/







































































































Текст
АКАДЕМИЯ НАУК
СОЮЗА СОВЕТСКИХ СОЦИАЛИСТИЧЕСКИХ РЕСПУБЛИК
ТРУДЫ
ПАЛЕОНТОЛОГИЧЕСКОГО
ИНСТИТУТА
ТОМ XIII
ВЫПУСК 2
Б. Б. РОДЕНДОРФ
ЭВОЛЮЦИЯ КРЫЛА И ФИЛОГЕНЕЗ
ДЛИННОУСЫХ ДВУКРЫЛЫХ
OLIGONEURA (DIPTERA, NEMATOCERA)
(С 97 рисунками в тексте и 16 таблицами фотографий)
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА
1946
ЛЕНИНГРАД
АКАДЕМИЯ НАУК
СОЮЗА СОВЕТСКИХ СОЦИАЛИСТИЧЕСКИХ РЕСПУБЛИК
ТРУДЫ
ПАЛЕОНТОЛОГИЧЕСКОГО
ИНСТИТУТА
ТОМ XIII
ВЫПУСК 2
Б. Б. РОДЕНДОРФ
ЭВОЛЮЦИЯ КРЫЛА И ФИЛОГЕНЕЗ
ДЛИННОУСЫХ ДВУКРЫЛЫХ
OLIGONEURA (DIPTERA, NEMATOCERA)
(С 97 рисунками в тексте и 16 таблицами фотографии)
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА
1946
ЛЕНИНГРАД
ПРЕДИСЛОВИЕ
Настоящее исследование филогении разных групп двукрылых и эво-
люции их крыльев явилось непосредственным следствием работ по ископае-
мой фауне Diptera из юры Кара-тау, с одной стороны, и результатом
исследования особенностей системы современных видов, с другой.
Такая связь палеонтологического изучения с исследованием системы
современных форм мне кажется единственно научным методом познания
эволюционных взаимоотношений. Необходимо лишь подчеркнуть, что
под «исследованием особенностей системы» мною понимается не только
формальное описание особенностей животных с целью их различения,
но ^всестороннее рассмотрение всех особенностей организмов — их морфо-
логии, развития, биологии и распространения, с непременным условием
охвата всей современной фауны. Отсутствие комплексного характера в ис-
следовании опасно не только для систематики, что очевидного и для мор-
фологии. Действительно, для эволюциониста-морфолога само понятие «эво-
люционной морфологии» заключает в себе обязательство изучать организмы
не тгкачестве чего-то постоянного, неизменного, а, наоборот, как продукт
долгой и разнообразной истории. Эволюция органического мира не может
быть отделена от понятия дивергенции: все современное многообразие
фауны и флоры есть следствие этого процесса. Само многообразие орга-
низмов есть одно из главнейших следствий их эволюции; большее или
меньшее многообразие есть один из важнейших показателей этапов в
Эволюции данной ветви. Морфолог, если он выхватывает для иссле-
дований ту или иную форму из современной фауны группы и при этом ни-
мало не интересуется общим развитием группы в настоящее время, — ко-
роче говоря, детально незнакомый с системой группы, — в значительной
мере обесценивает результаты своего исследования. Незнание системы груп-
пы, к которой относится исследуемый объект, ее распространения, объема?
лишает морфолога знания ряда важнейших моментов, освещающих фило-
генез и эволюцию группы. В результате односторонне морфологических
исследований мы получим в лучшем случае картины филогенезов отдель-
ных органов и весьма шаткие гипотетические схемы филогенезов крупных
систематических категорий.
Я сознаю трудность и непременное замедление темпа работы морфо-
лога при учете им всей системы группы. Но вместе с тем для меня оче-
видно, что только таким путем можно достичь вполне доброкачественных
результатов в области эволюционной морфологии; недоучет системы
Группы по существу равносилен недооценке значения дивергенции в эво-
люции.
В настоящем исследовании я делаю попытку анализа одной группы дву-
крылых насекомых, при использовании всех известных мне данных и до-
полнительно исследую ряд недостаточно известных деталей и& морфоло-
гии. В результате работы стало возможным осветить пути эволюции и вы-
яснить некоторые из ее закономерностей в этой группе, отчасти имею-
щие и более общее значение.
5
1. ЛИТЕРАТУРНЫЕ ДАННЫЕ
Двукрылые насекомые, объединенные в искусственную группу Nema-
tocera Oligoneura еще Шинером (Sehin er, 1864), разнообразны и рас-
пространены всесветно. К этим двукрылым принадлежат насекомые, имею-
щие важное хозяйственное значение в качестве вредителей, и к ним же от-
носятся одни из наиболее древних, первых представителей отряда Diptera,
появившихся в истории Земли еще в
нижнеюрское время.
Многочисленные семейства этой
группы объединяются в немногие бо-
лее или менее естественные надсемей-
ства. Нет особой нужды подробно из-
лагать историю развития наших зна-
ний по системе Oligoneura: необходи-
мо лишь уточнить границы и объем
принимаемых мною главнейших под-
разделений внутри этой искусствен-
ной группы.
Попытки исследования системы
современных Oligoneura делались уже
давно. Таковы схемы ряда авторов
(О s t е n‘-:S acken, Coquillet,
Johannsen и других, их после-
дователей); эти схемы ныне имеют
лишь исторический интерес, являясь
в значительной своей части искус-
ственными построениями. К началу те-
кущего столетия имела широкое рас-
Рис. 1. Fungivora punctata М е i'g. пространение схема деления изучае-
Общий вид насекомого (по jo h а пп- мого комплекса на три крупные семей-
sen, 1909). ства: Mycetophilidae (рис. 1), Cecido-
/ myiidae и Bibionidae; в дальнейшем
эти три семейства были разделены на меньшие: из первого семей-
ства было выделено семейство Sciaridae (—Lycoriidae), из послед-
него— семейство Scatopsidae. Последняя схема пяти семейств до сих
пор наиболее «популярна» и пользуется широким распространением
как в специальной литературе, так и в общих руководствах и учеб-
никах, хотя она явно не отражает действительных взаимоотношений
и структуры системы этих двукрылых. Большим шагом вперед, хотя и не
лишенным крупных ошибок, явилась классификация Моллока (М а 1-
1 о с h, 1917), выделившего из старого семейства Mycetophilidae в качестве
особых семейств Bolitophilidae, Macroceridae и Platyuridae (= Ceroplatidae).
Наконец, наиболее последовательной схемой явилась классификация Ген-
деля (Hen del, 1923, 1936), который правильно выделяет несколько
6
реликтовых групп в качестве особых семейств, правда, в ряде случаев про-
водя это разделение Лишь формально; так, им необоснованно отделены
Sciophilinae от остальных Fungivoridae. С другой стороны анализ проведен
недостаточно подробно: Penthetriidae им оставлены в рамках старого семей-
ства Bibionidae, Lestremiidae составляют лишь часть большого семейства
Itonididae. Кроме того, Гендель сгруппировал установленные им семейства
в три большие надсемейства —Fungivoroidea, Itonidoidea и Bibionoidea; эта
часть его схемы является уже чисто искусственной.
Я считаю необходимым попытаться объяснить существующую в спе-
циальной литературе «малую популярность» классификационных схем,
подобных системам Моллока и Генделя, схем, в которых произ-
ведено дробление старых, хорошо известных семейств, и установление
многочисленных мелких, бедных видами новых семейств.1
Основным в этом вопросе, на мой взгляд, является толкование и пони-
мание природы таких «крайних» форм, выделяемых в особые группы, иногда
характерно называемые «переходными»; такое определение, разумеется,
мало что объясняет, притом затемняя характерные черты этих форм,
всегда с легкостью обнаруживаемые. Действительно, главнейшими осо-
бенностями этих «крайних», «переходных» форм являются их реликтовые
черты, именно, бедность видами (и другими категориями) и своеобразное
Сочетание признаков специализации и примитивности. Мною принято, на мой
взгляд, правильное и всегда при ближайшем анализе доказуемое положе-
ние, что система и систематический состав группы отражают действи-
тельные филогенетические отношения. В свете этого положения вся-
кого рода «крайние» и тем более «переходные» группы не могут трактоваться
иначе как равноценные по таксономическому значению со сравниваемыми
соседними. Другое дело, если вовсе отбросить всякого рода соображения
о филогенетических отношениях групп и лишь формально оценивать сход-
ства и различия современных представителей данной группы, да еще к тому
же признавая признаки «равноценными». При этом последнем условии,—•
на мой взгляд, в корне неправильном,—конечно, возможно существова-
ние и «крайних» и «переходных» форм, так как необходимость их обособления
отпадает. Остается еще вопрос практического удобства: обилие системати-
ческих категорий очень громоздко и неудобно для справочных и учебных
целей. В этом случае очевидна неизбежность этого «неудобства», так как оно
отражает естественность системы, действительные взаимоотношения данной
группы.
Начиная с 900-х годов появляются первые опыты анализа родственных
отношений группы и попытки построения филогенетических схем. Боль-
шинство этих работ основывается на исследовании какой-либо одной си-
стемы органов или части тела (жилкования крыльев, скелета головы или
груди), или, наконец, особенностей метаморфоза; таковы, например, работы
,Л а м э р a (L a m е ё г е, 1906), Эндерлейна (Enderlein, 1911,
' 1916), Веше (Wesche, 1912), М е й е р э (М е i j е г е, 1916а, 1916b),
Моллока (М alloc h, 1917), Крэмптона (Crampton, 1925),
Эдвардса (Edwards, 1916). Наибольший интерес представляет
исследование Крэмптона, посвященное сравнительной морфологии скелета
груди Oligoneura, вернее, — всех длинноусых двукрылых, за исключением
Tipuloidea («Non-Tipuloid nematocerous Diptera» этого автора). В этой ра-
боте подробно исследовано строение склеритов груди, особенно плевр, и
установлены взаимоотношения между семействами по степени развития тех
1 Подобные случаи отнюдь не являются специфической особенностью изучаемой
мною группы двукрылых, будучи широко распространены в систематической литературе
по самым различным группам животных.
7
или иных склеритов. Несмотря на точность и сравнительно большое коли-
чество исследованного материала, заключения автора, хотя и строго аргу-
ментированные, все же являются односторонними: им почти не приняты во
внимание особенности жилкования и другие органы, не говоря уже о мета-
морфозе и биологии.
Работы Крэмптона и других авторов, несмотря на указываемую
мною односторонность, имеют все же крупное значение в качестве очень
ценной сводки описательного материала, позволяющего с большой полнотой
исследовать филогенетические отношения этих насекомых. Подобное же
значение имеют и другие работы, описывающие особенности двукрылых,
подчас даже совсем без попыток использования полученных данных в си-
стематике и тем более — в филогении; таковы, например, исследования
Питерсона (Peterson, 1916), K)Hra(Joung, 1921), Бишофа
(Bischoff, 1922—1924).
Немногие авторы пытались дать очерк филогенеза этой группы двукры-
лых при использовании всех известных им данных по морфологии и метамор-
фозу; мне известны две подобные попытки: это работы Эдвардса (1926)
иГен де ля (1936). Исследование Эдвардса представляет очень сжа-
тую статью, подытоживающую все литературные данные по морфологии и
превращению длинноусых двукрылых; в ней автор подробно разбирает осо-
бенности, используемые им в филогенетических построениях с точки зрения
их специализации. Но из поля зрения автора совершенно выпадает биоло-
гия; кроме того, при разборе особенностей их морфологии главное внимание
'им обращено на анализ отдельных признаков и не сделано попыток оценки
отдельных категорий подотряда в целом; основная цель автора заключается
лишь в установлении родственных отношений семейств.
Обзор филогенетических отношений современных групп двукрылых,
который дает Гендель, значительно интереснее, хотя и не столь точен,,
как исследование Эдвардса.В начале работы Гендель перечисляет
по пунктам те особенности, которые следует считать «примитивными» для
имаго и личинок порознь, а далее переходит к рассмотрению отдельных
«стволов» (Stamme) отряда. Автор пытается дать и «теоретическое» обосно-
вание закономерностей филогенеза двукрылых, но этот отдел очерка Г е н-
д е л я наименее интересен и ценен: в нем мы находим неправильные, но
многократно и разнообразно представленные в литературе рассуждения об
«ортогенезе» и ортогенетическом развитии (автор говорит о «прямолинейном
развитии некоторых систем органов», 1936, стр. 1872). В эту порочную схему
Гендель вводит и все многообразие личиночных форм и, что уже совсем
нетерпимо, настаивает на наличии у личинок «своего собственного филоге-
неза», добавляя при этом, что «естественная система может строиться лишь
на основании особенностей взрослых форм», а что «личиночные признаки
являются лишь к ней дополнением и контролем». Ошибочность такой точки
зрения очевидна: нельзя при исследовании системы группы рассматривать
раздельно особенности развития, с одной стороны, и строение взрослых форм
с другой. Естественная система нами мыслится как следствие и прямое произ-
водное истории —-филогенеза группы: особенности развития животного
есть один из важнейших моментов, позволяющих судить о его родственных
отношениях; вместе с тем эти особенности, так же как и признаки органи-
зации взрослых форм, неразрывно между собою связаны и характеризуют
целое—вид животного. Задачей систематика-филогенетика является изу-
чение филогенеза и построение современных систем групп животных,
но не их органов или стадий развития.
Появление подобных взглядов о «самостоятельности» систем личинок и
взрослых форм, по моему мнению, может быть естественно объяснено той
Крайней сложностью превращения, которая существует у двукрылых
8
Рис. 3. Plecia nigra L u n d. Голова самца спереди; Ус.
сур. край, колл, автора № 2734 (схема, ориг.).
Рис. 4. Plecia nigra Lund. Голова самки спереди;
Уссур. край, колл, автора № 2739 (схема, ориг,).
Голова спереди Рис. 5 Bibio japonicus Duda. Голова сам-
(стема, ориг.). ки спереди.( уССур. край, колл, автора
№ 2743 (схема, ориг.).
одного из наиболее молодых и специализованных отрядов насекомых. Раз-
витие двукрылого, как правило, проходит в совершенно иной среде, чем
жизнь взрослого насекомого. При этом личиночные и взрослые формы дву-
крылых в эволюции разных групп отряда играли, несомненно, очень
неравноценную роль: в некоторых случаях сроки развития очень сокра-
тились— личиночная фаза стала очень краткой и специализ званной,
в других же группах личинки оставались «примитивными» и, наоборот,
imagines крайне сократили срок своей жизни и так или иначе специали-
зовались (их афагия, бескрылость).
Кроме перечисленных выше работ, уместно указать на ценное исследо-
вание Мончадского (1936), хотя и не касающееся непосредственно
изучаемой мною группы, но представляющее несомненный шаг вперед в деле
изучения филогенеза длинноусых двукрылых. Этот автор, изучая личинок
кровососущих комаров, затрагивает вопрос о «несоответствии систем, по-
строенных по признакам личинок и взрослой фазы», и дает правильное
объяснение этому явлению в «различиях условий существования взрослых
комаров и личинок». К сожалению, он, описывая далее и объясняя различия
личиночной и взрослой фаз комаров, оперирует с неточными или ничего не
объясняющими понятиями. Так, неточны его положения о том, что «эволю-
ция каждой из этих фаз (личиночной и взрослой) шла разными путями,
часто в разных направлениях и разными темпами»; что «адаптивная способ-
ность обеих фаз неодинакова», что «у личинок она выражена значительно
сильнее» и что «связи между процессами эволюции обеих фаз обусловливают
каждую из них».
Действительно, наблюдающееся у комаров «несоответствие» систем,
построенных по разным фазам этих насекомых, как будто и наталкивает
исследователя на подобного рода заключение. Но вместе с тем очевидно,
что основная порочность этого положения заключается в недопустимости
раздельного рассмотрения личиночной и взрослой фаз; особенности разви-
тия животного есть один из главнейших характеризующих его признаков,
и «несовпадение» личиночной и имагинальных «систем» указывает только на
их несовершенство и неестественность. Лишь при таком понимании «несоот-
ветствия» систем личинок и комаров возможно плодотворное и правильное
исследование эволюции группы.
Далее тот же автор выясняет интереснейшие связи между характером
питания личинок и взрослых особей и различными специализациями этих
фаз, но, к сожалению, объяснение всех наблюденных фактов упирается
у него в загадочные «компенсаторные» (кавычки автора) изменения другой
фазы, наличие или появление которых никак не объяснены, хотя автор и
очень близко подходит к этому решению, указывая на значение белкового
питания. Мне кажется, что несогласованность эта произошла вследствие
недостаточной эволюционной целеустремленности исследователя, в задачу
которого не входило выяснение закономерностей эволюционного процесса.
Это и заставило его применить ничего не разъясняющий термин «компенса-
торный», поставив его поневоле в кавычки.
Можно кратко резюмировать современное состояние вопроса об эволю-
ции длинноусых двукрылых и, в частности, группы Oligoneura. Лишь в са-
мое последнее время сделаны попытки синтеза данных, существующих по
филогенезу этих насекомых, но при них большинство авторов старалось
выяснить лишь родственные отношения между современными группами
с целью улучшения их системы; биологические особенности и данные палеон-
тологии вовсе не были использованы. Подробные исследования филогенеза
разных групп в свете современных знаний закономерностей эволюционного
процесса при подобной неполноценности метода, естественно, вовсе отсут-
ствовали.
to
2. ОСОБЕННОСТИ СТРОЕНИЯ, РАЗВИТИЯ И БИОЛОГИИ
СОВРЕМЕННЫХ OLIGONEURA
Проведенное исследование современной системы группы Oligoneura,
изменчивости строения йх тела, биологии и развития показало большое
разнообразие в различных семействах. Весь большой отдел длинноусых
Oligoneura объединяет в современной фауне более 5700 видов (Г е н-
дель 1936), распределяющихся среди 20 семейств. Отдельные семей-
ства отличаются и характеризуются строением разных частей тела и органов,
причем жилкование крыльев является главнейшей особенностью, наиболее
точно указывающей взаимоотношения группы, «признаком-индикатором».
Строение головы и ее придатков, ног, груди,
брюшка и, наконец, биологические особенно-
сти,— все это позволяет характеризовать от-
дельные семейства и выясняет их взаимоотно-
шения и группировки. Строение же крыльев
наиболее точно определяет отношения группы.1
Кроме крыльев, как уже указано, в разных
группах Oligoneura изменчив ряд систем органов ;
Рис. 6. а — Ceroplatus sp.,
в — Platyroptilon sp. Антен-
ны (по Johannsen, 1909).
рассмотрим их по отделам тела.
Округлая голова и ее скелет, epicranium,
во всех семействах группы очень постоянны,
мало изменчивы, голова всегда более или менее
приближается к форме шара. Общую ’ее форму
'иногда нарушают удлиненные ротовые органы — у Lygistorrhinidae, неко-
торых Ceroplatidac, Fungivoridae-Sciophilinae и Bibionidae (рис. 2—5).
. Антенны являются, наоборот, очень изменчивыми
1 органами, позволяющими характеризовать семейства и
~ группы. Наиболее просто их строение у комплекса се-
(wW- мейсгв так называемых «грибных комариков», Fungivoridae,
и близких к ним; у них они имеют нитевидную или чет-
/ ковидную форму, с числом члеников до 17; в этих слу-
чаях все членики гомономны и лишь два основных всегда
более или менее увеличены и взаимно объединены (рис. 2).
IprRl Обычное среди Diptera направление специализации
антенн в виде их укорочения и слияния их отдельных
Ц. члеников в этой группе осуществляется- в ряде семейств.
W Простое укорочение антенны испытывают у Bibionidae,
W* Axymyiidae, Penthetriidae (рис. 3—5); у них число члени-
, ков уменьшено незначительно, и укорочение антенн дости-
PKCii^ktrtbv гается лишь сокращением размеров и относительной длины
' Антенна (ориг) члеников, без каких-либо вздутий или им подобных образо-
ваний. Несколько иного рода специализация антенн наблю-
дается у представителей Ceroplatidae(роды Ceroplatus и близкие—рис. 6); эти
насекомые обладают антеннами почти не укороченными, но зато резко утол-
щенными и расширенными в виде пластинок. Наибольшего укорочения,
1 Особое значение для филогении и систематики жилкование крыльев имеет во-
всем классе. Объяснение этого лежит, несомненно, в огромнолиразнообразии жилко-
вания, являющегося в подавляющем большинстве случаев у предковых форм более
сложным, чему потомков, вследствие того, что специализации крыльев входе эволюции
крылатых насекомых осуществляются, как правило, путем той или иной редукции жил-
кования. Способность к полету и органы непосредственно осуществляющие ее—крылья—
в эволюции насекомых играли главнейшую роль; это в конечном итоге, и-, объяс-
няет то особое значение, которое придается жилкованию крыльев в систематике.
II
и притом связанного с образованием комплексных крупных «члеников»,
антенны достигают у некоторых родов семейства Scatopsidae (Arthria, рис. 7)
и некоторых Fungivoridae (Polyxena); у них антенны имеют булавовидную
форму и на конце состоят из одного вздутого образования — продукта слия-
ния дистальных члеников (Arthria). Другого рода специализация наблю-
дается в семействах Macroceratidae, Bolitophilidae и Itonididae, у которых
антенны не претерпевают укорочения, а, наоборот, вторично удлиняются
за счет увеличения размеров члеников. Особенно длинны антенны у Macro-
ceratidae, у которых они часто значительно превышают длину тела, но со-
стоят при этом лишь из 16 сильно удлиненных члеников. Другое семейство
со специализованными длинными антеннами, это — галлицы, Itonididae;
они своеобразны вследствие появления на члениках их антенн особых и
сложно построенных гребней, щетинок и ребер (рис. 8 и 9); сами членики
антенн сильно удлинены и подразделены на несколько частей; общее число
их у этого семейства достигает иногда 30.
Ротовые органы, как уже отмечено, во всей группе развиты очень слабо,,
у некоторых семейств почти редуцированы; вероятно, афагия imagines имеет
.широкое распространение в
этой группе. Лишь у немногих
семейств ротовые органы силь-
но удлинены и образуют длин-
ный хоботок. Часто редук-
ция хоботка связана с полом
и наблюдается лишь у самцов.
Глаза -развиты хорошо;
иногда простые глазки реду-
цированы, например у Itoni-
didae, вследствие разрастания
сложных глаз, или же смеще-
ны к их краям, что наблю-
дается у Fungivoridae-Fungi vo-
rinae. Особенно сильно уве-
личены сложные глаза в се-
мействах Itonididae, Lycorii-
dae и Scatopsidae, у пред-
ставителей которых глаза сли-
1 2
Рис. 8. Contarinia tritici
Kirby. Самец. Части ан-
тенн: 7—1, 2,3 и часть 4
Алеников; 2 — 13 и 14 чле-
ники (по Venturi, 1938).
1 2
Рис. 9. Contarinia
tritici Kirby.
Самка. Части ан-
тенн ; обозначения
те же (по V е п-
t u г i, 1938).
ты один с другим посред-
ством мостика сзади основания антенн (рис. 2). Другого рода специализация
глаз наблюдается в семействах Bibionidae и Penthetriidae; у них глаза дости-
гают необычайного развития лишь у самцов, самки же обладают глазами уме-
ренной величины; при этом увеличение глаз самцов у Bibionidae сопро-
вождается обособлением и видоизменением части глаза (рис. 3 и 4).
Грудной отдел тела довольно изменчив, но недостаточно изучен. Важной
•собенностью его строения является разрастание и увеличение среднеспинки,
надвигающейся вперед, на голову, в виде горба, что связано с развитием
мускулатуры крыльев, с одной стороны, и с характерным образом жизни,
е другой. Подобное разрастание среднеспинки наблюдается,у Fungivoridae
и близких семейств и в меньшей степени у Lycoriidae, Itonididae} у Scatop-
sidae, Bibionidae и Penthetriidae, грудь не увеличивается в области
спинки. Особенности в строении переднегруди служат указателями
характера специализации отдельных групп этих насекомых. Таково разви-
тие плевральных склеритов эпистернитов, которые в ряде групп первично
обособлены от стернальных пластинок; таковы Fungivoridae и близкие к ним.
семейства, далее Scatopsidae, Axymyiidae. Иное наблюдается у Bibionidae,
Penthetriidae, Mycetobiidae, Bolitophilidae, далее у itonididae, Lycoriidae
12
Рис. 11. Leia winthemi L е h m
(Fungivoridae). Грудь, ос-
нования двух задних пар
ног и брюшка. Схема (по
Young, 1921). Сокраще-
ния см. рис. 10.
Рис. 12. Lycoria ochrolabis
L. (Lycoriidae). Грудь, осно-
вания двух задних пар ноги
брюшка. Схема (по Young,
1921), Сокращения см.
рис. 10.
Рис. 13. Rhabdophaga stro-
biloidea Walsh. (Itonidi-
dae). Грудь, основание
брюшка и двух задних пар
ног. Схема (по Young,
1921). Сокращения см.
рис. 10.
Рис. 10. Plecia heteroptera
Say (Penthetriidae). Грудь,
основания двух задних пар
ног и брюшка. Схема (по
Young, 1921), Сокраще-
ния: сх8, сх3 — тазики 2 и
3 пары ног; tr8, tr8 — верт-
луги 2 и 3 пары ног; fe8,
fe8 — бедра 2 и 3 пары ног;
sp8, sp3 — грудные дыхаль-
цы; 1 sp, 2sp, 3sp — брюш-
ные дыхальцы.
13
Рис. 14. Hesperinus brevipes (Hespe-
rintdae). Грудь и тазики 1 и 2 пары
ног; заднегрудь удалена. Схема (по
Crampton, 1925). Сокращения:ale,
sc, pic, bsi, emI; cf — части передне-
груди; aes2, spl2, em2, saf, m2, pt2, mt2,
psc2, p, pas2, sl2—части среднегруди;
cx2, ec2 — тазики передней и средней
пары ног.
Рис. 15. Plecia sp. (Penthetriidae).
Грудь и тазики 1 и 2 пары ног — зад-
негрудь удалена. Схема (по(С г а т-
р t о п, 1925). Сокращения см. рис. 19;
кроме того: арп1; ррп,—части перед-
неспинки; а — шов, делящий зпистер-
нит среднегруди; ptn, sal2 — склери-
ты среднегруди; sp3 — заднегрудное
дыхальце.
Рис. 16. Axymyia furcata Me
A tee (Axymylidae). Грудь и
тазики ног. Схема (по Cram-
pton, 1925). Сокращения см.
рис. 19 и 20; кроме того, р, пт,
s — борозды спинки среднегру-
ди; с, тр — шов между эпи-
стернитами и зпимеритами
среднегруди и углубление в
нем; рга — склерит между
спинкой и плеврой среднегру-
ди; tg — tegula; em3, es3 —
склериты плевр заднегруди;
сх2 — тазик 3 пары ног; аа —
прзанзпистернит; Ьр — пред-
крыловая ямка.
Рис. 17. Philia sp. (Bibionidae). Грудь
и тазики 1 и 2 пары ног; задне-
грудь удалена. Схема (по Cram-
pton, 1925). Сокращения см. рис, 14,
15, 16.
и особенно — у Pachyneuridae, у представителей которых эпистерниты пе-
реднегруди слиты со стернитом, чем, вероятно, достигается более прочная
основа для передней пары ног. Особенности строения плевр среднегруди
еще недостаточно изучены у представителей Oligoneura (рис. 10—24).
14
Большое значение в систематике группы имеет строение ног, которое
очень изменчиво. Большинство семейств обладает крепкими, сильными.
ногами, снабженными разного рода ши-
пами и щетинками; таковы бегатель-
ные ноги Fungivoridae, для которых,
кроме того, очень характерно мощное
Рис. 19. Bolitophila cinerea
Me i g. (Bolitophilidae), Грудь и
тазики ног. Схема (по Cram-
pton, 1925). Сокращения см,
рис. 14, 15, 16.
Рис. 18. Scatopse sp. <Scatopsidae).
Грудь и тазики 1 и 2 пары ног;
заднегрудь удалена. Схема (по
Crampton, 1925). Сокращения
см. рис. 14, 15, 16.]
развитие тазиков (coxae), достигающих необычайно крупных размеров.
Таковы также сильные «хватательные» ноги Bibionidae, снабженные шипами
Рис. 20. Asindulum montanum R о е d.
(Ceroplatidae). Грудь и тазики ног.
Схема (по Crampton, 1925). Сокра-
щения см. рис. 14, 15, 16.
Рис. 21. Mycetobia divergens Walk.
(Mycetobiidae). Грудь и тазики ног.
Схема (по С г a m р t о п, 1925). Со-
кращения см. рис. 14, 15, 16; кро-
ме того, aem, kem, Ьр — дополни-
тельные склериты или их части
среднегруди.
и хорошо развитыми коготками, пульвиллами и эмподием (рис. 25, 26).
Ряд семейств обладает умеренно развитыми ногами. Наконец, крайние члены
этого ряда обладают очень нежными, тонкими и длинными ногами, в большей
15
мере выполняющими сензорные, нежели моторные функции (Itonididae,
Bolitophilidae ).
Повидимому, есть связь между сильным развитием передней пары ног
и слиянием эпистернита и стернита переднегруди; Bibionidae и Penthetriidae,
em.
m
Lycoria
тазики
sp. (Lycoriidae).
ног. Схема (по
Рис. 22.
Грудь и
Crampton, 1925). Сокраще-
ния см. рис. 14, 15, 16, 21; кроме
того, pat — боковые участки спин-
ки среднегруди.
pi
sn,
I аа
aes.
№//> па! &
обладающие крепкими передними ногами,
и Itonididae и Bolitophilidae, хотя и имею-
щие тонкие, но зато очень длинные «сен-
зорные» передние ноги, обнаруживают опи-
санное выше слияние склеритов передне-
груди.
Брюшко у представителей рассматривае-
мых семейств длинноусых очень стабильно
и не обнаруживает большой изменчивости.
Генитальные придатки недостаточно изуче-
ны: самки обычно не обладают удлинен-
ными или более или менее сложными яй-
цекладами, и лишь в семействе Itonididae
и некоторых близких наблюдается силь-
ное развитие разного рода яйцекладов из
дистальных сегментов брюшка, часто до-
стигающих больших размеров и связан-
ных с особенностями биологии этих насе-
комых — специализованных паразитов
растений. Половые придатки самцов про-
сто устроены и лишь в семействе Fungi-
voridae сильно усложнены многочислен-
ными щетинками и подразделениями ос-
новных склеритов. Генитальные придатки самцов, как и у большинства
других двукрылых, имеют большое значение fi видовой
систематике.
Рис. 24. Rhabdophaga brassicoides
Walsh. (Itonididae). Грудь и
тазики ног. Схема (по Cram-
pton, 1925). Сокращения см. рис.
14, 15, 16.
Рис. 23. Со.tocha sp. (Lestremi-
idae). Грудь и тазики ног. Схе-
ма (по Crampton, 1925).
Сокращения см. рис. 14, 15,
16; кроме того, mtn — спинка
заднегруди.
Этими кратко перечисленными особенностями и ограничивается измен-
чивость различных частей тела, вернее его скелета, у взрослых фаз длинно-
усых двукрылых Oligoneura. Внутреннее строение тела—строение нервной,
16
2 Годендорф,
to
Рис. 25.Neosciara pagans Winn, (Lyco-
riidae). Ноги 1, 2 и 3 пары. Схема
(ориг.).
Рис. 27.Contarinia tritici Kirby. Ли^иицз сбоку /h<C>Ve n-
t u г i, 1938).
Рис. 26. Bibio clavipes M’e i'g. (Bibionidae). Ноги 1, 2 и 3 па-
ры. Схема (ориг.).
Рис. 28. Cantfiy-
loscelis antennata
Е d w.— личинка
снизу,; и сбоку; от-
дельно головной
сегмент личинки,
более увеличенный
(no Т о п п о i г,
1927). Сокраще-
ния: sp.—дыхаль-
ца; С — головной
сегмент; 1, II,
III.— грудные сег-
менты;;1—8 брюш-
ные сегменты.
пищеварительной, дыхательной, кровеносной и половой систем — до сих
пор не изучено достаточно подробно.
Прежде чем перейти к подробному описанию строения крыльев, необхо-
димо кратко описать метаморфоз, строение личинок этих двукрылых и их
биологию.
Строение яиц и характер яйцекладки известны лишь для немногих пред-
ставителей Oligoneura. В большинстве случаев яйца откладываются пооди-
ночке в питающий субстрат и обладают нежным хорионом. Лишь у некото-
рых групп (Mycetobiidae ) яйца откладываются в виде больших скоплений—
яйцевых масс.Личинки Oligoneura наиболее примитивны среди всех двукры-
лых, если понимать под примитивностью наличие хорошо развитой головы
с сильно склеротизованным epicranium, присутствие сложных ротовых ор-
ганов, построенных по плану, общему для всех насекомых с жующими рото-
выми частями, и хорошее развитие органов чувств на голове — антенн,
достигающих у Вolitophilidae особенно крупных размеров. Дыхательная
система построена по перипневстическому типу: трахеи сообщаются с внеш-
ним воздухом посредством парных дыхалец, расположенных почти на всех
сегментах тела. Куколка свободная, иногда лишь заключенная в слабый
кокон, сплетенный личинкой из шелковых нитей. У большинства семейств
группы специализации в строении личинок и куколок относительно незна-
чительны. Таковы, например, сильная склеротизация головной капсулы
у Fungivoridae и образование различного рода склеротизованных анасто-
мозов в шейной области головы у Lycoriidae, образование особых дыхатель-
ных трахейных выростов на заднем конце тела у Ceroplatidae.
Лишь два семейства из всей группы —• Itonididae и Corynoscelididae —
обладают резко отличными и высоко специализованными личинками, ли-
шенными головного скелета — epicranium и сложных жующих ротовых
органов (рис. 27, 28). У одного из этих семейств—галлиц (Itonididae),—
кроме того, наблюдается характерная специализация метаморфоза, именно
наличие пупария или ложной куколки. Эта особенность конвергентно
возникала в целом ряде несвязанных генетически стволов Diptera и, не-
сомненно, во всех случаях сыграла важную роль в эволюции этих групп.
Особенности биологии изучаемых двукрылых в достаточной мере разно-
образны и очень характерны для отдельных семейств. Личинки семейств
Fungivoridae, Ceroplatidae и близких живут на разных грибах; при этом ха-
рактер питательного субстрата личинок очень разнообразен: это ткани жи-
вых высших грибов — базидиомицетов и аскомицетов — и проросшая
мицелием древесина. Ныне наиболее богатое видами семейство Itonidi-
dae тесно связано с сосудистыми растениями: личинки галлиц явля-
ются одноядными паразитами живых растений, преимущественно покры-
тосемянных. Личинки других семейств, биология которых известна, живут
в гниющих растительных и животных веществах, экскрементах. В этой
последней группе, естественно, моно- или олигофагия выражена слабо.
Необходимо отметить, Фто биологические особенности и метаморфоз ряда
семейств — Ailactoneuridae, Hesperinidae, Lygistorrhinidae, Manotidae, Axy-
myiidae, Penthetriidae, т.е. почти треть всех современных семейств группы,
до сих пор остаются неизвестными, хотя как раз знание их очень важно
для понимания путей эволюции.
