























































































































































































































































































































Автор: Грин Н. Стаут У. Тейлор Д.
Теги: общая экология биоценология гидробиология биогеография общая биология биология экология издательство мир размножение учебник общей биологии опорно-двигательные системы гомеостаз осморегуляция
ISBN: 5-03-001602-3
Год: 1990




Н.ГРИН, У СТАУТ, Д.ТЕЙЛОР
БИОЛО ГИ Я
3
ИЗДАТЕЛЬСТВО «МИР»
BIOLOGICAL
SCIENCE
N.P.O. GREEN B.Sc., M.I.B10I
Headmaster
Sutton Manor High School for Boys,
Sutton, Surrey
G.W. STOUT B.Sc., M.Ed., M.I.Biol
Secretary for Biological Sciences
University of Cambridge Local Examinations Syndicate
DJ. TAYLOR B.Sc, Ph.D., M.I.Biol
Head of Biology
Strode's Sixth Form College, Egham
Editor
R. SOPER B.Sc., F.LBiol
Formerly Vice-principal and Head of Science
Collyers Sixth Form College, Horsham
CAMBRIDGE UNIVERSITY PRESS
Cambridge
London New York New Rochelle
Melbourne Sydney
H. ГРИН
У СТАУТ
Д.ТЕЙЛОР
Биология
3
В 3-х томах
ТОМ Под ред. P. Сопера
Перевод с английского под редакцией
E. P. Наумовой, д-ра биол. наук
канд. биол. наук Б. M. Медникова
M. С. Морозовой, и
канд. мед. наук Д"Ра биол· наУк
О. В. Протасовой Α· Α· Неифаха
Москва «Мир» 1990
ББК 28.0
Г85
УДК 574.577
Грин H., Стаут У., Тейлор Д.
Г85 Биология: В 3-х т. T. 3.: Пер. с англ./Под ред. P. Сопера.-
M.: Мир, 1990.-376 с, ил.
ISBN 5-03-001602-3
Современный учебник общей биологии, написанный английскими авторами. Весь материал
изложен на весьма высоком уровне, но простота изложения и удачное расположение тем делают
его доступным для неподготовленного читателя.
В т. 3 рассматриваются опорно-двигательные системы, гомеостаз, процессы выделения и
осморегуляции; размножение, рост и развитие; вопросы наследственности, изменчивости и
эволюции. В книгу включен также дополнительный материал по проблемам экологии из
последнего английского издания 1989 г.
Для студентов-биологов, преподавателей биологии в школе, биологов всех специальностей,
а также учащихся старших классов.
1901000000-007 ББК 28.0
Г 00-90, чЛ
041(01)-90
Редакция литературы по биологии
ISBN 5-03-001602-3 (русск.)
ISBN 5-03-001576-0
ISBN 0-521-26951-2 (англ.)
© Cambridge University Press 1985
This book was originally published in
the English language by Cambridge
University Press of Cambridge,
England
First published 1985
Ninth printing 1989
© перевод на русский язык,
коллектив переводчиков, 1990
Глава 17
Движение и опора
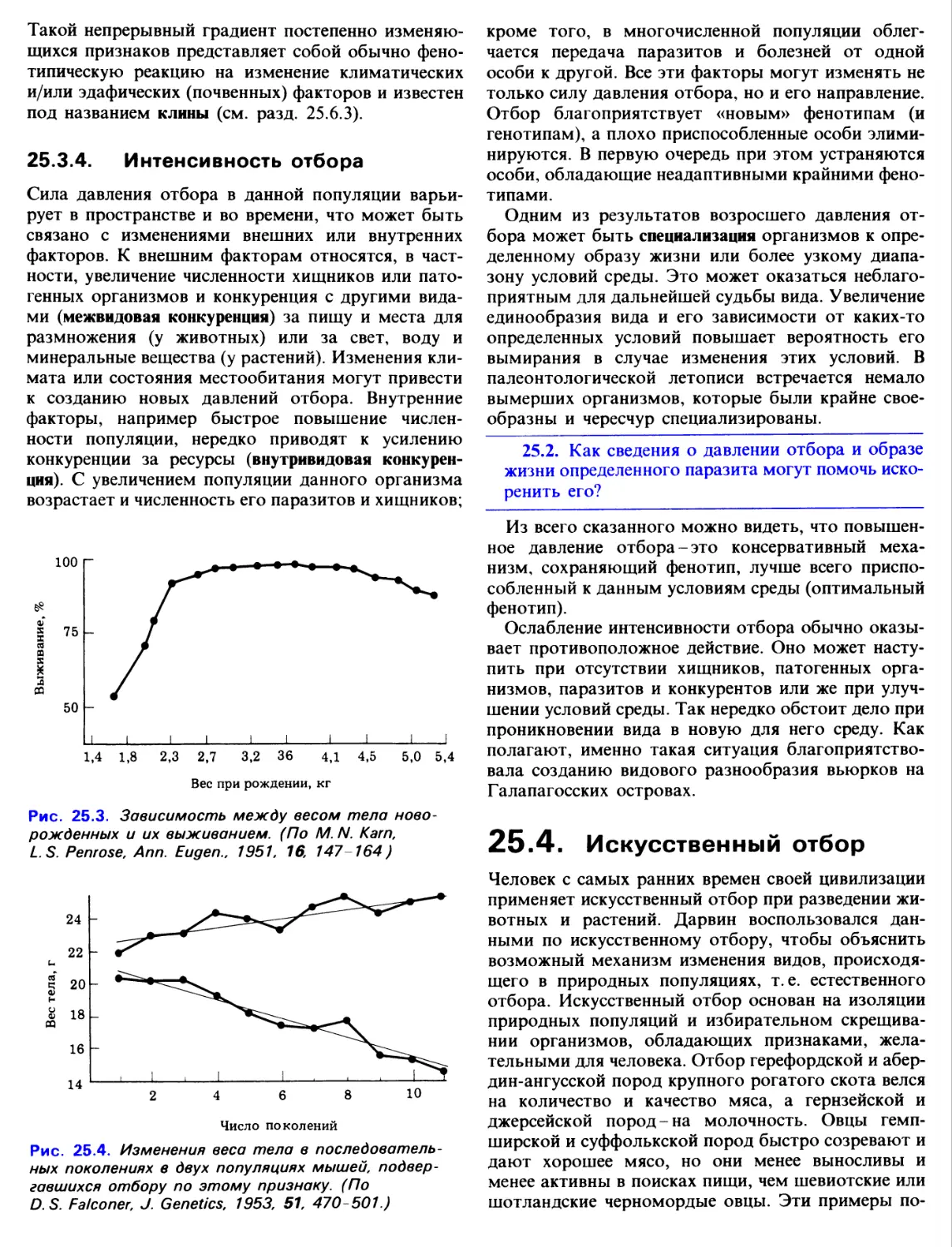
Движение может происходить на уровне клетки
(например, перетекание цитоплазмы или плавание
гамет), на уровне органа (сокращение сердца,
движение конечности и т. д.) или на уровне целого
организма. Передвижение всего ор1анизма с одного
места на другое называется локомоцией. Растениям
свойственно движение на клеточном и часто на
органном уровнях, локомоторная же активность,
т. е. перемещение всего организма в поисках пищи
или воды, у них отсутствует.
У огромного большинства животных в процессе
эволюции выработались сложные локомоторные
системы, позволяющие искать и добывать пищу.
Лишь немногие животные успешно приспособились
к сидячему образу жизни; однако даже у них
отдельные части тела отличаются большой
подвижностью.
Для некоторых животных локомоторная
активность-это не только способ поиска пищи, но и
средство для спасения от хищников. Кроме того,
перемещаясь, животные расселяются, осваивают
новые благоприятные местообитания, а также находят
себе половых партнеров.
Локомоция стала возможной в результате
формирования, взаимодействия и координированной
работы нервной, мышечной и скелетной систем.
Мышцы, участвующие в локомоции, прикреплены к
частям скелета, и их называют скелетными
мышцами (разд. 17.4.1). Они работают как машины,
преобразующие химическую энергию в механическую.
Мышцы способны сокращаться и при этом
приводить в движение системы рычагов, составляющие
часть скелета. Благодаря координированной работе
рычагов животное перемещается. Скелетно-мышеч-
ная система обеспечивает также поддержание позы
и находится под общим контролем центральной
нервной системы.
Другие мышцы не участвуют в передвижении всего
организма, но обеспечивают перемещение веществ
внутри тела. Сердечная мышца (разд. 8.5)
прогоняет по всему телу кровь, а сокращение или
расслабление гладкой мускулатуры (разд. 17.5.3) в
стенках различных кровеносных сосудов, изменяя их
просвет, регулирует кровоток. Гладкая мускулатура
кишечника своими волнообразными сокращениями
(перистальтика) проталкивает пищу по кишечному
тракту (разд. 10.4.11). Это лишь некоторые из
многочисленных процессов такого рода,
происходящих в организме.
В этой главе нас будет интересовать в первую
очередь локомоторная активность, и мы подробно
рассмотрим две системы организма-скелетную и
мышечную, а затем познакомимся с типами
локомоции, свойственными различным организмам.
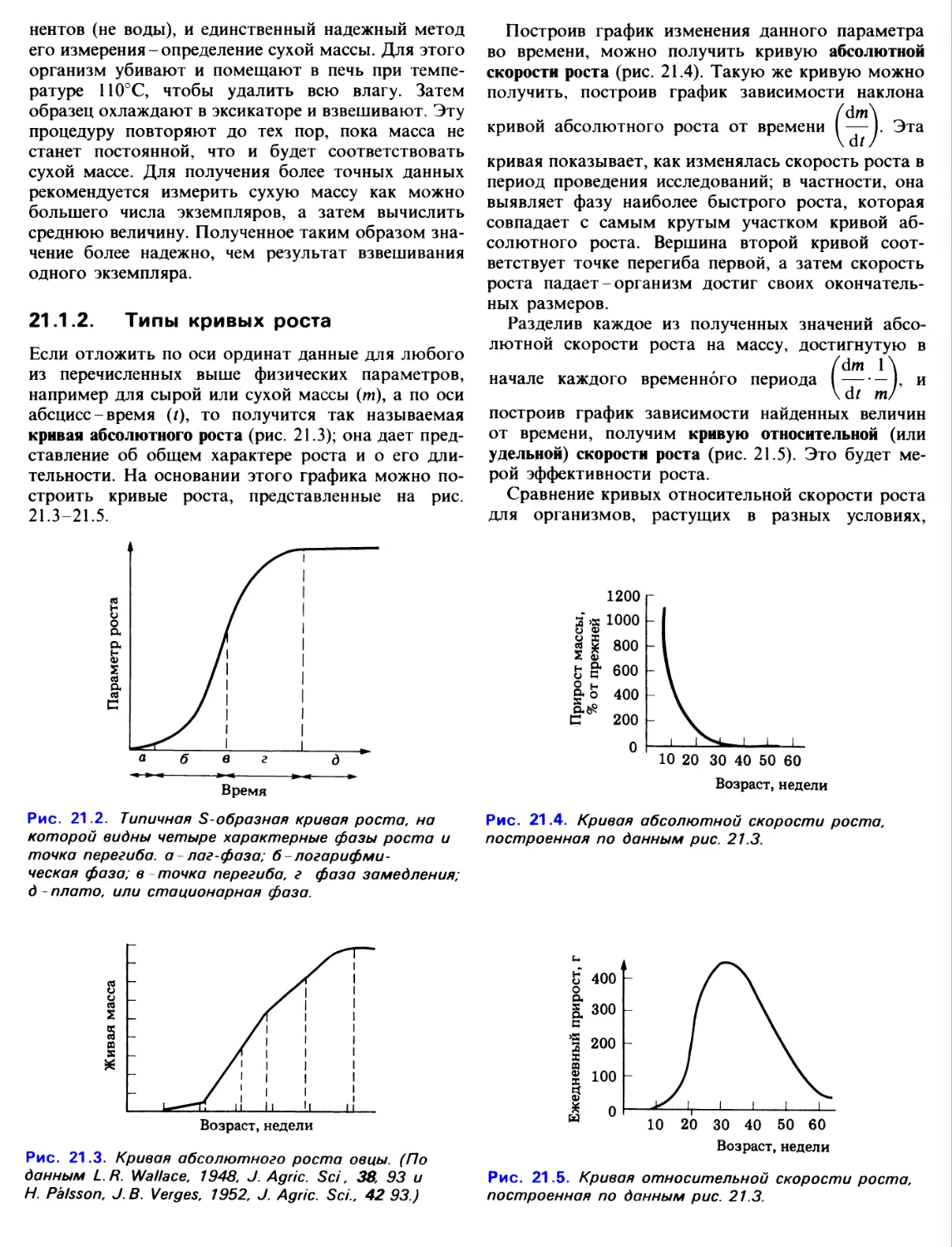
17.1 Скелетные системы
У огромного большинства животных имеются
разного рода опорные структуры, начиная от простых
плотных палочковидных образований у простейших
до сложно устроенного скелета у членистоногих и
позвоночных. Опорные системы, как правило,
обеспечивают характерную форму тела, а она в свою
очередь обусловлена определенными
потребностями организма. Поэтому такие системы у животных,
обитающих на суше или в воде, передвигающихся
на двух или четырех ногах, по земле или по воздуху,
неизбежно должны иметь разное строение.
Основные функции скелета следующие:
1. Опора. Скелет любого типа служит жестким,
устойчивым к сжатию каркасом тела. Он
помогает телу сохранять определенную форму. У
наземных организмов скелет обеспечивает опору
для всей массы тела, противодействуя силе
тяжести, и во многих случаях приподнимает тело
над землей. Это облегчает животным
передвижение по суше. Внутренние органы оказываются
закрепленными и подвешенными к скелету.
2. Защита. У некоторых животных (членистоногих)
имеется экзоскелет, или наружный скелет,
который защищает нежные внутренние органы. У
других животных эту функцию выполняют части
эндоскелета - внутреннего скелета. У человека,
например, черепная коробка обеспечивает
защиту головного мозга и органов чувств (зрения,
обоняния, равновесия и слуха),
позвоночник-защиту спинного мозга, а ребра и грудина-защиту
сердца, легких и крупных кровеносных сосудов.
3. Локомоция. Скелет, построенный из жесткого
материала, служит местом прикрепления мышц.
При сокращении мышц части скелета работают
как рычаги, и это приводит к различным
движениям. У мягкотелых животных при движении
опорой для сокращающихся мышц служит
полостная жидкость.
Различают три основных типа скелета:
гидростатический скелет, экзоскелет и эндоскелет.
17.1.1. Гидростатический скелет
Этот тип характерен для мягкотелых животных. У
них имеется полостная жидкость, заключенная
внутри мышечных стенок тела. Эта жидкость
оказывает давление на мышцы, а те в свою очередь
способны сокращаться, преодолевая это давление.
Мышцы не прикреплены к каким-либо структурам,
и поэтому при сокращении они тянут лишь друг
друга. Животное сохраняет определенные размеры
и форму тела благодаря давлению полостной
жидкости, с одной стороны, и сокращающихся мышц-с
другой. Обычно мышечные волокна образуют два
слоя - продольную и кольцевую мускулатуру.
Движение происходит благодаря тому, что эти слои
работают как антагонисты. У ^сегментированных
животных (таких, как нематоды) давление на
жидкость при сокращении мышц передается во все части
тела. У сегментированных животных (таких, как
Lumbricus terrestris - обыкновенный дождевой червь)
этот эффект локализован и лишь определенные
сегменты приводятся в движение или изменяют
форму. Подробно роль гидростатического скелета
при движении на примере дождевого червя будет
рассмотрена в разд. 17.6.5.
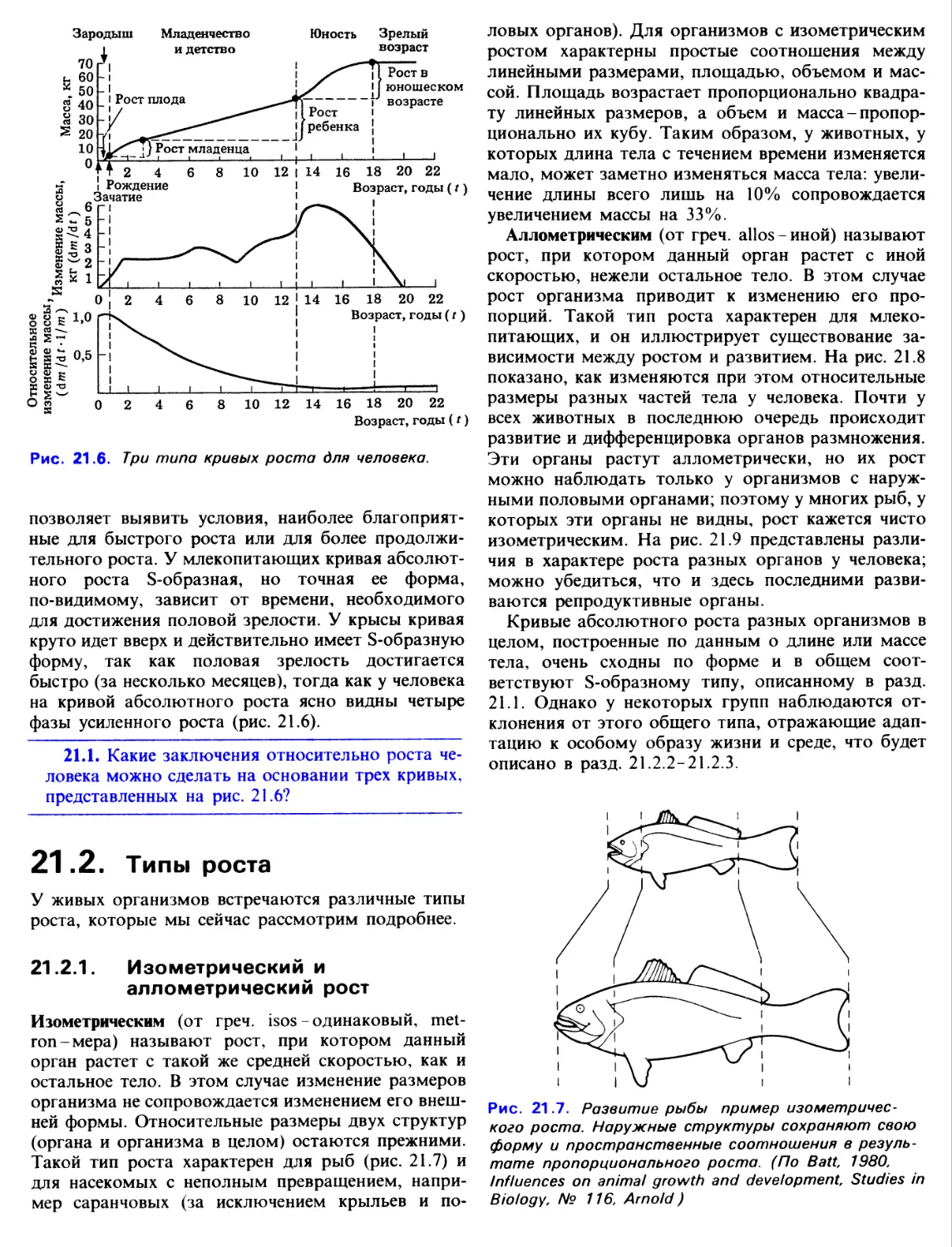
17.1.2. Экзоскелет
Скелет этого типа - характерная особенность
членистоногих. Экзоскелет (кутикула) секретируется
эпидермисом; он не содержит клеток и состоит в
основном из хитина. Это твердый наружный покров
тела, который построен из сочлененных между
собой пластинок или трубчатых образований. Трахеи,
передний (стомодеум) и задний (проктодеум)
отделы пищеварительного тракта тоже выстланы
хитином. Хитин-прочный, легкий материал, однако
он может приобретать твердость при встраивании в
него «задубленных» белков или при обызвествлении
(особенно у водных ракообразных). В участках
скелета, которые должны сохранять подвижность,
например в местах сочленения пластинок, хитин
остается неизмененным. Такая конструкция из
пластинок или трубочек, соединенных гибкими
пленками, обеспечивает одновременно и защиту, и
подвижность.
Членистоногие - это единственная группа
беспозвоночных, имеющая членистые конечности,
которые состоят из рычагов, соединенных подобием
шарниров. Эти рычаги приводятся в движение
мышцами-сгибателями и разгибателями,
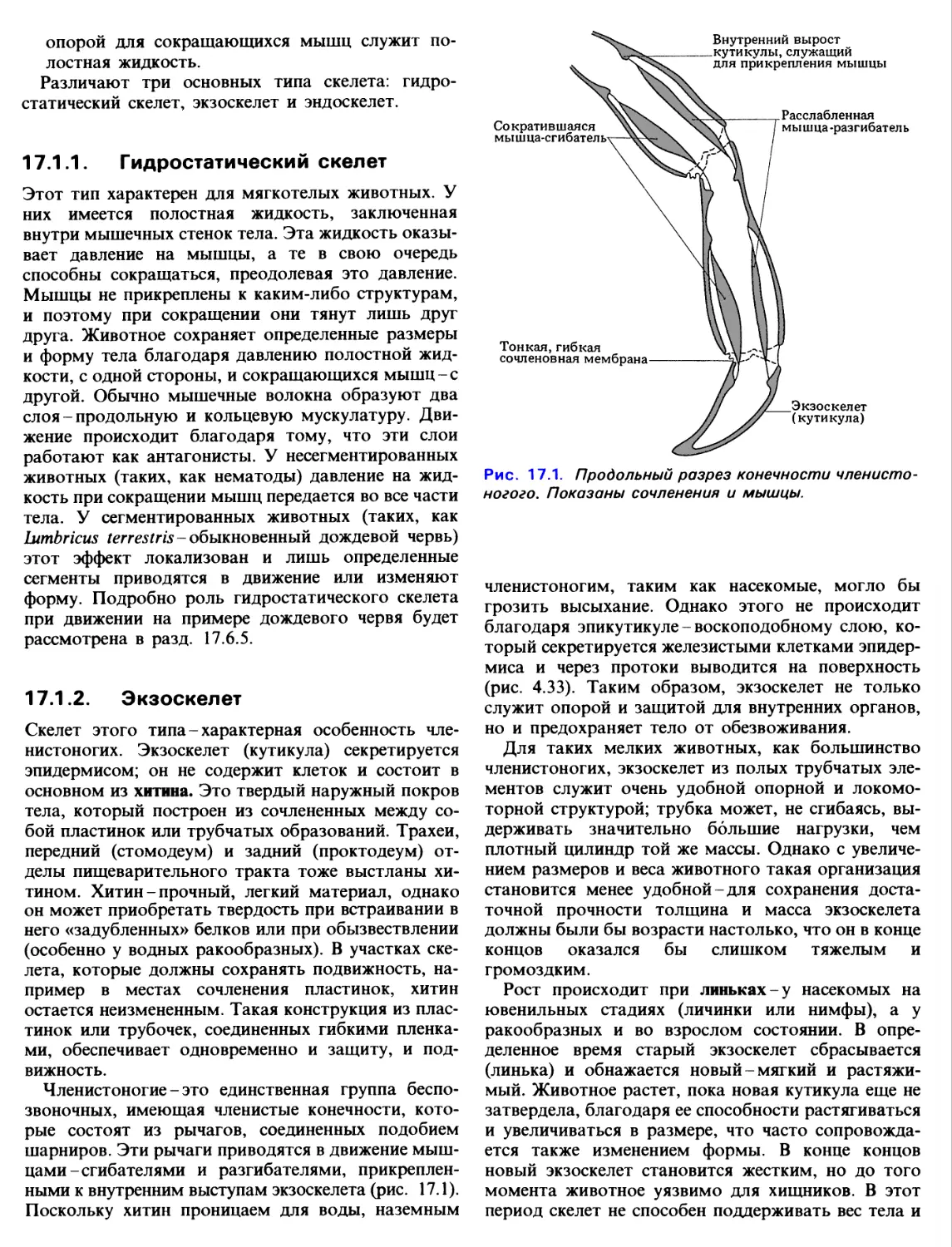
прикрепленными к внутренним выступам экзоскелета (рис. 17.1).
Поскольку хитин проницаем для воды, наземным
Сократившаяся
мышца-сгибатель
Внутренний вырост
кутикулы, служащий
для прикрепления мышцы
Расслабленная
мышца-разгибатель
Тонкая, гибкая
сочленовная мембрана
Экзоскелет
(кутикула)
Рис. 17.1. Продольный разрез конечности
членистоногого. Показаны сочленения и мышцы.
членистоногим, таким как насекомые, могло бы
грозить высыхание. Однако этого не происходит
благодаря эпикутикуле - воскоподобному слою,
который секретируется железистыми клетками
эпидермиса и через протоки выводится на поверхность
(рис. 4.33). Таким образом, экзоскелет не только
служит опорой и защитой для внутренних органов,
но и предохраняет тело от обезвоживания.
Для таких мелких животных, как большинство
членистоногих, экзоскелет из полых трубчатых
элементов служит очень удобной опорной и
локомоторной структурой; трубка может, не сгибаясь,
выдерживать значительно большие нагрузки, чем
плотный цилиндр той же массы. Однако с
увеличением размеров и веса животного такая организация
становится менее удобной-для сохранения
достаточной прочности толщина и масса экзоскелета
должны были бы возрасти настолько, что он в конце
концов оказался бы слишком тяжелым и
громоздким.
Рост происходит при линьках-у насекомых на
ювенильных стадиях (личинки или нимфы), а у
ракообразных и во взрослом состоянии. В
определенное время старый экзоскелет сбрасывается
(линька) и обнажается новый-мягкий и
растяжимый. Животное растет, пока новая кутикула еще не
затвердела, благодаря ее способности растягиваться
и увеличиваться в размере, что часто
сопровождается также изменением формы. В конце концов
новый экзоскелет становится жестким, но до того
момента животное уязвимо для хищников. В этот
период скелет не способен поддерживать вес тела и
всякого рода движения практически невозможны.
Для водных животных подобная проблема не столь
серьезна, так как вода помогает им поддерживать
вес тела, однако в период линьки и водные, и
наземные животные, как правило, прячутся в
укрытиях, чтобы уменьшить опасность быть
схваченными хищником. Линька-это процесс, требующий
значительных затрат энергии (в первую очередь на
построение нового экзоскелета) и сопряженный с
потерей материала при сбрасывании старой
кутикулы.
17.1.3. Эндоскелет
Внутренний скелет имеется у простейших из отряда
радиолярий (в виде кремневых спикул), у
головоногих моллюсков (например, у каракатицы есть
внутренняя раковина), но особенно характерен для
позвоночных, у которых он построен из хряща или
костной ткани и расположен в теле под слоем
мышц. Эндоскелет отличается от экзоскелета еще и
тем, что он образован живой тканью и может
непрерывно расти в теле животного; благодаря
этому нет необходимости в линьках. Существует
несколько типов суставов; образующие их кости
поддерживаются в определенном положении с
помощью эластичных связок.
Общий план строения скелета у четвероногих и
двуногих позвоночных практически одинаков,
однако есть некоторые различия в подвижности бедра и
плеча. Эти особенности, связанные с характерным
способом передвижения, мы рассмотрим позже.
17.1.4. Скелет позвоночных
животных
Скелет у позвоночных построен из костной или
хрящевой ткани. В обоих случаях он служит
внутренним каркасом для тела. Эндоскелет,
состоящий только из хряща, встречается лишь у пластино-
жаберных (к которым относятся акулы и скаты). Все
остальные позвоночные во взрослом состоянии
имеют костный скелет, но в отдельных его участках
сохраняется хрящ - например, в суставах или
межпозвоночных дисках. Скелет этих позвоночных в
эмбриогенезе закладывается в виде гиалинового
хряща (разд. 8.4.4). Эта особенность имеет важное
биологическое значение, так как хрящ способен
расти всей своей массой и в процессе развития
организма разные части скелета могут
пропорционально увеличиваться. В этом отношении костная
ткань отличается от хряща, поскольку рост ее
осуществляется путем откладки новых элементов на
поверхности. Если бы такой процесс происходил
при развитии организма, то пространственные
отношения между суставными поверхностями костей,
а также местами прикрепления мышц в результате
роста неизбежно нарушались бы.
17.2 Скелетные ткани
17.2.1. Хрящ
Выделяют три типа хряща: гиалиновый,
волокнистый и эластичный. Их гистологические
особенности подробно рассмотрены в разд. 8.4.4. Все типы
состоят из плотного матрикса (основного вещества),
пронизанного множеством соединительнотканных
волокон. Матрикс секретируют живые клетки-
хондробласты. Позднее эти клетки оказываются в
микроскопических полостях (лакунах),
разбросанных в матриксе. В этом состоянии они называются
хондроцитами. Наиболее распространенный тип
хряща - гиалиновый хрящ; им, например, покрыты
суставные поверхности костей. Его матрикс,
состоящий из хондроитинсульфата, сжимаем и эластичен,
он способен выдерживать большие нагрузки и
гасить резкие механические воздействия, которые
может испытывать сустав. Устойчивость к такого рода
нагрузкам матриксу придают пронизывающие его
тонкие коллагеновые волокна. На всей поверхности
такого хряща, кроме участков, обращенных внутрь
суставной сумки, находится плотная
соединительная ткань-надхрящница.
Волокнистый хрящ содержит плотную сеть кол-
лагеновых волокон; он образует межпозвоночные
диски и входит в состав сухожилий. Это очень
прочная, хотя и в известной степени гибкая ткань.
Эластичный хрящ содержит множество эластино-
вых волокон, из него образованы наружное ухо,
надгортанник и хрящи гортани.
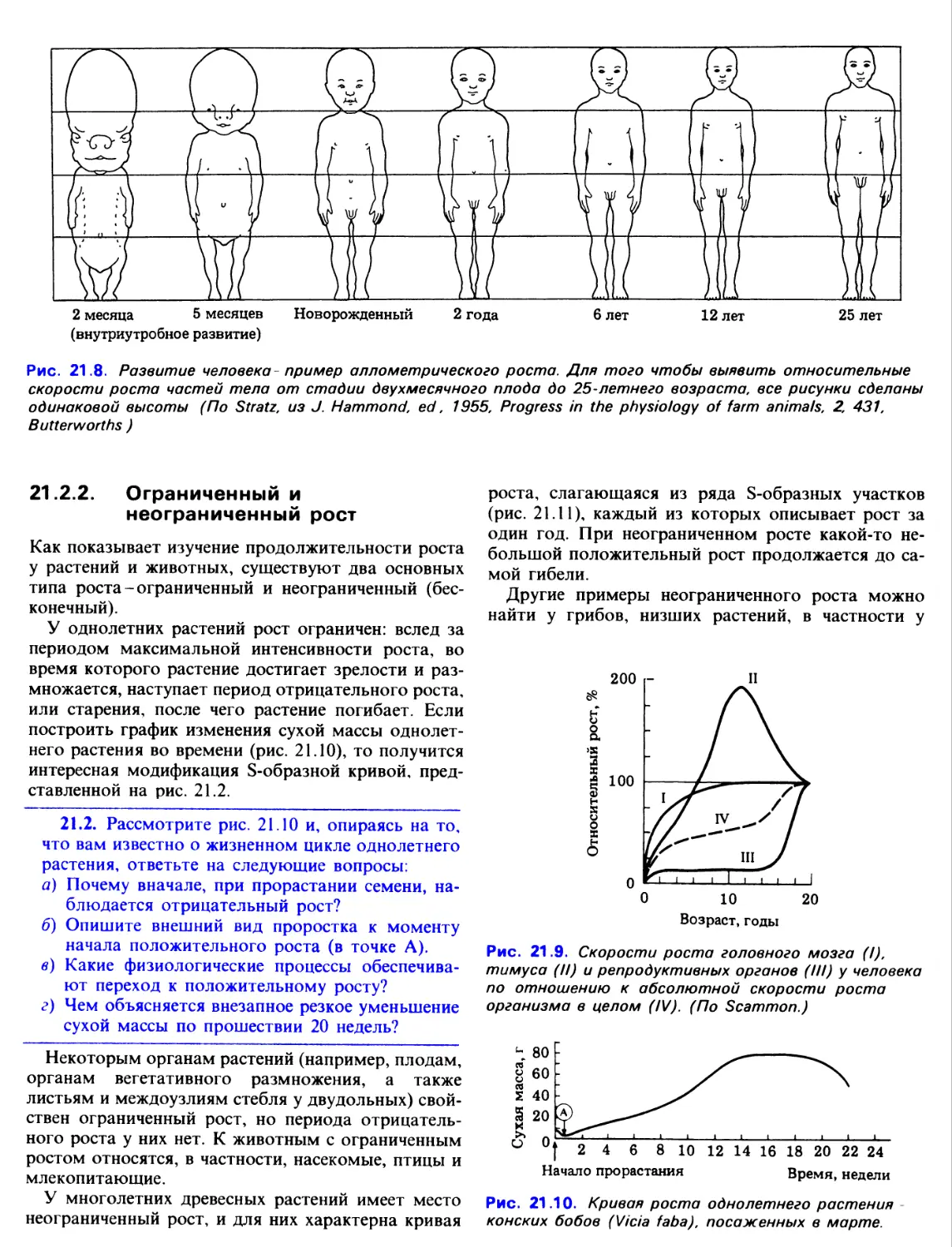
17.2.2. Костная ткань
Кость представляет собой плотную, твердую
соединительную ткань, в основном содержащую обыз-
вествленные элементы. Детали ее строения
приведены в разд. 8.4.4. На продольном разрезе длинной
кости (например, бедренной) хорошо различимы ее
части. Такая кость состоит из полого стержня-диа-
физа, на концах которого находятся две
расширенные головки - эпифизы. Снаружи вся кость покрыта
плотной соединительнотканной оболочкой -
надкостницей. Диафиз состоит из компактного
вещества, в то время как эпифизы образованы губчатой
костной тканью, окруженной тонким слоем плотной
костной ткани. Организация костной ткани придает
ей максимальную прочность в тех направлениях, в
которых на нее воздействуют нагрузки (рис. 17.2).
Полость диафиза занята желтым костным
мозгом, а красный костный мозг расположен в
эпифизах, между костными перекладинами (трабеку-
лами). Поверхность кости пронизана множеством
мелких отверстий, через которые костная ткань и
красный костный мозг снабжаются нервами и
кровеносными сосудами.
Помимо отмеченных ранее функций скелет
участвует в образовании эритроцитов и гранулоцитов.
.Суставной хрящ
на головке
(эпифизе)
-Трабекулы
Надкостница
.Компактное
вещество
Диафиз
Костный мозг
Рис. 17.2. Продольный разрез головки бедренной
кости. Показано расположение трабекул в губчатом
веществе.
Кроме того, он обеспечивает постоянство уровней
кальция и фосфора в крови (см. гл. 16): запасенные
в костях ионы кальция и фосфата могут
высвобождаться под действием кальцитонина и пара-
тиреоидного гормона, вырабатываемых
соответственно щитовидной и паращитовидными
железами.
17.1. Покажите на примере бедренной кости,
как ее строение, суставной хрящ, сухожилия
мышц и связки приспособлены для выполнения
своих функций.
17.2.3. Развитие костного скелета
у млекопитающих
Хрящевой скелет эмбриона постепенно по мере
роста животного замещается костным. Хрящ
распадается, и на его месте образуется костная ткань.
Этот процесс называют окостенением (оссификаци-
ей), и в длинной кости он начинается с диафиза.
Здесь в надхрящнице образуется грубоволокнистая
костная манжетка (разд. 8.4.4) и диафиз
оказывается в кольце плотной ткани. Сформировавшийся
таким образом слой называется надкостницей.
Находящиеся внутри этого кольца хондроциты
увеличиваются в размере, обызвествляется окружающий
их матрикс, и клетки, наконец, разрушаются. В
результате в хряще образуются полые ниши. Они
постепенно заполняются клетками эмбрионального
костного мозга и кровеносными сосудами, которые
тянутся от расположенной на поверхности диафиза
надкостницы. Часть клеток костного мозга
дифференцируется в остеобласты - клетки, образующие
кость. Располагаясь вокруг остального костного
мозга, они секретируют слой костного вещества.
Процесс завершается образованием прочной полой
трубки из костной ткани, окружающей
костномозговую полость.
Затем окостенение идет в направлении к
эпифизам. Этот процесс заканчивается только по
достижении зрелого возраста, и даже тогда
гиалиновый хрящ сохраняется на суставных поверхностях
костей. Эпифизы окостеневают вслед за диафизом и
независимо от него. Центров окостенения в
эпифизах может быть несколько.
Одновременно с этим процессом идет рост кости
в длину. Он происходит в обоих концах кости на
границе диафиза и эпифиза. Пока кость не достигнет
своей окончательной длины, хрящ в этой области
непрерывно образуется и окостеневает. Постепенно
окостеневают и эти зоны роста; в результате клетки
перестают делиться и удлинение кости
прекращается. Рост кости в толщину может продолжаться за
счет отложения костной ткани надкостницей.
17.2.4. Факторы, регулирующие
образование костной ткани
Даже после того как рост кости завершен,
происходят процессы ее разрушения (с помощью
особых клеток-остеокластов) и перестройки. Они
имеют большое значение, так как позволяют изменять
форму кости в зависимости от испытываемых ею
механических нагрузок; это особенно важно для тех
животных, у которых в процессе развития
существенно изменяется способ передвижения, что обычно
связано с перераспределением нагрузки на скелет.
Постоянная нагрузка на какой-либо участок кости
приводит к его разрушению, однако периодические
нагрузки на кость стимулируют откладку костной
ткани. Такие давления и нагрузки формируют
скелет определенным образом; в частности,
ритмические нагрузки приводят к появлению выступов и
гребней на поверхности кости, увеличивающих
площадь прикрепления мышц. При отсутствии
нагрузки кость атрофируется. Этой проблеме
посвящено сейчас много исследований, так как
подобные явления отмечаются при длительном
пребывании человека в условиях невесомости в космосе и
создают определенные трудности для
возвращающихся на Землю космонавтов и астронавтов.
Прочность костей может также уменьшаться при
недостатке в пище таких веществ, как витамины А и
D, или при дефиците гормона роста.
17.2.5. Опора при локомоции
у наземных позвоночных
Амфибии произошли от кистеперых рыб, и с
выходом амфибий из воды на сушу перед ними
возникла проблема, связанная с действием силы
тяжести и необходимостью поддерживать свое тело
над землей. В результате их позвонки
преобразовывались в сложные структуры, сочленяющиеся
друг с другом с помощью отростков. Все вместе они
образуют прочную, но достаточно гибкую балку-
позвоночник, который служит опорой для тела.
Рис. 17.3. Типы прикрепления конечностей у
позвоночных. А. У примитивных амфибий ноги
отходят от туловища в стороны, а затем вниз.
Б. У современных рептилий - промежуточное
положение между амфибиями и млекопитающими.
В. У млекопитающих ноги отходят от нижней
(брюшной) стороны тела.
У древних амфибий конечности отходили от
туловища вбок. Это давало возможность передвигаться,
почти не приподнимаясь над землей. Такой же тип
прикрепления конечностей и передвижения
отмечался и у примитивных рептилий (рис. 17.3, А). При
этом способе передвижения мышечная энергия
расходуется в основном на удержание тела над
поверхностью земли, и ее затраты настолько велики,
что большую часть времени на суше животное
проводит неподвижно, опираясь брюхом на землю.
В процессе дальнейшей эволюции у рептилий
отмечалась тенденция к смещению конечностей вниз,
так что туловище оказывалось уже заметно
приподнятым над землей (рис. 17.3, F). Такая поза
облегчала животному передвижение, и тяжесть тела
распределялась более равномерно между четырьмя
относительно прямыми конечностями.
Некоторые рептилии и млекопитающие перешли
к хождению на двух ногах. Задние конечности
служат им для ходьбы, бега или прыжков. Это
освобождает передние конечности для манипуляций,
связанных, например, с потреблением пищи,
строительной деятельностью, чисткой тела. Для
некоторых обезьян характерен особый способ
передвижения - брахиация. При этом животные
перебрасываются с дерева на дерево, раскачиваясь на длинных
руках и перехватывая ветви удлиненными кистями
рук. Небольшие размеры других обитателей
древесных крон не позволяют им перемещаться таким
способом; эти животные перескакивают с ветки на
ветку. Наиболее специализированный способ
передвижения по воздуху-полет. Он появился в юрский
период одновременно у летающих рептилий
(птеродактилей) и у первых птиц (которые произошли от
рептилий). Передние конечности, видоизменившись,
превратились в крылья. Летающие рептилии в конце
концов вымерли, а птицы в ходе дальнейшей
эволюции дали множество разнообразных форм.
17.3. Анатомическое строение
скелета млекопитающих
Основные отделы скелета у всех млекопитающих
одинаковы. Различают две главные части: осевой
скелет, состоящий из черепа, позвоночника и
грудины, и добавочный скелет, к которому относится
передний (плечевой) и задний (тазовый) пояса,
несущие по паре конечностей.
17.3.1. Осевой скелет
Череп состоит из мозговой коробки, с которой
сращена неподвижная верхняя челюсть и соединена
при помощи суставов нижняя челюсть.
Соответственно и мышцы связывают нижнюю челюсть с
другими костями черепа. Мозговой череп образован
несколькими плоскими костями, прочно
соединенными между собой неподвижными швами. Он
защищает не только находящийся внутри головной
мозг, но и органы обоняния, внутреннее и среднее
ухо и глаза. На задней поверхности черепа имеются
два округлых гладких выступа - затылочных
мыщелка, образующих сустав с верхним позвонком -
атлантом; благодаря этому животное может
поднимать и опускать голову.
Позвоночник является главной осью тела (см.
рис. 17.5). Он состоит из цепочки позвонков, между
которыми находятся хрящевые межпозвоночные
диски. Связки соединяют позвонки друг с другом и
не позволяют им смещаться, но все же некоторое
движение позвонков возможно, так что позвоночник
в целом обладает значительной гибкостью.
Позвоночник служит также для защиты спинного мозга.
Позвонки несут множество отростков для
прикрепления мышц. Мышцы, сокращаясь, могут изгибать
позвоночник вперед, назад или в сторону.
У разных млекопитающих общее число позвонков
варьирует. Тем не менее в позвоночнике у всех
млекопитающих выделяют пять отделов. В
табл. 17.1 приведено число позвонков у некоторых
представителей этого класса.
Таблица 17.1. Типы позвонков и их число у разных
млекопитающих
Типы
позвонков
Шейные
Грудные
Поясничные
Крестцовые
Хвостовые
Область
Шея
Грудная
клетка
Брюшная
область
Область
таза
Хвост
крыса
1
13
6
4
30 ±
Число позвонков
кролик
7
12-13
6-7
4
16
кошка
1
13
7
3
18-25
корова
1
13
6
5
человек
7
12
18-20
5
5
4
Остистый отросток
Невральная дуга
Поперечный
отросток
Передний суставной
отросток
Отверстие
Тело позвонка
Рис. 17.4. Типичный позвонок млекопитающего (вид
спереди).
Позвонки во всех отделах построены по одному
общему плану. Строение типичного позвонка
показано на рис. 17.4. Следует отметить, что при
горизонтальном положении позвоночника
(обычном у животных) две фасетки (сочленовные
поверхности) на передней стороне позвонка как бы
подогнаны к фасеткам на задней стороне другого,
расположенного перед ним. Такое строение
позвонков обеспечивает их взаимное сочленение, но при
этом конструкция не получается абсолютно
жесткой, так как благодаря гладким сочленовным
поверхностям позвонки могут слегка смещаться
относительно друг друга. Под каждой передней и
задней фасетками имеется небольшая вырезка.
Задняя вырезка одного позвонка лежит прямо против
передней вырезки позвонка, расположенного позади
него. Таким образом формируются
межпозвоночные отверстия, через которые проходят
спинномозговые нервы.
Для всех позвонков характерно также наличие
остистого и поперечных отростков, к которым
прикрепляются мышцы. Центральная плотная часть
позвонка представляет собой тело позвонка, а над
ним расположена невральная дуга, окружающая
спинной мозг.
Несмотря на значительное сходство всех
позвонков, все же их строение в разных отделах
позвоночника варьирует. Это связано с тем, что тяжесть
тела неравномерно распределяется по всей длине
позвоночника и что позвонки видоизменяются в
соответствии со специфическими функциями,
которые выполняют его отделы.
Когда кролик стоит, опорой для позвоночника
служат передние и задние лапы и вес тела
распределяется между ними. При этом на тела
позвонков действуют сжимающие силы, а на лежащие
над ними дорсальные связки и мышцы -
растягивающие силы (рис. 17.5).
17.3.2. Строение и функции позвонков
у кролика
Шейные позвонки
Шейные позвонки с 3-го по 7-й имеют одинаковое
строение (рис. 17.6, Л). Тело у них небольшое,
способное противостоять сжатию, а остистый отросток
короткий, к нему прикрепляются мышцы шеи.
Некоторые из них тянутся от шейных позвонков к
грудным, они удерживают шею в приподнятом
положении, в то время как другие идут к основанию
черепа и служат для поддержания головы. С каждой
стороны от тела позвонка имеется одно отверстие-
канал позвоночной артерии, который образуется в
результате слияния шейного ребра с поперечным
отростком. Как видно из названия, по этому каналу
проходит позвоночная артерия, снабжающая
кровью головной мозг. Таким образом, важный
кровеносный сосуд, расположенный в уязвимой
области шеи, оказывается защищенным.
Первые два шейных позвонка резко отличаются
по строению от остальных, так как они служат
опорой для головы и обеспечивают ее движение в
разных плоскостях. Первый шейный
позвонок-атлант (рис. 17.6,Б)-не имеет передних суставных
отростков и тела; остистый отросток редуцируется
до едва заметного выступа. На передней
поверхности атланта находятся два углубления - передние
суставные ямки, которые, сочленяясь с выпуклыми
затылочными мыщелками, образуют блоковидный
сустав. Таким образом, позвонок служит опорой
для черепа и позволяет животному поднимать и
опускать голову. Широкие уплощенные поперечные
отростки предоставляют большую площадь для
прикрепления мышц, участвующих в этих
движениях.
Второй шейный позвонок эпистрофей (рис. 17.6, В),
у него есть направленный вперед пальцеобразный
вырост - зубовидный отросток, образованный путем
слияния тел атланта и эпистрофея. Он заходит в
отверстие атланта, и расположенная там связка
отделяет отросток от позвоночного канала. При
этом создается цилиндрический сустав,
позволяющий поворачивать голову из стороны в сторону. На
боковых поверхностях шеи расположены мышцы,
которые производят эти движения; они тянутся от
остистого отростка эпистрофея вперед и
прикрепляются к поперечным отросткам атланта (рис. 17.7).
Передних суставных отростков у эпистрофея нет.
Грудные позвонки
У этих позвонков длинные направленные назад
остистые отростки и короткие поперечные отростки.
На последних расположены округлые реберные
фасетки (ямки), а на боковых поверхностях тела
позвонка - передние и задние по л у фасетки. Оба типа
фасеток служат для присоединения ребер (рис. 17.8, А).
На задних концах ребер, сочленяющихся с
грудными позвонками, имеются два ответвления-головка
и бугорок. Бугорок сочленяется с реберной фасеткой
поперечного отростка, а головка-с двумя
полуфасетками тела позвонка. Сустав устроен здесь
довольно сложно: передняя полу фасетка одного
позвонка вместе с задней полуфасеткой другого, лежа-
Сила.
создаваемая
передними
конечностями
Череп
Сила, создаваемая
задними конечностями
Шейные
позвонки
Грудные
позвонки
Нижняя
челюсть
^Поясничные
тюзвонки
Лопатка
Вес
Подвижные ребра
Реберный хрящ-
'Ребра
Метакромион.
Крестцовые
позвонки
(крестец)
Головка
плечевой
кости
Бедренная'
LKOCTb
Плечевая
кость
Хвостовые
позвонки
Локтевая
кость
Больше-
берцовая1
кость
Лобковая
кость
Грудина
Лучевая кость
Мало-
берцовая
кость
Седалищная
кость
Вес
Предплюсна
Плюсна
Запястье
Пясть
Фаланги
Вес
Рис. 17.5. Скелет кролика (вид слева). Тела позвонков работают на сжатие, в то время как связки и
мышцы, соединяющие позвонки между собой,-на растяжение. Мускулатура брюшной стороны
тела не позволяет поясам конечностей разойтись под действием силы тяжести.
Передний суставной
отросток
Позвоночный
канал
Поперечный
отросток
Шейное ребро
Остистый
отросток
Невральная дуга
Канал
позвоночной
артерии
Рис 17.6. А. Пятый шейный позвонок кролика (вид
спереди). Видны характерные отверстия для
позвоночных артерий. Б. Атлант кролика (вид
спереди). Обратите внимание на отсутствие тела
позвонка и наличие передних сочленовных
поверхностей. В. Эпистрофей кролика (вид сбоку).
Отличительные особенности: зубовидный отросток и
направленный вперед остистый отросток.
Тело позвонка
Позвоночный
канал
Остистый отросток
Невральная дуга
Поперечный
отросток
Канал
позвоночной
артерии
Передняя фасетка
для сочленения
с затылочным
мыщелком черепа
Шейные мышцы
Связка
Остистый
отросток
Задний суставной
отросток
Эпистрофей
Атлант-
Зубовидный
отросток
Шейное ребро
Тело позвонка
Зубовидный
отросток
Канал
позвоночной
артерии
Поверхности
для сочленения
с атлантом
Канал
позвоночной
артерии
К хвосту
К голове
Рис. 17.7. Расположение шейных мышц между
атлантом и эпистрофеем у кролика.
Реберная ямка
поперечного отроста
Поперечный
отросток
Передний
суставной
отросток
Передняя
реберная
полуямка
Остистый
отросток"
•К голове
Задний суставной
отросток
Задняя реберная
полуямка
Тело
позвонка
Остистый
отросток
Метапофиз
Задний суставной
отросток
К голове
Поперечный
отросток
К голове
•Тело позвонка
Поперечный отросток
Передний суставной
отросток
Область сращения
с тазовым поясом
Остистые отростки
четырех сросшихся
позвонков
Рис. 17.8. А. Грудной позвонок кролика (вид слева).
Видны длинный остистый отросток и
полуфасетки. Б. Поясничный позвонок кролика (вид
слева). Гипапофизы не показаны. Там. где они есть (у
первого и второго поясничных позвонков), они имеют
вид небольших выростов на вентральной
поверхности тела позвонка. В. Крестец кролика (вид со
спинной стороны).
щего впереди, образуют полную суставную ямку. В
это углубление и входит головка ребра, соединяясь
таким образом одновременно с двумя позвонками.
Грудные позвонки служат опорой для ребер, но
сложность сочленения ограничивает их
подвижность. Они могут несколько смещаться вперед и в
стороны, но в целом грудные позвонки самые
малоподвижные в позвоночнике.
Поясничные позвонки
Позвонки этой области позвоночника испытывают
наибольшие нагрузки под действием силы тяжести и
при передвижении. Они должны не только
обеспечивать опору для тела, но и позволять наклоняться
назад, вперед, в стороны и вращать туловищем.
Поэтому не удивительно, что именно в этой области
прикрепляются наиболее крупные мышцы спины и
именно здесь позвонки подверглись наибольшим
адаптивным видоизменениям. Хотя тела этих
позвонков довольно низкие, обычно они, как и их
невральные дуги, отличаются массивностью.
Строение поясничных позвонков делает их соединение
более гибким. Поперечные отростки длинные и
широкие; они направлены вперед и вниз. У
позвонков имеются дополнительные отростки для
прикрепления мышц-мета-, ана- и гипапофизы
(рис. 17.8, В); они тоже сочленяются между собой и
при больших нагрузках на эту часть позвоночника
обеспечивают правильное взаиморасположение
позвонков.
Крестцовые позвонки (крестец)
Крестцовые позвонки, срастаясь, образуют
широкую кость-крестец (рис. 17.8,5). У самых верхних
крестцовых позвонков имеются хорошо развитые
поперечные отростки, которые соединены с тазовым
поясом. Когда животное стоит неподвижно, именно
крестец передает тяжесть тела на тазовый пояс и
конечности. При перемещении животного вперед
сила, которую создают задние конечности, через
тазовый пояс и крестец передается другим отделам
осевого скелета.
Хвостовые позвонки
У разных видов млекопитающих число хвостовых
позвонков сильно варьирует (табл. 17.1) и зависит
от длины хвоста. Обычно по мере приближения к
его кончику поперечные отростки, невральные дуги
и сочленовные отростки уменьшаются в размерах и
постепенно исчезают, и в результате последние
хвостовые позвонки представлены лишь их
маленькими телами. У человека этот отдел состоит из
четырех позвонков, сросшихся в одну
кость-копчик. Снаружи он незаметен.
Ребра и грудина
Ребро представляет собой уплощенную изогнутую
кость. Его дорсальный конец, раздваиваясь,
образует головку и реберный бугорок, которые сочленяются
с грудными позвонками. При дыхании межреберные
мышцы приводят ребра в движение. Ребра вместе с
грудиной и грудным отделом позвоночника
образуют грудную клетку, которая защищает сердце,
легкие и главные кровеносные сосуды (рис. 17.9).
У кролика первые семь пар ребер с помощью
реберных хрящей прикрепляются своими
вентральными концами к грудине - плоской кости с
направленной вперед «рукояткой». Их называют
истинными ребрами. Следующие две пары ребер
вентральными концами тоже присоединены к хрящу
седьмой пары. Вентральные концы последних трех
или четырех пар остаются свободными - это так
называемые подвижные ребра (рис. 17.5).
17.3.3. Добавочный скелет (пояса
конечностей)
Пояса обеспечивают причленение конечностей к
осевому скелету. Шириной плечевого пояса
определяется расстояние между передними конечностями, а
тазового - между задними. От этого зависит и
устойчивость животного. Некоторые участки поясов
специально приспособлены для прикрепления к ним
мышц или сочленения с костями конечностей.
плечевой пояс. Он образован двумя
самостоятельными половинками, каждая из которых состоит из
лопатки с коракоидом (клювовидным отростком) и
ключицы. Пояс не срастается с осевым скелетом, а
присоединяется к нему подвижно с помощью мышц
и связок. Это обеспечивает подвижность передних
конечностей в разных плоскостях и под самыми
разными углами. Плечевой пояс достаточно прочен
и способен выдерживать основную массу тела,
когда четвероногое животное стоит неподвижно; он
также амортизирует удар в момент приземления
животного при прыжке.
Лопатка - плоская треугольная кость,
прилегающая к верхним ребрам (рис. 17.10, А). На ее
верхушке есть углубление-суставная впадина для
сочленения с головкой плечевой кости при помощи
шаровидного сустава. По наружной поверхности лопатки
тянется ость - гребень, на свободном конце
которого рядом с суставной впадиной находятся два
отростка: акромион (плечевой отросток) и метакро-
мнон; к ним прикрепляются мышцы. Клювовидный
отросток представляет собой рудимент маленькой
(вороновидной) косточки - коракоида, которая
приросла к лопатке над суставной впадиной.
Ключицы у разных млекопитающих различаются
по форме и размеру. У человека они хорошо
развиты. Ключица сочленяется одним концом с акро-
мионом, а другим-с грудиной. К ней
прикрепляются мышцы. Она позволяет совершать сложные
движения руками. У человека ее удаление не
приводит к серьезным последствиям. Еще менее важна
она для четвероногих зверей, и ее размеры у них
значительно меньше. У птиц ключицы срастаются в
так называемую «вилочку».
17.2. Какие преимущества дает
млекопитающим подвижное присоединение плечевого
пояса к позвоночнику?
тазовый пояс. Он тоже состоит из двух половин и
образован тремя парными костями: подвздошной,
лобковой и седалищной, которые срастаются в
единую тазовую (безымянную) кость. Подвздошные
кости прирастают к крестцу с обеих сторон. На
наружных краях тазовых костей имеется по
углублению - вертлужной впадине, которая служит для
образования шаровидного сустава с головкой бедренной
кости (рис. 17.10,5). Над этой впадиной
расположена подвздошная кость; на ее дорсальной стороне
Отросток
для прикрепления
связок
Бугорок
Тело
позвонка
Реберный
хрящ
Остистый
отросток
Передний
суставной
отросток
Поперечный
отросток
Головка
ребра
Ребро
Грудина
Рис. 179. Грудной позвонок кролика, сочлененный с
двумя ребрами (вид спереди).
Клювовидный
отросток
Суставная
впадина
Акромион
Ость
Надлопаточный
хрящ
Метакромион
А
Крестец
Подвздошная
кость
Лобковая,
кость
Седалищная
кость
Вертлужная
впадина
Запирательное
отверстие
Лобковое
сращение
(симфиз)
Рис. 17.10. А. Левая лопатка кролика.
Б. Тазовый пояс кролика (вид с брюшной стороны).
Обратите внимание на то, каким образом
подвздошная кость срастается с крестцом.
имеется большой гребень, к которому
прикрепляются мышцы бедра.
Между седалищной и лобковой костями имеется
большое запирательное отверстие. Оно затянуто
плотной неэластичной соединительнотканной
перепонкой, в которой есть лишь небольшое отверстие
для кровеносных сосудов и нервов, идущих к задним
(нижним) конечностям. Эта перепонка тоже служит
местом прикрепления мышц. Такое строение может
быть адаптацией, направленной на уменьшение веса
тазового пояса, а значит, и нагрузки на нижние
конечности.
На вентральной поверхности заметна граница
двух половин тазового пояса - лобковое сращение
(симфиз). Гибкий хрящ в этой области позволяет
отверстию таза увеличиваться при родах.
Конечности
Принцип строения задних и передних конечностей у
всех млекопитающих одинаков. В основе лежит
план пятипалой конечности, которая, как видно из
названия, оканчивается пятью пальцами (рис. 17.11),
однако у разных животных в связи с адаптацией к
разному образу жизни встречается множество
вариантов этого типа строения. Иногда в процессе
эволюции число пальцев уменьшалось (разд. 24.7).
передние конечности. Верхний отдел передней
конечности образован одной плечевой костью. Ее
проксимальный конец - головка, - входя в суставную
впадину лопатки, образует плечевой шаровидный
сустав, который позволяет совершать самые
различные движения. Рядом с головкой находятся два
шероховатых выроста - большой и малый бугорки,
между которыми расположена биципитальная
борозда (борозда двуглавой мышцы), по которой
проходит сухожилие бицепса. На дистальном конце
имеется блок плечевой кости, который сочленяется с
предплечьем локтевым блоковидным суставом. У
кролика плечевую кость прямо над блоком
прободает надблоковое отверстие. У человека такого
отверстия нет. На передней поверхности
проксимального отдела плечевой кости заметен
дельтовидный гребень (рис. 17.12, А). Расположенный ниже
отдел передней конечности - предплечье - состоит из
двух костей - локтевой и лучевой. Локтевая кость
более длинная. На ее проксимальном конце имеется
блоковидная вырезка, которая сочленяется с блоком
плечевой кости. Под локтевым суставом находится
локтевой отросток. Эта структура очень важна: она
ограничивает движение предплечья, после того как
передняя конечность уже выпрямлена, и таким
образом предотвращает вывих. На передней
поверхности плечевой кости над блоком имеется лучевая
(венечная) ямка, в которую входит конец лучевой
кости при сгибании конечности (рис. 17.12,5).
Лучевая кость уплощена, слегка изогнута и имеет
более простое строение. У человека она соединяется
Борозда
двуглавой
мышцы
Малый бугорок
Дельтовидный
гребень
Надблоковое
отверстие
Блок
Большой
бугорок
Головка
ВИД СПЕРЕДИ
ВИД СЗАДИ
Лучевая
кость
Ямка
локтевого отростка
Сигмоидная
вырезка
Дистальная
поверхность"
для сочленения
с костями запястья
Локтевая кость
Локтевой
отросток
Рис. 17.12. А. Левая плечевая кость кролика (вид
спереди и сзади). Б. Лучевая и локтевая
кости кролика (вид сбоку).
с локтевой костью относительно подвижно. В
результате сокращения мышц лучевая кость может
поворачиваться вокруг локтевой, при этом ладонь
руки поворачивается вверх или вниз, что позволяет
человеку совершать тонкие манипуляции руками. У
кролика обе кости жестко соединены друг с другом;
его кисть не обладает такой подвижностью и всегда
повернута тыльной стороной вверх. Это, однако, не
следует рассматривать как недостаток, поскольку
такое положение оказывается наиболее удобным
при рытье и беге. Дистальные концы локтевой и
лучевой костей сочленяются с несколькими
косточками, образующими запястье. Кости запястья
соединяются в свою очередь с удлиненными
костями пясти, к которым причленены пять пальцев.
Первый, обращенный внутрь палец состоит из двух
фаланг, а остальные пальцы-из трех фаланг.
задняя конечность. Верхний ее отдел образован
одной бедренной костью. Наверху кость
оканчивается круглой головкой, которая входит в вертлуж-
ную впадину таза, образуя шаровидный
тазобедренный сустав (рис. 17.13,/!). Под головкой бедренной
кости расположены три отростка - вертела. Они
служат местом прикрепления мышц бедра. На нижнем
конце бедра имеются два округленных мыщелка для
образования блоковидного коленного сустава с
большеберцевой костью. Мыщелки разделены
углублением, к которому спереди примыкает
надколенник (коленная чашка). Голень образована
большеберцовой и малоберцовой костями. Два
небольших углубления на верхнем конце больше-
берцовой кости представляют собой сочленовные
поверхности коленного сустава (рис. 17.13, Б).
Малоберцовая кость в образовании этого сустава не
участвует. Это тонкая кость, и у кролика она в
дистальном отделе сливается с большеберцовой. У
ПЕРЕДНЯЯ КОНЕЧНОСТЬ
1 задняя конечность]
Плечевая
кость
Лучевая
кость
Локтевая
кость
Запястье
Пясть
Пальцы -
(отдельные!
косточ ки -фаланги)
Бедренная
кость
Большеберцовая
кость
Малоберцовая
кость
Предплюсна
Плюсна
- Пальцы
Рис. 17.11. Пятипалая конечность позвоночных.
нижних концов большеберцовой и малоберцовой
костей расположен ряд косточек предплюсны. Две
самые большие из них-тарачная и пяточная
кости-образуют с большеберцовой и
малоберцовой костями голеностопный сустав. Кости
предплюсны в дистальной части сочленяются с
длинными костями плюсны, образующими стопу, а те в
свою очередь с фалангами пальцев. Следует
отметить, что задние конечности кролика имеют
только по четыре пальца.
17.3.4. Суставы
У позвоночных, имеющих костный скелет, в местах
соединения костей образуются суставы. Движение
смежных элементов скелета относительно друг
друга возможно лишь при наличии между ними
суставов. Суставы в скелете млекопитающих очень
разнообразны; сведения о них представлены в
табл. 17.2.
Истинные суставы (диартрозы) имеют сходную
«конструкцию». Сочленовные поверхности на
концах костей покрыты гладким гиалиновым хрящом.
И хотя хрящ-живая ткань, в ней нет нервов и
кровеносных сосудов. Нужные ему питательные
вещества и кислород диффундируют через
синовиальную оболочку и синовиальную жидкость. При
движении хрящ уменьшает трение между костями;
благодаря своей эластичности хрящ служит также
амортизатором при ударе.
Головка
Малый вертел
Поверхности
для сочленения'
с
большеберцовой костью
Большеберцовая·
кость
Сочленовная
поверхность
для предплюсны
Большой вертел
Третий вертел
Борозда
для надколенника
Гребень
большеберцовой
кости
Малоберцовая
кость
Рис. 17.13. А. Левая бедренная кость кролика (вид
спереди). Б. Левые большеберцовая и
малоберцовая кости кролика (вид спереди).
Связки, окружающие сустав, образуют плотную
волокнистую сумку и удерживают кости в
определенном положении. Связки прикреплены к обеим
костям и расположены таким образом, чтобы сустав
Таблица 17.2. Разнообразие суставов в эндоскелете млекопитающих
Тип сустава
Неподвижный (шов), син-
артроидный
Частично подвижный,
амфиартроидный
а. Эллипсовидный
б. Вращательный
(цилиндрический)
Подвижный
(синовиальный, диартроидный)
а. Блоковидный
б. Шаровидный
Общая характеристика
Тонкий слой волокнистой
соединительной ткани находится
между костями, жестоко
фиксируя их положение
Кости отделены друг от друга
хрящевыми прокладками
Сочленовные поверхности
костей покрыты хрящом и
разделены синовиальной полостью,
заполненной синовиальной
жидкостью
В движении участвует
относительно небольшое число
мышц
К костям, образующим сустав,
прикреплено множество
мышц
Примеры
Между костями
черепа; между крестцом
и подвздошной
костью тазового
пояса; между костями
тазового пояса
Суставы между
позвонками, костями
запястья и
предплюсны
Сустав между
атлантом и эпистрофеем
Локтевой и коленный
суставы, суставы
пальцев
Плечевой и
тазобедренный суставы
Функции
Обеспечивает опору тела,
придает прочность или
защищает нежные органы,
которым противопоказаны
любого рода деформации
Кости скользят относительно
друг друга, но подвижность
их ограниченна. Все вместе
они обеспечивают
разнообразные движения,
придают прочность конечности
Позволяет поворачивать
голову
Позволяет совершать
движения в одной плоскости
вокруг одной оси. Может
выдерживать большие нагрузки
Позволяет совершать
движения во всех плоскостях и
некоторое вращение. Не
способен выдерживать очень
большие нагрузки
Губчатое
вещество
Полость,
содержащая-
синовиальну]
жидкость
Гиалиновый
хрящ
Надкостница
Компактное
вещество
Суставная
сумка
Синовиальная
оболочка
Рис. 17.14. Диартроз (истинный сустав)
млекопитающего.
мог выдерживать приходящуюся на него нагрузку.
Внутренняя полость суставной сумки выстлана
синовиальной оболочкой, которая выделяет в эту
полость синовиальную жидкость (рис. 17.14).
Синовиальная жидкость содержит муцин и служит
смазкой для суставных поверхностей - уменьшает трение
между ними. Синовиальная оболочка является
водонепроницаемым барьером, предотвращающим
потерю синовиальной жидкости. Таким образом,
сустав фактически представляет собой
самоподдерживающуюся систему.
17.4. Мышечная система
Мышцы состоят из множества удлиненных
клеток - мышечных волокон, способных сокращаться и
расслабляться. Расслабленную мышцу можно
растянуть, но благодаря своей эластичности она после
растяжения способна возвратиться к исходным
размерам и форме. Мышцы хорошо снабжаются
кровью, которая доставляет им питательные
вещества и кислород и удаляет отходы метаболизма.
Приток крови к мышцам регулируется таким
образом, что в каждый данный момент мышца получает
Таблица 17.3. Типы движений, производимых парами мышц-
антагонистов.
Классификация
мышц
Сгибатель
Разгибатель
Приводящая
мышца
Отводящая
мышца
Протрактор
Ретрактор
Ротатор
Вид производимого движения
Сгибает конечность, притягивая два
скелетных элемента друг к другу
Распрямляет конечность, оттягивая
два скелетных элемента друг от
друга
Тянет конечность по направлению к
продольной оси тела
Отводит конечность от продольной
оси тела
Тянет дистальный отдел конечности
вперед
Оттягивает дистальный отдел
конечности назад
Поворачивает конечность целиком
или ее часть в одном из суставов
ее в необходимом количестве. Все мышцы имеют
самостоятельную иннервацию. У позвоночных
выделяют три гистологических типа мышц:
1. Скелетные мышцы (разд. 17.4.1) (их называют
также поперечнополосатыми или
произвольными). Это мышцы, прикрепляющиеся к костям.
Они обеспечивают локомоцию, с высокой
скоростью сокращаются и быстро утомляются;
иннервируются соматической нервной системой.
2. Гладкие мышцы (разд. 17.5.3) (их также
называют непроизвольными). Эти мышцы находятся в
стенках трубчатых органов тела и обеспечивают
передвижение содержимого этих органов; они
медленно сокращаются самопроизвольно и
иннервируются вегетативной нервной системой.
3. Сердечная мышца. Она имеется только в сердце,
сокращается самопроизвольно и не подвержена
утомлению. Иннервируется вегетативной
нервной системой.
17.4.1. Особенности скелетных мышц
Скелетные мышцы присоединяются к кости по
меньшей мере в двух местах, к одной неподвижной и
одной подвижной части скелета (первую из них
называют «началом» мышцы, а вторую -
«прикреплением»). Мышца прикрепляется с помощью
плотных, малорастяжимых сухожилий -
соединительнотканных образований, состоящих почти
исключительно из коллагеновых волокон (разд. 5.5). Один
конец сухожилия без перерыва переходит в
наружную оболочку мышцы, а другой-очень прочно
присоединен к надкостнице.
Поскольку мышцы способны развивать силу
только при укорочении (т. е. только тянуть, но не
толкать), ясно, что для того чтобы сместить кость, а
затем вернуть ее в прежнее положение, необходимы
по меньшей мере две мышцы или две группы мышц.
Пары мышц, действующих таким образом,
называются антагонистами. В табл. 17.3 приведена
классификация мышц по типу производимых ими
движений.
Очень редко в движении участвует лишь одна
пара мышц-антагонистов. Обычно каждое
отдельное движение обеспечивается группами мышц,
называемых синергистами.
17.4.2. Поперечнополосатые мышцы
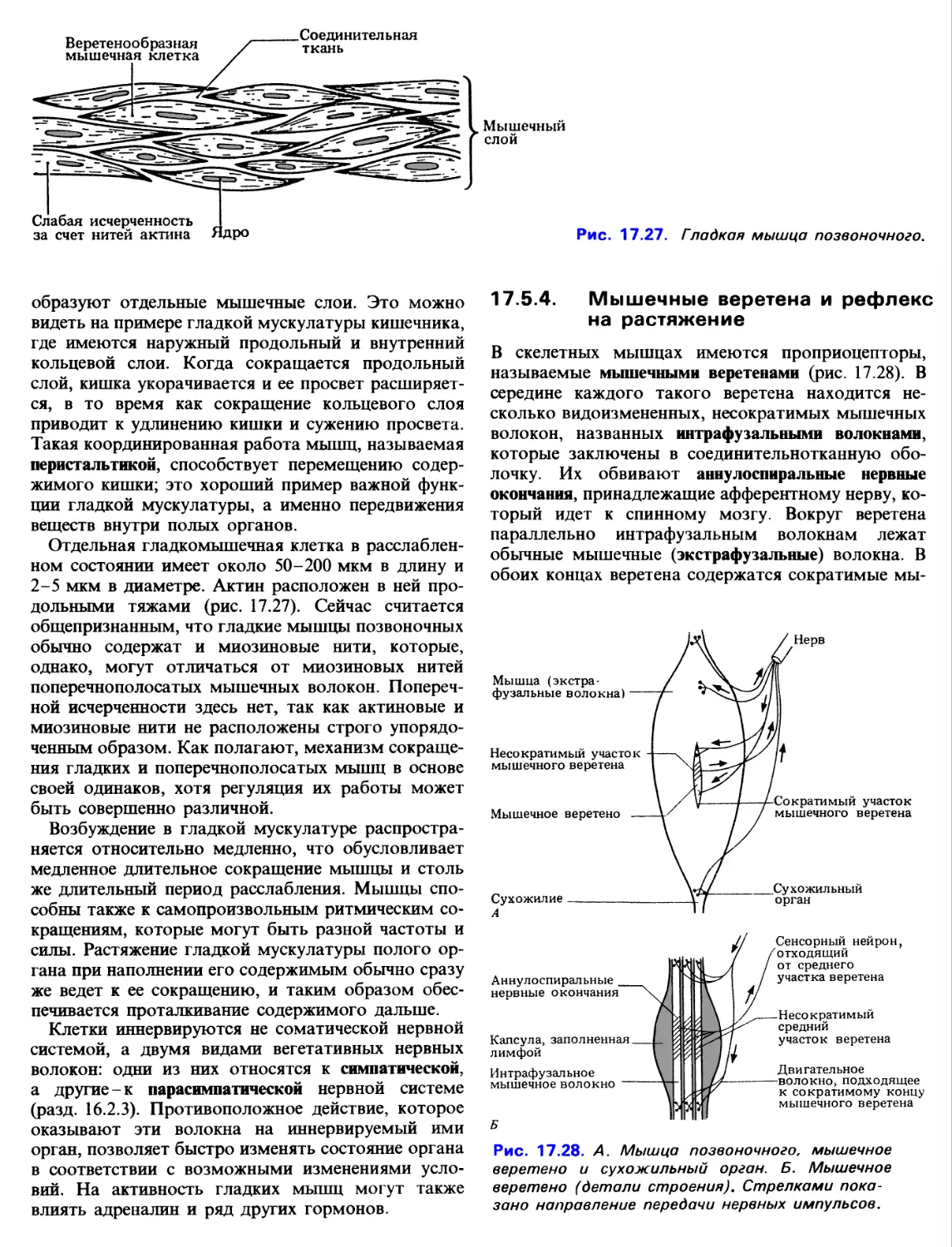
Поперечнополосатая мышца состоит из множества
функциональных единиц - мышечных волокон, или
мышечных клеток1. Они имеют цилиндрическую
форму и расположены параллельно друг другу. Это
многоядерные клетки 0,01-0,1 мм в диаметре,
достигающие нескольких сантиметров в длину. Ядра в
1 Строго говоря, мышечное волокно-не отдельная
клетка, а результат слияния многих клеток, т.е.
синцитий. -Прим. ред.
волокне расположены около его поверхности. Пучки
мышечных волокон окружены коллагеновыми
волокнами и соединительной тканью; между
волокнами тоже находится коллаген. На конце мышц
коллаген вместе с соединительной тканью образует
сухожилия, которые служат для прикрепления
мышц к разным частям скелета. Каждое волокно
окружено мембраной-сарколеммой, которая по
своему строению очень сходна с обычной
плазматической мембраной.
В мышечных волокнах содержится большое
количество миофибрилл (от греч. mys- мышца),
которые и создают характерную поперечную исчерчен-
ность. Каждая миофибрилла состоит из белковых
нитей двух типов -актиновых и миозиновых. Между
миофибриллами находится множество
митохондрий. Цитоплазма мышечного волокна называется
саркоплазмой и содержит сеть внутренних
мембран -саркоплазматический ретикулум. Поперек
волокна и между миофибриллами проходит система
трубочек, называемая Т-системой, которая связана с
сарколеммой (рис. 17.15). В определенных местах
трубочки Т-системы располагаются между двумя
цистернами саркоплазматического ретикулума.
Комплекс из одной Т-трубочки и двух цистерн
называется триадой. Трубочка и цистерны
соединены между собой поперечными мембранными
мостиками. Цистерны участвуют в захвате и
высвобождении ионов Ca2 + . В результате
концентрация этих ионов в саркоплазме снижается или
увеличивается, что в свою очередь влияет на активность
АТФазы, а значит, и на сократительную функцию
мышечного волокна.
В световой микроскоп видна лишь поперечная
исчерченность миофибрилл. Это выглядит как
правильное чередование светлых и темных полос,
названных соответственно зонами (или дисками) I и А.
По середине каждой зоны проходит тонкая темная
линия. Электронно-микроскопическое исследование
ясно показывает, что исчерченность обусловлена
определенным расположением нитей актина (тонких
филаментов) и миозина (толстых филаментов). Это
схематически представлено на рис. 17.16.
Зона I разделена на две половинки темной линией
Z. Участок миофибриллы между двумя линиями Z
называется саркомером. В обе стороны от линии Z
отходят актиновые нити, а в середине саркомера
находятся миозиновые нити. Все они лежат
параллельно друг другу, образуя, как видно на
поперечном срезе, гексагональную решетку (рис. 17.17. и
17.18). В определенных участках саркомера
актиновые и миозиновые нити перекрываются; в этих
участках вокруг одной миозиновой нити
размещаются шесть актиновых. Такое взаиморасположение
миофиламентов приводит к появлению в саркомере
нескольких дисков (полос). Там, где актиновые и
Миофибриллы
Триада
РИС 17.15. Саркоплазматический ретикулум и T-система.
Поперечная трубочка
Гликогеновые гранулы
Продольные цистерны
саркоплазматического
ретикулума
Наружные ("концевые")
цистерны
саркоплазматического
ретикулума
Мышца
Сухожилие
Мышечное
волокно
Миофиб-
рилла
Ядро
Видимая
исчерченность
Саркомер
Три саркомера
миофибриллы
Миозин
(толстая нить)
Зона H
Зона
I
Зона А
Поперечные через
разрезы зону H
саркомера.
Показано о о о
расположение о о о о
нитей о о о о о
в миофибрилле ° ° ° °
о о о
Актин
(тонкая нить)
через
зону I
+Тропомиозин
+Тропонин
Молекула
миозина
(с двумя головками)
Головки
lF-актин
G-актин
через зону
перекрывания нитей
ется при взаимодействии актина с миозином. Ионы
Mg2+ могут ингибировать этот процесс. Значение
описанных явлений станет ясным, когда мы
рассмотрим механизм сокращения саркомера.
Актин (тонкие филаменты)
Каждая актиновая нить образована двумя
спиральными тяжами из глобулярных молекул актина
(G-актина), закрученными один вокруг другого.
Весь комплекс актиновых молекул называется
F-актином (фибриллярным актином). Как
полагают, с каждой молекулой F-актина связана молекула
АТФ. Ни одна из форм актина не обладает
АТФазной активностью.
Актиновые филаменты состоят из F-актина и двух
вспомогательных белков - тропомиозина и тропони-
на. Тропомиозин - палочкообразный фибриллярный
белок, и эти палочки соединяются конец в конец и
образуют два тяжа, закрученные в виде растянутой
спирали вокруг F-актина. Тропомиозин служит для
включения и выключения сократительного
механизма. Тропонин - глобулярный белок, состоящий из
трех субъединиц. Каждая субъединица выполняет
определенную функцию. Тропонин T связывает
тропонин с тропомиозином; тропонин С чувствителен к
Рис. 17.16. Ультраструктура скелетного мышечного
миозиновые нити перекрываются, они образуют
диск А, в то время как в районе диска I находятся
только актиновые нити. Средняя часть зоны А более
светлая; она называется зоной Нив свою очередь
может подразделяться надвое линией M, которая
делит миозиновые нити на две равные части.
Миозин (толстые филаменты)
Молекула миозина состоит из двух частей:
длинного палочкообразного участка («хвоста») и
присоединенного к одному из его концов глобулярного
участка, который представлен двумя одинаковыми
«головками». Молекулы миозина расположены в
миозиновой нити таким образом, что головки
регулярно распределяются по всей ее длине, кроме
небольшого срединного участка, где их нет («голая»
зона). В тех местах, где нити актина и миозина
перекрываются, миозиновые головки могут
прикрепляться к соседним актиновым нитям, и в
результате этого взаимодействия может происходить
укорочение мышцы. Энергия для совершения такой
работы высвобождается при гидролизе АТФ; все
миозиновые головки обладают АТФазной
активностью. Как мы увидим дальше, прикрепление мио-
зиновых головок зависит от концентрации ионов
Ca2+ в саркоплазме. Миозиновая АТФаза активиру-
!Триада
!Саркомер,
Т-тру-|
бочка I
Рис. 17.17. Продольный срез мышечного волокна
рыбы (плотвы, Rutilus rutilus). Видны триады и
детали строения миофибрилл χ 7650.
Миозиновая
нить
Актиновая
нить
Поперечный
мостик
Рис. 17.18. Летательная мышца насекомого
(клопа белостомы). Поперечный срез волокна в
состоянии окоченения (х 137000).
Рис. 17.19. Изменение структуры актиновых
нитей. А. «Выключенное» состояние при
низком уровне Ca2+; тропомиозин блокирует участки
связывания миозина с актином. Б. «Включенное»
состояние при высоком уровне Ca2 + :
тропомиозин смещается, обнажая участки связывания с
миозином (указаны стрелками). А-актин; T-
тропомиозин. Тропонин не показан, он
расположен ближе к субъединицам актина, изображенным
серыми.
ионам Ca2+ и может обратимо присоединять их,
тогда как тропонин I способен ингибировать всякого
рода взаимодействие между актином и миозином.
Оба вспомогательных белка подавляют
взаимодействие актина с миозином в отсутствие ионов Ca2 +
(рис. 17.19).
17.4.3. Реакция типа «всё или ничего»
При раздражении скелетного мышечного волокна
оно будет сокращаться лишь в том случае, если
стимулирующий импульс достигнет определенной
пороговой величины или превысит ее. И это
сокращение будет для данных условий
максимальным. Даже значительное увеличение силы
раздражителя не приведет к большему укорочению волокна
или увеличению развиваемой им силы. Такое
явление называют реакцией типа «всё или ничего».
Слабый раздражитель, не способный вызвать
сокращение мышечного волокна, называется под-
пороговым.
После окончания ответа наступает период
абсолютной рефрактерности, когда волокно не способно
сокращаться. Затем следует период относительной
рефрактерности. В это время только сильный стимул
может вызвать сокращение. Период рефрактерности
представляет собой отрезок времени, в течение
которого происходит восстановление исходного
состояния мышечного волокна.
17.4.4. Типы сокращения скелетных
мышц позвоночных
В ответ на стимуляцию мышца совершает
механическую работу (т.е. сокращается), и это может
привести либо к укорочению мышцы, либо-если
оба конца мышцы неподвижно закреплены - к тому,
что мышца будет развивать напряжение без
изменения ее длины. Когда мышца, преодолевая
постоянную нагрузку, укорачивается, такое
сокращение называют изотоническим, а если при сокращении
мышцы не происходит изменения ее длины, его
называют изометрическим.
Мышечное сокращение можно изучать с
помощью кимографа. Такого рода исследования часто
проводят на икроножной мышце лягушки. Мышцу
выделяют из лапки только что забитой лягушки и
помещают в ванночку с хорошо оксигенируемым
физиологическим раствором. В этих условиях
мышца в течение нескольких часов сохраняет
способность адекватно реагировать на раздражение. Один
конец мышцы неподвижно закрепляют на
кронштейне, а другой-с сухожилием - прицепляют к
подвижному рычагу. Затем раздражают
электрическим током непосредственно мышцу или
подходящий к ней нерв. Когда мышца сокращается или в
ней возрастает напряжение, она смещает рычаг, и
след от этого смещения регистрируется на
вращающемся барабане кимографа. Полученная запись
таких событий называется миограммой.
Напряжение, возникающее в мышце,-это некоторая сила,
обычно измеряемая в граммах.
17.4.5. Сократительная реакция
одиночное раздражение. При воздействии
одиночного стимула мышца начинает сокращаться спустя
очень короткое время (около 0,05 с), называемое
латентным периодом. Затем мышца быстро
укорачивается, в ней развивается напряжение. Эта фаза
сокращения длится около 0,1 с. Вслед за ней
наступает более долгий период расслабления, когда
напряжение уменьшается и мышца возвращается в
исходное состояние; это занимает 0,2 с. Такое
сокращение мышцы называется одиночным
сокращением (рис. 17.20).
двукратное раздражение. Если интервал между
первой и второй стимуляцией значителен, миограм-
ма регистрирует два одинаковых ответа мышцы.
Однако если отрезок времени между двумя
стимулами укорачивается настолько, что второй из них
подается в тот момент, когда мышца еще
сокращается после первого, то второе сокращение
накладывается на первое. В результате получается
«бугристая» миограмма (рис. 17.21). При втором
сокращении развивается большее в сравнении с
первым напряжение. Этот эффект называется
механической суммацией.
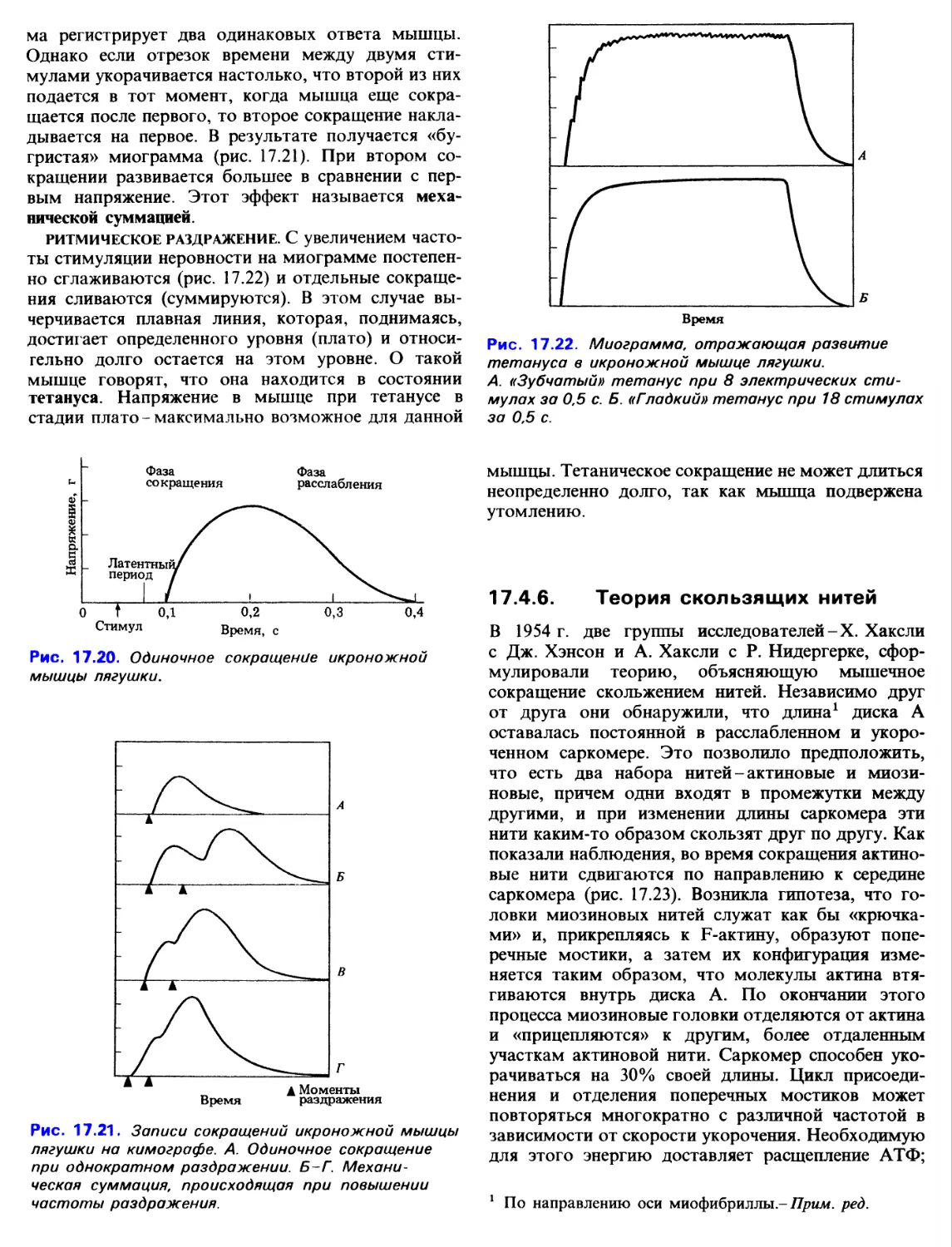
ритмическое раздражение. С увеличением
частоты стимуляции неровности на миограмме
постепенно сглаживаются (рис. 17.22) и отдельные
сокращения сливаются (суммируются). В этом случае
вычерчивается плавная линия, которая, поднимаясь,
достигает определенного уровня (плато) и
относительно долго остается на этом уровне. О такой
мышце говорят, что она находится в состоянии
тетануса. Напряжение в мышце при тетанусе в
стадии плато - максимально возможное для данной
Фаза
сокращения
Фаза
расслабления
Напряжение, г
Латентный^
период
Стимул
Время, с
Время
Рис. 17.22. Миограмма, отражающая развитие
тетануса в икроножной мышце лягушки.
А. «Зубчатый» тетанус при 8 электрических
стимулах за 0,5 с. Б. «Гладкий» тетанус при 18 стимулах
за 0,5 с.
Рис. 17.20. Одиночное сокращение икроножной
мышцы лягушки.
Время
А Моменты
раздражения
Рис. 17.21. Записи сокращений икроножной мышцы
лягушки на кимографе. А. Одиночное сокращение
при однократном раздражении. Б-Г.
Механическая суммация, происходящая при повышении
частоты раздражения.
мышцы. Тетаническое сокращение не может длиться
неопределенно долго, так как мышца подвержена
утомлению.
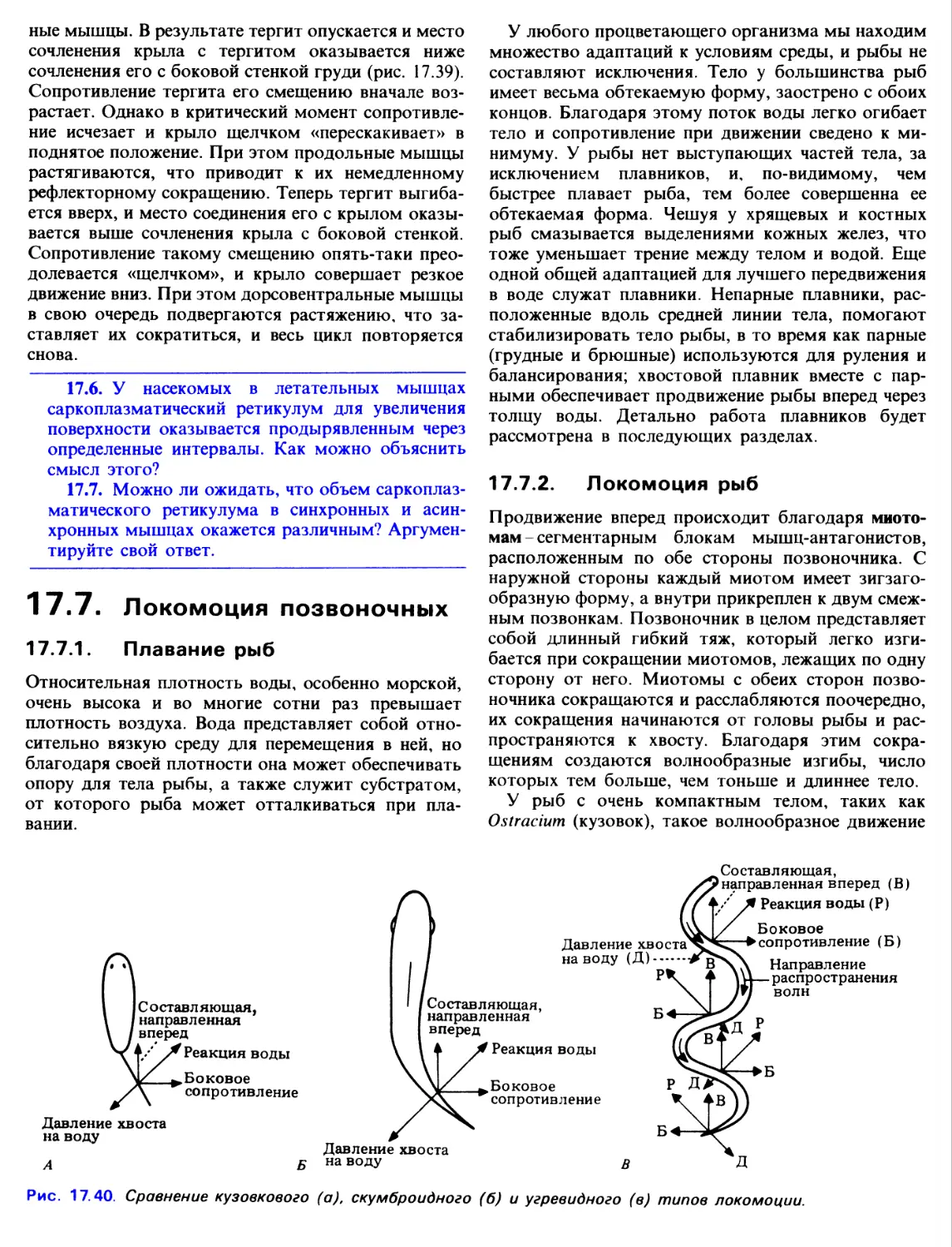
17.4.6. Теория скользящих нитей
В 1954 г. две группы исследователей-X. Хаксли
с Дж. Хэнсон и А. Хаксли с P. Нидергерке,
сформулировали теорию, объясняющую мышечное
сокращение скольжением нитей. Независимо друг
от друга они обнаружили, что длина1 диска А
оставалась постоянной в расслабленном и
укороченном саркомере. Это позволило предположить,
что есть два набора нитей - актиновые и миози-
новые, причем одни входят в промежутки между
другими, и при изменении длины саркомера эти
нити каким-то образом скользят друг по другу. Как
показали наблюдения, во время сокращения
актиновые нити сдвигаются по направлению к середине
саркомера (рис. 17.23). Возникла гипотеза, что
головки миозиновых нитей служат как бы
«крючками» и, прикрепляясь к F-актину, образуют
поперечные мостики, а затем их конфигурация
изменяется таким образом, что молекулы актина
втягиваются внутрь диска А. По окончании этого
процесса миозиновые головки отделяются от актина
и «прицепляются» к другим, более отдаленным
участкам актиновой нити. Саркомер способен
укорачиваться на 30% своей длины. Цикл
присоединения и отделения поперечных мостиков может
повторяться многократно с различной частотой в
зависимости от скорости укорочения. Необходимую
для этого энергию доставляет расщепление АТФ;
1 По направлению оси миофибриллы.- Прим. ред.
.Миозин
Z
Актин-
Рис. 17.23. Схема сокращения саркомера. Актиновые
нити скользят вдоль нитей миозина.
для осуществления полного цикла одного мостика
требуется одна молекула АТФ.
Сейчас эта гипотеза принята почти всеми. Однако
действительный процесс, приводящий при этом к
созданию механической силы (электромеханическое
сопряжение), еще мало изучен, и для физиологов,
занимающихся мышцами, он остается главной
проблемой, ждущей своего решения. Однако в этой
области достигнуты значительные успехи, и в
следующем разделе мы изложим современные
представления о механизме сокращения саркомера.
17.3. Как изменяется длина дисков A, H и I при
сокращении саркомера?
17.4. Как можно объяснить изменения дисков
A, H и I, исходя из того, что происходит с нитями
актина и миозина?
17.4.7. Электромеханическое
сопряжение
В состоянии покоя саркомер содержит
определенные количества ионов Mg2 + и АТФ, в то время как
концентрация свободных ионов Ca2+ очень мала. В
этих условиях актиновые нити находятся в
«нерабочем состоянии», так как тропомиозин,
располагаясь определенным образом на молекуле актина,
блокирует участки, к которым должны
прикрепляться головки миозина, и эти головки притянуты к
продольной оси миозиновой нити и удалены от
актина.
При стимуляции мышечного волокна нервным
импульсом по сарколемме распространяется волна
деполяризации, которая затем то Т-системе
переходит внутрь саркомера. Импульс достигает
цистерн триады и стимулирует высвобождение из них
ионов Ca2 + , концентрация которых в саркоплазме
соответственно увеличивается.
Ионы Ca2+ связываются с тропонином С.
который в свою очередь взаимодействует с
тропонином 1 и снимает обычный эффект системы тро-
понина, препятствующий образованию мостиков
между актином и миозином. Актин «включается»
при смещении расположенного на его поверхности
тропомиозина - места связывания актина с
миозином обнажаются. В мышцах некоторых
беспозвоночных животных ионы Ca2+ могут также
стимулировать АТФазную активность миозина,
которая усиливается в дальнейшем и под действием
актина. Во всех мышцах при активации актина и
миозина миозиновая головка выводится из
положения покоя и, связываясь с актином, образует
актомиозиновый поперечный мостик. Энергия для
образования таких мостиков высвобождается при
гидролизе АТФ, и за счет этой энергии изменяется
угол отхождения поперечных мостиков, что ведет к
подтягиванию актиновой нити по направлению к
середине саркомера. В результате такой активности
всех миофиламентов при раздражении саркомера в
нем создается сила (рис. 17.24), которая стремится
вызвать укорочение саркомера.
При затухании возбуждения в саркомере ионы
Ca2+ активно откачиваются назад в цистерны триад
с помощью АТФ-зависимого кальциевого насоса.
Концентрация этих ионов вскоре становится ниже
пороговой для сокращения, и саркомер
расслабляется. Комплекс тропомиозин-тропонин ингибирует
АТФазную активность; поперечные мостики
разрываются; актин, а возможно, и миозин переходят в
«выключенное» состояние, и напряжение саркомера
возвращается к обычному уровню покоя.
Актиновая нить
-G-актин
Головка
миозина
Образование
поперечного
мостика
Миозиновая
нить
Движение актина
Смена положения
миозиновой
головки
Разрушение
поперечного
мостика
Рис. 17.24. Электромеханическое сопряжение
(тропомиозин и тропонин не изображены).
Окисление
жирных кислот
Гликоген
в мышце
Кровь4
4/5 MK -♦
Печень
Ликвидация
кислородной
задолженности
Энергия
для мышечного
сокращения
Дополнительный
кислород, л/мин
»-» со
Потребность
в О,
Кислородная
задолженность;
ее ликвидация
Уровень покоя
Действительное
количество O2
- ι
Мышечная работа Восстановление
Рис. 17.25. А. Отношения между АТФ, креатинфосфатом и процессом дыхания при
сокращении мышцы. MK-молочная кислота; ПК - пировиноградная кислота; Kp-креатин; КрФ-креатинфосфат.
Б. Потребность в кислороде при физической роботе. Показано соотношение между потреблением
кислорода и созданием кислородной задолженности.
17.4.8. Снабжение энергией
Обычно первоисточником энергии для мышечного
сокращения служит гликоген, а иногда
используются жирные кислоты. В процессе расщепления этих
субстратов вырабатывается АТФ, гидролиз
которого доставляет энергию непосредственно для
самого сокращения:
АТФ -> АДФ + Фн + Энергия.
В покоящейся мышце количество АТФ невелико,
оно может обеспечить энергией лишь около восьми
одиночных сокращений. Такой уровень АТФ может
поддерживаться при обычном аэробном
метаболизме (дыхании). В процессе работы мышц АТФ
быстро расходуется, и его уровень должен быстро
восстанавливаться уже за счет других процессов.
В восстановлении уровня АТФ участвует
содержащийся в мышцах креатинфосфат (КрФ).
Образующийся при мышечном сокращении АДФ вновь фос-
форилируется в АТФ за счет креатинфосфата:
АДФ 4- КрФ
Креатинфосфотрансфераза
АТФ + Креатин.
Этот процесс непрерывно пополняет запасы АТФ,
которые могут использоваться непосредственно при
сокращении. Но в конце концов возникает
необходимость и в пополнении резервов креатинфосфата, и
оно происходит за счет окисления жирных кислот
или гликогена. Образующийся при этом АТФ
используется для ресинтеза креатинфосфата из
креатина (Kp):
Kp + АТФ -> КрФ + АДФ.
При очень интенсивной работе мышц
поступающего кислорода вскоре оказывается недостаточно
для поддержания на должном уровне
окислительного форфорилирования (разд. 11.3.6). В этих
условиях пировиноградная кислота - конечный продукт
гликолиза-при участии ионов H+ превращается в
молочную кислоту.
CH3COCOOH
Пировиноградная
кислота
+ H2- CH3-CHOH-COOH
Молочная кислота
Так происходит из-за нехватки кислорода,
связывающего ионы H + . В этом случае говорят, что
работа мышцы создает кислородную задолженность.
Образование молочной кислоты - процесс с
относительно малым энергетическим выходом:
высвобождаемая энергия составляет лишь 7% от энергии,
получаемой при полном окислении глюкозы.
Молочная кислота токсична и рано или поздно должна
быть выведена из организма или переработана. Так
и происходит при уменьшении или прекращении
мышечной работы. В этот период кислорода
становится достаточно для окисления молочной
кислоты и частичного превращения ее в гликоген. Такой
процесс обычно происходит в печени; здесь 1/5 часть
молочной кислоты полностью окисляется до
углекислоты и воды; высвобождаемая при этом энергия
расходуется на превращение остальной молочной
кислоты в глюкозу. Часть глюкозы возвращается в
мышцы, и там из нее в конце концов образуется
гликоген, а глюкоза, оставшаяся в печени,
превращается в гликоген и запасается в ней. Время полной
переработки молочной кислоты - это именно то
время, которое необходимо для ликвидации
кислородной задолженности после энергичной работы мышц
(рис. 17.25).
17.5. Иннервация
скелетной мышцы
К каждой мышце подходит много двигательных
аксонов, которые ветвятся и иннервируют группы
мышечных волокон. Каждая такая группа вместе с
иннервирующим ее аксоном называется двигатель-
Гликолиз
+ 1/2O2
.2H+
ПК
Аэрооные
процессы
Анаэробные
процессы у
Глюкоза
!Гликоген
Молочная
кислота
Кровь
ной единицей. При надлежащей стимуляции все
мышечные волокна в ней сокращаются
одновременно. Число этих волокон в двигательной единице
варьирует и зависит от того, насколько сложным
должно быть управление данной мышцей.
Например, в глазодвигательной мышце это число
составляет около 10, а в бицепсе - более 1000. Чем меньше
число волокон в двигательной единице, тем более
тонкий контроль осуществляется со стороны
нервной системы.
В месте контакта двигательного аксона с
мышечным волокном формируется нервно-мышечное
соединение, или двигательная концевая пластинка.
Здесь аксон не имеет миелиновой оболочки и его
ветвящиеся окончания погружены в разветвленные
бороздки концевой пластинки.
Сигнал для сокращения поперечнополосатых
мышц поступает из центральной нервной системы
(рис. 17.26). Импульсы по двигательному нерву
передаются на двигательную концевую пластинку.
Здесь в ответ на нервный сигнал в синаптическую
щель, отделяющую моторный аксон от сарколеммы
мышечного волокна, секретируется ацетилхолин.
События, происходящие в концевой пластинке и в
межнейронных синапсах (разд. 16.1.2), удивительно
сходны. Ацетилхолин диффундирует к сарколемме и
на короткое время повышает ее проницаемость для
ионов, особенно для Na+ и K + ; в результате
возникает потенциал концевой пластинки, в свою очередь
генерирующий потенциал действия; последний
быстро распространяется по всей длине и в глубь
мышечного волокна по Т-системе, вызывая
сократительную реакцию по принципу «всё или ничего».
В области сарколеммы в большом количестве
содержится ацетилхолинэстераза, которая быстро
расщепляет ацетилхолин на холин и уксусную
кислоту и таким образом предотвращает избыточную
стимуляцию мышечного волокна. Передав
возбуждение мышечному волокну, концевая пластинка
быстро возвращается в состояние покоя.
17.5.1. Регуляция силы сокращения
скелетных мышц
Для тонкого управления мышечной активностью
необходима регуляция напряжения, развиваемого
каждой отдельной мышцей. Такая регуляция
осуществляется одним из двух способов (или
одновременно обоими):
1. Может изменяться число мышечных волокон,
возбуждающихся в каждый данный момент.
Развиваемая мышцей сила будет тем больше, чем
больше будет число стимулированных волокон, и
наоборот. Так обычно обстоит дело в скелетных
мышцах позвоночных.
2. Может изменяться частота нервных импульсов,
приходящих к мышечным волокнам. Таким
образом, более частая стимуляция тоже будет
приводить к увеличению развиваемой мышцей силы.
Сокращение мышц в организме происходит
плавно и координирование. Это обеспечивается
асинхронным сокращением разных групп мышечных
волокон в мышцах-антагонистах.
17.5.2. Типы скелетных мышечных
волокон
Выделяют два типа скелетных мышечных волокон,
каждый из которых имеет свои физиологические
особенности. Это медленные (тонические) и быстрые
(фазические) волокна. Их строение, локализация и
общие свойства указаны в табл. 17.4. В некоторых
мышцах могут быть только быстрые или только
медленные волокна, в других - волокна обоих типов
в определенном соотношении.
Благодаря волокнам этих двух типов организм
способен передвигаться и поддерживать позу.
Быстрые волокна позволяют мышце сокращаться с
высокой скоростью. В большом количестве эти
волокна имеются у хищников; они обеспечивают
быстроту реакции при ловле добычи. С другой
стороны, потенциальная добыча, чтобы не стать
Складки
сарколемы
Двигательная
концевая пластинка
Синаптические пузырьки
с ацетилхолином
Аксон
Миелиновая оболочка
Саркоплазма
Ядро мышечного волокна
Митохондрия
Миофибриллы
Рис. 17.26. Двигательная концевая пластинка в месте контакта нерва и мышечного волокна.
Таблица 17.4 Строение, локализация и общие свойства быстрых и медленных мышечных волокон
жертвой хищников, тоже должна быть способна к
быстрому реагированию. В обоих случаях от
подвижности животного будут зависеть его шансы на
выживание. Когда животное находится в покое, оно
поддерживает определенную позу с помощью
тонических мышечных волокон. Им свойственно более
медленное и длительное сокращение и в то же время
энергетические затраты меньше, чем при
сокращении быстрых волокон. Сокращение их по своей
природе обычно изометрическое, при котором
мышцы, противодействуя силе тяжести и удерживая
конечности в определенном положении, сохраняют
постоянную длину.
У человека все мышцы тела состоят из волокон
обоих типов, но обычно один из них доминирует.
Это имеет физиологическое значение, поскольку
тонические мышцы способны к медленному и
длительному сокращению и их соответственно больше
в позных мышцах-разгибателях, тогда как в
сгибателях, предназначенных для быстрых реакций,
преобладают фазические волокна.
17.5.3. Гладкие мышцы
Гладкая мускулатура имеется в стенках многих
полых образований в теле животного, к которым
относятся кишечный тракт, мочевой пузырь,
кровеносные сосуды, мочеточники, матка, семявыно-
сящий проток. Гладкомышечные
клетки-одноядерные, веретенообразные; они скреплены
соединительной тканью, состоящей в основном из коллагена.
Клетки располагаются параллельно друг другу и
Строение
Расположение
Иннервация
Возбудимость
Тип ответа
Физиологическая
активность
Функция
Медленные (тонические) волокна
Много митохондрий
Саркоплазматический ретикулум развит слабо
Красные — из-за присутствия миоглобина и ци-
тохромовых пигментов
Содержание гликогена невелико
Тесный контакт волокон с капиллярами для
ускорения обмена веществами
В глубоких слоях мышц конечностей
Тонкие нервные волокна 5 мкм в диаметре
На одном мышечном волокне несколько
концевых пластинок (это называется мультитер-
минальной иннервацией)
Аксоны
Концевые
пластинки
Волокно
Скорость проведения импульса 2—8 м ■ с-'
Мембрана не обладает электрической
возбудимостью. Каждый импульс приводит к
высвобождению лишь небольшого количества аце-
тилхолина. Таким образом, степень
деполяризации мембраны зависит от частоты
стимуляции
Медленное градуальное сокращение. Медленное
расслабление (приблизительно в 100 раз
медленнее, чем у быстрых волокон)
Источник АТФ — аэробное дыхание
Многие волокна при недостатке O2 продолжают
работать за счет анаэробного гликолиза; в
этом случае образуется молочная кислота и
создается кислородная задолженность
По мере окисления дыхательного субстрата
мобилизуются резервные углеводы и жиры
Тепло отводится от мышцы по мере его
выработки
Устанавливается равновесие между работой
мышцы и ее потребностями
Обеспечивают длительное сокращение мышцы;
это используется для поддержания позы
Быстрые (фазические) волокна
Мало митохондрий
Саркоплазматический ретикулум хорошо развит
Белые — миоглобина и цитрохромовых пигментов
мало или нет вовсе
Обилие гликогеновых гранул
Ближе к поверхности
Толстые нервные волокна 10—20 мкм в диаметре
Обычно на одном мышечном волокне одна
концевая пластинка (могут быть две)
Скорость проведения 8—40 м · с-'
Мембрана обладает электрической
возбудимостью. Когда возникает потенциал действия,
развивается ответ типа «всё или ничего»
Быстрое сокращение (в три раза быстрее, чем у
медленных волокон). Довольно быстрое
утомление
Источник АТФ — анаэробные процессы (гликолиз)
Быстро создается кислородная задолженность
В качестве дыхательного субстрата интенсивно
используется гликоген
Тепло поглощается волокнами, так как
кровеносная система не обеспечивает его быстрого
отведения
Мышца некоторое время сокращается и тогда,
когда кровеносная система не успевает обеспечить
дополнительную доставку кислорода
Обеспечивают немедленное быстрое сокращение,
когда кровеносная система еще только
приспосабливается к новому уровню мышечной
активности; поэтому очень важны при локомоции
Веретенообразная
мышечная клетка
Соединител ьная
ткань
Мышечный
слой
Слабая исчерченность
за счет нитей актина
Ядро
Рис. 17.27. Гладкая мышца позвоночного.
образуют отдельные мышечные слои. Это можно
видеть на примере гладкой мускулатуры кишечника,
где имеются наружный продольный и внутренний
кольцевой слои. Когда сокращается продольный
слой, кишка укорачивается и ее просвет
расширяется, в то время как сокращение кольцевого слоя
приводит к удлинению кишки и сужению просвета.
Такая координированная работа мышц, называемая
перистальтикой, способствует перемещению
содержимого кишки; это хороший пример важной
функции гладкой мускулатуры, а именно передвижения
веществ внутри полых органов.
Отдельная гладкомышечная клетка в
расслабленном состоянии имеет около 50-200 мкм в длину и
2-5 мкм в диаметре. Актин расположен в ней
продольными тяжами (рис. 17.27). Сейчас считается
общепризнанным, что гладкие мышцы позвоночных
обычно содержат и миозиновые нити, которые,
однако, могут отличаться от миозиновых нитей
поперечнополосатых мышечных волокон.
Поперечной исчерченности здесь нет, так как актиновые и
миозиновые нити не расположены строго
упорядоченным образом. Как полагают, механизм
сокращения гладких и поперечнополосатых мышц в основе
своей одинаков, хотя регуляция их работы может
быть совершенно различной.
Возбуждение в гладкой мускулатуре
распространяется относительно медленно, что обусловливает
медленное длительное сокращение мышцы и столь
же длительный период расслабления. Мышцы
способны также к самопроизвольным ритмическим
сокращениям, которые могут быть разной частоты и
силы. Растяжение гладкой мускулатуры полого
органа при наполнении его содержимым обычно сразу
же ведет к ее сокращению, и таким образом
обеспечивается проталкивание содержимого дальше.
Клетки иннервируются не соматической нервной
системой, а двумя видами вегетативных нервных
волокон: одни из них относятся к симпатической,
а другие-к парасимпатической нервной системе
(разд. 16.2.3). Противоположное действие, которое
оказывают эти волокна на иннервируемый ими
орган, позволяет быстро изменять состояние органа
в соответствии с возможными изменениями
условий. На активность гладких мышц могут также
влиять адреналин и ряд других гормонов.
17.5.4. Мышечные веретена и рефлекс
на растяжение
В скелетных мышцах имеются проприоцепторы,
называемые мышечными веретенами (рис. 17.28). В
середине каждого такого веретена находится
несколько видоизмененных, несократимых мышечных
волокон, названных интрафузальными волокнами,
которые заключены в соединительнотканную
оболочку. Их обвивают аннулоспиральные нервные
окончания, принадлежащие афферентному нерву,
который идет к спинному мозгу. Вокруг веретена
параллельно интрафузальным волокнам лежат
обычные мышечные (экстрафузальные) волокна. В
обоих концах веретена содержатся сократимые мы-
Мышца (экстра -
фузальные волокна)
Несократимый участок
мышечного веретена
Мышечное веретено
Сухожилие
Нерв
Аннулоспиральные
нервные окончания
Капсула, заполненная
лимфой
Интрафузальное
мышечное волокно
Сократимый участок
мышечного веретена
Сухожильный
орган
Сенсорный нейрон,
отходящий
от среднего
участка веретена
Несократимый
средний
участок веретена
Двигательное
волокно, подходящее
к сократимому концу
мышечного веретена
Рис. 17.28. А. Мышца позвоночного, мышечное
веретено и сухожильный орган. Б. Мышечное
веретено (детали строения). Стрелками
показано направление передачи нервных импульсов.
Спинной мозг
Двигательный
аксон
Серое вещество
-Тормозный
промежуточный
нейрон
МЫШЦА
!СОКРАЩАЕТСЯ
МЫШЦА
РАССЛАБЛЯЕТСЯ
Пара мышц-
антагонистов
Рис. 17.29. Реципрокное торможение. При сокращении мышцы А импульсы направляются в спинной мозг, где
они передаются на промежуточный тормозный нейрон; торможение им соответствующего
двигательного нейрона, иннервирующего мышцу Б, заставляет эту мышцу расслабиться.
шечные волокна, которые иннервируются
эфферентными нервами, идущими от центральной нервной
системы.
При растягивании мышцы в аннулоспиральных
окончаниях веретена возникает напряжение,
вызывающее залп нервных импульсов, которые идут по
афферентным нейронам к спинному мозгу. Эти
нейроны образуют синапсы непосредственно с
эфферентными нейронами, которые посылают
импульсы к экстрафузальным мышечным волокнам,
вызывая их рефлекторное сокращение. Это рефлекс
на растяжение. Чем сильнее будет растянуто
веретено, тем больше импульсов пойдет в спинной мозг
и тем сильнее сократится мышца. Таким образом,
этот рефлекс направлен на поддержание постоянной
длины мышцы при изменении нагрузки на нее.
Мышечные веретена работают не все
одновременно: разные веретена возбуждаются в разное
время в зависимости от нужд организма. В любой
момент времени всегда есть несколько веретен,
стимулируемых непрерывным потоком сигналов из
центральной нервной системы. Это поддерживает
частично сокращенное состояние мышц, называемое
мышечным тонусом. Для хорошего мышечного
тонуса нужна систематическая работа мышц,
например физические упражнения.
Важную роль в поддержании мышечного тонуса
играют концевые участки веретена. При стимуляции
их импульсами эфферентного нерва они
сокращаются, и этим обеспечивается некоторое напряжение
ненагруженной мышцы. Если бы мышца не
поддерживалась в таком состоянии, то в случае
внезапной нагрузки она могла бы быть повреждена в
результате чрезмерного растяжения.
17.5.5. Тормозные рефлексы
Для перемещения конечности в том и другом
направлении нужны по меньшей мере две
противоположно действующие мышцы или группы мышц.
Когда одна из них сокращается, другая должна
расслабляться. Достигается это благодаря простым
тормозным рефлексам. Как мы уже видели,
сигналы, возникшие в мышечном веретене, поступают от
мышцы по афферентному нейрону в спинной мозг и
в конце концов возвращаются к той же мышце и
вызывают ее сокращение. Афферентный нейрон
образует также синапсы с промежуточными
нейронами в сером веществе спинного мозга (рис. 17.29).
При соответствующей стимуляции они блокируют
эфферентные нейроны, идущие к
мышце-антагонисту, и эта мышца, таким образом, оказывается
неспособной к сокращению и остается расслабленной.
Хорошим примером может служить механика
ходьбы. Сначала конечность сгибается, чтобы стопа
оторвалась от земли. Во время сгибания мышцы-
антагонисты - разгибатели - растягиваются, но
рефлекторное торможение делает их сокращение
невозможным. Затем согнутая конечность
выпрямляется и стопа вновь опускается на землю. По
окончании сокращения мышц-сгибателей торможение с
мышц-разгибателей снимается, и теперь становится
возможным рефлекс на растяжение, приводящий к
сокращению мышцы-разгибателя. Когда
конечность выпрямлена, мышечное веретено разгибателя
не подвергается больше растяжению и рефлекс на
растяжение исчезает. После этого весь процесс
может начаться снова.
17.5.6. Сухожильные рецепторы
Рецепторы растяжения есть и в сухожилиях, однако
они реагируют только на очень сильное растяжение.
Импульсы от них поступают в центральную
нервную систему и воздействуют на тормозные
промежуточные нейроны, которые затормаживают
сокращение мышцы и ее активное противодействие
растяжению. Этот процесс очень важен, так как он
предотвращает повреждение мышц, возможное при
внезапной перегрузке или при излишне сильном
сокращении в ответ на большую нагрузку.
17.5. Изучите рис. 17.28 и ответьте на вопросы:
а. Что происходит с мышечным веретеном при
растяжении всей мышцы (экстрафузальных
волокон)?
б. Что следует за сокращением мышечного
веретена?
в. Что произойдет, если мышца будет сильно
растянута, например при внезапной большой
нагрузке?
г. Что происходит при сокращении интрафузаль-
ных мышечных волокон?
17.6. Различные способы
локомоции у
беспозвоночных
17.6.1. Амебоидное движение
Для амебоидного движения характерно образование
в клетке временных выступов-так называемых
псевдоподии. Этим способом перемещаются
простейшие класса Rhizopoda и лейкоциты (белые
кровяные клетки) позвоночных. При движении в клетке
намечается «передний» конец - место, где
образуются новые псевдоподии. Изучение амебы показало,
что ее цитоплазма состоит из наружного вязкого
слоя - плазмагеля, или эктоплазмы, которым
окружен более жидкий слой-плазмазоль, или
эндоплазма. Как полагают, движение происходит благодаря
поочередным изменениям коллоидного состояния
цитоплазмы, обусловленным переходами золь τ±
гель, и перетеканию плазмазоля в псевдоподию
(разд. 4.1). В месте образования псевдоподии плаз-
магель разжижается, превращаясь в плазмазоль;
теперь плазмазоль перетекает в эту часть клетки и
дальше во вновь образующуюся псевдоподию. В
наружной зоне псевдоподии плазмазоль быстро
переходит в плазмагель, образующий вокруг
псевдоподии плотный воротничок. В заднем конце амебы
плазмагель быстро превращается в плазмазоль и
перетекает вперед (рис. 17.30)1.
Существует несколько гипотез, объясняющих
амебоидное движение. Согласно одной из них,
плазмазоль проталкивается вперед в формирующуюся
1 Эти представления имеют сейчас только
исторический интерес.- Прим. ред.
псевдоподию в результате сокращения заднего
конца амебы. Изменения вязкости цитоплазмы,
сопровождаемые небольшими изменениями давления в
разных частях клетки, по мнению исследователей,
приводят к перетеканию цитоплазмы внутри клетки.
Сторонники более распространенной гипотезы
«фонтанирования» утверждают, что клетка
перемещается вперед за счет сокращения цитоплазмы на
переднем ее конце. Белковые молекулы плазмазоля
находятся здесь в растянутом состоянии, а при
превращении плазмазоля в плазмагель они спирали-
зуются и укорачиваются. В это время молекулы
белка в заднем конце клетки автоматически
растягиваются, переходя в состояние плазмазоля, который
проталкивается к переднему концу.
Возможно, что вскоре эти гипотезы придется
пересмотреть, так как сейчас установлено, что во
всех эукариотических клетках имеются белки актин
и миозин, и показано, что актин, присутствующий в
виде тонких нитей, обратимо связывается с
миозином; эта реакция сопровождается гидролизом АТФ.
Таким образом, механизмы амебоидного движения
в целом, а может быть, и перетекания цитоплазмы в
клетке, возможно, окажутся в основе своей
сходными с механизмом мышечного сокращения.
17.6.2. Реснички и жгутики
Электронные микрофотографии показывают, что
реснички и жгутики имеют одинаковое внутреннее
строение. Реснички-это просто укороченный
вариант жгутиков, и в отличие от жгутиков они чаще
всего располагаются группами, а не по одиночке. На
поперечных срезах видно, что и другие органеллы
состоят из двух центральных фибрилл, окруженных
девятью периферическими (рис. 17.31),-так
называемая структура «9 + 2». Этот пучок фибрилл-
аксонема - окружен мембраной, которая является
продолжением плазматической мембраны.
Все периферические фибриллы построены из
белка тубулина и состоят из микротрубочек А и В.
Задний конец
Зона
разжижения
Направление
движения амебы
Плазмагель
Передний конец
Гиалиновый
колпачок
псевдоподии
Плазматическая
мембрана
Прозрачный жидкий слой,
перемешающийся назад
Зона
затвердевания
Движение
золя вперед
Гель-* Золь
Золь—►Гель
Плазмагель
Рис. 17.30. Образование псевдоподии у амебы. Согласно теории фонтанирования,
плазмазоль затвердевает, и образующийся плазмагель сокращается, проталкивая таким образом в псевдоподию еще
больше плазмазоля.
Дублет
Центральный
футляр
Центральная
микро-
трубочка
Нексиновая
связь
Плазматическа)
мембрана
Общие
протофиламенты
Динеин
Спица
Рис. 17.31. Строение реснички или жгутика. А. Схема расположения трубочек и сопутствующих
компонентов (вид сбоку). Б. Поперечный разрез. Каждый дублет состоит из микротрубочек AuB.
А-микротрубочка имеет пару отростков («ручек»). Обратите внимание, что на поперечном
разрезе А-трубочка представляется полым цилиндром, причем часть ее стенки становится общей для обеих
трубочек дублета. По всей длине трубочек через определенные интервалы отходят «спицыж
которые соединяют дублеты с «осевым футляром», окружающим центральные трубочки. В. Дублет микро-
трубочек в увеличенном виде.
Каждая А-микротрубочка несет по паре «ручек»,
образованных другим белком - динеином, который
обладает способностью гидролизовать АТФ, т.е.
действует как АТФаза. Центральные фибриллы
соединены с А-микротрубочками периферических
фибрилл при помощи радиальных перекладин.
В месте прикрепления к клетке ресничка или
жгутик оканчивается базальным тельцем, которое по
строению почти идентично аксонеме (имеет
структуру 9 + 0) и является производным центриоли. От
центриоли оно отличается лишь наличием у его
основания сложной структуры, названной «колесом
со спицами». Как полагают, базальное тельце в
процессе формирования ресничек и жгутиков служит
матрицей для сборки микротрубочек. Нередко от
базального тельца в цитоплазму отходят
волоконца, фиксирующие его в определенном положении.
Но хотя реснички и жгутики имеют одинаковый
план строения, способы их работы заметно
различаются.
Жгутик совершает симметричные движения, при
этом в каждый данный момент по нему проходит
несколько последовательных волн (рис. 17.32).
Биение жгутика может происходить в одной плоскости,
но (реже) может быть и спиралеобразным, что
приводит к вращению организма вокруг своей
продольной оси с одновременным продвижением
вперед по спиральной траектории (рис. 17.33). У
некоторых жгутиконосцев жгутик расположен на
переднем конце тела и как бы тянет животное за собой.
На жгутиках такого типа обычно есть мельчайшие
боковые выступы-мастигонемы, повышающие
эффективность такого способа локомоции. Чаще
жгутик располагается на заднем конце клетки (как,
например, у сперматозоида) и толкает ее вперед. На
рис. 17.34 обобщены сведения о типах движения,
осуществляемых с помощью жгутика.
Биение ресничек асимметрично (рис. 17.32); после
быстрого и энергичного удара прямой реснички она
изгибается и медленно возвращается в исходное
положение. При скоплении большого числа
ресничек необходим какой-то механизм,
координирующий их активность. У инфузории Paramecium эти
функции обычно приписывают нейрофанам - нитям,
соединяющим базальные тельца. Обычно биение
ресничек синхронизовано, так что вдоль тела
пробегают волны их активности в одном определенном
направлении. Это называется метахронным ритмом.
Было много споров относительно механизма
движения самих ресничек или жгутиков. Судя по
новейшим данным, этот механизм в основе своей
очень близок к взаимодействию актина и миозина
при мышечном сокращении. Как полагают,
изгибание жгутика связано с присоединением двух динеи-
Рис. 17.32. Последовательные стадии
движения жгутика (А) и реснички (Б). Рабочее
движение реснички начинается из положения 1.
Динеиновая
ручка
Радиальная
спица
Ручка
Рис. 17.33. Траектория движения эвглены.
Траектория переднего
конца клетки
Общее направление
движения
Направление
движения
изменяется
при участии
мионем
новых отростков А-микротрубочек к соседним
В-микротрубочкам в периферических фибриллах.
При этом происходит гидролиз АТФ, и
микротрубочки А и В скользят друг по другу, приводя в
движение жгутик. По-видимому, пять
периферических фибрилл на одной стороне инициируют
первоначальный изгиб, а остальные четыре фибриллы на
другой стороне включаются в работу позже, что
приводит к возвратному движению жгутика
(рис. 17.35). Радиальные перекладины, препятствуя
скольжению, сводят его к локальному изгибанию
жгутика. Возможно, что центральные фибриллы
проводят сигнал о начале скольжения от базального
тельца по всей длине реснички или жгутика. Было
показано, что реснички могут работать только в
присутствии ионов Mg2 + и что направление биения
определяется концентрацией Ca2+ внутри клетки.
Интересно, что реакция избегания контролируется у
парамеции следующим образом: когда инфузория
наталкивается на препятствие, направление биения
ресничек у нее меняется на противоположное, а
затем она возобновляет движение вперед. Такое
изменение стимулируется внезапным притоком
ионов Ca2+ в клетку в результате повышения
проницаемости для этих ионов.
У мелких эукариотических организмов реснички
или жгутики широко используются для
передвижения в воде, проталкивая тело сквозь окружающую
вязкую жидкость. Этот способ локомоции может
быть эффективным только при очень малых
размерах тела, когда отношение поверхности к объему
намного больше, чем у крупных животных. Для
последних создаваемая ресничками мощность была
Биение в одной плоскости
Движение
Жгутик
с боковыми
отростками
Движение
HPB
(например,
у сперм иев)
Жгутик тянет
клетку
HPB
(например,
у трипаносом)
(например,
у некоторых
одноклеточных
водорослей
и зооспор)
Спиральное биение
Жгутик на переднем
конце клетки,
но направлен назад
Движение
(например,
у эвглены)
Рис. 17.34. Типы движения, обусловленные работой
жгутика. HPB-направление распространения волн.
Эвглена вращается
вокруг своей оси
HPB
Движение
Отростки
мастигонемы
отталкивают
воду назад
HPB
Поперечный
разрез
на указанном
уровне
Рис. 17.35. Скольжение дублетов при работе
реснички в жабрах двустворчатого моллюска. Одна из
двух микротрубочек дублета выступает в кончике
реснички дальше другой. Поэтому на поперечных
срезах, сделанных на подходящем расстоянии от кончика
реснички на разных этапах ее изгибания, сначала
будут видны только дублеты, а позже наряду с
дублетами появятся одиночные микротрубочки.
Дублеты целиком скользят относительно друг друга,
однако взаиморасположение микротрубочек в
дублете остается неизменным. (По P. Satir, из Noviroff,
Holtzman.)
бы недостаточной. Однако реснички часто
встречаются внутри тела многоклеточных организмов,
где они выполняют ряд важных функций. Они могут
прогонять по протокам жидкость, как это
происходит у кольчатых червей в метанефридиях при
удалении отходов метаболизма. С помощью ресничек
перемещаются яйца в яйцеводах млекопитающих и
различные материалы по внутренней поверхности
органов, например слизь в дыхательных путях, где
работа ресничек позволяет удалять частички пыли и
другой «мусор». Реснички могут также создавать
поток наружной жидкости, из которой некоторые
организмы, в том числе парамеция,
отфильтровывают пищевые частицы (нередко с помощью ресничек
иного типа).
Жгутики есть и у некоторых бактерий, но они
существенно отличаются от жгутиков эукариот; они
более короткие, тонкие и весьма жесткие. Жгутик
бактерии-это внеклеточное образование, он не
покрыт плазматической мембраной и напоминает
отдельную микротрубочку в жгутике эукариот. Он
движется за счет силы, развиваемой в месте его
прикрепления к бактериальной клетке.
17.6.3. Локомоция эвглены
Локомоторный жгутик расположен на переднем
конце тела и тянет животное вперед. Волны
движения генерируются в самом жгутике, и по мере
прохождения этих волн от основания жгутика к его
концу их скорость и амплитуда увеличиваются.
Работа жгутика заставляет тело эвглены вращаться
вокруг своей оси со скоростью около одного оборота
в секунду и в то же время обеспечивает продвижение
в воде вперед по спирали. За 1 с эвглена может
продвинуться на полмиллиметра, что вчетверо
больше длины ее тела. С помощью сократимых
мионем, которые тянутся через всю клетку, эвглена
способна изменять форму своего тела и вместе с тем
направление его перемещения. Такое движение
называют эвгленоидным (рис. 17.33).
17.6.4. Передвижение парамеции
Реснички, расположенные на поверхности тела
рядами, бьют назад по диагонали слева направо; при
этом парамеция, продвигаясь вперед, вращается
вокруг своей продольной оси. В то же время сильное
биение ресничек околоротовой воронки заставляет
передний конец тела поворачиваться вокруг заднего
конца. Реснички работают в метахронном ритме и
их активность, возможно, координируется моториу-
мом-тельцем, связанным нейрофанами (нейроне-
мами) с базальными тельцами. Парамеция плавает
со скоростью около 1 мм/с, т.е. продвигается на
четыре длины собственного тела в секунду.
17.6.5. Локомоция кольчатых червей
Представитель класса
малощетинковых червей: Lumbricus
terrestris (обыкновенный дождевой
червь)
Полость тела (целом) дождевого червя окружена
стенками, состоящими из двух слоев
мышц-антагонистов: наружного кольцевого и внутреннего
продольного. Межсегментные перегородки - септы -
разделяют кольцевую мускулатуру животного на
самостоятельные единицы, в то время как
продольные мышцы тянутся через несколько сегментов.
Локомоция связана с координированной работой
обоих мышечных слоев, а также мышц, приводящих
в движение щетинки.
При передвижении дождевого червя вперед
сокращение кольцевых мышц начинается на переднем
конце тела и, захватывая сегмент за сегментом,
распространяется волной через все тело.
Сократившиеся мышцы в каждом сегменте давят на цело-
мическую жидкость, которая растягивает
расслабленные продольные мышцы и изменяет форму
сегмента-он становится длиннее и тоньше. Благодаря
этому передний конец червя продвигается вперед.
Щетинки, имеющиеся на всех сегментах тела, кроме
первого и последнего, при сокращении кольцевых
мышц втягиваются и таким образом не
препятствуют продвижению тела вперед (рис. 17.36).
По мере продвижения переднего конца тела
продольные мышцы в заднем его конце сокращаются, и
эта часть тела, утолщаясь, прижимается к
окружающей почве. Щетинки здесь выдвигаются и помогают
Передний конец А
Неподвижный
червь
Задний конец
Растяжение (P):
кольцевые мышцы
сокращаются,
продольные мышцы
расслабляются,
щетинки втянуты
Неподвижное
состояние (H):
оба мышечных слоя
расслаблены,
щетинки выдвинуты
и закрепляют
червя на месте
Рис. 17.36. Локомоция дождевого червя.
Сокращение (С):
кольцевые мышцы
расслабляются,
продольные мышцы
сокращаются,
щетинки втянуты
Движение
вперед
Ό
этому отделу закрепиться на месте. Это имеет
особое значение при рытье, так как, продвигаясь
вперед, червь может с силой давить на почву.
Вслед за кольцевыми мышцами быстро начинают
сокращаться продольные, и эта волна сокращения
тоже проходит по всей длине тела. Таким образом,
в разное время разные части тела могут
продвигаться вперед (при сокращении кольцевых мышц)
или оставаться неподвижными (при сокращении
продольных мышц). Суммарный эффект - плавная
перистальтическая волна сокращений, идущая вдоль
всего тела червя при его движении вперед. Изменив
последовательность сокращения мышц (т. е.
направление волны), червь способен ползти назад.
Работа мышц контролируется сложной сетью
внутри- и межсегментарных нейронов. Через
сегменты проходит брюшная нервная цепочка, и при
этом в каждом из них есть свои сегментарные
нервы. Это позволяет контролировать работу
каждого отдельного сегмента и всего организма в
Нервная цепочка
Кольцевая
мускулатура
Сенсорный
нейрон
Двигательный нейрон'
„Продольная
мускулатура
Промежуточные
нейроны
Септа
Целом
Рис. 17.37. Сегментарная иннервация продольной
мускулатуры дождевого червя (вид со спинной
стороны). Подобным же образом иннервируются и
кольцевые мышцы.
целом (рис. 17.37). В брюшной нервной цепочке
имеется гигантский аксон, который тянется вдоль
всего тела животного и проводит импульсы от
переднего конца к заднему. В брюшной цепочке есть
также два продольных латеральных (боковых)
волокна, проводящих импульсы от заднего конца к
голове. При раздражении чувствительных
рецепторов головы импульсы по гигантскому аксону идут к
продольным мышцам, вызывая их сокращение, и в
результате этого передний конец тела отдаляется от
раздражителя. Если раздражитель действует в
области заднего конца, то импульсы по латеральным
нервам передаются в головной конец и
обеспечивают подтягивание задней части тела вперед.
Такова основа реакций избегания у дождевого червя.
Представитель класса многощетин-
ковых червей: Nereis (нереис)
В отличие от дождевого червя у нереиса слой
продольных мышц разделен на две дорсальные и
две вентральные мышечные полоски (рис. 4.26).
Кольцевая мускулатура относительно слаба и в
каждом сегменте делится на группы косых мышц,
которые идут в параподии и приводят их (а также
связанные с ними структуры) в движение.
Параподии представляют собой полые выступы
тела, в которые заходят разветвления целома,
заполненные цел омической жидкостью. Параподия
подразделяется на спинной и брюшной отростки-
нотоподию и невроподию; каждый из них снабжен
палочкообразной ацикулой и пучком щетинок.
Совместная работа мышц стенки тела, параподии
и целомической жидкости делает возможными
движения нескольких типов. С помощью параподии,
которые действуют как рычаги, осуществляется
медленное ползание. При рабочем движении
параподии ацикула и щетинки в результате
повышения гидростатического давления в целоме
выдвигаются, обеспечивая зацепление за субстрат и
отталкивание от него. Затем параподия приподни-
мается над субстратом и переносится вперед, а
ацикула и щетинки при этом втягиваются.
Втягивание происходит путем сокращения
сегментарных косых мышц.
Работа правой и левой параподий в сегменте
скоординирована таким образом, что при движении
одной из них вперед другая перемещается назад.
При работе всех параподий создается несколько
перистальтических волн, проходящих вдоль всего
тела. В отличие от дождевого червя у нереиса эти
волны идут в направлении от хвоста к голове; при
этом группа параподий на одной стороне тела
движется вперед, а в то же самое время
симметрично расположенные параподий перемещаются назад.
При быстром ползании попеременно
сокращаются продольные мышцы по обеим сторонам тела и в
результате червь совершает волнообразные
движения в горизонтальной плоскости. В этом случае
параподий тоже опираются на субстрат.
Продольные мышцы, сокращаясь, эффективно продвигают
тело животного вперед относительно этих точек
опоры.
При свободном плавании в воде правая и левая
продольные мышцы сокращаются поочередно,
создавая волны горизонтальных изгибов тела,
которые распространяются от хвоста к голове. Широкие
веслообразные параподий во время рабочего
движения перемещаются назад; при этом они,
отталкиваясь от воды, продвигают животное вперед.
17.6.6. Локомоция членистоногих
Плавание и ходьба у речного рака
Astacus fluviatilis
У речного рака 8 грудных и 6 абдоминальных
сегментов, каждый из которых несет различные по
форме и величине придатки, выполняющие
определенные функции (см. рис. 4.34). К последним
четырем грудным сегментам причленяются
ходильные ноги (перейоподы), в то время как на брюшных
сегментах 2-5 у самок и 3-5 у самцов имеются
веслообразные разветвленные ножки - плеоподы.
Шестой брюшной сегмент-уроподит-видоизменен
в уплощенный хвостовой плавник, служащий в
основном для плавания.
При ходьбе клешненосные конечности
приподнимаются над субстратом, а брюшко выпрямляется
горизонтально. В любой данный момент только две
ноги (по одной с каждой стороны) оторваны от
субстрата, остальные шесть крепко цепляются за
субстрат. При этом ноги, расположенные на
грудных сегментах 5-7, тянут тело вперед, а на сегменте
8-толкают его.
Конечности приводятся в движение
поперечнополосатыми мышцами. Наиболее мощные мышцы
расположены в области брюшка;
мышцы-антагонисты сгибают брюшко вниз и выпрямляют его,
приподнимая вверх. При быстром подгибании
Тело-
Крыло опущено
мышца,
опускающая-
крыло,
сокращается
Мышца,
поднимающая
крыло,
расслаблена
Крыло поднято
Мышца,
опускающая-
крыло,
расслаблена
Мышца,
поднимающая
крыло,
сокращается
Рис. 17.38. Работа «прямых» летательных мышц у
насекомых с большими крыльями, таких как бабочка
или стрекоза
брюшка мощный толчок отбрасывает рака назад;
это движение используется как реакция избегания.
Локомоция насекомых
ходьба. Этот способ передвижения обеспечивает
координированная работа трех пар ног-по одной
паре на каждом из трех грудных сегментов. Каждая
нога представляет собой систему полых цилиндров
со стенками из твердого хитина. В местах
сочленений эти цилиндры связаны между собой гибкими
перепонками. Сочленение тазика (проксимального
сегмента ноги насекомого) с телом-это род
шаровидного сустава; все остальные суставы ноги блоко-
видные. Сгибание и выпрямление ног
осуществляется мышцами-антагонистами-сгибателями и
разгибателями, которые прикреплены к внутренней
поверхности экзоскелета с обеих сторон от сустава (см.
рис. 17.1).
Когда насекомое начинает ходьбу, три его ноги
опираются на землю и поддерживают тело, в то
время как три другие переступают вперед. При этом
первая нога с одного бока подтягивает тело, а
третья с того же бока подталкивает его, в то время
как вторая с другого бока служит просто опорой.
Затем весь процесс повторяется, только тройки
конечностей меняются ролями.
У многих насекомых на концах ног имеются пары
коготков и клейкие подушечки. Подушечки состоят
из мельчайших полых трубочек, выделяющих
клейкую жидкость, с помощью которой насекомое
прикрепляется к гладкой поверхности. Благодаря этому
такие насекомые способны ходить по вертикальным
поверхностям, а также вниз головой.
полет. Крылья насекомых представляют собой
плоские выросты экзоскелета, которые
поддерживаются сложной сетью жилок. Их движения
контролируются двумя главными группами
мышц-прямыми и непрямыми летательными мышцами. У
насекомых с большими крыльями (таких, как бабочки и
I вверх
Продольные мышцы расслаблены
Дорсовентральные мышцы сокращены
Нога
-Продольные
мышцы
сокращены
ВНИЗ
Дорсовентральные мышцы расслаблены
Передний край крыла
Рис. 17.39. Поперечный разрез через грудь
насекомого (вид спереди). Показаны отношения между
крыльями, стенками груди и мышцами при полете.
стрекозы) мышцы прикрепляются непосредственно
к основанию крыла (рис. 17.38). Это «прямые»
мышцы. Они поднимают и опускают крылья, а
также регулируют угол взмаха крыла при полете.
Когда крылья располагаются под определенным
углом друг к другу, насекомое совершает поворот в
воздухе. Такие мышцы участвуют и в складывании
крыльев после окончания полета.
У насекомых с меньшими крыльями главную
движущую силу при полете создают две группы
«непрямых» летательных мышц-антагонистов:
дорсовентральные мышцы тянутся от тергита (крыши) к
стерниту (основанию) груди, продольные - от
переднего отдела груди к заднему (рис. 17.39). Эти
мышцы не прикреплены непосредственно к крыльям.
Однако каждое крыло присоединено к тергиту и к
плевральной (боковой) поверхности груди. Такое
устройство представляет собой высокоэффективную
систему рычагов, так как сокращение какой-либо из
«непрямых» мышц приводит к большому
изменению формы груди, а в результате-к значительному
перемещению крыла. Поскольку амплитуда
изменений груди невелика, сокращение может быстро
повторяться. Это особенно важно для тех
насекомых, у которых частота взмахов крыльев очень
велика.
У крупных крылатых насекомых, таких как
бабочки или саранча, крылья совершают от 5 до 50
взмахов в секунду (табл. 17.5). У них отдельные
сокращения мышц вызываются одиночными
нервными импульсами. Импульсы здесь генерируются с
частотой, равной частоте взмахов. Летательные
мышцы, работающие таким образом, называются
синхронными. У комнатной мухи, у которой частота
взмахов составляет 120-200 в 1 с, летательные
мышцы должны работать слишком быстро для
того, чтобы каждое сокращение могло быть
ответом на отдельный нервный импульс. Такие мышцы
называются асинхронными, и к ним поступает один
нервный импульс приблизительно на 40 взмахов
крыла; эти импульсы нужны для поддержания
мышцы в активном состоянии во время полета. Такая
мышца может дольше сокращаться и развивать
большую мощность, чем синхронная мышца.
Асинхронная мышца способна автоматически
сокращаться в ответ на растяжение (так называемый
рефлекс на растяжение), не дожидаясь очередного
нервного импульса.
Асинхронный полет детально изучил Буттигер у
мухи Sacrophaga bullata. При полете по прямой
линии крылья у нее описывают в воздухе
«восьмерку». Тягу и подъемную силу в основном создает
движение крыльев вниз; при этом крылья
смещаются также вперед, и задний их край несколько
приподнят по отношению к переднему. При подъеме
крыло движется вверх и назад и его задний край
опущен по отношению к переднему. В таком
положении крыло испытывает наименьшее
сопротивление воздуха и в то же время обеспечивает большую
подъемную силу.
При подъеме крыла сокращаются дорсовентраль-
Таблица 17.5. Скорость работы крыльев у разных
насекомых"
Насекомые
Крупные бабочки,
например махаон
Саранча
Бражник
Комнатная муха
Пчела
Галлица
Число
взмахов
в 1 с
5
18
40
120
180
700—1000
Звук
Слышно жужжание
Слышен высокий
писк
" Как общее правило, чем меньше насекомое, тем быстрее оно
машет крыльями. У многих насекомых (у саранчи, стрекоз и др.)
две пары крыльев. B некоторых случаях (например, у пчел) обе пары
работают синхронно; в других случаях задняя пара несколько
опережает переднюю, как, например, у саранчовых. У других
насекомых (мухи, жуки) только одна пара крыльев. У комнатной мухи
задние крылья редуцированы и превращены в жужжальца —
булавовидные образования, выполняющие сенсорную функцию. Bo
время полета они быстро колеблются, воспринимают
аэродинамические силы и доставляют информацию, необходимую для
поддержания стабильности полета. У некоторых (очень немногих)
насекомых, например у блох, крыльев нет совсем.
ные мышцы. В результате тергит опускается и место
сочленения крыла с тергитом оказывается ниже
сочленения его с боковой стенкой груди (рис. 17.39).
Сопротивление тергита его смещению вначале
возрастает. Однако в критический момент
сопротивление исчезает и крыло щелчком «перескакивает» в
поднятое положение. При этом продольные мышцы
растягиваются, что приводит к их немедленному
рефлекторному сокращению. Теперь тергит
выгибается вверх, и место соединения его с крылом
оказывается выше сочленения крыла с боковой стенкой.
Сопротивление такому смещению опять-таки
преодолевается «щелчком», и крыло совершает резкое
движение вниз. При этом дорсовентральные мышцы
в свою очередь подвергаются растяжению, что
заставляет их сократиться, и весь цикл повторяется
снова.
17.6. У насекомых в летательных мышцах
саркоплазматический ретикулум для увеличения
поверхности оказывается продырявленным через
определенные интервалы. Как можно объяснить
смысл этого?
17.7. Можно ли ожидать, что объем саркоплаз-
матического ретикулума в синхронных и
асинхронных мышцах окажется различным?
Аргументируйте свой ответ.
17.7. Локомоция позвоночных
17.7.1. Плавание рыб
Относительная плотность воды, особенно морской,
очень высока и во многие сотни раз превышает
плотность воздуха. Вода представляет собой
относительно вязкую среду для перемещения в ней, но
благодаря своей плотности она может обеспечивать
опору для тела рыбы, а также служит субстратом,
от которого рыба может отталкиваться при
плавании.
У любого процветающего организма мы находим
множество адаптации к условиям среды, и рыбы не
составляют исключения. Тело у большинства рыб
имеет весьма обтекаемую форму, заострено с обоих
концов. Благодаря этому поток воды легко огибает
тело и сопротивление при движении сведено к
минимуму. У рыбы нет выступающих частей тела, за
исключением плавников, и, по-видимому, чем
быстрее плавает рыба, тем более совершенна ее
обтекаемая форма. Чешуя у хрящевых и костных
рыб смазывается выделениями кожных желез, что
тоже уменьшает трение между телом и водой. Еще
одной общей адаптацией для лучшего передвижения
в воде служат плавники. Непарные плавники,
расположенные вдоль средней линии тела, помогают
стабилизировать тело рыбы, в то время как парные
(грудные и брюшные) используются для руления и
балансирования; хвостовой плавник вместе с
парными обеспечивает продвижение рыбы вперед через
толщу воды. Детально работа плавников будет
рассмотрена в последующих разделах.
17.7.2. Локомоция рыб
Продвижение вперед происходит благодаря миото-
мам - сегментарным блокам мышц-антагонистов,
расположенным по обе стороны позвоночника. С
наружной стороны каждый миотом имеет
зигзагообразную форму, а внутри прикреплен к двум
смежным позвонкам. Позвоночник в целом представляет
собой длинный гибкий тяж, который легко
изгибается при сокращении миотомов, лежащих по одну
сторону от него. Миотомы с обеих сторон
позвоночника сокращаются и расслабляются поочередно,
их сокращения начинаются от головы рыбы и
распространяются к хвосту. Благодаря этим
сокращениям создаются волнообразные изгибы, число
которых тем больше, чем тоньше и длиннее тело.
У рыб с очень компактным телом, таких как
Ostracium (кузовок), такое волнообразное движение
Составляющая,
направленная
вперед
Реакция воды
Боковое
сопротивление
Давление хвоста
на воду
Давление хвоста
на воду (Д)
P
f Составляющая,
направленная
вперед
1 Реакция воды
Боковое
сопротивление
Давление хвоста
на воду
Составляющая,
направленная вперед (В)
Реакция воды (P)
Боковое
■►сопротивление (Б)
Направление
распро странения
волн
Рис. 17.40. Сравнение кузовкового (а), скумброидного (б) и угревидного (в) типов локомоции.
Боковое
сопротивление
Боковое
сопротивление
уравновешено
давлением
воды
на голову
и спинной
плавник
Реакция воды
Направление
движения хвоста
при отталкивании
от воды
Составляющая,
направленная
вперед
всего тела практически невозможно и продвижение
вперед приблизительно на 80% осуществляется за
счет боковых изгибов хвоста и хвостового плавника.
Этот способ локомоции называется кузовковым.
Для более удлиненных форм, например акул и
большинства костных рыб, характерна скумброид-
ная локомоция. У них волнообразные движения
совершает задняя половина тела. Угревидная
локомоция характерна для угрей, у которых тело
очень длинное и по нему пробегает несколько волн
сокращения, так что разные участки тела
изгибаются вправо и влево одновременно.
Перемещение тела вперед, как правило, является
результатом боковых движений хвоста с
прикрепленным к нему хвостовым плавником. При этом
хвостовой плавник слегка изгибается и с силой
отталкивается от относительно вязкой воды. Силу,
приложенную к телу, можно разложить на
составляющие, направленные вперед и вбок. Передняя
составляющая проталкивает рыбу сквозь толщу
воды, в то время как боковая заставляет голову рыбы
отклоняться в сторону, противоположную
движению хвоста; эта вторая сила создает боковое
сопротивление (рис. 17.41). Однако боковому
движению противодействует инерция воды, окружающей
относительно массивную (по сравнению с хвостом)
переднюю часть тела и имеющий большую площадь
спинной плавник. В воде для смещения тела вбок
потребовалось бы значительно большее усилие, чем
для проталкивания его вперед. Величина силы, с
которой хвост и хвостовой плавник отталкиваются
от воды, зависит от скорости их работы, площади
их поверхности и угла, под которым они
располагаются по отношению к потоку воды.
Рис. 17.41. Силы,
создаваемые хвостовым плавником акулы
при движении его из стороны
в сторону.
17.7.3. Локомоция хрящевых рыб
(акул)
Акула тяжелее морской воды, и если она перестанет
плавать, то начнет опускаться на дно. Чтобы этого
не произошло, парные грудные и брюшные
плавники работают как гидрокрылья. Они могут
поворачиваться под различным углом к длинной оси тела,
когда рыбе нужно подниматься или опускаться в
толще воды. Когда плавники расположены под
определенным углом, силу, которую они создают,
можно разделить на составляющие - подъемную
силу, направленную вверх, и лобовое сопротивление,
направленное назад (рис. 17.42). Однако тяга,
развиваемая хвостом, значительно превышает лобовое
сопротивление, и им можно пренебречь. У акулы
гетероцеркальиый хвостовой плавник-его брюшная
лопасть больше спинной. Когда хвост движется в
сторону, создаваемую им силу можно разделить на
вертикальную составляющую (отрицательную
подъемную силу) и боковую составляющую.
Положительная подъемная сила на переднем конце
тела уравновешивается отрицательной на заднем
конце.
При движении рыбы по горизонтали вперед на нее
могут действовать дестабилизирующие силы трех
типов, вызывающие отклонения в направлении
(рыскание), в поперечном равновесии (крен) и в
положении длинной оси тела (дифферент).
Различные плавники обычно поддерживают устойчивое
движение рыбы вперед; на рис. 17.43 показаны
способы их противодействия разного рода
отклонениям.
Плавники очень быстро реагируют на любую
дестабилизацию. Помогают им в этом системы из
трех полукружных каналов, расположенные на
обеих сторонах головы. Если возникает малейший
нежелательный поворот тела около одной из трех
Передний
спинной плавник
Задний
-спинной плавник
Подъемная
сила
Подъемная сила
Реакция
воды
Реакция воды
Реакция воды
Подъемная сила
/Лобовое
сопротивление
Лобовое
сопротивление
Тягаг направленная"
вперед
Вес тела
Давление парных грудных
и брюшных плавников на воду
Анальный плавник
Гетероцер кальный
хвост
Рис. 17.42. Работа парных грудных и брюшных плавников, а также гетероцеркального
хвостового плавника у акулы, создающая подъемную силу.
Рыскание
(ему противодействуют
^передний и задний
спинные плавники
и анальный плавник)
Крен
(контролируется
всеми плавниками)
Рыскание
Рыскание
Отклонение
по вертикали
Отклонение по вертикали
(контролируется грудными
и брюшными плавниками)
Крен
Рис. 17.43. Отклонения, возникающие при плавании (А), и противодействие им с помощью
плавников (Б).
осей, возбуждается соответствующий полукружный
канал и от него в головной мозг направляются
нервные импульсы. Головной мозг в свою очередь
посылает необходимые двигательные импульсы к
мышцам плавников, и эти мышцы должным
образом изменяют их положение.
17.7.4. Локомоция костистых рыб
У костистых рыб есть орган, называемый
плавательным пузырем. Хотя у предковых форм он
обеспечивал более эффективное использование
атмосферного воздуха для дыхания, у костистых рыб он
был превращен в гидростатическое устройство.
Плавательный пузырь представляет собой мешок,
расположенный между позвоночником и кишкой; он
обеспечивает рыбе так называемую «нейтральную
плавучесть». Благодаря ему средняя плотность тела
становится равной плотности окружающей воды;
поэтому рыбе не нужно расходовать энергию на
поддержание тела в воде-она может тратить
мышечные усилия только на движение.
Появившийся плавательный пузырь освободил
парные плавники от поддерживающей функции. Они
стали гораздо меньше, чем у акулы, и служат для
обеспечения устойчивости или для торможения; в
последнем случае они расправляются вертикально
под углом 90° к телу. Два грудных плавника могут
работать независимо один от другого, и благодаря
этому рыба может делать быстрый поворот вокруг
одного из них как оси вращения. Когда рыба плывет
по прямой, парные плавники плотно прижаты к
бокам тела, что делает его более обтекаемым.
Появление плавательного пузыря привело к развитию
симметричного гомоцеркального хвостового
плавника; при отталкивании им от воды основное усилие
направлено на продвижение вперед.
Плавательные пузыри бывают двух видов:
1. Открытый плавательный пузырь (например, у
золотой рыбки, сельдевых). Он соединен
протоком с глоткой, так что воздух может поступать
в пузырь или удаляться из него через рот; при
Плавательный
пузырь
Артерия
Область обратного
всасывания газа
Газовая железа
Рис. 17.44. Закрытый плавательный пузырь у костных рыб, его расположение (А) и кровоснабжение (Б).
этом средняя плотность тела соответственно
уменьшается или увеличивается.
2. Закрытый плавательный пузырь (например, у
трески). Такой пузырь полностью утратил связь с
глоткой. Рыба способна уравнивать плотность
тела с плотностью окружающей воды и
сохранять нейтральную плавучесть путем
автоматического увеличения или уменьшения количества
газа в пузыре.
Закрытый плавательный пузырь находится под
контролем нервной системы. На его переднем конце
расположена газовая железа, обильно снабженная
кровеносными капиллярами, которая выделяет в
полость пузыря газ, обогащенный кислородом. На
заднем конце пузыря находится участок,
пронизанный большим числом кровеносных сосудов, где
может происходить всасывание газов из пузыря
(рис. 17.44). В газовой железе артериальные и
венозные капилляры (рис. 17.44) расположены рядом
друг с другом. Именно здесь действует противо-
точная система для ускоренного выделения
кислорода в плавательный пузырь. В газовой железе при
секреции газа образуется молочная кислота,
которая, попадая в венозные капилляры, уменьшает
здесь сродство гемоглобина к кислороду (это
называется эффектом Рута). В результате этого в
венозных капиллярах, выходящих из газовой
железы, увеличивается напряжение кислорода - оно
становится выше, чем в артериальных капиллярах.
Поэтому кислород непрерывно диффундирует из
венозных капилляров в артериальные и переходит
из них в плавательный пузырь (рис. 17.45). Во время
секреции газа газовой железой кровеносные сосуды
в заднем участке плавательного пузыря
закрываются, так что газ из пузыря не выводится.
17.8. Перечислите адаптации, способствующие
эффективному плаванию, у рыб.
17.7.5. Локомоция лягушки
Парные конечности лягушки в отличие от
плавников не служат более стабилизаторами или рулями;
они приспособлены для работы в качестве
удлиненных членистых рычагов, приподнимающих тело
над землей, и обеспечивают передвижение
животного на суше. Передние конечности относительно
короткие со сросшимися локтевой и лучевой
костями. Задние конечности очень длинные; когда
животное неподвижно, они согнуты в коленном и
голеностопном суставах и поджаты под туловище.
Малоберцовая кость срослась с большеберцовой, и
кости предплюсны сильно удлинены. Пояса, к
которым причленяются конечности, намного крупнее,
чем у рыб, и способны, противодействуя силе
тяжести, выдерживать вес тела. Это в особенности
относится к тазовому поясу.
При медленном передвижении по субстрату у
лягушки одновременно выдвигаются вперед
конечности, расположенные по диагонали друг от друга.
10 мл O2 в 100 мл крови
Артериальный капилляр
Плавательный
пузырь
Ц)Д МПа O2
Венозный капилляр
9 мл O2 в 100 мл крови
Рис. 17.45. Противоточная система в газовой
железе закрытого плавательного пузыря у
костистой рыбы. В железе образуется молочная кислота,
которая, влияя на свойства гемоглобина,
повышает напряжение кислорода в венозных капиллярах.
Поэтому кислород диффундирует в
артериальные капилляры, оставаясь в петле. Оттекающая
от газовой железы венозная кровь содержит
меньше кислорода, чем приходящая артериальная
кровь. На схеме изображены только один
венозный капилляр и один артериальный. Газовая
железа содержит множество таких капилляров.
Спинная аорта
К сердцу
Воротная вена
^_ к печени
Газовая
железа
Рис. 17.46. «Ползание» лягушки (вид сверху). А.
Левая передняя нога сгибается, правая задняя нога и
правая передняя распрямляются, левая задняя
выносится вперед. Б. Левая передняя нога
выпрямляется, правая задняя выносится вперед, правая
передняя сгибается, левая задняя выпрямляется.
Если начать наблюдение над неподвижной
лягушкой, которая опирается на все четыре конечности, то
от момента начала ее движения можно проследить
следующие события. Левая передняя лапка
сгибается и одновременно выпрямляется правая задняя;
при этом обе они, опираясь на субстрат,
перемещают лягушку вперед. Вслед за этим сгибается
правая передняя лапка и разгибается левая задняя.
Весь этот цикл повторяется, пока животное
движется вперед (рис. 17.46).
При отрыве от земли во время прыжка благодаря
быстрому сокращению сильных мышц-разгибателей
Рис. 17.47. Отрыв от земли и прыжок лягушки.
все суставы задней конечности распрямляются.
Сила отталкивания от земли при этом передается от
конечностей через тазовый пояс на позвоночник.
Она по меньшей мере в три раза превосходит вес
животного и оказывается достаточной для того,
чтобы бросить тело вперед и вверх в воздух
(рис. 17.47). Во время приземления короткие
передние лапки, подвижно соединенные с плечевым
поясом, амортизируют удар.
17.7.6. Перья птиц и полет
Наличие перьев и значительное изменение скелета
привели к развитию у большинства птиц
структурных приспособлений, которые позволили им летать
и в значительной степени овладеть воздушной
стихией.
Перья птиц-это производные эпидермиса,
которые ведут свое происхождение от чешуи рептилий.
Перья служат изолирующим покровом тела, но
распределены на его поверхности неравномерно.
Они располагаются полосами, получившими
название птерилий, между которыми находятся лишенные
перьев участки - аптерии. Выделяют четыре типа
перьев, а именно стержневые (маховые и рулевые),
контурные, пуховые и нитевидные. Строение их
показано на рис. 17.48. В каждом крыле имеется по 23
маховых пера. Они направлены назад по
отношению к телу птицы и перекрывают друг друга. В
связи с этим создаются верхняя (выпуклая) и
нижняя (вогнутая) поверхности крыла, и благодаря
этому оно может функционировать как профильное
крыло, значение которого мы рассмотрим позже.
