ядрами блуждающих нервов и спинным мозгом,
общая функциональная схема которого не отличается от таковой у гомойотермных позво-
ночных. Однако особенности структурной организации и свойства нейронов дыхательного
центра разноцветной ящурки не выяснены.
5.7. ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Разноцветная ящурка, как и большинство ящериц, — насекомоядное животное. В основ-
ном питается жуками, перепончатокрылыми (муравьями), бабочками (гусеницами) ,дву-
крылыми, клопами и прямокрылыми [320]; пойманных насекомых, прежде чем заглот-
101
ну ть, разминает челюстями. Зубы пневродонтного типа, число их непостоянно, на каждой
челюсти от 35 до 41, имеются и птеригоидные (подробнее см. разд. 5Д .1.).
В преддверии рта и его полости располагаются многочисленные слизистые клетки и
многоклеточные железы. Язык ящурки относительно крупный, удлиненный, заметно раз-
двоен на конце. Многослойный эпителий слизистой оболочки, покрывающей язык, обра-
зует складки на дорсальной поверхности, ближе к* корню языка. Как и у других Sauria
[306, 356], в глубине складок располагаются слизистые клетки, а на их выпуклостях —
редкие вкусовые луковицы. Эпителий вентральной поверхности складок не образует;
здесь количество желез значительно меньше, чем на дорсальной. В мышечной толще органа
встречаются пигментные клетки (рис. 52). В области корня языка заложены симметрично
две язычные миндалины, имеющие вид крупных лимфатических узлов.
Значительная часть объема в толще языка занята поперечнополосатыми мышечными
волокнами, которые образуют переплетающиеся пучки. Среди них от корня к верхушке
языка Проходят крупные нервные стволики.Претерминальные нервные волокна подходят
как к мышечным волокнам, так и к слизистой языка. Среди мышц центральной части
языка залегает длинный прямой отросток подъязычной кости — entoglossum. Этот плот-
ный стержень вначале заметен на поверхности, затем погружается в глубь органа и его ко-
ническая концевая часть вновь выходит на вентральную поверхность языка (рис. 53).
Гистохимический анализ секрета, выделяемого железами языка ящериц, показал на-
личие в нем нейтральных и кислых мукополисахаридов, сиаломуцинов и небольшого коли-
чества белка [306], тх. компонентов, свойственных гомологичным секретам других на-
земных позвоночных. Возрастание числа компонентов, продуцируемых ротовыми желе-
зами, как и высокое содержание в секрете выполняющих защитные функции сульфому-
цинов, связывают с выходом рептилий на сушу; у них, в отличие от других позвоночни-
ков, преобладают мукосерозные железы [63].
Стенки пищеварительного тракта имеют обычное для позвоночных четырехслойное
строение: слизистая оболочка, подслизистый, мышечный и серозный слои. Пищевод выст-
лан изнутри многослойным плоским эпителием, расположенным на тонкой собственной
пластинке слизистой оболочки. Мышечная пластинка слизистой развита достаточно хоро-
шо. В подслизистом слое, а также в собственной пластинке, но уже в каудальном отделе
пищевода (ближе к желудку), встречаются отдельные железы. Мышечный слой неодина-
ков по всей длине пищевода. В наружном продольном слое поперечнополосатые волокна
постепенно замещаются гладкомышечными. Снаружи пищевод покрыт рыхлой соедини-
тельнотканной оболочкой — адвентицией.
Желудок простой, мешковидный, его кардиальный и пилорический сфинктеры вы-
ражены достаточно хорошо. На внутренней поверхности опорожненного желудка видны
глубокие складки, почти полностью закрывающие его просвет. Желудок такого же типа
встречается и у прыткой ящерицы [30]. После растягивания складок под действием ин-
трамуральной мускулатуры вместимость желудка значительно увеличивается; то же свой-
ственно и более отдаленным в систематическом отношении рептилиям, в частности змеям.
Кроме того, благодаря складчатости стенки желудка увеличивается и его переваривающая
способность, так как крупный пищевой ком или добычу больших размеров облегает сли-
зистая оболочка, изобилующая соковыделительными железами.
Слизистая оболочка покрыта однослойными цилиндрическими клетками, однотипны-
ми по строению, которые секретируют слизь. В толще ее располагаются собственно пище-
варительные железы — 5—8 секреторных клеток окружают просвет, формируя железу.
Секреторные клетки слизистого слоя двух типов (рис. 54). Преобладают пирамидаль-
ные клетки. Ядро сдвинуто к основанию, содержит более темное ядрышко. Верхние от-
делы желез открываются в желудочные ямки. Между железами располагается собственное
вещество слизистой: соединительнотканные волокна и клетки, кровеносные и лимфати-
ческие сосуды, нервные волокна. В слизистую проникают и гладкомышечные волокна из
двухслойной собственно мышечной пластинки. Подслизистая, образованная главным обра-
зом соединительнотканными волокнами, заходит глубоко в складки и формирует их ос-
нования. В складках часто встречаются кровеносные сосуды среднего и малого калибра.
Железы в подслизистой оболочке отсутствуют.
Мышечный слой желудка состоит из внутреннего, более развитого циркулярного и на-
ружного продольного слоев. Крупные кровеносные сосуды в этом слое редки. Снаружи
желудок покоыт серозной оболочкой.
102
Желудок разноцветной ящурки, как и других лацертид, в отличие от желудка птиц и
млекопитающих, не подразделяется на функционально, морфологически и гистологически
различимые отделы (у птиц — железистый и мускульный желудки, у млекопитаю-
щих — кардиальная, фундальная, пилорическая части или, более того, обособленные
камеры).
Железы пищеварительного канала ящурки, как и других Lacertidae, заслуживают даль-
нейшего исследования, если принять во внимание, что эти пресмыкающиеся, фактически
всеядные, способны переваривать разнообразную пищу.
Разделение кишечника на тонкий и толстый отделы достаточно отчетливое, особенно
по гистологическим данным. Переход желудка в 12-перстную кишку обозначен вполне
четко. К особенностям 12-перстной кйшки разноцветной ящурки относятся складчатость
слизистой оболочки, обилие кишечных ворсинок, наличие интрамуральных желез и хоро-
шо развитого мышечного слоя (рис. 55) . Наружная оболочка (адвентиция) на всем про-
тяжении кишечника в сравнении с другими слоями его стенки небольшой толшины.
Ворсинки, которые возвышаются над поверхностью складок слизистой оболочки, в
двенадцатиперстной кишке ящурки длиннее и шире, чем в более каудальных отрезках
кишечной трубки; они покрыты эпителиальными клетками двух типов: цилиндрическими
(резорбирующими) и бокаловидными, последние растянуты слизью. Вероятно, поверх-
ность ворсинки покрыта микроворсинками, образующими в совокупности щеточную кай-
му, поскольку она обнаружена уже у рыб. Щеточная кайма представляет собой своего род&
пористую поверхность, что, как установлено физиологическими опытами на других позво-
ночных, повышает эффективность ферментативных процессов в ходе пищеварения. Осно-
ва кишечной ворсинки образована собственным веществом слизистой оболочки. Основа-
ние кишечной складки сформировано подслизистым слоем.’
В дуоденальной стенке имеются железы, по строению соответствующие трубчатым.
Эти образования, которые могут рассматриваться как гомологи бруннеровых желез мле-
копитающих, располагаются преимущественно в основании слизистого слоя и подслизи-
стой оболочке.
В средней — мышечной — оболочке кишечной стенки обнаруживаются продольные
гладкомышечные волокна и мощный циркулярный слой. Более крупные кровеносные со-
суды проходят через мышечную оболочку транзитом, в апикальной части подслизистого
слоя делятся на ветви более мелкого калибра, которые проникают в слизистую оболоч-
ку, образуя в подслизистом слое сплетения. Среди рыхлой соединительной ткани этого
слоя встречаются и эластиновые волокна. В нем же имеются и нервные сплетения, состоя-
щие в основном из демиелинизированных волокон.
Адвентиция более каудальных отделов кишечника имеет типичное для серозных обо-
лочек строение. Мышечный и подслизистый слои развиты заметно слабее. Кишечные вор-
синки неодинаковой высоты, в целом они ниже дуоденальных. В собственно пластинке
слизистой оболочки встречаются лимфатические фолликулы (рис. 56) .
На границе тонкого и толстого кишечника хорошо заметен червеобразный отросток.
Поперечник толстого кишечника увеличивается постепенно. Складки слизистой оболочки
не выражены, отсутствуют кишечные ворсинки, толщина мышечного слоя уменьшена.
Общая длина кишечника у 5 особей E.a.arguta составила 139 ±6 % длины тела; длина тела
измеренных ящурок варьировала в пределах 8 %.
Особенности строения ротовой полости и желудочно-кишечного тракта позволяют вы-
сказать некоторые соображения об особенностях пищеварения разноцветной ящурки. Как
У других Squamata, пищеварительный тракт этого вида хорошо растяжим и может перева-
ривать добычу различного объема, требующую интенсивной ферментной обработки; зна-
чение ее особенно велико вследствие зависимости переваривания от внешней температуры.
Хотя эти процессы у ящурки не исследованы, но данные эксперимента по скорости перева-
ривания у прыткой ящерицы при колебаниях окружающей температуры, которые приве-
дены Г.ПЛукиной и др. [157, 158], являются достаточным подтверждением такой зави-
симости. Эффективность действия пищеварительных ферментов пойкилотермных живот-
ных [142] при недостаточно высоких температурах отчасти компенсируется высокой
каталитической активностью самих ферментов, которая у трипсина ящерицы не уступает
таковой лягушек и рыб — животных с выраженной пойкилотермией.
Печень разноцветной ящурки, как и других лацертид, — самый крупный из внутрен-
них органов. Она красная, консистенция очень мягкая, подвешена на двух тонких прозрач-
103
ных лентовидных связках к брюшной стенке (ligamentum suspensorium hepatis) и кишечни-
ку lig. hepatu-emericum). Вентральная поверхность печени выпуклая, дорсомедиальная не-
сколько вогнутая. Состоит из двух крупных долей — правой и левой, на свободном крае
которых имеются вырезки, а также небольшой центральной, выделяющейся на вентраль-
ной поверхности. С медиальной заметно разделение еще на несколько мелких долей. Круп-
ные доли асимметричны; левая — широкая, относительно плоская, правая - вытянутая,
больше по размеру. Ее удлиненная каудальная часть загибается дорсокаудально и охваты-
вает часть толстой кишки. На медиальной поверхности левой доли расположены ворота,
в которые вступают воротная вена печени и печеночная артерия и выходит общий желчевы-
носящий поток. Характерно, что через печень проходит задняя полая вена. Она вступает в
каудальный отдел правой лопасти печени, принимает печеночные вены, собирающие кровь
из воротной системы печени, и вновь выходит наружу по середине медиальной поверхности
органа.
Снаружи печень покрыта соединительнотканной капсулой. В поверхностном слое кап-
сулы проходят кровеносные сосуды мелкого калибра. Глубже волокна соединительной
ткани расположены плотнее, среди них встречаются клеточные элементы.
Основную массу клеток печени составляют гепатоциты, полифункциональные клетки,
выполняющие кроме метаболических и секреторных функций и опорные, так как коли-
чество соединительнотканных волокон в печени невелико. Они главным образом включе-
ны в стенки печеночных желчных протоков разного калибра и стенки сосудов. Среди не-
многочисленных соединительнотканных волокон залегает множество секреторных тру-
бок. В основном трубка образована 6 продольными рядами гепатоцитов. Это мелкие
клетки с ядром, смещенным к расширенному базальному краю, который через простран-
ство Диссе сообщается с синусоидами и кровеносными капиллярами. Секрет гепатоцитов
через узкий апикальный край попадает в просвет трубки, затем в мелкие желчные прото-
ки, выстланные плоским эпителием. По системе протоков, объединяющихся перед выхо-
дом из печени в один канал, желчь поступает в двенадцатиперстную кишку. По ходу об-
щего желчевыносящего протока располагается желчный пузырь (рис. 57) .
У разноцветной ящурки, как и у других рептилий, амфибий и рыб, в печени нет раз-
деления на дольки [68]. По этому признаку печень ящурки имеет строение, присущее низ-
шим позвоночным, что, однако, не означает функционального несовершенства органа.
Объемистый желчный пузырь свидетельствует об интенсивном образовании желчи.
Особенности энергетического обмена ящериц, их способность переносить продолжи-
тельное голодание, изменчивость массы печени в связи с фазой репродуктивного цикла
[373] указывают на высокую метаболическую лабильность печени лацертид, в том числе и
разноцветной ящурки.
Поджелудочная железа E^rguta компактная, листовидная, вытянутая. Располагается
в петле двенадцатиперстной кишки, только ее головка выступает за краниальную часть
левой лопасти печени, с брюшной стороны почти полностью прикрыта печенью. Ее проток
впадает в двенадцатиперстную кишку самостоятельно. По строению это альвеолярно-
трубчатый орган, типичный для всех рептилий. Концевые отделы экзокринной части желе-
зы сформированы 6—8 клетками, апикальная часть которых содержит гранулы зимогена;
ядро клеток смещено к основанию. Протоки железы выстланы однослойным эпителием.
Специальные исследования, проведенные на рептилиях, выявили тесную связь эндо-
кринной и экзокринной частей поджелудочной железы. Встречаются топографические ви-
довые особенности в расположении экзо- и эндокринных клеток, определяемые электрон-
но-микроскопическими и цитохимическими методами [93,172}.
5.8. МОЧЕПОЛОВАЯ СИСТЕМА
з 3 Л. ВЫДЕЛИТЕЛЬНАЯ СИСТЕМА
Важнейшая проблема для наземных рептилий — снижение потери воды, связанней с пере-
гревом. Проницаемость кожных покровов этих животных сопоставима с таковой некото-
рых амфибий [340, 438]. Существенное снижение потерь воды обеспечивается за счет
уменьшения локомоторной активности, использования убежищ, а также адаптаций ионо-
регулирующих механизмов почек. Преобладающим типом азотистого обмена у рептилий
104
является так называемый урикотелический, ведущий к образованию мочевой кислоты
При переходе канальцевой мочи в клоаку мочевая кислота, почти не растворимая в всас,
осаждается, выделяясь в виде сухого (или полужидкого) продукта; вода реабсорбирует-
ся в клоаке [143, 438]. Способность задерживать во внеклеточной жидкости натрий и то-
лерантность к повышению концентрации хлоридов в плазме крови также способствуют
снижению потерь воды [206, 233].
Изучение микроморфологии почек рептилий позволило выяснить то, что эволюция их
в сторону дегенерации клубочковой системы — преобразования эффекторных органов си-
стемы осмотической и ионной регуляции осуществлялись за счет приспособления послед-
них к использованию цикла мочевой кислоты в отличие от филогенетической линии двоя-
кодышащие — амфибии — млекопитающие, адаптированной к функционированию цикла
мочевины [215]. В нефронах почки рептилий выделяется 5 отделов, следующих за почеч-
ным тельцем: шейка, проксимальный каналец, промежуточный отдел, дистальный каналец
и соединительный отдел. Далее следует собирательная трубка, имеющая иной генезис. Тон-
кий сегмент петли Генле отсутствует, его замещает промежуточный отдел: короткий,
снабженный цилиарным эпителием [57,143, 177,178,414,449].
Почки Eremias представляют собой два удлиненных дольчатых органа, подве-
шенных на собственной брыжейке с дорсальной поверхности тазовой области, при-
крытых костями таза. Они сращены в каудальной части, образуя треугольную
(в плане) фигуру. При этом каудальные полюса их сращены в виде гребня. Ме-
диальные края почек, образующие собственно гребень, сведены вместе и отклонены вен-
тро дорсально. Вентральная поверхность почек сращена с дорсальной поверхностью клоаки
за счет капсулы почки и перитонеального листка, повторяя ее конфигурацию. В некоторых
случаях клоака расположена на поверхности правой почки, приобретающей вид желоба.
Сращение регистрируется только в каудальной части почек. Диаметр правой вены почки
в 3—4 раза превышает диаметр левой. Мочевые протоки собираются в короткий мочеточ-
ник, направляющийся к дорсальной поверхности клоаки. У самцов мочеточники впадаю;
в общее отверстие с семяпроводами, у самок разноцветной ящурки — открываются на
двух особых сосочках, расположенных на дорсальной поверхности клоаки.
На вентральной поверхности клоаки располагается белый плоский треугольный тонко-
стенный мочевой пузырь. В каудальной части его открывается круглый в сечении канал,
начинающийся из заднего отдела клоаки.
5 Я2. МИКРОАНАТОМИЯ ПОЧЕК
Почечные тельца (рис. 58), слабо выражены, немногочисленны. Капиллярная сеть в клу-
бочках уменьшена до немногих (в некоторых случаях до 2—3) капиллярных петель. Со-
хранение минимального числа этих структур, по-видимому, вызвано необходимостью на-
личия некоторого количества фильтрата для смывания мочевой кислоты и для продвиже-
ния ее по направлению к клоаке [143]. Шейка укорочена, выстлана кубическим реснит-
чатым эпителием, характерным также и для соединительного отдела. Вероятно, цилиар-
ный механизм способствует продвижению экскрета в канальцах, компенсируя низкое
фильтрационное давление. Извитые и прямые участки канальцев обладают однотипным
строением (рис. 59) Для клеток эпителия проксимального канальца в извитой части
характерно значительное развитие базальной мембраны. Имеется хорошо выраженная
плотная каемка.
Эпителий дистального канальца — высокий призматический, щеточной каемки нет.
Эпителий проксимального и дистального канальцев обладает базальными ъпячиваниями
(в дистальных канальцах эти образования выражены в большей степени) . Но мнению ряда
исследователей, описываемая структура может играть ощутимую роль в создании осмоти-
ческих градиентов, в конечном итоге образуя гиперосмотическую мочу [57]. Не ряде пре-
паратов в дистальных канальцах самцов разноцветной ящурки обнаружены небольшие
железистые участки. Нефронные системы и сосудистое русло почки фиксируются хоро-
шо развитым соединительнотканным остовом.
Мочевые канальцы впадают в разветвленные собирательные коллекторы. Собиратель-
ные трубки выстланы кубическим эпителием. Эпителиальная выстилка мочеточников об-
разована переходным эпителием. Нервные волокна проникают в паренхиму почек, следуя
к05
по ходу кровеносных сосудов. В стенках сосудов кровеносного русла обнаруживаются
тонкие периадвентициальные и адвентициальные волокна. На некоторых препаратах про-
слеживаются отдельные нервные волокна на нефронных системах.
5.8.3. МИКРОАНАТОМИЯ НАДПОЧЕЧНИКОВ
Топографические взаимоотношения, микроскопическое строение этих органов у рептилий
описаны в многочисленных оригинальных работах и обзорах [4, 236, 364,399,416 и др.].
К общим -чертам строения этих желез у представителей разных отрядов рептилий (кроме
ящериц и, частично • змей) относятся: сближение двух железистых типов тканей — супраре-
наловой (хромаффинной) и интерреналовой (корковой) . У ящериц и некоторых змей ад-
реналовые клетки лежат в основном на дорсальной поверхности надпочечника. У пред-
ставителей 2 других подклассов (крокодилы и черепахи) клеточные элементы адренало-
вой ткани рассеяны между интерреналовыми клетками [4,236].
Хромаффинные клетки исследованных ящурок обладают полигональной или кубиче-
ской неправильной формой; они лишены островков, содержат крупные или овальные
ядра. Клетки интерреналовой ткани более разнообразны по форме — преобладающий тип —
призматические клетки. Адреналовые клетки образуют компактную массу, которая в
несколько слоев располагается на дорсальной стороне железы и образует островки или
пальцевидные выросты, проникающие в глубь интерреналовой ткани.
Иннервация паренхимы надпочечников осуществляется за счет висцеральных ветвей
симпатического ствола и внутренностных нервов. Источник интрамуральных нервов —
капсулярное нервное сплетение, в составе которого встречаются отдельные нервные клет-
ки и мелкие микроганглии (рис. 60).
5ЗА. ПОЛОВАЯ СИСТЕМА
Семенники имеют вид овальных тел желтого цвета, расположенных впереди краниальных
долей почек, по бокам позвоночного столба. Правый семенник залегает значительно крани-
альнее левого. От этих органов отделяются многочисленные канальцы, образующие в сово-
купности придаток семенника (рис. 61, 62, 63). Продолговатый придаток продолжается
в семяпровод, открывающийся в мочеточник вблизи его впадения в клоаку.
Копулятивные органы самцов разноцветной ящурки достаточно своеобразны. Эти ор-
ганы имеют вид двух полых цилиндров, богато васкуляризованных. Копулятивные органы
сформированы за счет складки вентральной области каудального отдела клоаки (подроб-
нее о строении этих органов см. на с. 66, 67).
Эпителий кутикулы гемипениса имеет ’’шиповатый” вид. На наружной поверхности
кутикулярных клеток регистрируется ’’частокол” зубчиков, слабо загнутых к выпуклой
поверхности клеток. Такого рода ’’зубчатая” архитектура поверхности кутикулы сформи-
рована за счет изогнутых отростков клеток кутикулярного эпителия. По данным Л.АЛан-
тца [393] и В .Беме [342], в строении эпителия кутикулы гемипенисов у представителей
афроазиатской группы ящурок наблюдается значительное разнообразие, по-видимому,
имеющее систематическое значение.
Парные яичники (рис. 64) имеющие вид гроздевидных тел неправильной формы, под-
вешены к дорсальной стенке тела на собственной брыжейке. Правый яичник располагает-
ся несколько кражальнее левого. Зрелые органы принимают вид скоплений яиц разной
степени развития. Глубокий слой органа пронизан многочисленными кровеносными со-
судами и является сосудистой зоной яичника.
Яйцеводы также присоединены к дорсальной стенке тела, располагаясь с латерального
края яичников. Спереди яйцеводы уплощены в дорсовентральном направлении, сзади -
утончаются, принимают округлую форму, открываясь с дорсальной поверхности в полость
клоаки генитальными отверстиями; общие очертания этих органов имеют извитой вид.
Спереди яйцеводы соединяются с полостью тела удлиненными воронками, выстланными
мерцательным эпителием. В составе стенки их выделяются слизистая, мышечная и Сероз-
ная оболочки.
106
5.9 ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа половозрелых особей разноцветной ящурки имеет типичное для
представителей класса рептилий строение (рис, 65) .
Капсула щитовидной железы тонкая, представлена одним — двумя слоями упло-
щенных соединительнотканных клеток и коллагеновыми волокнами, В некоторых участ-
ках капсула отсутствует, и фолликулы щитовидной железы непосредственно контактиру-
ют с клетками белой жировой ткани, прилегающей к органу. В периферии ее нередко
встречаются крупные пигментные клетки.
На срезе щитовидной железы насчитывается до нескольких десяткой фолликулов, су-
щественно варьирующих по размерам и форме. Чаще всего фолликулы имеют размеры
более 100 мкм, округлую или овальную, нередко — неправильную форму. Стенка фолли-
кулов образована одним слоем призматических, иногда уплощенных тироцитов. Ядра ти-
роцитов занимают большую часть клетки, в них хорошо видны 1—2 крупных ядрышка и
мелкие глыбки хроматина.
Таблица 3. Морфометрическая характеристика некоторых структур щитовидной железы неполо-
возрелых и половозрелых разноцветных ящурок в процессе индивидуального развития
Параметр Неполово- зрелые (Хн±5г) Полово- зрелые х 100% t, норми- рованное отклонение Р, вероят- ность
Большой диаметр ядра, мкм 5,3 ±0,06 Тироциты 5,3 ±0,1' 0 0,00 >0.05
Меньший диаметр ядра, мкм 4,1 ±0.08 3,5 ±0,21 -15 2,6,7 < 0,05
Эксцентриситет ядра 34 ± 3.70 61 ± 8,5 + 79 2,91 < 0,05
Высота тироцита, мкм 5,2 ±0,18 5,7 ± 0,62 + 10 0,77 > 0,05
Ширина тироцита, мкм 7,0 ±0,35 6,2 ±0,55 -И 1,23 > 0,05
Большой диаметр фолликула, мкм 46 ± 2,40 Фолликулы 85 ±5,6 + 85 6,40 <0,001
Меньший диаметр фолликула, мкм 34 ± 2,70 63 ±4,6 + 85 5,44 <0,01
Эксцентриситет фолликула 40 ±5,60 35 ± 1,6 -12 0,86 >0,05
Количество тироцитов в фолликуле 18 ± 0,60 39 ± 3,2 + 117 6,44 <0,001
резорбционных вакуолей 6,0 ± 1,10 14 ±2£ + 133 2,66 <0,05
Довольно часто регистрируется слияние фолликулов, при котором контактирующие
участки стенок фолликулов претерпевают дистрофические изменения. В данных участках
располагаются тироциты с уплощенными, пикнотизированными ядрами. Внутри фоллику-
лов находится коллоид, плотность и окраска которого в различных фолликулах варьи-
руют; нередко регистрируются значительные различия в окрашивании отдельных участ-
ков коллоида одного фолликула.
Строма щитовидной железы представлена в основном расширенными элементами со-
судистого русла, которые заполнены эритроцитами, имеют неправильную форму и запол-
няют промежутки между фолликулами.
В процессе индивидуального развития существенно уменьшаются размеры ядер тиро-
цитов (табл. 3), в то же время размеры тиреоидных фолликулов значительно увеличива
ются (рис. 65.66).
ГЛАВА 6
ГАЗООБМЕН
Характер газообмена прямо зависит от колебания температуры тела рептилий. Проведен-
ные эксперименты [258, 260] по изучению потребления кислорода разноцветной ящуркой
в Центральном Предкавказье свидетельствуют, что с повышением температуры тела по-
требление кислорода увеличивается, а с понижением — уменьшается (табл. 4) , что объяс-
няется изменением скорости биохимических процессов в организме. Подобная закономер-
ность установлена и для других пресмыкающихся [103, 338, 339, 349, 350], в частности
показана нами для прыткой ящерицы из Центрального Предкавказья [258,260]. Наиболее
низкое потребление кислорода ящерицами мы отметили при 0°. При такой температуре
Таблица 4. Потребление кислорода
разноцветной ^щуркой при разной темпера-
туре ^ела (см /100 г массы в течение 1 ч
при 0 и 760 мм рт. ст.) [ 258, 260, с изме-
нениями]
Темпе- ратура. п М ± т Г
0 5 14,4 ±53 4^37
5 23 72,2 ± 123 1396
10 14 983 ± 113 1339
15 14 124,7 ± 113 3,891
20 18 195,0 ± 14,1 5364
25 14 347,0 ± 23,4 5303
30 11 11023 * Ш,7 5,522
31 4 264,0 ± 73,63 0,065
32 5 256,0 ± 98,65 0,489
33 3 340,0 ± 140,6 0333
34 4 243,0 ±61,0 0,480
35 26 273,0 ± 133 6323
40 3 1383 ±153 7,836
41 5 113 *43 —
47 4 озо
энергетический обмен у ящерицы очень низок,
что биологически оправдано в условиях зимов-
ки. Снижение температуры тела ящериц до ука-
занных В.М Родионовым [218] пределов приво-
дит их к гибели.
Данные табл .4 показывают, что количество по-
требляемого кислорода резко возрастает при
подъеме температуры тела до 5°. Далее более или
менее плавно повышается потребление кислорода
вплоть до подъема температуры тела до 20°
Затем, при 25—30°, вновь происходит резкий ска-
чок, а с 30 до 31° — такое же резкое падение.
Нам представляется, что в данном случае вкра-
лась ошибка в эксперимент, и с 25 до 35° вклю-
чительно потребление кислорода животным на-
ходилось на близком уровне. Дальнейшее повы-
шение температуры тела приводило к тому, что
потребление кислорода разноцветной ящуркой
прекращалось при температуре между 41 и 47°,
при 47,2° она погибала. Сопоставление этих дан-
ных с материалами о влиянии температуры на
активность ящурки (см. гл. 10) позволяет за-
тела между 25 и 35° для этих животных в Центральном
при температуре
ключить, что
Предкавказье существуют наиболее оптимальные условия, так называемая зона удобства
жизни [142]. Именно при этих температурах ящурка потребляет более или менее постоян-
ное количество кислорода. Это позволяет предположить наличие у разноцветной ящурки
химической терморегуляции. Однако уровень окислительных процессов невысок, посколь-
ку животное неспособно удерживать постоянную температуру тела при изменении темпе-
ратуры среды выше или ниже указанных величин.
Очевидно, ящерица способна повышать температуру тела за счет окислительных про-
цессов в организме; это возможно при температуре среды не ниже 14°
Эксперимент показал повышение газообмена от весны к осени и падение ко времени
перехода к спячке, а также более высокий уровень газообмена молодых особей сравни-
тельно с взрослыми. Последнее объясняется относительно большей поверхностью тела мо-
лодых. Различия в количестве потребляемого ящурками кислорода при разной освещенно-
сти, в разное время суток (при равных температурных условиях) не обнаружены.
ГЛАВА 7
НЕКОТОРЫЕ АСПЕКТЫ ЭНЕРГЕТИЧЕСКОГО МЕТАБОЛИЗМА
Сведения об энергетическом обмене тканей пойкилотермных животных, в частности реп*
тилий, скудны, а по разноцветной ящурке вообще отсутствуют. В связи с этим представля-
ет интерес изучение содержания энергетических метаболитов в ее тканях и его изменение
при зимней спячке, подготовке к ней, а также при длительной глубокой искусственной ги-
потермии, что необходимо для обнаружения сходства и различия между этими со-
стояниями.
Определяли содержание гликогена, глюкозы, незтерифицированных жирных кислот
(НЭЖК) , молочной кислоты, АТФ, креатинфосфата, неорганического фосфора в печени,
скелетных мышцах, мозге, а также глюкозы и НЭЖК в крови. Опыты по искусственной
гипотермии проведены в июне — июле, по зимней спячке — в октябре—ноябре. Темпера-
туру тела замеряли в полости рта термопарой. Животных умерщвляли, погружая в жид-
кий кислород или азот.
Глюкозу определяли глюкозооксидным методом [159], гликоген — по количеству
глюкозы, образовавшейся после 2,5 ч гидролиза в 1 н H2SO4 [209], НЭЖК — по методу
ВТДункомба [354], молочную кислоту — методом С.В.Баркера и В.Саммерсона [332] ,
креатинфосфат — по AM.Алексеевой [8] , неорганический фосфор — колориметрически
после осаждения магнезиальной смесью. О содержании АТФ судили по легкогидролизуе-
мому фосфору ртутного осадка.
В июне — июле содержание гликогена в печени разноцветной ящурки составляет
62 мкм/г, что в 3,2 раза выше уровня этого полисахарида в скелетных мышцах (табл. 5).
Полученные величины сопоставимы с данными, полученными для млекопитающих и
птиц, и с результатами наших экспериментов на полосатых ящерицах [162]. Высокое со-
держание гликогена (9 мкм/г) обнаружено в мозге разноцветной ящурки. Для полоса-
той ящерицы эта цифра составила 12 мкм/г.
Наличие значительных резервов гликогена в мозге холоднокровных позвоночных
отмечалось в литературе [204]. Это позволяет мозгу меньше зависеть от поступления глю-
козы с кровью, имеет большое значение для выживания вида, например при аноксии и
длительном голодании.
В исследованных тканях разноцветной ящурки отмечено также высокое содержание
глюкозы. Уровень этого энергетического субстрата в мозге, печени и мышцах ящурки со-
ответственно в 5,6; 3 и 3,5 раза выше, чем у полосатой ящерицы. Различий в содержании
глюкозы и НЭЖК в крови между разноцветной ящуркой и полосатой ящерицей не найде-
но, однако у ящурок в мышцах уровень НЭЖК достоверно выше.
Содержание молочной кислоты свидетельствует об использовании больших углевод-
ных запасов мозга ящурок преимущественно анаэробным путем (табл. 6) . Интенсивность
гликолиза в мозге ящурок выше, чем в других исследованных тканях, в том числе мышеч-
ной. Такая закономерность обнаружена и у полосатых ящериц, однако у них интенсив-
ность гликолиза ниже. В тканях разноцветной ящурки содержится много фосфатов: АТФ,
креатинфосфат и неорганический фосфат.
Осенью, в конце сентября, перед впадением ящурок к спячку, запасы гликогена в его
основном депо — печени — увеличиваются по сравнению с летом в 2,4 раза. Масса печени в
этот период максимальна. В мышцах также наблюдается повышение количества этого по-
лисахарида. Уроень гликогена в мозге существенно не изменяется. Причиной накопления
гликогена перед залеганием в спячку, по-видимому, является изменение активности эндо-
109
Таблица 5. Содержание гликогена, глюкозы (мкм/г) и жирных кислот (НЭЖК, мкэкв/г) в тканях разноцветной ящурки с чюня по ноябрь в норме, при ис
кусствеиной гипотермии в зимней спячке при температуре 2-4° (М ± т, п =6~ 10)
Состояние животных Печень Кровь Мозг Мышцы НЭЖК
Гликоген Глюкоза Глюкоза НЭЖК Гликоген Глюкоза Гликоген Глюкоза
Интактные животные Недельная гипотермия 3-недельная гипотермия 62 ±3,1 32 ± 2,8* 17 ± 0,7* 22 ± 3,3 18 ± 1,3 14 ± 1,1* 5,5 ± 0,6 4,7 ±0,1 7,2 ±0,2 Июнь - июль 0,86 ± 0,06 1,52 ±0,20* 7,80 ±0,45* 9 ±0,5 9 ±0,2 13 ± 1,3* 39 ± 5,9 18 ± 1,4* 43 ± 3,4 19 ± 1,0 11 ±0,1* 10 ±0,1* 14 ± 1,4 14 ± 2,0 7 ± 1,0* 4,6 ± 0,4 2,1 ±0,04* 4,3 ± 0,40
Интактные животные 153 ±5,1* 21 ± 1,0 6,5 ±0,2 Сентябрь 1,05 ± 0,09* 8,5 ± 0,2 26 ± 1,2 26 ± 3,7 7 ± 0,7* 6,0 ±0,54
Недельная зимняя спячка 3-недельная зимняя спячка 5-недельная зимняя спячка 112 ±17,8** 35 ± 1,4** 122 ±9,2** 14 ± 1,8** 9 ±0,6*» 4 ±0,3*» Октябрь — ноябрь 8,5 ±0,7 1,27 ±0,06 4,1 ±0,3 1,0 ±0,07 3,7 ±0,3 12 ± 1,0*» 12 ±0,2*» 18 ± 1,0*» 18 ± 2,1*» 18 ± 2,1*» 40 ± 4,6*» 34 ± 2,2 15 ± 1,0** 36 ± 1,4 5 ±0,4 4 ± 0,4*» 5 ±0,5 4,2 ±0,42** 4,0 ± 32 ** 4,0 ±0,48
* Здесь и в табл. 6 достоверные (Р < 0,05) различия данных для июня
** То же - для сентября.
Таблица б. Содержание молочной кислоты и фосфатов (мкм/г) в тканях разноцветной ящурки с июня по ноябрь в норме, при искусственной гипотермии и
зимней спячке при температуре 2-4 (ЛГ ± т, п = 6-10)
Состояние животных Печень Мозг Мышид
Молочная кислота Креатин- фосфат АТФ Молочная кислота Креатин- фосфат | АТФ 1 Молочная кислота Креатин- фосфат АТФ
Интактные животные 11 ±0,2 Июнь - июль 12 ± 1,6 27 ± 1,7 56 ±7,2 11 ±0,3 10 ± 0,7 12 ± 0,9 27 ± 1,9
Недельная гипотермия 30 ± 2,4* 16 ± 1,1 6 ± 0,7* 24 ± 4,0 62 ± 3,2 17 ± 1,8* 21 ± 1,7* 80 ± 2,2* 7 ± 1,1*
3-недельная гипотермия 19 ± 2,0* — 7 ± 0,8* 87 ± 3,8* — 36 ± 2,7* 18 ± 2,3* 56 ± 6,0 10 ± 0,7*
Интактные животные 17 ± 1,2* Сентябрь 7 ±0,7* 56 ±4,5* — 19 ± 1,9* 5 ±03* 57 ±5,4* 10 ± 1,1*
Недельная зимняя спячка 4 ± 0,2** 13 ± 1,1 Октябрь - ноябрь 7 ± 0,9 13 ± 03*» 4 ±0,6 13 ± 0,4*» 1 ± 0,2** 57 ±4,2 12 ±0,9
3-недельная зимняя спячка 8 ± 0,4** 7 ± 1,0 19 ± 2,0** 12 ± 1,1** 24 ± 2,4 27 ± 2,7*» 5 ±0,5 40 ± 3,6*» 25 ± 1,5**
5-недельная зимняя спячка 6 ± 0,6** 9 ±0,5 7 ±0,7 21 ±2,6** 55 ± 5,5 - 5 ±0,8 30 ± 2,3*» 11 ± 1,2
кринных желез, в частности наблюдаемое у холоднокровных усиление в этот пери-
од выработки инсулина [204], который стимулирует увеличение углеводных резер-
вов в печени.
Содержание глюкозы в тканях разноцветной ящурки осенью менялось со статистиче-
ской достоверностью лишь в мышцах, где ее уровень снижался в 1,8 раза. Одновременно в
мышцах в 2 раза снижалось содержание молочной кислоты, а концентрация неэтерифици-
рованных жирных кислот и макроэргических фосфатов достоверно увеличивалась. Вероят-
но, наблюдаемые в мышцах изменения обусловлены значительным снижением двигатель-
ной активности животных. Увеличение концентрации АТФ перед спячкой наблюдалось и
в мозге ящурок. Подготовка к зимней спячке сопровождалась накоплением запасных жи-
ров. Масса жировых тел у ящурки была минимальна в конце июня, в период размножения:
у самок — 0,18 г, у самцов — 0,055 г, что составляет соответственно 0,32 и 0,6 % массы те-
ла. К концу июля масса жировых тел увеличивалась в 3,3 и 4,7 раза, а в сентябре перед
спячкой у самок она составила 0,226 г, у самцов — 0.28 г, превышая июньский уровень со-
ответственно в 12,6 и 5,1 раза.
Следовательно, подготовка разноцветной ящурки к спячке сопровождается накопле-
нием энергетических ресурсов: гликогена в печени, жира в жировых телах, ростом концен-
трации микроэргических фосфатов в мышцах и мозге.
При недельной спячке (при температуре 2—4°) содержание глюкозы в крови несколь-
ко увеличивается. Резервы гликогена в печени понижаются при этом в 1,4 раза. Уровень
полисахарида в мышцах существенно не изменяется, а в мозге увеличивается на 48 % по
сравнению с осенним контролем. Резко тормозятся при спячке анаэробные процессы, со-
держание молочной кислоты в мозге, печени и мышцах ящурки снижается соответственно
в 43; 4,2 и 5 раз. В уровне макроэргических фосфатов заметных изменений не происхо-
дит. Распад резервных жиров и поступление жирных кислот в кровь также существенно
не изменяются. Таким образом, у ящурок на этом этапе спячки в качестве энергетического
субстрата используется в основном гликоген печени.
Продление спячки до 3 недель вело к дальнейшему расходованию запасов гликогена,
причем не только в печени, но и в мышцах. Содержание гликогена в печени при этом в
4 раза, в мышцах в 1,7 раза ниже, чем до спячки. Понижалось содержание глюкозы в тка-
нях, особенно в печени, а уровень молочной кислоты в печени и мышцах увеличивался в
2,4 раза по сравнению с недельной спячкой, хотя и не превышал контроля. Возрастало со-
держание макроэргических фосфатов в мозге. Следовательно, на этом этапе спячки усили-
вается утилизация углеводов, не компенсируется ресинтезом.
Однако при удлинении спячки до 5—6 недель в углеводном обмене наступали заметные
изменения: количество гликогена во всех исследованных тканях яшурки значительно воз-
растало по сравнению с предыдущим этапом спячки. В печени оно не достигало уровня
контроля, оставаясь в 1,2 раза ниже его, но в мышцах и мозге превышало контроль соот-
ветственно в 1,4 и 13 раза. По-видимому, это связано со значительным усилением глюко-
неогенеза. Литературные данные свидетельствуют, что у пойкилотермных животных этот
процесс интенсивно протекает в осенне-зимний период не только в печени [321], но и в
мышцах [160,221].
Результаты исследования показывают, что на начальных этапах (1—3 недели) спячки
разноцветной ящурки энергетические потребности организма, значительно пониженные в
результате снижения температуры тела, в основном удовлетворяются за счет использова-
ния резервов гликогена в печени, отчасти мышц. Использование липидов как энергетиче-
ских субстратов при зимней спячке ящурки ограничено.
При искусственной гипотермии (2—4°) разноцветной ящурки, как и при зимней спяч-
ке, используется гликоген печени и мышц. Так, при недельной гипотермии содержание
гликогена в печени и мышцах понижается соответственно в 2 и 1,7 раза, при 3-недельной
гипотермии — в 4 и 1,9 раза. Однако в отличие от спячки при длительной глубокой искус-
ственной гипотермии разноцветной ящурки наряду с углеводами в значительной мере ис-
пользуются и липиды: уровень НЭЖК в крови при недельной гипотермии увеличивается в
1,7 раза, при 3-недельной — в 9. Значительно интенсифицируются при длительной глубокой
гипотермии ящурок анаэробные процессы в тканях. Содержание молочной кислоты в пече-
ни и мышцах увеличивается при недельной гипотермии в 2,1—2,7 раза. При 3-недельной ги-
потермии наблюдается заметное увеличение уровня этого метаболита и в мозге.
111
Нервная система, видимо, не испытывает нехватки энергетических субстратов, по-
скольку содержание глюкозы в мозге не отличается от контроля, а гликогена —
превышает его.
Сказанное выше свидетельствует о наличии существенных различий в динамике изу-
ченных метаболитов при искусственной и физиологической гипотермии ящурок. При дли-
тельной глубокой гипотермии (в отличие от зимней спячки) наряду с углеводами в значи-
тельно большей мере используются и резервные жиры, а в тканях возникают гипоксиче-
ские явления,приводящие к усилению анаэробных процессов.
ГЛАВА 8
МИКРОЭЛЕМЕНТЫ В ОРГАНИЗМЕ
РАЗНОЦВЕТНОЙ ЯЩУРКИ
В целом в организме разноцветной ящурки обнаружено 24 химических элемента, в
яйцах — 15 (табл. 7) . Между самцами и самками по содержанию микроэлементов статисти-
чески достоверных различий не найдено, однако ящурки из разных мест ареала отличаются
по составу организма. Например, литий обнаружен только у ящурок из Астраханской обл.,
висмут — у ящурок из Крыма, галлий, иттрий и иттербий — из Казахстана, а бериллий — из
Калмыкии. Крымские разноцветные ящурки содержат свинца в среднем в 10, цинка в
Таблица 7. Содержание химических элементов (мг/кг сухого вещест-
ва (в организме и яйцах разноцветной ящурки
Элемент В организме В яйцах
п проб Um м т М
min max
Алюминий* 20 0,40 10,0 4,00 0,71 2,00
Кальций* 20 2,00 20,0 15,05 1,20 12,50
Магний* 20 2,00 10,0 6,80 0,61 5.00
Железо* 20 0,60 3,0 1,28 0,14 2,00
Титан 20 60,00 400,0 110,00 18,40 75,00
Марганец 20 1,60 160,0 29,80 9,35 2,00
Хром 20 6,00 1000,0 30,93 69,76 100,00
Никель 20 1,60 600,0 70,76 31,65 10,00
Медь 20 4,00 100,0 16,40 4,63 5,00
Серебро 20 0,01 1,0 0,11 0,04 0,02
Стронций 20 16,00 80,0 35,45 6,97 —
Олово 12 0,60 6,0 2,05 0,44 0,75
Свинец 10 0,60 6,0 1,46 0,55 0,75
Цинк 8 6,00 200,0 42,75 25,25 150,00
Цирконий 14 2,00 10,0. 4,85 1,46 250
Барий 9 16,00 160,0 38,20 15,57 —
Молибден 4 0,20 0,6 0,50 0,10 —
Бериллий 3 0,16 0,2 0,17 — —
Галлий 2 0,60 1,0 0,80 — —
Литий 2 6,00 6,0 6,00 — —
Висмут 2 2,00 20,0 11,00 — —
Иттрий 1 2,00 2,0 2,00 — —
Иттербий 1 0,40 0,4 0,40 — —
Фосфор* 20 > 10 > 10 > 10 — > 10
* В г/кг.
30 раз больше, чем ящурки из иных частей ареала. По сравнению с другими видами лацер-
тид у разноцветной ящурки в среднем в 2 раза больше титана. Она статистически достовер-
но отличается от прыткой ящерицы по содержанию 5 элементов, от крымской и скаль-
ной — 6, а от крымского геккона — 7 элементов. Разноцветная ящурка может служить био-
логическим индикатором загрязнения окружающей среды, так как у особей этого вида из
населенных пунктов и природных биотопов имеются достоверные отличия по содержанию
в организме 10 металлов,в том числе и тяжелых [301].
113
ГЛАВА 9
СТАЦИИ, ЧИСЛЕННОСТЬ, БИОМАССА!
Ящурка разноцветная относится к эврибионтным видам ящериц, обладающим высокой
пластичностью при выборе среды обитания. Однако при этом у данного вида наблюдаются
различия в выборе (предпочтении) стаций в разных зонах, что создает сложную картину
биотопического распределения. Здесь мы рассматриваем конкретные местообитания ви-
да — пространственно ограниченную совокупность условий абиотической среды, обеспечи-
вающую весь цикл развития особей, популяции и вида в целом [214].
Ареал разноцветной ящурки охватывает лесостепную, степную и полупустынную зоны,
поднимаясь в горы до высоты около 2000 м н.у.м. Определенные требования к субстрату
и освещенности обусловили мозаично-ленточный характер размещения животных, террито-
риальных группировок и популяций. В этой связи местообитания ящериц в разных природ-
ных зонах во многом схожи.
9.1. БИОТОПЫ, СТАЦИИ
Биотопы и стации ящурки разноцветной описаны на большей части ее ареала [5,7,15,
27, 35, 44, 49, 50, 64, 66, 75,78,86,94, 104, 110, 113, 118, 121, 125, 128, 144, 146, 147,
151, 170, 176, 188, 189, 192, 199,201, 202, 246,248,255,258, 266,273,278, 280,290,305,
313, 316, 320, 323, 324, 359]. Анализируя среду обитания ящурки в разных природных зо-
нах, ниже приводим классификацию ее биотопов в естественных и антропогенных ланд-
шафтах.
А, Естественные биогеоценозы
1. Песчаные массивы
Закрепленные и пол у закрепленные пески в долинах рек
Закрепленные и полузакрепленные пески в степных ландшафтах, с луговинами и зарослями
кустарников и полукустарников
Песчаные пляжи, морские косы с кумулятивными берегами
Верхняя часть полосы морского прибоя, с псаммофильной растительностью
Литоральный вал с псаммофильно-ксерофитной растительностью
2. Полупустыни
Полынные, солянковые, с другими растительными ассоциациями
3. Степи
Полынно-солянковые, разнотравные, разнотравно-ковыльные, ковыльно-солянковые, полынно-
солянковые с эфемерной растительностью и др.
Солончаки сухие
Сухие русла рек
4. Лесостепи
Разреженные колки сухие
Б, Антропогенные биогеоценозы
Отдельные необработанные участки на староосвоенных землях
Участки пара
Окраины посевов
Молодые или разреженные посадки из сосны, белой акации, лоха, тамарикса и др.
Молодые полезащитные лесонасаждения
Залежные земли
Террасированные склоны оврагов, балок
1 Материалы предоставили Т.Алиев, В.И.Бадмаева, З.К.Брушко, Э.В.Вашетко, А.Г.Высотин, В.И.Горо
вая, М.П.Зинякова, Н.Н.Иванова, В .А.Киреев, Т.И-Котенко, Е.Ю.Кудакина, В.В.Неручев, Н.М.Окуло
ва, В.Ф.Орлова, П.П.Руденко, А.М.Рудик, Х.Тербиш, Э.А.Шебзухова.
114
Окрестности колодцев в сухих степях
Заброшенные кошары
Обочины автомобильных дорог
Склоны железнодорожных насыпей
Грунтовые дороги в сухих степях и полупустынях
Склоны водохранилищ и оросительных каналов
Кладбища
Как отмечает Н.Н.Щербак [320], в европейской части ареала ящурки в целом обита-
ют на песчаных и мягких грунтах. Находки этого вида в степи с плотным грунтом очень
редки и известны для Восточного Предкавказья, низовий Волги,междуречья Волгин Ура-
ла, Закавказья. В Средней Азии и Казахстане он обитает преимущественно в местах с плот-
ной почвой и даже в стациях с твердым грунтом: в галечниковых и каменисто-щебнистых
пустынях. Все это, по мнению Н.НЩербака [320], указывает на смену стаций в направле-
нии с запада на восток и юг.
9.2. ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ БИОТОП И СТАЛИИ
Ящурки обитают преимущественно на горизонтальных поверхностях с песчаным, лесовым,
глинистым, щебнистым и каменисто-щебнистым грунтами с относительно разреженным
травостоем: до 50 % [35], 70 [131] и 95 % - по МЛ.Зиняковой и ПЛРуденко [устн.
сообщ.]. Растительность обычно псаммофильная и галофильная, а также ксерофитная.
Животное избегает крутых склонов, соров, сырых, развеваемых песков, отсутствует
обычно в населенных пунктах, очень редко встречается в антропогенных модификациях
пустынных ландшафтов, а также посевах, в разнотравно-злаковой степи, на пашнях, буграх
из чистой ракушки, территориях, подвергающихся значительной рекреационной нагрузке
(курорты на море).
Из приведенных данных следует, что выбор ящуркой того или иного биотопа, стации
в нем определяется рядом факторов. Очевидно, что выбор ящерицей места обитания зави-
сит от наличия корма, убежищ, возможности размножения. Такая же зависимость харак-
терна для других ящериц [262]. Для разноцветной ящурки Т.ИКотенко [131] обнаружи-
ла зависимость выбора биотопа от механического состава и влажности почвы, структуры
растительного покрова, степени освещенности и защитных свойств биотопа. Так, в Украи-
не животное распространено повсеместно на песчаных, песчано-ракушечных почвах, встре-
чается на сыпучих и легких супесях, но избегает илистых солончаковых и болотистых
почв. Ящурки предпочитают участки с сухим (0—1 балл)1, суховатым (1) увлажнением,
в меньшей степени — со свежеватым (1—2) и не встречаются в понижениях с высокой влаж-
ностью почвы (3—5 баллов) [131].
Кроме того, показана важная роль освещенности при выборе ящурками биотипа, ста-
ции, что определяется структурой растительного покрова. Так, высокая численность яще-
риц установлена в биотопах с проективным покрытием от нескольких процентов до 30-
40 %, очень низкое и даже полное отсутствие животных в биотопах с проективным покры-
тием выше 60 %. На участках, лишенных растительности, ящурки встречаются лишь в пе-
риод суточных перемещений.
Высота травяного покрова в стациях может варьировать от 5-10 см (песчаная степь
с чабрецом, низкорослым типчаком, лишайниками и пр.) до 1—2 м (кусты полыни мор-
ской , донника белого и др.) [131].
Благоприятные условия освещенности складываются также в насаждениях с освет-
ленной и полуосветленной структурой, состоящих из ажурнокронных и полуажурно-
кронных деревьев1 с сомкнутостью крон не более 0,5. В частности, подходящие условия
в посадках сосны, белой акации, гледичии, лоха, тамарикса, в сосновых посадках, где ши-
рокие междурядья с почти голым песком чередуются с рядами маленьких сосен, растущих
на полосах с сохранившейся степной растительностью. Наконец, ящурки обитают на участ-
ках с сильно разреженным древостоем любого возраста с единичными деревьями или рас-
тущими небольшими группами [131].
1 Использована классификация А.Л.Бельгарда [33].
115
Таким образом, наиболее предпочитаемы (типичны) группы биотопов с низкой влаж-
ностью, песчаным или каменисто-песчаным грунтом, с достаточным количеством убежищ,
позволяющих укрыться от перегрева и врагов и благополучно провести зимовку, где име-
ется высокая освещенность и возможность свободно передвигаться. Типы биотопов с ука-
занными условиями обычны для сухостепных, полупустынных и пустынных ландшафтов,
в меньшей мере встречаются в лесостепных ландшафтах. Типичные стации вида представле-
ны на рис. 67—70.
93. ЧИСЛЕННОСТЬ И БИОМАССА
Данных о численности и биомассе ящурки в территориальных группировках, популяциях,
биотопах и природных зонах до сих пор немного. Практически все имеющиеся сведения о
плотности населения в той или иной части ареала относительны, поскольку получены путем
экстраполяции результатов относительного учета на небольших участках, входящих в ту
или иную природную зону. За редким исключением, приводимые данные варьируют в ши-
роких пределах (табл. 8—12) - от нескольких единиц до нескольких сотен особей на гек-
Т а блица 8. Плотность и биомасса разноцветной ящурки в различных регионах
видового ареала
Регион Плотность, экз ./га Биомасса, г/га Источник информации
Украина Крым 20-100 134-670 [316] Степная эона Украины 2,5—640 10-3250 [386] Харьковская обл. 47—58 314,9—388,6 [Рудик, сообщ.] Юго-восток европейской части России Волгоградская обл. 6 40,2 [147] Калмыкия 4—163,3 26,8-1094,1 [26,114] Северный Кавказ Западное Предкавказье 150-200 1005-1340 [44,155] Юг Центрального Пред- кавказья (Кабарда) 48 321,6 [305] Центральное Предкав- казье 2-156 13,4-1045,2 [259] Предкавказье в целом 1,26 8,44 [Тертышников, со- общ.] Закавказье, Азербайджан 50-150 335-1005 [7; Алиев, сообщ. ] Казахстан и Средняя Азия Волго-Уральские пески 1,5—26,5 10,05-177,55 [192; Окулова, со- общ.] Хребет Нуратау 0,36 2,41 [НО] Джизакская обл. 1,2-3,0 8,04-20,1 [Вашетко, сообщ.] Западная Монголия 29 194,3 [Орлова, Тэрбиш, со-
общ.]
тар. В европейской части ареала численность и плотность населения оказываются значи-
тельно выше, чем в азиатской.
По-видимому, исследователи обращали внимание на типичные биотопы, где числен-
ность обычно значительно выше, чем в нетипичных. Однако, как показывают учеты на
больших площадях, средняя площадь, занятая ящерицей по всем биотопам, составляет не-
значительные размеры, Так, для стационарного участка в верховьях р.Томузловки (Став-
ропольская возв.) [258] средняя площадь, занятая ящуркой, составила всего 24 %, из 10
биотопов заселены были лишь 4 и только один из них на 10 %. Судя по этим данным, эк-
страполяция данных учета на большую площадь, в частности на незаселенные биотопы, при-
ведет к их усреднению и резкому снижению показателей плотности.
Однако отсутствие данных по площади населенных биотопов в каждой природной зо-
не не позволяет определить общую численность ящериц. В равной мере это относится и к
биомассе ящурок в разных зонах.
К этому следует добавить еще один момент: крайне ограниченные материалы, касаю-
щиеся колебания численности (периодических и апериодических), что также осложняет
116
Таблица 9. Плотность и биомасса разноцветной ящурки в Украине
в местах с высокой численностью [131]
Место учета Дата учета Масса ’’сред- ней” особи Плот- ность, экз./га Биомас- са^сг/га
Черноморский заповедник Ив ано-Рыб альчан- ский участок VHI.1975*r. 2,08 640,0 1331 О-в Тендра VII. 1975 г. 4,44 115,0 0,510 Потиевский участок V.1979 г. 4,84 153 3 0,742 О-в Джарылгач VII. 1976 г. 4,86 300,0 1,457 Буркуты VI. 1978 г. 3,89 250,0 0,973 Кирилловская коса VII. 1977* г. 5,90 550,0 3,245 * Максимальная плотность на отдельных небольших участках. Таблица 10. Плотность (экз^га) разноцветной ящурки в различных биотопах Калмыкии [25]
Биотопы 1973 г. 1974 г. 1975 г. 1976 г. 1977 г. 1978 г. 1979 г. 1980 г. 1981г.
Разнотравно-ко- выльные 32,0 32,0 26,0 24,0 20,0 11,0 9,0 8,0 83 Ромашко-тип- чаковые 34,0 32,0 39,0 40,0 39,0 32,0 26,4 29,4 30,2 Белополынно- тырсовые 90,0 90,0 85,0 85,0 80,0 80,0 80,0 75,0 60,0 Белополынно- прутняковые 90,0 95,0 90,0 90,0 88,8 79,9 82,6 81,5 82,4 Типчаково- тырсовые 813 803 80,4 79,6 72,4 733 703 72,8 72,4 Тамарик- совые 72,4 733 72,0 74,0 62,0 62,0 60,0 62,0 61,2 Кияковые 36,4 34,8 40,0 40,0 40,0 33,1 31,9 39,6 39,2 Рогозо-ка- мышовые 103 9,8 12,0 14,0 16,0 ИЗ 83 83 8,2 Тростниково- рогозовые 9,0 83 12,0 13,0 123 9,9 8,4 8,6 8,4 Песчано-по- лынные 493 493 43,4 413 41,0 41,0 40,9 40,0 40,0 Кмяково- джузгуновые 36,7 363 33,4 313 33,4 32,0 403 32,1 31,4 Полынно-злаковые 39,4 42,0 43,0 42 3 46,7 34,9 32 3 30,1 34,6 Закрепленные пески 42,6 42,6 41,4 41,2 46,4 42,0 40,0 41,4 40,0 Развеваемые пески 8,0 9,0 11,0 6,0 8,0 9,0 9,0 11,0 11,4 Участки, ли- шенные расти- тельности 4,0 6,0 9,1 8,4 83 8,0 7,6 73 8,1 Склоны оврагов, балок, ороси- тельных ка- налов 89,6 87,7 80,4 126,4 1293 131,9 121,4 120,0 123,6
попытку оценить численность и биомассу ящурок в природных зонах и в ареале в целом.
Известны, например, сезонные колебания, связанные с размножением и возрастной смерт-
ностью, прессом хищников, миграциями и др., в том числе и в связи с воздействием факто-
ров среды. А роль их порой немаловажна. По расчетам [259], например, на стационарном
участке в Центральном Предкавказье в 1969 г. погибло 61,6 % популяции в результате воз-
действия учтенных фоновых врагов из числа позвоночных. Кроме того, отход яиц и смерт-
ность сеголетков в первую зимовку составляют 87 %.
Осложняет расчеты численности и биомассы ящериц отсутствие точных сведений и
площади, занятой видом. Численность ящериц постоянно уменьшается в связи с корен-
ным изменением среды их обитания в связи с антропогенными воздействиями. Имеющие-
ся у нас данные свидетельствуют о том, что исчезли, например, популяции у х.Садки на по-
бережье Азовского моря [Зинякова, сообщ.], на плато Надреманное Ставропольского
117
Таблица 11. Численность и плотность разноцветной ящурки в Джунгарской Гоби
в различных местообитаниях ( Орлова, Тэрбиш, сообщ.]
Место учета Дата учета Биотоп Числен- ность (в сред- нем за 3 учета), зкз. Плот- ность, зкз ./га
ОкрАлтай- сомона 8.VI1982 Петрофитные ковыльково-солянковые пустынные степи на бурых супесчаных почвах 6,6 22,0
11.VI1982 Солончаковые бугристые песчаные гря- ды с селитрянкой и чием 0,3 3,0
10.VI1982 Терескеновые щебнисто-супесчаные пу- стыни с участием саксаула и солянки на серо-бурых засоленных почвах 8,7 29,0
Окр. колод- ца Загийн- Толгой 9.VI1983 Щебнисто-каменистые низкорослые сак- сауловые пустыни с участием терескена и парнолистника на серо-бурых супесча- ных высокогипсоносных почвах 0,6 6,0
Окр.Уенч- сомона 7.VI1984 Таровые пустыни со злаковыми и куста- ми на бурых суглинистых почвах 3,7 123
Таблица 12. Бмотопическое распределение, плотность, биомасса и числен-
ность разноцветной ящурки в Центральном Предкавказье [259]
Биотоп Площадь биото- па, га Плотность, экз ./га Б ио мае- са, кг/га Числен- ность, экз.
Пределы | Средняя
Разнотравно-злаковая степь 803 0-30 2ft 0,01 125
Типчаково-полынная степь 43
Полынная степь 43 — — — —
Опустененная степь 313 10-156 55 Д озз 1749
Посевы 123 0-30 3,0 0,02 36
Полезащитные лесные полосы 27,0 0-50 7,0 0,04 180
Толока 0,6 — — — —
Лес 83 — — — —
Лесные опушки 0,4 — — — —
Вырубки и просеки в лесу 0,2 — — — —
Всего 170,0 0-156 12Д 0,17 2090
края [Тертышников, сообщ.]; за счет распашки и обводнения целинных земель в Калмы-
кии [Киреев, сообщ.} ящурка исчезла на площади около 158 тыс. га и тд.
Очевидно, наиболее полные данные о численности и биомассе популяции разноцветной
ящурки имеются по Предкавказью. По расчетам М.Ф.Тертышникова [сообщ.],разноцвет-
ная ящурка занимает в этом регионе площадь около 1 374 800 га. При усредненной плот-
ности населения, равной 1,26 экз./га, общая численность ящериц в регионе достигает
1 732 250 экз. и биомассы 11,59 т. Плотность населения в пустынных ландшафтах варьиру-
ет от 0,67 до 13 экз Да (средняя 1,01) при биомассе 4,49—12,06 г/га (средняя 6/74)1.
Плотность населения в полупустынных ландшафтах варьирует от 0,6 до 187,5 экз./га
(средняя 5,4) при биомассе 4,01—1254,38 г/га (средняя 36,13) . В степных и лесостепных
ландшафтах плотность населения варьирует соответственно от 1,29 до 160 и от 0,1 до
54,6 экз^га при средних показателях 8,85 и 8,89, биомасса — от 8,63 до 1072 и от 0,67 до
365,82 г/га при средних показателях 59,25 и 59,56 (табл. 13).
Эти данные статичны и не учитывают изменения численности отдельных возрастных
групп. Поэтому правильней было бы производить расчеты до размножения, когда в попу-
1 В результате взвешивания 300 особей разных возрастных групп из популяций Предкавказья опреде-
лено, что средняя особь в популяции имеет массу 6,67 г. Эта величина и была использована в дальней-
ших расчетах. Приведенные цифры могут несколько изменяться в разных регионах.
118
Таблица 13. Плотность и биомасса разноцветной ящурки в различных ландшаф-
тах Предкавказья [ Тертышииков, сообщ.)
Ландшафт, биотоп Плотность, экз./га 1 Биомасса, г/га
Пустынные ландшафты
Рошинско-ЧервленобурунскиА ста- ционар Пески в бурунах 0,67(0,29-2,19) 4,49(1,94-14,67)
Межгрядовые понижения 13 8,71
Степь песчаная полынно-солян- ковая 1,8 12,06
В среднем 1,01 6,74
Полупустынные ландшафты
Махмуд-Мектебский стационар Пески в бурунах 0,86(0,45-2,86) 6,43(3,02-19,16)
Межгрядовые понижения 33 22,11
Степь песчаная полынная 0,6 4,02
Величаевский стационар Пески в бурунах 5,0 333
Степь глинистая полынная 13 8,71
Долина р.Чограйки Степь полынно-типчаковая 5,0 333
Пастбище 10,0 67,0
Посевы люцерны 0,6 4,02
Склоны Кумо-Манычского канала 5,0 333
Оз. Довсун, степь полынная 10,0 67,0
Пос. Б акр ес, степь полынная 187,5 125635
Оз. Дадынское Дорога 95,0 6363
Побережье с полынью и солянками 80,0 536,0
Пос.Турксад Пересохшее дно дренажного канала 1635 111,56
Степь полынная зззз 22331
Пос. Зимняя Ставка Степь типчаково-полынная 150,00 1005,0
Степь полынная с гребенчу ком 50,0 335,0
В среднем 5,4 36,13
Степные ландшафты
С.Казинка, склоны балок с террасиро- ванными склонами 5,78(2,97-1138) 38,73(19,9-76,25)
С Кочубеевское, разнотравные степи с участками песка 160 1072,0
Типчаково-полынные степи 139 8,63
В среднем 835 5935
Лесостепные ландшафты
Верховья р.Томузловки Разнотравно-злаковая степь 2,0 13,4
Типчаково-полынная степь 113 75,04
Полынная степь 0,1 0,67
Опустыненная степь 54,6 365,82
Посевы зерновых (края") 2,8 18,76
Полезащитные лесные полосы (края) 6,67 44,69
В среднем 8,89 5936
Усредненная по всем ландшафтам, рассчитанная по методу ’’объединен- ного гектара” 136 8,44
Примечание. В скобках указаны лимиты.
ляции имеются две возрастные группы — молодые и половозрелые, и после размноже-
ния, когда к указанным группам присоединяются сеголетки, учитывая при этом усред-
ненную биомассу особи каждой возрастной группы. Значительные коррективы были бы
внесены в расчеты, если бы принимались во внимание гибель яиц и общая смертность ящу-
рок разных возрастных групп. По крайней мере расчеты подобного рода (в среднем по
всем зонам) имеются для Центрального Предкавказья, а они важны, учитывая ключевую
роль разноцветной ящурки во многих типах биотопов степной зоны, полупустынных и
пустынных ландшафтов в пределах ареала.
ГЛАВА 10
ТЕРМОБИОЛОГИЯ1
ЮЛ. ОБЩИЕ ЗАМЕЧАНИЯ, ТЕРМИНОЛОГИЯ, ОЦЕНКА ДАННЫХ
Термобиологию разноцветной ящурки изучали в разное время и в различных местах, с
использованием различающихся методик и подходов, поэтому перед рассмотрением кон-
кретных материалов по E^rguta целесообразно бегло остановиться на основных понятиях
и показателях термобиологии (см. обзор [283]), что позволит более правильно оценить
полученные результаты. Термофизиологические показатели характеризуют возможные для
данного вида пределы температурной выносливости и активности и отражают норму реак-
ции вида на температуру. Конкретные экологические условия накладывают некоторые
ограничения на эти показатели, отражением чего являются термоэкологические показате-
ли. Термофизиологические показатели в чистом виде можно выявить, лишь исключив
влияние среды, а именно — в корректно поставленных опытах в термоградиент-приборс.
Термозкологические же показатели *- это то, что мы определяем в полевых условиях.
Основные термобиологические показатели жизнедеятельности рептилий следующие
[281—283]: критические минимум и максимум температуры тела, минимальная и макси-
мальная температуры тела активного состояния, или добровольный минимум и максимум,
и диапазон оптимальных (предпочитаемых) температур тела. В оптимальном диапазоне у
рептилий с наименьшим напряжением функционирует система физиологических и химиче-
ских механизмов терморегуляции, стабилизирован обмен веществ, оптимизированы
процессы переваривания пищи, гаметогенеза и др. Оптимальные температуры — это те,
которые рептилии активно избирают (предпочитают) при наличии широкого выбора
температурных условий и поддерживают специальными формами терморегуляционного
поведения.
Потенциально дневным видам рептилий, к которым относится и разноцветная ящурка,
свойственны следующие основные формы поведения, определяемые температурными
условиями среды и закономерно сменяющиеся в течение суток: ночной покой, нагревание,
термостабилизирующее поведение (рептилии ведут активный образ жизни в широком
диапазоне внешних температур, при этом температура их тела удерживается в пределах,
близких к оптимальному диапазону) , добровольный перегрев, или активное охлаждение
(температура тела выше оптимальных температур, при этом поведение и действие физио-
логических механизмов терморегуляции направлены на предотвращение перегрева) , днев-
ной отдых, термостабилизирующее поведение, остывание. Термостабилизирующее поведе-
ние, добровольный перегрев и дневной отдых соответствуют активному состоянию репти-
лий, ночной сон — неактивному, нагревание и остывание — переходные состояния.