3. ЗНАЧЕНИЕ ЖИЛКОВАНИЯ КРЫЛЬЕВ В СИСТЕМАТИКЕ
NEMATOCERA
При рассматривании системы семейств и подсемейств Nematocera Oligoneura
очевидно, что основными руководящими систематическими признаками,
позволяющими выяснить взаимоотношения различных групп этих насе-
18
комых, являются особенности строения крыла, именно особенности его
жилкования. Жилкование крыльев насекомых по существу определяет
главную функцию крыльев — их полет; жилки являются скелетными,
опорными элементами крыла. Поэтому всякого рода изменения жилкова-
ния— передвижения жилок, их слияние, редукция или, наоборот, ново-
образование их должны рассматриваться только в связи с изменением
функции крыла как органа полета. Короче говоря, поскольку жилкование
является приспособлением крыла для целей полета, постольку мы должны
каждое наблюдающееся различие жилкования у разных групп описывать
и оценивать как то или иное изменение качества органа полета — крыла.
Надо, однако, сразу же оговорить очень большие технические трудности,
которые возникают при попытках провести необходимый механический
анализ различных форм крыльев — «специализованных» и «неспециали-
зованных». Эти трудности с ясностью вытекают из ряда причин; в изучае-
мой группе Diptera, например, они таковы: неразработанность техники
механического анализа крыльев мелких объектов (от 1 до 5 мм величи-
ной), обилие материала (сотни объектов — отдельных родов, анализ ко-
торых необходим), часто незначительные отличия жилкования, учет меха-
ники которых, несомненно, потребует очень большой точности, наконец,
как последний момент, невозможность провести анализ ископаемого ма-
териала, когда по существу налицо лишь плоскостной отпечаток, часто
плохо сохранившийся. Поэтому полностью избежать упрека в неточности
и отсутствии научной объективности при оценке тех или иных изменений
жилкования не всегда возможно. ........
Остается единственный путь оценки жилкования, по которому обычно
и следуют, — чисто описательное исследование наблюдаемых редукций
и других превращений жилкования. Подобное исследование жилкования
в разных группах насекомых все же много дает для понимания направле-
ний эволюционного процесса. Так, с несомненностью выясняется путь
специализации крыльев, заключающийся, например, в изучаемой группе
двукрылых в редукции жилок, с одной стороны, и в их смещении к перед-
нему краю и к основанию крыла, с другой. Оба эти процесса всегда сопро-
вождают один другой, будучи неразрывно связаны взаимно; в результате
их мы наблюдаем те крайние формы специализации жилкования, которые
осуществились, например, у Scatopsidae.
Конечно, одними процессами усиления и укрепления переднего края
крыла — «костализацией» — не ограничиваются направления -эволюции
крыльев даже в этой одной группе длинноусых Diptera. Существуют и
иные процессы специализации крыльев; таково, например, удлинение
крыльев и смещение развилков R, RS и М в дистальную часть крыла у Ti-
puloidea и ряд других случаев; описывать все направления эволюции крыльев
Diptera не входит в мою задачу; я лишь хочу указать на плодотворность
чисто описательного метода изучения жилкования крыльев при обязатель-
ном и возможно более точном учете функциональных моментов. При по-
добном методе исследования наличие ископаемого материала представляет
выдающийся интерес, так как сразу же выясняет и объясняет намечаю-
щиеся на современном материале особенности жилкования: палеонтоло-
гические данные являются тем необходимым и безупречным коррективом,
который так нужен при всех филогенетических построениях, проводимых
на современном материале.
Литературные данные по строению крыла и полету насекомых в на-
стоящее время уже достаточно многочисленны. В мокИТастоящую задачу не
входят рассмотрение и изложение современного состояния вопроса о полете
насекомых: необходимо лишь кратко обрисовать те основные направления,
по которым велись исследования, и успехи, которые сейчас-уже достигнуты.
*
19
Работы морфологов (Janet, 1899; В е г 1 е s е, 1909, 1924; Snodgrassr
1909, 1927; Weber, 1933 и др.), в которых разбирается строение грудного
отдела насекомых — его скелет и мускулатура, позволили выяснить глав-
нейшие особенности тела насекомых и являются сейчас основными исследо-
ваниями, обеспечившими дальнейшее развитие изучения полета. С другой
стороны, полет насекомых изучался также и с точки зрения чисто механи-
ческой, без попыток подробного анализа особенностей насекомых и их
морфологии. В этом направлении исследованию подверглись не только
крылья насекомого, но и все его тело в целом: насекомые изучались как
летательные аппараты. Такова, например, работа Демоля (Demoll,
1918), в которой различные насекомые и птицы изучались чисто аэродина-
мически: исследовались вес насекомого и его частей, поверхность крыльев,
скорость полета. Особого совершенства этого рода исследования достигли
в работах Маньяна и его школы (Magnan, 1934; Magna n et
Saint e-L ague, 1933a, 1933b, Magnan et Girard, 1937 и др.).
Эти авторы применили для изучения движений крыльев метод сверхбыст-
рой киносъемки (до 25 000 снимков в 1 сек.), которая позволила разложить
на отдельные этапы движения крыльев в полете и выяснить направления
создаваемых крыльями потоков воздуха. Морфология насекомых в иссле-
дованиях Маньяна разрабатывается очень недостаточно: лишь форма
крыла (длина, ширина, его сечение) несколько затронута изучением, да
и то лишь в самых общих чертах, хотя и подверглась отчасти и матема-
тическому анализу (Magnan, 1934; Amans, 1901). Эти исследования
далее идут в направлении экспериментального изучения полета насекомых,
и более точного анализа всех общих механических моментов этого процесса
путем иостройки разного рода летающих моделей (Ш ва й к о в с к.и й, 1937).
Основной и характерной чертой в работах по изучению полета и осо-
бенностей летательных органов насекомых является стремление исследот
вателей охватить и анализировать все главные группы насекомых — стре-
коз, прямокрылых, жуков, перепончатокрылых, бабочек и мух. Особенно
это разительно в работах по механике полета, в которых авторы,
стремясь охватить все возможное разнообразие, всегда исследуют насеко-
мых самых разных отрядов. В результате подобные исследования дают
лишь самую общую картину особенностей полета насекомых, картину,
которая не позволяет судить о действительной эволюции полета и органов,
его осуществляющих. Эти данные ни в какой мере не могут удовлетворить
запросов биолога-эволюциониста и указывают лишь на те или иные меха-
нические особенности разных насекомых, что имеет общий интерес, и то
лишь для аэродинамика.
Но возможно наметить направление исследований по механике полета
и строению крыла, которые дадут действительно ценные результаты и для
эволюциониста. Так, несомненно, что точное исследование механики крыла
позволит подвести научный базис под все суждения морфолога о том или
ином характере летательной функции. Вместе с тем очевидно, что вместе
с большой сложностью изучения летательной функции крыла насекомого
сама методика этого исследования вполне выходит из сферы биологических
дисциплин: исследователь должен оценить структуры различных крыльев
и тел насекомых в целом как те или иные летательные аппараты, указать
на их особенности и взаимные различия. Роль биолога в этом исследовании
своеобразна: само Изучение объектов в своей методике почти не будет ну-
ждаться в обычных биологических приемах, основным же будет исследова-
ние аэродинамических и механических качеств насекомого и его крыльев
со всеми вытекающими из них особенностями этой работы.
Не следует думать, что биолог при этом останется только безучастным
зрителем, лишь доставившим нужный материал для аэродинамика и меха-
20
ника: наоборот, наиболее ответственным в подобных исследованиях яв-
ляется именно подбор материала и направление исследований механика
в нужную сторону. Только изучение механизма полета определенной и
большой группы близких форм насекомых сможет дать ответ как на во-
просы биолога-эволюциониста, так и на запросы аэродинамика. Необычай-
ное разнообразие насекомых и разнообразие типов их полета ведет к тому,
что исследования механики полета насекомых «вообще» не дают опреде-
ленных ответов на вопросы о генезисе того или другого типа крыльев:
обычно описываемые насекомые очень далеки между собою и малосравни-
мы одно с другим вследствие обладания очень различными летательными
особенностями. Изучая и описывая разные типы крыльев и полета насе-
комых, исследователь лишь статически регистрирует (возможно, и очень
точно) явления. Но моменты, выясняющие возможные пути филогенеза
летательных функций при подобных общих исследованиях над далекими
одна от другой формами насекомых, вовсе выпадают или страдают крайней
гипотетичностью.
Совсем иное мы получили бы при исследовании механики полета у пред-
ставителей одной определенной группы насекомых и достаточно при этом
разнообразной, например семейства. В этом случае вполне возможен был
бы ответ на вопрос: чем механически отличаются крылья, скажем, у пред-
ставителей двух соседних родов или семейств, отличающихся особенно-
стями жилкования? Конечно, в ряде случаев отличия окажутся малыми и
трудно регистрируемыми, но зато ценность подобных данных была бы очень
велика. Точное знание механических качеств крыльев многих различных,
но близких форм даст морфологу недвусмысленный ответ на вопрос о боль-
шем или меньшем адаптивном значении крыла; ценность подобных све-
дений не нуждается в пояснении.
4. СТРОЕНИЕ КРЫЛЬЕВ И НАПРАВЛЕНИЯ ИХ СПЕЦИАЛИЗАЦИИ
Как указывает само название изучаемой большой группы длинноусых
двукрылых Oligoneura, крыло их обладает сокращенным количеством жилок
по сравнению с другими семействами, объединяемыми под именем Poly-
пеига. Вместе с тем несомненно, что количество жилок крыла у этой группы
двукрылых не является главным признаком, отличающим их от соседних,
среди которых также есть формы с уменьшенным числом жилок; с другой
стороны, среди Oligoneura, особенно у архаичных семейств, наблюдается
более полное жилкование. Для Oligoneura гораздо более характерна форма
крыла, которое очень редко удлиняется и никогда не сужается в виде сте-
белька при основании.Также характерно строение тела и, в особенности,
брюшка, почти никогда не приобретающего удлиненной, параллельно-
крайней формы.
Крылья насекомых покрыты на обеих поверхностях короткими щетин-
ками, волосками или чешуйками. У представителей изучаемой нами группы
крыло снабжено очень короткими (в 5—20 р.) и тонкими, густо расположен-
ными микротрихиями, покрывающими, как правило, большую часть по-
верхности крыла. Кроме микротрихий, поверхность крыла снабжена у не-
которых форм более крупными, около 40—75 р, и резко отличимыми от ми-
кротрихий щетинками — макротрихиями. Микротрихии присутствуют на
крыльях у громадного большинства двукрылых, в том числе у всех Oligo-
neura; Наиболее густы и длинны, до 25 р, они у Bibionidae и Penthetriidae
(табл. I, фиг. 2 и 3), покрывая притом почти всю поверхность их крыльев.
У других семейств микротрихии более короткие и редко превышают 12—
15 р; некоторые участки поверхности крыльев лишены микротрихий. Боль-
шие щетинки-макротрихии расположены у большинства семейств лишь
21
на поверхности продольных жилок и отсутствуют, как правило,
на свободной перепонке: лишь у некоторых групп перепонка несет,
кроме микротрихий, также и менее многочисленные макротрихии (Согупо-
scelididae, Ditomyiidae и другие, табл. VII, фиг. 17). Наличие макротрихий
на свободной перепонке крыла рядом авторов (например Эдвардсом, 1924)
считается за признак более примитивный; исчезновение их является, таким
образом, явлением вторичным. Мне кажется, что это воззрение, в основе
правильное, все же нуждается в известном дополнении и разъяснении в том
отношении, что есть еще и третий случай, когда крыло снабжено много-
численными микротрихиями, покрывающими всю его поверхность, но до-
стигающими относительно большой величины. Подобный случай наблю-
дается у представителей Penthetriidae и Bibionidae; возможно, что эти
щетинки мы не имеем права трактовать в качестве микро- или макротрихий: /
они, вероятно, являются остатками первичного щетинистого покрова
крыльев, еще не дифференцированного на мелкие и крупные волоски.
Жилки на крыле Oligoneura, как и других насекомых, распадаются на
«верхние» и «нижние», расположенные на продольных выпуклостях или
в продольных бороздах крыла. Эта особенность жилкования, вызванная,
несомненно, механической потребностью крыла как органа полета, именно
Рис. 29. Zeltnira setnirufa М е i g. Разрез крыла на уровне середины основания —
через середину basianalis 1. Рисунок показывает относительные толщины мембраны
и жилок и их взаимное положение. Сокращения: ВАпх—basianalisl; ВАпа — ba-
sianalis 2; BSC—basisubcosta; ВС — basicosta; BR—basiradius (ориг.).
необходимостью его укрепления, вместе с тем дает в руки исследователя
верное средство для точного различения отдельных жилок, позволяя гомо-
логизовать их (рис. 29).
Прежде всего необходимо рассмотреть строение основания крыла, на
котором лежат базальные отрезки («рукоятки») главнейших стволов жилок.
Основание крыла естественно определяется как часть его, лежащая между
аксиллярными склеритами-эпидемами (epidemata), посредством которых
крыло причленяется к груди, и уровнем плечевой поперечной жилки,
с одной стороны, и выемкой между крылышком (alula) и анальной ло-
пастью (lobus analis), с другой. Основание крыла представляет сложный
орган, по форме очень далекий от плоскости, к которой приближается
большая часть крыла (рис. 29).
Передняя часть основания состоит из более или менее широкой, ди-
стально расширяющейся, плоской, почти горизонтально расположенной
пластины — костальной ячейки, спереди окаймленной костальной жилкой
и сзади постепенно понижающейся и переходящей в субкостальную бо-
розду, в которой лежит «нижняя», субкостальная жилка SC. Непосред-
ственно позади SC лежит «верхняя», очень крепкая радиальная жилка R.
Обе эти жилки, SC и R, ближе к основанию крыла, т. е. проксимально, по-
степенно утолщаются, образуя неправильной формы утолщения; их про-
ксимальные концы — basicosta, basisubcosta и basiradius, соединяющиеся
с эпидемами, опять заметно утончаются (рис. 30—40; табл. I—VII).
22
Задняя часть основания крыла наиболее изменчива, и в ее строении
у отдельных групп Oligoneura наблюдаются различные типы специализации
жилкования, сводящиеся в основном к редукции базальных частей аналь-
ных жилок. Верхняя жилка задней части основания крыла — анальная
•вторая (Ап2) хорошо развита у представителей семейств Bibionidae и близких
Рис. 30. Bibio japonicus Duda (Bibionidae). Основная часть
крыла, basiala. Схема. Сокращения обычные, кроме того Sq —
крыловая чешуйка; А1 — крылышко; La — анальная лопасть;
Ch — хетарии; знаками 4-и— обозначены выпуклые и вогнутые
жилки (ориг.).
(рис. 30); проксимальный конец этой жилки (basianalis 2) сильно увели-
чен и раздут, близко прилегая к основанию радиальной жилки (basiradius).
Места соприкосновения basiradius и basianalis 2 снабжены густораспо-
ложенными, короткими и крепкими щетинками в виде щеток, которые
можно назвать хетариями, анальным и радиальным (chaetarium ra-
diale, ch. anale); оба эти хетария (щетки) на основаниях~р?1диальной и аналь-
ной второй жилок могут тесно соприкасаться один с другим; при этом их
взаимное движение строго ограничивается определенным направлением
именно, «по ворсу»; в ином направлении щетинки их неподвижно
23
фиксируют. 1 Непосредственная связь Ап2 с описанным выше хетарием
хорошо выражена лишь у Bibionidae и Penthetriidae (рис. 30); у Fungivo-
roidea в узком смысле (Fungivoridae, Ceroplatidae и др.) анальный хетарий
прекрасно выражен, но зато утерял всякую связь со слабой Апа; чаще
же последняя вовсе редуцирована и от нее сохраняется лишь хетарий
(рис. 31—35).
Рис. 31. Bolitophila maculipennis Walk. (Bolitophilidae). Основная
часть крыла, Lbasiala. Схема. Сокращения обычные (ориг.).
JCoBceM особого рода устройство этой части крыла у Scatopsidae и Согу-
noscelididae (рис. 36, 37), у которых в построении радиоанального хетария
принимает участие и basianalis 1, обычно лежащий свободно в перепонке
крыла. Механическое значение радиоанального хетария, несомненно, очень
велико. Присутствие хетариев сопровождается всегда специализацией
крыла в направлении усиления переднего и ослабления заднего края крыла;
наоборот, хетарии отсутствуют или плохо выражены в тех случаях, когда
крыло не испытывает указанной специализации, например у Tipuloidea.
Короче говоря, хетарии присутствуют в случаях резко выраженного про-
пеллирующего полета и отсутствуют при гребном полете; имея в виду не-
достаточную определенность этих типов полета, можно сказать, что
1 Можно предположить действительное назначение описанного образования: ще-
тинки, покрывающие радиальный хетарий, направлены дистально, а щетинки аналь-
ного хетария — проксимально; при движении конца крыла вперед хетарии соприка-
саются, и анальная часть крыла со 2-й анальной жилкой фиксируется; при движении
крыла назад хетарии расходятся, и крыло смягчается и складывается.
24
La
Рис. 32. Boletina trilineata Meig. (Fungivttridae). Основная
часть крыла, basiala. Схема. Сокращения обычные (ориг.).
Рис. 33. Ze Im ira semirufa M e i g. (Ceroplatidae). Основная часть
крыла, basiala, Схема, Сокращения обычные (ориг,),
Рис. 34. Asindulum flavum M e i g. (Ceroplatidae). Основание
крыла, basiala. Схема. Сокращения обычные; см. рис. 30 (ориг.).
Zzz
Рис. 35. Symmerus annulatus М е i g. (Ditomyiidae). Основание
крыла, basiala. Схема. Сокращения обычные; см. рис. 30 (ориг.).
Рис. 36. Corynoscelis eximia Boh. (Corynoscelididae). Ос-
нование крыла, basiala. Схема, Сокращения обычные (ориг.).
Рис. 38. Puchyneura fasciata Z e 11. (Pachyneuridae). Основа-
ние крыла, basiala. Схема. Сокращения обычные (ориг.).
i
развитие костализации обусловливается увеличением числа взмахов в по-
лете, ускорением движения крыльев.
Жилки R и Ап2, т. е. те, рукоятки которых образуют описанные выше
радиоанальные хетарии, дистально резко расходятся; в углублении между
расходящимися стволами R и Ап2 расположена рукоятка Апп которая
является нижним опорным элементом задней половины основания крыла.
Рукоятка А^ (basianalis 1) проксимально лежит свободно в мембране крыла
и лишь у Bibionidae и Penthetriidae более или менее тесно прилегает к ba-
sianalis 2. В отличие от basianalis 2 рукоятка Апх присутствует всегда,
несомненно, являясь одной из важнейших частей основания крыла: даже
в случаях крайних редукций жилок, например у Lycoriidae и Itonididae,
basianalis 1 всегда может быть отмечена. В построении basianalis 1 прини-
мает участие, невидимому, в некоторых случаях также и’ прилегающая
спереди жилка СиР. В ряде случаев эта связь несомненна, например у Ly-
coriidae и Fungivoridae’, в других же случаях нельзя с определенностью ут-
верждать об участии СиР в построении basianalis 1 (у Penthetriidae, Bibioni-
dae, Pachyneuridae).
Последним опорным элементом основания крыла является рукоятка
СиА — basicubitus. В громадном большинстве случаев этот отрезок куби-
тальной жилки у представителей Oligoneura нацело редуцирован, невиди-
мому, в связи с общим направлением специализации крыла в сторону раз-
вития пропеллирующего полета. Лишь у некоторых немногих групп (Pachy-
neuridae, Bolitophilidae, рис. 38, 31) basicubitus развит в виде более или
менее длинного отростка, расположенного проксимальнее от места ответ-
вления М, лежащего под известным углом к дистальной части СиА и своим
проксимальным концом прилегающего к basianalis 1.
Заканчивая описание базальной части крыла, необходимо указать еще
на так называемый «базальный перелом» крыла. Приступая выше к опи-
санию особенностей строения основания крыла, я условно принял, что
дистальная граница основания крыла лежит на уровне плечевой попереч-
ной жилки и выемки между alula и lobus analis. Рассматривая строение
крыла разных двукрылых, в том числе и представителей Oligoneura, можно
легко заметить, что на уровне плечевой поперечной жилки наблюдаются
изменения в строении остального жилкования. Наиболее часты изменения
ствола R, заключающиеся по меньшей мере в легком его изгибе (у Сего-
platidae, Macroceratidae, Bibionidae, рис. 30, 33). В большинстве случаев
R в указанном месте крыла образует резкий изломили вырост назад ~(Phry-
jieidae, рис. 39), постепенно утончающийся и сливающийся с мембраной.
Сзади на этом же уровне относительно плечевой жилки лежит основание
жилки Мив случае обычной редукции basicubitus’а, основание СиА. Далее
назад на этом же уровне крыла располагается выемка — граница между
анальной лопастью и alula. В заключение необходимо отметить характер
поилок, проходящих или начинающихся на описанной зоне «перелома».
Три передние жилки, С, SC и R, проходя через зону, не меняют своего
положения верхних или нижних жилок. Нижняя жилка М начинается
лишь в зоне перелома. Жилки СиА, СиР и Апп переходя зону, ведут себя
очень своеобразно, меняя свое положение. СиА начинается нижним basi-
cubitus, переходящим в верхнюю СиА; СиР начинается верхней складкой,
постепенно превращающейся в нижнюю СиР; Ап1; начинаясь нижним
-basianalis 1, превращается в верхнюю Anv Все описанные особенности
этой части крыла с несомненностью указывают на особое, ближе еще не-
ясное значение зоны «перелома» крыла, вероятно, играющей роль своеобраз-
ного эластичного сочленения крыловой пластинки с основанием крыла.
Особой редукции подвергается основание крыла у Itonididae в связи
с выработкой своеобразного полета этих мелких насекомых: исчезают «ру-
29
коятки» многих жилок, редуцируются alula, анальная лопасть и крыловая
чешуйка, basiradius и basianalis 2 утолщаются, тесно прилегая друг к другу
(рис. 40).
Дистальная часть крыла двукрылых есть по существу большая часть
крыла, за исключением самого основания, лишь очень незначительного по
площади. Жилкование дистальной части крыла, в отличие от основной,
изучено с большой полнотой. Нам нет необходимости излагать длинную
историю развития знаний жилкования двукрылых. Наиболее точно описали
и интерпретировали жилкование крыла двукрылых Тильярд (Til-
lyard, 1918—1919) иАле к сан дер (Alexander, 1927). Тиль-
ярд, называя определенные ветви и жилки, учел и весь известный ему
палеонтологический материал по группе Mecopteroidea (его «Panorpoid-
Complex»). Совсем особо следует сказать о работах Виньона (Vignoa
Рис. 40. Contarinia tritici Kirby (Itonididae). Основание крыла,
basiala (по Venturi, 1938).
1932а, 1932b) иГенделя(Непбе1, 1936). В и н ь о н, исследуя жил-
кование самых различных двукрылых, учитывает все явно вторичные жил-
коподобные образования (например vena spuria у Syrphidae) и ставит их
в связь с истинными1 жилками; созданную таким образом искусственную
схему жилкования этот автор «подкрепляет» исследованием разных иско-
паемых групп Insecta и в заключение дает схему крыла «примитивного
двукрылого». Достаточно взглянуть на эту схему, чтобы стала ясной глу-
бокая ошибочность всех построений Виньона: жилкование его «примитива»
соответствует специализованным крыльям различных Palaeoptera, например
некоторых Paleodictyoptera, и никакого отношения к Mecopteroidea иметь,
конечно, не может. Эта схема’—яркий пример искусственного гипотети-
ческого предка, соединяющего в себе все возможные особенности потомков.
Очевидно, метод Виньона — простой «синтез» всех известных особен-
ностей жилкования различных двукрылых, и получение такой «примитив-
ной» синтетической формы в корне неправильно. Для того чтобы знать,
30
каким жилкованием обладали предки Diptera, совсем не нужно заниматься
подобной «синтетической» работой. Наиболее примитивным и мало специа-
лизованным крылом среди всех Mecopteroidea обладали некоторые предста-
вители отряда Mecoptera, например пермские Permocfwristidae (рис. 41);
их примитивность сказывается прежде всего в слабой выраженности «коста-
лизации» — укрепления переднего края, причем расположение жилок
крыла сохраняет свое первичное, симметричнее относительно продольной
оси крыла состояние, именно то, что наблюдается в крыльях Bl attoidea.
Рис. 41. Sylvopanorpa carpenteri Mart. (Permochoristidae,
Mecoptera). Крыло пермской скорпионницы; жилкование
имеет сходство с предполагаемым предком Diptera.
Подобные крылья с равномерно распределенными жилками, несомненно,
примитивны и с точки зрения их механики: отсутствие моментов укрепле-
ния переднего края крыла показывает незначительную разницу нагрузки
(давления) при полете между передним и задним краем; иначе говоря, ско-
рость поступательного движения подобного насекомого при полете была,
вероятно, очень невелика. Таким образом, с полной очевидностью можно
видеть несостоятельность схемы В и н ь о н а, крыло которой обладает
чертами уже большой специализации по сравнению хотя бы, например,
Рис. 42. Схема жилкования исходных, предковых форм,
Diptera (ориг.).
с современными Psychodidae. Не останавливаясь подробнее на высказанных
положениях о путях специализации крыла двукрылых (рис. 42), следует
лишь вкратце описать изменения Жилкования в изучаемой группе длинно-
усых — развитие и редукцию различных стволов жилок.
Костальная жилка (costa, С) идет по самому краю крыла и в громадном
большинстве случаев развита на протяжении лишь одного переднего края.
Наиболее крепка и толста эта жилка в проксимальной-лоловине крыла;
далее к вершине она постепенно утончается и внезапно оканчивается,
оставляя край крыла, свободным от жилки, перепончатым. Место окончания
С на краю крыла всегда очень определенно и колеблется у разных семейств
группы Oligoneura лишь между концом последней ветви R и серединой
расстояния между последней и концом передней ветви М.
Субкостальная жилка (subcosta, SC), расположенная непосредственно
позади костальной и обычно на конце соединяющаяся с ней, является пер-
вой «нижней» жилкой крыла. Эта- жилка всегда простая, неветвящаяся;
часто она очень слаба и оканчивается свободно в перепонке крыла (у Fun-
givoridae), притом сильно укорачиваясь (у Manotidae). Иногда присут-
ствует особая поперечная жилка, соединяющая дистальный конец SC со
стволом R; эту поперечную иногда ошибочно называют второй ветвью SC
(SC2, Johannsen).
Радиальные жилки (radius и sector radii, R и RS) наиболее изменчивы
в своем числе и строении. Передняя, неветвистая ветвь, собственно-ра-
диальная жилка (R) всегда очень крепкая и толстая, является на всем
своем протяжении резко выраженной «верхней» жилкой. От нее ответ-
вляется RS — радиальный сектор, который может ветвиться. Ряд семейств
обладает неразветвленной RS (Lycoriidae, Itonididae, Fungivoridae, Diado-
cidiidae, Bibionidae, Scatopsidae). У других семейств RS разветвляется на
две ветви (Ceroplatidae, Macroceratidae, Bolitophilidae, Ditomyiidae, Myce-
tobiidae, Penthetriidae, Hesperinidae. Наконец, у представителей двух релик-
товых семейств — Pachyneuridae и Axymyiidae — три ветви. Строение ра-
диальных жилок является одним из ведущих диагностических признаков
в группе Oligoneura, позволяющим различать семейства; вместе с тем эти
жилки, будучи расположены в передней части крыла, первые претерпевают
редукцию при его специализации — усилении переднего края. Поэтому
степень редукции радиальных жилок и число их ветвей может служить
одним из указателей меньшей или большей специализации крыла.
Медиальная жилка (media, М) очень постоянна в этой группе дву-
крылых и почти всегда присутствует в числе трех ветвей. Особенно харак-
терна вилка MiJ-Ma, хорошо выраженная у большинства семейств. Спе-
циализация жилкования отражается на этой системе жилок своеобразно:
жилки не перемещаются на крыле и не сливаются одна с другой; происхо-
дит лишь редукция медиальных жилок, притом идущая от основания крыла
к его вершине. Первым редуцируется базальный отрезок жилки М3 (вер-
нее М4), что наблюдается у Fungivoridae, Lycoriidae, Lestremiidae, Scatop-
sidae и некоторых других; в этом случае жилка М3 оказывается вполне
обособленной от остальной части системы М, что и дает повод некоторым
авторам (Н е n d е 1) считать кубитальную жилку двукрылых (СиА) ветви-
стой, в разрез всем данным палеонтологии. Иного рода редукция М наблю-
дается у Macroceratidae, Ceroplatidae, Bolitophilidae, Mycetobiidae и ряда
других: у них происходит ослабление и полная редукция общего ствола М;
при этом у двух первых семейств основной ствол жилок М4 и М2 сливается
на некотором протяжении с RS. Наконец, крайняя степень редукции М
наблюдается в семействах Lygistorrhinidae, Manotidae, некоторых Fungi-
voridae (Azaninae, Morioclonini), у которых редуцируются части ветвей М
вплоть до их дистальных концов или же нацело исчезает задняя ветвь М.
Особо следует описать характерную специализацию основной части-М,
RS и поперечной радиомедиальной жилки rm. Первичное положение rm,
лежащей поперечно между основаниями RS и М, у некоторых групп изме-
няется; так, у многих Fungivoridae rm приобретает косое положение, при-
чем угол между ней и дистальной частью RS резко уменьшается и в даль-
нейшем вовсе исчезает: rm становится непосредственным продолжением RS
к основанию крыла. Одновременно с этим процессом меняется и угол между
rm и основным стволом Mlt2. Крайняя стадия этой специализации наблю-
дается в семействе Lycoriidae, жилки MJ-rmJ-RS составляют одну тол-
стую и крепкую жилку, лежащую рядом с R. Основной отдел RS превра-
32
щается в короткую поперечную между ними. Образование этой крепкий
жилки M-J-rm-J-RS осуществляет сильное укрепление переднего края
крыла. ,
Кубитальные жилки (cubitus anterior и posterior, CuA, CuP) всегда
хорошо развиты, причем CuP, как обычно в отряде Diptera, имеет вид лишь
слабой, но весьма постоянной складки сзади CuA. . -
Анальные жилки (analis prima и secunda, Апг и Ап2) в большинстве
случаев рудиментарны, причем Апх развита в качестве настоящей жилки
лишь в основной части крыла и достигает края его в виде складки, Ап2
обычно вовсе отсутствует и различима лишь в виде дугообразной складки.
В заключение необходимо отметить различное строение заднего края
крыла, в частности степень развития анальной лопасти, крылышка (alula)
и чешуек (calyptrae). Эти части крыла имеют важное механическое значение
при полете; форма и строение заднего края крыла определяют, характер
полета насекомого. Отсутствие крылышка и чешуек наиболее характерно
из всех двукрылых для Tipuloidea, так же, как и для всех Mecoptera; по-
добное строение крыла справедливо может считаться первичным состоя-
нием. У большинства других двукрылых все эти образования явственно
развиты и связаны с развитием более совершенного типа полета этих насе-
комых. Среди изучаемых нами насекомых лишь у немногих семейств наблю-
дается первичное строение основания крыла, отсутствие крылышка и сла-
бое развитие чешуйки. Таково семейство Pachyneuridae, у которого, кроме
отсутствия лопастей, как мы видели выше, наблюдается и недоразвитие
хетариев на рукоятках R и Ап2. Также почти не выражена анальная лопасть
и крылышко у Bolitophilidae- (рис. 31). ;. .... .
^ другой стороны, у CewMcrB Bibionidae, Penthetriidae и Axymyiidae при
почти полной редукции alula наблюдается прекрасное развитие чешуйки.
Сильное увеличение всех этих задних перепончатых лопастей крыла по-
казывает большую степень развития пропеллирующего полетает, е. с боль-
шим числом взмахов в единицу времени.
Кроме того, следует вкратце упомянуть о явлениях ослабления и утол-
щения жилок, с одной стороны, и о развитии волосков и щетинок, с дру-
гой. Утолщение и ослабление жилок тесно связаны с общим процессом
специализации крыльев — со сдвигом и слиянием жилок. Как правило,
оба эти процесса сопровождают один другой: жилки, сдвинутые к перед-
нему краю, вместе с тем и заметно более утолщены. Ослабление испытывают,
наоборот, те жилки, которые не сдвигаются, а остаются на месте в нроцессе
специализации крыла; таковы медиальные, кубитальные и анальные жилки.
Все эти явления хорошо выражены у представителей семейств Bibionidae,
Scatopsidae и в меньшей степени у Lycoriidae. Значительно более неясно
значение волосков и щетинок крыла. До сих пор (см. Hend е 1, 1936)
считалось общепризнанным, что присутствие щетинок и волосков на крыльях
есть признак примитивный, а потеря опушения — явление вторичное.
‘В общих чертах эта закономерность, видимо, существует: таковы сильно
волосистые крылья примитивных Penthetriidae, Axytnyiidae и Ditomyiidae,
с одной стороны, и голые у специализованных Scatopsidae, с другой. Вместе
с тем, повидимому, утраты волосков не бывает у мелких форм, так как
волоски на крыльях этих насекомых, несомненно, вторично приобретают
очень важное значение в механике полета.1 Таковы, например, Itonididae,
1 Абсолютная величина представителей Oligoneura колеблется от 0,5 (некоторые
Itonididae) до 11 мм (Bibio pomonae L.), т. е. от 1 до 22; вес же, который является решаю-
щим моментом в механике полета, колеблется в гораздо более широких пределах, до-
стигающих третьей степени от 22 (221 * 3 = 10 648). Совершенно очевидно, что механизм
полета насекомых самым коренным образом зависит от абсолютных размеров их тела.
Увеличение размеров влечет за собой увеличение веса, который при этом растет в гео-
3 Родендорф, т. ХШ в. 2. 33
которых можно сравнить по их полету и строению крыльев с трипсами,
мелкими Hymenoptera (Mymaridae) и Coleoptera (у которых несущими опор-
ными поверхностями крыла являются волоски, а жилкование имеет второ-
степенное значение). С другой стороны, волоски на крыльях специализо-
ванных форм обыкновенно частично исчезают, укорачиваются и лишь
по краю перепонки остаются хорошо развитыми и длинными, играя, по-
видимому, в этом случае определенную аэродинамическую роль в полете.
Резюмируя эту главу, можно указать на те определенные направления
в эволюции крыла, которые выясняются при ближайшем рассмотрении.
