






























































































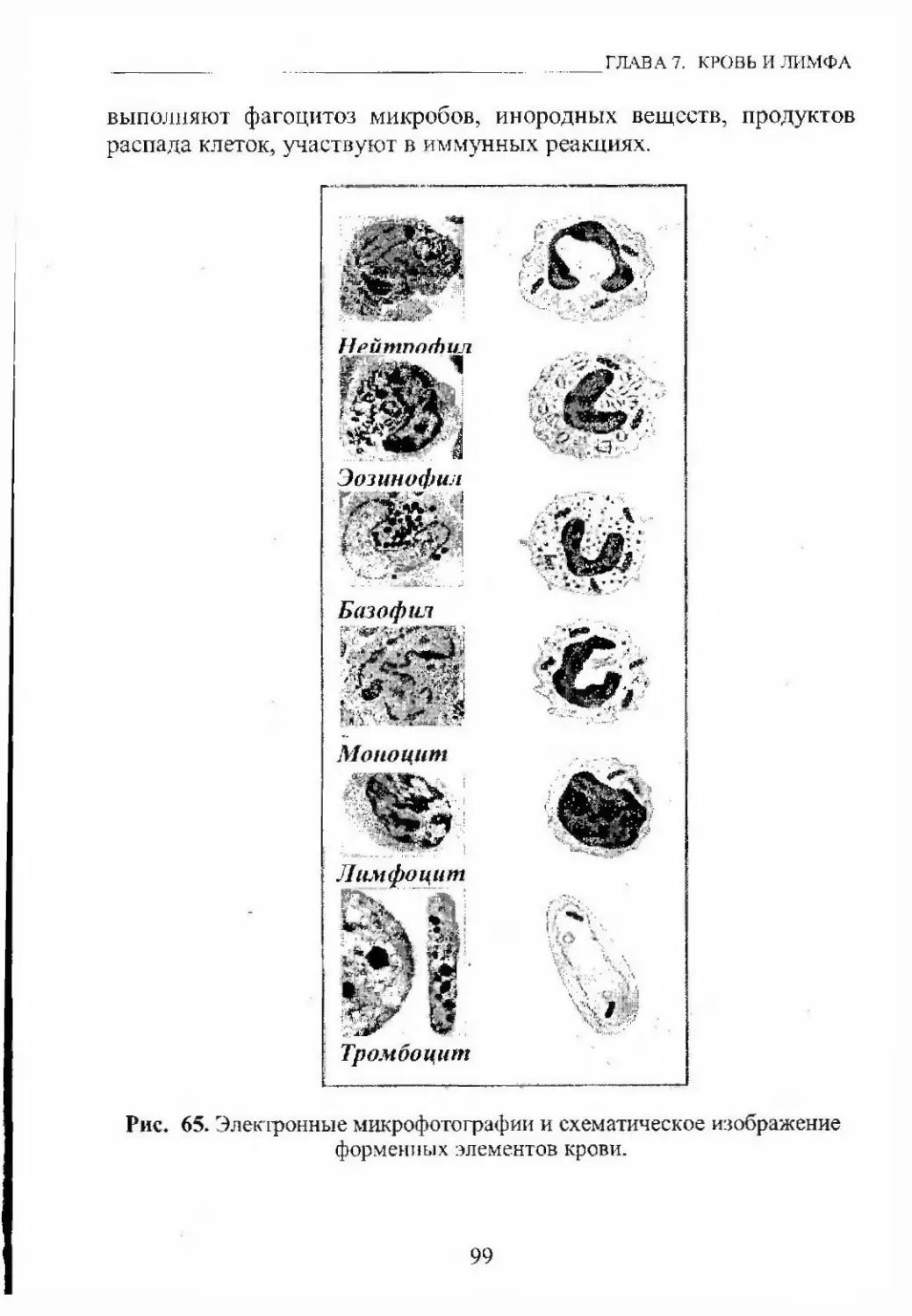
























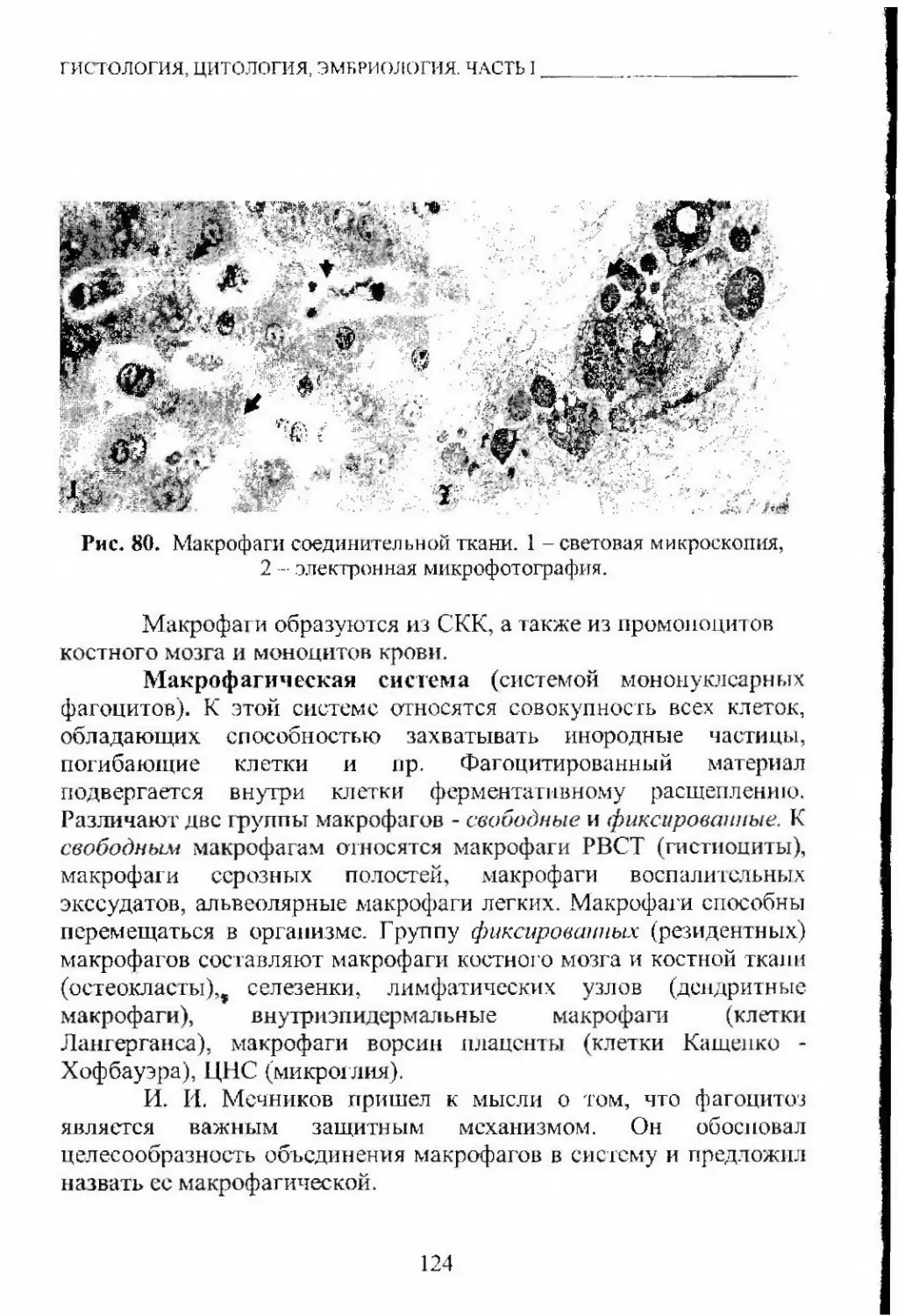


















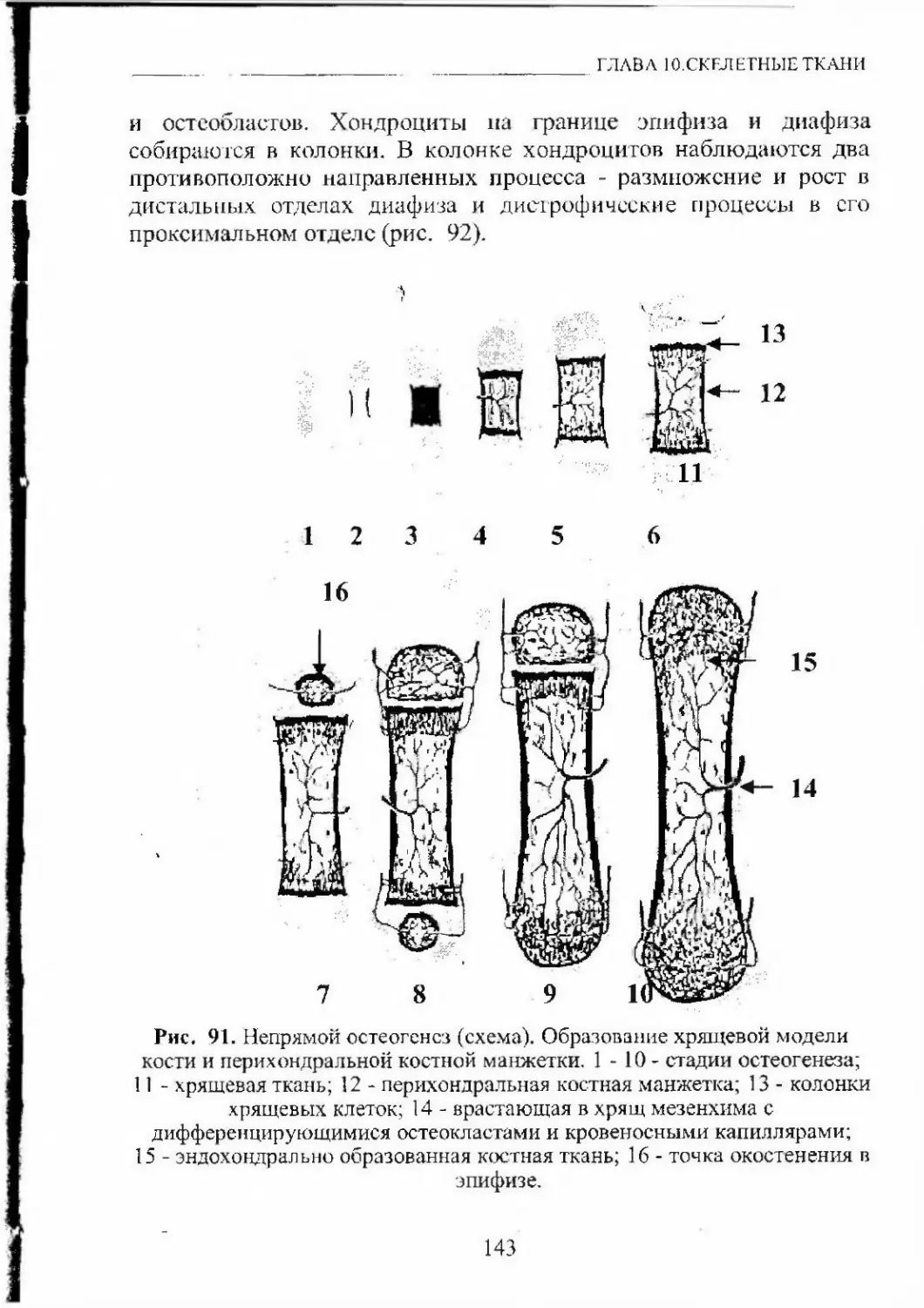

















































































Автор: Кащенко С.А. Бобрышева И.В.
Теги: анатомия анатомия человека сравнительная анатомия антропология медицина гистология цитология эмбриология
ISBN: 978-617-579-477-7
Год: 2012




1-ІядаiÄfлил'ЯЯГйЛШ'Я:И1
Ч D Л иГШ1“JiJli’-TJ:-» -С
ГЗ «ЛУГАНСКИЙ ГОСУДАРСТВЕННЫЙ
МЕДИЦИНСКИЙ УНИВЕРСИТЕТ»КАЩЕНКО С.А.
БОБРЫШЕВА И.В.ГИСТОЛОГИЯЦИТОЛОГИЯЭМБРИОЛОГИЯЧАСТЬ IЛУГАНСК
Издательство «Ноулидж»
2012
УДК 611.013+611.018
БЬК 28.705+2Н.706
Г51Рецензенты:
член - корреспондент АМН Украины, д.мед.н., профессор
Ю.Б. Чайкоаскийдоктор медицинских наук, профессор С.Ю. Мааювский;
доктор медицинских наук, профессор Э.Ф. Баринов.Кащенко С.А.Г 51 Гнсюлогня, цитология и :}мбриология. Часть I. (Учебник для
инос']ранных студентов высших медицинских учебных заведений
1И-]У уровней аккредитации, обучающихся на русском и;}ыке) /
С.Л. Кашенко, И.В. Бобрышева. - Луганск: Изд-во «Ноулидж»,
2012.-224 с.ISBN 978-617-579-477-7Кащенко С.А., доктор медицинских наук, профессор, зав. кафедрой
гистологии, цитологии эмбриологи!'! ГЗ «Луи'ански!! госуда]эственный
медицинский университет».Бобрышева И.В.., кандидаг медицинских наук, доцент кафедры
гистологии, цитологии эмбриологии ГЗ «Луганский государственный
медицинский университет»,в книге в сжатой форме изложен материал о сфоении и принципах
организации клетки, 1исгологическом строении тканей ор!'анизма и
особенностях их эмбрионального развития. Охарак['еризованы ранние этапы
эмбриогене:^а человека и развигие основных систем плода, Книз’а
иллю^рирована мнох очисленными рисунками и схемами.Материал изложен в соответствии с требованиями программы
учебной ДИСЦИШШНЫ «ГИСТ0Л01'ИЯ, ЦИТ0ЛО1ИЯ, ЭМбрИ0Л01'ИЯ» для студентов
высших учебных медицинских заведений ИМУ уровней аккредитации по
специальности «Лечебное дело» и «Педиатрия» (Киев, 2010), а также
ориентирован в соответствии с кредитно-модульной сис гемой обучения.Учебник предназначен для иностранных студентов медицинских
университетов,* академий, иЕтститутов, интернов, а 1'акже преподавателей
морфологических дисцшимн.УДК611.013+611.018
ББК 28.705+28.706£1С-А.Кащенко, И.В, Бобрышева, 2012
€) Издательство «Ноулидж», 2012
Предисловие к первому изданиюВ данном учебнике изложены основы гистологии,
цитологии, эмбриологии для иностранных студентов медицинских
специалыюстсй, которые обучаются на русском языке. Материал,
приведенный в данном издании, ориентирован на эффективное
освоение особенностей гистологического строения тканей и
органов. В основу положено содержание курса лекций по
гистологии, цитологии и эмбриологии, который читался студентам
в ГЗ «Луга1£ский государственный медицинский университет» с
2005 -2012 уч.г.г.При отборе материала в текст издания включена
информация, которая создаёт базис морфологических знаний,
необходимых для усвос11ия будущих клинических дисциплин.Издание предназначено для студентов медицинских
университетов, академий, ипстатутов, интернов, также
преподавателей морфологических дисциплин.Авторы будут весьма признательны за все замечания и
пожелания, высказанные в адрес содержания и оформления этой
книги, а также учтут их в дальнейшей работе.Заведующая кафедрой гистологии,
цитологии, эмбриологии
Государственного заведения
«Луга1£ский 1'осударствснный
медицинский университет»
доктор медицинских наук, профессор
С.А. Кащенко
ГЛАВА 1.
ПРЕДМЕТ И ЗАДАЧИ ГИСТОЛОГИИ И
цитологии, ИСТОРИЯ РАЗВИТИЯ,
МЕТОДЫ ИССЛЕДОВАНИЯГистология (от греч. histos - ткань, logos - учение) -
фундаментальная наука о строении, развитии и жизнедеятельности
тканей животных организмов. Гистология объединяет общую
гистологию И частн}то гистологию. Предметом общей гистологии
являются общие закономерности, присущие тканевому уровню
организации и особенности конкретных тканей организма; предметом
частной гистологии - закономерности жизнедеятельности и
взаимодействия различных тканей в органах на более высоких
уровнях организации.Курс гистологии включает в себя также цитологию (от феч.
kytos - клетка, logos - учение) ~ учение развитии, строении и
жизнедеятельности клеток и эмбриологию (от греч. embryon -
зародыш, logos - учение) - учение о пренатальном развитии
организма, о закономерностях развития зародыша.Цитология составляет необходимую часть гистологии, так как
клетки являются основой развития и строения тканей. Новые данные о
структуре ядра, его хромосомного аппарата легли в основу
цитодиагностики наследственных заболеваний, опухолей, болезней
крови. Раскрытие особенностей улыраструктуры и химического
состава клеточных мембран является основой для понимания
закономерностей взаимодействия клеток в тканевых системах,
защитных реакциях и др. В медицинской практике широко
использус’гся цитодиагностика.В курсе эмбриологии рассматриваются закономерности
эмбрионального развития человека, гистогенез - образование тканей
и их способность к регенерации.
ГЛАВА 1. НРКДМКТ И ЗА^ЧАМИ ] ИСТОЛОГИИ И ЦИТОЛОгаИ, ИСТОРИЯ РАЗВИТИЯметоды исслсдовлпия Методы исследования
в современной гистологии, цитологии и эмбриологии
применяются разнообразные методы исследования, позволяющие
ьсссгороиис изучать процессы развития, строения и функции клеток,
тканей и органов.1лавпыми этапами цитологического и гистологического
анализа являются вьтбор' объекта исследования, подготовка его для
изу^1ения в микроскопе, применение мегодов микроскопирования, а
также качественный и количссшенный а^1ализ изображений.Объектами исследования служат живые и мертвые
(фиксированные) клетки и ткани, и их изображения, полученные в
свсговььч и электронных микроскопах.Основным объектом исследования являются гистологические
препараты. Препарат можсг прсдсгавлять собой мазок (кровь,
коагпый мозг, слюна), отпечаток (селезенка, тимус, печень), пленку
из ткани (брюпшна, плевра, мягкая мозговая оболочка),
гистологический срез. Чаще всего применяю! специально
обрабоганпые препараты; фиксированные, заключенные в твердую
^еду и окрашенные.^ Процесс 1131 оюв.1ения 1 исгологическо! о препарата ддясветовой и элск'хронной микроскопии, которые используются в
патоморфологических лабораториях и на учебных занятиях можно
разделить на несколько этапов:1. Забор и фиксация кусочка ткани или органа размером 0,6 х 0,6 х0,6 см в одной из фиксируюищх жидкостей (спирт, раствор
формШ1ина, разнообразные фиксирующие смеси) в течение
нескольк'их суток.2. Промывка объекта от фиксатора в водонровод1Юй воде (1 сутки).3. Обезвоживание объекта в смесях спиртов восходягцей крепости
(60°, 70^ 80°, 90^ 96°, 100^-1, 100°-2).4. Просветление препарата в смесях ксилола со спиртом, а затем в
чисюм ксилоле.5. Пропитка препарата парафином в снегоподобной массе.6. Заливка препарата в парафин (56"^)Изюговлспис парафиновых блоков и наклеивание их на
деревянный блок.8* Изготовление срезов необходимой толпщпы с парафиновых
блоков, наклеивание срезов на предметные стёкла.
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I9. Окраска препаратов специальными гистологическими
красителями и заключение в канадский бальзам или синтсшческие
среды.Фиксация обеспечивает предотвращение процессов
разложения, что способствусг сохранению целостности ст|5укгур. Это
достигается тем, что взятый из органа образец погружают в фиксатор
(спирт, формалин, растворы солей тяжелых металлов, осмиевая
кислота, специальные фиксирующие смеси). Под действием
фиксатора в тканях и органах происходят сложные физико¬
химические изменения. Наиболее существенным из них является
процесс необратимой коагуляции белков, вследствие которого
жизнедеятельность прекращается, а структуры становятся
фиксированными. Фиксация приводит к уплотнению и уменьшению
объема кусочков, а также к улучшению последующей окрасюи клеток
и тканей.Уплотнение материала, необходимое для приготовления
срезов, производится путем пропитывания предварительно
обезвоженного материала парафином, цешюидином, органическими
смолами. Более быстрое уплотнение достигается применением метода
замораживания кусочков, например, в жидкой углекислоте.Приготовление срезов происходит на специальных приборах- микротомах (для световой микроскопии, рис. 1) и
ультрамикротомах (для электронной микроскопии).ращающиисяПриводдержатель блокаблок^,препаратмикротомныйножРис. 1. Ротационный микротом.
6
ГЛАВА Г ПРЕДМЕТ И ЗАДАЧИ ГИСТОЛОГИИ И ЦИТОЛОГИИ, ИСТОРИЯ РАЗВИТИЯ,МЕТОДЫ ИССЛНДОВАИИЯ Окрашивание срезов (в свеговой микроскопии) или
иапыленис их солями металлов (в электронной микроскопии)
применяют для увеличения контрастности изображения огдельньгх
структур при рассматривая™ их в микроскопе. Методы окраски
гистологических структур очень разнообразны и выбираюгся в
зависимости от задач иccJ(eдoвaния.Гистологическис * красители (по химической природе)
подразделякуг па кислые {эозин - окрашивает цитоплазму в розовый
цвет), основные {гематокстин - окрашивает ядра клеток в
фиолетовый цвет) и нейтральные. Структуры, хорошо
окрашивающиеся кислыми краситеу1ями, называются
оксифильными, а окрашивающиеся ос^ювными - базофильными.
Например, цигоплазма клеток чаще всего окрашивается оксифильно, а
ядра клеток - окрашиваются базофильно. Структуры,
воспринимаюпдие как кислые, так и основные красители, являются
нсйтрофильными (гетерофилышми). Метахромазия - изменение
цвега отдельных основных красителей при связывании их со
структурами, обладающими специфическими свойствами (например:
гранулы базофильных лейкоцитов и тучных клеток). Окрашенные
препараты ячя длительного сохранения заюшчают между
предметным и покфовным стеютами в канадский бальзам или
полиетирол. Готовый гистологический препарат может бьпь
использован дая изучения под микроскопом в течение многих лет.Для электронной микроскопии срезы, полученные на
ультрамшфотоме, помещают на специальные сетки, контраетируют
солями урана, свинца и других металлов, 1юсле чего просматривают в
микроскопе и фотограф1фуют. Полученные микрофотографии служат
объектом изучения наряду с гистологическими препаратами (рис. 2).Микроскопия может быть световая (с использованием
светового микроскопа) и электронная (с использованием
электронного микроскопа).- Световая микроскопия может
осуществляться в проходящем свете, когда свет проходит через
топкий прозрачньш гистологический препарат, или же в
отраженном свете, когда исследуют, например, толстый или
непрозрачный объект. Аналогичным образом, электронная
микроскопия может быть трансмиссионной, когда пучок
электронов проходит сквозь изучаемый ультратонкий срез, или же
растровой, или сканирующей, когда пучок электронов отражается
от поверхности исследуемого объекта. В первом случае
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I,электронный микроскоп называется трансмиссионным, а во втором- сканирующим.. Г\Рис. 2. Электронный микроскоп.Световая микроскопияМикросконирование - основной метод изучения препаратов- используется в практике уже более 300 лет. Современные
микроскопы - это сложные оптические системы, позволяющие
изучать тонкие детали строения клеток и тканей (рис. 3). Понятие
«увеличение микроскопа» относится к его оптической системе и
выражается в произведении увеличений объектива и окуляра.
Однако «разрешение» микроскопа зависит от характеристик
объектива и не зависит от окуляра.Ддя изучения гистологических препаратов чаще применяют
обычные свеювые микроскопы, в которых в качестве источника
освещения используют естественный или искусственный свет. Для
светового микроскопа обпхес увеличение достигает 2000 раз.
ГЛАВА 1. ПРЕДМЕТ И ЗАДАЧИ ГИСТОЛОГИИ И ЦИТОЛОГИИ, ИСТОРИЯ РАЗВИТИЯ,
МЕТОДЫ ИССЛЕДОВАНИЯокулнрревольверобъективПредмет¬
ный столикконденсордиафрагмаисточник.светаоснованиеколонкавинт
перемещениямалое
^увеличениебольшоеувеличениеРис. 3. Детали строения светового микроскопа.Размеры объектов и их деталей^ изучаемые с
использованием цитологических и гистологических методов,
обычно столь малы, что невидимы невооруженным глазом. Они
составляют преимущественно микрометры (мкм) в световой
микроскопии и нанометры (нм) в электронной микроскопии.
Широко используемая ранее единица ангстрем (А), равная 10’* нм, в
настоящее время болсс не применяется.
гистология, цитолог ия, ЭМБРИОЛОГИЯ. ЧАСТЬ I.Соотношения между величинами линейных единиц
измерения, наиболее часто используемых в гистологии и
цитологии:1 миллиметр (1мм) = 10"^ м = 10'^ мкм = 10^ нм = 10^ А
1 микрометр (1мкм) = 10'^ м - 10'^ мм = Ю^нм = 10^ А
1 нанометр (1нм) = 10'^ м = 10'^ мм = 10'^ мкм = 10 А1 ангстрем (1 А) = 10'^® м = 10'^ мм = 10"^ мкм = 10 * нмУльтрафиолетовая микроскопияВ ультрафиолетовом микроскопе используют
ультрафиолетовые лучи с длиной волны около 0,2 мкм. Полученное
в ультрафиолетовых лу*1ах невидимое глазом изображение
преобразуется в видимое с homoitimo регисфации на
фотопластинке, люминесцентном экране или электронно-
онтическом преобразователе.Флуоресцентная (люмипесцентпая) микроскопияЯвления флюоресценции заключаются в том, чго атомы и
молекулы ряда веществ, поглощая коротковолновые лу^ти,
переходят в возбужденное состояние. Обратный переход из
возбужденного состояния в нормальное происходит с испусканием
света. В флуоресцентном микроскопе в качестве источников света
применяют ртутные или ксеноновые лампы, обладающие высокой
яркостью в области спектра 0,25-0,4 мкм (ультрафиолетовые лу^ти)
и 0,4—0,5 мкм (сине-фиолстовые лучи).Любая клетка живого организма обладает собственной
флюоресценцией, однако она часто бывает чрезвычайно слабой.
Вторичная флюоресценция возникает при обработке препаратов
специальными красителями - флуорохромами (акридиновый
оранжевый). В этом случае ДНК и ее соединения в клетках имеют
ярко-зеленое, а РНК и ее производные - ярко-красное свечение.
Вариант метода флюоресцентной микроскопии, при котором
флюоресценция происходит в ультрафиолетовой области спектра,
получил название метода ультрафиолетовой флюоресцентной
микроскопии.Фазово-контрастная микроскопияЭтот метод служит для получс1Н1я контрастных
изображений прозрачных и бесцветных объектов за счет
специальной кольцевой диафрагмы, помещаемой в конденсоре, и10
ГЛЛВЛ 1. ПРЕДМЕТ И ЗАДАЧИ ГИСТОЛОГИИ И ЦИТОЛОГИИ, ИСТОРИЯ РАЗВИТИЯ,МНТОДЬ! ИССЛЕДОВАНИЯ так называемой фазовой пластинки, находящейся в объективе.
Повышение контраста позволяет видеть все структуры,
различающиеся по показателю преломления. Разновидностью
метода фазового контраста является метод фазово-темнопольного
контраста, дающий негативное изображение.Микроскопия в темпом поле используется при изучении
живых объектов, в падающем (отраженном) свете.Поляризационная микроскопия применяется для изучения
архите!сгоники гистологических структур ~ в поляризованном
свете.Электронная микроскопияВ элекгроппом микроскопе используется лоток электропоб.
С помо1Цью просвечивающего (трансмиссионного) электронного
микроскопа можно получить лишь плоскостное изображение
изучаемого объекта. Для получс£шя пространственного
представления о структурах используют растровые (сканирующие)
электронные микроскопы, способные создавать трехмерные
изображения. Для исследования выбранного участка микрозонд
двигается по его поверхности, такое исследование объекта
назьшасгся сканированием, а рисунок, по которому движется
микрозонд, -растром.Главными достоинствами электронной микроскопии
являются большая глубина резкости (в 100-1000 раз больше, чем у
световых микроскопов), широкий диапазон непрерывного
изменения увеличения (от десятков до десятков тысяч раз) и
высокая разрешающая способность.Цитохимические и гистох1шические методы исследованияПодобные методы направлены на выявление в клетках и
тканях определённых химических веществ (гликоген, липиды,
ферменты, железо и т.п.) или химических групп, а также оценивают
активность ферментов.Иммуноцитохимические и -гистохимические методы
исследованияВ основе метода_находится обработка мазков или срезов
маркированными специфическими антителами к выявляемому
веществу, которое служит антигеном.РадиоавтографияМетод основывается на выявлении в тканях введенных
извне веществ, меченых радиотопом. Меченое вещество вводят в11
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАС1 Ь Iорганизм животною, срезы материала в темноте покрывают
фотоэмульсией, После определённой экспозиции фотоэмульсия
засвечивается в местах расположения радиоактивного изотопа.Специачьиые методы окрашивания нефиксированных тканей
Витальная окраска. Некоторые красители в виде взвеси
вводят в организм животного, при этом они активно захватываются
фэ1 оцитами, накапливаются в них и маркируют эти к,тетки.Суправитаяьная окраска основана на связывании
некоторых красителей с компонентами живых юхеток, извлеченных
из ор1'анизма.Культивирование клеток^ тканей и органов вне органиша
Это метод используется для изучения влияния различных
факторов на рост, дифферснцировку и прочие процессы,
происходяпшс 13 клетках, тканях и opгaíJ[ax вне организма.Количественная оценка кчеток и тканей
Морфометрия позволяет определить высагу, толпщиу,
ширину, площадь сечения объекта на плоском срезе.Стереологические методы позволяют oпpeдcJПITЬ
трёхмерные параметры исследуемых объектов.Цитофотометрия
Этот метод позволяет оценить содержание исследуем01’0
вещества в структурных элементах клеток или тканей путём оценки
оптической плотности продукта гистохимической реакции по длине
поглощённой световой волны.проточная цитометрия и цитофлуориметрия
Эти методы дают возможность произвести оценку
содержания веществ в клетках, которые входят в состав суспензии.12
ГЛАВА 2. ЦИТОЛОГИЯКлетка - ограниченная мембраной, упорядоченная
система биополимеров, образующих ядро и цитоплазму,
участвующая в метаболических и энергетических процессах, в
поддержании и воспроизведении всей системы в целом.Основные компоненты клетки:❖ Плазмолемма❖ Ядро❖ Цитоплазма:* гиалоплазма (матрикс цитоплазмы - объединяет все
клеточные структуры и обеспечивает химическое взаимодействие
их друг с другом)- органеллы (постоянные структуры клетки, каждая из
которых выполняет определённую функцию)- включения (непостоянные структуры клетки) (рис.4).Гранулярная
\эндоплазматическая
сеть
Рибосом«Ядерная
/ пора
у "Ядрышко
-КариолеммаАграиулярпая
эндоплазмвтическая
1 сеткII МитохондрияЯдроКомплексГольджиЦентриолиЛизосомаЦитоп/шхиаПлазматическаямембранаРис. 4. Схема электронномикроскопического строения клетки.13
ГИС1ОЛОГИЯ, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ IПлазмолеммаЭго клеточная мембрана, ограничивающая клетку снаружи,
обеспечивающая её связь с внеклеточной средой, со вссми
веществами и стимулами, воздействующими на клетку.Наиболее признана жидкостно-мозаичная модель
биологической мембраны Синджера-Николсона (рис. 5).По принципу строения клеточных мембран построены:
плазмолемма, кариолемма, мембраны митохондрий,
эндоплазматической ссги, комплекса Гольджи, лизосом,
пероксисом.Химический состав клеточной мембраны:❖ липиды (40%),❖ белки (60%), (интегральные, периферические)зт’лсводы (5-10%) (преимущественно олигосахариды).НаружнаяповерхностьОлиго¬тетки иптеграль^'’^“^Перифери-Гпико- Гяико- ческий белок
протеин липиды \'■ скз ‘Г, \Гидрофобный] *•' фобиыеБислойхвосты. фосфо¬
липидов_ м -І ч ІІ І' 1І
■»> ‘і-і 44^т/IIII )ї І»,иГидрофобный 5' ^ к хбостВнутренняя Интегральный ' I Фосфолипидыповерхность белок Периферические Гидрофилъная .клетки белки головкаРис. 5. План строения кпеточной мембраны.Молекула фосфолипида состоит из полярной
гидрофильной части (головка) и гидрофобного углеводного хвоста.
Хвосты фосфолипидов направлены внутрь бислоя, а головки -
кнаружи.14
ІЛАВА2. ЦИТОЛОГИЯБелки:интегральные глобулярные белки встроены в билипидныйслой;периферические мембранные белки находятся на одной из
повсрхіюс гей юісточной мембраны (наружной или внутренней).С наружной поверхностью мембраны связаны рецепторные и
адгезионные белки. С внутренней поверхностью мембраны связаны
белки цитоскелета и белки системы вторых посредников.Углеводы входят в состав гликолипидов и гликопротеинов
мембраны. Цепи олигосахаридов выступают па наружной
поверхности мсдмбран клетки и образуют гликокаликс.К клеточной мембране прилегает❖ снаружи гликокаликс, образующий надмембранный
слой,*1* изнутри клетки - подмембранный слой - кортикальный
(узкий участок циюилазмы, который содержит компоненты
цитоскелета и участвует в процессе экзoт^итoзa).Функции плазмолеммы:❖ разграничительная,❖ рецепторная,<* Іране порт веществ,❖ образование мсжктюточных контактов,❖ передача сигналов от поверхности вглубь клетки.Проницаемость плазмолеммы обеспечивают:1-трансмембранные каналы,2-транспорт в мембранной упаковке (эндоцитоз:пиноцитоз и фагоцитоз). Транспорт с участием особыхферментов называется - пермеаз.Эндоцитоз - поглощение клеткой веществ, частиц и
макромолекул. Варианты эндоцитоза: пиноцитоз и фагоцитозПиноцитоз - захват и поглощение клеткой жидкости и
растворённых веществ (макротшоцитоз и микротшоцитоз).Фагоцитоз ~ поглошсеіис клеткой крупных частиц, при этом
образуются фагосомы.Экзоцитоз - внутриклеточные секреторные пузырьки и
секреторные гранулы сливаются с плазмолеммой, а их содержимое
высвобождается из клетки - секреция.15
гистология, ЦИТОЛОГИЯ. ЭМБРИОЛОГИЯ. ЧАСТЬ IМежклеточные контакты
Межклеточные контакты - специализированные структуры,
соединяющие клетки для формирования тканей.❖ Простые межклеточные контакты представляют собой
сближение плазмолемм соседних клеток на расстояние 15-
20 нм. При этом происходит icoirraicr гликокаликсов двух
соседних клеток. Примером являются индердигитации (рис.7).❖ Сложные межклеточные контакты: это небольшие парные
специализированные участки плазматических мембран двух
соседних клеток. Они подразделяются на замыкающие,
адгезионные и коммуникационные.- замыкающие -
Плотный контакт (zonula occludens) состоит из
непрерывных цепочек специальных белковых молекул,
соединяющих мембраны двух соседних клеток (рис. 6).Рис. 6. Плотный контакт (zonula occiudcns). Схема.- адгезионные -Адгезивный поясок {zonula adherens) - имеет вид ленты, которая по
периметру охватывает клетку в виде пояска. В зоне этого тина
контакта плазмолеммы утолщены, образуя пластинки16
_ГЛАВА 2. ЦИТОЛОГИЯприкрепления, в области пластинок содержатся связывающие
белки: а-актинин, винкулин, плакоглобин. Подобный контакт не
только связывает мембраны соседних клеток, но также и
стабилизирует их цитоскелет.Десмосома (полудесмосома) (macula adherens) - состоит из
утолщённых пластинок прикрепления (диаметр - 0,5 мкм) двух
соседних клеток, разделённых межклеточной щелью (25 нм).
Пластинки прикрепления содержат особые белки - десмоплакины,
плакоглобин, десмокальмин. Со стороны цитоплазмы в неё
вплетаются промежуточные филаменты. Десмосомы поддерживают
структурную целостность ткани, скрепляя клетки между собой
(рис. 7, 8). Полудесмосомы связывают клетки с базальной
мембраной.Фокальный контакт ~ соединение, характерное для
фибробластов.- коммуникационные -
Щелевой контакт (nexus) - мембраны соседних клеток разделены
щелью 2-3 нм, в плазмолеммах соседних клеток расположены
специальные белковые комплексы - коннексоны, которые образуют
каналы из одной клетки в другую.Рис. 7. Электронная микрофотография межклеточных контактов.
1 - ин'гердигитации, 2 - десмосомы.
ГИСТ0Л01’ИЯ, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I,Рис. 8. Электронная микрофотография десмосомы.Щелевой замыкательный контакт - образуется сближением
замыкательных пластин двух соседних клеток. Концы
интегральных белков мембран двух соседних клеток контактируют
друг с другом. Промежуток в области контакта уплотняется за счёт
фибрилл и ионов кальция.Синапс - специализированный контакт между нервными
клетками или нервной клеткой и мышечным волокном,
обеспечивающий передачу сигнала с одной клетки на другую с
помощью нейромедиатора.ЦитоплазмаЦитоплазма, отделенная от окружающей среды
плазмолеммой, включает в ссбя гиалоплазму, находящиеся в ней
обязательные клеточные компоненты - органеллы, а также
непостоянные структуры - включения.Гиалоплазма (от греч. ЬуаИпоз - прозрачный) или матрикс
цитоплазмы содержит органеллы и включения. Гиалоплазма
включает 11итозолъ (вода с расворёнными в ней органическими и
неорганическими веществами) и цитоматрикс (трабекулярная сеть
волокон белковой природы толщиной 2-3 нм). Роль гиалоплазмы
заключается в том, что она объединяет все клеточные структуры и
обеспечивает их взаимодействие друг с другом.18
_ГЛАВЛ2. ЦИТОЛОГИЯОрганеллы- это постоянные структуры клетки, которые имеют специфическое
строение и выполняют конкретную функцию.Классификация органелл клеткиТс органеллы, в структуру которых входит элементарная
биологическая мембрана, называют мембранными. К ним
относятся митохондрии, эндоплазматическая сеть (ЭПС), комплекс
Гольджи, лизосомы, пероксисомы, окаймлённые пузырьки.Немембранные органеллы: рибосомы, клеточный центр,
протеасомы, реснички, микроворсинки, жгутики, компоненты
цитоскелета.Органеллы подразделяются также на органеллы общего
значения и специальные органеллы.Органеллы общего значения имеются во всех клетках и
необходимы для обеспечения их жизнедеятаїьности. к ним
относятся митохондрии, рибосомы, протеасомы, ЭПС, комплекс
Гольджи, лизосомы, псроксисомы, клеточный центр, компоненты
цитоскелета.Специальные органеллы имеются лишь в некоторых
клсгках и обеспечивают выполнение специальных функций. К ним
относят миофибриллы, нейрофибриллы, реснички, жгутики,
микроворсинки, акросома (в сперматозоидах).Эндоплашатическая сетьГранулярная эндоплазматическая сеть (ГЭПС) - система
плоских мембранных цистерн, покрышх рибосомами.