На кисти (пясти и пальцах) находятся 11 маховых
перьев первого порядка, а к предплечью
(образуемому лучевой и локтевой костями) прикреплены
остальные 12 маховых перьев - маховые второго
порядка (рис. 17.49). Стержневые перья хвоста
называются рулевыми перьями.
Типичное стержневое перо состоит из жесткой
полой нижней части - очина, погруженного в кожу, и
верхней-плотного стержня. Стержень несет опахало,
которое образуют прикрепленные с двух сторон
Рис. 17.48. А. Маховое перо. Б. Контурное перо. В. Пуховое перо.
Маховые
1 -го порядка
Маховые
2-го порядка
Крылышко
Контурные
перья
Пух
Рис. 17.49.
Распрямленное крыло голубя.
Показаны маховые
перья первого и второго
порядка.
бородки первого порядка, а на них расположены в
два ряда бородки второго порядка. Последние с
помощью маленьких крючков и бороздок
сцепляются друг с другом (рис. 17.50), и благодаря этому
крыло сохраняет легкость и его поверхность
становится плотной, прочной и воздухонепроницаемой.
В том месте, где очин переходит в плотный
стержень, расположен небольшой пуховый участок,
который состоит из нескольких несцепленных
бородок.
Контурные перья по строению сходны с рулевыми
и маховыми, но они меньше рулевых и более гибки,
их бородки не так прочно сцеплены. Они образуют
основной покров тела и создают
ветронепроницаемый слой, а также определяют контур тела.
Пуховые перья у птенцов служат единственным
покровом. У более взрослых птиц они прижаты к
коже и располагаются между контурными перьями.
Бородки первого порядка у них длинные и мягкие,
бородки второго порядка не имеют крючков.
Нитевидные перья - мелкие волосовидные
образования; у них нет настоящего опахала, имеются лишь
небольшие пучки гибких бородок первого и второго
порядков, расположенные на кончике стержня. Эти
перья разбросаны по всему телу птицы, они
помогают удерживать воздушный слой около тела.
Помимо появления перьев у птиц произошло
множество других изменении, связанных с полетом.
Важнейшие из них перечислены ниже.
1. Плечевой пояс хорошо развит и служит
прочной опорой для крыльев. Широкая грудина с
развитым килем обеспечивает большую поверхность
для прикрепления мощных грудных мышц
(рис. 17.51). У таких сильных летающих птиц, как
почтовые голуби, вес летательной мускулатуры мо-
Стержень
Бородки
1-го порядка
Крючочки
на переднем ряду
бородок 2-го порядка
Задний ряд
бородок 2-го порядка-
Рис. 17.50. Опахало махового пера. Показаны
бородки первого и второго порядков.
Шарнир
Плечевой
пояс
Поднятие <ч
крыла
Опускание
крыла
Плечо
Малые
грудные
мышцы
Большие
грудные
мышцы
Грудина
(киль)
Рис. 17.51. Схематический разрез, показывающий
взаимодействие летательных мышц, костей крыльев
и плечевого пояса у птицы (голубя).
жет быть близок к половине веса всего тела.
2. Кости передних конечностей частично слились
и в значительной мере удлинились, что обеспечило
большой размах крыльев. Три косточки запястья
срослись с тремя пястными костями, образовав
carpometacarpus прочную опорную структуру для
прикрепления маховых перьев 1-го порядка. У птиц
осталось только три пальца. Некоторые части
скелета соединены очень жестко; например, срослись
между собой многие позвонки, а также тазовый пояс
с позвоночником. Это позволяет более эффективно
распределять силы, создаваемые крыльями, и
развивать большую подъемную силу. Задние
конечности удлинены, чтобы при взлете крылья не задевали
землю.
3. Форма тела обтекаемая, поэтому при полете
птицы она встречает лишь небольшое
сопротивление воздуха. У птиц компактный череп и обычно
обтекаемый, заостренный клюв. Расположение
перьев обеспечивает гладкую поверхность крыла. Хвост
короткий, что увеличивает маневренность и
уменьшает сопротивление. Ноги у птиц в полете прижаты
к телу.
4. Благодаря наличию нескольких губчатых
костей уменьшается вес тела. Многие полости тела
заполнены воздушными мешками. Мешки также
улучшают вентиляцию легких, снабжая их свежим
воздухом при вдохе и при выдохе (разд. 11.6.10). В
костях крупных птиц для прочности есть распорки.
Клюв представляет собой заостренные, лишенные
зубов челюсти, покрытые роговым чехлом. В связи с
отсутствием зубов нет надобности в тяжелых
челюстных мышцах для перемалывания пищи. Эту
функцию выполняет мускульный желудок,
расположенный недалеко от центра тяжести тела.
Подсчитано, что у пеликана, вес которого составляет 11 кг и
рост-1,5 м, скелет весит всего лишь 0,65 кг, а у
фрегата с размахом крыльев 2 м скелет весит ПО г.
Кстати, перья у него тяжелее скелета (1,4 -1,8 кг)!
Половые органы у птиц очень невелики и
полностью развиваются только в брачный период. Это
тоже уменьшает затраты энергии при полете, так
как облегчает тело.
5. Птицы обладают высокой температурой тела и
хорошей теплоизоляцией, которая препятствует
охлаждению тела воздухом во время полета; кроме
того, высокая температура повышает
эффективность сокращения мышц. У птиц большое, сильное
сердце, способное поддерживать высокое кровяное
давление.
6. Птицы обладают большой остротой зрения. У
них есть третье веко - мигательная перепонка,
которая удаляет инородные частицы, попадающие в
глаза во время полета. Шея у многих птиц длинная
и гибкая; она позволяет им доставать клювом до
земли при добывании пищи и вращать головой,
улучшая круговой обзор.
7. Как и следовало ожидать, всем летающим
птицам свойственна поразительная координация
работы мышц.
17.7.7. Полет птиц
Рассмотрим горизонтальный поток воздуха
относительно наклонной поверхности крыла в том случае,
когда его передняя кромка приподнята над задней.
В этом смысле крыло действует как несущая
плоскость. Поток воздуха над крылом встречает
меньшее сопротивление и развивает большую скорость,
чем под крылом (рис. 17.52). В результате давление
воздуха над крылом уменьшается, а под крылом-
увеличивается. Так возникает подъемная сила. Ее
величина зависит от размеров и формы крыла, угла
его наклона по отношению к длинной оси тела (угол
Крыло
птицы
Подъемная
сила
Пониженное
давление воздуха
(отрицательное
давление)
Воздушный
поток
Результирующая
Воздух над верхней
поверхностью крыла
движется быстрее,
чем под нижней
поверхностью
■► Лобовое
сопротивление
Повышенное
давление воздуха
(положительное Угол атаки
давление)
Рис. 17.52. Крыло как несущая плоскость.
Маховые 1-го порядка —
для продвижения вперед
-Маховые 2-го порядка-
для подъемной силы
(1) Начало опускания
крыльев
Мах вниз
Перо
Сопротивление воздуха
заставляет перья сомкнуться
Крылья наклоняются
и отталкиваются
от воздуха,
обеспечивая
продвижение вперед
Взмах вверх
Крылья изогнуты
для уменьшения
сопротивления
(3) Взмах вверх
Воздух проходит
между перьями,
сопротивление
уменьшается
Рис. 17.53. Работа крыльев и перьев во время машущего полета. А. Положение крыльев при машущем полете у
голубя. Б. Положение перьев при опускании крыльев вниз и взмахе вверх во время
машущего полета.
атаки) и скорости полета. В воздухе на тело птицы
действует еще одна сила, которая стремится отвести
крыло назад в направлении воздушного потока; она
называется лобовым, или аэродинамическим,
сопротивлением. Механическая эффективность крыла
зависит от его способности развивать большую
подъемную силу при небольшом относительном
росте лобового сопротивления.
Различают три основных типа полета: машущий,
парящий (планирующий) и зависание.
Машущий полет
У таких птиц, как голубь, у которых крылья делают
около двух взмахов в секунду, основная мощность
развивается при опускании крыльев. Это
происходит благодаря сокращению сильно развитых
больших грудных мышц, которые одним концом
прикреплены к плечевой кости, а другим-к килю
грудины. При отрыве от земли крыло в начале маха
опускается почти вертикально и его передняя
кромка располагается ниже задней. Маховые перья 1-го
порядка отклоняются вверх под давлением воздуха.
Они плотно сомкнуты, чтобы обеспечить
максимальное сопротивление воздуху, а значит, и
максимальную подъемную силу. Затем по мере опускания
крыло движется вперед и поворачивается таким
образом, что его передняя кромка отклоняется
вверх. В этом положении крыло создает силу,
поднимающую корпус. Воздух, проходящий между
маховыми перьями, стремится разделить их и
отогнуть кверху (рис. 17.53).
Подъем крыла начинается тогда, когда крыло
еще полностью не опущено. Внутренняя часть
предплечья резко поднимается вверх и назад, и при этом
передняя кромка крыла находится в наклонном
положении над задней. Это делают малые грудные
мышцы, прикрепленные к дорсальной поверхности
плечевой кости и к грудине. При движении крыла
вверх оно сгибается в запястье и кисть
поворачивается таким образом, что маховые 1-го порядка
резко отводятся назад и вверх до того момента,
пока все крыло в какой-то мере не выпрямится над
телом птицы. Во время этою движения маховые
1-го порядка разъединяются, так что воздух
проходит между ними и его сопротивление
уменьшается. Движением этих перьев назад в основном и
создается мощный толчок, который птица
использует для поступательного движения вперед. Еще до
того момента, как маховые 1-го порядка
поднимутся до высшей точки, снова начинают сокращаться
большие грудные мышцы, опускающие крылья, и
весь процесс повторяется.
При длительном машущем полете работа
крыльев заметно видоизменяется и требует гораздо
меньше энергии, чем при отрыве от земли. Взмахи при
этом не такие сильные, крылья не соприкасаются за
спиной, и нет движения вперед на заключительном
этапе опускания крыльев. Крылья обычно
выпрямлены, и махи вверх и вниз происходят в запястье (в
сочленении костей предплечья и запястья).
Активного отведения кисти вверх и назад не происходит-
крыло поднимается пассивно в результате давления
воздуха на его нижнюю поверхность.
По окончании полета птица приземляется,
опуская и распластывая хвост, который одновременно
служит тормозом и источником подъемной силы.
После создания этой силы ноги опускаются, и птица
прекращает движение. Хвост в полете служит также
рулем, и устойчивость птицы обеспечивается
нервным контролем при участии полукружных каналов.
В них возникают импульсы, которые стимулируют
вспомогательные мышцы, изменяющие форму и
положение крыльев и соотношение между их
взмахами.
Разные птицы летают с разными скоростями. Эти
различия обусловлены формой крыльев и ее
изменениями в полете, а также частотой взмахов.
Рис. 17.54 позволяет сравнить крылья быстрых
летунов (таких, как стрижи) и медленных (как
воробьи).
17.9. Перечислите характерные особенности
стрижа, позволяющие ему быстро летать.
Планирующий и парящий полет
При планировании крылья неподвижно
распластаны под углом 90° относительно тела, и птица
постепенно теряет высоту. Когда птица, планируя,
опускается, на нее действует сила тяжести, которую
можно разложить на две составляющие, одна из
которых (тяга) направлена вперед по линии полета,
а другая-вниз под прямым углом к первой
(рис. 17.55). С увеличением скорости планирования
эту вторую силу уравновешивает возрастающая
подъемная сила, а тягу уравновешивает лобовое
сопротивление, и с этого момента птица планирует с
постоянной скоростью. Скорость и угол скольжения
зависят от размеров, формы и угла атаки крыльев и
от веса птицы.
Птицы, обитающие на суше, используют при
планировании восходящие термальные потоки
воздуха, которые возникают, когда горизонтальный
поток, встретив преграду (например, гору),
отклоняется вверх или когда теплый воздух вытесняется
холодным и поднимается вверх; так происходит,
например, над городами. Птицы, имеющие легкое
тело и широкие крылья, такие как канюки и орлы,
Рис. 17.55. Силы, действующие на птицу при парении в безветренном воздушном пространстве.
А - направление воздушного потока относительно птицы, В тяга, С - вес тела; D - сила,
вызывающая снижение, E-лобовое сопротивление, F-результирующая аэродинамическая сила;
G-подъемная сила; H-направление движения (По Gray, с изменениями.)
Небольшой
размах
крыльев
Большая внутренняя
поверхность крыла
для подъемной силы
Широкое крыло
Сильно развитые
маховые
2-го порядка
Большой
размах
крыльев
Удлиненная кисть
для быстрого
пере дв ижения
вперед
Узкое уплощенное
крыло для уменьшения
лобового сопротивления
Развитые маховые
1-го порядка
Мало маховых
2-го порядка
Рис. 17.54. Сравнение строения крыльев у быстрых
(таких, как стриж) и медленных (таких, как воробей)
летунов.
искусно используют термальные потоки и могут
постепенно набирать высоту, делая небольшие
круги. Планирование без потери высоты и даже с
подъемом называется парением.
У морских птиц, например альбатросов, форма
тела и крыльев иная, и они парят по-другому
(рис. 17.56). У альбатроса большое тело и очень
длинные узкие крылья, и он использует порывы
Медленно парящие птицы
(грифы, орлы, ястребы и др.)
Маховых 2-го порядка_
много — для создания
подъемной силы
Хвост используется
для балансирования
Положение крыльев
при скольжении
с быстрой потерей высоты
Положение крыльев
при парений —
крылья распластаны
[Широкое крыло обеспечивает
большую подъемную силу
Маховые 1-го порядка
расправлены для увеличения
подъемной силы
Быстро парящие птицы
(альбатрос, чайка, баклан и др.)
Узкое
крыло
Большой размах крыльев
(например, у альбатроса 3,0 — 3,4 м)
Заостренные концы
крыльев уменьшают
лобовое сопротивление
Длинное узкое крыло
Рис. 17.56. Сравнение формы тела и крыльев у медленно и быстро парящих птиц. Медленно
парящие птицы - большей частью материковые; они могут использовать слабые ветры и
термальные потоки. Быстро парят преимущественно морские птицы; они могут использовать сильные ветры.
Кости передней
конечности
Рис. 17.57. Летучая мышь с расправленными крыльями. Показано прикрепление крыла к предплечью,
туловищу и лодыжке.
ветра над волнами. За время скольжения против
ветра вверх он поднимается на высоту около 7-10
метров. Затем он разворачивается по ветру и с
большой скоростью на отогнутых назад крыльях
спускается вниз. В конце скольжения вниз альбатрос
описывает дугу, возвращаясь во встречный поток
воздуха с крыльями, вынесенными несколько
вперед. Такое положение крыльев и быстрое движение
вперед относительно воздуха обеспечивают
подъемную силу, необходимую для набора высоты перед
очередным спуском. Альбатрос способен также
парить, покрывая большие расстояния параллельно
гребням волн; при этом он использует небольшие
восходящие потоки воздуха от волн, подобно тому
как сухопутные птицы используют потоки над
горными склонами.
Зависающий полет
При зависании птица машет крыльями, но при этом
остается на одном месте. Крылья совершают около
50 взмахов в секунду, и развиваемая ими тяга,
направленная вверх, уравновешивает вес тела.
Птицы, способные зависать, имеют очень сильно
развитые летательные мышцы (1/3 от веса тела). Их
крылья могут наклоняться почти под любым углом.
Большая часть маховых перьев -1 -го порядка
(маховых 2-го порядка только шесть), и они
используются для создания тяги.
17.7.8. Полет летучих мышей
Летучие мыши - единственные представители
млекопитающих, способные по-настоящему летать.
Каждое крыло представляет собой тонкую
перепонку из сильно васкуляризированной кожи,
растянутую между передней конечностью (предплечьем и
всеми пальцами, кроме большого), боковой
поверхностью тела и предплюсной задней конечности.
Пястные кости сливаются, придавая крылу
прочность, в то время как кости предплечья и кисти
значительно удлинены и служат опорным каркасом
крыла. Указательный и средний пальцы кисти
поддерживают переднюю кромку крыла, в то время как
остальные два пальца оттянуты назад к задней
кромке, которая в конце концов соединяется с
пятым пальцем стопы (рис. 17.57). Движения крыльев
у летучих мышей сходны с таковыми у птиц. Полет
может быть мощным и быстрым.
17.7.9. Локомоция четвероногих
(на примере собаки)
Ходьба
При ходьбе позвоночник собаки сохраняет
жесткость и продвижение вперед обеспечивается работой
задних конечностей. Они перемещаются вперед и
назад благодаря сокращению сгибателей и
разгибателей соответственно.
Когда сокращаются разгибатели, задняя лапа
работает как рычаг; она выпрямляется и
отталкивается от земли, продвигая тело животного вперед и
слегка приподнимая его. При сокращении
сгибателей лапа отрывается от земли и перемещается
вперед. В каждый момент времени приподнята только
одна лапа, остальные три служат опорой и
удерживают тело в равновесии. Если неподвижно
стоявшая собака начнет движение вперед с левой
передней ноги, то дальше ноги будут переступать в
такой последовательности: правая задняя, правая
передняя, левая задняя, снова левая передняя и т. д.
Бег
При беге собака перестает опираться на три ноги
одновременно; она переходит на другой аллюр,
когда одновременно перемещаются обе передние
лапы, а за ними обе задние. Лапы опираются на
землю гораздо меньше времени, чем при ходьбе, и
обычно одна передняя конечность на какую-то долю
секунды опережает другую. Так же обстоит дело и с
задними конечностями. Поэтому лапы касаются
земли в такой последовательности: левая передняя,
правая передняя, правая задняя, левая задняя.
Если собака развивает максимальную скорость,
движения ног ускоряются, и тогда все четыре ноги
после выпрямления могут одновременно оказаться
в воздухе. Сильные мышцы туловища изгибают
позвоночник дугой вверх, когда ноги согнуты, и
прогибают вниз, когда ноги полностью выпрямле-
Таз
Позвоночник
Лопатка
Плечевая
кость
Голеностопный
сустав
Бедро
Коленный
сустав
Локтевой
сустав
Лучезапястный
сустав
Рис. 17.58. Последовательность движений собаки
при беге (на примере борзой). А. Позвоночник изогнут
дугой, лапы находятся непосредственно под
туловищем. Б. Позвоночник полностью выпрямлен (даже
несколько прогибается вниз)г конечности
полностью вытянуты.
Рис. 17.59. Последовательная смена
положений правой ноги во время одного шага.
ны. Благодаря этому отталкивание получается
более сильным и шаг заметно длиннее. И то и другое
способствует быстроте бега (рис. 17.58).
17.7.10. Двуногое хождение: человек
Когда человек стоит, его вес распределен между
двумя ногами. Если правая нога делает шаг вперед,
то в первую очередь сокращается икроножная
мышца и поднимает пятку. Это приводит к
отталкиванию носком правой ноги от земли и появлению
движущей силы, направленной вперед. Правая нога
продолжает отталкиваться от земли, когда она
движется вперед, слегка сгибаясь в колене (рис. 17.59).
Тяжесть тела тем временем переносится на левую
ногу, которая все еще стоит на земле и служит
опорой для тела. Когда правая нога выпрямляется,
первой ставится на землю пятка. Вес тела
постепенно переносится с левой стопы на правую пятку, а
затем, по мере того как тело продолжает двигаться
вперед-на пальцы, и обычно большой палец правой
стопы отталкивается от субстрата. Теперь, когда вес
тела перенесен на правую ногу, левая пятка
поднимается и все последовательно повторяется. Так
чередуется работа правой и левой ноги, пятки и
пальцев, пока человек не остановится.
17.10. Почему спринтеры обычно бегут на
пальцах?
17.8. Опора у растений
У растений в опорной функции участвуют четыре
типа тканей-паренхима, колленхима, склеренхима
и ксилема. Их роль как опорных элементов
подробно рассмотрена в гл. 8.
Глава 18
Гомеостаз
Организм можно определить как
физико-химическую систему, существующую в окружающей среде
в стационарном состоянии. Именно эта способность
живых систем сохранять стационарное состояние в
условиях непрерывно меняющейся среды и
обусловливает их выживание. Для обеспечения
стационарного состояния у всех организмов - от
морфологически самых простых до наиболее
сложных-выработались разнообразные анатомические,
физиологические и поведенческие приспособления,
служащие одной цели - сохранению постоянства
внутренней среды.
Впервые мысль о том, что постоянство
внутренней среды обеспечивает оптимальные условия для
жизни и размножения организмов, была высказана
в 1857 г. французским физиологом Клодом Берна-
ром. На протяжении всей его научной деятельности
Клода Бернара поражала способность организмов
регулировать и поддерживать в достаточно узких
границах такие физиологические параметры, как
температура тела или содержание в нем воды. Это
представление о саморегуляции как основе
физиологической стабильности он резюмировал в виде
ставшего классическим утверждения: «Постоянство
внутренней среды является обязательным условием
свободной жизни».
Клод Бернар постоянно подчеркивал различие
между внешней средой, в которой живут организмы,
и внутренней средой, в которой находятся их
отдельные клетки (у млекопитающих это тканевая,
или интерстициальная, жидкость), и понимал, как
важно, чтобы внутренняя среда оставалась
неизменной. Так, например, млекопитающие способны
поддерживать температуру тела, несмотря на
колебания окружающей температуры. Если становится
слишком холодно, животное может переместиться в
более теплое или более защищенное место, а если
это невозможно, вступают в действие механизмы
саморегуляции, которые повышают температуру
тела и препятствуют теплоотдаче. Адаптивное
значение этого заключается в том, что организм как
целое функционирует более эффективно, так как
клетки, из которых он состоит, находятся в
оптимальных условиях. Системы саморегуляции
действуют не только на уровне организма, но и на
уровне клеток. Организм является суммой
составляющих его клеток, и оптимальное
функционирование организма как целого зависит от оптимального
функционирования образующих его частей.
В 1932 г. американский физиолог Уолтер Кэннон
ввел термин гомеостаз (от греч. homoios-τοτ же;
stasis-стояние) для определения механизмов,
поддерживающих, выражаясь словами Клода Бернара,
«постоянство внутренней среды». Функция гомео-
статических механизмов состоит в том, что они
поддерживают стабильность клеточного окружения
и тем самым обеспечивают независимость
организма от внешней среды-в той мере, в какой эти
механизмы эффективны. Независимость от условий
окружающей среды является показателем
жизненного успеха, и на этом основании млекопитающих
следует рассматривать как преуспевающий класс:
они способны поддерживать относительно
постоянный уровень активности, несмотря на колебания
внешних условий.
Для того чтобы обеспечить более или менее
стабильную активность организма, необходима
регуляция на всех уровнях-от молекулярного до по-
пуляционного. Это требует использования
различных биохимических, физиологических и
поведенческих механизмов, наиболее соответствующих
уровню сложности и образу жизни данного вида, и во
всех этих отношениях млекопитающие, очевидно,
лучше вооружены, чем простейшие.
Как показывают исследования, существующие у
живых организмов способы регуляции имеют много
общих черт с регулирующими устройствами в
неживых системах, таких как машины. И в том и в
другом случае стабильность достигается благодаря
определенной форме управления. Винер в 1948 г. дал
науке об управлении название кибернетики (от греч.
cybernos-рулевой). Кибернетика занимается, в
частности, общими закономерностями регулирования в
живых и неживых системах. Физиологи, изучающие
растения и животных, часто используют точные
математические модели теории управления для
объяснения механизмов действия биологических ре-
гуляторных систем; поэтому, прежде чем перейти к
изучению способов саморегуляции таких
параметров, как температура тела, газообмен, концентрации
различных веществ в крови и баланс электролитов,
следует получить хотя бы общее представление о
некоторых положениях теории управления.
18.1. Системы управления
в биологии
18.1.1. Введение в теорию управления
Строгое применение теории управления к
биологическим процессам позволило глубже понять
функциональные взаимоотношения между
компонентами многих физиологических механизмов и
прояснить многие вещи, которые ранее казались
запутанными. Так, например, живые системы
рассматриваются теперь как открытые системы,
поскольку они нуждаются в постоянном обмене
веществами с окружающей средой. В самом деле,
живые системы находятся в динамическом
равновесии со средой; нужен постоянный приток энергии,
чтобы предотвратить полное уравновешивание с
окружающим миром. Равновесие возможно только
после смерти организма, когда он становится
термодинамически стабильным по отношению к среде.
Основные компоненты любой системы управления
показаны на рис. 18.1.
Мерой эффективности всякой управляющей
системы является степень отклонения регулируемого
параметра от должного (оптимального) уровня и
скорость возвращения к этому уровню. Гомеостати-
ческие механизмы должны иметь свободу
колебаний, так как именно колебания активируют систему
управления и возвращают переменную к
оптимальной величине. Подобные системы основаны на
таком соединении их компонентов, при котором
выход может регулироваться входом, т.е. они
действуют по принципу обратной связи. В большинстве
систем с обратной связью выход служит
одновременно входом.
Для осуществления обратной связи необходимо,
чтобы результат работы данной системы
сравнивался с заданным значением («уставкой»),
являющимся оптимальным значением регулируемого
параметра (переменной), и в случае отклонения от
него соответствующим образом изменялся.
Существуют два вида обратной связи-отрицательная
и положительная. Первая более распространена в
гомеостатических системах живых организмов.
Отрицательная обратная связь повышает
стабильность системы (рис. 18.2). При нарушении
равновесия системы возникает ряд последствий,
которые приводят к устранению этого нарушения и к
возвращению системы в исходное состояние. Прин-
Вход ► Детектор-
РЕГУЛЯТОР
Заданная
величина
(уставка)
Эффектор —► Выход
Модулятор
Рис. 18.1. Основные компоненты системы
управления.
цип отрицательной обратной связи можно
рассмотреть на примере регулирования температуры в
электрической печи. Система управления в
электрической печи состоит из входа (электрический ток,
проходящий через нагревательный элемент), выхода
(температура печи) и термостата, который может
быть установлен на нужную температуру.
Термостат действует как модулятор. Если он настроен на
температуру 1500C, электрический ток будет идти
через нагревательный элемент до тех пор, пока
температура в печи не достигнет 1500C5 а затем
термостат выключится, и нагревание прекратится.
Когда температура упадет ниже 1500C, термостат
вновь включится и электрический ток опять повысит
температуру до заданного значения. В этой системе
термостат играет роль детектора ошибки. Ошибкой
является разница между фактическим выходом и его
заданным значением, и она ликвидируется за счет
увеличения входа. Это пример стабильной системы
с замкнутой цепью, типичной для многих
физиологических регуляторных механизмов.
Примером биологических механизмов с
отрицательной обратной связью может служить регуляция
напряжения дыхательных газов в крови, частоты
сердечных сокращений, артериального кровяного
давления, уровней гормонов и метаболитов в крови,
водного и электролитного баланса, регуляция рН и
температуры тела. Рис. 18.3 иллюстрирует роль
отрицательной обратной связи в регулировании
секреции тироксина щитовидной железой. В этом
случае модулятор состоит из трех компонентов-
детектора (гипоталамуса), регулятора (гипофиза) и
эффектора (щитовидной железы).
Положительная обратная связь редко встречается
в биологических системах, так как она приводит к
нестабильности системы и экстремальным
состояниям. В этих ситуациях возникшее возмущение
вызывает такие последствия, которые еще более его
усиливают (рис. 18.2). Например, во время
распространения нервного импульса деполяризация
мембраны нейрона повышает ее проницаемость для
Na+. Ионы Na+ входят в аксон через мембрану и
вызывают дальнейшую деполяризацию, которая
приводит к возникновению потенциала действия. В
этом случае положительная обратная связь
действует как усилитель ответа, величину которого
ограничивают другие механизмы, описанные в
разд. 16.1.
В организме существуют и более сложные ре-
гуляторные устройства, чем упомянутые выше. Эти
механизмы включают дополнительные детекторы
(физиологические системы раннего предупреждения)
или дополнительные эффекторы (на случай отказа
основных), действующие на разных уровнях. Так,
например, у гомойотермных животных детекторы
температуры, находящиеся внутри тела и на его
поверхности, обеспечивают почти постоянную
температуру внутренних областей тела.
Терморецепторы кожи, играющие роль детекторов окружающей
Дальнейшее увеличение
/
Положительная
обратная связь
Возмущение
Корректирующий
механизм
Увеличение
Отрицательная
обратная связь
Стабильное
состояние
Нормальная ситуация
Возмущения нет
Стабильное
состояние
Уменьшение,
Возмущение
Положительная
обратная связь
Дальнейшее уменьшение
Отрицательная
обратная связь
/
Корректирующий
механизм
Время
температуры, посылают импульсы в гипоталамус,
который выполняет функцию модулятора и вносит
коррективы раньше, чем успевает измениться
температура крови. В качестве других примеров
подобной системы может служить регуляция дыхания
при физической нагрузке, а также регуляция
аппетита и жажды. Аналогичным образом
множественные детекторы и эффекторы обеспечивают
дополнительную надежность регуляции таких
жизненно важных параметров, как артериальное давление:
рецепторы растяжения каротидного синуса и аорты
и барорецепторы продолговатого мозга реагируют
на изменения кровяного давления и вызывают
ответы различных эффекторов, включая сердце,
кровеносные сосуды и почки. Нарушение работы
одного из этих органов может компенсироваться
работой других.
18.1.2. Природа и регуляция
внутренней среды
Внутреннюю среду организма и ее регуляцию
можно рассматривать на двух уровнях-на уровне
клеток и на уровне тканей.
Клетка содержит цитоплазму, состав которой
модулируется избирательной проницаемостью кле-
Рис. 18.2. Гомеостатическая
система управления. Стрелками
показаны направления воздействий.
точной мембраны и активностью ферментов,
зависящей от синтеза белков. Плазматическая
мембрана позволяет проникать в клетки и выходить из
них лишь определенным молекулам, и скорость
обмена ими через мембрану строго регулируется
возможностями диффузии, осмотическими и
электрическими градиентами, активными механизмами,
включающими транспортные системы мембран, и
перемещениями мембранных структур, как,
например, при пиноцитозе и фагоцитозе (разд. 7.2.2).
Аналогичным образом природа и количества веществ,
синтезируемых внутри клетки, регулируются
скоростями синтеза белков. Внутриклеточным
метаболизмом управляют ферменты, образующиеся в
результате считывания последовательности
оснований ДНК и трансляции ее в первичную структуру
ферментных белков. Участки ДНК, кодирующие
специфические белки, называются генами. Как
полагают, «включение» и «выключение» генов
контролируется системами индукции и репрессии,
подробно описанными в разд. 22.7. С точки зрения теории
управления, поддержание стационарного состояния
в клетке зависит от скоростей поступления в клетку
и использования ею различных материалов, т. е. от
«входа», «выхода» и активности модуляторов
(рис. 18.4).
МОДУЛЯТОР
ДЕТЕКТОР
(гипоталамус)
Вход
(тироксин)
Отрицательная
обратная
связь
ТЛ
РЕГУЛЯТОР
(гипофиз)
(Тироксин в крови)
ΤΤΓ
Выход
(тироксин)
ЭФФЕКТОР
(щитовидная
железа)
Рис. 18.3. Пример простой биологической системы управления: регуляция секреции тироксина (см.
разд. 16.6). ТЛ-тиреолиберин; ТТГ -тиреотропный гормон.
вход
(метаболит)
Избирательное
поглощение
гМОДУЛЯТОР=
(клеточная^мембрана)
Избирательная
секреция
ВЫХОД
(метаболит)
ВХОД-
(ДНК)
► МОДУЛЯТОР-
(синтез белка)
.ВЫХОД 1-
(фермент)
Отрицательные обратные связи
►ВЫХОД2,
(продукт)
Рис. 18.4. Общая схема механизмов, участвующих в поддержании постоянства внутриклеточной среды.
Как у одноклеточных, так и у многоклеточных
организмов внутренняя среда клеток регулируется
по описанной выше схеме. В случае одноклеточных
организмов непосредственным окружением
является внешняя среда, которую они никак не могут
контролировать. Если они обладают какими-то
локомоторными механизмами, они могут
перемещаться в более благоприятные условия, но чаще
всего они целиком зависят от окружающей среды.
Насколько это возможно, они приспосабливаются к
условиям существования и приобретают
толерантность к ним. При этом само обилие этих
микроскопических существ указывает на их способность к
выживанию, несмотря на простоту строения.
Непосредственным окружением для клеток
многоклеточных растений и животных служит
межклеточная жидкость. У растений такой жидкостью
является сок, у насекомых - гемолимфа, у
иглокожих-вода, у большинства остальных животных-
тканевая жидкость. Состав внеклеточной жидкости
может регулироваться организмом в разной степени
в зависимости от совершенства и эффективности его
гомеостатических систем. У млекопитающих
ближайшее окружение для всех живых клеток
составляет тканевая жидкость. Механизм образования этой
жидкости, отличающейся удивительным
постоянством состава, описан в главе 14. Со времен Клода
Бернара состав тканевой жидкости и механизмы его
регуляции подвергались широкому изучению.
Птицы и млекопитающие обладают наиболее
совершенной регуляцией параметров тканевой жидкости-
содержания в ней воды, газов, ионов, питательных
веществ, гормонов, отходов метаболизма, рН и
температуры. Во всех случаях эти параметры
достаточно жестко регулируются при участии одной
или нескольких тканей, органов или систем органов.
У большинства животных механизм регуляции
включает реакции со стороны желез внутренней
секреции или нервной системы, которые
координируются регуляторными центрами головного и
спинного мозга, как показано на рис. 18.5.
В заключение следует еще раз подчеркнуть
адаптивное значение гомеостатических механизмов. Все
метаболические системы работают наиболее
эффективно лишь в узких пределах по обе стороны от
оптимальных условий. Роль органов и систем,
участвующих в гомеостазе, в том и состоит, чтобы,
работая порознь и сообща, препятствовать
отклонениям от оптимума, вызываемым изменениями
внешней и внутренней среды. Некоторые из этих
механизмов рассматриваются в последующих
разделах.
18.1.3. Регуляция содержания
дыхательных газов в крови
Для дыхания клеток тела необходимо постоянное
поступление в них кислорода из тканевой жидкости.
С другой стороны, образующаяся в процессе
дыхания углекислота не должна накапливаться в
клетках или тканевой жидкости, так как это привело бы
к нарушению равновесий участвующих в дыхании
реакций и к местным изменениям рН, которые
могли бы повлиять на скорость ферментативных
процессов. Организм осуществляет тонкую
регулировку концентрации (или напряжения) CO2 в крови,
внутренняя
СРЕДА
РЕГУЛЯТОР
СТИМУЛ
■Сенсорный вход -
Регулятор-
] ные центры
вЦНС
Нервы
периферические
вегетативные
Отрицательная обратная связь
ВХОД ► МОДУЛЯТОР
ВНУТРЕННЯЯ
СРЕДА
\
Железы -
Гормоны
Выход
РЕАКЦИЯ
Рис. 18.5. Гомеостатические механизмы, регулирующие внутреннюю среду.
внешняя
Двигательный выход
Аортальное
.-тельце
Высшие центры
головного мозга
Центр выдоха
\Дыхатель-
'ные центры)
продолговатого
мозга
Хемо- λ
рецептор
CO2
PH
Продолговатый
мозг
Ветвь
блуждающего
нерва
Трахея
Каротидное
тельце
Легкие
CO2 O2
PH
CO2
O2
Сердце
правое левое
Аорта
Тканевое
дыхание
Диафрагмальный нерв
Диафрагма
Наружные межреберные
™ ft- мь1ШцЫ|
Внутренние межреберные I
мышцы
Нейроны
Кровеносные
сосуды
Повышенный
уровень
Пониженный
уровень
Рис. 18.6. Общая схема механизмов, участвующих в
регуляции содержания дыхательных газов в крови.
и она остается относительно постоянной, несмотря
на колебания количества доступного кислорода и
потребности в нем, которая во время интенсивной
мышечной работы может увеличиваться в 20 раз.
Частота и глубина дыхания регулируются
дыхательными центрами, расположенными в варолие-
вом мосту и продолговатом мозге (у основания
головного мозга) (рис. 18.6). Эти центры посылают
к диафрагме и межреберным мышцам ритмические
импульсы, которые вызывают дыхательные
движения. В основе своей ритм дыхания является
непроизвольным, но может изменяться в некоторых
пределах высшими центрами головного мозга, о
чем свидетельствует способность к произвольной
задержке дыхания. Частота и глубина дыхания
непосредственно влияют на состав альвеолярного
воздуха, который в свою очередь определяет
напряжение O2 и CO2 в артериальной крови, снабжающей
ткани тела. У человека в покое парциальное
давление O2 и CO2 в альвеолярном воздухе и в
артериальной крови составляет на уровне моря в
среднем 100 и 40 мм рт. ст. соответственно.
Поддержание таких уровней O2 и CO2 обеспечивается
регуляцией активности дыхательных центров с
помощью отрицательной обратной связи. Эту
регуляцию осуществляют импульсы, поступающие от
рецепторов двух типов - механорецепторов и хемо-
рецепторов. К первым относятся рецепторы
растяжения, находящиеся в стенках трахеи и легких,
вторые - хеморецепторы - имеются в стенках аорты,
в каротидных тельцах (расположенных в стенках
сонных артерий) и в самом продолговатом мозге.
Этот механизм отрицательной обратной связи
может модифицироваться высшими центрами
головного мозга, что позволяет произвольно усиливать
или подавлять активность дыхательных центров,
например при задержке или форсировании дыхания,
при разговоре, пении, чихании или кашле.
Действие импульсов, поступающих от рецепторов
растяжения, связано в основном с механикой
дыхательных движений. Импульсы, возникающие в
дыхательных центрах, идут по эфферентным путям
спинного мозга. Некоторые аксоны, образующие
эти пути, выходят из спинного мозга в его шейном
отделе в виде диафрагмальных нервов,
направляющихся к диафрагме, тогда как аксоны других
нейронов выходят из грудного отдела в составе нервов,
направляющихся к наружным межреберным
мышцам. Импульсы, поступающие по этим нервам,
совместно вызывают вдох. Легкие наполняются
воздухом, и рецепторы растяжения, находящиеся в
стенках легких и трахеи, возбуждаются и активируют
афферентные нейроны блуждающего нерва.
Последний временно угнетает центры вдоха, и вдох
прекращается. В результате расслабления диафрагмы
объем грудной клетки уменьшается, эластичные
легкие спадаются и воздух выталкивается из них.
Поскольку рецепторы растяжения в легких и трахее
больше не стимулируются, снимается угнетение с
центра вдоха и дыхательный цикл повторяется. Во
время интенсивной физической нагрузки
повышенное напряжение CO2 в крови стимулирует центр
выдоха, и импульсы от него поступают к
внутренним межреберным мышцам; мышцы начинают
сокращаться сильнее, и это приводит к более
глубокому или частому дыханию.
Частота и глубина дыхания регулируются
импульсами от хеморецепторов, возникающими в
ответ на изменение напряжения O2 и CO2 в крови.
Опыты, в которых люди дышали воздухом с
различным содержанием O2 и CO2, показали, что в
стимуляции дыхания избыток CO2 играет более
важную роль, чем недостаток O2. При уменьшении
концентрации O2 с 20 до 5% частота дыхания
повышалась вдвое, и точно такой же эффект давало
увеличение концентрации CO2 всего лишь на 0,2%.
Регулирующее влияние CO2 и сниженного рН
крови на дыхание осуществляется почти всецело
через хеморецепторы аорты, каротидных телец и
самого продолговатого мозга. Хеморецепторы аор-
ты и каротидных телец чувствительны также к
изменениям концентрации O2, что имеет жизненно
важное значение при низком напряжении O2, так как
при этом падает активность продолговатого мозга.
Усиленная вентиляция облегчает выведение
углекислоты из крови путем ее диффузии в
альвеолярный воздух, где концентрация CO2 понижается. Это
же происходит в случае преднамеренного глубокого
дыхания - гипервентиляции.
18.1. Объясните, почему после гипервентиляции
частота дыхания уменьшается и человек
испытывает головокружение и слабость.
На больших высотах (свыше 3000 м) парциальное
давление O2 и CO2 в атмосферном воздухе снижено,
и соответственно снижено их содержание в
альвеолярном воздухе. В связи с уменьшением
концентрации CO2 в альвеолярном воздухе в него из крови
переходит больше углекислоты, ее содержание в
крови оказывается недостаточным для стимуляции
хеморецепторов, и она уже не может играть роль
стимулятора дыхания. Теперь эту роль играет
падение напряжения O2 в крови: оно вызывает
стимуляцию хеморецепторов в каротидных тельцах, а
последние активируют дыхательные центры.
Существует тесная взаимосвязь между
дыхательными и сердечно-сосудистыми центрами
продолговатого мозга. Изменение кровяного давления,
регулируемого сердечно-сосудистыми центрами,
влияет на дыхание; например, в случае падения
кровяного давления вентиляция легких усиливается, а
при его повышении она уменьшается. В свою
очередь изменения концентраций дыхательных газов в
крови, регистрируемые дыхательными центрами,
вызывают изменение кровяного давления (разд.
14.12.6).
18.1.4. Регуляция уровня метаболитов
в крови
Одним из наиболее важных метаболитов,
присутствующих в крови, является глюкоза. Уровень
глюкозы в крови должен находиться под строгим
контролем, поскольку она служит главным
субстратом тканевого дыхания и должна непрерывно
поступать в клетки. Особенно чувствительны к
нехватке глюкозы клетки головного мозга, которые не
могут использовать никакие другие метаболиты в
качестве источника энергии. Недостаток глюкозы
вызывает потерю сознания. Нормальный уровень
глюкозы в крови составляет около 90 мг на 100 мл
(90 мг%), но может колебаться от 70 мг% натощак
до 150 мг% после приема пищи. Источники
глюкозы и ее взаимоотношения с другими
метаболитами будут рассмотрены в разд. 18.5.2.
Регуляция уровня глюкозы в крови является
примером сложного гомеостатического механизма,
находящегося под контролем эндокринной системы и
включающего координированную секрецию по
меньшей мере шести различных гормонов и две
цепи отрицательной обратной связи. Повышение
уровня глюкозы в крови (гипергликемия)
стимулирует секрецию инсулина (разд. 16.6.7), а его падение
(гипогликемия) приводит к угнетению секреции
инсулина и к стимуляции секреции глюкагона
(разд. 16.6.7) и других гормонов, повышающих
уровень глюкозы в крови (гипергликемических
факторов). Общая схема этой регуляторной системы
представлена на рис. 18.7.
18.1.5. Регуляция ритма сердца и
кровяного давления
Изменения частоты сердечных сокращений и
кровяного давления косвенно влияют на состав
тканевой жидкости и участвуют в поддержании его на
относительно постоянном уровне, несмотря на
колебания внешних условий и меняющиеся потребности
тканей. Механизмы регуляции включают
деятельность вегетативной нервной системы и подробно
описаны в разд. 14.12.5.
Существуют две важные формы отклонений от
нормального ритма сердца-тахикардия и бради-
кардия. Тахикардия (греч. tachys - быстрый) - общее
название для всех случаев повышенной частоты
сердечных сокращений. Она может вызываться
разнообразными факторами, в том числе эмоциями
(тревога, ярость, и т. п.) и повышенной активностью
щитовидной железы. Сильная тахикардия часто
бывает результатом изменения электрической
активности сердца. Различные участки сердца, лежащие за
пределами синоатриального узла (определяющего
нормальный ритм сердца), могут становиться
очагами возникновения стимулов, вызывающих
сокращения сердечной мышцы, и это приводит к
тахикардии.
Брадикардия (греч. bradys - медленный) -
состояние, при котором сердце сокращается реже, чем в
норме. Характерна для спортсменов, у которых в
результате тренировок увеличен ударный объем
сердца и для поддержания постоянного минутного
объема в покое нужна меньшая частота сокращений.
Брадикардия может быть также связана с
гипофункцией щитовидной железы или с изменением
электрической активности синоатриального узла и
других проводящих участков сердца.
Устранению временной тахикардии или бради-
кардии и восстановлению нормального сердечного
ритма способствует антагонистическое воздействие
на синоатриальный узел со стороны симпатической
и парасимпатической нервной системы.
Кровяное давление - это давление крови на стенки
кровеносных сосудов. Обычно его измеряют в
плечевой артерии с помощью сфигмоманометра.
Систолическое давление возникает в результате
сокращения желудочков, а диастолическое отражает
давление крови в артериях при расслабленных желу-
мышцы
Поглощение глюкозы
клетками
Секретируемый инсулин
(регулятор) —
Островки Лангерганса
(детектор)
ПЕЧЕНЬ
Глюкоза—► CO2 + H2O
Глюкоза—► Гликоген
Глюкоза—► Жир
(Эффектор)
Пища .
(вход)
Повышение уровня глюкозы
Нормальный
уровень глюкозы
в крови
(заданная величина)
Снижение уровня глюкозы
Нормальный
уровень глюкозы
в крови
[(заданная величина)!
„ Ткани
(выход)
Ассимиляция _
(вход)
Снижение уровня глюкозы в крови
Островки Лангерганса,
кора и мозговое вещество
надпочечников, ч
гипоталамус \
(детекторы) χ
Секреция глюкагона,
глюкокортикоидов,
адреналина, гормона роста
и тироксина
(регулятор)
ПЕЧЕНЬ
Гликоген ►Глюкоза!
Белок ►Глюкоза
Повышение
уровня глюкозы
(эффектор)
Рис. 18.7. Общая схема двойной регуляции уровня глюкозы в крови.
дочках. Кровяное давление зависит от возраста,
пола и состояния здоровья, и у здоровых молодых
мужчин составляет 120 (систолическое) и 80 (диасто-
лическое) мм рт. ст. На кровяное давление влияют
как сила сердечных сокращений, так и сопротивление
периферических кровеносных сосудов; оно отражает
общее состояние сердца и сосудов. Условия,
вызывающие сужение сосудов и уменьшение их
эластичности (атеросклероз) или нарушение работы почек,
могут вызывать повышение кровяного давления
(состояние, называемое гипертонией) и тем самым
увеличивать нагрузку на сердце и артерии. В свою
очередь растяжение артериальных стенок может
привести к их ослаблению и разрыву или к
закупорке суженных сосудов сгустками крови
(тромбоз). Эти нарушения очень опасны, если происходят
в мозгу или в сердце и приводят к кровоизлияниям в
мозг или к тромбозу мозговых или коронарных
сосудов.
18.1.6. Защита от инфекции
Многие механизмы защиты от инфекции являются
гомеостатическими, так как они направлены на
поддержание постоянства внутренней среды
организма. Главной защитой служит кожа. Однако
вирусы, бактерии, простейшие, а также круглые и
плоские черви способны проникать в ткани через
естественные отверстия тела или даже
пробуравливаться через кожу. Присутствие патогенных
организмов и выделяемых ими токсичных веществ
приводит в действие ряд защитных механизмов, в том
числе активирует различные лейкоциты (фагоциты,
моноциты и лимфоциты). (Механизмы свертывания
крови, заживления ран и иммунитета подробно
были описаны в разд. 14.13.5 и 14.14.)
18.2. Терморегуляция
Тепло-форма энергии, имеющая очень важное
значение для поддержания живых систем. Все живые
системы нуждаются в непрерывном снабжении
теплом для предотвращения их деградации и гибели.
Главным источником тепла для всех живых
существ служит солнечная энергия. Солнечная
радиация превращается в экзогенный (находящийся вне
организма) источник тепла во всех случаях, когда
она падает на организм и им поглощается. Сила и
характер воздействия солнечного излучения зависят
от географического положения и являются важными
факторами, определяющими климат региона. В
свою очередь климат определяет наличие и обилие
видов растений и животных в данной местности. В
основном организмы обитают в таких областях
земного шара, где нормальная температура
варьирует от — 400C (в Арктике) до 500C (в пустынях). В
некоторых пустынных регионах температура на
поверхности земли может достигать 800C.
Большинство живых организмов существует в весьма
ограниченном диапазоне температур, например в
пределах от 10 до 35°С, однако многие виды обла-
Температура на поверхности Солнца
Точка кипения воды (100 °С)
Денатурация белков (50 0C)
Температура тела человека (37 0C)
Точка замерзания воды ( 0 вС )
Температурные пределы растительной жизни
Температурные пределы животной жизни
Всякое движение молекул
прекращается
Рис. 18.8. Шкала температур и пределы
переносимости для живых организмов.
дают приспособлениями, позволяющими им
заселять регионы с экстремальными температурами
(рис. 18.8). Солнечный свет-не единственный
источник тепла, доступный живым организмам.
Солнечная энергия видимой части спектра используется
автотрофами для фотосинтеза и аккумулируется в
химических связях синтезируемых ими органических
веществ (гл. 9). Молекулы этих веществ служат
эндогенными источниками тепловой энергии,
которая освобождается в процессе тканевого дыхания
(гл. 11).
Температура является показателем количества
тепловой энергии в системе и основным фактором,
определяющим скорость химических реакций как в
живых, так и в неживых системах. Как отмечалось в
разд. 6.4.3., тепловая энергия повышает скорость
движения атомов и молекул, благодаря чему
возрастает вероятность взаимодействия между ними.
Зависимость между температурой и скоростью
химических реакций выражается в виде
температурного коэффициента, Q10. В живых системах
температура влияет на структуру ферментов, которая в
свою очередь влияет на интенсивность обмена. От
температуры в большой степени зависит
распространение организмов и их активность.
18.2.1. Влияние температуры на рост и
распространение растений
Температура может быть фактором,
лимитирующим рост и развитие растений, так как влияет на
скорость деления клеток, интенсивность клеточного
метаболизма и фотосинтез. От температуры зависят
темновые реакции фотосинтеза, а они в свою
очередь контролируют различные метаболические
пути, описанные в гл. 9. Интенсивность фотосинтеза и
накопление достаточных количеств питательных
веществ, необходимых для завершения полного
жизненного цикла,- таковы факторы, определяющие
географическое распространение растений.
18.2.2. Адаптация растений к низким
температурам
Растительный мир северных областей умеренной
зоны и тундры демонстрирует множество
приспособлений, позволяющих максимально использовать
преимущества теплого периода во время короткого
лета. Единственные растения здесь-это мхи,
лишайники, некоторые травы и быстро растущие
однолетники. Растения высоких северных или южных
широт подвергаются длительному воздействию
таких неблагоприятных условий, как слабая
освещенность, низкие температуры и мерзлая почва.
Выживание в этих условиях обеспечивается
множеством разнообразных анатомических,
физиологических и поведенческих приспособлений, связанных с
определенными стадиями жизненного цикла.
Например, большинство многолетних деревянистых
растений в умеренных поясах относятся к
листопадным. Сбрасывание листвы (под действием
регулятора роста - абсцизовой кислоты) предотвращает
потерю воды в результате испарения через устьица в
те периоды, когда из-за низких температур
ограничено поглощение влаги из почвы (разд. 15.2.8).
Опадение листьев помогает также избежать
повреждения снегом и ветром в то время, когда фотосинтез
все равно был бы резко снижен из-за слабой
освещенности, низких температур и недостатка воды и
солей. В течение этих периодов зоны будущего
роста - почки - бывают прикрыты почечными
чешуйками, а их метаболическая активность
подавлена присутствием регулятора роста дормина. Такое
название этот регулятор получил за свое действие,
но недавно было показано, что он представляет
собой абсцизовую кислоту.
В более северных широтах среди деревянистых
растений преобладают хвойные. Они имеют
игольчатые листья, благодаря чему уменьшается
количество снега, который может скапливаться на них
зимой. Кроме того, эти листья покрыты толстой
кутикулой, уменьшающей потерю влаги летом.
Разнообразные виды однолетних растений имеют
короткие периоды роста и переносят зиму, образуя
холодостойкие семена или подземные органы.
Низшие растения переносят холодный период, образуя
устойчивые споры, которые могут сохранять
жизнеспособность сотни лет, как можно видеть на
примере спор, найденных в Сибири, которые после их
извлечения из вечной мерзлоты нормально
проросли.
Многие растения нуждаются в воздействии
низких температур, чтобы выйти из состояния покоя и
подвергнуться яровизации. Например, почки сирени
развиваются быстрее после выдерживания при
низкой температуре, чем при высокой. Другие примеры
влияния низкой температуры на рост растений
описаны в гл. 15 и разд. 21.6.
18.2.3. Адаптация растений к высоким
температурам
Во многих регионах мира высокие температуры
воздуха сочетаются с недостатком воды, и
адаптации, наблюдаемые здесь у растений, часто бывают
связаны, с одной стороны, со способностью
переносить засуху, а с другой-с необходимостью
испарять воду для охлаждения.
Растения не могут избегать высоких температур,
перемещаясь в затененные места, и для
предотвращения перегрева у них выработались
морфологические и физиологические приспособления.
Нагревающему воздействию солнечных лучей
подвергаются наземные части растения, а наибольшей
поверхностью обладают листья. Листья отличаются
малой толщиной и большим отношением площади
к объему, что облегчает газообмен и поглощение
света. Такое строение лучше всего предотвращает
повреждение от перегрева. С другой стороны,
тонкий лист обладает относительно небольшой
теплоемкостью и поэтому принимает температуру
окружающей среды (явление, известное как гетеротер-
мия). Связанную с этим опасность перегрева у
растений жарких стран уменьшает образование
глянцевой кутикулы, выделяемой эпидермисом. Кутикула
отражает значительную часть падающего на нее
света, препятствуя таким образом поглощению
тепла и перегреванию. Большая площадь поверхности
содержит многочисленные устьица, через которые
осуществляется транспирация. В жаркую сухую
погоду растение может терять за счет транспирации до
0,5 кг воды на 1 м2 за 1 ч, что соответствует потере
приблизительно 350 Вт тепловой энергии на 1 м2 и
составляет почти половину общего количества
энергии, поглощаемой растением. Благодаря этим
механизмам растения способны в значительной степени
регулировать свою температуру. Эта способность
тесно связана с относительной влажностью
воздуха.
18.2. Почему нарушается жизнедеятельность
растений при температуре воздуха выше 300C или
при высокой влажности воздуха?
В жаркие солнечные дни у многих растений,
подвергающихся опасности перегрева, наблюдается
явление «фотосинтетического кризиса».
Предполагают, что оно вызвано временным прекращением
метаболической активности вследствие изменения
структуры ферментов или закрытия устьиц в
результате накопления CO2 в палисадных клетках.
Возможно, однако, что причина закрытия устьиц-
просто отсутствие воды в листьях. Обычной
реакцией на высокую температуру и недостаток влаги
бывает увядание. Дисбаланс между скоростями
транспирации и поглощения воды приводит к
потере тургора в таких частях растений, как паренхима,
где клетки не имеют утолщенных клеточных стенок.
Адаптивное значение этого явления состоит в
уменьшении поверхности листа, подверженной
прямому действию света, благодаря чему
предотвращается перегрев. У растений, растущих в теплицах,
увядание может происходить под действием
слишком высокой температуры, которая создается в
листьях, несмотря на достаточное снабжение водой.
При снижении температуры растения начинают
очень быстро оправляться даже после сильного
ожога.
Растения, приспособленные к засушливым
условиям, называются ксерофитами. Они обладают
множеством разнообразных адаптации, позволяющих
им выживать (разд. 19.3.2). В большинстве случаев
эти адаптации связаны с характерными
игловидными листьями, позволяющими им максимально
рассеивать тепло. Механизмы устойчивости к
высоким температурам имеют в основном
физиологическую природу.
18.2.4. Влияние температуры на рост
и распространение животных
Температура влияет на метаболическую активность
животных и растений, воздействуя главным
образом на ферменты и подвижность атомов и молекул.
Это непосредственно сказывается на скорости роста
животных (гл. 21). Кроме того, температура может
определять географическое распространение и
экологию животных через свое влияние на растения-
первичных продуцентов в пищевых цепях. Границы
обитания большинства животных определяются
местными пищевыми ресурсами. Исключение
составляют некоторые насекомые, птицы и
млекопитающие, способные к миграциям. Другие
организмы из-за ограниченной способности к
передвижению вынуждены использовать легко доступные
источники пищи, которые определяются пищевыми
взаимоотношениями, существующими в экосистеме.
Разнообразие реакций животных на температуру
зависит от степени стабильности температуры
окружающей среды и от того, насколько данный вид
способен регулировать температуру тела. Как
полагают, жизнь возникла в морской среде, в которой
у живых организмов было очень мало проблем.
Колебания температуры в водной среде невелики, и
биологическое значение этого факта состоит в том,
что температурные условия жизни относительно
стабильны. У большинства водных организмов,
включая беспозвоночных и рыб, температура тела
колеблется вместе с температурой воды, хотя
некоторые очень активные рыбы, например тунцы,
способны поддерживать более высокую
температуру тела. Однако водные животные редко нуждаются
в том, чтобы проявлять физиологические и
поведенческие реакции, наблюдаемые у наземных видов,
так как физические свойства воды предохраняют их
от внезапных резких колебаний. Вода имеет
максимальную плотность при 4°С, поэтому при 00C она
замерзает только у поверхности - образующийся
лед не тонет. Благодаря этому многие водные
животные остаются активными в то время, когда
большинство наземных животных из-за
минусовых температур не могли бы сохранять
активность.
Воздух обладает относительно малой
теплоемкостью, поэтому его температура может сильно
колебаться в течение суток. Одна из проблем,
связанных с заселением животными суши, заключалась
в трудности адаптации к этим температурным
колебаниям. Необходимость такой адаптации привела к
появлению разнообразных физиологических,
поведенческих и экологических особенностей (а также
явилась основным фактором, определяющим
географическое распространение животных). Природа
этих адаптивных особенностей и их примеры будут
рассмотрены в последующих разделах.
18.2.5. Источники тепла для животных
Все животные получают тепло из двух
источников - непосредственно из внешней среды и из
химических субстратов, подвергающихся
расщеплению в клетках. Степень, в которой животные
способны генерировать и сохранять тепло, зависит от
физиологических механизмов, свойственных данной
филогенетической группе. Все беспозвоночные,
рыбы, амфибии и рептилии лишены способности
поддерживать температуру тела в узких границах.
Поэтому этих животных называют пойкилотермными
(греч. poikilos разный). Их называют также экто-
термными, так как они больше зависят от тепла,
поступающего извне, чем от того тепла, которое
образуется в обменных процессах. Раньше этих
животных обычно называли холоднокровными, но
этот термин неточен и может вводить в
заблуждение.
Птицы и млекопитающие способны
поддерживать достаточно постоянную температуру тела
независимо от окружающей температуры. Их
называют гомойотермными (греч. homoios подобный)
или, что менее правильно, теплокровными. Гомой-
отермные животные относительно мало зависят от
внешних источников тепла, так как благодаря
высокой интенсивности обмена у них вырабатывается
достаточное количество тепла, которое может
сохраняться. Поскольку эти животные существуют за
счет внутренних источников тепла, их называют
также эндотермными. У некоторых пойкилотермных
животных температура тела иногда может быть
выше, чем у гомойотермных; поэтому, чтобы
избежать путаницы, мы будем пользоваться
терминами «эктотермный» и «эндотермный». На рис. 18.9
Внутренняя температура тела, ° С
Кошка
Человек
Утконос
Ехидна
Ящерица
Внешняя температура, 0C
Рис. 18.9. Зависимость между окружающей
температурой и температурой тела у
позвоночных после двух часов пребывания при указанных
температурах.
показана способность некоторых позвоночных
регулировать температуру тела при различных
температурах окружающей среды.
18.2.6. Температура тела
Когда говорят о температуре тела животного,
обычно имеют в виду температуру внутренних областей
тела, т. е. тканей, лежащих глубже 2,5 см под
поверхностью кожи. Такую «внутреннюю»
температуру обычно измеряют в прямой кишке. В других
участках тела температура может сильно
варьировать в зависимости от их локализации и от
наружной температуры (рис. 18.10).
Хотя основные источники тепла у эктотермных и
у эндотермных животных различны, способы
теплообмена между организмом и окружающей средой у
них одинаковы и включают излучение, конвекцию,
теплопроводность и испарение. С помощью первых
трех способов тепло может передаваться в любом
направлении в зависимости от температурного
градиента (от более теплого к более холодному), но при
четвертом способе организм может только терять
тепло.
излучение. При излучении тепло переносится в
виде электромагнитных волн длинноволновой
инфракрасной части спектра, лежащей за пределами
видимой его части. Тела не просто излучают тепло в
окружающий воздух (так как он не может поглотить
много излученного тепла), а передают его другим
телам со скоростью, пропорциональной разности
температур между двумя телами. У человека на
излучение приходится около 50% общей
теплоотдачи, и этот способ служит главным регулируемым
путем теплоотдачи у животных.
конвекция. При этом способе тепло передается от
организма окружающей среде через воздух. У
эндотермных животных температура тела обычно бы-
34,2-
32,0-
27,2-
29,0-
27,0-
25,5-
20,5-
Температура в ротовой полости (37 0C)
Температура, 0C
Расстояние от верхнего края таза, см
Рис. 18.10. Температура кожи на разных расстояниях от точки К расположенной у верхнего края таза.
В течение всего периода исследования температура воздуха составляла 19°С. Низкая температура
в области коленной чашки отражает, очевидно, более слабое кровоснабжение этого участка.
вает выше окружающей температуры, поэтому
воздух, находящийся в контакте с телом, быстро
нагревается, поднимается вверх и замещается более
холодным воздухом. Скорость передачи тепла этим
способом зависит от скорости движения воздуха
около поверхности тела. Этот вид теплоотдачи
может быть уменьшен за счет материалов,
покрывающих кожу, таких, как перья, мех, волосы и
одежда.
теплопроводность. В этом случае тепло
передается при физическом контакте между телами,
например между организмом и землей. У большинства
наземных животных этот способ составляет
незначительную часть от общего теплообмена, но у
водных животных и у животных, живущих в земле,
его доля может быть весьма существенной.
испарение. При испарении тепло теряется с
поверхности тела в процессе превращения воды в
водяной пар. На испарение 1 мл (1 г) воды
затрачивается 2,45 кДж тепла. Вода непрерывно теряется
путем испарения через кожу и в легких с
выдыхаемым воздухом. Эта неощутимая потеря воды не
регулируется, а между тем она служит
лимитирующим фактором для распространения многих видов
растений и животных.
Существует также регулируемая форма
испарения, которая заключается в потере воды при
потоотделении и тепловой одышке.
18.3. Эктотермные животные
Большинство животных относится к эктотермным,
и их активность зависит от температуры
окружающей среды. Для эктотермных животных характерна
низкая интенсивность обмена и отсутствие
механизмов сохранения тепла. Водные эктотермные
животные живут в ограниченном интервале температур-в
зоне переносимости, определяемой размером
водоема, в котором они обитают. Например,
температура воды в пруду иногда значительно варьирует на
протяжении года, тогда как температура океана
может меняться всего лишь на несколько градусов.
Несмотря на широкие пределы температурных
колебаний в малых водоемах, стадии личинки и куколки
(или же нимфы) многих насекомых проходят в воде,
так как температура здесь более стабильна и в
зимние месяцы понижается не так сильно, как на
поверхности суши. У поденок, стрекоз, ручейников и
многих двукрылых насекомых диапауза проходит в
воде (разд. 21.10.1).
Благодаря относительной простоте строения и
физиологии водные беспозвоночные способны
переносить более сильные колебания температуры,
чем водные позвоночные. У рыб интенсивность
метаболизма выше, чем у водных беспозвоночных,
но большая часть эндогенного тепла быстро
рассеивается по всему телу и передается в окружающую
среду через жабры и кожу. В связи с этим рыбы
имеют обычно такую же температуру, как
окружающая вода. Температура тела у рыб не может быть
ниже температуры воды, но в некоторых случаях
бывает выше, как, например, у тунца. Тепло в этих
случаях удерживается в теле благодаря противоточ-
ной системе теплообмена, и температура «красной»
плавательной мускулатуры может подниматься на
12° выше температуры морской воды.
Наземным эктотермным животным приходится
сталкиваться с более резкими колебаниями
температуры, чем водным, но зато они имеют
возможность жить при более высокой температуре внешней
среды. Это позволяет им быть более активными и
осуществлять ряд сложных поведенческих реакций,
определяемых температурными условиями. Многие
из наземных эктотермных видов способны
поддерживать температуру несколько выше или ниже, чем
температура воздуха, и таким образом избегать
крайностей. Сравнительно малая теплопроводность
воздуха уменьшает у этих животных теплоотдачу, а
для охлаждения организма может использоваться
потеря воды за счет испарения.
18.3.1. Регуляция температуры тела
у наземных эктотермных
животных
В процессе поведенческой и физиологической
активности наземные эктотермные животные поглощают
и выделяют тепло. Главными источниками тепла
для них служат поглощаемое ими солнечное
излучение и соприкасающиеся с телом воздух и земля.
Количество поглощенного тепла зависит от цвета
покровов организма, величины его поверхности и
положения по отношению к солнечным лучам. Так,
например, один вид австралийских прямокрылых
имеет темную окраску, так что тело поглощает
солнечный свет и благодаря этому быстро
нагревается. Когда температура тела достигает
оптимума, кутикула светлеет и поглощение света
уменьшается. Как полагают, это изменение окраски
обусловлено прямой реакцией пигментных клеток на
температуру.
Изменение ориентации организма по отношению
к солнечным лучам приводит к изменению
обогреваемой поверхности тела. Такой способ
терморегуляции характерен для многих наземных
эктотермных животных, включая насекомых, паукообразных,
амфибий и рептилий; это так называемая
поведенческая терморегуляция. Например, пустынная
саранча Schistocerca относительно мало активна при
17 0C, но, расположившись под прямым углом к
направлению солнечных лучей, способна поглощать
тепловую энергию. Когда температура воздуха
повышается примерно до 400C, насекомое меняет
положение и располагается параллельно солнечным
лучам, чтобы уменьшить обогреваемую поверхность
тела. Дальнейшее повышение температуры тела,
которое может привести к гибели, предотвращается
тем, что саранча приподнимается над землей или
взбирается на растения. Температура воздуха
быстро снижается уже на небольшом расстоянии от
поверхности земли, и такое поведение помогает
насекомому обеспечить себе более благоприятный
микроклимат (разд. 12.4.4).
Крокодилы, находясь на суше, тоже регулируют
температуру тела, меняя положение по отношению
к солнцу и разевая пасть, чтобы увеличить
теплоотдачу путем испарения. Если становится слишком
жарко, они переходят в воду, которая остается
более прохладной, чем суша. Ночью же, наоборот,
крокодилы погружаются в воду для того, чтобы
избежать воздействия более прохладного теперь
воздуха.
Особенно широко изучался механизм
терморегуляции у ящериц. При изучении многих видов были
обнаружены разнообразные реакции на изменения
температуры. Ящерицы - наземные рептилии. Им
свойственно множество форм поведения, общих для
всех эктотермных животных; кроме того, некоторые
виды ящериц, например австралийский варан,
используют ряд физиологических механизмов,
позволяющих им разогреваться и поддерживать
температуру выше уровня окружающей среды. Другие
виды рептилий способны удерживать температуру
тела в ограниченных пределах, изменяя свою актив-
ноств, прячась в тени или вылезая на солнце. В
обоих этих случаях ящерицы предвосхитили многие
механизмы терморегуляции, существующие у птиц
и млекопитающих.
В пустынях температура земной поверхности
может повышаться до 70-800C днем и падать до 4°С к
концу ночи. В этих условиях большинство ящериц
ищет убежище, спасаясь от обеих крайностей в
норах или под камнями. Это поведение и
определенные физиологические реакции ярко выражены
у ящерицы Phrynosoma, обитающей в пустынях
юго-востока США и Мексики. Помимо того что эта
ящерица может зарываться в землю, она способна
менять положение тела и его окраску, а когда
температура становится высокой, может также
уменьшить поверхность тела, втягивая ребра. К
числу других реакций на высокую температуру
относятся тепловая одышка (при которой теплоотдача
увеличивается за счет испарения воды из ротовой
полости, глотки и легких), выпячивание глаз и
выведение мочи из клоаки.
У амфибий влажная кожа обеспечивает
идеальный механизм теплоотдачи путем испарения влаги.
Однако потеря воды не может у них регулироваться
физиологическими механизмами, как у
млекопитающих. В сухой среде амфибии сразу начинают
терять воду, и хотя это обстоятельство способствует
теплоотдаче, оно приведет к обезвоживанию, если
животное не сможет найти влажное и прохладное
место, где скорость испарения будет меньше. У
рептилии морской игуаны, когда она греется на
скалистых берегах Галапагосских островов,
температура тела поддерживается на уровне около 37°С.
Так как игуана питается морскими водорослями,
она вынуждена довольно долго находиться в море
при температуре около 25°С. Однако быстрое
охлаждение во время пребывания в воде
предотвращается у нее уменьшением кровотока между
поверхностными и глубокими областями тела в
результате замедления ритма сердца (брадикардии).
18.4. Эндотермные животные
К эндотермным животным относятся птицы и
млекопитающие, у которых активность мало зависит от
температуры окружающей среды. Для поддержания
постоянной температуры тела, которая обычно
выше окружающей температуры, этим животным
необходима высокая интенсивность обмена и эффек-
тивные способы регуляции теплоотдачи с
поверхности тела. Органом, который контактирует с
внешней средой и, следовательно, регистрирует
изменения ее температуры, является кожа. Регуляцию
температуры тела за счет различных метаболических
процессов осуществляет гипоталамус.
18.4.1. Кожа
Название «кожа» относится к наружному покрову
позвоночных животных, и его не следует применять
к наружным покровам червей, моллюсков и
членистоногих. Кожа-самый обширный орган тела.
Она состоит из соединительной ткани, кровеносных
сосудов, потовых желез и сенсорных клеток, что
позволяет ей выполнять множество разнообразных
функций. Строение кожи варьирует у разных групп
позвоночных; основные различия будут
рассмотрены после общего знакомства со строением кожи
человека.
Строение кожи человека
Кожа (рис. 18.11) состоит из двух основных слоев-
эпидермиса и дермы, или собственно кожи. Под
дермой лежит подкожная клетчатка, которая
содержит специализированные жировые клетки,
образующие жировую ткань. Толщина слоя
подкожной клетчатки варьирует в разных участках тела и у
разных людей.
эпидермис. Клетки эпидермиса имеют эктодер-
мальное происхождение и отделены от дермы ба-
зальной мембраной. Эпидермис состоит из многих
слоев клеток, образующих слоистый эпителий
(разд. 8.3.2.). Непосредственно на базальной
мембране лежат кубические эпителиальные клетки
(разд. 8.3.1), которые образуют зону активного
деления-так называемый мальпигиев слой. За счет
повторных делений клеток этого слоя идет
непрерывное обновление клеток эпидермиса. Мальпигиев
слой образует нижнюю часть зернистого слоя,
состоящего из живых клеток, которые по мере
перемещения к наружному слою
эпидермиса-роговому - становятся все более плоскими. Клетки
рогового слоя продолжают постепенно уплощаться, и
в них накапливается кератин - богатый серой
фибриллярный белок, который делает клетки
водонепроницаемыми. По мере накопления кератина
Мышца,
поднимающая
волос
Зернистый
слой!
Роговой
слой
Эпидермис
Зона
эластических и
коллагеновых
волокон
Мальпигиев
слой
Волосяной
фолликул
Дерма
Кровеносный
сосуд
Сальные
железы
Часть стержня
волоса
Секреторные
клетки
Подкожная
клетчатка
Подкожные
жировые
клетки
Потовая
f железа
Проток
железы
Волосяной
сосочек
Рис. 18.11. А Поперечный срез кожи человека.
Б. Схема структур, видимых на срезе.
клетки ороговевают, ядра их разрушаются, и они
отмирают. Толщина рогового слоя больше в тех
участках тела, которые подвергаются
значительному трению, например на подошвах ног и
подушечках пальцев. Поверхностный слой кожи
представляет собой полупрозрачный, тонкий, плотный,
эластичный водонепроницаемый покров с
многочисленными порами (отверстиями потовых желез) и
волосяными фолликулами. Самый наружный слой
ороговевших эпителиальных клеток непрерывно
слущивается в результате трения.
У многих позвоночных видоизмененный роговой
слой образует ногти, когти, рога, чешую, перья и
волосы. Основным компонентом всех этих структур
является кератин.
ДЕРМА. Большая часть клеток дермы имеет мезо-
дермальное происхождение. Дерма состоит из
плотной массы соединительной ткани, содержащей сеть
эластических волокон, кровеносные капилляры,
лимфатические сосуды, мышечные волокна,
сенсорные клетки, нервные волокна, хроматофоры
(пигментные клетки), потовые железы и волосяные
фолликулы.
Волосяные фолликулы представляют собой впя-
чивания эпидермиса. В основании фолликула
находится корень волоса, из которого развивается
стержень волоса. Волос состоит из кубовидных
эпителиальных клеток, которые ороговевают в
результате накопления в них кератина. Корковое
вещество волоса содержит различные количества
пигмента меланина, определяющего цвет волос.
Мозговое вещество волоса может содержать пузырьки
воздуха; с возрастом по мере увеличения количества
этих пузырьков и снижения выработки меланина
волосы седеют. Кровеносные капилляры
обеспечивают снабжение растущего волоса питательными
веществами и удаление продуктов обмена. Верхняя
часть волоса выходит за поверхность эпидермиса;
она остается эластичной и не смачивается благодаря
маслянистому секрету сальных желез,
открывающихся в волосяной фолликул. Секрет сальных желез
содержит жирные кислоты, воски и стероиды и
распределяется по волосу и поверхности кожи,
защищая фолликулы от пыли и микроорганизмов и
образуя на коже тонкий водонепроницаемый слой.
Этот слой препятствует не только потере воды с
поверхности кожи, но и проникновению воды через
кожу внутрь организма.
К основанию фолликула прикреплена гладкая
мышца, другой конец которой прикрепляется к ба-
зальной мембране. При сокращении таких мышц
уменьшается наклон волос по отношению к коже,
что увеличивает толщину неподвижного слоя
воздуха над ее поверхностью. Это используется как один
из способов терморегуляции, а также как
поведенческая реакция на опасность у некоторых
позвоночных. Когда волосы «встают дыбом», видимые
размеры тела как бы увеличиваются, и этого может
оказаться достаточно, чтобы отпугнуть возможного
врага. У человека волосяной покров развит
значительно слабее, чем у других млекопитающих.
Потовые железы представляют собой
закрученные клубочком трубчатые железы, расположенные в
дерме и соединенные выводным протоком с порами.
У человека они имеются на всей поверхности тела, а
у некоторых млекопитающих-только на
подушечках пальцев. У птиц потовые железы отсутствуют.
Кровеносные капилляры доставляют этим железам
воду, соли и мочевину. Секреторная активность
желез регулируется симпатическими нервными
волокнами. Существуют два типа потовых желез-
эккринные (мерокринные) и апокринные. Первые
наиболее распространены (у человека их около
2,5 · 106) и встречаются в большинстве участков
тела, а вторые имеются только в подмышечных
впадинах, вокруг сосков, на лобке, ладонях и
ступнях и вокруг анального отверстия. Железы
выделяют лишенную запаха жидкость, которая позднее
может издавать неприятный запах благодаря
деятельности бактерий. Некоторые соединения цинка и
алюминия способны подавлять активность желез и
уничтожать бактерий. Большинство дезодорантов и
препаратов против потливости содержит такие
вещества.
Кровеносные капилляры в дерме очень
многочисленны и снабжают кровью все имеющиеся в ней
структуры. Многие капилляры образуют петли и
анастомозы (рис. 18.12), позволяющие изменять
количество крови, протекающей через кожные
капилляры. Это один из многих механизмов
терморегуляции, описанных в разд. 18.4.3.
Мотонейроны иннервируют мышцы и железы
кожи, а сенсорные нейроны передают нервные
импульсы от многочисленных рецепторных клеток,
находящихся в коже. Эти клетки реагируют на
тепло, холод, прикосновение, давление и стимулы,
вызывающие боль. Некоторые из таких рецепторов
имеют очень простое строение и состоят из
свободных нервных окончаний, тогда как другие
окружены капсулой, как, например, тельца Пачини.
Строение кожи позвоночных
У многих позвоночных строение кожи отличается от
описанного выше. У рыб имеются хорошо развитые
чешуи или кожные пластинки и железы,
выделяющие слизь. У амфибий кожа гладкая и скользкая
благодаря секреции слизи многочисленными
железами, расположенными в дерме. Эта водянистая
слизь поддерживает влажность кожи, что облегчает
газообмен; у некоторых видов, например у жаб, ее
неприятный вкус служит защитой от хищников.
Кожа амфибий содержит многочисленные
пигментные клетки, позволяющие животным менять
окраску в соответствии с окружающим фоном, а также
поглощать или отражать солнечные лучи. Под
кожей имеется множество лимфатических полостей, и
кожа прикреплена к лежащим под ней мышцам
Поверхность кожи
Теряется
очень мало тепла
Мелкая артерия
-Капиллярная сеть
Артериовенозный анастомоз
(шунт)
Соединительная вена
Мелкая вена
Теряется
много тепла
Мелкая артерия
Мелкая вена
Рис. 18.12. Механизм регуляции кровотока через кожу. А. Распределение кровотока, уменьшающее
теплоотдачу. Сужение артериол приводит к уменьшению тока крови через капилляры и
анастомоз. В кожу поступает лишь столько крови, сколько необходимо для поддержания ее
жизнеспособности. Основная масса крови, притекающей из глубины тела, минует кожу, проходя через
соединительную вену, благодаря чему уменьшается теплоотдача. Б. Распределение кровотока, повышающее
отдачу тепла. Расширение артериол приводит к уменьшению тока крови через капиллярную
сеть и анастомозы. Капилляры расширяются под давлением протекающей в них крови. Согреваемая кровью
кожа отдает тепло путем излучения, конвекции и теплопроводности; кроме того, кровь в большем
количестве протекает через потовые железы.
лишь в отдельных местах, что делает кожу
чрезвычайно подвижной.
У рептилий тело покрыто роговыми чешуйками,
образующими непроницаемый слой. Кожа у них
всегда сухая, а многочисленные хроматофоры,
содержащиеся в эпидермисе, позволяют рептилиям
менять окраску. У птиц в процессе эволюции
образовались перья, представляющие собой выросты
эпидермиса. Существуют разные типы перьев,
выполняющие специфические функции (см. разд. 17.7.6.),
но все они уменьшают потерю тепла организмом. У
птиц поддерживается относительно высокая
температура тела (порядка 40-430C), а поскольку у них
велико отношение поверхности к объему, им
необходимы эффективные способы сохранения тепла.
Все млекопитающие имеют потовые железы и
волосы, количество и распределение которых
зависят от потребностей терморегуляции.
Функции кожи млекопитающих
Кожа выполняет четыре главные функции. Одна из
этих функций-терморегуляция-подробно описана
в разд. 18.4.3., а остальные функции будут
рассмотрены ниже.
защитная функция. Кожа предохраняет
внутренние органы от повреждения, обезвоживания и
инфекции. Ороговевший эпидермис защищает от
механических повреждений, особенно в тех участках,
которые подвергаются давлению или трению.
Дерма и подкожные ткани ослабляют воздействие
толчков и ударов. Темный пигмент меланин,
содержащийся в хроматофорах, защищает организм от
избытка ультрафиолетовых лучей. Различия в
пигментации волос определяют окраску меха животных и
позволяют им маскироваться в природной среде. У
некоторых видов, например у горностая и полярного
зайца, окраска полностью меняется в зависимости от
сезона, что помогает им укрываться от хищников
или оставаться незаметными для жертвы.
Ороговевший слой и выделения сальных желез препятствуют
неконтролируемой потере воды путем испарения.
Кроме того, секрет сальных желез предохраняет мех
таких водных животных, как выдра, от намокания,
которое нарушило бы теплоизоляцию, создаваемую
воздушными прослойками между волосками.
Строение кожи в большой степени затрудняет
проникновение в организм патогенных организмов - бактерий,
вирусов, грибов, простейших и плоских червей. Тем
не менее микроорганизмы могут проникать внутрь
через волосяные фолликулы и потовые железы, а
также при укусах насекомых. Личинки многих
паразитических червей, например Schistosoma, способны
пробуравливаться через кожу.
образование витаминов. Ультрафиолетовые лучи
солнца вызывают превращение некоторых
стероидов, называемых стеролами, в витамин D. Правда,
это лишь дополнительный источник витамина D,
так как главным его источником служит пища (см.
разд. 10.3.10).
ЗАПАСАНИЕ ЭНЕРГЕТИЧЕСКИХ МАТЕРИАЛОВ. В ЖИро-
вых клетках, находящихся в нижних слоях дермы и в
подкожной клетчатке, откладываются запасы жира.
Этот жир служит не только резервным питательным
веществом, но и теплоизолирующим материалом. У
многих животных жир запасается в подкожной
клетчатке для подготовки к неблагоприятному времени
года.
18.4.2. Источники тепла
У эндотермных животных основным источником
тепла служат экзотермические реакции,
протекающие в живых клетках. Тепло высвобождается при
расщеплении молекул веществ, получаемых с
пищей, главным образом углеводов и жиров.
Количество тепла, образующееся в организме
животного натощак в состоянии покоя, называют уровнем
основного обмена и используют как точку отсчета
при сравнении энергетических затрат во время
различных видов активности у разных организмов (см.
разд. 11.7.6). У мужчины среднего роста и веса
суточная потребность в энергии для поддержания
основного обмена составляет около 8000 кДж.
Точное количество энергии, необходимое каждому
индивидууму, зависит от его веса, возраста и пола и у
мужчин несколько больше, чем у женщин.
Большая часть метаболической энергии,
выделяющейся в виде тепла, поступает из таких активных
тканей, как печень и скелетные мышцы.
Теплопродукция регулируется рядом различных факторов,
таких как окружающая температура и гормоны.
Тироксин, выделяемый щитовидной железой,
повышает интенсивность метаболизма, а тем самым и
выработку тепла. Действие этого гормона бывает
продолжительным, тогда как эффекты адреналина
большей частью кратковременны. Другие
источники теплообразования включаются под действием
нервных импульсов. Периодическая стимуляция
скелетных мышц двигательными нейронами вызывает
дрожь, которая может повысить теплопродукцию
почти в 5 раз по сравнению с уровнем покоя. При
дрожи различные группы мышечных волокон одной
и той же мышцы сокращаются и расслабляются
неодновременно и суммарным результатом
оказываются некоординированные движения. Тепловой
эффект может быть усилен за счет мышечной
активности другого рода, например потирания рук,
притаптывания или легких физических упражнений.
У многих млекопитающих вокруг кровеносных
сосудов грудной полости имеются участки, богатые
бурой жировой тканью. Клетки этой ткани
содержат многочисленные жировые капельки, которые
быстро метаболизируются при стимуляции
симпатическими нервами. Для животных, впадающих в
зимнюю спячку, выработка тепла митохондриями
этих клеток имеет особенно важное значение, так
как позволяет быстро повысить температуру тела во
время пробуждения от спячки (см. разд. 21.10.3).
Для большинства описанных выше механизмов
теплообразования инициирующим центром служит
гипоталамус.
18.4.3. Теплоотдача
У эндотермных животных существуют четыре
механизма теплоотдачи, описанных в разд. 18.2.6,
а именно теплопроводность, конвекция, излучение
и испарение. Во всех случаях скорость отдачи
тепла зависит от разности температур между
внутренними областями тела и его поверхностью и между
кожей и окружающей средой. Теплоотдача может
расти или уменьшаться в зависимости от скорости
образования тепла и от внешней температуры.
Существуют три фактора, ограничивающих
теплоотдачу, которые мы сейчас рассмотрим.
скорость кровотока в коже. Потеря тепла через
кожу путем излучения, конвекции и
теплопроводности зависит от количества протекающей через
кожу крови. При слабом кровотоке температура
кожи приближается к температуре окружающей
среды, а при сильном-к температуре внутренних
областей тела. Кожа эндотермных животных обильно
снабжена кровеносными сосудами, и кровь может
протекать через нее любым из трех путей: через
капиллярные сети дермы, через анастомозы между
венами и артериолами в глубоких слоях дермы и
через мелкие подкожные соединительные вены,
связывающие кожные артериолы с венами.
Артериолы имеют относительно тонкие
мышечные стенки, которые могут сокращаться или
расслабляться, изменяя диаметр сосудов и скорость
кровотока через них. Степень сокращения
регулируется симпатическими сосудодвигательными
нервами, идущими от вазомоторного центра головного
мозга, а этот центр получает импульсы от гипо-
таламического центра терморегуляции. У человека
скорость кровотока в коже (на 100 г веса) может
варьировать от 1 мл/мин и менее на холоде до
100 мл/мин при высокой температуре среды,
благодаря чему теплоотдача может увеличиваться в
5-6 раз. Ниже уровня капиллярной сети в коже
лежат «шунты», называемые артерио-венозными
анастомозами. При сужении этих сосудов кровь
выталкивается в обладающие малым
сопротивлением «соединительные вены», которые связывают
артерии с венами, и основная масса крови минует
капилляры и анастомозы (рис. 18.12). Это типичная
реакция, уменьшающая теплоотдачу. При
расширении анастомозов ток крови направляется в
основном через капиллярное русло и анастомозы, а не
через соединительные вены. В результате кровоток в
коже увеличивается и отдача тепла возрастает.
ПОТООТДЕЛЕНИЕ И ИСПАРЕНИЕ ЧЕРЕЗ КОЖУ. ΠθΤ-
водянистая жидкость, содержащая 0,1-0,4%
хлористого натрия, лактата натрия и мочевины. По
отношению к плазме крови пот гипотоничен; он
образуется из тканевой жидкости благодаря
активности потовых желез, находящихся под контролем
особых нервных волокон. Эти волокна принадлежат
симпатической нервной системе, и по ним
поступают импульсы из гипоталамуса. У человека
потоотделение начинается всякий раз, когда температура
тела поднимается выше средней нормальной
температуры 36,7°. В условиях умеренного климата за
сутки выделяется около 900 мл пота, но при очень
сильной жаре и достаточном снабжении водой и
солями эта величина может возрасти до 12 л.
Температура, °С
Скорость
теплоотдачи,
Дж/с
Температура кожи
Температура гипоталамуса
г Теплоотдача путем испарения (потоотделения)
Время, мин
Рис. 18.13. Графики, показывающие зависимость
между температурой кожи, температурой
гипоталамуса и скоростью испарения воды у
человека в теплой камере (45еС). В точках а, б и в
испытуемый выпивал глоток ледяной воды.
18.3. Скрытая теплота испарения пота
составляет 2,45 кДж/мл. Рассчитайте, какая доля (%)
энергии расходуется на потоотделение у шахтера,
который за сутки теряет 4 л пота и потребляет
50 000 кДж энергии.
При испарении пота с поверхности кожи организм
теряет энергию в виде скрытой теплоты испарения,
что приводит к снижению температуры тела. При
низкой температуре среды, высокой влажности и
безветрии скорость испарения уменьшается.
У многих млекопитающих мех настолько густой,
что потоотделение может происходить только на не
покрытых им участках тела: у собак и кошек,
например, на подушечках лап, а у крыс-на ушах. Эти
млекопитающие увеличивают теплоотдачу путем
облизывания шерсти с последующим испарением
влаги, а также путем частого поверхностного
дыхания («тепловая одышка»), при котором тепло
отдает влажная поверхность слизистой рта и носа.
У человека, лошади и свиньи пот может свободно
выделяться через всю поверхность тела.
В настоящее время показано, что потоотделение
является результатом повышения температуры во
внутренних областях тела. В исследованиях,
проведенных на людях и животных, установлено, что
снижение температуры внутри тела после питья
ледяной воды или охлаждения сонных артерий
льдом, приложенным вокруг шеи, приводит к
уменьшению потоотделения, даже если кожа
подвергается воздействию тепла. В опытах с
противоположным распределением тепла и холода
наблюдается обратный эффект. Поскольку из сонных
артерий кровь попадает в гипоталамус, эти опыты
указывают на роль гипоталамуса в терморегуляции.
Помещая термометр в ухо рядом с барабанной
перепонкой, можно получить представление о
температуре, существующей в гипоталамусе. На
рис. 18.13 показано влияние температуры в этом
участке и температуры кожи на потоотделение.
Рассмотрите рисунок и ответьте на поставленные
ниже вопросы.
18.4. Почему температура и скорость
испарения пота остаются относительно постоянными в
течение первых 20 мин?
18.5. Опишите взаимоотношения между
температурой гипоталамуса и скоростью
потоотделения.
18.6. Попробуйте объяснить, почему сразу
после приема ледяной воды температура кожи
повышается.
Теплоизоляция между внутренней областью тела
и окружающей средой. Теплоизоляция тела
обеспечивается прослойкой неподвижного воздуха над
поверхностью кожи (пограничный слой), а затем
кожным и подкожным жиром. Перья, мех и одежда
сохраняют пограничный слой воздуха, а поскольку
воздух-плохой проводник тепла, он уменьшает
теплоотдачу. Степень достигаемой при этом
теплоизоляции зависит от толщины воздушной
прослойки. При низкой температуре в результате
рефлекторного сокращения кожной мускулатуры волосы
или перья приподнимаются и слой неподвижного
воздуха становится толще. У человека эта реакция
еще сохранилась, но так как волос на теле у него
очень мало, она дает лишь эффект, известный под
названием «гусиной кожи». Отсутствие волос
человек компенсирует теплоизолирующим действием
одежды. Для многих млекопитающих характерно
сезонное накопление толстого слоя подкожного
жира, особенно у тех видов, которые не впадают в
зимнюю спячку и переносят холода в активном
состоянии. У водных млекопитающих, особенно у
тех, которые живут в холодных водах (киты,
морские львы, моржи и тюлени), имеется толстый
слой жира, называемого ворванью, который
отлично защищает их от холода.
18.4.4. Тепловой баланс и роль
гипоталамуса
Тепловой баланс любого тела определяется
соотношением между теплом, которое оно получает, и
теплом, которое оно отдает.
Эндотермные животные способны вырабатывать
достаточное количество тепла и регулировать
теплоотдачу, поэтому равенство прихода и расхода
тепла у них всегда сохраняется (рис. 18.14). Это
свойство носит название гомойотермии. Как уже
говорилось в начале этой главы, любой механизм,
имеющий вход и выход, для поддержания своей
стабильности нуждается в регуляторной системе.
Земля
Небо
Температура воздуха
Солнце;
Атмосфера
Дерево
Конвекция
Испарение
Земля
Скала'
проводность
'Камни
Вода
Рис. 18.14. Схема теплообмена между телом лошади (температура 38°С) и окружающей средой в
жаркий солнечный день при температуре воздуха 300C. Прерывистыми линиями
показана передача тепла путем излучения.
У млекопитающих и птиц имеется хорошо
развитая система терморегуляции, включающая
рецепторы, эффекторы и чрезвычайно чувствительный ре-
гуляторный центр в гипоталамусе. Этот центр
следит за температурой крови, отражающей
температуру тех органов, через которые она протекает.
Чтобы гипоталамус мог регулировать температуру
внутренних областей тела (как у эндотермных
животных), он должен получать также информацию об
изменениях наружной температуры. Без такой
информации тело могло бы получить или потерять
массу тепла, прежде чем изменение внутренней
температуры вызовет активацию гипоталамуса и
заставит его принять надлежащие меры. Эту
информацию гипоталамус получает от периферических
терморецепторов, находящихся в коже; они
воспринимают изменения окружающей температуры и
посылают импульсы в гипоталамус еще до того, как
изменится температура внутренних областей тела.
Существуют терморецепторы двух типов - холодо-
вые и тепловые. При соответствующей стимуляции
эти рецепторы генерируют импульсы в отходящих
от них афферентных нервных волокнах. Одна часть
импульсов идет в гипоталамус, а другая-в
сенсорные зоны коры, где возникают температурные
ощущения, соответствующие силе и длительности
стимуляции, а также числу возбужденных
рецепторов. Подсчитано, что у человека имеется примерно
150 000 Холодовых и 16 000 тепловых рецепторов.
Это позволяет организму быстро и точно
регулировать температуру внутренних областей тела. С
точки зрения теории управления кожные рецепторы
действуют как детекторы внешнего возмущения,
«предсказывающие» вероятное изменение
температуры тела. В то же время другие факторы,
изменяющие эту температуру (такие, как интенсивность
обмена или болезненное состояние), будут
непосредственно влиять на нее, и ее изменения будут
обнаружены рецепторами гипоталамуса. В большинстве
случаев в поддержании постоянной температуры
тела участвуют и периферические, и гипоталами-
ческие рецепторы.
Изучение терморегуляторной активности
гипоталамуса показало, что здесь имеются два различных
центра терморегуляции, функции которых
суммированы в табл. 18.1. Взаимоотношения между корой
мозга, гипоталамическими центрами
терморегуляции, температурой внутренних областей тела и кожи
и температурой окружающей среды показаны на
рис. 18.15 и 18.16.
При некоторых заболеваниях температура тела
повышается в результате того, что «термостат»
оказывается настроенным на более высокую
температуру. Предполагается, что эту перенастройку
вызывают определенные вещества, называемые пиро-
генами, к числу которых относятся токсины
патогенных микробов и вещества, выделяемые нейтро-
филами крови. Эти вещества повышают «заданный»
уровень температуры, действуя, очевидно,
непосредственно на гипоталамус. Повышение температуры
тела стимулирует защитные реакции организма и
способствует разрушению патогенных факторов.
Жаропонижающие средства типа аспирина
понижают «заданное значение» и снимают тягостные
симптомы лихорадочного состояния, но при этом,
по-видимому, замедляют нормальные защитные
реакции. Однако в случаях чрезмерно высокой
температуры эти лекарственные препараты
оказываются необходимыми, так как позволяют
предотвратить необратимые изменения головного мозга.
Тепловые
рецепторы
кожи
Высокая
Окружающая
температура
Sr
Повышается
Температура
кожи
Понижается
Кора мозга
Холодов ые ι
рецепторы |
кожи ι
Передняя область
гипоталамуса
Центр теплоотдачи
Торможение
Торможение
Задняя область
гипоталамуса
Центр теплопродукции
Потоотделение
Артериолы
кожи
расширяются
Интенсивность
метаболизма
снижается
Отрицательная обратная связь
Дрожь
Понижается
Температура
крови
[
Повышается
Адреналин
Тироксин
Интенсивность |
метаболизма
повышается
Отрицательная^ обратная связь j
Волосы
поднимаются
Артериолы
кожи
сужаются
Рис. 18.15. Общая схема рефлекторной регуляции температуры тела у млекопитающих, включающая
окружающую среду, гипоталамус и температуру крови.
18.7. Развитие лихорадочного состояния часто
сопровождается дрожью и ощущением холода
(ознобом). Объясните эти симптомы на основе
представлений о механизме терморегуляции.
18.4.5. Адаптация к экстремальным
климатическим условиям
Размеры органов, а также физиологические и
поведенческие адаптации у животных могут быть
различными в разных частях географического ареала. У
эндотермных животных общая теплопродукция
зависит от объема метаболически активных тканей, а
скорость теплоотдачи - от величины поверхности
тела. Эти два параметра-поверхность и объем-не
пропорциональны друг другу. В связи с этим
животные, обитающие в холодных областях (например,
полярные медведи и киты), имеют, как правило,
крупные размеры, тогда как обитатели жарких
стран (например, многие насекомоядные
млекопитающие) обычно меньше по размерам. Это явление
носит название правила Бергмана и наблюдается у
многих видов, в том числе у тигров, размеры
которых уменьшаются по мере удаления от
полюсов. Существуют исключения из этого правила, но в
этих случаях организмы обладают особыми адап-
тациями для выживания в данном районе.
Например, мелкие млекопитающие умеренных или
арктических широт отличаются большим аппетитом,
позволяющим им поддерживать высокую
интенсивность метаболизма. У них небольшие конечности
(что уменьшает теплоотдачу), а зимой они
вынуждены впадать в спячку. Крупные млекопитающие
жарких стран, такие как слоны и бегемоты,
сталкиваются с противоположными проблемами. Слонам
помогают очень большие уши, которые хорошо
снабжаются кровью. Хлопая ушами, слон увеличивает
теплоотдачу путем излучения и конвекции. У
бегемота нет потовых желез, и он использует такую же
поведенческую стратегию, как
крокодил,-перемещается то на сушу, то в воду, чтобы уменьшить
влияние колебаний температуры. Размеры
выступающих частей тела тоже варьируют в соответствии
с температурой среды, так что у видов, живущих в
более холодном климате, различные придатки
меньше, чем у родственных видов из более теплых мест.
Таблица 18.1. Функции центров теплоотдачи и
теплопродукции, расположенных соответственно в переднем и заднем
отделах гипоталамуса и оказывающих противоположное
действие
Центр теплоотдачи
Активируется
повышением температуры в
гипоталамусе
Способствует
расширению кожных сосудов
Повышает теплоотдачу
путем излучения,
конвекции и
теплопроводности
Усиливает
потоотделение и тепловую
одышку
Снижает
метаболическую активность
Уменьшает толщину
слоя неподвижного
воздуха за счет
опускания волос или перьев
Центр теплопродукции
Активируется импульсами от
периферических Холодовых
рецепторов или
температурой в гипоталамусе
Способствует сужению кожных
сосудов
Уменьшает теплоотдачу путем
излучения, конвекции и
теплопроводности
Угнетает потоотделение и
тепловую одышку
Повышает метаболическую
активность, вызывая дрожь и
повышенную секрецию
тироксина и адреналина
Увеличивает толщину слоя
неподвижного воздуха за счет
поднятия волос или перьев
Выше 43 0C — смерть
Верхний предел
нормальной ^^
температуры 37,5°(j"
Средняя температура-
тела 36,70C
Нижний предел
нормальной
темпеоатуоы 35.8 0C
Температура тела
человека, 0C
Организм
■ выдерживает >
до 41,20C /
Повышение
температуры тела
Снижение
температуры тела
Организм
выдерживает
снижение до 32 ° С
Снижение метаболической активности;
расширение артериол кожи; увеличение
кровотока через кожу; повышение
излучения, конвекции и теплопроводности,
потоотделения и испарения; опускание
волос (уменьшение теплоизоляции);
увеличение поверхности тела,
снижение мышечной активности
ГИПЕРТЕРМИЯ
(в случае неэффективности
го мео стаза)
Снижение температуры тела
(отрицательная обратная связь)
Повышение метаболической активности;
сужение артериол кожи; уменьшение
кровотока через кожу; уменьшение
излучения, конвекции, теплопроводности,
потоотделения; поднятие волос (увеличение
теплоизоляции); поза, уменьшающая
открытую поверхность тела;
повышение мышечной активности
Средняя температура тела
Повышение температуры тела
(отрицательная обратная связь)
ГИПОТЕРМИЯ
(в случае неэффективности"
гомео стаза)
.Ниже 260C — смерть
Время
Рис. 18.16. Гомеостатический контроль температуры тела.
Это явление известно как правило Аллена, и его
удобно наблюдать у близких видов, например у
лисиц (рис. 18.17).
18.4.6. Адаптация к жизни при низких
температурах
Эктотермные и многие эндотермные животные
неспособны поддерживать в холодное время года
такую температуру тела, которая позволяла бы им
сохранять нормальную активность, и при низкой
температуре впадают в ту или иную форму спячки
(разд. 21.10). Известны совершенно удивительные
способы адаптации к холоду. Например, личинка
насекомого Вгасоп, паразитирующего на канадском
хлебном пилильщике, способна переносить
температуру ниже — 400C. Эта способность объясняется
тем, что в гемолимфе личинки накапливается гли-
церол, который действует как антифриз и
предотвращает образование кристаллов льда
благодаря процессу переохлаждения.
У многих животных избыточной отдаче тепла
через выступающие придатки тела препятствует
распределение в них кровеносных сосудов. Артерии,
несущие в эти придатки кровь, окружены венами, по
которым кровь течет обратно к телу. Теплая
артериальная кровь, поступающая от тела, охлаждается
холодной венозной кровью, текущей к телу. В свою
очередь холодная венозная кровь, оттекающая от
придатков, нагревается от теплой артериальной
крови, притекающей к ним. Поскольку в придатки
поступает уже охлажденная кровь, теплоотдача
значительно уменьшается. Такое устройство
называется противоточным теплообменником; оно имеется в
ластах тюленей и в хвостовых плавниках китов, в
конечностях птиц и млекопитающих и в системе
кровоснабжения семенников у млекопитающих
(рис. 18.18).
Принцип противоточного обмена используется
для переноса не только тепла, но и различных
веществ, в том числе дыхательных газов и ионов
(разд. 11.7 и 19.5.7.).
В настоящее время в сердечной хирургии широко
используется гипотермия, так как она позволяет
хирургу производить операции на сердце без риска
вызвать у больного повреждение головного мозга.
При понижении температуры тела до 15°С
метаболические потребности клеток мозга снижаются до
такой степени, что снабжение его кровью может
быть прервано без каких-либо вредных последствий
на время до 1 часа. При более продолжительных
Арктический вид
Температура тела 37 ° С
Средняя температура среды O0C
Европейский вид
Температура тела 37 0C
Средняя температура среды 12 0C
Африканский вид
Температура тела 37 ° С
Средняя температура среды 25 0C
Рис. 18.17. Различия β длине ушей у трех видов
лисиц, обитающих в разных
географических областях. Пример, иллюстрирующий правило
Аллена.
Артериальная _
кровь
Венозная „
кровь
Температура
окружающей
среды 40C
Капиллярная сеть :
Рис. 18.18. Схема кровотока между туловищем эндотермного животного, имеющим постоянную
температуру 35 С, и придатками при внешней температуре 4°С. Тепло передается от более
нагретого тела к менее нагретому, и скорость этой передачи пропорциональна разности
температур между телами. В приведенном примере противоточного теплообмена в
капиллярную сеть поступает кровь с температурой 5°С, а из сети собирается кровь с температурой 43C. Таким
образом, теплоотдача в окружающую среду соответствует разности температур в VC.
Аналогичным образом кровь, возвращающаяся во внутренние области тела, всего на 1° холоднее крови,
оттекающей от них. Этот механизм препятствует большой потере метаболического тепла и
способствует поддержанию внутренней температуры внутри тела на уровне 35°С.
операциях наряду с гипотермией применяют
аппарат «сердце - легкие» для поддержания
кровообращения в тканях.
18.4.7. Адаптация к жизни
при высоких температурах
Животные, обитающие в местах, где температура
воздуха превышает температуру кожи, поглощают
тепло, и единственным способом снижения
температуры тела служит испарение воды с его
поверхности. В силу климатических особенностей жаркие
страны могут быть засушливыми, или, наоборот,
очень влажными, что создает дополнительные
трудности. В районах с жарким сухим климатом
теплоотдача может осуществляться за счет свободного
испарения влаги, но обитающим здесь животным
нужно каким-то образом получать воду в
количестве, необходимом для охлаждения тела. В
условиях влажного жаркого климата воды вполне
достаточно, но малый градиент влажности между
организмом и окружающей средой нередко
препятствует испарению. В таких случаях к
физиологическим механизмам часто добавляются
поведенческие формы терморегуляции, например
использование тени и бризов, характерных для зоны
влажных лесов и джунглей.
Организм получает тепло за счет своей
метаболической активности и из окружающей среды.
Количество тепла, поступающею извне,
приблизительно пропорционально площади поверхности
тела. Большинство животных, обитающих в
пустынях, имеют небольшие размеры (как, например,
кенгуровая крыса) и испытывают меньше
трудностей, чем такие крупные животные, как верблюд.
К тому же мелкие животные могут жить в норах,
вырытых в песке или почве, где микроклимат более
благоприятный.
Верблюд в условиях жаркого и сухого климата,
но при наличии свободного доступа к воде может
поддерживать температуру тела в пределах 36-38°С
за счет испарения воды с поверхности тела. Если же
верблюд лишен воды, например во время
многодневных переходов через пустыню, то по мере того,
как он теряет воду, увеличивается разница в
температуре тела в утренние и вечерние часы; при этом
температура может колебаться от 34°С рано утром
до 4ГС после полудня. Верблюд накапливает тепло
в дневные часы и не нуждается в том, чтобы
отдавать его путем испарения. Шмидт-Ниельсен в
серии исследований установил, что верблюд весом
500 кг, способный переносить повышение
температуры тела на 7°, запасает около 12 000 кДж тепла.
Если бы он терял столько тепла путем испарения, то
для поддержания постоянной температуры тела ему
приходилось бы терять 5 л воды. Вместо этого
теплоотдача у верблюда осуществляется путем
излучения, теплопроводности и конвекции в ночные
часы. Важное преимущество такой временной
гипертермии, наблюдаемой у верблюда в дневные
часы, состоит также в том, что благодаря ей
уменьшается разность температур между горячим
воздухом пустыни и телом верблюда, а поэтому
уменьшается и приток тепла. К тому же шерсть верблюда
служит превосходным термоизолятором,
уменьшающим поглощение тепла и потерю воды. В
эксперименте было показано, что остриженный верблюд
теряет на 50% больше воды, чем контрольное
животное. Наконец, еще одно важное преимущество
верблюда заключается в том, что он может
переносить сильное обезвоживание. Большинство
млекопитающих не способно переносить потерю
воды, при которой вес тела уменьшается более чем
на 10-14%, тогда как верблюд может терять за счет
воды до 30% своего веса: даже в этом случае он
способен поддерживать нормальный объем плазмы
крови. Обычно смерть от перегрева при
обезвоживании связана с тем, что из-за уменьшения объема
плазмы кровеносная система не может
транспортировать тепло из внутренних областей тела к его
поверхности достаточно быстро, чтобы
предотвратить перегрев. Вопреки распространенному мнению,
верблюд не может заранее запасать воду на случай
ее дефицита. Вызывает также сомнение его
способность получать воду за счет метаболизма жира,
хранящегося в горбу. Но после периода сильного
обезвоживания верблюды могут выпивать огромное
количество воды. Известно, например, что однажды
верблюд весом 325 кг выпил 30 л воды менее чем за
10 минут. Это приблизительно равноценно тому,
как если бы человек среднего роста и веса выпил
около 7 л воды.
18.5. Печень млекопитающих
Печень-самый крупный из внутренних органов,
участвующих в гомеостазе. Она контролирует
многие обменные процессы, играющие важную роль в
поддержании постоянного состава крови.
Развивается печень из выпячивания энтодермальной
пищеварительной трубки, и многие ее функции
связаны с «обработкой» всасываемых компонентов
пищи. Печень обладает уникальным двойным
кровоснабжением, и благодаря обильной васкуляризации
регулирует многие процессы, связанные с кровью и
кровеносной системой (рис. 18.19). Удивительно то,
что несмотря на огромное многообразие
метаболических функций, выполняемых печенью, ее
гистологическая структура сравнительно проста и
однотипна.
18.5.1. Расположение и строение
печени
У человека печень имеет большие размеры и
составляет 3-5% общей массы тела. Расположена она
непосредственно под диафрагмой, к которой
прикреплена серповидной связкой. Она состоит из
нескольких долей, причем форма может изменяться в
зависимости от количества находящейся в ней
крови. Снаружи печень окружена оболочкой, состоящей
из двух слоев: наружный слой образует гладкая
влажная брюшина, а внутренний - фиброзная глис-
сонова капсула, которая одевает также все
структуры, входящие в печень и выходящие из нее.
Волокна глиссоновой капсулы образуют
«внутренний скелет» (строму) печени, который поддерживает
ее форму.
Клетки печени, называемые гепатоцитами, не
проявляют никакой структурной или функциональной
дифференциации. Кроме них, в печени имеются
только нервные элементы и клетки, связанные с
кровеносными и лимфатическими сосудами. Гепа-
тоциты содержат крупное ядро, хорошо развитый
аппарат Гольджи, многочисленные митохондрии и
лизосомы, а также множество гликогеновых гранул
Печеночная вена
Селезеночная вена
(от селезенки, желудка,
поджелудочной железы)
Передняя
брыжеечная вена
Нижняя
полая вена
Желчный
пузырь
ПЕЧЕНЬ
Желчный
проток
Двенадцатиперстная
кишка
Воротная
вена печени"'
Печеночная
артерия
Аорта
Тонкий
кишечник
Задняя брыжеечная вена
(от толстой и прямой кишок)
Рис. 18.19. Схема кровоснабжения печени.
и липидных капелек. Они плотно примыкают друг к
другу и на поверхности, обращенной к кровеносным
капиллярам, имеют микроворсинки, через которые
и происходит обмен веществами между
гепатоцитами и кровью.
В целом внутреннее строение печени довольно
сложно и до конца еще не изучено. Оно основано на
определенном взаимном расположении гепатоцитов
и двух систем кровеносных сосудов,
переплетающихся с желчными канальцами (желчными
капиллярами). Воротная вена печени многократно ветвится, и ее
многочисленные разветвления идут рядом с ветвями
печеночной артерии, желчными протоками, нервами
и соединительнотканными перегородками
глиссоновой капсулы. В совокупности все эти структуры
образуют портальный тракт (рис. 18.20).
Кровеносные сосуды, отходящие от воротной вены и
приносящие вещества, всосавшиеся в пищеварительном
тракте, и разветвления печеночной артерии, несущие
оксигенированную кровь, соединяются вместе,
образуя венулы, которые доставляют кровь к гепато-
цитам. Как полагают, такое расположение венул и
гепатоцитов образует функциональную единицу
печени, называемую ацинусом. От венул отходят более
мелкие кровеносные сосуды, называемые
синусоидами. Синусоиды образуют густую сеть капиллярных
сосудов, которые, соединяясь, впадают в ветви
печеночной вены. Соседние синусоиды отделены друг от
друга пластинками, или «балками», из гепатоцитов,
часто толщиной всего лишь в одну клетку. В
синусоидах происходит обмен веществами между
кровью и гепатоцитами. Этот обмен облегчается
благодаря наличию в эндотелии синусоидов пор
диаметром до 10 нм и микроворсинок на
поверхности гепатоцитов, обращенной к синусоидам.
Желчь, образующаяся в гепатоцитах, поступает не в
синусоиды, а в мельчайшие желчные капилляры,
Портальный тракт
Фиброзная
капсула
АЦИНУС
Ветвь
печеночной
вены
Лимфатический
сосуд
Ветвь
воротной вены
Эндотелиальная:
клетка
Гепатоцит
Венула
Купферовская
клетка
Синусоиды
Ток
желчи
Ветвь '
печеночного
нерва
Ветвь
печеночной
артерии
Желчный
проток
желчный
капилляр
Рис. 18.20. Упрощенная схема строения печеночного ацинуса. Портальный тракт показан в
поперечном разрезе, а ветвь печеночной вены-в продольном разрезе.
которые, перемежаясь с синусоидами, проходят
между соседними слоями гепатоцитов. В желчные
капилляры выступают микроворсинки гепатоцитов,
через которые желчь выводится в капилляры путем
активного транспорта. Желчные капилляры
образуют разветвленную сеть и сливаются в мелкие
желчные протоки. Последние соединяются вместе в
портальном тракте и образуют более крупные
протоки, сливающиеся в конце концов в общий
печеночный проток.
В целом, как видно из описания, печень
представляет собой густую сеть из гепатоцитов, кровеносных
сосудов и мелких желчных протоков. Такое
строение обеспечивает огромную поверхность, где
каждая клетка непосредственно контактирует с кровью,
и это обеспечивает максимальный обмен
веществами между клеткой и кровью и возможность
контролировать состав крови.
У свиней строение печени значительно проще, чем
у человека. Хотя оно не очень типично для других
млекопитающих, ниже приводится его описание, так
как на его примере легче понять многие
структурные и функциональные взаимоотношения, не столь
очевидные при типичном строении печени
млекопитающих, описанном выше. Печень свиньи состоит
из огромного числа отдельных единиц, называемых
дольками, которые на поперечном разрезе имеют
многоугольную форму и диаметр порядка 1 мм.
Дольки представляют собой наперстковидные
образования длиной 2-3 мм и окружены
соединительнотканными оболочками (продолжение глиссоновой
капсулы). Между дольками проходят междольковые
кровеносные сосуды, представленные одной арте-
риолой и одной веточкой воротной вены, и желчные
протоки (как в портальном тракте других
млекопитающих). В середине дольки проходит
центральная вена, в которую поступает кровь из синусоидов,
идущих от воротной вены и печеночной артерии. Из
центральных вен долек кровь поступает в
печеночную вену. Основную массу дольки составляют тяжи
(балки) из гепатоцитов, которые идут от периферии
и сходятся к центральной вене. Обмен веществами
между гепатоцитами и кровью осуществляется так,
как было описано выше. Синусоиды чередуются с
желчными капиллярами, по которым желчь
собирается во внутридольковый желчный проток. Этот
сравнительно простой и регулярный план строения
показан на рис. 18.21 и 18.22.
Функциональными единицами печени у свиньи
являются дольки, а регулярная гистологическая
Рис. 18.21. Срез печени свиньи; видны дольки.
Междольковые
кровеносные "
сосуды
Внутридольновая
вена
Ветвь воротной
вены печени
Ветвь
печеночной артерии
Печеночные струны
(синусоиды и желчные
капилляры)
Желчный
проток
Желчный
капилляр
Гепатоцит
Синусоид
Соединительнотканная капсула
дольки
Рис. 18.22. Участок печени свиньи
в поперечном разрезе. Показано
регулярное расположение долек,
обусловленное параллельным
расположением междольковых и вну-
тридольковых кровеносных сосудов.
Стрелки указывают направление
тока жидкости в синусоидах и
желчных канальцах.
структура печени обусловлена тем, что внутридоль-
ковые и междольковые сосуды идут параллельно
друг другу. Такое строение не свойственно другим
млекопитающим-у них нет деления на дольки или
оно выражено менее четко.
Лимфа, находящаяся в лимфатических сосудах
портального тракта, поразительно богата белками.
Она образуется в результате выхода плазмы через
поры в эндотелии венул и синусоидов.
В печени имеются клетки еще одного типа-куп-
феровские; они входят в состав широко
распространенной в организме ретикулоэндотелиальной
системы (см. разд. 14.14). Цитоплазматическими
выростами купферовские клетки прикреплены к стенкам
синусоидов. Они обладают способностью к
фагоцитозу и участвуют в разрушении старых,
изношенных эритроцитов и в поглощении патогенных
организмов.
18.5.2. Функции печени
Как полагают, печень выполняет несколько сотен
различных функций, включающих тысячи
различных химических реакций. Все эти функции связаны с
положением печени в кровеносной системе и с тем
огромным объемом крови, который протекает через
нее в единицу времени (приблизительно 20% от
общего объема крови). Печень и почки - главные
органы, ответственные за регуляцию содержания
метаболитов в крови и постоянство ее состава. Все
питательные вещества, всосавшиеся в
пищеварительном тракте, поступают прямо в печень, где они
либо хранятся, либо превращаются в другие
вещества, необходимые в данный момент организму.
Таким образом, все функции печени можно
разделить на две категории: 1) хранение питательных
веществ и синтез их производных и 2) расщепление
ненужных организму веществ перед их экскрецией.
Кроме того, благодаря ряду протекающих в
печени обменных реакций она может служить
источником тепла для животных, обитающих в холодных
зонах.
Углеводный обмен
Гексозы (шестиуглеродные сахара) поступают в
печень из кишечника по воротной вене печени-
единственному кровеносному сосуду, в крови
которого содержание сахара может очень сильно
колебаться. Это дает ключ к пониманию роли печени в
углеводном обмене как органа, поддерживающего
концентрацию глюкозы в крови на уровне около
90 мг% (90 мг на 100 мл). Печень препятствует
колебанию этого уровня в связи с режимом питания
и тем самым предотвращает повреждение таких
тканей, как мозг, которые не могут запасать
глюкозу. В печени все гексозы, включая галактозу и
фруктозу, превращаются в нерастворимый
полисахарид гликоген и в таком виде хранятся. Процесс
превращения глюкозы в гликоген называется глико-
генезом и стимулируется инсулином:
Инсулин
Глюкоза :
(фосфорилирование)
- Глюкозо-6-фосфат :
τ Глюкозо-1-фосфат —
(конденсация)
^Гликоген
Расщепляясь до глюкозы, гликоген препятствует
падению уровня глюкозы в крови ниже 60 мг%.
Процесс расщепления гликогена называется глико-
генолизом и включает активацию фермента фосфо-
рилазы под действием гормона глюкагона,
выделяемого поджелудочной железой. В момент опасности,
при стрессе или в условиях холода фосфорилазу
активируют также адреналин, выделяемый
мозговым слоем надпочечников, и норадреналин,
освобождаемый окончаниями симпатических нейронов
(разд. 15.2.3).
Гликоген:
(депо)
Фосфорилаза
: Глюкозо-1-фосфат
Фосфоглюкомутаза
Глюкозо-6-фосфат
Глюкозо-6-фосфатаза
; Глюкоза
(свободная)
Глицерол,
жирные кислоты
Аминокислоты
^Глюконеогенез/
Лглкжокорти/
коиды)
Гликогенолиз
(глюкагон, адреналин,^
глюкокортикоиды)
Гликоген
(печень)
Углевод
Переваривание
'Глюкоз;
крови
(Инсулин)
^ Глюкозо-б-фосфат~
(ткани + печень)
Глюкозо-1-
фосфат
Гликогенез
Гликоген
(мышцы)
Пировиноградная кислота
Аэробный путь
СС>2 + H2O + энергия
Анаэробный путь
Молочная кислота + энергия
Рис. 18.23. Общая схема углеводного обмена.
В мышцах нет фермента глюкозо-6-фосфатазы,
поэтому они не могут превращать гликоген в
глюкозу непосредственно через глюкозо-6-фосфат.
Вместо этого они превращают глюкозо-6-фосфат в
пировиноградную кислоту, которая используется в
процессе дыхания или анаэробного гликолиза с
образованием АТФ. Молочная кислота,
образующаяся в скелетных мышцах при гликолизе, может
затем превратиться в глюкозу, а последняя уже в
печени превращается в гликоген. Этот
биохимический путь известен как цикл Кори:
Молочная кислота -► Пировиноградная кислота ->
-> Глюкоза -> Гликоген
Когда в связи с раходованием глюкозы запасы
гликогена в печени истощаются, глюкоза может
синтезироваться из неуглеводных
предшественников. Этот процесс называется глюконеогенезом.
Низкий уровень глюкозы в крови (гипогликемия)
приводит к стимуляции симпатической нервной
системы и выбросу адреналина, который, как уже
отмечалось, способствует удовлетворению
сиюминутных потребностей организма в глюкозе. Низкий
уровень глюкозы в крови приводит также к
стимуляции гипоталамуса, который выделяет кортиколи-
берин (разд. 16.6.2.), стимулирующий секрецию
АКТГ передней долей гипофиза. Под действием
АКТГ усиливаются синтез и секреция глюкокорти-
коидных гормонов (в основном кортизола).
Последние стимулируют переход из тканей в кровь
аминокислот, глицерола и жирных кислот, а также синтез
в печени ферментов, катализирующих превращение
аминокислот и глицерола в глюкозу (жирные
кислоты превращаются в ацетил-КоА и
непосредственно используются в цикле Кребса). Углеводы,
которые не смогут быть использованы организмом или
запасены в виде гликогена, превращаются в жиры и
в такой форме депонируются. Общая схема
углеводного обмена, показывающая участие в нем печени,
мышц и других тканей, приведена на рис. 18.23.
Белковый обмен
Печень играет важную роль в белковом обмене,
который включает здесь дезаминирование,
образование мочевины, трансаминирование и синтез
белков плазмы.
дезаминирование. Организм не способен запасать
аминокислоты, и те из них, которые не могут быть
сразу использованы для биосинтеза белка,
подвергаются дезаминированию в печени. Этот процесс
состоит в ферментативном отщеплении
аминогруппы (—NH2) и одновременного окисления остатка
молекулы с образованием углевода, который
используется в процессе дыхания. Аминогруппа
отщепляется вместе с атомом водорода, поэтому
азотистым продуктом дезаминирования оказывается
аммиак (NH3):
Аминокислота
Кислород
Оксокислота
Аммиак
Конкретный пример:
CH3
I
2 С—COOH + 2NH3
Алании
Кислород
Пировиноградная
кислота
Аммиак
Образующийся аммиак может быть либо
использован для синтеза определенных аминокислот или же
азотистых оснований типа аденина или гуанина
(разд. 5.6), либо удален из организма.
образование мочевины. Аммиак, образующийся
при дезаминировании, превращается в мочевину-
растворимый продукт, подлежащий удалению:
Аммиак Углекислота Мочевина Вода
Образование мочевины происходит в орнитиновом
цикле, представленном на рис. 18.24.
трансаминирование. Это процесс синтеза
аминокислот путем ферментативного переноса
аминогруппы с аминокислоты на оксокислоту (гл. 5).
Например, глутаминовая кислота может быть
синтезирована следующим образом:
Алании
α - Оксоглутаровая
кислота
Глутаминовая
кислота
Пировиноградная
кислота
Общий принцип всех этих реакций - взаимный
обмен специфическими радикалами между
аминокислотой и оксокислотой:
Аминокислота А
Оксокислота В
Аминокислота В
Оксокислота А
Трансаминирование служит способом образования
тех аминокислот, которых не хватает в пищевом
рационе, т.е. это еще одна гомеостатическая
функция печени. «Незаменимые» аминокислоты,
описанные в разд. 10.3, не могут синтезироваться путем
трансаминирования в печени и должны поступать с
пищей.
синтез белков плазмы. Белки являются жизненно
необходимыми компонентами плазмы, и
большинство этих белков синтезируется из аминокислот
в печени. По количеству главным белком плазмы
является альбумин, концентрация которого в норме
составляет около 4 г на 100 мл. Он играет важную
роль в поддержании онкотического давления,
уравновешивающего гидростатическое давление в
кровеносных сосудах. Благодаря взаимному
противодействию этих двух факторов сохраняется баланс
жидкости, находящейся в кровеносных сосудах и вне
их (разд. 14.12.1.). Кроме того, альбумины
выполняют в кровеносной системе транспортную
функцию-служат переносчиками таких веществ, как
кальций, триптофан, билирубин, соли желчных
кислот, аспирин и некоторые стероидные гормоны.
Глобулины плазмы-очень крупные молекулы,
содержание которых в крови составляет около 34 г/л.
Альфа- и бета-глобулины транспортируют
гормоны, в том числе тироксин и инсулин, а также
холестерол, липиды, железо и витамины B12, A, D и
К. Гамма-глобулины образуются в лимфоцитах и
других клетках ретикулоэндотелиальных тканей и
участвуют в иммунном ответе (разд. 14.14.3.).
Важными белками плазмы являются также факторы
свертывания крови, протромбин и фибриноген,
функции которых описаны в разд. 14.13.5.
Жировой обмен
Печень больше участвует в метаболизме и
транспорте жиров, чем в их хранении. В жировом обмене
клетки печени выполняют следующие функции:
превращают в жиры избыток углеводов; поглощают
из крови и расщепляют холестерол и фосфолипиды,
а в случае надобности синтезируют их; образуют
глобулины для транспорта липидов.
Запасание витаминов в печени
В печени хранятся некоторые запасы
водорастворимых витаминов В и С, особенно такие витамины
группы В, как никотиновая кислота, витамин B12 и
фолиевая кислота. Витамин B12 (цианкобаламин) и
фолиевая кислота необходимы костному мозгу для
образования эритроцитов, и недостаток этих
витаминов приводит к различным формам анемии.
Однако главные витамины, запасаемые в печени,-это
жирорастворимые витамины A, D, E и К. Печень
некоторых рыб, например трески и палтуса, содер-
Дезаминирование
Цитруллин.
Аргинин
Орнитин
Мочевина
Рис. 18.24. Общая схема орнитинового цикла в
печени млекопитающего (орнитин и цитруллин-
аминокислоты, не получаемые с пищей, а
образующиеся в процессах обмена).
жит витамины AhDb высоких концентрациях.
Витамин К является незаменимым фактором
свертывания крови.
Запасание минеральных веществ
Наряду с железом и калием в печени хранятся
минеральные вещества, нужные в очень малых
количествах (микроэлементы), такие как медь, цинк,
кобальт и молибден. Железо содержится главным
образом в виде ферритина - комплекса железа с
бета-глобулином. У человека содержание железа в
печени составляет около 1 мг на Ir сухой массы.
Большая часть этого железа находится в печени
временно; оно освобождается при разрушении в
печени старых эритроцитов и хранится здесь, пока
не потребуется для образования новых эритроцитов
в костном мозге.
Депонирование крови
Кровеносные сосуды, отходящие от селезенки и
кишечника, соединяются, образуя воротную вену
печени, и вместе с кровеносными сосудами самой
печени содержат большой объем крови; это своего
рода кровяное депо, хотя и проточное, а не
инертное. Под действием симпатических нервов и
выделяемого мозговым веществом надпочечников
адреналина многие сосуды печени могут сужаться, и
тогда в общую систему циркуляции поступает
больше крови. И наоборот, при увеличении объема крови,
например при ее переливании, печеночные и другие
вены могут расширяться, вбирая излишки крови.
Образование эритроцитов
У плода образование эритроцитов (эритропоэз)
происходит в печени, но постепенно эту функцию берут
на себя клетки костного мозга (разд. 14.11.2.). После
этого печень начинает играть прямо
противоположную роль - участвует в разрушении эритроцитов и
гемоглобина.
Разрушение гемоглобина
Время жизни эритроцитов составляет около 120
дней. К концу этого периода они стареют и затем
разрушаются фагоцитирующими макрофагами ре-
тикулоэндотелиальной системы печени, селезенки и
костного мозга. Гемоглобин расщепляется на гем и
глобин. Глобин распадается на составляющие его
аминокислоты, которые поступают в общий фонд
свободных аминокислот печени и используются в
соответствии с потребностями. От гема
отщепляется железо, а остающиеся пиррольные кольца
образуют зеленый пигмент биливердин. Последний
превращается в билирубин-желтый пигмент,
входящий в состав желчи. Накопление билирубина в
крови служит признаком заболевания печени и
вызывает пожелтение кожи, называемое желтухой.
Образование желчи
Желчь-это вязкая зеленовато-желтая жидкость,
которую выделяют гепатоциты. Ее суточная секреция
составляет 500-1000 мл. Желчь содержит 98%
воды, 0,8% желчных кислот и их солей, 0,2% желчных
пигментов, 0,7%) неорганических солей и 0,6% хо-
лестерола. Она участвует в пищеварении,
способствуя всасыванию жиров, и в выведении желчных
пигментов.
Соли желчных кислот являются производными
стероида холестерола (холестерина), образующегося
в печени. Главные соли желчных кислот-это глико-
холат натрия и таурохолат натрия. Они секретиру-
ются вместе с холестеролом и фосфолипидами в
виде крупных частиц, называемых мицеллами. Холе-
стерол и фосфолипиды удерживают полярные
молекулы желчных солей вместе, так что все
гидрофобные концы молекул ориентированы одинаково.
Гидрофобные концы прикрепляются к липидным
каплям, а противоположные концы-к воде. Это
уменьшает поверхностное натяжение липидных
капель, благодаря чему они дробятся, образуя
эмульсию. Меньшие по размеру капли имеют большую
общую поверхность для воздействия
панкреатической липазы, расщепляющей липиды на глицерол и
жирные кислоты, которые затем всасываются в
кишечнике. Кроме того, соли желчных кислот
активируют фермент липазу, причем их действие носит
чисто физический характер. При недостатке солей
желчных кислот в желчи в ней повышается
концентрация холестерола, который может осаждаться в
желчном пузыре или в желчных протоках в виде
холестероловых желчных камней. Эти камни могут
вызывать закупорку желчного протока,
приводящую к весьма болезненным симптомам.
Желчные пигменты не выполняют никакой
функции-это просто продукты, подлежащие удалению
из организма.
Образующийся в печени холестерол
(холестерин) служит предшественником для синтеза других
стероидов. Главный источник холестерола-пища.
Многие молочные продукты богаты холестеролом
или жирными кислотами, из которых может
синтезироваться холестерол. Тироксин повышает как
синтез холестерола в печени, так и скорость его
выделения с желчью. Избыточное содержание холестерола
в крови может приводить к отложению его в стенках
артерий и развитию атеросклероза (сужению
артерий), что связано с повышенным риском
образования тромбов-сгустков крови, которые могут
закрывать просвет сосуда. Это явление называют
артериальным тромбозом. Если тромбоз происходит в
сердце или головном мозге, он часто приводит к
смерти. Нередко холестерол считают главной
причиной сердечно-сосудистых заболеваний, однако
многие из приводимых доказательств спорны.
Желчь запасается в желчном пузыре и
концентрируется в нем в результате всасывания ионов натрия
КИШЕЧНИК
Моносахариды —
Липиды
ПЕЧЕНЬ
-Гликоген-
КРОВЬ
Глюкоза
Жирные кислоты
и глицерол
Аминокислоты
Аминокислоты
Депо
-Жиры-
Аминокислоты.
Трансаминирование
. Дезаминирование
Оксо кислоты
NH3
Углеводы
Жирные кислоты
Мочевина
Мочевина
Белки плазмы
В и С
'Cu*+ Zn'+ Co2 +
Mb2+, Fe2* K +
Объем крови
Эритроциты
Fe2 +
Синтез белка
Витамины
Минеральные
соли
Кровь воротной
вены печени
^Запасы витаминов А, D, E и К|
-|Запасы микроэлементов Fe+2H К7}
Депо
•Образование эритроцитов (у плода)-
Расщепление гемоглобина
Биливердин
Билирубин
Холестерол
Желчь
ревращение в желчные кислоты
Холестерол
Гормоны
Токсины
Расщепление
Детоксикация
Безвредные соединения
Рис. 18.25. Общая схема функций печени.
и воды в кровеносные капилляры. Стимулятором
выведения желчи в двенадцатиперстную кишку
служит гормон холецистокинин (панкреозимин),
описанный в разд. 10.5.
Образование υ расщепление
гормонов
Печень не считают железой внутренней секреции,
но она синтезирует и выделяет под влиянием
гормона роста (продукта передней доли гипофиза)
ростовые факторы, называемые сом атоме динами. Более
детально регуляция процессов роста описана в разд.
21.8.1. В печени в той или иной степени
разрушаются почти все гормоны. Тестостерон и альдо-
стерон расщепляются довольно быстро, тогда как
инсулин, глюкагон, гормоны кишечника, женские
половые гормоны, гормоны надпочечников,
адиуретин и тироксин разрушаются не так быстро. Таким
образом печень осуществляет гомеостатический
контроль над действием многих гормонов.
Детоксикация
Этот термин относится к целому ряду гомеостати-
ческих функций печени, поддерживающих
постоянство состава крови. Бактерии и другие патогенные
организмы удаляются из крови синусоидов купфе-
ровскими клетками, а токсины, которые они
выделяют, обезвреживаются в результате биохимических
реакций, происходящих в гепатоцитах. К
обезвреживанию токсинов приводят такие реакции, как
окисление, восстановление, метилирование
(присоединение СН3-групп) или конденсация с другой
органической или неорганической молекулой. После
детоксикации эти вещества, теперь уже в виде
безвредных продуктов, выводятся почками. Самым
токсичным веществом, содержащимся в крови,
является аммиак, о судьбе которого уже говорилось в
этой главе. Детоксикации в печени подвергаются и
такие вещества, как алкоголь и никотин.
Избыточное потребление алкоголя в постепенно
возрастающих дозах может привести к разрушению печени-
заболеванию, называемому циррозом.
Некоторые виды метаболической активности
печени могут таить в себе потенциальную опасность
для организма. Так, например, накапливается все
больше данных в пользу того, что некоторые
пищевые добавки могут превращаться в печени в
токсичные или канцерогенные вещества. Даже
болеутоляющее средство парацетамол при избыточном его
употреблении превращается в вещество, которое
влияет на ферментные системы и может повреждать
печень и другие ткани.
Печень и теплопродукция
Накапливается все больше данных против широко
распространенного представления, что метаболи-
ческая активность печени-один из главных
источников тепла в организме. Многие метаболические
процессы, происходящие в печени, являются
эндотермическими, т.е. сопровождаются поглощением,
а не выделением энергии. При очень сильном
охлаждении тела гипоталамус действительно может
усиливать экзотермические процессы в печени,
стимулируя высвобождение адреналина симпатической
нервной системой и секрецию тироксина. Однако
при обычных температурах, как показано, печень
«термически нейтральна», хотя ее температура на
1-2° выше, чем в других внутренних областях тела.
Печень служит еще одним примером тесной
взаимосвязи между структурой и функцией
биологических систем. Но особенно примечательно то, что
многообразие функций этого органа сочетается с
относительно несложным, мало
дифференцированным гистологическим строением.
Глава 19
Экскреция и осморегуляция
Экскреция и осморегуляция-два важных гомеоста-
тических процесса, характерные для живых
организмов. Каждый из этих процессов в той или иной мере
способствует сохранению постоянства внутренней
среды организма в условиях изменяющейся внешней
среды.
Экскреция-это выведение из организма
ненужных ему продуктов обмена («отходов»), накопление
которых мешало бы поддержанию стационарного
состояния внутренней среды. Из организма
выводятся также многие вещества, которые не являются
отходами, и вещества, не образующиеся в самом
организме; в первом случае речь идет о секреции, а
во втором-о дефекации. Чтобы
отдифференцировать эти последние функции от экскреции, следует
дать им четкое определение.
Секреция-это пассивное или активное выделение
молекул из клеток во внеклеточное пространство
(кровеносное русло, пищеварительный тракт) или в
окружающую среду. Эти молекулы образуются в
самом организме, как, например, гормоны или
ферменты, поэтому их следует рассматривать как
продукты метаболизма, но при этом они не являются
отходами. Как мы увидим позже, секреция может
быть составной частью процесса экскреции.
Дефекация-это удаление из организма
балластных веществ (главным образом непереваренных
остатков пищи), которые никогда не участвовали в
метаболизме.
тивнои реакции
А + В
(реагирующие
вещества)
±C + D
(продукты)
19.1. Значение экскреции и
осморегуляции
Процессы экскреции и осморегуляции выполняют
ряд функций, которые можно описать следующим
образом.
1. Удаление отходов метаболизма, которые
нередко бывают побочными продуктами главных
метаболических путей. Оно необходимо для
сохранения нужного равновесия биохимических реакций.
Многие реакции обратимы, а по закону
действующих масс направление реакции определяется лишь
относительными концентрациями реагирующих
веществ и продуктов реакции. Например, в фермента-
непрерывное образование жизненно необходимого
метаболита С обеспечивается удалением побочного
продукта D, сдвигающим равновесие в сторону
прямой реакции.
2. Удаление таких отходов, которые в случае
накопления отрицательно влияли бы на
метаболическую активность организма. Многие из этих
веществ токсичны, так как подавляют активность
ферментов.
3. Регуляция ионного состава жидкостей тела. В
водных средах организма соли ведут себя как
электролиты и подвергаются диссоциации.
Например, поглощаемый с пищей NaCl в жидкостях тела
находится в виде ионов натрия (Na+) и хлора (Cl-).
Если концентрации этих и других ионов не будут
удерживаться в узких пределах, многие
физиологические и биохимические процессы могут быть
нарушены; например, снижение концентрации Na+
приводит к ухудшению нервной координации. Помимо
Na+ и Cl" важную роль в организме играют ионы
K+, Mg2+, Ca2+, Fe2+, H+, Г, РОГ и HCO3",
концентрация которых должна жестко
регулироваться, поскольку они участвуют во многих
процессах, включая работу ферментов, синтез белка,
образование гормонов и дыхательных пигментов,
проницаемость мембран, электрическую активность и
мышечное сокращение. Их влияние на содержание
воды, осмотическое давление и рН жидкостей тела
будет рассмотрено позже.
4. Регуляция содержания воды в жидкостях тела.
Содержание воды в жидкостях тела и его
регуляция-одна из основных проблем, с которыми
сталкивались организмы при колонизации многих
экологических ниш на планете. В ходе решения этих
проблем сформировался ряд важнейших
структурных и функциональных приспособлений.
Механизмы, обеспечивающие получение воды, ее сохранение
и выведение, весьма разнообразны, но все они, как
мы увидим позже, имеют огромное значение для
поддержания осмотического давления и объема
жидкостей тела на стабильном уровне. Прежде чем
перейти к рассмотрению этих механизмов,
необходимо подчеркнуть, что осмотическое давление
жидкостей тела зависит от количественного
соотношения между растворенными веществами и
растворителем, т.е. водой. Регулирование относительных
концентраций растворенных веществ и воды
называется осморегуляцией.
5. Регуляция концентрации водородных ионов в
жидкостях тела. Природа рН и методы его
измерения описаны в Приложении 1.1.5, но механизмы
экскреции ионов, оказывающих большое влияние на
рН (H+ и HCO^), будут рассмотрены в настоящей
главе. Например, рН мочи может колебаться от 4,5
до 8,0, благодаря чему рН жидкостей тела
поддерживается на достаточно постоянном уровне.
19.1.1. Продукты, подлежащие
экскреции
Важнейшие экскреты сведены в табл. 19.1. Как
видно из таблицы, далеко не все они являются
отходами, т. е. веществами, которые не могут быть
дальше использованы организмом. В
действительности многие из них до или после выведения из
организма еще могут служить определенным
целям.
Таблица 19.1. Основные продукты, подлежащие экскреции,
их источники, функции и дальнейшая судьба
Продукт
O2
CO2
Вода
Ионы
(соли)
Соли
желчных
кислот
Желчные
пигменты
Таннины и
другие
ганические
кислоты
Азотистые
соединения
Источник
Фотосинтез у ав-
тотрофов
Аэробное
дыхание у всех
организмов
Расщепление
мочевины
Аэробное
дыхание у всех
организмов
Реакции
конденсации
Пища
Жировой обмен
в печени
Расщепление
тема в печени
Азотистый и
углеводный
обмен у
некоторых
растений
Обмен белков и
нуклеиновых
кислот
Функция/судьба
Используется при
аэробном дыхании
Используется при
фотосинтезе
Снижает рН жидкостей
тела
Растворитель для всех
метаболических
реакций
Используются при
фотосинтезе и т. п.
Поддерживают
осмотическое давление.
Повторно используются в
экосистеме
Способствуют
эмульгированию жиров
Никакой функции не
выполняют
Горькие вещества,
защищают растение от
поедания животными
Расщепляются и
повторно используются в
экосистеме
19.1.2. Экскреция у растений
У растений экскреция не связана с таким
множеством проблем, как у животных. Это объясняется
фундаментальными различиями в физиологии и
образе жизни растений и животных. Растения
являются первичными продуцентами и синтезируют в
нужном количестве все необходимые им
органические соединения. Например, в растениях образуется
лишь столько белка, сколько его необходимо в
данный момент. Они никогда не синтезируют белок
в избытке и поэтому выделяют очень мало
азотистых отходов, образующихся при расщеплении
белков. Если же белки расщепляются до аминокислот,
то последние могут быть использованы для синтеза
новых белков. Три конечных продукта,
образующиеся в ходе определенных обменных процессов, -
O2, CO2 и вода-используются у растений как
исходные вещества для других реакций; это в особенности
относится к CO2 и воде. Единственный
газообразный продукт, выделяемый растениями в большом
количестве,-это молекулярный кислород. На свету
в растении образуется намного больше O2, чем ему
нужно для дыхания, и этот избыток кислорода
переходит в окружающую среду путем диффузии.
Многие органические отходы метаболизма
откладываются у растений в омертвевших постоянных
тканях (таких, как ядровая древесина), а также в
листьях или коре, которые периодически
сбрасываются. Многолетние растения состоят в основном
из мертвых тканей. Экскреты скапливаются в этих
тканях и уже не могут оказывать вредного
воздействия на активность живых тканей. Аналогичным
образом могут накапливаться многие минеральные
соли, поглощаемые растением в виде ионов,
накопление которых связано с разным использованием
катионов и анионов. Органические кислоты,
способные нанести вред растению, часто связываются с
избыточными катионами и выпадают в виде
нерастворимых кристаллов, которые могут
сохраняться в клетках растения, не причиняя им вреда.
Например, ионы кальция и сульфата одновременно
поглощаются растением, но сульфат сразу же
используется в синтезе аминокислот, а кальций
остается в избытке. Ионы Ca2+ легко реагируют со
щавелевой и пектиновой кислотами, образуя с ними
безвредные нерастворимые продукты - оксалат и
пектат кальция. Другие ионы, такие как железо и
марганец, а также органические кислоты вроде тан-
ниновой и никотиновой поступают в листья, где они
накапливаются, придавая листьям характерную
окраску перед их опадением. Подлежащие удалению
вещества элиминируются не только с листвой, но и с
лепестками, плодами и семенами, хотя экскреция не
является главной функцией этих образований. У
водных растений основная масса отходов
метаболизма переходит путем диффузии прямо в
окружающую воду.
19.1.3. Экскреция у животных
Потенциальным местом экскреции может быть
любая проницаемая поверхность, которая напрямую
связывает участок, содержащий экскрет, с внешней
средой. К таким поверхностям относятся наружная
мембрана одноклеточных организмов, эпидермис
низших беспозвоночных, трахеи членистоногих,
жабры и кожа рыб и амфибий, легкие и кожа
высших позвоночных. Клетки организмов,
имеющих относительно простое строение, обычно прямо
контактируют с окружающей средой, и их экскреты
сразу же удаляются путем диффузии. По мере
усложнения организации животных у них развиваются
выделительные органы, осуществляющие выведение
отходов из организма в окружающую среду через
протоки и поры прямым или непрямым путем. У
высших позвоночных имеются специализированные
органы выделения, к которым отходы метаболизма
доставляются из всех клеток кровеносной
системой. Наиболее важные органы выделения у
таких животных-это кожа, легкие, печень и почки.
В настоящем разделе мы рассмотрим только роль
первых трех органов.
кожа. Вода, мочевина и соли активно выводятся
из кожных капилляров в протоки потовых желез.
При выходе пота на поверхность кожи вода
испаряется, что приводит к потере тепла и способствует
терморегуляции.
легкие. CO2 и водяные пары диффундируют с
влажной поверхности легочных альвеол. Легкие у
млекопитающих - единственный орган выделения
CO2. Часть воды, испаряющейся в легких,
представляет собой метаболическую воду, т. е. продукт
клеточного дыхания, который можно было бы считать
экскретом, но истинное происхождение этой воды не
так уж важно ввиду большого общего объема воды,
содержащейся в организме.
печень. Учитывая многочисленные гомеостати-
ческие функции печени, описанные в разд. 18.5.2,
вряд ли следует удивляться тому, что в число этих
функций входит и экскреция. Выводимыми
продуктами являются желчные пигменты, образующиеся
при расщеплении гемоглобина старых эритроцитов.
В составе желчи эти пигменты поступают в
двенадцатиперстную кишку и выходят из организма
вместе с калом, которому они придают
характерный цвет. Но наиболее важная выделительная
функция печени состоит в переработке отходов
азотистого обмена, образующихся при дезаминировании
избыточных аминокислот (разд. 18.5.2).
19.2. Азотистые экскреты
Азотистые продукты, подлежащие экскреции,
образуются при расщеплении белков и нуклеиновых
кислот. Непосредственный продукт дезаминирова-
ния белков - аммиак (суть этой реакции описана в
разд. 18.5.2). Аммиак может выделяться прямо в
Рис. 19.1. Химическая структура трех важнейших
азотистых экскретов.
окружающую среду или превращаться в такие
продукты, как мочевина и мочевая кислота,
различающиеся по растворимости и токсичности
(рис. 19.1). Конкретно природа выделяемого
продукта определяется метаболическими
возможностями организма (наличием соответствующих
ферментов), доступностью для организма воды (т. е. средой
его обитания) и степенью контроля над ее потерей
организмом.
В соответствии с природой главного азотистого
экскрета всех животных можно разделить на три
основные группы. Как видно из табл. 19.2,
существует определенная корреляция между природой
выводимого вещества и средой обитания животного на
стадии эмбриона, личинки и взрослой особи. Эта
корреляция может быть суммирована следующим
образом:
Аммиак
Мочевина
Мочевая кислота
Экскрет
Аммиак
Мочевина
Мочевая
кислота
Тип экскреции
Аммониотелия
Уреотелия
Урикотелия
Животные
Водные
Водные/наземные
Наземные
19.2.1. Аммиак
Аммиак - чрезвычайно растворимое
низкомолекулярное соединение (мол. масса 17), легко
диффундирующее в воде. Он токсичен для животных и не
должен накапливаться в организме.
Млекопитающие очень чувствительны к аммиаку и не могут
переносить его в концентрациях выше 0,02 мг на
100 мл крови. У большинства водных организмов
(от простейших до амфибий) аммиак благодаря
своей высокой растворимости быстро выделяется в
виде ионов аммония (NH4), не успевая достигнуть
токсичной концентрации.
19.2.2. Мочевина
Мочевина образуется в печени позвоночных в
результате взаимодействия аммиака,
отщепляющегося при дезаминировании, и углекислоты,
образующейся в процессе дыхания. Синтез мочевины
происходит в ходе циклического процесса, называемого
орнитиновым циклом. Она менее растворима в воде
и менее токсична, чем аммиак, и является главным
азотистым экскретом у пластиножаберных и
некоторых костистых рыб, взрослых амфибий и
млекопитающих. Нормальное содержание мочевины в
крови млекопитающих составляет 2,5 -6,0 ммоль/л.
Превращение головастика в лягушку
сопровождается переходом от экскреции аммиака (аммонио-
телии) к экскреции мочевины (уреотелии).
19.2.3. Мочевая кислота
Мочевая кислота и ее соли - идеальные азотистые
экскреты для наземных животных, а для
организмов, откладывающих клейдоические яйца
(заключенные в оболочку, разд. 19.4.8), их образование
является обязательным, так как они сочетают
высокое содержание азота с низкой токсичностью. Эти
вещества могут храниться в клетках, тканях и
органах, не оказывая никаких токсических или вредных
осморегуляторных эффектов, и для их экскреции
требуется минимальное количество воды. При
увеличении содержания мочевой кислоты в тканях она
Таблица 19.2. Взаимоотношения между азотистым
экскретом и стадиями жизненного цикла у различных групп
животных
Группа животных
Простейшие
Губки
К ишечнопо л ост-
ные
Плоские черви
Водные
ракообразные
Наземные
насекомые
Брюхоногие
моллюски
Иглокожие
Пластиножабер-
ные
Пресноводные
костистые
рыбы
Морские
костистые рыбы
Головастики
бесхвостых
амфибий
Взрослые
бесхвостые амфибии
Пресмыкающиеся
Птицы
Млекопитающие
Экскрет
Аммиак
— » —
— » —
— » —
— » —
Мочевая
кислота
— » —
Аммиак
Мочевина
Аммиак
Мочевина, три-
метил-
амин-
оксид
Аммиак
Аммиак
Мочевая
кислота
— » —
Мочевина
Среда обитания
эмбриона
и IU
личинки
Водная
— » —
— » —
— » —
— » —
Клейдои-
ческое
яйцо
— » —
Водная
— » —
— » —
— » —
— » —
Клейдои-
ческое
яйцо
— » —
Водная
взрослого
животного
Водная
— » —
— » —
— » —
— » —
Наземная
— » —
Водная
— » —
— » —
— » —
Водная и
наземная
Наземная
— » —
— » —
выпадает в виде твердого осадка. Более подробно
биохимический механизм образования мочевой
кислоты описан в разд. 19.4.5. В виде мочевой кислоты
и урата аммония выводятся азотистые продукты
обмена у насекомых, ящериц, змей и птиц. Человек,
обезьяны и (в связи с особым дефектом почек)
далматский дог также выделяют небольшие
количества мочевой кислоты, однако у них она
образуется только при расщеплении нуклеиновых
кислот, но не белков. В норме содержание мочевой
кислоты в крови человека составляет 3 мг на
100 мл, а экскреция ее с мочой-около 1 г в сутки.
19.2.4. Другие азотистые экскреты
Помимо уже описанных продуктов у некоторых
морских рыб при расщеплении избыточного белка
образуется еще одно вещество - триметиламин-
оксид.
Триметиламиноксид
Синтез этого вещества происходит путем
присоединения метальных групп к аммиаку,
отщепляющемуся при дезаминировании, и последующего
окисления полученного продукта. Триметиламиноксид
придает рыбе ее характерный запах.
Единственным, кроме белков, источником
азотистых отходов являются нуклеиновые кислоты. Такие
продукты питания, как дрожжи, печень и почки,
содержат большое количество мелких клеток и
поэтому богаты клеточными ядрами. При
расщеплении компонентов этих пищевых продуктов
образуются значительные количества оснований
нуклеиновых кислот-пуринов и пиримидинов. У
паукообразных и некоторых млекопитающих пуриновые
основания - аденин и гуанин - выделяются в
неизмененном виде, но у других организмов они
расщепляются с образованием мочевой кислоты, как будет
описано дальше.
Аденин и гуанин сходны по строению с мочевой
кислотой, тогда как структура пиримидиновых
оснований - цитозина, тимина и урацила -
обусловливает их расщепление с образованием аммиака и
аминокислоты. Аминокислота подвергается затем
дезаминированию, и конечный азотистый продукт
выводится в форме, характерной для данного
организма. У насекомых, наземных рептилий, птиц,
человека, обезьян и далматского дога пурины
выделяются в виде мочевой кислоты. Однако у
большинства млекопитающих в печени есть фермент ури-
каза, катализирующий превращение мочевой
кислоты в аллантоин, который и выделяется. Двукрылые
насекомые также выделяют аллантоин. У
некоторых костистых рыб аллантоин окисляется в ал-
лантоевую кислоту, и последняя экскретируется.
К числу других азотистых продуктов обмена от-
носятся креатин и его производное креатинин.
Креатин образуется в печени позвоночных из
аминокислот аргинина, метионина и глицина. Около 2%
всего креатина, имеющегося в организме, за сутки
выводится в виде креатинина. Часть креатина фос-
форилируется в мышцах с образованием фосфо-
креатина (фосфагена), который служит запасным
источником энергии для регенерации АТФ путем
фосфорилирования АДФ (разд. 1.4.8).
Наконец, еще одна группа азотистых отходов
образуется в печени в процессах детоксикации.
Чаще всего при этом образуется гиппуровая
кислота, впервые обнаруженная в моче лошади (греч.
hippos-лошадь), которая синтезируется путем
связывания бензойной кислоты (поступающей с
растительной пищей) с аминокислотой глицином. При
помощи подобных реакций обезвреживаются и
многие другие фенольные соединения.
19.3. Экскреция азотистых
продуктов и
осморегуляция
Основной источник конечных продуктов азотистого
обмена-дезаминирование избытка аминокислот. В
результате дезаминирования образуется аммиак,
который чрезвычайно токсичен и подлежит
выведению из организма. Будучи растворимым, аммиак
может быть быстро и безопасно удален из
организма, если он растворится в достаточном объеме
воды. Для организмов, живущих в воде, это не
составляет никакой проблемы, но это относится
только к пресноводным организмам. Морские
животные сталкиваются с острой проблемой
получения воды, а наземные-с проблемой ее сохранения,
поэтому для выведения азотистых отходов они
могут использовать очень мало воды. Из табл. 19.2
видно, что у живущих в этих условиях организмов
выработались иные пути экскреции азота. Они
связаны с выработкой ряда анатомических,
биохимических, физиологических и поведенческих
механизмов, связанных с выведением азотистых веществ и
одновременным поддержанием стационарного
состава жидкостей тела. Так как проблемы экскреции и
осморегуляции здесь связаны между собой, эти два
процесса будут рассматриваться вместе.
Осморегуляция-это гомеостатический механизм,
с помощью которого у растений и животных
поддерживается постоянство концентрации
растворенных веществ в жидкостях внутренней среды.
Жидкости тела делятся на внутриклеточные и
внеклеточные. Например, у растений та жидкость, которая
находится в вакуолях клеток, является
внутриклеточной, а та, которая окружает клетки коры стебля
или корней,-внеклеточная. У многоклеточных
животных внутриклеточная жидкость распределена в
клетках довольно равномерно, а внеклеточная
представлена плазмой крови и тканевой жидкостью.
Последняя подразделяется у позвоночных на
собственно тканевую жидкость и лимфу. Для нормальной
метаболической активности клеток очень важно,
чтобы состав этих жидкостей оставался
постоянным. Природа внутри- и внеклеточных жидкостей и
регуляция их состава будут рассмотрены в
разд. 19.3.2.
Слово «осморегуляция» означает не просто
регуляцию водного баланса в организме, а регуляцию
состава жидких сред организма, которые во всех
случаях представляют собой растворы различной
сложности. Детали физических и химических
свойств растворов описаны в Приложении 1.4.
В живых системах даже в растворах с одинаковым
осмотическим давлением (т. е. изотоничных друг
другу) происходит перемещение молекул растворенных
веществ, если их относительные концентрации
различны. Перемещение молекул воды между двумя
растворами определяется относительным
осмотическим давлением этих растворов. Молекулы
растворенных веществ проходят через полупроницаемые
мембраны в направлении, зависящем от их
относительной концентрации по обе стороны мембраны и
от свойств самой мембраны. Это передвижение
может быть пассивным или активным. В первом
случае молекулы перемещаются путем диффузии
«вниз» по концентрационному градиенту, т.е. из
раствора с более высокой концентрацией в раствор с
более низкой концентрацией. Во втором случае
перемещение происходит против
концентрационного градиента и осуществляется специальными
механизмами, находящимися в мембране. Все
мембраны организма, включая плазмалемму (клеточную,
или плазматическую, мембрану), слои цитоплазмы,
поверхности тела и легких, способны действовать
как полупроницаемые мембраны, через которые
могут проходить вода и растворенные вещества.
Концентрацию осмотически активных веществ в
растворе мы будем называть в этой главе
осмотическим потенциалом или осмолярностью и выражать
в миллиосмолях на 1 л или на 1 кг воды.
В биологическом контексте при обсуждении
осморегуляции концентрацию растворенных веществ
можно выражать как депрессию точки замерзания
раствора. Чистая вода замерзает при 00C, но при
добавлении растворимых веществ точка замерзания
падает ниже 00C, и новая температура замерзания
показывает концентрацию растворенных веществ в
растворе. Например, осмолярность морской воды
составляет 1000 мосмоль/л, а ее депрессия точки
замерзания равна —1,7 0C.
19.3.1. Механизмы осморегуляции
Осмотические взаимоотношения между двумя
растворами, разделенными полупроницаемой
мембраной, можно выразить в виде тоничности или осмоляр-
ности. Между этими двумя понятиями есть важные
различия в отношении обстоятельств, при которых
они употребляются; однако для простоты мы будем
пользоваться словом «тоничность»: будем называть
раствор либо гипотоническим, либо
гипертоническим, либо изотоническим по отношению к какому-
то другому раствору.
У пресноводных организмов жидкости тела
обычно гипертоничны по отношению к их водному
окружению. Многие морские организмы, в том числе
позвоночные, гипотоничны по отношению к
морской воде, но многие морские беспозвоночные изо-
тоничны ей.
Если в водной среде повышается концентрация
растворенных веществ, животные отвечают на это
одним из двух способов. У животных, не способных
регулировать осмотическую концентрацию
жидкостей тела (осмоконформеров), она изменяется так же,
как в окружающей среде, тогда как у животных,
способных к осморегуляции (осморегуляторов), она
остается прежней при всех изменениях среды.
Применяя терминологию, связанную с гомеостазом,
первых все чаще называют теперь пойкилосмотичес-
кими, а вторых-гомойосмотическими животными,
используя те же приставки, что и в случае
терморегуляции.
19.3.2. Осморегуляция у растений
Ткани растений содержат больше воды, чем ткани
животных, и функционирование растительной
клетки, так же как и всего растения в целом, зависит от
постоянства содержания воды. У растений не
существует тех проблем осморегуляции, с которыми
встречаются животные, и растения можно
рассматривать просто в связи с местообитанием,
подразделив их на группы, описанные ниже.
Гидрофиты
Пресноводные растения типа элодеи (Elodea
canadensis), тысячелистника (Myriophyllum) и водяной
лилии (Nymphaea) относятся к гидрофитам и
меньше сталкиваются с проблемами осморегуляции, чем
растения всех остальных типов. В пресной воде
растения окружены гипотонической средой, и в сок,
содержащийся в вакуолях, вода поступает путем
осмоса. Она свободно проходит через клеточную
стенку и полупроницаемые плазматическую и ва-
куолярную мембраны. Когда за счет поглощения
воды объем вакуоли увеличивается, в ней создается
тургорное давление. Клетка становится тургесцент-
ной, и достигается равновесное состояние, когда
водный потенциал в ней повышается до той же
величины, что и в окружающей воде (почти
нулевой), и поступление воды прекращается (см. разд.
14.1.5). Это называют механической осморегуляцией.
Пресноводная водоросль нителла (Nitella clavata)
содержит чрезвычайно концентрированный сок, и
вся проблема осморегуляции у нее сводится к
поддержанию ионного состава этого сока, что
осуществляется путем активного поглощения ионов из
окружающей воды.
Процент от сырого веса
Fucus spiralis
Ascophyllum nodosum
Fucus vesicubsus
Fucus serratus
Время, ч
Рис. 19.2. Скорость отдачи воды на воздухе
четырьмя видами водорослей, растущих на
морском побережье.
Галофиты
Единственными растениями, способными
существовать в морской воде, являются
водоросли-основной источник растительной пищи на морском
побережье. Распределение видов водорослей по мере
удаления от берега определяется многими
факторами, в том числе устойчивостью к воздействию волн
и к обезвоживанию при отливе, а также составом
пигментов, участвующих в фотосинтезе. Во всех
случаях эти виды способны переносить повышение
солености, и главная задача осморегуляции
сводится у них к предотвращению потери воды в
результате испарения. Самую верхнюю зону
морской растительности на укрытых скалистых берегах
Британских островов занимает взморник
бороздчатый (Pelvetia canaliculata). Этой водоросли
помогают выживать толстые клеточные стенки, густой
слой слизи и ножка в форме желобка.
На рис. 19.2 показана скорость потери воды и
степень переносимости обезвоживания у четырех
видов распространенных у Британских островов
морских водорослей, которые располагаются по
зонам в зависимости от их способности удерживать
воду во время отлива.
Строго говоря, галофитами называются растения,
произрастающие на сильно засоленных почвах,
например в эстуариях рек и в соленых маршах около
морей, где соленость постоянно меняется и может
превышать соленость морской воды. Стебли этих
растений не всегда находятся в условиях высокой
солености, но их корневая система вынуждена
переносить повышенную соленость песка и ила, которая
создается во время отлива в жаркие ветреные дни.