Главный путь специализации — это направление, условно нами названное
как«костализация» крыла; при этом процессе крыло укрепляется на перед-
нем крае, а задний край ослабляется и превращается в тонкую перепонку;
такое крыло становится совершенным механизмом для пропеллирующего,
с частыми взмахами крыльями, полета. За исходную форму кры-
ла мною принимается то строение его, которое наблюдается у
пермских Mecoptera (рис. 41) и наиболее полно сохранилось у со-
временных Psychodidae", в нем отсутствуют какие бы то ни было
сдвиги жилок, которые равномерно распределены по всей поверхности
крыла. Это первичное жилкование в группе Oligoneura давно пережито,
и его непосредственные и малоизмененные . дериваты можно видеть в дру-
гих группах двукрылых. Таким мало изменившимся производным
первичного крыла является «гребное» крыло комаров-долгоножек Tipuloi-
dea: специализация в этом крыле заключается лишь в сильном, его удли-
нении, без образования эластичных «пропеллирующих» частей по заднему
краю (табл. III, фиг. 7, 8). В группе Oligoneura крылья этого типа почти
вполне отсутствуют, и лишь крылья одних Pachyneuridae (табл. III, фиг. 9)
и, может быть, Bolitophilidae (табл. II, фиг. 6) могут быть отнесены к кате-
гории «гребных». Совсем особое направление эволюции крыльев наблюдается
у. мелких Itonididae и Heteropezidae-. у них крылья обладают сокращенным
и, что особенно важно, «облегченным» жилкованием и снабжены хорошо
развитыми волосками; этот путь специализация» несомненно, обусловлен
вторичным уменьшением размеров тела.
5. СОСТАВ СОВРЕМЕННОЙ. ФАУНЫ
Современная фауна изучаемой группы длинноусых Oligoneura слагается
из более чем 5700 видов * 1 двадцати одного семейства; из них на долю трех
семейств—Itonididae, Fungivoridae и Lycoriidae — падает не менее пяти
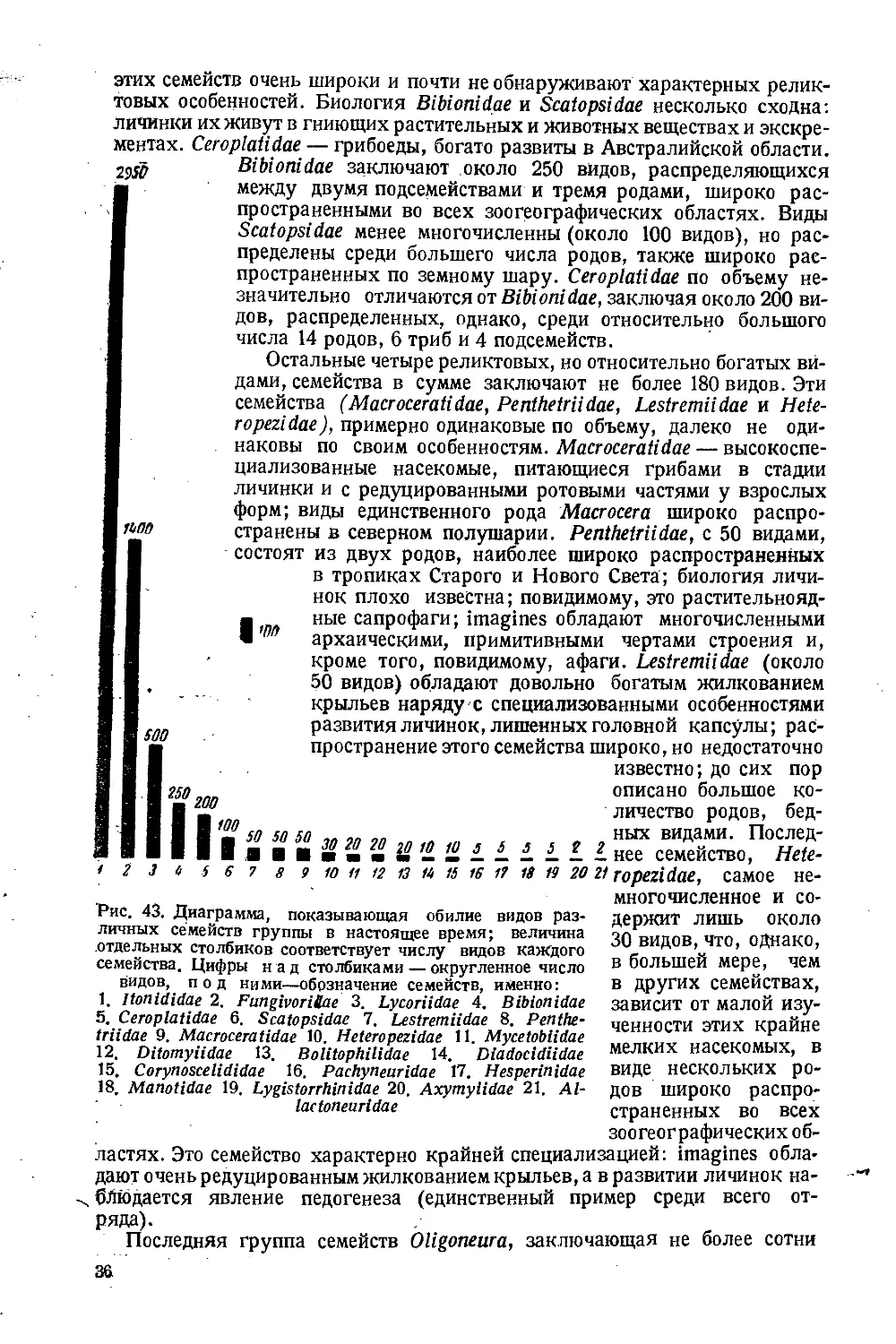
шестых (4850 видов) всего количества видов современной фауны (рис. 43).
Эти три семейства группы являются главными цветущими группами,
в настоящее время переживающими момент своего наибольшего развития.
Наибольшее по количеству видов семейство Itonididae, с его более чем 2900
метрической прогрессии. Ймея в виду, что плотность воздуха биосферы есть величина
практически постоянная, несомненно, что один и тот же тип крыла — летательного
приспособления насекомого — не может выполнять той же работы при различных аб-
солютных размерах его и тела насекомого. Иначе говоря, эволюция крыла двукрылых
должна была итти различными путями в зависимости от того, оставались ли размеры
животного постоянными, уменьшались или увеличивались. Оба эти процесса — изме-
нение размеров тела и специализация крыла в филогенезе группы являлись следствием
самых разнообразных причин; каждый отдельный случай в эволюции группы объясняется
своими специфическими особенностями.
1 Приводимые цифры взяты из последней сводки по отряду двукрылых, составлен-
ной Генделем( 1936), и, конечно, еще далеко не исчерпывают действительного бо-
гатства фауны этих двукрылых. Ценность этих цифр заключается, главным образом,
в том, что они позволяют выяснить относительный удельный вес в фауне каж-
дой группы, иначе говоря меньшее или большее развитие ее в настоящее время,
34
видов, широко распространено по всему земному шару, встречаясь во все*
зоогеографических областях; оно распадается на 6 вторичных категорий,
подсемейств или триб, с большим количеством отдельных родов и видов.
Itonididae, или галлицы, изо всех других семейств группы наиболее тесно
связаны с растительностью: личинки их живут в тканях живых растений,
будучи специализованными монофагами. Жизнь imagines-комариков очень
коротка; невидимому, они в громадном большинстве афаги.
Второе по объему семейство группы —Fungivoridae, или грибные кома-
рики,—хотя почти в два раза меньше предыдущего по числу видов (около
.1400 видов), но гораздо разнообразнее его, подразделяясь не менее чем на
8 вторичных категорий — подсемейств очень неравного объема; многие из
этих подсемейств носят явно реликтовый характер и бедны видами. Это
семейство обладает широким распространением во всех зоогеографических
областях; ареалы бедных видами реликтовых подсемейств очень узки или
разорваны. Название семейства точно передает черту его биологии: лй-
.чинки грибных комариков питаются разнообразными грибами: одни группы
живут в плодовых телах высших грибов, базидиальных и сумчатых; ли-
чинки других питаются гниющей древесиной, проросшей мицелием гри-
бов; одноядность у этих комариков почти вовсе не выражена. Наиболее
богато представлены Fungivoridae в лесных стациях; степные, а тем более
пустынные, зоны растительности очень бедны ими. Длительность имаги-
нальной жизни у большинства форм, повидимому, велика: зимуют взрослые
комарики.
Третье крупное семейство группы, Lycoriidae, значительно беднее ви-
дами первых двух, в нем около 500 видов, и горазд# однообразнее их: оно
подразделяется на немногие вторичные категории с небольшим числом ро-
дов, однако, заключающих относительно большое число Ридов. Это се-
мейство распределено, повидимому, равномерно по разным зоогеографи-
ческим областям; изучено оно гораздо хуже двух предыдущих. Личинки
Lycoriidae живут в разных гниющих или загнивающих растительных ве-
ществах; комарики, повидимому, не обладают длительной жизнью.
Остальные 18 семейств группы носят характер в большей или меньшей
степени выраженных реликтов, в настоящее время обладающих сокращен-
ными ареалами и крайне специализованными особенностями наряду с на-
личием многочисленных архаических признаков. Эта комбинация резко
контрастных черт строения — признаков большей специализации — на об-
щем фоне примитивных особенностей очень характерна' для реликтовых
групп и по сути дела определяет само существо реликта. Преобладание
специализованных или, наоборот, примитивных особенностей объясняет
современное состояние группы — ее реликтовый характер. Вместе с тем
последним и самым главным коррективом в решении вопроса о «реликто-
вости» является действительное состояние данной группы в настоящее
время — степень ее развития; последняя определяется числом видов, ре-
йдов и их распространением, широким или узким географически и эколо-
гически; только анализ каждого отдельного случая может дать ответ на
вопрос о факторах эволюции данной группы.
Вся группа реликтовых семейств Oligoneura может быть разбита на две
весьма неравные по объему, и разнообразию части; семь семейств — Bi-
bionidae, Ceroplatidae, Scatopsidae, Lestremiidae, Penthetriidae, Macrocera-
tidae и Heteropezidae — заключают в себе около 800 видов; на долю осталь-
ных одиннадцатй остается лишь около 100.
Среди первых семи семейств особенно сильным развитием отличаются
три — Bibionidae, Ceroplatidae и Scatopsidae — всего около 550 видов;
вместе с тем эти семейства объединены г аличием высокоспециализованных
особенностей строения крыльев и органов чувств; ареалы распространения
*
35
этих семейств очень широки и почти не обнаруживают характерных релик-
товых особенностей. Биология Bibionidae и Scatopsidae несколько сходна:
личинки их живут в гниющих растительных и животных веществах и экскре-
ментах. Ceroplatidae — грибоеды, богато развиты в Австралийской области.
295р Bibionidae заключают около 250 видов, распределяющихся
между двумя подсемействами и тремя родами, широко рас-
пространенными во всех зоогеографических областях. Виды
Scatopsidae менее многочисленны (около 100 видов), но рас-
I пределены среди большего числа родов, также широко рас-
пространенных по земному шару. Ceroplatidae по объему не-
I значительно отличаются от Bibionidae, заключая около 200 ви-
дов, распределенных, однако, среди относительно большого
числа 14 родов, 6 триб и 4 подсемейств.
I Остальные четыре реликтовых, но относительно богатых ви-
I дами, семейства в сумме заключают не более 180 видов. Эти
семейства (Macroceratidae, Penthetriidae, Lestremiidae и Hete-
I ropezidae), примерно одинаковые по объему, далеко не оди-
I каковы по своим особенностям. Macroceratidae — высокоспе-
I диализованные насекомые, питающиеся грибами в стадии
I личинки и с редуцированными ротовыми частями у взрослых
форм; виды единственного рода Масгосега широко распро-
Шгш странены в северном полушарии. Penthetriidae, с 50 видами,
I состоят из двух родов, наиболее широко распространенных
в тропиках Старого и Нового Света; биология личи-
' нок плохо известна; невидимому, это растительнояд-
I п ные сапР0Фаги> imagines обладают многочисленными
1 'т архаическими, примитивными чертами строения и,
' кроме того, невидимому, афаги. Lestremiidae (около
I 50 видов) обладают довольно богатым жилкованием
’ крыльев наряду с специализованными особенностями
Я развития личинок, лишенных головной капсулы; рас-
Iя пространение этого семейства широко, но недостаточно
известно; до сих пор
250 описано большое ко-
личество родов, бед-
I so so 5о „„ НЫХ видами. Послед-
I I I Я 1 £ 1 £ I нее семейство, Hete-
1 2 3 5 5 6? 8 9 io п is i3 м is 16 1? is 19 20 2t ropezidae, самое не-
Рис. 43. Диаграмма, показывающая обилие видов раз-
личных семейств группы в настоящее время; величина
отдельных столбиков соответствует числу видов каждого
семейства. Цифры над столбиками — округленное число
видов, под ними—обозначение семейств, именно:
1. Itonididae 2. Fungivoridae 3. Lycoriidae 4. Bibionidae
5. Ceroplatidae 6. Scatopsidae 7. Lestremiidae 8. Penthe-
triidae 9. Macroceratidae 10. Heteropezidae 11. Mycetobiidae
12. Ditomyiidae 13. Bolitophilidae 14. Diadocidiidae
15. Corynoscelididae 16. Pachyneuridae 17. Hesperinidae
18. Manotidae 19. Lygistorrhinidae 20. Axymyiidae 21. Al-
lactoneuridae
многочисленное и со-
держит лишь около
30 видов, что, оДнако,
в большей мере, чем
в других семействах,
зависит от малой изу-
ченности этих крайне
мелких насекомых, в
виде нескольких ро-
дов широко распро-
страненных во всех
зоогеографических об-
ластях. Это семейство характерно крайней специализацией: imagines обла-
дают очень редуцированным жилкованием крыльев, а в развитии личинок на-
блюдается явление педогенеза (единственный пример среди всего от-
ряда).
Последняя группа семейств Oligoneura, заключающая не более сотни
36
видов, но разделяющаяся вместе с тем на 11 семейств, обладает наиболее
резко выраженным реликтовым характером.
Три ее семейства (Mycetobiidae, Ditomyiidae и Bolitophilidae) относительно
«богаты» видами: Mycetobiidae состоят из 15—20 видов рода Mycetobia
и некоторых близких тропических родов; семейство плохо изучено: ли-
чинки живут в дуплах деревьев; более точно их биология неизвестна;
строение их мало специализовано; взрослые формы обладают богатым, но
сильно специализованным жилкованием. Семейство Ditomyiidae примерно
того же объема, что и предыдущее; распространение его немногих видов
недостаточно изучено; личинки сравнительно с Mycetobiidae значительно
более специализованы и живут в грибах; особенности imagines, наоборот,
явно более архаичны и жилкование их крыльев не костализованное. Семей-
ство Bolitophilidae заключает около 20 довольно редких видов одного
рода Bolitqphila, распространенных во всех зоогеографических областях;
личинки очень специализованы и живут в грибах; взрослые комарики обла-
дают рядом архаических, примитивных черт в жилковании и мало спе-
циализованным основанием крыльев, в котором почти не выражены
обычные редукции «рукояток» кубитальных и анальных жилок.
Последние восемь семейств являются наиболее резко выраженными
реликтами: все они редкие насекомые, обладающие широко разорванными
ареалами или, наоборот, ограниченные небольшой областью, притом обычно
обладающие различными примитивными особенностями, всегда сопрово-
ждаемыми признаками крайней специализации. Вследствие большой ред-
кости этих насекомых, особенности биологии их известны лишь для немно-
гих семейств. Таковы два семейства — Diadocidiidae и Corynoscelididae;
развитие остальных остается неизвестным.
Семейство Corynoscelididae заключает в себе немногие виды трех родов,
известные из Европы, Северной Америки и Новой Зеландии. Строение этих
насекомых представляет яркий пример комбинации крайне специализован-
ных и архаичных особенностей; архаичные особенности выражены в слож-
ном жилковании: крайняя специализация проявляется в строении,1 личи-
нок, лишенных головной капсулы и обладающих своеобразной биологией;
Немногие виды единственного рода Diadocidia, из одноименного се-
мейства, известны из лесных областей северного полушария; личинки их
живут в гниющем дереве; более точно превращение и особенности биологии
этих комариков не изучены; взрослые особи Diadocidiidae обладают слабо
специализованными крыльями, с мало выраженной костализацией^
Известные 3 или 4 вида семейства Pachyneuridae, пожалуй, наиболее
интересны по особенностям строения крыла, которое в этом семействе по-
строено наименее специализованно по сравнению с другими Oligoneura;
на нем вовсе отсутствуют всякие мягкие и эластичные перепонки у основа-
ния, которое не несет приспособлений к пройеллирующему полету; рукоятки
цочти всех жилок хорошо развиты и не редуцированы; специализация
крыла заключается лишь в редукции основной части медиальной жилки.
Это семейство известно лишь из немногих пунктов Европы, Дальнего
Востока и Северной Америки; превращение и биологий этих насекомых
остаются неизвестными.
Семейство Hesperinidae, известное по немногим видам из Европы и
Северной Америки, обладает смесью специализованных и архаичных при-
знаков. Крайней специализацией является резкий половой диморфизм:
самки лишены крыльев вовсе; крылья самцов обладают богатым не специа-
лизованным жилкованием; выражены лишь перепонки ГГо заднему краю;
развитие и биология неизвестны.
Два семейства, Manotidae и Lygistorrhinidae, обладающие немногими
единичными видами, могут быть охарактеризованы как примеры крайней
37
специализации крыльев. Manotidae распространены в Северной Америке
и в палеарктической области; характерно, кроме далеко зашедшей редук-
ции жилкования, также положение антенн, основания которых приближены
К темени. Lygistorrhinidae обладают сокращенным ареалом и распростра-
нены лишь в Австралии,; очень характерна морфологическая специализация
этих насекомых, обладающих сильно удлиненными ротовыми частями в виде
хоботка и очень редуцированным и резко костализованным жилкованием;
биология и развитие неизвестны.
• Два единственных и близких вида рода Axymyia известны лишь из
трех пунктов земного шара — Северной Америки, Венгрии и Приуралья.
Жилкование Axymylidae мало специализовано, без следов костализации;
более или менее видоизменено основание крыльев; превращение и биология
неизвестны.
Семейство Allactoneuridae очень плохо известно; оно описано мною
в 1938 г.; типом является род Allactoneura М е i j е г е, 1909, установленный
этим автором для единственного вида с острова Явы; он неизвестен мне
в натуре; биология и метаморфоз этой формы не исследованы. Для Allacto-
neuridae характерно архаичное ответвление RS от R и отсутствие костали-
зации крыла; большое сходство в жилковании этой современной яванской
формы с некоторыми юрскими родами и выяснило действительное таксоко-
мическое значение рода Allactoneura М е i j.
Прежде чем перейти к рассмотрению ископаемых представителей изу-
чаемой группы двукрылых, необходимо пояснить причины, побудившие
меня опустить данные, касающиеся ее третичных фаун, и посвятить все
внимание лишь древнейшим, мезозойским. Как ясно из дальнейшего, к на-
чалу третичной эры все основные группы Oligoneura в виде большинства
семейств уже сложились (середина эоценового времени), и лишь некоторые
семейства заметно отличались, по родовому составу и. иному удельному весу
в ископаемой фауне от современных. Относительно малые отличия третичной
фауны от современной и объясняют незначительную ценность третичных
материалов для решения вопросов филогении крупных систематических
категорий длинноусых двукрылых. Поэтому наличие коллекций по зна-
чительно более древней юрской фауне «двукрылых из Кара-тау явилось
счастливым обстоятельством, возможно более полно мною использованным.
6. ЮРСКАЯ ФАУНА OLIGONEURA
До сих пор ЯШИ знания о мезозойской фауне двукрылых, как и боль-
шинства другая отрядов Holometabola, были очень отрывочны и по существу
очень мало достоверны. Немногие описания лейасовых и среднеюрских видов
из Германии, верхнеюрских из Англии очень несовершенны; лишь послед-
ние описания Г а н<дл и р ш а (1925) более точны и могут дать материал
для суждения о взаимоотношениях видов. В особенности все это справед-
ливо по отношению к группе Oligoneura, состоящей в громадном своем боль-
шинстве из мелких насекомых, ископаемые остатки которых необходимо
исследовать при помощи сильной оптики. Те же изображения юрских
Oligoneura, которые опубликованы до сих пор, например из верхней юры
Англии, крайне дефектны (рис. 44), порою даже фантастичны, и не позволяют
делать каких-либо выводов б их действительных взаимоотношениях. Все
мои заключения о характере юрской фауны двукрылых основываются,
таким образом, на изучении богатой коллекции юрских двукрылых из
хребта Кара-тау в южном Казахстане, хранящейся в Палеонтологическом
институте Академии наук СССР.
'38
Всего известно 9 юрских семейств Oligoneura, из которых лишь одно,
Eopleciidae, отсутствует в фауне Кара-тау; остальные являются почти исклю-
чительно каратаускими и только одно, Protopleciidae, известно также и из
лейаса Германии. Впервые Diptera из Кара-тау начали обрабатываться
мною; в 1938 г. мною описано б видов из трех семейств; в настоящей работе
списывается 26 видов груп-
пы, принадлежащих 8 семей-
ствам, в большинстве своем
новым.
Необходимо отметить од-
ну особенность изученных ма-
териалов по фауне Oligoneura
Кара-тау; она заключается в
том, что все найденные виды
описаны по единичным, уни-
в
кальным экземплярам; описы-
ваемые роды в свою очередь, В
как правило, монотипичны.
Это, казалось бы, невероят-
ное обстоятельство естест-
венно объясняется случай-
ным характером захороне-
ния Oligoneura в юрском во-
доеме Кара-тау; те случаи,
когда ископаемые двукрылые
и других групп, например
Рис. 44. А — «Thimna defossa» Brodie (Хб,5);
Б — «Cecidomium grandaevurm Westwood
(X4,5); 'В — «Pseudadonia fittoni» Brodie (X5);
Г— «Bibionites priscus» G i e b e 1 (X5). Изобра-
жения ископаемых Oligoneura из верхней юры
Англии; слои Purbeck (из Handlirsch по
Brodie и Westwood).
из Tendipedoidea, обнаружи-
ваются в большом количестве экземпляров одного вида, всегда относятся
к нахождению насекомых большими скоплениями в одном горизонте карата-
уских сланцев. Находки Oligoneura всегда обнаруживались в виде единичных
остатков в разных микрогоризонтах сланцев: этим и объясняется большое
разнообразие найденной фауны, вместе с тем отражающее действительное бо-
гатство фауны того времени. Уместно вспомнить приблизительно сходное
положение с современной фауной Oligoneura', любой случайный сбор этих
комариков обычно заключает отдельных представителей целого ряда видов—
серии одного вида редки.
Список юрских Oligoneura
Семейство Protopleciidae, fam. nov.
Protoplecia Handlirsch, 1908
P. jurassica Handl. 1908, лейас Германии
Mesoplecia Rohdendorf, 1938
M. jurassica R о h d. 1938, Кара-тау
Mesopleciella, gen. nov.
M. minor, sp. nov., Кара-тау
Се мейство Eopleciidae, fam. nov.
Eoplecia Handlirsch, 1925
E. primitiva Handl. 1925, лейас Германии
Семейство Paraxymyiidae, fam. nov.
Paraxymyia, gen. nov.
P. quadriradialis, sp. nov., Кара-тау
Семейство Protobibionidae, fam. nov.
Protobibio, gen. nov.
P. jurassicus, sp. nov., Кара-тау
39
Семейство Protoscatopsidae, fam. nov.
Protoscatopse, gen. nov.
P. jurassica, sp. nov., Кара-тау,
Семейство Pleciofungivoridae, fam. nov.
Polyneurisca, gen. nov.
P. atavina, sp. nov., Кара-тау.
transversiplecia, gen. nov.
T. transversinervis, sp. nov., Кара-тау.
Pleciofungivorella, gen. nov.
P. binerva, sp. nov., Кара-тау.
Prohesperinus, gen. nov.
P. abdominalis, sp. nov., Кара-тау.
Eopachyneura, gen. nov.
E. trisectoralis, sp. nov., Кара-тау.
Pleciofungivora Ro hd. 1938
P. latipennis Ro h d. 1938, Кара-тау.
P. major, sp. nov., Кара-тау.
Eohesperinus, gen. nov.
E. martynovi, sp. nov., Кара-тау.
Allactoneurites Rohd. 1938
A. jurassicus Rohd. 1938, Кара-тау.
Семейство Pleciomimidae, fam. nov.
Подсемейство Plectomiminae, subfam. nov.
Lycoriomima, gen. nov.
' L. ventralis, sp. nov., Кара-тау.
Paralycoriomima, gen. nov.
P. sororcula, sp. nov., Кара-тау.
Lycoriop lecia, gen. nov.
L. elongata, sp. nov., Кара-тау.
Lycoriomimodes, gen. nov.
L. deformatus, sp. nov., Кара-тау.
L. sp., Кара-тау.
Lycoriomimella, gen. nov.
L. minor, sp. nov., Кара-тау.
Pleciomima Rohd. 1938
P. sepulta Rohd. 1938, Кара-тау.
P. secunda, sp. nov., Кара-тау.
Pleciomimella, gen. nov.
P. karatavica, sp. nov., Кара-тау.
Подсемейство Mimallactoneurinae, subfam. nov.
Mimallactoneura, gen. nov.
M. vetusta, sp. nov., Кара-тау.
Подсемейство Antefungivorinae Rohd. 1938
Antefungivora Rohd. 1938
A. prima Rohd. 1938, Кара-тау.
Antiquamedia Rohd. 1938
A. tenuipes Rohd. 1938, Кара-тау.
Подсемейство Paritonidinae, subfam. nov.
Paritonida, gen. nov.
P. brachyptera, sp. nov., Кара-тау.
Семейство Allactoneuridae Rohd. 1938
Подсемейство Mesosciophilinae, subfam. nov.
Mesosciophila, gen. nov.
M, venosa, sp. nov., Кара-тау.
Mesosciophilodes, gen. nov.
M. angustipennis, sp. nov., Кара-тау.
Подсемейство Mimalycoriinae, subfam. nov.
Mimalycoria, gen. nov.
M. allactoneuroides, sp. nov., Кара-тау.
40
Подсемейство Fungivoritinae, subfam. nov.
Eobeletina, gen. nov.
E. gracUis, sp. nov., Кара-тау.
Altactoneurisca, gen. nov.
A. indistlncta, sp. nov., Кара-тау.
Fungivorites R о h d. 1938
F. latimedius Ro h d, 1938, Кара-тау.
Определительная таблица юрских семейств длинноусых
Oligoneura (Fungi voroi de а и Bi bi onoi dea )
1 (18) Система жилок M не образует замкнутой ячейки; ноги умеренно
удлиненные; антенны многочлениковые, обычно тонкие; как правилог
мелкие, реже средней величины насекомые.
2 (17) Жилкование крепкое, хорошо различимое; ноги относительно ко-
роткие.
3 (6) Жилкование сильно специализованное, резко сдвинутое к перед-
нему краю.
4 (5) До вершины крыла доходит длинная жилка RS; SC крепкая; крыло
удлиненное.
Семейство Protobibionidae, fam. nov. (стр. 46)
Единственный род Protobibio, gen. nov. (стр. 47)
5 (4) Вершина крыла без жилок: все жилкование сконцентрировано на
переднем крае; SC отсутствует.
Семейство Protoscatopsidae, fam. nov. (стр. 48)
Единственный род Protoscatopse, gen. nov. (стр. 49}
6 (3) Жилки расположены по всему крылу; костализация выражена
слабо.
7 (14) RS всегда с ветвями, направленными вперед и соединяющимися
с С.
8 (11) Ветви RS всегда ответвляются дистальнее rm; поперечная rm уда-
лена от ветви RS.
9 (10) Поперечная rm расположена в отдалении от развилка R—RS;
базальный отрезок RS от развилка до rm не менее чем в 4 раза длин-
нее rm; средней величины или крупные виды, 5—8 мм.
Семейство Protopleciidae, fam. nov. (стр. 42}
2’рода из юры Кара-тау и 1 из лейаса Германии
10 (9) Поперечная жилка rm резко приближена к развилку R—"RS; ба-
зальный отрезок RS не более чем в 3 раза длиннее rm, часто гораздо
короче; мелкие, редко средней величины, насекомые в 1,75—5 мм.
Семейство Pleciofungivoridae, fam. nov. (стр. 51)
Ряд родов из юры Кара-тау
11 (8) RS с двумя передними ветвями, из которых первая ответвляется про-
I ксимальнее гт,т.е. между развилок R—RS и rm; мелкие насекомые
в 3—4 мм.
12 (13) SC значительно короче половины крыла; М слабая; общий ствол
этой жилки редуцирован; конец R очень удален от конца SC и резко
приближен к концу RS; есть стигмальное пятно; ноги короткие, не
особенно крепкие; антенны короче головы и груда, вместе взятых.
Семейство Paraxymyiidae, fam. nov. (стр. 44}
Единственный род Paraxymyia, gen. nov. (стр. 45)
13 (12) SC длинная, не менее половины крыла; М крепкая, не редуциро-
ванная; концы жилок SC, R и RS резко сближены; стигма неясная.
Семейство Eopleciidae, fam. nov. (стр. 43)
Единственный род Eoplecia Handl, из лейаса Германии
41
14 (7) RS простая, без ветвей; если же ветви есть, то они расположены
близ основания жилки и имеют вид поперечных между R и RS.
15 (16)’R короткая, не более 2/3 или 8/4 крыла; М часто слабая.
Семейство Pleciomimidae, fam. nov. (стр. 61)
4 подсемейства с 11 родами из Кара-тау
16 (15) R длинная, не менее 3/4 крыла, часто длиннее: М крепкая.
Семейство Allactoneuridae (стр. 75)
3 подсемейства с 6 родами из Кара-тау
17 (2) Жилкование слабое, неясное, сильно обедненное, но не сдвинутое
вперед; ноги очень тонкие и длинные.
Надсемейство Tendipedoidea
18 (I) Жилки М образуют замкнутую ячейку; если ячейка отсутствует,
то антенны очень коротки; антенны короткие или длинные; в послед-
нем случае насекомые крупные.
Все остальные юрские двукрылые.
НАДСЕМЕЙСТВО BIBIONOIDEA
Семейство Protopleciidae, fam. nov.
Тип семейства: р. Protoplecia Handlirsch, 1906 г.J
Рис. 45. Protoplecia
primitive Handl.;
крыло (no Hand-
lirsch).
дугообразная, равная 1{3—1{3 крыла; RS всегда имеет косую переднюю
ветвь; основная часть RS всегда значительно длиннее rm, которая распо-
ложена, примерно, на середине между развилком RS и ее ответвлением
от R; М4 и Ма образуют длинную вилку, значительно
более длинную, чем ее общий ствол; М3 отделена от
передних ветвей, но не слита с Си, отделяясь от по-
следней поперечной жилкой или даже совсем не сое-
диняясь с Си (Mesopleciella), mcu хорошо развита;
антенны умеренно удлиненные; ноги средней длины;
тазики слабые; брюшко очень массивное, состоящее
из 7 сегментов и генитального аппарата; брюшко бо-
лее чем в 2 раза длиннее головы и груди, вместе
взятых.
Тип семейства, описанный в 1908 г. Г а н д л и р-
ш е м, род Protoplecia (рис. 45) был им отнесен к
семейству Bibionidae (Bibionidae + Penthetriidae). Не-
сомненно, нельзя безоговорочно объединять роды
Protoplecia и близкие к нему с современными пред-
ставителями семейства Penthetriidae: эти две группы
(различаются рядом признаков, не позволяющих их
соединить. Для Protopleciidae характерны тонкие,
умеренно удлиненные антенны, короткая SC, не пре-
вышающая х/2 длины крыла (обычно короче!) и более мелкая величина.
Общий характер семейства заставляет считать его группой, очень близ-
кой к предкам современных Penthetriidae; счесть Protopleciidae прямыми
предками Penthetriidae не позволяет наличие у них более короткой SC,
т. е. признака, явно прогрессивного по сравнению с удлиненной SC у
Penthetriidae.
К семейству Protopleciidae, кроме Protoplecia, принадлежат еще следую-
щие юрские роды: Mesoplecia R о h d. (табл. VIII, фиг. 19) и описывае-
мая ниже Mesopleciella, gen. nov.
42
Род Mesopleciella, gen. nov.
Тип рода Mesopleciella minor, sp. nov.
SC хорошо развитая, равная x/3 крыла, впадающая в С,.на основании крыла
сильно приближенная к R; Нелегка волнистая, значительно тоньше С и SC;
передняя ветвь RS умеренно S-образно изогнутая, отходящая от ствола RS
под прямым углом, значительно ближе к основанию крыла, чем место соеди-
нения R с С. Передняя ветвь RS равна х/2 задней ветви; задняя ветвь RS
слабо и равномерно загнутая назад; концы RS2 и Мх сходящиеся; прокси-
мальный отрезок общего ствола RS почти в 1х/2 раза длиннее дистального;
поперечная rm расположена почти на середине крыла и при этом делит
общий ствол передних ветвей М пополам; ветви Мх и М2 резко расходятся
к краю крыла: Мх почти прямая, М2 слабо волнистая и является прямым
продолжением общего ствола М; М3 параллельная с М2 и лишь слабо рас-
ходится в дистальной четверти; поперечная meu расположена почти сразу
перед местом ответвления
М8 от общего ствола М,
Си правильно и резко за-
гнутая назад, Ап неясная.
Голова небольшая, с
умеренно выступающим
вперед хоботком; сохрани-
лись лишь 4 базальных
членика антенн.
Грудь резко и горбо-
образно выдается в своей
задней части.
Ноги умеренно удли-
ненные; тазики и бедра
неясные; голени длинные,
параллельно-крайние; зад-
ние голени с короткой,
но заметной шпорой на
вентральном крае дисталь-
ного конца; лапки равны
или короче голеней, пя-
тцмлениковые.
Брюшко из 8 сегмен-
тов, из которых послед-
ний, повидимому, состав-
ляет комплекс гениталь-
ных сегментов.
Рис. 46. Mesopleciella minor, sp. nov. Кара-тау,
ПИН № 2452/642; общий вид ископаемого (ориг.).
Mesopleciella minor, sp. nov.
(рис. 46 и 47; табл. VIII, фиг. 20).
Длина тела 5.0; крыла 3.75 Мм.
Кара-тау, сел. Галкино, А. В. Мартынов, 1925., Колл. ПИН,
№ 2452/642.
Семейство Eopleclldae, fam. nov.
Syn. Protofrhyphidae Edwards, 1928, pro parte
Тип семейства: род Eoplecia Handlirsch, 19254рис. 90).
SC более x/2 крыла; R почти параллельна на всем протяжении eSC; RS с
двумя передними ветвями, из которых проксимальная расположена ближе к
основанию крыла, чем rm; rm очень короткая, расположенная почти на сере-
43
дине крыла; М3 у основания с изломом: ее основная часть расположена
косо, под прямым углом к косой mcu; развиты две крепкие, хорошо
выраженные Ап; общий ствол М1+2 очень короткий; Си крепкая; СиР
неясная; строение тела неизвестно; величина тела мала; длина крыла
около 3.7 мм.
Рис. 47. Mesopleciella minor, sp. nov. Кара-тау, ПИН, № 2452/642;
крыло (ориг.).
Рис. 48. Eoplecia primitiva Handl,
крыло (по Handlirsch, 1925)
обозначения Гандлирша.
Семейство описывается по единственному роду. Eoplecia. из верхнего,
лейаса Германии (рис. 48); описание составлено по рисунку Н a n d 1 ir-
sch’a (1925).