Сведения, относящиеся к вопросам термобиологии разноцветной ящурки, довольно
многочисленны, но неравноценны. В некоторых работах приводятся лишь результаты от-
дельных измерений температур воздуха, почвы или тела ящурок в природных условиях
[48, 55, 56, 316; Бондаренко, сообщ.], в других есть попытки дать какие-либо термоэко-
логические характеристики активного состояния ящериц в природе [19, 20, 22, 24, 25,
27, 66, 97, ИЗ, 192, 203, 224, 225, 250, 258-260, 262, 266, 268, 315, 359, 436; Киреев,
Окулова, Рудик, Тэрбиш, Хонякина, сообщ. ]. Кроме того, имеются данные по критиче-
скому минимуму [218] и максимуму температур тела [ИЗ, 192, 210, 220, 258—260,
262], результаты единичных измерений температуры тела ящурок в неактивном состоянии
1 При участии МЛ Ахмедова, В Л Бадмаев ой, ДА .Бондаренко, В А .Киреева, НМ .Окуловой, AM Руди-
ка, Т.Сатторова, СБ.Таращука, М.Ф.Тертышникова, Х.Тэрбиша, 3 Л-Хонякиной.
120
[192,203] и температур в норах — убежищах этих рептилий [192; Киреев,Рудик, сообщ.].
Теплоустойчивость клеток разноцветных ящурок из Армении и Средней Азии изучал
Б Л .Ушаков [272], температуру одной особи в 17 точках поверхности тела измерил
ЛЛ.Хозацкий [277]. Особое место занимают исследования Н.НЛ1ербака [316, 318, 319],
выполненные в термоградиент-приборе. Наиболее подробно вопросы термобиологии разно-
цветной ящурки изучались в Западном Казахстане [192; Окулова, сообщ.], на Ставрополь-
ской возвышенности [210, 258, 260, 262, 266] и юге Украины [315, 316, 318, 319]. По-
следний регион в результате более поздних исследований [127, 129—131, 135, 232, 385,
386] оказался самым изученным. Сведения, полученные до начала 70-х годов, суммирова-
ны в сводке НН .Щербака [320] .
Даже беглое ознакомление с литературными данными показываем необычайный раз-
брос чисел по каждому из термоэкологических параметров. Местоположение региона мо-
жет накладывать отпечаток на те или иные показатели, но, учитывая, что многие из них
видоспецифичны, и что в определенных пределах температур среды рептилии в состоянии
очень точно поддерживать оптимальную температуру тела, такую разницу в температурах
тела активных ящурок, как 15 и 30° (минимум) , 35 и 40 (максимум) и 30 и 35 (сред-
няя оптимальная), или разницу в предпочитаемых температурах почвы (24-25 и 37-
40°) , минимальных (12 и 25°) и максимальных (40 и 56°) температурах субстрата и ми-
нимальных температурах воздуха (9 и 20°) в период активности ящурок, нельзя объяс-
нить климатическими различиями. Какие же факторы могут влиять на результаты термо-
биологических исследований?
1. Отсутствие единообразия в способах измерения температур воздуха и почвы. Раз-
ные исследователи измеряли температуру воздуха на высоте 2, 1,5 или 1 м, 10-15, 3—
5 см от поверхности почвы или ”в приземном слое воздуха”, при этом нередко вообще
не оговаривали, на каком уровне проводили замеры. Температуру поверхности почвы,
судя по полученным результатам, измеряли то на открытых, освещенных солнцем участ-
ках, то в тени, то в точке поимки ящурки, и это обычно также не оговаривают. Порой ис-
пользовали показания ближайшей метеостанции, иногда без соответствующих указаний,
Однако еще в работах ИД.Стрельникова [240], А «М .Сергеева [228] и И.В .Андреева [10]
отлично показаны большие различия в температурах среды в одном и том же биотопе в
конкретный момент времени.
2. Температура тела рептилий в разных его участках различна (данные по Е .arguta
см. [277]), поэтому важно единообразие и в этих измерениях. Обычно температуру тела
ящурок измеряли в ротовой полости; некоторые исследователи этот момент не оговари-
вали. Вообще же температуру тела рептилий лучше измерять в клоаке, т.е. ближе к поло-
сти тела.
3. Применение различных измерительных приборов также может влиять на резуль-
таты. Так, электротермометры дают показания очень быстро, тогда как на данных по тем-
пературе тела, полученных ртутным термометром, может сказаться температура субстрата
или рук исследователя. Например, температура тела разноцветной ящурки, помещенной на
субстрат с температурой 44 °, за 10 мин повысилась на 14° [192], температура тела дру-
гих ящериц могла повышаться в течение минуты на 4—5° [240].
4. Температуру тела целесообразно измерять только у быстро пойманного животного,
поскольку даже после непродолжительного бега температура тела ящериц существенно
возрастает [242], в том числе и у разноцветной ящурки [135].
5. Полученные в полевых условиях термоэкологические показатели могут значитель-
но различаться у разных исследователей еще и потому, что получены в разное время года
или суток, для разных возрастных групп или полов, для животных, находящихся в различ-
ных физиологических состояниях или формах поведения. По этим же причинам при вычи-
слении оптимальных температур тела или предпочитаемых температур среды нельзя усред-
нять все полученные в природе данные, так как это приводит к существенным искажени-
ям результатов.
6. При работе с термоградиент-прибором необходимо учитывать длительность содер-
жания животного в неволе. Как показали исследования.Н.Н±Цербака [318, 319], в уело
виях неволи со временем извращается присущее виду отношение к температуре (включая
средние оптимальные температуры) и характер его сезонной динамики.
7. Наконец, одной из важнейших причин существенных расхождений в термобиологи-
121
ческих показателях у разлитаых авторов является неполнота полученных данных. Оче-
видно, что отдельные, случайные измерения дадут и ’’случайные” результаты.
Перечисленные выше методические погрешности в большей или меньшей степени
присущи практически всем работам по термобиологии разноцветной ящурки, что сужает
возможности нашего анализа (в частности, делает невозможным выявление географиче-
ской изменчивости термоэкологических показателей). Поэтому дальнейший наш анализ
будет основываться преимущественно на нескольких наиболее полных исследованиях —
на работах НЛкОкуловой [192; сообщ.], М.Ф.Тертышникова [259, 262] и ТИКотенко
[129, 131, 135, 232, 386], выполненных в полевых условиях, и Н Л Щербака [316,319] —
в термоградиент-приборе. Статья, специально посвященная терморегуляции, суточной и
сезонной активности разноцветной ящурки [268], полностью компилятивна и содержит
ряд неточностей и опечаток, поэтому в дальнейшем изложении мы на нее не ссылаемся.
10.2. ТЕМПЕРАТУРНЫЕ ГРАНИЦЫ АКТИВНОСТИ,
ПРЕДПОЧИТАЕМАЯ ТЕМПЕРАТУРА СРЕДЫ
По данным различных исследователей для разных сезонов года и регионов, минимальные
температуры воздуха, при которых разноцветные ящурки покидают свои убежища, состав-
ляют 9—23°, минимальные температуры поверхности почвы — 12—25°. Поскольку формы
поведения рептилий ни один из авторов не учитывал, то эти цифры отражают не только гео-
графические и сезонные различия, но и состояние животных: в одних случаях отмечали
температуру среды при обнаружении первых особей на поверхности (т.е. в переходном
состоянии — в фазе нагревания), в других — регистрировали температурные параметры
первых активных ящурок (т.е. по сути отмечали добровольный минимум температур).
Температурные условия весеннего пробуждения разноцветных ящурок таковы. В Харь-
ковской обл. (Украина) первые особи встречены на поверхности в солнечную погоду при
температуре воздуха 15—16°. Минимальная температура поверхности почвы, на которой
отмечена активная ящурка, 23°. Критическим моментом для пробуждения животных яв-
ляется, видимо, переход температур через рубеж 5,5—7°, поскольку именно такие темпе-
ратуры были зарегистрированы в норах первых проснувшихся особей [Рудик, сообщ.].
(По данным для Калмыкии [Киреев, сообщ.], ящурки зимуют при температуре воздуха
в норах 4—9°.) В Волго-Уральских песках (Западный Казахстан) в период пробуждения
ящурок наблюдались следующие изменения погоды: переход через 0° среднесуточных тем-
ператур воздуха и почвы на глубине 20 см, подъем максимальной температуры воздуха
от 4° в конце марта до 16—18° в начале апреля. И хотя среднесуточная температура почвы
на глубине 20 см в начале апреля не превышала 2,8° (данные за 5 лет), надо полагать,
что максимальная температура достигала величин, необходимых для пробуждения репти-
лий [192].
В Ставропольском крае первые ящерицы появляются, когда температура воздуха
устанавливается в пределах 11—15° [259, 262]. В 1970 г. температура почвы и воздуха
в момент первого появления ящериц была 12°, в 1971 г. — 15, однако в течение несколь-
ких предыдущих дней почва прогревалась до 34°, что и вывело животных из зимней
спячки [259]. В Калмыкии первые особи отмечены ВИБадмаевой [22, 25] при темпера-
турах воздуха 10-15°, почвы - 14—15,2, В А Киреевым [ИЗ; сообщ.] - при 12,2-
15,5 и 14,1-18,7° соответственно. В Северном Таджикистане ящурки появляются при тем-
пературе воздуха 18—20°, почвы — 16—18 [225].
Массовый выход ящурок из зимних убежищ происходит обычно позже появления пер-
вых особей и при более высоких температурах среды: в Волго-Уральских песках он наблю-
дался при установлении максимальной температуры воздуха 21—28° [192], в Калмы-
кии — при температуре почвы 15,8—17,5° [25]. В Черноморском заповеднике (юг Украи-
ны) в последние дни марта — первые дни апреля обычно резко возрастает температура
среды. В течение нескольких дней, несмотря на высокие температуры воздуха (до 20—
25°) и поверхности почвы (до 30—40°), ящурки не выходили из нор. Только после доста-
точного прогрева почвы (до 11° на глубине 20 см и более 13° на глубине 10 см) появились
первые особи. В последующие дни ящурки выходили из своих убежищ при температурах
воздуха и поверхности почвы не ниже 11 и 17° соответственно [135]. На о. Обливной
в Каспийском море (Азербайджан) ящурки в марте появлялись при температуре воздуха
122
18—20е, в середине апреля - при температуре воздуха 16, почвы - 23.5° [20]. Наблюде-
ния за другими видами пресмыкающихся дали аналогичную картину: для того, чтобы жи-
вотные покинули свои зимние убежища, нужна температура на 2—3° (иной раз и более)
выше той, при которой они будут появляться на поверхности в последующие дни. В февра-
ле рептилии пробуждаются при более высокой температуре среды, чем в марте, в марте -
при более высокой температуре, чем в апреле [38,39].
Такой сдвиг температурного минимума характерен только для зимнего и ранневесен-
него периодов, когда оттепели чередуются с резкими похолоданиями, грозящими репти-
лиям замерзанием. Позже имеет место сезонный сдвиг температуры начала суточной актив-
ности в сторону повышения к жаркому периоду года. Так, летом пресмыкающиеся выхо-
дят на поверхность при более высоких температурах среды, чем весной [38, 39, 278,
318,319 и др.],это же отмечали и для разноцветной ящурки [113,192,315,316; Хоняки-
на, сообщ.]. В частности, в Западном Казахстане в первой декаде апреля ящурки наблюда-
лись при температуре субстрата 16—20°, уже в конце апреля и позже они не выходили из
нор до тех пор, пока температура не поднималась до 22-23° [192]. В Калмыкии в апреле
ящурки появлялись при температуре воздуха 13-15°, в мае - 14-15, июне - 19-20°
[22]. В Азербайджане [20] животные выходили на поверхность в мае—июне при темпера-
турах воздуха 20—21°,почвы — 24—25, в апреле — при более низких температурах (см. вы-
ше) . В Северном Таджикистане ящурки после зимовки появлялись при температуре воз-
духа 18—20°, в мае — 21—22, июле — 23° [225]. В Джунгарской Гоби (Монголия) они от-
мечались в первой декаде мая при минимальной температуре воздуха 10°, в первой дека-
де июня — 113,в третьей декаде июня - 15° [Тэрбиш, сообщ.].
В Нижнем Приднепровье данная закономерность нередко носит стертый характер
вследствие погодных особенностей весны. Высокие температуры апреля и мая позволяют
ящуркам в Черноморском заповеднике быть активными длительное время в течение суток
и появляться утром при температурах выше минимальных. В самый жаркий период года
ящурки обычно выходят на поверхность при более высоких температурах, но в отдельные
дни — при температурах, близких к весенним [135], что зависит прежде всего от темпера-
турного режима предыдущих дней.
В целом добровольный минимум температур среды в летний период примерно одина-
ков у ящурок из разных частей ареала (кроме Монголии) и составляет для воздуха 19-
23° (чаще 20-22°), почвы - 20-25 (обычно 22-23°) [20, 22, 66, 192, 225,232, 250;
наши данные]. В термоградиент-приборе яшурки, обследованные в июле, занимали места
с температурой не ниже 25° [316].
При указанных выше температурах ящурки появляются летом в сравнительно не-
жаркие периоды, а в особенно жаркие недели они выходят, как правило, при более высо-
ких температурах среды. В частности, в Дагестане в июне — июле 1962-1963 гг. первые
ящурки появлялись на поверхности при температуре почвы 26-29° в жаркие месяцы и
при 20—22° — в относительно прохладные [Хонякина, сообщ.]. Аналогичную картину
мы наблюдали и в Черноморском заповеднике в июле — августе 1975 г. По данным
В Л .Гончаровой [66], в мае — июне ящурки оставляли свои норы при температуре возду-
ха 23°, почвы — 22; при температурах соответственно 20 и 17° удавалось видеть
только молодых особей, при температуре воздуха 16°, почвы - 18° их на поверх-
ности не было.
Осенью ящерицы выходят из убежищ при более высоких температурах, чем весной
[19, 38, 39], уходят на зимовку также при более высоких температурах среды, чем пробу-
ждаются весной [25, 192, 258, 259, 315]. Так, на о.Обливной весной яшурки активны при
минимальной температуре воздуха 16°, почвы — 23,5, осенью — не ниже 19—21 и 23—26°
соответственно [19]. В Калмыкии, по данным 10-летних наблюдений, из зимней спячки
выходят при температуре почвы 14—15,2°, уходят на зимовку — при 17,6—19 [25]. В За-
падном Казахстане весной ящурки появляются на поверхности при среднесуточной темпе-
ратуре почвы на глубине 20 см 1—3°, осенью при 8—13° они уже не показываются. Это
вызвано, вероятно, тем, что осенью температура поверхности почвы становится неблаго-
приятной для ящурок, хотя на глубине она еще довольно высока; весной же к моменту'
перехода температуры на глубине 20 см через 0° поверхность почвы уже достаточно нагре-
та [192]. Однако в Харьковской обл. активные ящурки отмечались в октябре при тем-
пературе воздуха 12°, поверхности почвы — 12,5—23, температура в их норах была 6-9,5
[Рудик, сообщ.] ,что не выше температур весеннего пробуждения.
123
В Черноморском заповеднике мы наблюдали ящурок в октябре 1975 г. при темпера-
туре воздуха 8 , поверхности почвы — 15 [131, 135, 386], что ниже минимальных апрель-
ских показателей (11 и 17° соответственно) для этого же места и года. Такое различие
наших результатов и литературных данных мы объясняем следующими причинами. Во-пер-
вых, весна 1975 г. была бурной и теплой, с резким подъемом температуры среды, поэто-
му ящурки могли появляться при более высоких (благоприятных для них) температурах,
чем в годы с холодной затяжной весной. Во-вторых, длительность активного состояния
ящурок перед уходом на зимовку минимальна, крайне низок и уровень их активности
(подробнее см. в разд. 11.4), вследствие чего время (дата) залегания в спячку при недо-
статочно тщательном обследовании может быть определено как более раннее, чем было
на самом деле; отсюда возможность завышения температурных показателей прекращения
активности осенью в работах некоторых авторов. Наконец, самым главным моментом
является недоучет возрастной структуры популяции в осенний период и недооценка значе-
ния зависимости отношения рептилий к температуре от их возраста.
То, что сеголетки ящериц (в том числе разноцветной ящурки) позже взрослых особей
уходят на зимовку, давно известно [39,79,192,278,315,316 и др.],но на термобиологи-
ческую сторону вопроса внимания обращали мало. Исследования в Черноморском запо-
веднике показали, что для популяции ящурки в целом от августа к сентябрю и от сентяб-
ря к октябрю действительно наблюдается снижение температурного минимума. Но при
рассмотрении отдельно сеголеток и взрослых выясняется, что каждая возрастная группа
характеризуется своим специфическим отношением к температуре. Взрослые ящурки ока-
зались сравнительно постоянными в отношении к температуре, что осуществляется за счет
уменьшения периода активности [135, 232, 386]. Сеголетки активны при более низких,
по сравнению со взрослыми, температурах среды даже в летние месяцы, осенью же у них
наблюдается дополнительное снижение температурного минимума вследствие необходимо-
сти продлить активный период. Такое снижение особенно заметно у самых мелких (т.е.
поздно родившихся) сеголеток, у наиболее крупных температурный сдвиг практически от-
сутствует - они просто раньше залегают в спячку, избегая этим активности при низких
температурах среды.
Отношение ящурок к температуре зависит также от времени суток. В Калмыкии
[113] суточная активность Еarguta заканчивалась при более высокой температуре среды
по сравнению с ее началом. Это неоднократно мы наблюдали и на юге Украины. Примеры
неожиданно раннего окончания активности животных приводят 3 ЛХонякина и НЛ4 .Оку-
лова: активность ящурок вечером прекращалась в Дагестане при температуре поверхно-
сти почвы 28,2—28,5 , что на 2—3° выше температуры утреннего появления [Хонякина,
сообщ.], а в Западном Казахстане — при 29 и даже 38,5°, хотя утром этих же суток они
были активны при температуре субстрата 20—26° [192]. Такое поведение ящурок особен-
но удивляет в мае, в период наивысшей активности этих рептилий, когда они стараются
максимально использовать световой день. Подобные факты свидетельствуют о том, что ак-
тивность зависит не только от температурных условий.
Анализируя отношение ящурок к температуре в течение года и. суток, мы пришли к
выводу, что длительность периода суточной активности зависит от температур лишь тогда,
когда они низки (т.е. когда активность ограничена минимальными добровольными темпе-
ратурами) . При достаточно высоких температурах среды (в мае — сентябре для Черномор-
ского заповедника) продолжительность активного периода определяют другие причины
[135]: уровень половой активности, чувство голода, обилие корма, степень упитанности,
т.е. физиологическое состояние организма и ’’производственная необходимость9’. Наблю-
дения за мечеными ящурками одного пола и возраста показали, кроме того, наличие у
отдельных особей строго индивидуального отношения к факторам среды [127], что вызва-
но, вероятно, теми же причинами, но на более тонком уровне.
Зимой разноцветных ящурок отмечали на поверхности, насколько нам известно, лишь
в Каршннской степи (Узбекистан). Здесь вторая и третья декады декабря 1987 г. оказа-
лись очень теплыми (температура воздуха днем достигала 25°) , и 13.XII были обнаруже-
ны 2 особи при температуре воздуха 20,8 9 и поверхности почвы 30,5° [Бондаренко, со-
общ.] . На Ставропольской возвышенности, несмотря на частые оттепели, когда темпера-
тура воздуха может подниматься до 18°, снег стаивает, а земля достаточно прогревается,
М.Ф.Тертышников [259] не наблюдал выход ящурок на поверхность. В Черноморском
заповеднике в декабре - феврале неоднократно встречали змей (степную гадюку и обык-
124
новенного ужа) , но не видели ящериц, хотя в этом районе зимой часты сильные оттепели
и обычны длительные бесснежные периоды. Вероятно, зимой такая теплолюбивая ящерица,
как E.arguta, появляется редко и только в самых теплых частях ареала вида. Как
показали исследования в термоградиент-приборе [319], нарушение зимней спячки
в неволе нередко вызывает интересную реакцию — повышение предпочитаемых
температур.
Максимальная температура воздуха, при которой ящурки наблюдались на поверхно-
сти, для разных регионов и по данным различных авторов, составляет 28—40°, максималь-
ная температура поверхности почвы - 40—56° Необходимо учитывать, что установить
верхний температурный предел активности вида можно только в самой теплой части его
ареала и в самое жаркое время года, т.е. тогда, когда имеется широкий выбор темпера-
тур. В Черноморском заповеднике в 1974—1976 и 1987—1990 гг. в те дни, когда мы наблю-
дали за ящурками, температура воздуха не превышала 34° на высоте 1 м и 36 — на высоте
5 см над поверхностью песка. Обычно ящурки уходили в норы до того, как температу;
воздуха поднималась до таких величин, но в отдельные дни, когда жара начиналась с само-
го утра, некоторые особи оставались на поверхности и при достижении этих температурных
пределов. Вскоре, однако, ящурки уходили в убежища, так как продолжавшееся повыше-
ние температуры поверхности почвы на фоне мощной солнечной радиаций делали невоз-
можным дальнейшее пребывание рептилий на поверхности. Предельная для яшурок темпе-
ратура воздуха в 35—36° отмечена в Укараине, на Ставропольской возвышенности, в За-
падном Казахстане и Северном Таджикистане [192, 225,250, 259, 262], 38 — в Азербай-
джане [19,20] ,35—40° — в Калмыкии [22].
В Черноморском заповеднике при температуре поверхности почвы выше 44-45° коли-
чество активных ящурок резко сокращается, единичные особи отмечены при температуре
55°, при температуре выше 56° мы их на поверхности не встречали [135,386] (табл.16)
(максимальную температуру поверхности песка на ровных, освещенных солнцем участках
мы зарегистрировали в июле — 65°). По данным других исследователей, предельные тем-
пературы поверхности почвы для ящурок 40° — для Северного Таджикистана и Крыма
[225, 315], 46—48 — Азербайджана [19], 49 — Дагестана [Хонякина, сообщ.], 45-
52 — Западного Казахстана [192], 54-55 — Ставрополья [259, 262] и 56° - для Северо-
Западного Причерноморья [250]. Судя по всему, приведенные температуры почвы ниже
50° — результат недостаточно полных сведений, и верхние пределы температур лежат в диа-
пазоне 52—56° Однако в районах с особенно сильной солнечной радиацией рептилии во
избежание перегрева теоретически должны уходить с поверхности при более низких пре-
дельных температурах почвы и воздуха. В термоградиент-приборе [316] ящурки не за-
нимали места с температурой выше 50,6°
Предпочитаемые видом температуры воздуха, по данным разных авторов [20, 22,
97^113,192,225,250, 258, 259, 262, 266; Тэрбиш, сообщ.], лежат в диапазоне от 20 до
30° (в среднем около 24—26° ) . Предпочитаемые температуры поверхности почвы обычно
укладываются в диапазон 30—40° со средним оптимумом около 34—38° [97, 192, 224,
250, 315, 359]. (В статье В Л-Гончаровой [66] явно перепутаны оптимальные температу-
ры воздуха и почвы.) В термоградиент-приборе ящурки выбирали места с температурами
субстрата 35,1—40° [316], средние показатели в разных опытах колебались обычно в пре-
делах. 34,3—37,8° [319]. Вместе с тем М.Ф .Тертышников [258, 259, 262, 266], ВАКире-
ев [113], ВЛ Бадмаева [24] и МЛ Ахмедов [20] указывают значительно более низкие
температуры почвы — 24—26, 27, 22—32 и 25—35° соответственно (о возможных причи-
нах таких результатов см. разд. 10.1) . Проанализировав материалы (рис.71, Л, Б), полу-
ченные ВЛ Бадмаевой [25], мы пришли к выводу, что предпочитаемыми температурами
почвы для ящурок Калмыкии следовало бы считать не 22—32°, как делает автор [24],
а температуры в диапазоне 31—35° . Более низкие показатели наблюдали весной и осенью,
их не нужно учитывать, так как в эти периоды в природе отсутствовал достаточный выбор
температур. Зависимость активности ящурок от температуры воздуха и почвы в Ставро-
польском и Краснодарском краях показана на рис. 71, В, Г Обращает на себя внимание
большое сходство графиков 1 и 2 на рис. 71, В и фактическое равенство предпочитаемых
температур почвы и воздуха. Типична ситуация, отраженная на рис. 71,5
По нашим наблюдениям, наибольшее число активных ящурок встречается при тем-
пературе поверхности почвы 32—44° (рис. 72, Л) . Существенным моментом, однако, яв-
ляются возрастные различия в термопреферендуме. Известно [319], что сеголетки и мо-
125
Температура, °C
Рис. 71. Активность разноцветной ящурки в зависимо-
сти от температуры среды и тела:
А, Б - Калмыкия [25]: А — весна и осень,Б — ле-
то; 1 — март, 2 — апрель, 3 — сентябрь, 4 — октябрь,
5 - май, 6 - июнь, 7 — июль, 8 — август; В - Ставро-
польская возвышенность (Центральное Предкавказье)
[259]; Г — Кизилташская коса (Таманский п-ов, За-
падное Предкавказье) [97] \.а — температура почвы,
б — воздуха, в — тела
лодняк более пластичны по отношению к температуре, но предпочитаемые ими температу-
ры соответствуют таковым взрослых. Это заключение сделано на основании исследований
в термоградиент-приборе. Действительно, полученные результаты для сеголеток (37,85°)
и взрослых (37,83°) разноцветных ящурок практически идентичны. Поэтому вполне за-
кономерен вывод НЛДЦербака [319] о том, что при изучении средних оптимальных тем-
ператур в лаборатории (т.е. с применением использованных автором аппаратуры и мето-
дики) возраст подопытных рептилий можно во внимание не принимать.
126
Рис. 72. Активность разноцветной ящурки в зависимости от температуры поверхности почвы
(Л) и температуры тела (Б) в песчаной степи Ивано-Рыбальчанского участка Черноморского за-
поведника (юг Украины), апрель - октябрь 1975-1976 гг. (данные обрабртаны методом сколь-
зящей средней [135]);
1 — сеголетки, 2 — взрослые и полувзрослые особи
В природе наблюдается несколько иная картина. По нашим данным, заметные возраст-
ные различия имеются лишь между сеголетками и остальной частью популяции, причем
температурные границы активности сеголеток расширяются за счет низких температур.
Предпочитаемые температуры у сеголеток ниже, чем у взрослых: наибольшее число моло-
дых отмечалось при температуре почвы 32—39°, взрослых — при 37—44 [131, 135, 386]
(рис. 72, А) . На возрастные различия в термопреферендуме косвенно указывает и харак-
тер активности сеголеток и взрослых: в конце лета первые активны в основном утром и
вечером, вторые — в более жаркое дневное время (см. рис. 77). ВИ.Бадмаева [24] и
МИ Ахмедов [19] полагают, однако, что сеголетки по сравнению со взрослыми ящур-
ками толерантнее как к низким, так и к высоким температурам.
Исследования, проведенные ННДЦербаком в термоградиент-приборе, показали, что от
весны к лету средние оптимальные температуры повышаются, от лета к осени — снижают-
ся [319]. Необходимо, однако, помнить о влиянии содержания рептилий в неволе на получа-
емые результаты (см. разд. 10.1). Сезонную динамику оптимальных температур в природ-
ных условиях мы рассмотрим несколько позже на примере температуры тела ящурок.
Особо следует остановиться на температурных условиях активности разноцветных
ящурок в Монголии. Обращают на себя внимание низкие температуры начала, максимума
и конца суточной активности. Так, 10.V 1982 г. отдельные особи были еще активны в
1930 при температуре воздуха 10°. 7.VI 1983 г. первые ящурки вышли на поверхность
при температуре воздуха 11,5°, дневной пик активности наблюдался при 16—19,3 21.VI
1984 г. первые особи появились в 6.30 при температуре воздуха 15°, утренний пик был
при 183—23,9°, при более высокой температуре наблюдался спад активности; вечерний
пик отмечен при 22,1—20°. Единичные особи попадались даже в пасмурную погоду при
температуре воздуха 12—14° [Тэрбиш, сообщ.]. Столь низкие температурные показатели
активности разноцветных ящурок в Монголии объясняются, очевидно, тем, что Джунгар-
ская Гоби, где встречается этот вид, расположена на значительной высоте над уровнем
моря. По данным В.Ф.Орловой и Х.Тэрбиша [197], места обитания Е. arguta находятся на
высоте 1200—1800 м н.у.м., а, как известно [241], с высотой существенно возрастает ин-
тенсивность солнечной радиации, действие которой вполне заменяет недостающие граду-
сы тепла. В пределах ареала разноцветной ящурки имеются и другие высоко располо-
женные (до 2000—2100 м H.yjvi.) районы — Севанская котловина в Армении [76,82], Та-
ласская долина и Прииссыккулье в Кыргызстане [324] , где можно было бы наблюдать
аналогичные явления, но по этим регионам данные по термобиологии вида крайне скудны
[203] или отсутствуют. Таким образом, практически все рассмотренные ранее термоэко-
логические показатели активности Е.arguta относятся к равнинам.
127
103. ТЕМПЕРАТУРА ТЕЛА
Критические температуры тела разноцветной ящурки изучали В .Родионов [218], А Л Рю-
мин [220], Н.М.Окулова [192], М.Ф.Тертышников [210,258—260,262,266] и В. А Ки-
реев [113]. Установлено [218], что при температуре среды от —7 до —10° в организме
ящурок через 36—63 мин после переохлаждения начинается образование льда, что приво-
дит к гибели животных. Ящерицы другой группы погибали после содержания их при —7,8°
в течение 160—240 мин. Столь высокая продолжительность выживания в последнем слу-
чае вызвана, возможно, тем, что до опыта животные 4 мес находились без пищи в со-
стоянии неглубокой спячки. Одна из ящурок после 3-часового пребывания при темпера-
туре —5,0° жила, несмотря на то, что после первых 2 ч эксперимента ее дыхание останови-
лось. Другая ящерица погибла после пребывания в течение 160 мин при температуре
—3,5°, вероятно, вследствие сильного истощения. Те опыты, в которых кроме темпера-
туры воздуха в камере измерялась и температура тела, показали, что ящурки оставались
живыми после пребывания в течение 225 мин при температуре воздуха —4,0° и тела — 3,1
и в течение 85 мин — при —5,0 и —3,3° соответственно [218].
Заметный разброс данных по критическим температурам и по длительности выжива-
ния при низких температурах связан, на наш взгляд, с различным физиологическим со-
стоянием подопытных животных, разницей в их массе и индивидуальной изменчивостью
холодоустойчивости, а также с особенностями проведения опытов. Уровни критических
минимума и максимума зависят от той температуры, при которой рептилий содержали
до эксперимента [283]. Правда, влияние акклимации на критические температуры тела
краткосрочно.
Эксперименты по нагреванию ящурок, проведенные АЗ Рюминым [220] в октябре
над особями, добытыми в августе в уральских песках, показали следующее. При перегрева-
нии животные начинают вести себя беспокойно, возникает полипноэ. Частота дыхания все
время возрастает и при температуре тела 41—44° достигает максимума. При температуре
46—47,5° дыхание прекращается, при 47—47,5 появляются судороги конечностей, затем
век, и ящурки впадают в состояние теплового оцепенения. Некоторые находились в таком
состоянии более 24 ч и оживали. Если температура тела поднималась до 49°, тепловое око-
ченение становилось необратимым и заканчивалось смертью животного. В опытах М.Ф.Тер-
тышникова £210, 258—260, 262, 266] разноцветные ящурки погибали при температуре
тела 47—47,2 , ВАКиреев [ИЗ] летальной также назвал температуру 47,2°. По данным
Н.М.Окуловой, тепловой шок (длительностью 1—2 мин) наступал у ящурок при повыше-
нии температуры тела до 40—42°. При повторном нагревании только что вышедшей из шо-
ка ящерицы новый тепловой шок начинался значительно быстрее и при более низкой тем-
пературе [192]. В Черноморском заповеднике температура тела ящурок могла достигать
40г естественным путем, тепловое оцепенение наблюдалось при более высоких темпера-
турах.
Минимальные и максимальные температуры тела активного состояния разноцветных
ящурок, по данным различных авторов, существенно различаются. Так, А.Г.Банников
[27] для Дагестана указывает диапазон температур тела от 30 до 40°, НМ.Окулова [192]
для Западного Казахстана — от 25 до 38, М.Ф.Тертышников [258, 259, 262, 266] для
Ставропольской возвышенности — от 15 (18) до 35 .В Крыму у добытых в августе ящу-
рок температура тела достигала 39° [316], что также можно считать максимальным пока-
зателем. По нашим данным, максимальная температура тела активного состояния разно-
цветных ящурок равна 40° [129, 135, 386]. Температуру тела в пределах 39,5—40,0° мы
отмечали неоднократно, однако лишь в самое жаркое время года и только в конце утрен-
него пика активности, перед уходом ящериц на дневной отдых. Очевидно, это температура
тела при добровольном перегреве.
Таким образом, добровольный максимум, согласно приведенным данным, может раз-
личаться на 5°, добровольный минимум — на 15. Такие расхождения в максимальных
температурах вызваны, вероятно, неполнотой сведений, а различия в минимальных темпе-
ратурах связаны прежде всего с тем, что исследователи не разграничивали формы поведе-
ния рептилийу в результате чего в массив данных попали температуры тела и неактивных
особей.
По наблюдениям Н.М .Окуловой [192], при температуре тела 18—20° ящурки.уже не
могут передвигаться по поверхности земли, хотя еще, вероятно, способны закапываться
128
в грунт. Наблюдались случаи, когда при внезапном снижении температуры среды ящурка
теряла способность к передвижению и оставалась на поверхности почвы. Так, в апреле
при снижении за 10—15 мин температуры поверхности песка с 33 до 21° была обнаружена
малоподвижная особь с температурой тела 21° У другой не успевшей скрыться ящерицы
температура тела была 22,5° [192]. По данным С ЛЛерешкольника [203]. в неактивном
состоянии ящурки имели температуру тела в среднем 20°
Наши наблюдения в Черноморском заповеднике в апреле 1975-1976 гг. вследствие
специфики погодных условий и малого числа полученных данных не позволили выяснить
возможную для вида минимальную температуру тела взрослых активных особей: все до-
бытые ящурки имели температуру тела выше 30°. В конце сентября, т.е. непосредственно
перед уходом на зимовку, взрослые особи подолгу грелись у входов в норы, были актив-
ными очень короткое время и тоже при высоких температурах тела. И лишь сеголетки в
октябре проявляли активность при низких, температурах среды и тела. В частности, мини-
мальная температура тела ящурки, добытой на поверхности почвы в октябре, была 18,1°,
еще несколько пойманных сеголеток имели температуру тела от 18,7 до 21,2°. При этом
одни особи лежали неподвижно у входа в свою нору (форма поведения — нагревание),
другие были активными (табл. 15, группа 7). Таким образом, можно полагать, что сеголет-
ки разноцветной ящурки при температуре тела ниже 18° неактивны и находятся в убежи-
щах, при температуре выше 21° — активны, в диапазоне от 18 до 21° могут пребывать
либо в переходном состоянии, либо в активном.
Температуру тела ящурок, находящихся в норах, почти не измеряли. В Западном Ка-
захстане в конце апреля в норах утром находили неактивных особей с температурой тела
13°, равной температуре окружающей почвы [192]. В Черноморском заповеднике 14Л
1975 г., через 2 недели после залегания в спячку взрослых ящурок, при раскопке норы
была обнаружена спящая взрослая особь с температурой тела 20,4", температура песка
здесь была 13° [125, 131]. Большая разница в температурах тела и среды объяснялась,
очевидно, тепловой инерцией тела ящурки и действием механизмов физиологической тер-
морегуляции, поскольку за 1,5 ч до этого было еше тепло (температура воздуха - 17,2°
поверхности почвы — до 35).
Имеются также данные измерений (в лабораторных условиях) температуры в 17 точ-
ках поверхности тела одного экземпляра разноцветной ящурки из Восточного Казахста-
на [277]. Наиболее холодным местом оказалась ступня (22,6°), наиболее теплым - ухо
(28,6°) , т.е. разность температур достигала 6°. На нижней стороне тела температура была
заметно выше, чем на верхней: разница температур горла и пилеуса составила 4,1° ,низа и
верха шеи — 0,7° , брюха и спины — 0,9 , низа и верха хвоста в конце его передней трети -
0,2 (опыты проводились при температуре воздуха 22,5°). Наблюдавшиеся различия
кожных температур обусловлены особенностями строения покровов, степенью их васку-
ляризации, интенсивностью испарения с поверхности, и, прежде всего, степенью удален-
ности отдельных участков поверхности тела от внутренних органов и мышц, вырыбаты-
вающих тепловую энергию. В таких местах тела рептилий, как ноздри, утлы рта, ротовая
полость, клоакальное отверстие, температура (в связи с более интенсивным испарением)
относительно низка [277].
Температура тела ящурок, как и отношение их к температуре среды, зависит от возра-
ста. Это хорошо видно на материале из Черноморского заповедника. Здесь температура
тела была измерена у 293 особей разного возраста (129 сеголеток и 164 взрослых и полу-
взрослых) и в различное в^эемя года. У сеголеток она колебалась от 18,1 (у активных
особей — от 18,7°) до 37,5 , у остальных ящурок — от 30,8 до 40,0° [131, 135, 386]. Та-
ким образом, диапазон температур тела у сеголеток (19,4°) был примерно вдвое больше,
чем у прочих возрастных групп (9,2°) (рис. 72,Б) . Эта разница может быть существенно
меньше при других погодных условиях и более полном материале по взрослым ящуркам
весной, однако в любом случае сохранится несовпадение мод (рис. 72, Б), т.е. сдвиг кри-
вой по сеголеткам в сторону более низких температур.
Данные по оптимальным температурам тела вида малочисленны и противоречивы. Из-
вестно, что в Прииссыккулье в середине августа температура тела ящурок в период актив-
ности оставалась практически постоянной и составляла 35° [203]. МИАхмедов [19] для
сеголеток приводит оптимальную температуру тела, равную 34°. Однако эти и некоторые
другие сведения и обобщения по экологии E.arguta, содержащиеся в указанной работе,
почерпнуты из литературы, поэтому нами рассматриваются по первоисточникам. По
129
М.Ф.Тертышникову [258, 259, 262, 266], в Ставрополье наибольшая активность яшурок
проявляется при температуре тела 30,2° , которую и следует считать оптимальной (рис.71,
В). Его опыты по газообмену [210, 259, 260] показали, что количество потребляемого
ящурками кислорода по мере подъема температуры тела растет, при температуре 30° оно
максимально, а затем резко падает и в диапазоне температур 31—35° находится на относи-
тельно постоянном уровне. К сожалению, следующие опыты были проведены лишь при
температурах 40 и 47 , поэтому динамика газообмена при температурах выше 35“ неясна.
Известно, что именно при оптимальных температурах тела наиболее экономично работают
механизмы терморегуляции, стабилизирован обмен веществ. Поэтому по результатам изу-
чения газообмена оптимальной температурой тела следовало бы считать не 30° [210,262].
Таблица 14.Температура тела разноцветной ящурки в период максимумов ее суточной
активности на Ивано-Рыбальчанском участке Черноморского заповедника в 1975—1976 гг.
[ 135, с изменениями]
Возрастная группа Месяц, декада п xmln хшах X о Sx t
Взрослые и полу
взрослые IV, 3-я 6 32,3-35,3 34,08 1,01 0,41 с 75
V, 3-я 16 34,6-38,6 37,12 1,13 0,28 1 л л
VI, 3-я 23 33,1-39,4 36,56 1,25 0,26 1,44 1 1 Q
VII, 1-2-я 24 34,3-39,0 36,99 123 0,25 1,1^ □
Взрослые VIII, 2-я 43 32,2-39,5 35.60 1,55 0,24 о ,/ / Q Сй
Сеголетки VHI, 2-я 48 29,3-37,0 34,40 1,64 0,24 3 рс
Взрослые IX, 3-я 19 34,0-38,3 35,67 1,28 029 5.16
Сеголетки IX, 3-я 19 31,5-352 33.80 0,92 021 о ЙЛ
Х.2-я 25 20,0-305 26,55 3,49 0,70 О .OV
Взрослые V-VII 63 33,1-39.4 36,86 122 0,15 5 1 Я
vni-ix 62 32,2-39,5 35,62 1.46 0,19 V ,1 о с о л
Сеголетки VIII-IX 67 29,3-37,0 34,23 1,49 0,18 5 ,34
а температуру, лежащую, как минимум, в пределах 31—35° Однако мнение о величине
температурного оптимума вполне правомерно, если основывать его на результатах поле-
вых исследований (рис. 71, В). По данным НМ .Окуловой [192], в Западном Казахстане
наибольшее количество ящурок встречается на поверхности в активном состоянии при
температуре тела обычно в пределах 33-34° (у молодых несколько ниже) , что и принято
в качестве оптимальной температуры тела.
Нами в качестве температурного оптимума первоначально была принята средняя тем-
пература тела ящурок в период пиков их суточной активности [131, 135]. В дальнейшем
результаты полевых исследований были пересмотрены с учетом форм поведения рептилий,
при этом из общего массива данных были исключены температуры тела особей при добро-
вольном перегреве, в случае двух пиков активности рассматривался лишь утренний. Та-
ким образом, мы по сути учитывали температуру тела при первишгом термостабилизирую-
щем поведении, что позволило несколько уточнить средние оптимальные температуры тела
ящурок. Общие закономерности и соотношения, однако, не изменились, что видно при
сравнении данных прежней таблицы [135] и настоящей (табл. 14).
Установлено, что оптимальная температура тела ящурок в мае—июле практически по-
стоянна (различия недостоверны) и составляет для всех особей, кроме сеголеток, пример-
но 37°. В апреле, августе и сентябре она ниже, причем в августе - сентябре фактически
одинакова и несколько превышает апрельскую.У сеголеток оптимальные температуры тела
в августе и сентябре близки и достоверно ниже, чем у взрослых особей; в октябре сред-
няя температура тела заметно понижается (табл. 14) . Поскольку летом у ящурок имеется
больший выбор температур, мы считаем в действительности оптимальной температуру те-
ла, равную 36,9° для взрослых и 34,2° для сеголеток, а не более низкие весенние или осен-
ние показатели.
ВА.Черлин [281] отмечает, что нельзя усреднять температуру тела пресмыкающихся,
добытых в течение дня, поскольку такие данные не отражают предпочитаемых температур.
Поэтому он считает целесообразным характеризовать оптимальные температуры тела мо-
дой внутри диапазона наиболее часто встречающихся температур при термонейтральном
поведении [285]. Первое положение бесспорно. Что же касается последнего, то, на наш
взгляд, средняя температура тела рептилий в период максимумов их суточной активности
130
вполне удовлетворительно отражает температурный оптимум (во всяком случае, у днев-
ных видов) . По нашим данным, распределение температур тела у разноцветных ящурок в
период пиков их активности было близким к нормальному и, например, у взрослых осо-
бей в мае—июле мода равнялась 37° , а средняя арифметическая — 36,9. Преимущества же
последнего параметра с точки зрения статистики несомненны [135].
То, что сеголетки хуже взрослых переносят высокие температуры, может быть обу-
словлено, на наш взгляд, недостатком у них навыков поведенческой терморегуляции и
более мелкими размерами тела. У недавно родившихся ящурок поведенческие реакции
еше несовершенны — они не умеют в полной мере использовать растительность и различ-
ные убежища для охлаждения тела, значительно хуже взрослых особей знают свою терри-
торию, теряются в незнакомой обстановке, медленнее бегают. По мере роста ящурки при-
обретают опыт терморегуляционного поведения, что хорошо заметно при длительном на-
блюдении в природе за отдельными особями разного возраста. В неволе наблюдали [297]
выработку у яшериц сложных двигательных рефлексов при привыкании к термофилогра-
фу. Рептилии, помещенные в термоградиент-прибор, первые 2—3 недели имеют очень широ-
кий диапазон предпочитаемых температур тела, и лишь затем он устанавливается в преде-
лах, свойственных виду в природе [347], Таким образом, знакомство с местностью и
’’жизненный опыт” имеют немаловажное значение для успешной терморегуляции.
Роль массы тела в терморегуляции рептилий давно известна. При этом имеет значение
ряд моментов: увеличение тепловой инерции и уменьшение теплопродукции на единицу
массы тела с увеличением размеров (массы) тела, повышение роли испарительной влаго-
отдачи у крупных животных, увеличение теплоотдачи у мелких животных в связи с увели-
чением относительной поверхности тела; у мелких животных менее выражена разница ме-
жду внутренней и поверхностной температурами тела, так как у них скорее достигается
выравнивание этих температур [71,235, 242, 277 и др.]. Показано также влияние разме-
ров тела на терморегуляционные ’’способности” рептилий: установлено,что мелкие виды
ящериц обладают относительно небольшим арсеналом поведенческих приемов терморегу-
ляции по сравнению с крупными [173].
На наш взгляд [129, 135], сеголетки разноцветной ящурки (масса тела 0,4—0,9 г)
должны быстрее нагреваться, чем взрослые и полувзрослые особи (масса тела 3—8 ^дан-
ные по Черноморскому заповеднику). К тому же сеголетки находятся ближе к поверх-
ности почвы, чем взрослые ящурки, что усугубляет их положение при высоких темпера-
турах субстрата. В результате у сеголеток скорее наступает перегрев, поэтому активность
их при высоких температурах среды менее оправдана: период пребывания на открытом
солнце сокрашается настолько, что они не могут успешно охотиться. Наконец, энергетиче-
ские затраты организма на поддержание оптимальной температуры тела при высоких тем-
пературах среды столь значительны, что активность сеголеток оказывается энергетически
невыгодной.
Пол заметно не влияет на оптимальную температуру тела ящурок и предпочитаемые
температуры субстрата [319, 320] . Вместе с тем беременные ящерицы регулируют темпе-
ратуру тела менее успешно, чем яловые [283]. В состоянии беременности падает также и
теплоустойчивость нервной и мышечной тканей [272]. По нашим данным, уловимые раз-
личия в термопреферендуме имеются лишь у самок на последних стадиях беременности
(толстых). Такие особи по утрам и вечерам дольше греются на солнце у входов в норы,
а их оптимальная температура тела несколько выше (во всяком случае, не ниже) , чем у
остальных взрослых ящурок: для июня—июля 1975 г. соответственно 37,67±0,33 и 36,95±
±0,22°, t = 2,10 [135]. Возможно, какую-то роль в этих изменениях термопреферендума
играет увеличившаяся масса тела самок (как сам факт большей массы, так и возникаю-
щие сложности в поведенческой терморегуляции в связи с непривычной массой тела) .
По мнению НМ.Окуловой [192], самцы более устойчивы к воздействию низких темпера-
тур, чем самки: они первыми появляются весной или после сильных дождей.
Рассмотренные термофизиологические показатели (критические минимум и макси-
мум, диапазон добровольных и оптимальных температур тела) видоспецифичны, однако
могут несколько изменяться под действием ряда факторов. О влиянии температуры ак-
кламации уже упоминалось. Известны сезонно-циклические изменения теплоустойчивости
тканей. В частности, отмечены повышение теплоустойчивости клеток в активный период,
вызываемое усилением функции щитовидной железы, ее падение во время размножения
131
рептилий, происходящее под влиянием половых гормонов, и новое повышение теплоустой-
чивости в момент перехода ящериц в состояние зимней спячки, обусловленное гормональ-
ной перестройкой организма [272]. Во время пищеварения рептилии нуждаются в более
высокой, чем обычно, температуре, и их термофилия возрастает [295,297] , при этом чем
больше съедено корма, тем более высокую температуру избирают животные для ее пере-
варивания, но такие колебания оптимальных температур не превышают 2—3° [283]. Та-
ким образом, сезонные и суточные колебания термофизиологических показателей опреде-
ляются физиологическим состоянием рептилий и связаны прежде всего с гормональным
фоном и питанием.
Изменчивость термоэкологических показателей обусловлена действием различных
ограничителей, в качестве которых могут выступать климат и погода (рассмотрено ранее) ,
энергетика (как в случае с сеголетками), социальные и межвидовые взаимодействия
(у разноцветных ящурок практически не изучались). обилие корма в биотопе и др. (см.
обзор по вопросу [283]). Взаимодействия между различными особями в популяции мо-
гут, вероятно, влиять на точность терморегуляции: мы неоднократно наблюдали, как
взрослые особи занимали места, более выгодные, удобные для регулирования температу-
ры тела, прогоняя при необходимости из этих мест сеголеток. Самцы в мае—июне слиш-
ком заняты вопросами размножения, что должно сказываться на их терморегуляции (вы-
сокая половая активность самцов отражается даже на интенсивности их питания [131]).
Обилие корма также влияет на точность терморегуляции рептилий (подробнее см. разд.
10.4.6).
10.4. ВЛИЯНИЕ РАЗЛИЧНЫХ ФАКТОРОВ СРЕДЫ
НА ТЕМПЕРАТУРУ ТЕЛА И АКТИВНОСТЬ
Из внешних факторов, определяющих температуру тела рептилий и характер их активно-
сти, ведущими являются солнечная радиация и температура среды (прежде всего, суб-
страта) .
10.4.1. СОЛНЕЧНАЯ РАДИАЦИЯ
Действие солнечной радиации на рептилий подробно рассмотрено еще в статьях ИД.Стрель-
никова [240—242], И.ФАндреева [10, 11] и Л.ГДинесмана [88]. Степень воздействия
солнечных лучей на организм пресмыкающегося зависит от напряженности солнечной ра-
диации (следовательно, от времени года и суток, географической широты местности и ее
высоты над уровнем моря, а также физико-географических условий, определяющих режим
облачности и степень прозрачности атмосферы) , от угла падения лучей (значение имеют те
же временные и пространственные условия плюс положение тела животного относитель-
но оси луча) и поглощающей способности организма (зависит от окраски покровов тела и
степени пигментации перитонеума).
Перечисленные вопросы применительно к разноцветной ящурке практически не изуча-
ли, лишь в работе ЛГ Динесмана [88] в табл. 2 указано, что Earguta на территории бывше-
го СССР населяет 7 самых южных зон солнечной радиации (из 14, освоенных рептилиями
в пределах страны), которые характеризуются наличием в году 1—3 мес с летне-тропиче-
ским типом радиации, 0—2 мес с весенне-тропическим типом и 0—3 мес с переходным ти-
пом от первого ко второму. Автор, однако, ограничился рассмотрением зонального рас-
пределения рептилий на территории бывшего СССР между 48° вд.и Алтаем, т.е. не учиты-
вал районы западнее Волги и за пределами страны. Вместе с тем разноцветная ящурка
встречается и в лесостепной зоне бывшего СССР (например, в Украине и в Белгород-
ской обл. России [140], а также в Юго-Восточной Румынии, Западной Монголии и некото-
рых северных районах Китая, а также Ирана [197, 320], в результате чего список радиа-
ционных зон (по принятому Л.Г.Динесманом делению) должен быть расширен минимум на
одну — зону лесостепи, имеющую 1 мес с летне-тропическим типом радиации. Однако соглас-
но радиационным картам [18, 42], все перечисленные части ареала вида входят в те же радиа-
ционные зоны, в целом разноцветная ящурка распространена на территории, характери-
зующейся годовой суммой прямой солнечной радиации примерно 50—110 ккал/см, сум-
132
Таблица 15» Температура тела разноцветной ящурки в связи с поведением, темпера-
турой среды и солнечной радиацией на Ивано-Рыбальчанском участке Черноморского запо-
ведника в 1975 г. [ по данным ТЛ.Котенко]
Труп- па Дата Возраст Температура, °C Облач- ность, баллы Характеристика поведения
тела субстрата воздуха
1 28.V ad. 37,2 48,0 29,1 0 Активна
28 .V ad. 37,8 48,0 29,0 0
2 29 .V juv. 36,2 44,0 29,8 3
29 .V juv. 40.0 50,0 32,7 5
3 29 .VI ad. 39.4 56,0 35,0 0
4 13.VII ad. 3S 49,0 31,6 1
13.VI1 ad. 37,3 49,0 31,6 1
13.VII ad. 37,0 49,0 32,0 1
13.vn ad. 39,0 50,0 323 1
13.vn ad. 39,0 50,4 323 1
5 14.VDI juv. (сего- летки) 37,0 34,0 213 0
14.VTII juv. (сего- летки) 32,8 26,8 213 0
6 15 .VIII ad. 53,3 35,3 26,9 1
15.vm ad. 36,0 55,0 27,0 1
15 .VIII ad. 363 47,0 27,1 2
15 УШ ad. 35,8 51.0 273 2
15 .vm ad. 34,0 46,0 27,4 3
7 12Л juv. (сего- 20,7 253 н,о 0 Лежала у входа в
летки) нору
12Л juv. (сего- 20,0 22,5-24,0 11,2 0 Ходила вокруг
летки) норы
12Л juv. (сего- 21,0 22,0-22,8 11,4 0 Сидела у входа,
летки) 0 побежала
12Л juv. (сего- 18Д 26,0 11,7 Лежала у входа в
летки) нору
12X juv. (сего- 20.2 24,0-263 11,9 0 Бегала возле норы
летки
12.X juv. (сего- 213 24,0-283 123 0 Ходила вокруг
летки) норы
12Л juv. (сего- 18,7 в среднем 10,8 0 Бежала издалека в
летки) 19,0 нору (конец ак- тивности)
8 13.X juv. (сего- 28,3 15,0-20,0 9,3 0 Бегала
летки
13.X juv. (сего- летки) 27,8 22,0-24,0 9,6 0
9 16 JC juv. (сего- 25,6 21,0-22,0 16,7 10, пас- Сидела у входа в
летки) мурно нору
16.X juv. (сего- 25,3 19,0-19,6 16,7 То же Сидела у входа,
летки) побежала
16.X 27,3 193-21,0 16,8 Бегала
16.X 18,9 20,0 16,8 Лежала у входа в нору
10 16.X juv. (сего- 30,9 26,0 20,4 10, но Бегала
летки) солнце просве- чивает
16Л juv. (сего- летки) 293 25 3- 26,0 203 То же Ходила
16.X juv. (сего- 30,0 25,0 203 Ходила возле
летки) норы
16X juv. (сего- 30,9 25,0 203 То же
летки)
маркой (т.е. прямой и рассеянной) радиацией 100-160 ккал/см и общим радиационным
балансом приблизительно 40—60 ккал/см. В горных районах напряжение радиации с высо-
той увеличивается примерно на 10 % на каждый километр подъема [42], о чем необходи-
мо помнить при рассмотрении условий обитания и анализе термоэкологических показа-
телей рептилий.
133
Роль солнечной радиации в жизни разноцветной ящурки как гелиотермного пойкило-
термного животного очень велика. Косвенно об этом можно судить по результатам неко-
торых измерений температуры тела и среды. Так, в Черноморском заповеднике температу-
ра тела ящурок, добытых утром возле нор в фазе нагревания, часто была на 0,5-2вы-
ше, чем песка,на котором они находились [135] , и тем более выше температуры воздуха;
это обеспечивается за счет инсоляции. В активном состоянии превышение температуры
тела над таковой субстрата могло достигать 13 °, а над температурой воздуха — 19°
(табл. 15, группы 5, 8), что характерно для сравнительно холодных, но солнечных дней.
Здесь, однако, действие солнечной радиации в чистом виде выделить нельзя, так как на
температуру тела ящурок в той или иной степени влиял еще один существенный фак-
тор — движение животных (разогрев в результате мышечной работы) .
Пример губительного действия солнечной радиации на разноцветных ящурок привела
НЛ1.Окулова: при температуре субстрата 37—39° у особей с исходной температурой тела
26-28° через 8-10 мин наступал тепловой шок при температуре тела 40—42 [192].
В высокогорных районах, где температура среды низкая, напряженность солнечной ра-
диации высокая, действие последней выступает особенно ярко: температура тела ящериц
может превышать температуру воздуха на 25° на высоте 2200 м н.у.м. и почти на 30 — на
высоте 4000 м, где температура воздуха в момент наблюдений была всего 5° [241, для
Lacerta agilis]. Роль инсоляции в определении температуры тела рептилий пустыни хорошо
показана И.ФАндреевым [10, И] на различных видах ящурок и И.Д.Стрельниковым
[240] на ящурках и круглоголовках, однако в последней работе не учитывалась темпе-
ратура почвы.
Наиболее сильно действует на рептилий прямая солнечная радиация, но и рассеянная
имеет определенное значение. Она может обеспечить некоторое превышение температу-
ры тела над температурой среды даже в пасмурные дни. Влияние рассеянной радиации бы-
ло заметно в некоторых наших наблюдениях (табл. 15, группы 9, 10) , это же излучение
позволило ящуркам быть активными в пасмурную погоду в Монголии при температуре
воздуха 12—14°
1042. ТЕМПЕРАТУРА ВОЗДУХА И ПОЧВЫ
Из температурных факторов для наземных рептилий ведущим является температура суб-
страта, а не воздуха, что отмечалось и для разноцветной ящурки [135, 192] .По нашим на-
блюдениям в Черноморском заповеднике [131, 135], температура тела ящурок всегда
была существенно выше температуры воздуха и в общих чертах следовала за температу-
рой почвы (рис. 73, Л). На графике видно, что при температуре субстрата от 26 до 45°
средняя температура тела ящурок увеличивается с 28 до 37 й, причем сначала кривая идет
круто вверх, а в конце почти не растет; при температуре почвы ниже 26° температура те-
ла животных сильно варьирует. Это объясняется незначительным количеством данных на
конце температурной кривой, разным состоянием ящурок в интервале температур тела
от 18 до 21° (см. разд. 10.3) и поведенческой терморегуляцией. Именно малоблагоприятная
температура среды требует от рептилий наиболее активных мер по поддержанию темпера-
туры тела в пределах нормы.
Данные по Западному Казахстану [192] также свидетельствуют о ведущем значении
температуры субстрата в терморегуляции ящериц (рис. 73, Б). Вместе с тем выявлена
прямая линейная зависимость между температурой тела разноцветной ящурки и темпера-
турой воздуха в диапазоне 10—33°, при этом коэффициент корреляции составил 0,796
(Р = 0,999) , уравнение регрессии имело виду = 21,106 + 0,482 х (по результатам 70 наблю-
дений) . Линия регрессии почти полностью совпала с таковой для обитающей тут же круг-
логоловки-вертихвостки [Окулова, сообщ.]. По данным М.Ф.Тертышникова, зависи-
мость температуры тела ящурок от таковой воздуха и почвы выглядит следующим
образом (рис. 73, В, Г) , при этом при температуре среды 15,5—32,5° температура тела реп-
тилий оказывается или выше на 0—7, или ниже на 0—11° , при температуре среды 32,5—
55Р° температура тела ящериц становится ниже на 0—20° [259]. Хотя материал, представ-
ленный на рис. 73, В, Г, статистически не обработан, все же заметно, что связь между тем-
пературами тела и воздуха носит прямолинейный характер, между температурами тела и
почвы — криволинейный. Напрашивается вывод, что температура тела зависит прежде
всего от температуры воздуха. Но такая зависимость — скорее следствие, чем причина.
134
A
Температура тела, X
40V
30-
20
Рис. 73. Температура тела разноцветной ящурки в
связи с температурой почвы (Л-S) и температурой
воздуха (Г):
А - песчаная степь Ивано-Рыбальчанского участка
Черноморского заповедника, апрель - октябрь 1975-
1976 гг. Показаны эмпирические точки (диаметр про-
порционален числу данных, п = 293) и кривая функции
у = 1^04 х - 0,015 х2, удовлетворительно описывающая
эту зависимость на отрезке 20-50 [135,с исправления-
ми согласно 131]; Б - Волго-Уральские пески (Запад-
ный Казахстан) [ 192] : 1 - взрослые, 2 - молодые осо-
би; В, Г - Ставропольская возвышенность [ 259]: 1 -
взрослые самцы, 2 - взрослые самки, 3 - сеголетки
-J-------1------1_______I
20 30 40 50
Температура воздуха, X
Ведущее значение температуры субстрата (как и непосредственно солнечной радиации)
проявляется не в том, что температура тела строго следует за ее изменениями, а в том, что
она выступает лимитирующим фактором. Именно прогрев почвы побуждает рептилий к на-
чалу активности, а перегрев — к ее прекращению и уходу в убежища. Доказательством слу-
жит активность рептилий при низких температурах воздуха (для разноцветной ящурки -
при 8—10°) , лишь бы температура почвы была достаточной. Об этом свидетельствует и по-
ведение ящериц в холодные дни: они подолгу греются, плотно прижавшись к почве или
камням.
Ведущее значение температуры почвы по сравнению с температурой воздуха для актив-
ности ящурок выступает, на наш взгляд,.и при сопоставлении температур тела рептилий,
измеренных per os и на поверхности спины (брали одних и тех же особей в июле—августе
1975 г., п = 108) . Температура внутренняя и на поверхности спины составила в среднем
35^9±0,23 и 34,24±0,28° для взрослых (Г = 4,90) и 34,56±0,23 и 31,99±0,35о - для сего-
леток (t = 6,11) . При этом пероральная температура оказалась выше температуры поверх-
135
ности спины на 0,2—9,0° в 95,4 % случаев [131,135]. Превышение температуры поверхно-
сти спины рептилий над температурой тела в клоаке, наблюдавшееся АБ.Гражданкиным
[67], мы связываем с условиями опытов, при которых по сути устранялось влияние тем-
пературы субстрата на температуру тела пресмыкающихся, зато было сильным воздей-
ствие солнечной радиации.
1043. ВЕТЕР, ЗАПЫЛЕННОСТЬ АТМОСФЕРЫ
По наблюдениям в Черноморском заповеднике, часть популяции разноцветной ящурки
находилась на поверхности даже при скорости ветра 13,5 м/с, если другие погодные усло-
вия были благоприятными. Сам по себе ветер не препятствует активности ящурок, тем бо-
лее, что даже сильный ветер у поверхности почвы среди больших кустов полыни или ко-
лосняка заметно ослабевает. В частности, при скорости ветра до 2,3 м/с на высоте 03 м
движение воздуха в приземном слое анемометром не улавливалось. Однако ветер влияет
на отношение пресмыкающихся к температуре: он является одним из факторов, ограничи-
вающих действие солнечной радиации и высоких температур среды, поскольку усиливает
отдачу тепла животным путем конвекции. Поэтому ветер позволяет рептилиям быть ак-
тивными при более высоких температурах почвы и воздуха, снижая их активность при
низких температурах среды. Сильный ветер в теплый, но пасмурный день, или в солнечный,
но прохладный, может полностью прекратить активность ящурок.
Влияние ветра хорошо заметно в Черноморском заповеднике при сравнении уровня
активности рептилий в песчаной степи и на морском берегу при одинаковых температур-
ных условиях [135]: летом в первом биотопе в жаркие дневные часы ящурки на поверх-
ности практически отсутствуют, а во втором, где дует ветер с моря (бриз) , они довольно
многочисленны. В Ставрополье на защищенной от ветра площадке отметили 52 разноцвет-
ные ящурки, на другой, подверженной действию ветра, наблюдали всего 10 особей, хотя
плотность населения ящериц на обоих участках была одинаковой и составляла 55 экз У га
[259]. В Зайсанской котловине (Казахстан) ящурки переживают ветер в норах или рас-
пластавшись и прижав голову к субстрату; в июне на одном и том же участке в хорошую
погоду встречалось до 50 экз ./га, а во время ветра — всего 2—3 особи [48].
В Азербайджане наибольшее количество активных ящурок в июле отмечено при тем-
пературе воздуха 22°, почвы 25° и скорости ветра 3—7 м/с [20]. На Кизилташской косе
(Западное Предкавказье) за период наблюдений скорость ветра не превышала 15 м/с
(7 баллов по шкале Бофорта); при таком ветре ящурки оставались активными. Сильный
ветер подавляет активность этих ящериц при температуре воздуха ниже 22°, а также кос-
венно влияет на их активность, резко снижая таковую насекомых — объектов питания
ящурок [97]. В Дагестане [Хонякина, сообщ.] и Калмыкии [113, 115] при сильном вет-
ре ящурки обычно не покидают своих убежищ, при этом в Калмыкии они наиболее актив-
ны при силе ветра не более 2 баллов, в дни, когда сила ветра достигала 8 баллов и начи-
налась пыльная буря, они из нор не выходили [22]. В Монголии при сильном ветре ящур-
си на поверхности не появляются, но в какой-то мере активны в солнечную погоду при
скорости ветра до 4—6 м/с [Тэрбиш, сообщ.]. По другим данным для этого же региона
[197], при сильном ветре, когда другие виды уходят с поверхности, E^rguta остается у
основания кустов или у входа в нору.
Зависимость активности разноцветных ящурок от скорости ветра графически проде-
монстрировал М.Ф.Тертышников: из рис. 74, Л следует, что ящурки активны при скоро-
сти ветра 0—13 м/с, максимум активности приходится на 4 м/с [258,259,262]. Однако
ветер в 2—8 м/с (именно в таких условиях чаще всего отмечались ящурки в Ставрополье)
гипичен для погожих летних дней в степях европейской части бывшего СССР, он ежеднев-
но закономерно усиливается от практически полного штиля утром до средних величин в
тол день. Снижение или прекращение активности ящурок в середине дня вызвано
слишком высокими температурами среды и не связано с отрицательным отноше-
нием рептилий к ветру. Рисунок же, суммируя все подряд, не отражает реальную
картину.
Кроме обычного ветра охлаждающе действует на ящериц движение воздуха относи-
тельно тела животного, возникающее при беге. При этом ящерица находится как бы под
влиянием ветра такой силы, с какой скоростью она передвигается [240]. Действие солнеч-
аой радиации ослабляют также пылевые частицы, поднимаемые в воздух ветром. При
136
Рис, 74. Активность разноцветной ящурки в зави-
симости от скорости ветра (А) и относительной
влажности воздуха (Б) в условиях Ставрополь-
ской возвышенности [259]
значительной запыленности атмосферы на-
пряженность инсоляции может уменьшаться
в 2—3 раза против нормы и в середине
дня оказаться ниже, чем в утренние часы
[240]. Особенно сильно падает напряжен-
ность солнечной радиации во время пыль-
ных бурь: в степях Украины, например, от-
мечалось понижение суточной суммы радиа-
ции в 5 раз против нормы [274]. Сочетание
пыли и ветра сильно влияет на температуру
тела и активность рептилий: в частности,
активность ящурок резко упала в калмыц-
кой полупустыне с 20 по 26 .IX 1973 г., ког-
да была пыльная буря [22].
1044. ОСВЕЩЕННОСТЬ, ОБЛАЧНОСТЬ
Освещенность имеет большое значение в
жизни ящурок. Они ведут исключительно
дневной образ жизни и в сумерках или но-
чью находятся в убежищах при любых зна-
чениях температуры среды. По наблюдени-
ям в Черноморском заповеднике, наиболь-
шую активность ящурки проявляют в сол-
нечные дни при освещенности 30—60 тыс.
лк. Утром они появляются на поверхности
при 10—20 тыс. лк, вечером уходят в норы
при 1—20 тыс. лк и выше [135]. При этом
не наблюдается определенной зависимости
между началом или концом суточной актив-
ности ящурок и освещенностью, поскольку
основной лимитирующий фактор — температура среды. В Ставрополье в хорошую погоду
первые ящурки выглядывают из нор при освещенности 200 лк, вечером уходят с поверх-
ности при освещенности от 21 000 до 280 лк [259,262].