Считалось, что эти растения должны выдерживать
периоды «физиологической засухи», когда вода
бывает недоступна для тканей из-за гипертоничности
среды, окружающей корни. Однако дело,
по-видимому, обстоит не так, поскольку большая скорость
транспирации и высокое осмотическое давление в
клетках корневой системы обеспечивают
поглощение воды из почвы. В эстуариях рек и соленых
маршах растет галофит спартина (Spartina). Она
имеет обширную систему корневищ, с помощью
которой распространяется, и придаточные корни,
обеспечивающие ей закрепление в почве и
поглощение воды и солей. Другие галофиты, растущие в
эстуариях рек и маршах,-более мелкие растения,
запасающие воду, когда она имеется в изобилии.
Наиболее известные среди них-солерос (Salicornia),
сведа (Suaeda maritima) и морской портулак (HaIi-
mione). Некоторые виды, например глаукс морской
и спартина, способны регулировать содержание в
своих тканях солей, выделяя их через железы,
расположенные по краям листьев.
Мезофиты
Большинство покрытосеменных растений относится
к мезофитам и растет в местах, в достаточной мере
обеспеченных водой. Эти растения сталкиваются с
проблемой потери воды путем испарения со всех
надземных частей и имеют ряд морфологических
(ксероморфизм) и физиологических особенностей,
помогающих им уменьшать эту потерю. Из таких
особенностей следует упомянуть наличие кутикулы,
защищенные устьица, диаметр которых может регу-
Таблица 19.3. Способы преодоления недостатка воды у
различных растений (см. главы 14 и 16)
Основной
принцип
Уменьшение
скорости
транспирации
Запасание
воды
Поглощение
воды
Приспособления
Восковая кутикула;
мало устьиц
Погружение устьица
Устьица открыты
ночью и закрыты
днем; поверхность
покрыта мелкими
волосками
Курчавость листьев
Мясистые сочные
листья
Мясистый сочный
стебель
Мясистые подземные
части
Глубокая корневая
система ниже уровня
грунтовых вод
Поверхностная
корневая система,
поглощающая влагу
поверхностного слоя
Примеры
Опунция
(Opuntia)
Сосна (Pinus)
Хрустальная
травка (Me-
sembryanthe-
тит)
Песколюб
(Ammophila)
Бриофиллум
{Bryophyl-
lum)
Kleinia
Raphionacme
Акация,
олеандр
Кактусы
лироваться, разнообразную форму листьев,
сбрасывание листвы и экологическое распространение,
связанное с устойчивостью к обезвоживанию. Более
детальное описание и примеры этих механизмов
можно найти в разд. 14.3.
Ксерофиты
Ксерофитами называют растения сухих
местообитаний, способные переносить продолжительную
засуху. Они составляют типичную флору пустынь и
полупустынь и весьма обычны на морском
побережье и в песчаных дюнах. Некоторые из этих
растений переживают периоды экстремальных
условий в виде семян и спор, которые после
выпадения дождя могут прорастать; новые растения
иногда за четыре недели успевают вырасти, зацвести и
дать семена (как, например, эшшольция
калифорнийская), которые пребывают в состоянии покоя до
следующего дождливого периода.
С другой стороны, есть растения, обладающие
разнообразными анатомическими (ксероморфизм) и
физиологическими приспособлениями,
позволяющими им вегетировать во время сильной засухи.
Большинство ксерофитов Британских островов
растет вдоль береговой линии и на песчаных дюнах,
как, например, солянка (Salsola) и морская песчанка
(Нопкепуа), растущие на невысоких песчаных
холмах морского побережья. Песчаный пырей (Лдгору-
гоп) и песколюб (Ammophila) -доминирующие
растения на молодых дюнах, они обладают обширной
системой корневищ с придаточными корнями,
позволяющими добывать воду из водоносного слоя
ниже песка. Песчаный пырей способен переносить
концентрацию соли в песке, в 20 раз большую, чем в
морской воде. Оба растения имеют большое
значение как пионеры заселения песчаных дюн.
Ксерофиты, растущие в пустынях, обладают
рядом адаптации для уменьшения потери воды и для
ее запасания. Некоторые из этих адаптации
приведены в таблице 19.3.
19.3.3. Процессы, связанные с
экскрецией и осморегуляцией
Ультрафильтрация - процесс удаления из раствора
молекул растворителя и растворенных веществ в
соответствии с их способностью проходить через
поры фильтра. У большинства животных роль
фильтра выполняет слой, разделяющий циркулятор-
ную систему и орган осморегуляции или экскреции,
а движущей силой служит гидростатическое
давление крови. Раствор, образующийся при фильтрации,
называется фильтратом. При фильтрации крови из
нее удаляется большинство ее компонентов, и
остаются в ней только очень крупные молекулы, такие
как белки, и клетки, например эритроциты.
Избирательная реабсорбция представляет собой
избирательное обратное всасывание из фильтрата
молекул растворенных веществ и воды в нужных
организму количествах. Отходы метаболизма не
реабсорбируются. Растворенные вещества и вода
тоже не всасываются обратно, если это может
привести к избыточному их содержанию в
жидкостях тела, нарушающему нормальное стационарное
состояние. Вначале реабсорбция происходит путем
пассивной диффузии, продолжающейся до тех пор,
пока не уравняются концентрации, после чего
дальнейшая реабсорбция происходит путем активного
транспорта. По мере реабсорбции растворенных
веществ фильтрат становится все более
разбавленным и гитютоничным по отношению к жидкостям
тела; поэтому вслед за ионами будут двигаться
путем осмоса молекулы воды, и в конечном счете
образуется фильтрат, изотоничный жидкостям тела.
В том участке осморегуляторно-экскреторного
органа, который непроницаем для воды, фильтрат
опять становится гипотоничным в результате
поглощения из него ионов.
Секреция-это процесс активного переноса
растворенных веществ из жидкостей тела в фильтрат
или непосредственно в окружающую среду. Таким
образом, секреция действует в направлении,
противоположном реабсорбции. Этот процесс
приводит к дальнейшему повышению осмотического
давления фильтрата и к снижению осмотического
давления жидкостей тела.
Конечный результат действия этих трех
механизмов (ультрафильтрации, избирательной
реабсорбции и секреции) является гомеостатическим, так как
он поддерживает постоянство состава жидкостей
тела.
Существует большое многообразие осморегуля-
торных или экскреторных органов и органелл,
различных по своей локализации и строению, но в
основе функциональной активности всех этих
структур лежит один или несколько описанных выше
механизмов. Некоторые из этих структур
относительно мало специализированы и имеют ряд общих
черт в филогенетическом ряду организмов
(сократительные вакуоли, нефридии, почки), тогда как
другие относительно специализированы (жабры,
ректальные железы, солевые железы).
19.3.4. Влияние окружающей среды на
экскрецию и осморегуляцию
Для различных групп организмов характер
окружающей среды создает определенные проблемы осмо-
регуляции. Многие водные организмы, живущие в
гипертонической среде, теряют воду путем осмоса
и поглощают растворенные вещества путем
диффузии. Потеря воды возмещается разными способами,
включая питье и потребление пищи, но при этом
повышается также концентрация в организме солей
и возникает необходимость удаления их избытка
путем активного транспорта Организмы, живущие
в гипотонической среде, напротив, поглощают воду
путем осмоса и теряют растворенные вещества за
счет диффузии. Уменьшить этот обмен нередко
помогает организмам непроницаемый наружный
покров, а потерю солей приходится возмещать
путем их активного поглощения из окружающей
среды.
Все наземные организмы сталкиваются с
проблемой потери воды и растворенных веществ из
жидкостей тела в окружающую среду. Постоянство
состава внутриклеточной жидкости поддерживается
у этих организмов путем регуляции состава
внеклеточной жидкости специализированными осморе-
гуляторно-экскреторными органами, такими как
мальпигиевы сосуды и почки. Количество
получаемых и выделяемых молекул воды и ионов должно
быть сбалансировано. Проблемы водного баланса
будут рассмотрены в разд. 19.4.
Адаптация к сильнозасушливым
условиям
Кенгуровая крыса (Dipodomys) выделяется среди
млекопитающих своей удивительной способностью
переносить засушливые условия, существующие в
пустынях Северной Америки. Она прекрасно
чувствует себя в этих условиях благодаря уникальному
сочетанию морфологических, физиологических и
поведенческих адаптации. Потеря воды с выдыхаемым
воздухом снижается у нее за счет того, что
выдыхаемый воздух имеет более низкую температуру,
чем внутренние области тела. При вдохе воздух
забирает тепло в носовых ходах и охлаждает их. Во
время выдоха водяные пары, содержащиеся в
теплом воздухе, конденсируются на слизистой носа, и
таким образом вода задерживается. Питается
кенгуровая крыса сухими семенами и другой сухой
растительной пищей и совсем не пьет. Единственные
источники воды для нее - это вода, образующаяся в
организме в процессе тканевого дыхания, и те очень
малые количества воды, которые содержатся в
пище. Шмидт-Ниельсен в своих классических
исследованиях, суммированных в табл. 19.4, измерил
водный баланс у кенгуровой крысы, весящей 35 г и
съедающей 100 г ячменя, в экспериментальных
условиях (при 25 0C и относительной влажности 20%).
Единственным источником воды для нее в течение
исследуемого периода служили зерна ячменя.
Таблица 19.4. Водный обмен у кенгуровой крысы в
экспериментальных условиях. Животное получало только ту воду,
которая содержалась в пище
Источники воды
Тканевое дыхание
Пища
Всего
Количество, мл
54,0
6,0
60,0
Потери воды
С мочой
С фекалиями
Испарение
Всего
Количество, мл
13,5
2,6
43,9
60,0
Наконец, в природных условиях кенгуровая крыса
избегает потери воды за счет ее испарения, проводя
много времени в относительно влажной атмосфере
подземной норы.
Другой поразительный пример сохранения
воды водный баланс верблюда, физиологические
адаптации которого были рассмотрены в разд.
18.4.7.
19.4. Филогенетический обзор
органов и процессов,
участвующих в выделении
азотистых веществ и в
осморегуляции
При рассмотрении этого вопроса нужно иметь в
виду следующие моменты:
1) на характер выводимого продукта и на процесс
осморегуляции влияет окружающая среда;
2) в некоторых группах организмов имеются виды,
приспособленные к жизни в разных условиях;
3) некоторые организмы способны переносить
значительные колебания внешних условий.
Мембрана
вакуоли
Пузырьки
19.4.1. Простейшие
Простейшие обитают в пресной и морской воде, а
также в жидкостях тела других организмов.
Внутриклеточная жидкость у простейшего отделена от
внешней среды только полупроницаемой
плазматической мембраной. Выделение CO2 и аммиака
происходит путем простой диффузии через всю
поверхность клетки. Удалению отходов метаболизма
способствует высокое отношение поверхности к
объему
Все пресноводные виды простейших гипертонич-
ны по отношению к окружающей среде и обладают
специальными осморегуляторными органеллами -
сократительными вакуолями. Эти органе л лы
необходимы для удаления воды, поступающей в
клетку путем осмоса через клеточную мембрану из
гипотонической окружающей среды, и таким
образом участвуют в регуляции объема клетки,
предотвращая его увеличение. Строение и локализация
сократительной вакуоли сильно варьируют. У амёбы
Amoeba proteus такая вакуоль может образовываться
в любом участке клетки и освобождать
заключенную в ней жидкость во внешнюю среду в любом
участке поверхности тела (рис. 19.3). У инфузории
Paramecium aurelia имеются две сократительные
вакуоли, расположенные в определенных местах
(рис. 19.4). Однако способ функционирования
сократительных вакуолей, по-видимому, одинаков у всех
видов и состоит в том, что вода из цитоплазмы
поступает в мелкие пузырьки, которые сливаются с
сократительной вакуолью и передают ей свое
содержимое. Вокруг сократительной вакуоли
группируются митохондрии, которые, по-видимому,
Сократительная
вакуоль
Митохондрия
Сократи
тельная -
вакуоль
Мембрана
вакуоли
Пузырек
Рис. 19.3. А. Электронная микрофотография
сократительной вакуоли амебы, Б, Схема
функционирования вакуоли. Вода секретируется в мельчайшие
пузырьки, которые сливаются с мембраной вакуоли и
опорожняют в вакуоль содержащуюся в них воду.
доставляют энергию для выполняемой ею
осмотической работы.
Изучение функционирования сократительной
вакуоли у гигантской амебы (Chaos chaos) показывает,
что поступление в клетку воды путем осмоса,
рассчитанное по данным об осмотическом давлении
внутриклеточной жидкости, соответствует объему
воды, выводимой вакуолью. Содержимое
сократительной вакуоли гипотонично по отношению к
внутриклеточной жидкости, но гипертонично по
отношению к окружающей воде. Предложено
несколько гипотез для объяснения образования вакуоляр-
ной жидкости, и одно из вероятных объяснений
показано на рис. 19.5.
У многих морских корненожек сократительная
вакуоль отсутствует, так как их внутриклеточная
жидкость изотонична морской воде. Это позволяет
предполагать, что главная функция такой
вакуоли - осморегуляция.
19.4.2. Кишечнополостные
Кишечнополостные, по-видимому, не имеют
специальных органов или органелл для выделения и
осморегуляции, и механизм осморегуляции у них
неизвестен. Основные токсичные продукты обмена у
HHx-CO2 и аммиак, которые путем диффузии
выходят из клеток прямо в воду внеклеточной среды.
19.4.3. Плоские черви
У плоских червей основная масса отходов
метаболизма переходит в сильно разветвленный
кишечник и выводится из организма через ротовое
отверстие. Однако некоторая их часть поступает в
систему канальцев, выполняющих и
выделительную, и осморегуляторную функцию. Эти канальцы
представляют собой примитивный тип нефридиев и
называются протонефриднями. Главная функция
протонефридиев - осморегуляция. Протонефридии
характерны в основном для животных, лишенных
вторичной полости тела (целома), таких как плоские
черви и коловратки. У планарий имеется пара
протонефридиев, которые тянутся вдоль всего тела
и открываются наружу через многочисленные поры
(рис. 19.6, А). Отдельный протонефридии состоит из
множества ветвящихся канальцев, заканчивающихся
крупными клетками с просветом внутри. В этот
просвет выступают реснички; если имеется только
одна ресничка, клетку называют соленоцитом, а если
целый пучок ресничек, то пламенной клеткой (рис.
19.6,ZT). Реснички пламенных клеток колеблются, и
это движение напоминает колебание пламени свечи,
отсюда и их название. Биение ресничек способствует
постоянному току жидкости в канальцы и через
протоки-к выделительным порам. Находящаяся в
пламенных клетках жидкость состоит из воды и
конечных продуктов обмена, образующихся в
тканях. Предполагается, что одни из этих продуктов
Рис. 19.4. Микрофотография фиксированной
парамеции. Видны сократительные вакуоли. 1
-передняя сократительная вакуоль с канальцами;
2 - каналец задней вакуоли.
Пузырьки содержат
воду и изотоничны
по отношению
к цитоплазме
Ионы Na+
накачиваются в пузырьки в
обмен на ионы К*,
причем приток Na+
меньше, чем отток
К* (см. разд. 7 2 2
о Na+/K+= насосе)
Пузырек
Теперь пузырьки
в связи с потерей
K+ и меньшим
поглощением Na*
становятся гиперто-
ничными, но вода
удерживается в них,
так как мембрана
для нее непроницаема
Пузырек сливается
с сократительной
вакуолью, и его
содержимое
переходит в вакуоль.
Ионы Na+ выводят,
ся из цитоплазмы в
жидкость вакуоли
и замещаются
новыми благодаря их ак.
тивному поглощению
из окружающей среды
Сократительная
вакуоль
Рис. 19.5. Схема предполагаемого механизма поглощения воды сократительной вакуолью.
секретируются в канальцы путем активного
транспорта, а другие-путем ультрафильтрации через
цитоплазму пламенных клеток. Вода поступает в
просвет пламенных клеток путем осмоса.
Пламенные клетки имеются также у некоторых кольчатых
червей. Соленоциты характерны в основном для
головохордовых, представителем которых является
ланцетник.
19.4.4. Кольчатые черви
У кольчатых червей органами выделения и осмо-
регуляции, поддерживающими постоянный
химический состав жидкостей тела, служат метанефридии
(или просто нефридии). У представителей разных
классов круглых червей - у полихет, олигохет и
пиявок-строение и расположение нефридиев
варьируют, но основной структурный план у всех
одинаков. Нефридии представляют собой неразветвлен-
ные канальцы, соединяющие целом с внешней
средой. У некоторых форм, например у пескожила
(Arenicola), нефридий образуется путем слияния
эктодермального канальца, открывающегося порой
наружу, с мезодермальным канальцем,
открывающимся в целом. У дождевого червя (Lumbricus)
имеется пара нефридиев в каждом сегменте (кроме
трех первых и последнего), а у полихет и пиявок их
меньше.
Нефридий состоит из покрытой ресничками
воронки (нефростома), которая соединена длинным
канальцем, снабженным ресничками и мышечными
волокнами, с мочевым пузырем, где жидкость
накапливается и откуда выводится через наружное
отверстие - нефридиопор. Выделяемая жидкость
называется мочой и образуется путем
ультрафильтрации, избирательной реабсорбции и активной
секреции. Целомическая жидкость, содержащая и нужные
организму вещества, и отходы, загоняется реснич-
- Пламенная
клетка
- Выделительный
каналец
Выделительная
пора
Ядро
Цитоплазма
Пучок
ресничек
("пламя")
Нефриди-
альный
проток
Выделительный
каналец
Рис. 19.6. Строение выделительной системы
плоского червя Planaria, состоящей из
ηротонефридиев. А. Общее схематическое изображение.
Б. Пламенная клетка.
ками в нефростом (рис. 19.7) и, движимая биением
ресничек и сокращением мускулатуры, проходит по
узкому длинному канальцу, в котором еще не
происходит никакой реабсорбции ценных веществ.
Далее следуют короткий средний и более длинный
широкий канальцы, клетки которых всасывают
обратно нужные организму вещества и активно секре-
тируют их в кровеносные капилляры канальца, а из
капилляров в каналец дополнительно
секретируются отходы. По мере обратного всасывания нужных
веществ моча становится менее концентрированной,
но в ней растет содержание отходов метаболизма.
Эта гипотоничная по отношению к целомической
жидкости моча выводится через нефридиопор.
Способность к образованию гипотоничной мочи
Вода, глюкоза,
аминокислоты,
белки, соли,
аммиак,
мочевина
Аммиак,
мочевина
Глюкоза, белки,
аминокислоты,
некоторые
соли
Аммиак,
мочевина
Во
да
Соли
Вода,
аммиак,
мочевина
Концентрация мочевины,
% от концентрации в крови
Нефростом
Узкий
каналец
Средний
каналец
Широкий
каналец
Мышечный
каналец
Мочевой пузырь и
нефридиопор
Рис. 19.7. Стадии образования мочи у дождевого червя. Толстыми стрелками показаны участки, в
которых происходит активная секреция веществ. Известно, что белки имеются в нефростоме, но
отсутствуют в моче, выделяемой через нефридиопор; это означает, что на какой-то стадии они
извлекаются из мочи, но механизм их поглощения пока не ясен. (По Ramsey.)
указывает на то, что нефридий выполняет и осмо-
регуляторную функцию. Как показали
исследования, в отношении осморегуляции дождевой червь
сходен с пресноводными организмами, так как при
помещении его в солевые растворы он
поддерживает гипертоничность жидкостей своего тела по
отношению к окружающей среде и вместе с тем
выделяет гипотоничную мочу. Хотя дождевой
червь - казалось бы, сухопутное животное, он
находится в прямом контакте с водяной пленкой,
покрывающей частицы почвы в стенках подземного
хода; поэтому его можно рассматривать как
пресноводный организм. Осморегуляция у типичных
морских полихет вроде Nereis diversicolor описана в
разд. 19.4.6.
19.4.5. Членистоногие
Членистоногие приспособились к жизни в самых
разнообразных местообитаниях - от морских до
полностью наземных. Не удивительно поэтому, что
механизмы выделения и осморегуляции у
представителей этого типа весьма разнообразны. Для
иллюстрации этого разнообразия мы рассмотрим
адаптации насекомых к наземным условиям
существования и ракообразных - к жизни в морской воде, в
эстуариях рек и в пресной воде.
Одна из главных проблем жизни на
суше-предотвращение потери воды. У насекомых имеется
почти непроницаемая кутикула, уменьшающая
испарение воды с поверхности тела, и дыхальца,
уменьшающие потерю воды через систему
газообмена-трахеи и трахеолы. Прочная кутикула
состоит из хитиновой экзо- и эндокутикулы, покрытой
тонким водопроницаемым слоем эпикутикулой
толщиной 0,3 мкм (см. разд. 4.10.1) Потере воды
путем испарения препятствуют водонепроницаемые
свойства эпикутикулы, образованной
упорядоченным мономолекулярным слоем липидов, покрытым
несколькими слоями нерегулярно расположенных
липидных молекул. Если эти восковые или липид-
ные слои стираются острыми частицами, например
песком или кремнеземом, скорость испарения
возрастает и насекомому грозит обезвоживание.
Интересно, что по мере повышения окружающей
температуры скорость испарения растет постепенно, пока
температура не достигнет определенной величины,
после чего испарение быстро усиливается. Эту
температуру называют «критической». Построив
график зависимости испарения от температуры на
поверхности тела (рис. 19.8), эту критическую
температуру можно выявить более четко; она соответствует
той температуре, при которой нарушается
упорядоченное строение воскового монослоя.
Некоторые насекомые, питающиеся сухой пищей
и живущие в очень сухих местах, способны
поглощать влагу из воздуха, если относительная влаж-
Потеря воды, мг/ч
Температура, ° С
Рис. 19.8. Зависимость потери воды с
поверхности кутикулы таракана от температуры воздуха
(треугольники) и от температуры самой
кутикулы (кружочки). Видно, что отдача воды резко
возрастает при температуре кутикулы
около 29° С.
Средняя кишка
Прямая
кишка
Мальпигиева
трубочка
Рис. 19.9. Схема взаимного расположения маль-
пигиевых трубочек и пищеварительного тракта у
клопа Rhodnius prolixus.
ность воздуха превышает определенную величину,
например 90% для мучного червя (личинки жука
Tenebrio) и 70% для домового клеща (Dermatophagoi-
des).
У насекомых проблема предотвращения потери
воды при выведении экскретов решается с помощью
специализированных органов выделения,
называемых мальпигиевыми трубочками (или сосудами),
которые образуют и выделяют почти нерастворимый
продукт азотистого обмена - мочевую кислоту. Маль-
пигиевы трубочки представляют собой слепо
оканчивающиеся канальцы, расположенные в полости
брюшка и омываемые гемолимфой. Число трубочек
у насекомых варьирует: у некоторых всего одна
пара, у других может быть несколько сотен;
кровососущий клоп Rhodnius имеет четыре трубочки. Все
они открываются в заднюю кишку в месте ее
перехода в среднюю кишку и могут быть длинными и
тонкими или короткими и компактными (рис. 19.9).
Уиглсуорс, изучавший функцию мальпигиевых
трубочек в процессе образования мочевой кислоты у
Rhodnius, предполагает следующий механизм.
Трубочка состоит из двух разных по гистологическому
строению отделов-верхнего сегмента (дисталь-
ного по отношению к кишке), построенного из
одного слоя клеток и содержащего прозрачный
раствор, и нижнего сегмента. Клетки нижнего
сегмента имеют на внутренней поверхности
микроворсинки, на которых из раствора осаждаются
кристаллы мочевой кислоты (рис. 19.10). Содержимое
трубочек выводится в заднюю (прямую) кишку, где
смешивается с непереваренными частицами пищи.
Ректальные железы, расположенные в стенках
прямой кишки, осуществляют обратное всасывание
воды из экскрементов и из суспензии мочевой
кислоты, и в результате из организма выводятся
совершенно сухие экскременты в виде шариков.
Животные, ведущие исключительно наземный
образ жизни, не сталкиваются с теми проблемами
осморегуляции, с которыми встречаются водные
или земноводные организмы. Однако насекомым
приходится регулировать ионный состав
гемолимфы, и эта регуляция состоит в поддержании баланса
между ионами, получаемыми с пищей, и ионами,
теряемыми в процессах синтеза, дефекации и
экскреции. Больше всего адаптации к местообитанию и
образу жизни мы находим у органов осморегуляции
и выделения водных членистоногих, таких как
речной рак Astacus и представитель морских
ракообразных зеленый краб Carcinus maenas.
Речной рак живет в пресной воде - гипотонической
среде. У его нимф некоторая часть азотистых
продуктов обмена и CO2 (в виде карбоната)
откладывается в кутикуле и вместе с ней сбрасывается во
время линьки. Но у взрослого рака азотистые
отходы выводятся в виде аммиака через
специализированные органы выделения, называемые антенналь-
ными или зелеными железами.
Эти железы представляют собой слепо
оканчивающиеся мезодермальные образования, которые
лежат в гемолимфе перед ротовым отверстием и
открываются наружу порой, находящейся у
основания антенны. Каждая антеннальная железа состоит
из четырех отделов: слепого целомического
мешочка, зеленого канальца, называемого лабиринтом,
длинного белого нефридиального канальца и
тонкостенного мочевого пузыря, открывающегося
наружу (рис. 19.11).
Вода с растворенными в ней веществами
фильтруется в целомический мешочек из гемолимфы под
действием гидростатического давления крови. Ког-
Гемолимфа
H2U
Мочевая
кислота
Мальпигиева
трубочка
Мочевая
кислота
Верхний отдел I
Нижний отдел
Рис. 19.10. Схема предполагаемого механизма
выделения мочевой кислоты через мальпи-
гиевы трубочки. 1. Мочевая кислота (H2U),
образующаяся в клетках тела, выводится в гемолимфу,
где она вступает в реакцию с бикарбонатами натрия
и калия и водой с образованием уратов натрия и
калия (NaHU и KHU), CO2 и воды (на схеме показан
только калий). 2. Эти соли активно секретиру-
ются в просвет трубочки, а вслед за ними в него
путем осмоса поступает вода. 3. Во время
передвижения растворенных соединений по трубочке
образуются бикарбонаты, которые активно всасываются
в гемолимфу, а за ними выходит путем осмоса вода.
Так как в результате реабсорбции бикарбонатов в
проксимальной части трубочки снижается рН,
мочевая кислота выпадает в осадок в виде
кристаллов.
Глюкоза,
аминокислоты,
вода, аммиак,
мочевина, соли
Аммиак
Глюкоза,
аминокислоты
Соли
Вода,
аммиак,
мочевина,
аминокислоты
Цело ми
ческий Лабиринт
мешочек
Белый каналец
Мочевой
пузырь
Гипотоническая
моча
Концентрация Cl', мкМ
Рис. 19.11. Схема строения и функции антеннальной (зеленой) железы речного рока. На графике внизу
показаны изменения осмотической концентрации фильтрата. (По Peters, из Barrington )
да этот фильтрат проходит через лабиринт,
выстланный изнутри железистой тканью, и по нефридиаль-
ному канальцу, ценные для организма вещества
всасываются обратно в гемолимфу, а в фильтрат
секретируются новые порции отходов метаболизма,
в том числе азотистые соединения.
Внутренняя среда речного рака гипертонична по
отношению к окружающей среде, и вода поступает в
организм через любую проницаемую поверхность, в
первую очередь через жабры. Это поступление воды
путем осмоса уравновешивается выделением
большого количества мочи, гипотоничной по сравнению
с гемолимфой. Азотистые продукты обмена
выделяются в основном в виде аммиака, но образуется и
некоторое количество мочевины.
Зеленый краб живет в приливной зоне и большую
часть времени находится в морской воде. Он имеет
такие же антеннальные железы, как речной рак,
через которые выводятся азотистые продукты
обмена, главным образом аммиак. Как у многих других
морских животных, некоторые участки его покровов
проницаемы для воды и солей, а жидкости тела
изотоничны морской воде. Для морских организмов
это выгодно, так как им не приходится затрачивать
энергию на поддержание в жидкостях тела более
высокого или более низкого осмотического
давления по сравнению с окружающей средой. Однако,
несмотря на изотоничность жидкостей организма
окружающей среде, концентрация отдельных солей
в них может быть иной, чем в морской воде, и для
регуляции их ионного состава необходима затрата
энергии.
Зеленый краб обитает в весьма различных местах.
Он способен переносить гипертонические условия,
которые создаются в соленых озерцах,
образующихся на скалистом морском берегу или в затопляемых
морем низинах, когда в жаркое время из них
испаряется вода. В этих условиях он ведет себя как
осмоконформер - осмотическая концентрация
жидкостей тела повышается в результате поглощения
солей и ткани могут переносить это; моча в этом
случае гипертонична по отношению к жидкостям
внутренней среды. Краб способен также переносить
изменчивые условия солености в эстуариях рек, т.е.,
подобно большинству обитающих здесь животных,
является эвригалинным организмом. Если соленость
воды уменьшается вследствие разбавления ее
речной водой или в результате ливней, зеленый краб
становится гипертоничным по сравнению с
окружающей средой (рис. 19.12); вода теперь стремится
войти в организм путем осмоса, а растворенные
вещества, напротив, стремятся выйти из организма
путем диффузии. В этих условиях зеленый краб
становится «осморегулятором», или гомойосмоти-
ческим животным: состав жидкостей тела
стабилизируется у него за счет активной реабсорбции
натрия из мочи в гемолимфу в клетках антеннальных
желез. В связи с реабсорбцией натрия в организме
будет также удерживаться и некоторое количество
Содержание NaCI
во внутренней среде, %о
Абсолютная
пойки лосмотичность
Абсолютная
гомойосмотичность
Морская вода
Морская
вода
Palaemonetes
Nereis
Leander
Carcinus
Содержание NaCI в окружающей среде, %о"
Рис. 19.12. Взаимоотношения между
внутренней средой организма и внешней средой у пойки -
лосмотических и гомойосмотических форм и у
четырех видов морских
организмов-пелагических ракообразных Leander и Palaemonetes, червя
Nereis и краба Carcinus.
воды, но общий объем тела не будет увеличиваться
благодаря жесткой кутикуле. Выделяемая в этих
условиях моча изотонична жидкостям тела.
19.4.6. Адаптации к изменениям
окружающей среды у
беспозвоночных
Беспозвоночные, живущие в морской воде,
например актинии, изотоничны окружающей среде. Если
их поместить в разведенную морскую воду, они
немедленно начнут поглощать воду и терять соли.
То же самое при уменьшении солености среды
происходит с пескожилом. Вначале масса тела его
увеличивается, а затем возвращается к нормальной
величине. Объясняется это тем, что вода поступает
внутрь путем осмоса быстрее, чем выходят ионы.
Через короткий период времени условия
стабилизируются благодаря потере ионов, жидкости тела
становятся изотоничными окружающей среде и
масса тела возвращается к норме, если соленость среды
не падает слишком сильно. Многощетинковый
червь Nereis живет обычно в море, зарывшись в
песок или в ил, но он может осуществлять осморегу-
ляцию при уменьшении солености морской воды до
10% нормы и жить в эстуариях рек. Он является
гомойосмотическим организмом и способен
оставаться гипертоничным по отношению к среде
благодаря пониженной проницаемости наружных
покровов и способности поглощать ионы хлора из
окружающей воды (рис. 19.12).
Существует несколько видов морских
беспозвоночных, которые гипотоничны по отношению к
морской воде, как, например, ракообразные Palae-
monetes и Leander (рис. 19.12). Они успешно борются
с потерей воды и поступлением внутрь солей,
заглатывая морскую воду и активно секретируя соли
обратно во внешнюю среду через жабры. Эти
механизмы осморегуляции настолько эффективны, что
животные могут поддерживать стабильное
содержание воды в организме при колебаниях солености от
2 до 110% от обычной солености моря. Как
полагают, эта способность указывает на то, что эти
ракообразные произошли от пресноводных предков
и уже вторично поселились в морской среде. Соло-
новодный рачок Artemia гипотоничен по отношению
к чрезвычайно соленой среде его обитания и
поддерживает гипотоничность жидкостей тела путем
непрерывного удаления ионов Na+ и Cl",
поступающих в организм с тем концентрированным
раствором, который животное пьет. Эти ионы удаляются
из гемолимфы через жаберный эпителий. Рачок
способен также поддерживать стационарный состав
жидкостей тела при колебаниях солености среды от
10 до 100% по отношению к морской воде.
Некоторые животные, обитающие в солоноватой
воде, например мидия (Mytilus), могут
поддерживать состав внутриклеточной жидкости в довольно
стабильном состоянии, несмотря на изменения
состава внешней среды и внеклеточной жидкости.
Предполагают, что это связано с регуляцией
состава внутриклеточной жидкости самими клетками.
Полагают, что многие пресноводные животные,
например речной рак, произошли от обитателей
солоноватых вод, а те-от морских форм путем
постепенного совершенствования регуляции водно-
солевого обмена.
Для мелких пресноводных беспозвоночных
характерно сравнительно высокое отношение
поверхности к объему. Будучи гипертоничны по отношению к
среде, они поглощают воду и теряют соли, и это
происходит у них быстрее, чем у более крупных
беспозвоночных (вроде речного рака) с меньшим
отношением поверхности к объему. Эту проблему
мелкие беспозвоночные в какой-то мере разрешают
за счет того, что во внутренних жидкостях
концентрация ионов у них ниже, чем у более крупных форм.
19.4.7. Иглокожие
Все иглокожие - морские животные. Азотистые
отходы метаболизма выводятся у них в виде аммиака
через жабры и амбулакральные ножки. Иглокожие
имеют амбулакральную систему, содержащую
морскую воду, с которой клетки тела изотоничны.
Никаких проблем осморегуляции для них не
существует.
19.4.8. Позвоночные
У различных позвоночных встречается экскреция
всех трех азотистых продуктов, что, как правило,
зависит от доступности воды для того или иного
вида. Механизмы осморегуляции у позвоночных
более эффективны, чем у беспозвоночных,
благодаря малой проницаемости наружных покровов и
наличию почек. Биологи до сих пор спорят о том,
где возникли первые рыбы-в морской или пресной
воде. Многие биологи считают более вероятным
морское происхождение первых рыб и
рассматривают почки как более позднее приобретение,
необходимое для выживания в гипотонических
условиях пресных водоемов. В этих условиях почки
служат для удаления избытка воды и задержки
солей. Последующее развитие почек зависело от
характера окружающей среды и шло по линии все
большего усложнения в ряду позвоночных от рыб
до млекопитающих. Увеличение сложности
строения почек было связано с заселением суши.
Благодаря повышенной эффективности механизмов
выделения и осморегуляции состав внутренней среды у
позвоночных колеблется в более узких пределах,
чем у беспозвоночных.
Структурно-функциональной единицей почечной
ткани является нефрон. Нефроны - сегментарные
структуры, образовавшиеся из мезодермальных не-
фротомов (разд. 21.8), вступивших в тесный контакт
с кровеносными сосудами, отходящими от аорты, и
Приносящая
артериола
Выносящая
артериола
Клубочек
Капсула
нефрона
Примитивное почечное
(мальпигиево) тельце
Здесь образуется
клубочковый фильтрат
Каналец нефрона
Воронка с
ресничным
эпителием
Целом
Рис. 19.13. Строение примитивного нефрона.
Образующийся в клубочке фильтрат поступает
через воронку с ресничным эпителием в целом или
через соединительный каналец объединяется с
фильтратом из других нефронов.
связанных с целомом через ресничную воронку.
Нефроны у зародышей рыб имеют наиболее
примитивное строение, при котором несколько нефронов
открываются в перикардиальную полость и в
совокупности образуют структуру, называемую пронеф-
росом (рис. 19.13) или предпочкой. У всех взрослых
рыб и амфибий пронефрос утрачивается, а вместо
него развивается более компактное образование,
состоящее из значительно большего числа нефронов
и расположенное в брюшном и хвостовом отделах
тела. Это мезонефрос, или первичная почка. В мезо-
нефросе нефроны утратили связь с целомом и
объединены собирательным протоком, ведущим к
мочеполовому отверстию. Такое строение идеально
подходит для выделения разбавленной мочи,
образующейся в основном у обитателей пресных вод.
Рептилии, птицы и млекопитающие
приспособились к жизни на суше, где вместо проблемы
удаления воды, стоящей перед рыбами и амфибиями,
встает задача удержания воды в организме. У этих
животных орган выделения представляет собой еще
более компактную структуру - метанефрос, или
вторичную почку, которая состоит из еще большего
числа нефронов с еще более длинными канальцами.
В канальцах происходит реабсорбция воды и
образуется концентрированная моча, которая в конце
концов поступает в почечную лоханку, а из нее - в
мочевой пузырь. (Более подробно строение и
функции почки млекопитающего описаны в разд. 19.5.)
Образование мочи в почке позвоночных основано
на принципах ультрафильтрации, избирательной
реабсорбции и активной секреции. Моча
представляет собой жидкость, содержащую отходы
азотистого обмена, воду и те ионы, содержание которых
в организме превышает необходимый уровень.
Ультрафильтрации подвергаются также и ценные
для организма вещества, но они всасываются
обратно в кровь. Реабсорбируется 99% растворенных
веществ, и на этот процесс расходуется энергия. С
энергетической точки зрения такой механизм
кажется неэкономным, но он обеспечивает позвоночным
большую гибкость при освоении новых мест
обитания, так как позволяет выводить чужеродные или
«новые» вещества, как только они появляются в
организме, и для их удаления не нужно создавать
новый секреторный механизм.
Рыбы
У рыб органами выделения и осморегуляции служат
жабры и почки. Оба органа проницаемы для воды,
азотистых отходов и ионов и обладают большой
поверхностью, облегчающей обмен. Почки в
отличие от жабр отделены от окружающей среды
стенками тела, тканями и внеклеточной жидкостью, и
поэтому они могут контролировать состав
внутренней среды организма. Хотя все рыбы живут в
водной среде, механизм экскреции и осморегуляции
у пресноводных и морских рыб настолько
различен, что эти две группы следует рассмотреть
по отдельности.
пресноводные рыбы. У пресноводных костистых
рыб осмолярность жидкостей тела составляет около
300 мосмоль/л, и они гипертоничны по отношению
к внешней среде. Несмотря на относительную
непроницаемость наружного покрова из чешуи,
покрытых слизью, внутрь тела путем осмоса
поступает значительное количество воды через
высокопроницаемые жабры, и через них же теряются соли.
Жабры служат также органами выделения таких
азотистых продуктов, как аммиак. Для
поддержания стационарного состояния жидкостей внутренней
среды пресноводные рыбы должны постоянно
выводить много воды. Это происходит у них благодаря
образованию большого объема ультрафильтрата,
из которого растворенные вещества извлекаются
путем их избирательной реабсорбции в капилляры,
окружающие почечные канальцы. Почки образуют
большое количество сильно разведенной мочи (ги-
потоничной по отношению к крови), в которой
содержится и ряд других растворенных веществ.
Количество мочи, выделяемой за сутки, может
составлять до одной трети от всей массы тела. Потеря
солей с мочой возмещается за счет электролитов,
получаемых с пищей, и за счет активного
поглощения их из окружающей воды особыми клетками,
находящимися в жабрах.
морские рыбы. Полагают, что рыбы впервые
появились в морской среде, затем успешно заселили
пресноводные водоемы, и после этого некоторые из
них вторично переселились в море, дав начало
Осмотическая концентрация
крови (A0C)
моче-
вина
Морская
вода
Пластино-
жаберные
Костистые
рыбы
Рептилии
Птицы
Млекопитающие
Рис. 19.14. Приблизительные концентрации
растворенных веществ в жидкостях тела морских
позвоночных. Пластиножаберные рыбы -
единственные позвоночные, у которых жидкости тела
гипертоничны по отношению к окружающей
среде; но, как видно из диаграммы,
концентрация электролитов у них лишь ненамного выше, чем у
костистых рыб. Благодаря задержке мочевины
осмотическое давление у них такое же, как у
морской воды, о чем свидетельствует
депрессия точки замерзания (A0C).
пластиножаберным и морским костистым рыбам.
В процессе эволюции в пресной воде у рыб
сформировались многие физиологические механизмы,
приспособленные к такому осмотическому
давлению жидкостей тела, которое в 2-3 раза меньше,
чем у морской воды. После возвращения рыб в
морскую среду жидкости их тела сохранили
осмотическое давление, присущее их предкам, и в связи с
этим возникла проблема гомеостаза жидкостей тела
в условиях гипертонического окружения (рис. 19.14).
пластиножаберные рыбы. У этих рыб исходная ос-
молярность жидкостей тела примерно такова же,
что и у морских костистых рыб, т.е. эквивалентна
1%-ному раствору соли. Избыточная потеря воды в
гипертонической морской воде предотвращается за
счет синтеза и удержания мочевины в тканях и
жидкостях тела. По-видимому, большинство клеток
тела, за исключением клеток головного мозга,
способно синтезировать мочевину, и для своей
метаболической активности они не только нуждаются в
присутствии мочевины, но и обладают
толерантностью к высоким ее концентрациям. Исследования,
проведенные на изолированных сердцах акул,
показали, что сердце может сокращаться только при
перфузии сбалансированным солевым раствором,
содержащим мочевину. Жидкости тела у акул
содержат 2-2,5% мочевины, что в 100 раз превышает
концентрацию, переносимую другими
позвоночными. Как правило, высокая концентрация мочевины
приводит к разрыву водородных связей,
денатурации белков и тем самым к инактивации ферментов.
Однако у пластиножаберных рыб этого почему-то
не происходит. Мочевина в сочетании с
неорганическими ионами и другим азотистым продуктом
обмена - триметиламиноксидом (CH3)3N=0, менее
токсичным, чем аммиак,- создают в жидкостях тела
более высокое осмотическое давление, чем в
морской воде (Δ морской воды составляет 1.7°С, а у
жидкостей тела пластиножаберных-1,8°С) (рис. 19.14).
Будучи слегка гипертоничными по отношению к
окружающей среде, пластиножаберные рыбы
поглощают воду путем осмоса через жабры. Вода вместе
с избытком мочевины и триметиламиноксидом
выводится почками с мочой, которая слегка гипотонич-
на по отношению к жидкостям тела. Почки имеют
длинные канальцы, которые используются для
избирательной реабсорбции мочевины, а не для
выведения поступающих с пищей солей. Избыток же ионов
натрия и хлора удаляется из жидкой среды
организма путем активной секреции в прямую кишку
клетками ректальной железы - маленькой железки,
связанной протоком с прямой кишкой. Жабры
относительно непроницаемы для отходов азотистого
обмена, и их выведение целиком контролируется
почками. Таким путем осмотическое давление жидкостей
тела поддерживается на высоком уровне.
морские костистые рыбы . У морских костистых рыб
осмотическое давление жидкостей тела
поддерживается на уровне более низком, чем у морской воды
(рис. 19.14). Благодаря чешуе и слизи наружные
покровы рыб относительно мало проницаемы для
воды и ионов, но вода легко теряется из организма
(а ионы поглощаются) через жабры. Для регуляции
состава жидкостей тела костистые рыбы пьют
морскую воду, а особые секреторные клетки в
кишечнике извлекают из нее соли путем активного
транспорта и выделяют их в кровь. В жабрах имеются
хлоридные клетки, которые активно поглощают из
крови ионы хлора и выделяют их в окружающую
среду, а вслед за ионами хлора по принципу
сохранения электрохимической нейтральности выходят и
ионы натрия. Другие ионы, присутствующие в
морской воде в большом количестве,-ионы магния
и сульфата-удаляются с изотонической мочой,
образуемой в небольшом количестве почками. Почки
не имеют клубочков и поэтому не способны к
ультрафильтрации. Все компоненты мочи, такие как
азотистое соединение триметиламиноксид
(придающий рыбе характерный запах) и соли, секретируют-
ся в почечные канальцы, а за ними осмотическим
путем следует и вода.
эвригалинныерыбы. Существует ряд видов эврига-
линных рыб, которые не только переносят
небольшие изменения солености воды, но и могут
полностью адаптироваться к жизни в пресной и в
морской воде на протяжении длительных периодов
их жизни. В зависимости от того, куда эти рыбы
движутся на нерест, различают анадромных и ка-
тадромных рыб. Анадромные рыбы (греч. апа-
вверх, dromein - бежать), такие как лосось (Salmo
salar), выводятся из икры в пресной воде и
мигрируют в море; здесь они достигают зрелости и затем
возвращаются для нереста в реки. Катадромные
рыбы (греч. cata-вниз), к которым относится угорь
(Anguilla vulgaris), мигрируют в противоположном
направлении. Они выводятся в морской воде и
мигрируют в пресноводные водоемы, где достигают
зрелости, после чего возвращаются для нереста в
море. При переходе из реки в море угорь теряет за
10 часов около 40% своего веса. Чтобы
компенсировать эту потерю и сохранить гипотоничность
жидкостей тела, он пьет морскую воду, а соли выделяет
путем активной секреции через жабры. При
переходе угря из моря в реку его масса вначале
увеличивается за счет поступления воды путем осмоса, но
уже через два дня он достигает стабильного
осмотического состояния. В пресной воде угорь поглощает
соли через жабры путем активного транспорта.
На примере этих двух групп рыб мы видим, что
механизмы активного транспорта в жабрах могут
действовать в двух направлениях. Связано ли это с
переменой направления работы ионных насосов в
одних и тех же клетках или с функционированием
разных групп клеток, пока неизвестно.
Предполагается, что на эти механизмы влияют гормоны,
выделяемые гипофизом и корой надпочечников. У
рыб обеих групп при переходе в пресную воду
существует период «выжидания», позволяющий
механизмам осморегуляции приспособиться к новой
среде.
Амфибии
Считается, что амфибии произошли от
рыбообразных пресноводных предков и унаследовали от них
проблемы осморегуляции, связанные с тем, что их
кровь гипертонична по отношению к окружающей
среде. Кожа лягушек проницаема для воды, и
именно через кожу поступает из внешней среды в
организм основная масса воды. Избыток поглощаемой
организмом воды удаляется путем
ультрафильтрации в многочисленных крупных почечных
клубочках.
Почки амфибий широко использовались для
изучения физиологии этих органов, так как их крупные
клубочки расположены близко к поверхности. В эти
клубочки и канальцы можно вводить микрошприц и
извлекать из них фильтрат для анализа. Таким
путем можно определить эффективность
ультрафильтрации и избирательной реабсорбции.
Амфибии выделяют большое количество очень
разбавленной мочи, гипотоничной по отношению к
жидкостям тела. Моча содержит мочевину, которая
выводится путем ультрафильтрации и путем секреции в
канальцы. Преимущество этого механизма состоит
в том, что он позволяет амфибиям снижать
скорость клубочковой фильтрации в засушливых
условиях и таким образом уменьшать потерю воды с
мочой, тогда как канальцы продолжают получать
кровь из воротных сосудов почек, из которых в
канальцы активно секретируется мочевина. В этом
отношении данный механизм противоположен то-
Гипер-
Bo да
Соли
Большой объем сильно
разведенной мочи,
содержащей аммиак
Некоторое *
количество
воды
Изо-/Гипер-
Триметиламиноксид,
избыток мочевины, вода
Почка
Ректальная
железа
Морская
вода
Гипо-
Вода,
Соли
Активный перенос Cl
за которым пассивно
следует Na +
Небольшой объем
изотонической мочи,
содержащей
триметиламиноксид
Рис. 19.15. Экскреция и осморегуляция у
пресноводных костистых рыб (A)1 пластиножаберных
(Б) и морских костистых рыб (В). Сокращения
Гипо-, Изо- и Гипер- указывают тонич-
ность внутренней среды по отношению к внешней.
му, что имеет место у пластиножаберных рыб, у
которых мочевина в канальцах активно реабсорби-
руется.
Некоторая часть солей неизбежно теряется с
мочой и в результате диффузии через кожу, но эта
потеря возмещается за счет солей, поступающих с
пищей, а также активно поглощаемых из
окружающей воды кожей, которая служит у амфибий
главным органом осморегуляции. Личинка бесхвостой
амфибии - головастик - является полностью водным
организмом и выделяет аммиак через жабры, но при
метаморфозе состав азотистых экскретов и
механизм их выделения меняются и становятся такими,
как описано выше.
Лягушки способны накапливать воду в мочевом
пузыре и многочисленных подкожных
лимфатических пространствах. За счет этих запасов
возмещается потеря воды путем испарения в те периоды,
когда лягушка находится на суше. Жабы способны
находиться в сухих условиях более
продолжительное время, так как их почки могут реабсорбиро-
вать воду из клубочкового фильтрата и
образовывать более концентрированную мочу, а кожа менее
проницаема для воды. Известно, что проницаемость
кожи у амфибий регулируется антидиуретическим
гормоном, выделяемым задней долей гипофиза; как
полагают, механизм регуляции проницаемости
здесь тот же, что и в почечных канальцах
млекопитающих.
Водный баланс у сухопутных
организмов
Для нормального функционирования клеток в
организме животного необходимо стационарное
состояние внутриклеточной жидкости. Гомеостатический
обмен водой между клетками, тканевой жидкостью,
лимфой, плазмой крови и окружающей средой
представляет проблему и для водных, и для наземных
организмов. Водные формы получают или теряют
воду путем осмоса через все проницаемые участки
поверхности тела в зависимости от того, каково
окружение - гипотоническое оно или
гипертоническое. Наземные организмы сталкиваются с
проблемой потери воды и для поддержания устойчивого
водного баланса используют многочисленные
приспособления, приведенные в табл. 19.5. Это
устойчивое состояние водного обмена достигается за счет
баланса между отдачей воды и ее получением (табл.
19.6).
Рептилии
Эти животные первыми приспособились к наземной
жизни. Они обладают множеством
морфологических, биохимических и физиологических адаптации
для существования на суше. Однако во всех трех
отрядах (черепахи, ящерицы и змеи, крокодилы)
имеются виды, которые вторично приспособились к
жизни в пресной и морской воде. У всех этих
животных механизмы выделения и осморегуляции
адаптированы к соответствующим условиям.
У наземных пресмыкающихся потере воды
препятствует относительно непроницаемая кожа,
покрытая роговыми чешуйками. Органами
газообмена у них служат легкие, расположенные внутри тела,
что уменьшает потерю воды. В тканях образуется
нерастворимая мочевая кислота, которая может
выводиться без большой потери влаги. Для
удаления избытка ионов натрия и калия нужна вода, но
поскольку экономия воды имеет жизненно важное
значение, эти ионы соединяются с мочевой
кислотой, образуя нерастворимые ураты натрия и калия,
которые удаляются вместе с мочевой кислотой.
Почечные клубочки имеют малые размеры и
образуют лишь такое количество фильтрата, которое
необходимо для вымывания мочевой кислоты из
почечных канальцев в клоаку, где часть воды реаб-
сорбируется. У многих наземных рептилий
почечные клубочки вообще отсутствуют.
У сухопутных рептилий нет специальных
механизмов для выведения солей, а ткани способны
переносить повышение концентрации солей на 50% по
сравнению с обычным уровнем после приема их
Таблица 19.5. Механизмы сохранения воды у наземных
животных
Группа
животных
Насекомые
Рептилии
Птицы
Млекопитающие
Механизмы и приспособления
Непроницаемая кутикула; трахеи и
дыхальца; мальпигиевы трубочки;
мочевая кислота как азотистый экскрет;
клейдоические яйца
Чешуи и ороговевшая кожа, легкие; ме-
танефрос; мочевая кислота как
азотистый экскрет; реабсорбция в клоаке;
клейдоические яйца; поведенческие
реакции на избыток тепла;
физиологическая переносимость обезвоживания
Перья; легкие; метанефрос, мочевая
кислота как азотистый экскрет,
клейдоические яйца
Ороговевшая кожа и волосы, легкие;
метанефрос, гипертоническая моча,
содержащая мочевину; живорождение,
поведенческие реакции на избыток
тепла; ограниченная зона обитания;
физиологическая толерантность к
обезвоживанию
Таблица 19.6. Компоненты водного баланса у наземных
организмов
Потеря воды
Испарение с поверхности
тела
Испарение с поверхности
органов газообмена
Вода, содержащаяся в
фекалиях
Вода, выделяемая с мочой
Вода, выделяемая с
секретами, например со слезами
Поступление воды в организм
Питье
Вода, поступающая с
пищей, поглощение через
поверхность тела
Вода, образующаяся при
клеточном дыхании
(метаболическая вода)
с пищей или избыточной потери воды. Морские
пресмыкающиеся, такие как галапагосская игуана
и зеленая черепаха (Сhe lone my das), получают с пищей
большое количество соли. Их почки не способны
справиться с быстрым выведением этого избытка
соли из жидкостей тела, и им помогают особые
солевые железы, расположенные на голове. Эти
железы способны секретировать раствор хлористого
натрия, в несколько раз более концентрированный,
чем морская вода. Солевые железы находятся у
черепахи в глазницах, и протоки от них идут к
глазам; отсюда впечатление, что черепаха плачет. В
«слезах», выделяемых солевыми железами черепах,
концентрация солей очень высока.
Клейдоические яйца
Важной особенностью рептилий и птиц, благодаря
которой они могут существовать вне воды в течение
всего жизненного цикла, является наличие у них
клейдоических яиц (рис. 20.52). Яйцо заключено в
плотную оболочку, которая предохраняет
зародыша от обезвоживания. В процессе эмбриогенеза
вырост задней кишки образует мешковидную
структуру, называемую аллантоисом, в которой
откладывается мочевая кислота, выделяемая эмбрионом.
Поскольку мочевая кислота нерастворима и
нетоксична, это служит для эмбриона идеальным
способом депонирования экскретов. На более поздних
стадиях развития аллантоис васкуляризуется,
прижимается к оболочке и функционирует как орган
газообмена.
Птицы
Птицы, по-видимому, произошли от наземных
пресмыкающихся, таких как змеи и ящерицы, и
унаследовали те же проблемы. Кожа птиц относительно
непроницаема для воды, и благодаря наличию
перьев и отсутствию потовых желез скорость
испарения влаги у птиц очень мала. Однако
значительное количество воды теряется у них в дыхательных
путях в связи с очень активной вентиляцией легких и
сравнительно высокой температурой тела.
Вследствие большой интенсивности метаболизма
некоторые мелкие птицы могут терять за сутки до 35%
веса тела.
Азотистые продукты обмена удаляются в виде
мочевой кислоты с мочой, гипертоничной по
отношению к жидкостям тела. Моча поступает в клоаку,
где часть воды из мочи и фекальных масс
всасывается обратно, благодаря чему из организма выводятся
почти твердые экскременты.
Почки птиц содержат мелкие клубочки. Вся кровь,
снабжающая каналец, в котором происходит реаб-
сорбция воды и секреция солей, поступает от
клубочка, для эффективной работы которого
необходимо относительно высокое кровяное давление. Таким
образом осуществляется связь между образованием
большого объема клубочкового фильтрата и
последующим всасыванием большой части содержащихся
в нем воды и солей. Это всасывание облегчается
тем, что поверхность канальца увеличена за счет
образования петли Генле. В результате
деятельности этой структуры концентрация мочевой кислоты в
моче достигает 21%, что почти в 3000 раз выше ее
концентрации в жидкостях тела.
Некоторые морские птицы (пингвины, олуши,
бакланы, альбатросы), которые питаются рыбой и
пьют морскую воду, поглощают большие
количества солей. Соли выводятся из жидкостей тела
специализированными секреторными клетками
солевых, или носовых, желез. Эти железы сходны с
солевыми железами морских рептилий и тоже
расположены в глазницах. Они выделяют раствор
хлористого натрия, концентрация которого в 4 раза
выше, чем в жидкостях тела. Носовые железы
состоят из множества долек, содержащих большое
число секреторных трубочек, которые открываются
в центральный проток; этот проток ведет в носовую
полость, где раствор соли освобождается в виде
больших капель или выдувается в виде мельчайших
брызг.
19.5. Почки млекопитающих
У млекопитающих почки служат главными
органами выделения и осморегуляции. Их функции
включают: 1) удаление из организма ненужных
продуктов обмена и «чужеродных» веществ; 2) регуляцию
химического состава жидкостей тела путем
удаления веществ, количество которых превышает
текущие потребности; 3) регуляцию содержания воды в
жидкостях тела (и тем самым их объема) и рН этих
жидкостей.
Почки обильно снабжаются кровью и гомеоста-
тически регулируют состав крови. Благодаря этому
поддерживается оптимальный состав тканевой
жидкости, а следовательно, и внутриклеточной
жидкости омываемых ею клеток, что обеспечивает их
эффективную работу.
19.5.1. Расположение и строение
почек
У человека имеется пара почек, лежащих у задней
стенки брюшной полости по обе стороны
позвоночника на уровне грудных и поясничных позвонков.
Вес одной почки составляет около 0,5% общего веса
тела; левая почка слегка выдвинута вперед по
сравнению с правой.
Кровь поступает в почки через почечные артерии,
отходящие от аорты, а оттекает от них по почечным
венам, впадающим в нижнюю полую вену.
Образующаяся в почках моча стекает по двум
мочеточникам в мочевой пузырь, где накапливается до тех
пор, пока не будет выведена через
мочеиспускательный канал.
На поперечном разрезе почки видны две ясно
различимые зоны: лежащее ближе к поверхности
корковое вещество и внутреннее мозговое вещество.
Корковое вещество почки покрыто фиброзной
капсулой и содержит клубочки, едва видимые
невооруженным глазом. Мозговое вещество состоит из
канальцев, собирательных трубок и кровеносных
сосудов, собранных вместе в виде почечных пирамид.
Верхушки пирамид, называемые сосочками,
открываются в почечную лоханку, образующую
расширенное устье мочеточника (рис. 19.16). Через почки
проходит множество сосудов, образующих густую
сеть капилляров.
19.5.2. Общий план строения
и кровоснабжения нефрона
Основной структурной и функциональной единицей
почки является нефрон вместе с его кровеносными
сосудами. У человека в одной почке содержится
около миллиона нефронов, каждый из них длиной
около 3 см. Благодаря такому количеству нефронов
создается огромная поверхность для обмена
веществами.
Каждый нефрон включает шесть отделов, сильно
Фиброзная капсула
Юкстамедул
лярный нефрон
Корковое_
вещество
Мозговое,
вещество
Почечная
чашка
Корковый
нефрон
Собирательная
трубка
Сосочек
Отверстие
собирательной
Почечная
лоханка
Верхушка
пирамиды
Мочеточник
Рис. 19.16. Разрез почки млекопитающего.
Показано расположение кортикального и юкстамедул-
лярного нефронов.
различающихся по строению и физиологическим
функциям:
1) почечное тельце (мальпигиево тельце), состоящее
из боуменовой капсулы и клубочка;
2) проксимальный извитой каналец;
3) нисходящее колено петли Генле;
4) восходящее колено петли Генле;
5) дистальный извитой каналец;
6) собирательная трубка.
Структурные взаимоотношения между этими
отделами нефрона показаны на рис. 19.17.
Существуют нефроны двух типов-корковые и
юкстамедуллярные. Корковые нефроны
расположены в коре и имеют относительно короткие петли
Генле, которые лишь недалеко заходят в мозговое
вещество. В юкстамедуллярных нефронах почечные
тельца расположены около границы коркового и
мозгового вещества (лат. juxta-рядом). Они имеют
длинные нисходящие и восходящие колена петли
Генле, глубоко проникающие в мозговое вещество
(рис. 19.18). Значение этих двух типов нефронов
связано с различием их функций. При нормальном
количестве воды в организме объем плазмы
контролируют корковые нефроны, а при недостатке воды
происходит усиленная ее реабсорбция в
юкстамедуллярных нефронах.
Кровь поступает в почку по почечной артерии,
которая разветвляется сначала на междолевые, а
затем на дуговые и междольковые артерии; от
последних отходят приносящие артериолы, снабжающие
кровью клубочки. Из клубочков кровь, объем
которой уменьшился, оттекает по выносящим артерио-
лам. Далее она течет по сети перитубулярных
капилляров, находящихся в корковом веществе и
окружающих проксимальные и дистальные извитые
канальцы всех нефронов и петли Генле корковых
нефронов. От этих капилляров отходят прямые
сосуды, идущие в мозговом веществе параллельно
петлям Генле и собирательным трубкам. Функция
обеих описанных сосудистых сетей - возвращение
крови, содержащей ценные для организма вещества,
в общую кровеносную систему. Через прямые
сосуды протекает значительно меньше крови, чем через
перитубулярные капилляры, благодаря чему в ин-
терстициальном пространстве мозгового вещества
поддерживается высокое осмотическое давление,
необходимое для образования концентрированной
мочи.
19.5.3. Функции почек
Почки служат не только органами выделения, но
и гомеостатическими органами, поддерживающими
постоянный состав жидкостей тела на фоне больших
колебаний поступления воды и солей. Недостаток
воды или избыточное ее потребление, чрезмерное
Приносящая
артериола
Мальпигиево
тельце
Клубочек-
Боуменова
капсула
Проксимальный
извитой каналец
Выносящая артериола
МОЗГОВОЕ ВЕЩЕСТВО \ КОРКОВОЕ ВЕЩЕСТВО
Дистальный
извитой
каналец
Толстый
сегмент
восходящего
колена
Петля Генле
Тонкий
сегмент
" восходящего
колена
Почечный
сосочек
Собирательная
трубка
^Проток
Беллини
Тонкий
сегмент
нисходящего
колена
Рис. 19.17. Схема строения нефрона (масштаб отдельных частей не выдержан).
КОРКОВОЕ ВЕЩЕСТВО
МОЗГОВОЕ ВЕЩЕСТВО
Собирательная
трубка
Петля
Генле
Выносящая
артериола
Клубочек
Перитубу-
лярные
капилляры
Приносящая
артериола
Междолевая артерия
^7
Дуговая
артерия
Дуговая
вена
Междолевая
вена
Прямые сосуды
Рис. 19.18. А. Кортикальный нефрон (слева) и юкстамедуллярный нефрон (справа). Б.
Кровоснабжение нефронов этих двух типов.
потоотделение, нехватка или избыток солей - все это
имело бы серьезные последствия для организма,
если бы почка не могла приспосабливать свою
деятельность к этим изменениям. При этом только
в двух последних отделах нефрона-в дистальном
извитом канальце и собирательной
трубке-изменяется функциональная активность с целью
регуляции состава жидкостей тела. Остальная часть неф-
рона вплоть до дистального канальца
функционирует при всех физиологических состояниях
одинаково. Конечным продуктом деятельности почек
является моча, объем и состав которой варьируют
в зависимости от физиологического состояния
организма. В норме отделяется большое количество
разведенной мочи, но при недостатке в организме
воды образуется концентрированная моча.
19.5.4. Основные принципы работы
почек
Трудно дать логичное объяснение работы почек,
исходя лишь из пути фильтрата через нефрон, так
как процессы, происходящие в одном отделе нефро-
на, имеют важные последствия для процессов,
протекающих в других его отделах.
Образование мочи и поддержание постоянства
внутренней среды организма - динамичный процесс,
включающий перенос веществ из одной части неф-
рона в другую, например из собирательной трубки
в восходящее колено петли Генле и из нефрона
в окружающие его капилляры.
Задача этого раздела - описать общие принципы
работы нефрона, выделив те процессы и механизмы,
которые лежат в ее основе и которые будут
рассмотрены более подробно в последующих разделах.
Процессы
1. Ультрафильтрация. В клубочке все
низкомолекулярные вещества, такие как глюкоза, вода и
мочевина, переходят в фильтрат, заполняющий
боуменову капсулу и поступающий затем в
каналец нефрона.
2. Избирательная реабсорбция. Все вещества,
которые могут быть использованы организмом или
нужны для поддержания водно-солевого состава
жидкостей тела, всасываются из фильтрата в
кровеносные капилляры; например, глюкоза
всасывается в проксимальном извитом канальце.
3. Секреция. До того как фильтрат покинет нефрон
в виде мочи, в него могут секретироваться
ненужные организму вещества; например, избыток
ионов K + , H+ и NH^ секретируется клетками
дистального извитого канальца.
Механизмы
1. Активный транспорт. В описанных выше
процессах 2 и 3 молекулы и ионы активно секретируют-
ся в фильтрат или всасываются из него. Так,
например, осуществляется всасывание глюкозы
в перитубулярные капилляры, окружающие
проксимальный каналец, и NaCl-в толстом
восходящем колене петли Генле.
2. Избирательная проницаемость. Различные участки
нефрона обладают избирательной проницаемо-
стью для ионов, воды и мочевины. Например,
проксимальные канальцы относительно мало
проницаемы по сравнению с дистальными.
Проницаемость собирательной трубки может
регулироваться гормонами.
3. Концентрационные градиенты. В результате
действия механизмов 1 и 2 в интерстициальном
пространстве мозгового вещества
поддерживаются концентрационные градиенты; например, у
человека осмолярность интерстициальной
жидкости повышается с 300 мосмоль/л в корковом
веществе до 1200 мосмоль/л в сосочке.
4. Пассивная диффузия и осмос. Ионы натрия и
хлора и молекулы мочевины будут
диффундировать в фильтрат и из него по концентрационному
градиенту в тех участках нефрона, которые
проницаемы для них. А молекулы воды в
проницаемых для них участках нефрона будут выходить
путем осмоса из фильтрата в тканевую (интер-
стициальную) жидкость почки там, где эта
жидкость гипертонична.
5. Гормональная регуляция. Водный баланс
организма и экскрецию солей регулируют гормоны,
действующие на дистальные извитые канальцы
и собирательные трубки,-антидиуретический
гормон, альдостерон и другие гормоны.
ковой фильтрации) может быть непосредственно
вычислена по величине клиренса таких веществ, как
инулин (C6H10O5), которые фильтруются из крови
почками, но не подвергаются реабсорбции или
секреции. У человека общая скорость клубочковой
фильтрации в обеих почках составляет около
125 мл/мин.
19.5.6. Строение и функция нефрона
Мальпигиево тельце состоит из почечного клубочка
и окружающей его боуменовой капсулы. Эта капсула
образуется в результате впячивания слепого конца
эпителиального канальца и охватывает в виде
двуслойного мешочка почечный клубочек. Последний
состоит примерно из 50 собранных в пучок
капилляров, на которые разветвляется единственная
подходящая к клубочку приносящая артериола и которые
сливаются затем в выносящую артериолу (рис. 19.19).
Строение мальпигиева тельца целиком связано с его
функцией-фильтрацией крови. Стенки капилляров
состоят из одного слоя эндотелиальных клеток,
между которыми имеются поры диаметром 50-100 нм.
Эти клетки лежат на базальной мембране, которая
19.5.5. Методы изучения функции
почек
Работу почек можно исследовать как прямым
путем, например изучая процесс образования мочи
в самой почке, так и косвенным путем, сравнивая
состав плазмы и мочи.
Американский физиолог Ричарде, вводя
микропипетку в боуменову капсулу почки лягушки, собрал
некоторое количество клубочкового фильтрата и
показал, что он сходен по составу с плазмой крови.
Почки амфибий использовались в этих
исследованиях потому, что мальпигиевы тельца в них
относительно крупные и расположены близко к
поверхности почки. Этот метод почечной микропункции был
затем применен для получения жидкости из других
отделов нефрона, в том числе у млекопитающих;
с его помощью было получено много сведений
о функциях различных участков.
Несмотря на эти достижения, многие процессы,
протекающие в почках, еще недостаточно изучены
(например, неясна функция петли Генле). В этих
случаях наши представления - всего лишь гипотезы
о том, как могли бы осуществляться те или иные
функции.
Для определения скорости фильтрации и
реабсорбции в почках и скорости кровотока в них
использовались различные методы определения
почечного клиренса веществ, содержащихся в
плазме крови. Клиренс - один из показателей быстроты
удаления веществ из плазмы почками.
Скорость фильтрации плазмы в почках (клубоч-
Рис. 19.19. Клубочки и артериолы в почке собаки,
инъецированные силиконовой смолой. Ткани
растворены.
Приносящая
артериола
Фенестри рованн ые
капилляры
Базальная
мембрана
Подоцит
Вторичные
отростки
Щелевидные
поры
Выносящая
артериола
Капилляры
клубочка
Подоцит
Первичные
отростки
Плоские
эпителиальные
клетки
боуменовой
капсулы
Клетки
проксимального
извитого
канальца
Рис. 19.20. Схема строения мальпигиево тельца.
Показаны типичные клетки клубочка и боуменовой
капсулы.
полностью окружает каждый капилляр и образует
непрерывный слой, полностью отделяющий
находящуюся в капилляре кровь от просвета боуменовой
капсулы (рис. 19.20). Внутренний листок
боуменовой капсулы состоит из клеток, называемых подоци-
тами и напоминающих морскую звезду с ее лучами.
От тела подоцита отходят несколько больших
первичных отростков, а от них - вторичные мелкие
отростки, напоминающие амбулакральные ножки
морской звезды. Вторичные отростки поддерживают
базальную мембрану и окруженный ею капилляр.
Между первичными отростками соседних подоци-
тов имеются щелевидные поры шириной 25 нм,
которые облегчают процесс фильтрации. Клетки
наружного листка боуменовой капсулы
представляют собой плоские неспециализированные
эпителиальные клетки (рис. 19.21 и 19.22).
В результате ультрафильтрации, происходящей в
клубочках, из крови удаляются все вещества с
молекулярной массой менее 68 000 и образуется
жидкость, называемая клубочковым фильтратом.
Всего через обе почки проходит 1200 мл крови
в 1 мин, т.е. за 4-5 мин проходит вся кровь,
имеющаяся в кровеносной системе. В этом объеме
крови содержится 700 мл плазмы, из которых
125 мл отфильтровывается в мальпигиевых тельцах.
Вещества, фильтрующиеся из крови в клубочковых
капиллярах, проходят через их поры, базальную
мембрану и щелевидные поры под действием
давления в капиллярах. Основную часть этого давления
составляет гидростатическое давление крови, но
эффективное фильтрационное давление, под
действием которого происходит фильтрация жидкости из
Вторичные
отростки
подоцита
Поверхность
капилляра
Первичные
отростки
подоцита
Рис. 19.21. А. Электронная микрофотография, показывающая строение клубочкового капилляра и
стенки боуменовой капсулы. Плазма, находящаяся в просвете капилляра (К), проходит через поры капилляра
(указаны стрелками), BM-базальная мембрана, действующая как фильтр между
капилляром и полостью боуменовой капсулы (BK). Вторичные отростки (ВО) подоцита (П)
оканчиваются на базальной мембране, и между ними видны щелевидные поры (ЩП). Б. Микрофотография
клубочкового капилляра, полученная с помощью сканирующего электронного микроскопа.
Капсула
Базальная
мембрана
Капилляр
Вторичный
отросток
подоцита
Щелевидная
пора
ФИЛЬТРАТ
Эндотели-
альная
клетка
капилляра
Пути
фильтрации
ПЛАЗМА
Рис. 19.22. Схема путей, по которым происходит
фильтрация плазмы из клубочкового капилляра в
боуменову капсулу.
клубочков,-это равнодействующая трех видов
давления, показанных на рис. 19.23, и выражается
уравнением
Фильтрационное давление =
= Гидростатическое давление крови в клубочках —
— [Онкотическое давление -I- Гидростатическое
давление клубочкового фильтрата]
Кровяное давление в клубочковых капиллярах
может варьировать при изменениях диаметра
приносящей и выносящей артериол, находящихся под
нервным и гормональным контролем. Сужение
выносящей артериолы приводит к уменьшению оттока
крови из клубочка и повышению в нем
гидростатического давления. При таком состоянии в клубочко-
вый фильтрат начинают проходить и вещества с
молекулярной массой более 68000.
По химическому составу клубочковый фильтрат
сходен с плазмой крови. Он содержит глюкозу,
аминокислоты, витамины, некоторые гормоны,
мочевину, мочевую кислоту, креатинин, электролиты
и воду. Лейкоциты, эритроциты, тромбоциты и
такие белки плазмы, как альбумины и глобулины,
не могут выходить из капилляров - они
задерживаются базальной мембраной, которая выполняет
роль фильтра. Кровь, оттекающая от клубочков,
обладает повышенным онкотическим давлением,
поскольку в плазме повышена концентрация белков,
но ее гидростатическое давление снижено.
Почечные канальцы и процессы
избирательной реабсорбции и
секреции
Скорость ультрафильтрации довольно стабильна.
У человека образуется около 125 мл фильтрата
в минуту. Если бы весь этот фильтрат выводился
в виде мочи, то суточное количество мочи
достигало бы примерно 180 л. Поскольку на самом деле
выделяется в среднем лишь около полутора литров,
очевидно, что большая часть фильтрата должна
реабсорбироваться. И действительно, из 125 мл
фильтрата, образующихся за 1 мин, 124 мл
всасывается обратно. Главные места реабсорбции
показаны на рис. 19.24.
Ультрафильтрация - процесс совершенно
пассивный и неизбирательный (в отношении ценных для
организма веществ). Вместе с «отходами» из крови
удаляются и вещества, необходимые для
жизнедеятельности. Обратное всасывание веществ, которые
могут быть использованы организмом или нужны
для поддержания постоянства внутренней среды,
происходит в канальцах. В канальцах же может
происходить дополнительное активное выведение
ненужных продуктов из окружающих капилляров.
проксимальный извитой каналец. Это наиболее
длинная (14 мм) и широкая (60 мкм) часть нефрона,
по которой фильтрат поступает из боуменовой
капсулы в петлю Генле. Стенки этого канальца состоят
из одного слоя эпителиальных клеток с
многочисленными длинными (1 мкм) микроворсинками,
образующими щеточную каемку на внутренней
поверхности канальца. Наружная мембрана
эпителиальной клетки примыкает к базальной мембране,
и ее впячивания образуют базальный лабиринт.
Мембраны соседних эпителиальных клеток
разделены межклеточными пространствами, и через эти
Приносящая
артериола
Капиллярная
сеть клубочка
Боуменова
капсула
Выносящая
артериола
Знкотическ
давление
30 мм рт.ст.
-!Гидростатическое
давление- '
ч крови 70 мм4)
vV рт. ст. ^
Давление
клубочкового
„ фильтрата
20 мм рт.ст.
Эффективное
фильтрационное
давление =
= 70 - (30+20)
= 20 мм рт. ст.
Рис. 19.23. Направление и величины сил,
определяющих фильтрационное давление в почечных
клубочках человека.
пространства и лабиринт циркулирует жидкость
(рис. 19.25). Эта жидкость омывает клетки
проксимальных извитых канальцев и окружающую сеть
перитубулярных капилляров, образуя связующее
звено между ними. В клетках проксимального
извитого канальца около базальной мембраны
сосредоточены многочисленные митохондрии,
генерирующие АТФ, необходимый для активного транспорта
веществ.
избирательная реабсорбция. Большая поверхность
проксимальных извитых канальцев,
многочисленные митохондрии в них и близость перитубулярных
капилляров-все это приспособления для реабсорб-
ции веществ из клубочкового фильтрата. Здесь
всасывается обратно более 80% веществ, в том числе
вся глюкоза, все аминокислоты, витамины и
гормоны и около 85% хлористого натрия и воды.
Механизм всасывания можно описать следующим
образом:
1. Глюкоза, аминокислоты и ионы диффундируют
из фильтрата в клетки проксимального извитого
канальца, откуда активно переносятся
транспортными системами плазматической мембраны в
межклеточные пространства и щели лабиринта.
2. Из межклеточных пространств и лабиринта они
диффундируют в чрезвычайно проницаемые пери-
тубулярные капилляры и выводятся из нефрона.
3. В результате непрерывного удаления всех этих
веществ из клеток проксимального извитого
канальца создается диффузионный градиент между
находящимся в просвете канальца фильтратом и
клетками, и по этому градиенту в клетки переходят все
новые молекулы, которые затем активно
транспортируются из клеток в межклеточные пространства
и щели лабиринта, и весь процесс продолжается.
В результате активного поглощения натрия и
сопровождающих его анионов осмотическое
давление фильтрата снижается, и в перитубулярные ка-
125 мл
фильтрата
100 мл
24 мл
1 мл мочи
Рис. 19.24. Схематическое изображение главных
отделов нефрона, в которых происходит
реабсорбция веществ из почечного фильтрата (обведены
прерывистой линией).
пилляры путем осмоса переходит эквивалентное
количество воды. Основная масса растворенных
веществ и воды извлекается из фильтрата с довольно
постоянной скоростью. В результате этого процесса
в канальце образуется фильтрат, изотоничный
плазме крови перитубулярных капилляров.
Из фильтрата путем диффузии реабсорбируется
также около 50% мочевины, которая поступает
в перитубулярные капилляры и возвращается таким
образом в общую систему кровообращения;
остальная мочевина выводится с мочой.
Белки с мол. массой менее 68 000, поступающие
в процессе ультрафильтрации в просвет канальца,
Перитубулярный
капилляр
Базальный
лабиринт
Базальная
мембрана
со складками
Путь
диффузии
Активный
транспорт
Эпителиальная клетка
проксимального извитого канальца
Глюкоза
Плазма
Гл
ιοί коза
)Гпю
. коза
D Глюкоза,
Просвет канальца
Межклеточное
пространство
Микроворсинки
Фильтрат
Глюкоза
Плотное
соединение
Рис. 19.25. Схема, иллюстрирующая предполагаемый механизм реабсорбции растворенных веществ,
например глюкозы, из просвета проксимального извитого канальца в перитубулярные капилляры. Как
полагают, молекулы-переносчики, участвующие в активном транспорте, находятся в базальной мембране.
Цифры соответствуют описанным в тексте стадиям поглощения глюкозы.
извлекаются из фильтрата путем пиноцитоза,
происходящего у основания микроворсинок. Они
оказываются внутри пиноцитозных пузырьков, к
которым прикрепляются первичные лизосомы (разд.
7.2.8). Гидролитические ферменты лизосом
расщепляют белки до аминокислот, которые либо
используются самими клетками канальца, либо переходят
путем диффузии в перитубулярные капилляры. И
наконец, в этом отделе нефрона происходит
активная секреция креатинина и «чужеродных» веществ.
Эти вещества транспортируются из межклеточной
жидкости, омывающей канальцы, в канальцевый
фильтрат и выводятся с мочой.
19.5.7. Образование мочи
Моча образуется в результате обмена
растворенными веществами и водой между фильтратом,
покидающим проксимальный извитой каналец, и
всеми последующими отделами нефрона (табл. 19.7).
Способность к образованию гипертонической
мочи характерна только для тех позвоночных (птицы
и млекопитающие), у которых имеется петля Генле.
Концентрация образующейся мочи находится в
прямой зависимости от длины петли Генле и
толщины слоя мозгового вещества по сравнению с корой.
Оба показателя возрастают с увеличением дефицита
воды в зоне обитания. Например, у бобра (Castor) -
полуводного млекопитающего - имеется тонкий
слой мозгового вещества, состоящего из коротких
петель Генле, и выделяется большой объем
разведенной мочи (600 мосмоль/л), тогда как обитатели
пустынь-кенгуровая крыса (Dipodomys) и
тушканчик (Dipus)- обладают толстым слоем мозгового
вещества с длинными петлями Генле и выделяют
небольшое количество сильно концентрированной
мочи (6000 и 9000 мосмоль/л соответственно).
Таблица 19.7. Состав плазмы и мочи и изменения
концентрации в них растворенных веществ в процессе образования мочи
у человека
Компонент
Вода
Белок
Глюкоза
Мочевина
Мочевая кислота
Креатинин
Na+
NH;
K +
Mg'*
Cl-
ро34
S024
Содержание в
плазме, %
90
8
0,1
0,03
0,004
0,001
0,32
0,0001
0,02
0,0025
0,37
0,009
0,002
Содержание в моче,
%
95
0
0
2,0
0,05
0,075
0,35
0,04
0,15
0,01
0,60
0,27
0,18
Увеличение
—
—
67 χ
12 χ
75 χ
1 χ
400 χ
7 χ
4 χ
2 χ
30 χ
90 χ
Нисходящее и восходящее колена
петли Генле
Прежде чем подробно обсуждать строение и
функцию различных участков петли Генле, необходимо
рассмотреть ее функцию в целом. Петля Генле
вместе с капиллярами прямых сосудов и
собирательной трубкой создает и поддерживает
продольный градиент осмотического давления в мозговом
веществе по направлению от коркового вещества
к сосочку за счет повышения концентраций
хлористого натрия и мочевины. Благодаря этому
градиенту возможно удаление все большего количества
воды путем осмоса из просвета канальца в интер-
стициальные пространства мозгового вещества,
откуда она переходит в прямые сосуды. В конечном
счете в соединительной трубке образуется
гипертоническая моча.
Механизм, с помощью которого петля Генле
обеспечивает повышение концентрации хлористого
натрия в мозговом веществе по направлению к
сосочку, не вполне ясен. Не известна также степень
проницаемости нисходящего колена петли Генле
для хлористого натрия. Эта неясность не позволяет
полностью понять механизм образования
градиента, но не изменяет существенно того, что было
сказано выше и будет сказано ниже. Движение
ионов, мочевины и воды между петлей Генле,
прямыми сосудами и собирательной трубкой можно
описать следующим образом (см. соответствующие
цифры в кружках на рис. 19.26):
1. Короткий и относительно широкий (30 мкм)
верхний сегмент нисходящего колена непроницаем1
для солей, мочевины и воды. По этому участку
фильтрат переходит из проксимального извитого
канальца в более длинный тонкий (12 мкм) сегмент
нисходящего колена, свободно пропускающий воду.
2. Благодаря высокой концентрации хлористого
натрия и мочевины в тканевой жидкости мозгового
вещества создается высокое осмотическое давление,
вода отсасывается из фильтрата и поступает в
прямые сосуды.
3. В результате выхода воды из фильтрата его
объем уменьшается на 5% и он становится гиперто-
ничным. В верхушке мозгового вещества (в сосочке)
нисходящее колено петли Генле изгибается в виде
шпильки для волос и переходит в восходящее
колено, которое по всей своей длине проницаемо для
воды.
4. Нижний участок восходящего колена - тонкий
сегмент - проницаем для хлористого натрия и
мочевины, и хлористый натрий диффундирует из него,
тогда как мочевина, напротив, диффундирует внутрь.
5. В следующем, толстом сегменте восходящего
колена эпителий состоит из уплощенных кубовидных
клеток с рудиментарной щеточной каемкой и много-
1 Здесь и далее речь идет, конечно, о проницаемости
стенок.
Прямые
сосуды
Нисходящее
колено
Восходящее
колено
Собирательная
трубка
Гипотон1.
[Tool
I ^Моче-J
* вина'
'Моче>
вина
Гипертон.
Мочевина
Моче!
вина
Мочевина
Движение воды
Активный транспорт NaCl
Пассивный перенос Na+H СГ
Движение мочевины
Градиент
осмотического
давления
в тканевой
жидкости
11200|
Рис. 19.26. Общая схема, показывающая передвижение солей, мочевины и воды, в результате которого
создается градиент осмолярности в мозговом веществе. Цифры в прямоугольниках
показывают осмолярность (мосмоль/кг), а цифры в кружочках соответствуют пунктам текста. To-
ничность разных отделов нефрона указана по отношению к плазме крови за пределами почки.
численными митохондриями. В этих клетках
осуществляется активный перенос ионов натрия и хлора
из фильтрата.
6. Вследствие выхода ионов натрия и хлора из
фильтрата повышается осмолярность мозгового
вещества, а в дистальные извитые канальцы поступает
гипотоничный фильтрат.
Петля Генле- противоточный
концентрирующий механизм
Роль петли Генле как противоточного
концентрирующего механизма определяют следующие факторы:
1) близкое соседство нисходящего и восходящего
колен;
2) проницаемость нисходящего колена для воды;
3) непроницаемость нисходящего колена для
растворенных веществ;
4) проницаемость тонкого сегмента восходящего
колена для растворенных веществ;
5) наличие механизмов активного транспорта в
толстом сегменте восходящего колена.
Осмолярность фильтрата в обоих коленах петли
Генле тоже позволяет предположить, что петля действует
как противоточный концентрирующий механизм. В
такой системе разность концентраций между
жидкостями в восходящем и нисходящем коленах на
каждом уровне может быть небольшой, но поскольку эта
разность существует на большом протяжении (как
в петле Генле), она суммируется и конечная
концентрация фильтрата у изгиба петли оказывается
намного больше, чем у ее концов. Чем длиннее
петля, тем больше разность концентраций.
Выход хлористого натрия из фильтрата в
восходящем колене приводит к повышению его
концентрации в тканевой жидкости мозгового вещества,
в связи с чем из восходящего колена отсасывается
вода. Эта вода сразу поступает в прямые сосуды,
благодаря чему в тканевой жидкости и в фильтрате
сохраняется высокая концентрация растворенных
веществ, как показано на рис. 19.27. В процессе
продвижения фильтрата по петле противоточный
обмен продолжается. Непрерывная пассивная и
активная циркуляция растворенных веществ
обеспечивает постоянную высокую концентрацию
хлористого натрия в мозговом веществе. Параллельно
функционирует другая система - система противоточного
обмена в прямых сосудах (см. ниже), и эти два
противоточных механизма, работая сообща,
обеспечивают градиент осмотической концентрации в
мозговом веществе.
Прямые сосуды как противоточный
обменник
Узкий нисходящий и более широкий восходящий
капилляры прямых сосудов на всем протяжении
идут параллельно друг другу и образуют на разных
уровнях ветвящиеся петли. Эти капилляры проходят
очень близко к канальцам петли Генле, однако
прямого переноса веществ из фильтрата петли в
прямые сосуды не происходит. Вместо этого
растворенные вещества выходят сначала в интерстициаль-
ные пространства мозгового вещества, где мочевина
и хлористый натрий задерживаются из-за малой
скорости кровотока в прямых сосудах, и
осмотический градиент тканевой жидкости сохраняется.
Клетки стенок прямых сосудов свободно пропу-
Изотон.1
Нисходящее колено
Восходящее колено
Тканевая жидкость
Активный
транспорт NaCI
Пассивный перенос NaCl
Передвижение воды
КОРКОВОЕ
ВЕЩЕСТВО
Тканевая жидкость!
Градиент
осмотической
концентрации
Тканевая жидкость
МОЗГОВОЕ
ВЕЩЕСТВО
Рис. 19.27. Модель противоточного концентрирующего механизма, образуемого канальцами петли
Генле. Ионы Na+U Cl ~ активно транспортируются из толстого сегмента восходящего колена петли в
тканевую жидкость, и концентрация растворенных веществ в этой жидкости
повышается. Вода выходит из просвета нисходящего колена петли. Цифрами указана осмотическая
концентрация (мосмоль/кг). Градиент осмолярности между нисходящим и восходящим коленами петли Генле
произвольно принят за 200 мосмоль/кг.
екают воду, мочевину и соли, а поскольку эти
сосуды идут рядом, они функционируют как
система противоточного обмена. При вступлении
нисходящего капилляра в мозговое вещество из плазмы
крови вследствие прогрессирующего повышения
осмотического давления тканевой жидкости выходит
путем осмоса вода, а обратно входят путем
диффузии хлористый натрий и мочевина (рис. 19.28).
В восходящем капилляре происходит обратный
процесс: в связи с уменьшением осмотического
давления тканевой жидкости вода вновь переходит в
плазму, а хлористый натрий и мочевина выходят из
нее. Адаптивное значение этого механизма состоит
в том, что благодаря ему осмотическая
концентрация плазмы, выходящей из почек, остается
стабильной независимо от концентрации плазмы,
поступающей в них. Наконец, что особенно важно,
поскольку все перемещения растворенных веществ и
воды происходят пассивно, противоточный обмен
в прямых сосудах происходит без всяких затрат
энергии.
Дистэльный извитой каналец
Дистальный извитой каналец подходит обратно по
направлению к мальпигиеву тельцу и весь лежит
в корковом веществе. Клетки дистальных канальцев
имеют щеточную каемку и содержат много
митохондрий, и именно этот отдел нефронов ответствен
за тонкую регуляцию водно-солевого баланса и рН
крови. Гормональный контроль проницаемости
клеток дистального извитого канальца для воды связан
с таким же контролем собирательных трубок (оба
механизма описаны ниже). Регуляция баланса
электролитов и рН описана в разд. 19.7.
Собирательная трубка
Собирательная трубка начинается в корковом
веществе от дистального извитого канальца и идет
вниз через мозговой слой, где объединяется с
несколькими другими собирательными трубками в
более крупные протоки, называемые протоками
Беллини. Проницаемость стенок собирательной трубки
для воды и мочевины регулируется
антидиуретическим гормоном (АДГ), и благодаря этой регуляции
собирательная трубка участвует вместе с дисталь-
ным извитым канальцем в образовании
гипертонической или гипотонической мочи в зависимости от
потребности организма в воде.
19.6. Антидиуретический
гормон (АДГ) и
образование
гипертонической
или гипотонической мочи
Относительно стабильное осмотическое давление
крови поддерживается за счет баланса между
поступлением воды с питьем и пищей и потерей воды
с выдыхаемым воздухом, потом, калом и мочой.
Однако за тонкую регуляцию осмотического
давления ответственно в основном воздействие
антидиуретического гормона (АДГ) на проницаемость ди-
стальных извитых канальцев и собирательных
трубок.
При недостаточном потреблении воды, сильном
потоотделении или после приема большого
количества соли осморецепторы, находящиеся в
гипоталамусе, регистрируют повышение осмотического
давления крови. Возникают нервные импульсы,
которые передаются в заднюю долю гипофиза и
вызывают освобождение АДГ. АДГ повышает
проницаемость для воды стенок дистального извитого
канальца и собирательной трубки, вода выходит из
фильтрата в тканевую жидкость коркового и
мозгового вещества, и почки выделяют меньший объем
более концентрированной (1000 мосмоль/л) мочи.
АДГ повышает также проницаемость
собирательной трубки для мочевины, которая диффундирует из
мочи в тканевую жидкость мозгового вещества.
Здесь мочевина повышает осмолярность, что
приводит к увеличению выхода воды из тонкого
сегмента нисходящего колена петли Генле (рис. 19.29).
После приема большого количества воды,
напротив, осмотическое давление крови снижается и
секреция АДГ прекращается. Стенки дистального
извитого канальца и собирательной трубки
становятся непроницаемыми для воды, реабсорбция воды
при прохождении фильтрата через мозговое
вещество уменьшается и, как следствие, выводится боль-
Таблица 19.8. Изменения, вызываемые антидиуретическим
гормоном (АДГ) в эпителии дистального извитого канальца
и собирательной трубки
Осмотическое давление
крови
Повышается
Падает
АДГ
Освобождается
Не
освобождается
Эпителий
Проницаем
Непроницаем
Моча
Концентрированная
Разведенная
шой объем гипотонической мочи (рис. 19.29).
В табл. 19.8 суммированы процессы, участвующие
в регуляции водного баланса, а на рис. 19.30 дана
общая схема взаимодействий, регулирующих
водный и солевой баланс.
При недостаточности АДГ возникает
заболевание, называемое несахарным диабетом, при котором
выделяются очень большие количества
гипотонической мочи. Потерю жидкости с мочой приходится
при этом возмещать обильным питьем.
19.7. Регуляция содержания
ионов натрия в крови
Поддержание стабильной концентрации ионов
натрия в крови контролируется гормоном альдостеро-
ном, который вторично влияет также на реабсорб-
KOPKOBOE
ВЕЩЕСТВО
МОЗГОВОЕ ВЕЩЕСТВО
Хлористый натрий
Мочевина
Движение воды
Движение NaCl
Движение мочевины
Рис. 19.28. Пассивное
перемещение воды,
ионов и мочевины
между соседними
прямыми сосудами и
между этими сосудами
и тканевой жидкостью
мозгового вещества.
Прямые сосуды
образуют систему про-
тивоточного обмена.
Цифры - величины
осмолярности
(моемо ль 1кг).
КОРКОВОЕ
ВЕЩЕСТВО
МОЗГОВОЕ ВЕЩЕСТВО
В присутствии АДГ
Ионы Na+ и Cl ~
Движение воды
Движение
мочевины
В отсутствие АДГ
Рис. 19.29. Схема, показывающая действие антидиуретического гормона (АДГ) на
проницаемость дистального извитого канальца и собирательной трубки для воды и мочевины. МЧ-мочевина.
цию воды. Уменьшение объема крови, вызванное
потерей натрия, стимулирует группу клеток,
расположенных между дистальным извитым канальцем
и приносящей артериолой,-так называемый юкста-
гломерулярный аппарат. Эти клетки высвобождают
при стимуляции фермент ренин. Ренин воздействует
на содержащийся в плазме глобулин,
синтезируемый печенью, который при этом превращается в
активный гормон ангиотензин. Этот гормон
побуждает кору надпочечников выделять альдостерон,
а он в свою очередь стимулирует активный перенос
натрия из фильтрата в плазму перитубулярных
капилляров. Это поглощение ионов натрия
сопровождается также реабсорбцией осмотически
эквивалентного количества воды.
19.7.1. Гомеостатический контроль
содержания воды и натрия в
плазме
Когда организм имеет свободный доступ к воде
и получает нормальное количество натрия с пищей,
АДГ и альдостерон не секретируются; в этом случае
эпителий дистального извитого канальца и
собирательной трубки остается непроницаемым для солей,
мочевины и воды и образуется обильная, сильно
разведенная моча. Механизмы, поддерживающие
это нормальное состояние, схематически
представлены на рис. 19.30.
Снижение секреции АДГ
(эффектор)
ПОЧКА
Ocморецепторы в
гипоталамусе (детектор)
Повышенное
потребление
воды (вход)
Снижение осмотического
давления плазмы
Уменьшается
проницаемость дистального
извитого канальца и
собирательной трубки (эффек]
тор)
Выделяется обильная
разведенная моча (выход)
Нормальная осмо-
лярность плазмы
!(заданное значение)
Сниженное
потребление
воды (вход)
Увеличение
осмотического давления плазмы
Осморецепторы в
гипоталамусе (детектор)
Ч
Увеличение
секреции АДГ
(эффектор)
(Нормальная осмоляр
ность плазмы
(заданное значение)
Образуется меньший
объем концентрированной
мочи (выход)
ПОЧКА
Повышается
проницаемость дистального изви
|того канальца и
собирательной трубки (эффек-
Рис. 19.30. Общая схема регуляции осмотического давления плазмы.
19.8. Регуляция рН крови
Бикарбонатная и фосфатная буферные системы
крови препятствуют снижению рН из-за накопления
избытка водородных ионов (H+), образующихся
в процессах метаболизма. Эти системы регулируют
также содержание в крови углекислоты,
образующейся при дыхании, не позволяя ей изменять рН
плазмы (когда CO2 еще находится в крови).
Слишком сильным изменениям химического состава
крови, которые могли бы сдвинуть рН с его
нормальной величины, равной 7,4, препятствует дистальный
извитой каналец: при снижении рН он экскретирует
водородные ионы и задерживает ионы бикарбоната
(гидрокарбоната), а при повышении рН, напротив,
экскретирует ионы бикарбоната и задерживает ионы
водорода. В результате этого рН мочи может
колебаться в пределах от 4,5 до 8,5. Падение рН
стимулирует также клетки почек к образованию иона
аммония (NH^), который, обладая основным
характером, реагирует с поступающими в почки
кислотами и выделяется в виде аммонийных солей.
Глава 20
Размножение
Способность размножаться, т. е. производить новое
поколение особей того же вида,-одна из основных
особенностей живых организмов. В процессе
размножения происходит передача генетического
материала от родительского поколения следующему
поколению, что обеспечивает воспроизведение
признаков не только данного вида, но конкретных
родительских особей. Для вида смысл размножения
состоит в замещении тех его представителей, которые
гибнут, что обеспечивает непрерывность
существования вида; кроме того, при подходящих условиях
размножение позволяет увеличить общую
численность вида.
Каждая новая особь, прежде чем достигнуть
стадии, на которой она будет способна к размножению
(см. гл. 21), должна пройти ряд стадий роста и
развития. Некоторые особи погибают, не достигнув
репродуктивной стадии (или половозрелости) в
результате уничтожения хищниками, болезней и
разного рода случайных событий; поэтому вид
может сохраниться лишь при условии, что каждое
поколение будет производить больше потомков,
чем было родительских особей, принимавших
участие в размножении. Численность популяций
колеблется в зависимости от баланса между
размножением и вымиранием особей (разд. 12.6). Существует
ряд различных стратегий размножения, каждая из
которых имеет определенные преимущества и
недостатки; все они будут описаны в этой главе.
20.1. Бесполое и половое
размножение
Существуют два основных типа размножения -
бесполое и половое. Бесполое размножение происходит
без образования гамет, и в нем участвует лишь один
организм. При бесполом размножении обычно
образуются идентичные потомки, а единственным
источником генетической изменчивости служат
случайные мутации.
Генетическая изменчивость выгодна виду, так как
она поставляет «сырье» для естественного отбора, а
значит, и для эволюции. Потомки, оказавшиеся
наиболее приспособленными к среде, будут
обладать преимуществом в конкуренции с другими
представителями того же вида и будут иметь больше
шансов выжить и передать свои гены следующему
поколению. Благодаря этому виды способны
изменяться, т.е. возможен процесс видообразования
(разд. 25.7). Повышение изменчивости может быть
достигнуто путем смешения генов двух разных
особей - процесса, называемого генетической
рекомбинацией и составляющего важную особенность
полового размножения; в примитивной форме
генетическая рекомбинация встречается уже у некоторых
бактерий (разд. 2.2.4).
20.1.1. Бесполое размножение
При бесполом размножении потомки происходят от
одного организма, без слияния гамет. Мейоз в
процессе бесполого размножения не участвует (если
не говорить о растительных организмах с
чередованием поколений; см. разд. 3.3.1), и потомки
идентичны родительской особи. Идентичное потомство,
происходящее от одной родительской особи,
называют клоном. Члены одного клона могут быть
генетически различными только в случае
возникновения случайной мутации. Высшие животные не
способны к бесполому размножению, однако в
последнее время было сделано несколько успешных
попыток клонировать некоторые виды
искусственным образом; мы их рассмотрим в дальнейшем.
Существует несколько типов бесполого
размножения. Подробное описание этого процесса у
некоторых организмов, упоминаемых ниже, можно
найти в соответствующих разделах глав 2, 3 и 4.
Деление
Делением размножаются одноклеточные
организмы: каждая особь делится на две или большее число
дочерних клеток, идентичных родительской клетке.
Делению клетки предшествует репликация ДНК, а у
эукариот - также деление ядра. В большинстве
случаев происходит бинарное деление, при котором
образуются две идентичные дочерние клетки. Так
делятся бактерии, многие простейшие, такие как
амеба или парамеция, и некоторые одноклеточные
водоросли, например эвглена. При подходящих
условиях это приводит к быстрому росту популяции,
как было описано в разд. 2.2.4, где речь шла о
бактериях.
Множественное деление, при котором вслед за
рядом повторных делении клеточного ядра
происходит деление самой клетки на множество дочерних
клеток, наблюдается у споровиков - группы
простейших, к которой относится, в частности,
возбудитель малярии Plasmodium. Стадия, на которой
происходит множественное деление, называется шизон-
том, а сам этот процесс - шизогонией. У Plasmodium
шизогония непосредственно следует за заражением
хозяина, когда паразит проникает в печень. При
этом получается сразу около тысячи дочерних
клеток, каждая из которых способна инвазировать
эритроцит и произвести путем шизогонии еще 24
дочерние клетки. Такая высокая плодовитость
компенсирует большие потери из-за трудностей
успешной передачи паразита от одного хозяина другому,
а именно от человека организму-переносчику, т.е.
малярийному комару, и в обратном направлении.
Образование спор (споруляция)
Спора-это одноклеточная репродуктивная единица
обычно микроскопических размеров, состоящая из
небольшого количества цитоплазмы и ядра.
Образование спор наблюдается у бактерий, простейших,
у представителей всех групп зеленых растений и всех
групп грибов. Споры могут быть различными по
своему типу и функции и часто образуются в
специальных структурах. Например, у Rhizopus и
Dryopteris споры образуются в спорангиях;
микроспоры (пальцевые зерна) и мегаспоры (зародышевые
мешки) семенных растений образуются в особых
спорангиях, называемых соответственно пыльцевым
мешком и семязачатком.
Нередко споры образуются в больших
количествах и имеют ничтожный вес, что облегчает их
распространение ветром, а также животными,
главным образом насекомыми. Вследствие малых
размеров спора обычно содержит лишь минимальные
запасы питательных веществ; из-за того что многие
споры не попадают в подходящее место для
прорастания, потери спор очень велики. Главное
достоинство таких спор - возможность быстрого
размножения и расселения видов, в особенности грибов.
Споры бактерий служат, строго говоря, не для
размножения, а для того, чтобы выжить при
неблагоприятных условиях, поскольку каждая
бактерия образует только одну спору. Бактериальные
споры относятся к числу наиболее устойчивых: так,
например, они нередко выдерживают обработку
сильными дезинфицирующими веществами и
кипячение в воде.
Хотя этот раздел посвящен бесполому
размножению, следует указать, что существуют и половые
споры-они участвуют в половом размножении.
Таковы, например, зооспоры хламидомонады, иногда
выполняющие функции гамет, и зигоспоры Phizopus
и спирогиры. Зигоспоры-довольно крупные споры,
содержащие запасы питательных веществ и
окруженные защитной оболочкой; они образуются в
результате полового размножения, а в периоды
неблагоприятных условий могут переходить в
состояние покоя.
Необходимо отметить, что один организм может
производить споры более чем одного типа;
например, Rhizopus образует половые и бесполые споры, а
высшие растения производят бесполым путем
микро- и мегаспоры.
Почкование
Почкованием называют одну из форм бесполого
размножения, при которой новая особь образуется в
виде выроста (почки) на теле родительской особи, а
затем отделяется от нее, превращаясь в
самостоятельный организм, совершенно идентичный
родительскому. Почкование встречается в разных
группах организмов, особенно у кишечнополостных,
например у гидры (рис. 20.1), и у одноклеточных
грибов, таких как дрожжи. В последнем случае
почкование отличается от деления (которое тоже
наблюдается у дрожжей) тем, что две образующиеся
части имеют разные размеры.
Необычная форма почкования описана у сукку-
лентного растения Bryophyllum - ксерофита, часто
выращиваемого в качестве декоративного
комнатного растения: по краям его листьев развиваются
миниатюрные растеньица, снабженные маленькими
корешками (рис. 20.2); эти «почки» в конце концов
отпадают и начинают существовать как
самостоятельные растения.
Размножение фрагментами
(фрагментация)
Фрагментацией называют разделение особи на две
или несколько частей, каждая из которых растет и
образует новую особь. Фрагментация происходит,
например, у нитчатых водорослей, таких как
спирогира. Нить спирогиры может разорваться на две
части в любом месте.
Фрагментация наблюдается также у некоторых
низших животных, которые в отличие от более
высокоорганизованных форм сохраняют
значительную способность к регенерации из относительно
слабо дифференцированных клеток. Например, тело
немертин (группа примитивных червей, главным
образом морских) особенно легко разрывается на
много частей, каждая из которых может дать в
результате регенерации новую особь. В этом случае
регенерация - процесс нормальный и регулируемый;
однако у некоторых животных (например, у морских
звезд) восстановление из отдельных частей
происходит только после случайной фрагментации.
Животные, способные к регенерации, служат объектами
для экспериментального изучения этого процесса;
часто при этом используют свободноживущего
червя планарию. Такие эксперименты помогают
понять процесс дифференцировки (см. разд. 22.8).
Родительская
особь
Почка
Рис. 20.1. Почкование у гидры.
Вегетативное размножение
Вегетативное размножение представляет собой одну
из форм бесполого размножения, при которой от
растения отделяется относительно большая, обычно
дифференцированная, часть и развивается в
самостоятельное растение. По существу вегетативное
размножение сходно с почкованием. Нередко
растения образуют структуры, специально
предназначенные для этой цели: луковицы, клубнелуковицы,
корневища, столоны и клубни. Некоторые из этих
структур служат также для запасания питательных
веществ, что позволяет растению пережить периоды
неблагоприятных условий, таких как холода или
засуха. Запасающие органы позволяют растению
переживать зиму и давать в следующем году цветки
и плоды (двулетние растения) или выживать в
течение ряда лет (многолетние растения). К таким
органам, называемым зимующими, относятся луковицы,
клубнелуковицы, корневища и клубни.
Зимующими органами могут быть также стебли,
корни или целые побеги (почки), однако во всех
случаях содержащиеся в них питательные вещества
создаются главным образом в процессе
фотосинтеза, происходящего в листьях текущего года.
Образовавшиеся питательные вещества переносятся в
запасающий орган, а затем обычно превращаются в
Рис. 20.2. Бесполое размножение у Bryophyllum: по
краям листьев видны молодые растеньица.
какой-либо нерастворимый резервный материал,
например крахмал. При наступлении
неблагоприятных условий надземные части растения отмирают, а
подземный зимующий орган переходит в состояние
покоя. В начале следующего вегетационного
периода запасы питательных веществ мобилизуются с
помощью ферментов: почки пробуждаются, и в них
начинаются процессы активного роста и развития за
счет запасенных питательных веществ. Если
прорастает более одной почки, то можно считать, что
осуществилось размножение. Эта
последовательность событий тесно согласована со сменой времен
года, поскольку она регулируется такими внешними
факторами, как длина светового дня (фотопериод) и
температура, глубокое влияние которых на рост и
развитие было описано в гл. 15.
луковица. Это видоизмененный побег, имеющийся,
например, у лука (Allium), нарцисса (Narcissus) и
тюльпана (Tulipa). Она служит как зимующим
органом, так и органом вегетативного размножения.
Луковица состоит из очень короткого стебля
и мясистых листьев, содержащих запасные
питательные вещества. Снаружи она покрыта бурыми
пленчатыми листьями - остатками прошлогодних
листьев, запасные вещества которых были
израсходованы. Луковица содержит одну или несколько
дочерних луковиц (детки или зубки); каждая из них
может образовать побег, который к концу
вегетационного периода дает новую луковицу, и если их
будет несколько, то это уже вегетативное
размножение. Корни у луковиц придаточные, т.е. отходят
непосредственно от стебля, а главного корня нет.
Типичная луковица изображена на рис. 20.3.
клубнелуковица. Это короткий, вздутый
вертикальный подземный стебель, как, например, у
шафрана (Crocus) или гладиолуса (Gladiolus).
Клубнелуковицы служат как запасающими органами, так и
органами вегетативного размножения.
Клубнелуковица состоит из вздутого основания
стебля, окруженного защитными пленчатыми
листьями; в отличие от луковицы здесь нет
мясистых листьев. Пленчатые листья-это остатки
прошлогодних надземных листьев. Корни
придаточные. К концу вегетационного периода они
укорачиваются и втягивают новую клубнелуковицу в почву.
Клубнелуковица (так же как луковица) содержит
одну или несколько деток, которые могут
обеспечить вегетативное размножение.
Типичная клубнелуковица показана на рис. 20.4
корневище. Оно представляет собой подземный
стебель, растущий горизонтально. У одних
растений, например у ириса и соломоновой печати (PoIy-
gonatum), корневище короткое и вздутое,
содержащее запасные питательные вещества, а у других,
таких как пырей ползучий (Agropyron repens), мята
(Mentha) и астры (Aster sppj, оно длинное и тонкое.
Корневище обычно служит зимующим органом, а
также органом вегетативного размножения.
Корневище несет листья, почки и придаточные
Бурый пленчатый лист — остаток
прошлогоднего надземного листа,
выполянющий защитную функцию
Мясистые листья, содержащие пита
тельные вещества—вздувшиеся
основания надземных листьев
предшествующего вегетационного периода
Верхушечная почка с закладками стебля,
листьев и цветка для следующего
вегетационного периода
Верхушечная почка луковицы
следующего вегетационного периода
Пазушная почка, из которой также
может развиться новое растение
Короткий уплощенный стебель
Остатки придаточных корней
предшествующего вегетационного
периода
Рис. 20.3. Схематический продольный разрез через
покоящуюся луковицу.
Верхушечная почка с закладками стебля,
надземных листьев и цветка для
следующего вегетационного периода
Стебель и клубнелуковица
следующего вегетационного периода
Пазушная почка, из которой тоже
может развиться новое растение
Бурые пленчатые листья —
сморщенные остатки прошлогодних
листьев, несут защитную функцию
Клубнелуковица, образовавшаяся в
результате накопления в основании стебля
питательных веществ к концу
вегетационного периода
Клубнелуковица предшествующего года
Придаточные корни. Некоторые из них
способны сокращаться и втягивать новую
клубнелуковицу в землю
Рис. 20.4. Схематический продольный разрез через
покоящуюся клубнелуковицу.
корни. Листья могут быть либо пленчатые или
чешуйчатые (мелкие, тонкие, беловатого или
коричневатого цвета), как у пырея, либо только зеленые
надземные, как у ириса, либо того и другого типа,
как, например, у соломоновой печати, у которой
листья обоих типов растут и на надземных побегах.
Корневище ириса изображено на рис. 20.5.
столон. Это ползучий горизонтальный стебель,
стелющийся на поверхности почвы, как, например, у
ежевики (Rubus), крыжовника (Grossularia), черной
и красной смородины (Ribes sppj. Столон не служит
зимующим органом. Корни придаточные,
отходящие от узлов.
Схема строения типичного столона представлена
на рис. 20.6.
усы (плети). Это разновидность столонов,
которые быстро растут в длину, как, например, у
земляники (Fragaria) или у лютика ползучего (Ranunculus
repens).
Ус несет пленчатые листья с пазушными почками,
которые дают начало придаточным корням и
новым растениям. После укоренения новых растений
прежние усы в конце концов разрушаются. В роли
уса может выступать главный стебель; ус может
Остатки проводящих пучков -
Кольчатые листовые рубцы на узлах
Старые части у стебля не отмирают
так быстро, как у луковицы и
клубнелуковиц, а сохраняются в
течение нескольких лет
Придаточные корни отходят-
главным образом от узлов
Остатки листьев
Прирост
за один
год
Цветонос
Надземные листья
из верхушечной почки,
поднимающейся вверх
Новое корневище, растущее из
пазушной почки (вегетативное
размножение)
Рис. 20.5. Схема строения корневища ириса.
также расти от одной из нижних пазушных почек на
главном стебле, как показано на рис. 20.7. У
земляники пленчатые листья и пазушные почки есть у
каждого узла, но корни и надземные листья
образует лишь каждый второй узел. Все пазушные почки
могут дать начало новым усам.
клубень. Это подземный запасающий орган,
раздутый в результате накопления питательных
веществ и способный перезимовывать. Клубни живут
только один год, а затем, после того как их
содержимое будет использовано за время вегетационного
периода, они ссыхаются. К концу вегетационного
периода образуются новые клубни, но они
возникают не из старых клубней (в отличие от
клубнелуковиц, возникающих из прежних клубнелуковиц).
Стеблевые клубни представляют собой
структуры, образующиеся на концах тонких корневищ, как
у картофеля (Solarium tuberosum) и топинамбура
(Helianthus tuberosum). Ha их стеблевое
происхождение указывает наличие пленчатых листьев и почек в
их пазухах. В следующем вегетационном периоде
каждая такая почка может дать начало новому
растению.
Корневые клубни - это вздувшиеся придаточные
корни, как, например, у георгины (Dahlia) и у
Ranunculus ficaria. Новые растения развиваются из
пазушных почек у основания старого стебля.
Стеблевой клубень картофеля изображен на рис.
20.8, а корневые клубни георгины-на рис. 20.9.
мясистые стержневые корни. Стержневой корень-
это главный корень, развивающийся из первичного
корешка, т.е. первого корня зародыша. Корневая
система стержневого типа характерна для
двудольных растений. У некоторых растений, таких как
морковь (Daucus), пастернак (Pastinaca), брюква
(Brassica napus), репа (Brassica г ара) и редька (Ra-
phanus sativus), главный корень может утолщаться
за счет развития паренхимной ткани, содержащей
запасные питательные вещества. Вместе с почками,
расположенными у основания старого стебля
непосредственно над стержневым корневым, такие корни
образуют зимующие органы и органы вегетативного
размножения. Мясистые стержневые корни двух
типов показаны на рис. 20.10.
После цветения питательные вещества из листьев
возвращаются в корневище и поступают в пазушную
почку, обеспечивающую дальнейший рост корневища
Увядающие прошлогодние листья, побуревшие и
превратившиеся в чешуйки
Положение скрытой пазушной почки; она запасает
питательные вещества, поступающие из листьев, и
служит для продолжения роста корневища;
в конце последнего образуется верхушечная почка
В начале следующего сезона из нее будут
расти листья и цветки
Мясистые стержневые корни характерны для
двулетних растений, у которых в первый год
происходит вегетативный рост. Затем они переживают
зиму за счет подземного запасающего органа, а на
второй год образуют цветки и семена и к концу года
отмирают.
Помимо описанных выше специализированных
органов вегетативного размножения новое растение
могут регенерировать и некоторые другие,
неспециализированные органы, если отделить их от
родительского растения, например листья суккулента
Sedum. Одной из форм вегетативного размножения
можно считать также размножение черенками и
отводками, применяемое в плодоводстве и
цветоводстве.
черенки и отводки - это части растения, которые в
подходящих условиях пускают корни и образуют
листья, превращаясь в самостоятельные растения.
Таким образом можно искусственно размножать
нужные сорта, которые при этом не изменяются.
Нередко укоренение стимулируют добавлением
ростового гормона. Для вегетативного
размножения с успехом используют побеги пеларгонии и
колеуса, веточки ивы (Salix sppj и Forsythia, а также
листья бегонии и узамбарской фиалки (Saintpaulia
ionantha).
Другим важным и широко используемым
способом искусственного вегетативного размножения
служат прививки. Прививка состоит в пересадке
одного растения (побега или почки) на нижнюю
Родительское
растение
Лист
Пазушная-
почка
Стержневой _
корень
Боковые"
корни
Столон: побег изгиба
ется, от одного из
узлов отрастают
корни,а от почки
в этом узле берет
начало новый побег
Верхушечная почка
Новый побег
Придаточные корни
Рис. 20.6. Обобщенная схема строения столона.
Родительское растение
Стержневой
корень
Боковые
корни
Рис. 20.7. Строение уса земляники.
_Ус, растущий из
пазушной почки
пленчатого листа
Верхушечная почка
Новые растения,
вырастающие из
пазушных почек
Пленчатый лист
Придаточные корни
часть побега другого растения. Пересаженную часть
растения-донора называют привоем, а реципиента-
подвоем. Подвой обрезают над местом прививки.
Получающееся в результате растение обычно
обладает корневой системой подвоя и побегом (в том
числе цветками и плодами) привоя. Этот метод
применяется в широких масштабах для
размножения розовых кустов и плодовых деревьев, в
особенности яблонь. Он имеет два преимущества -
позволяет сочетать в одном растении желательные
признаки двух разных сортов или видов и дает
возможность быстро получать в больших количествах
новые комбинации привой/подвой для продажи.
Иногда удается получить прививки нескольких привоев
на данном подвое; примером служат яблони, у
которых на одном дереве вырастают десертные
плоды и плоды, предназначенные для варки варенья
и консервирования.
Клонирование высших растений и
животных
Как уже говорилось, получение идентичных
потомков при помощи бесполого размножения
называют клонированием. В начале шестидесятых годов
были разработаны методы, позволяющие успешно
клонировать некоторые высшие растения и
животных. Эти методы возникли в результате попыток
доказать, что ядра зрелых клеток, закончивших свое
развитие, содержат всю информацию, необходимую
для кодирования всех признаков организма, и что
специализация клеток обусловлена включением и
выключением определенных генов, а не утратой
некоторых из них (см. разд. 22.7). Первый успех был
достигнут проф. Стюардом из Корнелльского
университета, который показал, что, выращивая
отдельные клетки корня моркови (ее съедобной части)
в среде, содержащей нужные питательные вещества
и гормоны, можно индуцировать процессы
клеточного деления, приводящие к образованию новых
растений моркови.
Вскоре после этого Гёрдон, работавший в
Оксфордском университете, впервые сумел добиться
клонирования позвоночного животного.
Позвоночные в естественных условиях клонов не образуют;
однако, пересаживая ядро, взятое из клетки
кишечника лягушки, в яйцеклетку, собственное ядро
которой предварительно было разрушено путем
облучения ультрафиолетом, Гёрдону удалось вырастить
головастика, а затем и лягушку, идентичную той
особи, от которой было взято ядро (рис. 20.11).
Такого рода эксперименты не только
доказывают, что дифференцированные
(специализированные) клетки содержат всю информацию,
необходимую для развития целого организма, но и
позволяют рассчитывать, что подобные методы можно
будет использовать для клонирования позвоночных,
стоящих на более высоких ступенях развития, в том
числе и человека. Клонирование нужных животных,
например племенных быков, скаковых лошадей и
т.п., может оказаться столь же выгодным, как и
клонирование растений, которое, как было сказано,
уже производится. Однако применение методов
клонирования к человеку сопряжено с серьезными
проблемами нравственного порядка. Теоретически
можно создать любое число генетически
тождественных копий данного мужчины или данной жен-
Рубец
пленчатого
листа
Пазушная
( почка
"Глазок"; глазки расположены
на клубне по спирали и каждый
из них в следующем
вегетационном периоде может
дать начало новому побегу
Корневище растения
нынешнего года; несет мелкие
пленчатые листья с пазушными
почками. Корневище оставляет
рубец на клубне, отделяющемся
в конце вегетационного периода
Чечевички в наружном
пробковом слое клубня
Верхушечная почка
Стеблевой клубень,
запасающий крахмал
Рис. 20.8. Стеблевой клубень картофеля.
Стебель
Корневой клубень
— раздувшийся
придаточный корень,
содержащий инулин
(полимер фруктозы)
Рис. 20.9. Корневые луковицы георгины.
Кора
Боковой корень
Проводящая ткань
бокового корня
Стель:
паренхиматозные
клетки,
наполненные
питательными
веществами
Проводящая ткань
стержневого
корня
Морковь
(продольный разрез)
Рис. 20.10. Стержневые корни моркови и репы.
Почки
.(пазушные и верхушечные).
Черешки листьев
„нынешнего года;
надземные части
к концу вегетационного
периода отмирают
Стержневой корень,
содержащий питательные
вещества, синтезированные
листьями нынешнего года
Морковь
Мелкие боковые корни
Собственно корни
Репа
щины. На первый взгляд может показаться, что
таким образом можно было бы воспроизводить
талантливых ученых или деятелей искусства.
Однако надо помнить, что степень влияния,
оказываемого на развитие средой, еще не вполне ясна, а
между тем любая клонируемая клетка должна снова
пройти через все стадии развития, т. е. в случае
человека - стадии зародыша, плода, младенца и т.д.
20.1.2. Половое размножение
При половом размножении потомство получается в
результате слияния генетического материала
гаплоидных ядер. Обычно эти ядра содержатся в
специализированных половых клетках - гаметах; при
оплодотворении гаметы сливаются, образуя
диплоидную зиготу, из которой в процессе развития
получается зрелый организм. Гаметы гаплоидны-они
содержат один набор хромосом, полученный в
результате мейоза; они служат связующим звеном
между данным поколением и следующим (при
половом размножении цветковых растений сливаются не
клетки, а ядра, но обычно эти ядра тоже называют
гаметами.)
Мейоз- важный этап жизненных циклов,
включающих половое размножение, так как он ведет к
уменьшению количества генетического материала
вдвое. Благодаря этому в ряду поколений,
размножающихся половым путем, это количество остается
постоянным, хотя при оплодотворении оно каждый
раз удваивается. Во время мейоза в результате
случайного расхождения хромосом (независимое
распределение) и обмена генетическим материалом
между гомологичными хромосомами (кроссинговер)
возникают новые комбинации генов, попавших в
одну гамету, и такая перетасовка повышает
генетическое разнообразие (см. разд. 22.3). Слияние
содержащихся в гаметах гаплоидных ядер называют
оплодотворением или сингамией; оно приводит к
образованию диплоидной зиготы, т.е. клетки,
содержащей по одному хромосомному набору от
каждого из родителей. Это объединение в зиготе
двух наборов хромосом (генетическая рекомбинация)
представляет собой генетическую основу
внутривидовой изменчивости. Зигота растет и развивается
в зрелый организм следующего поколения. Таким
образом, при половом размножении в жизненном
цикле происходит чередование диплоидной и
гаплоидной фаз, причем у разных организмов эти фазы
принимают различные формы (см. рис. 20.13).
Гаметы обычно бывают двух типов - мужские и
женские, но некоторые примитивные организмы
производят гаметы только одного типа (см. в разд.
20.2 об изогамии, анизогамии и оогамии). У
организмов, образующих гаметы двух типов, их могут
производить соответственно мужские и женские
родительские особи, а может быть и так, что у одной и
той же особи имеются и мужские, и женские
половые органы. Виды, у которых существуют
отдельные мужские и женские особи, называются
раздельнополыми; таковы большинство животных и
человек. Среди цветковых растений тоже есть
раздельнополые виды; если у однодомных видов
мужские и женские цветки образуются на одном и том
же растении, как, например, у огурца (Cucumis) и
лещины (CorуIm), то у двудомных одни растения
несут только мужские, а другие - только женские
цветки, как у остролиста (Ilex) или у тиса (Taxus).
Гермафродитизм
Виды, у которых одна и та же особь способна
производить и мужские, и женские гаметы,
называют гермафродитными или двуполыми. К их числу
относятся многие простейшие, в том числе
парамеция, такие кишечнополостные, как Obelia, плоские
черви, например солитер (Taenia), олигохеты,
например дождевой червь (Lumbricus), ракообразные,
например морской желудь (Balanus), такие
моллюски, как улитка (Helix), некоторые рыбы и
ящерицы, а также большинство цветковых растений.
Гермафродитизм считается самой примитивной
формой полового размножения и свойствен многим
примитивным организмам. Он представляет собой
приспособление к сидячему, малоподвижному или
паразитическому образу жизни. Одно из
преимуществ гермафродитизма состоит в том, что он
делает возможным самооплодотворение, а это весь-
Мутанты-альбиносы — доноры
Самка
дикого
типа —
реципиент
Рис. 20.11. Клон шпорцевых лягушек (Xenopus laevis), полученный путем трансплантации ядер. А. От
скрещивания между двумя мутантными лягушками-альбиносами был получен зародыш (донор), на стадии
хвостовой почки его клетки были диссоциированы и выделенные ядра
пересажены в неоплодотворенные яйца самки дикого типа (реципиента), ядра которых были
разрушены УФ-облучением. Б. Группа из 30 лягушек, полученных в результате 54 таких
пересадок, все они самки-альбиносы.
ма существенно для некоторых эндопаразитов,
таких как солитер, ведущих одиночное существование.
Однако у большинства гермафродитных видов в
оплодотворении участвуют гаметы, происходящие
от разных особей, и у них имеются многочисленные
генетические, морфологические и физиологические
адаптации, препятствующие самооплодотворению и
благоприятствующие перекрестному
оплодотворению. Например, у многих простейших
самооплодотворение предотвращается генетической
несовместимостью, у многих цветковых растений -
строением андроцея и гинецея, а у многих животных-
тем, что яйца и спермии образуются у одной и той
же особи в разное время.
Партеногенез
Партеногенез-одна из модификаций полового
размножения, при которой женская гамета развивается
в новую особь без оплодотворения мужской
гаметой. Партеногенетическое размножение встречается
как в царстве животных, так и в царстве растений, и
преимущество его состоит в том, что в некоторых
случаях оно повышает скорость размножения.
Существует два вида партеногенеза - гаплоидный
и диплоидный, в зависимости от числа хромосом в
женской гамете. У многих насекомых, в том числе у
муравьев, пчел и ос, в результате гаплоидного
партеногенеза в пределах данного сообщества
возникают различные касты организмов. У этих видов
происходит мейоз и образуются гаплоидные
гаметы. Некоторые яйцеклетки оплодотворяются, и из
них развиваются диплоидные самки, тогда как из
неоплодотворенных яйцеклеток развиваются фер-
тильные гаплоидные самцы. Например, у
медоносной пчелы (Apis mellifera) матка откладывает
оплодотворенные яйца (2/7 = 32), которые, развиваясь,
дают самок (маток или рабочих особей), и не-
оплодотворенные яйца (п = 16), которые дают
самцов (трутней), производящих спермии путем
митоза, а не мейоза. Развитие особей этих трех типов у
медоносной пчелы схематически представлено на
рис. 20.12. Такой механизм размножения у
общественных насекомых имеет адаптивное значение, так
как позволяет регулировать численность потомков
каждого типа.
У тлей происходит диплоидный партеногенез, при
котором ооциты самки претерпевают особую
форму мейоза без расхождения хромосом (см. разд.
22.3)-все хромосомы переходят в яйцеклетку, а
полярные тельца не получают ни одной хромосомы.
Яйцеклетки развиваются в материнском организме,
так что молодые самки рождаются вполне
сформировавшимися, а не вылупляются из яиц. Такой
процесс называется живорождением. Он может
продолжаться в течение нескольких поколений,
особенно летом, до тех пор пока в одной из клеток не
произойдет почти полное нерасхождение, в
результате чего получается клетка, содержащая все пары
аутосом и одну Х-хромосому. Из этой клетки пар-
теногенетически развивается самец. Эти осенние
самцы и партеногенетические самки производят в
результате мейоза гаплоидные гаметы, участвующие
в половом размножении. Оплодотворенные самки
откладывают диплоидные яйца, которые
перезимовывают, а весной из них вылупляются самки,
размножающиеся партеногенетически и рождающие
живых потомков. Несколько партеногенетических
поколений сменяются поколением, возникающим в
результате нормального полового размножения,
что вносит в популяцию генетическое разнообразие
в результате рекомбинации. Главное преимущество,
которое дает тлям партеногенез,- это быстрый рост
численности популяции, так как при этом все ее
половозрелые члены способны к откладке яиц. Это
особенно важно в периоды, когда условия среды
благоприятны для существования большой
популяции, т.е. в летние месяцы.
Партеногенез широко распространен у растений,
где он принимает различные формы. Одна из них-
апомиксис -представляет собой партеногенез,
имитирующий половое размножение. Апомиксис
наблюдается у некоторых цветковых растений, у
которых диплоидная клетка семязачатка - либо
клетка нуцеллуса, либо мегаспора - развивается в
функциональный зародыш без участия мужской
гаметы. Из остального семязачатка образуется семя, а
из завязи развивается плод. В других случаях
требуется присутствие пыльцевого зерна, которое
стимулирует партеногенез, хотя и не прорастает;
пыльцевое зерно индуцирует гормональные изменения,
необходимые для развития зародыша, и на практике
такие случаи трудно отличить от настоящего
полового размножения.
Спермии
(п = 16)
Митоз
Матка
(фертильная самка)
(личинка получает
маточное молочко)"
(личинка получает мед
и пыльцу)
Матка
Оплодотворенное
яйцо (2 л =32)
Неоплодотворенное
яйцо ( η = 16)
(личинка получает
мед и пыльцу)
Партеногенез
Трутень
Рабочая особь
(стерильная самка)
Рис. 20.12. Схема, иллюстрирующая роль партеногенеза в жизненном цикле семьи
медоносной пчелы.
20.1.3. Происхождение полового
процесса
Любые рассуждения о событиях, происходящих на
неведомом нам этапе истории жизни, носят чисто
умозрительный характер, и вытекающие из них
выводы следует считать лишь
предположительными. Именно так обстоит дело с происхождением
полового процесса. Предполагается, что бесполое
размножение представляет собой примитивную
форму вопроизведения как способа «копирования»
генетически идентичных поколений. На более
позднем этапе возникли механизмы, делающие
возможным обмен нуклеиновыми кислотами между
особями. Быть может, при этом происходило
слияние целых организмов, за которым следовал мейоз;
и в самом деле, такой процесс мог предшествовать
образованию гамет и их слиянию. Принято считать,
что способы обмена генетическим материалом,
наблюдаемые у бактерий (см. разд. 2.2.4),
представляют собой ранний этап развития полового
процесса. Преимущество, которое дает фенотипи-
ческая изменчивость, обусловленная мейозом и
генетической рекомбинацией, несомненно, сыграло
важную роль в развитии более сложных форм
жизни и типов гамет - от идентичных гамет (изогамет) к
гетерогаметной стадии с подвижными мужскими
гаметами (спермиями и антерозоидами) и
неподвижными женскими гаметами (яйцеклетками). Тот
факт, что половое размножение мы находим во всех
крупных группах организмов, позволяет думать,
что оно обладает преимуществом перед бесполым,
которое, будучи редким у животных, стоящих по
своей организации выше плоских червей,
сохраняется у растений: в жизненных циклах большинства
растений наблюдается чередование бесполого и
полового способов размножения (разд. 20.2). Одна
из причин этого состоит в том, что у неподвижных
организмов обмен гаметами затруднен и они во
многих случаях предпочитают бесполое
размножение. Кроме того, при бесполом размножении
родительские особи, обосновавшиеся в определенном
месте, облегчают потомкам решение нелегкой
экологической проблемы - нахождение подходящего
местообитания. Однако при этом возникают другие
проблемы, создаваемые перенаселением, что
способствует ограничению размеров растительных
популяций.
Некоторые из наиболее существенных и типичных
черт бесполого и полового размножения
перечислены в табл. 20.1.
20.1.4. Разнообразие жизненных
циклов
Последовательность стадий развития, через
которые проходят представители данного вида от
зиготы одного поколения до зиготы следующего,
называют жизненным циклом. Сложность
жизненных циклов у разных организмов различна; в
некоторых случаях жизненный цикл включает два и
больше поколений, различающихся по своей морфоло-
Таблица 20.1. Сравнение бесполого и полового размножения
Бесполое размножение
Одна родительская особь
Гаметы не образуются
Мейоз отсутствует
Потомки идентичны
родительским особям
Характерно для
растений, некоторых
низших животных и
микроорганизмов. У
высших животных не
встречается
Часто приводит к
быстрому созданию
большого числа
потомков
Половое размножение (исключая
бактерий)
Обычно две родительские
особи
Образуются гаплоидные
гаметы, ядра которых
сливаются (оплодотворение),
так что получается
диплоидная зигота
На какой-либо стадии
жизненного цикла происходит
мейоз, что препятствует
удвоению хромосом в каждом
поколении1 >
Потомки не идентичны
родительским особям.У них
наблюдается генетическая
изменчивость, возникающая в
результате генетической
рекомбинации
Характерно для большинства
растений и животных
Менее быстрое увеличение
численности
" Обычный источник недоразумений — непонимание роли
митоза и мейоза в процессе размножения Для ее правильного
понимания необходимо изучить различные жизненные циклы и ясно
представлять себе, на какой стадии происходит каждый из этих двух
типов ядерного деления (см рис 20 13 и разд.20 1 4).
гии (внешнему виду) и способу
размножения,-явление, называемое иногда чередованием поколений.
Применение этого термина лучше ограничить
наземными растениями и некоторыми эволюционно
продвинутыми водорослями, у которых
наблюдается чередование диплоидного спорообразующе-
го поколения, называемого спорофитом, и
гаплоидного гаметофита, образующего гаметы (см. разд.
3.3.1). Чередование бесполого и полового поколений
имеет место также у некоторых кишечнополостных,
но в этом случае оба поколения диплоидны, а
гаплоидная стадия представлена только гаметами.
Такую форму чередования поколений иногда
называют метагенезом (см. разд. 4.4.3). В жизненном
цикле этих кишечнополостных разные формы
особей данного вида сменяют одна другую - явление,
известное под названием циклического
полиморфизма. Например, в жизненном цикле обелии
последовательно сменяются три различные формы-
две полипоидные и одна медузоидная (рис. 4.11).
Полиморфизм встречается и у некоторых других
организмов, например у первоцвета (Primula), обра-
Половой организм
С)
Гаметы
Зигота
(2")
Половое поколение
(2л)
медуза
Мейоз
Гаметы
Бесполое
поколение
(2п)
Размножающийся полип
(2")
Зигота
(2л)
Питающийся полип
(2л)
Половой организм
(2л) X \
Мейоз
Гаметы
/(я)
Зигота
(2л)
Споры
Половое поколение
(л)
гаметофит
Бесполое
поколение
(2л)
спорофит
Гаметы
Зигота
(2л)
Рис. 20.13. Схемы различных часто встречающихся жизненных циклов. Обратите внимание, что
в жизненном цикле каждого типа мейоз происходит только один роз. Но схемах А, Б и Г бесполое
размножение полового организма не показано, хотя у ряда видов оно возможно, п- гаплоид,
2п-диплоид.
A. Взрослый (половой) организм гаплоидный. Диплоидна только зигота. Первое деление ядра в прорастающей зиготе
происходит путем мейоза и приводит к возврату в гаплоидное состояние. Гаметы образуются не в результате мейоза.
Примеры: Chlamydomonas, Spirogyra, Rhizopus.
Б. Гаплоидны только гаметы, они образуются в результате мейоза. Примеры: Fucus, позвоночные и большинство других
животных.
B. В жизненном цикле участвуют три морфологически различные формы (полиморфизм). Все они диплоидные. Диплоидное
половое поколение чередуется с диплоидным бесполым поколением. Единственные гаплоидные клетки-это гаметы, они
образуются путем мейоза. Пример: Obelia.
Г. Чередование гаплоидных и диплоидных поколений. Гаметы образуются не в результате мейоза. Примеры: Laminaria (бурая
водоросль), все наземные растения, т. е. мхи, папоротники и семенные растения.
зующего цветки двух типов-длинностолбчатые и
короткостол бчатые.
Жизненные циклы паразитов часто бывают
весьма сложными и включают несколько поколений.
Каждое поколение приспособлено к определенной
жизненной ситуации -либо к существованию в
организме определенного хозяина, либо к переходу от
одного хозяина к другому (примером служит
жизненный цикл печеночной двуустки, см. разд. 4.5.3).
Добавочные поколения, возникающие путем
бесполого размножения (полиэмбриония), обеспечивают
увеличение численности данного вида. Некоторые
типичные жизненные циклы представлены на
рис. 20.13.
Ввиду того что вопрос об относительной роли
митоза и мейоза в бесполом и половом
размножении иногда вызывает затруднения, мы еще раз
рассмотрим здесь значение каждого из этих
процессов в жизненном цикле. Мейоз происходит только в
тех жизненных циклах, которые включают половое
размножение. Зигота, образующаяся в результате
слияния ядер двух гаплоидных гамет, содержит
диплоидное число хромосом и начинает следующий
жизненный цикл. Если на какой-либо стадии этого
цикла не произойдет мейоза, то ядра образующихся
гамет будут диплоидными, и новая зигота в таком
случае окажется тетраплоидной (будет содержать
четыре набора хромосом). Таким образом, мейоз
необходим для того, чтобы предотвратить удвоение
хромосом в каждом поколении в жизненных циклах,
включающих половое размножение. Мейоз не
обязательно происходит непосредственно перед
образованием гамет (см. рис. 20.13,,4 и 7"). Еще одно
преимущество мейоза-то, что он создает
изменчивость в результате независимого распределения
хромосом и кроссинговера (разд. 22.3). Эти
процессы непосредственно (жизненные циклы Б и В на рис.
22.3) или косвенным образом (циклы Λ и Г)
приводят к различию между гаметами. В особом случае
чередования поколений (жизненный цикл Г) мейоз
происходит при образовании спор, а не гамет.
Поэтому здесь создаются различия между спорами и
между развивающимися из них гаметофитами. Эта
изменчивость передается ядрам гамет,
образующихся в гаметофитах путем митоза. Во всех
случаях, однако, мейоз составляет основу полового
размножения. В жизненных циклах А, Б и В, т.е. в
циклах, не связанных с чередованием поколений, в
результате бесполого размножения образуются
генетически идентичные потомки, и это происходит
без мейоза.
20.1.А. На основании всего сказанного в разд.
20.1 укажите, какое из сформулированных ниже
утверждений верно.
а) Бесполое размножение всегда приводит к
образованию генетически идентичных
потомков.
б) Гаметы всегда гаплоидны.
в) Гаметы всегда образуются в процессе
мейоза.
г) В результате мейоза всегда образуются
гаплоидные, а в результате митоза -
диплоидные клетки.
д) Митоз происходит только в диплоидных
клетках.
Б. Приведите примеры, опровергающие
четыре остальных утверждения.
20.2. Половое размножение
у растений
У всех зеленых растений, обитающих на суше, а
также у некоторых продвинутых в эволюционном
отношении водорослей наблюдается смена
поколений, при которой гаплоидное гаметофитное
поколение чередуется с диплоидным спорофитным, как это
показано на рис. 20.13,Г. Половое размножение
происходит на стадии гаметофита, который
образует гаметы путем митоза.
Все прочие зеленые растения относятся к
водорослям, и у них можно встретить разнообразные
типы полового размножения (см. разд. 3.2.3). При
простейших типах (обычно их считают
примитивными) все гаметы идентичны, а процесс
размножения называют изогамией; при более сложных (или
эволюционно продвинутых) типах наблюдается
анизогамия-гаметы различаются по величине и
поведению; при крайней форме анизогамии - оогамии -
подвижная мужская гамета подплывает к
неподвижной женской.
На протяжении миллионов лет после выхода из
воды на сушу и у растений, и у животных
вырабатывались разнообразные приспособления,
необходимые для наземной жизни. Наиболее
продвинувшиеся в эволюционном отношении группы растений
и животных, а именно цветковые растения и
млекопитающие, в значительной степени обязаны своим
успехом адаптациям, касающимся полового
размножения, и здесь между этими двумя группами
можно усмотреть некоторые аналогии. Например, и
у тех и у других родительские особи обеспечивают
оплодотворение, не полагаясь на встречу гамет,
выпущенных в воду (как это делают, например, мхи
или амфибии), и, кроме того, обеспечивают
развивающемуся зародышу защиту и питание. Одно из
главных различий состоит в том, что у цветковых
растений гаметы производит гаметофитное
поколение путем митоза, тогда как у млекопитающих нет
чередования поколений и гаметы образуются у
диплоидных родительских особей в результате мейоза.
Как мы увидим, однако, гаметофит у цветковых
растений крайне редуцирован.
20.2.1. Жизненный цикл цветковых
растений (покрытосеменных)
Адаптации цветковых растений к жизни на суше
были описаны в гл. 3. Главные адаптации в области
размножения - это образование семян и плодов,
обеспечивающих питание и защиту зародышам и
способствующих расселению растений, отсутствие
свободноплавающих мужских гамет и крайняя
редукция полового гаметофитного поколения. Мужские
гаметы, заключенные в пыльцевых зернах,
переносятся на женские органы растения (процесс,
называемый опылением), а затем происходит
прорастание пыльцевой трубки, содержащей мужские ядра.
Жизненный цикл цветковых растений
схематически представлен на рис. 3.37, где он сопоставляется с
жизненными циклами, типичными для других групп
растений. В нем все еще сохраняется чередование
поколений, показанное в упрощенном виде на
рис. 20.13, Г. Поскольку гаметофитное поколение
почти исчезло и уже не представлено свободно-
живущей особью, было бы трудно заметить, что
чередование поколений имеет место, если бы не
возможность провести сравнение с более
примитивными предками. Жизненный цикл цветкового
растения подробно описан ниже; строго говоря, он
складывается из бесполого размножения
доминирующего спорофита (цветковое растение как таковое) и
полового размножения гаметофита.
20.2.2. Строение и функции цветка
Термин «цветковые растения», как обычно
называют покрытосеменные, указывает на признак,
присущий только этой группе,-наличие цветков.
Цветки-это репродуктивные органы, эволюционное
происхождение которых неясно; некоторые авторы
полагают, однако, что цветок построен из
высокоспециализированных листьев и аналогичен шишке
голосеменных, так как несет спорообразующие
структуры. Цветки можно считать органами как
бесполого, так и полового размножения - бесполого
потому, что они производят споры (пыльцевые
зерна и зародышевые мешки), а полового - потому что
в дальнейшем в спорах образуются гаметы. Обычно
цветки называют просто органами полового
размножения. В областях с четко выраженной сменой
времен года цветение приурочено к определенному
Один или несколько
плодолистиков
-* ГИНЕЦЕЙ
/Столбик
Завязь
Семязачаток
(один или несколько)
Прицветник
(маленькая
листовидная структура)
Рыльце
Цветок -
Пыльник Л
Тычиночная
-"— нить _,
Тычинка
АНДРОЦЕЙ
Нектарник
Лепесток —►ВЕНЧИК^
Цветоножка _
ОКОЛО-
Самый
молодой цветок
Цветок
Цветоножка.
Прицветник
Ось соцветия -
-Чашелистик-*ЧАШЕЧКА/ЦВЕТНИК
Цветоложе
Цветоножка
Модифицированные кисти
Одиночный цветок
(без цветоножки),
например
у тюльпана
Главная ось соцветия
продолжает расти,
например у
колокольчика,
желтофиоли
Платформа для насекомых
Щиток цветоножки
растут до одного и
того же уровня,
например у иберийки
Самый молодой цветок
Самый
старый о
цветок v
Цветоножка.]
Лучи
зонтика
Ось
соцветия
Цветоножка
Самый старый цветок
Самый
молодой цветок
Колосок: цветки
сидячие(без
цветоножек),
например у
подорожника
Простой зонтик:
все цветоножки
растут из одной
точки
Сложный зонтик
(встречается чаще,
чем простой зонтик)
оси соцветий растут из
одной точки, например
у моркови, борщевика
I
Завиток:на главной
оси образуется один
цветок, а затем рост
продолжается за
счет боковых почек,
например у лютика
Ложный зонтик:
рост продолжается за
счет двух боковых почек,
например у смолевки
Рис. 20.14. А. Схема строения цветка (продольный разрез). Собирательные термины указаны
стрелками, например, «лепесток -► ВЕНЧИК» означает, что совокупность лепестков называют венчиком.
Б. Расположение цветков по одному или группами (соцветия). Показаны некоторые обычные типы
соцветий, которые делятся на ботрические (или рацемозные кистевидные) и цимозные.
сезону. Регуляция цветения гормонами и факторами
среды описана в разд. 15.5.
Расположение частей цветка на цветоложе -
верхней части цветоноса (цветоножки) - бывает
спиральным или мутовчатым. На рис. 20.14 представлены
обобщенная схема строения цветка и различные
типы соцветий, т. е. групп объединенных цветков, а
ниже мы даем объяснение некоторых терминов,
используемых при описании цветков.
Части цветка
Соцветием называют совокупность цветков,
расположенных на конце одного стебля-ножке соцветия
(рис. 20.14,£). Вероятно, большое число цветков,
собранных вместе, лучше привлекает насекомых-
опылителей, чем маленький одиночный цветок. Дис-
тальную часть цветоножки, на которой
располагаются околоцветник, гинецей и андроцей,
называют цветоложем. Цветок называют подпестичным
(гипогинным), если тычинки и околоцветник
расположены под гинецеем (рис. 20.14,Л), надпес-
тичным (эпигинным), если они расположены над
гинецеем, и околопестичным (перигинным), если
гинецей находится в центре уплощенного или
вогнутого цветоложа, а тычинки и околоцветник
прикреплены к его верхнему краю (см. также описание
верхней и нижней завязи, данное ниже).
Околоцветник состоит из двух мутовок
листовидных частей, называемых чашелистиками и
лепестками. У однодольных обе мутовки обычно
одинаковые, как, например, у нарцисса (Narcissus),
тюльпана (Tulipa) и пролески Endymion (Scilla) non-scripta.
У двудольных мутовки часто отличаются одна от
другой: наружная, состоящая из чашелистиков,
называется чашечкой, а внутренняя, состоящая из
лепестков,- венчиком.
Чашелистики, образующие чашечку,-это обычно
листовидные образования зеленого цвета, которые
служат для защиты заключенным внутри
цветочным бутонам. Иногда они могут быть ярко
окрашены и похожи на лепестки; в таких случаях они
тоже участвуют в привлечении
насекомых-опылителей.
Свободнолистная чашечка состоит из свободных
(несросшихся) чашелистиков, а спайнолистная - из
чашелистиков, которые хотя бы частично слились в
трубку.
Венчиком называют совокупность лепестков
цветка. У растений, опыляемых насекомыми, лепестки
обычно крупные и яркоокрашенные-они
привлекают насекомых к цветкам. У ветроопыляемых
растений лепестки чаще мелкие и зеленые, а иногда
отсутствуют вовсе.
Свободнолепестный венчик состоит из свободных
(несросшихся) лепестков, как, например, у горошка
(Pisum), розы (Rosa) и лютика (Ranunculus); у
спайнолепестного венчика лепестки хотя бы частично
слиты в трубочку, как, например, у наперстянки
(Digitalis), первоцвета (Primula), одуванчика
(Taraxacum) и яснотки (lamium album).
Андроцей-это совокупность тычинок, образую-
щих мужские репродуктивные органы цветка.
Каждая тычинка состоит из пыльника и тычиночной
нити. Пыльник содержит пыльцевые мешки, в
которых образуется пыльца. Тычиночная нить содержит
проводящий пучок, по которому в пыльник
поступают питательные вещества и вода.
Гинецей, или пестик,-это совокупность
плодолистиков, или карпелей, образующих женские
репродуктивные органы цветка. Плодолистик состоит
из рыльца, столбика и завязи. Столбик
поддерживает рыльце в таком положении, чтобы на него
могла попадать пыльца во время опыления. Завязь
представляет собой вздутое полое основание
плодолистика, содержащее один или несколько
семязачатков. Семязачатки - это структуры, в которых
развиваются зародышевые мешки и которые после
оплодотворения превращаются в семена. Каждый
семязачаток прикреплен к стенке завязи короткой
ножкой - фуникулусом, а место его прикрепления
носит название плаценты.
Плодолистики цветка могут оставаться
свободными, т.е. не связанными друг с другом, образуя
апокарпный гинецей, как у лютика, или же слиты в
единую структуру-синкарпный гинецей, как у
яснотки. Завязь, образуемая слившимися
плодолистиками, может иметь одну камеру, или гнездо (одно-
гнездная завязь), или же несколько (многогнездная
завязь); в последнем случае каждый из
составляющих ее плодолистиков образует по одному гнезду.
Столбики синкарпного гинецея могут сливаться или
сохранять обособленность.
Верхняя завязь - завязь, расположенная на
цветоложе над другими частями цветка, как в подпестич-
ном или околопестичном цветке; нижняя завязь
расположена на цветоложе под другими частями
цветка, как в надпестичном цветке.
Нектарники - это железистые образования,
выделяющие нектар - сахаристую жидкость, которая
привлекает животных-опылителей (обычно это
насекомые, но в тропических областях опылителями
могут быть также птицы и летучие мыши).
Термины, перечисленные ниже, приложимы как
к растению в целом, так и к цветку.
Гермафродитные (обоеполые) растения: мужские и
женские половые органы образуются на одном и
том же растении.
Однодомные растения: мужские и женские цветки
растут по отдельности на одном и том же растении,
как, например, у дуба (Quercus), лещины (Corylus),
бука (Fagus) и белого клена (Acer pseudoplatanus).
Двудомные растения: мужские и женские половые
органы образуются на разных растениях, т.е.
каждое данное растение является либо мужским, либо
женским, как, например, у тиса (Тахш)% ивы (Salix),
тополя (Populus) и остролиста (Ilex).
Гермафродитные (обоеполые) цветки: мужские и
женские половые органы развиваются в одном и
том же цветке, как у лютика, яснотки, пролески
Endymion и гороха.
Однополые цветки: они несут либо только
тычинки, либо только плодолистики, как, например, у
дуба, лещины, тиса, тополя и остролиста.
Симметрия цветков
Если части цветка располагаются на цветоложе
радиально-симметрично, то цветок называют
правильным или актиноморфным (например, у лютика
или пролески). Если же цветок обладает только
двусторонней симметрией, то его называют
неправильным или зигоморфным (например, у яснотки
или гороха).
На рис. 20.15-20.18 изображено несколько
типичных цветков, опыляемых насекомыми. Морфология
цветка типичного злака - ветроопыляемого
однодольного - представлена на рис. 3.38.
Строение цветка удобнее всего показать, если
разрезать его по вертикали на две равные
половинки (в медиальной плоскости, т. е. так, чтобы
расщепить цветоножку надвое в продольном
направлении) и изобразить одну половинку, очертив
поверхность разреза непрерывной линией.
Развитие пыльцевых зерен
Каждая тычинка состоит из пыльника, который
содержит четыре пыльцевых мешка, производящих
пыльцу, и тычиночной нити, содержащей
проводящий пучок, по которому в пыльник поступают
питательные вещества и вода. На рис. 20.19
показано внутреннее строение пыльника с его четырьмя
пыльцевыми мешками, содержащими микроспоро-
циты, или материнские клетки микроспор. Каждый
микроспороцит претерпевает мейоз и образует
четыре пыльцевых зерна (рис. 20.20).
Непосредственно после мейоза можно видеть
тетрады (группы по четыре) молодых пыльцевых зерен.
У каждого пыльцевого зерна образуется толстая
стенка, поверхность которой часто бывает
причудливо скульптурирована, причем характер узоров
специфичен для вида или рода. Наружная стенка,
или экзина, состоит из спорополленина - материала,
близкого к кутину и суберину, но более стойкого,
чем оба этих вещества. Спорополленин - одно из
самых устойчивых веществ, существующих в
природе, и благодаря этому оболочки пыльцевых зерен
могут сохраняться, не изменяясь, на протяжении
длительного времени, иногда миллионов лет. Это
обстоятельство, а также возможность довольно
легко определять род или вид растения, которому
принадлежало пыльцевое зерно, привели к
созданию целой науки - палинологии, или пыльцевого
анализа. Изучая пыльцевые зерна, относящиеся к
определенному времени и собранные в
определенном месте, можно установить, какие там росли
растения, и получить информацию о
существовавших в этом месте экосистемах (включая и
животных) и климате. Особенно обильным источником
Тычинка (совокупность тычинок,
расположенных спирально
на цветоножке, образует андроцей
Тычиночная нить
(короткая, белая)
Пыльник
(желтый)
Рыльце
(изогнутое) ч
Столбик
(очень короткий)^
Чашелистик-
(пять чашелистиков образуют j
чашечку - зеленую,
опушенную, раздельнолистную)
Завязь
(верхняя, апокарпная, одногнезд-
ная; один плодолистик, один семя- '
зачаток; совокупность плодолисти-1
ков образует гинецей, они располо-^
жены на цветоложе по спирали )
Околоцветник
ЧТепесток;
пять лепестков |
I образуют ярко-
\ желтый раздель-|
\ нолепестный I
\ (свободнолепес(]
\ тный) венчик
[ с блестящей
верхней
стороной
Нектарник,
прикрытый маленькой
чешуйкой у основания
лепестка
-Цветоложе
-Цветоножка
Рис. 20.15. Схематический продольный разрез
цветка лютика (Ranunculus acris), отдел Sper-
matophyta, класс Angiospermae, подкласс Dicotyledonae,
семейство Ranunculaceae/ Это многолетнее
травянистое растение, часто встречающееся на
сырых лугах и пастбищах. Зимует благодаря
наличию корневища. Цветки актиноморфные, свобод -
нолепестные, с верхней завязью (подпестичные).
Цветет с апреля по сентябрь. Опылители -
двукрылые и мелкие перепончатокрылые. Каждый
плодолистик содержит одно семя и образует плод,
называемый семянкой. Специальных механизмов,
способствующих распространению семян, нет.
Нектарник
Одна свободная тычинка
Чашелистик
Цветоложе
Цветоножка.
_ Парус
Крылья (их два)
Лодочка
Рыльце
Столбик
Пыльник
Тычиночная
нить
Тычинки
(их десять)
Парус
ι Платформа
_для пчелы
Лодочка
1. Целый цветок
Желобок,
образованный
сросшимися '
нитями девяти
тычинок, окружает
завязь
Крыло
2. Удалено одно крыло
Завязь (один плодолистик),
после оплодотворения
превращается вбоб
■ - '(см. рис. 20.28)
Желобок
. Свободная тычинка
'Сросшиеся
тычинки
Девять сросшихся
тычиночных нитей
3. Удалена
половина лодочки
4. Удалены
сросшиеся тычинки
Рис. 20.16. Строение цветка душистого горошка (Lathyrus odoratus), отдел Spermatophyta, подкласс
Dicotyledonae, семейство Leguminosae. Цветок актиноморфный, свободнолепестный. Цветет в июле.
Чашечка состоит из пяти чашелистиков. Венчик пятилепестный: парус, два крыла и
примыкающая к ним лодочка, образованная двумя сросшимися нижними лепестками. Венчик белый или
окрашенный. Опыляется пчелами, которых привлекает окраска, запах и нектар. Особенно хорошо заметен парус.
Крылья служат платформой, на которую садится насекомое. Под тяжестью его тела
крылья опускаются вниз вместе с лодочкой, с которой они соединены. Столбик и рыльце выступают при
этом наружу и соприкасаются с нижней поверхностью тела пчелы, на которой может
находиться пыльца, захваченная с другого цветка. Пока пчела с помощью своего длинного хоботка добывает
нектар у основания завязи, пыльца из пыльников наносится на нижнюю поверхность столбика, откуда она
может попасть на пчелу. Иногда происходит и самоопыление. К душистому горошку близок горох
огородный (Pisum sativum), но у него самоопыление более обычно. Плод - боб, состоящий из одного
плодолистика со многими семенами. (Способ распространения семян-см. рис. 20.28.)
Семязачаток
^Гнездо
Стенка завязи
Верхняя губа венчика
(два лепестка); прикрывает
тычинки и столбик,
расположенные таким образом, что они
соприкасаются со спинкой
вошедшего в цветок шмеля
Лепесток
(пять лепестков образуют
двугубый венчик белого цвета)
Чашелистик-
(пять чашелистиков образуют
опушенную зеленую
сростнолистную чашечку)
Цветоложе -
Цветоножка-
Сочень короткая)
Пыльник
передней ты-,
чинки ι черный
с желтым вдоль
шва, опушенный)
Тычиночная нить
(белая)
. Рыльце
(двулопастное,
выступает под
пыльниками)
Столбик
(длинный и тонкий)
Нижняя губа венчика
(служит платформой для насекомых;
образована одним лепестком)
Кольцо волосков
на сужающейся части трубки
венчика преграждает мелким
насекомым доступ к нектарникам
Завязь
из двух плодолистиков, синкарпная,
верхняя, четырехгнездная
(первоначально гнезд два, а затем они
делятся "ложной" перегородкой),
по одному семязачатку на гнездо
Нектарник (два нектарника образуют
вздутия на передней стороне
завязи, у ее основания)
Рис. 20.17. Схематический продольный разрез цветка яснотки (Lamium album), отдел Spermatophyta, класс
Angiospermae, подкласс Dicotyledonae, семейство Labiatae. Травянистое многолетнее растение, часто
встречающееся у изгородей и на заброшенных землях. Перезимовывает благодаря наличию корневища. Цветки
сростнолепестные, зигоморфные, двусторонне-симметричные, подпестичные. Цветет с апреля по июнь и
осенью. Опыление происходит главным образом с помощью шмелей. Шмель садится на нижнюю губу венчика, и
когда он проникает в цветок, его спинка, на которой может находиться пыльца, соприкасается с рыльцем, что
приводит к перекрестному опылению, хотя у яснотки возможно и самоопыление. Плод-кар-
церула, состоящая из четырех орешков; каждый из них имеет твердую стенку и содержит по одному семени.
Андроцей
(шесть тычинок,
расположенных
двумя мутовками
по три в каждой ;
прикреплены к
элементам около
цветника; пыльники
кремового цвета)
Тычинки
наружной
мутовки
Тычинки
внутренней
мутовки
. Пыльники
(в наружной
мутовке мелкие)
!Тычиночные нити
прикреплены к
середине
L околоцветника
Пыльники —
(во внутренней
мутовке
крупные)
[Тычиночные
нити (прикреп
лены к основанию]
околоцветника)
Семязачатки
Гнездо завязи
Плацента
Шесть элементов околоцветника
(расположены двумя мутовками
по три в каждой; голубые или
фиолетовые, реже розовые или белые,
они сливаются своими основаниями,
образуя трубку)
Рыльце
(маленькое,
трехлопастное)
Столбик
(длинный и тонкий)
Завязь
(синкарпная, верхняя,
трехгнездная; три
плодолистика;
многочисленные семязачат -
ки на пазушных
плацентах)
Цветоложе
Цветоножка
ν Гинецей
Рис. 20.18. Схематический продольный разрез цветка пролески Endymion non-scripta, отдел
Spermatophyta, класс Angiospermae, подкласс Monocotyledonae, семейство Liliaceae. Травянистое
многолетнее растение, часто встречающееся в лесах и живых изгородях, предпочитает
затененные участки и легкие кислые почвы. Перезимовывает при помощи клубней. Цветок актино-
морфный, подпестичный Число частей цветка кратно трем, что типично для
однодольных. Цветет с апреля по май. Опыление производят насекомые, такие как медоносная пчела. Цветки
издают слабый аромат; верхняя часть завязи выделяет нектар. Плод коробочка,
растрескивающаяся с образованием трех вальв.
Тычинка
(четыре тычинки,
прикрепленные к
лепесткам, образуют
андроцей у двух
передних нити длиннее)
Линия разреза
Пыльник ■
Тычиночная
нить
Материнская клетка микроспоры
делится митотически
Тычинка
Волокнистые слои
(в стенках клеток
развиваются
утолщенные полосы,
участвующие в
растрескивании
пыльников)
. Наружная
стенка-
один слой крупу
ных клеток
Внутренняя·
стенка—
несколько
k слоев
уплощенных клеток
Тапетум·
клетки, снабжающие питательными
веществами материнские клетки
микроспор. По мере формирования,
пыльцевых зерен этот слой '
постепенно распадается
Эпидермис
Линия растрескивания-
Пыльцевые мешки
(по четыре на пыльник)
До растрескивания
Связник
(содержит проводящий
пучок из тычиночной нити)
Паренхима
После растрескивания
Рис. 20.19. Зрелый пыльник колокольчика в поперечном разрезе до и после растрескивания.
пыльцевых зерен служит торф, мощные слои
которого накапливаются на протяжении длительных
периодов времени в торфяных болотах.
20.2. Как можно использовать пыльцевые зерна
для определения а) климата данной области в
прошлом и б) деятельности человека в те
времена?
На этой стадии пыльцевое зерно эквивалентно
микроспоре (см. гл. 3). Его ядро делится надвое
путем митоза, образуя генеративное ядро и ядро
пыльцевой трубки (рис. 20.20). После этого
содержимое пыльцевого зерна можно рассматривать как
эквивалент мужского гаметофита, поскольку из
генеративного ядра в дальнейшем образуются
мужские гаметы.