Обеспечивает синтез белка в клетке, необходимого для построения
плазматической мембраны, а таклсе на «экспорт».Агранулярная эндоплазматическая сеть (АЭПС) - система
мембранных каналов, пузырьков, трубочек. АЭПС обеспечивает
синтез липидов, полисахаридов, стероидных гормонов, выполняет
роль депо кальция, а также деюксикационцую функп;ию (рис. 9).19
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЫ,Рис. 9. Электронная микрофотография гранулярной (а) и агранулярной
(б) эндоплазматической сети.Комплекс гольджиКомплекс Гольджи расположен около ядра, участвует в
образовании мукопротеидов, выведении веществ за пределы
клетки, образовании лизосом, концентрировании и упаковке
секреторных пузырьков (рисЛ 1).Комплекс Гольджи состоит из мембран, которые
налагаются друг на друга (диктиосома), на периферии
располагаются пузырьки.Различают: с18 (выпуклая) и 1гапз (вогнутая) поверхности
(рис. 10).20
ГЛАВА 2. ЦИТОЛОГИЯ) 3 •Vi?s| #v?0- ^ , ■■'“■ ‘ ^ ^' ' ^Рис. 10. Электронная микрофотография комплекса Гольджи.1 - диктиосома, 2 - eis поверхность, 3 - Irans поверхность,4 - секреторные пу:}ырьки.Рис. П. Комплекс Гольджи в нервных клетках спинального ганглия
(световая микроскопия).21
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ 1 Митохондрии
Митохондрии располагаются в местах усиленного
потребления АТФ. Митохондрии имеют собственный геном,
способны к делению, органелла функционирует около 10 суток.Митохондрии имеют две мембраны: наружную и
внутреннюю, внутренняя мембрана образуе-г кристы. Матрикс
содержит все ферменты цикла Кребса (рис. 12).Митохондрии у^таствуют в теплопродукции, синтезе
белков за счёт митохондриальной РНК, окислительных процессах,
транспорте электронов.Рис. 12. Электронная микрофотография митохондрии. 1 - матрикс,2 - кристы, 3 - гранулы.Митохондриальные болезниМутЬции, в митохопдриалънь^ ферментах приводят к
значительномуколичеетв^^^абощваний. ' Большинство . этих
мйтохондриальньгх^ нар)^^’Щ\. происходит из-за мутагщй в
Шерных ДНК белк£Т^^^$1ислёниость болезней .из-за
митохондриальной дисфункг^ии раетет год от года. Каждая из
болезнё^£!^омя)1утых ниже, отражает 'дефекты в различных
штоЩШриальных ферментах:миоратия скелетных мышц и
М1ю%^р^агНекоторые. ■ митохондритън ые мутации прингшаюгп22
ГЛАВА 2. ЦИТОЛОГИЯучастие в этиологии болезней старения, таких как, болезнь
Паркинсона и Альцгеймера.Лизосомы, пероксисомы
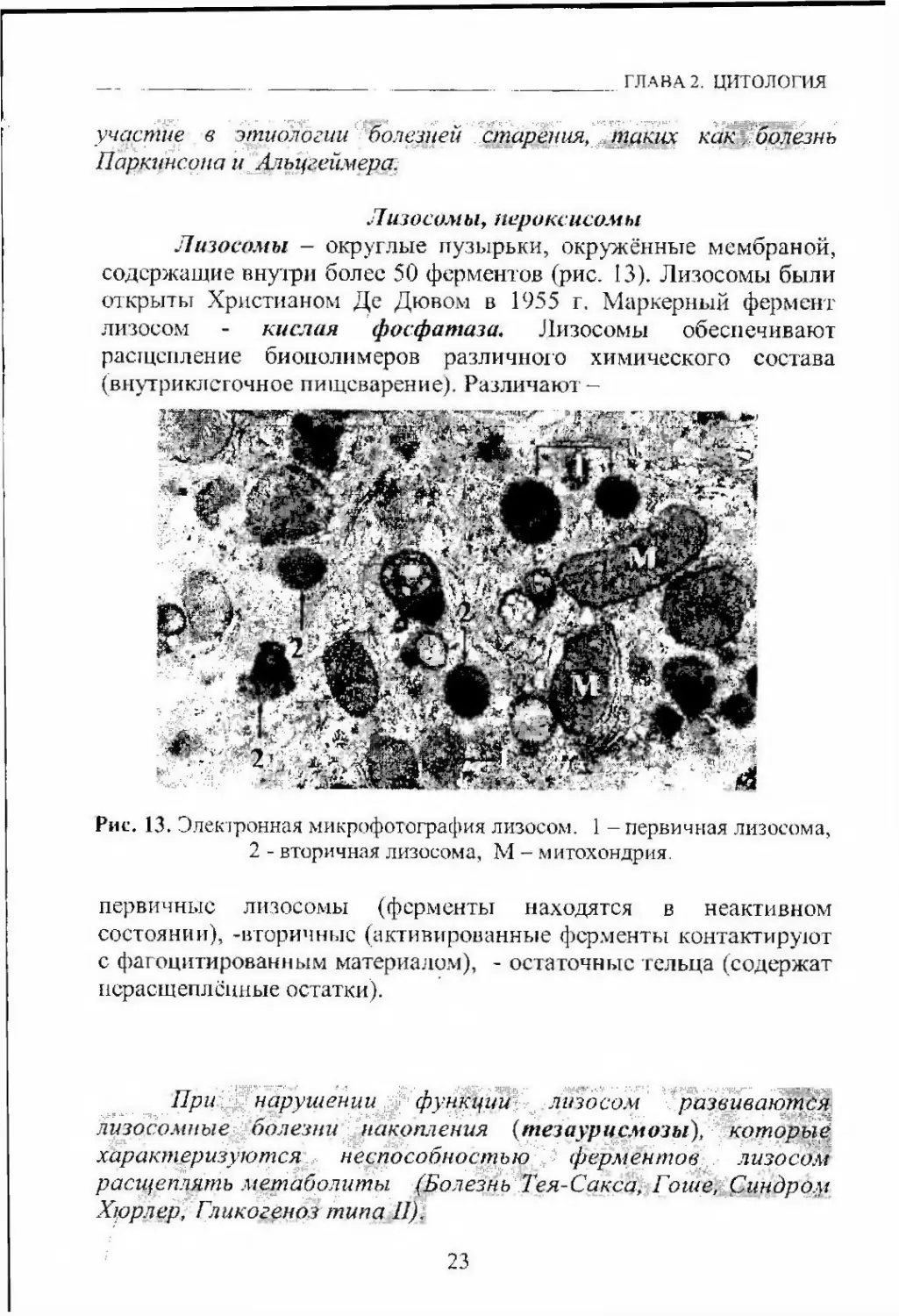
Лизосомы - округлые пузырьки, окружённые мембраной,
содержащие внуїри более 50 ферментов (рис. 13). Лизосомы были
открыты Христианом Де Дювом в 1955 г. Маркерный фермент
лизосом - кислая фосфатаза, Лизосомы обеспечивают
расщепление биополимеров различного химического состава
(внутриклсгочное пищеварение). Различают-Рис. 13. Элекфонная микрофотография лизосом. 1 - первичная лизосома,
2 - вторичная лизосома, М - митохондрия.первичные лизосомы (ферменты находятся в неактивном
состоянии), -вторичные (активированные ферменты контактируют
с фагоцитированным материалом), - остаточные тельца (содержат
нсрасщеплсиные остатки).При. /нарушении функции-. ^ лизосом развиваются
лизо'солтъге болезни накопления {тезаурцсМозы), котррщ
характ/^ризуются^ неепособностью ферментов, лизосат
расщеплять метаболиты (Болезнь Тея-Сакса, Гоше, Синдро^^
Х}ррлер; Гликогтоз типа И).23
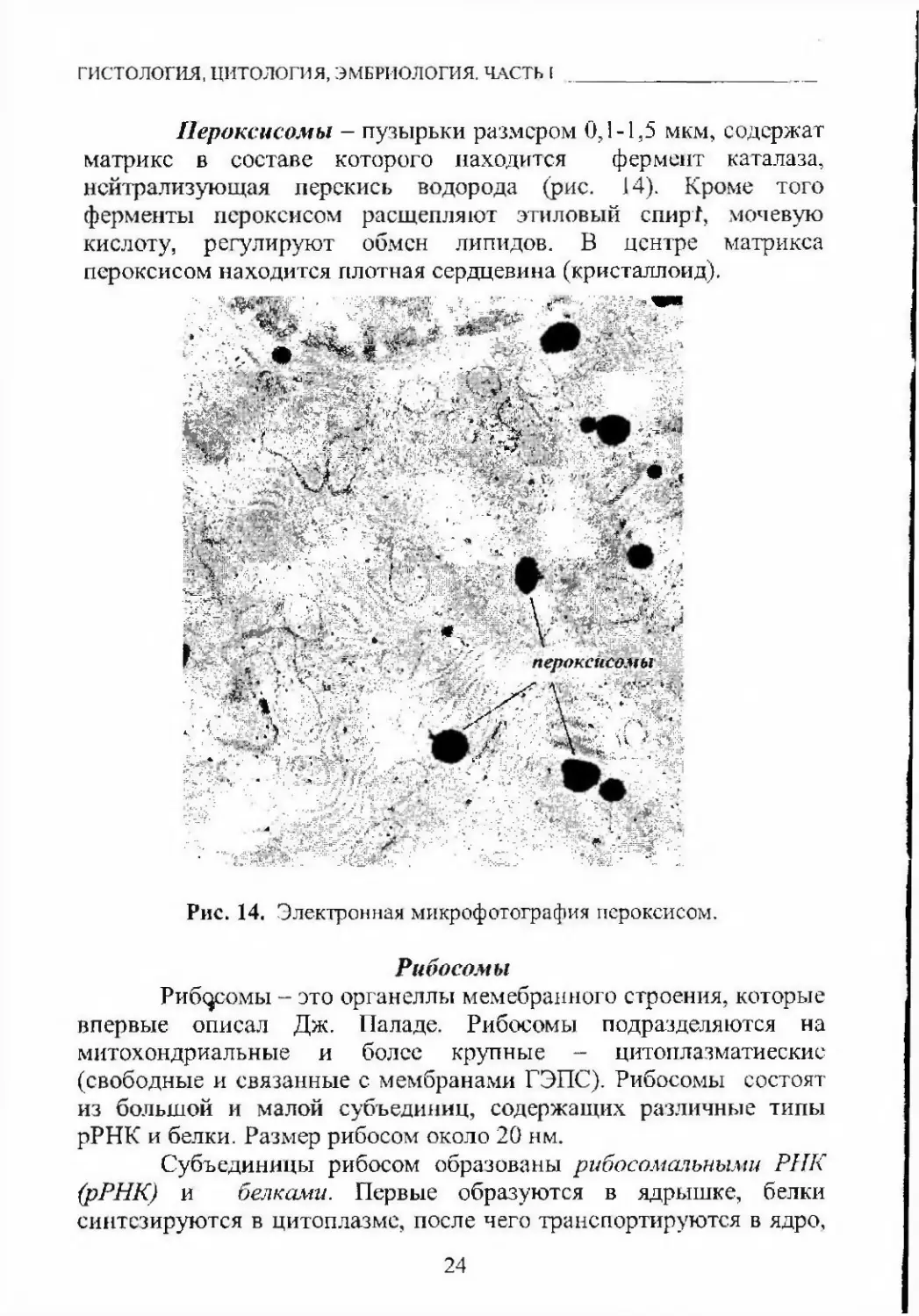
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ [Пероксисомы - пузырьки размером 0,1-1,5 мкм, содержат
матрикс в составе которого находится фермент каталаза,
нейтрализующая перекись водорода (рис. 14). Кроме того
ферменты пероксисом расщепляют этиловый спир1, мочевую
кислоту, регулируют обмен липидов, в центре матрикса
пероксисом находится плотная сердцевина (кристаллоид).р11№Я1йЛ|§^Шя«;:■■'';• •■■г.;Рис. 14. Электронная микрофотография пероксисом.РибосомыРиб^^омы - это органеллы мемебраиного строения, которые
впервые описал Дж. Паладе. Рибосомы подразделяются на
митохондриальные и болсс крупные -- цитоплазматиеские
(свободные И связанные с мембранами ГЭПС). Рибосомы состоят
из большой и малой субъединиц, содержащих различные типы
рРНК и белки. Размер рибосом около 20 нм.Субъединицы рибосом образованы рибосомстьными РНК
(рРНК) и белками. Первые образуются в ядрышке, белки
синтезируются в цитоплазме, после чего транспортируются в ядро,24
_ГЛАВЛ2. ЦИТОЛОП1Ягде связываются с рРНК. В дальнейшем субъединицы через
ядерные поры направляются из ядра в цитоплазму, где принимают
участие в синтезе белка.Рибосомы могуг располагаться свободно в гиалоплазме или
формировать ассоциации - полирибосомы (комплекс нескольких
рибосом, расположенных на одной молекуле мРНК), Также
рибосомы могуг быть связаны с мембранами эндоплазматической
сети (рис. 15).г—?"■-■ >.•^ гРис. 15. Схема строения рибосомы. 1 - малая субьединица; 2 -тРНК;3 - аминоацил-тРНК; 4 - аминокисл ага; 5 - большая субъединица;6 - мембрана эндоплазматической сети; 7 - полипептидная цепь.Основная функция рибосом - трансляция (считывание кода
мРНК и сборка полипептидов). Свободные полирибосомы
синтезируют белки для самой клетки, а полирибосомы ГЭПС
предназначены для синтеза белка на экспорт.ПротеасомыПротеасомы - это органеллы, которые наряду с
лизосомами обеспечивают разрушение белков в клетке.Каждая протсасома состоит из трубкоподобной и одной
или двух регуляторных частей, которые расположены на одном25
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iили обоих концах органеллы, В механизме распознавания белков,
которые в дальнейшем будут разрушены в протеасоме, ведущую
роль играет процесс присоединения белка убиквитина.Йа0шёниё этого процесса и блокирование разр^уЩепШ
белков в протеасомах лежит в основе патогенеза
нейродегенёративных заболеваний (болезнь 11аркш^о§1а,
болезцьЛльцгейм ера).Окаймлённые пузырькиОкаймленные пузырьки участвуют во внугрикл сточи ой
сортировке белков и образуют две разновидности - с клатрином и
без клатрина в оболочке. Окружённые клатрином пузырьки
переносят белки, поступившие в клетку посредством эндоцитоза, а
также белки со сторо11ы 1гапз-поверхности комплекса Г ольджи.Не содержащие клатрин пузырьки переносят белки из
ГЭПС в комплекс Гольд5КИ и из комплекса Гольджи в
плазматическую мембрану.ЦитоскелетЦитоскелет обеспечивает тургор клс1'ки, поддержание и
перемещение её в пространстве, изменение формы, двигательные
процессы внутри клетки.к цитоскелету относят микротрубочки, микрофиламенты,
промежуточные филаменты.Микротрубочки имеют диаметр 24 нм, толщина стенки
составляет* 5 нм (рис. 16). Микротрубочки состоят из 13
тубулиновых нитей, идухцих по спирали. Каждая нить собрана из
чередующихся димеров а- и )3-тубулина. Микротрубочки
обеспечивают расхождение хромосом при делении клетки.
Добавление колхицина приводит к нарушению сборки
микротрубочек или к разборке уже существующих.26
_ГЛАВЛ2. ЦИТОЛОГИЯРис. 16. Схема, объясняющая строение микротрубочек.Промежуточные филаменты имеют диаметр около 10 нм,
часто располагаются пучками, имеют разный белковый состав в
различных тканях, (кератин, виментин, десмин).Микрофиламенты имеют толщину 5-7 нм, встречаются во
всех видах клеток. Микрофиламенты часто образуют пучки,
которые направляются в клеточные отростки, В состав
микрофиламситов входят сократительные белки: актин, миозин,
тропомиозин, а-актинин. Таким образом, микрофиламенты
представляют собой сократительный аппарат клегки при её
перемещении, а также участвуют во внутриклеточных
передвижениях цитоплазмы, вакуолей, митохондрий, делении
клетки, эндоцитозе и экзоцитозе, а также в изменении
консистенции гиалоплазмы (рис. 17),27
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАС'ГЪ {Рис. 17. Электронная микрофотография цитоскслегга клетки.1 - микротрубочки, 2 - актиновые микрофиламенты.ЦентросомаЦентросома (клеточный астр) - является центром организации
митотического веретена при делении клетки. Она состоит из двух
центриолей, окружённых цапросферой. Центриоли,
расположенные в паре, называются диплосома, В неделящихся
клетках цсшриоли располагаются вблизи комплекса Гольджи.
Каждая центриоль состоит из 9 трицлсгов микротрубочек,
образующих полый цилиндр (рис. 18). Каждая центриоль имеет
следующую формулу: (9хЗ)-1-0. кроме микротрубочек в состав
центриоли входят «ручки», содержащие белок динеин. В
диплосомс центриоли располагаются под прямым углом друг- к
другу, вокруг них образуется особая зона - центросфера. При
подготовке клеток к митозу происходит удвоение центриолей.Центриоли - это место роста микротрубочек аксонсмы
ресничек иди жгутиков. Кроме того, сама центриоль индуцирует
полимеризацию тубулинов новой центриоли, возникающей при её
удвоении.28
_ГЛЛВА2. ЦИТОЛОГИЯРис. 18. Электронная микрофотография клеточного центра.
1 “ центриоли, 2 - триплеты микротрубочек.Реснички и жгутики
Это специальные органеллы движения, встречающиеся в
некоторых клетках различных организмов, В основании ресничек
(сШит) и жгутика (flagellum) видны хороню окрашивающиеся
мелкие гранулы, так называемые базальные тельца. Длина ресничек
5-10 мкм, а длина жгутиков может достигать 150 мкм.Ресничка представляет собой вырост цитоплазмы
диаметром 200 нм. Внутри реснички расположена аксонема
(«осевая нить»). Базальное тельце по своей структуре сходно с
центриолью. Аксонема в своем составе имеет 9 дублеров
микротрубочек с динеиновыми «ручками». В центре аксонемы
располагается также одна пара центральных микро1рубочек.
Систему микротрубочек реснички описывают формулой: (9x2) +
2.Жгутики обеспечивают передвижение отдельных видов
клсгок, нанримср, сперматозоидов. По своему строению идентичны
с ресничками, однако, имеют большую длину.карта^енера ^¡щ^иада (КартагМера синдром) /т-
щслёЬстветдя аномтт развития, Х(^актеризуето1; а^29
гистология. ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ Iбронхоэктазов, зЫту угБсетт туегзш' и пЩ(йоза слизистойобдд^^кй "носа; сШ1др^М^связан дефектом беЛНй0инеина и, какслед1^^^^е^-,--'-- ' сопровождается неподвижностью ■. реашчёк
дыхательного эпителия.ВключенияВключения - необязательные компоненты клетки,
возникающие и исчезающие в зависимости от метаболического
состояния клетки: пигментные (экзогет1Ыс - пылевые частицы,
красители, эндогенные - гемоглобин, липофусцин, меланин),
трофические (жиры, гликоген), секреторные (биологически
активные вещества, образующиеся в клетке), экскреторные
(продукты метаболизма) (рис. 19).Рис. 1^. Различные виды включений в цитоплазме клеток.1 - включения липофусцина в цитоплазме нервных клеток.1 - липофусцин, 2 - ядро нейрона, 3 - глиоциты.2 - включения гликогена в клетках печени. 1 - ядра,2 — гранулы гликогена.3 - жировые включения в клетках печени. 1 - ядра, 2 - капли жира.30
_ГЛЛВА2. ЦИТОЛОГИЯЯдроЯдро {nucleus) клетки - система генетической детерминации
и регуляции белкового синтеза. Ядро обеспечивает функцию,
связанную с хранением и передачей генетической информации, а
также синтез белка.В ядре происходит воспроизведение и удвоение молекул ДНК, а
также создание собственного аппарата белкового синтеза. Под Э'шм
понимается синтез и транскрипция на молекулах ДНК разных
информационных РНК и фанскрипция всех видов транспортных и
рибосомных РНК. В ядре происходит также образование
субъединиц рибосом.Структура и химический состав клеточного ядраЯдро состоит из хроматина, ядрышка, кариоплазмы
(нуклеоплазмы) и ядерной оболочки, отделяющей его от
цитоплазмы (рис. 20).Рис. 20. Электронная микрофотография ядра. 1 - зухроматин,2 - гетерохроматин, 3 - ядерная оболочка, 4 - ядрышко, 5 - цитошшма,6 - митохондрия.Хроматин, Внутри клеточного ядра выявляются зоны
плотного вещества, которые хорошо воспринимают основные31
гистология, цитология, ЭМБРИ0Л01 ИЯ. ЧАСТЬ Iкрасители. Этот компонент ядра получил название «хроматин» (от
греч. chroma - цвет, краска). В состав хроматина входиг ДНК в
комплексе с белком. Такими же свойствами обладают и
хромосомы, которые отчетливо видны во время митотического
деления клеток, (heterochromatinum) (рис. 21).Рис. 21. Участок клеточного ядра, содержащий хроматин. 1 - внутрегтияя
мембрана кариолем.мы. 2 - наружная мембрана кариол ем мы,3 - перинуклеарное пространство.В неделящихся (ицтерфазных) клетках хроматин
представлен деконденсированными хромосомами {еисИготаИпит)
или же располагается отделы1ыми глыбками конденсированного
хроматинаЧем диффузнее распределен хроматин в интерфазном ядре,
тем интенсивнее в нём протекают синтетические процессы.Максимально конденсированный хроматин во время
митотического деления клеток обнаруживается в виде плотных
хромосом (рис. 22). В этот период хромосомы НС выполняют
никаких синтетических функций, в них не происходит включения
предшественников ДНК и РНК.32
ГЛАВА 2. ЦИТОЛОГИЯЯдро\Две хроматизм/ *РОМ0С0теломср маРис. 22. Строение хромосомы. 1 - короткое плечо, 2 - длиппое плечо.Таким образом, хромосомы клеток могут находиться в двух
структурно-функциональных состояниях: в активном, частично или
полностью дскондснсированном, когда с их участием в интерфазном
ядре происходят процессы 1ра1Юкрипции и редупликации, и в
неактивном, в состоянии метаболического покоя при максимальной их
кондснсироваиности, в этот период они выполняют функцию
распределения и пера юса генетического матфиала и дочерние
клегки.В химическом отношении фибриллы хроматина представляют
собой комплексы дезоксирибонуклеопротеидов (ДНП), в состав
которых входят ДНК и специалыше хромосомные белки -
гистоновыс и псгистоновые. в составе хроматина обнаруживается
также РНК. Отнотенис ДНК, белка и РНК составляет 1:1,3:0,2.В хромосомах сушествуют места независимой репликации
ДНК - репликоны. Средний размер репликона около 30 мкм. В
составе генома человека встрсчаегся более 50 ООО репликонов,
участков ДНК, которые синтезируются как независимые единицы.Белки хроматина соегавляют 60-70% от его сухой массы
(гистоны и негистоновые белки). Негистоновые белки составляют
20% 01 количества гистонов. Гистоны - щелочные белки,
обогащенные основными аминокислотами, они расположены по
ДЛИ1Ю молекулы ДНК в виде блоков. В один такой блок входят 8
молекул гистонов, образуя нуклеосому (10 нм). При образовании33
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iнуклсосом происходит укорачиваиие длины хромосомной фибриллы
примерно в 5 раз. Сама же хромосошшя фибрилла имеет вид иитки
бус или четок, где каждая бусина - нуклсосома. Такие фибриллы
толщиной 10 нм дополнительно продольно конденЬируются и
образуют основную элементарную фибриллу хроматина толщиной 25
нм. Нс1истоновые белки интерфазных ядф образуют внутри ядра
структурную сеть, которая носит название ядерный белковый
матрикс.В ядрах также встречаются пqшxpoмaтинoвьíe фибриллы,
перихроматиновые и интерхроматин овыс гранулы. Они содержат РНК
и встречаются во всех активных ядрах, представляют собой
информационные РНК, связанные с бажами. Особый тип матричной
ДНК, а именно ДНК для синтеза рибосомной РНК, собран обычно в
нескольких компактных участках, входящих в состав ядрышек
интффазных ядф.Для каждого вида харакгфен свой хромосомный набор по их
количеству (например, у человека 46 хромосом, у дрозофилы - 8, у
лошади - 66). Хромосомы в ядрах соматических клсгок могут быть
сгруппированы пapa^ш, получившими название гомологичных
хромосом. В ядрах половых клеток (гамет) из каждой пары
гомологичных хромосом содержится только одна, т. с. обшлй набор
хромосом вдвое меньше, чем в соматических клс11<ах;Одинарный набор хродмосом в половых клегках называется
гатоидньш и обозначается буквой п, а в соматических - дипчоыдньш
(2п).В результате образования гамет выявляется- большое
количертво дефектных гашт^ Значительное количество дефен^тнш
гххмет не жизнеспособно. В противном сл^ае рШсдаются^^ети с
генетическим дефектом. Иаруше^тя м(§ущ.прошойтщ^ клетках вр
Ъремя митоза, что можещ^ пфвести к синтезу полностью цли
частично дефектного белка (серповидноклетонная анемщц,
кистозный фиброз). Микроудац^енш или перемещения могут привести
к синтезу усеченных .белков. Могут .возникнуть мутации ти
перемещения хро.мосомы, которые регулирует проявление гена. Их
называют , ,апруктурными трщлиями хромосом. Прш1ером
хромосомных бо.пезней является трисомия ПЩ:21 паре хромосом
(сшндром Дс^на}^: при которой наблюдаетсяг47¡щюмосом. СиндрОм
Тернера - при котором жтщыны тдлько: имеют одну X хромосому и
гщ^тф]щ^ общ^ сюжности сд^ерэ^ат 0 хромосом.34
ГЛАВА 2. 11ИТ0Л0ГИЯЯдрышкоВо всех живых клетках эукариотических организмов в ддрс
содержится ядрышко {п1(с1ео(и.ч). Ядрышко является производным
хромосомы. Оио ИС является самостоятельной струкгурой Ш1И
органеллой. Ядрышко - это место образования рибосомных РНК
(рРНК) и рибосом, иа которых происходит си1пез полипептидных
цепей улче в цитоплаз1\те. Образовагшс ядрышек и их количссшо
связаны с ак^гивностью определенных участков хромосом -
ядрышковых органитпгоров, которые расположены в зоне вторичной
исрстя:ж1ш. Вокруг ядрышка обычно выявляется зона
конденсированного хроматина -околоядрышковый хроматин.
Ядрышко неоднородно по своему строению, в его структуре
выявляются два основных компонента: гранулярный и фибриллярный.
Фибриллярный компонент сосредоточен в центральной части
ядрышка, а гранулярный - по периферии (рис. 23). Гранулярный
компонент образует нитчатые структуры - нуклеолонемы.
Фибр1шлярный комгюпепт ядрышек представлясг собой
рибоиуклеопротеидные тяжи предшссгвенников рибосом, а гранулы “
созреваюище субъединицы рибосом. В зоне фибрилл можно выявить
участки ДНК ядрьшжовых организаторов.Рис. 23. Электронная микрофотография ядрышка. 1 - ядрышковый
организатор, 2 - фибриллярный компонент, 3 - гранулярный компонент, 4
- околоядрышковый хроматин, 5 - ядерная оболочка, 6 - цитош1азма,
Ядерная оболочка
Ядро ограничено ядерной оболочкой, отделяющей его
содержимое (кариоплазму) от цитоплазмы. Оболочка состоит из35
г истология, гистология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iдвух мембран (наружной и внутренней ядерной мембраны),
разделенных перинуклеарным пространством (рис. 24). Обе они
пронизаны многочисленными порами, благодаря которым
возможен обмен вen^ecтвaми между ядром и цитоплазмой.“Наружная мембрана ядерной оболочки, непосредственно
контактирующая с цитоплазмой клетки может прямо переходить в
мембраны эндоплазматической сети. Внутренняя мембрана связана
с хромосомным материалом ядра.Ядерные поры образуются за счег слияния двух ядерных
мембран и имеют диаметр около 80-90 нм. Эти отверстия в
ядерной оболочке заполнены глобулярными и фибриллярными
структурами. Совокупность всех компонентов поры называют
комплексом поры (рис. 24). По границе ядерной поры
располагается три ряда гранул по 8 в каждом: один ряд лсжиг со
стороны ядра, другой - со стороны цитоплазмы, третий расположен
между ними в центральной части поры. Размер гранул около 25 н.м.
От этих гранул отходят фибриллярные отростки. Фибриллы,
отходящие от периферических гранул, могут сходиться в центре и
создавать как бы перегородку, диафрагму поперек поры.Тонкие филаментыНаружные V
гранулыПромежуточныегранулыВнутренниегранулыП ром еж>т очные
гранулы' л. .Наружная ядерная
мембранаеринуклсарнос
пространстпо''‘Вп)’трснняя
ядерная мембранаФиламенты,
присоединяющиеся
к центральной
грануле.Рис. 24, Ядерный поровый комплекс (схема).36
ГЛАВА 2. цитологияЧисленность ядерных пор зависит от мсгаболичсской
активности клеток: чем интенсивнее синтетические процессы в
клетках, тем больше пор приходится па сди1пщу поверхности
клеточного ядра. Ядерная оболочка ограничивает свободный
доступ в ядро кру1П1ых агрегатов биополимеров, регулирует
транспорт макромолекул между ядром и цитоплазмой, учас'1'вусг в
фиксации хромосомного материала в трехмерном пространстве
ядра.Половой хроматин дкенский (ттьце Барра , Х-хроматип) -
интенсивно окрашиваемый участок хромосомы треугольной,
круглой или палочковидной формы, расположенный на
периферии ядра соматической клетки Ыенскдго организма,
представляет собой конденсированную Х-хромосому^Хроматин половой мужской (У-хроматин) - крупная глыбка
хромосомы в ядре соматической клетки мужского организма,
представляет собой конденсировиннмйучасток длинного плеча
Т-хромосомы.Клеточный цикл и его регуляцияКлеточный цикл - это период жизни клетки от одного
деления до другого или от деления до смерти. Клеточный цикл
состоит из интерфазы (период вне деления) и самого клеточного
делеиим. Интерфаза состоит из трех периодов. Сразу после выхода
из митоза клетка вступает в пресинтетический или С] период,
далее переходит в синтетический или 8 период и потом “ в
постсинтетический или Сг период. После этого клетка вступает в
следующий митоз (рис. 25).Нели клетка не делится, то она как бы выходит из
клсгочного цикла и вступает в период покоя, или Со. При
необходимости клетка выходит из Со периода и вступает в С1
период клеточного цикла. Длительность периодов клеточного
цикла различна. Так, С1 период может продолжаться от 2-4 ч до
нескольких недель или даже месяцев, 8"Период длится от 6 до 8 ч,
длительность Сз периода - до получаса. Длительность митоза
составляет в среднем от 40 до 90 минут.37
гистология, цитология. ЭМБРИОЛОГИЯ. ЧАСТЬ I.Рис. 25. Клеточный цикл.GJ период характеризуется высокой синтетической
активностью. Начинается рост клеток главным образом за счет
накопления клеточных белков, что определяется увеличением
количества РНК. В этот период 11ачинается подготовка клетки к
синтезу ДНК (8-периоду).В S периоде происходит удвоение или редупликация ДНК.
Дополнительно синтезируются ГИСТО£ЮВЫе белки, ферменты,
регулирующие и обеспечивающихесинтез нуклеотидов и
образование новых нитей ДНК.в 6г2 периоде происходит образование веществ,
необходимых для самого процесса митоза (синтез белков, из
которых об^^азуются микротрубочки веретена деления (тубулин,
динеин, нексин, спектрин), синтез АТФ).За прохождение 1С(1еткой определённых фаз клеточного
цикла отвечают специальные белки-ферменты. Они называются
циклии-зависимыми протсинкиназами (cdc).38
ГЛАВА 2. ЦИТОЛОГИЯРЕГУЛЯЦИЯ КЛЕТ()ЧНОГО ЦИКЛАicdk2 + циклин
¡DiIcdkS и циклин
ЬзGi периодR-пункт период?^ Gi
переход из G| в S период
переход из S в Ог периодicdc2 + циклин С
|cdk2 + цикли Е!cdk2 + циклин Апереход из Сг периода в митоз + циклин В ‘период) [ циклин Н I cdk7 необходим для фосфорилирования
и активациии cdc2 в комплексе с циклином ВАпоптозАпоптоз - запрограммированная юісгочная гибель, которая
происходит в нормальных физиологических условиях. Клетки,
вступаюіцие в апоптоз, имеют специфические морфологические и
биохимические характфистики. При этом в ядре наблюдается
копдеіюация хроматина в виде «полумесяца», Затем нроисходиг
расиїепление цитоплазмы и ядра на несколько частей, окруженных
мембраной (аиоитогические тельца), которые содержат
морфологически интактные органеллы и ядерный материал (рис. 26).ІП vivo эти тельца быстро фагоцитируются макрофагами или
другими рядом лежащими клетками. Апоптоз протекает без
возникновения воспалительной реакции. Процесс апоптоза
регулируется специальными генами.Рис. 26. Апоптоз в животной клетке. 1 -- апоптотические тельца.39
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ IНекрозНекроз ~ гибель клсгки, которая наступает в результате
повреждения.МитозМитоз (кариокинез, непрямое деление) - универсальный
способ деление эукариотических клеток. Митоз следует за Gi
периодом. Митоз длится 1-3 часа и сопровождается равномерным
распределением генетического материала между дочерними
клетками.Митоз вк;ночает 4 следующие друг за другом фазы:
профазу, метафазу, анафазу и телофазу (рис. 27, 28).Профаза сопровождается конденсацией хромосом, каждая
хромосома состоит из двух сестринских хроматид, связанных в
области центромеры. Ядрышко и ядерная оболочка исчезают,
цитоплазма и кариоплазма смешиваются, центриоли мигрируют к
полюсам клетки, появляются нити митотического веретена. В
области цешромеры образуются кинетохоры - центры -
организаторы микротрубочек.Метафаза - хромосомы выстраиваются в области экватора
клетки, образуя мстафазную пластинку (материнскую звезду).Анафаза - хромосомы расщепляются на сестринские
хроматиды, дочфние хромосомы направляются к
противоположным полюсам клетки, в конце анафазы по
окружности клетки начинает образовываться клеточная перетяжка,
которая углубляясь, приводит к цитотомии в телофазе.Телофаза - завершается реконструкция ядер дочерних
клеток, ядра постепенно увеличиваются, хромосомы
деспирализуются. Цитоплазма клеток перешнуровывается с
образованием двух дочерних клеюк.Эндомитоз (эндорепродукция)- разновидность митоза, при
котором происходит удвоение числа хромосом внутри ядерной
оболочки без её разрушения и образования веретена деления.Полиплоидия - образование клеток с повышенным
содержанием ДНК. Отмечастся при повторных эндомитозах, может
явиться следствием неоконченных обычных митозов. Смысл
полиплоидии заключается в усилении функциональной активности
клеток.40
ГЛАВА 2. ЦИТОЛОГИЯРис. 27. Фазы мито'за: 1 - профаза, 2 - ■ метафаза, З - анафаза,
4 - телофаза.Рис. 28. Митотітческое деление ютеток в корешке лука.41
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ \ МейозМсЙОЗ - это деление половых клеток. Мейоз СОСТОИТ из двух
после-довательных делений с короткой иитсрфазой между иими
(рис. 29).Профаза первого деления очень сложная и состоит из 5
стадий, в зиготеие профазы первого деления происходит
соединение гомологичных хромосом (образуются биваленты или
тсфады), которые остаются связанными между собой до анафазы
первого деленияВ анафазе первого деления происходит разделение и
расхождение к полюсам целых хромосом, состоящих из двух
хроматид, а не разделение хромосом на отдельные хроматиды, как
в митозе.В интерфазе между первым и вторым делениями мейоза
отсутствует период и перед вторым делением НС происходит
редупликации ДНК.в процессе мейоза образующиеся дочерние клетки
полностью не разделяются между собой, а остаются связанными
тонкими цитоплазматическими мостиками.Сперматидыинтерф(иа V Мейоз IХромосомы*1*ис. 29. Схема мейотического деления.42
ГЛАВА 3.
ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙТкаии - это исторически сложивтииеся системы Ю1СГ0К и
исклсточных структур, обладаютцие общностью строения,
происхождения и специализированные на выполнении определенных
функций.Ведущими элементами гкалсвой системы являются клетки.
Кроме клеток, различают клеточные производные и меисклсточное
вещество.Производные клеток:- с1штасты (мышечные воло1а1а, симпластотрофобласт),- стщитий (развивающиеся мужские половые клетки, пульпа
эмалевого органа),- постклеточпые структуры (эритроцигы, тромбоциты,
роговые ЧСШуЙЕСИ эпидермиса и т. д.).Межклеточное вещество подразделяют на основное вещество
и на волокна. Оно может быть находиться с состоянии золя, геля или
быть минерализованным.Развитие тканейТкани как системы, состоящие из клеток и их производных,
возникли с появлением MHOJOKЛ сточных организмов. По мере
развития животною мира совершалось закрепление свойств
отдельных гкансй, количество тканей постепенно увеличивалось в
соответствии со все более возрастающей специализацией.Детерминация и коммитированиеРазвитие организма начинается с зиготы. В ходе дробления
возникают бласгомерьт, но совокупность бластомеров - это еще не
ткань. Бластомеры на начальных этапах дробления еще не
детерминированы. Нели отделить их один от другого, - каждый может
дать начало полноценному самостоятельному организму, таков
механизм В03НИ1Ш0ВСНИЯ монозиготных близнецов. Постепенно на
следующих стадиях происходит огршшчсние потенций, в основе его
лежат процессы, связанные с блокированием отдельных компонентов
генома клеток и детерминацией.Детерминации - эго процесс определения дальнейшего пути
развития клеток на основе блоюгровшшя отдельных генов.43
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАС’ТЬ [Коммитирование - это ограничение возможных нугсй
развития вследствие детерминации. Вначале детерминируются
наиболее общие свойства клеток, а затем и более чаш ные.Па этапе іаструляции возникают эмбриональные зачаї ки.
Клетки, которые входят в их состав, еще не окончательно
детерминированы. Следовательно, один эмбриональный .зачаток
может служить источником развития нсскольюіх ткаисй.Теория эволюции тканейПоследовательная детерминация и коммитирование потенций
однородных клеточных іругишровок - дивергентный процесс.
Концепция дивергентного развития тканей в филогенезе и в
онтогенезе была сформулирована Н.Г.Хлопиным, который ввел
понятие о генетичесюїх тканевых типах. Концепция Хлопина отвечает
на вопрос, как и какими иуіями ироисходшіо развигие и становление
тканей.Причинные аспекты развития тканей раскрывает теория
ггараллслизмов А.А.Заварзина. Он обратил внимание на сходство
строения тканей, которые вьяіолііяют одиііакоЕзьіе функции у
животных, принадлежащих даже к весьма удаленным друг от друї'а
эволюционным группировкам. Следовательно, в ходе эволюции в
разных вегвях фююгспсгичсского древа самостоятельно, как бы
параллельно, возникали одинаково организованные ткани,
выполняющие сходную функцию.Концепции А.А.Заварзина и П.Г.Хлопина, разрабшанмые
независимо одна от другой, дополняют друг друга.Развитие тканей в эмбриогенезе происходит в результате
дифференцировки клеток. Под дифференцировкой іюнимаюг
изменения в структуре клеток в результате их функциональной
специализации, обусловленные активностью их генетического
аппарата. Различают четыре основных периода дифференцировки
клеток зародыпш - оотипичсскую, бласгомсріїую, зачатковую и
тканевую дифферснцировку. Проходя через эти период>л клспш
зародьила образуют ткани (гистогенез).у Юшссификация тканейНаиболее распространенной является морфофуикциональная
классификация, по которой ткани подразделяют на четыре груины;1. эпителиальные ткани;2. ткаии виуїренней среды;3. мышечные ткани;4. нервная ткань,К тканям внутренней среды относятся соединительные ткани,
кровь и лимфа.44
ГЛАВАЗ ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙЭпителиальные ткаи и хараіасризуются объединением
клсгок в пласты. Через эти ткани совершается обмен веществ между
организмом и внешней срсдой. Эпителиальные ткани выполняют
функции защиты, всасывания и зкскрсции. Источниками
формирования эпителИШ1Ы1ЫХ тканей являются все три зародьппевых
листка - эктодерма, мезодерма и энтодерма.Ткани внутренней среды ^соединительныс ткани, скелетные,
КРОВЬ и лимфа) развивакугся из мезенхимы. Ткани внутренней среды
харакгеризуются наличием большого количества межклеточного
іісїцества и содержаг различные клеікл. Они специализируются на
выполнении трофической, пластической, опорной и защитной
функциях.Мышечные ткани специализированны на вьшолнении
функции движения. Они развиваегся в основном из мезодермы
(поперечнополосатая мышечная ткань) и мезенхимы (гладкая
мышечная гкань).Нервная ткань развивается из эктодер.мы и специализируется
на выполнении восприятия, проведения и передачи информации.Основы кинетики клеточных популяцийКаадая ткань имеет шш имела в эмбриогенезе стволовые
юіегки (СК) - наименее дифференцированные и наименее
комлштированные. СК образую"г самоподдерживаютцуюся
популяцию, их потомки способны дифференцировагься в нескольких
направлениях под влиянием микроокружения (факторов
дифференцировки), образуя клегки-предшественішки и
функционирующие дифференцированные клетки. Таким образом,
стволовые клегки П0ЛИП0ТеНТ11Ы. Они делятся редко, пополнение
зрелых клеток ткани осуществляется за счет клсток-
предшественников. По сравнению со всеми другими клетками данной
гкани стволовые клетки наиболее устойчивы к повреждающим
воздействиям. Хотя в состав ткани входят не только к^легки, именно
клетю! являются ведущими элементами системы, т. с. определяют ее
основные свойства. Их разрушение приводит к деструкции системы и,
как правило, их гибель делает ткань неясизнеспособной, особенно если
были затронуты сгволовые клетки.Нели одна из СК вступает на путь дифференциации, то в
результате последовательного ряда митозов возникают сначала
полустволовые, а затем и дифферетщированные клегки со
специфической функцией. Выход стволовой клетки из попу.ляции
служит сигнало.м для деления другой стволовой клетки. Обиіая
численность стволовых клеток в итоге восстанавливается. В условиях
іюрмальной жизнедеятельности она является постоянной величиной.45
гистология, 11.ИТ0Л0ГИЯ, Г^М КРИОЛОГИ я, ЧАСТЬ I,Совокупность клеток, развивающихся из одного вида
сгволовых клеток, составляет стволовой диффсроіі. Часто в
образовании ткани участвуют различіїьіе диффероны.Дифференцированные клегки наряду с вьшолнеїЛіем своих
специфических функций способны синтезировать кейлоны,
тормозящие интенсивность размножения клеток-прсдшссівснников и
стволовых клеток. Если в силу каких-либо причин количество
дифффенцированных функционирующих клеток уменьшается
(например, ІЮСЛЄ травмы), тормозящее действие кейлонов ослабевает
и 'шсленность популяции восстанавливается. Кроме кейлонов,
клеточное размножение контролирусі ся гормонами.Выбор пути дифференциации юісгок определяется
мсжклсгочными взаимодействиями. Влияние микроокружения
изменяет активность генома дифффенцирующейся клетки, активируя
одни и блокируя другие гены, у Ю1С10К, уже дифференцированных и
утративших способность к дальнейшему размножению, строение и
функция тоже могут изменяться. Такой процесс не приводит к
возникновению различий среди потомков клетки и для него больше
подходит название «специачизация».Регенерация тканейСоответственно уровням организации живого организма
различают клеточную (или внутриклеточную), тканевую и органную
регенфацию.Также различают регенерацию физиологическую, которая
совфшается постоянно в здоровом организме, и рспаративную -
вследствие повреждения, у разных тканей возможности регенерации
неодинаковы.В ряде тканей гибель клеток генетически запрограммирована и
совершается постоянно. За счет непрфывного размножения, в пфвую
очередь полустволовых клеток-предшсственников, количество клеток
в іюпуляции пополняется и постоянно находіггся в состоянии
равновесия. Хотя в ряде тканей запрограммированной і ибели нет, но в
течение всей жизни в них сохраняются стволовые и полустволовые
клетки, в отвїгг на случайную і ибель возникает их размножение и
популяция восстанавливается.У взрослого человека в тканях, где стволовых клеток не
остается, регенерация на ткаїісвом уровне невозможна, она
происходит лишь на клеточном уровне.46
ГЛАВА 4.
ОСНОВЫ ЭМБРИОЛОГИИЭмбриология - это наука о закономерностях, развития
зародыша.Медицинская эмбриология изучает закономерности
развития зародыша человека.Знания в этой области необходимы врачам в акушерско-
гинекологической практике (анализ и ликвидация причин
бесплодия, экстракорпоральное оплодотворение (1УР), разработка
специфических лекарственных препаратов, анализ механизмов
тератогенеза, коррекция нарушений системы «мать-плацента-
плод»). Знание особенностей эмбриогенеза человека необходимо
врачу для его практической деятельности.Сравнительная эмбриология даёт материал для
понимания развития человека.Эмбриогенез (пренатальный онтогенез, внутриутробныйпериод)“ период от момента оплодотворения до рождения, тесно связан с
прогенезом и ранним постэмбриональным периодом(посчнатальным, первый год после рождения).Ирогенез - это период развития половых клеток —
женских и мужских (яйцеклетка и сперматозоид).В основе биологических прот^ессов, определяющих
развитие зародыша лежат: индукция, детерминация, деление,
миграция, рост, дифферснцировка, взаимодействие клеток,
разрушение. Морфогенез формирование пространственной
организации организма и е10 частей. Многоклеточный организм
развивается из оплодотворённой яйцеклетки (зигота) не только
путём увеличения количества клеюк (пролиферация) и увеличения
массы зародыша (рост).Индущия - влияние индуктивных факторов одних участков
зародыша на другие участки, которые развиваются в определенном
направлении. Например, организатор первого порядка индуцирует
развитие нервной пластинки из эктодермы. В нервной пластинке47
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ [возникает организатор второго порядка, способствующий
превращению у^тастка нервной пластинки в глазной бокал и т.д.