Облачность, как и ветер, влияет на отношение ящурок к температуре, ослабляя дей-
ствие солнечной радиации. Поэтому в пасмурные дни они покидают свои убежища при бо-
лее высокой температуре среды, чем в солнечные, а характер их активности с двухпиково-
го меняется на относительно однородный в течение дня. Так, в Узбекистане в мае в пасмур-
ные дни ящурки встречались с 11.00 при температуре воздуха 20,5° и почвы 35,5°, в яс-
ные дни — с 9.00 при температуре воздуха 19° [56]. В Черноморском заповеднике в пас-
мурную погоду они выходили на поверхность на 1 —3 ч позже и при температуре воздуха на
несколько градусов выше, чем в солнечную. В целом ящурки были активны при облачно-
сти от 0 до 10 баллов, однако явно предпочитали безоблачную или малооблачную погоду
(имели более высокий уровень активности). В отдельные дни при резком увеличении об-
лачности (и связанным с этим похолоданием и уменьшением освещенности) их активность
быстро прекращалась [135]. В Калмыкии, по данным В.И.Бадмаевой [22], активность
ящурок в сентябре при пасмурной погоде длилась весь световой день, в Краснодарском
крае АБЪочарникова [44] этих рептилий в пасмурные дни не встречала. По наблюдениям
МЛ.Зиняковой и ПЛ .Руденко на Кизилташской косе (Краснодарский край) , облачность
существенно не влияет на активность ящурок. Лишь тогда, когда весь день облачность со-
ставляла 10 баллов, активность ящериц немного снижалась, но причиной этого был слабый
лет насекомых в пасмурные дни [97]. В Прииссыккулье [96] в пасмурную погоду ящурки
иногда совсем не появлялись.
1045. АТМОСФЕРНОЕ ДАВЛЕНИЕ, ВЛАЖНОСТЬ , ОСАДКИ
Зависимость активности ящурок от атмосферного давления мы не обнаружили [131,
135]: они активны при различных показаниях барометра (в том числе непосредственно
137
перед сильной грозой), если другие погодные условия благоприятны — тепло, солнечно,
отсутствуют осадки. М.Ф.Тертышников [259] также не отметил какой-либо зависимости
поведения ящериц от изменения давления, хотя за время его наблюдений атмосферное дав-
ление колебалось от 709 до 742 мм рт. ст.
Влияние влажности на температуру тела животных состоит из двух противоположных
по результатам воздействий: во влажном воздухе увеличивается отдача тепла путем тепло-
проводности, но уменьшается испарение, снижающее температуру тела. На Ставропольской
возвышенности разноцветная ящурка активна при относительной влажности воздуха от
50 до 95 %,проявляя наибольшую активность при влажности 60 % [258, 259,262] (рис.74,
Б). В песках Черноморского заповедника ящурки были активными при относительной
влажности воздуха от 22 до 91 %, в период максимальной активности влажность составля-
ла 22-72, чаще 25-50 % [131,135].
Таким образом, влажность сама по себе мало влияет на активность ящурок: они могут
быть активными при любых ее значениях, отмечаемых в период их сезонной активности,
если другие метеорологические факторы благоприятны. Более высокие величины относи-
тельной влажности, сопровождающие активность вида на Ставропольской возвышенности,
обусловлены в целом более высоким уровнем влажности этого района по сравнению с Чер-
номорским заповедником. В Монголии разноцветная ящурка населяет район, отличающий-
ся крайней сухостью воздуха, ничтожным количеством осадков (50—60 мм в год) и часты-
ми ураганными ветрами [197]. В пустынях Средней Азии и Казахстана годовая сумма
осадков колеблется от 75—80 до 200—300 мм (чаще составляет около 100 мм) , относи-
тельная влажность воздуха летом — 20—30 % [211].
Во время и после дождя ящурки находятся в убежищах [113, 115]. В Монголии после не-
большого дождя они вновь появились на поверхности через 1,5 ч после его окончания, когда
почва была еще влажной, а температура воздуха составляла 17—18° [Тэрбиш, сообщ.]. На Ки-
зилташской косе 5.VI и 3.VII во время учета начинался сильный дождь, и в течение 15 мин
после его начала ящурки все еще оставались на поверхности; в последнем случае во время
дождя было отмечено 11 особей, или 19 % общего количества учтенных в этот день на
маршруте [97].
По нашим наблюдениям, во время сильного дождя ящурки находятся в норах незави-
симо от температуры среды. Однако в жаркие летние дни они некоторое время могут оста-
ваться на поверхности и после того, как начнется небольшой дождь. Совсем незначитель-
ные и непродолжительные осадки в жаркий солнечный день почти не отражаются на уровне
активности ящурок. Если до и после дождя (даже сильного) светит солнце и жарко, жи-
вотные уходят в убежища лишь с началом дождя, а выходят через 30—60 мин после его
окончания. Обычно утром их активность начинается после того, как спадет роса, но в дни
с высокой влажностью воздуха или особенно большой росой и безветрием — до того, как
полностью обсохнет растительность [135].
В террариуме эти ящерицы охотно пили воду, а когда песок долго не увлажнялся и во-
да в поилке отсутствовала, они с явным удовольствием сидели под искусственным дождем
и жадно слизывали с себя, с др утих особей и предметов капли воды [131; Хонякина, со-
обш.]. Родившаяся в неволе ящурка от пищи отказывалась, но с удовольствием лакала
сок с ломтиков огурца и помидора [Окулова, сообщ.]. ННДПнитников наблюдал за ящур-
кой, которая лакомилась сочными кусочками дыни, выжимая из них сок и оставляя клет-
чатку [313]. Очевидно, и в природе после особенно жарких и сухих дней ящурки употреб-
ляют воду (капли дождя, росы); обычно же они довольствуются влагой, содержащейся
в пище. Там,где имеются водоемы, расположенные вблизи мест обитания ящурок, в самое
жаркое и сухое время года эти рептилии периодически посещают водоемы и пьют из них
воду (наблюдения 3 Л-Хонякиной в Дагестане) .
Разноцветные ящурки хорошо плавают и во время затопления мест обитания или в
случае опасности могут спасаться вплавь. Плавающих ящурок наблюдали Н Л .Щербак
[314] в соленой воде оз. Сасык (западное побережье Крыма) и Г Л Лукина [156] в канале
близ Новотроицкой ГЭС (Ставропольский край) ; о способности разноцветных ящурок пе-
реплывать небольшие водоемы упоминала и 3 Л.Хонякина [278]. Мы неоднократно на-
блюдали плывших по мелководью и даже нырявших и затаивавшихся у основания куста
травы ящериц на о-вах Тендра, Джарылгач (Черное море) и Бирючий (Азовское море) ,
т.е. там, где многочисленны постоянные и временные солоноводные и пресные водо-
138
емы и обычны кратковременные затопления участков под действием нагонных ветров.
По наблюдениям М.Ф.Тертышникова [259], периодические ливни в районе Прикалаус-
ских высот (Ставрополье) весной и в первой половине лета 1968—1969 гг. приводили к ги-
бели часть популяций прыткой ящерицы и разноцветной ящурки, а незначительная толщи-
на снежного покрова в 1969 г. на некоторых участках не могла предохранить ящериц от
гибели при действии низких зимних температур (в местах, лишенных снежного покрова,
почва промерзла на глубину до 100 см).
10.4.6. ОСОБЕННОСТИ БИОТОПА
Характер микрорельефа, почв, растительности, убежищ и другие характеристики биотопа
играют существенную роль в терморегуляции рептилий и накладывают заметный отпечаток
на динамику суточной и сезонной активности животных.
Микрорельеф создает большие перепады температур в пределах биотопа. Так, в песча^
ной степи Ив ано-Рыб альчанск ого участка Черноморского заповедника (типичная стация
разноцветной ящурки в Украине) на освещенном солнцем голом песке мы в одно и то же
время регистрировали разницу температур до 10—15° [131, 135], вызванную исключитель-
но разнообразием микро- и. нанорельефа (дюны и понижения между ними, более мелкие
элементы эолового рельефа, выбросы слепышей, следы от копыт домашних и диких ко-
пытных, мельчайшие бугорки и углубления от деятельности мышевидных грызунов, ему-
ранчиков, насекомых и др.). С учетом же различий в освещенности (на солнце - в тени) ,
характере почвы и растительности перепады температур субстрата будут еще значительнее.
Таблица 16. Температура поверхности почвы и тела взрослых разноцветных ящурок в разных био-
топах Западного Казахстана в 1959-1960 гг. [ по данным НА!.Окуловой]
1 Биотоп г — —— — —— — — Температура, ° С
почвы тела
п 1 ?±5х Г | С. V. % (п) п | х Г I C.V % (и)
Глинистая пустыня 26 28Д±0,9 Апрель 2 0 7,3(20) 22,4(21) 25 29,0 ±0,6 2,75 10,9(20)
Пески 32 273 ±1,0 43 26,0 ±0,9 19,6(36)
Глинистая пустыня 17 40,2 ±0,7 Май — начало июня ,о, 73(17) 17 343 ±03 3,20 6,0(17)
Пески 72 37,0 ±0,8 17,4(55) 34 32,0 ±0,6 11,7(27)
Разные почвы характеризуются различным температурным режимом, поскольку по-
глощение солнечной радиации почвой (в частности, теплоемкость, теплопроводность и от-
ражающая способность) зависит от ее механического состава, структуры, цвета и влажно-
сти. Глинистые почвы холодные, плохо прогреваемые, тепловой режим в биотопах с таки-
ми почвами отличается более низкой температурой самого субстрата и приземного слоя
воздуха, что значительно снижает тепловой эффект солнечной радиации. Песчаные почвы -
теплые, кроме того, они дают в одно и то же время очень пеструю картину температур
[10, 11]. Темные почвы нагреваются сильнее, чем светлые, влажные нагреваются медлен-
нее, а охлаждаются быстрее, чем сухие [65]. Таким образом, из почв, на которых встреча-
ются разноцветные ящурки, самыми теплыми являются сухие темноокрашенные песчаные
почвы. В Западном Казахстане на участке, где глинистая пустыня соседствует с песчаной,
температура поверхности почвы в апреле—мае, по наблюдениям HJ4.Окуловой [сообщ.],
а также температура тела активных взрослых Е arguta были выше в глинистой пустыне,
чем в песках, но в последних значительно выше была степень изменчивости обеих темпера-
тур (табл. 16) .
Растительность и убежища определяют защитные свойства биотопа, они играют исклю-
чительно важную роль в поведенческой терморегуляции рептилий. От структуры раститель-
ного покрова (густоты и высоты травяного покрова, проективного покрытия, наличия
древесно-кустарниковой растительности, ее структуры и сомкнутости крон) зависят свето-
вой и температурный режимы мест обитания и, следовательно, отношение к температуре,
139
дремя суток,ч
Рис. 75. Изменение температуры тела разноцветной
ящурки в связи с изменениями температуры и относи-
тельной влажности среды в условиях Ставропольской
возвышенности [259].'
1 - температура тела, 2 — температура поверхности
почвы, 3 — температура почвы в норе, 4 — температура
воздуха в норе, 5 — температура воздуха над поверхно-
стью почвы, 6 — относительная влажность воздуха над
поверхностью почвы; 7 — количество ящериц на поверх-
ности, 8 - относительная влажность воздуха в норе
длительность и характер активности
разноцветной ящурки. Тень от расте-
ний и сами растения ящерицы широко
используют для регулирования темпе-
ратуры тела.
Особая роль в терморегуляции
рептилий принадлежит норам [284].
В частности, норы в зависимости от
времени года и суток могут выпол-
нять функции нагревания или охла-
ждения тела рептилий, температура
нор определяет нижнюю границу тем-
пературы тела в нежаркое время го-
да, наконец, норы в аридных усло-
виях выступают важнейшим факто-
ром экономии воды. В течение суток
температура почвы в норе достигает
минимума на 4—8 ч, максимума —
на 6—12 ч позже, чем на поверхности;
с глубиной резко уменьшается суточная амплитуда колебаний температур, и в пустынях,
где на поверхности почвы она может достигать 40° , на глубине 50 см она уже практически
не ощущается [284]. Менее резкие, чем снаружи, суточные колебания температуры почвы,
воздуха и относительной влажности воздуха в норах (рис. 75) позволяют ящуркам пере-
живать неблагоприятные условия среды.
Определенное значение для успешной терморегуляции имеют и другие убежища (тре-
щины в почве, щели и ниши в твердых породах, пространства под корнями кустов и др.).
Наличие камней позволяет ящерицам быстро нагреваться от контакта с субстратом, рых-
лые почвы (с пылевой структурой или песчаные) дают возможность животным быстро
закапываться и изменять температуру тела. Зарывание разноцветных яшурок в пыль у
основания кустов для снижения температуры тела наблюдала НЛ1.Окулова [сообщ.] в За-
падном Казахстане, закапывание в песок или рыхлый лесс отмечали в различных частях
ареала [92,359].
Обилие корма может влиять на длительность и характер суточной активности репти-
лий, в том числе и разноцветной ящурки [232]. При низкой численности насекомых яще-
рицы вынуждены больше времени находиться на поверхности почвы и не всегда при самых
благоприятных температурных условиях среды, к тому же им приходится охотиться на
большей по площади территории, т.е. в менее знакомой обстановке и на значительном уда-
лении от своих нор. Все это не может не сказаться на точности терморегуляции рептилий.
10.5. ТЕРМОРЕГУЛЯЦИОННОЕ ПОВЕДЕНИЕ
Для повышения температуры тела до уровня добровольного минимума разноцветные
ящурки обогреваются в лучах солнца, используют тепло нагретого субстрата или активно
увеличивают теплопродукцию организма путем мышечных сокращений. В Азербайджане,
Калмыкии и Ставропольском крае ящурки затрачивают на обогрев до 40—45 мин [20,
22, ИЗ, 259, 262; Киреев, сооб.], в Украине — до 60 мин (обычно не более 30—40) Дече-
ром также нередко подолгу греются на солнце у входа в нору [131,135). Столько времени
тратится на обогрев обычно осенью и в прохладные летние дни, а в жаркие дни этот период
заметно сокращается. Так, в Азербайджане в марте ящурки грелись у входов в норы 30—
45 мин, в мае — 20—25, в июне — 10—15 мин [20]; в Калмыкии и Казахстане время обо-
грева составляло летом 10—20 мин [22,48; Киреев, сообщ.].
По нашим наблюдениям, в жаркие летние дни ящурки не обязательно греются на по-
верхности у входов в норы: для начала активности им бывает достаточно посидеть в норе
возле ее выхода, выставив на солнце лишь голову. Этим создается впечатление, что ящур-
ки как бы минуют стадию нагревания, сразу из норы появляясь на поверхности в актив-
ном состоянии. Как показала А.Ф.Баженова [26], температурный коэффициент метаболиз-
140
ма Сю разных видов дневных ящериц в мае — июне имеет хорошо выраженный градиент
мозг > сердце > печень > мышцы. При этом интенсивность дыхания тканей скелетных
мышц практически не зависит от температуры (210 = 1, реже 1.1) , зависимость же мета-
болизма мозга от этого фактора велика (£?10 для разных видов колеблется от 1,7 до 3,1) .
Поэтому для наступления двигательной активности достаточно нагревания головы ящери-
цы. Это физиологическое свойство позволяет рептилиям нагреваться, не выходя из норы
на поверхность, и, очевидно, помогает им успешно избегать врагов тогда, когда они осо-
бенно уязвимы из-за малой подвижности (низкой температуры тела) . Осенью наблюдает-
ся исчезновение градиента температурного коэффициента и стабилизация его величины на
относительно высоком уровне. Это означает, что параллельно уменьшению температуры
среды равномерно снижается жизнедеятельность всех систем организма, что и обеспечивает
переход от активного состояния к зимнему оцепенению [26,235]
При невысоких температурах среды ящурки искусно используют особенности микро-
и нанорельефа, выбирая наиболее освещенные и нагретые участки субстрата. Плотно при-
жавшись телом к почве или камню и приняв положение, обеспечивающее наибольшее воз-
действие прямых солнечных лучей, ящерица в минимальные сроки повышает температуру
тела до уровня добровольного минимума, затем усиливает и поддерживает достигнутый
эффект за счет бега. Влияние движения на температуру тела разноцветных ящурок хорошо
заметно в полевых условиях при низких температурах среды и невысокой инсоляции
(табл. 15, группы 8—10). При содержании ящурок в стеклянных банках в одинаковых
условиях температура тела при отсутствии активных движений лишь немного превышала
температуру среды. Но стоило ящерицам сделать несколько резких движений или пробе-
жаться, как температура тела существенно возрастала и, например, при температуре среды
9,7° могла подняться до 18—24 [131, 135]
Достигнув оптимальных величин, температура тела ящурок некоторое время удержива-
ется животными в пределах диапазона оптимальных температур (термостабилизируюшее по-
ведение) . Это достигается чередованием уже рассмотренных способов нагревания с приема-
ми охлаждения тела — использованием тени от растений и различных предметов, быстрым
перемещением по горячему субстрату с приподнятым над ним телом, влезанием на стебли
растений и др. В частности, в Черноморском заповеднике на стационарной площадке
яшурки часто сидели в тени от деревянных колышков, которыми были помечены их
норы, а некоторые особи влезали на колышек и, зацепившись за его верх передними ла-
пами, свисали с теневой стороны.
На морском берегу они охотно прячутся под выброшенные морем доски и деревян-
ные ящики. Так, на о.Тендра во время учетов в июле на 100 м маршрута отмечено 7 яшу-
рок, из них 6 в качестве убежиш выбрали доски; на 12 км маршрута учтено 1ь особей,
из них 4 спрятались под доски и ящики, 12 — в кусты полыни. Как убежиша большие
кусты песчаной полыни настолько хороши, что это влияет на поведение ящурок: на о.Тен-
дра они, спасаясь от жары или опасности, редко уходят в норы, а укрываются обычно под
полынными кустами, откуда их очень трудно выгнать [131]. Неоднократно наблюдали
ящурок, спасавшихся от жары на приподнятых над землей стеблях растений [97. 131. 135].
При слишком высокой температуре среды у ящурок может быть короткий период
добровольного перегрева. В это время они ведут себя беспокойно, стремительно перебега-
ют с места на место; температура тела может повышаться до 40°. В таких экстремальных
условиях заметно действие физиологических механизмов терморегуляции: увеличивается
частота дыхания, соответственно возрастает и испарение с поверхности ротовой полости и
легких; наблюдается некоторая задержка в подъеме температур в оптимальном диапазо-
не [131,192, 220]. Испарительное охлаждение усиливается также за счет изменения кожно-
легочной проницаемости, при этом функцию рецептора выполняет теменной орган, реаги-
рующий на инфракрасное излучение [89]. В дальнейшем во избежание теплового шока
ящурки уходят в норы, где температура тела быстро снижается.
Подводя итог, отметим следующее. Отношение разноцветных ящурок к температуре сре-
ды зависит прежде всего от климатических и погодных условий (уровня солнечной радиа-
ции, ветра, облачности, осадков и др.) , возраста и физиологического состояния животных
141
(степени упитанности, стадии полового цикла, режима питания), особенностей биотопа
(микрорельефа, почв, растительности и убежищ, обилия корма) и связано с сезонностью
и временем суток в соответствии с динамикой перечисленных факторов.
Температура тела ящурок в каждый конкретный момент определяется, в первую оче-
редь, такими факторами, как солнечная радиация, температура поверхности почвы и дви-
гательная активность, и зависит от возраста, физиологического состояния организма и ха-
рактера биотопа. На нее также влияют абиотические факторы, снижающие уровень солнеч-
ной радиации или увеличивающие теплоотдачу (ветер, запыленность атмосферы, облач-
ность, осадки). Зависимость терморегуляции от возраста связана с размерами (массой те-
ла) рептилий и степенью совершенства поведенческих реакций. Исключительно велика
роль убежищ (прежде всего нор), а также растительности в регулировании температуры
тела разноцветных ящурок.
Сеголетки, как правило, активны при температуре поверхности почвы не ниже 15°
воздуха — не ниже 8, остальные — при температуре не ниже 17 и 11° соответственно. Верх-
ние температурные пределы активности ящурок 55—56° (температура почвы) и 36—38°
(температура воздуха) , у сеголеток несколько ниже. Температура тела активных ящурок
может колебаться от 18 (обычно от 20—21°) до 40°, оптимальной является температура
в диапазоне 34—38 (в среднем около 37° для взрослых и 34 — для сеголеток) . Темпера-
тура тела ящурок в природе может быть выше температуры поверхности почвы на 10—
13° (весной и осенью) или ниже ее на 15—20° (в летнюю жару) . Первое достигается за счет
обогрева в лучах солнца и последующего бега, в результате чего животные некоторое вре-
мя не зависят от низкой температуры среды, второе,— за счет перемещения по горячему
субстрату с приподнятым над ним телом, использования тени и различных убежищ, влеза-
ния на стебли растений и др. Таким образом, хорошо развитая поведенческая терморегу-
ляция позволяет ящуркам быть активными в широком диапазоне температур среды —
не менее 40° для поверхности почвы и около 30 — для воздуха.
ГЛАВА 11
СУТОЧНЫЙ ЦИКЛ АКТИВНОСТИ1
11.1. ОБЩИЕ ЗАМЕЧАНИЯ, ОЦЕНКА ДАННЫХ
Сведения о суточной активности разноцветной ящурки содержатся примерно в полусотне
работ, однако в большинстве случаев они отрывочны или скудны [19, 27, 48, 55, 56, 77,
96, 115, 120, 121,125,179, 203,324,359] либо носят самый общий характер [35,107,118,
155, 198, 266, 310, 315, 385] . В ряде источников имеются более обстоятельные материалы
по данному вопросу [49, ИЗ, 122, 130,197, 225,258. 271,316,386]. Сведения, опублико-
ванные до начала 70-х годов, суммированы в сводках ННЩербака [320, 436]. Наиболее
подробно суточный цикл активности E^rguta рассмотрен в работах НМ.Окуловой [192]
по Западному Казахстану, М.Ф.Тертышникова [259, 262] по Ставрополью, В .И .Бадмаев ей
[22. 25] по Калмыкии, М Л .Зиняковой и ПЛ Руденко [97] по Краснодарскому краю,
МИ Ахмедова [20] по Азербайджану и ТИКотенко [127, 131, 232] по югу Украины.
В итоге Южная Украина [121, 122. 125, 127, 130, 131, 135, 232, 315, 316, 385,'386] в дан-
ном плане стала самым изученным регионом.
В связи с тем что начало и конец суточной активности популяций разноцветной яшур-
ки большинство исследователей отмечало по первым и последним зарегистрированным на
поверхности почвы особям, то в категорию активных (и соответственно в понятие ’’пе-
риод активности”) вошли не только действительно активные особи, но и те, которые на-
ходились в переходных состояниях (нагревания и остывания, см. разд. 10.1). Время на-
чала и окончания фаз суточного цикла и время пиков активности у разных авторов сильно
различаются. Происходит это не только вследствие объективной географической и сезон-
ной изменчивости данных параметров, но и по иным причинам: во-первых, накладывает от-
печаток специфика сетки часовых поясов1 2 , во-вторых, большую путаницу вносит практи-
куемый ежегодный перевод стрелок часов на 1 ч в ту или иную сторону. В результате осло-
жняется сопоставление данных, полученных до 1981 г. и позже, либо тех, что получены в
1981—1989 гг. в апреле-сентябре и в другое время года. Поскольку большинство работ
по суточной активности E.arguta выполнено до учреждения летнего и зимнего времени, а
также во избежание ошибок, мы в дальнейшем везде даем время, указанное самими ав-
торами.
11.2. ИНДИВИДУАЛЬНАЯ, ГРУППОВАЯ
И ПОПУЛЯЦИОННАЯ АКТИВНОСТЬ
Необходимо различать индивидуальную, групповую и популяционную активность. Попу-
ляционная слагается из активности отдельных групп (возрастных, половых) , групповая -
из активности отдельных особей. Популяционная активность всегда продолжительнее ин-
дивидуальной. Так, в Черноморском заповеднике 19.VIII 1978 г. взрослая разноцветная
ящурка находилась на поверхности с 9.00 до 13.00 и с 16.00 до 18.00 (т.е. всего 6 ч) ,в то
время как популяция в целом была активна с 8.00 до 18.00 (10 ч подряд) . В третьей дека-
де сентября и в первой половине октября 1975 г. одна из сеголеток покидала свое убежи-
1 При участии М И Ахмедова, В.И.Бадмаевой,И.С.Даревского, Т.Сатторова, Х.Тэрбиша, 3 ЛХонякиной.
2 Для сопоставимости данных декретное время следует переводить в местное солнечное, однако различ-
ные авторы это не делают и/или не оговаривают.
143
ще обычно после 14.00, и период ее активности длился 2,5—4 ч. Популяционная же актив-
ность начиналась в 830—1030 и заканчивалась в 1630—18.00, т.е. продолжалась 6—9,5 ч.
Наблюдая за мечеными ящурками, мы обнаружили, что у отдельных особей тип актив-
ности может не совпадать с типом популяционной активности. Так, в летние жаркие дни,
когда популяционная активность носит дифазный характер, отдельные особи могут быть
активными только утром, только вечером или, с небольшими перерывами, весь день. Ин-
тересно, что определенный тип активности может быть присущ какой-либо особи доволь-
но длительное время (1—3 недели) , либо ящурка часто меняет его в зависимости от погод-
ных и других условий. Отсюда понятно, почему в 10.00—12.00 можно поймать ящурку с
большим наполнением желудка, в 16.00—17.00 - с малым. Поскольку особи с индиви-
дуальной активностью, резко отличающейся от схемы популяционной активности, состав-
ляют меньшинство, суммарная активность популяции носит все же довольно упорядочен-
ный характер [122, 127, 386]. Активность разных групп (прежде всего, возрастных) мо-
жет существенно разниться между собой и отличаться от общей популяционной актив-
ности.
113. СТРУКТУРА СУТОЧНОЙ АКТИВНОСТИ
И ЕЕ СЕЗОННАЯ ДИНАМИКА
В течение года закономерно изменяются условия жизни и физиологическое состояние реп-
тилий. Последнее определяет потребности организма и термофизиологические характери-
стики. Факторы среды обусловливают термоэкологические характеристики животных. Все
это вместе непосредственно или косвенно влияет на формирование структуры активности
рептилий. Суточный цикл активности и его изменения по сезонам у разноцветной ящурки,
как гелиотермного пресмыкающегося, зависят, прежде всего, от изменяющихся темпера-
турных условий среды. Влияние температуры и других метеорологических факторов на
активность ящурок подробно рассмотрено в предыдущей главе, поэтому здесь мы оста-
новимся лишь на анализе кривых суточной активности.
Как показали исследования в различных частях ареала [19, 20, 22, 24,25.35,97,107,
113, 115, 122, 127, 130, 131, 155, 192, 197, 198, 225, 232, 258, 259, 262, 266, 271,310,
315,316,386;Тэрбиш, Хонякина, сообщ.], для разноцветной ящурки весной и осенью свой-
ствен монофазный тип активности с максимумом в самое теплое время дня, летом — ди-
фазный с перерывом в жаркое время. Поскольку сезонное изменение характера суточной
активности обусловлено динамикой температуры среды, то в разных по климатическим
условиям частях ареала переход от монофазного к дифазному типу активности и обратно
происходит в разное время. Так, на Кизилташской косе (Западное Предкавказье) актив-
ность ящурок имеет 2-вершинный характер в июне—августе [97], в Черноморском запо-
веднике (юг Украины) — с конца мая до середины — конца июля, иногда до середины ав-
густа (для сеголеток — до начала сентября) [127], в Волго-Уральских песках (Западный
Казахстан) — с третьей декады мая до третьей декады августа [192], на Ставропольской
возвышенности (Центральное Предкавказье) — во второй декаде мая — августе [262], в
Калмыкии — в мае — начале сентября [22], на о.Обливной и мысе Бендован (Восточный
Азербайджан) — в мае—августе [20], в Джунгарской Гоби (Монголия) — в третьей дека-
де июня — августе [197, 291; Тэрбиш, сообщ.].
В течение года закономерно изменяются также продолжительность фаз активности и
(соответственно) время начала и окончания активного периода. В первое время после весен-
него пробуждения разноцветные ящурки появляются на поверхности только к 11.00—12.00
[77], активны в полуденные часы [35] и очень непродолжительное время — не более 2—
3 ч в Западном Казахстане [192] или примерно с 11.00 до 15.00 — в Азербайджане [20].
В конце апреля в Дагестане ящурки при хорошей погоде активны обычно с 9.00—10.00
до 16.00 с максимумом в 11.00—13.00 [Хонякина, сообщ.]. В Северном Таджикистане они
весной активны с 1030—11.00 до 17.30—18.00 [225], на юге Узбекистана в апреле — пос-
ле 10.00 [55].
В мае ящурки появлялись на поверхности на Кизилташской косе в 730—11.15 (в зави-
симости от погодных условий), уходили на ночевку с 16.00 до 17.00 [97]; в Прииссык-
кулье активность этих ящериц начиналась в 9.00—10.00 [96], в Ферганской долине —
144
Месяц
и декада
Март щ
I
Апрель ц
_______
1
Май п
______П1
I
Июнь и
_______ш
1
Июль п
_______ш
I
Август и
\1и
Сентябрь.
Октябрь
Время суток
1 -L I J. J I
Март
Апрель
Май
Июнь
Июль
Август
Месяц
декада
UI
п
т
I
п
т
11
in
I
Сентябрь II
в
9
п
7
19 20
Время суток
~ю
iiuiiiiiuiinminniui
IlMiiinwuinmiimiiiimiNmmiiiiiiiiiuiMl
ЯН.
liiiiHimuHiHUiimiiiimilumiiii
r^iillllllllllilllilllllllllin I
Октябрь
П
Рис, 76, Суточная и сезонная активность разноцветной ящурки:
А — Волго-Уральские пески (Западный Казахстан) [ 192] : а — только взрослые самцы, б — взрослые
самцы и самки, в — только молодые, г — вся популяция; Б — Ставропольская возвышенность (Цен-
тральное Предкавказье) [ 259, 262] : а — вся популяция, б - сеголетки; В, Г - песчаная степь Ивано-Ры-
бальчанского участка Черноморского заповедника, 1975 г. [ 127] : В — взрослые и полувзрослые, Г - се-
голетки
п
т
п
ш
ш
с 9.00 [56], в Дагестане — с 7.00—8.00 [Хонякина, сообщ.]. В Калмыкии она длилась с
6.00—630 до 1830 и долее [22, 113],в Восточном Азербайджане — с 930 до 18.00 [20],в
Монголии — с 8.00—9.00 до 17.00—1930 [197; Тэрбиш, сообщ.]. Сезонная динамика вре-
менной структуры суточной активности ящурок Западного Казахстана, Ставрополья и
Южной Украины представлена на рис. 76.
145
Рис. 77. Суточные циклы активности раз-
ноцветной ящурки в песчаной степи Ивано-
Рыбальчанского участка Черноморского за-
поведника, 1975 г., постоянная площадка
[127]:
А: 1 - 6 XV, 2 - 17.IV, 3 - 22.V; Б: 1 -
27.V, 2 - 6.УП, 3 - 14.УП; В - 19.УП;
Г - 12.УШ; Д - 20.УШ; Е: 1 - 20.IX,
2 — 11.Х. Сплошные линии - взрослые
и полувзрослые особи, пунктирные - се-
голетки
Таким образом, длительность ак-
тивного периода разноцветной ящур-
ки с момента весеннего пробуждения
постепенно увеличивается и к началу
мая для самых теплых частей ареала,
к середине — концу мая для юга
Украины, Западного Казахстана и
Центрального Предкавказья [127, 192,
262] достигает максимальных вели-
чин, составляя для всей популяции
около 11—12 ч/сут (рис. 76) . В июне-
июле временные границы активности
разноцветных ящурок расширяются.
Так, утром первые особи появляются
в Северном Таджикистане, Западном
Казахстане и Крыму с 8.00, в Восточ-
ном Азербайджане и на Ставрополь-
ской возвышенности — с 730 (по
[266] — с 5.00—6.00) , на юге Украи-
ны — с 7.00, в Западном Предкав-
казье и Монголии — с 630, в Чуйской
долине (Кыргызстан) и Дагестане —
с 6.00, в Калмыкии — с 5.00. Послед-
ние ящурки на поверхности в это вре-
мя года наблюдались в Кыргызстане
и Западном Предкавказье в 18.00, на
юге Украины, в Ставрополье, Дагеста-
не и Восточном Казахстане — в 20.00,
в Калмыкии, Восточном Азербайджа-
не и Северном Таджикистане — в 2030,
в Западном Казахстане и Монголии —
до 21.00 [20,22,48,97,113,115,121,
127, 192, 225, 262, 271,316,324; Тэр-
биш, Хонякина, сообщ.]. Выше приве-
дены максимальные из зарегистриро-
ванных границы суточной активности
ящурок в июне — июле. В указанные
сроки животные утром появляются
лишь в жаркие безоблачные и безвет-
ренные дни, уходят же на ночевку
обычно на час (в отдельные дни даже
на 2—3 ч) раньше максимально воз-
можного времени. В частности, в За-
падном Предкавказье в июле—августе ящурки на ночь уходят обычно с 17.00 до 18.00
[97]. Случаи неожиданно раннего окончания активности ящурок приводились для Запад-
ного Казахстана [192] и Дагестана [Хонякина, сообщ.]; неоднократно наблюдали мы это
явление и в Черноморском заповеднике. Причины прекращения активности ящурок ранее
того времени, когда температурные условия среды становятся неблагоприятными, рас-
смотрены в предыдущей главе.
146
7 3 11 В « 17 a 21 s 7 9 71 13 13 17 19 21
Время суток, у
Рис. 78. Суточные циклы активности разноцветной ящурки:
А — Кизилташская коса (Западное Предкавказье) [97] : 1 — май, 2 — июнь, 3 — июль-август;
Б — о.Обливной в Каспийском море (Азербайджан) [20]: 1 - апрель, 2 — май-август, 3 - сен-
тябрь — ноябрь; В — Калмыкия [ 25] : 1 — май, 2 — июнь, 3 — июль, 4 — август; Г — Джунгарская
Гоби (Монголия) [271; Тэрбиш, сообщ.] : 1 - 10.V 1982 г., 2 - 7.V] 1983 г., 3 - 21.VI 1984 г.
Количество особей: Л - %,Б~Г - экз.
Несмотря на раннее начало и поздний конец, общая продолжительность активного
периода в июне—июле не увеличивается, а остается прежней или несколько сокращает-
ся (рис. 76) , поскольку в самое жаркое время дня наступает фаза дневного отдыха, когда
все или большая часть ящурок находятся в убежищах. Такой перерыв в активности яще-
риц начинается (в зависимости от климата и погоды) в 11.00—13.00 и заканчивается в
14.00—18.00, но чаще всего наблюдается с 12.00 до 15.00 [20, 22, 97, 120, 127, 192, 225,
232, 262, 271; Тэрбиш, сообщ.]. Сезонная динамика продолжительности фаз активности
и дневного отдыха хорошо видна на рис. 76.
Через 1—3 ч после появления первых особей наступает утренний пик активности. В са-
мые жаркие дни максимальное количество ящериц наблюдается в 8.00—10.00,как,напри-
мер, в Черноморском заповеднике [127], на Кизилташской косе [97] и в Калмыкии [22,
25, 113, 115] (рис. 77, 78, А, Б) . Но в большинстве регионов и при не очень высоких тем-
пературах среды утренний пик активности приходится на 9.00—11.00. Вечерний пик актив-
ности в норме всегда ниже, чем утренний, и имеет место обычно в 16.00—18.00, сдвигаясь
в ту или Иную сторону на 1 ч в зависимости от региона и погоды (конкретные сведения
см. [ 20, 22, 25, 97, 113, 120, 127, 179,232,271; Тэрбиш, Хонякина, сообщ.]). Данные по
Румынии [359] (в 9.00 — начало активности, в 11.00—13.00 — максимум) относятся
ими не к июню—июлю или к прохладным дням.
В августе (для Украины и Западного Казахстана) или в сентябре (для Калмыкии) ак-
тивность разноцветных ящурок снова приобретает одновершинный характер с максиму-
мом в 12.00—14.00 [49, 127, 232]. При этом длительность активности взрослых ящурок
в августе сокращается до 5—6 ч в Днепропетровской обл. Украины и в Прииссыккулье
[49, 203] или до 8—9 ч — в Черноморском заповеднике (Херсонская обл. Украины)
[127,232].
Осенью ящурки в Узбекистане активны во второй половине дня [35], в Северном
Таджикистане отмечались на поверхности с 1230—13.00 до 16.00 [225], в Восточном
Азербайджане в сентябре — с 12.00 до 17.00, в октябре — начале ноября — с 13.00 до 16.30,
хотя единичные особи наблюдались в течение всего дня [20] . В Калмыкии в первой поло-
чине сентября ящурки были активны с 10.00 до 18.00 с некоторым спадом активности в
1130—1430, а с середины месяца активность приобрела 1-вершинный характер [22].
147
В Крыму в сентябре активность ящурок еще сохраняла 2-вершинный характер, хотя и в
сглаженном виде, при этом наибольшее число особей наблюдалось между 11.00—12.00 и
16.00—17.00 [316]. В Западном Казахстане период активности сокращается до 4—6 ч/сут
в конце сентября и до 2—3 ч — в октябре [192]. В Черноморском заповеднике длитель-
ность активной фазы у взрослых особей изменяется от 6 ч в начале сентября до 3,5—
4,5 ч в конце месяца [127, 232]. Сезонная динамика структуры суточной активности раз-
ноцветных ящурок в этом регионе, а также в Центральном Предкавказье и Западном Ка-
захстане хорошо видна на рис. 76 и 77, особенности активности Earguta в других частях
ареала отражены на рис. 78.
11.4. ЗАВИСИМОСТЬ СУТОЧНОЙ АКТИВНОСТИ ОТ ВОЗРАСТА,
ПОЛА И ФИЗИОЛОГИЧЕСКОГО СОСТОЯНИЯ ЯЩУРОК
Рассмотренные выше структура и динамика суточной активности касаются взрослых ящу-
рок. Активности сеголеток в литературе уделяли мало внимания. Отмечали, в частности,
что в Среднем Приднепровье длительность суточной активности молодых ящурок 4—5 ч,
взрослых — 5—6 [49], в Прииссыккулье в августе соответственно 4 и 5 ч [203]. В Чуй-
ской долине в июле молодые встречались в основном утром, а взрослые были малоактив-
ны [324]. В Калмыкии с появлением молоди длительность суточной активности популя-
ции несколько увеличивается, поскольку сеголетки выходят на поверхность раньше, а
уходят позже взрослых [25].
По нашим наблюдениям в Черноморском заповеднике [127, 131, 386], появление се-
голеток усложняет общую картину популяционной активности, поскольку возраст (и свя-
занные с этим специфика терморегуляции и степень упитанности) заметно влияет на ха-
рактер активности. Если в августе—сентябре (в отдельные годы даже во второй полови-
не июля [232]) у взрослых ящурок наблюдается один пик активности, приходящийся
на жаркое время дня, то у сеголеток активность носит 2-вершинный характер с максиму-
мами в утренние и вечерние часы. При этом взрослые на 0,5—3,5 ч позже сеголеток покида-
ют свои убежища утром и на 0—2 ч раньше уходят в них вечером. Интересно, что в сентяб-
ре даже при высокой температуре среды (воздуха — до 30°, почвы — до 50) у взрослых
ящурок сохраняется монофазный тип активности.
Таким образом, периоды максимальной активности взрослых и сеголеток в значитель-
ной мере разобщены во времени (рис. 76, В, Г; 77, В—Е) . Эго позволяет более полно ис-
пользовать кормовую базу и приводит к ослаблению пищевой конкуренции у разных воз-
растных групп. Последнему способствует также заметное понижение уровня активности
взрослых ящурок при высоком значении этого показателя у сеголеток [122]. Все это, на
наш взгляд, имеет значение для нагула сеголеток: за весь-период активности разноцветных
ящурок именно в августе—сентябре численность и биомасса беспозвоночных минимальны
[139], численность популяции ящурок — максимальна (см. разд. 20.1). Осенью актив-
ность сеголеток приобретает одновершинный характер, а период активности постепенно
сужается (рис.76,Г; 77,В).
Упомянутый выше показатель ’’уровня активности” мы используем для оценки степе-
ни активности популяции в конкретный период времени. Такой показатель можно опре-
делить как отношение количества особей, находящихся на поверхности какой-либо пло-
щадки в период пика активности, к абсолютной численности вида на этой площадке [127].
Уровень активности популяции зависит от длительности индивидуальной активности и от
того, ежедневно или нет выходят отдельные особи из убежищ, т.е. наряду с погодными
условиями он в сильной степени определяется физиологическим состоянием животных.
Ориентировочно этот показатель можно оценить по высоте графиков суточной активности.
Уровень активности ящурок со времени весеннего пробуждения постепенно увеличи-
вается, достигая наивысшего значения в конце мая — начале июня (для юга Украины) , а
затем начинает снижаться. В целом период наибольшей активности ящурок на юге Украины
приходится на конец апреля (в годы с ранней и теплой весной) , май и июнь. В это время
они подолгу находятся на поверхности, встречаются на значительных расстояниях от нор
(особенно самцы) , что объясняется повышенной половой активностью ящериц. Значитель-
ное понижение уровня активности взрослых ящурок в конце лета создается за счет того,
что во второй половине августа и особенно в сентябре — начале октября взрослая особь
148
появляется на поверхности далеко не каждый день (через 1,2,3 и даже более дней). Не-
которые ящурки выходят по-прежнему ежедневно (хотя бы несколько дней подряд),
отдельные особи начинают появляться не ежедневно уже в начале июля. Данное явление
мы объясняем разной упитанностью ящурок и, следовательно, различной потребностью
в корме. Такой характер индивидуальной активности резко понижает уровень популя-
ционной активности и создает впечатление раннего залегания в спячку части взрослых
ящурок [127, 386]. Аналогичная закономерность наблюдалась также в других частях
ареала вида: максимальная активность ящурок была приурочена к брачному периоду,
а в июле (для Чуйской долины в Кыргызстане) или во второй половине июня — авгу-
сте (для нижнего Поволжья) встречаемость этих рептилий значительно понижалась [118,
324]. На Ставропольской возвышенности уровень активности снижался постепенно, начи-
ная с третьей декады июня [259], в бывшей Харьковской губернии — с середины авгу-
ста [239].
Степень упитанности (масса жировых тел) непосредственно связана со сроками раз-
множения, а последние определяются, прежде всего, погодными условиями. Так, в Чер-
номорском заповеднике в 1975 г. (необычайно ранняя и теплая весна, жаркое лето) ос-
новная масса самок отложила яйца в июне — начале июля и успела накопить значительные
жировые запасы за июль — начало августа. Поэтому в августе у взрослых ящурок чаще от-
мечался ’’осенний” тип активности — один пик в 11.00—14.00, позднее появление утром и
ранний уход вечером, низкий уровень активности, малая подвижность. В 1978 г. (поздняя
весна, холодное дождливое лето) период размножения затянулся, захватив первую дека-
ду августа, в результате чего в этом месяце наблюдалась необычно высокая активность
взрослых ящурок,часто по ’’летнему” типу [127].
Чувство голода оказывается одним из основных факторов, определяющих продолжи-
тельность и характер суточной активности [127, 386]. После нескольких дней непогоды
(когда ящурки совсем не появлялись на поверхности) в первый солнечный и теплый день
они необычайно дружно выходят из убежищ и активно питаются. Уровень их активности
в это время резко увеличивается, достигая даже в августе (сравниваются только взрослые
особи) показателей мая. Последнее обстоятельство позволяет использовать такие дни для
проведения учетов с целью определения численности рептилий (если нельзя выполнить
эту работу в мае—июне). В этот первый погожий день ящурки активны, как правило, в
первую половину дня (следствие дружного выхода и интенсивного питания) , 2-вершин-
ный характер активности (если это лето) восстанавливается лишь на второй, иногда — на
третий день. Влияние чувства голода на активность ящурок особенно наглядно демонстри-
рует следующий пример: в дни с малоблагоприятной для ящериц погодой (пасмурно,
прохладно) на опытной площадке активными были лишь особи, выпущенные сюда нака-
нуне вечером после содержания в течение нескольких дней в неволе без пищи [127]. На-
блюдения 3 Л Хонякиной [сообщ.] также показали, что продолжительность фазы дневной
активности у сытых ящурок заметно короче, чем у голодных (установлено путем
эксперимента в природе). В связи со сказанным понятно, почему утром ящурки
обычно дружно выходят из убежищ и утренний пик активности, как правило, выше
вечернего.
По данным 3 Л Хонякиной [сообщ.], активность несколько снижается у линяющих
особей и у самок в период откладки яиц.
115. ВЛИЯНИЕ РАЗЛИЧНЫХ ФАКТОРОВ СРЕДЫ
НА СУТОЧНУЮ АКТИВНОСТЬ
Активность ящурок в наибольшей степени зависит (прямо и косвенно) от климата и по-
годных условий — температуры, осадков, ветра, облачности и др. Подробно влияние этих
факторов рассматривалось в предыдущей главе. Здесь же мы приведем лишь некоторые
примеры. Так, влияние микроклимата на суточную активность ящурок отмечено в Восточ-
ном Азербайджане [19, 20]: на ©.Обливной в Каспийском море активность рептилий пре-
кращалась на 1—2 ч позже, чем на рядом (в 7 км) расположенном побережье материка, а
начиналась либо на 1 ч раньше (весной и осенью) ,либо на 1 ч позже (летом) . В Черномор-
ском заповеднике наблюдались различия в характере и уровне активности ящурок в пес-
чаной степи и на морском берегу (см. разд. 10.43)
149
Рис. 79. Активность разноцветной ящурки в раз-
личных ландшафтах и при разной погоде (Дне-
пропетровская обл. Украины, август 1970 г.)
[49]:
1 — открытые пространства, ясный день; 2 — об-
лесенные пространства, ясный день; 3 — открытые
пространства, пасмурный день; 4 — облесенные
пространства, пасмурный день
Влияние погоды на суточный цикл ак-
тивности наглядно продемонстрировано для
Среднего Приднепровья в августе [49]: в
пасмурный день активность начиналась поз-
же, заканчивалась раньше с разницей при-
мерно в 2 ч по сравнению с ясным днем
(рис. 79). По наблюдениям в Центральном
Предкавказье, в хорошую погоду у ящурок
больше длина перемещений и величина
дневного индивидуального участка, чем в
плохую [255,259].
Особенности биотопа также могут вли-
ять на продолжительность и характер ак-
тивности, что связано со спецификой мик-
роклимата и защитными свойствами био-
топа. Влияние (через термальный режим
биотопа) характера субстрата и раститель-
ности на структуру суточной активности
ящурок отметил еще А.Г.Банников для Да-
гестана [27]. Наличие большого количества
нор и кустов растений облегчает рептилиям
как процесс терморегуляции, так и обеспе-
чение безопасности, что отражается на дли-
тельности пребывания ящурок в активном состоянии на поверхности почвы (наши наблю-
дения) . В Л Зул ахов [49] показал, что на облесенных участках начало активности разно-
цветной ящурки сдвинуто на 1 ч по сравнению с открытыми участками (рис. 79) .
Рис. 80. Экспозиция входных отвер-
стий нор, вырытых разноцветными
ящурками (стационарная площадка
на Ивано-Рыбальчанском участке Чер-
номорского заповедника, п = 150)
На длительность и характер суточной активности может влиять обеспеченность кор-
мом. Так, монофазный тип активности с пиком в первой половине дня, наблюдавшийся у
ящурок в Черноморском заповеднике в июле 1974 г., мы связываем с высокой числен-
ностью прямокрылых в это время, благодаря чему ящерицы могли удовлетворять свои
потребности в пище в утренние часы и не нуждались в продолжительной вечерней
охоте [232]. Исследованиями М.Ф.Тертышникова установлено, что размер инди-
видуальной территории (следовательно, и длительность активности) разноцветной
ящурки зависит от обилия корма [255, 259].
На суточную активность E.arguta некоторым об-
разом влияет экспозиция входа в нору [125]. Это осо-
бенно заметно на ящурках, обитающих в норах ему-
ранчиков (Scirtopoda telum) (наблюдали летом в Чер-
номорском заповеднике). Особи, живущие в норах с
выходом на север, северо-восток и восток, появляют-
ся утром, как правило, раньше других. У них же рань-
ше заканчивается период активности. Вечером дольше
всех остаются на поверхности ящурки, норы которых
имеют западную экспозицию входа и освещены до за-
хода солнца. Норы, вырытые самими ящурками, мо-
гут иметь вход, направленный в любую сторону, од-
нако предпочтение отдается южным, восточным и, от-
части, северным румбам (рис. 80). Еще большее вли-
яние на сроки начала и окончания активного периода
оказывают особенности рельефа — в частности, эк-
спозиция склона.
Подводя итог, можно заключить, что продолжи-
тельность и характер суточной активности разноцвет-
ной ящурки зависят от климатических и погодных
условий, сезона (И связанного с сезонностью физио-
логического состояния животных), характера биото-
па (рельефа, почв, растительности, кормовых усло-
вий и др.), возраста и индивидуальных особенностей
ящериц.
ГЛАВА 12
СЕЗОННЫЙ ЦИКЛ АКТИВНОСТИ1
Конкретные сведения о сроках начала, окончания и длительности активного периода раз-
ноцветной ящурки, условиях ее зимовки или наличии летней спячки имеются не менее
чем в 46 публикациях (не учитывая рукописей диссертаций, а также тех работ, что содер-
жат лишь данные литературы) . Ареал вида в этом плане изучен неравномерно. В частности,
из регионов, находящихся за пределами бывшего СССР, данные имеются лишь по Монго-
лии [Тэрбиш, сообщ.]. На территории Российской федерации хорошо изучены Ставрополь-
ский край [144, 258, 259, 262, 266, 270, 273], Калмыкия [22, 24, 25, 113, 115; Киреев,
сообщ.] и Дагестан [Хонякина, сообщ.], а данные по другим районам республики отры-
вочны и касаются Ростовской, Волгоградской и Саратовской области [70, 118,310], а так-
же Кабардино-Балкарии [304].
В Закавказье сезонная активность разноцветной ящурки наиболее подробно исследо-
валась в Азербайджане [7, 19, 20, 86] ,в меньшей степени — в Армении [77]; отсутствуют
данные по Грузии. Довольно много сведений по Казахстану,в том числе по южным, север-
ным, северо-западным и юго-восточным районам [35, 48, 192,199,313]. Хорошо изучены
Кыргызстан [96, 323,324] и юго-восток Узбекистана [35,92,107,198; Вашетко, сообщ.],
есть дашые по Северному Таджикистану [222, 224, 225] отсутствуют по Туркменистану.
Наконец, наибольшее количество работ со сведениями о сезонной активности разноцвет-
ной ящурки имеется по Украине [49,66, 121—125, 127, 130, 131,151,201,239,249,250,
316, 386; Рудик, сообщ.], в том числе по разным областям республики, вследствие чего
украинская часть ареала E^rguta изучена наиболее основательно. Данные литературы,
опубликованные до 70-х годов, обобщены в сводках ННЩербака [320,436].
12.1. НАЧАЛО И КОНЕЦ
СЕЗОННОЙ АКТИВНОСТИ ПОПУЛЯЦИЙ
Длительность активного периода рептилий определяется, прежде всего, климатом региона
и погодными условиями. Поэтому сроки начала и окончания сезонной активности законо-
мерно изменяются в зависимости от географической широты местности и высоты над
уровнем моря, демострируя при этом значительную изменчивость по годам. Ориентируясь
в основном на средние или часто встречающиеся сроки весеннего пробуждения разноцвет-
ной ящурки в том или ином регионе, получаем следующую картину. Наиболее рано, во
второй половине февраля — начале марта, ящурки появляются в Ферганской долине, в
Ташкентской обл. и Голодной степи, тх. на юго-востоке Узбекистана, юго-западе Кыргыз-
стана и крайнем юге Казахстана, несколько позже (в конце февраля — первой половине
марта) — в Чуйской долине Кыргызстана [35,107,198,199,323,324]. Во второй—третьей
декадах марта пробуждаются обычно ящурки в Таджикистане и Азербайджане [7, 19, 20,
86, 222, 224, 225], в Джизакской обл. Узбекистана [Вашетко, сообщ.] и на юге Калмыкии
[24, 25; Киреев, сообщ.], а в годы с ранней весной — также на юге Волгоградской обл.
[118].
Конец марта — начало апреля — обычное время появления ящурок на юго-востоке Ка-
захстана, в Калмыкии, Дагестане и Ставропольском крае [113, 115, 199,258, 259; Хонякина,
1 При участии МЛ Ахмедова, В Л Бадмаевой, Э .В .Вашетко, И.С.Даревского, В.А.Киреева, АМТудика,
СБ .Таращука, М .Ф.Тертышникова, Х.Тэрбиша, 3 Л Хонякиной.
151
сообщ.]. В первой декаде апреля обычно начинается активность ящурок в ряде районов
Казахстана (в Волго-Уральский песках, на севере и юго-востоке республики) , в Прииссык-
кулье,Кабардино-Балкарии,на юге Украины [48,96, 127,192,199,201,313;кол.ИЗ АНУ].
В середине апреля появляются ящурки в Джунгарской Гоби (Монголия) [Тэрбиш, со-
общ.] и Николаевской обл. Украины [250] , наиболее поздно они пробуждаются в Лесосте-
пи и на севере Степи Украины [49, 66, 151,201], а также в Армении [77] - после 15.IV,
чаще в третьей декаде месяца.
На зимовку первыми (обычно во второй половине сентября) уходят ящурки в Сред-
нем Приднепровье (Украина) и, возможно, в северных районах Казахстана [151, 199,
201], в конце сентября — первой декаде октября — в Кабардино-Балкарии, Армении, Вол-
гоградской обл.России,Харьковской, Днепропетровской и Николаевской областях Украи-
ны [49,77,118, 239, 250, 304; Рудик, сообщ.]. В первой половине октября исчезают ящур-
ки в Ставропольском крае, Волго-Уральских песках,на юго-востоке Казахстана [144, 192,
258, 259, 262], в конце октября, иногда в начале ноября — на юге Украины (Черномор-
ский заповедник и Крым) , в Ростовской обл. России, в Калмыкии, Дагестане, Прииссык-
кулье и Северном Таджикистане [24, 25, 70,96, 113, 122, 123, 127, 131, 222, 224, 225, 324;
Хонякина, сообщ.].
Дольше всего ящурки активны в Азербайджане — до начала — середины ноября [7,
19, 20], Чуйской долине (Кыргызстан) и в Ферганской долине и Голодной степи (юго-
восток Узбекистана, юго-запад Кыргызстана и самые южные районы Казахстана) ,где этих
ящериц встречали в течение первой половины ноября, вплоть до начала третьей декады
месяца [35, 107, 198, 199, 324; Вашетко, сообщ.]. Следует отметить, что процесс залега-
ния в спячку происходит постепенно, сроки ухода на зимовку разных возрастных групп
и популяции в целом значительно растянуты, поэтому выше рассматривались сроки исчез-
новения с поверхности последних активных особей (т.е. сеголеток) .
Работы, в которых сроки начала и окончания периода сезонной активности разноцвет-
ной ящурки малоправдоподобны для того или иного региона (т.е. отмечены явно не пер-
вые и не последние особи либо указано окончание активности лишь взрослых ящурок) ,
мы здесь не приводим. Сроки, указанные для Саратовской обл. России [310: активна с
конца марта — начала апреля до октября—ноября], не совсем понятны.
12.2. ОСОБЕННОСТИ ВЕСЕННЕГО ПРОБУЖДЕНИЯ
И ПОРЯДОК УХОДА НА ЗИМОВКУ ОТДЕЛЬНЫХ ГРУПП ЯЩУРОК
Сроки первого появления разноцветных ящурок для одного и того же региона могут
сильно отличаться в зависимости от погодных условий. Так, в Калмыкии в окр. пос. Хо-
мутниково в разные годы первые ящурки на поверхности отмечались с общим интервалом
в 50 дней [Киреев, сообщ.). Чаще всего разброс дат первого появления ящурок не столь
велик и обычно составляет 1—2 недели, реже - до 1 мес (конкретные данные см. [20, 25,
66, 127, 131, 151, 192, 199, 222, 259, 324]). В годы с ранней, но неустойчивой весной от-
дельные особи могут отмечаться задолго до тех сроков, которые являются собственно
началом активности популяции. В той же Калмыкии, где ящурки обычно активны с конца
марта — начала апреля, в 1966 г. ВАКиреев [113] первую особь на поверхности отметил
27Л. В Волгоградской обл. в год с неустойчивой весенней погодой (частые заморозки, хо-
лодные ветры) первая ящурка наблюдалась 10.III, а массовое появление этих рептилий от-
мечено лишь 20.IV [118]. Обычно же сроки первого и массового выхода ящурок из убе-
жищ разделены 1—3 неделями, что хорошо показано для Западного Казахстана [192] и
Калмыкии [25]. В тех регионах и в те годы, где и когда для весны характерен резкий и
устойчивый переход от низких температур к высоким (как, например, нередко бывает в
Черноморском заповеднике), первое появление ящурок может почти совпадать с мас-
совым [131].
По вопросу о последовательности весеннего пробуждения ящурок разного пола и воз-
раста единого мнения нет. Так, по данным НЛ1 .Окуловой [192], первые 7—10 дней (иног-
да дольше) на поверхности встречаются почти исключительно взрослые самцы, затем по-
являются самки, последними — молодые особи. ИД.Яковлева [324] также наблюдала
сначала выход самцов и лишь через 10—15 дней — самок. Напротив, 3 ЛХонякина [со-
152
общ.] и В J4 Бадмаева [22] отметили, что первыми покидают свои убежища молодые
ящурки, затем взрослые самцы и несколько позднее (или через неделю) - взрослые сам-
ки. На такую же закономерность указал О Л Богданов [39] для рептилий Средней Азии.
В то же время М.Ф.Тертышников определенной очередности появления особей разного по-
ла и возраста не выявил. В 1966 и 1968 гг. первыми были обнаружены взрослые самцы и
самки, в 1967 и 1969 гг. — взрослые самцы, в 1970 и 1971 гг. — молодые особи и взрослые
самцы [258, 259, 262]. Из этого сделан вывод, что сроки пробуждения зависят от глубины
залегания ящерицы во время спячки: чем глубже она находится в норе, тем позже прогре-
ется и выйдет на поверхность.
Наши наблюдения в Черноморском заповеднике [127] также не позволяют вычленить
какую-либо определенную очередность пробуждения ящурок. Вероятно, это обусловлено
тем, что в годы наших исследований весна начиналась бурно, и резкий подъем температу-
ры среды пробуждал к активности всех ящурок. Имеет значение, очевидно, и то, что раз-
ные особи одной половой или возрастной группы могут зимовать на различной глубине
(например, в глубоких норах емуранчика, менее глубоких норах мышевидных грызунов
и в совсем мелких, вырытых ящурками); поэтому весеннее появление ящурок не носило
здесь определенной закономерности. Напротив, в регионах с плотным субстратом, где зи-
мовальными убежищами ящуркам служат достаточно глубокие норы грызунов, очеред-
ность появления особей разного пола и возраста вполне возможна, особенно в годы с хо-
лодной весной. При этом первыми должны появляться взрослые самцы и молодые ящур-
ки (из-за повышенной половой активности первых и необходимости усиленно питаться и
расти — у вторых) , последними — взрослые самки.
В отличие от весеннего пробуждения на зимовку ящурки уходят по вполне определен-
ному порядку: первыми исчезают с поверхности взрослые особи, последними — сеголет-
ки [19, 20, 25, 39, 77, 113, 115,121—125,127,151,192,239,258,262,316; Тэрбиш,Хоня-
кина, сообщ.]. Среди взрослых первыми в спячку залегают взрослые самки [192], что хо-
рошо согласуется с физиологическим состоянием животных: самки после периода раз-
множения успевают накопить достаточное количество жировых запасов, у самцов же
осенью наступает новый цикл сперматогенеза, требующий энергетических затрат (подроб-
нее см. гл. 14) . По ОЛБогданову [39], в Средней Азии первыми на зимовку уходят
взрослые самцы рептилий, позже — самки. Зависимость сроков залегания в спячку ящериц
разных видов от степени упитанности хорошо показал И.СДаревский [79]. Он установил,
что скорость роста жировых тел и степень их развития перед уходом ящериц на зимовку
вырабатывается в процессе приспособления к конкретным условиям существования и спе-
цифичны для каждого вида. Ко времени залегания в спячку взрослых разноцветных ящу-
рок жировые тела сеголеток развиты еще ниже того минимума, который необходим ор-
ганизму для поддержания обмена веществ в зимнее время, и недостающее количество ре-
зервных веществ сеголетки накапливают за счет более продолжительной, чем у взрослых,
осенней активности.
В группе сеголеток уход на зимовку, как показали исследования в Черноморском за-
поведнике, также происходит в определенной очередности. В этом районе Украины взрос-
лые особи залегают в спячку с середины сентября до начала октября, сеголетки встречают-
ся до середины — конца октября, очень редко — и в начале ноября. Поскольку залегание
в спячку происходит по мере достижения ящурками определенной степени упитанности, а
величина жировых накоплений зависит у сеголеток, в первую очередь, от сроков рожде-
ния, устанавливается порядок ухода молодых особей на зимовку в зависимости от их
возраста. Сеголетки, родившиеся в июле — начале августа, к осени успевают подрасти и
накопить жир, что позволяет им залегать в спячку в первой половине октября. У поздно
родившихся сеголеток жировые тела в сентябре еще малы, поэтому такие ящурки остают-
ся активными до конца октября — начала ноября, что возможно вследствие их большей,
чем у взрослых,толерантности к низким температурам среды. Так, в 1975 г. на Ивано-
Рыбальчанском участке Черноморского заповедника наиболее крупные сеголетки залегли
в спячку в конце сентября — первых числах октября (т.е. одновременно с последними
взрослыми ящурками) , массовый уход на зимовку сеголеток средних размеров проходил
во второй декаде октября, а самые мелкие оставались активными до конца октября [122,
124, 127]. Сроки исчезновения с поверхности последних взрослых ящурок и последних
сеголеток могут отличаться на 1—4 недели (рис. 76).
153
123. ЗИМНЯЯ СПЯЧКА
Данных по условиям зимовки разноцветных ящурок немного. Ясно, что глубина залега-
ния рептилии зависит от климата местности и должна быть ниже уровня промерзания поч-
вы. Поэтому в районах с теплой зимой ящурки зимуют сравнительно неглубоко — на глу-
бине 29—45 см в Ставрополье [259, 262], 13—50 см — в Черноморском заповеднике (на-
ши наблюдения). В последнем районе глубже других зимовали ящурки, укрывшиеся в
норах емуранчика; близко к поверхности находились особи, зимовавшие в собственных
норах. Так, 14Х 1975 г., через 2 недели после исчезновения с поверхности последних
взрослых ящурок, в горизонтальном колене норы, вырытой ящуркой, находилась спя-
щая взрослая особь на глубине 13 см и в 15—20 см от входа в нору [125].
В холодные зимы ящурки, зимующие неглубоко, могут погибнуть (в первую очередь,
мелкие сеголетки в вырытых ими норах) . В Волго-Уральских песках осенью обнаружива-
лись норы ящурок, идущие почти вертикально вниз на глубину до 70 см. Поскольку на
глубине ВО см температура почвы зимой падала до —7°, предположили, что ход норы
углубляется ящуркой по мере охлаждения верхних слоев почвы, при этом осенью живот-
ное движется в глубь земли, а весной — к ее поверхности [192].
В Калмыкии при раскопке 200 нор малого суслика и 40 нор полуденной и тамариско-
вой песчанок зимующие разноцветные ящурки найдены в состоянии оцепенения на глубине
40—110 см, где температура воздуха колебалась от 4 до 9° . При этом в норах песчанок вы-
явлена всего одна ящурка, в 17 норах суслика — 27, в том числе в одной норе зимовало
4 особи, в двух — по 3 и в трех — по 2 ящурки [Киреев, сообщ.]. В Кыргызстане 2.П1
при раскопках обнаружены 3 ящурки (2 самца и самка) , зимовавшие в одной норе [323,
324]. В JB Дубинин зимующих ящурок находил в октябре и ноябре в глубине нежилых
нор песчанок, сусликов и черепах, где ящерицы лежали, зарывшись в мягкий лесс и свер-
нувшись обычно в кольцо [92].
Для успешной зимовки рептилиям необходим достаточный запас энергетических ве-
ществ. Определенное значение имеют жировые тела (см. разд. 14.2), часть резервных ве-
ществ откладывается в печени. Сезонную динамику массы печени'у самцов и самок разно-
цветной ящурки в двух пунктах Азербайджана изучал МИ Ахмедов [20], некоторые
из полученных им результатов представлены нами графически на рис. 87,Г. Однако опре-
деленные закономерности на материале из Азербайджана не прослеживаются — видимо,
вследствие того, что в разные месяцы вскрытию подвергались особи различных размеров
и возраста.
Роль печени как депо резервных питательных веществ, расходуемых во время зимней
спячки, отмечалась для других видов ящурок [171]. Как показали исследования С Л Льво-
вой [161], подготовка разноцветной ящурки к зимовке сопровождается накоплением
энергетических ресурсов: гликогена в печени, липидов в жировых телах, макроэргических
фосфатов в мозгу и мышцах. В начале спячки используются в основном запасы гликогена
в печени и мышцах, а при длительной гипотермии начинают активнее расходоваться резерв-
ные жиры (см. гл. 7).
12.4. ЛЕТНЯЯ СПЯЧКА
На большей части ареала летняя спячка у разноцветной ящурки не отмечалась. В частно-
сти, не наблюдалась она в Центральном Предкавказье, Западном Казахстане, Дагестане и на
юге Украины [127, 131, 192, 258, 259, 262, 386; Рудик, Хонякина, сообщ.], т.е. в тех ре-
гионах, где экология ящурки изучена наиболее подробно. Исследования в Черноморском
заповеднике, проведенные на стационарной площадке с применением мечения животных,
показали, что в августе — сентябре резко снижается активность взрослых ящурок (см.
разд. 11.4), что создает впечатление раннего ухода их на зимовку и может трактоваться
как наличие летней спячки у части взрослых особей [386] .МИАхмедов [19] указал, что
в Азербайджане у разноцветной ящурки на побережье (мыс Б ендов ан) в конце июля —
154
августе имеется 2-недельная пауза в активности, отсутствующая на рядом расположенном
о.Обливной,
По данным В .В Дубинина [92], на юго-Востоке Узбекистана взрослые особи на равни-
не перестали встречаться в середине мая, в предгорьях — в середине июня, а вновь они
появились лишь в сентябре. В Кыргызстане ИД Яковлева [324] наблюдала крайне низкую
активность взрослых ящурок в Чуйской долине в июле, в то время как в Прииссыккулье
эти ящерицы встречались все лето, что автор связывает с более низкой температурой воз-
духа в последнем районе. Наконец, СБ.Таращуком [249] ’’путем раскопок установлен
факт летней спячки у взрослых особей разноцветной ящурки” в окр. г Николаев (юг
Украины) . Он отметил [250], что взрослые ящерицы отсутствовали на поверхности со вто-
рой декады августа по первую декаду сентября 1985 г. в связи с жарой и засухой, длив-
шейся больше месяца. Удивление, однако, вызывает то, что сеголетки, появившиеся в этом
году лишь в середине августа (вследствие поздней затяжной весны и холодной первой по-
ловины лета) , позже не наблюдались и отмечались вновь только в первой декаде сентября
(см. [250]) , т^. вели себя аналогично взрослым особям.