Этот род был впервые помещен описавшим его Гандлиршем (1925)
в семействе Bibionidae; Edwards (1928) поместил его вместе с Protor-
rhyphas в семейство Protorrhyphidae
Handl. Ошибочность этого положения
несомненна; Protorrhyphas принадлежит
к группе^Phryneoidea, для которой ха-
рактершГприсутствие не менее четырех
ветвей М и наличие между ними интер-
медиальных -поперечных жилок. Род
Eoplecia обладает лишь тремя ветвями
М, притом не образующими интерме-
диальной ячейки. Все строение жилко-
. вания этого рода очень близко К се-
мействам Axymyiidae и Penthetriidae;
Eopleciidae, несомненно, были близки к исходным формам названных
групп: присутствие трех ветвей RS, длинная SC, две крепкие Ап — все
это признаки примитивные, подлежащие редукции.
Семейство Paraxymyiidae, fam. nov.
i Тип семейства: род Paraxymyia, gen. nov.
' SC умеренно короткая, тонкая, но ясная, впадающая в С; RS ответ-
вляется от R под острым углом и состоит из трех крепких ветвей, соединяю-
щихся с С; rm расположена на середине крыла между ответвлениями RS
и RS2 от общего ствола RS. М слабая, трехветвистая; общий ствол М реду-
цирован; Си тонкая; Ап неясные; крыло довольно широкое. Грудь
очень слабо выпуклая, почти не нависающая над головой. Антенны*
в 2—3 раза длиннее головы, невидимому 15-члениковые. Ноги довольно
тонкие, умеренно удлиненные; тазики средней величины. Брюшко кони-
ческое, 8-члениковое. Размеры тела около 4 мм.
44
Описываемая груййа своеобразна и обнаруживает как черты большой
примитивности,- тате и Некоторые, несомненно, специализованные особен*
Ноам. Признаки примитивные следующие: присутствие трехветвистого RS,
все ветвикоторого крепки и хорошо развиты; слабое развитие спинки
груди, почти плоской; слабое развитие ног, тазики которых лишь
умеренно увеличены. Специализация описываемой формы ясна из ряда
признаков строения крыла и тела: редукции основного ствола М, слабого
развития SC, более сильного развития (утолщения) жилок системы R и RS
и ослабления М и Си, короткости антенн.
Каковы же отношения Paraxymyiidae к другим группам Oligoneura? Наи-
более характерная особенность специализации крыла описываемой формы
заключается в редукции основной части медиального ствола, именно то,
что наблюдается у современных семейств Mycetobiidae, Macroceratidae,
Ceroplatidae, Ditomyiidae, Diadocidiidae, Axymyiidae, Pachyneuridae и некото-
рых других. Другая важная черта строения крыла Paraxymyiidae заклю-
чается в расположении ветвей RS, из которых передняя ветвь RSj ответ-
вляется от общего ствола значительно проксимальнее rm; эта последняя
поперечная жилка оказывается расположенной между двумя ветвями RS.
Подобное смещение ветви RSr среди различных групп Nematocera мы наблю-
даем у очень немногих семейств: таковы Axymyiidae, Phryneidae, отчасти
Mycetobiidae и Pachyneuridae. Вместе с тем этот процесс специализации
жилкования является у названных семейств, невидимому, уже более поздним
этапом: в первую очередь происходит редукция ветвей RS; у Mycetobiidae и
Phryneidae отсутствуют всякие остатки RS 2+3, у Axymyiidae и Pachyneu-
fidae обе ветви RS сливаются в одну вильчатую жилку. Ископаемые из
верхнелейасовых групп длинноусых, именно Protorrhyphidae и Eopleciidae,
обнаруживают большое сходство с описываемой группой в строении си-
стемы R; в особенности замечательна общность ветвления RS у всех назван-
ных семейств Nematocera с Brachycera и даже Cyclorrhapha: у всех этих дву-
крылых rm расположена дистальнее ответвления первой ветви RS.
Вместе с тем высокая специализация жилкования — редукция системы
М, ее общего ствола, ветви М3 и SC не позволяет видеть в Paraxymyiidae
предков каких-либо современных групп, близких по характеру ветвления
RS. Как Axymyiidae, так и Paraxymyiidae произошли от предков, близких
К лейасовымЕор/есп'Дйе; Paraxymyiidae приобрели ряд черт высокой специа-
лизации (редукции), сохранив вместе с тем некоторые примитивные особен-
ности.
Род Paraxymyla, gen. nov.
Тип рода: Paraxymyla quadriradialis, sp. nov.
С достигает лишь конца последней ветви RS, не доходя до вершины крыла;
SC: нежная и тонкая, но ясная, равная примерно х/3 крыла, приближенная
и параллельная с R вплоть до ответвления RS; дистальнее уровня ответвле-
ния RS SC довольно круто загибается вперед, впадая в С; R крепкая, почти
прямая, равная, примерно, 3'5 длины крыла; RS ответвляется от R прокси-
мальнее конца базальной трети крыла; ствол RS дает две крепкие ветви,
отходящие вперед и впадающие в C; RSX ответвляется от ствола RS почти
под прямым углом на уровне середины 2-го отрезка С, проксимальнее fm; RSM
ясно S-образно изогнутая; RS24.3 почти прямая и отходит от RS4 под острым
углом; конец этой жилки RS4 слабо загибается назад, не достигая вершины
крыла; rm крепкая, расположенная почти точно на середине крыла, попе-
речная; М и Си слабее и тоньше жилок R и RS; ветви Mj и М2 в несколько
раз длиннее общего ствола М1+2 от rm до развилка, почта прямые, слабо
расходящиеся; общий ствол М редуцированный и заметный в виде слабого
следа, лежащего вдоль большой базальной ячейки; лишь часть этой жилки
45
и основание М3 хорошо заметны в виде изогнутого «стебелька», выходящего
из М3; mcu имеет вид непосредственного основания жилки М3, которая почти
параллельна с М2; СиА в основной части прямая, в дистальной не сохрани-
лась; вершина субкостальной и радиальной ячеек вокруг конца R затем-
нена, образуя хорошо заметную птеростигму; крыло умеренно удлиненное,
с закругленной тупой вершиной.
Голова не особенно крупная; антенны короче головы и груди, вместе
взятых; глаза, глазки и ротовые органы неясны.
Грудь слабо выпуклая спереди, на спинке почти плоская, с заметно
выступающим кверху щитком.
.Тазики передних ног не менее г/а длины бедер, средние и задние плохо
видны; передние бедра прямые и толстые, довольно резко утончающиеся
к концам; передние голени прямые, в 2г/а—3 раза тоньше бедер и несколько
их длиннее; средние ноги плохо сохранились; средние бедра заметно утол-
щены в основной половине и постепенно утончаются к дистальному концу;
задние ноги длиннее двух других пар; задние бедра веретеновидной формы;
голени в Р/а раза толще бедер и едва их длиннее; лапки всех пар ног плохо
сохранились.
Брюшко коническое, сильно сплющенное и растянутое; сегменты 2-й,
3-й и 4-й короткие, 5-й, б-й и 7-й более длинные; последний, 8-й сегмент, ко-
нический, плохо видимый.
Paraxymyia quadriradialis, sp. nov.
(Рис. 49 и 50; табл. IX, фиг. 21)
Остатки насекомого, лежащего на правой стороне, со сложенными нс*
гами, откинутыми крыльями и вытянутыми антеннами; сохранность хоро-
Рис. 49. Paraxymyia quadriradialis, sp. nov. Кара-тау,
ПИН, № 2452/584; общий вид ископаемого (ориг.).
шая: частично разрушены голова и ее придатки, грудь, ноги и одно крыло;
окраска темнокоричневая, ноги светлые; длина крыла 2.25, длина насеко-
мого с растянутым брюшком 3.75, истинная длина около 3.25 мм.
1 самец. Сел. Галкино, А. В. Мартынов, Колл. ПИН, № 2452/584.
Семейство Protobibtonidae, fam. nov.
Тип семейства: род Protobibio, gen. nov.
SC хорошо развитая, несколько короче г/2 крыла, впадающая в С; RS
делится на две ветви, из которых длинная (задняя) достигает вершины крыла,
а короткая (передняя) соединяется с R далеко до конца R; пн косолежащая,
-46
длинная; М и Си слабые; их строение вследствие плохой сохранности отпе-
чатка неясно; есть поперечно лежащая mcu; С, R и RS крепкие, значительно
толще и темнее М и Си; крыло удлиненное, почти в 3 раза длиннее ширины.
Строение головы неясно: ноги очень тонкие и длинные, длиннее тела; бедра и
голени не вздутые, без шипов; тазики увеличенные; брюшко умеренно взду-
тое, утончающееся к концу. Размеры тела несколько менее 2 мм.
Описываемое по единственному роду семейство Protobibionidae очень
интересно: оно сочетает в себе признаки чисто «фунгивороидного» характера
с особенностями специализованного крыла современного семейства Bibio-
Рис. 50.Paraxytnyia quadriradialis, sp. nov. Кара-тау, ПИН
№ 2472/584; крыло (ориг.).
nidae. Длинные, тонкие ноги, слабая склеротизация их и брюшка, первичный
характер ответвления RS от R — все это ставит род Protobibio в непосред-
ственную близость к семействам Allactoneuridae и Pleciofnngivoridae. С другой
стороны, величина и косое положение rm, сильное утолщение жилок С, R
и RS сближает это новое семейство с Bibionidae, именно с родом Philia (==Dilo-
phus). Признаки жилкования, отличающие Protobibionidae от рецентных Bibio-
nidae, заключаются в присутствии передней, правда, короткой ветви RS,
более крепкой, но. короткой SC, в иной форме основных частей М и Си и при-
сутствии настоящей mcu; некоторые из этих признаков явно более прогрес-
сивного характера по сравнению с Bibionidae (такова, например, укорочен-
ная SC) и не позволяют считать Protobibionidae за непосредственных пред-
ков названной современной группы. Но очевидно, что Protobibionidae ушли
от предков Bibionidae очень недалеко и должны быть поставлены в непосред-
ственной близости к этому стволу.
Род Protobiblo, gen. nov.
Тип рода: Protobiblo jurassicus, sp. nov.
С слегка заходит за конец RS, достигая вершины крыла, крепкая; SC
тоньше С, заметно короче 1/.2 крыла, соединяется с С и короткой попереч-
ной с R; R почти прямая, крепкая,
примерно равная 2/з крыла; RS ответ-
вляется от R сразу дистальнее уровня
. поперечной между SC и R; ответ-
вление RS от R нормальное, под ост-
рым углом; RS очень слабо и посте-
пенно дугообразно изогнутая, резко
загнутая назад на самом конце, по- Рис. 51. Protobiblo jurassicus, sp. nov.
чти достигающая вершины крыла; есть Кара-тау, ПИН №2452/50; крыло (ориг.).
короткая передняя ветвь RS, впадаю-
щая в середину R; rm косо лежащая, почти в 3 раза длиннее проксимального
отрезка общего ствола RS; М видна на отпечатке лишь в проксимальной
47
части, как и Си; обе эти жилки значительно слабее радиальных; хорошо
заметна поперечно расположенная mcu; крыло узкое, в 2.85 раза длиннее
ширины,
Голова плохо сохранилась; антенны и ротовые органы не видны.
Грудь массивная; характер отпечатка не позволяет судить о степени
вздутости ее спинки; ноги тонкие и длинные; тазики сильно увеличенные;
вытянутая задняя нога превышает длину тела.
Брюшко умеренно массивное, к концу суженное; число его сегментов
плохо различимо.
Protobibio jurassicus, sp. nov.
(Рис. 51)
Отпечаток самца, лежащего на спинной поверхности, с головой, вдавлен-
ной в грудной отдел; антенны не сохранились; ноги и крылья расправлены,
но плохо сохранились; окраска отпечатка темная, коричнево-черная. Длина
тела 1,9, крыла 1,5 мм.
Сел. Галкино, А. В. Мартынов, 1925, Колл. ПИН, №^2452/50.
Семейство Protoscatopsidae, fam. nov.
Тип семейства: род Protoscatopse, gen. nov.
Жилкование крыла сильно сокращено и сдвинуто на передний край;
большая часть крыла лишена жилок, перепончатая; SC отсутствует; R почти
прямая; RS с тремя ветвями; С несколько заходит за конец последней ветви
RS, не достигая вершины крыла; М нежная и тонкая, двухветвистая; С почти
прямая, крепкая лишь до середины крыла, далее—в виде складки. Крылья
довольно узкие. Грудь умеренно вздутая. Антенны короткие, булавовидные,
едва длиннее головы, 12-члениковые. Брюшко коническое, 7-члениковое.
Строение ног не видно. Размеры тела 3.5 мм.
Описываемая новая форма наиболее близка к представителям современ-
ного семейства Scatopsidae, но отличается рядом явно более примитивных
признаков. Такими примитивными признаками являются: присутствие вет-
вей RS (развитые короткие ветви RSj и RS2+3), расположенные в ячейке
между R и основным стволом RS, доказывают вторичное редуцированное
состояние жилкования системы RS у современных Scatopsidae); характер
соединения М с RS (эти жилки у Protoscatopsidae расположены своеобразно,
будучи слиты на некотором протяжении); место ответвления М от общего
ствола М -f- RS расположено значительно дистальнее, чем развилок R — RS,
и напоминает жилкование крыльев у Corynoscelididae.
Эти особенности жилкования описываемой юрской формы позволяют
внести серьезные поправки в существующую трактовку жилкования совре-
менных Scatopsidae, с одной стороны, а с другой, выясняют взаимные гене-
тические связи этих групп. До сих пор (Duda, 1928) жилкование у Sca-
topsidae сравнивалось с (таковым у группы настоящих Oligoneura, именно
у Lycoriidae, и поэтому считалось, что RS у своего основания превратился
в поперечную жижу, а истинная радиомедиальная жилка (rm) вошла в си-
стему прямой крепкой жилки М — RS. Положение жилок у Corynosceli-
didae никак не укладывалось в эту схему, поэтому считалось, что эта группа
является аберративной ветвью; явно реликтовый, весьма древний характер
Corynoscelididae недоучитывался большинством авторов, проходивших мимо
очень интересного и поучительного жилкования этих редких насекомых.
Проводя сравнение жилкования Scatopsidae, Corynoscelididae и новой юрской
формы, сразу же можно отметить разительное сходство в ветвлении жилок
RS и М между Corynoscelididae и Protoscatopsidae: это сходство заключается
в слиянии общих стволов RS и М1+2, как у Ceroplatidae и близких к ним
48
групп. Вместе с тем общий характер специализации жилкования, его сдвиг
и усиление к переднему краю, — в частности, сближение стволов RS и
M-J-RS у Scatopsidae и Protoscatopsidae, — с очевидностью указывают, что
основание RS — ответвление этой жилки от R — у Scatopsidae исчезло,
что так называемое «основание RS» (br8, D u d а) у Scatopsidae есть лишь
сместившаяся проксимально передняя ветвь RS — RSX.
Теперь уместно разобрать вопрос, является ли описываемая юрская
группа непосредственным предком рецентных Scatopsidae и Corynoseetidi-
dae. С самого начала следует ответить на этот вопрос резко отрицательно
в отношении семейства Corynoscelididae; эта последняя группа обладает
рядом примитивнейших черт строения, не позволяющих считать ее потомком
уже высокоспециализованных Protoscatopsidae; такова слабая механиче-
ская специализация крыльев, многочленистость антенн и их длина. Cory-
noscelididae, несомненно, очень древняя реликтовая группа, отделившаяся
ют общего ствола «скатопсоидных» форм гораздо раньше Protoscatopsidae.
Совсем иное наблюдается в отношениях Scatopsidae и Protoscatopsidae;
по существу лишь короткие антенны и отсутствие следов SC могут считаться
признаками более высокой специализации Protoscatopsidae по сравнению
с современным семейством; все остальное описанное выше жилкование застав-
ляет меня считать эту юрскую группу очёнь близки стоящей, к исходным
формам современных Scatopsidae.
Род Protoscatopse, gen. nov.
Тип рода: Protoscatopse jurassica, sp. nov.
С крепкая, не достигающая вершины крыла, на месте соединения жилок С
и R на краю крыла образуется заметная выпуклость; SC отсутствует; R тоньше
С, параллельная с ней, на
конце равномерно дугооб-
Рис. 53. Protoscatopse ju-
rassica, sp. nov. Кара-тау,
ПИН, № 2452/351; го-
лова (ориг.).
Рис. 52. Protoscatopse jurassica, sp. nov.
Кара-тау, ПИН, № 2452/351; общий
вид ископаемого (ориг.).
разно загибающаяся вперед; RS ответвляется от R под острым углом, при-
мерно в базальной х/5 крыла; непосредственно после ответвления от R, RS
сливается на некотором протяжении со стволом М1+2. продолжаясь
параллельно и в непосредственной близости от R; в конце базаль-
ной трети крыла ответвляется под сравнительно-—большим углом
ствол М1+а, слабый и нежный, дающий развилок перед своей середи-
ной; RS дает две передние, довольно тонкие ветви, из которых пер-
вая RSi располагается параллельно с концом R, вторая ветвь RS
4 Розендорф, т. XIII, в. 2. 49
лежит резко поперечно, впадая в.С почти в одной точке с RSV Основной
ствол RS (=RS4) почти совсем прямой; СиА в основной половине крепкая
и толстая; дистальная ее часть слабая и нежная. Крыло умеренно удли-
нённое, в 2.4 раза длиннее своей ширины. -
Голова плохо сохранилась; глаза крупные, повидимому сближенные на
темени, позади антенн; антенны булавовидные, 12-члениковые, в Р/з раза
длиннее головы; последний членик их заостренный, конический; членики
от 8-го до 11-го наиболее толстые; 1-й и 2-й членики более тонкие и длин-
ные; ротовые органы, повидимому, короткие и мягкие; грудь и брюшко
плохо сохранились.
Protoscatopse jurassica, sр. nov.
(Рис. 52, 53 и 54)
Отпечаток насекомого, лежащего на брюшной поверхности, с неправильно
распростертыми в стороны крыльями и антеннами; антенны раздвинуты
в стороны; ноги не сохранились; сохранность плохая; брюшко сильно фаз-
Рис. 54. Protoscatopse jurassica, sp. nov. Кара-тау, ПИН,
№ 2452/351; крыло (ориг.).
рушено; окраска тела темная, черно-коричневая; длина 3.5, длина крыльев
2.0 мм.
Самка. Сел. Галкино, А. В. М а р т ы н о в, 1925. Колл. ПИН, № 2452/351.
НАДСЕМЕЙСТВО FUNGIVOROIDEA
В 1937 г. при описании первых представителей двукрылых юрской фауны
Кара-тау мною установлено семейство Allactoneuridae с типичным родом
Allactoneura М е i j е г е и тремя подсемействами — Allactoneurinae, s. str.,
Antefungivorinae, и Antiquamediinae. Обработка дальнейшего материала
выяснила преждевременность установления этих семейственных и подсе-
мейственных категорий. Подробное исследование жилкования крыльев и
других особенностей морфологии этих двукрылых с ясностью показало
гетерогенный характер установленного семейства Allactoneuridae. Первые
два рода Pleciojungivora и Allactoneurites должны быть выделены в особую
группу — семейство Pleciofungivoridae, описываемое ниже; род Pleciomima
является представителем особого, также самостоятельного семейства Р1е-
ciomimidae; наконец, последний род, Fungivorites, лишь один относится к се-
мейству Allactoneuridae, правда,, к особому, юрскому подсемейству, вместе
с рядом вновь описываемых родов. Подсемейства Antejungivorinae и Anti-
quamediinae должны быть объединены в одну группу, принадлежащую в ка-
честве, подсемейства к семейству Pleciomimidae-, взаимоотношения этих трех
семейств излагаются ниже.
50
Семейство Pleciofungivoridae, fam. nov.:
Тип семейства: Pleciofungivora Rohd., 1938
SC умеренно удлиненная, равна r/4 или */3 крыла, всегда хорошо развитая,
впадающая в С; RS всегда с одной, двумя или большим числом передних
ветвей, впадающих частично в С; основная часть RS, лежащая проксималь-
нее гт, очень короткая, равна или лишь не более чем в 4 раза длиннее rrti,
которая резко приближена к месту ответвления RS от R; вилка М всегда
значительно длиннее общего ствола М, между rm и развилком Мг+М2.
М8 отдалена от передних ветвей, но всегда ответвляется от общего ствола М,
не соединяясь с Си вовсе или же лишь посредством поперечных жилок (Trans-
versiplecia). Антенны не превышают головы и груди, вместе взятых, 14-
или 15-члениковые; ноги тонкие и довольно длинные; тазики слабо разви-
тые; брюшко массивное, более чем в 2 раза длиннее головы и груди, вместе
взятых, состоящее из 8 или 9 сегментов; размеры тела 3.75—5.75 мм.
К описываемому семейству принадлежит ряд юрских родов, перечислен-
ных ниже в определительной таблице. Для семейства особенно характерно
строение- радиальной системы, именно присутствие передних ветвей RS
и укорочение общего ствола RS и SC. Наиболее близко семейство Plecio-
fiingivorldae к современным им Allactoneuridae и Pleciomimidae и, вероятно,
является генетически очень близким к вторым. В настоящее время Plecio-
fungivoridae нацело отсутствуют в фауне и, невидимому, представлены
своими почти непосредственными потомками — семействами Ditomyiidae,
Macroceratidae и Ceroplatidae.
Определительная таблица родов Pleciofnngivoridae
fam. ftbv.
1 (2) Субкостальное поле с одной поперечной жилкой, расположенной
сразу за вершиной SC; единственная передняя ветвь RS мало изогну-
тая; проксимальный отрезок общего ствола RS в 2 раза длиннее rm.
(рис. 57) Род Transversiplecia,gen. nov.
Т. transversinervis, sp. nov. (стр. 53)
2 (1) Субкостальное поле без поперечных жилок.
3 (10) Развита лишь одна передняя ветвь RS, притом крепкая, как пра-
вило, хорошо заметная.
4 (5) Проксимальный отрезок общего ствола RS не менее как в 3 раза
длиннее rm; последняя расположена на уровне конца SC; место ответ-
вления RS от R лежит на уровне середины крыла (рис. 61)
Род Prohesperinus, gen.nov.
Р. abdominalis, sp. nov. (стр. 57)
-5 (4) Проксимальный отрезок RS равен или в Р/2 раза длиннее rm; RS
ответвляется от R на уровне конца SC; rm расположена проксималь-
нее конца SC.
6 (7) Передняя ветвь RS равна или лишь незначительно короче отрезка
RS между rm и развилком RS; развилок М расположен примерно на
середине между уровнями концов SC и R; передняя ветвь RS крепкая,
косая (рис. 67) Род. Eohesperinus, gen. nov.
Е-. mdrtynovi,' sp. nov. (стр. 61)
7 (6) Передняя ветвь RS гораздо короче среднего отрезка RS; развилок
М1+2 гораздо ближе к уровню конца R, чем SC; ветвь более тонкая.
8 (9) Конец RS удален от вершины крыла; передняя-ветвь RS почти пря-
мая; крыло широкое и притупленное (рис. 65)
Род. Pleciofungivora Rohd.
Р. latipennis R о h d. u P. major, sp. nov. (стр. 58)
* 5J
9 (8) Конец RS почти на самой вершине крыла; передняя ветвь RS до-
вольно резко изогнутая, неравномерно утолщенная; крыло удлинен-
ное и на конце не притупленное. / Род. Allactoneurites R о h d.
A. jttrassicus R о h d. (стр. 61).
10 (3) Передних ветвей RS не менее двух, часто нежных и слабых.
И (12) Проксимальный отрезок RS очень короткий, короче пи; передние
ветви RS косые, впадающие в С; rm расположена значительно прокси-
мальнее конца SC (рис. 63) Род. Eopachyneura, .gen. nov.
Е. trisector alls, sp. nov. (Стр. 57)
12 (11) Проксимальный отрезок RS всегда длиннее rm; передние ветви RS
более поперечные, частично впадающие в R; rm расположена на уровне
самого конца SC или дистальнее.
13 (14) Основные задние стволы RS и Mj резко сходятся дистально; rm
лежит на уровне конца SC; передних ветвей RS не менее трех; заметны
еще следы ветвей в дистальном конце радиального поля; основная
(задняя) ветвь RS на конце сильно загнута назад и оканчивается на
самой вершине (рис. 56) Род. Polyneurisca, gen. nov.
Р. atavina, sp. nov. (стр. 52)
14 (13) RS и параллельные; rm расположена значительно дистальнее
конца SC; RS лишь с двумя передними ветвями; дистальный конец ради-
ального поля без поперечных жилок; основная ветвь RS почти прямая,
очень слабо загнутая назад (рис. 59) Род Pleciofungivorella, gen. not*.
Р. binerva, sp. nov. (стр. 55)
Род Polyneurisca, gen. nov.
Тип рода: Polyneurisca atavina, sp. nov.
достигает вершины крыла, слегка заходя за конец Мх; SC тонкая,
но хорошо-развитая, заметно короче */з крыла; R прямая, дашь слабо загну-
тая, на самом конце RS состоит из крепкой задней ветви, довольно сильно
загнутой назад, и нескольких передних, более слабых ветвей; основной отре-
зок RS примерно в Р/2 раза длиннее rm, равномерно изогнутый; передние
ветви RS в числе трех S-образно изогнутых жилок, из которых самая прокси-
мальная соединяется с R, остальные впадают в С; в дистальном конце радиаль-
ного поля еще заметны 1 или 2 слабых ветви RS, носящие характер попереч-
ных жилок; rm расположена на уровне конца SC, притом почти строго попе-
речно к оси крыла и делит основной ствол передних ветвей М почти на рав-
ные половины; RS и М, заметно сходящиеся к вершине; общий ствол М,
и Mg от rm до развилка почти в 3 раза короче Мг; М3 ответвляется от общего
ствола М проксимальнее развилка R и RS, довольно тонкая и слабая; развиты
нежные, но заметные поперечные жилки между М3, М1+2и Си; Си хорошо раз-
витая, слабо загнутая на конце; Ап и CuP неясные; крыло короткое и при-
тупленное; отношение егр длины к ширине равно примерно 1:2; крыло
заметно длиннее брюшка1.
Голова плохо сохранилась; антенны и щупики не сохранились.
Грудь вздутая посредине, умеренно нависающая над головой.
Брюшко в 1г/2 раза длиннее головы и груди, вместе взятых; заметны
8 его сегментов, которые значительно короче своей высоты; конец брюшка
притуплен.
Polyneurisca atavina, sp. nov.
(Рис. 55, 56, табл. IX, фиг. 22) .
Отпечаток самца, лежащего на правой стороне. Длина тела 2.25, крыла
.L75 мм; окраска светлая, коричневато-желтая; стигма на крыле слабо
развитая, но хорошо заметная, охватывающая конец R и две ветви RS.
52
Самец. Кара-тау, горы Кочкар-ата, ур. Ак-тас, Терс-джейлау, Б.Ро-
дсндорф, 1937. Колл. ПИН, № 340/167.
Род Polyneurisca наиболее своеобразен по присутствию многочисленны*
ветвей, принимающих характер поперечных жилок; также замечательно
присутствие поперечных жилок между М1+2 и М3 и между М3 и Си. Вместе
с тем могут быть отмечены и черты явной х
специализации; такова укороченная SC, f \
основание RS (вернее, сдвиг к основа-
нию крыла rm), утолщенные радиальные
жилки: все это заставляет считать род
Polyneurisca сохранившим ряд примитив-
ных черт и при этом уже сильно специ-
ализованным, приближающимся к таким
формам семейства, как роды Plecio/un-
givora и Pleciofungivorella; к послед-
нему и наиболее близка Polyneurisca.
Род Traltsversiplecla, gen. nov.'
Тип рода: Transverslplecia transverstnervis,
sp. nov.
С достигает вершины крыла, заходя
до середины расстояния между концами
RS и Мх; SC хорошо развитая, равная
>/3 крыла; R прямая, резко загнутая
на вершине; развита крепкая попереч-
Рис. 55.Polyneurisca atavina, sp. nov.
Кара-тау, ПИН,№ 340/167; общий вид
ископаемого (ориг.).
ная жилка, расположенная сразу ди-
стальнее конца SC между С и R; RS равномерно изогнутая, ответ-
вляющаяся от R примерно на уровне середины SC и обладающая одной перед-
ней ветвью, расположенной на уровне конца R; проксимальный отрезок
Рис. 56. Polyneurisca atavina, sp_nov. Кара-тау,'ПИН,
№ 340/167; крыло [(ориг.).
RS примерно в 2 раза длиннее rm; передняя ветвь слабо изогнутая, умеренно
косо лежащая, впадающая в С несколько дистальнее конца R; конец RS
загнут назад и заметно приближен к Мх; общий ствол М очень короток и
резко загнут назад, соединяясь с Си; проксимальный отрезок общего ствола
иередних ветвей М короткий, почти в два раза короче дистального
отрезка; последний в свою очередь почти в два раза короче Мх; rm таким
образом расположена почти на границе проксимальной трети ствола Мх+2;
ветви Мх и М2 в своей дистальной половине параллельною; М3 почти прямая,
слабо дугообразная; различимы две mcu, поперечные; Си тонкая, но хо-
рошо заметная, равномерно загнутая; СиР и Ап неясные. Крыло короткое,
|фитупленное, в 2 раза длиннее ширины.
53
Антенны тонкие, слегка длиннее головы и груди; членистость их плох»
заметна вследствие плохой сохранности; 1-й .членик сильно вздутый, шаро-
видный.
Грудь небольшая; вздутие ее неясно; отпечаток насекомого виден сверху;
ноги плохо сохранились, тонкие; бедра умеренна вздутые.
Брюшко заостренное к концу; заметны лишь 4 дистальных его сегмента.
Transversiplecia transversinervis, sp. nov.
(Рис. 57, табл. X, фиг. 23)
Отпечаток самки, лежащей на брюшной стороне, с отодвинутым: слегка
в сторону левым крылом, протянутыми вперед передними лапками и голе-
нями, расправленными в стороны антеннами и подогнутыми под тело осталь-
ными ногами; окраска темная, коричнево-черная; птеростигма на крыле
Рис. 57. Transversiplecia transversinervis, sp. n>v. Кара-тау,
ПИН, № 2452/358; крыло (ориг.).
хороню выраженная, лишь расплывчатая по краям и охватывающая концы
R и ветви RS. Длина тела 4.5, крыла 3.5 мм
1 самка. Кара-тау, сел.Галкино, А.В. М а р ты н о в, 1925. Колл. ПИН,
№ 2452/358.
Этот род, как и Polyneurisca, обнаруживает ряд примитивных черт;
таково присутствие поперечной жилки в субкостальном поле, а также и
двух поперечных mcu. Другие признаки — длина SC, основного отрезка
RS, положение rm и характер вилки М1+2— сближают род Transversiple-
cia с Plecicfungivora; этот последний, несомненно, потомок ближайших
предков Transversiplecia.
Род Pleciofangivorella, gen. nov.
Тип рода: Pleciofungivorella binerva, sp. nov.
SC слабая и тонкая, но заметная, не более х/4 крыла; С почти достигает
вершины крыла, слегка заходя за конец RS; R почти прямая, слабо загну-
тая на вершине; RS крепкая, очень слабо загнутая, ответвляющаяся от
R несколько проксимальнее конца SC; развиты две косо лежащие, почти
прямые передние ветви, из которых проксимальная соединяется с R, а дисталь-
ная с С; rm примерно в два раза короче основного отрезка общего ствола
RS и расположенная дистальнее уровня конца SC; М и Си плохо видны,
строение их неясно; повидимому, дистальный отрезок общего ствола М1+2
почти равен по длине Мх или М2; крыло умеренно удлиненное и притуплен-
ное на конце.
Глаза крупные, почти в Р/2 раза выше своей длины. Антенны равны
по длине голове и груди, вместе взятым, и, повидимому, состоят из 15 чле-
ников, из которых основной сильно вздут; ротовые органы укороченные.
54
Грудь равномерно и довольно сильно вздутая, сильно выетупайЙЗЯ
вперед и нависающая над головой; ноги тонкие, но не особенно удлиненный;
тазики умеренно увеличенные; бедра слегка короче голеней и лишь немного
толще их, почти параллельно-крайние, очень слабо расширяющиеся к дисталь-
ным концам; лапки незначительно тоньше голеней, пятичлениковые; про-
порции между отдельными члениками не могут быть даны с достовер-
ностью вследствие плохо заметных границ члеников.
Брюшко, невидимому, ссохлось при
захоронении и поэтому ненормально
укорочено; по этой же причине плохо
заметны границы между его сегмен-
тами; оно примерно в Р/а раза длин-
нее головы и труди и на конце за-
остренное.
Pleciofungivorella binerva, sp. nov.
(Рис. 58 и 59)
Отпечаток, вероятно, самки, лежа-
щей на правой стороне с несколько
подтянутыми ногами, расправленными
антеннами и крыльями, из которых
правое сложено вдоль; окраска черная;
ноги более светлые, коричневатые;
крылья слабо окрашенные; птеростигма
не резкая, захватывающая конец R и
часть проксимальной ветви RS; длина
Рис. 58. Pleciofungivorella binerva,
sp. nov. Кара-тау, ПИН, №22/167;
общий вид ископаемого (ориг.).
тела 1.75, крыла Л.50 мм.
Самка (?). Кара-тау, горы Кочкар-ата, ур. Ак-тас, Терс-джейлау.
Б. Р о д е п д о р ф, 1937. Колл. ПИН, № 22/167.
Род Pleciofungivorella, gen. nov., невидимому, наиболее близок к роду
Pleciofungivora. Плохая сохранность крыльев,—именно жилок, лежащих
сзади RS, — не позволяет с точностью установить взаимоотношения этого
рода с соседними. Сравнение лишь одной передней области крыла все же
Рис. 59. Pleciofungivorella binerva, sp. nov. Кара-тау,
ПИН, № 22/167; крыло (ориг.).
позволяет сделать некоторые заключения о своеобразии Pleciofungivorella.
В первую очередь характерно присутствие двух ветвей RS, притом распо-
ложенных более поперечно, чем это наблюдается у родов Eopachyneura,
Prohesperinus и Eohesperinus, и по положению приближающихся к жилко-
ванию рода Polyneurisca, а особенно Pleciofungivora. (Гдругой стороны, SC
у Pleciofungivorella очень слаба и коротка, что не может не рассматриваться
как признак специализации. Таким образом, этот род обладает, наряду
с примитивными признаками в ветвях RS, также и рядом признаков явной
55
специализации; строение RS все же в общем близко к роду Pleciojungivora
я» вероятно, описываемый род является параллельной, близкой ветвь»
общих исходных форм этих родов.
Род Prohesperinus, gen. nov.
Тип рода: Prohesperimis abdominalis, sp. nov.
С едва заходит за конец задней ветви RS; SC не особенно тонкая, короче
Vs длины крыла; R почти прямая, очень слабо загнутая на протяжении
дистальной трети; RS ответвленная от R на уровне середины SC и обладает
единственной передней ветвью; ветвь RS расположена косо, ответвляется
на уровне последней трети R и соединяется с С дистальнее конца R; перед-
няя’ ветвь RS несколько тоньше задней основной ветви; последняя равно-
мерно дугообразно изогнутая и достигает самой вершины крыла; rm рас-
положена на уровне конца SC; проксимальный отрезок RS в 4 раза длиннее
rm; rm делит общий ствол М1+2 на две равные части; дистальный отрезок
общего ствола М1+2 и RS заметно расходятся; М2 и Мг в своих дистальных
половинах параллельны; М3 ответ-
вляется от общего ствола М не-
сколько проксимальнее развилка
f R—RS; М прямая и резко дуго-
образно изогнутая на своем конце;
Си слабая, равномерно изогнутая;
mcu и; An f неясные; крыло
Рис. 60. Prokesperinus abdominalis, sp. nov.