20.2.3. Развитие зародышевого мешка
и женской гаметы
Каждый плодолистик состоит из рыльца, столбика
и завязи. В завязи образуются один или несколько
семязачатков; каждый из них прикреплен к стенке
завязи в месте, называемом плацентой, с помощью
короткой ножки - фуникулуса, по которому к
развивающемуся семязачатку поступают питательные
вещества и вода. Часто семязачаток в процессе своего
развития загибается вниз, так что его кончик
поворачивается к основанию фуникулуса; в таких
случаях семязачаток называют анатропным. Если же
фуникулус и семязачаток остаются прямыми, то
семязачаток называют ортотропным.
Главную часть семязачатка составляет нуцеллус,
окруженный двумя защитными покровами -интегу-
ментами. На одном конце семязачатка слева имеется
маленькая пора-микропиле. Другой его конец, где
фуникулус сливается с нуцеллусом и интегументами,
называется халазой.
В нуцеллусе, у его микропилярного конца,
начинает развиваться одна материнская клетка
мегаспоры, известная под названием материнской
клетки зародышевого мешка. Эта диплоидная клетка
делится путем мейоза и образует гаплоидную
мегаспору, или зародышевый мешок (рис. 20.21).
Зародышевый мешок растет, его ядро делится путем
митоза, и теперь его содержимое можно
рассматривать как женский гаметофит. В результате
дальнейших митозов образуется восемь ядер; одно из
них-ядро женской гаметы.
Два полярных ядра перемещаются к центру
зародышевого мешка и сливаются, превращаясь в одно
диплоидное ядро (в некоторых случаях их слияние
происходит позднее, при оплодотворении).
Остальные шесть ядер, по три на каждом конце,
разделяются тонкими клеточными стенками; по-видимо-
Мейоз I
Материнская клетка
микроспоры (диплоидная)
Мейоз II
Тетрада — четыре
гаплоидные клетки
Образование
оболочек
Отдельные
пыльцевые зерна
Генеративное ядро
Ядро пыльцевой трубки
Экзина -
толстая скульптурированная
наружная оболочка
Интина -тонкая
внутренняя оболочка
Пора
Зрелое пыльцевое зерно
Рис. 20.20. Развитие пыльцевых
зерен.
Рыльце
Столбик
Интегументы
Нуцеллус
Зародышевый мешок
(см. схему внизу)
Микропиле
Клетки-
антиподы (три)
Завязь
Молодой семязачаток
Фуникулус
Молодой семязачаток
Плодолистик
Клетка растет
Мейоз
Митоз
Митоз
Митоз
Два
полярных
ядра
Яйцеклетка
(женская гамета)
Слияние
ядер
Диплоидное
ядро
Материнская
клетка
зародышевого
мешка
(диплоидная)
Клетки
дегене·
рируют
Рост
за счет
нуцеллуса
Четыре
гаплоидные
клетки
Мегаспора,
или
зародышевый
мешок (диплоидная)
. Конец,
ближайший
к микропиле
Женский гаметофит,
или зародышевый
мешок
Две синергиды
Зрелый зародышевый мешок
(женский гаметофит)
непосредственно перед
оплодотворением; из семи его
ядер одно диплоидное,
а шесть гаплоидные
Рис. 20.21. Развитие зародышевого мешка и женской гаметы (в анатропном семязачатке).
му, лишь одно из них-ядро женской
гаметы-принимает участие в дальнейшем развитии.
Окончательный вид зрелого плодолистика перед
оплодотворением показан на рис. 20.22.
20.2.4. Опыление
После того как в пыльцевых мешках сформируются
пыльцевые зерна, клетки, образующие стенки
пыльника, начинают подсыхать и сморщиваться;
возникает напряжение, которое в конце концов приводит
к растрескиванию (раскрыванию) пыльников
продольными щелями, образующимися на боковых
поверхностях (рис. 20.19), что ведет к освобождению
пыльцы.
Перенос пыльцевых зерен из пыльника на рыльце
называется опылением. Опыление необходимо для
того, чтобы мужские гаметы, развивающиеся в
пыльцевых зернах, могли встретиться с женскими
гаметами; в процессе эволюции возникли особые,
нередко очень замысловатые, механизмы,
обеспечивающие успешное опыление.
Перенос пыльцы с пыльника на рыльце того же
самого цветка или другого цветка того же
экземпляра растения называют самоопылением. Перенос
пыльцы с пыльника одного экземпляра растения на
рыльце другого называют перекрестным опылением.
Перекрестное опыление ведет к перекрестному
оплодотворению и способствует генетической
изменчивости. Таким образом, это особого рода аут-
бридинг. У многих растений имеются специальные
особенности, благоприятствующие перекрестному
опылению; некоторые из них будут описаны ниже.
Преимущество самоопыления, ведущего к
самооплодотворению, состоит в том, что оно более
надежно, особенно в тех случаях, когда
представители данного вида встречаются относительно редко
и на больших расстояниях один от другого. Это
связано с тем, что самоопыление не зависит от
внешних агентов, таких как ветер или насекомые.
Однако самооплодотворение, будучи крайней
формой инбридинга, может приводить к уменьшению
жизнеспособности потомков (см. разд. 25.4).
Примерами самоопыляющихся растений могут служить
крестовник (Senecio) и звездчатка (Stellaria); их
цветки не образуют нектара и лишены запаха.
Как перекрестное опыление, так и самоопыление
Прорастающее
пыльцевое зерно
Диплоидная
ткань
Гаплоидная
ткань
Рыльце
Столбик
Пыльцевая трубка
Стенка завязи
Гнездо
Халаза
Интегументы
Нуцеллус
Три клетки-
антиподы
^Диплоидное
[ядро — результат
' слияния
полярных ядер
•Яйцеклетка
Две синергиды
Микропиле
Мужские ядра
Ядро пыльцевой
трубки
Семязачаток
Зародышевый
мешок
Фуникулус
Проводящий
пучок
на конце
пыльцевой
трубки
Рис. 20.22. Плодолистик в момент
оплодотворения (в продольном разрезе). Обратите внимание,
что семязачаток (анатропный). который после
оплодотворения превращается в семя, содержит как
диплоидную родительскую ткань, так и
гаплоидную ткань зародышевого мешка. Содержимое
пыльцевого зерна и пыльцевой трубки
эквивалентно мужскому гаметофиту, а зародышевый
мешок эквивалентен женскому гаметофиту.
имеют свои преимущества и свои недостатки, и у
многих растений выработались приспособления,
которые, благоприятствуя перекрестному
опылению, вместе с тем дают возможность, если оно
почему-либо не удалось, прибегнуть к
самооплодотворению. Например, у фиалки (Viola) и у кислицы
(Oxalis) некоторые бутоны не раскрываются, так
что для них неизбежно самоопыление.
Особенности, благоприятствующие
самоопылению
двудомные растения. У двудомных растений
самоопыление невозможно. Однодомные виды, у
которых на одном и том же гермафродитном растении
есть и мужские, и женские цветки, тоже
предпочитают перекрестное опыление, но у них может
происходить и самоопыление.
20.3. Двудомные растения встречаются редко,
несмотря на преимущества перекрестного
опыления. Приведите две возможные причины этого.
20.4. «Двудомность» (раздельнополость)
широко распространена у животных. Почему эта
система оказалась более подходящей для животных,
чем для цветковых растений?
дихогамия. Иногда созревание пыльников и рылец
происходит в разные сроки - явление, называемое
дихогамией. Если пыльники созревают раньше, чем
рыльца, то говорят о протандрии, а если сначала
созревают рыльца-о протогинии. Протандрия
встречается гораздо чаще; примерами служат
яснотка, одуванчик, иван-чай (Epilobium angustifolium),
а также шалфей (Salvia, рис. 20.23).
Протогиния свойственна пролеске и норичнику
(Scrophularia). При дихогамии в большинстве
случаев имеется такой период, когда и пыльники, и
рыльца находятся в состоянии зрелости, что делает
возможным самоопыление, если перекрестное
опыление не произошло. Сходный механизм,
обеспечивающий перекрестное оплодотворение, существует у
некоторых гермафродитных животных, например у
гидры, у которой мужские гонады созревают
раньше, чем женские.
самонесовместимость (самостерильность). Даже
если самоопыление произошло, пыльцевые зерна
часто не развиваются или развиваются очень
медленно, что препятствует самооплодотворению или
уменьшает его вероятность. Во всех таких случаях
происходит специфическое взаимодействие, ингиби-
рующее процесс прорастания пыльцевой трубки в
столбик, причем это ингибирование определяется
генами самонесовместимости.
В случаях самонесовместимости частота
совместимых перекрестных комбинаций варьирует и
опять-таки генетически детерминирована. Для
максимально эффективного использования пыльцы
значительная доля скрещиваний должна быть
совместимой. Крайним примером служит клевер, у
которого все растения самонесовместимы, но
перекрестная несовместимость встречается менее чем в одной
из 22000 пар. Менее эффективны системы, в
которых совместимые типы различаются по морфологии
цветков. Примером служит первоцвет (см. ниже).
20.5. Самонесовместимость контролируется
множественными аллелями. Если принять, что
а) имеется три аллеля-Sj, S2 и S3 и б)
самонесовместимость возникает в тех случаях, когда
пыльцевое зерно и клетки рыльца имеют один
общий аллель, то какая доля пыльцевых зерен от
растения с генотипом S1S2 будет способна
успешно прорасти на растении с генотипом S2S3I
структурные адаптации. У большинства
гермафродитных цветков имеются структурные особенности,
способствующие перекрестному опылению.
У энтомофильных (опыляемых насекомыми)
цветков рыльца обычно выступают над
пыльниками, что не позволяет пыльцевым зернам падать
прямо на рыльца того же цветка. Когда насекомое,
несущее на себе пыльцу с другого растения,
посещает такой цветок, оно сначала прикасается к его
рыльцу. Затем, когда насекомое ищет нектар, оно
либо обмазывается пыльцой, либо стряхивает ее на
себя. Так обстоит дело, например, при опылении
яснотки (рис. 20.17). Более примитивный механизм
состоит в том, что рыльце прикасается к насекомому,
когда оно садится на цветок, как при опылении
душистого горошка (см. рис. 20.16). Такие
механизмы обычно сочетаются с дихогамией, и цветки
при этом часто бывают сложными и
зигоморфными.
Цветки привлекают насекомых, предоставляя им
пищу (нектар или пыльцу) и стимулируя их зрение и
обоняние. Это возможно благодаря наличию у
цветков особых признаков, которые будут рассмотрены
ниже.
У многих ветроопыляемых цветков тычинки,
цветок в целом или все соцветие свисают вниз, так что
пыльца высыпается, а затем уносится прочь (как,
например, у лещины).
20.6. На рис. 20.24 изображены цветки
первоцвета двух типов, встречающихся в природе
примерно в равном количестве и различающихся
длиной столбиков (гетеростилия) и
расположением тычинок, а) Известно, что пчелы
высасывают нектар из нижней части трубки венчика;
объясните, почему перекрестное опыление
происходит главным образом между длинно- и ко-
роткостолбчатыми цветками, а не между
цветками одного типа? б) В чем преимущество такой
системы?
Хотя гетеростилия (см. вопрос 20.6 и рис. 20.24),
очевидно, способствует аутбридингу, гораздо более
эффективен в этом смысле механизм
самонесовместимости, существующий у длинно- и коротко-
Рис. 20.23. Пчела, входящая в цветок лугового шалфея. Голова пчелы надавливает на
пластинку, к которой прикреплены тычинки, и они опускаются, стряхивая пыльцу на
брюшко пчелы. Если пчела посетит теперь более старый цветок (по мере
старения цветка его рыльце удлиняется), то ее брюшко соприкоснется с рыльцем и приставшая к брюшку пыльца
будет перенесена на рыльце. В результате всего этого произойдет
перекрестное опыление.
Рыльце
Пыльники
Нектарник
Пыльники
Рыльце
Нектарники
Рис. 20.24. Гетеростилия у первоцвета (Primula).
А. Длинностолбчатый цветок. Б. Коротко -
столбчатый цветок.
столбчатых первоцветов, благодаря которому
перекрестное оплодотворение происходит только между
цветками разных типов. Гены, контролирующие
несовместимость, длину столбика и высоту
расположения тычинок, локализованы очень близко друг к
другу в одной и той же хромосоме и ведут себя как
одна наследуемая единица.
Опыление ветром и опыление
насекомыми
Пыльцевые зерна-это споры, но в отличие от спор,
образуемых споровыми растениями, они не могут
прорастать на суше и должны быть перенесены на
женские половые структуры шишек (у
голосеменных) или цветков (у покрытосеменных).
Первоначально споры распространялись ветром, но такой
механизм переноса пыльцы очень неэффективен, так
как при этом попадание пыльцевых зерен на шишки
или цветки всецело зависит от случая. Все хвойные и
многие цветковые растения (злаки и большинство
деревьев умеренного пояса, такие как дуб и лещина)
по-прежнему опыляются с помощью ветра, но им
приходится производить огромные количества
пыльцы, что требует больших затрат вещества и
энергии. После того как в процессе эволюции
возникли цветки, растения очень быстро начали
использовать для опыления насекомых, так как это
гораздо более надежный способ переноса пыльцы.
Насекомое может переносить небольшое количество
пыльцы с тычинок одного цветка точно на рыльце
другого. В результате в процессе эволюции между
цветками и насекомыми сложились особые
взаимоотношения: наградой, которую насекомые
получают от цветков, служит пища в виде нектара, а
иногда пыльцы. Насекомые, специализированные
для питания на цветках, появились одновременно с
цветковыми растениями; к их числу относятся
пчелы, шмели, осы и бабочки. В некоторых случаях
насекомое и опыляемое им растение настолько
взаимозависимы, что ни одно из них не может
существовать без другого, как, например, юкка и
юкковая моль (Pronuba yuccasella). Энтомофилия
обладает еще одним преимуществом: она
благоприятствует перекрестному опылению, а тем самым
и перекрестному оплодотворению; поэтому
описанные ниже модификации цветков,
облегчающие опыление насекомыми, можно добавить к
перечню признаков, способствующих
перекрестному опылению.
У энтомофильных растений цветки обычно
крупные, с ярко окрашенными лепестками, что
привлекает насекомых; если же цветки мелкие, то они
собраны в соцветия. Насекомые воспринимают лучи
ультрафиолетовой области спектра, невидимые для
человека, а поэтому цветки, кажущиеся человеку
белыми, насекомые могут воспринимать как
окрашенные. Часто на лепестках имеются полоски,
пятнышки или более интенсивно окрашенные участки,
указывающие насекомым путь к нектарникам; они
есть, например, у фиалок, включая анютины глазки
(Viola), орхидей (Orchis и другие роды) и
наперстянки (Digitalis).
Более специфичны, чем окраска, запахи,
издаваемые цветками; некоторые из них, например лаванда
и роза, используются в парфюмерной
промышленности. Некоторые растения обладают запахом
гниющего мяса, который привлекает насекомых,
питающихся падалью; арум (Arum maculatum)
привлекает навозных мух. Специфическим
опознавательным признаком может служить также форма цветка.
Один из самых сложных и странных механизмов
для обеспечения перекрестного опыления обнаружен
у некоторых орхидей, цветки которых по форме,
окраске и даже запаху подражают самкам ос-сфек-
сов и притом настолько убедительно, что сфексы-сам-
цы пытаются копулировать с цветками (рис. 20.25).
При этих попытках насекомое стряхивает с себя
пыльцу на цветок, а затем, покидая его, уносит на
себе его пыльцу, которую передает потом другому
цветку.
Типичные различия между цветками,
опыляемыми ветром и опыляемыми насекомыми, приведены в
табл. 20.2.
20.2.5. Оплодотворение
Как только на рыльце попадает пыльцевое зерно,
эпидермальные клетки рыльца выделяют раствор
сахарозы, который стимулирует прорастание
пыльцевого зерна и, возможно, используется для его
питания (рис. 20.26). Сквозь одну из пор,
имеющихся в стенке пыльцевого зерна, выходит
пыльцевая трубка, которая быстро растет вниз внутри
столбика, направляясь к завязи. Ее рост
сопровождается секрецией пищеварительных ферментов и
регулируется полученным от пыльцевого зерна
ядром пыльцевой трубки, которое находится у ее
растущего кончика. Рост пыльцевой трубки
стимулируют ауксины, вырабатываемые гинецеем, а к
завязи ее направляют определенные вещества
(пример хемотропизма). Возможно также, что здесь
имеет место отрицательный аэротропизм, т. е. рост
Таблица 20.2. Типичные различия между цветками,
опыляемыми ветром и цветками, опыляемыми насекомыми
Характерные признаки ветро-
опыляемых цветков
Мелкие лепестки, не
имеющие яркой окраски
(обычно зеленые), или же
лепестков нет, так что
цветки малозаметны
Лишены запаха
Нектарников нет
Крупное многолопастное и
перистое рыльце
свешивается из цветка наружу,
чтобы захватывать пыль-
цу
Тычинки свешиваются из
цветка наружу, так что
пыльца высыпается
Пыльники подвижные: они
прикреплены к кончикам
тычиночных нитей
только в своих средних
точках, так что свободно
раскачиваются ветром
Производят большие
количества пыльцы, так как
потери ее очень велики
Пыльцевые зерна
относительно легкие и мелкие, с
сухими, часто гладкими
стенками
Строение цветка
относительно простое
Цветки расположены
значительно выше листьев
благодаря длинным стеблям
(например, у злаков) или
появляются до развития
листьев (например, у
многих деревьев
Британских островов)
Характерные признаки цветков,
опыляемых насекомыми
Крупные, ярко окрашенные
лепестки делают цветки
хорошо заметными. Если
цветки относительно
невзрачны, то они могут
быть собраны в соцветия
Издают запах
Нектарники имеются
Маленькое рыльце, не
выступающее из цветка и се-
кретирующее клейкое
вещество, к которому
прилипает пыльца
Тычинки заключены в
цветке
Пыльники неподвижные:
они прикреплены к
тычиночным нитям своим
основанием или сращены с
нитями дорсальной
поверхностью
Производят меньше
пыльцы
Пыльцевые зерна
относительно тяжелые и
крупные. Шипики на стенках
и клейкость
способствуют прикреплению к
телу насекомого (рис. 20.26)
Строение цветка часто
усложненное,
приспособлено к переносу пыльцы
определенными
насекомыми
Расположение и время
появления цветков по
отношению к листьям
варьируют, хотя часто цветки
возвышаются над
листьями, что делает их более
заметными
в направлении от воздушной среды. Как уже
говорилось, рост зависит от совместимости между
пыльцой и тканью рыльца.
Во время роста пыльцевой трубки генеративное
ядро пыльцевого зерна делится митотически,
образуя два мужских ядра, представляющие собой
мужские гаметы (рис. 20.22). В отличие от спермиев
низших растений они неподвижны и могут
добраться до женской гаметы, находящейся в
зародышевом мешке семязачатка, только с помощью
пыльцевой трубки. Пыльцевая трубка проникает в
семязачаток через микропиле, ее ядро дегенерирует,
а кончик трубки разрывается, освобождая мужские
гаметы вблизи зародышевого мешка, в который они
и проникают. Одно ядро сливается с женской
гаметой, образуя диплоидную зиготу, а другое - с двумя
полярными ядрами (или с диплоидным ядром, если
эти последние уже слились), образуя триплоидное
ядро, называемое первичным ядром эндосперма.
Такое двойное оплодотворение свойственно
только цветковым растениям.
Если, как это часто бывает, гинецей содержит
несколько семязачатков, то каждый из них, чтобы
превратиться в семя, должен быть оплодотворен
отдельным пыльцевым зерном. Поэтому может
оказаться, что один цветок даст семена,
образовавшиеся в результате оплодотворения пыльцевыми
зернами от разных растений.
Опыт. 20.1. Изучение роста пыльцевых
трубок
Рыльца выделяют раствор, содержащий от 2 до 45%
сахарозы. Этот раствор способствует прилипанию
пыльцевых зерен к рыльцу и их прорастанию.
Добавив к экспериментальному раствору борат натрия,
можно предотвратить осмотический разрыв
кончиков пыльцевых трубок и стимулировать рост.
Материалы
Микроскоп
Предметное стекло с лункой
Цветки с растрескивающимися пыльниками, например
цветки яснотки или лакфиоли
10-20%-ный (масса/объем) раствор сахарозы,
содержащий также борат натрия (0,01%)
Ацетокармин или нейтральный красный
Методика
Каплю раствора сахарозы помещают в лунку
предметного стекла и вносят в нее пыльцевые зерна,
прикасаясь к капле поверхностью растрескавшегося
пыльника. Препарат рассматривают под
микроскопом спустя различные промежутки времени на
протяжении 1-2 часов. Ядра, находящиеся в кончике
растущих пыльцевых трубок, можно окрасить,
добавив к раствору каплю ацетокармина или
нейтрального красного.
20.2.6. Развитие семени вплоть до
стадии покоя
После оплодотворения семязачаток называют уже
семенем, а завязь-плодом.
Зигота растет путем многократных митотических
делений и превращается в многоклеточный
зародыш, состоящий из первичного побега, или плю-
мулы, первичного корешка и одной (у однодольных)
или двух (у двудольных) семядолей. Семядоли
имеют более простую структуру, чем первые
настоящие листья; иногда они наполнены
питательными веществами и служат запасающей тканью,
как, например, у гороха и у конских бобов (Vicia
faba). Плюмула состоит из стебля, первой пары
настоящих листьев и терминальной почки. Трип-
Рис. 20.25. Роющая оса Argogorytes mystaceus, пытающаяся копулировать с цветком офриса пчелоносного
(Ophrys insectifera).
лоидное ядро первичного эндосперма многократно
делится (путем митоза), образуя эндосперм
скопление триплоидных ядер, отделенных друг от друга
тонкими клеточными стенками. У некоторых
растений, например у кукурузы (Zea mays), эндосперм
сохраняет роль запасающей ткани.
Если запасающей тканью служат семядоли, то
они растут за счет эндосперма, который может при
этом совершенно исчезнуть. У некоторых семян
запасы питательных веществ содержатся и в
эндосперме, и в семядолях.
В развивающемся семени рост зародыша, а
иногда и эндосперма происходит в зародышевом мешке.
По мере продолжения роста окружающий их нуцел-
лус дезорганизуется и разрушается, доставляя
необходимые для роста питательные вещества. В
дальнейшем снабжение питательными веществами
обеспечивает проводящий пучок фуникулуса (ножки
семязачатка), подобно тому как развивающийся
зародыш млекопитающего получает питание от
материнского организма через плаценту. Как уже
отмечалось, место прикрепления фуникулуса к стенке
Поверхность рыльца.
покрытая мелкими выступами
Пыльцевые зерна,
покрытые шипами
Рис. 20.26. Микрофотография пыльцевых зерен на рыльце цветка, сделанная с помощью сканирующего
электронного микроскопа. Шиповатая поверхность пыльцевых зерен характерна для
цветков, опыляемых насекомыми.
Таблица 20.3. Изменения, происходящие у цветковых растений после оплодотворения
Семяза- <
чаток
Зигота
Первичное
ядро
эндосперма
H у цел л ус
Интегументы —
Стенка завязи —
Плюмула I
/^ Первичный ко-
- Зародыш^— решок
^^ Одна или две се-
""* мядоли
(содержащие или
не
содержащие запасы ,
питательных
веществ)
- Эндосперм (содержащий или не
содержащий запасы
питательных веществ)
- Исчезает
- Семенная кожура
- Перикарпий J
Семя —
оплодотворенный семязачаток (один
рубчик — от ножки се-
► мязачатка)
J
Плод — оплодотворенная
завязь. Содержит одно
или более семян (два
► рубчика — от
цветоложа и столбика)
завязи тоже называется плацентой.
Микропиле сохраняется в виде маленькой поры в
тесте (семенной кожуре), через которую поступают
кислород и вода при прорастании семени. Теста-
это тонкий, но прочный защитный слой,
образующийся из интегументов. На завершающих стадиях
созревания семени содержание в нем воды
снижается от обычного для растительных тканей -
примерно 90% (по массе)-до 10-15%. Это
сопровождается заметным снижением метаболической
активности и представляет собой важный шаг к переходу
семени в состояние покоя.
По мере развития семени завязь превращается в
зрелый плод и стенка ее получает название
перикарпия. Происходящие при этом изменения у разных
видов различны, но обычно направлены на то,
чтобы плод мог защищать семена и способствовать
их распространению (см. разд. 20.2.8).
Остальные части цветка увядают, отмирают и
сбрасываются, подобно листьям у листопадных
растений. В некоторых случаях такие части, как
цветоложе, столбик или чашелистики сохраняются и
участвуют в распространении семян; цветоложе в
таких случаях, например у земляники (Fragaria),
получает название ложного плода.
Гормональная регуляция развития плода
рассмотрена в разд. 15.3.5.
Некоторые изменения, происходящие после
оплодотворения, суммированы в табл. 20.3.
Некоторые стадии развития зародыша показаны
на рис. 20.27.
20.2.7. Преимущества и недостатки
размножения семенами
Семя - типичный продукт полового размножения у
семенных растений; согласно определению, это
оплодотворенный семязачаток. Семя содержит
зародыш растения и одну или более семядолей, а иногда
эндосперм, и окружено защитной кожурой.
Разрастающийся
зародышевый мешок
Эндосперм
Собственно
- зародыш
Суспензор
Питательные вещества
от родительского
растения
Семенная кожура
Семядоли,
наполненные
питательными
веществами
Остатки
эндосперма
I Первое
1митотическое\
[деление зиготы
Микропиле
Зародыш
Растущие
семядоли
Плюмула
Зародышевый
корешок
Микропиле
Зародыш
Суспензор
Базальная клетка
Семядоли
Развивающиеся
плюмула и
зародышевый
корешок
Рис. 20.27 Развитие зародыша в
лишенном эндосперма семени
двудольного растения - пастушьей
сумки (Capsella bursa -pastoris).
Преимущества
1. Растение не нуждается в водной среде для
полового размножения и поэтому лучше
приспособлено к наземным условиям.
2. Семя защищает зародыш.
3. Семя содержит питательные вещества,
необходимые зародышу (либо в семядолях, либо в
эндосперме).
4. Семя обычно имеет приспособления для
распространения.
5. Семя способно оставаться в состоянии покоя и
переживать неблагоприятные условия.
6. Семя реагирует на благоприятные условия и
иногда должно пройти период дозревания, т.е.
прорастает не сразу (см. гл. 15).
7. Семена образуются в результате полового
размножения, а поэтому обеспечивают виду
преимущества, связанные с генетической
изменчивостью.
Недостатки
1. Семена имеют относительно крупные размеры
из-за находящихся в них больших запасов
питательных веществ. Это затрудняет их
распространение по сравнению со спорами.
2. Животные часто поедают семена ради
содержащихся в них питательных веществ.
3. Опыление часто зависит от внешних факторов
(ветер, насекомые, вода). Поэтому оно (а значит,
и оплодотворение) не всегда гарантировано,
особенно если производится ветром.
4. Потери семян велики, так как вероятность
выживания каждого данного семени незначительна.
Поэтому для обеспечения успеха родительский
спорофит должен вкладывать в производство
семян большие количества вещества и энергии.
5. Запасы питательных веществ в семени
ограниченны, тогда как при вегетативном размножении
родительское растение доставляет дочернему
растению питательные вещества до тех пор, пока
это второе не станет вполне самостоятельным.
6. У двудомных видов в размножении участвуют
два растения, что повышает вероятность неудачи
по сравнению с размножением при участии одной
родительской особи. Напомним, что
двудомность-явление относительно редкое.
Приведенные выше сведения можно использовать
для сопоставления выгодных или невыгодных
сторон организации семенных растений по сравнению
со споровыми или относительных преимуществ
полового и вегетативного размножения у семенных
растений.
20.2.8. Распространение плодов и
семян
После образования семян либо весь плод, либо
содержащиеся в нем семена (или семя) отделяются
от родительского спорофита. Если перикарпий
(стенка плода) становится твердым и сухим, плод
называют сухим, если же он становится мясистым,
то плод называют сочным. Сухой плод может быть
вскрывающимся или невскрывающимся в
зависимости от того, связано ли освобождение семян с
разрывом его стенки или нет. Сухие плоды могут
также распадаться на несколько односеменных
члеников; такие плоды называют схизокарпными.
Чем дальше разносятся семена, тем менее
вероятна конкуренция со стороны родительского растения.
Это дает также больше шансов колонизировать
новую территорию, что со временем приводит к
увеличению размеров популяции в целом. Однако
при этом существует опасность, что семя, будучи
занесено слишком далеко от родительского
растения, не найдет подходящего места для прорастания.
Существуют три главных внешних фактора,
распространяющих семена,-ветер, животные и вода.
Кроме того, имеются механизмы автохории, часто
связанные со взрывным освобождением семян из
плода. Примеры всех этих способов
распространения семян представлены на рис. 20.28.
Рис. 20.28. Разные типы плодов и семян и способы их
распространения.
РАСПРОСТРАНЕНИЕ ЖИВОТНЫМИ
Распространение семян животными в общем более надежно,
так как животные обычно посещают плодородные
участки, подходящие для прорастания семян. Плоды,
снабженные колючками или крючочками, прииепляются к
коже или шерсти проходящих мимо животных и могут быть
перенесены на некоторое расстояние, прежде чем
отпадут или будут содраны. Прииепки развиваются из
разных частей плода или окружающих структур.
Примерами служат плоды подмаренника цепкого (Galium sparine),
гравилата (Geum), лопуха (Arctium), моркови (Daucus),
лютика (Ranunculus), череды (Bidens), репешка (Agrimonia). У
гравилата крючочки имеются на столбике. У лопуха
скопление мелких плодов окружено крючковидными
листочками обертки (видоизмененные листья), образующими
единую структуру; кроме того, здесь имеются мелкие
жесткие волоски, которые проникают в кожу,
вызывая раздражение и расчесывание, в результате чего плоды
отпадают. У подмаренника, моркови и лютика
перикарпий покрыт выростами (прицепками). У череды плод
снабжен летучкой, как у одуванчика, но с
прочными шипами. Цветоложе репешка покрыто жесткими
загнутыми колючками.
Подмаренник цепкий
(Galium aparine )
Две
односеменных
части плода
Перикарпий
с крючками
Плодоножка
Жесткие загнутые
колючки по верхнему
краю цветоложа
Цветоложе покрыто
мелкими волосками
Репешок
Колючки
перикарпия
Плодоножка
Плод подмаренника с
многочисленными крючочками, развивающимися
из околоплодника.
Микрофотография, полученная с помощью
сканирующего электронного
микроскопа.
Примерами сочных плодов служат плоды сливы (Prunus),
ежевики (Rubus), томата (Lycopersicum), яблони
(Malus), земляники (Fragaria). Мясистые части этих
плодов служат пищей животным, в том числе птицам. При
созревании они обычно приобретают яркую окраску и
аромат, привлекающие животных. Животное съедает и
переваривает плод, но семена, устойчивые к
пищеварительным ферментам, проходят невредимыми через его
кишечник и выходят с испражнениями, часто попадая на
плодородную почву. Содержащиеся в экскрементах
питательные вещества могут повышать плодородие
почвы вокруг семени.
Слива
Слива служит примером плода, называемого костянкой; ее
перикарпий состоит из трех слоев: экзокарпия
(защитной кожицы), сочного мезокарпия и
внутреннего каменистого эндокарпия («косточки»). Эти слои
защищают семя, предохраняя его от переваривания. Другие
примеры костянок-плоды персика, вишни и миндаля (также
принадлежащих к роду Prunus), бузины (Sambucus) и
кокосовой пальмы (Cocos nucifera; плод с волокнистым
мезокарпием). Миндаль, поступающий в продажу, обычно
очищают от экзо- и мезокарпия.
Иногда цветоложе разбухает, становится мясистым и само
напоминает плод. Такие плоды называют ложными.
Примерами служат плоды яблони и земляники.
Яблоко
Ложный плод яблони относится к типу яблока, или
тыквины. У плодов этого типа мякоть образуется из полого
цветоложа, окружающего заключенные в нем
плодолистики. Перикарпий настоящего плода превращается в
«сердцевину», содержащую семена. Другими примерами
служит груша (Pyг us) и боярышник (Crataegus).
Слива (в продольном разрезе)
Гнездо—
Фуникулус
Семенная
кожура
Остатки
столбика
Экзокарпий -
(кожица)
Мезокарпий
(мякоть)
Эндокарпий.
(косточка)
Одно семя
Чашелистик
Плодоножка
Перикарпий
* из одного
плодолистика
Яблоко ( в продольном разрезе)
Остатки чашелистиков
(первоначально по краю
.вогнутого цветоложа)
Цветоложе
(разросшееся и мясистое)
Перикарпий (твердый
наружный слой
"сердцевины")
Семя ("зернышко")
Фуникулус
Плацента
Плодоножка
Плодолистики; 3—5, сросшиеся,
по два семени на плодолистик
Ягода ежевики в продольном разрезе - совокупность
костянок, образовавшихся из одного цветка
Отдельные плоды, каждый
из одного плодолистика
Семя
Цветоложе
Плодоножка
.Увядшие тычинки
-Чашелистики
Гнездо
Фуникулус.
Семенная кожура
Остатки
столбика
Экзокарпий
Мезокарпий
Эндокарпий
(зернышко")
Семя
Один плод (костянка) в продольном разрезе
Томат
Плод томата, образующийся в результате слияния двух,
трех или четырех плодолистиков, относится к типу
ягодовидных. Он сходен с костянкой, но
эндокарпий у него мясистый, а не каменистый. К ягодовидным
принадлежат также плоды черной смородины и
крыжовника (Ribes sppj, тыквы (Cucurbita), апельсина и лимона (Citrus
sppj, банана (Musa), винограда (Vitis), финика (Phoenix
dactylifera; содержит одно деревянистое семя).
Томат (в продольном разрезе)
.Остатки столбика
Экзокарпий (кожица")
Мезокарпий (мякоть)
Эндокарпий (мягчей
сочнее мезокарпия)
Семя
Фуникулус
Плацента
Чашелистик
- Плодоножка
Земляника
У земляники мелкие зеленые плоды, называемые семянками и
расположенные на разросшемся цветоложе; каждая семянка
содержит по одному семени.
Ягода земляники (в продольном разрезе)
Отдельные плоды
Цветоложе
(разросшееся и мясистое)
Чашелистик
Плодоножка
Перикарпий_
Семенная кожура
Плацента
Остатки
столбика
Гнездо
Фуникулус
Один плод
(в продольном разрезе)
Ореховидные плоды, например у дуба (Quercus),
бука (Fagus), каштана (Castanea),-относительно крупные
сухие плоды, которые не расщепляются и не выбрасывают
семена. Распространяются ореховидные плоды целиком,
часто при помощи животных, делающих запасы, в
особенности таких грызунов, как белки. На рисунке изображен
плод дуба, называемый желудем.
Плод дуба — желудь
Остатки столбика
Ореховый плод
(желудь)
Плюска-чашеобразная
структура, образованная
прицветниками (моди
фицированные листья)
.Плодоножка
Рис. 20.28 (продолжение)
Желудь в продольном
разрезе (образован
тремя слившимися
плодолистиками, но семя
развивается только одно)
Деревянистый
перикарпий
Гнездо
Семя
Семенная кожура
Плюска
РАСПРОСТРАНЕНИЕ ВЕТРОМ
Рис. 20.28 (продолжение)
Многие растения, распространяемые ветром, имеют
специальные приспособления. К их числу относятся летучки,
которыми снабжены, например, семена кипрея (Epilobium),
ивы (SaHx), хлопчатника (Gossypium hirsutum), плоды
одуванчика (Taraxacum), ломоноса (Clematis), и
крылатки, как, например, у сосны (голосеменные), вяза
(UImus), ясеня (Fraxinus), клена (Acer), граба (Carpinus).
Цветок
Структуры, I
расположенные
над летучкой,
отпадают после
оплодотворения
Одуванчик
Венчик
Рыльце
Пыльники
Летучка'
Завязь
Плод (два слившихся
плодолистика)
из нижней завязи,
маленький и легкий
Летучка (белые волоски,
образующиеся из чашелистиков
после оплодотворения; их большая
поверхность облегчает
распространение ветром)
Плод в продольном разрезе
Семенная кожура-
Ножка летучки
(вырастает после
оплодотворения)
Маленькие крючки
Шов между
плодолистиками,
виден на перикарпии
-Рубец на месте
прикрепления к цветоложу
Семя.
Плацента
Гнездо
Перикарпий
-Фуникулус
Длинные волоски -
Семя
Клен A cer pseudoplatanum
Кипрей
Опушенные семена,
расположенные.
четырьмя вертикаль
ными рядами
Шов между
плодолистиками
Перикарпий-
Коробочка,
вскрывающаяся
по четырем швам
Плодоножка
Два крыла,
образующиеся
из одного перикарпия i
(два плодолистика)/
Рубец
от столбика
Шов между
плодолистиками
Рубец от элементов
цветка
Плодоножка-
Гнездо "
Семя
внутри
перикарпия
Семенная
кожура
-Фуникулус
-Плацента
Перикарпий
|Продольный
разрез
f правой
стороны
плода —
видно семя
Плод клена до или после того, как он будет перенесен
ветром на новое место, распадается на две части по шву
между плодолистиками (такой плод называют схизо-
карпным); опадающий плод вращается и подхватывается
ветром.
У ряда растений, например у мака
(Pa г aver), чернушки (Nigella),
наперстянки (Digitalis), лихниса
(Lichnis), плод -коробочка,
сидящая на конце длинного
стебелька, которую колышет
ветер, так что многочисленные
мелкие семена высыпаются через
поры в верхней части плода.
Мак
Рыльца, от 4 до 20,
по одному на плодо -
листик; это гребни,
выступающие над
рыльцевым диском
(столбиков нет)
Рыльцевый диск —
"крышка" завязи
Пора
Швы между
плодолистиками
Перикарпий
Рубцы цветка
Плодоножка
Рыльце
_Лора
Фуникулус
Плацента
Гнездо
(только одно)
Семена
Коробочка (в продольном разрезе)
АВТОХОРИЯ
Рис. 20.28. (продолжение)
Семена некоторых растений, например орхидей, настолько
мелкие и легкие, что разносятся ветром, не имея никаких
специальных придатков, которые бы увеличивали их
поверхность. Такое распространение, называемое обычно
автохорией, наблюдается у вскрывающихся плодов.
Вскрывание у них часто происходит путем
растрескивания по шву между плодолистиками, как, например, у бобов.
Семена при этом либо выбрасываются из плодов более или
менее сильным толчком, либо просто выпадают из них.
Некоторые из таких семян имеют приспособления для
распространения ветром (например, у кипрея). Коробочки
тоже бывают вскрывающимися (например, у мака и кипрея),
причем у некоторых растений семена
выбрасываются из них с силой (например, у фиалки).
Бобы-плоды, характерные для представителей сем.
бобовых (LeguminosaeJ, например для гороха (Pisum), конских
бобов (Vic/a), фасоли (Phaseolus), утесника (Ulex), ракитника
(Cytisus), золотого дождя (Laburnum). Боб состоит
из одного плодолистика, содержащего много семян.
Перикарпий, высыхая, скручивается, так как его наружные и
внутренние волокнистые слои сокращаются в разных косых
направлениях; в результате создается напряжение, и боб
может внезапно раскрыться по двум боковым швам, с силой
выталкивая наружу несколько семян. Другие семена
выбрасываются менее энергично по мере дальнейшего
скручивания (ранние стадии развития плода гороха показаны на
рис. 20.16).
-Остатки
столбика
РАСПРОСТРАНЕНИЕ ВОДОЙ
Лишь немногие плоды и семена специально
приспособлены для распространения с помощью воды. Они содержат
воздушные полости, удерживающие их на поверхности воды.
Кокосовый орех (плод Cocos nucifera )-костянка (ср. со
сливой) с волокнистым мезокарпием и многочисленными
воздухоносными полостями. У кувшинки (Nymphaea) семя
снабжено губчатой оболочкой (ариллусом), происходящей из
ножки семязачатка.
СЛУЧАЙНОЕ РАСПРОСТРАНЕНИЕ
Разделение семян и плодов на категории в зависимости от
способа распространения никак нельзя считать
строгим, и степень специализации к распространению ветром,
животными или автохории сильно варьирует.
Очевидно, во многих случаях большую роль играет элемент
случайности, и данный плод или семя может
распространяться двумя из названных трех способов или даже
всеми тремя. Один из главных факторов
случайного распространения является человек; семена могут
прицепляться или прилипать к его одежде и т. п. или
перевозиться вместе с разными грузами на транспортных
средствах. Засорение зерновых культур семенами
сорняков-обычное явление, наблюдаемое по всему
земному шару. Орехи, припрятанные про запас грызунами, могут
остаться и прорасти следующей весной. Наводнения,
ураганы и т. п. могут занести семена дальше, чем обычно.
Горох или ракитник
Шов-
Гнездо-
Семя-
Фуникулус
Плацента-
Плодоножка
Чашечка
Перикарпий
Два шва вдоль
одного плодолистика
Впадина для семени,
лежащего на
против опол ожной
стороне
Кувшинка
Семенная_
кожура
Семя
Губчатый
ариллус
Фуникулус
20.3. Половое размножение
у человека
20.3.1. Мужская половая система
Мужская половая система состоит из парных
семенников (яичек), семявыносящих протоков, ряда
придаточных желез и полового члена (пениса).
Семенник-сложная трубчатая железа яйцевидной формы;
она заключена в капсулу-белочную оболочку-и
состоит примерно из тысячи сильно извитых
семенных канальцев, погруженных в соединительную
ткань, в которой содержатся интерстициальные (лей-
диговы) клетки. В семенных канальцах образуются
гаметы-спермин (сперматозоиды), а
интерстициальные клетки вырабатывают мужской половой
гормон тестостерон. Семенники расположены вне
брюшной полости, в мошонке, а поэтому спермин
развиваются при температуре, которая на 2-3 0C ниже
температуры внутренних областей тела. Более
низкая температура мошонки частично определяется ее
положением, а частично - сосудистым сплетением,
образуемым артерией и веной семенника и
действующим как противоточный теплообменник.
Сокращения особых мышц перемещают семенники ближе
или дальше от тела в зависимости от температуры
воздуха, чтобы поддерживать температуру в
мошонке на уровне, оптимальном для образования
спермы. Если мужчина достиг половой зрелости, а
семенники не опустились в мошонку (состояние,
называемое крипторхизмом), то он навсегда
остается стерильным, а у мужчин, носящих слишком
тесные трусы или принимающих очень горячие ванны,
образование спермиев может так сильно понизиться,
что это приведет к бесплодию. Лишь у немногих
млекопитающих, в том числе у китов и слонов,
семенники всю жизнь находятся в брюшной полости.
Семенные канальцы достигают 50 см в длину и
200 мкм в диаметре и расположены в участках,
называемых дольками семенника. Оба конца
канальцев соединяются с центральной областью
семенника-сетью семенника (rete testis) - короткими
прямыми семенными канальцами. Здесь сперма собирается
в 10-20 выносящих канальцах; по ним она
переносится в головку придатка (эпидидимиса), где
концентрируется в результате обратного всасывания
жидкости, выделяемой семенными канальцами. В
головке придатка спермин созревают, после чего они
проходят по извитому 5-метровому выносящему
канальцу к основанию придатка; здесь они остаются
в течение короткого времени, прежде чем попадают
в семявыносящий проток. Семявыносящий проток-
это прямая трубка длиной около 40 см, которая
вместе с артерией и веной семенника образует
семенной канатик и переносит сперму в уретру
(мочеиспускательный канал), проходящую внутри
полового члена. Взаимоотношения между этими
структурами, мужскими придаточными железами и
половым членом показаны на рис. 20.29 и 20.30.
Образование гамет (гаметогенез)
Клетки зачаткового эпителия в мужских и женских
половых железах (гонадах) претерпевают ряд
последовательных митотических и мейотических делений,
называемых в совокупности гаметогенезом, в
результате чего образуются зрелые мужские гаметы
(сперматогенез) и женские гаметы (оогенез). В обоих
случаях процесс делится на три фазы - фазу
размножения, фазу роста и фазу созревания. Фаза
размножения включает многократные митотические
деления, приводящие к образованию множества сперма-
тогоний или оогоний. Каждая из них проходит
период роста в порядке подготовки к первому
делению мейоза и последующему цитокинезу. Затем
начинается фаза созревания, во время которой
происходят первое и второе деления мейоза с
последующей дифференцировкой гаплоидных клеток и
формированием зрелых гамет.
Развитие сперматозоидов
(сперматогенез)
Сперматозоиды (спермин) образуются в результате
ряда последовательных клеточных делений, назы-
Мочевой пузырь
Семяизвергательный
канал
Предстательная
железа
Перепончатая часть
Половой член
Губчатая часть
уретры
Пещеристые тела
Крайняя плоть
Головка полового
члена
Мочеточник
Ампула
семявыносящего протока
Семенной пузырек
Бульбоуретральная
железа
Семявыносящий проток
Придаток яичка
Яичко
Мошонка
Рис. 20.29. Схема строения мужской половой системы.
Долька яичка
Сеть яичка (rete testis)
Прямой семенной
каналец
Семенной каналец
Белочная оболочка
К уретре
Головка придатка
Выносящие канальцы
Основание придатка
Семявыносящий проток
Рис. 20.30. Упрощенная схема
строения семенника человека и
протоков, по которым сперма посту-
пает из семенных канальцев в
уретру.
ваемых в совокупности сперматогенезом, за
которыми следует сложный процесс дифференцировки,
называемый спермиогенезом (рис. 20.31). Процесс
образования спермия занимает примерно 70 дней;
на 1 г веса яичка образуется 107 спермиев в сутки.
Эпителий семенного канальца состоит из наружного
слоя клеток зачаткового эпителия и примерно шести
слоев клеток, образовавшихся в результате
многократных делений клеток этого слоя (рис. 20.32 и
20.33); эти слои соответствуют последовательным
стадиям развития сперматозоидов. Сначала деление
клеток зачаткового эпителия дает начало
многочисленным сперматогониям, которые увеличиваются
в размерах и становятся сперматоцитами первого
порядка. Эти сперматоциты в результате первого
деления мейоза образуют гаплоидные сперматоциты
второго порядка, после чего претерпевают второе
деление мейоза и превращаются в сперматиды.
Между «тяжами» из развивающихся клеток
расположены крупные клетки Сертоли, или трофические
клетки, расположенные во всем пространстве от
наружного слоя канальца до его просвета.
20.7. Клетки Сертоли богаты элементами
гладкого эндоплазматического ретикулума, содержат
обширный аппарат Гольджи и много
митохондрий и лизосом. Что можно сказать о функции этих
клеток на основании такой их структуры?
Фаза
развития
Механизм деления
Размножение
Митоз и цитокинез
Тип половых клеток
Клетка зачаткового
. эпителия
Число
хромосом
46
Повторные
деления
Созревание
Рост
Первое деление мейоза
Второе деление мейоза
I Спермиогенез
I Сперматогонии 46
Сперматоцит 4g
первого порядка
Сперматоциты
второго порядка
Сперматиды
(претерпеваю- 23
щие
спермиогенез)
Сперматозоиды 23
(спермин)
Рис. 20.31. Схема процессов сперматогенеза и спермиогенеза у человека.
Сперматогонии
Сперматиды
Лейдиговы
клетки
Базальная
мембрана
Сперматоцит|
первого
порядка
Спермин
Капилляры
Сперматиды
Спермин
Семенной
каналец
Ядро клетки
Сертоли
Рис. 20.32. Микрофотография среза семенника.
Видны семенные канальцы и лейдиговы (интерсти-
циальные) клетки.
Сперматоциты располагаются в многочисленных
впячиваниях на боковых поверхностях клеток
Сертоли; здесь они превращаются в сперматиды, а
затем переходят на тот край клетки Сертоли,
который обращен к просвету семенного канальца, где
они созревают, образуя спермин (рис. 20.33). По-
видимому, клетки Сертоли обеспечивают
созревающим спермиям механическую опору, защиту и
питание. Все питательные вещества и кислород,
доставляемые развивающимся гаметам по окружающим
семенные канальцы кровеносным сосудам, и
выделяемые в кровь отходы метаболизма проходят через
клетки Сертоли. Эти клетки секретируют и
жидкость, с которой спермин проходят по канальцам.
тительных механизмов, обеспечивающих движения
жгутика. Главная и хвостовая части сперматозоида
имеют строение, характерное для жгутиков: на
поперечном срезе видна типичная структура «9 + 2» -
9 пар периферических микротрубочек, окружающих
пару центральных микротрубочек.
Если смотреть на головку человеческого спермия
сверху, то она кажется округлой, а при взгляде
сбоку-уплощенной. Одного только жгутикового
движения недостаточно, чтобы сперматозоид мог
пройти расстояние от влагалища до места, где
происходит оплодотворение. Главная
локомоторная задача спермиев состоит в том, чтобы скопиться
вокруг ооцита и ориентироваться определенным
Сперматозоиды
Сперматозоиды, или спермин,-это очень мелкие
подвижные мужские гаметы, образуемые мужскими
гонадами-семенниками; их количество исчисляется
миллионами. Форма спермиев у разных животных
различна, однако строение их однотипно. Каждый
сперматозоид можно подразделить на пять
участков (рис. 20.34). В головке сперматозоида находится
ядро, содержащее гаплоидное число хромосом и
прикрытое акросомой Акросома - особая структура,
ограниченная мембраной,- содержит
гидролитические ферменты, способствующие проникновению
спермия в ооцит непосредственно перед
оплодотворением; таким образом, функционально ее можно
рассматривать как увеличенную лизосому. В
короткой шейке спермия расположена пара центриолей,
лежащих под прямым углом друг к другу.
Микротрубочки одной из центриолей удлиняются, образуя
осевую нить жгутика, проходящую вдоль всей
остальной части сперматозоида. Средняя часть
расширена за счет содержащихся в ней многочисленных
митохондрий, собранных в спираль вокруг жгутика.
Эти митохондрии доставляют энергию для сокра-
Развивающиеся
спермин (5)
Клетка
Сертоли
Ядро клетки
Сертоли
Сперматогония
в процессе -—
митоза
Базальная
мембрана \
Волокнистая
соединительная'
ткань
Капилляр
Сперматида
в процессе
спермиогенеза
Ранняя
сперматида (4)
Сперматоцит
второго порядка (3)
Сперматоцит
первого порядка (2)
Сперматогонии
(D
Лейдигова клетка
Рис. 20.33. Участок стенки семенного канальца
(схема). Показаны лейдиговы клетки и
различные стадии сперматогенеза и спермиогенеза.
, 25 MKMi
Головка.
(5 мкм)
Шейка
Промежуточный]
отдел
(5 мкм)
Жгутик
(хвост)
(50 мкм)
Акросома
Ядро
Спермий
(продольный
разрез)
Центриоль
Митохондрии
(расположены
по спирали)
Головка спермин
(в разрезе)
Парные
периферические
микро-
I трубочки
f Центральные
^икротрубочки
Жгутик
(поперечный разрез)
Рис. 20.34. Схема строения зрелого спермин
человека.
образом, прежде чем проникнуть сквозь мембраны
ооцита.
Эндокринная функция яичек
у человека
Рост, развитие и поддержание яичек у человека
контролируются гонадотропными гормонами
гипофиза - фолликулостимулирующим (ФСГ) и лютеини-
зирующим (ЛГ). ФСГ стимулирует развитие спер-
миев, а ЛГ-синтез стероидного гормона
тестостерона лейдиговыми клетками яичка. Л Г оказывает
влияние на эти клетки, заставляя их синтезировать
циклический аденозинмонофосфат (цАМФ); цАМФ
переходит в ядро, где воздействует на генетический
аппарат и таким образом стимулирует синтез
ферментов, участвующих в образовании тестостерона
из холестерола. (У мужчин Л Г иногда называют по
месту его действия гормоном, стимулирующим ин-
терстициальные клетки).
Тестостерон - это главный андрогенный гормон,
влияющий как на первичные, так и на вторичные
половые признаки. Для успешного образования
спермиев необходимы и тестостерон, и ФСГ, тогда
как развитие вторичных половых признаков в
период полового созревания и поддержание их в
течение всей взрослой жизни контролируется только
тестостероном. К этим признакам относятся
развитие наружных мужских половых органов и
придаточных желез репродуктивной системы, усиленное
развитие мускулатуры, рост гортани и
соответственно более низкий голос, рост и распределение
волосяного покрова, особенности поведения,
связанные с половой жизнью и родительскими
заботами.
20.3.2. Женская половая система
Роль женщины в процессе размножения гораздо
больше, чем роль мужчины, и она сопряжена с
взаимодействиями между гипофизом, яичниками,
маткой и плодом. Женская половая система состоит
из парных яичников и фаллопиевых труб, матки,
влагалища и наружных половых органов (рис.
20.35).
Яичники прикреплены к стенке брюшной полости
с помощью складки брюшины и выполняют две
функции: производят женские гаметы и секретируют
женские половые гормоны. Яичник имеет
миндалевидную форму, состоит из наружного коркового и
внутреннего мозгового слоев и заключен в
соединительнотканную оболочку, называемую белочной
оболочкой. Наружный слой коркового вещества
состоит из клеток зачаткового эпителия, из которого
образуются гаметы. Корковое вещество образовано
развивающимися фолликулами, а мозговое
вещество стромой, содержащей соединительную ткань,
кровеносные сосуды и зрелые фолликулы (см.
рис. 20.38).
Фаллопиева труба это мышечная трубка длиной
около 12 см, по которой женские гаметы выходят из
яичника и попадают в матку. Отверстие
фаллопиевой трубы заканчивается расширением, край
которого образует бахромку, приближающуюся во
время овуляции к яичнику. Просвет фаллопиевой
трубы выстлан ресничным эпителием; перемещению
женских гамет к матке способствуют
перистальтические движения мышечной стенки фаллопиевой
трубы.
Матка представляет собой толстостенный
мешок длиной примерно 7,5 см и шириной 5 см,
состоящий из трех слоев. Наружный слой называется
серозной оболочкой. Под нею находится самый
толстый средний слой - миометрий; его образуют пучки
гладких мышечных клеток, которые во время родов
чувствительны к окситоцину. Внутренний слой-
Фаллопиева
труба
Бахромк;
Отверстие фаллопиевой
трубы
Правый яичник
Складка брюшины
Тело матки
Полость матки
Шейка матки
Влагалище
Вульва
tБольшая губа
Малая губа
Клитор
Отверстие
уретры
Отверстие влагалища
Анальное отверстие
Рис. 20.35. Упрощенная схема строения женской
половой системы. Матка и влагалище изображены в
разрезе, а наружные половые органы, отверстие
мочеиспускательного канала и анальное
отверстие со стороны поверхности, причем половые губы
раздвинуты.
Фаза
развития
Механизм
деления
Тип половых
клеток
Стадия
жизненного
цикла
Число
хромосом
Клетка
зачаткового
эпителия
46
Размножение
Митоз
Первичная
половая
клетка
Оогонии
Внутриутробное
развитие
Рост
Профазное-
ядро
Первое
деление
мейоза
Ооцит
первого порядка
— Первичный
фолликул
Первое
полярное тельце
Ооцит
второго порядка
Рождение
46
До овуляции
23
Овуляция
23
Созревание
Спермий-
Второе
деление мейоза,
стимулируемое
оплодотворением
Оплодотворение
Второе
деление
мейоза
Второе
полярное тельце
46
Яйцо (содержащее
мужской и женский
пронуклеусы)
Рис. 20.36. Схема процесса оогенеза у человека.
эндометрий - мягкий и гладкий; он состоит из
эпителиальных клеток, простых трубчатых желез и
спиральных артериол, снабжающих клетки кровью. Во
время беременности полость матки может
увеличиться в 500 раз-от 10 см3 до 5000 см3. Нижним
входом в матку служит ее шейка, соединяющая
матку с влагалищем. Вход во влагалище, наружное
отверстие мочеиспускательного канала и клитор
прикрыты двумя кожными складками - большими и
малыми половыми губами, образующими вульву.
Клитор-это небольшое способное к эрекции
образование, гомологичное мужскому половому члену.
В стенках вульвы находятся бартолиниевы железы,
выделяющие при половом возбуждении слизь,
которая увлажняет влагалище во время полового акта.
Развитие яйца у человека (оогенез)
В отличие от образования спермиев, которое
начинается у мужчин только при половом созревании,
образование яйцеклеток у женщин начинается еще
до их рождения и завершается для каждой данной
яйцеклетки только после ее оплодотворения. Стадии
оогенеза показаны на рис. 20.36. Во время развития
плода первичные половые клетки многократно
делятся путем митоза, образуя много крупных клеток,
называемых оогониями. Оогонии снова
претерпевают митоз и образуют ооциты первого порядка,
которые остаются на стадии профазы почти до
самой овуляции. Ооциты первого порядка
окружены одним слоем клеток - гранулезной оболочкой-и
образуют так называемые примордиальные
фолликулы. Плод женского пола непосредственно перед
рождением содержит около 2· 106 этих фолликулов,
но лишь примерно 450 из них достигают стадии
ооцитов второго порядка и выходят из яичника
(овуляция). Перед овуляцией ооцит первого порядка
проделывает первое деление мейоза, образуя
гаплоидный ооцит второго порядка и первое полярное
тельце. Второе деление мейоза доходит до стадии
метафазы, но не продолжается дальше до тех пор,
пока ооцит не сольется со сперматозоидом. При
оплодотворении ооцит второго порядка совершает
второе деление мейоза, образуя крупную клетку-
яйцо, а также второе полярное тельце. Все полярные
тельца представляют собой мелкие клетки; они не
играют никакой роли в оогенезе и в конечном счете
разрушаются.
Менструальный цикл
У мужчин образование и выделение гамет - процесс
непрерывный, начинающийся с наступлением
половой зрелости и продолжающийся в течение всей
жизни. У женщин же это циклический процесс,
повторяющийся примерно через каждые 28 дней
и связанный с изменениями в строении и функциях
всей репродуктивной системы. Процесс этот
называется менструальным циклом, и его можно
разделить на четыре стадии (рис. 20.37). В событиях,
происходящих во время менструального цикла,
участвуют яичники (овариальный цикл) и матка
(маточный цикл), и они регулируются гормонами
яичников, секреция которых в свою очередь
регулируется гипофизарными гонадотропинами.
Овариальный цикл
У взрослой женщины овариальный цикл (рис. 20.38)
начинается с развития нескольких первичных
фолликулов (содержащих ооциты первого порядка) под
(14) Овуляция
<-(ЛГ и ФСГГ
выделение ооцита
(7)'
Фолликулярная
фаза к
(ФСГ, ЛГ и
эстроген) развитие
фолликула
(прогестерон)
развитие
желтого тела
(21)Лютеальная
фаза
(Снижение уровня
прогестерона)
инволюция
желтого тела
(5)
Менструация
(1)1(28)
Рис. 20.37. Главные фазы менструального цикла
(цифры в скобках-дни, на которые
приходится данная фаза).
Первичный
^ фолликул
Вторичный
) фолликул
Тека
фолликула
Примордиальные
фолликулы
Зачатковый
эпителий
Белочная
оболочка
_ Полость
Третичный
фолликул'
Клетки
гранулозы
Граафов пузырек
Тека фолликула
Гранулоза
Полость
Corona radiata
Zona pellucida
Строма
с соединительной тканью и
кровеносными сосудами
Инволюция
желтого тела
Функционирующее
желтое тело
Разорвавшийся
граафов пузырек
Corona radiata
Zona pellucida
Ооцит
второго порядка
Рис. 20.38. Схема яичника женщины (в разрезе). Показаны последовательные стадии
(обозначены цифрами) развития граафова пузырька, овуляции, образования желтого тела и его
инволюции. На самом деле нельзя наблюдать все эти стадии одновременно.
действием фолликулостимулирующего гормона
(ФСГ), выделяемого передней долей гипофиза. Из
этих фолликулов только один продолжает расти,
тогда как остальные разрушаются в результате
дегенеративного процесса (атрезия фолликулов).
Клетки гранулезной оболочки растущего фолликула
пролиферируют, образуя наружный волокнистый
слой толщиной в несколько клеток, называемый
theca externa, и внутренний слой, богатый
кровеносными сосудами,- theca interna. Клетки
гранулезной оболочки секретируют фолликулярную
жидкость, которая накапливается в полости фолликула
(antrum). Лютеинизирующий гормон (ЛГ),
выделяемый гипофизом, стимулирует клетки обеих thecae,
побуждая их вырабатывать стероиды, главным
образом 17р-эстрадиол. Возрастание уровня эстрадио-
ла во время фолликулярной фазы действует на
гипофиз по принципу отрицательной обратной
связи, вызывая понижение уровней ФСГ в крови
(4-й-11-й дни); уровень ЛГ остается неизменным.
Уровень эстрогена достигает максимума примерно
за три дня до овуляции и действует в это время
на гипофиз по принципу положительной
обратной связи, стимулируя выделение как ФСГ, так
и ЛГ.
Как полагают, ФСГ необходим для стимуляции
роста фолликулов, но дальнейшее развитие
фолликулов контролируется главным образом Л Г.
Гранулезные клетки располагаются по периферии, яйцо
смещено к одной стороне фолликула, но все еще
окружено слоем гранулезных клеток. Зрелый
фолликул, называемый граафовым пузырьком,
достигает примерно 1 см в диаметре и выступает над
поверхностью яичника в виде бугорка (рис. 20.39).
Механизм овуляции в точности неизвестен, но
предполагается, что в нем участвуют Л Г, ФСГ и
простагландины.
Во время овуляции ооцит второго порядка
отделяется от стенки лопнувшего фолликула, выходит
в брюшную полость и попадает в фаллопиеву трубу.
Обычно каждый месяц освобождается только один
ооцит одним из яичников, так что овуляция
происходит поочередно то в одном, то в другом
яичнике. Овулировавший ооцит представляет собой
клетку, ядро которой находится в метафазе I мейо-
за; он окружен слоем клеток, называемым zona
pellucida, и слоем гранулезных клеток (corona
radiata), который создает защиту ооциту вплоть до
оплодотворения.
После овуляции уровень ЛГ падает до уровня,
характерного для фолликулярной фазы, и под
действием другого гонадотропина - пролактина - клетки
лопнувшего фолликула изменяются, образуя желтое
тело. Желтое тело начинает секретировать еще один
женский гормон - прогестерон - и небольшие
количества эстрогена. Эти два гормона поддерживают
нормальное строение эндометрия, выстилающего
матку, и ингибируют выделение ФСГ и ЛГ,
воздействуя по принципу отрицательной обратной связи
на гипоталамус. Если оплодотворения не
произошло, то под действием факторов, которые будут
описаны позднее, желтое тело подвергается
инволюции и от него остается лишь небольшой рубец-
corpus albicans; это сопровождается снижением
уровней прогестерона и эстрогена, в результате чего
прекращается ингибирование секреции ФСГ, его
уровень повышается и начинается новый цикл
развития фолликула. Соотношения между активностью
гипофиза и яичников и изменениями в яичниках и в
матке показаны на рис. 20.40.
20.8. При вскрытии трупа 22-летней женщины
оказалось, что яичники имеют неодинаковые
размеры. При исследовании яичников в них было
обнаружено:
Меньший яичник
17 000 фолликулов
4 желтых тела
10 рубцов от желтых
тел
Больший яичник
25 000 фолликулов
5 желтых тел
48 рубцов от желтых тел
Фолликулы были по большей части мелкие, но
219 имели свыше 100 мкм в диаметре. Пять
фолликулов содержали по два ооцита каждый и 2%
ооцитов содержали по два ядра. Если
предположить, что каждый фолликул образует одно желтое
тело, то
\.а) В каком возрасте у этой женщины начались
овуляции?
б) Примерно сколько лет у нее могли бы
продолжаться овуляции?
в) Сколько пар близнецов могла бы она родить
и какие это были бы близнецы?
2. Чем можно объяснить наличие в некоторых
ооцитах двух ядер (см. разд. 22.3)?
Маточный цикл
Маточный цикл делится на три фазы, связанные с
определенными структурными и функциональными
изменениями эндометрия.
ι. менструальная фаза. В этой фазе происходит
отторжение эпителиального слоя эндометрия. Перед
самой менструацией кровоснабжение этой области
уменьшается в результате сужения спиральных ар-
териол в стенке матки, вызываемого падением
уровня прогестерона в крови после инволюции желтого
тела. Недостаточное кровоснабжение приводит к
гибели эпителиальных клеток. Затем сужение
спиральных артериол сменяется их расширением, и под
действием усилившегося притока крови эпителий
отторгается и его остатки выводятся вместе с
кровью в виде менструальных выделений.
2. пролиферативная фаза. Эта фаза совпадает с
фолликулярной фазой овариального цикла и состоит в
быстрой пролиферации клеток эндометрия,
приводящей к его утолщению под контролем эстрогена,
выделяемого развивающимся фолликулом.
3. секреторная фаза. Во время этой фазы
прогестерон, выделяемый желтым телом, стимулирует
секрецию слизи трубчатыми железами; это
поддерживает выстилку матки в состоянии, при котором
возможна имплантация в нее оплодотворенного
яйца.
Гранулезная
оболочка
Полость
Corona radiata
Zona pellucida
Ооцит
Тека
фолликула
Zona glomerulosa
Рис. 20.39. Микрофотография зрелого граафова пузырька.
Что происходит,
если оплодотворение
не состоялось?
Если в течение суток после овуляции
оплодотворения не произойдет, то ооцит второго порядка
претерпевает автолиз в фаллопиевой трубе; то же
самое происходит со спермиями, оставшимися в
половых путях женщины. Желтое тело сохраняется
в течение 10-14 дней после овуляции (обычно до
26-го дня цикла), но затем перестает секретировать
прогестерон и эстроген из-за недостаточного уровня
JIГ в крови и подвергается автолизу. Как показали
недавние исследования, у некоторых видов стенка
матки, не содержащая оплодотворенного яйца,
по-видимому, выделяет фактор, названный лютео-
лизином; этот фактор представляет собой проста-
гландин (простагландин F2a). Как полагают, лю-
теолизин поступает с кровью в яичник, где он
вызывает инволюцию желтого тела, разрушая лизо-
сомы в гранулезных клетках последнего, что
приводит к их автолизу.
Результаты оплодотворения
Если происходит оплодотворение, то из
образовавшейся зиготы развивается бластоциста, которая
через 8 дней после овуляции погружается в стенку
матки. Наружные клетки бластоцисты, образующие
трофобласт, начинают затем секретировать
гормон - хорионический гонадотропин, по своей функции
сходный с лютеинизирующим гормоном. Эта
функция включает предотвращение автолиза желтого
тела и побуждение его к секреции больших
количеств прогестерона и эстрогенов, которые
вызывают усиленный рост эндометрия. Отторжение
эпителиальной выстилки эндометрия подавляется, и
очередная менструация не наступает, что служит
самым ранним признаком беременности.
(Хорионический гонадотропин стимулирует также интерсти-
циальные клетки семенников у зародыша мужского
пола и заставляет их вырабатывать тестостерон,
который индуцирует рост мужских половых
органов.) Роль плаценты возрастает примерно на 10-ю
неделю беременности, когда она начинает сама
Менструация
Содержание
ФСГиЛГ
в крови
.ЛГ
ФСГ
Содержание
170-эстра-
диола и
прогестерона в крови
Яичник
Новый цикл
|17р-эстрадиол
! (эстроген)
Время, дни
Прогестерон
Ооцит
Время, дни
Матка
Первичный
фолликул
Желтое тело
ПРОГЕСТЕРОН
Толщина
эндометрия
Развивающаяся
тека фолликула^
Менструальная фаза
ЭСТРОГЕН
Овуляция
Пролиферативная
фаза
Секреторная
фаза
Время, дни
Рис. 20.40. Общая схема взаимоотношений между гонадотропными гормонами гипофиза (А),
стероидными гормонами яичников (Б), развитием фолликулов и желтого тела (В) и
толщиной эндометрия (Г) на протяжении менструального цикла женщины.
секретировать большую часть прогестерона и
эстрогена, необходимых для нормального течения
беременности. Преждевременное прекращение
активности желтого тела (до того как плацента в полной
мере проявит свою секреторную способность) -
частая причина выкидыша на 10-й-12-й неделях
беременности.
Во время беременности хорионический гонадо-
тропин можно обнаружить в моче, и на этом
основаны тесты на беременность. В настоящее время
применяется тест, основанный на подавлении
агглютинации: частицы латекса, покрытые хориони-
ческим гонадотропином, добавляют к смеси мочи с
антисывороткой, агглютинирующей этот гормон.
Если он присутствует в моче, то он будет
реагировать с агглютинирующей антисывороткой, а не с
частицами латекса. Отсутствие агглютинации
латекса служит показателем беременности; этот тест
можно применять уже через 14 дней после
несостоявшейся менструации.
20.3.3. Копуляция
Внутреннее оплодотворение составляет важную
часть репродуктивного цикла у наземных
организмов, и у многих из них, в том числе у человека, оно
облегчается благодаря совокупительному органу-
мужскому половому члену, который вводится во
влагалище и вносит гаметы как можно глубже в
половые пути самки. Эрекция полового члена
происходит вследствие локального повышения
кровяного давления в его эректильной ткани в результате
сужения вен и расширения артерий. Эта реакция
обусловлена активацией парасимпатической
нервной системы при половом возбуждении. В таком
состоянии половой член может быть введен во
влагалище, где трение, создаваемое ритмическими
движениями во время полового акта, производит
тактильное раздражение сенсорных клеток в головке
полового члена. Это активирует симпатические
нейроны, которые вызывают сокращение внутреннего
сфинктера мочевого пузыря, гладкой мускулатуры
эпидидимиса, семявыносящего протока и
придаточных желез (семенного пузырька, простаты и бульбо-
уретральных желез). В результате всего этого
будущие компоненты семенной жидкости
выталкиваются в проксимальную часть уретры, где они
смешиваются. Возросшее давление в проксимальной
части уретры вызывает рефлекторную активность
двигательных нейронов, иннервирующих мышцы у
основания полового члена. Ритмические
волнообразные сокращения этих мышц проталкивают
семенную жидкость через дистальную часть уретры, и
происходит эякуляция (выбрасывание спермы)-
кульминационный момент полового акта.
Ощущения, связанные с этим моментом как у мужчины, так
и у женщины, объединяются под названием оргазма.
Смазку во время полового акта обеспечивают
главным образом секреты желез влагалища и вульвы, а
частично также прозрачная слизь, выделяемая буль-
боуретральными железами мужчины после эрекции.
Секреты придаточных мужских желез имеют
щелочную реакцию и содержат слизь, фруктозу,
витамин С, лимонную кислоту, простагландины и
различные ферменты; они повышают рН среды во
влагалище, которая обычно бывает кислой, до
6-6,5, что является оптимальным для подвижности
спермиев после эякуляции. Количество семенной
жидкости, извергаемой при эякуляции, составляет
около 3 мл, из которых лишь 10% составляют
спермии. Несмотря на такой низкий процент,
сперма содержит примерно 108 спермиев на 1 мл.
20.3.4. Оплодотворение
Сперма изливается глубоко во влагалище, вблизи
шейки матки. Как показали исследования,
сперматозоиды проходят из влагалища в матку и доходят
до верхнего конца фаллопиевых труб за пять минут
благодаря сокращениям матки и труб. Как
полагают, эти сокращения инициируются окситоцином,
выделяющимся во время полового акта, и
локальным воздействием простагландинов, содержащихся
в семенной жидкости, на матку и фаллопиевы
трубы. Спермии сохраняют жизнеспособность в
женских половых путях в течение 24 - 72 ч, но остаются
высокофертильными лишь 12-24 ч. Сперматозоид
способен оплодотворить ооцит только после того,
как он проведет в половых путях несколько часов,
претерпевая процесс так называемой капацитации;
при этом свойства мембраны, покрывающей акро-
сому, изменяются, что делает возможным
оплодотворение, которое обычно происходит в верхней
части фаллопиевой трубы.
Когда спермии приближается к ооциту (рис.
20.41), его наружная мембрана, покрывающая
область акросомы, и мембраны самой акросомы
разрываются, и находившиеся в акросоме ферменты-
гиалуронидаза и протеаза - «переваривают»
окружающие ооцит клеточные слои, в том числе zona
pellucida. Эти изменения в головке спермия
называют акросомальной реакцией. В результате
дальнейших изменений в головке спермия внутренняя
мембрана акросомы выворачивается наизнанку, что
позволяет спермию проникнуть сквозь zona
pellucida и плазматическую мембрану в ооцит; у
человека сперматозоид входит в ооцит целиком.
После того как в яйцеклетку проник один спермии,
расположенные под ее плазматической мембраной
кортикальные гранулы разрываются, начиная с
места вхождения спермия, и освобождают вещество,
под действием которого zona pellucida утолщается и
отделяется от плазматической мембраны
яйцеклетки. Эта так называемая кортикальная реакция
распространяется по всей клетке, в результате чего
zona pellucida образует непроницаемую преграду-
оболочку оплодотворения, которая препятствует
Рис. 20.41. А. Спермии человека, скопившиеся вокруг ооцита второго порядка.
Микрофотография, полученная с помощью сканирующего электронного микроскопа. Б. Момент
оплодотворения у морского ежа - проникновение спермия сквозь мембрану яйца. Темный клин-головка спермия, в которой
заключен генетический материал. Серое образование позади головки доставляет энергию, необходимую
хвосту. Спермий переварил наружную оболочку яйца, состоящую из глико-
протеина, и проник внутрь. Содержащаяся в яйце жидкость вышла наружу, покрыла спермий и теперь
втягивает внутрь, чтобы завершилось оплодотворение. Каким-то неизвестным образом проникновение в яйцо
одного спермия закрывает доступ в него для других, вероятно, это связано с быстрыми
изменениями наружной оболочки яйца.
проникновению в ооцит других спермиев, т. е.
явлению полиспермии.
Проникновение сперматозоида служит стимулом
для завершения второго деления мейоза, и ооцит
второго порядка становится зрелым яйцом. При
этом образуется также второе полярное тельце,
которое тотчас же дегенерирует, а хвост сперматозоида
рассасывается в цитоплазме яйца. Ядра обеих гамет
превращаются в пронуклеусы и сближаются.
Мембраны пронуклеусов разрушаются, а отцовские
и материнские хромосомы прикрепляются к
образовавшимся нитям веретена. К этому времени оба
гаплоидных набора, содержащих у человека по 23
хромосомы, уже реплицировались, и возникшие в
результате 46 пар хроматид выстраиваются по
экватору веретена, как в метафазе митоза. Слияние
пронуклеусов называется кариогамией. На этой
стадии восстанавливается диплоидное число хромосом,
и оплодотворенное яйцо получает название зиготы.
Зигота проходит стадии анафазы и телофазы и
завершает свое первое митотическое деление.
Следующий за этим цитокинез приводит к образованию
двух диплоидных дочерних клеток. Важнейшие
события, связанные с оплодотворением,
суммированы на рис. 20.42.
20.3.5. Имплантация
Проходя по фаллопиевым трубам в матку, зигота
дробится (т.е. претерпевает ряд ядерных и
клеточных делений) и образует комочек из клеток-
морулу. Дробление не приводит к увеличению
размеров морулы, так как клетки продолжают
оставаться внутри zona pellucida. Получающиеся в
результате дробления клетки называют бластомерами;
в дальнейшем они располагаются по периферии,
образуя стенку вокруг центральной полости-
Corona radiata
Zona pellucida
Кортикальные гранулы
Ооцит второго порядка
Спермин
Второе полярное
тельце
Женский пронуклеус
Мужской пронуклеус
Хроматин второго
деления мейоза
Оболочка
оплодотворения
Плазматическая
мембрана
Перивителлиновое
пространство
Материнские
хромосомы
Волокна веретена
Отцовские хромосомы
Рис. 20.42. Упрощенная схема оплодотворения. А. Спермий после процесса капацитации способен
проникнуть сквозь zona pellucida ооцита второго порядка и вызвать разрушение кортикальных
гранул. Б. В результате кортикальной реакции zona pellucida превратилась в оболочку оплодотворения, которая
отходит от плазматической мембраны, так что между ними образуется перивителлиновое
пространство. В. При втором делении мейоза образуется второе полярное тельце, а затем формируются
женский и мужской пронуклеусы. Г. Ядерные оболочки пронуклеусов разрушаются; материнские и отцовские
хромосомы, прикрепленные к нитям веретена, выстраиваются на экваторе яйца. Это метафаза первого
митотического деления зиготы.
бластоцеля, наполненного жидкостью из яйцевода.
Наружный слой бластомеров, называемый трофо-
бластом, в одном участке дифференцируется,
образуя внутреннюю клеточную массу. Эта стадия
развития, достигаемая спустя 4-5 дней после овуляции,
называется бластоцистой (рис. 20.43).
Попав в матку, бластоциста в течение двух дней
остается в ее полости; за это время zona pellucida
исчезает, так что клетки трофобласта могут
вступить в контакт с клетками эндометрия. Клетки
трофобласта размножаются, получая питательные
вещества из эндометрия, и на 6-й -9-й день после
овуляции бластоциста погружается в стенку матки,
располагаясь между клетками эндометрия. Этот
процесс носит название имплантации. Клетки
трофобласта дифференцируются на два слоя; из клеток
наружного слоя образуются ворсинки трофобласта,
которые врастают в эндометрий (рис. 20.44).
Участки эндометрия между этими ворсинками образуют
сообщающиеся друг с другом полости, называемые
лакунами, что придает этой области эндометрия
сходство с губкой. Под действием гидролитических
ферментов, выделяемых этой многоядерной
структурой, артериальные и венозные сосуды эндометрия
разрываются и вытекающая из них кровь заполняет
лакуны. На ранних стадиях развития бластоцисты
обмен питательными веществами, кислородом и
отходами метаболизма между клетками
бластоцисты и материнской кровью в стенке матки
происходит через ворсинки трофобласта. На более
поздних стадиях развития эту функцию принимает на
себя плацента.
20.3.6. Ранние стадии эмбрионального
развития
Наружные клетки бластоцисты, из которых состоит
трофобласт, растут и развиваются, образуя
наружную оболочку, называемую хорионом; хорион
играет важную роль в питании развивающегося
зародыша и удалении ненужных продуктов обмена. Во
внутренней клеточной массе появляются две
полости; клетки, выстилающие эти полости, дают
начало двум оболочкам - амниону и желточному
мешку. Амнион представляет собой тонкую
оболочку, которая покрывает зародыша и несет
защитные функции; его клетки выделяют амниотическую
жидкость, заполняющую амниотическую полость,
расположенную между амнионом и зародышем. По
мере роста зародыша амнион расширяется, так что
он всегда бывает прижат к стенке матки. Амнио-
Трофобласт (каждая
его клетка — бластомер)
Бластоцель
Внутренняя клеточная масса
Рис. 20.43. Упрощенная схема строения
бластоцисты человека спустя четыре дня после
оплодотворения.
Стенка
матки
Полость матки
.Венула матки
_ Эрозированная венула
.Ворсинки трофобласта
(образовались из наружного
трофобласта)
Лакуны
(пространства, заполненные кровью)
Зародышевый диск
Желточный мешок
Амниотическая полость
Бластоцель
Внутренний слой трофобласта
Эпителий стенки матки
Рис. 20.44. Бластоциста человека вскоре после имплантации в эндометрий (упрощенная схема).
Ферменты, вырабатываемые синцитием наружного слоя трофобласта, разрушают
кровеносные сосуды эндометрия, в результате чего образуются наполненные кровью лакуны, через которые
бластоциста получает питательные вещества и выводит продукты, подлежащие экскреции.
тическая жидкость поддерживает зародыша и
защищает его от механического повреждения. У
человеческого эмбриона желточный мешок не играет
существенной роли, однако у рептилий и птиц он
поглощает запасенные в желтке питательные
вещества и переносит их в среднюю кишку
развивающегося зародыша.
Клетки, составляющие внутреннюю клеточную
массу, и желточный мешок образуют структуру,
называемую зародышевым диском, из которой
развивается эмбрион. Клетки этого диска на ранней
стадии, когда его диаметр не достигает 2 мм,
дифференцируются на два слоя - эктодерму (наружный
слой) и энтодерму (внутренний). На более поздней
стадии образуется мезодерма, и эти три
зародышевых листка дают начало всем тканям
развивающегося эмбриона, как это описано в разд. 21.8
На ранних стадиях развития обмен веществами
между зародышем и материнским организмом
происходит через ворсинки трофобласта, однако до-
Ворсинки
трофобласта
Хорион
Амнион
Пуповина
Ворсинки
хориона
Амниотическая
полость
Зародыш
Желточный
мешок
В этой области
разовьется
плацента
Аллантоис
Хорион
Рис. 20.45. Упрощенная схема взаимоотношений
между человеческим эмбрионом и зародышевыми
оболочками примерно через 5 недель после
овуляции. В области хориона и аллантоиса в дальнейшем
образуется плацента.
вольно скоро ворсинки уже не могут справляться с
этими функциями и из задней кишки зародыша
развивается четвертая оболочка - аллантоис.
Хорион, амнион, желточный мешок и аллантоис
называют зародышевыми, или позднее плодными,
оболочками (рис. 20.45). Аллантоис растет в наружном
направлении, пока не приходит в соприкосновение с
хорионом, образуя богатую сосудами структуру-
хориоаллантоис, который участвует в формировании
плаценты - органа, более эффективно
осуществляющего обмен между зародышем и материнским
организмом.
Плацента
Плацента - временный орган, имеющийся только у
плацентарных млекопитающих; у животных это
единственный орган, состоящий из клеток,
происходящих от двух разных организмов - матери и
плода. В плаценте кровеносные сосуды
материнского организма и плода вступают в тесный контакт
между собой, что облегчает перенос питательных
веществ, кислорода и отходов метаболизма.
Плацента представляет собой обособленное дисковид-
ное образование, расположенное в одном из
участков стенки матки; по мере своего развития она
принимает на себя функции ворсинок трофобласта и
по прошествии 12 недель становится главной
структурой, обеспечивающей обмен веществами между
матерью и плодом.
Часть плаценты, происходящая от плода, состоит
из соединительнотканных клеток хориона, которые
инвазируют ворсинки трофобласта и образуют
более крупные выросты - ворсинки хориона.
Внутренние участки этих ворсинок заполняются сложными
сетями капилляров, отходящих от двух кровеносных
сосудов плода - пупочной артерии и пупочной вены.
Эти сосуды образуются в аллантоисе и идут от
плода к стенке матки, находясь в пупочном канатике,
или пуповине,- плотном тяже длиной около 40 см,
ПЛАЦЕНТА
Пространство, заполненное
материнской кровью
К ПЛОДУ
Пупочные артерии
Лакуна
Лакуна.
Лакуна
Лакуна:
Пупочная вена
Амниотическая полость
Амнион
Артерии плода
Вены плода
Хорион
Место отторжения -
эндометрия
после родов
Децидуальный эндометрий
Перманентный эндометрий
Миометрий
Ворсинка хориона
Капилляры хориона
Венула матки
Артериола матки
Рис. 20.46. Упрощенная схема взаимоотношений между пупочными кровеносными сосудами,
капиллярами ворсинок хориона и наполненными кровью лакунами в плаценте человека. Плацента служит
связующим звеном между кровеносными системами плода и материнского организма.
покрытом клетками, происходящими из амниона и
хориона (рис. 20.46).
Материнская часть плаценты состоит из выростов
поверхностных слоев эндометрия - децидуальной
оболочки. Между ними и ворсинками хориона
расположены лакуны, снабжаемые артериальной
кровью по артериолам матки и отдающие венозную
кровь по венулам в маточную вену. Направление
тока крови через лакуны определяется разницей
давления в артериальных и венозных сосудах.
Мембраны клеток в стенках ворсинок хориона,
омываемых материнской кровью, образуют
микроворсинки, что увеличивает их поверхность, через
которую происходит обмен веществами путем
диффузии и иными способами. В этих мембранах
имеются молекулы-переносчики, служащие для
поглощения веществ ворсинками путем активного
транспорта; клетки содержат множество
митохондрий. То, что в клетках ворсинок много мелких
пузырьков, позволяет предполагать, что
поглощение веществ из материнского организма происходит
путем пиноцитоза (см. разд. 7.2.2). Вероятно,
необходимы различные механизмы, так как расстояние,
отделяющее кровь плода от крови матери, довольно
велико (в десять раз больше, чем, например,
расстояние, которое приходится преодолевать при
газообмене в легких). Через «плацентарный барьер»
от матери плоду передаются вода, глюкоза,
аминокислоты, простые белки, липиды, неорганические
соли, витамины, гормоны, антитела и кислород, а в
обратном направлении - вода, мочевина и другие
азотсодержащие отходы, гормоны и CO2. Плоду
могут передаваться от матери потенциально
вредоносные факторы, такие как бактерии, вирусы,
токсины и лекарственные препараты, однако их
воздействие нейтрализуется передачей в том же
направлении некоторых антител, глобулинов,
антибиотиков и антитоксинов; благодаря этому рожденный
младенец обладает пассивным иммунитетом к
некоторым болезням (см. разд. 14.14.5).
Плацентарный барьер не только защищает плод
от последствий многих вредных воздействий,
которым может подвергнуться мать, но и экранирует
его кровеносную систему от высокого кровяного
давления в сосудах матери. Он не может, однако,
служить иммунологическим барьером, а плод,
поскольку он несет отцовские гены, будет
вырабатывать антигены, чужеродные для организма
матери, который будет вырабатывать против них
антитела. Чем обусловлена удивительная способность
плода (в иммунологическом отношении, по-сущест-
ву, гомотрансплантата) на протяжении 40 недель
сопротивляться отторжению, неизвестно;
предполагается, однако, что она связана с выработкой имму-
носупрессивных веществ, циркулирующих в плазме
матери.
Для жизни и развития плода необходимо, чтобы
он непрерывно получал от материнского организма
кислород; это обеспечивается различным сродством
к кислороду гемоглобинов плода и матери (см.
разд. 14.13.1).
Плацента представляет собой эндокринный
орган, секретирующий главным образом хориониче-
ский гонадотропин, эстрогены, прогестерон и
плацентарный лактоген (соматомаммотропный
гормон). Последний стимулирует развитие молочных
желез при подготовке к лактации. Все эти гормоны
образуются в соединительной ткани хориона.
Половое развитие зародыша
Генетический пол зародыша определяется половой
хромосомой, содержащейся в отцовском
сперматозоиде: если это Х-хромосома, то родится девочка,
если Y-мальчик. Однако создается впечатление, что
человеческий организм исходно имеет тенденцию к
развитию в сторону женского пола, в основном
из-за присутствия в клетках обоих полов Х-хро-
мосом. На ранних стадиях онтогенеза у зародыша
развивается пара недифференцированных зачатков
гонад-половых складок-и образуются зачатки
элементов как женской, так и мужской репродуктивной
системы. В результате все эмбрионы вплоть до
шестой недели развития являются потенциально
обоеполыми.
В недавних исследованиях был выявлен
возможный механизм, с помощью которого половые
хромосомы определяют, какая из двух систем будет
активирована и достигнет полного развития у
данного индивидуума.
Х-хромосома несет ген Tfm (ген тестикулярной
феминизации), нормальный аллель которого
детерминирует синтез в клетках развивающейся
репродуктивной системы белкового рецептора для андрогена.
Поскольку зародыш как мужского, так и женского
пола несет по крайней мере по одной Х-хромосоме,
это вещество имеется у обоих полов.
Y-хромосома несет особый ген,
детерминирующий синтез определенного белка - H-Y-антигена,
который стимулирует дифференцировку клеток
половых складок в семенные канальцы и интерстициаль-
ные клетки. Тестостерон, выделяемый в
кровеносную систему зародыша, взаимодействует с
рецептором андрогенов в клетках-мишенях
потенциальной репродуктивной системы. Образующийся в
результате комплекс рецептора с тестостероном
переходит в клеточное ядро, где он активирует гены,
влияющие на развитие тканей. Тестостерон
стимулирует развитие только тех тканей, которые дают
Генетический
пол
Ген
ι /
Сцепленный с /
Y-хромосомой ген, /
Tfm детерминирующий /
I развитие семенника
Tfm
Белковый
рецептор
андрогена
H-Y-антиген
Белковый
рецептор
андрогена
Семенник
Яичник
Синтезируемый
белок
Половая
складка
зародыша
Гонада
зародыша
Тестостерон
Комплекс рецептора
андрогена с тестостероном
Мужская/женская
Зачаток
репроду ктивной
системы
Плод Мужского пола
Мужская/женская
Женского пола
Рис. 20.47. Общая схема событий, с которыми
связана дифференцировка половых складок зародыша
и зачатков репродуктивной системы в мужские или
женские гонады и репродуктивную систему плода.
начало мужской репродуктивной системе, и поэтому
из зародыша XY развивается плод мужского пола.
Ткани потенциальной женской репродуктивной
системы не активируются и не развиваются. У
зародыша XX отсутствие тестостерона позволяет
репродуктивной системе развиваться в присущем ей
направлении - женском.
Таким образом, можно сделать вывод, что
плацентарные влияния направляют развитие в сторону
женского пола, если механизм, приводимый в
действие Y-хромосомой, не направит его в
противоположную сторону. Все это схематически
представлено на рис. 20.47.
Таблица 20.4. Основные функции эстрогена и прогестерона
во время беременности
Эстроген
Стимулирует рост
молочных желез
Подавляет выделение фол-
ликулостимулирующего
гормона
Подавляет выделение про-
лактина
Предохраняет матку от
инфекций
Увеличивает размеры
мышечных клеток матки
Усиливает синтез АТФ и
креатинфосфата
Повышает
чувствительность миометрия к окси-
тоцину
Прогестерон
Стимулирует рост
молочных желез
Подавляет выделение фол-
ликулостимулирующего
гормона
Подавляет выделение про-
лактина
Подавляет сокращения
миометрия
20.3.7. Роды
С начала третьего месяца беременности
человеческий зародыш называют плодом; в общей сложности
беременность обычно продолжается 40 недель, по
истечении которых наступают роды. На 12-й неделе
беременности у плода уже сформированы все
основные органы, а остальное время занимает рост.
На всем протяжении беременности секретируются
все большие количества эстрогена и прогестерона;
сначала их источником служит желтое тело, а затем
главным образом плацента. В последние три месяца
беременности секреция эстрогена усиливается
быстрее, чем секреция прогестерона, а перед самыми
родами уровень прогестерона снижается, а уровень
эстрогена возрастает. Функции этих гормонов в
период беременности представлены в табл. 20.4.
Раньше считали, что сроки наступления родов
Гипоталамус и
!задняя доля ги-1
!пофиза материн-Г
ского организма]
Пониженное
выделение
прогестерона (5)
Окситоцин (6)
Кора надпочечников
Повышенное
выделение
простагландинов (7)у
Плацента
-Вегетативные.
сенсорные и
(моторные
Пнейронь!
Спинной
мозг ,
Кортикостероидь1
АКТГ
Неизвестный начальный стимул
детальный стресс"?
Иммунологическое отторжение?
Созревший гипоталамус плода?
Созравшие надпочечники плода?
Расширение шейки матки
Рис. 20.48. Гормональные и нервные механизмы, возможно связанные с родами. Из всех
приведенных на схеме механизмов твердо установлено только воздействие окситоцина на матку. Цифры 1-7
указывают возможную последовательность событий, вызывающих сокращение стенки матки, а 8, 9а и
96-возможные рефлекторые реакции, участвующие в регуляции этих сокращений.
регулируются гормонами; однако, как показали
результаты недавних исследований на ряде
млекопитающих, значительную роль, видимо, играет и
плод. Это, вероятно, имеет большой адаптивный
смысл, обеспечивая рождение плода на такой стадии
развития, когда он способен к относительно
самостоятельному существованию. Как полагают,
начальные этапы родов вызываются какими-то еще не
установленными факторами, заставляющими
гипоталамус плода стимулировать секрецию адренокор-
тикотропного гормона (АКТГ) гипофизом.
Согласно одному из современных представлений, сигналом
для начала родов служит «фетальный стресс»,
который создается в результате иммунологического
отторжения зрелого плода материнским организмом.
Выделяющийся АКТГ побуждает надпочечники
плода секретировать кортикостероиды, которые,
проникая через плацентарный барьер в кровоток
матери, вызывают снижение синтеза прогестерона и
повышение секреции простагландинов. Снижение
уровня прогестерона делает возможным секрецию
октапептида окситоцина задней долей гипофиза
матери и снимает ингибирующее воздействие на
сокращение миометрия. В то время как окситоцин
вызывает сокращение гладкой мускулатуры матки,
простагландины увеличивают силу этих
сокращений. Окситоцин выделяется во время родов
«волнами» и облегчает изгнание плода из матки. Начало
сокращений миометрия - так называемых «родовых
схваток»-сопровождается расширением шейки
матки, разрывом амниона и хориона, вытеканием
амниотической жидкости через шейку матки и
стимуляцией рецепторов растяжения в матке и ее
шейке. Импульсы от этих рецепторов возбуждают
вегетативную нервную систему, что приводит к
рефлекторному сокращению матки. Другие импульсы
передаются вверх по спинному мозгу и стимулируют
гипоталамус, который побуждает заднюю долю
гипофиза к секреции окситоцина. Давление
проходящей через таз головки плода раздражает шейку и
вызывает более сильные сокращения миометрия.
Сокращения матки распространяются сверху
вниз, усиливаясь по мере приближения к шейке и
проталкивая ребенка книзу. При этом шейка
постепенно расширяется, а перерывы между
приступами сокращений уменьшаются. Это первая стадия
родов, которая завершается, когда шейка матки
достигнет того же диаметра, что и головка плода.
На второй стадии головка и тельце проходят через
влагалище и младенец рождается на свет. Пуповину
перевязывают в двух местах и перерезают между
этими местами, полностью выводя ребенка из-под
прямой физиологической опеки со стороны матери.
В течение 10-45 мин после рождения ребенка матка
резко сокращается, плацента отделяется от стенки
матки и выходит через влагалище. Это третья
стадия родов. Кровотечение при этом ограничивается
благодаря сокращению гладких мышечных воло-
Рис. 20.49. А. Врач, принимающий роды, направляет
головку, которая целиком вышла из влагалища.
В. Тельце ребенка наполовину вышло наружу, ножки
еще находятся внутри. В. Ребенка уже обтерли, и он
лежит на груди у матери. Пуповина пока не
перерезона, так что ребенок все еще связан с
плацентой. Г. Врач перерезает пуповину, и процесс родов
завершается.
Рис. 20.49. (продолжение)
кон, которые окружают все кровеносные сосуды,
снабжающие плаценту. В среднем потеря крови
составляет около 350 мл.
20.3.8. Лактация
Молочные, или грудные, железы состоят из тканей
двух типов - железистой ткани и поддерживающей
ткани, или стромы. Клетки железистого эпителия
выстилают небольшие мешочки, называемые
альвеолами и сгруппированные в молочной железе в 15
долек. Альвеолы окружены слоем сократительной
миоэпителиальной ткани, участвующей в выведении
молока. Молоко выделяется железистыми клетками
и проходит по ряду протоков и синусов, в которых
оно собирается к отдельным отверстиям в соске.
В период полового созревания молочные железы
увеличиваются в результате развития стромы под
влиянием эстрогена и прогестерона. Внутренние
изменения характера стромы и молочных протоков, а
также количества жира в молочных железах,
происходящие на протяжении менструального цикла,
приводят лишь к небольшому увеличению молочных
желез во время лютеальной фазы, однако во время
беременности происходит значительное увеличение
размеров желез и их функциональной активности.
Увеличение молочных желез во время
беременности связано с развитием их секреторных областей
под влиянием эстрогена, прогестерона, кортикосте-
роидов, гормона роста, пролактина и
плацентарного лактогена. Однако на всем протяжении
беременности наличие в крови прогестерона
подавляет образование молока (лактогенез). При родах,
когда уровень прогестерона снижается, подавление
действия пролактина прекращается и он
стимулирует альвеолы к секреции молока.
Женское молоко содержит жир, лактозу
(молочный сахар) и белки лактальбумин и казеин; все эти
вещества легко перевариваются. Молоко
синтезируется из метаболитов, содержащихся в крови:
лактоза - из глюкозы под действием фермента лактозо-
синтетазы, белки- из аминокислот, а жиры-из
жирных кислот, глицерола и ацетата. Одного
материнского молока достаточно, чтобы младенец
ежедневно прибавлял в весе 25-30 г.
В промежутках между кормлениями грудью про-
лактин стимулирует образование молока для
следующего кормления. Для непрерывной секреции
Грудная мышца
Альвеолы молочной железы
Молочный проток
Молочные синусы
Отверстия протоков на соске
.Пигментированный
участок (ареола)
.Поддерживающая строма
. Жировая ткань
Рис. 20.50. Упрощенная схема строения женской
груди. Видны дольки, где секретируется молоко, а
также протоки и синусы, по которым молоко
направляется к соску.
молока необходим эстроген; однако с помощью
искусственно создаваемого избытка эстрогена
можно подавлять лактогенез у матерей, не желающих
кормить ребенка грудью (в настоящее время для
этого вместо эстрогена используют бромокриптин).
Выделение молока из соска связано с простой
рефлекторной реакцией - активным выведением
молока. Когда младенец сосет грудь, он стимулирует
рецепторы соска, которые посылают импульсы
через спинной мозг к гипоталамусу; гипоталамус
секретирует окситоцин, выделяющийся через
заднюю долю гипофиза. Это вызывает сокращение
миоэпителиальной ткани, окружающей альвеолы, и
молоко через протоки и синусы выталкивается из
сосков.
После родов молочные железы вначале выделяют
не молоко, а молозиво. Молозиво желтого цвета,
содержит клетки альвеол и богато белком
(глобулином), но в нем мало жира. Как полагают, с
молозивом ребенок получает от матери антитела, в
частности YgA (см. разд. 14.14.3).
20.3.9. Изменения в системе
кровообращения плода
при рождении
В период внутриутробного развития легкие и
пищеварительная система плода не функционируют,
поскольку газообмен и питание обеспечивает
материнский организм через плаценту. Большая часть
насыщенной кислородом крови, возвращающейся к
плоду по пупочной вене, обходит печень по
венозному протоку, из которого по нижней полой вене
идет в правое предсердие (рис. 20.51). Некоторое
количество крови из пупочной вены проходит и
через печень; поэтому в правое предсердие
поступает смесь оксигенированной (насыщенной O2) и
неоксигенированной крови. Из правого предсердия
большая часть крови проходит через овальное
отверстие (в перегородке между предсердиями) в левое
предсердие. Некоторое количество крови идет из
правого предсердия в правый желудочек, а из
него - в легочную артерию, но не поступает в легкие.
Вместо этого кровь направляется по артериальному
(боталлову) протоку прямо в аорту, обходя легкие,
легочную вену и всю левую половину сердца
(предсердие и желудочек). Кровь из левого предсердия
поступает в левый желудочек и в аорту, которая
снабжает кровью голову, верхние конечности,
туловище, нижние конечности и пупочную артерию.
Давление крови в кровеносной системе плода
достигает наибольшей величины в легочной артерии,
чем и определяется направление тока крови через
плод и плаценту.
20.9. Опишите главное изменение, которое
произошло бы в кровообращении плода, если бы
кровяное давление оказалось наивысшим в аорте.
После родов внезапное расправление легких
снижает сопротивление току крови через легочные
капилляры, и в них направляется больше крови, чем в
артериальный проток; это снижает давление в
легочных артериях. Одновременно с этим перевязка
Артериальный проток
Легочная-1
артерия
Нижняя полая-
вена
ЛЕГКИЕ
Овальное отверстие
Венозный-
проток
ПП
ПЖ
ЛП
ЛЖ
ТКАНИ
ПЕЧЕНЬ
_ Воротная
вена печени
4КИШЕЧНИК1
-Легочная
вена
_ Аорта
Пупочная вена-
- Пупочная
артерия
ПЛАЦЕНТА
Функционирует:
только до рождения
только после рождения
и до, и после рождения
Рис. 20.51. Изменения в кровеносной системе,
происходящие при рождении (упрощенная схема),
Π Π -правое предсердие; ЛП-левое предсердие;
ПЖ- правый желудочек; ЛЖ-левый желудочек.
пуповины препятствует току крови через плаценту;
это увеличивает объем крови, протекающей через
тело ребенка, и ведет к внезапному повышению
кровяного давления в аорте, левом желудочке и
левом предсердии. В результате изменения давления
маленькие клапаны, прикрывающие овальное
отверстие и открывающиеся в сторону левого
предсердия, закрываются, препятствуя току крови по
сокращенному пути - из правого предсердия в левое.
Через несколько месяцев эти клапаны срастаются с
перегородкой между предсердиями и овальное
отверстие полностью закрывается. Если отверстие не
зарастает, то возникает необходимость
хирургического вмешательства для устранения этого
дефекта.
Повышенное давление в аорте и пониженное в
легочной артерии проталкивает кровь назад по
артериальному протоку в легочную артерию и далее в
легкие, резко повышая их кровоснабжение. Спустя
несколько часов мышцы в стенке артериального
протока сокращаются под действием
возрастающего парциального давления кислорода в крови и
отключают этот кровеносный сосуд. Аналогичный
механизм, основанный на мышечном сокращении,
отключает венозный проток и усиливает ток крови
через печень. Непосредственный стимул,
вызывающий отключение венозного протока, неизвестен, но
он играет важную роль в преобразовании прена-
тальной (дородовой) системы кровообращения в
постнатальную (послеродовую).
20.4. Филогенез
полового размножения
у позвоночных
Половое размножение встречается у представителей
всех типов животных. Лежащие в его основе
процессы гаметогенеза, оплодотворения и образования
зиготы в принципе одинаковы во всех группах,
однако морфологические, физиологические и
поведенческие аспекты полового размножения весьма
разнообразны. Многие примеры различий,
наблюдаемых среди беспозвоночных, были описаны в
гл. 4.
У позвоночных тоже существуют огромные
различия во многих аспектах полового размножения;
некоторые примеры представлены в табл. 20.5. В
эволюции позвоночных можно проследить
постепенную адаптацию к жизни на суше. Одна из
главных трудностей, которую было необходимо
преодолеть при переходе от водного образа жизни к
наземному, была связана с размножением.
Большинство рыб выбрасывает свои гаметы прямо в
воду, оплодотворение у них наружное; яйца со-
Таблица 20.5. Особенности полового размножения у некоторых позвоночных
Организм
Треска
Колюшка
Кошачья
акула (Scylio-
rhinus canicu-
Ia)
Травяная
лягушка (Rana
temporaria)
Прыткая
ящерица (Lacer-
ta agilis)
Зарянка (En-
thacus rube-
cula)
Число яиц,
производимых
самкой за год
3 7 млн.
60
144
2000
6
6
Диаметр
яйца, мм
0,13
2,0
15,0
2
8,0
20,0
Оплодотворение
Наружное
— » —
Внутреннее
Наружное
Внутреннее
— » —
Откладка яиц и развитие
зародыша
Яйцекладущие;
личинки развиваются в
море
Яйцекладущие; гнездо
строит самец
Самка откладывает
яйца в твердой
капсуле; личиночной
стадии нет
Массовая откладка
яиц в воду; личинки
(головастики)
претерпевают
метаморфоз
Амниотические яйца;
откладываются в
ямки в почве
Амниотические яйца;
откладываются в
гнездо, которое
строит самка
Брачное и родительское поведение
Заботы о потомстве нет; яйцо
содержит очень мало желтка
Самец занимает территорию;
ухаживает за самкой; яйца
откладываются и оплодотворяются в
гнезде; молодь охраняется в
гнезде
Спаривание происходит у
поверхности воды; яйца богаты
желтком
Несложный ритуал ухаживания,
затем спаривание; умеренное
количество желтка в яйце; нет
заботы о потомстве
Несложный ритуал ухаживания,
затем спаривание; большое
количество желтка в яйце; нет
заботы о потомстве
Самец занимает территорию, на
которой происходит
спаривание; самка насиживает яйца в
течение 14 дней; птенцов кормят
оба родителя;
самостоятельными птенцы становятся через
6 недель
Скорлупа
Хорион
Амнион
Амниотическая полость
Зародыш
Аллантоис
Желточный мешок
Рис. 20.52. Упрощенная схема строения яйца у
амниот.
держат значительные количества желтка, у многих
видов имеются личиночные стадии, а забота о
потомстве встречается редко. У амфибий известен
ряд примеров адаптации к наземному образу жизни,
однако лишь немногие из них связаны с механизмом
размножения. Для размножения амфибии должны
возвращаться в воду, и ранние стадии их развития
также проходят в воде. Однако у многих видов
амфибий наблюдаются весьма сложные формы
поведения, связанные с заботой о потомстве.
Например, у пипы самец распределяет оплодотворенную
икру по спине самки, к которой икринки прилипают,
попадая в имеющиеся на ней ячейки, где
развиваются головастики. Примерно через три недели
молодые потомки соскакивают с материнской
спины и переходят к самостоятельной жизни.
Рептилии были первыми позвоночными,
разрешившими проблему оплодотворения и развития на
суше. Очевидно, что на суше нельзя просто
выбрасывать гаметы в окружающую среду, так что
первым условием для полного перехода к наземной
жизни было введение мужских гамет в тело самки,
т.е. внутреннее оплодотворение. Именно такое
оплодотворение свойственно рептилиям, и
поскольку при этом вероятность слияния гамет возрастает,
число их может быть уменьшено. Образующаяся
в результате оплодотворения зигота развивается
в специализированной структуре - амниотическом
(клейдоическом) яйце; в таком яйце эмбрион
развивается в полости, наполненной жидкостью.
Снаружи яйцо покрыто скорлупой, предохраняющей от
механических повреждений. Под скорлупой
имеются еще четыре оболочки, окружающие зародыш:
желточный мешок, амнион, хорион и аллантоис,
происходящие из эктодермы, энтодермы и
мезодермы; они защищают зародыш и облегчают
осуществление ряда метаболических функций -
питания, дыхания, выделения. Желточный мешок
закладывается как вырост эмбрионального кишечника;
этот мешок обрастает желток, который постепенно
поглощается желточными кровеносными сосудами.
После того как весь желток использован,
желточный мешок спадается и втягивается в кишку. Амнион
развивается из клеток, лежащих под зародышем,
которые растут вверх и полностью смыкаются над
зародышем, образуя амниотическую полость; клетки
амниона секретируют амниотическую жидкость,
которая заполняет эту полость. Таким образом,
эмбрион оказывается в жидкой среде (замена
водной среды амфибий), в которой он может
развиваться. Амнион имеется у всех рептилий, птиц и
млекопитающих, и их называют амниотами. По
мере роста зародыша амнион оттесняется кнаружи,
пока он не сливается с третьей зародышевой
оболочкой-хорионом, которая лежит непосредственно
под скорлупой и препятствует чрезмерной потере
воды амнионом. Четвертая оболочка - аллантоис-
образуется в виде выроста задней кишки; у рептилий
и птиц аллантоис быстро разрастается, образуя
слой, подстилающий хорион. Он выполняет
главным образом функцию «мочевого пузыря», в
котором накапливаются отходы метаболизма, и
осуществляет газообмен между внешней средой и
амниотической жидкостью через пористую
скорлупу яйца. Строение яйца амниот схематически
показано на рис. 20.52.
Яйца такого типа откладывают также птицы и
примитивные яйцекладущие млекопитающие,
называемые однопроходными, как, например, утконос
(Ornithorhynchus); у высших млекопитающих яйцо
утратило скорлупу, но сохранило четыре
зародышевые оболочки; у плацентарных млекопитающих две
из них, хорион и аллантоис, образуют плаценту (см.
разд. 20.3.6).
Глава 21
Рост и развитие
Способность к росту-одна из главных
особенностей всех живых организмов. Нередко рост
представляют себе просто как увеличение в размерах;
однако, внимательно рассмотрев такое определение,
приходится признать его неверным. Так, например,
размеры растительной клетки могут увеличиться за
счет осмотического поглощения воды, но этот
процесс иногда бывает обратимым и, следовательно,
его нельзя считать истинным ростом. Во время
дробления зиготы и на ранних стадиях развития
зародыша происходит увеличение числа клеток без
увеличения общих размеров (объема или массы).
Это результат клеточных делений без
последующего роста дочерних клеток. Процесс этот
представляет собой морфогенетическое событие, и поэтому
его, возможно, следует считать ростом, хотя
увеличения размеров при этом не происходит1.
Развитие так тесно связано с ростом, что для
описания процессов, которые обычно считают
ростом, часто пользуются выражением «рост и
развитие». Рост многоклеточного организма,
начинающийся с одной клетки, можно разделить на три
стадии:
1) деление клеток (гиперплазия) - увеличение числа
клеток в результате митозов;
2) рост клеток (гипертрофия) - необратимое
увеличение размеров клеток в результате поглощения
воды или синтеза протоплазмы;
3) дифференцировка клеток - их специализация (рост
в широком смысле включает также и эту стадию
развития клетки).
Каждый из этих процессов может, однако, быть
отделен от других во времени. Мы уже приводили
пример с дроблением. Возможно также увеличение
объема клеток без изменения их числа, как,
например, в зоне растяжения клеток позади кончиков
корня и побегов у высших растений. У
одноклеточных организмов, таких как бактерии, клеточное
деление приводит к репродукции (но не к росту)
данной особи и к росту популяции.
Все стадии роста сопряжены с биохимической
активностью. Особенно важная роль принадлежит
1 Обычно все же ростом называют увеличение
размеров (клетки, органа, организма) за счет процессов
биосинтеза.- Прим. ред.
белковому синтезу, так как в результате этого
процесса заключенная в ДНК информация реализуется
в форме синтезируемых клеткой ферментов.
Ферменты регулируют функциональную активность
клетки. Изменения, происходящие на клеточном
уровне, приводят к изменению общей формы и
структуры как отдельных органов, так и организма
в целом - процесс, называемый морфогенезом.
Определение понятия «рост» должно
удовлетворять критерию увеличения размеров, которое
происходит у всех организмов - от одноклеточных до
самых высокоорганизованных растений и
животных, а также учитывать связанную с ростом
метаболическую активность. Поэтому рост можно
определить как необратимое увеличение сухой массы
протоплазмы. Это в свою очередь отражает увеличение
количества синтезированного белка и тот факт, что
процесс белкового синтеза составляет основу роста.
Рост может быть положительным или
отрицательным. Положительным рост бывает тогда, когда
анаболизм преобладает над катаболизмом, а
отрицательным - когда катаболизм преобладает над
анаболизмом (см. гл. 11). Например, в процессе
прорастания семени и образования проростка многие
физические параметры - число клеток, их размеры,
сырая масса, длина, объем, сложность
формы-возрастают, а другие, такие как сухая масса,
уменьшаются. Согласно определению, прорастание в
этом случае представляет собой, строго говоря,
период отрицательного роста.
21.1. Измерение роста
Рост происходит на многих уровнях биологической
организации - от уровня сообщества до
молекулярного уровня. Во всех случаях график зависимости
измеряемого параметра от времени представляет
собой S-образную кривую роста. На рис. 21.1
приведено несколько кривых роста, полученных путем
откладывания по оси ординат различных
параметров (длина, высота, масса, площадь поверхности,
объем, численность), а по оси абсцисс-времени.
Такие кривые называют сигмоидными или S-образ-
ными.
S-образную кривую можно разделить на четыре
участка (рис. 21.2). Начальная фаза, или лаг-фаза,
Длина, мм
Самка дафнии
Время, дни
Параметр растения
Средняя высота
Средняя сухая
масса
Средняя площадь
листьев
Zea mais
Время, дни
Средний объем, м 3
Pinus sylvestris
Время, годы
Число организмов
Дрожжи
Время, часы
Рис. 21.1. Кривые роста четырех разных организмов, основанные на шести различных параметрах. Во всех
случаях кривые имеют S-образную форму (Б по Kreusler; В по Tischendorf; Г - по Knopf.)
характеризуется незначительным ростом. Она
переходит во вторую, логарифмическую, фазу
интенсивного роста, в которой рост происходит по
экспоненте. Во время этой фазы скорость роста достигает
максимума и в любой момент пропорциональна
количеству материала или числу уже имеющихся
клеток или организмов.
Во всех случаях экспоненциальный прирост
наконец убывает, и скорость роста начинает
снижаться. Точку, в которой это происходит, называют
точкой перегиба. Третья фаза-это фаза замедления
(самозамедления), во время которой рост
ограничивается в результате влияния какого-либо
внутреннего или внешнего фактора или взаимодействия
того и другого. Конечная фаза-фаза плато, или
стационарная фаза. В этот период общий рост уже
прекратился и рассматриваемый параметр остается
постоянным. Характер кривой в этой фазе может
несколько варьировать в зависимости от природы
данного параметра, вида организма и внутренних
факторов. В некоторых случаях может
продолжаться незначительное восхождение кривой, до тех
пор пока организм не погибнет; это наблюдается у
листьев однодольных, у некоторых беспозвоночных,
у рыб и у ряда рептилий и свидетельствует о
положительном росте. У некоторых
кишечнополостных кривая роста уплощается, что отражает
отсутствие изменений в скорости роста, тогда как у
других организмов кривые роста отклоняются вниз,
что указывает на отрицательный рост. Последнее
характерно для многих млекопитающих, в том
числе для человека, и служит признаком
физического старения.
21.1.1. Способы измерения роста
Рост можно измерять на разных уровнях
биологической организации, например измерять рост
клетки, организма или популяции. Изменения
численности популяции во времени отражает кривая
роста, представленная на рис. 21.1, Г (см. также
разд. 12.6.3). На уровне организма можно измерять
множество различных параметров; часто измеряют
длину, площадь поверхности, объем или массу. У
растений нередко нужно бывает получать кривые
роста корней, стеблей, междоузлий и листовой
поверхности, и измеряемыми параметрами служат
длина и площадь. При построении кривых роста
целого животного или растения обычно используют
длину и массу. Массу можно оценивать по двум
показателям - по сырой и сухой массе. Сырую массу
измерять легче, так как это не требует специальной
обработки и не причиняет организму вреда, что
позволяет многократно измерять один и тот же
организм на протяжении длительного времени.
Главный недостаток использования сырой массы
в качестве показателя роста состоит в том, что
получаемые при этом данные могут варьировать
из-за колебаний содержания влаги в тканях.
Истинный рост отражается в изменении других компо-
нентов (не воды), и единственный надежный метод
его измерения - определение сухой массы. Для этого
организм убивают и помещают в печь при
температуре 1100C, чтобы удалить всю влагу. Затем
образец охлаждают в эксикаторе и взвешивают. Эту
процедуру повторяют до тех пор, пока масса не
станет постоянной, что и будет соответствовать
сухой массе. Для получения более точных данных
рекомендуется измерить сухую массу как можно
большего числа экземпляров, а затем вычислить
среднюю величину. Полученное таким образом
значение более надежно, чем результат взвешивания
одного экземпляра.
21.1.2. Типы кривых роста
Если отложить по оси ординат данные для любого
из перечисленных выше физических параметров,
например для сырой или сухой массы (т), а по оси
абсцисс - время (/), то получится так называемая
кривая абсолютного роста (рис. 21.3); она дает
представление об общем характере роста и о его
длительности. На основании этого графика можно
построить кривые роста, представленные на рис.
21.3-21.5.
Построив график изменения данного параметра
во времени, можно получить кривую абсолютной
скорости роста (рис. 21.4). Такую же кривую можно
получить, построив график зависимости наклона
fdm\
кривой абсолютного роста от времени 1 — 1· Эта
кривая показывает, как изменялась скорость роста в
период проведения исследований; в частности, она
выявляет фазу наиболее быстрого роста, которая
совпадает с самым крутым участком кривой
абсолютного роста. Вершина второй кривой
соответствует точке перегиба первой, а затем скорость
роста падает - организм достиг своих
окончательных размеров.
Разделив каждое из полученных значений
абсолютной скорости роста на массу, достигнутую в
/dm 1\
начале каждого временного периода I J, и
Vd/ т/
построив график зависимости найденных величин
от времени, получим кривую относительной (или
удельной) скорости роста (рис. 21.5). Это будет
мерой эффективности роста.
Сравнение кривых относительной скорости роста
для организмов, растущих в разных условиях,
Параметр роста
Время
Рис. 21.2. Типичная S-образная кривая роста, на
которой видны четыре характерные фазы роста и
точка перегиба, а-лаг-фаза;
б-логарифмическая фаза; в-точка перегиба, г фаза замедления;
д - плато, или стационарная фаза.
Прирост массы,
% от прежней
Возраст, недели
Рис. 21.4. Кривая абсолютной скорости роста,
построенная по данным рис. 21.3.
о
Σ
§
OQ
S
Возраст, недели
Рис. 21.3. Кривая абсолютного роста овцы. (По
данным LR. Wallace, 1948, J. Agric. Sci, 38, 93 и
H. Palsson, J. В. Verges, 1952, J. Agric. Sci., 42 93.)
Ежедневный прирост, г
Возраст, недели
Рис. 21.5. Кривая относительной скорости роста,
построенная по данным рис. 21.3.
Младенчество
и детство
Юность
Зрелый
возраст
Возраст, годы (/)
Зародыш
Масса, кг
Рост плода
Рост младенца
Изменение массы,
кг (dm/df)
Относительное
изменение массы,
(dm /df-1/m)
Рост в
юношеском
возрасте
Возраст, годы (t)
Возраст, годы (t)
Рис. 21.6. Три типа кривых роста для человека.
позволяет выявить условия, наиболее
благоприятные для быстрого роста или для более
продолжительного роста. У млекопитающих кривая
абсолютного роста S-образная, но точная ее форма,
по-видимому, зависит от времени, необходимого
для достижения половой зрелости. У крысы кривая
круто идет вверх и действительно имеет S-образную
форму, так как половая зрелость достигается
быстро (за несколько месяцев), тогда как у человека
на кривой абсолютного роста ясно видны четыре
фазы усиленного роста (рис. 21.6).
21.1. Какие заключения относительно роста
человека можно сделать на основании трех кривых,
представленных на рис. 21.6?
21.2. Типы роста
У живых организмов встречаются различные типы
роста, которые мы сейчас рассмотрим подробнее.
21.2.1. Изометрический и
аллометрический рост
Изометрическим (от греч. isos - одинаковый, met-
гоп-мера) называют рост, при котором данный
орган растет с такой же средней скоростью, как и
остальное тело. В этом случае изменение размеров
организма не сопровождается изменением его
внешней формы. Относительные размеры двух структур
(органа и организма в целом) остаются прежними.
Такой тип роста характерен для рыб (рис. 21.7) и
для насекомых с неполным превращением,
например саранчовых (за исключением крыльев и
половых органов). Для организмов с изометрическим
ростом характерны простые соотношения между
линейными размерами, площадью, объемом и
массой. Площадь возрастает пропорционально
квадрату линейных размеров, а объем и масса -
пропорционально их кубу. Таким образом, у животных, у
которых длина тела с течением времени изменяется
мало, может заметно изменяться масса тела:
увеличение длины всего лишь на 10% сопровождается
увеличением массы на 33%.
Аллометрическим (от греч. alios-иной) называют
рост, при котором данный орган растет с иной
скоростью, нежели остальное тело. В этом случае
рост организма приводит к изменению его
пропорций. Такой тип роста характерен для
млекопитающих, и он иллюстрирует существование
зависимости между ростом и развитием. На рис. 21.8
показано, как изменяются при этом относительные
размеры разных частей тела у человека. Почти у
всех животных в последнюю очередь происходит
развитие и дифференцировка органов размножения.
Эти органы растут аллометрически, но их рост
можно наблюдать только у организмов с
наружными половыми органами; поэтому у многих рыб, у
которых эти органы не видны, рост кажется чисто
изометрическим. На рис. 21.9 представлены
различия в характере роста разных органов у человека;
можно убедиться, что и здесь последними
развиваются репродуктивные органы.
Кривые абсолютного роста разных организмов в
целом, построенные по данным о длине или массе
тела, очень сходны по форме и в общем
соответствуют S-образному типу, описанному в разд.
21.1. Однако у некоторых групп наблюдаются
отклонения от этого общего типа, отражающие
адаптацию к особому образу жизни и среде, что будет
описано в разд. 21.2.2-21.2.3.
Рис. 21.7. Развитие рыбы пример
изометрического роста. Наружные структуры сохраняют свою
форму и пространственные соотношения в
результате пропорционального роста. (По Batt, 1980,
Influences on animal growth and development Studies in
Biology. № 116, Arnold)
Рост
ребенка
ι Рождение
Зачатие
2 месяца
5 месяцев
(внутриутробное развитие)
Новорожденный
2 года
6 лет
12 лет
25 лет
Рис. 21.8. Развитие человека- пример аллометрического роста. Для того чтобы выявить относительные
скорости роста частей тела от стадии двухмесячного плода до 25-летнего возраста, все рисунки сделаны
одинаковой высоты (По Stratz, из J. Hammond, ed, 1955, Progress in the physiology of farm animals, 2, 431,
Butterworths )
21.2.2. Ограниченный и
неограниченный рост
Как показывает изучение продолжительности роста
у растений и животных, существуют два основных
типа роста-ограниченный и неограниченный
(бесконечный).
У однолетних растений рост ограничен: вслед за
периодом максимальной интенсивности роста, во
время которого растение достигает зрелости и
размножается, наступает период отрицательного роста,
или старения, после чего растение погибает. Если
построить график изменения сухой массы
однолетнего растения во времени (рис. 21.10), то получится
интересная модификация S-образной кривой,
представленной на рис. 21.2.
21.2. Рассмотрите рис. 21.10 и, опираясь на то,
что вам известно о жизненном цикле однолетнего
растения, ответьте на следующие вопросы:
а) Почему вначале, при прорастании семени,
наблюдается отрицательный рост?
б) Опишите внешний вид проростка к моменту
начала положительного роста (в точке А).
в) Какие физиологические процессы
обеспечивают переход к положительному росту?
г) Чем объясняется внезапное резкое уменьшение
сухой массы по прошествии 20 недель?
Некоторым органам растений (например, плодам,
органам вегетативного размножения, а также
листьям и междоузлиям стебля у двудольных)
свойствен ограниченный рост, но периода
отрицательного роста у них нет. К животным с ограниченным
ростом относятся, в частности, насекомые, птицы и
млекопитающие.
У многолетних древесных растений имеет место
неограниченный рост, и для них характерна кривая
роста, слагающаяся из ряда S-образных участков
(рис. 21.11), каждый из которых описывает рост за
один год. При неограниченном росте какой-то
небольшой положительный рост продолжается до
самой гибели.
Другие примеры неограниченного роста можно
найти у грибов, низших растений, в частности у
Относительный рост, %
Возраст, годы
Рис. 21.9. Скорости роста головного мозга (I),
тимуса (II) и репродуктивных органов (III) у человека
по отношению к абсолютной скорости роста
организма в целом (IV). (По Scammon.)
Сухая масса, г
Начало прорастания
Время, недели
Рис. 21.10. Кривая роста однолетнего растения
конских бобов (Vic/a faba), посаженных в марте.
ев
ί-
Ο
и
3
Таблица 21.1. Факторы, влияющие на рост и развитие
Время, годы
Рис. 21.11. Кривая роста многолетнего
древесного растения- березы.
S
о
ссГ
X
S
5
Нимфы:
1-го/возраста
2-го возраста.
3-го возраста
4-го BOSPaCTa1
5-го возраста
Имаго
Время, дни
Рис. 21.12. Кривая роста длины тела у саранчи (По
R. Soper, T. Smith, 1979, Modern Biology, Macmillan.)
водорослей, и у многих животных, особенно у
беспозвоночных, рыб и рептилий. Неограниченный
рост характерен также для листьев однодольных
растений.
21.2.3. Рост у членистоногих
Очень своеобразный тип роста наблюдается у
ракообразных и у других членистоногих, таких как
насекомые с неполным превращением и личинки
насекомых с полным превращением. Поскольку
наружный скелет у этих животных неэластичен, рост у
них происходит как бы отдельными рывками,
разделенными рядом линек (прерывистый рост).
Картина роста, типичная для насекомых с неполным
превращением, показана на рис. 21.12. Такие кривые
роста, основанные на общей длине тела, не дают
верного представления о росте. Если построить
кривую роста для того же насекомого, используя в
качестве меры сухую массу, то получится S-образ-
ная кривая; это показывает, что истинный рост,
оцениваемый по увеличению массы протоплазмы,
непрерывен.
21.3. Регуляция роста и
развития
Процесс роста и развития данного организма
определяется в конечном счете информацией, содержа-
Внешние факторы
Свет (интенсивность,
спектральный состав,
продолжительность
воздействия)
Коротковолновое
электромагнитное
излучение (рентгеновские
лучи, γ-лучи и т. п.)
Питательные вещества
Температура
Кислород
Вода
Сезонные явления
Отходы метаболизма
Внутренние факторы
Гены
Гормоны и ростовые
вещества
Процесс, на который они влияют
Фотосинтез — источник
энергии (гл.9)
Фотоморфогенез (гл. 15)
Фототропизм (гл. 15)
Позеленение (гл. 15)
Фотопериодизм (гл. 15 и 16)
Синтез витамина D у человека
Мутагенез (гл. 23)
Автотрофное питание —
потребность в CCh, воде,
неорганических солях; отсутствие
этих веществ вызывает
болезни недостаточности (гл. 9)
Гетеротрофное питание —
потребность в органических
веществах, воде и
неорганических солях (гл. 10); при
отсутствии этих веществ —
болезни недостаточности
(гл. 9 и 10)
Воздействует на рост через
посредство ферментов (гл. 6
и рис. 21.13); термоперио-
дизм (гл. 15)
Необходим аэробным
организмам для дыхания; особенно
важен для активного
поглощения ионов корнями
растений и для развития плода у
животных (гл. 12)
Играет важную роль во
многих процессах (гл. 5)
Состояние покоя почек, семян
и зимующих органов (гл. 15)
Листопад (гл. 15)
Размножение (гл. 15, 16 и 20)
Фотопериодизм и термоперио-
дизм (гл. 15 и 16)
Обычно не создают проблем,
но иногда могут оказывать
ингибирующее воздействие
(например, в колониях
бактерий)
Процесс, на который они влияют
Белковый синтез (гл. 23), а
следовательно, синтез
ферментов и регуляция химизма
клетки
Многие процессы (гл. 15 и 16)
Масса, кг
Потребление пищи,
кДж/кг ♦ день
Среднемесячная
температура, 0C
Рис. 21.13. Влияние количества потребляемой пищи
и внешней температуры на массу тела у лайки. (По
данным J. L. Durrer, J. P. Наппоп. 1962, Am J. Physio/,
20Z 375.)
щейся в его ДНК. Рост, однако, представляет собой
результат взаимодействия между ДНК и факторами
внутренней и внешней среды организма. К важным
внешним факторам относятся пища, свет, тепло и
вода. Внутренняя среда создается такими
веществами, как гормоны, и открытыми недавно цитоплаз-
матическими белками (факторами транскрипции),
которые прямо или косвенно влияют на экспрессию
генов. Внешние факторы могут оказывать влияние и
на внутреннюю среду; например, при отсутствии в
пищевом рационе иода организм человека не
способен вырабатывать гормон тироксин, что
приводит к замедлению роста.
В недавних экспериментах по пересадке тканей
между видами с разными типами роста была
обнаружена регуляция роста, обусловленная
взаимодействием между тканями. Если бы каждая ткань росла
со своей обычной скоростью, то возникли бы
несогласованности и структура органа оказалась бы
нарушенной. Но благодаря тому, что скорости
роста двух тканей приспосабливаются одна к
другой, рост органа протекает нормально. Из этих
исследований можно сделать вывод, что одна ткань
влияет на скорость роста другой.
Некоторые из факторов, влияющих на рост и
развитие, перечислены в табл. 21.1 (см. также
рис. 21.13).
21.3. Используя данные, приведенные на
рис. 21.13, объясните, почему масса тела
снижается до минимума в периоды максимального
потребления пищи.
21.4. Развитие
В самом широком смысле рост-это не только
необратимое увеличение сухой массы как результат
клеточного деления и увеличения размеров клеток,
но также и последующий процесс развития. В
процессе развития клетки специализируются для
выполнения определенных функций в организме - это
называется дифференцировкой. Другими словами,
между клетками происходит разделение труда.
Кроме того, как уже говорилось, при этом
осуществляется также морфогенез. Как видно из табл. 21.2,
степень дифференцировки клеток связана с уровнем
филогенетической организации.
Одна из интригующих проблем
дифференцировки-это вопрос о ее механизме. Главным фактором
роста служит клеточное деление, которое у всех
многоклеточных организмов совершается путем
митоза. Это означает, что все клетки, происходящие от
исходной зиготы или споры, идентичны по своему
генотипу и поэтому должны были бы иметь
идентичную структуру и функцию. Очевидно, однако,
что у взрослого организма это не так. Какие же
механизмы создают различия между клетками,
тканями, органами и системами органов? Структура и
функции клеток определяются активностью генов, и
если клетки различаются по своей структуре и
функциям, то это, по-видимому, обусловлено
различиями в экспрессии их генов. Это могло бы достигаться
двумя способами: либо клетки при специализации
утрачивают некоторые гены, сохраняя только гены,
необходимые для своей специальной функции, либо
специализация обусловлена «включением» и
«выключением» разных генов в разных клетках. В
начале 60-х годов стало очевидно, что действует второй
механизм, во всяком случае у растений. Проф.
Стьюард из Корнелльского университета показал,
что если поместить дифференцированные клетки,
например клетки флоэмы моркови, в подходящую
культуральную среду, то из них может вырасти
новое растение моркови; это означает, что клетки
все еще содержат всю необходимую информацию. В
1967 г. Гёрдон, работая в Оксфордском
университете, установил, что так же обстоит дело и у
животных. Гёрдон пересаживал ядра из клеток кишечного
эпителия шпорцевой лягушки (Xenopus) в яйца того
же вида, ядра которых были предварительно
разрушены ультрафиолетом. Небольшое число таких яиц
нормально развивалось, прошло через стадию
головастика и достигло половозрелости; это позволяет
думать, что даже в дифференцированной клетке
возможна реактивация генов, если они окажутся в
подходящей среде. При использовании менее
дифференцированных клеток, находившихся на такой
ранней стадии развития, как бластула, частота случа-
Таблица 21.2. Приблизительное число клеток и клеточных
типов как отражение степени дифференцировки,
наблюдаемой на разных ступенях филогенетического развития
Организмы
Простейшие
Губки
Гидра
Кольчатые черви
Насекомые
Человек
Число клеток
1
>103
108
1012
10'2
1015
Число
клеточных типов
1
>10
10- 20
102
102
103
Месяцы
ев успешного развития была выше. С помощью
этого метода можно получать любое число
генетически идентичных лягушек. Идентичное
потомство, полученное от одной родительской особи,
называют клоном, а сам метод - клонированием (см.
разд. 20.1.1).
Выращивать в культуре изолированные клетки
млекопитающих трудно, а заставить их
дифференцироваться-еще труднее. В этом сказывается
высокая специализация клеток млекопитающих.
Уровень сложности, достигнутый в результате
дифференцировки, может быть таким высоким, что
он препятствует даже росту или делению клеток.
Так обстоит дело с нейронами. В отличие от них
клетки печени остаются сравнительно
неспециализированными и способны выполнять множество
различных функций (см. разд. 18.5). Если удалить у
взрослого млекопитающего две трети печени, то она
вскоре восстанавливает прежние размеры и форму.
Это свидетельствует не только о высокой
способности к регенерации, но и о том, что размеры
органов каким-то образом предетерминированы.
Если ядра дифференцированных клеток
полностью сохраняют свои генетические потенции, то это
означает, что в регуляции дифференцировки
участвует цитоплазма. Шпеман и Мангольд изучали
влияние цитоплазматических факторов на ход
развития. Пересаживая ткань от зародыша-донора
зародышу-реципиенту, они сумели показать, что
донорская ткань способна изменить процессы
развития у реципиента. Такое воздействие называют
индукцией, а участки эмбриона-донора,
вызывающие индукцию,-организаторами. Более подробно
процесс индукции и эксперименты Шпемана и
Мангольд будут описаны в разд. 22.8.2. Каким именно
образом осуществляется индукционное воздействие,
не вполне ясно, однако полагают, что в этом
участвует дифференциальная репрессия и активация
генов в различных клетках (подробнее см. разд.
22.8.3).
На дифференцировку оказывают также влияние
гормоны. Типичные примеры влияния гормонов на
рост у растений и у различных животных описаны в
этой главе, а также в гл. 15 и 16. Хотя действие
ростовых гормонов у растений варьирует,
имеющиеся данные указывают на то, что в некоторых
отношениях оно сходно у всех организмов.
Гормоны могут оказывать прямое или косвенное
воздействие на гены, «включая» или «выключая» их в
определенной последовательности,
детерминирующей ход развития. При изучении гигантских
хромосом из слюнных желез личинок двукрылых в
разных участках этих хромосом были обнаружены
утолщения - «пуфы» (см. разд. 22.8.5 и 23.5.1).
Местоположение пуфов характерно для тех или
иных стадий развития личинки. Эти участки - места
активного синтеза мРНК; они соответствуют генам,
которые включаются на определенных стадиях
жизненного цикла и обеспечивают в конечном счете
синтез веществ, необходимых на данной стадии.
Введение личинке соответствующего гормона
может вызвать образование пуфов у личинки, у
которой были удалены железы, вырабатывающие этот
гормон. Если же ввести личинке сверхоптимальную
дозу гормона, нормальная последовательность
включения генов ускорится и особь раньше
достигнет взрослой стадии. Результаты этих
исследований показывают, что гормоны нередко действуют
путем включения генов, связанных с развитием.
21.5. Морфогенез
Во время развития организма дифференцированные
клетки постепенно занимают предназначенные им
участки, где происходит их дальнейший рост и
деление. Этот процесс составляет часть
морфогенеза, так как он приводит к образованию клеток,
тканей и органов, которые, сформировавшись в
надлежащих местах, придают зрелому организму
свойственные ему форму и структуру. Детали этих
морфогенетических движений у разных организмов
различны, но тем не менее в двух последующих
разделах будут описаны некоторые характерные
примеры морфогенеза у растений и животных. У
растений главные морфогенетические события
связаны с развитием стебля, корня, листьев и цветка.
У животных можно найти много удивительных
примеров морфогенеза, но мы ограничимся здесь
теми, на которых можно продемонстрировать
резкие изменения формы, происходящие при
метаморфозе, и все возрастающее усложнение тканей в
процессе развития позвоночных.
21.6. Рост и размножение
цветковых растений
21.6.1. Покой семян
Для развития находящегося в семени зародыша
необходимы определенные условия среды, а именно:
влажность, оптимальная температура и кислород.
Однако даже при наличии этих условий некоторые,
казалось бы, зрелые семена не могут прорасти, пока
в них не произойдут известные внутренние
изменения, совокупность которых можно назвать
дозреванием. Необходимость дозревания предотвращает
преждевременное прорастание. Например, если бы
семена, высыпавшиеся летом или осенью, тотчас же
прорастали, то проростки, возможно, не пережили
бы зиму. Другими словами, имеются механизмы,
обеспечивающие совпадение сроков прорастания с
наступлением сезона, благоприятного для роста.
В этих механизмах часто участвуют наружные
слои семени, которые могут содержать ингибиторы
роста, непроницаемы для воды или для кислорода
или же обладают достаточной физической прочно-
стью, чтобы не допускать рост зародыша, как,
например, у многих бобовых растений. Иногда
механическое повреждение (скарификация) оболочки
семян устраняет эти препятствия; такое
повреждение можно нанести искусственно, удалив семенную
кожуру или просто проколов ее булавкой. В
природных условиях аналогичный эффект могут
вызвать бактерии. Чаще, однако, подавление
прорастания снимается при участии факторов,
рассмотренных ниже.
ингибиторы роста. Многие плоды или семена
содержат ингибиторы роста, препятствующие
прорастанию. В этой роли часто выступает абсцизовая
кислота, например в семенах ясеня. При достаточно
длительном вымачивании семян ингибитор из них,
возможно, вымывается или же действие его
ослабевает в результате увеличения количества такого
стимулятора роста, как гиббереллин.
свет. Покой семян прерывается при воздействии
света после вымачивания в воде - реакция,
контролируемая фитохромом. Она связана с повышением
содержания гиббереллина в семени. Реже свет
подавляет прорастание, как у Phacelia и Nigella.
(Соответствующий эксперимент описан в табл. 15.5,
разд. 15.4.2.)
температура. Как отмечалось в разд. 15.5.1, для
того чтобы семена могли прорасти, они обычно
должны подвергнуться действию низких температур
(стратификации). Это характерно для
представителей сем. Rosaceae и для злаков. При стратификации
возрастает активность гиббереллинов, а иногда
снижается содержание ингибиторов роста.
Каков конкретный механизм действия света и
низких температур, неясно; возможно, что он связан
не только с изменениями количеств ростовых
веществ, но и с повышением проницаемости семенных
оболочек.
21.4. Семена, для прорастания которых
необходима стимуляция светом, обычно бывают
относительно мелкими. Какое это может иметь
значение?
21.5. Свет, прошедший сквозь листья,
обогащается зеленым и дальним красным
компонентами по сравнению со светом, падающим на
листовую поверхность.
а) Чем это обусловлено?
б) Какое это может иметь экологическое значение
для семян таких растений, как салат,
прорастание которых контролируется фитохромом? (В
случае необходимости обратитесь к разд.
15.4.2.)
21.6.2. Прорастание
Прорастание знаменует собой начало развития
зародыша, обычно по прошествии периода покоя.
Строение семени к моменту прорастания описано в
разд. 20.2.6.
Условия среды, необходимые
для прорастания
вода. Первоначальное поглощение воды семенем
происходит путем всасывания. Вода всасывается
через микропиле и семенные оболочки в результате
чисто физического процесса абсорбции воды
находящимися в семени коллоидами; к ним относятся
белки, крахмал и вещества, входящие в состав
клеточных стенок, такие как гемицеллюлозы и
пектины. Набухание этих веществ может создать
большую силу, достаточную для разрыва семенной
кожуры или околоплодника, окружающих семя. В
дальнейшем вода движется от клетки к клетке под
действием осмотических сил. Вода необходима для
активации биохимических процессов, связанных с
прорастанием, поскольку эти процессы протекают в
водном растворе. На этой стадии вода участвует
также в гидролизе (переваривании) запасных
питательных веществ.
МИНИМАЛЬНАЯ ИЛИ ОПТИМАЛЬНАЯ ТЕМПЕРАТУРА. ДЛЯ Ce-
мян каждого типа существует известный диапазон
температур, за пределами которого они не
прорастают. Такие диапазоны зависят от условий
среды, нормальных для данного растения, и все они
лежат в интервале температур 5-400C. Температура
влияет на скорость ферментативных реакций (см.
разд. 6.4.3).
кислород. Кислород необходим для аэробного
дыхания, которое в случае нужды может дополняться
анаэробными процессами.
Физиология прорастания
Типичное семя содержит запасы углеводов, липидов
и белков либо в своем эндосперме, либо в семядолях
зародыша. Главным резервом обычно служат ли-
пиды, представленные маслами; заметное
исключение составляют Leguminosae (бобовые) и Grami-
пеае (злаки, в том числе хлебные), семена которых
содержат в основном крахмал. Почти все
сельскохозяйственные культуры, выращиваемые человеком,
относятся к этим двум группам, и от них он
получает большую часть необходимых ему углеводов.
Бобовые, в частности соя, богаты также белками;
поэтому сою используют в качестве источника
белка во многих новых пищевых продуктах. Кроме
того, семена богаты минеральными веществами, в
особенности фосфором, а также содержат обычные
компоненты цитоплазмы - нуклеиновые кислоты и
витамины. В результате процессов всасывания и
осмоса зародыш гидратируется, что приводит к
активации определенных ферментов, в частности
дыхательных. Другие ферменты должны быть еще
синтезированы, возможно с использованием
аминокислот, освобождающихся при переваривании
запасенных белков.
Грубо говоря, в прорастающем семени
существуют две зоны активности: область запасных веществ
и область роста (зародыш). Главные события,
происходящие в первой из них, носят (за исключением
синтеза ферментов) катаболический характер, т.е.
связаны с процессами распада.
Переваривание запасов питательных веществ
происходит главным образом путем гидролиза:
Растворимые продукты переваривания
переносятся затем в зоны роста зародыша. Сахара, жирные
кислоты и глицерол могут служить субстратами для
дыхания как в зоне запасных веществ, так и в
области роста; в последней они могут также
использоваться для анаболических реакций, т. е. для
реакций синтеза. Особенно важное значение для этих
реакций имеют глюкоза и аминокислоты. Глюкоза
используется главным образом для синтеза
целлюлозы и других веществ, образующих клеточные
стенки, а аминокислоты - для синтеза белков, как
ферментных, так и структурных, входящих в состав
протоплазмы. Кроме того, для многих процессов,
перечисленных в табл. 9.10, необходимы
минеральные вещества.
И область расщепления запасных веществ, и зона
роста получают необходимую им энергию за счет
дыхания. В процессе дыхания происходит окисление
какого-либо субстрата (обычно сахара) до CO2 и
воды. Это сопровождается уменьшением сухой
массы семени, так как двуокись углерода улетучивается
(что же касается воды, то она в сухую массу вообще
не входит). Такое уменьшение массы продолжается
до тех пор, пока у проростка не появятся зеленые
листья и он не начнет сам синтезировать для себя
пищу (см. разд. 21.2.2).
Хорошо изученным примером прорастания
семени, богатого полисахаридами, служит ячменное
зерно; было показано, что у ячменя синтез а-амила-
зы и других ферментов происходит в наружных
слоях эндосперма под воздействием гиббереллина,
выделяемого зародышем. Наружные слои
эндосперма содержат резервный белок, служащий
источником аминокислот для белкового синтеза. Этот
процесс и соответствующие экспериментальные
исследования описаны в разд. 15.2.6. На рис. 15.21
приведен пример, иллюстрирующий роль гормонов на
ранних стадиях прорастания. Появление амилазы в
прорастающих зернах ячменя можно также изучать,
размалывая зерна в воде, фильтруя суспензию,
центрифугируя фильтрат и определяя затем амилаз-
ную активность полученного прозрачного экстракта
Белки -
Протеазы
Аминокислоты
Полисахариды
Карбогидразы
Сахара
Например,
Крахмал
Амилаза
Мальтоза
Мальтаза
Глюкоза,
Липиды
Липазы
Жирные кислоты -I- Глицерол
4 Общая масса
Сухая масса, г
Эндосперм
Зародышу
Время после посева, дни
Рис. 21.14. Относительные изменения сухой массы
эндосперма и зародыша при прорастании ячменя.
с помощью раствора крахмала. Если брать пробы
зерен ячменя в разные сроки после начала
прорастания, можно определить повышение амилазной
активности в расчете на одно зерно на протяжении
недели.
21.6. Объясните результаты, представленные на
рис. 21.14.
Семена, запасающие жиры, превращают их в
жирные кислоты и глицерол. Каждая молекула
жира дает три молекулы жирных кислот и одну-
глицерола (см. разд. 5.3.1). Жирные кислоты либо
непосредственно окисляются в процессе дыхания,
либо превращаются в сахарозу, которая затем
переносится в зародыш. В этом превращении участвует
глиоксилатный цикл (разд. 11.5.5).
21.7. (Этот вопрос служит для проверки знания
основ химии вообще и химии липидов в
частности. Химия липидов изложена в разд. 5.3.)
Предположим, что в 51,2 г (по сухой массе)
семян, содержащих 50% (по массе) жирных
кислот, все жирные кислоты превратились в сахар в
результате реакции
Ci6H32O2 + HO2 -+ Ci2H22On 4- 4CO2 + 5H2O +
Жирная Сахар + Энергия
кислота
а) Считая, что не произошло никаких других
превращений, которые могли бы изменить сухую массу,
вычислите увеличение или уменьшение сухой массы
семян.
(Относительные атомные массы: С = 12, H = I,
О = 16.)
б) Какое другое важное изменение могло бы повлиять
на сухую массу?
в) Вычислите объем двуокиси углерода, выделяющейся
из семян при нормальных температуре и давлении.
(1 моль газа при нормальных температуре и
давлении занимает 22,4 л.)
г) Как можно получить из жира жирные кислоты?
Какой другой компонент содержит молекула жира?
д) Сколько атомов углерода должна была содержать
одна молекула исходного жира, если единственной
полученной из него жирной кислотой была
Ci6H32O2?
е) Какой сахар образуется в результате приведенной
выше реакции?
ж) Как кислород попадает в запасающую ткань?
Дыхание прорастающих семян
Интенсивность дыхания как запасающей ткани, так
и зародыша высока, что связано с высокой
метаболической активностью этих двух участков
семени. Дыхательные субстраты в этих участках
могут быть различными; кроме того, они могут
изменяться в процессе прорастания. Это выявляется
по изменениям дыхательного коэффициента (см.
разд. 11.7.7).
21.8. Анализ семян клещевины на содержание
липидов и сахара при проращивании в темноте
дал результаты, приведенные на рис. 21.15, При
измерении дыхательного коэффициента
проростков на 5-й день оказалось, что у зародыша он
равен примерно 1,0, а у остатков семядолей-
примерно 0,4-0,5.
а) Дайте как можно более полное объяснение
этим результатам (воспользуйтесь данными
разд. 21.6.2).
б) Чему будет равен дыхательный коэффициент
проростка в целом на 11-й день? Объясните
очень кратко ход ваших рассуждений.
21.9. Дыхательный коэффициент семян гороха
в первые семь дней прорастания лежит между 2,8
и 4, но если удалить у них кожуру, то он
понижается до 1,5-2,4. В обоих случаях в семенах
накапливается этиловый спирт, но при удалении
семенной кожуры-в значительно меньших
количествах. Объясните эти наблюдения.
Масса 100 проростков, г
Общая сухая масса
Сахар
Липиды
Время после посева, дни
Рис. 21 15. Изменения содержания липидов и
сахара в семенах клещевины при прорастании в
темноте (По данным R. Desveaux, M. Kogane-
Charles, 1952, Annls. Inst. Nath Rech. Agron. Paris,
3. 385-416)
Рост зародыша
Рост зародыша происходит путем деления клеток,
увеличения их размеров и дифференцировки.
Количество белков, целлюлозы, нуклеиновых кислот и
других веществ в растущих участках зародыша
постепенно увеличивается, а сухая масса запасов
питательных веществ уменьшается. Первый видимый
признак роста - появление зародышевого корешка.
Корешок обладает положительным геотропизмом,
т.е. растет вниз, закрепляя семя в почве. Затем
появляется зачаток побега - плюмула, или почечка,
проявляющая отрицательный геотропизм (и
положительный фототропизм, если она находится над
землей) и растущая вверх.
Различают два типа прорастания, в зависимости
от того, остаются ли семядоли под землей или
выносятся на поверхность. У двудольных, если
вытягивается междоузлие, расположенное
непосредственно под семядолями (гипокотиль), семядоли
выносятся на поверхность (эпигеальное прорастание);
если же вытягивается междоузлие, находящееся
непосредственно над семядолями (эпикотиль), то
семядоли остаются под землей (гипогеальное
прорастание).
При эпигеальном прорастании гипокотиль,
пробиваясь сквозь почву, остается искривленным
(рис. 21.16,Б), т.е. именно он преодолевает
сопротивление почвы, а не тоненький кончик плюмулы,
который дополнительно защищен прикрывающими
его семядолями. При гипогеальном прорастании у
двудольных искривлен эпикотиль, т.е. кончик
плюмулы опять-таки защищен (рис. 21.16,5). В обоих
случаях искривленный участок, оказавшись на свету,
тотчас же выпрямляется - реакция, контролируемая
фитохромом.
У злаков, относящихся к однодольным, плюмула
защищена чехликом, так называемым колеоптилем,
который обладает положительным фототропизмом
и отрицательным геотропизмом (см. разд. 15.1.1).
Первый лист пробивается через колеоптиль и
разворачивается, оказавшись на свету. Под действием
света быстро возникает ряд реакций,
контролируемых фитохромом; они известны под названием
фотоморфогенеза и приводят к переходу от
этиолированного роста (разд. 15.4.1) к нормальному.
Главные изменения, которые при этом происходят,
приведены в табл. 15.5; они включают, в частности,
разрастание и развертывание семядолей или первых
настоящих листьев и синтез хлорофилла
(«позеленение»). На этой стадии начинается фотосинтез и
начинает увеличиваться сухая масса проростка;
теперь он становится независимым от запаса
питательных веществ и переходит к автотрофному
существованию. Оказавшись на свету, побег тоже
проявляет фототропные реакции, хотя они не
контролируются фитохромом.
Эпикотиль
Семядоля
Корешок
Эпигеальное прорастание
(удлиняется гипокотиль)
Искривленный
гипокотиль
Эндосперм
Плюмула
Семенная кожура
Гипокотиль
Гипогеальное прорастание
(удлиняется эпикотиль)
Две семядоли
Семенная кожура
Первые зеленые
листья
•Плюмула
■Фотосинте-
зирующие
,семядоли
Искривленная
плюмула
Выпрямление,
контролируемое*,
фитохромом'г
Выпрямление,
} контролируемое.
W фитохромом*1
Боковые корни
Боковые корни
Рис. 21.16. А. Строение семени. Б. Эпигеальное (надземное) прорастание, например у
подсолнечника (эндосперма нет) или у клещевины (эндосперм имеется). В. Гипогеальное (подземное)
прорастание, например у огородных бобов (эндосперма нет).
21.6.3. Первичный рост растения
Меристемы
У многоклеточных растений в отличие от животных
рост (за исключением ранних стадий развития
зародыша) происходит лишь в определенных участках,
называемых меристемами. Меристема - это группа
клеток, сохраняющих способность к митотическому
делению; в результате этого деления образуются
дочерние клетки, которые растут и формируют
постоянную ткань из клеток, уже не способных
делиться. Существуют три типа меристемы, описанные
в табл. 21.3. В этой таблице упоминаются также два
типа роста, а именно первичный и вторичный рост,
которые приводят соответственно к увеличению
длины и толщины растения.
Таблица 21.3. Типы меристем и их функции
Сначала происходит первичный рост. В результате
первичного роста может сформироваться целое
растение, и у большинства однодольных и
травянистых двудольных это единственный тип роста.
В нем участвуют апикальная, а иногда и интер-
калярная меристемы. Строение зрелых первичных
корней и стеблей описано в разд. 14.3.9.
У некоторых растений за первичным ростом
следует вторичный рост, в котором участвуют
латеральные меристемы. Он в наибольшей степени
выражен у кустарников и деревьев, которые иногда
называют древесными растениями в отличие от
травянистых, или трав, у которых значительного
вторичного роста не бывает. У некоторых
травянистых растений наблюдается некоторое вторичное
утолщение стебля, например развитие добавочных
проводящих пучков у подсолнечника (Helianthus).
Тип меристемы
Апикальная
Латеральная
(камбий)
Интеркалярная
Местоположение
В кончиках корней и побегов
В более старых частях растения;
лежит параллельно длинной оси
органа [например, пробковый
камбий (феллоген), васкулярный
камбий]
Между участками постоянных
тканей, например в узлах у многих
однодольных (в основании
листьев у злаков)
Роль
Обеспечивает первичный рост, образуя
первичное тело растения
Обеспечивает вторичный рост.
Васкулярный камбий дает начало
вторичным проводящим тканям; из фел-
логена образуется перидерма,
которая замещает эпидермис и содержит
пробку
Делает возможным рост в длину в
промежуточных участках. Это
существенно для тех растений, у которых
апикальные участки часто
подвергаются повреждению или
разрушению, например объедаются
травоядными животными (у злаков) или
повреждаются волнами (у бурых
водорослей); при этом отпадает
необходимость в ветвлении
Результат
Удлинение
Утолщение
Апикальные меристемы и
первичный рост
Для апикальной (верхушечной) меристемы типичны
относительно мелкие кубовидные клетки с тонкой
целлюлозной стенкой и густой цитоплазмой. В ней
имеется несколько небольших вакуолей (в отличие
от крупных вакуолей клеток паренхимы), а
цитоплазма содержит мелкие недифференцированные
пластиды, называемые пропластидами. Меристема-
тические клетки плотно упакованы, так что между
ними нет заметных воздухоносных межклетников.
Клетки апикальной меристемы называют инициа-
лями. При митотическом делении этих клеток одна
из дочерних клеток остается в меристеме, а другая
увеличивается в размерах и дифференцируется, с
тем чтобы стать частью постоянного тела растения.
21.6.4. Первичный рост побега
Строение типичной апикальной меристемы побега
представлено на рис. 21.17 и 21.18. На рис. 21.18
показано примерное подразделение верхушки
побега на зоны деления, роста и дифференцировки
клеток. У побега эти зоны перекрываются сильнее,
чем в корне. В направлении от куполообразной
апикальной меристемы назад клетки становятся все
старше, так что в одной верхушке побега можно
одновременно видеть разные этапы роста. Поэтому
изучать последовательные стадии развития
растительной ткани относительно несложно.
В побегах различают три основные зоны меристе-
матической ткани: 1) протодерму, дающую начало
эпидермису; 2) прокамбий, образующий проводящие
ткани, т.е. перицикл, флоэму, васкулярный камбий
и ксилему; 3) основную меристему, образующую
паренхимные основные ткани (у двудольных это
кора и сердцевина). Эти меристематические ткани
закладываются в результате деления инициалей в
верхушке побега. В зоне роста дочерние клетки,
образующиеся путем деления инициалей,
увеличиваются в размерах - главным образом за счет
осмотического поглощения воды, поступающей в
цитоплазму, а из нее-в вакуоли. Рост стеблей и
корней в длину достигается в основном за счет
Зона
клеточного
деления
апикальной
меристемы
Эпидер-
мальный
волосок
Основная
меристема
Лист
Листовой
примордий
Протодерма
Прокамби-
альный тяж —
вытянутые
клетки,
образующие
ксилему и
флоэму
Лист (на срезе
только
часть его)
Рис. 21.17. Апикальная меристема.
происходящего на этой стадии удлинения клеток
(рис. 21.19).
Мелкие вакуоли увеличиваются в размерах и
сливаются в конце концов в одну большую вакуоль.
Развивающееся в клетке тургорное давление
растягивает ее тонкие стенки, а относительное
расположение целлюлозных фибрилл в них определяет
окончательную форму клетки. Объем цитоплазмы
на конечном этапе может быть лишь немногим
больше, чем в исходной меристематической клетке,
но теперь вакуоль оттесняет ее на периферию. Когда
рост почти завершается, у многих клеток
происходит дополнительное утолщение стенок за счет
целлюлозы или лигнина, в зависимости от типа
клетки. Это может ограничить дальнейший рост, но
не обязательно ведет к его полному прекращению.
Например, удлинение колленхимных клеток в коре
может продолжаться, в то время как на внутренней
стороне их первичных стенок откладывается
целлюлоза в виде столбиков. Таким образом, клетки
Апикальная меристема
Зона
клеточного
деления
Зона
деления и
роста
клеток
Зона
дифференцировки
клеток
Постоянные
ткани
Листовой примордий,
образующийся в узле
Протодерма -*· эпидермис
Основная меристема -*■ основные ткани
Прокамбий -* первичные
проводящие
ткани
Протофлоэма
Протоксилема
Междоузлие — участок между
двумя узлами
Узел
Эпидермис ]
Камбий
Ксилема
Кора
Сердцевина
Флоэма
паренхима
Рис. 21.18. Кончик побега
двудольного растения (в продольном разрезе,
схема). Видны апикальная меристема
и зоны первичного роста. Для
простоты проводящие ткани, идущие
в листья и почки, не показаны.
зачатки проводящих
тканей
-Пазушная почка
Перицикл - склеренхима
колленхимы еще в период своего роста могут нести
опорные функции. В отличие от этого на стенках
развивающихся клеток склеренхимы
откладываются толстые слои лигнина, и клетки вскоре отмирают,
т.е. их дифференцировка начинается лишь после
того, как рост фактически завершен.
Прокамбий образует ряд продольных тяжей,
состоящих из более узких и длинных клеток, чем
основная меристема. Первыми в прокамбии
начинают дифференцироваться клетки протоксилемы во
внутренней его части и клетки протофлоэмы - в
наружной. Это элементы первичной ксилемы и
первичной флоэмы, которые образуются до завершения
роста клеток в длину. В протоксилеме лигнин, как
правило, образует только кольцевидные или
спиральные утолщения на трахеидах (см. разд. 8.2.1),
которые благодаря своей прерывистости делают
возможным рост и растяжение целлюлозы между
утолщениями по мере роста в длину окружающей
ткани. Элементы как протоксилемы, так и
протофлоэмы вскоре отмирают и по мере роста
окружающих тканей сминаются, растягиваются и наконец
спадаются. Их функции принимают на себя ксилема
и флоэма, развивающиеся позднее в зоне дифферен-
цировки.
В зоне дифференцировки каждая клетка
полностью специализируется для выполнения своей
собственной особой функции в соответствии со своим
положением в данном органе относительно других
клеток. Самые крупные изменения происходят в
протокамбиальных тяжах, которые
дифференцируются в проводящие пучки. Это связано с лигнифика-
цией стенок склеренхимных волокон и элементов
ксилемы, а также с развитием трубок, характерных
для сосудов ксилемы и ситовидных трубок флоэмы.
Эти ткани в их окончательном виде описаны в
разд. 8.2. Теперь склеренхима и ксилема тоже несут
опорные функции, выполнявшиеся ранее только
колленхимой и тургесцентной паренхимой. Между
ксилемой и флоэмой имеются клетки, сохраняющие
способность к делению. Они образуют васкулярный
камбий, деятельность которого будет рассмотрена
позже, при описании вторичного утолщения.
Листовые зачатки и боковые почки
Развитие побега включает также рост листьев и
боковых почек. Листья закладываются в виде
мелких вздутий или складок, называемых листовыми
зачатками; они хорошо видны на рис. 21.17. Эти
вздутия состоят из групп меристематических клеток
и распределены с равномерными интервалами;
места их возникновения называются узлами, а
промежутки между ними - междоузлиями. Способ
расположения листьев может быть различным и носит
название филлотаксиса. Листья могут располагаться
мутовками-от каждого узла отходит два или
большее число листьев; или же от каждого узла отходит
по одному листу, причем листья либо образуют два
Цитоплазма
Ядро
Клеточная ^
стенка
Большая
центральная-
вакуоль
Развивающиеся вакуоли
Рис. 21.19. Стадия растяжения в росте меристе-
матической клетки.