(теория организационных центров Шпсмана), Индукция клеток
осуществляется особыми веществами белковой ‘ природы;
мезодермальным М - агентом и нейральным N - агс1ггом.
Эмбриональная индукция - влияние одних зачатков на другие.Детерминация - генегичсски запрограммированный путь
развития клегок и тканей.Дифференцировка - изменения в структуре клеток,
связанные с их функциональной специализацией, обусловленные
активностью определенных генов. Детерминирующими в
эмбриогенезе являются: факторы наследственности (гены),внутренние, внешние факторы.Этапы дифференцировки:1 - оотипическая: материал будущих зачатков представлен
презумптивными участками цитоплазмы яйцеклеток или зиготы;2 - бластомер11ая: различия наблюдаются уже в первых
бластомерах;3 - зачатковая: появление обособленных угастков -
зародышевых листков;4 - гисхогенетическая: в пределах одного зародышевого
листка появляются зачатки различных тканей.Разрушение - внутриутробное развитие сопровождается
образованием огромного количества клеток, которые своевременно
уничтожаются путем апоптоза, Апоптоз - физиоло1ическая
запрограммированная гибель клеток. Например, в нервной системе
или же в тимусе способом аноптоза погибает до 85% клегок.ПрогенезПрогенез - созревание и развитие мужских и женских
половых клеток, к половым клеткам (гаметам) относятся
сперматозоид! и яйцеклетки.Сперматозоид имеег головку и хвостовой отдел (рис. 30).Головка содержит небольшое шютное ядро с гаплоидным
набором хромосом (22 + X, 22 + У у человека) (1п) и
акросомальный чехлик. В нем находится акросома, содержащая
набор ферментов: гиалуронидаза и протсаза, которые способны
растворять оболочки яйцеклетки при оплодотворении. А1фосома -
производное комплекса Гольджи.48
ГЛЛВЛ 4 ОСНО ВЫ ЭМБРИОЛОГИИХвостовой отдел имеет:1 - связующую часть (шейку), которая содержит проксимальную
центриоль;2 - промежуточную часть, которая содержит 2 центральные и 9 пар
периферических микротрубочек, окруженных митохондриальным
влагалищем;3 - главную часть, которая по строению напоминает ресничку
(9х2)+2, окруженную фибриллами;4 - конечную часть, которая содержит единичные сократительные
филаменты.Анросома¡1::лЯдроУОм Цектриоль^ Митохондрии' ІІІІI_ФибриллярныйфутлярРис. 30. Строение сперматозоида. 1 - головка, 2 - связующая часть хвоста,3 - промежуточная часть хвоста, 4 - главная часть хвоста, 5 - конечнаячасть хвоста.49
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ IЖенские половые клетки (яйцеклетки)Яйцеклетки, или овоциты (о\а1П1 - яйцо), со:^ревают в
неизмеримо меньшем количестве, чем сперматозоиды.Овоцит - это крупная неподвижная клетка. Яйцеклетка
имеет шаровидную форму, больший объем цитоплазмы,
неподвижна.Оболочки яйцеклетки (рис. 31):- первичная (оолемма),- вторичная (прозрачная зона, фолликулярная оболочка),- третичная (подскорлуповая у птиц),- четвертичная (екорлуповая у птиц).Рис. 31. Стр^>ег1ие овоцита. 1 - ядро, 2 - оолемма, 3 - лучистый венец,4 - кортикальные гранулы, 5 - желточные включения, 6 - фолликулярные
* клетки, 7 - блестящая зона.Ядро яйцеклетки имеет гаплоидный набор хромосом (22 + х).Цитоплазма содержит ЭПС, КГ, рибосомы, митохондрии,
кортикальные 1'ранулы и желточные включения. Желточные
гранулы содержат протеины, фосфолипиды и углеводы.Яйцеклетка человека - вторичная олиголецигальная
изолецитальная, имеет диаметр около 130 мкм.50
ГЛАВА 4 ОСНОВЫ ЭМБРИОЛОГИИЦитоплазма содфжит в периферических отделах
кортикальные гранулы число которых достигас'г около 4000,
которые включают ферменты, участвующие в кортикалыюй
реакции, защищая яйцею1саку от полиспермии.К оолемме прилежит прозрачная или блестящая зона (zona
pcllucida), которая состоит из гликозоаминогликанов (запщтная
функция), а далее - слой фолликулярных клеток. Отростки
фолликулярных Ю1СТ0К проникают через прозрачную зону,
нанравляясь к цитолеммс яйцеклетки. Цитолемма яйцеклетки имеет
микроворсинки, располагающиеся между отростками
фолликулярных клегок. Фолликулярные клетки выполняют
трофическую и защитную функции.в образовании прозрачной зоны принимают участие овоцит
и фолликулярная оболочка. Зернистая оболочка состоит из
фолликулярных клеток.Классификация яйцеклеток у живых организмов
основывается на признаках наличия, количества и распределения
желтка (Iccithos)» представляющего собой белков о-липидное
включение в цитоплазме, используемое для питания зародыша.Различают следующие типы яйцеклеток:- безжслтковые (алецитальные),- маложсл'1кивыс (олиголецитальные)первичные (ланцетник)
вторичные (человек)- многожелтковые (полилецитальные)ценпгролег{итапьные (желток находится в центре)
теяолецитальные (желток находится навегетативном полюсе)
умеренно телолещталъные
(мезолецитачьные) (амфибии)
резко телолециталыше (птицы)В маложелтковых яйцеклшках желточные включения
расгюлолсены равномерно в цитоплазме, поэтому они называются
изолецитальнымм
гистология, ЦИТ0Л01'ИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ IОплодотворен неОплодотворение {fertUisatio)- слияние мужской и женской
половых клеток, в результате чего восстанавливается диплоидный
набор хромосом, характерный для данного вида ячивотных, и
возникает качественно новая клетка - зигота (оплодотворенная
яйцеклетка, или одноклеточный зародыш). Оплодотворение у
человека происходит в просвете маточной трубы. В процессе
оплодотворения различают три фазы: 1) дистантное
взаимодействие и сближение гамет; 2) контактное взаимодействие
и активизация яйцеклетки; 3) вхождение сперматозоида в яйцо и
последующее слияние - сингамия.Первая фаза обеспечивается хемотаксисом. Важную роль в
этом играют гамоны - химические вещества, вырабатываемые
половыми клетками. Яйцеклетки выделяют пептиды,
способствующие привлечению сперматозоидов. После эякуляции
спермин неспособны к проникновению в яйцеютетку до тех пор,
пока не произойдсг канацитация - приобретение снермиями
оплодотворяющей способности под действием секрета женских
половых путей. В процессе капацитации в области акросомы
удаляются гликопротеины, что способствует акросомальной
реакции.Вторая фаза оплодотворения - контактное взаимодействие,
во время которого сперматозоиды вран^ают яйцеклетку. Яйцсклсгка
вращаетсельные движения со скоростью 4 оборота в Nmnyry в
течение 12 часов. В это время происходит акросомальная реакция,
Она заключасгся в слиянии наружной мембраны акросомы с
плазмолеммой спармия. Затем ферменты акросомы выходят в
окружающую среду. Ферменты разрушают блecтян^yю зону,
спермин проходит через нес и входит в неривителлиновое
пространство, расположенное между блестящей зоной и
плазмолеммой яйцеклетки. Через несколько секунд изменяются
свойства плазмолеммы яйцеклетки и начинается кортикальная
реакция, а через несколько минут изменяются свойсгва блестян1ей
зоны. У млекопитающих при оплодотворении в яйцсклсгку
проникает лингь один сперматозоид.Третья фаза. В ооплазму проникают головка и
промежуточная часть хвостового отдела. После вхождения
сперматозоида в яйцеклетку на периферии ооплазмы происходит
уплотнение и образуется оболочка оплодотворения.52
ГЛАВА 4 ОСНОВЫ ЭМБРИОЛОГИИКортикальная реакция - слияние плазмолеммы яйцеклетки
с мембранами кортикальных гранул, в результате чего содержимое
из гранул выходит в неривителлиновое пространство и
воздсйсгвусг на молекулы глн-конротеидов блестящей зоны.
Образуется оболочка оплодотворения толщиной 50 нм,
препятствующая полиснермии.После сближения^ женскою и мужского пронуклеусов
образуется зигота - одноклеточный зародыш. Уже на этой стадии
выявляются презумптивные зоны (лат. рге8итр11о - вероятность,
предположение) как источники развития соответствующих
участков бластулы, из которых в дальнейшем формируются
зародышевые листки.Дроблениедробление - вариант митотического деления, при котором
образуются дочерние клетки (бластомеры) без дальнейшего роста
до размера материнской. Бластомеры не расходятся и образуют
морулу (рис. 32). Вследствие отсугствия 0|-периода интерфазы, во
время которого происходит рост клеток до размеров материнской,
масса зародыша не превышаег массу зиготы.Дробление продолжается до тех пор, пока соотношения
объемов ядра и цитоплазмы бластомеров не станет характерным
для соматических клеток данного организма. После этого
растормаживается 01-преиод и начинается обычный митоз.г \ ■: :Г'У'"'. ■ ч..Л.'Ш'Рис. 32. Дробление зиготы.53
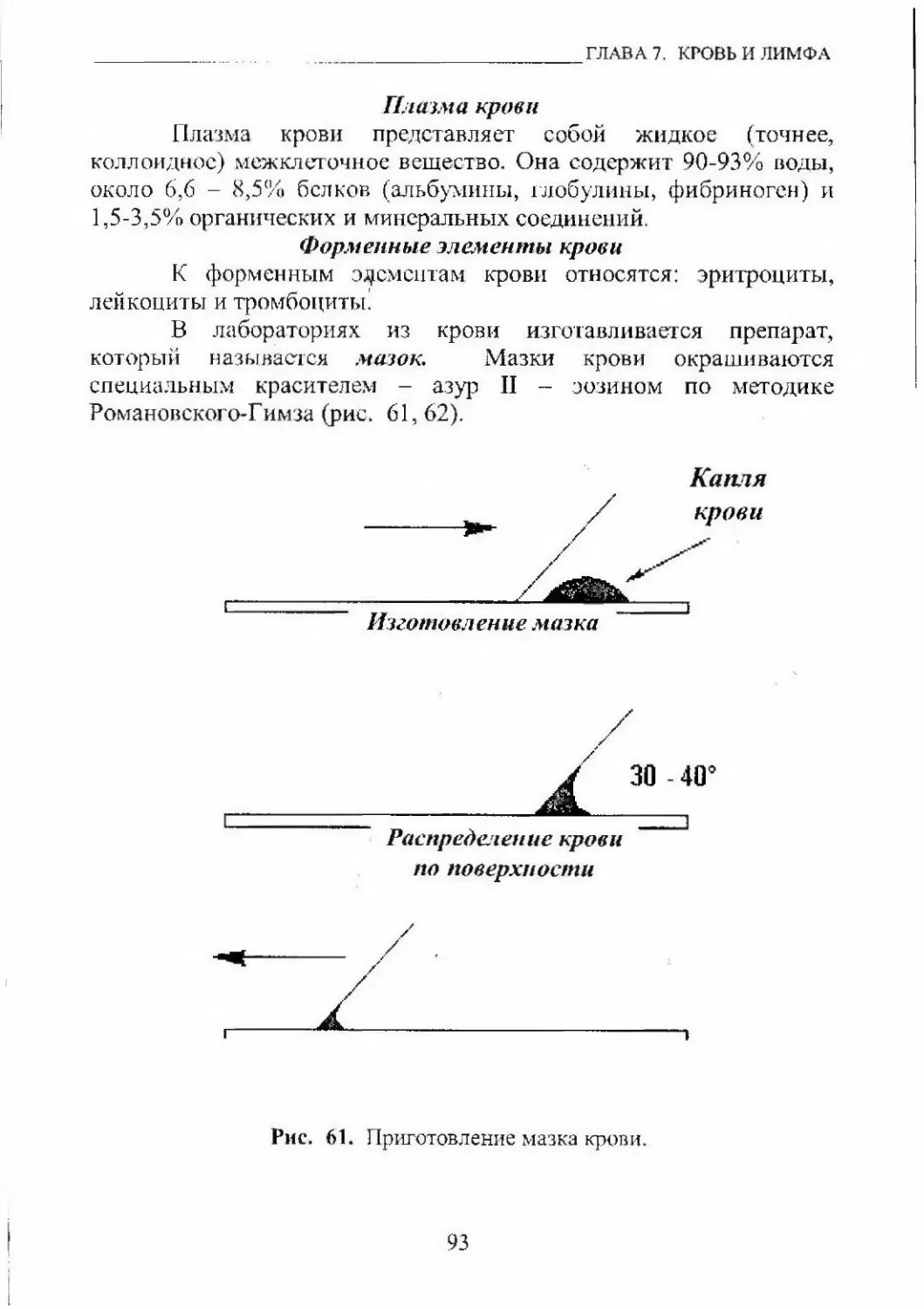
гистология, цитология, ЭМБРИОЛОгаЯ. ЧАСТЬ I...Классификация дробленииПо полноте деления зиготы различают следующие виды дробления:• полное - ЗНГ0Тс1 делится полностью и образуются две
отдельные клегки• неполное - борозда деления не гюлноетью разделяет дочерние
Ю1СПСИ и не происходит образования агдельиых юхетокпо размерам образующихся блас гомеров• равномерное - бластомеры имеют одинаковые размеры• неравномерное - бластомеры имеют разные размеры
по временным интервалам между делениями• синхронное - интервалы между дeлcния^ш всех бластомеров
одинаковые• асинхронное - ннтфвалы между делениями всех бластомеров
различные.Первичные олиголецитальные яйцеклетки (ла1щсгник)
дробятся полно и равномерно. Дробление - голобластическое.
Бластула имссг вид пузырька с центрально расположенной пoJюcтью
(33 -1).Мезолецитальные яйцеклетки дробятся полно, но дробление
на вегетативном полюсе, где находится желток, более медленное, чем
на противоположном - анимальном. Такой тип дробления называется
полное неравномерное.. Бластула ~ амфибластула имеет эксцентрично
расположенную полость, дно содержит более крупные бластомфы (33
-2).У организмов с резко телолецитальными яйцск-испсами
дробление частичное или меробластическое. У птиц дробится часть
яйцеклетки на анимальном полюсе. Бластула - дискобластула (33 - 3).Для вторично олиголецитальных яйцеклеток характф1Ю
полное (голобластические) неравномерное асинхронное дробление.
Бластула - бластоциста (33 - 4).Рис. 33. Основные типы бластул: 1 - целобластула, 2-амфибластула,3 - дискобластула, 4 - бластоциста.54
ГЛАВА4 ОСНОВЬІ ЭМБРИОЛОГИИВ результате дробления вначале образуется морула -
МН010КЛсточный зародыш, не имеющий полости, а затем бластула,
в которой появляется полость.Бластула имеет еіспку - бластодерму, полость - бластоцель.
В С1СНКС различают крышу, дно и краевые зоны (рис. 34).Рис. 34. Строение амфибластулы. 1 - анимальный полюс,2 - вегетативный полюс, 3 - краевая зона, 4 - бластоцель (полость).ГаструляцияГаструляция - сложный процесс химических и
морфогенетических изменений, сопровождающийся размножением,
ростом, направленным перемещением клеток, в результате чего
образуются зародышевые листки: наружный - эктодерма,
внутренний - энтодерма и средний - мезодерма, а также комплекс
осевых 0рга1юв {нервная трубка, хорда, кишечная трубка).
Промежутки между зародышевыми листками заполнены клетками
звёздчатой формы, которые образуют мезенхиму.Способы гаструляции :Инвагинация - часть стенки зародыша прогибается в
полость бластулы.55
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ IЭпиболия (обрастание) - рост и перемещение быстро
делящихся клеток аиималыюх’О полюса бластулы на медленно
делящиеся клетки вегетативного полюса.Дел амил ация ~ расслоение стенки бластулы на два слоя.Иммиграция - перемещение части бластомсров сгонки
бластулы внутрь. В результате гаструляции формируется зародыш,
назьгоасмый - гаструла.Дифференцировки зародышевых листковДифферснцировка зародышевых листков - эю изменения в
строении клеток, которые связаны с их функциональной
специализацией.Зародышевые листки являются источниками развития
тканей (гистогенез). Из тканей в дальнейшем образуются органы
(органогенез)^ из органов - системы органов (системогенез).
Результатом гистогсиегических процессов является формирование
основных групп тканей - эпителиальных, крови и лимфы,
сосдинитахьных, мышечных и нервных. Их формирование
начинается в эмбриональном периоде и заканчивается после
рождения.Дифферснцировка эктодермыЭктодерма подразделяется на: внезародышевую
(выстилающая амнион) и зародышевую.Зародышевая эктодерма в свою очередь имеет такие части:- кожная - из нсс формируется эпидермис кожи, эпителий
полости рта, эмаль и кутикула зубов, роговица, анальный отдел
прямой кишки, влагалище;- нейроэктодерма - нервная трубка и ганглиозная пластинка;- нервный гребень - нейроны спинальных и вегетативных
ганглиев, мозговое вещество надпочечников;- плакоды - внутрс1шсе ухо;- прехорд1льная пластинка - эпителий трахеи, бронхов, легких.Дифференцировки энтодермыЭнтодерма подразделяется на; внезародышевую (источник
образования жслточ1Юго мешка и аллантоиса) и зародышевую.
Зародышевая энтодерма даёт начало эпителию желудка, тонкой и
толстой кишки, эпителию печени и поджелудочной железы.56
ГЛАВА4 ОСНОВЫ ЭМБРИОЛОГИИДифферснцировка мезодермыМезодерма подразделяется на внезародышевую (эпителий
желточного мешка и аллантоиса) и зародышевую. Этот процесс
начлиастся на 3-й педеле эмбриогенеза. Дорсальные участки
мезодермы разделяются на сомиты. На 22-е сутки развития у
эмбриона имеется 7 пЬр сегментов, на 35-е - 43-44 пары.
Вентральные отделы мезодермы {спланхнотом) не сегментируются,
а расщепляются на два листка - висцеральный и париетальный.
Участок мезодермы, связывающий сомиты со спланхнотомом,
называется - сег.ментная ножка (нефрогонотом). Из мезодермы
зародыша развивается также парамезонефралъный канал.Сомиты дифференцируются на 3 части: миотом^ дающий
начало поперечнополосатой скелетной мышечной ткани,
склеротом, являющийся источником развития костных и хрящевых
тканей, а 1акже дерматом, формирующий соединительнотканную
основу кожи - дерму.Из сегментных ножек (нефрогонотом) развиваются
эпителий почек, гонад и семявыводящих путей, а из
парамезонефра [ьного канала - эпителий матки, маточных труб
(яйцеводов) и лпггелий первичной выстилки влагалища.Париетальный и висцеральный листки спланхнотома
образуют эпителиальную выстилку серозных оболочек
мезоте.гшй. Из части висцерального листка мезодермы
{миоэпикардиальная пластинка) развиваются средняя и наружная
oбoJЮчки сердца - миокард и эпикард, а также корковое вещество
надпочечников.МезенхимаМезенхима - скопление отростчатых клеток,
pacnoJЮжcllныx между зародышевыми листками. Мезенхима в теле
зародыша является источником формирования многих структур -
клеток крови и кроветворных органов, соединительной ткани,
сосудов, гладкой мышечной ткани, микроглии. Из в незародышевой
мезодермы развивается мезенхима, дающая начало соединительной
ткани внезародышевых органов, - амниона, хориона, желточного
мешка.В незародышевые органыВнезародышсвыс органы (провизорные), развивающиеся в
процессе эмбриогенеза вне тела зародыша, выполняют57
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ !,многообразные функции, обеспечивающие рост и развитие самого
зародыша. Некоторые из этих органов, окружающих зародыш,
называют также зародышевыми оболочками. К этим органам
огносятся амнион, желточный мешок, аллантоис, хорион,
пупочный канатик, тацента.АмнионАмнион - временный орган, обеспечивающий водную среду
для развития зародыша. Стенка пузырька образусг внезародышевую
эктодерму, которая соединяется с внезародышевой мезодермой,
разрастается и окружает зародыш тонкой полупрозрачной
амниотической оболочкой. Амнион заполнен амниотической
жидкостью, которая обеспечивает защиту плода от механических
повреждений и вредных факторов.Желточный мешокЖелточный мешок образован внезародышевой энтодермой
и внезародышевой мезодермой. У человека в питании зародыша
принимает участие до 8 недели эмбриогенеза. Является
кроветворным органом, в стенке которого образуются первые
клетки крови и предшественники половых клеток (I 01юбласты).АллантоисАллантоис - пальцевидный вырост, является производным
желточного мешка и из внезародышевой энтодермы и
внезародышевой мезодермы. Аллантоис выполняет функцию
газообмена и выделения.ХорионХорион (ворсинчатая оболочка) развивается из трофобласта
бластоцисты и внезародышевой мезодермы. Выделяет
протеолитические ферменты, с помощью которых разрушается
слизистая оболочка матки и осуществляется имплантация.%Пупочный канатикПупочный канатик - упругий тяж, соединяющий зародыш с
плацентой. Внутри находится слизистая соединительная ткань,
покрытая амниотической оболочкой. Слизистая соединительная
ткань (Вартонов студень) предохраняет пупочные сосуды от
сдавления.58
ГЛАВА4 ОСНОВЫ ЭМБРИОЛОГИИКлассификация плацент по строению
Эпителиохориальная - в плацентах такого типа ворсинки
хориоиа контактируют с эпителием маточных желез (верблюд,
лошадь, свинья).Десмохориальиая - ворсинки хориона разрушают эпителий
маточных желез, контактируют с подлежащей соединительной
тканью (корова, овца). >Эндотелиохориапышя - ворсинки хориоиа разрушают
эпителий и соединительную ткань (десму) , контактируют с
эндотелием кровеносньж сосудов (кошка, собака).Гемохориальная ~ ворсиики хорио11а свободно омываются
материнской кровью (крыса, человек) (рис. 35).Рис. 35. Типы плацент. 1 - эпителиохориальная, 2 - десмохориальиая,3 - эвдотелиохориапьная, 4 - гемохориальная.Юшссификация плацент по способу питания1. В плацентах 1 типа хорион поглощает белки из тканей
матери и расщспляст их до аминокислот, образование
специфических для эмбриона белков происходит в печени
эмбриона (лошадь, корова).2. В плацентах 2 типа хорион получает из материнских
тканей аминокислоты, синтез эмбриоспецифических белков
происходит в плаценте.Эмбрион получает готовые белки для строительства собственных
тканей. Такие организмы после рождения длительное время
питаются только молоком матери и не способны питаться
самостоятельно (человек).59
ГЛАВА 5.
ЭМБРИОЛОГИЯ ЧЕЛОВЕКАВ процессе эмбрионального развития человека сохраняются
общие закономерности и стадии, характерные для позвоночных
животных. Эмбриональное (внутриутробное) развитие
продолжается 280 суток, 40 недель или 10 лунных месяцев.Эмбриональное развитие можно разделить на 3 нсриодГ:> начальный (1 неделя развития),> зародышевый (2-8 неделя развития ),> плодный (9-40 неделя развития).Гаметы (половые клетки)Сперматозоид - мужская половая клетка, состоит из
головки, шейки и хвоста. В головке имеется ядро и акросома, в
шейке - центриоли, а хвост состоит из 3 отделов; промежуточного
(в нем имеются митохондрии и аксонема жгутика (9 дунлегов
микротрубочек по периферии и 1 дуплет - в центре), главного и
терминального (представляют собой аксон ему жгутика), в
терминальной части - неполный набор микротрубочек (рис. 36).Терминальная частьСвязующая
ча^ть хвост>головкаПромежуто чная
часть хвостачасть хвостаРис. 36. Строение сперматозоида.60
ГЛАВА 5 ЭМБРИОЛОГИЯ ЧЕЛОВЕКАЯйцеклетка - женская половая клетка, крупная клетка, в
ооплазме (цитоплазме) имеется большой запас РНК, кортикальные
гранулы, бслково-липидно-углеводііьіе (желточные) включения.
Кортикальные гранулы являются производным комплекса Гольджи,
представляют собой мембранные пузырьки, содержащие
протеолитические ферменты, которые способны разрушать
редсторы для сперматс^зоидов на дитомембране и блестящей
оболочке яйцеклетки с образованием оболочки оплодотворения. На
дитомембране яйцеклетки есть рецепторы для сперматозоидов.
Яйцеклетка человека имеет 2 дополнительные оболочки;
блестящую, состоящую из гликозамингликаїюв, и лучистый венец,
образовагшый из фолликулярных клеток, которые прилипают к
яйцеклетке пока она находится в фолликуле яичника. Яйцеклетка
человека являегся изолецитальной, вторично олиголецитальной
(рис. 37).ІЇШШ:
т1 3ГЖ''"с 'Щ4 Рис. 37. Строение овоцита человека. 1 - оолемма, 2 - лучистый венец,3 - желточные включения, 4 - фолликулярные клетки, 5 - блестящая зона.Начальный период (1 неделя развития)Этот период харак1'сризустся оплодотворением и
образованием зиготы, а таюке последуюпщм дроблением и
образованием бластулы, соверпгаегся в просвете маточной трубы
(рис. 38).61
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ I,Рис. 38. Овоцит в просвете маточной трубы.ОПЛОДОТВОРЕНИЕ - слияние мужской и женской
половых клеток с образованием одноклсгочного зародыша -
зиготы. В процессе оплодотворения различают несколько фаз:1. Дистантное взаимодействие - сближение
сперматозоидов с яйцеклеткой под действием веществ, выделяемых
яйцеклеткой, в эту фазу сперматозоид начинает направленно
двигаться к яйцеклетке (хемотаксис), а также происходит его
активация (капацитация),2. Контактное взаимодействие - происходит
акросомальная реакция сперматозоида, при которой
высвобождаются ферменты из акросомы и разрушают небольшой
у^шсток блестящей оболочки.3* Проникновение головки и шейки сперматозоида в
ооплазму. В эту фазу происходит взаимодействие между
рецепторамй сперматозоида и яйцеклетки, после чего их мембраны
сливаются, и головка и шейка сперматозоида оказываются в
ооплазме. После проникновения одного сперматозоида в
яйцеклетке возникает кортикальная реакгшя. В ооплазму входят
ионы натрия, в результате чего меняется заряд цитомембраны
яйцеклетки (с отрицательного на положительный), резко
повышается концентрация ионов кальция. Кортикальные i ранулы
начинают двигаться к цитомембранс яйцеклетки и их мембрана62
ГЛАВА5 ЗМКРИОЛОГИЯ ЧЕЛОВЕКАсливается с ци гомембраной яйцеклетки, т.е. происходит экзоиитоз
кортикальных гранул. Ферменты кортикальных гранул разрушают
рецепторы для сперматозоидов, в результате чего другие
сперматозоиды уже не могут проникнуть в ооплазму.
Цигомсмбрапа и блестящая оболочка яйцеклетки превращаются в
оболочку оплодотворения. После проникновения сперматозоида в
яйцсклсгку ядра этих клсгой сначала располш аются ио отдельности
(стадия двух пронуклеусов), а потом сливаются (синкарион) (рис.
39.1).ДРОБЛЕНИЕ - последовательное деление зиготы без роста
образуюп1ихся клеток - бластомеров. Дробление у человека полное,
неравномерное, асинхронное. После первого деления образуются 2
блас'гомсра (39.2). Один из них болсс гсмиый и крупный
эмбриобласт, другой более мелкий и светлый - трофобласта. Из
эмбриобласга развивасгся зародыш и почти все провизорные
органы (хорион, плодная часть плаценты, амнион, желточный
мешок, аллантоис). В процессе дробления клетки трофобласта
делятся быстрее эмбриобласга. В результате этого клетки
трофобласта обрастают снаружи клетки эмбриобласта. Поэтому
образующаяся клеточная масса - морула состоит из двух групп
клеток. Внутри располагаются клетки эмбриобласта, а снаружи -
клетки трофобласта (рис. 39.6).В процессе деления клеток трофобласта и эмбриобласта
объем морулы увеличивается, клетки зародыша начинают
секретировать жидкость, внутри зародыша образуется полость,
заполненная этой жидкостью. Образующаяся бластула называется
бластоцистой (39.7).Она состоит из 1) трофобласта, образующего стенку
бласгулы; 2) клеток эмбриобласга, располагающихся внутри; 3)
полости бластулы, заполненной жидкостью.Поверхность бластоцисты неровная, так как трофобласт
образует вьфосты. Эти выросты называются первичными
ворсинками трофобласта, они состоят только из клс^юк самого
трофобласта. Трофобласт является первым провизорным
органом, образующимся у зародыша человека (рис. 40).63
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСЛЪ 1»ШтФ'-шш:А:Г-''^¿жгМгШз;Рис. 39. Стадия двух пронуклеусов (1), дробление зиготы (2-5),
морула (6), бластоциста (7),64
ГЛАВА5 ЭМБРИОЛОГИЯ ЧЕЛОВЕКАРис. 40. Бластоциста человека.ИмплантацияИмплантация - внедрение зародыша в слизистую оболочку
матки. Различают две стадии имплантации: адгезию (прилиианис),
когда зародыш прикрепляется к внутренней поверхности матки, и
инвазию - внедрение зародыша в ткани слизистой оболочки матки.Имплантация продолжается около 40 час. Одновременно с
имп.иан'шцией происходит и начало гаструляции (образование
зародышевых листков).Б первой стадии трофобласт прикрепляется к слизистой
оболочке матки и в нем начинают дифференцироваться два слоя -
цитотрофобласт и симпластотрофобласт. Во второй фазе
симпластотрофобласт, продуцируя протеолитические ферменты,
разрушает слизистую оболочку матки. Образуется
ымплантациониая ямка. Трофобласт первые 2 недели потребляет
продукты распада материнских тканей (гистиотрофпый тип
питания), затем питание зародыша осуществляется из материнскай
крови (гсматотрофный тип питания). Из крови матери зародыш
кислород необходимый для дыхания: Одновременно в слизистой
оболочке матки усиливается образование децидуальных клеток65
ГИСТОЛОГ'ИЯ, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I.богатых гликогеном. После полного погружения зародыша в
имплантационную ямку дефект слизистой оболочки покрывается
регенерирующим эпителием,1 ематотрофный тип пихания, сменяющий гистиофофный,
сопровождается переходом ко второй фазе гаструляции и заюшдке
внезародышевых органов.ГАСТРУЛЯЦИЯ - это процесс образования зародышевых
листков. Гаструляция у человека происходит в два этапа. Первый
этап идет пугем дсляминадии (расщепления), а второй - путем
миграции.В процессе первого этапа (7 сут.) образуются два
зародышевых листка (экто- и энтодерма), два провизорных органа
(амнион и желточный мешок). Перед нaчaJЮм первого этапа
происходит образование такого провизорного органа, как хорион.Из клеток эмбриобласта образуется зародышевый диск
(щиток) и в незародышевая мезенхима. Внезародышевые
мезенхима подрастает к трофобласту, при эхом образуются
вторичные ворсинки хориона.Клетки зародышевого диска расщепляются (деляминация)
на два слоя. Верхний слой клеток - это эктодерма (эпибласт),
нижний слой - энтодерма (гипобласт).Клетки эктодермы разрастаются за пределы зародышевого
диска и обрастают поверхность амниотического пузырька.
Получается амнион. Стенка амниона состоит из внезародышевой
эктодермы и внезародышевой мезенхимы (рис. 41),Энтодерма разрастается за пределы зародышевого диска,
обрастает поверхность желточного пузырька и получается
желточный мешок. Его стенка состоит из внезародышевой
энтодермы и внезародышевой мезенхимы.Во время второго этапа гаструляции (14-15 сут.)
образуются мезодерма и мезенхима, провизорный орган -
аллантоис и происходит образование плаценты: формируются
тре1'ичные ворсинки хориона, которые в последующем
соединяются с decidua basalis и формирзтот плаценту. Образуются
осевые органы - хорда, нервная трубка, кишечная трубка.66
ГЛАВА 5 ЭМБРИОЛОГИЯ ЧЕЛОВЕКАРис. 41. Ранняя стадия развития зародыша. 1 - амниотический пузырёк,2 - эктодерма, 3 - желточный пузырёк, 4 - энтодерма. 5 - внезародышевая
мезенхима, 6 - цитотрофобласт, 7 - синцитиотрофобласт, 8 - лакуны,9 - кровеносные сосуды.В эктодерме клетки начинают мигрировать от головного к
хвоотовому концу зародыша. В срединной части образуется
первичная полоска, в сс средней части ~ первичный узелок
(рис.42).Образование мезодермы происходит из клеток первичной
полоски, которые прорастают в пространство между экто- и
энтодермой и там разрастаются, образуя мезодерму. Образование
хорды происходит из клеток первич£юго узелка.В срединной части эктодермы образуется yглyбJЮHиe -
нфвная пластинка. Нервная пл^ютинка погружается под эктодерму,
а края эктодермы смыкаются и образуется нерв11ая трубка.Образование кишечной трубки происходит в процессе
^ формирования туловищной складки. В результате этого кишечная
(зародышевая) энтодерма отделяется от желточного мешка
(желточная энтодерма).67
гистология, ЦИТОЛО]'ИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ I.IIРис. 42. Вторая стадия гаструляции (схема). I - амниотический пузырёк,
II - желточный пузырёк, 1 - зародышевая эктодерма,2 - внезародышевая эктодерма, 3 - зародышевая энтодерма,4 - внезародышевая энтодерма, 5 - внезародышевая мезенхима,6 - первичная полоска, 7 - головной узелок, 8 - прехордальный отросток,9 - мезодерма,Дифферснцировка мезодермы и образование мезенхимы.Сразу после своего образования мезодерма подразделяется на два
главных отдела: сомиты - спинной отдел и снлахнотом - брюшной
отдел. Между сомитами и сплахнотомом имеется сегмснгная
ножка, с помощью которой они соединяются. Сомиты разделяются
на три части: дерматом, склеротом, миотом. ДермаIом дает
начало дерме - собственно кожа. Миотом является источником
поперечнополосатых мышц. Из склеротома образуются кости и
хрящи (рис. 43). Сплахнотом делится на висцеральный и
париетачъный листки, между которыми находится полость тела -
целом. Висцеральный и париетальный листки дают нача.10
висцеральным и париетальным серозным оболочкам. Мезенхима
образуется из трех источников - висцерального листка68
ГЛАВА 5 ЭМБРИОЛОГИЯ ЧЕЛОВЕКАспланхнотома, дерматома, склеротома. Из мезенхимы развивасгся
вся соединительная ткань.Из сегментных ножек (нефрогонотом) закладываегся
эпителий П04СК, гонад и семявыводяпщх путей, а из
парамезонефрального канала - эпигслий матки, маточных труб
(яйцеводов) и эпителий первичиой выстилки влагалища.Оплодотворённая
яйцеклетка (1 сут*)Бластоциста
(5-6 сут»)трофобласт
эмбриобластГаструла
(14-16 сут.) кЗародышевые
листки и
формирую щ иеся
из них тканиЭНТОДЕРМА
(виутреипин
листок)
Поджелудочн
ая железа
Печень,- - ПтМЕЗОДЕРМАЭКТОДЕРМА
(промежуточный (наружны й
листок) листок)Хрящевая и Эпидермис,
костная ткань, Нервная
дерма^ШМТ, [^трубка.Рис. 43. Дифференцировка зародышевых листков.
69
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I.Аллантоис образуется как всшральный вырост передней части
задней кипжи. Он растет кперед!^, дорастасг до желточного стебелька и
в этой области выходит из xeaia зародыша. Стенка аллантоиса состоит
из энтодермы и внезародышевой мезенхимы.Образование плаценты. По аллантоису ко вгоричлым
ворсинкам хориона подрастают кровеноснью сосуды и такие ворсинки,
состоящие уже из трофобласта, внезародышевой мезенхимы и
кровеносных сосудов назьшаются третичными ворсинками хориона.
Хорион прикрепляется к подлежащему участку слизистой оболочхси
матки - decidua basalis и вместе с ней формируег плаценту. Таким
образом, плацента представляет собой соединение фсгичпых ворсинок
хориона (плодная часть) и decidua basalis (материнская часть).Внезщюдмшевые органы
Внезародышевые органы, развиваюпщеся в процессе
эмбриогенеза, вьшолняют многообразные функцш!, обеспечивающие
рост и развитие самого зародыша. К ним относятся aNfflHOH, желточный
меиюк, аллаетоис, хорион, плацента, пупочный канатик (рис. 44).дййіїМ ■ й ^МаточнаятрубаСосудыпуповиныПупочныйкаиа-|икDeciduaparíetaJisГладкийхориоиПолостьхорионаDeciduacapsularisDecidua
Лакуны с
материнской
кровьюВорсиныхорилиаЖелточныймешок-ЖелточныйсгсбслёкNfHOMcrpHHїШШггСлнзнсганпробкаРис. 44. В;»аимоотношение зародыша, внезародышевых органов и
оболочек матки.70Í
ГЛАВА5 ЭМБРИОЛОГИЯ ЧЕЛОВЕКААмниол - временный орган, обеспечивающий водную среду
для развития зародыша. Стенка амниотического пузырька состоит
из юшгок внезародышевой эктодермы и из внезародышевой
мезенхимы. Образуется амнион па 7-8 сутки эмбрионального
развития. Основная функция амниотической оболочки - выработка
околоплодных вод, обесп/рчивающих среду для развивающегося
организма и предохраняютцих его от механического повреждения.
Амнион выполняет также защитную функцию, предупреждая
попадание в плод вредоносных агентов.Желточный мешок - депонируег питательные вещества
(желток), необходимые ДЛЯ развития зародыша. Он образован
внезародышевой энтодермой и внезародышевой мезодермой
(мезенхимой). Появляется на 2-й неделе эмбрионального развития.
В стенке желточного мешка развиваются первые клетки крови,
первые кровегюсные сосуды и предшественники половых клеток -
гоиобласты. Желточный мешок функционирует до 7-8-й недели, а
затем подвергается обратному развитию и остается в составе
пупочного канатика в виде узкой трубочки, служащей проводником
кровеносных сосудов к плаценте.Аллантоис - небольшой пальцевидный отросток в
каудальном отделе зародыша, врастающий в амниотическую
ножку. Он состоит из внезародышевой энтодермы и висцерального
листка мезодермы. У человека с помощью аллантоиса к хориону
растут сосуды, располагающиеся в пупочном канатике. Это орган
газообмена и выделения. По сосудам аллантоиса доставляется
кислород, а в аллантоис выделяются продукты обмена веществ
зародыша. На 2-м месяце эмбриогенеза аллантоис редуцируется и
превращается в тяж клегок, который вместе с редуцированным
желточным мешком входит в состав пупочного канатика.Пупочный канатик (пуповина) - упругий тяж,
соединяющий зародыш (плод) с плацентой. Он покрыт
амниотической оболочкой, окружающей слизисгую
соединительную ткань с кровеносными сосудами (две пупочные
артерии и одна вена) и рудиментами желточного мешка и
аллантоиса. Слизистая соединительная ткань (вартонов студень)
обеспечивает упругость канатика, предохраняет пупочные сосуды
от сжатия.71
гистология, цитология. ЭМБРИОЛОГИЯ. ЧАСТЬ I \ ’ кШРис. 45. Пупочный канатик. 1 - пупочная артерия, 2 - пупочная вена,3 * слизистая соединительная ткань (вартонов студень).Хорион, или ворсинчатая оболочка развивается из
трофобласта и внезародышевой мезодермы. Пфвоначально
трофобласт представлен слоем клеток, образующих первичные
ворсинки. На 2-й неделе трофобласт приобрсхает двухслойное
строение {цитотрофобласт и симпластотрофобласт).
Внезародышевая мезодерма на 2-3»й неделе развития подрастает к
трофобласту и образует вместе с ним вторичные ворсинки. С этого
времени трофобласт превращастся в хорион, или ворсинчатую
оболочку. В начале 3-й недели в ворсинки хориона врастают
кровеносные капилляры и формируются третичные ворсинки. Это
совпадаег с началом гематотрофного питания зародыша.
Дальнейшее развития хориона связано развитием плаценты.Плацента (детское место) человека относится к типу
дискоидальных гсмохориальных ворсинчатых плацент. Это важный
временный орган с многообразными функциями, которые
обеспечивают связь плода с материнским организмом. Плацента
создает барьер между кровью матери и плода. Плацента состоит из
двух частей: зародышевой, или плодной (pars fetalis) и материнской
(pars materna). ПJЮднaя часть представлена ветвистым хорионом и
амниотической оболочкой, а материнская - видоизмененной
слизистой оболочкой матки, отторгающейся при родах (decidua
basalis) (рис. 4б).Развитие плаценты начинается на 3-й неделе и
заканчивается к концу 3-го месяца беременности. На 6-8-й неделе
вокруг сосудов дифференцируются элементы соединительной
ткани, которые содержат значительное количество гиалуроновой и
хо1щроитинсерной кислот, с которыми связана регуляция
проницаемости плаценты. При развитии плаценты происходит
разрушение слизистой оболочки матки и смена гистио1рофного72
ГЛАВА 5 ЭМБРИОЛОгаЯ ЧЕЛОВЕКАпитания Неї гсматотрофнос, Ворсины хориона омываются кровью
матфи, излившейся из разрушенных сосудов эндометрия в лакуны
(рис. 47). Однако кровь матери и плода в нормальных условиях
никогда ПС смешивасгся.Материнская
часть
плаценты
- гкотиледоРис. 46. Плацента человека (макропрепарат).АмнионПупочная
артерия
Хориальная
п.іас і ина сПупочные веныкровеноснымиСОСУЛЯМН1'И ■Кропепосные
сосуды ворсипБазальная пластина....Плацентарныесспты Вены мкометрия Миометрий
Артерии миометрияРис. 47. Гемохориальная дискоидальная плацента (схема).73
гистология, ЦИ'І’ОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ 1,Гсматохориальный барьер, разделяющий оба кровотока,
состоит из эндотелия сосудов плода, окружающей сосуды
соединительной ткани, эпителия хориальных ворсин
(цитотрофобласт и симпластотрофобласт), а кроме foro, из
фибриноида, который местами покрывает ворсины снаружи (рис.