Кара-тау, ПИН, № 2452/580; общий вид
ископаемого (ориг.).
Рис. 61. Prohesperinus abdominalis,
sp. nov. Кара-тау, ПИН, № 2452/580,
крыло (ориг.).
слегка удлиненное, не притупленна; длина его более чем в 2 раза больше
ширины.
Антенны равны по длине голове и груди.и, повидимому, имеют 16 чле-
ников, из которых 1-й сильно увеличен и вздут; форма члеников плохо
различима; повидимому, они незначительно длиннее своей толщины; дисталь-
ные членики более тонкие; щупики хорошо заметные, но точное число их
члеников установить невозможно; хоботок короткий, не выступающий.
Грудь сильно вздутая, нависающая над головой; ноги не особенно тон-
кие, длинные; бедра умеренно вздутые в середине; голени тонкие, постепенно
расширяющиеся к дистальному концу; средние равны по длине бедрам,
задние почти в Р/2 раза их длиннее; лапки не особенно.тонкие, длиннее
голеней; на передних лапках 2-й членик наиболее длинный, 1-й и 5-й —
самые короткие; на средних лапках 1-й членик очень длинный, равный
всем остальным, вместе взятым; эти последние почти равной величины;
на задних лапках 1-й членик равен примерно 2/в длины лапки, размеры
остальных члеников плохо различимы.
Брюшко очень массивное, состоящее из 8 сегментов и генитального
аппарата; оно более чем в 2 раза длиннее головы и груди; сегментация его
хорошо выражена; конец брюшка заостренный.
56
Profiesperinus abdominalis, sp. nov.
(Рис. 60 и 61)
Отпечаток самки, лежащей на правом боку, с расправленными антеннами,
Крыльями и ногами; окраска коричнево-черная, равномерная; птеростиг-
малъное пятно небольшое, но резко обособленное, охватывающее конец R,
приближенное, но не заходящее дистальнее конца передней ветви RS. Длина
тела 5.75, крыла 3.25 мм.
Самка. Кара-тау, сел. Галкино, А. В. Мартынов, 1925, Колл. ПИН,
№ 2452/580.
Род Profiesperinus, gen. nov., близок к родам Eohesperinus, gen. nov., и
Eopachyneura, gen. nov., образуя с ними ясную группу, для которой харак-
терно положение передней ветви RS; эта жилка у перечисленных родов
крепкая, не обнаруживающая тенденции к редукции и, кроме того, располо-
женная очень косо, почти параллельно с концом R. Среди этих родов род
Profiesperinus характерен по удлиненному основанию RS и положению rm.
отдаленной от развилка R—RS; эти особенности носят явно примитивный
характер; с другой стороны, короткая SC й крепкая передняя ветвь RS
показывают уже достаточную степень специализации. В общем все же надо,
повидимому, считать род Profiesperinus по сравнению с двумя другими наибо-
лее близким к исходным формам; связи с другими родами семейства оста-
ются еще недостаточно выясненными.
Род Eopachyneara, gen. nov.
Тип рода: Eopachyneura trisectoralis, sp. nov.
Рис, 62. Eopa-
chyneura trisec-
toralis, sp. nov,
Кара-тау,ПИН,
№ 2452/673;
общий вид ис-
копаемого
(ориг.).
С достигает вершины крыла и далеко заходит за конец основной ветви
RS; в месте соединения с SC, С образует хорошо заметную выпуклость; SC
ддвольно тонкая, но хорошо развитая, короткая, менее */з
крыла; R прямая н лишь в дистальной трети слабо изогну-
тая; RS очень характерного строения и обладает двумя пе-
редними ветвями; обе передние ветви RS соединяются с С
и расположены косо, параллельно с концом R; дистальная
ветвь в два раза короче и заметно тоньше проксимальной;
основная задняя ветвь RS равномерно изгибается назад;
место ответвления RS от R расположено на уровне пример-
но середины SC; проксимальный отрезок общего ствола
RS очень короткий вследствие сильно сдвинутой жилки
rm; длина этого отрезка меньше rm; последняя распо-
ложена значительно проксимальнее конца SC; дистальный
отрезок М1+2 из-за положения rm очень велик, более
чем в 2 раза короче Мх; обе ветви М—Мх слабо расхс-
дящиеся, почти параллельные, как и концы RS и Мх; М3
цлохо заметная, прямая и тонкая; Си, шеи и Ап не видны;
крыло умеренно удлиненное, несколько более чем в
2 раза длиннее своей ширины.
Голова, грудь и брюшко сохранились очень плохо, и
детали их строения не могут быть выяснены; брюшко
сильно заостренное; заметны 5 или б дистальных его
сегментов.
Eopachyneura trisectoralis, sp. nov.
(Рис. 62 и 63)
Отпечаток самки, лежащей на спинной стороне. Голова, грудь с ногами
и брюшко сжаты в комок и их детали неразличимы; крылья откинуты на
голову и налегают при этом одно на другое; окраска темная, корйчнево-
57
черная; птеростигмальное пятно небольшое и плохо отграниченное. Длина
тела 4.0, крыла 3.0 мм.
Самка. Кара-тау, сел. Галкино, А. В. Мартынов, 1925. Колл. ПИН,
№2452/673.
Этот род обладает своеобразным мозаичным характером. Так прими-
тивно строение RS, несущего две передние ветви, притом косые и довольно
длинные; проксимальная ветвь крепкая, не отличающаяся по толщине от
R и собственно RS. С другой стороны, есть черты большой специализации,
выражающиеся в далеком сдвиге жилки rm к основанию крыла, укороче-
Рис. 63. Eopachyneura trisectoralis, sp. nov. Кара-тау, ПИН,
№ 2452/673; крыло (ориг.).
нии проксимального отрезка RS, ослаблении SC и своеобразии С — обра-
зовании выступа у SC. Все это заставляет считать род Eopachyneura осо-
бой уклонившейся формой, сохранившей наряду с общим достаточно
высокоспециализованным характером некоторые черты большой прими-
тивности. Будучи в общем близок к родам Eohesperinus и Prohesperinus,
этот род не обнаруживает ясно намечающихся связей с каким-либо одним
из них.
Род Pleciofungivora, R о h d.
Родендорф, 1938, Тр. ПИН, т. VII, вып. 3, стр. 42
Тип рода: Pleciofungivora latipennis Rohd.
Этот род довольно специализованный; это выражается в сдвиге rm,
коротком проксимальном отрезке общего ствола RS, в тонкой и слабо на-
клонной передней ветви RS, в сдвиге главного ствола RS к переднему краю
крыла и в относительна коротких антеннах.
Pleciofungivora, невидимому, является потомком форм, близких к родам
Polyneurisca и Pleciofungivorella, но утерявших те черты примитивности,
которые еще сохранились у этих двух названных родов.
Pleciofungivora major, sp. nov.
(Рис. 64 и 65)
^Отпечаток самки, лежащей на правой стороне, с расправленными антен-
нами, ногами и крыльями; обе передние й одна средняя нога не сохрани-
лись; левое крыло смято; окраска темнокоричневая, местами черная. Ан-
тенны состоят из умеренно вздутого базального членика и 13 члеников более
тонких; форма члеников неодинаковая; 4 или 5 дистальных длиннее своей
58
Рис. 64. Pleciofungivora major, sp. nov. Кара-тау,
ПИН, № 2452/659; общий вид ископаемого (оригГ).
толщины; базальные толще своей длины. Грудь умеренно вздутая; крылья
не длиннее брюшка, довольно широкие; SC оканчивается на уровне пи;
передняя ветвь RS нежная, лежащая почти поперечно относительно оси
крыла; проксимальный отрезок общего ствола RS лишь в Р/2—2 раза длин-
нее rm; последняя слегка наклонная; задняя ветвь RS изогнутая я
своей дистальной половине;
дистальный отрезок общего
'ствола М1+2 меньше х/2 вет-
ви Мх; ветви Мх и М2 очень
слабо сходящиеся. Ноги не
особенно тонкие; бедра рав-
номерно утолщающиеся к
середине; передние равны го-
леням; средние и задние бо-
лее короткие; голени почти
па ра лле льно-кра йние; задние
заметно утолщающиеся к
концу; передние лапки равны
голени, с наиболее коротки-
ми 3-м и 4-м члениками; сред-
ние лапки короче голени,
с очень большим 1-м члени-
ком, достигающим почти */2
длины всей лапки, задние
лапки несколько более удли-
ненные. Брюшко в 2 раза
длиннее груди и головы, за-
остренное. Длина тела 3.75,
крыла 3.00 мм.
Самка. Кара-тау, сел.
Галкино, А. В. Марты-
н о в, 1925. Колл. ПИН,
№ 2452/659.
Описанный второй вид ро-
да Pleciofungivora довольно
резко отличается от ранее из-
вестного ; эти отличия даны
в следующей таблице.
1 (2) Передняя ветвь RS рас-
положена косо; место
ее ответвления от RS
проксимальнее уровня
конца R; проксималь-
ный отрезок общего ство-
ла RS не менее чем в 2 ра-
за длиннее rm; дисталь-
ный отрезок общего ство-
ла М1+2 заметно больше
J/2 Мх; длина тела; 2.5, Рис. 65. Pleciofungivora major, sp. nov. Кара-тау,
. крыла 2.0 ММ. пин. № 2452/659; крыло (ориг.).
Р. tatipennis R о h d.
2 (1) Передняя ветвь RS более поперечная и ответвляется" от RS едва ди-
стальнее уровня конца R; проксимальный отрезок RS очень корот-
кий, лишь в Р/2 раза длиннее rm; общий ствол М1+2 (дистальнее rm)
меньше х/2 Мх; длина тела 3.95, крыла 3.0 мм.............Р. major, sp. nov.
5»
Род Eohesperinus, gen. nov.
Тип рода: Eohespertnus martynovi, sp. nov.
С достигает вершины крыла, лишь слегка заходя за конец RS; SC лишь
незначительно короче 1/3 крыла, нежная, но хорошо заметная; R почти
прямая, слабо загнутая на конце; RS имеет крепкую, резко изогнутую перед-
нюю ветвь, отчасти параллельную с концом
R; место ответвления передней ветви RS
расположено значительно проксимальнее
уровня конца R: эта ветвь лежит очень
косо; проксимальный отрезок общего ство-
ла RS очень короткий, лишь едва длиннее
rm; основная задняя ветвь RS равномерно
изогнутая; rm довольно короткая, располо-
женная косо; дистальный отрезок общего-
ствола . М1+2 в 2х/2 раза короче Mt; М3:
почти совершенно прямая; Си, mcu и Ап
неясные; заметна лишь тонкая, дугообраз-
ная жилка Си; место соединения rm с RS
расположено точно на уровне конца SC;
форма крыла умеренно удлиненная, отно-
шение ширины крыла к длине 1:2.
Голова небольшая; антенны сохрани-
лись не целиком; повидимому, они рав-
Рис. 66. Eohesperinusmartynovi, sp. ны длине голове и груди, вместе взя-
nov. Кара-тау, ПИН, № 2452/289; тым; основной их членик резко вздутый,
общий вид ископаемого (ориг.). остальные членики лишь в P/j—2 раза
длиннее своей толщины; хоботок неясен.
Грудь умеренно вздутая, слабо нависающая над головой; ноги тонкие
и длинные; тазики удлиненные; бедра слегка утолщенные, короче голеней;
голени тонкие и длинные, слабо расширенные на конце, с короткими, но
хорошо заметными шпорами; лапки тонкие, равные по длине голеням; 1-й
Рис. 67. Eohesperinus martynovi, sp. nov. Кара-тау, ПИН,
№ 2452/289; крыло (ориг.).
их членик наиболее длинен; на груди заметны короткие булавовидные жуж-
жальца.
Брюшко в 2 раза длиннее головы и груди, довольно толстое, на конце
заостренное, из 10 сегментов; последний видимый «сегмент», вероятно,
Является церками.
М
Eohesperinus martynovi, sp. nov. ... t
(Рис. 66 и 67; табл. XI, фиг. 27)
Отпечаток самки, лежащей на левой стороне; одно крыле продольно
сложено; ноги более или менее расправлены; антенны частично обломаны;
окраска темнокоричневая; птеростигма окружает конец R и не заходит ди-
стально за переднюю ветвь RS; конец R и передняя ветвь RS ясно сходя-
щиеся; последняя не изогнутая на конце; основной отрезок общего ствола
RSb Р/граза длиннее rm; SC прямая, лишь на конце дугообразно изогнута.
Длина тела 4.0, крыла 3.0 мм.
Самка. Кара-тау, сел. Галкино, А. В. Мартынов, 1925. Колл. ПИН,
№ 2452/289 и № 2452/308 (негатив).
Род Allactoneurites R о h d.
Родендорф, 1938. Тр. ПИН, т. VII, вып. 3, стр. 43
Тип рода: Allactoneurites jurassicus Rohd-, 1938
Этот род наиболее близок к роду Pleciofungivora. Отличия его от послед-
него заключаются в более удлиненной форме крыла, более длинного и вы-
прямленного, в задней ветви RS и изгибе и неравномерном утолщении перед-
ней ветви RS; все эти признаки носят черты меньшей специализации по
сравнению с тем, что наблюдается у Pleciofungivora. Признаки специали-
зации выражаются, так же как и у Pleciofungivora, в сдвиге rm к основанию
крыла и в слабой SC.
Известен лишь один вид, тип рода из юры Кара-тау.
Семейство Pteclomimidae, fam. nov.
Тип семейства: род Pleciomima Rohd., 1938.
SC умеренно удлиненная, равная х/<—2/s крыла, почти всегда слабая,.
иногда очень нежная и тонкая, впадающая в С; RS простая, изогнутая или
прямая (у Mimallactoneurinae, Lycoriomimella, Pleciomimella); расположе-
ние rm изменчиво: оно очень близко от ответвления RS от R (у Pleciomi-
mitiae) или дистальнее; общий ствол М1+2 обычно короче ветвей Мх и М2,
реже равен им; М3 изменчивой длины, обычно соединенная с М1+2; антенны
12—17-члениковые, умеренно тонкие, равные голове и груди, вместе взя-
тым; грудь умеренно вздутая; ноги утолщенные, довольно короткие,.реже
удлиненные и тонкие (у Antefungivoriпае); крылья обычно довольно корот-
кие, закругленные на вершине; брюшко крупное; размеры тела от 1.6 до
4.25 мм.
Это семейство является наиболее крупным изо всех известных кара-
тауских Oligoneura; к нему принадлежат не менее 20 видов, большая часть
которых описывается ниже. Будучи наиболее близким к Pleciofuagivoridae
и> Allactoneuridae, семейство Pleciomimidae отличается гораздо большей
специализацией, наиболее резко проявленной в строении крыльев, в их
редуцированном жилковании. Для Pleciomimidae характерна простая,
Лишенная ветвей жилка RS, далее — укорочение жилки R, резкое ослаб-
ление жилок SC и М.
Определительная таблица подсемейств и родов
Pleciomimidae, fam. nov. _
1 (14) Базальный отрезок RS очень короткий, не-более чем в 2 раза длин-
нее rm; ноги более или менее утолщенные.
Подсемейство Pleciomiminae, subfam,nov.
61
2 (5) RS прямая или очень слабо изогнутая в дистальной половине; R на
! конце прямая, соединяющаяся с С под очень острым углом; SC, по
крайней мере в основной части, крепкая.
3 (4) RS совершенно прямая, в 2 раза длиннее дистальной части R
(рис. 77) Род Ljcoriomimella, gen. nov. (стр. 69}
'4 (3) RS в дистальной половине слабо изогнутая, в Р/2 раза длиннее
дистальной части R (рис. 80). Род Pleciomimella, gen. nov. (стр. 71)
5 (2) RS равномерно и иногда довольно сильно дуговидно изогнутая;.
R на конце обычно изогнутая, впадающая в С под большим углом;
SC обычно слабая.
6 (7) SC и М крепкие, мало отличающиеся от других жилок; rm распо-
ложена значительно проксимальнее конца SC; R прямая (рис. 79)
Род Pleciomima R о h d. (стр. 70)
7 (6) SC и М слабее других жилок; R изогнутая.
8 (9) rm расположена проксимальнее места впадения SC в С; М3 очень
длинная, непосредственно соединенная с М1+2; 3-й отрезок С короче
2-го; М очень слабая и плохо заметная (рис. 72)
Род Paralycoriomima, gen. nov. (стр. 64}
9 (8) rm расположена на уровне или дистальнее конца SC; М обычно
. гораздо короче; 3-й отрезок С длиннее или равен 2-му.
10 (И) RS и М ясно сходящиеся; есть слабое стигмальное пятно. (рис* 74)
Род Lycorioplecia, gem nov. (стр. 66)
И (10) RS и Мх параллельные; стигмальное пятно неясное.
12 (13) СиР ясная; ноги короткие (рис. 70)
Род Lycoriomitna, gen. nov. (стр. 62)
13 (12) СиР неясная; ноги более длинные (рис. 75)
Род Lycoriomimodes, gen. nov. (стр. 67)
14 (1) Базальный отрезок RS длинный, не менее чем в 5 раз длиннее rm;
ноги тонкие.
15 (16) М3 обособленная от М142 и ответвляется от общего стволаМз+СиА;
ветви Мг и М3 очень слабые; крылья короткие; спинка груди в про-
филь плоская (рис. 85—87). Подсемейство Paritonidinae, subfam. nov.
Единственный род Paritonida, gen. nov. (стр. 74)
16 (15) M1+a ответвляется от М3; ветви М более крепкие; крылья
длинные.
17 (18) Крыло широкое; SC короткая; R не более 8/6 длины крыла; ответ-
вление М3 от СиА происходит значительно проксимальнее развилка
R—RS; оба отрезка ствола М1+2 почти равные (рис. 83 и 84)
Подсемейство Antefungivorinae Rohd. emend.
Роды Antefungivora Rohd. и Antiquamedia Rohd.
18 (17) Крыло более узкое; SC длинная, более 1/3 крыла; R тоже значи-
тельно длиннее» не менее 3/4 крыла; ответвление М3 от СиА происхо-
дит несколько ‘дистальнее развилка R—RS; дистальный развилок
ствола М1+2 более чем в 2 раза длиннее проксимального (рис. 81 и 82)
Подсемейство Mimallactoneurinae, subfam. nov.
Единственный род Mimallactoneura, gen. nov. (стр. 72)
Подсемейство Pleciomimlnae, s. str.
Род Lycorlomima, gen. nov.
Тип рода.' Lycoriomitna ventralis, sp. nov.
С доходит лишь до RS; SC тонкая, равномерно изогнутая, впадающая
в край крыла дистальнее уровня rm; R почти прямая, соединяющаяся с С
на границе 2-й и 3-й третей крыла; RS ответвляется от R на уровне конца
62
[Рис. 68. Lycoriomima ventralis, sp. nov. Кара-тау Д ПИН,
№ 2452/293; общий вид ископаемого (ориг.).
Рис. 69. Lycoriomima ventralis, sp. nov. Кара-тау, ПИН,
№ 2452/293; части ног: а — передняя лапка и конец го*
лени, б — среднйя нога; в — заднее бедро, голень и осно-
вание лапки (ориг.).
основной трети крыла; rm расположена близко от ответвления RS от R
и лишь в 2—2х/2 раза короче базального отрезка RS; М нежная и тонкая;
развилок Мх4-М2, повидимому, значительно короче общего ствола М1+3;
М3 крепче передних ветвей, правильно дугообразно изогнутая; СиА
изогнутая; СиР хорошо заметная, параллельная с СиА; крылья короче
брюшка, довольно широкие.
Грудь умеренно вздутая, слабо нависающая над головой; щиток мало
выдается; ноги крепкие, слабо удлиненные; тазики умеренно удлиненные;
задние голени лишь достигают 7-го сегмента брюшка. Голова округлая
в профиль; антенны 15-члениковые; бедра и голени покрытые нежными
прилегающими волосками и более крепкими и длинными щетинками;
шпоры на концах голеней отсутствуют или слабо развиты.
Этот род наиболее близок к описываемым ниже родам Lycoriomimodes
и Paralycoriomima и отличается от них более короткими ногами, слабым
развитием (редукцией) некоторых жилок (SC, М, СиР) и их пропорциями.
Lycoriomima ventralis, sp. nov.
(Рис. 68, 69, и 70; табл. XIII, фиг. 29)
Отпечаток самца, лежащего на левой стороне; хорошо сохранились
ноги, антенны, брюшко и крылья; последние разрушены в середине и нале-
гают одно на другое; хорошо сохранились ноги; передние бедра тонкие,
слабо вздутые, передние голени равны бедрам, более тонкие, на конце
с короткой щетинкой; передние лапки слегка длиннее голеней, пятичленн-
ковые; 1-й членик самый длинный, притом лишь незначительно длиннее
2-го, который слегка длиннее 3-го; 4-й членик короче 3-го и почти в 2 раза
длиннее 5-го, который несет слабо изогнутые коготки; бедро средней пары
ног умеренно вздутое, веретеновидное, покрытое, кроме коротких волосков,
хорошо заметными, но .короткими щетинками; голень длиннее бедра, равно-
мерно, но слабо утолщается к дистальному концу, на конце обрубленная;
на голени расположены хорошо заметные щетинки; лапка пятичлениковая;
1-й членик наибольший, длиннее 2-го и 3-го, вместе взятых; эти последние
почти равной длины; 4-й членик второй по величине, несколько короче
суммы 2-го и 3-го члеников; 5-й членик равен 3-му, округлых очертаний
и несет довольно длинный коготок; задняя пара ног длиннее передних и
средних; строение похоже на среднюю пару, лишь бедро более вздутое,
а голень более удлиненная и более расширенная дистально; на конце с за-
кругленными щетинками, на голени с укороченными, в виде шипиков.
Брюшко сильно вздутое, из трех сегментов, на конце со сложно построен-
ным копулятивным аппаратом; строение последнего плохо различимо;
все стерниты и задние края тергитов, особенно последних, с торчащими
щетинками. Длина тела 3.25, крыла 1.75 мм.
Позитивный отпечаток самца. Колл. ПИН, № 2452/293. Кара-тау, сел.
Галкино. А. В. Мартынов. 1924.
Род Paralycoriomima, gen. nov.
Тип рода: Paralycoriomima sororcula, sp. nov.
С едва заходит за RS; SC тонкая и нежная, равномерно дугообразно
изогнутая, впадающая в край крыла значительно дистальнее уровня rm;
R почти прямая, достигающая 2/3 длины крыла; RS ответвляется от R на
уровне конца х/5 длины крыла; rm расположена очень близко от ответвле-
ния RS от R, лишь в Р/г раза короче основного отрезка RS; М нежная и
тонкая: развилок короче общего ствола М1+2; Мх очень слабая и плохо
видимая; М3 тонкая, неравномерно изогнутая; общий ствол М1+2 соеди-
64
няется с М8; место соединения Mg с Си неясно; СиР хорошо видна; СиА
резко загнутая назад в последней четверти;крылья короче брюшка, широкие.
Грудь резко вздутая; спинка в профиль округлая, нависающая над
головой; щиток очень мал; ноги крепкие, довольно короткие, тазики уме
ренно удлиненные; задние голени не достигают конца брюшка; голова
в профиль почковидная; антен- х-ч. /Х\
ны 15-члениковые, шпоры на / \ \
концах голеней плохо развитые. / \ / I
Род Paralycoriomima наибо- / / /
лее приближается к родам Ly- / /
coriomimella и Lycoriomi modes, / /
особенно к первому, отличаясь / / /
иными пропорциями жилок и 'х/ / /// /
другими признаками, указанны-
мн в определительной табли- \
це- L
Г //</
Paralycoriomima sororcula, х\
sp. nov. »
(.рис. 71 и 72; табл. XIII, фиг. 30) t *
Отпечаток самки, лежащей на **
левой стороне; хорошо сохрани- рис. 71. Paralycoriomima sororcula, sp. nov.
лось тело, крыло И отчасти ноги; Кара-тау, ПИН, № 2452/287; общий вид
ископаемого (ориг
голова и ее придатки плохой
сохранности.
Передние бедра очень слабо утолщенные, мало отличимые от голени,
значительно короче их; членистость передних лапок плохо различима;
бедра средних и задних ног явственно вздутые; средние бедра и голени
почти равные по длине; голени тонкие, почти параллельно-крайние; сред-
Рис. 72. Paralycoriomima sororcula, sp. nov. Кара-тау, ПИН,
№ 2452/287; крыЛо (ориг,).
ние лапки равны голеням; первые два членика их наиболее длинны, почти
равной длины, на основании заметно суженные; 3-й, 4-й и 5-й более корот-
кие; их строение плохо различима; задние ноги наиболее длинные; задаие
голени заметно длиннее бедер, очень слабо расширенные к концу, покрытые
щетинками; задние лапки длинные; 2-й их членик наибольший; все осталь-
ные почти равной длины; коготки хорошо развитые. Брюшко, невидимому,
конической формы с восемью сегментами, на конце с придатками, церками;
щетинки неясные. Длина тела 2.75, крыла 1.75 мм.
Позитивный отпечаток самки. Колл. ПИН, № 2452/287. Кара-тау, сел.
Галкино. А. В. М а р т ы н о в, 1924.
5 Родендорф, т. ХШ, в. 2.
65
Род Lycorioplecia, gen. nov.
Тип рода: Lycorioplecia elongata^sp. nov.
С доходит до RS; SC нежная, но хорошо заметная; R слабо изогнутая,
оканчивающаяся примерно на уровне 2-й и 3-й третей крыла; RS ответ-
вляется от R на уровне конца пер-
вой трети крыла, оканчиваясь пе-
ред концом крыла; rm располо-
жена вблизи от развилка R — RS
и незначительно короче основного
отрезка RS; М слабая; развилок
М примерно равен общему ство-
лу М; место ответвления М1+2 от
ствола Си неясно и, повидимому,
расположено близ основания кры-
ла; М3 почти прямая, более.креп-
кая, чем М2 и Мх, ответвляется
от СиА едва проксимальнее уров-
ня rm; СиА крепкая, равномерно
дугообразно изогнутая; СиР и Ап
неясные, крылья значительно ко-
роче брюшка, широкие.
Грудь слабо вздутая, почти не
нависающая над головой; щиток
небольшой, но заметно торчащий;
ноги слегка удлиненные, покрытые
волосками и щетинками; шпоры
на голенях очень короткие; та-
Рис. 73. Lycorioplecia elongata, sp. nov. зики удлиненные, примерно рав-
Кара-тау, ПИН, № 2452/91; общий вид цые высоте головы; задние голени
ископаемого (ориг.). почти достигают конца брюшка.
Голова в профиль округлая; рото-
вые органы, видимо, короткие и мягкие; антенны умеренно удлиненные;
число их члеников неясное.
Рис. 74. [Lycorioplecia elongata, sp. nov. Кара-тау, ПИН,
№ 2452/91; крыло (ориг.).
Описываемый род наиболее близок к роду Pleciomima R o h d., но
хорошо от него отличается более короткими крыльями, слегка сдвинутыми
вперед жилками RS и Мх и более слабой SC. Сильное удлинение брюшка
объясняется как следствие раздавливания при захоронении.
66
Lycorioplecia elongata, sp. nov.
(Рис. 73 и 74; табл. XIII, фиг. 31)
Отпечаток самки, лежащей на левой стороне; тело и ноги удовлетво-
рительной сохранности; голова, ее придатки и крылья сохранились хуже*
Крыло широкое и короткое; отношение его ширины к длине равно
1 ; 1.8; окраска крыла темная, желтовато-коричневая; хорошо заметна
птеростигма, охватывающая конец R; RS и Мх заметно сходятся. Ноги
покрыты многочисленными короткими щетинками, особенно передние и
средние бедра и задние голени; передние бедра умеренно вздутые и лишь
в Г/2 раза длиннее тазиков: передние голени тонкие, в 1*/8 раза длиннее
бедер, параллельно-крайние, на конце с короткими шпорами; передние
лапки длинные, лишь незначительно тоньше и почти в 2 раза длиннее
голеней; первые три их членика длиннее, равные по величине; членики
4-й и 5-й в 2 раза короче 2-го; коготки неясные; средние ноги длиннее перед-
них; бедра в два раза длиннее тазиков, умеренно вздутые; голени тонкие,
в Р/з раза длиннее бедер; лапки целиком не сохранились; остались лишь
два базальных членика; задние ноги наиболее велики; бедра их более
чем в 2 раза длиннее тазиков и почти равны (лишь немного менее) толстым
голеням; задние лапки равны голеням; 1-й их членик очень велик и длин-
нее суммы всех остальных; 2-й равен сумме 3-го, 4-го и 5-го; 3-й, самый
малый, не сохранился полностью; 4-й и 5-й членики равные. Брюшко
удлиненное, из восьми сегментов, на конце последнего стернита несет
короткий густой гребень щетинок; все сегменты по заднему краю с торча-
щими, не особенно длинными щетинками. Длина тела 4.25, крыла 2.5 мм.
Отпечаток самки. Колл. ПИН, №2452/91. Кара-тау, сел. Галкино.
А. В. Мартынов, 1924.
Род Lycoriomimodes, gen. nov.
Тип рода: Lycoriomimodes deformatus, sp. nov.
С достигает лишь конца RS; SC слабая и нежная; место ее соединения
с С неясное; R прямая, на конце внезапно загнутая к краю крыла; RS
волнисто изогнутая, ответвляющаяся от R проксимальнее конца SC; rm
слегка короче базального отрезка RS, расположенная, повидимому, на
уровне конца SC; система жилок М хорошо развитая; развилок и общий
ствол Мх+3 примерно равный общему стволу М1+3, который ответвляется
от М3 на уровне ответвления RS от R; М3 более крепкая, чем передние
ветви, слабо и равномерно дугообразно изогнутая, ответвляющаяся от СиА
лишь едва проксимальнее ответвления Мх+2; СиА крепкая, сильно, но
равномерно дуговидно изогнутая; СиР и Ап неясные; относительные раз-
меры крыльев неясны вследствие деформации брюшка.
Грудь умеренно вздутая; щиток резко выступающий. Ноги крепкие,
умеренно удлиненные; шпоры на задних голенях заметные; тазики, пови-
Димому, удлиненные; пропорции размеров ног и брюшка неясные. Строение
головы неизвестно. Брюшко деформировано.
Род Lycoriomimodes наиболее близок к родам Lycoriomima и Lycorio-
mimella, но отличается от них длинными ногами и хорощо развитой
системой М.
Lycoriomimodes deformatus, sp. nov.
(Рис. 75; табл. XV, фиг. 33)
Отпечаток самца, лежащего на правой стороне; голова не сохранилась;
правое крыло сложено вдоль; левое загнуто вперед и сохранилось удовле-
творительно; ноги сохранились довольно хорошо.
67
Отношение длины и ширины крыла равно 1: 1.77; крыло неокрашен-
ное; бедра умеренно вздутые; голени значительно более тонкие, длиннее
бедер; лапки плохо сохранились; брюшко смятое; конец брюшка глубоко
расщепленный: повидимому, это генитальный аппарат; щетинки тела плохо
заметны.
Длина тела 2.0, крыла 1.6 мм.
Позитивный отпечаток самца. Колл. ПИН, № 2452/651. Кара-тау, сел.
Галкино. А. В. Мартынов, 1925.
Lycoriomimodes sp.
(Рис. 76)
Отпечаток насекомого плохой сохранности; лучше всего сохранились
остатки груди, брюшка и основной части антенны; остались фрагменты
головы и крыльев; ноги не сохрани-
лись.
Плохая сохранность отпечатка не
позволяет определить точно видовой
принадлежности этой формы; повидимо-
му, все же это насекомое следует отнести
к роду Lycoriomimodes; за это говорит
укороченная R, изгиб RS, четкая М;
иное строение гт неосновной части RS,
Рис. 76. Lycoriomimodes sp. Кара-тау,
ПИН, № 12/167; общий вид ископае
мото (ориг.).
Рис. 75. Lycoriomimodes de/ormatus, sp. nov.
Кара-тау, ПИН, № 2452/651; крыло (ориг.).
форматразвилка и основной части М заставляют предполагать принадлеж-
ность/этого вида к особому роду, устанавливать который я считаю преж-
девременным вследствие неполной сохранности объекта.
R, повидимому, правильной дугообразной формы; RS резко волнооб-
разно изогнутая; ее основная часть более чем в два раза длиннее rm и соеди-
няется с последней под тупым, а не острым углом; М2 является прямым
продолжением общего ствола М1+2, так что Мх довольно резко выступает
вперед на своем основании; развилок М1+2 едва длиннее общего ствола М1+2
от ггп до развилка; место соединения М3 с СиА неясно; М3 резко загнутая
назад; СиА плохо заметна.
Грудь умеренно вздутая; щиток выпуклый, округлый; ноги не сохра-
нились. Голова разрушена; от антенны сохранились 8 базальных члеников,
из которых 2-й наиболее велик. Брюшко довольно широкое; сохранились
6‘ сегментов И остатки 7-го и 8-го. Длина остатков тела 2.8 мм.
Позитивный отпечаток. Колл. ПИН, № 12/167. Кара-тау, горы Кочкар-
ата, ур. АК’ТЭС, Терс-джейлау. Б. Родендорф, 1937.
Род LycoriomimeUa, gen. nov.
Тип рода: Lycoriomimella minor, sp. nov.
С несколько заходит за вершину RS; SC крепкая, лишь на конце более
тонкая, резко загнутая вперед и впадающая в С на уровне ответвления
RS от R; R слабо изогнутая, на конце загнутая вперед, короткая, лишь
незначительно заходящая за середину крыла; RS почти совершенно пря-
мая, кроме основания; rm лишь в l1^ раза короче основного отрезка RS,
расположенная несколько дистальнее уровня конца SC; строение М и Си
полностью неизвестно вследствие плохой сохранности крыла; основная
часть ствола М1+2 крепкая, ответвляющаяся от М8 (или М8 + СиА?) зна-
чительно проксимальнее конца SC и развилка R—RS, при этом идущая на
некотором протяжении параллельно с М8 (или М8 -]-CuA?); крылья ясно
короче брюшка.
. Грудь очень слабо вздутая, лишь едва выпуклая в профиль; щиток
очень слабо выступающий. Ноги довольно крепкие и короткие; задние
голени лишь едва достигают конца брюшка; тазики, видимо, умеренно раз-
Рис, 77. Lycoriomimella minor, sp. nov, Кара-тау, ПИН
№ 2452/364; остаток крыла (ориг.).
витые; шпоры неясные. Строение головы и ее придатков неясно вследствие
плохой сохранности объекта. Брюшко умеренно вздутое, коническое; за-
метны хорошо 2-й и 7-й сегменты и два конечных сегмента, 8-й и 9-й, обра-
зующие генитальный аппарат.
Этот род хорошо отличается от всех ближайших родов группы совер-
шенно прямой RS, крепкой SC, короткой R, слабо вздутой грудью и мень-
шей величиной. Недостаточная сохранность крыльев не позволяет с точ-
ностью выяснить взаимоотношения этой формы; повидимому, все.,же род
наиболее близок к роду Lycoricmima.