ряда, направленные в противоположные стороны,
либо расположены по спирали. Как правило, они
обычно располагаются таким образом, чтобы
перекрывание между ними, а значит, и взаимное
затенение было минимальным (мозаичное
расположение).
Листовые зачатки быстро удлиняются, так что
вскоре они окружают апикальную меристему,
защищая ее как чисто механически, так и тем теплом,
которое они выделяют при дыхании. Позднее они
растут, площадь их увеличивается и они образуют
листовые пластинки. Деление клеток постепенно
прекращается, хотя может продолжаться до тех пор,
пока листья не достигнут половины своей
окончательной величины.
Вскоре после того, как листья начинают расти, в
пазухах между ними и стеблем образуются почки.
Они представляют собой небольшие группы
меристематических клеток, обычно пребывающих в
состоянии покоя, но способных приступить к делению
и росту на более поздней стадии. Из этих почек
могут развиваться ветки или такие
специализированные органы, как цветки, а также подземные
структуры - корневища и клубни. Полагают, что
пазушные почки находятся под контролем
апикальной меристемы (см. разд. 15.3.3 об апикальном
доминировании).
21.6.5. Первичный рост корня
Строение апикальной меристемы типичного корня
представлено на рис. 21.20.
На самом кончике апикальной меристемы
имеется покоящийся центр-группа инициалей
(меристематических клеток), от которых в конечном счете
происходят все другие клетки корня, но которые
делятся значительно медленнее, чем их потомки в
окружающей апикальной меристеме. На наружной
стороне образуются клетки корневого чехлика. Они
превращаются в крупные паренхимные клетки,
которые защищают апикальную меристему при
углублении корня в почву. Эти клетки непрерывно
снашиваются и замещаются новыми. Они несут, кроме
того, важную дополнительную функцию - служат
рецепторами гравитации: в них содержатся крупные
Появление бокового
корня.
Корневой чехлик
Постоянные ткани
Зрелый корневой "
волосок
Зона дифференцировки
клеток
Зона клеточного
деления
(апикальная
меристема)
Корневой чехлик
Зона растяжения
клеток
Примордий бокового корня,
выходящий из перицикла
Эпидермис
Коря
Эндодерма
Перицикл
Молодой корневой волосок
Протоксилема
Метаксилема
Начало дифференцировки
элементов сосудов ксилемы;
протоксилема снаружи
Сердцевина (иногда отсутствует)
Первые зрелые ситовидные трубки
Начинают дифференцироваться
ситовидные элементы флоэмы
Прокамбий -*· перицикл, первичная
флоэма, васкулярный камбий,
первичная ксилема, сердцевина (если
имеется); образует сплошной
центральный
цилиндр
- Основная меристема -> кора,
включая эндодерму
- Протодерма -* эпидермис
■ Покоящийся центр —
апикальные инициали
Рис. 21.20. Апикальная меристема
типичного корня (в продольном
разрезе). Справа показана
дифференциация ксилемы, а слева
флоэмы. На самом деле ксилема и
флоэма чередуются по окружности
корня и расположены на разных
радиусах (см рис 14.16). Кроме
того, в корне зона растяжения в
действительности длиннее.
крахмальные зерна, играющие роль статолитов, т. е.
опускающиеся на «дно» клетки под действием силы
тяжести (подробнее роль этих зерен описана в
разд. 15.2.2).
Позади покоящегося центра можно видеть
правильные ряды клеток; в этой области различимы
зоны меристемы, уже описанные для побега, а
именно протодерма, основная меристема и
прокамбий (рис. 21.20). В корне прокамбием называют
весь центральный цилиндр, хотя в зрелом состоянии
в нем находятся непроводящие ткани перицикла и
сердцевины, если она имеется.
Зона клеточного деления обычно занимает
1-2 мм позади кончика корня и слегка
перекрывается с зоной растяжения клеток. Кончики корней-
удобный объект для изучения митоза; один из
применяемых для этого методов описан в разд. 22.2.
Позади зоны деления рост происходит главным
образом за счет растяжения клеток; клетки
увеличиваются в размерах так же, как это описано для
побега (рис. 21.19). Граница зоны растяжения
клеток находится на расстоянии примерно 10 мм от
кончика корня, и удлинение этих клеток
проталкивает кончик корня вниз, заставляя его углубляться в
почву.
Некоторая дифференцировка клеток начинается
еще в зоне деления, с развитием первых ситовидных
элементов флоэмы (рис. 21.20). На продольных
срезах кончика корня видны четкие продольные ряды
ситовидных элементов, достигающие все большей
зрелости по мере удаления от кончика, пока они не
превращаются в полностью сформированные
ситовидные трубки. Развитие флоэмы идет
центростремительно, т. е. по направлению снаружи внутрь,
к центральной оси корня.
Еще дальше от кончика корня в зоне растяжения
начинают дифференцироваться сосуды ксилемы,
тоже по направлению внутрь (экзархная ксилема) в
отличие от центробежной дифференцировки в
стебле (эндархная ксилема). Первыми, как и в стебле,
образуются сосудистые элементы протоксилемы, и
они таким же образом лигнифицируются и
растягиваются по мере роста окружающих клеток. Затем
их функции переходят к метаксилеме, которая
развивается позднее и созревает в зоне
дифференцировки после того, как прекратится растяжение
клеток. Ксилема часто доходит до центральной оси
корня, и в таких случаях сердцевина не образуется.
Развитие легче изучать в корнях, чем в побегах. В
побегах тяжи прокамбия, идущие к листьям,
усложняют распределение развивающихся тканей.
Развитие ксилемы особенно хорошо видно на давленых
препаратах апикальных участков тонких корешков
(например, проростков кресс-салата) при
надлежащей окраске.
Дифференцировка заканчивается, когда все клетки
перестают удлиняться. К этому времени
завершается и развитие корневых волосков из эпидермиса.
Боковые корни
Главный корень может ветвиться, но в отличие от
побега-не при помощи почек. Вместо этого
небольшие группы клеток перицикла в зоне
дифференцировки возобновляют меристематическую
активность и образуют новую корневую апикальную
меристему. Она начинает расти, пробиваясь сквозь
эндодерму, кору и эпидермис (рис. 21.20). Такое
развитие называется эндархическим в отличие от
экзархического развития боковых почек
Адвентивный рост
Адвентнвными, или придаточными, называют
структуры, растущие в нетипичных местах. Адвентивные
корни и почки могут возникать в разнообразных
ситуациях в результате возобновления меристема-
тической активности некоторых клеток.
Адвентивные, или придаточные, корни развиваются
независимо от первичного корня и образуют основную
корневую систему у однодольных, возникая из
узлов на стебле. Корневища и столоны - стеблевые
структуры, из которых придаточные корни
вырастают непосредственно. Придаточные корни играют
также важную роль в размножении растений при
помощи черенков. Плющ использует придаточные
корни для того, чтобы цепляться за субстрат.
Адвентивные почки могут развиваться на корнях,
побегах или листьях. Например, узамбарские
фиалки можно размножать листьями, образующими
адвентивные почки и корни. У деревьев могут
развиваться новые ветки из адвентивных почек,
образующихся в стволе.
21.6.6. Латеральные меристемы и
вторичный рост
Вторичный рост происходит после первичного роста
в результате активности латеральных меристем. Он
приводит к утолщению стебля или корня. Обычно
вторичный рост связан с отложениями больших
количеств вторичной ксилемы - древесины, которая
совершенно изменяет первичную структуру и
составляет характерную особенность деревьев и
кустарников.
Известно два типа латеральной меристемы-
васкулярный (сосудистый) камбий, из которого
образуется новая проводящая ткань, и пробковый
камбий, или феллоген, который развивается позднее,
замещая эпидермис, разрывающийся при
утолщении тела растения.
Клетка
делится в
тангенциальной
плоскости
Клетка-спутница-
Веретеновидная
инициаль
Сосуд вторичной
ксилемы
Одна
веретеновидная
инициаль
Клеточное
деление/
Клетка
делится в
радиальной
плоскости
Ситовидная пластинка
ситовидной трубки
Вторичная флоэма
Веретеновидная
инициаль
Вторичная ксилема
Рост
Две веретеновидные
инициали
Рис. 21.22. А. Два последовательных деления вере-
теновидной инициали, в результате которых
образуется ксилема и флоэма (в поперечном разрезе).
На самом деле дифференцировка ксилемы и флоэмы
до представленных здесь стадий занимает довольно
много времени, и за это время образуется много
клеток. Б. Деление веретеновидной инициали с
образованием новой веретеновидной инициали
(в поперечном разрезе).
внутри-вторичную ксилему (обычно в большем
количестве, чем флоэму). Последовательные деления
схематически показаны на рис. 21.22. Вторичная
флоэма содержит ситовидные трубки,
клетки-спутницы, склеренхимные волокна и склереиды, а также
паренхиму.
Лучевые инициали имеют почти шаровидную
форму и делятся митотически, образуя паренхим-
ные клетки, которые накапливаются и образуют
лучи между соседними проводящими пучками,
состоящими из флоэмы и ксилемы.
Васкулярный камбий
Этот камбий состоит из клеток двух типов -ве-
ретеновидных инициалей и лучевых инициалей
(рис. 21.21). Веретеновидные инициали-это узкие
вытянутые клетки, которые делятся путем митоза,
образуя кнаружи от себя вторичную флоэму, а ко-
Две веретеновидные
инициали
(продольный разрез)
Две веретеновидные
инициали
(поперечный разрез)
Лучевые инициали —
почти сферические
клетки,
одинаковые
в поперечном
и продольном
разрезе
Толстые
целлюлозные
стенки
- Сужающийся кончик
Рис. 21.21. Веретеновидные и лучевые инициали.
Вторичный рост стебля
у деревянистых двудольных
Васкулярный камбий вначале располагается между
первичной ксилемой и первичной флоэмой
проводящих пучков (его образование из апикальной
меристемы показано на рис. 21.18). Он становится
активным почти сразу же после завершения
первичной дифференцировки клеток. На рис. 21.23
показаны ранние стадии вторичного утолщения стебля
типичного деревянистого двудольного.
На рис. 21.23, Л представлена первичная
структура стебля (для простоты перицикл не показан). На
рис. 21.23,/> можно видеть сплошной камбиальный
цилиндр. На рис. 21.23, В изображено сплошное
кольцо вторичного утолщения. Здесь
веретеновидные инициали образовали большое количество
вторичной ксилемы и меньшее количество вторичной
флоэмы, а лучевые инициали - лучевую паренхиму.
С утолщением стебля должна увеличиваться и
окружность слоев камбия. Это становится
возможным благодаря делению камбиальных клеток в
Эпидермис
Кора
Первичная флоэма
Пучковый камбий
Первичная ксилема
Сердцевина
Первичный сердцевинный луч,
связывающий кору с древесиной
Пучковый камбий
(в проводящем пучке)
Межпучковый камбий (между
проводящими пучками)
Вторичная ксилема
Вторичная флоэма
Первичная флоэма
Вторичная флоэма
Васкулярный камбий
Вторичная ксилема
Первичная ксилема
Сердцевина
Первичный сердцевинный луч
(идет от коры до сердцевины)
Вторичный сердцевинный луч
Рис. 21.23. Ранние стадии вторичного
утолщения ствола у типичного деревянистого
двудольного. А. Первичное строение стебля. Б. Камбий
образует сплошной цилиндр, по мере того как
сердцевинные лучи становятся меристематичес-
кими и растут кнаружи от проводящих пучков.
Тем временем уже происходит образование
вторичной ксилемы и вторичной флоэмы из имеющегося
камбия. В. Сплошное кольцо вторичного
утолщения уже сформировалось. Утолщение особенно
сильно выражено на местах первых проводящих
пучков, где начиналась камбиальная
активность.
радиальном направлении (рис. 21.22).
Первоначально возникшие лучевые инициали образуют
первичные сердцевинные лучи, которые тянутся от
сердцевины до коры, в отличие от вторичных
сердцевинных лучей, образующихся из лучевых инициалей,
возникших позднее. Эти лучи служат живым
связующим звеном между сердцевиной и корой. Они
обеспечивают радиальный перенос внутри стебля
воды и минеральных солей из ксилемы, а
питательных веществ-из флоэмы. Кроме того, через
межклеточные пространства может происходить
газообмен путем диффузии. Лучи могут также
служить для запасания питательных веществ, что имеет
особенно важное значение в периоды покоя,
например зимой. В трех измерениях они имеют вид
продольных радиальных пластов, так как лучевые
инициали лежат друг над другом, образуя
вертикальные колонки (рис. 21.24). На рис. 21.24
схематически представлено строение древесины
(вторичной ксилемы) и содержащиеся в ней лучи показаны в
поперечном, тангенциальном и продольном
радиальном разрезе.
На рис. 21.25 изображена часть стебля
деревянистого двудольного растения на третий год роста;
видны большие количества вторичной ксилемы. На
рис. 21.26 представлены срезы стволов липы (TiUa) в
возрасте трех и пяти лет.
Годичные кольцо (слои прироста)
В областях с умеренным климатом каждую весну
рост возобновляется. Первые образующиеся при
этом сосуды - широкие и тонкостенные, способные
проводить большие количества воды. Вода
необходима для инициации роста, особенно для
увеличения в размерах новых клеток, например клеток
развивающихся листьев. Позднее сосудов
образуется меньше, они более узкие, а стенки у них толще. В
течение зимы камбий пребывает в состоянии покоя.
Осенняя древесина, образующаяся в конце
вегетационного периода перед самым окончанием роста,
резко отличается от непосредственно примыкающей
к ней весенней древесины следующего года. В
результате получаются так называемые годичные
кольца, ясно видимые на рис. 21.26. Если сосуды
сконцентрированы в ранней древесине, то древесину
называют кольцепоровой; если же они распределены
равномерно, так что годичные кольца выражены
менее четко, древесину называют рассеянно-поровой.
В тропиках сезонные засухи могут вызвать сходные
колебания камбиальной активности.
Ширина годичного кольца частично зависит от
климата: благоприятный климат способствует
образованию большого количества древесины,
поэтому расстояния между кольцами бывают больше.
Эта зависимость используется в двух областях
науки-в дендроклиматологии и дендрохронологии.
Дендроклиматология занимается изучением климата
по данным о годичных кольцах, например выявляет
корреляции между результатами метеорологических
наблюдений и ростом деревьев для той или иной
местности или же исследует климатические события,
происходившие несколько сотен или даже тысяч лет
назад. Возраст самых древних из ныне живущих
деревьев - остистых сосен - достигает примерно 5000
лет, а среди ископаемых остатков деревьев известны
еще более древние.
Дендрохронология занимается датировкой
древесины путем изучения годичных колец, рисунок
которых позволяет установить, в какое время росло
дерево. Это дает возможность определять возраст
бревен, найденных при археологических раскопках,
возраст старых строений, кораблей и т.п.
Первичный сердцевинный луч
Вторичный сердцевинный луч
Вторичная флоэма
Васкулярный камбий
Вторичная ксилема
Сердцевина
Кора
Веретеновидные
инициалы
Лучевые
инициалы
Васкулярный
камбий
Столбик
лучевых
инициалей
На поперечном срезе сердцевинный
луч имеет вид радиального ряда
клеток лучевой паренхимы; если
это первичный луч, то он тянется
от сердцевины до коры. Некоторые
лучи на поперечном срезе имеют
толщину более чем в одну клетку
Кора
На тангенциальном срезе
сердцевинный луч имеет вид столбика
лучевой паренхимы. Некоторые
лучи на таком срезе имеют толщину
более чем в одну клетку
На продольно-радиальном срезе сердцевинный
луч имеет вид пласта клеток; если это
первичный луч, то он тянется от сердцевины
до коры
Рис. 21.24. Схематическое изображение первичных и вторичных сердцевинных лучей в стволе
типичного деревянистого двудольного. Справа показан первичный, а слева -вторичный луч.
Ядро и заболонь
По мере того как дерево стареет, древесина в центре
ствола перестает выполнять проводящие функции и
заполняется темноокрашенными отложениями,
такими как таннины. Эту внутреннюю часть называют
ядром, или ядровой древесиной, а более влажную
наружную проводящую часть-заболонью.
Пробка и чечевички
По мере роста вторичной ксилемы в направлении
кнаружи прилегающие к ней с этой стороны ткани
все больше спрессовываются, а также
растягиваются из-за увеличения длины окружности. Это
оказывает влияние на эпидермис, кору, первичную
флоэму и почти на всю вторичную флоэму, кроме
только что образовавшегося слоя. Эпидермис в
конце концов разрывается и замещается пробкой в
результате активности второй латеральной
меристемы - пробкового камбия (феллогена), который
обычно образуется под самым эпидермисом. Пробка
(феллема) откладывается кнаружи от пробкового
камбия, тогда как с внутренней стороны он
формирует один или два слоя паренхимы. Эти слои
неотличимы от первичной коры и образуют фелло-
дерму, или вторичную кору. Феллоген, пробка и
феллодерма в совокупности образуют перидерму
(рис. 21.25).
При созревании пробковых клеток их стенки
пропитываются жировым веществом суберином,
непроницаемым для воды и газов. Клетки постепенно
отмирают, утрачивая живое содержимое, и
наполняются либо воздухом, либо смолой или таннина-
ми. Старые, мертвые пробковые клетки смыкаются
друг с другом и окружают ствол, предохраняя его
от высыхания, инфекций и механических
повреждений. По мере утолщения ствола эти клетки спрессо-
Местоположение первичной флоэмы
(смята до неузнаваемости)
Перицикл (склеренхима)
Полоса ситовидных трубок.
и клеток-спутниц
Полоса флоэмных волокон
Мелкие осенние сосуды
(поздняя древесина)
Широкие весенние сосуды
(ранняя древесина)
Вторичные сердцевинные лучи
Штрихами показано продолжение
рядов сосудов ксилемы
Прирост
первого года
Остатки эпидермиса
Пробка (феллема)
Пробковый камбий (феллоген)
Вторичная кора (феллодерма)
Первичная кора (паренхима)
Вторичная флоэма
Васкулярный камбий
Годичное кольцо
Первичные сердцевинные лучи
Вторичная ксилема (древесина)
в приросте второго года
Метаксилема
Прото ксилема
Перидерма
Первичная ксилема
Рис. 21.25. Ствол типичного деревянистого двудольного, такого как липа, на третьем году
роста (возраст два года) в поперечном разрезе. Детали строения вторичной флоэмы, вторичной ксилемы и
вторичных сердцевинных лучей показаны только в левом секторе.
вываются и могут в конце концов быть замещены
лежащими под ними более молодыми клетками.
Если бы пробковый слой был сплошным,
дыхательный газообмен между живыми клетками стебля
и наружной средой был бы невозможен и эти клетки
погибли бы. Однако в пробке имеются чечевички-
щелевидные отверстия, расположенные случайным
образом. Чечевички образует пробковый камбий, и
они представляют собой рыхлую массу
тонкостенных мертвых клеток, не содержащих суберина, с
обширными межклеточными пространствами,
делающими возможным газообмен (рис. 21.27).
Кора
В конце концов деревянистый стебель покрывается
слоем, известным под названием коры. «Кора»-
неточный термин, которым обозначают либо все
ткани, лежащие кнаружи от проводящей системы,
либо - в более узком смысле - ткани, лежащие
кнаружи от пробкового камбия. При сдирании коры с
дерева обычно удаляются все ткани вплоть до
васкулярного камбия - тонкого слоя клеток,
который легко разрывается.
Пробковый камбий обычно возобновляется
каждый год, по мере увеличения ствола в обхвате.
Часто пробковый камбий образуется во вторичной
флоэме; в таких случаях кора с течением времени
становится слоистой, что обусловлено
чередованием слоев вторичной флоэмы и коры.
Вторичный рост корней двудольных
У двудольных, у которых происходит вторичный
рост стебля, в большинстве случаев наблюдается
также вторичный рост корней. У растений с
запасающими корнями, таких как морковь или репа,
этот рост может быть более заметным, чем в стебле,
хотя в приведенных примерах в ксилеме и флоэме
преобладает паренхима. Процесс вторичного роста
схематически показан на рис. 21.28; он в основном
сходен с вторичным ростом в стеблях.
Развитие в корне вторичной ксилемы, вторичной
флоэмы и сердцевинных лучей в основе своей сходно
с их развитием в стебле; в корнях, как и в стеблях,
обычно образуются годичные кольца. Из перицикла
развивается пробковый камбий, выполняющий те
же функции, что и в стебле. Иногда образуется
феллодерма, но она неотличима от перицикла. Так
же как и в стебле, первоначальный пробковый
камбий может замещаться пробковым камбием,
возникающим в глубине корня. На рис. 21.29 представлен
поперечный срез корня с вторичным утолщением.
Годичные кольца,
обозначающие окончание
первого и второго года роста
Вторичная ксилема Остатки эпидермиса
(древесина) второго
года роста
Широкие
весенние сосуды
третьего года роста
,Пробка
тглг»я Вторичная
флоэма
Полоса
волокон
Маленькая
чечевичка
Первичная
ксилема
Мелкие осенние
сосуды второго
года роста _
г Васкулярныи
камбий
Первичные
сердцевинные
лучи
Вторичные
сердцевинные
лучи
Рис. 21.26. А. Поперечный срез ветки Tilia vulgaris в
возрасте двух лет (третий год роста) (х 2,2). Б.
Часть поперечного среза ветки Tilia vulgaris в
возрасте пяти лет (шестой год роста) ( χ 11,5).
Рис. 21.27. Чечевичка в вертикаль -
ном разрезе (содержимое клеток не
показано)
Воздушные полости, делающие
возможным газообмен
Радиальные ряды
спрессованных
пробковых
клеток
Чечевичка
Перидерма
Остатки
эпидермиса
Рыхлая
паренхима
Пробка
(феллема)
Пробковый камбий
'(феллоген)
Феллодерма
(вторичная кора)
Паренхима
первичной коры
Полоса
ситовидных
трубок и
клеток-спутниц
Ситовидные трубки
и клетки- спутницы
Кора
Волокна
Вторичная
флоэма
Васкулярный
камбий
Вторичный
• сердцевинный
луч
Первичный
сердцевинный
луч
Первичный
сердцевинный
луч
Широкие сосуды
весенней древесины
(ранняя древесина)
Годичное
кольцо
Вторичная
ксилема
(древесина),
прирост
за один год
Мелкие сосуды
осенней
древесины
(поздняя
древесина)
21.7. Метаморфоз
Термином «метаморфоз» (от греч. meta-между,
morphe - форма) обозначают быстрые изменения,
происходящие при переходе от личиночной стадии к
взрослой форме. Это процесс постэмбрионального
созревания, который встречается у многих
беспозвоночных, особенно у насекомых, иглокожих,
полухордовых, оболочников, головохордовых, а также у
некоторых рыб и амфибий. В табл. 21.4 перечислены
различные группы животных и названия их
личиночных стадий.
21.7.1. Адаптивное значение
личиночных стадий
Личинки обычно служат стадией, предназначенной
для расселения, т.е. обеспечивают распространение
вида. Это очень важно для сидячих организмов, так
как позволяет избежать перенаселения, которое
могли бы создать потомки в местообитании
родителей. Перенаселение повлекло бы за собой
усиление конкуренции за пищу и другие ресурсы, угро-
Таблица 21.4. Личиночные стадии некоторых животных
Группа животных
Кишечнополостные
Сосальщики
Ленточные черви
Полихеты
Ракообразные
Насекомые
Моллюски
Иглокожие
Оболочники
Миноги (кругло-
ротые)
Амфибии
Личиночная стадия
Планула
Мирацидий, редия, церкария
Онкосфера, цистицерк
Трохофора
Науплиус, циприсовидная личинка,
зоэа
Червеообразная личинка (гусеница и
др), нимфа
Трохофора, велигер
Диплеврула, плутеус
(Особого названия нет)
Пескоройка
Головастик
Эпидермис
Кора
Эндодерма
Перицикл
Первичная ксилема
Васкулярный камбий
Первичная флоэма
Эндодерма
Первичная флоэма и часть
перицикла
Вторичная флоэма
Широкий первичный
сердцевинный луч, часто
образующийся против рукава
Васкулярный камбий ксилемы
Вторичная ксилема
Первичная ксилема
Вторичный сердцевинный луч
Первичный сердцевинный луч
Остатки первичной флоэмы
Вторичная флоэма
Вторичный сердцевинный луч
Вторичная ксилема
Первичная ксилема
Васкулярный камбий
Первичный сердцевинный луч
Пробка
Пробковый
камбий
Перидерма
Рис. 21.28. Стадии вторичного роста двудольного
растения. А. Развитие васкулярного камбия из клеток
прокамбия (см. рис. 21.23), оставшихся меристе-
матическими. Камбий продолжает расти, образуя
сплошное кольцо (см. стрелки), по мере того как
клетки перицикла за пределами рукавов ксилемы
становятся меристематическими. Б. Камбий
образует ряды сосудов вторичной ксилемы ковнутри и
вторичной флоэмы - кнаружи. Рост вторичной
ксилемы начинается раньше и протекает быстрее
между рукавами ксилемы, пока волнообразный
контур камбия не выравнивается, превратившись в
круг. В. Рост продолжается, и корень, утолщаясь,
вызывает растяжение, разрывы и слущивание
эндодермы, коры и эпидермиса. Из перицикла
образуется пробковый камбий. (Схема В дана в
меньшем масштабе, чем А и Б)
жающей выживанию вида. Например, многие
морские организмы, ведущие сидячий образ жизни,
такие как морские желуди и мидии, производят
огромное число планктонных личинок,
распространяющихся с морскими течениями. На определенной
стадии они прикрепляются к твердым субстратам,
после чего продолжают свое развитие.
Личинки обычно отличаются от взрослых особей
по своему местообитанию, по биологии питания,
способу локомоции и особенностям поведения;
благодаря этому данный вид может использовать на
протяжении своего жизненного цикла возможности,
предоставляемые двумя экологическими нишами,
что повышает его шансы на выживание в период
между яйцом и взрослой стадией. Многие виды,
например стрекозы, питаются и растут только на
личиночных стадиях, составляющих самый
длительный период их жизни.
Другая особенность личинок заключается в том,
что они играют роль как бы переходного этапа, во
время которого вид может приспособиться к новым
условиям, ожидающим его во взрослой жизни.
Кроме того, личинки обладают иногда физиологической
выносливостью, благодаря которой они при
неблагоприятных условиях выступают в роли покоящейся
стадии. Например, некоторые насекомые
перезимовывают в почве в форме личинок, но в большинстве
случаев это происходит на другом этапе
метаморфоза-на стадии куколки; происходящие на этих
стадиях физиологические изменения описаны в
разд. 21.7.3.
И наконец, личиночные стадии обладают иногда
тем преимуществом, что на этих стадиях возможно
увеличение числа личинок. Это обычное явление в
жизненных циклах некоторых плоских червей (см.
разд. 4.5.3).
Не следует, однако, считать личинок непременно
какими-то недоразвитыми формами. Во многих
случаях они достигают весьма высокой организации,
как, например, личиночные стадии насекомых, у
которых недоразвитыми остаются только
репродуктивные органы. В разд. 21.7.4 будет описан
аксолотль-интересный пример вполне развитой
личинки, достигшей половой зрелости.
21.7.2. Характерные черты
метаморфоза
Структурные и функциональные изменения,
происходящие во время метаморфоза, подготавливают
организм к взрослой жизни в новом местообитании.
В некоторых случаях необходимые новые адаптации
могут появляться в то время, когда организм еще не
вышел из личиночной стадии и занимает
личиночное местообитание. Иными словами,
метаморфоз-это не просто реакция на изменение среды, а
подготовка к ее изменению. Например, головастики,
обитающие в воде, где им не грозит высыхание,
выделяют примерно 80% азотистых продуктов
обмена в виде аммиака. На более поздних стадиях
метаморфоза, но все еще оставаясь в воде,
головастики начинают выделять мочевину. А к тому
времени, когда молодые лягушки готовы выйти из
воды, они выделяют уже 80% азотистых отходов в
виде мочевины.
Если на рост и дифференцировку гормоны могут
оказывать влияние, то метаморфоз всецело
контролируется и регулируется эндокринными факторами.
В одних случаях метаморфоз представляет собой
реакцию на повышение уровня гормонов, а в дру-
Перидерма
Остатки коры
Пробковый
камбий
Пробка
Расположение
васкулярного
камбия
Вторичная
флоэма
Вторичная!
ксилема j
(широкие
сосуды и
волокна)
Первичная ксилема
(четыре рукава)
Первичный
сердцевинный
луч
Рис. 21.29. Вторичное утолщение корня Vicia faba (x 120).
гих-это результат различных реакций разных
органов-мишеней на один и тот же уровень какого-то
гормона. Эти реакции могут выражаться либо в
гибели клеток и уменьшении размеров какого-либо
органа (инволюция), либо в делении клеток, их
дифференцировке и росте органа.
21.7.3. Гормональный контроль
линьки у насекомых
Участие гормонов в линьке впервые описал Уиггл-
суорс, работавший в Кембридже. В результате ряда
экспериментов он установил, что линька
контролируется гормоном линьки (ГЛ), который выделяется в
ответ на определенный стимул. Результат линьки
(т.е. следующая за ней стадия жизненного цикла)
может модифицироваться другим гормоном - юве-
нильным, который препятствует появлению имаги-
нальной (взрослой) формы.
Уигглсуорс изучал гормон роста насекомых у
южноамериканского кровососущего клопа Rhodnius
prolixus. У этого насекомого с неполным
превращением пять личиночных (нимфальных) стадий, а
стадия куколки отсутствует. На каждой нимфальной
стадии клопу необходимо один раз насосаться
крови, и этот акт стимулирует линьку. Уигглсуорс
показал, что при наполнении брюшка кровью
находящиеся в его стенке рецепторы реагируют на
растяжение, генерируя нервные импульсы, передающиеся в
головной мозг. Эти импульсы стимулируют
координированную секрецию ряда гормонов, что и
вызывает линьку. Линька, приводящая к превращению
личинки 4-го возраста в личинку 5-го возраста,
наступает (при 280C) через 14 дней после кормежки.
Если декапитировать личинку (т. е. удалить у нее
голову) в первые четыре дня после кормежки и
залить рану воском, то она может прожить 18
месяцев без каких-либо признаков дальнейшего
роста или развития. Если же произвести декапитацию в
более поздний срок, личинка продолжает расти, но
линька не наступает. Эти результаты позволяют
предположить, что передача импульсов от брюшка
в головной мозг и выделение гормонов, влияющих
на рост и дифференцировку тканей-мишеней, в том
числе эпидермальных клеток, занимает четыре дня.
В своих экспериментах Уигглсуорс пересаживал
головной мозг от личинки, готовой к линьке, в
брюшко голодавшей декапитированной личинки, и
последняя начинала линять. При пересадке ткани
головного мозга от линяющей личинки другой, у
которой были отрезаны голова или грудь, линька не
наступала. Очевидно, линьку вызывает какое-то ве-
- Эпитикула
-Экзокутикула
Эндокутикула
Эпидермис
Утолщение
эпидермальных
клеток
Эпикутикула
Экзокутикула
Эндокутикула
Новая
эпикутикула
Эпи кути кула
-Экзокутикула
Эндокутикула
Новая
эпикутикула
Новая
экзокутикула
^Эпидермис
Среднеспинная линия
Рис. 21.30. А. Кутикула клопа Rhodnius в
поперечном разрезе. До того как клоп насосется крови,
эпидермис тонкий и уплощенный. После кормежки
эпидермис начинает утолщаться и отстает от
лежащей над ним эндокутикулы. Б. Над эпидермисом
образуется новая эндокутикула. Ферменты хитиназа
и протеиназа, секретируемые эпидермисом,
разрушают старую кутикулу. Новая кутикула защищена
от действия этих ферментов благодаря
образованию новой эпикутикулы. Продукты переваривания
прежней эндокутикулы всасываются
эпидермисом и включаются в состав новой эндокутикулы.
В. Rhodnius вбирает в себя воздух, раздувается,
разрывает старую кутикулу и сбрасывает ее. Новая
кутикула вначале бывает мягкой, но затем твердеет.
Г. Старая кутикула разрывается по среднеспинной
линии, где она тоньше, чем в других местах.
щество, содержащееся в головном мозге и
воздействующее на определенный участок груди.
Проводившиеся ранее работы на гусеницах тутового
шелкопряда показали, что в первом грудном сегменте у
них имеется проторакальная железа, которая
способна индуцировать линьку, если пересадить ее
гусенице в брюшко, отделенное лигатурой от
грудного отдела. Это наводило на мысль, что у клопа
Rhodnius линька тоже контролируется гормоном
проторакальной железы.
Гормон линьки был выделен в 1953 г.; это
стероид, сходный по структуре с холестеролом (см.
разд. 5.3.6). Его назвали экдизоном; он содержит
больше гидроксильных групп, чем холестерол, и
поэтому растворим в воде. Впоследствии этот
гормон был синтезирован; при введении насекомым
синтетического гормона у них возникают все
реакции, обычно связанные с линькой.
Как полагают, экдизон активирует гены,
контролирующие синтез ферментов, участвующих в
процессах роста. Эта реакция особенно сильно
выражена в эпителиальных клетках, которые
претерпевают ряд изменений, связанных с линькой (рис.
21.30).
При введении экдизона личинкам насекомых,
обладающим «гигантскими хромосомами», у них
возникают «пуфы» в определенных участках
хромосом. Поскольку это те участки, в которых
происходит синтез мРНК, можно думать, что экдизон
действует непосредственно на уровне генетической
транскрипции.
В ряде дальнейших экспериментов Уигглсуорс
получил данные, указывающие на то, что в
головной области личинок Rhodnius синтезируется еще
один гормон, который способствует сохранению
личиночных признаков, препятствуя тем самым
превращению во взрослую форму. Как показали
исследования, этот гормон вырабатывают органы,
лежащие позади мозга,-corpora allata (прилежащие
тела). Если пересадить личинке 5-го возраста такой
орган, взятый у личинки 3-го или 4-го возраста, то
она превратится не во взрослого клопа, а в
гигантскую личинку 6-го возраста. Аналогичная
пересадка от личинки 5-го возраста такого результата
не дает.
Corpora allata вырабатывают так называемый
ювенильный гормон. Теперь считают, что рост и
метаморфоз у членистоногих контролируется
взаимодействием между экдизоном и ювенильным
гормоном. Как полагают, ювенильный гормон
обусловливает сохранение личиночных (или нимфаль-
ных) признаков, активируя гены, определяющие эти
признаки, и подавляя гены, детерминирующие
развитие кутикулы и строения тела, характерного для
имаго
Выделение экдизона и ювенильного гормона
контролируется еще одним гормоном, который
вырабатывает головной мозг. Механизм секреции
этого гормона будет описан на примере саранчи
(рис. 21.31).
Тела нейросекреторных клеток, расположенные в
интерцеребральной железе, выделяют проторакотроп-
ный гормон, который по аксонам нейросекреторных
клеток поступает в нервные волокна, идущие к
кардиальным телам (corpora cardiaca), возможно, с
током аксоплазмы (см. разд. 16.6.2). В кардиальных
телах, расположенных непосредственно позади
головного мозга, этот гормон накапливается. Фактор,
стимулирующий линьку, каким бы он ни был,
возбуждает в мозгу нервные импульсы, которые
передаются по нервным волокнам в кардиальные
тела и вызывают выделение в кровь проторакотроп-
ного гормона (рис. 21.32). Этот гормон поступает в
Нейросекреторные
клетки интерцеребральной
железы
Нервные волокна
кардиальных тел
Кардиальное тело
(corpus cardiacum)
-Corpus allatum
Пищевод
Проторакальная железа
Рис. 21.31. Железы, участвующие в процессе линьки
у саранчи.
Пища
Свет
Температура
Мозг
Нейросекреторные
клетки
Проторакотропный гормон
J Corpus cardiacum
Проторакотропный гормон
Corpus allatum
Проторакальные железы
Ювенильный гормон
Экдизон
Имагинальные признаки
Личиночные признаки
Рис. 21.32. Гормональная регуляция линьки у
насекомых.
проторакальные железы, стимулируя выделение
экдизона, который в свою очередь стимулирует
эпидермальные клетки к образованию новой
кутикулы.
С аналогичной последовательностью событий
связана регуляция линьки у Rhodnius, но парные
кардиальные тела, так же как и парные прилежащие
тела, слились у него в единые структуры,
расположенные по средней линии тела.
Насекомые с полным превращением проходят
стадию куколки при очень низком содержании юве-
нильного гормона в крови, а в отсутствие этого
гормона сразу превращаются во взрослую форму.
Если у насекомого с неполным превращением
удалить на второй, третьей или четвертой нимфальной
стадии прилежащее тело, то нимфа, перелиняв,
превращается в миниатюрную взрослую особь.
Возможно поэтому, что увеличение размеров и
достижение половозрелости у насекомых вызывается
снижением уровня ювенильного гормона. Регуляция
роста и метаморфоза у насекомых с полным и
неполным превращением схематически
представлена на рис. 21.33.
У крылатых насекомых по достижении стадии
имаго проторакальные железы атрофируются и
исчезают, так что дальнейшие линьки становятся
невозможными. У бескрылых насекомых этого не
происходит. У всех взрослых особей прилежащее
тело вновь активируется и выделяет ювенильный
гормон. Самкам это необходимо для того, чтобы в
яйцах откладывался желток, а самцам-для
нормального функционирования придаточных желез,
образующих сперматофор - капсулу, в которой
сперматозоиды вводятся в половое отверстие самки.
21.7.4. Метаморфоз у амфибий
При метаморфозе у амфибий личинка, обитающая в
воде, превращается в полуназемную взрослую
форму. Механизмы метаморфоза контролируются
секретами эндокринных желез, подобно тому как это
было описано для насекомых. В 1911 г. немецкий
биолог Гудернач обнаружил, что если скармливать
головастикам вытяжку из щитовидной железы
млекопитающего, то они быстро превращаются в
миниатюрных лягушек. Этот преждевременный
метаморфоз указывал на то, что гормон щитовидной
железы - тироксин - является фактором, ответственным
за многообразные морфологические,
физиологические, биохимические и поведенческие изменения,
происходящие в жизненном цикле лягушки. В
дальнейших исследованиях у одних головастиков
удаляли щитовидную железу, а у других - гипофиз; в
обоих случаях наблюдалось подавление
метаморфоза, которое снималось скармливанием тироксина.
Насекомое с полным превращением — капустница Pieris brassicae
г л +юг
-гл + юг
гл
Яйцо ► 1 —► 2—► 3—► 4 —► 5 —► Куколка -► Имаго
Личиночные стадии
Насекомое с неполным превращением — Rhodnius prolixus
Яйцо-
гл+юг
гл+юг
гл+юг
ГЛ -г юг
гл
♦> Имаго
Нимфальные стадии
Рис. 21.33. Влияние гормонов на жизненные циклы
насекомых с полным и неполным превращением.
ГЛ-гормон линьки, ЮГ— ювенильный гормон. Высота
букв ЮГ отражает относительную концентрацию
гормона.
Полагают, что первоначальным стимулом для
метаморфоза служат такие изменения условий
среды, как повышение температуры или увеличение
длины дня. Нервные сигналы об этих внешних
стимулах передаются в головной мозг и активируют
гипоталамус, который начинает выделять тиреоли-
берин. Этот гормон попадает в кровеносные сосуды
гипоталамо-гипофизарной воротной системы (см.
разд. 16.6.2) и поступает с кровью в переднюю долю
гипофиза. Здесь он вызывает высвобождение тирео-
тропного гормона, который переходит в кровяное
русло и побуждает щитовидную железу к секреции
тироксина (тетраиодтиронина) и трииодтиронина. Эти
гормоны оказывают непосредственное влияние на
все ткани, участвующие в метаморфозе, вызывая
разнообразные реакции - атрофию хвоста и жабр
головастика, ускорение роста конечностей и языка,
дифференцировку кожи с образованием железистых
клеток и хроматофоров (клеток, содержащих
пигменты). Прямое местное воздействие тироксина
на метаморфоз было продемонстрировано в
следующем эксперименте: нескольким головастикам
прикладывали к левому глазу и к левой стороне
Таблица 21.5. Изменения внешних признаков лягушки при
метаморфозе. Время и размеры тела можно указать только
приблизительно, так как скорость метаморфоза зависит от
количества пищи, температуры и внутренних факторов
Время,
недели
0
1
2
4
7
9
11—12
13
14
16
Длина
тела,
MM
—
7
9
12
28
35
35
25
20
15
Стадия развития
Оплодотворение
Вылупляется из слизистой капсулы.
Имеются наружные жабры, хвост,
рот с роговыми челюстями; под
ротовым отверстием слизистые
железы
Наружные жабры начинают
атрофироваться, над внутренними
жабрами образуется жаберная крышка.
Глаза хорошо развиты
Утрата наружных жабр и слизистых
желез. Развивается брызгальце.
Хвост расширяется и помогает
плавать
Появляются почки задних
конечностей
Задние конечности вполне
сформированы, но не используются при
плавании Голова начинает
расширяться
Левая передняя конечность выходит
через брызгальце, а правая
прикрыта жаберной крышкой Задние
конечности используются при
плавании
Глаза увеличиваются, рот
расширяется
Хвост начинает рассасываться
Все внешние личиночные признаки
исчезли. Лягушка выходит на сушу
хвоста препарат тироксина на жировой основе,
используя правую сторону в качестве контроля (к ней
прикладывали только чистую жировую основу);
через несколько дней на левой стороне можно было
наблюдать регрессию хвоста и замену личиночного
зрительного пигмента порфиропсина на родопсин-
пигмент, характерный для взрослых особей. В
контрольных тканях никаких изменений не
происходило.
На ранних стадиях метаморфоза контроль
осуществляется по принципу положительной обратной
связи, т.е. увеличение количества тироксина,
выделяемого растущей щитовидной железой,
стимулирует дальнейшую секрецию самого тироксина. При
повышении уровня тироксина в крови различные
органы-мишени реагируют на него гибелью клеток,
их делением или дифференцировкой, как это было
описано в приведенных выше примерах. После
достижения имагинальной стадии секреция тироксина
контролируется по принципу отрицательной
обратной связи (см. разд. 16.6.5). Последовательность
внешних морфологических изменений,
наблюдаемых у головастиков при метаморфозе, представлена
в табл. 21.5.
Аксолотль {Ambystoma mexicanum) (рис. 21.34) в
обычных условиях не претерпевает метаморфоза,
так как его гипофиз не способен вырабатывать
тиреотропный гормон. Поэтому животное
достигает половозрелости, сохраняя личиночные признаки:
наружные жабры, хвостовой плавник и светлую
окраску; это явление называют неотенией. Как
полагают, неотения сыграла существенную роль в
эволюции многих групп животных, например самых
ранних позвоночных, которые, вероятно,
произошли от неотенических личинок асцидий.
У аксолотля можно вызвать метаморфоз,
добавив к воде аквариума надлежащее количество
тироксина. При этом он теряет жабры и большую
часть хвостового плавника, становится взрослой
амфибией, внешне очень похожей на своего
близкого родича - мраморную амбистому.
21.8. Развитие позвоночных
В предыдущем разделе были описаны примеры
морфологических изменений, происходящих при
превращении личиночной формы во взрослую. Эти
изменения, конечно, впечатляют; они имеют
огромное адаптивное значение, подготавливая организм к
тем условиям, в которых ему предстоит жить во
взрослом состоянии. Однако они кажутся
незначительными по сравнению с изменениями,
происходящими после оплодотворения яйца. Так, например,
из 47 клеточных поколений, которые, по
имеющимся оценкам, сменяются на протяжении жизни
человеческого организма, 42 приходятся на период
внутриутробного развития. Изучение
эмбрионального (зародышевого) развития составляет предмет
эмбриологии, и события, происходящие в этот пе-
Рис. 21.34. Аксолотль
риод, в основе своей сходны у всех животных. Они
помогают нам лучше понять происхождение тканей,
органов, систем органов и функциональные связи
между ними. Эмбриональное развитие удобно
разбить на три главные стадии; следует помнить,
однако, что процесс развития в целом непрерывен и одна
его стадия незаметно переходит в следующую.
Ниже рассмотрены главные черты этих стадий и их
значение.
дробление яйца. Дроблением называют ряд ми-
тотических делений зиготы, которые происходят
после оплодотворения. У лягушки точка, в которой
спермий проник в яйцо, определяет плоскость
симметрии будущего зародыша и положение будущей
дорсальной губы бластопора. Оплодотворение
служит стимулом для дробления, в результате
которого получаются дочерние клетки, или бластомеры,
образующие полый шар - бластулу, внутреннюю
полость которой называют бластоцелем.
Вводя в клетки разноцветные красители (метод
прижизненного, или витального, окрашивания),
можно проследить за судьбой клеток бластулы на
протяжении двух дальнейших процессов - гаструляции и
органогенеза. Фогт с помощью этого метода создал
карты зачатков, на которых указана проспективная,
или презумптивная, судьба клеток разных участков
бластулы (рис. 21.35).
гаструляция. Этот процесс начинается с
образования в бластуле круглого отверстия-бластопора. У
лягушки клетки бластулы быстро делятся,
дифференцируются и в результате ряда морфогенетических
движений впячиваются внутрь (инвагинируют) через
бластопор; при этом бластоцель исчезает и
образуется новая полость - полость первичной кишки
(архентерон). Клетки перераспределяются,
располагаясь в виде трех отдельных зародышевых листков в
трехслойной структуре, называемой гаструлой.
Зародышевые листки - эктодерма, мезодерма и
энтодерма - приобретают таким образом определенную
пространственную организацию, что служит
подготовкой к органогенезу.
органогенез. Происходят дальнейшие клеточные
деления, рост и дифференцировка - сложные морфо-
генетические процессы, приводящие к образованию
хорды, центральной нервной системы и развитию
мезодермы. У позвоночных при этом образуются
ткани, органы и системы органов, представленные
на рис. 21.36 и 21.37. Органогенез завершается вы-
луплением из яйца или рождением детеныша.
Эктодерма-
Бластоцель-
Энтодерма
Нервная пластинка
Полость первичной
кишки
Хорда
Дорсальная губа
бластопора
Желточная пробка
— Мезодерма
Рис. 21.35. Схематическое изображение
развивающейся га cm рулы (разрез по срединной плоскости) с
указанием будущей судьбы отдельных участков.
Стрелки показывают направление дальнейшего
роста и миграции клеток эктодермы и
мезодермы. Мезодерма перемещается внутрь и
располагается под эктодермой.
Нервный гребень
Нервная трубка
Дерматом
Миотом
Хорда
Склеротом
Нефротом
Эктодерма
Соматическая мезодерма
Спланхническая мезодерма
Кишечник
Целом
Желточные клетки
энтодермы
Рис. 21.36. Схематическое изображение
поперечного среза только что вылупившегося головастика
лягушки. Показаны производные зародышевых
листков (см рис. 21.37). Головастик продолжает
питаться за счет желтка, так как эктодерма еще
недостаточно дифференцировалась, чтобы могло
завершиться формирование пищеварительного
тракта.
21.8.1. Гормоны и рост
млекопитающих
На рост и развитие влияют гормоны, выделяемые
четырьмя железами: щитовидной железой, печенью,
корой надпочечников и половыми железами. У всех
этих желез процесс секреции происходит под
контролем других гормонов, выделяемых гипофизом. В
свою очередь на гипофиз оказывают влияние
особые стимулирующие и ингибирующие факторы,
вырабатываемые гипоталамусом (см. разд. 16.6.2).
Гормональная регуляция на этих трех уровнях
будет описана на примере секреции печенью гормона
роста соматомедина.
Многочисленные эксперименты по удалению
гипофиза и заместительной терапии, проводившиеся в
начале нашего века, выявили роль гипофиза в
определении скорости и степени роста у животных.
Из гипофиза было экстрагировано вещество,
которое при введении животным вызывает увеличение
массы тела. Это вещество было названо «гормоном
роста». Дальнейшие исследования показали, что это
один из тропных гормонов гипофиза - соматотропин,
секрецию которого регулируют два других гормона,
вырабатываемые гипоталамусом: соматостатин
(пептид, состоящий из 40 аминокислот) и сомато-
либерин. Считается, что эти два гормона обладают
противоположным действием: соматостатин
подавляет, а соматолиберин стимулирует секрецию со-
матотропина, причем влияние соматостатина
доминирует над влиянием соматолиберина.
Соматотропин человека - это белок, состоящий из
190 аминокислот. Он воздействует на рецепторные
участки в клетках печени (гепатоцитах), стимулируя
синтез и выделение соматомедина в печеночную
вену. Соматомедин усиливает процессы роста во
всем организме, ускоряя синтез нуклеиновых кислот
для подготовки к митозу и способствуя
поглощению аминокислот хрящевой и мышечной тканями.
Он вызывает также повышение содержания в
клетках калия, фосфора, кальция и натрия и изменяет
уровни глюкозы и липидов в крови; в результате
создаются дополнительные количества
метаболитов, необходимых для процессов биосинтеза и для
роста клеток.
21.9. Восстановление и
регенерация
Когда организм достигает своих окончательных
размеров, рост клеток не прекращается. Утрата или
повреждение клеток и тканей в результате старения,
Эктодерма <
Бластомеры -4—Мезодерма
Энтодерма <
' Эктобласт
. Нервная
" пластинка
» Нервный
гребень
Эпидермис, эпидермальные железы, волосы, перья, чешуя, эмаль,
конъюнктива,хрусталик, уши, эпителиальная выстилка носа и ротовой
полости, анального отверстия и влагалища, передняя доля гипофиза
Центральная нервная система, симпатические и парасимпатические
ганглии, соматические моторные нейроны, сетчатка глаза, задняя
доля гипофиза
Челюсти, ганглии спинных корешков, соматические сенсорные
нейроны, мозговое вещество надпочечников
Скелетные мышцы, диафрагма
Позвонки, оболочка хорды
Дерма, дентин
Мышцы стенки тела
Почечные канальцы, мочеточники, яйцеводы, матка, части
яичников и семенников, кора надпочечников
Гладкая мускулатура, сердце, кровеносные сосуды, кровь,
лимфатическая система, легкие, соединительная ткань (хрящи, кости,
брыжейки и др.) склера, сосудистая и роговая оболочки глаза
- Большая часть пищеварительного тракта
- Выстилка кишечника, мочевого пузыря, легких; евстахиевы трубы
Печень, поджелудочная железа, щитовидная железа, паращитовидные
железы, тимус
" Первичные половые клетки
Сомиты
Миотомы-
Склеротомы
Дерматомы-
.Нефротомы-
^Спланхнотомы
(мезенхима)
4 Хорда
у Кишечник
-Эпителии-
- Железы—
N Гонады—
Рис. 21.37. Судьба трех зародышевых листков в процессе развития позвоночного.
заболеваний, несчастных случаев или нападения
других организмов может стимулировать деление и
дифференцировку клеток, что приводит к
заживлению ран, восстановлению и замещению
поврежденных или недостающих тканей.
У млекопитающих многие ткани, например кожа,
клетки кишечного эпителия и клетки крови,
непрерывно замещаются на протяжении всей жизни
организма с постоянной скоростью -
приблизительно такой, с какой они обычно отмирают. В других
органах, например в печени, щитовидной железе и
поджелудочной железе, клетки у взрослой особи при
нормальных обстоятельствах почти не делятся.
Если, однако, часть такого органа по той или иной
причине будет утрачена, то клетки остальной его
ткани начинают быстро делиться и
дифференцироваться, с тем чтобы восстановить нормальные
размеры органа. Этот процесс носит название
компенсаторной гипертрофии. В отличие от тканей таких
органов клетки центральной нервной системы не
способны к регенерации.
Регенерацией называют замещение частей
организма, утраченных по одной из перечисленных выше
причин. Она очень часто наблюдается у растений,
особенно у покрытосеменных, у которых из одного
листа с черешком можно иногда вырастить целое
растение (например, у традесканции). Способность
растений к регенерации лежит в основе
вегетативного размножения (см. разд. 20.1.1). У животных
способность к регенерации утраченных структур,
по-видимому, зависит от сложности организации. У
губок, кишечнополостных и плоских червей она
достигает поразительной степени. Губок и
некоторых кишечнополостных, таких как гидра, можно
сильно измельчить, после чего каждый кусочек
регенерирует и образует целый организм. Планарию
можно разрезать пополам вдоль или поперек, и
каждая половинка регенерирует недостающую
часть. Другой плоский червь - Dugesia - способен
регенерировать целую особь из маленького кусочка,
отрезанного от взрослого червя. Большинство
кольчатых червей восстанавливают недостающие
участки тела; например, дождевой червь может
регенерировать первые пять сегментов и значительную
часть заднего конца тела в случае их утраты. У
ракообразных, насекомых и иглокожих возможна
регенерация конечностей или лучей, подобно тому
как у личинок амфибий регенерируют конечности, а
у взрослых ящериц - хвосты, утраченные в
результате нападения других особей своего вида или
хищников. Ящерицы способны также в случае нападения
сами отламывать собственный хвост; отделившийся
от тела извивающийся хвост отвлекает внимание
врага, что позволяет ящерице спастись бегством.
Способность самопроизвольно отбрасывать
придатки тела, известная под названием аутотомии,
свойственна также крабам и омарам.
21.10. Состояние покоя
В жизненном цикле многих растений и животных
наблюдаются периоды, когда метаболическая
активность снижается до минимума и рост
прекращается. Это периоды покоя, представляющие собой
физиологически детерминированные реакции и
наблюдаемые у взрослых организмов на одной из
стадий развития, например у куколки, или у
специализированных репродуктивных структур, таких
как споры. Период покоя наступает под влиянием
изменений, происходящих во внешней среде; его
биологическая роль состоит в том, чтобы дать
возможность виду пережить время, когда снабжение
энергией недостаточно для нормального роста и
метаболизма.
Большинство растений тропических и умеренных
поясов и все арктические растения реагируют на
изменение освещенности, температуры и влажности
переходом в состояние покоя. Механизмы
управления периодами покоя у растений при помощи
регуляторов роста описаны в разд. 15.5.
У животных известны три формы покоя, в
которых участвуют физиологические адаптации,
позволяющие переносить экстремальные внешние
условия. Однако некоторые животные благодаря
своей высокой подвижности могут избегать
неблагоприятных условий среды, мигрируя в более
подходящие области. Эти реакции, хотя они и
координируются с помощью физиологических механизмов,
носят чисто поведенческий характер и описаны в
разд. 16.8.
21.10.1. Диапауза
Диапауза-это своего рода остановка развития,
наблюдаемая у насекомых, но этим термином
обозначают также период покоя, характерный для цист
простейших и для зимующих яиц многих
пресноводных организмов. Жизненные циклы насекомых
организованы во времени таким образом, чтобы
потомство появлялось в периоды изобилия корма
для личиночных или нимфальных стадий. Диапауза
создает возможность для координирования этих
двух факторов. Остановка развития возможна на
любой стадии жизненного цикла-на стадии яйца,
личинки, куколки или имаго, а когда она
происходит у данного вида-это детерминировано
генетически. У некоторых насекомых (например, у
саранчи Nomadacris) развитие непременно
сопровождается диапаузой, которую в этом случае называют
облигатной диапаузой, тогда как у других диапауза
наблюдается только при возникновении
неблагоприятных условий (как у саранчи Schistocerca)- это
факультативная диапауза.
Главный фактор, индуцирующий диапаузу у всех
видов,-длина дня (фотопериод), однако могут
Температура, °С
-Куколка-
Температура ■
Мозг
Кровь
высокий
Имаго
низкий
Уровень
проторако-
тропного
гормона
Время, недели
Рис. 21.38. Зависимость между уровнями протора-
котропного гормона в мозге и крови Bombyx топ при
3°С и при 25°С.
иметь значение также наличие пищи, температура и
влажность. Для того чтобы диапауза была
эффективна, действие индуцирующего фактора должно
опережать наступление неблагоприятных условий.
Поэтому механизм синхронизации часто
воздействует на стадию, предшествующую той, на
которой наступает диапауза. Например, диапауза на
стадии яйца у тутового шелкопряда (Bombyx mori)-
результат фотопериода, действию которого
подвергалась самка во время развития яйца. Диапауза у
взрослых особей индуцируется условиями
освещения, имевшими место на ранних личиночных
стадиях.
Свет, как полагают, оказывает прямое влияние на
мозг, который в свою очередь воздействует на
активность нейросекреторной системы. У тлей свет,
по-видимому, непосредственно влияет на нейро-
секреторные клетки. У шелкопряда Hyalophora
cecropia короткий осенний фотопериод инактиви-
рует нейросекреторные клетки мозга,
вырабатывающие проторакотропный гормон. Это препятствует
секреции экдизона торакальными железами,
развитие приостанавливается, и насекомое
перезимовывает на стадии куколки. Нормальная секреция
и дальнейшее развитие возобновляются лишь
после продолжительного пребывания на холоде.
Парадоксальным образом именно те
неблагоприятные условия, которых диапауза позволяет избе-
жать,стимулируют прерывание периода покоя
(рис. 21.38).
21.10. Используя экспериментальные данные,
представленные на рис. 21.38, объясните
механизмы, приводящие к окончанию диапаузы.
Большинство насекомых, например бобовая тля
(Aphis fabae), если содержать их в условиях длинного
дня, растут и развиваются непрерывно и дают
несколько поколений в год. У таких видов в
условиях длинного дня диапауза не наступает. Это
«организмы длинного дня», и они вступают в диа-
паузу только в том случае, если периодически
содержать их в условиях короткого дня. Другие виды,
например тутовый шелкопряд (Bombyx mori), у
которого диапауза происходит на стадии яйца
(эмбриональная диапауза), в условиях короткого дня
растут и развиваются нормально, а диапауза
наступает у них только после содержания в течение
некоторого времени в условиях длинного дня. Это
«организмы короткого дня». Диапаузирующие яйца
тутового шелкопряда начинают расти и развиваться
в период весенних коротких дней, а взрослые особи
появляются в начале лета. Поскольку развитие
приходится на период коротких дней, из яиц,
отложенных этими особями, гусеницы вылупляются и
развиваются летом, без диапаузы. Однако у яиц,
отложенных этими летними шелкопрядами,
диапауза наступает в порядке подготовки к зиме. Так как у
насекомых этого типа диапаузу индуцируют
длинные фотопериоды, тем самым число поколений в
год ограничивается; у тутового шелкопряда их два
(рис. 21.39).
У Bombyx обнаружен гормон диапаузы, от
которого зависит наличие или отсутствие диапаузы в
развитии яйца. Длинные летние дни побуждают
мозг в секреции этого гормона, вырабатываемого
парой нейросекреторных клеток, расположенных в
подглоточном ганглии. Этот гормон оказывает
прямое воздействие на яичник, в котором
образуются темноокрашенные яйца, вступающие в диа-
Фотопериод:
короткий
длинный
короткий
короткий
Стадия
жизненного
цикла
Яйцо -► Личинка -♦ Имаго -► Яйцо ->· Личинка -► Имаго -¥- Диапауза на стадии -►
яйца
Время
года
Зима Весна Лето Осень Зима
Рис. 21.39. Зависимость между длиной дня, диапаузой и стадиями жизненного цикла Bombyx топ. Это
пример «организма короткого дня».
Мозг
Короткий фотопериод
Отсутствие стимула·
Длинный фотопериод
Подглоточный ганглий-
Гормона диапаузы нет
- Стимул
- Окологлоточная
комиссура
Гормон диапаузы
- Яичник
Непигментированные
яйца
Без диапаузы
■ Пигментированные
яйца
Диапауза
Рис. 21.40. Схема, иллюстрирующая влияние фото-
периода на продукцию яиц у Bombyx топ.
паузу. Особи, развивающиеся в конце зимы и
весной, т.е. в условиях короткого дня, не
вырабатывают гормона диапаузы и откладывают
непигментированные яйца, которые развиваются без
диапаузы (рис. 21.40).
21.10.2. Летняя спячка (эстивация)
Летняя спячка-это особое состояние покоя,
наблюдаемое у некоторых рыб и амфибий в периоды
жаркой сухой погоды. В такие периоды эти
организмы зарываются в ил, пока он еще остается
мягким, и снижают интенсивность метаболизма до
минимума, что дает им возможность пережить
условия острого дефицита влаги. Во время летней
спячки они дышат атмосферным воздухом.
Нормальная метаболическая активность возобновляется
только тогда, когда река, пруд или озеро после
периода засухи вновь наполняется водой. Два из
трех родов двоякодышащих рыб - Lepidosiren в
Южной Америке и Protopterus в Африке -
способны проводить в спячке не менее шести месяцев,
когда реки, в которых они живут, временно
пересыхают.
21.10.3. Зимняя спячка (гибернация)
Зимняя спячка-это состояние относительно низкой
метаболической активности, связанное с периодом
низких температур. Такая спячка дает возможность
многим видам амфибий, рептилий, птиц и
млекопитающих переживать периоды нехватки пищи,
снижая энергетические потребности до минимального
уровня.
Это имеет особенно важное значение для го-
мойотермных животных, которым для поддержания
высокой температуры тела обычно необходима
высокая интенсивность метаболизма. При
настоящей спячке температура тела падает почти до
температуры окружающей среды, а сокращения
сердца, дыхание, общий метаболизм, рост и
процессы развития резко замедляются, так что
животное становится почти пойкилотермным. Среди
организмов, впадающих в настоящую спячку, можно
назвать насекомоядных (землеройки, ежи),
грызунов (сурки, суслики, сони, хомяки), летучих мышей,
питающихся насекомыми, и нескольких птиц.
Большинство других животных, у которых наблюдается
та или иная форма спячки, на самом деле находятся
в состоянии оцепенения, сна или псевдоспячки. К
таким животным относятся змеи, ящерицы,
черепахи, саламандры, жабы, тритоны и лягушки; в случае
повышения температуры среды они могут
просыпаться, с тем чтобы подкормиться имеющимися в
норе запасами, или же для мочеиспускания и
дефекации.
Начало спячки связано с понижением
температуры среды и укорочением фотопериода, а также с
накоплением соответствующих запасов пищи. Судя
по имеющимся экспериментальным данным, в
крови содержится какой-то метаболит, способный
стимулировать спячку, поскольку введение крови,
взятой от суслика, находящегося в спячке,
бодрствующему животному, уже через 48 ч вызвало у
него спячку. Подобным же образом была
индуцирована спячка у сусликов в такое время года, когда
они обычно активны: им вводили кровь, взятую от
животного в период спячки. Некоторые грызуны,
например Arctomys, впадают в спячку за несколько
часов, тогда как у других это занимает несколько
ночей, в течение которых температура внутренних
областей тела у них постепенно понижается, пока
не достигнет уровня, характерного для зимней
спячки.
По мере снижения температуры тела до уровня,
превышающего температуру среды примерно на
1-2°С, частота сердечных сокращений может
понизиться с 400 до 8 в 1 мин, как у суслика, а
основной обмен-до менее чем 2% от нормального
уровня.
Спячка не спасает животных от гибели при очень
сильном охлаждении, но они избегают замерзания,
проводя спячку в гнездах или норах, где
температура значительно выше, чем снаружи. Свертываясь в
комок, животное уменьшает открытую поверхность
тела, и это, так же как погружение морды в мех,
способствует сохранению тепла. Если наступают
очень сильные морозы и температура тела падает
до 2°С, возникает опасность повреждения тканей. В
таких случаях животное просыпается и начинает
двигаться, чтобы повысить температуру тела, или
же повышает интенсивность метаболизма, не
просыпаясь.
Многие зимнеспящие животные запасают
энергию в виде липидных капелек, накапливаемых в
особой жировой ткани-буром жире. Клетки этой
ткани содержат много сферических митохондрий,
которые и придают ей характерную темную
окраску. Липиды рассеяны по всей цитоплазме в виде
капелек, а не в твердом состоянии. Животное рас-
ходует эти резервные липиды понемногу на
протяжении всего периода спячки, но как только оно
просыпается, метаболическая активность клеток
жировой ткани резко возрастает. Используя
освобождающуюся при этом энергию, животное
начинает сильно дрожать и генерирует тепло, которое
разносится по организму кровью благодаря
учащению сердечного ритма. В первую очередь тепло
поступает в сердце, головной мозг и легкие; это
достигается благодаря сужению сосудов, идущих к
другим частям тела, под воздействием
симпатической нервной системы. Постепенно весь организм
разогревается и его нормальные метаболические и
физиологические функции в конце концов
возобновляются.
Некоторые мелкие птицы и млекопитающие, у
которых отношение поверхности тела к его объему
велико, отличаются крайне интенсивным
метаболизмом; поэтому по ночам, когда они не могут
потреблять корм, им приходится, для того чтобы
избежать гибели, понижать температуру своего
тела. Это явление, наблюдаемое у колибри и у мелких
насекомоядных летучих мышей, известно под
названием оцепенения.
Глава 22
Непрерывность жизни
В начале главы 7 говорилось о создании клеточной
теории Шлейденом и Шванном. Рудольф Вирхов
расширил эту теорию, провозгласив в 1855 г.:
"omnis cellula e cellula" («каждая клетка из клетки»).
Признание непрерывности живого побудило ученых
второй половины XIX века заняться исследованием
строения клетки и механизмов клеточного деления.
Совершенствование гистологических методов и
создание микроскопов с более высокой разрешающей
способностью позволило выявить важную роль
ядра и в особенности заключенных в нем хромосом
как структур, обеспечивающих преемственность
между последовательными поколениями клеток.
В 1879 г. Бовери и Флемминг описали
происходящие в ядре события, в результате которых
образуются две идентичные клетки, а в 1887 г. Вейсман
высказал мысль о том, что при образовании гамет
происходит деление иного типа. Эти два типа
деления называют соответственно митозом и мейо-
зом. Происходящие при этом процессы почти
идентичны, однако они приводят к совершенно разным
результатам.
Митоз-это такое деление клеточного ядра, при
котором образуются два дочерних ядра с наборами
хромосом, идентичными наборам родительской
клетки. Вслед за ядерным делением обычно сразу же
происходит деление цитоплазмы на две равные
части, восстановление клеточной
(плазматической) мембраны и клеточной стенки (у растений) или
одной только клеточной (плазматической)
мембраны (у животных) и разделение возникших таким
образом двух дочерних клеток. Весь этот процесс и
называют клеточным делением. Митотическое
деление клеток приводит к увеличению их числа,
обеспечивая процессы роста, регенерации и
замещения клеток у всех высших животных и растений.
У одноклеточных организмов митоз служит
механизмом бесполого размножения, ведущего к
увеличению их численности.
Мейоз-это процесс деления клеточного ядра с
образованием четырех дочерних ядер, каждое из
которых содержит вдвое меньше хромосом, чем
исходное ядро. Его называют также редукционным
делением (от лат. reductio - уменьшение): число
хромосом в клетке уменьшается с диплоидного (2п) до
гаплоидного (п). Значение мейоза состоит в том, что
он обеспечивает сохранение в ряду поколений
постоянного числа хромосом у видов с половым
размножением. Мейоз происходит только при
образовании гамет у животных и при образовании спор у
тех растений, которым свойственно чередование
поколений (см. разд. 20.2). В результате мейоза
получаются гаплоидные ядра, слияние которых при
оплодотворении ведет к восстановлению
диплоидного числа хромосом.
Хромосомы играют главную роль в процессе
клеточного деления, так как они обеспечивают
передачу наследственной информации от одного
поколения другому (см. разд. 23.2) и участвуют в
регуляции клеточного метаболизма. В состав
хромосом эукариотических клеток входят ДНК, белки и
небольшие количества РНК (см. разд. 5.6). В не-
делящихся клетках хромосомы представлены
чрезвычайно длинными тонкими нитями,
распределенными во всем объеме ядра. Отдельные хромосомы
неразличимы, но хромосомный материал
окрашивается некоторыми основными красителями (см.
Приложение 4.2) и поэтому был назван хроматином.
В начале клеточного деления хромосомы
укорачиваются и окрашиваются более интенсивно, так что
становятся видимыми по отдельности. В
диспергированном растянутом состоянии хромосомы
участвуют в регуляции всех процессов биосинтеза,
протекающих в клетке, но во время клеточного деления
эта их функция прекращается.
При всех формах клеточного деления ДНК
каждой хромосомы реплицируется, так что образуются
две идентичные двойные полинуклеотидные цепи
ДНК (см. разд. 22.4.2). Эти цепи окружаются
белковой «оболочкой» и в начале клеточного деления
имеют вид двух идентичных нитей, лежащих бок о
бок. Каждая нить носит название хроматиды и
соединена со второй нитью неокрашивающимся
участком -центромерой (кинетохором).
22.1. Клеточный цикл
Последовательность событий, происходящих между
образованием данной клетки и ее делением на
дочерние клетки, называют клеточным циклом. Этот
цикл состоит из трех главных стадий.
I. Интерфаза. Период интенсивного синтеза и
ПРОФАЗА
МЕТАФАЗА
АНАФАЗА
Процессы синтез;
Ядерное деление
Деление цитоплазмы
ТЕЛОФАЗА
ДЕЛЕНИЕ
ЦИТОПЛАЗМЫ
Фаза
G1
S
G2
M
С
События, происходящие в клетке
Интенсивные процессы биосинтеза.
Образование митохондрий, хлоропластов (у
растений), эндоплазматического ретику-
лума, лизосом, аппарата Гольджи, вакуолей
и пузырьков. Ядрышко продуцирует рРНК,
мРНК и тРНК; образуются рибосомы;
клетка синтезирует структурные и
функциональные белки. Интенсивный клеточный
метаболизм, контролируемый ферментами.
Рост клетки. Образование веществ,
подавляющих или стимулирующих начало
следующей фазы
Репликация ДНК. Синтез белковых молекул,
называемых гистонами, с которыми
связывается каждая нить ДНК. Каждая
хромосома превращается в две хроматиды.
Интенсивные процессы биосинтеза. Деление
митохондрий и хлоропластов. Увеличение
энергетических запасов. Репликация цент-
риолей (в тех клетках, где они имеются) и
начало образования веретена деления
Деление ядра, состоящее из четырех стадий
Равномерное распределение органелл и
цитоплазмы между дочерними клетками
Рис. 22.1. Клеточный цикл
роста. В клетке синтезируется много веществ,
необходимых для ее роста и осуществления всех
свойственных ей функций. Во время интерфазы
происходит репликация ДНК.
2. Митоз (кариокинез). Это процесс деления ядра,
при котором хроматиды отделяются одна от другой
и перераспределяются в виде хромосом между
дочерними клетками.
3. Цитокинез - процесс разделения цитоплазмы
между двумя дочерними клетками1.
1 Обычно под названием «митоз» цитологи
объединяют стадии 2 и 3-деление ядра (кариокинез) и
цитоплазмы (цитокинез).- Прим. ред.
Весь цикл показан на рис. 22.1.
Продолжительность клеточного цикла зависит от
типа клетки и от внешних факторов, таких как
температура, питательные вещества и кислород.
Бактериальные клетки могут делиться каждые
20 мин, клетки кишечного эпителия - каждые 8-10 ч,
клетки в кончике корня лука - каждые 20 ч, а многие
клетки нервной системы не делятся никогда.
Опыт 22.1. Изучение фаз митоза
Обычно хромосомы можно наблюдать только во
время деления ядра. Подходящим материалом для
этого служит апикальная меристема кончиков корня
чеснока (2п = 16), лука (2п = 16) и конских бобов
(2п = 12). Этот материал помещают в такие
условия, чтобы началось развитие корешков, кончики
корешков срезают, фиксируют, окрашивают и ма-
церируют, после чего хромосомы можно изучать
под микроскопом.
Материалы
Булавки
Пробирка с водой
Скальпель
Маленькие пробирки с пробками
Пинцет
Две чашки Петри
Водяная баня и пробирки
Предметное стекло
Покровное стекло
Пара тонких иголок
Несколько листков фильтровальной бумаги
Зубчик чеснока
Дистиллированная вода
Уксусная кислота
Одномолярный раствор соляной кислоты
Реактив Фёльгена
Метод
1. Проткните зубчик чеснока булавкой и подвесьте
его вверху пробирки с водой так, чтобы
основание зубчика находилось в воде. Оставьте на 3-4
дня в покое, так как любое постороннее
воздействие может временно подавить клеточное
деление.
2. После образования нескольких корешков
длиной 1-2 см отрежьте от них концевые участки
длиной 1 см.
3. Поместите отрезанные участки корешков в
небольшую пробирку с уксусной кислотой,
заткните ее пробкой и оставьте на ночь при комнатной
температуре для фиксации.
4. Ухватив корешки пинцетом за верхний конец,
перенесите их в чашку Петри с
дистиллированной водой и отмывайте в течение нескольких
минут для удаления фиксатора.
5. Перенесите кончики корешков в пробирку,
содержащую одномолярный раствор соляной
кислоты, и выдержите 3 мин при 600C (для
корешков лука, горошка или бобов-6-10 мин).
При этом срединные пластинки, удерживающие
клетки вместе, разрушаются, а ДНК хромосом
гидролизуется с образованием альдегидных
форм дезоксирибозы, способных
взаимодействовать с красителем (реактивом Фёльгена).
6. Кислоту вместе с кончиками корешков вылейте
в чашку Петри. Перенесите корешки в другую
чашку Петри, содержащую дистиллированную
воду, и отмойте кислоту. Оставьте на 5 мин.
7. Перенесите корешки в маленькую пробирку с
реактивом Фёльгена и заткните ее пробкой.
Поставьте в прохладное темное место (лучше в
холодильник) минимум на 2 ч.
8. Выньте один кончик и поместите его в капле
уксусной кислоты на чистое предметное стекло.
9. Отрежьте концевой участок длиной 1-2 мм и
отбросьте остальное.
10. Растреплите кончик корешка с помощью двух
тонких иголок и накройте покровным стеклом.
Поместите препарат на плоскую поверхность,
накройте несколькими листками
фильтровальной бумаги и сильно нажмите через нее на
покровное стекло подушечкой большого пальца
Не допускайте смещения покровного стекла в
стороны.
11. Изучите препарат под микроскопом при малом
и большом увеличении и найдите клетки,
находящиеся на разных стадиях митоза.
12. Зарисуйте ядра в разных фазах митоза и
надпишите рисунки.
22.2. м итоз
События, происходящие в ядре во время митоза,
обычно наблюдают на фиксированных и
окрашенных клетках (см. Приложение 2.4.2). Такие
препараты позволяют увидеть фазы, через которые
проходят хромосомы при клеточном делении, но не
выявляют их последовательность. Методы фазово-
контрастной микроскопии и цейтраферной съемки
дали возможность наблюдать, как происходит
деление ядра в живой клетке. При быстром
прокручивании пленки митоз предстает как непрерывный
процесс, включающий четыре стадии. Изменения,
происходящие на этих стадиях в животной клетке,
показаны на рис. 22.2.
22.2.1. Центриоли и образование
веретена
Центриоли - это органеллы, расположенные в
цитоплазме около ядерной оболочки; они имеются в
животных клетках и в клетках низших растений. Две
центриоли, образующие пару, лежат
перпендикулярно друг другу.
Каждая центриоль имеет примерно 500 нм в
длину и 200 нм в диаметре; она состоит из девяти
групп микротрубочек, по три в каждой группе.
Полагают, что соседние тройки микротрубочек
соединены между собой фибриллами (рис. 22.3).
Структуры типа центриолей имеются также у
оснований ресничек и жгутиков; в этом случае их
называют базальными тельцами (см. разд. 17.6.2).
Нити веретена имеют трубчатую форму и
диаметр около 25 нм. Они образуются во время митоза
и мейоза и построены из микротрубочек, состоящих
из тубулина и других белков. Прежде считалось, что
центриоли играют роль организаторов нитей
веретена, но теперь от этого мнения отказались: в
большинстве растительных клеток центриолей нет,
однако у них образуются нити веретена, состоящие
из таких же микротрубочек, как и в животных
клетках. Некоторые нити веретена идут от одного
полюса к другому, тогда как другие образуют
пучки, прикрепленные к центромерам хромосом. Как
полагают, расхождение дочерних хромосом в
анафазе митоза обусловлено движением нитей веретена
относительно друг друга. Как показывают
результаты электронно-микроскопических исследований,
между нитями двух типов имеются поперечные
мостики; это наводит на мысль, что относительное
смещение нитей сходно по своему механизму со
скольжением миофиламентов в мышечных волокнах
(см. разд. 17.4.6).
Добавление колхицина (см. разд. 23.9.1) к активно
делящимся клеткам подавляет образование
веретена, так что пары хроматид остаются там, где они
находились в метафазе. Этот метод позволяет
производить подсчет хромосом и изучать их структуру
под микроскопом.
22.2.2. Деление цитоплазмы
Деление цитоплазмы называют цитокинезом. Оно
обычно следует за телофазой и ведет к периоду G1
интерфазы. При подготовке к делению клеточные
органеллы вместе с хромосомами равномерно
распределяются по двум полюсам телофазной клетки.
В животных клетках плазматическая мембрана во
время телофазы начинает впячиваться внутрь на
том уровне, где прежде располагался экватор
веретена. Как полагают, это происходит под
действием находящихся здесь микрофиламентов. В
результате этого впячивания образуется непрерывная
борозда, опоясывающая клетку по экватору. В конце
концов клеточные мембраны в области борозды
смыкаются, полностью разделяя две клетки.
В растительных клетках нити веретена во время
телофазы начинают исчезать, сохраняясь лишь в
области экваториальной пластинки. Здесь они
сдвигаются к периферии клетки, число их увеличивается,
и они образуют боченковидное тельце - фрагмо-
пласт. В эту область перемещаются также
микротрубочки, рибосомы, митохондрии, эндоплазмати-
Интерфаза
Ее часто неправильно называют стадией покоя.
Продолжительность интерфазы различно и зависит от функции
данной клетки. Это период, во время которого клетка
обычно синтезирует органеллы и увеличивается в
размерах. Ядрышки хорошо видны и активно синтезируют
рибосомный материал. Непосредственно перед
клеточным делением ДНК и гистоны каждой хромосомы
реплицируются. Каждая хромосома представлена теперь парой
хроматид, соединенных друг с другом центромерой.
Вещество хромосом окрашивается и носит название
хроматина, но сами эти структуры увидеть трудно.
Профаза
Обычно самая продолжительная фаза клеточного
деления. Хроматиды укорачиваются (до 4% своей
первоначальной длины) и утолщаются в результате их спирали -
зации и конденсации При окрашивании хроматиды четко
видны, но центромеры не выявляются В разных парах
хроматид центромера располагается по-разному. В
животных клетках и у низших растений центриоли расходятся к
противоположным полюсам клетки. От каждой центриоли в
виде лучей расходятся короткие микротрубочки,
образующие в совокупности звезду. Ядрышки уменьшаются, так как
их нуклеиновая кислота частично переходит в
определенные пары хроматид. К концу профазы ядерная
мембрана распадается и образуется веретено деления.
Метафаза
Пары хроматид прикрепляются своими центромерами к
нитям веретена (микротрубочкам) и перемещаются вверх и
вниз по веретену до тех пор, пока их центромеры не
выстроятся по экватору веретена перпендикулярно его оси.
Анафаза
Это очень короткая стадия. Каждая центромера
расщепляется на две, и нити веретена оттягивают дочерние
центромеры к противоположным полюсам. Центромеры
тянут за собой отделившиеся одна от другой
хроматиды, которые теперь называются хромосомами.
Твлофаза
Хромосомы достигают полюсов клетки, деспирализуются,
удлиняются, и их уже нельзя четко различить. Нити
веретена разрушаются, а центриоли реплицируются. Вокруг
хромосом на каждом из полюсов образуется ядерная
оболочка. Вновь появляется ядрышко. За телофазой
может сразу следовать цитокинез (разделение всей клетки на
две).
Рис. 22.2. Схемы и краткие описания
последовательных стадий митоза в животной клетке.
Ядерная оболочка
Нити хроматина
Ядрышко
Центриоли
Цитоплазма
Клеточная мембрана
Интерфаза
Звезда
Центриоль
Ядерная оболочка
Ядрышко
Пара хроматид
Центромера
Профаза
- Нити веретена
-Центромера на
экваторе веретена
Метафаза
Дочерние хромосомы
растаскиваются в
стороны своими
центромерами
Анафаза
Пара центриолей
Ядрышко
Нити хроматина
Ядерная оболочка
Телофаза
Рис. 22.2. (продолжение). Микрофотографии и основанные на них схемы, изображающие четыре
стадии митоза в растительных клетках на примере клеток кончика корня Crocus balansae (2n = 6).
(Любезно предоставлены д-ром S. A. Henderson, Dep. of Genetics, Univ. of Cambridge.) На этих давленых
препаратах хорошо видны только хромосомы, которые были окрашены. Ядрышки, веретена и
цитоплазма не окрашены и не видны. В начале деления (1) хромосомы длинные, тонкие, но уже явно двойные. Они
спирализуются и укорачиваются. После полного укорочения хромосом ядерная мембрана
разрушается и хромосомы выстраиваются на экваторе веретена (2, вид с полюса). Спустя
некоторое время центромеры (изображены на схемах в виде точек) под действием нитей веретена отделяют
сестринские хроматиды друг от друга (3). Нити веретена растаскивают центромеры,
а вместе с ними и хроматиды к полюсам (4), где образуются две идентичные группы
хромосом. После разделения цитоплазмы из них формируются ядра дочерних клеток.
Триплеты микротрубочек
Соединительные
фибриллы
Радиальные фибриллы
("спицы")
• 200 нм-
Рис 22.3. А. Электронная
микрофотография поперечного среза
центриоли из клетки
поджелудочной железы куриного эмбриона.
Б. Схематический рисунок,
сделанный на основе микрофотографии.
ческий ретикулум и аппарат Гольджи; последний
образует множество мелких пузырьков,
наполненных жидкостью. Пузырьки появляются сначала в
центре клетки, а затем, направляемые
микротрубочками, перемещаются и сливаются друг с другом,
образуя клеточную пластинку, расположенную в
экваториальной плоскости (см. рис. 7.23).
Содержимое пузырьков идет на построение срединной
пластинки и стенок дочерних клеток, а из их мембран
образуются новые клеточные мембраны. Клеточная
пластинка, разрастаясь, в конце концов сливается со
стенкой родительской клетки и полностью разделяет
две дочерние клетки. Новообразованные клеточные
стенки называют первичными; в дальнейшем они
могут дополнительно утолщаться за счет отложения
целлюлозы и других веществ, таких как лигнин и
суберин, образуя вторичную клеточную стенку (рис.
22.4). В определенных участках клетки пузырьки
клеточной пластинки не сливаются, так что между
цитоплазмами дочерних клеток сохраняется контакт.
Эти цитоплазматические мостики покрыты
клеточной (плазматической) мембраной и образуют
структуры, называемые плазмодесмами.
22.2.3. Митоз в животных
и растительных клетках
Самое важное событие, происходящее во время
митоза,-это равное распределение удвоившихся
хромосом между двумя дочерними клетками.
Митоз протекает в животных и растительных клетках
почти одинаково, но имеется и ряд различий
(табл. 22.1).
У разных организмов и в разных тканях митоти-
ческое деление клеток протекает с различной
скоростью-с наибольшей у бактерий и у зародышей
многоклеточных организмов и с наименьшей в вы-
сокодифференцированных тканях. Очень быстро
могут делиться изолированные растительные и
животные клетки при росте на питательных средах в
условиях, оптимальных для деления. Клеточную
популяцию, полученную от одной родительской
клетки, называют клоном (см. разд. 20.1.1). Клетки,
входящие в состав данного клона, не обязательно
должны быть идентичны по строению и функции.
Отдельные клетки, взятые из какого-либо
организма, могут дать начало новой особи или новой ткани,
идентичной той, из которой они были выделены;
например, одна клетка, взятая из легкого, может
дать начало легочной ткани с ее альвеолами и
бронхиолами.
22.2.4. Значение митоза
генетическая стабильность. В результате митоза
получаются два ядра, содержащие каждое столько же
хромосом, сколько их было в родительском ядре.
Эти хромосомы происходят от родительских
хромосом путем точной репликации ДНК, поэтому
гены их содержат совершенно одинаковую
наследственную информацию. Дочерние клетки генетически
идентичны родительской клетке, так что никаких
Таблица 22.1. Особенности митоза у растений и у животных
Растительная клетка
Центриолей нет
Звезды не образуются
Образуется клеточная
пластинка
При цитокинезе не
образуется борозды
(перетяжки)
Митозы происходят
главным образом в
меристемах
Животная клетка
Центриоли имеются
Звезды образуются
Клеточная пластинка не
образуется
При цитокинезе образуется
борозда
Митозы происходят в
различных тканях и участках
организма
Дочерняя клетка
Срединная пластинка
Плазмодесма
Первичные клеточные
стенки
Вторичная клеточная
стенка родительской
клетки
.Первичная клеточная
стенка родительской
клетки
.Межклеточное
пространство
-Срединная пластинка
Рис. 22.4. Строение клеточной стенки,
образовавшейся в результате деления родительской
растительной клетки.
Клеточная.
мембрана
Дочерняя клетка
изменений в генетическую информацию митоз
внести не может. Поэтому клеточные популяции
(клоны), происходящие от родительских клеток,
обладают генетической стабильностью.
рост. В результате митозов число клеток в
организме увеличивается (процесс, известный под
названием гиперплазии), что представляет собой один из
главных механизмов роста (см. гл. 21).
БЕСПОЛОЕ РАЗМНОЖЕНИЕ, РЕГЕНЕРАЦИЯ И ЗАМЕЩЕНИЕ
КЛЕТОК. Многие виды животных и растений
размножаются бесполым путем при помощи одного лишь
митотического деления клеток. Способы
вегетативного размножения описаны более подробно в разд.
20.1.1. Кроме того, митоз обеспечивает регенерацию
утраченных частей (например, ног у ракообразных)
и замещение клеток, происходящее в той или иной
степени у всех многоклеточных организмов.
22.3. Мейоз
Мейоз (от греч. meiosis-уменьшение)-форма
ядерного деления, сопровождающаяся уменьшением
числа хромосом с диплоидного (2п) до гаплоидного
(л). Не вдаваясь в подробности, можно сказать, что
при этом в родительской клетке происходит
однократное удвоение хромосом (репликация ДНК, как
при митозе), за которым следуют два цикла
клеточных и ядерных делений (первое деление мейоза и
второе деление мейоза). Таким образом, одна
диплоидная клетка дает начало четырем гаплоидным
клеткам, как это схематически показано на рис. 22.5.
Мейоз происходит при образовании спермиев и
яйцеклеток (гаметогенез) у животных (см. разд. 20.3.1
и 20.3.2) и при образовании спор у большинства
растений (у тех, у которых имеет место чередование
поколений; см. разд. 20.2.2). У некоторых низших
растений чередования поколений нет, и мейоз у них
происходит при образовании гамет. Стадии мейоза
удобно наблюдать на ядрах сперматоцитов из
мужских гонад прямокрылых или на ядрах незрелых
пыльцевых мешков крокуса.
Подобно митозу, мейоз-процесс непрерывный,
но его тоже можно подразделить на профазу, мета-
фазу, анафазу и телофазу. Эти стадии имеются в
первом делении мейоза и еще раз повторяются во
втором. Поведение хромосом во время этих стадий
представлено на рис. 22.6, где показано деление
ядра, содержащего четыре хромосомы (2п = 4), т. е.
два пары гомологичных хромосом.
22.3.1. Значение мейоза
половое размножение. У организмов,
размножающихся половым путем, в результате мейоза
образуются четыре дочерние клетки, каждая из которых
содержит половинное число хромосом по
сравнению с родительской клеткой. При оплодотворении
ядра двух половых клеток (гамет) сливаются,
образуя зиготу, которая содержит постоянное для
Диплоидная родительская клетка ( с одной
парой гомологичных
хромосом)
От матери
От отца
Каждая хромосома удваивается (синтез ДНК), образуя
две пары хроматид
'Первое деление мейоза
(Мейоз I)
Второе деление мейозау
(Мейоз
Гаплоидные гаметы
Рис. 22.5. Схема основных этапов мейоза
(дупликация одной хромосомы и два последующие ядерные и
клеточные деления).
каждого вида число хромосом. У всех организмов
это число соответствует диплоидному (2л)
состоянию (исключение из этого правила составляет поли-
плоидизация, см. разд. 23.9). Если бы не было
мейоза, слияние гамет приводило бы к удвоению
числа хромосом в каждом новом поколении,
полученном половым способом. У всех организмов с
половым размножением это предотвращается
благодаря происходящему на какой-либо стадии
жизненного цикла особому клеточному делению, при
котором диплоидное число хромосом (2п)
уменьшается до гаплоидного (п).
22.1. На рис. 22.10 показаны изменения
количества ДНК в клетке при нескольких делениях
ядра.
а) Какой тип деления представлен на рис. 22.10?
б) Каким стадиям соответствуют прерывистые
линии W, X и Y?
в) Какой тип клеток представлен линией Z?
генетическая изменчивость. Мейоз создает также
возможности для возникновения в гаметах новых
генных комбинаций. Это ведет к изменениям в
генотипе и фенотипе потомства, получаемого в
результате слияния гамет. Механизмы мейоза,
участвующие в создании этой изменчивости, сводятся
к следующему:
1. Уменьшение числа хромосом от диплоидного до
гаплоидного сопровождается расхождением (раз-
Интерфаза
Продолжительность различна у разных видов.
Происходит репликация органелл, и клетка увеличивается в
размерах. Репликация ДНК и гистонов в основном
заканчивается в премейотической интерфазе, но частично
захватывает и начало профазы I. Каждая хромосома
представлена теперь парой хроматид, соединенных
центромерой. Хромосомный материал окрашивается, но из всех
структур четко выявляются только ядрышки (ср. с рис. 22.2,
где показан митоз).
Профаза I
Самая продолжительная фаза. Ее часто делят но пять
стадий (лептотена, знготена, пахитена, диплотена и
диакинез), но здесь она будет рассматриваться как
непрерывная последовательность изменений хромосом.
А. Хромосомы укорачиваются и становятся видимыми как
обособленные структуры. У некоторых организмов они
выглядят как нитки бус: участки интенсивно
окрашивающегося материала - хромомеры-чередуются у них с
неокрашивающимися участками. Хромомеры-это те места,
где хромосомный материал сильно спирализован.
Б. Гомологичные хромосомы, происходящие из ядер
материнской и отцовской гамет, приближаются одна к другой
и конъюгируют. Эти хромосомы одинаковой длины, их
центромеры занимают одинаковое положение, и они
обычно содержат одинаковое число генов,
расположенных в одной и той же линейной
последовательности. Хромомеры гомологичных хромосом лежат бок о бок.
Процесс конъюгации называют также синапсисом он
может начинаться в нескольких точках хромосом, которые
затем соединяются по всей длине (как бы
застегиваясь на «молнию»). Пары конъюгировавших гомологичных
хромосом часто называют бивалентами. Биваленты
укорачиваются и утолщаются. При этом происходит как более
плотная упаковка на молекулярном уровне, так и внешне
заметное закручивание (спирализация). Теперь каждая
хромосома с ее центромерой ясно видна.
В. Гомологичные хромосомы, составляющие бивалент,
частично разделяются, как будто отталкиваясь друг
от друга. Теперь видно, что каждая хромосома состоит
из двух хром эти д. Хромосомы все еще соединены между
собой в нескольких точках. Эти точки называют хиазмами
(от греч chiasma-перекрест). В каждой хиазме происходит
обмен участками хроматид в результате разрывов и
воссоединений, в которых участвуют две из четырех
имеющихся в каждой хиазме нитей. В результате гены из
одной хромосомы (например, отцовской-А, В, С)
оказываются связанными с генами из другой хромосомы
(материнской а, Ь, с), что приводит к новым генным комбинациям в
образующихся хроматидах. Этот процесс называют
кроссинговером. Гомологичные хромосомы после кроссин-
говера не расходятся, так как сестринские хроматиды
(обеих хромосом) остаются прочно связанными вплоть до
анафазы.
Рис. 22.6. Схемы и краткие описания
последовательных стадий мейоза в животной клетке.
Ядерная оболочка
Хромомеры
- Ядрышко
А. Ранняя профаза 1
Пара гомологичных хромосом
— бивалент
Центромеры
Бивалент
Б. Профаза I
Пара сестринских
хроматид
Хиазма
Пара сестринских
хроматид
В. Кроссинговер во время профазы I
Г. Хроматиды гомологичных хромосом продолжают
отталкиваться друг от друга, и биваленты
приобретают определенную конфигурацию в зависимости от числа
хиазм. Биваленты с одной хиазмой имеют
крестообразную форму, с двумя хиазмами- кольцевидную, а с тремя и
более-образуют петли, лежащие перпендикулярно друг
другу. К концу профазы I все хромосомы полностью
уплотнены и интенсивно окрашиваются. В клетке
происходят и другие изменения миграция центриолей (если они
имеются) к полюсам, разрушение ядрышек и ядерной
мембраны, а затем образование нитей веретена.
Поворот
Г (I). Бивалент с одной хиазмой
Хиазмы
Мвтафаза I
Биваленты выстраиваются в экваториальной плоскости,
образуя метафазную пластинку. Их центромеры ведут себя
как единые структуры (хотя часто выглядят
двойными) и организуют прикрепленные к ним нити веретена,
каждая из которых направлена только к одному из полюсов.
В результате слабого тянущего усилия этих нитей каждый
бивалент располагается в области экватора, причем обе
его центромеры оказываются на одинаковом
расстоянии от него - одна снизу, а другая сверху.
Г (II) Бивалент с двойной хиазмой. В результате поворота
плеч хромосом образуется кольцевая структура
Нити веретена
Клеточная мембрана
Д. Поздняя метафаза I
Рис. 22.6. (продолжение)
Анафаза I
Имеющиеся у каждого бивалента две центромеры еще не
делятся, но сестринские хроматиды уже не
примыкают одна к другой. Нити веретена тянут центромеры,
каждая из которых связана с двумя хроматидами, к
противоположным полюсам веретена. В результате
хромосомы разделяются на два гаплоидных набора, попадающих в
дочерние клетки.
E. Анафаза I
Телофаза I
Расхождение гомологичных центромер и связанных с ними
хроматид к противоположным полюсам означает
завершение первого деления мейоза. Число хромосом в
одном наборе стало вдвое меньше, но находящиеся на
каждом полюсе хромосомы состоят из двух хроматид.
Вследствие кроссинговера при образовании хиазм эти
хроматиды генетически неидентичны, и при втором
делении мейоза им предстоит разойтись. Веретена и их
нити обычно исчезают.
У животных и у некоторых растений хроматиды деспи-
рализуются, вокруг них на каждом полюсе формируется
ядерная мембрана и образовавшееся ядро вступает в
интерфазу. Затем начинается деление цитоплазмы (у
животных) или формирование разделяющей клеточной стенки
(у растений), как при митозе. У многих растений не
наблюдается ни телофазы, ни образования
клеточной стенки, ни интерфазы, и клетка прямо переходит из
анафазы I в профазу II.
Интерфаза Il
Эта стадия обычно наблюдается только в
животных клетках, продолжительность ее варьирует. Фаза S
отсутствует, и дальнейшей репликации ДНК не происходит.
Процессы, участвующие во втором делении мейоза, по
своему механизму сходны с происходящими в митозе. Они
включают разделение хроматид в обеих дочерних
клетках, получившихся в результате первого деления мейоза.
Второе деление мейоза отличается от митоза главным
образом двумя особенностями: 1) в метафазе Il
мейоза сестринские хроматиды часто сильно обособляются
друг от друга, 2) число хромосом гаплоидное.
Профаза Il
В клетках, у которых выпадает интерфаза II, эта стадия
тоже отсутствует. Продолжительность профазы Il
обратно пропорциональна продолжительности телофазы I.
Ядрышки и ядерные мембраны разрушаются, а
хроматиды укорачиваются и утолщаются. Центриоли, если они
есть, перемещаются к противоположным полюсам
клеток; появляются нити веретена. Хроматиды
располагаются таким образом, что их длинные оси перпендикулярны
оси веретена первого деления мейоза.
Рис. 22.6 (продолжение)
Ж. Телофаза I в животной клетке
3. Профаза II
Метафаза Il
При втором делении центромеры ведут себя как
двойные структуры. Они организуют нити веретена,
направленные к обоим полюсам, и таким образом выстраиваются по
экватору веретена.
И. Метафаза II
К. Анафаза II
Л. Стадии, следующие за телофазой II ,
разделение цитоплазмы (в случае животной
Анафаза Il
Центромеры делятся, и нити веретена растаскивают их к
противоположным полюсам. Центромеры тянут за
собой отделившиеся друг от друга хроматиды, которые
теперь называются хромосомами.
Телофаза Il
Эта стадия очень сходна с телофазой митоза.
Хромосомы деспирализуются, растягиваются и после этого
плохо различимы. Нити веретена исчезают, а центриоли
реплицируются. Вокруг каждого ядра, которое
содержит теперь половинное (гаплоидное) число хромосом
исходной родительской клетки, вновь образуется
ядерная мембрана. В результате последующего деления
цитоплазмы (у животных) или образования клеточной
стенки (у растений) из одной исходной
родительской клетки получается четыре дочерних клетки.
Рис. 22.6 (продолжение)
делением) аллелей, так что каждая гамета несет
только один аллель по данному локусу (см.
разд. 23.2).
2. Расположение бивалентов в экваториальной
пластинке веретена в метафазе I и хромосом в мета-
фазе II определяется случайным образом.
Последующее их разделение в анафазах I и II
соответственно создает новые комбинации аллелей в
гаметах. Этот процесс, называемый независимым
распределением, приводит к случайному
распределению материнских и отцовских хромосом
между дочерними ядрами. Он лежит в основе
второго закона Менделя (разд. 23.1.3).
3. В результате образования хиазм между гомо-
Рис. 22.7. А. Восемь стадий
мейоза на примере сперматоцитов
пустынной саранчи (Schisto-
cerca gregaria, 2п = 22, +Xy самцов).
Как и на рис 22.2.
микрофотографии сделаны с давленых
препаратов, на которых видны только
окрашенные хромосомы. Ядрышки,
веретена и цитоплазма не
окрашены, и детали их строения не
видны. В начале первого деления
мейоза (1) хромосомы длинные,
тонкие, как бы растрепанные и
плохо различимы по
отдельности, кроме интенсивно
окрашивающейся Х-хромосомы. В
результате конъюгации гомологичных
(материнской и отцовской)
хромосом образуется набор из
гаплоидного числа (11) бивалентов и
одной Х-хромосомы (2, 3). Между
тесно соприкасающимися
материнской и отцовской хромосомами
происходит обмен генетическим
материалом. Затем они
отделяются одна от другой по всей своей
длине, за исключением тех точек,
где произошли обмены (4). Эти
точки носят название хиазм
Рис. 22.7 (продолжение). Б. В
результате дальнейшего
уплотнения образуются короткие толстые
биваленты, которые после
разрушения ядерной мембраны
прикрепляются к веретену своими
центромерами (5; вид с полюса). Нити
веретена тянут хромосомы,
составляющие биваленты, в разные
стороны (S; вид сбоку).
X-хромосома, которая теперь
окрашивается слабо, направляется, не
разделившись, к одному полюсу.
При втором делении мейоза
хромосомы (в гаплоидном числе)
располагаются в экваториальной
плоскости (7; вид с полюса) и в
результате активности нитей
веретена хроматиды каждой
хромосомы расходятся к разным
полюсам (8; вид сбоку).
(Микрофотографии и схемы любезно
предоставлены д-ром S A. Henderson,
Dер of Genetics, Univ. of Cambridge.)
Рис. 22.8. Хромосомы из одной клетки саранчи Locusta migrator/a на стадии диплотены (профаза I). Видны
биваленты с одной или двумя хиазмами. На схеме слева материнские и отцовские
хромосомы изображены соответственно сплошными и прерывистыми линиями. В каждой хиазме
произошел обмен генетическим материалом. Форма бивалентов варьирует от палочковидной до
крестообразной или кольцевидной, в зависимости от числа хиазм и их расположения.
Рис. 22.9. Мейоз в живых клетках.
Конъюгация и клеточное деление в
живых сперматоцитах саранчи
Locusta migratoria. Препараты
сфотографированы методом,
известным под названием
интерференционного контраста Hо
морского; этот метод с
использованием поляризованного света
позволяет получать удивительно
объемные картины живых неокрашенных
клеток. В двух клетках можно
видеть конъюгацию хромосом в
ранней профазе I с совмещением
хром о мер (стрелка). Две другие
клетки заканчивают первое
деление мейоза. После того как
образовались две полярные группы,
начинается деление всей клетки с
образованием двух дочерних клеток
примерно одинаковой величины.
Нитевидные структуры, тянущиеся
от клетки к клетке между двумя
полярными группами и
перехваченные посередине подобно снопу
пшеницы,-это митохондрии.
Содержание
ДНК на одну
клетку
Время
Рис. 22.10. График, который следует использовать
при выполнении задания 22.1.
логичными хромосомами в профазе I часто
происходит кроссинговер, ведущий к возникновению
новых комбинаций аллелей в хромосомах
половых клеток. При этом распадаются
существовавшие ранее группы сцепления и возникают
новые.
Значение этих трех механизмов в процессах
наследования и в создании изменчивости будет
подробно описано в разд. 23.8.4.
22.3.2. Сходства и различия между
митозом и мейозом
Главные черты сходства между митозом и мейозом
касаются механизмов, с помощью которых
хромосомы и другие клеточные органеллы
реплицируются и перемещаются в клетке перед ее делением
и во время самого деления. Механизмы цитокинеза
при митозе и мейозе тоже сходны.
Различия между этими двумя процессами
перечислены в табл. 22.2.
Особенности, связанные с полом, и различия между
животными и растениями. Приведенное выше
описание мейоза в общем приложимо ко всем животным
и растениям обоего пола, но между ними
существуют также некоторые важные различия.
А. Мужской пол. У животных разделение клетки
происходит в конце как первого, так и второго
делений мейоза, дочерние клетки тотчас же теряют
связь друг с другом и становятся независимыми. Все
четыре продукта мейоза выживают, превращаются
в сперматиды, а затем в спермин. У растений
происходит формирование клеточных стенок, и
дочерние клетки остаются связанными, образуя к
концу первого деления мейоза диаду, а к концу
второго - тетраду. Все продукты мейоза тоже выживают и
превращаются в пыльцевые зерна.
Б. Женский пол. Как у растений, так и у животных
из четырех продуктов мейоза выживает только
один, образующий ядро яйцеклетки. У животных
первое деление мейоза асимметричное - оно
приводит к образованию ооцита второго порядка и
полярного тельца. Второе деление мейоза тоже
асимметричное: ооцит второго порядка делится на
яйцеклетку и второе полярное тельце (первое полярное
тельце также может делиться на два, но все
полярные тельца дегенерируют); таким образом, в
результате мейоза образуется только один
функциональный ооцит. У растений мейоз приводит к
образованию четырех ядер, заключенных в зародышевом
мешке. Три из них дегенерируют, а четвертое дает
начало ядрам зародышевого мешка и ядру
яйцеклетки.
Таблица 22.2. Различия между стадиями митоза и мейоза
Стадия
Профаза
Метафаза
Анафаза
Телофаза
Где
происходит деление
данного
типа
Митоз
Хромомеры не видны
Гомологичные хромосомы обособлены
Хиазмы не образуются
Кроссинговера не происходит
Пары хроматид располагаются на экваторе
веретена
Центромеры выстраиваются в одной плоскости
на экваторе веретена
Центромеры делятся
Хроматиды расходятся
Расходящиеся хроматиды идентичны
Число хромосом в дочерних клетках то же, что и
в родительских клетках
Дочерние клетки содержат обе гомологичные
хромосомы (у диплоидов)
Возможно в гаплоидных, диплоидных и
полиплоидных клетках
Происходит при образовании соматических
клеток и некоторых спор, а также при образовании
гамет у растений, у которых имеет место
чередование поколений
Мейоз
Хромомеры видны
Гомологичные хромосомы конъюгируют
Хиазмы образуются
Кроссинговер может иметь место
Пары хроматид располагаются на экваторе
веретена только во втором делении мейоза
Центромеры в первом делении мейоза
располагаются над и под экватором на одинаковых
расстояниях от него
Центромеры делятся только во втором делении
мейоза
Хроматиды расходятся во втором делении мейоза.
В первом делении расходятся целые хромосомы
Расходящиеся хромосомы могут оказаться
неидентичными в результате кроссинговера
Число хромосом в дочерних клетках вдвое меньше,
чем в родительских
Дочерние клетки содержат только по одной из
каждой пары гомологичных хромосом
Только в диплоидных и полиплоидных клетках
При гамето- или спорогенезе
22.4. Структура хромосом
Гистохимическое и цитологическое изучение
хромосом эукариотических клеток показало, что они
состоят из дезоксирибонуклеиновой кислоты (ДНК)
и белка с небольшой примесью хромосомной РНК.
«Хромосомы» прокариотических клеток (бактерий и
сине-зеленых водорослей) состоят из одной только
ДНК и, строго говоря, их не следует называть
хромосомами. Молекула ДНК несет отрицательные
заряды, распределенные по всей ее длине, а
присоединенные к ней белки, так называемые гистоны
(относящиеся к основным белкам), заряжены
положительно. Этот комплекс ДНК с белком
называют хроматином1. Белковые молекулы увеличивают
толщину хромосомы и, возможно, служат наружной
защитной оболочкой для ДНК. Структурные
взаимоотношения между ДНК и гистонами в хромосоме
еще не выяснены с полной достоверностью, однако
создается впечатление, что хромосома-это не
просто единичная молекула ДНК, окруженная слоем
гистонов, а нечто гораздо более сложное.
Предложено несколько моделей строения хромосом.
Толщина одной двойной спирали ДНК равна 2
нм, тогда как самые тонкие хромосомы, видимые в
световой микроскоп, имеют толщину от 100 до
200 нм. Это может означать, что одна хромосома
содержит либо много двойных спиралей ДНК, либо
одну двойную спираль, еще дополнительно
закрученную крупными витками. На электронных
микрофотографиях хромосом типа ламповых щеток
(названных так за их сходство со щетками, которыми в
прежние времена чистили стекла керосиновых ламп)
из ооцитов амфибий видно, что каждая хроматида
состоит из плотно скрученной осевой нити с
отходящими от нее боковыми петлями,
образованными одной двойной спиралью ДНК (рис. 22.11).
Эти петли, возможно, представляют собой ДНК,
освобожденную от белков для осуществления
транскрипции (см. разд. 22.6.6). Недавние исследования
структуры хромосом позволяют думать, что
спираль ДНК соединяется с группами из восьми гисто-
новых молекул, образующих нуклеосомы - частицы,
имеющие вид нанизанных на нитку бусинок. Эти
нуклеосомы и соединяющие их участки ДНК плотно
упакованы в виде спирали толщиной 36 нм, на
каждый виток которой приходится примерно 6
нуклеосом и которая по своим размерам и
другим признакам соответствует хромосоме.
Предполагаемая структура такой хромосомы показана
на рис. 22.12.
В хромосомах спермиев у некоторых видов,
например у лосося и сельди, белками, связанными с
ДНК, служат не гистоны, а протамины.
1 Кроме гистонов, в состав хроматина входит и много
других белков.- Прим. ред.
Хромомеры одной
"двойной спирали ДНК
Одна двойная.
спираль ДНК
Хромомера
Несколько закрученных
нитей ДНК
Рис. 22.11. А. Хромосомы типа ламповых щеток из
ооцита амфибии, видны центромера и три хиазмы.
Б и В. Хромосомы растянуты, чтобы
показать центральную нить ДНК и петли ДНК, в которых
происходит синтез мРНК. Как полагают,
уплотненные участки представляют собой хромомеры, а
каждая хромомера и связанная с ней петля
соответствуют определенному генному локусу.
(H. G. Callan, Int. Rev. Cytology, 1963, 151.)
22.4.1. Данные, указывающие на роль
ДНК в наследственности
Еще в начале XX века Саттон и Бовери высказали
верную мысль, что именно хромосомы передают
генетическую информацию от одного поколения
другому (см. разд. 23.2), однако потребовалось еще
много лет, для того чтобы выяснить, что служит
генетическим материалом-ДНК или белок
хромосом. В ряде экспериментов Альфред Мирский
показал, что у особей данного вида все соматические
клетки содержат одинаковое количество ДНК,
которое вдвое больше количества ДНК в гаметах. Но то
же самое относится и к содержанию в хромосомах
белка, так что эти данные мало способствовали
выяснению природы генетического материала.
Ученые склонны были думать, что белок-это
единственное вещество, молекулы которого облада-
ют достаточным структурным разнообразием,
чтобы служить генетическим материалом.
В 1928 г. английский микробиолог Фредерик
Гриффит сделал наблюдение, которое впоследствии
оказалось важным для решения этой проблемы. Во
времена, когда еще не было антибиотиков, Гриффит
пытался приготовить вакцину против
пневмококка-возбудителя одной из форм пневмонии. Были
известны две формы этой бактерии, из которых
одна обладает студенистой капсулой и вирулентна
(вызывает заболевание), а другая не имеет капсулы
и невирулентна. Способность этих бактерий
вызывать пневмонию, по-видимому, была связана с
наличием капсулы. Граффит надеялся, что если
ввести больному бескапсульную или убитую
нагреванием инкапсулированную форму, то его
организм начнет вырабатывать антитела, которые
смогут предохранить от заболевания пневмонией. В
ряде экспериментов Гриффит вводил мышам обе
формы бактерий и получил результаты,
представленные в табл. 22.3. При вскрытии погибших
мышей в них были обнаружены живые
инкапсулированные формы. На основании этих результатов
Гриффит сделал вывод, что от убитых нагреванием
инкапсулированных форм к живым бескапсульным
формам, очевидно, передается какой-то фактор,
заставляющий их вырабатывать капсулы и
становиться вирулентными. Однако природа этого
трансформирующего фактора оставалась неизвестной вплоть
до 1944 г., когда его удалось выделить и
идентифицировать. На протяжении 10 лет Эвери, Мак-Карти
Спираль,
образуемая '
нуклеосомами!
Одна молекула ДНК-
"Сердцевина" из восьми
гистоновых молекул
5,5 нм
-11 нм-
Двойная спираль ДНК
2 нм
Рис. 22.12. Предполагаемая структура нуклеосом и
их соотношение с хромосомой и молекулой ДНК
(в метафазной хромосоме) (E. J. Du Praw, School of
Medicine, Univ. of Maryland.)
Таблица 22.3. Результаты экспериментов Гриффита
В какой форме инъецирован пневмококк
Живой бескапсульный
Живой инкапсулированый
Инкапсулированный, убитый
нагреванием
Инкапсулированный, убитый
нагреванием + живой бескапсульный
Действие на мышей
Выживают
Гибнут
Выживают
Гибнут
и Мак-Леод занимались выделением и очисткой
молекул, входящих в состав убитых нагреванием
инкапсулированных клеток пневмококка, и изучали
их способность трансформировать бескапсульные
клетки. Удаление полисахаридной капсулы и
белковой фракции из клеточных экстрактов не оказывало
влияния на эту способность, но добавление
фермента дезоксирибонуклеазы (ДНКазы), гидролизу-
ющей ДНК, препятствовало трансформации.
Способность высокоочищенных экстрактов ДНК из
инкапсулированных клеток вызывать трансформацию
показала, что фактором Гриффита была ДНК.
Несмотря на эти результаты, многие ученые все еще
отказывались признать, что генетическим
материалом служит ДНК, а не белок. В начале пятидесятых
годов множество неоспоримых данных, полученных
при изучении вирусов, наконец,
продемонстрировали универсальность ДНК как носителя генетической
информации.
В сороковые годы вирусы стали одним из главных
объектов экспериментальных генетических
исследований; проведенные на них эксперименты считаются
теперь такими же классическими, как эксперименты
на горохе, плодовой мушке и, как будет описано
позже, хлебной плесени Neurospora. Вирусные
частицы имеют очень простое строение; они состоят из
оболочки, образованной в основном из белка, и
заключенной внутри нее молекулы нуклеиновой
кислоты ДНК или РНК (см. разд. 2.5.2). Это
делает их идеальным материалом для изучения вопроса
о том, что служит генетическим материалом - белок
или нуклеиновая кислота. В 1952 г. Херши и Чейс
приступили к ряду экспериментов на вирусах
особого типа, заражающих бактериальные клетки и
называемых бактериофагами. Бактериофаг T2
проникает в клетку кишечной палочки (Escherichia coli) и
заставляет ее за очень короткий срок образовать
множество фаговых частиц. Херши и Чейс
выращивали частицы фага T2 в клетках E. coli, которые
росли на среде с радиоактивными изотопами серы
(35S) или фосфора (32P). Фаговые частицы,
образующиеся в клетках E. coli на среде с радиоактивной
серой, включали 35S в свои белковые оболочки, а
частицы, образующиеся в E. coli на среде с
изотопом фосфора, содержали ДНК, меченную 32Р.Такое
избирательное распределение изотопов связано с
тем, что белки не содержат фосфора, а нуклеиновые
кислоты не содержат серы. Мечеными частицами
Заражают E coli,
выращенную на среде с P
ФАГ T2
Белковая оболочка
ДНК
ЗаражаютE coli
выращенную на среде с 35S
Радиоактивность /.
только в ДНК
Радиоактивность
только в белковой
оболочке
ДНК этой
клетки
радиоактивна
Фаговые частицы прикрепляются
к клеточной стенке E coli
Фаговые частицы "впрыскивают"
в клетку свою ДНК
Фаговые частицы удаляют с
E coli механическим путем
Обе бактериальные клетки
продуцируют новые фаговые
частицы
В клетке нет
никаких следов
радиоактивности
Рис. 22.13. Схема экспериментов Херши и Чейза на фаге T2 и E. coli.
фага T2 заражали немеченые клетки E. coli, и спустя
несколько минут эти клетки встряхивали в
смесителе, чтобы отделить фаговые частицы от клеточных
стенок. Затем бактерий инкубировали и проверяли
на радиоактивность. Результаты представлены на
рис. 22.13.
Из полученных данных Херши и Чейз сделали
вывод, что в бактериальную клетку проникает
фаговая ДНК, которая и дает начало многочисленному
фаговому потомству. Эти эксперименты показали,
что наследственным материалом служит ДНК,
которая определяет в новообразованных фаговых
частицах структуру не только фаговой ДНК, но и
фаговых белков. Результаты
электронно-микроскопических исследований и более полные данные о
жизненном цикле вирусов подтверждают, что в
бактериальную клетку проникает только ДНК фага.
(Жизненный цикл вирусных и фаговых частиц
описан в разд. 2.5.3 и 2.5.4.)
22.4.2. Репликация ДНК
Модель структуры ДНК в виде двойной спирали,
предложенная Уотсоном и Криком, описана в
разд. 5.6.3. Одна из самых привлекательных
особенностей этой модели состоит в том, что она
одновременно подсказывает, каким способом могла бы
происходить репликация ДНК. Уотсон и Крик
высказали предположение, что две цепи, образующие
спираль, могут раскручиваться и разделяться, после
чего каждая из них служит матрицей, к которой
путем спаривания оснований пристраивается
комплементарная цепочка нуклеотидов. Таким
образом, из каждой исходной молекулы ДНК
получаются две копии с идентичной структурой.
В 1956 г. Корнбергу удалось продемонстрировать
in vitro синтез молекулы ДНК, используя в качестве
матрицы одиночную цепь ДНК. (Этот метод был
затем использован другими учеными для выяснения
природы генетического кода; см. разд. 22.5.) Корн-
берг выделил из E. coli и очистил фермент, который
способен связывать друг с другом свободные нукле-
отиды в присутствии АТФ как источника энергии с
образованием комплементарной цепи ДНК. Он
назвал этот фермент ДНК-полимеразой. В своих
последующих экспериментах Корнберг использовал
вместо нуклеотидов и АТФ дезоксинуклеозидтрифосфа-
ты (дАТФ, дГТФ, дТТФ, дЦТФ), из которых тоже
строилась комплементарная цепь ДНК.
Дальнейшие данные подтвердили, что именно в этой форме
нуклеотиды легко присоединяются к ДНК-матрице
и друг к другу. Когда нуклеозидтрифосфаты
связываются между собой, две концевые фосфатные
группы отщепляются. Оставшаяся фосфатная группа
нуклеотида и освобождающаяся энергия
используются для образования эфирной связи с углеродными
атомами 5 и 3 остатков сахара соседних
нуклеотидов (рис. 22.14).
Позднее, в 1967 г., Корнберг показал, что ДНК-
полимераза присоединяет нуклеотиды только в
направлении 5 -► 3. Поскольку две цепи ДНК анти-
параллельны, т. е. направление 5 -> 3 у них
противоположно, ДНК-полимераза может непрерывно
строить лишь одну новую цепь молекулы ДНК.
Другая дочерняя молекула ДНК синтезируется
отдельными короткими участками под действием
ДНК-полимеразы, движущейся в противоположном
направлении1. Эти короткие участки новосинтези-
руемой полинуклеотидной цепи связываются
воедино другим ферментом-ДНК-лигазой (рис. 22.15).
Молекулы ДНК, синтезируемые in vitro в
присутствии ДНК-лигазы, биологически активны и могут
использоваться в системе синтеза белка. Молекулы
ДНК, полученные в 1956 г. Корнбергом, хотя они
имели в целом правильную структуру, были
лишены биологической активности, так как участки
одной из синтезируемых цепей не могли соединиться
друг с другом.
Способ репликации ДНК, предложенный Уотсо-
ном и Криком и показанный на рис. 22.16, известен
под названием полуконсервативной репликации, так
как при этом каждая новая двойная спираль
сохраняет одну цепь исходной двойной спирали ДНК.
Этот механизм основан на данных, полученных
Мезелсоном и Сталем в серии классических
экспериментов в 1958 г. Клетки E. coli содержат одну
кольцевую хромосому; при выращивании этих
клеток в течение многих поколений на среде,
содержащей тяжелый изотоп азота (15N), вся их ДНК
метилась этим изотопом. Клетки с меченой ДНК
переносили на среду, содержавшую обычный изотоп
азота 14N. По прошествии срока, соответствующего
времени генерации E. coli (50 мин при 36°С), брали
пробы клеток, выделяли из них ДНК и
центрифугировали 20 ч при 40 000 g в растворе хлористого
цезия (CsCl). При этом тяжелые молекулы CsCl
начинали осаждаться, образуя градиент плотности,
которая возрастала от верхушки к дну пробирки.
ДНК собиралась на том уровне, на котором
плотность раствора CsCl была равна ее плотности. При
исследовании в ультрафиолетовых лучах ДНК в
центрифужной пробирке имела вид узкой полосы.
Расположение полос ДНК, выделенной из клеток,
выращенных на средах с 15N и с 14N, и
интерпретация их структуры представлены на рис. 22.16. Эти
эксперименты убедительно показали, что
репликация ДНК происходит полуконсервативным
способом.
22.2. Были выдвинуты три гипотезы для
объяснения процесса репликации ДНК. Один вариант
получил название полуконсервативного; он
описан выше и представлен на рис. 22.14. Две другие
гипотезы-о консервативной репликации и о дис-
персивной репликации-представлены на рис. 22.17.
Нарисуйте схемы распределения различных
типов ДНК в градиенте плотности, которые обнару-
1 На рис. 22.14 этот факт не отображен: показано,
будто обе цепи реплицируются одной молекулой ДНК-
полимеразы, движущейся в одном направлении. В этом
отношении приведенная на рисунке схема неправильна.-
Прим. ред.
жили бы Мезелсон и Сталь для двух первых
поколений клеток, если бы были верны
консервативная и дисперсивная гипотезы.
22.5. Природа генов
Изучение наследственности уже давно было связано
с представлением о ее корпускулярной природе. В
1866 г. Мендель высказал мнение, что признаки
организмов определяются наследуемыми
единицами, которые он назвал «элементами». Позднее их
стали называть «факторами» и, наконец, генами;
было показано, что гены находятся в хромосомах, с
которыми они и передаются от одного поколения
другому (см. разд. 23.2).
Двойная спираль родительской ДНК
ДНК- полимераза
Свободные
нуклеозид-
три фосфаты
Освобождающийся
фосфат
Новая цепь
Пара идентичных двойных спиралей ДНК
Направление
движения
ДНК-полимеразы
Свободные
нуклеозид-
три фосфаты
Цепь,
служащая
матрицей
Рис. 22.14. Репликация двойной спирали ДНК.
Спираль родительской ДНК раскручивается, и
комплементарные цепи отделяются одна от другой в
результате разрыва слабых водородных связей
между комплементарными основаниями. Полинук-
леотидные цепи ДНК не разрываются благодаря
прочности их фосфодиэфирных связей.
Происходит спаривание между основаниями матричной цепи
и свободными нуклеозидтрифосфатами, и последние
связываются друг с другом, образуя новую
полинуклеотидную цепь. Таким образом получаются
две идентичные молекулы ДНК. В разделении
родительских цепей и образовании новой
полинуклеотидной цепи участвует фермент ДНК-полиме-
раза, но, по всей вероятности, на репликацию
оказывают влияние также и другие ферменты.
Рис. 22.15. Роль ДНК-лигазы в репликации ДНК. Стрелками показаны направления, в которых
перемещается ДНК-полимераза при образовании новых двойных спиралей ДНК. ДНК-лигаза обеспечивает
соединение коротких отрезков нуклеотидной цепи в точках, помеченных звездочками.
Несмотря на все то, что нам теперь известно о
хромосомах, а также о структуре и функциях ДНК,
дать точное определение гена все еще очень трудно.
В результате изучения природы гена пока удалось
сформулировать три возможных определения гена;
мы кратко рассмотрим их, так как они важны для
понимания некоторых вопросов генетики и
эволюции.
ген как единица рекомбинации. На основании своих
работ по построению хромосомных карт
дрозофилы (см. разд. 23.3) Морган постулировал, что
ген-это наименьший участок хромосомы, который
может быть отделен от примыкающих к нему
участков в результате кроссннговера. Согласно этому
определению, ген представляет собой крупную
единицу, специфическую область хромосомы,
определяющую тот или иной признак организма.
ген как единица мутирования. В результате
изучения природы мутаций было установлено, что
изменения признаков возникают вследствие случайных
Клетки,
выросшие
на среде C15N ι
Клетки из пробирки А,
выросшие на среде с 14N
' 1-е
поколение
2-е
поколение
3-е Λ
поколение
Градиент
плотности
хлористого
цезия
ДНК, с 14N
ДНК, с 14N15N
ДНК, с is N
Все цепи
содержат
тяжелую
(15ЮДНК
Все цепи
содержат
гибридную
(14Ν/15Ν)
ДНК
Все цепи
содержат
только ι*ν
14N-цепь
15N-uem>
Рис. 22.16. Результаты экспериментов Мезелсона
и Сталя по изучению репликации ДНК и их
интерпретация. Ширина полос ДНК в центрифужных
пробирках отражает относительные количества
различных типов молекул ДНК, указанных справа. В
пробирке В отношение ширины полос 1:1, а в
пробирке Г-3 1
спонтанных изменении в структуре хромосомы, в
последовательности оснований или даже в одном
основании (разд. 23.9). В этом смысле можно было
сказать, что ген-это всего-навсего одна пара
комплементарных оснований в нуклеотидной
последовательности ДНК, т. е. наименьший участок
хромосомы, способный претерпеть мутацию.
ген как единица функции. Поскольку было извест-
но, что от генов зависят структурные,
физиологические и биохимические признаки организмов, было
предложено определять ген как наименьший участок
хромосомы, обусловливающий синтез определенного
продукта.
Самое приемлемое определение - третье, но в нем
не указано, какого рода продукт синтезируется.
Известны случаи, когда один «ген» воздействует на
несколько признаков (см. разд. 23.1.1), а в других
случаях несколько «генов» (полигены) могут
определять один специфический признак (разд. 23.7.6).
Третье определение получило признание в
результате исследований Бидла и Татума.
Сформулированная ими гипотеза, известная под названием
«один ген - один фермент», нашла много
подтверждений с развитием новой области биологии -
молекулярной генетики. Впоследствии эта гипотеза была
преобразована в функциональную концепцию «один
цистрон-один полипептид» (разд. 22.5.1).
22.5.1. Гены и ферменты
Предположение о связи между генами и
ферментами впервые высказал, хотя и без употребления этих
слов, английский врач Арчибальд Гаррод в 1908 г.
Гаррод постулировал, что некоторые «врожденные
ошибки метаболизма» - результат неспособности
тела вырабатывать определенные химические
вещества, что в свою очередь обусловлено
наследственными механизмами. Прошло почти 40 лет, прежде чем
первые работы по молекулярной генетике
подтвердили эту гипотезу и во всей полноте
продемонстрировали ее значение.
Исследования в области молекулярной генетики
начались с выявления различных веществ и фермен-
Половина
цепей
содержит
легкую
(14Ы)ДНК,
а половина
-
"гибридную*'
Двойные спирали
родительской ДНК
Первое
поколение
дочерних
двойных
спиралей ДНК
Консервативная
репликация
Дисперсивная
репликация
Рис. 22.1 7. Схема, поясняющая две другие гипотезы
о способе репликации ДНК.
тов, участвующих в метаболических процессах.
Некоторые нарушения метаболизма связаны с
дефектами, которые наследуются так, как если бы они
определялись единичными генами. Эти нарушения
возникают спонтанно, как в случае мутаций, или же
передаются по наследству в соответствии с
обычными законами генетики. Рассмотрим, например,
обмен аминокислот фенилаланина и тирозина; обычно
они используются для синтеза клеточных белков и
других веществ, выполняющих структурные и
физиологические функции, а их избыток распадается с
образованием CO2, воды и азотистых отходов
метаболизма. Во всех случаях судьбу этих
аминокислот определяет простой метаболический путь, в
котором участвует несколько ферментов.
Дефектный фермент или отсутствие фермента в одной из
четырех точек на этом пути приводит к таким
нарушениям метаболизма, как фенилкетонурия,
кретинизм, альбинизм и алкаптонурия. Характер
наследования этих четырех заболеваний указывает на
то, что каждое из них контролируется каким-то
одним рецессивным геном.
Дальнейшие важные данные в пользу гипотезы
«один ген - один фермент» были получены в работах
Бидла и Татума, начатых в 1941 г. и посвященных
механизмам наследования ферментов у розовой
хлебной плесени (Neurospora crassa). Подобно
Менделю и Моргану, эти авторы очень тщательно
выбирали объект для своих экспериментов. Нейро-
спора-сумчатый гриб (аскомицет), обладающий
следующими ценными для генетика качествами:
1) его легко выращивать;
2) его можно получать в больших количествах;
3) у него короткое время генерации (10 дней);
4) вегетативная стадия гаплоидная.
Последняя особенность очень важна. Поскольку
на протяжении большей части жизненного цикла
клетки этой плесени содержат только один набор
хромосом, проявление рецессивных генов не
маскируется доминантными аллелями. Если в результате
мутации возникает рецессивный ген, то его действие
сказывается немедленно. Нейроспора обычно
образует бесполые споры (конидии), которые
прорастают и дают начало мицелию. Кусочки мицелиев,
принадлежащих к двум противоположным типам
скрещиваемости, могут сливаться, образуя
диплоидную зиготу. Эта зигота тотчас же проходит мейо-
тическое деление, за которым следует одно митоти-
ческое деление, и образуется сумка (аск) - плодовое
тело, содержащее восемь гаплоидных аскоспор;
четыре из них происходят от одного из родительских
штаммов, а четыре-от другого. Каждая аскоспора,
прорастая, дает начало новому мицелию. Таким
образом, мицелий может происходить либо от
конидии, возникшей бесполым путем, либо от аско-
споры, образовавшейся половым путем (рис. 22.18).
Нейроспора способна расти на культуральной
среде, содержащей только агар, сахара, соли и
витамин биотин. Такую среду называют
минимальной. Если нейроспора может расти на ней, то это
означает, что она способна синтезировать все
необходимые для роста углеводы, жиры,
аминокислоты и витамины с помощью ферментов,
вырабатываемых собственными клетками.
Эксперименты Бидла и Татума состояли в
следующем.
Аскоспора
Рис. 22.18. Аскоспоры в сумке.
Рентгеновские лучи
Конидии
(2) Полная среда
(мутантный штамм)
Минимальная
среда
(отсутствие роста,I
т. е. здесь мутантный
штамм)
[Образцы от
каждого мицелия переносят
на минимальную среду; наличие
роста указывает на
принадлежность к дикому типу, отсутствие ^J
роста — на мутантный штамм
(3) Полная среда
(дикий тип)
Сумка
Аскоспоры
, Полная среда
(все аскоспоры образовали
мицелий)
Минимальная
среда
(наличие роста,
т. е здесь
дикий тип)
Мутантные штаммы переносят наI
20 различных минимальных сред
в каждой из которых только
одна аминокислота
+ Алании
(отсутствие роста)
У этого штамма отсутствует
фермент (или ферменты),
необходимый для синтеза
валина
+ Валин
(наличие роста)
Рис. 22.19. Схема, иллюстрирующая главные этапы в идентификации мутантных аллелей,
которые контролируют синтез ферментов, участвующих в образовании аминокислот. Подробности см. в
тексте (цифры на схеме соответствуют цифрам в тексте).
1. Конидии подвергали рентгеновскому облучению,
чтобы повысить частоту мутаций.
2. Облученные конидии переносили на полную
среду (содержащую все аминокислоты и витамины,
необходимые для нормального роста) и
выращивали на ней.
3. Образовавшиеся мицелии скрещивали с мицелия-
ми, развившимися из конидий, не подвергнутых
рентгеновскому облучению («дикий тип»).
4. Образующиеся аски содержали по четыре
аскоспоры от каждого из родительских штаммов
(мутантного и дикого типа).
5. Аскоспоры извлекали из асков и выращивали по
отдельности на полной питательной среде.
6. Пробы мицелиев помещали на обогащенную
витаминами минимальную среду. В некоторых
случаях роста не наблюдалось.
7. Штаммы, которые не росли, не были способны
синтезировать некоторые аминокислоты. Для
того чтобы определить, какие это были
аминокислоты, эти штаммы переносили на ряд
минимальных сред, к каждой из которых
добавлялась какая-либо одна аминокислота.
8. Если данный штамм рос на определенной среде,
то, следовательно, эта среда содержала ту
аминокислоту, которую данный штамм синтезировать
не способен. Таким образом были
идентифицированы мутантные штаммы нейроспоры. Анализ
полученных результатов показал, что во всех
случаях, в которых половина аскоспор из данного
аска давала мутантный штамм, другая половина
была способна расти на минимальной среде.
Эти результаты указывали на то, что мутантный
ген ведет себя как единичный рецессивный ген и
наследуется в соответствии с законами Менделя.
Бидл и Татум пришли к выводу, что в каждом
случае рентгеновские лучи вызывали мутацию
одного гена, контролирующего синтез одного фермента,
необходимого для синтеза одной аминокислоты.
Это легло в основу их гипотезы «один ген-один
фермент» (рис. 22.19).
В этих ранних исследованиях был разработан
экспериментальный метод с использованием
минимальных сред, применяемый в генетике
микроорганизмов во многих модификациях и давший
огромное количество информации о роли генов.
В настоящее время твердо установлено, что гены
контролируют происходящие в клетке процессы
путем синтеза ферментов и других белков. Эти
ферменты в свою очередь определяют синтез всех
прочих веществ клетки.
С течением времени, однако, определение гена
было видоизменено. Работы по строению генов
бактериофага T4, проведенные Бензером в 1955 г.,
привели к созданию концепции цистрона как
единицы функции. Цистрон-это участок ДНК, несущий
информацию, необходимую для синтеза одной
полипептидной цепи; такая цепь может
функционировать самостоятельно как биологически активная
белковая молекула или становится частью более
крупной макромолекулы. В настоящее время
концепцию «один ген-один фермент» сменила
концепция «один цистрон-один полипептид».
Каждую аскоспору
переносят на полную среду,
где все они растут
22.5.2. Генетический код
Когда Уотсон и Крик в 1953 г. предложили модель
структуры ДНК в виде двойной спирали, они
высказали также предположение, что генетическая
информация, передаваемая из поколения в поколение
и определяющая метаболизм клетки, заключена
в последовательности оснований молекулы ДНК.
После того как было установлено, что ДНК
кодирует синтез белковых молекул, стало ясно, что
последовательность оснований в нуклеотидах ДНК
должна определять аминокислотную
последовательность белков. Эта зависимость между
основаниями и аминокислотами известна под названием
генетического кода. Оставалось показать, что ну-
клеотидный код в самом деле существует,
расшифровать его и выяснить, каким образом он
переводится в аминокислотную последовательность
белковой молекулы.
22.5.3. Структура кода
Молекула ДНК построена из нуклеотидов четырех
типов, в состав которых входят четыре разных
основания: аденин (А), гуанин (Г), тимин (T) и
цитозин (Ц) (см. разд. 5.6). Нуклеотиды соединены в
полинуклеотидную цепь; их обозначают
начальными буквами названий соответствующих оснований.
С помощью этого четырехбуквенного алфавита
записаны инструкции для синтеза потенциально
бесконечного числа различных белковых молекул. Если
бы одно основание определяло положение одной
аминокислоты в первичной структуре какого-то
белка, то этот белок мог бы содержать только четыре
вида аминокислот. Если бы каждая аминокислота
кодировалась двумя основаниями, то с помощью
такого кода можно было бы определить 16
аминокислот.
22.3. Составьте список 16 возможных попарных
сочетаний из оснований А, Г, T и Ц.
Только код, состоящий из троек оснований
(триплетов), мог бы обеспечить включение в белковые
молекулы всех 20 аминокислот. В такой код входит
64 разных триплета.
22.4. Четыре основания, используемые по
одному, позволяют закодировать четыре
аминокислоты; используемые попарно-16 аминокислот;
используемые по три-64 аминокислоты.
Выведите математическое выражение, объясняющее это.
Доказательства триплетности кода представил
Фрэнсис Крик в 1961 г., получив у фагов T4
мутации, вызванные добавлением или выпадением
оснований. Эти добавления и выпадения, приводящие к
сдвигам рамки при «чтении» кода (рис. 22.20),
проявились у фагов T4 в изменениях фенотипа. В
результате сдвига рамки получались такие
последовательности триплетов оснований, которые не
могли обеспечить синтез белковых молекул с прежней
последовательностью аминокислот (прежней
первичной структурой). Только добавление или
удаление одного основания в определенных точках могло
привести к восстановлению правильного кода.
Восстановление первоначальной последовательности
оснований предотвращало появление мутантов. Эти
эксперименты показали также, что триплеты не
перекрываются, т.е. каждое основание может
принадлежать только одному триплету. Ни одно из
Нормальный триплетный код
ΓΑΤίΓΑΤίΓΑΤ',ΓΑΤ'ΓΑΤΙΓΑΤ'ΓΑΤ'ΓΑΤ'ΓΑΤ
Добавлено одно основание (Ц)
ГАТ;ГАТ!ЦГА!тГА!ТГа!ТГА!ТГАТГА!ТГА
Удалено одно основание (А)
γατ!Γατ!γατ!γατ;γτγ!ατγιατγ|ατγ;ατγ
Добавлено одно (Ц) и удалено одно (А) основание
гат!гат!цга!тга!тгт.гат1ГаТ'Гат'Гат
Код снова в фазе
Рис. 22.20. Схема, поясняющая результаты добавления или удаления оснований в триплет -
ном коде. Добавление основания Ц приводит к сдвигу рамки, так что первоначальное сообщение ГА T, ГА T,
превращается в TГА, TГА, ... Выпадение основания А вызывает сдвиг рамки,
приводящий к замене первоначального сообщения ГAT, ГAT, ... на ATГ, ATГ, ... Добавление
основания Ц и удаление основания А в точках, указанных на схеме, приводят к восстановлению
первоначального сообщения ГAT, ГAT, ... (По F H. С Crick, 1962, The genetic cod I, Scientific American,
Offprint N123, Wm. Saunders and Co.)
ц а т ! г τ ц !
Первоначальная
последовательность
триплетов
(неперекрывающаяся)
A T Г ! T Ц
Первая перекрывающаяся
последовательность
T Г T ! Ц
Вторая перекрывающаяся
последовательность
Рис. 22.21. Триплеты оснований в
неперекрывающемся и перекрывающемся кодах
основании, входящих в данный триплет, не является
частью другого триплета (рис. 22.21).
22.5. Используя повторяющиеся триплеты ГТА
и основание Ц, покажите, что исходную
последовательность триплетов можно восстановить
только путем добавления или удаления трех
оснований. (Представьте свой ответ в такой форме, как
показано на рис. 22.20.)
22.5.4. Расшифровка кода
Для того чтобы понять ход экспериментов,
проводившихся с целью установить, какие триплеты
соответствуют тем или иным аминокислотам (т.е.
расшифровать генетический код), нужно иметь
представление о механизме, с помощью которого три-
плетный код переводится в структуру белковой
молекулы.
В синтезе белка участвуют нуклеиновые кислоты
двух типов-дезоксирибонуклеиновая кислота (ДНК)
и рибонуклеиновая кислота (РНК),
взаимодействующие друг с другом. Существует три главных типа
РНК: информационная, или матричная, РНК (мРНК),
рибосомная РНК (рРНК) и транспортная РНК
(тРНК)1. ДНК содержится и в таких органеллах, как
митохондрии и хлоропласты, но в основном она
сосредоточена в ядре, где ее нуклеотидная
последовательность копируется (транскрибируется) с
образованием матричной РНК (мРНК), переходящей из
ядра в цитоплазму. Оказавшись в цитоплазме, нить
мРНК прикрепляется к рибосомам, где
нуклеотидная последовательность мРНК транслируется в
аминокислотную последовательность белка. Каждая
аминокислота связывается с соответствующей тРНК,
которая присоединяется к комплементарному
триплету оснований мРНК. Аминокислоты, оказавшиеся
в результате этого друг подле друга, соединяются,
образуя полипептидную цепь. Таким образом, для
белкового синтеза необходимы ДНК, мРНК, рибо-
1 Сейчас известны еще предшественники мРНК и
рРНК-пре-мРНК и пре-рРНК, а также
низкомолекулярные РНК (помимо транспортных) нмРНК. Прим. ред.
сомы, тРНК, аминокислоты, АТФ и ГТФ как
источники энергии и различные ферменты и кофакторы,
катализирующие каждую стадию этого процесса.
Ниренберг использовал эти данные и различные
методы, созданные в конце пятидесятых годов, для
разработки ряда экспериментов, поставленных с
целью расшифровки кода. Суть его экспериментов
сводилась к тому, чтобы, используя мРНК с заранее
известной последовательностью оснований,
выяснить последовательность аминокислот в
полипептидной цепи, синтезируемой в присутствии этой
мРНК. Ниренбергу удалось синтезировать РНК (по-
лирибонуклеотид), состоящую из многократно
повторяющихся триплетов УУУ. Это соединение,
названное полиуридиловой кислотой (поли-У), было
использовано в качестве мРНК. В каждую из 20
пробирок поместили бесклеточный экстракт E. coli,
содержавший рибосомы, тРНК, АТФ, ГТФ,
ферменты и какую-либо одну меченую аминокислоту.
Затем в каждую пробирку добавили поли-У и
оставили на некоторое время, чтобы произошел синтез
полипептидов. Анализ содержимого пробирок
показал, что полипептид образовался только в той
пробирке, которая содержала аминокислоту
фенил ал анин. Это было первым шагом в расшифровке
генетического кода: выяснилось, что входящий в
мРНК триплет оснований, или ко дон, УУУ
определяет включение в полипептидную цепь фенилалани-
на. Затем Ниренберг и его сотрудники занялись
получением синтетических полинуклеотидов,
соответствующих всем 64 возможным кодонам, и к
1964 г. расшифровали коды для всех 20 аминокислот
(табл. 22.4).
Как видно из табл. 22.4, для большинства
аминокислот имеется по нескольку кодонов. Код, в
котором число аминокислот меньше числа кодонов,
называют вырожденным. Кроме того, можно
видеть, что для многих аминокислот существенное
значение имеют только первые буквы. Три из
представленных в табл. 22.4 кодонов не кодируют
аминокислот («нонсенс-кодоны») и действуют как
«стоп-сигнал» - означают конец закодированного
сообщения. По-видимому, стоп-кодон-это концевая
точка функциональной единицы ДНК-цистрона.
Во всех экспериментах, проводившихся с целью
расшифровки генетического кода, в качестве
источника триплетов использовалась мРНК. Однако от
одной клетки другой и от одного поколения
другому «генетический текст» передается
последовательностью триплетов в ДНК. Поскольку мРНК
образуется непосредственно на полинуклеотидной цепи
ДНК путем комплементарного спаривания
оснований, запись наследуемого генетического «текста»
ДНК комплементарна его записи в мРНК. Код
ДНК можно получить, заменяя основания,
содержащиеся в РНК, комплементарными им основаниями
ДНК в соответствии с табл. 22.5.
22.6. Выпишите последовательность оснований
в мРНК, образованной на цепи ДНК с такой
последовательностью:
АТГТТЦГАГТАЦЦАТГТААЦГ
Одна из примечательных особенностей
генетического кода состоит в том, что он, по-видимому,
универсален. У всех живых организмов имеются
одни и те же 20 аминокислот и одни и те же пять
азотистых оснований (А, Г, T, Ц и У). Ниренберг
показал, что если ввести мРНК, взятую от вида А, в
бесклеточную систему от вида Б, то в ней начнется
синтез того же полипептида, который образовался
бы у вида А. Например, в бесклеточных экстрактах
E. coli, в которые добавляли мРНК, кодирующую
гемоглобин млекопитающего, синтезировались
молекулы гемоглобина, свойственного этому
млекопитающему1.
Некоторые кодоны служат «стартовыми»
(инициирующими) сигналами-означают начало
полипептидной цепи (как, например, АУГ-кодон ме-
тионина), тогда как другие, такие как У А А,-не
кодируют ни одну аминокислоту, а служат «стоп-
1 Оказалось, однако, что код митохондриальной ДНК
несколько отличается от универсального кода.- Прим.
ред.
сигналами», т.е. означают конец синтеза
полипептидной цепи.
В настоящее время успехи молекулярной
биологии достигли такого уровня, что становится
возможным определять последовательности оснований
в целых генах и удалось даже расшифровать весь
генетический «текст» одного организма - фага φχ 174.
Это серьезная веха в развитии науки, поскольку
теперь можно искусственно синтезировать целые
гены, что уже нашло применение в генной
инженерии (см. разд. 2.3.6).
Главные черты генетического кода можно вкратце
сформулировать следующим образом.
1. Кодом, определяющим включение
аминокислоты в полипептидную цепь, служит триплет
оснований в полипептидной цепи ДНК.
2. Код универсален: одни и те же триплеты
кодируют одни и те же аминокислоты у всех
организмов.
3. Код является вырожденным: данная
аминокислота может кодироваться более чем одним
триплетом.
4. Код неперекрывающийся: например,
последовательность мРНК, начинающаяся с нуклеотидов
АУГАГЦГЦА, не считывается как АУГ/УГА/
ГАГ... (перекрывание по двум основаниям) или
АУГ/ГАГ/ГЦГ... (перекрывание по одному ос-
Таблица 22.4. Последовательности оснований в триплетах и кодируемые ими аминокислоты. Приведены кодоны,
т. е. триплеты оснований в мРНК, а не в ДНК. В ДНК содержатся комплементарные основания, а У заменен на T
2-е основание
1-е основание
У
ц
А
Г
У
УУУ
УУЦ.
ууа'
УУГ ,
ЦУУ
ЦУЦ
ЦУА
ЦУГ
АУУ
АУЦ
АУА
АУГ
ГУУ
ГУЦ
ГУА
[ ГУГ
гФен
Лей
Лей
► Иле
Мет
у Вал
ц
УЦУ
УЦЦ
УЦА
УЦГ
ЦЦУ
ццц
ЦЦА
ццг(
АЦУ
АЦЦ
АЦА
АЦГ
ГЦУ
ГЦЦ
I ГЦА
| ГЦГ
► Сер
► Про
►Тре
Ала
А
УАУ]
УАЦ]
УАА
УАГ
ЦАУ^
ЦАЦ.
ЦАА'
ЦАГ.
ААУ'
ААЦ.
AAA
ААГ.
ГАУ
ГАЦ.
ГАА^
ГАГ,
.Тир
Стоп1
Стоп!
^Гис
► Глн
у Асн
► Лиз
► Асп
► Глу
Г
УГУ1
угц]
УГА
УГГ
ЦГУ ]
ЦГЦ
ЦГА
ЦГГ J
АГУ 1
АГЦ,
ΑΓΑ'
АГГ ,
ГГУ
ГГЦ
ГГА
[ ггг
Цис
Стоп1
Трп
Apr
Сер
► Apr
I Гли
у|
ц
А
Г
У
ц
А
Г
У
ц
А
Г
ц
А
Г
3-е основание
" Кодон, означающий конец синтеза полипептидной цепи
нованию). Недавно было обнаружено, однако,
перекрывание некоторых генов у бактериофага
φχ174 и ряда других вирусов. Эти случаи, по-
видимому, составляют исключение, возможно,
связанное с экономным использованием
нуклеиновой кислоты (количество которой у вирусов
очень невелико)1.
22.6. Синтез белка
Из всего того, что было сказано в предыдущих
разделах этой главы, можно видеть, что
единственные молекулы, которые синтезируются под
прямым контролем генетического материала клетки,-
это белки (если не считать РНК). Белки могут быть
структурными (кератин, коллаген) или играть
функциональную роль (инсулин, фибриноген и, главное,
ферменты, ответственные за регуляцию клеточного
метаболизма). Именно набор содержащихся в
данной клетке ферментов определяет, к какому типу
клеток она будет относиться. «Инструкции»,
необходимые для синтеза этих ферментов и всех других
белков, заключены в ДНК, которая почти вся
находится в ядре; однако, как было показано в начале
пятидесятых годов, синтез белка фактически
происходит в цитоплазме и в нем участвуют рибосомы.
Стало ясно, что должен существовать какой-то
механизм, переносящий генетическую информацию из
ядра в цитоплазму. В 1961 г. два французских
биохимика Жакоб и Mоно, исходя из теоретических
соображений, постулировали существование особой
формы РНК, выполняющей в синтезе белка роль
посредника; впоследствии этот посредник получил
название мРНК.
22.6.1. Роль РНК
РНК содержится во всех живых клетках в виде
одноцепочечных молекул. Она отличается от ДНК
тем, что содержит в качестве пентозы рибозу
(вместо дезоксирибозы), а в качестве одного из пирими-
диновых оснований - урацил (вместо тимина).
Анализ РНК, содержащейся в клетках, показал, что
существует три типа РНК, участвующих в синтезе
белковых молекул. Это матричная, или
информационная, РНК (мРНК), транспортная РНК (тРНК) и
рибосомная РНК (рРНК). Все три типа РНК
синтезируются непосредственно на ДНК, которая
служит матрицей для этого процесса. Количество РНК
в каждой клетке находится в прямой зависимости от
количества вырабатываемого этой клеткой белка.
1 В этих случаях перекрывание состоит не в том, что
соседние аминокислоты в белке кодируются
перекрывающимися триплетами, а в том, что на одном участке
ДНК закодированы два различных белка, имеющие
разные точки инициации.- Прим. ред.
22.6.2. Матричная, или
информационная, РНК
Как показали исследования, мРНК составляет 3-5%
всей содержащейся в клетке РНК. Это одноцепочеч-
ная молекула, образующаяся на одной из цепей
ДНК в процессе так называемой транскрипции. При
синтезе мРНК копируется только одна цепь
молекулы ДНК. Механизм, определяющий, какая
именно цепь будет при этом копироваться, до конца
еще не выяснен. Возможно, что в этом процессе
участвуют промотор и оператор (см. разд. 22.7.1).
Нуклеотиды, из которых синтезируется мРНК,
присоединяются к ДНК в соответствии с правилами
спаривания оснований и при участии фермента
РНК-полимеразы связываются между собой,
образуя полинуклеотидную цепь мРНК.
Последовательность оснований в мРНК представляет собой
комплементарную копию цепи ДНК-матрицы; длина ее
может быть различной в зависимости от длины
полипептидной цепи, которую она кодирует. Самая
короткая молекула мРНК состоит примерно из 300
нуклеотидов. Большинство мРНК существует в
клетке лишь в течение короткого времени; в
бактериальных клетках это время измеряется минутами, тогда
как в эритроцитах млекопитающих синтез
гемоглобина может продолжаться в течение нескольких
дней после утраты ими ядра (значит, мРНК все это
время сохраняется).
22.6.3. Рибосомная РНК
Рибосомная РНК, составляющая более 80% всей
РНК клетки, была идентифицирована раньше
других типов РНК. Она кодируется особыми генами,
находящимися в нескольких хромосомах и
расположенными в участке ядрышка, известном под
названием ядрышкового организатора.
Последовательность оснований в рРНК сходна у всех организмов-
от бактерий до высших растений и животных; рРНК
содержится в цитоплазме, где она связана с
белковыми молекулами, образуя вместе с ними
клеточные органеллы, называемые рибосомами (см.
разд. 7.2.6).
На рибосомах происходит синтез белка. Здесь
«код», заключенный в мРНК, транслируется в
аминокислотную последовательность строящейся
полипептидной цепи. Рибосомы часто образуют группы,
соединяясь друг с другом одной цепочкой мРНК.
Такие группы-полирибосомы, или
полисомы,-делают возможным одновременный синтез
нескольких молекул полипептида при участии одной
молекулы мРНК.
22.6.4. Транспортные РНК
Существование транспортных РНК (тРНК) было
постулировано Криком и продемонстрировано Хог-
лендом в 1955 г. Для каждой аминокислоты имеется
Водородные
связи между
комплементарными
основаниями
Антикодон
Рис. 22.22. Модель строения транспортной РНК
(тРНК). Молекула состоит из 80 нуклеотидов,
но пар, образованных за счет
комплементарного связывания, в молекуле только 20.
специфическая тРНК, и все они доставляют
содержащиеся в цитоплазме аминокислоты к
рибосомам. Таким образом, тРНК играют роль
связующих звеньев между триплетным кодом,
содержащимся в мРНК, и аминокислотной
последовательностью полипептидной цепи. На долю тРНК
приходится примерно 15% всей клеточной РНК; у этих
РНК самая короткая полинуклеотидная цепь-в нее
входит в среднем 80 нуклеотидов. Так как многие
аминокислоты кодируются несколькими
триплетами, число различных тРНК, переносящих
определенные аминокислоты, значительно больше
двадцати (идентифицировано уже 60). Все молекулы
тРНК имеют сходную основную структуру,
показанную на рис. 22.22.
На 5'-конце молекулы тРНК всегда находится
гуанин, а на З'-конце-группировка ЦЦА.
Последовательность нуклеотидов в остальной молекуле
варьирует и может содержать «необычные»
основания, такие как инозин (И) и псевдоурацил (ψ).
Последовательность оснований в триплете антико-
дона (рис. 22.22) строго соответствует той
аминокислоте, которую переносит данная молекула тРНК.
Каждая аминокислота присоединяется к одной из
своих специфических тРНК при участии своей
особой формы фермента аминоацил-тРНК-синтетазы.
В результате образуется комплекс аминокислоты с
тРНК - аминоацил-тРНК, в котором энергия связи
между концевым нуклеотидом А (в триплете ЦЦА)
и аминокислотой достаточна для того, чтобы в
дальнейшем могла образоваться пептидная связь с
карбоксильной группой соседней аминокислоты.
Таким образом синтезируется полипептидная цепь.
В экспериментах с введением в бесклеточные
экстракты E. coli рибосом из клеток крысиной печени
наблюдали синтез белков E. coli, несмотря на «чуже-
родность» рибосом. Эти эксперименты
продемонстрировали универсальность механизмов,
осуществляющих белковый синтез с участием мРНК, тРНК
и рРНК.
22.6.5. Механизм синтеза белка
Данные, полученные с помощью различных
методов в экспериментах на самых разнообразных
организмах-от вирусов до млекопитающих, показали,
что процесс синтеза белка состоит из двух этапов,
схематически представленных на рис. 22.23.
22.6.6. Транскрипция
Транскрипцией называют механизм, с помощью
которого последовательность оснований в одном из
цистронов цепи ДНК «переписывается» в
комплементарную ей последовательность оснований мРНК.
Как полагают, в области этого цистрона гистоны,
связанные с двойной спиралью ДНК, отделяются,
обнажая полинуклеотидные последовательности
молекулы ДНК. Относительно слабые водородные
связи между комплементарными основаниями поли-
нуклеотидных цепей разрываются, что приводит к
раскручиванию двойной спирали ДНК и
освобождению одиночных цепей. С помощью механизма,
который пока не выяснен, одна из этих цепей
избирается в качестве матрицы для построения
комплементарной одиночной цепи мРНК. Молекула
мРНК образуется в результате связывания друг с
другом свободных рибонуклеотидов под действием
РНК-полимеразы в соответствии с правилами
спаривания оснований ДНК и РНК (табл. 22.5 и
рис. 22.24).
Таблица 22.5. Комплементарное™ между основаниями РНК
и ДНК
Основания ДНК
А (аденин)
Г (гуанин)
T (тимин)
Ц (цитозин)
Основания РНК
У (урацил)
Ц (цитозин)
А (аденин)
Г (гуанин)
Как именно происходит транскрипция оснований
ДНК в основания РНК, было продемонстрировано
в опытах с синтетической ДНК, состоящей из
нуклеотидов только одного вида - тимидина (TTT...).
При введении этой ДНК в бесклеточную систему,
содержавшую РНК-полимеразу и все четыре рибо-
нуклеотидтрифосфата (А, У, Г и Ц), синтезирова-
ЯДРО
днк-
Клеточная мембрана
ЦИТОПЛАЗМА
тРНК/ Аминокислоты
Транскрипция
-►мРНК-
*Рибосома
Трансляция
Полипептид!
Ядерная оболочка
Рис. 22.23. Схема главных этапов в процессе
белкового синтеза.
Нетранскриби-
руемая цепь
Двойная спираль ДНК
Транскрибируемая
цепь
Направление
к транскрипции
РНК.
-полимераза!
Одиночная цепь
мРНК, транскри
бируемая с
молекулы
ДНК
Свободные
нуклеозидтрифосфаты
Транскрибируемая и
нетранс крибиру емая
цепи ДНК вновь
соединяются
Рис. 22.24. Схема механизма транскрипции. В
присутствии РНК-полимеразы двойная спираль ДНК
раскручивается в результате разрыва
водородных связей между комплементарными основаниями, и
из свободных рибонуклеозидтрифосфатов строится
полинуклеотидная цепь мРНК. Она
комплементарна транскрибируемой цепи ДНК, которая служит
матрицей. (По E. J. Ambrose, D. M. Easty, Cell
Biology, 1977, 2nd ed., Nelson.)
этом на одной молекуле мРНК становится
возможным одновременный синтез нескольких
полипептидных цепей (см. разд. 22.6.3). Каждая
рибосома состоит из двух субъединиц-малой и большой
(рис. 7.18). Как полагают, мРНК обратимо
присоединяется к поверхности малой субъединицы в
присутствии ионов магния (Mg2+). При этом два ее
первых транслируемых кодона оказываются
обращенными к большой субъединице рибосомы.
Первый кодон связывает молекулу тРНК, содержащую
комплементарный ему антикодон и несущую
первую аминокислоту (обычно это метионин)
синтезируемого полипептида. Затем второй кодон
присоединяет комплекс аминоацил-τΡΗΚ, содержащий
антикодон, комплементарный этому кодону (рис.
22.26, А и Б). Функция рибосомы заключается в том,
чтобы удерживать в нужном положении мРНК,
тРНК и белковые факторы, участвующие в процессе
трансляции, до тех пор пока между соседними
аминокислотами не образуется пептидная связь.
Как только новая аминокислота присоединилась
к растущей полипептидной цепи, рибосома
перемещается по нити мРНК с тем, чтобы поставить на
надлежащее место следующий кодон. Молекула
тРНК, которая перед этим была связана с
полипептидной цепью, теперь, освободившись от
аминокислоты, покидает рибосому и возвращается в
цитоплазму, чтобы образовать новый комплекс амино-
ацил-τΡΗΚ (рис. 22.26,5).
лась мРНК, содержавшая один лишь аденин.
Синтезированные молекулы мРНК, несущие
генетическую информацию, выходят из ядра через
ядерные поры и направляются к рибосомам. После того
как образовалось достаточное число молекул мРНК,
транскрибированных с данного цистрона,
транскрипция прекращается и две цепи ДНК на этом участке
вновь соединяются, восстанавливая двойную
спираль («молния застегивается»), и опять связываются
с гистонами.
22.6.7. Трансляция
Трансляцией называют механизм, с помощью
которого последовательность триплетов оснований в
молекулах мРНК переводится в специфическую
последовательность аминокислот в полипептидной
цепи. Этот процесс происходит на рибосомах.
Несколько рибосом могут прикрепиться к молекуле
мРНК подобно бусинам на нитке, образуя
структуру, называемую полисомой Вся эта структура
показана на рис. 22.25. Входящие в ее состав рибосомы
связаны общей нитью толщиной 1,5 нм, что
соответствует толщине одной цепи мРНК.
Преимущество такого комплекса состоит в том, что при
Начало сообщения
0,5 мкм
Неактивный участок
хромосомы
Рибосома
Активный участок
хромосомы
РНК-полимераза
Полирибосома
Направление синтеза белка
мРНК
Рис. 22.25. Процесс транскрипции и образование
полисомы у бактерий. А. Электронная
микрофотография участка хромосомы, на которой можно
видеть последовательные стадии образования мРНК
и присоединения рибосом. Б. Схематическое
изображение структуры вроде показанной на
микрофотографии.
Метионин
τΡΗΚι-
τΡΗΚ2
Антикодон
Кодон
5'-конец
.Большая субъединица
рибосомы
3' -конец
мРНК
Малая субъединица рибосомы
Метионин
2-я аминокислота
Между двумя аминокислотами
образуется пептидная связь
тРНК1-
тРНК2
^Вторая молекула тРНК с
аминокислотой присоединяется
к мРНК
У А Ц
А У Г
Метионин
[2-я аминокислота|
тРНК2
JPHK3-
А У Г
Третий комплекс тРНК/амино кислота
приближается
кмРНК
тРНК отделилась
от рибосомы, чтобы
присоединить новую
молекулу метионина
Комплекс продвинулся на один триплет
вперед относительно рибосомы
Рис. 22.26. А и Б. Последовательные стадии
прикрепления комплексов тРНК - аминокислота их
антикодонами к кодонам мРНК и образования
пептидной связи между соседними аминокислотами.