48).Гемохориалъный барьер проницаем для ряда возбудитедец
заболеваний (краснуха, цитомегаловирус, ветряная оспа,
простой герпес, вирус иммунодефицита человека, токсоплазмоз,
сифилис) химических веществ и лекарственных., препаратов
(стрептомицин (глухота), имипрамин(деформация конечностей), тетрациклин (аномалии костей и
зубов), кокаин (гипоксия плода), курение табака (задержщ
внутриутробного роста плода), алкоголь (нарушает развитие
нервной и других систем плода), гипертермия).te""■‘■'І- «itu A“ft' ‘ •> '"'4 ''-^J-.V я- чРис. 48. Строение ворсин хориона. 1 - межворсинчатое пространстпо,2 ~ ворсины }?ориона, 3 - симпластотрофобласт и цитотрофобласт,4 - кровеносные сосуды, 5 - соединительная ткань ворсины, 6 - макрофаги
(клетки Кащенко-Гофбауэра).Плодная часть плаценты к концу 3-го месяца
представлена хориальной пластинкой, покрытой цито- и
симпластотрофобластом (рис. 49.1), Ветвящиеся ворсины хориона
(стволовые, якорные) хорошо развиты лишь со стороны,Iі74 j'І
ГЛЛВЛ5 ЭМБРИОЛОГИЯ ЧЕЛОВЕКАобращенной к миометрию. Здесь они проходят через всю толщу
плаценты и своими вершинами погружаются в базальную часть
разруигснного эндомегрия. Во второй половине беременности и
особенно в конце се трофобласт сильно истончается и ворсины
покрываются фибриноподобной оксифильной массой, являющейся
продук1'Ом свфтывания плазмы и распада трофобласта
("фибриноид Лаигханса").Структурно-функциональной единицей плаценты является долька
{котиледон), Общее количество котиледонов в плаценте - 200.Материнская часть плаценты представлена базальной
пластинкой и соедииитсльнотканными септами, отделяющими
котиледоны друг от друга, а также лакунами, заполненными
материнской кровью, в которые свободно свисают ворсины
хориона (рис. 49.2). В местах контакта cтвo^ювыx ворсин с
отпадающей оболочкой встречаются также трофобластичсские
клетки (периферический трофобласт). Глубокие части отпадающей
оболочки вместе с ^рофобластом образуют базальную пластинку.Рис. 49. Гистоструктура плодной (1) и материнской (II) частей плаценты.1 - хориальная пластина, 2 - амниотический эпителий,3 - сосуд ребёнка, 4 - ворсины плодной части плаценты,5 - лакуны с материнской кровью, 6 - цито- и симпластотрофобласт,7 - ііорсиньї хориона, 8 - сосуды с материнской кровью, 9 - дещщуальные
клетки, 10 - базальная пластина.75
гистологая, ЦИТОЛОГИЯ, эмбриология, часть iБазальный слой эндометрия (lamina basalis)
соединительная ткань слизистой оболочки матки, содержащая
óeijudyanhHMe клетки. Эти крупные, богатые гликогеном клетки
соединительной ткани расположены в глубоких слоях слизистой
оболочки матки. В течение 2-го месяца беременности
децидуальные клетки значительно укрупняются. В базальной
пластинке, чаще в месте прикрепления ворсин к материнской части
плаценты, встречаются скопления клеток псрифсрическохо
цитотрофобласта. Они напоминают децидуальные клетки, но
отличаются более интенсивной базофилией цитоплазмы. Аморфная
субстанция (фибриноид Popa) находится на поверхности базальной
пластинки, обращенной к хориальным ворсинам. Фибриноид
играет существенную роль в обеспечении иммунологического
г омеостаза в системе мать - плод.На границе ветвистого и гладкого хориона, т.е. по краю
плацентарного диска, образус'гся замыкающая пластинка,
препятствующая истечению крови из лакун плаценты.Кровь в лакунах непрерывно циркулирует. Она поступает из
маточных артерий, а оттекает от плаценты по венам, берущим
начало от лакун крупными отверстиями.Формирование плаценты заканчивается в конце 3-го месяца
беременности. Плацента обеспечивает питание, тканевое дыхание,
рост, регуляцию образовавшихся к этому времени зачатков органов
плода, а также его защиту.Функции плаценты. Основные функции плаценты: 1)
дыхательная, 2) транспорт питательных веществ, воды,
электролитов и иммуноглобулинов, 3) выделительная, 4)
эндокринная, 5) участие в регуляции сокращения миометрия.Одним из первых шацепта синтезирует хорионический
i:OHadomponuHy концентрация которого быстро, нарастает на 2-3-
и неделе беременности, достигая максимума на S-lO-i} неделе,
причем в крови плода она в 10-20 раз вышесчем в крови матери. На
определены}!^наличия этого гормона в .\юче основывается тест на
беременность. Среди гормонов плаценты следует упомянуть:
АКТГ гипофиза, плацентарный лакгоген, прогестерон и
прсгнандиол, эстрогены ■ (эстрадиол, эстрон, эстриол),
меланоцитостимудируюащЙ, и адренокортикртропный гормоны^
соматостатин и др.76
ГЛАВА 5 ЭМБРИОЛОГИЯ ЧЕЛОВЕКАКритические периодыКритические периоды - периоды короткой качсствсииой
перестройки и поиьтшенной чувствительности организма к
действию факторов внешней среды (Норман Грег-1944,
П,Г.Светлов-1960). В Э'50 время эмбрион наиболее восприимчив к
повреждаютцим воздействиям.В онтогенезе человека выделяют критические периоды
развития: в прогенезе, эмбриогенезе и постнатальной жизни. К ним
относят:- прогенез- оилодотворение,- имплантация (7-8 сут.),- формирование плаценты и осевых органов (3-8 нед.),- стадия усшюнного роста г/м (15-20 нед.),- формирование систем организма (20-24 нед.),- рождение,- период новорожденности (до 1 года),- половое созревание (11-16 лег).77
ГЛАВА 6.
ЭПИТЕЛИАЛЬНЫЕ ТКАНИЭпителиальные ткани предстаиляют совокупі^ть
дифферонов клеток на границе с внешней и внутренней срсдой, а
таїслсе образующих больпшнство желез орі анизма.Эпителиальная ткань покрывает поверхность тела и выстилает'
слизистые и серозные оболочки внутренних органов (покровный
энитслий), а таюке образует паренхиму большинства желез
(железистый эпителий).функции эпителиальных тканейЭпителий обеспечивает защиту подлежащих тканей
организма ш механических, физических и химических
воздействий.Разграничительная, барьерная - основная функция
эпителисв.Транспортная функция характеризуется переносом
веществ сквозь пласты эпителиальных клегок.Всасываи)Н|ая - многие эпителии активно всасывают
вещества.Посредством эпителиев происходит связь между
организмом и окружающей средойЧасть клеток специализируется на синтезе и выделении
(секреции) специфических вeп^ecтв, необходимых для
деятельности других клеток и организма в целом.Экскреторная - эпителии участвуют в удалении из
организма конечных продуктов обмена веществ.Сенсорная (рецепторная) - эпителиальные ткани с
помощью расположенных в них рецепторов воспринимаіот
сигналы внутренней и внешней среды.Общий план строения эпителиальных тканей
Имеется пять основных особеніюстей эпителиев:78
ГЛАВА 6 ЭПИТЕЛИАЛЬНЫЕ ТКАНИ1. Эпителии представляют собой пласты клеток -
эпителиоцитов. Между ними исг межклеточного вещества,
Ю1СТКИ тесно связаны друг с другом с помощью различных
контактов.2. Эпителии рас1юлагаются на базалыюй мембране.3. Эпителиальные клетки обладают полярностью. Различают
базальный (ло1^ащий в основании) и апикальный
(верхушечный) отделы ютеток, которые имеют разное
строение.4. Эпителий ПС содержит кровеносных сосудов. Питание
^питслиопитов осуществляется диффузно через базальную
мембрану со стороны подлежащей соединительной ткани.5. Эпителиям присупш высокая способность к регенерации.
Базальная мембрана, на которой расположенэпителиальный пласт имеет толщину около 1 мкм и состоит из двух
пластинок: светлой {lamina lucida) и темной {lamina densa). Светлая
пластинка включает аморфное вещество, относительно бед1юе
белками, но богатое ионами кальция. Темная пластинка
представляет аморфный матрикс, в который впаяны фибриллярные
структуры, обеспечивающие механическую прочность мембраны.
Гликопротеины базальной мембраны - фибронектин и ламинин -
выполняют роль адгезивного субстрата, к которому прикрепляются
эпителиоциты. Ионы кальция обеспечивают связь между
адгезивными хликопротеинами базальной мембраны и
полудесмосомами эпител иоцитов. Клетки эпителия связаны с
базальной мембраной с помощью полудесмосом. Здесь от
плазмолеммы эпителиоцитов к темной пластинке базальной
мембраны проходят «якорные» филаменты, В этой же области, но
со стороны подлежащей соединительной ткани в темную пластинку
базальной мембраны вплетаются пучки «заякоривающих» фибрилл
коллагена VII типа, обеспечивающих прочное прикрепление
эпителиального пласта к подлежащей ткани.Функции базальной мембраны:1. механическая (закрепление эпителиоцитов),2. трофическая и барьерная (избирательный транспорт
вещесгв),3. морфогенетическая (обеспечение процессов регенерации и
ограничение возможности инвазивного роста эпителия).79
гистология, цриология, эмбриология, часть I.Морфол оги ческая юшссифика ция эпител и ев
Различают две основные группы эпителиев: однослойные и
многослойные, в однослойных эпитслиях всс клетки связаны с
базальной мсмбра1юй, а в многослойных с ней нсхюсрсдсгвенно
связан лишь один нижний слой клсгок.I. Однослойный эпителий по форме клегок подразделяют на:
плоский, кубический, призма! ический (столбчатый или
цилиндрический).Однослойный эпителий может быть: однорядным и
многорядным, у однорядного эпителия все клетки имеют
одинаковую форму, а их ядра лежат на одном уровне, т.е. в один
ряд. Однослойный эпителий, имеющий клетки различной формы и
высогы, ядра которых лежат на разных уровнях, т.е. в несколько
рядов, носит название многорядного, или псевдомногослойною.
п. Многослойный эпителий бывает ороговеваюшим,
неороговевающим и переходным. Эпителий, в котором
протекают проттессы ороговения, называют многослойным плоским
ороговсвагощим. При отсутствии ороговения эпителий является
многослойным неороговевающим. Переходный эпителий
выстилает мочевыводящис пути, подверженные сильному
растяжению.Онтофидогепетическая классификация (Н.Г. Хлопин)Она включает 5 типов эпителиев: эпидермальный (или
кожный), энтсродермальный (или кишечный),цслонефродермальный, эпендимоглиальный и ангиодсрмальный
типы эпителиев.Эпидермальный тип эпителия образуется из эктодермы
(многослойный плоский ороговевающий эпителий кожи).
Энтсродермальный тип эпителия развиваегся из энтодермы,
является по строению однослойным призматическим (однослойный
каемчатый эпителий тонкой кишки), выполняет железистую
функцию (однослойный эпителий желудка).Целонефродерйальный тип эпителия развивается из мезодермы,
по строегтю однослойный; выполняет барьерную или
экскреторную функцию (мезотелий, кубический и призматический
эпителий в канальцах почек).Эпендимоглиальный тип выстилаег полости мозга. Источником
его образования является нервная трубка.80
ГЛАВА 6 ЭПИТЕЛИАЛЬНЫЕ ТКАНИК ацгиодсрмальному типу эпителия относят эндотелиальиую
выстилку кровеносных СОСУДОВ. имеющую мезенхимное
происхождение.Однослойный эпителий
Однослойный плоский эпителий представлен в организме
мсзотслием и эндотелием,Мезотелий покрь^вас!' серозные оболочки (плевра,
брюпшна, околосердечная сумка). Клетки мезотслия плоские, на
свободной поверхности имеются микроворсинки (рис. 50). Через
мезогслий происходят выделение и всасывание серозной жидкости.
Благодаря его гладкой поверхности легко осуществлясася
скольжение внутренних органов. Мезотелий препятствует
образованию спаек между органами брю1иной или грудной
полостей.W ¡?^ y# ШЖШРис. 50. Мезотелий сальника. 1 - ядра эпителиальных клеток,2 - клеточные границы.Эндотелий выстилает кровеносные и лимфатические
сосуды, а таюке камеры сердца (рис. 51). Эндотелий участвует в
обмене веществ между кровью и другими тканями. При его
повреждении возможно образование тромбов.81
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ IРис. 51. Однослойный плоский эпителий (эндотелий), выстилающий
кровеносные сосуды.Однослойный кубический эпителий выстилает часті»
почечных канальцев (рис. 52). Эпителий почечных канальцев
выполняет функцию обратного всасывания (реабсорбии) ряда
веществ из первичной мочи в кровь. Рис. 52. ОднослойньЕЙ кубический эпителий, выстилающий почечныеканальцы.82
ГЛАВА 6 ЭПИТЕЛИАЛЬНЫЕ ТКАНИОднослойный призматический эпителий выстилает внутреннюю
поверхность желудка, тонкой и толстой кишки, желчного пузыря,
ряда протоков печени и поджелудочной железы. Эпителиальные
клетки связаны между собой с помощью десмосом, щелевых
coeдинcílий, плотных замыкающих соединений.в желудке все эпителиальные клетки являются
железистыми, продуциругЬщими слизь, которая защищает стенку
желудка от грубого влияния комков пищи и переваривающего
действия желудочного сока. За счет гкамбиальных клеток каждые 5
сут. происходит полное обновление эпителия желудка.Рис. 53. Однослойный призматический железистый эпителий слизистойоболочки желудка.В тонкой кишке эпителий однослойный призматический
каемчагый. Он покрывает поверхность кишечных ворсинок и
состоит из каемчатых эпителиоцитов, среди которых
располагаются железистые бокаловидные югетки (рис. 54). Каемка
эпителиоцитов образована микроворсинками, покрытыми
гликокаликсом. В нем и микроворсинках находятся ферменты,
которые осуществляют мембранное пищеварение - расщепление
пи1цевых веществ и всасывание их в кровеносные и лимфатические
капилляры соединительной ткани.Благодаря камбиальным (бескаемчатым) клеткам
каемчатые эпителиоциты ворсинок полностью обновляются в
течение 5-6 сут. Бокаловидные клетки выделяют слизь на
поверхность эпителия. Эндокринные клетки нескольких видов,83
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iтакже входящие в состав эпителиальной выстилки кишечника,
секрстируют в кровь гормоны, которые осуществляют местную
регуляцию функции органов пищеварительного аппарата.Рис. 54. Однослойный [1ризматический каемчатый эпителий слизистой
оболочки тонкой кишки.Однослойные многорядные эпителии
Многорядные (псевдомногослойныс) .эпителии выстилают
воздухоносные пути - носовую полость, трахею, бронхи, а также
ряд других органов (рис. 55). В воздухоносных путях многорядный
эпителий является реснитчатым, и содержит клетки, различные но
форме и функции. Базальные клетки лежат на базальной
мембране, являются камбиальными клетками, которые делятся и
дифференцируются в реснитчатые и бокаловидные клегки.
Реснитчатые клетки высокие, призматической формы. Их
апикальная поверхность покрыта ресничками. В воздухоносных
путях они очишают вдыхаемый воздух от частиц пыли, выталкивая
их в направлении носоглотки, Бокаловидные клетки секретируют
на поверхность эпителия слизь. Вставочные клетки. Все эти виды
клеток имеют разную форму и размеры, поэтому их ядра
располагаются на разных уровнях эпителиаль1Юго пласта. Нередко
всгречаются также эндокринные клетки.84
ГЛЛВЛ 6 ЭПИТЕЛИАЛЬНЫЕ ТКАНИ..І'ітшшРис. 55. Однослойный миогорядный (псевдомногослойный) эпителий
слизистой оболочки трахеи. Стрелками указаны мерцательные реснички ибокаловидная клетка.Однослойный двурядный призматический эпителий
встрсчаегся в:протоке придатка яичка, ссмявыносящем протоке, концевых
отделах предстательной железы и семенных пузырьков.Многослойные эпителии
В таких эпител ИЯХ лишь часть клегок располагается на
базальной мембране. Клетки, входящие в состав остальных слоев
утрачивают с ней связь. Форма клеток в различных слоях
неодинакова, форму всего эпителиального пласта оценивают по
форме клеток поверхностного слоя.Миогислойный плоский неоро! овевающий эпителийпокрывает снаружи роговицу глаза, выстилает полость рта и
пищевода (рис. 56). В нем различают три слоя: базальный,
шиповатый (промежуточный) и плоский (поверхностный).
Базальный слой состоит из эпителиоцитов призматической формы.
Среди них имеются стволовые клегки, способные к митотическому
делению. Шиповатый слой состоит из клеток неправильной
многоугольной формы, в базальном и птиповатом слоях в
эпиташоцитах хорошо развиты тонофибриллы (пучки
тонофиламентов из белка кератина), а между эпителиоцитами -85
гистология, 1 (ИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ {десмосомы и другие виды контактов. Верхние слои эпителия
образованы плоскими клетками. Заканчивая свой жизненный цикл,
последние отмирают и слущиваются с поверхности эпителия.;:Г ::;аШЛй*ШвЯа.'Иг-Рис. 56. Многослойный плоский неороговевающий эпителий.1 - базальный слой, 2 - шиповатый, 3 - поверхностный.Многослойный плоский ороговевашщий эпителийпокрывает поверхность кожи, образуя ее эпидермис, в котором
происходит процесс ороговения, или кератинизации, связанный с
дифференцировкой эпителиальных клсгок - кератииоцитов в
роговые чешуйки наружного слоя эпидермиса. В эпидермисе
различают несколько слоев клеток - базальный, шиповатый
(остистый), зернистый, блестящий и роговой (рис.57),Кератиноциты - основные клетки эпидермиса по мере
дифференцир(^ки перемещаются из базального слоя в
вышележащие слои. Базальный слой эпидермиса состоит из
призматических кератииоцитов, в цитоплазме которых
синтезируется кератиновый белок, формирующий тонофиламенты.
Здесь же находятся стволовые клетки дифферона кератииоцитов.
Поэтому базальный слой называют ростковым, или
герминативным.86
ГЛАВА6 ЭПИТЕЛИАЛЬНЫЕ ТКАІІИКроме кератииоцитов, в эпидермисе находятся меланоциты
(пигментные клетки), виутриэпидермальные макрофаги (клетки
Лаигерганеа), лимфоциты и некоторые другие.Меланоциты препятствуюют воздействиюультрафиолетовых лучей на ядра базальных кератииоцитов. Клетки
Лангерганса являются разновидностью макрофагов, участвуют в
защитных иммунных peг^кцияx и регулируют размножение
кератииоцитов, образуя вместе с ними «пролиферативные
единицы».Роговой слой эпидермиса состоит из плоских роговых
чешуек, заполненных ксратиновыми фибриллами. Между
чешуйками находится цементирующее вещество. Наружные
роговые чешуйки утрачивают связь друг с друїом и отпадают с
поверхности эпителия. На смену им приходят новые из
нижележащих слоев. Благодаря этим процессам в эпидермисе
полностью обновляегся состав кератииоцитов через каждые 3^
недели.5421Рис. 57. Мно1'ослойный плоский ороговсвающий эпителий (эпидермис).
1 - базальный, 2 - шиповатый, 3 - зернистый, 4 - блестящий, 5 - роговой
слои эпидермиса кожи пальцев человека.87
гистология, цитология, ЭМБРИОЛОГИЯ, ЧАСТЬ I.Переходный эпителий
Этот вид эпителия типичен для МОЧеОТВОДЯТПИХ органов “
лоханок почек, мочеточников, мочевого пузьфя, стенки которых
подвержены значительному растяжению при заполнении точой. В
нем различают базальный, промежуточный и поверхностный слои
клеток (рис. 58).Базальный слой образован мелкими камбиальными
клетками. В промежуточном слое располагаются клетки
полигональной формы. Поверхностный слой состоит из очень
крупных, нередко дву- и трехъядерных клеток, имеющих
куполообразную или уплощенную форму в зависимости от
состояния стенки органа. При растяжении стснки вследствие
заполнения органа мочой эпителий становится более тонким и его
поверхностные клетки уплощаются. Во время сокращения стенки
органа толщина эпителиального пласта возрастает.||Ь,■irЧт-:А.-.. М, ■Рис. 58. Переходный эпителий слизистой оболочки мочевого пузыря,1 - базальный, 2 - промежуточный, 3 - поверхностный слои клеток.* Железистый эпителийЖелезистый эпителий состоит из железистых клеток -
гландулоцитов. Они осуществляют синтез и выделение
специфических продуктов - секретов на поверхность: кожи,
слизистых оболочек и в полости ряда внутренних органов
(экзокринная секреция) или же в кровь и лимфу (эндокринная
секреция).88
ГЛАВА 6 ЭПИТЕЛИ АЛЫ 1ЫЕ ТКАН ИГлапдулоииты содержат секреторные включения в
цитоплазме, развитую ЭПС и аппарат Гольджи. Гландулоциты
лежат на базальной мембране. Форма их весьма меняется в
зависимости от фазы секреции, В цитоплазме гландулоцитов,
которые вырабатывают секреты белкового характера хорошо
развита гранулярная эндоплазматическая сеть, В клетках,
синтезирующих небелковЬе секреты (липиды, стероиды),
выражена агранулярная эндоплазматическая сеть. На базальной
поверхности клеток цитолсмма образует небольшое число складки,
апикальная поверхность клеток покрыта микроворсинками.Синтез секрета в гландулоците и его выделение протекают
практически непрерывно. Механизм выделения секрета в
различных железах неодинаковый, различают три типа секреции:• мерокриновый (или ЭККрИНОБЫЙ),• апокриновый• голокриновый.При мсрокриновом типе секреции железистые клетки
полностью сохраняют’ свою структуру (клетки слюнных желез).При апокриновом тиие секреции происходит частичное
разрушение железистых клеток (клетки молочных желез), вместе с
секреторными продуктами отделяются апикальная часть
цитоплазмы железистых 1слеток.Г олокри новый тип секреции сопровождается похшым
разрушением железистых клеток (клегки сальных желез кожи) (рис.
59),:.:'Л СркпанияРазрушениеИ 'О ц оШ^ ММ ШЗ /о\0о Апикальная \частьклетки1Рис. 59. Типы секреции гландулоцитов. 1 - голокриновый,2 - мерокриновый, 3 - апокриновый.89
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I.ЖелезыЖелезы ВЫПОЛНЯЮТ секреторную функцию, вырабатывают и
выделяют секреты, обеспечивающие различные функции
организма.Эндокринные железы - продуцируют гормоны,
циркулирующие в крови в низких концентрагтиях и обладающие
высокой биологической активностью. Выводные протоки в таких
железах отсутствуют (гипофиз, эпифиз, гипоталамус, щитовидная
железа, надпочечники)Экзокринныс железы вырабатывают секрегы, которые
выделяются на поверхность тела (потовые железы) или в полости
организма (железы желудка, экзокринная часть поджелудочной
железы, слюнные железы) (рис. 60).ВыводнойпротокСекреторныйотделРис. 60. План строения экзокринной железыПо форме концевых отделов выделяют железы;
трубчатые,
альвеолярные,90
ГЛАВА 6 ЭПИТЕЛИАЛЬНЫЕ ТКАНИтруочато-альвсолярные.По ветвлению контгевых отделов:
разветвлённые,
неразветвленные.По ветвлению выводных протоков:простые (с неразветвлённым протоком),
сложные (с развС^гвлснным протоком),Регенерация тканейПокровный эпителий, занимая пограничное положение,
постоянно испытывает влияние внешней среды, поэтому
эпителиальные клетки сравнительно быстро изнашиваются и
погибают. Источником их восстановления являются стволовые
клетки эпителия.Процесс, обеспечивающий обновление тканей в ходе
нормальной жизнедеятельности (физиологическая регенерация)
или восстановление после поврежде1шя (репаративиая
регенерация) называется регенерация.Различают: внутриклеточный, клеточный, тканевой и
органный уровни регенерации.91
ГЛАВА 7.
КРОВЬ И ЛИМФАВсе элементы системы крови имеют общие сіруктурно-
функциональные особенности, они происходят из мезенхимы,
подчиняются общим законам нейрогуморальной регуляции,
объединены тесным взаимодействием всех звеньев. Постоянный
состав периферической крови поддерживается сбалансированными
процессами новообразования и разрушения клеток крови.Кровь и лимфа вместе с соединительной тканью образуют
внутреннюю среду организма, в них происходит постоянный обмен
форменными элементами, а также веществами, находящимися в
плазме. Как в крови, так и в лимфе различают плазму и взвешенные
в ней форменные элементы. Все клетіоі крови развиваются из
общей полипотентной стволовой клетки крови (СКК) в
эмбриогенезе и после рождения.Кровь (sangшsi Ігаіта)Масса крови в организме человека составляет 4-9% от
массы тела, объем - 5-5,5 л.Плазма составляет 55-60% объема крови, форменные
элементы - 40-45%. Отношение объема форменных элементов ко
всему объему крови называется гематокритным показателем - и
составляет в норме 0,40 - 0,45.Основные функции крови❖ дьіхатаїьная функция (перенос кислорода из легких во всс
органы и углекислоты из органов в легкие);❖ трофическая функция (доставка органам питательных веществ);❖ защитная функция (обеспечение гуморального и клеточного
иммуніїгета, свертывание крови при травмах);❖ вьщелительная функция (удаление и транспортировка в почки
продуктов обмена веществ);❖ гомеостатическая функция (поддержание постоянства
внутренней среды организма);❖ транспортная (перенос гормонов, биолоіически активных
веществ, лекарственных препаратов).92
ГЛАВА 7. КРОВЬ И ЛИМФАПлата кровиПлазма крови представляет собой жидкое (точнее,
коллоидное) межклеточное вещество. Она содержит 90-93% воды,
около 6,6 - 8,5% белков (альбу'мипы, глобулины, фибриноген) и
1,5-3,5% органических и минеральных соединений.Форменные элементы крови
К форменным эд]1смситам крови относятся: эритроциты,
лейкоциты и тромбоциты!в лабораториях из крови изготавливается препарат,
который назьшасгся мазок. Мазки крови окрашиваются
специальным красителем - азур II ~ эозином по методике
Романовского-Гимза (рис. 61,62).Капля
кровиИзготовление мазкаРаспределение крови
по поверхностиРис. 61. Приготовление мазка крови.93
П1СТОЛОГИЯ, цитология, ЭМБРИОЛОГИЯ. ЧАСТЫч-''ШРис. 62. Мазок крови человека. 1 - эритроциты, 2 - лейкоциты.Ни одна ткань организма не исследуется в диагностических:
целях так часто, как кровь. Результаты анализа записываются в
виде гемограммы, отражающей содержание отдельных форменных
элементов и некоторые биохимические показатели.Большое значение для характеристики состояния организма
имеет подсчег лейкоцитов, процентные соотношения разных видов
лейкоцитов называются дейкощтарной формулой.Гемограмма взрослого человекаЭРИТРОЦИТЫГЕМО¬ГЛОБИН(г/л)ЦВЕТНОЙПОКАЗАТЕЛЬТРОМБО¬ЦИТЫСОЭ(мм/ч)м4,5-’5,0х10'^л130-1600,85-1,05180-320х10^/л2-10ж3 9-
4,7х10*^л120-1400,85-1,05180-320х10’/л2-1594
ГЛАВА 7. КРОВЬ И ЛИМФАЛейкоцитарная формулаЛейко¬цитыБазо-филы(%)Эозиио-филы(%)Нейтрофилы (%)Юн. Пал СсгмЛим-фоц.(%)Моноц.(%)3,9-9,0x10%0-10,5-50,5-51-647-7219-173-11ЭритроцитыЭритроциты являются высокодифференцированными
П0СТЮ1СТ0ЧНЫМИ структурами, несиособными к делению.
Эритроцигы у человека не имеют ядра. Основная функция
эритроцитов - дыхательная - транспортировка кислорода и
углекислоты. Эта функция обеспечивается дыхательным пигментом- гемоглобином. Кроме того, эритроциты участвуют в транспорте
аминокислот, антител, токсинов и ряда лекарственных веществ,
адсорбируя их на поверхности плазмолеммы.В крови человека основную массу составляют эритроциты
двояковогнутой формы - дискоциты (80-90%) (рис. 63), Кроме
того, имеются планоциты (с плоской поверхностью) и стареющие
формы эритроцитов - эхиног^иты, куполообразные, или
стоматоциты, и шаровидные, или сфероциты.Рис. 63. Эритроциты. 1 - дискоцит, 2 - сканирующая электронОфамма
эритроцитов, 3 - электронная микрофотография эритроцитов в
кровеносном сосуде.95
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ 1Одним из проявлений процессов старения эрифоцитов
является их гемолиз, сопровождающийся выхождением
гемоглобина. В этом случае крови обнаруживаются оболочки
эритроцитов - «тени». Молодые формы эритроцитов называются
ретикулоцитами (1 - 5%) (рис. 64). В них сохраняются рибосомы и
ЭПС^ формирующие зернистые и сетчатые структуры. При
обычной гематологической окраске (азур И - эозином) они
проявляют полихроматофилию и окрашиваются в серо-голубой
цвет.г•'V. =;=• •• ■Рис. 64. Ретшсулоциты в мазке крови человека. Окраска азур II- эозин.При заболеваниях могут появляться аномальные формы
эритроцитов, что чаще всего обусловлено изменением структуры
гемоглобина (№). Процесс нарушения формы эритроциюв при
заболеваниях называется пойкилоцитоз.Большинство эритроцитов имеют диаметр около 7,5 мкм и
назьшаются нормоцитами (75%). Микроциты имеют диамсф <7, а
макроциты >8 мкм (по 12,5%). Изменение размеров эритроцитов
назьшасгся акизоцитозом.В плазмолемме эритроцита содержит белок спектрин,
гликофорип и т.н. полосу 5. Наружная поверхность мембраны
эритроцита несет отрицательный заряд.Спектрин - белок цитоскелета эритроцита, связанный с
внутренней стороной плазмолеммы, участвусг в поддержании
двояковогнутой формы эритроцита. На вну1ренней поверхности
плазмолеммы образуется сетевидная структура, которая96
ГЛЛВЛ 7. КРОВЬ И ЛИМФАподдерживает форму эритроцита и прогивостоит давлению при
прохождении его через тонкий капилляр.Соединение спектрипового цитоскелета с плазмолеммой
обеспечивает внугрикл сточный белок аикерин. Анкирии связывает
спектрин с трансмсмбранным белком плазмолеммы (полоса 3).Гликофорш - трансмсмбранный белок, выполняющий
рецепторные функции. Г^икофорииы обнаружены только в
эритроцитах.Полоса 3 - трансмсмбранный гликопротеид, который
у^шствует в обмене кислорода и углекислоты.Олигосахариды гликолипидов и гликопротсидов образуют
гликокаликс. Они определяют антигенный состав эритроцитов.
При связывании этих антигенов соответствующими антителами
происходит склеивание эритроцитов - агглютинация. Антигены
эритроцитов получили название агглютиногены, а
соответствующие им антитела плазмы крови - агглютинины. В
норме в плазме крови нет агглютининов к собственным
эритроцитам. В настоящее время выделяют 4 группы крови по
антигенным свойствам эригроцитов. По системе ABO выявляют
агглюгиногены А и В. Этим антигенам эритроцитов соответствуют
а- и р-агглютинины плазмы крови.Скорость оседания эритроцитов (СОЭ) в 1 час у здорового
человека составляет 2-10 мм у мужчин и 2 - 15 мм у женщин.
Этот показатель может значительно изменяться при заболеваниях и
поэтому служит важным диагностическим признаком. Цитоплазма
эритроцита состоит из воды (60%) и сухого остатка (40%),
содержащего, в основном, гемоглобин.Гемоглобин - это сложный пигмент,, состоящий из 4
полипептидных цепей глобина и гелш (железосодержащего
порфирина), обладающий высокой способностью связывать
кислород (02), углекислоту (С02), угарный газ (СО).Гемоглобин способен связывать кислород в легких, - при
этом в эритроцитах образуется оксигемоглобш. В тканях
выделяемая углекислота (конечный продукт тканевого дыхания)
поступает в эритроциты и соединяясь с гемоглобином образует
карбоксигемоглобин.Утилизация старых или поврежденных эритроцитов
производится макрофагами главным образом в селезенке, а также в
печени и костном мозге, при этом гемоглобин распадается, а
высвобождающееся из гема железо используется для образования97
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I.новых эритроцитов. Эритроциты участвуют в транспорте
аминокислот и полипептидов, регулируют их концентрацию в
плазме крови. Средняя продолжительность жизни эритроцитов
составляет около і20 дней. В организме ежедневно разрешается (и
образуется) около 200 млн. эритроцитов.ЛейкоцитыЛейкоциты в свежей крови бесцветны. Численноть их
составляет в среднем 4 - 9 х10^ в 1 литре крови. Лейкоциты
способны к активным движениям, могут переходить через стенку
сосудов в соединительную ткань органов, где они вьшолняют
основные зашдтные функции. По морфологическим признакам
лейкоциты подразделяют на две группы: зернистые лейкоциты, или
гранулоцити, и незернистые лейкоциты, или аграиулоциты.При стандартной гематологической окраске по
Романовскому - Гимза используются два красителя: кислый эозин
и основной азур-П. Структуры, окрашиваемые эозином (в розовый
цвет) называют эозинофильными, оксифильными или
ацидофильными. Структуры, окрашиваемые красителем азур-П (в
фиолетово-красный цвет) называют базофильными шш
азурофильными (рис. 65).У зернистых лейкоцитов при окраске азур-П - эозином, в
цитоплазме выявляется зернистость. В соответствии с окраской
специфической зернистости различают нейтрофилъные,
эозинофильные и базофильныс гранулоциты.Группа незернистых лейкоцитов (лимфоциты и моноциты)
характеризуется отсутствием специфической зернистости и
несегментированными ядрами.Процентное соотношение основных видов лейкоцитов
называется лейкоцитарной формулой. Обш,ее число лейкоцитов и
их процентное соотношение у человека могут изменяться как в
норме, так и при различных заболеваниях. Исследование
показателей крови является необходимым для установления
диагноза и наз&ачсиия лечения.Все лейкоциты способны к активному перемещению путем
образования псевдоподий, при этом у них изменяются форма тела и
ядра. Они способны проходить между клетками эндотелия сосудов
и клетками эпителия, через базальные мембраны и перемещаться
по основному веществу соединительной ткани. Направление
движения лейкоцитов определяется хемотаксисом. Лейкоциты98
ГЛАВА 7. КРОВЬ И ЛИМФАВЫПОЛНЯЮТ фагоцитоз ми1фобов, инородных веществ, продуктов
распада клеток, участвуют в иммунных реакциях.БазофилМоноцитТромбоцит/м|%'|||Рис. 65. Элек'фонные микрофотофафии и схематическое изображение
форменных элементов крови.99
гистология, цитология, ЭМКРИОЛОГИЯ. ЧАСТЬ IГранулоциты (зернистые лейкоциты)К граиулоцитам относятся нейтрофильные, эозинофильные
и базофильные лейкоциты. Они образуются в красном костном
мозге, содержат специфическую зернистость в цитоплазме и имеют
сегментированные ядра.Нейтрофилъные гралулоциты (нейтрофилы) — самая
многочисленная группа лейкоцитов, составляющая (65-75% от
общего числа лейкоцитов. Среди нейтрофилов крови находятся
клетки различной степени зрелости - юные, папочкоядерные и
сегментоядерные. Юные юхетки составляют 0,5%, они имеют
бобовидное ядро. Палочкоядерные (1-6%о) имеют
нессгментированное ядро в форме буквы 8, изогнутой пшючки или
подковы. Увеличение в крови количества юных и палочкоядерных
форм нейтрофилов {сдвиг лейког{ытарной формулы влево)
свидетельствует о наличии кровопотери иш острого
воспалительного процесса в организме, сопровождаемых
усилением гемопоэза в костном мозге). В зрелом ссхментоядерном
нейтрофиле ядро содержит 3-5 сегментов, соединенных тонкими
перемычками. Для женщин характерно наличие в ряде
нейтрофилов половою хроматина (Х-хромосома) в виде
барабанной паиючки - тельце Барра (рис. 66).:: нейтрофш^ л..... г ■
тромр^итгГ.шщ н ^итрофилРис. 66. Нешрофилы периферической крови. Окраска азур П~ эозин.
1 - тельце Барра, 2 - псевдоподия.100
ГЛАВА 7. КРОВЬ И ЛИМФАВ цитоплазме нейтрофилов можно различить два типа гранул:
специфичес1а1с и азурофильные. Специфические гранулы содержат
лшоцим и щелочную фосфатазу, а также белок лактоферрин,
Азурофильные гранулы являются первичными лизосомами, содержат
лйзосомальные ферменты и миелопсроксидазу. Азурофильиые
гранулы называются первичными в отличие от вторичных -
специфических. лОснов! {ая функция пейфофилов - фагоцитоз
микроорганизмов, поэтому их называют микрофагами. В очаге
воспаления убитые бактерии и погибшие нейтрофилы образуют гной.Среди нейтрофилов здоровых людей фагоцитирующие клетки
составляют 69-99%. Этот показатель называют фагоцитарной
актгтностью. Фагоцитарный индекс -число частиц, поглощенных
одной клеткой. Для нейтрофилов он равен 12-23. Продолжительность
жизни нейтрофилов составляет 5-9 суток.Эозинофильные гранулоциты (эозинофилы). Количество
эозинофилов в крови составляет от 1,0 до 5 % от общего числа
лейкоцитов. Ядро эозинофилов имеег 2 сегмента, соединенных
перемычкой, в цитоплазме расположены орга]Ю1Шы общего
назначения и азурофильиые (первичные) и эозинофильные
(вторичные). Специфические эозинофильные гранулы заполняют
почти 13СЮ цитоплазму. В центре гранулы находится кристаллоид,
кагорый содержит глаи11ый основной белок, богатый аргинином,
лизосомные ферменты, пероксидазу, эозинофильный катионный
белок, а также гистаминазу (рис. 67).Главный оаювной белок обеспечивает антипаразитарную
фуикцию эозинофилов, Гисташтаза - разрушающая гистамин, -
один из основных медиаторов воспаления. Эозинофилы - подвижные
клетки, способные к фагоцитозу. Эозиио4)ильт участвуют в реакциях
на чужеродный белок. Гистамин повышает проницаемость сосудов,
вызывает развитие отека тканей. .Эозинофилы обладают специфической антипаразитарной
функцией. При паразиг'арньгч заболеваниях наблюдается резкое
увеличение числа эозинофилов. Эозинофшгы убивают личинки
паразитов, поступивише в кровь или органы. Они прилипают к
паразитам благодаря наличию на них обволакиваюгцих компонентов
комплемента. При этом происходят дегрануляция эозинофилов и
выделение главного основного белка, оказывающего
антипаразитарное дейегвие. Эозинофилы находятся в периферической
крови 12 час. и потом переходят в ттсани. Уменьшение содержания101
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iэозинофилов при стресс-реакции связано с увеличением содержания
г ормонов надпочечников.Шэозинофил■«■¡.-.ж«1 розипофшііі'эозинофилРис. 67. Эозинофилы. 1, 2 - световая микроскопия. Ол:/?ас/с« ¿т/)II—эозин. 3 - электроппая микроскопия, 4 - вторичная зернистость,
содержащая кристаллоид,Базофшьпые гранулоциты (базофилы). Количество
базофилов в крови составляет до 0,5 % от общего числа
лейкоцитов. Ядра базофилов сегментированы, содержат 2-3
дольки. Характерно наличие специфических крупных
метахроматических гранул (рис, 68). Метахромазия обусловлена
наличием гепарина. Гранулы базофилов содержат протсогликаны,
гликозаминогликаны, вазоактивный гистамин, нейтральные
протеазы. Дегрануляция базофилов происходит в реакциях
гиперчувствительности немедленного типа (при астме,
анафилаксии, сыпи).базофилРис. 68. Базофилг,!. Окраска азур II - эозин.102
ГЛАВА 7. КРОВЬ И ЛИМФАБазофилы образуются в кост^юм мозге. Они так же, как и
нейтрофилы, находятся в периферической крови около 1-2 суток.
Помимо спсцифичесю^х гранул, в базофил ах содержатся и
а,зурофильные гранулы. Базофилы, выделяя гепарин и гистамин,
участвую!’ в регуляции процессов свертывания крови и
проницаемости сосудов. Базофилы у^1аствуют в иммунологических
реакциях организма, в ^ частности в реакциях аллергического
характера.Аграиулоциты (незернистые лейкоциты)К этой группе лейкоцитов относятся лимфоциты и моноциты.
Эти клетки не содержат в цитоплазме специфической зернистости, а
их ядра не сегментированы.Лгшфогщты в крови взрослых людей составляют 19-37 % от
общего числа лейкоцитов. Среди лимфоцитов различают малые
лимфоциты, средние и большие. Большую часть всех лимфоцитов
крови человека составляют малые лимфоциты. Для всех видов
.'шмфоцигов харакгфно наличие интенсивно окрашенного ядра
округлой или бобовидной формы (рис. 69). В цитоплазме лимфоцитов
содержится небольшое количество азурофильных гранул (лизосом).Основная функция лимфохщтов - участие в 11\шунных
реакциях. Среди лимфоцитов различают три основных
функциональных класса: В-лимфониты, Т-лимфоциты и нулевые
лимфоциты.лимфоцитлимфоцитРис. 69. Яимфоцты. Окраска азур II - эозин.В-лгшфоциты впервые были обнаружены в специальном
органе у птиц -фабрициевой сумке, {bursa Fabricius), поэтому и
получили соответствующее название. Ofin образуются в костном
мозге. В-лимфоциты составляют около 30 % циркулирующих103
гистологии, ЦИТОЛОГИЯ, ЭМЬРИОЛОГИЯ. ЧАСТЬ глимфоцитов. При действии антигенов В-лимфоциты способны к
пролиферации и дифференцировке в плазмоциты - клетки, способные
синтезировать и секретировать защитные белки - антитела, или
иммуноглобулины, которые поступают в кровь, обеспечивая
гуморальный иммунитет.Т-лимфогщты образуются из стволовых клеток костного
мозга, а созревают в тимусе, что и обусловило их назва11ис. Они
составляют около 70% циркулирующих лимфоцитов. Т-клетки
имеют специфические рецепторы, способные распознавать и
связывать антигены, участвовать в иммунных реакциях.
Основными функциями Т-лимфоцитов являются обеспечение
реакций клеточного иммунитета и регуляция гуморального
иммунитета). Т-лимфоциты способны к выработке сигнальных
веществ - лимфокинов, которые регулируют деятельность В-
лимфоцитов и других клсгок в иммунных реакциях. Среди Т-
лимфоцитов выявлено несколько функциональных групп: Т-
хелперы, Т-супрессоры, Т-киллеры,Нулевые лимфог^иты не имеют поверхностных маркеров на
плазмолемме, характерных для В- и Т-лимфоцитов. Их
расценивают как резервную популяцию недифференцированных
лимфоцитов.Продолжительность жизни лимфоцитов варьирует от
нескольких недель до нескольких лет, Т-лимфоциты являются
«долгоживугцими» (месяцы и годы) клетками, а В-лимфоциты
относятся к «короткоживущим» (недели и месяцы). Для Т-
лимфоцитов характерно явление рециркуляции, т.е. выход из крови
в ткани и возвращение по лимфатическим путям снова в кровь.