Исследования, проведенные нами в Черноморском заповеднике [126, 127,131,386 и
др.], расположенном в 60 км от г Николаев и характеризующемся более жарким и сухим
климатом, показали, что даже в 1975 г., когда засуха здесь длилась с июня по октябрь,
а температ^уа воздуха достигала 27° уже в первых числах апреля и держалась на высоком
уровне (30 ) до начала октября, активность не прекращалась даже у взрослых ящурок.
Из-за необходимости усиленно питаться, чтобы расти и накапливать жировые запасы к зи-
мовке, летняя спячка у сеголеток лишена всякого смысла и даже представляет опасность
для жизни, так как при высоких температурах (т.е. интенсивном обмене веществ) отсут-
ствие пище в течение 15—20 дней может привести к гибели молодых ящериц. Неблагопри-
ятные высокие температуры среды сеголетки (см. разд. 102 и 11.4) успешно избегают,
сдвигая активность на утренние и вечерние часы, низкая же (порядка 25-30 %) относи-
тельная влажность воздуха (см. разд. 10.4.5) не является препятствием для активности
разноцветной ящурки.
Относительно летней спячки в целом можно сделать следующее заключение. Половой
цикл разноцветной ящурки обусловливает динамику накопления резервных питательных
веществ, что определяет целесообразность той или иной длительности активности ящериц.
Климатические и погодные условия позволяют реализоваться вполне определенной схеме
активности. В результате летняя спячка у E.arguta возможна, но лишь у взрослых
особей и в тех частях ареала (или в те годы), для которых, кроме жаркого лета,
характерны раннее весеннее пробуждение ящериц и ранние сроки окончания раз-
множения.
*
В заключение отметим следующее. Длительность сезонной активности разноцветной яшур-
ки определяется прежде всего климатическими условиями местности и погодными особен-
ностями года, из других факторов наибольшее значение имеют возраст и физиологиче-
ское состояние ящериц (в первую очередь, степень их упитанности) . Особенности биотопа
также влияют на сроки начала и окончания сезонной активности, что обусловлено специ-
фикой микроклимата и связано с особенностями рельефа (значение имеют, прежде всего,
экспозиция склона и защита от холодных ветров), характером растительности и почв
(определяют световой и тепловой режимы) , близостью крупных водоемов (в первую оче-
редь, моря) . Так, на о.Обливной (Азербайджан ) разноцветные ящурки пробуждались вес-
ной на 5—10 дней позже и позже уходили на зимовку,чем на ближайшем побережье мате-
рика [20]. Влияние характера почвы на сроки весеннего пробуждения ящериц рода Eremi-
as показал И.ФАндреев [10] : на песках массовое появление ящурок наблюдалось между
20 и 30JII, рядом на глинистых почвах лишь 16.IV появились первые одиноко ползающие
особи.
В целом длительность периода сезонной активности разноцветной ящурки (определяе-
155
мая как отрезок времени между первым появлением этих ящериц весной и исчезновением
последних активных особей осенью) заметно варьирует в пределах ареала (следствие
климатических условий) и по годам (результат влияния погоды) и равна 167—176 дням
на Днепропетровщине (Украина) [49], 169—210 — в Центральном Предкавказье [259,
262], около 210 - в Калмыкии [22, 24,113], 185 — 220 дней (обычно 200—210) — на юге
Украины [123], а на юго-востоке Узбекистана и в прилегающих районах Казахстана и
Кыргызстана, судя по данным ряда авторов [35, 107, 198, 324; Вашетко,сообщ.],может
достигать 8,5—9 мес (без учета возможной летней спячки). Необходимо подчеркнуть,
что ни одна группа ящериц в той или иной популяции не имеет столь длительных перио-
дов активности, какие указаны выше. В частности, взрослые ящурки прекращают актив-
ность обычно за 2—4 недели до окончания популяционной активности, сеголетки, уходя
на зимовку последними, в первый сезон жизни активны, в зависимости от региона и сро-
ков рождения (см. разд. 14.10), всего 1—6 мес.
ГЛАВА 13
ПИТАНИЕ’
Имеющиеся в литературе сведения о питании разноцветной ящурки в целом немногочис-
ленны и приведены для большей части ареала: Крыма [316], Левобережной Украины [66,
131], Западного [44], Центрального [255—257, 259] и Восточного [278] Предкавказья,
Калмыкии [25,113], Волгоградской обл. [118], Казахстана и Средней Азии [35,94, 298,
313,324] Азербайджана [7] и Восточной Грузии [176].
В перечисленных работах с разной степенью -полноты приведены сведения о составе
пищи, иногда о сезонных и биотопических изменениях рациона. Еще реже встречаются ма-
териалы, касающиеся массы потребляемой в сутки пищи, интенсивности сезонного пита-
ния, длительности переваривания пищи, ее калорийности, избирательности питания и др.
За редким исключением [131, 259], состав пищи их рассматривается вне связи с числен-
ностью ящериц и беспозвоночных в биоценозах.
Мы обобщаем имеющиеся в литературе сведения об особенностях питания ящурки
разноцветной, дополнив их материалами, полученными в последние годы.
13.1. ОБЩАЯ ХАРАКТЕРИСТИКА ПИТАНИЯ
Как показывают существующие материалы, в содержимом кишечно-желудочного тракта
ящурки разноцветной встречаются разнообразные беспозвоночные животные. Практиче-
ски во всех частях ареала основой рациона этого вида следует считать жесткокрылых, пе-
репончатокрылых (муравьев), чешуекрылых (гусениц), двукрылых, кожистокрылых,
прямокрылых, клопов. Реже ящурка питается другими группами насекомых, а также мол-
люсками, ракообразными (мокрицы), паукообразными и др. Отмечены, хотя и крайне
редко, случаи поедания позвоночных животных (канибализм) .
В пищеварительных трактах ящурок в разных частях ареала обнаружены также части-
цы растительности. Так, у ящурок из Левобережной Украины найдены остатки листьев и
стеблей кохии, молочая, полыни, мха, а также семена крестоцветных [131], в Западном
Казахстане — молочая, солянок, изеня, цветов цмина и семена злаков [Окулова,сообщ.],
в Восточном Дагестане — листьев полыни песчаной и семена Польши песчаной и колосни-
ка гигантского, ягоды шелковицы [Хонякина, сообщ.]. Количество желудков ящурок с
остатками растительности неодинаково в разных частях ареала. В частности, в пределах
Предкавказья от 0,5 % в центральной части [Горовая, сооб.] до 17,5 % на западе (Кизил-
ташская коса) [Зинякова, сообщ.].
Эти обстоятельства позволили Г.Ф.Сухову [246] даже связывать биотопическое рас-
пределение ящурок с наличием или отсутствием молочая как объекта питания этого вида.
Однако многие исследователи [131, 255, .256, 320] считают, что растительность попала в
желудок ящериц случайно вместе с отлавливаемыми беспозвоночными и не служит объек-
том питания.
В желудке и кишечнике ящурок находят и различные неорганические остатки, кото-
рые, возможно, играют роль гастролитов [142,257,259]. ТЛКотенко [131] находила не-
органические остатки в 84,6 % исследованных ящурок. Это были песчинки, мелкие ка-
мешки, обломки раковин морских моллюсков, кусочки глины, древесного и каменного
1 Материалы предоставили В Я Бадмаева, ЗКБрушко, МЛ.Зинякова, Н.НИванова, ВАКиреев,
Т И Котенко, ЕЮКудакина, ВБ.Неручев, Н-М.Окулова’ В.Ф .Орлова, AM .Рудик, Х.Тэрбиш, ЗЛХо-
някина.
157
угля. По ее данным, неорганические частицы в изобилии встречаются в желудках вместе с
муравьями, личинками муравьиного льва, чернотелками и жужелицами. По мнению Т Л Ко-
тенко [131,386], неорганические остатки не являются гастролитами и захватываются яще-
рицами случайно при отлове беспозвоночных.
13.2. СООТНОШЕНИЕ КОМПОНЕНТОВ В ПИЩЕ ЯШУРОК
В РАЗНЫХ ЧАСТЯХ АРЕАЛА
Имеющиеся материалы показывают, что в рацион ящурки входят самые разнооб-
разные объекты питания. Причем набор животных кормов зависит от географи-
ческой точки. Сравнение рациона ящурок из разных частей ареала показывает сле-
дующее.
В Крыму по числу съеденных экземпляров основную роль в питании играют перепон-
чатокрылые (31,07 %), жесткокрылые (28,27 %), двукрылые (11,92 %), чешуекрылые
(8,64 %) , паукообразные (5,14 %) , полужесткокрылые (4,91 %) и прямокрылые (3,50 %)
[316].
В Черноморском заповеднике [131] в желудках ящурок были обнаружены перепон-
чатокрылые (27,86 %) , жесткокрылые (17,39 %) , равнокрылые хоботные (16,41 %) , че-
шуекрылые (9,66 %) , прямокрылые (9,15 %) , полужесткокрылые (6,27 %) , паукообраз-
ные (6,20 %), двукрылые (3,66 %) и др.
В Запорожской обл. [49] основу рациона ящерицы составляют жесткокрылые
(43,18 %), чешуекрылые (13,92 %) , перепончатокрылые и двукрылые (каждые по 10,13%) ,
паукообразные (8,86 %).
В Западном Предкавказье (Кизилташский лиман) [Зинякова, сообщ.] в пище ящериц
обычны двукрылые (56,15 %) , паукообразные (15,39 %), жесткокрылые (13,26 %) , пе-
репончатокрылые (6,86 %) и чешуекрылые (4,91 %) .
В Центральном Предкавказье [255, 256] основу рациона ящурок составляют жестко-
крылые (43,5 %), перепончатокрылые (26,2 %) , двукрылые (7,3 %) , мокрицы (55 %) ,
многоножки (5,3 %), чешуекрылые (3,8 %), прямокрылые (3,6 %) и паукообразные
В Восточном Предкавказье [Хонякина, Костенко, сообщ.] в пище ящурок преобла-
дают перепончатокрылые (24,47 %), чешуекрылые (15,4 %), равнокрылые хоботные
(13,85 %), двукрылые (8,75 %) , паукообразные (5,76 %) , жесткокрылые (3,95 %) и пря-
мокрылые (3 ,53 %).
В Калмыкии [25] в питании ящериц преобладают жесткокрылые (46,73 %) , перепон-
чатокрылые (15,15 %), чешуекрылые (9,67 %), двукрылые (7,78 %), прямокрылые
(7,15 %) , паукообразные (6,31 %) и равнокрылые хоботные (4,84 %) .
В Западном Казахстане ящурки поедают преимущественно жесткокрылых (43,22 %) ,
чешуекрылых (21,23 %) , прямокрылых (10,23 %) , паукообразных (7,03 %) и перепонча-
токрылых (6,27 %) [Окулова, сообщ.].
На юго-востоке Туркмении [298] в пище ящериц обнаружены преимущественно жест-
кокрылые (51,2 %) , термиты (32,5 %) , перепончатокрылые (14,2 %) и мокрицы (2,3 %) .
В Азербайджане [7] в пище ящурок преобладают жесткокрылые (78,6 %), перепон-
чатокрылые (10,7 %) и прямокрылые (4,2 %) .
По Узбекистану имеются следующие данные [Брушко, сообщ. ]. В Ферганской доли-
не (июль, 1971 г.) в желудках ящурок найдены преимущественно саранчевые (40,4 %) ,
перепончатокрылые (30,4 %), жесткокрылые (15,2 %) и клопы (4,6 %), чешуекрылые
(4,0 %), в Джизакской обл. — перепончатокрылые (60,1 %) и жесткокрылые (30,5 %)
(март — сентябрь, 1968—1969 гт.).
В Западной Монголии [Орлова, Тэрбиш, сообщ.] ящурки питаются главным обра-
зом жесткокрылыми (47,76 %), чешуекрылыми (15,42%), перепончатокрылыми
(13,43 %) и паукообразными (9,45 %) .
Таким образом, основу пищевого рациона ящурки разноцветной в пределах ареала со-
ставляют прежде всего жесткокрылые и перепончатокрылые, далее в порядке уменьше-
ния располагаются чешуекрылые, двукрылые, прямокрылые, паукообразные, равнокры-
лые хоботные, ракообразные (мокрицы) и др. (табл. 17).
158
6SI
Примечание. (+)- наличие компонента в желудках ящурок.
Oligochaeta - — _ _ _ - 0,02 _ _ _ _ _ Mollusc^ Gastropoda — 0,07 0,27 - 1,02 - 0,43 0,21 _ _ _ _ Crustacea 1,87 — — 1,09 5,25 — -1,65 — 1,8 1,02 — 1,11 Arachnoidea 5,14 6,20 11,16 15,39 1,78 4,16 5,62 6,52 1,2 7,03 0,58 1,66 Myriapoda - - 0,27 - 5,08 - - 0,21 - - 1,17 0,28 Insecta Podura - 0,03 - - Ephemeroptera - 0,07 - - - - ~ ~ ~ ~ ” Odonatoptera 1,17 0,03 - 0,09 - - 0,39 7,14 - - - 22,61 Blattoptera 0,23 - - 0,12 _ _ - - 3,0 ,9 Mantoptera 0,23 0,24 - 0,03 - - 3,45 - - - 1,36 Orthoptera 3,50 9,15 3,36 0,12 3,56 - - 4.2 10.23 °>19 Dermaptera 1,17 — — 0,25 0,68 — 0,47 0,42 — — Psocoptera — 0,07 — — — — — — — — ~ Homoptera 0,93 16,41 5,91 0,71 0,17 - 13,51 4,84 - 1,79 - - Hemiptera 4,91 6,27 5,91 0,06 1,27 - 1,27 - 1,8 1,02 0,58 3,05 ColeoptCTa”3 28,27 17^39 46,91 13,26 42,76 50,00 25,91 46,73 78,6 43,22 31,19 23,82 Neurontera 0 47 0 75 2.42 0,59 - - 0,25 1,05 - - ~ 0,11 Lepidoptera 8*64 9*66 6,05 4,91 4,40 15,00 9,67 1,8 21,23 9,36 3,05 Hymenoptera 31,07 27,86 16,13 6,86 24,72 41,7 23,84 15,15 10,7 6,27 50,88 43 77 Dlptera 11,9 2 3,66 6,05 56,13 7,37 4,66 8,5 2 7,78 1,8 1,79 - 0,55 Vertebrata, Reptilia _ _ _ 0 39 Eremias arguta _ _ _ - 0,08 0,02 _ Частицы растительности + + - + 0,51 + - » Компоненты
Крым (316] (л = 75)
Песчаная степь Черно- морского заповедника [131] (л =227)
Харьковская область (Рудик, сообщ.] (л =69)
Западное Предкавказье [ Зинякова, Руденко, сообщ.] (л =63)
Центральное Предкав- казье [259] (л = 172)
Север Восточного Пред- кавказья [ 170] (л =7)
Юг Восточного Предкав- казья [ Хонякина, Костен- ко, сообщ.] (л = 719)
Калмыкия [25] (л =338)
Азербайджан [ 7] (л=21)
Уральская область, Ка- захстан [Окулова, со- общ.] (л =461)
Волго-Уральское между- речье [ Неручев, Кудаки- на, Иванова, сообщ.] (л = 100)
Ферганская, Самарканд- ская и Джизакская об- ласти [ Вашетко, сообщ.] (и =71)
Таблица 17. Соотношение основных компонентов питания (% съеденных объектов) разноцветной ящурки в разных частях ареа.
133. БИОТОПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ КОРМОВОГО РАЦИОНА
Ящерицы поедают беспозвоночных, обитающих с ними в общих стациях, поэтому их ра-
цион зависит от качественного и количественного состава биоценотических комплексов
беспозвоночных и доступности отдельных видов.
В рационе ящурок со слабо закрепленных песков Каспия чаще встречались жесткокры-
лые, двукрылые и перепончатокрылые, у ящурок с солонцеватых и суглинистых пустынь-
моллюски, ракообразные (мокрицы) и клопы [Хонякина, сообщ.]. В окультуренных
ландшафтах в составе кормов появляются тараканы, что свидетельствует о расширении на-
бора доступных кормов в мозаике фации антропогенного ландшафта [Неручев,Кудакина,
Иванова, сообщ.].
Таблица 18. Пищевой рацион разноцветной ящурки в разных биотопах Центрального Предкав-
казья [ 257]
Беспозвоночные Опустыненная Степь Разнотравно-зла- ковая Степь Лесная полоса Посевы
Количество беспозвоночных, %
учтен- съеден- ных ных учтен- съеден- ных ных учтен- съеден- ных ных учтен- съеден- ных ных
Mollusca. Gastropoda Pulmonata 1,42 0,80 1,26 1,71 Anhropoda. Arachnoidea Araneae 2,84 1,60 2,52 5,13 - 7,70 Insecta (всего) 93,72 96,80 95,76 91,77 100,0 93,30 100,00 100,00 Orthoptera (всего) 4,26 2,40 3,78 9,12 8,80 7,70 Grillodea - - 0,84 0,57 - Tettigoniodea 4,26 2,40 2,94 8,55 8,80 7,70 Dermaptera 1,42 2,40 ______ Homoptera — — 0,42 0,57 - - — - Hemiptera, Penta tomidae, 2,84 1,60 2,94 1,14 - - - - Coleoptera (всего) 35,50 40,00 6,72 29,07 22,30 38,50 23,56 40,00 Carabidae 2,84 1,60 0,84 7,41 6,90 7,70 8,82 10,00 Coccinellidae 1,42 3,20 - — - — - - Chrysomelidae 9,94 24,00 1,26 3,99 _ _ - - Scarabaeidae 7,10 4,80 0,84 2,85 3,40 7,70 2,94 10,00 AnisopHa 1,42 1,60 - - 1,70 7,70 2,94 10,00 Aphodius, Geotrupes 5,68 3,20 0,84 2,28 - - - - Coleoptera sp. 14,20 6,40 3,78 14,82 12,00 23,10 20,58 30,00 Diptera (всего) 12,78 17,60 61,32 6,27 31,00 23,10 32,34 40,00 Muscidae 4,26 2,40 61,32 6,27 17,20 23,10 20,58 30,00 Diptera, sp. 8,52 15,20 0,88 0,39 13,80 - 1.1,76 10,00 Hymenoptera (всего) 31,24 27,20 14,28 31,35 15,50 16,30 35,28 20,00 Formicidae 21,30 26,40 14,28 28,50 15,50 16,30 35,28 20,00 Hymenoptera, sp. 9,94 1,60 2,52 2,85 - - - - Lepidoptera (imago, larv.) 5,68 5,60 3,36 14,25 22,40 7,70 8,82 —
В глинистой пустыне Западного Казахстана ящурка чаще поедает саранчевых (24,2 %)
и паукообразных (14,6 %) , в мелкобугристых песках — муравьев (9,5 %) и равнокры-
лых хоботных (4,7 %) [Окулова, сообщ.].
Анализ состава пищи ящурок, обитающих в разных биотопах Центрального Предкав-
казья, показывает, что они поедают наиболее часто встречающихся там беспозвоночных
[255, 256, 320]. В опустыненной степи — это жесткокрылые, перепончатокрылые, дву-
крылые и чешуекрылые, в разнотравно-злаковой степи — жесткокрылые, перепончатокры-
лые, чешуекрылые, прямокрылые, двукрылые и паукообразные, в полезащитных лесона-
саждениях — жесткокрылые, двукрылые и перепончатокрылые, на посевах зерновых —
жесткокрылые, двукрылые и перепончатокрылые (табл. 18) .
Очень показательны также материалы, полученные Т.ИКотенко [131] на юге Украи-
ны. Так, на о. Тендра ящурки охотно поедали бокоплавов, выбегая в прибойную зону.
В желудках 33,3 % особей обнаружено 2—17 экз. бокоплавов.
160
13.4. СЕЗОННАЯ ИЗМЕНЧИВОСТЬ КОРМОВОГО РАЦИОНА
В связи с изменением численности беспозвоночных в биоценозах по сезонам изменяется
и состав кормов ящурки разноцвеАной в разные месяцы активности. К такому мнению
пришли многие исследователи, изучая питание ящурки в разных частях видового ареала.
Так, аг ал из рациона ящурки в различные месяцы активности показал, что на дагестан-
ском побережье Каспия в пище ящурок в мае чаще встречались жесткокрылые (63 $ %),
двукрылые (443 %) и чешуе- крылые (32,8 %), в июне — че- шуекрылые (59,2 %), жестко- крылые (493 %) и перепончато- крылые (32,1 %), в июле — че- шуекрылые (57,7 , жестко- Таблица 19. Пищевой рацион (% съеденных объектов) разноцветной ящурки в разные сезоны активности [755]
Компоненты Весна Лето Осень
Апрель — май Июнь - август Сентябрь - октябрь
крылые (46,1), прямокрылые Mollusca. Gastropoda
(263 %) и перепончатокрылые Pulmonata — 1,10 2,70
(263 %) , в августе — сентяб- Arthropods. Crustacea 21,60 8,10
ре — чешуекрылые (63,6 %), Isopoda Arachnoidea, Araneae 1,30 5,45 1,60
перепончатокрылые (39,4 %) , па- Myriapods, Dlplopods,
укообразные (303 %) и жестко- Juhformia — 6,10 —
крылые (303 %) [Хонякина, Insects (всего) 97,93 83,05 67,65
сообщ.]. Orthoptera — 4,25 —
В Западном Казахстане зна- Dermaptera Homoptera 1,95 0,50 о,ю- 2,70
чительную роль в питании имели Hemiptera, Penta-
в мае саранчевые (61,5 %) и че- tomidae — 1,50 —
шуекрылые (523 %) 9 в июне — Coleoptera (всего) 50,66 42,25 2435
июле — муравьи (17,0 %) и па- укообразные (14,0%), осенью Carabidae Cocdnellidae Chrysomelidse 18,82 0,65 3,90 0,80 4,75 3,15 5,40
вновь увеличивается встречае- Scarabaeidae 14,95 23,35 —
мость в пище ящурок жестко- Coleoptera, sp. 12,34 10,20 18,95
крылых и чешуекрылых [Оку- лова, сообщ.] . В Калмыкии весной в пище Diptera (всего) Muscidae Diptera, sp. Hymenoptera (всего) 2,60 1,30 1,30 37,65 8,10 5,65 2,45 22,40 8,10 8,10 32,45
ящурок преобладали жесткокры- Formlcidae 36,35 21,40 32,45
лые (36,2 %), перепончатокры- Hymenoptera, sp. 1,30 1,00 —
лые (16,6 %), двукрылые и че- шуекрылые (соответственно по Lepidoptera (imago, L) Vertebrata, Reptilia, Squamata, Lacertilia 5,20 4,45 0,10 —
10,4 %) , в июне — жесткокрылые Частицы раститель-
(343 %) 9 перепончатокрылые ности 0,65 озо —
(25,6 %), двукрылые (18,8 %), Неорганические остатки — 1,60 —
чешуекрылые (11,6 %) и прямо-
крылые (9,3 %) , в июле — жесткокрылые (28,0 %) , прямокрылые и паукообразные (по
19,2 %),чешуекрылые (10,1 %) ’[23].
В Центральном Предкавказье [255, 259] весной в пище ящурок преобладают жестко-
крылые (58,1 %), перепончатокрылые (31,6 %) , летом — жесткокрылые (42,2 %) , пере-
пончатокрылые (23,0 %), двукрылые (8,1 %), осенью — перепончатокрылые (32,5 %),
жесткокрылые (243 %) , мокрицы (21,6 %) и паукообразные (8,1 %) . Кроме того, весной
и летом в содержимом желудков находили частицы растительности (0,65,0,50 %) , а летом
и осенью — наземных моллюсков и мокриц (соответственно 1,10—2,70 и 5,45—21,60 %
и др.) (табл. 19).
Таким образом, в весеннем рационе ящерицы основное значение имеют жесткокры-
лые, чешуекрылые (гусеницы), перепончатокрылые и двукрылые, основу летнего рациона
составляют жесткокрылые, перепончатокрылые, чешуекрылые и прямокрылые, в осен-
нем — перепончатокрылые, жесткокрылые, паукообразные. Видимо, закономерно умень-
шение к осени поедания жесткокрылых и увеличение встречаемости перепончатокрылых и
паукообразных.
161
13.5. ИЗМЕНЕНИЕ КОРМОВОГО РАЦИОНА ПО ГОДАМ
Количественный и качественный состав беспозвоночных в биоценозах изменяется доволь-
но быстро. Смена отдельных фаз развития у некоторых насекомых делает их недоступны-
ми для охотящихся за ними ящерицами. Некоторые насекомые мигрируют с одного вида
растений на другой, из одного яруса в другой и др. Под воздействием биотических и абио-
тических факторов среды происходят более глубокие изменения фауны беспозвоночных,
когда некоторые семейства и виды могут исчезнуть в биоценозах, следовательно, выпасть
из рациона ящериц на сезон, год и более [100] . Качественное и количественное изменение
фауны беспозвоночных по годам в стациях ящериц сказывается и на их рационе. Это под-
тверждается в исследованиях ряда герпетологов. Так, наблюдениями ЗТГХонякиной в те-
чение 4 лет у ст. Ал мало и г Махачкалы за питанием ящурок установлено следующее.
У Махачкалы в пище ящурок преобладали по встречаемости в 1957 г. двукрылые (85,05%) .
в 1958 и 1963 гг — жесткокрылые (соответственно 192 % и 68,1 %) ,в 1972 г. — перепон-
чатокрылые (86^2 %), у ст. Алмало в 1958 г. — перепончатокрылые (823 %) , в 1960 и
1962 гг. — жесткокрылые (соответственно 82,2 и 81,2 %), в 1961 г. — жесткокрылые и
пауки (каждые по 70.6 %).
В Центральном Предкавказье [255, 259] также отмечены изменения рациона ящурок
в течение ряда лет (табл. 20). Так, в 1966 г. основу рациона составляли жесткокрылые
(44,5 %), перепончатокрылые (25,77 %), кивсяки и мокрицы (каждые по 4,12 %) , в
1967 г. — прямокрылые (3330 %) , жесткокрылые (7,40 %), перепончатокрылые (3330%)
и чешуекрылые (7,40 %), в 1968 г. — жесткокрылые (44,20 %), перепончатокрылые
(22,73 %), многоножки и мокрицы (каждые по 6,80 %) , наконец в 1969 г. — жесткокры-
лые и перепончатокрылые (каждые по 30,70 %) и чешуекрылые (13,90 %) .
Таблица 20. Пищевой рацион (% съеденных объектов) разноцветной ящур-
ки в разные годы [ 255,259]
Компоненты 1966 г. 1967 г. 1968 г. 1969 г.
Mollusca, Gastropoda
Pulmonata 1,05 — 1,00 1,55
Arthropoda, Crustacea
Isopoda 4,12 — 6,80 1,55
Arachnoidea, Araneae 2,10 — 1,70 1,55
Myriapoda, Diplopoda,
Juliformia 4,12 — 6,60 —
Insecta
Orthoptera 3,30 33,30 2,60 1,55
Homoptera 0,21 — — 1,55
Dermaptera 1,05 — 0,50 —
Hemiptera, Pentatomidae 2,10 — озо —
Coleoptera 44,55 7,40 44,20 30,70
Carabidae 3,50 7,40 2,80 1,55
Coccinellidae 4,12 — 4,50 1,55
Chrysomelidae 2,40 — 4,70 1,55
Scarabaeidae 21,63 — 23,90 6,12
Coleoptera sp. 13,10 — 8,30 19.90
Diptera 6,22 18,60 1,50 3,10
Muscidae 4,12 — 6,30 4,60
Diptera, sp. 2,10 18,60 1,50 3,10
Hymenoptera 25,77 33,00 22,73 30,70
Formicidae 24,72 33,30 21,93 27,60
Hymenoptera, sp. 1,05 — 0,80 3,10
Lepidoptera (imago, larv.) 3,74 7,40 3,80 13,90
Venebrata, Squamata, Lacer-
tilia — — 0,17 —
Частицы растительности 0,62 — озо —
Неорганические остатки 1,05 — озо 9,25
162
13.6. ПОЛОВЫЕ И ВОЗРАСТНЫЕ ОСОБЕННОСТИ
В ПИТАНИИ ЯЩУРОК
Исследования многих герпетологов позволяют говорить о некоторых особенностях пита-
ния ящурок разных возрастных и половых групп. В частности, в пище молодых ящурок
преобладают мелкие объекты, взрослых — более крупные. Кстати, подобная закономер-
ность была установлена и для других ящериц. Имеющиеся на сегодня материалы позволя-
ют оценить половые и возрастные особенности питания разноцветной ящурки в разных час-
тях ареала.
Таблица 21. Половозрастные особенности питания разноцветной ящурки
в Западном Казахстане в конце мая - начале июня 1958-1960 гг. (п = 235)
[ Окулова, сообщ.]
Количе- ство желудков Встречаемость корма в желудках, %
Жуки Саранче- вые Гусе- ницы Муравьи Пауки Остатки раститель- ности
Самки
109 63,5 14,7 38,5 Самцы 12,9 9,2 4,5
126 53,1 18,2 35,0 Взрослые особи 9,5 11,5 3,2
189 59,5 15,9 39,6 Молодые особи 10,6 10,1 3,9
46 52,0 19,6 26,1 13,1 10,9 4,4
Так, НМ.Окулова [сообщ.], изучавшая особенности питания ящурок разных возраст-
ных и половых групп, установила, что в пище взрослых особей чаще встречаются жуки и
гусеницы и реже саранчовые и муравьи, чем у молодых. Кроме того, пиша самцов и бере-
менных самок состоит из одних и тех же компонентов, но их количественное соотношение
неодинаково (табл. 21).
В Дагестане 3 ЛХонякина [сообщ.] также находит ряд отличий в составе пищи взрос-
лых и молодых особей. При сходстве качественного состава пищи сравниваемых животных
она находит отличия в том, что молодые не поедают прямокрылых, редко употребляют
в пищу жесткокрылых и клопов, но в большем количестве поедают пауков, двукрылых
и равнокрылых хоботных. И кроме того, беременные самки по сравнению с самцами очень
редко поедают быстро перемещающихся прямокрылых, предпочитая животных с нежными
хитиновыми покровами. Однако отло-
жившие яйца самки поедают те же
группы беспозвоночных, что и самцы.
В Центральном Предкавказье [255,
259] рацион взрослых самцов и са-
мок представлен однородными по ка-
честву и количеству объектами добы-
чи (рис. 81). Мелкие и малоподвиж-
Рис. 81, Соотношение основных компонентов пиши раз-
ноцветных ящурок разного пола и возраста [259]:
а — взрослые самцы, б - взрослые самки, в - молодые
самцы и самки; 1 — жесткокрылые, 2 — кивсяки,5 -
паукообразные, 4 — двукрылые, 5- прочие объекты,
6 - мокрицы, 7 — чешуекрылые (имаго и гусеницы) ,
8 — прямокрылые, 9 - перепончатокрылые
ные молодые самцы и самки поедают
значительно меньше жесткокрылых,
но больше мелких двукрылых и пере-
пончатокрылых, чем взрослые.
В этом плане интересны материа-
лы Т И Котенко [31] из Южной
Украины о возрастных и половых от-
личиях питания разноцветной ящурки. Автор показал, что весной в рационе взрослых
ящурок преобладают жесткокрылые, перепончатокрылые и равнокрылые (цикадки), летом
они в большом количестве поедают прямокрылых, осенью — жесткокрылых и двукрылых
(рис. 82, а—е), В питании сеголетков с июля по октябрь возрастает доля равнокрылых и
жесткокрылых (рис. 82, ж—к) . Сеголетки поедают более мелких и менее хитинизирован-
ных беспозвоночных — равнокрылых, перепончатокрылых (муравьи) и чешуекрылых
(гусеницы) . В это же время в пище взрослых ящурок преобладают прямокрылые и жест-
163
Рис. 82. Соотношение основных компонентов пищи раз-
ноцветной яшурки на Ивано-Рыбальчанском участке
Черноморского заповедника [131] :
а—г — взрослые и полувзрослые особи, 1975 г.; ж—к -
сеголетки, 1974-1977 гг.: а - апрель; б - май; в -
июнь; г, ж — июль; д,з- август; е, и — сентябрь; к -
октябрь. Здесь и на рис. 83, 84: 1 - пауки, 2-прямо-
крылые, 3 - равнокрылые, 4 - полужесткокрылые,
5 - жесткокрылые, 6 - чешуекрылые, 7 - перепончато-
крылые, 8 - двукрылые, 9 - прочие
кокрылые (рис. 83). Также различно питание взрослых самок и самцов (рис. 84) : самки
чаще поедают жесткокрылых и перепончатокрылых, самцы — прямокрылых и двукры-
лых. Беременные самки (с яйцами в яйцеводах) поедают более мелких и менее подвиж-
ных беспозвоночных (муравьи), что сближает их по характеру питания с сеголетками.
Таким образом, приведенные материалы подтверждают ранее высказанное мнение
О Л Богданова [39] о том, что общая закономерность для ящериц — преобладание в пище
молодых мелких объектов. Одинаковый (или сходный) качественный состав пищи взрос-
лых и молодых ящериц свидетельствует о быстрой выработке реакции на внешний вид
объекта добычи,которую ВД.Черномордиков [296] отмечал, например, у прыткой ящерицы
уже в конце первой недели жизни. Справедливо и то, что беременность снижает подвижность
самок, что приводит к увеличению в ее рационе малоподвижных форм беспозвоночных.
13.7. ИЗБИРАТЕЛЬНАЯ СПОСОБНОСТЬ К ОПРЕДЕЛЕННЫМ КОРМАМ
Список кормовых объектов разноцветной ящурки в пределах ареала крайне обширен.
Среди животных, поедаемых ящуркой, встречаются виды с разнообразной окраской тела,
кусающиеся, жалящие, ядовитые, выделяющие неприятный резкий запах и др.
164
Судя по набору бес-
позвоночных, потребляемых
ящур кой, она добывает
корм на поверхности поч-
вы, низко над ней. Как пра-
вило, это малоподвижные
формы (муравьи, гусени-
цы, некоторые жесткокры-
лые и др.) , которые встре-
чаются в содержимом же-
лудков ящурок чаше, чем
быстро передвигающиеся
(прямокрылые, осы, пче-
лы, мухи и др.). Нередко
в пище ящурок встречают-
ся куколки, личинки жест-
Рис. 83. Соотношение основных компонентов пиши разноцветной
ящурки на Ивано-Рыбальчанском участке Черноморского заповед-
ника в июле - сентябре 1974-1977 гг. [ 131]:
а - взрослые и полувзрослые особи» б - сеголетки
кокрылых и некоторые двукрылые, которых они, каК считает НМ.Окулова [сообщ.].
могли отловить в норах тушканчика, суслика. Есть наблюдения о том, что ящурка может
выкапывать беспозвоночных из песка [Хонякина, сообщ.], иногда в поисках корма взби-
Рис. 84. Соотношение основных компонентов пищи разноцветной ящурки на Ивано-Рыбальчан-
ском участке Черноморского заповедника в июне - июле 1974-1978 гг. [ 131 ]
а — самки с яйцами в яйцеводах, б — прочие самки, в - самцы
рается на растения [Горовая, Хонякина, сообщ.]. На такыровидных почвах ящурка отыс-
кивает корм под приподнявшимися верхними слоями почвы, проделывая порой путь под
коркой земли на расстояние до 30 м [Брушко, сообщ.]. Любопытны в этом отношении
материалы табл. 22, из которых следует, что основное количество корма собирается на
растениях и поверхности почвы.
Как показывают наблюдения [Орлова, Тэрбиш,
сообщ.], длина съеденных объектов колеблется от 3,5
до 21,3 мм, основная часть добычи (83,9 %) имеет дли-
ну 4.5—12,5 мм. Добыча размером 3,5—4.5 мм состав-
ляет 9,5 %, более 12,5 мм — 6,5 % количества съеден-
ных объектов. Сильно хитинизированные животные в
нишевом рационе составляют 39,4 %, слабохитинизи-
рованные — 37,95, мягкие — 22,6 %.
Говоря об избирательности питания, следует от-
метить разные точки зрения по этому вопросу.
НМ.Окулова [сообщ.] считает, например, что в глини-
стой полупустыне Западного Казахстана ящурки пред-
почитают поедать гусениц и саранчовых, несмотря на
большую численность в биоценозах мокриц, муравьев
и жуков. М.Ф.Тертышников [257, 259]., изучая пита-
ние ящурок в Центральном Предкавказье, установил,
что многие объекты добычи встречаются в содержи-
мом желудков пропорционально их обилию в биоце-
Таблица 22. Ярусная приурочен-
ность пищевых объектов разноцвет-
ной ящурки [Рудик, Ширяева, со-
общ.]
Т
Местонахожде- ние пищевого объекта Количество съеденных объ- ектов
абсо- лютное %
На растении 225 55,8
На поверхности почвы, реже на растении 116 28,8
На поверхно- сти почвы 46 11,4
В верхнем слое почвы 16 4,0
165
нозах: при увеличении численности беспозвоночных в биоценозах заметно возрастает их
поедание ящерицами, при уменьшении поедание их также уменьшается (табл. 18).
Видимо, избирательная способность к объектам пищи все же существует, но она под-
вержена большим изменениям и зависит от набора пищевых компонентов. Пока доказа-
тельно можно говорить об избирательности в.питании лишь по отношению к размерам,
подвижности и степени хитинизации жертв [131,386] . Основными факторами, определяю-
щими состав пищевого рациона ящурок, являются количественный и качественный состав
беспозвоночных в биоценозах, доступность, подвижность и величина кормовых объектов.
13.8. КОЛИЧЕСТВО И МАССА ПОЕДАЕМОГО КОРМА
Ящурки в период дневной активности поедают разнообразный корм. Взвешивание содер-
жимого желудочно-кишечного тракта ящериц разного пола и возраста позволило оценить
массу поедаемого корма в течение суток. Так, по данным ВАКиреева [сообщ.], ящурки
различного пола и возраста поедают 0,08—0,95 г пищи: взрослые самцы — до 0,95, самки —
до 0,7, сеголетки и молодые — 0,08 — 0,3 г. В Западном Казахстане средняя масса пищи
взрослых особей весной равна 0,58 г [Окулова,сообщ.] .В Дагестане [Хонякина,сообщ.]
масса пиши ящурок варьирует в пределах 0,01—1,35 г. По данным М.Ф.Тертышникова
[257, 259; Горовая, сообщ.], взрослые особи съедают в сутки ОД-0,98, сеголетки —
0,06—0,29 г пищи. Количество съеденных животных - от 1 до 11 (в среднем 4,18) [Орло-
ва, Тэрбиш, сообщ.].
Рис. 85. Зависимость массы содержимого желудка разноцветной ящурки от времени суток (в часовой
□робе усредненная масса пищи 20 особей) [ 259]
Рис. 86. Сезонные изменения интенсивности питания разноцветных ящурок [257]:
1 - взрослые самцы, 2 — взрослые самки, 3 — молодые самки и самцы
В процессе охоты в течение дня желудок ящурки постепенно наполняется проглочен-
ными беспозвоночными. Наибольшая масса пищи и наименьшее количество пустых желуд-
ков наблюдаются к вечеру. Рис. 85 показывает характер наполнения желудка взрослого
самца в конце июня [259] в течение светового дня при нормальных погодных условиях.
Установлено, что интенсивность питания ящурок изменяется в течение активного пе-
риода и зависит от погодных условий, возраста и, как отметила ТЛКотенко [131,386],
от степени упитанности и уровня половой активности. На рис. 86 показано изменение
массы съедаемого корма ящурками разного возраста и пола в течение сезона активности.
Как показывают данные ЗЛХонякиной [сообщ.], средний индекс наполнения желуд-
ка ящурок в течение сезона активности колеблется в широких пределах — от 1,7 до 5,6
(табл. 23). Средний индекс наполнения желудка у взрослых самцов колеблется от 2,5
до 3,3, у взрослых самок — от 1,7 до 4,7, у неполовозрелых особей — от 3,8 до 5,6. Самый
высокий индекс (5,6 %) отмечен у сеголетков, что, по мнению ЗЛЛонякиной, связано
с относительно быстрым темпом роста молодых животных и, следовательно, большей по-
требностью в пище.
Анализируя данные рис. 86 и табл. 23 мы можем отметить, что интенсивность питания
166
ящурки начинает увеличиваться вскоре
после весеннего пробуждения. В период
размножения у самцов интенсивность пи-
тания возрастает. У самок заметно воз-
растает интенсивность питания после от-
кладки яиц — в июне — июле. Со второй
половины июля — в августе индекс на-
полнения желудка понижается, что мож-
но объяснить повышением дневных тем-
ператур и уменьшением активности яще-
риц и беспозвоночных в местах обитания
ящериц и тем, что к этому времени ящур-
ки восстанавливают затраченную орга-
низмом энергию на размножение.
Молодые особи, появляющиеся в
июле — августе, в первые дни поедают
небольшое количество пищи. Но уже
через неделю интенсивность питания
возрастает очень быстро, что связано
с интенсивным ростом. Максимальное
количество пищи они поедают в авгус-
те. В дальнейшем, по мере
Таблица 23. Индекс наполнения желудка разно- цветной ящурки в равнинном Дагестане [Хонякина, сообщ.]
Пол, возраст Количе- ство особей, экз. Индекс наполнения желудка
min max среднее
Май Взрослые самцы 42 0,1 8,1 3,08 Взрослые самки 18 0,4 8,6 3.4 Беременные самки 5 0,5 3,9 1,7 Полувзрослые 21 0,5 9,3 4,5 Июнь Взрослые самцы 154 0,0-0,1 16,7 3,3 Взрослые самки 107 0.0-0,1 16,3 4,7 Беременные самки 52 0,0-0,1 12,3 2,6 Полувзрослые 54 0,0—0,3 11,7 3,8 Июль Взрослые самцы 13 0,0-0,2 8,0 2,5 Взрослые самки 16 0,0-0,1 9,2 3,7 Август Взрослые самцы 8 0,5 6,3 3,3 Взрослые самки 14 0,6 12,7 4,6 Сеголетки 18 0,3 11,5 5,6
накопления питательных веществ, интенсивность питания
постепенно уменьшается вплоть до ухода в спячку.
13.9. ПРОДОЛЖИТЕЛЬНОСТЬ ПЕРЕВАРИВАНИЯ ПИЩИ
Как показали эксперименты в лаборатории, в ходе которых подопытным животным
скармливали беспозвоночных, окрашенных пищевыми красителями, полная эвакуация пи-
щи из желудка наблюдалась через 21—22 ч у 60 % ящурок, у 40 % сохранялись остатки на-
секомых и пауков [Хонякина, сообщ.]. По данным М.Ф.Тертышникова [257, 259] желу-
док от пищи освобождается через 24 ч. Установлено также, что продолжительность пере-
варивания беспозвоночных с хитиновым покровом (жесткокрылые преимущественно)
длится 18—19 ч, мягкой пищи (гусеницы и др.) —8—11.
Говоря о скорости переваривания добычи, следует иметь в виду и образ жизни ящу-
рок. Наблюдения показывают, что в плохую погоду ящерицы не охотятся и при затяжном
ненастье могут оказаться без пищи. По сообщению 3 ЛХонякиной, за 20 дней голодания
(весна, взрослые самцы) ящерицы потеряли около 1/4 массы тела и половину массы жи-
ровых тел, после чего погибли. При нормальной погоде ящурка охотится практически пол-
ный световой день и всякий раз после приема пищи греется в лучах солнца. Такое поведе-
ние ящерицы объясняется повышением скорости ферментативных процессов при повыше-
нии температуры тела под воздействием солнечной радиации.
Максимальная ферментативная деятельность отмечена при оптимальной температуре
тела [142]. Видимо, у разноцветной ящурки оптимальные условия деятельности пищевари-
тельных ферментов будут при оптимальной температуре тела, которая, например, для Цен-
трального Предкавказья находится в пределах 30—35° (см. гл. 10) . Отсюда следует, что
в весенне-осенний период сезона активности при значительном колебании температуры сре-
ды скорость переваривания съеденной добычи также колеблется. В данной связи интерес-
ны материалы Р.Скокциласа [441], подтверждающие нашу мысль. Из них следует, что при
5° у ужа пищеварение не отмечено, при 15° добыча переваривается медленно, 25 — опти-
мальная температура для пищеварения. Аналогичный механизм действия пищеварительных
ферментов при перепаде температуры среды был отмечен нами ранее для прыткой ящери-
цы [157,259].
13.10. СПОСОБЫ ДОБЫВАНИЯ ПИЩИ
Разнообразие объектов питания характеризует ящурку как активного энтомофага, поедаю-
щего двигающихся, а иногда неподвижных мертвых беспозвоночных. Среди объектов ее
167
добычи встречаются медленно передвигающиеся (моллюски, некоторые жуки, муравьи,
гусеницы и др.) , быстро передвигающиеся (многие жуки, пчелы, осы, бабочки, прямокры-
лые, двукрылые и др.) формы, имеющие критическую и апосематическую окраску. Кро-
ме того, среди них немало ядовитых, жалящих и неприятно пахнущих животных.
Свою добычу ящурка отыскивает с помощью зрения1, слуха1 2, обоняния. В период
активности она начинает охотиться с раннего .утра до наступления сумерек с перерывами
в самое жаркое время дня. После выхода из убежищ она некоторое время греется в лучах
солнца, затем приступает к поиску пищи. Быстро передвигаясь зигзагами по поверхно-
сти почвы, она внимательно осматривает различные объекты, наклоняя голову вниз, пово-
рачивая ее по оси в бок. Заметив двигающееся насекомое, она на короткое время замира-
ет на месте, затем бежит к нему и схватывает резким броском. Можно часто видеть, как
она подпрыгивает и ловит пролетающих над ней мух, мелких молей и др.
Возможно, разноцветная ящурка способна отыскивать зарывшихся в почву личинок
насекомых, как это было установлено, например, для быстрой и сетчатой ящурок, а также
для ящурки Штрауха [53]. Интересны наблюдения ЗКЗрушко (сообщ.) о способности
ящурок отыскивать корм под коркой земли на такырах (см. выше) .
Наряду с активным поиском пищи ящерица добывает его, находясь в засаде. При этом
она располагается на небольшом возвышении, затаивается невдалеке от какого-либо невы-
сокого цветущего растения, прижав тело к земле и приподняв голову. Заметив появивше-
еся на растении или проползающее мимо насекомое, ящерица, сделав несколько шагов,
бросается на него и старается схватить.
Объекты добычи ящурки обычно невелики. Мелких животных она глотает быстро,
крупных — медленно, около 2—3 мин. Охватив крупный объект, ящерица сильно сжимает
челюсти, иногда трясет головой из стороны в сторону, выпускает ее и вновь схватывает и
заглатывает. При этом ящерица как бы ’’поправляет” положение добычи в ротовой поло-
сти так, чтобы она легче проходила через пищевод. Во время глотания она делает паузы.
После этого ящурка облизывает края рта языком и ’’обтирает” их о землю, а иногда сма-
хивает (или обтирает) остатки пищи передней или задней конечностью. ЗКЪрушко
[сообщ.] наблюдала, как во время кормежки (Зайсанская котловина, май — июнь 1976 г.)
закручивала кончик хвоста так, как это делают круглоголовки.
1 Многие ящерицы воспринимают 5—8 цветов, но реагируют только на движущиеся .объекты [54,
381].
2 Изучение акустических способностей ящериц показало, что некоторые ящурки воспринимают звуки
от 200 до 6000 Гц [53].
ГЛАВА 14
РАЗМНОЖЕНИЕ1
Сведения о разноцветной ящурке по вопросам, рассматриваемым в этой главе, имеются
примерно в 90 работах (без тех, что содержат лишь данные литературы, а также рукопи-
сей диссертаций). Во многих публикациях приводятся фрагментарные или немногочис-
ленные данные [6, 12,19,24,55,56,66,70,74,85,86,91,92,94,97,98,104, 107,111, 114,
115, 118, 122, 125, 127, 144, 145, 151,155,161,168,179,192-194,197,201,222,224,227
253, 257, 266, 271, 273, 276, 290, 291, 294, 304, 305,310, 313,.315, 328, 359, 367, 385]
В ряде работ содержатся более обстоятельные сведения [7, 27, 35, 49, 77, ПО, 121, 124ч
156, 191, 198, 199, 223, 225, 250, 293, 316; Зинякова, Колбинпев, сообщ.]. Отдельно сле-
дует выделить публикации, содержащие небольшой материал по размножению конкретно
разноцветной ящурки, зато носящие обобщающий характер [37—39, 79, 278] Данные
по размножению вида, опубликованные до 70-х годов, суммированы в сводках
НЯЩербака [320,436].
Наиболее обширные сведения по рассматриваемому вопросу содержатся в работах
11 авторов [20,21,25,48,113, 123, 126, 130, 131,258,259,263,323,324,386; Вашетко,
Киреев, Окулова, Хонякина, сообщ.]. При этом исследования НМ.Окуловой, ЗЛХоняки-
ной и Т И Котенко наиболее детальны и охватывают широкий круг вопросов, связанных
с размножением разноцветной ящурки, вследствие чего Западный Казахстан, Дагестан и
Южная Украина — наиболее изученные регионы в рассматриваемом плане.
Работы, выполненные в разные годы и различными исследователями, неравноценны не
только по объему материала, но и по степени обработки данных и характеру выводов.
Некоторые авторы приводят взаимоисключающие данные или выводы, противоречащие
собственным материалам. В таких случаях мы ориентировались на фактические данные и
обычно учитывали те, которые казались наиболее правдоподобными. В дальнейшем мы
ссылаемся лишь на сведения, полученные разными авторами на основании собственных
исследований.
14.1. НАСТУПЛЕНИЕ ПОЛОВОЙ ЗРЕЛОСТИ
Половая зрелость в разных популяциях разноцветной ящурки может наступать при различ-
ных размерах, массе тела и возрасте животных, что связано с особенностями климата, по-
годными условиями и другими факторами, влияющими на скорость роста и созревания
рептилий. Наступление половой зрелости разноцветной яшурки объективно можно опре-
делить лишь по состоянию гонад в репродуктивный период: у самок — по наличию в яични-
ках овоцитов желтого цвета диаметром более 2 мм или желтых тел, у самцов — по увели-
ченным семенникам и присутствию в них зрелых сперматозоидов.
Менее надежными показателями половой зрелости ящурок являются состояние жиро-
вых тел и поведение. Наши наблюдения на юге Украины показали, что попытки спаривать-
ся бывают и у явно неполовозрелых особей. При этом у молодых самок в задней части ту-
ловища и на хвосте остаются следы от челюстей взрослых самцов, а полувзрослые самцы
не только имеют следы укусов, но и могут выдвигать наружу копулятивные органы,
если надавливать пальцем на основание хвоста. Развитие жировых тел тесно связано с раз-
1 При участии Т.Р Алиева, М.ИАхмедова, В J4Ъадмаевой, ЭЗЗашетко, И.СДаревского, М ЛЗиняко-
вой, В А.Киреева, ВЛ Колбинцева, Г Л-Лукиной, НМ.Окуловой, Т.Сатторова, С.В.Таращука, М.Ф.Тер-
тышникова, ЗЛХонякиной,
169
витаем половых продуктов (подробнее ниже) , поэтому если во второй половине периода
размножения, когда жировые запасы у ящурок истощены, встречаются небольшие самцы
с жировыми телами, резко выделяющимися крупными размерами на общем фоне, таких
особей следует признать неполовозрелыми. Поскольку определение половозрелое™ сам-
цов по наличию сперматозоидов сравнительно трудоемко и редко используется, то в по-
давляющем большинстве работ наступление половой зрелости отмечалось по состоянию
гонад самок. При этом предполагают, что самцы становятся взрослыми в том же возрасте
и при тех же размерах, что и самки. В целом это вполне приемлемо, но в тех регионах, где
сеголетки появляются очень рано и ко времени ухода на зимовку успевают дораста до раз-
меров взрослых животных, половая зрелость у самцов (в силу специфики их полового
цикла) может наступить и осенью, т.е. в более раннем возрасте, чем у самок (хотя и при
сходных размерах тела, поскольку во время спячки ящерицы не растут) .
В связи с тем что размеры ящурок, как и других рептилий, зависят от возраста, длина
туловища (в меньшей степени — масса тела) служит довольно хорошим критерием на-
ступления половой зрелости — при условии, однако, что предварительно размеры живот-
ных были сопоставлены с состоянием гонад, поскольку минимальные размеры половозре-
лых ящериц в разных популяциях и подвидах отличаются. Более надежным, чем длина ту-
ловища, критерием является длина пилеуса, поскольку последняя более фиксирована при
измерениях. Однако этот показатель обычно не используется. В случае его применения
необходимо учитывать, что для абсолютной и относительной длины пилеуса ящурок харак-
терен половой диморфизм, выраженный нередко еще сильнее, чем по абсолютной и относи-
тельной длине хвоста (см. 131).
Во многих частях ареала разноцветные ящурки становятся половозрелыми после пер-
вой зимовки в возрасте около года, точнее — в 9—12,чаще 11 мес [25, ИЗ, 114, 123,126,
191, 258—260, 266, 278, 293, 386; Киреев, Окулова, Хонякина,сообщ.]. В Западно-Казах-
станской обл. ящурки созревают, по предположению АМ.Сергеева [227], на вторую вес-
ну, в Казахском мелкосопочнике и Джамбулской обл. [12; Колбинцев, сообщ.] также в
конце 2-го года жизни, в Кыргызстане [324] — на 3-й год (очевидно, имеется в виду тре-
тий сезон активности, т.е. возраст около 2 лет) . Для Западного Предкавказья М Л .Зиня-
кова и ПЛ Руденко [97] приняли разделение ящурок на размерно-возрастные группы, со-
гласно которому в категорию полувзрослых попадают особи, перезимовавшие один раз и
имеющие длину туловища 42—59 мм, массу тела — 2,0—4,0 г.
В большей части публикаций не указывается, все ли особи новой генерации созревают
в одинаковом возрасте. По предположению С А .Чернова [293], в Волго-Уральском между-
речье после первой зимовки половозрелыми становится подавляющее большинство осо-
бей. По НМ .Окуловой [сообщ.], в том же регионе половая зрелость ящурок, вышедших
из яиц первой кладки, наступает весной следующего года, а для тех, что вышли из яиц вто-
рой и последующих кладок, — летом следующего года, т.е. в любом случае в возрасте 9—
10 мес. По М.Ф.Тертышникову [259], в Ставрополье все ящурки становятся взрослыми
в 11 мес. В Дагестане большинство ящурок созревает в возрасте около 11 мес и лишь
некоторые особи, по-видимому, около 22—23 мес [Хонякина, сообщ.].
На юге Украины половая зрелость у ящурок наступает обычно на 10—12-м месяце
жизни, т.е. в мае — июле. В большинстве случаев это особи, родившиеся в июле — начале
августа предыдущего года. Часть годовиков (в основном родившихся в конце августа —
сентябре) не успевает созреть к июлю и дает потомство лишь после второй зимовки, т.е.
на 21—23-м месяце жизни. В разные годы соотношение рано или поздно созревающих ящу-
рок различно (как правило, преобладают первые) и зависит прежде всего от погодных
условий, определяющих (через продолжительность суточной и сезонной активное™) тем-
пы роста рептилий. Неодновременное созревание ящурок приводит к растягиванию перио-
дов спаривания, откладки и инкубации яиц, появления сеголеток [123, 124, 126 , 386].
В различных частях ареала половая зрелость разноцветных ящурок наступает при раз-
ных размерах и массе тела. Однако на Данные различных авторов влияют не только есте-
ственные причины (географическая изменчивость размеров тела, различные темпы роста в
разные по погодным условиям годы), но и объем полученного материала. Только на боль-
ших выборках (лучше — при целенаправленном обследовании особей ’’промежуточных”
размеров) можно выявить действительную нижнюю границу длины туловища и массы тела
половозрелых особей. Наибольшие значения минимальной длины туловища взрослых ящу-
170
рок отмечены для Зайсанской котловины (64 мм) [48] и для других районов Казахста-
на (60 мм) — Южного Прибалхашья [48], Казахского мелкосопочника [12], Волго-
Уральского междуречья [227; иные данные см. ниже], а также для Западного Предкав-
казья [97] ,в последнем случае — при массе тела более 5,0 г.
В Крыму ящурки становятся половозрелыми при длине туловища 592 мм [316],
Азербайджане — при 58 мм, по данным AM Алекперова [7], или при 55 мм для самок
и 60 мм для самцов — по данным М.И Ахмедов а, при этом сеголетки достигают размеров
половозрелых особей к осени [20]. В Волго-Уральском междуречье минимальная длина
туловища взрослых ящурок составляла 57 мм [293] или 55—56 мм [191, 193; Окулова,
сообщ.], в Монголии — 562 мм [197], в Калмыкии, Ставрополье и Дагестане — 55 мм
[25, 258, 259, 263, 266; Хонякина, сообщ.]. Интересно, что в Калмыкии тем же автором
6.05 [21] была добыта самка с длиной туловища 53 см и массой тела 4,6 г с развивающи-
мися фолликулами диаметром 3,1 мм, 4.07 — самка длиной 45 мм и массой 5,4 г с фолли-
кулами до 5,1 мм. Мелкие размеры начинающих размножаться ящурок (51 мм) отмечали
и для Румынии [359].
На юге Украины половая зрелость у разноцветных ящурок наступает обычно при дли-
не туловища 50—54 мм (зависит от популяции) , длине пилеуса 11-12 мм для самок и
12—13 мм для самцов, массе тела около 3 г (2,90—3,17) . На Нижнеднепровских песках
отдельные особи приступают к размножению при длине туловища 47—49 мм. Так, 20.VII
1975 г. была добыта самка с 2 готовыми к откладке яйцами с длиной туловища 47,4 мм
и массой тела 3,10 г (без массы яиц и содержимого пищеварительного тракта - 2,15 г)
[123,126,386].
142.ЖИРОВЫЕ ТЕЛА И ИХ ЗНАЧЕНИЕ
На протяжении периода сезонной активности и в течение жизни ящериц масса жировых тел
закономерно изменяется (рис. 87—89, 91) . При этом обнаруживается тесная связь между
динамикой массы жировых тел и половыми циклами самцов и самок. Объсняется это тем,
что жировые запасы ящериц расходуются на развитие половых продуктов. Поэтому к
концу периода размножения, который у самцов заканчивается раньше, чем у самок, жиро-
вые тела разноцветной ящурки достигают минимальной массы (у части особей могут рас-
ходоваться полностью) , в самом конце периода покоя гонад или чуть позже, т.е. в авгус-
те или сентябре (в зависимости от региона) , они имеют максимальную массу. Ко времени
залегания ящурок в зимнюю спячку жировые тела несколько уменьшаются, особенно у
самцов, что связано с подготовкой животных к новому циклу размножения (рис. 87, 89,
91) . Внешний вид жировых тел E.arguta дан в статье И.С Даревского [79].
Исследования в Дагестане [Хонякина, сообщ.], Западном Казахстане [Окулова, сообщ.]
и на юге Украины (наши данные) показали следующее. Масса жировых тел у самцов в сред-
нем ниже, чем у самок. У крупных особей жировые тела крупнее. Средняя и максималь-
ная массы жировых тел колеблются по годам в зависимости от кормовой базы и погоды,
определяющих продолжительность суточной и сезонной активности. Межпопуляционные
различия в массе жировых тел обусловлены различиями в размерах ящурок, кормовых и
погодно-климатических условиях среды. Так, на юге Украины в первой декаде июня
1979 г. на о.Бирючий (Азовское море) у всех самок были необычайно мелкие жировые
тела, весившие в среднем 20 мг при диаметре овоцитов 3,1—3,5 мм. В то же время на Поти-
евском участке Черноморского заповедника во второй половине мая того же года масса
жировых тел самок составляла 130—320 мг при диаметре овоцитов 5,3—82 мм [126].
Наибольшая зарегистрированная нами на Ивано-Рыбальчанском участке этого заповедника
масса жировых тел достигала 365 мг, в Дагестане — 820 [Хонякина, сообщ.], в Арме-
нии перед уходом ящурок на зимовку средняя масса жировых тел составляла 900 мг -
для самок и 570 мг — для самцов, или 9,0 и 72 % массы тела животных [79]. В Казахстане
жировые запасы ящурок достигали максимальных величин в сентябре и составляли 4,7—
5,9 % массы тела [191], сразу после весеннего пробуждения ящериц жировые тела у са-
мок весили 500-600 мг, у самцов — 200—220 [199] .По данным МИ.Фоминой [276], изу-
чавшей изменчивость массы тела и внутренних органов разноцветной ящурки, наибольшая
вариабельность характерна для индекса жировых тел.
171
Рис. 87. Сезонная динамика массы жировых тел и печени, массы и размеров семенников разно-
цветной ящурки:
А - динамика массы жировых тел самок: а — Восточный Азербайджан [ 20] t б - Дагестан
[Хонякина, сообщ.], в — Западный Казахстан, % массы тела [ Окулова, сообщ.]; Б - динамика
массы жировых тел самцов, массы и средней длины семенников (Дагестан) [ Хонякина, сообщ.];
В — динамика индекса (относительной массы) жировых тел самцов и средней длины семенников
(Западный Казахстан) [ Окулова, сообщ.]; Г — динамика массы жировых тел, семенников и пе-
чени самцов (о.Обливной, Каспийское море) [ 20]; 1 — масса жировых тел, 2 — длина семенни-
ков, 3 — масса семенников, 4 — масса печени
Рис. 88. Накопление жировых запасов у
сеголеток разноцветной ящурки в Черно-
морском заповеднике (юг Украины) [126] :
А - петом (июль - август) (п = 46,
г = 0,66 (Р > 99 %), у = 0,28х-7,35); Б-
осенью (сентябрь - октябрь): (п = 41,
г = 0,85 (Р > 99 %),у = 1,34х-38,83).Диа-
метр точек пропорционален количеству
данных
Прослежена также связь между динамикой
накопления жировых запасов и темпами роста
ящурок. У недавно родившихся сеголеток жиро-
вые тела практически отсутствуют, затем проис-
ходит накопление резервных веществ, идущее
вначале медленно. Распределения массы жиро-
вых тел в зависимости от размеров тела в июле —
августе и сентябре — октябре существенно отли-
чаются (рис. 88) и свидетельствуют о том, что
летом поступающие с пищей питательные веще-
ства расходуются прежде всего на рост сеголе-
ток, а осенью преимущественно используются
для накопления жировых запасов [126]. Это со-
гласуется с данными по росту сеголеток (см.
[124]). Связь между массой жировых тел и рос-
том ящериц отмечала и 3ЛХонякина [сообщ.]
для Дагестана: после первой зимовки у неполо-
возрелых особей, несмотря на более интенсив-
ное питание по сравнению со взрослыми, мас-
са жировых тел была невелика и во второй поло-
вине мая составляла 0—150 мг (в среднем
32,1 мг), в июне — 0—110 (26,2 мг); незначи-
тельные жировые запасы обусловливались быс-
трыми темпами роста молодых особей.
Некоторая часть жировых запасов расходуется во время зимней спячки, поэтому мас-
са жировых тел после весеннего пробуждения ящурок ниже, чем во время ухода их на зи-
мовку (рис. 89, А, некоторые графики на рис. 87) . Случаи, когда авторы получали иные
соотношения — слишком большое превышение осенних показателей над весенними (1)
или большие велич1Ыы весной по сравнению с осенью (2) — мы объясняем неполнотой
172
данных, а именно тем, что жировые тела ящурок анализировались не вскоре после весен-
него пробуждения, позже, когда они уже частично были израсходованы на развитие го-
над (в первом случае) , либо в осеннюю выборку попали мелкие взрослые или вообще мо-
лодые особи, что исказило общую картину (во втором случае). Как показал И.С Дарев-
ский [79] на разных видах ящериц Армении, расходование жировых тел во время зимов-
ки у самок значительно меньше, чем у самцов, поскольку яичники зимой находятся в со-
стоянии покоя. Из двух совместно обитающих видов ящурок у вида с более длинным пе-
риодом зимней спячки жировые тела истощаются за зиму сильнее.
Кроме жировых тел, большое значение как депо питательных резервных веществ име-
ет печень (подробнее см. в гл. 7 и 123) .
14.3.ПОЛОВОЙ ЦИКЛ САМЦОВ
У взрослых самцов масса и размеры семенников максимальны в апреле (рис.89, Л ), сра-
зу после весеннего пробуждения. Семенники в это время желтые и покрыты густой сетью
кровеносных сосудов, жировые тела мелкие. В мае — первой половине июня размеры и
масса семенников постепенно уменьшаются и в июле достигают минимальных значений. За-
тем они увеличиваются и достигают в сентябре, ко времени ухода взрослых особей на зи-
7
I \ / Рис. 89. Половые циклы самцов и самок разноцветной
40 ~ \ /Ч/ ящурки в различных частях ареала:
/ ) I 2 Л — Ивано-Рыбальчанский участок Черноморского
/ \ / запрведника [126]: 1 — максимальная длина семенни-
। 1 । ! i t ! ка, 2 — масса семенника, 3 — масса жирового тела, 4 -
IV V VI VII Ш И X масса придатка семенника; Б — Ставропольская возвы-
МвСЯЦ шенность (Центральное Предкавказье) [259] 1 - мас-
са жирового тела, 2 - масса семенника, 5 — нахождение зрелых сперматозоидов в семенниках;
В — Калмыкия [25] 1 — масса жировых тел, 2 - масса семенников; Г-Кыргызстан[324].отно-
шение длины наибольшего семенника к длине туловища: а - Чуйская долина, б - Прииссык-
кулье, в — приферганские районы; Д- Южный Узбекистан! Вашетко, сообщ.] .индексы (относи-
тельная масса) : 2 — семенников, 6 — половых продуктов самок, 7 — жировых тел самок
мовку, примерно таких же значений, как в апреле. Придатки семенников наиболее круп-
ные в мае—июне, в период наибольшей брачной активности самцов (рис. 89,Л) . Динамика
массы жировых тел имеет противоположный характер, поскольку жировые запасы расхо-
дуются на развитие половых продуктов. Поэтому масса жировых тел минимальна в июне,
достигает максимальных величин в августе, в течение сентября существенно уменьшает-
173
ся, однако ко времени ухода самцов на зимовку все же заметно превышает таковую пос-
ле зимней спячки [126].
Таков половой цикл самцов у ящурок, заканчивающих зимнюю спячку к началу апре-
ля и активных до конца сентября — начала октября (взрослые особи) и конца октября
(сеголетки), что характерно для большинства хорошо изученных популяций вида. В са-
мых жарких районах Средней Азии и Южного ’ Казахстана ящурки примерно на месяц
раньше появляются весной и месяцем позже исчезают осенью, в результате размеры и мас-
са семенников самцов максимальны в марте — начале апреля и в октябре — ноябре [222,
323, 324; Вашетко, сообщ.] (89, Г, Д). Соответственно этому изменяется и динамика
массы жировых тел. Некоторые исследователи отмечали иной характер динамики разме-
ров и массы семенников: после выхода самцов из зимней спячки семенники начинали уве-
личиваться и достигали максимальных размеров и массы в мае [25; Киреев, сообщ.] и
даже в июне [Зинякова, сообщ.]. В.И£адмаева [25] отметила два пика увеличения мас-
сы семенников — в середине мая и в августе (рис. 89, В) , что трудно увязать с имеющими-
ся данными различных авторов по динамике сперматогенеза (см. ниже). ЗЛЛонякина
[сообщ.] наблюдала в Дагестане следующую картину: после пробуждения от спячки у
самцов несколько увеличиваются семенники по сравнению с осенью, причем у наиболее
крупных особей гонады достигают максимальной величины в конце апреля, а у самых мел-
ких — в конце июня. Последнее, на наш взгляд, можно объяснить просто более поздним
созреванием мелких самцов.