Lycoriomimella minor, sp. nov.
(Рис. 77;'табл. XVI, фиг. 35)
Отпечаток самки, лежащей на правей стороне; сохранность груди,
'• брюшка и части крыльев хорошая; строение головы и ног плохо разли-
чимы.
Крыло описано в родовом описании; общая его форма неясна вследствие
де формации;, ноги плохо сохранились, частично смяты и поломаны; зад-
ние бедра равны голеням, но значительно толще их, веретенообразные;
задние голени прямые, параллельно-крайние; строение остальных ног и их
частей неясно. Брюшко с широкой плевральной перепонкой на сегментах
от 2-го до 7-го; 8-й сегмент цельный, без ясной перепонки; 9-й сегмент
в виде узких вытянутых пластинок, церков; 1-й сегмент неясен. Длина
тела 2.5, крыла 1.25 мм.
Отпечаток колл. ПИН, № 2452/364. Кара-тау, сел. Галкино.
А. В. Мартынов, 1924.
69
Род Pleciomima Rohd.
Родендорф, 1938, Tp. ПИН, т.VII, вып. 3, стр. 44.
Pleciomima secunda, sp. nov.
(Рис. 78 и 79; табл. XIV, фиг. 32)
Отпечаток самца, лежащего на левой стороне с расправленными крыль-
ями, ногами и головой; сохранность довольно хорошая.
Крыло довольно широкое; отношение его длины к ширине 1 : 1.91;
С едва заходит за конец RS, SC крепкая, косолежащая, впадающая в край
Рис.С, 78. Pleciomima secunda, sp. nov. Кара-тау,
ПИН, № 987/2465; общий вид ископаемого (ориг.).
крыла дистальнее уровня
rm; R почти совершенно
прямая до конца, впадаю-
щая в край крыла на гра-
нице 2-й и 3-й третей; RS
ответвленная под острым
углом от R в базальной
четверти крыла; RS равно-
мерно дугообразно изогну-
тая, оканчивающаяся ров-
но на вершине крыла; раз-
вилок М1+2 короткий, но
хорошо выраженный, рав-
ный общему стволу М1+2
до rm; rm расположенная
поперечно, почти в 2 раза
короче основного отрезка
RS; ветвь Мх параллельная
RS и М2; М3 слитая с СиА;
более слабая, чем Мх и Ма,
параллельная с последней;
место развилка М3 + СиА
расположено на уровне се-
редины базального отрез-
ка RS; свободная часть
СиА довольно тонкая, не-
равномерно загнутая назад.
Грудь умеренно выпук-
лая, больше всего в задней
части, перед щитком; те-
sonotum заметно выдается
в виде валика над головой;
тазики, видимо, массивные
и крупные, но плохо раз-
личимые; средние и задние
бедра, равныеголеням,уме-
Рис. 79. Pleciomima secunda, sp. nov. Кара-тау, ренно вздутые, веретено-
ПИН, № 287/2465; крыло (ориг.). видные; голени топкие, па-
раллельно-крайние; шпоры
неясные; лапки длинные; членистость их неясная; концы задних бедер
достигают заднего края 4-го тергита; голени, видимо, заходят за ко-
нец брюшка.
Голова небольшая, с выступающим лицом и сильно вздутой задней
поверхностью; антенны толстые, равные по длине голове и груди, вместе
взятым; членистость антенн неясная.
го
Брюшко продолговато-яйцевидной формы, на конце закругленное;
хорошо различимы 3-й и 7-й тергиты; границы между тергитами и стерни-
тами неясные.
Длина тела 2.75, крыла 2.25 мм.
Позитивный отпечаток самца. Колл. ПИН, № 987/2465. Кара-тау, ур.
Чугурчак. А. И. Турутанова-Кетова, 1930.
Род Pleciomimella, gen. nov.
Тип рода: Pleciomimella karatavica, sp. nov.
С заходит за конец RS, не достигая, однако, вершины крыла; SC тон-
кая, приближенная и параллельная с R, на конце неясная, загибающаяся
к С; SC равная */* длины крыла; R почти совершенно прямая, несколько
менее 2/з крыла; RS слабо изогнутая, ответвляющаяся от R несколько
далее конца базальной трети; базальный отрезок PS лишь в 1.75 раза
длиннее rm; конец RS далеко не достигает вершины крыла, на которой
оканчивается Мх; М более тонкая, чем Яли RS; общий ствол М1+8 короче
ветвей, из которых Ма является непосредственным продолжением общего
ствола, а М, отходит под углом в виде дуги; М1+а непосредственно перед
Рис. 80. Pleciomimella karatavica, sp. nov. Кара-тау, ПИН № 2452/549; общий вид
ископаемого (ориг.).
rm ответвляется от М, которая в свою очередь несколько проксимальнее
ответвляется от СиА; общий ствол М, повидимому, редуцирован; СиА тон-
кая, но ясная; Ап не видны; крыло умеренно удлиненное; отноше1ще..вьо^.
длины к ширине равно 1 : 2.4.
Тело не сохранилось; брюшко, повидимому, было короч^’:
Этот род резко отличен от других родов подсемейства п$'
жилкования: очень характерны прямые, неизогнутые жилки^К
Тальных признаках можно отметить лишь некоторые черты сходства с дру- -
гими формами; так, длинная R сближает этот род с
lycoriomima и особенно с Mimallactoneura, т. е. с представителем/0уже *
другого подсемейства.
Pleciomimella karatavica, sp. nov.
(Рис. 80; табл. XV, фиг. 34)
Отпечаток насекомого, лежащего с расправленными крыльями; тело
очень плохой сохранности в виде разрушенных фрагмент® склеритов груди
и брюшка; голова с придатками и ноги не сохранились. Длина остатков
тела 1.6, длина крыла 2.1, ширина крыла 0.95 мм.
1 экземпляр — колл. ПИН, №2452/549, Галкино, А. В. Мартынов.
71
: П о д с е м е й с т в о МIm a llacto neuri п ае, subfam. nov.
Тип подсемейства: род Mimallactoneura, gen. nov.
Это подсемейство устанавливается по вновь описываемому роду Mimal-
lactoneura из юры Кара-тау. Прямые, совсем не изогнутые жилки R и RS
резко отличают этот род от всех других Pleciomimidae и напоминают совре-
менный^род Allactoneura, от которого он, однако, хорошо отличается
большей специализацией — более укороченными жилками RS и особенно
R, слабым развитием Мх. Вероятно, описываемый род является особой
боковой ветвью, сохранившей ряд примитивных черт жилкования в виде
длинного базального отрезка RS и прямых жилок наряду с признаками
специализованности. Наиболее близко это подсемейство к группе Ante-
fungivorinae, что, кроме признаков жилкования, сказывается еще в строе-
нии ног, именно в довольно тонких бедрах.
Род Mimallactoneura, gen. nov.
Тип рода: Mimallactoneura vetusta, sp. nov.
С не достигает вершины крыла, оканчиваясь ровно у конца RS; SC
слабая, заметная в виде следа, впадающая в С, длинная, более1/» крыла;
R почти прямая, несколько больше а/8 крыла; RS с длинным базальным
отрезком, отходящим подострым
углом от R; rm расположена
поперечно, крепкая, почти в5раз
короче базального отрезка RS;
RS имеет слабый, но заметный
излом в месте соединения с rm;
Рис. 81. Mimallactoneura vetusta, sp. nov. Рис. 82. Mimallactoneura vetusta, sp.
Кара-тау, ПИН, № 2452/611; общий вид nov. Кара-тау, ПИН, № 2452/611; крыло
ископаемого (ориг.). (ориг,).
М слабо выпуклая спереди; ствол и ветви М1+2 слабее R и RS; базаль-
ный отрезок М1+2 короткий, ответвляющийся от М8, в 3 раза длиннее
rm; проксимальный отрезок М1+2 длинный, длиннее М2, но короче
Мх; М2 является прямым продолжением общего ствола М1+2; Мх отходит
Под углом вперед; М3 значительно крепче передних ветвей, так же
как М1+2, слабо дугообразно изогнутая и ответвляющаяся от CuA
несколько дистальнее ответвления М1+2; этот базальный отрезок Ма
есть не что иное, как косо расположенная mcu; CuP хорошо замет-
ная на уровне средней части CuA; эта последняя жилка крепкая, резко
загнутая в дистальной четверти; крыло несколько более чем в 2 раза длин-
нее своей ширины; вершина его умеренно заостренная; крылья длиннее
брющка.
Тело плохо сохранилось; детали его неясны; антенны несколько короче
головы и груди, вместе взятых; брюшко длиннее головы и груди, на основа-
72
нии, повидимому, суженное, с 7 или 8 сегментами, вздутое; бедра задник
ног равномерно расширенные, толще голеней. Размеры насекомого около-
3 мм.
Mimallactoneura vetusta, sp. nov.
(Рис. 81 и 82)
Отпечаток насекомого, лежащего на вентральной поверхности тела,,
с распростертыми в стороны крыльями; тело и его придатки, кроме левого
крыла, плохой сохранности. Длина тела 3.25, крыла 3.00 мм.
1 самка (?). Колл. ПИН, № 2452/611. Галкино, А. В. Мартынов.
Подсемейство Antefunglvorlnae Rohd. 1938]
Это подсемейство, установленное мною в 1938 г., с типичным родом
Antefungivora Rohd. 1938, должно быть объединено вместе с родом
Antiquamedia Rohd. 1938 (для которого мною ошибочно было установ-
Рис. 83. Antefungivora prima? Rohd. Кара-тау, ПИН,
№ 2452/316 — тип; общий вид ископаемого (ориг.).
Рис. 84. Antejungi-
vora prima Rohd.
Кара-тау, ПИН,
№ 2452/316-
тип; крыло (ориг,).
лено также особое подсемейство) в особую группу, входящую в семейство
Pleciomimidae в качестве особого подсемейства, название которого сохра-
няется по роду Antefungivora, описанного первым в упомянутой работе
1938 г. (фиг. 83 и"84; табл. XVI, фиг. 36).
Подсемейство Р ar it onidinae, ubfam. nov.
<’ Тип подсемейства: род Paritonida, genZnov.
этом роде характерно мозаичное сочетание признаков большой спе-
циализации с наличием явно примитивных особенностей. Общий характер
жилкования приближает род Paritonida к видам группы Pleciomiminae-,
таковы изогнутые и укороченные R и RS, слабые жилки системы М, корот-
кие крылья; вместе с тем положение пп вдали от развилка RS—R, слабо
вздутая грудь и, повидимому, тонкие ноги, резко отличают Paritonidinae
от всех других Pleciomimidae’ последние перечисленные признаки мы вправе
считать примитивными. Взаимоотношения описываемого рода Paritonida
более или менее ясны: он является потомком предков, общих с группой
Pleciomiminae, сохранившим наряду с высокой специализацией ряд при-
митивных черт.
73
Род Paritonida, gen. nov.
Тип рода: Paritonida brachyptera, sp. nov.
С крепкая, заходящая за конец RS, но не достигающая вершины крыла;
SC бледная, но не особенно тонкая, свободно оканчивающаяся в перепонке
костальной ячейки; эта жилка короткая, менее’/3 крыла; R крепкая,
Рис. 85. Paritonida brachyptera, sp. nov.
Кара-тау, ПИН, № 2452/262; общий вид
ископаемого — схема (ориг.).
прямая, лишь на самом конце
крыла загнутая вперед, равная
примерно 3/5 крыла; RS креп-
кая, ответвляющаяся от R под
острым углом на уровне конца
базальной четверти крыла; R
волнообразно, довольно сильно
изогнутая; rm расположенная
примерно на середине крыла,
лишь несколько проксимальнее
ее; базальный отрезок RS более
чем в б раз длиннее rm; ствол М1+а
заметно отдален от М3; его ба-
зальный отрезок, повидимому,
длиннее дистального; пропорции
жилок М1; Ма и дистального от-
резка общего ствола М1+2 неяс-
ны вследствие плохой сохранно-
сти; место соединения М1+а и М3 неясно; М3 крепче М1+а; ответвление этой
жилки от CuA расположено несколько дистальнее развилка R — RS; CuA
Рис. 86. Paritonida bra-
chyptera, sp. nov. Кара-
тау, ПИН, № 2452/262;
голова (ориг.).
Рис. 87. Paritonida brachyptera, sp. nov.
Кара-тау, ПИН, № 2452/262; крылья и
брюшко (ориг.).
Крепкая, такая же, как и R и RS, довольно правильно дугообразно
изогнутая; CuP тонкая и ясная; крыло довольно широкое и притуплен-
ное на вершине; отношение его ширины к длине равно 1 : 1.9; крылья
короче брюшка.
74
Голова и профиль равной длины и ширины; глаза неправильно круг-
лые; глазки неясные; лицо выступающее в виде тупого, широкого бугра;
антенны длинные, равные голове и груди, вместе взятым, 17-члениковые;
два их основных членика сильно сближенные и более толстые чем остальные;
членики от 3-го до 5-го боченковидные, незначительно длиннее своей тол-
щины, членики от б-го до 12-го такой же формы, но меньше; членики от
13-го до 16-го более тонкие и вытянутые; последний, 17-й, членик заострен-
ный, более или менее конусовидный. Грудь слабо выпуклая, с хорошо
заметным, но слабо выступающим щитком; вентральная часть груди и
основания ног не сохранились; остатки ног передней и средней пары пока-
зывают, что бедра незначительно отличаются по толщине от тонких голе-
ней. Брюшко широкое в основании, постепенно утончающееся к концу;
заметны 8 его сегментов, из которых 2 базальных узкие; 3-й и 6-й средней
длины, 7-й самый длинный и 8-й, плохо сохранившийся, довольно корот-
кий и несет половые придатки; брюшко покрыто неособенно густыми
короткими волосками. Насекомые менее 3 мм.
Paritonida brachyptera, sp. nov.
(Рис. 85,86 и 87; табл. XVI, фиг. 37)
Позитивный отпечаток насекомого, лежащего на левой стороне; хоро-
шей сохранности; не сохранилась часть груди и крыльев; окраска темная;
ноги и крылья светлые. Длина тела 2.75, крыла 1.75 мм.
Самка. Колл. ПИН,№2452/262.Сел. Галкино, А. В. М а р т ы н о в, 1925.
Семейство Allactoneuridae, Rohd. 1938
Основным и наиболее характерным признаком этой группы являются,
кроме первичного положения RS, ответвления его от R под острым углом,
особенно размеры и сильное развитие R, достигающей всегда не менее
3/4 крыла, а часто гораздо больше. Эта особенность в жилковании не может
оцениваться иначе, как состояние более примитивное по сравнению с уко-
роченной и изогнутой R у Pleciomimidae и Pleciofnngivoridae; вместе с тем
длинную R .Mbi наблюдаем и у ряда современных Fungivoridae и в особен-
ности— у вымершего третичного рода Мусе! о phi Hies Scudder из
семейства Mycetophilitidae, fam. nov. Все эти семейства генетически, несо-
мненно, близко связаны друг с другом; подробнее вопрос о филогенезе груп-
пы разобран ниже. Юрская фауна семейства Allactoneuridae заключает
небольшое число родов, которые, однако, обнаруживают очень богатое его
развитие в юрское время и дают ряд вторичных группировок — подсемейств.
Подсемейство Allactoneurinae, заключающее единственный и притом
современный род Allactoneura, характерно по ряду примитивных черт
строения. Не располагая в наличности материалом по этой интересной
форме, описанной de Meijere в 1904 г. с острова Явы, я пользуюсь,
кроме оригинального описания, также и описанием Johannsen (1909).
' Строение крыла обладает многими явно примитивными признаками;
таково положение rm, удаленной от развилка RS — R, почему базальный
отрезок RS очень велик; R и RS прямые, не изогнутые; очень длинная М3,
сближенная с общим стволом М1+2, присутствие SC2. С другой стороны,
признаки специализации у рода Allactoneura в строении крыла немного-
численный заключаются лишь в некотором укорочении R и RS; большую
специализацию мы наблюдаем: в строении ног — в очень сильном развитии
тазиков, головы — в редукции среднего глазка. Однако все черты спе-
циализации по сравнению с примитивными особенностями Allactoneurinae
явно менее значительны; род Allactoneura следует считать ярко выраженной
реликтовой формой, сохранившей до наших дней ряд древних юрских
черт строения.
75
Определительная таблица подсемейств и родов
Allactoneuridae
' 1 (10) RS равномерно загнутая назад; rm всегда ближе к основанию
крыла, чем к его вершине, равная или в 3 раза короче базального
отрезка RS; брюшко тонкое, по крайней мере на основании.
2 (5) Между R и RS лежит поперечная жилка, рудимент RS; изгиб RS
очень слабый, rm равная базальному отрезку RS, косая, расположен-
ная к нему под прямым углом и на уровне конца SC.
Подсемейство Mesosci ophili пае, subfam. nov.
3 (4) Рудимент передней ветви RS, поперечная между R и RS изогнутая;
Ап крепкая, изогнутая и длинная; крыло широкое (рис. 89).
Род Mesosciophila, gen. nov. (стр. 76).
4 (3) Поперечная между R и RS прямая; Ап прямая, более слабая;
крыло суженное (рис. 90). Род Mesosciophilodes, gen. nov. (стр. 77).
5 (2) Между R и RS нет поперечных жилок; rm расположена поперечно,
под тупым углом к основной части RS.
6 (7) R несколько укороченная; rm в 3 раза короче базальной части RS;
RS слабо изогнутая, общий ствол М1+2 лишь в Р/а раза короче Mg.
(рис. 91). Подсемейство Mimalycoriinae, subfam. nov. (стр. 78).
Единственный род из Кара-тау — Mimalycoria, gen. nov.
7 (6) R длинная; rm больше, равна или лишь в 2 раза короче базального
отрезка RS; общий ствол М1+2 очень короткий, не менее чем в 2 или
больше раз короче М3. Подсемейство Fungivorilinae, subfam. nov,
8 (9) RS на конце резко загибается назад; крыло на конце притупленное
(рис. 94). Род Fungivorites Rohd. (стр. 80).
9 (8) RS на конце лишь умеренно загнутая назад; крыло заостренное на
конце (рис. 93). Род Eoboletina, gen. nov. (стр. 79).
10 (1) RS совершенно прямая; rm расположенная на середине крыла и во
много раз короче базального отрезка RS; SC прямая, около 1/3 крыла.
Современное подсемейство Allactoneurinae с единственным родом
Allactoneura Meij.
Подсемейство Mesosciophilinae, subfatn. nov.
Тип подсемейства: род Mesosciophila, gen. nov.
Это подсемейство наиболее близко к группе Fungivorilinae и хорошо от
нее отличается присутствием поперечной жилки между R и RS — руди-
ментом ветви RS; основной отрезок RS очень мал и иногда принимает харак-
тер почти поперечной жилки, как это мы наблюдаем у некоторых современ-
ных Sciophilinae из семейства Fungivoridae, именно у Boletinini-, изгиб RS
незначителен. Строение тела плохо известно: ноги, повидимому, не утол-
щены. Несомненно, подсемейство Mesosciophilinae является исходной груп-
пой современных Sciophilinae (см. ниже).
Род Mesosciophila, gen. [nov.
Тип’рода: Mesosciophila venosa, sp. nov.
С не особенно крепкая, достигающая вершины крыла и конца RS; SC
тонкая, но хорошо выраженная, правильно дугообразно изогнутая и впадаю-
щая в С; длина ее несколько превышает г/л крыла; R очень длинная, пре-
вышающая 8/10 длины крыла, слабо загнутая назад; RS ответвляется от R
под углом примерно в 45° и после соединения с rm расположена параллельно
с R, лишь в своей дистальной четверти загнута назад; развита поперечная,
неправильно изогнутая жилка, передняя ветвь RS, расположенная на сере-
76
дине крыла, несколько дистальнее развилка М; пи лежит несколько косо/
лишь в Р/а Раза короче базального отрезка RS; жилки М лишь незначи-
тельно слабее других; развилок М1+2 очень длинный; проксимальный отре-
зок общего ствола М1+а в ---------------- —
2 раза короче дистально- S' "\Л Дхд
го, который в свою оче- (
редь в 23/4 раза короче М2 X. А. Лл |
и в З1/^ короче Мх; обе I)
ветви М1+2 постепенно рас- yf / XkX/z \
ходится; М3 длинная, ела- г~-^
бо и равномерно изогнутая /SS!a!^vy —-<
дугой, отходящая от обще-
го ствола М, который про- 7)
ксимально сливается с CuA.
эта последняя жилка креп-
кая и резко загнутая на
мому, на конце загибаю-
щаяся назад параллельно с СнА; крыло в 2 раза длиннее своей ширины,
короче брюшка, на конце заостренное и расширенное на середине.
Рис. 89. Mesosciophila venosa, sp. nov. Кара-тау, № 26. Геол, ин-т
САГУ; крыло (ориг.). ________
Тело недостаточно сохранилось; голова с большими овальными глазами;
придатки ее неясные; грудь выпуклая; брюшко суженное на основании,
вздутое дистально; заметны 7 его сегментов; ноги, невидимому, тонкие.
Mesosciophila venosa, sp. nov.
(Рис. 88 и 89; табл. XVI, фиг. 38)
Положительный отпечаток насекомого, лежащего на брюшной и отчасти
на левой стороне, с распростертыми крыльями; антенны и большая часть
ног не сохранились; окраска светлая, коричнево-желтая. Длина тела 6.0,
крыла 4.5 мм.
Самка (?). Колл. Геол. инет. Среднеазиатского гос. университета, № 26.
Род MesosciophUodes, gen. nov.
Тип рода: MesosciophUodes angustipennis, sp. nov.
l.C не достигает R; SC равная около 1/3 крыла, доволыЯТ слабая, но ясная,
почти прямая, косо идущая от основания крыла к С; R равная ®/10 крыла,
почти прямая и лишь на конце слегка загнутая назад, соединяясь с С под
очень небольшим углом; RS волнообразно изогнутая, ответвляющаяся от
R под острым углом; ее базальный отрезок резко обособлен от дисталь-
ной части; rm расположена косо и почти приобретает характер ложного
основания RS (то, что наблюдается у современных Fungivoridae); между
R и RS лежит крепкая, прямо расположенная поперечная жилка, рудимент
ветви RS; жижи М заметно
слабее R и RS; проксималь-
ный отрезок М1+а несколько
менее чем в 2 раза короче ди-
стального; этот последний в
2.6 раза короче и в 2.3
раза короче Ма; обе ветви
М1+а слабо расходящиеся; М3
очень слабо загнутая назад,
отходящая от так называемо-
Рис. 90. Mesosciophilodes angustipennis, sp. nov. ГО общего ствола M, вероятно,
Кара-тау, ПИН, № , 2452/290; крыло (ориг.). mcu; СиА равномерно дуго-
„ образная: СиР в виде прямой
тонкой черты у основания жилки СиА между этой жилкой и Ап; Ап более ко-
роткая и почти прямая; крыло на конце округлое, довольно узкое и длин-
ное, почти в 2!/а раза длиннее своей ширины. Строение тела неизвестно.
Mesosciophilodes angustipennis, sp. nov.’
(Рис. 90; табл. XVI, фиг. 39)
Положительный отпечаток правого крыла, местами разрушенный;
остатков тела не сохранилось; окраска крыла светлая. Длина крыла 4.0 мм.
Колл. ПИН, № 2452/290. Сел. Галкино, А. В. Мартынов, 1925.
Подсемейство Mima ly cori inae, sub fam. nov.
Тип подсемейства: род Mimalycoria, gen. nov.
Род Mimalycoria обладает рядом своеобразных черт в жижовании,
заставляющих считать его представителем особой группы, провизор но
принимаемой мною за подсемейство. Наряду с признаками специализации,
заключающимися в удлинении крыла, укорочении R, ослаблении конца SC,
укорочении ветвей М1+а, наблюдаются некоторые относительно примитив-
ные особенности; таково положение rm вдали от развилка R — RS. Наи-
более близок род Mimalycoria к современному подсемейству Allactoneurinae
и отличается от него деталями жилкования, притом более специализован-
ными; вероятно, эти две группы обладали одной исходной формой, к кото-
рой род Mimalycoria был довольно близок.
Род Mimalycoria, gen. nov.
Тип рода: Mimalycoria allactoneuroides, sp. nov.
С достигает вершины крыла, заметно заходя за конец RS; SC тонкая г
но ясная, слабо изопфтая, около */3 крыла, на конце слабая, почти сво-
бодная; R почти совершенно прямая, лишь на самом конце слабо загнутая
назад и соединяющаяся с С под очень небольшим углом, равная 8/ю
крыла; RS ответвляется от R под острым углом и в большей своей части
лежит параллельно с R, лишь едва расходясь с ней в дистальной половине;
RS равномерно загибается назад и достигает вершины крыла; rm распо-
ложена поперечно, в 3 раза короче базального отрезка RS; М1+а более нежна,
чем rm, R и RS; проксимальный отрезок М1+а очень длинный, длиннее
дистального; последний в 2.2 раза короче Mj и в 1.6 раза Ма; строение жи-
лок М3, СиА, Ап и СиР неизвестно вследствие плохой сохранности ископае-
мого; крыло относительно узкое, в 2’/4 раза длиннее своей ширины, на
вершине незначительно заостренное.
78
Mimalycoria allactoneuroides, sp. nov.
(Рис. 91).
Отпечаток крыла и остатков тела.
Длина тела 3.75, крыла 2.75 мм.
Колл. ПИН, № 2452/172. Сел. Галкино, А. В. Мартынов, 1925.
Подсемейство Fangivoritinae, subfam. nov.
Тип подсемейства: род Fungivorites Rohd. 1938.
Недостаточно известный род Fungivorites R о h d. и описываемый ниже
род Eoboletina gen. nov. обладают рядом признаков отчасти примитивного,
отчасти специализованного
характера; жилкование этих
форм близко к подсемейству
Mesosciophilinae, но отличает-
ся большей специализованно-
стью. Оценивая весь ком-
плекс особенностей Fungivo-
ritinae, особенно строение
тела, следует признать эти
оба рода представителями осо-
бой группы, провизорно счи-
таемой за подсемейство.
Примитивными признаками Fungivorilinae, несомненно, являются удли
ненная R, поперечное положение rm, крепкая и длинная SC и слабое раз-
витие груди — ее малая выпуклость.
Из отдельных моментов в специализации следует упомянуть об отсут-
ствии поперечной RSj и изогнутости RS. Филогенетические отношения
группы разобраны ниже.
Род Eoboletina, gen. nov.
Тип рода: Eoboletina gracilis, sp. nov.
С едва не достигает вершины крыла, не доходя до RS и оканчиваясь на
середине между R и RS; SC крепкая и длинная, правильно, но слабо дуго-
образно изогнутая; отношение ее к длине крыла равно 5 : 11; R очень длин-
ная, в дистальной половине слабо, но равномерно загибающаяся назад,
лишь незначительно короче крыла, равна 0,93 его; RS ответвляется от
R под острым углом значительно проксимальнее конца SC; после места
соединения RS с rm первая жижа расположена почти параллельно R,
лишь едва заметно расходясь дистально; конец RS загибается равномерно
назад, впадая в край крыла за его вершиной; жилки системы М крепкие,
лишь незначительно ослабленные по сравнению с R и RS; оба отрезка об-
щего ствола М]+а почти равной длины; дистальный отрезок М1+а в 2.6 раза
короче и в 2.1 раза короче Ма; М3 крепкая, ответвляющаяся от общего
ствола М, который проксимальнее направлен прямо к корню крыла и, по-
степенно сходясь с СиА, соединяется с ней; СиА крепкая, довольно резко
загнутая назад, СиР неясная; Ап очень тонкая, заметна в виде прямой
линии в своей основной половине; крыло ровно в 2 раза длиннее своей
ширины, на вершине умеренно закругленное, длиннее брюшка.
Тело тонкое; голова умеренной величины, с большими, в профиль высо-
кими, овальными глазами, лицо резко вздутое; антенны и ротовые органы
не сохранились. Грудь равномерно выпуклая, с небольшим* торчащим щит-
ком; ноги плохо сохранились: заметны лишь бедро и голень задней пары ног
и остаток голени (или бедра) средней пары; бедро слабо и равномерно взду-
тое на середине, лишь незначительно толще длинной голени; тазики не сохра-
7J
нились, вероятно, умеренной величины. Брюшко тонкое и длинное, в 2 раза
длиннее головы и груди, из 8 сегментов, на основании (1-м и 2-м сегментах)
заметно суженное; 1-й сегмент неясно отчлененный от груди, 2-й, 3-й,
4-й и 5-й примерно равной длины, б-й и 8-й укороченные, 7-й несколько
удлиненный.
Eoboletina gracilis, sp. nov.
(Рис. 92 и 93)
Позитивный отпечаток насекомого, лежащего на правой стороне; при-
датки головы не сохранились, равно как и большая часть ног; крылья ле-
жат одно на другом, незначительно сдвину-
тые; окраска темная. Длина тела 5.75, кры-
ла 4.5 мм.
Рис. 92. Eoboletina gracilis, sp. nov.
•Кара-тау, ПИН, ^s 2452/249; общий
вид ископаемого—схема (ориг.).
Рис. 93. Eoboletina gracilis, sp. nov.
Кара-тау, ПИН, № 2452/249;
крыло (ориг.).
1 самец. Колл. ПИН, № 2452/249. Сел. Галкино. А. В. Марты-
нов, 1926.
Род Fungivorites Rohd. 1938.
Этот род, описанный мною в 1938 г. из Кара-тау по недостаточно хорошо
сохранившемуся ископаемому, резко отличается от рода Eoboletina gen.
nov., почему и объединяется с последним лишь провизорно. Для рода Fun-
givorites характерна резко загибающаяся назад RS, короткий базальный
отрезок RS и, повидимому, более крепкие ноги, обладающие резко взду-
тыми задними бедрами (строение ног известно лишь у одного вида).
Род Fungivorites по сравнению с Eoboletina явно более специализован,
что проявляется как в жилковании, так и в строении ног.
Отличия видов рода Fungivorites Rohd.
1 (2) Базальный отрезок RS заметно длиннее rm; дистальный конец RS
резко загибается назад, на некотором протяжении будучи параллель-
ным с С; длина крыла 3 мм............F. latimedius Rohd.
2 (1) Базальный отрезок RS равен rm; дистальный конец менее загнут и не
параллелен с С; длина"крыла 5 мм.......F. indistinctus, sp. nov.
Fungivorites indistinctus, sp- nov.
(Рис. 94)
(Отпечаток насекомого, сильно смятого и сжатого; выдается лишь часть
крыла, которая и описывается.
С достигает вершины крыла, слегка заходя за конец RS; SC крепкая,
слабо изогнутая на конце и впадающая в С; R очень слабо и равномерно
«о
загнутая назад, равная 0.38 крыла; RS ответвляется от R; под острым
углом, на всем своем протяжении довольно правильно S-образно изогнутая,
особенно на конце; rm длинная, поперечно расположенная, равная базаль-
ному отрезку RS; строение жилок М, СиА и Ап неизвестно вследствие пло-
хой сохранности ископаемого.
Тело плохо сохранилось, и детали его строения неясны. Длина крыла
5 мм.
Гэкземпляр; колл. ПИН, № 2335/31, Михайловка. А. И.Турута-
нова-Кетова.
ОСОБЕННОСТИ ЮРСКОЙ ФАУНЫ OLIGONEURA
Необходимо сделать несколько замечаний об общих чертах юрской
фауны насекомых Кара-тау. Общий состав захороненной фауны с очевид-
ностью выясняет ее случайный характер: в ее состав входят как водные
насекомые, так и насекомые чисто сухопутные, не связанные с водоемами;
с другой стороны, найдены ра-
стительноядные виды, хищники — -—
и, наконец, виды, живущие в ". "X. '
гниющих веществах; далее, най- -----—
дены исключительно крылатые
формы,—бескрылые почти вовсе - ' ’ " 1
отсутствуют; при этом немногие /
бескрылые насекомые, которые у
захоронились, например некою- _ .
рые Homoptera, могли быстро _______ ________
двигаться посредством сильных “ "
прыжков. Рис, 94. Fungivorites indistinctus, sp. nov. Kapa-
Все эти данные заставляют тау, ПИН, № 2323/31; часть крыла (ориг.).
предполагать чисто пассивный
занос сильным ветром летающих или прыгающих насекомых в очень мел-
кий водоем (может быть, глубиной в 2—5 см—прибрежные лужи?), тцпа ли-
манов, периодически пересыхающий и наполняющийся водой, богатой легко
выпадающими карбонатами.Заносимые ветром на поверхность такого рода
водоема насекомые не испытывали каких-либо дальнейших переносов, а фи-
ксировались тут же, на месте падения, может быть прилипая даже к влаж-
ной поверхности ила высыхающей лужи. За это говорит исключительная
сохранность насекомых. Вместе с тем эти обстоятельства позволяют
считать захороненную фауну насекомых своего рода «пробной группой»,
более или менее точно отражающей действительный состав существо-
вавшей в то время фауны.
В составе юрской фауны двукрылых группа Oligoneura имеет по срав-
нению с современной гораздо больший «удельный вес»: число видов совре-
менных Oligoneura достигает 42% всех Nematocera (по данным Н е и d е I,
,1936); число же юрских видов составляет 58% ко всем до сих пор зареги-
строванным длинноусым двукрылым из Кара-тау.1
Вся большая группа Oligoneura в юрское время подразделялась на ряд
семейств, из которых на долю трех — Pleciomimidae, Pleciojungivoridae
и Allactoneuridae — приходилось, повидимому, подавляющее большинство
видов группы: 40 видов из 48, т. е. ®/6 всего количества.
Семейство Pleciomimidae заключает в себе около 20 видов, распределяю-
щихся между 4 подсемействами и 11—12 родами. Для него характерна до-
вольно высокая степень специализации жилкования? заключающаяся
1 Возможно, конечно, что это преобладание вызвано отчасти более мелкими раз-
мерами,
6 Родендорф, т. XIII, в. 2.
в редукции SC, ветвей RS, укорочении R и ослаблении М. Тело мало сиециа-
лизовано, ноги довольно слабые. Остается архаичным тип ответвления RS
от R, также первично слабое развитие грудного отдела и длина антенн.
Различные подсемейства Pleciomimidae обнаруживают связи с другими
семействами группы Fungivoroidea и различаются деталями жилкования,
развитием ног и вздутием спинки груди. Наиболее. архаично строение
представителей подсемейства Mimallactoneurinae, обладающих относительно
длинными крыльями, тонкими ногами, длинным и прямым основным отрез-
ком RS и R. Жилки R и SC также длинные у Antefungivorinae-, представи-
тели этого подсемейства вместе с тем обладают специализованными ногами.
Наиболее специализовано главное подсемейство — Pleciomiminae, для ко-
торого характерна редукция жилкования: укорочение большинства жилок,
равно как и относительных размеров крыла; ноги обычно заметно утолщен-
ные. Последнее подсемейство — Paritonidinae— характерно по смеси
архаичных и специализованных признаков; наряду с редуцированным
жилкованием наблюдается плоская спинка и слабые ноги.