Таким образом, они осуществляют иммунологический надзор за
состоянием всех органов.Моноциты крупнее других лейкоцитов. В крови человека
количество моноцитов от 3 до 11 % от общего числа лейкоцитов.
Ядра моноци’^ов обычно бобовидные, подковообразные, редко -
дольчатые. Цитоплазма моноцитов имеет бледно-голубой цвет. В
цитоплазме содержится различное количество мелких
азурофильных зерен (лизосом), расположенных чаще около ядра
(рис. 70).104
ГЛАВА 7. КРОВЬ И ЛИМФАКомплексГольджимоноцит' моноцит.:i¿4tSíí 'ЩAV»«.Гольджи\Рис. 70. Ыояоши^^. Окраска шур и - эозин.Характерно наличие пальцеобразных выростов цитоплазмы
и образование фагоцитарных вакуолей. В цитоплазме расположено
множество пиноцитозных везикул. Моноциты относятся к
макрофагической системе организма, шш к мононуклеарной
фагоцитарной системе. Клетки этой системы происходят из
промоноцитов костного мозга, способны прикрепляться к
поверхности стекла, осуществлять иммунный фагоцитоз.
Моноциты циркулирующей крови представляют собой подвижный
пул относительно незрелых клеток, находящихся на пуги из
костного мозга в ткани. Время пребывания моноцитов в
периферической крови - от 36-104 часов. Моноциты,
выселяющиеся в ткани, превращаются в макрофаги, при этом у них
появляются большое количество лизосом, фагосом, фаголйзосом.Кровяные пластинкиКровяные пластинки (тромбоциты) в свежей крови
человека имеют вид мелких бесцветных телец округлой или
вереген ОБИДНОЙ формы (рис, 72). Они могут объединяться
(агглютинировать) в мелкие или большие группы. Количество их
колеблется от 200 до 400 х 10^ в 1 литре крови. Кровяные
пластинки представляют собой безъядерные фрагменты
цитоплазмы, отде^швшиеся от мегакариоцитов - гигантских клеток
костного моз1'а (рис. 71).В тромбоцитах выявляются более светлая периферическая
часть - гиаюмер и более темная, зернистая часть - грануломер.
1’иаломер в молодых пластинках окрашивается в голубой цвет, а в
зрелых - в розовый. Молодые формы тромбоцитов крупнее старых.
Плазмолемма тромбоцитов имеет толстый слой rnHKOKajmKca и105
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ Iсодержит гликопротсины, которые BЫnOJUIЯЮT функцию
поверхностных рецепторов, участвующих в процессах адгезииРис. 71. Мегакариоцит в костном мозге. Окраска азур II- эозин.и агрегации кровяных пластинок (процессах свертывания крови).
Элементы цитоскелета обеспечивают поддержание формы
кровяных пластинок, участвуют в образовании их отростков.
Актиновые филаменты участвуют в сокращении объема
(ретракции) образующихся кровяных тромбов. В гранул ом ере
выявлены органеллы, включения и специальные гранулы.
Органеллы представлены рибосомами, элементами
эндоплазматической сети аппарата Гольджи, митохондриями,
лизосомами, пероксисомами. Имеются включения гликогена и
ферритина в виде мелких гранул.К'.:- ■ .Ш .ШЯЙЬ-[у | ) тромбоцит , .'.І" " траябоцитд/г _ ^ I * .41%■ .і':. 'Рис. 72. Тромдощпы. Окраска азур ІГ'эозин.106
ГЛАВА 7. КРОВЬ И ЛИМФАОсновная функция кровяных нластинок - участае в процессе
свертывания, или коагуляции, крови. В фомбоцитах содержится
около 12 факторов, участвующих в свертывании крови. При
повреждении стснки сосуда пластинки быстро агрегируют, прилипают
к образующимся нитям фибрина, в результате чего формируется
фомб, закрывающий дефект (рис. 73).Рис. 73. Формирование тромба. Сканирующая микрофотография.В популяции тромбоцитов различают; юные, зрелые,
старые, дегенеративные, гигантские.Важной функцией тромбоцитов является их участие в
метаболизме серотонина. Тромбоциты - это практически
единственные элементы крови, в которых из плазмы
накапливаются резервы серотонина. В процессе свертывания крови
из разрушающихся тромбоцитов высвобождается серотонин,
который действует на сосудистую проницаемость и сокращение
гладкомышечных клеток сосудов. Продолжительность жизни
тромбоцитов составляет 9-10 дней. Стареющие тромбоциты
фагоцитируются в селезенке.Возрастные изменения крови
Число эритроцитов в момент рождения и в первые часы
жизни выше, чем у взрослого человека, и достигает 6,0-7,0 х 10^^ в
] литре крови, к 10-14 сут. оно равно тем же цифрам, что и во
взрослом организме. В последующие сроки происходит снижение
числа эритроцитов с минимальными показателями на 3-6-м месяце107
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ I,жизни (фи:^иологическая аиемия). Численность эритроциюв
возвращается к нормальным значениям в период полового
созревания. Для новорожденных характерно наличие анизоиитоза с
преобладанием макроцитов, увеличенное содержание
ретикулоцитов, а также присутствие незначительного числа
ядросодержащих предшественников эритроцитов.Численность лейкоцитов у новорожденных достигает 30 X
10^ в 1 литре крови, в течение 2 недель после рождения число их
падает до 9-15 х 10^ в 1 литре. Количество лейкоцитов достигает к
14-15 годам уровня, который сохраняется у взрослого.Гeмoгpa^fма ребенкаВИДФОРМЕННЫХЭЛЕМЕНТОВ1СУТКИу10-14СУТКИ6МЕСЯЦЕВ14ЛЕТЭРИТРОЦИТЫ/С6-7х10’^л4,5-
5,0x1 о’1Т|1п4,5-5,0х10'^лСоотношение числа нейтрофилов и лимфоцитов у
новорожденных такое же, как и у взрослых. В последуюпше сроки
содержание лимфоцитов возрастает, а нейтрофилов падает, и к
четвертым-пятым суткам количество этих видов лейкоцитов
сравнивается (первый физиологический перекрсст). Дальнейший рост
числа лимфоцитов и снижение нейтрофилов приводят к тому, что на
1-2-м году жизни ребенка лимфоциты составляют 65%, а нейтрофилыВид лейкоцитов
(содержание в Уо)Возраст1 день4 день1 год4 года14 летНейтрофильныегранулоциты65%45%25%45%65%Лимфоциты25%45%65«/;,45%25%108
ГЛАВА 7. КРОВЬ И ЛИМФА- 25%. Новое снижение числа лимфоцитов и гювытпение нейтрофилов
приводят к вьіравниБаїшю обоих показателей у 4-летних детей (второй
физиологический перекрест). Постепенное снижение содержания
лимфошттов и поБЬ]шспис нейтрофилов продолжаются до полового
созревания, когда количество этих видов лейкоцитов достигает нормы
взрослого.)ЛимфаЛимфа представляет собой слегка желтоватую жидкую
ткапь, протекающую в лимфатических капиллярах и сосудах. Она
состоит из лимфоплазмы (plasma lymphae) и форменных элементов.
По химическому составу лимфоплазма близка к плазме крови, но
содержит меньше белков. Лимфоплазма содержит также
нейтральные жиры, простые сахара, соли (NaCl, Na2C03 и др.), а
также различные соединения, в состав которых входят кальций,
магний, железо.Форменные элементы лимфы представлены главным
образом лимфоцитами (98%), моноцитами и другими видами
лейкоцитов. Лимфа фильтруется из тканевой жидкости в
лимфатические капилляры. Из капилляров лимфа перемещается в
периферические лимфатические сосуды, по ним - в лимфатические
узлы, затем в крупные лимфатические сосуды и вливается в
венозную кровь.Состав лимфы постоянно меняется. Различают лимфу
периферическую (т.е. до лимфатических узлов), промежуточную
(после прохождения через лимфатические узлы) и центральную
(лимфу грудного и правого лимфатического протоков).109
ГЛАВА 8.
КРОВЕТВОРЕНИЕГсмопоэзом (haemopoesis) называют развитие крови.
Различают эмбриональный гемопоэз, который происходит в
эмбриональный период и приводит к развитию крови как ткани, и
постэмбриональный 1емопоэз, который представляет собой процесс
физиологической регенерации крови.Развитие эритроцитов называют эритропоэзом, развитие
гранул оци-тов - гранулоцитопоэзом, тромбоцитов
тромбоцитопоэзом, развитие моноцитов - моноцитопоэзом,
развитие лимфоцитов и иммуноцитов - лш^фоцито- и
ильчуноцитопоэзом.Эмбриональный гемопоэз
В развитии крови как ткани в эмбриональный период можно
выделить 3 этапа, c^lfeняющиx друг друга: 1) мезобластичсский,
когда начинается развитие клеток крови во внезародышевых
органах; 2) печеночный, который начинается в печени с 5-6-й
недели развития плода; 3) медуллярный ( костномозговой) -
появление СКК в костном мозге.Кроветворение в стенке желточного мешка, у чаювека
оно начинается в конце 2-й - начале 3-й недели эмбрионального
развития. В мезенхиме стенки желточного мешка появляются
кровяные островки. В них мезенхимные клетки преобразуются в
стволовые клетки крови (СКК). Клетки, ограничиваюище кровяные
островки, образуют эндотелиальную выстилку сосуда. Часть СКК
превращается в мегалобласты. Такой тип кроветворения
называется мегалобластическим. Он характерен для
эмбрионального периода, но может появляться в посшатальном
периоде при некоторых заболеваниях (злокачественное
малокровие).При нормобластическом кроветворении из бластов
образуются вторичные эритробласты; из них образуются
вторичные эритроциты (нормоциты). Развитие эритроцитов в
стенке желточного мешка происходит внутри первичных
кровеносных сосудов, т.е. интраваскулярно (рис. 74).110
ГЛАВА 8.КРОВЕТВОГЕНИЕОдновременно экстраваскулярно дифференцируется небольшое
количество неЙ1рофилов и эозинофилов. Часть СКК разносится
током крови по различным органам зародыша, где происходит их
дальнейшая дифференцировка в клетки крови. После редукции
желточного мешка основным кроветворным органом временно
становится печень.Рис. 74. Эмбриональный гемопоэз. Поперечный срез кровяного островка:
1,3- эндотелий; 2 - интраваокулярные кровяные клетки;4 - полость сосуда, 5 - энтодерма; 6 - формирование первичной кровяной
клетки, 7 - клетки мезенхимы, 8 - делящаяся кровяная клетка.Кроветворение в печени. Печень с 5-й недели становится
центром зкстраваскулярного кроветворения. Одновременно с
развитием эритроцитов в печени образуются зернистые лейкоциты.
В цитоплазме бласта появляется специфическая зернистость, после
чего ядро приобретает неправильную форму. Кроме гранулоцитов,
печени формируются гигантские клетки - мегакариоциты. К111
гистология, 11И ГОЛОГИЯ, ЭМЬРИОЛОГИЯ. ЧАСТЬ I.концу внутриутробного периода кроветворение в печени
прекращается.кроветворение в тимусе. Тимус на 7-8-й неделе, ¡«ачинает
заселяться стволовыми клетками крови, которые
дифференцируются в лимфоциты тимуса. Увеличиваю1цееся число
лимфоцитов тимуса дает начало Т-лимфоиитам, заселяющим Т-
зоны периферических органов иммунопоэза.Кроветворение в селезенке, в селезёнке происходит
экстраваскулярное образование всех видов форменных элементов
крови, т.е. сслсзенка в эмбриональном периоде представляет собой
универсальный кроветворный орган. Образование эритроцитов и
гранулоцитов в селезенке достигает максимума на 5-м месяце
эмбриогенеза. После этого в ней начинает преобладать
лимфоцитопоэз,Кроветворение в лимфатических узлах. На 9-10-й неделе
начинается проникновение в лимфатические узлы СКК, из которых
дифференцируются эритроциты, гранулоциты и мегакариоциты.
Однако формирование этих элементов быстро подавляется
образованием лимфоцитов. Массовое "заселение" лимфатических
узлов предш^твенниками Т- и В-лимфоцитов начинается с 16-й
недели. Дифференцировка Т- и В-лимфоцитов происходит в Т-и В-
зависимых зонах лимфатических узлов.Кроветворение в костном мозге. Заклад1са костного мозга
осуществляется на 2-м месяце эмбрионального развития. На 12-й
неделе основную массу гемопоэтичсских клеток составляют
эритробласты и предшественники гранулоцитов. Из СКК в костном
мозге формируются все форменные э^юменты крови, развитие
которых происходит экстраваскулярно. Часть СКК сохраняется в
костном мозге, они могут расселяться по другим органам и тканям
и являться источником развития клеток крови. Костный мозг
становится центральным органом, осуществляюшцм
универсальнь!й гемопоэз в течение постнатальной жизни.Постэмбриональный гемопоэз
Постэмбриональный гемопоэз представляет процесс
физиологической регенерации крови. Миелопоэз происходит в
миелоидной ткани (1ех1и& туе1о1(1еик), расположенной в эпифизах
трубчатых и полостях многих 1 убчатых костей. Здесь развиваются
форменные элементы крови: эритроциты, гранулоциты, моноциты,
кровяные пластинки, предшественники лимфоцитов.112
ГЛАВА 8. КРОВЕТВОРЕНИЕПредшественники лимфоцитов постепенно мигрируют и заселяют
такие органы, как тимус, селезенка, лимфатические узлы и др.Лимфопоэз происходит в лимфоидной ткани (1ех1и5
1утрЬо{(1еи8) тимуса, селезенки, лимфатических узлов. Она
выполняет функции образования Т- и В-лимфоцитов и
плазмоцитов.В миелоидной лимфоидной тканях ретикулярные,
жировые, тучные и остеогенные клетки вместе с межклеточным
веществом формируют микроокружение для гемопоэтичсских
элементов. Микроокружение оказывает воздействие на
дифференцировку клеток крови.в тимусе имеется сложная строма, представленная
соединительнотканными и ретикулоэпителиальными клетками.
Эпителиальные клетки секретируют тимозины, оказывающие
влияние на дифферснцировку из СКК Т-лимфоцитов. В
лимфатических узлах и селезенке ретикулярные клетки создают
микроокружение, необходимое для пролиферации и
дифференцировки в специальных зонах Т- и В-лимфоцитов и
плазмоцитов.СКК являются плюрипотсптными (полипотентными)
предшественниками всех клеток крови и относятся к
самоподдерживающейся популяции.Каждая СКК в селезенке образует одну колонию и
называется селезеночной колониеобразующей единицей (КОЕ-С).
Подсчет колоний позволяет судить о количестве стволовых клеток.
Установлено, что у мышей на 10^ клеток костного мозга
приходится около 50 стволовых клеток, в сслсзенке - 3,5 клетки,
среди лейкоцитов крови - 1,4 клегки. Ультраструктура СКК близка
к малым темным лимфоцитам.В клеточном составе колоний выявлено две линии их
дифферсгщировки. Одна линия даст начало мультипотентной
клсгкс - родоначальнице гранулоцитарного, эритроцитарного,
моноцитарного и мсгакариоцитарного рядов гемопоэза (КОЕ-
ГЭММ).Вторая линия дает начало мультипотентной клсгкс -
родоначальнице лимфопоэза (КОЕ-Л). Из мультипотентных клеток
дифференцируются олигопотентные (КОЕ-ГМ) и унипотентные
клетки, из которых образуются клетки-прсдшсственники
(прекурсорныс). в лимфопоэтическом ряду выделяют113
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iунипотентньте клетки - предшественницы для В-лимфоцитов и Т-
лимфоцитов.Дифферснцировка полипотентных клеток в унипотентныс
опреде;шется эритропоэтинами (для эритрббластов),
гранулопоэтинами (для миелобластов), лимфопоэтинами (для
лимфобластов), тромбопоэтинами (для мегакариобласгов) и др.Процессы гемопоэза представлены на схеме (рис. 77),
ЭритропоэзНачалом эритроидного ряда является взрывообразующая
(бурстообразующая) единица эритроцитов (БОЕ-Э). При
интенсивном размножении она может быстро образовать крупную
колонию клеток.В дальнейшем из неё образуется унипотентный
предшественник эритроцитов (КОЕ-Э), Под влиянием
эритропоэтина, который образуется в юкстагломсрулярном
аппарате почки (90 %) и печени (10 %), происходит дальнейшее
образование последовательных клеток дифферона эритроцитов.Проэритробласт (18 мкм) имеет большое круглое ядро, 1-2
ядрышка, базофильную цитоплазму.Б^офильный эрытробласт - диаметр до 13-16 мкм.
Цитопла^а клетки обладает выражен! юй базофильностью в связи с
накоплением в ней рибосом, в которых начинается синтез НЬ.Полихроматофильиый эрытробласт (10-12 мкм), В
цитоплазме клетки накапливается НЬ, окрашиваемый оксифильно.Нормобласт (8-10 мкм), имеющий маленькое пикнотичное
ядро. В цитоплазме содержится много НЬ, обеспечивающего
окрашивание эозином в ярко-розовый цвет. Пикнотическое ядро
выталкивается из клетки, клсгка утрачивает способность к
делению.Pemuкyлolium - безъядерная клетка с небольшим
содержанием рибосом, преобладанием НЬ, определяющим
оксифилию, что дает многоцветную (полихромную) окраску
(поэтому эта клетка получила название "полихроматофильный
эритроцит”) (рис. 75).114
ГЛАВА 8. КРОВЕТВОРЕНИЕПроэритробластЕазофильпый эрйтробластПолихроматофилъный эрйтробластОртохроматофилъный эрйтробластРетикулоцитЭритроцит*Рис. 75. Дифференцировка эритроцитов в красном костном мозге.При выходе в кровь ретикулоцит созревает в эритроцит в
течение 1-2 суток.Эритро11ит (7-8 мкм) имеет форму двояковогнутого диска,
ацидофильиую цитоплазму, насыщенную НЬ. Период образования
эритроцита занимает 7 дней, продолжительность его жизни в крови
-120 дней.Из одной СКК в течещ0 7‘1О д^ней в результате 12Эритропоэз у млекопитающих и человека протекает в
костном мозге в эритробластических островках. В норме из
костного мозга в кровь поступают только эритроциты и
ретикулоциты.115
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ !Гранулоцитопоэз
Источником для гранулоцитопоэза являются также СКК и
мультипо-тентные КОЕ-ГЭММ (рис. 76), одновременно
начинающие дифференцироваться через ряд промежуточных
стадий в трех различных направлениях и образующие гранулоциты
трех видов: нейтрофилы, эозинофилы и базофилы.МиелобластПромиелоцитМиелоцитНейтрофильныйметамиелоцитПалочкоядерный нейтрофилСегмептоядерный нейтрофилРис. 76. Дифференцировка нейтрофильного гранулоцита в костном мозге.Основные ряды для каждой из групп гранулоцитов
слагаются из следующих клеточных форм: СКК ~> КОЕ-ГЭММ —>
КОЕ-ГМ —унипотентньте предгаественники (КОЕ-Б, КОЕ-Эо,
КОЕ-Гн) - миелобласт - промиелоцит - миелоцит - метамиелоцит -
палочкоядерный гранулоцит -ссі ментоядерньш гранулоцит.По мере созревания гранулоцитов клетки уліеньпшются в
размерах, изменяется форма их ядер от округлой до
сегментированной, в цитоплазме накапливается специфическая
зернистость.116
ГЛАВА 8. КРОВЕТВОРЕНИЕЭозинофильные миелоциты (14-16 мкм) по строению ядра
мало отличаются от нейтрофильных миелоцитов. Миелоциты
митотичсски делятся, а ядро приобрегает подковообразную форму.
Постепенно ядро становится двудольчатым, в цитоплазме
увеличивается количество специфических гранул. Клетка
утрачивает способность к делению. Среди зрелых форм различают
палочкоядерные и сегменто^дериые лейкоциты.Базофильные миелоциты) содержат специфические
базофильные зерна неодинаковых размеров, которые проявляют
мстахромазию при окрашивании азуром. По мере созревания
базофильный миелоцит превращается в базофильный
метамиелоцит, а затем в зрелый базофильный лейкоцит.Все миcJЮциты, особенно нейтрофильные, обладают
способностью фагоцитировать, а начиная с метамиелоцита,
приобретают подвижность.Мегакариоцитопоэз. ТромбоцитопоэзКровяные пластинки образуются в костном мозге из
мегакариоцитов - гигантских клеток, которые дифференцируются
из СКК, проходя ряд стадий. Последовательные стадии
дифференцировки можно представить следующим рядом клеток:
СКК —> КОЕ-ГЭММ КОЕ'МГЦ —>■ мегакариобласт —»
промегакариоцит мегакариоцит —> тромбоциты (кровяные
пластинки). Весь период образования пластинок составляет около10 дней.Мегакариобласт (15-25 мкм), имеет ядро с инвагинациями
и ободок базофильной цитоплазмы. Клетка способна к делению
митозом, иногда содержит два ядра.Промегакариоцит (30-40 мкм), содержит полиплоидные
(4п, 8п), несколько пар центриолей. Объем цитоплазмы возрастает,
Б ней начинают накапливаться азурофильные гранулы,Мегакарио1{ыт - дифференцированная форма. Резервные
мегакариоциты (50-70 мкм), имеют большое, дольчатое ядро (16-32
п) Зрелый, активированный мегакариоцит (до 100 мкм). Содержит
крушюе, сильно дольчатое полиплоидное ядро (64 п). В ее
цитоплазме накапливасгся много азурофильных гранул, которые
объединяются в группы. В цитоплазме мегакариоцита наблюдается
скопление линейно расположенных микровезикул, которые
формируют демаркационные мембраны, разделяющие цитоплазму
мегакариоцита на участки диаметром 1-3 мкм, содержащие по 1-3117
гистология, цитология,ЭМБРИОЛОГИЯ. ЧАСТЫ.гранулы (будущие кровяные пластинки. После отделения
пластинок остается клетка - резидуалълый мегакариоцит, который
затем подвергается разрушению.Мопоцитопоэз
Образование моноцитов происходит из стволовых клеток
костного мозга по схеме: СКК - КОЕ-ГЭММ - КОЕ-ГМ -
унипотентный предшественник моноцита (КОЕ-М) - мои облает -
промоноцит -моноцит. Моноциты из крови поступают Б ткани, где
являются источником развития различных видов макрофагов.Лимфоцитопоэз и гшмуноцитопоэз
Лимфоцитопоэз проходит следующие стадии: СКК - КОН-Л- унипотентныс предшественники лимфоцитов - лимфобласг -
пролимфоцит - лимфоцит. Особенностью лимфоцитопоэза является
способность дифференцированных клеток дедифференцироваться в
бластные формы.Процесс дифференцировки Т-лимфоцитов в тимусе
приводит к образованию из унипотентных предшественников Т-
бластов, из которых формируются эффекторныс лимфоциты -
киллеры, хелперы, супрессоры.Дифференцировка унипотентных предшественников В-лимфоцитов
в лимфоидной ткани ведет к образованию плазмобластов, затем
проплазмоцитов, плазмог1итов.Регуляция гемопоэза
Кроветворение регулируется факторами роста,
обеспсчиваюищми пролиферацию и дифференцировку СКК и
последующих стадий их развития, факторами транскрипции,
влияющими на экспрессию генов, определяющих направление
дифференцировки гемопоэтичсских клсгок, а также витаминами,
гормонами, интерлейкинами.118
ГЛАВА 8, КРОВЕТВОРЕНИЕРис. 77. Схема кроветворения119
ГЛАВА 9.
СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ
ТКАНЬСоединительные ткани - это комплекс мезенхимных
производных, состоящий из клеточных дифферонов и
межклеточного вещества, у^шствующих Б поддержании гомеостаза
внутренней среды. Соединительная ткань составляет более 50 %
массы тела человека. Она участвует в формировании стромы
органов, прослоек между другими тканями, дермы кожи, скелета.Соединительные ткани выполняют различные функции:
трофическую, защитную, опорную, пластическую,
морфогенетическую.Классификация соединительных тканей.Соединительные ткани подразделяются па собственно
соединительную ткань (волокнистые соединительные ткани
(рыхлые и плотные) и соединительные ткани со специальными
свойствами) и скелетные ткани. Последние в свою очередь
подразделяются на гиалиновую, эластическую, волокнистую
хрящевые ткани и две разновидности костной ткани
(ретикулофиброзная и пластинчатая), а также цемент и дентин зуба
(табл. 1).Общие принципы организации соединительных тканей.Главными компонентами соединительных тканей являются клетки
и межклеточное вещество (волокна коллагенового, эластического
или ретикулярного типов и основное аморфное вещество^.»ВОЛОКНИСТЫЕ СОЕДИНИТЕЛЬНЫЕ ТКАНИ
Рыхлая волокнистая соединительная тканьВ рыхлой волокнистой соединительной ткани (РВСТ)
превалируют клетки и аморфное вещество над волокнами. РВСТ
встречается во всех органах, так как она сопровождает кровеносные
и лимфатические сосуды и образует строму многих органов (рис.
78).120
ГЛАВА 9.СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ ТКАНЬРис. 78. Рыхлая волокнистая соединительная ткань. 1 - основное
аморфное вещество, 2 - коллагсиовые волокна, 3 - эластические волокна,4 - клетки.У121
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ IКлетки. Основными клетками соединительной ткани
являются фибробласты, макрофаги, тучные, адвентициальные,
плазматические клетки, перициты, жировые клетки, ‘ а также
лейкоциты, мигрирующие из крови; иногда встречаются
пигментные клетки.Фибробласты - клетки, синтезирующие компоненты
межклеточного вещества: белки (коллаген, эластин),
протеогликаны, гликопротсины.Дифферон фибробластов: стволовые клетки, полустволовые
клетки-предшественники, малоспециализированные,дифференцированные фибробласты (зрелые, активно
функционируюпше), фиброциты (дефинитивные формы клеток), а
также миофибробласты и фиброкласты. Главная функция
фибробластов - образование основного вещества и волокон,
заживление ран, развитие рубцовой ткани, образование
соединительнотканной капсулы вокруг инородного тела и др.Мачоспщиализированиые фибробласты (20-25 мкм). В
цитоплазме этих клеток обнаруживается большое количество
свободных рибосом, Эндоплазматическая сеть и митохондрии
развиты слабо. Аппарат Гольджи представлен скоплениями
коротких трубочек и пузырьков.Дифферетщированные зрелые фибробласты (40-50 мкм),
Это активно функционирующие клетки. Биосинтез белков и
протеогликанов, необходимых для формирования основного
вещества и волокон, осуществляется интенсивно. Фибробласты
способны к движению.Фиброциты - дефинитивные формы развития
фибробластов. Эти клетки веретенообразные с крыловидными
отростками. Они содержат небольшое число органелл, вакуолей,
липидов и гликогена. Синтез коллагена и других веществ в
фиброцитах ]>сзко снижен.Мифибробласты - клетки, сходные морфологически с
фибробластами. Фибробласты могут превращаться в
миофибробласты, функционально сходные с гладкими мышечными
клетками. Такие клетки наблюдаются в грануляционной ткани в
условиях раневого процесса и в матке при развитии беременности.122
.ГЛАВА 9.СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ ТКАНЬФиброкласты - принимают участие в "рассасывании”
межклсгонного вещества в период инволюции органов (в матке
после окончания беременности)..уфибробластГк ^ I.'І/Рис. 79. Фибробласты и фиброциты в рыхлой волокнистой
соединитсяг^ной ткани, а - световая микроскопия, б - схема.Макрофаги. Макрофаги имеют одно ядро. Цитоплазма
базофильна, богата лизосомами, фагосомами и пиноцитозными
пузырьками, содержит митохондрии, ГЭПС, аппарат Г ольджи,
включения гликогена, липидов и др.. В цитоплазме макрофагов
выделяют "клеточную периферию”, обеспечивающую макрофагу
способность передвигаться, осуществлять эндо- и экзоцитоз (рис.
80 а, б). На поверхности плазмолеммы имеются рецепторы для
опухолевых клеток и эритроцитов, Т-и В-лимфоцитов, антигенов,
иммуноглобулинов, гормонов. Наличие рецепторов к
иммуноглобулинам обусловливает' их участие в иммунных
реакциях.Функции макрофагов: 1) поглощение и расщепление
чужеродного материала; 2) обезвреживание его при контакте; 3)
передача информации о чужеродном материале
иммунокомпсгентным клеткам. Макрофаги содержат протеазы,
кислые гидролазы, нироген, интерферон, лизоцим и др. Количество
макрофагов возрастает при воспалительных процессах. Макрофаги
вырабатывают факторы, активирующие выработку
иммуноглобулинов В-лимфоцитами, дифференцировку Т- и В-
лимфоцитов; противоопухолевые факторы, а также факторы роста,123
гистология, цитология. ЭMБPИOJ[OГИЯ. ЧАСТЬ I,* л~* /" /<#Рис. 80. Макрофаги соединительной ткани. 1 - световая микроскопия,2 - электронная микрофотография.Макрофаги образуются из СКК, а также из промоноцитов
костного мозга и моноцитов крови.Макрофаги ческая сисіема (системой моноыуклсарных
фагоцитов). К этой системе относятся совокупность всех клеток,
обладающих способностью захватывать инородные частицы,
погибающие клетки и пр. Фагоцитированный материал
подвергается внутри клетки ферментативному расщеплению.
Различают две группы макрофагов - свободные и фиксировапные. К
свободным макрофагам шносятся макрофаги РВСТ (гистиоциты),
макрофаги серозных полостей, макрофаги воспалительных
экссудатов, альвеолярные макрофаги легких. Макрофаі и способны
перемещаться в организме. Группу фиксированных (резидентных)
макрофагов составляют макрофаги костного мозга и костной ткапи
(остеокласты),^ селезенки, лимфатических узлов (дендритные
макрофаги), внутриэпидермальные макрофаги (клетки
Лангерганса), макрофаги ворсин плаценты (клетки Кащенко -
Хофбауэра), ЦНС (микрої лия).И. И. Мечников пришел к мысли о том, что фагоцитоз
является важным защитным механизмом. Он обосіювал
целесообразность объединения макрофагов в систему и предложил
назвать ее макрофагической.124
ГЛАВА 9.СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ ТКАНЬТучные клетки (тканевые базофилы, лаброциты) являются
регуляторами местного гомеостаза соединительной ткани (рис. 81
а, б). Они принимают участие в понижении свертывания крови,
повышении проницаемости гсматотканевого барьера, в процессе
воспаления, имму1югенеза и др.Особенно много тканевых базофилов в стенке органов
желудочно-кишечного тракта, матке, молочной железе, тимусе
(вил очковая железа), миндалинах. В цитоплазме имеются
многочисленные гранулы, большинство из них отличается
мегахромазией, содержит гепарин, хондроитинсерные кислоты,
гиалуроновую кислоту, гистамин, серотонин.Ту^шыс клетки способны выбросу гранул, содержащих
биологически активные вещества, что изменяет местный или
общий гомеостаз. Гистамин вызывает расширение кровеносных
капилляров и повышает их проницаемость, что проявляется в
локальных отеках. Он обладает также выраженным гипотензивным
действием и является важным медиатором воспаления. Гепарин
снижает проницаемость межклегочного вещества и свертываемость
крови, оказывает противовоспалительное влияние.Рис. 81. Тканевые базофилы соединительной ткани. 1 - световая
микроскопия, 2 - электронная микрофотография.Плазматические клетки (плазмоциты) обеспечивают
^фаботку антител - у-глобулинов при гюявлении в организме125
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iантигена. Они образуются в лимфоидных органах из В-
лимфоцитов, встречаются в РВСТ собственного слоя слизистых
оболочек полых ор1'анов, сальнике, соединительной ткани
различных желез (молочных, слюнных и др.), лимфатических
узлах, селезенке, костном мозге и др.Величина плазмоцитов колеблется от 7 до 10 мкм.
Цитоплазма содержит хорошо развитую концентрически
расположенную гранулярную эндоплазматическую сеть, в которой
синтезируются антитела (рис. 82 б). Небольшая светлая зона
цитоплазмы около ядра образует так называемую сферу или дворик
(рис,82 а). Количество плазмоцитов увеличивается при различных
инфекционно-аллергических и воспалительных заболеваниях.Рис. 82. Плазмоциты соединительной ткани, 1 - световая микроскопия,2 - электронная микрофотография,Адипоциты (жировые клетки) обладают способностью
накапливать в больших количествах резервный жир, принимающий
участие в трофике, энергообразовании и метаболизме воды.
Адипоциты располагаются группами, реже поодиночке и, как
правило, около кровеносных сосудов. Форма жировых клеток
шаровидная, ^нутри содержится одна большая капля нейтрального
жира, окруженная тонким цитоплазматическим ободком.Расходование жира, депонированного в адипоцитах,
происходит под действием гормонов (адреналин, инсулин) и
тканевого липолитического фермента (липаза), расщепляющего
триглицериды до глицерина и жирных кислот. Новые жировые
клетки в соединительной ткани взрослого организма могут
развиваться при усиленном питании из адвентициальных клеток,
прилегающих к кровеносным капиллярам. Скопления жировых126
ГЛАВА 9.СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ ТКАНЬклеток, встречающихся во многих органах, называют жировой
тканью.Адвентициальные клетки. Это малоспециализированные
клетки, сопровождающие кровеносные сосуды, в процессе
дифференцировки эти клетки могут, по-видимому, превращаться, в
фибробласты, миофибробласты и адипоциты.Перициты - клетки; окружающие кровеносные капилляры
и входящие в состав их стенки.Пигментоциты (пигментные клетки, меланоциты). Эти
клетки содержат в своей цитоплазме пигмент меланин. Их много в
родимых пятнах, а также в соединительной ткани людей черной и
желтой рас. Меланоциты образуются из нервного гребня, а не из
мезенхимы. Межююточное веществоМежклеточное вещество (матрикс)Матрикс соединительной ткани состоит из коллагеновых и
эластических волокон, а также из основного (аморфного) вещества.
Межклеточное вещество образуется за счёт секреции,
осуществляемой соединительнотканными клетками, а с другой - из
плазмы крови, поступающей в межклеточные пространства. В
течение жизни межклеточное вещество постоянно обновляется -
резорбируется и восстанавливается.Коллагеновые волокна в составе соединительной ткани
определяют их прочность, в рыхлой неоформленной волокнистой
соединительной ткани они располагаются в различных
направлениях в виде волнообразно изогнутых тяжей толщиной 1-3
мкм и более. Длина их различна. Белок коллаген синтезируется на
рибосомах qDaнyляpнoй эндоплазматической сети фибробластов.
Различают 14 типов коллагена, отличающихся молекулярной
организацией, органной и тканевой принадаежностыо.Коллаген / типа естреча^й^ся ' главным ^брагрт в
соединительной ткани цотщ сухожилиях косгпщ ^ раговице
глазау ^склере, стенке артерий и др.; коллаген Л типа »состав гиалиновых и фиброзных хряи^ей, стекловидного'телЩ
роговицы' коллаген Ш типа находитсАг$ дерме кожи плода,
стенках /срупных кровеносных сосудов, вретикулярньрс^врлокнщс
органов кроветворения;^ ГУ, типа - в -базацьны^. мембрщах^
капсуле хрусталика (в отличие от другЕштипоё^ колл^ена &н
содержит гораздо больше ботвых углеводных цепей,127
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iгидрооксилизшёа^.щ^ гидроокси-пролина). V тип коллагена
прифщщеует ^ориЬт, амнионе, эндомиэии, пёримизии, кдже,
во^фуг . ^^¡№юк (фибробластов, эндотелиальных
^фйдкомышечных), синтезирующих коллаген. Коллаген IV и V
%ипа не, образует выраженных фибрилл. Коллагены У1^ХУ1
типа{тученыШёиее полно.Различают 4 уровня организации коллагенового волокна.
Первый уровень организации - молекулярный. Второй -
надмолекулярный. Третий - фибриллярный. Четвертый
волоконный уровень организации.Волокна могут складываться в пучки толщиной до 150 мкм
(рис. 83). Коллагеновые волокна отличаются малой растяжимостью
и большой прочностью на разрыв. Под электронным микроскопом
можно различить поперечнополосатую исчерченность волокон. В
воде толщина сухожилия в результате набухания увеличивается на
50 %. При термической обработке в воде коллагеновые волокна
образуют клейкое вещество.Эластические волокна. Наличие эластических волокон в
соединительной ткани определяет ее эластичность и растяжимость,
В РВСТ они анастомозируют друг с другом. Толщина эластических
волокон 0,2-1 мкм. В составе эластических волокон различают
микрофибргтлярный и аморфный компоненты.Основой эластических волокон является эластин.,
синтезируемый фибробластами и гладкими мышечными клетками
(первый уровень организации). Молекулы эластина соединяются в
цепочки толщиной 3-3,5 им (второй, надмолекулярный, уровень
организации). Эластин овые протофибриллы в сочетании с
гликопротеином (фибриллином) образуют микрофибрнллы (третий,
фибриллярный, уровень организации). Четвертый уровень
организации волоконный. Зрелые эластические волокна содержат
белок эластин в центре, а по периферии - микрофибриллы. В
эластическщ волокнах в отличие от коллагеновых нет структур с
поперечной исчерченностью на их протяжении.кроме зрелых эластических волокон, различают
элауииноеые и окситалановые волокна. В элауниновых волокнах
соотношение микрофибрилл и аморфного компонента примерно
равное, а окситалановые волокна состоят только из микрофибрилл.128
.ГЛЛВЛ 9.С0БСТВЕІИІ0 СОЕДИНИТЕЛЬНАЯ ТКАНЬРис. 83. Коллагеновые и эластические волокна в рыхлой волокнистой
соединительной ткани. 1 - коллагеновые волокна, 2 - эластическиеволокна.Основное аморфное вещество. Клетки и волокна
соединительной ткани заключены в аморфный компонент или
основное всщсство, Эта гелеобразная субстанция окружает
клеточные и волокнистые структуры сосдинитсльной ткани,
нервные и сосудистые элементы, в состав компонентов основного
вещества входят белки плазмы крови, вода, неорганические ионы,
продукты метаболизма, растворимые предшественники коллагена и
эластина, протеогликаны, гликопротеины и комплексы,
образованные ими.Протеогликаны - белково-углеводные соединения,
содержащие 90-95 % углеводов (ПГ). Гликозаминогликаны (Г4Г)- полисахаридные соединения, содержащие обычно гексуроновугд
кислоту 'с''аминосахарами (N^‘aцemuлглuкoзaмuнy 1Ч~ацет1{л-
галактозамин). Молекулы ГАГ легко присоединяют молекулы
воды и ионы N0+ и поэтому определяют гидрофильные свойства
ткани. Гликозаминогликаны участвуют ,ж^фо^^
во.щкнистых _ структур соединительной ткапи и их
механичесш1х сеойстваху"" репаративньос процессйх
сочинительной ткани, в регуляции роста и дифференцировке129
гистология, ТЩТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ ]кчеток, среди этих соединений наиболее распространена
гиалуроновая кислота^ а также хондроитиисульфаты й
коже, роговице), дерматансульфат (в коже, сухожилиях, в
стенке кровеносных сосудов и др,), кератансульфат,
гепаринсульфат /в составе многих ба1ольных мембран).Гепарин - гликоюминогликан, состоящий уу из
глюкуронрщоц кислоты и^&чи-козамина. В организме человека и
живот^ЩгЪт^в^ тучными клетками, являетсяестест(№ёцнь1АШротиво.свертывающим фактором крови.Гликопротеины (ГП) - класс соединений белков с
олигосахаридами (гексозамины, гексозы, фукозы, сиаловые
кислоты), входящими в состав как волокон, так и аморфного
вещества. К ним относятся растворимые ГП, связанные с
%рртеогликанами; ГП кальцинированных тканей; ГП,
связанные с коллагеном. Гликопротеины играют большую роль в
формировании структуры межклеточного вещества
соединительной ткани и определяют его функциональные
особенности (фибронектин, хондронектин, фибриллин,
ламинин и др.).Фибронектин - главный поверхностный гликопротеин
фибробласта. Полагают, что фибронектин обусловливает
липкость, подвижность, рост и специализацию клеток и др^Фибриллин формирует микрофибриллы, усиливает связь
между внеклеточными компонентами.Даминин - компонент базальной мембраны, состоящий
из трех полипептидных цепочек, связанных между собой
дисульфидными соединениями, а также с коллагеном V типа и
поверхностными рецепторами кле^к^Плотные волокнистые соединительные ткани
Плотные волокнистые соединительные ткани (1ех1и8
солпсейуиз collagenosus compactus) характеризуются большим
количество?^» плотно расположенных волокон и незначительным
количеством клеточных элементов и ОАВ между ними. Эта ткань
подразделяется на плотную пеоформлеппую и плотную
оформленную соединительную ткань.Плотная неоформленная соединительная ткань
характеризуется неупорядоченным расположением волокон. ^В плотной оформленной волокнистой соединительной
ткани расположение волокон строго упорядочено. Оформленная130
ГЛЛВЛ 9.С0БСТВЕ[1И0 СОЕДИШПЕЛЬНАЯ ТКАНЬволокнистая соединительная ткань встречается в сухожилиях,
связках и фиброзных мембранах.Сухожилие состоит из толстых, плотно лежащих
параллельных пучков коллагеновых волокон между которыми
располагаются фиброциты, фибробласты и основное аморфное
вещество (рис. 84). Фиброциты сухожильных пучков Нс1зываются
сухижш1ьпыми клеткалЬл. Каждый пy^юк коллагеновых волокон,
отделенный от соседнего слоем фиброцитов, называется пучком
первого порядка. Несколько пучков первого порядка, окруженных
тонкими прослойками рыхлой волокнистой соединительной ткани,
составляют пучки второго порядка. Прослойки РВСТ, разделяющие
пучки второго порядка, называются эидотенонием. Из пучков
второго порядка слагаются пучки третьего порядка, разделенные
перитеиолием. В крупных сухожилиях могут быть и пучки
чегвсртого порядка. В перитенонии и эндотенонии проходят
кровеносные сосуды, питающие сухожилие, нервы и
[{роприоцсптивпыс нервные окончания, посылающие в
центральную нервную систему сигналы о состоянии натяжения
ткани СУХОЖИЛИЙ.■с.-'• Г-,:.яёш Рис. 84. Сухожилие телёнка. 1 - пучок первого порядка,
2 - эндотеноний.131
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iк ПЛОТНОЙ оформленной ВОЛОКНИСТОЙ соединительной
ткани ОТНОСИТСЯ и выйная связка, только пу*1ки ее образованы
эластическими волокнами и нечетко подразделены.Фиброзные мембраны, фасции, апоневрозы, сухожильный
центр диафрагмы, капсула некоторых органов, твердая мозговая
оболочка, склера, надхрящница, надкостница, белочная оболочка
яичника и яичка образованы плотной оформленной соединительной
тканью. Фиброзные мембраны трудно растяжимы, пучки
коллагеновых волокон располагаются в определенном порядке в
несколько слоев друг над другом.Соединительные ткапи со специальньши свойствами
К таким тканям относят ретикулярную, жировую,
пигментную и слизистую.Ретикулярная ткань состоит из отростчатых
ретикулярных, клеток и ретикулярных (аргирофильных) волокон
(рис. 85), образующих трёхмерную сеть. Ретикулярная ткань
образует строму кроветворных органов и микроокружение для
развивающихся в них клеток крови. Ретикулярные волокна
(диаметр 0,5-2 мкм) обнаруживаются при импрегнации солями
серебра, поэтому называются еще аргирофильными.Рис. 85. Ретикулярные волокна в строме мозгового вещесгв4
лимфатического узла (указаны стрелками).132
.ГЛАВА 9.СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ ТКАНЬЖировая тканьЖировая ткань представляет скопления жировых клсгок,
встречающихся во многих органах. Различают - бедую и бурую
жировую ткань. Белая жировая гкаиь широко распространена в
организме человека, а бурая встречается главным образом у
новорожденных дсгсй и у некоторых животных (грызунов и
зимоснящих) в течение всей жизии.Белая жировая ткань у человека располагасгся под кожей в
нижней части брюшной стенки, на ягодицах и бедрах, где она
образует подкожный жировой слой, в сальнике, брыжейке и
ретроперитонсальной области. Жировая ткань делится
прослойками рыхлой волокнистой соединительной ткани на дольки
различных размеров и формы (рис. 86 а). Между ними
располагаются фибробласты, лимфоидные элементы, тканевые
базофилы и тонкие коллагеновые волокна. Кровеносные и
лимфатические капилляры, охватывают своими псглями группы
жировых клеток. При распаде жиров высвобождается большое
количество воды и выдслясгся энергия. Во время голодания
подкожная и oкoJЮПoчeчнaя жировая ткань, жировая ткань
сальника и брыжейки быстро теряют запасы жира. В области
орбиты глаз, в коже ладоней и подошв жировая ткань теряет лишь
небольшое кoJшчccтвo липидов даже во время продолжительного
голодагсия.Бурая жировая гкань встречается у новорожденных детей
и у некогорых животных на шее, около лопаток, за грудиной, вдоль
позвоночника, под кожей и между мышцами.Она состоит из жировых клеток, густо оплетенных
гемокапиллярами. Эти клетки принимают утастие в процессах
теплопродукции. Адипоциты бурой жировой ткани имеют
множество мелких жировых включений в цитоплазме (рис. 86 б).