Максимальные размеры и масса семенников, по данным различных авторов, состави-
ли 5,9x3,3 мм для Западного Предкавказья [Зинякова, сообщ.], 6,4 мм и 44 мг (масса
обоих семенников 72 мг) — для Ивано-Рыбальчанского участка Черноморского заповед-
ника (наши данные), 7x3 мм [21] или 7x4 мм и 140 мг [Киреев,сообщ.] — для Калмы-
кии, 8 мм и 210 мг [Хонякина, сообщ.] — для Дагестана, 9x4 мм [323] — для Кыргыз-
стана, 11x3,5 мм [225] и 12x3,1 мм [222] — для Таджикистана, 11x6 мм [110] и 13х
хб мм [Вашетко, сообщ.] — для Узбекистана. Чем крупнее особи в популяции, тем, есте-
ственно, крупнее семенники у самцов, что, в частности, наглядно показано для Казахстана
[48, табл. 2]. Но большие различия в массе семенника по данным разных авторов объяс-
няются не только этими причинами, но и тем, что в одних работах приведена масса одного
семенника, в других — двух (часто это не оговаривается) , а иные исследователи взвешива-
ли семенники явно вместе с придатками. Правый семенник обычно бывает крупнее лево-
го (см. [21, 48]). По результатам вскрытия 48 самцов с Ивано-Рыбальчанского участка
Черноморского заповедника, длина и масса правого семенника были выше, чем левого,
в 85 % случаев. У недавно родившихся сеголеток масса семенника менее 0,5 мг, его длина
0,5-0,8 мм [126].
Динамика развития семенников и их придатков становится понятной, если учесть,
что зрелые спермин у разноцветных ящурок образуются еще в сентябре, до наступления
зимней спячки, а весной, после пробуждения, их продуцирование продолжается. Таким об-
разом, осеннее и весеннее состояния гонад фактически являются вершинами одного пика,
разделенного зимовкой. Такой цикл развития гонад у ящериц впервые установил И.С Да-
ревский [79]. Он также показал, что в весенне-летний период продуцирование спермиев
продолжается и тогда, когда семенники начинают уменьшаться. В придатках семенников
зрелые спермин сохраняются до окончания брачного периода. Столь длительная поло-
вая активность самцов приводит к тому, что практически все половозрелые самки в по-
пуляции оказываются неоднократно оплодотворенными и процент яловости ничтожен
[79,80].
Для разноцветной ящурки явление осенней активизации сперматогенеза впервые от-
метил М.Ф.Тертышников [258, 259, 263], изучая ящериц Ставрополья. В частности, он уста-
новил, что в этом регионе зрелые спермин у Е. arguta формируются во второй половине
сентября, весной их продуцирование возобновляется; в мазках из семенников сперма-
тозоиды обнаруживались до начала июля [259] (рис. 89, Б), Динамику сперматогенеза
разноцветной ящурки исследовали также в Узбекистане, Восточном Казахстане и (в какой-
то мере) в Калмыкии, Дагестане. Так, в Калмыкии из 24 исследованных в начале мая сам-
цов активный сперматогенез обнаружен у 83 % особей [Киреев, сообщ.]. В Дагестане у неко-
торых самцов сперматозоиды в семенниках находили в конце июля [Хонякина, сообщ.].
В Восточном Казахстане спермин в семенниках и придатках обнаруживали в июне, августе
174
и сентябре, хотя оба органа к этому времени уменьшаются в размерах и становятся дряб-
лыми [48].
Наиболее подробно сперматогенез изучен у Earguta из южных районов Узбекистана
[55, 56, 223; Вашетко, сообщ.]. Здесь период активного сперматогенеза приходится на
март — апрель. В это время в стенках семенника наблюдаются все стадии деления спермио-
генного эпителия, в просветах канальцев — большое количество сперматозоидов. Размеры
последних (измерения на мазках) следующие: длина головки 6,3—10,0 (7,60±0,14) мкм,
длина хвостовой нити 25—60 (35,50±0,75) мкм. Стенки канальцев придатка состоят из
однослойного высокого цилиндрического эпителия с крупными ядрами, расположенными
в базальной части клеток. Цитоплазма апикальной части клетки заполнена секретом, круп-
ные капли его имеются и в просветах канальцев, содержащих также большое количество
спермиев. Состояние придатка свидетельствует о готовности к спариванию.
В конце апреля на части препаратов видно, что в семенниках еще продолжается актив-
ное деление спермиогенного эпителия (рис. 90, л) , в придатках часть канальцев уже пустая,
т.е. произошло извержение спермы; в других канальцах капли секрета и сперматозоиды
еще имеются. В мае в семенных канальцах присутствует умеренное количество спермиев,
а в канальцах придатка — остатки спермиев и секрета. В июне семенники и придатки боль-
шинства самцов находятся в состоянии покоя, тх. спермиогенный эпителий не активен,
канальцы меньшего диаметра, просветы их малы. Отмечается усиленное развитие симпла-
ста Сертоли и утолщение соединительнотканной оболочки семенника. Таким образом, к
концу июня процесс сперматогенеза у основной массы ящурок затухает. Вместе с тем в
это же время попадались самцы, в семенниках которых шел активный процесс образова-
ния спермиев, и канальцы придатков имели большое количество секрета и сперматозои-
дов. В июле — сентябре семенники и придатки находятся в состоянии покоя, а в конце сен-
тября — ноябре наблюдается новая волна сперматогенеза (рис. 90,6) . Осенняя активность
семенного эпителия приводит к образованию умеренного количества спермиев,но каналь-
цы придатка пустые [Вашетко, сообщ.].
По данным В .И .Бадмаевой для Калмыкии, у самцов разноцветной ящурки имеются
два пика увеличения массы семенников, что соответствует периодам усиленного спермато-
генеза. Первый пик приходится на вторую половину мая, второй — на первую половину
июля. Во второй половине июля семенники находятся в состояний покоя [21]. Однако на
рис. 89,В ТЦ1Я того же региона представлена несколько иная картина.
14.4. ПОЛОВОЙ ЦИКЛ САМОК
У половозрелых самок после весеннего пробуждения гонады (в зависимости от региона)
могут находиться на разных стадиях развития. Так, в Черноморском заповеднике в апреле
в яичниках ящурок содержатся беловатые овоциты диаметром до 2,2—2,4 мм, и лишь в
конце месяца — начале мая (в зависимости от года) начинается рост и пожелтение овоци-
тов. ВЛХончарова [66] на юго-востоке Украины встречала беременных самок с конца
апреля. В Дагестане [Хонякина, сообщ.] развитие гонад у самок начинается обычно также
с конца апреля, т.е. почти через месяц после окончания зимней спячки, а в Северном Уз-
бекистане на Устюрте [35] — лишь в первых числах мая.
В Западном Казахстане [Окулова, сообщ.] сразу после пробуждения ящурок в их яич-
никах не удается обнаружить развивающиеся овоциты; они начинают появляться, в зависи-
мости от погоды, через 5—10 дней. В Центральном Предкавказье [259] и Калмыкии [113]
у проснувшихся самок в яичниках уже имеется несколько желтых и крупных овоцитов
диаметром до 3—5 мм. В окр. Новой Сардобы (Южный Казахстан) самки, добытые в пер-
вой декаде марта, имели желтые овоциты диаметром 3—6 мм [35], на юго-востоке Узбе-
кистана сразу после выхода ящурок из зимних убежищ (в марте) желтые овоциты дости-
гали 3—9 мм, и индекс гонад был уже высоким [Вашетко, сообщ.] (рис. 89, Д) .
Развитие яиц в теле самки продолжается примерно месяц [35, 126, 191; Окулова,
сообщ.], что также следует из рис. 92 и схематически отражено на рис. 91,Б. Поскольку
сроки наступления половой зрелости и спаривания растянуты, самки с ярко-желтыми
круглыми овоцитами, находящимися на разных.стадиях развития, встречаются в течение
примерно 3 мес, что отмечалось для разных регионов [66, 324; Хонякина, сообщ.; наши
175
500
Рис. 91. Половой цикл самок разноцветной ящурки:
А, Б — популяция Ивано-Рыбальчанского участка Черноморского заповедника [ 126] : А — по ре-
зультатам вскрытий, Б — схема для одной особи (пояснения в тексте; В — популяция из Центрального
Предкавказья [263]; Г — популяция из Калмыкии [25] : 2 — максимальные размеры овоцитов или
яиц, 2 - масса яитаика и яиц, 5 — масса жирового тела. Точкой отмечен момент перехода овоцита из
яичника в яйцевод
наблюдения]. Кроме крупных желтых овоцитов в яичнике содержатся мелкие белые: нес-
колько штук диаметром 1,8—2,4 мм (следующая генерация) и до десятка совсем мел-
ких — последующих генераций (по нашим данным для юга Украины). В Кыргызстане диа-
летр белых овоцитов первой генерации составлял 1—3 мм [324], в Калмыкии — 2—2^2
[21], в Азербайджане — 1,5—2,0 мм [20]. Пожелтение овоцитов наступает при диаметре
2,5 мм и более, поскольку для желтых'овоцитов указывались минимальные размеры
2,5 мм [35] и 2,7 мм [126, 156]; в Кыргызстане минимальный диаметр желтых овоци-
тов составлял 2—3 мм [324].
176
16182022» 262850 2 4 6 8 101214 1618 20222426 2830 1 3 57 9
Апрель май Июнь
Месяца число
Рис. 92. Развитие овоцитов и формирование яиц у самок разноцветной
ящурки (Волго-Уральские пески, Западный Казахстан, 1959 г.) [ Окулова,
сообщ.)
1 - первая кладка, 2 - вторая кладка, 3 — отложены первые яйца
Развивающиеся фолликулы по достижении определенных размеров лопаются, и яйце-
клетки выпадают в полость тела, откуда переходят в яйцеводы. Это обычно происходит
при диаметре яйцеклеток 8—9 мм, так как именно такие размеры указывались в качестве
максимальных для фолликулов в яичнике и минимальных — для яиц в яйцеводах (кон-
кретные данные см.: 20, 35, 48, 113, 223, 250, 290, 293, 324; Зинякова, Колбинцев, Тэр-
биш) . При этом минимальный зарегистрированный диаметр яйца в яйцеводе равен 8 мм
[290; Колбинцев, Тэрбиш, сообщ.], максимальные размеры фолликула в яичнике —9,6 мм
[126J . По данным НМ.Окуловой, яйцеклетки из яичника в яйцевод переходят при диамет-
ре 10—12 мм [191 ]. Оплодотворение происходит в верхних отделах яйцеводов.
В июне — июле (для северных и центральных частей ареала) E.arguta откладывает яйца,
поэтому, наряду с самками с развивающимися в яичниках овоцитами, попадаются особи, в
яйцеводах которых содержатся готовые к откладке яйца. У таких самок в яичниках име-
ются желтые тела (по числу находящихся в яйцеводах яиц) и овоциты диаметром до 1,6—
2,8 мм, которые будут развиваться в следующем году [126]. В тех популяциях, где наблю-
даются две кладки, у части самок кроме яиц в яйцеводах имеются еще и крупные желтые
овоциты в яичниках — будущая вторая кладка. Жировые запасы расходуются на развитие
овоцитов и формирование яиц, поэтому масса жировых тел минимальна в период отклад-
ки яиц и максимальна в конце лета — в начале осени, когда ящерицы успевают накопить
необходимое количество резервных веществ.
При наличии двух кладок жировые тела используются большей частью или полностью
на развитие первой кладки яиц, вторая кладка формируется обычно за счет поступления
питательных веществ с пищей. По данным В.ИЬадмаевой [25], яйца обеих кладок форми-
руются в яйцеводах при полном истощении жировых тел. Динамика массы жировых тел
и размеров овоцитов (для популяции с одной кладкой в году) показана на рис. 91. По-
скольку кривые на рис. А, В, Г построены по усредненным данным, они отражают не толь-
ко общую закономерность развития трех показателей, но и значительную их изменчивость,
обусловленную прежде всего различной плодовитостью самок и неодновременностью от-
кладки яиц. Для одной особи графики приняли бы примерно такой вид (рис. 91, Б}. Ис-
ходными данными для построения этих кривых послужили средние величины плодовито-
сти, размеров и массы яиц, массы жирового тела и яичника самок популяции Ивано-Ры-
бальчанского участка Черноморского заповедника, т.е. взята ’’средняя самка”, отложив-
шая яйца 10.VI. Кривые реальных самок будут в той или иной степени отличаться от дан-
ной схемы. Так, пики кривых 1 и 2 на рис. 91, Б в годы с различными погодными усло-
виями могут смещаться до 10 дней влево или до 60 дней вправо (для этой же популяции)
Соответственно этому изменится и кривая 3. В зависимости от кормовых и погодных
условий будет меняться упитанность ящурок и соответственно высота графика 3. С изме-
нением величины кладки и размеров и массы яиц будут меняться максимальные значения
кривых 1 и 2 [126].
В августе, после окончания периода размножения, в яичниках самок содержатся ово-
циты диаметром до 1,6—2,2 мм, и ко времени ухода ящурок на зимовку размеры овоци-
тов почти не увеличиваются (126, для юга Украины]. В Дагестане после завершения пе-
риода откладки яиц масса 2 яичников ящурок составляет в среднем 20 мг, а размер фол-
ликулов равен 0^—1,5 мм, причем заметно желтеют они только следующей весной [Хо-
някина, сообщ,] .В Восточном Казахстане в сентябре и в Центральном Предкавказье перед
залеганием в спячку у самок имеются овоциты до 3 мм в диаметре [48,263].
Для юго-востока Узбекистана осенью отмечено постепенное увеличение размеров ово-
цитов: в сентябре их диаметр достигал 2 мм, в октябре - 25, в ноябре — 3 мм, при этом
они приобретали желтый цвет [Вашетко, сообщ,]. О ШБогданов у самок из этого района,
вскрытых в ноябре, обнаружил развивающиеся желтые фолликулы диаметром до 3 мм, а
у некоторых — даже яйпа в яйцеводах [35]. Тот же автор показал, что на развитие поло-
вых продуктов заметно влияют осадки. В частности, у разноцветной ящурки в централь-
ных районах Узбекистана в годы с сухой осенью развитие овоцитов перед уходом на зи-
мовку обычно не наблюдается, если же в конце августа — октябре идут частые дожди, а
после них стоит теплая погода, то в октябре начинается рост и пожелтение овоцитов [37,
39].
У недавно родившихся сеголеток масса яичника не превышает 0,5 мг, диаметр овоци-
тов не более 0,5 мм [126].
145. СРОКИ СПАРИВАНИЯ
Спаривание разноцветных ящурок начинается вскоре после пробуждения — обычно через
2—3 недели после начала их массового появления на поверхности. Поэтому первы-
ми к спариванию приступают ящурки на юго-востоке Узбекистана, последними — в лесо-
степи Украины. О начале брачного периода в популяции при отсутствии непосредственно
наблюдавшихся случаев спаривания можно судить по появлению на теле самок следов уку-
сов сампов. Подковообразные следы от челюстей самцов хорошо видны на брюхе, хвосте
и особенно многочисленны в области таза самок. У самцов также имеются (в основном
на хвосте и на туловище выше таза) следы укусов, нанесенных другими самцами, а также
самками во время спаривания. По наблюдениям в Южном Прибалхашье [48], такие следы
встречаются у особей обоего пола по июль (спаривание здесь наблюдалось 10.IV). По на-
шим данным, на юге Украины следы укусов заметны у самцов до конца июля — середины
августа, у самок — до середины августа — начала сентября, т.е. сохраняются в течение 1-
15 мес после окончания брачного периода.
Общая длительность периода спаривания на большей части ареала вида, насколько
можно судить по обычно неполным данным разных авторов, составляет примерно 2 мес.
На юго-востоке Узбекистана спаривание происходит в марте — апреле (однажды отмечено
даже 25.П) [55, 198; Вашетко, сообщ.], в других районах Средней Азии (в Таджикистане,
Джизакской степи, Зеравшанской долине, на хребте Нуратау) , в Казахстане (Южном При-
балхашье, Джамбулской обл.) и в Азербайджане — в апреле — мае, иногда с конца марта
[7, 20, 48, 104,223,225; Колбинцев, сообщ,]. С конца апреля начинают спариваться ящур-
ки в Волгоградской обл., Калмыкии и на юге Украины [ИЗ, 118, 126; Киреев, сообщ.].
В .И .Бадмаева [24] для начала спаривания в Калмыкии указывает середину марта, что не
согласуется с данными о сроках весеннего пробуждения ящурок (см. [25].) В мае -
июне наблюдалось спаривание у ящурок в Волго-Уральских песках, Прииссыккулье, на
Ставрополье и в Дагестане [192, 259,263,324; Хонякина, сообщ.].
У других видов ящериц [210] период спаривания в северных частях ареала характери-
зуется более сжатыми сроками, чем в южных. Это вызвано тем, что благоприятный кли-
матический период, обеспечивающий оптимальные условия для развития яиц, в северных
районах гораздо короче, чем в южных. Поскольку основная часть данных по срокам спа-
ривания фрагментарна (отмечены либо только время начала периода спаривания, либо
отдельные даты, когда спаривание наблюдалось), определенные выводы по разноцветной
ящурке сделать сложно. Во всяком случае на юге Украины период спаривания длится не
менее 2 мес (с конца апреля по конец июня, иногда немного дольше), в Дагестане пример-
но столько же (май — начало июля), в Калмыкии 2,5-3 мес (май — июль или конец апреля -
первая половина июля, по данным разных авторов), в Узбекистане опять 2 мес (март —
апрель или апрель — май, в зависимости от региона) [25,110, 198, 223; Вашетко, Киреев,
Хонякина, сообщ.; наши данные].
178
Повторное спаривание отмечалось для Зеравшанской долины и хребта Нуратау (Узбе-
кистан) в первой половине июля, не менее чем через месяц после окончания первого пе-
риода спаривания [104, ПО]. В Прииссыккулье массовое спаривание ящурок наблюда-
лось в мае, отдельные случаи — в июне — июле [324]. Поскольку даже в тех регионах, где
у ящурок имеется всего одна кладка в году, период спаривания растянут на 2—2,5 мес (на-
пример, в Черноморском заповеднике) из-за неодновременного созревания ящурок, гово-
рить о реальном существовании или отсутствии повторного спаривания, как правило, сло-
жно. Вероятно, в тех районах, где у самок 2 кладки в году, повторное спаривание присут-
ствует, хотя, возможно, и не является обязательным условием для оплодотворения яиц
второй кладки (в связи с предполагаемой способностью сперматозоидов к длительному
хранению в половых путях самок) . М JU .Ахмедов предположил наличие осеннего спарива-
ния у ящурок Азербайджана [20].
Судя по многочисленным следам челюстей самцов на теле самки, одна самка спарива-
ется несколько раз. Это подтверждается также тем, что спаривание наблюдалось задолго
до овуляции, в период овуляции и даже после нее. В частности, у самок, добытых в мо-
мент спаривания в различных частях ареала, фолликулы в яичниках достигали в диаметре
2—4 мм [324], 3,5 [48], 3—6 [35], 4—5 [Окулова, сообщ.], 4—7 мм [Хонякина, сообщ.]
По мнению В А Киреева [113], спаривание и оплодотворение имеют место в период пере-
хода овоцитов из полости тела в яйцеводы. По нашим наблюдениям, спаривание чаще все-
го происходит до момента овуляции, но отмечались также самки, спаривающиеся в период
беременности. Первыми к спариванию приступают крупные взрослые особи, последними -
прошлогодки, успевшие созреть к концу периода размножения. Так. в Калмыкии [Кире-
ев, сообщ.] спаривание у крупных ящурок начинается в конце апреля и продолжается до
середины июня, а у мелких особей происходит во второй половине июня — первой поло-
вине июля.
14.6. ПРОДОЛЖИТЕЛЬНОСТЬ БЕРЕМЕННОСТИ
Непосредственных наблюдений в природе нет, поэтому о продолжительности беременно-
сти судят по времени между началом спаривания и началом откладки яиц в популяции
[210, 259] . На основании таких расчетов М.Ф.Тертышников считает, что беременность у
разноцветных ящурок длутся около месяца [258]. Такой же срок указывают и другие ав-
торы [35, 126, 191; Окулова, сообщ.], однако не как длительность беременности, а как
продолжительность периода развития половых продуктов самок (от начала роста овоци-
тов до откладки яиц) , что не одно и то же. Строго говоря, мы имеем дело с тремя различ-
ными периодами: сроком развития половых продуктов самок, периодом между временем
спаривания и временем откладки яиц и периодом между моментом оплодотворения яйце-
клетки и временем откладки яйца. В зависимости от обстоятельств могут совпадать во
времени первый и второй, второй и третий периоды, но лишь третий является собственно
периодом беременности. Определить его можно приблизительно по времени между появле-
нием в популяции первых самок с минимальными размерами яиц в яйцеводах (8—9 мм) и
первых особей с готовыми к откладке яйцами. Длительность такого периода составляет
примерно 2 недели (наши наблюдения на юге Украины, в популяции одна кладка) или нес-
колько меньше (следует из данных НМ .Окуловой, представленных на рис. 92, и относит-
ся ко второй беременности) . По данным В И Бадмаевой [25: диаграмма в приложении],
первая беременность у ящурок Калмыкии длится 35 дней, вторая 20. На чем основаны
эти и другие сроки в указанной схеме — неясно.
14.7. СРОКИ ОТКЛАДКИ ЯИЦ И КОЛИЧЕСТВО КЛАДОК
Сроки откладки яиц обусловлены, как и другие фенологические явления, погодно-клима-
тическими условиями региона. Во второй половине апреля начинают откладывать яйца
ящурки на юго-востоке Узбекистана [35, 56, 107; Вашетко, сообщ.], по некоторым дан-
ным [291] — даже во второй половине февраля, что явно ошибочно. Очень ранние сроки
указаны для ящурок из Таджикистана: конец апреля [225] и даже первая половина его
179
[222, 294] . В первой половине мая откладка яиц начинается в Кызылкуме [199] и Азер-
байджане [7], в Чуйской долине и на склоне Киргизского хребта [324], во второй поло-
вине мая — в тех же районах, но по данным других авторов [20, 85, 94], а также в Даге-
стане, Волго-Уральских песках, Муюнкуме [199; Окулова, Хонякина, сообщ.] . С конца
мая к откладке яиц приступают ящурки Волгоградской и Саратовской областей России
[118, 310], в мае размножаются также ящурки Кабардино-Балкарии [304]. В послед-
них числах мая — первой половине июня начинают откладывать яйца самки на юге Украи-
ны [123, 126, 250] , во второй половине июня — в Зайсанской котловине, Южном Прибал-
хашье и на Ставропольской возвышенности [48, 144, 258, 263]. Июнь указан также для
Армении, Монголии и предгорий в Джамбулской обл. Казахстана [77, 197, 272; Колбин-
цев, сообщ.] , июль - для Румынии [359] . Позже всех размножаются ящурки в Прииссык-
кулье - с третьей декады июля до начала сентября [324]. Имеющиеся сведения по Калмы-
кии противоречивы: откладка яиц начинается в мае и продолжается по июль [21] либо
происходит в июне — июле [25]; по В.А Кирееву, ящурки начинают откладывать яйца во
второй половине июня, реже — в июле [113]. В целом можно считать, что в жарких низи-
нах Средней Азии откладка яиц у разноцветной ящурки происходит в апреле — мае, в дру-
гих районах зоны пустынь, в том числе в Азербайджане, — мае — июне, в зоне степей —
июне — июле, а в лесостепи и некоторых высокогорных районах — в июле — августе.
Сроки начала и окончания периода размножения существенно зависят от конкрет-
ных погодных условий [20, 24, 37, 113, 126, 127, 224; Окулова, Хонякина, сообщ.] и в
различные годы могут отличаться на 10—20 дней на юге Украины и в Дагестане и даже бо-
лее чем на месяц — в глинистых пустынях Средней Азии [39; набл. Котенко, Хонякиной].
Влияние микроклимата продемонстрировано МИ Ахмедовым [20] на о. Обливной от-
кладка яиц и появление сеголеток происходят примерно на неделю позже, чем на ближай-
шем побережье материка, что обусловлено сильными ветрами на острове.
Отмечалось также [37, 39, 98] влияние рельефа местности и характера почвы на сро-
ки размножения рептилий. В частности, по данным О Л Богданова [37,39] ,на песках раз-
множение ящериц начинается позже, чем на твердом субстрате, и период откладки яиц бо-
лее растянут. Вместе с тем МЛ.Зинякова и ГJC .Плотников [98] отметили, что в Восточном
Приазовье разноцветная ящурка размножается раньше, чем прыткая ящерица, именно в
связи с тем, что первый вид населяет биотопы с ракушечно-песчаным хорошо прогревае-
мым субстратом. По нашим наблюдениям, на юге Украины прыткая ящерица всегда начи-
нает откладку яиц на 10—20 дней раньше, чем разноцветная ящурка, как на плотных чер-
ноземах, так и в песках в местах совместного обитания этих видов. Что касается отмечен-
ного О Л Богдановым более позднего размножения ящериц на песках, то это не увязыва-
ется с наблюдениями И.Ф Андреева [10] — более ранними сроками пробуждения ящурок
на песках по сравнению с глинистыми почвами (см. гл. 12) .
Период размножения у разноцветной ящурки сильно растянут даже в пределах одной
популяции [49, 66, 113, 126, 130, 278, 324, 386 и др.]. Это вызвано тем, что вначале к от-
кладке яиц приступают более старые (крупные) особи [20, 35, 37, 114, 126, 156, 191;
Окулова, сообщ. ], позже подключаются впервые размножающиеся животные, созреваю-
щие нередко лишь к концу периода размножения популяции. На материале из Дагестана
(табл. 24) хорошо видна динамика развития овоцитов в зависимости от размеров (возра-
ста) самок. 3ЛХонякина [278] последовательность участия ящериц в размножении объ-
ясняет тем, что у крупных особей темп роста ниже, чем у мелких, в связи с чем энергети-
ческие затраты организма невелики, и жировые запасы расходуются в основном на обеспе-
чение репродуктивной функции.
НЛ1 .Окулова | сообщ. 1 по поводу сроков размножения ящурок в Западном Казахста-
не отметила, что дружно происходит только начало репродуктивного периода в популяции
и откладка яиц первой кладки. При этом весной в размножение вступают главным обра-
зом крупные особи, в июне включаются мелкие ящурки. Так, среди самок с развивающи-
мися овоцитами или яйцами в яйцеводах в апреле — мае (и = 120) 40,1 % приходилось на
особей длиной 66—75 мм, 50,8 % - на самок длиной 61—65 мм и лишь 9,1 % — на особей
с длиной туловища 56—60 мм; в июне (п = 37) распределение размножающихся самок по
этим размерным группам было следующим,: 27,0; 43,2 и 29,8 %. Таким образом, состав
размножающихся самок изменялся во времени, но основу всегда составляли особи дли-
ной 61—65 мм.
180
Таблица 24. Развитие яиц у разноцветной ящурки в зависимости от длины туловища
(возраста) самок (по данным З.П.Хонякинои для Дагестана)
Длина туло- вища, мм Показатель Время отлова
16-30.V 1-15, VI 1б-з а Vi 23-27, VII
55-59 Количество вскрытых самок, экз. 6 26 26
с овоцитами крупнее 4 мм, % 16,7 383 26,9
с яйцами в яйцеводах, % — 30.8 15,4 —
60-64 Количество вскрытых самок, экз. 12 61 40 11
с овоцитами крупнее 4 мм, % 50,0 52,5 50,0 9Д
с яйцами в яйцеводах, % 50,0 27,9 20,0 9,1
65-69 Количество вскрытых самок, экз. 9 43 16 3
с овоцитами крупнее 4 мм, % 33,3 533 623 ззз
с яйцами в яйцеводах,% 66,7 32,6 373 —
70-74 Количество вскрытых самок, экз. 3 14 5 1
с овоцитами крупнее 4 мм, % 66,7 28,6 20,0
с яйцами в яйцеводах, % 33,3 35,7 60,0
Всего Количество вскрытых самок, экз. 30 144 87 17
с овоцитами крупнее 4 мм, % 40,0 47,9 43,7 11,8
с яйцами в яйцеводах, % 43,3 30,6 24,1 5,9
В целом период откладки яиц длится, по данным разных авторов, примерно месяц
[6,20,48, 77, 107, 118, 197, 223, 359], до 1,5 мес [85, 191,310; Зинякова, Окулова,со-
общ.] , около 2 мес. [25, 49, 123, 124, 126, 225, 291,304,324,386] и даже до 2,5 мес [21,
278, 293; Вашетко, Хонякина, сообщ.]. По нашему мнению, длительность периода размно-
жения обычно составляет около 2 мес, и указанные выше значительно более сжатые сроки
либо являются следствием неполноты сведений, либо могут быть присущи лишь отдель-
ным популяциям, обитающим в наиболее суровых условиях (например, в высокогорьях) .
Интересно, что длительность периода откладки яиц практически не зависит (или зависит
незначительно) от числа кладок в году. Связано это, по-видимому, с тем, что поздно со-
зревающие (мелкие) ящурки в популяциях, характеризующихся наличием одной кладки,
удлиняют период размножения популяции, а там, где имеются 2 кладки, участвуют лишь
в одной, совпадающей по срокам со второй кладкой.
По вопросу о количестве кладок в году единого мнения нет. Одни авторы [49, 293]
растянутость сроков размножения объясняют наличием второй кладки, другие [124,
126] считают, что это может быть вызвано неодновременным созреванием ящурок. Много-
летние исследования, проведенные на Ивано-Рыбальчанском участке Черноморского запо-
ведника [126; наши неопубликованные материалы], показали отсутствие второй кладки,
хотя сроки размножения популяции в целом были велики (июнь — июль, в годы с поздней
весной и холодным летом — вторая декада июня — первая половина августа) . Поэтому го-
ворить о наличии второй кладки можно лишь в тех случаях, когда при вскрытии самок об-
наруживают одновременно присутствие яиц в яйцеводах (или желтых тел в яичниках, если
яйца недавно были отложены) и развивающихся овоцитов — в яичниках.
Достоверно 2 кладки бывает у ящурок в Дагестане [278; Хонякина, сообщ.], Волго-
Уральском междуречье [191,293; Окулова, сообщ.], Армении [77] и Узбекистане [107;
Вашетко, сообщ.], а также у части самок с побережий Черного и Азовского морей в Украи-
не, в частности в Одесской обл., — на Будакской косе [250] и в окр. г.Вилково и в Донец-
кой обл. — в окр. гМариуполь и на Кривой косе [386; наши неопубликованные данные].
О 2 кладках для ящурок юга Казахстана упомянул К Л Лараскив [199], но на чем основа-
но такое утверждение, неясно.
181
Двукратная кладка возможна, по НН .Щербаку [316], в Крыму и Северном Причерно-
морье. По предположению Т.Сатторова [225], в Северном Таджикистане у ящурок одна,
возможно, 2 кладки, происходящие в конце апреля — мае и в июне. ВЛБулахов [49]
для Днепропетровской обл. Украины также предполагает наличие 2 кладок — на основа-
нии растянутости периода размножения (с июня по август) и того, что у вскрытых самок
видны две генерации овоцитов. Учитывая, что указанное автором количество овоцитов в
каждой генерации (в среднем 9,2 и 6,9) заметно превышает средние (и даже максимальные)
размеры кладок разноцветной ящурки в Украине, а также, что результаты анализа только
размеров фолликулов в яичниках не могут определенно свидетельствовать о наличии 2 кла-
док (неоднозначен также факт растянутости периода размножения) , вывод В Л Булахова
представляется нам неубедительным.
Противоречивы данные по Калмыкии и Азербайджану. Так, В Л Бадмаева [21] сооб-
щила, что в Калмыкии готовые к откладке яйца появились в яйцеводах самок в начале
июня, а в первой половине июля наблюдалась вторая кладка. Из приведенной таблицы
[21] следует, что средняя масса половых органов самок крупных размеров (65—75 мм)
была максимальной во второй половине мая, затем шел резкий спад, а в первой половине
июля — новое возрастание массы. У мелких самок (55—65 мм) пик массы яиц и овоци-
тов также пришелся на вторую половину мая, а затем шло постепенное понижение этого
показателя. Во второй половине июля яичники всех самок находились в состоянии покоя.
Следовательно, период откладки яиц длился примерно со второй половины мая до середи-
ны июля. В более поздней работе ВИБадмаева [25] также сообщает о двух пиках роста
массы яичников, но уже в середине июня и начале августа (рис. 91,Г) , отмечая тут же, что
к июлю в яичниках прекращается развитие овоцитов. Откладка яиц на диаграмме [25, при-
ложение] показана с середины мая до конца второй декады июня и во второй декаде июля.
В А Киреев [113; сообщ.] в том же регионе наблюдал откладку первых яип у ящурок
7.VI. последнюю кладку зарегистрировал 3.VII и полагает, что у самок здесь имеется
1 кладка. В Азербайджане на о. Обливной и мысе Бендован, по данным МИ Ахмедова
[19, 20], у разноцветной ящурки, в отличие от других местных ящериц, имеется лишь
1 кладка, приходящаяся на вторую половину мая — первую половину июня. По Т Алиеву
[сообщ.] для тех же пунктов и годов, у ящурок 2 кладки — в мае — июне и в июле. Нако-
нец, АТ Банников [27] для Дагестана указал предположительно 2—3 (и даже 4) кладки
за сезон.
Опираясь на безуловно достоверные данные, можно в целом считать, что в ряде райо-
нов Средней Азии, Казахстана и в Дагестане у разноцветной ящурки обычно 2 кладки, в
западной части ареала вида — 1, при этом часть самок в отдельных популяциях может здесь
также давать 2 кладки. В высокогорных районах [77,324] бывает либо 2 кладки (Армения),
либо 1 (Прииссыккулье) . По мнению М.Ф.Тертышникова [258,263], ящурки Ставрополья
в течение года откладывают яйца только раз, что является приспособлением к перенесе-
нию ухудшающихся условий среды во второй половине лета. Согласно Н Л Щербаку
[320], на всей территории ареала, кроме Прииссыккулья, у Еarguta обычны 2 кладки; на
Восточном Кавказе возможны 3 кладки, в Средней Азии этому препятствует, по-видимо-
му, наступление очень сухого и жаркого лета.
Наиболее подробные сведения о второй кладке имеются для Дагестана и особенно для
Волго-Уральских песков (Западный Казахстан) . Установлено, что овоциты второй кладки
начинают развиваться в яичниках сразу после того, как яйца первой кладки выйдут в яйце-
воды [Окулова, сообщ.]. Во второй кладке участвует меньше самок, чем в первой: ящур-
ки, ставшие половозрелыми к концу периода размножения, успевают отложить за лето
липа одну кладку [Окулова, Хонякина, сообщ.].
В Дагестане в годы с теплой весной первая массовая откладка яиц у разноцветных
ящурок приходится на первую половину июня,вторая — на первую половину июля. Вслед-
ствие неодновременного созревания яиц у самок разных размеров, происходит перекрыва-
ние сроков первой и второй кладок, что усложняет анализ картины размножения [Хоня-
кина, сообщ.]. В Западном Казахстане первая кладка начинается в середине мая, вторая —
под конец месяца, и сроки обеих кладок существенно перекрываются (рис. 92) . Массовая
откладка яиц здесь происходит на рубеже мая и июня. В этом плане показателен следую-
щий пример [Окулова, сообщ.]: в течение июня 1960 г. из 20 самок впервые участвовали
в размножении 4, отложили яйца, но не содержали в яичниках развивающиеся ово-
циты второй кладки — 4 особи, одновременно имели в организме яйца 2 кладок —
8 самок.
182
В Украине вторая кладка достоверно из- вестна лишь у ящурок с морских побережий, при этом на Кривой косе признаки второй кладки имели 25,0 % из 8 самок, добытых 19.VI 1983 г., в окр. г Мариуполь — 23,8 % из 21 самки,отловленной 16—18.VI 1984 г., у с Приморское (окр. гБилково) — 42,9 % из Таблица 25. Интенсивность размножения самок разноцветной ящурки в разше годы (по данным НАС .Окуловой для Волго-Ураль- ских песков)
Год Размножаю- щиеся особи среди взрос- Среднее коли- чество яиц в кладке,
лых самок, % шт.
14 самок, добытых 1—3.VI 1985 г. (наши данные). Указанные популяции характеризу- ются крупными размерами взрослых особей, сравнительно большими кладками и высокой плотностью населения. Судя по состоянию по- ловых продуктов самок, откладка яиц первой кладки начинается, в зависимости от года, в 1958 79,0 (43) 3,90 (28) 1959 91,0 (83) 3,97 (62) 1960 70,1 (60) 3,50 (25) В среднем за 3 года 81,1 (186) 3,85 (115) Примечание. В скобках - количество вскрытых самок, экз.
последних числах мая — в первой половине июня, второй кладки — во второй половине
июня — первой половине июля.
По данным НМ.Окуловой [сообщ.] для Западного Казахстана, не все самки размно-
жаются ежегодно; интенсивность размножения популяции колеблется по годам (табл. 25)
и может зависеть от погоды (в годы с холодной весной этот показатель несколько снижа-
ется) и биотопа (в группе крупных самок в степи размножалось 80 % из 35 особей, а в пес-
ках — 98 % из 44) .
14.8. КОЛИЧЕСТВО ЯИЦ В КЛАДКЕ И ИХ ХАРАКТЕРИСТИКА
Плодовитость разноцветной ящурки различна в разных регионах и в различных популя-
циях одного региона (табл. 26) . В северных и западных частях ареала число яиц в кладке
колеблется от 1 до 7 (для Саратовской обл. указано до 10) , обычно составляя 3-4 яйца.
В южных и восточных районах-, в частности в Армении, Кабардино-Балкарии, Средней
Азии, на юге и юго-востоке Казахстана, ящурки могут откладывать до 11-12 яиц [145,
199. 222, 304, 313; Вашетко, сообщ.], в Голодной степи (юго-восток Узбекистана) в
яйцеводах самок обнаруживали до 14—15 яиц [198].
Таблица 26. Количество яиц в кладке и их характеристика в различных частях ареала разноцветной
ящурки
Регион Количество яиц в кладке Длина яйца, мм Ширина яйца, мм Масса яйца, мг Источник инфор- мации
min - max обычно или в среднем
Румыния (Дрбруджа) Украина 3-4 — 14 8 — [359]
различные регионы 2-6 2-3 — — — [49,66,201]
юго-западная часть 3-7 — — — — [328]
Среднее Приднепровье 1-3 — — — — [Ш]
Одесская обл. 2-7 — 9,8-12,4 73-8,8 — [250]
Крым Черноморский запо- 3-5 4,09* 10,5—16,0 7,0-93 — [316]
ведник 2-6 3-4 11,4-16,0 7,9-9,0 231-588 [121,123]
степная зона 2-7 3,57 11,4—18,9 6,8—9,2 231-685 [126, 386]
Кривая коса 3-7 5,13 12Д-15.7 7,6-10,1 280-580 1 Данные Т.И Ко- тенко
окр. г Мариуполь пересыпь Днестровско- 2-8 4,52 — — — То же
го лимана 3-5 4,00 12Д>-14Д 8Д—9,0 320-400
окр. г Бил ков о 3-7 4,62 13,9-16,5 8,1-10,0 433-601
Западное Предкавказье 2-7 — 11,0-15,0 6,0-8,6 — [Зинякова, со- общ.]
2-8 — 12,0—15,0 8,0-8,1 — [156]
3-4 — 12,0—15,0 8,0-8,1 — [70,155]
Центральное Предкавказье Др 5 — До 15 До 8 — [144]
2-5 3,60 14,0 8,0 520-600 [258,259,263]
3-7 3,60 14 8 — [266]
183
Окончание табл. 26
Регион Количество яиц в кладое Длина яйца, мм Ширина яШщ, мм Масса яйца, мг Источник инфор- мации
min ~ max обычно или в среднем
Северный Кавказ (Кабар-
диио-Балкария) 6—12 — До 132 ДР 7,0 — 004]
2-14 — — — — [179]
Армения (Севанская
котловина) 4-6 — 13-14 — — [77]
1-12 — — — — [145]
Азербайджан
разлитаые регионы 4-7 — — — — [86]
3-9 — До 15 7—9,5 — [7]
Апшерон 3-5 — Размером 7—9 мм — [85]
4-7 — — — [6]
о. Обливной и мыс
Беидован 2-8 — 13-14,5 8-83 — [20]
Дагестан 2-6 4 12,4 6,4 — [27]
2-7 4.14* 10-16 5,9-9,9 306-508 [278; Хонякина,
сообщ.]
Калмыкия 2-6 3-5 — — — [21]
Др9 — До 18 До 13 353 [25]
3-5 — 10,4-20,0 6,0-93 [113, 115; Кире-
ев, сообщ.]
Саратовская обл. 3-10 — — — — [310]
Волгоградская обл. — 5,0 и 4,0 — — — [168]
— — 14 7 420 [118]
Казахстан
Волго-Уральское
междуречье 1-7 11-16,5 — — [293]
1-7 3,7 16-18 — — [191]
2-6 3,73* — — — [ Окулова, со-
общ.]
Зяйсанская котловюа 4-6 4,75* 9,0—15,5 8,0-11,0 — [48]
Юго-Восточное При-
балхашье 4—11 — 16 10 — [313]
Джамбулская обл. 3-9 — 1 j Тб 438 [Колбинцев, со-
общ.]
Голодная степь 3-11 — 12 8,5 450 [199]
Узбекистан
Устюрт 3-8 — — — — [35]
Нуратжу 3-9 — 19-20 9,2-10 — [94]
До 8 — 10 82 — [НО]
Зеравшаяская долина €-10 — — — — 104]
Ддызакская стет» 7-9 — 13-14,5 9-93 — 223]
Голодная степь 11-15 13* — — — 198]
3-12 — 16-20 8-11 — [40]
6-15 — 13 6 — [35]
8 — 17-20 9 Д-11 — [92]
Ферганская долина 8-10 — 13 9 — 55, 56]
юго-востошкя часть 5-11 — 10Л*17 4,2-9,8 275-580 Вашетко, сообщ.]
Кыргызстан
Чуйская долина 2-10 4,91 10-15Д 6-93 — [323, 324]
Прииссыккулье 3-6 — 10 8-9 — [323, 324]
3-4 — 10,5-12 7-8 — {313]
другие районы 5-6 — 13Д-15Д 8,2-9,5 — [324]
Таджикистан 3— 11 7 ДО* — — — [222]
6-10 13-17 8-10 — [225]
Средняя Азия 4-12 — Размером 10-15 мм — [107]
Бывший СССР (эона пустынь) 3—7 — — — — [291]
Монголия (Джунгарская
Гоби) 2-6 — — — — [197,271]
Весь ареал вида 3-11 — До 16 — — 253]
1-12 3-4 10,5—20 6-10 306-508 320,436]
Примечание: Величина кладки определялась по количеству яиц в яйцеводах или крупных овоци-
тов в катанках, реже — по количеству отложенных яиц. * Числа, рассчитанные нами по данным указан-
ных авторов. Средине размеры или масса яиц, если это оговорено в источнике, подчеркнуты. Все чис-
ла даны с точностью, с которой они приведены в источниках.
184
Рис, 93, Зависимость плодовитости разноцветной
ящурки от длины туловища самок:
А: I - пески речных террас (Ивано-Рыбаль-
чанский участок Черноморского заповедника)
(п = 34, г = 0,55, Р > 99 %, у = 0,12х - 3,86);
Б: 2 — пески морских побережий, островов и кос
(о. Тевдра и Потиевский участок Черноморского
заповедника, оДжарылгач на Черном море, Ара-
батская стрелка и Кирилловская коса на Азов-
ском море) (л = 40, г = 0,62,Р > 99 %,у = 0,17х-
- 6,84); В - те же выборки, обработанные путем
разбивки на классы с усреднением количества
яиц [126, 131]; Г: 3 — низменный Дагестан
[ Хонякина, сообщ.], 4 - Волго-Уральские пес-
ки [ Окулова, сообщ.]
56~64 65-72
Длина туловища, мм
Во второй кладке яиц обычно нес-
колько меш»ше, чем в первой [191, 293;
Окулова, сообщ.], В частности, в Западном
Казахстане в 1960 г. среднее количество
яиц на одну самку составляло для первой
кладки 3,6, для второй — 3,2 [Окулова,
:ообщ.]. На юге Украины, согласно нашим
данным, такая закономерность не прояв-
пяется.
Даже близко расположенные популяции
разноцветной ящурки могут существенно
отличаться по плодовитости. Так, в Черно-
морском заповеднике ящурки на Ивано-
Рыбальчанском участке (пески речной тер-
расы) откладывают 2—6 яиц (3,26±0,15),
а на Потаевском (морское побережье) —
4—7 (5ДО±ОЗТ) , и вообще в Украине пло-
довитость популяций с морских побережий
и кос выше [126, табл. 1] (табл. 26) . Во
всех указанных случаях более высокая
плодовитость связана с более крупными
размерами ящурок в популяции. Имеются
указания на различия в плодовитости
E«arguta в зависимости от условий обитания (при сходных размерах самок) .В частности,
число-овоцитов разных генерации выше у особей из популяций, населяющих открытие про-
странства Приднепровья, по сравнению с таковыми из облесенных районов [49]. В Запад-
ном Казахстане среднее количество яиц на одну размножавшуюся самку в песках оказа-
лось ниже, чем в глинистой пустыне, — 3,5 и 4,2 соответственно [Окулова, сообщ.]. Напро-
тив, интенсивность размножения была выше в песках (см. разд. 14.7) .
По данным НМ .Окуловой [сообщ.], в популяции разноцветной ящурки интенсифика-
ция размножения может происходить как за счет заметного увеличения числа яиц в клад-
ке, так и в результате усиленного размножения более мелких ящурок. Например, в год
наиболее интенсивного размножения ящериц (1959 г., самый влажный за период 1958-
1960 гг.) количество яиц в кладке и процент размножавшихся самок были максимальны-
ми (табл. 25) , в 1960 г. оба показателя существенно снизились, причем последний — за
счет мелких особей. Так, если в 1959 г. среди самок длиной 55—64 мм размножалось 85 %,
то в 1960 г. — всего 56,5 %; более крупные самки в оба года размножались практически
с равной интенсивностью (97 и 91 % соответственно) .
В пределах популяции наблюдается положительная связь между размерами (возрас-
том) самок и количеством откладываемых ими яиц. Данная закономерность отмечалась
многими авторами [21,24, 25, 37—39,113,115,126,130,191,278,316,386; Киреев, Оку-
лова, Хонякина, сообщ.] (рис. 93) . Об этом же свидетельствуют и материалы ВИБадмае-
вой [21]: у мелких самок (размером до 65 мм) количество откладываемых яиц 2—4 (в
среднем 3) , у крупных (длиной 70—75 мм) — от 4 до 6 (в среднем 5) . Однако М.Ф.Тер-
185
Количество яиц в кладке
Рис. 94. Зависимость между количеством яиц
в кладке и средней массой яйца у разноцветной
ящурки (Хонякина, сообщ,]
тыпшиков такой зависимости не установил
и полагает, что малое количество яиц в
кладках разноцветной ящурки компенсиру-
ется быстрым созреванием и одинаковой пло-
довитостью самок разного возраста, что яв-
ляется приспособлением для сохранения ви-
да при низкой продолжительности жизни жи-
вотных [258, 259, 263]. По ВЛ£улахову
[49], абсолютная плодовитость возрастает
с увеличением длины туловища самок, но
относительная (т.е, количество яиц на 1 г
массы тела) почти всегда остается постоян-
ной и лишь у старых ящурок несколько
снижается. По нашим данным для Ивано-
Рыбальчанского участка Черноморского за-
поведника, относительная плодовитость
примерно на одном уровне (около 0,7) дер-
жится в течение всей жизни самок.
Левый яичник, как правило, продуцирует меньше яиц, чем правый; в левом яйцеводе
количество яиц обычно также меньше, чем в правом (48,126,130]. Так, на юге Украины в пра-
вом яичнике самок отмечалось до 4 развивающихся фолликулов, в левом — до 3, что в
среднем составило 2,00 и 1,75 (по 52 самкам), количество яиц в разных яйцеводах также
заметно различалось [126]. Вместе с тем данные ВИ£адмаевой [21,25], С А.Саид-Алиева
[222] и М Я Ахмедова [сообщ.] свидетельствуют об обратной закономерности; возмож-
но, это связано с иным определением сторон тела у животного. Изредка встречается пере-
Таблица 27. Характеристика зрелых яиц разноцветной ящурки из южных областей
Укравши [ 126]
Показатель Размер кладки Коли- чество яиц X mm шах X St t
Дшваяйце,мм I 41 11,4-18,9 14Д8 0,22 71 П 42 11,4-15,8 13Д8 0Д6 Все 83 11,4-18,9 13Д7 0Д4 Арика яйца, мм I 41 7,4-9Д 8ДЗ 0,06 7 П 42 63-9Д 7,86 0,09 Все 83 6,8-9,2 7,98 0,06 Индекс яйца I 41 1,49-2Д0 1,744 0,022 700 (отешмнедаи- II 42 1,38-1,96 1,687 0JJ18 ныкширпе) Все 83 1,38—2Д0 1,715 0,015 Масса яйца,мг I 31 268-685 406,8 20,6 7 17 П 38 231-568 352,4 15Д • Все 69 231-685 376 Л 12,8
Примечание. I - малые кладаш (2-3 яйввьр среднем 2^6±О,13), П - большие кладки
(4—5 среднем 4 ДО±ОДЗ) .
крестный переход овоцитов из одного яичника в противоположный яйцевод [48; Вашет-
ко, сообщ.]. По нашим наблюдениям, на юге Украины это довольно обьгшое явление,по-
скольку нередко добывали самок, у которых распределение желтых тел в яичниках и яиц
в яйцеводах было различным.
Масса и размеры готовых к откладке или уже отложенных яиц заметно варьируют
в пределах ареала, конкретной гюпуляции (табл. 26) , одной особи и даже кладки. Опреде-
ленная закономерность в географической изменчивости размеров яиц не просматривается:
наиболее крупные (длиной до 19—20 мм) яйца обнаружены в Узбекистане (Голодная
Стеш»и предгорья хребта Нуратау), Калмыкии и на юге Украины [40, 92, 94, 113, 126],
самые мелкие — в Прииссыккулье в Кыргызстане [313, 324] (табл. 26) . Интересный при-
мер, демонстрирующий изменчивость величины кладки и размеров и массы яиц у одной и
той же особи, а также потенциальную способность к частому размножению, реализовавшу-
юся в условиях неволи, привела Э .В .Вашетко [сообщ.]: разноцветная ящурка с длиной ту-
ловища 81,4 мм,добытая в конце мая у пос. Кассансай Наманганской обл. Узбекистана и
1К
содержавшаяся в террариуме вместе с самцами, отложила 14X1 5 яиц размером 10,0—
13,5 х 7—8 мм и массой 203—396 мг, 26.1 — 8 яиц величиной 10,0—13,8 х 7,5—83 мм и мас-
сой 190—380 мг, 17Л — еще 5 яиц размером 13,4—15,0 х 7,8—8,5 мм; погибла самка
15JV, при этом в яйцеводах у нее содержалось по одному яйцу размером 15 х 9 мм.
По данным В.А Киреева [113, 115; сообщ.], размеры яиц у старых (крупных) самок
обычно больше, чем у молодых. Другие исследователи такой закономерности не обнаружи-
ли, зато отметили зависимость размеров и массы яиц от величины кладки [126,130,179,386;
Хонякина, сообщ.] . Так, на юге Украины у самок с небольшим количеством (2—3) яиц в
яйцеводах яйца крупнее, чем у особей с 4—5 яйцами, при этом яйца в малых кладках бо-
лее вытянуты (табл. 27) . Поскольку малые кладки характерны для молодых ящурок, то
именно у этих особей яйца более крупные. Обратная связь между количеством яиц в
кладке и средней массой яйца хорошо показана для Дагестана (рис. 94).
Данные по соотношению длины и ширины яиц немногочисленны: в Восточном Казах-
стане [48] этот индекс составлял в среднем 1,6 (1,1—2,0) , на юге Украины [126, 131] —
1,7 (1,4-2,1) (табл. 27).
14.9. МЕСТО И ПРОЦЕСС ОТКЛАДКИ ЯИЦ
Разноцветные ящурки откладывают яйца в норах или в специально вырытых ямках [27,
86, 113, 115, 125, 126, 258, 259, 266; Зинякова, Киреев, Окулова, Хонякина, сообщ.].
В Калмыкии на плотных почвах ящурки откладывают яйца в заброшенных норах грызу-
нов, а на бурых песчаных и супесчаных почвах (пустынная зона республики) — в специаль-
но вырытых норках глубиной 5—8 см [113, 115; Киреев, сообщ.]. В Азербайджане яйца
помещаются в небольшие норки [86], на Ставропольской возвышенности [258, 259] —
в норы или ямки в местах, достаточно увлажненных и хорошо прогреваемых солнцем, при
этом яйца самка размещает в горизонтальном положении и засыпает почвой так, что они
оказываются на глубине 4—6 см.
В Дагестане [27] яйца откладываются в песок на глубину 6—8 см, где влажность суб-
страта составляет около 10 %. На Кизилташской косе в Западном Предкавказье [Зиняко-
ва, сообщ.] 5 выявленных кладок (4—7 яиц) разноцветной ящурки располагались в рых-
лом песке на глубине 10—15 см на южных склонах дюн ближе к гребню. В Черноморском
заповеднике на юге Украины [125] при раскопке жилой норы ящурки на глубине 13 см
обнаружена кладка из 3 яиц, слегка зарытых в песок в стенке норы. В Западном Казахста-
не ящурки охотно закапывают свои яйца в рыхлую, хорошо прогреваемую землю курган-
чиков малого суслика [194] . В частности, 2 кладки были найдены на таких курганчиках
в глинистой полынной пустыне. Первая находилась в ямке глубиной 14 см, выкопанной
в рыхлой почве, вторая — в ямке глубиной 6 см и диаметром 3 см, вырытой в плотном
грунте. Обе кладки были засыпаны землей вровень с поверхностью, температура почвы на
дне ямок в конце июня составляла 24 и 26° [Окулова, сообщ.] . В Дагестане [Хонякина,
сообщ.] наблюдались миграции самок разноцветной ящурки, связанные с откладкой
яиц: в июне на песках, граничащих с глинистыми, солончаковыми и суглинистыми почва-
ми, отмечалось резкое увеличение численности самок. Песок привлекает этих ящериц
рыхлостью, влажностью, хорошей прогреваемостью.
По данным В А Киреева [сообщ.] , у ящурок из восточных районов Калмыкии наблю-
далась порционная откладка яиц, что, вероятно, характерно и для других частей республи-
ки. При этом яйца откладываются в двух, а иногда и в трех местах. СБ.Таращук [250]
факт, что у большинства более крупных самок количество яиц при вскрытии было мень-
ше, чем у мелких особей, объясняет тем, что разноцветная ящурка откладывает яйца пор-
ционно. Учитывая, однако, что разница в размерах самок двух групп составляла немногим
более 1 мм, а объем выборок был 6 и 8 особей, вывод трудно признать убедительным. В Л .Бу-
лахов [49] также заключает, что откладка яиц происходит в нескольких местах, — на осно-
вании того, что в яичниках самок обнаруживалось в среднем 16,1 овоцита (первой гене-
рации — 9,2, второй — 6,9), а в кладках отмечалось 2—6 яиц. Наиболее обосновано,
на наш взгляд, предположение ЗЛХонякиной [сообщ.] о наличии порционной отклад-
ки яиц, поскольку она не раз добывала самок,.у которых желтых тел было больше, чем
яиц в яйцеводах.
187
В других работах данный вопрос не обсуждался, но обычно подразумевалась отклад-
ка яиц одной порцией, поскольку в разных частях ареала обнаруживали кладки разноцвет-
ной ящурки с количеством яиц, близким к характерной для конкретного региона величи-
не всей кладки, определяемой по числу яиц в яйцеводах или крупных фолликулов в яич-
никах. В частности, были найдены кладки с количеством яиц до 5 [250; Окулова, сообщ.],
7 [Зинякова, сообщ.] и даже 11 [313]. Нам на юге Украины не попадались самки, у кото-
рых количество желтых тел не совпадало с количеством яиц в яйцеводах. Наблюдения
ВХКолбинцева [сообщ.] над ящурками, содержавшимися в террариуме, также свидетель-
ствуют в пользу однопорционной кладки: одна из самок отложила 9 яиц в течение 2 ч с ми-
нимальными интервалами в 10 мин, после чего сгребла яйца в более компактную кучку и
засыпала песком.
Приведенные выше факты противоречивы и не позволяют сделать окончательный вы-
вод о процессе откладки яиц разноцветной ящуркой.
14.10. ДЛИТЕЛЬНОСТЬ ИНКУБАЦИИ ЯИЦ
И ПОЯВЛЕНИЕ МОЛОДЫХ ОСОБЕЙ
О сроках инкубации в природе судят по времени откладки яиц и появления сеголеток,
так как прямых наблюдений за мечеными кладками нет. В частности, ИД Яковлева [324]
отметила, что в Кыргызстане от времени спаривания до появления молодых особей прохо-
дит не менее 2 мес. М.Ф.Тертышников [258] указал для Ставрополья, что инкубационный
период длится около 2 мес, однако конкретный материал [259] не позволяет сделать опре-
деленный вывод. В Азербайджане длительность инкубации яиц составила 40—45 дней [Ах-
медов, сообщ.] . Наблюдения на стационаре в Черноморском заповеднике (юг Украины)
показали [123, 124, 126, 386], что продолжительность инкубационного периода значитель-
но варьирует даже в пределах одной популяции и зависит от сроков откладки яиц (связа-
но с сезонными изменениями факторов среды) и погодных особенностей года. Так, из
яиц, отложенных в первой половине июня, сеголетки вывелись через 30—40 дней в 1975 г.
(ранняя теплая весна, жаркое лето) и через 45—55 дней — в 1974 г. (менее благоприятные
погодные условия), а из отложенных во второй половине июля — спустя 55—70 дней.
ВТКолбинцев [сообщ.] инкубировал кладку из 9 яиц на влажном простерилизованном
песке при температуре 29°, и через 37 дней из всех яиц появились молодые ящурки. По
данным В А Киреева [сообщ.}, ящурки из яиц одной кладки вылупляются в течение 2—3
дней в различное время суток.
Сроки появления первых сеголеток существенно отличаются дня разных частей ареа-
ла, поскольку тесно связаны со сроками начала откладки яиц. Раньше всего молодые
ящурки появляются в юго-восточной части Узбекистана — в различных числах июня [92,
104, 107, 110, 198] и даже в середине мая [92]. В Чуйской долине (Кыргызстан) первые
сеголетки отмечались в июне [324], в Джамбулской обл. Казахстана — с конца июня
[Копбшщев, сообщ.], в других частях Казахстана — с 10.VII по 5.УШ [12,199], в Азер-
байджане — в начале июля [20] (по Б А Домбровскому [91] - в первой декаде августа) ,
в Дагестане — во второй половине июля [Хонякина, сообщ.], в Северном Таджикистане —
в конце этого месяца [225], в Калмыкии и Саратовской обл. России — в конце июля
[25, 113, 310; Киреев, сообщ.], в Монголии — в конце июля — начале августа [271].
В Крыму первые сеголетки появляются в начале августа [316], в Одесской обл. —
с 15.VII до середины августа, в зависимости от года [250], в Херсонской обл. Украины
(в Черноморском заповеднике) — обычно в третьей декаде июля, но в отдельные годы —
гораздо раньше или позже (в 1975 г. — 6.УП,в 1978 г. — 5УШ, т.е.разница в сроках мо-
жет достигать 1 мес), и продолжают вылупляться до конца сентября — начала октября
[124, 131, 386]. Позже всего молодые ящурки появляются на северо-западной границе
ареала вида (в Черкасской обл. Украины) и в горах Прииссыккулья — во второй декаде
августа и в августе — сентябре соответственно [151,324].
Большой разброс сроков появления первых сеголеток для одного и того же региона
нередко объясняется не только, зависимостью сроков вылупления от погодных условий,
но и неполнотой сведений: часто пропускается начало появления сеголеток и указывает-
ся лишь массовое появление, которое обычно имеет место через 2—4 недели после появле-
188
ния первых особей. Последние сеголетки вылупляются в различных регионах примерно
в одно и то же время — в конце сентября, реже в начале октября. Растянутость сроков
рождения молодых ящурок в популяции обусловлена как растянутостью периода отклад-
ки самками яиц, так и разной продолжительностью их инкубации (см. выше) .
3 ЛХонякина [сообщ.] предположила, что из некоторых отложенных в конце лета
яиц при благоприятных погодных условиях зимы вылупление молоди может происходить
и весной, так как очень маленьких ящурок она иногда встречала в середине мая. Учитывая,
однако, поздние сроки рождения последних сеголеток и особенности роста молодых ящу-
рок осенью (см. разд. 14.10 и 15.1.) , данные 3 Л Хонякиной можно объяснить и без пере-
зимовки яиц.
Размеры и масса только что родившихся ящурок, судя по данным различных авторов
[12, 20, 25, 27,49,77,92, ИЗ, 124, 131,227; 259,263,316,324; Зинякова, Колбиниев,
Окулова, Хонякина, сообщ.] , сильно варьируют не только в пределах всего ареала, но и
внутри отдельных популяций. Наименьшая длина туловища указана для новорожденных
из юго-восточной части Узбекистана (20—25 мм) и Волго-Уральского междуречья (22 мм),
наибольшая — для молоди из Херсонской обл. Украины, Джамбулской обл. Казахстана
(до 33,7 мм) и Западного Предкавказья (до 35 мм) [92,124; Зинякова,Колбинцев, Оку-
лова, сообш.]. Крупные размеры для сеголеток Казахстана указал также КЛЛараскив
[199] — 37 мм для 5-дневной особи; но в большинстве регионов ящурки рождаются дли-
ной 27—30 мм. В популяции длина туловища и масса тела недавно родившихся особей мо-
гут колебаться в пределах, например, 27,5—33,7 мм и 0,45—0,97 для ящурок с Ивано-Ры-
бальчанского участка Черноморского заповедника [124, 131] или 28—35 мм и 0,66-
0,89 г — для сеголеток с Кизилташской косы [Зинякова, сообщ.] Особи (9 экз.), вылу-
пившиеся из яиц одной кладки, имели длину туловища 32,0—33,7 мм, длину хвоста 40,0-
45,0 мм,массу тела 0,73—0,80 г [Колбинцев, сообщ.]
ГЛАВА 15
РАЗВИТИЕ И РОСТ. СТРУКТУРА ПОПУЛЯЦИЙ1
15.1.ТЕМПЫ РОСТА
По постэмбриональному росту ящериц имеются отдельные данные в литературе [113,
156,199,228,229,259,266,278, 293,316,320,324].
При длине туловища 25—28 мм масса новорожденных ящериц варьирует от 0,3 до 0,4 г
(0,35±0,04; п = 178) [Тертышников, сообщ.] . В Черноморском заповеднике (юг Украи-
ны) недавно вылущившиеся сеголетки имели длину туловища 27,5—33,7(30,69±0,19) мм
[124]. В связи с неодновременностью откладки яиц и, следовательно, неодновременностью
появления новорожденных эта возрастная группа в популяциях ящериц представлена осо-
бями, имеющими перед уходом в спячку различную длину туловища и хвоста, разную мас-
су. Так, по данным НШЩербака [316, 320], у сеголеток разноцветной ящурки из Крыма
при длине туловища 27 мм в первый месяц размер его увеличивается на 8,7 мм, во вто-
рой — 5,3, в третий — на 3,2 мм, т.е. за 3 мес до зимовки они выросли на 17,2 мм.М.Ф.Тер-
тышников [259, 263] установил, что в Центральном Предкавказье тело новорожденных
ящурок за вторую половину июля, август, сентябрь и первую половину октября увеличи-
вается на 15 мм при средней скорости роста, равной почти 5 мм в месяц.
Рис. 95. Изменение длины туловища разноцветной ящурки с возрастом по данным мечения
на постоянной площадке (Ивано-Рыбальчанский участок Черноморского заповедника)
[131]:
1—3 — ящурки 1974 г. рождения (J — родившиеся в июле, 2 — родившиеся в августе,
3 — особи с замедленными темпами роста) ; 4, 5 - ящурки 1975 г. рождения (4 — родив-
шиеся в июле, 5 — родившиеся в августе) . Пунктиром даны кривые на тех отрезках, где из-
мерения ящурок не производились
ТЯКотенко при наблюдениях за ящурками Черноморского заповедника установила,
что родившиеся в июле — начале августа сеголетки достигают до ухода в спячку длины ту-
ловища 37—49 мм (средний прирост 10 мм), а у родившихся в конце августа — сентябре
туловище увеличивается до 32—36 мм (средний прирост 2,7 мм). Сеголетки,родившиеся
в конце сентября — начале октября, почти не растут [124,386].
По данным В А Киреева [сообщ.], у сеголеток, вылупившихся в августе, длина туло-
вища достигает 28 мм и к уходу в спячку увеличивается до 45 мм. Следовательно, прирост
достигает 17 мм при средней скорости роста 5,4 мм/мес.
1 Материалы предоставили В .И .Бадмаева, В А Киреев, ТИКотенко, АН Рудик.
190
Во время зимней спячки ящерицы практически не растут, но после выхода из спячки
размеры животных продолжают увеличиваться. По-видимому, прежние темпы роста со-
храняются по достижении половозрел ости, после чего снижаются. При рассмотрении имею-
щихся в литературе данных [113,124,131,239,263,316,320); Киреев, сооб.; идр,] о ско-
рости роста ящурок и времени наступления половозрелости видно, что в первые месяцы
после спячки средняя скорость роста ящериц лежит в прежних пределах, в частности в
Центральном Предкавказье — около 5 3 мм/мес, на юге Украины в различные годы — 4,0—
8р [124, 386]. Затем скорость роста (после размножения) снижается. По данным ТЯКо-
тенко, в июле скорость роста ящурок снижается (рис. 95) , с конца августа и до ухода на
вторую зимовку они почти не растут и в спячку залегают раньше очередных сеголетков
[124,131].
Возможно, ящурки, родившиеся из поздно отложенных
яиц (в августе — сентябре) , в группу половозрелых особей
входят на вторую весну, т.е. на 3-м году жизни [Киреев,
сообщ.]. После второй зимовки скорость роста ящурок со-
ставляет уже 1—3 мм/мес. По нашим данным [259, 263],
наступление половозрелости не ограничивает увеличение раз-
меров животных. В то же время Т И Котенко [124] показа-
ла, что в возрасте 2 лет часть ящурок (в основном самки)
перестает расти, остальные растут, хотя и медленно, до кон-
ца жизни.
Как показывают материалы исследований различных
герпетологов, в разных частях ареала половозрелость насту-
пает при различной длине туловища (см. разд. 14.1) .
НЯДЦербак [320] , анализируя темпы роста и время на-
Рис. 96. Кривая роста и продол-
жительность жизни ящериц
1259]:
7 - кривая роста, 2 - вре-
мя наступления половозрелости
ступления половозрелости у ящурок в пределах ареала, от-
метил, что по сравнению с особями из равнинной части ареала у ящурок из высо-
когорья темп роста более медленный, однако они достигают более крупных размеров и
характеризуются большей продолжительностью жизни.