Второе по объему семейство Pleciofungivoridae, заключающее около
14 видов и примерно 8 родов, характерно по сравнению с предыдущим своей
цельностью: все известные роды Pleciofungivoridae, хотя и четко отграничи-
ваются один от другого, но не могут быть объединены в какие-либо вторич-
ные, подсемейственные категории. Семейство характеризуется ветвистым
строением жилки RS, образующей стойкие ветви, направленные вперед;
в большинстве случаев RS несет лишь одну, притом крепкую изогнутую
ветвь (в родах Prohesperinus, Eohesperinus, Allactoneurites и др.); реже этих
ветвей бывает две (у Eopachyneura, Pleciofungivorella) или даже много сла-
бых, похожих на поперечные жилки веточек (у Polyneurisca). Другим архаич-
ным признаком жилкования является присутствие крепкой и длинной SC
и у некоторых родов поперечных жилок между С и R и основными частями
М и Си (у Transversiplecia и Polyneurisca). Признаком специализации крыла
в этом семействе является сдвиг к основанию поперечной радиомедиальной
жилки и образование пигментного пятна — птеростигмы. Тело этих ко-
мариков довольно сильно специализовано; на это указывает хорошее раз-
витие спинки груди, у некоторых родов образующей настоящий, нависаю-
щий над головой горб, например у Pleciofungivorella binerva, gen. nov., sp.
nov. Ноги умеренно развитые; антенны укороченные.
Семейство Allactoneuridae, названное по ныне живущему роду, хотя
и уступает по объему двум предыдущим, заключая всего лишь 8 видов,
принадлежащих к б родам, но далеко не столь цельно и однообразно. Юр-
ские роды семейства с ясностью распределяются среди трех обособленных
подсемейств. Для юрских Allactoneuridae очень характерно архаичное
строение жилки R, а именно ее большая длина, незначительно отличаю-
щаяся от длины всего крыла; кроме того, всегда хорошо развита SC; перед-
няя ветвь RS иногда (у Mesosciophilinae) также развита, но всегда прибли-
жена к основанию ‘RS и вследствие большой длины R соединяется с этой
жилкой, а не с С. Из признаков специализации на крыле следует отметить
смещенное положение поперечной rm, которая очень приближена к осно-
ванию RS; в ряде случаев наблюдается симметричное положение rm и
основного отрезка RS, именно то положение, которое ныне существует
у большинства Fungivoridae; лишь у одного подсемейства (Mimalycoriinae)
rm расположена «нормально», т. е. архаично. Строение тела плохо известно;
невидимому, грудь умеренной величины, но с ясно выраженным вздутием
спинки. Брюшко довольно длинное и тонкое у основания. Три подсемейства—
Mesosciophilinae, Fungivoritinae и Mimalycoriinae различаются деталями
жилкования. Современный род Allactoneura—тип семейства — резко отли-
чается от своих юрских родственников своеобразным и более примитивным
«2
положением поперечной жилки rm, которая значительно удалена от развилка
R—RS. Этот признак не позволяет считать ни одного изо всех до сих пор
известных юрских Allactoneuridae непосредственным предком современ-
ного рода Allactoneura. Все юрские роды в жилковании специализованы,
и лишь один род Mimalycoria несколько приближается к современному;
эти взаимоотношения выражены мною в обособлении рода Allactoneura
в отдельное подсемейство, отличное от юрских.
Семейства Paraxymyiidae, Protopleciidae, Protobibionidae и Protoscatop-
sidae известны по одному-трем видам монотипических родов; все они нахо-
дятся в определенной связи с некоторыми современными; степень сходства
очень различна у разных семейств.
Семейство Paraxymyiidae, известное по единственному виду рода Para-
xymyia, очень характерно по строению жилкования, именно по ветвлению
радиальных жилок и редукции основного ствола М. Архаичное строение R
сочетается с прогрессивными моментами — ослаблением М, усилением
переднего края крыла посредством мощно развитой системы R и птеростиг-
мой. Строение тела мало своеобразно; грудь уплощенная, брюшко кони-
ческое, ноги умеренно развитые. Характерны укороченные антенны и не-
большая голова. Величина насекомого не достигает 4 мм.
Семейство Protopleciidae впервые установлено мною для лейасового рода
Protoplecia Handl., к нему же относятся два рода из юры Кара-тау.
Protopleciidae характерны рядом архаичных признаков жилкования: их
крылья почти лишены костальной специализации, широкие и с хорошо раз-
витой медиальной системой жилок. Грудь умеренно вздутая, с хо-
рошо развитыми цепкими ногами. Размеры тела средней величины — 5—7 мм.
Семейство Protobibionidae известно лишь по одному виду единственного
рода Protobibio. Для него характерно высокоспециализованное жилкова-
ние с резко выраженными явлениями костализации и ослабления задних
жилок. Размеры тела небольшие; его особенности неясны вследствие плохой
сохранности. Это и следующее семейство являются наиболее специализо-
ванными юрскими длинноусыми двукрылыми.
Семейство Protoscatopsidae также известно лишь по единственному
монотипическому роду и обладает высокоспециализованными крыльями и
антеннами; в крыльях резко проявляется костализация; антенны укорочены
и слегка булавовидно вздуты; грудь резко вздута; строение ног. неизвестно.
Размеры тела очень невелики — около 3 мм.
Оба последних семейства по облику очень напоминают представителей
семейств Pleciofungivoridae и Pleciomimidae — у них вздутая груды и не-
большие размеры; и лишь строение крыльев и антенн указывает на более
далеко зашедшую специализацию и связи с совсем иными современными
группами. J
♦ * "1
I *
В заключение этого обзора юрской фауны следует указать, .что^богатей-
шее каратауское юрское местонахождение еще далеко не исчерпано. Уже
после окончания составления настоящей статьи были исследованы новые
материалы по двукрылым из Кара-тау, которые обнаружили новых пред-
ставителей Oligoneura', фотография одной из этих новых форм приводится
на таблице Х(фиг. 24)— Tipuloplecia breviventris gen. sp.^noу., являющегося
представителем нового своеобразного семейства.
S3
7. СВЯЗЬ ЮРСКОЙ ФАУНЫ С СОВРЕМЕННОЙ И ФИЛОГЕНЕЗ
ОТДЕЛЬНЫХ ГРУПП
Критически рассматривая особенности юрских двукрылых и сравнивая
их с современными группами, можно наметить и изобразить графически
родственные связи отдельных семейств, иначе говоря, показать родословное
древо этих насекомых (рис. 95).
Соврепенм» зпага
Рис. 95. Схема филогенетических отношений семейств Oligoneura. Сокращения: I —
Oligoneura, II—Phryneoidea, III — Brachycera, IV — Tendipedoidea, V — Culicoidea,
VI — Psychodoidea, VII — Pachyneuroidea, VIII — Tipuloidea-, 1 —Fungivoridae, 2 —
Manotidae, 3 — Lygistorrhinidae, 4 — Necromyzidae, 5 — Mycetophilitidae, 6 — Allac-
toneuridae, 6a — Mesosciophilinae, 6b—Mimalycoriinae, 6c—Fungivoritinae, 7 —
Pleciomimidae, 7a—Pleciomiminae; 7b — Mimallactoneurinae, 7c — Paritonidinae, 7d —
Antefungivorinae, 8 — Pleciofungivoridae, 9 — Diadocidiidae, 10 — Ditomyiidae, 11 —
Macroceratidae, 12 — Ceroplatidae, 13 — Bolitophilidae, 14 — Lycoriidae, 15—Heterope-
zidae, 16—Itonididae, 17 — Lestremiidae, .18 — Mycetobiidae, 19 — Hesperinidae,20—
Pachyneuridae, 21 — Axymyiidae, 22 — Paraxymyiidae, 23 — Protopleciidae, 24 — Eople-
ciidae, 25 — Penthetriidae, 26 — Protobibionidae, 27—Bibionidae, 28 — Protoscatopsidae,
29 — Scatopsidae, 30 — Corynoscelididae (ориг.). — На схеме толщина фигур возможно
более точно показывает обилие видов и разнообразие группы в % к общему коли-
честву видов фауны данной эпохи, именно юрской (Каратауской), эоценовой (балтий-
ский янтарь) и современной, показанных черными отрезками, в отличие от предпола-
гаемых, заштрихованных.
Общее сравнение юрских Oligoneura с современными сразу же показы-
вает отсутствие резких отличий в разнообразии той и другой фауны: нельзя
сказать, что юрские Oligoneura были менее разнообразны и более однородны.
Несмотря на небольшое число зарегистрированных видов (48) каратауские
комарики принадлежат не менее чем к 8 семействам. Любой случайный
84
современный сбор этих насекомых даст примерно подобные же цифры:
будут собраны многочисленные виды Itonididae, Fungivoridae и Lycoriidae
и более редкие виды 4 или 5 семейств — Bibionidae, Ceroplatidae, Scatop*
sidae, Macroceratidae; в тропиках прибавится еще семейство Penthetriidae.
Подобное «сходство» ископаемой и современной фаун может свидетельство-
вать лишь об одном, именно о значительно большем богатстве группы в про-
шедшем времени, в течение юрского периода. Несомненно, нам стали из-
вестны остатки пока лишь очень незначительной части юрской фауны,
только представителей наиболее богато развитых групп и притом самых
обычных форм. Наиболее естественно предположить, поэтому, что это ка-
жущееся сходство юрской и современной фаун в их объеме и разнообразии
есть лишь следствие недоучета особенностей палеоэнтомологического ма-
териала—его бедности. Вероятнее всего, что юрская фауна Oligoneura
была значительно богаче современной.
На прилагаемой схеме (рис. 95) я изобразил филогенетические взаимо-
отношения групп Oligoneura, основываясь на результатах исследования
юрских каратауских двукрылых и учете всех особенностей современных
представителей группы.
Все юрские Oligoneura распадаются на четыре близкие, но ясно обособлен-
ные группы. Наиболее крупной является семейство Pleciomimidae (отме-
ченное на схеме цифрами 7a-d), сильно притом специализованное и со-
стоящее из четырех отдельных подсемейств. Оно, вероятно, в начале мело-
вого периода отщепило от себя ствол галлиц — Itonidiae (на схеме 16),
достигших расцвета лишь ныне. Подсемейство Antefungivorinae дошло до
настоящего времени лишь в виде реликтовой группы Diadocidiidae. Подсе-
мейство Paritonidinae уже в юре было высокоспециализованной группой
и в дальнейшем дало начало крайне специализованным Heteropezidae, с одной
стороны^ и реликтовым Lestremiidae, с другой, сохранившим многие арха-
ичные черты. Подсемейство Mimallactoneurinae, несомненно, было слепой
ветвью, не давшей потомства.
Вторая группа, семейство Pleciofungivoridae (8), довольно специали-
зована, но обладает вместе с тем рядом архаических черт. Вторичные под-
разделения в нем едва намечены. Непосредственные потомки Pleciofungi-
voridae ныне почти отсутствуют; вместе с тем ряд современных реликтовых
групп обнаруживает сходство с этим семейством. Наиболее близки к Pie*
eiofungivoridae современные Ditomyiidae, отщепившиеся от форм, близких
к родам Eohesperinus, Transversiplecia, вероятно, в конце юры или-начале
мела. Более далеки от Pleciofungivoridae три современных семейства —
Ceroplatidae, Macroceratidae и Mycetobiidae, вместе с тем, несомненно, род-
ственные одно с другим, и, несомненно, происходящие с Pleciofungivoridae
из общего корня и притом обладающие некоторыми более архаичными чер-
тами жилкования. Отщепление комплекса Ceroplatidae A- Macroceratidae А-
Mycetobiidae от ствола Pleciofungivoridae произошло, вероятно, в конце
Ариаса или в начале юры. Еще более далека от Pleciofungivoridae современ-
ная группа, до сих пор не обнаруженная в юрской ископаемой фауне, но,
несомненно, уже существовавшая в то время — Hesperinidae. Это ярко
выраженное реликтовое семейство .обособилось от предков описываемого
выше комплекса, вероятно, уже в начале или середине триасового периода.
Примерно в это же время или, может быть, несколько позже, исходные
стволы Pleciomimidae и Pleciofungivoridae разделились; еще ранее отщепился
ствол, давший начало третьей главной группе — семейству Allactoneuridae:
Последнее семейство, дожившее до настоящего времени""!? виде единствен-
ного тропического рода на Яве, в юрское время было, повидимому, богато
представлено и, сочетая в организации архаичные и специализованные осо-
бенности; состояло не менее чем из трех, вернее, четырех подсемейств;
85
Наиболее крупное подсемейство, Fungivoritinae, обладает рядом специади-
зованных черт, не позволяющих считать его предковой группой по отно-
шению R каким-либо современным семействам; то же следует сказать и о
Mimalycoriinae: оба они были слепыми, не давшими потомства ветвями.
Лишь подсемейство Mesosciophilinae, несомненно, дало начало всем нынеш-
ним Fungivoridae; отщепление последних произошло, вероятно, в середине
мелового периода.
Неясна история Lycoriidae: высокая специализация этих комариков
очень затрудняет решение вопроса о их филогенезе; повидимому, вероятнее
всего, что это семейство отделилось от предков Allactoneuridae в начале
юры; за это говорит присутствие у архаичных родов Lycoriidae ряда при-
митивных признаков: длинная R или даже присутствие ветвей на RS;
последний признак при прочих особенностях указывает на большую древ-
ность, не уступающую юрским группам.
Кроме описанных трех больших групп в юрской фауне открыто несколько
семейств специализованных Oligoneura, обнаруживающих очень ясные
связи с современными реликтовыми группами; таковы Protopleciidae —
несомненные предки современных Penthetriidae, Protobibionidae — предки
Bibibionidae, Protoscatopsidae — предки Scatopsidae, и Paraxymyiidae, очень
близкие к исходным формам современных вымирающих Axymyiidae. Все
эти группы, может быть, за исключением Protoscatopsidae, родственны одна
другой и имеют общих предков, от которых в свою очередь отщепились
предки «настоящих», т. е. всех остальных Fungivoridae; произошло это,
вероятно, еще в верхней или даже средней перми. Положение Scatopsidae
и близких к ним Protoscatopsidae и Corynoscelididae еще неясно; возможно,
что они окажутся не столь родственными комплексу Bibionoidea, как это
кажется сейчас.
Следует отметить совсем особые филогенетические отношения двух релик-
товых семейств Oligoneura — Pahyneuridae (20) и Bolitophilidae (13). Первое
из них обладает в основной части крыла настолько архаическими особен-
ностями— отсутствием радиоанальных хетариев, хорошим развитием basi-
cubitus, basianales 1 и 2, отсутствием перепончатых лопастей по краю
основания крыла — что заставляет противопоставлять его всем остальным
группам Oligoneura; семейство Pachyneuridae до сих пор по существу со-
хранило первичный тип строения крыла, именно «гребной», и не «пропел-
лирующий»; мне кажется несомненным, что Pachyneuridae отщепились от
общего ствола Diptera в числе первых групп еще в всфхнепермские времена.
Эти двукрылые, несомненно, очень далеки от остальных Oligoneura, как
Bibionoidea, так и Fungivoroidea и Должны быть обособлены системати-
чески в качестве своеобразного надсемейства Pachyneuroidea. Что касается
Bolitophilidae, то уровень специализации его крыла — развитие анальной
лопасти и зачатков alula и другие особенности, — несомненно, выше, чем
у Pachyneuridae; отщепление этого семейства от ствола Oligoneura произо-
шло, вероятно, одновременно с дивергенцией его на Bibionoidea и Fungivoroidea.
Из этого наброска филогенетических отношений Oligoneura можно
сделать ряд общих заключений о ходе эволюционного процесса в этой группе.
Состав современной и юрской фаун и анализ их взаимоотношений позво-
ляют вывести заключение о различных сроках формирования групп и о не-
скольких периодах большей интенсивности эволюционного процесса Oli-
goneura. Особенности групп, близких к современным Bibionidae -J- Pen-
thetriidae и Scatopsidae, и нахождение их в юре в сравнительно мало отлич-
ном виде с очевидностью свидетельствуют о реликтовом характере назван-
ных семейств еще в юрское время: Bibionoidea пережили свой расцвет рань-
ше, в «доюрское» время, вероятно в верхней перми или раннем триасе;
тогда сложились все основные стволы семейства, которые в дальнейшем
86
лишь более специализовались, приобретая все больший реликтовый
характер.
Все остальные группы Oligoneura можно вполне противопоставить на-
званным выше Bibionoidea (понимая под этим названием условное объеди-
нение более древних групп) под термином надсемейства Fungivoroidea. Три
крупнейших семейства юры Кара-тау по существу близки одно к другому;
формирование и дивергенция их начались, вероятно, в самых верхах триаса
или низах юры; дальнейшие же пути эволюции этих главных групп очень
различны. Наиболее архаичное изо всех трех семейств — Allactoneuridae,
обладающее наименее специализованным жилкованием, не отличалось
особым разнообразием в юрское время и достигло максимального расцвета
лишь к началу или середине кайнозоя; об этом свидетельствует богатая
фауна Fungivoridae балтийского янтаря, т. е. верхнего эоцена, вероятно
не уступавшая в разнообразии современной; дивергенция отдельных се-
мейств и подчиненных им категорий в этом стволе продолжалась, вероятно,
в течение всего мела, достигнув к концу палеогена расцвета.
Второй комплекс семейств — Pleciofungivoridae и близкие — обладал
совсем иной судьбой. Юрская фауна этих комариков состоит из крупного
семейства Pleciofungivoridae, находящегося в цветущем состоянии с еще
почти не намеченными вторичными подразделениями; это семейство, вернее
его непосредственные предки, является несомненными исходными формами
для ряда современных семейств — Ceroplatidae и близких (см. выше).
€ другой стороны, кайнозойская, в частности верхнеэоценовая фауна этих
семейств, уже обладает чертами реликтовости, вполне ясно выраженными
у современных групп. Очевидно, что расцвет этого комплекса произошел
в середине или во второй половине мезозоя;1 начало расцвета мы наблюдаем,
несомненно, на примере разнообразного семейства Pleciofungivoridae в сред-
неюрском времени Кара-тау; своего апогея развитие этого ствола^ вероятно,
достигло на границе юрского и мелового периодов.
Рассматривая отношения наиболее обильной и разнообразной юрской
группы Pleciomimidae и близких к ней современных семейств, мы видим
примерно ту же судьбу отдельных специализованных ветвей, какая была
и у потомков предыдущей группы; дивергенция лишь в семействе Plecio-
mimidae началась, повидимому, несколько раньше, приблизительно с начала
юры; расцвета этот ствол достиг к концу юрского периода. Основным отли-
чием от предыдущей группы является отщепление от этого ствола прогрессив-
ного семейства Itonididae, достигшего наибольшего расцвета лишь в совре-
менную эпоху и появившегося еще в балтийском янтаре — верхнем эоцене.
8. ПУТИ И ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИОННОГО ПРОЦЕССА
В ГРУППЕ OLIGONEURA
Как показано в предыдущей главе, обособление от древнейшей ветви
Bibionoidea — всех остальных Oligoneura (Fungivoroidea, собственно) про-
изошло не позднее конца перми или начала триаса. В задачу настоящего
исследования не входило решение вопроса о связях всего комплекса Oligo-
neura с другими группамиDiptera-, но очевидно, что отщепление этого ствола
от остальных двукрылых, вероятнее всего, случилось не позднее середины
пермского периода. За всю свою долгую историю, за время, следовательно,
с середины пермского периода до современной эпохи, со стволом Oligoneura
произошли важные изменения. Исследуя эволюцию всей группы, можно
•с ясностью выделить ее этапы, носящие резко выраженный прогрессивный
1 За это, между прочим, говорит богатство семейства Ceroplatidae в Австралийской
области, обычном убежище мезозойских реликтов.
87
характер, именно то, что Северцев (1939) называет ароморфозами;
такие изменения могут быть охарактеризованы как резко повышающие
всю организацию животного, значительно расширяя ареал его обитания
и давая ему преимущества перед остальными.
Отчетливо выясняются также Изменения, не носящие характер аромор-
фозов, именно всякого рода адаптации и специализации. Эти изменения
распространены гораздо более широко по сравнению с моментами прогрес-
сивной эволюции, ароморфозами. Дегенерация, т. е. упрощение и узкая
специализация, на нашем примере двукрылых Oligoneura почти не имеет
места за исключением частных случаев.1
Первый период появления ароморфозов в эволюции Oligoneura, несо-
мненно, был на рубеже перми и триаса, когда от бибионоидных предков
отщепился ствол настоящих Fungivoroidea; полезным приобретением этой
новой группы (Fungivoroidea) было усиленное развитие крыльев и ног при
резком увеличении грудного отдела. Сильное развитие органов движения
дало большое преимущество сформировавшейся группе и позволило ей
широко расселиться, захватив новые экологические стации и географи-
ческие районы. После этой «пермской радиации», давшей в результате аро-
морфоза ствол Fungivoroidea, последний претерпел разнообразные идио-
адаптивные изменения.
Этот процесс продолжался до конца триаса и начала юры и за-
ключался в приобретении полезных изменений крыльев, осуществлявшихся
при наличии крепких ног. и мощного грудного отдела. Изменения крыльев
уже происходили и ранее в истории основного ствола бибионоидных
Предков группы, но там они не сопровождались одновременным усилением
всех органов движения насекомого и потому не сыграли столь большой
роли как в этом случае. У предков Bibionidae, именно в семействе Proto-
bibionidae, костализация крыльев пошла очень далеко, но по существу
являлась лишь случаем частной адаптации органа: этот процесс не был
♦поддержан» соответствующими изменениями остальных моторных органов
Тела, и поэтому его значение не вышло из рамок простой идиоадаптацйи.
Совсем иное произошло при эволюции главного ствола Oligoneura, именно
Fungivoroidea-. за время этой эволюции одновременное усиление претерпели
все органы передвижения и в первую очередь сам грудной отдел тела; в этом
случае изменения жилкования имели гораздо более серьезное значение
в филогенезе и они должны быть оценены именно как прогрессив-
ное изменение (ароморфоз Северцова). Биологическое значение
этих изменений несомненно: укрепление крыльев и ног значительно повы-
шает активность взрослых насекомых и позволяет им не только быть чисто
«воздушными» животными, но и свободно передвигаться среди растительных
остатков на почве, скрываться среди них при неблагоприятной температуре
и влажности или от хищников. Усиление органов Передвижения — груд-
ного отдела с крыльями и ногами — было важным шагом вперед в эволю-
ции ствола Fungivoripidea, давшим большие преимущества его представите-
лям перед их предками, значительно расширив их экологические границы.
Прежде чем перейти к рассмотрению путей эволюции ствола Fungi-
voroidea, полезно вкратце рассмотреть особенности филогенеза основных
ветвей Oligoneura, от которых сохранились до настоящего времени лишь
реликты.
f Точно представить себе пути эволюции древней группы Oligoneura,
именно Bibionoidea, при существующих данных палеонтологии нельзя;
1 Как будет ясно из дальнейшего, нельзя считать эволюцию группы галлиц (Itoni-
diidae) прошедшей по этому пути. Особенности развития этой группы представляют
явно прогрессивные изменения.
Л
современные семейства далеки одно от другого и в большинстве, какука-
зано выше, обладают или резко выраженным реликтовым характером или
несут далеко зашедшие черты адаптивных изменений.
Эволюция Bibionidae и Penthetriidae, групп, лучше всего представлен-
ных ныне, проходила в направлении «половой специализации»; разви-
вался резкий половой диморфизм; у самцов увеличились и усложнились
глаза, усилились крылья и ноги, у самок увеличилось брюшко, крылья
мало изменялись. Сравнение названных двух семейств этого ствола даёт
очень многое для понимания причин и путей их эволюции. Семейство
Bibionidae обладает сильно измененными вследствие «костализации» кры-
льями, ротовые органы у самцов развиты слабо, у самок крупные и функ-
ционирующие, ноги у обоих полов с многочисленными шипами и вырос-
тами; функция этого вооружения неизвестна, но почти несомненно, что
оно служит для раскапывания, рытья или, может быть, схватывания.
Соседнее семейство, Penthetriidae, не обнаруживает в крыльях ясно-
выраженной костализации; ноги у них тонкие, довольно крепкие, но лишен-
ные вооружения; ротовые органы рудиментарные, вероятно не функциони-
рующие. Очевидно, что обе эти группы резко отличаются тем, что у Bibio-
nidae имеет большое значение питание взрослых фаз, преимущественно
самок, в то время как у Penthetriidae imagines не принимают пищи.1 На-
прашивается само собой заключение о связи афагии у Penthetriidae с от-
сутствием специализации крыльев и ног. Imagines Bibionidae нуждаются
в питании и должны искать пищу; усиленные органы передвижения —
крылья и ноги, — несомненно, при этой полезны. Imagines Penthetriidae —
афаги, которым не надо передвигаться для поисков пищи, почему их могут
удовлетворить и мало костализованные крылья.
Возможно сделать заключение о предполагаемых больших адаптивных из-
менениях личинок Penthetriidae по сравнению с Bibionidae. Вероятно,
личинки Penthetriidae обладают такими полезными особенностями, обес-
печивающими, например, обилие пищи, которые обусловили афагию ima-
gines, что заставляет нас приписывать им во всяком случае более прогрес-
сивный характер по сравнению с Bibionidae.
Аналогичный случай наблюдается, в Отношениях между семействами
Scatopsidae и Corynoscelididae. Первое семейство о бла дает целым рядом дале ко-
зашедших адаптивных изменений органов imago и заключает сравнительно
многочисленные роды и виды. Личинки его представителей живут в навозе
и гниющих веществах, где ведут хищный образ жизни; морфология их
довольно примитивна, не специализована. Второе семейство;— Coryno-
scelididae — являет собой характернейший пример примитивного по има-
гинальным особенностям вымирающего реликта и состоит всего из 5—8
видов (см. стр. 36); биология европейско-азиатских видов до сих пор
неизвестна; новозеландский вид рода Canthyloscelis, очень близкий к европей-
скому, изучен Тонуаром (Tonnoir, 1927), причем оказалось, что
'его личинки обладают необычайно специализованным строением. Несо-
мненно, что только особенности превращения и биологии личинки и позво-
лили этой группе сохраниться до настоящего времени. Imagines Cory-
noscelididae обладают архаичным жилкованием и, вероятно, так же как и
Penthetriidae, афаги.
Представители реликтового и известного лишь по трем экземплярам
насекомых из далеких точек земного шара семейства Axymyiidae также не
питаются во взрослом состоянии, обладая редуцированными ротовыми
1 Личинки обоих семейств живут в почве, гниющей древесине и навозе; более точно
биология и превращение Penthetriidae почти неизвестны.
89-
органами. И в этом случае справедливо ожидать узкой специализации
в личиночной биологии.
Рассмотрение особенностей эволюции двух групп, пожалуй, наиболее
удаленных от остальных ветвей — семейств Bolitophilidae и Pachyneuridae —
дает мало; оба они еще очень недостаточно изучены. Pachyneuridae известны
лишь по редким в музеях экземплярам imagines, биология которых до
сих пор не выяснена; особенности их крыльев заставляют их считать вовсе
чуждой группой и противопоставлять остальным Oligoneura. Сохранение
в основании крыла (basiala) базальных отделов всех жилок и отсутствие
развитых радиоанальных хетариев вполне оправдывают обособление Pachy-
neuridae и вместе с тем показывают очень большую древность этой реликто-
вой группы. Очень досадно отсутствие каких бы то ни было данных о мета-
морфозе Pachyneurw, вероятнее всего, особенности превращения и эколо-
гии этих насекомых заключают в себе такие полезные приспособления,
наличие которых и объясняет сохранение до настоящего времени этой релик-
товой группы, по существу верхнепалеозойской.
Взаимоотношения Bolitophilidae с другими Oligoneura более ясны; эта
группа не обнаруживает столь резкого обособления, как Pachyneuridae,
и явно тяготеет к стволу Fungivoroidea, правда, отличаясь архаич-
ными особенностями крыльев. Наряду с архаической природой imagi-
nes у Bolitophilidae необходимо отметить особенности личинки: хорошее
развитие органов чувств (антенн), наряду с относительно измененным
скелетом головы, приближающим эту группу к более молодому семейству
Fungivoridae.
Переходя к рассмотрению путей эволюционного развития основного
ствола Fungivoroidea, в первую очередь следует анализировать особенности
бедных видами, в своем большинстве реликтовых, ныне вымирающих групп.
Как уже указано, большинство реликтовых групп этого ствола плохо
известны биологически; поэтому, не зная особенностей превращения и эколо-
гии Diodocididae, Manotidae, Lygistorrhinidae, Allactoneuridae, Hesperinidae,
можно лишьс очень большой осторожностью судить о их эволюции и ее путях.
В развитии Macroceratidae наблюдается улучшение органов чувств
в виде увеличения антенн и глаз; с другой стороны, эти насекомые обладают
сильно редуцированными ротовыми органами, следовательно, имагинальной
афагией. Несмотря на относительно малый объем семейства, заключающий
около 50 видов единственного рода, эти насекомые не являются редкостью
и распространены широко и в большом количестве индивидов. Поэтому
весьма вероятно, что Macroceratidae отнюдь не являются реликтовой, выми-
рающей группой. Особенности близкого семейства — Ceroplatidae — напо-
минают черты предыдущего семейства. Реликтовая природа этой группы
выражена менее отчетливо — в группе около 200 видов 15 родов; морфо-
логически семейство Ceroplatidae приближается к Macroceratidae, обладая
также умеренно измененным жилкованием. Строение антенн также является
главным систематическим признаком; однако характер специализации
этих органов совсем иной: антенны увеличивают свою площадь путем не
удлинения члеников, а расширения и разрастания их в ширину (рис. 11).
С другой стороны, в этом семействе афагия не является правилом, если даже
вовсе не отсутствует, так как многие роды обладают хорошо развитым
удлиненным хоботком. Развитие немногих изученных форм Ceratoplatidae
очень своеобразно: личинки их обладают высокоспециализованной дыха-
тельной системой в виде редуцированного числа стигм и образования ды-
хательных выростов на заднем конце тела, и живут в гниющей древесине
и грибах-трутовиках.
Исследования над Ceroplalus testaceus D а 1 m (Stammer, 1932),
как и мои наблюдения над превращением и экологией Ceroplatus sesioides
so
Wahl, показали, что личинки этих видов живут под прикрытием пены И»
секрета слюнных желез, обладают способностью свечения и питаются
исключительно спорами грибов-трутовиков, на нижней поверхности ко-
торых живут. Тонкие выросты на заднем конце тела, погруженные во
влажную пену, несомненно, играют роль своеобразных жабер. Imago не
нуждается в питании и откладывает яйца через 1 или 2 дня после окры-
ления. Столь далеко зашедшая специализация личинок очень примечательна.
Можно очень пожалеть о неизученности биологии Macroceratidae: в ко-
нечном итоге лишь точное знание этих особенностей позволило бы ответить
на вопрос о причинах и путях эволюции группы. В общих чертах уже оче-
видно, что и в случае этих двух семейств первенствующее значение в ка-
честве фактора эволюции имела специализация личинки.
Особый интерес представляет рассмотрение эволюции современной
реликтовой группы Mycetobiidae. Немногие группы двукрылых претерпели
столь разнообразную «историю», как род Mycetobia: его то соединяли в одну
группу с родами Phryne или Petaurista, или только с последним и лишь
недавно было выяснено окончательно его истинное положение в системе
среди настоящих Fungivoroidea. Причиной разногласий во взглядах на
систематическое положение Mycetobia явилось сходство в строении
личинок трех названных групп — Phryneidae, Petauristidae и Mycetobii-
dae; очевидно, наблюдаемое сходство действительно отражает родственные
отношения; но, с другой стороны, большие различия во взрослых фазах
этих насекомых никак не могут подтвердить действительной близости этих
групп. Объяснением этого интереснейшего факта может быть лишь то
обстоятельство, что сходство личинок в данном случае указывает на род-
ственные, но очень далекие отношения: личинки сохранили свое
архаичное внешнее строение и не специализовались с давних времен, с сере-
дины пермского времени. Эволюционировали эти насекомые в направлении
дифференциации лишь особенностей imagines, личинки же обладали та-
кими выгодными особенностями в биологии питания и стаций, которые по-
зволили им сохранить архаичные черты и до сих пор.1
. Исследование путей эволюции семейств, не обладающих в современную
эпоху реликтовыми особенностями, а, наоборот, многочисленных и широко-
распространенных, именно Fungivoridae и Itonididae, очень плодотворно.
Эти исследования указали на большие различия в направлении эволюции
грибных комариков, с одной стороны, и галлиц, с другой.
Эволюция грибных комариков—Fungivoridae, развившихся из юрской
группы Mesosciophilinae семейства Allactoneuridae, шла по пути изменения
особенностей зрелой фазы насекомого, личинки же претерпели умеренную
специализацию. Действительно, эвцефальные личинки Fungivoridae не обла-
дают выраженными морфологическими признаками узкой специализации:
ротовые органы их хорошо развиты, дыхательная система перипневстиче-
бкая. Лишь головная капсула склеротизована, да и слабое развитие антенн
достойно внимания. Биологическая особенность в виде мицетофагии очень
характерна, причем выражена далеко не одинаково у разных групп; так,
трибы и роды более архаичные, например Sciophilini, Boletinini,
1 Следует вспомнить биологические особенности Petauristidae и Phryneidae. Первые
являются одним из немногих представителей Nematocera, летающих при низкой темпе-
ратуре (зимой) и распространенных преимущественно в умеренных зонах Земли, на
север до 82° северной широты (Lindner, 1930), в высокогорной зоне, в пещерах и
глубоких шахтах, Phryneidae, более широко распространенная группа, личинки которой
питаются гниющими растительными веществами, обнаруживая ясно выраженное тяго-
тение к постройкам человека—синантропию. Оба случая, особенно с Petauristidae, очень
хорошо указывают на своеобразие биологии описываемых групп, заключающееся в за-
нятии данным видом животного определенной экологической стации — «ниши», менее
заселенной по сравнению с любыми другими.
&
Monocionini, Tetragoneurini имеют личинок, живущих большей частью
в гниющей древесине или древесных грибах, в то время как более специа-
лизованные группы, например большая часть Fungivorinae, Leiinae, живут
за счет шляпочных грибов.
Гораздо более характерны морфологические и биологические особен-
ности взрослых Fungivoridae; они заключаются в очень сильном развитии
ног, груди и в «обтекаемой» форме головы и груди (голова глубоко «вдавлена»
в грудь), с одной стороны, и в большой длительности имагинальной жизни
и распространенности зимней диапаузы у imagines, с другой. Несомненно,
что развитие мощных бегательных и прыгательных ног, равно как и форма
головы и груди, очень полезны для жизни комариков в среде, состоящей
из растительных остатков — частей коры, древесины, трухи из гниющей
древесины, сухих листьев; в такой среде эти двукрылые очень оживленно-
передвигаются, забиваясь в щели и пустоты, где они проводят и зимовку.
Крылья в этом семействе испытали сравнительно умеренную специ-
ализацию: костализация выражена слабо и лишь у некоторых групп
вполне ясна.
На основе этого краткого очерка выясняется общее направление эволю-
ции Fungivoridae. Эта группа, отщепившись от юрских Allactoneuridae,
пошла по пути усиления бегательных и прыгательных способностей комаров
наряду с удлинением имагинальной жизни; это дало возможность насе-
комым активно искать места убежищ для зимовки, более надежных и защи-
щенных, чем те, которых Могли бы достигнуть лишь летающие комарик#
со слабыми ногами. Каковы же причины такой эволюции группы? Мне
кажется, что семейство Fungivoridae могло сложиться в условиях умерен-
ного и холодного климата с резкими колебаниями летних и зимних темпера-
тур, притом в лесах, богатых растительными остатками на поверхности почвы.