В. Перемещение мРНК относительно рибосомы,
приводящее к тому, что в надлежащем положении
оказывается новый триплет, к которому теперь
может присоединиться новый комплекс тРНК-
аминокислота. Первая молекула тРНК отделилась
от рибосомы и возвращается в цитоплазму, где
она реактивируется ферментами, с тем чтобы
образовать комплекс с аминокислотой.
Такое последовательное «считывание» рибосомой
заключенного в мРНК «текста» продолжается до
тех пор, пока процесс не доходит до одного из
стоп-кодонов (терминирующих кодонов). Такими
кодонами служат триплеты У А А, УАГ или У ГА.
На этом этапе полипептидная цепь, первичная
структура которой была детерминирована цистро-
ном ДНК, покидает рибосому, и трансляция
завершена. Перечислим главные этапы процесса
трансляции:
1) присоединение мРНК к рибосоме;
2) активация аминокислоты и ее присоединение к
тРНК;
3) инициация (начало синтеза) полипептидной
цепи;
4) элонгация (удлинение) цепи;
5) терминация (окончание синтеза) цепи;
6) дальнейшее использование мРНК (или ее
разрушение).
Процесс трансляции схематически представлен на
рис. 22.27.
После того как полипептидные цепи отделились
от рибосомы, они могут тотчас же приобретать
свойственную им вторичную, третичную или
четвертичную структуру (см. разд. 5.5.3).
Доказательства того, что включение
аминокислоты в полипептидную цепь определяется
комплементарным спариванием оснований между кодоном
мРНК и антикодоном тРНК, а не самой
аминокислотой, были получены в следующем
эксперименте. Комплекс тРНК-цистеин обычно спаривается
при помощи своего антикодона АЦА с кодоном
УГУ мРНК. При воздействии на этот комплекс
катализатора - никеля Ренея - цистеин превращается
в аминокислоту аланин. Когда такой новый
комплекс тРНК-аланин (несущий антикодон цистеиновой
тРНК) помещали в бесклеточную систему,
содержавшую в качестве мРНК поли-УГУ,
синтезированная полипептидная цепь состояла из одного только
аланина. Этот эксперимент подтвердил важную
роль взаимодействия кодонов мРНК с
антикодонами тРНК в процессе трансляции.
Вся последовательность событий, из которых
складывается процесс белкового синтеза,
схематически представлена на рис. 22.28.
22.7. Регуляция активности
генов
В этой главе мы описали механизмы, с помощью
которых клетки передают генетический материал из
поколения в поколение. Структура и функции
генетического материала в настоящее время изучены
достаточно подробно, однако в генетике остается
еще ряд областей, где вопросов много, а ответов
мало. За последние 30 лет генетические
исследования сильно продвинулись вперед, и на многие
фундаментальные вопросы ответы уже получены. Самые
главные достижения - это, несомненно, открытие
структуры ДНК и расшифровка генетического кода.
Эти два открытия вдохновили других ученых и
побудили их попытаться глубже проникнуть в
тайны молекулярной генетики. Большая часть
неразрешенных проблем, стоящих перед молекулярными
генетиками, связана с механизмами, которые
регулируют активность генов, участвующих в процессах
метаболизма, развития и дифференцировки.
Классической генетикой установлено, что все
соматические клетки организма несут один и тот же
набор генов, т. е. содержат одинаковое число
хромосом, несущих одни и те же аллели. И тем не менее
клетки многоклеточного организма очень
разнообразны по структуре и функциям. Даже в одной и той
же клетке скорость синтеза белковых молекул
может варьировать в зависимости от обстоятельств и
потребностей. Данные о механизмах, регулирующих
Растущая полипептидная цепь
Метионин
Большая субъединица рибосомы
В алии
Алании
{Фенил ал анин]
Лизин
Молекула тРНК возвращается
в цитоплазму
Аминокислота
Ί ,
Лейцин Глицин
Тирозин!
Антикодон
5 -конец -
УАА. АЦУУГГГУАУГЦУА
Кодон 4
Кодон 5
Кодон 6
Кодон 7
Кодон 8
3 -конец
Молекула
тРНК
Ц Г А
vy у у
Г А А Ц Ц Ц
А У А'
^. Направление
считывания
Одноцепочечная мРНК
Положение рибосомы
Малая субъединица рибосомы
Рис. 22.27. Схема процесса трансляции. Антикодон каждого специфического комплекса
тРНК-аминокислота спаривается с комплементарным ему кодоном мРНК на рибосоме. В
приведенном здесь примере пептидная связь должна образоваться между лейцином и глицином, в результате чего к
растущей полипептидной цепи добавится еще одна аминокислота.
активность генов в клетке, были впервые получены
при изучении регуляции синтеза ферментов у E. coli.
В 1961 г. Жакоб и Моно провели ряд
экспериментов, желая понять природу индукции синтеза
ферментов у E. coli. Полагают, что в клетках E. coli
синтезируется около 800 ферментов. Синтез
некоторых из них происходит непрерывно, и их называют
конститутивными ферментами; другие же
образуются только в присутствии надлежащего индуктора,
который может и не быть субстратом данного
фермента. Такие ферменты, примером которых
служит β-галактозидаза, называют индуцибельными
ферментами.
E. coli быстро растет на культуральной среде,
содержащей глюкозу. При перенесении клеток на
среду, содержащую вместо глюкозы лактозу, рост
начинается не сразу, а после короткой задержки, но
затем идет с такой же скоростью, как и на среде с
глюкозой. Проведенные исследования показали, что
для роста на лактозной среде необходимо наличие
двух веществ, которые E. coli обычно не
синтезирует: β-галактозидазы, гидролизующей лактозу до
Трансляция
мРНК
Двойная спираль ДНК
РНК-полимераза
Ядерная оболочка
Ядерная пора
ДНК-матрица
Кодон
Аминоацил-тРНК-синтетаза _
[Вал
Ала |
Аминокислота
Комплекс
тРНК-амино-
кислота (
Рибосомы
(рРНК + белок)
Антикодон
тРНК
мРНК
Транскрипция
мРНК (вместе с
рибосомами
образует
полисому)
Полипептидная цепь
сходит с рибосомы
Рис. 22.28. Упрощенная схема основных структур и процессов, участвующих в белковом
синтезе.
ΟΠΕΡΟΗ
Двойная
спираль ДНК
Ген-регулятор
^)ооооооооо(^
Оператор
Транскрипция
мРНК
Трансляция
Может связаться или
не связаться
с оператором
Трансляция
Молекула-репрессор
Структурные гены
) Транскрипция
'Синтез полипептидов
мРНК
Рис. 22.29. Основные структуры и процессы, участвующие в регуляции белкового синтеза
согласно гипотезе Жакоба - Моно Цифры указывают последовательность событий.
глюкозы и галактозы, и лактозопермеазы,
делающей клетку способной быстро поглощать лактозу из
среды. Это служит примером того, как изменение в
условиях среды-замена глюкозы лактозой -
индуцирует синтез определенного фермента. Другие
эксперименты с E. coli показали, что высокое
содержание в среде аминокислоты триптофана подавляет
выработку триптофансинтетазы - фермента,
необходимого для синтеза триптофана. Синтез β-галакто-
зидазы служит примером индукции, а подавление
синтеза триптофансинтетазы - примером репрессии
фермента. На основании этих наблюдений Жакоб и
Моно предложили механизм, объясняющий
индукцию и репрессию,-механизм «включения» и
«выключения» генов.
22.7.1. Гипотеза Жакоба-Моно
Генетические инструкции, определяющие
аминокислотную последовательность упомянутых выше
белков, заключены в структурных генах, причем
инструкции для β-галактозидазы и лактозопермеазы
тесно сцеплены в одной хромосоме. Активность
этих генов регулируется еще одним геном, который
называют геном-регулятором и который
препятствует переходу структурных генов в активное
состояние. Ген-регулятор может находиться на некотором
расстоянии от структурных генов. Доказательства
его существования получены при изучении мутант-
ных клеток E. coli, лишенных этого гена и поэтому
вырабатывающих β-галактозидазу непрерывно. Ген-
регулятор содержит генетическую информацию для
синтеза репрессора, который препятствует
активности структурных генов. Репрессор действует на
структурные гены не прямо, а опосредованно, оказывая
влияние на участок, примыкающий к структурным
генам и называемый оператором. Оператор и
управляемые им структурные гены в совокупности
называют опероном (рис. 22.29).
Репрессор представляет собой особый аллостери-
ческий белок, который либо связывается с
оператором, подавляя его активность («выключает» его),
либо не связывается с ним, позволяя ему проявлять
активность (оставляет его включенным). Когда
оператор включен, на структурных генах
осуществляется транскрипция и происходит образование мРНК,
которую рибосомы и тРНК транслируют в
полипептиды; а когда оператор выключен, мРНК не
образуется и кодируемые ею полипептиды не
синтезируются (рис. 22.29).
Механизм, от которого зависит, присоединится
ли аллостерический белок к оператору или нет,
прост и при этом чувствителен к изменениям
условий внутри клетки. В молекуле репрессора имеется
по меньшей мере два активных участка; к одному из
них может присоединиться молекула индуктора, а
другой служит для присоединения к оператору,
выключающего весь оперон.
22.7.2. Индукция ферментов
Присоединение молекулы индуктора к активному
участку молекулы репрессора изменяет третичную
структуру репрессора (аллостерический эффект; см.
разд. 6.6) так, что он не может связаться с геном-
оператором и репрессировать его; в результате
оператор оказывается в активном состоянии и
включает структурные гены.
При выращивании E. coli на среде с глюкозой
ген-регулятор продуцирует белок, обладающий
свойствами репрессора, который связывается с геном-
оператором и выключает его. Структурные гены
при этом не активируются, и ни β-галактозидаза, ни
лактозопермеаза не синтезируются. При переносе
бактерий на среду с лактозой последняя действует
как индуктор, присоединяясь к молекуле репрессора
и препятствуя ее соединению с оператором.
Структурные гены переходят в активное состояние и
продуцируют мРНК для синтеза ферментов,
ответственных за поглощение и расщепление лактозы.
Таким образом, лактоза индуцирует собственное
расщепление (рис. 22.30).
Ген-регулятор
Оператор
ι
Структурный ген
Молекула-репрессор
Лактоза действует
как индуктор
Транскрипция
мРНК
Комплекс индуктора с
репрессором, неспособный
связаться с оператором
0-Галактозидаза
(фермент)
Лактоза
Трансляция
„Глюкоза +
Галактоза
Рис. 22.30. Индукция синтеза β-галактозидазы согласно гипотезе Жакоба-Моно. Цифры указывают
последовательность событий.
22.7.3. Репрессия ферментов
Если молекула корепрессора присоединяется к
соответствующему активному участку репрессора, то
это усиливает способность репрессора связываться с
оператором; при этом происходит инактивация
оператора и тем самым предотвращается включение
структурных генов.
E. coli может синтезировать аминокислоту
триптофан при участии фермента триптофансинтетазы.
Если клетка содержит избыток триптофана,
некоторая его часть действует как корепрессор, связываясь
с молекулой репрессора. Молекулы корепрессора и
репрессора присоединяются к оператору и
подавляют его активность. Структурные гены
выключаются, мРНК не образуется, и синтез
триптофансинтетазы прекращается. Это пример ингибирования по
типу обратной связи на генном уровне (рис. 22.31)1.
22.7.4. Регуляция метаболических
путей
Описанный выше двойной механизм делает
возможным взаимодействие между внутриклеточной
средой и генетическим аппаратом для обеспечения
1 В настоящее время показано, что действительный
механизм более сложен.- Прим. ред.
тонкой регуляции клеточного метаболизма. На
рис. 22.32 показан простой метаболический путь, в
котором исходный субстрат и конечный продукт
могут играть роль соответственно индуктора и
корепрессора. Благодаря этому клетка может
синтезировать фермент в таком количестве, которое
необходимо в данное время для того, чтобы
поддерживать на нужном уровне количество конечного
продукта. Такой способ регуляции метаболизма
чрезвычайно экономен. Отрицательная обратная
связь, осуществляемая путем инактивации первого
фермента (а) при его связывании с конечным
продуктом (E), быстро блокирует данный
метаболический путь, но не приостанавливает синтез других
ферментов (Ь, с и d). B модели, предложенной
Жакобом и Моно, конечный продукт (E),
присоединяясь к репрессору и тем самым усиливая его
ингибирующее действие на оператор, подавляет
синтез всех ферментов (а, Ъ, с и d) и выключает
данный метаболический путь. Индуцирующая
функция исходного субстрата на схеме не показана.
22.7.5. Видоизмененная гипотеза
оперона
После того как в 1961 г. Жакоб и Моно предложили
механизм, с помощью которого гены могут
включаться и выключаться, были получены другие
данные, позволившие прояснить различные аспекты
Ген-регулятор
Оператор
Структурный ген
Неактивная
молекула-
репрессор
Активная
молекула-
репрессор
Связывается
с оператором
Триптофан
действует как
корепрессор
Предшественник -
Транскрипция
чмРНК
Трансляция
Триптофансинтетаза
Триптофан
Рис. 22.31. Механизм репрессии синтеза триптофанситетазы согласно гипотезе Жакоба Моно.
Цифры указывают последовательность событий. Прерывистыми стрелками обозначены
репрессированные стадии.
Структурные гены
Ген-регулятор
Оператор
Молекула-репрессор
Репрессия
Фермент
Субстрат
Отрицательная обратная связь
Конечный
продукт
Рис. 22.32. Механизм регуляции метаболического пути А -> E. Сплошными линиями показаны механизмы,
действующие по принципу отрицательной обратной связи, прерывистыми
линиями-механизмы, действующие во время репрессии. Крестом( χ ) обозначено блокирование синтеза
данного фермента.
этого механизма. Исходя из генетических данных,
было высказано предположение о существовании
промотора, расположенного рядом с оператором и
действующего между ним и геном-регулятором.
Предполагается, что промотор выполняет две
функции. Во-первых, промотор-это то место, к
которому присоединяется РНК-полимераза, прежде чем
начать перемещаться вдоль ДНК, транскрибируя
структурные гены в соответствующую мРНК. Это
перемещение, конечно, зависит от того, находится
ли оператор в активном состоянии или нет.
Во-вторых, последовательность оснований в промоторе
определяет, какая из цепей двойной спирали ДНК
присоединит к себе РНК-полимеразу. Таким
образом, от промотора зависит, какая из цепей
двойной спирали ДНК будет служить матрицей для
синтеза мРНК.
22.8. Генетическая регуляция
развития
Большинство многоклеточных животных и
растений начинает свой жизненный цикл с одной клетки -
зиготы. Из этой клетки в результате многократных
митотических делений получается сложный, высоко-
дифференцированный организм. Процесс этот
называют ростом и развитием, и он включает также
дифференцировку. В результате дифференцировки
каждая клетка приобретает определенную структуру,
позволяющую ей выполнять ряд специфических
функций более эффективно, и это является одним из
важнейших событий, происходящих в процессе
развития. Почему клетки, принадлежащие одному
организму, образовавшиеся путем повторных
клеточных делений и содержащие один и тот же
генетический материал, отличаются таким широким
разнообразием, типичным для многоклеточных
организмов? Причина этого далеко не ясна, однако она,
несомненно, связана с индукцией и репрессией генов
при участии механизмов, вероятно, сходных с
описанными в предыдущем разделе. Судя по
имеющимся данным, дифференцировка связана с различными
взаимодействиями трех факторов - ядра,
цитоплазмы и окружающей среды.
22.8.1. Роль ядра
Значение ядра как хранилища генетического
материала и его главная роль в определении фенотипи-
ческих признаков были установлены давно.
Немецкий биолог Хаммерлинг одним из первых
продемонстрировал важнейшую роль ядра. Он выбрал в
качестве объекта своих экспериментов необычайно
крупную одноклеточную (или неклеточную) морскую
водоросль Acetabularia. Существуют два близко
родственных вида Л. mediterranea и Л. crenulata,
различающиеся только по форме «шляпки» (рис. 22.33).
В ряде экспериментов, в том числе таких, в
которых «шляпку» отделяли от нижней части
«стебелька» (где находится ядро), Хаммерлинг показал,
что для нормального развития шляпки необходимо
ядро. В дальнейших экспериментах, в которых
соединяли нижнюю, содержащую ядро часть одного вида
с лишенным ядра стебельком другого вида, у таких
химер всегда развивалась шляпка, типичная для
того вида, которому принадлежало ядро.
При оценке этой модели ядерного контроля
следует, однако, учитывать примитивность организма,
использованного в качестве объекта. Метод
пересадок был применен позднее в экспериментах,
проведенных в 1952 г. двумя американскими
исследователями, Бриггсом и Кингом, с клетками лягушки
Rana pipiens. Эти авторы удаляли из неоплодотво-
ренных яйцеклеток ядра и заменяли их ядрами из
клеток поздней бластулы, уже проявлявших
признаки дифференцировки. Во многих случаях из яиц-
реципиентов развивались нормальные взрослые
лягушки.
22.7. О чем свидетельствуют результаты
описанных выше экспериментов?
22.8.2. Роль цитоплазмы
Дальнейшие сведения о роли цитоплазмы дали
эмбриологические исследования. У очень многих
организмов цитоплазма яйца выглядит
неоднородной уже на самой ранней стадии зародышевого
развития: в ней можно различить слои и зоны,
"Шляпка"-
'Стебелек"
-Основание
Ядро
A.mediterranea
A.mediterranea
A.crenulata
A.crenulata
'.'Шляпка/
/ срезана/
'по линии
W-W ·'
Вырезан участок
стебля между
W-Wh Y-Y
^ "Шляпка*
срезана
по линии
X-X
Основание
отрезано
по линии
/Z-Z
Развивается новая
"шляпка" как у
A.crenulata
Рис. 22.33. Морская водоросль Acetabularia,
использованная Хаммерлингом для того, чтобы
продемонстрировать роль ядра. А. Два вида Acetabularia.
Б. Эксперименты, проведенные Хаммерлингом.
создаваемые неравномерным распределением
зернистого или по-разному окрашенного материала.
У тех видов, у которых первые деления дробления
происходят в вертикальной плоскости, как,
например, у амфибий, каждый из образующихся при этом
бластомеров, если отделить его от других, обычно
дает начало целому нормальному эмбриону. У
других видов, например у моллюсков, изолированные
бластомеры не способны к нормальному развитию
и образуют только часть зародыша. В первом
случае разные зоны цитоплазмы распределяются
между обоими бластомерами поровну, тогда как во
втором они распределены неравномерно. Яйца, в
которых цитоплазма дифференцирована, так что
разные их участки всегда дают начало
определенным частям зародыша, называют мозаичными. Тем
не менее во всех случаях (за редкими исключениями)
ядра всех клеток содержат одинаковые наборы
генов. Следует поэтому полагать, что различие в
дальнейшей судьбе эмбриональных клеток -
результат какого-то влияния на гены со стороны
цитоплазмы.
В 1924 г. в ряде эмбриологических экспериментов,
ставших теперь классическими, Шпеман и Мангольд
показали, что дифференцировка в значительной
степени контролируется влиянием цитоплазмы клеток
одного типа на клетки другого типа. В одном из
своих экспериментов эти авторы брали гаструлу
амфибии, вырезали кусочек ткани из дорсальной
губы бластопора (рис. 22.34) и пересаживали ее в
вентральную область другой гаструлы. Клетки
дорсальной губы при нормальном развитии образуют
хорду и мезодермальные сомиты (миотомы). В этом
опыте у гаструлы-реципиента из тканей
трансплантата (донора) развивались вторая хорда и миотомы.
Над ними из эктодермы реципиента возникала
новая, дополнительная нервная трубка, что в итоге
приводило к образованию органов второго
головастика (рис. 22.34). Структуры, образовавшиеся в
эксперименте на необычном для них месте,
называют атопическими.
На основании этих данных Шпеман и Мангольд
выдвинули гипотезу о механизме дифференцировки,
получившем название эмбриональной индукции.
Согласно этой гипотезе, определенные клетки
действуют как организаторы на другие, подходящие для
этого клетки. Организатор способен побуждать
такие клетки к развитию в направлении, отличном от
того, в котором они развивались бы в отсутствие
организаторов.
Согласно представлениям Шпемана, в процессе
эмбрионального развития организма определенный
участок, называемый первичным организатором,
детерминирует все дальнейшее течение развития.
У амфибий это дорсальная губа бластопора, а
у птиц-область, называемая первичной полоской.
Эти первичные организаторы определяют
положение оси зародыша и побуждают другие ткани
действовать как вторичные и третичные организаторы - и
Бластопор
Нервная пластинка
-Хорда
^Вырезанный
/ кусочек ткани
\ Бластопор
Энтодерма
Сюда пересажена
ткань от зародыша Б
Эктодерма
Нервная трубка
Хорда
Мезодермальные сомиты
Хорда
Нервная трубка
Зародыш, развившийся
из гаструлы-реципиента
Зародыш, развившийся
из пересаженного
кусочка дорсальной
губы бластопора
Рис. 22.34. Эксперименты Шпемана и Мангольд по
эмбриональной индукции. А. Развивающаяся гаструла
(вид сверху), виден бластопор. Б. Гаструла
амфибии в продольном разрезе; видна область
дорсальной губы бластопора, кусочек которого
вырезали и пересадили в га cm рулу А. В. Поперечный
разрез развивающегося зародыша, у которого
образовалось два идентичных набора эмбриональных
тканей. Г. Сросшиеся зародыши, возникшие в
результате пересадки ткани из дорсальной губы
бластопора.
так далее, пока не завершится дифференцировка
всех органов и систем органов эмбриона на
нормальных для них местах1.
22.8.3. Природа организатора
В ряде экспериментов у зародышей вырезали
участки ткани, обладающие свойствами индуктора, и
помещали их на несколько часов на кусочки агара.
Когда затем эти кусочки агара имплантировали в
области зародыша, обладавшие соответствующей
компетенцией, эти области дифференцировались так
же, как и при воздействии самого индуктора. Таким
образом было показано, что организатор выделяет
химическое вещество, способное диффундировать в
1 Эмбриональная индукция (действие
организаторов) является лишь одним из механизмов развития.
Многие явления развития и, в частности, развитие мозаичных
яиц требуют иных механизмов, которые активно
изучаются, но пока еще не понятны. Прим. ред.
агар. Попытки выделить это вещество пока не дали
достаточно ясных результатов. Высказывались
предположения, что оно может быть стероидом, белком
или нуклеиновой кислотой. Пол и Гилмур обратили
внимание на то, что в разных тканях гистоновые и
негистоновые белки распределены по поверхности
молекул ДНК неравномерно. По их мнению, гисто-
ны «прикрывают» нефункционирующие участки
ДНК, т.е. те участки, в которых гены необратимо
репрессированы, тогда как негистоновыми белками
«прикрыты» гены, которые должны
транскрибироваться.1
22.8.4. Роль внешней среды
Большую роль в выяснении степени влияния
внешней среды на развитие сыграли исследования,
подобные экспериментам Жакоба и Моно. Оказалось,
например, что лактоза - это внешний фактор,
оказывающий прямое влияние на функционирование
генов у E. coli. Ha развитие и рост растений и
животных влияют такие факторы, как свет,
температура, снабжение водой, питательными
веществами, CO2 и кислородом (см. гл. 9, 10, 15, 16 и 21).
Факторы среды, вероятно, обычно оказывают
влияние на дифференцировку, воздействуя через
цитоплазму, которая в свою очередь влияет на гены.
22.8.5. Роль генов в развитии
Связь между определенными участками молекулы
ДНК и морфологическим развитием усиленно
изучалась на организмах, обладающих гигантскими
(политенными) хромосомами. Такие хромосомы
имеются, например, в клетках слюнных желез у личинок
многих двукрылых, в том числе дрозофилы. Как эти
политенные хромосомы достигают таких размеров,
объясняется в разд. 23.5.1. Гигантские хромосомы
относительно легко изучать с помощью светового
микроскопа, и окраска по Фёльгену выявляет в них
специфическую для каждого участка поперечную
исчерченность. В процессе развития личинки
дрозофилы проходят несколько стадий («возрастов»),
разделенных линьками - периодами интенсивной
активности эпидермиса, за которой следует сбрасывание
старой кутикулы. Две последние линьки
сопровождаются особенно резкими изменениями; одна из
них ведет к образованию куколки, а другая - к выходу
взрослой особи (имаго). Для этих стадий
характерны интенсивный метаболизм и процессы дифферен-
цировки. Ход развития регулируется гормонами
(см. разд. 21.4). На определенных стадиях некото-
1 В последние годы выделен ряд веществ белковой
природы, обладающих индукторной активностью.
Представления же Пола и Гил мора сейчас уже не
рассматриваются: хорошо известно, что гистоны равномерно
распределены вдоль всей длины хромосом. Однако
негистоновые белки действительно играют важную роль в
регуляции работы генов.- Прим ред.
рые из поперечных полос на хромосомах
увеличиваются и образуют структуры, известные под
названием пуфов или колец Бальбиани (по имени
ученого, впервые обнаружившего их в 1890 г.).
Специфическое окрашивание, выявляющее РНК, и
радиоавтографические исследования с
использованием меченых предшественников РНК показали, что в
пуфах происходит синтез РНК. Было также
установлено, что величина пуфов находится в прямой
зависимости от скорости синтеза РНК. Полагают, что
пуфы образуются в результате раскручивания
молекул ДНК, разделения комплементарных цепей и
синтеза мРНК в процессе транскрипции.
В процессе развития особи наблюдается
определенная последовательность образования пуфов,
регулируемая гормоном линьки-экдизоном. На
разных личиночных стадиях и на стадии куколки пуфы
возникают в различных участках хромосом; это
позволяет предположить, что они соответствуют
структурным генам. Доказательства того, что это
участки генетической активности, представил Беер-
ман, изучавший личинок двух видов комаров,
принадлежащих к роду Chironomus. Некоторые клетки
слюнных желез у личинок одного вида зернистые, а
у другого вида-нет. Генетические карты,
построенные по данным скрещиваний между этими двумя
видами, показали, что аллели, определяющие эти
признаки, расположены вблизи центромеры одной
из хромосом, причем аллель зернистости
доминирует. Исследование хромосом этих видов показало,
что у вида, обладающего зернистыми клетками,
пуфы образуются только в том участке, где
локализован этот ген. Кроме того, у личинок,
гетерозиготных по признаку зернистости, пуф в этом
локусе возникает только в хромосоме, несущей до-
Рис. 22.35. Радиоавтографы
четвертой хромосомы личинки
Chironomus tentans, демонстрирующие
связь между пуфингом и синтезом
мРНК. Черные
точки-радиоактивный уридин, включенный в процессе
синтеза мРНК А. Как показывает
радиоавтограф, синтез РНК
сосредоточен в области пуфа. Б. После
добавления небольших количеств
актиномицина D, подавляющего
синтез мРНК, как пуфинг, так и
включение уридина выражены
слабо.
минантный аллель.
Дальнейшим подтверждением связи между
хромосомными пуфами и синтезом мРНК служат
результаты введения организмам, обладающим
гигантскими хромосомами, актиномицина D. Актино-
мицин D подавляет транскрипцию, препятствуя
синтезу мРНК, и у особей, которым вводили это
вещество, пуфы не образуются (рис. 22.35).
На рост и развитие растений и животных
оказывают также влияние гормоны. Во многих случаях их
действие осуществляется на уровне транскрипции,
т.е. синтеза мРНК. Конкретные механизмы
воздействия различных гормонов на транскрипцию и
на синтез белка весьма разнообразны. Некоторые
гормоны присоединяются к рецепторным участкам
клеточной мембраны. После связывания гормона
рецептором, который связан с лежащим под ним
ферментом аденилатциклазой, образуется
циклический АМФ, который переходит в цитоплазму и
действует как «второй посредник», индуцирующий
транскрипцию. Подробнее об этом механизме
говорится в разд. 16.6.1.
В этой главе мы попытались описать некоторые
процессы, связанные с непрерывностью жизни.
Живым системам, очевидно, необходима как
генетическая стабильность в малых временных
масштабах, так и генетическая пластичность в
долговременном плане. Стабильность обеспечивают механизмы
митоза, тогда как мейоз создает огромную
генетическую изменчивость, позволяющую организмам
приспосабливаться к меняющейся среде.
В следующей главе будут описаны особенности
наследования признаков, правила, которым оно
подчиняется, и пути распространения генетических
изменений в популяциях.
Глава 23
Изменчивость и генетика
Генетика по праву может считаться одной из самых
важных областей биологии. На протяжении
тысячелетий человек пользовался генетическими методами
для улучшения домашних животных и
возделываемых растений, не имея представления о механизмах,
лежащих в основе этих методов. Судя по
разнообразным археологическим данным, уже 6000 лет
назад люди понимали, что некоторые физические
признаки могут передаваться от одного поколения
другому. Отбирая определенные организмы из
природных популяций и скрещивая их между
собой, человек создавал улучшенные сорта растений
и породы животных, обладавшие нужными ему
свойствами.
Однако лишь в начале XX в. ученые стали
осознавать в полной мере важность законов
наследственности и ее механизмов. Хотя успехи микроскопии
позволили установить, что наследственные
признаки передаются из поколения в поколение через
сперматозоиды и яйцеклетки, оставалось неясным,
каким образом мельчайшие частицы протоплазмы
могут нести в себе «задатки» того огромного
множества признаков, из которых слагается каждый
отдельный организм.
Первый действительно научный шаг вперед в
изучении наследственности был сделан австрийским
монахом Грегором Менделем, который в 1866 г.
опубликовал статью, заложившую основы
современной генетики. Мендель показал, что
наследственные задатки не смешиваются, а передаются от
родителей потомкам в виде дискретных
(обособленных) единиц. Эти единицы, представленные у особей
парами, остаются дискретными и передаются
последующим поколениям в мужских и женских
гаметах, каждая из которых содержит по одной
единице из каждой пары. В 1909 г. датский ботаник
Иогансен назвал эти единицы генами, а в 1912 г.
американский генетик Морган показал, что они
находятся в хромосомах. С тех пор генетика
достигла больших успехов в объяснении природы
наследственности и на уровне организма, и на уровне гена.
23.1. Исследования Менделя
Грегор Мендель родился в Моравии в 1822 г.
В 1843 г. он поступил в монастырь августинцев в
Брюнне (ныне Брно, Чехословакия), где принял
духовный сан. Позже он отправился в Вену, где
провел два года, изучая в университете
естественную историю и математику, после чего в 1853 г.
вернулся в монастырь. Такой выбор предметов,
несомненно, оказал существенное влияние на его
последующие работы по наследованию признаков у
гороха. Будучи в Вене, Мендель заинтересовался
процессом гибридизации растений и, в частности,
разными типами гибридных потомков и их
статистическими соотношениями. Эти проблемы и
явились предметом научных исследований Менделя,
которые он начал летом 1856 г.
Успехи, достигнутые Менделем, частично
обусловлены удачным выбором объекта для
экспериментов - гороха огородного (Pisum sativum).
Мендель удостоверился, что по сравнению с другими
этот вид обладает следующими преимуществами:
1) имеется много сортов, четко различающихся по
ряду признаков;
2) растения легко выращивать;
3) репродуктивные органы полностью прикрыты
лепестками, так что растение обычно
самоопыляется; поэтому его сорта размножаются в чистоте,
т. е. их признаки из поколения в поколение
остаются неизменными;
4) возможно искусственное скрещивание сортов, и
оно дает вполне плодовитых гибридов.
Из 34 сортов гороха Мендель отобрал 22 сорта,
обладающие четко выраженными различиями по
ряду признаков, и использовал их в своих опытах со
скрещиванием. Менделя интересовали семь главных
признаков: высота стебля, форма семян, окраска
семян, форма и окраска плодов, расположение и
окраска цветков.
И до Менделя многие ученые проводили
подобные эксперименты на растениях, но ни один из них
не получил таких точных и подробных данных;
кроме того, они не смогли объяснить свои
результаты с точки зрения механизма наследственности.
Моменты, обеспечившие Менделю успех, следует
признать необходимыми условиями проведения
всякого научного исследования и принять их в качестве
образца. Условия эти можно сформулировать
следующим образом:
1) проведение предварительных исследований для
ознакомления с экспериментальным объектом;
2) тщательное планирование всех экспериментов, с
тем чтобы всякий раз внимание было
сосредоточено на одной переменной, что упрощает
наблюдения;
3) строжайшее соблюдение всех методик, с тем
чтобы исключить возможность введения
переменных, искажающих результаты (подробности
см. ниже);
4) точная регистрация всех экспериментов и запись
всех полученных результатов;
5) получение достаточного количества данных,
чтобы их можно было считать статистически
достоверными.
Как писал Мендель, «достоверность и полезность
всякого эксперимента определяются пригодностью
данного материала для тех целей, в которых он
используется».
Следует, однако, отметить, что в выборе
экспериментального объекта Менделю кое в чем и просто
повезло: в наследовании отобранных им
признаков не было ряда более сложных особенностей,
открытых позднее, таких как неполное
доминирование (разд. 23.7.1), зависимость более чем от одной
пары генов (разд. 23.7.6), сцепление генов (разд. 23.3).
23.1.1. Наследование при
моногибридном скрещивании
и закон расщепления
Для своих первых экспериментов Мендель выбирал
растения двух сортов, четко различавшихся по
какому-либо признаку, например по расположению
цветков: цветки могут быть распределены по всему
стеблю (пазушные) или находиться на конце стебля
(верхушечные). Растения, различающиеся по одной
паре альтернативных признаков, Мендель
выращивал на протяжении ряда поколений. Семена от
пазушных цветков всегда давали растения с
пазушными цветками, а семена от верхушечных цветков-
растения с верхушечными цветками. Таким
образом, Мендель убедился, что выбранные им растения
размножаются в чистоте (т.е. без расщепления
потомства) и пригодны для проведения опытов по
гибридизации (экспериментальных скрещиваний).
Его метод состоял в следующем: он удалял у ряда
растений одного сорта пыльники до того, как могло
произойти самоопыление (эти растения Мендель
называл «женскими»); пользуясь кисточкой, он
наносил на рыльца этих «женских» цветков пыльцу из
пыльников растения другого сорта; затем он
надевал на искусственно опыленные цветки
маленькие колпачки, чтобы на их рыльца не могла
попасть пыльца с других растений. Мендель
проводил реципрокные скрещивания - переносил
пыльцевые зерна как с пазушных цветков на верхушечные,
так и с верхушечных на пазушные. Во всех
случаях из семян, собранных от полученных гибридов,
вырастали растения с пазушными цветками. Этот
признак - «пазушные цветки», наблюдаемый у
растений первого гибридного поколения, Мендель
назвал доминантным; позднее, в 1902 г., Бэтсон и
Сондерс стали обозначать первое поколение
гибридного потомства символом F1. Ни у одного из
растений F1 не было верхушечных цветков.
На цветки растений F1 Мендель надел колпачки
(чтобы не допустить перекрестного опыления) и дал
им возможность самоопылиться. Семена,
собранные с этих растений F1, были пересчитаны и
высажены следующей весной для получения второго
гибридного поколения, F2 (поколение F2-это всегда
результат инбридинга в поколении F1, в данном
случае самоопыления). Во втором гибридном
поколении у одних растений образовались пазушные
цветки, а у других-верхушечные. Иными словами,
признак «верхушечные цветки», отсутствовавший в
поколении F1, вновь появился в поколении F2.
Мендель рассудил, что этот признак присутствовал
в поколении F1 в скрытом виде, но не смог
проявиться; поэтому он назвал его рецессивным. Из 858
растений, полученных Менделем в F2, у 651 были
пазушные цветки, а у 207-верхушечные. Мендель
провел ряд аналогичных опытов, используя всякий
раз одну пару альтернативных признаков.
Результаты экспериментальных скрещиваний по семи парам
таких признаков приведены в табл. 23.1. Во всех
случаях анализ результатов показал, что отношение
доминантных признаков к рецессивным в поколении
Таблица 23.1. Результаты экспериментов Менделя по наследованию семи пар альтернативных признаков.
(Наблюдаемое соотношение доминантных и рецессивных признаков приближается к теоретически ожидаемому 3:1)
Признак
Высота стебля
Семена
Окраска семян
Форма плодов
Окраска плодов
Положение цветков
Окраска цветков
Итого
Родительские растения
доминантный
признак
Высокий
Гладкие
Желтые
Плоские
Зеленые
Пазушные
Красные
рецессивный признак
Низкий
Морщинистые
Зеленые
Выпуклые и с
перетяжками
Желтые
Верхушечные
Белые
Поколение Fi
доминантные
787
5474
6022
882
428
651
705
14 949
рецессивные
277
1850
2001
299
152
207
224
5010
Отношение
2,84 : 1
2,96 : 1
3,01 : 1
2,95 : 1
2,82 : 1
3,14: 1
3,15: 1
2,98 : 1
А-пазушные цветки (доминантный признак)
а-верхушечные цветки (рецессивный признак)
Фенотипы родительских
особей
Генотипы родительских
особей (2п)
Мейоз
Гаметы (п)
Случайное
оплодотворение
Генотипы F1 (2п)
Гомозиготы с
пазушными цветкам*
Фенотипы F.
Все растения гетерозиготные с пазушными
цветками (аллели А и а остаются обособленными,
несмотря на доминирование А)
Самоопыление потомков F
Фенотипы F1
Генотипы F1 (2п)
Мейоз
Гаметы (п)
Случайное
оплодотворение
Генотипы F2(2n)
Пазушные цветки
Пазушные цветки
Гомозиготы
Гетерозиготы
Гомозиготы
Фенотипы F2
3 с пазушными цветками : 1 с верхушечными
цветками
Рис. 23.1. Полное генетическое объяснение одного
из моногибридных скрещиваний, проводившихся
Менделем. А-пазушные цветки (доминантный
признак), а-верхушечные цветки (рецессивный признак);
2п-диплоидное состояние; η-гаплоидное состояние
(см. разд. 22.3).
F2 составляло примерно 3:1.
Приведенный выше пример типичен для всех
экспериментов Менделя, в которых изучалось
наследование одного признака (моногибридные
скрещивания). Вкратце он сводится к следующему:
Наблюдения
Родительские
растения
F1
F2
Соотношение
BF2
С пазушными
цветками
χ С
верхушечными цветками
Все с пазушными цветками
С пазушными
цветками 651
3
С
верхушечными цветками 207
1
На основании этих и аналогичных результатов
Мендель сделал следующие выводы:
1. Поскольку исходные родительские сорта
размножались в чистоте (не расщеплялись), у сорта с
пазушными цветками должно быть два
«пазушных» фактора, а у сорта с верхушечными
цветками -два «верхушечных» фактора.
2. Растения F1 содержали по одному фактору,
полученному от каждого из родительских растений
через гаметы.
3. Эти факторы в F1 не сливаются, а сохраняют
свою индивидуальность.
4. «Пазушный» фактор доминирует над
«верхушечным» фактором, который рецессивен.
Разделение пары родительских факторов при
образовании гамет (так что в каждую гамету попадает
лишь один из них) известно под названием первого
закона Менделя, или закона расщепления. Согласно
этому закону, признаки данного организма
детерминируются парами внутренних факторов. В одной
гамете может быть представлен лишь один из
каждой пары таких факторов.
Теперь мы знаем, что эти факторы,
детерминирующие такие признаки, как расположение цветка,
соответствуют участкам хромосомы, называемым
генами.
Описанные выше эксперименты, проводившиеся
Менделем при изучении наследования одной пары
альтернативных признаков, служат примером
моногибридного скрещивания. Его можно описать при
помощи символов и связать с современными
представлениями о формировании гамет и
оплодотворении. Генотип принято обозначать первой
буквой слова, описывающего доминантный признак,
причем большой буквой обозначают доминантный
аллель, а маленькой-рецессивный. Все
используемые здесь генетические термины и символы
объяснены в табл. 23.2.
На рис. 23.1 показан правильный способ описания
моногибридного скрещивания или решения
генетических задач по наследованию одной пары
альтернативных признаков.
Отношение доминантных фенотипов к
рецессивным 3:1 характерно для расщепления при
моногибридном скрещивании. Выводы Менделя о
передаче каждой гаметой одного аллеля и о его фено-
типическом проявлении соответствуют
вероятностным законам. Вероятность того, что гамета,
образованная гетерозиготной родительской особью
F1, будет нести доминантный аллель А или
рецессивный аллель а, равна 50%, или 1/2. Если среди
гамет данной родительской особи гаметы каждого
из двух типов встречаются с вероятностью 1/2,
то вероятность каждой из четырех возможных
комбинаций гамет при оплодотворении составит
1/2 χ 1/2= 1/4. Статистическая вероятность встречи
гамет, содержащих аллели А и а, при случайном
оплодотворении показана на рис. 23.2. Вследствие
доминирования отношение фенотипов среди
потомков составит 3 доминантных фенотипа на 1
рецессивный фенотип. Как видно из табл. 23.1,
результаты, полученные Менделем в его экспериментах по
скрещиванию, подтверждают это теоретически
выведенное отношение.
23.1. При скрещивании чистой линии мышей с
коричневой шерстью с чистой линией мышей с
серой шерстью получаются потомки с коричневой
шерстью. В F2 от скрещивания между этими
χ Гомозиготы с
верхушечными цветками
Вероятность появления аллелей А и а в гетерозготах равна IJ
поэтому Л =1/2
а =1/2
Пользуясь этими оценками, вероятность появления в поколении F2 каждого
генотипа и фенотипа можно определить следующим образом:
Генотипы F\ (2п)
Мейоз
Гаметы (п)
Вероятность появления
Случайное оплодотворение
Генотипы F2(2n)
Фенотипы F,
3/4 доминантных :1/4 рецессивных,
т.е. 3 доминантных : 1 рецессивный
Рис. 23.2. Вероятностное объяснение менделевского
отношения 31 при моногибридном скрещивании.
мышами F1 получаются коричневые и серые
мыши в отношении 3:1.
а) Дайте полное объяснение этим результатам.
б) Каким будет результат скрещивания гетеро-
зиготы с коричневой шерстью из поколения F2
с серой особью из чистой родительской линии?
23.1.2. Возвратное, или
анализирующее, скрещивание
Организм из поколения F1, полученного от
скрещивания между гомозиготной доминантной и
гомозиготной рецессивной особями, гетерозиготен по
своему генотипу, но обладает доминантным
фенотипом. Для того чтобы проявился рецессивный
фенотип, организм должен быть гомозиготным по
рецессивному аллелю. В поколении F2 особи с
доминантным фенотипом могут быть как
гомозиготами, так и гетерозиготами. Если селекционеру
понадобилось выяснить генотип такой особи, то
единственным способом, позволяющим сделать это,
служит эксперимент с использованием метода,
называемого анализирующим (возвратным)
скрещиванием. Скрещивая организм неизвестного генотипа с
организмом, гомозиготным по рецессивному
аллелю изучаемого гена, можно определить этот
генотип путем одного скрещивания. Например, у
плодовой мушки Drosophila длинные крылья доминируют
над зачаточными. Особь с длинными крыльями
может быть гомозиготной (LL) или гетерозиготной
(L/). Для установления ее генотипа надо провести
анализирующее скрещивание между этой мухой и
мухой, гомозиготной по рецессивному аллелю (//).
Если у всех потомков от этого скрещивания будут
длинные крылья, то особь с неизвестным
генотипом-гомозигота по доминантному аллелю.
Численное соотношение потомков с длинными и с
зачаточными крыльями 1:1 указывает на гетеро-
зиготность особи с неизвестным генотипом (рис. 23.3).
23.2. Почему в анализирующем скрещивании
нельзя использовать особь, гомозиготную по
доминантному аллелю (например, TT)9 чтобы
определить генотип особи, обладающей доминантным
фенотипом? Проиллюстрируйте свой ответ
схемой скрещивания с применением надлежащих
генетических символов.
23.1.3. Дигибридное скрещивание
и закон независимого
распределения
Установив возможность предсказывать результаты
скрещиваний по одной паре альтернативных
признаков, Мендель перешел к изучению наследования
двух пар таких признаков. Скрещивания между
Таблица 23.2. Перечень наиболее употребительных генетических терминов (с пояснением на примере скрещиваний,
представленных на рис. 23.1)
Термин
Ген
Аллели
Локус
Гомозигота
Гетерозигота
Фенотип
Генотип
Доминантный
Рецессивный
Поколение F,
Поколение F2
Объяснение
Основная единица наследственности для данного
признака
Альтернативные формы одного и того же гена,
определяющие альтернативные признаки
Местоположение аллеля в хромосоме
Диплоид, содержащий два идентичных аллеля данного
гена
Диплоид, содержащий два разных аллеля данного гена
Физическое или химическое проявление исследуемого
признака
Имеющиеся у особи аллели в локусе, определяющем
данный признак
Аллель, определяющий фенотип как в гомозиготном,
так и в гетерозиготном состоянии
Аллель, определяющий фенотип только в
гомозиготном состоянии
Первое гибридное поколение
Второе гибридное поколение, полученное от
скрещивания двух особей из F,
Пример
Ген, определяющий положение
цветка
А или а
AA или аа
Aa
Пазушный цветок,
верхушечный цветок
AA, Aa, аа
А
а
L- длинные крылья (доминантный признак)
1 - зачаточные крылья (рецессивный признак)
Длиннокрылые родительские
особи гомозиготные
Фенотипы участников
анализирующего
скрещивания
Генотипы участников
анализирующего
скрещивания (2п)
Мейоз
Гаметы (п) (^
Случайное ^-
оплодотворение
Генотипы Tj
потомков (2η)
Фенотипы потомков
Длинные крылья
(гомозигота)
Зачаточные крылья
Все длиннокрылые (гетерозиготы)
Длиннокрылые родительские особи
гетерозиготные
Длинные крылья
(гетерозиготы)
Зачаточные крылья
Длинные крылья
(гетерозиготы)
Зачаточные крылья
(гомозиготы)
Рис. 23.3. Полное генетическое объяснение метода анализирующего скрещивания, с помощью
которого можно определить генотип организма с доминантным признаком. Указаны
фенотипы получающихся при этом потомков.
особями, различающимися по двум признакам,
называют дигибридными.
В одном из своих экспериментов Мендель
использовал растения гороха, различающиеся по форме и
окраске семян. Применяя метод, описанный в
разд. 23.1.1, он скрещивал между собой
чистосортные (гомозиготные) растения с гладкими желтыми
семенами и чистосортные растения с
морщинистыми зелеными семенами. У всех растений F1 (первого
поколения гибридов) семена были гладкие и
желтые. По результатам проведенных ранее
моногибридных скрещиваний Мендель уже знал, что эти
признаки доминантны; теперь, однако, его
интересовали характер и соотношение семян разных
типов в поколении F2, полученном от растений F1
путем самоопыления. Всего он собрал от растений
F2 556 семян, среди которых было
гладких желтых 315
морщинистых желтых 101
гладких зеленых 108
морщинистых зеленых 32
Соотношение разных фенотипов составляло
примерно 9:3:3:1 (дигибридное расщепление). На
основании этих результатов Мендель сделал два
вывода:
1. В поколении F2 появилось два новых сочетания
признаков: морщинистые и желтые; гладкие и
зеленые.
2. Для каждой пары аллеломорфных признаков
(фенотипов, определяемых различными аллелями)
получилось отношение 3:1, характерное для
моногибридного скрещивания-среди семян было
423 гладких и 133 морщинистых, 416 желтых и
140 зеленых.
Эти результаты позволили Менделю утверждать,
что две пары признаков (форма и окраска семян),
наследственные задатки которых объединились в
поколении F1, в последующих поколениях
разделяются и ведут себя независимо одна от другой. На
этом основан второй закон Менделя-принцип
независимого распределения, согласно которому
каждый признак из одной пары признаков может
сочетаться с любым признаком из другой пары.
Приведенный выше эксперимент можно описать с
помощью известных нам генетических символов
так, как это показано на рис. 23.4, Л. В результате
разделения (сегрегации) аллелей (R, г, Y и у) и их
независимого распределения (рекомбинации) в
каждой из мужских и женских гамет возможно одно из
четырех сочетаний аллелей. Для того чтобы
показать все возможные сочетания гамет,
возникающие при случайном оплодотворении, используют
запись в виде решетки Пённета, названной так
по имени кембриджского генетика; она позволяет
свести к минимуму ошибки, которые могли бы
возникнуть при составлении списка всех возможных
сочетаний гамет. При заполнении решетки Пённета
рекомендуется сначала внести все мужские гаметы в
клеточки по вертикальным столбцам, а затем все
«женские»-в клеточки горизонтальных строк.
Кроме того, определяя фенотипы особей F2, полезно
обозначать идентичные фенотипы какими-нибудь
хорошо различимыми значками (как это сделано на
рис. 23.4,5"). Как показывают рис. 23.4, Л и 2>,
основанные на первом и втором законах Менделя, при
каждом мужском или женском генотипе F1
возможно образование гамет со следующими
сочетаниями аллелей:
R может встречаться только в сочетании с Y или у
(но не с г), т. е. в виде RY или Ry;
г может встречаться только в сочетании с Y или у
(но не с R), т. е. в виде rY или гу.
Таким образом, для любой гаметы шанс получить
какое-то одно из четырех указанных здесь сочетаний
аллелей равен 1 из 4. Поскольку при
моногибридном скрещивании у 3/4 потомков F2 проявляется
доминантный аллель, а у 1/4-рецессивный,
вероятности проявления четырех рассматриваемых нами
R-гладкие семена (доминантный признак)
г-морщинистые семена (рецессивный признак)
Y — желтые семена (доминантный признак)
У — зеленые семена (рецессивный признак)
Фенотипы
родительских
особей
Генотипы родительских
особей (2п)
Мейоз
Гаметы ( η )
Случайное оплодотворение
Генотипы F1 (2 η )
Фенотипы F.
Гладкие желтые семена (гомозиготы)
Морщинистые зеленые семена (гомозиготы)
Все (ry)
Все (ту)
Все RrYy
Все семена гладкие и желтые
(гетерозиготы)
Гладкие желтые семена
RrYy
Фенотипы F1
Генотипы Fx(2n)
Мейоз
Гаметы ( η ) -
обозначены значками О и о
Гладкие желтые семена
RrYy
Случайное оплодотворение
(по решетке Пеннета)
Генотипы F2 (2п) указаны в
каждой клетке
Фенотипы F0
9 гладк. желт. : 3 гладк. зел.: 3 морщ. желт.: 1 морщ. зел.
Рис. 23.4. А. Формирование фенотипов F1 от скрещивания между гомозиготными родительскими
особями. Это пример дигибридного скрещивания, поскольку рассматриваются две пары
контрастирующих признаков. Б. Использование решетки Пеннета с целью показать все возможные сочетания гамет при
образовании генотипов в F2.
аллелей в F2 равны:
гладкие семена (доминантный 3/4
аллель)
желтые семена » » 3/4
морщинистые семена (рецессив- 1/4
ный аллель)
зеленые семена » » 1/4
Отсюда вероятности проявления соответствующих
возможных сочетаний аллелей у потомков F2
равны:
гладкие и желтые = 3/4-3/4 = 9/16
гладкие и зеленые = 3/4· 1/4 = 3/16
морщинистые и желтые = 1/4-3/4 = 3/16
морщинистые и зеленые = 1/41/4= 1/16
Результаты экспериментов Менделя со
скрещиванием сортов, различающихся по двум парам
альтернативных признаков, близки к результатам этих
теоретических расчетов.
23.3. У морской свинки (Cavia) имеются два
аллеля, определяющих черную или белую окраску
шерсти, и два аллеля, определяющих короткую
или длинную шерсть. При скрещивании между
гомозиготами с короткой черной шерстью и
гомозиготами с длинной белой шерстью у всех
потомков F1 шерсть была короткая и черная.
а) Какие аллели являются доминантными?
б) Каким будет соотношение различных
фенотипов в F2?
23.4. Окраска цветков у душистого горошка
определяется двумя парами аллелей, R, г и S, s.
При наличии по крайней мере одного
доминантного гена из каждой пары аллелей цветки
пурпурные; при всех других генотипах цветки белые.
Каким будет соотношение разных фенотипов в
потомстве от скрещивания двух растений RrSs с
пурпурными цветками?
23.1.4. Краткое изложение сути
гипотез Менделя
В сформулированных ниже положениях
используются термины, принятые в современной генетике.
1. Каждый признак данного организма
контролируется парой аллелей.
2. Если организм содержит два различных аллеля
для данного признака, то один из них
(доминантный) может проявляться, полностью подавляя
проявление другого (рецессивного).
3. При мейозе каждая пара аллелей разделяется
(расщепляется) и каждая гамета получает по
одному из каждой пары аллелей (принцип
расщепления).
4. При образовании мужских и женских гамет в
каждую из них может попасть любой аллель из
одной пары вместе с любым другим из другой
пары (принцип независимого распределения).
5. Каждый аллель передается из поколения в
поколение как дискретная неизменяющаяся
единица.
6. Каждый организм наследует по одному аллелю
(для каждого признака) от каждой из
родительских особей.
Описанный здесь механизм наследования при ди-
гибридном скрещивании, приведенные примеры и
типичное отношение 9:3:3:1 относятся только к
тем признакам, которые контролируются генами,
лежащими в разных хромосомах. Если же гены
лежат в одной и той же хромосоме, такое
независимое распределение наблюдается не всегда
(см. разд. 23.3).
23.2. Хромосомная теория
наследственности
Мендель опубликовал результаты своих
исследований и свои гипотезы в 1866 г. в журнале «Труды
Брюннского общества естествоиспытателей»,
который рассылался в научные общества многих стран.
Однако ученые не сумели оценить значение его
открытий; возможно, это объяснялось тем, что в то
время нельзя было связать данные Менделя с
какими-то конкретными структурами в гаметах, с
помощью которых наследственные факторы могли
бы передаваться от родителей потомкам.
К концу XIX в. в результате повышения
оптических качеств микроскопов и совершенствования
цитологических методов возможно стало наблюдать
поведение хромосом в гаметах и зиготах. Еще в 1875 г.
Гертвиг обратил внимание на то, что при
оплодотворении яиц морского ежа происходит слияние
двух ядер-ядра спермия и ядра яйцеклетки. В 1902 г.
Бовери продемонстрировал важную роль ядра в
регуляции развития признаков организма, а в 1882 г.
Флемминг описал поведение хромосом во время
митоза.
В 1900 г. законы Менделя были вторично
открыты и должным образом оценены почти
одновременно и независимо друг от друга тремя учеными -
Таблица 23.3. Соответствие между событиями,
происходящими при мейозе и оплодотворении, и гипотезами Менделя
Мейоз и оплодотворение
Диплоидные клетки
содержат пары гомологичных
хромосом
Гомологичные хромосомы
расходятся во время
мейоза
В каждую гамету попадает
одна из гомологичных
хромосом
Только ядро мужской
гаметы сливается с ядром
яйцеклетки
При оплодотворении пары
гомологичных хромосом
восстанавливаются;
каждая гамета (мужская или
женская) вносит одну из
гомологичных хромосом
Гипотезы Менделя
Признаки контролируются
парами факторов
Парные факторы
разделяются при образовании
гамет
Каждая гамета получает
один из пары факторов
Факторы передаются из
поколения в поколение как
дискретные единицы
Каждый организм
наследует по одному фактору
от каждой из
родительских особей
Рис. 23.5. Клетка с двумя парами гомологичных
хромосом. Положение двух разных генных локусов
показано черными кружками. В данном случае два
локуса расположены в разных парах гомологичных
хромосом и каждый ген представлен двумя аллелями.
де Фризом, Корренсом и Чермаком. Корренс
сформулировал выводы Менделя в привычной нам
форме двух законов и ввел термин «фактор», тогда как
Мендель для описания единицы наследственности
пользовался словом «элемент». Позднее американец
Уильям Сэттон заметил удивительное сходство
между поведением хромосом во время образования
гамет и оплодотворения и передачей менделевских
наследственных факторов (табл. 23.3).
На основании изложенных выше данных Сэттон и
Бовери высказали мнение, что хромосомы являются
носителями менделевских факторов, и
сформулировали так называемую хромосомную теорию
наследственности. Согласно этой теории, каждая пара
факторов локализована в паре гомологичных
хромосом, причем каждая хромосома несет по одному
фактору. Поскольку число признаков у любого
организма во много раз больше числа его хромосом,
видимых в микроскоп, каждая хромосома должна
содержать множество факторов.
В 1909 г. Иогансен заменил термин фактор,
означавший основную единицу наследственности,
термином ген. Альтернативные формы гена,
определяющие его проявление в фенотипе, назвали
аллелями. Аллели - это конкретные формы, которыми
может быть представлен ген, и они занимают одно и
то же место - локус - в гомологичных хромосомах
(рис. 23.5).
Менделевский закон расщепления можно теперь
объяснить расхождением гомологичных хромосом,
происходящим в анафазе I мейоза и случайным
распределением аллелей между гаметами. Эти
события схематически представлены на рис. 23.6.
23.2.1 Поведение хромосом
как основа независимого
распределения
Менделевский принцип независимого распределения
тоже можно объяснить особенностями
передвижения хромосом во время мейоза. При образовании
гамет распределение между ними аллелей, находя-
Гаплоидные
клетки
Профаза II
Хроматиды расходятся
Гаплоидные клетки, в каждой из
которых по одному аллелю от
исходной пары
Анафаза I
Гомологичные
хромосомы расходятся
Хроматиды расходятся
Гомологичные хромосомы
(каждая хромосома представлена
парой хроматид, несущих аллели А и а)
Рис. 23.6. Объяснение менделевского закона расщепления факторов (аллелей) А и а расхождением
гомологичных хромосом, происходящим в мейозе.
Профаза I *. две пары
гомологичных хромосом,
несущих аллели А, а, В, b
Во время метафазы I хромосомы
могут выстроиться на экваториальной
пластинке либо так, как показано вверху,
либо так, как внизу — в этом и состоит
независимое распределение
В результате последующего
деления ядер образуются
изображенные здесь гаметы
четырех типов
Рис. 23.7. Объяснение менделевского закона независимого распределения факторов (аллелей) A, я, B,
b независимым расхождением разных пар гомологичных хромосом в мейозе.
щихся в данной паре гомологичных хромосом,
происходит совершенно независимо от распределения
аллелей из других пар (рис. 23.7). Именно случайное
расположение пар гомологичных хромосом на
экваторе веретена в метафазе I мейоза и их последующее
разделение в анафазе I ведет к разнообразию
сочетаний аллелей в гаметах. Число возможных
комбинаций аллелей в мужских или женских гаметах можно
определить по общей формуле 2й, где η - гаплоидное
число хромосом. У человека η = 23, а возможное
число различных комбинаций равно 223 = 8 388 609.
23.5. Отложение крахмала в пыльцевых зернах
кукурузы контролируется наличием одного из
аллелей определенного гена. При наличии другого
аллеля этого гена отложения крахмала не
происходит. Объясните, почему половина пыльцевых
зерен, образуемых на данном растении кукурузы,
содержит крахмал, а в другой половине его нет.
23.6. Рассчитайте число возможных сочетаний
хромосом в пыльцевых зернах шафрана {Crocus
balansae), у которого диплоидное число хромосом
равно шести (2л = 6).
23.3. Сцепление
Все ситуации и примеры, обсуждавшиеся до сих пор
в этой главе, относились к наследованию генов,
находящихся в разных хромосомах. Как выяснили
цитологи, у человека все соматические клетки
содержат по 46 хромосом. Поскольку человек обладает
тысячами различных признаков-таких, например,
как группа крови, цвет глаз, способность секретиро-
вать инсулин,-в каждой хромосоме должно
находиться большое число генов.
Гены, лежащие в одной и той же хромосоме,
называют сцепленными. Все гены какой-либо одной
хромосомы образуют группу сцепления; они обычно
попадают в одну гамету и наследуются вместе.
Таким образом, гены, принадлежащие к одной
группе сцепления, обычно не подчиняются менделевско-
му принципу независимого распределения. Поэтому
при дигибридном скрещивании они не дают
ожидаемого отношения 9:3:3:1. В таких случаях
получаются самые разнообразные соотношения,
которые теперь, когда нам известна цитологическая
основа закономерностей, открытых Менделем,
можно довольно легко объяснить. (Здесь следует еще раз
подчеркнуть, что Менделю посчастливилось выбрать
для изучения наследования пары признаков,
определяемых генами разных хромосом.) У дрозофилы
гены, контролирующие окраску тела и длину крыла,
представлены следующими парами аллелей
(назовем соответствующие признаки): серое
тело-черное тело, длинные крылья - зачаточные (короткие)
крылья. Серое тело и длинные крылья доминируют.
Ожидаемое отношение фенотипов в F2 от
скрещивания между гомозиготой с серым телом и длинными
крыльями и гомозиготой с черным телом и
зачаточными крыльями должно составить 9:3:3: 1. Это
указывало бы на обычное менделевское
наследование при дигибридном скрещивании, обусловленное
случайным распределением генов, находящихся в
разных, негомологичных хромосомах. Однако
вместо этого в F2 были получены в основном
родительские фенотипы в отношении примерно 3:1. Это
можно объяснить, предположив, что гены окраски
тела и длины крыла локализованы в одной и той же
хромосоме, т.е. сцеплены (рис. 23.8).
Практически, однако, соотношение 3:1 никогда
не наблюдается, а возникают все четыре фенотипа.
Это объясняется тем, что полное сцепление
встречается редко. В большинстве экспериментов по
скрещиванию при наличии сцепления помимо мух с
родительскими фенотипами обнаруживаются особи с
новыми сочетаниями признаков. Эти новые
фенотипы называют рекомбинантными. Все это позволяет
дать следующее определение сцепления: два или
более генов называют сцепленными, если потомки
с новыми генными комбинациями (рекомбинанты)
встречаются реже, чем родительские фенотипы.
События, приведшие американского генетика
Томаса X. Моргана к открытию сцепления, можно
проиллюстрировать одним из его экспериментов на
дрозофиле, в котором он предсказывал результаты
возвратного скрещивания между серой
длиннокрылой гетерозиготой (из поколения F1 от
скрещивания, представленного на рис. 23.8) и рецессивной
G-cepoe тело (доминантный признак)
g-черноетело (рецессивный признак)
L-длинные крылья (доминантный признак)
1-зачаточные крылья (рецессивный признак)
Фенотипы
родительских
особей
Генотипы
родительских
особей (2п)
Серое тело,
длинные крылья
Черное тело,
зачаточные крылья
Мейоз
Гаметы (п)
Случайное
оплодотворение
Генотипы F1 (2п)
Фенотипы F. Все потомки — гетерозиготы с серым
телом и длинными крыльями
Скрещивание между потомками F.
Фенотипы F1
Серое тело,
длинные крылья
χ Серое тело,
длинные крылья
Генотипы F1 (2η )
Мейоз
Гаметы ( η )
Случайное
оплодотворение
Генотипы F (2η )
Фенотипы Fn
3 серое тело,
длинные крылья
1 черное тело,
зачаточные крылья
Рис. 23.8. Генетическое объяснение отношения
фенотипов 3:1, получающегося в F2 в результате
сцепления.
гомозиготой с черным телом и зачаточными
крыльями. Было предсказано два возможных результата:
1. Если две пары аллелей, определяющие серую
или черную окраску тела и длинные или зачаточные
крылья, лежат в разных парах хромосом (т.е. не
сцеплены), то они должны распределяться
независимо и давать следующее отношение фенотипов:
1 серое тело, длинные крылья
1 серое тело, зачаточные крылья
1 черное тело, длинные крылья
1 черное тело, зачаточные крылья
2. Если аллели, определяющие окраску тела и
длину крыльев, лежат в одной и той же паре
хромосом (т. е. сцеплены), то отношение фенотипов
будет иным:
1 серое тело, длинные крылья
1 черное тело, зачаточные крылья
Объяснение этих предсказаний представлено на
рис. 23.9.
Морган провел такое возвратное скрещивание
несколько раз и ни разу не получил ни один из
предсказанных результатов. Всякий раз он получал
следующие результаты:
41,5%-серое тело, длинные крылья
41,5%-черное тело, зачаточные крылья
8,5%-серое тело, зачаточные крылья
8,5%-черное тело, длинные крылья
На основании этих результатов Морган
постулировал, что
1) изучаемые гены находятся в хромосомах;
2) оба гена находятся в одной хромосоме, т.е.
сцеплены;
3) аллели каждого гена расположены в
гомологичных хромосомах;
4) во время мейоза между гомологичными
хромосомами происходил обмен генами.
Появление рекомбинантных сочетаний аллелей у
17% потомков было объяснено на основании
пункта 4. Это явление получило название кроссинговера
(перекреста).
23.7. Гомозиготное растение с пурпурными
цветками и коротким стеблем скрестили с
гомозиготным растением с красными цветками и
длинным стеблем; у гибридов F1 были пурпурные
цветки и короткий стебель. При анализирующем
скрещивании растения F1 с двойной гомозиготой
по рецессивным генам были получены следующие
потомки:
52 с пурпурными цветками и коротким стеблем
47 с пурпурными цветками и длинным стеблем
49 с красными цветками и коротким стеблем
45 с красными цветками и длинным стеблем.
Дайте полное объяснение этим результатам.
А Если все четыре аллеля находятся в разных парах хромосом:
Фенотипы
участников
анализирующего
скрещивания
Серое тело,
длинные крылья
(гетерозиготы)
χ Черное тело,
зачаточные крылья
(гомозиготы)
Генотипы участников
анализирующего
скрещивания (2 η )
Мейоз
Гаметы ( η )
(обозначены
сГи j )
Случайное
оплодотворение
(по решетке
Пеннета)
gi
GL
GL
gl
Gl
Gl
sl
gL
gL
gl
gl
gl
gl
Фенотипы потомков (2п)
(указаны в каждой клетке)
Фенотипы потомков
1 серое тело, длинные крылья:
1 серое тело, зачаточные крылья:
1 черное тело, длинные крылья:
1 черное тело, зачаточные крылья
Б Если четыре аллеля находятся в одной паре хромосом:
Фенотипы участников
анализирующего
скрещивания
Серое тело,
длинные крылья
(гетерозиготы)
Черное тело,
зачаточные крылья
(гомозиготы)
Генотипы участников
анализирующего
скрещивания (2п)
Мейоз
Гаметы (п )
Случайное
оплодотворение
Генотипы потомков (2п)
Фенотипы потомков
1 серое тело,
длинные крылья
: 1 черное тело,
зачаточные крылья
Рис. 23.9. Генетическое объяснение предсказаний,
сделанных Морганом.
23.3.1. Кроссинговер и частота
рекомбинаций
В 1909 г. бельгийский цитолог Янссенс наблюдал
образование хиазм во время профазы I мейоза (см.
разд. 22.3). Генетическое значение этого процесса
разъяснил Морган, высказавший мнение, что
кроссинговер (обмен аллелями) происходит в результате
разрыва и рекомбинации гомологичных хромосом
во время образования хиазм. В дальнейшем
сопоставление цитологических данных с данными о
соотношениях рекомбинантных фенотипов
подтвердило, что обмен генетическим материалом в мейозе
происходит почти буквально между всеми
гомологичными хромосомами. Аллели, входящие в
группы сцепления у родительских особей, разделяются и
образуют новые сочетания, которые попадают в
гаметы,-процесс, называемый генетической
рекомбинацией. Потомков, которые получаются из таких
гамет с «новыми» сочетаниями аллелей, называют
рекомбинантами. Таким образом, кроссинговер
представляет собой важный источник генетической
изменчивости, наблюдаемой в популяциях.
Для иллюстрации принципа кроссинговера можно
рассмотреть поведение пары гомологичных
хромосом дрозофилы, несущих аллели серой окраски тела
и длинных крыльев (оба аллеля доминантные) и
черной окраски тела и зачаточных крыльев (оба
аллеля рецессивные), во время образования хиазм.
Скрещивание между гомозиготным серым
длиннокрылым самцом и гомозиготной черной самкой с
зачаточными крыльями дало в F1 гетерозиготных
потомков с серым телом и длинными крыльями
(рис. 23.10).
При возвратном скрещивании мух из поколения
F1 с гомозиготными двойными рецессивами были
получены следующие результаты:
Родительские |
фенотипы <
Рекомбинант- /
ные фено- <
типы '
' серое тело, длинные крылья
, черное тело, зачаточные
крылья
' черное тело, длинные крылья
, серое тело, зачаточные
крылья
965
944
206
185
Как показывают эти результаты, гены,
определяющие окраску тела и длину крыльев, сцеплены.
(Вспомните, что если бы эти гены находились в
разных хромосомах и поэтому распределялись
случайным образом, то при дигибридном скрещивании
гетерозиготы F1 с гомозиготой по двум
рецессивным признакам получилось бы соотношение
фенотипов 1:1:1:1.) Из приведенных цифр можно
вычислить частоту рекомбинации генов,
определяющих окраску тела и длину крыльев.
Частота рекомбинаций вычисляется по формуле
Число рекомбинантов
Общее число потомков
•100.
В нашем примере частота рекомбинации равна
(206 + 185)
(965 + 944) + (206 + 185)
100 =
391
2300
100= 17%.
Это значение соответствует числу рекомбинаций,
происходящих при образовании гамет. Один из
учеников Моргана, А. X. Стертевант, высказал мысль,
что частоты рекомбинаций свидетельствуют о ли-
Серое тело,
длинные крылья
(гетерозиготы)
Черное тело,
зачаточные крылья
(гомозиготы)
Фенотипы участников
анализирующего
скрещивания
Генотипы участников
анализирующего
скрещивания (2п)
Мейоз
(имеет место
кроссинговер)
Гаметы (п)
Генотипы
потомков (2η )
Рекомбинантныегенотипы (х)
Все генотипы (у)
Рис. 23.10. Генетическое объяснение процесса
кроссинговера и появления рекомбинантных генотипов.
Подсчитав число особей, у которых выявляется
рекомбинация (х), и общее число особей (у), можно
вычислить частоту рекомбинаций по формуле:
χ
Частота рекомбинации (%) = -· 100.
У
нейном расположении генов вдоль хромосомы. Еще
более важное предположение Стертеванта состояло
в том, что частота рекомбинаций отражает
относительное расположение генов в хромосоме: чем
дальше друг от друга находятся сцепленные гены, тем
больше вероятность того, что между ними
произойдет кроссинговер, т.е. тем выше частота
рекомбинантов (рис. 23.11).
23.8. На приведенной ниже схеме изображены
локусы двенадцати аллелей, расположенные в
паре хромосом. Показаны их относительные
расстояния от центромеры.
Коричневое тело
Волосатое тело
Толстое тело
Слияние жилок
Рубиновые глаза
Миниатюрные крылья
Желтое тело
Гладкое тело
Тонкое тело
Сплетение жилок
Розовые глаза
Миниатюрные
крылья
Центромера
Центромера
а) Как называют изображенные здесь
хромосомы?
б) Между какими двумя локусами чаще всего
будет происходить кроссинговер?
в) Произойдет ли кроссинговер между генами,
определяющими цвет глаз и форму антенн?
Обоснуйте свой ответ.
Рис. 23.11. А, В и С-три генных локуса.
расположенных в одной хромосоме. Вероятность крос-
синговера и разделения для генов AuC выше, чем для
генов BuC или AuB, так как частота крос-
синговера зависит от расстояния между генами.
23.4 Генетические карты
Данные о частотах рекомбинаций важны прежде
всего потому, что дают генетикам возможность
составлять карты относительного расположения
генов в хромосомах. Хромосомные карты строятся
путем прямого перевода частоты рекомбинаций
между генами в предполагаемые расстояния на
хромосоме. Если частота рекомбинации между
генами А и В равна 4%, то это означает, что они
расположены в одной и той же хромосоме на
расстоянии 4 морганид друг от друга (морганида-
единица расстояния на генетической карте); если
частота рекомбинации между генами А я С равна
9%, то они разделены расстоянием 9 морганид.
Однако эти данные еще не позволяют сказать,
в каком порядке расположены гены А, В и С
(рис. 23.12).
Рис. 23.12. Определение возможного положения
генов А, В и С в хромосоме, основанное на
данных, изложенных в тексте.
На практике обычно определяют частоту
рекомбинации по меньшей мере для трех генов
одновременно; этот метод, называемый триангуляцией,
позволяет определить не только расстояния между
генами, но и их последовательность. Рассмотрим,
например, частоты рекомбинации, установленные в
результате ряда экспериментальных скрещиваний
при участии четырех генов:
P-Q = 24%
R-P = 14%
R-S = 8%
S-P = 6%
Для того чтобы установить последовательность
генов и расстояния между ними, вычерчивают
линию, изображающую хромосому, и производят
следующие действия.
1. В середину хромосомы помещают гены с
наименьшей частотой рекомбинации, т. е. S-P =
= 6% (рис. 23.13,/).
2. Выбирают следующую по величине частоту
рекомбинации, т.е. R-S = 8%, и указывают два
возможных положения R в хромосоме по от-
Рис. 23.13. Установление положений генов P, Q, R и
S в хромосоме с помощью метода
триангуляции.
ношению к 5 (рис. 23.13,77).
3. Проделывают то же самое со следующей
частотой рекомбинации, т.е. R-P= 14%. При этом
выясняется, что R не может находиться вправо от
P (рис. 23.13,777).
4. Проделывают то же самое для P-Q = 24%
(рис. 23.13,7F). Положение Q не может быть
установлено без дополнительной информации.
Если, например, окажется, что частота
рекомбинации Q-R= 10%, то это подтвердит
расположение гена Q в левом конце хромосомы.
При построении хромосомных карт могут
возникнуть затруднения, создаваемые двойным кроссинго-
вером; особенно это касается тех случаев, когда
изучаемые гены разделены большими
расстояниями, так как число выявляемых рекомбинантов
бывает при этом меньше фактического числа
перекрестов. Если, например, произойдет кроссинговер в
двух местах-между А и В и между В и С (рис. 23.14),
то А и С внешне проявят себя как сцепленные, но
хромосома будет нести теперь рецессивный аллель Ъ.
23.9. У кукурузы гены окрашенного семени и
гладкого эндосперма доминируют над генами
неокрашенного семени и сморщенного
эндосперма. Линию, гомозиготную по обоим
доминантным признакам, скрещивали с линией,
гомозиготной по рецессивным признакам; при
анализирующем скрещивании F2 с растениями F1 были
получены следующие результаты:
окрашенные семена, гладкий эн- 380
досперм
неокрашенные семена, сморщен- 396
ный эндосперм
окрашенные семена, сморщенный 14
эндосперм
неокрашенные семена, гладкий 10
эндосперм
Рис. 23.14. /. Пара гомологичных хроматид, одна из
которых несет доминантные аллели Af B и С,
а другая рецессивные аллели а, Ь и с. Кроссинго-
вер происходит в двух точках-*1 и *2. //.
Результат разделения хроматид: последовательности
аллелей в них иные, хотя последовательность
генных локусов и расстояния между ними
остаются прежними.
Вычислите расстояние в морганидах между
генами окраски семян и характера эндосперма.
23.5. Группы сцепления
и хромосомы
Изложенные в этой главе данные демонстрируют
постепенный рост наших знаний о механизмах
наследственности. Генетические исследования,
проводившиеся в начале нашего века, в основном были
направлены на выяснение роли генов в передаче
признаков. Работы Моргана с плодовой мушкой
Drosophila melanogaster показали, что большинство
фенотипических признаков объединено у нее в
четыре группы сцепления и признаки каждой группы
наследуются совместно. Было замечено, что число
групп сцепления соответствует числу пар хромосом.
Изучение других организмов привело к сходным
результатам. При экспериментальном скрещивании
разнообразных организмов обнаружилось, что
некоторые группы сцепления больше других (т.е. в
них больше генов). Изучение хромосом этих
организмов показало, что они имеют разную длину.
Морган доказал наличие четкой связи между этими
наблюдениями. Они послужили дополнительными
подтверждениями локализации генов в хромосомах.
клетках. По каким-то причинам эти хромосомы
многократно удваиваются, но не отделяются друг
от друга, до тех пор пока их не наберется несколько
тысяч, лежащих бок о бок. Окрасив хромосомы и
изучая их с помощью светового микроскопа, можно
увидеть, что они состоят из чередующихся светлых
и темных поперечных полос (рис. 23.15). Для
каждой хромосомы характерен свой особый рисунок
полос. Первоначально предполагали, или скорее
надеялись, что эти полосы представляют собой
гены, но оказалось, что дело обстоит не так просто.
У дрозофилы можно искусственным путем
вызывать различные фенотипические аномалии, которые
сопровождаются определенными изменениями в
рисунке поперечных полос, видимых под
микроскопом. Эти фенотипические и хромосомные аномалии
коррелируют в свою очередь с генными локусами,
обозначенными на хромосомных картах, которые
были построены по данным о частоте рекомбинаций
в экспериментальных скрещиваниях. Это позволяет
сделать вывод, что полосы на хромосомах
действительно как-то связаны с генами, но
взаимоотношения между теми и другими остаются пока неясными.
23.5.1. Гигантские хромосомы и гены
В 1913 г. Стертевант начал свою работу по
картированию положения генов в хромосомах дрозофилы,
но это было за 21 год до того, как появилась
возможность связать различимые в хромосомах
структуры с генами. В 1934 г. было замечено, что в
клетках слюнных желез дрозофилы хромосомы
примерно в 100 раз крупнее, чем в других соматических
Рис. 23.15. Гигантские хромосомы из клеток
слюнных желез Drosophila melanogaster. Видны четыре пары
хромосом, соединенных своими центромерами.
Самка
Самец
Рис. 23.16. Хромосомные наборы самца и самки
D. melanogaster. Они состоят из четырех пар
хромосом (пара I-половые хромосомы)
Рис. 23.17. Вид половых хромосом человека в мета-
фазе митоза.
23.6. Определение пола
Особенно четким примером описанного выше
метода установления зависимости между фенотипи-
ческими признаками организмов и строением их
хромосом служит определение пола. У дрозофилы
фенотипические различия между двумя полами явно
связаны с различиями в хромосомах (рис. 23.16).
При изучении хромосом у самцов и самок ряда
животных между ними были обнаружены некоторые
различия. Как у мужских, так и у женских особей во
всех клетках имеются пары одинаковых
(гомологичных) хромосом, но по одной паре хромосом
они различаются. Это половые хромосомы (гетеро-
сомы). Все остальные хромосомы называют аутосо-
мами. Как можно видеть на рис. 23.16, у дрозофилы
четыре пары хромосом. Три пары (II, III и IV)
идентичны у обоих полов, но пара I, состоящая из
идентичных хромосом у самки, различается у самца.
Эти хромосомы называют X- и Y-хромосомами;
генотип самки XX, а самца-XY. Такие различия по
половым хромосомам характерны для большинства
животных, в том числе для человека (рис. 23.17), но
у птиц (включая кур) и у бабочек наблюдается
обратная картина: у самок имеются хромосомы XY,
а у самцов-XX. У некоторых насекомых, например
у прямокрылых, Y-хромосомы нет вовсе, так что
самец имеет генотип XO.
При гаметогенезе наблюдается типичное менде-
левское расщепление по половым хромосомам.
Например, у млекопитающих каждое яйцо содержит
одну Х-хромосому, половина спермиев - одну X-
хромосому, а другая половина - одну Y-хромосому
(рис. 23.18). Пол потомка зависит от того, какой
спермий оплодотворит яйцеклетку. Пол с
генотипом XX называют гомогаметным, так как у него
образуются одинаковые гаметы, содержащие
только Х-хромосомы, а пол с генотипом XY-гетерога-
метным, так как половина гамет содержит X-, а
половина-Y-хромосому. У человека генотипичес-
кий пол данного индивидуума определяют, изучая
неделящиеся клетки. Одна Х-хромосома всегда
оказывается в активном состоянии и имеет обычный
вид. Другая, если она имеется, бывает в покоящемся
состоянии, в виде плотного темно-окрашенного
тельца, называемого тельцем Барра. Число телец Барра
всегда на единицу меньше числа наличных Х-хромо-
сом, т.е. у самца (XY) их нет вовсе, а у самки
(XX)-только одно. Функция Y-хромосомы,
очевидно, варьирует в зависимости от вида. У человека
Y-хромосома контролирует дифференцировку
семенников, которая в дальнейшем влияет на
развитие половых органов и мужских признаков (см.
разд. 20.3.1). У большинства организмов, однако,
Y-хромосома не содержит генов, имеющих
отношение к полу. Ее даже называют генетически инертной
или генетически пустой, так как в ней очень мало
генов. Как полагают, у дрозофилы гены,
определяющие мужские признаки, находятся в аутосомах, и их
фенотипические эффекты маскируются наличием
пары Х-хромосом; в присутствии одной Х-хромосомы
мужские признаки проявляются. Это пример
наследования, ограниченного полом (в отличие от
наследования, сцепленного с полом), при котором,
например, у женщин подавляются гены,
детерминирующие рост бороды.
Морган и его сотрудники заметили, что
наследование окраски глаз у дрозофилы зависит от пола
родительских особей, несущих альтернативные
аллели. Красная окраска глаз доминирует над белой.
При скрещивании красноглазого самца с белоглазой
самкой в F1 получали равное число красноглазых
самок и белоглазых самцов (рис. 23.19, Л). Однако
при скрещивании белоглазого самца с красноглазой
самкой в F1 были получены в равном числе
красноглазые самцы и самки (рис. 23.19,2>). При
скрещивании этих мух F1 между собой были получены
красноглазые самки, красноглазые и белоглазые
Фенотипы родительских
особей
Генотипы родительских
особей (2 η )
Мейоз
Гаметы (п) *
Случайное
оплодотворение
Генотипы потомков(2η)
Фенотипы потомков
Самка
Самец
Отношение полов 1 самка : 1 самец
Рис. 23.18. Генетическое объяснение соотношения
полов у человека.
R- красные глаза (доминантный признак)
г - белые глаза (рецессивный признак)
XX-самка (9)
XY-самец (d")
Красноглазые
) XRY
Фенотипы родителей
Генотипы родителей
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы F{(2n)
Фенотипы F.
Красноглазые
Белоглазые
Белоглазыес
Белоглазые
Красноглазые
Красноглазые 9
Красноглазые cf
Фенотипы F.
Генотипы Fy(2n)
Мейоз
Гаметы (η)
Случайное оплодотворение
Генотипы FJ2 п)
Фенотипы F2
Красноглазые <
Красноглазые
Белоглазые*
Красноглазые
Краен.
Бел.
Краен .
Краен.
Бел.
Краен.
Бел.
Рис. 23.19. AuB. Реципрокные экспериментальные скрещивания между красноглазой и
белоглазой дрозофилами, проведенные Морганом. Обратите внимание на низкую частоту появления белых глаз.
В. Инбридинг между красноглазым самцом F1 и красноглазой (гетерозиготной) самкой F1
подтвердил полученные результаты. Г. Скрещивание между белоглазым самцом и
красноглазой (гетерозиготной) самкой F1. Обратите внимание на то, что признак белоглазости
наблюдается только у самок, гомозиготных по этому аллелю.
самцы, но не было ни одной белоглазой самки
(рис. 23.19, В). Тот факт, что у самцов частота
проявления рецессивного признака была выше, чем у
самок, наводил на мысль, что рецессивный аллель,
определяющий белоглазость, находится в Х-хромо-
соме, а Y-хромосома лишена гена окраски глаз.
Чтобы проверить эту гипотезу, Морган скрестил
исходного белоглазого самца с красноглазой
самкой из F1 (рис. 23.19, Г). В потомстве были
получены красноглазые и белоглазые самцы и самки.
Из этого Морган справедливо заключил, что только
Х-хромосома несет ген окраски глаз. В Y-хромосо-
ме соответствующего локуса вообще нет. Это
явление известно под названием наследования,
сцепленного с полом.
23.10. У дрозофилы гены длины крыльев и
окраски глаз сцеплены с полом. Нормальная
длина крыла и красные глаза доминируют над
миниатюрными крыльями и белыми глазами.
а) Каким будет потомство F1 и F2 при
скрещивании между самцом с миниатюрными крыльями
и красными глазами и гомозиготной самкой с
нормальными крыльями и белыми глазами?
Объясните ожидаемые результаты.
б) Скрещивание между самкой из полученного
выше поколения и белоглазым самцом с
миниатюрными крыльями дало следующие
результаты:
Самцы и самки с нормальными
крыльями и белыми глазами 35
Самцы и самки с нормальными
крыльями и красными глазами 17
Самцы и самки с миниатюрными
крыльями и белыми глазами 18
Самцы и самки с миниатюрными
крыльями и красными глазами 36
Объясните появление этих фенотипов и их
численные соотношения.
23.6.1. Наследование, сцепленное
с полом
Гены, находящиеся в половых хромосомах,
называют сцепленными с полом. В Х-хромосоме имеется
участок, для которого в Y-хромосоме нет гомолога
(рис. 23.20). Поэтому у особей мужского пола
признаки, определяемые генами этого участка,
проявляются даже в том случае, если они рецессивны.
Эта особая форма сцепления позволяет объяснить
наследование признаков, сцепленных с полом,
например цветовой слепоты, раннего облысения и
гемофилии у человека. Гемофилия - сцепленный с
полом рецессивный признак, при котором
нарушается образование фактора VIII, ускоряющего
свертывание крови. Ген, детерминирующий синтез
фактора VIII, находится в участке Х-хромосомы, не
имеющем гомолога, и представлен двумя
аллелями - доминантным нормальным и рецессивным му-
тантным. Возможны следующие генотипы и фе-
нотипы:
Генотип
ХНХН
XHXh
Хну
XhY
Фенотип
Нормальная женщина
Нормальная женщина
(носитель)
Нормальный мужчина
Мужчина-гемофилик
Особей женского пола, гетерозиготных по любому
из сцепленных с полом признаков, называют
носителями соответствующего рецессивного гена. Они
фенотипически нормальны, но половина их гамет
несет рецессивный ген. Несмотря на наличие у отца
нормального гена, сыновья матерей-носителей с
вероятностью 50% (-J будут страдать гемофилией.
От брака женщины-носителя с нормальным
мужчиной могут родиться дети с различными фенотипами
(рис. 23.21).
Один из наиболее хорошо документированных
примеров наследования гемофилии мы находим в
родословной потомков английской королевы
Виктории. Предполагают, что ген гемофилии возник в
результате мутации у самой королевы Виктории или
у одного из ее родителей. На рис. 23.22 показано,
как этот ген передавался ее потомкам.
23.11. Окраска тела у кошек и у крыжовниковой
пяденицы (Abraxas grossulariata) контролируется
сцепленным с полом геном, локализованным в
Х-хромосоме. В двух экспериментальных
скрещиваниях, в которых гомогаметный пол в
родительском поколении был гомозиготным по
окраске тела, получены следующие разультаты:
Фенотипы
родительских
особей
Соотношение
фенотипов в
потомстве
Пяденица
(нормальная
окраска
доминирует над
бледной)
Бледный
самец χ
Нормальная самка
1 нормальный
самец
1 нормальная
самка
Кошка
(черная
окраска
доминирует над рыжей)
Черный самец
χ Рыжая
самка
1 рыжий
самец
1 черная
самка
Какой пол является гетерогаметным у каждого из
этих организмов?
23.7. Взаимодействие между
генами
До сих пор в этой главе рассматривались
относительно простые аспекты генетики: доминирование,
моногибридное и дигибридное скрещивание,
сцепление, определение пола и наследование, сцепленное с
Участок, не имеющий гомолога
в Y-хромосоме
Гомологичные
участки
половых хромосом
Рис. 23.20. Гомологичные и негомологичные участки
половых хромосом.
Н— нормальный аллель (доминантный)
h - аллель гемофилии (рецессивный)
XX-женские хромосомы
XY-мужские хромосомы
Фенотипы родителей
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Здоровая женщина
(носитель)
Здоровый мужчина
Генотипы потомков (2п)
Фенотипы потомков
Здоровая
женщина
Здоровый
мужчина
Здоровая
женщина
(носитель)
Мужчина с
гемофилией
Рис. 23.21. Механизм наследования аллеля
гемофилии, сцепленного с полом.
полом. Известны, однако, и другие взаимодействия
между генами, и возможно, что именно они
определяют большую часть фенотипических признаков
организма. Ниже будут описаны некоторые из этих
взаимодействий.
23.7.1. Неполное доминирование
Известны случаи, когда два или более аллелей не
проявляют в полной мере доминантность или
рецессивность, так что в гетерозиготном состоянии ни
один из аллелей не доминирует над другим. Это
явление неполного доминирования, или кодоминант-
ность, представляет собой исключение из
описанного Менделем правила наследования при
моногибридных скрещиваниях. К счастью, Мендель выбрал
для своих экспериментов признаки, которым не
свойственно неполное доминирование; в противном
случае оно могло бы сильно осложнить его первые
исследования.
Неполное доминирование наблюдается как у
растений, так и у животных. В большинстве случаев
гетерозиготы обладают фенотипом, промежуточ-
Королева Виктория
Принц Альберт
Фридрих X Виктория
Король
Эдуард
Королева Александра
Алиса
Людвиг IV Гессенский
Леопольд,
герцог Олбани
Елена
Беатриса
Генрих Баттенбургский
София
Георг V
Вальдемар Сигизмунд
Ирен
Гессенская
Принц
Фредерик
Виктория
Принц Луи
Баттенбу pre кий
Царица
Александра I
Царь
Николай
Алиса
Александр Леопольд Морис
Виктория,
королева
Испании
Альфонс XHI ,
король Испании
Георг VI
Алиса
Альфонс
Гонсало
Александр
Елизавета II
Генрих,
умер 4 лет
Вальдемар,
дожил до 56 лет
Царевич
Алексей
Морис, умер
в раннем
детстве
Руперт
(лорд Трематон)
Принц Филип
Эндрю Эдуард
Диана Чарлз
Анна | Марк Филлипс
Уильям
Генрих
Питер
Зара
Истекли кровью
после несчастных
случаев
Здоровая женщина
Женщина-носитель
Здоровый мужчина
Мужчина- возможный
гемофилик
Мужчина-гемофил и к
Рис. 23.22. Наследование гемофилии у потомков королевы Виктории. На схеме указаны только те потомки, которые участвовали в передаче
гемофилии или были поражены ею. Родословная британского королевского дома продолжена, чтобы показать, почему гемофилия не проявилась
здесь ни у одного из потомков королевы Виктории на протяжении семи поколений.
ным между фенотипами доминантной и рецессивной
гомозигот. Примером служат андалузские куры,
полученные в результате скрещивания
чистопородных черных и «обрызганных белых» (splashed white)
кур. Черное оперение обусловлено наличием аллеля,
определяющего синтез черного пигмента меланина.
У «обрызганных» кур этот аллель отсутствует. У
гетерозигот меланин развивается не в полной мере,
создавая лишь голубоватый отлив на оперении.
Поскольку общепринятых символов для
обозначения аллелей с неполным доминированием не
существует, нам необходимо ввести для генотипов
такие символы, чтобы сделать понятными
приведенные ниже схемы получения андалузских кур.
Возможны, например, такие обозначения: черные-
В; «обрызганные»-6, W1 В™ или Z?BW. Результаты
скрещивания между гомозиготными черными и
«обрызганными» курами представлены на рис. 23.23.
При скрещивании между собой особей F1
отношение фенотипов в F2 отличается от менделевско-
го отношения 3:1, типичного для моногибридного
скрещивания. В этом случае получается отношение
1:2:1, где у половины особей F2 будет такой же
генотип, как у F1 (рис. 23.24). Отношение 1:2:1
характерно для результатов скрещиваний при
неполном доминировании. Другие примеры
приведены в табл. 23.4.
23.12. У кошек гены, контролирующие окраску
шерсти, находятся в Х-хромосоме и проявляют
неполное доминирование (кодоминантность). При
скрещивании черной кошки с рыжим котом в F1
были получены черные самцы и черепаховые
самки. Каково ожидаемое отношение фенотипов в
F2? Объясните полученные результаты.
23.7.2. Множественные аллели
Во всех рассмотренных до сих пор случаях каждый
признак контролировался одним геном, который
мог быть представлен одной из двух аллельных
форм. Известно, однако, немало примеров, когда
один признак проявляется в нескольких различных
формах, контролируемых тремя и более аллелями,
из которых любые два могут находиться в
соответствующих локусах гомологичных хромосом. В
Таблица 23.4. Примеры неполного доминирования
Объект
Львиный зев
(Antirrhinum)
Mirabilis
Скот шортгорн-
ской породы
Кролики породы
реке и
ангорские
Гомозиготные
фенотипы
Красные цветки;
белые цветки
Красные цветки;
белые цветки
Рыжая масть;
белая масть
Длинная шерсть;
короткая
шерсть
Гетерозиготный
фенотип
Розовые цветки
Розовые цветки
Чалая масть
Промежуточная
шелковистая
шерсть
таких случаях говорят о множественных аллелях.
Такие признаки, как окраска шерсти у мышей, цвет
глаз у мышей и группы крови у человека,
контролируются множественными аллелями.
Наследование групп крови
Группа крови контролируется аутосомным геном.
Локус этого гена обозначают буквой / (от слова
«изогемагглютиноген»), а три его аллеля буквами А,
В и О. Аллели А и В доминантны в одинаковой
степени, а аллель О рецессивен по отношению к ним
обоим. Генотипы, представленные в табл. 23.5,
определяют фенотипическое проявление групп
крови. При наличии одного доминантного аллеля в
крови образуется вещество, называемое
агглютинином, которое действует как антитело. Например,
при генотипе /А/° на мембране эритроцитов
образуется агглютиноген А, а в плазме содержится
агглютинин анти-В (кровь соответствует группе А).О
методе определения групп крови см. в разд. 14.14.6.
23.13. а. Используя надлежащие генетические
символы, укажите возможные группы крови у
детей, оба родителя которых гетерозиготны,
причем у отца группа крови А, а у матери-В.
б. Если эти родители произведут на свет разно-
яйцовых близнецов, то какова вероятность
того, что у обоих детей кровь будет относиться
к группе В?
23.7.3. Летальные гены
Известны случаи, когда один ген может оказывать
влияние на несколько признаков, в том числе на
жизнеспособность. У человека и других
млекопитающих определенный рецессивный ген вызывает
образование внутренних спаек легких, что приводит
к смерти при рождении. Другим примером служит
ген, который влияет на формирование хряща и
вызывает врожденные уродства, ведущие к смерти
плода или новорожденного.
У кур, гомозиготных по аллелю, вызывающему
«курчавость» перьев, неполное развитие перьев
влечет за собой несколько фенотипических
эффектов. У таких кур теплоизоляция недостаточна, и они
страдают от охлаждения. Для компенсации потери
тепла у них появляется ряд структурных и физиоло-
Таблица 23.5. Генотипы групп крови у человека
Генотип
JA /А
/A JO
JB /В
JB JO
JA JB
JO JO
Группа крови (фенотип)
А
А
В
В
AB
О
B - аллель черной окраски
Bw- аллель "обрызганной" окраски
Фенотипы родителей
Генотипы родителей (2η )
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы F1 (2п)
Фенотипы F1
Черные >
(гомозиготы)
BB >
0 0 >
BBW BBW
с "Обрызганные" белые
(гомозиготы)
( RWRW
с (£w) (g)
BBW BBW
Все куры — "голубые" гетерозиготы
Рис. 23.23. Скрещивание кур андалузской породы:
гибриды F1
Фенотипы F1
Генотипы F1 (2 η)
Мейоз
Гаметы (п)
"Голубые" >
BBW >
0 @ >
Случайное оплодотворение
Генотипы F2 (2п)
Фенотипы F2
BB BBW
Черные
1
< "Голубые"
< BBW
< (В) (BW)
BBW Bw Bw
Голубые
: 2
"Обрызганные"
белые
: 1
Рис. 23.24. Скрещивание кур андалузской породы:
гибриды F2
гических адаптации, но эти адаптации
малоэффективны и среди таких кур высока смертность.
Воздействие летального гена ясно видно на
примере наследования окраски шерсти у мышей.
У диких мышей шерсть обычно серая, типа агути;
но у некоторых мышей шерсть желтая. При
скрещиваниях между желтыми мышами в потомстве
получаются как желтые мыши, так и агути в
отношении 2:1. Единственное возможное объяснение таких
результатов состоит в том, что желтая окраска
шерсти доминирует над агути и что все желтые
мыши гетерозиготны. Атипичное менделевское
отношение объясняется гибелью гомозиготных
желтых мышей до рождения (рис. 23.25). При вскрытии
беременных желтых мышей, скрещенных с желтыми
же мышами, в их матках были обнаружены мертвые
желтые мышата. Если же скрещивались желтые
мыши и агути, то в матках беременных самок не
оказывалось мертвых желтых мышат, поскольку
при таком скрещивании не может быть потомства,
гомозиготного по гену желтой шерсти (YY).
23.7.4. Генные комплексы
В некоторых случаях в каком-то генном локусе
присутствует пара аллелей, полностью
контролирующая один фенотипический признак. Однако
такие случаи сравнительно редки. Большинство при-
γ—желтая шерсть (доминантный признак)
у—агути (рецессивный признак)
Фенотипы родителей
Генотипы родителей (2п)
Мейоз
Гаметы (п )
Случайное оплодотворение
Генотипы потомков (2η )
Фенотипы потомков
Желтая ι
шерсть )
Yy
® Θ :
YY .Yy Yy,
1 желтые
(погибают
в утробе
матери)
х Желтая шерсть
< Yy
® ©
УУ
2 желтые : 1 агути,
Рис. 23.25. Генетическое объяснение наследования
окраски шерсти при скрещивании желтых мышей друг
с другом (YY-летальный генотип)
знаков определяется взаимодействием нескольких
генов, образующих генный комплекс. Например,
признак может контролироваться двумя или
большим числом взаимодействующих генов,
находящихся в разных локусах. Так, наследование формы
гребня у петухов контролируется генами,
находящимися в двух локусах и притом в разных
хромосомах; в результате взаимодействия этих генов
возникают четыре разных фенотипа, известных под
названиями гороховидного, розовидного, ореховидного и
простого гребня (рис. 23.26). Развитие
гороховидного и розовидного гребня определяется
соответственно наличием доминантного аллеля P или R при
отсутствии другого доминантного аллеля.
Ореховый гребень образуется в результате
модифицированной формы неполного доминирования, когда
имеется по меньшей мере по одному аллелю
гороховидного и розовидного гребня (т. е. при генотипе
PR). Простой гребень развивается только у
гомозигот по обоим рецессивным генам (т. е. рргг). Эти
фенотипы и генотипы представлены в табл. 23.6.
Генотипы F2 и отношение фенотипов в F2 от
скрещивания между гомозиготной курицей с
гороховидным гребнем и гомозиготным петухом с розо-
видным гребнем показаны на рис. 23.27.
23.14. У кур аллель белого оперения (W)
доминирует над аллелем черного оперения (w). Аллель
P дает гороховидный, а аллель /?-розовидный
гребень; при одновременном наличии этих двух
аллелей образуется ореховидный гребень, а их
рецессивные аллели в гомозиготном состоянии
обусловливают развитие простого гребня.
При скрещивании черного петуха с розовидным
гребнем и белой курицы с ореховидным гребнем
были получены потомки со следующими
фенотипами: 3 белых с ореховидным гребнем, 3 черных с
ореховидным, 3 белых с розовидным, 3 черных с
розовидным, 1 белый с гороховидным, 1 черный с
гороховидным, 1 белый с простым и 1 черный с
простым гребнем.
Укажите генотипы родительских особей.
Объясните подробно, каким образом могли получиться
указанные фенотипы.
23.7.5. Эпистаз
Ген называют эпистатическим (от греч. epi-над),
если его присутствие подавляет эффект какого-либо
гена, находящегося в другом локусе. Эпистати-
ческие гены иногда называют ингибирующими
генами, а те гены, действие которых ими
подавляется,-гипостатическими (от греч. hypo-под).
Окраска шерсти у мышей контролируется парой
генов, находящихся в разных локусах. Эпистати-
ческий ген определяет наличие окраски и имеет два
аллеля: доминантный, определяющий окрашенную
шерсть, и рецессивный, обусловливающий
альбинизм (белая окраска). Гипостатический ген
определяет характер окраски и имеет два аллеля: агути
(доминантный, определяющий серую окраску) и
черный (рецессивный). Мыши могут иметь серую
или черную окраску в зависимости от своих
генотипов, но наличие окраски возможно только в том
случае, если у них одновременно имеется аллель
окрашенной шерсти. Мыши, гомозиготные по
рецессивному аллелю альбинизма, будут альбиносами
даже при наличии у них аллелей агути и черной
шерсти. Возможны три разных фенотипа: агути,
черная шерсть и альбинизм. При скрещивании
можно получить эти фенотипы в различных
соотношениях в зависимости от генотипов скрещиваемых
особей (рис. 23.28 и табл. 23.7).
23.15. У кур-белых леггорнов - окраска
оперения контролируется двумя группами генов:
W (белая окраска) доминирует над w (цветная)
В (черная окраска) доминирует над b
(коричневая)
Гетерозиготное потомство F1 имеет генотип
WwBb и белую окраску.
Объясните происходящее в этом случае
взаимодействие между генами и укажите численные
отношения фенотипов в поколении F2.
23.7.6. Полигенное наследование
Многие из самых заметных признаков организма
представляют собой результат совместного дейст-
Таблица 23.6. Возможные генотипы кур с различной формой
гребня
Фенотип
Гороховидный гребень
Розовидный гребень
Ореховидный гребень
Простой гребень
Возможные генотипы
РРгг Рргг
RRpp Rrpp
PPRR, PpRr, PPRr, PpRR
рргг
Рис. 23.26. Изменчивость формы гребня у кур.
А-простой гребень. Б-гороховидный, В—розовидный, Г—ореховидный.
Ρ— наличие гороховидного гребня (доминантный аллель)
р—отсутствие гороховидного гребня (рецессивный аллель)
R— наличие розовидного гребня (доминантный аллель)
г — отсутствие розовидного гребня (рецессивный аллель)
Фенотипы родителей
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы F1 (2п)
Фенотипы Fx
Фенотипы F1
Мейоз
Гаметы (п) (обозначены
О* HJ )
Случайное оплодотворение
Генотипы F2 (2 η) (указаны
в клетках решетки Пеннета)
Гороховидный
РРгг
Розовидный
RRpp
Все PpRr
У всех потомков
гребень ореховидный
Ореховидный гребень
Ореховидный гребень
Фенотипы F2
Символы
9 ореховидный : 3
гороховидный : 3 розовидный : 1 простой
Рис. 23.27. Генетическое объяснение наследования формы гребня у кур.
вия многих различных генов; эти гены образуют
особый генный комплекс, называемый полигенной
системой. Хотя вклад каждого отдельного гена,
входящего в такую систему, слишком мал, чтобы
оказать сколько-нибудь значительное влияние на
фенотип, почти бесконечное разнообразие,
создаваемое совместным действием этих генов (полигенов),
составляет генетическую основу непрерывной
изменчивости (см. разд. 23.8.2).
23.8. Изменчивость
Изменчивостью называют всю совокупность
различий по тому или иному признаку между
организмами, принадлежащими к одной и той же природной
популяции или виду. Поразительное
морфологическое разнообразие особей в пределах любого вида
привлекло внимание Дарвина и Уоллеса во время их
путешествий. Закономерный, предсказуемый
характер передачи таких различий по наследству
послужил основой для исследований Менделя. Дарвин
установил, что определенные признаки могут
развиваться в результате отбора (см. разд. 24.4.2), тогда
как Мендель объяснил механизм, обеспечивающий
передачу из поколения в поколение признаков, по
которым ведется отбор.
Мендель описал, каким образом наследственные
факторы определяют генотип организма, который в
процессе развития проявляется в структурных,
физиологических и биохимических особенностях
фенотипа. Если фенотипическое проявление любого
признака обусловлено в конечном счете генами,
контролирующими этот признак, то на степень развития
Таблица 23.7. Некоторые соотношения фенотипов,
получающиеся в потомстве при эпистатическом взаимодействии генов
(см. также рис. 23.28)
Фенотипы
родительских особей
Агути χ агути
Агути χ
черная
Агути χ
альбинос
Агути χ
альбинос
Агути χ
альбинос
Агути χ
черная
Альбинос χ
черная
Альбинос χ
черная
Генотипы
родительских особей
AaCc χ
AaCc
AaCc χ
ааСс
AaCc χ
Аасс
AaCc χ
аасс
AACc χ
аасс
AaCc χ
ааСс
ААсс χ
ааСС
ААсс χ
ааСс
Отношение фенотипов
9 агути : 3 черных : 4
альбиноса
3 агути: 3 черных : 2
альбиноса
3 агути : 1 черная : 4
альбиноса
1 агути: 1 черная : 2
альбиноса
1 агути: 1 альбинос
1 агути : 1 черная
Все агути
1 агути : 1 альбинос
А— окраска агути (доминантный признак)
а—черная (рецессивный признак)
C- окрашенная шерсть (доминантный признак)
с— альбинизм (рецессивный признак)
Фенотипы родителей
Генотипы родителей
Мейоз
Гаметы (п)
(обозначены о*и ρ)
Случайное оплодотворение
Генотипы потомков (2п)
(указаны в клетках решетки
Пеннета)
Фенотипы потомков
Символы
Агути
Альбиносы
3 агути : 4 альбиносы : 1 черные
Рис. 23.28. Генетическое объяснение необычных соотношений фенотипов в случае эпистоза.
определенных признаков может оказывать влияние
среда.
Изучение фенотипических различий в любой
большой популяции показывает, что существуют две
формы изменчивости-дискретная и непрерывная.
Для изучения изменчивости какого-либо признака,
например роста у человека, необходимо измерить
этот признак у большого числа индивидуумов в
изучаемой популяции. Результаты измерений
представляют в виде гистограммы, отражающей
распределение частот различных вариантов этого
признака в популяции. На рис. 23.29 представлены
типичные результаты, получаемые при таких
исследованиях, и они наглядно демонстрируют различие
между дискретной и непрерывной изменчивостью.
23.8.1. Дискретная изменчивость
Некоторые признаки в популяции представлены
ограниченным числом вариантов. В этих случаях
различия между особями четко выражены, а
промежуточные формы отсутствуют; к таким
признакам относятся, например, группы крови у человека,
длина крыльев у дрозофилы, меланистическая и
светлая формы у березовой пяденицы (Biston be-
tularia), длина столбика у первоцвета (Primula) и пол
у животных и растений. Признаки, для которых
характерна дискретная изменчивость, обычно
контролируются одним или двумя главными генами, у
которых может быть два или несколько аллелей, и
внешние условия относительно мало влияют на их
фенотипическую экспрессию.
Поскольку дискретная изменчивость ограничена
некоторыми четко выраженными признаками, ее
называют также качественной изменчивостью в
отличие от количественной, или непрерывной,
изменчивости.
23.8.2. Непрерывная изменчивость
По многим признакам в популяции наблюдается
полный ряд переходов от одной крайности к другой
без всяких разрывов. Наиболее яркими примерами
служат такие признаки, как масса (вес), линейные
размеры, форма и окраска организма в целом или
отдельных его частей. Частотное распределение по
признаку, проявляющему непрерывную
изменчивость, соответствует кривой нормального
распределения (см. Приложение 2.7.3, т. 1). Большинство
членов популяции попадает в среднюю часть
кривой, а на ее концах, соответствующих двум крайним
значениям данного признака, находится примерно
одинаковое (очень малое) число особей. Признаки,
для которых характерна непрерывная изменчивость,
обусловлены совместным воздействием многих
генов (полигенов) и факторов среды. Каждый из этих
генов в отдельности оказывает очень небольшое
влияние на фенотип, но совместно они создают
значительный эффект.
Число особей
Число особей
Признак
Признак
Рис. 23.29. Гистограммы, отражающие
распределение частот в случае прерывистой (А) и
непрерывной (Б) изменчивости.
23.8.3. Влияние среды
Главный фактор, детерминирующий любой фено-
типический признак,-это генотип. Генотип
организма определяется в момент оплодотворения, но
степень последующей экспрессии этого генетического
потенциала в значительной мере зависит от
внешних факторов, воздействующих на организм во
время его развития. Так, например, использованный
Менделем сорт гороха с длинным стеблем обычно
достигал высоты 180 см. Однако для этого ему
необходимы были соответствующие условия -
освещение, снабжение водой и хорошая почва. При
отсутствии оптимальных условий (при наличии
лимитирующих факторов) ген высокого стебля не мог в
полной мере проявить свое действие. Эффект
взаимодействия генотипа и факторов среды
продемонстрировал датский генетик Иогансен. В ряде
экспериментов на карликовой фасоли он выбирал из
каждого поколения самоопылявшихся растений
самые тяжелые и самые легкие семена и высаживал их
для получения следующего поколения. Повторяя
эти эксперименты на протяжении нескольких лет, он
обнаружил, что в пределах «тяжелой» или «легкой»
селекционной линии семена мало различались по
среднему весу, тогда как средний вес семян из
разных линий сильно различался. Это позволяет
считать, что на фенотипическое проявление
признака оказывают влияние как наследственность, так
и среда. На основании этих результатов можно
определить непрерывную фенотипическую
изменчивость как «кумулятивный эффект варьирующих
факторов среды, воздействующих на вариабельный
генотип». Кроме того, эти результаты показывают,
что степень наследуемости данного признака
определяется в первую очередь генотипом. Что касается
развития таких чисто человеческих качеств, как
индивидуальность, темперамент и интеллект, то, судя
по имеющимся данным, они зависят как от
наследственных, так и от средовых факторов, которые,
взаимодействуя в различной степени у разных
индивидуумов, влияют на окончательное выражение
признака. Именно эти различия в тех и других
факторах создают фенотипические различия между
индивидуумами. Мы пока еще не располагаем
данными, которые твердо указывали бы на то, что
влияние каких-то из этих факторов всегда
преобладает, однако среда никогда не может вывести
фенотип за пределы, детерминированные
генотипом.
23.8.4. Источники изменчивости
Необходимо ясно представлять себе, что
взаимодействие между дискретной и непрерывной
изменчивостью и средой делает возможным существование
двух организмов с идентичным фенотипом.
Механизм репликации ДНК при митозе столь близок к
совершенству, что возможности генетической
изменчивости у организмов с бесполым
размножением очень малы. Поэтому любая видимая
изменчивость у таких организмов почти наверное
обусловлена воздействиями внешней среды. Что же касается
организмов, размножающихся половым путем, то у
них есть широкие возможности для возникновения
генетических различий. Практически
неограниченными источниками генетической изменчивости
служат два процесса, происходящие во время мейоза:
1. Реципрокный обмен генами между хромати-
дами гомологичных хромосом, который может
происходить в профазе I мейоза. Он создает новые
группы сцепления, т. е. служит важным источником
генетической рекомбинации аллелей (см. разд. 23.3
и 22.3).
2. Ориентация пар гомологичных хромосом
(бивалентов) в экваториальной плоскости веретена в
метафазе I мейоза определяет направление, в
котором каждый член пары будет перемещаться в
анафазе I. Эта ориентация носит случайный
характер. Во время метафазы II пары хроматид опять-
таки ориентируется случайным образом, и этим
определяется, к какому из двух противоположных
полюсов направится та или иная хромосома во
время анафазы II. Случайная ориентация и
последующее независимое расхождение (сегрегация)
хромосом делают возможным большое число
различных хромосомных комбинаций в гаметах; число это
можно подсчитать (см. разд. 23.2.1).
Третий источник изменчивости при половом
размножении-это то, что слияние мужских и женских
гамет, приводящее к объединению двух гаплоидных
наборов хромосом в диплоидном ядре зиготы,
происходит совершенно случайным образом (во всяком
случае, в теории); любая мужская гамета
потенциально способна слиться с любой женской
гаметой.
Эти три источника генетической изменчивости и
обеспечивают постоянную «перетасовку» генов,
лежащую в основе происходящих все время
генетических изменений. Среда оказывает воздействие на
весь ряд получающихся таким образом фенотипов,
и те из них, которые лучше всего приспособлены к
данной среде, преуспевают. Это ведет к изменениям
частот аллелей и генотипов в популяции (см. гл. 25).
Однако эти источники изменчивости не порождают
крупных изменений в генотипе, которые
необходимы, согласно эволюционной теории, для
возникновения новых видов. Такие изменения возникают
в результате мутаций.
23.9. Мутации
Мутацией называют изменение количества или
структуры ДНК данного организма. Мутация
приводит к изменению генотипа, которое может быть
унаследовано клетками, происходящими от мутант-
ной клетки в результате митоза или мейоза. Мути-
Родительская клетка
(2п),гдеп = 1
Нерасхождение
Пара
гомологичных
хромосом
Гаметы (η)
Слияние
гамет
/Нормальная
гаплоидная
гамета
Нормальная
гаплоидная
гамета
Трисомия
Моносомия
Рис. 23.30. Нерасхождение хромосом при гамето-
генезе и результаты слияния образующихся при этом
аномальных гамет с нормальными гаплоидными
гаметами. Это приводит к различным формам
полисомии, при которой число хромосом может
быть равно 2п+1 (трисомия), 2п + 2 (тетрасомия),
2п + 3 (пентасомия) и т. д. или же 2п-1
(моносомия)-в зависимости от числа гомологичных
хромосом, не разошедшихся нормальным образом.
рование может вызывать изменения каких-либо
признаков в популяции. Мутации, возникшие в
половых клетках, передаются следующим поколениям
организмов, тогда как мутации в соматических
клетках наследуются только дочерними клетками,
образовавшимися путем митоза, и такие мутации
называют соматическими.
Мутации, возникающие в результате изменения
числа или макроструктуры хромосом, известны под
названием хромосомных мутаций или хромосомных
аберраций (перестроек). Иногда хромосомы так
сильно изменяются, что это можно увидеть под
микроскопом. Но термин «мутация» используют
главным образом для обозначения изменения
структуры ДНК в одном локусе, когда происходит так
называемая генная, или точечная, мутация.
Представление о мутации как о причине
внезапного появления нового признака было впервые
выдвинуто в 1901 г. голландским ботаником Гуго де
Фризом, изучавшим наследственность у энотеры
Oenothera lamarckiana. Спустя 9 лет T. Морган начал
изучать мутации у дрозофилы, и вскоре при участии
генетиков всего мира у нее было идентифицировано
более 500 мутаций.
23.9.1. Частота мутаций и их причины
Мутации возникают случайным образом и
спонтанно, т.е. любой ген может мутировать в любой
момент. Чатота возникновения мутаций у разных
организмов различна, но, по-видимому, связана с
продолжительностью жизненного цикла: у
организмов с коротким жизненным циклом она выше.
В результате работ Г. Дж. Мёллера в двадцатые
годы нашего века было установлено, что частоту
мутаций можно повысить по сравнению с их
спонтанным уровнем, воздействуя на организмы
рентгеновскими лучами. В дальнейшем выяснилось, что
частоту мутаций можно также значительно
повысить с помощью ультрафиолетовых лучей и гамма-
лучей. Частицы высокой энергии, такие как а- и
β-частицы, нейтроны и космическое излучение, тоже
мутагенны, т. е. способны вызывать мутации.
Мутагенным действием обладают и разнообразные
химические вещества, в частности иприт, кофеин,
формальдегид, колхицин, некоторые компоненты
табака и все возрастающее число лекарственных
препаратов, пищевых консервантов и пестицидов.
23.9.2. Хромосомные мутации
Хромосомные мутации включают изменения числа
хромосом и их макроструктуры. Некоторые типы
хромосомных мутаций изменяют действие
некоторых генов и оказывают на фенотип гораздо более
глубокое влияние, чем генные мутации. Изменения
числа хромосом обычно происходят в результате
ошибок при мейозе, но они возможны и при
нарушениях митоза. Эти изменения выражаются либо
в анэуплоидии - утрате или добавлении отдельных
хромосом, либо в полиплоидии-добавлении целых
гаплоидных наборов хромосом.
Анэуплоидия
Анэуплоидия может выражаться, например, в
наличии добавочной хромосомы (п + 1, 2л + 1 и т. п.)
или в нехватке какой-либо хромосомы (п — 1, 2л — 1
и т.п.). Анэуплоидия может возникнуть, если в
анафазе I мейоза гомологичные хромосомы одной
или нескольких пар не разойдутся. В этом случае
оба члена пары направляются к одному и тому же
полюсу клетки, и тогда мейоз приводит к
образованию гамет, содержащих на одну или несколько
хромосом больше или меньше, чем в норме (рис. 23.30).
Это явление известно под названием нерасхождения.
Когда гамета с недостающей или лишней
хромосомой сливается с нормальной гаплоидной гаметой,
образуется зигота с нечетным числом хромосом:
вместо каких-либо двух гомологов в такой зиготе их
может быть три или только один.
Зигота, в которой число хромосом меньше
диплоидного, обычно не развивается, но зиготы с
лишними хромосомами иногда способны к развитию.
Однако из таких зигот в большинстве случаев
развиваются особи с резко выраженными аномалиями.
Одна из наиболее часто встречающихся
хромосомных мутаций, возникающих у человека в результате
нерасхождения,-трисомия-21, или синдром Дауна
(2л = 47). Эта аномалия, названная так по имени
врача, впервые описавшего ее в 1866 г., вызывается
нерасхождением хромосом 21 (рис. 23.31). К числу
ее симптомов относятся задержка умственного раз-
Таблица 23. 8. Фенотипические отклонения, обусловленные
нерасхождением половых хромосом у человека
Синдром, генотип
Синдром,
Клайнфельте-
ра, XXY
Синдром
Тернера, XO
XXX
XYY
Симптомы
Мужчина обладает
некоторыми вторичными
женскими половыми
признаками; бесплоден;
яички слабо развиты,
волос на лице мало;
иногда развиваются
молочные железы; обычно
низкий уровень
умственного развития
У женщин отсутствуют
обычные вторичные
половые признаки; низкий
рост, соски сближены
Женщины внешне
нормальны, плодовиты, но
отмечается умственная
отсталость
Мужчины высокого роста
с различным уровнем
умственного развития;
иногда обладают
психопатическими чертами
или проявляют
склонность к мелким
правонарушениям
Частота
среди населения
в странах
Запада, %
0,02
0,03
0,12
0,1
вития, пониженная сопротивляемость болезням,
врожденные сердечные аномалии, короткое
коренастое туловище и толстая шея, а также
характерные складки кожи над внутренними углами глаз, что
создает внешнее сходство с представителями
монголоидной расы. Синдром Дауна и другие сходные
хромосомные аномалии чаще встречаются у детей,
рожденных немолодыми женщинами. Точная
причина этого неизвестна, но, по-видимому, она как-то
связана с возрастом яйцеклеток матери.
Возможно также нерасхождение мужских и
женских половых хромосом, которое приводит к анэу-
плоидии, влияющей на вторичные половые
признаки и плодовитость, а иногда и на умственное
развитие (табл. 23.8)1.
Полиплоидия
Гаметы и соматические клетки с увеличенным
числом хромосом, кратным гаплоидному числу,
называют полиплоидными; приставки три-, тетра- и
т.д. указывают, во сколько раз увеличено число
хромосом, т.е. степень плоидности: Ъп-триплоид,
1 Возникает, очевидно, и трисомия по другим
хромосомам, но ее носители нежизнеспособны и погибают в
раннем эмбриогенезе. Прим. ред.
4л - тетраплоид, 5л-пентаплоид и т. д. У растений
полиплоидия встречается гораздо чаще, чем у
животных. Например, из 300 тысяч известных видов
покрытосеменных примерно
половина-полиплоиды. Относительная редкость полиплоидии у
животных объясняется тем, что увеличенное число
хромосом значительно повышает вероятность
ошибок при мейозе во время гаметогенеза. Что касается
растений, то большинство из них способно к
вегетативному размножению, поэтому они могут
эффективно воспроизводиться и в полиплоидном
состоянии. Полиплоидные организмы часто обладают
благоприятными признаками - более крупными
размерами, выносливостью, устойчивостью к
заболеваниям. Большинство наших культурных растений
полиплоиды, образующие крупные плоды,
запасающие органы, цветки или листья.
Существуют две формы полиплоидии-аутополи-
плоидия и аллополиплоидия.
Аутополиплоидия может возникать как
естественным, так и искусственным путем в результате
увеличения числа хромосомных наборов у данного
вида. Например, если произойдет репликация
хромосом (в интерфазе) и хроматиды нормально
разойдутся в (анафазе), но цитоплазма не разделится,
то образуется тетраплоидная (4я) клетка с большим
ядром. Такая клетка затем делится и дает тетра-
плоидные дочерние клетки. Количество цитоплазмы
Рис. 23.31. Хромосомы женщины с синдромом
Дауна. Нерасхождение хромосом G21 в одной из
гамет привело к трисомии по этой хромосоме.
Полный набор хромосом индивидуума, подобный
представленному на этой фотографии,
называют кариотипом.
в этих клетках возрастает, чтобы сохранилось
постоянство ядерно-плазменного отношения, и это
ведет к увеличению общих размеров растения или
какой-либо его части. Аутополиплоидизацию
можно вызвать с помощью алкалоида колхицина,
выделяемого из клубнелуковицы безвременника (Colchi-
сит). В концентрации порядка 0,01% колхицин
подавляет образование веретена деления, разрушая
микротрубочки, так что хромосомы не расходятся
во время анафазы. Колхицин и близкие к нему
вещества использовали для выведения
определенных сортов хозяйственно ценных культур, таких как
табак, томаты и сахарная свекла. Аутополиплоиды
обычно менее плодовиты, чем диплоиды, так как во
время мейоза гомологичные хромосомы должны
объединяться более чем по две.
У животных иногда наблюдается видоизмененная
форма полиплоидии, при которой образуются
отдельные полиплоидные клетки и ткани. Это обычно
происходит при удвоении хромосом, не
сопровождающемся разделением клетки. Гигантские
хромосомы в клетках слюнных желез дрозофилы
возникают в результате многократной репликации
ДНК без перехода к митозу.
Аллополиплоидией называют удвоение числа
хромосом у стерильного гибрида, в результате чего он
становится плодовитым. Гибриды F1, получаемые
при скрещиваниях между разными видами, обычно
стерильны, так как их хромосомы не могут
образовать гомологичные пары во время мейоза. Если,
однако, число хромосом становится кратным
исходному гаплоидному их числу, например равным
2(/I1 + л2), 3(W1 + л2) и т.д. (где п1 и л2-гашюидные
числа хромосом у родительских видов), то
возникает новый вид, который дает плодовитых гибридов
при скрещивании с такими же полиплоидами, но
стерилен при скрещивании с любым из
родительских видов.
Для большинства аллополиплоидных видов
характерно диплоидное число хромосом, которое
представляет собой сумму диплоидных чисел их
родительских видов; например, Spartina anglica
(2л = 122)-плодовитый гибридный аллополиплоид,
полученный в результате скрещивания Spartina mari-
tima (stricta) (2л = 60) со Spartina alterniflora (2п = 62).
(Гибрид F1 от скрещивания между этими двумя
видами стерилен и получил название Spartina town-
sendii.) Большинство аллополиплоидных растений
отличается по своим признакам от обоих
родительских видов; к ним относятся многие из весьма
ценных культур, выращиваемых человеком.
Например, вид пшеницы Triticum aestivum (2л = 42), из
которого получают муку, был выведен путем
скрещивания и отбора за 5000 лет. В результате
скрещивания дикорастущей пшеницы-однозернянки (2л =
= 14) с неидентифицированным диплоидом (2л =
= 14) был получен новый вид пшеницы - эммер
(2л = 28). Пшеница эммер скрещивалась еще с
одним дикорастущим диплоидным видом (2л = 14), в
результате чего получилась форма Triticum aestivum
(2л = 42), которая представляет собой гекса-
плоид (6л) первоначальной пшеницы-однозернянки.
Другой пример межвидовой гибридизации -
скрещивание редьки с капустой - будет описан в разд. 25.9.
У животных аллополиплоидия почти неизвестна,
так как у них межвидовые скрещивания происходят
редко. В результате полиплоидии генофонд не
получает новых генов (см. разд. 25.1.1), но создаются
новые генные комбинации.
Структурные изменения хромосом
(хромосомные перестройки)
При кроссинговере во время профазы I мейоза
происходит реципрокный обмен генетическим
материалом между гомологичными хромосомами. Это
ведет к изменению последовательности аллелей в
родительских группах сцепления, и в результате
получаются рекомбинанты, но без потери
каких-либо генных локусов. Сходные эффекты возникают
при таких хромосомных перестройках, как инверсии
и транслокации. При перестройках других типов-
делециях и дупликациях - число генных локусов в
хромосомах изменяется, и это может оказать
глубокое влияние на фенотипы. Структурные изменения в
хромосомах, связанные с инверсиями, делециями,
дупликациями, а в некоторых случаях и
транслокациями, можно наблюдать под микроскопом, когда в
профазе I мейоза гомологичные хромосомы
приступают к конъюгации. Гомологичные хромосомы
конъюгируют (см. разд. 22.3), а в тех участках,
которые затронула перестройка, одна из
гомологичных хромосом образует петлю или же
перекручивается. Какая из хромосом образует петлю и как
расположатся ее гены, зависит от типа перестройки.
Инверсия возникает в результате вырезания
участка хромосомы, который поворачивается на 180°, а
затем вновь встраивается на прежнем месте. При
этом никаких изменений генотипа не происходит, но
возможны фенотипические изменения (рис. 23.32, А).
Это показывает, что последовательность генов в
данной хромосоме может быть небезразлична для
их действия (так называемый эффект положения).
При транслокации от одной из хромосом
отрывается участок и присоединяется либо к другому
концу той же хромосомы, либо к другой,
негомологичной хромосоме (рис. 23.32, Б). И в этом случае в
фенотипе может проявиться эффект положения.
Реципрокная транслокация между
негомологичными хромосомами приводит к образованию двух
новых пар гомологичных хромосом. В тех случаях,
когда при синдроме Дауна сохраняется нормальное
диплоидное число хромосом, причиной этого
служит транслокация сверхкомплектной хромосомы 21
на одну из более крупных хромосом, обычно 15.
Самая простая форма хромосомной мутации - это
делеция, или нехватка, т.е. утрата хромосомой
какого-нибудь участка, промежуточного или концево-
Нормальная хромосома
Инверсия
Транслокация
Нормальная
хромосома
Хромосома-
с инверсией,
образовавшая
петлю
Рис. 23.32, Инверсия и транслокация (схемы) и их
влияние на расположение генов. А.
Образование петли в профазе мейоза, вызванное
инверсией. Б. Часть хромосомы, несущая
гены CDuE, оторвалась и
присоединилась к хромосоме, несущей гены К,
LuM.
Нормальная хромосома
Делеция
Дупликация
Нормальная
- хромосома -
-Хромосома
с делецией
Хромосома
с дупликацией
Нормальная
хромосома
Рис. 23.33. Делеция и дупликация (схемы) и их
влияние на расположение генов. В обоих случаях
можно видеть образование петель.
го. При этом в хромосоме уже недостает некоторых
генов (рис. 23.33). Делеция может произойти в
одной из двух гомологичных хромосом; в таком
случае аллели, находящиеся в другой, нормальной,
хромосоме, будут экспрессироваться, даже если они
рецессивны. Если же делеция затронула одни и те же
генные локусы в обеих гомологичных хромосомах,
это обычно ведет к летальному исходу.
Иногда какой-либо участок хромосомы
удваивается, так что возникает дупликация - повторение
набора генов, локализованных в этом участке. Этот
дополнительный набор может оказаться внутри той
же хромосомы или на одном из ее концов, а иногда
присоединяется к какой-нибудь другой хромосоме
(рис. 23.33).
23.9.3. Генные мутации
Внезапные спонтанные изменения фенотипа,
которые нельзя связать с обычными генетическими
явлениями или микроскопическими данными о
наличии хромосомных аберраций, можно объяснить
только изменениями в структуре отдельных генов.
Генная, или точечная (поскольку она относится к
определенному генному локусу), мутация -
результат изменения нуклеотидной последовательности
молекулы ДНК в определенном участке
хромосомы. Такое изменение последовательности
оснований в данном гене воспроизводится при
транскрипции в структуре мРНК и приводит к изменению
последовательности аминокислот в полипептидной
цепи, образующейся в результате трансляции на
рибосомах (см. разд. 22.6).
Существуют различные типы генных мутаций,
связанных с добавлением, выпадением или
перестановкой оснований в гене. Это дупликации, вставки,
делеции, инверсии или замены оснований. Во всех
случаях они приводят к изменению нуклеотидной
последовательности, а часто-и к образованию
измененного полипептида. Например, делеция
вызывает сдвиг рамки, последствия которого описаны в
разд. 22.5.
Генные мутации, возникающие в гаметах или в
будущих половых клетках, передаются всем
клеткам потомков и могут влиять на дальнейшую
судьбу популяции. Соматические генные мутации,
происходящие в организме, наследуются только теми
клетками, которые образуются из мутантной клетки
путем митоза. Они могут оказать воздействие на
тот организм, в котором они возникли, но со
смертью особи исчезают из генофонда популяции.
Соматические мутации, вероятно, возникают очень
часто и остаются незамеченными, но в некоторых
случаях при этом образуются клетки с повышенной
скоростью роста и деления. Эти клетки могут дать
начало опухолям-либо доброкачественным,
которые не оказывают особого влияния на весь
организм, либо злокачественным, что приводит к
раковым заболеваниям.
Эффекты генных мутаций чрезвычайно
разнообразны. Большая часть мелких генных мутаций фе-
нотипически не проявляется, поскольку они
рецессивны, однако известен ряд случаев, когда
изменение всего лишь одного основания в определенном
гене оказывает глубокое влияние на фенотип.
Одним из примеров служит серповидноклеточная
анемия заболевание, вызываемое у человека заменой
основания в одном из генов, ответственных за
синтез гемоглобина. Молекула дыхательного пигмента
гемоглобина у взрослого человека состоит из
четырех полипептидных цепей (двух а- и двух β-цепей), к
которым присоединены четыре простетические
группы гема. От структуры полипептидных цепей
зависит способность молекулы гемоглобина
переносить кислород. Изменение последовательности ос-
нований в триплете, кодирующем одну
определенную аминокислоту из 146, входящих в состав β-це-
пей, приводит к синтезу аномального гемоглобина
серповидных клеток (HbS). Последовательности
аминокислот в нормальных и аномальных β-цепях
различаются тем, что в одной точке аномальных
цепей гемоглобина S глутаминовая кислота
замещена вал ином. В результате такого, казалось бы,
незначительного изменения гемоглобин S
кристаллизуется при низких концентрациях кислорода; а это в
свою очередь приводит к тому, что в венозной
крови эритроциты с таким гемоглобином
деформируются (из округлых становятся серповидными) и
быстро разрушаются. Физиологический эффект
мутации состоит в развитии острой анемии и снижении
количества кислорода, переносимого кровью.
Анемия не только вызывает физическую слабость, но и
может привести к нарушениям деятельности сердца
и почек и к ранней смерти людей, гомозиготных по
мутантному аллелю. В гетерозиготном состоянии
этот аллель вызывает значительно меньший эффект:
эритроциты выглядят нормальными, а
аномальный гемоглобин составляет только около
40%. У гетерозигот развивается анемия лишь в
слабой форме, а зато в тех областях, где широко
распространена малярия, особенно в Африке и
Азии, носители аллеля серповидноклеточности
невосприимчивы к этой болезни. Это объясняется тем,
что ее возбудитель - малярийный плазмодий-не
может жить в эритроцитах, содержащих
аномальный гемоглобин (см. разд. 25.1.5).
23.9.4. Значение мутаций
Хромосомные и генные мутации оказывают
разнообразные воздействия на организм. Во многих
случаях эти мутации летальны, так как нарушают
развитие; у человека, например, около 20%
беременностей заканчиваются естественным выкидышем в
сроки до 12 недель, и в половине таких случаев
можно обнаружить хромосомные аномалии. В
результате некоторых хромосомных мутаций
определенные гены могут оказаться вместе, и их общий
эффект может привести к появлению какого-либо
«благоприятного» признака. Кроме того, сближение
некоторых генов друг с другом делает менее
вероятным их разделение в результате кроссинговера, а в
случае благоприятных генов это создает
преимущество.
Генная мутация может привести к тому, что в
определенном локусе окажется несколько аллелей.
Это увеличивает как гетерозиготность данной
популяции, так и ее генофонд, и ведет к усилению
внутрипопуляционной изменчивости. Перетасовка
генов как результат кроссинговера, независимого
распределения, случайного оплодотворения и
мутаций может повысить непрерывную изменчивость, но
ее эволюционная роль часто оказывается
преходящей, так как возникающие при этом изменения
могут быстро сгладиться вследствие «усреднения».
Что же касается генных мутаций, то некоторые из
них увеличивают дискретную изменчивость, и это
может оказать на популяцию более глубокое
влияние. Большинство генных мутаций рецессивны по
отношению к «нормальному» аллелю, который,
успешно выдержав отбор на протяжении многих
поколений, достиг генетического равновесия с
остальным генотипом. Будучи рецессивными, мутантные
аллели могут оставаться в популяции в течение
многих поколений, пока им не удастся встретиться,
т. е. оказаться в гомозиготном состоянии и
проявиться в фенотипе. Время от времени могут возникать
и доминантные мутантные аллели, которые
немедленно дают фенотипический эффект (см. разд. 25.5).
Материал, изложенный в этой главе, дает
представление о происхождении внутрипопуляционной
изменчивости и о механизмах наследования
признаков, но он не объясняет, каким образом могло
возникнуть поразительное разнообразие живых
организмов, описанных в главах 2-4. Попытка
ответить на этот вопрос составляет содержание двух
следующих глав.
Глава 24
Эволюция-история жизни
Природа жизни, ее происхождение, разнообразие
живых существ и объединяющая их структурная
и функциональная близость занимают одно из
центральных мест в биологической проблематике.
В этой главе мы попытаемся изложить и обсудить
различные теории возникновения жизни и
возможные способы видообразования. В изучении истории
жизни всегда проявлялась склонность к
доктринерству. Доктринерство можно определить как
сознательные попытки внедрения слепой приверженности
к какому-либо убеждению или учению. Подобный
подход не только далек от научного, но и
интеллектуально нечестен, и мы постараемся избегать его
в этой книге. В настоящей главе будут кратко
изложены наиболее распространенные теории
происхождения жизни, с тем чтобы учащиеся получили
представление о различии разных точек зрения на
характер этого события. Большая часть
соображений, на которых основываются эти теории,
умозрительны, так как воспроизвести в сколько-нибудь
наглядном виде события, происходившие при
возникновении жизни, невозможно. Это относится как
к научным, так и к теологическим построениям.
Однако одна теория, теория эволюции, все больше
и больше воспринимается не как некая отдельная
метафизическая теория, а как совокупность ряда
научных гипотез, каждая из которых поддается
проверке (см. Приложение 2.1, т. 1).
В этой главе и в главе 25 изложены научные
данные, отобранные так, чтобы создать связную
картину процессов, лежащих в основе
происхождения живых организмов и их разнообразия. Ввиду
неизбежности такого отбора картина не может быть
абсолютно объективной, как, впрочем, и при
изложении любой другой проблемы,-относится ли она
к истории, естественным наукам или метафизике.
Однако, подчеркивая ограничения, присущие
фактическим данным и принимаемым допущениям,
авторы старались придать представленной здесь картине
ту степень объективности и гипотетичности,
которая должна быть свойственна доброкачественному
научному обзору. Следует особо отметить, что
собранные в этой главе данные и выведенные из них
заключения соответствуют сегодняшним взглядам:
эти взгляды постоянно пересматриваются, и их
достоверность ограничена уровнем наших знаний
в каждый данный момент.
24.1. Теории возникновения
жизни
Теории, касающиеся возникновения Земли, да и всей
Вселенной, разнообразны и далеко не достоверны.
Согласно теории стационарного состояния,
Вселенная существовала извечно. Согласно другим
гипотезам, Вселенная могла возникнуть из сгустка
нейтронов в результате «Большого взрыва», родилась
в одной из черных дыр или же была создана
Творцом. Вопреки бытующим представлениям, наука не
в состоянии опровергнуть идею о божественном
сотворении первозданной Вселенной, так же как
теологические взгляды не обязательно отвергают
возможность того, что жизнь в процессе своего
развития приобрела черты, объяснимые на основе
законов природы.
Среди главных теорий возникновения жизни на
Земле следует упомянуть следующие:
1) жизнь была создана сверхъестественным
существом в определенное время (креационизм);
2) жизнь возникала неоднократно из неживого
вещества (самопроизвольное зарождение);
3) жизнь существовала всегда (теория стационарного
состояния);
4) жизнь занесена на нашу планету извне
(панспермия);
5) жизнь возникла в результате процессов,
подчиняющихся химическим и физическим законам
(биохимическая эволюция).
24.1.1. Креационизм
Согласно этой теории, жизнь возникла в результате
какого-то сверхъестественного события в прошлом;
ее придерживаются последователи почти всех
наиболее распространенных религиозных учений. В
1650 г. архиепископ Ашер из г. Арма (Ирландия)
вычислил, что бог сотворил мир в октябре 4004 г. до
нашей эры и закончил свой труд 23 октября в
9 часов утра, создав человека. Ашер получил эту
дату, сложив возрасты всех людей, упоминающихся
в библейской генеалогии, от Адама до Христа
(«кто кого родил»). С точки зрения арифметики это
разумно, однако при этом получается, что Адам
жил в то время, когда, как показывают
археологические находки, на Ближнем Востоке существовала
хорошо развитая городская цивилизация.
Традиционное иудейско-христианское
представление о сотворении мира, изложенное в Книге Бытия,
1.1.-26, вызывало и продолжает вызывать споры.
Хотя все христиане признают, что Библия-это
завет господа людям, по вопросу о длине «дня»,
упоминающегося в Книге Бытия, существуют
разногласия. Некоторые считают, что мир и все
населяющие его организмы были созданы за шесть дней
продолжительностью по 24 часа. Они отвергают
любые другие точки зрения и целиком полагаются
на вдохновение, созерцание и божественное
откровение. Другие христиане не относятся к Библии как
к научной книге и считают, что в Книге Бытия
изложено в понятной для людей всех времен форме
теологическое откровение о сотворении всех живых
существ всемогущим творцом. Для них описание
сотворения живых существ скорее относится к
ответу на вопрос «почему?», а не «каким образом?» Если
наука в поисках истины широко использует
наблюдение и эксперимент, то богословие постигает
истину через божественное откровение и веру.
«Вера же есть осуществление ожидаемого и
уверенность в невидимом... Верою осознаём, что вещи
устроены словом Божьим, так что из невидимого
произошло видимое» (послание к евреям, 11, 1-3).
Вера признает вещи, которым нет доказательств
в научном смысле слова. Это означает, что
логически не может быть противоречия между научным
и богословским объяснением сотворения мира, так
как эти две сферы мышления взаимно исключают
одна другую. Для ученого научная истина всегда
содержит элемент гипотезы, предварительности, но
для верующего теологическая истина абсолютна.
Процесс божественного сотворения мира
мыслится как имевший место лишь единожды и потому
недоступный для наблюдения; этого достаточно,
чтобы вынести всю концепцию божественного
сотворения за рамки научного исследования. Наука
занимается только теми явлениями, которые
поддаются наблюдению, а поэтому она никогда не
будет в состоянии ни доказать, ни отвергнуть эту
концепцию.
24.1.2. Самопроизвольное (спонтанное)
зарождение
Эта теория была распространена в древнем Китае,
Вавилоне и Египте в качестве альтернативы
креационизму, с которым она сосуществовала.
Аристотель (384-322 до н.э.), которого часто
провозглашают основателем биологии, придерживался
теории спонтанного зарождения. На основе
собственных наблюдений он развивал эту теорию дальше,
связывая все организмы в непрерывный
ряд-«лестницу природы» (scala naturae):
«Ибо природа совершает переход от безжизненных
объектов к животным с такой плавной
последовательностью, поместив между ними существа, которые живут,
не будучи при этом животными, что между соседними
группами, благодаря их тесной близости, едва можно
заметить различия» (Аристотель).
Этим утверждением Аристотель укрепил более
ранние высказывания Эмпедокла об органической
эволюции. Согласно гипотезе Аристотеля о
спонтанном зарождении, определенные «частицы»
вещества содержат некое «активное начало», которое
при подходящих условиях может создать живой
организм. Аристотель был прав, считая, что это
активное начало содержится в оплодотворенном
яйце, но ошибочно полагал, что оно присутствует
также в солнечном свете, тине и гниющем мясе.
«Таковы факты—живое может возникать в результате
не только спаривания животных, но и разложения почвы...
Так же обстоит дело и у растений: некоторые развиваются
из семян, а другие как бы самозарождаются под действием
сил природы, возникая из разлагающейся земли или из
определенных частей растений» (Аристотель).
С распространением христианства теория
спонтанного зарождения жизни оказалась не в чести; ее
признавали лишь те, кто верил в колдовство и
поклонялся нечистой силе, но эта идея все же
продолжала существовать где-то на заднем плане в
течение еще многих веков.
Ван Гельмонт (1577-1644), весьма знаменитый и
удачливый ученый, описал эксперимент, в котором
он за три недели якобы создал мышей. Для этого
нужны были грязная рубашка, темный шкаф и
горсть пшеницы. Активным началом в процессе
зарождения мыши ван Гельмонт считал
человеческий пот.
24.1. Какое упущение допустил ван Гельмонт в
своем эксперименте?
В 1688 г. итальянский биолог и врач Франческо
Реди, живший во Флоренции, подошел к проблеме
возникновения жизни более строго и подверг
сомнению теорию спонтанного зарождения. Реди
установил, что маленькие белые червячки, появляющиеся
на гниющем мясе,-это личинки мух. Проведя ряд
экспериментов, он получил данные,
подтверждавшие мысль о том, что жизнь может возникнуть
только из предсуществующей жизни (концепция
биогенеза).
«Убежденность была бы тщетной, если бы ее нельзя
было подтвердить экспериментом. Поэтому в середине
июля я взял четыре больших сосуда с широким горлом,
поместил в один из них змею, в другой - немного рыбы, в
третий-угрей из Арно, в четвертый - кусок молочной
телятины, плотно закрыл их и запечатал. Затем я поместил
то же самое в четыре других сосуда, оставив их
открытыми... Вскоре мясо и рыба в незапечатанных сосудах
зачервивели; можно было видеть, как мухи свободно залетают в
сосуды и вылетают из них. Но в запечатанных сосудах я не
видел ни одного червяка, хотя прошло много дней, после
того как в них была положена дохлая рыба» (Реди).
24.2. В чем состояло, по вашему мнению,
исходное представление Реди?
Эти эксперименты, однако, не привели к отказу от
идеи самозарождения, и хотя эта идея несколько
отошла на задний план, она продолжала оставаться
главной теорией в неклерикальной среде.
В то время как эксперименты Реди, казалось бы,
опровергали спонтанное зарождение мух, первые
микроскопические исследования Антона ван Левен-
гука усилили эту теорию применительно к
микроорганизмам. Сам Левенгук не вступал в споры между
сторонниками биогенеза и спонтанного зарождения,
однако его наблюдения под микроскопом давали
пищу обеим теориям и в конце концов побудили
других ученых поставить эксперименты для решения
вопроса о возникновении жизни путем спонтанного
зарождения.
В 1765 г. Ладзаро Спалланцани провел
следующий опыт: подвергнув мясные и овощные отвары
кипячению в течение нескольких часов, он сразу же
их запечатал, после чего снял с огня. Исследовав
жидкости через несколько дней, Спалланцани не
обнаружил в них никаких признаков жизни. Из
этого он сделал вывод, что высокая температура
уничтожила все формы живых существ и что без них
ничто живое уже не могло возникнуть.
24.3. Какая другая причина могла
препятствовать росту микроорганизмов в экспериментах
Спалланцани?
В 1860 г. проблемой происхождения жизни
занялся Луи Пастер. К этому времени он уже многое
сделал в области микробиологии и сумел разрешить
проблемы, угрожавшие шелководству и виноделию.
Он показал также, что бактерии вездесущи и что
неживые материалы легко могут быть заражены
живыми существами, если их не простерилизовать
должным образом.
24.4. В чем состояли главные предположения
Пастера относительно возникновения жизни?
В результате ряда экспериментов, в основе
которых лежали методы Спалланцани, Пастер доказал
справедливость теории биогенеза и окончательно
опроверг теорию спонтанного зарождения.
Однако подтверждение теории биогенеза
породило другую проблему. Коль скоро для возникновения
живого организма необходим другой живой
организм, то откуда же взялся самый первый живой
организм? Только теория стационарного состояния
не требует ответа на этот вопрос, а во всех других
теориях подразумевается, что на какой-то стадии
истории жизни произошел переход от неживого к
живому. Было ли это первичным самозарождением?
Опыт 24.1. Изучение вопроса о
возникновении микроорганизмов:
спонтанное зарождение или
биогенез?
Цель этого опыта - повторить исследования
Спалланцани и Пастера, провести дальнейшие
эксперименты с учетом критики применявшихся ими
методов и дать объективную оценку теориям
самозарождения и биогенеза.
Материалы
8 термостойких пробирок на 30 мл
Штатив для этих пробирок
120 мл питательного бульона
5 ватных пробок
Алюминиевая фольга
Водяная баня
Прямая стеклянная трубка 0,5 χ 6 см
S-образная стеклянная трубка 0,5 χ 10 см
Автоклав; термостат (32°С)
Метод
1. Простерилизуйте в автоклаве 8 термостойких
пробирок
2. Налейте в каждую из этих пробирок
(пронумерованных от 1 до 8) по 15 мл питательного бульона.
3. Проведите с пробирками 1-8 следующие
процедуры.
Пара А
Пробирка 1:
Пробирка 2:
Пара Б
Пробирка 3:
Пробирка 4:
Пара В
Пробирка 5:
Пробирка 6:
Пара Г
Пробирка 7:
Пробирка 8:
оставьте открытой, не нагревайте.
заткните ватной пробкой и
оберните верхний конец алюминиевой
фольгой; не нагревайте.
оставьте открытой; нагревайте на
кипящей водяной бане 10 мин.
заткните ватной пробкой и
оберните верхний конец алюминиевой
фольгой; нагревайте на кипящей
водяной бане 10 мин.
оставьте открытой; поместите на
20 мин в автоклав при давлении
0,8 атм.
заткните ватной пробкой и
оберните сверху алюминиевой
фольгой; поместите на 20 мин в
автоклав при давлении 0,8 атм.
оберните прямую стеклянную
трубку ватой и заткните ею пробирку;
поместите на 20 мин в автоклав
при давлении 0,8 атм.
оберните S-образную стеклянную
трубку ватой и заткните ею
пробирку; поместите на 20 мин в
автоклав при давлении 0,8 атм.
4. Поместите все пробирки в термостат (32°С).
5. Осматривайте все пробирки каждые 2 дня на
протяжении 10 дней. Записывайте результаты
в виде таблицы.
6. Спустя 10 дней возьмите из каждой пробирки,
соблюдая стерильность, по капле бульона и
исследуйте под микроскопом при большом
увеличении; запишите результаты в виде таблицы.
7. Сделайте вывод из проведенных наблюдений.
24.5. Четко сформулируйте гипотезу, которая
могла бы объяснить появление микроорганизмов
в питательном бульоне.
24.6. Перечислите переменные (факторы),
которые могут влиять на появление в бульоне
микроорганизмов.
24.7 По какому фактору различаются пробирки 1
и 2, 3 и 4, 5 и 6, 7 и 8?
24.8. По какому фактору различаются пары А, Б,
В и Г?
24.9. В каких пробирках воспроизводятся
эксперименты Спалланцани и Пастера?
24.10. Какие пробирки служат контролем?
24.11. Считаете ли вы, что описанные выше
эксперименты отвечают всем требованиям,
предъявляемым к научному исследованию? Насколько
достоверными вы считаете сделанные вами выводы?
24.1.3. Теория стационарного
состояния
Согласно этой теории, Земля никогда не возникала,
а существовала вечно; она всегда способна
поддерживать жизнь, а если и изменялась, то очень мало;
виды также существовали всегда.
Оценки возраста Земли сильно варьировали - от
примерно 6000 лет по расчетам архиепископа Ашера
до 5000· 106 лет по современным оценкам,
основанным на учете скоростей радиоактивного распада.
Более совершенные методы датирования
(Приложение 5) дают все более высокие оценки возраста
Земли, что позволяет сторонникам теории
стационарного состояния полагать, что Земля
существовала всегда. Согласно этой теории, виды также
никогда не возникали, они существовали всегда и у
каждого вида есть лишь две возможности-либо
изменение численности, либо вымирание.
Сторонники этой теории не признают, что
наличие или отсутствие определенных ископаемых
остатков может указывать на время появления или
вымирания того или иного вида, и приводят в качестве
примера представителя кистеперых рыб-латиме-
рию. По палеонтологическим данным кистеперые
вымерли в конце мелового периода, 70 млн. лет
назад. Однако это заключение пришлось
пересмотреть, когда в районе Мадагаскара были найдены
живые представители кистеперых. Сторонники
теории стационарного состояния утверждают, что
только изучая ныне живущие виды и сравнивая их с
ископаемыми остатками, можно делать вывод о
вымирании, да и в этом случае весьма вероятно, что
он окажется неверным. Используя
палеонтологические данные для подтверждения
теории стационарного состояния, ее
немногочисленные сторонники интерпретируют
появление ископаемых остатков в экологическом
аспекте. Так, например, внезапное появление
какого-либо ископаемого вида в определенном
пласте они объясняют увеличением численности его
популяции или его перемещением в места,
благоприятные для сохранения остатков. Большая
часть доводов в пользу этой теории связана с
такими неясными аспектами эволюции, как значение
разрывов в палеонтологической летописи, и она
наиболее подробно разработана именно в этом
направлении.
24.1.4. Теория панспермии
Эта теория не предлагает никакого механизма для
объяснения первичного возникновения жизни, а
выдвигает идею о ее внеземном происхождении.
Поэтому ее нельзя считать теорией возникновения
жизни как такового; она просто переносит проблему
в какое-то другое место во Вселенной.
Теория панспермии утверждает, что жизнь могла
возникнуть один или несколько раз в разное время и
в разных частях Галактики или Вселенной. Для
обоснования этой теории используются
многократные появления НЛО («неопознанных летающих
объектов»), наскальные изображения предметов,
похожих на ракеты и «космонавтов», а также
сообщения якобы о встречах с инопланетянами.
Советские и американские исследования в космосе
позволяют считать, что вероятность обнаружить жизнь в
пределах нашей Солнечной системы ничтожна,—
однако они не дают никаких сведений о возможной
жизни вне этой системы. При изучении материала
метеоритов и комет в них были обнаружены многие
«предшественники живого»-такие вещества, как
цианогены, синильная кислота и органические
соединения, которые, возможно, сыграли роль «семян»,
падавших на голую Землю. Появился ряд
сообщений о нахождении в метеоритах объектов,
напоминающих примитивные формы жизни, однако
доводы в пользу их биологической природы пока не
кажутся ученым убедительными.
24.1.5. Биохимическая эволюция
Среди астрономов, геологов и биологов принято
считать, что возраст Земли составляет примерно
4,5-5 млрд. лет.
По мнению многих биологов, в далеком прошлом
состояние нашей планеты было мало похоже на
нынешнее: по всей вероятности, температура ее
поверхности была очень высокой (4000-80000C), и
по мере того, как Земля остывала, углерод и более
тугоплавкие металлы конденсировались и
образовали земную кору; поверхность планеты была, ве-
К вакуумному насосу
и газовому баллону
( для удаления
атмосферного
воздуха
и введения
газов)
Кипящая вода
(поставляет
водяные пары
и обеспечивает
циркуляцию)
Направление!
потока
Газовая камера
Метан (CH4)
Аммиак (NH3)
Водяные пары (H2O)
Водород (H2)
Электрический разряд
высокого напряжения
(источник энергии)
Водяная рубашка
(охлаждение)
Отделитель жидкости
(накапливает продукты
химических реакций)
(предполагаемый
состав
первичной
атмосферы)
Подогрев
Рис. 24.1. Установка Стэнли
Миллера, в которой он синтезировал
аминокислоты из газов, создав
условия, предположительно
существовавшие в атмосфере
первобытной Земли. Газы и водяные
пары, циркулировавшие в установке
под высоким давлением, подвергали
в течение недели воздействию
высокого напряжения. После этого
вещества, собранные в «ловушке»,
исследовали методом
хроматографии на бумаге. В общей
сложности было выделено 15
аминокислот, в том числе глицин, аланин
и аспарагиновая кислота.
роятно, голой и неровной, так как на ней в
результате вулканической активности, непрерывных
подвижек коры и сжатия, вызванного охлаждением,
происходило образование складок и разрывов.
Полагают, что в те времена атмосфера была
совершенно не такая, как теперь. Легкие газы-
водород, гелий, азот, кислород и аргон-уходили из
атмосферы, так как гравитационное поле нашей еще
недостаточно плотной планеты не могло их
удержать. Однако простые соединения, содержащие
(среди прочих) эти элементы, должны были
удерживаться; к ним относятся вода, аммиак, двуокись
углерода и метан. До тех пор пока температура
Земли не упала ниже 1000C, вся вода, вероятно,
находилась в парообразном состоянии. Атмосфера
была, по-видимому, «восстановительной», о чем
свидетельствует наличие в самых древних горных
породах Земли металлов в восстановленной форме,
таких как двухвалентное железо. Более молодые
горные породы содержат металлы в окисленной
форме, например трехвалентное железо. Отсутствие
в атмосфере кислорода было, вероятно,
необходимым условием для возникновения жизни;
лабораторные опыты показывают, что, как это ни
парадоксально, органические вещества (основа живых
организмов) гораздо легче создаются в
восстановительной среде, чем в атмосфере, богатой кислородом.
В 1923 г. А. И. Опарин высказал мнение, что
атмосфера первичной Земли была не такой, как
сейчас, а примерно соответствовала сделанному
выше описанию. Исходя из теоретических
соображений, он полагал, что органические вещества,
возможно углеводороды, могли создаваться в океане
из более простых соединений; энергию для этих
реакций синтеза, вероятно, доставляла интенсивная
солнечная радиация (главным образом
ультрафиолетовая), падавшая на Землю до того, как
образовался слой озона, который стал задерживать
большую ее часть. По мнению Опарина, разнообразие
находившихся в океанах простых соединений,
площадь поверхности Земли, доступность энергии и
масштабы времени позволяют предположить, что в
океанах постепенно накопились органические
вещества и образовался тот «первичный бульон», в
котором могла возникнуть жизнь. Эта идея была не
нова: в 1871 г. сходную мысль высказал Дарвин:
«Часто говорят, что все необходимые для создания живого
организма условия, которые могли когда-то существовать,
имеются и в настоящее время. Но если (ох, какое это
большое «если») представить себе, что в каком-то
небольшом теплом пруду, содержащем всевозможные
аммонийные и фосфорные соли, при наличии света, тепла,
электричества и т. п. образовался бы химическим путем белок,
готовый претерпеть еще более сложные превращения, то в
наши дни такой материал непрерывно пожирался бы или
поглощался, чего не могло случиться до того, как
появились живые существа».
В 1953 г. Стэнли Миллер в ряде экспериментов
моделировал условия, предположительно
существовавшие на первобытной Земле. В созданной им
установке (рис. 24.1), снабженной источником
энергии, ему удалось синтезировать многие вещества,
имеющие важное биологическое значение, в том
числе ряд аминокислот, аденин и простые сахара,
такие как рибоза. После этого Орджел в Институте
Солка в сходном эксперименте синтезировал нук-
леотидные цепи длиной в шесть мономерных
единиц (простые нуклеиновые кислоты).
Позднее возникло предположение, что в
первичной атмосфере в относительно высокой
концентрации содержалась двуокись углерода. Недавние
эксперименты, проведенные с использованием
установки Миллера, в которую, однако, поместили смесь
CO2 и H2O и только следовые количества других
газов, дали такие же результаты, какие получил
Миллер. Теория Опарина завоевала широкое
признание, но она оставляет нерешенными проблемы,
связанные с переходом от сложных органических
веществ к простым живым организмам. Именно в
этом аспекте теория биохимической эволюции
предлагает общую схему, приемлемую для большинства
современных биологов. Однако они не пришли к
единому мнению о деталях этого процесса.
Опарин полагал, что решающая роль в
превращении неживого в живое принадлежала белкам.
Благодаря амфотерности белковых молекул они
способны к образованию коллоидных гидрофильных
комплексов - притягивают к себе молекулы воды,
создающие вокруг них оболочку. Эти комплексы могут
обособляться от всей массы воды, в которой они
суспендированы (водной фазы), и образовывать
своего рода эмульсию. Слияние таких комплексов
друг с другом приводит к отделению коллоидов от
водной среды - процесс, называемый коацервацией
(от лат. coacervus - сгусток или куча). Богатые
коллоидами коацерваты, возможно, были способны
обмениваться с окружающей средой веществами
и избирательно накапливать различные соединения,
в особенности кристаллоиды. Коллоидный состав
данного коацервата, очевидно, зависел от состава
среды. Разнообразие состава «бульона» в разных
местах вело к различиям в химическом составе
коацерватов и поставляло таким образом сырье для
«биохимического естественного отбора».
Предполагается, что в самих коацерватах
входящие в их состав вещества вступали в дальнейшие
химические реакции; при этом происходило
поглощение коацерватами ионов металлов и образование
ферментов. На границе между коацерватами и
внешней средой выстраивались молекулы липидов
(сложные углеводороды), что приводило к
образованию примитивной клеточной мембраны,
обеспечивавшей концерватам стабильность. В результате
включения в коацерват предсуществующей
молекулы, способной к самовоспроизведению, и
внутренней перестройки покрытого липидной оболочкой
коацервата могла возникнуть примитивная клетка.
Увеличение размеров коацерватов и их
фрагментация, возможно, вели к образованию идентичных
коацерватов, которые могли поглощать больше
компонентов среды, так что этот процесс мог
продолжаться. Такая предположительная
последовательность событий должна была привести к
возникновению примитивного самовоспроизводящегося
гетеротрофного организма, питавшегося
органическими веществами первичного бульона.
Хотя эту гипотезу происхождения жизни
признают очень многие ученые, астроном Фред Хойл
недавно высказал мнение, что мысль о
возникновении живого в результате описанных выше
случайных взаимодействий молекул «столь же нелепа и
неправдоподобна, как утверждение, что ураган,
пронесшийся над мусорной свалкой, может привести
к сборке Боинга-747»1.
1 Самое трудное для этой теории объяснить
появление способности живых систем к самовоспроизведению.