Как показали исследования ТЯКотенко [124, 131,386] в Черноморском заповедни-
ке, скорость и характер роста ящурки зависят от времени рождения, условий года, пола и
обнаруживают значительную индивидуальную изменчивость. Скорость роста ящериц даже
одного пола, возраста и примерно одинаковых размеров в отдельные периоды жизни мо-
жет отличаться почти в 2 раза. В результате этого к концу мая — началу июня происходит
частичное перекрывание размеров разновозрастных ящурок предыдущего года рождения,
а к концу июня перекрываются размеры многих особей, отличающихся по возрасту на год,
и уже в августе невозможно отличить ящурок разных возрастных групп по длине тулови-
ща [124]. По мере роста у ящурок изменяются пропорции тела: с возрастом относительная
длина головы и хвоста уменьшается, причем в большей степени у самок, что с возрастом
ведет к усилению полового диморфизма [386].
О продолжительности жизня ящурок в пределах ареала единого мнения нет. Так, для
Кыргызстана предельный срок жизни составляет 5 лет [324], для Калмыкии — 2-3,изред-
ка 4 года [Киреев, сообщ.]. По данным Т Я Котенко [123,124, 386], ящурки на юге Укра-
ины живут обычно 3 года, реже — дольше. Мы считаем [259, 263] , что в Центральном Пред-
кавказье ящурка живет 3 года, а некоторые, возможно, доживают до 4-летнего возраста
(рис. 96). Видимо, к этому времени они достигают окончательных размеров: дли-
на туловища самок — 72 мм, самцов — 80. Так же постепенно с возрастом изменяется
и масса ящурок: при длине туловища 30, 35, 40, 45, 50, 55, 60, 65, 70, 75 и 80 мм соответ-
ственно 0,7; 1,1; 1,8; 3,1; 3,7; 5,1; 7,0; 8,2; 9,5; 12,0и15г.
15.2. СТРУКТУРА ПОПУЛЯЦИЙ
Сведений о структуре популяций (возрастном и половом составе) и их пространственном
распределении крайне мало, в излагаемом ниже материале использованы данные исследова-
ний по Центральному Предкавказью [259] и Калмыкии [25].
Возрастная структура популяции. Имеющиеся данные о темпах роста позволяют выде-
191
Т а б л i ц I 28. Возрастные группы соотношение иолов в популяции разноцветной ящурки в Цен-
тральном Предкавказье (верховья р.Томузловки) в 1966-1971 гг. [ 259]
Возрастная группа Самки Самцы Всего
Количе- ство, жз. % в по- пуляции % в во- зраст- ной группе Количе- ство, ЭКЗ.' % в по- пуляции % в воз- растной группе Количе- ство, экз. % в по- пуляции % в воз- растной группе
Сеголетки 48 27,0 50,0 48 24,0 50,0 96 25,6 100,0
Полувзрослые Взрослые 19 10,8 36,6 33 17,0 64,4 52 14,0 100,0
1-я группа 93 52,6 50,2 92 46,0 49,8 185 49.2 100,0
2-я группа 17 9,6 40,0 25 13,0 60,0 42 11,2 100,0
Всего 177 100,0 47,0 198 100,0 53,0 375 100,0 100.0
40-
20-
Ю-
Рис, 97, Структура популяции разноцветной ящур-
ки в Калмыкии в разные месяцы [ 25]:
1 - особи генерация первого года, 2 — особи
генерации второго года, 3 - особи генерации пре-
дыдупмх лет (3-4 года)
пять в популяции несколько возрастных
групп: особи этого года рождения (сеголет-
ки) , перезимовавшие и не достигшие поло-
вой зрелости (полувзрослые) и более стар-
шие по возрасту животные, включающие
только половозрелых особей. Мы [259] в
популяции яшурок из верховьев р.Томуз-
ловки среди половозрелых особей выдели-
ли две группы: взрослых 1-й группы и взро-
слых 2-й группы (табл. 28) . По этим мате-
риалам к сеголеткам отнесены особи данно-
го года рождения с длиной туловища до
40 мм, к группе полувзрослых — перезимо-
вавшие особи и не вступившие в размноже-
ние с длиной туловища до 55 мм (данная
возрастная группа существует 2—4 мес и,
достигнув указанных размеров, вступает в
размножение), взрослые 1-й группы с дли-
ной тела 55—70 мм, вступающие в размно-
жение сразу после выхода из спячки, и, на-
конец, взрослые 2-й группы, вступающие в
размножение весной повторно.
Близкие по содержанию материалы
приводит ВМБадмаева [25], описывая воз-
растную структуру популяции ящурки раз-
ноцветной в Калмыкии.
Описанная выше структура популяции
характерна для нюня, когда полувзрослые
переходят в группу половозрелых, но еще
не начавших размножение, и одновременно
появляются сеголетки, вышедшие из яиц
первых кладок. В другие месяцы структура популяций иная. Так, В Я Бадмаева [25],
изучая возрастную структуру популяций ящурки разноцветной в Калмыкии, установила,
что в апреле (вскоре после выхода из зимней спячки) популяция состоит из молодых
и половозрелых двух групп, в мае молодых меньше, поскольку часть их переходит в 1-ю
группу половозрелых, в июне в популяции только 1-я и 2-я группы половозрелых, в июле к
ним добавляется группа сеголеток, в августе — сентябре популяция состоит из 30—50 %
особей генерации текущего года, 45 % в августе и 35 % в сентябре — из особей генерации
2-го года и 20 % особей 3-го и, возможно, 4-го годов (рис. 97). Видимо, в конце сентября
— в октябре в популяции должны быть встречены в активном состоянии только сеголетки,
поскольку ож дольше других задерживаются на поверхности перед зимовкой.
Приведенная схема-модель возрастного состава популяции пригодна лишь для равнин-
ных популяций западной части ареала, где продолжительность жизни ящериц находится в
пределах 3—4 лет, но не годна для высокогорий, где продолжительность жизни этих живот-
192
Таблица 29. Биотопическое ра»
пределение разноцветной ящурки hl
стационаре в верховьях р.Томузлов-
ки в Центральном Предкавказье
[259]
Биотоп Пло- щадь биотопа, га Площадь, занятая видом,%
Разнотравно-зла- ковая степь Типчаково-по- 80,3 3,0
лынная степь 4,5 —
Полынная степь Опустыненная 4,5 -
степь 31,8 100.0
Посевы Полезащитные 12,5 24,5
лесные полосы 27,0 16,0
Толока 0,6 —
Лес 8,2 —
Лесные опушки Вырубки и про- 0,4 —
секи в лесу 0,2 —
Всего 170,0 24,0
ных больше. Возрастной состав популяции на той или
иной части видового ареала может быть усложнен
разнообразными факторами: увеличением количества
кладок в сезон, порционностью откладки яиц, что при-
водит к увеличению приплода, и растянутостью его по-
явления. низкой плодовитостью старых самок, значи-
тельным колебанием численности самок и эмбриональ-
ной смертностью, что может повлиять на присутствие
в популяции той или иной возрастной группы и др.
Половая структура популяции. Сведений, отражаю-
щих соотношение полов в популяции ящурок, немно-
го. Так, НН Щербак [316] указывает для популяций
из Крыма отношение самцов и самок, равное 1,08:1.
ВИБадмаева [125] приводит сведения для Калмы-
кии, из которых следует, что среди молодых особей
соотношение самцов и самок равно 1:1,7, среди взрос-
лых 1-й и 2-й групп — 1:7,3. Разрозненные данные
ЭБВашетко [сообщ.] показывают, что соотношение
самцов и самок в Ферганской долине в апреле близко
1:1, в Самаркандской обл. в мае — 1,4:1, в Нуратау
в ноябре — 2:1.
Приведенные данные не отражают истинного соот-
ношения особей разного пола б популяциях, посколь-
ку в них не учтена, например, активность животных разного пола в разные периоды сезон-
ной активности, пресс хищников на животных разного пола и возраста, возрастная струк-
тура популяции в разные периоды весенне-осенней активности и др. По данным Т И Котен-
ко [126], соотношение полов в выборке существенно зависит от сезона, что связано с раз-
ным уровнем активности самцов и самок в отдельные месяцы. В частности, в апреле — мае
самцы наиболее активны и преобладают в выборках за этот период года, в июне — авгус-
те могут превалировать самки (в Черноморском заповеднике весной в выборках было
73 % самцов и 27 % самок, летом соответственно 38,5 и 61,5 %) ,в то время как истинное
соотношение полов в популяции было близко 1:1.
Имеется ряд материалов, показывающих, что действительное соотношение полов в по-
пуляции примерно 1:1 для сеголеток. Однако в дальнейшем под влиянием различных фак-
торов среды процентное соотношение полов в популяции может измениться (табл. 28)
Так, у сеголеток соотношение самцов и самок равно 1:1, в группе полувзрослых (моло-
дых, еще не размножавшихся после выхода из зимовки) — 1:1,76, в группе 1-й взрослой —
1:1,01. во 2-й взрослой группе — 1:1,5. К сожалению, механизм этих процессов изучен сла-
бо. а приведенный пример показывает, что во всех возрастных группах, кроме сеголеток,
смертность самцов несколько выше, чем самок.
Пространственная структура популяции. В пределах ареала разноцветная ящурка оби-
тает в определенных типах биотопов в степных, полупустынных и пустынных ландшафтах.
В связи с неоднородностью занимаемого пространства в плане ’’удобств” для жизни ящу-
рок, а также из-за некоторых особенностей их биологии прослеживается неравномерное
размещение членов популяций. Ящерицы в редком случае занимают всю территорию
биотопа, ландшафтной зоны. В качестве примера мы приводим данные о размещении
ящурки разноцветной на одном из стационарных участков в Центральном Предкавказье
(табл., 29) . Анализируя эти материалы, можно сделать вывод о том, что наиболее ’’удоб-
ный” биотоп — опустыненная степь, в меньшей степени — разнотравно-злаковая степь. По-
севы и полезащитные лесные полосы заселяются ящуркой только по границе с опустынен-
ной степью; поселения возникли в результате расселения мигрантов.
Исследование биотопического и специального размещения ящурки показывает, что не-
равномерное размещение членов популяции проявляется в разных точках ареала. Обычно
размещение животных может быть мозаичным, соответствуя аналогичному распределению
типичных местообитаний в биотопе, типичных биотопов в ландшафте. Примером такого
типа размещения животных могут служить обширные пространства сухих степей на При-
каспийской низменности, имеющие примерно равные условия для обитания на огромной
территории. Не менее распространенным типом размещения является ленточный. Приме-
193
Рис. 98. Поселения разноцветной ящурки в опустыненной степи в Центральном Предкавказье
[259]/
А — 1969 г., Б - 1971 г.; 1 — взрослый самец, 2 — взрослая самка, 3 — молодая особь, 4 -
камни, 5 - травянистый покров, 6 - обрывчик, 7 - границы индивидуальной территории,£ -
нора
ром его могут быть песчано-каменистые степи по склонам Прикалаусских высот в Цен-
тральном Предкавказье [259] или песчаные террасы вдоль небольших речек, просек, дорог
в Харьковской обл. Украины [Рудик, сообщ.].
Между указанными типами размещения животных имеются переходные варианты,
определяемые системой взаимоотношений между членами популяций в различные периоды
активности.
Внутри популяций ящерицы также распределены неравномерно [259]. Они. как прави-
ло, концентрируются у различных убежищ — нор мелких млекопитающих, камней, неболь-
ших кустов травянистых растений и др., в связи с чем формируются небольшие грушы
особей — внутршюпуляционные территориальные группировки (рис. 98) . Имеются наблю-
дения, что такие группировки состоят из 5—6 особей и даже 8 [Рудик,сообщ.] .Ящерицы
в такой группировке нередко прячутся при опасности в одни и те же убежища. Они ведут
одиночный образ жизни (см. гл. 17), переходящий, по-видимому, в одиночно-семейный в
период размножения. При этом некоторые особи занимают норы в непосредственной бли-
зости друг от друга, ’’соседи” обычно успешно избегают частых встреч, активная защита
территорю! проявляется самцами по отношению к мигрантам из других внутрипопуляци-
онных территориальных группировок и в меньшей степени — к своим, чаше в период раз-
множения и меньше вне его.
Каждая территориальная группировка отделена от соседней какой-либо нетипичной
стацией, что исключает между ними частый контакт, но поддерживается кочующими осо-
бями: самцами в период размножения, молодыми (новорожденными) в период расселе-
ния (рис. 98) . Изменение условий обитания может привести как к слиянию территориаль-
ных группировок, так и к исчезновению их.
ГЛАВА 16
ЛИНЬКА1
16.1. СРОКИ И КОЛИЧЕСТВО ЛИНЕК
Сведения о линьке разноцветной ящурки сравнительно немногочисленны и порою проти-
воречивы.. Так, на юго-востоке Украины 1—13 .IX наблюдались как уже перелинявшие осо-
би, так и находившиеся в процессе линьки или еще не линявшие [66]. В окр. городов Фео-
досия и Керчь (Крым) линька отмечена 20 .IV 1959 г., 12 и 16 JX 1958 г. [316]. В Дагеста-
не линяющих ящурок встречали с мая по сентябрь включительно, особенно часто — в кон-
це мая, в июне и начале августа, при этом у большинства взрослых особей в году бывает
2—3 линьки [Хонякина, сообщ.].
В Зайсанской котловине (Казахстан) первые линяющие ящурки встречались со второй
декады июня [48], в Казахском нагорье линька наблюдалась с середины июля до середи-
ны августа, причем только у взрослых особей [12]. На юге Джамбулской обл.Казахстана
ящурки, по мнению ВТКолбинцева [сообщ.), линяют не менее 3 раз в год — после спари-
вания, летом и перед уходом на зимовку, поскольку линяющие особи наблюдались 6.V,
8.VH, 19 JX, 14 и 21X. В Западном Казахстане первая линька происходила во второй поло-
вине мая — июне, вторая — в конце августа — начале сентября, когда линяли и взрослые, и
молодые ящурки [Окулова, сообщ,].
На Ставропольской возвышенности первые линяющие особи отмечались в июне, а
повторная лидока — в июле — сентябре, при этом неодновремённость протекания этого
процесса у разшпшых особей в популяции создавала ложное впечатление значительно боль-
шего <шсла линек у одного животного [259]. На Кизилташской косе (Западное Предкав-
казье) массовая линька тоже бывает 2 раза в год — весной и осенью. Во второй половине
мая встречались только перелинявшие и линяющие ящурки, в сентябре 86 % осмотренных
особей линягш. Кроме того, линяющие ящурки отмечены и в июне [Зинякова, сообщ.].
ВЛ .Бадмаева [24, 25] также сообщает о двукратной линьке яшурок в Калмыкии:
первая массовая линька в популяции происходит с 20JV по 10.V (из добытых 209 особей
169 находились в процессе линьки или недавно перелиняли), вторая — с 10 VIII по 10JX,
при этом в промежутке между указанными сроками в 1973—1982 гг. взрослых линяющих
особей не встречали. Сеголетки линяют с конца июля по конец сентября и даже в ноябре:
из 12 сеголеток, добытых 2X1 1981 г, в окр. г, Элиста, 4 были с признаками диньки [25].
Вместе с тем В А Киреев для той же Калмыкии сообщает о наличии у разждаетной ящур-
ки 3—4 линек в году [114], не менее 4 линек [113] и даже 4—5 в течение лета (для моло-
дых еще чаще) , при этом первые линяющие особи встречались через 15—20 дней после вы-
хода из зимних убежищ [115]. Первую линьку этот автор наблюдал в конце апреля, вто-
рую — в мюле, третью — в конце сентября, перед уходом на зимовку [113]. Массовая линь-
ка отмечалась в первой половине мая, в некоторые годы ее сроки могут сдвигаться в ту
или иную сторону на 5—10 дней. В годы с холодной весной ящурки дольше остаются в
старом покрове, а в теплую и дружную весну линяют в более сжатые сроки. Осенью линя-
ют, как правило, не все взрослые особи. Большая часть сеголеток перед уходом на зимов-
ку линяет [Киреев, сообщ.]. Ящурки 2-го года жизни линяют не менее 4 раз. Аналогичные
данные получены и в условиях неволи [113].
Для Азербайджана также указаны 3 линьки: на о.Обливной (Каспийское море) первая
1 При участии МЛ Ахмедова, В Л .Бадмаевой, И.СЛаревского, МЛЗинжковой, В А Киреев а , ВТ Кол-
бинцева, НМ .Окуловой, Т.Сатторова, М .Ф .Тертышникова, ХТзрбиша, 3 ЛХоыякиной.
195
линька начинается в конце апреля и продолжается до первой половины июня, в прибреж-
ной полосе материка она происходит несколько раньше — со второй декады апреля до кон-
ца мая. Вторая линька протекает летом, с начала июля до второй декады августа, третья на-
чинается в конце сентября и продолжается до ухода животных в зимнюю спячку как на
острове, так и на материке. Сеголетки, появившиеся в начале июля, впервые линяют в
августе [20]. (По другим источникам [19], сеголетки здесь линяют спустя 10-15 дней
после вылупления из яиц.) AM Алекперов [7] отметил лишь одну линьку у ящурок из
Азербайджана, длившуюся с 10—15.V по 3— 10.V1.
В Узбекистане линяющие особи часто попадались в мае [35], в Ферганской долине на-
блюдались в июне и июле [55, 56]. В Кыргызстане линных ящурок встречали в окр. г. Ош —
17.V, в Прииссыккулье — в конце июля — начале августа, в Чуйской долине — 5.III, 6—
14.V и 26X. В неволе ящурки линяли 4 раза: 3JV, 8.VI, 1 .VIII и ЮХ [324]. В Северном
Таджикистане ящурки с признаками линьки попадались 7—10.V и 3—8.VIII 1978 г. [225].
У ящериц Средней Азии массовая линька происходит вскоре после выхода с зимовки и
продолжается с марта до конца мая. Единичные линяющие особи встречаются в течение
всего лета, а в середине лета и осенью их несколько больше, что позволяет говорить о
3 линьках пресмыкающихся в году. Наиболее дружно и в сжатые сроки происходит весен-
няя линька, несколько растянута и не такая массовая осенняя и наименее выражена лет-
няя. Сеголетки до ухода в спячку успевают перелинять несколько раз [38, 39]. В Джун-
гарской Гоби (Монголия) массовая линька разноцветных ящурок наблюдалась в первой
половине мая: из 21 особи линяли 66,6 %. В середине июня из 9 пойманных животных
линяющие составили 44,4 %, в начале сентября из 14 экз. — 35,7 %. В середине июля из 7
добытых ящурок ни одна не линяла [Тэрбиш, сообщ.].
Отмечены половые различия в сроках линьки. Так, летняя линька у самок растянута
во времени, а у самцов происходит в более сжатые сроки [Киреев, сообщ.]. Обычно сам-
цы линяют раньше самок [20; Киреев, Хонякина, сообщ.]. Это же наблюдала й НМ.Оку-
лова [сообщ.]: в 1969 г. первые линяющие самцы стали попадаться с 16.V, самки — в нача-
ле июня, по окончании первой кладки; в 1960 г. линька у самца отмечена 15 .VI, у самки —
29.V1.
Сопоставление перечисленных фактов дает пеструю картину, что объясняется, на наш
взгляд, следующими причинами: существует географическая изменчивость сроков линьки,
обусловленная климатическими различиями регионов, поскольку климат определяет сро-
ки всех жизненных процессов рептилий; в пределах каждой популяции периоды линьки
растянуты, так как разные особи линяют неодновременно; данные по линьке, приведен-
ные для большинства регионов, в той или иной степени отрывочны и не позволяют полу-
чить полную картину, а сравнительно массовый материал по Калмыкии носит противоречи-
вый характер; о количестве линек у пресмыкающихся в природе нельзя судить по резуль-
татам наблюдений в неволе, поскольку число линек зависит от условий содержания живот-
ного.
Рассмотревдые выше данные все же позволяют сделать некоторые выводы: разноцвет-
ная ящурка линяет не менее 2 раз в год — в апреле — июне и в августе — октябре; возмож-
но, имеет место третья линька — в июне — августе, после окончания периода размножения;
сеголетки линяют чаще и до ухода на зимовку успевают перелинять несколько раз; самцы,
по-видимому, линяют раньше самок. Более полную информацию о линьке можно получить
лишь при тщательных стационарных исследованиях на большом материале и-с применени-
ем мечения. Такая работа была проведена на юге Украины в Черноморском заповеднике
[123,126,130,131,386].
По нашим данным, линяющих ящурок можно встретить почти в любое время на про-
тяжении периода сезонной активности.Первые линяющие особи отмечены 26.IV 1976 г.,
последние — 20 X 1975 г. Суммируя все случаи линек у ящурок, отловленных на Ивано-
Рыбальчанском участке Черноморского заповедника, получаем следующую картину
(рис. 99, А-В) . Взрослые и полувзрослые особи начинают линять в последней декаде апре-
ля, т^. через 3—4 недели после весеннего пробуждения. В мае — первой половине июня
ваблюдается максимальное количество линяющих ящурок, что совпадает с периодом их
наиболее быстрого роста. В июле линяет небольшое число особей, а в августе — сентябре
отмечается второй пик, что также совпадает с ростом и нагулом ящурок после периода
размножения. Самки линяют обычно на 1—4 недели позже самцов (во всяком случае,
196
Рис. 99. Линька разноцветной ящурки (процент линяющих особей в выборках из различных по-
пуляций) [126,131; Котенко, сообщ.)
А—В — Ивано-Рыбальчанский участок Черноморского заповедника (Л - меченые особи,
1975 г., Б — все особи, 1975 г.,В — все особи. 1974—1978 гг.) : а — взрослые и полувзрослые осо-
би (п = 559), б - сеголетки (п = 298); Г - юг Украины, июль 1977 г. (взрослые и полувзрослые
особи) : 1 — Ивано-Рыбальчанский участок, 2 — окр. г. Голая Пристань Херсонской обл., 3 -
о. Тендра, 4 - Кирилловская коса (пересыпь Молочного лимана), 5 - Арабатская стрелка; Д -
европейская часть бывшего СССР, июнь 1983-1988 гг. (взрослые и полувзрослые особи) J -
берег Черного моря у с Приморское Килийского р-на Одесской обл. (л =36); 2 - Каролине-Бу-
газская коса (пересыпь Днестровского лимана) (п = 28); 3 - Ивано-Рыбальчанский участок Чер-
номорского заповедника (и = 212) ; 4 — окр. г. Мариуполь (п = 39); 5 — Кривая коса, Азовское
море (п = 43) ; 6 - Дрно-Арчединские пески (и =68) ; 7 - левый берег Цимлянского водохрани-
лища (и =6)
Месяц
во время весенней линьки). Сеголетки начинают линять через 2—3 недели после появле-
ния на свет (некоторые даже через неделю) , и с каждым месяцем число линяюших особей
увеличивается. Последние линяющие сеголетки встречались за неделю до залегания в
лтячку.
Анализ данных по меченым яшуркам показал, что сеголетки линяют обычно 2—3 раза
(т.е. примерно раз в месяц) — в зависимости от сроков рождения. Так, в 1975 г. особи,
родившиеся в первой половине июля, линяли в конце июля, в августе и сентябре; родив-
шиеся позже — в августе, сентябре и октябре или сентябре и октябре. Слишком поздно
родившиеся сеголетки до залегания в спячку линяют, очевидно, всего 1 раз, а родившие-
ся рано и отличавшиеся наиболее быстрыми темпами роста могут, вероятно, линять 4 раза.
После зимовки молодые ящурки линяют 3—4 раза за период активности: чаще всего в
конце апреля — начале мая, в июне и августе — сентябре. На следующий год эти же ящурки
линяют 2 раза, реже — 3: в мае — июне, иногда в июле, и августе — сентябре. После очеред-
ной (третьей) зимовки ящерицы линяют 2 раза, примерно в то же время. За 3 года (обыч-
ная продолжительность жизни Earguta в Черноморском заповеднике) ящурки линяют
9—12 (в среднем 10) раз.
16.2. ФАКТОРЫ, ВЛИЯЮЩИЕ НА ЛИНЬКУ
В целом количество линек в году и то, в какое время они происходят, зависит непосред-
ственно от скорости роста ящурок, а следовательно, от погодных и климатических условии,
времени рождения, пола и индивидуальных особенностей рептилий. Именно закономерности
постэмбрионального роста ящурок (см. [ 124]) определяют сезонную динамику линьки, а воз-
растная структура популяции в конкретный момент — процент линяющих в это время особей.
Поэтому кривые на рис. 99, А-В - это как бы результат сложения кривых отдельных возра-
стных групп (точнее, групп особей, родившихся примерно в одно время и растущих с оди-
197
наковой скоростью). В частности, первый пик кривых (май — июнь) — результат нало-
жения первой линьки взрослых ящурок (май — июнь) на первую (конец апреля - нача-
ло мая) и вторую (июнь) линьки молодых особей. Отсюда ясно, что обычно применяв-
шиеся способы — простая регистрация фактов линьки или вычисление процента линяю-
щих особей от общего объема выборки — не позволяют определить истинное количество
линек у особи за сезон активности или за всю жизнь и (тем более) объяснить сроки и ко-
личество линек. Для этого необходимы наблюдения за отдельными мечеными особями,
В разные годы (т.е. в зависимости от погодных условий) сроки линьки в одной и той
же популяции могут несколько сдвигаться, вследствие чего изменяется процент линяющих
особей (рис. 99). Влияние климатического фактора в наиболее чистом виде проявляется
при сравнении выборок ящурок, взятых в одном и том же месяце одного года в различных
пунктах юга Украины (рис. 99, Г, половозрастная структура популяций примерно одина-
кова) ; некоторое представление могут дать и материалы, полученные нами в другое вре-
мя и в иных районах (рис. 99, Д) .
Большое значение для линьки имеет также физиологическое состояние животного. Из-
вестно. что при содержании в террариуме рептилии часто линяют, и это является своеобраз-
ной реакцией на ухудшение условий существования. Вместе с тем у истощенных особей
процесс линьки сильно растянут, а ороговевший слой сбрасывается клочьями. Нормально
и в сжатые сроки линяют упитанные пресмыкающиеся [38, 39].
163. ПРОЦЕСС ЛИНЬКИ
По наблюдениям И.С Даревского [77], кожа у разноцветных ящурок нередко линяет не
клочьями, а целиком: часто приходилось находить под камнями почти целые шкурки, вы-
вернутые наизнанку. МЛАхмедов [20] встречал ящурок, у которых старые покровы
снимались целиком, только на о. Обливной (Каспийское море) ,на материке же они схо-
дили клочьями. По данным ИД Яковлевой [324], верхний слой кожи слезает целиком с
нижней стороны головы (изредка и с верхней), со всего туловища и основания хвоста,
при этом лишь в одном случае кожа на спине была разорвана вдоль. На голове, ногах и
хвосте остается бахрома, которая сходит кусками через 2—3 дня. Кожа, слезшая с лапок,
имеет форму перчаток.
ЗКДрушко и РАКубыкин [48] сообщают, что эпидермис может сходить как отдель-
ными кусками, так и единым пластом с головы, спины, боков туловища и основания хвос-
та; хвост и конечности линяют последними. Напротив, по наблюдениям ВАЛСиреева, от
старых покровов вначале освобождаются передние и задние лапки, затем остальные части
конечностей и хвост, позже — спина и голова, и в последнюю очередь сбрасываются покро-
вы с брюха. При этом верхний слой эпидермиса со спины сходит лоскутами, с брюшной
стороны целиком, а с лапок в виде перчаток. Весь процесс линьки длится от 5 до 9 дней
[115; Киреев, сообщ.]. Чтобы освободиться от старых покровов, ящурки многократно
проползают между кустами растительности, где и оставляют кусочки эпидермиса [48]
Дчя успешного завершения процесса линьки большое значение имеет влажность, поэтому
количество выползков рептилий всегда увеличивается после дождя [38,39].
По нашим наблюдениям, разноцветные ящурки линяют клочьями, при этом с тулови-
ща покровы сходят нередко одним куском (обычно сильно изорванным) , голова, хвост
и конечности линяют отдельно. Типичная картина процесса линьки такова. Первым линяет
туловище: вначале появляется разрыв рогового слоя кожи между пилеусом и тулови-
щем, затем трещина проходит вдоль спины (может и отсутствовать) , и верхний слой эпи-
дермиса с туловища сходит (наиболее целые шкурки состоят из покровов брюха, боков,
основания хвоста и, в той или иной степени, покровов спины, а нередко и горла). В это
же время или чуть позже очищается пилеус. Затем линяют конечности (шкурка сходит с
лап в виде перчаток), хвост, остальные части головы. Дольше всего старый роговой слой
держится на самых кончиках пальцев и хвоста, а также на щитках морды — носовых и
подбородочном. Линька туловища длится обычно 2—3 дня (от начала отслоения старых
покровов на шее до полного сбрасывания),весь процесс занимает, как правило, 5—7 дней
Ослабленные животные линяют дольше, а эпидермис сходит более мелкими клочками
(как бы шелушится).
198
Ьыползки ящурок чаще всего попадаются у входов в норки (непосредственно у от-
верстий или рядом у основания кустика травы) . Поскольку в западной части ареала E.ar-
guta населяет вполне определенные биотопы с разреженной травянистой растительностью
и роет характерные норы (т.е. поиск шкурок облегчен), а также отличается от других
местных ящериц своеобразным фолидозом (кроме того, сброшенный роговой слой яще-
риц хорошо сохраняет характерный для вида рисунок) , эта ящурка может быть легко об-
наружена даже по небольшим обрывкам сброшенных покровов [134]. Этим способом
мы часто пользовались при изучении распространения разноцветной ящурки в Украине,
особенно в плохую погоду и при недостатке времени, что позволяло за один день обследо-
вать несколько различных пунктов даже при отсутствии активных рептилий на поверхно-
сти почвы.
ГЛАВА 17
ПОВЕДЕНИЕ, ПЕРЕМЕЩЕНИЯ И МИГРАЦИИ. УБЕЖИЩА1
Рис. 100. Ориентация разноцветных ящурок при
возвращении к месту первоначального отлова
[2591:'
1 — место поимки и мечения ящерицы, 2 -
место выпуска меченых ящериц, 3 - место по-
вторной поимки меченых ящерии, 4 - направле-
ние движения ящериц
Под перемещениями мы понимаем передвижения ящериц в пределах индивидуальной
территории, а под миграциями — длительные перемещения на значительное расстояние,
обычно совершаемые за пределы индивидуальной территории. Сведений о перемещениях и
миграциях, убежищах и поведении ящурки разноцветной немного [120, 254, 259, 264,
265, 316, 320]. Известно, что они, как и другие ящерицы, имеют индивидуальные участки,
в пределах которых перемещаются, у них отмечены местные миграиии, обусловленные
временными изменениями факторов окружающей среды [255, 264]. Сведения о переме-
щениях и миграциях важны, так как позволяют глубже понять образ жизни отдельных
особей, популяции в целом, установить место и роль вида в биогеоценозах,
Ящурка разноцветная — животное со строго дневным циклом активности (см. гл. 11) .
В течение светового дня на протяжении сезона активности она непрерывно перемещается
в поисках корма, особей другого пола
(взрослые) в период размножения, при
переселении или по другим причинам.
Протяженность перемещений такого рода
и скорость их варьируют в широких преде-
лах и определяются влиянием разнообраз-
ных факторов среды, физиологическим со-
стоянием организма. Наблюдения за ящур-
кой в пределах ареала однозначно свиде-
тельствуют о ее привязанности к опреде-
ленному участку, на котором находятся
постоянные и временные убежища, ’’пред-
почитаемые” места кормления, наконец, что
она всю жизнь или часть времени ведет
оседлую жизнь, а поэтому, возможно, хо-
рошо знает топографию местности. Наши
наблюдения [259] показали, что отловлен-
ные и выпущенные на некотором расстоя-
нии от места поимки ящерицы стремятся
обычно возвратиться назад, сохраняя в
большинстве случаев правильную ориента-
цию: 64 особи сохранили 100%-, 14—75%-,
14—50 %-ную ориентацию. Установлено
также, что не вернулись на прежнее место те
ящурки, которые были унесены на рас-
стояние свыше 125 м от места первоначального отлова (рис. 100). Выяснено и то, что
с расстояния 30 м ящурки возвращались за 1—2 дня, с расстояния 50 м — через 3 дня.
Если предположить, что радиус ’’знакомой” ящерипе территории равен 125 м, то площадь
•е будет равна примерно 49 000 м2 Возможно, животные в проведенном эксперименте
могли достичь знакомых мест в результате поисков ’’наудачу”
L Материалы предоставили Т Алиев, МЛ Ахмедов, 3 КВрушко, Э .В .Вашетко, МЛ.Зинякова, В А Ки-
реев, Т Л Котенко, НМ .Окулова, В.Ф .Орлова, ПЛТуденко, АМТудик, Х.Тэрбиш, ЗЛХонякина.
zoo
17,1. ТЕРРИТОРИАЛИЗМ
Ящурки территориально консервативны, радиус индивидуальной активности не превыша-
ет 14—22 м. Нахождение меченых ящериц длительное время на одних и тех же участках
подтверждает высказанную мысль. Исследования, проведенные в Центральном Предкав-
казье [255, 259], показали, что размеры индивидуального участка яшурки в разных био-
топах не превышают 34—178 (80,0±24,2) м2: в опустыненной степи — 85—178— (122,3±
±28,4), а в разнотравно-злаковой — 34—45 (37,6±3,6) м2 На Таманском полуострове
[Зинякова, Руденко, сообщ.] площадь индивидуального участка яшурки варьирует от 25
до 80 м2, а по наблюдениям в Харьковской обл. [Рудик, сообщ.] — от 4—7 до 20 м2
Можно считать установленным, что размер индивидуального участка зависит от оби-
лия корма [259; Зинякова, Руденко, сообщ.], форма его, довольно разнообразная, от ми-
крорельефа, распределения растительности и связанного с ней корма ящурок (беспозво-
ночных) , обилия его (рис. 101) . Индивидуальные участки обнаружены у взрослых и моло-
дых особей, размеры их примерно одинаковы.
Рис. 101. Индивидуальная территория и дневные участки разноцветных ящурок [255]
I - опустыненная степь, II - разнотравно-злаковая степь. Границы участка ящерицы :
1 — за первый день, 2 - за второй день, 3 — за третий день, 4 - за последующие дни; 5 -
нора ящурки
В разные по погодным условиям дни ящурки неравномерно используют индивидуаль-
ную территорию. Размеры их дневных участков варьируют в разнотравно-злаковой степи
от 5,5 до 32 (16,7±2,2) м2 в опустыненной степи — от 5,5 до 104 (48,4± 13,3) , т.е. ящурки
разного пола и возраста в разных биотопах Центрального Предкавказья используют свою
индивидуальную территорию до 88,2 %.
Как показывают наблюдения, индивидуальные участки [255,265; Зинякова,Руденко,
сообщ.] могут быть изолированными или перекрываться индивидуальными территориями
других особей. AM Рудик [сообщ.] предполагает, что часто приходится говорить не об ин-
дивидуальном участке, а об основной норе.
На индивидуальном участке ящерицы располагаются постоянные (на плотном грунте)
и временные убежища. Последние характерны для песков, так как быстро разрушаются,
заносятся песком, и ящурки практически каждый день роют новые норы в разных точ-
ках своего участка. Убежища чаще располагаются группами, расстояние между ними ред-
ко превышают 3 м (обычно 0,3—1). Территория индивидуального участка неравноценная
по количеству и расположению убежищ, количеству и распределению растительности, объ-
ектов питания. По-видимому, поэтому ящурки чаще находятся у ’’предпочитаемых”
мест кормления и убежищ, которые приурочены к обрывчикам, кучам почвы, камней,
растительности и др., и редко используют всю территорию своего участка каждый день.
Бродяжничая в поисках корма, ящерица выедает его на каком-либо участке за день,
на следующий день выедает на другом участке. За день ящерица осматривает 5,5—32,0
(16,7±2,2) м2 в пределах индивидуального участка в разнотравно-злаковой степи и
5,5-104 (48,4± 13,3) м2 в опустыненной степи. При хорошей погоде средняя величина
осмотренного участка больше, чем при плохой. Размеры дневных участков закономерно
201
изменяются в связи с изменением суточной активности ящериц: постепенно увели-
чиваются с весны до середины июня и также постепенно уменьшаются с июня до
осени по мере накопления ящерицами жировых запасов и приближения осенних
холодов.
Анализ имеющихся материалов показывает, что ящурка — маломобильное животное.
При поисковых и терморегуляционных передвижениях она ведет себя не всегда одинако-
во. В одном случае ящерица придерживается индивидуального участка, а в другом широ-
ко передвигается за его пределами. Но и тогда мы сталкиваемся не с явлением мигра-
ции в общепринятом смысле слова. Однако для систематизации типов передвижений
ящериц по протяженности мы выделяем: перемещения (передвижения ящериц в пределах
индивидуального участка); миграции, кочевки (передвижения ящериц на большей терри-
тории, обычно вне пределов индивидуальной территории) .
17.2. ПЕРЕМЕЩЕНИЯ
Ящурка в течение светового дня непрерывно перемещается в поисках корма, партнера в
период размножения, участков с подходящим микроклиматом, убежищ при преследова-
нии врагами и др. По известным данным, ящурка передвигается в пределах индивидуаль-
ной территории на расстояние до 80—100 м рЗинякова, Руденко, сообщ.] и даже 150
[255, 259]. Длина дневных перемещений варьирует в широких пределах, завися от погод-
ных условий, характера распределения корма, присутствия врага и др. В разные сезоны и
при разных погодных условиях продолжительность суточной активности ящерки колеб-
лется от 0 до 12 ч, но в движении на поверхности она находится не более 5 ч 30 мин. Ос-
тальное время затрачивается на обогрев, отдых, нахождение в укрытии и др. В нормальных
условиях средняя дневная скорость передвижения около 0,14-0,47 м/с.
Исследования в Центральном Предкавказье [255] позволили установить, что в раз-
ных биотопах ящурка передвигается в течение светового дня на различное расстояние.
Например, в опустыненной степи она перемещается на 21—150 (135,6± 12,0) м, в разно-
травно-злаковой степи — на 21—82 (76,4±7,0). Наблюдения за перемещением ящурки в
межбарханных понижениях (урочище Яман-Кую, Ставрополье, 1982—1987 гг.) показали,
что длина дневных перемещений на оголенных песках достигает 120—200 (175,0±21,0) м
[Тертышников, Горовая, Высотин, сообщ.]. Видимо, на участках со скудной раститель-
ностью и низкой численностью объектов питания ящерицы передвигаются на большее рас-
стояние, чем в биотопах с хорошо развитым растительным покровом.
По-видимому, к группе перемещений следует также отнести передвижение ящериц,
связанное с изменением освещенности той или иной части биотопов. Так, по сообщению
АМРудика, ящурки, оказавшиеся на теневой стороне холма к концу лета, постепенно сме-
стились на освещенную его сторону.
173. МИГРАЦИИ, КОЧЕВКИ
Данные об этом типе передвижений единичны. Их характеристика была отражена в пе-
чати [265]. Причины миграций сложны, недостаточно изучены, и о некоторых из них мож-
но говорить пока лишь предположительно. Другие можно объяснить влиянием климатиче-
ских и погодных факторов, особенностью кормовой базы, поиском мест для зимовки, от-
кладки яиц, конкурентными отношениями и др. [Хонякина, Алиев, сообщ.].
Миграции, связанные с размножением и расселением молоди. В период размножения
самцы широко кочуют в поисках самок и за короткий срок перемещаются на многие
десятки метров (до 450 м). Самки более консервативны, их перемещения менее значи-
тельны. По наблюдениям М.Ахмедова [сообщ.], самцы в поисках самок обходят участок
площадью около 70—80 м2. Самки в поиске подходящих мест для откладки яиц смещают-
ся на расстояние до 400 м, и, видимо, этой причиной З.ПЛонякина [сообщ.] объясняет
увеличение их количества на песках между городами Махачкалой и Каспийском в период
откладки яиц.
Описаны и миграции сеголеток. По сообщению AM Рудика, сеголетки переселяются на
расстояние 11 м от места вылупления. Наши исследования [259] показывают, что сеголет-
ки после вылупления некоторое время задерживаются в пределах ’’родительской” терри-
202
Рис, 102. Предполагаемые пути расселения сеголеток ящурок на стационарном участке в верховь-
ях р. Томузловки в Центральном Предкавказье [259]
1 - место откладки яиц самкой, взрослые самки; 2 — взрослые самцы; 3 — сеголетки после
миграции; 4 — норы; 5 - выходы известняка на поверхность; 6 - овраг; 7 - предполагаемые
пути расселения сеголеток; 8 — границы индивидуальной территории взрослого животного
тории, т.е. на том месте, где были отложены яйца, затем мигрируют на участки, пригодные
для обитания и свободные oi посещения взрослых ящериц (рис. 102), поскольку к этому
их вынуждают столкновения с взрослыми особями. Видимо,сеголетки изредка мигрирую!
на значительное расстояние и в процессе переселения задерживаются там, где имеются
свободные убежища и достаточная кормовая база. Так, ТИКотенко [сообщ.] обнаружи-
ла меченую сеголетку в 200 м от места рождения.
Миграции, связанные с изменением влажности. По наблюдениям ЗЛХонякиной
[сообщ.], в засушливые годы ящурки мигрировали на участки с повышенной влажно-
стью к небольшим водоемам, расположенным на расстоянии до 400 м в другом типе био-
ценоза. По ее же наблюдениям [Хонякина, сообш.] , часть прибрежных песков Каспия, рас-
положенных между городами Махачкалой и Каспийском, после высадки молодых деревь-
ев вскоре были заселены ящурками. перекочевавшими сюда из ближайших суглинистых и
солончаковых участков. Сведения о миграциях ящурок к каналам, рекам в период засух
(до 150 м) приводит также Т Алиев [сообщ.].
17.4. УБЕЖИЩА
При опасности ящурка использует разнообразные убежища: трещины и пустоты в почве,
зарывается в песок, прячется среди кустиков травы, у стволов кустарника, под опавшей
листвой, под лепешками высохшего помета крупного рогатого скота [Киреев, сообщ.],
в пустоты, оставшиеся на месте высохшей и сгнившей или выгоревшей корневой системы
тростника [Брушко, сообщ.], под выброшенные морем коряги, доски, бревна, кучи му-
сора, оставшиеся среди дюн после сильных штормов [125; Зинякова, Руденко, сообщ.],
пустоты под камнями и др., в том числе и норы различных животных. Так, в глинистой пу-
стыне Алма-Атинской обл. ящурки нередко прячутся в норы среднеазиатской черепахи
[Брушко. сообщ,], норы песчанок и мышевидных грызунов в Калмыкии и Западном Ка-
захстане [Киреев, Окулова, сообщ.], на юге Украины [125, 131] — в норы мышевидных
грызунов, емуранчиков, сусликов, слепышей и даже лисиц, но предпочитают норы мыше-
видных грызунов и емуранчиков. При этом последние в Нижнеднепровских песках заселе-
ны ящурками обычно на 90—100%.
Немаловажное значение в жизни ящурки имеют собственные норы. Способность рыть
норы — характерная черта поведения ящурки разноцветной [292] . Исследования НН Щер-
бака [320] показали, что ящурки охотно роют норы в мягком грунте, на твердой или ка-
менистой почве. На песках ящурка роет норы почти ежедневно, поскольку они быстро раз-
рушаются и недолговечны. Норы в плотном грунте — на черноземе, каштановых почвах —
более долговечны и долго используются ящерицами.
203
Рис. 103. Схемы основных типов {А —Г) нор разноцветной ящурки
в Предкавказье (Увел. 1:10) [ 259] Объяснения в тексте
При выборе места для
норы отдается предпочте-
ние мягкому грунту — пес-
ку, выбросам слепыша,
емуранчика, кучам навеян-
ного песка и др. Нередко
норы устраиваются под кор-
нями кустарников, полу-
кустарников, в дерновинах
злаковых, под камнями,
досками и другими пред-
метами [125, 131; Орло-
ва, Тэрбиш, сообщ.]. Так,
по данным Т.Н.Котенко
[131], например, в песча-
ной степи Ивано-Рыбальчан-
ского участка на площадке
0,25 га из 175 нор 33 выры-
ты в выбросах слепыша,
11 — в покопах емуранчи-
ка, 3 — в следах копыт,
20 — под кустиками полы-
ни. На участке пустыни в
Западной Монголии 100 %
нор выкопано ящуркой у
основания кустарников и
травянистых растений (Ор-
лова.Тэрбиш,сообщ.].
Нередко норы концентрируются у обрывчиков, возникших в песках в результате вы-
дувания, останцев. Так, на стационарном участке в Калмыкии на одном из останцев пло-
щадью 3,5 м2 и высотой до 60 см размещалось 29 нор [Киреев, сообщ.]. Во внутренней
части его располагался ’’грот” площадью 7x11 см и высотой около 3 см, в который выхо-
дило 11 отверстий нор.
Вход в нору может иметь любую экспозицию. Так, на Ивано-Рыбальчанском участке
степи 21,3 % нор имело южную экспозицию, 10,7 — юго-восточную, 12,7 — восточную,
15,3 — северо-восточную и 13,3 % — северную [125]. В Калмыкии большинство нор имело
южную экспозицию [Киреев, сообщ.]. То же отмечено и для Харьковской обл. [Рудик,
сообщ.]. В Западной Монголии 23,2 % нор имело южную экспозицию, 16,1 — юго-восточ-
ную, 17,85 — северо-восточную, 14,3 — северную и 12,5 — восточную, 1,78 — северо-запад-
ную, а западную и юго-западную — по 7,1 % [Орлова, Тэрбиш, сообщ.]. Таким образом,
определенной закономерности направления входного отверстия не отмечается.
Входное отверстие норы взрослой ящурки овальное. Его размер 1,2—1,5 — 2,3—3,0 х
х 2,5—2,8 — 3,7—4,0 см [125, 131, 259; Высотин. Горовая, Орлова, Тэрбиш,сообщ.] .Ход
норы идет в глубь почвы под углом 5—85° в зависимости от рельефа. Ход чаще прямой, ре-
же изогнутый или же с несколькими ответвлениями (отнорками) от основного хода [125,
131,259,278,316,324]. Нередко отнорки расположены рядом с отверстием.
Мы изучили строение более 750 нор разноцветной ящурки на разных типах почв в Цен-
тральном Предкавказье [259], выделили 4 типа нор (рйс. 103). Норы первого типа (Л)
имеют вид прямых коротких ходов длиной до 27 см, опускающихся вглубь на 7—27 см.
Норы второго типа (Б) также просты по строению, по форме напоминают булеву ”у”. Об-
щая протяженность хода такой норы не превышает 72 см, оканчивается она на глубине 7—
40 с. Несколько сложнее норы третьего типа (В) . Они обычно имеют несколько отнорков.
Общая длина хода норы 10—35 см, а глубина расположения нижнего тупика — 10—16 см.
И наконец, норы четвертого типа (Г) имеют несколько входных отверстий, несколько от-
норков. Общая длина хода редко превышает 75 см, а глубина расположения нижнего ту-
пика — 20—30 см.
Бытующее подразделение нор, используемых ящуркой, на летние и зимние, по-видимо-
му, условно. ТЯКотенко [125], например, считает, что специальных летних и зимних убе-
204
жшц у яшурок нет, одна и та же нора (если она не будет разрушена) служит им в качестве
укрытия от врагов, жары, осадков, как место ночевок и зимовок и даже для откладки
яиц. Свои норы ящурки роют в течение всего периода активности по мере необходимости.
Норы могут быть временными (легко разрушаемые, вырытые ящурками на песках, не-
глубокие) и постоянными (служат ящерицам один или более сезонов, чаще расположе-
ны на плотном грунте, первоначальными хозяевами были мышевидные грызуны или дру-
гие млекопитающие).
17.5. ПОВЕДЕНИЕ
Активные и пассивные способы защиты от врагов. При опасности ящурка чаще спасается
бегством. Увидев приближающегося врага, она настораживается и при дальнейшем продви-
жении его устремляется к ближайшему укрытию, приподнимая при беге брюхо и хвост.
Короткими перебежками от укрытия к укрытию ящурка пробегает в один прием до SO-
TO м. Взрослые самцы иногда останавливаются, принимают угрожающую позу, раскрыв
пасть и приподняв голову. Скорость бега взрослого животного достигает 7 м/с, но с такой
скоростью ящерица может бежать всего лишь 15—25 м, после чего становится малопод-
вижной [Зинякова,Руденко, сообщ.].
Нередко убегающая ящерица часто кружит вокруг куста, находясь все время с проти-
воположной стороны его, и не выпускает врага из поля зрения. Изредка способна взби-
раться на ветки. На открытой местности (в местах с очень редкой растительностью) „за-
гнанная” ящурка может зарываться в песок при помощи боковых движений тела, подоб-
но круглоголовке-вертихвостке. Наконец, убегающая ящурка иногда бросается в воду
(оросительный канал, ручей) и переплывает на другой берег [Хонякина,сообщ.].
Другим защитным приспособлением является аутотомия, когда ящурка отбрасывает
хвост, схваченный врагом. Он извивается и отвлекает внимание врага от животного. Труд-
но оценить действенность аутотомии при отсутствии материалов, дающих представление
о количестве съеденных и оставшихся в живых ящурок в популяции. Однако данные из
разных частей ареала показывают порой значительное количество ящурок, утративших
хвост. Так, в Краснодарском крае количество аутотомированных животных в выборках
достигает 20,6 % [Зинякова, Руденко, сообщ.], Зайсанской котловине — 8,3 [Брушко,
сообщ.], Дагестане — 23,6 [Хонякина, сообщ.], на юге Украины — 5,9—30,0 [131], в Цен-
тральном Предкавказье — 1,9—3,2 % [259]. При этом замечено, что пойманная ящурка ста-
рается вырваться, извиваясь всем телом и выделяя экскременты, пытается укусить врага.
Как упоминали, застигнутая врагом ящурка старается укрыться в ближайшем убежи-
ще. Нередко забегает в небольшие углубления в почве, и если они не укрывают надежно,
то убегает дальше, мечется в поисках убежища, делает попытку зарыться. Роют нору
ящурки довольно быстро. Взрослая самка за 2 мин вырыла в песчаной почве норку, рав-
ную длине тела (Зинякова, Руденко, сообщ.]. Сначала она сделала углубление передними
лапками, затем уперлась в него головой, дальнейшие ее движения напоминали ”бег на
месте”.
Скрывшись в убежище, ящурка долго не показывается. Затем через 1—3 мин выгля-
дывает и рассматривает преследователя. Нередко ящурки прячутся в чужие норы, где мо-
жет находиться 2—6 других ящурок, молодые ящурки, прыткая ящерица, чесночница,
жаба. Отмечались случаи [131; Киреев, сообщ.], когда ящурка, спасаясь от преследования,
попадала в нору с находившейся там змеей. Одни моментально выскакивали из нее, дру-
гие становились жертвой змеи. Нахождение 2 и более ящурок в одной норе приводит ряд
исследователей к мысли, что они могут жить парами [131; Зинякова, Руденко, сообщ.;
и др.].
Поведение во время питания. Утром ящурка покидает убежище не сразу, некоторое
время находится у выходного отверстия. Затем понемногу высовывает голову и выпол-
зает на площадку перед норой, греясь в лучах солнца. Если заметит опасность, то возвра-
щается в нору. Предварительный обогрев в лучах солнца отмечен в разные сезоны — от
ранней весны до поздней осени. После согревания ящурка приступает к активному поис-
ку пищи, реже подстерегает. В поисках пищи иногда раскапывает песок, обследует норы и
норки, взбирается на небольшие растения или осматривает их с земли. Нередко наблюдает
за летящими насекомыми, замечает место их приземления и ползет по направлению к
205
насекомому, стараясь схватить его в броске. Иногда бежит за летящими насекомыми,
хватая их или на лету, или в момент приземления [131,259, и др.].
Пищу добывают по-разному: медленно ползающих подбирают, а быстро бегающих
настигают прыжком. Поймав добычу, ящурка треплет ее, мнет челюстями, прижимает к
земле. Часто при таких манипуляциях отламываются конечности, надкрылья. Мелких по
размерам насекомых ящурка глотает быстро, а крупных заглатывает долго — около 1—
3 мин с некоторыми паузами. После проглатывания пищи ящурка облизывает челюсти
языком, иногда вытирает их о песок.
Поведение во время размножения. В брачный период самец в течение светового дня
разыскивает самку. При встрече с другим самцом хозяин территории пытается прогнать
конкурента, однако драк между ними не происходит. Встретившиеся самцы принимают
характерные позы: они выгибают спину и мотают головой из стороны в сторону. Как пра-
вило, пришелец убегает, а ’’хозяин” затем некоторое время гонится за ним [Хонякина,
сообщ.].
По имеющимся данным [Хонякина, Высотин, сообщ.], возбужденный самец при встре-
че с самкой ведет себя по-разному. На неполовозрелых он не реагирует. Взрослые самки,
готовые к размножению, демонстрируют определенные позы: они поднимают хвост и кру-
жатся на одном месте. Настигнув самку, самец хватает ее ртом за заднюю часть туловища
ближе к основанию хвоста, за бок брюха и, сильно изгибаясь, подтягивает свое тело к сам-
ке, а свою клоаку к клоаке самки. В этот период ящурки мало реагируют на окружающее
и не убегают, позволяют взять себя в руки. Впервые сведения в литературе — описание и
рисунок спаривающихся ящурок — были приведены Е.Эверсманном [355].
Самка ящурки откладывает яйца в неглубокую ямку, вырытую в песке. Животные,
обитаюпдее на шютном грунте (каштановые почвы, черноземы), яйца откладывают в но-
ре: по туловищу самки проходит волнообразное движение и из клоаки выходит яйцо, ко-
торое скатывается на дао ямки. Интервал между яйцами составляет 2—3 мин. По заверше-
нию откладки самка засылает их землей.
Наблюдение показывает [Хонякина, сообщ.], что вылупляющаяся молодая ящурка,
прорвав головой оболочку и высунув ее наружу, некоторое время осматривается, затем
постепенно выползает на почву. Некоторое время она лежит неподвижно, потом присту-
пает к рытью норы.
Поведение в неволе. Ящерицы, помещенные в террариум, непрерывно бегают в попыт-
ке найти выход. После многократных неудачных попыток они роют норы или же прячутся
под предметы, находящиеся в террариуме. Первые дни они настороженно относятся к под-
ходящему наблюдателю, но в дальнейшем привыкают к нему. Периодически пьют воду,
начинают разыскивать корм, постепенно становятся более доверчивыми и при появлении
человека начинают суетиться в ожидании корма и даже некоторые берут его из рук. Как
отметил АЛиудшс [сообщ.], у ящурок в террариуме не наблюдали позу угрозы.
ГЛАВА 18
ВРАГИ1
Сводные материалы с территории ареала позволяют считать, что среди врагов, уничтожа-
ющих ящурку, известны пресмыкающиеся, птицы и млекопитающие.
Пресмыкающиеся. Известны случаи поедания ящурки разноцветной прыткой ящери-
цей [25, 266, 320], удавчиками западным и песчаным [25, 320] , желтопузиком [25, 35,
39], полозами желтобрюхим [35, 131, 266, 320], четырехполосым [131, 225, 266, 316,
320; Высотин, сообщ.], узорчатым [25, 35,39, 266],разноцветным [35,39,320], ящерич-
ной змеей [25, 320], медянкой [131, 259], Иногда ящурку поедает степная гадюка [14,
110, 131, 164,266; Высотин, сообщ.], палласов щитомордник [320] и стрела-змея [320]
С учетом канибализма можно считать, что 14 видов рептилий поедают ящурку. По данным
В Л .Бадмаевой [25], в Калмыкии встречаемость ящурки в содержимом желудков пре-
смыкающихся достигает 33 3—40 % у ящериц и 33 3—57,1 % у змей..
Птицы. Среди птиц у ящурки врагов больше. Так, в разных частях ареала ящурку по-
едают цапли серая [25], рыжая [237, 238] и желтая [25,237, 238] ,выпь [25], луни болот-
ный [25, 200], луговой и степной [200], аисты [320], осоед [25], коршун черный [266,
320], сарычи рыжий [35,39] и обыкновенный [35, 39, 286], кобчик [25, 131,320], пус-
тельги обыкновенная и степная [25,131,320] , дрофа-красотка [25,35,39,320], орел степ-
ной [286, 320], крачка чайконосая [131, 134, 286, 307], сова ушастая [25]. сыч домовой
[131], сорокопуты чернолобый ижулан [25], скворец [131],ворона серая [320; Высотин,
Г оровая, сообщ.]
Таким образом, в пределах ареала ящурка разноцветная входит в состав корма 26 ви-
дов птиц. Однако встречается она в желудках птиц реже, чем у пресмыкающихся. Как
показывают материалы В .И .Бадмаевой, для Калмыкии встречаемость ящурки в содержи-
мом желудков ряда птиц (цапли, выпь, луни, осоед, кобчик, пустельги, совы и сорокопу-
ты) колеблется от 1,3 до 32,6 %. Видимо, в этом регионе постоянно ими питаются только
хищные птицы и сорокопуты, а для остальных ящурка — сопутствующий корм.
Млек отстающие. Из этой группы животных ящурку разноцветную поедают ежи обык-
новенный и ушастый [278,320],корсак,лисица [25,278,286,320],барсук [320],домаш-
ние собаки и кошки [131], енотовидная собака [117], перевязка, хорь степной [25]. Та-
ким образом, среди врагов зарегистрировано 9 видов млекопитающих. Из имеющихся в
нашем распоряжении материалов следует, что много ящурок поедают в Калмыкии, напри-
мер, корсак (встречаемость 75 %) и хорь степной (встречаемость 88,8 %) [25] .Большую
опасность для ящурок представляет человек, который, уничтожая их или изменяя среду
обитания, в пределах ареала снижает численность популяций во многих точках.
В заключение следует сказать, что приведенные сведения о врагах ящурки разноцвет-
ной, за редким исключением, не дают представления о прессе хищников на популяции яще-
рицы . Существующие материалы о проценте особей с регенерированными хвостами не по-
зволяют не только определить конкретных врагов, но и установить также процент изъя-
тых из популяций ящурок. Поэтому, приводя их В тексте, мы лишь указываем на интен-
сивность преследования ящурок со стороны всего комплекса хищных позвоночных.
1 Материалы предоставили ВИБадмаева, ЭВБашетко, АГВысотин, ВЛТоровая и ТИЛотенкс.
ГЛАВА 19
ПАРАЗИТЫ
19.1. ЭНДОПАРАЗИТЫ
Разноцветная ящурка — один из достаточно полно исследованных в паразитологическом
отношении видов пресмыкающихся. Имеющиеся данные о паразитах этого вида относятся,
однако, лишь к территории бывшего Советского Союза — европейской части страны, Ка-
захстану и Средней Азии и касаются в основном 2 наиболее распространенных подвидов:
Eremias arguta deserti и Еа. arguta. Сведения о паразитах этой ящерицы за пределами этой
территории (в Румынии, Турции, Иране, Китае) нам неизвестны.
Всего у разноцветной ящурки зарегистрировано 24 вида паразитов, представленных
простейшими (жгутиконосцы, гемогрегарины) и многоклеточными — гельминтами (тре-
матоды, цестоды, акантоцефалы и нематоды). Паразитофауна разноцветной ящурки не
имеет каких-либо специфических особенностей: все известные у нее виды паразитов заре-
гистрированы и у других видов ящурок. Несмотря на значительное количество обнаружен-
ных паразитов, вызываемые ими заболевания, которые, несомненно,имеют место при вы-
сокой зараженности разноцветной ящурки, в литературе не описаны. В связи с этим судить
о влиянии паразитарных заболеваний на ее жизнедеятельность нельзя.
Ниже приводим данные о паразитах, зарегистрированных к настоящему времени у раз-
ноцветной ящурки в пределах ее ареала. В основу обзора положены оригинальные данные
автора, материалы, содержащиеся в опубликованных работах, большинство из которых
можно найти в монографии ВЛДПарпило [300] и А.Овезмухамедова ”Протистофауна реп-
тилий”1 . В нашем распоряжении были также материалы, переданные на определение
НМ.Окуловой.
Простейшие
Класс Zoomastigophorea
Proteromonas lacertae Grassi, 1879
Паразит задней части кишечника.
ChSomastix wenyom Markov et al., 1961
Паразит задней части кишечника.
Monocercomonas colubrorum Hammerchmidt, 1884
Паразит задней части кишечника.
Класс Sporozoea
Baemogregarina eremiae Zmeev, 1936
Паразит крови.
Наряду с перечисленными видами у разноцветной ящурки обнаружен также Pirhemocy-
don eremiasi Blanc et Asione, 1958, товдое систематическое положение которого остается
неустановленным.
Гельминты. Эта группа паразитов представлена в составе паразитофауны разноцветной
ящурки 19 видами и включает облигатные и факультативные виды. Наряду с имагинальны-
ми формами гельминтов значительное место принадлежит также личиночным формам, для
которых разноцветная ящурка выступает в роли промежуточного и резервуарного хозяи-
1 А.Овезмухамедев. Протистофаунарептилий. - Ашхабад : Ылым, 1987. - 372 с.
208
на. Как промежуточный и резервуарный хозяин эта ящурка может служить источником
заражения хищных, врановых и других птиц, а также хищных млекопитающих.
Класс Trematoda
Plagiorchis elegans (Rud., 1802)
Паразит кишечника. Принадлежит к редким факультативным паразитам разноцветной
ящурки.
Plagiorchis molini Lent et Freitas, 1940
Паразит кишечника. Как и предыдущий вид, принадлежит к редким факультативным
паразитам разноцветной ящурки.
Класс Cestoda
Oochoristica tuberculata (Rud., 1819)
Паразит кишечника. Один из наиболее обычных и широко распространенных парази-
тов разноцветной ящурки, зараженность которой может достигать 60—70 %.
Cladotaenia circi Yamaguti, 1935 (larvae)
Паразит печени. Разноцветная ящурка служит для этого вида промежуточным хозя-
ином.
Класс Acanthocephala
Centrorhynchus aluconis (Muller, 1780) (larvae)
Паразит полости тела, печени, стенок пищеварительного тракта. Разноцветная ящурка
служит для этого вида резервуарным хозяином.
Centrorhynchus sp. (larvae)
Паразит полости тела и стенок кишечника. Разноцветная яшурка служит для этого ви-
да резервуарным хозяином.
Sphaerirostris teres (Rud., 1819) (larvae)
Паразит различных внутренних органов. Разноцветная ящурка служит для этого вида
резервуарным хозяином.
Oligacanthorhynchus sp. (larvae)
Паразит печени и полости тела. Разноцветная ящурка служит для этого вида резервуар-
ным хозяином.
Класс Nematoda
Spauligodon eremiasi Markov et Bogdanov, 1961
Паразит задней части кишечника. Один из наиболее обычных паразитов разноцветной
ящурки, зараженность которым может достигать 50—60 %.
Spaunligodon pseuderemiasi Sharpilo, 1976
Паразит задней части кишечника. Обычный паразит разноцветной ящурки, заражен-
ность которой может достигать 60 %.
Ascarops strongylina (Rud., 1819) (larvae)
Паразит печени, стенок пищеварительного тракта, подкожной клетчатки. Разноцветная
ящурка служит для этого вида резервуарным хозяином.
Spirocerca lupi (Rud., 1819) (larvae)
Паразит печени, стенок пищеварительного тракта. Разноцветная ящурка служит для
этого вида резервуарным хозяином.
Habronema sp. (larvae)
Паразит печени и стенок пищеварительного тракта. Разноцветная ящурка служит для
этого вида резервуарным хозяином.
Physaloptera lausa (Rud., 1819) (larvae)
Паразит желудка. Разноцветная ящурка служит для этого вида резервуарным хозя-
ином.
Abbreviate abbreviate (Rud., 1819)
Паразит желудка. Один из наиболее обычных паразитов разноцветной ящурки, зара-
женность которым может достигать 80 %.
Thubunaea schukurovi Annaev, 1973
Паразит желудка. Редкий паразит разноцветной ящурки, зарегистрирован по единич-
ным находкам.
209
Agamospinira maaacanthis Sharpilo, 1963 (larvae)
Паразит стенок пищеварительного тракта. Разноцветная ящурка служит для этого ви-
да резервуарным хозяином.
Agamospirura magna Sharpilo, 1963 (larvae)
Паразит печени, стенок пищеварительного тракта. Разноцветная ящурка служит для
этого вида резервуарным хозяином.
Agamospinira punctata Sharpilo, 1971
Паразит стенок пищеварительного тракта. Разноцветная ящурка служит для этого ви-
да резервуарным хозяином.
19.2. ЭКТОПАРАЗИТЫ
Эта группа паразитов разноцветной ящурки представлена клещами и изучена хуже, чем
предыдущая. Сводка по эктопаразитам рептилий или ящериц, включающая Е.arguta, отсут-
ствует, и сведения о клещах, паразитирующих на данном виде, разрозненны, сравнительно
немногочисленны и содержатся нередко в труднодоступной литературе [7, 19, 20, 35,39, 48,
95 , 96, 130, 131, 136, 165, 169, 175, 205, 212, 222, 258, 259, 269, 309, 316]. В некоторых
работах имеются лишь указания на обнаружение клещей [278] ,иксодовых клещей [Зиня-
кова, сообщ.] или гамазид [Окулова, сообщ.] в качестве паразитов разноцветной ящурки.
Всего на E^rguta выявлено 8 видов клещей, из них 2 — гамазовых, 3 — иксодовых и
3 — клещей-краснотелок. Обычно экстенсивность и интенсивность заражения разноцвет-
ных ящурок эктопаразитами крайне низки. В ряде популяций E.arguta из Казахстана и
южных областей европейской части России клещи вообще не были обнаружены [51, 156,
166, 168, 167 и др.], несмотря на тщательное обследование больших выборок ящурок.
Вместе с тем на юге Украины трудно найти популяцию Е.arguta, не зараженную гамазовы-
ми клещами. Скорее всего, это обусловлено высокой плотностью населения разноцветной
ящурки. На результаты некоторых исследований могла повлиять методика сбора клещей.
В частности, осмотр ящурок и мешочков через 8—20 ч после умерщвления рептилий, как
это делала HJH.Окулова [сообщ.], дает очень большие потери по такой группе, как гама-
зовые клещи, и наилучшим методом, как мы убедились, является снятие клещей с живых
ящериц сразу после их поимки. Автор искренне признателен Г «И Щербак (Киевский уни-
верситет) и АА.Стекольникову (ЗИН РАН) за определение материала.
Отряд Acariformes
Семейство Trombicididae
Leptotrombidium (Ericotrombidium) caucasicum Schluger, 1967
Обнаружен в Ставропольском крае (с. Безопасное Труновского р-на) и в Одес-
ской обл. (с. Зеленое) на прыткой ящерице и разноцветной ящурке [309]. Поскольку в
последнем пункте Еarguta не обитает, факт паразитирования этого клеща на данном виде
ящериц следует относить к Ставрополью. Паразитируют личинки.
Neotrombicula tragardhiana (Feider, 1953)
Найден нами у ящурок из окрестностей Мариуполя (пос. Виноградное) Донецкой обл.