По сравнению с клетками белой жировой ткани в них значите;пэНО
больше митохондрий. Бурый цвет жировым клеткам придают
железосодержащие пигменты - цитохромы митохондрий.
Окислительная способность бурых жировых клеток примерно в 20
раз выше белых. При понижении температуры окружающей среды
вьщеляется тепловая энергия, обогревающая кровь в кровеносных
капиллярах. При голода!ши бурая жировая ткань изменяется
меньше, чем белая.133
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. Ч.АСТЬ I'и.■"¿Щ. 1'.*■:,V V' --.’..г.г^т" . Кровеносные--••■"А I :Й г '. >’Рис. 86. Строоше белой (а) и бурой (б) жировой ткани. 1 - адипоциты
белой жировой ткани с крупной каплей липидов; 2 - адипоциты б>рой
жировой ткани с множественными мелкими каплями липидов;3 - коллагеновые волокна.Слизистая ткань встречается только у зародыша в
пупочном канатике. Слизистая соединительная ткань пупочного
канатика ("вартонов студень") синтезирует коллаген IV типа,
характерный для базальных мембран. Между клетками этой ткани в
первой половине берсмшносги в большом количестве
обнаруживается гиалуроновая кислота, что обусловливает
желеобразную консистенцию основного вещества.Пигментная соединительная ткань содержит клетки -
пигментоциты и встречается в околоареолярной области молочной
железы, коже мошонки и околоанальной области, склере,
пигментных и родимых пятнах.134
ГЛАВА 10.
СКЕЛЕТНЫЕ ТКАНИСкелетные ткани - это соединительные ткани с выраженной
опорной, механической функцией, обусловленной наличием
плотного межклеточного вещества: хрящевые, костные ткани,
дентин и цемент зуба.Хрящевые ткаииХрящевые ткани выполняют опорную функцию и входят в
состав органов дыхагельной системы, суставов, межпозвоночных
дисков и др. Хрящевые ткани состоят из клеток и большого
количества межклеточного вещества, oтJшчaющeгocя упругостью.
В свежей хрящевой ткани содержится около 70-80 % воды, 10-15 %
органических веществ и 4-7 % солей. От 50 до 70 % сухого
вещества хрящевой ткани составляет коллаген. Собственно
хрящевая ткань не имеет кровеносных сосудов, а питательные
вещества диффундируют из окружаюгцей ее надхрящницы.PaзJШчaют три вида хрящевой ткани: гиаишювую,
эластическую, волокнистую.Развитие хрящевой ткани осущесчвлястся как у эмбриона,
так и в постэмбриональном периоде при регенерации. В процессе
развития образуется хрящевой дифферон: стволовые клетки,
пoлycтвoJювыe, хондробласты, хондроциты.Хондробласты - это молодые клетки, способные к
пролиферации и синтезу межклеточного вещества хряща
(протеогликанов). При участии хондробластов происходит
периферический (аппозиционный) рост хряща. Эти клетки в
процессе развития хряща превращаются в хондроциты,Хондроциты расположены в особых полостях (лакунах) в
межклеточном веществе поодиночке или изогенными группами, в
изогенных группах различают три типа хондроцитов. Хондроциты I
типа преобладают в молодом, развивающемся хряще. Хондроциты
II типа обеспечивают образование и секрецию
гликозаминогликанов и протеогликанов в межклсгочное вещество.135
гастология, цитология, эмбриология, часть 1.Хондроциты III типа сохраняют способность к образованию и
сскрсции белка, но в них снижается синтсз гликозаминогликанов.Эм брионал ьны й хондрогистогепезИсточником развития хрящевых тканей является мезенхима.
В участках тела зародыша, где образуется хрящ, клетки мезенхимы
ТфЯЮТ свои отростки и плотно прилегают друг к другу ~ это
хондрогсииые зачатки. Находящиеся в их сосгаве стволовые к.четки
дифференцируются в хондробласты, из которых затем образуется
хрящевая ткань.Затем первичные хондроциты увеличиваются в размере и
приобретают способность синтезировать гликозамииогликаны,
хондроитиисульфаты и протсогликаны.По периферии хряща формируется надхрящница, состоятцая
из наружного волокнистого и внутреннего камбиального слоев. Во
внутренней зоне клетки интенсивно делятся и дифференцируются в
хондробласты. В процессе секреции продуктов синтеза и
наслаивания на уже имеющийся хрящ по его периферии сами
клетки "замуровываются" в продукты своей деятельности. Так
происходит рост хряща способом наложения, или аппозиционный
рост.Хрящевые клетки, лежащие в центре молодого хряща,
сохраняют способность делиться, оставаясь в одной лакуне. За счет
увеличения количества этих клеток происходи!’ увеличение массы
хряг!!;а изнутри, что назьшается интерстициальным ростом.По мерс роста хряща С10 центральные участки все более
отдаляются от близлежащих сосудов и начинаю г испытывать
затруднения в питании. Вследствие этого хондроциты теряют
способность размножаться, некоторые из них подвергаются
разрушсгшю, а протеогликаны превращаются в более простой
оксифильный белок - альбумоид.Гиалиновая хрящевая ткапьГиалиновая хрящевая ткань встречается в местах
соединения ребер с грудиной, 1'ортани, воздухоносных путях, на
суставных поверхностях костей.Большая часть гиациновой хрящевой ткани покрыта
надхрящницей. В надхрящнице вьщеляют два слоя: наружный,
состоящий из волокнистой соединительной ткани с кровеносными
сосудами; внутрс!!ний, содержащий хо!щробласты и
прехондробласты. Под !1адхрящницсй располагаются ^юлодыс136
ГЛЛВЛ Ю.СКЕЛЕТПЫЕ ТКЛПИхондроциты вфсгспообразлой формы, в глубоких слоях хрящевые
клетки далеко не расходятся и образуют изогснные группы из 2-4
хондроцитов.Межклеточное всщсство (ма'фикс) окружает хрящевые
клетки или их группы (рис. 87). В его составе различают
коллагеновые волокна П типа и протеогликаны. В структурной
организации межклето^1^ного вспгества хряща большую роль играет
хондронектин. Высокая гидрофильность межклеточного вещества
способствусг диффузии 1штательиых веществ и солей.Рис. 87. Гиалиновый хрящ. 1-надхрящница, 2-изогенные группы
клеток, 3 - матрикс.Структурной особенностью гиалинового хряща суставной
поверхности является отсутствие надхрящницы на поверхности,
обращенной в полость сустава. Суставной хрящ состоит из трех
нечетко очерченных зон; поверхностной, промежуточной и
базальной. Питание суставного хряща осутцествляется лишь за счет
синовиальной жидкости полости сустава.Эластическая хрящевая ткань
Эластическая хрящевая ткань встречается в ушной
раковине, рожковидных и кгшновидных хряшдх гортани. В свежем,
нефиксированном препарате эластическая хрящевая ткань имеет
желтоватый цвег и невысокую прозрачность. Снаружи
эластический хрящ покрыт надхряъцницей. Хрящевые клетки137
гистология, цитология, эмбриология, часть 1располагаются иоодиаочкс или образую'г изогенные группы. Одним
из главных отличительных признаков эластического хряща
является наличие в его мслскл сточном веществе наряду с
коллагеновыми волокнами эластических волокон (рис. 88).^ ■ ■ --''ЯРис. 88. Эластический хрящ. 1 - - изогенные группы клеток, 2 - матрикс,
содержащий эластические подокна.Волокнистая хрящевая тканьВолокнистая хрящевая ткань находится в мсжиозво1ючных
дисках, полуподвижных сочленениях, в местах перехода
волокнистой соединительной ткани (сухожилия, связки) в
гиалиновый хрящ. Межклеточное веп1ССтво содержит параллельно
направленные коллагеновые пучки, постепенно переходящие в
гиалиновый хрящ. Хрящевые клетки распола1'аются поодиночке
или образуют небольшие изогенные группы. Цитоплазма клеток
часто бывает вакуолизировашюй. По направлению от гиалинового
хряща к сухожилию волокнистый хрящ становится все более
похожим иа с^^хожилие (рис. 89).Возрастные [1зменепия. По мере старения организма в
хрящевой ткаии уменьшаются концентрация протсогликахюв и
гидрофильность. Ослабляются процессы размножения
хондробластов и молодых хондроцитов.В резорбции дистрофически измененных клеток и
межклеточного вещества участвуют хондрокласты, Часть лакун
после гибехш хондроцитов заполняется аморфным вещ^^ством и138
ГЛАВА Ю.СКЕЛЕТНЫН ТКАНИколлагеновыми фибриллами, Местами в межклеточном ветцестве
обнаруживаются отложения солей кальция, хрящ становится
мутным, непрозрачным, приобретает твердость и ломкость. Может
произойти врастание кровеносных сосудов с последующим
костеобразованием.Рис. 89. Волокнистый хрящ. 1 - и;зогснные группы клеток, 2 - матрикс.Регенерация хрящевой ткани осуществляется за счс'г
ма]юспепиализированных клсгок надхрящницы и хряща путем
размножения и дифференцировки прехондробластов и
хо1?дробластов.Факторы регуляции метаболизма хрящевых тканей.Регуляция метаболизма хрящевой ткани происходит под действием
механической нагрузки, нервных и гормональных факторов (СТГ,
гироксин, инсулин, глюкокортикоиды, эстрогены).Костные тканиКостные ткани содержат около 70 % неорганических
соединений, главным образом фосфатов кальция. В костной ткани
обнаружено более 30 микроэлементов (медь, стронций, цинк,
барий, магний и др.).Матрикс костной ткани представлен бeJíкaми коллагенового
типа и липидами. Костная ткань обладает опорной, механической,
защитной функциями для внутренних органов, а также является
депо солей кальция, фосфора и др. Морфофункциональные139
гистология, ЦИТ0Л0П1Я, ЭМБРИОЛОГИЯ, ЧАСТЬ I.свойства костной ткани ^feняютcя в зависимости от возраста,
мышечной деятельности, условий питания, а также под влиянием
деятельности желез внутренней секреции, иннервации и Д[1.Классификации. Существует два основных типа костной
ткани: ретикулофиброзная (грубоволокнистая) и пластинчатая. К
костной ткапи относятся также дентин и цемент зуба.Костный дифферон и остеогистогепезРазвитие костной ткани у эмбриона осуществляется двумя
способами: 1) из мезенхимы {прямой остеогенез), 2) на месте
хряшевой модели кости {непрямой остеогенез).
Постэмбриональное развитие костной ткани происходит при
физиологической и рспаративной регенерации.В процессе развития костной ткаии образуется костный
дифферон: стволовые, полустволовые клетки, остеобласты,
остеоциты.Остеокласты развиваются из стволовых клеток крови.Остеобласты (15-20 мкм) - это молодые клегки, создающие
костную ткань. Они встречаются в глубоких слоях надкостницы и в
местах регенерации костной ткани после се травмы. Остеобласты
способны к пролиферации, форма клеток - кубическая,
пирамидальная или угловатая. В цитоплазме определяется врлсокая
активность щелочной фосфатазы (рис. 90).Остеоциты (22 до 55 мкм) дефинигив11ые клетки костной
ткани, утратившие способность к делению. Они имеют
отростчатую форму, компактное, крупное ядро и базофильную
цитоплазму, Костные клетки лежат в костных лакунах. Канальцы
костных полостей заполнены тканевой жидкостью и
анастомозируют между собой (рис. 90).Остеокласты (90 мкм) С1Юсобиы разрушать
обызвествленный хрящ и кость, располагаются на поверхности
костных перекладин. Та сторона остеокласта, которая прилежит к
разрушаемой поверхности, (гофрированная каемка), является
областью синтеза и секреции гидролитических ферментов. По
периферии остеокласта находится зона плотного прилегания клетки
к костной поверхности. Остеокласты выделяют СО2 в окружающую
среду, который преобразуется в Н2СО3 и растворяет соли кальция,
В том месте, где остеокласт соприкасается с костным веществом,Л140
IГЛАВА Ю.СКЕЛЕТНЫЕ ТКЛ11Иобразуется лакуна. Один остеокласт может разрушить столько
кости, сколько создают 100 остеобластов за это же время (рис. 90),Рис. 90. Остеобласты, остеоциты и остеокласты.Межклеточное вещество состоит из основного аморфного
вегцества, импрспшрованного неорганическими солями, в котором
располагаются коллагеновые волокна (коллахен I и V типов).
Волокна могут иметь беспорядочное (в регикулофиброзной
костной ткапи) или строго ориентированное (в пластапчатой
костной ткани) направление.Прямой остеогистогенез характерен для развитая
грубоволокнистой костной ткани при образовании плоских костей.
Эгот процесс наблюдается в основном в течение первого месяца
внутриутробного развития. В первой стадии происходит
образование скелетогсшюго островка и его васкуляризация. Во
второй - образуется оксифилыюе межклеточное вещество. В
основном веществе появляются мукопротеиды (оссеомукоид),141
гистология, цитология, ЭМБРИОЛОГИЯ, ЧАСТЬцементирующие волокна в одну прочную массу. Клетки внутри
островка дифференцируются в остеоциты, располагающиеся по
поверхности - в остеобласты. Из окружаюи^ей мезенхимы
образуются новые генерации остеобластов, которые на^)ащиваюг
кость снаружи (аппозиционный рост). Трсгья стадия
кальцифи1сация мсжк.^с! очного вещества. При этом остеобласты
выделяют фермент щелочную фосфатазу. Последняя всгупаст в
реакцию с солями кальция, в дальнейшем образуются кристаллы
гидроксиапатита [Caíо (Р04)б (ОН)2]. Часть волокнистой ткаш!
превращасгся в периост, который обеспечивает трофику и
регенерацию кости, Такая кость, состоящая из перекладин
ретикулофиброзной костной ткани, назьшается первичной губчатой
костью, в болсс поздних стадиях развития она заменяется
вторичной губчатой костью взрослых, которая огличасгся от
первой тем, что построена из пластинчатой костной ткаии
(четвертая стадия остеогснсза).Развитие пластинчатой костной ткани тесно связано с
врастанием кровепоспых сосудов в толщу ретикулофиброзной
кости, в этом процессе принимают у^шстие остеокласты. Костные
пластинки обычно образуются вокруг кровеносных сосудов. Над
такими пластинками образуется слой новых остеобластов и
возникают новые пластинки, Коллагеновые во.'юкна в каждой
пластинке ориентированы под углом к волокнам предыдущей
пластинки. Вокруг сосуда формируются как бы костные цилиндры,
вставленные один в другой (первичные остеоны). С момента
появления остеонов ретикулофиброзная костная ткань заменяется
пластинчатой костной тканью. Перестройка кости продолжается
всю жизнь.Непрямой остсогистогснсз. На 2-м месяце эмбрионального
развития в местах будущих трубчатых костей закладывается из
мезенхимы хрящевой зачаток, который принимасг форму будущей
кости (хрящевая модель). Зачаток состоит из гиалинового хряща,
покрытого надхрящницей.В области диафиза образуется костная манжетка, что
нарушает питание хряща (рис. 91). Вследствие этого в центре
диафизарной части возникают дистрофические изменения.
Хондроциты вакуолизируются, их ядра пикнотизируются,
образуются так называемые пузырчатые хондроциты. Появившиеся
остеокласты очищают пути для врастающих кровеносных сосудов142
ГЛЛВЛ Ю.СКГЛЁТНЫЕ ТКАНИи остеобластов, Хондроциты на границе эпифиза и диафиза
собиршотся в колонки. В колонке хондроцитов наблюдаются два
противоположно направленных процесса - размножение и рост в
дистальных отделах диафиза и дисгрофичсские процессы в его
проксимальном отделе (рис. 92).I м1 2
161514Рис. 91. Непрямой остеогенсз (схема). Образование хрящевой модели
кости и перихондральной костной манжетки, 1 - 10 - стадии остеогенеза;11 - хрящевая ткань; 12 - перихондральная костная манжетка; 13 - колонки
хрящевых клеток; 14 - врастающая в хрящ мезенхима с
дифференцирующимися остеокластами и кровеносными капиллярами;15 - эндохондрально образованная костная ткань; 16 - точка окостенения вэпифизе.143
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ 1.Рис. 92. Эпифизарный хрящ (7). 1 - зона неизменённого хряща, 2 - зона
пролиферащти, 3 - зона гипертрофированного хряща, 4 - зона остеогенеза,5 зона резорбции.Надхряищица перестраивастся и превращается в
надкостницу. Диафизарный хрящ разрушасгся, в нем возникают
пространства, в которых "поселяются" остеоциты, образующие на
поверхности оставшихся участков обызвествленного хряща
костную ткань. *Первичный, или диафизарный, центр окостенения.
Процесс отложения кости внутри хрятцевого зачатка получил
название энхондрального окостенения. Внутри возникает
костномозговая полость.Вокруг сосудов, которые идут по длинной оси зачатка
кости, начинают образовываться концентрические пластинки,
состояпще из параллельно ориентированных тонких коллагеновых144
ГЛЛВЛ Ю.СКЕЛЕТНЫЕ ТКАІІИВОЛОКОН - первичные остеоны. Вслед за диафизом центры
окостенения появляются Б эпифизах. Оссификация сопровождается
врастанием в эпифизы сосудов.В промсжугочной области между диафизом и эпифизами
сохраняется хрящевая ткань - метафизарный хрящ, являющийся
зоной росга костей в длину.Различают 2 вида ^костной ткани: ретикулофиброзная и
пластинчатая.Ретикулофиброзная костная ткань встречается главным
образом у зародышей, У взрослых она встречается на месте
заросших черепных швов, в местах прикрепления сухожилий к
костям. Беспорядочно расположенные коллагеновые волокна
образуют в ней толстые пучки. В основном веществе
ретикулофиброзной костной ткани находятся удлиненные костные
полости, с длинными анастомозирующими канальцами, в которых
лежат костные клетки - остеоциты с их оіростками. С поверхности
грубоволокнистая кость покрыта надкостницей.Пластинчатая костная ткань - наиболее распространенная
разновидность костной ткани во взрослом организме. Она состоит
из костных пластинок. Из этой ткаїш построены компактное и
губчатое вещества в большинстве плоских и трубчатых костей
скелета.Гистологическое строение трубчатой кости как органаТрубчатая кость как орган в основном построена из
пластинчатой костной ткани. Снаружи кость покрыта
надкостницей, за исключением суставных поверхностей эпифизов,
покрытых разновидностью гиалинового хряща.В надкостнице различают два слоя: наружный
(волокнистый) и внутренний (юіеточньїй). Наружный слой
образован в основном волокнистой соединительной тканью.
Внутренний слой содержит осіеогенньїе камбиальные клетки,
преостеобласты и остеобласты различной степени
дифференцировки.Камбиальные клетки имеют веретеновидную форму.
Остеобласты характеризуются сильно развитымбелоксинтезирующим аппаратом. Через надкостницу проходят
питающие косгь сосуды и нервы. Надкостница связывает кость с
окружающими тканями и принимает участие в ее трофике,
развитии, росте и регенерации.145
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЪ IКомпактное вегцество, образующее диафнз кости, состоит
из костных пластинок, толщина которых колеблется от 4 до 12-15
мкм. Костные пластинки образуют сложные структуры (гаверсовы
системы). В диафизе различают три слоя: наружный слОй общих
пластинок, средний, образованный остсонами, и внутренний слой
общих пластинок (рис. 93).Наружные
■ снсральные
иластмиыНаружнымслойнадкостницыВнутренний слои
нядкостикныКонцснгричсскис
костные пластины
остеонаСосудынадкостницыКомпактнаяК0С1ЬРис. 93. Строение фубчатой кости (схема).146
ГЛАВА 1 «.СКЕЛЕТНЫЕ ТКАНИВ наружных общих пластинках залегают прободающие
(фолькмановы) каналы, по которым из надкостницы внутрь кости
входят сосуды. Со стороны надкостницы в косгь под разными
углами проникают коллагеновые пpoбoдaюп^иe (шарпеевы)
волокна.В среднем слое наблюдаются вставочные пластинки,
лежащие между остеонами. Остеоны (гаверсовы системы) являются
структурными единицами компактного вещества трубчатой кости.
Они представляют собой цилиндрические образования, состоящие
из костных пластинок, как бы вставленных друг в друга. В костных
пластинках и между ними располагаются тела остеопитов и их
офостки, замурованные в костном межклеточном веществе (рис,
94). Каждый остеон отграничен от соседних остеонов спайной
линией, образованной основным веществом. В центральном канале
остеона проходят кровеносные сосуды с остеогенными клетками.Рис. 94. Сфоение пластинчатой костной ткани. 1 - остеон; 2 - канал
остеона; 3 - кровеносный сосуд внутри канала остеона; 4 -остеоциты,
5 вставочные пластинки, 6 - концентрические костные пластинки.147
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ I,Каналы остеонов анастомозируют друг с другом, в местах
анастомозов прилежащие к ним пластинки изменяют свое
направление. Такие каналы называют прободающими, или
питательными. На впутреиней поверхности диафиза, граничащей с
костномозговой полостью, пластинчатая костная ткань образует
костные перекладины губчатого вещества кости. Полость диафиза
трубчатых костей заполнена костным мозгом (красным и желтым).Эндост - оболочка, покрывающая кость со стороны
костномозговой полости (рис. 95). В областях активного
формирования кости толщина эндоста возрастает в 10-20 раз за
счет повышения синтетической активности остеобластов и их
предшествешшков. в эндостс стареющей кости уменьшается
популяция остеобластов и клеток-предшествспников, но возрастает
активность остеокластов, что ведет к истончению компактного слоя
и перестройке губчатого вещества кости.Васкуляризация костной ткани. Кровеносные сосуды
образуют во внутреннем слое надкостницы густую сеть. Отсюда
берут начало тонкие артериальные веточки, которые, помимо
кровоснабжения остеонов, проникают в костный мозг через
питательные отверстия и принимают участие в образовании
питающей его сети капилляров. Лимфатические сосуды
располагаются главным образом в наружном слое надкостницы.Рис. 95. Расположение периоста (1) и эндоста (2) в диафизе бедренЕюйкости.148
ГЛЛВЛ Ю.СКБЛЕТНЫЕ ТКАНИИннервация костной ткани« В надкостнице миелиновые и
бсзмислиновыс нервные волокна образуют сплетение. Часть
волокон сопровождает кровеносные сосуды и проникает с ними
через питательные отверстия в одноименные каналы, а затем в
каналы остеонов и далее достигает костного мозга. Другая часть
волокон заканчивается в надкостнице свободными нервными
разветвлениями, а также участвусг в образовании
инкапсулированных телец.Рост костей начинается у человека с ранних
эмбриональных стадий и кончается в среднем к 20-летнему
возрасту. В течение всего периода роста кость увеличивается как в
длину, так и в ширину. Рост трубчатой кости в длину
обеспечивается наличием метаэпифизарной хрящевой пластинки
роста, в которой проявляются два противоположных
гисто1'снетических процесса.Рос г 1рубчатой кости в ширину осуществляется за счет
лсриоста.Физиологическая регенерация костных тканей происходит за счет
остеогенных клсгок надкостницы, эндоста и остеогенных клеток в
канале остеона.149
ГЛАВА 11.
МЫШЕЧНЫЕ ТКАНИМышечные ткани - это ткани, различные по строению и
происхождению, но сходные по способности к сокращению. Они
обеспечивают перемещения организма в пространстве, его частей и
движение органов внуїри организма.Классификации мышечных тканей
В соответствии с морфофункциональным принципом
мышечные ткани подразделяют на две поді руппьі.Первая - поперечнополосатые (исчерчсішьіе) мышечные
ткани. В цитоплазме их злсмсіітов миозиновые филаменты
образуют с актиновыми нитями миофибриллы. Последние
организованы в специальные комгшексы - сарком еры, в соседних
миофибриллах структурные субъединицы саркомеров расположены
на одинаковом уровне и создают поперечную исчерчсниость.Вторая - гладкие (неисчсрченные) мышечные ткаїш.
Миофибриллы в этом виде ткани не имеют поперечной
исчерчсиности.в соотвегсгвии с источником развития мышечные ткани
подразделяются на: мезенхимные, эпидермачьиые, нейральнї>іе,
целомические и соматические. Первые три типа относятся к подгруппе
гладких мышечных тканей, четвертый и пятый - к по,тгрупгю
попсречіюполосатьіх.Гладкие мышечные ткани
Гладкая мышечная ткань мезенхимного происхождения
находится в стенках кровеносных сосудов (рис. 96) и многих
трубчатых ^іутренних органов, а также образует отдельные мелкие
мышцы (цилиарные).Структурной единицей мышечной ткани мезенхимного
происхождения является гладкий миоцит - веретеновидная клсгка
длиной 20-500 мкм, шириной 5-8 мкм. Ядро палочковидное,
находится в центре. Когда миоцит сокращается, его ядро изгибается
и даже закру^швается. Органеллы общего значения сосредоточены
около полюсов ядра.150
ГЛАВА И.МЫШЕЧНЫЕ ТКАНИ’р Рис. 96. Гладкие миоциты в составе стенки кровеносных сосудов.1 - в состоянии расслабления, 2 - в состоянии констрикции.Актиновые филаменты образуют в цитоплазме трехмерную
сеть. Концы филаментов скреплены между собой и с плазмолеммой
специальными сшивающилш белками. Сигнал к сокращению
поступает по нервным волокнам, Медиатор, который выделяется из
их тсрмипалсй, изменяет состояние плазмолеммы. Она образует
впячивания - кавсолы, в которых концентрируются ионы кальция.
Кавеолы отшнуровываются в сторону цитоплазмы в виде
пузырьков, здесь освобождается кальций.' Актиновые филаменты смещаются друг другу навстречу,
плотные пятна сближаются, усилие псрсдасгся на плазмолемму, и
вся клсгка укорачивается (рис. 97). Когда поступление сигналов со
стороны нервной системы прекращается, ионы кальция
эвакуируются из кавеол, ’ миозин дсполимеризуется и
"миофибриллы" распадаются. Сокращение прекращается. Таким
образом, актино-миозиновые комплексы существуют в гладких
миоцитах только в период сокращения.Гладкие миоциты разделены базальной мембраной. На
отдельных участках в ней образуются "окна", поэтому
плазмолеммы соседних миоцитов сближаются. Здесь формируются
нексусы и между клетками возникают не только механические, но и
метаболические связи. Поверх базальной мембраны между151
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I.миодитами находятся эластические и ретикулярные волокна,
объединяющие клетки Б единый тканевой комплекс.IРис. 97. Гладкие миоциты в расслабленном состоянии (1) и в период
сокращения (2). 3 - ядро, 4 - цитопла:5ма, 5 - нити актина,6 - снгавающие белки.Физиологическая регенерация гладкой мышечной ткаии
осуществляется не столько на тканевом, сколько на клеточном
уровне: миоциты растут, в цитоплазме активизируются
синтетические процессы, количество миофиламентов
увеличивается.Мышечная ткань эпидермального происхождения
формируется из эпидермального зачатка. Такая ткань встречается в
потовых, молочных, слюнных и слезных железах.
Миоэпители^льные клетки непосредственно прилежат к
эпителиальным и имеют общую с ними базальную мембрану. Эти
клетки имеют отростчатую форму: их отростки охватывают
концевые отделы и мелкие протоки желез (рис. 98).152
ГЛАВА И.МЫШЕЧНЫЕ ТКАНИРис. 98. Миоэш1телиоциты, окружаюш;ие секреторные отделы
околоушной слюнной железы.Миоциты мышечной ткани нейрального происхождения
развиваются из клеток нейральног о зачатка в составе внутренней
С1С11КИ глазного бокала. Каждая из них имеет отросток, который
направляется в толщу радужки и ложится параллельно сс
поверхности. В отростке находится сократительный аппарат,
организованный так же, как и во всех гладких миоцитах. Подобные
миоциты образуют две мышцы - суживающую и расширяющую
зрачок.Поперечнополосатые мышечные ткани
Скелетная мышечная ткань. Источником развития
элементов скелетной (соматической) поперечнополосатой
мышечной ткапи являются клетки МИОТОМОВ, в ходе
дифференцировки возникают две клеточные линии. Клетки одной
из линий образуют симпласты. Миофибри;шы заполняют большую
часть миотубы, ядра смещаются к периферии. Клеточные центры и
микротрубочки полностью исчезают. Такие дефинитивные
структуры называют миосимпластами. Клетки другой линии153
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ [,дифференцируются в миосателлитоциты, они располагаются на
поверхности миосимпластов.Структурной единицей скелетной мышечной ткани является
мышечное волокно, состоящее из ми оси МП л а ста и
миосатсллитоцитов, покрытых общей базальной мембраной (рис.
99). Длина всего волокна достигает нескольких сантиметров при
толщине 50-100 мкм. Плазмолемма миосимпласта и базальная
мембрана называются сарколеммой. Цитоплазма называется
саркоплазмой.Миосимпласт имеет несколько десятков тысяч ядер,
расположенных под сарколеммой. У полюсов ядер располагаются
органеллы общего значения. Миофибриллы заполняют основную
часть миосимпласта и расположены продольно.Рис. 99.^Поперечнополосатое скелетное мышечное волокно.1 - миосимпласт, 2 - миофибрилла, 3 - ядра миосимпласта,4 - ядра миосатсллитоцитов,5 - соединительная ткань между мыпгечными волокнами.Структурная единица миофибриллы называется с ар ком ер.
Каждая миофибрилла имеег поперечные темные и светлые диски,
имеющие неодинаковое лу^іепреломлсние (анизотропные А-диски и
изотропные 1-диски). Каждая миофибрилла окружена петлями154
ГЛАВА 11 .МЫШЕЧНЫЕ ТКАНИагранулярной эндоплазматической сети - саркоплазматическои
СС1И. Границей саркомера служит 2-линия (рис. 100, 101). Она
построена в виде сети из белковых фибриллярных молекул, среди
которых существенную роль играет (1-актинин. С этой сетью
связаны концы актиновых филаментов. От соседних 7-линий
актиновые филамигш наиравляются к центру саркомера, но не
доходят до его середины, филаменты актина объсди}1ены с Ъ-
линией и нитями миозина молекулами нсбулина. Посередине
темного диска саркомсра располагается сеть^ построенная из
миомсзина. Она образует М-линию. Другие их концы
направляются в сторону 2-линий и располагаются между
филаментами актина, но до самих 7-линий тоже не доходят. Вместе
с тем эти концы фиксированы по отношению к 2-линиям
растяжимыми гигaнтcки^ш белковыми молекулами титина.саркомерIIПиск ИПиск АПиск ИМиозинАктинРис. 100. Схема строения саркомера. Z - линия, образованная
телофрагмой, Н - полоса.155
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I^11 V.Саркоплаша-
тический
ретикулумТ- трубочкаТриадаРис. 101. Электронная микроскопия скелетной мышечной ткани.Молекулы миозина имеют длинный хвост и две головки,
при повышении концентрации ионов кальция в области
присоединения головок молекула изменяет свою конфигурацию.
При этом головки миозина связываются с актином с помощью
тропомиозина и тропонина. Затем головка миозина наклоняется и
тянет за собой актиновую молекулу в сторону М-линга. 2-линии
сближаются, саркомер укорачивается.Саркомеры всех миофибрилл располагаются на одном
уровне. Это и создает при наблюдении в микроскоп впечатление
поперечной исчерченности всего волокна.Для обеспечения сокращения необходимы ионы кальция,
источником его поступления служат цистерны АЭПС. Они
вытянуты вдоль миофибрилл и образуют саркоилазматическую156
ГЛЛВЛ 11 МЫШЕЧНЫЕ ТКАНИсеть. На границе А- и 1-дисков канальцы сети образуют
герминальные (Ь) цистерны (рис. 102).Саркоплазма- |
тический ретикулумДиск А
Саркоплазма- \ |
тический ретикулум0^"Т~ трубочкиТ- трубочки
ТпиадаСаркоплазма-
тический ретикулумСаркоплаша-
т--. тический ретикулумУД Триада/гйТ- трувочк» 4''411Э-- 1'етикул.р,шеРис. 102. Трёхмерное изображение ультраструктурной организации
скелетного мышечного волокна.157
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ [с поверхности миосимпласта плазмолемма образует трубочки,
идущие поперечно в глубину клетки (Т-трубочки) между темными и
светлыми дисками. Сигнал о начале сокращения распространяется на
мембрану Т-трубочек, кальций освобождае1х;я из цистерн ссги и
взаимодействует с актино-миозиновыми комплексами. Когда
потенциал действия исчезает, кальций снова аккумулируется и
сокращение миофибрил-т прекрапщегся. Источником энергии (АТФ)
служат образом митохондрии, расположенные между
миофибриллами.Гликогет! служит источником энергии, необходимой для
поддержания теплового баланса всего организма. Миоглобин
связывает кислород, во время сокращения мышцы и сосуды
сдавливаются, а запасенный кислород освобождается и участвусг в
биохимических реакциях.Миосателлитоциты ябляются источником регенерации
мьш1ечной ткани. Они прилежат к поверхности миосимпласта, так что
их плазмолеммы соприкасаются.Типы мышечных волокон. Различают белые, красные и
промежуточные волокна. По функциональным особсниосгям
мышечные волокна подразделяют на быстрые, медлент1ые и
промежуточные.Свойства мыги<0чЫгх волокон меняются при ишенении
нагрузок - спортивных, проф'ессионачьных, а также в экстрефлы{ых
условйт (невесомоапь). При возврате к обычной деятелыюсШ 7пакие
изменения обратимы. При некоторых заболеваниях (мышечные
атрофии, дистрофии, последствия денерващи) мышечные волокна й
разными исходными свойстваушъизменяются. неодинаково. Это
позволяет уточнять диагноз, 'для чего исследуют биоптаты
скелетйШмъшщРегенерация скелетной мышечной ткани. Камбиальными
элементами являются миосателлитоциты. После повреждения
мьииечного волокна па некотором протяжении от места травмы оно
разрушается и его фрагменты фагоцитируются макрофагами.
Восстановлени^г тканей осуществляется за счсг компенсаторной
гипертрофии самого симпласта и пролиферации миосатсллитоцитов.
Поврежденный конец миосимпласта yгoJш^aeтcя, образуя мышечную
почку, Миосателлитоциты, сохранившиеся рядом с повреждением,
делятся. Од1ш из них мигрируют к мышечной почке и встраиваются в
нее, другие сливаются (так же, как миобласты при гисгогенезе) и
образуют миотубы, которые затем входят в состав вновь
образованных мышечных волокон.158
ГЛЛВЛ 11 .МЫШЕЧНЫЕ ТКАНИСкелешая мышца как органПередача усилий сокращения на скелет осуществляется
посредством сухожилий или прикрепления мышц непосредственно к
надкостнице. Между мышечными волокнами находятся тонкие
прослойки рыхлой волокнистой соединительной ткани - эндомизий.
Более толстые прослойки рыхлой соединительной ткани окружают по
нескольку мышечных волокон^ образуя неримизий и разделяя мышцу
11а пучки, Несколько пучков объединяются в более к-рупные группы,
разделенные болсс толстыми соединительнотканными прослойками -
эпими'^ий (рис. 103).Сарколе.чмаКатилярыПери.мишйМиофибриллыРис. 103. Трёхмерная реконструкция мь11!1цы как органа.159
П1СТ0Л0ГИЯ, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ 1Васкуляризация. Артерии вступают в мьттцу и
распространяются по прослойкам соединительной ткани,
постепенно истончаясь. В перимизии расположены артериолы, в
эндомизип - капилляры. Рядом с сосудами много тканевых
базофилов, принимающих участие в регуляции проницаемости
сосудистой стенки.Иннервация. В мышцах выявлены миелипизировапньте
эфферентные (двигательные), афферентные (чувствительные), а
также немиелинизированные вегетативные нервные волокна. При
поступлении нервного импульса из терминалсй вьщеляется
ацетилхолин - медиатор, который вызывает возбуждение,
распространяющееся отсюда по плазмолемме миосимпласта.Каждое мышечное волокно иннервируется самостоятельно
и окружено сетью гемокапилляров, образуя комплекс, именуемый
мионом.группа мышечных волокон, иннервируемых одним
мотонейроном, называется нервно-мышечной единицей.Чувствительные нервные окончания располагаются не на
рабочих (экстрафузальных) мышечных волокнах, а связа11ы со
специализированными мышечными во;юкнами в мышечных
веретенах (с интрафузальными мышечными волокнами), которые
расположены в перимизии.Интрафузальные мышечные волокна. Существует два
вида - волокна с ядерной сумкой и волокна с ядерной цепочкой
(рис. 104). Ядра расположены в толще симпласта, а не у его
поверхности. В волокнах с ядерной сумкой ядра симпласта
образуют скопления в его средней части. В волокнах с ядерной
цепочкой ядра лежат продольно одно за другим. Миофибриллы
находятся в концах симпластов. Каждое мышечное волокно
веретена спирально обвито терминалью чувствительного нервного
волокна. В результате сокращения или расслабления мышечных
волокон изъясняется натяжение капсулы веретена. Вследствие этого
возбуждаются чувствительные нервные окончания, обвивающие
их, и возникают афферентные нервные импульсы. На каждом
миосимпласте располагается также своя моторная бляшка.160
ГЛАВА И.МЫ[[1НЧНЫКТКАНИРис. 104. Схема строения нервно-мышечного веретена.1 - иитра-фугзальпьге мы[иечт1ые подокна, 2 - экстрафу^альные
мышечные волокна, 3 - поперечнополосатая мышечная ткань, 4 - волокно
с ядерной сумкой, 5 - волокно с ядерной цепочкой,6 - чувствительное нервное волокно.Сердечная мышечная ткань
Гистогенез. Источники развития сердечной
поперечнополосатой мышечной ткани - участки висцерального
листка спланхнотома в шейной части зародыша
миоэпикардиальные пластинки. Из них дифференцируются также
клетки мезотелия эпикарда, В ходе гистогенеза возгшкают161
г истология, цитология, ЭМБРИОЛОП1Я, ЧАСТЬ Iследующие виды кардиомиоцигов: рабочие (сократительные),
синусные (псйсмекерные), переходные, проводящие, секреторные.Сократительные кардиомиоциты образуют цепочки.
Рабочие кардиомиоциты способны передавать сигиалы друг друх у.
Синусные (псйсмекерные) кардиомиоциты способны сменять
состояние сокращения на состояние расслабления, они
воспринимают управляющие сигналы от нервных волокон, в ответ
на что изменяют ритм сократительной деятельности. Синусные
кардиомиоциты передают управляющие сигналы переходным
кардиомиоцитам, а последние - проводящим. Проводящие
кардиомиоциты образуют цепочки клеток и передают имиульс
рабочим. Секреторные кардиомиоциты вырабатывают
натрийуретический фактор (гормон), у^шствующий в процессах
регуляции мочеобразования и в некоторых других процессах. Все
кардиомиоциты покрыты базальной мембраной,Сократительные кардиомиоциты имеют удлиненную (100-
150 мкм) форму, близкую к цилиндрической. Их концы
соединяются друг с другом с помощью вставочных дисков, так что
цепочки клеток составляют функциональные волокна (рис. 105).
Кардиомиоциты могут ветвиться и образуют пространственную
сеть. Их поверхности покрыты базальной мембраной, в которую
снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро
кардиомиоцита (иногда их два) овальное и лежит в центральной
части клетки. У полюсов ядра сосредоточены органеллы общего
значения, за исключением АЭПС. Миофибриллы кардиомиоцитов
сходны со строением миофибрилл миосимпласта скелетного
мышечного вoJЮKнa. Каждая митохондрия располагается на
протяжении всего саркомера. От поверхности плазмолеммы в глубь
кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-
линии. Их мембраны контактируют с мембранами АЭПС. Петли
последней вытянуты вдоль поверхности миофибрилл и имеют
латеральные утолщения (L-системы), формирующие вместе с Т-
трубочками тp^йlды или диады. В цитоплазме имеются включения
гликогена и липидов, особенно много включений миоглобин а.