Гипотезы по этому вопросу пока мало убедительны.-
Прим. ред.
24.2. Природа самых первых
организмов
Данные, которыми мы сейчас располагаем,
позволяют думать, что первые организмы были гетеро-
трофами, так как только гетеротрофы могли
использовать имевшиеся в среде запасы энергии,
заключенные в сложных органических веществах первичного
бульона. Химические реакции, необходимые для
синтеза питательных веществ, слишком сложны,
поэтому они вряд ли могли возникнуть у самых
ранних форм жизни.
Предполагается, что по мере образования в ходе
«биохимической эволюции» более сложных
органических веществ некоторые из них оказались
способными использовать солнечную радиацию как
источник энергии для синтеза новых клеточных
материалов. Возможно, что включение этих веществ в уже
существующие клетки позволило последним
синтезировать новые клеточные материалы, так что им
больше не надо было поглощать органические
вещества-клетки стали автотрофными. Возрастание
численности гетеротрофов должно было привести к
уменьшению пищевых ресурсов первичного
бульона, и возникшая конкуренция ускорила появление
автотрофов.
Самые первые фотосинтезирующие организмы
хотя и использовали в качестве источника энергии
солнечную радиацию, были лишены
метаболического пути, ведущего к образованию молекулярного
кислорода. Полагают, что на более позднем этапе
возникли организмы, способные к фотосинтезу с
выделением кислорода, подобные современным
сине-зеленым водорослям (см. разд. 3.2), и это
привело к постепенному накоплению кислорода в
атмосфере. Увеличение количества O2 в атмосфере и его
ионизация с образованием озонового слоя
уменьшили количество ультрафиолетовой радиации,
достигающей Земли. Это привело к замедлению
синтеза новых сложных веществ, но одновременно
повысило устойчивость преуспевающих форм жизни.
Изучение физиологии современных организмов
выявило большое разнообразие биохимических путей,
участвующих в связывании и освобождении энергии,
которые, возможно, отражают первые
эксперименты, проводившиеся природой на живых организмах.
Несмотря на все сказанное выше, проблема
возникновения жизни остается нерешенной, и при всех
огромных успехах биохимии ответы на вопросы
носят умозрительный характер. Читателю был
предложен в упрощенном виде сплав из имеющихся
гипотез. Гипотезы, которая могла бы стать
«руководящей» и превратиться во всеобъемлющую
теорию, пока еще нет (см. Приложение, 2.1).
Подробности перехода от сложных неживых веществ к
простым живым организмам покрыты тайной.
Возникновение
А. Стационарное
состояние
Не было
Б. Креационизм
В определенные
эпохи
В. Спонтанное
зарождение
В любое время
Виды
Г. Панспермия
Д. Биохимическая
эволюция
Возникновение в бесконечности / D
В определенное время В определенное время
Эволюция
Эволюция
Виды
Возникновение
E. Креационизм
В определенные эпохи
(или)
. Панспермия
В бесконечности /
В определенное время
(подразумевается)
Спонтанное зарождение
(в определенное время)
Биохимическая эволюция
Эволюция
Виды
Рис. 24.2. Схемы различных теорий о
происхождении жизни и видообразовании.
24.3. Общие выводы о
«теориях» возникновения
жизни
Многие из этих «теорий» и предлагаемые ими
объяснения существующего разнообразия видов
используют одни и те же данные, но делают упор на
разные их аспекты. Научные теории могут быть
сверхфантастическими, с одной стороны, и
сверхскептическими - с другой. Теологические
соображения тоже могут найти себе место в этих рамках
в зависимости от религиозных взглядов их авторов.
Одним из главных пунктов разногласий, даже еще
в додарвиновские времена, был вопрос о
соотношении между научными и теологическими взглядами
на историю жизни.
Схемы А-Д на рис. 24.2 просто отображают
теории, гипотезы или верования относительно
происхождения жизни, тогда как схемы E и Ж
представляют собой попытку объединить некоторые аспекты
теорий Б, В и Г в нечто целостное, приемлемое для
многих людей. Наука и религия на практике не
обязательно взаимно исключают друг друга, о чем
свидетельствует число ученых, придерживающихся
религиозных убеждений.
24.4. Теория эволюции
Теория эволюции занимает особое место в изучении
истории жизни. Она стала той объединяющей
теорией, которая служит фундаментом для всей
биологии. Эволюция подразумевает всеобщее
постепенное развитие, упорядоченное и последовательное.
Применительно к живым организмам эволюцию
можно определить как «развитие сложных
организмов из предсуществующих более простых организмов
с течением времени».
Представление об эволюции ведет свое начало не
от Дарвина с его книгой «Происхождение видов».
Еще задолго до Дарвина попытки человека
объяснить очевидное разнообразие окружающих его
живых организмов парадоксальным образом
привлекли его внимание к чертам структурного и
функционального сходства между ними. Выдвигались
различные эволюционные гипотезы, чтобы объяснить
это сходство, и такие идеи сами
«эволюционировали» по мере развития науки со времен Дарвина.
История развития эволюционной теории (табл.
24.1) показывает, что концепция непрерывности или
постепенного развития более сложных видов из
предсуществующих более простых форм возникла у
ряда философов и естествоиспытателей еще до
формального провозглашения в начале XIX в.
эволюционных гипотез.
24.4.1. Теория эволюции Ламарка
Французский биолог Ламарк в 1809 г. выдвинул
гипотезу о механизме эволюции, в основе которой
лежали две предпосылки: упражнение и
неупражнение частей организма и наследование
приобретенных признаков. Изменения среды, по его мнению,
могут вести к изменению форм поведения, что
вызовет необходимость использовать некоторые
органы или структуры по-новому или более
интенсивно (или, наоборот, перестать ими пользоваться).
В случае интенсивного использования
эффективность и (или) величина органа будет возрастать, а
при неиспользовании может наступить дегенерация
и атрофия. Эти признаки, приобретенные
индивидуумом в течение его жизни, согласно Ламарку,
наследуются, т.е. передаются потомкам.
С точки зрения ламаркизма, длинная шея и ноги
жирафа - результат того, что многие поколения его
некогда коротконогих и короткошеих предков
питались листьями деревьев, за которыми им
приходилось тянуться все выше и выше. Незначительное
удлинение шеи и ног, происходившее в каждом
поколении, передавалось следующему поколению,
пока эти части тела не достигли своей нынешней
длины. Перепонки между пальцами у
водоплавающих птиц и форму тела камбалы объясняли таким
же образом. Перепонки возникли в результате
постоянного раздвигания пальцев и растягивания ко-
Таблица 24.1. История эволюционных идей
Древний Китай
Конфуций
Жизнь возникла из одного источника путем постепенного развертывания и разветвления
Античная эпоха и Средневековье
Диоген
Эмпедокл
Демокрит
Анаксагор
Фалес
(640—546 до н. э.)
Анаксимандр
Аристотель
(384—322 до н. э.)
Средневековье
(400—1400 н.э.)
Все вещи — результат дифференциации одной и той же вещи и подобны ей
Воздух, земля, огонь и вода — четыре корня всего сущего. Жизнь возникла в результате
действия сил притяжения и отталкивания между этими четырьмя элементами. Пытался
объяснить происхождение Вселенной, растений, животных и человека (впервые
высказал зачатки идеи об эволюции органического мира)
Живые существа возникли на Земле путем самозарождения из ила
Организмы возникли из «зародышей», носящихся в воздухе
Все живое происходит из воды
Растения, затем животные и, наконец, человек возникли из тины на формировавшейся
Земле
Сформулировал теорию непрерывного и постепенного развития живого из неживой
материи, основанную на его наблюдениях над животными. Создал представление о
«лестнице природы» применительно к миру животных
Теории, основанные на более ранних концепциях, перечисленных выше, или признание
креационизма
Время умозрительных построений
(1400—1790)
Джон Рей
(1627—1705)
Карл Линней
(1707-1778)
Бюффон
(1707-1788)
Джеймс Хаттон
(1726—1797)
Время создания теорий
(1790-1900)
Эразм Дарвин
(1731 — 1802)
Жан-Батист Ламарк
(1744—1829)
Жорж Кювье
(1769-1832)
Уильям Смит
(1769-1838)
Чарлз Лайель
(1797—1875)
Чарлз Дарвин
(1809—1882)
Альфред Рассел Уоллес
(1823—1913)
Гуго де Фриз
(1848—1935)
Август Вейсман
(1834—1914)
Грегор Мендель
(1822-1884)
Создал концепцию вида
Ввел биномиальную систему номенклатуры. Считал, что роды были созданы по
отдельности, а виды представляют собой варианты родов
Высказал мнение, что разные типы животных имеют различное происхождение и возникли
в разное время. Признавал влияние внешней среды и наследование приобретенных
признаков
Теория униформизма. Исчислял возраст Земли миллионами лет
Жизнь возникла из одной «нити», созданной богом. Не признавал отдельного сотворения
человека. «Нить» эволюционировала в результате наследования приобретенных
признаков
Наследование приобретенных признаков (воздействие внешней среды на организмы и
передача фенотипических признаков потомству). Концепция упражнения и
неупражнения органов
Использование палеонтологической летописи. Ископаемые остатки — результаты
«катастроф», после которых создавались новые виды
Возражал против теории катастроф Кювье, основываясь на непрерывном
распространении сходных видов в близких по возрасту слоях
Продемонстрировал прогрессивные изменения ископаемых остатков
Находился под влиянием идей Лайеля и Мальтуса. Сформулировал теорию эволюции
в результате естественного отбора
Сформулировал теорию, сходную с дарвиновской, но не включал в нее человека
Открыл существование наследуемых мутаций, составляющих основу дискретной
изменчивости; считал, что виды возникают в результате мутаций
Показал, что половые клетки обособлены от остального организма и поэтому не
подвержены влияниям, действующим на соматические ткани
Генетические исследования (опубликованы в 1865 г.), получившие признание только после
1900 г. Открыл законы наследственности
Двадцатый век (неодарвинизм)
В. Л. Иогансен
T. Хант Морган
Г. Дж. Мёллер (1927)
Фенотипические признаки определяются генотипом и факторами среды
Создал хромосомную теорию наследственности, основанную на генетических и
цитологических данных
Генотип может изменяться под действием рентгеновских лучей: индуцированные мутации
Продолжение табл. 24.1
P. А. Фишер (1930)
Дж. У. Бидл и Э. Jl. Татум
(1941)
Дж. Ледерберг и
А. Д. Херши (1951)
Дж. Уотсон и Ф. Крик
(1953)
Ф. Жакоб, Ж. Моно(1961)
Изменения, изучаемые генетиками, аналогичны изменениям, выявляемым в
палеонтологической летописи
Продолжая работы А. Геррода (1909) и Дж. Холдейна (1935), выявили генетическую
основу процессов биосинтеза
Продемонстрировали ценность бактерий как объектов для изучения генотипических
изменений
Предложили модель молекулярной структуры ДНК и механизма ее репликации
Открыли механизм регуляции генной активности
жи между ними при плавании в поисках пищи или
для спасения от хищников, а уплощенное тело-
из-за лежанья на боку на мелководье. Хотя теория
Ламарка способствовала подготовке почвы для
принятия эволюционной концепции, его взгляды на
механизм изменения никогда не получили широкого
признания.
Однако Ламарк был прав, подчеркивая роль
условий жизни в возникновении фенотипических
изменений у данной особи. Например, занятия
физкультурой увеличивают объем мышц, но хотя эти
приобретенные признаки затрагивают фенотип, они не
являются генетическими и, не оказывая влияние на
генотип, не могут передаваться потомству. Для того
чтобы доказать это, Вейсман на протяжении многих
последовательных поколений отрезал мышам
хвосты. По теории Ламарка вынужденное
неупотребление хвостов должно было бы привести к их
укорочению у потомков; однако этого не произошло.
Вейсман постулировал, что признаки, приобретаемые
сомой (телом) и приводящие к изменению
фенотипа, не оказывают прямого воздействия на половые
клетки (гаметы), с помощью которых признаки
передаются следующему поколению. Его теория
непрерывности зародышевой плазмы» была
исторически необходима, для того чтобы стало
возможным признание наследования генетических
особенностей при половом размножении.
24.4.2. Дарвин, Уоллес и
происхождение видов в
результате естественного
отбора
Чарлз Дарвин родился в 1809 г. Он был сыном
состоятельного врача и, подобно многим великим
людям, вначале как ученый ничем особенно не
выделялся. В 1831 г. он принял предложение
отправиться в качестве натуралиста (без жалованья) в
путешествие на военном корабле «Бигль», который
уходил на пять лет в море для проведения
топографических съемок у восточного побережья Южной
Америки. «Бигль» возвратился в Фалмут в октябре
1836 г., проделав путь вдоль берегов Чили, через
Галапагосские острова, Таити, Новую Зеландию,
Тасманию и Южную Африку. Большую часть этого
времени Дарвин занимался геологическими
исследованиями; однако во время пятинедельного
пребывания на Галапагосских островах его внимание
привлекло сходство между флорой и фауной этих
островов и материка. Особенно его заинтересовало
распространение черепах и вьюрков (см. разд. 24.7.2).
Он собрал множество данных об изменчивости
организмов, которые убедили его в том, что виды
нельзя считать неизменяемыми. После возвращения
в Англию Дарвин занялся изучением практики
разведения голубей и других домашних животных, что
привело его к концепции искусственного отбора,
однако он все еще не мог представить себе, каким
образом отбор мог бы действовать в природных
условиях. В 1778 г. священник Томас Мальтус
опубликовал свой труд «Трактат о народонаселении»,
в котором ярко обрисовал, к чему мог бы привести
рост населения, если бы он ничем не сдерживался.
Дарвин перенес его рассуждения на другие
организмы и обратил внимание на то, что несмотря на их
высокий рупродуктивный потенциал численность
популяций остается относительно постоянной.
Сопоставляя огромное количество сведений, он начал
понимать, что в условиях интенсивной конкуренции
между членами популяции любые изменения,
благоприятные для выживания в данных условиях,
повышали бы способность особи размножаться и
оставлять плодовитое потомство, а
неблагоприятные изменения, очевидно, невыгодны, и у
обладающих ими организмов шансы на успешное
размножение понижались бы. Эти соображения послужили
остовом для теории эволюции путем естественного
отбора, сформулированной Дарвином в 1839 г. В
сущности, наибольший вклад Дарвина в науку
заключается не в том, что он доказал существование
эволюции, а в том, что он объяснил, как она может
происходить.
Тем временем другой естествоиспытатель,
Альфред Рассел Уоллес, много путешествовавший по
Южной Америке и островам Юго-восточной Азии
и тоже читавший Мальтуса, пришел к тем же
выводам о естественном отборе, что и Дарвин.
В 1858 г. Уоллес изложил свою теорию на 20
страницах и послал их Дарвину. Это стимулировало
и ободрило Дарвина, и в июле 1858 г. Дарвин
и Уоллес выступили с докладами о своих идеях на
заседании Линнеевского общества в Лондоне.
Спустя год с небольшим, в ноябре 1859 г., Дарвин
опубликовал «Происхождение видов путем
естественного отбора». Все 1250 экземпляров книги были
проданы в первый же день, и говорят, что по своему
воздействию на человеческое мышление она
уступала только Библии.
24.5. Естественный отбор
Согласно Дарвину и Уоллесу, механизмом, с
помощью которого из предсуществовавших видов
возникают новые виды, служит естественный отбор.
Эта гипотеза (или теория) основана на трех
наблюдениях и двух выводах, которые можно
сформулировать следующим образом:
Наблюдение 1. Особи, входящие в состав
популяции, обладают большим репродуктивным
потенциалом.
Наблюдение 2. Число особей в каждой данной
популяции примерно постоянно.
Вывод 1. Многим особям не удается выжить
и оставить потомство. В популяции происходит
«борьба за существование».
Наблюдение 3. Во всех популяциях существует
изменчивасть.
Вывод 2. В «борьбе за существование» те особи,
признаки которых наилучшим образом
приспособлены к условиям жизни, обладают
«репродуктивным преимуществом» и производят больше
потомков, чем менее приспособленные особи.
Вывод 2 содержит гипотезу о естественном
отборе, который может служить механизмом эволюции.
24.5.1. Доказательства существования
естественного отбора
Наблюдение 1. Мальтус привлек внимание к
репродуктивному потенциалу человека и отметил, что
численность народонаселения возрастает по
экспоненте (см. разд. 12.6.3). Способность к размножению
свойственна всему живому и представляет собой ту
основную силу, которая обеспечивает сохранение
вида. Это относится к самым разным организмам
(табл. 24.2). Если бы каждая женская гамета была
оплодотворена и развилась в половозрелую особь,
Земля через несколько дней оказалась бы
перенаселенной.
Таблица 24. 2. Репродуктивный потенциал некоторых видов
Crassostrea
virginica
Ly coper don sp.
Papaver rhoeas
Carcinus maenas
Виргинская
устрица
Дождевик
Мак-самосейка
Зеленый краб
1,0 · 106 яиц за
сезон
7,0- 10" спор
6,0 · 103 семян
4,0 · 106 яиц за
сезон
Наблюдение 2. Численность всех популяций
ограничивается или контролируется различными
факторами среды, такими как пищевые ресурсы,
пространство и свет. Размеры популяции возрастают до тех
пор, пока среда еще может выдерживать их
дальнейшее увеличение, после чего достигается некое
равновесие. Численность колеблется вблизи этого
равновесного уровня (см. разд. 12.6.3). Таким
образом, величина популяции обычно остается
относительно постоянной в течение периода времени,
продолжительность которого зависит от
продолжительности жизненного цикла данного организма.
Вывод 1. Непрерывная конкуренция между
индивидуумами за факторы среды приводит к «борьбе за
существование». Происходит ли конкуренция в
пределах одного вида (внутривидовая конкуренция) или
между представителями разных видов (межвидовая
конкуренция), не имеет значения в смысле ее влияния
на численность популяции (см. разд. 12.6.5), но
в любом случае некоторые организмы не смогут
выжить или оставить потомство.
Наблюдение 3. Изучение жуков в студенческие
годы в Кембридже, наблюдения, сделанные во
время путешествия на «Бигле», и знания,
приобретенные при разведении и селекции голубей с целью
выработки определенных признаков, убедили
Дарвина в важном значении внутривидовой
изменчивости. А адаптивное значение межвидовой
изменчивости, обнаруженной у галапагосских вьюрков (род
Geospiza), дало Дарвину ключ к его второму выводу.
Материалы, собранные Уоллесом на Малайском
архипелаге, подтверждали существование межпопу-
ляционных различий. Однако Дарвин и Уоллес не
смогли выявить источники всех этих форм
изменчивости. Вопрос оставался неясным до тех пор, пока
Мендель не открыл корпускулярную природу
наследственности и не показал, каким образом
сохраняются генетические различия.
Вывод 2. Поскольку все особи в пределах данной
популяции подвержены изменчивости и поскольку
ясно, что неизбежна «борьба за существование», из
этого следует, что особи, обладающие
определенными признаками, будут более приспособлены к тому,
чтобы выжить и оставить потомство. Решающий
фактор, определяющий выживание,-это
приспособленность к среде. Любое, пусть самое
незначительное физическое, физиологическое или поведенческое
изменение, дающее одному организму
преимущество перед другим, будет действовать в «борьбе за
существование» как селективное преимущество.
(Термин «селективное преимущество» имеет не столь
эмоциональную окраску, как термин «выживание
наиболее приспособленных», созданный философом
и социологом Гербертом Спенсером.)
Благоприятные изменения будут передаваться следующим
поколениям, а неблагоприятные отметаются
(элиминируются) отбором, так как они невыгодны
организму. Действуя таким образом, естественный
отбор ведет к повышению «мощности» вида, а в
филогенетическом плане - обеспечивает его
выживание (если условия среды остаются постоянными).
Вся суть теории естественного отбора Дарвина и
Уоллеса в наиболее сжатом виде выражена самим
Дарвином:
«Так как рождается гораздо больше особей каждого
вида, чем может выжить, и так как между ними поэтому
часто возникает борьба за существование, то из этого
следует, что любое существо, если оно хотя бы
незначительно изменится в направлении, выгодном для него
в сложных и нередко меняющихся условиях его жизни,
будет иметь больше шансов выжить и, таким образом,
будет сохраняться естественным отбором. В силу действия
закона наследственности всякая сохраненная отбором
разновидность будет размножаться в своей новой,
видоизмененной форме». (Дарвин, 1859).
Теория эволюции, сформулированная Дарвином,
обросла множеством неверных представлений;
поэтому необходимо сделать следующие замечания:
1. Дарвин не пытался объяснить возникновение
жизни на Земле; его интересовало, каким образом из
существующих видов могут возникать новые виды.
2. Естественный отбор - это не просто негативная,
разрушающая сила; он может быть механизмом,
с помощью которого в популяцию вносятся
позитивные новшества (см. разд. 25.5). В процессе
популяризации идеи «борьбы за существование»
распространились неудачные выражения, такие как «выживание
наиболее приспособленных» и «устранение
неприспособленных», введенные философом Гербертом
Спенсером и подхваченные тогдашней прессой.
3. Упрощенная, слишком прямолинейная
трактовка прессой концепции о «происхождении человека от
обезьяны» болезненно задевала чувства как
клерикальных, так и мирских слоев общества. Церковь
рассматривала это как оскорбление ее учения о том,
что бог создал человека «по своему образу и
подобию», а общество было возмущено принижением
«высокого» положения человека в царстве
животных.
4. Явное противоречие между описанным в Книге
Бытия сотворением Вселенной за шесть дней и
концепцией постепенного формирования все новых
видов еще более обострилось после заседания
Британской ассоциации по распространению научных
знаний, состоявшегося в июне 1860 г. На этом
заседании епископ Оксфордский Сэмюэль Уилбер-
форс неистово обрушился на выводы Дарвина,
изложенные в «Происхождении видов», но, поскольку
он не был биологом, его выступление страдало
неточностями. Заключая свою речь, епископ
обратился к одному из защитников дарвиновской
теории-профессору Томасу Гексли-с вопросом,
считает ли он себя связанным с обезьяньими предками
через дедушку или бабушку. В ответ на это Гексли
изложил основные идеи Дарвина и указал на
искажения, допущенные Уилберфорсом. В заключение
он дал понять, что предпочел бы иметь своим
предком обезьяну, чем «находиться в родстве с
человеком, который использует большие таланты
для того, чтобы затемнять истину». Эта досадная
дискуссия продолжалась и известна под названием
«Книга Бытия против Эволюции». Профессор
P. Дж. Берри охарактеризовал крайние группировки
в этих дебатах следующим образом:
а) те, кто преклоняется перед учеными и считает,
что библейская история опровергнута;
б) те, кто остается под влиянием Священного
писания и собственной его интерпретации и
закрывает глаза на тот факт, что творение господа бога
можно изучать научными методами.
24.6. Современные
представления об
эволюции
Теория эволюции, предложенная Дарвином и
Уоллесом, была расширена и разработана в свете
современных данных генетики, палеонтологии,
молекулярной биологии, экологии и этологии и получила
название неодарвинизма. Неодарвинизм можно
определить как теорию органической эволюции путем
естественного отбора признаков,
детерминированных генетически.
Термин «эволюция» может означать как сам этот
процесс, так и его результат; соответственно разные
аспекты неодарвинизма опираются на
доказательства разного типа. Для того чтобы признать
сформулированную выше неодарвинистскую эволюционную
теорию, необходимо:
1) установить факт изменения форм жизни во
времени (эволюция в прошлом);
2) выявить механизм, производящий эволюционные
изменения (естественный отбор генов);
3) продемонстрировать эволюцию, происходящую в
настоящее время («эволюция в действии»).
Свидетельствами эволюции, происходившей в
прошлом, служат ископаемые остатки организмов и
стратиграфия. Данные о механизме эволюции
получают путем экспериментальных исследований и
наблюдений, касающихся естественного отбора
наследуемых признаков (например, отбора по окраске
раковин у Сераеа; см. разд. 25.5.1) и механизма
наследования, демонстрируемого классической
генетикой (работы Менделя на горохе и др). Наконец,
сведения о действии этих процессов в наше время
дает изучение популяций современных организмов,
например исследование видообразования у
серебристой чайки (см. разд. 25.8.4), а также результаты
искусственного отбора и генной инженерии, такие
как создание новых сортов пшеницы или получение
моноклональных антител.
Не существует пока твердо установленных
законов эволюции; у нас есть лишь хорошо
подкрепленные фактическими данными гипотезы, которые в
совокупности составляют достаточно
обоснованную теорию. Преждевременное принятие
современных концепций как неких догматических истин на
достигнутом сейчас уровне, равно как и на любом
уровне научного исследования, может подавить
интеллектуальный рост и поиски истины.
Некритическое принятие ряда представлений эволюционной
теории служит тому примером. Некоторые из
событий, приводимых в качестве доказательств
эволюционной теории, могут быть воспроизведены в
лаборатории, однако это не значит, что они
действительно имели место в прошлом; они просто
свидетельствуют о возможности таких событий. В
современных научных дискуссиях об эволюции
обсуждается не самый факт ее существования, а то, что
она происходит путем естественного отбора
случайно возникающих мутаций.
24.7. Подтверждения теории
эволюции
Сведения, подтверждающие современные
представления об эволюции, поступают из разных
источников, среди которых главное место занимают
палеонтология, биогеография, систематика, селекция
растений и животных, морфология, изучение
адаптивной радиации, сравнительная эмбриология и
сравнительная биохимия.
Большая часть данных, приводимых в этой главе,
не была доступна Дарвину и Уоллесу в то время,
когда они публиковали свои статьи о
происхождении видов путем естественного отбора. Многие
великие ученые обладают большими способностями
к индукции, чем к дедукции, основанной на
наблюдении и эксперименте, однако у Дарвина и Уоллеса,
по-видимому, имелось удачное сочетание того и
другого. Дарвин так пишет о совмещении этих двух
подходов:
«В октябре 1838 г., т.е. спустя 15 месяцев после того,
как я начал свое систематическое исследование, я взялся
читать просто для развлечения работу Мальтуса о
народонаселении, и поскольку длительные наблюдения над
жизнью животных и растений вполне подготовили меня
к тому, чтобы оценить происходящую повсеместно борьбу
за существование, я вдруг понял, что в таких условиях
благоприятные изменения должны сохраняться, а
неблагоприятные - отметаться. Результатом всего этого должно
быть формирование новых видов. Итак, я, наконец,
получил теорию, с которой можно работать».
Представленные здесь данные в значительной
мере подтверждают теорию эволюции путем
естественного отбора, изложенную в разд. 24.5, хотя и не
могут быть бесспорными доказательствами. Эти
данные получены в разных областях знания, и их
интерпретация во всех случаях исходит из
предположения о достоверности принятой здесь концепции
эволюции. Большая часть этих данных
подтверждает доводы в ее пользу, полученные из других
источников. Среди них немало также доказательств,
в свою очередь нуждающихся в подтверждении, а
также исключений или сведений, которым можно
дать иную интерпретацию; однако концепция
эволюции в широком смысле основана на огромном
количестве научных сведений, которые на нашем
уровне изложения трудно представить в понятной
и притом совершенно свободной от догматизма
форме.
24.7.1. Палеонтология
Палеонтология занимается изучением ископаемых
остатков, т. е. любых сохранившихся в земной коре
остатков, предположительно принадлежавших
каким-либо живым организмам. Это могут быть
целые организмы, твердые скелетные структуры,
наружные и внутренние ядра, окаменелости,
отпечатки, следы и копролиты (окаменевшие
экскременты) (табл. 24.3).
Одних лишь ископаемых данных недостаточно
для обоснования какой-либо эволюционной теории,
однако они подтверждают факт прогрессивного
возрастания сложности организмов и отвергают
представление о неизменяемости видов. Ископаемые
остатки были хорошо известны еще до того, как
мысль об эволюции получила всеобщее признание.
Их считали либо остатками существ, сотворенных
раньше других, либо артефактами, помещенными
в горные породы богом. Большую часть найденных
до сих пор остатков можно распределить по тем же
самым таксономическим группам (типам и
классам), что и ныне живущие виды, однако вопрос
о том, являются ли они предками современных
форм, можно обсуждать, но доказать с помощью
одной лишь палеонтологии ничего нельзя.
В самых древних породах, содержащих
ископаемые остатки, встречаются организмы очень
немногих типов, и все они имеют простое строение. Более
молодые породы содержат более разнообразные
ископаемые остатки со все более сложным
строением. Во всей палеонтологической летописи многие
виды, появляющиеся на каком-либо
стратиграфическом уровне, на более позднем уровне исчезают. В
эволюционном смысле это истолковывают как
возникновение и вымирание видов в соответствующие
эпохи.
Геофизические данные указывают на то, что
географические области и климатические условия
изменялись на всем протяжении истории Земли.
Поскольку каждый организм приспособлен к
определенной среде, непрерывно изменяющиеся условия
могли благоприятствовать возникновению некоего
механизма эволюционного изменения, что
позволяет объяснить прогрессивные изменения в
строении организмов, отраженные в палеонтологической
летописи. Экологические соображения тоже
согласуются с палеонтологическими данными: так,
например, растения появились на суше раньше, чем
животные, а насекомые - раньше чем опыляемые
ими растения.
Одно из главных возражений против
использования ископаемых остатков для доказательства эво-
Таблица 24. 3. Типы ископаемых остатков, их образование и примеры
Тип остатков
Целые организмы
Твердые скелетные
структуры
Наружные и внутренние
ядра
Окаменелости
Отпечатки
Следы
Копролиты
Процесс фоссилизации
Вмерзшие в лед в период оледенения
Заключенные в янтарь — отвердевшую смолу
хвойных деревьев
Заключенные в асфальт
Захороненные в кислых торфяных болотах,
где отсутствие бактерий и грибов
препятствует полному разложению
Захороненные в осадочном песке и глинах,
образующих осадочные породы, например
известняк, песчаник и др.
Твердые объекты, захороненные как указано
выше. При литификации осадка части
скелета растворяются, оставляя отпечаток
своей наружной или внутренней
поверхности, получающаяся полость может
заполняться тонкозернистым материалом,
который затвердевает, образуя слепок
(возможно сохранение тонких деталей)
Постепенное замещение приносимыми водой
минеральными веществами, такими как
кремнезем, пириты, карбонат кальция или
углерод. Медленное их внедрение в
организм по мере его разложения обеспечивает
сохранение тонких структур
Отпечатки организмов или их частей на
поверхности тонкозернистых отложений, на
которых они погибли
Следы ног животных, следы ползания и ходы,
оставшиеся в иле, быстро высыхают и
заполняются песком, а затем покрываются
другими отложениями
Экскременты животных, избежавшие
разложения, а затем включенные в осадочные
породы. Часто содержат остатки съеденной
пищи, например зубы или чешуи
Примеры
Мамонты, найденные в вечной мерзлоте
в Сибири
Экзоскелеты насекомых в олигоценовых
отложениях на Балтийском побережье
"Мумии", найденные в асфальтовых озерах
Калифорнии
"Мумии", найденные в болотах и
торфяниках на Скандинавском полуострове
Кости, раковины и зубы (очень часто
встречаются на Британских островах)
Брюхоногие из портландского камня (юра).
Отпечатки гигантских хвощей
(каламитов), росших в лесах каменноугольного
периода. Отпечатки внутренних
поверхностей раковин моллюсков, на которых
видны места прикрепления мышц
Иглокожее Micraster, структуры тела
которого были замещены кремнеземом
Перья археоптерикса из верхней юры.
Медузы из кембрия Британской
Колумбии. Отпечатки листьев из карбона
Следы ног и хвоста динозавра дают
представление о размерах и позе животного
Экскременты кайнозойских
млекопитающих
люции - отсутствие непрерывности в
палеонтологической летописи. Разрывы в ней («недостающие
звенья») считают веским доводом против теории
образования новых форм путем постепенного
изменения. Однако существует ряд соображений,
позволяющих объяснить неполноту палеонтологических
данных:
1) мертвые организмы быстро разлагаются;
2) мертвые организмы поедаются животными,
питающимися падалью;
3) животные с мягким телом плохо поддаются
фоссилизации;
4) лишь небольшая часть особей погибла в
условиях, благоприятных для фоссилизации;
5) обнаружена только часть ископаемых остатков.
Данные в пользу эволюционного процесса
пополняются по мере нахождения все большего числа
возможных «недостающих звеньев»-либо
ископаемых остатков, таких как сеймурия (амфибии -*
рептилии), археоптерикс (рептилии -► птицы) и циног-
натус (рептилии -► млекопитающие), либо ныне
живущих, как, например, Peripatus (см. рис. 24.18)
и латимерия, близкие по своему строению к
вымершим формам.
Возможно также, что новые виды возникали
внезапно и что промежуточных форм не существовало.
Элдредж и Гоулд описали процесс, позволяющий
объяснить внезапное появление некоторых видов
в палеонтологической летописи. По их мнению,
скорости эволюции варьируют и некоторые новые
виды возникают очень быстро, что и приводит к
неполноте палеонтологической летописи. Эти
«скачки» в эволюционной последовательности породили
термин «скачкообразная эволюция». Сам Дарвин
обсуждал такую возможность и написал в
«Происхождении видов»: «Я не думаю, что этот процесс
[видообразование]... происходит непрерывно;
гораздо более вероятно, что каждая форма долгое
время остается неизменной, а затем вновь
претерпевает модификацию».
Палеонтологическая история
лошади
Эволюция лошади служит одним из лучших
примеров филогении, основанной на почти полном
палеонтологическом материале из
североамериканских осадочных отложений от нижнего эоцена до
настоящего времени.
Самые первые непарнокопытные появились 54
млн. лет назад; к современным представителям
непарнокопытных относятся лошади, тапиры и
носороги. Самое древнее животное, напоминающее
лошадь, остатки которого обнаружены в
палеонтологической летописи, принадлежало к роду Нугасо-
therium, широко распространенному по всей
Северной Америке и Европе в раннем эоцене. К началу
олигоцена оно сохранилось только в Северной
Америке. Это было небольшое субтильное животное,
приспособленное к бегу. Его короткие тонкие ноги
оканчивались вытянутыми ступнями, а пальцы
располагались почти вертикально. На передних ногах
имелось по четыре, на задних-по три пальца. Резцы
были небольшими, а коренные зубы имели
низкую коронку с округлыми бугорками, покрытыми
эмалью.
В предполагаемый эволюционный ряд лошадей
от Hyracotherium до Equus входит не менее 12 родов
и несколько сот видов. Главные направления в
эволюции лошадей касались передвижения и
питания. Они отражали адаптацию к изменявшимся
условиям среды и включали следующие тенденции:
1) увеличение размеров тела;
2) удлинение ног и, в частности, стопы;
3) редукцию боковых пальцев;
4) удлинение и утолщение третьего пальца;
5) выпрямление спины и увеличение ее жесткости;
6) совершенствование органов чувств;
7) увеличение и усложнение головного мозга
(связанное с развитием органов чувств);
8) увеличение резцов в ширину;
9) замещение ложнокоренных зубов коренными;
10) удлинение зубов;
11) увеличение высоты коронки коренных зубов;
12) укрепление зубов благодаря развитию цемента
между складками;
13) увеличение поверхности зубов благодаря
развитию складок эмали.
На рис. 24.4 представлено прогрессивное развитие
лошади в кайнозое; в качестве примеров выбраны
роды, доминирующие в данной геологической
эпохе. Однако нет данных, которые позволяли бы
утверждать, что эти формы связаны прямым
родством.
Значение последовательного ряда форм,
показанного на рис. 24.4, состоит в том, что он служит
подтверждением теории прогрессивного развития
гомологичных структур, в данном случае
конечностей и зубов. Каждый из видов на рис. 24.4
соответствует некой стадии развития, которая процветала
в течение нескольких миллионов лет (судя по
обилию ископаемых остатков), прежде чем вымерла.
Вымирание одного вида не означало, однако,
исчезновения линии всего семейства. Как показывают
ископаемые данные, место вымершего вида всегда
занимал другой, близко родственный ему вид.
Поскольку все виды, составляющие
последовательность, обладают структурным и экологическим
сходством, это подкрепляет теорию об эволюции
путем видоизменения. Другие ископаемые остатки,
найденные в тех же слоях, свидетельствуют об
изменявшихся климатических условиях; это вместе с
другими данными позволяет считать, что каждый
вид был приспособлен к преобладавшим в его время
условиям.
Эволюция предков лошади-это не постепенный
переход, равномерно распределенный во времени
Рис. 24.3. Фотография ископаемого трилобита из
кембрийских отложений.
Эпоха и возраст
ее древнейших
горных пород
Род
Внешний вид и рост
(от холки до земли)
Плейстоцен
Ы06лет
Equus
Кости правой передней ноги
Пятка
(поджилка)
Кости <
запястья
3-й палец
-, Пястные
-Бабка
Копыто
До 1,6 м
Плиоцен
7-Ю6 лет
Pliohippus
1,0 м
Миоцен
26-106 лет
Meryhippus
До 1,0 м
Олигоцен
38-106лет
Mesohippus
До 0,6 м
Эоцен Hyracotherium
54-106лет
Примерно 0,4 м
Пястные кости
(с номерами
соответствующих
пальцев)
Образ жизни, климат
и изменения строения тела
Адаптированы к жизни в сухих
степях. Очень быстро бегают.
Пястные и плюсневые кости удлинены.
Расширенная 3-я фаланга покрыта
роговым копытом (видоизмененный
коготь). Зубы с широкой
жевательной поверхностью Под стирающимся
цементом обнажается эмаль. Ложно-
коренные замещены коренными.
Животное перетирает пищу
Все больше полагается на быстроту
бега Второй и четвертый пальцы
сильно редуцированы. Утолщение
пястных и плюсневых костей для
большей устойчивости. На 3-й
фаланге образуется копыто. Зубы с
высокой коронкой — для питания травой
Очень сухая среда — прерии.
Быстрота бега становится важнее.
Редукция 2-го и 4-го пальцев.
При беге опора на 3-й палец
Удлинение оставшихся пястных и
плюсневых костей Удлинение зубов,
развитие цемента на коронках
Сухая среда — леса и прерии
Быстрота передвижения важна для
бегства от врагов. Хорошо
различимы только три пальца
Третий палец сильно утолщен
Размером с лисицу. Жил на мягкой
почве вблизи рек По четыре пальца
на передних и по три на задних ногах.
Коренные зубы с низкими
коронками, адаптированные к тому,
чтобы щипать мягкую траву
Рис. 24.4. Эволюция предков современной лошади
и пространстве, да и палеонтологическую летопись
нельзя считать полной. По-видимому, от линии,
представленной на рис. 24.4, отходило несколько
боковых ветвей, но все они вымерли. Все
современные лошади, очевидно, происходят от Pliohippus.
Современный род Equus возник в Северной Америке
в плейстоцене и мигрировал в Евразию и Африку,
где он дал начало зебрам и ослам, а также
современной лошади. Удивительно то, что лошади,
просуществовав в Северной Америке миллионы лет,
вымерли там несколько тысяч лет назад, в период,
совпавший с появлением человека. Рисунки,
сохранившиеся на стенах пещер в других частях земного
шара, указывают на то, что в самом начале
лошадей, по-видимому, использовали в пищу. Лошади
были вторично интродуцированы в Северную
Америку испанцами почти 500 лет назад.
24.7.2. Географическое
распространение
Все организмы в большей или меньшей степени
приспособлены к своей среде. Если абиотические
и биотические факторы (см. разд. 12.2), имеющиеся
в определенном местообитании, могут обеспечить
существование какого-то вида в одной
географической области, то можно было бы ожидать, что этот
вид будет обнаружен в аналогичном местообитании
и в другой сходной географической области,
например львы - в африканских саваннах и в
южноамериканской пампе. Однако на самом деле это не так.
Распространение растений и животных на земном
шаре носит прерывистый характер. Это нередко
обусловлено экологическими факторами, однако
данные об успешной колонизации новых
местообитаний растениями и животными, интродуцирован-
ными в них человеком, позволяют думать, что в
этом участвуют наряду с экологической адаптацией
и какие-то иные факторы. Кроликов в Австралии не
было, но быстрое увеличение их численности после
того, как они были завезены туда человеком,
указывает на то, что австралийские местообитания им
подходят. Сходными примерами служат факты
распространения человеком домашних животных и
культурных растений, таких как овцы, кукуруза,
картофель и пшеница. Рациональное объяснение
прерывистого распространения организмов
основано на концепции, согласно которой виды возникают
в какой-то данной области, а затем расселяются из
нее. Степень расселения зависит от того, насколько
успешно сможет обосноваться данный организм
в новых местах, от эффективности механизма его
расселения и от наличия или отсутствия
естественных преград, таких как океаны, горные хребты и
пустыни. Наиболее приспособлены для
распространения через сушу и моря, по-видимому, споры
и семена, переносимые ветром, и летающие
насекомые.
В противоположность этому несмотря на то, что
организмы, как правило, ограничены в своем
распространении определенными частями земного
шара, во многих случаях близко родственные формы
обитают в сильно удаленных друг от друга
областях; например, три современных вида
двоякодышащих рыб (Dipnoi) встречаются порознь в
тропических областях Южной Америки (Lepidosiren),
Африки {Protopterus) и Австралии (Neoceratodus);
верблюды и ламы (сем. Camelidae) живут в Северной
Америке, Азии и Южной Америке; а еноты широко
распространены в Северной и Южной Америке, но
обитают также в небольшом районе
Юго-Восточной Азии. Судя по ископаемым данным, в прошлом
эти животные были распространены гораздо шире,
чем в наши дни.
Все эти сведения не имеют непосредственного
значения для эволюционной теории, однако они
указывают на то, что распределение суши на земном
шаре не всегда было таким, как сейчас.
Прежде считалось, что материки и океаны всегда
занимали такое положение, какое они занимают
теперь. Ранние геологи, такие как Хаттон и Лайель
(см. табл. 24.1), полагали, что осадочные породы
образовались в результате периодических поднятий
и опусканий уровня моря. Позднее возникло
представление о том, что в прошлом на земном шаре
существовали два больших континента: Лавразия - в
Северном полушарии и Гондвана-в Южном; эти
два массива были соединены обширными мостами
суши, по которым могли мигрировать и расселяться
растения и животные. Позже в результате
геологических исследований эта точка зрения несколько
изменилась, и в настоящее время общепринятой
стала теория дрейфа континентов, основанная на
тектонике литосферных плит. Гипотезу о дрейфе
континентов впервые выдвинул Снайдер в 1858 г.,
а затем в конце XIX в. развили Тэйлор в Америке
и Вегенер в Германии. Вегенер считал, что в карбоне
Лавразия и Гондвана составляли единый массив
суши, названный им Пангеей (греч. «вся земля»),
который «плавал» в более плотном расплавленном
внутреннем веществе Земли. В настоящее время,
однако, принято считать, что континенты
раздвинулись под действием глубинных конвективных
течений, направленных вверх и в стороны и тянущих за
собой плиты, на которых плавают континенты. Эта
гипотеза позволяет объяснить непрерывное
перемещение массивов суши и современное
распространение таких животных, как, например,
двоякодышащие рыбы (рис. 24.5).
Что касается верблюдов и лам, то, как полагают,
они происходят от общего предка, который, судя по
ископаемым находкам, возник в Северной Америке.
В плейстоцене этот предок распространился на юг,
перейдя по Панамскому перешейку в Южную
Америку, и на север, проникнув в Азию до того, как в
результате изменений уровня моря она была
отделена от Северной Америки (рис. 24.6).
Предполагается, что на протяжении всего этого времени в группе
Африка
Южная
Америка
Австралии
Ант арктида
Lepidosiren
Protopterus
Neoceratodus
Рис. 24.5. А. Относительное расположение Южной
Америки, Африки и Австралии на ранних стадиях
дрейфа континентов, указывающее на близость
областей, в которых, возможно, возникли
двоякодышащие рыбы Б. Современные ареалы
двоякодышащих.
Camelidae происходили прогрессивные изменения,
приведшие к образованию двух родов -Camelus и
Lama-в краевых областях их плейстоценовой
миграции. В ископаемых остатках, находимых по всей
Северной Америке, в Азии и в северных областях
Африки, были обнаружены формы, промежуточные
между современными верблюдами и ламами. Как
показывают палеонтологические данные, другие
представители Camelidae в других частях земного
шара вымерли в конце последнего оледенения.
Другим примером прерывистого
распространения, возникшего в результате географической
изоляции, служат однопроходные и сумчатые
австралийской области. Как полагают, Австралия откололась
от остального массива суши в конце юры, вскоре
после появления примитивных млекопитающих.
Млекопитающих делят на три подкласса:
однопроходных (Monotremata) сумчатых (Marsupialia) и
плацентарных (Eutheria). B австралийской области
развивались только две первые группы. Здесь они
существуют, претерпев адаптивную радиацию, в
результате которой сложилась характерная
австралийская фауна, представленная однопроходными
Tachyglossus и Zaglossus (ехидны и проехидны) и
Omithorhynchus (утконос) и 24 родами сумчатых. В
других областях земного шара возникли также
более высокоразвитые плацентарные млекопитающие.
Видимо, они по мере своего распространения по
континентам вытесняли более примитивных
однопроходных и сумчатых из их экологических ниш и
остановились лишь там, где их дальнейшему рассе-
Считается, что Camelidae
возникли здесь
в плейстоцене
Рис. 24.6. Распространение современных представителей Camelidae- верблюдов в Азии и в
Северной Африке и лам в Южной Америке. Судя по ископаемым находкам, в
плейстоцене Camelidae были распространены по всей Северной и всей Южной Америке, в значительной части Азии и на
севере Африки. Толстыми черными стрелками указаны возможные пути миграции. (По
Matthews, Climate of evolution, 1938, vol. 1, 2nd ed., NY Acad of Sci.)
лению помешали географические преграды, как на
пути в Австралию.
Все изложенное можно вкратце свести к
следующему:
1) виды возникали в какой-то определенной
области;
2) они расселялись за пределы этой области;
3) большинство видов могло расселяться только в
том случае, если массивы суши располагались
достаточно близко один от другого;
4) отсутствие в какой-либо области более
высокоорганизованных форм обычно указывает на то,
что она отделилась от родины этих форм до
возникновения последних.
Ни один из приведенных выше фактов не позволяет
объяснить механизм возникновения видов, однако
все они указывают на то, что разные группы
возникали в разное время и в разных областях.
Палеонтологическая летопись дает представление о
направлениях, в которых происходило постепенное
изменение этих организмов, но она опять-таки не содержит
никаких указаний на возможный механизм такого
изменения.
О вероятном механизме возникновения видов в
результате естественного отбора можно говорить на
основании распространения растений и животных
на океанических островах. И Уоллеса, и Дарвина
поразило огромное разнообразие видов,
обитающих на таких группах островов, как Гавайские и
Галапагосские острова. Как показывают
геологические данные, эти острова образовались в результате
вулканической активности, поднявшей их со дна
океана, так что они никогда не имели прямой
географической связи ни с каким массивом суши.
Найденные на этих островах растения были, по всей
вероятности, занесены туда ветром в виде спор
и семян или по воде в виде плавучих семян и
растительной массы. Водные и полуводные животные,
как полагают, были принесены морскими
течениями, а наземные организмы приплыли на бревнах
или на плавучих скоплениях растений. Птицам,
летучим мышам и летающим насекомым заселить
эти острова было значительно проще.
Галапагосские острова находятся в Тихом океане,
на экваторе, на расстоянии почти 1000 км от
Эквадора и образуют архипелаг, описанный более
подробно в разд. 25.8.3. В 1835 г., когда Дарвин впервые
посетил эти острова, его внимание привлекло
сходство обитающих на них видов с видами,
населяющими ближайший континент; аналогичное сходство
островных и материковых видов он обнаружил на
островах Зеленого мыса, лежащих вблизи
побережья Западной Африки. Однако на океанических
островах растения и животные были в большинстве
случаев заметно крупнее. Это можно объяснить
отсутствием здесь крупных, более
высокоорганизованных доминантных видов, с которыми более
мелким родичам островных видов приходилось
конкурировать на континенте. Например, слоновая
Рис. 24.7. Гигантская морская игуана
Галапагосских островов.
черепаха (Geochelone elephantopus), которая питается
имеющейся на островах обильной растительностью,
достигла, вероятно, своих гигантских размеров-
почти 2 м длины и 260 кг веса - благодаря
отсутствию конкуренции со стороны различных
млекопитающих, населяющих континент. Дарвин заметил
также обилие на Галапагосских островах игуан,
которые опять-таки были значительно крупнее
своих континентальных родичей. Ящерицы обычно
живут на суше, однако на Галапагосских островах
из обнаруженных там двух видов игуан один
обитает в море. Водная форма-галапагосская игуана
(Amblyrhyncus cristatus) - питается морскими
водорослями и приспособлена к передвижению в воде
благодаря уплощенному с боков хвосту и хорошо
развитым перепонкам между пальцами на всех
четырех конечностях (рис. 24.7). Предполагают, что
конкуренция между наземными игуанами за пищу,
пространство и брачного партнера создала давление
отбора в пользу тех особей, у которых намечались
изменения, благоприятные для жизни в водной
среде. Этот механизм воздействия факторов среды
на изменчивый генотип называют естественным
отбором, и он уже был нами описан. Возможно, что
именно в результате такого отбора постепенно
сформировался морской вид игуан. Однако самое
сильное влияние на мысли Дарвина о механизме
возникновения видов оказало разнообразие
адаптивных признаков у 13 видов вьюрков, обитающих
на островах. В Эквадоре, т.е. на континенте,
имелись вьюрки только одного типа, у которых клюв
был приспособлен для раздробления семян. На
Галапагосских островах обнаружено шесть главных
типов клюва, каждый из которых приспособлен для
определенного способа питания. Различные группы
вьюрков, особенности их питания и число видов в
Типы вьюрков
Большой земляной вьюрок
(предковая форма)
Земляные вьюрки
Кактусовые земляные вьюрки
Насекомоядные древесные
вьюрки
Растительноядный
древесный вьюрок
Славковый вьюрок
Дятловые вьюрки
Форма клюва
Типичный для материковых
видов — короткий и прямой
Различный, но короткий
и прямой, как у материковых
видов
Длинный, несколько изогнутый,
язык расщепленный
Похожий на клюв попугая
Загнутый, похожий на клюв
попугая
Тонкий
Большой, прямой (использует
кактусовые иглы или веточки,
чтобы выковыривать
насекомых из дырок в коре)
Пища
Семена
Семена, насекомые
Нектар опунции
Семена, насекомые
Плоды, почки;
мягкие плоды
Насекомые
(ловит налету)
Личинки насекомых
Местообитание
Прибрежная зона
Прибрежная зона
и низменности
Низменности
Лес
Лес
Лес
Лес
Число видов
1
3
2
3
1
1
2
каждой группе представлены на рис. 24.8.
Дарвин предположил, что острова были
колонизированы группой вьюрков с континента. Здесь
они хорошо прижились, и неизбежная конкуренция,
возникшая в результате роста их численности, а
также наличие незанятых экологических ниш
благоприятствовали освоению разнообразных ниш теми
формами, у которых были соответствующие
адаптивные вариации. Видовые особенности сводятся к
небольшим различиям в размерах тела, окраске
оперения и форме клюва. Несколько видов
встречаются на всех крупных островах. Земляные и славко-
вые вьюрки, которых считают самыми
примитивными, живут на большей части островов.
Насекомоядных древесных вьюрков и растительноядного
древесного вьюрка нет на периферийных островах, а
дятловые вьюрки обитают только на островах
центральной группы. Современное распространение
видов представляет большой интерес; согласно Лэку,
его можно объяснить адаптивной радиацией и
географической изоляцией. Например, на центральных
островах обитает много видов, принадлежащих к
разным группам (земляные, древесные, славковые и
дятловые), а не к одной какой-то группе. Даже там,
где обитают несколько видов из одной группы (как
на периферийных островах), все они различаются по
своим экологическим требованиям. Это
соответствует принципу конкурентного исключения Гаузе
(см. разд. 12.6), согласно которому два или большее
Рис. 24.8. А. Адаптивная радиация дарвиновых
вьюрков (по Lack). Б. Самец кактусового вьюрка
(Geospiza scandens).
число близко родственных видов могут занимать
одну и ту же область только в том случае, если их
экологические требования различны.
24.7.3. Классификация
Система классификации, описанная в Приложении
3, была создана Линнеем задолго до Дарвина и
Уоллеса, но тем не менее она содержит кое-какие
намеки в связи с проблемой происхождения видов и
эволюции. Конечно, можно представить себе, что
все виды, как ныне живущие, так и вымершие, были
сотворены каждый в отдельности в какой-то
отдельный момент времени или существовали всегда,
однако структурное сходство между организмами,
составляющее основу естественной
филогенетической классификации, наводит на мысль о
существовании эволюционного процесса. Черты сходства и
различия между организмами можно объяснить как
результат прогрессивной адаптации организмов в
пределах каждой таксономической группы к
определенным условиям среды на протяжении
некоторого периода времени.
Специалисты по нумерической таксономии,
занимающиеся главным образом сравнением фено-
типических признаков, нашли возможным
разработать фенетическую классификацию (см. Приложение
3), которая согласуется, насколько позволяют
судить современные знания, с концепцией
эволюции. Обе системы классификации могут иметь и
самостоятельное значение как способы описания
множества живых организмов, но в то же время они
явно подсказывают мысль об эволюционном
процессе.
24.7.4. Селекция растений и животных
Одним из самых ранних достижений человеческой
цивилизации было выведение сортов растений и
пород домашних животных от диких предков.
Отбирая тех особей, которые обладали каким-либо
желательным отклонением, например более
крупными размерами или более приятным вкусом и
запахом, человек сохранял эти признаки путем
искусственного разведения с помощью избирательного
размножения или опыления. В результате
непрерывной селекции человек создал породы домашних
животных и сорта культурных растений, которыми
мы располагаем сейчас. Как показывают
археологические данные, человек проявил большое
искусство в разведении крупного рогатого скота, свиней и
домашней птицы и в выращивании зерновых
культур и некоторых овощей. До того как стали
известны работы Менделя, теоретические основы генетики
и селекции животных и растений оставались
неясными, однако это не ограничивало практические
усилия человека. Если перейти на генетическую
терминологию, человек сохраняет гены,
желательные для его целей, и элиминирует те, которые его не
устраивают. Производя отбор, он использует
существующую в природе генетическую изменчивость, а
также возникающие время от времени случайные
мутации.
Наибольшее внимание растениеводы и
животноводы уделяли выведению хозяйственно ценных
пород животных (рис. 24.9) и сортов растений, хотя
создавались также разнообразные породы собак,
кошек, птиц, рыб и сорта декоративных растений.
Некоторые примеры фенотипических признаков, по
которым вели искусственный отбор, перечислены в
табл. 24.4. Недавно возникла новая форма
искусственного отбора-неумышленный отбор на
устойчивость к антибиотикам, пестицидам и гербицидам,
которому подвергаются соответственно патогенные
микроорганизмы, вредители и сорняки. Создается
порочный круг: все возрастающее число химических
веществ, изобретаемых для борьбы с вредными
организмами, приводит к появлению новых форм,
устойчивых к этим веществам.
Способность человека «создавать» признаки
путем направленного отбора, как, например, при
разведении собак и голубей, Дарвин использовал
как довод в пользу того, что с помощью
аналогичного механизма виды могут возникать и в
естественных условиях: он полагал, что в природе
вместо человека в роли фактора отбора выступает
внешняя среда. Формы, созданные в результате
искусственного отбора, вероятно, не могли бы
возникнуть в природе: в большинстве случаев они были
бы неспособны успешно конкурировать с близкими
к ним неодомашненными формами.
24.7.5. Сравнительная анатомия
При сравнительном изучении анатомии
(морфологии) групп животных или растений становится ясно,
что по ряду структурных особенностей они в основе
Таблица 24.4. Некоторые фенотипические признаки, по
которым ведется селекция
Признак
Холодостойкость
Размеры
Увеличение
продуктивности
Раннее созревание
Более длительный период
плодоношения
Вкусовые качества
Облегчение уборки урожая
Длительность хранения
Повышенная пищевая
ценность
Устойчивость к болезням
Примеры
Сахарная кукуруза в
Англии
Картофель, капуста
Молоко, яйца, шерсть,
плоды
Зерновые культуры (два
урожая за сезон)
Земляника
Яблоки, виноград без
косточек
Горох
Фасоль и горох для
быстрого замораживания
Соя и другие растения
(содержание белка)
Пшеница, устойчивая к
ржавчине и к милдью
Рис. 24.9. Результаты использования отбора в
животноводстве. Из дикого кабана
(А)-коренного обитателя Европы, Азии и Африки-путем
отбора было выведено много домашних пород, в
том числе английская крупная белая (Б), дающая
превосходное мясо.
своей сходны. Например, у всех цветков имеются
чашелистики, лепестки, тычинки, рыльце, столбик и
завязь; однако каждый отдельный вид отличается от
других по размерам, окраске, числу этих частей и
деталям их строения. Точно так же скелет
конечностей у всех четвероногих, от амфибий до
млекопитающих, построен по единому плану-по плану
пятипалой конечности (рис. 17.11). Эта основная
структура у разных животных различным образом
модифицирована (рис. 24.10) и в каждом случае
приспособлена для определенного способа
передвижения в соответствующей среде обитания.
Органы, построенные по одному плану,
занимающие сходное положение в организме животного,
сходные по гистологическому строению и
развивающиеся из одних и тех же зачатков, называют
гомологичными (термин, введенный в 1843 г. Ричардом
Оуэном).
Гомологичные структуры, обладающие адапта-
циями к разным условиям среды и к разному образу
жизни, служат примерами адаптивной радиации.
Экологическое значение этих процессов
рассмотрено в разд. 24.7.6. Специфические функции,
выполняемые гомологичными структурами, могут
различаться у разных организмов; их различия отражают
особые способы адаптации каждого организма к его
среде и образу жизни. Ниже приведено несколько
примеров гомологии.
Жаберные дуги - слуховые косточки. Можно
проследить, как некоторые челюстные кости рыбы,
преобразующиеся у других позвоночных в подвесок
для нижней челюсти, у млекопитающих превращены
в слуховые косточки—молоточек, наковальню и
стремя (рис. 24.11).
Жужжальца. Задняя пара крыльев, имеющаяся у
большинства насекомых, у двукрылых
преобразована в небольшие палочковидные структуры-
жужжальца (гальтеры), которые служат органами
равновесия при полете.
Околоплодник. У цветковых растений стенка
завязи после оплодотворения семязачатков
различным образом видоизменяется, чтобы
способствовать рассеиванию семян (см. рис. 20.28 и 24.12).
Явление гомологии само по себе не может
служить доказательством эволюции, однако
существование гомологии в пределах данной группы
организмов расценивается как свидетельство их
происхождения от общего предка и указание на близкое
филогенетическое родство.
Линней использовал гомологию как основу для
своей системы классификации. Чем более
специфичны общие для двух данных организмов
гомологичные признаки, тем теснее родство между ними и тем
ниже ранг таксономической группы, к которой их
обоих следует отнести. Например, дневные и ночные
бабочки принадлежат к одному отряду (Lepido-
ptera), а осы и пчелы-к другому (Hymenoptera).
Некоторые структуры у многих видов,
по-видимому, не несут никакой функции, и их называют
рудиментарными органами. Червеобразный отросток
человека, не участвующий в пищеварении,
гомологичен отростку, имеющему функциональное
значение у растительноядных млекопитающих. Точно
так же некоторые явно нефункционирующие кости
змей и китов считаются гомологами тазовых костей
и костей задних конечностей четвероногих
позвоночных. Копчиковые позвонки у человека считают
рудиментами хвоста, имевшегося у наших предков и
Плечевая кость
Лучевая кость
Локтевая кость -
Летучая мышь
(летает)
Плечевая кость
Лучевая
кость
Запястье
Пясть
Фаланги
Локтевая
кость
Запястье
Пясть
Фаланги
Лошадь
(бегает)
Плечевая
кость
Лучевая
кость
Локтевая
кость
Увеличенная
смещенная
запястная кость
"Колено"
Одна развитая J
пястная кость *
Мослак
Бабка -
Копыто
Запястье
Пясть
Фаланги
Тюлень (плавает)
Человек
(манипулирует)
Фаланги
Крот(роет)
Плечевая кость
Локтевая кость
(слившаяся с лучевой)
Лучевая кость
Запястье
Рудиментарные
пястные кости
Фаланга
Плечевая кость
Лучевая кость
Локтевая кость
Запястье
Пясть
Рис. 24.10. Адаптивные изменения пятипалой конечности у некоторых млекопитающих.
Череп
Верхняя
челюсть
Нижняя
челюсть
А. Рыбы
Верхняя
челюсть
приросла
к черепу —
Квадрат-"
ная кость
Сочленовная KOCTbJ
Нижняя
челюсть
Б. Амфибии
Молоточек
Наковальня
Гиомандибулярный
хрящ, причленяющий
челюсти к черепу
Квадратная
Сочленовная ->
Кости
челюстей
Стремя — передает
звуковые колебания
во внутреннее ухо
Причленение
нижней
челюсти
Слуховой]
проход
В. Млекопитающие
Стремя
Овальное окно
Круглое окно
Евстахиева труба
Высохшее
рыльце
Семя
Перикарпий,
напоминающий
перечницу
Мак (распространение
ветром)
Крыло
перикарпия
Семя
Клен (распространение ветром)
Высыхаю;
щий боб
Семена
Утесник (механическое
распространение)
Стенка w
оплодотворенной
завязи
Сочный
мезокарпий
Каменистый
эндокарпий
Семя
-Эпикарпий
■ Мезокарпий
Эндокарпий
Семя
Кокосовый орех
(распространение водой)
Вишня
(распространение
животными)
Подмаренник
(распространение
животными)
Рис. 24.11. Костные структуры рыб и амфибий,
превратившиеся у млекопитающих в
слуховые косточки.
Рис. 24.12. Изменение строения околоплодника при
разных способах распространения семян.
Сосущие ротовые части
на примере бабочки
верхняя губа редуцирована,
жвалы утрачены; максиллы
длинные, образуют хоботок;
нижняя губа редуцирована
Лакающие ротовые
части на примере пчелы,
длинная нижняя губа
служит для лакания
нектара, жвалы —
для пережевывания
пыльцы и для постройки
ячеек из воска
Грызущие ротовые части
(примитивное состояние)
на примере кузнечика
сильные жвалы; максиллы
приспособлены для
захватывания пищи
Колющие и сосущие
ротовые части
на примере самки
комара верхняя губа
и максиллы образуют
хоботок, жвалы
образуют колющий
стилет, нижняя губа
превращена в желоб,
в котором находятся
остальные части
сохранившегося у зародышей. Существование
рудиментарных органов было бы трудно объяснить вне
связи с процессом эволюции.
24.7.6. Адаптивная радиация
Адаптивной радиацией называют развитие какой-
либо гомологичной структуры у разных
представителей данной группы в различных направлениях, в
соответствии с выполняемыми ею различными
функциями. Примеры адаптивной радиации можно
найти в любой таксономической группе выше
видового ранга. У всех организмов, принадлежащих к
определенному классу, имеется ряд диагностических
признаков; при этом различия между разными
видами в пределах этого класса дают им возможность
вести различный образ жизни, приспособленный к
определенным местообитаниям. Так, структура
ротовых частей насекомых в своей основе одинакова:
верхняя губа, пара жвал, гипофаринкс (подглоточ-
ник), пара максилл и нижняя губа (слившаяся
вторая пара жвал). Насекомые способны
использовать разнообразную пищу, так как у разных видов
одни из перечисленных выше структур увеличены и
видоизменены, а другие редуцированы или вовсе
утрачены (рис. 24.13). В результате ротовой аппарат
насекомых очень разнообразен.
Относительно высокая степень адаптивной
радиации, наблюдаемая у насекомых, отражает высокую
приспособляемость и полезность основных
особенностей этой группы. Именно такая «эволюционная
пластичность» позволила им занять столь широкий
диапазон экологических ниш. Это один из ряда
связанных между собой критериев, на основании
которых судят о биологическом успехе
таксономической группы, например типа или класса. К другим
критериям относятся эволюционный возраст
группы и число относящихся к ней видов.
Наличие у предкового организма какой-либо
структуры или физиологической функции, которая
имеется в сильно модифицированной форме у более
высокоразвитых, по-видимому родственных,
организмов, можно истолковать как указание на
происхождение последних путем видоизменения
первого; это и составляет основу эволюционной теории,
согласно определению, данному в разд. 24.4.2.
Значение адаптивной радиации состоит в том, что
она указывает на возможность дивергентной
эволюции, основанной на модификации гомологичных
структур.
Рис. 24.13. Адаптивная радиация на примере
ротовых частей насекомых 1-антенны; 2-сложный
глаз, 3-нижняя губа, 4-верхняя губа; 5—жвалы,
6-максиллы.
Таблица 24.5. Примеры параллельной эволюции у сумчатых
и плацентарных млекопитающих
Сумчатые (Австралия)
Сумчатый крот
Сумчатая мышь
Сумчатый муравьед
Вомбат
Кенгуру
Бандикут
Сумчатая летяга
Коала
Сумчатый волк
Плацентарные (другие области
земного шара)
Крот
Мышь
Муравьед
Луговая собачка
Антилопа
Кролик
Летяга
Ленивец
Гиена
Сходные структуры, физиологические процессы
или особенности образа жизни, наблюдаемые у
организмов, явно не связанных близким
филогенетическим родством, но обладающих адаптациями
для выполнения одних и тех же функций, называют
аналогичными. Примерами могут служить глаза
позвоночных и головоногих моллюсков, крылья
насекомых и летучих мышей, членистые ноги
насекомых и высших позвоночных, колючки растений и
иглы животных, обнаружение нейрогормонов
позвоночных (ацетилхолина, 5-гидрокситриптамина и
гистамина) в жгучих волосках крапивы. Сходство
между аналогичными структурами чисто внешнее.
Так, крылья насекомых поддерживаются жесткими
жилками, состоящими из кутикулы, а у птиц и
летучих мышей-полыми костями. Подобным же
образом глаза головоногих моллюсков и
позвоночных развиваются по-разному. У головоногих
сетчатка не инвертирована и фоторецепторы обращены
в сторону падающего на них света, тогда как у
позвоночных сетчатка инвертирована и
фоторецепторы отделены от падающего света нейронами (см.
рис. 16.33). Поэтому в глазу позвоночных имеется
слепое пятно, а у головоногих его нет.
Существование аналогичных структур говорит о
возможности конвергентной эволюции.
Конвергентную эволюцию можно объяснить как результат
действия среды путем естественного отбора,
благоприятствующего тем изменениям, которые
сообщают организмам повышенную выживаемость и
репродуктивный потенциал.
Значение дивергенции, прямо наводящей на
мысль об эволюционном процессе, и конвергенции,
указывающей на механизм эволюционных
изменений, особенно хорошо видно на примере
параллельной эволюции сумчатых и плацентарных
млекопитающих. Как полагают, обе группы претерпели
конвергентную эволюцию и в результате заняли
идентичные экологические ниши в разных частях
замного шара (рис. 24.14 и табл. 24.5).
24.7.7. Сравнительная эмбриология
Фон Бэр (1792-1867), изучая эмбриональное
развитие у представителей разных групп позвоночных,
обнаружил удивительное структурное сходство во
всех этих группах, особенно на стадиях дробления,
гаструляции и ранних стадиях дифференцировки
(см. разд. 21.5). Геккель (1834-1919) высказал
мысль, что это сходство имеет эволюционное
значение. Он сформулировал закон рекапитуляции,
согласно которому «онтогенез повторяет филогенез», т. е.
стадии, через которые проходит организм в
процессе своего развития, повторяют эволюционную
историю той группы, к которой он относится. Хотя этот
принцип слишком упрощает действительное
положение дела, он привлекателен и в известной мере
справедлив. Изучение одних только ранних
зародышей любых позвоночных показывает, что
определить группу, к которой они принадлежат,
невозможно. Как видно из рис. 24.15, только на
относительно поздних стадиях развития эмбрион
начинает приобретать некоторое сходство с
соответствующей взрослой формой. На определенных
сравнимых стадиях эмбрионы всех позвоночных
имеют следующие структуры:
1. Карманообразные впячивания эктодермы и
растущие им навстречу выпячивания стенок
глотки. У рыб они соединяются и срастаются, а
затем прорываются; в результате образуются
жаберные щели, которые служат для газообмена. У
других групп позвоночных сохраняется одна
такая щель, из которой образуются евстахиева
труба и слуховой проход.
2. Сегментированные миотомы в хвостообразной
структуре, сохраняющейся не у всех видов.
3. Один круг кровообращения с двухкамерным
сердцем, не разделенным на правую и левую
половины; во взрослом состоянии такое строение сердца
сохраняется только у рыб.
По мере развития зародыша позвоночного у него
происходят изменения, в результате которых он
приобретает признаки рыбы, амфибии, рептилии,
птицы или млекопитающего в соответствии со своей
видовой принадлежностью. Изначальное сходство
между эмбрионами объясняют тем, что все они, а
следовательно, и классы, к которым они относятся,
имели общего предка. Если у организма в
эмбриональном периоде образуются зачатки, из которых
не будут развиваться функционирующие органы, то
их можно интерпретировать лишь как остатки
структур, имевшихся у его предков. Закон
рекапитуляции, однако, не может быть принят
безоговорочно, так как ни у одного из ныне живущих
организмов нельзя обнаружить всех признаков его
предполагаемых эволюционных предков. Но
кажется вероятным, что организмы сохраняют
механизмы развития, унаследованные от предков. Поэтому
возможно, что на разных стадиях развития у
данного организма будут черты структурного сходства
с зародышами предковых форм. Последующие
адаптации к иным условиям среды и образу жизни
изменяют дальнейший ход развития. Как
показывают наблюдения, чем ближе группы, к которым
относят два данных организма на основании общих
Сумчатая
летяга
Карликовый
летучий кускус'
Древесный
кенгуру
Коала
Примитивные сумчатые
Сумчатый волк
Намбат
Гигантский
красный кенгуру
Сумчатый черт
Сумчатая мышь
Бандикут
Сумчатый крот
Вомбат
Рис. 24.14. Адаптивная радиация сумчатых в Австралии (По различным источникам.)
Черепаха
Курица
Кролик
Рис. 24.15. Сравнение стадий эмбрионального
развития на примере представителей трех классов
позвоночных."
гомологичных дефинитивных структур, тем дольше
сохраняется их сходство на эмбриональных стадиях.
Организмы, приспособленные к определенному
образу жизни и определенному местообитанию, не
типичному для крупной группы, к которой они
принадлежат, менее сходны с другими членами этой
группы и в процессе эмбрионального развития. Это
видно на примере двух паразитических плоских
червей Fasciola и Taenia: у них имеется ряд
личиночных стадий с адаптациями к вторичным хозяевам,
отсутствующих в жизненном цикле свободноживу-
щих плоских червей Planaria. Точно так же у дожде-
} Данный материал, также как и рисунок на
обложке, основан на фальшивке,
сделанной Э.Геккелем в 1874 г.
Ниже приведены рисунки Геккеля в
сопоставлении с подлинными фотографиями.
Видно, что эмбрионы разных видов
сильно различаются. (См. Richardson,
M. К., J. Hanken, L. Selwood, G. M.
Wright, R. J. Richards and C. Pieae, 1998.
"Haeckel, embryos, and evolution".
Science 280: 983-986.)
вого червя Lumbricus, ведущего наземный образ
жизни, нет стадии трохофоры-личинки, типичной
для более примитивных кольчецов. На этих
примерах ясно видна ограниченность геккелевского
закона рекапитуляции.
Изучение индивидуального развития
представителей крупных групп организмов выявляет черты
структурного сходства, очевидные на
эмбриональных и личиночных стадиях, но отсутствующие у
взрослых особей. Эти наблюдения
интерпретируются как признаки филогенетического родства между
различными группами организмов, что
подразумевает существование эволюционного процесса. По
характеру дробления зиготы и судьбе бластопора
организмы с трехслойным зародышем можно
разделить на две группы - первичноротых и вторично-
ротых. У первичноротых дробление спиральное и
бластопор превращается в ротовое отверстие
взрослой особи. Такой тип развития свойствен
кольчецам, моллюскам и членистоногим. Для вторично-
ротых характерно радиальное дробление, и
бластопор превращается у них в анальное отверстие
взрослой особи. Так происходит развитие иглокожих и
хордовых. Различия между этими двумя группами
показаны на рис. 24.16. Именно такого рода данные
помогли выяснить вопрос о филогенетических
родственных связях иглокожих. Строение взрослых
иглокожих заставляет рассматривать их как один из
типов беспозвоночных, но эмбриональное развитие,
протекающее у них по типу, характерному для
вторичноротых, подтверждает их близость к линии
хордовых. Этот пример показывает, что вопрос о
филогенетических взаимоотношениях нельзя решать
на основании одних лишь сведений о гомологичных
структурах взрослых особей.
В результате изучения эмбриогенеза растений
тоже получены данные, свидетельствующие о
прогрессивной эволюции различных групп, однако
изученные примеры не так хорошо
документированы, как в царстве животных. У мхов и папоротников
рыба саламандра черепаха курица кролик человек
Радиальное
дробление
Зигота
Рот
Бластопор
Анус
Реснички
Стадия бластулы
Типичная личинка
вторичноротых
Рот
Спиральное
дробление
Бластопор
Реснички
Анус
Типичная личинка
первичноротых
Рис. 24.16. Ранние стадии развития вторичноротых и первичноротых.
Зрелое
пыльцевое
зерно
Ядро пыльцевой трубки·
■Вегетативная клетка-
Антеридиальная
клетка
Проталлиальная
клетка
Жгутики-
Прорастающая
пыльцевая
трубка
Зрелый
антерозоид
Рис. 24.17. Развитие подвижных мужских гамет из
пыльцевых зерен у Cycas.
гаметофит (на ранних стадиях развития
представленный протонемой, образующейся в результате
прорастания спор) по своей структуре, физиологии и
типу роста сходен с нитчатыми зелеными
водорослями; поэтому принято считать, что они
происходят от этих водорослей. Чередование
поколений в жизненных циклах растений и различные его
варианты, отражающие адаптацию к разным
условиям среды, можно интерпретировать как примеры
гомологии и рассматривать как дополнительные
доводы в пользу эволюционных связей между
различными группами растений.
Голосеменные - это группа, промежуточная по
своим признакам между формами,
приспособленными к жизни на суще, и формами, у которых для
встречи гамет все еще необходима вода. У
саговников (порядок Cycadales) мужской гаметофит похож
на легкую сухую микроспору (пыльцевое зерно)
покрытосеменных тем, что он тоже
распространяется ветром. В процессе развития мужского гамето-
фита одна из его клеток образует, как у
покрытосеменных, пыльцевую трубку, однако она не
доставляет неподвижную мужскую гамету к архего-
нию; вместо этого из концевой (антеридиальной)
клетки мужского гаметофита образуются два
снабженных жгутиками антерозоида (спермия), которые
подплывают к семязачатку и осуществляют
оплодотворение (рис. 24.17). Поэтому можно полагать, что
саговники-группа, промежуточная между низшими
растениями и покрытосеменными, а это позволяет
предположить, что в царстве растений существует
филогенетическая непрерывность. Существование
группы организмов, обладающих чертами, общими
для двух других групп, находящихся на разных
уровнях сложности или адаптированных к
различным местообитаниям, можно интерпретировать как
указание на филогенетическую непрерывность
между этими тремя группами - на происхождение одной
группы (такой, как покрытосеменные) от другой
(низшие растения) через промежуточную группу
(саговники). Многие из таких промежуточных форм
вымерли, и только изучая палеонтологическую
летопись, можно вывести эволюционную
последовательность. Во многих случаях промежуточные
формы обнаружить не удалось. Эти формы
эквивалентны «недостающим звеньям», которые
выглядят как разрывы в ископаемой летописи. Возможно
также, что эти звенья и не существуют (в
соответствии с гипотезой прерывистого равновесия,
описанной в разд. 24.7.1). Если, однако, согласиться с
изречением «Natura non facit saltum» («природа не
делает скачков»), то отсутствие таких звеньев
можно объяснить тем, что их остатки либо не
сохранились, либо еще не найдены. Промежуточным
звеном между кольчатыми червями и
членистоногими служит группа Onychophora, представленная
родом Peripatus, который обладает признаками и
кольчецов, и членистоногих (рис. 24.18). С
кольчецами перипатуса сближают строение стенки тела, в
которой имеются кольцевые и продольные мышцы,
нечленистые конечности, напоминающие параподии,
Рис. 24.18. Примитивное членистоногое Peripatus.
Таблица 24.6. Аминокислотные последовательности цитохрома с у 21 вида (по Dayhoff M. О., Eck R. V (1967—1968) Atlas
of protein sequence and structure, National Biomedical Research Foundation, Silver Spring, Md)
Вид
Человек
Макак-резус
Лошадь
Свинья, корова, овца
Собака
Серый кит
Кролик
Кенгуру
Курица, индейка
Пингвин
Утка (пекинская)
Каймановая черепаха
Лягушка-бык
Тунец
Муха Chrysomyia
Тутовый шелкопряд
Пшеница
Нейроспора
Пекарские дрожжи
Гриб Candida
Бактерия Rhodospirillum
Обозначения аминокислот
А — аланин
С — цистеин
D — аспарагиновая
кислота
E — глутаминовая
кислота
F — фенилаланин
G — глицин
70
0
D
D
E
E
E
E
D
D
D
D
D
E
D
D
D
D
N
N
N
P
А
1
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
N
T
N
2
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
M
M
L
3
M
M
M
M
M
M
M
M
M
M
M
M
M
N
F
F
Y
F
S
S
А
4
E
E
E
E
E
E
E
E
E
E
E
E
E
E
E
E
D
E
E
D
А
5
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
6
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
V
Аминокислотные последовательности
7
E
E
E
E
E
E
E
E
E
E
E
E
E
E
E
E
L
E
T
E
К
8
N
N
N
N
N
N
N
N
N
N
N
N
N
N
N
N
N
N
N
N
N
H — гистидин
I — изолейцин
К — лизин
L — лейцин
M — метионин
N — аспарагин
P — пролин
9
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
80
0
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
1
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
А
2
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
Y
F
3
1
I
I
1
I
I
I
I
I
I
I
I
I
1
I
1
I
I
1
I
V
4
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
P
L
5
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
E
6
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
T
I
T
S
7
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
к
8
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
M
Q — глутамин
R — аргинин
S — серии
T — треонин
V — валин
W — триптофан
Y — тирозин
9
I
I
I
I
I
I
I
I
I
I
I
I
1
I
I
I
V
V
А
А
T
90
0
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
F
1
V
V
А
А
А
А
А
А
А
А
А
А
А
А
А
А
А
P
G
G
К
2
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
G
—
выделительные канальцы, сходные с нефридиями,
имеющиеся в каждом сегменте, и двойная брюшная
нервная цепочка. К признакам сходства с
членистоногими относятся хитиновая кутикула, трахеи,
дыхальца и незамкнутая система кровообращения.
Другая группа «живых ископаемых», служащая
промежуточным звеном между рыбами и
амфибиями,-это двоякодышащие (Dipnoi).
Хотя большая часть этих данных указывает на
существование той или иной формы эволюционного
процесса, следует помнить, что твердыми данными
о том, что он имел место, мы не располагаем.
24.7.8. Сравнительная биохимия
По мере разработки более точных методов
биохимического анализа эта область исследований
стала источником новых данных в пользу
эволюционной теории. Наличие одинаковых веществ у всех
организмов указывает на возможную
биохимическую гомологию, подобную морфологической
гомологии на уровне органов и тканей. Следует снова
подчеркнуть, что эти данные лишь подкрепляют
другие доводы в пользу существования эволюции,
но сами по себе не могут считаться бесспорными
доказательствами. Большая часть сравнительно-
биохимических исследований касалась первичной
структуры широко распространенных белков, таких
как цитохром с и гемоглобин, а позднее -
нуклеиновых кислот, в особенности рибосомной РНК.
Незначительные изменения в генетическом коде ДНК,
связанные с генными мутациями, приводят к
тонким изменениям в общей структуре
соответствующих белков или РНК. Это создает основу для
выяснения филогенетических связей, если
предположить, что чем меньше различия в структуре того
или иного вещества, тем меньше число
произошедших мутаций и тем ближе родство между
организмами, содержащими это вещество. Крупные
различия в молекулярной структуре отражают крупные
различия в ДНК, и можно предсказать, что такие
различия будут обнаружены у организмов со
сравнительно небольшим числом морфологических
гомологии.
Цитрохромы представляют собой белки
(дыхательные пигменты), содержащиеся в митохондриях
клеток и участвующие в переносе электронов по
дыхательной цепи, т.е. в той последовательности
реакций, которая приводит к высвобождению
энергии, необходимой для синтеза АТФ (см. разд.
11.5.4). Цитохром с-один из белков, входящих в эту
цепь. Это сложный белок, состоящий из
железосодержащей простетической группы, которую
окружает полипептидная цепь из 104-112 аминокислот
(в зависимости от вида организма). Современные
методы масс-спектрометрических исследований с
применением ЭВМ позволили установить
первичную структуру полипептидной цепи цитохрома с у
многих организмов, в том числе некоторых
бактерий, грибов, пшеницы, мухи Chrysomyia, тутового
шелкопряда, тунца, пингвина, кенгуру и приматов.
У исследованного таким образом 21 организма
аминокислотные последовательности оказались
удивительно сходными. У 20 из этих организмов-
от гриба Candida до человека - аминокислоты в
положениях 78-88 идентичны (табл. 24.6).
Аминокислотные последовательности цитохрома с у
человека и шимпанзе одинаковы, а от
последовательности макака-резуса они отличаются лишь по одной
аминокислоте. Филогенетические деревья растений
и животных, основанные на современных данных об
аминокислотных последовательностях цитохрома с,
близки к деревьям, построенным на основании
морфологических гомологии.
Такие же результаты были получены при изучении
глобинов-гемоглобина и миоглобина,
участвующих в переносе и запасании кислорода. Степень
сходства между молекулами гемоглобина у четырех
видов приматов показаны в табл. 24.7.
Эволюционные связи между различными глобинами,
предполагаемые на основании аминокислотных
последовательностей (с указанием организмов, у которых они
встречаются), представлены на рис. 24.19. Различия
в последовательностях цитохрома с и этих глобинов
возникли, по-видимому, в результате генных
мутаций.
Иммунологические исследования тоже
свидетельствуют об эволюционном родстве между
организмами. Если белки, содержащиеся в сыворотке
крови, ввести в кровь животным, у которых этих
белков нет, то они действуют как антигены, т.е.
побуждают организм вырабатывать соответствую-
Таблица 24.7. Различия между полипептидными цепями
гемоглобина четырех видов приматов"
Вид
Человек
Шимпанзе
Горилла
Гиббон
Полипептидные цепи
а-Цепь
(141
аминокислота)
1
3
$-Цепь
(146
аминокислот)
—
1
3
у-Цепь
—
1
1
2
" Гемоглобин состоит из четырех полипептидных цепей,
построенных из α-, β- и γ-полипептидов Минус означает отсутствие
отличий в аминокислотной последовательности данного вида от
таковой у человека, а цифры — число отличий в аминокислотах
Миоглобин
а · Цепь
(позвоночные,
обладающие
челюстями)
I
γ-Цепь
(млекопитающие на стадии
утробного
плода)
0 -Цепь
(приматы)
δ -Цепь
(человек
и
человекообразные
обезьяны)
/З-Цепь млекопитающих
Пред ковая цепь β-типа
Предковая цепь гемоглобина
Предковая цепь глобина
Рис. 24.19. Предполагаемое происхождение
миоглобина и полипептидных цепей глобина у
позвоночных. У человека имеются цепи всех пяти типов
(По V M. Ingram, Haemoglobin in genetics and
evolution, 1963, Columbia Univ. Press.)
щие антитела; в результате возникает реакция
антиген—антитело (см. разд. 14.14). Эта иммунная
реакция обусловлена способностью
животного-реципиента распознавать присутствие в сыворотке
чужеродных белков. Человеческая сыворотка, введенная
кроликам, сенсибилизирует их и вызывает у них
образование антител к сывороточным белкам
человека. Если спустя некоторое время к пробе
сенсибилизированной сыворотки кролика добавить
человеческую сыворотку, то произойдет образование
комплексов антиген-антитело, выпадающих в
осадок (преципитат), который можно измерить. Если
добавлять к пробам кроличьей сыворотки,
содержащей антитела против сыворотки человека,
сыворотку различных животных, образуются разные
количества преципитата. Предполагая, что количество
преципитата находится в прямой зависимости от
сходства между белками сравниваемых сывороток,
этот метод можно использовать для оценки степени
родства между разными группами животных (табл.
24.8).
Сравнительно-серологический метод широко
используется для подтверждения филогенетических
связей. Например, зоологам оставалось неясным, к
какой систематической группе следует отнести
мечехвоста (Limulus). Когда к сыворотке против
антигенов мечехвоста добавляли антигены различных
членистоногих, образование наибольшего
количества преципитата вызывали антигены паукообразных.
Этот результат подкрепил имевшиеся
морфологические данные, и мечехвоста теперь уверенно
относят к классу Arachnida. Аналогичные исследования
позволили устранить многие неясности в
филогенетических взаимоотношениях млекопитающих.
Таблица 24.8. Количества преципитата, образующиеся при
добавлении сыворотки разных млекопитающих к кроличьей
сыворотке, содержащей антитела к сыворотке крови
человека (количество преципитата, образующегося с человеческой
сывороткой, принято за 100%)
Организм
Человек
Шимпанзе
Горилла
Гиббон
Павиан
Паукообразная обезьяна
Лемур
Еж
Свинья
Количество преципитата, %
100
97
92
79
75
58
37
17
8
Таблица 24.9. Действие пролактина у позвоночных
Группа
животных
Костные
рыбы
Амфибии
Рептилии
Птицы
Млекопитающие
Размножение
Стимулирует
выделение слизи
кожей
Стимулирует
секрецию студенистой
капсулы яиц
Подавляет
продукцию яиц
Стимулирует
выделение зобного
молочка
Стимулирует
развитие молочных
желез и
лактацию
Осморегуляция
Усиливает
образование мочи
Повышает
проницаемость кожи для
воды
Стимулирует потерю
воды у черепах
Повышает
потребление воды
Действует наподобие
антидиуретического гормона
Справедливость разделения животных на первич-
норотых и вторичноротых, основанного на
различиях в эмбриональным развитии, подтвердило и
изучение фосфатсодержащих веществ мышечной
ткани, участвующих в синтезе АТФ. У первично-
ротых, к которым относятся кольчатые черви,
моллюски и членистоногие, в мышцах содержится арги-
нинфосфат, а у вторичноротых, т.е. иглокожих и
хордовых,- креатинфосфат.
Еще один пример биохимической гомологии - тот
факт, что у позвоночных имеются сходные или
идентичные гормоны, выполняющие целый ряд
различных функций. Например, выделяемый
гипофизом гормон, сходный с пролактином
млекопитающих, найден у представителей всех групп
позвоночных. По имеющимся данным, пролактин вызывает
множество разнообразных эффектов, которые
можно разбить на две большие группы: одни эффекты
связаны с размножением, а другие-с осморегуля-
цией (табл. 24.9).
24.7.9. Заключение
Неодарвинистская теория эволюции основана на
данных из обширного круга источников и
подтверждается множеством совершенно независимых
наблюдений. Для ученого такие данные служат самым
убедительным доказательством достоверности этой
теории. Эволюционная теория завоевала широкое
признание, однако предстоит еще много работы по
ее уточнению и приложению ко всем наблюдаемым
ситуациям.
Все научные объяснения, гипотезы и теории
относительно истории жизни носят предварительный
характер и до тех пор пока человек в своих поисках
истины сохраняет объективность, будут оставаться
такими.
Поскольку проблема эволюции занимает
центральное место в изучении биологии, было бы
непростительно окончить эту главу, не указав ее
место в общей системе наших знаний о природе. Это
лучше всего сделать, процитировав фразу, которой
Дарвин заключает «Происхождение видов»:
«Есть некое величие в представлении о том, что жизнь с ее
различными проявлениями Творец первоначально вдохнул
всего лишь в несколько форм или даже в одну; и в то
время, как наша планета продолжает вращаться,
подчиняясь раз и навсегда установленному закону тяготения,
из такого простого начала развивалось и развивается
бесконечное число прекраснейших и удивительнейших
форм».
Глава 25
Механизмы видообразования
В предыдущей главе было описано, каким образом
Дарвин пришел к выводу о существовании у
растений и животных наследственной изменчивости как
при искусственном разведении, так и в природных
популяциях. Он понимал, что наследственные
изменения должны играть важную роль в эволюции,
но не мог предложить механизм, который объяснял
бы их возникновение при сохранении дискретности
признаков. Лишь после того, как были вторично
открыты работы Менделя о наследственности и
оценено их значение для эволюционной теории,
появилась возможность разрешить многие из этих
проблем. Современное объяснение изменчивости
живых организмов - это результат синтеза
эволюционной теории, основанной на работах Дарвина и
Уоллеса, и теории наследственности, основанной на
законах Менделя. Сущность изменчивости,
наследственности и эволюции можно теперь объяснить с
помощью данных, полученных в одной из областей
биологии, известной под названием популяционной
генетики.
25.1. Популяционная генетика
Популяция-это группа организмов,
принадлежащих к одному и тому же виду и занимающих
обычно четко ограниченную географическую
область. Дарвина интересовало, каким образом
естественный отбор, действуя на уровне отдельного
организма, вызывает эволюционное изменение.
После вторичного открытия работ Менделя,
доказавших корпускулярную природу
наследственности, большое внимание при изучении изменчивости,
наследственности и эволюционных изменений стали
уделять генотипу. Бэтсон, который в 1905 г. ввел
термин «генетика», видел задачу этой науки в
«освещении явлений наследственности и изменчивости».
Основу современной эволюционной теории,
которую называют неодарвинизмом или синтетической
теорией эволюции, составляет изучение
популяционной генетики. Гены, действуя независимо или
совместно с факторами среды, определяют фенотипи-
ческие признаки организмов и обусловливают
изменчивость в популяциях. Фенотипы,
приспособленные к условиям данной среды или «экологическим
рамкам», сохраняются отбором, тогда как
неадаптивные фенотипы подавляются и в конце концов
элиминируются. Естественный отбор, влияя на
выживание отдельных особей с данным фенотипом,
тем самым определяет судьбу их генотипа, однако
лишь общая генетическая реакция всей популяции
определяет выживание данного вида, а также
образование новых видов. Только те организмы,
которые, прежде чем погибнуть, успешно произвели
потомство, вносят вклад в будущее своего вида. Для
истории данного вида судьба отдельного организма
не имеет существенного значения.
25.1.1. Генофонд
Генофонд слагается из всего разнообразия генов и
аллелей, имеющихся в популяции, размножающейся
половым путем; в каждой данной популяции состав
генофонда из поколения в поколение может
постоянно изменяться. Новые сочетания генов
образуют уникальные генотипы, которые в своем
физическом выражении, т. е. в форме фенотипов,
подвергаются давлению факторов среды, производящим
непрерывный отбор и определяющим, какие гены
будут переданы следующему поколению.
Популяция, генофонд которой непрерывно
изменяется из поколения в поколение, претерпевает
эволюционное изменение. Статичный генофонд
отражает отсутствие генетической изменчивости среди
особей данного вида и отсутствие эволюционного
изменения.
25.1.2. Частоты аллелей
Любой физический признак, например окраска
шерсти у мышей, определяется одним или несколькими
генами. Каждый ген может существовать в
нескольких различных формах, которые называют
аллелями (см. табл. 23.2). Число организмов в данной
популяции, несущих определенный аллель,
определяет частоту данного аллеля (которую иногда
называют частотой гена, что менее точно).
Например, у человека частота доминантного аллеля,
определяющего нормальную пигментацию кожи, волос
и глаз, равна 99%. Рецессивный аллель,
детерминирующий отсутствие пигментации - так называемый
альбинизм,-встречается с частотой 1%. В
популяционной генетике частоту аллелей или генов чаще
выражают не в процентах или простых дробях, а в
десятичных дробях. Таким образом, в данном
случае частота доминантного аллеля равна 0,99, а
частота рецессивного аллеля альбинизма-0,01.
Общая частота аллелей в популяции составляет 100%,
или 1,0, поэтому
Частота
нантного
аллеля
+ Частота
рецессивного
аллеля
= 1
0,99 + 0,01 = 1.
Как это принято в классической генетике, аллели
можно обозначить буквами, например
доминантный аллель (нормальная пигментация) - буквой N, а
рецессивный (альбинизм) - буквой п. Для
приведенного выше примера частота N = 0,99, а частота
« = 0,01.
Популяционная генетика заимствовала у
математической теории вероятностей два символа, ρ и q,
для выражения частоты, с которой два аллеля,
доминантный и рецессивный, встречаются в
генофонде данной популяции. Таким образом,
p + q= I,
где ρ - частота доминантного, a q - частота
рецессивного аллеля.
В примере с пигментацией у человека ρ = 0,99, а
Я = 0,01;
p + q = 1
0,99 + 0,01 = 1
Значение этого уравнения состоит в том, что, зная
частоту одного из аллелей, можно определить
частоту другого. Пусть, например, частота
рецессивного аллеля равна 25%, или 0,25. Тогда
p + q = 1
ρ + 0,25 = 1
ρ = 1-0,25
ρ = 0,75
Таким образом, частота доминантного аллеля равна
0,75, или 75%.
25.1.3. Частота генотипов
Частоты отдельных аллелей в генофонде позволяют
вычислять генетические изменения в данной
популяции и определять частоту генотипов. Поскольку
генотип данного организма - главный фактор,
определяющий его фенотип, вычисление частоты
генотипа используют для предсказания возможных
результатов тех или иных скрещиваний. Это имеет
важное практическое значение в сельском хозяйстве
и медицине.
Математическая зависимость между частотами
аллелей и генотипов в популяциях была установлена
в 1908 г. независимо друг от друга английским
математиком Дж. Харди и немецким врачом
В. Вайнбергом. Эту зависимость, известную под
названием равновесия Харди-Вайнберга, можно
сформулировать так: частоты доминантного и
рецессивного аллелей в данной популяции будут
оставаться постоянными из поколения в поколение при
наличии определенных условий. Условия эти
следующие:
1) размеры популяции велики;
2) спаривание происходит случайным образом;
3) новых мутаций не возникает;
4) все генотипы одинаково плодовиты, т.е. отбора
не происходит;
5) поколения не перекрываются;
6) не происходит ни эмиграции, ни иммиграции, т. е.
отсутствует обмен генами с другими
популяциями.
Поэтому любые изменения частоты аллелей
должны быть обусловлены нарушением одного или
нескольких из перечисленных выше условий. Все эти
нарушения способны вызвать эволюционное
изменение; и если такие изменения происходят, то
изучать их и измерять их скорость можно с
помощью уравнения Харди-Вейнберга.
25.1.4. Уравнение Харди-Вайнберга
Это уравнение дает простую математическую
модель, которая объясняет, каким образом в
генофонде сохраняется генетическое равновесие; но
главное применение его в популяционной генетике-
вычисление частот аллелей и генотипов.
Если имеется два организма, один гомозиготный
по доминантному аллелю Л, а другой-по
рецессивному аллелю я, то все их потомки будут
гетерозиготными (Aa):
Фенотипы родителей
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы Fi (2п)
Фенотипы F1
А = доминантный аллель
а = рецессивный аллель
Доминантный χ Рецессивный
Все доминантные
Если наличие доминантного аллеля Л обозначить
символом р, а рецессивного аллеля а-символом q,
то картину скрещивания между особями F1,
возникающие при этом генотипы и их частоты можно
представить следующим образом:
Фенотипы Fχ
Генотипы F1 f2 η)
Мейоз
Гаметы (η)
Случайное
оплодотворение
Доминантный
Aa
χ Доминантный
х Aa
Генотипы F2(Z")
Фенотипы F2
Доминантные
(гомозиготы)
Доминантные
(гетерозиготы)
Рецессивные
(гомозиготы)
Однако для большинства популяций частоту
обоих аллелей можно вычислить только по доле особей,
гомозиготных по рецессивному аллелю, так как это
единственный генотип, который можно распознать
непосредственно по его фенотипическому
выражению.
Например, один человек из 10 000 - альбинос, т.е.
частота альбинотического генотипа составляет 1 на
10 000. Поскольку аллель альбинизма рецессивен,
альбинос должен быть гомозиготным по
рецессивному гену, т.е. на языке теории вероятности
Поскольку аллель А доминантный, отношение
доминантных генотипов к рецессивным составляет
3:1 -это менделевское отношение при
моногибридном скрещивании. Используя символы ρ и q,
результаты приведенного выше скрещивания можно
представить следующим образом:
р2 - доминантные
гомозиготы;
2pq - гетерозиготы;
q2 - рецессивные
гомозиготы.
Такое распределение возможных генотипов носит
статистический характер и основано на
вероятностях. Три возможных генотипа, образующихся при
таком скрещивании, представлены со следующими
частотами:
AA
0,25
2Aa
0,50
аа
0,25
Сумма частот трех генотипов, представленных в
рассматриваемой популяции, равна единице;
пользуясь символами ρ и q, можно сказать, что
вероятности генотипов следующие:
/72 + 2ОТ + <72 = 1,
На математическом языке ρ + q = 1 представляет
собой уравнение вероятности, тогда как р2 + 2pq +
+ q2 = 1 является квадратом этого уравнения [т. е.
(P + яП
Поскольку
ρ - частота доминантного аллеля;
q - частота рецессивного аллеля;
р2 - гомозиготный доминантный
генотип;
2pq - гетерозиготный генотип;
q2 - гомозиготный рецессивный
генотип,
можно вычислить частоты всех аллелей и
генотипов, пользуясь выражениями
для частот аллелей: ρ + q = 1;
для частот генотипов: р2 + 2pq + q2 = 1 ·
Зная, что q2 = 0,0001, можно определить частоты
аллеля альбинизма (#), доминантного аллеля
нормальной пигментации (/?), гомозиготного
доминантного генотипа (р2) и гетерозиготного генотипа (2pq).
Так как
т.е. частота аллеля альбинизма в популяции равна
0,01 или 1%. Поскольку
p + q= I,
ρ =1-0=1- 0,01 = 0,99,
частота доминантного аллеля в популяции равна
0,99, или 99%. А если
ρ = 0,99 и q = 0,01, то
2/><? = 2·(0,99)·(0,01) = 0,0198,
т.е. частота гетерозиготного генотипа составляет
0,0198; иными словами, примерно 2%
индивидуумов в данной популяции несут аллель альбинизма
либо в гетерозиготном, либо в гомозиготном
состоянии.
Как показывают эти вычисления, частота
рецессивного аллеля в популяции неожиданно велика при
малом числе индивидуумов с гомозиготным
рецессивным генотипом.
Гетерозиготных индивидуумов, нормальных по
фенотипу, но обладающих рецессивным геном,
который в гомозиготном состоянии может вызвать
нарушение метаболизма, называют носителями. Как
показывают вычисления с использованием
уравнения Харди-Вайнберга, частота носителей в
популяции всегда выше, чем можно было бы ожидать на
основании оценок частоты фенотипического
проявления данного дефекта. Это ясно видно из табл.
25.1.
25.1. Кистозный фиброз поджелудочной
железы встречается среди населения с частотой 1 на
2000. Вычислите частоту носителей.
25.1.5. Следствия уравнения
Харди-Вайнберга
Из уравнения Харди-Вайнберга следует, что
значительная доля имеющихся в популяции рецессивных
аллелей находится у гетерозиготных носителей.
Фактически гетерозиготные генотипы служат
важным потенциальным источником генетической
изменчивости. Это приводит к тому, что в каждом
поколении из популяции может элиминироваться
лишь очень малая доля рецессивных аллелей.
Только те рецессивные аллели, которые находятся в
гомозиготном состоянии, проявятся в фенотипе и
тем самым подвергнутся селективному воздействию
факторов среды и могут быть элиминированы.
Многие рецессивные аллели элиминируются
потому, что они неблагоприятны для фенотипа -
обусловливают либо гибель организма еще до того, как
он успеет оставить потомство, либо «генетическую
смерть», т.е. неспособность к размножению.
Однако не все рецессивные аллели
неблагоприятны для популяции. Например, у человека из всех
групп крови чаще всего встречается группа О,
соответствующая гомозиготности по рецессивному
аллелю. Другим примером служит серповиднокле-
точная анемия. Это наследственное заболевание
крови, широко распространенное в ряде областей
Африки и Индии, в некоторых средиземноморских
странах и у негритянского населения Северной
Америки. Индивидуумы, гомозиготные по
соответствующему рецессивному аллелю, обычно умирают, не
достигнув половой зрелости и элиминируя таким
образом из популяции по два рецессивных аллеля.
Что касается гетерозигот, то они не гибнут.
Установлено, что во многих частях земного шара
частота аллеля серповидноклеточности остается относи-
Таблица 25.1. Некоторые наследственные метаболические
дефекты и частоты рецессивных гомозиготных и
гетерозиготных генотипов
Нарушение
Альбинизм (отсутствие
пигментации)
Алкаптонурия (моча на
воздухе чернеет)
Семейная амавротическая
идиотия (ведет к слепоте
и смерти)
Сахарный диабет
(неспособность секретировать
инсулин)
Фенилкетонурия (может,
если не будет вовремя
выявлена, привести к
задержке умственного
развития)
Приблизительная частота
рецессивного
гомозиготного
генотипа (q2)
1 на 10 000
(в Европе)
1 на 1 000 000
1 на 40 000
1 на 200
1 на 10 000
(в Европе)
Частота
терозиготного
генотипа (2 pq)
1 на 50
1 на 503
1 на 100
1 на 7,7
1 на 50
тельно стабильной. У некоторых африканских
племен частота гетерозиготного фенотипа достигает
40%. Раньше думали, что этот уровень
поддерживается за счет появления новых мутантов. Однако в
результате дальнейших исследований выяснилось,
что дело обстоит иначе: оказалось, что во многих
частях Африки, где среди факторов, угрожающих
здоровью и жизни, важное место занимает малярия,
люди, несущие аллель серповидноклеточности,
обладают повышенной резистентностью к этой
болезни. В малярийных районах Центральной Америки
это селективное преимущество гетерозиготного
генотипа поддерживает частоту аллеля
серповидноклеточности среди населения на уровне 10-20%. У
североамериканских негров, которые уже 200-300
лет не испытывают на себе селективного эффекта
малярии, частота аллеля серповидноклеточности
упала до 5%. Это снижение можно частично отнести
на счет обмена генами в результате браков между
представителями черной и белой расы, однако
важным фактором служит отсутствие в Северной
Америке малярии, устраняющее селективное давление в
пользу гетерозигот; в результате рецессивный
аллель медленно элиминируется из популяции.
Этот пример эволюции в действии ясно
демонстрирует селективное влияние среды на частоту
аллелей - механизм, нарушающий генетическое
равновесие, предсказываемое законом Харди-Вейн-
берга. Именно такого рода механизмы вызывают в
популяциях сдвиги, ведущие к эволюционному
изменению.
25.2. Факторы, вызывающие
изменения в популяциях
Принцип равновесия Харди-Вайнберга гласит, что
при наличии определенных условий частота аллелей
остается постоянной из поколения в поколение. При
этих условиях популяция будет находиться в
состоянии генетического равновесия и никаких
эволюционных изменений происходить не будет. Однако
принцип Харди-Вайнберга носит чисто теоретический
характер. Очень немногие популяции находятся в
условиях, при которых сохраняется равновесие (см.
разд. 25.1.3).
В разд. 23.8.4 были подробно описаны четыре
главных источника генетической изменчивости: крос-
синговер во время мейоза, независимое
распределение хромосом при мейозе, случайное оплодотворение
и мутационный процесс. Первые три источника часто
объединяют под общим названием половой
рекомбинации; они обусловливают перетасовку генов,
лежащую в основе происходящих изо дня в день
непрерывных изменений. Но хотя эти процессы и
приводят к образованию новых генотипов и
изменяют частоты генотипов, они не вызывают
никакого изменения имеющихся аллелей, так что
частоты аллелей в популяции остаются постоянными.
Многие эволюционные изменения, однако,
происходят вслед за появлением новых аллелей, а
главным источником последних служат мутации.
Условия, необходимые для равновесия Харди-
Вайнберга, нарушаются и в ряде других случаев:
когда скрещивание носит неслучайный характер;
когда популяция мала, что ведет к дрейфу генов;
когда генотипы обладают различной фертильно-
стью, что создает генетический груз; при наличии
обмена генами между популяциями. Ниже
рассматривается каждая из этих ситуаций.
25.2.1. Неслучайное скрещивание
В большинстве природных популяций спаривание
происходит неслучайным образом. Во все тех
случаях, когда наличие одного или нескольких
наследуемых признаков повышает вероятность успешного
оплодотворения гамет, имеет место половой отбор.
У растений и животных существует много
структурных и поведенческих механизмов, исключающих
чисто случайный подбор родительских особей.
Например, цветки, у которых лепестки крупнее и
нектара больше, чем обычно, вероятно, будут привлекать
больше насекомых, что повысит вероятность
опыления и оплодотворения. Характер окраски
насекомых, рыб и птиц и особенности их поведения,
связанные с постройкой гнезда, охраной территории
и брачными церемониями, повышают
избирательность при скрещивании.
Влияние неслучайного скрещивания на генотип и
на частоту аллелей демонстрируют, например,
эксперименты, проведенные на дрозофиле. В
культуре мух, содержавшей вначале равное число
красноглазых и белоглазых самцов и самок, через 25
поколений исчезли все белоглазые особи. Как
показали наблюдения, и красноглазые, и белоглазые
самки предпочитали спариваться с красноглазыми
самцами. Таким образом, половой отбор как
механизм избирательного скрещивания обеспечивает
некоторым особям более высокий репродуктивный
потенциал, в результате чего вероятность передачи
генов этих особей следующему поколению
повышается. Репродуктивный потенциал особей с менее
благоприятными признаками понижен, и передача
их аллелей последующим поколениям происходит
реже.
25.2.2. Дрейф генов
О дрейфе генов говорят в тех случаях, когда
изменения частоты генов в популяциях бывают
случайными и не зависят от естественного отбора.
Случайный дрейф генов, или эффект Сьюэлла Райта
(названный по имени американского генетика, который
понял его роль в эволюции), может служить
важным механизмом эволюционных изменений в
небольших или изолированных популяциях. В
небольшой популяции могут быть представлены не все
аллели, типичные для данного вида. Случайные
события, например, преждевременная гибель особи,
бывшей единственным обладателем какого-то ал-
леля, приведут к исчезновению этого аллеля в
популяции. Если данный аллель встречается в популяции
из 1000000 особей с частотой, скажем, 1% (т.е.
q = 0,01), то им будут обладать 10000 особей; а в
популяции, состоящей из 100 особей, этот аллель
будет иметься только у одной особи, так что
вероятность его случайной утраты в малой популяции
гораздо выше.
Точно так же, как некий аллель может исчезнуть
из популяции, частота его может и повыситься
чисто случайным образом. Случайный дрейф генов,
как показывает само его название, непредсказуем.
Небольшую популяцию он может привести к
гибели, а может сделать ее еще более приспособленной к
данной среде или усилить ее дивергенцию от
родительской популяции. С течением времени
возможно образование из нее нового вида под
действием естественного отбора. Дрейф генов считают
существенным фактором в возникновении новых
видов в островных и других репродуктивно
изолированных популяциях.
С дрейфом генов связано явление, известное под
названием принципа основателя. Оно состоит в том,
что при отделении от родительской популяции
небольшой ее части последняя может случайно
оказаться не вполне типичной по своему аллельному
составу. Некоторые аллели в ней могут
отсутствовать, а другие будут представлены с
непропорционально высокой частотой. Постоянное скрещивание
внутри такой пионерной популяции приведет к
созданию генофонда, отличающегося по частотам
аллелей от генофонда исходной родительской
популяции. Дрейф генов обычно снижает генетическую
изменчивость в популяции, главным образом в
результате утраты тех аллелей, которые встречаются
редко. Длительное скрещивание особей внутри
малой популяции уменьшает долю гетерозигот и
увеличивает долю гомозигот. Примеры действия
принципа основателя были выявлены при изучении
небольших популяций, образованных в Америке
религиозными сектами, эмигрировавшими из Германии
в XVIII веке. В некоторых из этих сект браки
заключались почти исключительно между членами
данной секты. В таких случаях частота ряда аллелей
здесь сильно отличается от их частоты среди
населения как ФРГ, так и Америки. Например, изученные
общины данкеров (религиозная секта,
обосновавшаяся в Пенсильвании) состояли примерно из 100
семей каждая; это такие маленькие популяции, что в
них должен был происходить дрейф генов.
Определение групп крови дало следующие результаты:
Частота группы А
Население Пенсильвании
Население ФРГ
Община данкеров
42%
45%
60%
Эти данные, по-видимому, отражают результаты
дрейфа генов, происходящего в малых популяциях.
Дрейф генов может вести к уменьшению
изменчивости в пределах популяции, но он может также
увеличить изменчивость в пределах вида в целом.
В небольших изолированных популяциях могут
возникать нетипичные для основной популяции
признаки, которые в случае изменения среды могут дать
селективное преимущество. Таким образом, дрейф
генов может участвовать в процессе
видообразования.
25.2.3. Генетический груз
Существование в популяции неблагоприятных
аллелей в составе гетерозиготных генотипов называют
генетическим грузом. Как отмечалось в разд. 25.1.5,
некоторые рецессивные аллели, вредоносные в
гомозиготном состоянии, могут сохраняться в
гетерозиготных генотипах и при некоторых
условиях среды доставлять селективное преимущество;
примером служит аллель серповидноклеточности в
местах распространения малярии. Любое
повышение частоты рецессивных аллелей в популяции в
результате вредных мутаций увеличивает ее
генетический груз.
25.2.4. Поток генов
В генофонде скрещивающейся внутри себя
популяции происходит непрерывный обмен аллелями
между особями. Если частоты аллелей не изменяются в
результате мутаций, происходящая при таком
обмене перетасовка генов ведет к генетической
стабильности или равновесию генофонда. В случае
возникновения мутантного аллеля он
распространится по всему генофонду в результате случайного
оплодотворения.
Такое перемещение аллелей в пределах популяции
часто не вполне правильно называют «потоком
генов». Строго говоря, этот термин относится к
перемещению аллелей из одной популяции в другую
в результате скрещивания между членами этих двух
популяций. Случайное внесение новых аллелей в
популяцию-реципиента и удаление их из
популяции-донора изменяет частоту аллелей в обеих
популяциях и ведет к повышению генетической
изменчивости. Несмотря на то что поток генов вносит в
популяции генетическую изменчивость, в смысле
эволюционного изменения его действие оказывается
консервативным. Распространяя мутантные аллели
по всем популяциям, поток генов приводит к тому,
что все популяции данного вида приобретают
общий генофонд, т.е. различия между популяциями
уменьшаются. Поэтому прерывание потока генов
между популяциями представляет собой одну из
предпосылок для образования нового вида.
Интенсивность обмена генами между двумя
популяциями зависит от их пространственной близости и
от легкости, с которой организмы или гаметы могут
переходить из одной популяции в другую.
Например, две популяции могут находиться так близко
друг к другу, что скрещивание между ними
происходит непрерывно, и тогда в генетическом смысле
их можно считать одной популяцией, поскольку они
обладают общим генофондом; пример - две
популяции улиток, обитающие в соседних садах,
разделенных живой изгородью.
Летающим животным и пыльцевым зернам
относительно легко активно или пассивно
распространяться в новые места. Здесь они могут скрещиваться
между собой или с местной популяцией, внося в нее
при этом генетическую изменчивость.
25.3. Отбор
Действие отбора можно рассматривать на двух
связанных между собой уровнях-на уровне
организма и на уровне аллелей.
Отбор-это процесс, благодаря которому
организмы, по своей морфологии, физиологии и
проведению лучше приспособленные к данной среде,
выживают и размножаются а менее приспособленные
гибнут или им не удается оставить потомков.
Первые передают свои удачные признаки следующему
поколению, а вторые-нет. Таким образом, можно
видеть, что действие отбора достигается благодаря
дифференциальной смертности и дифференциальному
размножению. Адаптивное значение отбора состоит
в том, что он обеспечивает сохранение тех
организмов, которые с наибольшей вероятностью могут
обеспечить выживание данного вида, а его
эффективность зависит от наличия в популяции фенотипи-
ческой изменчивости.
При увеличении численности популяции
некоторые факторы среды, такие как пища (для животных)
или свет (для растений), становятся
лимитирующими. Это приводит к конкуренции за ресурсы
между членами популяции. Организмы,
индивидуальные особенности которых дают им
преимущество в конкуренции, будут легче овладевать
этими ресурсами, выживут и оставят потомство.
Организмы, лишенные таких признаков, окажутся в
невыгодном положении и могут погибнуть, прежде
чем успеют произвести потомство. Совместное
действие лимитирующих факторов среды и
численности популяции создает давление отбора,
интенсивность которого может быть различной.
Таким образом, отбор-это процесс,
определяющий, какие аллели будут переданы следующему
поколению благодаря дифференциальным
преимуществам, которые они доставляют, экспрессируясь в
фенотипах. Давление отбора можно поэтому
рассматривать как способ увеличить или уменьшить
распространенность какого-либо аллеля в
генофонде, и эти изменения частоты аллелей могут вести
к эволюционному изменению. Крупные изменения в
генотипе возникают в результате распространения в
генофонде мутантных аллелей.
Степень отбора и его ход во времени зависят от
характера мутантного аллеля и его влияния на
данный фенотипический признак. Доминантный
аллель сразу проявляется в фенотипе, и тут же
начинается его положительный или отрицательный
отбор. Если же аллель рецессивен, как это обычно для
большинства мутаций, то он не подвергается отбору
до тех пор, пока не появится в гомозиготном
состоянии; вероятность немедленного появления таких
рецессивных гомозигот невелика, и новый аллель
может исчезнуть из генофонда, прежде чем они
возникнут. Аллель, рецессивный в данных условиях,
может сохраниться в популяции и дождаться таких
изменений среды, при которых его эффект будет
доминантным; если он при этом окажется
выгодным для организма, то отбор будет
благоприятствовать распространению аллеля в популяции, как,
например, в случае серповидноклеточной анемии.
Рецессивный мутантный аллель может быстро
распространиться в популяции, если его локус в
хромосоме сцеплен (находится в тесной близости) с
локусом какого-либо доминантного аллеля,
имеющего важное функциональное значение и
подвергающегося сильному положительному отбору. При
таком сцеплении шансы мутантного аллеля
соединиться с другим мутантным аллелем и оказаться в
гомозиготном состоянии сильно возрастают
(рис. 25.1).
Фенотипические эффекты мутантных аллелей
могут быть различными. Мутации генов,
контролирующих важные функции, вероятнее всего, окажутся
летальными и будут тотчас же элиминированы из
популяции. Эволюционные сдвиги происходят
обычно в результате постепенного появления
многих мутантных аллелей, производящих небольшие
прогрессивные изменения в фенотипических
признаках.
Существует три типа отбора: стабилизирующий,
направленный и дизруптивный. Смысл этих
терминов легче всего пояснить, построив кривые
нормального распределения для непрерывной фенотипичес-
кой изменчивости, наблюдаемой в природных
популяциях (рис. 25.2).
25.3.1. Стабилизирующий отбор
Стабилизирующий отбор происходит в тех случаях,
когда фенотипические признаки оптимально
соответствуют условиям среды и конкуренция
относительно слабая. Такой отбор действует во всех
популяциях, элиминируя особей с крайними
отклонениями признаков. Например, существует некая
оптимальная длина крыла для хищной птицы
определенных размеров с определенным образом жизни в
данной среде. Стабилизирующий отбор, действуя
благодаря дифференциальному размножению,
будет элиминировать тех птиц, у которых размах
Появление аллеля
ι результате мутации
Случайное оплодот
ворение гамет
Генофонд
Отбор сильно благо
приятствует аллелю
В, так что его частота
в следующем
поколении возрастает
Две особи, несущие
мутантный аллель,
спариваются
Мутантный аллель в
гомозиготном состо -
янии проявляется в
фенотипе
Рис. 25.1. Ускорение распространения в
популяции мутантного аллеля (а), сцепленного с
доминантным аллелем (В), которому сильно
благоприятствует отбор.
крыльев больше или меньше оптимального.
Карн и Пенроуз изучали корреляцию между
весом новорожденных и постнатальной смертностью
у 13 730 младенцев, родившихся в Лондоне в период
1935-1946 гг. Среди них 614 родились мертвыми
или умерли в первый месяц после рождения. Как
показывает рис. 25.3, оптимальный для
новорожденных вес равен примерно 3,6 кг. Младенцы с
большим или меньшим весом оказываются в
смысле отбора в невыгодном положении, и смертность
среди них несколько повышена. На основании этих
данных можно вычислить интенсивность отбора.
Если в момент рождения или в первый месяц 614
младенцев (из 13 730) умерли, то смертность
составляет 4,5%. Даже среди младенцев с оптимальным
весом смертность равна 1,8%. Следовательно,
интенсивность отбора на вес при рождении, равный
3,6 кг, составляет 4,5% - 1,8% = 2,7%, или 0,027.
Среди новорожденных, весящих 1,8 кг, смертность
равна 34%, что соответствует интенсивности отбора
примерно 30%, или 0,3. Следует указать, однако,
что в результате успехов, достигнутых педиатрией
после 1946 г., постнатальная смертность
значительно снизилась.
Стабилизирующий отбор не способствует
эволюционному изменению, а поддерживает фенотипичес-
кую стабильность популяции из поколения в
поколение.
Стабилизирующий
Направленный
Дизруптивный
Частота
Поколения
Частота
Поколения
Частота
Давление
отбора
Давление
отбора
Фенотипический признак
Фенотипический признак
Фенотипический признак
Поколения
Рис. 25.2. Три типа отбора, действующего в популяциях. О-первоначальное положение точки
соответствия оптимального фенотипа оптимальным условиям среды; N-новое положение точки
соответствия. Особи, признаки которых попадают в заштрихованные участки, оказываются в
невыгодном положении и элиминируются отбором. Цифрами 1-3 указана последовательность
поколений.
25.3.2. Направленный отбор
Эта форма отбора возникает в ответ на постепенные
изменения условий среды. Направленный отбор
влияет на диапазон фенотипов, существующих в
данной популяции, и оказывает селективное
давление, сдвигающее средний фенотип в ту или другую
сторону. После того как средний фенотип придет в
оптимальное соответствие с новыми условиями
среды, вступает в действие стабилизирующий отбор.
Направленный отбор приводит к эволюционному
изменению, оказывая на популяцию такое давление,
которое благоприятствует увеличению в ней
частоты новых аллелей. Направленный отбор лежит в
основе искусственного отбора, при котором
избирательное скрещивание особей, обладающих
желательными фенотипическими признаками, повышает
частоту этих признаков в популяции (см. разд. 25.4).
В ряде экспериментов Фальконер выбирал из
популяции шестинедельных мышей самых тяжелых
особей и давал им спариваться между собой. То же
самое он проделывал с самыми легкими мышами.
Такое избирательное скрещивание по признаку
массы тела привело к созданию двух популяций, в
одной из которых масса возрастала, а в другой-
уменьшалась (рис. 25.4).
После прекращения селекции ни та, ни другая
группа не вернулась к первоначальной массе
(примерно 22 г). Это показывает, что искусственный
отбор по фенотипическим признакам привел к
некоторому генотипическому отбору и частичной
утрате обеими популяциями каких-то аллелей. В
настоящее время непосредственно изучено много
примеров классического направленного отбора; они
описаны в разд. 25.5.
25.3.3. Дизруптивный отбор
Эта форма отбора, вероятно, встречается реже
всего, но она может играть очень важную роль в
возникновении эволюционных изменений.
Колебания условий среды, связанные, например, со сменой
времен года и климатическими факторами, могут
благоприятствовать наличию в данной популяции
не одного, а двух или нескольких фенотипов.
Давление отбора, действующее в самой популяции, может
привести к отклонению фенотипов от среднего для
данной популяции к обоим крайним типам. В
результате популяция окажется разделенной на две
субпопуляции. Если поток генов между этими
субпопуляциями почему-либо нарушится, каждая из
них может дать начало новому виду. В некоторых
случаях такая форма отбора приводит к появлению
в одной популяции нескольких различных
фенотипов - явление, называемое полиморфизмом,
которое будет рассмотрено в разд. 25.5.1. В пределах
одного вида популяции с различными фенотипами,
или экотипы, могут быть адаптированы к
определенным условиям среды (см. разд. 25.6.2). Если вид
занимает очень обширный географический ареал, у
населяющих этот ареал популяций могут
наблюдаться локальные изменения фенотипических
признаков, в результате чего они становятся
промежуточными между популяциями краевых областей.
Такой непрерывный градиент постепенно
изменяющихся признаков представляет собой обычно
стенотипическую реакцию на изменение климатических
и/или эдафических (почвенных) факторов и известен
под названием клины (см. разд. 25.6.3).
25.3.4. Интенсивность отбора
Сила давления отбора в данной популяции
варьирует в пространстве и во времени, что может быть
связано с изменениями внешних или внутренних
факторов. К внешним факторам относятся, в
частности, увеличение численности хищников или
патогенных организмов и конкуренция с другими
видами (межвидовая конкуренция) за пищу и места для
размножения (у животных) или за свет, воду и
минеральные вещества (у растений). Изменения
климата или состояния местообитания могут привести
к созданию новых давлений отбора. Внутренние
факторы, например быстрое повышение
численности популяции, нередко приводят к усилению
конкуренции за ресурсы (внутривидовая
конкуренция). С увеличением популяции данного организма
возрастает и численность его паразитов и хищников;
6S
αί
s
X
CQ
OQ
S
А
CQ
Вес при рождении, кг
Рис. 25.3. Зависимость между весом тело
новорожденных и их выживанием. (По M. N. Кат,
L. S. Penrose, Ann. Eugen., 1951, 16, 147-164)
§
H
υ
CU
CQ
Число поколений
Рис. 25.4. Изменения веса тела в
последовательных поколениях в двух популяциях мышей,
подвергавшихся отбору по этому признаку. (По
D. S. Falconer, J. Genetics, 1953, 51, 470-501.)
кроме того, в многочисленной популяции
облегчается передача паразитов и болезней от одной
особи к другой. Все эти факторы могут изменять не
только силу давления отбора, но и его направление.
Отбор благоприятствует «новым» фенотипам (и
генотипам), а плохо приспособленные особи
элиминируются. В первую очередь при этом устраняются
особи, обладающие неадаптивными крайними
фенотипами.
Одним из результатов возросшего давления
отбора может быть специализация организмов к
определенному образу жизни или более узкому
диапазону условий среды. Это может оказаться
неблагоприятным для дальнейшей судьбы вида. Увеличение
единообразия вида и его зависимости от каких-то
определенных условий повышает вероятность его
вымирания в случае изменения этих условий. В
палеонтологической летописи встречается немало
вымерших организмов, которые были крайне
своеобразны и чересчур специализированы.
25.2. Как сведения о давлении отбора и образе
жизни определенного паразита могут помочь
искоренить его?
Из всего сказанного можно видеть, что
повышенное давление отбора-это консервативный
механизм, сохраняющий фенотип, лучше всего
приспособленный к данным условиям среды (оптимальный
фенотип).
Ослабление интенсивности отбора обычно
оказывает противоположное действие. Оно может
наступить при отсутствии хищников, патогенных
организмов, паразитов и конкурентов или же при
улучшении условий среды. Так нередко обстоит дело при
проникновении вида в новую для него среду. Как
полагают, именно такая ситуация
благоприятствовала созданию видового разнообразия вьюрков на
Галапагосских островах.
25.4. Искусственный отбор
Человек с самых ранних времен своей цивилизации
применяет искусственный отбор при разведении
животных и растений. Дарвин воспользовался
данными по искусственному отбору, чтобы объяснить
возможный механизм изменения видов,
происходящего в природных популяциях, т.е. естественного
отбора. Искусственный отбор основан на изоляции
природных популяций и избирательном
скрещивании организмов, обладающих признаками,
желательными для человека. Отбор герефордской и абер-
дин-ангусской пород крупного рогатого скота велся
на количество и качество мяса, а гернзейской и
джерсейской пород-на молочность. Овцы гемп-
ширской и суффолькской пород быстро созревают и
дают хорошее мясо, но они менее выносливы и
менее активны в поисках пищи, чем шевиотские или
шотландские черномордые овцы. Эти примеры по-
называют, что в одной породе невозможно
объединить все признаки, необходимые для максимального
экономического эффекта в любых условиях, так что
программы селекции часто разрабатывают таким
образом, чтобы улучшить количественно и
качественно главный ценный признак данной породы.
При искусственном отборе человек создает
направленное селективное давление, которое ведет к
изменениям частот аллелей или генотипов в
популяции. Это эволюционный механизм, приводящий к
созданию новых пород, линий, сортов, рас и
подвидов. Генофонды всех этих групп изолированы, но
они сохраняют основную генную и хромосомную
структуру, характерную для вида, к которому они
все еще принадлежат.
Существует две формы искусственного отбора-
инбридинг и аутбридинг.
25.4.1. Инбридинг (родственное
разведение)
Инбридинг состоит в избирательном скрещивании
между близко родственными особями (например,
между потомками от одних и тех же родительских
особей) с целью сохранения и распространения
особенно желательных признаков. Инбридинг очень
часто применяют при разведении таких животных,
как кошки и собаки. Прежде животноводы
использовали инбридинг для получения крупного рогатого
скота, свиней, домашней птицы и овец, дающих
много молока, мяса, яиц и шерсти соответственно,
но по причинам, изложенным ниже, инбридинг не
находит теперь широкого применения.
Длительный инбридинг может привести к
снижению плодовитости, что создает особенно серьезные
проблемы в животноводстве. Интенсивный
инбридинг уменьшает изменчивость генома (общее число
всех аллелей данного вида), повышая число
гомозиготных генотипов за счет гетерозиготных. Для
того чтобы преодолеть эти проблемы, животноводы
после нескольких поколений инбридинга
прибегают к аутбридингу. Например, владелец молочной
фермы может проводить случку своего быка с
коровами из последовательных поколений собственного
стада для получения высокоудойных коров. Однако
еще до того, как фермер обнаружит у своего скота
признаки ослабления устойчивости к заболеваниям
и снижения плодовитости, он приводит для случки
другого быка или искусственно осеменяет коров
спермой, приобретенной в животноводческом
центре. Таким образом он вводит в стадо новые аллели
и повышает гетерозиготность скрещивающейся
популяции.
25.4.2. Аутбридинг (неродственное
разведение)
Аутбридинг особенно полезен в растениеводстве,
однако его все шире используют также в
производстве мяса, яиц и шерсти. Этот метод состоит в
скрещивании особей из генетически различных
популяций. Аутбридинг обычно производят между
представителями различных сортов или линий, а у
некоторых растений - между близко родственными
видами. Потомков от таких скрещиваний называют
гибридами, и они превосходят по ряду признаков обе
родительские формы - явление, называемое
гибридной мощностью или гетерозисом. Гибридов,
получаемых при скрещивании гомозиготных родительских
линий из различных популяций, называют
гибридами F1; они превосходят родительские формы по
величине и количеству плодов, устойчивости к
заболеваниям и скорости созревания. В случае
кукурузы гибридизация привела к повышению урожаев
зерна у гибридов F1 до 250% по сравнению с
родительскими линиями (рис. 25.5). При двойной
гибридизации гибридов, полученных от
скрещивания двух инбредных линий, скрещивают между
собой. Качество початков и урожайность таких
гибридов с избытком окупает расходы, связанные с
двухгодичной программой скрещиваний (рис. 25.6).
Увеличение мощности растений обусловлено
повышением гетерозиготности в результате смешения
генов. Например, если каждая гомозиготная
родительская особь обладает некоторыми, но не всеми
доминантными аллелями, определяющими мощный
рост, то полученная при скрещивании таких особей
гетерозигота может нести все доминантные аллели
(рис. 25.7). У некоторых сортов повышение
мощности может быть обусловлено не просто
увеличением числа доминантных аллелей, но и каким-то
взаимодействием между определенными
сочетаниями аллелей в гетерозиготе. При продолжительном
инбридинге гибридов F1 их мощность понижается
по мере возрастания доли гомозигот (рис. 25.8).
Иногда гибридизация может сопровождаться
изменением числа хромосом - полиплоидизацией,
которая может приводить к возникновению новых видов
(один пример этого описан в разд. 23.9.2).
25.4.3. Искусственный отбор у
человека
Достигнутые за последнее время успехи в изучении
структуры гена, генетического кода, механизмов
наследственности и дородовой диагностики
генетических дефектов создали возможности для
закрепления или элиминации некоторых признаков у
человека. Евгеника давно уже занимается вопросом, как
можно было бы «улучшить» род человеческий путем
избирательного скрещивания определенных
индивидуумов. Это весьма волнующая тема, и в связи с ней
возникают самые разнообразные возражения. Ол-
Рис. 25.5. Пример гибридной мощности. А. Две родительские линии кукурузы (слева и справа) и
гибридная кукуруза (в середине), полученная в результате скрещивания между ними. Б. Початки этих
родительских линий (вверху) и гибрида (внизу) (Фото D. F Janes, Connecticut Agricultural
Experiment Station.)
Рис. 25.6. Фенотипы, образующиеся при двойной гибридизации кукурузы. Растения на правой
стороне - результат скрещивания между гибридами инбредных линий (слева).
Генотипы родителей (2η)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы F^(2n)
Фенотипы Fy
FFgghhlljj χ FFGGHHiUJ
FfGgHhIiJj
Потомки несут доминантные
аллели по каждому гену
Рис. 25.7. Простое генетическое объяснение
увеличения мощности у гибридов F1.
дос Хаксли в своем романе «Дивный новый мир»,
опубликованном в 1932 г., описал то воображаемое
время, когда евгеника достигнет вершины своих
возможностей и будет создавать индивидуумов,
соответствующих потребностям общества. Подобные
идеи противоречат морали любого общества,
ставящего на первое место свободу и права личности;
однако можно привести немало аргументов в
пользу ограниченного применения в этой области
некоторых достижений генетики. В медицине получают
все большее признание генетические консультации,
где супругам, у которых в роду имеются
генетические аномалии, разъясняют, с каким риском связано
для них рождение детей. С помощью уравнения
Харди-Вейнберга можно вычислить частоту
носителей таких нарушений метаболизма, как фенилке-
тонурия, или таких болезней крови, как талассемия,
серповидноклеточная анемия или гемофилия.
Носителям генов того или другого из этих заболеваний
можно разъяснить, какова вероятность того, что
они вступят в брак с другим носителем тех же генов,
и каковы шансы на то, что их дети в этом случае
окажутся больными. В таких формах
профилактическая медицина дает советы, а не диктует
требования. Любые успехи науки, позволяющие уменьшить
страдания, следует приветствовать. Такой подход
становится опасным лишь в том случае, если им
злоупотребляют.
Рис. 25.8. Растения кукурузы из восьми последовательных поколений. 1 - исходный гибрид. У
растений 2-8 видна постепенная утрата гибридной мощности в результате инбридинга. В трех
последних поколениях, достигших гомозиготности, утрата мощности замедлилась. (Фото D. F. Jones, Connecticut
Agricultural Experiment Station.)
25.5. Естественный отбор
Естественный отбор, постулированный Дарвином и
Уоллесом, представлял собой гипотезу, основанную
на исторических данных. Для Дарвина промежуток
времени, необходимый для эволюционного
изменения популяции, был слишком велик, чтобы такое
изменение можно было наблюдать
непосредственно. Происходящие в последние десятилетия
изменения, связанные с промышленной, технической и
медицинской революцией, создают столь сильные
факторы направленного и дизруптивного отбора,
что мы можем теперь наблюдать резкие изменения
в генотипе и фенотипе популяций, происходящие
достаточно быстро. Открытие в сороковых годах
антибиотиков создало сильное давление отбора в
пользу бактериальных штаммов, обладающих
генетической устойчивостью к антибиотикам. Бактерии
очень быстро размножаются и дают ежедневно
много поколений и миллионы индивидуумов.
В результате случайной мутации может появиться
устойчивая клетка, потомки которой будут
процветать благодаря отсутствию конкуренции со стороны
других бактерий, уничтожаемых данным
антибиотиком. В ответ на это приходится создавать новые
антибиотики для уничтожения устойчивых бакте-
Рис. 25.9. Полиморфизм у березовой пяденицы Biston
betиIaria. А. Нормальная форма. В. betularia typica.
Б. Меланистическая форма. В. betularia carbonaha.
(E. В. Ford, Evolution studied by observation and
experiment, Oxford Biology Readers, 1973, 55. Oxford Univ.
Press )
рий, и цикл продолжается. Селективное давление
создается также при использовании таких веществ,
как ДДТ, для борьбы с платяной вошью и
комарами или антикоагулянта уорферина для уничтожения
крыс. После возникновения устойчивости она
быстро распространяется во всей популяции.
Классическим примером эволюционного
изменения служит реакция некоторых бабочек на
направленное селективное давление, создаваемое
загрязнением атмосферы в результате промышленной
революции. За последние 100 лет у более чем 80 видов
бабочек появились темные формы, встречающиеся
теперь с различной частотой по всей
Великобритании. Это явление известно под названием
индустриального меланизма. До 1848 г. все описанные формы
березовой пяденицы (Biston betularia) имели
бледно-кремовую окраску с черными точками и
отдельными темными пятнышками (рис. 25.9,Л). В 1848 г.
в Манчестере была обнаружена черная форма этой
бабочки, а к 1895 г. популяция березовой пяденицы
в Манчестере состояла на 98% из черных бабочек.
Эта черная «меланистическая» форма появилась в
результате повторных случайных мутаций, причем
фенотип мутантных особей обладал большим
селективным преимуществом в промышленных районах
по причинам, выдвинутым и проверенным д-ром
Кеттлуэллом.
Березовые пяденицы активны в ночное время, а
днем они отдыхают на стволах деревьев.
Нормальная форма имеет очень эффективную
покровительственную окраску, которая сливается с
лишайниками, покрывающими стволы. С развитием
промышленной революции двуокись серы,
образующаяся при сжигании угля, вызвала в промышленных
районах гибель лишайников, и в результате
обнажилась темная кора деревьев, которая сделалась
еще темнее из-за покрывающей ее сажи (рис. 25.10).
В пятидесятых годах нашего века Кэттлуэлл
пометил определенное число светлых и темных
бабочек и выпустил их в двух местах: в загрязненном
районе вблизи Бирмингема, где популяция на 90%
состояла из темной формы, и в незагрязненном
районе в Дорсете, где темная форма встречалась
редко. С помощью световой ловушки он провел
отлов меченых бабочек и получил следующие
результаты:
Меченые темные
бабочки, %
Меченые светлые
бабочки, %
Бирмингем
34,1
15,9
Дорсет
6,3
12,5
С помощью киносъемки Кэттлуэлл показал, что
малиновки и дрозды питаются этими бабочками,
осуществляя форму естественного отбора,
называемую избирательным выеданием, которая в данном
случае оказывает селективное давление на
распространение меланистической и светлой форм.
Рис. 25.10. Мелонистическая и нормальная формы
В/ston betularia на стволах деревьев вблизи
Бирмингема (А) и в Дорсете (Б). (С любезного
разрешения д-ра HBD Kettlewell, Dep of Zoology
Uη/ν of Oxford.)
Как показывают полученные результаты, мела-
нистическая форма, Biston betularia carbonaria, в
промышленных районах обладает селективным
преимуществом перед светлой формой, Biston betularia
typica, тогда как светлая форма обладает
преимуществом в незагрязненных районах.
Позднее было установлено, что окраска темной
формы определяется доминантным меланистичес-
ким аллелем. На рис. 25.11 показано
распространение этих двух форм на Британских островах.
Распространение темных бабочек в
непромышленных районах восточной части Англии
объясняется преобладанием западных ветров. После того
как в 1956 г. был введен в действие Акт о чистоте
воздушного бассейна, доля светлых форм слегка
возросла, так как в промышленных районах
элиминация их отбором несколько снизилась.
25.5.1. Полиморфизм
Полиморфизм играет значительную роль в процессе
естественного отбора. На примере полиморфных
групп можно продемонстрировать ряд описанных в
этой главе принципов, касающихся соотношения
частот генотипов в популяции и изменений давления
отбора. Полиморфизм можно определить как
существование в пределах одной популяции двух или
нескольких форм данного вида, различающихся по
биохимическим, морфологическим или
поведенческим признакам. Различают две формы
полиморфизма: переходный и сбалансированный, или
стабильный, полиморфизм.
Классическое количественное изучение
сбалансированного полиморфизма провели Кейн, Карри и
Шеперд на обыкновенной наземной улитке Сераеа
nemoralis. У этих улиток раковина бывает желтая
(если внутри находится живая улитка, раковина
выглядит зеленой), различных оттенков
коричневого (в том числе светло-палевая), розовая,
оранжевая или красная. Устье раковины может быть
темно-коричневым, розовым или белым, а на самой
раковине может быть до пяти темных полос,
идущих по ее контурам (рис. 25.12). Как цвет раковины,
так и характер ее полос детерминированы
генетически. Окраска определяется множественными
аллелями, причем коричневая окраска доминирует над
розовой, а обе они доминируют над желтой. Поло-
сатость - рецессивный признак.
Улиток поедают дрозды, которые переносят их на
близлежащий камень и, используя его в качестве
«наковальни», разбивают на нем раковину, чтобы
добраться до улитки. Изучая соотношение раковин
разных типов вокруг «наковален» и в самих
местообитаниях улиток, Кейн, Карри и Шеперд показали,
что в популяции действуют селективные факторы.
В тех местах, где фон был довольно однородным,
например на траве и на лесной подстилке,
селективным преимуществом обладали желтые и
коричневые раковины без полос, так как вблизи наковален
таких раковин было меньше (рис. 25.13). Более
темные полосатые раковины имели селективное
преимущество там, где фон был пестрым, как,
например, на пастбищах с грубыми травами и в живых
изгородях. На любых участках дрозды больше всего
истребляют те формы, которые хорошо заметны.
Обширная популяция полиморфных улиток может
занимать несколько участков, различающихся по
фону. Кроме того, характер и окраска фона могут
изменяться со сменой времен года. Хотя выедание
хорошо заметных форм происходит непрерывно, ни
одна форма не обладает абсолютным селективным
преимуществом; поэтому доля каждой формы в
популяции из года в год остается довольно
постоянной.
Баланс численностей разных форм не обязательно
определяется только окраской и характером полоса-
тости. Судя по некоторым данным, сохранению
равновесия при полиморфизме способствуют также
физиологические факторы. В некоторых областях с
сухой известковой почвой и светлой поверхностью
не всегда доминируют формы с наименее
заметными окраской и характером полос. Полагают, что
полиморфизм у Сераеа определяется генетическим
сцеплением особого типа. Гены окраски и полоса-
тости сцеплены и образуют суперген, который
действует и наследуется как одна генетическая
единица. Входящие в него гены определяют признаки,
обладающие такими селективными
преимуществами, благодаря которым они сохраняются в
популяции. Именно разнообразие аллельных форм этих
генов, охраняемое благодаря широко
распространенной гетерозиготности, составляет основу
полиморфизма. К этому добавляется сцепление генов,
детерминирующих некоторые физиологические
функции, что, как полагают, тоже способствует
поддержанию сбалансированного полиморфизма.
Наличие в одной и той же популяции нескольких
обособленных форм, доля которых слишком велика,
чтобы их можно было отнести за счет повторных
мутаций, называют генетическим полиморфизмом;
примером служит Сераеа.
Переходный полиморфизм
Переходный полиморфизм возникает в тех случаях,
когда различные формы, или морфы, существуют в
популяциях, испытывающих сильное давление
отбора. Частота фенотипического проявления каждой
формы определяется интенсивностью отбора, как в
случае меланистической и светлой форм березовой
пяденицы. Переходный полиморфизм обычно
наблюдается при постепенном замещении одной
формы другой.
Сбалансированный полиморфизм
Сбалансированный полиморфизм создается при
сосуществовании в одной популяции различных форм
в стабильных условиях среды. Наиболее ярким
примером служит наличие двух полов у животных и
растений. Частоты генотипов разных форм
сбалансированы, так как обе формы обладают
равноценными селективными преимуществами. Примером
сбалансированного полиморфизма служат группы
крови А, В, AB и О у человека. Частоты разных
генотипов в разных популяциях могут варьировать,
однако в данной популяции они остаются
постоянными из поколения в поколение. Это объясняется
тем, что ни один генотип не обладает селективным
преимуществом перед другими. Как показывают
статистические данные, у мужчин белой расы с
группой крови О ожидаемая продолжительность
жизни выше, чем у мужчин с другими группами
крови; однако у обладателей группы О чаще, чем у
других, развивается язва двенадцатиперстной
кишки, которая в случае прободения может привести к
смерти. Другие примеры полиморфизма -
нормальное зрение и цветовая слепота у человека, касты
рабочих, трутней и маток у общественных
насекомых и длинно- и короткостолбчатые формы у
первоцвета.
Biston betularia typica
Bistort betularia carbonaria
Рис. 25.11. Распространение
меланистической и нормальной форм
Biston betularia на Британских
островах в 1958 г. (H. В. D. Kett-
lewell. Heredity, 1978, 1Z 51-72.)
Рис. 25.12. Изменчивость окраски и рисунка полос у раковины Cepaea nemoralis. A-вид сверху, Б-
вид сбоку. Показан постепенный переход от желтой раковины без полос
(верхний ряд) до темно-коричневой полосатой (нижний ряд справа). (По Tribe, Ta IIa η,
Er ant, Basic Biology Course, Book 12, Cambridge Univ Press. 1978.)
Рис. 25.13. Раковины Cepaea
nemoralis без полос на фоне
листовой подстилки. Раковина,
лежащая справа,- желтая,
вверху розовая, а две раковины
слева - коричневые. (По Ford,
Evolution studied by observation
and experiment, Oxford Biology
Reader, 1973, 55, Oxford Univ.
Press, 1973.)
25.6. Концепция вида
Вид представляет собой самую низкую
таксономическую категорию, которой еще можно дать
сколько-нибудь точное определение. Существует
несколько определений вида; некоторые из них приведены в
табл. 25.2.
Особи, принадлежащие к одному виду, редко
существуют в природе в форме одной обширной
популяции. Обычно вид представлен рядом
небольших скрещивающихся популяций, называемых де-
мами; каждый дем имеет собственный генофонд.
Такие популяции могут занимать примыкающие
друг к другу или сильно разобщенные
географические области. Пространственная разобщенность
популяций означает, что данный вид может
сталкиваться с разнообразными условиями среды и разной
интенсивностью отбора. Мутационный процесс и
отбор в изолированных популяциях могут
создавать различную степень фенотипической
изменчивости в пределах вида, как это описано ниже.
25.6.1. Географические расы
Популяции, занимающие обширный
географический ареал или в течение длительного времени
разбросанные по сильно разобщенным местообитани-
Таблица 25.2. Некоторые определения вида
Биологический аспект,
положенный
в основу
Размножение
Экология
Генетика
Эволюция
Определение
Группа особей, способных скрещиваться
между собой и давать плодовитое
потомство
Группа особей, занимающих одну и ту же
экологическую нишу; два разных вида
никогда не могут занимать одну
экологическую нишу
Группа особей, очень сходных по кариотипу
Группа особей, обладающих единственным
в своем роде набором структурных и
функциональных признаков
ям, могут обладать значительными фенотипичес-
кими различиями. В основе этих различий обычно
лежат приспособления к климатическим факторам.
Например, непарный шелкопряд (Lymantria dispar)
распространен во всей Японии и восточной части
Азии. На этом пространстве существуют самые
разнообразные климатические условия-от
субарктических до субтропических. Различают десять
географических рас шелкопряда, различающихся по
срокам вылупления из яиц. Северные расы
вылупляются позднее, чем южные. Как полагают, феноти-
пические особенности этих рас - результат влияния
климатических факторов на частоты генов в их
генофондах. О генетической основе такой
изменчивости свидетельствует тот факт, что при помещении
представителей этих рас в одинаковые условия они
продолжают вылупляться в разные сроки.
25.6.2. Экологические расы (экотипы)
Популяции, приспособленные к экологически
несходным местообитаниям, могут занимать смежные
географические области; например, в прибрежных
областях Калифорнии растение Gilia achilleaefolia
представлено двумя расами. Одна раса -
«солнечная» - растет на открытых травянистых южных
склонах, тогда как «теневая» раса встречается в
тенистых дубовых лесах и рощах секвойи. Эти расы
различаются по величине лепестков - признаку,
детерминированному генетически.
25.6.3. Клины
Клиной называют постепенное изменение фенотипи-
ческих признаков данного вида не протяжении его
ареала. У одного вида может быть несколько клин,
и они могут иметь разное направление (рис. 25.14).
Виды с выраженной фенотипической изменчи-
Крупные
W Светлые
Темные E
Мелкие
Рис. 25.14. Фенотипическая изменчивость размеров
тела и окраски шерсти у лесных мышей (Apodemus) в
Шотландии.
востью, связанной с частичной географической
изоляцией, называют политипическими видами.
Классическим примером такого вида служат чайки рода
Larus (см. разд. 25.8.4).
Все описанные выше случаи фенотипической
изменчивости отражают различную степень
генетической несхожести, которая может оказывать
влияние на репродуктивный потенциал членов
соответствующих популяций при их встрече.
25.7. Видообразование
Видообразованием называют процесс
возникновения одного или нескольких новых видов из видов,
существовавших ранее. Новые виды могут
возникнуть из одного вида (внутривидовое
видообразование) или, как это нередко происходит у многих
цветковых растений, два вида дают начало новому
виду (межвидовая гибридизация). Если
внутривидовое видообразование происходит в условиях
пространственной разобщенности популяций, то его
называют аллопатрическим видообразованием. Если
же во время этого процесса популяции занимают
одну и ту же географическую область, его называют
симпатрическим видообразованием.
25.8. Внутривидовое
видообразование
Во внутривидовом видообразовании участвует
несколько факторов, однако во всех случаях
непременным условием является прекращение обмена
генами между популяциями; в результате каждая
популяция становится генетически изолированной.
Изменение частоты аллелей и генотипов в
отдельных популяциях, обусловленное действием
естественного отбора на диапазон фенотипов,
создававшихся в результате мутаций и половой
рекомбинации, ведет к образованию рас и подвидов. Если
генетическая изоляция сохраняется в течение
длительного времени, а затем подвиды вновь
встречаются в одной и той же области, то они либо снова
скрещиваются, либо это оказывается невозможным.
В случае успешного скрещивания их все еще можно
считать принадлежащими к одному виду.
Невозможность скрещивания означает, что произошло
видообразование и прежние подвиды следует теперь
рассматривать как самостоятельные виды.
Полагают, что именно таким образом могут
происходить эволюционные изменения.
Начальным фактором в процессе
видообразования может служить снижение интенсивности отбора
в популяции. Это может привести к повышению
внутривидовой изменчивости. Если новые фенотипы
обладают адаптациями к условиям среды,
имеющимся в краевых участках ареала, то популяция
получает возможность расширить область своего
распространения. В тех случаях, когда обмен генами
внутри популяции не ослабевает, данный вид,
проявляя локализованную фенотипическую
изменчивость (экотипы), все еще сохраняет общий генофонд
и продолжает оставаться единым видом. В таких
ситуациях обычно создаются клины.
Видообразование может происходить только в
результате возникновения преград, ведущих к
репродуктивной изоляции между членами данной
популяции. Репродуктивная изоляция создается
изолирующими механизмами (термин, введенный генетиком
T. Добржанским) того или иного типа.
25.8.3. Симпатрическое
видообразование
Наследственные различия могут накапливаться и в
таких аллопатрических популяциях, которые были
географически изолированы на протяжении гораздо
более коротких периодов времени. Если эти
популяции затем встречаются, то в зонах перекрывания
могут образоваться гибриды. Например, на
Британских островах обитают черная ворона (Corvus co-
rone) и серая ворона (Corvus corone comix).
У черной окраска целиком черная, эта форма
Таблица 25.3. Изолирующие механизмы (по Добржанскому)
Презиготические механизмы (преграды для возникновения
гибридов)
25.8.1. Изолирующие механизмы
Изолирующий механизм - это фактор, создающий и
поддерживающий репродуктивную изоляцию.
Репродуктивная изоляция может быть обусловлена
механизмами, действующими до оплодотворения или
после него. В табл. 25.3 приведена в несколько
модифицированном виде классификация
изолирующих механизмов, предложенная Добржанским.
25.8.2. Аллопатрическое
видообразование
Для аллопатрического видообразования характерно
то, что на какой-либо его стадии существенную роль
играет пространственное разобщение. Такое
разобщение, создаваемое географическими барьерами
(например, горными хребтами, морями или реками)
или же различиями в предпочитаемых
местообитаниях, может препятствовать потоку генов;
организмы или их гаметы теряют возможность встретиться,
и это ведет к репродуктивной изоляции. Адаптация
к новым условиям и случайный дрейф генов в
небольших популяциях приводят к изменениям
частоты аллелей и генотипов. В результате
длительного разобщения популяций между ними может
возникнуть генетическая изоляция, сохраняющаяся
даже в том случае, если они вновь окажутся вместе.
Таким образом могут возникать новые виды.
Например, разнообразие видов вьюрков семейства
Goespizidae и их распространение на островах
Галапагосского архипелага считают результатом
аллопатрического видообразования. Как полагает
Дэвид Лэк, первоначально вьюрки прибыли на острова
с южноамериканского материка и здесь в отсутствие
конкуренции со стороны местных видов (слабое
давление отбора) произошла адаптивная радиация,
приведшая к возникновению разнообразных видов,
приспособленных к различным экологическим
нишам; эволюция разных видов, происходившая в
условиях географической изоляции, зашла так
далеко, что когда впоследствии они снова встречались
на некоторых островах, то могли уже
сосуществовать как самостоятельные виды.
Сезонная
изоляция
Экологическая
изоляция
Поведенческая
изоляция
Механическая
изоляция
Брачный период или сроки цветения
двух видов приходятся на разное
время года. Например, в Калифорнии
Pinus radiata цветет в феврале, а
P. attenuata — в апреле
Два вида, обитающие в одной и той
же области, предпочитают различные
местообитания. Например, Viola аг-
vensis растет на известковых почвах, а
V tricolor предпочитает кислые почвы
Сложные, специфичные для каждого
вида ритуалы ухаживания, на
которые реагируют только особи того же
вида, предотвращают спаривание
между особями разных видов.
Характерны для определенных видов рыб,
птиц и насекомых
Различия в строении половых
органов у животных, делающие
невозможной успешную копуляцию, и у
цветковых растений, у которых
близко родственные виды опыляются
различными животными
Постзиготические механизмы (определенные особенности
гибридов)
Нежизнеспособность
гибридов
Стерильность
гибридов Fi
Неполноценность
гибридов F2
Образование гибридов возможно, но
они не достигают зрелости, как,
например, гибриды между северной и
южной расами леопардовой лягушки
(Rana pipiens) в Северной Америке
Гибриды не способны производить
функциональные гаметы, как,
например, мул (2/1 = 63), получающийся от
скрещивания между лошадью (Equus
equus, 2λι = 60) и ослом (E.hemionus,
In = 66)
Гибриды Fi фертильны, но поколение
F2 и потомки от возвратных
скрещиваний между гибридами Fi и
родительскими формами либо не
развиваются, либо стерильны, как,
например, у гибридов между разными
видами хлопчатника (Gossypium)
Серая ворона
Зона гибридизации
Черная ворона
Вид
Рис. 25.15. Зона гибридизации как барьер,
препятствующий обмену генами между двумя
популяциями. Благодаря наличию этой зоны, тянущейся
поперек Шотландии, два вида ворон сохраняют
обособленность. Между соседними популяциями
часто существуют такие «гибридные» преграды,
действующие следующим образом: в тех местах,
где ареалы видов AuB перекрываются, в
результате межвидовых скрещиваний появляются
гибриды с пониженной фертильностью; в дальнейшем
вид А свободно скрещивается с гибридом AB,
a AB с видом В, но существование гибрида AB
препятствует свободному скрещиванию между
популяциями AuB.
распространена в Англии и на юге Шотландии. Серая
ворона - черная с серой спинкой и брюшком и
обитает в северной части Шотландии. Гибриды между
черной и серой воронами занимают узкую полосу в
центральных районах Шотландии (рис. 25.15). У
этих гибридов низкая фертильность, и это
эффективно препятствует обмену генами между
популяциями черной и серой ворон.
Со временем может возникнуть отбор,
направленный против скрещивания между обеими формами,
что приведет к образованию двух отдельных видов.
Поскольку такое видообразование происходит в
конечном счете в одной географической области, его
называют симпатрическим видообразованием.
Симпатрическое видообразование не связано с
территориальным разобщением популяций в период
создания генетической изоляции. Для него
необходимо развитие какого-либо механизма
репродуктивной изоляции, возникающего в результате отбора в
пределах географически изолированной области.
Этот механизм может быть структурным,
физиологическим, поведенческим или генетическим.
Симпатрическое видообразование чаще
рассматривают как механизм, позволяющий объяснить,
каким образом близкие виды, которые, вероятно,
произошли от общего предка в результате
временной изоляции, могут сосуществовать как
обособленные виды в одной и той же географической области.
Например, на Галапагосских островах вьюрок Ca-
marhyncus pauper обитает только на острове Чарлз,
где он сосуществует с родственной формой С.
psittacula, широко распространенной на всех
центральных островах (рис. 25.16). Виды вьюрков при
выборе брачного партнера руководствуются,
по-видимому, размерами клюва. Диапазоны размеров
клюва у С. pauper на острове Чарлз и С. psittacula на
острове Альбемарль примерно одинаковы, но на
острове Чарлз у С. psittacula клюв длиннее. Такое
отличие уже достаточно велико для того, чтобы эти
два вида, которые различаются по своему питанию,
не привлекали друг друга во время брачного
периода. Таким образом, виды сохраняют
обособленность и способны к сосуществованию.
25.8.4. Кольцевые клины
Симпатрическое видообразование особого типа
наблюдается в местах, где две популяции,
занимающие краевые участки клины, встречаются и
обитают совместно в одной области, замыкая таким
Джеймс
Нарборо
Альбемарль
Чарлз
Экватор
Индефатигейбл
Четем
Распространение
Camarhyncus psittacula
Худ
Распространение
Camarhyncus pauper
Рис. 25.16. Распространение двух видов вьюрков на
Галапагосских островах как пример
сосуществования видов после аллопатрического
видообразования.
Капуста
(Brassica oleracea)
2л = 18
х Редька
(Raphanus sativus)
2«= 18
Мейоз
Гаметы
Гибриды F1 2/f=18
В мейозе у гибридов F1 хромосомы редьки и капусты не
могут конъюгировать, так как они не гомологичны
Поэтому гибриды F1 стерильны. Иногда в результате
нерасхождения гибриды Fj дают гаметы с диплоидным
набором хромосом (2я=18).
Гаметы 2л= 18 х 2л= 18
Гибриды Fi
An = 36 (тетраплоид)
(Raphanobrassica)
Гибриды F2 плодовиты. Bo время мейоза может
происходить конъюгация гомологичных хромосом, поскольку в
наличии имеются два набора родительских хромосом.
Образуются диплоидные гаметы (2л = 18), содержащие 9
хромосом капусты и 9 хромосом редьки
Рис. 25.17. Этапы создания межвидового гибрида
капусты с редькой.
образом кольцо. Например, чайки рода Larus
образуют непрерывную популяцию, окружающую
Северный полюс кольцом между 50 и 80 ° с. ш. В это
кольцо входит 10 рас или подвидов, различающихся
главным образом по общим размерам тела и
окраске ног, спинки и крыльев. Между всеми этими
расами происходит свободный обмен генами, если
не считать того места, где на Британских островах
концы кольца смыкаются. Здесь, в крайних точках
ареала, чайки ведут себя как два отдельных вида-
серебристая чайка (Larus argentatus) и клуша (L
fuscus). Они различаются по внешнему виду, по
крику и по характеру миграций и скрещиваются
между собой лишь в редких случаях. Отбор,
направленный против их гибридизации, происходит в
условиях симпатрии.
У видов с половым размножением симпатричес-
кое видообразование без географической изоляции
маловероятно. Однако у организмов,
размножающихся бесполым путем, в том числе у высших
растений с вегетативным размножением, один
мутант, достаточно отличающийся от родительской
популяции, чтобы быть генетически
изолированным, может симпатрически дать начало новому
виду. Примером служит полиплоидия у Spartina (см.
разд. 23.9.2).
25.3. Десять подвидов популяции Larus
argentatus-fuscus образуют непрерывное кольцо,
которое тянется от Британских островов через
Скандинавию, СССР, Берингов пролив, Аляску и
Канаду и возвращается к Британским островам с
другой стороны. Если бы подвиды, обитающие в
районе Берингова пролива и Аляски, исчезли, то
какие это имело бы последствия для популяции?
25.9. Межвидовая
гибридизация
Это одна из форм симпатрического
видообразования, при которой новый вид возникает в результате
скрещивания между особями двух разных видов.
Плодовитые гибриды обычно получаются при этом
только в тех случаях, когда межвидовая
гибридизация приводит к хромосомной мутации, в результате
которой образуется аллополиплоид (см. разд. 23.9.2).
Примером служит гибрид между капустой и
редькой, полученный Г. Д. Карпеченко. Происходящие
при такой гибридизации генетические изменения
показаны на рис. 25.17.
Ответы и обсуждение
Глава 17
17.1. Бедренная кость в основном полая. Когда
при сопротивлении изгибу на одну ее сторону
действует сжимающая сила, другая сторона
подвергается растяжению. Вдоль
центральной оси кости эти силы взаимно
компенсируются. Поэтому материал внутренней
области не выполняет существенной
механической функции, и для животного выгодно
иметь здесь полость, чтобы вес бедренной
кости был меньше. Компактное вещество,
которое испытывает воздействие сил сжатия
и растяжения, расположено по периметру
кости. Губчатая ткань в головке бедренной
кости представляет собой сеть связанных
между собой костных пластинок (трабекул).
Такая структура обеспечивает прочность
кости при минимальном весе.
Хрящ служит амортизирующей
прокладкой между двумя сочлененными костями.