Украины на песчаном берегу Азовского моря: в выборке из 39 особей, взятой 16—18.VI
1984 г., зараженность личинками этого вида краснотел ок составила 82,05 % при интенсив-
ности заражения 1—300 экз. (в среднем 32 клеща на одну ящурку) . Данная выборка отли-
чалась и высокой зараженностью (84,62%) гамазовым клещом Sauronyssus saurarum
(рис. 104). Осмотр ящурок в этом же пункте 25.VIII 1983 г., а также в других точках
азовского побережья летом 1983-1984 гг., находок N.tragardhiana не дал. Вид впервые об-
наружен на прыткой ящерице в Румынии [462], данные литературы по паразитированию
этой краснотелки на рептилиях фауны бывшего СССР нам неизвестны.
Geckobia sp.
Выявлен у одной из 113 ящурок, добытых в Узбекистане на плато Устюрт [165], и
указан в сводке ОЛ .Богданова [39] для Средней Азии.
210
Отряд Parasitiformes
Семейство Ixodidae
Haemaphysalis punctata Can. et Fanz., 1877
Найден на ящурках в Ставропольском
крае и, возможно, в Азербайджане [95,169,
212, 258, 259,266,269]. Зараженность этим
клещом очень низкая: 0,4 и 1,8 % в выбор-
ках из 256 и 397 ящурок. Паразитируют ли-
чинки и нимфы. Треххозяинный паразит.
Может передавать возбудителей клещевого
сыпного тифа и бруцеллеза.
Haemaphysalis sulcata Can. et Fanz., 1877
Обнаружен на ящурках из Азербайджа-
на [7], предгорий Нуратау в Узбекистане
[35], из Таджикистана [222] и Иссыккуль-
ской котловины в Кыргызстане [96], а так-
же приведен без указания места [205]. Па-
разитируют личинки и нимфы. Для вида
характерна четко выраженная специфич-
ность паразитирования молодых фаз на реп-
тилиях. Треххозяинный клещ. Заражен-
ность может быть довольно высокой: экс-
тенсивность 14,3 % (п = 21) в Азербайджа-
не и 30,6 % (п = 49) — в Кыргызстане, при
Популяции.
Рис. 104. Зараженность (%) разноцветной ящурки
клешем Sauronyssus saurarum в различных популя-
циях (июнь 1983-1988 гг.) (по материалам
Т Я Котенко)
1 - берег Черного моря у с Приморское Ки-
лийского р-на Одесской обл., 2-6.VI 1985 г.
(л = 36); 2 — Каролино-Бугазская коса (пере-
сыпь Днестровского лимана), 23.VI 1985 г.
(л = 28); 3 — Ивано-Рыбальчанский участок Чер-
номорского заповедника, 13-23.VI 1988 г. (л=
= 212); 4 - окр. гМариуполь, 16-18.VI
1984 г. (л = 39); 5 — Кривая коса, Азовское мо-
ре, 19. VI 1983 г. (л = 43) и 15.VI 1984 г. (л =
= 10) ; 6 - Доно-Арчединские пески, 13.VI 1986 г.
(л = 68); 7 - левый берег Цимлянского водохра-
нилища, 16.VI 1986 г. (л =6)
интенсивности инвазии 1—4 экз. Может передавать животным возбудителя бруцеллеза.
Hyalomma plumbeum (Panz., 1795)
Зарегистрирован на ящурках в Узбекистане [35]. Паразитируют личинки и нимфы.
Двуххозяинный паразит. Может передавать человеку вирус геморрагической лихорадки.
Иксодовые клещи (ближе не определены) выявлены также на ящурках с Кизилташ-
ской косы в Краснодарском крае [Зинякова, сообщ.]
Семейство Dermanyssidae
Ophionyssus eremiades Naglov et Naglova
Выявлен в подмышечных впадинах разноцветной ящурки из Зайсанской котловины,
Казахстан [48].
Sauronyssus saurarum (О ud emails, 1902)
Специфичный убежищный паразит ящериц, отмечался для многих видов рода Lacerta
и других ящериц. Является переносчиком кровепаразитов рептилий — Schellackia bolivari и
Karyolysus lacertarum. Паразитируют в основном протонимфы. В качестве паразита разно-
цветной ящурки указан для Крыма [316] и Азербайджана [19, 20, 95, 175]. Мы [130,
131, 136; неопубликованные данные] обнаружили его практически по всей украинской
части ареала E^rguta (в Донецкой, Запорожской, Крымской, Николаевской, Одесской,
Полтавской, Харьковской и Херсонской областях), а также в Волгоградской обл. Рос-
сии — в Доно-Арчединских и Цимлянских песках. Экстенсивность заражения в различных
популяциях разноцветной ящурки составила 1,5—89,0 % (всего обследовано более 1200
особей), при этом максимальная зараженность наблюдалась в местах с высокой плотно-
стью населения ящериц (рис. 104) . Интенсивность инвазии — 1—28 экз. Клещи локализу-
ются на брюхе, нижней поверхности хвоста, горле, реже — на нижней поверхности бедер и
голеней, располагаясь по 1—3 экз. под чешуйкой или щитком; иногда они сидят на бара-
банной перепонке по ее краю.
Исследования, проведенные на стационарной плошадке Ивано-Рыбальчанского участ-
ка Черноморского заповедника, показали, что данным видом клещей взрослые самцы и
самки заражены примерно в равной степени, зараженность полувзрослых обычно ниже,
сеголеток — существенно ниже, чем взрослых. Максимальная зараженность отмечена в
сентябре, довольно высока она и весной, в апреле — мае [131, 136] (табл. 30) . Накопле-
211
Таблица 30. Сезонная динамика зараженности разноцветной ящурки гамазовым клещом Sauronys-
sus sauiarum на Ивано-Рыбальчанском участке Черноморского заповедника, по результатам обследова-
ния меченых особен на стационарной площадке в 1975—1977 гг. (по данным Т Л .Котенко)
Месяц Взрослые самцы Взрослые самки „ I Полувзрослые Взрослые и полу-, взрослые в целом' Сеголетки
Всего Заражен- ные, % Всего Заражен- ные, % Всего Заражен- ные, % Всего Заражен- ные, % Всего Заражен- ные, %
Апрель 15 33,33 3 33,33 8 37,50 26 34,62
Май 20 40.00 22 31,82 20 20,00 62 30,65 — —
Июнь 21 0 22 22,73 31 323 74 8,11
Июль 10 20,00 12 8,33 1 0 23 13,04 5 0
Август 18 27,78 31 25.fl — — 49 26,53 69 4,35
Сентябрь 17 41,18 10 50,00 — — 27 44,44 74 28,38
Октябрь — — — — — — — — 25 8,00
Таблица 31. Зараженность разноцветной ящуркн гамазовым клещом Sauronys-
sus sauiarum на Ивано-Рыбальчанском участке Черноморского заповедника в разные
годы (по данным ТЛ .Котенко)
Год Взрослые самцы Взрослые самки Полувзрослые Все ящурки
Всего Заражен- ные, % Всего Заражен- ные, % Всего Заражен- ные, % Всего Заражен- ные, %
1975 21 0 22 22,73 31 323 74 8,11
1987 41 14,63 47 12,77 73 9,59 161 11,80
1988 61 2235 87 13,79 64 12,50 212 16,04
1989 26 92,31 39 89,74 18 77,78 82 89,02
Примечание. Все данные относятся ко второй половине июня.
ние клещей к сентябрю идет параллельно с увеличением плотности популяций яшурок и на
фоне благоприятных температурных условий среды [136]. Колебания зараженности
E^rguta по годам могут быть очень большими. В частности, заЗ года зараженность наблю-
давшейся популяции возросла почти в 8 раз, а зараженность отдельных половозрастных
групп в разные годы колебалась от 0 до 92 % (табл. 31).
В Азербайджане S^aurarum встречается на ящерицах во все времена года, особенно
высока зараженность этим клещом осенью [95, 175]. Данный вид — наиболее многочис-
ленный эктопаразит рептилий (в том числе и разноцветной ящурки) полупустынных низ-
менностей Азербайджана — он составил 97,4 % от сборов всех клещей [95].
Gamasoidea sp.
Обнаружены в Волго-Уральских песках (Западный Казахстан) на 2 особях из несколь-
ких сотен просмотренных ящурок [Окулова, сообщ.].
ГЛАВА 20
ПРАКТИЧЕСКОЕ И БИОЦЕНОТИЧЕСКОЕ ЗНАЧЕНИЕ. ОХРАНА
20.1. БИОЦЕНОТИЧЕСКИЕ СВЯЗИ РАЗНОЦВЕТНОЙ ЯЩУРКИ1
Значение животного в экосистеме определяется прежде всего его численностью и местом
в системе трофических отношений. Как показано в главе 9, в степных, полупустынных и
пустынных экосистемах разноцветная ящурка зачастую является фоновым видом, а в не-
которых биотопах достигает значительной плотности популяций (табл. 8). В отдельных
биотопах юга Украины этот вид является самым многочисленным позвоночным животным
и занимает важнейшее место в соответствующих биоценозах [386}.
Из всех звеньев биоценотических связей разноцветной ящурки наиболее существенна
ее роль как консумента. Анализ питания E.arguta и данные учетов численности этого вида и
беспозвоночных позволили нам определить количество и биомассу изымаемых ящерица-
ми объектов питания и выяснить степень воздействия этих рептилий на кормовую базу в
некоторых биотопах юга Украины [123. 130. 131, 386] и Центрального Предкавказья
[258,259,261,270].
Таблица 32. Воздействие разноцветных ящурок на беспозвоночных в песчаной степи Ивано-Рыбаль-
чанского участка Черноморского заповедника в июне 1975 г. [ 131 ]
Группа беспозво- ночных Абсолютное количество беспозвоночных Степень воздей- ствия, % Соотношение, % Изби- ратель- ность питания
В 10 про- бах На пло- щади 1 га В 12 же- лудках Съедено яще- рицами за месяц на 1 га В степи В пище
Паукообразные 12 48 000 16 16 480 34ДЗ 5,43 10,96 +034
Прямокрылые 32 128 000 23 23 690 18,51 14,48 15,75 +0,04
Равнокрылые Полужестко- 61 244 000 26 26 780 10,98 27,60 17,81 -032
крылые 15 60 000 10 10 300 17,17 6,79 6,85 0,00
Жесткокрылые 11 44 000 42 43 260 98,32 4,98 28,77 +0,70
Чешуекрылые Перепончато- 7 28 000 8 8240 29,43 3,17 5,48 +037
крылые 64 256 000 18 18 540 734 28,96 12,33 -0,40
Двукрылые 13 52 000 2 2060 3,96 5,88 1,37 -0,62
Прочие 6 24 000 1 1030 439 2,71 0,68 -0,60
Всего 221 884 000 146 150 380 17,01 100,00 100,00 —
Прим ечание. Проанализировано содержимое желудков 8 взрослых и 4 молодых ящурок. По-
скольку в июне 1975 г. взрослые и молодые особи съедали в сутки почти одинаковое количество
беспозвоночных (см. табл. 34), суточный рацион 1 ящурки принимается равным в среднем 12,2 экз.
беспозвоночных. Учет беспозвоночных проводили биоценометром (1 проба - 0,25 м2), то же в
табл. 34 и 35.
Как видно из приводимых таблиц, ящурки, уничтожая большое количестве беспозво-
ночных, оказывают неодинаковое давление на различные их группы в разных биотопах и
регионах (все данные приведены для июня) . Так, в песчаной степи Черноморского запо-
ведника (табл. 32) это жесткокрылые, пауки и чешуекрылые; в опустыненной степи на
Ставропольской возвышенности (Центральное Предкавказье) — жесткокрылые и пауки,
там же в разнотравно-злаковой степи -т жесткокрылые и перепончатокрылые, а в лесной
полосе — пауки, жесткокрылые и моллюски (табл. 33) . При этом степень изъятия беспоз-
1 При участии 3 ЛЛонякиной.
213
Таблица 33. Воздействие разноцветных ящурок на беспозвоночных в различных биотопах Цен-
трального Предкавказья в июне 1971 г. [ 259]
r*nvnna Абсолютное количество беспозвоночных Степень воздей- ствия, % Соотношение, % Изби-
Л yjj ШЛ4Х беспозвоночных В про* бах На пло- щади 1 га В желуд- ках Съедено ящерицами за месяц на 1 га В степи В пище -ратель- ность питания
Опустынення степь (6 проб,21 желудок)
Паукообразные Полужестко- 1 1700 3 248 14,5 1,0 2,1 +0,3
крылые 1 1700 1 82 4,8 1,0 0,7 -0,1
Жесткокрылые 20 34 000 84 6930 203 20,0 57,1 +0,4
Чешуекрылые Перепончато- 6 10 200 11 908 8,8 6,0 73 +0,1
крылые 65 110 500 38 3135 2,8 65,0 25,8 “0,4
Двукрылые 3 5 100 4 330 6,4 з,о 2,7 -0,05
Прочие 4 6 800 6 495 73 4,0 4,1 + 0,01
Всего 100 170 000 147 Разнотравно-злаковая 12 128 степь 7,1 100,0 (3 пробы, 5 желудков) 100,0 —
Паукообразные 1 3 300 1 12 036 2,7 23 +0,03
Прямокрылые Полужестко- 3 10 000 <> 24 0,24 8,1 5,7 “0,1
крылые 4 13 000 1 12 0,09 10,8 23 -03
Жесткокрылые 5 16 500 9 108 0,65 13,5 253 +0,3
Чешуекрылые Перепончато- 2 7 000 1 12 0,17 5,4 2,9 “03
крылые 12 40 000 18 216 034 24,4 513 +0,3
Двукрылые 7 23 300 1 12 0,05 19,0 23 -0,7
Протае 3 10 000 2 24 0,24 8,1 5,7 “0,1
Всего 37 122 100 Лесная 35 полоса 420 0,34 (6 проб, 17 желудков) 100,0 100,0 —
Моллюски 1 1 700 2 24 1,4 2,1 0,6 1,7 +03
Паукообразные 1 1700 3 37 0,6 2,6 +0,6
Прямокрылые Полужестко- 13 22 100 3 37 0,1 8,4 2.6 “0,5
крылые 2 3400 1 12 03 1,2 03 “0,1
Жесткокрылые 28 47 600 65 793 13 18.0 553 +0,5
Чешуекрылые Перепончато- 13 22 100 12 146 0,6 8,4 10,4 +0,1
крылые 40 68 000 24 292 0,4 26,0 20,5 -од
Двукрылые 29 49 300 2 24 0,04 18,7 1,7 -0,8
Прочие 28 47 600- 5 60 0,1 18,1 43 -0.6
Всего 155 263 500 117 1425 034 100,0 100,0 —
Примечание. Учет беспозвоночных проводили методом энтомологического кошения 10 взмахов на площади 1 м2) . (1 проба -
Таблица 34. Численность, масса и суточный рацион разноцветных ящурок на стационарной площад-
ке (0,25 га) в песчаной степи Ивано-Рыбальчанского участка Черноморского заповедника в 1975 г.
[131]
“! Месяц Количество ящу- рок на площадке, экз. Средняя мас- са ящурки, г Масса сред- ней особи, г Суточный раци- он ящурок I возрастной группы Суточный ра- цион ящурок П возрастной группы Съедено беспозво- ночных за сутки всеми ящурками на площадке
I воз- раст- ная груп- па П воз- раст- ная груп- па Все- го I воз- раст- ная груп- па П воз- раст- ная груп- па
экз. г экз. г экз. г
Апрель 38 40 133 5,13 4,94 16,0 0,079 7,0 0,197 298,0 7,644
Май 49 62 111 1,61 4,91 3,45 7,8 0,083 9Д 0,134 946,4 12375
Июнь 40 63 103 2,22 4,73 3,76 12,8 0,137 11,9 0,288 1261,7 23,624
Июль 13 91 104 0,66 4,11 3,68 8,0 0,056 6,6 0324 704,6 30,212
Август Сен- 114 46 160 1,13 4,44 2,08 18,0 0,071 5,2 0,229 2291,2 18,628
тябрь 106 27 133 1,27 434 1.87 20,4 0,086 73 0,162 23593 13,490
Октябрь 34 — 34 0,94 — 0,94 25,4 0,057 — — 863,6 1,938
Примечание. I возрастная группа в апреле — июне включает молодых и полу взрослых, в июле -
октябре — сеголеток, П возрастная группа в апреле — июне — взрослых, в июле — сентябре — взрослых
и полувзрослых.
214
Рис. 105. Схема биоценстических связей разноцветной ящурки [ 2591:
1 — пищевые связи; 2 — паразитарные связи, 3 — враги
воночных из биоценоза (степень воздействия ящурок на кормовую базу) варьировала в
широких пределах как по отдельным группам беспозвоночных (0,04—98.32 %) , так и в пе
лом (0,34—17,01 %, для июня) .
Изучение кормовой базы разноцветной ящурки в Черноморском заповеднике показа-
ло [130, 131, 139], что плотность и биомасса беспозвоночных сильно варьируют в зависи-
мости от биотопа и в значительной степени определяются густотой и разнообразием расти-
тельного покрова. В одном и том же биотопе в течение года закономерно изменяются чис-
ленность и биомасса беспозвоночных как в целом, так и по отдельным группам, а также
видовой состав подвижных фаз насекомых (сезонную динамику видового состава проб,
плотности и биомассы беспозвоночных в местах обитания разноцветной ящурки см, на гра-
фиках [139]) . В соответствии с этими изменениями кормовой базы и в связи с динамикой
потребностей организма в пище суточный рацион разноцветных ящурок и воздействие их
на энтомофауну в разных биотопах и в разное время года различны (табл. 34, 35), при
Таблица 35. Воздействие разноцветных ящурок на беспозвоночных в различных биотопах Черно-
морского заповедника в 1975 г.[131]
Месяц Биотоп Ящурки Беспозвоночные Изъято ящур ками за месяц на 1 га
экз ./га кг/га экз./га кг/га . экз. % кг %
Ивано-Рыбальчанский участок Апрель Песчанаяраэ- 160 0,790 3 483 333 7,031 35 760 1,03 0,917 13,04 нотр авно-злако вая степь реч- ной террасы Май Тоже 444 1,532 2 543 529 13,465 117 354 4,61 1,535 11,40 Июнь ”• ” 412 1,549 884 000 5,918 151 404 17,13 2,835 47,90 Июль 416 1,531 588 000 11,083 87 370 14.86 3,746 33,80 Август 640 1Д31 316 000 1,294 284 109 89,91 2.310 178,52 Сентябрь 532 0,995 908 000 3,045 283 140 31,18 1,619 53,17 Октябрь 136 0,128 542 000 4,473 107 086 19,76 0,240 5,37 Потиевскии участок Июль Песчаная при- 32 0,124 1 016 000 4,097 9 920 0,98 0,288 7,03 морская степь Июль Заросли ко- 35 0,136 2 640 000 7,771 10 850 0,41 0315 4,05 лосняка на бе- реговом валу о. Тендра Июль Песчаная при- 35 3,155 732 000 1329 9 042 1,24 0,354 26,64 морская степь Июль Заросли колос- 65 0,289 544 000 0,653 16 792 3,09 0,658 100,77 няка и полыни на береговом валу
215
Таблица 36. Сравнительный анализ питания разноцветной ящурки и насекомоядных птиц в Цен-
тральном Предкавказье [ 259 ]
Вид Коли- чество иссле- дован- ных же- лудков Общее количество животных в пище, %
Мол- люски Пауки Много- ножки Прямо- кры- лые Хо- ботные Жуки Дву- кры- лые Пере- пон- чато- кры- лые Чешуе кры- лые Прочая пища
Ящурка разно- цветная 172 1,0 1,8 5,0 3,5 0,1 42,2 73 24,5 4,4 10,0
Удод 5 2,0 — 4,0 3,о 40,0 20,0 10,0 3,0 7,0 14,0
Жав~эонок по- левом 20 з,о 2,0 713 16,2 63 1,0
Конек лесной 10 2,0 з,о — 4,0 15,0 25,0 1,0 15,0 35,0 —
Трясогузка белая 5 20,0 5,0 30,0 5,0 20,0 20,0
Жулан 20 — 1,0 — 10,0 9,0 42,0 2,0 10,0 20,0 6,0
Дрозд певчий 10 1,0 5,0 — 10Д — 20,0 50,0 10,0 — 4,0
Дрозд черный 20 6,0 2,0 5,0 — юд 37,0 4,0 10,0 25,0 1,0
Каменка обык- новенная 5 24,5 36,0 393
Пеночка-тень- ковка 5 10,0 35,0 35,0 10,0 5,0 5,0
Славка серая 5 — 5,0 — — — 35,0 10,0 5,0 45,0 —
Синиту» большая 10 1,0 6,0 — — 10,0 32,0 2,0 2,0 45,0 —
Просянка 5 — 10,0 10,0 — — 40,0 — 40,0 —
Овсянка черного- ловая 5 зл зз 883 33 1,0
Овсянка садовая 15 ол 0.5 — 5,0 33 55,0 — 0,5 30,0 5,0
Зяблик 15 1,0 13 1,0 1,0 10Д 4,0 5,0 10,0 30,0 1,0
Сойка 15 — 5,0 — — 30,0 20,0 5,0 10,0 30,0 —
Таблица 37. Сравнительным анализ штания разноцвет-
ной ящурки и некоторых позвоночных в низменном Даге-
стане [ Хонякина, сообщ.]
Пища Встречаемость в желудках, %
Ящурка раз- ноцветная (л = 719) Ящурка быстрая (л = 228) Жаба зеленая (л = 331)
Дождевые черви 0,14 0,88 1037
Моллюски 234 0,88 634
Ракообразные 332 — 1037
Паукообразные 21,70 1бзз 1934
Многоножки 0,44 1239
Стрекозы 2,09 1,75 —
Уховертки 237 132 937
Прямокрылые 1430 3,94 7,25
Равнокрылые 539 7,89 3,02
Клопы 536 6,14 5536 11,48
Жесткокрылые 62,87 73,72
Сетчатокрылые 0,97 0,88 —
Перепончатокрылые 3338 57,45 4531
Двукрылые 22,25 22,86 18,13
Чешуекрылые 4533 2335 25,08
Земноводные (сего- летки жабы зеленой) 030
Пресмыкающиеся (се- голетки ящурки раз- ноцветной) 0,14
Примечание. Животные отвловлены в общих стациях.
этом воздействие ящериц на беспозвоночных оказывается максимальным в биотопах с от-
носительно невысокой плотностью беспозвоночных и высокой — рептилий. По биомассе
ящурки изымают в несколько раз большую часть беспозвоночных, чем по числу экземпля-
216
Таблица 38. Питание некоторых амфибий и рептилий на ста-
ционарном участке в Центральном Предкавказье [ 259]
Пища Количество съеденных хищником животных, %
Жаба зеленая (л=30) Верете- ница (л = 17) Медянка (л = 20) Гадюка степная (л =7) Полоз четырех- полосый (и = 10)
Насекомые Другие беспозво- 80,0 10,0 - 83 Д 9,0
ночные 20,0 90,0 — 2,0 —
Ящерица прыткая Ящурка разноцвет — — 80,0 1,0 1,0
ная — — 20,0 0,8 1,0
Птицы — — — — 9,0
Млекопитающие — — — 5,0 80,0
Прочая пища — — — 8,0 —
Таблица 39. Питание некоторых хищных птиц, хищных и насекомоядных млекопитающих на ста-
ционарном участке в Центральном Предкавказье [ 259}
Пища Количество съеденных хищником животных, %
Подор- лик ма- лый (Л=6) Канюк обык- новен- ный (л=30) Кор- шун чер- ный (л =15) Лунь поле- вой (л=4) Пус- тельга обык- новен- ная 1 («=Ю) Лиси- ца (и=130) Бар- сук (л=20) Соба- ка ено- товид- ная (п=20) Еж обык- новен- ный (л=2) Еж ушас- тый (и=2)
Насекомые — 3,0 10,0 5,0 70,0 16,0 25,1 22,1 38,0 56,0
Другие беспозвоночные — — — — — — 1,1 3,0 42,0 35.0
Ящурка разноцветная — 0,5 2,0 2,0 4,0 1,5 — — 1,и
Птицы — 7,5 10,0 15,0 0,5 10,0 3,0 ИЗ — —
Млекопитающие 69,0 81,5 54,0 56,0 11,0 51,0 57,0 33,8 —
Прочая пища 30,0 5,0 20,0 — 8,5 20,0 19,8 29,7 20,0 7,0
ров. Это свидетельствует о том, что данные пресмыкающиеся предпочитают питаться отно-
сительно крупными объектами, что энергетически оправдано [131].
Из табл, 35 видно, что ящурки в песчаной степи Черноморского заповедника в июле
изымали наибольшую биомассу, а в августе — сентябре — наибольшее количество беспозво-
ночных. При этом самое сильное воздействие на энтомофауну ящурки оказывали в авгус-
те, когда их плотность была максимальной (за счет массового появления сеголеток), а
плотность насекомых — минимальной [130,131,386]. В это время ящурки изымали в сут-
ки 2,90 % беспозвоночных по количеству экземпляров и 5,76 % — по биомассе. В пересчете
на месяц, как принято делать (см. [100, 210, 259]), это составило соответственно 89,91 и
178,52 % (табл. 35) . Очевидно, что такой способ подсчетов условен, поскольку не учитыва-
ет постоянного пополнения беспозвоночных за счет их высокой репродуктивной способ-
ности и миграций из почвы на поверхность (почвенные раскопки мы не проводили) , и не
оправдывает себя в биотопах с большой плотностью рептилий и сравнительно низкой —
беспозвоночных [131].
Как показали исследования, проведенные в Центральном Предкавказье на стационаре
в верховьях р.Томузловка [261], доля 2 видов ящериц (разноцветной ящурки и прыткой
ящерицы) в уничтожении беспозвоночных составила 61,30 %, доля птиц — 31,03,млекопи-
тающих — 0,24%.
Сложные трофические связи разноцветной ящурки в биоценозах (рис. 105) обуслови-
ли разнообразие ее пищевых конкурентов. Среди беспозвоночных к трофическим конку-
рентам следует отнести различных энтомофагов — пауков, хищных многоножек, прямо-
крылых, клопов, жужелиц, кокцинеллйд, перепончатокрылых и двукрылых, различных
сетчатокрылых, богомолов, стрекоз и др. Поскольку многие из них многочисленны,мож-
но предположить, что они существенно влияют на кормовую базу ящурки. Из позвоночных
трофическими конкурентами E.arguta являются некоторые амфибии и рептилии, многие
виды птиц и млекопитающих (табл. 36, 37) Впрочем, конкуренция за пищу между ящур-
217
кой и другими насекомоядными позвоночными часто оказывается ослабленной ьы прак-
тически отсутствует за счет разной избирательности в питании и разобщенности видов в
пространстве и времени (в силу различной биотопической приуроченности и различий в ха-
рактере суточной активности).
Анализ трофических связей показал, что разноцветная ящурка представляет собой
преимущественно консумента II—III порядков, изредка — I или IV порядков. При этом она
не замыкает ряд консументов в трофических цепях, поскольку и сама является объектом
питания многих позвоночных животных (см. гл. 18) . В частности, в некоторых экосисте-
мах юга Украины, Центрального Предкавказья и междуречья Волга — Урал эта ящурка явля-
ется одним из основных элементов кормовой базы степной гадюки, медянки, желтобрюхо-
го полоза, обыкновенной пустельги, степного орла, чайконосой крачки, корсака и некото-
рых других позвоночных [131, 194] (табл. 38, 39) . При этом, как считает ВИБадмаева
[24], пресс хищников никогда не вызывал в популяциях ящериц Калмыкии катастрофи-
ческих последствий. Наконец, разноцветная ящурка является хозяином паразитических
клещей и гельминтов и принимает участие с обмене паразитарными формами с другими
группами животных (рис. 105).
202. ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ1
Рис. 106. Содержание вредных, полезных и
аейтральных беспозвоночных в пище разно-
цветной ящурки в разные годы [259].*
1 - вредные беспозвоночные; 2 - по-
лезные беспозвоночные, 3 — нейтральные
беспозвоночные
С практической точки зрения наиболее важным является уничтожение разноцветной ящур-
кой беспозвоночных — вредителей сельского и лесного хозяйств. В разных регионах и
биотопах количество уничтожаемых этой ящерицей вредителей варьирует. Так, в Украине
в Днепропетровской обл. [49] доля вредных беспозвоночных в питании ящурки составля-
ет 68 % (по массе) , в Калмыкии [24] — 67,6 % (здесь и далее — по количеству экземпля-
ров) , в Краснодарском крае [43] равна 57,4 %, на Ставропольской возвышенности [259]
в разные годы колеблется от 54 до 62 % (рис .106) .
По данным Б АКрасавцева [144] для Предкав-
казья, коэффициент полезности разноцветной
яшурки составил 80 % по числу съеденных насе-
комых и 87 % — по их биомассе. Материал по пита-
нию ящурки собран в степях, но неподалеку от
посевов, и содержал таких вредителей сельско-
го хозяйства, как прус Calliptamus italicus, личин-
ки Elateridae, хлебный жук Anisoplia leucaspis,
гусеницы бабочек Noctuidae [144].
Исследования в Черноморском заповеднике
показали [131], что процент вредных насекомых
в питании разноцветных ящурок постоянно из-
меняется во времени и зависит от возраста реп-
тилий (табл. 40) . При этом доля вредителей в ра-
ционе вида особенно велика в период их массово-
го размножения. Так, у сеголеток ящурок в од-
ном желудке отмечалось до 8 личинок листоеда
Chrysomela camifex (октябрь 1975 г.), до 20 червецов и до 50 тлей (сентябрь 1975 г.). Во вре-
мя вспышки численности лугового мотылька Pyrausta sticticalis, нанесшего большой ущерб
вельскому хозяйству юга Украины в 1975 г., ящурки в массе поедали гусениц и бабочек
этого вредителя (в июле отмечалось до 72 головных капсул этих гусениц в одном пищева-
рительном тракте ящурок).
По данным В Л Бул ахова [49], ящурка в большом количестве уничтожает пыльце-
едов, долгоносиков, листоедов, чернотелок, зерновок, тлю, клопов, листоверток, древо-
точцев, сосновых пядениц, совок, саранчевых, и ее трофический пресс почти в равной сте-
пени направлен на все стадии развития вредителей (данные не совсем понятны) . Ящурки,
заселяющие молодое сосновое насаждение; за месяц на площади 100 га истребляют до
28 кг беспозвоночных, в том числе 19,04 кг опасных вредителей, чем способствуют защите
формирующегося лесного биоценоза. Вокруг этого насаждения на пограничной полосе
1 При участии М.Ф.Тертышникова.
218
Таблица 40. Вредные насекомые в питании разноцветных ящурок на Ивано-Ры-
бальчанском участке Черноморского заповедника [131]
Пища Год Месяц Возрастная группа ящурок Коли- чество ящурок Съеденные насекомые, %
Коли- чество Встречае- мость
Acrididae 1974 VI ad. 7 19,50 71,43
1974 VII ad. 24 16,46 70,83
1974 VIII ad. 7 29,41 85,71
1975 VI ad. 19 14,67 89,47
1975 vn ad. 6 6136 100,00
1975 VIII juv. 24 4,48 66,67
1975 IX iuv. 17 4 Д9 76,47
Aphidinea 1975 VIII juv. 24 15,48 6230
1975 IX juv. 17 30,48 64,71
1975 X juv. 8 17,65 100,00
Homoptera 1975 VIII juv. 24 19,35 66,67
1975 1У juv. 17 4337 8235
1975 X juv. 8 30,72 100,00
Scarabaeidae 1974 VII ad. 24 31,65 70,83
Chrysolina carnifex,
larv. (Chrysome-
lidae) 1975 X juv. 8 11,11 8730
Curculionidae 1974 V ad. 26 2236 57,69
1974 IX juv. 8 8,65 50,00
1975 V ad. 15 21,74 66,67
1975 IX juv. 17 2,14 47,06
Pyrausta stictica-
lis larv. (Lepidop-
tera) 1975 VII juv. 12 58,59 8333
Lepidoptera 1975 VII juv. 12 59,38 91,67
1975 VIII ad. 7 13,73 57,14
1975 vin juv. 24 2,65 41,67
1975 IX juv. 17 439 3539
1975 X juv. 8 9,80 100,00
1977 IX juv. 9 4,59 77,78
шириной 200 м за месяц ящурки уничтожают еще 26 кг вредителей, что создает защитный
барьер в проникновении вредных насекомых из открытых пространств в молодую сосно-
вую посадку [49]. На площади 1 км2 эти ящерицы за сутки поедают 0,63 кг насекомых,
что за весь период их сезонной активности составляет 0,13 т вредителей [50].
По данным М.Ф.Тертышникова [257], в Ставропольском крае на площади около
150 га все разноцветные ящурки (примерно 5550 особей) в течение суток съедали 2 кг
беспозвоночных (суточный рацион ящурки принят в среднем равным 0,36 г) , за месяц —
60, за весь период активности (200 дней) — около 400 кг.
Некоторую пользу ящурки приносят тем, что являются объектом питания ряда ценных
видов птиц, млекопитающих и пресмыкающихся.
При оценке практического значения разноцветной ящурки следует, разумеется, учиты-
вать и то, что она может приносить вред, поедая полезных беспозвоночных, являясь про-
кормителем клещей и промежуточным хозяином гельминтов. Однако, судя по данным
глав 13, 18 и 19, значение ее в уничтожении полезных насекомых невелико, а в распростра-
нении паразитов, опасных для домашних и промысловых животных, ничтожно, поэтому
в целом разноцветную ящурку следует признать полезным животным. Поскольку роль ви-
да как консумента и продуцента местами велика и Earguta — неотъемлемый элемент мно-
гих биоценозов, придающий своеобразие ряду ландшафтов, необходимо всемерно охра-
нять это полезное и интересное животное.
20.3.БИОЭНЕРГЕТИЧЕСКАЯ ОЦЕНКА
РОЛИ РАЗНОЦВЕТНОЙ ЯЩУРКИ В БИОГЕОЦЕНОЗАХ
При решении этой задачи мы использовали имеющиеся рекомендации [60,348] для пере-
вода биомассы ящериц, яиц в кладках, поедаемой ими пищи в энергетический эквивалент
(в Дж) . Подобный подход к биоэнергетическим возможностям ящурки имел место толь-
219
Таблица 41. Биоэнергетическая оценка роли разноцветной ящурки в биогеоценозах
Предкавказья [ Тертышников, сообщ.]
На гектар, кДж На усредненный гектар биогео- ценоза в Пред- кавказье, кДж/год На территории ареала в преде- лах Предкав- казья, МДж/год
в пустыне и полу- пустыне в степи на равнине в степи на возвышенности
Биоэнергетика населения весной до размножения
30,34 0,39 41,2 7,08 Биомасса пищи 8,73
12,25-13,27 0,16-2,17 16,56—17,94 2,86-3,10 Биоэнергетика размножения Яйцекладки 3,93-4,26
2,20 0,29 2,98 0,52 Сеголетки в первые сутки жизни 0,71
1,71 0,022 2,32 0,40 0,55
ко в работе С.Л.Перешкольника [203] по Прииссыккулью. Хотя приводимые нами мате-
риалы не учитывают расход энергии в популяции ящурок на прокормление различных па-
разитов, построены на ряде допущений и являются статичными, однако более или менее
реально отражают какие-то определенные моменты и закономерности в жизни конкретных
популяций разноцветной ящурки на примере Центрального Предкавказья как одного из
модельных участков для всего ареала.
Усредненная многолетняя среднегодовая биомасса ящурки разноцветной (табл. 41)
составляет 7,08 кДж/га. Ее максимум отмечен для опустыненных степей возвышенно-
стей (в частности, Ставропольской возвышенности и Прикалаусских высот) —41,02 кДж/га,
а также для полупустынных и пустынных ландшафтов Восточного Предкавказья —
30,4 кДж/га. В течение сезона активности биомасса ящурки колеблется. Первое изменение
ее в связи с яйцекладкой (+0,52 кДж/га/год) и развитием сеголеток (+ 0,40 кДж/га/год) .
В разных биоценозах биомасса пресмыкающихся в связи с яйцекладкой увеличивается
неодинаково: на 7,25—7,44 % и более всего в степях равнин.
После завершения инкубации яиц биомасса популяции ящурки несколько снижается
(1£9—2,19 %) в связи с меньшей массой сеголеток по сравнению с массой яйца. По-види-
мому, имеет место и гибель яиц. Нагул сеголеток до их ухода в спячку, гибель их по ряду
причин также сказываются на изменении количества энергии, проходящей через популя-
цию в связи с указанными процессами, но такие данные у нас отсутствуют.
Ящурка извлекает из биоценозов большое количество биопродуктов — 2,86—
3,10 кДж/га на одном усредненном гектаре. Наибольшее количество энергии проходит че-
рез популяции ящурки в опустыненных степях возвышенностей (16,56—17,94 кДж/га)
и пустынных и полупустынных биоценозах (12^25—13^7 кДж/га), наименьшее — в рав-
нинных степях (0,16—2,17 кДж/га).
По нашим расчетам, ящурка разноцветная при усредненной плотности 7,1 экз У га в
Предкавказье занимает площадь около 1 374 800 га. Через ее популящш в регионе прохо-
дит примерно до 14,7 МДж/год, что указывает на немаловажную роль ящурки в энергети-
ке функционирующих биогеоценозов.
20.4. ОХРАНА
Во многих местах, особенно на западе ареала, разноцветная ящурка быстро и резко со-
кращает численность и ареал под влиянием антропогенных факторов. На всем побережье
моря от гМамайи (Румыния) до г.Одесса и далее до г Мариуполь, где еще недавно этот
вид был доминирующим на морских пляжах, он уже исчез во многих местах в результате
строительства курортов, санаториев, пансионатов и др. В других местах — Харьковская
и Херсонская области, Севанская котловина — местообитания ящурки изменяются в ре-
зультате лесопосадок, что также приводит к их исчезновению. Строительство водохрани-
лищ, каналов, развитие орошаемого земледелия (долина Днепра, Таманское побережье
Азовского моря, долина Маныча, Калмыкия, западная часть Терско-Кумской низменно-
220
сти и др.) дают тот же конечный результат. Все это приводит к мысли о необходимости
вьючения данного вида (по крайней мере в Украине) в Красную книгу Украины.
Однако положение не столь трагическое вследствие того, что на территории обита-
ния большинства форм ящурок существуют заповедники, которые пока удовлетворитель-
но сохраняют генофонд данного вида (см. Амфибии и рептилии 1987) 1. Так, Е.а. deser-
ti охраняется в Украине в Черноморском заповеднике, Азово-Сивашском заповедно-охот-
ничьем хозяйстве, встречается в Луганском заповеднике, в Российской федерации — на
участках Дагестанского заповедника (Кумторкала и Северном) , Воронежском (?) /Жигу-
левском и Хоперском; Ел. transcaucasica — в Грузии в Сагурамском и Сатанлийском запо-
ведниках; в Азербайджане — в Караязском, Кызылагачском (?) , Пиркулинском,Туркап-
чайском и Ширванском заповедниках. К сожалению, в Армении, в Севанской котловине
(типовое местообитание) пока нет заповедных участков, а это необходимо сделать, пока,
может быть, сохранилась там хоть небольшая популяция. Тем более, что эта форма вклю-
чена в Красную книгу Армянской ССР [145]. Ел. arguta в Казахстане охраняется в Алма-
Атинском, Кургальджинском, Е.а. potanini — в Маркакольском заповедниках,,Ел. da-
rewskii в Кыргызстане — в Иссыккульском заповеднике, Ел.uzbekistanica — в Заамин-
ском и Нуратинском заповедниках. Разноцветная ящурка включена также в Красную кни-
гу Туркменистана.
1 Амфибии и рептилии заповедных территорий : Сб. науч.тр. - М., 1987. - 160 с.
СПИСОК ЛИТЕРАТУРЫ
1. Абашидзе В.С. К устройству пояснично-крестцового сплетения земноводных, рептилий и птиц /7
Тр. Ин-та эксперим. морфологии. - 1957. - 6. - С. 27—35,
2. Абашидзе В.С. К устройству спинномозговых сплетений и периферических нервов позвоночных
животных // Там же. - 1958. - 7. - С. 111-128.
3. Абашидзе В.С. К сравнительной анатомии спинномозговых сплетений и симпатических стволов. -
Тбилиси : Изд-во АН ГССР, 1962. - 135 с.
4. Агарков Г.Б. Эволюция и адаптация нервного аппарата надпочечных желез позвоночных. — Киев :
Наук, думка, 1985,- 136 с.
5. Алекперов А.М. Земноводные и пресмыкающиеся Азербайджанской ССР Автореф. дис. д-ра
биол. наук. - Баку, 1957. — 19 с.
6. Алекперов AM., Логинов А А. Очерки по биологии пресмыкающихся Апшерона // Тр. Азерб. ун-та.
Сер. биол. - 1953.- 5. - С. 106-127.
7. Алекперов AM, Земноводные и пресмыкающиеся Азербайджана. — Баку: Элм, 1978. - 264 с.
8. Алексеева AM. К вопросу о превращении креатинфосфата в креатин-и о новом методе определе-
ния креатинфосфата // Биохимия. - 1951. - 16, выл. 2. — С. 93-103.
9. Алисов Б.П., Берлин ИА., Михель ВМ. Курс климатологии. — Л. Гидрометеоиздат, 1954. —
Ч. 1/2.-487 с; Ч.З.- 320с.
10. Андреев И.Ф. К экологии рептилий района г«КзылЮрда // Учен. зап. Черновиц. ун-та. Сер. биол. на-
ук. - 1948. - 1, вып. 1.-С. 94—106.
11. Андреев И.Ф. Приспособления рептилий к высоким температурам пустыни // Там же. — С.109—118.
12. Андрушко AM. Пресмыкающиеся Казахского нагорья и их хозяйственное значение // Учен. зап.
Ленингр. ун-та.Сер. биол. наук. — 1955. — Вып. 38,№ 181. - С. 19—43.
13. Антипчук ЮЛ., Соболева А Д. Эволюция респираторных систем. - Новосибирск Наука, 1976. —
207 с.
14. Ардаматская Т.Б. К биологии степной гадюки района Черноморского заповедника // Тр. Черномор,
заповедника. - 1958,- Вып. 2.- С. 107-109.
15. Аренс ЛТ. К вопросу о северной границе распространения разноцветной ящурки (Eremias arguta
Pall.) в Восточной Европе // Докл. АН СССР. - 1928. — 13. - С. 189—290.
16. Асатиани Д.Л. Материалы к сравнительной анатомии плечевого сплетения : Автореф. дис. ... канд.
биол. наук. — Тбилиси, 1968. — 18 с.
17. Атаев ЧА. О нахождении разноцветной ящурки в Юго-Западном Туркменистане // Изв. АН ТССР.
Сер. биол.наук,- 1988.-№ 1, —С.66-67,
18. Атлас теплового баланса / Под ред. М Л Будыко. — Л., 1955 .-41с.
19. Ахмедов ММ. Герпетологическая фауна островов Апшеронского и Бакинского архипелагов Кас-
пийского моря : Автореф. дис.... канд. биол. наук. - Бзасу, 1988. - 19 с.
20. Ахмедов ММ. Герпетологическая фауна островов Апшеронского и Бакинского архипелагов Кас-
пийского моря : Дне.... канд. биол. наук. — Баку, 1988, - 198 с.
21. Бадмаева ВМ. Размножение разноцветной ящурки на территории Калмыцкой АССР // Герпетоло-
гия. - Краснодар, 1976. - С. 114—117.
22. Бадмаева ВМ. Суточная активность разноцветной ящурки в Калмыцкой АССР // Там же. — С. 109—
113.
23. Бадмаева ВМ., Онаева Н.С., ИнжиноваГА. О питании разноцветной ящурки в Калмыцкой АССР //
Животный мир Калмыкии, его охрана и рациональное использование. — Элиста, 1977. — С. 45—
48.
24. Бадмаева ВМ. Ящерицы Калмыкии : Автореф. дис. ... канд. биол. наук. - Киев, 1983. - 24 с.
25. Бадмаева ВМ. Ящерицы Калмыкии : Дис.... канд. биол. наук. - Киев, 1983. - 227 с.
26. Баженова А.Ф. Температурные зависимости тканевого метаболизма у пустынных рептилий // Тем-
пературная компенсация и поведенческий гомеостазис. - Л. : Наука, 1980. - С. 89-95.
27. Банников АТ. Материалы по биологии земноводных и пресмыкающихся Южного Дагестана //
Учен. зап. Моск. гор. дед. ин-та им. Потемкина. - 1954. — 28, вып. 2.— С. 75-88.
28. Банников АТ., Даревский И.С., Рустамов AJC. Земноводные и пресмыкающиеся СССР. - М.
Мысль, 1971. - 303 с.
29. Банников АТ., Даревский И.С., Ищенко В.Г., Рустамов AJC., Щербак НМ. Определитель земновод-
ных и пресмыкающихся фауны СССР. — М.: Просвещение, 1977. — 414 с.
30. Баранов А.С., Яблоков А Б. и др. Численность и ее динамика // Прыткая ящерица : Моногр. описа-
ние вида. — М.: Наука, 1976. - С. 303—321.
31, Баранов А.С., ВалецкийАД., Яблоков АД. Морфология // Там же, - С. 131-134.
222
32. Белехова МТ. Таламо-телеэнцефальная система рептилий. — Л.: Наука, 1977. — 216 с.
33. Белъгард АЛ. Введение в типологию искусственных лесов степной зоны // Искусственные леса
степной зоны Украины. — Харьков : Изд-во Харьк. ун-та, 1960. — С. 33—55.
34. Бескровный МА. Практикум по экологии животных. — Харьков Изд-во Харьк. ун-та, 1953. —
4.1. - 232 с.
35. Богданов ОЛ. Земноводные и пресмыкающиеся. — Ташкент: Изд-во АН УзССР, 1960. - 260 с. -
(Фауна Узбекской ССР; Т. 1) .
36. Богданов ОЛ. Пресмыкающиеся Туркмении. — Ашхабад: Изд-во АН ТССР, 1962. — 234 с.
37. Богданов ОЛ. Факторы, влияющие на размножение пресмыкающихся Средней Азии // Вопросы
герпетологии. - Л.: Изд-во Ленингр. ун-та, 1964. - С. 11-12.
38. Богданов О Л. Экология пресмыкающихся Средней Азии Автореф. дис.... д-ра биол. наук. - Ал-
ма-Ата, 1964. - 33 с.
39. Богданов О Л. Экология пресмыкающихся Средней Азии. — Ташкент : Фан, 1965. - 260 с.
40. Богданов ОЛ. Животные Узбекистана (Позвоночные) . - Ташкент Средн, и высш, шк., 1961, -
316 с.
41. Бодемер Ч. Современная эмбриология. — М.: Мир, 1971. — 446 с.
42. Борисов А А. Климаты СССР в прошлом, настоящем и будущем. - Л. Изд-во Ленингр. ун-та,
1975.-432с.
43. Бочарникова АВ. Значение ящериц Краснодарского края для сельского и лесного хозяйства //
П науч. конф, зоологов пед, ин-тов РСФСР. — Краснодар, 1964. - С. 136-138.
44. Бочарникова А.В. Экология ящурки разноцветной и ящерицы луговой в Краснодарском крае //
Материалы IV науч. конф, зоологов пед. ин-тов. - Горький, 1970. - С. 304—305.
45. Браунер А. Третье предварительное сообщение о пресмыкающихся и земноводных губерний Сува-
лковской, Минской, Подольской. Черниговской, Бессарабской, Херсонской, Екатеринославской и
Днепровского уезда Таврической. - Одесса, 1906. - 17 с. (Зап. Новорос. о-ва естествоиспыта-
телей; Т. 28).
46. Браунер А. Гады Бессарабии // Тр. Бессараб, о-ва естествоиспытателей. — 1907. — 1, ч. 2. — С.1—25.
47. Браунер А А. Сельскохозяйственная зоология. — Одесса : Госиздат Украины, 1923. — 435 с.
48. Брушко ЗК., Кубыкин РА. К биологии ящурки разноцветной восточной в Казахстане // Изв. АН
КазССР. Сер.биол. - 1988.- 147,№ 3. - С. 31-35.
49. Булахов ВЛ Морфологическая характеристика и роль Eremias arguta (Pall.) в биогеоценозах степ-
ной зоны УССР // Вопросы степного лесоведения и охраны природы. - Днепропетровск, 1975. -
Вып. 5 - С. 217-228.
50. Булахов ВЛ., Бобылев ЮЛ., Константинова Н.Ф. Земноводные и пресмыкающиеся и их роль в
жизни степных лесов // Там же. - Днепропетровск, 1977. — Вып. 8. — С. 124—130.
51. Ваккер ВЛ. Паразитофауна рептилий юга Казахстана и их роль в циркуляции некоторых гельмин-
тов человека и животных : Автореф. дис.... канд. биол. наук. - Алма-Ата, 1970. - 27 с.
52. Василевская ГЛ., Веселовский МВ. Морфологические особенности нервной системы разноцветной
ящурки // Вопросы ’ герпетологии. - Киев : Наук, думка, 1989. - С. 42-43.
53. Васильев Б.Д. Морфофункциональная характеристика акустического анализатора амфибий и реп-
тилий : Дис. ... канд. биол. наук. — М., 1969. - 261 с.
54. Васильев БД., Симкин ГЛ. О пространственной ориентации рептилий и амфибий // Вопросы гер-
петологии. - Л.: Изд-во Ленингр. ун-та, 1964. — С. 13-15.
55. Вашетко ЭВ. Распространение и экология ящурок юга Средней Азии : Автореф. дис.... канд. биол.
наук. - Краснодар, 1974. - 20 с.
56. Вашетко ЭЛ., Камалова ЗЯ. Черепахи, ящерицы // Позвоночные животные Ферганской долины. -
Ташкент : Фан, 1974. — С. 60-74.
57. Вельш У., Шторх Ф. Введение в цитологию и гистологию животных. - М. : Мир, 1976. - 256 с.
58. Винников Я А., Титова JLK. Морфология органа обоняния. - М.: Медгиз, 1957. - 296 с.
59. Воробьева ЭЛ., Чугунова TJ0. Морфофункциональные особенности строения зубной системы яше-
риц // Морфология и эволюция животных. — М. : Наука, 1986. - С. 187-211.
60. Второв ПЛ. Пути познания места амфибий и рептилий в потоке энергии экосистем // Вопросы гер-
петологии. — Л. : Наука, 1973. - С. 53-55.
61. Гаранин ВЛ. Земноводные и пресмыкающиеся Волжско-Камского края. - М. Наука, 1983. -
175 с.
62. Гассо В Я. Сравнительная характеристика микроэлементного состава различных видов пресмыкаю-
щихся степного Приднепровья // Вопросы герпетологии. — Киев Наук, думка, 1989. — С. 55-56.
63. Герловин EJ11. Гистогенез и дифференцировка пищеварительных желез. - М. Медицина, 1978.-
263 с.
64. Гожее АД.О некоторых представителях животного мира, встреченных в пределах западной части
Терско-Дагестанского песчаного массива // Изв. Гос. рус. геогр. о-ва. - 1930. - 57, вып. 3. -
С. 269-286.
65. Голубев И.Ф. Почвоведение с основами геоботаники. - М. : Колос, 1970. - 440 с.
66. Гончарова ВЛ. Некоторые данные по биологии разноцветной ящурки (Eremias arguta deserti Gmell.)
на юго-востоке УССР // Науч. зап. Днепропетр. ун-та. - 1955. - 51. - С. 91-98.
67. Гражданкин АВ. Реакции рептилий аридных территорий на высокие температуры и инсоляцию //
Зоол.журн. — 1973. — 52, вып. 4. — С. 552—561.
68. Григорьев Н.И. Строение и регенерация печени после ее местного повреждения. — Л. Медицина,
1975,- 190 с.
223
69. Гуртовой Н.Н., Матвеев Б.С., Дзержинский Ф.Я. Практическая зоотомия позвоночных, Зем-
новодные. Пресмыкающиеся / Под ред. Б.С.Матвеева, Н.Н.Гуртового. — М.: Высш, шк.. 1978 —
408 с.
70. Гуськов ЕЛ., Лукина ГЛ., Конева В А. Определитель земноводных и пресмыкающихся Ростов-
ской области. - Ростов-н/Д : Изд-во Ростов, ун-та, 1983. - 50 с.
71. Гэнн ДЛ. Температура тела пойкидотермных животных // Успехи соврем, биологии. - 1944. -
17, вып. 1.-С. 87—107.
72. Даль СК. Позвоночные низовьев реки Зеравшан // Тр. Узб. гос. ун-та. - Самарканд, 1936 - 7 -
С.142.
73. Даль СК. Позвоночные животные прибрежной полосы озера Севан и изменение их группировок в
связи со спуском водоема // Зоол. сб. АН АрмССР. - 1950. - Вып. 7. - С. 5-53.
74. Даль СК. Позвоночные животные. - Ереван Изд-во АН АрмССР, 1954. - 415 с. - (Животный
мир Армянской ССР ; Т. 1).
75. Даль СК. Ландшафтно-экологический очерк позвоночных животных мест возможной природной
очаговости бруцеллеза в Ставропольском крае // Тр. Н-и. противочум. ин-та Кавказа и Закав-
казья. - Ставрополь, 1959. - Вып. 2. - С. 93-114.
76. Даревский И.С. О систематическом положении закавказских разноцветных ящурок Eremias arguta
(Pall.) (Reptilia, Sauna) // Докл. АН АрмССР. - 1953. - 16, № 4. - С. 117-121.
77. Даревский И.С. Фауна пресмыкающихся Армении и ее зоогеографический анализ Дис. ... канд.
биол.наук. - Ереван. 1957,— 457, 26 с.
78. Даревский И.С. Зоогеографические особенности герпетофауны бассейна озера Севан и вероятные
причины их возникновения // Изв. АН АрмССР. - 1959. - 12, № 10. - С. 15-22.
79. Даревский И.С. Сезонные изменения жировых тел и гонад у некоторых ящериц долины реки
Араке в Армении // Зоол. журн. - 1960. - 39, вып. 8. - С. 1209-1217.
80. Даревский И.С. Скальные ящерицы Кавказа. - Л.: Наука, 1967, - 214 с.
81. Даревский И.С. Исчезающие и редкие виды земноводных и пресмыкающихся Закавказья // Мате-
риалы конф. ’’Фауна и ее охрана в республиках Закавказья”. - Ереван, 1975. - С. 64—67.
82. Даревский И.С. Сохранилась ли высокогорная популяция разноцветной ящурки (Eremias arguta
Pall.) в бассейне озера Севан // Материалы V Всесоюз.совещ. ”Вид и его продуктивность в ареале”,—
Вильнюс, 1988. - С. 130-131.
83. Даниелян ФД. Некоторые данные по сравнительному эмбриогенезу бисексуальных и партеногене-
тических форм скальных ящериц Армении // Биол. журн. Армении. - 1971. - 24, №10.- С8-12.
84. Даниелян ФД., Симонян А А. Развитие и рост // Прыткая ящерица : Моногр. описание вида. — М.:
Наука, 1976. - С.227-245.
85. Джафаров РД. Пресмыкающиеся Апшеронского полуострова. Herpetologia Арscheronica // Тр. ес-
теств.-истор. музея им. Зар даб и. - 1946. - Вып. 1. - С. 5-25.
86. Джафаров РД. Пресмыкающиеся Азербайджанской ССР. - Herpetologia Azerbajdzhanica // Там
же. - 1949. - Вып. 3. - С. 3-85.
87. Дидусенко ЛМ. Ящерицы Молдавии // Вопросы герпетологии. - Л.: Наука, 1964. - С. 20-21.
88. Динесман ЛЛ. О распределении и экологии рептилий в связи с зонами солнечной радиации //
Проб л. физ. географии. - 1949.- 14. - С. 153-165.
89. Долинин В А. Исследование основного обмена и водных потерь у пустынных видов рептилий //
Видовые и природно-климатические адаптации организма животных : Реф. докл. к III Всесоюз.
совещ. по экол. физиологии, биохимии и морфологии. — Новосибирск, 1967. - С. 33—36.
90. Дольник ВР. Энергетический метаболизм и размеры животных: физические основы соотношения
между ними // Журн. общ. биологии. - 1978. - 39, № 6. - С. 805-816.
91. Домбровский БА. К герпетофауне Апшеронского полуострова. — Изв. Кавказ. Музея. - Тифлис,
1913.- 7, вып. 3/4.-С. 295-302.
92. Дубинин ВЛ. Эколого-фаушстический очерк земноводных и пресмыкающихся Хавастского
района Ташкентской области УзССР // Тр. Ин-та зоологии и паразитологии АН УзССР. - 1954. -
З.-С. 159-170.
93. Елецкий ЮК. Некоторые итоги и перспективы изучения пищеварительных желез в условиях нару-
шения иннервации // Регуляция морфогенеза и регенерация пищеварительных желез. - Л.: ЛГСМИ,
1974.-С. 6-8.
94. Захидое Т.З. Биология рептилий Южных Кызыл-Кумов и хребта Нура-Тау // Тр. Среднеазиат. ун-
та. Сер. 8. Зоология. - 1938. — Вып. 54. - С. 1-52.
95. Гаджиев А.Т.,Мустафаева ЗА., Джафаров С Л. Эктопаразиты ящериц (Sauria, Reptilia) Азербайджа-
на // Паразитологические исследования в Азербайджане. - Баку : Зим, 1982. - С. 155—161.
96. Зимина Р.П. О пресмыкающихся Иссык-Кульской котловины // Тр. Ин-та географии АН СССР. —
1959. - Вып. 75. - С. 156-167.
97. ЗиняковаМ.П.,РуденкоП.П. Особенности экологии разноцветной ящурки в Краснодарском крае //
Фауна и экология амфибий и рептилий : Сб. науч. тр. — Краснодар, 1984. — С. 55-64.
98. Зинякова МЛ., Плотников ГК. К герпетофауне побережья Восточного Приазовья // Актуальные
вопросы экологии и охраны природы Азовского моря и Восточного Приазовья. — Краснодар,
1989.-Ч. 1.-С. 153-155.
99. Иванова НЛ. К изучению популяций разноцветной ящурки Eremias arguta в северном Прикас-
пии // Вопросы герпетологии. - Л.: Наука, 1985. - С. 83—84.
100. Иноземцев АА. Трофические связи бурых лягушек в хвойных лесах Подмосковья // Зоол. журн.-
1969. - 48, вып. 11. - С. 1687-1694.
224
101. Иорданский НЛ. Кинетизм черепа ящериц. О функциональном значении кинетизма черепа // Там
же. - 1966. - 45, вып. 9. - С. 1398-1410.
102. Иорданский НЛ. К функциональному анализу черепа ящериц (Lacertilia) . Особенности строения,
связанные с кинетизмом // Там же. - 1971. - 50, вып. 5. - С. 724-733.
103. Калашников ЕЛ. Физиология газообмена у ядовитых змей Средней Азии // Вопросы герпетоло-
гии. - Л. : Изд-во Ленингр. ун-та, 1964. - С. 31-32.
104. Калужина МЛ. Морфология и биология отряда ящериц Зеравшанской долины // Тр. биол.-почв,
ф-та Узб. ун-та. Н.С. - 1951. - Вып. 46. - С. 75-97.
105. Кольбере В А. Анатомия большого чревного нерва // Сб. работ Риж. мед. ин-та. - 1954. - 3. -
С.5-12.
106. Калъберг В А. Анатомия пограничного симпатического ствола человека // Кауч, тр.высш. учеб,
заведений ЛитССР. - Вильнюс Минтис, 1964. - С. 23-25.
107. Камалова ЗЯ. Ящерицы Средней Азии. - Ташкент : Узбекистан. - 1978. - 48 с.
1№.К арамян АЛ. Эволюция межинтегративной деятельности мозга домлекопитающих позвоночных //
Эволюционная физиология. - Л. : Наука, 1979. - С. 81-143.
109. Карамян АЛ., Веселкин НЛ., Белехова МГ О некоторых общих закономерностях морфофунк-
циональной эволюции конечного мозга позвоночных // IX съезд Всесоюз. физиол. о-ва им.И Л .Пав-
лова. - Л., 1970. - 2. - С. 353-354.
110. Карпенко ВЛ. Ящерицы // Экология позвоночных животных хребта Нуратау. - Ташкент Фан,
1970.- С. 12-35.
111. Кесслер К.Ф. Естественная история губерний Киевского учебного округа. Животные земновод-
ные. - Киев, 1853. - 94 с.
112. Кесслер К.Ф. Путешествие с зоологической целью к северному берегу Черного моря и в Крым в
1858 г.-Киев, 1861.- 160 с.
113. Киреев В А. К изучению экологии и биологии разноцветной ящурки Eremias arguta Pallas в Калмы-
кии // Животный мир Калмыкии, его охрана и рациональное использование. - Элиста, 1977. -
С.70-81.
114. Киреев В А. Земноводные и пресмыкающиеся Калмыкии Автореф. дис. ... канд. биол. наук. -
Киев, 1982.- 20 с.
115. Киреев В А. Земноводные и пресмыкающиеся. - Элиста Калм. кн. изд-во, 1983. - 112 с. - (Жи-
вотный мир Калмыкии).
116. Корууев ПА. Эволюция, гравитация, невесомость. - М.: Наука, 1971. - 152 с.
117. Корнеев АЛ. Енотовидная собака Nyctereutes procyonoides на Украине // Тр. зоол. музея Киев,
ун-та. - 1954. - № 4. - С. 13-72.
118. Косарева Н. Рептилии юга Сталинградской области // Учен. зап. Сталингр. пед. ин-та - 1950. -
Вып. 2.-С. 227-240.
119. Костин ВД. Материалы к фауне млекопитающих левобережья низовьев Аму-Дарьи и Устюрта и
очес к распределения видов позвоночных животных // Тр. Ин-та зоологии и паразитологии АН
УзСС!.- 1956.- 8.-С. 5-77.
120. Костин ВЛ. Заметки по распространению и экологии земноводных и пресмыкающихся древней
дельты Амударьи и Кара-Калпакского Устюрта // Там же. - 1956. - 5. - С. 47-66.
121,Котенко ТЛ. Герцетофауна Черноморского заповедника и прилежащих территорий // Вести,
зоологии. - 1977. - № 2. - С. 55-66.
122. Котенко ТЛ. Некоторые особенности поведения и суточной активности разноцветной ящурки //
Управление поведением животных: Докл. участников II Всесоюз. конф, по поведению живот-
ных. - М.: Наука, 1977.- С. 154-155.
123. Котенко ТЛ. О экологии разноцветной ящурки в Нижнем Заднепровье // Вопросы герпетоло-
гии. - Л.: Наука, 1977. - С. 120-121.
124. Котенко Т.И. К изучению постэмбрионального роста разноцветной ящурки (Eremias arguta desert.
Gm. ) // Эколого-морфологические особенности животных и среда их обитания. - Киев : Наук,
думка, 1981.- С. 25-28.
125. Котенко ТЛ. Материалы по экологии разноцветной ящурки на юге Украины // Там же. - С.29-32.
126, Котенко ТЛ. Разноцветная ящурка - Eremias arguta deserti (Reptilia, Sauria) на Украине. Сообщение
i // Вести, зоологии. - 1981. - № 4, - С. 15-21.
127. Котенко ТЛ. Разноцветная ящурка Eremias arguta deserti (Gmelin, 1788) на Украине. Сообщение 2.
Суточный и сезонный циклы активности // Там же. - 1981. - № 5. - С. 47-52.
128. Котенко Т.И. Влияние особенностей биотопа на активность и поведение некоторых пресмыкающих-
ся юга Украины // Прикладная этология : Материалы III Всесоюз. конф, по поведению животных.-
М.: Наука, 1983. - 3. - С. 251-252.
129. Котенко ТЛ. Влияние температуры среды на активность, поведение и температуру тела разноцвет-
ной ящурки Eremias arguta // Механизмы поведения: Материалы III Всесоюз. конф, по поведению
животных. - М. : Наука, 1983. - 1. - С. 175-176.
130. Котенко ТЛ. Пресмыкающиеся левобережной степи Украины Автореф. дис. канд. биол. на-
ук. - Киев, 1983.- 24 с.
131. Котенко ТЛ. Пресмыкающиеся левобережной степи Украины Дис. ... канд. биол. наук. - Киев,
1983,- 555 с.
132. Котенко ТЛ. Влияние антродических факторов на распространение и численность разноцветной
ящурки // Вид и его продуктивность в ареале Материалы 4-го Всесоюз. совещ. - Свердловск,
1984.-4.5.-С. 21.
225
133. Котенко ТЛ. Изменения в герпетофауне левобережной степи Украины // VIII Всесоюз. зоогеогр.
конф.: Тез. докл. - М., 1984. - С. 391-392.
134. Котенко ТЛ. О регистрации редких и малочисленных видов пресмыкающихся на Украине // Все-
союз. совещ. по пр об л. кадастра и учета животного мира Тез. докл. - М„ 1986. - Ч. 2. - С 429-
431.
135. ХЬгенко ТЛ. Разноцветная ящурка на Украине. Сообщение 3. Отношение к погодным факторам;
температура тела // Веста, зоологии. - 1986. - № 4. - С. 45-52.
136. Котенко ТЛ. Sauronyssus sauiarum - обычный паразит ящериц степной Украины // X конф. Укр.
о-ва паразитологов : Материалы конф. - Киев : Наук, думка, 1986. - Ч. 1. - С. 305.
137. Котенко ТЛ. Земноводные и пресмыкающиеся Украины с точки зрения их охраны. - Киев,1987 -
67 с. - Деп. в ВИНИТИ, № 8014-В 87.
138. Котенко ТЛ. Охрана амфибий и рептилий в заповедниках Украины // Амфибии и рептилии запо-
ведных территорий : Сб. науч. тр. - М., 1987. - С. 60-80.
139. Котенко ТЛ. К изучению энтомофауны Черноморского заповедника: некоторые вопросы методи-
ки и результаты исследования // Веста, зоологии. - 1988. - № 1. - С. 44-50.
140. Котенко ТЛ. О северной границе ареала разноцветной ящурки на Украине // Там же. - №6,-
С.67-71.
141. Котенко Т.Н Роль Черноморского заповедника в охране герпетофауны степной зоны Украины и
некоторые проблемы малых заповедников // Заповедники СССР - их настоящее и будущее. -
Новгород, 1990. - Ч. 3. - С. 262-265.
142. Коштоянц Х.С. Основы сравнительной физиологии. - М.; Л: Изд-во АН СССР, 1950. - Т.1 - 524 с.
143. Кравчинский БД. Современные основы физиологии почек. - Л.: Медгиз, 1958. - 364 с.
144. Красавцев БА. Герпетологические наблюдения в Предкавказье // Тр. Ворошилов, пед. ин-та. —
1941. - 3, вып. 2. - С. 25-38.
145. Красная книга Армянской ССР. - Ереван : Айастан, 1987. - 124 с.
146. Красовский Д. Материалы к познанию фауны Reptilia et Amphibia Хасавъюртовского окр. Даге-
станской ССР // Изв. Горек, пед. ин-та. - Владикавказ, 1929. - 6. - С. 217-228.
147. Кубанцев Б.С., Косарева НА. Новые данные о распространении и численности некоторых видов жи-
вотных в Волго-Донском междуречье Волгоградской области // Учен. зап. Волгоград, пед. ин-та. -
1964. - Вып. 16. - С. 90—98.
148. Кудакина EJ0. О географической изменчивости разноцветной ящурки Eremias arguta в северном
Прикаспии // Вопросы герпетологии. - Л. : Наука, 1981. - С. 75.
149. Кулик ИЛ. Изучение индивидуальных участков пустынных ящериц // Зоол. журн. - 1971. -
50, вып. 2.-С. 300-302.
150. Куприянова ЛА. О возможных путях эволюции кариотипа ящериц // Тр, зоол, ин-та АН СССР. -
1986. - 157,—С. 86-100.'