Механизм сокращения кардиомиоцитов такой же, как у
миосимпласта.162
ГЛАВА 11.МЫШЕЧНЫЕ ТКАНИ•is* •'•=;!!¡,i---. -i:... i ■.,ДТ - ‘"'У'>..:ж^^~1^ _ ЛШШвк'''^- "Ф Л‘ *..ai.Рис. 105, Световая микроскопия сердечной мышцы. 1 - вставочные
диски, 2 ~ ядра кардиомиоцитов, 3 - соединительная ткань.Кардиомиоциты соединяются друг с другом вставочными
дисками. Соприкасающиеся noeqpxHOCTH кардиомиоцитов
неровные, поэтому соседние Ю1СТКИ соединены друг с другом
интердигитациями и десмосомами. К каждой десмосоме со стороны
цитоплазмы подходит миофибрилла, закрепляющаяся концом в
дссмоплакиновом комплексе. При сокращении тяга одного
кардиомиоцита передается другому. Боковые поверхности
выступов кардиомиоцитов объединяются нексусами. Это создает
между ними метаболические связи и обеспечивает синхронность
сокращений.При длительной усиленной работе происходит рабочая
гипертрофия кардиомиоцитов. Стволовых клеток ИЛ1И клсток-
предшсетвснииков в сердечной мышечной ткани нет, поэтому
погибающие кардиомиоциты (в частности, при инфаркте миокарда)
не восстанавливаются.163
ГЛАВА 12.
НЕРВНАЯ ТКАНЬНервная ткань обеспечивает восприятие раздражений,
возбуждения, выработки импульса, его передачу, интеграцию
тканей и органов и связь с окружающей средой.Нервные клетки (нейроны, нсйроциты) - основные
структурные компоненты нервной ткани.Нейроглия обеспечивает существование ифункционирование нервных клеток, осуществляя опорную,
1рофическую, разграничительную, секреторную и защитную
функции.Нервная ткань развивас'гся из дорсальной эктодермы. На 18
сутки у эмбриона дифференцируется нервная пластинка, края
которой приподнимаются, образуя нервные валики, а затем
нервный желобок. Передний конец нервной пластинки образует
головной мозг. Латеральные края сливаются но средней линии в
нервную трубку. Полость нервной трубки образует желудочки
X оловного мозга и центральный каххал спинного мозх а. Часть клеток
нервной пластинки образует скопления хю бокам от нервххой трубки
“ нервный гребень (ганглиозная пластинка) (рис. 106). Из нервххой
трубхш в дальнейхнем формируются нейроны и макроглия
центральной нервной системы. Нервный гребень дасг начало
нейронам чувствительных (сенсорххых) и автономных ганглиев,
клеткам мягкой мозговой и паутинной оболочек мозга и некоторым
видам глии: ххейролеммоцитам (шванновским клеткам), клехкам-
сателлитам ганглиев, хслеткам мозгового вещества ххадпочечников,
меланоцитам кожи, части клеток АРUD-системы, сенсорным
юхеткам кароти^ххых телец и др.Из плакод, представляющих собой утолщения эктодермы по
бокам нервной трубки в краниальххом отделе зародыша, образуются
ганглии V, VII, IX и X череххххых нервов. В нервной трубхсе
дифференцируется вентрикулярная (энсххдимная), промежуточная
(плащевая, мантийная) и краевая (маргинальная) зоны.
Цилиндрические клетки вентрикулярной зоны являются164
ГЛАВА 12.НЕРВ1ІЛЯ ТКАНЬПредшественниками нейронов и клсгок макроглии. Промежуточная
зона состоит из -исйробластов и глиобластов.Из клеток плаш;евого слоя образуются серое вещество
спинного и часть серого вещества головного мозга. Маргинальная
зона дает начало белому всшсству. В некоторых областях
головного мозіа клетки плащевого слоя образуют кортикальные
пластинки - скопления ісле'гок, из которых формируется кора
большого мозга и мозжечка.Рис. 106. Формирование нервной tpyбки зародыша. А - стадия нервной
Ш1астинки; Б - замыкание нервной трубки; В - обособление нервной
трубки и ганглиозной пластинки от эктодермы: 1 - нервный желобок;2 - нервные валики; 3 - кожная эктодерма; 4 - хорда; 5 - мезодерма;6 - ганглиозная пластинка; 7 - нервная трубка; 8 - мезенхима;9 - энтодерма.НейроныНейроны (нейроциты) - клетки нервной системы. Нейрон
является морфологически и функционально самостоятельной165
гистология, ЦИТОЛОГИЯ, ЭМБРИ0Л01ИЯ. ЧАСТЫ.единицси, но с помощью своих отростков осуществляет контакт с
другими нейронами, образуя рефлекторные дуги. В зависимости от
функции в рефлекторной дуге различают рецепторные
(чувствительные, афференгные), ассоциативные и эфферентные
(эффекторныс) нейроны. Афферентные нейроны воспринимают
импульс, эфферентные передают его на ткани рабочих органов,
побуждая их к действию, а ассоциативные осуществляют связь
между нейронами. Нейроны отличаются разнообразием форм и
размеров. Диаметр тел клеток-зерен коры мозжечка 4-6 мкм, а
гигантских пирамидных нейронов коры большого мозга - 130-150
мкм. Нейроны состоят из тела (перикариона) и отростков: аксона и
различного числа ветвящихся дендритов.По количеству отростков различают униполярные нейроны,
имеющие только аксон, биполярные, имеющие аксон и один
дендрит, и мультиполярные, имеющие аксон и много дендритов
(рис. 107). Псевдоуниполярные - от тела отходит один общий
отросток, разделяющийся затем на дендрит и аксон.
Псевдоуниполярные нейроны присутствуют в спинальных
ганглиях, биполярные - в органах чувств. Большинство нейронов
мультиполярные.телоБиполяоиый нейоонатеолгиналдендрит I аксон теш\коллатерали——^.Псевдоуыиполятшйтело _ 0^;
нейрона
дендрит аксонМхгльтиполярпый%д^дритыÁ irРис. 107. Типы нервнЕ>1х клеток.
166
ГЛАВА 12.НЕРВНЛЛ ТКАПЬВ псйрояс различают тело - перикариои, аксон, передающий
импульс, и дентриты, воспринимающие информацию. За счет
дендритов рецепторная поверхность нейрона увеличивается в 1000
и более раз. Аксон - огросток, по которому импульс передастся от
тела клетки. Он содержит митохохщрии, нейротубулы и
нсйрофиламенты, а также АЭПС. Нейроны в большинстве
содержаг 0Д1Ю ядро (рис. Ш8).дендритыол игодерндроцитдендритысинапсыперикариончое веществоаксональпыи холмик
миелиновая оболочкаколлатераль аксонаЦентральная
нервная системаШванновские клеткимиелиновая оболочка
моторная бляшкаРис. 108. Нейрон. Схема строения.
167
гистология, цитология, ЭМБРИОЛОГИЯ, ЧАСТЬ 1,в ядре имеется 1, а иногда 2-3 крупных ядрышка. Усиление
функциональной активности нейронов сопровождается
увеличением количества ядрышек.Плазмолемма нейрона обладает способностью генерировать
и проводить импульс.При окрашивании нервной ткани анилиновыми
красителями в цитоплазме нейронов выявляется в виде
базофильных глыбок и зерен различных размеров и форм
хроматоф ильная субстанция (вещество Ниссля) (рис. 109).
Базофильные глыбки локализуются в перикарионах и дендритах
нейронов, но никогда не обнаруживаются в аксонах и аксональных
холмиках. Базофилия глыбок объясняется высоким содержанием
рибонуклеопротеидов. Электронная микроскопия показала, что
каждая глыбка хроматофильной субстанции состоит из цистерн
гранулярной эндоплазматической сети, свободных рибосом и
полисом.Ргапх (1860-1919)ІІК;-- і-»«- .ґ-Рис. 109. Структурные компоненты нейрона. 1 - дендриты, 2 - ядрышко,
З - тельца Ниссля, 4 - аксональный холмик, 5 - аксон.»Аппарат Гольджи в нейронах хорошо развит. При световой
микроскопии он выявляется в виде различных нитей и зернышек.
Его ультраструктура обычна.Митохондрии обеспечивают энергией транспорт ионов и
синтсз белков. Нейроны нуждаются в постоянном притоке глюкозы
и кислорода с кровью, и прекращение кровоснабжения і оловного
мозга вызывает потерю сознания. Лизосомы участвуют в168
ГЛАВА 12.НЕРВИ АЛ ТКАНЬферментативном расщеплении компонентов клетки рецепторов и
мембран.С возрастом в нейронах накопливается липофусцин -
"пигмент старения" (рис. 110).аксоиРис. 110. Ультраструктурная организация іїервной клетки коры головногомозга' Из элементов цитоскелета в цитоплазме нейронов
присутствуют нейрофиламенты диаметром 12 нм и нейротубулы
диаметром 24-27 нм. Пучки нсйрофиламентов на препаратах видны
в виде нейрофибрилл (рис. 111). Нейрофибриллы образуют сеть в
теле нейрона, а в отростках расположены параллельно.
Нейротубулы и нейрофиламенты участвуют в поддержании формы
клеток, росте oipocTKOB и аксональном транспорте.Аксональный транспорт - это перемещение веществ от тела
в агростки и от отростков в тело нейрона. Транспорт веществ от
тела клетки в отростки называется антероградным, к телу -
ретроградным. Аксональный транспорт представлен двумя1'лавными компонентами: быстрым компонентом (400-2000 мм в169
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ 1день) и медленным (1-2 мм в день). Обе транспортные системы
присутствуют как в аксонах, так и в дендритах.і?*,
■fXV-Рис. 111. Нейрофибриллы в нервных клетках спинного мозга.Однако ретроградный транспорт имеет отрицательное
с^дстео, С ним пейротропиые &щусь^^ такие кат вирус
бешенства, доставляются в центральную нервную систему.
Дефект нейротубул может быть причиной ij№
неврологических нарушений у человеШ!Существуют отдельные нейроциты, специализироваццыс на
синтезе и секреции биологически активных веществ, - секреторные
нейроны, например, клетки нсйросскреториых ядер
гипоталамической области головного мозга. В цитоплазме
нейронов находятся гранулы нейросекрета, содержащие белок,
которые выводятся в кровь или мозговую жидкость.НсйроглияНейрогл1|я выполняет опорную, 'фофичсскую,
разграничительную, поддержание постоянства среды вокруг
нейронов, защитную и секреторную функции. Различают глию
центральной и периферической нервной системы.Глия ценгральной нервной системы. Клетки глии
цстральиой нервной системы делятся на макроглию и микроглию.
Макроглия развивается из глиобластов нервной трубки. К170
ГЛАВА 12.ИЕРВІ1ЛЯ ТКАНЬмакроглии относятся эпеидимоциты, астроциты иОЛИГОДеНДрОі'ЛИОЦИТЬІ.Эпендимоциты выстилают желудочки головііоіо мозга и
центральный канал спиилого мозга (рис. !!2). Эти клетки
цилиндрической формы. Большинство эпсидимоцитов имеют
подвижиыс реснички, вызывающие ток цереброспинальной
жидкости. Базальная поверхность большинства эпендимоцитов
имеет длинный ОфОСТОК, идуттіий глубоко в нервную ткань.
Эпендимный эпителий сосудистых сплегений желудочков
продуцирует цереброспиналы1ую жидкость.Спишіо^озгі
. кипал '■ШШ-ресничкаСпинномозговойканал«г¡МтШЙШШШШэпендимоцитыРис. 112. Эпендимоциты спинномозгового канала.Астроциты - клетки отростчатой формы. Они выполняют
опорн>тю и разграничительную функции (рис. 113). Различают
протоплазматическис астроциты, локализующиеся в сером
всщсстве ЦНС, и волокнистые астроциты, присутствующие в бсіюм
веп^естве.Протоплазматическис астроциты имеют короткие сильно
ветвящиеся отростки. Волокнистые астроциты имеют 20-40
длинных, слабо ветвящихся отростков, в которых много фибрилл.
Отростки ас'гроцитов тянутся к базальным мембранам капилляров,
к телам и дендритам нейронов. Подходя к капиллярам, их отростки
образуют расширенные "ножки", полностью окружающие сосуд.
Астроциты накапливают и передают вещества от капилляров к171
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГ ИЯ. ЧАСТЬ ]нейронам, захватывают избыток экстрацеллюлярного калия и
других всщсств, таких как нейромедиаторы, из экстрацеллюлярного
пространства.Волокнисты й астра ц ит Астроцатфпі
ножкаПротоплататический астроцитЛстроцатарнаячожка/ШШ\ V'.Аксон^^УИ^риневропальпая
ножкаПротоплазматический астроцитРис. 113. Прстоплазматические и волокнистые астроциты в ЦНС.* 1 - кровеносный капилляр.Олигодендроциты имеют немногочисленные отростки, эти
глиоциты присутствуют 1сак в ссром, гак и в белом всщсствс. В
сером веществе они локализуются вблизи перикарионов. В белом
веществе их ОТрОСТКЗ! образуют мислиновый слой Б мислиновых
нервных волокнах.172
ГЛАВА 12.11ЕРВПАЯ ТКАНЬ14 икрогли ЯФагоцигирующие клетки микроглии относятся к системе
мононуклеарных фагоцитов и происходят из стволовой
кроветворной Ю1сгк11, Ес функция - защита от инфекции ы
новрслсдсния и удаление продуктов разрушения нервной ткани
(рис. !14).Реактивная лшкро^лыя появляется после травмы в любой
области мозга. В цитоплазме юде-гок реактивной микроглии
присутствуют плотные тельца, липидные включения, лизосомы.' \Г’' .Рис. П4. Микроглиопит.Глия периферической нервной системы происходит из
нервного гребня, к периферической нейроглии относятся
нейролеммоциты (шванновские клетки) и глиоциты ганглиев
(мантийные глиоциты).Нейролеммоциты формируют оболочки отростков нервных
клеток в нервных волокнах периферической нервной системы.
Глиоциты ганглиев (окружают тcJJa нсйр01юв в нервных узлах и
участвуют в обмене веществ нейронов.Рост опухолей (неоплазмы), происходящих из нервной, ткагш,
щчяется 'Ж сожалению нередким событием. Опухоли могут быть
классифицированы на две категории: мягкие (неинвазивные) и
^(олигнитрующиеся Рост всех опухолей отраокает173
гистология, ЦИТОЛОГИЯ. ЭМБРИОЛОГИЯ. ЧАСТЬ I.ндрущение в регуляции клеточного деления.. Существует нескоящо
факторов определяющих, станет ли опухоль инвазивной:• если рпухоль растет больше определенного размера.• если клетки опухоли продуцируют протеазы, г^дторше
разрушают базальную мембрану.Когда . протеазы опухолевъ^>^ клеток разрушают бтальяую
мембрану, клетки опухоли свободно рассеиваются в отдалённые
области организма, где они дают начало поврщ отдалённым
очагам (метастазам)^Нервные волокнаОтростки нервных клеток, покрытые оболочками,
называются нервными волокнами. По строению оболочек
различают миелиновые и безмиелиновые нервные волокна.
Отросток нервной клетки в нервном волокне называют осевым
цилиндром, или аксоном.В центральной нервной системе оболочки отростков нейронов
образуют отростки олигодендроглиоцитов, а в периферической -
цейролсммоциты,Безмиелиновые нервные волокна находятсяпреимущественно в составе вегетативной нервной системы. В
нервных волокнах имеется не один, а несколько (10-20) осевых
цилиндров, принадлежащих различным нейронам. Такие волокна,
содержащие несколько осевых цилиндров, называются волокнами
кабельного тина. По мере погружения осевых цилиндров в тяж
нейролемоцитов оболочки последних прогибаются и плотно
охватывают осевые цилиндры. Сближенные в области складки
участки оболочки нейролеммоцита образу 1от сдвоенную мембрану- мезаксон (рис. 115).Миелиновые нервные волокна встречаются как в
центральной, так и в периферической нервной систсмс. Они также
состоят из усевого цилиндра, "одетого" оболочкой из
нейролеммоцитов (шванновских клеток). Различают два слоя
оболочки: внутренний, более толстый, - миелиновыи слой (рис. 116
и наружный, тонкий, состоящий из цитоплазмы, ядер
нейролеммоцитов и нейролсммы, в миелиновом слое периодически
встречаются насечки миелина (Шмидта - Лантсрмана). Через
интервалы 1-2 мм видны участки волокна, лишенные миелинового
слоя^ - узловатые перехваты или перехваты Ралвье.174
ГЛАВА 12.ИЕРВПЛ^ ТКАНЬРис. 115. Схема строения безмиелинового нервного волокна.ПерехватРачвьеШеанновскиеклеткиПерехеатРапвьеНасечки Шмидта-Ппит^п И лн/7ЦитоташаРис. 116. Продольное сечение миелиновос'о нервного волокна.в процессе развития аксон погружается в цитоплазму
нейролемм оцита. При этом образуется двойная складка
плазмолеммы нейролсммоцита - мезаксон. Мезаксо1£ удлиняется,
концентрически наслаивается на осевой цилиндр и образует вокруг
него миелиновый слой (рис. 117). Отсутствие миелинового слоя в
области узловых перехватов объясняется тем, что в этом участке
волокна кончаегся один нейролеммоцит и начинается другой.
Снаружи от нейролемм оцита располагается базальная мембрана.175
гистология. ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ IЯдроЦитоп.чашаРис. 117. Развитие и строение миелинового волокнаСкорость передачи импульса мислиновыми волокнами
больше, чем бсзмиелиновьтми. Безмиелиновые волокна проводят
нервный импульс со скоростью 1-2 м/с, тогда как толстые
миелиновые - со скоростью 5-120 м/с.Реакция нейронов и их вс^.іокрН: на травіЦу. Шререзка
нервного волокна вызывает различные реакции в телемейриицу в
участке волокна между телом нейрона и местом перерезки
(проксимальный сегмент) и в отрезке^ рцсположенном
дистальнеє от места травмы^ и не связанном с телом нейрона
(дистальныйтегмепт). Изменения в теле нейрона (перикарионе)
выражаются в его набухании, тигролизе ■•растворении глубок
тигроида и в перемещении ядра нц периферию Шела клетки.
Дегенеративные изменения в центральном отрезке
ограничиваются распадом миелинового слоя и осевого цилиндра
вблизи травмы. В дистальном отрезке миелиновый слой и
осевой цилиндр фрагментируются и продукты распада
удаляются макрофагами в течение 1 недели.Полноцеї^ной регенерации нервных волокон в
центральной нервной системе обычно не происходит, но
первые волокна^ в составе периферических^: нервов обычно
хорошо рі^фЩируют. При этом кейролеммоццпт
періІф^рического отрезка и ближайшего к ojmacmu травмы
участка центрального отрезка пролиферируют и
выстраиваются компактными " тяжами. Осевые цилиндры
цепщршіьного отрезкаЦ дают мно^численные коллатералщ
которш растут со скоростью- мм в сутки вдоль176
ГЛАВА 12.НЕРВНАЯ ТКАНЬнейроленмтьных тяжей, Вьижтают только те волокнщ
которые^достигают^ соответствующих окончаний* Остальные
дегенерируют^ Если существует препятствие длЛ врастания
аксонов центрального отрезка нерва м тяжи нейролелтоципща
периферического отрезка (обширная травма^ воспалительный
процесс^ наличие рубца), аксоны центрального отрезка растущ
беспорядочно и могут ^ образовать кчубок, называемый
ампутационной невромой. При ее раздражении возникает
сщшная больу которая воспрцнимается как ¿производящая из
первоначально иннервируемой области, например как боль в
ш^^йировшщц конечности (фантомные боли).Нервные окончанияНервные волокна заканчиваются концевыми аппаратами -
нервными окончаниями. Различают 3 группы нервных окончаний;
мсжнейрональные синапсы; эффекторные окончания (эффекторы);
рецепторные (аффск1’орные, или чувствительные).СинапсыСинапсы предназначены для передачи импульса с одного
нейрона на другой или на мышечные и железистые структуры. В
зависимости от способа передачи импульса синапсы могут быть
химическими или электрическими.в заиисимости от локализации окончаний терминальных
веточек аксона первого нейрона различают аксодендритические,
аксосоматические и аксоаксональныс синапсы.Химические синапсы передают импульс на другую юхетку
с помощью биологически активных веществ “ нейромедиаторов,
находящихся в синаптических пузырьках (рис. 118). Тсрминаль
аксона представляет собой пресинаптическую часть, а область
второго нейрона, или другой клегки, с которой она контактирует, -
постсинаптическую часть. В пресинаптической части находятся
синаптические пузырьки, митохондрии и отдельные
нейрофиламенты. Холинергическими являются парасимпатические
и преганглионарные симпатические синапсы, аксо-мышечные
синапсы и некоторые синапсы ЦНС. Адетилхолин и норадреналин- наиболее распространенные медиаторы, но существует и
множество других. Различают низкомолекулярпые нейромедиа юры
(ацетилхолин, норадреналин, дофамин, серотонин, глицин, гамма-
аминомасляная кислота, серотонин, гистамин, глютамат) и
нейропептиды: опиоидныс (эндорфины, энксфалины, динорины),177
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ 1 _ вещество Р и др. Дофамин, глицин и гамма-аминомасляная кислота
ЯВЛЯЮТСЯ медиаторами тормозящих синапсов. Вырабатывающиеся
в головном мозге эндорфины и энксфалины являются
ингибиторами восприятия боли. Однако большинство мед1и(аторов и
соответственно синапсов являются возбуждающими.ДеиАрит[Тресииапмическан
частьAкcвнa^íьuь^й
холмикАксоиРис. 118. Синаптические контакты нейрона. 1 - схема, 2 - конфокальнаямикроскопия.Пресинаптическая мембрана - это мембрана клетки,
передающей импульс. В этой области локализованы кальциевые
каналы, способствующие слиянию синаптических пузырьков с
пресинаптической мембраной и выделению медиатора в
синаптическую щель (рис. 119, 120).Синоптическая щель имеет ширину 20-30 им.Постсинаптическая мембрана - это участок плазмолеммы
клетки, воспринимающий медиаторы генерирующий импульс. Она
снабжена рецепторными зонами для восприятия соотвсгсгвующего
нейромедиатора.178
ГЛАВА 12.НЕРВНАЯ ТКАНЬРис. 119. Строение синапсов (схема).Рис. 120. Электронная микрофотофафия синапса.179
гистология, [ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ IЭлектрические синапсы
млекопитающих встречаются редко.нервнойсистемеЭффекторныс нервные окончанияЭффекторные нервные окончания бывают двух типов -
двигательные и секреторные.Двигательные нервные окончании - это концевые
аппараты аксонов двигательных клеток соматической, или
вегетативной, нервной системы. При их участии нервный импульс
передается на ткани рабочих органов. Двигательные окончания в
поперечнополосатых мышцах называются нервно-мышечными
окончаниями. Нервно-мышечное окончание состоит из ко1щевого
ветвления осевого цилиндра нервного волокна и
специализированного участка мышечного волокна (рис. 121).
Миелиновое нервное волокно, подойдя к мышечному волокну,
теряет мислиновый слой и 1югружается в него.Рис. 121.1 - 121.2. Нервно-мышечное окончание (моторная бляшка).180
ГЛЛВЛ 12.НЕРВНАЯ ТКАНЬПлазм0ле1ммы терминальных ветвей аксона и мышечного
волокна раздел СИ bj синоптической щелью шириной около 50 нм.
Мембрана мышечного волокна образует складки, формирующие
вторичные синаптические щели эффекторного окончания (рис.
122). В области окончания мышечное волокно не имеет типичной
поперечной нсчсрченност]и и характеризуется обилием
митохондрий. Саркоплазма с митохондриями и ядрами в
совокупности образует постсинаптическую часть синапса.
Постсинаптическая мембрана моюрного нервного окончания
содержит фермент ацетилхолинэстеразу, разрушающий медиатор
ацетилхолин.' - Осевой/■ цшшпдр... . ‘. .1ЯдрошеапиовскойкпеткиСшштпииесниепузырьки:;,4bНаружнаямембранаЦитоплашашваииовскойклеткиПресииаптическая
мембрана Постсшшптическаяшштf.меморапа
■ Ядро мышечногоРис. 122. Ультрамикроскопическое строение нервно-мышечного
окончания (схема).Нарушения в нервно-мышечных окончаниях еышвают
развитие неизлечимого Ш заболевания .miastenia gravis^
характеризуюш,ег0ся прогрессирующей мышечной слабостью и
часто заканчивающегося параличом дыхательной мускулатуры
(межреберных мышц и диафрагмй). При этом заболтщии в
крови циркулируют антитела против ацетилхошновых
рецепторов сар^рлрнмы. Этц~; антитела связываются с181
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iхщипорецепторами постсииапШичеЬкой "ШембраньГ и
инактивируют их, нарушаядервно^ышечнбё взаимодействие.Двигательные нервные окончания в гладкой мышечной
ткани представляют собой чсгкообразиые утолщения нервного
волокна, идущего среди гладких миоцитов.Сходное строение имеют нейрожелез истые секрсгорные
нервные окончания. Они представляют собой концевые утолщения
терминален или утолщения по ходу нервного волокна, содержащие
пресинаптические пузырьки.Рецепторные нервные окончания
Эти нервные окончания рассеяны по всему организму и
воспринимают различные раздражения как из внешней среды, так и
от внутренних органов. Выделяют две группы рецепторов:
экстерорецепторы (слуховые, зрительные, обонятельные, вкусовые
и осязательные) и интерорецепторы (висцерорецепторы и
нроприорецепторы). в зависимости от специфичности
раздражения, воспринимаемого данным видом рецептора, все
чувствительные окончания делят 11а механорецепторы,
барорецепторы, хеморецепторы, терморецепторы и др.Чувствительные окончания подразделяют li^d свободные
нервные окончания, состоящие только из конечных ветвлений
осевого цилиндра, и несвободные, содержащие в своем составе все
компоненты нервного волокна. Несвободные окончания могут быть
покрыты соединительнотканной капсулой (инкапсулированные) и
не иметь её (неинкапсулированные).Свободные нервные окончания в эпителии воспринимают
холод, тепло и боль, в этом случае миелиновые нервные волокна
подходят к эпителиальному пласту, теряют миелин, а осевые
цилиндры проникают в эпителий и распадаются там между
клетками на тонкие терминалы1ые ветви (рис. 123).182
ГЛЛВЛ 12.НЕРВНАЯ ТКАНЬЩШШ:■ ■ ■■ I ■!¥г': *|Л-Ь' ■ - . ф^-. ,ч\.,<Л^ ■■ :й’б" ■*'■?х. ..;■ &'Н^1Р•Й!'.. Ш "}■ Лй-Рис. 123. Свободные нервные окончания.Большинство рецепторов сосди1штельной ткани
представлясг собой ветвления оссвого цилиндра. В состав таких
концевых аппаратов входят нсйролсммоциты, которые
сопровождают все всгвления волокна (это несвободные
нсиикаисулированные рецепторы).Инкапсулированные рецепторы соединительной ткани
состоят из ветвления осевого цилиндра и глиальных клеток.
Снаружи такие рецепторы покрыты соединительнотканной
капсулой (пластинчатые тельца Фатера-Пачиии). Их размеры 0,5х I-2 мм. В центре такого тельца располагается внутренняя луковица,
образованная видоизмененными JЮммoцитaми (рис. 124.2).
Миелиновое нервное волокно теряет мислиновый слой, проникает'
во внугреннюю луковицу и разветвляется. Снаружи тельце
окружено слоистой капсулой, состоящей из фибробластов и
спирально ориентированных волокон. Давление на капсулу
передается на внутреннюю луковицу и воспринимасгся
безмиелиновыми волокнами. Пластинчатые тельца воспринимают
давление и вибрацию. Они присутствуют в глубоких слоях дермы
(особенно в коже пальцев), в брыжейке и внутренних органах.К чувствительным инкапсулированным окончаниям
относятся осязательные тельца - тельца Мейснера, они183
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iрасполагаются в верхушках соединительно1’каиных сосочков кожи.
Осязательные тельца состоят из видоизмененных
нейролеммоцитов, расположенных перпендикулярно длинной оси
тельца (рис. 124.1).К инкапсулированным нервным oкoнчaния^f относятся
та1сже рецепторы мышц и сухожилий: нервно-мышечные веретена и
нервно-сухожильныс веретена.О»;.Рис. 124.1-124.2. Строение инкапсулированных нервных окончаний.1 - осязательное тельце Мейсснера, 2 - пластинчатое тельце Фатера-Пачипи.Понятие о рефлекторной дугеРефлекторная дуга представляет собой цепь нейронов,
связанных друг с другом синапсами и обеспечиваюпщх проведение
нервного импульса от рецептора чувствительного нейрона до
эфферентного окончания в рабочем органе.Самая простая рефлекторная дуга состоит из двух нейронов- чувствительного и двигательного. В подавляющем большинстве
случаев между чувствительными и двигательными нейронами
включены вставочные, или ассоциативные, нейроны.
ГЛАВА 13.
НЕРВНАЯ СИСТЕМАНервная сисгема обеспечивает регуляцию всех жизненных
процессов в организме и его взаимодействие с внешней средой,
Нфвпую систему делят на центральную {головной и спинной мозг) и
пфиферическую {нервные узлы, стволы и окончания).С физиологической точки зрения нервная система делится на
соматическую, иннервирующую все тело, кроме внутренних органов,
сосудов и желез, и автономную, или вегетативную, регулирующую
деятельность перечисленных Орга1ЮВ,Развитие. Нфвная система развивается из нервной трубки и
ганглиозной пластинки. Из краниальной части нервной трубки
дифференцируются головной мозг и органы »1увств. Из туловшцного
отдела нервной фубки и ганглиозной пластинки формируются
сттной люзг, сттномозговые и вегетативные узлы и хромаффинная
ткань.Стенки нервной трубки делятся на три зоны: эпендиму,
плащевой слой и краевую вуачъ. Из плащевого аюя развивается серое
вещество спинного мозга, а из краевой вуали - его белое вещество.
Нейробласты передашх столбов дифференцируются в двигательные
нейроны ядер передних рет ов. Их аксоны выходяг из спинного мозга
и образуют а о передние корешки. В задних столбах и промежуточной
зоне развиваются различные ядра вставочных (ассоциативных) клсток.
Их аксоны, поступая в белое вещество спинного мозга, образуют
проводящие пути. В задние ро1а входят нейриты чувствительных
1спеток спинномозговых ганглиев.Сгшшюмозговые и всгсштивные ’ нервные узлы образуются из
ганглиозной пластинки.СПИННОМОЗГОВОЙ УЗЕЛ
Спинномозговой узел окружен капсулой, в паренхиму узла
проникают тонкие прослойки соединительной ткани, в которой
расположены кровеносные сосудьгПссвдоуниполярные нейроны спинномозгового узла
располагаются группами по периферии органа. В центре находятся
отросгкц этих клсток. Дсндриты идут в составе чувствительной части185
1'ИСТОЛОГИЯ, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ Iсмешанных спинномозговых нервов на периферию и заканчиваются
там рецепторами. Нейриты в совокупноеги образуют задние корешки,
несущие нервные импульсы в серое вещество спинного мозга (рис.
106). Нервные клетю! СПИННОМОЗ! ОВЬ[Х узлов окружены слоем
мантийных глиоцитов..-.л#:'::., ,
.м .• . ^ ¥-4 ї!.V4IРис. 106. Спиниомозговой узел. 1 - задиий корешок, 2 - передний
корешок, 3 - спинномозговой нерв, 4 “ псевдоуниполярные нейроны,5 - капсула узла, 6 - нервные волокна.Периферические нервыПериферические нервы образован нервными волокнами,
которые собраны в пучки и отграничены друг от друга
прослойками соединительной ткани. Между отдельными нервными
волокнами располагаются тонкие прослойки соединительной ткали- эпдоневрий. Пучки нервных волокон окутаны перииеврием.
Наружная оболочка нервного ствола - этшеврий - представляет
собой оболочку, объединяющую несколько пучков нервных
волокон (рис. 107).Рис. 107. Строение периферического нерва.
186
ГЛАВА 13.НЕРВНАЯ СИСТЕМАСпиииой MOJrСпинной мозг состоит из двух симметричных половин,
отграниченных друг от друга спереди щелью, а сзади -
соединительнотканной перегородкой (рис. 108). На периферии
спишюго мозга располагасгся более светлое белое вещество.
Внутренняя часть ор1ана -Ьто серое вещество. Различают передние
(вентральные), задние (дорсальные) и боковые (латеральные) рога.Серое вещество спинного мозга состоит из тел нейронов,
безмиелиновых и тонких миелиновых волокон и нейроглии.Белое вещество спинного мозга представляет собой
совокупность продольно ориентированных волокон. Пучки
нервных волокон называются проводящими путями спинного
мозга.Нейроны, сходные по строению и функциональному
значению, лежат в сером веществе группами, которые называются
ядрами. Различают: корешковые клетки, нейриты которых
образуют передние корешки, внутренние клетки, отростки которых
заканчиваются в сером веществе спинного мозга, и пучковые
клетки, аксоны которых образуют проводящие пути.В задних рогах различают губчатый слой, желатинозное
вещество, собственное ядро заднего рога и грудное ядро. Между
задними и боковыми рогами различают рсгикулярную формацию.В н ром ежу точной зоне различают медиальное и
латеральное промежуточные ядра. В промежуточной зоне
расположены центры вегетативной нервной системы.В передних рогах расположены крупные нейроны спинного
мозга (100-150 мкм). Их нейриты составляют основную массу
вoJюкoн передних корешков. В составе смешанных
спинномозговых нервов они образуют моторные окончания в
скелетной мускулатуре. Медиальная группа моторных клеток
иннервирует мьинцы туловища и хорошо развита на всем
протяжении спинного мозга. Латеральная группа находится в
области шейного и поясничного утолщений и иннервирует мышцы
конечностей (рис. 108).Аксоны пучковых нейронов выходят в белое вещество и
делятся на длинную восходящую и более короткую нисходящую
ветви, в совокупности эти BOjmKHa образуют собственные, или
основные, пучки белого вещества.187
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I .Глиоциты спинною мозга. Спинномозговой канал
вьісі'лан эпендимоцитами, участвующими в выработке
спинномозговой жидкости, в сером всщсстве находятся
протоплазматические и волокнистые астроциты.“ Отростки
волокнистых астроцитов принимают участие в образовании
перегородок в белом веществе и глиальных мембран вокруг
кровеносных сосудов и на поверхности спинного мозга.
Олигодендроглиоциты входят в состав оболочек нервных волокон.
Микроглия встречается в сером и белом веществе.Задняя серединная
7 бдроздаБелое вещество f.Передняя середин пая
гцельРис. 108. Спинной мозг. 1 - передний рог, 2 - задний рог, 3 - боковой
рог, 4 - спинномозговой канал, 5 - передний канатик, 6 - боковой канатик,7 - задний канатик.188
ГЛАВА 13.НЕРВНАЯ СИСТЕМАСТВОЛ МОЗГАВ состав ствола мозга входят продолговатый мозг, мост,
мозжечок и структуры среднего и промежуточного мозга (рис. 109).
Все ядра серого вещества ствола мозга состоят из мультиполярных
нейронов. Различают ядра черепных нервов и переключательные
ядра.В продолговатом л1озге паходятся ядера черепных нервов,
которые концентрируются преимущественно в его дорсальной части,
образующей дно IV желудочка. В нижние оливы поступают волокна
от мозжечка, красного ядра, ретикулярной формации и спищ10Г0
мозга. В центральной области продолговатого мозга располагается
ретикулярная формация (сетчатое образование). Ретикулярная
форма1щя представляет собой сложный рефлекторный центр и
принимает участие в контроле над тонусом мышц и стереотипными
движениями. Белое вещество в продолговатом мозге занимает
преимущественно вентролатеральное положение.Мост делится па дорсальную и вентральную части.
Дорсальная часть содержит волокна проводящих путей
продолговатого мозга, ядра У-УШ черепных нервов, ретикулярную
формацию моста. В вентральной части располагаются собственные
ядра моста и волокна пирамидных путей, идущие продольно.Средний мозг состоит из крьппи среднего мозга
(четверохолмия), покрышки среднего мозга, черного вещества и
ножек мозга. Четверохолмие состоит из пластшжи крыши, двух
верхних (звено зрительного анализатора) и двух нижних холмиков
(часть слухового анализатора). В покрышке среднего мозга
находится до 30 ядер. Ножки мозга образованы мислиновыми
волокнами, идущими от коры большого мозга.Промежуточный мозг, в промежуточном мозге
преобладает зрительный бугор. Вентрально от него рас1юлагаегся
бохатая мелкими ядрами гипоталамыческая область. Зрительный
бугор содержит множество ядер, отграпичс1шых друг от друга
прослойками белого вещества. В каудальной группе ядер
заканчиваются волокна зрительного пути.Гипоталамическая область - вегетативный центр
головного мозга, регулирующий температуру, кровяное давление,
водный, жировой обмен и др.189
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I.средний мозгмостПродолговатыймозг^промежуточныймозгВерхние бугоркиНижние бугорки
четверохолмияНожкимозжечкаРис. 109. Ствол мозг (схема)., МОЗЖЕЧОКМозжечок представляег собой центральный орган
равновесия и координации движений. Он связан со стволом мозга
тремя парами ножек. На поверхности мозжечка много извилин и
бороздок, которые создают на разрезе картину "древа жизни".
Основная масса серого вещества в мозжечке располагается на
поверхности и образует его кору. Меньшая часть серого вещества
лежит глубоко в белом веществе в виде ядер.190
ГЛАВА 13.НЕРВНАЯ СИСТЕМАВ коре мозжечка различают три слоя: наружный -
молекулярный, средний - ганглионарный и внутренний - зернистый
слои (рис. ПО). Гаиглиопариый слой содержит грушевидные
нейроны (клетки ГТуркинье), клетки располагаются строго в один
ряд. От их крупного (60 X 35 мкм) грушевидного тела в
молекулярный слой отходят 2-3 дендрита. От основания тел этих
клегок огходят нейриты, чзаканчиваюш;исся на клетках ядер
мозжечка.Молекулярный слой содержит корзинчатые и звездчатые
нейроны. Корзинчатые нейроны (10-20 мкм) находятся в нижней
трети слоя. Их длинные дендриты ветвятся поперечно к извилине.
Длинные нейриты идут поперек извилины, они отдают коллатерали
к телам грушевидных нейронов и формируют корзинки нервных
волокон.Звездчатые нейроны бывают двух типов: мелкие и крупные.
Ветви их нейритов соединяются с дендритами грушевидных клеток
или входят в состав корзинок. Корзинчатые и звездчатые нейроны
передают тормозные импульсы к грушевидным клеткам.В зернистом слое различают клетки-зерна (5-8 мкм). Клетка
имеет 3-4 коротких дендрита, заканчивающихся в виде лапки
птицы. Моховидные (возбуждающие) волокна и дендриты клеток-
зерен образуют клубочки мозжечка. Нейриты клеток-зерен
проходят в молекулярный слой и в нем Т-образно делятся. Эти
волокна образуют с дендритами грушевидных, корзинчатых и
звездчатых нейронов синапсы. Нейриты клеток-зерен передают
возбуждение, полученное ими от моховидных волокон. Тормозные
большие звездчатые нейроны с короткими нейритами лежат вблизи
ганглионарного слоя. Их разветвленные дендриты находятся в
молекулярном слое. Нейриты направляются в зернистый слой к
юіубочкам мозжечка и заканчиваются синапсами на дендритах
клеток-зерен проксимальнеє синапсов моховидных волокон.
Возбуждение звездчатых нейронов может блокировать импульсы,
поступающие по моховидным волокнам. Дендриты и нейриты
звездчатых нейронов с длинными нейритами ветвятся в зернистом
слое. Веретеновидные горизонтальные клетки встречаются между
зернистым и ганглионариым слоями, их нейриты уходят в белое
вещество. Афферентные волокна, поступающие в кору мозжечка
(моховидные и лазящие), они контактируют с дендритами клеток-
зерен. Таким образом, возбуждаюпще импульсы в кору мозжечка
поступают по лазящим волокнам, или по параллельным волокнам191
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I,клеток-зфсн. Торможение - функция звездчатых нейронов
молекулярного слоя, корзинчатых нейронов, а также больших
звездчатых нейронов зернистого слоя.Кора мозжечка содержит волокнистые,протоплазматические астроциты и олигоде*1дроциты. Особенно
богаты этими клетками зернистый слой и белое вещество мозжечка.
Микроглия содержится в молекулярном и ганглионарном слоях.Рис. 110. Мозжечок, а - древо жизни мозжечка, б - кора мозжечка.1 - молекулярный слой, 2 - ганглионарный слой, 3 -зермистый слой,4 - белое вещество.КОРА ГОЛОВНОГО МОЗГАВ развитии коры больших полушарий человека принимают
участие малоспсциа/шзированные пролиферирующие клетки, из
которых дифференцируются нейроциты нсокортекса. в корковую
пластинку поступают нейроциты будущих Т и VÍ слоев. Затем в нсс
встраиваются нейроны V, IV, III и II слоев. Кора большого мозга
(серое вещество) имеет толщину 3-5 мм. Поверхность мозга
содержит борозды и извилины. Основные клетки коры имеют
пирамиднук* форму (10 - 140 мкм). От вершины и боковых
поверхностей тела клеток отходят дендриты. От основания
пирамидных клеток берут начало нейриты.Мелкие по размерам клетки представляют собой
вставочные нейроны, нейриты которых связывают отдельные
участки коры одного полушария (ассоциативные нейроны) или
двух полушарий (комиссуральные нейроны). Нейриты крупных
пирамид принимают участие в образовании пирамидных путей.192
ГЛАВА 13.НЕРВНАЯ СИСТЕМАВ коре головного мозга различают 6 основных слоев: I -
молекулярный, II - наружный зернистый, TTI - пирамидный, IV -
внутренний зернистый, V - ганглионарный, VI - слой полиморфных
клеток (рис. 111).Молекулярный слой коры содержит мелкие ассоциативные
клетки верстсиовидной фо^)мы. Их аксоны проходят параллельно
поверхности мозга и образуют тангенциатьное сплетение нервных
вoJЮкoн молекулярного слоя.Наружный зернистый слой образован мелкими нейронами,
имеющими округлую, звёздчатую и пирамидальную форму.
Дендриты этих клеток поднимаются в молекулярный слой.