При сжатии его матрикс может
деформироваться, но после снятия нагрузки
возвращается в исходное состояние. Кроме того,
хрящ уменьшает трение между гладкими
суставными поверхностями при движении.
Сухожилия образованы белой
нерастяжимой волокнистой тканью и служат для
прикрепления мышц к бедренной кости. Тянущая
сила мышц оказывается приложенной к
небольшой поверхности. Наличие сухожильных
рецепторов позволяет предотвратить
повреждение при внезапном воздействии большой
нагрузки.
Связки тоже состоят из нерастяжимой
волокнистой ткани; они соединяют
бедренную кость с другими костями, образующими
с ней суставы. Связки позволяют
сочлененным костям двигаться лишь в определенном
направлении и таким образом обеспечивают
эффективную работу суставов. Кроме того,
они укрепляют сустав.
17.2. а) Обеспечиваются свободные движения
грудной клетки млекопитающего.
б) Упругая «подвеска» позволяет животному
амортизировать удар, который испытывают
его передние конечности во время
приземления в конце прыжка.
в) Передние конечности приобретают
широкий диапазон движений, необходимый для
таких действий, как лазание, «умывание»,
манипулирование с пищей, рытье.
17.3. Длина полосы А не изменяется, полосы H и I
укорачиваются.
17.4. Миозиновые нити (образующие полосу А)
сохраняют постоянную длину, а актиновые
нити скользят вдоль них навстречу друг
другу, что фактически приводит к укорочению
(сужению) полос H и I.
17.5. а) Центральная несократимая часть
мышечного веретена растягивается. Аннулоспираль-
ные окончания посылают импульсы в
центральную нервную систему. Оттуда по
эфферентным нейронам поступают ответные
импульсы к экстрафузальным мышечным
волокнам, и эти волокна сокращаются (рефлекс
на растяжение).
б) Мышечное волокно теперь уже не
растянуто, и число импульсов, поступающих из его
центрального несократимого участка,
уменьшается.
в) В сухожильных рецепторах генерируются
импульсы, которые поступают к тормозным
нейронам спинного мозга. Активация
последних рефлекторно подавляет сокращение
растянутой мышцы, и активное сопротивление
растягивающей силе уменьшается. Этот
механизм обеспечивает защиту мышцы от
возможного разрыва при внезапной ее
перегрузке.
г) Центральный участок веретена
растягивается, и импульсы от него поступают в
спинной мозг. Двигательные нейроны
активируются, импульсы передаются по ним к
экстрафузальным волокнам мышцы и вызывают ее
сокращение. После того как сокращение
интрафузальных волокон приводит к
соответствующему сокращению экстрафузаль-
ных волокон, центральный участок веретена
уже не подвергается дальнейшему
растяжению и наступает состояние равновесия. Этот
механизм очень важен, так как он регулирует
тонус мышц. Он не позволяет мышце
полностью расслабиться: в ответ на более
сильное растяжение веретена тонус мышцы вновь
возрастает. Если бы мышца была полностью
расслаблена, внезапное воздействие большой
нагрузки могло бы привести к ее
повреждению.
17.6. Это обеспечивает больший поток ионов
Ca2 + , необходимых для мышечного
сокращения.
17.7. В мышцах синхронного типа саркоплазма-
тический ретикулум развит сильнее, так как
для регуляции их работы необходимо
большее число нервных импульсов, а каждый
импульс связан с высвобождением Ca2+ из
ретикулума.
17.8. а) Обтекаемая форма тела.
б) Гладкая поверхность тела-чешуи
перекрывают друг друга в соответствующем
направлении, а слизистая или маслянистая
смазка уменьшает трение.
в) Плавники различных типов,
обеспечивающие продвижение вперед и устойчивость при
плавании.
г) Развитая мускулатура тела.
д) Боковая линия (см. табл. 4.12).
е) Плавательный пузырь у костистых рыб.
ж) Четко координированная работа нервно-
мышечного аппарата.
17.9. а) Мощные грудные мышцы.
б) Обтекаемая форма тела.
в) Узкие заостренные крылья,
обеспечивающие малое лобовое сопротивление.
г) Развиты главным образом маховые перья
1-го порядка (для продвижения вперед).
д) Быстрота взмахов (около 10 в 1 с).
е) Тяга, направленная вперед, создается как
при подъеме крыла, так и при его опускании.
17.10. Это увеличивает полезную длину
конечностей. В результате каждый шаг получается
длиннее и тело переносится вперед на
большее расстояние. Таким образом, при равной
быстроте перестановки ног скорость бега
возрастает.
Глава 18
18.1. При гипервентиляции воздух в легких
обновляется быстрее, углекислота усиленно
выводится из легких и ее парциальное давление
в альвеолярном воздухе понижается. Когда
кровь проходит через легочные капилляры,
уровень CO2 в ней приходит в новое
равновесие с альвеолярным воздухом. В результате
этого напряжение CO2 в крови уменьшается
и рН крови возрастает. Защелачивание крови
может приводить к головокружению и даже
обмороку. Импульсы от хеморецепторов
перестают поступать в дыхательный центр, и
это затормаживает вентиляцию легких.
18.2. Скорость транспирации и испарения воды
обратно пропорциональна влажности
воздуха. При высокой влажности эти процессы
идут медленно, поэтому растение не может
таким путем отдавать тепло и понижать
свою температуру.
18.3. 19,6%.
18.4. В этот период испытуемый имел
возможность прийти в равновесие с окружающей
средой.
18.5. Существует прямая зависимость между
этими двумя переменными; это позволяет
предполагать, что потоотделение регулируется
гипоталамусом.
18.6. Прямая зависимость между температурой
кожи и испарением влаги в первые 20 мин
показала, что между ними устанавливается
равновесие. Когда под влиянием
гипоталамуса, реагирующего на прием ледяной воды,
испарение замедляется, кожа начинает терять
за счет испарения меньше тепла; этим и
объясняется наблюдаемое повышение ее
температуры.
18.17. Лихорадка обусловлена изменением
настройки гипоталамического «термостата»: он
стремится теперь поддерживать более высокую
температуру. Пока температура не
поднимется до этого нового уровня, организм
реагирует на более низкую «нормальную»
температуру как на охлаждение. Возникает дрожь,
и мы чувствуем, что нам холодно, пока
температура внутренних областей тела не
придет в соответствие с настройкой
гипоталамического «термостата».
Глава 20
20.1. Л. Верно утверждение б: гаметы всегда
гаплоидны. Но даже оно не всегда справедливо,
так как полиплоидные родительские клетки
дают начало гаметам, в которых число
хромосом больше гаплоидного; например,
гаметы гексаплоидной пшеницы будут три-
плоидными.
Б. Исключения для утверждений а, в, г и д:
а) В бесполом размножении растений,
которым свойственно чередование поколений,
участвует мейоз; поэтому среди
образующихся гаметофитов наблюдается изменчивость.
б) В жизненных циклах А и Г (рис. 20.13)
гаметы образуются в результате митоза.
в) В результате мейоза всегда образуются
гаплоидные клетки (исключение -
полиплоидные организмы). В результате митоза могут
образоваться гаплоидные клетки [у таких
гаплоидных организмов, как спирогира
(рис. 20.13,/ί) и при росте гаметофита
(рис. 20.13, Г)]. Митоз может также давать
диплоидные клетки-при росте
многоклеточных диплоидных организмов.
г) Митоз может происходить в гаплоидных
клетках (как в примерах, приведенных в
пункте в).
20.2. а) Если есть возможность определить
растения, которым принадлежат пыльцевые зерна,
то можно сделать некоторые выводы о
климате, который в то время существовал.
б) Любое вмешательство человека в
естественную растительность будет отражено в
палинологической летописи. Например, пыльца
сорняков и сельскохозяйственных культур,
таких как пшеница, будет указывать на
уничтожение природной растительности и
возделывание земли, отсутствие в какой-то
области пыльцы древесных пород - на
вырубку лесов.
20.3. У двудомных растений половина
экземпляров не производит семян. Кроме того,
большое количество пыльцы расходуется зря, что
невыгодно с точки зрения использования
материальных и энергетических ресурсов.
20.4. У животных раздельнополость более
целесообразна, чем у растений, так как перенос
мужских гамет сопряжен у них с меньшим
риском благодаря подвижности этих
организмов и особенностям их поведения.
20.5. Половина (50%). Вспомните, что пыльцевые
зерна гаплоидны:
Генотип
родительского
растения
S1S2
S2
Возможные
генотипы
пыльцы
в равном
числе
Пыльцевые зерна S1
совместимы со столбиками S2S3
Пыльцевые зерна S2
совместимы со столбиками S1S3
Обратите внимание на то, что ни пыльцевые
зерна S1, ни зерна S2 не будут совместимы со
столбиком родительского растения (S1S2), так
что самоопыление невозможно.
20.6. а) Больше всего пыльцы попадет на ту часть
тела пчелы, которая соприкасается с
пыльниками, когда пчела высасывает нектар.
Поэтому перекрестное опыление обычно
происходит между цветками, у которых
пыльники и рыльца расположены в цветке на
одинаковой высоте, т. е. между длинно- и
короткостолбчатыми.
б) Она способствует аутбридингу.
20.7. Функции органелл, содержащихся в клетках
Сертоли, указывают на то, что эти клетки
синтезируют вещества, используемые в них
самих. Сырье для синтезов они получают
путем расщепления материалов,
поступающих в клетку, используя для этого ферменты,
находящиеся в лизосомах. Синтезированные
продукты запасаются в аппарате Гольджи
для последующего использования.
20.8. 1. а) Складывая число желтых тел и число
рубцов от желтых тел, можно установить
общее число имевших место овуляций; в
данном случае их было 67. Считая, что из
каждого фолликула выходит по одному ооциту в
месяц, получим репродуктивный период,
равный 5 годам и 7 месяцам. Следовательно, у
этой 22-летней женщины овуляция началась в
16,5 лет.
б) Из 42000 фолликулов только 219 имели
более 100 мкм в диаметре, т. е. содержали
ооциты первого порядка. Предполагая снова,
что в месяц высвобождается по одному
ооциту, находим, что запас ооцитов был бы
достаточен еще на 18 лет и 3 месяца.
в) Пять фолликулов содержали по два ооци-
та. Поэтому у женщины потенциально могло
бы родиться 5 пар разнояйцовых близнецов.
2. Нерасхождением, имевшим место в ана-
фезе I.
20.9. Кровь стала бы течь по боталлову протоку в
обратном направлении.
Глава 21
21.1. На основании этих кривых можно сделать
три заключения:
а) относительный рост достигает максимума
во время эмбрионального развития;
б) скорость роста максимальна в
младенческом и подростковом возрасте;
в) максимальный результат роста, если
оценивать его по одному параметру, например
по длине тела, достигается у взрослого
человека.
21.2. а) Происходит потеря массы, обусловленная
расходованием запасенных питательных
веществ в процессе дыхания.
б) Появились и распустились зеленые листья.
в) Процессы фотосинтеза; их скорость
должна быть выше скорости дыхания.
г) Оно объясняется сбрасыванием семян и
плодов.
21.3. С ноября по март масса тела составляет
в среднем 33,5 кг; в это время потребление
пищи соответствует 320 кДж в сутки. На
протяжении всего этого периода
среднемесячная температура не бывает выше
— 100C. Для того чтобы поддерживать
постоянную температуру крови на уровне
около 35°С, несмотря на разность температур в
45°С, лайке (гомойотермное животное)
необходима очень высокая интенсивность
метаболизма; а это в свою очередь требует
соответствующего снабжения пищей, чтобы
обеспечить источники энергии для дыхания.
Поскольку приведенные на рис. 21.13 данные
показывают, что масса тела в зимний период
меньше, чем летом, это дополнительно
свидетельствует о том, что для поддержания
постоянной температуры тела нужны
большие затраты энергии.
21.4. В мелких семенах запасы питательных
веществ невелики; поэтому растущий побег
должен как можно быстрее оказаться на
свету, чтобы фотосинтез начался раньше, чем
эти запасы будут исчерпаны.
21.5. а) Хлорофилл интенсивно поглощает свет в
красной и синей, но не в зеленой и дальней
красной областях (см. спектр поглощения
хлорофилла на рис. 9.9).
б) Красный компонент света стимулирует
прорастание семян латука, а дальний
красный подавляет (см. разд. 15.4.2). Поэтому
прорастание семян, находящихся под
пологом листьев, где свет обогащен дальним
красным компонентом, может быть подавлено до
тех пор, пока между листьями не появится
просвет, так что семена получат достаточно
света для фотосинтеза и роста.
21.6. При прорастании ячменного зерна
мобилизуются содержащиеся в нем питательные
вещества (главным образом крахмал и
некоторое количество белка). Крахмал
превращается в сахара, а белки - в аминокислоты;
продукты переходят в зародыш, где
используются в процессах роста. Поэтому с
увеличением сухой массы зародыша сухая масса
эндосперма уменьшается.
В то же самое время сухая масса в целом в
течение первой недели уменьшается. Это
обусловлено аэробным дыханием как
эндосперма, так и зародыша (хотя в большей
степени зародыша), при котором расходуется
сахар. Примерно на 7-й день появляется
первый лист и начинается фотосинтез. Это
приводит к увеличению сухой массы, которое
более чем компенсирует ее потери за счет
дыхания, так что в итоге общая сухая масса
возрастает. Одновременно ускоряется рост
зародыша, который теперь превратился в
проросток.
21.7. а) При этом сухая масса увеличивается на
8,6 г. Вычисление проводится следующим
образом:
Масса семян = 51,2 г
Масса жирных кислот = 51,2/2 = 25,6 г
Мол. масса жирных кислот = 256
Отсюда 1 моль = 256 г, а 25,6 =
= 0,1 моль
Из уравнения реакции следует:
0,1 моль жирных кислот -> 0,1 моль
сахара + 0,5 моль воды + 0,4 моль CO2
Мол. масса сахара = 342
Поэтому: 25,6 г жирных кислот -► 34,2 г
сахара 4- вода + CO2. Вода не входит в сухую
массу, a CO2 улетучивается, поэтому прирост
сухой массы =(34,2 — 25,6) г = 8,6 г.
б) Дыхание должно приводить к
уменьшению сухой массы. На самом деле сухая масса
все же возрастает.
в) Объем CO2, выделившейся из
семян, = 8,96 л при нормальных температуре и
давлении; это вычисляется следующим
образом: согласно уравнению, 0,1 моль жирных
кислот -► 0,4 моль CO2. 0,4 моль CO2 при
нормальных температуре и давлении
занимает 0,4-22,4 л = 8,96 л.
г) Жирные кислоты можно получить из жира
путем гидролиза, катализируемого липазой.
Другим компонентом липида является гли-
церол.
д) 51 атом углерода (липидом был трипаль-
митин; жирная кислота-это пальмитиновая
кислота). Каждая молекула липида содержит
три молекулы жирной кислота, по 16 атомов
С в каждой, плюс одну молекулу глицерола с
тремя атомами С.
ё) Сахароза или мальтоза.
ж) Кислород диффундирует в запасающую
ткань через семенную кожуру и микропиле.
21.8. а) Среди запасных веществ преобладают ли-
пиды, составляющие около 70% сухой массы
семян до начала прорастания. К 4-му дню
масса липидов начинает уменьшаться, а
масса Сахаров-увеличиваться: липиды
превращаются в сахара и переносятся в зародыш.
Образования Сахаров путем фотосинтеза
происходить не может, так как семена
прорастают в темноте. На 5-й день дыхательный
коэффициент (ДК) зародыша равен 1; это
указывает на то, что зародыш дышит за счет
сахара, образующегося из липидов. Между
тем семядоли (ДК от 0,4 до 0,5) получают
энергию за счет превращения липидов в сахар
и, возможно, за счет окисления сахара и
жирных кислот.
С18Н34°3
Рицинолеи-
новая
кислота (жирная
кислота,
получающаяся
из липида)
+ 13O2 -► C12H22O11
Сахароза
+ 6CO2 +
+ 6H2O + Энергия
ДК = 6Дз = 0,46.
Превращение липида в сахар сопровождается
увеличением сухой массы, так что суммарная
сухая масса проростков возрастает вплоть до
6-го или 7-го дня. Затем запасы липидов
истощаются, так что скорость использования
сахара начинает превышать скорость его
образования. После этого масса сахара и
общая масса проростков начинают
уменьшаться. Сахар расходуется в процессе
дыхания и в анаэробных реакциях.
б) На 11-й день ДК целых зародышей должен
быть, вероятно, чуть меньше 1,0. Эта
величина складывается из результатов двух
процессов; главный из них - окисление сахара при
дыхании (ДК =1), но возможен также
небольшой вклад превращения липида в сахар
(ДК от 0,4 до 0,5).
21.9. Обычно количества кислорода, проникающе-
го сквозь семенную кожуру, недостаточно
для обеспечения полностью аэробного
дыхания; ДК складывается из ДК для аэробного
дыхания (вероятно, около 1,0) и ДК для
анаэробного дыхания, который равен
бесконечности. Удаление семенной кожуры
создает возможность для более быстрого
проникновения кислорода путем диффузии, что
ведет к усилению аэробного дыхания и
снижению ДК. Этиловый спирт представляет
собой продукт анаэробного дыхания, поэтому
при удалении семенной кожуры он
накапливается в меньших количествах.
21.10. Во время 6-недельного воздействия на
куколку постоянной низкой температуры 3°С ней-
росекреторные клетки мозга вырабатывают и
запасают торакотропин. Последующее
содержание куколки при постоянной температуре
25°С приводит к выведению торакотропина в
кровь; этот гормон побуждает торакальные
железы к секреции гормона линьки, диапауза
прерывается и из куколки выходит имаго.
Глава 22
22.1. а) Мейоз
б) W-интерфаза
Х-телофаза I
Y-телофаза II
в) Половые клетки
22.2. См. рис. 22.2 (отв.).
22.3.
Основания
А
Г
T
Ц
А
AA
ГА
ТА
ЦА
Г
АГ
ГГ
ТГ
ЦГ
T
AT
ГТ
TT
ЦТ
Ц
АЦ
ГЦ
ТЦ
цц
22.4. 4 основания, используемые поодиночке: 4 1 =
= 41 =4; 4 основания, используемые парами:
4·4 = 42 = 16; 4 основания, используемые
тройками: 4 · 4 · 4 = 43 = 64. Математическое
выражение имеет вид ху, где χ-число
различных оснований, а у- число оснований в
кодоне.
22.5. См. рис. 22.5 (отв.).
Клетки из А, растущие в 14N
15N -ДНК
Первое
поколение
Второе
поколение
14
ДНК, содержащая N
-ДНК, содержащая 5N
Консервативная репликация
-ДНК с 14N и 15N
-ДНК с 15N
Дисперсивная репликация
Рис. 22.2. (отв). Схемы, поясняющие гипотезы о
консервативной и дисперсивной репликации ДНК.
Показано распределение ДНК в градиенте плотности
хлористого цезия в соответствии с теориями,
проиллюстрированными на рис. 22.14.
22.6. УАГ ААГ ЦУЦ АУГ ГУА ЦАУ УГЦ.
22.7. Результаты этих экспериментов показывают,
что процесс дифференцировки связан не с
утратой или повреждением генетического
материала (ДНК), а с выключением
определенных генов. ДНК сохраняла способность
функционировать, оказавшись в новой для
себя среде недифференцированной клетки.
Дальнейшие эксперименты по
трансплантации ядер, взятых из клеток на более поздней
стадии онтогенеза, не привели к развитию
жизнеспособных зародышей. Это было
сочтено доказательством того, что в процессе
развития в генетическом материале
происходят какие-то необратимые изменения,
воздействующие на дифференцировку путем не-
Рис. 22.5. (отв.). Общий принцип, на
котором основано восстановление
нормальной рамки считывания
триплетов путем добавления или удаления
оснований, состоит в том, чтобы внести
в любой участок нуклеотидного кода или
удалить из него три основания.
Исходная последовательность
Три основания добавлены
Три основания удалены
|гТа[ ГТА| ГТА ГТА| ГТА ГТА ГТА ГТА|
ΓΤΑΙЦГТ АГТ|АЦГ ΤΑΓΙТАЦГТА ГТА|
Рамка ι
считывания!
',восстановлена
I ГТА ГТГI ТАГ! TTT|АГТ | ГТА ГТА| ГТА
Рамка считывания
восстановлена
обратимого выключения генов. Однако в
1960-х годах Гёрдон провел в Оксфорде
эксперименты с пересадкой ядер из клеток
кишечного эпителия головастиков в энуклеирован-
ные яйцеклетки от половозрелой лягушки. Во
многих случаях из таких яиц развивались
нормальные лягушки с признаками,
соответствующими признакам донора ядра. Этот
метод известен под названием клонирования
особей; полученные результаты позволяют
думать, что хотя ядру принадлежит
доминирующая роль в процессе развития,
цитоплазма (в данном случае цитоплазма яйца) играет
важную роль в определении избирательной
экспрессии ядерного материала, т.е. в диф-
ференцировке. В данном случае она
побуждает ядро к полной реализации его генетических
потенций и созданию целого организма.
Глава 23
23.1. Пусть:
В-коричневая шерсть (доминантный
признак)
Ь-серая шерсть (рецессивный признак)
Фенотипы родителей
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы Fl(2n)
Фенотипы F1
Коричневая
шерсть
BB
Серая
шерсть
Все коричневые
Фенотипы F2
Генотипы F](2n)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы F2(2 п)
Фенотипы F2
Коричневая
шерсть
Bb :
® ® >
BB Bb
3 коричневые
Коричневая
к шерсть
< Bb
< ® Θ
Bb bb
1 серая
(б)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы потомков (2п)
Фенотипы потомков
Коричневая
шерсть
Bb
Серая
шерсть
bb
1 коричневая
1 серая
В случае моногибридного скрещивания
гетерозиготной особи с особью, гомозиготной по
рецессивному аллелю, среди потомков будет
равное число особей того и другого фенотипа,
в данном случае 50% с коричневой и 50% с
серой шерстью.
23.2. Если провести анализирующее скрещивание
особи неизвестного генотипа с особью,
гомозиготной по доминантному аллелю
изучаемого гена, то у всех потомков в фенотипе
проявится доминантный признак, как это
показано ниже. Обозначим доминантный
аллель T1 а рецессивный t.
Гомозигота
Гомозигота Гетерозигота
Гомозигота
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы потомков (2 η)
Все с доминантным фенотипом (гомозиготные)
Все с доминантным
фенотипом (половина гомозиготных,
половина гетерозиготных)
23.3. а) Если в F1 у всех морских свинок была
короткая черная шерсть, то это означает, что
короткая шерсть доминирует над длинной, а
черная окраска над белой.
б) Рассмотрим приведенную ниже схему.
Примем следующие обозначения:
В-черная шерсть
Ь-белая шерсть
S-короткая шерсть
s-длинная шерсть
Фенотипы F1
Генотипы F1 (2п)
Мейоз
Гаметы ( п)
Случайное оплодотворение
(по решетке Пеннета)
Генотипы F2(2n)
(указаны в каждой клетке)
С короткой
черной шерстью *
С короткой
' черной шерстью
Фенотипы F2
9 с короткой:
черной
шерстью
. 3 с корот-:
кой белой
шерстью
: 3 с длинной :
черной шерстью
_
1 с длинной
белой шерстью
ι—ι
(Символы)
23.4. Пусть: R, г и S, s-две пары аллеломорфных
генов, контролирующих окраску цветков.
Фенотипы родителей
Генотипы родителей (2 η )
Мейоз
Гаметы ( п)
Случайное оплодотворение
(по решетке Пеннета)
Генотипы потомков (2п)
(указаны в каждой клетке)
Фенотипы потомков
(Символы)
Пурпурные цветки х Пурпурные цветки
9 пурпурных. 7 белых
23.5. В метафазе I и анафазе I происходит
расщепление по этим двум аллелям.
23.6. Число различных комбинаций хромосом в
пыльцевых зернах (и мужских гаметах)
вычисляется по формуле 2я, где «-гаплоидное
число хромосом. У шафрана 2п = 6,
следовательно η = 3. Поэтому число комбинаций
равно 23 = 8.
23.7. Фенотипы F1 свидетельствуют о том, что
пурпурные цветки и короткий
стебель-доминантные признаки, а красные цветки и
длинный стебель - рецессивные. Примерное
соотношение 1:1:1:1 при дигибридном
скрещивании означает, что два гена,
контролирующие окраску цветка и длину стебля, не
сцеплены и что четыре соответствующих ал-
леля находятся в разных парах хромосом.
Поясним это следующим образом:
P-пурпурные цветки
ρ-красные цветки
S-короткий стебель
s-длинный стебель
Поскольку оба родительских растения были
гомозиготны по обоим признакам, растения
F1 должны иметь генотип PpSs.
23.8. а) Гомологичные хромосомы.
б) Между локусами окраски тела и длины
крыльев.
в) На основании имеющихся данных
ответить на этот вопрос нельзя, так как на схеме
не указано положение гена, определяющего
форму антенн.
23.9. Из 800 полученных семян только у 24
наблюдаются результаты кроссинговера между
генами окраски семян и характера
эндосперма. У остальных 776 семян аллели этих
генов остаются сцепленными, о чем
свидетельствует отношение, примерно равное 1:1.
Таким образом, частота рекомбинации равна
(24/800)· 100 = 3%. Значит, расстояние между
генами окраски семян и характера
эндосперма равно 3 морганидам.
3.10. а) Пусть:
N-нормальные крылья (доминантный
признак)
η-миниатюрные крылья (рецессивный
признак)
R-красные глаза (доминантный признак)
г-белые глаза (рецессивный признак)
XX - самка
XY-самец
Фенотипы родителей
Миниатюрные крылья,
красные глаза
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы F\(2n)
Фенотипы F1
XnRY
Нормальные крылья,
белые глаза
X
XNrXN,
(χ**) (V) χ (х^) (х")
vnRyNr YnRYNr
Нормальные крылья,
красные глаза
XNry XNry
Нормальные крылья,
белые глаза
Пурпурный цветок, >
короткий стебель
< Пурпурные цветок,
длинный стебель
Фенотипы родителей
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
(по решетке Пеннета)
Генотипы потомков (2п)
(указаны в каждой клетке)
Фенотипы потомков
1 с пурпурными цветками и коротким стеблем :
1 с пурпурными цветками и длинным стеблем :
1 с красными цветками и коротким стеблем :
1 с красными цветками и длинным стеблем
(2) Если между генами длины крыльев и
цвета глаз не происходит перекрестов, то
можно ожидать следующих результатов:
(2)
Фенотипы F1
Генотипы F1 (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы F2 (2п)
Фенотипы F2
Нормальные крылья
белые глаза
Нормальные крылья,
красные глаза
XNrXnR
Нормальные
крылья,
белые глаза
ΧΝγΧΝγ
Нормальные
крылья,
белые глазг
XnRY
Миниатюрные
крылья,
ι красные глаза
ΧΝΎ
Нормальные
крылья,
белые глаза
б) Отсутствие отношения 1:1:1:1 между
фенотипами особей, полученных от этого
скрещивания, указывает на кроссинговер между
генами длины крыльев и цвета глаз у самки.
Фенотипы родителей
Генотипы родителей (2 η)
Мейоз
Гаметы (п)
Случайное оплодотворение
(по решетке Пеннета)
Генотипы потомков (2 п)
(указаны в каждой клетке)
Фенотипы потомков
Число потомков в опыте
Нормальные крылья, χ Миниатюрные крылья,
красные глаза О белые глаза cf
xnRxNr х Хпгу
Крылья:
Глаза:
миниатюрные
красные
36
нормаль- мини-
нормальные атюрные ные
белые
35
красные
18
белые
17
Аллели, определяющие длину крыльев и цвет
глаз, указаны выше для двух «женских» (X)
хромосом в F1. В результате кроссинговера
между этими аллелями получаются
указанные рекомбинантные генотипы. Из 106 мух
35 (18 + 17) являются рекомбинантными;
таким образом, частота рекомбинации равна
35/106 * 30%.
23.11. а) Пяденица
N-нормальная окраска (доминантный
признак)
η-бледная окраска (рецессивный признак)
Фенотипы родителей
Бледно
окрашенный самец *
Нормально
< окрашенная самка
Генотипы родителей (2 п)
ЛИБО
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы потомков (2п)
ЛИБО
ЛИБО
ЛИБО
Фенотипы потомков
Нормальная
ίο краска
Нормаль-
I ная j
ν окраска
На основании фенотипов потомства можно
сделать вывод, что гетерогаметным полом у
пяденицы является самка.
б) Кошка
В-черная окраска (доминантный признак)
Ь- рыжая окраска (рецессивный признак)
ЛИБО
Фенотипы родителей
Генотипы родителей (2 п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы потомков (2п)
ЛИБО
Черный самец
Рыжая самка
ЛИБО
ЛИБО
Фенотипы потомков
Черная
окраска
Рыжая
окраска
Черная
окраска
j
Черная
окраска
Нормаль-J
ная
окраска
f
Бледная
о крас ю
На основании фенотипов потомства можно
сделать вывод, что у кошки гетерогаметным
полом является самец.
23.12. Пусть:
В-черная шерсть
G-рыжая шерсть
XX - кошка
ΧΥ-κοτ
Фенотипы родителей
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы Fi (2п)
Фенотипы F1
Рыжий кот
X0Y
© Θ
XGXB XGXB
Черепаховая
окраска
9
х Черная кошка
χ ХВХВ
XBY XBY
Черная окраска,
(Родительская особь женского пола должна
быть гомозиготна по черной окраске шерсти,
так как только в этом случае могут получиться
потомки с черной шерстью.)
Черный кот
XBY
Фенотипы F1
Генотипы F1 (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы F2 (2 п)
Фенотипы F2
Черепаховая кошка
XGXB
XBXG
Черепаховые
ХВХВ
Черные
XGY
Рыжие,
XBY
Черные,
Cf
23.13. а) Пусть:
I-ген, детерминирующий группу крови
А-аллель Al (доминантны в одинаковой
В-аллель BJ степени)
О-аллель О (рецессивен)
Фенотипы родителей
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
Генотипы потомков (2п)
Фенотипы потомков
Группа крови А >
JA1O ,
© ® >
,А,В JA1O
Группы AB А
крови:
< Группа крови В
< 1в1°
< © ©
JO1B !O1O
В О
б) Для каждого из детей вероятность того,
что кровь будет принадлежать к группе А,
равна 1/4 (25%). Поэтому вероятность того,
что у обоих будет группа А, равна 1/41/4 =
= 1/16 (6,25%).
23.14. Пусть:
P - гороховидный гребень
R-розовидный гребень
Один аллель P и один аллель R вместе дают
ореховидный гребень. Гомозигота по обоим
рецессивным аллелям имеет простой гребень.
W-белое оперение (доминантный признак)
w-черное оперение (рецессивный признак).
Если среди потомков от скрещивания
получено восемь различных фенотипов, то каждая
из родительских особей должна быть
гетерозиготна по максимально возможному числу
аллелей. Поэтому они должны иметь
генотипы, указанные ниже:
Фенотипы родителей
Генотипы родителей (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
(по решетке Пеннета)
Генотипы потомков (2п)
(указаны в каждой клетке)
Фенотипы потомков
и символы
Черное оперение, розовидный гребень X
wwRrpp X
Белое оперение, ореховидный гребень
WwRrPp
Белые с ореховидным
гребнем
Черные с
ореховидным гребнем
Белые с розо-
видным гребнем
Черные с
розовид-
ным
Белые с
гороховидным гребнеы
Черные с
гороховидным гребнем
Белые с простым
гребнем
Черные с простым
гребнем
23.15. Поскольку в гетерозиготном генотипе
поколения F1 содержатся оба доминантных алле-
ля, W (белая окраска) и В (черная окраска), а
фенотипически куры белые, можно сделать
вывод, что эти аллели эпистатически
взаимодействуют друг с другом, причем белый
аллель эпистатичен, а черный - гипостатичен.
Ниже показаны соотношения фенотипов в
F2 (использованы символы, приведенные в
задаче):
Фенотипы F1
Генотипы F1 (2п)
Мейоз
Гаметы (п)
Случайное оплодотворение
(по решетке Пеннета)
Генотипы F2 (2п)
(указаны в каждой клетке)
Белый петух
WwBb х
Белая курица
WwBb
Фенотипы
(Символы)
Белые
12
Черные
3
Коричневые
1
Глава 24
24.1. Ван Гельмонту следовало поставить
контрольные эксперименты, в которых
исключалась бы по одной каждая из переменных.
24.2. Реди исходил из предположения, что наличие
«червяков» было связано с мухами, свободно
залетавшими в открытые сосуды.
24.3. Запечатывание бульона могло
препятствовать попаданию организмов в сосуды.
Отсутствие же в сосудах воздуха могло лишить
организмы необходимого для дыхания кислорода.
24.4. Главное предположение Пастера
заключалось в том, что каждое поколение
организмов происходит от предшествующего
поколения, а не возникает спонтанно.
24.5. Микроорганизмы развиваются в питательном
бульоне в результате его загрязнения
организмами из атмосферы.
24.6 Плотное закрывание пробирок, кипячение, ав-
токлавирование, прямой контакт с
атмосферой, непрямой контакт с атмосферой.
24.7. По контакту с атмосферой.
24.8. Пробирки пары А не нагревали, пары Б
кипятили 10 мин, пары В - автоклавировали
20 мин при давлении 0,8 атм, пары Г
различались в отношении контакта с атмосферой.
24.9. Эксперименты Спалланцани - в пробирках 3 и
4, Пастера-в пробирках 7 и 8.
24.10. Пробирки 1, 3, 5 и 7.
24.11. Проведение экспериментов соответствует всем
требованиям, предъявляемым к научному
методу. Они основаны на определенной
гипотезе, сопровождаются соответствующим
контролем, и в них последовательно исключаются
переменные факторы. Если идентичные
эксперименты дают одинаковые результаты, то
полученные данные можно считать
достоверными. Достоверность выводов, сделанных на
основании этих (или любых других) данных,
зависит от того, насколько правильно эти
данные интерпретируются.
Глава 25
25.1. Носители являются гетерозиготами. Частоты
генотипов вычисляются по уравнению
Харли-Вайнберга
р2 + 2pq + q2 = 1,
где ρ2 - частота доминантного гомозиготного
генотипа, 2pq-частота гетерозиготного
генотипа и q2-частота рецессивного
гомозиготного генотипа.
Кистозный фиброз поджелудочной железы
поражает индивидуумов с рецессивным
гомозиготным фенотипом; следовательно, q2 =
= 1 на 2000, или 1/2000 = 0,0005. Отсюда
q = ^/0,0005 = 0,0224.
Поскольку ρ + q = \, ρ = \ — q = \ — 0,0224 =
= 0,9776.
Таким образом, частота гетерозиготного
фенотипа (2pq) = 2 · (0,9776) · (0,0224) = 0,044 =
= 1 на 23 % 5%, т.е. носители рецессивного
гена кистозного фиброза поджелудочной
железы составляют около 5% популяции.
25.2. Печеночная двуустка (Fasciola
hepatica)-паразит, заражающий овец. У него есть
промежуточный хозяин-малый прудовик (Ытпаеа
truncutula), живущий в пресных водоемах и на
сырых лугах. Осушение прудов и сырых
участков приведет к изменению условий среды и
создаст давление отбора, направленное на
элиминацию улитки. С уменьшением
численности улитки сократится число подходящих
хозяев, что повлечет за собой снижение
численности паразита.
25.3. Пониженная интенсивность отбора в краевых
областях ареала каждой новой популяции
будет благоприятствовать увеличению
изменчивости. Новые фенотипы могут оказаться
приспособленными к областям, которые
прежде были заняты элиминированными
подвидами, и будут распространяться внутрь ареала,
занимая освободившуюся экологическую
нишу. Географическое разделение клины могло
привести к аллопатрическому
видообразованию. В случае восстановления кольца обмен
генами может оказаться невозможным
вследствие генетической изоляции, и тогда обе
субпопуляции будут дивергировать еще сильнее
на пути к образованию самостоятельных
видов, подобно нынешнему положению на
Британских островах, где два вида сосуществуют
симпатрически. Если генетическая изоляция
между двумя субпопуляциями зашла не
слишком далеко, то при воссоединении
субпопуляций между ними возможна гибридизация.
Такая зона гибридизации может играть роль
репродуктивного барьера, как в случае
черной и серой ворон.
Дополнения к гл. 12
12.4.1. Нарушенные биогеохимические
циклы и загрязнение среды
Круговорот углерода: диоксид
углерода и парниковый эффект в
планетарном масштабе
Основные пути, по которым происходит круговорот
углерода, показаны на рис. 9.2 (т. 1).
Дополнительный, индуцированный человеком цикл
(мощность его оценивается в 5 1012 кг год-1) связан с
использованием ископаемого топлива. Некоторые
ученые полагают, что сведение лесов также является
важным дополнительным источником
атмосферного диоксида углерода (углекислого газа), а скорость
антропогенного поступления углерода в атмосферу
приближается к 10· 1012 кг год"1.
12.14. Как сведение лесов приводит к
увеличению содержания диоксида углерода в атмосфере?
В норме диоксид углерода содержится в нижних
слоях атмосферы (так называемой тропосфере) в
очень небольшом количестве - около 300 частей на
миллион или 0,03% в объемных единицах. Важность
этого газа определяется его вкладом в парниковый
эффект, проявляющийся в масштабе всей планеты.
Диоксид углерода прозрачен для коротковолнового
излучения, поступающего от Солнца, но он сильно
поглощает энергию в длинноволновой части
спектра, которая переизлучается Землей обратно в
космос. Таким образом, диокид углерода «ловит»
уходящее излучение, согревая нижние слои атмосферы,
в свою очередь излучающие энергию обратно к
поверхности Земли (а также вверх и в стороны).
Естественно, что в конце концов любая «порция»
входящей энергии будет рассеяна и потеряна в
космосе, но энергообмена между атмосферой и
поверхностью Земли, обусловленного наличием
диоксида углерода и других парниковых газов,
достаточно, чтобы повысить температуру поверхности
Земли примерно на 400C по сравнению с той, что
наблюдалась бы в отсутствие такого обмена. Очень
важно осознать, что без некоторого основного
уровня парникового эффекта, мало изменявшегося на
протяжении миллионов лет, живые системы, во
всяком случае в той форме, в какой мы их знаем,
существовать бы не могли. Имеющиеся в настоящее
время данные ясно свидетельствуют о том, что
содержание в атмосфере диоксида углерода и
других газов, обеспечивающих парниковый эффект, в
частности моноксида углерода, метана и хлорфтор-
углеродов, увеличивается со скоростью,
беспрецедентной для недавней истории Земли; это
увеличение по логике вещей должно способствовать
повышению температуры окружающей среды (см. рис.
12.12). В свою очередь такое повышение
температуры усиливает испарение и соответственно
увеличивает концентрацию водяных паров в
атмосфере. Поскольку водяной пар также действует как
мощный поглотитель длинноволнового излучения,
это приведет к дальнейшему повышению
температуры в приповерхностных слоях атмосферы.
Достигнутое в результате всех этих процессов
повышение температуры вызовет изменение в
распределении и интенсивности основных воздушных и
водных потоков, определяющих погоду на планете,
что в свою очередь может существенным образом
повлиять на хозяйственную деятельность человека.
Некоторые ученые-метеорологи и
правительственные чиновники не придавали большого
значения угрозе парникового эффекта, полагая, что он
будет компенсирован за счет возросшей
запыленности атмосферы i (из-за усилившейся эрозии почвы
в результате сведения лесов, интенсификации
сельского хозяйства, уничтожения живых изгородей,
перевыпаса и т.д.) и, таким образом,
результирующее воздействие окажется незначительным.
Однако необычные (порой катастрофические для
человека) погодные явления, наблюдавшиеся в
планетарном масштабе летом 1988 г., в частности сильная
засуха в юго-западной части США, наводнения в
Китае и Судане, частые случаи понижения давления
и выпадения осадков в Англии (обычно такое
происходит в летнее время значительно севернее), а также
не вызывающее уже сомнений воздействие хлор-
фторуглеродов на озоновый слой, привели к
гораздо более широкому признанию реальной воз-
1 Наличие в атмосфере пыли в большей степени
приводит к сокращению поступающего
коротковолнового излучения, чем к задержке исходящего от
поверхности земли длинноволнового излучения
можности возникновения глобальных
климатических изменений при загрязнении среды и к осознанию
угрожающих последствий этих изменений.
Независимо от того, вызваны ли описываемые события
парниковым эффектом или нет, надо
приветствовать усилия правительств, направленные на
контроль за поступлением в атмосферу загрязняющих ее
парниковых газов, так как природные механизмы,
обеспечивающие их удаление, работают слишком
медленно, чтобы изменить ситуацию (такое
наблюдается, например, в случае диоксида углерода), или
же создают новые проблемы, как, например, в
случае разрушения озона стратосферы хлорфтор-
углеродами (см. разд. 12.9).
Диоксид углерода изымается из атмосферы
посредством фотосинтеза, а также в результате
обмена с океаном и захоронения в виде карбонатов.
Главный из этих процессов - обмен с океаном.
Верхние 75 метров океанической толщи представляют
собой слой воды, хорошо перемешиваемый ветром
и согреваемый солнцем. Указанные особенности
данного слоя в сочетании с химическими
свойствами карбонатов обеспечивают довольно быстрое
поглощение им диоксида углерода. В конце концов
скорость поступления диоксида углерода в океан
определяется процессами обмена между его
поверхностными и глубинными слоями, так как именно в
результате этого обмена восстанавливается
способность поверхностных вод к дальнейшему
поглощению диоксида углерода. Обмен поверхностных вод с
глубинными в большинстве районов океана
протекает очень медленно. Громадные массы холодной (с
температурой менее 5°) океанической воды
изолированы от поверхностных перемешиваемых вод
зоной стагнации - так называемым океаническим
термоклином. Эта зона, характеризующаяся тем, что по
мере возрастания глубины в ней снижается
температура и повышается плотность воды, простирается
до глубины порядка 1000 м. По имеющимся
оценкам за сотни лет к поверхности подходит не более
10% общего объема воды в океане. Таким образом,
хотя океаны потенциально могут поглотить весь
образовавшийся в результате деятельности человека
излишек атмосферного диоксида углерода, на
практике реализации этой возможности (во всяком
случае во временных масштабах, сопоставимых с
временем интенсивной хозяйственной деятельности
человека) препятствует медленный перенос от
поверхностных вод к глубинным. Возможно также,
что, по мере того как вода у поверхности будет
насыщаться диоксидом углерода, способность
океана к дальнейшему поглощению этого газа
уменьшится, в результате чего вся проблема повышения
содержания его в атмосфере станет еще более
серьезной.
Предсказать будущую концентрацию диоксида
углерода в атмосфере довольно трудно. Отчасти это
объясняется недостаточным пониманием всех
тонкостей химического процесса абсорбции (видимо, в
океане может быть абсорбировано большее
количество диоксида углерода, чем это следует из
химической модели), а отчасти также тем, что возросшая
озабоченность общественности состоянием
окружающей среды может затормозить процесс сведения
лесов и ускорить развитие альтернативных
источников энергии; это в свою очередь сократит скорость
потребления ископаемого топлива. Тем не менее
широко распространено мнение о том, что к
середине двадцать первого века концентрация диоксида
углерода в атмосфере увеличится до 600 частей на
миллион, превысив, таким образом, более чем вдвое
уровень, зарегистрированный для конца
девятнадцатого века. Трудно также предсказать
климатические последствия, поскольку существует большое
число весьма вероятных обратных связей,
включенных в рассматриваемые процессы. В соответствии с
предсказаниями большинства моделей к середине
двадцать первого века средняя для планеты
температура увеличится на 3 + 1,5°. В высоких широтах
может сложиться еще более серьезная ситуация
из-за наличия механизмов обратной связи. По мере
того как снег и лед тают, альбедо земли (т.е. ее
отражательная способность) снижается, вследствие
чего усиливается поглощение поверхностью Земли
солнечного излучения. Это повлечет за собой
интенсификацию процессов испарения, в результате чего в
конце концов возрастет содержание паров воды в
атмосфере. Поскольку водяной пар сам по себе
является газом, усиливающим парниковый эффект,
скорость потепления в высоких широтах при
удвоении концентрации атмосферного диоксида углерода
окажется в два-три раза выше, чем в тропиках.
Последствия такого потепления могут быть самыми
разными-от затопления обширных участков
прибрежных низменностей (что может угрожать юго-
восточной Англии, Нидерландам, Бангладеш и
тропическим островам, возникшим на коралловых
рифах, например Мальдивским), до крупных сдвигов в
распределении мировых климатических зон. Все эти
события существенным образом повлияют на
производство пищи и на сложившийся баланс сил в
сфере торговли и политики. Некоторая
незначительная по масштабам компенсация процессов
увеличения содержания в воздухе CO2 может быть
достигнута за счет возрастания интенсивности
фотосинтеза в наземных экосистемах (обычно диоксид
углерода-это фактор, лимитирующий фотосинтез).
Может также возрасти продукция морского
фитопланктона, особенно в прибрежных водах, где за
счет поступления загрязняющих веществ увеличена
концентрация нитратов и фосфатов - основных
биогенных элементов, ограничивающих развитие
фитопланктона. Эти процессы в конечном счете могут
способствовать изъятию диоксида углерода из
атмосферы, однако желательность их вызывает
большие сомнения. Отмеченное возрастание
продуктивности прибрежных вод благоприятствует
увеличению численности быстро растущих видов,
уменьшению разнообразия фитопланктона и
появлению в нем доминантов, слабо поедаемых консу-
ментами. Все это не способствует возрастанию
продукции консументов-фитофагов, а массовое
отмирание водорослей приводит к снижению концентрации
кислорода и увеличению смертности консументов.
Обзор по проблеме парникового эффекта
опубликован в журнале „New Scientist" от 22 октября
1988 г.
Эвтрофизация
Эвтрофизация - это обогащение водных экосистем
биогенными элементами. В течение длительного,
измеряемого тысячелетиями, периода времени
озерные экосистемы испытывают постепенное
превращение, переходя из олиготрофного состояния
(характеризующегося малым содержанием биогенных
элементов) в эвтрофное (характеризующееся
высоким содержанием биогенных элементов) или даже
дистрофное (характеризующееся высоким
содержанием биогенных элементов, находящихся в
трудноусвояемой форме) (табл. 12.4). Однако в XX в. во
многих озерах (среди которых Эри в Америке,
Цюрихское озеро в Швейцарии, Лох-Ней в Северной
Ирландии), во внутренних морях (например, в
Балтийском и Черном) и во многих речных системах,
разбросанных по всему свету, наблюдалась очень
быстрая эвтрофизация, что обусловлено
интенсификацией хозяйственной деятельности человека. К
числу главных факторов, непосредственно
воздействующих на это явление, следует отнести
массированное использование азотных удобрений на
сельскохозяйственных угодьях и увеличившийся сток
фосфатов в сточных водах. Проблемы, связанные с
избыточным поступлением фосфатов в водоемы,
отражают не только возросшую численность
населения, но и существующую тенденцию его
скопления в городах, что приводит к развитию крупных
систем сброса сточных вод. Таким образом,
происходит не только увеличение абсолютного
количества фосфорсодержащих сточных вод, но и
возрастание (причем значительно быстрее, чем раньше) их
локальной концентрации в местах сброса в реки,
озера и моря. Особенно серьезные проблемы
возникают при сбросе стоков в озера. Примером может
служить ситуация, возникшая в середине
пятидесятых годов в оз. Вашингтон (США). Наблюдавшееся
здесь бурное «цветение» фитопланктона
сопровождалось существенным снижением концентрации
растворенного кислорода в глубинных водах озера
(так называемом гиполимнионе). Произошло это
вследствие сильно возросшего сброса сточных вод,
поступавших от быстро развивающегося города
Сиэтл. Ситуацию удалось поправить, отведя
сточные воды города непосредственно в Тихий океан
через залив Пьюджет-Саунд. В то же время удалось
улучшить очистку сбрасываемых вод и избежать
таким образом сильного загрязнения самого залива.
Эвтрофизация озерных систем в настоящее время
наблюдается повсеместно по всему миру. Везде, где
только возможно, следует избегать сброса стоков в
озера и другие более или менее замкнутые водоемы,
например в бухты.
Процесс эвтрофизации, быстро развивающийся в
Таблица 12.4. Основные черты олиготрофных и эвтрофных озер
Глубина
Содержание кислорода в гиполимнионе
летом
Фитопланктон
«Цветение» фитопланктона
Приток биогенных элементов
Животная продукция
Рыбы
Олиготрофные
Глубокие
Содержится
Высокое видовое разнообразие,
небольшая численность, низкая
продуктивность; часто доминируют
зеленые водоросли
Наблюдается редко
Значительный
Низкая
Нередко преобладают ценные
промысловые виды (представители
лососевых и сиговых)
Эвтрофные
Мелкие
Не содержится
Низкое видовое разнообразие,
большая численность, высокая
продуктивность; часто доминируют сине-
зеленые водоросли (цианобактерии)
Наблюдается часто
Незначительный
Высокая
Обычно доминируют «сорные» рыбы
(например, окунь, плотва, карп и
др)
Примечание В классическом варианте процесса естественной эвтрофизации недавно образовавшееся глубокое озеро
(например, при отступлении ледника) содержит мало питательных веществ, поскольку вещества эти еще не
высвободились из материнских и осадочных пород территории водосбора. Первичная и вторичная продукция в таком озере низка,
прозрачность высокая, а кислородный режим благоприятный. Со временем при разрушении и выщелачивании пород
приток биогенных элементов увеличивается, первичная и вторичная продукция возрастают, на дне накапливаются
органические и неорганические осадки и озеро мелеет. Прозрачность снижается, а в гиполимнионе к концу лета
наблюдается дефицит кислорода. Дистрофным называют озеро, которое основное количество органических веществ,
синтезированных растениями, получает с суши Этим обусловливается коричневый цвет воды в дистрофных озерах.
Располагаются такие озера обычно среди торфяных болот и постепенно сами превращаются в болота
результате хозяйственной деятельности человека,
правильнее называть «культурной» (или
«антропогенной») эвтрофизацией. Возникает он обычно в
результате нарушений в круговоротах азота и
фосфора. Антропогенные воздействия приводят к тому,
что количество вещества, вовлекаемого в активную
фазу цикла, начинает превышать то, которое
соответствует саморегулирующимся (гомеостатическим)
возможностям системы по крайней мере тем, что
имеются у них в настоящее время. Антропогенная
эвтрофизация создает серьезные экономические и
экологические проблемы. Вода хорошего качества
необходима для многих технологических процессов,
для питья человеку и домашним животным, для
коммерческого и любительского рыболовства, а
также для поддержания водных путей в
удовлетворительном состоянии (см. табл. 12.5).
Нитраты и фосфаты наиболее часто выступают в
качестве питательных веществ, ограничивающих
увеличение первичной продуктивности водных
экосистем. Добавление нитратов и фосфатов
способствует, таким образом, увеличению численности
быстро растущих и являющихся более сильными
конкурентами водорослей, например Oscillatoria ги-
bescens, и уменьшению общего видового
разнообразия фитопланктона. Подобные процессы в озерах
нередко приводят к непропорциональному
возрастанию доли несъедобных водорослей, т.е.
водорослей, неподходящих в качестве пищи для консу-
ментов. Эта особенность доминирующих
водорослей, а также более длительный жизненный цикл
консументов (и соответственно - большее время
реакции их на изменения в среде), приводят к тому,
что не вся возросшая первичная продукция
потребляется консументами. Отсюда и включение
излишка образовавшегося органического вещества
непосредственно в процессы разложения
(минерализации). Поскольку для разрушения сложных
органических веществ (и превращения их в простые
минеральные) требуется кислород, содержание этого
газа, растворенного в толще воды, может снизиться
ниже уровня, необходимого для успешного роста и
размножения таких чувствительных видов, как
лосось или форель (Salmo spp.). B некоторых
крайних случаях гибель рыб и последующее разложение
их трупов приводит в свою очередь к дальнейшему
снижению концентрации кислорода и ухудшению
всей ситуации (по принципу положительной
обратной связи). Непосредственные эффекты такой
ситуации могут наблюдаться в географически
отдаленных областях. Наличие зон пониженного
содержания кислорода в реке бывает достаточно для того,
чтобы даже при отсутствии других
неблагоприятных воздействий нарушилось размножение таких
мигрирующих видов, как лососи или угри.
Проблема снижения концентрации
растворенного кислорода при эвтрофизации глубоких озер
усугубляется наличием сезонной термической
стратификации (т.е. расслоением водной толщи на эпи-
Таблица 12.5. Основные проявления эвтрофизации в
экосистемах и связанные с этим проблемы водопользования
Проявления
1. Уменьшается видовое разнообразие и меняется состав
доминирующих в биоте видов
2. Возрастают биомасса растений и животных
3. Уменьшается прозрачность водоема
4. Возрастает скорость седиментации, укорачивается
продолжительность жизни озера
5. Могут формироваться бескислородные зоны
Проблемы
1. Очистка питьевой воды усложняется, и поступающая
к потребителю вода может иметь неприятный вкус или
запах
2. Вода может быть опасна для здоровья
3 Вода утрачивает благоприятные для потребителя
качества
4. Увеличившаяся масса растительности уменьшает
проточность и препятствует навигации
5. Важные в коммерческом отношении виды (например,
лососевые и сиговые рыбы) исчезают
лимнион - хорошо перемешиваемый и
прогреваемый поверхностный слой, термоклин, в пределах
которого наблюдается резкое падение температуры
с глубиной, и гиполимнион-глубинные воды,
сохраняющие низкую температуру с весны до осени).
В умеренных широтах термическая стратификация,
препятствующая проникновению кислорода в
гиполимнион, устанавливается обычно в начале лета и
держится до осени, а затем нарушается в результате
охлаждения воды у поверхности и усиления
ветрового перемешивания. Вода впадающих в озеро рек
обычно теплая (так как реки мелкие), поэтому
растворенные в речной воде питательные вещества и
кислород попадают только в эпилимнион.
Животные же, чувствительные к изменениям температуры,
обычно обитают только в гиполимнионе. Масса
фитопланктона, образовавшаяся в прогреваемом и
богатом питательными веществами эпилимнионе,
при отмирании попадает в гиполимнион, где в
процессе ее разложения потребляются и без того
ограниченные здесь запасы кислорода. Продувание
воздухом гиполимниона может компенсировать
возникший дефицит кислорода и тем самым
сохранить обитающих там животных. Однако, если не
следить внимательно за складывающейся
ситуацией, то в конце лета может произойти внезапная
катастрофическая гибель рыб вследствие
наступившего дефицита кислорода.
Анализ конкретного случая
В бродах * Норфолка содержание нитратов и
особенно фосфатов очень сильно возросло по сравне-
1 «Броды» (broads)-это заброшенные торфяные
выборки, затопленные водой в конце XIII-начале XIV
веков. Многие «броды» соединены узкими неглубокими
протоками с речной системой. Отсюда и частая
практика изолирования их дамбами.
Таблица 12.6. Уровень фосфатов в озерах и «бродах» Норфолка начиная с 1800 г.
Суммарные данные
Чистое озеро около истока
Естественным образом обогащенное нижележащее озеро
Броды Норфолка до 1800 г.
Броды Норфолка в XIX в. после изолирования их дамбами
Броды Норфолка в XX в. после возрастания численности населения
и перехода к современным методам сброса стоков
Концентрация фосфатов, мкг л i
5
10-30
10-20
до 80
макс. 2000
обычно 150-300
По В. Moss (1979) Alarm call for the Broads (Geographical Magazine)
Изменение содержания фосфатов в Бартон-Брод (Норфолк) и основные их источники
Дата
1800
1900
1920
1940
1975
Концентрация в воде, мкг л i
13,3
52,0
72,0
119,0
361,0
За счет сброса сточных вод, мкг л 1
0
(раздельных данных нет)
(раздельных данных нет)
72
287
Сток с суши, мкг л i
13,3
47
74
По В. Moss, 1980, Ecology of fresh waters, Blackwell Scientific Publications
нию с уровнем конца прошлого века, причем
наиболее заметные изменения произошли за последние
40 лет (табл. 12.6). В то же время чистые прозрачные
воды, характеризующиеся низкой продуктивностью
фитопланктона и развитием прикрепленной
растительности (например, водорослей Chara spp.),
сменились водами непрозрачными, богатыми
фитопланктоном, в которых произрастали только немногие
прикрепленные растения. В 1973 г. прозрачность
воды в Бартон-Брод, измеренная с помощью диска
Секки1, составляла только 11см. Хотя большая
часть обитавшей здесь до 1800-х годов фауны
(включая ценные виды промысловых рыб) позднее
исчезла, к середине двадцатого века местность эта
снова приобрела славу мирового центра
спортивного рыболовства. Продолжающийся, однако,
быстрый рост количества поступающих биогенных
элементов, особенно фосфора, а также воздействие
целого комплекса других факторов, означают
реальную угрозу рыболовству. Дополнительные
проблемы возникают из-за периодического
затопления территории морской водой, которое к тому же
усугубляется эрозией берега, нарушением структуры
донных отложений и уничтожения тростниковых
зарослей, не только защищающих речные берега, но
и создающих удобные места для нереста рыб.
Разрушение поясов прибрежной полупогруженной
растительности (тростник, рогоз, камыш и т. д.)
происходит вследствие расширяющейся и меняющей свой
1 Диск, разделенный на 4 сектора, выкрашенных в
черный и белый цвета. Опуская диск Секки на
размеченной веревке, отмечают ту глубину, на которой
нельзя уже более различить его контрастных секторов.
Данная глубина используется как показатель
прозрачности.
характер рекреационной нагрузки, в частности все
большим вытеснением парусного спорта катанием
на различного рода моторных лодках.
Недавние эксперименты, проведенные на
«бродах» Англии, свидетельствуют о том, что эвтрофи-
зация может быть приостановлена и даже как бы
обращена вспять. Так, например, когда два брода-
Алдерфен и Кокшут-отгородили от речной системы
запрудами, поступление в них воды, богатой
биогенными элементами, прекратилось. В Алдерфене
ничего более не предпринималось и вслед за
наблюдавшимся вначале улучшением ситуации
(увеличением прозрачности и т. п.) состояние водоема снова
ухудшилось. Произошло это вследствие сложного
взаимодействия процессов, протекающих, с одной
стороны, в донных отложениях, а с другой - в толще
воды при разложении фитопланктона, давшего
весной вспышку численности («цветение»).
Результатом этого взаимодействия было высвобождение
фосфатов из богатых биогенными элементами
донных отложений. Если сразу после запруживания
протоков рост водорослей в Алдерфене
лимитировали нитраты, то позднее в нем развились формы,
фиксирующие атмосферный азот, и летом 1985 г.,
спустя шесть лет после начала эксперимента,
прозрачность воды снова стала крайне низкой из-за
очень большого количества содержащихся в ней
водорослей и цианобактерий. При отделении
запрудой другого брода-Кокшута-было проведено
драгирование и очистка водоема от богатых
фосфором донных отложений. Постепенно здесь
сформировались заросли прикрепленной водной
растительности и возросла численность рыб. Стоимость
драгирования Кокшута оценивалась в 75 тыс. фунтов
стерлингов, но для большинства бродов, учитывая
их размеры, она должна составлять примерно
250 тыс. фунтов стерлингов. К сожалению, во
многих случаях действующие правила судоходства
запрещают сооружение дамб и изоляцию бродов,
особенно если это не временная мера.
Последствия эвтрофизации и меры
борьбы с ней
Отделение водоемов от питающей их речной
системы далеко не всегда осуществимо практически, но
описанные выше эксперименты показывают, что
улучшение сложившейся в таких водоемах ситуации
реально возможно, хотя и достижимо порой только
довольно сложным путем. Эти эксперименты
помогают также лучше понять суть процесса
эвтрофизации и возможных способов борьбы с ним. Изыма-
ние фосфатов из сточных вод сейчас нередко
осуществляется в местах сброса стоков. Основывается
оно на химическом осаждении фосфора известью и
солями железа. Подобная процедура, особенно если
она сопровождается предварительным
драгированием, может заметно облегчить очистку протоков.
Вынутые из водоемов донные отложения могут с
успехом использоваться в качестве удобрения, по
крайней мере при установлении на них льготных
цен. В результате драгирования озёра и каналы
углубляются, поэтому проточность всей системы
улучшается, а толща воды обильно снабжается
кислородом. В более глубоких стратифицированных
озерах при этом увеличивается объем гиполимниона
и соответственно растут запасы растворенного
кислорода. Углубление водоемов приводит также к
установлению в них стратификации, к ограничению
тем самым притока биогенных элементов в эпилим-
нион, а следовательно, и к сокращению продукции
фитопланктона. Содержание биогенов в воде может
быть уменьшено и такими способами, как
изолирование дна от толщи воды (например,
полиэтиленовой пленкой), или же химическое осаждение
(например, с помощью сульфата алюминия).
«Цветение» водоемов может также
непосредственно подавляться альгицидами - химическими
веществами, убивающими водоросли, но способ этот
отнюдь не бесспорный, так как нельзя исключить
возникновения трудно предсказуемых
отрицательных последствий для других видов организмов.
Серьезную экологическую проблему
представляет собой эвтрофизация водохранилищ. Так,
например, водоросли могут забивать фильтры
водозаборных сооружений, что сильно ограничивает
пропускную способность станций очистки и порой даже
требует их временной остановки для смены
фильтров. Мелкие водоросли могут некоторое время
существовать в водопроводных системах; в
результате разложения этих водорослей потребляемая
вода порой приобретает неприятный вкус и запах, что,
однако, еще не свидетельствует о ее вредности.
Большая часть усилий по борьбе с эвтрофиза-
цией направляется на сокращение количества
сбрасываемых в воду фосфатов. Делается это главным
образом потому, что фосфаты попадают в водоемы
в легко выявляемых местах («точечных источниках»
загрязнения1). Что же касается избыточного
количества нитратов, то их источник-это по сути дела
весь сток с суши, охваченной сельскохозяйственным
производством. Таким образом, поступление азота
в водные системы рассредоточено по большой
площади и контроль за его поступлением может
осуществляться не иначе как через радикальные
перемены во всей системе ведения сельского хозяйства.
Уровень нитратов в питьевой воде вызывает,
однако, серьезную озабоченность. Европейские
санитарные нормы устанавливают пороговую
концентрацию нитратов в питьевой воде, равную 50 мг-л"1,
хотя и допускают ее подъем до 100 мг-л"1. В
низменной части Англии речная вода обычно
содержит нитраты в концентрации, большей чем
100 мг-л"1, и поэтому для нужд питьевого
водоснабжения ее необходимо предварительно
смешивать с водой, содержащей малое количество
нитратов.
Мониторинг эвтрофизации
Связанные с эвтрофизацией изменения в водоеме
могут быть подвергнуты биологическому и
химическому мониторингу. Полученные при этом данные
позволяют предпринять определенные
оздоровительные меры, не допуская возможности
катастрофического разрушения экосистемы. Изменения
видового состава фитопланктона могут
свидетельствовать о начавшейся эвтрофизации. Характерными
примерами являются случаи «цветения»
сине-зеленых водорослей (цианобактерий): Oscillatoria rubes-
cens в оз. Вашингтон и Anabaena flos-aquae в оз.
Лох-Ней. В табл. 12.7 приведены группы
планктонных организмов (водорослей и цианобактерий),
типичных для олиготрофных и эвтрофных озер.
Обычно для эвтрофных вод характерна высокая
численность фитопланктона и малое видовое
разнообразие. Ссылки на руководства, позволяющие
определить планктонные организмы, можно получить от
Philip Harris Ltd.2 При другом варианте
мониторинга прослеживаются во времени изменения содержа-
1 Фосфатам (и фосфору вообще) при борьбе с
эвтрофизацией в пресных водах уделяется также
первоочередное внимание потому, что важная группа
продуцентов-цианобактерий (или, как их раньше
называли, «сине-зеленые водоросли») - обладают
способностью фиксировать для своих нужд азот воздуха, т. е.
использовать практически неограниченные его запасы.
Фосфор оказывается при этом единственным ресурсом,
лимитирующим рост численности цианобактерий -
Прим. ред.
2 Нашему читателю можно предложить,
например, такое руководство как «Определитель
пресноводных беспозвоночных Европейской части СССР»,/Л.:
Гидрометеоиздат, \911.-Прим. ред.
Таблица 12.7. Ассоциации водорослей (и цианобактерий),
характерных для планктона олиготрофных и эвтрофных
озер
Олиготроф-
ные озера
Эвтрофные
озера
Характерная группа
Десмидиевые
Хризофитные
Диатомовые
Динофлагеллаты
Хлорококковые
Диатомовые
Динофлагеллаты
Хлорококковые
Миксофиты
(цианобактерий)
Примеры
Staurodesmus,
Staurastrum
Dinobryon
Cyclotella, Tabellaria
Peridiniwn, Ceratium
Oocystis
Asterionella, Fra-
gillaria crotonensis,
Stephanodiscus
astraea, Melosira
granulata
Peridiniwn bipes,
Ceratium, Glenodi-
nium
Pediastrum, Scene-
desmus
Anacystis, Aphanizo-
menon, Anabaena
Из: Mason C F. (1981), Biology of freshwater pollution,
Longman
ния хлорофилла а как показателя общей биомассы
водорослей. В летнее время средние значения
биомассы фитопланктона в олиготрофных озерах
составляют 0,3-2,5 μγ·μ-3, а в эвтрофных-5-
140 мгм"3.
Полезным индикатором качества воды в реках
может быть видовой состав крупных
беспозвоночных животных. Простая пятиуровневая схема
такого анализа приведена в табл. 12.8. При сборе
проб, необходимых для проведения подобного
анализа, следует позаботиться о стандартизации всей
процедуры и обеспечении достаточно
репрезентативной выборки из каждого обследуемого
местообитания. В Великобритании специалисты водного
хозяйства используют чаще всего две принятые
системы оценки качества воды: индекс Ф. Вудевиса
(известный также как «биотический индекс реки
Трент»; табл. 12.9) и показатель Чендлера (или
«баллы Чендлера»). Индекс Вудевиса учитывает
наличие или отсутствие «ключевых» видов и общее
число видов, но не требует знания численности
разных видов. Подобный способ расчета индекса
позволяет быстро определить его величину, но
вместе с тем величина эта может существенно
зависеть от случайного попадания (или непопадания) в
пробу того или иного чувствительного к
загрязнению вида. При расчете показателя Чендлера обилие
каждого отдельного вида оценивается по
5-балльной шкале. Недостаток этого индекса в том, что
оценка самого обилия в каждом случае довольно
произвольна. Но поскольку в принципе все
организмы в пробе могут быть не только определены, но и
подсчитаны, реальные возможности показателя
Чендлера больше, чем у индекса Вудевиса. Полное
описание обоих методов, включая разбор примеров,
можно найти в книге: Mason CF. (1981), Biology of
Freshwater Pollution, Longman.
Полезным химическим показателем степени
эвтрофизации может быть величина биохимического
потребления кислорода (БПК). Величина эта,
оценивающая скорость поглощения кислорода
организмами, отражает активность микроорганизмов,
которые разлагают содержащееся в воде органическое
вещество (как уже упоминалось, содержание
органического вещества обычно возрастает при
обогащении воды биогенными элементами в
минеральной форме). Потребление кислорода в процессе
дыхания водорослей также неизбежно должно
включаться в итоговую величину БПК. В большинстве
случаев оно практически не имеет особого значения,
но иногда на дыхание водорослей приходится до
50% общего БПК. Величина БПК, следовательно,
довольно приблизительный показатель качества
воды, использовать который наиболее целесообразно
в комбинации с другими индексами. Стандартная
оценка БПК дается обычно как количество
кислорода (в мг), поглощенного из пробы воды объемом
1 л, помещенной в темноту на 5 сут при температуре
200C. Чистая речная вода характеризуется, как
правило, величиной БПК порядка Змг-л"1, вода из
сильно загрязненной реки-10 мгл-1, а типичные
значения БПК для бытовых стоков равны 250-
350 мгл-1 (см. табл. 12.10).
Уменьшение содержания кислорода в реке при
сбросе в нее богатых органическим веществом сточ-
Таблица 12.8. Пятиуровневая шкала для оценки загрязнения вод по данным о присутствии или отсутствии индикаторных
видов
Уровень загрязнения
А. Чистая вода или очень низкий уровень
загрязнения
Б. Низкий уровень загрязнения
В. Высокий уровень загрязнения
Г Очень высокий уровень загрязнения
Д. Крайне высокий уровень загрязнения
Концентрация кислорода
Высокая
Низкая
Кислород отсутствует
Индикаторные организмы
Нимфы веснянок,
нимфы поденок
Нимфы ручейников.
пресноводные креветки
Водяной ослик,
мотыль (личинки хирономид)
Трубочник (черви тубифициды),
«крыска» (личинка мухи-журчалки)
Заметных признаков жизни нет
Таблица 12.9. Индекс Вудевиса («Биотический индекс реки Трент»)
Итоговая таблица
Чистая вода
; в по-
iepe воз-
нения
Организмы, перечисленные
рядке их исчезновения по м
растания степени загряз!
Сильно
загрязненная вода
Виды-индикаторы
Присутствуют нимфы
веснянок (Plecoptera)
Присутствуют личинки
поденок (Ephemeroptera)
Присутствуют личинки
ручейников (Trichoptera)
Присутствует Gammarus
Присутствует Asellus
Присутствуют тубифициды
и(или) личинки мотыля
Все перечисленные выше
группы отсутствуют
Более одного вида
Только один вид
Более одного вида1*
Только один вид1*
Более одного вида2)
Только один вид2)
Все перечисленные выше
виды отсутствуют
То же
» »
Некоторые организмы,
например личинки
Eristalis tenax, не требующие
растворенного в воде
кислорода, могут присутствовать
Общее число присутствующих
групп
0-1
2-5
6-10
11-15
16 +
Значения индекса
—
—
—
—
4
3
2
1
0
7
6
6
5
5
4
4
3
2
1
8
7
7
6
6
5
5
4
3
2
9
8
8
7
7
6
6
5
4
—
10
9
9
8
8
7
7
6
—
—
u Исключая Baetis rhodani
2) Baetic rhodani (Ephemeroptera) учитывается в этой графе
Перечень групп
Термин «группа» в данном случае просто уровень, до
которого ведется определение.
Группы эти следующие:
Любой известный вид плоских червей (Platyhelminthes)
Кольчатые черви (Annelida) за исключением рода Nais
Род Nais
Любой известный вид пиявок (Hirudinae)
Любой известный вид моллюсков (Mollusca)
Любой известный вид ракообразных (Asellus, креветки)
Любой известный вид веснянок (Plecoptera)
Любой известный вид поденок (Ephemeroptera), за
исключением Baetis rhodani
Поденка Baetis rhodani
Любое семейство ручейников (Trichoptera)
Любой вид сетчатокрылых (Neuroptera)
Личинки хирономид (Chironomidae), за исключением
мотыля Chironomus thummi
Личинки Chironomus thummi (мотыль)
Семейство мошки (Simuliidae)
Личинки любых других видов двукрылых
Любые виды жуков и их личинки (Coleoptera)
Любые виды водяных клещей (Hydracarina)
Максимальная величина индекса равна 10. Более низкие
значения соответствуют некоторому загрязнению.
Наибольшая степень загрязненности-0.
Процедура подсчета:
1. Животные из взятой пробы сортируются по группам
(согласно приведенному списку). Подсчитывается
общее число групп.
2. Отмечаются виды-индикаторы, начиная с верхней
части списка.
3. Для того чтобы определить величину индекса, берется
вид наибольшего ранга из присутствующих, например
Trichoptera, и отмечается число групп (из приведенного
списка). Таким образом, индикаторные виды
определяют строку таблицы, а общее число обнаруженных
групп-столбец. Считываемая на пересечении цифра
и есть значение индекса.
Пример: Вид-индикатор, занимающий наивысший ранг,
относится к ручейникам (Trichoptera); ручейников более
чем один вид в пробе;
Общее число групп-7.
Соответствующее значение индекса в таблице равно 6.
Таблица 12.10. А. Руководство по определению качества
воды по величине БПК. Б. Сравнение оценок величины БПК
в некоторых типичных промышленных стоках
А.
БПК,
мгл
1
2
3
5
10
20
Качество реки
Очень хорошее
Хорошее
Довольно
хорошее
Сомнительное
Плохое
Очень плохое
БПК
стоков,
мг ·л ~'
600
350
200
20
Качество стоков
Сильное загрязнение
Умеренное
загрязнение
Слабое загрязнение
Максимальный
уровень БПК в стоках,
допускаемый
Королевской комиссией и
требующий
10-кратного разбавления в
реке
Б. Тип стока
(вид хозяйственной деятельности)
Хлопчатобумажная
Кожевенная
Прачечная
Пивоваренная
Винокуренная
Молочная
Консервная:
переработка бобовых
переработка фруктов
(цитрусовых)
Стоки ферм
Отходы при силосовании
Бумажная
Коксовая печь
Маслоочистка
5-суточн БПК,
мг-л~х
200-1000
1000-2000
1600
850
7000
600-1000
570
2000
1000-2000
50000
100-450
780
100-500
ных вод-процесс медленный, поэтому то место, где
наблюдается наиболее выраженный дефицит
кислорода, может находиться значительно ниже по
течению от места сброса сточных вод.
12.15. Перечислите факторы, определяющие
степень дефицита кислорода.
В р. Темза в 1967 г. в сравнительно маловодный
осенний период зона пониженного содержания
кислорода начиналась сразу за Лондонским мостом и
тянулась на 40 км, тогда как весной при высокой
скорости течения зона эта начиналась только в
22 км ниже моста, а протяженность ее составила
12 км.
12.8. Земледелие
Выдающиеся успехи в сельском хозяйстве были
достигнуты за последние 20 лет благодаря
широкому внедрению научно обоснованных
высокоразвитых технологий. Ряд примеров использования
таких технологий приводится ниже.
12.8.1. Вспашка
Цель этой процедуры - дробление комков почвы и
создание удобных мест для прорастания семян
посредством перевертывания или закапывания
дерновины. Природный травянистый покров уступает при
этом место искусственно создаваемым агроценозам.
В Великобритании вспашка проводится обычно
осенью, а зимние морозы способствуют
разрыхлению почвы.
Изогнутые стальные лезвия плуга
протаскиваются трактором через толщу почвы. Устанавливаемые
сейчас на тракторах гидравлические устройства
автоматически контролируют глубину вспашки.
Затем используются хорошо разламывающие
твердую почву дисковые плуги, а окончательная
подготовка пашни включает также боронование
(дисковыми или рамочными боронами).
12.8.2. Сев
После уборки стерня обрабатывается гербицидами,
таким, например, как паракват, с целью
уничтожения всех сорняков. Используя сеялку, способную
взламывать ненарушенную почву, можно
непосредственно производить посев в нераспаханную землю
и тем самым избежать расходов на вспашку
(сэкономив, например, горючее).
12.8.3. Севооборот
Исторически сложилось так, что земледелие в
Великобритании основывается на трехгодичном цикле:
озимая пшеница сменяется яровой, а затем поле
оставляют на год невозделанным, чтобы сохранить
или восстановить утраченное плодородие. По мере
развития производства корнеплодов и осознания
важности бобовых (как фиксаторов азота и
создателей «зеленого удобрения») начал формироваться
четырехгодичный цикл, включающий
последовательное выращивание на одном поле корнеплодов,
пшеницы дикорастущих трав (или бобовых) и
ячменя. Иногда этот цикл модифицируют, включая в
него более широкий спектр возделываемых культур,
например капусту или сахарную свеклу.
Чередование культур не только поддерживает плодородие
почвы, но также препятствует развитию вредителей.
Однако с введением в практику сельского хозяйства
пестицидов стало возможным возделывание
монокультур. При этом достигается очень интенсивное
сельскохозяйственное производство за счет
успешного выращивания из года в год одной культуры на
том же самом участке земли.
На впервые осваиваемых (а в развивающихся
странах и на давно освоенных) территориях может
использоваться система, известная под названием
подсечно-огневого земледелия. Такая система
основывается на вырубке и выжигании всей
существующей на данной территории растительности, что
приводит к возвращению в почву важных
минеральных компонентов питания. Расчищенная подобным
образом площадь используется для земледелия
непрерывно в течение нескольких лет, до тех пор пока
плодородие почвы не падает. Затем участок этот
забрасывается по крайней мере на несколько лет, а
выращивание сельскохозяйственных культур
переносится в новые места. Земледелие такое обычно
смешанное, т. е. несколько разных культур
выращиваются бок о бок на одном участке.
12.8.4. Сбор урожая
Способ сбора урожая определяется типом
выращиваемой культуры и может варьировать от жатвы с
помощью самых современных комбайнов до
ручного сбора фруктов. Урожай собирается тогда, когда
по опыту он достигает необходимой стадии
зрелости. При этом в случае зерна учитывается
содержание в нем влаги, а в случае сбора фруктов
необходимо предусмотреть те изменения, которые
произойдут в плодах до момента попадания их на
рынок. При современной технологии зерно, однако,
может быть искусственно высушено до
необходимого уровня. Собранный урожай отправляется либо
непосредственно покупателю или на
перерабатывающую фабрику (так поступают, например, с
сахарной свеклой), либо помещается в хранилище.
12.8.5. Хранение
Основные задачи, возникающие при хранении
урожая, связаны с предотвращением (или хотя бы
сокращением масштабов) его порчи в ходе
ожидания момента, удобного для продажи. Главные
потери определяются вредом, наносимым насекомыми и
грибами. Особенности хранения зависят от вида
продукции; это могут быть силосные башни и
элеваторы для злаковых, бурты для картофеля и
корнеплодов, специальные хранилища для фруктов.
Было выяснено, что продолжительность
хранения фруктов находится в обратной зависимости от
интенсивности их дыхания, которую можно снизить,
помещая фрукты в холодильник или в некоторых
случаях продувая хранилище воздухом,
обогащенным диоксидом углерода (10%). Ускорить же
созревание и сократить время хранения можно, используя
этилен, который, кстати сказать, выделяется самими
зреющими фруктами. В воздух, циркулирующий в
картофелехранилищах, добавляются также
различные летучие вещества, препятствующие
прорастанию клубней, а в хранилищах фруктов
используются фунгициды - вещества, подавляющие рост
грибов.
12.8.6. Контроль за сорняками
Сорняк можно определить как растение,
произрастающее «не там, где надо». Для сорняков
характерно быстрое прорастание и рост, кроме того, они
продуцируют большое число семян, способных
долго находиться в покоящемся состоянии и сохранять
при этом хорошую всхожесть. Взрослые сорняки
бывает трудно уничтожить, поскольку наличие
длинного стержневого и боковых придаточных
корней, а также быстрая регенерация нарушенных
тканей позволяют им эффективно противостоять
обработке мотыгой, выеданию фитофагами и действию
контактных гербицидов. Благодаря таким
свойствам сорняки способны успешно конкурировать с
культурными растениями (особенно на стадии
проростков) за влагу, биогенные элементы и свет, что
приводит к значительному снижению урожая. На
стадии уборки урожая сорняки, кроме того,
засоряют зерно своими собственными семенами.
12.8.7. Гербициды
Одной из задач, давно стоявшей перед
сельскохозяйственной наукой, было увеличение урожая
зерновых культур с помощью растительных гормонов.
Использование ауксина привело к открытию герби-
цидного действия феноксиуксусных кислот, таких,
например, как метилхлорфеноксиуксусная кислота.
В настоящее время разработано много различных
гербицидов, значительно более совершенных, чем
старые «контактные» гербициды вроде хлората
натрия. Эти новые вещества, называемые «системными
гербицидами» (поскольку они переносятся по
проводящим системам растений), обладают нередко
селективным действием. Так, например, паракват и
дикват, проникая внутрь растения, воздействуют на
процесс фотосинтеза и могут быть использованы
для уничтожения сорняков, находящихся среди
древесных насаждений. Другие гербициды, например
атразин и симазин, остаются в почве и
препятствуют прорастанию семян.
12.8.8. Инсектициды
Действие инсектицидов, как и других пестицидов,
чаще всего основано на ингибировании каких-либо
метаболических процессов. Инсектициды весьма
значительно различаются по своему составу,
эффективности, способу и скорости воздействия, а также
по применяемым дозам. В идеальном варианте
диапазон их действия должен быть ограничен
строго определенной группой организмов (табл. 12.21).
12.8.9. Пестициды и окружающая
среда
Пестициды - это химические вещества,
используемые для борьбы с вредителями, т. е. организмами,
которые, по мнению человека, непосредственно за-
Таблица 12.21. Примеры трех основных типов инсектицидов и характеристика их использования
Тип
1. Контактные
(проникают через кутикулу
насекомых)
2. Системные
(проникают через
пищеварительный тракт. Часто яды,
действующие на кишечник.
Некоторые препятствуют
прохождению нервных импульсов)
3. Фумиганты
(вещества, которые, будучи
распыленными в воздухе,
уничтожают насекомых)
Примеры
а) Природные
Никотин (из листьев табака)
Пиретрум (экстракт из цветков)
Chrysanthemum sp.
Ротенон (из корней Derris)
б) Синтетические
Вещества, в настоящее время
широко разрабатываемые и
постоянно модифицируемые по мере того,
как возникают в результате отбора
новые устойчивые к ним расы
насекомых; примеры: ДДТ, гаммаксан,
тиофос, карбофос
Некоторые из контактных ядов
используются как системные
инсектициды, например тиофос и ДДТ
Формальдегид, оксид этилена
Примечания
Применяются для уничтожения
насекомых (в частности, тлей), которые
прокалывают покровные ткани
растений для того, чтобы сосать из
них соки
Первые использованные вещества -
это хлорированные углеводороды
(ДДТ), но они оказались очень
токсичными и устойчивыми во
внешней среде. В настоящее время
в ряде стран запрещены и заменены
фосфорорганическими
соединениями (например, такими, как тиофос)
Используются против насекомых с
кусающими ротовыми частями.
Могут применяться в кронах против
листогрызущих насекомых или в
в составе ядов против саранчовых
Используется в теплицах
трагивают его интересы. Таким образом,
пестициды-широкое понятие, включающее в себя
гербициды, уничтожающие растения; инсектициды,
уничтожающие насекомых; фунгициды, уничтожающие
грибы и т.д. Большинство пестицидов - яды,
вызывающие гибель нежелательных видов, но, как
правило, к пестицидам относят также ингибиторы
роста, и вещества, приводящие к стерилизации
организмов. В Великобритании пестициды обычно
используют в сельском хозяйстве при выращивании
урожая; однако широко используются они и для
защиты запасов продуктов питания, а также
древесины, шерсти и других природных материалов. Во
многих странах пестициды широко применяются в
лесном хозяйстве и при борьбе с переносчиками
различных заболеваний человека и животных (см.,
например, разд. 12.3.8). Пестициды используют по
всему свету в самых разнообразных
местообитаниях. Наиболее стойкие из них распространяются по
всему земному шару, и их обнаруживают даже там,
где они никогда не применялись, например в
открытом океане или в высоких широтах. Таким образом,
большая часть живых существ на земле
контактирует с пестицидами. Конечно, человек давно уже
употреблял природные органические пестициды
(примером может служить пиретрум, получаемый
из растения с таким же названием) или некоторые
неорганические вещества, например CuSO4
(входящий в состав бордосской жидкости, используемой в
виноградарстве), но наиболее широкое
распространение новых синтезированных органических веществ
(а к ним относится примерно 90% всех известных
пестицидов) - это уже завоевание эпохи,
наступившей после второй мировой войны. Понятно, что по
всей планете организмы любого вида не
сталкивались в ходе своей эволюции с такими веществами,
которые теперь фактически стали новым фактором
среды. Экологические последствия применения
пестицидов предсказать трудно, что делает задачу
изучения их чрезвычайно важной и интересной. Пока, к
сожалению, мы не можем до конца оценить
побочные результаты широкого применения пестицидов,
такие, например, как уничтожение видов, с
которыми не предполагалось вести борьбу, выработка
устойчивости к пестицидам у видов-вредителей и
хроническое воздействие пестицидов на
человеческие популяции.
Наиболее важные экологические характеристики
пестицидов - это токсичность, устойчивость,
неспецифичность и независимость силы воздействия от
плотности контролируемой популяции.
Токсичность и устойчивость взаимосвязаны; проявляется
это в том, что вещество, вызывающее летальный
исход, но неустойчивое во внешней среде, со
временем может оказаться менее эффективным, чем
вещество, обладающее сублетальным действием, но
являющееся более устойчивым. Объясняется это
тем, что у такого устойчивого, но не очень
ядовитого вещества больше шансов включиться в
пищевые цепи и превратиться в ходе метаболизма в более
токсичную форму, или же (что более вероятно) -
накопиться в верхних звеньях пищевых цепей (см.
разд. 12.3.8). Применение пестицидов в форме
распыляемых порошков и разбрызгиваемых жидкостей
приводит к их широкому распространению в воде и
в воздухе, а тем самым и к включению в пищевые
цепи там, где их применение даже и не
предполагалось. Хлорорганические соединения (например,
ДДТ), отличающиеся высокой устойчивостью и
растворимостью в жирах, пожалуй, наиболее
распространенные из всех пестицидов. Следы хлор-
органических соединений обнаружены не только в
организмах всех видов птиц, обитающих на
Британских островах, но и, что гораздо более
знаменательно,-в телах пингвинов, обитающих в
Антарктиде.
Устойчивость рассматривается обычно как
нежелательное свойство пестицидов, особенно если
они попадают в продукты питания, однако в
некоторых случаях, например при борьбе с паразитами
или болезнетворными организмами, обитающими в
почве, определенная степень устойчивости
оказывается абсолютно необходимой по ряду чисто
технических и экономических соображений.
Токсичность для какого-либо конкретного вида
организмов обычно определяется как 50%-ная летальная
доза (ЛД50). Это - однократная доза введенного per
os вещества, достаточная для того чтобы погибло
50% экспериментальной лабораторной популяции.
В природных условиях та же доза может привести к
гибели большей доли популяции, поскольку
организмы испытывают дополнительный стресс от
воздействия различных внешних факторов. Тем не
менее по определению некоторые особи выживают и в
этом случае. (Задачу использования пестицидов в
сельском хозяйстве кратко можно сформулировать
как снижение потерь урожая до некоторого уровня,
определяемого простым экономическим балансом
затрат и выгод.) К сожалению, выжившие особи
служат основой для становления устойчивых к
пестицидам популяций вредителей. Во всяком случае
среди насекомых с их коротким жизненным циклом
формирование способности противостоять
пестицидам и выходить из-под их контроля наблюдается
очень часто. Типичная реакция на подобные
явления-это разработка новых пестицидов, что
является делом дорогостоящим и, по мнению многих
экологов, довольно бессмысленным, так как
приводит к тем же самым результатам.
Большинство пестицидов открыто чисто
эмпирически методом проб и ошибок или же в результате
случайных наблюдений. Не всегда поэтому
известно, каким именно способом вызывают пестициды
гибель тех или иных организмов. Токсичность
определенного пестицида может сильно варьировать для
организмов, относящихся к разным
систематическим группам, а иногда существенно различаться
даже по отношению к близкородственным видам.
Так например, в случае ДДТ LD50 для овцы (при
расчете на единицу массы тела) больше, чем для
крысы. У некоторых видов представители разных
полов обладают различной чувствительностью к
пестицидам: так, для самцов мышей LD50 по ДДТ
составляет 500 мг-кг-1, а для самок-550 мг-кг"1.
Эти различия в реакции на пестициды у отдельных
организмов, особенно у представителей разных
таксонов, могут быть использованы практически: к
примеру, произрастающие среди посевов хлебных
злаков сорняки двудольных могут быть уничтожены
без нанесения заметного вреда возделываемым
растениям и потребляющим их людям. Блохи и другие
паразитические насекомые могут быть истреблены
пестицидами так, чтобы не пострадали при этом
хозяева-млекопитающие. Однако к заключению о
безвредности того или иного пестицида надо
подходить с крайней осторожностью. Недавние
сообщения о заболеваниях рабочих, имевших
многолетние контакты с фосфорорганическими
соединениями1, используемыми, например, в жидкостях
для уничтожения паразитов на овцах, заставляют
радикально пересмотреть установленные ранее
санитарные нормы. Пока, в силу недавнего появления
большинства пестицидов, еще крайне слабо изучены
долгосрочные последствия длительного
употребления даже малых доз пестицидов и, возможно, син-
эргические эффекты при контактах их с другими
загрязнителями или с переносчиками заболеваний.
В настоящее время все более возрастает
озабоченность по поводу того, что «безвредные» следовые
количества пестицидов (или продуктов их
переработки), остающиеся в продуктах питания, хотя и не
оказывают прямого токсического (не говоря уж о
летальном) воздействия, могут тем не менее
снижать устойчивость организмов к различным
заболеваниям или же, переходя по пищевым цепям,
накапливаться до опасных концентраций. В частности,
именно присутствие пестицидов (особенно так
называемых ПХБ - полихлорбифенилов) в Северном
море способствовало, по мнению многих ученых,
быстрому распространению летом 1988 г.
вирусного заболевания в популяции обыкновенного тюленя.
В практике сельского хозяйства серьезные
экономические проблемы могут возникать при
воздействии пестицидов не только на виды-вредители, но и
на виды, являющиеся хищниками по отношению к
данным вредителям. Хорошим примером может
быть история применения ДДТ для борьбы с
бабочкой репницей (Pieris rapae), гусеницы которой
наносят вред брюссельской капусте. Первоначально
использование пестицида казалось успешным, но
затем на обработанных территориях численность
гусениц стала даже превышать ту, что отмечалась
на контрольных (не обработанных) участках.
Данный эффект еще более усиливался при повторных
применениях ДДТ с целью регулирования новых
вспышек численности вредителя. Изучение всей этой
агроэкосистемы показало, что концентрация
пестицидов на листьях быстро снижалась, благодаря
1 Фосфорорганические соединения -это
воздействующие на нервную систему яды, находящие все
большее практическое применение вместо более устойчивых
хлорорганических соединений.
росту уже существующих листьев и образованию
новых. Однако содержание пестицидов в почве
оставалось высоким, особенно при запашке остатков
возделываемых растений. В результате яйца,
отложенные после распыления пестицидов взрослыми
особями, прилетевшими с других территорий,
оказывались вне опасности, тогда как главные
хищники, нападающие на гусениц репницы, а именно
обитающий в почве жук Harpalus rufipes и сенокосец
Phalangium opilio, существенно снижали свою
численность и пищевую активность. Таким образом,
пресс хищников значительно ослабевал, а
численность гусениц начинала превышать уровень,
наблюдавшийся до обработки пестицидами. Дальнейшее
использование ДДТ еще более усугубило эту
ситуацию.
Хищные виды обычно оказываются наиболее
уязвимыми по отношению к применяемым
пестицидам. Происходит это потому, что численность этих
видов ниже численности жертв-вредителей, и
соответственно затруднено восстановление их
популяций после каких-нибудь нарушений. Особенно плохо
положение тех хищников, которые питаются
животными, устойчивыми к пестицидам, но содержащими
небольшие количества их в жировой ткани. К тому
же, отравленные, умирающие, больные и
характеризующиеся нарушенным поведением особи гораздо
чаще становятся добычей хищников, чем здоровые
особи с нормальным поведением. Обычный
результат применения пестицидов-снижение видового
разнообразия. Продуктивность нижних
трофических уровней экосистемы при этом нередко
увеличивается, а более высоких-уменьшается. О том, как
пестициды влияют на редуцентов, известно пока еще
крайне мало. Почти не исследованным остается и
вопрос о воздействии пестицидов на круговорот
биогенных элементов и на плодородие почвы.
Серьезные проблемы, связанные с
использованием пестицидов, особенно проблемы, касающиеся
выхода вредителей из-под их контроля, а также
риска для здоровья человека, приводят к тому, что
очень быстро растет заинтересованность в развитии
альтернативных подходов к борьбе с вредителями.
Ниже кратко рассматриваются главнейшие из них;
это - биологический контроль вредителей и
интегрированная борьба с ними (осторожное, строго
направленное применение пестицидов в сочетании с
методами биологического контроля).
Биологический контроль
Под биологическим контролем за вредителями
традиционно понимается регуляция численности их
популяций естественными врагами: хищниками,
паразитами и различными патогенными организмами.
По сути дела такой контроль предполагает
управление популяциями, не допускающее бесконтрольного
экспоненциального роста численности видов
вредителей. Некоторые исследователи рассматривают
биологический контроль в более широком
понимании, включая в это понятие и другие методы,
например генетические. К биологическому
контролю можно отнести и методы контроля
сельскохозяйственных культур, а именно поддержание
севооборота, соответствующая обработка почвы,
использование смешанных культур, уборка
растительных остатков и установление сроков
сельскохозяйственных работ таким образом, чтобы они
оказались благоприятными для возделываемых культур и
популяций врагов вредителей, но не для самих
вредителей. В своем классическом варианте
биологический контроль наиболее успешно применяется
по отношению к завезенным видам, у которых в
новой среде нередко отсутствуют естественные
ограничивающие факторы, как биологические, так и
физические, связанные, например, с сезонными
изменениями климата.
Борьба с червецом Icerya purchasi, начавшаяся в
конце XIX в. на плантациях цитрусовых в
Калифорнии,-первый пример действительно удачного,
научно обоснованного биологического контроля за
видом-вредителем. Червец этот был завезен
случайно из Австралии, являющейся его родиной. В
результате полевых исследований, проведенных в
Австралии, были выявлены два природных врага
червеца, а именно паразитирующая на нем муха
Cryptochetum iceryae и хищная божья коровка Ro-
dolia cardinalis. Эти враги червеца были интродуци-
рованы в Калифорнию и после тщательного
изучения (в том числе - после контролируемого выпуска в
большие садки с цитрусовыми) выпущены в
насаждения цитрусовых. Оба вида-паразит и хищник-
быстро размножились и, распространившись, уже
через несколько месяцев стали эффективно
контролировать численность червеца Icerya purchasi.
Далеко не всегда, однако, удается так быстро
достичь успеха. Во всяком случае скрупулезный
анализ климатических условий и внимательное
наблюдение за взаимоотношениями завезенного вида
с аборигенными абсолютно необходимы. Так, при
попытках борьбы с ореховой тлей Chromaphis juglan-
dicola в Калифорнию было завезено из Канн
(Франция) паразитическое перепончатокрылое Trioxys
pallidus. Паразит этот прижился в прибрежных
районах Калифорнии, но в более жарких
континентальных районах (где как раз и достигала наибольшей
численности ореховая тля) он вымер в течение
одного сезона. Спустя десятилетие, потраченное на
напряженные поиски паразитов тли в жарких и
сухих местообитаниях, была ввезена в Калифорнию
новая линия Trioxys, на сей раз из Ирана. Паразиты
новой линии успешно перезимовали, и в течение
следующего года уже более 50 тыс. квадратных
миль (1 кв. миля = 2,59 кв. км) посадок цитрусовых
были очищены от тлей, зараженность которых
паразитами превышала порой 90%.
Биологический контроль может быть особенно
эффективен, если он направлен на экзотические (не
аборигенные) виды вредителей, угрожающие
местной природной растительности или окультуренным
пастбищам. Классическим примером может
служить ситуация с кактусом опунцией в Австралии.
Опунция, происходящая из Нового Света, стала
серьезным вредителем целого ряда пастбищ в
теплых районах Старого Света и Австралии. В поисках
естественных врагов опунции энтомологи
обнаружили 150 возможных кандидатов на интродукцию.
Из них 51 был завезен в Австралию, где содержался
в условиях карантина, 19 видов затем были
выпущены в природную среду и из них 12 прижились.
Однако долгосрочный эффективный контроль был
достигнут с помощью только одного вида -Cacto-
blastis cactorum. При использовании в качестве
агентов биологического контроля растительноядных
насекомых необходимо выбирать
высокоспециализированные виды (предпочтительнее даже, чтобы эти
виды были монофагами). Не следует забывать, что
в любом случае выпущенного в природу хищника
(или фитофага) назад уже не вернешь. Поэтому
абсолютно необходимо проведение очень
тщательных предварительных исследований в местах
исконного обитания предполагаемого вселенца, чтобы
удостовериться в безвредности интродукции для
экономически важных видов растений.
Биологический контроль широко применяется
также в умеренных широтах в теплицах, где нередко
выращивается основной урожай помидоров и
огурцов. Два главных тепличных вредителя - это двух-
пятнистый паутинный клещ Tetranychus urticae и
парниковая белокрылка Trialeurodes vaporariorum.
Стерилизация почвы в зимний период убивает всех
возможных хищников, а вместе с новыми
растениями, высаживаемыми в теплицу весной, обычно
попадают новые белокрылки. Что касается
паутинных клещиков, то они зимуют, забившись где-
нибудь в щелях теплицы, а весной нападают на
высаженные в теплице растения. Поскольку никаких
врагов у этих вредителей нет, их популяции растут
очень быстро вплоть до момента применения
пестицидов или же восстановления контроля со стороны
естественных врагов. Биологический контроль, для
того чтобы быть эффективным, должен вступать в
действие достаточно быстро. Поэтому естественных
врагов белокрылки - паразита Encarsia formosia и
хищника Phytoseiulus persimilis специально разводят
в коммерческих масштабах. Запускают их обычно
через неделю после преднамеренного выпуска
вредителей, для того чтобы те, размножившись, могли
обеспечить поддержание популяций паразита и
хищника. Существует, правда, в общем понятное
сопротивление растениеводов акту сознательного выпуска
вредителей в теплицу. Другой возможный метод-
это повторяющийся (обычно с 10-дневным
интервалом) выпуск естественных врагов в тот период,
когда вероятно появление в теплице вредителей.
Для успешного контроля жизненно важно
использовать естественных врагов уже на самых первых
этапах становления популяций вредителей. Если
критическое время упущено, то данный метод
может не сработать - вредители выйдут из-под
контроля.
Интегрированная борьба
Под интегрированной борьбой понимают обычно
осуществление контроля над популяцией
организма-вредителя, основанное на комплексном,
интегрированном применении как биологических, так и
химических методов ограничения роста
численности. Пестициды при этом используются, но только
так, чтобы не нарушать систему биологического
контроля. Цель программы интегрированной
борьбы-это управление популяцией вредителя
экономически обоснованными и экологически
оправданными способами. Популяция вредителя при этом
должна поддерживаться на уровне, более низком,
чем тот, при котором проявляется экономический
ущерб, или же развитие ее вообще не допускается.
Используемые методы в свою очередь должны
причинять минимальный вред агроэкосистемам (или
природным экосистемам), но благоприятствовать
популяциям естественных врагов вредителя. В
большинстве программ интегрированной борьбы
предусматривается или полный отказ от дорогих и
опасных для окружающей среды химических
веществ или же крайне ограниченное и строго
направленное их применение. Один из способов
достижения этой цели - разработка пестицидов селективного
действия. Так, например, появление афицида пири-
мор-карбаматного инсектицида, который строго
избирательно действует на тлей, но почти не
причиняет вреда их естественным врагам, положило
начало программе интегрированной борьбы с
персиковой тлей Myzus persicae и паутинным клещом
Tetranychus- основными вредителями хризантем,
выращиваемых в теплицах Англии. Раньше с этими
вредителями боролись с помощью фосфороргани-
ческих инсектицидов, которые не обладали
селективным действием, и к которым у тлей быстро
вырабатывалась устойчивость. Теперь же
паутинный клещ контролируется хищным клещом
Phytoseiulus, а тля-инсектицидом.
По чисто экономическим причинам более
удобным подходом является, однако, не столько
разработка новых строго специализированных
пестицидов (невыгодное для производителя соотношение
затрат на исследования и внедрение в сравнении со
спросом на рынке), сколько более прицельное
использование пестицидов широкого действия. Такое
узко направленное применение пестицидов может
быть более ограниченным как в пространстве, так и
во времени. Открытие и получение феромонов1
1 Вещества, используемые насекомыми как
сигналы, свидетельствующие о той или иной форме
поведенческой активности, например о готовности к
размножению или о формировании скоплений.
намного сократило пространство, обрабатываемое
пестицидами. Используя, например, половые ат-
трактанты, можно заманить насекомых в какое-то
определенное место, где их легко уничтожить
пестицидами или стерилизовать с помощью специальных
веществ, воздействующих на половую систему.
Очевидно, что при этом используется значительно
меньше пестицидов, а к тому же сокращается
воздействие их на виды, не являющиеся вредителями, и
вообще на все местообитание. Другой
многообещающий путь использования феромонов - это
подавление поведенческих реакций (обеспечивающих,
например, эффективное спаривание) путем
насыщения приземного слоя воздуха соответствующими
феромонами (для чего достаточна очень низкая их
концентрация). Насекомые привыкают к постоянно
действующим стимулам, и необходимые для
спаривания реакции подавляются. Частота применения
пестицидов может быть уменьшена путем
тщательного выбора такого момента, когда популяции
вредителя причиняется наибольший ущерб (например,
в период его спаривания) и в то же время
минимизируется воздействие на другие обитающие в том же
месте виды. Понятно, что подобная стратегия
борьбы с вредителями требует тщательного изучения
жизненных циклов всех затрагиваемых видов и
глубокого понимания процессов, происходящих во всей
экосистеме.
Программы интегрированной борьбы были с
успехом осуществлены во многих
сельскохозяйственных экосистемах, где ранее сплошная обработка
посевов ядохимикатами широкого действия
разрушила естественные взаимосвязи вредителей с их
исконными врагами. Так, например,
комбинированное использование феромонов и пестицидов
позволило восстановиться популяциям естественных
врагов бабочки репницы и соответственно перейти к
другим методам ограничения численности этого
опасного вредителя, способного нанести серьезный
ущерб посевам всех видов капусты (Brassica).
Методы интегрированной борьбы в настоящее
время широко применяются при выращивании
хлопка, посевы которого ранее во всем мире
обрабатывали пестицидами широкого действия с целью
уничтожения рано появляющихся вредителей,
например таких, как долгоносик Anthonomus grandis.
Неудивительно, что подобная обработка вела к
уничтожению естественных врагов вредителей,
способствуя тем самым появлению новых вредителей
(зачастую устойчивых к пестицидам), и вместе с тем
требовала больших денежных затрат. В долине
Канет (Перу) сплошная обработка монокультуры
хлопка пестицидами практически уничтожила всю
обитавшую там фауну и привела к экономической и
экологической катастрофе. В середине пятидесятых
годов законодательным путем было ограничено
использование ядохимикатов широкого действия и
одновременно стали поощряться такие методы
борьбы с вредителями, которые основывались на
внедрении смешанных культур, повторной
интродукции полезных насекомых и использовании не
столь устойчивых инсектицидов. К концу
пятидесятых годов урожаи хлопка значительно
возросли. Сходные системы мер были проведены и на
хлопковых плантациях в других местах. В
настоящее время методы интегрированной борьбы широко
применяются при выращивании люцерны,
цитрусовых, яблок, масличной пальмы, гевеи (каучукового
дерева), какао и др. Очень широко используются эти
методы и в тепличном хозяйстве.
12.9. Зеленая революция
Первоначально этот термин употребляли тогда,
когда речь шла об успешном введении в культуру
высокопродуктивных сортов риса и пшеницы с целью
повышения производства пищи в развивающихся
странах.
В конце сороковых годов низкорослые сорта
пшеницы из Японии были скрещены с
полукарликовыми американскими сортами. Выведение
низкорослых форм злаковых культур в значительной
степени решало проблему полегания стеблей с
тяжелыми колосьями, что в свою очередь открыло
возможности гораздо более широкого использования
азотных удобрений и искусственного орошения с
целью повышения урожайности. В результате
проведенных мер урожай зерновых удалось увеличить в
два раза.
Успех, достигнутый при выращивании пшеницы,
подтолкнул к организации Международного центра
по изучению риса на Филиппинах. Удачное
скрещивание высоких мощных сортов из Индонезии и
карликовых форм с Тайваня дало очень хороший
сорт 1R8, который при использовании азотных
удобрений созревает очень рано, что позволяет
собирать два урожая за один сезон.
Описанные нововведения принесли, однако, и
некоторое разочарование, поскольку выяснилось,
что использование их связано с внесением большого
количества дорогостоящих удобрений. Поэтому
крестьянам эти нововведения особого
преимущества не давали. Кроме того, из-за необычного вкуса и
консистенции рис новых сортов часто оказывался
непопулярным среди местного населения.
На выбор основной сельскохозяйственной
культуры, используемой для пропитания, могут влиять
различные факторы, порой не связанные с
производством пищи. В обычно возделываемых зерновых
злаковых может не хватать некоторых незаменимых
аминокислот; например, в кукурузе очень низко
содержание лизина и триптофана. Жители развитых
стран предпочитают обычно получать необходимые
аминокислоты из белков животного, а не раститель-
ного происхождения (содержащихся, например, в
бобовых и злаковых). Зерно при этом скармливается
животным, что добавляет лишнее звено консумен-
тов в пищевую цепь и создает, таким образом,
довольно неэффективную систему, превращающую
растительный белок в высококлассные животные
белки.
Решение названных проблем может заключаться
в:
1) развитии программ, направленных на отбор
мутантов и выведение новых сортов, лучше
приспособленных к климатическим условиям, или
способных противостоять временному
ухудшению погоды, например засухе;
2) развитии генной инженерии, способной перенести
благоприятные гены в организм
растения-хозяина, для того чтобы придать ему определенные
качества, например устойчивость к какому-то
заболеванию. В настоящее время уже
предпринята попытка перенести N/F-гены (гены азотфикса-
ции), обусловливающие взаимодействие бобовых
растений с бактерией Rhizobium, в
сельскохозяйственные культуры, не относящиеся к бобовым.
Если данная попытка окажется успешной, это
значительно сократит затраты на дорогостоящие
азотные удобрения.
Для аридных зон выводятся сорта, способные
выносить высокое содержание солей в почве.
Оказывается, что некоторые виды обладают генами,
определяющими возможность накапливать ионы солей в
вакуолях (в качестве механизма детоксикации).
Другие виды растений, видимо, способны переносить
высокие концентрации ионов натрия в листьях без
какого-либо отрицательного воздействия на рост и
развитие.
У ряда растений устойчивость к засухе
определяется, по-видимому, наличием в цитоплазме
специальных веществ, которые ограничивают вредные
последствия высыхания и которые способны быстро
реактивироваться при наличии воды. По мнению
некоторых исследователей, способность растения
переносить засуху зависит прежде всего от
структуры клеточных стенок, а также от размеров и формы
клеток (или вакуолей).
Выражение «зеленая революция» в настоящее
время используется в более широком смысле и
обозначает просто успешное применение научно
обоснованных методов растениеводства в
тропических условиях.
Сельскохозяйственные технологии, позволившие
увеличить производство органических веществ,
обусловили продолжающийся быстрый рост мирового
народонаселения. Тем не менее в Европе
возможности получения пищи на базе совершенных
технологий пересматриваются (например, в рамках
сельскохозяйственной политики ЕЭС) в связи с
наблюдающимся перепроизводством, уже приведшим к
образованию «гор продуктов». Однако в мире в
целом проблема нехватки пищи остается
по-прежнему актуальной, так же как и проблема
зависимости урожая от погодных условий, особенно
количества выпадающих осадков.
По скорости прироста населения все страны
можно разделить на три основные группы:
Группа L Страны, где скорость прироста населения
меньше, чем 1,5% в год. Сюда относятся
вся Северная Америка, Западная Европа
и СССР, на долю которых в сумме
приходится примерно 26% всего мирового
населения.
Группа 2. Страны, где скорость роста составляет
1,5-2,5% в год. Это Китай, Япония,
Южная Африка, на долю которых
приходится около 28% мирового населения.
Группа 3. Страны, где скорость ежегодного
прироста более 2,5%. Сюда относятся
Индия, Пакистан, Индонезия и большая
часть государств Африки. В сумме на эту
группу приходится примерно 46%
мирового населения. Было подсчитано,
например, что Индия должна ежегодно
увеличивать производство зерна примерно
на 3 млн. τ просто для того, чтобы
прокормить увеличивающееся население. К
тому же именно эти районы наиболее
подвержены внезапным стихийным
бедствиям (наводнениям или засухам), и
следовательно, именно здесь наиболее
вероятны предпосылки для возникновения
голода.
Все сельскохозяйственные технологии
направлены на то, чтобы повысить эффективность
фотосинтетических систем и в конечном итоге увеличить
продукцию сухого органического вещества,
используемого человеком. Эффективность эта базируется
на превращении солнечного излучения в прирост
растительной массы. Выбор основной
сельскохозяйственной культуры определяется, однако, не
только желанием получить наибольший выход
полезного продукта и не только ограничениями,
накладываемыми климатическими условиями (прежде
всего температурой) и активностью вредителей, но
и разными привходящими обстоятельствами, а
именно вкусовыми предпочтениями, легкостью
возделывания, религиозными и социальными
моментами. С целью удовлетворения пищевых
потребностей растущего народонаселения (особенно в
развивающихся странах) отмечается стремление
расширить географические (определяемые
возможностями адаптации растений к тому или иному
климату) пределы возделывания различных культур, для
чего осваиваются новые районы и вместе с тем
выводятся новые сорта, способные успешно
произрастать в пограничных (маргинальных) районах
основного ареала.
Сельское хозяйство, в той форме, в какой оно
развивается на Западе, зависит от больших вкладов
энергии (механизация) и от интенсивного
использования азотных удобрений. Производство таких
удобрений связано не только с потреблением
атмосферного азота, но также требует большого
количества энергии. Интенсивное земледелие возможно,
таким образом, только на малой части
сельскохозяйственных угодий. Поэтому в мировом
масштабе основное внимание уделяется селекции новых
сортов, способных дать больше высококачественной
пищи, но вместе с тем требующих при
культивировании меньшего вклада энергии и меньшего
количества азотных удобрений. Подобная целевая
установка сочетается с необходимостью использовать
для выращивания сельскохозяйственной продукции
новые местообитания и расширять сам круг
возделываемых культур. Так, например, в
Великобритании обычной культурой стал теперь рапс и изучены
возможности выращивания подсолнечника (для
производства масла).
Программы интенсивного земледелия,
основывающиеся на искусственной оптимизации внешних
условий, позволили добиться высоких урожаев.
Использование теплиц и гидропоники дает
возможность особенно четко выявить основные принципы,
связанные с такими программами.
12.9.1. Теплицы
В полевых условиях скорость фотосинтеза
определяется не только уже упомянутыми выше
физиологическими и генетическими факторами, но зависит
также от:
1) минерального питания растений;
2) водного режима;
3) наличия патогенных для растений
микроорганизмов;
4) теплового стресса;
5) ветра;
6) возраста растения и его листьев: факторы,
препятствующие быстрому наступлению старения,
увеличивают интенсивность фотосинтеза.
Например, цитокинины задерживают старение листьев.
В листьях, обработанных цитокининами (бензи-
ладенином и кинетином), интенсивность
фотосинтеза заметно повышается по сравнению с
интенсивностью фотосинтеза в листьях
контрольных растений.
Для того чтобы получить высокий урожай,
необходимо контролировать все указанные факторы,
поддерживая значения их на наиболее оптимальном
для растений уровне. Выращивая растения под
стеклом или дешевым пластиком (например, под
полиэтиленовой пленкой) можно достичь гораздо более
строгого контроля над окружающими условиями,
что позволяет получить урожай вне обычного для
данной культуры сезона и соответственно с выгодой
сбыть товар на рынке. Теплицы очень удобны также
для выращивания рассады.
Технология выращивания сельскохозяйственных
культур в теплицах во многом определяется
конкретными особенностями тех или иных растений, но
общие принципы сводятся к следующим:
1. Контроль за температурой. У большинства
растений хорошо выражен определенный диапазон
оптимальных температур. Поддержание
оптимальной температуры достигается подогревом
зимой, и затенением и вентиляцией летом. В
больших коммерческих теплицах температура
тщательно контролируется и регулируется
посредством автоматического управления системой
вентиляции.
2. Вентиляция. Посредством вентиляции
достигается необходимый приток свежего воздуха,
возобновление запаса диоксида углерода, а также
поддержание соответствующей температуры и
влажности.
3. Поливной режим и влажность. Растения очень
сильно различаются по необходимому для них
уровню увлажненности. Кактусы, например,
лучше всего чувствуют себя в сухом воздухе, а
широколиственные растения требуют порой
значительной влажности. Поддержание
определенной влажности необходимо и для того, чтобы
контролировать распространение грибковых
заболеваний. Повысить уровень влажности можно,
разбрызгивая воду или используя специальные
генераторы тумана. Многие из перечисленных
выше операций контролируются теперь с
помощью компьютеров. Предпринимались также
попытки искусственного увеличения
концентрации CO2 в закрытых теплицах. Это приводило к
увеличению урожая, но требовала
дорогостоящего оборудования. Иногда используют различные
колпачки из пластика, которыми закрывают
растения, высаженные в открытом грунте; тем
самым повышают температуру и сохраняют
необходимую влажность.
12.9.2. Гидропоника
Данный метод получения сельскохозяйственной
продукции основывается на принципах, выявленных
в ходе давно проводимых экспериментов по
изучению минерального питания растений (что
достигалось выращиванием растений на питательных
растворах разного состава). Системы гидропоники
устанавливают теперь в теплицах при непрерывной
циркуляции питательного раствора, за счет которой не
только приносятся необходимые биогенные
элементы, но и осуществляется аэрация корней. Состав
питательной среды находится под постоянным
наблюдением и регулируется с помощью
компьютеров. Корневая система находится в песке или в
гравии. Гидропоника используется обычно там, где:
а) земля бедна биогенными элементами; б)
лимитирована вода; в) есть потребность в некоторой сель-
Таблица 12.22. Отмеченное увеличение урожая при
выращивании его методом гидропоники
Томаты, кг на 1 куст
Картофель, τ та-1
Рис, кг та-1
Обычные
сельскохозяйственные
методы
5,4
12,1
551,0
Гидропоника
7,3-10,2
26,3
1652
скохозяйственной продукции, выращиваемой вне
обычного сезона и потому реализуемой по высоким
рыночным ценам.
12.10. Охрана природы
Основную цель охраны природы можно
сформулировать следующим образом: это поиск таких
методов эксплуатации биологических и минеральных
ресурсов Земли, которые позволят создать условия,
оптимальные для всех связанных общей судьбой
живых существ планеты, в том числе и человека.
Для достижения этой цели необходима не только
практическая деятельность по защите среды и
сохранению ее в состоянии пригодном для обитания,
но и разъяснение важности этих проблем и
мероприятий. Накопленные экологические данные
позволяют надеяться на то, что при бережном отношении
к воде, земле и природе вообще человечество
сможет и в будущем обеспечить себя всем
необходимым: пищей, жильем, промышленной продукцией,
сохранить физическое и интеллектуальное здоровье
и возможности для нормального отдыха.
Охрана природы это совсем не то же самое, что
охрана памятников старины, когда объект
поддерживается в неизменном виде. Процессы,
происходящие в биосфере чрезвычайно сложны и изменчивы, а
значит и охрана природы должна быть гибкой и
динамичной. Для эффективного управления
природой необходимо знать, что происходит в
окружающем нас мире; эти знания дает экология и другие
науки о Земле.
Вся деятельность по охране природы в неявном
виде определена нашим отношением к миру, в
котором мы живем. Это своего рода этика охраны
природы; вкратце ее можно выразить следующими
высказываниями:
«Мы унаследовали мир от наших родителей, но
гораздо больше взяли взаймы у собственных
детей» (лозунг «зеленых»).
«Обмани Землю, и она обманет тебя»
(старинная китайская пословица).
«Вымирание необратимо!» (надпись на
рекламной майке, агитирующей за спасение китов).
Таким образом, проблема охраны природы часто
затрагивает нравственные и политические аспекты.
Чтобы охрана природы была эффективна,
необходима интеграция задач, которые ставит перед
собой наука и общество. Длительное процветание
человеческого общества возможно лишь при
благополучном состоянии биосферы. Но на небольших
отрезках времени цели охраны природы часто
находятся в противоречии с прогрессом в развитии
общества и промышленности. Таким образом,
результаты охраны природы часто бывают
чрезвычайно противоречивы и многогранны. Ниже
приводятся некоторые примеры того, какова практическая
роль охраны природы.
Сохранение генетического разнообразия. Это один
из важнейших доводов в пользу охраны влажного
тропического леса, обладающего высоким видовым
разнообразием. Культурные растения и животные
также являют собой источник генетического
материала, а значит, и они нуждаются в охране. Этот
материал хранится в многочисленных,
разбросанных по всему миру, банках семян и спермы.
Предположим, что в окружающей среде произойдут
значительные изменения абиотических или биотических
факторов, например наступит глобальное
потепление климата, обусловленное парниковым эффектом,
или под влиянием спонтанных или индуцированных
мутаций появятся новые паразиты и болезни. Даже
в этом случае накопленный наследственный
материал окажется бесценным источником новых пород
домашних животных и сортов
сельскохозяйственных культур, которые будут приспособлены к
новым условиям среды.
Восстановление заброшенных земель, таких, как
отвалы шахт и каменоломни и создание на их месте
лесов, парков, или сельскохозяйственных угодий, а
также создание новых участков суши-все это
уменьшает пресс цивилизации на мало нарушенные
ландшафты сельской местности.
Вторичное использование отходов значительно
снижает расход сырья. Так, если бумагу не сжигать
безо всякой пользы и не выбрасывать, а
перерабатывать вновь, то потребности бумажной
промышленности в древесине резко уменьшатся. Бытовой
мусор можно использовать для получения энергии,
сжигая его в системах отопления небольших
районов. При этом необходимо принять меры по
снижению загрязнения воздуха.
Сохранение лесов на водоразделах
предотвращает эрозию, слишком быстрый отток дождевой воды
по склонам и паводки, и в то же время позволяет
сберечь древесину, которую можно будет
использовать в будущем.
Сохранение ресурсов моря и суши.
12.10.1. Земля как космический
корабль
В течение многих тысяч лет человеческая
деятельность не наносила природе заметного ущерба. Если
в какой-либо местности истощались ресурсы, люди
откочевывали в другие районы. Там они выжигали
лес и возделывали освободившиеся участки, или
находили иное пропитание. В сообществах
охотников-собирателей существовала полная гармония
между потребностями человека и возможностями
природы; такой уклад жизни сохранился и по сей
день у бушменов Калахари, аборигенов Австралии
и эскимосов.
Серия технологических революций, которые
претерпела история человечества, нарушили равновесие
между человеком и природой. Возникновение 10000
лет назад земледелия и животноводства привело к
быстрому росту населения, в результате которого
постепенно появились первые крупные поселения.
Затем произошли дальнейшие революционные
сдвиги в технологии добывания пищи, здравоохранении
и промышленности, превратившие мизерное по
началу население земного шара в огромное технически
оснащенное общество, которому требуется все
больше сырья и энергии. В шестидесятые годы XX
века, когда люди впервые покинули планету,
появилась первая возможность взглянуть на Землю из
космоса, после чего все ясно осознали, что
возможности для роста населения и ресурсы Земли не
беспредельны.
Итак, экологи пришли к заключению, что
Земля-это космический корабль, оснащенный всем
необходимым для длительного полета, но не
имеющий никаких иных источников энергии, кроме
собственных, а также лучистой энергии ближайшей
звезды-Солнца. Считается, что жизнь на Земле
существует примерно 3,5 млрд. лет и нет никаких
оснований опасаться, что она не просуществует по
крайней мере еще столько же, если мы сами не
уничтожим ее.
Как сказано в разд. 12.3, энергия, необходимая
для жизни, поступает на Землю в основном в виде
солнечного излучения, которое используется
зелеными растениями для фотосинтеза, а от них
поступает далее в пищевые цепи, и следовательно,
управляет биогеохимическими циклами. Кроме
того, солнечная энергия определяет климатическую
зональность планеты и океанические течения, т.е.
непосредственно влияет на среду обитания живых
существ.
В 1980 г. три всемирные организации (UNEP,
IUCN, WWF) разработали Международную
программу охраны природы, основу которой
составляет экологический анализ того, как управлять
космическим кораблем-Землей. В этом документе
указывается, что сохранение физических свойств среды и
охрана растительных сообществ планеты-это тот
минимум, который необходим для поддержания
системы жизнеобеспечения. Атмосфера, океан и
почва дают жизнь растениям, а они обеспечивают
существование всех животных, и в то же время
представляют собой важнейший фактор,
препятствующий эрозии почвы.
Кроме того, Международная программа охраны
природы постулирует необходимость сохранения
генетического разнообразия. В процессе эволюции
возникло великое множество разнообразных живых
существ, появившихся в результате естественного
или искусственного отбора. При вымирании видов
происходит непрерывная потеря биологических
ресурсов, и это обязательно скажется на будущем
благополучии человечества. В Международной
программе охраны природы впервые выдвинута
концепция оптимального использования экосистем
Земли. Согласно этой концепции, все население
земного шара может рассчитывать на
удовлетворительные условия существования до тех пор, пока
система жизнеобеспечения используется нормально
и не происходит подавления других обитателей
«космического корабля», ведущего к их гибели.
Многие представители флоры и фауны Земли могут
оказаться в будущем более полезными, чем это
представляется сейчас.
12.10.2. Загрязнение-результат
ошибок в эксплуатации системы
жизнеобеспечения
До самого последнего периода истории Земли
живые системы планеты эволюционировали почти в
полной гармонии с атмосферой, гидросферой и
литосферой, не испытывая влияния человеческой
деятельности. Но по мере развития сельского
хозяйства и промышленности воздействие человека на
среду стало все заметнее. Повсеместная
индустриализация, особенно развернувшаяся за последние два
столетия, привела к потенциально опасным уровням
загрязнения среды.
Можно сказать, что загрязнение - это
поступление в окружающую среду каких-либо веществ или
энергии в таких больших количествах или в течение
столь длительного времени, что эти вещества или
энергия начинают наносить ущерб людям и
окружающей их природе. Легко распространяясь от
одних компонентов системы жизнеобеспечения к
другим, загрязнение в той или иной степени влияет
на все параметры среды - антропогенные и
природные, физические и биотические. Например,
поступление в атмосферу чрезмерного количества
некоторых газов ведет к возникновению «кислотных
дождей» (гидросфера), которые в свою очередь
приводят к закислению почвы (литосфера), а также к
глубоким изменениям в лесных и водных
экосистемах (биосфера), разрушению зданий и других
объектов человеческой деятельности.
Загрязнение атмосферы
Еще в начале шестидесятых годов считали, что
загрязнение атмосферы - это локальная проблема
больших городов и индустриальных центров, но
позже стало ясно, что атмосферные загрязнители
способны распространяться по воздуху на большие
расстояния, оказывая неблагоприятное воздействие
на районы, находящиеся на значительном удалении
от места выброса этих веществ. Таким образом,
загрязнение атмосферы - это глобальное явление, и
для контроля за ним необходимо международное
сотрудничество. К числу наиболее
распространенных загрязнителей атмосферы относятся такие газы,
как хлорфторуглероды, диоксид серы (SO2),
углеводороды и оксиды азота (NOx).
Загрязнение может привести к значительному
снижению естественной концентрации газов,
входящих в состав атмосферы, например озона (O3) в
стратосфере. Но поразительно другое:
концентрация озона выше в тех районах, где среднемесячное
содержание загрязнителей в нижних слоях
атмосферы составляет 200 млн.-1, чем там, где
максимальная величина загрязнения не превышает
0,04 млн.-1. Подобные аномалии высоких
концентраций озона наносят ущерб многим
сельскохозяйственным культурам, например томатам: суммарные
потери урожая за счет избытка O3 составляют в
Калифорнии около 1 млрд. долларов в год. В
сочетании с углеводородами и соединениями группы NOx
озон может представлять собой прямую опасность
для здоровья людей, и кроме того, является одной из
важных составных частей фотохимического смога.
Пыль, шум, чрезмерное количество тепла, радиация
и электромагнитные поля, все это - загрязнение
атмосферы. Детальное обсуждение полного перечня
загрязняющих веществ, эффектов их воздействия и
методов контроля за ними выходит за рамки
настоящего изложения (см. D. Elson, 1987. Atmospheric
pollution, Blackwell). Все возрастающая
концентрация диоксида углерода, вклад этого и других газов в
создание планетарного парникового эффекта
обсуждались в разд. 12.4.1. Ниже речь пойдет о двух
других актуальных глобальных проблемах; одна из
них - обеднение озонового слоя (образование
озоновых дыр), другая - «кислотные дожди».
обеднение озонового слоя. Атмосфера - это
термостатический и радиационный щит Земли. В
верхних слоях атмосферы на высоте 15-50 км от
земной поверхности кислород и озон поглощают
большую часть поступающей извне
коротковолновой радиации. Это ультрафиолетовое излучение
(УФ), рентгеновские и гамма-лучи, которые по своей
физической природе пагубны для живых существ,
так как разрушают их генетический аппарат. По
оценкам, полученным в США, при уменьшении
концентрации озона стратосферы на 5% интенсивность
УФ-излучения у поверхности Земли возрастет на
7,5-15%, что может повлечь за собой увеличение
заболеваемости раком кожи на десятки, а то и сотни
тысяч случаев в год.1 При поглощении излучения
озоном происходит прогрев стратосферы, возникает
хорошо выраженный слой температурной инверсии
1 УФ-излучение определенной длины волны
оказывает важный целебный эффект, способствуя
превращению стероидов кожи в витамин D
(в отличие от температуры стратосферы,
температура тропосферы, т. е. нижнего слоя атмосферы, по
мере возрастания высоты падает). Существование
этого слоя заметно ограничивает интенсивность и
масштабы конвективного перемешивания
атмосферы, поэтому любые нарушения слоя инверсии
приведут к резкой глобальной смене погодных условий,
а значит и к изменению климата на Земле.
В верхних слоях атмосферы молекулы кислорода
(O2) диссоциируют под действием излучения на
атомы (О), которые соединяются с молекулами,
образуя озон (O3). Под действием солнечного света
эта реакция обратима (O3 + О ^ 2O2). В «озоновом
слое» концентрация озона поддерживается на
некотором равновесном уровне в 1 млн.-1.
Хлорфторуглероды представляют собой группу
химических соединений, куда входят, в частности,
четыреххлористый углерод и хлороформ. Обычно
хлорфторуглероды используют в качестве
растворителей, распылителей аэрозолей и фреонов в
холодильниках. Эти вещества сравнительно устойчивы в
нижних слоях атмосферы, где часто способствуют
возникновению парникового эффекта (см. разд.
12.4.1). Они очень летучи и поэтому в конечном
счете оказываются в стратосфере. На высоте более
25 км хлор- и фторсодержащие соединения
распадаются под влиянием солнечного света, высвобождая
атомы хлора или фтора, которые реагируют с
озоном, причем каждый такой атом разрушает 105
молекул озона, т.е. диссоциация озона на
молекулярный и атомарный кислород происходит с
большей скоростью, чем обратная реакция.
Таким образом, примесь хлорфторуглеродов
нарушает равновесие между кислородом и озоном.
При современном уровне загрязнения этими
соединениями через 20 лет количество озона уменьшится
на 10%, а за полвека может разрушиться около двух
его третей. В 1987 г. впервые наблюдалось полное,
хотя и носившее сезонный характер, разрушение
озонового слоя над Антарктидой.
кислотные дожди. Кислотные дожди - не
простое и не единичное явление. При сжигании
ископаемых горючих материалов образуется диоксид
серы (SO2) и оксиды азота (NOx), а при неполном
сгорании образуются еще и углеводороды. Все эти
вещества поступают в атмосферу в газообразном
состоянии, но иногда «вымываются» оттуда,
выпадая на землю с осадками. Во всех промышленных
районах, таких, как восточная часть США, Западная
Европа и северовосточные районы Китая и Японии,
иногда выпадают дожди, рН которых значительно
ниже 4,0 (в норме нижний предел кислотности
дождевой воды равен 5).1
1 При естественной концентрации диоксида
углерода в атмосфере рН дождевой воды составляет 5,6 В
присутствии оксидов серы, поступающих в атмосферу
из океана и при извержении вулканов, эта величина
иногда понижается до 4,7.
Частое выпадение кислотных дождей (рН < 5)
нередко приводит к крупным изменениям в
экосистемах и повреждению зданий. Подобные явления
обычны для государств, граничащих с теми, в
которых находятся наиболее мощные источники
загрязнения. Так, кислотные дожди, выпадающие в
Норвегии и Швеции,- это результат загрязнения воздуха
в Великобритании и Западной Европе, причем
вредные вещества распространяются господствующими
ветрами, дующими на большой высоте. В
Центральной Швеции и Северной Норвегии под
действием кислотных дождей снизились уловы лосося и
форели, пострадали леса. Повреждение деревьев,
связанное с кислотным загрязнением,-явление,
очень распространенное сейчас в Европе, и есть
данные о том, что в Англии от такого загрязнения
страдают бук и тисе.
Нередко обнаруживается, что там, где почва
бедна карбонатами (такие почвы образуются,
например, на подстилающих гранитных породах), и не
в состоянии нейтрализовать воздействие кислотных
осадков, страдает фауна озер и рек. Особенно
чувствительными оказываются молодь и икра рыб. В
частности, в Скандинавии проблема осложняется
тем, что килотные загрязнители, накопившиеся за
зиму в снежном покрове, быстро высвобождаются
весной при таянии снега и сразу попадают в
большом количестве в талую воду в самый разгар
нереста и выклева мальков.
Под влиянием кислотных дождей магний и
кальций вымываются из почвы и листового опада, а
алюминий, марганец и тяжелые металлы
постепенно проникают в почвенные воды, где могут
достигать токсических концентраций, приводящих к
гибели корней деревьев и к разрушению микоризы. В
результате этого снижается способность деревьев
получать из почвы воду и минеральные соли.
Болезни, связанные с недостатком минерального питания,
стали весьма обычными, причем в условиях
пониженной влажности заболеваемость деревьев
увеличивается. К категории кислотных дождей следует
отнести озонно-кислотные туманы, служащие,
видимо, одной из важнейших причин суховершинности
древостоя в Шварцвальде (ФРГ), а также выпадение
сухих кислотных осадков. Озон образуется при
взаимодействии углеводородов и оксидов азота в
самых нижних слоях атмосферы под влиянием
солнечного света. Даже в том случае, когда
концентрация каждого отдельного загрязнителя безвредна,
смесь таких веществ, как озон, диоксид серы, оксиды
азота, другие фотооксиданты и тяжелые металлы,
может привести к сильному угнетению
растительности. В сочетании с климатическими стрессами,
особенно засухой, такой «коктейль» из
загрязняющих веществ часто бывает причиной гибели
деревьев.
Таким образом, выражение «кислотные дожди»
фактически обозначает несколько разнообразных
явлений. Методы борьбы с закислением, например
внесение извести в озера (Швеция) или лесную почву
(ФРГ) дают лишь кратковременный положительный
эффект. Серьезных успехов в этом направлении
можно достигнуть лишь одним
способом-снижением количества газообразных загрязнителей,
поступающих в атмосферу. Особое внимание следует
сосредоточить на уменьшении выбросов диоксида
серы, который поступает в атмосферу в особенно
больших количествах, а источники выбросов
сравнительно легко обнаружить: это промышленные
предприятия, главным образом работающие на угле
электростанции. Кроме того, существуют
эффективные и доступные, но дорогостоящие технологии
десульфуризации \ при использовании которых
диоксид серы в атмосферу не выбрасывается. В
долгосрочной перспективе, видимо, также важно будет
ограничить поступление в атмосферу углеводородов
и оксидов азота.
Загрязнение воды
До последнего времени загрязнение воды было
локальным явлением, касавшимся преимущественно
промышленно развитых стран. Проблема эвтрофи-
зации, вызванной применением чрезмерного
количества удобрений при интенсивном ведении
сельского хозяйства, а также сточными водами,
обогащенными фосфатами, уже обсуждалась в разд.
12.4.1. Такие ситуации все чаще возникают то в
одном, то в другом уголке земного шара,
затрагивая не только пресноводные, но и морские
экосистемы. Например, летом 1988 г. огромное,
протяженностью в несколько миль пятно «цветения»
планктонных водорослей покрыло часть Северного
моря и достигло Балтики. Сброс плохо очищенных
сточных вод из населенных пунктов побережья
приводит к возникновению прямой угрозы для
здоровья купающихся в море людей и для морских
животных.
Городские стоки и крупные свалки часто бывают
причиной загрязнения вод тяжелыми металлами и
углеводородами. Поскольку тяжелые металлы
накапливаются в морских пищевых цепях, их
концентрация может достигнуть летальных доз, что и
произошло после большого промышленного
выброса ртути в прибрежные воды Японии близ г. Мини-
мата. Повышенная концентрация этого металла в
тканях рыбы привела к гибели многих людей и
животных, съевших зараженный продукт.
Сублетальные дозы тяжелых металлов, пестицидов и
продуктов переработки нефти могут заметно
ослаблять защитные свойства организмов. Концентрация
полихлорбифенилов (известны еще и как
канцерогены) в Северном море достигает в настоящее время
1 Стоимость таких технологий велика при
внедрении их на уже действующих предприятиях, но она
значительно снижается при включении их в
первоначальный проект.
0,000002 млн.-1, а в тканях дельфинов,
представляющих собой конечное звено пищевой цепи, их
концентрация достигает уже 16 млн.1. Страны,
расположенные на побережье Северного моря с
недавнего времени проводят комплекс мероприятий,
направленных на снижение, а в перспективе и на
полное прекращение сброса в море и сжигания
токсических отходов. Что же касается
законодательных мер по контролю за нефтяным загрязнением
при работе нефтяных терминалов и слива
балластной воды из нефтеналивных судов, то они были
приняты гораздо раньше, после того как стала
очевидной опасность больших разливов нефти из
танкеров (как, например, это было при аварии
Торри Каньон в 1967 г.). Результаты исследований
показали, однако, что малоизвестные небольшие по
объему, но постоянно повторяющиеся разливы и
утечки нефти наносят экосистемам даже больший
ущерб, чем нашумевшие катастрофы. Детергенты,
применяемые для уничтожения нефтяных пятен с
поверхности моря, часто оказываются более
опасными для среды, чем сама нефть. Настоятельно
необходимо дальнейшее изучение проблемы и
дальнейшее совершенствование мониторинга нефтяного
загрязнения моря.
На пути юридического контроля загрязнений
морской, а особенно открытой океанической среды
возникают большие трудности, поскольку проблема
уничтожения отходов стала сейчас для многих
государств весьма острой, а незаконный сброс их в море
чрезвычайно привлекателен. Вредные химические
соединения и радиоактивные отходы, попавшие в
море, постепенно разносятся течениями и волнами,
но если они попадают в мелководные приконтинен-
тальные моря, и особенно на литораль, то
немедленно наносят вред экосистеме, еще до того, как
удается принять эффективные меры борьбы с этим
воздействием. В этом отношении малые моря
особенно уязвимы. Воды Ирландского моря имеют
более высокую радиоактивность, чем в других при-
континентальных морях, а объясняется это
утечками на атомной станции в Селлафилде.
Еще одна серьезная проблема связана с эрозией
почвы. В результате эрозии увеличивается
количество взвеси в речных и морских прибрежных
водах. Есть данные о том, что наличие взвеси в воде
благоприятно для рыболовства, однако это не
доказано. Вместе с тем совершенно очевидно, что
осаждение чрезмерного количества взвеси ведет к
разрушению коралловых рифов. Так, на Большой
барьерный риф неблагоприятное воздействие
оказало уничтожение лесов на континенте.
Уничтожение лесных богатств Земли
Лес-это естественная устойчивая формация
растительного покрова обширных областей земного
шара, занимавшая до последнего времени одну треть
поверхности суши. Если площадь, занятая лесами
умеренной зоны, уменьшается незначительно, то
тропические леса исчезают с такой скоростью, что к
2000 г. будут занимать лишь 30 млн. га или 7%
суммарной поверхности суши, тогда как в 1950 г.
эта цифра составляла 15%. Каждый год в мире
уничтожается лес на 12 млн. га (что равно площади
Англии), а еще 10 млн. га лесов деградируют, так
как вырубка ценных пород на этих площадях не
сопровождается соответствующим лесоустройством
и природоохранными мероприятиями. А между тем
тропические леса крайне нуждаются в охране. В
древесине ценных пород заинтересованы
практически все страны мира, и следовательно, все они
должны отвечать за состояние тропических лесов.
Страны Третьего мира, где эти леса произрастают,
слабо развиты и не в состоянии самостоятельно
справиться с этой задачей.
Традиционная практика расчистки лесов под
пашню с помощью подсечно-огневого метода
долгое время не приносило лесу ощутимого вреда, так
как плотность населения была невелика.
Необходимо также подчеркнуть, что времени, на которое
поле забрасывали и оно снова зарастало, хватало
для того, чтобы восстанавливалась естественная
лесная экосистема и плодородие лесных почв.
Позже, когда численность населения возросла и
появились такие конкурентные виды землепользования,
как постоянные сельскохозяйственные плантации
или затопление при строительстве ГЭС, периода
«отдыха» земли стало не хватать, и для
освобождения новых посевных площадей потребовалась
дополнительная вырубка лесов. Но развитие сельского
хозяйства-не единственная причина уничтожения
тропического леса: там заготавливают дрова и
ценную древесину тропических пород (например тисе, и
красное дерево), кроме того, лес вырубают и
выжигают, расчищая пространства под пастбища. За
сложившуюся ситуацию ответственны не только
страны Третьего мира. Крупномасштабное
уничтожение тропических лесов является результатом как
роста численности местного населения (мелким
фермерам нужно все больше земли и топлива), так и
эксплуататорских интересов богатых стран в
получении по низким ценам ценнейшей древесины и
мяса.
Уничтожение лесов чревато серьезными
последствиями.
а) Оно сопровождается уменьшением
количества сырья и продуктов, которые всегда давал
лес, а местное население использовало для
своих нужд. Это - ценная древесина,
стройматериалы, топливо, мед, дикие фрукты,
охотничья дичь и дикорастущие растения.
б) Во всем мире неуклонно возрастает
потребление лесоматериалов (строительство),
древесной массы (производство бумаги) и
ценной древесины тропических пород
(изготовление мебели). Однако длительная добыча
этого сырья в столь больших объемах может
нанести тропическим лесам невосполнимый
ущерб.
в) Леса, произрастающие на горных склонах и
водоразделах, задерживают большое
количество дождевой воды и медленно отдают ее
в ручьи и реки. Уничтожение лесов -
важнейшая причина наводнений в долинах. Индия
на борьбу с наводнениями ежегодно тратит
1 млрд. долларов. Летом 1988 г. в Бангладеш
происходили небывалые наводнения,
охватившие всю страну. Основная причина
бедствия - уничтожение горных лесов вблизи
северных границ государства - в Индии и Непале.
Чтобы возместить нанесенный наводнениями
ущерб, необходимы международная помощь
и сотрудничество разных государств.
г) Уничтожение лесов ведет к эрозии почвы, а
эрозия в свою очередь-к экономическому
спаду и голоду. Источники чистой воды
загрязняются, возникают заболевания, как это
имело место в Бангладеш. Заиление
водохранилищ сокращает срок их службы, а порты и
эстуарии необходимо постоянно расчищать,
чтобы они оставались судоходными.
д) В результате сведения лесов усиливается
возрастание концентрации диоксида углерода
(см. разд. 12.4.1) и увеличивается альбедо
земной поверхности. Оба этих фактора могут
стать причиной глобальных и длительных
изменений климата.
е) Уменьшение площади лесов может повлечь
за собой изменение количества атмосферных
осадков и частоты их выпадения, в
результате чего могут быть утеряны весенние запасы
воды, необходимые для нормального
развития природных сообществ.
ж) В лесах обитают наиболее богатые по числу
видов и их разнообразию природные
сообщества. При их нарушении исчезает масса
малоизвестных и редких живых существ,
происходит значительная потеря генетического
разнообразия и потенциальных ресурсов.
Эрозия почвы и потеря
плодородных земель
Уничтожение лесов-не единственная причина
эрозии почвы: это явление наблюдается и при
чрезмерной эксплуатации пашни и пастбищ.
Формирование почвы - длительный процесс (разд. 12.5) и
сейчас на планете в целом разрушение почвы
происходит гораздо интенсивнее, чем ее образование.
Особенно значительная эрозия возникает в холмистых
местностях, где выпадает много осадков и
происходит активная вспашка земли. Во многих районах
Юго-Восточной Азии издавна существует система
террасного земледелия, которая позволяет хорошо
удерживать почву даже там, где нет леса.
Применение этого агроприема наряду с безотвальной
вспашкой и сооружением на полях перемычек,
препятствующих вымыванию почвы во время дождей -
основные меры борьбы с эрозией. Примеры такой
сберегающей почву культуры земледелия имеются на
Бали, тогда как на соседней Яве быстрый рост
населения и преобладание мелких единоличных
хозяйств привели к значительному оскудению запасов
плодородных земель. Вследствие эрозии во всем
мире ежегодно теряется 5 млн. га плодородной
пахотной земли.
На пастбищах, где происходит перевыпас скота,
часто уничтожается растительный покров,
удерживающий плодородный слой почвы. Растения
выедаются целиком и погибают, в результате чего
дождевая вода беспрепятственно размывает поверхность
почвы, вызывая обширную эрозию, что в конечном
итоге ведет к возникновению глубоких оврагов.
Ежегодно в мире из-за перевыпаса и последующей
эрозии теряется около 7 млн. га пастбищных
угодий, многие из которых превращаются в пустыню.
Пустыни обычно возникают естественным
образом, однако этот процесс, называемый
опустыниванием, часто ускоряется в результате деятельности
человека. Треть поверхности суши располагается в
засушливых и полузасушливых областях, а 700 млн.
человек, населяющие эти области, находятся
буквально на грани выживания. Хозяйственная
деятельность этих малоимущих людей вызывает
значительную эрозию почв, но у них нет иного выхода.
Размеры Сахеля, полузасушливого пояса,
расположенного в Африке южнее Сахары, увеличиваются
из-за перенаселенности и слишком интенсивного
выпаса скота. Процесс опустынивания можно
остановить и снова превратить пустыню в оазис. Для
этого необходимо переселить население в другие
районы и насадить леса. В силу административных,
социальных, экономических и политических причин
эта задача чрезвычайно трудна, однако ее решение
привело бы к увеличению количества и
регулярности выпадения осадков, что в свою очередь через
изменение величины альбедо влияло бы на климат.
Вопросы для обдумывания
1. а) Для любой из тем, обсуждавшихся в этом
разделе, или разд. 12.4.1 (парниковый эффект или
эвтрофизация) составьте итоговую таблицу, из
которой было бы видно, кто расплачивается за
загрязнение среды, а кто получает прибыль.
б) Когда владельцу предприятия,
загрязняющего среду, приходится оплачивать мероприятия,
направленные на борьбу с загрязнением, эта сумма
включается в себестоимость продукции (принцип
«нарушитель платит за все»). Чем выгоден такой
подход, и в чем трудности его применения?
12.10.3. Сохранение генетического
разнообразия
Эволюционный процесс всегда сопровождался
гибелью видов, не выдержавших борьбы за
существование. Вымирание необратимо - дронта и
динозавров не возродить. В настоящее время скорость
вымирания видов все увеличивается, и это связано с
нарушением местообитаний организмов,
увеличением степени эксплуатации природных популяций,
сокращением природных ландшафтов под влиянием
расширяющейся деятельности человека. По
существующим оценкам сейчас на Земле ежедневно
исчезает один вид. Многие виды, особенно обитатели
влажных тропических лесов, вымерли еще до того,
как были описаны систематиками. Больше всего
страдают насекомые. Каждый десятый из живущих
сейчас видов высших растений или животных
находится под угрозой вымирания. Большая панда (AiIu-
ropoda melanoleuca)- один из подобных примеров.
Этот вид, изображение которого служит
символом Всемирного фонда охраны природы, обитает в
Восточном Тибете и юго-западных районах Китая.
Косолапое существо с грустной мордой завоевало
симпатию миллионов людей. Был момент, когда
вид этот подвергался очень большой опасности, так
как его местообитание - бамбуковый лес, стали все
чаще посещать люди. Было решено сохранить
большую панду, лес стал заповедником, и с тех пор
популяция этого вида увеличилась примерно до
1000 особей. Большая панда питается почти
исключительно молодыми проростками бамбука,
растения с циклической сменой периодов высокого и
низкого обилия. В 1983 г. в провинции Сычуань на
большой площади неожиданно вымер бамбуковый
лес, в результате чего по меньшей мере 59 панд
погибли от голода. Без помощи человека этот
угрожаемый вид мог в подобной ситуации вообще
исчезнуть с лица Земли.
Только в самые последние годы достаточно
широко развернулись исследования по экологии и
физиологии большой панды в естественных условиях.
Сейчас стало совершенно очевидно, сколь уязвим
этот вид и как трудно ему при современных
условиях устоять под натиском цивилизации. Панды ведут
одиночный образ жизни и встречаются между собой
только в период размножения. 95% активного
времени взрослое животное тратит на поедание
бамбука, и хотя эта пища низкокалорийна, не ест почти
ничего другого. Попытки добиться размножения
панды в зоопарках Китая оказались неудачными, но
создание в Волонге (провинция Сычуань) «ферм» по
разведению животных увенчалось успехом. Здесь
были использованы новейшие достижения в области
разведения животных, в том числе - искусственное
осеменение, банк спермы, и в результате в неволе
родилось несколько детенышей. Сейчас стало
возможным предсказывать начало течки у самок и
обеспечивать своевременное спаривание. При
сохранении среды обитания вида и небольшой помощи со
стороны человека большую панду, видимо, удастся
сохранить.
Имеется много хорошо известных примеров
угрожаемых видов и существует целый ряд
программ их спасения. Некоторые виды, например
калифорнийский кондор, оказались под угрозой
вымирания из-за отравления пестицидами;
численность других, таких, как филиппинский орел,
нападающий на обезьян, снижается, потому что
сокращается число подходящих местообитаний. Каждой
из этих птиц необходимо по меньшей мере 50 км2
леса и сейчас во всей Юго-Восточной Азии осталось
менее 300 особей данного вида.
Уменьшение численности некоторых видов
происходит потому, что они плохо уживаются с
человеком. Бенгальский тигр - исконный враг человека-
некоторое время назад оказался на грани
вымирания, и только благодаря созданию нескольких
больших тигровых заповедников его удалось сохранить.
Браконьерская добыча слоновой кости и рогов
носорога привела к значительному сокращению
поголовья этих животных, и это стало особенно заметно
с появлением автоматического оружия. Другие
редкие животные, например горные гориллы,
чрезвычайно чувствительны к нарушениям среды,
вызванным человеческой деятельностью. Все угрожаемые
виды животных и растений заносятся в Красную
книгу, издаваемую Международным комитетом
охраны природы и ресурсов. Для успешной охраны
фауны и флоры необходимо образование, гласность,
проведение экологических исследований и оказание
финансовой поддержки заповедникам, зоопаркам и
ботаническим садам.
Генетические ресурсы на службе
человека
Теперь стало совершенно ясно, что значительное
сокращение генофондов ныне существующих видов
приведет к исчезновению многих ценных аллелей и
это уже наблюдается у культурных растений и
домашних животных. В мире существует около
80 тыс. видов съедобных растений, но только 20 из
них возделывается для пищевых нужд в
значительных масштабах, причем в большинстве развитых
стран выращиваются лишь очень немногие сорта.
Обычно в практике земледелия используются
только тщательно отобранные и апробированные
высокоурожайные сорта (например, половина всей
пшеницы на полях Канады относится к одному сорту
«Neepawa»). Селекционеры иногда сталкиваются с
явлением генетической эрозии, которая возникает в
монокультуре зерновых. Поэтому они стремятся
"собрать как можно больше старых сортов, и
сохраняют их либо в коллекциях семян, либо в состоянии
вегетации путем ежегодного пересева. Крупнейший
селекционный семенной банк находится в Научно-
исследовательском институте риса на Филиппинах;
там собраны тысячи известных сортов этой
культуры (см. разд. 12.9). Многие из них дают небольшой
урожай, который почти не увеличивается даже после
внесения удобрений, но зато обладают генетически
обусловленной резистентностью к пестицидам и
болезням.
В Великобритании Организация по охране
редких пород имеет в своих прекрасных коллекциях
великое множество разновидностей домашних
животных. Редкая корнишская порода кур была
недавно использована при выведении новых пород как
источник генов быстрого роста. Свиньи тамуорт-
ской породы оказались хорошо приспособленными
для пастбищного содержания в Австралии.
Дикие виды в перспективе также могут оказаться
чрезвычайно полезными, поэтому их
охрана-единственная разумная стратегия. Так, розовый
барвинок, произрастающий на Мадагаскаре, используется
для изготовления медикаментов, применяемых для
лечения рака. Недавно в Вест-Индии найдена еще
одна разновидность этого растения, которая дает
десятикратный выход противоракового препарата.
Становится все более очевидным тот факт, что
наилучшую гарантию сохранения диких видов
животных и растений дает охрана среды их обитания, а
это требует экосистемного подхода. Сейчас в
каждой стране проводится выявление характерных
ландшафтов, которым в дальнейшем будет
присвоен статус особо охраняемых территорий, что будет
способствовать лучшему сохранению генетического
разнообразия растений и животных во всем мире
(см. разд. 12.10.6).
Вопросы для обдумывания
2. Каким образом можно использовать банки
спермы для увеличения генетического разнообразия
таких видов, как африканский носорог, популяции
которого будут становиться все более
изолированными?
12.10.4. Образ мыслей «зеленых»
Экологические познания и понимание принципов
функционирования и эволюции систем
жизнеобеспечения Земли заставило многих критически взглянуть
на то, как используются ресурсы планеты. Если мы
хотим, чтобы эксплуатируемые виды растений и
животных восстанавливались, а среда обитания
была пригодна для жизни, нам необходимо
руководствоваться концепциями «зеленых».
Использование и возобновление
животных и растительных ресурсов
Методы рациональной эксплуатации экосистем
обсуждались в разд. 12.3.7. Применимость данных о
динамике популяций рыб для достижения
оптимального промысла и воспроизводства
рассмотрены в разд. 12.7. В долгосрочной перспективе
экономическое процветание и экологическое
благополучие неотделимы друг от друга.
Практика лесного хозяйства, рассчитанная на
длительное время, показывает, что лес,
используемый в настоящий момент, должны были посадить
люди предыдущего или более старшего поколения.
Иногда потребности общества меняются, и от земли
хотят получить совсем не то, что требовалось
раньше, например вырубают лес для расширения
посевных площадей. В таких случаях необходимо, чтобы
новая система землепользования оказалась не менее
стабильной и долговечной, чем существовавшая
ранее. В некоторых районах Амазонки значительные
на первых порах достижения сельского хозяйства,
развивавшегося на месте вырубленной сельвы,
оказались недолговечными. Даже серьезные
программы, подкрепленные экспертизой и солидной
финансовой поддержкой других стран, были
приостановлены через 10-15 лет. Вторжение сорняков,
проблемы, связанные с использованием пестицидов и
болезнями растений, а главное-уменьшение
плодородия почвы, все это делает подобные проекты
экономически нежизнеспособными. В силу целого ряда
причин влажный тропический лес редко
восстанавливается в первоначальном виде там, где он был
вырублен. Почвы на вырубках теряют запас
питательных веществ и структуру, там уже нет запаса
семян. Но даже в том случае, когда на вырубке
сажают семена деревьев, которые обитали там
раньше, они крайне плохо приживаются на открытом
пространстве: деревья влажного тропического леса
прорастают только при высокой влажности под
пологом сомкнутых крон. В лучшем случае на этом
месте вырастает вторичный лес; он значительно
менее продуктивен, чем изначальный, и хотя здесь
используется подсечно-огневое земледелие,
возможности его крайне ограничены. Ясно, что развитие
интенсивного сельского хозяйства на месте
влажного тропического леса не выгодно. Оно ведет лишь к
деградации лесных ресурсов, тогда как многие
виды, свойственные зрелому лесу, постоянно
исчезают. Таким образом, важнейшая задача, стоящая
перед мировым сообществом,-это поиск таких
путей оптимального использования тропических
лесов, которые учитывали бы особенности
экономического развития прилегающих районов.
Использование вторичного сырья
Эффективная охрана природы будет возможна
только тогда, когда удастся смоделировать
происходящие в природе циклические процессы. Многие
вещества используются в экосистемах многократно,
и возникает вопрос, почему этого не происходит с
продуктами человеческой деятельности. Если речь
идет о каких-то необычных материалах, например
редких металлах, то они иногда используются пов-
торно, но в целом мы представляем собой
«общество транзита».
Во время второй мировой войны
Великобритания перерабатывала значительное количество
макулатуры (слаборазвитые страны Третьего мира
занимаются этим и по сей день). После окончания войны
в стране появились средства на приобретение
древесной массы, и переработка макулатуры как здесь,
так и в других промышленно развитых странах,
отошла на второй план. А между тем технология
этого процесса стала совершенной, что позволяет
сократить производство новой бумаги по крайней
мере вдвое.
Технология обработки сточных вод основана на
использовании процессов жизнедеятельности
гнилостных бактерий; в природе они присутствуют и в
чистой воде, и в почве, где разлагают различные
отходы. Этот естественный процесс и лежит в
основе очистной технологии, которая дает побочный
продукт - активный ил, богатый солями и
органическим веществом. Если он не загрязнен тяжелыми
металлами (например, свинцом из нефтяных
отходов), то его можно вносить в почву в качестве
удобрения. Кроме того, активный ил часто
используется для рекультивации почвы, так как улучшает
ее структуру и содержит много микроэлементов.
Энергию, выделяющуюся при очистке сточных вод,
можно использовать для получения метана (биогаз),
а он в свою очередь дает энергию для работы
очистных сооружений.
Новые источники энергии
Поскольку ископаемое топливо расходуется с
большой скоростью, а эксплуатация атомных
электростанций сопряжена с серьезными проблемами,
многие стали задумываться над идеей создания каких-
либо других источников энергии.
Можно сказать, что Солнце-это термоядерный
реактор, расположенный в 150 млн. км от Земли.
Под воздействием Солнца происходит движение
атмосферы и океана, а также круговорот воды на
Земле; оно служит единственным источником
энергии для фотосинтеза, происходящего в биосфере.
Ветровые, прибойные, гидроэлектрические станции
и солнечные батареи-вот источники энергии
будущего. Для получения топлива можно также
использовать и материалы растительного происхождения.
В Бразилии построены большие заводы по
переработке сахарного тростника в этиловый спирт (газо-
хол), используемый в качестве автомобильного
горючего. Процессы получения энергии из
«биомассы» оказываются экологически более чистыми и
менее трудоемкими, чем добыча ископаемого
топлива. А поскольку запасы ископаемого горючего не
бесконечны, их следует рассматривать как
неприкосновенный запас и заменить альтернативными
энергоресурсами.
Рекультивация промышленных и
городских пустырей
Охрана природы-это, кроме всего прочего, и
восстановление пространств с нарушенной средой,
чтобы сделать их опять пригодными для получения
биологической продукции, отдыха, или
дальнейшего использования в промышленных целях.
Отработанные отвалы шахт и заброшенные
промышленные пустыри иногда представляют собой
значительную опасность: так, например, в 1962 г.
близ г. Аверфен (Ю. Уэльс) произошел большой
оползень. Но если создать в таких местах
растительный покров, то он воспрепятствует чрезмерному
осушению или переувлажнению почвы, улучшит
качество воды, укрепит склоны.
Основная проблема, связанная с
восстановлением растительного покрова на заброшенных
территориях - создание почвы, пригодной для прорастания
и дальнейшего развития растений. При
неблагоприятной температуре и влажности ухудшается
всхожесть растений, а при недостатке нитратов и
фосфатов резко замедляется их дальнейшее развитие.
Терриконы шахт и промышленные отходы могут,
кроме того, содержать тяжелые металлы; в таком
случае их рекультивация практически невозможна:
если разместить в таких местах лесные или
сельскохозяйственные угодья, возникнет серьезная
опасность бионакопления тяжелых металлов в пищевых
цепях. Если почва имеет слишком высокую
щелочность, либо наоборот слишком закислена, как это
бывает в отвалах, содержащих железный колчедан,
появления устойчивого растительного покрова
можно ожидать лишь после предварительной обработки
почвы. Однако на таких не поддающихся
рекультивации пространствах можно снова разместить
промышленные объекты, которые тем самым не будут
распространяться за пределы уже нарушенного
ландшафта.
Терриконы шахт в средней и северной Англии
были засеяны травами, устойчивыми к засолению
почвы, и сейчас они выглядят, как обыкновенные
холмы. Во многих заброшенных каменоломнях
также обычно удается восстановить нормальный
сухопутный ландшафт или превратить их в водоемы,
пригодные либо для рыболовства, либо для катания
на лодках и водных лыжах. В Нортхэмптоншире
5700 га открытых угольных копей были засеяны
злаками и клевером. Через 20 лет почвенный покров
полностью восстановился и эта местность снова"
была отдана под пахоту, а окружающий ландшафт
удалось сохранить нетронутым.
Научное издание
Найджел Грин, Уилф Стаут, Деннис Тейлор
БИОЛОГИЯ
Под ред. P. Сопера
В трех томах
Том 3
Заведующий редакцией чл.-корр. АН СССР T. M. Турпаев
Зам. зав. редакцией M. Д. Гроздова
Ст. научный редактор Ю. И. Лашкевич
Мл. редакторы О. В. Шагинян, И. А. Деменцова
Художник В. Перлин
Художественный редактор А. Мусин, Л. M. Аленичева
Технический редактор 3. И. Резник
Корректор В. И. Киселева
ИБ № 7297
Сдано в набор 31.03.89. Подписано к печати 27.09.90. Формат
84 χ 108/16. Бумага офсетная № 2. Печать офсетная. Гарнитура
тайме. Объем 11,75 бум. л. Усл.печ.л. 39,48. Усл.кр.-отт. 79,81.
Уч.-изд.л. 44,81. Изд. № 4/6698. Тираж 100000 экз. Зак. 450. Цена
5 руб.
Издательство «Мир»
В/О «Совэкспорткнига» Государственного комитета СССР по
делам издательств, полиграфии и книжной торговли
129820, ГСП, Москва, 1-й Рижский пер. 2.
Можайский полиграфкомбинат В/О «Совэкспорткнига»
Государственного комитета СССР по делам издательств, полиграфии и
книжной торговли,
г. Можайск, ул. Мира, 93.
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформлении,
качестве перевода и другие просим присылать по адресу:
129820, Москва,
1-й Рижский пер., д. 2,
издательство «Мир»