151. Куцевй ПЛ. Попередш даш про рептилий Черкаського району КшвськоГ обласп // Наук. зап. Чер-
кас. пед.ш-ту. - 1941. - Вил. I. - С. 173-182:.
152. Ланге НО. Взаимоотношение между редукцией конечностей и развитием брюшной мускулатуры
рептилий // Памяти акад. АЛ.Северцова: Сб. ст. в 2 т. - М. ; Л. Изд-во АН СССР, 1940. - Т. 2,
ч.1.-С. 349-374.
153. Ленинджер. Биохимия. - М,: Мир, 1974. - 597 с.
154. Лукин ЕЛ. Различия в скорости эволюции разных систем органов и приспособлений к размноже-
нию и развитию животных//Зоол. журн. - 1964. - 43, вып. 8. - С. 1105—1119.
155. Лукина ГЛ. Пресмыкающиеся Западного Предкавказья Автореф. дис. ... канд. биол. наук. -
Ростов-н/Д, 1966. - 19 с.
156. Лукина ГЛ. Пресмыкающиеся Западного Предкавказья : Дис. ... канд. биол. наук. - Ростов-н/Д,
1966.- 218с.
157. Лукина ГЛ. Питание // Прыткая ящерица. - М.: Наука, 1976. - С. 179—213.
158. Лукина ГЛ., Гуськов ЕЛ., Конева В А. Определитель земноводных и пресмыкающихся Ростов-
ской области. - Ростов-н/Д, 1983. - 50 с.
159. Лукомская И.С., Городецкий BJC. Применение глюкозооксидазы для специфического количест-
венного определения глюкозы в крови // Вопр. мед. химии. — 1969. — 6, вып. 6. — С. 641—644.
160. Лызлова ЕМ., Вержбинская НА. Фосфоэнолпируваткарбоксикиназа з тканях миноги//Журн. эво-
люц. биохимии и физиологии. - 1976. - 12. - С. 75-77.
161. Львова СЛ. Особенности энергетического метаболизма ящурки Eremias arguta при зимней спячке //
Там же. - 1980. - 16, № 5. - С. 524-526.
162. Львова СЛ., Плотников ВЛ. Содержание гликогена и глюкозы в тканях лягушки и некоторых
рептилий // Там же. - 1978. - 14, № 4. - С. 400—402.
163,Майр Э. Зоологический вид и эволюция. - М.: Мир, 1968. - 598 с.
164. Макеев ВМ. К экологии степной гадюки в окрестностях ст. Отар // Позвоночные животные Сред-
ней Азии. - Ташкент : Фан, 1966. - С. 18-20.
165. Марков Г.С., Богданов ОЛ. Паразитофауна среднеазиатских ящурок // Учен. зап. Сталинград, пед.
ин-та. — 1961. - Вып. 13. — С. 101 — 123.
166. Марков Г.С., Иванов ВЛ., Крючков Б Л. и др. Простейшие и клеши - паразиты пресмыкающихся
Прикаспия // Учен. зап. Волгоград, пед. ин-та. - 1964. - Вып. 16. - С. 106-110,
161 .Марков Г.С., Лукина ГЛ., Маркова ЛЛ. и др. К паразитофауне пресмыкающихся Северного Кав-
каза // Там же. - С. 99-105.
168, Маркое Г.С., Косарева НА., Кубанцев Б.С. Материалы по экологии и паразитологии ящериц и змей
226
в Волгоградской области // Паразитические животные Волгоградской области. - Волгоград,
1969.- С. 198-220.
169.Марков Г.С., Тертышников М.Ф., Резник ПА. Материалы по паразитофауне массовых видов яще-
риц в Центральном Предкавказье // Вопросы паразитологии животных юго-востока СССР. - Вол-
гоград, 1974. - С. 54-63.
170.Медведев CJf., Петров В.С. Заметки о питании круглоголовок и ящурок в песках Восточного
Предкавказья // Учен. зап. Харьк. ун-та. - 1959. - 106, вып. 28. - С. 65.
Ш.Мелкумян Л.С. Степень физиологической готовности и время ухода на зимовку некоторых видов
ящериц // Экология. - 1972. - № 1. - С. 105-107.
112. Михеева ЕА. Цитохимический и авторадиографический анализ реактивных изменений в поджелу-
дочной железе с сохраненным и нарушенным оттоком секрета при действии хлористого кобальта :
Автореф. дис. ... канд. биол. наук. — Л., 1974. - 24 с.
113. Морев БРО. Предпочитаемые температуры среды и поведение ящериц в пустыне // Температурная
компенсация и поведенческий гомеостазис.— Л.: Наука, 1980, —С. 101-105.
174. Мунхбаяр X. Описание Eremias arguta Pall. // Науч, метод, зап. Монгол, пед. ин-та. — 1969. - № 4. -
С. 103-106.
175. Мустафаева ЗА., Гаджиев А.Т., Джафарова С.К., Ахмедов М.И. Рептилии - прокормители парази-
тических клещей в Азербайджане // Тез. докл. III Всесоюз. совещ. по теорет. и прикл. акароло-
гии. - Ташкент, 1976. - С. 177.
176. Мусхелишвили ТА. Пресмыкающиеся Восточной Грузии. — Тбилиси Мецниереба, 1970. - 320 с.
177. Наточин ЮЛ. Ионорегулирующая функция почки. - Л.: Наука, 1976. - 268 с.
118. Наточин ЮЛ. Проблемы эволюционной физиологии водносолевого обмена. - Л. : Наука, 1984. -
40 с.
179. Неемченко МТ., Темботов АК. К герпетофауне Кабардино-Балкарской АССР // Учен. зап. Кабард.-
Балкар.. ун-та. Сер. сельхоз. и биол. наук. - 1959. - Вып. 6. - С. 23-24.
180. Неручев ВЛ., Васильев Н.Ф. Фауна рептилий (Reptilia) Северо-восточного Прикаспия // Вести, зоо-
логии. - 1978. - №6. - С. 36-41.
181, Неручев ВЛ., Козырева О А., Накаренок ЕГ. К феногеографической характеристике популяций
Eremias arguta в Северном Прикаспии // Вопросы герпетологии, — Киев Наук, думка, 1989. -
С. 171-172.
182. Яедучев ВЛ., Кудакина EJ0., Васильев Н.Ф. и др. Исторические и антропогенные факторы в со-
временном распространении рептилий в северном*Прикаспии // VII Всесоюз. зоогеогр, конф.
Тез. докл. — М.: Наука, 1979. - С. 212—214.
183. Неручев В.В., Кудакина Е.Ю., Васильев Н.Ф. Влияние выпаса на фауну и население рептилий в
пустынях северного Прикаспия // Наземные и водные экосистемы. - Горький, 1981. - С.56-61.
184. Яер7чев ВЛ., Шатилович О А. О некоторых закономерностях формирования герпетофауны северо-
восточного Прикаспия // Вопросы герпетологии. - Л.: Наука, 1985. - С. 147-148,
185. #еру*ев ВЛ., Ширяев А.Ф., Капустина С.Ф., Кудакина ЕРО. Фауна и ландшафтные комплексы реп-
тилий юга урало-эмбинского междуречья // Наземные и водные экосистемы. - Горький, 1984. -
С. 36-39.
186. Никольский AM. Пресмыкающиеся и земноводные Российской империи Herpetologia Rossica //
Зап. Акад.Наук, Эсер, - 1905,- 17,№ 1. - 515 с.
181 .Никольский AM. Определитель пресмыкающихся и земноводных Российской империи. - Харьков,
1907,- 182 с.
188. Никольский AM. Пресмыкающиеся и земноводные Кавказа (Herpetologia caucasica). - Тифлис:
Изд-во Кавказ.музея, 1913. - 272 с.
189. Никольский AM. Фауна России и сопредельных стран. Пресмыкающиеся. - Пг.: Изд-во Рос. АН. -
1915.- 1, —464с.
190. Ньюсхолм Э., Старк К. Регуляция метаболизма. - М.: Мир, 1977. - 344 с.
191. Окулова НМ. Некоторые черты биологии разноцветной ящурки в Западном Казахстане // Вопро-
сы герпетологии. - Л.: Изд-во Ленингр. ун-та, 1964. - С. 50-51.
192. Окулова НМ. Влияние внешних температур на активность, поведение и температуру тела разно-
цветной ящурки (Eremias arguta) // Зоол. журн. - 1969. - 48, вып. 10. - С. 1500-1506.
193. Окулова НМ. Популяционный полиморфизм разноцветной ящурки на северо-восточной окраине
Волго-Уральских песков // Вопросы герпетологии. - Л.: Наука, 1973. - С. 135-137.
194. Окулова НМ. Биоценотические связи разноцветной ящурки междуречья Волга-Урал // Там же,
1977.-С. 160-161.
195,-Ордели ЛА. Эволюционный принцип в применении к физиологии центральной нервной системы //
Избр. тр. - М.; Л.: Изд-во АН СССР, 1961. - Т. 1. - С. 166-192.
196. Орлова В.Ф. Распространение и изменчивость ящурок Монголии // Вопросы герпетологии. - Киев :
Наук, думка, 1989.-С. 181-183.
191 .Орлова В.Ф., Тзрбиш X. Материалы по герпетофауне Джунгарской Гоби // Герпетологические ис-
следования в Монгольской Народной Республике: Сб. науч. тр.-М.,1986.-С. 95-110.
198. Павленко ТА. Позвоночные животные Голодной степи // Животный мир Голодной степи. - Таш-
кент : Изд-во АН УзССР, 1962. - С. 127-175.
199. Параскив КЛ. Пресмыкающиеся Казахстана. - Алма-Ата: Изд-во АН КазССР, 1956. - 228 с.
200. Пачосский ИРС. Материалы по вопросу о сельско-хозяйственном значении птиц. - Херсон, 1909-
680 с.
201. Пащенко ЮЛ. Визначник земноводних та плазуюв УРСР. - К.: Рад. шк., 1955. - 148 с.
227
202. Пащенко Ю.И., Межжерин В А. К распространению и экологии разноцветной ящурки (Eremias
arguta Pall.) на Украине // Тр. зоол. музея Киев.ун-та. - 1954. - № 4. - С. 133-134.
203. Перешколъник СЛ. Опыт определения суммарного метаболизма популяции разноцветной ящурки
из горной пустыни Прииссыккулья // Вопросы герпетологии. - Л. : Наука, 1973. - С. 145-146.
204, Плисецкая ЭМ. Гормональная регуляция углеводного обмена у низших позвоночных. — Л, : На-
ука, 1975. -С, 47-51.
205. Поспелова-Штром М3. О распространении, экологии и динамике клещей рода На ema physalis, преи-
мущественно в пределах СССР // Патогенные животные Тр. отд. паразитологии. - 1936. - 2. -
С. 97-104.
206. Проссер Л. Сравнительная физиология животных, - М.: Мир, 1977. - 1. - 608 с.
207. Проссер Л. Температура // Сравнительная физиология животных. - М. : Мир, 1977. - 3. - С.84-
209,
208. Проссер Л. Дыхательные функции крови // Там же. - С. 5-83.
209. Прохорова MM.t Тупикова ЗЯ. Большой практикум по углеводному и липидному обмену. — Л.:
зд-во Ленингр. ун-та, 1965. - 327 с.
210. Прыткая ящерица : Моногр. описание вида. - М.: Наука, 1976. - 374 с.
211. Пустыни / АТ.Бабаев, Н.Н.Дроздов, И.С.Зонн, ЗХ.Фрейкин. - М. Мысль. - 1986. - 318 с. -
(Природа мира) .
212,Радченко_НМ., Тертышников М.Ф., Шарпило ВЛ. О паразитофауне амфибий и рептилий Ставро-
полья // Фауна и экология амфибий и рептилий : Сб. науч. тр. - Краснодар, 1984. — С. 84—88.
213. Роль ЮМ. Некоторые методы экологического учета грызунов // Вопр. экологии и биоценологии. —
1936. - Вып. 3. - С. 140-157.
214. Реймерс Н.Ф. Природопользование. - М.: Мысль, 1990. - 637 с.
215. Рефф Р.,Кофман Т. Эмбрионы, гены, эволюция. - М. Мир, 1986. - 402 с,
216. Риклефс Р. Основы общей экологии. - М.: Мир, 1979. - 424 с.
217 .Рихтер В., Кранц Д. Авторадиографическое изучение возрастных особенностей нейрогенеза мозга
низших позвоночных // Материалы III Груз. респ. совещ. по проблеме ”Нейрогенез, реактивные и
регенеративные процессы”. - Тбилиси, 1974.- С. 111-113,
218. Родионов В. Некоторые данные по газообмену у рептилий в состоянии переохлаждения // Бюл.
Моск, о-ва испытателей природы. Отдел, биол. - 1938. — 47, вып. 2. - С. 182-187.
219. Руководство по изучению земноводшлх и пресмыкающихся. - Киев Наук, думка. 1989. - 172 с.
220. Рюмин АЗ. Температурная чувствительность позвоночных животных и биологический путь про-
исхождения теплокровных форм // Сб. науч. студ. работ. Биология. - М. Изд-во Моск. унн:а,
1939. - Вып.6. - С. 55-84.
221, Сави на М3., Вилкова В А. Глюконеогенез из глицерина в органах миноги в преднерестовый пери-
од// Жури, эволюц, биохимии и физиологии, - 1976. - 12, № 2. - С. 189-192,
222. Саид-Алиев С А. Земноводные и пресмыкающиеся Таджикистана. - Душанбе Дрниш, 1979. -
146 с.
223. Сергеева ХМ., Акрамов MJU. О размножении некоторых видов ящериц Джизакской степи // Во-
просы герпетологии, - Л.: Наука, 1985. - С. 186.
224. Сатторов Т. Пресмыкающиеся Северного Таджикистана: Автореф. дис.... канд. биол. наук. - Киев,'
1981,-22 с.
225. Сатторов Т. Пресмыкающиеся Северного Таджикистана Дис. ... канд. биол. наук. - Душанбе,
1981.- 289 с.
226. Conn ЕЛ. История развития нервной системы позвоночных. - М. *. Медицина, 1959. - 428 с.
227 .Сергеев AM. Материалы по постэмбриональному росту рептилий // Зоол. ж урн. - 1939. - 18,
вып. 5.-С. 888-903.
228. Сергеев А. Температура пресмыкающихся в естественных условиях // Докл. АН СССР. - 22,№ 1.-
С. 49-52.
229. Сергеев AM. Материалы к познанию популяций у рептилий // Вопр. экологии и биоценологии. -
1939. - Вып. 4, - С. 276-280.
230. Сергеев AM. Эволюция эмбриональных приспособлений рептилий. — М, Сов. наука, 1943. -
203 с.
231. Силантьев А А. Фауна ”Падов” имения В А Лар ыш кина, Балашовского уезда. Саратовской губ. //
Пады. - Спб, 1984. - 213 с.
232. Ситко ТМ. Суточный цикл активности ящурки разноцветной (Eremias arguta deserti (Gmelin ,
1789) на лесостепных участках Черноморского государственного заповедника // Некоторые вопро-
сы экологии и морфологии животных : Материалы IX науч. конф. Ин-та зоологии АН УССР. - Ки-
ев : Наук, думка, 1975. - С. 48-50.
233. Слоним А Л- Физиологические адаптации к аридной зоне // Экологическая физиология животных.-
Л.; Наука, 1982. - Ч. 2. - С. 236-259.
234. Слоним АЛ Среда и поведение. - Л.: Наука, 1976. - 209 с.
235, Слоним А Л- Эволюция терморегуляции. - Л.: Наука, 1986. - 74 с.
236. Смиттен НА. Симпато-адреналовая система в фило- и онтогенезе позвоночных. - М. Наука,
1972.- 346 с.
237. Смогоржевсъкий Л.О. Рибощш птахи Украши. - К.: Вид-во Кшв. ун-ту, 1959. - 122 с.
238. Смогоржевсъкий Л.О. Гагари, норш, трубконосу вес л оногупяпнасту фламшго. - К:: Наук, думка,
1979.- 187 с.
239. Сомов НН. Орнитологическая фауна Харьковской губернии. - Харьков, 1897. - 680 с.
228
240. Стрельников ИЛ, Свет как фактор в экологии животных. Статья первая. Действие солнечной ра-
диации на температуру тела некоторых пойкилотермных животных // Изв. науч, ин-та им Л.Ф Лес-
гафта. - 1934. - 17/18. - С. 313-372.
241. Стрельников ИЛ. Значение солнечной радиации в экологии высокогорных рептилий // Зоол.
журн. - 1944. - 23, вып. 5. - С. 250-257.
242. Стрельникое ИЛ. О терморегуляции у современных и вероятном тепловом режиме мезозойских
рептилий // Вопросы палеобиологии и биостратиграфии. - М. Госгеолтехиздях, 1959. - С. 129-
144.
243. Суханов ВЛ. Некоторые особенности глубокой тазобедренной мускулатуры ящериц (Lacertilia) //
Сб. науч. студ. работ МГУ (биол.). - 1957. - С. 7-13.
244. Суханов ВЛ. Некоторые вопросы филогении и системы Lacertilia (Sauria) // Зоол. журн. — 1961. -
40, вып. 1.- С. 73-83.
245. Суханов ВЛ. Общая систематика симметричной локомоции наземных позвоночных и особенно-
сти пере движения низших тетрапод. - Л.: Наука, 1968. - 228 с.
746. Сухов Г.Ф. Нотатки про спепиф!чшсть грунту та рослинносп тих м!сць на Украпп, де перебувае
Eremias arguta (Pall.) // 36. праш» Зоол. музею. - 1927. - № 3. - С. 179-183.
247. Основы сенсорной физиологии. - М.: Мир, 1976. — 520 с.
248. Таращук В Г. Земноводш та плазуни. — К.: Вид-во АН УРСР, 1959. — 191 с. -(Фауна Украши;Т.7;.
249. Таращук СЛ. Герпетофауна северо-западного Причерноморья и ее изменения под действием антро-
пических факторов : Автореф.... дис. канд. биол. наук. - Киев, 1987. - 25 с.
250, Таращук СЛ. Герпетофауна северо-западного Причерноморья и ее изменения под действием антро-
пических факторов : Дис.... канд. биол. наук. - Киев, 1987. - 309 с.
251. Терентьев П.В., Чернов С.А. Краткий определитель земноводных и пресмыкающихся СССР. - М.;
Л. : Учпедгиз, 1936. - 96 с.
252. Терентьев П.В., Чернов С.А. Краткий определитель земноводных и пресмыкающихся СССР. - Л.
Учпедгиз, 1940. - 184 с.
253. Терентьев ПЛ., Чернов СА. Определитель пресмыкающихся и земноводных. - М. Сов. наука,
1949. — 340 с.
254. Тертышников М.Ф. Материалы по экологии двух видов рептилий в Ставрополье // Материалы
III зоол. конф. пед. ин-тов РСФСР. - Волгоград, 1967. - С. 396-397.
255. Тертышников М.Ф. Индивидуальная территория прыткой ящерицы (Lacerta agilis) и разноцветной
ящурки (Eremias arguta) и особенности ее использования // Зоол. журн. - 1970. - 49, вып. 9. -
С. 1377-1385.
256. Тертышников М.Ф. О питании разноцветной ящурки (Eremias arguta deserti Gmehn, 1788) в Пред-
кавказье // Материалы IV науч. конф, зоологов пед. ин-тов. - Горький, 1970. - 7.316-318.
257. Тертышников М.Ф. Питание разноцветной ящурки (Eremias arguta deserti Gmel., 1788) в Централь-
ном Предкавказье // Экология. - 1970. - № 4. - С. 92-96.
258. Тертышников М.Ф. Экологический анализ и биоценологическое значение популяций прыткой яще-
рицы (Lacerta agilis exigna Eichw., 1831) и разноцветной ящурки (Eremias arguta deserti Gmelin,
1789) в условиях Ставропольской возвышенности Автореф. дис. канд. биол. наук. - Киев.
1972.- 30 с.
259. Тертышников М.Ф. Экологический анализ и биоценологическое значение популяций прыткой яще-
рицы (Lacerta agilis exigua Eichw., 1831) и разноцветной ящурки (Eremias arguta deserti Gmelin,
1789) в условиях Ставропольской возвышенности Дис. ... канд. биол. наук. - Киев, 1972.
247 с.
260. Тертышников М.Ф. К вопросу об особенностях газообмена прыткой ящерицы и разноцветной
ящурки // Вопросы герпетологии. - Л. Наука, 1973. - С. 178-179.
261. Тертышников М.Ф. Значение амфибий и рептилий в биоценозах Северного Кавказа // Географиче-
ские проблемы изучения, охраны и рационального использования природных условий и ресурсов
Северного Кавказа. - Ставрополь, 1973. - С. 134-135.
262. Тертышников М.Ф. Влияние погоды и климата на активность прыткой ящерицы и разноцветной
ящурки // Экология. - 1976. - № 3. - С. 57-61.
263. Тертышников М.Ф. О размножении прыткой ящерицы и разноцветной ящурки в Предкавказье //
Там же. - 1978. - № 2. - С. 94-96.
264. Тертышников М.Ф. О перемещениях и миграциях разноцветной ящурки // Сб. материалов III со-
вещ. "Вид и его продуктивность в ареале". - Вильнюс, 1980. - С. 73-74.
265. Тертышников М.Ф. О перемещениях и миграциях разноцветной ящурки // Вид и его продуктив-
ность в ареале. - М.: Наука, 1983. - С. 72-76.
266. Тертышников М.Ф., Горовая ВЛ. Пресмыкающиеся Ставропольского края. Сообщ. 1. Черепаха,
ящерицы // Фауна Ставрополья. - Ставрополь, 1984. - Вып. 3. - С. 48-91.
267. Тертышников М.Ф., Горовая ВЛ.,Игропуло В.С, и др. Биомасса и продуктивность массовых видов
бесхвостых амфибий в центральной части Северного Кавказа // Тр. Зоол. ин-та АН СССР. - 1987. -
158.-С. 155-159.
268. Тертышников М.Ф., Котенко ТЛ. Об особенностях терморегуляции разноцветной ящурки // Про-
блемы региональной зоологии. - Ставрополь, 1986. — С. 73—84.
269. Тертышников М.Ф., Шарпило ВЛ., Радченко НМ. Паразитофауна амфибий и рептилий Ставро-
полья // Болезни с природной очаговостью на Кавказе Тез. конф., посвящ. 60-летию образова-
ния СССР. — Ставрополь, 1982. - С. 144-145.
270. Тертышников М.Ф., Щербак НЛ. Роль прыткой яшерицы и разноцветной яшурки в биоценозах
229
Ставропольской возвышенности // Вопросы герпетологии. - Л. Наука, 1973. - С. 179-181.
271. Тэрбиш X. I ерпетофауна юго-западной части МНР и ее природно-хозяйственное значение Авто-
реф. дис. ...канд. биол.наук.- Улан-Батор, 1989. - 25 с.
272, Ушаков БД. Анализ теплоустойчивости клеток и белков пойкилотермных животных в связи с
проблемой вида. - Л„ 1964. - 71 с.
273. Федоров СМ. Пресмыкающиеся, земноводные, круглоротые и рыбы Ставропольского края // Ма-
териалы по изучению Ставропольского края. - 1956. - Вып. 8. - С. 203-220.
274. Фвична география УкрайнськоГ РСР / О.М.Маринич, АДЛанько, МДДербань. ПД?ДНишенко. - К. :
Вища шк„ 1982. - 207 с.
275. Филимонов ИЛ. Сравнительная анатомия большого мозга рептилий. — М. Изд-во АН СССР
1963.- 243 с.
276. Фомина МЛ. Коэффициенты вариации индексов некоторых органов разноцветной ящурки из
Калмыкии и Казахстана// Вопросы герпетологии. - Л.: Наука, 1973, - С. 188—189.
277. Хозацкий ЛЛ. Температура поверхности тела некоторых земноводных и пресмыкающихся // Вести.
Ленингр. ун-та.Сер. биол. - 1959. - № 21. - С. 92-105.
278. Хонякина ЗЛ. Ящерицы Дагестана Автореф. дис. ... канд. биол. наук. - Махачкала, 1964. -
20 с.
279. Царук СЛ. Изменчивость фолидоза головы и внутривидовая систематика разноцветной ящурки
Eremias arguta (Pallas, 1773) // Систематика и экология амфибий и рептилий. - Л., 1986. - С.203-
214.
280. Цемш I.O. Герпетолопчш замггки // Тр. Зоол. музею Ки1в. ун-ту. — 1941. — 1. - С. 313—322.
281. Черлин В А. Поведение, температура тела и терморегуляция у рептилий // Механизмы поведения :
Материалы Ш Всесоюз. конф, по поведению животных. - М. Наука, 1983. - 1.-С. 173-175.
232. Черлин В А. Способы адаптации пресмыкающихся к температурным условиям среды // Журн.
общ. биологии. - 1983. - 44, № 6. - С. 753-764.
283. Черлин В А. Популяционные аспекты термальных адаптаций пресмыкающихся // Проблемы попу-
ляционной экологии земноводных и пресмыкающихся. - М., 1989. - С. 135-172. - (Итоги науки
и техники; Зоология позвоночных; Т. 17) .
284. Черлин В А., Музыченко ИЛ. Использование нор в терморегуляции рептилий // Прикладная этоло-
гия : Материалы III Всесоюз. конф, по поведению животных. - М.: Наука, 1983. - Т. 3. - С.172-
174.
235. Черлин В А., Музыченко ИЛ. Термобиология сетчатой ящурки (Eremias grammica), ушастой (Phry-
nocephalus mystaceus) и песчаной (Ph. inter scapularis) круглоголовок в Восточных Каракумах // Зо-
ол. журн. - 1983. - 62, вып. 6. - С. 897-908.
286. Чернай А. Фауна Харьковской губернии и прилежащих к ней мест ... - Харьков, 1852. - 49 с.
287. Чернов С А. О подвидах и распространении разноцветной ящурки // Док. АН СССР. - 1934. -
3.-С. 666-671.
288. Чернов СА. Определитель змей, ящериц и черепах Армении. - М. : Изд-во АН СССР, 1937. - 54 с.
289. Чернов С А. Герпетофауна Армянской ССР и Нахичеванской АССР // Зоол. сб. - Арм. фил. АН
СССР, 1939. - 3. - С. 79-194.
290. Чернов С А. Материалы к герпетофауне Казахского нагорья, северного побережья Балхаша и гор
Кан-Тау // Изв. АН КазССР.Сер. зоол. - 1947. - Вып. 6. — С. 120-124.
291. Чернов СА. Пресмыкающиеся - Reptilia // Зона пустынь. - М.; Л.; Изд-во АН СССР, 1948. -
С. 127-161.- (Животный мир СССР; Т. 2).
292. Чернов С А. Пресмыкающиеся // Степи. — М.; Л.: Изд-во АН СССР, 1950. - С. 140-153. - (Жи-
вотный мир СССР; Т. 3) .
293. Чернов СА. Эколого-фаунистический обзор пресмыкающихся юга междуречья Волга-Урал // Тр.
Зоол. ин-та АН СССР. - 1954.- 16.-С. 137-158.
294. Чернов СА. Пресмыкающиеся // Тр. Ин-та зоологии и паразитологии АН ТаджССР. - 1959. - 98 —
204 с. - (Фауна Таджикской ССР; Т. 18) .
295. Черномордиков ВЛ. О температурных реакциях пресмыкающихся. Сообщение 1-е, термофилия //
Зоол.журн. - 1943. - 22, вып. 5. - С. 274-279.
296. Черномордиков ВЛ. О врожденных и приобретенных пищевых реакциях пресмыкающихся //
Докл. АН СССР. - 1944.-43, №4.-С. 181-184.
297. Черномордиков ВЛ. Эколого-физиологические особенности пресмыкающихся, птиц и млекопи-
тающих : Автореф. дис.... д-ра биол. наук. — М., 1965. — 39 с.
293. Шаммаков С. Пресмыкающиеся равнинного Туркменистана. — Ашхабад : Ылым, 1981. — 312 с.
299. Шарлеманъ М. По заповедниках швдня Укршни // Охорона пам’яток природа на Украли.- X.,
1928.- 36.2.-С. 1-15.
300. Шарпило ВЛ. Паразитические черви пресмыкающихся фауны СССР. - Киев : Наук, думка, 1976.-
287 с.
301. Шарыгин С А. Микроэлементы в организме разноцветной ящурки // Вид и его продуктивность
в ареале. - Свердловск, 1984, — Ч. 5. — С. 48.
302. Шарыгин С А. Приложимость данных по химическому элементарному составу к систематике не-
которых амфибий и рептилий // Журн. общ. биологии. - 1984. - 45, № 5. - С. 624—630.
303. Шварц С.С., Смирнов В.С., Добринский Л Л. Метод морфофизиологических индикаторов в эко-
логии наземных позвоночных. — Свердловск, 1968. — 388 с. — (Тр. Ин-та экологии растений и
животных; Вып. 58).
304. Шебзухова ЭА. Данные по биологии и распространению некоторых видов якцериц в центральной
230
части Северного Кавказа // Сб. науч, работ аспирантов / Кабард.-Балкар. ун-т. - Нальчик, 1968. -
Вып. 2. -С. 416-423.
305. Шебзухова ЭЛ. Пресмыкающиеся центральной части Северного Кавказа Автореф. дис. ... канд.
биол. наук. - Нальчик, 1969. - 16 с.
306. Шевлюк НМ. К гистохимической характеристике слюнных желез языка некоторых классов позво-
ночных // Арх. анатомии, гистологии и эмбриологии. - 1976. - № 3. - С. 58-62.
307. Шевченко ВИ. До екологп чайок i крячкгв Присивашшя // Праш н.-д. зоол.-бюл. iH-ту Харк. ун-ту.
Сектор екологп. - 1937. - 4. - С. 127-154.
308. Шимкевич ВМ. Курс сравнительной анатомии позвоночных животных. - М.; Пг.: Гос. изд-во,
1923.- 620 с.
309 .Шлугер ЕГ. Новые краснотелки юга Европейской части СССР // Вести, зоологии. - 1967. - № 3. -
С.41-50.
310. Шляхтин Г И., Голикова ВЛ. Методика полевых исследований экологии амфибий и рептилий:
Учеб, пособие. - Саратов : Изд-во Саратов, ун-та, 1986. - 78 с.
311. Шмалъгаузен ИИ. Основы сравнительной анатомии. - М. : Учпедгиз, 1938. - 488 с.
312. ШмидтЛиелъсен К. Размеры животных: почему они так важны ? - М.: Мир, 1987. - 259 с.
313. Шнитников ВМ. Пресмыкающиеся Семиречья // Тр. О-ва изучения Казахстана. - 1928. - 8, вып.З-
85 с.
314,Ztfep6flK НН. Плавающие ящурки // Природа. - 1959. - № 7. - С. 110-111.
315. Щербак НМ. Герпетофауна Крыма и ее зоогеографический анализ (Herpetologia Taurica) : Автореф.
дис. ... канд. биол. наук. — Л., 1962. — 16 с.
31Ь. Щербак НМ. Земноводные и пресмыкающиеся Крыма : Herpetologia Taurica. - Киев : Наук, думка,
1966.- 240с.
317. Щербак НМ. Есть ли в Турции ящурка разноцветная Eremias arguta (Pall.)? // Вести, зоологии. -
1970, — № 5. — С. 31.
318. ЛГер(мк НМ. Зависимость предпочитаемых температур от различных факторов у пустынно-степных
ящериц р. Eremias // Адаптация к аридной зоне. Зимняя спячка. - Новосибирск. 1970. - С. 136-
138. - (Адаптация организма человека и животных к экстремальным природным факторам сре-
ды : Материалы симп j .
319. Щербак ММ. Методика вивчення в!дношення до температуря плазушв за матер!алами досл!дження
реакшГ на градент температуря у представниюв роду Eremias (Sauria, Reptilia) фауни СРСР // 36.
праць Зоол. музею, - 1971. - № 34. - С. 76-88.
320. Щербак НМ. Ящурки Палеарктики. - Киев Наук, думка, 1974. - 296 с.
321. Щербина МЛ., Мукосева ЗЛ. Глюконеогенез как один из источников энергетического обеспечения
карпа в период зимнего голодания // В о пр. ихтиологии. - 1978. - № 3. - С 557-561.
322. Яблоков AJB., Тертышников М.Ф. Биомасса вида и отдельных его популяций // Прыткая ящерица :
Моногр. описание вида. - М.: Наука, 1976. - С. 322-328.
323. Яковлева ИД. Определитель пресмыкающихся Киргизии. - Фрунзе Изд-во АН КиргССР, 1961-
111 с.
324. Яковлева ИД. Пресмыкающиеся Киргизии. - Фрунзе : Илим, 1964. - 272 с.
325. Adams W.E. The cervical region of the Lacertilia. A critical review of certain aspects of its anatomy J.
Anat. - 1939. - 74. - P. 57 -71.
326. Adams W.E. Observations on the lacertilian sympathetic system // Ibid. - 1942. - 77. - P. 6-11.
327. Xdams ЖЕ*. The carotid sinus complex and epithelial body of Varanus varius // Anat. Rec. - 1952. -
113. - P.1-27.
323. Andrzefowski A. Reptilia imprimis Volhyniae, Podoliae et Gubernii Chersonensis // Nouv. Mem. Soc.
Nat. Moscou. - 1832. - 2. - P. 319-346.
329. Ariens-Kappers C.U. The corpus striatum, its phylogenetic and ontogenetic development and functions //
Acta psychiat. and neurol. - 1928. - 13. -P. 93-113.
330. Ariens-Kappers C.U., Huber G.C., Crosby E.C.The comparative anatomy of the nervous system of Vertebra-
tes including man. - New York : Macmilan Co., 1936. - 2. - P. 856-1845.
331. Balfur K. On the Early Development ol Lacertilia // Quart. J. Mier. Soc. - 1879. - P. 44.
332. Barker S.B., Summerson W. The colorimetric determination of lactic acid in biological material // J. Biol.
Chem. - 1941. - 138, N 2. -P. 535-554.
333. Bartels H., Hiller G., Reinhardt И'. Oxigen affinity of chicken blood // Respirat. Physiol. - 1966. - 1,
N3. -P. 345-356.
334. Basoglu M., Baran J. Turkije surungenleri. - Bor nova; Izmir : ilk г; matbaasi, 1977. - К. 1. - S. 272.
335. De Beer G.R. The early development of the chondrocranium of the lizard // Quart. J. Mier. Sci. - 1930. -
73, N 292. -P. 707-739.
336. De Beer G.R. The development of the vertebrate skull. - Oxford : Oxford Univ, press, 1937.
337. Bellairs A. d’A, Boyd G.D. The lachrymal Apparatus in Lizards and Snakes // Proc. Zool. Soc. - 1947. -
117, N 1. — P. 81-108,
338. Benedict F.G. The physiology of large Reptilies. - Washington, 1932.-613p.
339. Benedict F.G., Lee R.S. Lipogenesis in the animal body, with special reference to the physiology of the
goose. - Washington, 1938. - 286 p.
340. Bentley P.J., Schmidt—Nielsen K. Cutaneus water loss, reptiles // Science. - 1966. - 151. - P. 1547-
1549.
341. Berger-Dell’mour H. Der Ubergang von Eches zu Schleiche in der Gattung Tetradactylus Menem. Ver-
gleichend anatomische studien, biometrische Untersuchungen und Bewegungsanalysen an 3 sudafrikani-
231
schen Eidechsenarten (Fam. Gerrhosauridae) // Zool. Jahrb. Ait. - 2. - 1983. - 110, N 1. - S. 1-152.
342. Bohme W. tlber das stachelepithel am hemipenis lacertides eicechsen und seine systematische Bedeutung
Diss. Doktor math.-naturwiss. - 1971. - 60 S.
343. Boyd J.D. The nerve supply of the branchial arch arteries in Vipera berus // J. Anat. — 1942. — 76. -
P. 248- 257.
344. Brattstrom B.H. The evolution of reptilian social behavior // Amer. Zool. - 1974. - 14. - P. 35-
49.
345. Camp Ch. Classification of the lizards // Bull. Amer. Mus. Nat. Hist. - 1923. - 48. - P. 289-482.
346. Cope E.D. On the characters of the higher Groups of Reptilias Squamata, and especialy of the Diploglos-
sa // Proc. Acad. Natui. Sci. Philadelphia. - 1864. - 16. - P. 224-237.
347. De Witt C.B. Precision on thermoregulation and its relation to environmental factors in the desert iguana,
Dipsosaurus dorsalis // Physiol. Zool. - 1967. - 40, N 1. - P. 49-66.
348. Darevskij I.S., Terentev P. V. Estimation of energy flow through amphibian and reptile population // Secon-
dary productivity of terrestris ecosystems. - Warszawa ; Krakow, 1967. - P. 181-197.
349. DtfWon W.R., Bartholomew G.A. Metabolic and cardiac responses to temperature in the lizard Dipsosaurus
dorsalis // Physiol. Zool. - 1958. - 31, N 2. - C. 14.
350. Dawson W.R., Templeton J.R. Physiological responses to temperature in the lisard Crototaphytus colla-
ris // Ibid. - 1963. - 36, N 3. - C. 21.
351. Deraniyagala P.E.P. Reproduction in the monitor lizard Varanus bengalensis (Daudin) // Sopolia Zeylani-
ca. - 1958. - 28. - P. 11.
352, Du faure J.P., Hubert J. Table de developpement du lezard vivipare Lacerta (Zootoca) vivipara jacquin /./
Arch. anat. microsc. et moiphol. exp. - 1961. - 50, N 3. - C. 6.
353. Duke-Elder System of ophthalmology. The eye in evolution. - London Rimpton, 1958. - 1. -
843 p.
354. Dunkombe W. G. The colorimetric micro-determination of nonesterified fatty acids in plasma // Clin. chim.
acta. - 1964. - 9, N 2. - P. 122-124.
355. Eversmann E. Lacertae Imperii Rossici variis in itineribus meis observate // Nouv. Mem. Soc. Nat. Mos-
cou. - 1834. - 3. - P. 339-370.
356. Fahrenholz C. Drusen der Mundhohle // Hand. d. Vergl. Anat. d. Wirbelthiere.Berlin; Wien, 1937. -
3. - S. 115.
357. Foxon Y. Problems ot the double circulation in vertebrates // Biol. Rev. - 1955. - 30, N 3. - P. 61.
358. Frazzetta Т.Н. A functional consideration of cranial kinesis in lizards // J. Morphol. - 1962. - 111, N 3.-
P. 287-319.
359. Fuhn I.E. Vancea St. Eidonomische und okologische Untersuchungen an Eremias arguta (Gmelin) aus
Rumanien // Vestn. Cs. spolec. zool. - 1961. - 25, N 1.- S. 5-11.
360. Furbringer M. Die Knochen und Muskeln der Extremitaten bei des schlangenalnlichen Sauriem. - Leipzig,
1870. - 58 S.
361. Furbringer M. Zur vergleichenden Anatomie der Schultermuskeln. Th. 3, Ch. 4: Saurier und Crocodilia //
Morphol. Jahrb. - 1876. - 1. - S. 636-816.
362. Furbringer M. Zur vergleichenden Anatomie das Brustschulter apparatus und Schulter-muskein // Jenai-
sche Z. - 1900. - 34. - S. 215-718.
363. Gabe M., Marotja M., Saint-Girons H. Etat actuel des connaissances sur la glaude surrenale des reptiles //
Annee biol. - 1964. - 3. - P. 303-376.
364. Gabe M., Saint-Girons H. Contribution a la morphologic compares des fosses nasales et de leurs annexes
chez les lepidosaurieus. // Mem. Mus. nat. hist, natur. A. - 1976. - 98. - 87 p.
365. Gadow H. Beitrage zur Myologie der hint er Extremitaten der Reptilien // Morphol. Jahrb. - 1881. - 7,
Drittes Heft. - S. 329-466.
366. Gadow H. Amphibia and Reptiles. - London, 1901. - 8, p. 11, ch. 12. - P. 491-580.
367. Fuhn I.E., Vancea S. Reptilia (Testoase, Sopirle, Serpi). - Bucurejti, 1961. - 352 p. - (Fauna Republicii
Populare Ronfine; 14, f. 2).
368. Garus G. Traite elementaire d’anatomie comparee. - Paris, 1838. - 201 p.
369. Gaupp E. Das Chondrocranium von Lacerta agilis // Anat. Hefne. - 1900. - 14, N 1. - S. 333-
594.
370. Gnananuthu C.P. Comparative study of the hyoid and tongue of some typical genera of Reptiles // Proc.
Zool. Soc. London. Sor. B. - 1937. - 107, pt. 1. - P. 1-64.
371. Gregory W.K. Evolution emerging. A survey of changing patterns from primeval life to man. - New York,
1951. - Vol. 1-2.
372. Gregory W.K., Camp C.L. Studies in comparative myology and osteology. Note on the origin and evolu-
tion of certain adaptations for forward locomotion in the pectoral and pelvic girdles of reptiles and mam-
mals // Bull. Amer. Mus. Nat. Hist. - 1918. - 38, N 3. - P. 447-563.
373. Guibe J. Reptiles d’lran recoltes par M. Fransis Petter. Description d’un viperide nouveau: Pseudoceras-
tes latirostris // Bull. Mus. Nat. Hist. Natur. - 1957. - 29, N 2. - P. 96.
374. Guibe J. Traite do Zoologie. Publ. sons la director de Pierre.P. Crasse. - Paris; Masson, 1970. - T. 14:
Reptiles. Caracteres generaux et anatomie, fasc. 2. - S. 144-180.
375. Guieyss-Pellissier A. Les appareils respirotoires dans la serie animal. - Paris, 1945. - 245 p.
376. Gundy G.C., Ralph Ch,L.t Wurst G.Z. Parietal eyes in lizards zoogeographical correlates // Science. -
1975.- 190, N 4215. - P. 671-673.
377. Gundy G.C. , Wurst G.Z. Parietal eye - pineal morphology in lizards and its physiological implications. //
Anat. Rec. - 1976. - 185, N 4. - P. 419-431.
378. Hofer R.t Ladumer H., Gattrunger A. et al. Relationship between the temperature preferenda of fishes,
232
ampraoians ana reptuies, ana me suostrate ariiniues oi tneir trypsins // u. uomp. rnysioi. - 1У75. -
92, N 2. -P. 345-355.
379. Hoffman C.K. In: Dr H.G.Bronn’s: Klassen und Ordnungen des Thier-Reichs. Reptilien. 11. Eidechsen
und Wasserechsen. - Leipzig, 1890. - S. 610-707.
380. Hoffman C.K. Zur Eutwicklungsgeschichte des Sympathieus bei den Urodelen // Z. wiss. Zool. - 1902. -
74. -S. 71-73.
381. Holkenov C. Synssansen i dyreriget // Vor viden. - 1962 - 1963. - 26. - P. 801-810.
382. Holmgren W. Points of view concerning forebrain morphology in hieher vertebrates // Acta zool. - 1925 —
6. - p. 413-477.
383. Hutchinson V.H., Kosh P.G.Termoreeulatory function of the parietal eye in the lizard Anolis carolinensis //
Oecologia. - 1974. - 16, N 2. - Р. 173-177.
384. Kaushiva B.S. The arterial system of the pond-turtle, Lissemys vunctata (Bonnaterra) // Proc. Indian Acad.
Sci. - 1940. - 12. - P. 84-94.
385. Kotenko T.I. Eremias arguta deserti in Ukraine // Third ordinary general meeting of Societas Herpetologica
Europaea. — Prague, 1985. - P. 7'7.
386. Kotenko T. Eremias arguta deserti (Reptilia: Sauria) in the Ukraine // Studies in Herpetology. - Prague,
1986. -P. 479—482.
387. Kratzing G.E. The fine structure of the olfactory and vomeronasal organs of a lizard (Tiligna scincoides
scincoides) // Cell and Tissue Res. - 1975. - 156, N 2. - P. 239-252.
388. Kriegler M. Zur Myologie des Beckens und der Hinterextremitat der Reptilien // Gegenbaurs Morphol.
Jahrd. -1961. - 101, N 4. - S. 541-625.
339. Kumar Sh.t Majupuria T.Sh. The respiratory mechanism in Calotes versicolor Daud. (Agamidae, Lacer-
tilia) // Zool. Aux. - 1973. - 191, N 1/2. - S. 50-61.
390. Lakjer T. Studien fiber die Gaumenregion bei Saurien im Vergleich mit Anamniern und primitive Saurop-
siden // Zool. Jahrb. Anat. - 1927. - 49, N 1/2. - S. 57-356.
391. Lente L.A. Revision des reptilies decrits dans le ’’Journal de vojage” d’ Iwan Lepechin Par ЬЛ.Lantz //
Bull. Soc. zool. France. — 1922. - 47. - P. 193.
392. Lantz LA. Les Eremias de 1’Asie occidentale // Bull. Mus. de Ceorgie. - 1928. - 4. - P. 1-72; 5. -
P. 1-64.
393. Lantz LA. Les Eremias de 1’Asie occidentale // Bull. Soc. zool. France. - 1928/1930. - 6, N 4. - P.228-
243.
394. Larsell O. Morphogenesis and evolution of the cerebellum // Arch. Neurol, and Psych. - 1934. - 31. -
P. 357-435.
395. Lecuru S. Myologie et innervation du membre anterieur des Lacertiliens // Mem. Mus. nat. hist, natur.
A. - 1968. - 48, N 3. - P. 128-215.
396. Luppa H. Histologie, Histogeneze und Topochemie der Druzen des sauropsidenmagens. 1. Reptilie//Acta
histochem. - 1961. - 12. - C. 23.
397. Mertens R. Tierleben im Donau-Delta. 1. Lurche und Kriechtiere // Natur. und Volk. - 1957. -87, \T 5‘ -
S. 160-168.
398. Miller M.R. The normal histology and experimental alteration of the adrenal of the viviparous lizard. Xan-
tusia vigilis // Anat. Rec. - 1952. - 113. - P. 309-323,
399. Mivart S.G. Notes on the Myologie of Iguana tuberculata // Proc. Zool. Soc. of London. - 1967. - P.766-
797.
400. Mivart S.G. On the Myoloeie of Chameleon Parsonii // Proc. Zool. Soc. - 1870. - P. 850-890.
401. Myhre K., Hammel H.T. Behavioral regulation of the temperature in lizard Tiligua // Amer. J. Physiol. -
1969. -217,N 6. -P. 1490-1495.
4O2. Aegy K.A., Schoemaker V.H. Energy and nitrogen budgets in the free living desert lizard Sauromalus obe-
sus // Physiol. Zool. - 1975. - 48, N 3. - P. 252-262.
МУЗ. Northcutt R.G. The telencephalon of the western painted turtle (Chrysemys picta belli). - Urbana a. o.
Univ, press, 1970. - 113 p.
404. Oelrich T.M. The anatomy of the head of Ctenosaura pectinata (Iguanidae) // Misc. Pubis. Mus. Zool.
Univ. Mich. -1956. - 94. - P. 1-122.
405. Osawa G. Beitrage zur Anatomie der Hatteria punctata // Arch. Mikrobiol. Anat. - 1898. - 51. - S. 481-
691.
M)6. Parker W.K. On the structure and development of the skull in the Lacertilia. 1. Lacerta agilis, L. viridis,
Zootoca vivipara // Phill. Trans. Roy. Soc. London. - 1879. - 170. — P. 595-640.
407. Pasteels J. Une table analytique du developpemeut des Reptiles. 1. Stades de gastrulation chez les Lacerti-
lians // Ann. Soc. roy zool. Belg. - 1957. - 87. - P. 168-174.
MM. Pastor L.M., Ballesta G., Perez-Tomas R., Murine G. Immunocytochemical localization of serotonin in the
reptilian lung // Cell and Tissue Res. — 1987. - 248, N 3. - P. 713-715.
409. Pedreira de Oliveira A., Chequer Chemas R. Data on the pineal eye of some brazilian lizards // Rev. brazil,
pesquisas med. ebiol. - 1975. - 8, N 3/4. - P. 249-254. ,
410. Perrin A. Contribution a 1’etude de la myologie comparee // Bull. Sci. France et Belg. - 1892. - 24. -
S. 372-552.
411. Perrin A. Recherches sur les affinites zoologiques de 1’Hatteria punctata // Ann. Sci. natur. Zool. - 1895. -
7, N 20.-P. 33-102.
412. Perrin A. Contributions a 1’etude de la myologie comparee; membre anterieur chez un certain membre de
batraciens et de sauriens // Bull. Sci. France et Belg. - 1899. - 32. - P. 220—282.
413. Peter K, Normentafeln zur Entwicklunggeschichte der Zauneidechse (Lacerta agilis) // Keibel’s Normen-
tafeln. - 1904. - S. 63-69.
233
414. Peter К. Untersuchungen uber Bau und Entwicklung der Niere - Jena, 1909. - S. 100—107.
415. A>AwntoW7 The structure of the interstitial space of pulmonary alveolar septa in vertebrates // Anat. Anz. -
1986. - 160, N 8. - P. 373-375.
416. Reese A.M. The ductless glands of Alligator mississipiensis // Smiths. Misc. СоП. — 1931. — 82. — P. 1—14.
417. Renous S. Discussion sur la valeur phylogenique de divers caract^re empruntes a la myologie du membre
anterier des Lacertiliens // Gegenbaurs morph, Jahrb., Leipzig. - 1974. - 120, N 4. - P. 519-528.
418. Renous S. Retentissement de la rudimentation dux membre pelvien sur le complexe caudo-femoral des Squ-
amates // Bull, du Mus. Nat. D’Hist. Natur. - 1977. - 3, N 458, Zool 321. - P. 661-672.
419. Ribbing L. Die distale Armmuskulatur der Amphibien, Reptilien und Saugetiere // Zool. Jahrb. Abt. 2. -
1907. — 2, N 23. -P. 587-682.
420. Richter H. Das Zungenbein und seine Muskulatur bed den Lacertilia vera // Jen. Z. Natur. — 1933. - 66,
N2.-S. 395-480.
421. RieppelO. Die orbitotemporale Region im Schadel von Chelydra serpentina Linnaeus (Chelonia) und Lacer-
ta sicula Rafinesque (Lacertilia) // Acta Anat. - 1976. - 96. - S. 309-320.
422. Rieppel O. Uber die Entwicklung des Basicranium bei Chelydra serpentina Linnaeus (Chelonia) und Lacerta
sicula Rafinesque (Lacertilia) // Verh. Natur. Ges. Basel. - 1977. - 86, N 1/2. - S. 153-170.
423. Romer AA. The locomotor apparatus of certain primitive and mammal-like reptiles // BulL Amer. Mus.
Natur. Hist. - 1922. - 46. - P. 519-606.
424. Romer A.S. Crocodilian pelvic muscles and their avian and reptilien homologues // Ibid. - 1923. — 48. —
P. 533-552.
425. Romer A.S, The development of the thigh musculature of the chic // J. Morphol. — 1927. — 43. — P.347—
385.
426. Romer A.S, The development of Tetrapod limb musculature. The thigh of Lacerta // Ibid. - 1942. - 71. -
P. 251-298.
427. Romer A.S. The development of Tetrapod limb musculature - the schoulder region of Lacerta //Ibid. —
1944.-74. -P. 1—41.
428. Romer A.S. Osteologie of the Reptiles. - Chicago : Chicago univ. press, 1956. - 772 p.
429. Romer A.S. The Vertebrate Body, - Chicago : Chicago Univ, press, 1962. - 592 p.
430. Rudinger Dr. Die Muskeln der vorderen Extremit&ten der Reptilien und Vogel mit besonderer rucksicht
auf die analogen und homologen muskeln bei den Saugethieren und dem Menschen // Naturk. und verhan-
delingen van de Hollandsche Maatschappij der Wetenschappen te Haarlem. - Haarlem : De Erven Loosjes,
1868. - S. 1-187.
43X. Ruhner M. Uber den Einfluss der Korpergrosse auf Stoff ur;d Kraftwechsel // Zo BioL - 1883. - 19. -
S. 24-35.
432. Saint Girons H. Developpement respectif de I’epithelius sensoriel du cavum et de 1’organe de jacobson
chez les lepidosauriens // C.r. Acad. sci. D. - 1975. - 286, N 6. - P. 721 -724.
433. Sanders A. Notes of the Myology of Platydactylus japonicus // Proc. Zool. Soc. - 1870. - P. 413-426.
434. Sanders A. Notes on the Myology of Phrynosoma coronatum // Ibid. — 1874. - P. 71.
435. Stive-S&derbergh G. Notes on the brain-case in Sphenodon and certain Lacertilia // Zool. bidr. Uppsala. -
1947. -25.-P. 489-516.
436, SHerbak Nikolaj N. Eremias arguta (Pallas 1773) - Steppenrenner // Handb. der Reptilien und Amphibien
Europas. - 1981. - Bd. 1. - S. 432-446.
437. Schauinsland SJP. Entwicklung der Eihaute der Reptilien und der Vogel // Heiberg, VergL exper. Entwick.
Wirbeltiere. - 1906. - S. 145-160.
438. Schmidt-Nielsen B., Skadhauqe E. Renal function in crocodile // Amer. J. Phys. - 1967. - 212. — P.973-
980.
439. Шагдарсурэн О., Мунхбаяр X. Eremias arguta Pall. Монголоос олдов // Улсык тов нувбнн бутээл. -
1968. - № 1. — С. 32Zt
440. Schwarz-Karsten Н. Uber die Orbitaidrtfsen von Lacerta agilis, L. muralis, Ophisops elegans, Tarentola
mauretanica und Tropidonotus natrix // Morphol. Jahrb. - 1937. - 80. - S. 248-279.
441. Scoczylas R. Influence of temperature on gastric digestion in the grass snake Natrix natrix L. // Comp.
Biochem. and PhysioL - 1970. - 33, N 4. - P. 793-804.
442. Sewertzoff A .N. Die Entwicklung der pentadaktylen Extremitat der Wirbeltiere // Anat. Anz. - 1904. -
25.-S. 472-494.
443. Sewertzoff A.N. Studien uber die Entwickelung der Muskeln, Nerven und des Skeletts der Extremitaten
der niederen Tetrapoda // Bull, Soc. Imper. Natur. Moscow. (N.S.). - 1908. - 21. — P. 1—430.
444. Shoemaker V.H., Licht P., Dawson D.R. Water and salt excretion in lizards // Comp. Biochem. and Phy-
siol. - 1967. - 23. - 255-262.
445. Siebenrock F. Das Skelett der Lacerta symonyi Steind. und der Lacertidenfamilie uberhaupt // Sitzb.
kais. Akad. Wiss. Wien, Math. - Naturw. KI. - 1894. - 103, N 1. - S. 205-292.
446. Sieglbauer F. Zur Anatomie der Schildkroten - extremitat // Arch. Anat. Phys., Anat. Abt. - 1909. -
S. 183-280.
447. Smimowsky B. Zur Morphologic der respiratorischen Musculatur einiger Lacertilien // Anat. Anz. - 1930. -
70. - S. 58-77.
448. Smith R.V., Sotchell D.G. Histochemistry of the lung of the australian snake-nered tortoise Chelodina
longicallis // J. MorphoL - 1987. - 192, N 3. - P. 257 -268.
449. Starling EJL, Ferney E.B. The secretion of urine as saudied on the isolated kidney /f Proc. Roy. Soc. —
1925.-97.-P. 321.
450. Stebbins R.C., Eakin R.M. The role of the *4hird eye” m reptilian behaviour // Amer. Mus. Novit. - 1958.-
1876. -P. 1-16.
234
451. Sullivan B.f Riggs A, Structure, function and evolution of turtle hemoglobin // Comp. Biochem. and Phv-
sioL - 1967. - 23, N 3. - P. 437-474.
452. Terentiev P. V. Miscellanea Herpeto-Batrachologica // ZooL Anz. - 1923. - 5/6. - S. 136.
453. Terni T. И simpatico cervicale degliamnioti (Ricerche di morphologia comparata) // Z. Anat. und. Entwic-
klungsgesch. -r-1931. - 96. - P. 289—426.
454. Texik I. Contribution of phylogenesis of tracheal glans of vertebrates // Anat. Anz. - 1986. - 160, N 1. -
S. 393-394.
455. Tretjakoff D, Die orbitalen Sinusse bei den Amphibien, Reptilien und Vogcln // Morphol. Jahrb. - 1930. -
64, N 1. -S. 133-17'.
456. Tinkle D.W., Hadley N.F. Lizard reproductive effort: Caloric estimates and comments on its evolution //
Ecology. - 1975. - 56, N 2. - P. 427-434.
457. Underwood A., MenakerM. Extraretinal photoreception in lizards // Photochem. and Photobiol. - 1976. -
23, N 4. -P. 227-243.
458. Walls G.L. The vertebrate eye and its adaptive radiation. - Bloomfield hills (Mich.), 1942.- 785 p.>
459. Watson D.M.S. lhe evolution of the tetrapod shoulder girdle and fore-limb // J. Anat. - 1971. - 52.
P. 1-63.
460. Weldon J, Note on the Early Development of Lacerta muralis // Quart. J. Mier. Soc. - 1883. - P. 11л
461. Willnow L Bauplan der Lunge von Sphenodon punctatus. Eine vergleichend-anatomische Betrachtung // Ac-
ta Anat. - 1976. - 94, N 4. - S. 504-519.
462. Feider Z, Citeva larve ale genului Trombicula (Acarieni) si descrierea unui caz de trombidioza la sopiila
Lacerta agilis // Bull, stinnt., Sec. Biol. - 1953. - 5, N 4. - 775 -806.
463. Wright A., Chester Jones I. The adrenal gland in lizards and Snakes // J. Endocrinol. - 1957. - 15. - P.83-
99.
464. Boettger O. Die Reptilien und Batrachien Transcaspien // Zool. Jahrb. Abt 3. - 1888. - P. 871-1014.
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ (Щербак Н.Н.)......................................................... 3
ГЛАВА 1. ИСТОРИЯ ИЗУЧЕНИЯ ВИДА (Щербак Н.Н.) . . ............................. 4
ГЛАВА 2. МАТЕРИАЛ И МЕТОДИКА ИССЛЕДОВАНИЙ (Щербак Н.Н.)........................... 6
ГЛАВА 3. АРЕАЛ (ЩЕРБАК НЯ.)....................................................... 9
3.1. Распространение (места находок)........................................ 9
32. Общая характеристика ареала 20
ГЛАВА4. СИСТЕМАТИКА, ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ И ВНУТРИВИДОВАЯ СТРУК-
ТУРА (Щербак Н.Н^ Неручев В.В., Окулова Н.М., Орлова В.Ф.)....................... 22
4.1. Описание вида......................................................... 22
42. Географическая изменчивость.........................'.................. 23
4 2. Половой диморфизм..................................................... 30
4.4.Внутривидовая структура.............................................. 30
Г ЛАВА 5. МОРФОЛОГ ИЯ............................................................ 35
5.1. Скелет................................................................ 35
5.1.1. Череп (Иорданский НЯЭ............................................... 35
5.U. Осевой скелет (Коток В.С.) ........................................... 41
5.12. Скелет конечностей (Коток В .С.).................................... 46
52. Мускулатура (Коток В.С.) .......................................... . 53.
5.2.1. Мышцы шеи........................................................... 53
522. Мышцы туловища..........................................,............. 61
5,2,3, Мыппр.т хвоста...................................................... 66
5,2.4. Мышцы грудной конечности.......................................... 67
5.2.5. Мышцы тазовой конечности............................................ 73
52. Нервная система.................................................... 81
52.1. Головной мозг (Василевская Г ЛЭ..................................... 81
5.3 2. Черепномозговые нервы (Василевская Г J4J........................... 87
523. Периферическая нервная система (Веселовский МЯ.) ................... 88
5.4. Орган зрения (Василевская Г ЛЭ ...................................... 91
5.4.1. Глазное яблоко ..................................................... 91
5.4 2. Вспомогательные структуры глаза..................................... 92
5.5. Кровеносная система (Веселовский МЯ.).............. .............. 94
5.5.1. Сердце............................................................ 94
5.52. Артериальная система................................................ 94
5.5.3 . Дорсальная аорта и ее ветви ..................................... 96
5ЗА.Венозная система..................................................... 96
53. Система органов дыхания (Василевская ГЛ.) ............................ 96
5.7. Пищеварительная система (Василевская ГЛ.) ............................. 101
5.8. Мочеполовая система (Веселовский МД.).............................. 104
5.8.1. Выделительная система.............. .............................. 104
5.8.2. Мккроанатомия почек................................................ 105
5.8.3. Микроанатомия надпочечников...................................... 106
5.8.4. Половая система .................................................. 106
5Э. Щитовидная железа (Гербильский Л.В., Усенко Б.СЭ...................... 107
ГЛАВА 6. ГАЗООБМЕН (Тертышников М.Ф.).......................................... 108
ГЛАВА7. НЕКОТОРЫЕ АСПЕКТЫ ЭНЕРГЕТИЧЕСКОГО МЕТАБОЛИЗМА (Львова СП.) . . 109
236
ГЛАВА8.МИКРОЭЛЕМЕНТЫ В ОРГАНИЗМЕ РАЗНОЦВЕТНОЙ ЯЩУРКИ (Шарыгин СА.) . ... цз
Г ЛАВА 9. СТАЦИИ, ЧИСЛЕННОСТЬ, БИОМАССА (Тертышников М.Ф.)...................... 114
9.1. Биотопы, стации....................................................... 114
9 2. Факторы, определяющие биотоп и стации................................. 115
9 3. Численность и биомасса................................................ 116
ГЛАВА 10. ТЕРМОБИОЛОГИЯ (Котенко Т.И.)............................................ 120
10.1. Общие замечания, терминология, оценка данных......................... 120
103. Температурные границы активное™, предпочитаемая температура среды..... 122
103. Температура тела ..................................................... 128
10.4. Влияние различных факторов среды на температуру тела и активность.... 132
10.4.1. Солнечная радиация................................................. 132
10.4.2. Температура воздуха и почвы........................................ 134
10.4.3. Ветер, запыленность атмосферы ..................................... 136
10.4.4. Освещенность, облачность........................................... 137
10.4.5, Атмосферное давление, влажность, осадки ........................... 137
10.4.6 . Особенности биотопа............................................... 139
103. Терморегуляционное поведение ........................................ 140
ГЛАВА 11. СУТОЧНЫЙ ЦИКЛ АКТИВНОСТИ (Котенко Т.И.)................................. 143
11,1. Общие замечания, оценка данных ...................................... 143
113. Индивидуальная,групповая и популяционная активность................... 143
113. Структура суточной активности и ее сезонная динамика.................. 144
11,4. Зависимость суточной активности от возраста, пола и физиологического состояния
ящурок . ..............'................................................... 148
11.5. Влияние различных факторов среды на суточную активность.............. 149
ГЛАВА 12. СЕЗ ОННЫЙ ЦИКЛ АКТИВНОСТИ (Котенко Т.И.)................................ 151
12.1, Начало и конец сезонной активности популяций......................... 151
123. Особенности весеннего пробуждения и порядок ухода на зимовку отдельных групп
ящурок..................................................................... 152
123. Зимняя спячка........................................................... 154
12.4. Летняя спячка.......................................................... 154
ГЛАВА 13. ПИТАНИЕ (Тертышников М.Ф., Горовая В.И.).................................. 157
13.1. Общая характеристика питания......................................... 157
133. Соотношение компонентов в пище ящурок в разных частях ареала ........ 158
133. Биотопическая изменчивость кормового рациона ........................ 160
13.4. Сезонная изменчивость кормового рациона.............................. 161
13.5, Изменение кормового рациона по годам................................. 162
13.6, Половые и возрастные особенности в питании ящурок.................... 163
13.7. Избирательная способность к определенным кормам . .............. 164
13.8. Количество и масса поедаемого корма.................................. 166
133. Продолжительность переваривания пищи................................... 167
13.10. Способы добывания пищи................................................ 167
ГЛАВА 14. РАЗМНОЖЕНИЕ (Котенко Т.И.).............................................. 169
14.1. Наступление половой зрелости........................................... 169
143. Жировые тела и их значение...........................................
14.3. Половой цикл самцов.................................................... 173
14.4. Половой цикл самок..................................................... 175
14.5. Сроки спаривания....................................................... 178
14.6. Продолжительность беременности......................................... 179
14,7. Сроки откладки яиц и количество кладок................................. 179
14.8. Количество яиц в кладке и их характеристика .......................... 183
14.9. Место и процесс откладки яиц.......................................... 187
14.10. Длительность инкубации яиц и появление молодых особей............... 188
ГЛАВА 15. РАЗВИТИЕ И РОСТ.СТРУКТУРА ПОПУЛЯЦИЙ (Тертышников М.Ф.).................. 190
15.1. Темпы роста ......................................................... 190
153. Структура популяций................................................... 191
ГЛАВА 16. ЛИНЬКА (Котенко Т.И.)................................................... 195
16.1, Сроки и количество линек............................................. 195
237
16 Факторы, влияющие на линьку............................................ 197
163. Процесс линьки . . . ............................................... 198
ГЛАВА 17. ПОВЕДЕНИЕ» ПЕРЕМЕЩЕНИЯ И МИГРАЦИИ. УБЕЖИЩА (Тертышников М.Ф.) .... 200
17.1. Территориалиэм...................................................... 201
17 2, Перемещения.......................*.................................. 202
173. Миграции, кочевки..................................................... 202
17.4. Убежища.............................................................. 203
173. Поведение............................................................. 205
ГЛАВА 18.ВРАГИ (Тертышников М.Ф.)................................................ 207
ГЛАВА 19. ПАРАЗИТЫ............................................................... 208
19.1. Эндопаразиты (Шарпило В.П.) 208
192. Эктопаразиты (Котенко Т.И.) .......................................... 210
ГЛАВА 20. ПРАКТИЧЕСКОЕ И БИОЦЕНОТИЧЕСКОЕ ЗНАЧЕНИЕ. ОХРАНА........................ 213
20.1. Биогенетические связи разноцветной ящурки (Котенко ТЛ., Тертышников М.Ф J 213
202. Практическое значение (Котенко ТЛЭ.................................... 218
203. Биоэнергетическая оценка роли разноцветной ящурки в биогеоценозах (Тертышни-
ков М.Ф J ............................................................... 2W
20.4. Охрана (Щербак НЯ J ................................................. 220
СПИСОК ЛИТЕРАТУРЫ.............................................................. 222
Наукове видання
АКАДЕМ1Я НАУК УКРА1НИ
Тнститут зоологиям. Ы.Шмальгаузена
ЩЕРБАК Микола Миколайович
КОТЕНКО Тетяна leantвна
ТЕРТИШН1КОВ Михайло Федорович та in.
Р13НОБАРВНА ЯЩУРКА
Кшв, видавництво ’’Наукова думка*’, 199?
РосЙською мовою
Художнй редактор А.В.Косяк
Технмний редактор Н.ТКусъик
Коректори ЕЛ.МазнЛченко, Н.Б.Кудрявцева
Оператор Л.I.Про копчу к
Здано до набору 05.03.92. Пщп. до друку 26.02.93. Формат 70x108/16. Ilanip офс. № 1. Гарн. Прее
Роман. Офс. друк. Ум.-друк. арк. 22,40. Ум. фарбо-вздб. 22,75. Обл.-вид.арк. 25,61 + 1,05 вкл. =26,66.
Зам. 2.-6V7.
Орипнал-макет пщготовлено у видавництв! ’’Наукова думка” 252601, Киш 4, в ул. Терещенювська, 3.
Ки1вська книжкова друкарня науково! книги. 252004, Ки1в 4, вул. Терещенк1вська, 4.
НАУ КО ВА ДУМКА