Нейриты уходят в белое вещество.Пирамидный слой особенно хорошо развит в
прсцсп фальной извилине. От верхушки пирамидной клетки
отходит главный дендрит, который располагается в молекулярном
слое. Дендриты, берущие начало от боковых поверхностей
пирамиды и ес основания, образуют синапсы с нейронами в
пределах этого слоя. Нейрит крупной пирамидной клетки обычно
формирует миелиновое ассоциативное или комиссуральное
волокно, идущее в белое вещество.Внутренний зернистый слой образован мелкими
звездчатыми нейронами. В его состав входит большое количество
горизонтальных волокон.Г англионарный слой коры образован крупными
пирамидными нейронами (клетки Беца, 80 х 120 мкм). Гигантские
пирамиды характеризуются наличием крупных глыбок
хроматофильного вещества. Нейриты клеток этого слоя образуют
главную часть кортико-спинальных и кортико-нуклеарных путей и
оканчиваются синапсами на клетках моторных ядер.Слой полиморфных Ю1СТОК образован нейронами различнойформы.Сентаготаи и представители его школы установили, что
структурно-функционалыюй единицей нсокортекса является
модуль - вертикальная колонка диаметром около 300 мкм, В центре
находится кортико-кортикальное волокно - ассоциативное либо
комиссуральное. В модуль входят два таламо-кортикальных
волокна. Всего в нсокортексе человека насчитывается примерно 3
млн. модулей. Помимо специфических афферентных волокон на
пирамидные нейроны возбуждающее влияние оказывают
шипиковые звездчатые нейроны фокального и диффузного типа,193
ГИСТ0Л0П1Я, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ IТормозная система модуля представлена следующими
типами нейронов: 1) клетки с аксональной кисточкой; 2)
корзинчатые нейроны; 3) аксоаксональные нейроны; 4)^клетки с
ДВ0Й11ЫМ букетом дендритов.■п|1ЯШ, ^■ \?'-1Ш1Ш1is;sisiisa;isiBKsщизшiPIM«Шч:■ -i'plsSщ>ьшtftiiliРис. 111. Кора головного мозга. I - молекулярный слой, II-наружный
зернистый слой, III - пирамидный слой, IV - внутренний зернистый
слой, V - ганглионарный слой, VI - слой полиморфных клегок.Фокальные шипиковые звездчатые юютки одновременно
возбуждают пирамидные нейроны и клетку с двойным букетом
дендритов. Система тормозных нейронов играет роль фильтра,
тормозящего часть пирамидных нейронов коры.194
ГЛАВА 13.НЕРВНАЯ СИСТЕМААгранулирный тип коры встречается в моторных центрах
(передняя центральная извилина). В коре такого типа хорошо
развиты III, V и VI и плохо выражены И и IV слои.Гранулярный тип коры находится в ’^тувствитсльпых
корковых центрах, здесь хорошо развиты II и 1У слои.Мислоархитектоника коры. Ассоциативные волокна
связывают отдельные ^участки коры одного полушария,
ко.миссуральиые, соединяют кору различных полушарий, и
проекционные волокна, связывают кору с ядрами низших отделов
центральной нервной системы.ГЕМАТО-ЭНЦЕФАЛИЧЕСКИЙ БАРЬЕРСосуды проходят в мягкую мозговую оболочку, отсюда мелкие
веточки следуют в вещество мозга. Капилляры мозга имеют
непрерывную эндотелиальную высгилку. Здесь пpoиcxoд^rг обмен
веществ между нервной тканью и кровью, в котором принимает участие
гематоэнцефачический барьер. Отростки астроцитов образуют на
поверхности капилляров слой, агграничивающий нейроны от
соприкосновения с сосудистой стенкой (рис. 112).АстроцитарныеЭритроцитНейронСоединениеумдотелиоцитовЭндотепио цитБазальная мембрана
Рис, 112. Строение гематоэнцефалического барьера (схема).195
гистология, цитология, ЭМБРИОЛОГИ я. ЧАСТЬ I,ОВОЛОЧКИ МОЗГАМягкая мозговая оболочка - наиболее близко прилежит к
мозгу, она образована тонким слоем соединительной ткани,
покрыта непрерывным слоем плоского эпителия (рис. 113).Паутинная мозговая оболочка - располагается снаружи от
мягкой мозговой оболочки, образована рыхлой соедини!ель^юй
тканью, между мягкой и паугинной оболочками имеется
субарахноидальное пространство, заполненноецереброспинальной жидкостью.Твердая мозговая оболочка - находится снаружи от
паутинной и состоит из плотной волокнистой соединительной
ткани. Между паутинной и твердой оболочками есть субдуральное
пространство. В черепе твердая мозговая оболочка сращена с
надкостницей, а между твердой мозговой оболочкой снинного
мозга и надкостницей позвонков имеется эпидуралыюе
пространство.Кожа
Кость черепаТвёрдая мозговая
оболочкаС};6дуральпое
пространство^. _ПаутиннаяоболочкаВенаАртерияСу барахпоидальное
про страчст во ^Мягкая мозговая
оболочка—Рис. 113. Оболочки головного мозга (схема).196
ГЛ/\ВЛ 13.НЕРВНАЯ СИСТЕМААВТОНОМНАЯ НЕРВНАЯ СИСТЕМАЧасть нервной системы, контролирующая висцеральные
функции организма (моторика и секреция органов
пищеварительной системы, кровяное давление, потоотделение,
температура тела, обменные процессы) называется автономной^
или вегетативной, нервной системой. Вегетативная нервная
система делится на симпэтичсскую и парасимпатическую.Вегетативная нервная система состоит из центральных
отделов (ядра головного и спинного мозга), и периферических
(нервные сгволы, узлы и сплетения).Ядра центрального отдела вегегативпой нсрв1ЮЙ системы
находятся в среднем и продолговатом мозге, в боковых рогах
грудных, 1ЮЯСНИ4НЫХ и крестцовых сегментах спинного мозга, к
симпатической нервной системе относятся вегетативные ядра
боковых рогов грудного и верхнепоясничного отделов спинного
мозга, к парасимпатической - вегетативные ядра III, VII, IX и X пар
черепных нервов и вегетативные ядра крестцового отдела спинного
мозга (рис. 114). Нейриты покидают центральную нервную систему
через передние корешки спинного мозга или черепные нервы и
оканчиваются синапсами на нейронах периферических
вегетативных ганглиев. Это преганглионарные волокна
вегетативной нервной системы, обычно миелиновые.
Преганглионарные волокна симпатической и парасимпатической
вегетативной нервной системы - холинергичсскис.Периферические узлы вегетативной нервной системы лежат
как вне орга1Юв (симпатические паравертебральные и
превертебральные ганглии, парасимпатические узлы головы), так и
в стенке органов в составе интрамуральных нервных сплетений
пищеварительного тракта, сердца, матки, мочевого пузыря и др.
Вегетативные ганглии состоят из мультиполярных нервных клсток.Г англ ИИ парасимпатического отдела вегетативной нервной
системы лежат или вблизи иннервируемого органа или в его
ип1рамуралы1ых нервных сплетениях (в пищеварительном тракте,
сердце, мочевом пузыре и др.),Силтатический отдел ве1'стативной нервной системы.
Перикарионы первого нейрона расположены в боковых столбах
спинного мозга. Их аксоны проходят в составе передних корешков
и белой соединительной ветви и заканчиваются на перикарионах
второго нейрона в пара вертебральных ганглиях симпатической
цепочки, превертебральных ганглиях и ганглиях, расположенных197
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I.вблизи иннервируемых ими органов. Аксоны второго нейрона
паравсртебральных, прсвертебральных и терминальных ганглиев
заканчиваются во внутренних органах, коже, стенке кровеносных
сосудов. Аксоны некоторых нейронов паравсртебральных ганглиев
проходят через серую соединительную ветвь.Рис. 114. Схема строения вегетативной нервной системы.ЛарпсимпатическийотделСимпатическийотдел198
ГЛАВА 13.НЕРВНАЯ СИСТЕМАЗадний корешоктрубка железа сосудь(Рис. 115. Простая вегетативная рефлекторная дз^а.199
ГЛАВА 14,
ОРГАНЫ ЧУВСТВОрганы чувстіз обеспечивают восприятие раздражителей,
действующих на организм, передачу нервного импульса в
подкорковые и корковые центры, где происходят анализ
информации.Сенсорная система - это анализаторы внешней и внутренней
среды, которые обеспечивают адаптацию организма к
определённым условиям. В анализаторе различают 3 части;
периферическую, промежуточную и центральную.Периферическая часть - это органы, в которых находятся
рецепторные клетки.Промежуточная часть - цепь вставочных нейронов, по
которым нервный импульс от рецепторных клеток персдасгся к
корковым центрам.Центральная часть сенсорной системы представлена
участками коры больших полушарий.Классификация органов чувствК первому типу относятся органы чувств, у которых
рецепторами являются иейросенсорные клетки (орган зрения, орган
обоняния).Ко второму типу относятся органы чувств, у которых
рецепторами являются сенсоэпителиальные клетки (органы слуха,
равновесия, вкуса).К третьему типу относятся инкапсулировапные и
неинкапсулированные рецепторы.ОРГАН ЗРЕНИЯОрган ^зрения представляет собой периферическую часть
зрительного анализатора.Глаз развивается из различных эмбриональных зачатков.
Сетчатка и зрительный нерв формируются из нервной трубки.
Вначале образуются глазные пузырьки, сохраняющих связь с
эмбриональным мозгом. Затем передняя часть глазного пузырька
приобретает форму двустенного глазного бокала. Часть эктодермы200
ГЛАВА М ОРГАНЫ ЧУВСТВнапротив отверстия глазного бокала даст начало зачатку
хрусталика (рис. 114).Рис. 114. Ра:^витие глаза. 1, 2, 3 - сагиттальные разрезы органа зрения
эмбрионов на различных стадиях развития.Из внутренней стенки глазного бокала образуемся сетчатка
глаза, а из наружной - ей пигментный слой. Все виды нейронов
сетчатки формируются из внутренней стенки глазного бокала.
Аксоны ганглиозных клеток образуют зрительный нерв,
направляющийся в мозг.Из окружающей мезенхимы формируются сосудистая
оболочка и склера, которая переходит впереди в прозрачную
роговицу, Сосуды и мезенхима принимают участие в образовании
стекловидного тела и радужки. Мышцы радужной оболочки201
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I.(суживающая и расширяющая зрачок) по своему происхождению
являются нейральными.СТРОЕНИЕ ГЛАЗА
' Стенка глазного яблока состоит из трех оболочек (рис,115). Наружная (фиброзная) оболочка глазного яб.'юка имеет 2
отдела: роговица (передняя часть) и склера (задняя часть). Средняя
(сосудистая) оболочка имеет три части: радужная оболочка,
цилиарное тело и собственно сосудистая. Внутренняя оболочка -
сетчатка - сенсорная, рецепторная часть зрительного анализатора.
Оболочки глаза образуют три функциональных аппарата;
светопреломляющий или диоптрический (роговица, жидкость
передней и задней камер глаза, хрусталик и стекловидное тело);
аккомодационный (радужка, ресничное тело с ресничными
отростками); рецепторный аппарат (сетчатка).Наружная фиброзная оболочка - склера - образована
плотной оформленной волокнистой соединительной тканью.роговица Передняя камералимбШлемов капалЦилиарныемышцыСклераСклераПигментны й /эпителииРис. 115, Строение глазного яблока (схема).
202
ГЛАВА 14.0РГА11Ы ЧУВСТВСветопреломляющий аппарат глазаРоговица (comea) пропускает и преломляет световые лучи.
Толщина роговицы 0,8 - 1,1 мкм, радиус кривизны 7,8 мкм. В
роговице выделяют 5 слоев; 1) многослойный плоский
неороговевающий эпителий; 2) переднюю пограничную мембрану
(боуменову); 3) собственное вещество роговицы; 4) заднюю
пограничную мсмбраиу\(десцеметову); 5) задний эпителий (рис.116).Клетки переднего эпителия роговицы располагаются в 5
слоев, содержат свободные нервные окончания, что обусловливает
её высокую чувствительность. Поверхность роговицы увлажнена
секретом слезных и конъюнктивальных желез.Боуменова оболочка имеет толщину 6-9 мкм.Собственное вещество роговицы состоит из параллельно
расположенных пучков коллагеновых волокон, которые погружены
в аморфное вещество, богатое гликозаминогликанами. Между
волокнами располагаются клетки, являющиеся разновидностями
фибробластов. Собственное вещество роговицы не имеет
кровеносных сосудов.У'Рис. 116. Роговица глаза. 1 - передний эпителий роговицы; 2 - боуменова
мембрана; 3 - собственное вещество; 4 - десцеметова мембрана; 5 - заднийэпителий роговицы.Десцеметова оболочка представлена коллагеновыми
волокнами диаметром 10 нм, погруженными в аморфное вещество.203
гистология, цитология, ЭМКРИОЛОГИЯ. ЧАСТЬ I.Задний эпигелий состоит из одного слоя ПЛОСІСИХ
полигональных клсток.Лимфатическая система роговицы состоит из узких
лимфатических щслсй, сообщающихся с ресничным венозным
сплетением.При воспалительных процессах кровеноаше капилляры и
клетки (лейкоциты, макрофаги и др,) проникают: из обла<:ти
лимба в собственное вещество роговицы, что рриводит к щ
помутнению и ороговению, дбразованиюШтшо.Передняя камера находится между роговицей (наружная
стейка) и радужной оболочкой (задняя стсика). В углу передней
камеры различают радужно-роговичиый угол, который граничит с
дренажным аппаратом - шлсм.мовым каналом.Трабекулярный аппарат состоит из 2 частей:
склерокорнеальной и увеальной. Склерокорнеальная часть состоит
из сети переплетающихся трабекул. Между ними остаются
щелевидные отверстия - фонтановы пространство. Фонтановы
пространства направлены к стснкс ишеммова канала, который
выстлан эндотелием.Хрусталик (lens). Это прозрачное двояковыпуклое і ело,
форма которого меняется. Хрусталик покрыт прозрачной капсулой.
Его передняя стсика состоит из однослойного плоского эпителия
(рис. 1П^/1Рис. 117. Хрусталик. 1 - капсула хрусталика,По направлению к экватору эпителиоциты образуют
ростковую зону хрусталика. Новые эпителиоциты преобразуются в
хрусталиковые волокна. Каждое волокно представляет собой
прозрачную шестиугольную призму. BoJюкнa склеиваются друг с204
ГЛАВА 14.0РГЛ11Ы ЧУВСТВДругом особым веществом. ТДентрально расположенные волоиха
теряют свои ядра и образуют ядро хрусталика.Хрусталик поддерживается в глазу с помощью ресничного пояска,
образовашю1'о пучками волокон, прикрепленных с одной стороны к
цилиарному телу, а с другой - к капсуле хрусталика.Стекловидное тело - желеобразное всщсство, заполняющее
полость между хрусталикрм и сетчаткой. Через стекловидное тело
проходит канал - остаток эмбриональной сосудистой системы
глаза. Стекловидное тело содержит белок витреин и гиалуроновую
кислоту.Аккомодационный аппарат глазаАккомодационный аппарат глаза (радужка, ресничное тело
с ресничным пояском) обеспечивает изме1гс1шс формы и
преломляющей силы хрусталика, фокусировку изображения на
сетчатке, а также приспособлепие глаза к интенсивиости
освещения.Радужка представляет собой дисковидное образование с
отверстием в центре (зрачок). Она является производным
сосудистой и ссгчатой оболочек. Строма радужки состоит из
рыхлой во.’юкнистой соединительной ткани, богатой пигментными
клетками. Здесь располагаются гладкие миоциты, образующие
мышцы, суживающие или расширяющие зрачок.В радужке различают 5 слоев: передний эпителий,
наружный пограничный слой, сосудистый слой, внутренний
пограничный слой и пигментный эпителий.Ресничное тело является производным сосудистой и
сетчатой оболочек. Выполняет функцию фикcaщiи хрусталика и
изменения его кривизны. Цилиарное тело имеет вид трезтольника и
подразделяется на две части: внутреннюю - цилиарную корону и
наружную - цилиарное кольг^о. От цилиарной короны по
направлению к хрусталику отходят цилиарные отростки, к
которым прикрепляется ресничный поясок. Основная часть
цилиарного тела образована ресничной мышцей. Между
мышечными пучками расположена соединительная ткань с
пигментными клетками. Сокращение цилиарной мышцы приводит
к расслаблению волокон круговой связки, вследствие чего
хрусталик становится выпуклым и его преломляющая сила
увеличивается.Сосудистая оболочка богата меланоцитами, в ней
различают иадсосудистую, сосудистую, сосудисто-капиллярную
пластинки и базальный комплекс.205
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ I.Рецепторный аппарат глазаРецепторный аппарат глаза представлен зрительной частью
сетчатки. Сетчатка состоит из наружного пигментного слоя и
внутреннего светочувствительного. Выделяют зрительную,
цилиарную и радужковую части сетчатки.Свет входит в глаз через роговицу, жидкость передней
камеры, хрусталик, жидкость задней камеры, стекловидное тело и
попадает на отростки фоточувствительных нервных клеток.
Сетчатка глаза человека относится к типу инвертированных
органов, в которых фоторецепторы направлены от света и образуют
глубокие слои сетчатки, обращенные к слою пигментного эпителия.
Сетчатка состоит из трех типов радиально расположенных
нейронов. Первый тип нейронов - это фоторецепторные нейроны
(палочковые и колбочковые), второй тип - биполярные нейроны и
третий тип - ганглионарные нейроны. Кроме того, имеются
горизонтальные и амакриновые клетки.Слои сетчатки (рис. 118):1 пигментный эпителий отделяет сепатку от сосудистой
оболочки;2 слой палочек и колбочек (светочувствительный)
арсдставлсн дендритами нервных клеток палочек и колбочек;3 наружная глиальная мембрана образована глиальными
клетками;4 наружный зернистый (ядерный) слой содержит тела
палочек и колбочек;5 наружный сетчатый слой - состоит из аксонов
светочувствительных клеток и дендритов биполярных клеток;6 внутренний зернистый (ядерный) слой образован телами
биполярных клеток;7 внутренний сетчатый слой - состоит из аксонов
биполярных и дендритов ганглионарных нейронов;а ганглионарный слой содержит те/ш мультиполяных
га^плионарных клеток;9 слой ^1ервных волокон состоит из аксонов ганглионарных
нейронов, которые образуют зрительный нерв;10. внутренняя глиальная мембрана образована глиальными
клегками.На поверхности сетчатки имеется зона длиной 1,7 мм, в
которой отсутствуют фоторецепторы сетчатки - слепос пятно, а
аксоны ганглиозных нсйроцитов формируют зрительный нерв (рис.
119.1). На внуфсннсй поверхности сетчатки имеется желтое
пятно диаметром 2 мм. Углубленный центр этого образования206
ГЛАВА 14.0РГАНЫ ЧУВСТВназывается центральной ямкой (рис. 119.2). Центральная ямка -
место наилучшего восприятия зрительных раздражений. В этой
области внутренний ядерный и ганглиозный слои истончаются,
утолщенный наружный ядерный слой представлен главным
образом телами колбочковых нейросенсорных клеток.'“¿ги111Рис, 118. Строение задней стенки глаза. I - сетчатка, И - сосудистая
оболочка, 111“ склера. 1 - пигментный CJЮЙ, 2 - слой палочек и
колбочек, 3 - наружная глиальная пограничная мембрана, 4 - наружный
зернистый слой, 5 - наружный сетчатый слой, 6 - внутренний зернистый
слой, 7 - внутренний сетчатый слой, 8 - ганглионарный, 9 - слой нервных
волокон, 10 - кнуфенняя глиальная пограничная мембрана.Рис. 119. Диск зрительного нерва (I) и центральная ямка (II)Фоторег^епторы сетчатки делятся на два типа: палочковые и
колбочковые. Палочковые клетки являются рецепторами сумеречного207
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ 1(нотаого зрения), колбочковые - дневн010 зрения. Периферические
отростки фоторецеториых юкток состоят из наружного и
вну'феннего сегментов, соединённых ресничкой. Дистальная часть
фоторецепторов - наружный сегмент - содержит мембраны, здесь
происходит поглощение света и начинается зрительное возбуждение.
Во внутреннем сегменте находятся митохондрии, рибосомы,
комплекс Гольджи и ЭПС (рис. 120).Аксонтщ1Внутреппийсе,гм<хпшМитохондрияРесничка::Ларуж1шйсегментКолбочкаПалочкаРис. 120 . Сіроение фотосенсорных клеток (палочек и колбочек).208
ГЛАВА 14.0РГАНЫ ЧУВСТВУ колбочек в наружном сегменте диски не замкнуты, во
ВИуїрСННСМ СС1МСИТС колбочек имеется эллипсоид, состоящий из
липидной капли и митохондрий. От ядросодсржащей части отходит
аксоп^ который образует синапсы с дендриташі биполяров и
горизонтальных клеток.Колбочки содержат три типа зрительных пигментов, один из
них — гюдопсин. ^.Горизонтальные нервные клетки контактируют с аксонами
фоторецепторных клеток. Аксоны горизонтальных нейронов вступают
в контаїа с аксонами шж палочковых, так и колбочковых клеток.Биполярные нервные клетки соединяют палочковые и
колбочковые нейроны с ганглиозными клетками сетчатки. В
центральной части сетчатки несколько палочковых нейронов
соединяются с одной биполяріюй, а колбочковые нейроны
контактируют в соотношении 1:1 или 1:2. Такое сочетание
обеспечивает более высокую остроту цветового видения по
сравнению с черно-белым.Амакриниые клетки относятся к нейронам, которые
осуществляют связь: фоторецептор - биполяр - ганглионарная клетка.Ганглионарные клетки собирают информацию от всех слоев
сетчатки и передают ее в мозг. Аксоны ганглионарных клсток (более
миллиона волокон) формируют внутренний слои нервных волокон,
іюрсходящий в зрительный нерв.Нейроглия сетчатки представлена клетками микро- и
макроглии.Пигментный эпителий - наружный слой сетчатки - состоит из
пигментоцйтов (4-6 млн.). Апикальная поверхность пигментоцитов
имеет отростки, в цитоплазме содержатся меланосомы. Наличие
меланосом обусловливает поглощение 85-90 % света, попадающего в
глаз. Под воздействием свста мсланосомы перемещаются в
апикальные отростки пип^тентоцитов, а в темноте возвращаются в
цитоплазму.Фшиологмческая регенерация палочковых и колбочковых
нейронов происходит в течение всей жизни, процесс обновления
каждой палочковой клетки длится 9-12 дней.Особенности кровоснабжения. Ветви глазничной артерии
образуют ретинальную сосудистую систему сетчатки, а также
цилиарную систему, снабжающую кровью сосудистую оболочку,
цилиарное тело, радужку и склеру. Лимфатические капилляры
располагаются только в склеральной конъюнктиве.209
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ 1Вспомогательный аппарат глазаК вспомогательному аппарату глаза относятся глазные
мышцы, веки и слезный аппарат. Глазные мышцы представлены
поперечнополосатыми мьшхечными волокнами.Веки. В них разгптчают переднюю кожную поверхность и
задшою - конъюнктиву, которая продолжается в конъюнктиву глаза,
покрытую многослойным эпителием.Конъюнктива - тонкая соединительнотканная пластинка с
многослойш>ш плоским неороговеваюш,им эпителием, которая
покрывает заднюю поверхность век и переднюю часть глазного
яблока.Слезный аппарат глаза. Он состоит из слезопродуцирующей
слезной железы и слезоотводящих путей - слезное мясцо, слезные
канальцы, слезный мешок и слезно-носовой канал.Возрастные изменения. С возрастом ослабляется функция
всех аппаратов глаза. В связи с изменением общего метаболизма в
организме в хрусталике и роговще часто происходят уплотнение
межклеточного вещества и помутнение, у пожилых людей
откладываются липиды в роговице и склере, что обусловливает их
потемнение. Снижается аккомодационная возможность хрусталика.
Склеротические процессы в сосудистой системе глаза приводят к
изменению структуры и функции рецепторного аппарата.Орган обонянияОбонятельный анализатор представлен основ1юй и
вомсроназальной системами, каждая из которых имеет три части:
периферическую (орган обоняния), промежуточтгую, состоящую из
проводников (аксоны нейросенсорных обонятельных клегок и
нервных клеток обонятельных луковиц) и центральную.Основной орган обоняния представлен ограниченным
участком слизистой оболочки носа желтоватого цвета - обонятельной
областью, покрывающей верхнюю и частичтю среднюю раковины
носовой полости, а также верхнюю часть носовой перегородкл!.Вомероназальный орган (якобсонов) расхюложен в
соединителАюй ткани передней третти носовой перегородки и имеет
вид парных эпителиальных трубок. Его функция связана с фyнкция^ffl
половых органов и эмоциональной сферой. Вомероназальпая система
включает в себя вомероназальный нерв, терминальный нерв и
добавочную обонятельную луковицу.Орган обоняния развивается из плакод, в области которых
формируются обонятельные ямки, поддерэ/атающие эпителиогщиты и
иейросенсорные обонятельные тетки. Аксоны обонятельных клеток
образуют 20-40 нервных пучков, направляющихся обонятельным210
ГЛАВА 14.ОРГАНЫ ЧУВСТВлуковицам головного мозга. Орган обоііялия состоит из многорядного
эпителия, покрытого слоем слизи, в котором различают обонятельные
нейросенсорные (рецепторные) клетки, поддерживающие и базальные
эпителиоциты (рис. 121).Рис. 121, Строение обонятельного эпителия. I - обонятельный эпителий.
II - собственная пластинка слизистой оболочки; 1 - обонятельные клетки;
2 - обонятельные реснички; 3 - поддерживающие эпителиоциты;4 - базальные эпителиоциты; 5 - базальная мембрана; 6 - слизистые
железы, 7 - обонятельные нити.Рецепторные клетки располагаются между
поддерживающими эпителиоцитами. Дендриты обонятельных клеток
заканчиваются обонятельными булавами с 10-12 подвижными211
гистология, Г^ИТОЛОГИЯ, ЭМБРИОЛОГИЯ ЧАСТЬ 1ресничками. Цитоплазма периферических афоетков содержиг
митохондрии и лшкротрубочки. От базальной части клетки отходит
аксои, их совокупности составляЕот пучки обонятельного нерва,
которые объединяются в 20-40 обонятельных нитей и найравляютея в
обонятельные луковицы.Поддерживающие эпитечиоциты форм1фуют многорядпый
эпителиальный 1шаст. На апикальной поверхности этих клеток
имеются многочисленные микроворсинки длиной до 4 мкм, в
цитоплазме содержится коричнсво-жс;ггый ПИГМСП'Г,
Поддерживающие эпителиоциты проявляют признаки апокриновой
секреции.Базальные эпнтелгюгщты служат иегочииком регенерации
рецепторных клеток.Центральный отдел обонятельной сенсорной системы
локализуется в гиппокампе и гиппокамповой извилине, здесь
происходит аначиз обонятельной информации.В соединительной ткани обонятельной области располагаются
концевые отделы трубчато-альвеолярных желез, секрет которых
вместе с секретом поддерживающих эпителиоцитов увлажняет
поверхность обонятельного эпителия. Слизистая оболочка полости
носа обильно снабжена лимфатическими и кровеносными сосудалш,
которые способны депонировать кровь. При действии различных,
раздражителей слизистая оболочка носа сильно набухаст и
покрываст'ся слоем слизи, что затрудняет восприятие запахов.
Обновление обонятельных клеток происходит в течение 30 суток.Орган вкусаОрган вкуса - это периферическая часть вкусового
анализатора, которая представлена рецепторными эпителиальными
клетками вкусовъи: почек (2000). Вкусовые почки располагаются в
многослойном плоском эпителии боковых СТС1ЮК жслобовахых,
листовидных и грибовидных сосочков языка человека. У дсгсй
вкусовые почки могут находиться на губах, задней стенке глотки,
небных дужках, надгортаннике.Вкусовая почка состоит из 40-60 клеток, среди которых
различают: сенсоэпителиалъные, поддерживаюп^ие и базальные
клегки. Вершина почки сообщается с поверхностью слизистой
оболочки при помощи вкусовой поры. Вкусовая пора ведет во
вкусовую ямку (рис. 122).212
ГЛАВА 14.0РГАНЫ ЧУВСТВВкусовая пораМикроворсинкиСепсоэпи-телиальиыеклеткиРис, 122. Вкусовая почка (схема).Сенсоэпителиальные клетки содержат микроворсинки. С
цитолеммой базальной части клеток контактируют дендриты
чувствительных нейронов, которые передают импульс в
промежуточную часть анализатора, а также афферентные нервные
волокна Во вкусовых почках передней части языка обнаружен
сладкочувствительный рецепторный белок, задней части
горькочувствительный.Поддержнваюгцт эпителиоциты содержат много
митохондрий, мембран ГЭПС и свободных рибосом. Сенсорные и
поддерживающие :>пителиоциты вкусовой почки обновляются в
течение 10 суток.Статоакустическая сенсорная система. Орган слуха и
равновесияНаружное ухо включает ушную раковину, наружный
слуховой проход и барабанную перепонку.Ушная раковина состоит из тонкой пластинки
эластического хряща, покрытой кожей с немногочисленными
тонкими волосами и сальными железами (рис. 123). Наружный
слуховой проход образован хрящевой и костной частью.
Поверхность прохода покрыта тонкой кожей, содержащей вoJЮcы и
сальные железы. Глубже сальных желез расположены
церуминозные железы, выделяющие ушную серу. Барабанная
перепонка состоит из пу^шов коллагеновых и эластических волокон213
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ, ЧАСТЬ I.и имеет овальную, слегка вогнуїую форму. Одна из слуховых
косточек среднего уха - молоточек - сращена с внутренней
поверхностью барабанной перепонки. На наружной поверхности
перепонки располагасгся эпидермис, на внутренней поверхности -
слизистая оболочка.Среднее ухо состоит из барабанной полости, слуховых
косточек и слуховой трубы. На медиальной стенке барабанной
полости имеется овачьное окно, в нем располагается основание
стремечка. Овальное окно отделяет барабанную поіюсть от
вестибулярной лестницы улитки. Второе окно круглое, закрыто
мембраной. Круглое окно отделяс'г барабанную пoJЮCть от
барабанной лестницы улитки (рис. 124).Слуховые косточки - молоточек, наковальня, стремечко
передают колебания барабанной перепоііки наружного уха к
овальному окну, от которого начинается вестибулярная лестница
внутреннего уха.НаружноеухоСреднее ухоВнутреннее ухо
ухоРис. 123. Наружное, среднее и внуфеннее ухо.214
ГЛЛВЛ 14.0РГАНЫ ЧУВСТВСлуховая труба, соединяющая барабанную полость с
носовой частью глотки, имеет просвет диаметром 1-2 мм. Чqpcз
слуховую трубу регулируется давление воздуха в полости среднего
уха.124, Внутренне ухо. 1- улитковый канал перепопчагого лабиринта,2 - вестибулярная лестница, 3 - барабанная лестница, 4 - спиральный узел,5 - костный стержень. Стрелками указано направление распространениязвуковой волны.Внутреннее ухоВн утреннее ухо состоит из костного лабиринта и
pacпoJЮжcннoгo в нем перепончатого лабиринта. Рецепторные
клетки органа слуха расположены в спиральном органе улитки, а
рецепторные клетки органа равновесия - в эллиптическом и
сферическом мешочках и ампулярных гребешках полукружных
каналов.Перепончатый лабиринт образуется из эктодермы первой
жаберной щели, которая образует слуховой пузырек. В дальнейшем
пузырек делится на две части; вестибулярная часть превращасгся в
эллиптический мешочек с полукружными каналами и сферический
мешочек. Спиральной орган (кортисв) развивается из эпителия
базальной стснки перепончатого канала.Улитковый каналУлитковый канал представлясг собой спиральный мешок
длиной 3,5 см, заполненный эндолимфой и окруженный снаружи
перилимфой. Улитковый канал и окружаюпще его полости
барабанной и вестибулярной лестницы заключены в костную
улитку, образующую 2 /2 завитка.215
гистология, ЦИТОЛОГИЯ, ЭМБРИОЛОГИЯ. ЧАСТЬ I.На поперечном разрезе улитковый канал имеет треуголы!ую
форму, его стенки образованы вестибулярной мембраной,
сосудистой полоской и базилярной пластинкой.Сосудистая полоска (наружная стенка) расположена на
спиральной связке. В многорядном эпителии находятся капилляры,
эпителиальные клетки продуцируют эндолимфу. Базилярная
пластинка с внутренней стороны прикрепляется к спиральной
костной пластинке. Надкостница (лимб) делится на вестибулярную
барабанную губу. Барабанная губа продолжается в базилярную
пластинку, которая прикрепляется к спиральной связке. В основе
базилярной пластинки находятся коллагеновые волокна разной
длины. Длинные волокна (505 мкм) находятся на вершине улитки,
короткие (105 М1ш) - в се основании. Волокна сосюят из фибрилл
(30 нм). Со стороны барабанной лестницы базилярная пластинка
покрыта эндотелием, с противоположной стороны - базальной
мембраной эпителия кортиевого органа (рис. 125),Рис. 125. Строение перепончатого канала улитки. 1 - перепончатый канал
улитки; 2 - вестибулярная лестница; 3 - барабанная лестница;4 “ спиральная костная пластинка; 5 - спиральный узел; 6 - вестибулярная
мембрана; 7 - базилярная мембрана; 8 - спиральная связка; 9 - сосудистая
полоска; 10 - покровная пластинка; 11 - туннель.216
ГЛАНА 14.0РГАНЫ ЧУВСТВПоверхность спирального лгшба покрыта плоским
эпителием. Клетки спиральной бороздки переходят в
поддерживающие эпителиоциты, примыкающие к внутренним
волоскоЕым клеткам спирального органа. Покровная
(текториалъная) мембрана имеет желеобразную консистенцию и
располагается над вершинами волосковых клеток.\Спиральный органСпиральный (кортиев) орган расположен на базилярной
пластинке исреиончатого лабиринта улитки. В его составе
различают сенсоэпителиальные и поддерживающие клсаки,
которые подразделяется на внутренние и наружные (рис. 126).
Разграничивает эти клетки туннель.Внутренние сенсоэпителиальные клетки (3500) имеют
кувтинообразную форму и расположены в один ряд на
по,ддерживающих эпителиоцитах.На апикальной поверхности находится ретикулярная
пластинка и 30-60 коротких отростков - стереоцилий. В цитоплазме
клеток содержатся митохондрии, эндоплазматическая сеть,
актиновые и миозиновые миофиламенты. К основанию клсіки
подходят афферентные и эфферентные нервные окончания.Рис. 126. Кортиев орган. 1 — нокровная мембрана, 2 - туннель,3 - базилярная пластинка, 4 - эпителиальные клетки, 5 ~ внутренние
сенсоэпителиальные клетки, 6 - внутренние клетки-сголбы, 7 - наружные
сенсоэпителиальные клетки, 8 - наружные поддерживающие клетки,9 - наружные клетки-столбы.217
гистология, цитология, ЭМБРИОЛОГИЯ. ЧАСТЬ IНаружные сенсоэпителиальные клетки (12000-20000) имеют
цилиндрическую форму, образуют 3-5 рядов на поддерживающих
наружных фалашовых зпитслиоциіах. На апикальной поверхности
имеется кугикулярпая плаиипка со сгсреоцилиями, которые образуют
щеточку в виде буквы V (рис. 127). Стереоцилии наружных
волосковых клеток вершинами прикасаются к внутренней
гюверхпосш покровной мсмбраиы. в облас^ги стереоцилии
присутствует фермент ацстилхо;шпэсгсраза. Высокие звуки
раздражают волосковые клетки, расположенные в нижних завитках
улитки, а низкие звуки - волосковые клетки вершины улитки. Ув:-:-' ^Рис. 127. Электронная микроскопия наружных (А) и внутренних (Б)
сенсоэпителиальных клеток. 1-стереоцилии.при воздействии звука барабанная перепонка передаёт
колебания на молоточек, наковальню и стремечко, через овальное
окно на перилимфу, базилярную и текториальную мембраны. При
этом происходят отклонение стереоцилий, возбуждение
рецепторных клеток, Б последствие возникает микрофонный
эффект'. Далее информация по слуховому нерву передается в
центральные части слухового анализатора.Поддерживающие эпителиоциты спирального органа
располагаЯотся на базальной мембране. Внутренние фаланговые
эпителиоциты на апикальной поверхности имеют тонкие
пальцевидные отростки (фаланги). Этими отростками вершины
рецепторных клсток отделены друг от друга. Наружные
фаланговые клетки призматической формы залегают в 3-4 ряда.Внутренние и наружные столбовые эпителиоциты сходятся
под острым углом друг к другу и образуют треугольный туннель,
заполненный эндолимфой. Через туннель проходят безмиелиновые218
ГЛАВА 14.0РГАНЫ ЧУВСТВнервные волокна, идущие от нейронов спирального ганглия к
сенсорным клеткам.Вестибулярная часть перепончатою лабиринтаЗдесь располагаются рецепторы органа равновесия. Она
состоит из двух мещрчков - эллиптического (utriculus) и
сферического (sacculus), связанных с тремя полукружными
каналами и выстланных однослойным плоским эпителием. Каналы
на месте соединения с эллиптическим мешочком имеют
расширения - ампулы. В мешочках есть участки, содержащие
чувствительные (сенсорные) клетки, эти участки назьшаются
пятнами, или макулами. В ампулах различают гребешки (кристы).Пятна мешочков (макулы) выстланы эпителием, состоящим
из сенсорных и опорных клегок (рис, 128). На поверхности
эпителия находится отолитов ая мембрана, в составе которой
находятся кристаллы карбоната кальция - отолиты. В области
макулы эллиптического мешочка происходит восприятие линейных
ускорений и земного притяжения. Макула сферического мешочка,
являясь рецептором гравитации, дополнительно воспринимает
вибрационные колебания.-.'■,.'■*'’4 ' MaKvnaШ "" "’-ЩРис. 128. Макула. 1 - поддерживающие эпителиоциты;2 - сенсоэпителиальные клетки; 3 - волоски; 4 - отолитовая мембрана;5 - отолиты.Сенсорные волосковые клетки в области базальной части
контактирует с афферентными и эфферентными нервными
окончаниями. Различают клетки первого типа (грушевидные) с219
гистология, цитология, ЭМБРИОЛОГИЯ. Ч.ЛСТЬ Iшироким основанием, к которому примыкает нервное окопчаиис,
образующее вокруг него футляр в виде чаши. Клетки второго типа
(столбчатые) имеют призматическую форму, в основании
различают точечные афферентные и эфферентные нервные
окончания. Апикальная поверхность клеток имеет кутикулу, от
которой отходят 60-80 неподвижных стереоцилий (40 мкм) и одна
подвижная киноцилия (рис. 129). При смещении стереоцилий в
сторону киноцилии клетка активизируется. Клетки макул образуют4 группы, во время скольжения отолитовой мембраны
стимулируется только одна группа, рех'улирующая тонус
определенных мышц туловища. Поддерживаюгцие эпителиоциты
на апикальной повсрхносги содержат множество микроворсинок.Тип 1 Тип IIКиноцилияСтеиеоцилииБазальноетельцеАфферент¬
ное нервное
окончаниеАфферентныйнервЭфферентное нервное окончаниеРис. 129. Сенсорные волосковые клетки макулы I и II типов.220
ГЛАВА 14.0РГАНЫ ЧУВСТВАмпулярные гребешки (кристы) находятся в ампулярных
распгирениях полукружных каналов. Сенсорные волосковые и
поддерживающие эпителиоциты покрыты прозрачным куполом
высотой 1 мм (рис. 130). Строение волосковых клеток сходно со
строением сенсорных клеток мешочков. Желатинозный купол
является рецептором угловых ускорений. Отклонение купола при
движении оидолимфы ^ в полукружных каналах раздражает
волосковые клетки. Их возбуждение вызывает рефлекторный ответ
той части скелетной мускулатуры, которая корригирует положение
тела и движение глазных мышц.♦ \ 'V i-, iV:>.Рис. 130. Ампулярные гребешки. 1 - поддерживающие эпителиоциты;2 - сенштптелиальиыс кгаетки; 3 - купол.Иннервация. К базалвной части сенсорных эпителиоцитов
спирального и вестибулярного органов подходят нервные
окончания биполярных нейронов, тела которых образуют
спиральный гаш’лий. lío аксонам биполярных клеток через ядра
продолговатого мозга звуковое раздражение передаётся в верхнюю
височную извилину.Во'Зрастныс изл1снения. С возрастом у человека нарушения
органа слуха связаны с тем, что в области овального окна костного221
гистологая, цитология, эмбриология, часть Iлабиринта появляется усиленное отложение солей кальция.
Стремечко, вставленное в овальное окошко, теряет подвижность,
что резко снижает порог слышимости. С возрастом гибнет
значительное количество сенсорных клеток, что отражается на
качестве звуковых восприятий.222
СОДЕРЖАНИЕПредисловие к первому изданию 3ГЛАВА 1.ПРЕДМЕТ И ЗАДАЧИ^ГИСТОЛОГИИ И ЦИТОЛОГИИ,ИСТОРИЯ РАЗВИТИЯ, МЕТОДЫ ИССЛЕДОВАИИЯ 4ГЛАВА 2.ЦИТОЛОГИЯ 13ГЛАВА 3.ПРИНЦИПЫ ОРГАНИЗАЦИИ ТКАНЕЙ 43ГЛАВА 4.ОСНОВЫ ЭМБРИОЛОГИИ 47ГЛАВА 5.ЭМБРИОЛОГИЯ ЧЕЛОВЕКА 60ГЛАВА 6.ЭПИТЕЛИАЛЬНЫЕ ТКАНИ 78ГЛАВА 7.КРОВЬ И ЛИМФА 92ГЛАВА 8.КРОВЕТВОРЕНИЕ 110ГЛАВА 9.СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ ТКАНЬ 120ГЛАВА 10.СКЕЛЕТНЫЕ ТКАНИ 135ГЛАВА П.МЫШЕЧНЫЕ ТКАНИ , 150ГЛАВА 12.НЕРВНАЯ ТКАНЬ 164ГЛАВА 13.НЕРВНАЯ СИСТЕМА 185ГЛАВА 14.ОРГАНЫ ЧУВСТВ 200223
Учебное изданиеКАЩЕНКО С.А.
БОБРЫШЕВА И.В.ГИСТОЛОГИЯЦИТОЛОГИЯЭМБРИОЛОГИЯЧАСТЬ IУчебник
В авторской редакцииПодписано в печать 01.08.2012 г.Формат 60x84 1/16. Бумага офсетная. Гарнитура Times.
Печать офсетная, Условн. печ. л. 13,02. Уч. изд. л. 13,87.
Тираж 900 экз. Иад. №647. Зак. №115.Цепа договорная.Издательство "НОУЛИДЖ”Свидетельство о регистрации ДК №2884 от 26,06.2007
91051, г. Луганск, кв, Якира, 3/316,
тел, (050) 475-35*13, e-maü: пickvnu@gmai 1.соmПечать МЧП «Глобус», г. Луганск, ул. Лермонтова, 16, оф. 314
тел,: 711-777