Постепенное похолодание, богатая пища личинок в виде грибов и места
убежищ в форме растительного мусора — вот основные моменты, опреде-
лившие эволюцию группы. Особенности развития в этом случае не играли
ведущей роли в отличие от других семейств группы.
Совсем иное мы видим в истории ствола галлиц Itonididae, более моло-
дой группы, эволюция которой шла другими путями. Наиболее характерны в
эволюции галлиц изменения личинок, лишившихся жующего аппарата и став-
ших ацефальными; эти изменения явились, несомненно, отражением точно
неизвестных, но предполагаемых физиологических особенностей пищеваре-
ния. Повидимому, в течение эволюционного процесса у личинок Itonididae
выработались такие полезные особенности пищеварительного аппарата, кото-
рые сделали жующие ротовые органы излишними; вероятно, деятельность пи-
щеварительных желез, в первую очередь слюнных, изменилась в сторону
образования секретов, которые позволили не размельчать пищу челюстями
и заглатывать твердые ее частицы, а лишь пить жидкости — продукты
растворения пищи секретом слюнных Желез, подобно тому как это про-
исходит при питании (личинок круглошовных двукрылых, Cyclorrhapha;
в этом нас убеждают особенности строения кишечника и его желез у личи-
нок видов Itonididae, Heteropezidae и Lestremiidae (W е h г m е i s t е
1925). В кишечнике у различных видов этих семейств наиболее сложное его
строение наблюдено у Lestremiidae и Heteropezidae; оно заключается в при-
сутствии обособленного зоба и полного для Diptera числа из четырех маль-.
пигиевых сосудов (рис. 96 а); более измененные Itonididae обладают уко-
роченным и упрощенным кишечником и лишь одной парой мальпигиевых
сосудов (рис. 96 с); наоборот, слюнные железы в этом ряде форм услож-
нены и увеличены в размерах, будучи простыми и небольшими у Lestre-
miidae и сложными и крупными у Itonididae. Этот пример, хотя иллюстри-
рованный и очень небольшим числом форм, все же позволяет утверждать^
«2
что среди этой группы Oligoneura мы действительно наблюдаем выработку
явления «внекишечного», экстраинтестинального, пищеварения (L е п-
g е г к е п, 1924). Вместе с тем очевидно прогрессивное значение этого про-
цесса, несмотря на то, что он имеет внешние проявления в виде ацефализа-
ции личинок, заставляющей предполагать дегенерацию или, по крайней
мере, узкую специализации). Значительность этого пути эволюции доказы-
вается не только необычайно широким географическим распространением
и богатством семейства в настоящее время, но и присутствием среди галлиц
наряду с настоящими фитофагами также и свободно живущих хищников
на тлях, равно как и сапрофагов; последнее,
вероятно, было первичным. Наличие столь раз-
личных биологических особенностей отнюдь,
конечно, не указывает на узкую специализацию
группы.
Своеобразны также и те изменения, кото-
рые претерпевают imagines Itonididae. Основ-
ным и характерным процессом в эволюции взрос-
лой фазы галлиц явилась, с одной стороны,
редукция имагинального питания — развилась
в той или иной степени афагия, а с другой, раз-
ного рода половые специализации в антеннах
самцов, яйцекладах самок. Эти явления тесно
связаны с особенностями развития этих насе-
комых; образ жизни их личинок в условиях
избытка пищи, как и у очень многих других па-
разитных двукрылых, стимулировал редукцию
имагинального питания; с другой стороны, раз-
витие личинок й тканях разных растений и хоро-
шо выраженные их моно- или олигофагия стиму-
лировали выработку развитых органов чувств
и яйцекладов. При анализе изменений imagines
по сравнению с личинками с очевидностью мож-
но отметить отсутствие прогрессивного характера
в строении комариков, являющихся, в отличие
от личинок, узко специализованными насеко-
мыми.
На основе всего изложенного можно сделать
Рис. 96. Кишечник и слюн-
ные железы Lestremiidae,
Heteropezidae и Itonididae.
а—кишечник Lestremia sp.;
b — слюнная железа Mias-
tor metroloas W a g п.; c —
кишечник Lestodiplosis sp.;
d—слюнная железа tested i-
предположение о причинах эволюции этой группы ptesis sp. (по w е h г m е i s-
длинноусых. Ацефализация могла возникнуть у ter> 1925)-
личинок, живших в среде, очень богатой пи-
тательными веществами, поедание которых не требовало предвари-
тельного раздробления; питательный субстрат такого рода мог быть,
например, вытекающим соком растений, богатым сахарами, или бел-
ковой полужидкой средой, вроде тел слизистых грибов (М у хо-
лл у с е t е s), или, наконец, жидкими продуктами гниения, напри-
мер, тел высших грибов. Возможно и иное представление о по-
добных питательных средах, позволивших выработать экстраинтестинальное
пищеварение: пищей могли оказаться твердые, но легко растворимые тела,
например, выкристаллизовавшиеся на поверхности растений сахаристые
вещества, которые с легкостью растворялись слюной.
Эволюция питания ацефальных личинок нам представляется в следую-
щем виде. (См. таблицу на следующей странице).
Наибольшее развитие Itonididae в дальнейшем совпадает по времени
с развитием в меловом периоде покрытосемянных растений; совпадение
это далеко не случайно. Как известно, современные Itonididae биологи-
93
Строение личинки
Питательный субстрат
Первая, исходная стадия
Эвцефальиые личинки с жующими рото-1 Твердый субстрат, преимущественно и»
выми органами. | гниющих растительных веществ.
Вторая, переходная стадия
Различные стадии редукции ротовых
органов и усложнение ферментативных ка-
честв слюны.
Жидкий, или, наоборот, очень твердый,
но легко растворимый питательный суб-
страт.
Третья, конечная стадия
Ацефальные личинки с полностью реду-
цированными ротовыми органами и слож-
ными слюнными железами, обильный се-
крет которых обладает разнообразными пи-
щеварительными свойствами.
Самый разнообразный субстрат: ткани
живых растений и животных и гниющие
вещества.
чески очень тесно связаны с цветковыми растениями, и их многообразие,
конечно, объясняется именно богатством в кайнозойскую эру покрыто-
семянных.
Рассмотрением филогенеза семейства Itonididae я закончу очерк путей
эволюции группы Oligoneura. Далеко не все семейства этого комплекса
длинноусых могли быть сколько-нибудь подробно разобраны; некоторые,
притом довольно крупные семейства, например Lycoriidae и Scatopsidae*
или небольшие реликтовые группы, например Ditomyiidae, вследствие не-
достаточной изученности вовсе не смогли быть материалом для общих вы-
водов о ходе их эволюции*.
* *
*
~~ Можно подвести итоги и формулировать некоторые намечающиеся зако-
номерности эволюционного процесса. В мою задачу отнюдь не входят под-
робный анализ и классификация различнейших путей эволюции и способов
их осуществления. Посвященные этому исследования эволюционистов-
морфологов достаточно иллюстрируют эту область эволюционного
учения, например Северцова (1939), Кр ыжан о вского (1938),
Шмальгаузена (1939). Вместе с тем, как мною уже указано в предис-
ловии, исследования морфологов страдают фактическим отрывом от систе-
мы животных, т. е. от той действительной картины животного мира,
которая создана современной фауной, ее большим или меньшим многообра-
зием. Пренебрежение к систематике с ее статистическими данными, к фау-
нистике и экологии форм^значительно обесценивает результаты чисто морфо-
логического изучения, которые в результате рисуют лишь эволюцию органов
или их систем, а вовсе не групп организмов, что очевидно являлось главной
задачей. Исследование конкретных фило генезов организмов, несомненно,
есть основной метод, могущий дать материал для выяснения закономерно-
стей эволюционного Процесса; но до сих пор этот естественный источник
подменялся односторонними сравнительно-морфологическими исследова-
ниями, позволившими, правда, точно анализировать частные закономер-
ности эволюции, например значение онтогенеза, но зато очень мало разъяс-
нявшими вопросы причинности путей эволюции, связи организма со средой
в его эволюции и взаимную обусловленность различных изменений орга-
низмов в процессе эволюции. Очевидно, в моем столь кратком и частном
64
исследовании трудно достаточно конкретизовать высказанные положения
и совершенно лишить их неизбежного декларативного характера.
Лишь в работах А. Северцева (1939, стр. 267—343) — хотя этот
автор и не касается вовсе данных современной системы" животных, опери-
руя исключительно со сравнительно-морфологическими материалами, —
начата разработка вопросов о переживании примитивных и архаических
групп животных и о роли в их эволюции различного рода специализаций.
Указания Северцоваи натолкнули меня на исследования особенностей
архаических форм.
Прежде чем перейти к изложению тех особенностей и закономерностей
эволюционного процесса, которые наметились в результате моих исследо-
ваний, необходимо остановиться на определении понятия «реликт» или «оста-
точная форма». Реликты наиболее четко характеризуются статистическими
данными систематики и зоогеографии — редкостью индивидов, или бед-
ностью родов видами, семейств родами, узкими или разорванными ареа-
лами распространения и, наконец, историческими свидетельствами па-
леонтологии. Эта статистическая характеристика реликта, несомненно,
является отражением существующих отношений между реликтовыми груп-
пами и их средой обитания, — точнее, той или иной экологической нишей,
которую они стали занимать в процессе эволюции. Основной особенностью
экологии реликта — остатка некогда хорошо развитой группы — должно
быть, очевидно, ее крайнее своеобразие, которое и обусловило сохра-
нение реликтовой формы; на первый взгляд подобное утверждение зву-
чит парадоксально, и тем не менее справедливо, как я поясню Далее.
С другой стороны, для меня очевидно, что особенности остаточной формы,
особенности реликта, могут быть выражены во всех случаях далеко не
в одинаковой, крайней форме, и что в таких случаях приходится говорить
лишь о той или иной, более или менее ярко проявленной остаточной, ре-
стантной природе. Само определение реликта одновременно обусловливает
определение и противоположного понятия — формы нереликтовой; в ко-
нечном итоге очевидно, что точное решение может быть получено лишь при
широком охвате всего вопроса о качестве и равноценности различных систе-
матических категорий, о строении системы как продукта филогенетического
развития группы. В мою задачу не входят обсуждение и анализ всего во-
проса, и я считаю нужным затронуть особенности лишь остаточных форм.
При изучении Oligoneura мне пришлось столкнуться с многочисленными
формами, обладающими в разной мере выраженными особенностями реликта.
Подробный анализ некоторых реликтовых групп показал, что бесспорная
реликтовая, остаточная природа этих семейств отнюдь не подкрепляется их
обязательной примитивностью. В большинстве случаев мы видим обратное:
для реликтовой группы обязательна та или иная степень специали-
зации, часто ушедшей очень далеко; примитивные, архаичные особенности
могут присутствовать, но их отсутствие отнюдь не изменяет основную при-
роду реликта как такового. В конечном итоге важна лишь степень утраты
примитивных качеств реликтом, степень его специализации. Таким образом,
правильно притти к выводу, что все реликты специализованы и что термины
«примитивные» и «специализованные» реликты имеют лишь условный смысл.
Но рассмотрим, чем являются эти «примитивные» и «специализованные»
реликты с эволюционной точки зрения. Наличие так называемых «при-
митивных», или «архаических», особенностей указывает на близость и
сходство их носителей с исходными для них вымершими формами, предками
многочисленных новых групп. Очевидно, эти примитивные реликты яв-
ляются непосредственными дериватами древних предковых форм,
сохранившихся до данного геологического времени. Чем же являются
реликты с мало выраженной примитивностью или вовсе лишенные этих
95-
Рис. 97. Схема, поясняющая филогенетические
отношения реликтов. Обозначения А, В, С, D,
Е, F — геологические отрезки времени; RA —
реликты с хорошо выраженными архаичными
особенностями; RSj, RS,, RS3 — реликты с
резко выраженными признаками специализа-
ции, степень которой обозначается цифровыми
указателями (ориг.).
фактической части моей работы; вполне
особенностей, реликты специализованные? Бедность и пи отсутствие архаи-
ческих особенностей у реликтов естественно указывает на случай крайней
специализации; происхождение этих форм, особенно при отсутствии палеон-
тологических данных, обычно неясно. Вместе с тем реликты этого рода,
в меньшей или большей мере специализованные, несомненно, являются
остатками некогда богато развитой, обособленной ветви. Это
последнее положение обычно подтверждается данными палеонтологии,
в других же случаях очевидно и без этих документальных свидетельств,
лишь на основании особенностей самих реликтовых групп (рис. 97).
После этих замечаний о природе реликтовых форм я перейду к изложе-
нию тех частных закономерностей, которые мною отмечены при анализе
реликтов и их особенностей. Повидимому, для реликтов является правилом
постоянное сопровождение приз-
наков примитивных, архаичных,
признаками высокой специали-
зации. Возможно итти даже и
дальше и утверждать, что чем
примитивнее мы на-
ходим у реликта при-
знаки, тем более край-
нюю степень измене-
ний мы наблюдаем у
него в иных, немно-
гих, особенн ос т я х
его организации или
экологии. Это явление,
очевидно, выражено наиболее
резко и ясно у реликтов, сохра-
нивших многочисленные архаи-
ческие признаки; в этих случа-
ях особенно отчетливо выража-
ются крайние изменения от-
дельных особенностей. Примеры,
данные ниже в виде таблицы,
лишь отчасти суммированы из
возможно иллюстрировать вы-
сказанные выше положения также и примерами из других отделов жи-
вотного мира, что я попытался сделать.
Приведенных примеров достаточно, чтобы Пояснить высказанное поло-
жение. По существу ничего нового в этих примерах нет: специализации
реликтовых форм давно известны. Но до сих пор сам-факт сохранения ре-
ликтовых форм объяснялся лишь самыми общими причинами; таковы
«изоляции» всякого рода — географические, в случае, например, австра-
лийской фауны или островных фаун, экологические «ниши». Эти объяснения
страдают неконкретностью: сама по себе изоляция, например географиче-
ская,притом на «островах» такого масштаба, как Австралия в особенности,
мало что объясняет. Более понятна и ясна роль экологических изоляций,
«ниш», которые, действительно, часто играют главнейшую роль в сохра-
нении реликтов.
Моей целью является уточнение условий сохранения реликтовых форм
и объяснение причины «замедления темпов эволюции» у реликтов.
-Отмеченная выше закономерность, заключающаяся в обязательном сопрово-
ждении признаков примитивных особенностями специализованными, при-
том, так сказать, «обратно пропорционально», т. е. так, что чем примитивнее
характер реликта, тем более крайними изменениями в какой-либо черте
56-
Реликтовая группа Примитивные особенности Особенности специализации
Из группы Diptera Oligoneura
Corynoscelididae Строение имаго: крылья, Ацефалия личвнки - признак край-.
(см. стр. 44) грудь и голова обладают примитивным строением. и ей специализации.
Penthetriidae и Аху- Жилкование, тело, ноги Имаго ие принимает пищи; афаги,
myiidae (см. стр. 89) примитивного строения. плохо летают; предполагаются высо- коспециализованные личинки.
Из других групп жи вотных
Мечехвосты (Xypho- Развитие и скелет при- Крайняя централизация нервной си-
sura). митивны. стемы и экология — признаки вы- сокой специализации.
Двоякодышащие Скелет и другие черты Экология: пресные воды, способ-
рыбы (Dipnoi). организации примитив- ны. вость анабиотического переживания при неблагоприятных условиях.
Виды рода Nautilus. Вся организация близка к таковой вымерших, Nautiloidea. Высокоспециализованное развитие: крупные яйца, в которых проходит развитие первых двух оборотов ра- ковины.
Ластовогие раки- Метамерия тела и гомо- Необычайно быстрое развитие и стой-
щитни (например р. Triops). номия коиечвостей очевь примитивны. кость яиц — полезная специализация.
Неполнозубые: ленив- Примитивен ряд черт Крайняя специализация в экологии,
ды, муравьеды, броне- носцы и т. д. (Eden- внутренней организа- ции — скелета, нервной покровах, зубах.
iota) [С е в е р ц о в, 1939] системы. •
Акуловые рыбы (Se- Скелет и сосудистая си- Развитие в виде крупных яиц, вну-
lachii). стема очевь примитивны. тревнее оплодотворение — высокая степень специализации.
Хамелеоны (Chamae- Покровы,короткая шея Крайняя специализация в ряде осо-
leontes) [Север- цев, 1939] примитивны. бенностей, связанная с экологией.
Пингвины (Impen- Покровыидругие черты Экология: способность совершенно
nes). примитивны: свободный крестец, строение костей. плавать в воде, пользуясь крыльями; распространевие.
Круглоротые рыбы Скелет и дыхательные Специализован рот; своеобразны
(Cyclostomata). органы архаичны. экологические особенности.
Поденки (Ephemero- Две крыл атые фазы, три Афагия и сокращение жизни имаго,
ptera). длинных церка, жилко- вание крыльев. узкая специализация личинок.
7 Розендорф, т. XIII, в. 2.
97
организации он обладает, — имеет естественное объяснение. Действительно,
черты примитивные, или архаичные, так нами называются потому, что они
давно пережиты другими формами и являются характерными для предковых,
исходных форм. Чем же может быть объяснена их консервация у реликтов?
Почему реликты до сих пор обладают особенностями организации древних
предков и почему они их не утеряли? Совершенно очевидно, что причиной
Этого явилось приобретение таких новых полезных черт,
которые позволили законсервироваться особен-
ностям предков. Эти новые признаки оказались столь по-
лезными для организма, что дали возможность
е Му эволюционировать в ином направлении, по
новому пути, по пути, который определился этим
новым приобретением.
Эта закономерность в связи примитивных и крайне измененных особен-
ностей, думается мне, поможет объяснить и другие особенности эволюции.
Так, несомненно, что связь примитивности и специализации, притом тесная,
существует не только у форм остаточных, реликтовых: она проявляется
и у групп, не обладающих реликтовыми особенностями. Я не буду развивать
этой стороны вопроса: это составляет предмет уже особого исследования.
Высказанные мною положения, касающиеся особенностей реликтовых
форм, возможно, помогут несколько изменить современные воззрения на
эти явления, придав им большую значимость и вместе с тем стимулируют
более продуктивное исследование закономерностей эволюционного про-
цесса.
SUMMARY
1. There are practically no published data on the evolution of Oligoneura,
the few papers dealing with the structure or the development of the members
of this group do not elucidate the problem of the phylogeny of different
stocks. They give only more or less detailed descriptions of the structure and
examine the interrelations of different recent groups. The noticeable discre-
pancy between the «systems» built on characters of adult or larval stages is
due to the fundamental fault of these schemes: they are artificial construc-
tions based on one developmental stage only, but not on a full consideration
of all the features of each form.
2. An examination of the principal, taxonomically important characters
of the structure, development, and bionomics of recent Oligoneura allows
to find out the different directions of specialization of organs or systems.of
organs. Usually the specialization proceeds by way of reduction.of different
organs. Such are, e. g., the evolution of antennae from long multisegmented
ones, composed of hemonomous segments, to shortened antennae consisting
of a few unequally developed segments; the development of the aphagy of
adult insects, and reduction of their mouth - parts; the reduction of dif-
ferent parts of wing-venation. These reductions are usually accompanied by
the development and differentiation of other organs, as the complication and
strengthening of legs, of sense organs, antennae and eyes, strengthening of
the anterior edge of wing. Besides a simple stating of different ways of evo-
lution of organs, it is quite possible to make out the origin and the functional
value of the observed changes. The reduction of the number of segments of
the antennae, of the mouth parts, and of certain parts of wing-venation,
are manifestations of specialization-processes resulting in improvement and
greater concentration of functions (antennae and wings), or substitution and
shifting of some functions to the larval stage (aphagy of the imagines). In
98
consequence a certain adaptive value of main taxonomic characters becomes
clear; they are not characters sui generis, distinct from the rest.
3. The structure of wings is of a special value in the evolution of Oli-
goneura, as well as of many other insects. It is clear from their importance
as chief organs of locomotion and expansion, besides the great complexity
of structure. Also, the obvious, mechanical significance of all the details of
wing-structure calls for an exact study of the aerodynamic properties of dif-
ferent insects, of their wings and bodies. The mechanical analysis is of course
to be carried out under the direction of a biologist-systematicist; most
important is the selection of the material, such selection being without the
means of an experimenting mechanician.
A comparative morphological analysis of the wings of Oligoneura permit-
ted to trace only the main paths of the evolution, consisting in the develop-
ment of another evidently more perfect mode of flight in the later groups.
An examination of the base of wing (up to the present time hardly investi-
gated) confirmed with certainty the reality of this process.
4. The composition and geographic distribution of the recent fauna of
Oligoneura is peculiar; 5700 species belong to this group of Diptera (Hendel,
1936), assigned to 21 families of a very unequal compass. Only 3 families,
Itonididae (gall-gnats), Fungivoridae (fungus-gnats), and Lycoriidae have
a world-wide spreading and are flourishing; 4800 species, or 5/6 of the total
number of species of the group, belong to these families. The remaining 18
families have mostly a more or less distinct character of relicts: of these, 7
families have a less pronounced character of relicts and comprise about 800
species. The remaining 100 species belong to 11 families, representing the
most isolated groups subject to extinction.
5. The earliest Mesozoic fauna of Oligoneura, comprising about 50 species,
and being described in the present paper from the Jurassic of Kara-tau, is
of an outstanding interest. It is sharply distinct from the recent fauna, as
well as from any of the known Tertiary ones. Only three of the 8 Jurassic
families of Oligoneura, present in the Kara-tau fauna, viz. the Pleciofungi-
voridae, Pleciomimidae, and Allactoneuridae, are rich in species. The other
families are represented by a few species and genera only. A single family,
Allactoneuridae, survives in Oriental Region (Javal), the rest being exclusi-
vely Mesozoic groups.
6. The investigation of the Jurassic fauna supplied chief, in some cases
decisive evidence in the elucidation of the phylogeny of different Oligorieuran
families; as a result, a well-based scheme of phyletic relations of these families
became attainable.
7. The main recent families are direct descendants of some Jurassic ones:
Itonididae descend from Pleciomimidae; Fungivoridae, from Allactoneuridae;
Bibionidae, from Protobibionidae; Scatopsidae, from Protoscatopsidae.
! The descent of Lycoriidae remains unsufficiently clear. Some families,
as Ceroplatidae, Macroceratidae, etc., have doubtlessly common ancestors
with Pleciofungivoridae.
8. There were scarcely any specialized, now extinct, families in the Juras-
sic fauna, and only some groups of minor taxonomic value, subfamilies, undoub-
tedly became extinct since that period.
9. At the beginning of the Tertiary there existed already practically all
now living families. The European Upper Eocene fauna from Baltic amber
is strikingly different from the Jurassic one, and nearly related to the recent.
The main groups of Oligoneura were formed duringthe Mesozoic. The Bibio-
noidea branched off not later than the end of the Permian; the forming of
the fungus-gnats (families Fungivoridae, Ceroplatidae, and their allies) occur-
red without doubt during the Cetaceous, and merely the gall-gnats, Itoni-
♦ 99
didae, the latest, now flourishing group, branched off at the junction of the
Cretaceous and the Tertiary.
10. It is possible to approach an explanation of the paths and causes of
evolution of some families. The gall-gnats, Itonididae, attained their recent
flourishing condition, owing to the acquisition of a complete metamorphosis,
an extra-intestinal digestion of the larvae, and-the close connection with angio-
sperm plants and the development of the latter. The fungus-gnats, Fungi-
voridae, could attain their diversity and abundance by the middle of the
Tertiary period thanks to the development of perfect means of locomotion and
the concealment in the plant detritus of woods. Along with the development
of cursory capacity the lengthening of the adult life played an important part,
allowing a considerable enlargement of the distribution area of the family.
11. The study of the phyletic relations of different groups of Oligoneura
shows the necessity to consider for this purpose, the statistical data of syste-
matics: the compass of groups, the number of subordinate taxonomic units,
as well as the evidence of geographic distribution. Habitually these facts
are not taken into consideration in evolutionary investigations of any kind,
which are based only upon the study of morphology, development, paleontolo-
gy, and at best also of ecology. Yet, this evidence of systematics clearly re-
flects and proves the divergence in evolution. Indeed, the indications of the
systematics as to scarcity or abundance of forms, of subordinate taxonomic
units, the narrow or wide distribution, point with certainty at the flourishing
or declining condition of a given group, at a definite stage of evolution of
the latter. The absence of systematical testimony in evolutinary studies of
a morphologist weakens considerably the value of the results attained. These
latter explain at best only the evolution of certain organs or systems there of,
but speak nothing about the real evolution of organisms and its laws.
12. The study of the evolutionary laws owes to be based upon as complete
investigations as possible of morphology, development, ecology, facts of
systematics and geographic distribution. Only such a thorough consideration
of all the features of organisms can yield sufficient evidence on the real phy-
logeny and its laws.
13. The statistical facts of systematics and geographic distribution indi-
cate most exactly the vestigial, restant nature of a group. These indications
of the nature of relicts are: the small absolute number of species of the whole
group, a relatively large number of genera and major units, the scarcity of
. species in genera, and the restricted or disrupted areas of distribution.
14. The examination of relicts reveals some evolutionary laws. The main
./ . law of the evolution of relicts, usually characterized by archaic features, is
the appearance in these forms of some few, but extreme and deep changes in
the structure, or development, or bionomics. The development of these extreme
changes just accounts for the nature of a relict, being essentially a reflection
of special relation^' between the organism and the environment. In the evolu-
tion of relicts the development of such peculiar ecologic connections («niches»)
is observed which called forth the appearance of the mentioned specializa-
tion characters. Owing to these peculiarities the relicts might survive con-
siderably longer than their nearest relatives. This statement is illustrated
by facts from the descriptive part of the present paper, as well as by numerous
examples from different groups of animals (Molluscs, Arthropods, Vertebrates).
ЛИТЕРАТУРА
Крыжановский С. Г. 1938. Принцип рекапитуляции и условия исторического
понимания развития. — Сборник памяти А. Н. Северцова, т. I, стр. 281—381.
Мончадский А. С. 1936. Личинки комаров (Culicidae) СССР и сопредельных стран,—
Определители по фауне СССР. Изд. Зоологического ин-та АН СССР, 24.
Родендорф В. 1937. Двукрылые насекомые юры Кара-тау. I. Brachycera и часть
Nematocera. Труды ПИН АН СССР, т. VII, вып. 3.
Швайковский, 1937. Аэродинамические особенности насекомых. Докл. АН СССР,
т. XVII, № 1—2, стр. 77—80.
СеверцовА. Н. 1939. Морфологические закономерности эволюции. Москва.
ШмальгаузенИ. И. 1939. Пути и закономерности эволюционного процесса. Москва.
Alexander Ch. Р. 1937. The interpretation of the radial field of the wing in the Ne-
matocerous Diptera. — Proc. Linn. Soc, N. S. Wales, 52, 42—72.
♦ Ama ns P. 1901 Geometrie descriptive et comparde des ailes rigides.—C. R. Assoc.
Fr. p. avan, d. Sciences.
В e r 1 e s e A. 1909—1924. Gli insetti. Milano.
Bischoff W. 1922. Ueber die Kopfbildung der Dipterenlarven. 1. Teil. Arch. f. Na-
turgesch. A. 6, 1—51. — 1924. Ueber die Kopfbildung d. Dipterenlarven. 3. Teil.
Arch. f. Naturgesch. A. 8, 1—105.
♦ С о q u i 11 e t D. W. 1901. A systematic arrangement of the families of the Diptera.—
Proc. U. S. Nat. Mus. XXXIII, 653.
Crampton О. C. 1925a. Phylogeny of Nematocera, Psyche, 31, 238.— 1925b. A phylo-
genetic study of the thoracic sclerites of the non-tipuloid nematocerous Diptera.—
Ann. Entom. Society of America, XVIII, 49—74.
D e m о 11. 1918. Flug d. Insekten und Vogel. Jena.
Duda O. 1928. Scatopsidae; in: Lindner. Die Fliegen d. paldarktisch. Region. Lief. 26.—
1930. Bibionidae; ibidem, Lief. 45.
Edwards. 1916. Systematic position of Mycetobia.—Ann. Mag. Nat. Hist. Ser. 9,
17, 108. — 1926. The Phylogeny of Nematocerous Diptera: a critical review of some
suggestions. — Verhandl. d. Ill, Internation. Entom. Kongresses. Weimar. Ill—
130. — 1928, Protorrhyphidae, Anisopodidae, Pachyneuridae, Trichocejidae, in
Genera Insectorum, fasc. 190, Bruxelles. 1
E n d e r 1 e i n G. 1911. Die phyletischen Beziehungen der Lycoriiden zu denJFuf&iwMMesfc
und ihre systematische Gliederung. — Arch. Naturgesch. I, 3. Sup^f. 116—201. —
1912. Zur Kenntnis d. Zygophthalmen. Zool. Anzeiger. XL, 26Я* [f ffgJWnn
Feuerborn H. J. 1922. Dersexuelle Reizapparat (Schmuck-, Duft- utW Bgrfihrungsoi*
gane der Psychodiden nach bioiogischen und physioiogischen Gesifl^ri^punkten un-
tersucht. — Arch. f. Naturg. 8. Jahrg., Abt. A., 4 Heft, 1—137.
J a n e t Ch. 1899. Sur le mecanisme du vol chez les insectes. — C. R. hebd. АэдОЙ^Ра^
ris. CXXVIII, 249—253. ’
Johannsen O. A. 1909. Diptera, fam. Mycetophilidae. — Genera Insectorum,fasc.
93, Bruxelles.
Handlirsch A. 1908. Die fossilen Insekten und die Phylogenie derrezenten Formen,
Leipzig.— 1925. In: Handbuch d. Entomologie, hrsg. v. Ch. Schroder. Bd. Ill,
Jena.
H e n de 1 F. 1928, Zweiflflgler, Oder Diptera; Allgemeiner Teil; in: Dahl. Die Tierwelt
Deutschlands usw, II Teil. Jena.— 1936. Diptera; Handbuch d. Zoologie v. W.
Kiikenthal, 4 Bd. 2. Halfte. 8—11 Lief., 1729—1998.
La me i re A, 1906. Notes pour la classification des Dipteres. — Mem. Soc. Ent. Belg.
XII, 106.
Landrock. 1927. Fungivoridae; in: Lindner. Die Fliegen d. palaarkt. Region.
Stuttgart.
* L e n ge r k e n H. 1924. Biolog. Zentralblatt, XLIV, 275—295.
Lengersdorf. 1930. Lycoridae: in: Lindner. Die Fliegen d. palaarkt. Region.
Stuttgart.
1QI
Lindner E. 1930. Die Fliegen d. palaarkt. Region, Lief. 50. Stuttgart.
Magnan A. 1934. La locomotion chez les animaux. I. Le vol des insectes, 1—186. Paris,
Hermann & C-ie.
Magnan A., Ma gna n Cl, 1933. Stir la structure des ailes d’insectes et son role dans
le vol par battement. C. R. Ac. Sc. (S6ance 29 mai).
Magnan A., Girard H. 1937. Essais de cin6matographie d’ailes d’oiseaux en mouve-
ment, etc. — C. R. Ac. Sc. Paris. 204. 723—726,
Magnan A. et S a i n t-L a g u e A. 1933a. De quelques methodes en morphologic.—
Annales d. Sc. Nat. Zool. Ser X.— 1933b. Le vol au point fixe. Exp. de Morphol.
dyn. et de Mec. du mouvement, № 4. Paris, Hermann & C-ie.
M a 1 1 о c h 1917. A preliminary classification of Diptera excL of Pupipara, based upon
larval and pupal characters. Part 1. Bull. Ulin, State Labor.of Nat. Hist. Vol. XII,
161—410.
M e i j e r e I. C. H.,de. 1916a. Beitrage zur Kenntnis d. Dipterenlarven u. Puppen. Zool.
Jahrb. (Anatomie). Bd. 40, 177—322. — 1916b, Zur Kenntnis d. Kopfbaues der
Dipterenlarven u. Imagines. Zool. Anzeiger, Bd. 46, 241—251.
О s t e n-S а с к e n R. 1893. On the characters of the three divisions of Diptera. Berl.
Ent. Ztschr. XXXVII, 417.
Peterson A. 1916. The head capsule and mouthparts of Diptera. Ulin. Biol. Monograph,
vol. II1-2, 1—2.
Sc hiner. 1863. Die Fliegen Oesterreichs (Fauna Austriaca), Wien.
Snodgrass. 1909. Thorax of Insects and the articulation of the wing.—Proc. U. S.
Nat. Mus. 36, № 1687, 511—595. — 1927. Morphology and Mechanism of the insect
thorax. Smithson. Misc. Coll., 80, 1—108.
Stammer H. J. 1932. Zur Biologie und Anatomie der leuchtenden Pilzmflckenlarve von
Ceroplatus testaceus D a 1 m. — Zeitschr. f. Morph, u. Oekologie, 26 Bd. */a H.,
135—146.
T i 11 у a r d R. J. 1916—1919. The «Panorpoid» Complex. Part 1. — Proc. Linn. Soc.
N. S. Wales 43, 1916, 286—319; Part II, idem 43, 1918, 626—637; Part III, idem,
1919. 533—718.
T о n n о i r A. L. 1937. An acephalous larva of a nematocerous Dipteron: The early stages
of Canthylosceiis (Diptera Scatopsidae). Annals Mag, Nat. Hist. Ser. 9, XIX,
№ 110, 291—300.
Venturi F. 1938. Contributi alia conoscenza dell Entomofauna deile Graminaceae,
V. — Contarinia triiici Kirby. — Redia. Zool. XXIV, 57—95.
V i g n о n P. 1932a. Nomenclature des veines de I’aile chez les Dipteres.— Encyclopedic
entomol., Serie B. Diptera, VI, 133—142. — 1932b. L’Aile primitive des Dipte-
res. — Ibidem, 142—143.
We b e r. 1933. Lehrbuch d. Entoniologie, Jena.
Wehrmeister H. 1925. BeitrSge zur Kenntnis der Cecidomyidenlarven mit besonde-
rer Berflcksichtigung des Darmes. Zoolog. Jahrbflcher. Bd. 49, 299—334.
* W e s c h e W. 1912. The phylogeny of the Nematocera with notes on the leg bristles,
hairs and certain mouth glands of Diptera. Biol. Bull. 23, 250—270.
Young В. P. 1921. Attachment of the Abdomen to the Thorax in Diptera. Cornell.
Univ. Agric. Exp. Station Memoir., 44, 256—282.
Примечание. Статьи, обозначенные звездочкой, остались мне неизвестны
а оригинале.
СОДЕРЖАНИЕ
Стр.
Предисловие....................................................... 5
1. Литературные данные............................................. 6
2. Особенности строения, развития и биологии современных Oligoneura. 11
3. Значение жилкования крыльев в систематике Nematocera .......... 18
4. Строение крыльев и направления их специализации................. 21
5. Состав современной фауны........................................ 34
6. Юрская фауна Oligoneura .............................. 38
7. Связь юрской фауны с современной и филогенез отдельных групп.... 84
8, Пути и закономерности эволюционного процесса в группе Oligoneura.SI
Summary............................................................ 98
Литература...................................................... 101
Таблицы